Настоящее изобретение в целом относится к антителам, которые связываются с CD3, включая мультиспецифические антитела, например, для активации Т-клеток. Кроме того, настоящее изобретение относится к полинуклеотидам, кодирующим такие антитела, и к векторам и клеткам-хозяевам, содержащим такие полинуклеотиды. Изобретение дополнительно относится к способам получения антител и к способам их применения в лечении заболевания.
УРОВЕНЬ ТЕХНИКИ
CD3 (кластер дифференцировки 3) представляет собой белковый комплекс, состоящий из четырех субъединиц - цепи CD3γ, цепи CD3δ и двух цепей CD3ε. CD3 связывается с Т-клеточным рецептором и ζ-цепью, генерируя сигнал активации в Т-лимфоцитах.
CD3 был всесторонне изучен как лекарственная мишень. Моноклональные антитела, нацеленные на CD3, использовали в качестве иммуносупрессирующей терапии при аутоиммунных заболеваниях, таких как диабет I типа, или при лечении отторжения трансплантата. Антитело к CD3, муромонаб-CD3 (ОКТ3), было первым моноклональным антителом, одобренным для клинического применения на людях в 1985 г.
Более позднее применение антител к CD3 относится к форме биспецифических антител, с одной стороны связывающих CD3, а с другой стороны - антиген опухолевой клетки. Одновременное связывание такого антитела со своими обеими мишенями будет способствовать временному взаимодействию между клеткой-мишенью и Т-клеткой, приводя к активации любой цитотоксической Т-клетки и последующему лизису клетки-мишени.
В терапевтических целях важным требованием, которому должны удовлетворять антитела, является достаточная стабильность как in vitro (для хранения лекарственного средства), так и in vivo (после введения пациенту).
Такие модификации, как дезамидирование аспарагина, являются типичными вариантами деградации для рекомбинантных антител и могут влиять как на in vitro стабильность, так и на in vivo биологические функции.
Учитывая огромный терапевтический потенциал антител, в частности биспецифических антител, для активации Т-клеток, существует необходимость в антителах к CD3 с оптимизированными свойствами.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В настоящем изобретении предложены антитела, включая мультиспецифические (например, биспецифические) антитела, которые связываются с CD3 и являются устойчивыми к деградации путем дезамидирования аспарагина и, таким образом, исключительно стабильными, что требуется в терапевтических целях. Предложенные (мультиспецифические) антитела дополнительно характеризуются комбинацией хорошей эффективности и технологичности с низкой токсичностью и благоприятными фармакокинетическими свойствами.
Как показано в данном документе, антитела, такие как мультиспецифические антитела, которые связываются с CD3, предложенные в настоящем изобретении, сохраняют более чем около 90% активности связывания с CD3 через 2 недели при рН 7,4, 37°С, по сравнению с активностью связывания через 2 недели при рН 6, -80°С, по определению методом поверхностного плазмонного резонанса (ППР).
В одном аспекте в настоящем изобретении предложено антитело, которое связывается с CD3, причем антитело содержит первый антигенсвязывающий домен, содержащий вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область тяжелой цепи (HCDR) 1 с SEQ ID NO: 2, HCDR 2 с SEQ ID NO: 3 и HCDR 3 с SEQ ID NO: 5, и вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область легкой цепи (LCDR) 1 с SEQ ID NO: 8, LCDR 2 с SEQ ID NO: 9 и LCDR 3 с SEQ ID NO: 10. В одном аспекте VH содержит аминокислотную последовательность, которая по меньшей мере на около 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 7, и/или VL содержит аминокислотную последовательность, которая по меньшей мере на около 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 11.
В дополнительном аспекте в изобретении предложено антитело, которое связывается с CD3, причем антитело содержит первый антигенсвязывающий домен, содержащий последовательность VH с SEQ ID NO: 7 и последовательность VL с SEQ ID NO: 11.
В одном аспекте первый антигенсвязывающий домен представляет собой молекулу Fab.
В одном аспекте антитело содержит Fc-домен, состоящий из первой и второй субъединиц.
В одном аспекте антитело содержит второй и, необязательно, третий антигенсвязывающий домен, который связывается со вторым антигеном.
В одном аспекте второй и/или, в случае наличия, третий антигенсвязывающий домен представляет собой молекулу Fab.
В одном аспекте первый антигенсвязывающий домен представляет собой молекулу Fab, причем вариабельные домены VL и VH или константные домены CL и СН1, в частности, вариабельные домены VL и VH легкой цепи Fab и тяжелой цепи Fab замещены друг другом.
В одном аспекте второй и, в случае наличия, третий антигенсвязывающий домен представляет собой стандартную молекулу Fab.
В одном аспекте второй и, в случае наличия, третий антигенсвязывающий домен представляет собой молекулу Fab, причем в константном домене CL аминокислота в позиции 124 независимо замещена лизином (К), аргинином (R) или гистидином (Н) (нумерация в соответствии с Kabat) и аминокислота в позиции 123 независимо замещена лизином (К), аргинином (R) или гистидином (Н) (нумерация в соответствии с Kabat), а в константном домене СН1 аминокислота в позиции 147 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация в соответствии с индексом EU по Kabat) и аминокислота в позиции 213 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация в соответствии с индексом EU по Kabat).
В одном аспекте первый и второй антигенсвязывающие домены слиты друг с другом, необязательно посредством пептидного линкера.
В одном аспекте каждый из первого и второго антигенсвязывающих доменов представляет собой молекулу Fab и либо (i) второй антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первого антигенсвязывающего домена, либо (ii) первый антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab второго антигенсвязывающего домена.
В одном аспекте каждый из первого, второго и, в случае наличия, третьего антигенсвязывающих доменов представляет собой молекулу Fab, и антитело содержит Fc-домен, состоящий из первой и второй субъединиц; и при этом либо (i) второй антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первого антигенсвязывающего домена, а первый антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом первой субъединицы Fc-домена, либо (ii) первый антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab второго антигенсвязывающего домена, а второй антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом первой субъединицы Fc-домена; и третий антигенсвязывающий домен, в случае наличия, слит на С-конце тяжелой цепи Fab с N-концом второй субъединицы Fc-домена.
В одном аспекте Fc-домен представляет собой Fc-домен IgG, в частности IgG1. В одном аспекте Fc-домен представляет собой Fc-домен человека. В одном аспекте Fc содержит модификацию, способствующую ассоциации первой и второй субъединиц Fc-домена. В одном аспекте Fc-домен содержит одну или более аминокислотных замен, которые уменьшают связывание с Fc-рецептором и/или эффекторную функцию.
В одном аспекте второй антиген представляет собой антиген клетки-мишени, в частности антиген опухолевой клетки.
В одном аспекте второй антиген представляет собой TYRP-1. В одном аспекте второй и, в случае наличия, третий антигенсвязывающий домен содержит VH, содержащую HCDR 1 с SEQ ID NO: 15, HCDR 2 с SEQ ID NO: 16 и HCDR 3 с SEQ ID NO: 17, и VL, содержащую LCDR 1 с SEQ ID NO: 19, LCDR 2 с SEQ ID NO: 20 и LCDR 3 с SEQ ID NO: 21. В одном аспекте второй и, в случае наличия, третий антигенсвязывающий домен содержит VH, содержащую аминокислотную последовательность, которая по меньшей мере на около 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 18, и/или VL, содержащую аминокислотную последовательность, которая по меньшей мере на около 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 22.
В одном аспекте второй антиген представляет собой EGFRvIII. В одном аспекте второй и, в случае наличия, третий антигенсвязывающий домен содержит VH, содержащую HCDR 1 с SEQ ID NO: 85, HCDR 2 с SEQ ID NO: 86 и HCDR 3 с SEQ ID NO: 87, и VL, содержащую LCDR 1 с SEQ ID NO: 89, LCDR 2 с SEQ ID NO: 90 и LCDR 3 с SEQ ID NO: 91. В одном аспекте второй и, в случае наличия, третий антигенсвязывающий домен содержит VH, содержащую аминокислотную последовательность, которая по меньшей мере на около 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 88, и/или VL, содержащую аминокислотную последовательность, которая по меньшей мере на около 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 92.
В соответствии с дополнительным аспектом изобретения предложены выделенный полинуклеотид, кодирующий антитело по изобретению, и клетка-хозяин, содержащая выделенный полинуклеотид по изобретению.
В другом аспекте предложен способ получения антитела, которое связывается с CD3, включающий этапы (а) культивирования клетки-хозяина по изобретению в условиях, подходящих для экспрессии антитела, и, необязательно, (б) выделения антитела. Изобретение также включает антитело, которое связывается с CD3, полученное способом по изобретению.
В изобретении дополнительно предложена фармацевтическая композиция, содержащая антитело по изобретению и фармацевтические приемлемый носитель.
Также в изобретение включены способы применения антитела и фармацевтической композиции по изобретению. В одном аспекте в изобретении предложены антитело или фармацевтическая композиция в соответствии с изобретением для применения в качестве медикамента. В одном аспекте предложены антитело или фармацевтическая композиция в соответствии с изобретением для применения в лечение заболевания. В конкретном аспекте заболевание представляет собой рак.
Также предложено применение антитела или фармацевтической композиции в соответствии с изобретением при производстве лекарственного средства, применение антитела или фармацевтической композиции в соответствии с изобретением при производстве лекарственного средства для лечения заболевания, в частности рака. В изобретении также предложен способ лечения заболевания у индивида, включающий введение указанному индивиду эффективного количества антитела или фармацевтической композиции в соответствии с изобретением.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1. Типовые конфигурации (мультиспецифических) антител по изобретению. (А, Г) Иллюстрация молекулы «1+1 CrossMab». (Б, Д) Иллюстрация молекулы «2+1 IgG Crossfab» с альтернативным порядком компонентов Crossfab и Fab («инвертированная молекула»). (В, Е) Иллюстрация молекулы «2+1 IgG Crossfab». (Ж, Л) Иллюстрация молекулы «1+1 IgG Crossfab» с альтернативным порядком компонентов Crossfab и Fab («инвертированная молекула»). (3, М) Иллюстрация молекулы «1+1 IgG Crossfab». (И, Н) Иллюстрация молекулы «2+1 IgG Crossfab» с двумя Cross Fab. (К, О) Иллюстрация молекулы «2+1 IgG Crossfab» с двумя CrossFab и альтернативным порядком компонентов Crossfab и Fab («инвертированная молекула»). (П, У) Иллюстрация молекулы «Fab-Crossfab». (Р, Ф) Иллюстрация молекулы «Crossfab-Fab». (С, X) Иллюстрация молекулы «(Fab)2-Crossfab». (Т, Ц) Иллюстрация молекулы «Crossfab-(Fab)2». (Ч, Щ) Иллюстрация молекулы «Fab-(Crossfab)2». (Ш, Ы) Иллюстрация молекулы «(Crossfab)2-Fab». Черная точка: необязательная модификация в Fc-домене, способствующая гетеродимеризации. ++, --: аминокислоты с противоположными зарядами, необязательно внесенные в домены СН1 и CL. Молекулы Crossfab на изображении содержат взаимную замену областей VH и VL, но могут в аспектах, в которых зарядовые модификации внесены в домены СН1 и CL, - в альтернативном варианте содержать взаимную замену доменов СН1 и CL.
Фиг. 2. Относительная активность связывания оригинальных и оптимизированных связывающих CD3 фрагментов CD3orig и CD3opt с рекомбинантным CD3 по данным измерений методом ППР в бесстрессовых условиях через 14 сут. при 40°С, рН 6, или через 14 сут. при 37°С, рН 7,4 (формат IgG).
Фиг.3. Связывание оригинальных и оптимизированных связывающих CD3 фрагментов CD3orig и CD3opt с клетками Jurkat NFAT по данным измерений методом проточной цитометрии (формат IgG). Обнаружение антител, связанных с клетками Jurkat NFAT, проводили с помощью флуоресцентно меченного античеловеческого Fc-специфического вторичного антитела.
Фиг. 4. Схематическая иллюстрация анализа активации CD3, применяемого в примере 3.
Фиг. 5. Активация Jurkat NFAT оригинальными и оптимизированными связывающими CD3 фрагментами CD3orig и CD3opt (формат IgG). Репортерные клетки Jurkat NFAT инкубировали совместно с экспрессирующими анти-PGLALA клетками СНО (CHO-PGLALA) в присутствии CD3orig или CD3opt IgG PGLALA или CD3opt IgG дт в качестве отрицательного контроля. Количественную оценку активации CD3 проводили путем измерения люминесценции через 24 ч.
Фиг. 6. Схематическая иллюстрация молекул Т-клеточного биспецифического антитела (ТСВ), полученного в примерах. Все тестируемые молекулы антител ТСВ были получены в виде «2+1 IgG CrossFab, инвертированного» с зарядовыми модификациями (взаимная замена VH/VL в связывающей CD3 молекуле, зарядовые модификации в связывающий целевой антиген молекулах, ЕЕ=147Е, 213Е; RK=123R, 124К).
Фиг. 7. Относительная активность связывания TYRP1 ТСВ, содержащих оригинальные или оптимизированные связывающие CD3 фрагменты CD3orig или CD3opt, с рекомбинантным CD3 по данным измерений методом ППР в бесстрессовых условиях через 14 сут. при 40°С, рН 6, или через 14 сут. при 37°С, рН 7,4.
Фиг. 8. Относительная активность связывания TYRP1 ТСВ, содержащих оригинальные или оптимизированные связывающие CD3 фрагменты CD3orig или CD3opt, или соответствующего TYRP1 IgG с рекомбинантным TYRP1 по данным измерений методом ППР в бесстрессовых условиях через 14 сут. при 40°С, рН 6, или через 14 сут. при 37°С, рН 7,4.
Фиг. 9. Связывание TYRP1 ТСВ, содержащих оригинальные или оптимизированные связывающие CD3 фрагменты CD3orig или CD3opt, с клетками Jurkat NFAT по данным измерений методом проточной цитометрии. Обнаружение ТСВ, связанных с клетками Jurkat NFAT, проводили с помощью флуоресцентно меченного античеловеческого Fc-специфического вторичного антитела.
Фиг. 10. Активация Jurkat NFAT TYRP1 ТСВ, содержащими оригинальные или оптимизированные связывающие CD3 молекулы. Репортерные клетки Jurkat NFAT инкубировали совместно с линией клеток меланомы М150543 в присутствии TYRP1 ТСВ CD3orig или TYRP1 ТСВ CD3opt. Количественную оценку активации CD3 в присутствии ТСВ проводили путем измерения люминесценции через 24 ч.
Фиг. 11. Уничтожение опухолевых клеток и активация Т-клеток TYRP1 ТСВ, содержащими оригинальные или оптимизированные связывающие CD3 фрагменты. Уничтожение линии клеток меланомы M150543 после обработки TYRP1 ТСВ CD3orig и TYRP1 ТСВ CD3opt МКПК от трех разных здоровых доноров (А-Е: донор 1, Ж-М: донор 2, Н-Т: донор 3) определяли по высвобождению ЛДГ через 24 ч (А, Ж, Н) и 48 ч (Б, 3, О). Параллельно, как маркер активации Т-клеток, измеряли повышение регуляции CD25 (В, Д, И, Л, П, С) и CD69 (Г, Е, К, М, Р, Т) на CD8 (Д, Е, Л, М, С, Т) и CD4 (В, Г, И, Л, П, Р) Т-клетках в МКПК методом проточной цитометрии через 48 ч.
Фиг. 12. Специфическое связывание EGFRvIII IgG PGLALA. Специфическое связывание антител EGFRvIII IgG PGLALA с EGFRvIII без перекрестной реактивности с EGFRwt исследовали методом проточной цитометрии на CHO-EGFRvIII (А), EGFRvIII-положительных DK-MG (Б) и экспрессирующих EGFRwt MKN-45 (В). Цетуксимаб был включен в качестве положительного контроля для экспрессии EGFRwt.
Фиг. 13. Активация CAR J с помощью EGFRvIII IgG PGLALA. Репортерные клетки Jurkat NFAT, экспрессирующие анти-PGLALA CAR, совместно инкубировали с экспрессирующими EGFRvIII клетками DK-MG и антителами EGFRvIII IgG PGLALA или DP47 IgG PGLALA в качестве отрицательного контроля. Количественную оценку клеток Jurkat NFAT проводили путем измерения люминесценции через 22 ч.
Фиг. 14. Связывание EGFRvIII IgG PGLALA и соответствующих ТСВ с EGFRvIII. Специфическое связывание связывающих EGFRvIII молекул, в виде IgG PGLALA и преобразованных в ТСВ, с клетками CHO-EGFRvIII (А) и MKN-45 (Б) измеряли методом методом проточной цитометрии.
Фиг.15. Активация Jurkat NFAT с помощью ТСВ к EGFRvIII. Активацию Jurkat NFAT определяли как маркер взаимодействия CD3 с EGFRvIII ТСВ в присутствии EGFRvIII-положительных клеток DK-MG. DP47 ТСВ был включен в качестве отрицательного контроля.
Фиг. 16. Лизис опухолевых клеток EGFRvIII ТСВ. Индукцию специфического лизиса опухолевых клеток EGFRvIII ТСВ определяли после совместного культивирования со свежевыделенными МКПК и EGFRvIII-положительными клетками DK-MG (А, Б) или EGFRwt-положительными клетками MKN-45 (В, Г) для 24 ч (А, В) или 48 ч (Б, Г).
Фиг. 17. Активация Т-клеток EGFRvIII ТСВ. Индукцию активации Т-клеток EGFRvIII ТСВ определяли после совместного культивирования со свежевыделенными МКПК и EGFRvIII-положительными клетками DK-MG (А, Б, Д, Е) или EGFRwt-положительными клетками MKN-45 (В, Г, Ж, 3), используя маркеры активации CD25 (А, В, Д, Ж) или CD69 (Б, Г, Е, 3) на CD4 Т-клетках (А Г) или CD8 Т-клетках (Д-З).
Фиг. 18. Высвобождение цитокинов с EGFRvIII ТСВ. Индукцию высвобождения IFNγ (А, Г), TNFα (Б, Д) и гранзима В (В, Е) EGFRvIII ТСВ определяли после совместного культивирования со свежевыделенными МКПК и EGFRvIII-положительными клетками DK-MG (А В) или EGFRwt-положительными клетками MKN-45 (Г-Е).
Фиг. 19. Специфическое связывание EGFRvIII IgG PGLALA с созревшей аффинностью. Специфическое связывание антител к EGFRvIII с созревшей аффинностью с EGFRvIII сравнивали с родительской связывающей EGFRvIII молекулой на клетках U87MG-EGFRvIII (А) и на линии EGFRwt-положительных клеток MKN-45 (Б).
Фиг. 20. Активация Jurkat NFAT с помощью EGFRvIII ТСВ. Активацию Jurkat NFAT определяли как маркер взаимодействия CD3 с EGFRvIII ТСВ в присутствии EGFRvIII-положительных клеток DK-MG (А), клеток U87MG-EGFRvIII (Б) и клеток MKN-45 (В). DP47 ТСВ был включен в качестве отрицательного контроля.
Фиг. 21. Относительная активность связывания EGFRvIII ТСВ, содержащих оригинальные или оптимизированные связывающие CD3 фрагменты CD3orig или CD3opt, с рекомбинантным CD3 по данным измерений методом ППР в бесстрессовых условиях через 14 сут. при 40°С, рН 6, или через 14 сут. при 37°С, рН 7,4.
Фиг. 22. Относительная активность связывания EGFRvIII ТСВ, содержащих оригинальные или оптимизированные связывающие CD3 фрагменты CD3orig или CD3opt, с рекомбинантным EGFRvIII по данным измерений методом ППР в бесстрессовых условиях через 14 сут. при 40°С, рН 6, или через 14 сут. при 37°С, рН 7,4.
Фиг. 23. Связывание EGFRvIII ТСВ, содержащих оригинальные или оптимизированные связывающие CD3 фрагменты CD3orig или CD3opt, с клетками Jurkat NFAT по данным измерений методом проточной цитометрии. Обнаружение ТСВ, связанных с клетками Jurkat NFAT, проводили с помощью флуоресцентно меченного античеловеческого Fc-специфического вторичного антитела.
Фиг. 24. Связывание EGFRvIII ТСВ, содержащих связывающий EGFRvIII фрагмент Р063.056 или Р056.021, с клетками U87MG-EGFRvIII по данным измерений методом проточной цитометрии. Обнаружение ТСВ, связанных с клетками U87MG-EGFRvIII, проводили с помощью флуоресцентно меченного античеловеческого Fc-специфического вторичного антитела.
Фиг. 25. Лизис опухолевых клеток и активация Т-клеток EGFRvIII ТСВ. Индукция специфического лизиса опухолевых клеток (А, Б) и активации Т-клеток (В, Г) EGFRvIII ТСВ определяли после совместного культивирования со свежевыделенными МКПК и клетками U87MG-EGFRvIII для 24 ч (А, В) или 48 ч (Б, Г). DP47 ТСВ был включен в качестве отрицательного контроля.
Фиг. 26. Активация Jurkat NFAT со сравнением формата 2+1 и формата 1+1 EGFRvIII ТСВ. Активацию Jurkat NFAT определяли как маркер взаимодействия CD3 с EGFRvIII ТСВ в инвертированном формате 2+1 и формате голова-к-хвосту 1+1 в присутствии EGFRvIII-положительных клеток U87MG-EGFRvIII.
Фиг. 27. Лизис опухолевых клеток и активация Т-клеток со сравнением формата 2+1 и формата 1+1 EGFRvIII ТСВ. Индукцию специфического лизиса опухолевых клеток (А, Б) и активации Т-клеток (В, Г) EGFRvIII ТСВ в инвертированном формате 2+1 и формате голова-к-хвосту 1+1 определяли после совместного культивирования со свежевыделенными МКПК и клетками U87MG-EGFRvIII для 24 ч (А, В) или 48 ч (Б, Г).
Фиг.28. Активация и пролиферация Т-клеток с помощью EGFRvIII ТСВ. Индукцию пролиферации Т-клеток (А, В) и активации Т-клеток CD4 (А, Б) и Т-клеток CD8 (В, Г) ТСВ к EGFRvIII определяли после совместного культивирования U87MG-EGFRvIII и МКПК, выделенных от здоровых доноров.
Фиг. 29. Лизис опухолевых клеток, активация Т-клеток и высвобождение цитокинов с помощью ТСВ к EGFRvIII. Индукцию лизиса опухолевых клеток (А, Б), активации Т-клеток (В, Г) и высвобождения IFNγ и TNFα (Д, Е) EGFRvIII ТСВ определяли после совместного культивирования клеток U87MG-EGFRvIII с МКПК. Лизис опухолевых клеток определяли через 24 ч и 48 ч после обработки, активацию Т-клеток и высвобождение цитокинов определяли через 48 ч.
Фиг. 30. Лизис опухолевых клеток, активация Т-клеток и высвобождение цитокинов с помощью TYRP-1 ТСВ. Индукцию лизиса опухолевых клеток (А, Б), активации Т-клеток (В, Г) и высвобождения IFNγ и TNFα (Д, Е) TYRP-1 ТСВ определяли после совместного культивирования полученной от пациента линии клеток меланомы M150543 с МКПК. Лизис опухолевых клеток определяли через 24 ч и 48 ч после обработки, активацию Т-клеток и высвобождение цитокинов определяли через 48 ч.
Фиг. 31. In vivo эффективность TYRP-1 ТСВ. Линию клеток меланомы человека IGR-1 подкожно инъецировали гуманизированным мышам NSG, чтобы исследовать ингибирование роста опухоли в модели с подкожным ксенотрансплантатом меланомы. Значительное ингибирование роста опухоли (ИРО) наблюдали в группе TYRP-1 ТСВ (68% ИРО, р=0,0058*) по сравнению с группой носителя.
Фиг. 32. In vivo эффективность EGFRvIII ТСВ. Линию клеток глиобластомы человека U87-huEGFRvIII подкожно инъецировали гуманизированным мышам NSG, чтобы исследовать ингибирование роста опухоли в модели с подкожным ксенотрансплантатом глиобластомы. Значительный контроль опухоли наблюдали в группе EGFRvIII ТСВ, в которой все мыши достигли полной ремиссии.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
I. ОПРЕДЕЛЕНИЯ
В данном документе термины используются так, как это общепринято в данной области техники, если ниже не приведено иное определение.
В контексте данного документа термины «первый», «второй» или «третий» в отношении антигенсвязывающих доменов и т.д. используются для того, чтобы облегчить различение в случае наличия более чем одного из каждого типа фрагментов. Использование этих терминов не подразумевает конкретного порядка или ориентации фрагмента, если явным образом не указано иное.
Термины «антитело к CD3» и «антитело, которое связывается с CD3» относятся к антителу, которое способно связывать CD3 с достаточной аффинностью, чтобы антитело можно было применять в качестве диагностического и/или терапевтического агента для нацеливания на CD3. В одном аспекте степень связывания анти-CD3 антитело с неродственным отличным от CD3 белком составляет менее чем около 10% от связывания антитела с CD3 по данным измерения, например, методом поверхностного плазмонного резонанса (ППР). В определенных аспектах антитело, которое связывается с CD3, имеет константу диссоциации (KD)≤1 мкМ, ≤500 нМ, ≤200 нМ или ≤100 нМ. Говорят, что антитело «специфически связывается» с CD3, если антитело имеет KD 1 мкМ или менее по данным измерения, например, ППР. В определенных аспектах анти-CD3 антитело связывается с эпитопом CD3, который является консервативным для CD3 от разных видов.
Термин «антитело» в данном документе используется в самом широком смысле и включает различные структуры антител, включая, но не ограничиваясь этим, моноклональные антитела, поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела) и фрагменты антител, при условии, что они проявляют необходимую антигенсвязывающую активность.
«Фрагмент антитела» относится к молекуле, отличной от интактного антитела, которая содержит часть интактного антитела, которая связывает антигеном, с которым связывается интактное антитело. Примеры фрагментов антител включают, но не ограничиваются этим, Fv, Fab, Fab', Fab'-SH, F(ab')2, диатела, линейные антитела, молекулы одноцепочечных антител (например, scFv и scFab), однодоменные антитела и мультиспецифические антитела, образованные из фрагментов антител. Обзор некоторых фрагментов антител смотрите в Hollinger and Hudson, Nature Biotechnology 23:1126-1136 (2005).
В контексте данного документа термины «полноразмерное антитело», «интактное антитело» и «цельное антитело» используются взаимозаменяемо для обозначения антитела, имеющего структуру, по существу сходную со структурой нативного антитела.
В контексте данного документа термин «моноклональное антитело» относится к антителу, полученному из популяции по существу гомогенных антител, т.е. отдельные антитела, составляющие популяцию, являются идентичными и/или связывают один и тот же эпитоп, за исключением возможных вариантных антител, например, содержащих естественные мутации или возникающих во время получения препарата моноклональных антител, причем такие варианты в общем случае присутствуют в незначительных количествах. В отличие от препаратов поликлональных антител, которые обычно содержат разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело препарата моноклональных антител направлено против одной детерминанты на антигене. Таким образом, определение «моноклональный» указывает на особенность антитела, как полученного из по существу гомогенной популяции антител, и его не следует интерпретировать как требование получения антитела каким-либо конкретным методом. Например, моноклональные антитела можно получать различными методами, включая, но не ограничиваясь этим, метод гибридомы, методы рекомбинантных ДНК, методы фагового дисплея и методы с использованием трансгенных животных, содержащих все или часть локусов человеческого иммуноглобулина, причем такие методы и другие типовые методы получения моноклональных антител описаны в данном документе.
«Выделенное» антитело представляет собой антитело, которое было отделено от компонентов его естественного окружения. В некоторых аспектах антитело является очищенным до более чем 95% или 99% чистоты по определению, например, электрофоретическими (например, ДСН-ПААГ, изоэлектрическое фокусирование (ИЭФ), капиллярный электрофорез) или хроматографическими (например, ионообменная или обращенно-фазовая ВЭЖХ, аффинная хроматография, эксклюзионная хроматография) методами. Обзор методов оценки чистоты антител смотрите, например, в Flatman et al., J. Chromatogr. В 848:79-87 (2007). В некоторых аспектах предложенные в настоящем изобретении антитела являются выделенными антителами.
Термин «химерное» антитело относится к антителу, в котором часть тяжелой и/или легкой цепи получена из конкретного источника или от конкретного вида, тогда как остаток тяжелой и/или легкой цепи получен из другого источника или от другого вида.
«Гуманизированное» антитело относится к химерному антителу, содержащему аминокислотные остатки из нечеловеческих CDR и аминокислотные остатки из человеческих FR. В определенных аспектах гуманизированное антитело содержит по существу все из по меньшей мере одного и, как правило, двух вариабельных доменов, в которых все или, по существу все CDR соответствуют таковым из нечеловеческого антитела, а все или по существу все FR соответствуют таковым из человеческого антитела. Такие вариабельные домены называются в данном документе «гуманизированной вариабельной областью». Гуманизированное антитело, необязательно, может содержать по меньшей мере часть константной области антитела, полученную из человеческого антитела. В некоторых аспектах некоторые остатки FR в гуманизированном антителе замещены соответствующими остатками нечеловеческого антитела (например, антитела, из которого получены остатки CDR), например, с целью восстановления или улучшения специфичности или аффинности антитела. «Гуманизированная форма» антитела, например, нечеловеческого антитела, относится к антителу, которое было подвергнуто гуманизации.
«Человеческое антитело» представляет собой антитело, которое имеет аминокислотную последовательность, соответствующую последовательности антитела, вырабатываемого человеком или клеткой человека, или полученную из источника, отличного от человека, в котором используются репертуары человеческих антител или другие последовательности, кодирующие человеческие антитела. Это определение человеческого антитела явным образом исключает гуманизированное антитело, содержащее нечеловеческие антигенсвязывающие остатки. В определенных аспектах человеческое антитело получено от отличного от человека трансгенного млекопитающего, например, мыши, крысы или кролика. В определенных аспектах человеческое антитело получено из линии клеток гибридомы. Антитела или фрагменты антител, выделенные из библиотек человеческих антител, также считаются в данном документе человеческими антителами или фрагментами человеческих антител.
Термин «антигенсвязывающий домен» относится к части антитела, которая содержит участок, связывающийся и комплементарный с частью антигена или со всем антигеном. Антигенсвязывающий домен может быть образован, например, одним или более вариабельными доменами антитела (также называемыми вариабельными областями антитела). В предпочтительных аспектах антигенсвязывающий домен содержит вариабельный домен легкой цепи антитела (VL) и вариабельный домен тяжелой цепи антитела (VH).
Термин «вариабельная область» или «вариабельный домен» относится к домену тяжелой или легкой цепи антитела, который участвует в связывании антитела с антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL, соответственно) нативного антитела в общем случае имеют сходные структуры, причем каждый домен содержит четыре консервативные каркасные области (FR) и определяющие комплементарность области (CDR). Смотрите, например, Kindt et al., Kuby Immunology, 6th ed., W.H. Freeman & Co., страница 91 (2007). Одного домена VH или VL может быть достаточно для придания антигенсвязывающей специфичности. Кроме того, антитела, которые связывают конкретный антиген, можно выделять, используя домен VH или VL из антитела, которое связывает антиген, для скрининга библиотеки комплементарных доменов VL или VH, соответственно. Смотрите, например, Portolano et al., J. Immunol. 750:880-887 (1993); Clarkson et al., Nature 352:624-628 (1991). В контексте данного документа в связи с последовательностями вариабельных областей «нумерация Kabat» относится к системе нумерации, описанной в Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991).
В контексте данного документа аминокислотные позиции всех константных областей и доменов тяжелой и легкой цепи пронумерованы в соответствии с системой нумерации Kabat, описанной в Kabat, et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991), называемой в данном документе «нумерацией в соответствии с Kabat» или «нумерацией Kabat». В частности, систему нумерации Kabat (смотрите страницы 647-660 в Kabat, et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991)) используют для константного домена легкой цепи CL изотипа каппа или лямбда, а систему нумерации по индексу EU по Kabat (смотрите страницы 661-723) используют для константных доменов тяжелой цепи (СН1, шарнирная область, СН2 и СН3), что в таком случае дополнительно подчеркивается в данном документе названием «нумерация в соответствии с индексом EU по Kabat» или «нумерация по индексу EU по Kabat».
В контексте данного документа термин «гипервариабельная область» или «HVR» относится к каждой из областей вариабельного домена антитела, которые являются гипервариабельными по последовательности и которые определяют антигенсвязывающую специфичность, например, к «определяющим комплементарность областям» («CDR»). В общем случае антитела содержат шесть CDR: три в VH (HCDR1, HCDR2, HCDR3) и три в VL (LCDR1, LCDR2, LCDR3). Типовые CDR по данному документу включают:
(а) гипервариабельные петли, находящиеся в аминокислотных остатках 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (H1), 53-55 (Н2) и 96-101 (H3) (Chothia and Lesk, J. Mol. Biol. 196:901-917 (1987));
(б) CDR, находящиеся в аминокислотных остатках 24-34 (LI), 50-56 (L2), 89-97 (L3), 31-35b (H1), 50-65 (H2) и 95-102 (Н3) (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991)); и
(в) антигенные контакты, находящиеся в аминокислотных остатках 27 с-36 (L1), 46-55 (L2), 89-96 (L3), 30-35b (H1), 47-58 (Н2) и 93-101 (H3) (MacCallum et al. J. Mol. Biol. 262: 732-745 (1996)).
Если не указано иное, CDR определены в соответствии с Kabat et al., выше. Специалисту в данной области техники известно, что отнесение CDR также можно определить в соответствии с Chothia, выше, McCallum, выше, или любой другой научно принятой системой номенклатуры.
«Каркасная область» или «FR» относится к остаткам вариабельного домена, отличным от определяющих комплементарность областей (CDR). FR вариабельного домена в общем случае состоит из четырех FR-доменов: FR1, FR2, FR3 и FR4. Соответственно, последовательности HVR и FR в общем случае расположены в VH (или VL) в следующем порядке: FR1-HCDR1(LCDR1)-FR2-HCDR2(LCDR2)-FR3-HCDR3(LCDR3)-FR4.
Если не указано иное, остатки CDR и другие остатки в вариабельном домене (например, остатки FR) пронумерованы в данном документе в соответствии с Kabat et al., выше.
В целях данного документа «человеческая акцепторная каркасная область» представляет собой каркасную область, содержащую аминокислотную последовательность каркасной области вариабельного домена легкой цепи (VL) или каркасной области вариабельного домена тяжелой цепи (VH), полученную из каркасной области человеческого иммуноглобулина или человеческой консенсусной каркасной области, определенных ниже. Человеческая акцепторная каркасная область, «полученная из» каркасной области человеческого иммуноглобулина или человеческой консенсусной каркасной области, может содержать такую же самую аминокислотную последовательность или может содержать изменения в аминокислотной последовательности. В некоторых аспектах число аминокислотных изменений составляет 10 или менее, 9 или менее, 8 или менее, 7 или менее, 6 или менее, 5 или менее, 4 или менее, 3 или менее, или 2 или менее. В некоторых аспектах человеческая акцепторная каркасная область VL идентична по последовательности с последовательностью каркасной области VL человеческого иммуноглобулина или последовательностью человеческой консенсусной каркасной области.
«Человеческая консенсусная каркасная область» представляет собой каркасную область, которая представляет наиболее часто встречающиеся аминокислотные остатки в наборе последовательностей каркасных областей VL или VH человеческого иммуноглобулина. В общем случае набор последовательностей VL или VH человеческого иммуноглобулина получен из подгруппы последовательностей вариабельных доменов. В общем случае подгруппа последовательностей представляет собой подгруппу согласно Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, NIH Publication 91-3242, Bethesda MD (1991), тома. 1-3.
В данном документе термин «молекула иммуноглобулина» относится к белку, имеющему структуру антитела природного происхождения. Например, иммуноглобулины класса IgG представляют собой гетеротетрамерные гликопротеины массой около 150000 Дальтон, состоящие из двух легких цепей и двух тяжелых цепей, которые связаны дисульфидными связями. В направлении от N-конца к С-концу каждая тяжелая цепь содержит вариабельный домен (VH), также называемый вариабельным доменом тяжелой цепи или вариабельной областью тяжелой цепи, за которым следуют три константных домена (CH1, СН2 и CH3), также называемых константной областью тяжелой цепи. Аналогично, в направлении от N-конца к С-концу каждая легкая цепь содержит вариабельный домен (VL), также называемый вариабельным доменом легкой цепи или вариабельной областью легкой цепи, за которым следует константный домен легкой цепи (CL), также называемый константной областью легкой цепи. Тяжелая цепь иммуноглобулина может быть отнесена к одному из пяти типов, называемых α (IgA), δ (IgD), ε (IgE), γ (IgG) или μ (IgM), некоторые из которых могут быть дополнительно поделены на подтипы, например, γ1 (IgG1), γ2 (IgG2), γ3 (IgG3), γ4 (IgG4), α1 (IgA1) и α2 (IgA2). Легкая цепь иммуноглобулина может быть отнесена к одному из двух типов, называемых каппа (κ) и лямбда (λ), на основании аминокислотной последовательности ее константного домена. Иммуноглобулин состоит главным образом из двух молекул Fab и Fc-домена, связанных посредством шарнирной области иммуноглобулина.
«Класс» антитела или иммуноглобулина относится к типу константного домена или константной области, содержащихся в его тяжелой цепи. Существует пять основных классов антител: IgA, IgD, IgE, IgG и IgM, а некоторые из них могут быть дополнительно поделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, соответствующие разным классам иммуноглобулинов, называются α, δ, ε, γ и μ, соответственно.
«Молекула Fab» относится к белку, состоящему из доменов VH и СН1 тяжелой цепи («тяжелая цепь Fab») и доменов VL и CL легкой цепи («легкая цепь Fab») иммуноглобулина.
Под «кроссоверной» молекулой Fab (также называемой «Crossfab») подразумевается молекула Fab, в которой обменены (т.е. замещены друг другом) вариабельные домены или константные домены тяжелой и легкой цепи Fab, т.е. кроссоверная молекула Fab содержит пептидную цепь, состоящую из вариабельного домена легкой цепи VL и константного домена тяжелой цепи 1 CH1 (VL-CH1, в направлении от N- к С-концу), и пептидную цепь, состоящую из вариабельного домена тяжелой цепи VH и константного домена легкой цепи CL (VH-CL, в направлении от N- к С-концу). Для ясности, в кроссоверной молекуле Fab, в которой обменены вариабельные домены легкой цепи Fab и тяжелой цепи Fab, пептидная цепь, содержащая константный домен тяжелой цепи 1 СН1, называется в данном документе «тяжелой цепью» (кроссоверной) молекулы Fab. В то же время, в кроссоверной молекуле Fab, в которой обменены константные домены легкой цепи Fab и тяжелой цепи Fab, пептидная цепь, содержащая вариабельный домен тяжелой цепи VH, называется в данном документе «тяжелой цепью» (кроссоверной) молекулы Fab.
В противоположность этому, под «стандартной» молекулой Fab подразумевается, что молекула Fab имеет свой естественный формат, т.е. содержит тяжелую цепь, состоящую из вариабельных и константных доменов тяжелой цепи (VH-CH1, в направлении от N- к С-концу), и легкую цепь, состоящую из вариабельных и константных доменов легкой цепи (VL-CL, в направлении от N- к С-концу).
В данном документе термин «Fc-домен» или «Fc-область» используют для определения С-концевой области тяжелой цепи иммуноглобулина, которая содержит по меньшей мере часть константной области. Этот термин включает Fc-области с нативной последовательностью и вариантные Fc-области. В одном аспекте Fc-область тяжелой цепи человеческого IgG распространяется от Cys226 или от Pro230 до карбокси-конца тяжелой цепи. При этом антитела, вырабатываемые клетками-хозяевами могут подвергаться посттрансляционному расщеплению одной или более, в частности, одной или двух аминокислот из С-конца тяжелой цепи. Следовательно, антитело, вырабатываемое клеткой-хозяином посредством экспрессии конкретной молекулы нуклеиновой кислоты, кодирующей полноразмерную тяжелую цепь, может содержать полноразмерную тяжелую цепь или может содержать расщепленный вариант полноразмерной тяжелой цепи. Такое может происходить, когда двумя последними С-концевыми аминокислотами тяжелой цепи являются глицин (G446) и лизин (К447, нумерация в соответствии с индексом EU по Kabat). Следовательно, С-концевой лизин (Lys447) или С-концевые глицин (Gly446) и лизин (Lys447) Fc-области могут присутствовать или нет.Аминокислотные последовательности тяжелых цепей, включая Fc-область (или субъединицу Fc-домена по определению в данном документе) обозначены в данном документе без С-концевого глицин-лизинового дипептида, если не указано иное. В одном аспекте тяжелая цепь, включая Fc-область (субъединицу) по определению в данном документе, содержащаяся в антителе в соответствии с изобретением, содержит дополнительный С-концевой глицин-лизиновый дипептид (G446 и К447, нумерация в соответствии с индексом EU по Kabat). В одном аспекте тяжелая цепь, включая Fc-область (субъединицу) по определению в данном документе, содержащаяся в антителе в соответствии с изобретением, содержит дополнительный С-концевой остаток глицина (G446, нумерация в соответствии с индексом EU по Kabat). Если в данном документе не указано иное, нумерация аминокислотных остатков в Fc-области или константной области соответствует системе нумерации EU, также называемой индексом EU, описанной в Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991 (смотрите также выше). В контексте данного документа «субъединица» Fc-домена относится к одному из двух полипептидов, образующих димерный Fc-домен, т.е. полипептиду, содержащему С-концевые константные области тяжелой цепи иммуноглобулина, способные к стабильной самоассоциации. Например, субъединица Fc-домена IgG содержит константные домены СН2 IgG и CH3 IgG.
Под «слитым» подразумевается, что компоненты (например, молекула Fab и субъединица Fc-домена) связаны пептидными связями, как напрямую, так и посредством одного или более пептидных линкеров.
Термин «мультиспецифическое» означает, что антитело способно специфически связываться с по меньшей мере двумя отличными антигенными детерминантами. Мультиспецифическое антитело может, например, представлять собой биспецифическое антитело. Как правило, биспецифическое антитело содержит два антигенсвязывающих сайта, каждый из которых является специфическим к отличной антигенной детерминанте. В определенных аспектах мультиспецифическое (например, биспецифическое) антитело способно одновременно связывать две антигенные детерминанты, в частности, две антигенные детерминанты, экспрессируемые на двух разных клетках.
В контексте данного документа термин «валентный» обозначает наличие определенного числа антигенсвязывающих сайтов в антигенсвязывающей молекуле. Следовательно, термин «одновалентное связывание с антигеном» обозначает наличие одного (и не более одного) антигенсвязывающего сайта, специфического в отношении антигена, в антигенсвязывающей молекуле.
«Антигенсвязывающий сайт» относится к сайту, т.е. одному или более аминокислотным остаткам антигенсвязывающей молекулы, который обеспечивает взаимодействие с антигеном. Например, антигенсвязывающий сайт антитела содержит аминокислотные остатки из определяющих комплементарность областей (CDR). Нативная молекула иммуноглобулина, как правило, имеет два антигенсвязывающих сайта, молекула Fab, как правило, имеет один антигенсвязывающий сайт.
В контексте данного документа термин «антигенная детерминанта» или «антиген» относится к сайту (например, непрерывной последовательности аминокислот или конформационной конфигурации, состоящей из разных областей несмежных аминокислот) в макромолекуле полипептида, с которым связывается антигенсвязывающий домен с образованием комплекса антигенсвязывающий домен - антиген. Подходящие антигенные детерминанты могут находиться, например, на поверхности опухолевых клеток, на поверхностях инфицированных вирусом клеток, на поверхностях других пораженных заболеванием клеток, на поверхности иммунных клеток, в свободном виде в сыворотке крови и/или во внеклеточном матриксе (ВКМ). В предпочтительном аспекте антиген представляет собой белок человека.
«CD3» относится к любому нативному CD3 из любого источника, относящегося к позвоночным, включая млекопитающих, таких как приматы (например, люди), отличные от человека приматы (например, яванские макаки) и грызуны (например, мыши и крысы), если не указано иное. Этот термин охватывает «полноразмерный» непроцессированный CD3, а также любую форму CD3, полученную в результате процессинга в клетке. Данный термин также включает варианты CD3 природного происхождения, например, сплайс-варианты или аллельные варианты. В одном аспекте CD3 представляет собой CD3 человека, в частности эпсилон-субъединицу CD3 человека (CD3ε). Аминокислотная последовательность CD3ε человека приведена в SEQ ID NO: 112 (без сигнального пептида). Также смотрите номер доступа UniProt (www.uniprot.org) №Р07766 (версия 189) или NCBI (www.ncbi.nlm.nih.gov/) RefSeq NP_000724.1. В другом аспекте CD3 представляет собой CD3 яванского макака (Масаса fascicularis), в частности CD3ε яванского макака. Аминокислотная последовательность CD3ε яванского макака приведена в SEQ ID NO: 113 (без сигнального пептида). Также смотрите NCBI GenBank №ВАВ71849.1. В определенных аспектах антитело по изобретению связывается с эпитопом CD3, который является консервативным для антигенов CD3 от разных видов, в частности, CD3 человека и яванского макака. В предпочтительных аспектах антитело связывается с CD3 человека.
В контексте данного документа «антиген клетки-мишени» относится к антигенной детерминанте, представленной на поверхности клетки-мишени, например, клетки в опухоли, такой как раковая клетка или клетка опухолевой стромы (в этом случае - «антиген опухолевой клетки»). Предпочтительно, антиген клетки-мишени не представляет собой CD3 и/или экспрессируется на другой клетке, чем CD3. В предпочтительном аспекте антиген клетки-мишени представляет собой TYRP-1, в частности, TYRP-1 человека. В другом предпочтительном аспекте антиген клетки-мишени представляет собой EGFRvIII, в частности, EGFRvIII человека.
«TYRP1» или «TYRP-1» обозначает тирозин-родственный белок 1, который представляет собой фермент, участвующий в синтезе меланина. Зрелая форма TYRP1, также оригинально названная gp75, представляет собой 75 кДа трансмембранный гликопротеин. Последовательность TYRP1 человека приведена в SEQ ID NO: 114 (без сигнального пептида). Также смотрите запись UniProt №Р17643 (версия 185). В контексте данного документа «TYRP1» относится к любому нативному TYRP1 из любого источника, относящегося к позвоночным, включая млекопитающих, таких как приматы (например, люди), отличные от человека приматы (например, яванские макаки) и грызуны (например, мыши и крысы), если не указано иное. Этот термин охватывает «полноразмерный» непроцессированный TYRP1, а также любую форму TYRP1, полученную в результате процессинга в клетке. Данный термин также включает варианты TYRP1 природного происхождения, например, сплайс-варианты или аллельные варианты. В одном аспекте TYRP1 представляет собой TYRP1 человека.
«EGFRvIII» обозначает рецептор эпидермального фактора роста, вариант III, который представляет собой мутант EGFR, образованный за счет удаления экзонов 2-7 в рамке считывания, что приводит к удалению 267 аминокислот с заменой глицина в соединении. Последовательность EGFRvIII человека приведена в SEQ ID NO: 115 (без сигнального пептида). Последовательность EGFR человека дикого типа приведена в SEQ ID NO: 116 (без сигнального пептида). Также смотрите запись UniProt №Р00533 (версия 258). В контексте данного документа «EGFRvIII» относится к любому нативному EGFRvIII из любого источника, относящегося к позвоночным, включая млекопитающих, таких как приматы (например, люди), отличные от человека приматы (например, яванские макаки) и грызуны (например, мыши и крысы), если не указано иное. Этот термин охватывает «полноразмерный» непроцессированный EGFRvIII (но не EGFR дикого типа), а также любую форму EGFRvIII, полученную в результате процессинга в клетке (например, EGFRvIII без сигнального пептида). В одном аспекте EGFRvIII представляет собой EGFRvIII человека.
«Аффинность» относится к силе суммарных нековалентных взаимодействий между одним связывающим сайтом молекулы (например, антитела) и ее партнером по связыванию (например, антигеном). Если не указано иное, в контексте данного документа «аффинность связывания» относится к характерной аффинности связывания, которая отражает 1:1 взаимодействие между членами пары связывания (например, антителом и антигеном). Аффинность молекулы X в отношении ее партнера Y в общем случае можно представить константой диссоциации (KD). Аффинность можно измерять хорошо отработанными методами, известными в данной области техники, включая методы, описанные в данном документе. Предпочтительным методом измерения аффинности является поверхностный плазмонный резонанс (ППР).
Антитело с «созревшей аффинностью» относится к антителу с одним или более изменениями в одной или более определяющих комплементарность областях (CDR) по сравнению с родительским антителом, которое не содержит такие изменения, при этом такие изменения приводят к улучшению аффинности антитела в отношении антигена.
«Сниженное связывание», например, сниженное связывание с Fc-рецептором, относится к снижению аффинности для соответствующего взаимодействия по определению, например, ППР. Для ясности, этот термин также включает снижение аффинности до нуля (или ниже предела обнаружения аналитического метода), т.е. полное устранение взаимодействия. И наоборот, «повышенное связывание» относится к повышению аффинности связывания для соответствующего взаимодействия.
В контексте данного документа «активация Т-клеток» относится к одному или более клеточным ответам лимфоцита, в частности, цитотоксического Т-лимфоцита, выбранным из: пролиферации, дифференцировки, секреции цитокинов, высвобождения цитотоксических эффекторных молекул, цитотоксической активности и экспрессии маркеров активации. Подходящие анализы для определения активации Т-клеток известны в данной области техники и описаны в данном документе.
«Модификация, способствующая ассоциации первой и второй субъединиц Fc-домена» представляет собой манипуляцию с пептидным остовом или посттрансляционные модификации субъединицы Fc-домена, которые уменьшают или предотвращают ассоциацию полипептида, содержащего субъединицу Fc-домена, с идентичным полипептидом с образованием гомодимера. В контексте данного документа модификация, способствующая ассоциации, предпочтительно включает отдельные модификации, проведенные в каждой из субъединиц Fc-домена, которые должны ассоциировать (т.е. первой и второй субъединицах Fc-домена), причем модификации являются комплементарными по отношению друг к другу так, чтобы способствовать ассоциации двух субъединиц Fc-домена. Например, модификация, способствующая ассоциации, может изменять структуру или заряд одной или обеих субъединиц Fc-домена так, чтобы сделать их ассоциацию стерически или электростатически выгодной, соответственно. Таким образом, (гетеро)димеризация происходит между полипептидом, содержащим первую субъединицу Fc-домена, и полипептидом, содержащим вторую субъединицу Fc-домена, которые могут быть неидентичными в том смысле, что дополнительные компоненты, слитые с каждой из субъединиц (например, антигенсвязывающие домены), не являются идентичными. В некоторых аспектах модификация, способствующая ассоциации первой и второй субъединиц Fc-домена, включает аминокислотную мутацию в Fc-домене, в частности аминокислотную замену. В предпочтительном аспекте модификация, способствующая ассоциации первой и второй субъединиц Fc-домена, включает отдельную аминокислотную мутацию, в частности аминокислотную замену, в каждой из двух субъединиц Fc-домена.
Термин «эффекторные функции» относится к видам биологической активности, обеспечиваемым Fc-областью антитела, которые варьируются в зависимости от изотипа антитела. Примеры эффекторных функций антител включают: связывание Clq и комплементзависимую цитотоксичность (КЗЦ), связывание Fc-рецептора, антителозависимую клеточноопосредованную цитотоксичность (АЗКЦ), антителозависимый клеточный фагоцитоз (АЗКФ), секрецию цитокинов, опосредованное иммунным комплексом поглощение антигена антигенпрезентирующими клетками, понижающую регуляцию рецепторов клеточной поверхности (например, В-клеточного рецептора) и активацию В-клеток.
«Активирующий Fc-рецептор» представляет собой Fc-рецептор, который после взаимодействия с Fc-доменом антитела вызывает события сигнализации, которые стимулируют несущую рецептор клетку осуществлять эффекторные функции. Человеческие активирующие Fc-рецепторы включают FcγRIIIa (CD16a), FcγRI (CD64), FcγRIIa (CD32) и FcαRI (CD89).
Антителозависимая клеточноопосредованная цитотоксичность (АЗКЦ) представляет собой иммунный механизм, приводящий к лизису покрытых антителом клеток-мишеней иммунными эффекторными клетками. Клетки-мишени представляют собой клетки, с которыми специфически связываются антитела или их производные, содержащие Fc-область, в общем случае посредством белковой части, N-терминальной относительно Fc-области. В контексте данного документа термин «сниженная АЗКЦ» определяется как снижение числа клеток-мишеней, лизированных за заданное время при заданной концентрации антитела в среде, окружающей клетки-мишени, посредством определенного выше механизма АЗКЦ, и/или как повышение концентрации антитела в среде, окружающей клетки-мишени, необходимое для обеспечения лизиса заданного числа клеток-мишеней за заданное время посредством механизма АЗКЦ. Снижение АЗКЦ определяется относительно АЗКЦ, опосредованной таким же антителом, вырабатываемым таким же типом клеток-хозяев, с использованием таких же стандартных методов получения, очистки, составления и хранения (которые хорошо известны специалистам в данной области техники), но которое не было сконструировано. Например, снижение АЗКЦ, опосредованное антителом, содержащим в Fc-домене аминокислотную замену, которая снижает АЗКЦ, определяется относительно АЗКЦ, опосредованной таким же антителом без аминокислотной замены в Fc-домене. Подходящие анализы для измерения АЗКЦ хорошо известны в данной области техники (смотрите, например, публикацию РСТ №WO 2006/082515 или публикацию РСТ №WO 2012/130831).
В контексте данного документа считается, что термины «конструировать, сконструированный, конструирование» включают любую манипуляцию с пептидным остовом или посттрансляционные модификации встречающегося в природе или рекомбинантного полипептида или его фрагмента. Конструирование включает модификации аминокислотной последовательности, профиля гликозилирования или групп боковых цепей отдельных аминокислот, а также комбинации этих подходов.
В контексте данного документа подразумевается, что термин «аминокислотная мутация» включает аминокислотные замены, делеции, вставки и модификации. Можно осуществлять любую комбинацию замены, делеции, вставки и модификации, чтобы получить конечную конструкцию, при условии, что конечная конструкция обладает необходимыми характеристиками, например, сниженным связыванием с Fc-рецептором или повышенной ассоциацией с другим пептидом. Делеции и вставки в аминокислотной последовательности включают амино- и/или карбокси-концевые делеции и вставки аминокислот. Предпочтительными аминокислотными мутациями являются аминокислотные замены. В целях изменения, например, характеристик связывания Fc-области, в особенности предпочтительными являются неконсервативные аминокислотные замены, т.е. замещение одной аминокислоты другой аминокислотой, имеющей отличные структурные и/или химические свойства. Аминокислотные замены включают замещение не встречающимися в природе аминокислотами или встречающимися в природе аминокислотными производными двадцати стандартных аминокислот (например, 4-гидроксипролином, 3-метилгистидином, орнитином, гомосерином, 5-гидроксилизином). Аминокислотные мутации можно создавать, используя хорошо известные в данной области техники генетические или химические методы. Генетические методы могут включать сайт-направленный мутагенез, ПЦР, генный синтез и т.п. Подразумевается, что также могут быть применимы методы изменений групп боковых цепей аминокислот методами, отличными от генной инженерии, такими как химическая модификация. Для указания одной и той же аминокислотной мутации в данном документе можно использовать различные обозначения. Например, замена пролина в позиции 329 Fc-домена на глицин может быть обозначена как 329G, G329, G329, P329G или Pro329Gly.
«Процент (%) идентичности аминокислотной последовательности» относительно референсной полипептидной последовательности определяется как процентная доля аминокислотных остатков в кандидатной последовательности, которые являются идентичными с аминокислотными остатками в референсной полипептидной последовательности, после выравнивания последовательностей и внесения, в случае необходимости, гэпов для достижения максимального процента идентичности последовательностей, и без учета каких-либо консервативных замен в качестве части идентичности последовательностей. Выравнивание в целях определения процента идентичности аминокислотной последовательности можно осуществлять различными способами, которые известны в данной области техники, например, используя общедоступное компьютерное программное обеспечение, такое как программное обеспечение BLAST, BLAST-2, Clustal W, Megalign (DNASTAR) или пакет программ FAS ТА. Специалисты в данной области техники могут определить подходящие параметры для выравнивания последовательностей, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. В альтернативном варианте значения процента идентичности можно получить, используя компьютерную программу для сравнения последовательностей ALIGN-2. Компьютерная программа для сравнения последовательностей ALIGN-2 была разработана Genentech, Inc., а исходный код был подан вместе с пользовательской документацией в Бюро регистрации авторских прав США, Вашингтон, округ Колумбия, 20559, где он зарегистрирован под номером регистрации авторского права США TXU510087, и описан в WO 2001/007611.
Если не указано иное, в целях данного документа значения % идентичности аминокислотной последовательности получают, используя программу ggsearch из пакета FASTA версии 36.3.8 с или более поздней с матрицей сравнения BLOSUM50. Авторство пакета программ FASTA принадлежит W. R. Pearson и D. J. Lipman ("Improved Tools for Biological Sequence Analysis", PNAS 85 (1988) 2444-2448), W. R. Pearson ("Effective protein sequence comparison" Meth. Enzymol. 266 (1996) 227- 258) и Pearson et. al. (Genomics 46 (1997) 24-36), а сам пакет находится в открытом доступе на www.fasta.bioch.virginia.edu/fasta_www2/fasta_down.shtml or www.ebi.ac.uk/Tools/sss/fasta. В альтернативном варианте для сравнения последовательностей можно использовать открытый сервер, доступный по адресу fasta.bioch.virginia.edu/fasta_www2/index.cgi, используя программу ggsearch (global protein:protein) и опции по умолчанию (BLOSUM50; открытие: -10; продление: -2; Ktup=2), чтобы гарантировать проведение глобального, а не локального выравнивания. Процент аминокислотной идентичности выводится в заголовочном файле вывода выравнивания.
Термин «полинуклеотид» или «молекула нуклеиновой кислоты» включает любое соединение и/или вещество, которое содержит полимер нуклеотидов. Каждый нуклеотид состоит из основания, в частности пуринового или пиримидинового основания (т.е. цитозина (С), гуанина (G), аденина (А), тимина (Т) или урацила (U)), сахара (т.е. дезоксирибозы или рибозы) и фосфатной группы. Часто молекула нуклеиновой кислоты описана с помощью последовательности оснований, причем указанные основания представляют первичную структуру (линейную структуру) молекулы нуклеиновой кислоты. Последовательность оснований, как правило, представлена от 5' к 3'. В данном документе термин молекула нуклеиновой кислоты охватывает дезоксирибонуклеиновую кислоту (ДНК), включая, например, комплементарную ДНК (кДНК) и геномную ДНК, рибонуклеиновую кислоту (РНК), в частности матричную РНК (мРНК), синтетические формы ДНК или РНК и смешанные полимеры, содержащие две или более из этих молекул. Молекула нуклеиновой кислоты может быть линейной или кольцевой. Кроме того, термин «молекула нуклеиновой кислоты» включает как смысловую, так и антисмысловую цепи, а также одноцепочечные и двухцепочечные формы. Кроме того, описанная в данном документе молекула нуклеиновой кислоты может содержать встречающиеся в природе или не встречающиеся в природе нуклеотиды. Примеры не встречающихся в природе нуклеотидов включают модифицированные нуклеотидные основания с дериватизированными сахарами, или фосфатными остовными связями, или химически модифицированными остатками. Молекулы нуклеиновых кислот также включают молекулы ДНК и РНК, подходящие в качестве вектора для прямой экспрессии антитела по изобретению in vitro и/или in vivo, например, в организме-хозяине или организме пациента. Такие ДНК (например, кДНК)- или РНК (например, мРНК)-векторы могут быть немодицифированными или модифицированными. Например, мРНК может быть химически модифицирована для повышения стабильности РНК-вектора и/или экспрессии кодируемой молекулы так, чтобы мРНК можно было вводить субъекту для генерации антител in vivo (смотрите, например, Stadler et al. (2017) Nature Medicine 23:815-817, или ЕР 2101823 B1).
«Выделенная» молекула нуклеиновой кислоты относится к молекуле нуклеиновой кислоты, которая была отделена от компонента своего естественного окружения. Выделенная молекула нуклеиновой кислоты включает молекулу нуклеиновой кислоты, содержащуюся в клетках, которые обычно содержат эту молекулу нуклеиновой кислоты, но при этом молекула нуклеиновой кислоты присутствует внехромосомно или в хромосомном положении, которое отличается от ее естественного хромосомного положения.
«Выделенные полинуклеотид (или нуклеиновая кислота), кодирующие антитело» относятся к одной или более полинуклеотидным молекулам, кодирующим тяжелые и легкие цепи антитела (или их фрагменты), включая такие полинуклеотидные молекулы в одном векторе или отдельных векторах и такие полинуклеотидные молекулы, присутствующие в одной или более локациях в клетке-хозяине.
В контексте данного документа термин «вектор» относится к молекуле нуклеиновой кислоты, способной увеличивать число копий другой нуклеиновой кислоты, с которой она связана. Этот термин включает вектор в виде самореплицирующейся структуры нуклеиновой кислоты, а также вектор, включенный в геном клетки-хозяина, в которую его внесли. Некоторые векторы способны управлять экспрессией нуклеиновых кислот, с которыми они функционально связаны. Такие векторы называют в данном документе «экспрессионными векторами».
Термины «клетка-хозяин», «линия клеток-хозяев» и «культура клеток-хозяев» используются взаимозаменяемо и относятся к клеткам, в которые была внесена экзогенная нуклеиновая кислота, включая потомство таких клеток. Клетки-хозяева включают «трансформантов» и «трансформированные клетки», которые включают первично трансформированные клетки и полученное от них потомство вне зависимости от числа пассажей. Потомство может не быть полностью идентичным родительской клетке по содержанию нуклеиновой кислоты и может содержать мутации. Сюда включено мутантное потомство, которое имеет такую же функцию или биологическую активность, в отношении которых проводится скрининг или отбор изначально трансформированных клеток. Клетка-хозяин представляет собой любой тип клеточной системы, который можно использовать для генерации антител по настоящему изобретению. Клетки-хозяева включают культивируемые клетки, например, культивируемые клетки млекопитающих, такие как клетки НЕК, клетки СНО, клетки ВНК, клетки NS0, клетки SP2/0, клетки миеломы YO, клетки миеломы мышей Р3Х63, клетки PER, клетки PER.C6 или клетки гибридомы, клетки дрожжей, клетки насекомых и клетки растений, если называть только некоторые, но также клетки, присутствующие в организме трансгенного животного, трансгенного растения или культивируемой растительной или животной ткани. В одном аспекте клетка-хозяин по изобретению представляет собой эукариотическую клетку, в частности клетку млекопитающего. В одном аспекте клетка-хозяин не является клеткой в организме человека.
Термин «фармацевтическая композиция» или «фармацевтический состав» относится к препарату, который находится в форме, обеспечивающей эффективность биологической активности содержащегося в нем активного ингредиента, и который не содержит дополнительные компоненты, которые являются неприемлемо токсичными для субъекта, которому будет введена композиция.
«Фармацевтически приемлемый носитель» относится к ингредиенту в фармацевтической композиции или фармацевтическом составе, отличному от активного ингредиента, который нетоксичен для субъекта. Фармацевтически приемлемый носитель включает, но не ограничивается этим, буфер, вспомогательное вещество, стабилизатор или консервант.
В контексте данного документа термин «лечение» (и его грамматические вариации, такие как «лечить» или «лечащий») относится к клиническому вмешательству с целью изменения естественного течения заболевания у индивида, которого лечат, и может проводиться как для профилактики, так и во время клинической патологии. Необходимые эффекты лечения включают, но не ограничиваются этим, предотвращение возникновения или рецидива заболевания, облегчение симптомов, уменьшение любых прямых или косвенных патологических последствий заболевания, предотвращение метастазирования, снижение скорости прогрессирования заболевания, улучшение или временное облегчение состояния болезни и ремиссию или улучшенный прогноз. В некоторых вариантах осуществления антитела по изобретению используют для задержки развития заболевания или для замедления прогрессирования заболевания.
«Индивид» или «субъект» представляет собой млекопитающее. Млекопитающие включают, но не ограничиваются этим, одомашненных животных (например, коров, овец, кошек, собак и лошадей), приматов (например, людей и отличных от человека приматов, таких, как обезьяны), кроликов и грызунов (например, мышей и крыс). В определенных аспектах индивид или субъект представляет собой человека.
«Эффективное количество» агента, например, фармацевтической композиции, относится к количеству, эффективному в дозах и в течение периодов времени, необходимых для достижения необходимого терапевтического или профилактического результата.
Термин «вкладыш в упаковку» используют для обозначения инструкций, обычно включаемых в коммерческие упаковки терапевтических продуктов, которые содержат информацию о показаниях, применении, дозировке, введении, комбинированной терапии, противопоказаниях и/или предупреждениях относительно использования таких терапевтических продуктов.
II. КОМПОЗИЦИИ И СПОСОБЫ
В изобретении предложены антитела, которые связывают CD3, включая мультиспецифические антитела, которые связывают CD3 и второй антиген. Эти антитела демонстрируют превосходную стабильность в сочетании с другими благоприятными свойствами для терапевтического применения, например, в отношении эффективности и безопасности, фармакокинетики, а также возможности производства. Антитела по изобретению применимы, например, для лечения заболеваний, таких как рак.
A. Анти-CD3 антитела
В одном аспекте в изобретении предложены антитела, которые связываются с CD3. В одном аспекте в предложены выделенные антитела, которые связываются с CD3. В одном аспекте в изобретении предложены антитела, которые специфически связываются с CD3. В определенных аспектах анти-CD3 антитела сохраняют более чем около 90% активности связывания с CD3 через 2 недели при рН 7,4, 37°С, по сравнению с активностью связывания через 2 недели при рН 6, -80°С, по определению методом поверхностного плазмонного резонанса (ППР).
В одном аспекте в настоящем изобретении предложено антитело, которое связывается с CD3, причем антитело содержит первый антигенсвязывающий домен, содержащий вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область тяжелой цепи (HCDR) 1 с SEQ ID NO: 2, HCDR 2 с SEQ ID NO: 3 и HCDR 3 с SEQ ID NO: 5, и вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область легкой цепи (LCDR) 1 с SEQ ID NO: 8, LCDR 2 с SEQ ID NO: 9 и LCDR 3 с SEQ ID NO: 10.
В одном аспекте антитело представляет собой гуманизированное антитело. В одном аспекте антигенсвязывающий домен представляет собой гуманизированный антигенсвязывающий домен (т.е. антигенсвязывающий домен гуманизированного антитела). В одном аспекте VH и/или VL является гуманизированной вариабельной областью.
В одном аспекте VH и/или VL содержит акцепторную человеческую каркасную область, например, каркасную область человеческого иммуноглобулина или человеческую консенсусную каркасную область.
В одном аспекте VH содержит одну или более каркасных последовательностей тяжелой цепи (т.е. последовательность FR1, FR2, FR3 и/или FR4) последовательности вариабельной области тяжелой цепи SEQ ID NO: 7. В одном аспекте VH содержит последовательность, которая является по меньшей мере на около 95%, 96%, 97%, 98% или 99% идентичной аминокислотной последовательности SEQ ID NO: 7. В одном аспекте VH содержит последовательность, которая является по меньшей мере на около 95% идентичной аминокислотной последовательности SEQ ID NO: 7. В одном аспекте VH содержит последовательность, которая является по меньшей мере на около 98% идентичной аминокислотной последовательности SEQ ID NO: 7. В определенных аспектах последовательность VH, имеющая по меньшей мере 95%, 96%, 97%, 98% или 99% идентичности, содержит замены (например, консервативные замены), вставки или делеции по сравнению с референсной последовательностью, но антитело, содержащее эту последовательность, сохраняет способность связываться с CD3. В определенных аспектах была проведена замена, вставка и/или делеция в общей сложности 1-10 аминокислот в аминокислотной последовательности SEQ ID NO: 7. В определенных аспектах замены, вставки или делеции находятся в областях за пределами CDR (т.е. в FR). В одном аспекте VH содержит аминокислотную последовательность SEQ ID NO: 7. Необязательно, VH содержит аминокислотную последовательность SEQ ID NO: 7, включая посттрансляционные модификации этой последовательности.
В одном аспекте VL содержит одну или более каркасных последовательностей легкой цепи (т.е. последовательность FR1, FR2, FR3 и/или FR4) последовательности вариабельной области легкой цепи SEQ ID NO: 11. В одном аспекте VL содержит последовательность, которая является по меньшей мере на около 95%, 96%, 97%, 98% или 99% идентичной аминокислотной последовательности SEQ ID NO: 11. В одном аспекте VL содержит последовательность, которая является по меньшей мере на около 95% идентичной аминокислотной последовательности SEQ ID NO: 11. В одном аспекте VL содержит последовательность, которая является по меньшей мере на около 98% идентичной аминокислотной последовательности SEQ ID NO: 11. В определенных аспектах последовательность VL, имеющая по меньшей мере 95%, 96%, 97%, 98% или 99% идентичности, содержит замены (например, консервативные замены), вставки или делеции по сравнению с референсной последовательностью, но антитело, содержащее эту последовательность, сохраняет способность связываться с CD3. В определенных аспектах была проведена замена, вставка и/или делеция в общей сложности 1-10 аминокислот в аминокислотной последовательности SEQ ID NO: 11. В определенных аспектах замены, вставки или делеции находятся в областях за пределами CDR (т.е. в FR). В одном аспекте VL содержит аминокислотную последовательность SEQ ID NO: 11. Необязательно, VL содержит аминокислотную последовательность SEQ ID NO: 11, включая посттрансляционные модификации этой последовательности.
В одном аспекте VH содержит аминокислотную последовательность, которая по меньшей мере на около 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 7, a VL содержит аминокислотную последовательность, которая по меньшей мере на около 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 11. В одном аспекте VH содержит аминокислотную последовательность SEQ ID NO: 7, a VL содержит аминокислотную последовательность SEQ ID NO: 11.
В дополнительном аспекте в изобретении предложено антитело, которое связывается с CD3, причем антитело содержит первый антигенсвязывающий домен, содержащий VH, содержащую аминокислотную последовательность SEQ ID NO: 7, и VL, содержащую аминокислотную последовательность SEQ ID NO: 11.
В дополнительном аспекте в изобретении предложено антитело, которое связывается с CD3, причем антитело содержит первый антигенсвязывающий домен, содержащий последовательность VH с SEQ ID NO: 7 и последовательность VL с SEQ ID NO: 11.
В другом аспекте в изобретении предложено антитело, которое связывается с CD3, причем антитело содержит первый антигенсвязывающий домен, содержащий VH, содержащую последовательности CDR тяжелой цепи VH с SEQ ID NO: 7, и VL, содержащую последовательности CDR легкой цепи VL c SEQ ID NO: 11.
В дополнительном аспекте первый антигенсвязывающий домен содержит аминокислотные последовательности HCDR1, HCDR2 и HCDR3 VH с SEQ ID NO: 7 и аминокислотные последовательности LCDR1, LCDR2 и LCDR3 VL с SEQ ID NO: 11.
В одном аспекте VH содержит последовательности CDR тяжелой цепи VH с SEQ ID NO: 7 и каркасную область с по меньшей мере 95%, 96%, 97%, 98% или 99% идентичности последовательности с каркасной последовательностью VH с SEQ ID NO: 7. В одном аспекте VH содержит последовательности CDR тяжелой цепи VH с SEQ ID NO: 7 и каркасную область с по меньшей мере 95% идентичности последовательности с каркасной последовательностью VH с SEQ ID NO: 7. В другом аспекте VH содержит последовательности CDR тяжелой цепи VH с SEQ ID NO: 7 и каркасную область с по меньшей мере 98% идентичности последовательности с каркасной последовательностью VH с SEQ ID NO: 7.
В одном аспекте VL содержит последовательности CDR легкой цепи VL с SEQ ID NO: 11 и каркасную область с по меньшей мере 95%, 96%, 97%, 98% или 99% идентичности последовательности с каркасной последовательностью VL с SEQ ID NO: 11. В одном аспекте VL содержит последовательности CDR легкой цепи VL с SEQ ID NO: 11 и каркасную область с по меньшей мере 95% идентичности последовательности с каркасной последовательностью VL с SEQ ID NO: 11. В другом аспекте VL содержит последовательности CDR легкой цепи VL с SEQ ID NO: 11 и каркасную область с по меньшей мере 98% идентичности последовательности с каркасной последовательностью VL с SEQ ID NO: 11.
В одном аспекте в изобретении предложено антитело, которое связывается с CD3, причем антитело содержит первый антигенсвязывающий домен, содержащий последовательность VH, соответствующую любому из вышеприведенных аспектов, и последовательность VL, соответствующую любому из вышеприведенных аспектов.
В одном аспекте антитело содержит человеческую константную область. В одном аспекте антитело представляет собой молекулу иммуноглобулина, содержащую человеческую константную область, в частности молекулу иммуноглобулина класса IgG, содержащую человеческий домен CH1, СН2, CH3 и/или CL. Типовые последовательности человеческих константных доменов приведены в SEQ ID NO: 120 и 121 (человеческие каппа и лямбда CL-домены, соответственно) и SEQ ID NO: 122 (константные домены СН1-СН2-CH3 тяжелой цепи человеческого IgG1). В одном аспекте антитело содержит константную область легкой цепи, содержащую аминокислотную последовательность, которая является по меньшей мере на около 95%, 96%, 97%, 98%, 99% или 100% идентичной аминокислотной последовательности SEQ ID NO: 120 или SEQ ID NO: 121, в частности аминокислотной последовательности SEQ ID NO: 120. В одном аспекте антитело содержит константную область тяжелой цепи, содержащую аминокислотную последовательность, которая является по меньшей мере на около 95%, 96%, 97%, 98%, 99% или 100% идентичной аминокислотной последовательности SEQ ID NO: 122. В частности, константная область тяжелой цепи может содержать аминокислотные мутации в Fc-домене, как описано в данном документе.
В одном аспекте первый антигенсвязывающий домен содержит человеческую константную область. В одном аспекте первый антигенсвязывающий фрагмент представляет собой молекулу Fab, содержащую человеческую константную область, в частности, человеческий домен СН1 и/или CL. В одном аспекте первый антигенсвязывающий домен содержит константную область легкой цепи, содержащую аминокислотную последовательность, которая является по меньшей мере на около 95%, 96%, 97%, 98%, 99% или 100% идентичной аминокислотной последовательности SEQ ID NO: 120 или SEQ ID NO: 121, в частности аминокислотной последовательности SEQ ID NO: 120. В частности, константная область легкой цепи может содержать аминокислотные мутации, описанные в данном документе как «зарядовые модификации», и/или может содержать делецию или замены одной или более (в частности двух) N-концевых аминокислот, если она принадлежит кроссоверной молекуле Fab. В некоторых аспектах первый антигенсвязывающий домен содержит константную область тяжелой цепи, содержащую аминокислотную последовательность, которая является по меньшей мере на около 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности домена СН1, содержащейся в аминокислотной последовательности SEQ ID NO: 122. В частности, константная область тяжелой цепи (в частности домен СН1) может содержать аминокислотные мутации, описанные в данном документе как «зарядовые модификации».
В одном аспекте антитело представляет собой моноклональное антитело.
В одном аспекте антитело представляет собой IgG, в частности антитело IgG1. В одном аспекте антитело представляет собой полноразмерное антитело.
В другом аспекте антитело представляет собой фрагмент антитела, выбранный из группы из молекулы Fv, молекулы scFv, молекулы Fab и молекулы F(ab')2; в частности молекулы Fab. В другом аспекте фрагмент антитела представляет собой диатело, триатело или тетратело.
В одном аспекте первый антигенсвязывающий домен представляет собой молекулу Fab. В предпочтительном аспекте первый антигенсвязывающий домен представляет собой молекулу Fab, причем вариабельные домены VL и VH или константные домены CL и СН1, в частности вариабельные домены VL и VH легкой цепи Fab и тяжелой цепи Fab замещены друг другом (т.е. первый антигенсвязывающий домен представляет собой кроссоверную молекулу Fab).
В дополнительном аспекте антитело по любому из вышеприведенных аспектов может иметь любые из признаков, по отдельности или в комбинации описанных в разделах П. А. 1.-8.ниже.
В предпочтительном аспекте антитело содержит Fc-домен, в частности Fc-домен IgG, конкретнее Fc-домен IgG1. В одном аспекте Fc-домен представляет собой Fc-домен человека. В одном аспекте Fc-домен представляет собой Fc-домен IgG1 человека. Fc-домен состоит из первой и второй субъединиц и может иметь любые из признаков, по отдельности или в комбинации описанных ниже в данном документе в отношении вариантов Fc-доменов (раздел II. А. 8.).
В другом предпочтительном аспекте антитело содержит второй и, необязательно, третий антигенсвязывающий домен, который связывается со вторым антигеном (т.е. антитело представляет собой мультиспецифическое антитело, как дополнительно описано ниже в данном документе (раздел II. А. 7.).
1. Фрагменты антител
В определенных аспектах предложенное в данном документе антитело представляет собой фрагмент антитела.
В одном аспекте фрагмент антитела представляет собой молекулу Fab, Fab', Fab'-SH или F(ab')2, в частности молекулу Fab, как описано в данном документе. «Молекула Fab'» отличается от от молекулы Fab добавлением остатков на карбокси-конце домена СН1, в том числе одного или более остатков цистеина из шарнирной области антитела. Fab'-SH представляют собой молекулы Fab', в которых остаток(ки) цистеина константных доменов несет(ут) свободную тиольную группу. Обработка пепсином приводит к получению молекулы F(ab')2, которая содержит два антигенсвязывающих сайта (две молекулы Fab) и часть Fc-области.
В другом аспекте фрагмент антитела представляет собой диатело, триатело или тетратело. «Диатела» представляют собой фрагменты антител с двумя антигенсвязывающими сайтами, которые могут быть двухвалентными или биспецифическими. Смотрите, например, ЕР 404,097; WO 1993/01161; Hudson et al., Nat. Med. 9:129-134 (2003); и Hollinger et al., Proc. Natl. Acad. Sci. USA 90: 6444-6448 (1993). Триатела и тетратела также описаны в Hudson et al., Nat. Med. 9:129-134 (2003).
В дополнительном аспекте фрагмент антитела представляет собой одноцепочечную молекулу Fab. «Одноцепочечная молекулы Fab» или «scFab» представляет собой полипептид, состоящий из вариабельного домена тяжелой цепи антитела (VH), константного домена 1 тяжелой цепи антитела (СН1), вариабельного домена легкой цепи антитела (VL), константного домена легкой цепи антитела (CL) и линкера, причем указанные домены антитела и указанный линкер имеют один из следующих порядков в направлении от N-конца к С-концу: а) VH-CH1-линкер-VL-CL, б) VL-CL-линкер-VH-CH1, в) VH-CL-линкер-VL-CH1 или г) VL-CH1-линкер-VH-CL. В частности, указанный линкер представляет собой полипептид из по меньшей мере 30 аминокислот, предпочтительно от 32 до 50 аминокислот. Указанные одноцепочечные молекулы Fab стабилизированы за счет естественной дисульфидной связи между доменом CL и доменом СН1. Кроме того, одноцепочечные молекулы Fab могут быть дополнительно стабилизированы за счет создания межцепочечных дисульфидных связей посредством вставки остатков цистеина (например, в позиции 44 вариабельного домена тяжелой цепи и позиции 100 вариабельного домена легкой цепи в соответствии с нумерацией Kabat).
В другом аспекте фрагмент антитела представляет собой одноцепочечный вариабельный фрагмент (scFv). «Одноцепочечный вариабельный фрагмент» или «scFv» представляет собой слитый белок из вариабельных доменов тяжелой (VH) и легкой цепей (VL) антитела, соединенных линкером. В частности, линкер представляет собой короткий полипептид из 10-25 аминокислот и обычно обогащен глицином для гибкости, а также серином или треонином для растворимости, и может соединять N-конец VH с С-концом VL или наоборот.Этот белок сохраняет специфичность исходного антитела несмотря на удаление константных областей и внесение линкера. Обзор фрагментов scFv смотрите, например, в Pluckthun, in The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., (Springer-Verlag, New York), стр. 269-315 (1994); также смотрите WO 93/16185; и патенты США Ms 5571894 и 5587458.
В другом аспекте фрагмент антитела представляет собой однодоменное антитело. «Однодоменные антитела» представляют собой фрагменты антител, содержащие весь вариабельный домен тяжелой цепи антитела или его часть, или весь вариабельный домен легкой цепи антитела или его часть. В определенных аспектах однодоменное антитело представляет собой человеческое однодоменное антитело (Domantis, Inc., Waltham, MA; смотрите, например, патент США №6248516 В1).
Фрагменты антител можно получать различными способами, включая, но не ограничиваясь этим, протеолитическое расщепление интактного антитела, а также рекомбинантную выработку рекомбинантными клетками-хозяевами (например, Е. coli), как описано в данном документе.
2. Гуманизированные антитела
В определенных аспектах предложенное в данном документе антитело представляет собой гуманизированное антитело. Как правило, антитело нечеловеческого происхождения гуманизируют с целью снижения иммуногенности для человека, сохраняя при этом специфичность и аффинность родительского антитела нечеловеческого происхождения. В общем случае гуманизированное антитело содержит один или более вариабельных доменов, в которых CDR (или их части) получены из антитела нечеловеческого происхождения, a FR (или их части) получены из последовательностей человеческого антитела. Гуманизированное антитело, необязательно, также содержит по меньшей мере часть человеческой константной области. В некоторых аспектах некоторые остатки FR в гуманизированном антителе замещены соответствующими остатками нечеловеческого антитела (например, антитела, из которого получены остатки CDR), например, с целью восстановления или улучшения специфичности или аффинности антитела.
Гуманизированные антитела и способы их получения рассмотрены, например, в Almagro and Fransson, Front. Biosci. 13:1619-1633 (2008), и дополнительно описаны, например, в Riechmann et al., Nature 332:323-329 (1988); Queen et al., Proc. Nat'l Acad. Sci. USA 86:10029-10033 (1989); патентах США Ms 5821337, 7527791, 6982321 и 7087409; Kashmiri et al., Methods 36:25-34 (2005) (где описано прививание определяющей специфичность области (SDR)); Padlan, Mol. Immunol. 28:489-498 (1991) (где описано «изменение поверхности»); Dall'Acqua et al., Methods 36:43-60 (2005) (где описана «перетасовка FR»); и Osbourn et al., Methods 36:61-68 (2005) и Klimka et al., Br. J. Cancer, 83:252-260 (2000) (где описан подход «направленного отбора» при перетасовке FR).
Человеческие каркасные области, которые можно использовать для гуманизации, включают, но не ограничиваются этим, каркасные области, выбранные методом «наилучшего соответствия» (смотрите, например, Sims et al. J. Immunol. 151:2296 (1993)); каркасные области, полученные из консенсусной последовательности человеческих антител конкретной подгруппы вариабельных областей легких или тяжелых цепей (смотрите, например, Carter et al. Proc. Natl. Acad. Sci. USA, 89:4285 (1992); и Presta et al. J. Immunol, 151:2623 (1993)); человеческие зрелые (соматически мутированные) каркасные области или каркасные области зародышевой линии человека (смотрите, например, Almagro and Fransson, Front. Biosci. 13:1619-1633 (2008)); и каркасные области, полученные при скрининге библиотек FR (смотрите, например, Baca et al., J. Biol. Chem. 272:10678-10684 (1997) и Rosok et al., J. Biol. Chem. 271:22611-22618 (1996)).
3. Варианты no гликозилированию
В определенных аспектах предложенное в данном документе антитело изменяют с целью повышения или снижения степени гликозилирования антитела. Добавление или удаление сайтов гликозилирования к антителу удобно осуществлять путем изменения аминокислотной последовательности так, чтобы создать или удалить один или более сайтов гликозилирования.
Если антитело содержит Fc-область, можно изменять присоединенный к ней олигосахарид. Нативные антитела, вырабатываемые клетками млекопитающих, обычно содержат разветвленный биантеннарный олигосахарид, который обычно присоединен посредством N-связи к Asn297 домена СН2 Fc-области. Смотрите, например, Wright et al. TIBTECH 15:26-32 (1997). Олигосахарид может включать различные углеводы, например, маннозу, N-ацетилглюкозамин (GlcNAc), галактозу и сиаловую кислоту, а также фукозу, присоединенную к GlcNAc в «стволе» биантеннарной олигосахаридной структуры. В некоторых аспектах модификации олигосахарида в антителе по изобретению можно осуществлять с целью создания вариантов антитела с определенными улучшенными свойствами.
В одном аспекте предложены варианты антител, содержащие нефукозилированный олигосахарид, т.е. олигосахаридную структуру, в которой отсутствует фукоза, присоединенная (прямо или непрямо) к Fc-области. В частности, такой нефукозилированный олигосахарид (также называемый «афукозилированным» олигосахаридом) представляет собой N-связанный олигосахарид, в котором отсутствует остаток фукозы, присоединенный к первому GlcNAc в стволе биантеннарной олигосахаридной структуры. В одном аспекте предложены варианты антител, имеющие повышенную долю нефукозилированных олигосахаридов в Fc-области по сравнению с нативным или родительским антителом. Например, доля нефукозилированных олигосахаридов может составлять по меньшей мере около 20%, по меньшей мере около 40%, по меньшей мере около 60%, по меньшей мере около 80% или даже около 100% (т.е. фукозилированные олигосахариды отсутствуют). Процентное содержание нефукозилированных олигосахаридов представляет собой (среднее) количество олигосахаридов, в которых отсутствуют остатки фукозы, по отношению к сумме всех олигосахаридов, присоединенных к Asn 297 (например, сложных, гибридных структур и структур с высоким содержанием маннозы), по результатам измерения методом масс-спектрометрии ВП-МАЛДИ, например, как описано в WO 2006/082515. Asn297 относится к остатку аспарагина, расположенному в позиции 297 в Fc-области (согласно нумерации ЕС остатков Fc-области); при этом Asn297 также может быть расположен на около ±3 аминокислот выше или ниже позиции 297, т.е. между позициями 294 и 300, вследствие незначительных вариаций последовательностей в антителах. Такие антитела, имеющие повышенное процентное содержание нефукозилированных олигосахаридов в Fc-области, могут демонстрировать улучшенное связывание FcγRIIIa-рецептора и/или улучшенную эффекторную функцию, в частности улучшенную функцию АЗКЦ. Смотрите, например, US 2003/0157108; US 2004/0093621.
Примеры линий клеток, способных вырабатывать антитела со сниженным фукозилированием, включают клетки Lecl3 СНО с дефицитом фукозилирования белка (Ripka et al. Arch. Biochem. Biophys. 249:533-545 (1986); US 2003/0157108; и WO 2004/056312, в особенности в примере 11), и нокаутные линии клеток, такие как клетки СНО с нокаутом гена альфа-1,6-фукозилтрансферазы, FUT8 (смотрите, например, Yamane-Ohnuki et al. Biotech. Bioeng. 87:614-622 (2004); Kanda, Y. et al., Biotechnol. Bioeng., 94(4):680-688 (2006); и WO 2003/085107), или клетки со сниженной или устраненной активностью синтеза ГДФ-фукозы или белка-переносчика (смотрите, например, US2004259150, US2005031613, US2004132140, US2004110282).
В дополнительном аспекте предложены варианты антител с раздвоенными олигосахаридами, например, в которых биантеннарный олигосахарид, присоединенный к Fc-области антитела, разделен пополам GlcNAc. Такие варианты антител могут иметь сниженный уровень фукозилирования и/или улучшенную функцию АЗКЦ, как описано выше. Примеры таких вариантов антител описаны, например, в Umana et al., Nat Biotechnol 17, 176-180 (1999); Ferrara et al., Biotechn Bioeng 93, 851-861 (2006); WO 99/54342; WO 2004/065540, WO 2003/011878.
Также предложены варианты антител с по меньшей мере одним остатком галактозы в олигосахариде, присоединенном к Fc-области. Такие варианты антител могут иметь улучшенную функцию КЗЦ. Такие варианты антител описаны, например, в WO 1997/30087; WO 1998/58964; и WO 1999/22764.
4. Цистеин-сконструированные варианты антител
В определенных аспектах может существовать необходимость в создании антител, сконструированных с использованием цистеина, например, антител ТНIOМАВ™, в которых один или более остатков замещены остатками цистеина. В предпочтительных аспектах замещенные остатки находятся в доступных сайтах антитела. За счет замещения этих остатков цистеином реакционноспособные тиольные группы располагаются в доступных сайтах антитела и могут использоваться для конъюгации антитела с другими фрагментами, такими как лекарственные фрагменты или фрагменты линкер лекарственное средство, для создания иммуноконъюгата, как дополнительно описано в данном документе. Антитела, сконструированные с использованием цистеина, можно создавать, как описано, например, в патентах США №№7521541, 830930, 7855275, 9000130 или WO 2016040856.
5. Производные антител
В определенных аспектах предложенное в данном документе антитело можно дополнительно модифицировать так, чтобы оно содержало дополнительные небелковые фрагменты, которые известны в данной области техники и легко доступны. Фрагменты, подходящие для дериватизации антитела, включают, но не ограничиваются этим, водорастворимые полимеры. Неограничивающие примеры водорастворимых полимеров включают, без ограничений, полиэтиленгликоль (ПЭГ), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпирролидон, поли-1, 3-диоксолан, поли-1,3,6-триоксан, сополимер этилена/малеинового ангидрида, полиаминокислоты (гомополимеры или статистические сополимеры), декстран или поли(п-винилпирролидон)полиэтиленгликоль, гомополимеры пропропиленгликоля, сополимеры оксида/этиленоксида, полиоксиэтилированные полиолы (например, глицерин), поливиниловый спирт и их смеси. Пропиональдегид полиэтиленгликоля может иметь преимущества при производстве благодаря своей стабильности в воде. Полимер может иметь любую молекулярную массу и может быть разветвленным или неразветвленным. Количество молекул полимера, присоединенных к антителу, может варьироваться и, в случае присоединении более чем одного полимера, они могут представлять собой одинаковые или разные молекулы. В целом, количество и/или тип полимеров, применяемых для дериватизации, можно определить на основании соображений, которые включают, но не ограничиваются этим, конкретные свойства или функции антитела, которые нужно улучшить, то, будет ли производное антитела применяться в терапии в определенных условиях и т.д.
6. Иммуноконъюгаты
В изобретении также предложены иммуноконъюгаты, содержащие анти-CD3 антитело по данному документу, конъюгированное (химически связанное) с одним или более терапевтическими агентами, такими как цитотоксические агенты, химиотерапевтические агенты, лекарственные средства, ингибиторы роста, токсины (например, белковые токсины, ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения или их фрагменты) или радиоактивные изотопы.
В одном аспекте иммуноконъюгат представляет собой конъюгат антитело - лекарственное средство (ADC), в котором антитело конъюгировано с одним или более терапевтическими агентами, упомянутыми выше. Как правило, антитело соединено с одним или более терапевтическими агентами с помощью линкеров. Обзор технологии ADC, включая примеры терапевтических агентов, лекарственных средств и линкеров, приведен в Pharmacol Review 68:3-19 (2016).
В другом аспекте иммуноконъюгат содержит антитело по изобретению, конъюгированное с ферментативно активным токсином или его фрагментом, включая, но не ограничиваясь этим, цепь А дифтерийного токсина, несвязывающиеся активные фрагменты дифтерийного токсина, цепь А экзотоксина (из Pseudomonas aeruginosa), цепь А рицина, цепь А абрина, цепь А модецина, альфа-сарцин, белки Aleurites fordii, диантиновые белки, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор из момордики харантской, курцин, кротин, ингибитор из Saponaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены.
В другом аспекте иммуноконъюгат содержит антитело по изобретению, конъюгированное с радиоактивным атомом с образованием радиоконъюгата. Для получения радиоконъюгатов доступен ряд радиоактивных изотопов. Примеры включают At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu. Если радиоконъюгат используют в целях обнаружения, он может содержать радиоактивный атом для сцинтиграфических исследований, например, Tc99m или I123, или спиновую метку для ядерного магнитного резонанса (ЯМР) (также известного как магнито-резонансная томография, МРТ), такую как I, I131, In111, F19, С13, N15, О17, гадолиний, марганец или железо.
Конъюгаты антитела и цитотоксического агента можно создавать, используя ряд бифункциональных агентов, связывающих белок, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные сложных имидоэфиров (такие как диметиладипимидат HCl), активные сложные эфиры (такие, как дисукцинимидилсуберат), альдегиды (такие, как глутаральдегид), бис-азидо соединения (такие, как бис(п-азидобензоил)-гександиамин), производные бис-диазония (такие, как бис(п-диазоний бензоил)-этилендиамин), диизоцианаты (такие, как толуол 2,6-диизоцианат) и бис-активные соединения фтора (такие, как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин на основе рицина можно получать, как описано в Vitetta et al., Science 238:1098 (1987). Меченная углеродом-14 1-изотиоцианатобензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) представляет собой типичный хелатирующий агент для конъюгации радионуклида с антителом. Смотрите WO 94/11026. Линкер может представлять собой «расщепляемый линкер», облегчающий высвобождение цитотоксического лекарственного средства в клетке. Например, можно использовать кислотно-лабильный линкер, чувствительный к пептидазе линкер, фотолабильный линкер, диметиловый линкер или дисульфид-содержащий линкер (Chari et al., Cancer Res. 52:127-131 (1992); патент США №5208020).
В контексте данного документа под иммуноконъюгатами или ADC прямо подразумевают, без ограничения конъюгатами, полученными с помощью перекрестно-сшивающих реагентов, помимо прочего, BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, МРВН, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB, а также SVSB (сукцинимидил-(4-винилсульфон)бензоат), которые являются коммерчески доступными (например, от Pierce Biotechnology, Inc., Rockford, IL., U.S.A).
7. Мультиспецифические антитела
В определенных аспектах предложенное в данном документе антитело представляет собой мультиспецифическое антитело, в частности биспецифическое антитело. Мультиспецифические антитела представляют собой моноклональные антитела, имеющие специфичность связывания в отношении по меньшей мере двух антигенных детерминант (например, двух разных белков или двух разных эпитопов на одном белке). В определенных аспектах мультиспецифическое антитело имеет три или более специфичности связывания. В определенных аспектах одна специфичность связывания направлена на CD3, а другая - на любой другой антиген. В определенных аспектах мультиспецифические антитела могут связываться с двумя (или более) разными эпитопами CD3. Мультиспецифические (например, биспецифические) антитела также можно использовать для локализации цитотоксических агентов или клеток в клетках, которые экспрессируют CD3. Мультиспецифические антитела можно получать в виде полноразмерных антител или фрагментов антител.
Методики для создания мультиспецифических антител включают, но не ограничиваются этим, рекомбинантную коэкспрессию двух пар тяжелая цепь легкая цепь иммуноглобулина, имеющих разную специфичность (смотрите Milstein and Cuello, Nature 305: 537 (1983)), и конструирование по типу «выступ-во-впадину» (смотрите, например, патент США №5731168 и Atwell et al., J. Mol. Biol. 270:26 (1997)). Мультиспецифические антитела также можно создавать путем конструирования с использованием эффекта электростатического взаимодействия для создания Fc-гетеродимерных молекул антител (смотрите, например, WO 2009/089004); перекрестного сшивания двух или более антител или фрагментов (смотрите, например, патент США №4676980 и Brennan et al., Science, 229: 81 (1985)); применения лейциновых молний для получения биспецифических антител (смотрите, например, Kostelny et al., J. Immunol., 148(5): 1547-1553 (1992) и WO 2011/034605); применения технологии общей легкой цепи для преодоления проблемы неправильного спаривания легкой цепи (смотрите, например, WO 98/50431); применения технологии «диател» для получения биспецифических фрагментов антител (смотрите, например, Hollinger et al., Proc. Natl. Acad. Sci. USA, 90:6444-6448 (1993)); и применения одноцепочечных димеров Fv (sFv) (смотрите, например, Gruber et al., J. Immunol., 152:5368 (1994)); и получения триспецифических антител, как описано, например, в Tutt et al. J. Immunol. 147: 60 (1991).
Сконструированные антитела с тремя или более антигенсвязывающими сайтами, включая, например, «антитела-осьминоги» или DVD-Ig, также включены в данный документ (смотрите, например, WO 2001/77342 и WO 2008/024715). Другие примеры мультиспецифических антител с тремя или более антигенсвязывающими сайтами можно найти в WO 2010/115589, WO 2010/112193, WO 2010/136172, WO 2010/145792 и WO 2013/026831. Мультиспецифические антитела или их антигенсвязывающие фрагменты также включают «FAb двойного действия» или «DAF», содержащие антигенсвязывающий сайт, который связывается с CD3, а также другим отличным антигеном, или с двумя разными эпитопами CD3 (смотрите, например, US 2008/0069820 и WO 2015/095539).
Мультиспецифические антитела могут быть предоставлены в асимметрической форме с кроссовером доменов в одном или более связывающих плечах с одинаковой антигенной специфичностью (так называемая технология «CrossMab»), т.е. посредством обмена доменов VH/VL (смотрите, например, WO 2009/080252 и WO 2015/150447), доменов CH1/CL (смотрите, например, WO 2009/080253) или полных плеч Fab (смотрите, например, WO 2009/080251, WO 2016/016299, также смотрите Schaefer et al, PNAS, 108 (2011) 1187-1191 и Klein at al., MAbs 8 (2016) 1010-20). Асимметрические плечи Fab также можно сконструировать путем внесения заряженных или не заряженных аминокислотных мутаций в поверхности контакта доменов для обеспечения правильного спаривания Fab. Смотрите, например, WO 2016/172485.
Различные другие молекулярные форматы для мультиспецифических антител известны в данной области техники и включены в данный документ (смотрите, например, Spiess et al., Mol Immunol 67 (2015) 95-106).
Конкретным типом мультиспецифических антител, также включенным в данный документ, являются биспецифические антитела, предназначенные для одновременного связывания с поверхностным антигеном на клетке-мишени, например, опухолевой клетке, и с активирующим инвариантным компонентом комплекса Т-клеточного рецептора (ТКР), таким как CD3, для перенацеливания Т-клеток на уничтожение клеток-мишеней. Следовательно, в предпочтительных аспектах предложенное в данном документе антитело представляет собой мультиспецифическое антитело, в частности биспецифическое антитело, причем одна из специфичностей связывания направлена на CD3, а другая - на антиген клетки-мишени.
Примеры форматов биспецифических антител, которые можно применять в этих целях, включают, но не ограничиваются этим, так называемые молекулы «BiTE» (привлекающий Т-клетки биспецифический активатор), в которых две молекулы scFv сшиты гибким линкером (смотрите, например, WO 2004/106381, WO 2005/061547, WO 2007/042261 и WO 2008/119567, Nagorsen and Bauerle, Exp Cell Res 317, 1255-1260 (2011)); диатела (Holliger et al., Prot Eng 9, 299-305 (1996)) и их производные, такие как тандемные диатела («TandAb»; Kipriyanov et al., J Mol Biol 293, 41-56 (1999)); молекулы «DART» (перенацеливание с двойной аффинностью), которые основаны на формате диатела, но имеют С-концевой дисульфидный мостик для дополнительной стабилизации (Johnson et al., J Mol Biol 399, 436-449 (2010)), и так называемые triomab, которые представляют собой полностью гибридные молекулы IgG мыши/крысы (обзор в Seimetz et al., Cancer Treat Rev 36, 458-467 (2010)). Конкретные включенные в данный документ форматы Т-клеточных биспецифических антител описаны в WO 2013/026833, WO 2013/026839, WO 2016/020309; Bacac et al., Oncoimmunology 5(8) (2016) е1203498.
Предпочтительные аспекты мультиспецифических антител по настоящему изобретению описаны ниже.
В одном аспекте в изобретении предложено антитело, которое связывается с CD3, содержащее первый антигенсвязывающий домен, который связывается с CD3, как описано в данном документе, и содержащее второй и, необязательно, третий антигенсвязывающий домен, который связывается со вторым антигеном.
В соответствии с предпочтительными аспектами изобретения антигенсвязывающие домены, содержащиеся в антитела, представляют собой молекулы Fab (т.е. антигенсвязывающие домены, состоящие из тяжелой и легкой цепи, каждая из которых содержит вариабельный и константный домен). В одном аспекте первый, второй и/или, в случае наличия, третий антигенсвязывающий домен представляет собой молекулу Fab. В одном аспекте указанная молекула Fab является человеческой. В предпочтительном аспекте указанная молекула Fab является гуманизированной. В другом аспекте указанная молекула Fab содержит константные домены тяжелой и легкой цепи человека.
Предпочтительно по меньшей мере один из антигенсвязывающих доменов представляет собой кроссоверную молекулу Fab. Такая модификация снижает неправильное спаривание тяжелых и легких цепей из разных молекул Fab, тем самым улучшая выход и чистоту (мультиспецифического) антитела по изобретению при рекомбинантном получении. В предпочтительной кроссоверной молекуле Fab, применимой для (мультиспецифического) антитела по изобретению, обменены вариабельные домены легкой цепи Fab и тяжелой цепи Fab (VL и VH, соответственно). Однако даже при таком обмене доменов препарат (мультиспецифических) антител может содержать некоторые побочные продукты вследствие так называемого взаимодействия Бене-Джонса между неправильно спаренными тяжелой и легкой цепями (смотрите Schaefer et al, PNAS, 108 (2011) 11187-11191). Чтобы дополнительно снизить неправильное спаривание тяжелых и легких цепей из разных молекул Fab и, таким образом, повысить чистоту и выход необходимого (мультиспецифического) антитела, заряженные аминокислоты с противоположными зарядами можно вносить в определенных аминокислотных позициях в доменах СН1 и CL молекулы Fab, связывающейся с первым антигеном (CD3), или молекулы (молекул) Fab, связывающейся(ихся) со вторым антигеном (например, антигеном клетки-мишени, таким как TYRP-1 или EGFRvIII), как дополнительно описано в данном документе. Зарядовые модификации осуществляют в стандартных молекулах Fab, содержащихся в (мультиспецифическом) антителе (как показано, например, на Фиг. 1 А-В, Ж-К), или в VH/VL кроссоверных молекулах Fab, содержащихся в (мультиспецифическом) антителе (как показано, например, на Фиг. 1 Г-Е, Л-О) (но не в обеих). В предпочтительных аспектах зарядовые модификации осуществляют в стандартных молекулах Fab, содержащихся в (мультиспецифическом) антителе (которое в предпочтительных аспектах связывается со вторым антигеном, например, антигеном клетки-мишени, таким как TYRP-1 или EGFRvIII).
В предпочтительном аспекте в соответствии с изобретением (мультиспецифическое) антитело способно одновременно связываться с первым антигеном (т.е. CD3) и вторым антигеном (например, антигеном-мишенью, таким как TYRP-1 или EGFRvIII). В одном аспекте (мультиспецифическое) антитело способно перекрестно связывать Т-клетку и клетку-мишень посредством одновременного связывания с CD3 и антигеном клетки-мишени. В даже более предпочтительном аспекте такое одновременное связывание приводит к лизису клетки-мишени, в частности, экспрессирующей антиген клетки-мишени (например, TYRP-1 или EGFRvIII) опухолевой клетки. В одном аспекте такое одновременное связывание приводит к активации Т-клеток. В других аспектах такое одновременное связывание приводит к клеточному ответу Т-лимфоцита, в частности, цитотоксического Т-лимфоцита, выбранному из группы из пролиферации, дифференцировки, секреции цитокинов, высвобождения цитотоксических эффекторных молекул, цитотоксической активности и экспрессии маркеров активации. В одном аспекте связывание (мультиспецифического) антитела с CD3 без одновременного связывания с антигеном клетки-мишени не приводит к активации Т-клеток.
В одном аспекте (мультиспецифическое) антитело способно перенаправлять цитотоксическую активность Т-клетки на клетку-мишень. В предпочтительном аспекте указанное перенаправление не зависит от опосредованной ГКГС презентации пептидного антигена клеткой-мишенью и/или специфичности Т-клетки.
Предпочтительно Т-клетка в соответствии с любыми аспектами изобретения представляет собой цитотоксическую Т-клетку. В некоторых аспектах Т-клетка представляет собой CD4+ или CD8+ Т-клетку, в частности CD8+ Т-клетку.
а) Первый антигенсвязывающий домен
(Мультиспецифическое) антитело по изобретению содержит по меньшей мере один антигенсвязывающий домен (первый антигенсвязывающий домен), который связывается с CD3. В предпочтительных аспектах CD3 представляет собой CD3 человека (SEQ ID NO: 112) или CD3 яванского макака (SEQ ID NO: 113), в частности CD3 человека. В одном аспекте первый антигенсвязывающий домен является перекрестно-реакционноспособным в отношении CD3 человека и яванского макака (т.е. специфически связывается с ним). В некоторых аспектах CD3 представляет собой эпсилон-субъединицу CD3 (CD3 эпсилон).
В предпочтительном аспекте (мультиспецифическое) антитело содержит не более одного антигенсвязывающего домена, который связывается с CD3. В одном аспекте (мультиспецифическое) антитело обеспечивает одновалентное связывание с CD3.
В одном аспекте антигенсвязывающий домен, который связывается с CD3, представляет собой фрагмент антитела, выбранный из группы из молекулы Fv, молекулы scFv, молекулы Fab и молекулы F(ab')2. В предпочтительном аспекте антигенсвязывающий домен, который связывается с CD3, представляет собой молекулу Fab.
В предпочтительных аспектах антигенсвязывающий домен, который связывается с CD3, представляет собой кроссоверную молекулу Fab, описанную в данном документе, т.е. молекулу Fab, в которой вариабельные домены VH и VL или константные домены СН1 и CL тяжелой и легкой цепей Fab заменены/замещены друг другом. В таких аспектах антигенсвязывающий домен, который связывается со вторым антигеном (например, антигеном клетки-мишени, таким как TYRP-1 или EGFRvIII), предпочтительно представляет собой стандартную молекулу Fab. В аспектах, в которых присутствует более одного антигенсвязывающего домена, в частности молекулы Fab, которая связывается со вторым антигеном, содержащихся в (мультиспецифическом) антителе, антигенсвязывающий домен, который связывается с CD3, предпочтительно представляет собой кроссоверную молекулу Fab, а антигенсвязывающие домены, которые связываются со вторым антигеном, представляют собой стандартные молекулы Fab.
В альтернативных аспектах антигенсвязывающий домен, который связывается с CD3, представляет собой стандартную молекулу Fab. В таких аспектах антигенсвязывающий домен, который связывается со вторым антигеном (например, антигеном клетки-мишени, таким как TYRP-1 или EGFRvIII), представляет собой кроссоверную молекулу Fab, описанную в данном документе, т.е. молекулу Fab, в которой вариабельные домены VH и VL или константные домены СН1 и CL тяжелой и легкой цепей Fab заменены/замещены друг другом. В аспектах, в которых присутствует более одного антигенсвязывающего домена, в частности молекулы Fab, которая связывается с CD3, содержащихся в (мультиспецифическом) антителе, антигенсвязывающий домен, который связывается со вторым антигеном, предпочтительно представляет собой кроссоверную молекулу Fab, а антигенсвязывающие домены, которые связываются с CD3, представляют собой стандартные молекулы Fab.
В предпочтительных аспектах первый антигенсвязывающий домен представляет собой молекулу Fab, причем вариабельные домены VL и VH или константные домены CL и СН1, в частности вариабельные домены VL и VH легкой цепи Fab и тяжелой цепи Fab замещены друг другом (т.е. в соответствии с таким аспектом первый антигенсвязывающий домен представляет собой кроссоверную молекулу Fab, причем вариабельные или константные домены легкой цепи Fab и тяжелой цепи Fab обменены). В одном таком аспекте второй (и, в случае наличия, третий) антигенсвязывающий домен представляет собой стандартную молекулу Fab.
В одном аспекте в (мультиспецифическом) антителе присутствует не более одного антигенсвязывающего домена, который связывается с CD3 (т.е. антитело обеспечивает одновалентное связывание с CD3).
б) Второй (и третий) антигенсвязывающий домен
В определенных аспектах (мультиспецифическое) антитело по изобретению содержит по меньшей мере один антигенсвязывающий домен, в частности молекулу Fab, который связывается со вторым антигеном. Второй антиген предпочтительно не является CD3, т.е. отличается от CD3. В одном аспекте второй антиген представляет собой антиген, экспрессируемый на другой клетке, чем CD3 (т.е. экспрессируемый на клетке, отличной от Т-клетки). В одном аспекте второй антиген представляет собой антиген клетки-мишени, в частности антиген опухолевой клетки. В конкретном аспекте второй антиген представляет собой TYRP-1. В другом конкретном аспекте второй антиген представляет собой EGFRvIII. Второй антигенсвязывающий домен способен направлять (мультиспецифическое) антитело к сайту-мишени, например, к конкретному типу опухолевой клетки, которая экспрессирует второй антиген.
В одном аспекте антигенсвязывающий домен, который связывается со вторым антигеном, представляет собой фрагмент антитела, выбранный из группы из молекулы Fv, молекулы scFv, молекулы Fab и молекулы F(ab')2. В предпочтительном аспекте антигенсвязывающий домен, который связывается со вторым антигеном, представляет собой молекулу Fab.
В определенных аспектах (мультиспецифическое) антитело содержит два антигенсвязывающих домена, в частности две молекулы Fab, которые связываются со вторым антигеном. В таком предпочтительном аспекте каждый из этих антигенсвязывающих доменов связывается с одной и той же антигенной детерминантой. В даже более предпочтительном аспекте все из этих антигенсвязывающих доменов являются идентичными, т.е. они имеют одинаковый молекулярный формат (например, стандартной или кроссоверной молекулы Fab) и содержат одинаковые аминокислотные последовательности, содержащие одинаковые аминокислотные замены в доменах СН1 и CL, как описано в данном документе (в случае наличия). В одном аспекте (мультиспецифическое) антитело содержит не более двух антигенсвязывающих доменов, в частности двух молекул Fab, которые связываются со вторым антигеном.
В предпочтительных аспектах антигенсвязывающий(ие) домен(ы), который(ые) связывается(ются) со вторым антигеном, представляет(ют) собой стандартную молекулу Fab. В таких аспектах антигенсвязывающий домен (домены), который связывается с CD3, представляет собой кроссоверную молекулу Fab, описанную в данном документе, т.е. молекулу Fab, в которой вариабельные домены VH и VL или константные домены СН1 и CL тяжелой и легкой цепей Fab заменены/замещены друг другом.
В альтернативных аспектах антигенсвязывающий домен (домены), который связывается со вторым антигеном, представляет собой кроссоверную молекулу Fab, описанную в данном документе, т.е. молекулу Fab, в которой вариабельные домены VH и VL или константные домены СН1 и CL тяжелой и легкой цепей Fab заменены/замещены друг другом. В таких аспектах антигенсвязывающий домен (домены), который связывается с CD3, представляет собой стандартную молекулу Fab.
В одном аспекте второй (и, в случае наличия, третий) антигенсвязывающий домен содержит константную область человека. В одном аспекте второй (и, в случае наличия, третий) антигенсвязывающий домен представляет собой молекулу Fab, содержащую человеческую константную область, в частности, человеческий домен СН1 и/или CL. типовые последовательности человеческих константных доменов приведены в SEQ ID NO: 120 и 121 (человеческие каппа и лямбда CL-домены, соответственно) и SEQ ID NO: 122 (константные домены СН1-СН2-CH3 тяжелой цепи человеческого IgG1). В одном аспекте второй (и, в случае наличия, третий) антигенсвязывающий домен содержит константную область легкой цепи, содержащую аминокислотную последовательность, которая является по меньшей мере на около 95%, 96%, 97%, 98%, 99% или 100% идентичной аминокислотной последовательности SEQ ID NO: 120 или SEQ ID NO: 121, в частности аминокислотной последовательности SEQ ID NO: 120. В частности, константная область легкой цепи может содержать аминокислотные мутации, описанные в данном документе как «зарядовые модификации», и/или может содержать делецию или замены одной или более (в частности двух) N-концевых аминокислот, если она принадлежит кроссоверной молекуле Fab. В некоторых аспектах второй (и, в случае наличия, третий) антигенсвязывающий домен содержит константную область тяжелой цепи, содержащую аминокислотную последовательность, которая является по меньшей мере на около 95%, 96%, 97%, 98%, 99% или 100% идентичной последовательности домена СН1, содержащейся в аминокислотной последовательности SEQ ID NO: 122. В частности, константная область тяжелой цепи (в частности домен СН1) может содержать аминокислотные мутации, описанные в данном документе как «зарядовые модификации».
TYRP-1
В предпочтительных аспектах второй антиген представляет собой TYRP-1, в частности TYRP-1 человека (SEQ ID NO: 114).
В одном аспекте второй (и, в случае наличия, третий) антигенсвязывающий домен содержит вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область тяжелой цепи (HCDR) 1 с SEQ ID NO: 15, HCDR 2 с SEQ ID NO: 16 и HCDR 3 с SEQ ID NO: 17, и вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область легкой цепи (LCDR) 1 с SEQ ID NO: 19, LCDR 2 с SEQ ID NO: 20 и LCDR 3 с SEQ ID NO: 21.
В одном аспекте второй (и, в случае наличия, третий) антигенсвязывающий домен представляет собой гуманизированное антитело (или получен из него). В одном аспекте второй (и, в случае наличия, третий) антигенсвязывающий домен представляет собой гуманизированный антигенсвязывающий домен (т.е. антигенсвязывающий домен гуманизированного антитела). В одном аспекте VH и/или VL второго (и, в случае наличия, третьего) антигенсвязывающего домена представляет собой гуманизированную вариабельную область.
В одном аспекте VH и/или VL второго (и, в случае наличия, третьего) антигенсвязывающего домена содержит акцепторную человеческую каркасную область, например, каркасную область человеческого иммуноглобулина или человеческую консенсусную каркасную область.
В одном аспекте VH второго (и, в случае наличия, третьего) антигенсвязывающего домена содержит одну или более каркасных последовательностей тяжелой цепи (т.е. последовательностей FR1, FR2, FR3 и/или FR4) с SEQ ID NO: 18. В одном аспекте VH содержит последовательность, которая является по меньшей мере на 95%, 96%, 97%, 98% или 99% идентичной аминокислотной последовательности SEQ ID NO: 18. В одном аспекте VH содержит последовательность, которая является по меньшей мере на около 95% идентичной аминокислотной последовательности SEQ ID NO: 18. В одном аспекте VH содержит последовательность, которая является по меньшей мере на около 98% идентичной аминокислотной последовательности SEQ ID NO: 18. В определенных аспектах последовательность VH, имеющая по меньшей мере 95%, 96%, 97%, 98% или 99% идентичности, содержит замены (например, консервативные замены), вставки или делеции по сравнению с референсной последовательностью, но антитело, содержащее эту последовательность, сохраняет способность связываться с TYRP-1. В определенных аспектах была проведена замена, вставка и/или делеция в общей сложности 1-10 аминокислот в аминокислотной последовательности SEQ ID NO: 18. В определенных аспектах замены, вставки или делеции находятся в областях за пределами CDR (т.е. в FR). В одном аспекте VH содержит аминокислотную последовательность SEQ ID NO: 18. Необязательно, VH содержит аминокислотную последовательность SEQ ID NO: 18, включая посттрансляционные модификации этой последовательности.
В одном аспекте VL второго (и, в случае наличия, третьего) антигенсвязывающего домена содержит одну или более каркасных последовательностей легкой цепи (т.е. последовательностей FR1, FR2, FR3 и/или FR4) с SEQ ID NO: 22. В одном аспекте VL содержит последовательность, которая является по меньшей мере на 95%, 96%, 97%, 98% или 99% идентичной аминокислотной последовательности SEQ ID NO: 22. В одном аспекте VL содержит последовательность, которая является по меньшей мере на около 95% идентичной аминокислотной последовательности SEQ ID NO: 22. В одном аспекте VL содержит последовательность, которая является по меньшей мере на около 98% идентичной аминокислотной последовательности SEQ ID NO: 22. В определенных аспектах последовательность VL, имеющая по меньшей мере 95%, 96%, 97%, 98% или 99% идентичности, содержит замены (например, консервативные замены), вставки или делеции по сравнению с референсной последовательностью, но антитело, содержащее эту последовательность, сохраняет способность связываться с TYRP-1. В определенных аспектах была проведена замена, вставка и/или делеция в общей сложности 1-10 аминокислот в аминокислотной последовательности SEQ ID NO: 22. В определенных аспектах замены, вставки или делеции находятся в областях за пределами CDR (т.е. в FR). В одном аспекте VL содержит аминокислотную последовательность SEQ ID NO: 22. Необязательно, VL содержит аминокислотную последовательность SEQ ID NO: 22, включая посттрансляционные модификации этой последовательности.
В одном аспекте VH второго (и, в случае наличия, третьего) антигенсвязывающего домена содержит аминокислотную последовательность, которая по меньшей мере на около 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 18, и VL второго (и, в случае наличия, третьего) антигенсвязывающего домена содержит аминокислотную последовательность, которая по меньшей мере на около 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 22. В одном аспекте VH содержит аминокислотную последовательность SEQ ID NO: 18, a VL содержит аминокислотную последовательность SEQ ID NO: 22.
В дополнительном аспекте второй (и, в случае наличия, третий) антигенсвязывающий домен содержит VH, содержащую последовательность SEQ ID NO: 18, и VL, содержащую последовательность SEQ ID NO: 22.
В дополнительном аспекте второй (и, в случае наличия, третий) антигенсвязывающий домен содержит последовательность VH SEQ ID NO: 18 и последовательность VL SEQ ID NO: 22.
В другом аспекте второй (и, в случае наличия, третий) антигенсвязывающий домен содержит VH, содержащую последовательности CDR тяжелой цепи VH с SEQ ID NO: 18, и VL, содержащую последовательности CDR легкой цепи VL с SEQ ID NO: 22.
В дополнительном аспекте второй (и, в случае наличия, третий) антигенсвязывающий домен содержит аминокислотные последовательности HCDR1, HCDR2 и HCDR3 VH с SEQ ID NO: 18 и аминокислотные последовательности LCDR1, LCDR2 и LCDR3 VL с SEQ ID NO: 22.
В одном аспекте VH второго (и, в случае наличия, третьего) антигенсвязывающего домена содержит последовательности CDR тяжелой цепи VH с SEQ ID NO: 18 и каркасную область с по меньшей мере 95%, 96%, 97%, 98% или 99% идентичности последовательности с каркасной последовательностью VH с SEQ ID NO: 18. В одном аспекте VH содержит последовательности CDR тяжелой цепи VH с SEQ ID NO: 18 и каркасную область с по меньшей мере 95% идентичности последовательности с каркасной последовательностью VH с SEQ ID NO: 18. В другом аспекте VH содержит последовательности CDR тяжелой цепи VH с SEQ ID NO: 18 и каркасную область с по меньшей мере 98% идентичности последовательности с каркасной последовательностью VH с SEQ ID NO: 18.
В одном аспекте VL второго (и, в случае наличия, третьего) антигенсвязывающего домена содержит последовательности CDR легкой цепи VL с SEQ ID NO: 22 и каркасную область с по меньшей мере 95%, 96%, 97%, 98% или 99% идентичности последовательности с каркасной последовательностью VL с SEQ ID NO: 22. В одном аспекте VL содержит последовательности CDR легкой цепи VL с SEQ ID NO: 22 и каркасную область с по меньшей мере 95% идентичности последовательности с каркасной последовательностью VL с SEQ ID NO: 22. В другом аспекте VL содержит последовательности CDR легкой цепи VL с SEQ ID NO: 22 и каркасную область с по меньшей мере 98% идентичности последовательности с каркасной последовательностью VL с SEQ ID NO: 22.
EGFRvIII
В предпочтительных аспектах второй антиген представляет собой EGFRvIII, в частности EGFRvIII человека (SEQ ID NO: 115).
В одном аспекте второй (и, в случае наличия, третий) антигенсвязывающий домен содержит вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область тяжелой цепи (HCDR) 1 с SEQ ID NO: 85, HCDR 2 с SEQ ID NO: 86 и HCDR 3 с SEQ ID NO: 87, и вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область легкой цепи (LCDR) 1 с SEQ ID NO: 89, LCDR 2 с SEQ ID NO: 90 и LCDR 3 с SEQ ID NO: 91.
В одном аспекте второй (и, в случае наличия, третий) антигенсвязывающий домен представляет собой гуманизированное антитело (или получен из него). В одном аспекте второй (и, в случае наличия, третий) антигенсвязывающий домен представляет собой гуманизированный антигенсвязывающий домен (т.е. антигенсвязывающий домен гуманизированного антитела). В одном аспекте VH и/или VL второго (и, в случае наличия, третьего) антигенсвязывающего домена представляет собой гуманизированную вариабельную область.
В одном аспекте VH и/или VL второго (и, в случае наличия, третьего) антигенсвязывающего домена содержит акцепторную человеческую каркасную область, например, каркасную область человеческого иммуноглобулина или человеческую консенсусную каркасную область.
В одном аспекте VH второго (и, в случае наличия, третьего) антигенсвязывающего домена содержит одну или более каркасных последовательностей тяжелой цепи (т.е. последовательностей FR1, FR2, FR3 и/или FR4) с SEQ ID NO: 88. В одном аспекте VH содержит последовательность, которая является по меньшей мере на 95%, 96%, 97%, 98% или 99% идентичной аминокислотной последовательности SEQ ID NO: 88. В одном аспекте VH содержит последовательность, которая является по меньшей мере на около 95% идентичной аминокислотной последовательности SEQ ID NO: 88. В одном аспекте VH содержит последовательность, которая является по меньшей мере на около 98% идентичной аминокислотной последовательности SEQ ID NO: 88. В определенных аспектах последовательность VH, имеющая по меньшей мере 95%, 96%, 97%, 98% или 99% идентичности, содержит замены (например, консервативные замены), вставки или делеции по сравнению с референсной последовательностью, но антитело, содержащее эту последовательность, сохраняет способность связываться с EGFRvIII. В определенных аспектах была проведена замена, вставка и/или делеция в общей сложности 1 10 аминокислот в аминокислотной последовательности SEQ ID NO: 88. В определенных аспектах замены, вставки или делеции находятся в областях за пределами CDR (т.е. в FR). В одном аспекте VH содержит аминокислотную последовательность SEQ ID NO: 88. Необязательно, VH содержит аминокислотную последовательность SEQ ID NO: 88, включая посттрансляционные модификации этой последовательности.
В одном аспекте VL второго (и, в случае наличия, третьего) антигенсвязывающего домена содержит одну или более каркасных последовательностей легкой цепи (т.е. последовательностей FR1, FR2, FR3 и/или FR4) с SEQ ID NO: 92. В одном аспекте VL содержит последовательность, которая является по меньшей мере на 95%, 96%, 97%, 98% или 99% идентичной аминокислотной последовательности SEQ ID NO: 92. В одном аспекте VL содержит последовательность, которая является по меньшей мере на около 95% идентичной аминокислотной последовательности SEQ ID NO: 92. В одном аспекте VL содержит последовательность, которая является по меньшей мере на около 98% идентичной аминокислотной последовательности SEQ ID NO: 92. В определенных аспектах последовательность VL, имеющая по меньшей мере 95%, 96%, 97%, 98% или 99% идентичности, содержит замены (например, консервативные замены), вставки или делеции по сравнению с референсной последовательностью, но антитело, содержащее эту последовательность, сохраняет способность связываться с EGFRvIII. В определенных аспектах была проведена замена, вставка и/или делеция в общей сложности 1 10 аминокислот в аминокислотной последовательности SEQ ID NO: 92. В определенных аспектах замены, вставки или делеции находятся в областях за пределами CDR (т.е. в FR). В одном аспекте VL содержит аминокислотную последовательность SEQ ID NO: 92. Необязательно, VL содержит аминокислотную последовательность SEQ ID NO: 92, включая посттрансляционные модификации этой последовательности.
В одном аспекте VH второго (и, в случае наличия, третьего) антигенсвязывающего домена содержит аминокислотную последовательность, которая по меньшей мере на около 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 88, и VL второго (и, в случае наличия, третьего) антигенсвязывающего домена содержит аминокислотную последовательность, которая по меньшей мере на около 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности SEQ ID NO: 92. В одном аспекте VH содержит аминокислотную последовательность SEQ ID NO: 88, a VL содержит аминокислотную последовательность SEQ ID NO: 92.
В дополнительном аспекте второй (и, в случае наличия, третий) антигенсвязывающий домен содержит VH, содержащую последовательность SEQ ID NO: 88, и VL, содержащую последовательность SEQ ID NO: 92.
В дополнительном аспекте второй (и, в случае наличия, третий) антигенсвязывающий домен содержит последовательность VH SEQ ID NO: 88 и последовательность VL SEQ ID NO: 92.
В другом аспекте второй (и, в случае наличия, третий) антигенсвязывающий домен содержит VH, содержащую последовательности CDR тяжелой цепи VH с SEQ ID NO: 88, и VL, содержащую последовательности CDR легкой цепи VL с SEQ ID NO: 92.
В дополнительном аспекте второй (и, в случае наличия, третий) антигенсвязывающий домен содержит аминокислотные последовательности HCDR1, HCDR2 и HCDR3 VH с SEQ ID NO: 88 и аминокислотные последовательности LCDR1, LCDR2 и LCDR3 VL с SEQ ID NO: 92.
В одном аспекте VH второго (и, в случае наличия, третьего) антигенсвязывающего домена содержит последовательности CDR тяжелой цепи VH с SEQ ID NO: 88 и каркасную область с по меньшей мере 95%, 96%, 97%, 98% или 99% идентичности последовательности с каркасной последовательностью VH с SEQ ID NO: 88. В одном аспекте VH содержит последовательности CDR тяжелой цепи VH с SEQ ID NO: 88 и каркасную область с по меньшей мере 95% идентичности последовательности с каркасной последовательностью VH с SEQ ID NO: 88. В другом аспекте VH содержит последовательности CDR тяжелой цепи VH с SEQ ID NO: 88 и каркасную область с по меньшей мере 98% идентичности последовательности с каркасной последовательностью VH с SEQ ID NO: 88.
В одном аспекте VL второго (и, в случае наличия, третьего) антигенсвязывающего домена содержит последовательности CDR легкой цепи VL с SEQ ID NO: 92 и каркасную область с по меньшей мере 95%, 96%, 97%, 98% или 99% идентичности последовательности с каркасной последовательностью VL с SEQ ID NO: 92. В одном аспекте VL содержит последовательности CDR легкой цепи VL с SEQ ID NO: 92 и каркасную область с по меньшей мере 95% идентичности последовательности с каркасной последовательностью VL с SEQ ID NO: 92. В другом аспекте VL содержит последовательности CDR легкой цепи VL с SEQ ID NO: 92 и каркасную область с по меньшей мере 98% идентичности последовательности с каркасной последовательностью VL с SEQ ID NO: 92.
В альтернативных аспектах второй (и, в случае наличия, третий) антигенсвязывающий домен содержит последовательность VH по любому из аспектов, приведенных выше в этом разделе в связи с EGFRvIII, и последовательность VL по любому из аспектов, приведенных выше в этом разделе в связи с EGFRvIII, но на основании следующих последовательностей (упорядоченных строками) вместо SEQ ID NO 85 (HCDR1), 86 (HCDR2), 87 (HCDR3), 88 (VH), 89 (LCDR1), 90 (LCDR2), 91 (LCDR3) и 92 (VL):

В одном аспекте второй (и, в случае наличия, третий) антигенсвязывающий домен содержит последовательность VH по любому из аспектов, приведенных выше в этом разделе, и последовательность VL по любому из аспектов, приведенных выше в этом разделе.
Анти-TYRP-1 и анти-EGFRvIII антитела
В изобретении также предложено антитело, которое связывается с TYRP-1, содержащее последовательность VH по любому из аспектов, приведенных выше в этом разделе в связи с TYRP-1, и последовательность VL по любому из аспектов, приведенных выше в этом разделе в связи с TYRP-1 (например, антитело, которое связывается с TYRP-1, содержащее вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область тяжелой цепи (HCDR) 1 с SEQ ID NO: 15, HCDR 2 с SEQ ID NO: 16 и HCDR 3 с SEQ ID NO: 17, и вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область легкой цепи (LCDR) 1 с SEQ ID NO: 19, LCDR 2 с SEQ ID NO: 20 и LCDR 3 с SEQ ID NO: 21; или антитело, которое связывается с TYRP-1, содержащее VH, содержащую последовательность SEQ ID NO: 18, и VL, содержащую последовательность SEQ ID NO: 22).
В изобретении также предложено антитело, которое связывается с EGFRvIII, содержащее последовательность VH по любому из аспектов, приведенных выше в этом разделе в связи с EGFRvIII, и последовательность VL по любому из аспектов, приведенных выше в этом разделе в связи с EGFRvIII (например, антитело, которое связывается с EGFRvIII, содержащее вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область тяжелой цепи (HCDR) 1 с SEQ ID NO: 85, HCDR 2 с SEQ ID NO: 86 и HCDR 3 с SEQ ID NO: 87, и вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область легкой цепи (LCDR) 1 с SEQ ID NO: 89, LCDR 2 с SEQ ID NO: 90 и LCDR 3 с SEQ ID NO: 91; или антитело, которое связывается с TYRP-1, содержащее VH, содержащую последовательность SEQ ID NO: 88, и VL, содержащую последовательность SEQ ID NO: 92).
В дополнительном аспекте антитела, которые связываются с TYRP-1 или EGFRvIII, в соответствии с любым из вышеприведенных аспектов могут иметь другие характеристики, отдельно или в комбинации, описанные в связи с антителом, которое связывается с CD3 (если они явно не относятся к анти-CD3 антителу, например, связывающие последовательности).
в) Зарядовые модификации
(Мультиспецифическое) антитело по изобретению может содержать аминокислотные замены в содержащихся в нем молекулах Fab, которые являются в особенности эффективными для снижения неправильного спаривания легких цепей с несовпадающими тяжелыми цепями (побочные продукты типа Бенс-Джонса), которое может происходить при получении мультиспецифических антител на основе Fab с обменом VH/VL в одном (или более в случае молекул, содержащих более двух антигенсвязывающих молекул Fab) из их связывающих плеч (также смотрите публикацию РСТ №WO 2015/150447, в полном объеме включенную в данный документ посредством ссылки, в частности, примеры в ней). Соотношение необходимого (мультиспецифического) антитела в сравнении с нежелательными побочными продуктами, в частности побочными продуктами типа Бенс-Джонса, возникающими в мультиспецифических антителах с обменом доменов VH/VL в одном из их связывающих плеч, можно улучшить путем внесения заряженных аминокислот с противоположными зарядами в определенные аминокислотные позиции в доменах СН1 и CL (что иногда называется в данном документе «зарядовыми модификациями»).
Соответственно, в некоторых аспектах, в которых первый и второй (и, в случае наличия, третий) антигенсвязывающие домены (мультиспецифического) антитела, оба, представляют собой молекулы Fab, и в одном из антигенсвязывающих доменов (в частности, в первом антигенсвязывающем домене) вариабельные домены VL и VH легкой цепи Fab и тяжелой цепи Fab замещены друг другом,
i) в константном домене CL второго (и, в случае наличия, третьего) антигенсвязывающего домена аминокислота в позиции 124 замещена положительно заряженной аминокислотой (нумерация в соответствии с Kabat), и при этом в константном домене СН1 второго (и, в случае наличия, третьего) антигенсвязывающего домена аминокислота в позиции 147 или аминокислота в позиции 213 замещена отрицательно заряженной аминокислотой (нумерация в соответствии с индексом EU по Kabat); или
ii) в константном домене CL первого антигенсвязывающего домена аминокислота в позиции 124 замещена положительно заряженной аминокислотой (нумерация в соответствии с Kabat), и при этом в константном домене СН1 первого антигенсвязывающего домена аминокислота в позиции 147 или аминокислота в позиции 213 замещена отрицательно заряженной аминокислотой (нумерация в соответствии с индексом EU по Kabat).
(Мультиспецифическое) антитело не содержит обе модификации, указанные в i) и ii). Константные домены CL и СН1 антигенсвязывающего домена, содержащего обмен VH/VL, не замещены друг другом (т.е. остаются необмененными).
В более конкретном аспекте
i) в константном домене CL второго (и, в случае наличия, третьего) антигенсвязывающего домена аминокислота в позиции 124 независимо замещена лизином (К), аргинином (R) или гистидином (Н) (нумерация в соответствии с Kabat), а в константном домене СН1 второго (и, в случае наличия, третьего) антигенсвязывающего домена аминокислота в позиции 147 или аминокислота в позиции 213 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация в соответствии с индексом EU по Kabat); или
ii) в константном домене CL первого антигенсвязывающего домена аминокислота в позиции 124 независимо замещена лизином (К), аргинином (R) или гистидином (Н) (нумерация в соответствии с Kabat), а в константном домене СН1 первого антигенсвязывающего домена аминокислота в позиции 147 или аминокислота в позиции 213 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация в соответствии с индексом EU по Kabat).
В одном таком аспекте в константном домене CL второго (и, в случае наличия, третьего) антигенсвязывающего домена аминокислота в позиции 124 независимо замещена лизином (К), аргинином (R) или гистидином (Н) (нумерация в соответствии с Kabat), а в константном домене СН1 второго (и, в случае наличия, третьего) антигенсвязывающего домена аминокислота в позиции 147 или аминокислота в позиции 213 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация в соответствии с индексом EU по Kabat).
В дополнительном аспекте в константном домене CL второго (и, в случае наличия, третьего) антигенсвязывающего домена аминокислота в позиции 124 независимо замещена лизином (К), аргинином (R) или гистидином (Н) (нумерация в соответствии с Kabat), а в константном домене СН1 второго (и, в случае наличия, третьего) антигенсвязывающего домена аминокислота в позиции 147 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация в соответствии с индексом EU по Kabat).
В предпочтительном аспекте в константном домене CL второго (и, в случае наличия, третьего) антигенсвязывающего домена аминокислота в позиции 124 независимо замещена лизином (К), аргинином (R) или гистидином (Н) (нумерация в соответствии с Kabat) и аминокислота в позиции 123 независимо замещена лизином (К), аргинином (R) или гистидином (Н) (нумерация в соответствии с Kabat), а в константном домене СН1 второго (и, в случае наличия, третьего) антигенсвязывающего домена аминокислота в позиции 147 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация в соответствии с индексом EU по Kabat) и аминокислота в позиции 213 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация в соответствии с индексом EU по Kabat).
В более предпочтительном аспекте в константном домене CL второго (и, в случае наличия, третьего) антигенсвязывающего домена аминокислота в позиции 124 замещена лизином (К) (нумерация в соответствии с Kabat) и аминокислота в позиции 123 замещена лизином (К) (нумерация в соответствии с Kabat), а в константном домене СН1 второго (и, в случае наличия, третьего) антигенсвязывающего домена аминокислота в позиции 147 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat) и аминокислота в позиции 213 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat).
В даже более предпочтительном аспекте в константном домене CL второго (и, в случае наличия, третьего) антигенсвязывающего домена аминокислота в позиции 124 замещена лизином (К) (нумерация в соответствии с Kabat) и аминокислота в позиции 123 замещена аргинином (R) (нумерация в соответствии с Kabat), а в константном домене СН1 второго (и, в случае наличия, третьего) антигенсвязывающего домена аминокислота в позиции 147 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat) и аминокислота в позиции 213 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat).
В предпочтительных аспектах, если аминокислотные замены в соответствии с вышеприведенными аспектами проведены в константном домене CL и константном домене СН1 второго (и, в случае наличия, третьего) антигенсвязывающего домена, константный домен CL второго (и, в случае наличия, третьего) антигенсвязывающего домена относится к изотипу каппа.
В альтернативном варианте аминокислотные замены в соответствии с вышеприведенными аспектами могут быть проведены в константном домене CL и константном домене СН1 первого антигенсвязывающего домена вместо константного домена CL и константного домена СН1 второго (и, в случае наличия, третьего) антигенсвязывающего домена. В предпочтительных таких аспектах константный домен CL первого антигенсвязывающего домена относится к изотипу каппа.
Соответственно, в одном аспекте в константном домене CL первого антигенсвязывающего домена аминокислота в позиции 124 независимо замещена лизином (К), аргинином (R) или гистидином (Н) (нумерация в соответствии с Kabat), а в константном домене СН1 первого антигенсвязывающего домена аминокислота в позиции 147 или аминокислота в позиции 213 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация в соответствии с индексом EU по Kabat).
В дополнительном аспекте в константном домене CL первого антигенсвязывающего домена аминокислота в позиции 124 независимо замещена лизином (К), аргинином (R) или гистидином (Н) (нумерация в соответствии с Kabat), а в константном домене СН1 первого антигенсвязывающего домена аминокислота в позиции 147 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация в соответствии с индексом EU по Kabat).
В другом аспекте в константном домене CL первого антигенсвязывающего домена аминокислота в позиции 124 независимо замещена лизином (К), аргинином (R) или гистидином (Н) (нумерация в соответствии с Kabat) и аминокислота в позиции 123 независимо замещена лизином (К), аргинином (R) или гистидином (Н) (нумерация в соответствии с Kabat), а в константном домене СН1 первого антигенсвязывающего домена аминокислота в позиции 147 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация в соответствии с индексом EU по Kabat) и аминокислота в позиции 213 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация в соответствии с индексом EU по Kabat).
В одном аспекте в константном домене CL первого антигенсвязывающего домена аминокислота в позиции 124 замещена лизином (К) (нумерация в соответствии с Kabat) и аминокислота в позиции 123 замещена лизином (К) (нумерация в соответствии с Kabat), а в константном домене СН1 первого антигенсвязывающего домена аминокислота в позиции 147 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat) и аминокислота в позиции 213 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat).
В другом аспекте в константном домене CL первого антигенсвязывающего домена аминокислота в позиции 124 замещена лизином (К) (нумерация в соответствии с Kabat) и аминокислота в позиции 123 замещена аргинином (R) (нумерация в соответствии с Kabat), а в константном домене СН1 первого антигенсвязывающего домена аминокислота в позиции 147 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat) и аминокислота в позиции 213 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat).
В предпочтительном аспекте (мультиспецифическое) антитело по изобретению содержит
(а) первый антигенсвязывающий домен, который связывается с CD3, причем первый антигенсвязывающий домен представляет собой молекулу Fab, в которой вариабельные домены VL и VH легкой цепи Fab и тяжелой цепи Fab замещены друг другом, и содержит вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область тяжелой цепи (HCDR) 1 с SEQ ID NO: 2, HCDR 2 с SEQ ID NO: 3 и HCDR 3 с SEQ ID NO: 5, и вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область легкой цепи (LCDR) 1 с SEQ ID NO: 8, LCDR 2 с SEQ ID NO: 9 и LCDR 3 с SEQ ID NO: 10, и
(б) второй и, необязательно, третий антигенсвязывающий домен, который связывается со вторым антигеном;
причем в константном домене CL второго (и, в случае наличия, третьего) антигенсвязывающего домена аминокислота в позиции 124 независимо замещена лизином (К), аргинином (R) или гистидином (Н) (нумерация в соответствии с Kabat) (в предпочтительном аспекте - независимо лизином (К) или аргинином (R)) и аминокислота в позиции 123 независимо замещена лизином (К), аргинином (R) или гистидином (Н) (нумерация в соответствии с Kabat) (в предпочтительном аспекте - независимо лизином (К) или аргинином (R)), а в константном домене СН1 второго (и, в случае наличия, третьего) антигенсвязывающего домена аминокислота в позиции 147 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация в соответствии с индексом EU по Kabat) и аминокислота в позиции 213 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация в соответствии с индексом EU по Kabat).
г) Форматы мультиспецифических антител
(Мультиспецифическое) антитело в соответствии с настоящим изобретением может иметь различные конфигурации. Типовые конфигурации проиллюстрированы на Фиг. 1.
В предпочтительных аспектах антигенсвязывающие домены, содержащиеся в (мультиспецифическом) антителе, представляют собой молекулы Fab. В таких аспектах первый, второй, третий и т.д. антигенсвязывающий домен может называться в данном документе первой, второй, третей и т.д. молекулой Fab, соответственно.
В одном аспекте первый и второй антигенсвязывающие домены (мультиспецифического) антитела слиты друг с другом, необязательно посредством пептидного линкера. В предпочтительных аспектах каждый из первого и второго антигенсвязывающих доменов представляет собой молекулу Fab. В одном таком аспекте первый антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab второго антигенсвязывающего домена. В другом таком аспекте второй антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первого антигенсвязывающего домена. В аспектах, в которых (i) первый антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab второго антигенсвязывающего домена или (ii) второй антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первого антигенсвязывающего домена, легкая цепь Fab первого антигенсвязывающего домена и легкая цепь Fab второго антигенсвязывающего домена дополнительно могут быть слиты друг с другом, необязательно посредством пептидного линкера.
(Мультиспецифическое) антитело с одним антигенсвязывающим доменом (таким как молекула Fab), способным специфически связываться со вторым антигеном, например, антигеном клетки-мишени, таким как TYRP-1 или EGFRvIII (например, как показано на Фиг 1А, Г, Ж, З, Л, М), применимо, в частности, в случаях, когда после связывания с высокоаффинным антигенсвязывающим доменом можно ожидать интернализации второго антигена. В таких случаях наличие более одного антигенсвязывающего домена, специфического в отношении второго антигена, может усилить интернализацию второго антигена, тем самым снижая его доступность.
При этом в других случаях преимущественным является (мультиспецифическое) антитело, содержащее два или более антигенсвязывающих доменов (таких как молекулы Fab), специфическим в отношении второго антигена, например, антигена клетки-мишени (смотрите примеры, проиллюстрированные на Фиг. 1Б, 1 В, 1Д, 1Е, 1И, IK, 1Н или 1О), например, для оптимизации нацеливания на сайт-мишень или для обеспечения возможности перекрестного связывания антигенов клетки-мишени.
Соответственно, в предпочтительных аспектах (мультиспецифическое) антитело в соответствии с настоящим изобретением содержит третий антигенсвязывающий домен.
В одном аспекте третий антигенсвязывающий домен связывается со вторым антигеном, например, антигеном клетки-мишени, таким как TYRP-1 или EGFRvIII. В одном аспекте третий антигенсвязывающий домен представляет собой молекулу Fab.
В одном аспекте третий антигенсвязывающий домен идентичен второму антигенсвязывающему домену.
В некоторых аспектах каждый из третьего и второго антигенсвязывающих доменов представляет собой молекулу Fab, и третий антигенсвязывающий домен идентичен второму антигенсвязывающему домену. Таким образом, в этих аспектах второй и третий антигенсвязывающие домены содержат одинаковые аминокислотные последовательности тяжелой и легкой цепи и имеют одинаковое расположение доменов (т.е. стандартное или кроссоверное). Кроме того, в этих аспектах третий антигенсвязывающий домен содержит такие же аминокислотные замены, в случае их наличия, что и второй антигенсвязывающий домен. Например, аминокислотные замены, описанные в данном документе как «зарядовые модификации» проводят в константном домене CL и константном домене СН1 каждого из второго антигенсвязывающего домена и третьего антигенсвязывающего домена. В альтернативном варианте указанные аминокислотные замены можно проводить в константном домене CL и константном домене СН1 первого антигенсвязывающего домена (который, в предпочтительных аспектах, также представляет собой молекулу Fab), но не в константном домене CL и константном домене СН1 второго антигенсвязывающего домена и третьего антигенсвязывающего домена.
Как и второй антигенсвязывающий домен, третий антигенсвязывающий домен предпочтительно представляет собой стандартную молекулу Fab. При этом также предусмотрены аспекты, в которых второй и третий антигенсвязывающие домены представляют собой кроссоверные молекулы Fab (а первый антигенсвязывающий домен представляет собой стандартную молекулу Fab). Таким образом, в предпочтительных аспектах каждый из второго и третьего антигенсвязывающих доменов представляет собой стандартную молекулу Fab, а первый антигенсвязывающий домен представляет собой кроссоверную молекулу Fab, как описано в данном документе, т.е. молекулу Fab, в которой вариабельные домены VH и VL или константные домены CL и СН1 тяжелой и легкой цепей Fab заменены/замещены друг другом. В других аспектах каждый из второго и третьего антигенсвязывающих доменов представляет собой кроссоверную молекулу Fab, а первый антигенсвязывающий домен представляет собой стандартную молекулу Fab.
В случае наличия третьего антигенсвязывающего домена, в предпочтительном аспекте первый антигенсвязывающий домен связывается с CD3, а второй и третий антигенсвязывающие домены связываются со вторым антигеном, в частности антигеном клетки-мишени, таким как TYRP-1 или EGFRvIII.
В предпочтительных аспектах (мультиспецифическое) антитело по изобретению содержит Fc-домен, состоящий из первой и второй субъединиц. Первая и вторая субъединицы Fc-домена способны к стабильной ассоциации.
(Мультиспецифическое) антитело в соответствии с изобретением может иметь отличные конфигурации, т.е. первый, второй (и, необязательно, третий) антигенсвязывающий домен может быть слит с каждым другим и с Fc-доменом разными способами. Компоненты могут быть слиты друг с другом напрямую или, предпочтительно, посредством одного или более подходящих пептидных линкеров. Если молекула Fab слита с N-концом субъединицы Fc-домена, как правило, слияние происходит посредством шарнирной области иммуноглобулина.
В некоторых аспектах каждый из первого и второго антигенсвязывающих доменов представляет собой молекулу Fab, а первый антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом первой или второй субъединицы Fc-домена. В таких аспектах второй антигенсвязывающий домен может быть слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первого антигенсвязывающего домена или с N-концом другой субъединицы Fc-домена. В таких предпочтительных аспектах второй антигенсвязывающий домен представляет собой стандартную молекулу Fab, а первый антигенсвязывающий домен представляет собой кроссоверную молекулу Fab, как описано в данном документе, т.е. молекулу Fab, в которой вариабельные домены VH и VL или константные домены CL и СН1 тяжелой и легкой цепей Fab заменены/замещены друг другом. В других таких аспектах второй антигенсвязывающий домен представляет собой кроссоверную молекулу Fab, а первый антигенсвязывающий домен представляет собой стандартную молекулу Fab.
В одном аспекте каждый из первого и второго антигенсвязывающих доменов представляет собой молекулу Fab, первый антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом первой или второй субъединицы Fc-домена, а второй антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первого антигенсвязывающего домена. В конкретном аспекте (мультиспецифическое) антитело состоит преимущественно из первой и второй молекул Fab, Fc-домена, состоящего из первой и второй субъединиц, и, необязательно, одного или более пептидных линкеров, причем вторая молекула Fab слита на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первой молекулы Fab, и первая молекула Fab слита на С-конце тяжелой цепи Fab с N-концом первой или второй субъединицы Fc-домена. Такая конфигурация схематически изображена на Фиг. 1Ж и 1Л (в этих примерах первый антигенсвязывающий домен представляет собой VH/VL кроссоверную молекулу Fab). Необязательно, легкая цепь Fab первой молекулы Fab и легкая цепь Fab второй молекулы Fab могут быть дополнительно слиты друг с другом.
В другом аспекте каждый из первого и второго антигенсвязывающих доменов представляет собой молекулу Fab и каждый из первого и второго антигенсвязывающих доменов слит на С-конце тяжелой цепи Fab с N-концом одной из субъединиц Fc-домена. В конкретном аспекте (мультиспецифическое) антитело состоит преимущественно из первой и второй молекул Fab, Fc-домена, состоящего из первой и второй субъединиц, и, необязательно, одного или более пептидных линкеров, причем каждая из первой и второй молекул Fab слита на С-конце тяжелой цепи Fab с N-концом одной из субъединиц Fc-домена. Такая конфигурация схематически изображена на Фиг. 1А и 1Г (в этих примерах первый антигенсвязывающий домен представляет собой VH/VL кроссоверную молекулу Fab, а второй антигенсвязывающий домен представляет собой стандартную молекулу Fab). Первая и вторая молекулы Fab могут быть слиты с Fc-доменом напрямую или посредством пептидного линкера. В предпочтительном аспекте каждая из первой и второй молекул Fab слита с Fc-доменом посредством шарнирной области иммуноглобулина. В конкретном аспекте шарнирная область иммуноглобулина представляет собой шарнирную область IgG1 человека, в частности, когда Fc-домен представляет собой Fc-домен IgG1 человека.
В некоторых аспектах каждый из первого и второго антигенсвязывающих доменов представляет собой молекулу Fab, а второй антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом первой или второй субъединицы Fc-домена. В таких аспектах первый антигенсвязывающий домен может быть слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первого антигенсвязывающего домена или (как описано выше) с N-концом другой субъединицы Fc-домена. В таких предпочтительных аспектах указанный второй антигенсвязывающий домен представляет собой стандартную молекулу Fab, а первый антигенсвязывающий домен представляет собой кроссоверную молекулу Fab, как описано в данном документе, т.е. молекулу Fab, в которой вариабельные домены VH и VL или константные домены CL и СН1 тяжелой и легкой цепей Fab заменены/замещены друг другом. В других таких аспектах указанный второй антигенсвязывающий домен представляет собой кроссоверную молекулу Fab, а первый антигенсвязывающий домен представляет собой стандартную молекулу Fab.
В одном аспекте каждый из первого и второго антигенсвязывающих доменов представляет собой молекулу Fab, второй антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом первой или второй субъединицы Fc-домена, а первый антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab второго антигенсвязывающего домена. В конкретном аспекте (мультиспецифическое) антитело состоит преимущественно из первой и второй молекул Fab, Fc-домена, состоящего из первой и второй субъединиц, и, необязательно, одного или более пептидных линкеров, причем первая молекула Fab слита на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab второй молекулы Fab, и вторая молекула Fab слита на С-конце тяжелой цепи Fab с N-концом первой или второй субъединицы Fc-домена. Такая конфигурация схематически изображена на Фиг. 13 и 1М (в этих примерах первый антигенсвязывающий домен представляет собой VH/VL кроссоверную молекулу Fab, а второй антигенсвязывающий домен представляет собой стандартную молекулу Fab). Необязательно, легкая цепь Fab первой молекулы Fab и легкая цепь Fab второй молекулы Fab могут быть дополнительно слиты друг с другом.
В некоторых аспектах третий антигенсвязывающий домен, в частности третья молекула Fab, слит на С-конце тяжелой цепи Fab с N-концом первой или второй субъединицы Fc-домена. В таких предпочтительных аспектах каждый из указанных второго и третьего антигенсвязывающих доменов представляет собой стандартную молекулу Fab, а первый антигенсвязывающий домен представляет собой кроссоверную молекулу Fab, как описано в данном документе, т.е. молекулу Fab, в которой вариабельные домены VH и VL или константные домены CL и СН1 тяжелой и легкой цепей Fab заменены/замещены друг другом. В других аспектах каждый из указанных второго и третьего антигенсвязывающих доменов представляет собой кроссоверную молекулу Fab, а первый антигенсвязывающий домен представляет собой стандартную молекулу Fab.
В таком предпочтительном аспекте каждый из первого и третьего антигенсвязывающих доменов слит на С-конце тяжелой цепи Fab с N-концом одной из субъединиц Fc-домена, а второй антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первой молекулы Fab. В конкретном аспекте (мультиспецифическое) антитело состоит преимущественно из первой, второй и третьей молекул Fab, Fc-домена, состоящего из первой и второй субъединиц, и, необязательно, одного или более пептидных линкеров, причем вторая молекула Fab слита на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первой молекулы Fab, первая молекула Fab слита на С-конце тяжелой цепи Fab с N-концом первой субъединицы Fc-домена, а третья молекула Fab слита на С-конце тяжелой цепи Fab с N-концом второй субъединицы Fc-домена. Такая конфигурация схематически изображена на Фиг. 1Б и 1Д (в этих примерах первый антигенсвязывающий домен представляет собой VH/VL кроссоверную молекулу Fab, а второй и третий антигенсвязывающие домены представляют собой стандартную молекулу Fab) и Фиг. 1К и 1О (в этих примерах первый антигенсвязывающий домен представляет собой стандартную молекулу Fab, а второй и третий антигенсвязывающие домены представляют собой VH/VL кроссоверную молекулу Fab). Первая и третья молекулы Fab могут быть слиты с Fc-доменом напрямую или посредством пептидного линкера. В предпочтительном аспекте каждая из первой и третьей молекул Fab слита с Fc-доменом посредством шарнирной области иммуноглобулина. В конкретном аспекте шарнирная область иммуноглобулина представляет собой шарнирную область IgG1 человека, в частности, когда Fc-домен представляет собой Fc-домен IgG1 человека. Необязательно, легкая цепь Fab первой молекулы Fab и легкая цепь Fab второй молекулы Fab могут быть дополнительно слиты друг с другом.
В другом таком аспекте каждый из второго и третьего антигенсвязывающих доменов слит на С-конце тяжелой цепи Fab с N-концом одной из субъединиц Fc-домена, а первый антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab второго антигенсвязывающего домена. В конкретном аспекте (мультиспецифическое) антитело состоит преимущественно из первой, второй и третьей молекул Fab, Fc-домена, состоящего из первой и второй субъединиц, и, необязательно, одного или более пептидных линкеров, причем первая молекула Fab слита на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab второй молекулы Fab, вторая молекула Fab слита на С-конце тяжелой цепи Fab с N-концом первой субъединицы Fc-домена, а третья молекула Fab слита на С-конце тяжелой цепи Fab с N-концом второй субъединицы Fc-домена. Такая конфигурация схематически изображена на Фиг. 1 В и 1Е (в этих примерах первый антигенсвязывающий домен представляет собой VH/VL кроссоверную молекулу Fab, а второй и третий антигенсвязывающие домены представляют собой стандартную молекулу Fab) и Фиг. 1И и 1Н (в этих примерах первый антигенсвязывающий домен представляет собой стандартную молекулу Fab, а второй и третий антигенсвязывающие домены представляют собой VH/VL кроссоверную молекулу Fab). Вторая и третья молекулы Fab могут быть слиты с Fc-доменом напрямую или посредством пептидного линкера. В предпочтительном аспекте каждая из второй и третьей молекул Fab слита с Fc-доменом посредством шарнирной области иммуноглобулина. В конкретном аспекте шарнирная область иммуноглобулина представляет собой шарнирную область IgG1 человека, в частности, когда Fc-домен представляет собой Fc-домен IgG1 человека. Необязательно, легкая цепь Fab первой молекулы Fab и легкая цепь Fab второй молекулы Fab могут быть дополнительно слиты друг с другом.
В конфигурациях (мультиспецифического) антитела, в которых молекула Fab слита на С-конце тяжелой цепи Fab с N-концом каждой из субъединиц Fc-домена посредством шарнирной области иммуноглобулина, две молекулы Fab, шарнирные области и Fc-домен преимущественно образуют молекулу иммуноглобулина. В предпочтительном аспекте молекула иммуноглобулина представляет собой иммуноглобулин класса IgG. В еще более предпочтительном аспекте иммуноглобулин представляет собой иммуноглобулин подкласса IgG1. В другом аспекте иммуноглобулин представляет собой иммуноглобулин подкласса IgG4. В дополнительном предпочтительном аспекте иммуноглобулин представляет собой иммуноглобулин человека. В других аспектах иммуноглобулин представляет собой химерный иммуноглобулин или гуманизированный иммуноглобулин. В одном аспекте иммуноглобулин содержит человеческую константную область, в частности, человеческую Fc-область.
В некоторых из (мультиспецифических) антител по изобретению легкая цепь Fab первой молекулы Fab и легкая цепь Fab второй молекулы Fab слиты друг с другом, необязательно, посредством пептидного линкера. В зависимости от конфигурации первой и второй молекул Fab легкая цепь Fab первой молекулы может быть слита на С-конце с N-концом легкой цепи Fab второй молекулы Fab или легкая цепь Fab второй молекулы может быть слита на С-конце с N-концом легкой цепи Fab первой молекулы Fab. Слияние легких цепей Fab первой и второй молекул Fab дополнительно снижает неправильное спаривание несовпадающих тяжелых и легких цепей Fab и также снижает число плазмид, необходимое для экспрессии некоторых из (мультиспецифических) антител по изобретению.
Антигенсвязывающие домены могут быть слиты с Fc-доменом или друг с другом напрямую или посредством пептидного линкера, содержащего одну или более аминокислот, как правило, около 2-20 аминокислот.Пептидные линкеры известны в данной области техники и описаны в данном документе. Подходящие неиммуногенные пептидные линкеры включают, например, пептидные линкеры (G4S)n, (SG4)n, (G4S)n или G4(SG4)n. «n» в общем случае является целым числом от 1 до 10, как правило, от 2 до 4. В одном аспекте указанный пептидный линкер имеет длину по меньшей мере 5 аминокислот, в одном аспекте - длину от 5 до 100, в дополнительном аспекте - от 10 до 50 аминокислот. В одном аспекте указанный пептидный линкер представляет собой (GxS)n или (GxS)nGm, где G=глицин, S=серии, а (х=3, п=3, 4, 5 или 6 и m=0, 1,2 или 3) или (х=4, n=2, 3, 4 или 5 и m=0, 1, 2 или 3), в одном аспекте х=4 и n=2 или 3, в дополнительном аспекте х=4 и n=2. В одном аспекте указанный пептидный линкер представляет собой (G4S)2. В особенности подходящим пептидным линкером для слияния легких цепей Fab первой и второй молекул Fab друг с другом является (G4S)2. Типовой пептидный линкер, подходящий для соединения тяжелых цепей Fab первого и второго Fab-фрагментов, содержит последовательность (D)-(G4S)2 (SEQ ID NO 118 и 119). Другой такой подходящий линкер содержит последовательность (G4S)4. Помимо этого, линкеры могут содержать шарнирную область иммуноглобулина (или ее часть). В частности, если молекула Fab слита с N-концом субъединицы Fc-домена, она может быть слита посредством шарнирной области иммуноглобулина или ее части с или без дополнительного пептидного линкера.
В определенных аспектах (мультиспецифическое) антитело в соответствии с изобретением содержит полипептид, в котором вариабельная область легкой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab первой молекулы Fab (т.е. первая молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи), которая в свою очередь имеет общую карбокси-концевую пептидную связь с субъединицей Fc-домена (VL(1)-CH1(1)-CH2-CH3(-CH4)), и полипептид, в котором тяжелая цепь Fab второй молекулы Fab имеет общую карбокси-концевую пептидную связь с субъединицей Fc-домена (VH(2)-CH1(2)-СН2-CH3(-СН4)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид, в котором вариабельная область тяжелой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab первой молекулы Fab (VH(1)-CL(1)), и полипептид легкой цепи Fab второй молекулы Fab (VL(2)-CL(2)). В определенных аспектах полипептиды ковалентно связаны, например, дисульфидной связью.
В определенных аспектах (мультиспецифическое) антитело в соответствии с изобретением содержит полипептид, в котором вариабельная область тяжелой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab первой молекулы Fab (т.е. первая молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой константная область тяжелой цепи замещена константной областью легкой цепи), которая в свою очередь имеет общую карбокси-концевую пептидную связь с субъединицей Fc-домена (VH(1)-CL(1)-CH2-CH3(-СН4)), и полипептид, в котором тяжелая цепь Fab второй молекулы Fab имеет общую карбокси-концевую пептидную связь с субъединицей Fc-домена (VH(2)-СН1(2)-СН2-CH3(-СН4)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид, в котором вариабельная область легкой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab первой молекулы Fab (VL(1)-CH1(1)), и полипептид легкой цепи Fab второй молекулы Fab (VL(2)-CL(2)). В определенных аспектах полипептиды ковалентно связаны, например, дисульфидной связью.
В некоторых аспектах (мультиспецифическое) антитело содержит полипептид, в котором вариабельная область легкой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab первой молекулы Fab (т.е. первая молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи), которая в свою очередь имеет общую карбокси-концевую пептидную связь с тяжелой цепью Fab второй молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с субъединицей Fc-домена (VL(1)-CH1(1)-VH(2)-CH1(2)-CH2-CH3(-СН4)). В других аспектах (мультиспецифическое) антитело содержит полипептид, в котором тяжелая цепь Fab второй молекулы Fab имеет общую карбокси-концевую пептидную связь с вариабельной областью легкой цепи Fab первой молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab первой молекулы Fab (т.е. первая молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи), которая в свою очередь имеет общую карбокси-концевую пептидную связь с субъединицей Fc-домена (VH(2)-CH1(2)-VL(1)-CH1(1)-CH2-CH3(-СН4)). В некоторых из этих аспектов (мультиспецифическое) антитело дополнительно содержит полипептид кроссоверной легкой цепи Fab первой молекулы Fab, в котором вариабельная область тяжелой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab первой молекулы Fab (VH(1)-CL(1)), и полипептид легкой цепи Fab второй молекулы Fab (VL(2)-CL(2)). В других аспектах (мультиспецифическое) антитело дополнительно содержит полипептид, в котором вариабельная область тяжелой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab первой молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с полипептидом легкой цепи Fab второй молекулы Fab (VH(1)-CL(1)-VL(2)-CL(2)), или полипептид, в котором полипептид легкой цепи Fab второй молекулы Fab имеет общую карбокси-концевую пептидную связь с вариабельной областью тяжелой цепи Fab первой молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab первой молекулы Fab (VL(2)-CL(2)-VH(1)-CL(1)), в зависимости от ситуации. (Мультиспецифическое) антитело в соответствии с этими аспектами может дополнительно содержать (i) полипептид субъединицы Fc-домена (СН2-CH3(-СН4)) или (ii) полипептид, в котором тяжелая цепь Fab третьей молекулы Fab имеет общую карбокси-концевую пептидную связь с субъединицей Fc-домена (VH(3)-CH1(3)-CH2-CH3(-CH4)), и полипептид легкой цепи Fab третьей молекулы Fab (VL(3)-CL(3)). В определенных аспектах полипептиды ковалентно связаны, например, дисульфидной связью.
В некоторых аспектах (мультиспецифическое) антитело содержит полипептид, в котором вариабельная область тяжелой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab первой молекулы Fab (т.е. первая молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой константная область тяжелой цепи замещена константной областью легкой цепи), которая в свою очередь имеет общую карбокси-концевую пептидную связь с тяжелой цепью Fab второй молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с субъединицей Fc-домена (VH(1)-CL(1)-VH(2)-CH1(2)-CH2-CH3(-СН4)). В других аспектах (мультиспецифическое) антитело содержит полипептид, в котором тяжелая цепь Fab второй молекулы Fab имеет общую карбокси-концевую пептидную связь с вариабельной областью тяжелой цепи Fab первой молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab первой молекулы Fab (т.е. первая молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой константная область тяжелой цепи замещена константной областью легкой цепи), которая в свою очередь имеет общую карбокси-концевую пептидную связь с субъединицей Fc-домена (VH(2)-CH1(2)-VH(1)-CL(1)-CH2-CH3(-CH4)). В некоторых из этих аспектов (мультиспецифическое) антитело дополнительно содержит полипептид кроссоверной легкой цепи Fab первой молекулы Fab, в котором вариабельная область легкой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab первой молекулы Fab (VL(1)-CH1(1)), и полипептид легкой цепи Fab второй молекулы Fab (VL(2)-CL(2)). В других аспектах (мультиспецифическое) антитело дополнительно содержит полипептид, в котором вариабельная область легкой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab первой молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с полипептидом легкой цепи Fab второй молекулы Fab (VL(1)-CH1(1)-VL(2)-CL(2)), или полипептид, в котором полипептид легкой цепи Fab второй молекулы Fab имеет общую карбокси-концевую пептидную связь с вариабельной областью тяжелой цепи Fab первой молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab первой молекулы Fab (VL(2)-CL(2)-VH(1)-CL(1)), в зависимости от ситуации. (Мультиспецифическое) антитело в соответствии с этими аспектами может дополнительно содержать (i) полипептид субъединицы Fc-домена (СН2-CH3(-СН4)) или (ii) полипептид, в котором тяжелая цепь Fab третьей молекулы Fab имеет общую карбокси-концевую пептидную связь с субъединицей Fc-домена (VH(3)-CH1(3)-CH2-CH3(-CH4)), и полипептид легкой цепи Fab третьей молекулы Fab (VL(3)-CL(3)). В определенных аспектах полипептиды ковалентно связаны, например, дисульфидной связью.
В определенных аспектах (мультиспецифическое) антитело не содержит Fc-домен. В таких предпочтительных аспектах каждый из указанных второго и, в случае наличия, третьего антигенсвязывающих доменов представляет собой стандартную молекулу Fab, а первый антигенсвязывающий домен представляет собой кроссоверную молекулу Fab, как описано в данном документе, т.е. молекулу Fab, в которой вариабельные домены VH и VL или константные домены CL и СН1 тяжелой и легкой цепей Fab заменены/замещены друг другом. В других таких аспектах каждый из указанных второго и, в случае наличия, третьего антигенсвязывающих доменов представляет собой кроссоверную молекулу Fab, а первый антигенсвязывающий домен представляет собой стандартную молекулу Fab.
В одном таком аспекте (мультиспецифическое) антитело преимущественно состоит из первого и второго антигенсвязывающих доменов и, необязательно, одного или более пептидных линкеров, причем первый и второй антигенсвязывающие домены, оба, представляют собой молекулы Fab, а второй антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи первого антигенсвязывающего домена. Такая конфигурация схематически изображена на Фиг. 1П и 1У (в этих примерах первый антигенсвязывающий домен представляет собой VH/VL кроссоверную молекулу Fab, а второй антигенсвязывающий домен представляет собой стандартную молекулу Fab).
В другом таком аспекте (мультиспецифическое) антитело преимущественно состоит из первого и второго антигенсвязывающих доменов и, необязательно, одного или более пептидных линкеров, причем первый и второй антигенсвязывающие домены, оба, представляют собой молекулы Fab, а первый антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи второго антигенсвязывающего домена. Такая конфигурация схематически изображена на Фиг. 1Р и 1Ф (в этих примерах первый антигенсвязывающий домен представляет собой VH/VL кроссоверную молекулу Fab, а второй антигенсвязывающий домен представляет собой стандартную молекулу Fab).
В некоторых аспектах вторая молекула Fab слита на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первой молекулы Fab, а (мультиспецифическое) антитело дополнительно содержит третий антигенсвязывающий домен, в частности третью молекулу Fab, причем указанная третья молекула Fab слита на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab второй молекулы Fab. В определенных таких аспектах (мультиспецифическое) антитело состоит преимущественно из первой, второй и третьей молекул Fab и, необязательно, одного или более пептидных линкеров, причем вторая молекула Fab слита на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первой молекулы Fab, и третья молекула Fab слита на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab второй молекулы Fab. Такая конфигурация схематически изображена на Фиг. 1С и 1X (в этих примерах первый антигенсвязывающий домен представляет собой VH/VL кроссоверную молекулу Fab, а каждый из второго и третьего антигенсвязывающих доменов представляет собой стандартную молекулу Fab) и Фиг. 1Ш и 1Ы (в этих примерах первый антигенсвязывающий домен представляет собой стандартную молекулу Fab, а каждый из второго и третьего антигенсвязывающих доменов представляет собой VH/VL кроссоверную молекулу Fab).
В некоторых аспектах первая молекула Fab слита на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab второй молекулы Fab, а (мультиспецифическое) антитело дополнительно содержит третий антигенсвязывающий домен, в частности третью молекулу Fab, причем указанная третья молекула Fab слита на N-конце тяжелой цепи Fab с С-концом тяжелой цепи Fab второй молекулы Fab. В определенных таких аспектах (мультиспецифическое) антитело состоит преимущественно из первой, второй и третьей молекул Fab и, необязательно, одного или более пептидных линкеров, причем первая молекула Fab слита на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab второй молекулы Fab, и третья молекула Fab слита на N-конце тяжелой цепи Fab с С-концом тяжелой цепи Fab второй молекулы Fab. Такая конфигурация схематически изображена на Фиг. 1Т и 1Ц (в этих примерах первый антигенсвязывающий домен представляет собой VH/VL кроссоверную молекулу Fab, а каждый из второго и третьего антигенсвязывающих доменов представляет собой стандартную молекулу Fab) и Фиг. 14 и 1Щ (в этих примерах первый антигенсвязывающий домен представляет собой стандартную молекулу Fab, а каждый из второго и третьего антигенсвязывающих доменов представляет собой VH/VL кроссоверную молекулу Fab).
В определенных аспектах (мультиспецифическое) антитело в соответствии с изобретением содержит полипептид, в котором тяжелая цепь Fab второй молекулы Fab имеет общую карбокси-концевую пептидную связь с вариабельной областью легкой цепи Fab первой молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab первой молекулы Fab (т.е. первая молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи) (VH(2)-CH1(2)-VL(1)-CH1(1)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид, в котором вариабельная область тяжелой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab первой молекулы Fab (VH(1)-CL(1)), и полипептид легкой цепи Fab второй молекулы Fab (VL(2)-CL(2)).
В определенных аспектах (мультиспецифическое) антитело в соответствии с изобретением содержит полипептид, в котором вариабельная область легкой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab первой молекулы Fab (т.е. первая молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи), которая в свою очередь имеет общую карбокси-концевую пептидную связь с тяжелой цепью Fab второй молекулы Fab (VL(1)-CH1(1)-VH(2)-СН1(2)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид, в котором вариабельная область тяжелой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab первой молекулы Fab (VH(1)-CL(1)), и полипептид легкой цепи Fab второй молекулы Fab (VL(2)-CL(2)).
В определенных аспектах (мультиспецифическое) антитело в соответствии с изобретением содержит полипептид, в котором тяжелая цепь Fab второй молекулы Fab имеет общую карбокси-концевую пептидную связь с вариабельной областью тяжелой цепи Fab первой молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab первой молекулы Fab (т.е. первая молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой константная область тяжелой цепи замещена константной областью легкой цепи) (VH(2)-CH1(2)-VH(1)-CL(1)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид, в котором вариабельная область легкой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab первой молекулы Fab (VL(1)-CH1(1)), и полипептид легкой цепи Fab второй молекулы Fab (VL(2)-CL(2)).
В определенных аспектах (мультиспецифическое) антитело в соответствии с изобретением содержит полипептид, в котором вариабельная область тяжелой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab первой молекулы Fab (т.е. первая молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой константная область тяжелой цепи замещена константной областью легкой цепи), которая в свою очередь имеет общую карбокси-концевую пептидную связь с тяжелой цепью Fab второй молекулы Fab (VH(1)-CL(1)-VH(2)-CH1(2)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид, в котором вариабельная область легкой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab первой молекулы Fab (VL(1)-CH1(1)), и полипептид легкой цепи Fab второй молекулы Fab (VL(2)-CL(2)).
В определенных аспектах (мультиспецифическое) антитело в соответствии с изобретением содержит полипептид, в котором тяжелая цепь Fab третьей молекулы Fab имеет общую карбокси-концевую пептидную связь с тяжелой цепью Fab второй молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с вариабельной областью легкой цепи Fab первой молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab первой молекулы Fab (т.е. первая молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи) (VH(3)-CH1(3)-VH(2)-CH1(2)-VL(1)-CH1(1)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид, в котором вариабельная область тяжелой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab первой молекулы Fab (VH(1)-CL(1)), и полипептид легкой цепи Fab второй молекулы Fab (VL(2)-CL(2)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид легкой цепи Fab третьей молекулы Fab (VL(3)-CL(3)).
В определенных аспектах (мультиспецифическое) антитело в соответствии с изобретением содержит полипептид, в котором тяжелая цепь Fab третьей молекулы Fab имеет общую карбокси-концевую пептидную связь с тяжелой цепью Fab второй молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с вариабельной областью тяжелой цепи Fab первой молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab первой молекулы Fab (т.е. первая молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой константная область тяжелой цепи замещена константной областью легкой цепи) (VH(3)-CH1(3)-VH(2)-CH1(2)-VH(1)-CL(1)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид, в котором вариабельная область легкой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab первой молекулы Fab (VL(1)-CH1(1)), и полипептид легкой цепи Fab второй молекулы Fab (VL(2)-CL(2)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид легкой цепи Fab третьей молекулы Fab (VL(3)-CL(3)).
В определенных аспектах (мультиспецифическое) антитело в соответствии с изобретением содержит полипептид, в котором вариабельная область легкой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab первой молекулы Fab (т.е. первая молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи), которая в свою очередь имеет общую карбокси-концевую пептидную связь с тяжелой цепью Fab второй молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с тяжелой цепью Fab третьей молекулы Fab (VL(1)-CH1(1)-VH(2)-CH1(2)-VH(3)-CH1(3)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид, в котором вариабельная область тяжелой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab первой молекулы Fab (VH(1)-CL(1)), и полипептид легкой цепи Fab второй молекулы Fab (VL(2)-CL(2)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид легкой цепи Fab третьей молекулы Fab (VL(3)-CL(3)).
В определенных аспектах (мультиспецифическое) антитело в соответствии с изобретением содержит полипептид, в котором вариабельная область тяжелой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab первой молекулы Fab (т.е. первая молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой константная область тяжелой цепи замещена константной областью легкой цепи), которая в свою очередь имеет общую карбокси-концевую пептидную связь с тяжелой цепью Fab второй молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с тяжелой цепью Fab третьей молекулы Fab (VH(1)-CL(1)-VH(2)-CH1(2)-VH(3)-CH1(3)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид, в котором вариабельная область легкой цепи Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab первой молекулы Fab (VL(1)-CH1(1)), и полипептид легкой цепи Fab второй молекулы Fab (VL(2)-CL(2)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид легкой цепи Fab третьей молекулы Fab (VL(3)-CL(3)).
В определенных аспектах (мультиспецифическое) антитело в соответствии с изобретением содержит полипептид, в котором тяжелая цепь Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с вариабельной областью легкой цепи Fab второй молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab второй молекулы Fab (т.е. вторая молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи), которая в свою очередь имеет общую карбокси-концевую пептидную связь с вариабельной областью легкой цепи Fab третьей молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab третьей молекулы Fab (т.е. третья молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи) (VH(1)-CH1(1)-VL(2)-CH1(2)-VL(3)-СН1(3)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид, в котором вариабельная область тяжелой цепи Fab второй молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab второй молекулы Fab (VH(2)-CL(2)), и полипептид легкой цепи Fab первой молекулы Fab (VL(1)-CL(1)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид, в котором вариабельная область тяжелой цепи Fab третьей молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab третьей молекулы Fab (VH(3)-CL(3)).
В определенных аспектах (мультиспецифическое) антитело в соответствии с изобретением содержит полипептид, в котором тяжелая цепь Fab первой молекулы Fab имеет общую карбокси-концевую пептидную связь с вариабельной областью тяжелой цепи Fab второй молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab второй молекулы Fab (т.е. вторая молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой константная область тяжелой цепи замещена константной областью легкой цепи), которая в свою очередь имеет общую карбокси-концевую пептидную связь с вариабельной областью тяжелой цепи Fab третьей молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab третьей молекулы Fab (т.е. третья молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой константная область тяжелой цепи замещена константной областью легкой цепи) (VH(1)-CH1(1)-VH(2)-CL(2)-VH(3)-CL(3)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид, в котором вариабельная область легкой цепи Fab второй молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab второй молекулы Fab (VL(2)-CH1(2)), и полипептид легкой цепи Fab первой молекулы Fab (VL(1)-CL(1)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид, в котором вариабельная область легкой цепи Fab третьей молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab третьей молекулы Fab (VL(3)-CH1(3)).
В определенных аспектах (мультиспецифическое) антитело в соответствии с изобретением содержит полипептид, в котором вариабельная область легкой цепи Fab третьей молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab третьей молекулы Fab (т.е. третья молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи), которая в свою очередь имеет общую карбокси-концевую пептидную связь с вариабельной областью легкой цепи Fab второй молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab второй молекулы Fab (т.е. вторая молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой вариабельная область тяжелой цепи замещена вариабельной областью легкой цепи), которая в свою очередь имеет общую карбокси-концевую пептидную связь с тяжелой цепью Fab первой молекулы Fab (VL(3)-CH1(3)-VL(2)-CH1(2)-VH(1)-CH1(1)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид, в котором вариабельная область тяжелой цепи Fab второй молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab второй молекулы Fab (VH(2)-CL(2)), и полипептид легкой цепи Fab первой молекулы Fab (VL(1)-CL(1)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид, в котором вариабельная область тяжелой цепи Fab третьей молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab третьей молекулы Fab (VH(3)-CL(3)).
В определенных аспектах (мультиспецифическое) антитело в соответствии с изобретением содержит полипептид, в котором вариабельная область тяжелой цепи Fab третьей молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab третьей молекулы Fab (т.е. третья молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой константная область тяжелой цепи замещена константной областью легкой цепи), которая в свою очередь имеет общую карбокси-концевую пептидную связь с вариабельной областью тяжелой цепи Fab второй молекулы Fab, которая в свою очередь имеет общую карбокси-концевую пептидную связь с константной областью легкой цепи Fab второй молекулы Fab (т.е. вторая молекула Fab содержит кроссоверную тяжелую цепь Fab, в которой константная область тяжелой цепи замещена константной областью легкой цепи), которая в свою очередь имеет общую карбокси-концевую пептидную связь с тяжелой цепью Fab первой молекулы Fab (VH(3)-CL(3)-VH(2)-CL(2)-VH(1)-CH1(1)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид, в котором вариабельная область легкой цепи Fab второй молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab второй молекулы Fab (VL(2)-CH1(2)), и полипептид легкой цепи Fab первой молекулы Fab (VL(1)-CL(1)). В некоторых аспектах (мультиспецифическое) антитело дополнительно содержит полипептид, в котором вариабельная область легкой цепи Fab третьей молекулы Fab имеет общую карбокси-концевую пептидную связь с константной областью тяжелой цепи Fab третьей молекулы Fab (VL(3)-CH1(3)).
В одном аспекте в изобретении предложено (мультиспецифическое) антитело, содержащее
а) первый антигенсвязывающий домен, который связывается с CD3, причем первый антигенсвязывающий домен представляет собой молекулу Fab, в которой вариабельные домены VL и VH или константные домены CL и СН1 легкой цепи Fab и тяжелой цепи Fab замещены друг другом, и содержит вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область тяжелой цепи (HCDR) 1 с SEQ ID NO: 2, HCDR 2 с SEQ ID NO: 3 и HCDR 3 с SEQ ID NO: 5, и вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область легкой цепи (LCDR) 1 с SEQ ID NO: 8, LCDR 2 с SEQ ID NO: 9 и LCDR 3 с SEQ ID NO: 10;
б) второй антигенсвязывающий домен, который связывается со вторым антигеном, в частности антигеном клетки-мишени, конкретнее TYRP-1 или EGFRvIII, причем второй антигенсвязывающий домен представляет собой (стандартную) молекулу Fab;
в) Fc-домен, состоящий из первой и второй субъединиц;
причем
(i) первый антигенсвязывающий домен по а) слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab второго антигенсвязывающего домена по б), а второй антигенсвязывающий домен по б) слит на С-конце тяжелой цепи Fab с N-концом одной из субъединиц Fc-домена по в), или
(ii) второй антигенсвязывающий домен по б) слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первого антигенсвязывающего домена по а), а первый антигенсвязывающий домен по а) слит на С-конце тяжелой цепи Fab с N-концом одной из субъединиц Fc-домена по в).
В предпочтительном аспекте в изобретении предложено (мультиспецифическое) антитело, содержащее
а) первый антигенсвязывающий домен, который связывается с CD3, причем первый антигенсвязывающий домен представляет собой молекулу Fab, в которой вариабельные домены VL и VH или константные домены CL и СН1 легкой цепи Fab и тяжелой цепи Fab замещены друг другом, и содержит вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область тяжелой цепи (HCDR) 1 с SEQ ID NO: 2, HCDR 2 с SEQ ID NO: 3 и HCDR 3 с SEQ ID NO: 5, и вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область легкой цепи (LCDR) 1 с SEQ ID NO: 8, LCDR 2 с SEQ ID NO: 9 и LCDR 3 с SEQ ID NO: 10;
б) второй и третий антигенсвязывающие домены, которые связываются со вторым антигеном, в частности антигеном клетки-мишени, конкретнее TYRP-1 или EGFRvIII, причем каждый из второго и третьего антигенсвязывающих доменов представляет собой (стандартную) молекулу Fab; и
в) Fc-домен, состоящий из первой и второй субъединиц;
причем
(i) первый антигенсвязывающий домен по а) слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab второго антигенсвязывающего домена по б), а второй антигенсвязывающий домен по б) и третий антигенсвязывающий домен по б), каждый, слиты на С-конце тяжелой цепи Fab с N-концом одной из субъединиц Fc-домена по в), или
(ii) второй антигенсвязывающий домен по б) слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первого антигенсвязывающего домена по а), а первый антигенсвязывающий домен по а) и третий антигенсвязывающий домен по б), каждый, слиты на С-конце тяжелой цепи Fab с N-концом одной из субъединиц Fc-домена по в).
В другом аспекте в изобретении предложено (мультиспецифическое) антитело, содержащее
а) первый антигенсвязывающий домен, который связывается с CD3, причем первый антигенсвязывающий домен представляет собой молекулу Fab, в которой вариабельные домены VL и VH или константные домены CL и СН1 легкой цепи Fab и тяжелой цепи Fab замещены друг другом, и содержит вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область тяжелой цепи (HCDR) 1 с SEQ ID NO: 2, HCDR 2 с SEQ ID NO: 3 и HCDR 3 с SEQ ID NO: 5, и вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область легкой цепи (LCDR) 1 с SEQ ID NO: 8, LCDR 2 с SEQ ID NO: 9 и LCDR 3 с SEQ ID NO: 10;
б) второй антигенсвязывающий домен, который связывается со вторым антигеном, в частности антигеном клетки-мишени, конкретнее TYRP-1 или EGFRvIII, причем второй антигенсвязывающий домен представляет собой (стандартную) молекулу Fab;
в) Fc-домен, состоящий из первой и второй субъединиц;
причем
(i) первый антигенсвязывающий домен по а) и второй антигенсвязывающий домен по б), каждый, слиты на С-конце тяжелой цепи Fab с N-концом одной из субъединиц Fc-домена по в).
Во всех из разных конфигураций (мультиспецифического) антитела в соответствии с изобретением описанные в данном документе аминокислотные замены («зарядовые модификации»), в случае наличия, могут находиться в доменах СН1 и CL второго и (в случае наличия) третьего антигенсвязывающего домена/молекулы Fab, или в доменах СН1 и CL первого антигенсвязывающего домена/молекулы Fab. Предпочтительно они находятся в доменах СН1 и CL второго и (в случае наличия) третьего антигенсвязывающего домена/молекулы Fab. В соответствии с концепцией изобретения, если описанные в данном документе аминокислотные замены проводят во втором (и, в случае наличия, третьем) антигенсвязывающем домене/молекуле Fab, в первом антигенсвязывающем домене/молекуле Fab такие аминокислотные замены не проводят.И наоборот, если описанные в данном документе аминокислотные замены проводят в первом антигенсвязывающем домене/молекуле Fab, во втором (и, в случае наличия, третьем) антигенсвязывающем домене/молекуле Fab такие аминокислотные замены не проводят.Аминокислотные замены предпочтительно проводят в (мультиспецифических) антителах, содержащих молекулу Fab, в которой вариабельные домены VL и VH1 легкой цепи Fab и тяжелой цепи Fab замещены друг другом.
В предпочтительных аспектах (мультиспецифического) антитела в соответствии с изобретением, в частности, в которых описанные в данном документе аминокислотные замены проводят во втором (и, в случае наличия, третьем) антигенсвязывающем домене/молекуле Fab, константный домен CL второй (и, в случае наличия, третьей) молекулы Fab принадлежит к изотипу каппа. В других аспектах (мультиспецифического) антитела в соответствии с изобретением, в частности, в которых описанные в данном документе аминокислотные замены проводят в первом антигенсвязывающем домене/молекуле Fab, константный домен CL первого антигенсвязывающего домена/молекулы Fab принадлежит к изотипу каппа. В некоторых аспектах константный домен CL второго (и, в случае наличия, третьего) антигенсвязывающего домена/молекулы Fab и константный домен CL первого антигенсвязывающего домена/молекулы Fab принадлежат к изотипу каппа.
В одном аспекте в изобретении предложено (мультиспецифическое) антитело, содержащее
а) первый антигенсвязывающий домен, который связывается с CD3, причем первый антигенсвязывающий домен представляет собой молекулу Fab, в которой вариабельные домены VL и VH легкой цепи Fab и тяжелой цепи Fab замещены друг другом, и содержит вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область тяжелой цепи (HCDR) 1 с SEQ ID NO: 2, HCDR 2 с SEQ ID NO: 3 и HCDR 3 с SEQ ID NO: 5, и вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область легкой цепи (LCDR) 1 с SEQ ID NO: 8, LCDR 2 с SEQ ID NO: 9 и LCDR 3 с SEQ ID NO: 10;
б) второй антигенсвязывающий домен, который связывается со вторым антигеном, в частности антигеном клетки-мишени, конкретнее TYRP-1 или EGFRvIII, причем второй антигенсвязывающий домен представляет собой (стандартную) молекулу Fab;
в) Fc-домен, состоящий из первой и второй субъединиц;
причем в константном домене CL второго антигенсвязывающего домена по б) аминокислота в позиции 124 замещена лизином (К) (нумерация в соответствии с Kabat) и аминокислота в позиции 123 замещена лизином (К) или аргинином (R) (нумерация в соответствии с Kabat) (наиболее предпочтительно аргинином (R)), и при этом в константном домене СН1 второго антигенсвязывающего домена по б) аминокислота в позиции 147 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat) и аминокислота в позиции 213 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat); и
причем
(i) первый антигенсвязывающий домен по а) слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab второго антигенсвязывающего домена по б), а второй антигенсвязывающий домен по б) слит на С-конце тяжелой цепи Fab с N-концом одной из субъединиц Fc-домена по в), или
(ii) второй антигенсвязывающий домен по б) слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первого антигенсвязывающего домена по а), а первый антигенсвязывающий домен по а) слит на С-конце тяжелой цепи Fab с N-концом одной из субъединиц Fc-домена по в).
В предпочтительном аспекте в изобретении предложено (мультиспецифическое) антитело, содержащее
а) первый антигенсвязывающий домен, который связывается с CD3, причем первый антигенсвязывающий домен представляет собой молекулу Fab, в которой вариабельные домены VL и VH легкой цепи Fab и тяжелой цепи Fab замещены друг другом, и содержит вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область тяжелой цепи (HCDR) 1 с SEQ ID NO: 2, HCDR 2 с SEQ ID NO: 3 и HCDR 3 с SEQ ID NO: 5, и вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область легкой цепи (LCDR) 1 с SEQ ID NO: 8, LCDR 2 с SEQ ID NO: 9 и LCDR 3 с SEQ ID NO: 10;
б) второй и третий антигенсвязывающие домены, которые связываются со вторым антигеном, в частности антигеном клетки-мишени, конкретнее TYRP-1 или EGFRvIII, причем каждый из второго и третьего антигенсвязывающих доменов представляет собой (стандартную) молекулу Fab; и
в) Fc-домен, состоящий из первой и второй субъединиц;
причем в константном домене CL второго антигенсвязывающего домена по б) и третьего антигенсвязывающего домена по б) аминокислота в позиции 124 замещена лизином (К) (нумерация в соответствии с Kabat) и аминокислота в позиции 123 замещена лизином (К) или аргинином (R) (нумерация в соответствии с Kabat) (наиболее предпочтительно аргинином (R)), и при этом в константном домене СН1 второго антигенсвязывающего домена по б) и третьего антигенсвязывающего домена по б) аминокислота в позиции 147 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat) и аминокислота в позиции 213 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat); и
причем
(i) первый антигенсвязывающий домен по а) слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab второго антигенсвязывающего домена по б), а второй антигенсвязывающий домен по б) и третий антигенсвязывающий домен по б), каждый, слиты на С-конце тяжелой цепи Fab с N-концом одной из субъединиц Fc-домена по в), или
(ii) второй антигенсвязывающий домен по б) слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первого антигенсвязывающего домена по а), а первый антигенсвязывающий домен по а) и третий антигенсвязывающий домен по б), каждый, слиты на С-конце тяжелой цепи Fab с N-концом одной из субъединиц Fc-домена по в).
В другом аспекте в изобретении предложено (мультиспецифическое) антитело, содержащее
а) первый антигенсвязывающий домен, который связывается с CD3, причем первый антигенсвязывающий домен представляет собой молекулу Fab, в которой вариабельные домены VL и VH легкой цепи Fab и тяжелой цепи Fab замещены друг другом, и содержит вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область тяжелой цепи (HCDR) 1 с SEQ ID NO: 2, HCDR 2 с SEQ ID NO: 3 и HCDR 3 с SEQ ID NO: 5, и вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область легкой цепи (LCDR) 1 с SEQ ID NO: 8, LCDR 2 с SEQ ID NO: 9 и LCDR 3 с SEQ ID NO: 10;
б) второй антигенсвязывающий домен, который связывается со вторым антигеном, в частности антигеном клетки-мишени, конкретнее TYRP-1 или EGFRvIII, причем второй антигенсвязывающий домен представляет собой (стандартную) молекулу Fab;
в) Fc-домен, состоящий из первой и второй субъединиц;
причем в константном домене CL второго антигенсвязывающего домена по б) аминокислота в позиции 124 замещена лизином (К) (нумерация в соответствии с Kabat) и аминокислота в позиции 123 замещена лизином (К) или аргинином (R) (нумерация в соответствии с Kabat) (наиболее предпочтительно аргинином (R)), и при этом в константном домене СН1 второго антигенсвязывающего домена по б) аминокислота в позиции 147 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat) и аминокислота в позиции 213 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat); и
первый антигенсвязывающий домен по а) и второй антигенсвязывающий домен по б), каждый, слиты на С-конце тяжелой цепи Fab с N-концом одной из субъединиц Fc-домена по в).
В соответствии с любым из вышеприведенных аспектов компоненты (мультиспецифического) антитела (например, молекулы Fab, Fc-домен) могут быть слиты напрямую или посредством различных линкеров, в частности, пептидных линкеров, содержащих одну или более аминокислот, как правило, около 2-20 аминокислот, которые описаны в данном документе или известны в данной области техники. Подходящие неиммуногенные пептидные линкеры включают, например, пептидные линкеры (G4S)n, (SG4)n, (G4S)n или G4(SG4)n, где n в общем случае является целым числом от 1 до 10, как правило, от 2 до 4.
В предпочтительном аспекте в изобретении предложено (мультиспецифическое) антитело, содержащее
а) первый антигенсвязывающий домен, который связывает CD3, причем первый антигенсвязывающий домен представляет собой молекулу Fab, в которой вариабельные домены VL и VH легкой цепи Fab и тяжелой цепи Fab замещены друг другом, и содержит вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область тяжелой цепи (HCDR) 1 с SEQ ID NO: 2, HCDR 2 с SEQ ID NO: 3 и HCDR 3 с SEQ ID NO: 5, и вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область легкой цепи (LCDR) 1 с SEQ ID NO: 8, LCDR 2 с SEQ ID NO: 9 и LCDR 3 с SEQ ID NO: 10;
б) второй и третий антигенсвязывающие домены, которые связываются с TYRP-1, причем каждый из второго и третьего антигенсвязывающих доменов представляет собой (стандартную) молекулу Fab и содержит вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область тяжелой цепи (HCDR) 1 с SEQ ID NO: 15, HCDR 2 с SEQ ID NO: 16 и HCDR 3 с SEQ ID NO: 17, и вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область легкой цепи (LCDR) 1 с SEQ ID NO: 19, LCDR 2 с SEQ ID NO: 20 и LCDR 3 с SEQ ID NO: 21;
в) Fc-домен, состоящий из первой и второй субъединиц;
причем
в константном домене CL второго и третьего антигенсвязывающего домена по б) аминокислота в позиции 124 замещена лизином (К) (нумерация в соответствии с Kabat) и аминокислота в позиции 123 замещена лизином (К) или аргинином (R) (нумерация в соответствии с Kabat) (наиболее предпочтительно аргинином (R)), и при этом в константном домене СН1 второго и третьего антигенсвязывающего домена по б) аминокислота в позиции 147 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat) и аминокислота в позиции 213 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat);
и при этом дополнительно
второй антигенсвязывающий домен по б) слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первого антигенсвязывающего домена по а), а первый антигенсвязывающий домен по а) и третий антигенсвязывающий домен по б), каждый, слиты на С-конце тяжелой цепи Fab с N-концом одной из субъединиц Fc-домена по в).
В дополнительном предпочтительном аспекте в изобретении предложено (мультиспецифическое) антитело, содержащее
а) первый антигенсвязывающий домен, который связывается с CD3, причем первый антигенсвязывающий домен представляет собой молекулу Fab, в которой вариабельные домены VL и VH легкой цепи Fab и тяжелой цепи Fab замещены друг другом, и содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 7, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 11;
б) второй и третий антигенсвязывающие домены, которые связываются с TYRP-1, причем каждый из второго и третьего антигенсвязывающих доменов представляет собой (стандартную) молекулу Fab и содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 18, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 22;
в) Fc-домен, состоящий из первой и второй субъединиц;
причем
в константном домене CL второго и третьего антигенсвязывающего домена по б) аминокислота в позиции 124 замещена лизином (К) (нумерация в соответствии с Kabat) и аминокислота в позиции 123 замещена лизином (К) или аргинином (R) (нумерация в соответствии с Kabat) (наиболее предпочтительно аргинином (R)), и при этом в константном домене СН1 второго и третьего антигенсвязывающего домена по б) аминокислота в позиции 147 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat) и аминокислота в позиции 213 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat);
и при этом дополнительно
второй антигенсвязывающий домен по б) слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первого антигенсвязывающего домена по а), а первый антигенсвязывающий домен по а) и третий антигенсвязывающий домен по б), каждый, слиты на С-конце тяжелой цепи Fab с N-концом одной из субъединиц Fc-домена по в).
В предпочтительном аспекте в изобретении предложено (мультиспецифическое) антитело, содержащее
а) первый антигенсвязывающий домен, который связывает CD3, причем первый антигенсвязывающий домен представляет собой молекулу Fab, в которой вариабельные домены VL и VH легкой цепи Fab и тяжелой цепи Fab замещены друг другом, и содержит вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область тяжелой цепи (HCDR) 1 с SEQ ID NO: 2, HCDR 2 с SEQ ID NO: 3 и HCDR 3 с SEQ ID NO: 5, и вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область легкой цепи (LCDR) 1 с SEQ ID NO: 8, LCDR 2 с SEQ ID NO: 9 и LCDR 3 с SEQ ID NO: 10;
б) второй и третий антигенсвязывающие домены, которые связываются с EGFRvIII, причем каждый из второго и третьего антигенсвязывающих доменов представляет собой (стандартную) молекулу Fab и содержит вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область тяжелой цепи (HCDR) 1 с SEQ ID NO: 85, HCDR 2 с SEQ ID NO: 86 и HCDR 3 с SEQ ID NO: 87, и вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область легкой цепи (LCDR) 1 с SEQ ID NO: 89, LCDR 2 с SEQ ID NO: 90 и LCDR 3 с SEQ ID NO: 91;
в) Fc-домен, состоящий из первой и второй субъединиц;
причем
в константном домене CL второго и третьего антигенсвязывающего домена по б) аминокислота в позиции 124 замещена лизином (К) (нумерация в соответствии с Kabat) и аминокислота в позиции 123 замещена лизином (К) или аргинином (R) (нумерация в соответствии с Kabat) (наиболее предпочтительно аргинином (R)), и при этом в константном домене СН1 второго и третьего антигенсвязывающего домена по б) аминокислота в позиции 147 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat) и аминокислота в позиции 213 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat);
и при этом дополнительно
второй антигенсвязывающий домен по б) слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первого антигенсвязывающего домена по а), а первый антигенсвязывающий домен по а) и третий антигенсвязывающий домен по б), каждый, слиты на С-конце тяжелой цепи Fab с N-концом одной из субъединиц Fc-домена по в).
В дополнительном предпочтительном аспекте в изобретении предложено (мультиспецифическое) антитело, содержащее
а) первый антигенсвязывающий домен, который связывается с CD3, причем первый антигенсвязывающий домен представляет собой молекулу Fab, в которой вариабельные домены VL и VH легкой цепи Fab и тяжелой цепи Fab замещены друг другом, и содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 7, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 11;
б) второй и третий антигенсвязывающие домены, которые связываются с EGFRvIII, причем каждый из второго и третьего антигенсвязывающих доменов представляет собой (стандартную) молекулу Fab и содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 88, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 92;
в) Fc-домен, состоящий из первой и второй субъединиц;
причем
в константном домене CL второго и третьего антигенсвязывающего домена по б) аминокислота в позиции 124 замещена лизином (К) (нумерация в соответствии с Kabat) и аминокислота в позиции 123 замещена лизином (К) или аргинином (R) (нумерация в соответствии с Kabat) (наиболее предпочтительно аргинином (R)), и при этом в константном домене СН1 второго и третьего антигенсвязывающего домена по б) аминокислота в позиции 147 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat) и аминокислота в позиции 213 замещена глутаминовой кислотой (Е) (нумерация в соответствии с индексом EU по Kabat);
и при этом дополнительно
второй антигенсвязывающий домен по б) слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первого антигенсвязывающего домена по а), а первый антигенсвязывающий домен по а) и третий антигенсвязывающий домен по б), каждый, слиты на С-конце тяжелой цепи Fab с N-концом одной из субъединиц Fc-домена по в).
В одном аспекте в соответствии с этими аспектами изобретения в первой субъединице Fc-домена остаток треонина в позиции 366 замещен остатком триптофана (T366W), а во второй субъединице Fc-домена остаток тирозина в позиции 407 замещен остатком валина (Y407V) и, необязательно, остаток треонина в позиции 366 замещен остатком серина (T366S), а остаток лейцина в позиции 368 замещен остатком аланина (L368A) (нумерация в соответствии с индексом EU по Kabat).
В дополнительном аспекте в соответствии с этими аспектами изобретения в первой субъединице Fc-домена дополнительно остаток серина в позиции 354 замещен остатком цистеина (S354C) или остаток глутаминовой кислоты в позиции 356 замещен остатком цистеина (Е356С) (в частности остаток серина в позиции 354 замещен остатком цистеина), а во второй субъединице Fc-домена дополнительно остаток тирозина в позиции 349 замещен остатком цистеина (Y349C) (нумерация в соответствии с индексом EU по Kabat).
В дополнительном аспекте в соответствии с этими аспектами изобретения в каждой из первой и второй субъединиц Fc-домена остаток лейцина в позиции 234 замещен остатком аланина (L234A), остаток лейцина в позиции 235 замещен остатком аланина (L235A) и остаток пролина в позиции 329 замещен остатком глицина (P329G) (нумерация в соответствии с индексом EU по Kabat).
В дополнительном аспекте в соответствии с этими аспектами изобретения Fc-домен представляет собой Fc-домен IgG1 человека.
В предпочтительном конкретном аспекте (мультиспецифическое) антитело содержит полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 23, полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 24, полипептид (в частности два полипептида), содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 25, и полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 27. В дополнительном предпочтительном конкретном аспекте (мультиспецифическое) антитело содержит полипептид, содержащий аминокислотную последовательность SEQ ID NO: 23, полипептид, содержащий аминокислотную последовательность SEQ ID NO: 24, полипептид (в частности два полипептида), содержащий аминокислотную последовательность SEQ ID NO: 25, и полипептид, содержащий аминокислотную последовательность SEQ ID NO: 27.
В одном аспекте в изобретении предложено (мультиспецифическое) антитело, которое связывается с CD3 и TYRP-1, содержащее полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 23, полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 24, полипептид (в частности два полипептида), содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 25, и полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 27. В одном аспекте в изобретении предложено (мультиспецифическое) антитело, которое связывается с CD3 и TYRP-1, содержащее полипептид, содержащий аминокислотную последовательность SEQ ID NO: 23, полипептид, содержащий аминокислотную последовательность SEQ ID NO: 24, полипептид (в частности два полипептида), содержащий аминокислотную последовательность SEQ ID NO: 25, и полипептид, содержащий аминокислотную последовательность SEQ ID NO: 27.
В дополнительном предпочтительном конкретном аспекте (мультиспецифическое) антитело содержит полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 109, полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 110, полипептид (в частности два полипептида), содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 111, и полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 27. В дополнительном конкретном аспекте (мультиспецифическое) антитело содержит полипептид, содержащий аминокислотную последовательность SEQ ID NO: 109, полипептид, содержащий аминокислотную последовательность SEQ ID NO: 110, полипептид (в частности, два полипептида), содержащий аминокислотную последовательность SEQ ID NO: 111, и полипептид, содержащий аминокислотную последовательность SEQ ID NO: 27.
В одном аспекте в изобретении предложено (мультиспецифическое) антитело, которое связывается с CD3 и EGFRvIII, содержащее полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 109, полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 110, полипептид (в частности два полипептида), содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 111, и полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 27. В одном аспекте в изобретении предложено (мультиспецифическое) антитело, которое связывается с CD3 и EGFRvIII, содержащее полипептид, содержащий аминокислотную последовательность SEQ ID NO: 109, полипептид, содержащий аминокислотную последовательность SEQ ID NO: 110, полипептид (в частности два полипептида), содержащий аминокислотную последовательность SEQ ID NO: 111, и полипептид, содержащий аминокислотную последовательность SEQ ID NO: 27.
8. Варианты Fc-доменов
В предпочтительных аспектах (мультиспецифическое) антитело по изобретению содержит Fc-домен, состоящий из первой и второй субъединиц.
Fc-домен (мультиспецифического) антитела состоит из пары полипептидных цепей, содержащих домены тяжелой цепи молекулы иммуноглобулина. Например, Fc-домен молекулы иммуноглобулина G (IgG) представляет собой димер, каждая субъединица которого содержит константные домены тяжелой цепи СН2 и CH3 IgG. Две субъединицы Fc-домена способны к стабильной ассоциации друг с другом. В одном аспекте (мультиспецифическое) антитело по изобретению содержит не более одного Fc-домена.
В одном аспекте Fc-домен (мультиспецифического) антитела представляет собой Fc-домен IgG. В предпочтительном аспекте Fc-домен представляет собой Fc-домен IgG1. В другом аспекте Fc-домен представляет собой Fc-домен IgG1. В более конкретном аспекте аспекте Fc-домен представляет собой Fc-домен IgG4, содержащий аминокислотную замену в позиции S228 (нумерация в соответствии с индексом EU по Kabat), в частности аминокислотную замену S228P. Эта аминокислотная замена снижает in vivo обмен плеч Fab антител IgG4 (смотрите Stubenrauch et al., Drug Metabolism and Disposition 38, 84-91 (2010)). В дополнительном предпочтительном аспекте Fc-домен представляет собой Fc-домен человека. В даже более предпочтительном аспекте Fc-домен представляет собой Fc-домен IgG1 человека. Типовая последовательность Fc-области IgG1 человека приведена в SEQ ID NO: 117.
а) Модификации Fc-домена, способствующие гетеродимеризации
(Мультиспецифические) антитела в соответствии с изобретением содержат разные антигенсвязывающие домены, которые могут быть слиты с одной или другой из двух субъединиц Fc-домена, таким образом, две субъединицы Fc-домена, как правило, содержатся в двух неидентичных полипептидных цепях. Рекомбинантная коэкспрессия этих полипептидов и последующая димеризация приводят к нескольким возможным комбинациям двух полипептидов. Следовательно, для улучшения выхода и чистоты (мультиспецифических) антител при рекомбинантном получении будет целесообразно внести в Fc-домен (мультиспецифического) антитела модификацию, способствующую ассоциации необходимых полипептидов.
Соответственно, в предпочтительных аспектах Fc-домен (мультиспецифического) антитела в соответствии с изобретением содержит модификацию, способствующую ассоциации первой и второй субъединиц Fc-домена. Сайтом наиболее интенсивного белок-белкового взаимодействия между двумя субъединицами Fc-домена IgG человека является СН3-домен Fc-домена. Таким образом, в одном аспекте указанная модификация находится в СН3-домене Fc-домена.
Существует несколько подходов для модификации в СН3-домене Fc-домена с целью стимуляции гетеродимеризации, которые хорошо описаны, например, в WO 96/27011, WO 98/050431, ЕР 1870459, WO 2007/110205, WO 2007/147901, WO 2009/089004, WO 2010/129304, WO 2011/90754, WO 2011/143545, WO 2012058768, WO 2013157954, WO 2013096291. Как правило, во всех таких подходах СН3-домен первой субъединицы Fc-домена и СН3-домен второй субъединицы Fc-домена сконструированы комплементарным образом так, чтобы каждый СН3-домен (или содержащая его тяжелая цепь) больше не мог образовывать гомодимер сам с собой, но был бы вынужден образовывать гетеродимер с комплементарно сконструированным другим СН3-доменом (так, чтобы происходила гетеродимеризация первого и второго СН3-доменов и не происходило образование гомодимеров между двумя первыми или двумя вторыми СН3-доменами). Эти разные подходы для улучшения гетеродимеризации тяжелой цепи предусмотрены как разные альтернативные варианты в комбинации с модификациями тяжелой-легкой цепи (например, обмен/замена VH и VL в одном связывающем плече и внесение замен из заряженных аминокислот с противоположными зарядами в поверхность контакта CH1/CL) в (мультиспецифическом) антителе, что снижает неправильное спаривание тяжелой/легкой цепи и количество побочных продуктов Бенс-Джонса.
В конкретном аспекте указанные модификации, способствующие ассоциации первой и второй субъединиц Fc-домена, представляют собой так называемую модификацию «выступ-во-впадину», включающую модификацию «выступа» в одной из двух субъединиц Fc-домена и модификацию «впадины» в другой из двух субъединиц Fc-домена.
Технология «выступ-во-впадину» описана, например, в US 5731168; US 7695936; Ridgway et al., Prot Eng 9, 617-621 (1996) и Carter, J Immunol Meth 248, 7-15 (2001). Как правило, это способ включает внесение выпуклости («выступа») на поверхности контакта первого полипептида и соответствующей полости («впадины») на поверхности контакта второго полипептида, так, чтобы выпуклость могла располагаться в полости, чтобы способствовать образованию гетеродимера и затруднять образование гомодимера. Выпуклости конструируют путем замещения небольших боковых цепей аминокислот на поверхности контакта первого полипептида более крупными боковыми цепями (например тирозина или триптофана). Компенсирующие полости идентичного или сходного размера с выпуклостями создают на поверхности контакта второго полипептида путем замены крупных боковых цепей аминокислот на более мелкие (например, аланина или треонина).
Соответственно, в предпочтительном аспекте в СН3-домене первой субъединицы Fc-домена (мультиспецифического) антитела аминокислотный остаток замещают аминокислотным остатком, имеющим больший объем боковой цепи, с созданием, таким образом, выпуклости в СН3-домене первой субъединицы, которая может размещаться в полости в СН3-домене второй субъединицы, а в СН3-домене второй субъединицы Fc-домена аминокислотный остаток замещают аминокислотным остатком, имеющим меньший объем боковой цепи, с созданием, таким образом, полости в СН3-домене второй субъединицы, в которой может размещаться выпуклость в СН3-домене первой субъединицы.
Предпочтительно указанный аминокислотный остаток, имеющий больший объем боковой цепи, выбран из группы, состоящей из аргинина (R), фенилаланина (F), тирозина (Y) и триптофана (W).
Предпочтительно указанный аминокислотный остаток, имеющий меньший объем боковой цепи, выбран из группы, состоящей из аланина (А), серина (S), треонина (Т) и валина (V).
Выпуклость и полость можно создавать путем изменения нуклеиновой кислоты, кодирующей полипептиды, например, с помощью сайт-специфического мутагенеза, или путем пептидного синтеза.
В конкретном аспекте в (СН3-домене) первой субъединице Fc-домена (субъединице «выступов») остаток треонина в позиции 366 замещен остатком триптофана (T366W), а во (СН3-домене) второй субъединице Fc-домена (субъединице «впадин») остаток тирозина в позиции 407 замещен остатком валина (Y407V). В одном аспекте во второй субъединице Fc-домена дополнительно остаток треонина в позиции 366 замещен остатком серина (T366S), а остаток лейцина в позиции 368 замещен остатком аланина (L368A) (нумерация в соответствии с индексом EU по Kabat).
В дополнительном аспекте в первой субъединице Fc-домена дополнительно остаток серина в позиции 354 замещен остатком цистеина (S354C) или остаток глутаминовой кислоты в позиции 356 замещен остатком цистеина (Е356С) (в частности остаток серина в позиции 354 замещен остатком цистеина), а во второй субъединице Fc-домена дополнительно остаток тирозина в позиции 349 замещен остатком цистеина (Y349C) (нумерация в соответствии с индексом EU по Kabat). Внесение двух остатков цистеина приводит к образованию дисульфидного мостика между двумя субъединицами Fc-домена, дополнительно стабилизируя димер (Carter, J Immunol Methods 248, 7-15 (2001)).
В предпочтительных аспектах первая субъединица Fc-домена содержит аминокислотные замены S354C и T366W, а вторая субъединица Fc-домена содержит аминокислотные замены Y349C, T366S, L368A и Y407V (нумерация в соответствии с индексом EU по Kabat).
В предпочтительном аспекте антигенсвязывающий домен, который связывается с CD3, слит (необязательно посредством второго антигенсвязывающего домена, который связывается со вторым антигеном, и/или пептидного линкера) с первой субъединицей Fc-домена (содержащей модификацию «выступа»). Не ограничиваясь теорией, слияние антигенсвязывающего домена, который связывается с CD3, с содержащей выступ субъединицей Fc-домена будет (дополнительно) сводить к минимуму создание антител, содержащих два антигенсвязывающих домена, которые связываются с CD3 (вследствие стерического несоответствия двух содержащих выступ полипептидов).
Другие методики СН3-модификации для стимуляции гетеродимеризации предусмотрены как альтернативные варианты в соответствии с изобретением и описаны, например, в WO 96/27011, WO 98/050431, ЕР 1870459, WO 2007/110205, WO 2007/147901, WO 2009/089004, WO 2010/129304, WO 2011/90754, WO 2011/143545, WO 2012/058768, WO 2013/157954, WO 2013/096291.
В одном аспекте подход гетеродимеризации, описанный в ЕР 1870459, используют в качестве альтернативы. Этот подход основан на внесении заряженных аминокислот с противоположными зарядами в определенных аминокислотных позициях в поверхности контакта доменов CH3/CH3 между двумя субъединицами Fc-домена. Конкретным аспектом для (мультиспецифического) антитела по изобретению являются аминокислотные мутации R409D; К370Е в одном из двух СН3-доменов (Fc-домена) и аминокислотные мутации D399K; Е357К в другом из двух СН3-доменов Fc-домена (нумерация в соответствии с индексом EU по Kabat).
В другом аспекте (мультиспецифическое) антитело по изобретению содержит аминокислотную мутацию T366W в СН3-домене первой субъединицы Fc-домена и аминокислотные мутации T366S, L368A, Y407V в СН3-домене второй субъединицы Fc-домена, и дополнительно аминокислотные мутации R409D; К370Е в СН3-домене первой субъединицы Fc-домена и аминокислотные мутации D399K; Е357К в СН3-домене второй субъединицы Fc-домена (нумерация в соответствии с индексом EU по Kabat).
В другом аспекте (мультиспецифическое) антитело по изобретению содержит аминокислотные мутации S354C, T366W в СН3-домене первой субъединицы Fc-домена и аминокислотные мутации Y349C, T366S, L368A, Y407V в СН3-домене второй субъединицы Fc-домена, или указанное (мультиспецифическое) антитело содержит аминокислотные мутации Y349C, T366W в СН3-домене первой субъединицы Fc-домена и аминокислотные мутации S354C, T366S, L368A, Y407V в СН3-доменах второй субъединицы Fc-домена и дополнительно аминокислотные мутации R409D; К370Е в СН3-домене первой субъединицы Fc-домена и аминокислотные мутации D399K; Е357К в СН3-домене второй субъединицы Fc-домена (вся нумерация в соответствии с индексом EU по Kabat).
В одном аспекте подход гетеродимеризации, описанный в WO 2013/157953, используют в качестве альтернативы. В одном аспекте первый СН3-домен содержит аминокислотную мутацию Т366К, а второй СН3-домен содержит аминокислотную мутацию L351D (нумерация в соответствии с индексом EU по Kabat). В дополнительном аспекте первый СН3-домен содержит дополнительную аминокислотную мутацию L351K. В дополнительном аспекте второй СН3-домен содержит дополнительную аминокислотную мутацию, выбранную из Y349E, Y349D и L368E (в частности L368E) (нумерация в соответствии с индексом EU по Kabat).
В одном аспекте подход гетеродимеризации, описанный в WO 2012/058768, используют в качестве альтернативы. В одном аспекте первый СН3-домен содержит аминокислотные мутации L351Y, Y407A, а второй СН3-домен содержит аминокислотные мутации Т366А, K409F. В дополнительном аспекте второй СН3-домен содержит дополнительную аминокислотную мутацию в позиции Т411, D399, S400, F405, N390 или К392, например, выбранную из а) T411N, T411R, T411Q, Т411К, T411D, Т411Е или T411W, б) D399R, D399W, D399Y или D399K, в) S400E, S400D, S400R или S400K, г) F405I, F405M, F405T, F405S, F405V или F405W, д) N390R, N390K или N390D, е) K392V, К392М, K392R, K392L, K392F или К392Е (нумерация в соответствии с индексом EU по Kabat). В дополнительном аспекте первый СН3-домен содержит аминокислотные мутации L351Y, Y407A, а второй СН3-домен содержит аминокислотные мутации T366V, K409F. В дополнительном аспекте первый СН3-домен содержит аминокислотную мутацию Y407A, а второй СН3-домен содержит аминокислотные мутации Т366А, K409F. В дополнительном аспекте второй СН3-домен дополнительно содержит аминокислотные мутации К392Е, Т411Е, D399R и S400R (нумерация в соответствии с индексом EU по Kabat).
В одном аспекте подход гетеродимеризации, описанный в WO 2011/143545, используют в качестве альтернативы, например, с аминокислотной модификацией в позиции, выбранной из группы, состоящей из 368 и 409 (нумерация в соответствии с индексом EU по Kabat).
В одном аспекте подход гетеродимеризации, описанный в WO 2011/090762, в котором также используется описанная выше технология «выступы-во впадины», используют в качестве альтернативы. В одном аспекте первый СН3-домен содержит аминокислотную мутацию T366W, а второй СН3-домен содержит аминокислотную мутацию Y407A. В одном аспекте первый СН3-домен содержит аминокислотную мутацию T366Y, а второй СН3-домен содержит аминокислотную мутацию Y407T (нумерация в соответствии с индексом EU по Kabat).
В одном аспекте (мультиспецифическое) антитело или его Fc-домен относится к подклассу IgG2, и в качестве альтернативы используют подход гетеродимеризации, описанный в WO 2010/129304.
В альтернативном аспекте, модификация, способствующая ассоциации первой и второй субъединиц Fc-домена, включает модификацию, которая опосредует эффекты электростатического взаимодействия, например, как описано в публикации РСТ WO 2009/089004. В общем случае этот способ включает замещение одного из аминокислотных остатков на поверхности контакта субъединиц Fc-домена заряженным аминокислотным остатком так, чтобы образование гомодимера становилось электростатически невыгодным, но гетеродимеризация была бы электростатически выгодной. В одном таком аспекте первый СН3-домен содержит аминокислотную замену К392 или N392 отрицательно заряженной аминокислотой (например, глутаминовой кислотой (Е) или аспарагиновой кислотой (D), в частности K392D или N392D), а второй СН3-домен содержит аминокислотную замену D399, Е356, D356 или Е357 положительно заряженной аминокислотой (например, лизином (К) или аргинином (R), в частности D399K, Е356К, D356K или Е357К, и конкретнее D399K и Е356К). В дополнительном аспекте первый СН3-домен дополнительно содержит аминокислотную замену К409 или R409 отрицательно заряженной аминокислотой (например, глутаминовой кислотой (Е) или аспарагиновой кислотой (D), в частности K409D или R409D). В дополнительном аспекте первый СН3-домен дополнительно или в качестве альтернативы содержит аминокислотную замену К439 и/или К370 отрицательно заряженной аминокислотой (например, глутаминовой кислотой (Е) или аспарагиновой кислотой (D)) (вся нумерация в соответствии с индексом EU по Kabat).
В дополнительном аспекте подход гетеродимеризации, описанный в WO 2007/147901, используют в качестве альтернативы. В одном аспекте первый СН3-домен содержит аминокислотные мутации К253Е, D282K и K322D, а второй СН3-домен содержит аминокислотные мутации D239K, Е240К и K292D (нумерация в соответствии с индексом EU по Kabat).
В другом аспекте подход гетеродимеризации, описанный в WO 2007/110205, можно использовать в качестве альтернативы.
В одном аспекте первая субъединица Fc-домена содержит аминокислотные замены K392D и K409D, а вторая субъединица Fc-домена содержит аминокислотные замены D356K и D399K (нумерация в соответствии с индексом EU по Kabat).
б) Модификации Fc-домена, снижающие связывание Fc-рецептора и/или эффекторную функцию
Fc-домен придает (мультиспецифическому) антителу благоприятные фармакокинетические свойства, включая продолжительное сывороточное время полужизни, которое способствует хорошему накоплению в целевой ткани и благоприятному соотношению распределения в тканях и крови. Однако в то же время это может привести к нежелательному нацеливанию (мультиспецифического) антитела на клетки, экспрессирующие Fc-рецепторы, а не на предпочтительные несущие антиген клетки. Кроме того, совместная активация путей сигнализации Fc-рецепторов может приводить к высвобождению цитокинов, что в комбинации со свойствами активации Т-клеток и продолжительным временем полужизни (мультиспецифического) антитела приводит к чрезмерной активации цитокиновых рецепторов и тяжелым побочным эффектами после системного введения. Активация (несущих Fc-рецепторы) иммунных клеток, отличных от Т-клеток, может даже снизить эффективность (мультиспецифического) антитела вследствие потенциального разрушения Т-клеток, например, NK-клетками.
Соответственно, в предпочтительных аспектах Fc-домен (мультиспецифического) антитела в соответствии с изобретением демонстрирует сниженную аффинность связывания с Fc-рецептором и/или сниженную эффекторную функцию по сравнению с нативным Fc-доменом IgG1. В одном таком аспекте Fc-домен (или (мультиспецифическое) антитело, содержащее указанный Fc-домен) демонстрирует менее 50%, в частности менее 20%, конкретнее менее 10% и наиболее конкретно менее 5% аффинности связывания с Fc-рецептором по сравнению с нативным Fc-доменом IgG1 (или (мультиспецифическим) антителом, содержащим нативный Fc-домен IgG1), и/или менее 50%, в частности менее 20%, конкретнее менее 10% и наиболее конкретно менее 5% эффекторной функции по сравнению с нативным Fc-доменом IgG1 (или (мультиспецифическим) антителом, содержащим нативный Fc-домен IgG1). В одном аспекте Fc-домен (или (мультиспецифическое) антитело, содержащее указанный Fc-домен) практически не связывается с Fc-рецептором и/или не индуцирует эффекторную функцию. В предпочтительном аспекте Fc-рецептор представляет собой Fcγ-рецептор. В одном аспекте Fc-рецептор представляет собой Fc-рецептор человека. В одном аспекте Fc-рецептор представляет собой активирующий Fc-рецептор. В конкретном аспекте Fc-рецептор представляет собой активирующий Fcγ-рецептор человека, конкретнее FcγRIIIa, FcγRI или FcγRIIa человека, наиболее конкретно FcγRIIIa человека. В одном аспекте эффекторная функция представляет собой одну или более функций, выбранных из группы из КЗЦ, АЗКЦ, АЗКФ и секреции цитокинов. В предпочтительном аспекте эффекторная функция представляет собой АЗКЦ. В одном аспекте Fc-домен демонстрирует практически аналогичную аффинность связывания с неонатальным Fc-рецептором (FcRn) по сравнению с нативным Fc-доменом IgG1. Практически аналогичное связывание с FcRn достигается, когда Fc-домен (или (мультиспецифическое) антитело, содержащее указанный Fc-домен) демонстрирует более чем около 70%, в частности более чем около 80%, конкретнее более чем около 90% аффинности связывания нативного Fc-домена IgG1 (или (мультиспецифического) антитела, содержащего нативный Fc-домен IgG1) с FcRn.
В определенных аспектах Fc-домен конструируют так, чтобы он имел сниженную аффинность связывания с Fc-рецептором и/или сниженную эффекторную функцию по сравнению с не сконструированным Fc-доменом. В предпочтительных аспектах Fc-домен (мультиспецифического) антитела содержит одну или более аминокислотных мутаций, которые уменьшают аффинность связывания Fc-домена с Fc-рецептором и/или эффекторную функцию. Как правило, в каждой из двух субъединиц Fc-домена присутствуют одинаковые одна или более аминокислотных мутаций. В одном аспекте аминокислотная мутация снижает аффинность связывания Fc-домена с Fc-рецептором. В одном аспекте аминокислотная мутация снижает аффинность связывания Fc-домена с Fc-рецептором по меньшей мере в 2 раза, по меньшей мере в 5 раз или по меньшей мере в 10 раз. В аспектах, в которых присутствует более одной аминокислотной мутации, которая снижает аффинность связывания Fc-домена с Fc-рецептором, комбинация этих аминокислотных мутаций может снижать аффинность связывания Fc-домена с Fc-рецептором по меньшей мере в 10 раз, по меньшей мере в 20 раз или даже по меньшей мере в 50 раз. В одном аспекте (мультиспецифическое) антитело, содержащее сконструированный Fc-домен, демонстрирует менее 20%, в частности менее 10%, конкретнее менее 5% аффинности связывания с Fc-рецептором по сравнению с (мультиспецифическим) антителом, содержащим не сконструированный Fc-домен. В предпочтительном аспекте Fc-рецептор представляет собой Fcγ-рецептор. В некоторых аспектах Fc-рецептор представляет собой Fc-рецептор человека. В некоторых аспектах Fc-рецептор представляет собой активирующий Fc-рецептор. В конкретном аспекте Fc-рецептор представляет собой активирующий Fcγ-рецептор человека, конкретнее FcγRIIIa, FcγRI или FcγRIIa человека, наиболее конкретно FcγRIIIa человека. Предпочтительно связывание с каждым из этих рецепторов снижено. В некоторых аспектах также снижена аффинность связывания с компонентом комплемента, в частности аффинность связывания с Clq. В одном аспекте аффинность связывания с неонатальным Fc-рецептором (FcRn) не снижена. Практически аналогичное связывание с FcRn, т.е. сохранение аффинности связывания Fc-домена с указанным рецептором, достигается, когда Fc-домен (или (мультиспецифическое) антитело, содержащее указанный Fc-домен) демонстрирует более чем около 70% аффинности связывания не сконструированной формы Fc-домена (или (мультиспецифического) антитела, содержащего указанную не сконструированную форму Fc-домена) с FcRn. Fc-домен или (мультиспецифические) антитела по изобретению, содержащие указанный Fc-домен, могут демонстрировать более чем около 80% и даже более чем около 90% такой аффинности. В определенных аспектах Fc-домен (мультиспецифического) антитела конструируют так, чтобы он имел сниженную эффекторную функцию по сравнению с не сконструированным Fc-доменом. Сниженная эффекторная функция может включать, но не ограничивается этим, одно или более из следующего: сниженной комплементзависимой цитотоксичности (КЗЦ), сниженной антителозависимой клеточноопосредованной цитотоксичности (АЗКЦ), сниженного антителозависимого клеточного фагоцитоза (АЗКФ), сниженной секреции цитокинов, сниженного опосредованного иммунным комплексом поглощения антигена антигенпрезентирующими клетками, сниженного связывания с NK-клетками, сниженного связывания с макрофагами, сниженного связывания с моноцитами, сниженного связывания с полиморфноядерными клетками, сниженной прямой сигнализации, индуцирующей апоптоз, сниженного перекрестного связывания связанных с мишенью антител, сниженного созревания дендритных клеток или сниженного примирования Т-клеток. В одном аспекте сниженная эффекторная функция представляет собой одну или более функций, выбранных из группы из сниженной КЗЦ, сниженной АЗКЦ, сниженного АЗКФ и сниженной секреции цитокинов. В предпочтительном аспекте сниженная эффекторная функция представляет собой сниженную АЗКЦ. В одном аспекте сниженная АЗКЦ составляет менее 20% АЗКЦ, индуцируемой не сконструированным Fc-доменом (или (мультиспецифическим) антителом, содержащим не сконструированный Fc-домен).
В одном аспекте аминокислотная мутация, которая снижает аффинность связывания Fc-домена с Fc-рецептором и/или эффекторную функцию, представляет собой аминокислотную замену. В одном аспекте Fc-домен содержит аминокислотную замену в позиции, выбранной из группы из Е233, L234, L235, N297, Р331 и Р329 (нумерация в соответствии с индексом EU по Kabat). В более конкретном аспекте Fc-домен содержит аминокислотную замену в позиции, выбранной из группы из L234, L235 и Р329 (нумерация в соответствии с индексом EU по Kabat). В некоторых аспектах Fc-домен содержит аминокислотные мутации L234A и L235A (нумерация в соответствии с индексом EU по Kabat). В одном таком аспекте Fc-домен представляет собой Fc-домен IgG1, в частности Fc-домен IgG1 человека. В одном аспекте Fc-домен содержит аминокислотную замену в позиции Р329. В более конкретном аспекте аминокислотная замена представляет собой Р329А или P329G, в частности P329G (нумерация в соответствии с индексом EU по Kabat). В одном аспекте Fc-домен содержит аминокислотную замену в позиции Р329 и дополнительную аминокислотную замену в позиции, выбранной из Е233, L234, L235, N297 и Р331 (нумерация в соответствии с индексом EU по Kabat). В более конкретном аспекте дополнительная аминокислотная замена представляет собой Е233Р, L234A, L235A, L235E, N297A, N297D или P331S. В предпочтительных аспектах Fc-домен содержит аминокислотные замены в позициях Р329, L234 и L235 (нумерация в соответствии с индексом EU по Kabat). В более предпочтительных аспектах Fc-домен содержит аминокислотные мутации L234A, L235A и P329G («P329G LALA», «PGLALA» или «LALAPG»). В частности, в предпочтительных аспектах каждая субъединица Fc-домена содержит аминокислотные замены L234A, L235A и P329G (нумерация в соответствии с индексом EU по Kabat), т.е. в каждой из первой и второй субъединиц Fc-домена остаток лейцина в позиции 234 замещен остатком аланина (L234A), остаток лейцина в позиции 235 замещен остатком аланина (L235A) и остаток пролина в позиции 329 замещен остатком глицина (P329G) (нумерация в соответствии с индексом EU по Kabat).
В одном таком аспекте Fc-домен представляет собой Fc-домен IgG1, в частности Fc-домен IgG1 человека. Комбинация аминокислотных замен «P329G LALA» практически полностью устраняет связывание Fcγ-рецептора (а также комплемента) Fc-домена IgG1 человека, как описано в публикации РСТ №WO 2012/130831, которая в полном объеме включена в данный документ посредством ссылки. В WO 2012/130831 также описаны способы получения таких мутантных Fc-доменов и способы определения их свойств, таких как связывание Fc-рецептора или эффекторные функции.
Антитела IgG4 демонстрируют сниженную аффинность связывания с Fc-рецепторами и сниженные эффекторные функции по сравнению с антителами IgG1. Следовательно, в некоторых аспектах Fc-домен (мультиспецифических) антител по изобретению представляет собой Fc-домен IgG4, в частности Fc-домен IgG4 человека. В одном аспекте Fc-домен IgG4 содержит аминокислотную замену в позиции S228, в частности аминокислотную замену S228P (нумерация в соответствии с индексом EU по Kabat). Для дополнительного снижения аффинности связывания с Fc-рецептором и/или эффекторной функции в одном аспекте Fc-домен IgG4 содержит аминокислотную замену в позиции L235, в частности аминокислотную замену L235E (нумерация в соответствии с индексом EU по Kabat). В другом аспекте Fc-домен IgG4 содержит аминокислотную замену в позиции Р329, в частности аминокислотную замену P329G (нумерация в соответствии с индексом EU по Kabat). В предпочтительном аспекте Fc-домен IgG4 содержит аминокислотные замены в позициях S228, L235 и Р329, в частности аминокислотные замены S228P, L235E и P329G (нумерация в соответствии с индексом EU по Kabat). Такие мутантные Fc-домены IgG4 и их свойства связывания с Fcγ-рецептором описаны в публикации РСТ №WO 2012/130831, в полном объеме включенной в данный документ посредством ссылки.
В предпочтительном аспекте Fc-домен, демонстрирующий сниженную аффинность связывания с Fc-рецептором и/или сниженную эффекторную функцию по сравнению с нативным Fc-доменом IgG1, представляет собой Fc-домен IgG1 человека, содержащий аминокислотные замены L234A, L235A и, необязательно, P329G, или Fc-домен IgG4 человека, содержащий аминокислотные замены S228P, L235E и, необязательно, P329G (нумерация в соответствии с индексом EU по Kabat).
В определенных аспектах было устранено N-гликозилирование Fc-домена. В одном таком аспекте Fc-домен содержит аминокислотную мутацию в позиции N297, в частности аминокислотную замену с замещением аспарагина аланином (N297A) или аспарагиновой кислотой (N297D) (нумерация в соответствии с индексом EU по Kabat).
Помимо Fc-доменов, описанных выше в данном документе и в публикации РСТ №WO 2012/130831, Fc-домены со сниженными связыванием Fc-рецепторов и/или эффекторной функцией также включают домены с заменой одного или более остатков Fc-домена 238, 265, 269, 270, 297, 327 и 329 (патент США №6737056) (нумерация в соответствии с индексом EU по Kabat). Такие Fc-мутанты включают Fc-мутантов с заменами в двух или более аминокислотных позициях 265, 269, 270, 297 и 327, включая так называемый Fc-мутант «DANA» с заменой остатков 265 и 297 аланином (патент США №7332581).
Мутантные Fc-домены можно получать с помощью удаления, замены, вставки или модификации аминокислот, используя генетические или химические методы, хорошо известные в данной области техники. Генетические методы могут включать сайт-специфический мутагенез кодирующей последовательности ДНК, ПЦР, генный синтез и т.п. Правильные нуклеотидные изменения можно верифицировать, например, с помощью секвенирования.
Связывание с Fc-рецепторами можно легко определить, например, с помощью ELISA или поверхностного плазмонного резонанса (ППР), используя стандартное оборудование, такое как инструмент BIAcore (GE Healthcare), и Fc-рецепторы, которые можно получить с помощью рекомбинантной экспрессии. В альтернативном варианте аффинность связывания Fc-доменов или (мультиспецифических) антител, содержащих Fc-домен, в отношении Fc-рецепторов, можно оценивать, используя линии клеток, которые, как известно, экспрессируют конкретные Fc-рецепторы, такие как NK-клетки, экспрессирующие FcγIIIa-рецептор.
Эффекторную функцию Fc-домена или (мультиспецифического) антитела, содержащего Fc-домен, можно определить известными в данной области техники методами. Примеры in vitro анализов для оценки АЗКЦ-активности представляющей интерес молекулы, описаны в патенте США №5500362; Hellstrom et al. Proc Natl Acad Sci USA 83, 7059-7063 (1986) и Hellstrom et al., Proc Natl Acad Sci USA 82, 1499-1502 (1985); патенте США №5821337; Bruggemann et al., J Exp Med 166, 1351-1361 (1987). В альтернативном варианте можно использовать нерадиоактивные анализы (смотрите, например, нерадиоактивный анализ цитотоксичности для проточной цитометрии ACTI™ (CellTechnology, Inc. Mountain View, CA); и нерадиоактивный анализ цитотоксичности CytoTox 96® (Promega, Madison, WD). Эффекторные клетки, подходящие для такого анализа, включают мононуклеарные клетки периферической крови (МКПК) и естественные клетки-киллеры (NK). В альтернативном или дополнительном варианте АЗКЦ-активность представляющей интерес молекулы можно оценить in vivo, например, в животной модели, такой как описана в Clynes et al., Proc Natl Acad Sci USA 95, 652-656 (1998).
В некоторых аспектах снижено связывание Fc-домена с компонентом комплемента, в частности с Clq. Соответственно, в некоторых аспектах, в которых Fc-домен сконструирован так, чтобы иметь сниженную эффекторную функцию, указанная сниженная эффекторная функция включает сниженную КЗЦ. Анализы связывания Clq можно проводить, чтобы определить, способен ли Fc-домен или (мультиспецифическое) антитело, содержащее Fc-домен, связывать Clq, и, следовательно, обладает ли он активностью КЗЦ. Смотрите, например, ELISA связывания Clq и С3с в WO 2006/029879 и WO 2005/100402. Для оценки активации комплемента можно проводить анализ КЗЦ (смотрите, например, Gazzano-Santoro et al., J Immunol Methods 202, 163 (1996); Cragg et al., Blood 101, 1045-1052 (2003); и Cragg and Glennie, Blood 103, 2738-2743 (2004)).
Определение связывания FcRn и in vivo клиренса/времени полужизни также можно проводить, используя методы, известные в данной области техники (смотрите, например, Petkova, S.B. et al., Int'l. Immunol. 18(12):1759-1769 (2006); WO 2013/120929).
Б. Полинуклеотиды
В изобретении дополнительно предложен выделенный полинуклеотид, кодирующий антитело по изобретению. Указанный выделенный полинуклеотид может представлять собой один полинуклеотид или множество полинуклеотидов.
Полинуклеотиды, кодирующие (мультиспецифические) антитела по изобретению, можно экспрессировать в виде одного полинуклеотида, который кодирует все антитело, или в виде нескольких (например, двух или более) коэкспрессируемых полинуклеотидов. Полипептиды, кодируемые коэкспрессируемыми полинуклеотидами, могут объединяться посредством, например, дисульфидных связей или других способов с образованием функционального антитела. Например, часть легкой цепи антитела может кодироваться отдельным полинуклеотидом от части антитела, содержащей тяжелую цепь антитела. При коэкспрессии полипептиды тяжелой цепи будут объединяться с полипептидами легкой цепи с образованием антитела. В другом примере часть антитела, содержащая одну из двух субъединиц Fc-домена и, необязательно, одну или более молекул Fab (их части), может кодироваться отдельным полинуклеотидом от части антитела, содержащей другую из двух субъединиц Fc-домена и, необязательно, молекулу Fab (ее часть). При коэкспрессии субъединицы Fc-домена будут объединяться с образованием Fc-домена.
В некоторых аспектах выделенный полинуклеотид кодирует всю молекулу антитела в соответствии с изобретением, как описано в данном документе. В других аспектах выделенный полинуклеотид кодирует полипептид, содержащийся в антителе в соответствии с изобретением, как описано в данном документе.
В определенных аспектах полинуклеотид или нуклеиновая кислота представляет собой ДНК. В других аспектах полинуклеотид по настоящему изобретению представляет собой РНК, например, в форме матричной РНК (мРНК). РНК по настоящему изобретению может быть одноцепочечной или двухцепочечной.
В. Рекомбинантные методы
Антитела по изобретению можно получать, например, путем твердофазного пептидного синтеза (например, твердофазного синтеза Меррифильда) или рекомбинантного получения. Для рекомбинантного получения один или более полинуклеотидов, кодирующих антитело, например, описанных выше, выделяют и вставляют в один или более векторов для последующего клонирования и/или экспрессии в клетке-хозяине. Такой полинуклеотид можно легко выделять и секвенировать, используя традиционные процедуры. В одном аспекте предложен вектор, в частности экспрессионный вектор, содержащий полинуклеотид (помимо прочего один полинуклеотид или множество полинуклеотидов) по изобретению. Методы, хорошо известные специалистам в данной области техники, можно использовать для конструирования экспрессионных векторов, содержащих кодирующую последовательность антитела наряду с соответствующими транскрипционными/трансляционными регуляторными сигналами. Эти методы включают in vitro технологии рекомбинантных ДНК, технологии синтеза и in vivo рекомбинации/генетической рекомбинации. Смотрите, например, методики, описанные в Maniatis et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, N.Y. (1989); и Ausubel et al., Current Protocols in Molecular Biology, Greene Publishing Associates and Wiley Interscience, N.Y (1989). Экспрессионный вектор может быть частью плазмиды, вируса или может быть фрагментом нуклеиновой кислоты. Экспрессионные векторы включают экспрессионную кассету, в которую клонируют полинуклеотид, кодирующий антитело (т.е. кодирующую область) в функциональной связи с промотором и/или другими транскрипционными или трансляционными регуляторными элементами. В контексте данного документа «кодирующая область» представляет собой часть нуклеиновой кислоты, которая состоит из кодонов, транслируемых в аминокислоты. Хотя «стоп-кодон» (TAG, TGA или ТАА) не транслируется в аминокислоту, он может считаться частью кодирующей области, в случае наличия, но любые фланкирующие последовательности, например, промоторы, сайты связывания рибосомы, терминаторы транскрипции, интроны, 5' и 3' нетранслируемые области и т.п., не являются частью кодирующей области. Две или более кодирующих областей могут присутствовать в одной полинуклеотидной конструкции, например, в одном векторе, или в отдельных полинуклеотидных конструкциях, например, в отдельных (разных) векторах. Кроме того, любой вектор может содержать одну кодирующую область или может содержать две или более кодирующих областей, например, вектор по настоящему изобретению может кодировать один или более полипептидов, которые разделяются после или во время трансляции на конечные белки посредством протеолитического расщепления. Кроме того, вектор, полинуклеотид или нуклеиновая кислота по изобретению могут кодировать гетерологичные кодирующие области, слитые или не слитые с полинуклеотидом, кодирующим антитело по изобретению или его вариант или производное. Гетерологичные кодирующие области включают, без ограничения, специализированные элементы или мотивы, такие как секреторный сигнальный пептид или гетерологичный функциональный домен. Функциональная связь имеет место, когда кодирующая область генетического продукта, например, полипептида, связана с одной или более регуляторными последовательностями так, чтобы экспрессия генного продукта находилась под влиянием или управлением этих регуляторных последовательностей. Два фрагмента ДНК (такие как кодирующая полипептид область и связанный с ней промотор) являются «функционально связанными», если индукция промотора приводит к транскрипции мРНК, кодирующей необходимый генный продукт, и если природа связи между двумя фрагментами ДНК не препятствует способности экспрессионных регуляторных последовательностей управлять экспрессией генного продукта или не препятствует способности транскрибировать ДНК-матрицу. Таким образом, промоторная область является функционально связанной с нуклеиновой кислотой, кодирующей полипептид, если промотор способен влиять на транскрипцию этой нуклеиновой кислоты. Промотор может быть клеточно-специфическим промотором, который обуславливает достаточную транскрипцию ДНК только в определенных клетках. Помимо промотора, другие транскрипционные регуляторные элементы, например, энхансеры, операторы, репрессоры и сигналы терминации транскрипции, могут быть функционально связаны с полинуклеотидом с целью управления клеточно-специфической транскрипцией. Подходящие промоторы и другие транскрипционные регуляторные области описаны в данном документе. Специалистам в данной области техники известны различные транскрипционные регуляторные области. Они включают, без ограничения, транскрипционные регуляторные области, которые являются функциональными в клетках позвоночных, таких как, без ограничения, промоторные и энхансерные сегменты из цитомегаловирусов (например, немедленно ранний промотор в сочетании с интроном-А), вируса обезьян 40 (например, ранний промотор) и ретровирусов (таких как, например, вирус саркомы Рауса). Другие транскрипционные регуляторные области включают полученные из генов позвоночных, таких как гены актина, белка теплового шока и кроличьего β-глобина, а также другие последовательности, способные регулировать экспрессию генов в эукариотических клетках. Дополнительные подходящие транскрипционные регуляторные области включают тканеспецифические промоторы и энхансеры, а также индуцибельные промоторы (например, промоторы, индуцируемые тетрациклинами). Аналогично, специалистам в данной области техники известны различные трансляционные регуляторные элементы. Они включают, но не ограничиваются этим, сайты связывания рибосомы, кодоны инициации и терминации трансляции и элементы, полученные из вирусных систем (в частности, внутренний сайт посадки рибосомы или IRES, также называемый последовательностью CITE). Экспрессионная кассета также может включать другие элементы, такие как точка начала репликации и/или элементы интеграции в хромосому, такие как ретровирусные длинные концевые повторы (ДКП) или инвертированные концевые повторы (ИКП) аденоассоциированного вируса (AAV).
Кодирующие области полинуклеотида и нуклеиновой кислоты по настоящему изобретению могут быть связаны с дополнительными кодирующими областями, которые кодируют секреторные или сигнальные пептиды, которые управляют секрецией полипептида, кодируемого полинуклеотидом по настоящему изобретению. Например, если необходима секреция антитела, ДНК, кодирующую сигнальную последовательность, можно размещать выше нуклеиновой кислоты, кодирующей антитело по изобретению или его фрагмент.В соответствии с сигнальной гипотезой, белки, секретируемые клетками млекопитающих, имеют сигнальный пептид или секреторную лидерную последовательность, которые отщепляются от зрелого белка после инициации экспорта растущей белковой цепи через шероховатый эндоплазматический ретикулум. Специалистам в данной области техники известно, что полипептиды, секретируемые клетками позвоночных в общем случае имеют сигнальный пептид, слитый с N-концом полипептида, который отщепляется от транслируемого полипептида с образованием секретируемой или «зрелой» формы полипептида. В определенных аспектах используется нативный сигнальный пептид, например, сигнальный пептид тяжелой цепи или легкой цепи иммуноглобулина, или функциональное производное такой последовательности, которое сохраняет способность управлять секрецией полипептида, который с ним функционально связан. В альтернативном варианте можно использовать гетерологичный сигнальный пептид млекопитающих или его функциональное производное. Например, лидерная последовательность дикого типа может быть замещена лидерной последовательностью человеческого тканевого активатора плазминогена (TPA) или мышиной β-глюкуронидазой.
ДНК, кодирующая короткую белковую последовательность, которую можно использовать для облегчения последующей очистки (например, гистидиновый тег) или мечения антитела, может быть включена внутри или в концах кодирующего антитело (фрагмент) полипептида.
В дополнительном аспекте предложена клетка-хозяин, содержащая полинуклеотид (т.е. один полинуклеотид или множество полинуклеотидов) по изобретению. В определенных аспектах предложена клетка-хозяин, содержащая вектор по изобретению. Полинуклеотиды и векторы могут включать любые элементы, по отдельности или в комбинации, описанные в данном документе в связи с полинуклеотидами и векторами, соответственно. В одном таком аспекте клетка-хозяин содержит (например, была трансформирована или трансфицирована) один или более векторов, содержащих один или более полинуклеотидов, которые кодируют антитело (или его часть) по изобретению. В контексте данного документа термин «клетка-хозяин» относится к любому типу клеточной системы, которую можно сконструировать для генерации антитела по изобретению или его фрагментов. Клетки-хозяева, подходящие для репликации и для поддержания экспрессии антител, хорошо известны в данной области техники. Такие клетки, при необходимости, могут быть трансфицированы или трансдуцированы конкретным экспрессионным вектором, а большие количества содержащих вектор клеток можно выращивать для засевания крупномасштабных ферментеров для получения достаточных количеств антител для клинических применений. Подходящие клетки-хозяева включают прокариотические микроорганизмы, такие как Е. coli, или различные эукариотические клетки, такие как клетки яичника китайского хомяка (СНО), клетки насекомых и т.п. Например, полипептиды могут вырабатываться в бактериях, в частности, если отсутствует необходимость гликозилирования. После экспрессии полипептид можно выделять из бактериальной клеточной пасты в растворимую фракцию и дополнительно очищать. Помимо прокариот, эукариотические микробы, такие как нитевидные грибы или дрожжи, являются подходящими хозяевами для клонирования или экспрессии кодирующих полипептид векторов, включая штаммы грибов и дрожжей, пути гликозилирования которых были «гуманизированы», что приводит к получению полипептида с частично или полностью человеческим профилем гликозилирования. Смотрите Gerngross, Nat Biotech 22, 1409-1414 (2004), и Li et al., Nat Biotech 24, 210-215 (2006). Клетки-хозяев для экспрессии (гликозилированных) полипептидов также получают из многоклеточных организмов (беспозвоночных и позвоночных). Примеры клеток беспозвоночных включают клетки растений и насекомых. Были идентифицированы многочисленные бакуловирусные штаммы, которые можно использовать в сочетании с клетками насекомых, в частности для трансфекции клеток Spodoptera frugiperda. Культуры клеток растений также можно применять в качестве хозяев. Смотрите, например, патенты США №№5959177, 6040498, 6420548, 7125978 и 6417429 (в которых описана технология PLANTIBODIES™ для получения антител в трансгенных растениях). Клетки позвоночных также можно использовать в качестве хозяев. Например, можно применять линии клеток млекопитающих, которые адаптированы для роста в суспензии. Другими примерами применимых линий клеток-хозяев млекопитающих являются линия клеток почки обезьяны CV1, трансформированная SV40 (COS-7), линия клеток почки эмбриона человека (клетки 293 или 293Т, описанные, например, в Graham et al., J Gen Virol 36, 59 (1977)), клетки почки новорожденного хомяка (ВНК), клетки Сертоли мышей (клетки ТМ4, описанные, например, в Mather, Biol Reprod 23, 243-251 (1980)), клетки почки обезьяны (CV1), клетки почки африканской зеленой мартышки (VERO-76), клетки карциномы шейки матки человека (HELA), клетки почки собаки (MDCK), клетки печени серой крысы (BRL ЗА), клетки легкого человека (W138), клетки печени человека (Hep G2), клетки опухоли молочной железы мышей (ММТ 060562), клетки TRI, описанные, например, в Mather et al., Annals N.Y. Acad Sci 383, 44-68 (1982)), клетки MRC 5 и клетки FS4. Другие подходящие линии клеток-хозяев млекопитающих включают клетки яичника китайского хомяка (СНО), включая dhfr- клетки СНО (Urlaub et al., Proc Natl Acad Sci USA 77, 4216 (1980)); и линии клеток миеломы, такие как YO, NSO, РЗХ63 и Sp2/0. Обзор некоторых линий клеток-хозяев млекопитающих, подходящих для выработки белка, смотрите, например, в Yazaki and Wu, Methods in Molecular Biology, Vol.248 (B.K.C. Lo, ed., Humana Press, Totowa, NJ), pp.255-268 (2003). Клетки-хозяева включают культивируемые клетки, например, культивируемые клетки млекопитающих, клетки дрожжей, клетки насекомых, клетки бактерий и клетки растений, если называть лишь некоторые, но также клетки, находящиеся в организме трансгенного животного, трансгенного растения или в культивируемой растительной или животной ткани. В одном аспекте клетка-хозяин представляет собой эукариотическую клетку, в частности клетку млекопитающего, такую как клетка яичника китайского хомяка (СНО), эмбриональная клетка почки человека (НЕК) или лимфоидная клетка (например, клетка YO, NSO, Sp20). В одном аспекте клетка-хозяин не является клеткой в организме человека.
Стандартные технологии для экспрессии чужеродных генов в этих системах известны в данной области техники. Клетки, экспрессирующие полипептид, содержащий либо тяжелую, либо легкую цепь антигенсвязывающего домена, такого как антитело, можно конструировать так, чтобы они также экспрессировали другую из цепей антитела, чтобы экспрессируемый продукт представлял собой антитело, которое имеет как тяжелую, так и легкую цепь.
В одном аспекте предложен способ получения антитела в соответствии с изобретением, включающий культивирование клетки-хозяина, содержащей полинуклеотид, кодирующий предложенное в данном документе антитело, в условиях, подходящих для экспрессии антитела, и, необязательно, выделение антитела из клетки-хозяина (или культуральной среды клеток-хозяев).
Компоненты (мультиспецифического) антитела по изобретению могут быть генетически слиты друг с другом. (Мультиспецифическое) антитело можно сконструировать так, чтобы его компоненты были напрямую слиты друг с другом или ненапрямую посредством линкерной последовательности. Композицию и длину линкера можно определять в соответствии со способами, хорошо известными в данной области техники, и можно тестировать в отношении эффективности. Примеры линкерных последовательностей между разными компонентами (мультиспецифических) антител предложены в данном документе. Также при необходимости могут быть включены дополнительные последовательности для включения сайта расщепления для разделения отдельных компонентов слияния, например, последовательность распознавания эндопептидазой.
Антитела, полученные как описано в данном документе, можно очищать известными в данной области техники методами, такими как высокоэффективная жидкостная хроматография, ионообменная хроматография, гель-электрофорез, аффинная хроматография, эксклюзионная хроматография и т.п. Фактические условия, используемые для очищения конкретного белка, будут зависеть частично от факторов, таких как суммарный заряд, гидрофобность, гидрофильность и т.д., и будут очевидны для специалистов в данной области техники. В случае очистки методом аффинной хроматографии можно использовать антитело, лиганд, рецептор или антиген, с которыми связывается антитело. Например, для очистки методом аффинной хроматографии антител по изобретению можно использовать матрицу с протеином А или протеином G. Последовательные этапы аффинной хроматографии с протеином А или G и эксклюзионной хроматографии можно использовать для выделения антитела, по существу так, как это описано в примерах. Чистоту антитела можно определять любым из ряда хорошо известных аналитических методов, включая гель-электрофорез, жидкостную хроматографию высокого давления и т.п.
Г. Методы анализа
Идентификацию, скрининг или изучение характеристик предложенных в данном документе антител в отношении их физических/химических свойств и/или биологической активности можно проводить с помощью различных методов анализа, известных в данной области техники.
1. Анализы связывания
Связывание (аффинность) с Fc-рецептором или антигеном-мишенью можно определить, например, с помощью поверхностного плазмонного резонанса (ППР), используя стандартное оборудование, такое как инструмент BIAcore (GE Healthcare), и рецепторы или белки-мишени, которые можно получить с помощью рекомбинантной экспрессии. В альтернативном варианте связывание антител с разными рецепторами или антигенами-мишенями можно оценивать, используя линии клеток, экспрессирующие конкретный рецептор или антиген-мишень, например, методом проточной цитометрии (FACS). Конкретный иллюстративный и типовой аспект для определения активности связывания с CD3 описан ниже.
В одном аспекте активность связывания с CD3 определяют с помощью ППР следующим образом:
ППР проводят на инструменте Biacore Т200 (GE Healthcare). Захватывающее анти-Fab антитело (GE Healthcare, №28958325) иммобилизуют на сенсорном чипе СМ5 серии S (GE Healthcare), используя стандартную химическую реакцию аминного сопряжения, при поверхностной плотности 4000-6000 резонансных единиц (РЕ). В качестве подвижного буфера и буфера для разведения используют HBS-P+ (10 мМ ГЭПЭС, 150 мМ NaCl, рН 7,4, 0,05% поверхностно-активного вещества Р20). Антитела к CD3 в концентрации 2 мкг/мл (в 20 мМ His, 140 мМ NaCl, рН 6,0) вводят в течение приблизительно 60 с при скорости потока 5 мкл/мин. Используемым антигеном CD3 является гетеродимер эктодоменов CD3 дельта и CD3 эпсилон, слитых с Fc-доменом человека с модификациями типа «выступ-во-впадину» и С-концевым Avi-тегом (смотрите SEQ ID NO 28 и 29). Антиген CD3 вводят в концентрации 10 мкг/мл в течение 120 с и отслеживают диссоциацию при скорости потока 5 мкл/мин в течение около 120 с. Поверхность чипа восстанавливают посредством двух последовательных введений 10 мМ глицина, рН 2,1, приблизительно по 60 с каждый. Разницу объемного показателя преломления корректируют путем вычитания холостых введений и путем вычитания ответа, полученного от холостого контроля с проточной ячейкой. Для оценки ответ связывания определяют через 5 секунд после окончания введения. Для нормализации сигнала связывания связывание CD3 делят на ответ анти-Fab (сигнал (РЕ), полученный после захвата антитела к CD3 на иммобилизованном анти-Fab антителе). Активность связывания антитела с CD3 после определенной обработки по сравнению с активностью связывания антитела с CD3 после другой обработки (также называемую относительной активной концентрацией (OAK)) рассчитывают путем сопоставления активности связывания образца антитела после определенной обработки с активностью связывания соответствующего образца антитела после другой обработки.
2. Анализы активности
Биологическую активность (мультиспецифических) антител по изобретению можно измерять с помощью различных анализов, как описано в примерах. Биологическая активность может, например, включать индукцию пролиферации Т-клеток, индукцию сигнализации в Т-клетках, индукцию экспрессии маркеров активации в Т-клетках, индукцию секреции цитокинов Т-клетками, индукцию лизиса клеток-мишеней, таких как опухолевые клетки, и индукцию регрессии опухоли и/или улучшения выживаемости.
Д. Композиции, составы и пути введения
В дополнительном аспекте в изобретении предложены фармацевтические композиции, содержащие любое из предложенных в данном документе антител, например, для применения в любом из нижеописанных терапевтических способов. В одном аспекте фармацевтическая композиция содержит антитело в соответствии с изобретением и фармацевтически приемлемый носитель. В другом аспекте фармацевтическая композиция содержит антитело в соответствии с изобретением и по меньшей мере один дополнительный терапевтический агент, например, как описано ниже.
Дополнительно предложен способ получения антитела по изобретению в форме, подходящей для введения in vivo, включающий (а) получение антитела в соответствии с изобретением и (б) составление антитела с по меньшей мере одним фармацевтически приемлемым носителем, при этом препарат антитела составляют для введения in vivo.
Фармацевтические композиции по настоящему изобретению содержат эффективное количество антитела, растворенного или диспергированного в фармацевтически приемлемом носителе. Выражение «фармацевтически приемлемый» относится к молекулярным веществам и композициям, которые в общем случае являются нетоксичными для реципиентов в применяемых дозировках и концентрациях, т.е. не вызывают побочную, аллергическую или другую нежелательную реакцию при введении животному, такому как, например, человек, в зависимости от ситуации. Способ получения фармацевтической композиции, которая содержит антитело и, необязательно, дополнительный активный ингредиент, должен быть понятен специалисту в данной области техники в свете настоящего изобретения и как проиллюстрировано в Remington's Pharmaceutical Sciences, 18th Ed. Mack Printing Company, 1990, включенной в данный документ посредством ссылки. Кроме того, в случае введения животному (например, человеку) следует понимать, что препараты должны соответствовать стандартам стерильности, апирогенности, общей безопасности и чистоты, согласно требованиям Отдела по биологическим стандартам FDA или соответствующих органов в других странах. Предпочтительными композициями являются лиофилизированные составы или водные растворы. В контексте данного документа «фармацевтически приемлемый носитель» включает любые и все растворители, буферы, дисперсионные среды, покрытия, поверхностно-активные вещества, антиоксиданты, консерванты (например, антибактериальные агенты, противогрибковые агенты), изотонические агенты, агенты, замедляющие всасывание, соли, консерванты, антиоксиданты, белки, лекарственные средства, стабилизаторы лекарственных средств, полимеры, гели, связующие вещества, вспомогательные вещества, разрыхлители, лубриканты, подсластители, ароматизаторы, красители, подобные материалы и их комбинации, как должно быть известно специалисту в данной области техники (смотрите, например, Remington's Pharmaceutical Sciences, 18th Ed. Mack Printing Company, 1990, pp.1289-1329, включенную в данный документ посредством ссылки). За исключением случаев, когда какой-либо традиционный носитель несовместим с активным ингредиентом, предусмотрено его использование в фармацевтических композициях.
Антитело по изобретению (и любой дополнительный терапевтический агент) можно вводить любыми подходящими способами, включая парентеральное, внутрилегочное и интраназальное, и, при необходимости местного лечения, внутриочаговое введение. Парентеральные инфузии включают внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение. Введение дозы можно проводить любым удобным способом, например путем инъекций, таких как внутривенные или подкожные инъекции, частично в зависимости от того, является ли введение кратковременным или постоянным.
Парентеральные композиции включают разработанные для введения путем инъекции, например, подкожной, интрадермальной, внутриочаговой, внутривенной, внутриартериальной, внутримышечной, интратекальной или внутрибрюшинной инъекции. Для инъекции антитела по изобретению можно составлять в водных растворах, в частности физиологически совместимых буферах, таких как раствор Хенкса, раствор Рингера или физиологический солевой буфер. Раствор может содержать вспомогательные агенты, такие как суспендирующие, стабилизирующие и/или диспергирующие агенты. В альтернативном варианте антитела могут находиться в форме порошка для смешивания с подходящим носителем, например, стерильной апирогенной водой, перед использованием. Стерильные инъекционные растворы готовят путем внесения антител по изобретению в необходимом количестве в подходящий растворитель с различными перечисленными ниже другими ингредиентами, в случае необходимости. Стерильность можно легко обеспечить, например, путем фильтрации через мембраны для стерильной фильтрации. В общем случае дисперсии готовят путем внесения различных стерилизованных активных ингредиентов в стерильный носитель, который содержит основную дисперсионную среду и/или другие ингредиенты. В случае стерильных порошков для приготовления стерильных инъекционных растворов, суспензий или эмульсий предпочтительными способами приготовления являются технологии вакуумной сушки или сублимационной сушки, которые позволяют получать порошок активного ингредиента плюс любой дополнительный необходимый ингредиент из предварительно стерильно-профильтрованной жидкой среды. При необходимости жидкую среду следует должным образом забуферить, а жидкому растворителю перед инъекцией сначала придать изотоничность с помощью достаточного количества солевого раствора или глюкозы. Композиция должна быть стабильной в условиях производства и хранения и защищенной от загрязняющего действия микроорганизмов, таких как бактерии и грибы. Следует понимать, что загрязнение эндотоксинами следует свести к минимальному безопасному уровню, например, менее 0,5 нг/мг белка. Подходящие фармацевтически приемлемые носители включают, но не ограничиваются этим: буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония; хлорид бензетония; фенол; бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (содержащие менее 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; металлсодержащие комплексы (например, комплексы Zn-белок) и/или неионные поверхностно-активные вещества, такие как полиэтиленгликоль (ПЭГ). Водные инъекционные суспензии могут содержать соединения, которые увеличивают вязкость суспензии, такие как карбоксиметилцеллюлоза натрия, сорбит, декстран и т.п. Необязательно, суспензия может также содержать подходящие стабилизаторы или вещества, которые повышают растворимость соединений, для возможности получения высококонцентрированных растворов. Кроме того, суспензии активных соединений можно готовить в виде подходящих масляных инъекционных суспензий. Подходящие липофильные растворители или носители включают жирные масла, такие как кунжутное масло, или синтетические сложные эфиры жирных кислот, такие как этилолеат или триглицериды, или липосомы.
Активные ингредиенты также могут быть заключены в микрокапсулах, полученных, например, с помощью технологий коацервации или путем межфазной полимеризации, например, гидроксиметилцеллюлозные или желатиновые микрокапсулы и полиметилметакрилатные микрокапсулы, соответственно, в коллоидных системах для доставки лекарственных средств (например, липосомах, альбуминовых микросферах, микроэмульсиях, наночастицах и нанокапсулах) или в макроэмульсиях. Такие методики описаны в Remington's Pharmaceutical Sciences (18th Ed. Mack Printing Company, 1990). Можно получать препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие полипептид, причем матрицы находятся в виде формованных изделий, например, пленок или микрокапсул. В конкретных аспектах длительное всасывание инъекционной композиции можно обеспечивать путем использования в композициях агентов, замедляющих всасывание, таких как, например, моностеарат алюминия, желатин или их комбинации.
Помимо описанных ранее композиций, антитела также можно составлять в виде депо-препаратов. Такие составы пролонгированного действия можно вводить путем имплантации (например, подкожной или внутримышечной) или путем внутримышечной инъекции. Таким образом, например, антитела можно составлять с подходящими полимерными или гидрофобными материалами (например, в виде эмульсии в приемлемом масле) или ионообменными смолами, или в виде слаборастворимых производных, например, в виде слаборастворимой соли.
Фармацевтические композиции, содержащие антитела по изобретению, можно получать посредством традиционных процессов смешивания, растворения, эмульгации, инкапсуляции, заключения или лиофилизации. Фармацевтические композиции можно составлять традиционным образом, используя один или более физиологически приемлемых носителей, разбавителей, вспомогательных веществ, которые облегчают обработку белков в препараты, которые можно использовать в фармацевтических целях. Надлежащий состав зависит от выбранного пути введения.
Антитела можно составлять в виде композиции в форме свободных кислоты или основания, нейтральной или солевой форме. Фармацевтически приемлемыми солями являются соли, которые по существу сохраняют биологическую активность свободных кислоты или основания. Они включают соли присоединения кислот, например, образуемые со свободными аминогруппами белковой композиции или образуемые с неорганическими кислотами, таким как, например, хлористоводородная или фосфорная кислоты, или такими органическими кислотами, как уксусная, щавелевая, винная или миндальная кислота. Соли, образуемые с карбоксильными группами, также можно получать из неорганических оснований, таких как, например, гидроксиды натрия, калия, аммония, кальция или железа; или таких органических оснований, как изопропиламин, триметиламин, гистидин или прокаин. Фармацевтические соли, как правило, более растворимы в водных и других протонных растворителях, чем соответствующие формы свободных оснований.
Е. Терапевтические способы и композиции
Любые из предложенных в данном документе антител можно применять в терапевтических способах. Антитела по изобретению можно использовать в качестве иммунотерапевтических агентов, например, при лечении раков.
Для применения в терапевтических способах антитела по изобретению следует составлять, дозировать и вводить способом, соответствующим надлежащей медицинской практике. Факторы, которые необходимо учитывать в этом контексте, включают конкретное нарушение, подлежащее лечению, конкретное млекопитающее, подлежащее лечению, клиническое состояние отдельного пациента, причину нарушения, место доставки агента, способ введения, схему введения и другие факторы, известные практикующим врачам.
В одном аспекте предложены антитела по изобретению для применения в качестве лекарственного средства. В дополнительных аспектах предложены антитела по изобретению для применения в лечения заболевания. В определенных аспектах предложены антитела по изобретению для применения в способе лечения. В одном аспекте в изобретении предложено антитело по изобретению для применения в лечении заболевания у нуждающегося в этом индивида. В определенных аспектах в изобретении предложено антитело для применения в способе лечения индивида, имеющего заболевание, включающем введение индивиду эффективного количества антитела. В определенных аспектах подлежащее лечению заболевание представляет собой пролиферативное нарушение. В предпочтительном аспекте заболевание представляет собой рак. В определенных аспектах способ дополнительно включает введение индивиду эффективного количества по меньшей мере одного дополнительного терапевтического агента, например, противоракового агента, если подлежащее лечению заболевание представляет собой рак. В дополнительных аспектах в изобретении предложено антитело по изобретению для применения в индукции лизиса клетки-мишени, в частности опухолевой клетки. В определенных аспектах в изобретении предложено антитело по изобретению для применения в способе индукции лизиса клетки-мишени, в частности опухолевой клетки, у индивида, включающем введение индивиду эффективного количества антитела для индукции лизиса клетки-мишени. «Индивид» в соответствии с любым из вышеприведенных аспектов представляет собой млекопитающее, предпочтительно человека.
В дополнительном аспекте в изобретении предложено применение антитела по изобретению в производстве или приготовлении лекарственного средства. В одном аспекте лекарственное средство предназначено для лечения заболевания у нуждающегося в этом индивида. В дополнительном аспекте лекарственное средство предназначено для применения в способе лечения заболевания, включающем введение индивиду, имеющему заболевание, эффективного количества лекарственного средства. В определенных аспектах подлежащее лечению заболевание представляет собой пролиферативное нарушение. В предпочтительном аспекте заболевание представляет собой рак. В одном аспекте способ дополнительно включает введение индивиду эффективного количества по меньшей мере одного дополнительного терапевтического агента, например, противоракового агента, если подлежащее лечению заболевание представляет собой рак. В дополнительном аспекте лекарственное средство предназначено для индукции лизиса клетки-мишени, в частности опухолевой клетки. В дополнительном аспекте лекарственное средство предназначено для применения в способе индукции лизиса клетки-мишени, в частности опухолевой клетки, у индивида, включающем введение индивиду эффективного количества лекарственного средства для индукции лизиса клетки-мишени. «Индивид» в соответствии с любым из вышеприведенных аспектов может представлять собой млекопитающее, предпочтительно человека.
В дополнительном аспекте в изобретении предложен способ лечения заболевания. В одном аспекте способ включает введение индивиду, имеющему такое заболевание, эффективного количества антитела по изобретению. В одном аспекте указанному индивиду вводят композицию, содержащую антитело по изобретению в фармацевтически приемлемой форме. В определенных аспектах подлежащее лечению заболевание представляет собой пролиферативное нарушение. В предпочтительном аспекте заболевание представляет собой рак. В определенных аспектах способ дополнительно включает введение индивиду эффективного количества по меньшей мере одного дополнительного терапевтического агента, например, противоракового агента, если подлежащее лечению заболевание представляет собой рак. «Индивид» в соответствии с любым из вышеприведенных аспектов может представлять собой млекопитающее, предпочтительно человека.
В дополнительном аспекте в изобретении предложен способ индукции лизиса клетки-мишени, в частности опухолевой клетки. В одном аспекте способ включает приведение клетки-мишени в контакт с антителом по изобретению в присутствии Т-клетки, в частности цитотоксической Т-клетки. В дополнительном аспекте предложен способ индукции лизиса клетки-мишени, в частности опухолевой клетки, у индивида. В одном таком аспекте способ включает введение индивиду эффективного количества антитела по изобретению для индукции лизиса клетки-мишени. В одном аспекте «индивид» представляет собой человека.
В определенных аспектах подлежащее лечению заболевание представляет собой пролиферативное нарушение, в частности рак. Неограничивающие примеры раков включают рак мочевого пузыря, рак головного мозга, рак головы и шеи, рак поджелудочной железы, рак легкого, рак молочной железы, рак яичника, рак матки, рак шейки матки, рак эндометрия, рак пищевода, рак толстой кишки, колоректальный рак, ректальный рак, рак желудка, рак предстательной железы, рак крови, рак кожи, плоскоклеточную карциному, рак кости и рак почки. Другие клеточно-пролиферативные нарушения, которые можно лечить с помощью антитела по настоящему изобретению, включают, но не ограничиваются этим, новообразования, расположенные в: брюшной полости, кости, молочной железе, пищеварительной системе, печени, поджелудочной железе, брюшине, эндокринных железах (надпочечнике, паращитовидной железе, гипофизе, семенниках, яичниках, вилочковой железе, щитовидной железе), глазе, голове и шее, нервной системе (центральной и периферической), лимфатической системе, органах таза, коже, мягкой ткани, селезенке, области грудной клетки и мочеполовой системе. Также включены предраковые состояния или поражения и раковые метастазы. В определенных аспектах рак выбран из группы, состоящей из рака почки, рака мочевого пузыря, рака кожи, рака легкого, колоректального рака, рака молочной железы, рака головного мозга, рака головы и шеи и рака предстательной железы. В одном аспекте, в частности, в котором антитело представляет собой мультиспецифическое антитело, связывающееся с TYRP-1 в качестве второго антигена, рак представляет собой рак с экспрессией TYRP-1. В одном аспекте, в частности, в котором антитело представляет собой мультиспецифическое антитело, связывающееся с TYRP-1 в качестве второго антигена, рак представляет собой рак кожи, в частности меланому. В одном аспекте, в частности, в котором антитело представляет собой мультиспецифическое антитело, связывающееся с EGFRvIII в качестве второго антигена, рак представляет собой рак с экспрессией EGFRvIII. В одном аспекте, в частности, в котором антитело представляет собой мультиспецифическое антитело, связывающееся с EGFRvIII в качестве второго антигена, рак представляет собой рак головного мозга, в частности глиобластому. Специалисту в данной области техники понятно, что во многих случаях антитело может не обеспечивать излечение, но может только обеспечивать частичный благоприятный эффект.В некоторых аспектах физиологическое изменение, имеющее некоторый благоприятный эффект, также считается терапевтически благоприятным. Таким образом, в некоторых аспектах количество антитела, которое обеспечивает физиологическое изменение, считается «эффективным количеством». Субъект, пациент, индивид, нуждающийся в лечении, как правило, представляет собой млекопитающее, в частности человека.
В некоторых аспектах эффективное количество антитела по изобретению вводят индивиду для лечения заболевания.
Для профилактики или лечения заболевания соответствующая доза антитела по изобретению (используемого отдельно или в комбинации с одним или более другими дополнительными терапевтическими агентами) будет зависеть от типа заболевания, подлежащего лечению, пути введения, массы тела пациента, типа антитела, тяжести и течения заболевания, того, вводится ли антитело с профилактической или терапевтической целью, предыдущих или одновременных терапевтических вмешательств, анамнеза пациента и его ответа на антитело, а также решения лечащего врача. Практикующий врач, ответственный за введение, в любом случае определит концентрацию активного(ых) ингредиента(ов) в композиции и соответствующую(ие) дозу(ы) для каждого отдельного субъекта. В данном документе предусмотрены различные схемы дозирования, включая, но не ограничиваясь этим, однократное или многократное введение в различные моменты времени, болюсное введение и импульсную инфузию.
Антитело удобно вводить пациенту однократно или в течение серии курсов лечения. В зависимости от типа и тяжести заболевания, начальная кандидатная дозировка для введения пациенту может составлять от около 1 мкг/кг до 15 мг/кг (например, 0,1 мг/кг - 10 мг/кг) антитела, например, за одно или более отдельных введений или в виде непрерывной инфузии. Одна типичная суточная дозировка может находиться в диапазоне от около 1 мкг/кг до 100 мг/кг или более, в зависимости от факторов, упомянутых выше. В случае повторных введений в течение нескольких суток или более, в зависимости от состояния, лечение, как правило, проводят до достижения необходимой степени подавления симптомов заболевания. Одна типовая дозировка антитела находится в диапазоне от около 0,005 мг/кг до около 10 мг/кг. В других неограничивающих примерах доза также может составлять от около 1 микрограмм/кг массы тела, около 5 микрограмм/кг массы тела, около 10 микрограмм/кг массы тела, около 50 микрограмм/кг массы тела, около 100 микрограмм/кг массы тела, около 200 микрограмм/кг массы тела, около 350 микрограмм/кг массы тела, около 500 микрограмм/кг массы тела, около 1 миллиграмм/кг массы тела, около 5 миллиграмм/кг массы тела, около 10 миллиграмм/кг массы тела, около 50 миллиграмм/кг массы тела, около 100 миллиграмм/кг массы тела, около 200 миллиграмм/кг массы тела, около 350 миллиграмм/кг массы тела, около 500 миллиграмм/кг массы тела до около 1000 мг/кг массы тела или более на введение, а также любой получаемый из этих значений диапазон. В неограничивающих примерах получаемого из перечисленных в данном документе значений диапазона можно вводить дозу в диапазоне от около 5 мг/кг массы тела до около 100 мг/кг массы тела, от около 5 микрограмм/кг массы тела до около 500 миллиграмм/кг массы тела и т.д. с учетом вышеприведенных числовых значений. Таким образом, пациенту можно вводить одну или более доз, составляющих около 0,5 мг/кг, 2,0 мг/кг, 5,0 мг/кг или 10 мг/кг (или любую их комбинацию). Такие дозы можно вводить с интервалами, например, раз в неделю или раз в три недели (например, чтобы пациент получал от около двух до около двадцати или, например, около шести доз антитела). Можно вводить начальную более высокую нагрузочную дозу, за которой следует одна или более меньших доз. В то же время можно использовать другие схемы введения доз. Эффективность такой терапии легко отслеживать с помощью традиционных методик и методов анализа.
Антитела по изобретению в общем случае используют в количестве, эффективном для достижения предполагаемой цели. В случае применения для лечения или предотвращения болезненного состояния антитела по изобретению или их фармацевтические композиции вводят или применяют в эффективном количестве.
В случае системного введения эффективную дозу можно сначала оценить из in vitro анализа, такого как анализ клеточной культуры. После этого можно составить дозу для применения в животных моделях для достижения диапазона циркулирующей концентрации, который включает IC50, определенную в клеточной культуре. Такую информацию можно использовать для более точного определения доз, применимых для людей.
Начальные дозировки также можно оценить из in vivo данных, например, животных моделей, используя методики, которые хорошо известны в данной области техники.
Количество и интервал дозировки можно индивидуально корректировать, чтобы обеспечить плазменные уровни антител, достаточные для поддержания терапевтического эффекта. Обычные дозировки для пациентов при введении путем инъекции находятся в диапазоне от около 0,1 до 50 мг/кг/сутки, как правило, от около 0,5 до 1 мг/кг/сутки. Терапевтически эффективные плазменные уровни можно обеспечить путем введения нескольких доз каждые сутки. Уровни в плазме можно измерять, например, с помощью ВЭЖХ.
Эффективная доза антител по изобретению в общем случае обеспечит терапевтический благоприятный эффект без существенной токсичности. Токсичность и терапевтическую эффективность антитела можно определить посредством стандартных фармацевтических процедур на культуре клеток или экспериментальных животных. Анализы клеточных культур и исследования на животных можно использовать для определения LD50 (дозы, летальной для 50% популяции) и ED50 (дозы, терапевтически эффективной в 50% популяции). Соотношение доз между токсическим и терапевтическим эффектами представляет собой терапевтический индекс, который может быть выражен как соотношение LD50/ED50. Антитела, которые демонстрируют высокие терапевтические индексы, являются предпочтительными. В одном аспекте антитело в соответствии с изобретением демонстрирует высокий терапевтический индекс.Данные, полученные из анализов клеточных культур и исследований на животных, можно использовать при составлении диапазона дозировок, подходящего для применения человеком. Дозировка предпочтительно находится в пределах диапазона циркулирующих концентраций, которые включают ED50 с небольшой или отсутствующей токсичностью. Дозировка может варьироваться в пределах этого диапазона в зависимости от ряда факторов, например, применяемой дозированной формы, используемого пути введения, состояния субъекта и т.п. Точные состав, путь введения и дозировку может выбирать личный лечащий врач с учетом состояния пациента (смотрите, например, Fingl et al., 1975, в: The Pharmacological Basis of Therapeutics, Ch. 1, p.1, в полном объеме включенную в данный документ посредством ссылки).
Лечащий врач пациентов, которых лечат антителами по изобретению, должен знать, как и когда прекращать, прерывать или корректировать введение вследствие токсичности, органной дисфункции и т.п. И наоборот, лечащий врач также должен знать, как скорректировать лечение до более высоких уровней, если клинический ответ был недостаточным (не допуская токсичности). Величина вводимой дозы при лечении представляющего интерес нарушения будет варьироваться в зависимости от тяжести состояния, подлежащего лечению, пути введения и т.п. Тяжесть состояния можно, например, оценить частично с помощью стандартных методов оценки. Кроме того, доза и, возможно, частота введения дозы также будут варьироваться в соответствии с возрастом, массой тела и ответом каждого отдельного пациента.
Антитела по изобретению можно вводить в комбинации с одним или более другими агентами в терапии. Например, антитело по изобретению можно вводить совместно с по меньшей мере одним дополнительным терапевтическим агентом. Термин «терапевтический агент» включает любой агент, вводимый для лечения симптома заболевания у индивида, нуждающегося в таком лечении. Такой дополнительный терапевтический агент может включать любые активные ингредиенты, подходящие для конкретного заболевания, лечение которого проводят, предпочтительно те, которые имеют дополняющую активность и не оказывают негативного влияния друг на друга. В определенных аспектах дополнительный терапевтический агент представляет собой иммуномодулирующий агент, цитостатический агент, ингибитор клеточной адгезии, цитотоксический агент, активатор апоптоза клеток или агент, который повышает чувствительность клеток к индукторам апоптоза. В предпочтительном аспекте дополнительный терапевтический агент представляет собой противораковый агент, например, разрушитель микротрубочек, антиметаболит, ингибитор топоизомеразы, интеркалятор ДНК, алкилирующий агент, гормональную терапию, ингибитор киназы, антагонист рецептора, активатор апоптоза опухолевых клеток или антиангиогенный агент.
Такие другие агенты приемлемо присутствуют в комбинации в количествах, которые эффективны для предполагаемой цели. Эффективное количество таких других агентов зависит от используемого количества антитела, типа нарушения или лечения и других факторов, обсуждаемых выше. Антитела в общем случае используют в таких же дозировках и вводят путями, которые описаны в данном документе, или в дозировках, составляющих от около 1 до 99% дозировок, описанных в данном документе, или в любой дозировке и любым путем, которые эмпирически/клинически определены как подходящие.
Такие комбинированные виды терапии, отмеченные выше, включают комбинированное введение (когда два или более терапевтических агентов включены в одну или в отдельные композиции), а также раздельное введение, в случае чего введение антитела по изобретению может происходить до, одновременно и/или после введения дополнительного терапевтического агента и/или адъюванта. Антитела по изобретению также можно использовать в комбинации с лучевой терапией.
Ж. Готовые изделия
В другом аспекте изобретения предложено готовое изделие, содержащее материалы, предназначенные для лечения, предотвращения и/или диагностики вышеописанных нарушений. Готовое изделие содержит контейнер и этикетку или вкладыш в упаковку, находящиеся на нем или связанные с ним. Подходящие контейнеры включают, например, бутылки, флаконы, шприцы, пакеты для в/в раствора и т.д. Контейнеры могут быть выполнены из ряда материалов, таких как стекло или пластик. Контейнер содержит композицию, которая сама по себе или в комбинации с другой композицией эффективна для лечения, предотвращения и/или диагностики патологического состояния, и может иметь стерильное входное отверстие (например, контейнер может представлять собой пакет для внутривенного раствора или флакон с пробкой, прокалываемой гиподермической иглой для инъекции). По меньшей мере один активный агент в композиции представляет собой антитело по изобретению. На этикетке или вкладыше в упаковку указано, что композицию применяют для лечения выбранного патологического состояния. Кроме того, готовое изделие может содержать (а) первый контейнер с содержащейся в нем композицией, причем указанная композиция содержит антитело по изобретению; и (б) второй контейнер с содержащейся в нем композицией, причем указанная композиция содержит дополнительный цитотоксический или иной терапевтический агент.Готовое изделие в этом аспекте может дополнительно содержать вкладыш в упаковку, на котором указано, что композиции можно применять для лечения конкретного патологического состояния. В альтернативном или дополнительном варианте готовое изделие может дополнительно содержать второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (БВДИ), фосфатно-солевой буферный раствор, раствор Рингера и раствор декстрозы. Оно также может содержать другие материалы, необходимые с коммерческой и пользовательской точки зрения, включая другие буферы, разбавители, фильтры, иглы и шприцы.
3. Способы и композиции для диагностики и обнаружения
В определенных аспектах любые из предложенных в данном документе антител применимы для обнаружения наличия их мишени (например, CD3, TYRP-1, EGFRvIII) в биологическом образце. В контексте данного документа термин «обнаружение» включает количественное или качественное обнаружение. В определенных аспектах биологический образец включает клетку или ткань, такую как ткань предстательной железы.
В одном варианте предложено антитело по изобретению для применения в способе диагностики или обнаружения. В дополнительном аспекте предложен способ обнаружения наличия CD3, TYRP-1 или EGFRvIII в биологическом образце. В определенных аспектах способ включает приведение биологического образца в контакт с антителом по настоящему изобретению в условиях, допускающих связывание антитела с CD3, TYRP-1 или EGFRvIII, и обнаружение того, образовался ли комплекс между антителом и CD3, TYRP-1 или EGFRvIII. Такой способ может представлять собой in vitro или in vivo способ. В одном варианте антитело по изобретению используют для отбора субъектов, подлежащих терапии антителом, которое связывает CD3, TYRP-1 и/или EGFRvIII, например, если CD3, TYRP-1 и/или EGFRvIII является биомаркером для отбора пациентов.
Типовые нарушения, которые можно диагностировать, используя антитело по изобретению, включают рак, в частности рак кожи или рак головного мозга.
В определенных аспектах предложено антитело в соответствии с настоящим изобретением, причем антитело является меченным. Метки включают, но не ограничиваются этим, метки или фрагменты, обнаруживаемые прямым образом (например, флуоресцентные, хромофорные, электронноплотные, хемилюминесцентные и радиоактивные метки), а также фрагменты, такие как ферменты или лиганды, обнаруживаемые непрямым образом, например, посредством ферментативной реакции или молекулярного взаимодействия. Типовые метки включают, но не ограничиваются этим, радиоактивные изотопы 32Р, 14С, 125I, 3Н и 131I, флуорофоры, такие как редкоземельные хелаты или флуоресцеин и его производные, родамин и его производные, дансил, умбеллиферон, люциферазы, например, люциферазу светляков и бактериальную люциферазу (патент США №4737456), люциферин, 2,3-дигидрофталазиндионы, пероксидазу хрена (HRP), щелочную фосфатазу, β-галактозидазу, глюкоамилазу, лизозим, сахаридооксидазы, например, глюкозооксидазу, галактозооксидазу и глюкозо-6-фосфат дегидрогеназу, гетероциклические оксидазы, такие как уриказа и ксантиноксидаза, сопряженные с ферментом, который использует пероксид водорода для окисления предшественника красителя, такой как HRP, лактопероксидазу или микропероксидазу, биотин/авидин, спиновые метки, бактериофаговые метки, стабильные свободные радикалы и т.п.
II. ПОСЛЕДОВАТЕЛЬНОСТИ









IV. ПРИМЕРЫ
Ниже приведены примеры способов и композиций по изобретению. Следует понимать, что на практике можно реализовать ряд других аспектов с учетом общего описания, приведенного выше.
Пример 1 - Создание оптимизированного связывающего CD3 фрагмента
Начиная с ранее описанного (смотрите, например, WO 2014/131712, включенную в данный документ посредством ссылки) связывающего CD3 фрагмента, называемого в данном документе «CD3orig» и содержащего последовательности VH и VL SEQ ID NO 6 и 11, соответственно, мы поставили цель оптимизировать свойства этого связывающего фрагмента путем удаления двух аспарагиновых мотивов последовательности дезамидирования в позициях 97 и 100 по Kabat CDR3 тяжелой цепи.
С этой целью мы создали подходящую для фагового дисплея библиотеку антител тяжелой цепи с удаленными обоими аспарагинами в позициях 97 и 100 по Kabat и дополнительно рандомизированными H1, Н2 и H3 CDR с целью компенсации потери аффинности, вызванной замещением Asn97 и Asn100 посредством процесса созревания аффинности.
Эту библиотеку применяли к нитевидному фагу посредством слияния с минимальным белком оболочки р3 (Marks et al. (1991) J Mol Biol 222, 581-597) и проводили отбор в отношении связывания с рекомбинантным CD3ε.
Во время начального скрининга идентифицировали 10 клонов-кандидатов, демонстрирующих приемлемое связывание на рекомбинантном антигене по данным измерения ППР, в виде Fab-фрагментов (вырабатываемых Е. coli).
Однако только один из этих клонов демонстрировал приемлемую активность связывания с экспрессирующими CD3 клетками по данным измерений методом проточной цитометрии после преобразования в формат IgG.
Отобранный клон, называемый в данном документе «CD3opt» и содержащий последовательности VH и VL SEQ ID NO 7 и 11, соответственно, дополнительно оценивали и преобразовывали в биспецифический формат, как описано ниже.
Пример 2 - Связывание оптимизированного связывающего CD3 фрагмента с CD3
Связывание с рекомбинантным CD3
Связывание с рекомбинантным CD3 определяли методом поверхностного плазмонного резонанса (ППР) для оптимизированного связывающего CD3 фрагмента «CD3opt» и оригинального связывающего CD3 фрагмента «CD3orig», обоих в формате человеческого IgG1 с мутациями P329G L234A L235A («PGLALA», нумерация EU) в Fc-области (SEQ ID NO 12 и 14 (CD3orig) и SEQ ID NO 13 и 14 (CD3opt)).
Чтобы оценить эффект удаления сайта дезамидирования и его влияние на стабильность антител, исследовали связывание оригинального и оптимизированного связывающего CD3 фрагмента с рекомбинантным CD3 после температурного стресса в течение 14 суток при 37°С или 40°С. Образцы, которые хранили при -80°С, использовали в качестве референса. Референсные образцы и образцы, подвергнутые стрессу при 40°С, находились в 20 мМ His, 140 мМ NaCl, рН 6,0, а образцы, подвергнутые стрессу при 37°С - в ФСБ, рН 7,4, все в концентрации 1,2-1,3 мг/мл. После стрессового периода (14 суток) проводили обратный диализ образцов в ФСБ в 20 мМ His, 140 мМ NaCl, рН 6,0 для дополнительного анализа.
Относительную активную концентрацию (OAK) образцов определяли методом ППР следующим образом.
ППР проводили на инструменте Biacore Т200 (GE Healthcare). Захватывающее анти-Fab антитело (GE Healthcare, №28958325) иммобилизовали на сенсорном чипе СМ5 серии S (GE Healthcare), используя стандартную химическую реакцию аминного сопряжения, что приводило к поверхностной плотности 4000-6000 резонансных единиц (РЕ). В качестве подвижного буфера и буфера для разведения использовали HBS-P+(10 мМ ГЭПЭС, 150 мМ NaCl, рН 7,4, 0,05% поверхностно-активного вещества Р20). Антитела к CD3 в концентрации 2 мкг/мл вводили в течение 60 с при скорости потока 5 мкл/мин. Антиген CD3 (смотрите ниже) вводили в концентрации 10 мкг/мл в течение 120 с и отслеживали диссоциацию при скорости потока 5 мкл/мин в течение 120 с. Поверхность чипа восстанавливали посредством двух последовательных введений 10 мМ глицина, рН 2,1, по 60 с каждый. Разницу объемного показателя преломления корректировали путем вычитания холостых введений и путем вычитания ответа, полученного от холостого контроля с проточной ячейкой. Для оценки ответ связывания определяли через 5 секунд после окончания введения. Для нормализации сигнала связывания связывание CD3 делили на ответ анти-Fab (сигнал (РЕ), полученный после захвата антитела к CD3 на иммобилизованном анти-Fab антителе). Относительную активную концентрацию рассчитывали путем сопоставления каждого подвергнутого температурному стрессу образца с соответствующим не подвергнутым стрессу образцом.
Используемым антигеном был гетеродимер эктодоменов CD3 дельта и CD3 эпсилон, слитых с Fc-доменом человека с модификациями типа «выступ-во-впадину» и С-концевым Avi-тегом (смотрите SEQ ID NO 28 и 29).
Результаты этого эксперимента проиллюстрированы на Фиг. 2. Как можно видеть, оптимизированный связывающий CD3 фрагмент CD3opt демонстрировал заметно улучшенное связывание с CD3 после температурного стресса (2 недели при 37°С, рН 7,4) по сравнению с оригинальным связывающим CD3 фрагментом CD3orig. Этот результат демонстрирует, что удаление сайта дезамидирования было успешным и позволило получить антитело с превосходными свойствами стабильности, относящимися к его in vivo времени полужизни, а также составу антитела при нейтральном рН.
Связывание с CD3 на клетках Jurkat
Связывание с CD3 на репортерной линии Т-клеток человека Jurkat NFAT определяли методом FACS для оптимизированного связывающего CD3 фрагмента «CD3opt» и оригинального связывающего CD3 фрагмента «CD3orig», обоих в формате человеческого IgG1 с мутациями P329G L234A L235A («PGLALA», нумерация EU) в Fc-области (SEQ ID NO 12 и 14 (CD3orig) и SEQ ID NO 13 и 14 (CD3opt)).
Репортерные клетки Jurkat-NFAT (GloResponse Jurkat NFAT-RE-luc2P; Promega №CS 176501) представляют собой репортерную линию клеток острого лимфолейкоза человека с промотором NFAT, экспрессирующую CD3 человека. Клетки культивировали в RPMI1640, 2 г/л глюкозы, 2 г/л NaHCO3, 10% ФТС, 25 мМ ГЭПЭС, 2 мМ L-глутамина, 1 х NEAA, 1 х пирувата натрия при 0,1-0,5 млн клеток на мл. Каждый раз, когда клетки пассировали, добавляли конечную концентрацию 200 мкг на мл гигромицина В.
Для анализа связывания клетки Jurkat NFAT собирали, промывали в ФСБ и ресуспендировали в буфере FACS. Окрашивание антитела проводили в 96-луночном круглодонном планшете. Следовательно, высевали от 100000 до 200000 клеток на лунку. Планшеты центрифугировали в течение 4 мин при 400 х g и удаляли супернатант.исследуемые антитела разводили в буфере FACS и добавляли в лунки по 20 мкл раствора антитела в течение 30 мин при 4°С. Для удаления несвязанного антитела клетки дважды промывали буфером FACS перед добавлением разведенного вторичного антитела (РЕ-конъюгированный AffiniPure Р(ab')2-фрагмент козьего античеловеческого IgG, специфический к Fcg-фрагменту; Jackson Immuno Re search №109-116-170). Через 30 мин инкубации при 4°С несвязанное вторичное антитело смывали. Перед измерением клетки ресуспендировали в 200 мкл буфера FACS, после чего анализировали методом проточной цитометрии, используя устройство BD Canto П.
Как видно на Фиг. 3, оптимизированный связывающий CD3 фрагмент «CD3opt» и оригинальный связывающий CD3 фрагмент «CD3orig» сопоставимо хорошо связывались с CD3 на клетках Jurkat.
Пример 3 - Функциональная активность связывающего CD3 фрагмента
Функциональную активность оптимизированного связывающего CD3 фрагмента «CD3opt» исследовали в анализе с репортерными клетками Jurkat и сравнивали с активностью оригинального связывающего CD3 фрагмента «CD3 orig». Для исследования функциональной активности IgG, экспрессирующие анти-PGLALA клетки СНО инкубировали совместно с репортерными клетками Jurkat NFAT в присутствии возрастающих концентраций IgG1 PGLALA человека к CD3opt или IgG1 PGLALA человека к CD3orig. Активация CD3 на репортерных клетках Jurkat NFAT после перекрестного связывания Т-клеток индуцирует выработку люциферазы, и можно измерять люминесценцию в качестве маркера активации. IgG1 человека к CD3orig дт был включен в качестве отрицательного контроля, который не может связываться с экспрессирующими анти-PGLALA клетками СНО и, следовательно, не может перекрестно связываться на клетках Jurkat NFAT. Схематическая иллюстрация этого анализа приведена на Фиг. 4.
Экспрессирующие анти-PGLALA клетки СНО представляют собой клетки СНО-К1, сконструированные для экспрессии на своей поверхности антитела, которое специфически связывается с IgG1 Fc(PGLALA) человека (смотрите WO 2017/072210, включенную в данный документ посредством ссылки). Эти клетки культивировали в среде DMEM/F12, содержащей 5% ФТС+1% GluMax. Репортерные клетки Jurkat NFAT соответствуют описанным в примере 2.
После одновременного связывания CD3 hulgG1 PGLALA с анти-PGLALA, экспрессируемый на СНО, и CD3, экспрессируемый на репортерных клетках Jurkat-NFAT, активируется промотор NFAT и приводит к экспрессии активной люциферазы светляков. Интенсивность сигнала люминесценции (получаемая после добавления субстрата люциферазы) пропорциональна интенсивности активации и сигнализации CD3. Репортерные клетки Jurkat-NFAT выращивали в суспензии и культивировали в RPMI1640, 2 г/л глюкозы, 2 г/л NaHCO3, 10% ФТС, 25 мМ ГЭПЭС, 2 мМ L-глутамина, 1 х NEAA, 1 х пирувата натрия при 0,1 0,5 млн клеток на мл, 200 мкг на мл гигромицина. Для анализа собирали клетки СНО и определяли жизнеспособность с помощью ViCell. 30000 клеток-мишеней/лунка высевали в плоскодонный 96-луночный планшет с белыми стенками (Greiner bio-one №655098) в 100 μл среды и добавляли к клеткам СНО по 50 мкл/лунка разведенных антител или среды (для контролей). После этого собирали репортерные клетки Jurkat-NFAT и определяли жизнеспособность с помощью ViCell. Клетки ресуспендировали при 1,2 млн клеток/мл в клеточной культуральной среде без гигромицина В и добавляли к клеткам СНО при 60000 клеток/лунка (50 мкл/лунка) для получения конечного отношения эффектора к мишени (Э:М) 2:1 и конечного объема 200 мкл на лунку. Потом в каждую лунку добавляли 4 μл GloSensor (Promega №Е1291) (2% от конечного объема). Клетки инкубировали в течение 24 ч при 37°С в увлажненном инкубаторе. В конце времени инкубации регистрировали люминесценцию, используя TECAN Spark 10М.
Как показано на Фиг. 5, оптимизированный связывающий CD3 фрагмент CD3opt имел активность на клетках Jurkat NFAT после перекрестного связывания, аналогичную с CD3orig.
Пример 4 - Создание Т-клеточного биспецифического антитела, содержащего оптимизированный связывающий CD3 фрагмент
TYRP1 ТСВ
Оптимизированный связывающий CD3 фрагмент, идентифицированный в примере 1 («CD3opt», SEQ ID NO 7 (VH) и 11 (VL)), использовали для создания Т-клеточного биспецифического антитела (ТСВ), нацеленного на CD3 и TYRP1 («TYRP1 ТСВ»).
Связывающий TYRP1 фрагмент, содержащийся в этом ТСВ, создавали путем гуманизации связывающего TYRP1 фрагмента «ТА99» (смотрите записи GenBank AXQ57811 и AXQ57813 в отношении тяжелой и легкой цепи, соответственно), и он содержал последовательности вариабельных областей тяжелой и легкой цепи, приведенные в SEQ ID NO 18 и 22, соответственно.
Схематическая иллюстрация молекулы ТСВ приведена на Фиг. 6, а ее полные последовательности приведены в SEQ ID NO 23, 24, 25 и 27.
Также получали аналогичную молекулу с оригинальными связывающими CD3 последовательностями (SEQ ID NO 23, 24, 25 и 26).
Биспецифические молекулы создавали путем временной трансфекции клеток НЕК293 EBNA. Клетки трансфицировали соответствующими экспрессионными векторами в соотношении 1:2:1:1 («тяжелая цепь вектора (VH-CH1-VL-CH1-CH2-CH3)»: «легкая цепь вектора (VL-CL)»: «тяжелая цепь вектора (VH-CH1-CH2-CH3)»: «легкая цепь вектора (VH-CL)»). Клетки центрифугировали и заменяли среду предварительно нагретой средой CD СНО (Thermo Fisher, №10743029). Экспрессионные векторы смешивали в среде CD СНО, добавляли ПЭИ (полиэтиленимин, Polysciences, №23966-1), раствор перемешивали на вортексе и инкубировали в течение 10 минут при комнатной температуре. После этого клетки (2 млн/мл) смешивали с раствором вектор/ПЭИ, переносили в колбу и инкубировали в течение 3 часов при 37°С во встряхивателе-инкубаторе с атмосферой 5% СО2. После инкубации добавляли среду Excell с добавками (80% от общего объема). Через одни сутки после трансфекции добавляли добавки (Feed, 12% от общего объема). Клеточные супернатанты собирали через 7 суток путем центрифугирования и последующей фильтрации (0,2 мкм фильтр).
Белки очищали из профильтрованных клеточных культуральных супернатантов стандартными методами. Вкратце, Fc-содержащие белки очищали из клеточных культуральных супернатантов с помощью аффинной хроматографии с протеином A (MabSelect Sure, GE Healthcare: уравновешивающий буфер: 20 мМ цитрата натрия, 20 мМ фосфата натрия, рН 7,5; элюирующий буфер: 20 мМ цитрата натрия, 100 мМ NaCl, 100 мМ глицина, рН 3,0). Элюирование проводили при рН 3,0 с последующей немедленной нейтрализацией рН образца. Белок концентрировали путем центрифугирования (Millipore Amicon® ULTRA-15 (№UFC903096)) и отделяли агрегированный белок от мономерного белка с помощью эксклюзионной хроматографии (Superdex 200, GE Healthcare) в 20 мМ гистидина, 140 мМ хлорида натрия, рН 6,0.
Концентрацию очищенных белков определяли путем измерения поглощения на 280 нм, используя массовый коэффициент экстинкции, рассчитанный на основании аминокислотной последовательности в соответствии с Расе et al. (1995), Protein Science 4, 2411-23. Чистоту и молекулярную массу белков анализировали с помощью КЭ-ДСН в присутствии или отсутствие восстанавливающего агента, используя LabChipGXII (Perkin Elmer) (таблица 1). Определение содержания агрегатов проводили с помощью ВЭЖХ-хроматографии при 25°С, использую аналитическую эксклюзионную колонку (TSKgel G3000 SW XL или UP-SW3000), уравновешенную в подвижном буфере (25 мМ К2НРО4, 125 мМ NaCl, 200 мМ моногидрохлорида L-аргинина, рН 6,7, или 200 мМ KH2PO4, 250 мМ KCl, рН 6,2, соответственно) (таблица 2).
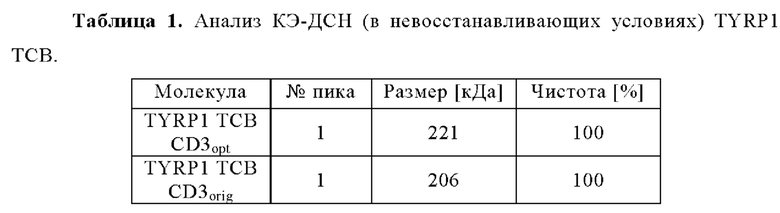

Пример 5 - Связывание Т-клеточного биспецифического антитела, содержащего оптимизированный связывающий CD3 фрагмент, с CD3 и TYRP1
Связывание с рекомбинантным CD3
Связывание TYRP1 ТСВ с рекомбинантным CD3 оценивали методом ППР, используя ТСВ с оптимизированными (TYRP1 ТСВ CD3opt) или оригинальными (TYRP1 ТСВ CD3orig) связывающими CD3 последовательностями.
Эксперименты ППР проводили на Biacore Т200 с HBS-EP в качестве подвижного буфера (0,01 М ГЭПЭС, рН 7,4, 0,15 М NaCl, 0,05% (об./об.) поверхностно-активного вещества Р20 (GE Healthcare)).
TYRP1 ТСВ захватывали на поверхности сенсорного чипа СМ5 с иммобилизованным антителом, которое специфически связывается с IgG1 Fc(PGLALA) человека (смотрите WO 2017/072210, включенную в данный документ посредством ссылки). Захватывающее антитело связывали с поверхностью сенсорного чипа путем прямой иммобилизации около 8700 резонансных единиц (РЕ) при рН 5,0, используя набор для стандартной реакции аминного сопряжения (GE Healthcare). Захват молекул ТСВ проводили в течение 30 с при 5 нМ со скоростью потока 10 мкл/мин.
Антигены человека и яванского макака проводили в концентрации 12,35 3000 нМ со скоростью потока 30 мкл/мин через проточные ячейки в течение 240 с. Фазу диссоциации отслеживали в течение 240 с и инициировали путем переключения с раствора для образца на HBS-EP. Поверхность чипа восстанавливали после каждого цикла, используя одно введение 10 мМ глицина, рН 2,0, в течение 30 с.
Используемым антигеном были гетеродимеры эктодоменов CD3 дельта и CD3 эпсилон человека или яванского макака, слитых с Fc-доменом человека с модификациями типа «выступ-во-впадину» и С-концевым Avi-тегом (смотрите SEQ ID NO 28 и 29) (CD3 человека) и SEQ ID NO 30 и 31 (CD3 яванского макака)).
Разницу объемного показателя преломления корректировали путем вычитания ответа, полученного на референсной проточной ячейке (без захваченного ТСВ). Константы аффинности получали из кинетических констант скорости путем аппроксимации 1:1 моделью связывания Ленгмюра, используя программное обеспечение BIAeval (GE Healthcare).
Значения KD для связывания с CD3 человека и яванского макака были определены как 50 нМ и 20 нМ, соответственно, для TYRP1 ТСВ CD3opt и были аналогичными для TYRP1 ТСВ CD3orig (50 нМ и 40 нМ, соответственно).
Это показывает, что в бесстрессовых условиях оба ТСВ, содержащие CD3opt или CD3orig, сопоставимо хорошо связывались с рекомбинантным CD3.
Связывание TYRP1 ТСВ с рекомбинантным CD3 человека также оценивали после температурного стресса в течение 14 суток при 37°С или 40°С, используя ТСВ с оптимизированными или оригинальными связывающими CD3 последовательностями. Этот эксперимент проводили, как описано в примере 2 выше, используя ТСВ вместо молекул IgG.
Результаты этого эксперимента проиллюстрированы на Фиг. 7.
Как можно видеть на Фиг. 7, ТСВ, содержащие оптимизированный связывающий CD3 фрагмент CD3opt, демонстрировал заметно улучшенное связывание с CD3 после стресса (2 недели при 37°С, рН 7,4) по сравнению с ТСВ, содержащими оригинальный связывающий CD3 фрагмент CD3orig. Этот результат подтверждает, что улучшенные свойства оптимизированного связывающего CD3 фрагмента (смотрите пример 2) сохраняются на уровне ТСВ.
Связывание с рекомбинантным TYRP1
Связывание с рекомбинантным TYRP1 анализировали методом ППР, используя Fab-фрагменты TYRP1, полученные путем расщепления плазмином соответствующего антитела.
Эксперименты ППР проводили на Biacore Т200 с HBS-EP в качестве подвижного буфера (0,01 М ГЭПЭС, рН 7,4, 0,15 М NaCl, 0,05% (об./об.) поверхностно-активного вещества Р20 (GE Healthcare)).
Антитело, которое специфически связывается с IgG1 Fc(PGLALA) человека (смотрите WO 2017/072210, включенную в данный документ посредством ссылки), напрямую связывали на сенсорном чипе СМ5 при рН 5,0, используя набор для стандартной реакции аминного сопряжения (GE Healthcare). Захват антигенов (смотрите ниже) проводили при скорости потока 10 μл/мин в течение 30 с. 3-Кратные серийные разведения Fab-фрагментов TYRP1 проводили через проточные ячейки при 30 μл/мин в течение 180 с для записи фазы ассоциации. Фазу диссоциации отслеживали в течение 180 с или 1200 с и инициировали путем переключения с раствора для образца на HBS-EP. Поверхность чипа восстанавливали после каждого цикла, используя одно введение 10 мМ глицина, рН 2, в течение 30 с при 30 мкл/мин.
Используемыми антигенами были мономерные слияния внеклеточного домена (ВКД) TYRP1 человека, яванского макака или мыши с Fc-доменом человека с модификациями типа «выступ-во-впадину» (и PG LALA) и С-концевым Avi-тегом (смотрите SEQ ID NO 32 и 35) (TYRP1 человека), SEQ ID NO 33 и 35 (TYRP1 яванского макака) или SEQ ID NO 34 и 35 (TYRP1 мыши)).
Разницу объемного показателя преломления корректировали путем вычитания ответа, полученного на референсной проточной ячейке (без захваченного антигена). Константы аффинности (KD) получали из кинетических констант скорости путем аппроксимации 1:1 моделью связывания Ленгмюра, используя программное обеспечение BIAeval (GE Healthcare).
Значения KD для связывания с TYRP1 человека, яванского макака и мыши были определены как 130 пМ, 180 пМ и 530 пМ, соответственно, и были аналогичными с родительским антителом ТА99 (90 пМ, 120 пМ и 310 пМ, соответственно).
Связывание TYRP1 ТСВ с рекомбинантным TYRP1 человека также оценивали после температурного стресса в течение 14 суток при 37°С или 40°С, используя ТСВ с оптимизированными или оригинальными связывающими CD3 последовательностями.
Этот эксперимент проводили, как описано выше для связывания с CD3, используя рекомбинантный TYRP1 (Sino Biologicals) в качестве антигена.
Результаты этого эксперимента проиллюстрированы на Фиг. 8. Они подтверждают, что на связывание с TYRP1 человека в случае обоих ТСВ (а также в случае соответствующего связывающего TYRP1 фрагмента в формате IgG) не влияют стрессовые условия.
Связывание с CD3 на клетках Jurkat
Связывание с CD3 на человеческой репортерной Т-клеточной линии Jurkat NFAT определяли методом FACS для TYRP1 ТСВ, содержащих оптимизированный связывающий CD3 фрагмент «CD3opt» или оригинальный связывающий CD3 фрагмент «CD3orig», как описано выше в примере 2.
Как показано на Фиг. 9, ТСВ, содержащие оптимизированный связывающий CD3 фрагмент «CD3opt», связывается с CD3 на клетках Jurkat по меньшей мере сопоставимо хорошо с ТСВ, содержащим оригинальный связывающий CD3 фрагмент «CD3orig».
Пример 6 - Функциональная активность Т-клеточного биспецифического антитела, содержащего оптимизированный связывающий CD3 фрагмент
Активация CD3
TYRP1 ТСВ, содержащие оптимизированный связывающий CD3 фрагмент CD3opt или оригинальный связывающий CD3 фрагмент CD3orig (пример 4), исследовали в анализе с репортерными клетками Jurkat NFAT (смотрите пример 3) в присутствии TYRP1-положительных клеток меланомы Ml50543 (линия клеток первичной меланомы, полученная от дерматологического банка клеток Университета Цюриха).
После одновременного связывания TYRP1 ТСВ с TYRP1-положительными клетками-мишенями и антигеном CD3 (экспрессируемым на репортерных клетках Jurkat-NFAT) активируется промотор NFAT и приводит к экспрессии активной люциферазы светляков. Интенсивность сигнала люминесценции (получаемая после добавления субстрата люциферазы) пропорциональна интенсивности активации и сигнализации CD3. Этот анализ проводили, как описано в примере 3, используя М150543 вместо экспрессирующих анти-PGLALA клеток СНО.
Как видно для IgG (пример 3) оба ТСВ, содержащих CD3opt или CD3orig, имели аналогичную функциональную активность на репортерных клетках NFAT и индуцировали активацию CD3 зависимым от концентрации образом (Фиг. 10).
Уничтожение клеток-мишеней
На следующем этапе обе молекулы ТСВ исследовали в анализе уничтожения опухолевых клеток со свежевыделенными МКПК человека от трех разных доноров, которые инкубировали совместно с линией клеток меланомы человека M150543. Лизис опухолевых клеток определяли по высвобождению ЛДГ через 24 ч и 48 ч. Активацию CD4 и CD8 Т-клеток анализировали по повышению регуляции CD69 и CD25 в обеих подгруппах клеток через 48 ч.
Вкратце, клетки-мишени собирали с трипсином/ЭДТА, промывали и высевали при плотности 30000 клеток/лунка, используя плоскодонные 96-луночные планшеты. Клетки оставляли на ночь для прикрепления. Мононуклеарные клетки периферической крови (МКПК) подготавливали путем центрифугирования в градиенте плотности Histopaque свежей крови, полученной от здоровых доноров-людей. Свежую кровь разводили в стерильной ФСБ и расслаивали в градиенте Histopaque (Sigma №Н8889). После центрифугирования (450 х g, 30 минут, комнатная температура) плазму выше МКПК-содержащей промежуточной фазы сливали и переносили МКПК в новую пробирку Falcon, которую после этого наполняли 50 мл ФСБ. Смесь центрифугировали (400 х g, 10 минут, комнатная температура), супернатант сливали, а осажденные МКПК дважды промывали стерильным ФСБ (этапы центрифугирования 350 х g, 10 минут). Полученную в результате популяцию МКПК автоматически подсчитывали (ViCell) и хранили в среде RPMI1640, содержащей 10% ФТС и 1% L-аланил-L-глутамина (Biochrom №К0302), при 37°С, 5% CO2 в клеточном инкубаторе до дальнейшего использования (не дольше 24 ч). Для анализа уничтожения антитела добавляли в указанных концентрациях в трех повторностях. МКПК добавляли к клеткам-мишеням при конечном отношении эффектора к мишени (Э:М) 10:1. Уничтожение клеток-мишеней анализировали через 24 ч инкубации при 37°С, 5% CO2 путем количественной оценки ЛДГ, высвобождаемой в клеточные супернатанты апоптотическими/некротическими клетками (набор для обнаружения ЛДГ, Roche Applied Science №11 644 793 001). Максимальный лизис клеток-мишеней (=100%) достигался путем инкубации клеток-мишеней с 1% Тритон Х-100. Минимальный лизис (=0%) относится к клеткам-мишеням, которые инкубировали совместно с эффекторными клетками без биспецифической конструкции.
Активацию CD8 и CD4 Т-клеток после уничтожения Т-клетками клеток-мишеней, опосредованного ТСВ, анализировали методом проточной цитометрии, используя антитела, распознающие маркеры активации Т-клеток CD25 (поздний маркер активации) CD69 (ранний маркер активации). Через 48 ч инкубации МКПК переносили в круглодонный 96-луночный планшет, центрифугировали при 350 х g в течение 5 мин и дважды промывали буфером FACS. Поверхностное окрашивание для CD4 АРС (BioLegend №300514), CD8 FITC (BioLegend №344704), CD25 BV421 (BioLegend №302630) и CD69 РЕ (BioLegend №310906) проводили в соответствии с указаниями поставщика. Клетки дважды промывали 150 мкл/лунка буфера FACS и фиксировали в течение 15 мин при 4°С, используя 100 мкл/лунка буфера для фиксации (BD №554655). После центрифугирования образцы ресуспендировали в 200 мкл/лунка буфера FACS. Образцы анализировали на BD FACS Fortessa.
В случае всех трех доноров оба ТСВ, содержащие оптимизированный или оригинальный связывающий CD3 фрагмент, индуцировали активацию Т-клеток и лизис опухолевых клеток сопоставимым образом (Фиг. 11). Значения ЕС50 лизиса опухолевых клеток для всех трех доноров через 48 ч приведены в таблице 3.

Пример 7 - Исследование ФК с Т-клеточными биспецифическими антителами на мышах
Фармакокинетику (ФК) TYRP1 ТСВ с разными связывающими CD3 фрагментами (CD3orig и CD3opt) исследовали после внутривенного болюсного введения при 1 мг/кг трансгенным в отношении человеческого FcRn (линия 32, гомозиготные) и имеющим нокаут FcRn мышам (номера линий Jackson Laboratory 003982 и 014565) (n=3/линия/исследуемое соединение). Серийные микрообразцы крови брали у трансгенных (tg) в отношении человеческого FcRn мышей до 672 ч (9 образцов на мышь от 5 мин до 672 ч после дозы) и до 96 ч у мышей с нокаутом (ко) FcRn (8 образцов на мышь от 5 мин до 96 ч после дозы). Получали сыворотку и хранили замороженной до анализа. Образцы мышиной сыворотки анализировали с помощью общего метода ECLIA, специфического в отношении Ig/Fab CH1/каппа домена человека, используя инструмент cobas® е411 (Roche), в условиях без GLP. Фармакокинетическую оценку проводили, используя стандартный некомпартментный анализ.
Результаты этого исследования проиллюстрированы в таблице 4. Они показывают, что конструирование CDR не приводит к другим свойствам последовательностей, которые повлияли бы на клиренс антитела. CD3opt является одинаково эффективным с CD3orig в терминах сывороточного времени полужизни, но имеет дополнительное преимущество повышенной стабильности CDR.

Пример 8 - Создание дополнительного Т-клеточного биспецифического антитела, содержащего оптимизированный связывающий CD3 фрагмент
Оптимизированный связывающий CD3 фрагмент, идентифицированный в примере 1 («CD3opt», SEQ ID NO 7 (VH) и 11 (VL)), использовали для создания Т-клеточного биспецифического антитела (ТСВ), нацеленного на CD3 и EGFRvIII («EGFRvIII ТСВ»).
Связывающий EGFRvIII фрагмент, содержащийся в этом ТСВ (Р063.056), был получен из фагового дисплея после созревания аффинности (смотрите ниже) и содержал последовательности вариабельных областей тяжелой и легкой цепи, приведенные в SEQ ID NO 88 и 92, соответственно.
Схематическая иллюстрация молекулы ТСВ приведена на Фиг. 6, а ее полные последовательности приведены в SEQ ID NO 109, 110, 111 и 27.
Также получали аналогичную молекулу с оригинальными связывающими CD3 последовательностями (SEQ ID NO 109, 110, 111 и 26).
Биспецифические молекулы создавали путем временной трансфекции клеток НЕК293 EBNA, очищали и анализировали, как описано выше в примере 4.
Кроме того, антитела к EGFRvIII, полученные из фагового дисплея, получали в формате IgG1 человека аналогичным образом (трансфекция клеток НЕК293 EBNA экспрессионными векторами для тяжелой и легкой цепей IgG в соотношении 1:1) для применения, как описано ниже.
Все конструкции IgG и ТСВ очищали до сопоставимого качества с содержанием мономеров более 95% по определению методом эксклюзионной хроматографии.
Отбор антитела к EGFRvIII
Антитела к EGFRvIII получали из фагового дисплея и обеспечивали созревание аффинности. Антитела, демонстрирующие высокую аффинность связывания и специфичность в отношении EGFRvIII (Р056.021 (SEQ ID NO 40 и 44), P056.052 (SEQ ID NO 48 и 52), P047.019 (SEQ ID NO 56 и 60), P057.012 (SEQ ID NO 64 и 68), P057.011 (SEQ ID NO 72 и 76), P056.027 (SEQ ID NO 80 и 84)), исследовали в отношении связывания с EGFRvIII, экспрессируемый на клеточной поверхности, используя клетки СНО, стабильно экспрессирующие EGFRvIII, и EGFRvIII-положительную линию клеток глиобластомы человека DK-MG. Для подтверждения специфичности и исключения перекрестной реактивности с EGFR дикого типа (EGFRwt) отобранные антитела исследовали в отношении связывания с EGFRwt-положительной опухолевой линией клеток человека MKN-45 (Фиг. 12). Цетуксимаб был включен в качестве положительного контроля для связывания с EGFRwt, а ненацеленный DP47 IgG - в качестве отрицательного контроля. Все отобранные антитела специфически связывались с EGFRvIII без перекрестной реактивности с EGFRwt и рассматривались для дополнительного изучения.
На следующем этапе анализировали функциональную активность этих антител к EGFRvIII в виде IgG1 PGLALA (формат человеческого IgG1 с мутациями P329G L234A L235A («PGLALA», нумерация EU) в Fc-области) на клетках DK-MG, которые инкубировали совместно с репортерными клетками Jurkat NFAT, экспрессирующими анти-PGLALA химерный антигенный рецептор (CAR), путем измерения люминесценции (анализ CAR J, смотрите заявку РСТ №РСТ/ЕР2018/086038, в полном объеме включенную в данный документ посредством ссылки). DP47 IgG1 PGLALA был включен в качестве отрицательного контроля. Все исследуемые антитела к EGFRvIII индуцировали сильную активацию CAR-экспрессирующих репортерных клеток Jurkat NFAT (Фиг. 13). Все исследуемые антитела к EGFRvIII за исключением Р047.019, которое демонстрировало самые слабые связывание и активацию, были отобраны для преобразования в формат ТСВ (с CDorig в качестве связывающего CD3 фрагмента).
Связывание выбранных антител к EGFRvIII, преобразованных в формат ТСВ, с клетками CHO-EGFRvIII сравнивали со связыванием соответствующих IgG (Фиг. 14), чтобы подтвердить, что преобразование в формат ТСВ не влияет на связывающую способность антител к EGFRvIII. Большинство из исследуемых клонов EGFRvIII сохраняли свою способность связываться с EGFRvIII после преобразования в формат ТСВ; только клон Р057.011 демонстрировал немного сниженное связывание с EGFRvIII в формате ТСВ по сравнению с соответствующим IgG (таблица 5).

После этого исследовали функциональную активность EGFRvIII ТСВ в анализе с репортерными клетками Jurkat NFAT на EGFRvIII-положительных клетках DK-MG (Фиг. 15). Все исследуемые EGFRvIII ТСВ обладали активностью в анализе с репортерными клетками Jurkat NFAT, при этом Р056.021 был наиболее активным, за ним следовали Р056.027, Р056.052 и Р057.012, которые имели сходную активность, и Р057.011, который имел самую низкую активность. После этого EGFRvIII ТСВ исследовали в анализе лизиса опухолевых клеток с МКПК, которые культивировали вместе с клетками DK-MG или MKN-45, чтобы исключить перекрестную реактивность EGFRvIII ТСВ с EGFRwt (Фиг. 16). В этом анализе помимо лизиса опухолевых клеток в качестве дополнительных результатов измеряли активацию Т-клеток (Фиг. 17) и высвобождение цитокинов (Фиг. 18). Как видно в проведенном ранее анализе с репортерными клетками, EGFRvIII ТСВ Р056.021 имел наибольшую активность на EGFRvIII-положительных клетках без какой-либо активности на EGFRwt-положительных клетках. EGFRvIII ТСВ Р057.011 демонстрировал неспецифическую активность на клетках EGFRwt и, следовательно, был исключен. EGFRvIII TCBs Р056.027, Р056.052 и Р057.012 имели сопоставимую активность. На основании этих результатов связывающие EGFRvIII фрагменты Р056.021 и Р057.012 были отобраны для дополнительного раунда созревания аффинности.
Из Р057.012 невозможно было получить хорошие связывающие фрагменты (результаты не показаны). Аффинность и специфичность к EGFRvIII отобранных связывающих EGFRvIII фрагментов, полученных из Р056.021, определенные методом ППР, приведены в таблице 6.
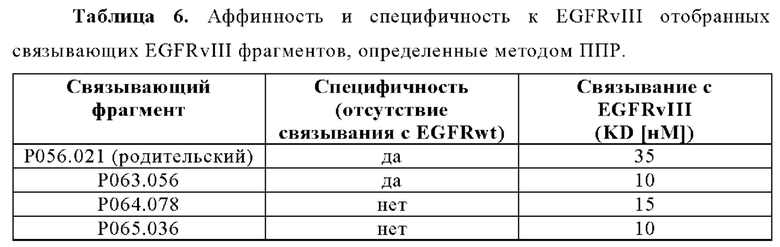
Связывающие EGFRvIII фрагменты с созревшей аффинностью (Р063.056 (SEQ ID NO 88 и 92), Р064.078 (SEQ ID NO 96 и 100), P065.036 (SEQ ID NO 104 и 108)) также сравнивали с родительским связывающим фрагментом в отношении специфического связывания с EGFRvIII на клетках U87MG-EGFRvIII 5 и MKN-45 (Фиг. 19). Наилучший связывающий EGFRvIII фрагмент в терминах аффинности и специфичности к EGFRvIII, Р063.056, был отобран для преобразования в формат ТСВ с CD3orig или CD3opt в качестве связывающего CD3 фрагмента.
Функциональную активность EGFRvIII ТСВ Р063.056 (с CD3opt или 10 CD3orig) сравнивали с родительским EGFRvIII ТСВ Р056.021 в анализе с репортерными клетками Jurkat NFAT на клетках U87MG-EGFRvIII, DK-MG и MKN-45 (Фиг. 20). Все три ТСВ индуцировали специфическую активацию Jurkat NFAT только в присутствии EGFRvIII-положительных клеток. EGFRvIII ТСВ Р063.056 имел немного большую активность, чем родительский EGFRvIII ТСВ 15 Р056.021.
Методы
Поверхностный плазмонный резонанс
Аффинность антитела к EGFRvIII к EGFRvIII определяли методом поверхностного плазмонного резонанса на Biacore Т200 с HBS-EP в качестве подвижного буфера (0,01 М ГЭПЭС, рН 7,4, 0,15 М NaCl, 0,005% (об./об.) поверхностно-активного вещества Р20; GE Healthcare) при 25°С. Анти-EGFRvIII PGLALA IgG захватывали в течение 30 с при 25 нМ с помощью антитела, которое специфически связывает человеческий IgG1 Fc(PGLALA) (смотрите WO 2017/072210, включенную в данный документ посредством ссылки), иммобилизованным на чипе СМ5. Антиген EGFRvIII-ECD avi his (смотрите ниже, пример 9) проводили в концентрации 12,4-1000 нМ со скоростью потока 30 мкл/мин через все проточные ячейки в течение 200 с. Фазу диссоциации отслеживали в течение 300 с и инициировали путем переключения с раствора для образца на HBS-EP. Поверхность чипа восстанавливали после каждого цикла, используя два введения 10 мМ глицина, рН 2,0, в течение 30 с. Разницу объемного показателя преломления корректировали путем вычитания ответа, полученного на референсной проточной ячейке. Константы аффинности получали из кинетических констант скорости путем аппроксимации 1:1 моделью связывания Ленгмюра, используя программное обеспечение BIAeval (GE Healthcare).
Для определения специфичности проводили захват антигенов EGFRvIII и ВКД EGFRwt с помощью анти-his (Penta His, Qiagen), иммобилизованного на чипе СМ5, в течение 40 с при 100 нМ. Проводили одно введение анти-EGFRvIII антител при 500 нМ в течение 60 с перед восстановлением 10 мМ глицина, рН 2,0, в течение 60 с. Наблюдали значения единиц ответа выше 50 для связывания EGFRvIII. Ответы выше 5 единиц ответа (ЕО) считали положительными для связывания EGFRwt, a IgG относили к категории специфических при ответе ниже 5 ЕО для EGFRwt.
Линии клеток
Репортерные клетки Jurkat-NFAT (GloResponse Jurkat NFAT-RE-luc2P; Promega №CS 176501) представляют собой репортерную линию клеток острого лимфолейкоза человека с промотором NFAT, экспрессирующую CD3 человека. Клетки культивировали в RPMI1640, 2 г/л глюкозы, 2 г/л NaHCO3, 10% ФТС, 25 мМ ГЭПЭС, 1% GlutaMAX, 1 х NEAA, 1 х пирувата натрия при 0,1-0,5 млн клеток на мл. Каждый раз, когда клетки пассировали, добавляли конечную концентрацию 200 мкг на мл гигромицина В.
Клетки Jurkat NFAT с PGLALA CAR создавали в лаборатории. Оригинальная линия клеток (Jurkat NFAT; Signosis) представляет представляют собой репортерную линию клеток острого лимфолейкоза человека с промотором NFAT, который приводит к экспрессии люциферазы после активации с помощью CD3 человека. Их конструировали для экспрессии химерного антигенного рецептора, способного распознавать мутацию P293G LALA. При культивировании клетки выращивали в суспензии в RPMI1640, дополненной 10% ФТС и 1% глутамина, и поддерживали при 0,4-1,5 млн клеток на мл.
Клетки CHO-EGFRvIII создавали в лаборатории. Клетки СНО-К1 стабильно трансдуцировали EGFRvIII. Клетки культивировали в среде DMEM/F12, содержащей 5% ФТС, 1% GlutaMAX и 6 μг/мл пуромицина.
DK-MG (DSMZ №АСС 277) представляет собой линию клеток глиобластомы человека. Клетки DK-MG обогащали путем сортировки клеток в отношении экспрессии EGFRvIII. Клетки культивировали в RPMI 1860, 10% ФТС и 1% GlutaMAX.
U87MG-EGFRvIII (АТСС НТВ-14) представляет собой линию клеток глиобластомы человека, которую стабильно трансдуцировали EGFRvIII. Клетки культивировали в DMEM, 10% ФТС и 1% GlutaMAX.
MKN-45 (DSMZ АСС 409) представляет собой клетки аденокарциномы желудка человека, экспрессирующие высокие уровни EGFRwt. Клетки культивировали в расширенной RPMI1640, содержащей 2% ФТС и 1% GlutaMAX.
Связывание мишени по данным проточной цитометрии
Клетки, используемые для экспериментов по связыванию, собирали, промывали ФСБ и ресуспендировали в буфере FACS. Окрашивание антитела проводили в 96-луночном круглодонном планшете. Клетки собирали, подсчитывали и высевали от 100000 до 200000 клеток на лунку. Планшеты центрифугировали в течение 4 мин при 400 х g и удаляли супернатант. Исследуемые антитела разводили в буфере FACS и добавляли в лунки по 20 мкл раствора антитела в течение 30 мин при 4°С. Для удаления несвязанного антитела клетки дважды промывали буфером FACS перед добавлением разведенного вторичного антитела, РЕ-конъюгированного AffiniPure F(ab')2-фрагмента козьего античеловеческого IgG, специфического к Fcg-фрагменту (Jackson ImmunoResearch №109-116-170 или №109-116-098). Через 30 мин инкубации при 4°С несвязанное вторичное антитело смывали. Перед измерением клетки ресуспендировали в 200 мкл буфера FACS и анализировали методом проточной цитометрии, используя BD Canto II или BD FACS Fortessa.
Анализ с репортерными клетками CAR J NFAT с EGFRvIII PGLALA IgG
Способность EGFRvIII PGLALA IgG индуцировать активацию Т-клеток оценивали, используя анализ с репортерными клетками CAR J NFAT. Принцип этого анализа состоит в совместной культивации сконструированных эффекторных клеток Jurkat-NFAT с раковыми клетками, экспрессирующими опухолевый антиген. Только после одновременного связывания IgG с CAR за счет мутации PGLALA и антигеном-мишенью EGFRvIII активируется промотор NFAT и приводит к повышению экспрессии люциферазы в эффекторных клетках Jurkat. После добавления достаточного количества субстрата активная люцифераза светляков приводит к испусканию люминесценции, которую можно измерять как сигнал CAR-опосредованной активации. Вкратце, собирали клетки-мишени и определяли жизнеспособность. 30000 клеток-мишеней/лунка высевали в плоскодонный 96-луночный планшет с белыми стенками (Greiner bio-one №655098) в 100 μл среды за сутки до начала анализа. На следующие сутки среду удаляли и добавляли к клеткам-мишеням 25 мкл/лунку разведенных антител или среды (для контролей). После этого собирали репортерные клетки Jurkat-NFAT и определяли жизнеспособность с помощью ViCell. Клетки ресуспендировали при 1,5 млн клеток/мл в клеточной культуральной среде и добавляли к опухолевым клеткам при 75000 клеток/лунка (50 мкл/лунка) для получения конечного отношения эффектора к мишени (Э:М) 2,5:1 и конечного объема 75 мкл на лунку. Потом в каждую лунку добавляли 4 μл GloSensor (Promega №E1291) (2% от конечного объема). Клетки инкубировали в течение 24 ч при 37°С в увлажненном инкубаторе. В конце времени инкубации планшеты адаптировали к комнатной температуре (около 15 мин). Потом добавляли 25 цл/лунка люциферазы One-Glo (Promega, №Е6120) и инкубировали планшет в течение 15 мин в темноте перед регистрацией люминесценции с помощью TEC AN Spark.
Анализ с репортерными клетками Jurkat NFAT с EGFRvIII ТСВ
Способность EGFRvIII ТСВ с улучшенным связывающим CD3 или оригинальным связывающим CD3 фрагментом индуцировать перекрестное связывание Т-клеток и последующую активацию Т-клеток оценивали, используя EGFRvIII-положительные клетки и репортерные клетки Jurkat-NFAT. После одновременного связывания EGFRvIII ТСВ с EGFRvIII-положительными клетками-мишенями и антигеном CD3 (экспрессируемым на репортерных клетках Jurkat-NFAT) активируется промотор NFAT и приводит к экспрессии активной люциферазы светляков. Интенсивность сигнала люминесценции (получаемая после добавления субстрата люциферазы) пропорциональна интенсивности активации и сигнализации CD3. Для анализа собирали клетки-мишени и определяли жизнеспособность. 30000 клеток-мишеней/лунка высевали в плоскодонный 96-луночный планшет с белыми стенками (Greiner bio-one №655098) в 100 μл среды и добавляли к клеткам-мишеням по 50 мкл/лунка разведенных антител или среды (для контролей). После этого собирали репортерные клетки Jurkat-NFAT и определяли жизнеспособность с помощью ViCell. Клетки ресуспендировали при 1,2 млн клеток/мл в клеточной культуральной среде без гигромицина В и добавляли к опухолевым клеткам при 60000 клеток/лунка (50 мкл/лунка) для получения конечного отношения эффектора к мишени (Э:М) 2:1 и конечного объема 200 мкл на лунку. Потом в каждую лунку добавляли 4 μл GloSensor (Promega №Е1291) (2% от конечного объема). Клетки инкубировали в течение 24 ч при 37°С в увлажненном инкубаторе. В конце времени инкубации регистрировали люминесценцию, используя TECAN Spark.
Опосредованное Т-клетками уничтожение опухолевых клеток
Клетки-мишени собирали с трипсином/ЭДТА, промывали и высевали при плотности 30000 клеток/лунка, используя плоскодонные 96-луночные планшеты. Клетки оставляли на ночь для прикрепления. Мононуклеарные клетки периферической крови (МКПК) подготавливали путем центрифугирования в градиенте плотности Histopaque свежей крови, полученной от здоровых доноров-людей. Свежую кровь разводили в стерильной ФСБ и расслаивали в градиенте Histopaque (Sigma, №Н8889). После центрифугирования (450 х g, 30 минут, комнатная температура) плазму выше МКПК-содержащей промежуточной фазы сливали и переносили МКПК в новую пробирку Falcon, которую после этого наполняли 50 мл ФСБ. Смесь центрифугировали (400 х g, 10 минут, комнатная температура), супернатант сливали, а осажденные МКПК дважды промывали стерильным ФСБ (этапы центрифугирования 350 х g, 10 минут). Полученную в результате популяцию МКПК автоматически подсчитывали (ViCell) и хранили в среде RPMI1640, содержащей 10% ФТС и 1% GlutaMAX, при 37°С, 5% CO2 в клеточном инкубаторе до дальнейшего использования (не дольше 24 ч). Для анализа уничтожения антитело добавляли в указанных концентрациях в трех повторностях. МКПК добавляли к клеткам-мишеням при конечном отношении эффектора к мишени (Э:М) 10:1. Уничтожение клеток-мишеней анализировали через 24 ч инкубации при 37°С, 5% СО2 путем количественной оценки ЛДГ, высвобождаемой в клеточные супернатанты апоптотическими/некротическими клетками (набор для обнаружения ЛДГ, Roche Applied Science, №11 644 793 001). Максимальный лизис клеток-мишеней (=100%) достигался путем инкубации клеток-мишеней с 1% Тритон Х-100. Минимальный лизис (=0%) относится к клеткам-мишеням, которые инкубировали совместно с эффекторными клетками без биспецифической конструкции.
Активацию CD8 и CD4 Т-клеток после уничтожения Т-клетками клеток-мишеней, опосредованного ТСВ, анализировали методом проточной цитометрии, используя антитела, распознающие маркеры активации Т-клеток CD25 (поздний маркер активации) CD69 (ранний маркер активации). Через 48 ч инкубации МКПК переносили в круглодонный 96-луночный планшет, центрифугировали при 350 х g в течение 5 мин и дважды промывали буфером FACS. Поверхностное окрашивание для CD4 АРС (BioLegend, №300514), CD8 FITC (BioLegend, №344704), CD25 BV421 (BioLegend, №302630) и CD69 РЕ (BioLegend, №310906) проводили в соответствии с указаниями поставщика. Клетки дважды промывали 150 мкл/лунка буфера FACS и фиксировали в течение 15 мин при 4°С, используя 100 мкл/лунка буфера для фиксации (BD, №554655). После центрифугирования образцы ресуспендировали в 200 мкл/лунка буфера FACS. Образцы анализировали на BD FACS Fortessa.
Секрецию цитокинов в супернатанте измеряли методом проточной цитометрии, используя цитометрическую гранульную матрицу (СВА) в соответствии с инструкциями производителя, но вместо 50 мкл гранул и образца использовали только 25 мкл супернатанта и гранул. Использовали следующие наборы СВА (BD Biosciences): Flex Set СВА с интерфероном гамма человека (IFNγ), Flex Set СВА с гранзимом В человека и Flex Set СВА с TNF человека. Измерения образцов проводили, используя BD FACS Canto II или BD FACS Fortessa, а анализ проводили, используя программное обеспечение Diva (BD Biosciences).
Пример 9 - Связывание Т-клеточного биспецифического антитела, содержащего оптимизированный связывающий CD3 фрагмент, с CD3 и EGFRvIII
Связывание с рекомбинантным CD3
Связывание EGFRvIII ТСВ с рекомбинантным CD3 оценивали методом ППР, используя ТСВ с оптимизированными (EGFRvIII ТСВ CD3opt) или оригинальными (EGFRvIII ТСВ CD3orig) связывающими CD3 последовательностями, как описано для TYRP1 ТСВ в примере 5 выше. Захватывающее антитело связывали с поверхностью сенсорного чипа путем прямой иммобилизации около 5200 резонансных единиц (РЕ) при рН 5,0, используя набор для стандартной реакции аминного сопряжения (GE Healthcare), а захват молекул ТСВ проводили в течение 30 с при 20 нМ со скоростью потока 10 мкл/мин.
Значения KD для связывания с CD3 человека и яванского макака были определены как 30 нМ и 20 нМ, соответственно, для TYRP1 ТСВ CD3opt и были аналогичными для TYRP1 ТСВ CD3orig (40 нМ и 30 нМ, соответственно).
Это показывает, что в бесстрессовых условиях оба ТСВ, содержащие CD3opt или CD3orig, сопоставимо хорошо связывались с рекомбинантным CD3.
Связывание EGFRvIII ТСВ с рекомбинантным CD3 человека также оценивали после температурного стресса в течение 14 суток при 37°С или 40°С, используя ТСВ с оптимизированными или оригинальными связывающими CD3 последовательностями. Этот эксперимент проводили, как описано в примере 2 выше, используя ТСВ вместо молекул IgG.
Результаты этого эксперимента проиллюстрированы на Фиг. 21.
Как можно видеть на Фиг. 21, ТСВ, содержащие оптимизированный связывающий CD3 фрагмент CD3opt, демонстрировал заметно улучшенное связывание с CD3 после стресса (2 недели при 37°С, рН 7,4) по сравнению с ТСВ, содержащими оригинальный связывающий CD3 фрагмент CD3orig. Этот результат снова подтверждает, что улучшенные свойства оптимизированного связывающего CD3 фрагмента (смотрите пример 2) сохраняются на уровне ТСВ.
Связывание с рекомбинантным EGFRvIII
Связывание EGFRvIII ТСВ с рекомбинантным EGFRvIII анализировали методом ППР.
Эксперименты ППР проводили на Biacore Т200 с HBS-EP в качестве подвижного буфера (0,01 М ГЭПЭС, рН 7,4, 0,15 М NaCl, 0,05% (об./об.) поверхностно-активного вещества Р20 (GE Healthcare)).
Анти-Fc антитело (GE Healthcare) напрямую связывали на сенсорном чипе СМ5 при рН 5,0, используя набор для стандартной реакции аминного сопряжения (GE Healthcare). Захват EGFRvIII ТСВ (5 нМ) проводили при скорости потока 10 μл/мин в течение 30 с. 3-Кратные серийные разведения антигена EGFRvIII проводили через проточные ячейки при 30 μл/мин в течение 200 с для записи фазы ассоциации. Фазу диссоциации отслеживали в течение 300 с и инициировали путем переключения с раствора для образца на HBS-EP. Поверхность чипа восстанавливали после каждого цикла, используя одно введение 3 М MgCL2 в течение 30 с при 20 мкл/мин.
Используемый антиген содержал внеклеточный домен EGFRvIII человека, слитый с Avi-тегом и His-тегом на С-конце (EGFRvIII-ECD avi his; SEQ ID NO: 36).
Разницу объемного показателя преломления корректировали путем вычитания ответа, полученного на референсной проточной ячейке (без захваченного ТСВ). Константы аффинности (KD) получали из кинетических констант скорости путем аппроксимации 1:1 моделью связывания Ленгмюра, используя программное обеспечение BIAeval (GE Healthcare). Кажущуюся константу авидности KD аппроксимировали с помощью кинетического анализа через константы скорости, используя 1:1 аппроксимацию связывания для этого 2:1 взаимодействия.
Значения KD (аффинности) для связывания с EGFRvIII человека были определены как 6 нМ для EGFRvIII ТСВ, содержащих как CD3opt, так и CD3orig.
Связывание EGFRvIII ТСВ с рекомбинантным EGFRvIII также оценивали после температурного стресса в течение 14 суток при 37°С или 40°С, используя ТСВ с оптимизированными или оригинальными связывающими CD3 последовательностями. Этот эксперимент проводили, как описано выше в примере 5, используя EGFRvIII-ECD avi his в качестве антигена (смотрите выше).
Результаты этого эксперимента проиллюстрированы на Фиг. 22. Они подтверждают, что на связывание с EGFRvIII в случае обоих ТСВ не влияют стрессовые условия.
Связывание с CD3 на клетках Jurkat
Связывание с CD3 на человеческой репортерной Т-клеточной линии Jurkat NFAT определяли методом FACS для EGFRvIII ТСВ, содержащих оптимизированный связывающий CD3 фрагмент «CD3opt» или оригинальный связывающий CD3 фрагмент «CD3orig», как описано выше в примере 2.
Как видно на Фиг. 23, ТСВ, содержащие оптимизированный связывающий CD3 фрагмент «CD3opt» и оригинальный связывающий CD3 фрагмент «CD3orig» сопоставимо хорошо связывались с CD3 на клетках Jurkat.
Связывание с EGFRvIII на клетках U87MG-EGFRvIII
Связывание с EGFRvIII на линии клеток глиобластомы человека U87MG-EGFRvIII определяли методом FACS для EGFRvIII ТСВ, содержащих связывающий EGFRvIII фрагмент Р063.056 с CD3opt или CD3orig, или клон EGFRvIII Р056.021 с CD3orig. Связывающий EGFRvIII фрагмент Р063.056 был включен также в формате IgG.
Как показано на Фиг. 24, все три ТСВ с высокой аффинностью связываются с EGFRvIII, экспрессируемым на клетках U87MG-EGFRvIII, а преобразование в формат ТСВ не влияет на связывание.
Пример 10 - Функциональная активность Т-клеточного биспецифического антитела, содержащего оптимизированный связывающий CD3 фрагмент
Активация CD3
EGFRvIII ТСВ, содержащие выбранный связывающий EGFRvIII фрагмент (Р063.056) и оптимизированный связывающий CD3 фрагмент CD3opt или оригинальный связывающий CD3 фрагмент CD3orig исследовали в анализе с репортерными клетками Jurkat NFAT в присутствии EGFRvIII-положительных клеток глиобластомы DK-MG, U87MG-huEGFRvIII и EGFRwt-положительных клеток MKN45, как описано выше в примере 8.
Как видно для IgG (пример 3) оба ТСВ, содержащих CD3opt или CD3orig, имели аналогичную функциональную активность на репортерных клетках NFAT и индуцировали активацию CD3 зависимым от концентрации образом (Фиг. 20).
Уничтожение клеток-мишеней
EGFRvIII ТСВ, содержащие выбранный связывающий EGFRvIII фрагмент (Р063.056) и оптимизированный связывающий CD3 фрагмент CD3opt или оригинальный связывающий CD3 фрагмент CD3orig, сравнивали с EGFRvIII ТСВ, содержащими родительский связывающий EGFRvIII фрагмент Р056.021 и связывающий CD3 фрагмент CD3orig в эксперименте по уничтожению опухолевых клеток в присутствии линии клеток глиобластомы U87MG-EGFRvIII и МКПК, как описано выше в примере 8. Как видно на репортерных клетках Jurkat NFAT, функциональная активность ТСВ с CD3opt или CD3orig является сходной в отношении индукции лизиса опухолевых клеток и активации CD4 и CD8 Т-клеток по измерению повышения регуляции CD69 (Фиг. 25).
Кроме того, функциональную активность EGFRvIII ТСВ со связывающим EGFRvIII фрагментом Р063.056 и оптимизированным связывающим CD3 фрагментом CD3opt в «формате 2+1» (как проиллюстрировано на Фиг. 6) сравнивали с EGFRvIII ТСВ с такими же связывающими EGFRvIII и CD3 фрагментами в «формате 1+1 голова-к-хвосту» (схематически изображенном на Фиг. 1Ж). Эти два EGFRvIII ТСВ исследовали в анализе с репортерными клетками Jurkat NFAT и анализе уничтожения опухолевых клеток с линией клеток глиобластомы U87MG-EGFRvIII, как описано в примере 8. EGFRvIII ТСВ в формате 2+1 имел превосходящую функциональную активность как в отношении активации CD3, измеряемой в анализе с репортерными клетками Jurkat NFAT (Фиг. 26), так и индукции уничтожения опухолевых клеток и активации Т-клеток в анализе уничтожения с МКПК (Фиг. 27).
Пример 11 - Функциональные характеристики Т-клеточных биспецифических антител, содержащих оптимизированный связывающий CD3 фрагмент
Пролиферация и активация Т-клеток EGFRvIII ТСВ
Функциональную активность EGFRvIII ТСВ, содержащих выбранный связывающий EGFR фрагмент (Р063.056) и оптимизированный связывающий CD3 фрагмент CD3opt или оригинальный связывающий CD3 фрагмент CD3orig, сравнивали с EGFRvIII ТСВ, содержащими родительский связывающий EGFRvIII фрагмент Р056.021 и связывающий CD3 фрагмент CD3orig в анализе пролиферации Т-клеток на клетках U87MG-EGFRvIII (Фиг. 28). Все три ТСВ индуцировали сильную пролиферацию и активацию CD4 Т-клеток и CD8 Т-клеток. Р063.056 EGFRvIII ТСВ с CD3opt обладал более высокой активностью, чем другие два EGFRvIII ТСВ.
Лизис опухолевых клеток EGFRvIII ТСВ
После этого EGFRvIII ТСВ, содержащие выбранный связывающий EGFR фрагмент (Р063.056) и оптимизированный связывающий CD3 фрагмент CD3opt, и EGFRvIII ТСВ содержащие родительский связывающий EGFRvIII фрагмент Р056.021 и связывающий CD3 фрагмент CD3orig, исследовали в анализе лизиса опухолевых клеток с МКПК, которые культивировали совместно с клетками DK-MG (Фиг. 29). В этом анализе помимо лизиса опухолевых клеток в качестве дополнительных результатов измеряли активацию Т-клеток и высвобождение цитокинов. Как можно было видеть ранее, Р063.056 EGFRvIII ТСВ с CD3opt обладал более высокой активностью, чем Р056.021 EGFRvIII ТСВ с CD3orig в отношении лизиса опухолевых клеток, активации Т-клеток и высвобождения IFNγ и TNFα.
Активация Т-клеток и лизис опухолевых клеток TYRP1 ТСВ
Функциональное свойство TYRP1 ТСВ индуцировать высвобождение цитокинов исследовали путем совместного культивирования линии клеток первичной меланомы М150543 с МКПК, выделенными от здорового донора. Лизис опухолевых клеток, опосредованный Т-клетками через TYRP1 ТСВ, анализировали через 24 ч и 48 ч после обработки (Фиг. 30). Высвобождение IFNγ и TNFα в супернатант, а также активацию CD4 и CD8 Т-клеток анализировали через 48 ч после обработки. TYRP1 ТСВ был способен индуцировать активный лизис опухолевых клеток уже через 24 ч. Это сопровождалось сильной активацией CD4 и CD8 Т-клеток, определяемой по повышению регуляции CD25, а также значительному высвобождению IFNγ и TNFα.
Методы
Выделение МКПК
Мононуклеарные клетки периферической крови (МКПК) подготавливали путем центрифугирования в градиенте плотности Histopaque свежей крови, полученной от здоровых доноров-людей. Свежую кровь разводили в стерильной ФСБ и расслаивали в градиенте Histopaque (Sigma, №Н8889). После центрифугирования (450 х g, 30 минут, комнатная температура) плазму выше МКПК-содержащей промежуточной фазы сливали и переносили МКПК в новую пробирку Falcon, которую после этого наполняли 50 мл ФСБ. Смесь центрифугировали (400 х g, 10 минут, комнатная температура), супернатант сливали, а осажденные МКПК дважды промывали стерильным ФСБ (этапы центрифугирования 350 х g, 10 минут). Полученную в результате популяцию МКПК автоматически подсчитывали (ViCell) и хранили в среде RPMI1640, содержащей 10% ФТС и 1% GlutaMAX, при 37°С, 5% CO2 в клеточном инкубаторе до дальнейшего использования (не дольше 24 ч) или замораживали и хранили в жидком азоте для дальнейшего использования. За сутки до использования замороженные МКПК размораживали и культивировали в течение ночи при 37°С.
Пролиферация Т-клеток
Вкратце, клетки-мишени собирали, подсчитывали и дважды промывали ФСБ. Клетки ресуспендировали при 5 млн клеток на мл в ФСБ. Клетки окрашивали красителем для анализа пролиферации клеток eFluor 670 (eBioscience, №65-0840-85) с конечной концентрацией 5 мкМ в течение 10 мин при 37°С. Чтобы остановить реакцию окрашивания, к клеткам добавляли 4 объема полной клеточной культуральной среды и инкубировали в течение 5 мин при 4°С, а потом три раза промывали средой. Меченные клетки-мишени подсчитывали и доводили до 0,1 млн клеток на мл в RPMI1640, 10% ФТС и 1% GlutaMax. 10000 клеток-мишеней на лунку высевали в 96-луночный планшет.Потом добавляли обработку в указанных концентрациях, а в конце добавляли 100000 МКПК, выделенных от здорового донора, на лунку. Клетки инкубировали в течение 5 суток при 37°С, потом собирали МКПК и окрашивали CD3 BUV395 (BioLegend, №563548), CD4 РЕ (BioLegend, №300508), CD8 АРС (BioLegend, №344722), CD25 РЕ/Су7 (BioLegend, №302612). Пролиферацию определяли путем разведения красителя eFluor 670 в CD4 Т-клетках и CD8 Т-клетках, измеряемого методом проточной цитометрии (FACS Fortessa, BD Bioscience), а активацию CD4 Т-клеток и CD8 Т-клеток - путем измерения повышения регуляции CD25.
Опосредованное Т-клетками уничтожение опухолевых клеток
Клетки-мишени собирали с трипсином/ЭДТА, промывали и высевали при плотности 30000 клеток/лунка, используя плоскодонные 96-луночные планшеты. Клетки оставляли на ночь для прикрепления. Для анализа уничтожения антитела добавляли в указанных концентрациях в трех повторностях. МКПК добавляли к клеткам-мишеням при конечном отношении эффектора к мишени (Э:М) 10:1. Уничтожение клеток-мишеней анализировали через 24 ч инкубации при 37°С, 5% CO2 путем количественной оценки ЛДГ, высвобождаемой в клеточные супернатанты апоптотическими/некротическими клетками (набор для обнаружения ЛДГ, Roche Applied Science, №11 644 793 001). Максимальный лизис клеток-мишеней (=100%) достигался путем инкубации клеток-мишеней с 1% Тритон Х-100. Минимальный лизис (=0%) относится к клеткам-мишеням, которые инкубировали совместно с эффекторными клетками без биспецифической конструкции.
Активация Т-клеток
Активацию CD8 и CD4 Т-клеток после уничтожения Т-клетками клеток-мишеней, опосредованного ТСВ, анализировали методом проточной цитометрии, используя антитела, распознающие маркеры активации Т-клеток CD25 (поздний маркер активации) CD69 (ранний маркер активации). Через 48 ч инкубации МКПК переносили в круглодонный 96-луночный планшет, центрифугировали при 350 х g в течение 5 мин и дважды промывали буфером FACS. Поверхностное окрашивание для CD4 АРС (BioLegend, №300514), CD8 FITC (№344704, BioLegend), CD25 BV421 (BioLegend, №302630) и CD69 РЕ (BioLegend, №310906) проводили в соответствии с указаниями поставщика. Клетки дважды промывали 150 мкл/лунка буфера FACS и фиксировали в течение 15 мин при 4°С, используя 100 мкл/лунка буфера для фиксации (BD, №554655). После центрифугирования образцы ресуспендировали в 200 мкл/лунка буфера FACS. Образцы анализировали на BD FACS Fortessa.
Секреция цитокинов
Секрецию цитокинов в супернатанте измеряли методом проточной цитометрии, используя цитометрическую гранульную матрицу (СВА) в соответствии с инструкциями производителя, но вместо 50 мкл гранул и образца использовали только 25 мкл супернатанта и гранул. Использовали следующие наборы СВА (BD Biosciences): Flex Set СВА с интерфероном гамма человека (IFNγ) и Flex Set СВА с TNF человека. Измерения образцов проводили, используя BD FACS Canto II или BD FACS Fortessa, а анализ проводили, используя программное обеспечение Diva (BD Biosciences).
Пример 13 - In vivo эффективность Т-клеточных биспецифических антител, содержащих оптимизированный связывающий CD3 фрагмент
TYRP1 ТСВ (содержащий оптимизированный связывающий CD3 фрагмент, идентифицированный в примере 1) исследовали в отношении его противоопухолевой эффективности в мышиной модели с ксенотрансплантатом опухолевой линии клеток человека, модели с ксенотрансплантатом меланомы IGR-1.
Клетки IGR-1 (меланомы человека) культивировали в среде DMEM, содержащей 10% ФТС (Sigma). Клетки культивировали при 37°С в насыщенной влагой атмосфере при 5% СО2. Для трансплантации использовали пассаж 6. Жизнеспособность клеток составляла 96,7%. 2×106 клеток на животное вводили подкожно в 100 мкл клеточной культуральной среды RPMI (Gibco) в бок мышей, используя 1 мл туберкулиновый шприц (BD Biosciences, Germany).
Полностью гуманизированных самок мышей линии NSG (Roche Glycart AG, Switzerland) содержали в специальных свободных от патогенов условиях с суточными циклами 12 ч день/12 ч ночь в соответствии с обязательными руководствами (GV-Solas; Felasa; TierschG). Протокол экспериментального исследования был рассмотрен и одобрен местным органом управления (ZH223/2017). Непрерывный мониторинг состояния здоровья проводили на постоянном основании.
На 0 сутки исследования мышам подкожно вводили 2×106 клеток IGR-1, а потом рандомизировали и взвешивали. Через двадцать суток после инъекции опухолевых клеток (объем опухоли >200 мм3), мышам дважды в неделю вводили в/в 10 мкг (0,5 мг/кг) TYRP1 ТСВ в течение пяти недель. Всем мышам вводили в/в по 200 мкл соответствующего раствора. Мышам в группе носителя вводили гистидиновый буфер, в группе обработки - конструкцию TYRP1 ТСВ. Для получения надлежащего количества антитела на 200 мкл исходные растворы разводили при необходимости гистидиновым буфером. Размер опухолей измеряли три раза в неделю с помощью штангенциркуля и, используя программное обеспечение GrahPad Prism, строили график зависимости объема в мм3 +/- СПС. Статистический анализ проводили с помощью программного обеспечения JMP12.
На Фиг. 31 показано, что TYRP1 ТСВ опосредовал значительную эффективность в терминах ингибирования роста опухоли по сравнению с группой носителя (68% ИРО, р=0,0058*).
EGFRvIII ТСВ (содержащий оптимизированный связывающий CD3 фрагмент, идентифицированный в примере 1) аналогично исследовали в отношении его противоопухолевой эффективности в мышиной модели с ксенотрансплантатом опухолевой линии клеток человека, модели с ксенотрансплантатом глиобластомы U87-EGFRvIII.
Клетки U87 (глиобластома человека) изначально получали от АТСС (Manassas, USA) и стабильно трансфицировали для экспрессии белка EGFRvIII человека (Roche Glycart AG, Switzerland). После размножения клетки депонировали во внутренний банк клеток Roche Glycart. Линию клеток U87-EGFRvIII культивировали в среде DMEM, содержащей 10% ФТС (Sigma) и 0,5 мкг/мл пуромицина (Invitrogen). Клетки культивировали при 37°С в насыщенной влагой атмосфере при 5% CO2. Для трансплантации использовали пассаж 8. Жизнеспособность клеток составляла 94,7%. 5×105 клеток на животное вводили подкожно в 100 мкл клеточной культуральной среды RPMI (Gibco) в бок мышей, используя 1 мл туберкулиновый шприц (BD Biosciences, Germany).
Полностью гуманизированных самок мышей линии NSG (Roche Glycart AG, Switzerland) содержали в специальных свободных от патогенов условиях с суточными циклами 12 ч день/12 ч ночь в соответствии с обязательными руководствами (GV-Solas; Felasa; TierschG). Протокол экспериментального исследования был рассмотрен и одобрен местным органом управления (ZH223/2017). Непрерывный мониторинг состояния здоровья проводили на постоянном основании.
На 0 сутки исследования мышам подкожно вводили 5×105 клеток U87-EGFRvIII, а потом рандомизировали и взвешивали. Через две недели после инъекции опухолевых клеток (объем опухоли >200 мм3), мышам дважды в неделю вводили в/в 10 мкг (0,5 мг/кг) EGFRvIII ТСВ в течение пяти недель. Всем мышам вводили в/в по 200 мкл соответствующего раствора. Мышам в группе носителя вводили гистидиновый буфер, в группе обработки - конструкцию EGFRvIII ТСВ. Для получения надлежащего количества антитела на 200 мкл исходные растворы разводили при необходимости гистидиновым буфером. Размер опухолей измеряли три раза в неделю с помощью штангенциркуля и, используя программное обеспечение GrahPad Prism, строили график зависимости объема в мм3 +/- СПС.
На Фиг. 32 показано, что EGFRvIII ТСВ опосредовал значительную эффективность в терминах контроля роста опухоли, при этом все мыши достигли полной ремиссии.
Пример 12 - ФК-исследование с EGFRvIII ТСВ на мышах
Фармакокинетику (ФК) EGFRvIII ТСВ, содержащих оптимизированный связывающий CD3 фрагмент CD3opt, исследовали после внутривенного болюсного введения при 1 мг/кг мышам, трансгенным в отношении человеческого FcRn (линия 32, гомозиготные), и мышам NOD-SCID. Серийные микрообразцы крови брали у трансгенных (tg) в отношении человеческого FcRn мышей и мышей NOD-SCID до 672 ч (9 образцов на мышь от 5 мин до 672 ч после дозы). Образцы мышиной сыворотки, обработанные EGFRvIII ТСВ, анализировали, используя специфический ферментный иммуносорбентный анализ (ELISA) в условиях без GLP. Захват EGFRvIII ТСВ проводили с биотинилированным антигеном EGFRvIII (huEGFRvIII his биотин) на покрытых стрептавидином микротитровальных планшетах (SA-MTP). Обнаружение связанного EGFRvIII ТСВ проводили с использованием меченного дигоксигенином моноклонального антитела против IgG1 Fc(PGLALA) человека (смотрите пример 3) с последующим добавлением анти-дигоксигенин-POD вторичного антитела для обнаружения. Сигнал генерировали путем добавления субстрата пероксидазы (ABTS). Диапазон калибровки составляли от 2,35 нг/мл до 150 нг/мл, а 2,5 нг/мл соответствовало нижнему пределу количественного обнаружения (НПКО).
Результаты этого исследования проиллюстрированы в таблице 7. ФК-профиль EGFRvIII ТСВ соответствует ожидаемому диапазону для обеих исследуемых линий мышей. Это показывает, что конструирование CDR или связывающего CD3 фрагмента не приводит к другим свойствам последовательностей, которые повлияли бы на клиренс антитела.
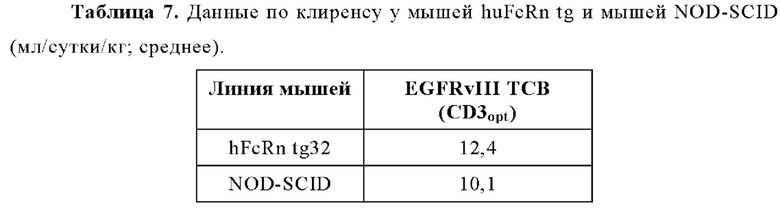
Хотя вышеизложенное изобретение было подробно описано посредством иллюстрации и примеров в целях ясности понимания, описания и примеры не следует воспринимать как ограничивающие объем данного изобретения. Раскрытие всех патентов и научной литературы, цитируемых в данном документе, в полном объеме и явным образом включено посредством ссылки.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Ф. ХОФФМАНН-ЛЯ РОШ АГ
<120> АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3
<130> P35214
<150> EP18214994.8
<151> 2018-12-21
<160> 122
<170> PatentIn version 3.5
<210> 1
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD3orig HCDR1
<400> 1
Thr Tyr Ala Met Asn
1 5
<210> 2
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD3opt HCDR1
<400> 2
Ser Tyr Ala Met Asn
1 5
<210> 3
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD3orig / CD3opt HCDR2
<400> 3
Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser
1 5 10 15
Val Lys Gly
<210> 4
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD3orig HCDR3
<400> 4
His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe Ala Tyr
1 5 10
<210> 5
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD3opt HCDR3
<400> 5
His Thr Thr Phe Pro Ser Ser Tyr Val Ser Tyr Tyr Gly Tyr
1 5 10
<210> 6
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD3orig VH
<400> 6
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 7
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD3opt VH
<400> 7
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Gln Phe Ser Ser Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Thr Thr Phe Pro Ser Ser Tyr Val Ser Tyr Tyr
100 105 110
Gly Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 8
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD3orig / CD3opt LCDR1
<400> 8
Gly Ser Ser Thr Gly Ala Val Thr Thr Ser Asn Tyr Ala Asn
1 5 10
<210> 9
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD3orig / CD3opt LCDR2
<400> 9
Gly Thr Asn Lys Arg Ala Pro
1 5
<210> 10
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD3orig / CD3opt LCDR3
<400> 10
Ala Leu Trp Tyr Ser Asn Leu Trp Val
1 5
<210> 11
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD3orig / CD3opt VL
<400> 11
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Glu Lys Pro Gly Gln Ala Phe Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 12
<211> 453
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD3orig IgG HC
<400> 12
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro
450
<210> 13
<211> 453
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD3opt IgG HC
<400> 13
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Gln Phe Ser Ser Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Thr Thr Phe Pro Ser Ser Tyr Val Ser Tyr Tyr
100 105 110
Gly Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro
450
<210> 14
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD3orig / CD3opt IgG LC
<400> 14
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Glu Lys Pro Gly Gln Ala Phe Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 15
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> TYRP1 HCDR1
<400> 15
Asp Tyr Phe Leu His
1 5
<210> 16
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> TYRP1 HCDR2
<400> 16
Trp Ile Asn Pro Asp Asn Gly Asn Thr Val Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 17
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> TYRP1 HCDR3
<400> 17
Arg Asp Tyr Thr Tyr Glu Lys Ala Ala Leu Asp Tyr
1 5 10
<210> 18
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> TYRP1 VH
<400> 18
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Asn Ile Lys Asp Tyr
20 25 30
Phe Leu His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Asp Asn Gly Asn Thr Val Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Ala Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Arg Asp Tyr Thr Tyr Glu Lys Ala Ala Leu Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 19
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> TYRP1 LCDR1
<400> 19
Arg Ala Ser Gly Asn Ile Tyr Asn Tyr Leu Ala
1 5 10
<210> 20
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> TYRP1 LCDR2
<400> 20
Asp Ala Lys Thr Leu Ala Asp
1 5
<210> 21
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> TYRP1 LCDR3
<400> 21
Gln His Phe Trp Ser Leu Pro Phe Thr
1 5
<210> 22
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> TYRP1 VL
<400> 22
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gly Asn Ile Tyr Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Lys Thr Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln His Phe Trp Ser Leu Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 23
<211> 674
<212> Белок
<213> Искусственная последовательность
<220>
<223> TYRP1 VH-CH1(EE) - CD3orig/CD3opt VL-CH1 - Fc (выступ, PGLALA)
<400> 23
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Asn Ile Lys Asp Tyr
20 25 30
Phe Leu His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Asp Asn Gly Asn Thr Val Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Ala Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Arg Asp Tyr Thr Tyr Glu Lys Ala Ala Leu Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ala Val Val Thr
225 230 235 240
Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly Thr Val Thr Leu Thr
245 250 255
Cys Gly Ser Ser Thr Gly Ala Val Thr Thr Ser Asn Tyr Ala Asn Trp
260 265 270
Val Gln Glu Lys Pro Gly Gln Ala Phe Arg Gly Leu Ile Gly Gly Thr
275 280 285
Asn Lys Arg Ala Pro Gly Thr Pro Ala Arg Phe Ser Gly Ser Leu Leu
290 295 300
Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Ala Gln Pro Glu Asp Glu
305 310 315 320
Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn Leu Trp Val Phe Gly
325 330 335
Gly Gly Thr Lys Leu Thr Val Leu Ser Ser Ala Ser Thr Lys Gly Pro
340 345 350
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
355 360 365
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
370 375 380
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
385 390 395 400
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
405 410 415
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
420 425 430
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
435 440 445
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
450 455 460
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
465 470 475 480
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
485 490 495
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
500 505 510
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
515 520 525
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
530 535 540
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro
545 550 555 560
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
565 570 575
Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val
580 585 590
Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
595 600 605
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
610 615 620
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
625 630 635 640
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
645 650 655
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
660 665 670
Ser Pro
<210> 24
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> TYRP1 VH-CH1(EE) -Fc (впадина, PGLALA)
<400> 24
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Asn Ile Lys Asp Tyr
20 25 30
Phe Leu His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Asp Asn Gly Asn Thr Val Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Ala Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Arg Asp Tyr Thr Tyr Glu Lys Ala Ala Leu Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro
<210> 25
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> TYRP1 VL-CL(RK)
<400> 25
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gly Asn Ile Tyr Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Lys Thr Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln His Phe Trp Ser Leu Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Arg Lys Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 26
<211> 232
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD3orig VH-CL
<400> 26
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val Ser Trp Phe
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Val
115 120 125
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
130 135 140
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
145 150 155 160
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
165 170 175
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
180 185 190
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
195 200 205
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
210 215 220
Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 27
<211> 232
<212> Белок
<213> Искусственная последовательность
<220>
<223> CD3opt VH-CL
<400> 27
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Gln Phe Ser Ser Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Thr Thr Phe Pro Ser Ser Tyr Val Ser Tyr Tyr
100 105 110
Gly Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Val
115 120 125
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
130 135 140
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
145 150 155 160
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
165 170 175
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
180 185 190
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
195 200 205
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
210 215 220
Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 28
<211> 360
<212> Белок
<213> Искусственная последовательность
<220>
<223> Ствол CD3 эпсилон человека - Fc(выступ) - Avi
<400> 28
Gln Asp Gly Asn Glu Glu Met Gly Gly Ile Thr Gln Thr Pro Tyr Lys
1 5 10 15
Val Ser Ile Ser Gly Thr Thr Val Ile Leu Thr Cys Pro Gln Tyr Pro
20 25 30
Gly Ser Glu Ile Leu Trp Gln His Asn Asp Lys Asn Ile Gly Gly Asp
35 40 45
Glu Asp Asp Lys Asn Ile Gly Ser Asp Glu Asp His Leu Ser Leu Lys
50 55 60
Glu Phe Ser Glu Leu Glu Gln Ser Gly Tyr Tyr Val Cys Tyr Pro Arg
65 70 75 80
Gly Ser Lys Pro Glu Asp Ala Asn Phe Tyr Leu Tyr Leu Arg Ala Arg
85 90 95
Val Ser Glu Asn Cys Val Asp Glu Gln Leu Tyr Phe Gln Gly Gly Ser
100 105 110
Pro Lys Ser Ala Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
115 120 125
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
130 135 140
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
145 150 155 160
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
165 170 175
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
180 185 190
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
195 200 205
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
210 215 220
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
225 230 235 240
Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys
245 250 255
Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
260 265 270
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
275 280 285
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
290 295 300
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
305 310 315 320
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
325 330 335
Leu Ser Leu Ser Pro Gly Lys Ser Gly Gly Leu Asn Asp Ile Phe Glu
340 345 350
Ala Gln Lys Ile Glu Trp His Glu
355 360
<210> 29
<211> 325
<212> Белок
<213> Искусственная последовательность
<220>
<223> Ствол CD3 дельта человека - Fc (впадина) - Avi
<400> 29
Phe Lys Ile Pro Ile Glu Glu Leu Glu Asp Arg Val Phe Val Asn Cys
1 5 10 15
Asn Thr Ser Ile Thr Trp Val Glu Gly Thr Val Gly Thr Leu Leu Ser
20 25 30
Asp Ile Thr Arg Leu Asp Leu Gly Lys Arg Ile Leu Asp Pro Arg Gly
35 40 45
Ile Tyr Arg Cys Asn Gly Thr Asp Ile Tyr Lys Asp Lys Glu Ser Thr
50 55 60
Val Gln Val His Tyr Arg Met Cys Arg Ser Glu Gln Leu Tyr Phe Gln
65 70 75 80
Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
85 90 95
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
100 105 110
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
115 120 125
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
130 135 140
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
145 150 155 160
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
165 170 175
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
180 185 190
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
195 200 205
Val Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
210 215 220
Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
225 230 235 240
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
245 250 255
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr
260 265 270
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
275 280 285
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
290 295 300
Ser Pro Gly Lys Ser Gly Gly Leu Asn Asp Ile Phe Glu Ala Gln Lys
305 310 315 320
Ile Glu Trp His Glu
325
<210> 30
<211> 351
<212> Белок
<213> Искусственная последовательность
<220>
<223> Ствол CD3 эпсилон яванского макака - Fc (выступ) - Avi
<400> 30
Gln Asp Gly Asn Glu Glu Met Gly Ser Ile Thr Gln Thr Pro Tyr Gln
1 5 10 15
Val Ser Ile Ser Gly Thr Thr Val Ile Leu Thr Cys Ser Gln His Leu
20 25 30
Gly Ser Glu Ala Gln Trp Gln His Asn Gly Lys Asn Lys Glu Asp Ser
35 40 45
Gly Asp Arg Leu Phe Leu Pro Glu Phe Ser Glu Met Glu Gln Ser Gly
50 55 60
Tyr Tyr Val Cys Tyr Pro Arg Gly Ser Asn Pro Glu Asp Ala Ser His
65 70 75 80
His Leu Tyr Leu Lys Ala Arg Val Ser Glu Asn Cys Val Asp Glu Gln
85 90 95
Leu Tyr Phe Gln Gly Gly Ser Pro Lys Ser Ala Asp Lys Thr His Thr
100 105 110
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
115 120 125
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
130 135 140
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
145 150 155 160
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
165 170 175
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
180 185 190
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
195 200 205
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
210 215 220
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
225 230 235 240
Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val
245 250 255
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
260 265 270
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
275 280 285
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
290 295 300
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
305 310 315 320
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Ser Gly
325 330 335
Gly Leu Asn Asp Ile Phe Glu Ala Gln Lys Ile Glu Trp His Glu
340 345 350
<210> 31
<211> 334
<212> Белок
<213> Искусственная последовательность
<220>
<223> Ствол CD3 дельта - Fc (впадина) - Avi
<400> 31
Phe Lys Ile Pro Val Glu Glu Leu Glu Asp Arg Val Phe Val Lys Cys
1 5 10 15
Asn Thr Ser Val Thr Trp Val Glu Gly Thr Val Gly Thr Leu Leu Thr
20 25 30
Asn Asn Thr Arg Leu Asp Leu Gly Lys Arg Ile Leu Asp Pro Arg Gly
35 40 45
Ile Tyr Arg Cys Asn Gly Thr Asp Ile Tyr Lys Asp Lys Glu Ser Ala
50 55 60
Val Gln Val His Tyr Arg Met Ser Gln Asn Cys Val Asp Glu Gln Leu
65 70 75 80
Tyr Phe Gln Gly Gly Ser Pro Lys Ser Ala Asp Lys Thr His Thr Cys
85 90 95
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
100 105 110
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
115 120 125
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
130 135 140
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
145 150 155 160
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
165 170 175
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
180 185 190
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
195 200 205
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser
210 215 220
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys
225 230 235 240
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
245 250 255
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
260 265 270
Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
275 280 285
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
290 295 300
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Ser Gly Gly
305 310 315 320
Leu Asn Asp Ile Phe Glu Ala Gln Lys Ile Glu Trp His Glu
325 330
<210> 32
<211> 699
<212> Белок
<213> Искусственная последовательность
<220>
<223> ВКД TYRP1 человека - Fc (выступ) - Avi
<400> 32
Gln Phe Pro Arg Gln Cys Ala Thr Val Glu Ala Leu Arg Ser Gly Met
1 5 10 15
Cys Cys Pro Asp Leu Ser Pro Val Ser Gly Pro Gly Thr Asp Arg Cys
20 25 30
Gly Ser Ser Ser Gly Arg Gly Arg Cys Glu Ala Val Thr Ala Asp Ser
35 40 45
Arg Pro His Ser Pro Gln Tyr Pro His Asp Gly Arg Asp Asp Arg Glu
50 55 60
Val Trp Pro Leu Arg Phe Phe Asn Arg Thr Cys His Cys Asn Gly Asn
65 70 75 80
Phe Ser Gly His Asn Cys Gly Thr Cys Arg Pro Gly Trp Arg Gly Ala
85 90 95
Ala Cys Asp Gln Arg Val Leu Ile Val Arg Arg Asn Leu Leu Asp Leu
100 105 110
Ser Lys Glu Glu Lys Asn His Phe Val Arg Ala Leu Asp Met Ala Lys
115 120 125
Arg Thr Thr His Pro Leu Phe Val Ile Ala Thr Arg Arg Ser Glu Glu
130 135 140
Ile Leu Gly Pro Asp Gly Asn Thr Pro Gln Phe Glu Asn Ile Ser Ile
145 150 155 160
Tyr Asn Tyr Phe Val Trp Thr His Tyr Tyr Ser Val Lys Lys Thr Phe
165 170 175
Leu Gly Val Gly Gln Glu Ser Phe Gly Glu Val Asp Phe Ser His Glu
180 185 190
Gly Pro Ala Phe Leu Thr Trp His Arg Tyr His Leu Leu Arg Leu Glu
195 200 205
Lys Asp Met Gln Glu Met Leu Gln Glu Pro Ser Phe Ser Leu Pro Tyr
210 215 220
Trp Asn Phe Ala Thr Gly Lys Asn Val Cys Asp Ile Cys Thr Asp Asp
225 230 235 240
Leu Met Gly Ser Arg Ser Asn Phe Asp Ser Thr Leu Ile Ser Pro Asn
245 250 255
Ser Val Phe Ser Gln Trp Arg Val Val Cys Asp Ser Leu Glu Asp Tyr
260 265 270
Asp Thr Leu Gly Thr Leu Cys Asn Ser Thr Glu Asp Gly Pro Ile Arg
275 280 285
Arg Asn Pro Ala Gly Asn Val Ala Arg Pro Met Val Gln Arg Leu Pro
290 295 300
Glu Pro Gln Asp Val Ala Gln Cys Leu Glu Val Gly Leu Phe Asp Thr
305 310 315 320
Pro Pro Phe Tyr Ser Asn Ser Thr Asn Ser Phe Arg Asn Thr Val Glu
325 330 335
Gly Tyr Ser Asp Pro Thr Gly Lys Tyr Asp Pro Ala Val Arg Ser Leu
340 345 350
His Asn Leu Ala His Leu Phe Leu Asn Gly Thr Gly Gly Gln Thr His
355 360 365
Leu Ser Pro Asn Asp Pro Ile Phe Val Leu Leu His Thr Phe Thr Asp
370 375 380
Ala Val Phe Asp Glu Trp Leu Arg Arg Tyr Asn Ala Asp Ile Ser Thr
385 390 395 400
Phe Pro Leu Glu Asn Ala Pro Ile Gly His Asn Arg Gln Tyr Asn Met
405 410 415
Val Pro Phe Trp Pro Pro Val Thr Asn Thr Glu Met Phe Val Thr Ala
420 425 430
Pro Asp Asn Leu Gly Tyr Thr Tyr Glu Ile Gln Trp Pro Ser Arg Glu
435 440 445
Phe Ser Val Pro Glu Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys
450 455 460
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
465 470 475 480
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
485 490 495
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
500 505 510
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
515 520 525
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
530 535 540
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
545 550 555 560
Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
565 570 575
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu
580 585 590
Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr
595 600 605
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
610 615 620
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
625 630 635 640
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
645 650 655
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
660 665 670
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Ser Gly Gly Leu Asn Asp
675 680 685
Ile Phe Glu Ala Gln Lys Ile Glu Trp His Glu
690 695
<210> 33
<211> 698
<212> Белок
<213> Искусственная последовательность
<220>
<223> ВКД TYRP1 яванского макака - Fc (выступ) - Avi
<400> 33
Gln Phe Pro Arg Glu Cys Ala Thr Val Glu Ala Leu Arg Ser Gly Met
1 5 10 15
Cys Cys Pro Asp Leu Ser Pro Met Ser Gly Pro Gly Thr Asp Arg Cys
20 25 30
Gly Ser Ser Ser Gly Arg Gly Arg Cys Glu Ala Val Thr Ala Asp Ser
35 40 45
Arg Pro His Ser Pro Arg Tyr Pro His Asp Gly Arg Asp Asp Arg Glu
50 55 60
Val Trp Pro Leu Arg Phe Phe Asn Arg Thr Cys His Cys Asn Gly Asn
65 70 75 80
Phe Ser Gly His Asn Cys Gly Thr Cys Arg Pro Gly Trp Arg Gly Ala
85 90 95
Ala Cys Asp Gln Arg Val Leu Val Val Arg Arg Asn Leu Leu Asp Leu
100 105 110
Ser Lys Glu Glu Lys Asn His Phe Val Arg Ala Leu Asp Met Ala Lys
115 120 125
Arg Thr Thr His Pro Leu Phe Val Ile Ala Thr Arg Arg Ser Glu Glu
130 135 140
Ile Leu Gly Pro Asp Gly Asn Thr Pro Gln Phe Glu Asn Ile Ser Ile
145 150 155 160
Tyr Asn Tyr Phe Val Trp Thr His Tyr Tyr Ser Val Lys Lys Thr Phe
165 170 175
Leu Gly Ala Gly Gln Glu Ser Phe Gly Glu Val Asp Phe Ser His Glu
180 185 190
Gly Pro Ala Phe Leu Thr Trp His Arg Tyr His Leu Leu Arg Leu Glu
195 200 205
Lys Asp Met Gln Glu Met Leu Gln Glu Pro Ser Phe Ser Leu Pro Tyr
210 215 220
Trp Asn Phe Ala Thr Gly Lys Asn Val Cys Asp Ile Cys Thr Asp Asp
225 230 235 240
Leu Met Gly Ser Arg Ser Asn Phe Asp Ser Thr Leu Ile Ser Pro Asn
245 250 255
Ser Val Phe Ser Gln Trp Arg Val Val Cys Asp Ser Leu Glu Asp Tyr
260 265 270
Asp Thr Leu Gly Thr Leu Cys Asn Ser Thr Glu Ser Gly Pro Ile Arg
275 280 285
Arg Asn Pro Ala Gly Asn Val Ala Arg Pro Met Val Gln Arg Leu Pro
290 295 300
Glu Pro Gln Asp Val Ala Gln Cys Leu Glu Val Gly Leu Phe Asp Thr
305 310 315 320
Pro Pro Phe Tyr Ser Asn Ser Thr Asn Ser Phe Arg Asn Thr Val Glu
325 330 335
Gly Tyr Ser Asp Pro Thr Gly Lys Tyr Asp Pro Ala Val Arg Ser Leu
340 345 350
His Asn Leu Ala His Leu Phe Leu Asn Gly Thr Gly Gly Gln Thr His
355 360 365
Leu Ser Pro Asn Asp Pro Ile Phe Val Leu Leu His Thr Phe Thr Asp
370 375 380
Ala Val Phe Asp Glu Trp Leu Arg Arg Tyr Asn Ala Asp Ile Ser Thr
385 390 395 400
Phe Pro Leu Glu Asn Ala Pro Ile Gly His Asn Arg Gln Tyr Asn Met
405 410 415
Val Pro Phe Trp Pro Pro Val Thr Asn Thr Glu Met Phe Val Thr Ala
420 425 430
Pro Asp Asn Leu Gly Tyr Thr Tyr Glu Val Gln Trp Pro Ser Arg Glu
435 440 445
Phe Ser Val Pro Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro
450 455 460
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
465 470 475 480
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
485 490 495
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
500 505 510
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
515 520 525
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
530 535 540
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
545 550 555 560
Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
565 570 575
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu
580 585 590
Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro
595 600 605
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
610 615 620
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
625 630 635 640
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
645 650 655
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
660 665 670
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ser Gly Gly Leu Asn Asp Ile
675 680 685
Phe Glu Ala Gln Lys Ile Glu Trp His Glu
690 695
<210> 34
<211> 699
<212> Белок
<213> Искусственная последовательность
<220>
<223> ВКД TYRP1 мыши - Fc (выступ) - Avi
<400> 34
Gln Phe Pro Arg Glu Cys Ala Asn Ile Glu Ala Leu Arg Arg Gly Val
1 5 10 15
Cys Cys Pro Asp Leu Leu Pro Ser Ser Gly Pro Gly Thr Asp Pro Cys
20 25 30
Gly Ser Ser Ser Gly Arg Gly Arg Cys Val Ala Val Ile Ala Asp Ser
35 40 45
Arg Pro His Ser Arg His Tyr Pro His Asp Gly Lys Asp Asp Arg Glu
50 55 60
Ala Trp Pro Leu Arg Phe Phe Asn Arg Thr Cys Gln Cys Asn Asp Asn
65 70 75 80
Phe Ser Gly His Asn Cys Gly Thr Cys Arg Pro Gly Trp Arg Gly Ala
85 90 95
Ala Cys Asn Gln Lys Ile Leu Thr Val Arg Arg Asn Leu Leu Asp Leu
100 105 110
Ser Pro Glu Glu Lys Ser His Phe Val Arg Ala Leu Asp Met Ala Lys
115 120 125
Arg Thr Thr His Pro Gln Phe Val Ile Ala Thr Arg Arg Leu Glu Asp
130 135 140
Ile Leu Gly Pro Asp Gly Asn Thr Pro Gln Phe Glu Asn Ile Ser Val
145 150 155 160
Tyr Asn Tyr Phe Val Trp Thr His Tyr Tyr Ser Val Lys Lys Thr Phe
165 170 175
Leu Gly Thr Gly Gln Glu Ser Phe Gly Asp Val Asp Phe Ser His Glu
180 185 190
Gly Pro Ala Phe Leu Thr Trp His Arg Tyr His Leu Leu Gln Leu Glu
195 200 205
Arg Asp Met Gln Glu Met Leu Gln Glu Pro Ser Phe Ser Leu Pro Tyr
210 215 220
Trp Asn Phe Ala Thr Gly Lys Asn Val Cys Asp Val Cys Thr Asp Asp
225 230 235 240
Leu Met Gly Ser Arg Ser Asn Phe Asp Ser Thr Leu Ile Ser Pro Asn
245 250 255
Ser Val Phe Ser Gln Trp Arg Val Val Cys Glu Ser Leu Glu Glu Tyr
260 265 270
Asp Thr Leu Gly Thr Leu Cys Asn Ser Thr Glu Gly Gly Pro Ile Arg
275 280 285
Arg Asn Pro Ala Gly Asn Val Gly Arg Pro Ala Val Gln Arg Leu Pro
290 295 300
Glu Pro Gln Asp Val Thr Gln Cys Leu Glu Val Arg Val Phe Asp Thr
305 310 315 320
Pro Pro Phe Tyr Ser Asn Ser Thr Asp Ser Phe Arg Asn Thr Val Glu
325 330 335
Gly Tyr Ser Ala Pro Thr Gly Lys Tyr Asp Pro Ala Val Arg Ser Leu
340 345 350
His Asn Leu Ala His Leu Phe Leu Asn Gly Thr Gly Gly Gln Thr His
355 360 365
Leu Ser Pro Asn Asp Pro Ile Phe Val Leu Leu His Thr Phe Thr Asp
370 375 380
Ala Val Phe Asp Glu Trp Leu Arg Arg Tyr Asn Ala Asp Ile Ser Thr
385 390 395 400
Phe Pro Leu Glu Asn Ala Pro Ile Gly His Asn Arg Gln Tyr Asn Met
405 410 415
Val Pro Phe Trp Pro Pro Val Thr Asn Thr Glu Met Phe Val Thr Ala
420 425 430
Pro Asp Asn Leu Gly Tyr Ala Tyr Glu Val Gln Trp Pro Gly Gln Glu
435 440 445
Phe Thr Val Ser Glu Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys
450 455 460
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
465 470 475 480
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
485 490 495
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
500 505 510
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
515 520 525
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
530 535 540
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
545 550 555 560
Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
565 570 575
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Asp Glu
580 585 590
Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr
595 600 605
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
610 615 620
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
625 630 635 640
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
645 650 655
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
660 665 670
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Ser Gly Gly Leu Asn Asp
675 680 685
Ile Phe Glu Ala Gln Lys Ile Glu Trp His Glu
690 695
<210> 35
<211> 225
<212> Белок
<213> Искусственная последовательность
<220>
<223> Fc (впадина)
<400> 35
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn Arg Phe Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro
225
<210> 36
<211> 393
<212> Белок
<213> Искусственная последовательность
<220>
<223> ВКД EGFRvIII - Avi - His
<400> 36
Leu Glu Glu Lys Lys Gly Asn Tyr Val Val Thr Asp His Gly Ser Cys
1 5 10 15
Val Arg Ala Cys Gly Ala Asp Ser Tyr Glu Met Glu Glu Asp Gly Val
20 25 30
Arg Lys Cys Lys Lys Cys Glu Gly Pro Cys Arg Lys Val Cys Asn Gly
35 40 45
Ile Gly Ile Gly Glu Phe Lys Asp Ser Leu Ser Ile Asn Ala Thr Asn
50 55 60
Ile Lys His Phe Lys Asn Cys Thr Ser Ile Ser Gly Asp Leu His Ile
65 70 75 80
Leu Pro Val Ala Phe Arg Gly Asp Ser Phe Thr His Thr Pro Pro Leu
85 90 95
Asp Pro Gln Glu Leu Asp Ile Leu Lys Thr Val Lys Glu Ile Thr Gly
100 105 110
Phe Leu Leu Ile Gln Ala Trp Pro Glu Asn Arg Thr Asp Leu His Ala
115 120 125
Phe Glu Asn Leu Glu Ile Ile Arg Gly Arg Thr Lys Gln His Gly Gln
130 135 140
Phe Ser Leu Ala Val Val Ser Leu Asn Ile Thr Ser Leu Gly Leu Arg
145 150 155 160
Ser Leu Lys Glu Ile Ser Asp Gly Asp Val Ile Ile Ser Gly Asn Lys
165 170 175
Asn Leu Cys Tyr Ala Asn Thr Ile Asn Trp Lys Lys Leu Phe Gly Thr
180 185 190
Ser Gly Gln Lys Thr Lys Ile Ile Ser Asn Arg Gly Glu Asn Ser Cys
195 200 205
Lys Ala Thr Gly Gln Val Cys His Ala Leu Cys Ser Pro Glu Gly Cys
210 215 220
Trp Gly Pro Glu Pro Arg Asp Cys Val Ser Cys Arg Asn Val Ser Arg
225 230 235 240
Gly Arg Glu Cys Val Asp Lys Cys Asn Leu Leu Glu Gly Glu Pro Arg
245 250 255
Glu Phe Val Glu Asn Ser Glu Cys Ile Gln Cys His Pro Glu Cys Leu
260 265 270
Pro Gln Ala Met Asn Ile Thr Cys Thr Gly Arg Gly Pro Asp Asn Cys
275 280 285
Ile Gln Cys Ala His Tyr Ile Asp Gly Pro His Cys Val Lys Thr Cys
290 295 300
Pro Ala Gly Val Met Gly Glu Asn Asn Thr Leu Val Trp Lys Tyr Ala
305 310 315 320
Asp Ala Gly His Val Cys His Leu Cys His Pro Asn Cys Thr Tyr Gly
325 330 335
Cys Thr Gly Pro Gly Leu Glu Gly Cys Pro Thr Asn Gly Pro Lys Ile
340 345 350
Pro Ser Val Asp Gly Gly Ser Pro Thr Pro Pro Thr Pro Gly Gly Gly
355 360 365
Ser Gly Leu Asn Asp Ile Phe Glu Ala Gln Lys Ile Glu Trp His Glu
370 375 380
Ala Arg Ala His His His His His His
385 390
<210> 37
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.021 HCDR1
<400> 37
Ser Tyr Trp Ile Ala
1 5
<210> 38
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.021 HCDR2
<400> 38
Val Ile His Pro Tyr Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe Gln
1 5 10 15
Gly
<210> 39
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.021 HCDR3
<400> 39
Val Ser Arg Ser Ser Tyr Ala Phe Asp Tyr
1 5 10
<210> 40
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.021 VH
<400> 40
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Asp Ser Tyr
20 25 30
Trp Ile Ala Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Val Ile His Pro Tyr Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Val Ser Arg Ser Ser Tyr Ala Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 41
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.021 LCDR1
<400> 41
Lys Ser Ser Gln Ser Val Leu Tyr Ser Ser Asn Asn Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 42
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.021 LCDR2
<400> 42
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 43
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.021 LCDR3
<400> 43
Gln Gln Val His Ser Gly Pro Pro Val Thr
1 5 10
<210> 44
<211> 114
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.021 VL
<400> 44
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Val His Ser Gly Pro Pro Val Thr Phe Gly Gln Gly Thr Lys Val Glu
100 105 110
Ile Lys
<210> 45
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.052 HCDR1
<400> 45
Asn Tyr Trp Ile Gly
1 5
<210> 46
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.052 HCDR2
<400> 46
Thr Ile Tyr Pro Gly Asp Ser Asp Arg Arg Tyr Ser Pro Ser Phe Gln
1 5 10 15
Gly
<210> 47
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.052 HCDR3
<400> 47
Val Ser Arg Ser Ser Tyr Ala Phe Asp Tyr
1 5 10
<210> 48
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.052 VH
<400> 48
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Thr Phe Met Asn Tyr
20 25 30
Trp Ile Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Thr Ile Tyr Pro Gly Asp Ser Asp Arg Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Leu Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Val Ser Arg Ser Ser Tyr Ala Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 49
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.052 LCDR1
<400> 49
Lys Ser Ser Gln Ser Val Leu Tyr Ser Ser Asn Asn Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 50
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.052 LCDR2
<400> 50
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 51
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.052 LCDR3
<400> 51
Gln Gln Val His Ser Gly Pro Pro Val Thr
1 5 10
<210> 52
<211> 114
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.052 VL
<400> 52
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Val His Ser Gly Pro Pro Val Thr Phe Gly Gln Gly Thr Lys Val Glu
100 105 110
Ile Lys
<210> 53
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P047.019 HCDR1
<400> 53
Ser Ile Trp Ile His
1 5
<210> 54
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P047.019 HCDR2
<400> 54
Thr Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe Gln
1 5 10 15
Gly
<210> 55
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P047.019 HCDR3
<400> 55
Thr Gly Pro Gly Leu Ala Phe Asp Tyr
1 5
<210> 56
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P047.019 VH
<400> 56
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Pro Ser Ile
20 25 30
Trp Ile His Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Thr Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Thr Gly Pro Gly Leu Ala Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 57
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P047.019 LCDR1
<400> 57
Lys Ser Ser Gln Ser Val Leu Tyr Ser Ser Asn Asn Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 58
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P047.019 LCDR2
<400> 58
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 59
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P047.019 LCDR3
<400> 59
Gln Gln Ser Tyr Ser Thr Pro Ile Thr
1 5
<210> 60
<211> 113
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P047.019 VL
<400> 60
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Ser Tyr Ser Thr Pro Ile Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
100 105 110
Lys
<210> 61
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P057.012 HCDR1
<400> 61
Asn Tyr Trp Ile Ala
1 5
<210> 62
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P057.012 HCDR2
<400> 62
Ile Ile Tyr Pro Asp Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe Gln
1 5 10 15
Gly
<210> 63
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P057.012 HCDR3
<400> 63
Ala Thr Asn Ile Ala Ser Gly Gly Tyr Phe Asp Tyr
1 5 10
<210> 64
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P057.012 VH
<400> 64
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Ala Asn Tyr
20 25 30
Trp Ile Ala Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Tyr Pro Asp Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Ala Thr Asn Ile Ala Ser Gly Gly Tyr Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 65
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P057.012 LCDR1
<400> 65
Lys Ser Ser Gln Ser Val Leu Trp Asn Ser Asn Asn Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 66
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P057.012 LCDR2
<400> 66
Trp Ala Ser Lys Arg Glu Ser
1 5
<210> 67
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P057.012 LCDR3
<400> 67
Gln Gln Ser Tyr Ser Ala Pro Ile Thr
1 5
<210> 68
<211> 113
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P057.012 VL
<400> 68
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Trp Asn
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Lys Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Ser Tyr Ser Ala Pro Ile Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
100 105 110
Lys
<210> 69
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P057.011 HCDR1
<400> 69
Arg Arg Trp Ile Ala
1 5
<210> 70
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P057.011 HCDR2
<400> 70
Ile Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe Gln
1 5 10 15
Gly
<210> 71
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P057.011 HCDR3
<400> 71
Ala Thr Asn Ile Ala Ser Gly Gly Tyr Phe Asp Tyr
1 5 10
<210> 72
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P057.011 VH
<400> 72
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Asn Phe Gly Arg Arg
20 25 30
Trp Ile Ala Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Tyr Pro Gly Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Ala Thr Asn Ile Ala Ser Gly Gly Tyr Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 73
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P057.011 LCDR1
<400> 73
Lys Ser Ser Gln Ser Val Leu Trp Asn Ser Asn Asn Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 74
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P057.011 LCDR2
<400> 74
Trp Ala Ser Lys Arg Glu Ser
1 5
<210> 75
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P057.011 LCDR3
<400> 75
Gln Gln Ser Tyr Ser Ala Pro Ile Thr
1 5
<210> 76
<211> 113
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P057.011 VL
<400> 76
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Trp Asn
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Lys Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Ser Tyr Ser Ala Pro Ile Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
100 105 110
Lys
<210> 77
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.027 HCDR1
<400> 77
Asn Asn Trp Ile Ala
1 5
<210> 78
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.027 HCDR2
<400> 78
Val Ile Tyr Pro Gly Asp Ser Asp Lys Arg Tyr Ser Pro Ser Phe Gln
1 5 10 15
Gly
<210> 79
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.027 HCDR3
<400> 79
Val Ser Arg Ser Ser Tyr Ala Phe Asp Tyr
1 5 10
<210> 80
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.027 VH
<400> 80
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Thr Phe Gly Asn Asn
20 25 30
Trp Ile Ala Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Val Ile Tyr Pro Gly Asp Ser Asp Lys Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Val Ser Arg Ser Ser Tyr Ala Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 81
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.027 LCDR1
<400> 81
Lys Ser Ser Gln Ser Val Leu Tyr Ser Ser Asn Asn Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 82
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.027 LCDR2
<400> 82
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 83
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.027 LCDR3
<400> 83
Gln Gln Val His Ser Gly Pro Pro Val Thr
1 5 10
<210> 84
<211> 114
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P056.027 VL
<400> 84
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Val His Ser Gly Pro Pro Val Thr Phe Gly Gln Gly Thr Lys Val Glu
100 105 110
Ile Lys
<210> 85
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P063.056 HCDR1
<400> 85
Ser Tyr Trp Ile Ala
1 5
<210> 86
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P063.056 HCDR2
<400> 86
Val Ile His Pro Tyr Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe Gln
1 5 10 15
Gly
<210> 87
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P063.056 HCDR3
<400> 87
Val Ser Arg Ser Ser Tyr Ala Phe Asp Tyr
1 5 10
<210> 88
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P063.056 VH
<400> 88
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Asp Ser Tyr
20 25 30
Trp Ile Ala Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Val Ile His Pro Tyr Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Val Ser Arg Ser Ser Tyr Ala Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 89
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P063.056 LCDR1
<400> 89
Lys Ser Ser Gln Ser Val Leu Tyr Ser Ser Asn Asn Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 90
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P063.056 LCDR2
<400> 90
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 91
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P063.056 LCDR3
<400> 91
Gln Gln Gln Arg Asp Gly Pro Pro Val Thr
1 5 10
<210> 92
<211> 114
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P063.056 VL
<400> 92
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Gln Arg Asp Gly Pro Pro Val Thr Phe Gly Gln Gly Thr Lys Val Glu
100 105 110
Ile Lys
<210> 93
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P064.078 HCDR1
<400> 93
Ser Tyr Trp Ile Ala
1 5
<210> 94
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P064.078 HCDR2
<400> 94
Val Ile His Pro Tyr Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe Gln
1 5 10 15
Gly
<210> 95
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P064.078 HCDR3
<400> 95
Val Ser Arg Leu Ser Tyr Ala Leu Asp Tyr
1 5 10
<210> 96
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P064.078 VH
<400> 96
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Asp Ser Tyr
20 25 30
Trp Ile Ala Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Val Ile His Pro Tyr Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Val Ser Arg Leu Ser Tyr Ala Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 97
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P064.078 LCDR1
<400> 97
Lys Ser Ser Gln Ser Val Leu Tyr Ser Ser Asn Asn Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 98
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P064.078 LCDR2
<400> 98
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 99
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P064.078 LCDR3
<400> 99
Gln Gln Val His Ser Gly Pro Pro Val Thr
1 5 10
<210> 100
<211> 114
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P064.078 VL
<400> 100
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Val His Ser Gly Pro Pro Val Thr Phe Gly Gln Gly Thr Lys Val Glu
100 105 110
Ile Lys
<210> 101
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P065.036 HCDR1
<400> 101
Ser Tyr Trp Ile Ala
1 5
<210> 102
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P065.036 HCDR2
<400> 102
Val Ile His Pro Tyr Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe Gln
1 5 10 15
Gly
<210> 103
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P065.036 HCDR3
<400> 103
Val Ser Arg Ser Ser Tyr Ala Leu Asp Tyr
1 5 10
<210> 104
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P065.036 VH
<400> 104
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Asp Ser Tyr
20 25 30
Trp Ile Ala Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Val Ile His Pro Tyr Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Val Ser Arg Ser Ser Tyr Ala Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 105
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P065.036 LCDR1
<400> 105
Lys Ser Ser Gln Ser Val Leu Tyr Ser Ser Asn Asn Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 106
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P065.036 LCDR2
<400> 106
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 107
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P065.036 LCDR3
<400> 107
Gln Gln Val Tyr Ser Gly Pro Pro Val Thr
1 5 10
<210> 108
<211> 114
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII P065.036 VL
<400> 108
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Val Tyr Ser Gly Pro Pro Val Thr Phe Gly Gln Gly Thr Lys Val Glu
100 105 110
Ile Lys
<210> 109
<211> 672
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII VH-CH1(EE) - CD3orig/CD3opt VL-CH1 - Fc (выступ,
PGLALA)
<400> 109
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Asp Ser Tyr
20 25 30
Trp Ile Ala Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Val Ile His Pro Tyr Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Val Ser Arg Ser Ser Tyr Ala Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Gly
210 215 220
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ala Val Val Thr Gln Glu
225 230 235 240
Pro Ser Leu Thr Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly
245 250 255
Ser Ser Thr Gly Ala Val Thr Thr Ser Asn Tyr Ala Asn Trp Val Gln
260 265 270
Glu Lys Pro Gly Gln Ala Phe Arg Gly Leu Ile Gly Gly Thr Asn Lys
275 280 285
Arg Ala Pro Gly Thr Pro Ala Arg Phe Ser Gly Ser Leu Leu Gly Gly
290 295 300
Lys Ala Ala Leu Thr Leu Ser Gly Ala Gln Pro Glu Asp Glu Ala Glu
305 310 315 320
Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn Leu Trp Val Phe Gly Gly Gly
325 330 335
Thr Lys Leu Thr Val Leu Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
340 345 350
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
355 360 365
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
370 375 380
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
385 390 395 400
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
405 410 415
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
420 425 430
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
435 440 445
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
450 455 460
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
465 470 475 480
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
485 490 495
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
500 505 510
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
515 520 525
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
530 535 540
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu
545 550 555 560
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
565 570 575
Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
580 585 590
Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
595 600 605
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
610 615 620
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
625 630 635 640
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
645 650 655
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
660 665 670
<210> 110
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII VH-CH1(EE) -Fc (впадина, PGLALA)
<400> 110
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Asp Ser Tyr
20 25 30
Trp Ile Ala Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Val Ile His Pro Tyr Asp Ser Asp Thr Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Val Ser Arg Ser Ser Tyr Ala Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser
355 360 365
Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 111
<211> 221
<212> Белок
<213> Искусственная последовательность
<220>
<223> EGFRvIII VL-CL(RK)
<400> 111
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Gln Arg Asp Gly Pro Pro Val Thr Phe Gly Gln Gly Thr Lys Val Glu
100 105 110
Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser
115 120 125
Asp Arg Lys Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn
130 135 140
Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala
145 150 155 160
Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys
165 170 175
Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp
180 185 190
Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu
195 200 205
Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<210> 112
<211> 186
<212> Белок
<213> Homo sapiens
<400> 112
Gln Asp Gly Asn Glu Glu Met Gly Gly Ile Thr Gln Thr Pro Tyr Lys
1 5 10 15
Val Ser Ile Ser Gly Thr Thr Val Ile Leu Thr Cys Pro Gln Tyr Pro
20 25 30
Gly Ser Glu Ile Leu Trp Gln His Asn Asp Lys Asn Ile Gly Gly Asp
35 40 45
Glu Asp Asp Lys Asn Ile Gly Ser Asp Glu Asp His Leu Ser Leu Lys
50 55 60
Glu Phe Ser Glu Leu Glu Gln Ser Gly Tyr Tyr Val Cys Tyr Pro Arg
65 70 75 80
Gly Ser Lys Pro Glu Asp Ala Asn Phe Tyr Leu Tyr Leu Arg Ala Arg
85 90 95
Val Cys Glu Asn Cys Met Glu Met Asp Val Met Ser Val Ala Thr Ile
100 105 110
Val Ile Val Asp Ile Cys Ile Thr Gly Gly Leu Leu Leu Leu Val Tyr
115 120 125
Tyr Trp Ser Lys Asn Arg Lys Ala Lys Ala Lys Pro Val Thr Arg Gly
130 135 140
Ala Gly Ala Gly Gly Arg Gln Arg Gly Gln Asn Lys Glu Arg Pro Pro
145 150 155 160
Pro Val Pro Asn Pro Asp Tyr Glu Pro Ile Arg Lys Gly Gln Arg Asp
165 170 175
Leu Tyr Ser Gly Leu Asn Gln Arg Arg Ile
180 185
<210> 113
<211> 177
<212> Белок
<213> Macaca fascicularis
<400> 113
Gln Asp Gly Asn Glu Glu Met Gly Ser Ile Thr Gln Thr Pro Tyr Gln
1 5 10 15
Val Ser Ile Ser Gly Thr Thr Val Ile Leu Thr Cys Ser Gln His Leu
20 25 30
Gly Ser Glu Ala Gln Trp Gln His Asn Gly Lys Asn Lys Glu Asp Ser
35 40 45
Gly Asp Arg Leu Phe Leu Pro Glu Phe Ser Glu Met Glu Gln Ser Gly
50 55 60
Tyr Tyr Val Cys Tyr Pro Arg Gly Ser Asn Pro Glu Asp Ala Ser His
65 70 75 80
His Leu Tyr Leu Lys Ala Arg Val Cys Glu Asn Cys Met Glu Met Asp
85 90 95
Val Met Ala Val Ala Thr Ile Val Ile Val Asp Ile Cys Ile Thr Leu
100 105 110
Gly Leu Leu Leu Leu Val Tyr Tyr Trp Ser Lys Asn Arg Lys Ala Lys
115 120 125
Ala Lys Pro Val Thr Arg Gly Ala Gly Ala Gly Gly Arg Gln Arg Gly
130 135 140
Gln Asn Lys Glu Arg Pro Pro Pro Val Pro Asn Pro Asp Tyr Glu Pro
145 150 155 160
Ile Arg Lys Gly Gln Gln Asp Leu Tyr Ser Gly Leu Asn Gln Arg Arg
165 170 175
Ile
<210> 114
<211> 513
<212> Белок
<213> Homo sapiens
<400> 114
Gln Phe Pro Arg Gln Cys Ala Thr Val Glu Ala Leu Arg Ser Gly Met
1 5 10 15
Cys Cys Pro Asp Leu Ser Pro Val Ser Gly Pro Gly Thr Asp Arg Cys
20 25 30
Gly Ser Ser Ser Gly Arg Gly Arg Cys Glu Ala Val Thr Ala Asp Ser
35 40 45
Arg Pro His Ser Pro Gln Tyr Pro His Asp Gly Arg Asp Asp Arg Glu
50 55 60
Val Trp Pro Leu Arg Phe Phe Asn Arg Thr Cys His Cys Asn Gly Asn
65 70 75 80
Phe Ser Gly His Asn Cys Gly Thr Cys Arg Pro Gly Trp Arg Gly Ala
85 90 95
Ala Cys Asp Gln Arg Val Leu Ile Val Arg Arg Asn Leu Leu Asp Leu
100 105 110
Ser Lys Glu Glu Lys Asn His Phe Val Arg Ala Leu Asp Met Ala Lys
115 120 125
Arg Thr Thr His Pro Leu Phe Val Ile Ala Thr Arg Arg Ser Glu Glu
130 135 140
Ile Leu Gly Pro Asp Gly Asn Thr Pro Gln Phe Glu Asn Ile Ser Ile
145 150 155 160
Tyr Asn Tyr Phe Val Trp Thr His Tyr Tyr Ser Val Lys Lys Thr Phe
165 170 175
Leu Gly Val Gly Gln Glu Ser Phe Gly Glu Val Asp Phe Ser His Glu
180 185 190
Gly Pro Ala Phe Leu Thr Trp His Arg Tyr His Leu Leu Arg Leu Glu
195 200 205
Lys Asp Met Gln Glu Met Leu Gln Glu Pro Ser Phe Ser Leu Pro Tyr
210 215 220
Trp Asn Phe Ala Thr Gly Lys Asn Val Cys Asp Ile Cys Thr Asp Asp
225 230 235 240
Leu Met Gly Ser Arg Ser Asn Phe Asp Ser Thr Leu Ile Ser Pro Asn
245 250 255
Ser Val Phe Ser Gln Trp Arg Val Val Cys Asp Ser Leu Glu Asp Tyr
260 265 270
Asp Thr Leu Gly Thr Leu Cys Asn Ser Thr Glu Asp Gly Pro Ile Arg
275 280 285
Arg Asn Pro Ala Gly Asn Val Ala Arg Pro Met Val Gln Arg Leu Pro
290 295 300
Glu Pro Gln Asp Val Ala Gln Cys Leu Glu Val Gly Leu Phe Asp Thr
305 310 315 320
Pro Pro Phe Tyr Ser Asn Ser Thr Asn Ser Phe Arg Asn Thr Val Glu
325 330 335
Gly Tyr Ser Asp Pro Thr Gly Lys Tyr Asp Pro Ala Val Arg Ser Leu
340 345 350
His Asn Leu Ala His Leu Phe Leu Asn Gly Thr Gly Gly Gln Thr His
355 360 365
Leu Ser Pro Asn Asp Pro Ile Phe Val Leu Leu His Thr Phe Thr Asp
370 375 380
Ala Val Phe Asp Glu Trp Leu Arg Arg Tyr Asn Ala Asp Ile Ser Thr
385 390 395 400
Phe Pro Leu Glu Asn Ala Pro Ile Gly His Asn Arg Gln Tyr Asn Met
405 410 415
Val Pro Phe Trp Pro Pro Val Thr Asn Thr Glu Met Phe Val Thr Ala
420 425 430
Pro Asp Asn Leu Gly Tyr Thr Tyr Glu Ile Gln Trp Pro Ser Arg Glu
435 440 445
Phe Ser Val Pro Glu Ile Ile Ala Ile Ala Val Val Gly Ala Leu Leu
450 455 460
Leu Val Ala Leu Ile Phe Gly Thr Ala Ser Tyr Leu Ile Arg Ala Arg
465 470 475 480
Arg Ser Met Asp Glu Ala Asn Gln Pro Leu Leu Thr Asp Gln Tyr Gln
485 490 495
Cys Tyr Ala Glu Glu Tyr Glu Lys Leu Gln Asn Pro Asn Gln Ser Val
500 505 510
Val
<210> 115
<211> 919
<212> Белок
<213> Homo sapiens
<400> 115
Leu Glu Glu Lys Lys Gly Asn Tyr Val Val Thr Asp His Gly Ser Cys
1 5 10 15
Val Arg Ala Cys Gly Ala Asp Ser Tyr Glu Met Glu Glu Asp Gly Val
20 25 30
Arg Lys Cys Lys Lys Cys Glu Gly Pro Cys Arg Lys Val Cys Asn Gly
35 40 45
Ile Gly Ile Gly Glu Phe Lys Asp Ser Leu Ser Ile Asn Ala Thr Asn
50 55 60
Ile Lys His Phe Lys Asn Cys Thr Ser Ile Ser Gly Asp Leu His Ile
65 70 75 80
Leu Pro Val Ala Phe Arg Gly Asp Ser Phe Thr His Thr Pro Pro Leu
85 90 95
Asp Pro Gln Glu Leu Asp Ile Leu Lys Thr Val Lys Glu Ile Thr Gly
100 105 110
Phe Leu Leu Ile Gln Ala Trp Pro Glu Asn Arg Thr Asp Leu His Ala
115 120 125
Phe Glu Asn Leu Glu Ile Ile Arg Gly Arg Thr Lys Gln His Gly Gln
130 135 140
Phe Ser Leu Ala Val Val Ser Leu Asn Ile Thr Ser Leu Gly Leu Arg
145 150 155 160
Ser Leu Lys Glu Ile Ser Asp Gly Asp Val Ile Ile Ser Gly Asn Lys
165 170 175
Asn Leu Cys Tyr Ala Asn Thr Ile Asn Trp Lys Lys Leu Phe Gly Thr
180 185 190
Ser Gly Gln Lys Thr Lys Ile Ile Ser Asn Arg Gly Glu Asn Ser Cys
195 200 205
Lys Ala Thr Gly Gln Val Cys His Ala Leu Cys Ser Pro Glu Gly Cys
210 215 220
Trp Gly Pro Glu Pro Arg Asp Cys Val Ser Cys Arg Asn Val Ser Arg
225 230 235 240
Gly Arg Glu Cys Val Asp Lys Cys Asn Leu Leu Glu Gly Glu Pro Arg
245 250 255
Glu Phe Val Glu Asn Ser Glu Cys Ile Gln Cys His Pro Glu Cys Leu
260 265 270
Pro Gln Ala Met Asn Ile Thr Cys Thr Gly Arg Gly Pro Asp Asn Cys
275 280 285
Ile Gln Cys Ala His Tyr Ile Asp Gly Pro His Cys Val Lys Thr Cys
290 295 300
Pro Ala Gly Val Met Gly Glu Asn Asn Thr Leu Val Trp Lys Tyr Ala
305 310 315 320
Asp Ala Gly His Val Cys His Leu Cys His Pro Asn Cys Thr Tyr Gly
325 330 335
Cys Thr Gly Pro Gly Leu Glu Gly Cys Pro Thr Asn Gly Pro Lys Ile
340 345 350
Pro Ser Ile Ala Thr Gly Met Val Gly Ala Leu Leu Leu Leu Leu Val
355 360 365
Val Ala Leu Gly Ile Gly Leu Phe Met Arg Arg Arg His Ile Val Arg
370 375 380
Lys Arg Thr Leu Arg Arg Leu Leu Gln Glu Arg Glu Leu Val Glu Pro
385 390 395 400
Leu Thr Pro Ser Gly Glu Ala Pro Asn Gln Ala Leu Leu Arg Ile Leu
405 410 415
Lys Glu Thr Glu Phe Lys Lys Ile Lys Val Leu Gly Ser Gly Ala Phe
420 425 430
Gly Thr Val Tyr Lys Gly Leu Trp Ile Pro Glu Gly Glu Lys Val Lys
435 440 445
Ile Pro Val Ala Ile Lys Glu Leu Arg Glu Ala Thr Ser Pro Lys Ala
450 455 460
Asn Lys Glu Ile Leu Asp Glu Ala Tyr Val Met Ala Ser Val Asp Asn
465 470 475 480
Pro His Val Cys Arg Leu Leu Gly Ile Cys Leu Thr Ser Thr Val Gln
485 490 495
Leu Ile Thr Gln Leu Met Pro Phe Gly Cys Leu Leu Asp Tyr Val Arg
500 505 510
Glu His Lys Asp Asn Ile Gly Ser Gln Tyr Leu Leu Asn Trp Cys Val
515 520 525
Gln Ile Ala Lys Gly Met Asn Tyr Leu Glu Asp Arg Arg Leu Val His
530 535 540
Arg Asp Leu Ala Ala Arg Asn Val Leu Val Lys Thr Pro Gln His Val
545 550 555 560
Lys Ile Thr Asp Phe Gly Leu Ala Lys Leu Leu Gly Ala Glu Glu Lys
565 570 575
Glu Tyr His Ala Glu Gly Gly Lys Val Pro Ile Lys Trp Met Ala Leu
580 585 590
Glu Ser Ile Leu His Arg Ile Tyr Thr His Gln Ser Asp Val Trp Ser
595 600 605
Tyr Gly Val Thr Val Trp Glu Leu Met Thr Phe Gly Ser Lys Pro Tyr
610 615 620
Asp Gly Ile Pro Ala Ser Glu Ile Ser Ser Ile Leu Glu Lys Gly Glu
625 630 635 640
Arg Leu Pro Gln Pro Pro Ile Cys Thr Ile Asp Val Tyr Met Ile Met
645 650 655
Val Lys Cys Trp Met Ile Asp Ala Asp Ser Arg Pro Lys Phe Arg Glu
660 665 670
Leu Ile Ile Glu Phe Ser Lys Met Ala Arg Asp Pro Gln Arg Tyr Leu
675 680 685
Val Ile Gln Gly Asp Glu Arg Met His Leu Pro Ser Pro Thr Asp Ser
690 695 700
Asn Phe Tyr Arg Ala Leu Met Asp Glu Glu Asp Met Asp Asp Val Val
705 710 715 720
Asp Ala Asp Glu Tyr Leu Ile Pro Gln Gln Gly Phe Phe Ser Ser Pro
725 730 735
Ser Thr Ser Arg Thr Pro Leu Leu Ser Ser Leu Ser Ala Thr Ser Asn
740 745 750
Asn Ser Thr Val Ala Cys Ile Asp Arg Asn Gly Leu Gln Ser Cys Pro
755 760 765
Ile Lys Glu Asp Ser Phe Leu Gln Arg Tyr Ser Ser Asp Pro Thr Gly
770 775 780
Ala Leu Thr Glu Asp Ser Ile Asp Asp Thr Phe Leu Pro Val Pro Glu
785 790 795 800
Tyr Ile Asn Gln Ser Val Pro Lys Arg Pro Ala Gly Ser Val Gln Asn
805 810 815
Pro Val Tyr His Asn Gln Pro Leu Asn Pro Ala Pro Ser Arg Asp Pro
820 825 830
His Tyr Gln Asp Pro His Ser Thr Ala Val Gly Asn Pro Glu Tyr Leu
835 840 845
Asn Thr Val Gln Pro Thr Cys Val Asn Ser Thr Phe Asp Ser Pro Ala
850 855 860
His Trp Ala Gln Lys Gly Ser His Gln Ile Ser Leu Asp Asn Pro Asp
865 870 875 880
Tyr Gln Gln Asp Phe Phe Pro Lys Glu Ala Lys Pro Asn Gly Ile Phe
885 890 895
Lys Gly Ser Thr Ala Glu Asn Ala Glu Tyr Leu Arg Val Ala Pro Gln
900 905 910
Ser Ser Glu Phe Ile Gly Ala
915
<210> 116
<211> 1186
<212> Белок
<213> Homo sapiens
<400> 116
Leu Glu Glu Lys Lys Val Cys Gln Gly Thr Ser Asn Lys Leu Thr Gln
1 5 10 15
Leu Gly Thr Phe Glu Asp His Phe Leu Ser Leu Gln Arg Met Phe Asn
20 25 30
Asn Cys Glu Val Val Leu Gly Asn Leu Glu Ile Thr Tyr Val Gln Arg
35 40 45
Asn Tyr Asp Leu Ser Phe Leu Lys Thr Ile Gln Glu Val Ala Gly Tyr
50 55 60
Val Leu Ile Ala Leu Asn Thr Val Glu Arg Ile Pro Leu Glu Asn Leu
65 70 75 80
Gln Ile Ile Arg Gly Asn Met Tyr Tyr Glu Asn Ser Tyr Ala Leu Ala
85 90 95
Val Leu Ser Asn Tyr Asp Ala Asn Lys Thr Gly Leu Lys Glu Leu Pro
100 105 110
Met Arg Asn Leu Gln Glu Ile Leu His Gly Ala Val Arg Phe Ser Asn
115 120 125
Asn Pro Ala Leu Cys Asn Val Glu Ser Ile Gln Trp Arg Asp Ile Val
130 135 140
Ser Ser Asp Phe Leu Ser Asn Met Ser Met Asp Phe Gln Asn His Leu
145 150 155 160
Gly Ser Cys Gln Lys Cys Asp Pro Ser Cys Pro Asn Gly Ser Cys Trp
165 170 175
Gly Ala Gly Glu Glu Asn Cys Gln Lys Leu Thr Lys Ile Ile Cys Ala
180 185 190
Gln Gln Cys Ser Gly Arg Cys Arg Gly Lys Ser Pro Ser Asp Cys Cys
195 200 205
His Asn Gln Cys Ala Ala Gly Cys Thr Gly Pro Arg Glu Ser Asp Cys
210 215 220
Leu Val Cys Arg Lys Phe Arg Asp Glu Ala Thr Cys Lys Asp Thr Cys
225 230 235 240
Pro Pro Leu Met Leu Tyr Asn Pro Thr Thr Tyr Gln Met Asp Val Asn
245 250 255
Pro Glu Gly Lys Tyr Ser Phe Gly Ala Thr Cys Val Lys Lys Cys Pro
260 265 270
Arg Asn Tyr Val Val Thr Asp His Gly Ser Cys Val Arg Ala Cys Gly
275 280 285
Ala Asp Ser Tyr Glu Met Glu Glu Asp Gly Val Arg Lys Cys Lys Lys
290 295 300
Cys Glu Gly Pro Cys Arg Lys Val Cys Asn Gly Ile Gly Ile Gly Glu
305 310 315 320
Phe Lys Asp Ser Leu Ser Ile Asn Ala Thr Asn Ile Lys His Phe Lys
325 330 335
Asn Cys Thr Ser Ile Ser Gly Asp Leu His Ile Leu Pro Val Ala Phe
340 345 350
Arg Gly Asp Ser Phe Thr His Thr Pro Pro Leu Asp Pro Gln Glu Leu
355 360 365
Asp Ile Leu Lys Thr Val Lys Glu Ile Thr Gly Phe Leu Leu Ile Gln
370 375 380
Ala Trp Pro Glu Asn Arg Thr Asp Leu His Ala Phe Glu Asn Leu Glu
385 390 395 400
Ile Ile Arg Gly Arg Thr Lys Gln His Gly Gln Phe Ser Leu Ala Val
405 410 415
Val Ser Leu Asn Ile Thr Ser Leu Gly Leu Arg Ser Leu Lys Glu Ile
420 425 430
Ser Asp Gly Asp Val Ile Ile Ser Gly Asn Lys Asn Leu Cys Tyr Ala
435 440 445
Asn Thr Ile Asn Trp Lys Lys Leu Phe Gly Thr Ser Gly Gln Lys Thr
450 455 460
Lys Ile Ile Ser Asn Arg Gly Glu Asn Ser Cys Lys Ala Thr Gly Gln
465 470 475 480
Val Cys His Ala Leu Cys Ser Pro Glu Gly Cys Trp Gly Pro Glu Pro
485 490 495
Arg Asp Cys Val Ser Cys Arg Asn Val Ser Arg Gly Arg Glu Cys Val
500 505 510
Asp Lys Cys Asn Leu Leu Glu Gly Glu Pro Arg Glu Phe Val Glu Asn
515 520 525
Ser Glu Cys Ile Gln Cys His Pro Glu Cys Leu Pro Gln Ala Met Asn
530 535 540
Ile Thr Cys Thr Gly Arg Gly Pro Asp Asn Cys Ile Gln Cys Ala His
545 550 555 560
Tyr Ile Asp Gly Pro His Cys Val Lys Thr Cys Pro Ala Gly Val Met
565 570 575
Gly Glu Asn Asn Thr Leu Val Trp Lys Tyr Ala Asp Ala Gly His Val
580 585 590
Cys His Leu Cys His Pro Asn Cys Thr Tyr Gly Cys Thr Gly Pro Gly
595 600 605
Leu Glu Gly Cys Pro Thr Asn Gly Pro Lys Ile Pro Ser Ile Ala Thr
610 615 620
Gly Met Val Gly Ala Leu Leu Leu Leu Leu Val Val Ala Leu Gly Ile
625 630 635 640
Gly Leu Phe Met Arg Arg Arg His Ile Val Arg Lys Arg Thr Leu Arg
645 650 655
Arg Leu Leu Gln Glu Arg Glu Leu Val Glu Pro Leu Thr Pro Ser Gly
660 665 670
Glu Ala Pro Asn Gln Ala Leu Leu Arg Ile Leu Lys Glu Thr Glu Phe
675 680 685
Lys Lys Ile Lys Val Leu Gly Ser Gly Ala Phe Gly Thr Val Tyr Lys
690 695 700
Gly Leu Trp Ile Pro Glu Gly Glu Lys Val Lys Ile Pro Val Ala Ile
705 710 715 720
Lys Glu Leu Arg Glu Ala Thr Ser Pro Lys Ala Asn Lys Glu Ile Leu
725 730 735
Asp Glu Ala Tyr Val Met Ala Ser Val Asp Asn Pro His Val Cys Arg
740 745 750
Leu Leu Gly Ile Cys Leu Thr Ser Thr Val Gln Leu Ile Thr Gln Leu
755 760 765
Met Pro Phe Gly Cys Leu Leu Asp Tyr Val Arg Glu His Lys Asp Asn
770 775 780
Ile Gly Ser Gln Tyr Leu Leu Asn Trp Cys Val Gln Ile Ala Lys Gly
785 790 795 800
Met Asn Tyr Leu Glu Asp Arg Arg Leu Val His Arg Asp Leu Ala Ala
805 810 815
Arg Asn Val Leu Val Lys Thr Pro Gln His Val Lys Ile Thr Asp Phe
820 825 830
Gly Leu Ala Lys Leu Leu Gly Ala Glu Glu Lys Glu Tyr His Ala Glu
835 840 845
Gly Gly Lys Val Pro Ile Lys Trp Met Ala Leu Glu Ser Ile Leu His
850 855 860
Arg Ile Tyr Thr His Gln Ser Asp Val Trp Ser Tyr Gly Val Thr Val
865 870 875 880
Trp Glu Leu Met Thr Phe Gly Ser Lys Pro Tyr Asp Gly Ile Pro Ala
885 890 895
Ser Glu Ile Ser Ser Ile Leu Glu Lys Gly Glu Arg Leu Pro Gln Pro
900 905 910
Pro Ile Cys Thr Ile Asp Val Tyr Met Ile Met Val Lys Cys Trp Met
915 920 925
Ile Asp Ala Asp Ser Arg Pro Lys Phe Arg Glu Leu Ile Ile Glu Phe
930 935 940
Ser Lys Met Ala Arg Asp Pro Gln Arg Tyr Leu Val Ile Gln Gly Asp
945 950 955 960
Glu Arg Met His Leu Pro Ser Pro Thr Asp Ser Asn Phe Tyr Arg Ala
965 970 975
Leu Met Asp Glu Glu Asp Met Asp Asp Val Val Asp Ala Asp Glu Tyr
980 985 990
Leu Ile Pro Gln Gln Gly Phe Phe Ser Ser Pro Ser Thr Ser Arg Thr
995 1000 1005
Pro Leu Leu Ser Ser Leu Ser Ala Thr Ser Asn Asn Ser Thr Val
1010 1015 1020
Ala Cys Ile Asp Arg Asn Gly Leu Gln Ser Cys Pro Ile Lys Glu
1025 1030 1035
Asp Ser Phe Leu Gln Arg Tyr Ser Ser Asp Pro Thr Gly Ala Leu
1040 1045 1050
Thr Glu Asp Ser Ile Asp Asp Thr Phe Leu Pro Val Pro Glu Tyr
1055 1060 1065
Ile Asn Gln Ser Val Pro Lys Arg Pro Ala Gly Ser Val Gln Asn
1070 1075 1080
Pro Val Tyr His Asn Gln Pro Leu Asn Pro Ala Pro Ser Arg Asp
1085 1090 1095
Pro His Tyr Gln Asp Pro His Ser Thr Ala Val Gly Asn Pro Glu
1100 1105 1110
Tyr Leu Asn Thr Val Gln Pro Thr Cys Val Asn Ser Thr Phe Asp
1115 1120 1125
Ser Pro Ala His Trp Ala Gln Lys Gly Ser His Gln Ile Ser Leu
1130 1135 1140
Asp Asn Pro Asp Tyr Gln Gln Asp Phe Phe Pro Lys Glu Ala Lys
1145 1150 1155
Pro Asn Gly Ile Phe Lys Gly Ser Thr Ala Glu Asn Ala Glu Tyr
1160 1165 1170
Leu Arg Val Ala Pro Gln Ser Ser Glu Phe Ile Gly Ala
1175 1180 1185
<210> 117
<211> 225
<212> Белок
<213> Homo sapiens
<400> 117
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro
225
<210> 118
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> linker
<400> 118
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 119
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> linker
<400> 119
Asp Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 120
<211> 107
<212> Белок
<213> Homo sapiens
<400> 120
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 121
<211> 105
<212> Белок
<213> Homo sapiens
<400> 121
Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
1 5 10 15
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
20 25 30
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
35 40 45
Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys
50 55 60
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser
65 70 75 80
His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu
85 90 95
Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 122
<211> 328
<212> Белок
<213> Homo sapiens
<400> 122
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2832669C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ АНТИ-FAP КЛОН 212 | 2019 |
|
RU2799429C2 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2837821C1 |
| МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2018 |
|
RU2837770C2 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2808030C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2020 |
|
RU2841523C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩИЕСЯ С PD1 И LAG3 | 2018 |
|
RU2778805C2 |
| БЕЛКИ УПРАВЛЕНИЯ, НАВИГАЦИИ И КОНТРОЛЯ И СПОСОБ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2019 |
|
RU2811457C2 |
| БИСПЕЦИФИЧЕСКИЙ АНТИГЕНСВЯЗЫВАЮЩИЙ БЕЛОК EGFR/CD16 | 2019 |
|
RU2792240C2 |
| HER-2-НАПРАВЛЕННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ 4-1BBL | 2019 |
|
RU2815451C2 |
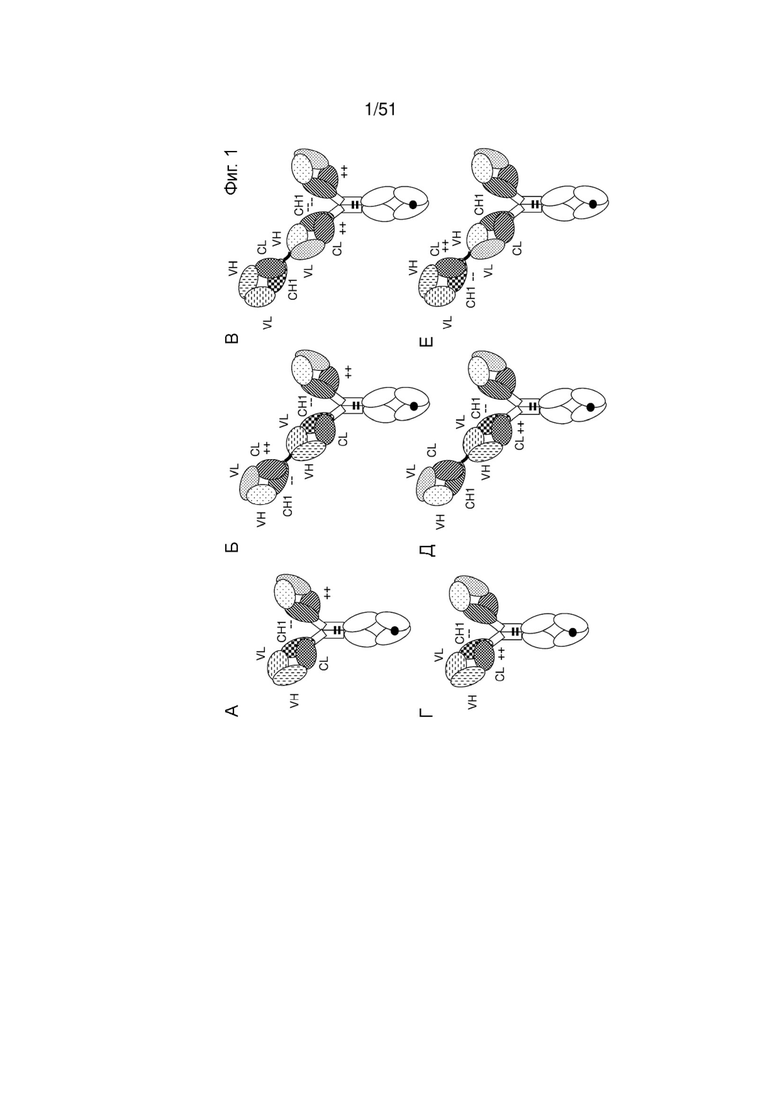
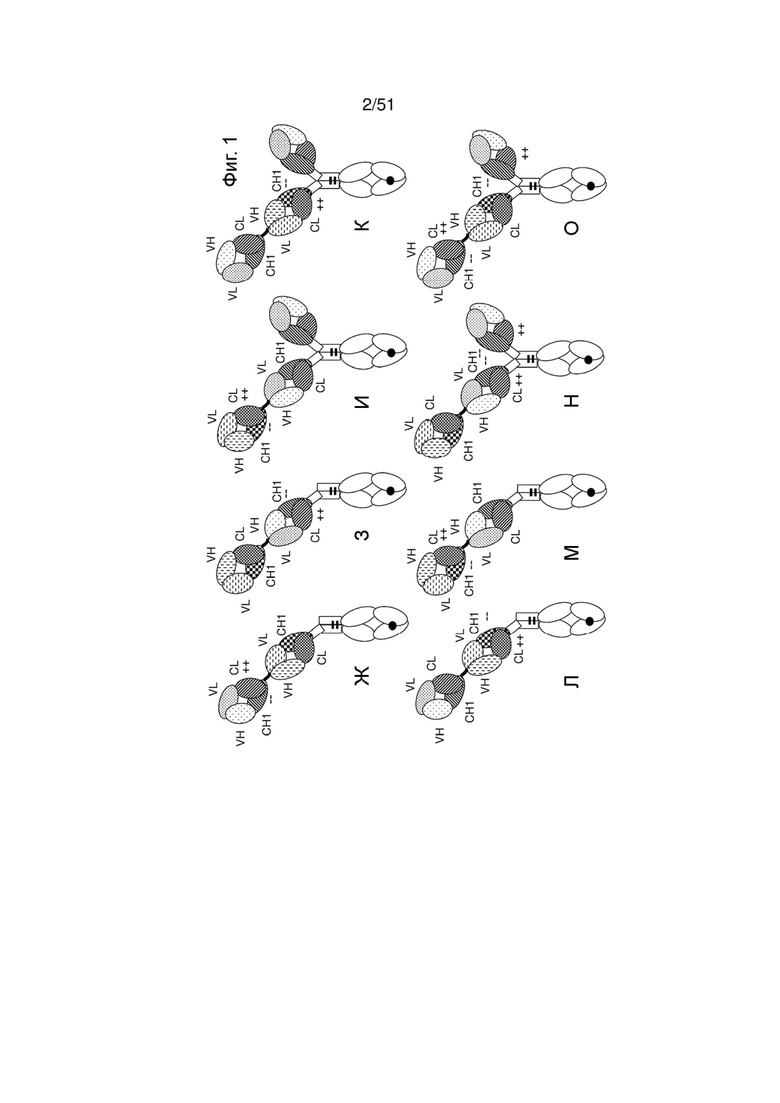




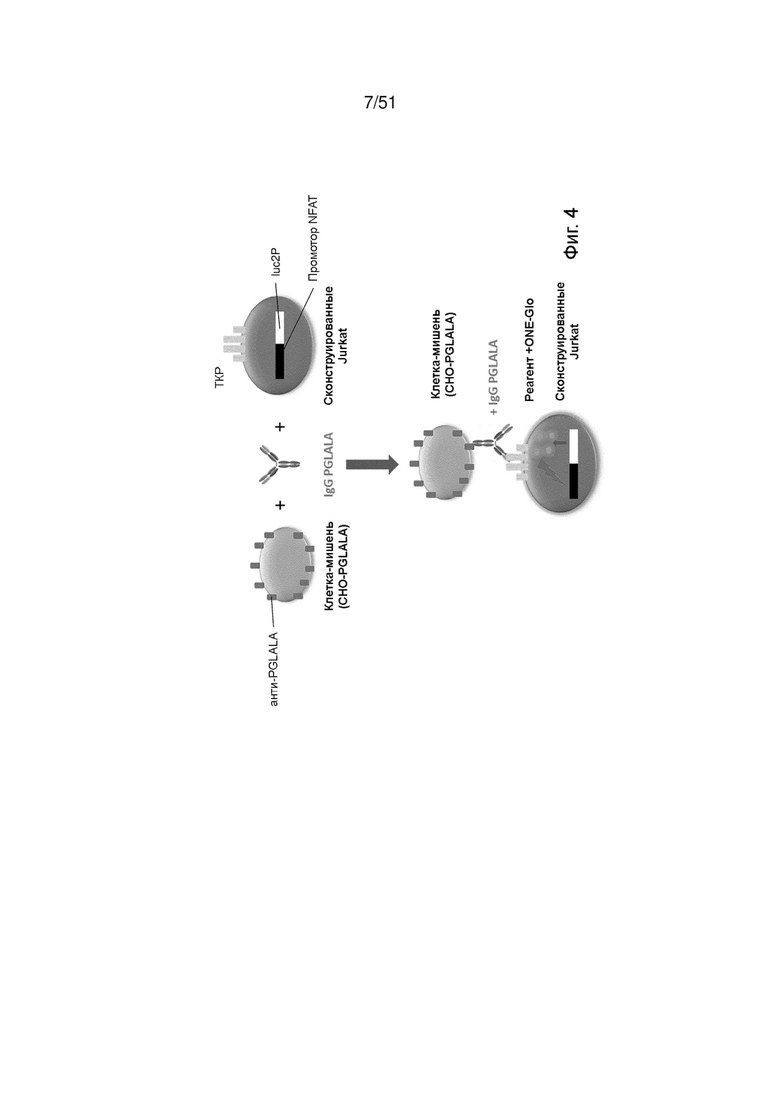


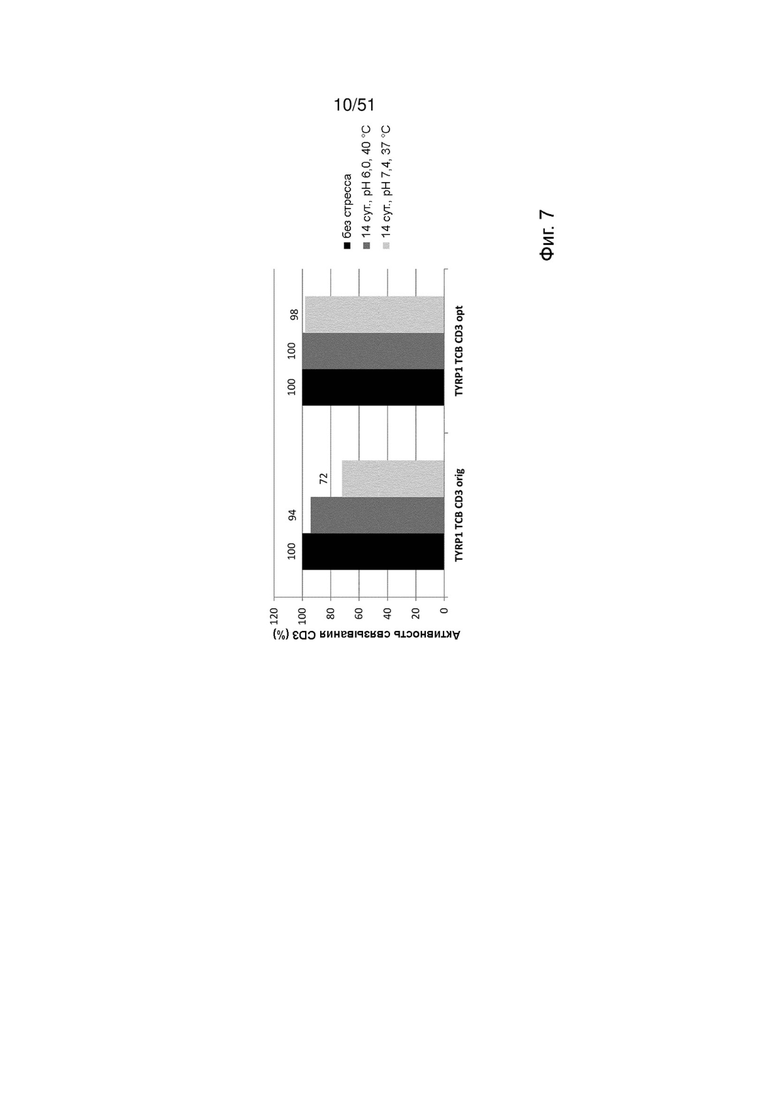

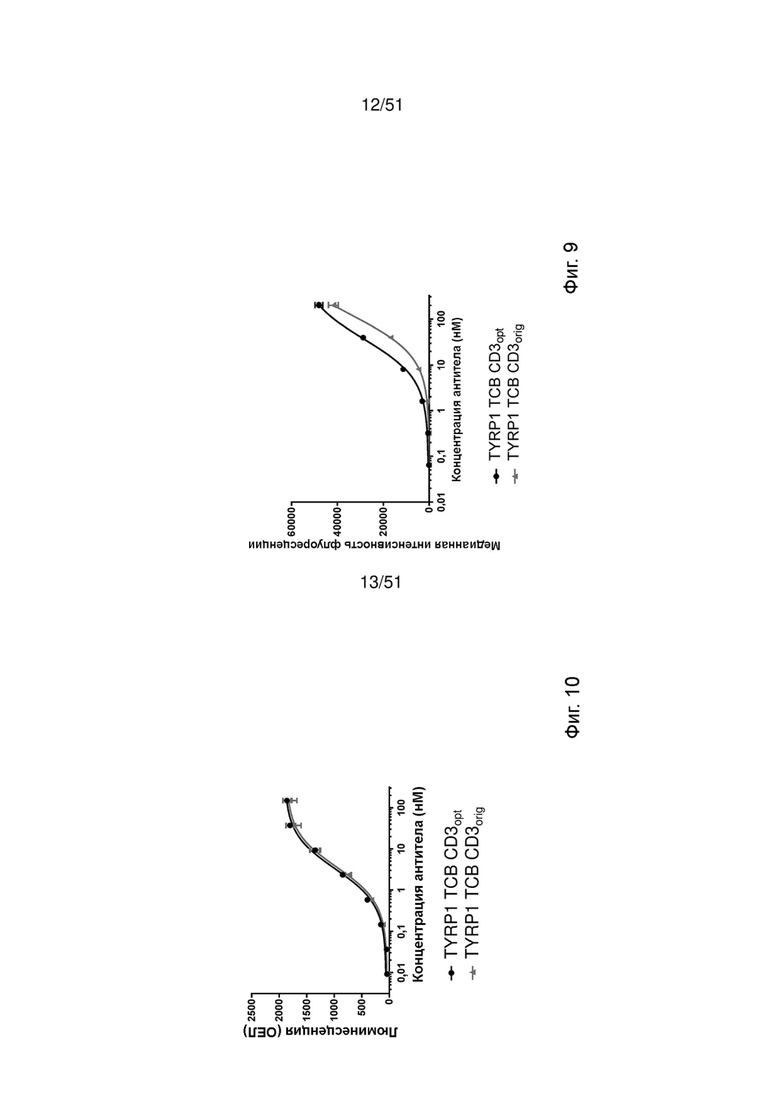




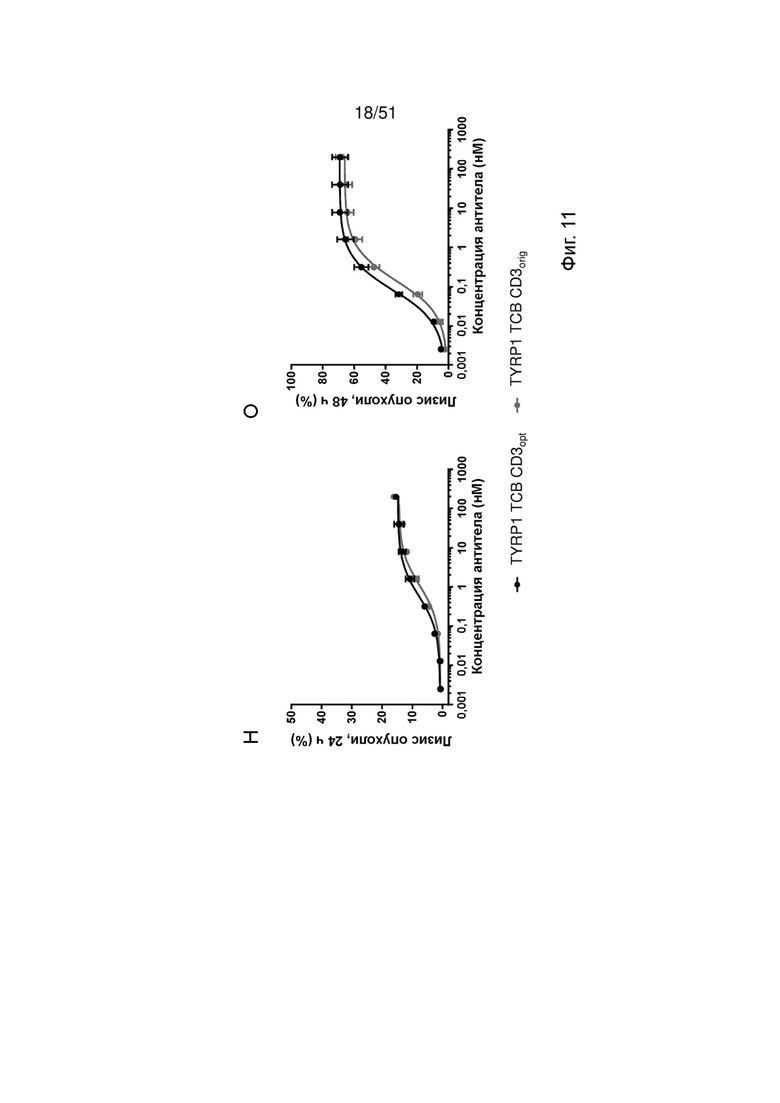

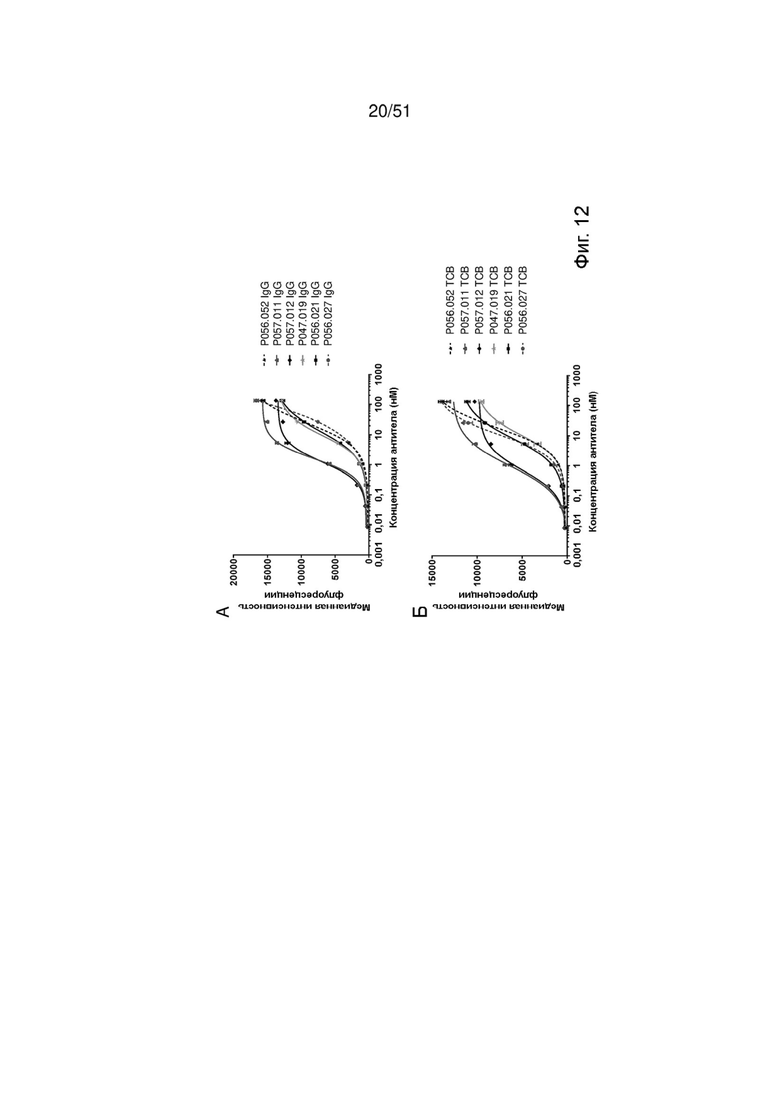




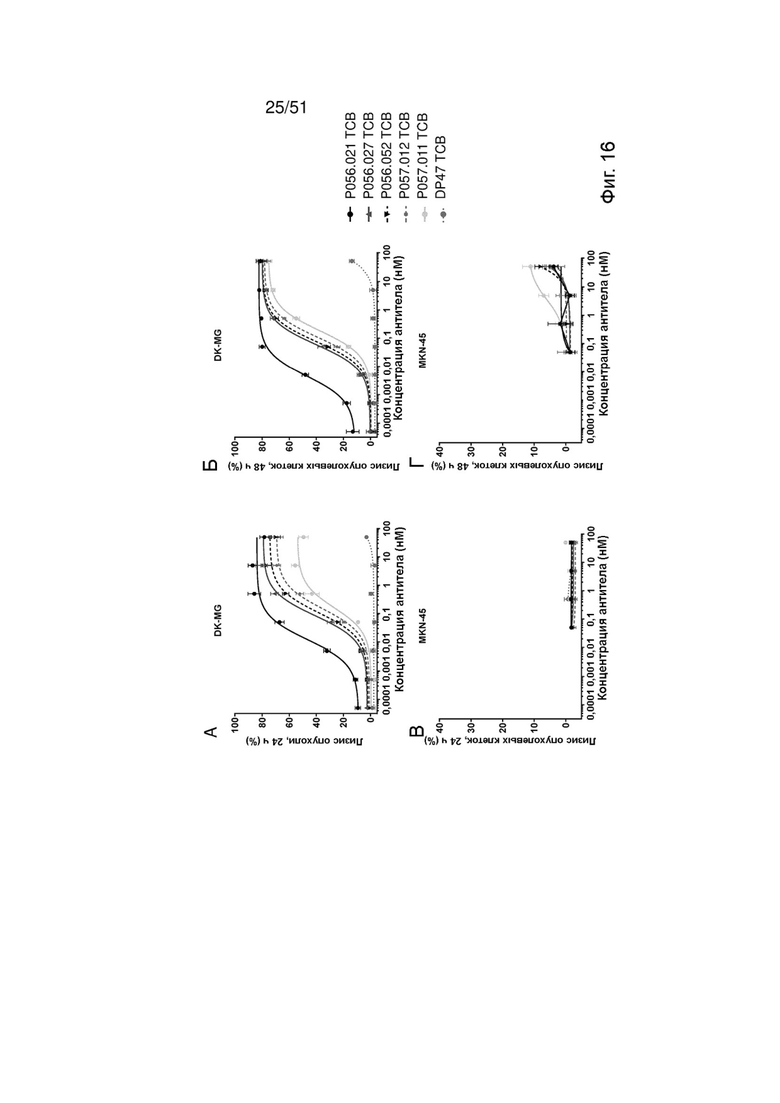

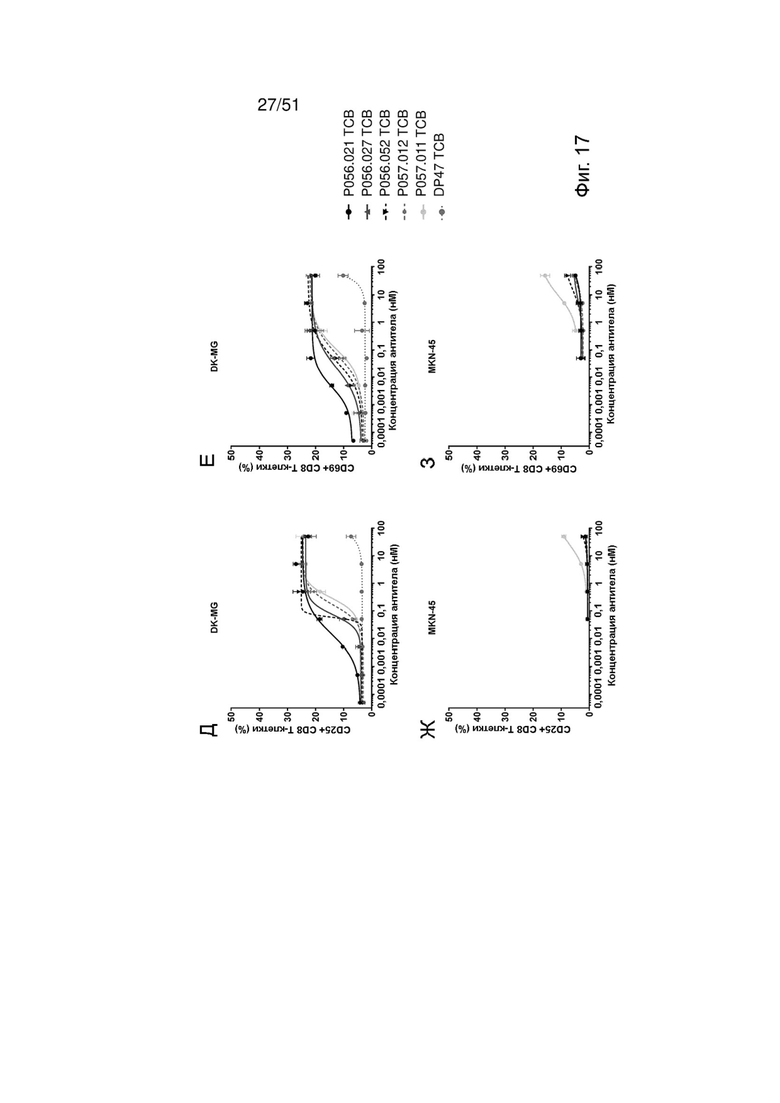




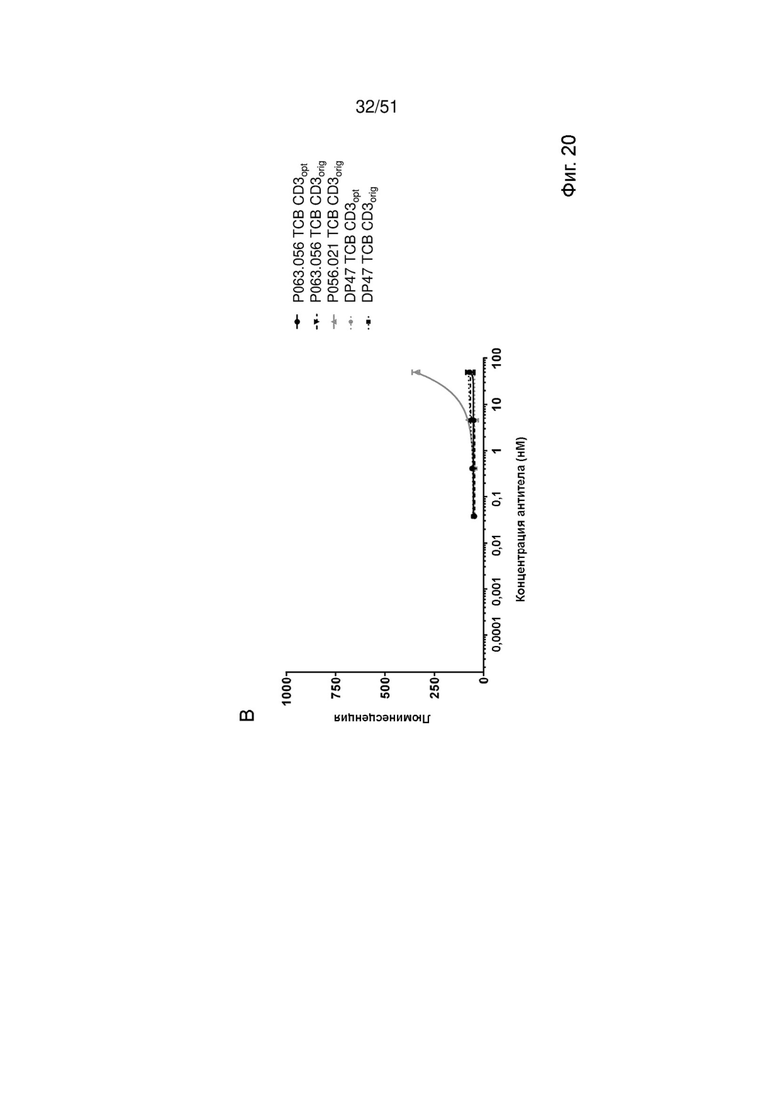
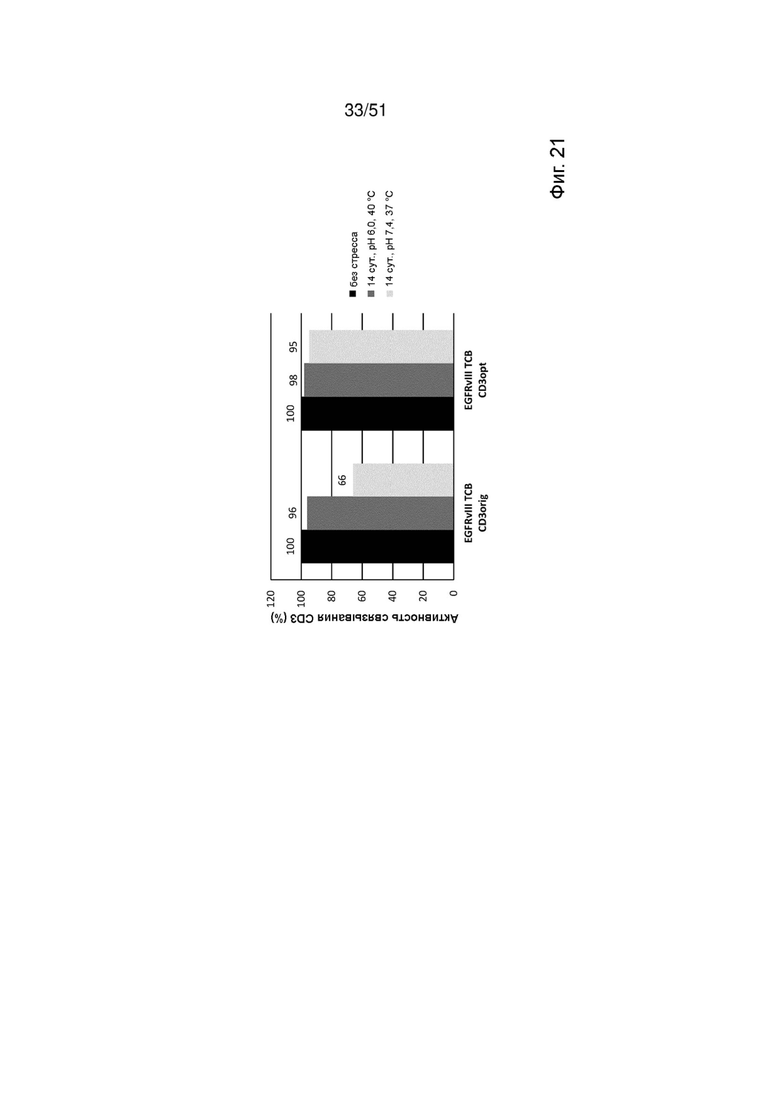
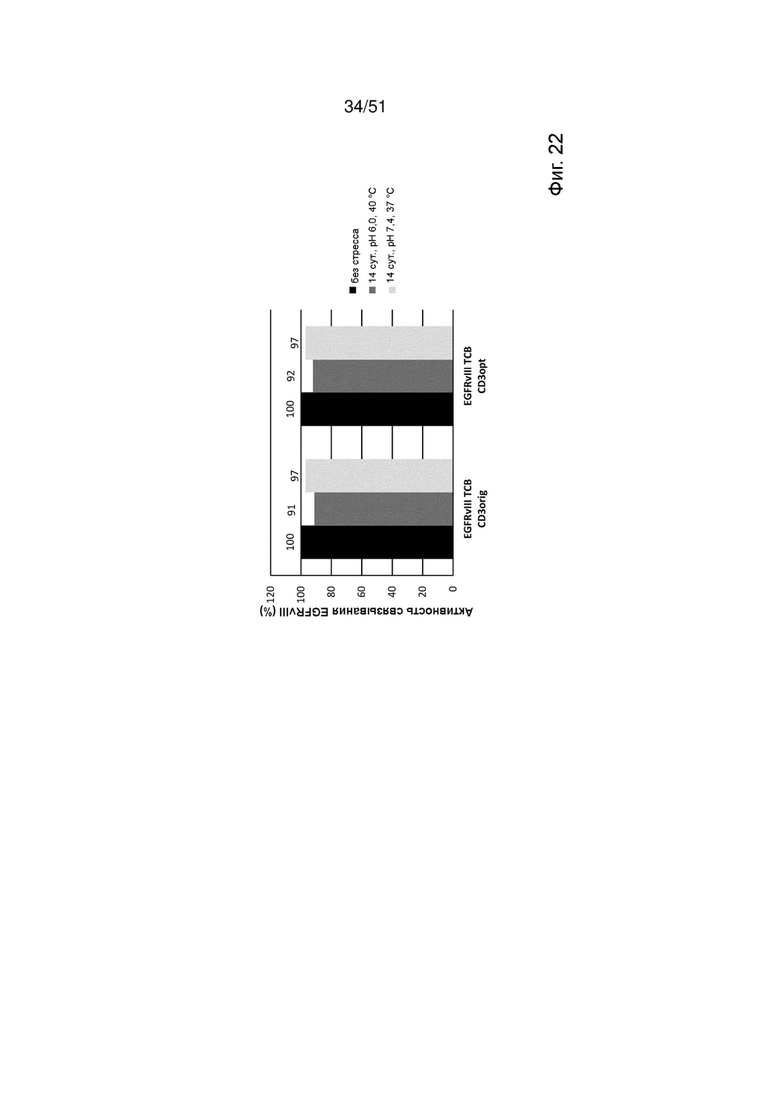



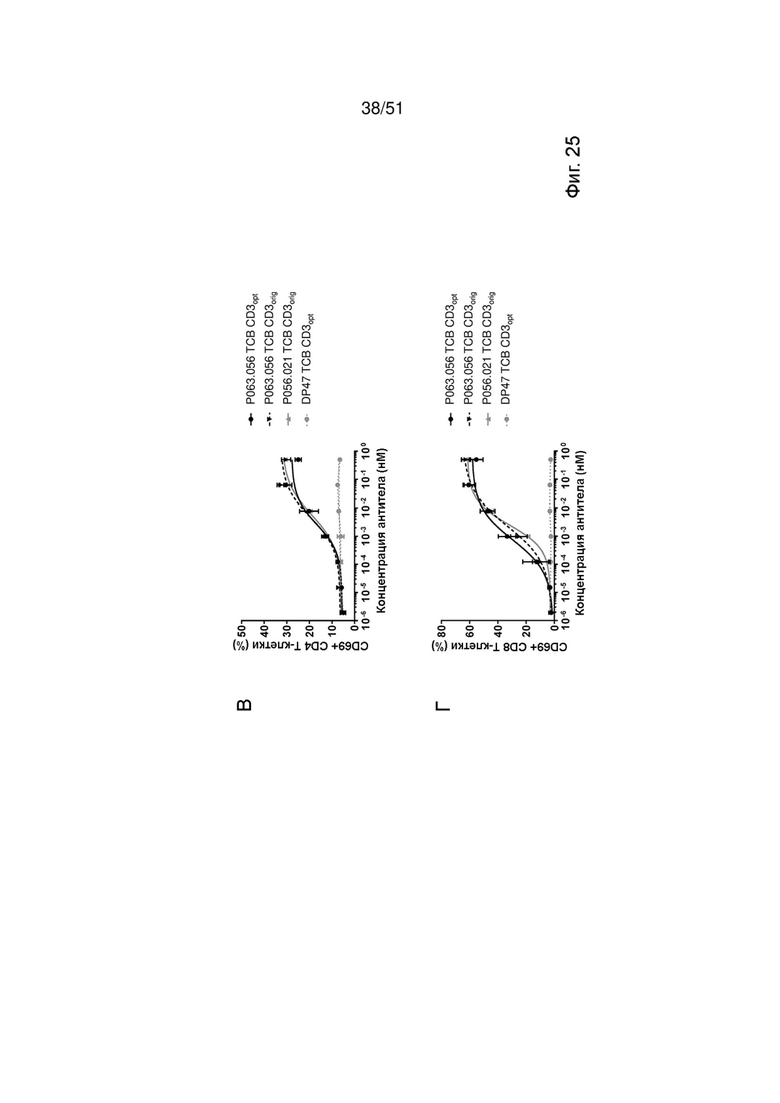


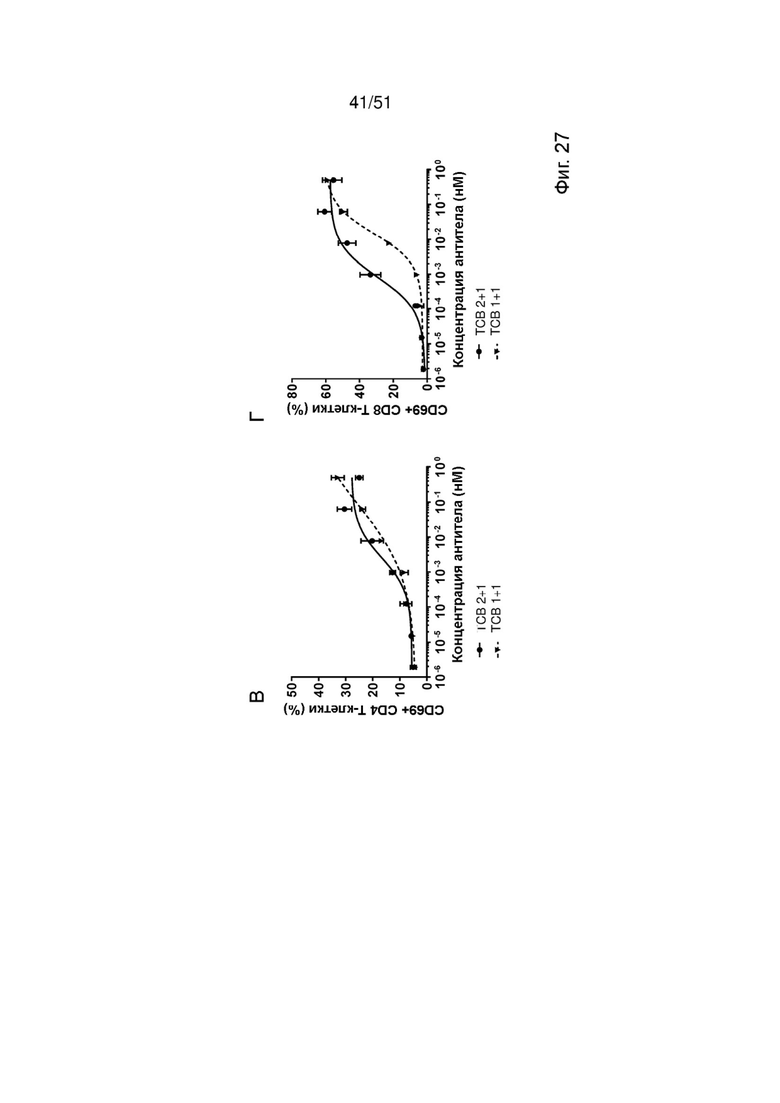
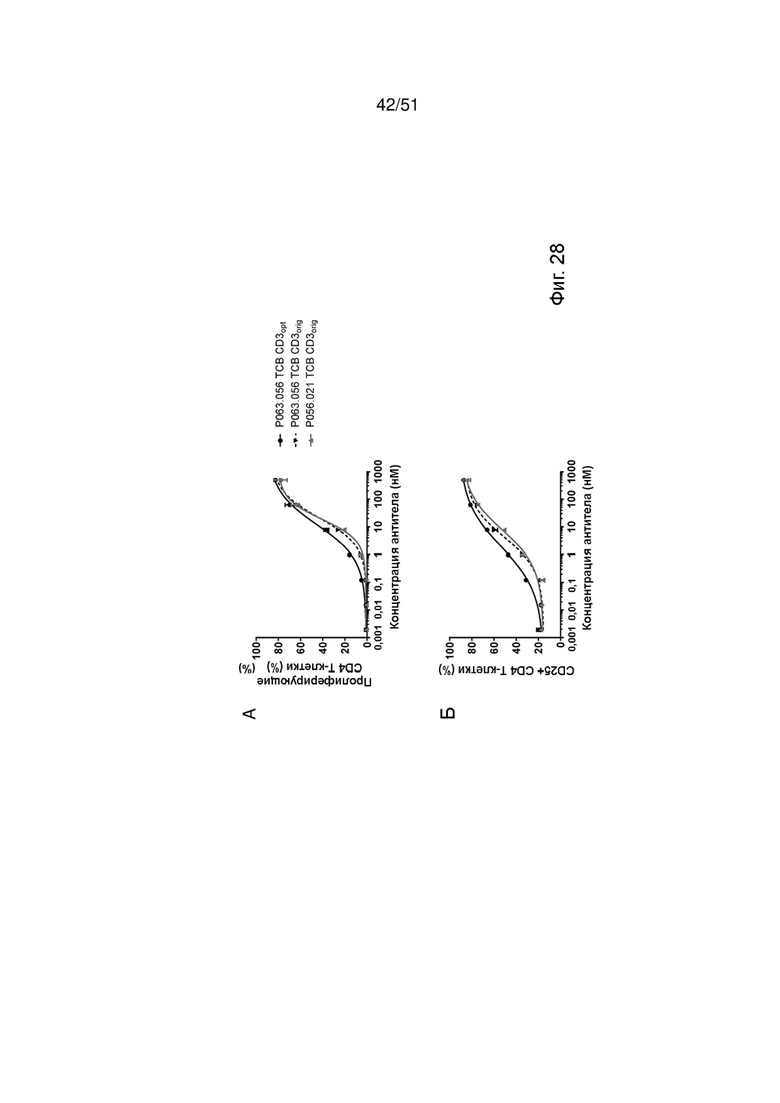
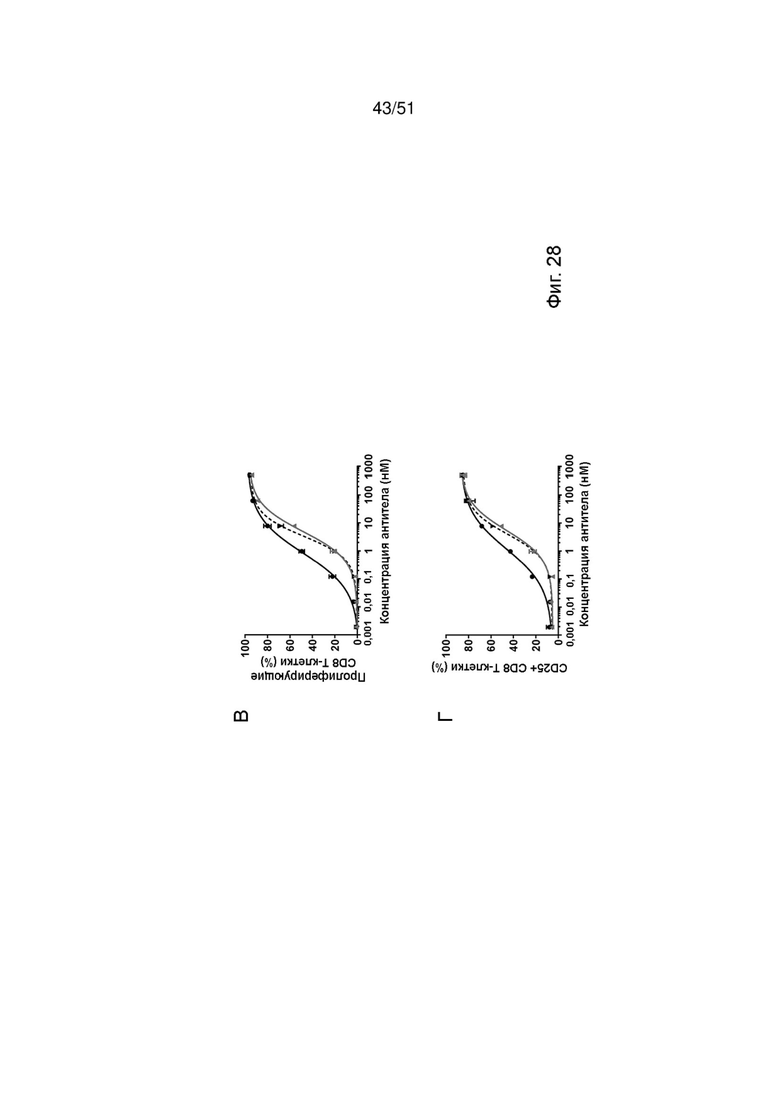
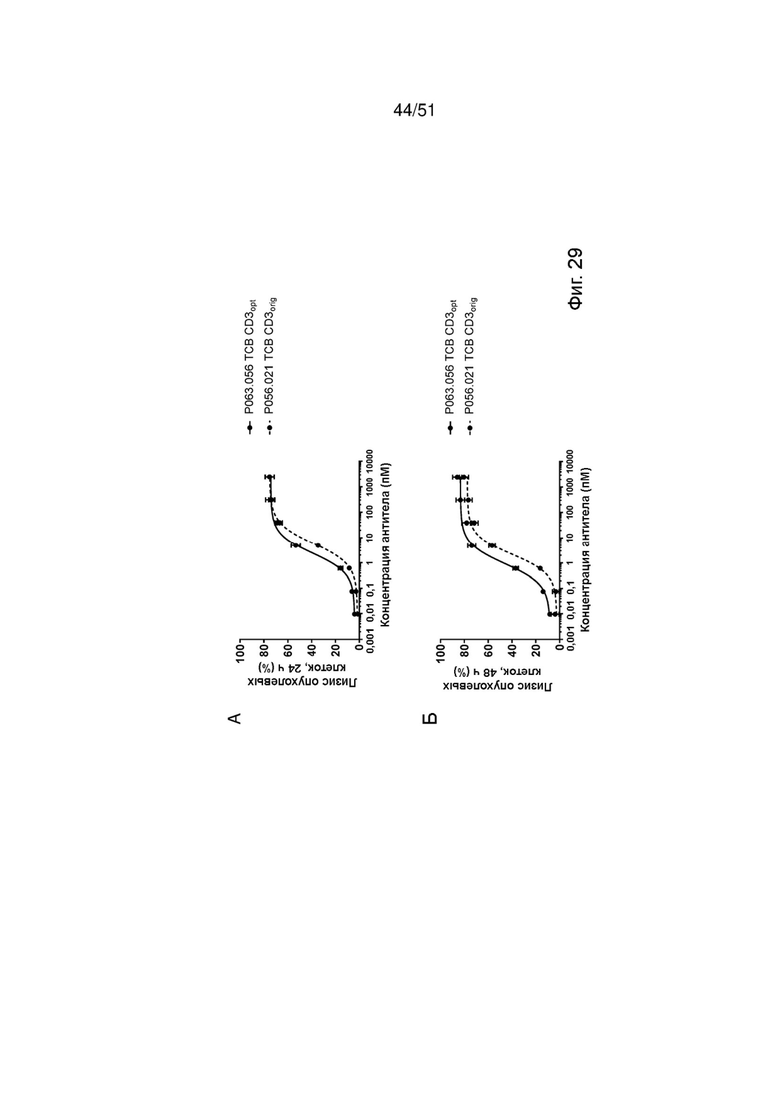
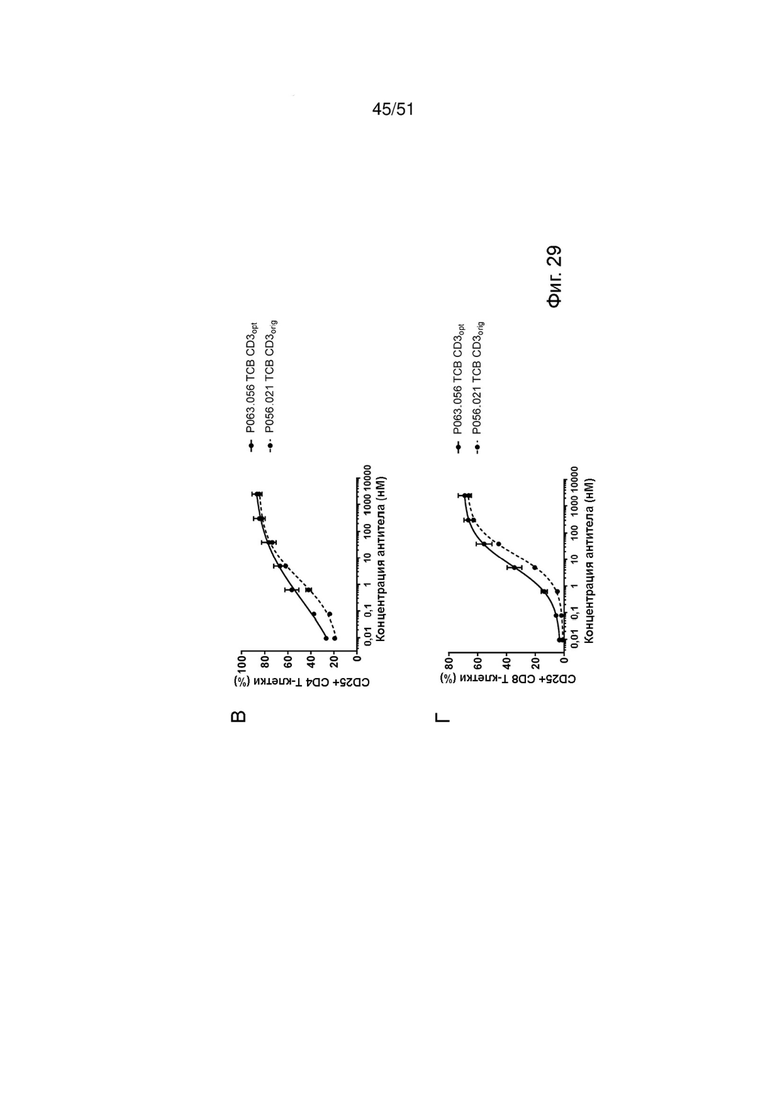

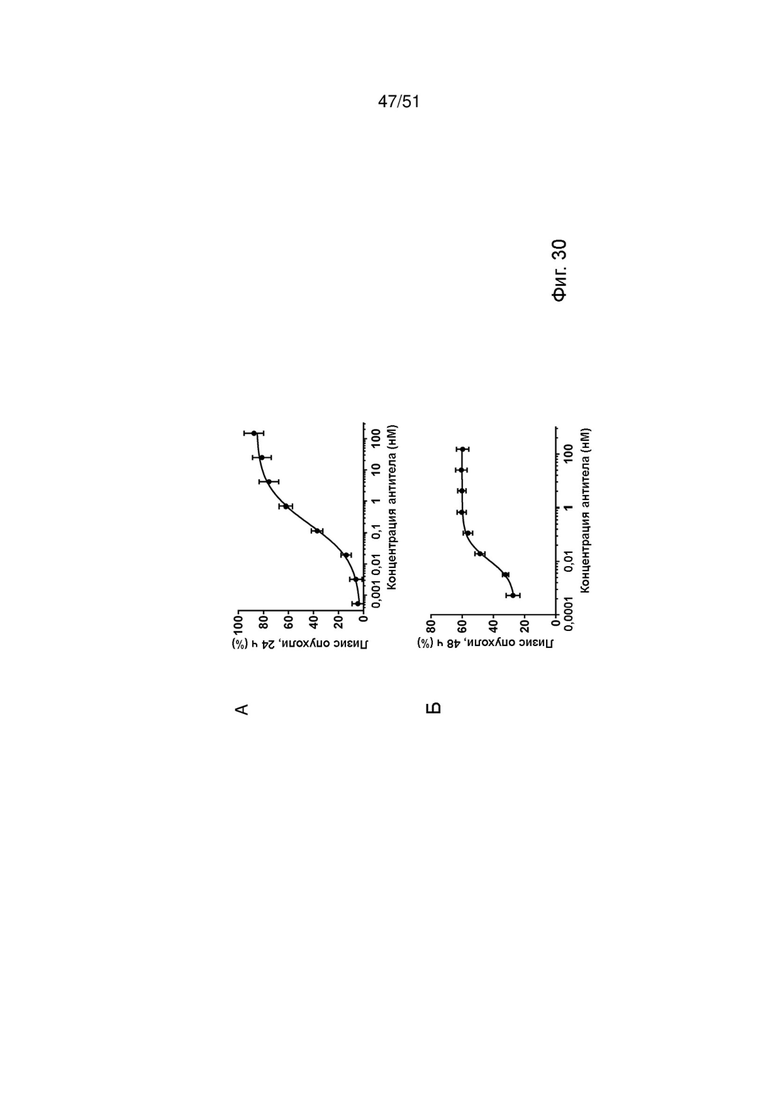


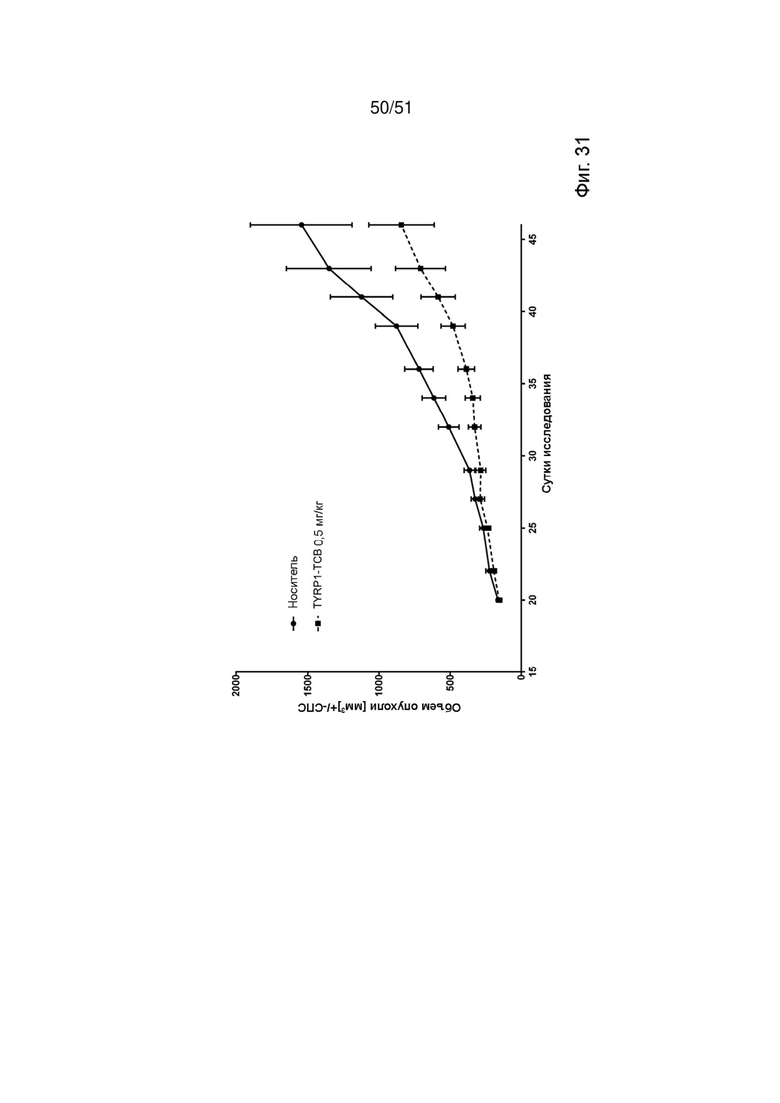
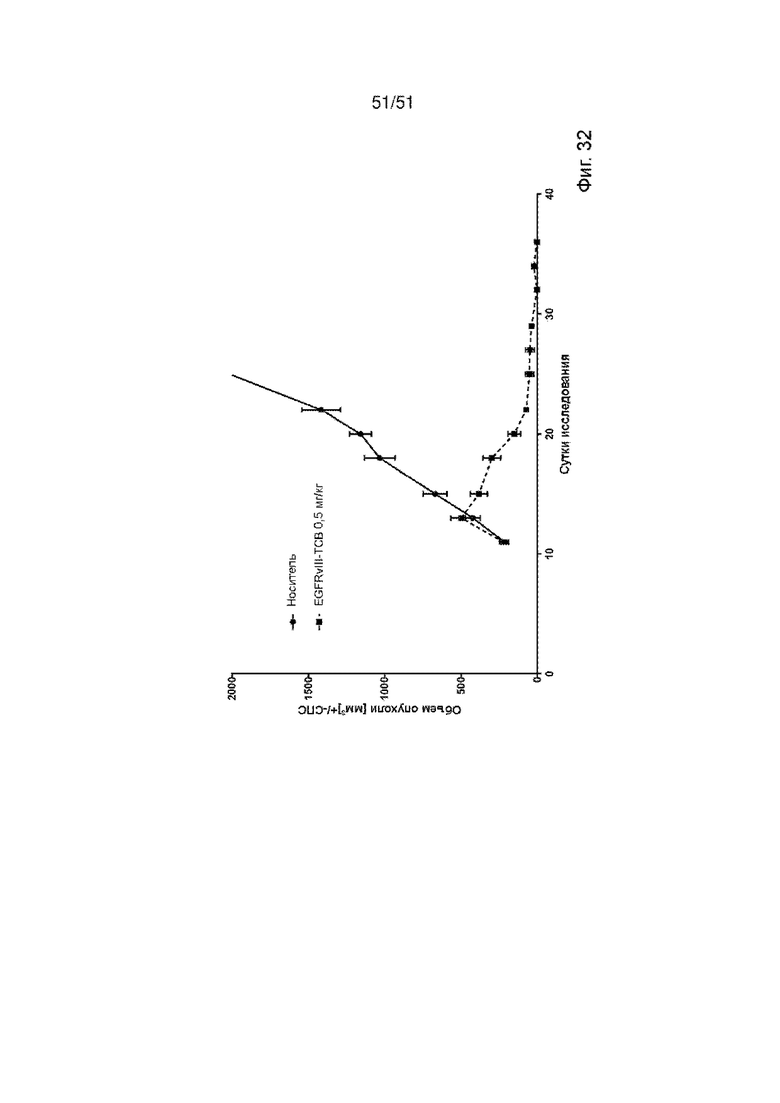
Изобретение относится к области биотехнологии, в частности к антителу, которое связывается с CD3 и EGFRvIII, способу его получения, а также к композиции, его содержащей. Также раскрыты выделенный полинуклеотид, кодирующий вышеуказанное антитело, а также клетка, его содержащая. Изобретение эффективно для лечения рака. 13 н. и 30 з.п. ф-лы, 32 ил., 7 табл., 13 пр.
1. Антитело, которое связывается с CD3 и EGFRvIII, причем антитело содержит (а) первый антигенсвязывающий домен, который связывается с CD3, содержащий вариабельную область тяжелой цепи (VH), содержащую определяющую комплементарность область тяжелой цепи (HCDR) 1 с SEQ ID NO: 2, HCDR 2 с SEQ ID NO: 3 и HCDR 3 с SEQ ID NO: 5, и вариабельную область легкой цепи (VL), содержащую определяющую комплементарность область легкой цепи (LCDR) 1 с SEQ ID NO: 8, LCDR 2 с SEQ ID NO: 9 и LCDR 3 с SEQ ID NO: 10; и (б) второй и, при необходимости, третий антигенсвязывающий домен, который связывается с EGFRvIII, содержащий VH, содержащий HCDR1 с SEQ ID NO: 85, HCDR 2 с SEQ ID NO: 86 и HCDR 3 с SEQ ID NO: 87, и VL, содержащий LCDR 1 с SEQ ID NO: 89, LCDR 2 с SEQ ID NO: 90 и LCDR 3 с SEQ ID NO: 91.
2. Антитело по п. 1, в котором VH первого антигенсвязывающего домена содержит аминокислотную последовательность, которая по меньшей мере на около 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 7, и/или VL первого антигенсвязывающего домена содержит аминокислотную последовательность, которая по меньшей мере на около 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 11.
3. Антитело по п. 1 или 2, в котором VH второго и, где присутствует, третьего антигенсвязывающего домена содержит аминокислотную последовательность, которая по меньшей мере на около 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 88, и/или VL второго и, где присутствует, третьего антигенсвязывающего домена содержит аминокислотную последовательность, которая по меньшей мере на около 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности SEQ ID NO: 92.
4. Антитело, которое связывается с CD3 и EGFRvIII, причем антитело содержит (а) первый антигенсвязывающий домен, который связывается с CD3, содержащий последовательность VH с SEQ ID NO: 7 и последовательность VL с SEQ ID NO: 11; и (б) второй и, при необходимости, третий антигенсвязывающий домен, который связывается с EGFRvIII, содержащий последовательность VH с SEQ ID NO: 88 и последовательность VL с SEQ ID NO: 92.
5. Антитело по любому из пп. 1-4, в котором первый антигенсвязывающий домен представляет собой молекулу Fab.
6. Антитело по любому из пп. 1-5, содержащее Fc-домен, состоящий из первой и второй субъединиц.
7. Антитело по любому из пп. 1-6, в котором второй и/или, в случае наличия, третий антигенсвязывающий домен представляет собой молекулу Fab.
8. Антитело по любому из пп. 1-7, в котором первый антигенсвязывающий домен представляет собой молекулу Fab, причем вариабельные домены VL и VH или константные домены CL и СН1, в частности вариабельные домены VL и VH легкой цепи Fab и тяжелой цепи Fab, замещены друг другом.
9. Антитело по любому из пп. 1-8, в котором второй и, в случае наличия, третий антигенсвязывающий домен представляет собой стандартную молекулу Fab.
10. Антитело по любому из пп. 1-9, в котором второй и, в случае наличия, третий антигенсвязывающий домен представляет собой молекулу Fab, причем в константном домене CL аминокислота в позиции 124 независимо замещена лизином (К), аргинином (R) или гистидином (Н), нумерация в соответствии с Kabat, и аминокислота в позиции 123 независимо замещена лизином (К), аргинином (R) или гистидином (Н), нумерация в соответствии с Kabat, а в константном домене СН1 аминокислота в позиции 147 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D), нумерация в соответствии с индексом EU по Kabat, и аминокислота в позиции 213 независимо замещена глутаминовой кислотой (Е) или аспарагиновой кислотой (D), нумерация в соответствии с индексом EU по Kabat.
11. Антитело по любому из пп. 1-10, в котором первый и второй антигенсвязывающие домены слиты друг с другом, необязательно посредством пептидного линкера.
12. Антитело по любому из пп. 1-11, в котором каждый из первого и второго антигенсвязывающих доменов представляет собой молекулу Fab и либо (i) второй антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первого антигенсвязывающего домена, либо (ii) первый антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab второго антигенсвязывающего домена.
13. Антитело по любому из пп. 1-12, в котором каждый из первого, второго и, в случае наличия, третьего антигенсвязывающих доменов представляет собой молекулу Fab и антитело содержит Fc-домен, состоящий из первой и второй субъединиц; и при этом либо (i) второй антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первого антигенсвязывающего домена, а первый антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом первой субъединицы Fc-домена, либо (ii) первый антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab второго антигенсвязывающего домена, а второй антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом первой субъединицы Fc-домена; и третий антигенсвязывающий домен, в случае наличия, слит на С-конце тяжелой цепи Fab с N-концом второй субъединицы Fc-домена.
14. Антитело по п. 13, в котором антитело состоит из первого, второго и третьего антигенсвязывающего домена, Fc-домена и необязательно одного или более пептидных линкеров, где второй антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом тяжелой цепи Fab первого антигенсвязывающего домена, а первый антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом первой субъединицы Fc-домена и третий антигенсвязывающий домен слит на С-конце тяжелой цепи Fab с N-концом второй субъединицы Fc-домена.
15. Антитело по любому из пп. 6-14, в котором Fc-домен представляет собой Fc-домен IgG, в частности IgG1.
16. Антитело по любому из пп. 6-15, в котором Fc-домен представляет собой Fc-домен человека.
17. Антитело по любому из пп. 6-16, в котором Fc-домен является Fc-домен IgG1.
18. Антитело по любому из пп. 6-17, в котором Fc-домен содержит модификацию, способствующую ассоциации первой и второй субъединиц Fc-домена.
19. Антитело по любому из пп. 6-18, в котором в СН3-домене первой субъединицы Fc-домена аминокислотный остаток замещают аминокислотным остатком, имеющим больший объем боковой цепи, таким образом, с созданием выпуклости в СН3-домене первой субъединицы, которая может размещаться в полости в СН3-домена второй субъединицы, а в СН3-домене второй субъединицы Fc-домена аминокислотный остаток замещают аминокислотным остатком, имеющим меньший объем боковой цепи, таким образом, с созданием полости в СН3-домене второй субъединицы, в которой может размещаться выпуклость в СН3-домене первой субъединицы.
20. Антитело по любому из пп. 6-19, в котором
в первой субъединице Fc-домена дополнительно остаток треонина в позиции 366 замещен остатком триптофана (T366W), а во второй субъединице Fc-домена остаток тирозина в позиции 407 замещен остатком валина (Y407V) и где необязательно
(а) во второй субъединице Fc-домена дополнительно остаток треонина в позиции 366 замещен остатком серина (T366S), а остаток лейцина в позиции 368 замещен остатком аланина (L368A), нумерация в соответствии с индексом EU по Kabat, и/или
(б) в первой субъединице Fc-домена дополнительно остаток серина в позиции 354 замещен остатком цистеина (S354C) или остаток глутаминовой кислоты в позиции 356 замещен остатком цистеина (Е356С), а во второй субъединице Fc-домена дополнительно остаток тирозина в позиции 349 замещен остатком цистеина (Y349C), нумерация в соответствии с индексом EU по Kabat.
21. Антитело по любому из пп. 6-20, в котором первая субъединица Fc-домена содержит аминокислотные замены S354C и T366W, а вторая субъединица Fc-домена содержит аминокислотные замены Y349C, T366S, L368A и Y407V, нумерация в соответствии с индексом EU по Kabat.
22. Антитело по любому из пп. 6-21, в котором Fc-домен содержит одну или более аминокислотных замен, которые уменьшают связывание с Fc-рецептором и/или эффекторную функцию.
23. Антитело по любому из пп. 6-22, где Fc-домен содержит аминокислотную замену в положениях, выбранных из группы Е233, L234, L235, N297, Р331 и Р329, в частности в положении, выбранном из группы L234, L235 и Р329, нумерация в соответствии с индексом EU по Kabat.
24. Антитело по любому из пп. 6-23, в котором каждая субъединица Fc-домена содержит аминокислотные замены L234A, L235A и P329G, нумерация в соответствии с индексом EU по Kabat.
25. Антитело по любому из пп. 1-24, где антитело включает полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 109, полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 110, полипептид, в частности два полипептида, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 111, и полипептид, содержащий аминокислотную последовательность, которая по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 27.
26. Выделенный полинуклеотид, кодирующий антитело по любому из пп. 1-25.
27. Клетка-хозяин для получения антитела, содержащая выделенный полинуклеотид по п. 26.
28. Способ получения антитела, которое связывается с CD3 и EGFRvIII, включающий этапы (а) культивирования клетки-хозяина по п. 27 в условиях, подходящих для экспрессии антитела, и, необязательно, (б) выделения антитела.
29. Фармацевтическая композиция для лечения рака, содержащая антитело по любому из пп. 1-25 и фармацевтически приемлемый носитель.
30. Применение антитела по любому из пп. 1-25 в качестве лекарственного средства для лечения рака.
31. Применение по п. 30, где рак представляет собой рак, экспрессирующий EGFRvIII и/или где рак представляет собой рак мозга, в частности глиобластому.
32. Применение фармацевтической композиции по п. 29 в качестве лекарственного средства для лечения рака.
33. Применение по п. 32, где рак представляет собой рак, экспрессирующий EGFRvIII и/или где рак представляет собой рак мозга, в частности глиобластому.
34. Применение антитела по любому из пп. 1-25 для лечения рака.
35. Применение по п. 34, где рак представляет собой рак, экспрессирующий EGFRvIII и/или где рак представляет собой рак мозга, в частности глиобластому.
36. Применение фармацевтической композиции по п. 29 для лечения рака.
37. Применение по п. 36, где рак представляет собой рак, экспрессирующий EGFRvIII и/или где рак представляет собой рак мозга, в частности глиобластому.
38. Применение антитела по любому из пп. 1-25 для получения лекарственного средства для лечения рака.
39. Применение по п. 38, где рак представляет собой рак, экспрессирующий EGFRvIII и/или где рак представляет собой рак мозга, в частности глиобластому.
40. Применение фармацевтической композиции по п. 29 для получения лекарственного средства для лечения рака.
41. Применение по п. 40, где рак представляет собой рак, экспрессирующий EGFRvIII и/или где рак представляет собой рак мозга, в частности глиобластому.
42. Способ лечения рака у индивида, включающий введение указанному индивиду эффективного количества антитела по любому из пп. 1-25 или фармацевтической композиции по п. 29.
43. Способ по п. 42, где рак представляет собой рак, экспрессирующий EGFRvIII и/или где рак представляет собой рак мозга, в частности глиобластому.
| WO 2014131712 A1, 04.09.2014 | |||
| WO 2016079076 A1, 26.05.2016 | |||
| WO 2017021370 A1, 09.02.2017 | |||
| XIAOJUN LU et al., Deamidation and isomerization liability analysis of 131 clinical-stage antibodies, MABS, published online: 10 Dec 2018, VOL | |||
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА ПРОТИВ CD3*CD19 | 2015 |
|
RU2651776C2 |

