ОБЛАСТЬ ТЕХНИКИ:
В настоящем изобретении предложены фармацевтические и пищевые композиции, вызывающие насыщение и продлевающие чувство сытости у нуждающихся в этом субъектов.
УРОВЕНЬ ТЕХНИКИ:
Состав кишечной микробиоты связывают с метаболическими фенотипами хозяина (Ley et al., 2006), и перенос микробиоты, «соответствующей ожирению», может вызывать ожирение (Turnbaugh et al., 2006) и гипрефагию (Vijay-Kumar et al., 2010), что дает основания предполагать, что кишечная микробиота может влиять на пищевое поведение хозяина. Несмотря на то что механизмы, лежащие в основе влияния кишечных бактерий на аппетит хозяина неизвестны, вероятно, что данные бактерии могут использовать молекулярные пути хозяина, контролирующие потребление пищи.
Существующая сегодня модель контроля потребления пищи включает передачу сигналов гормонами чувства голода и насыщения от кишечника к некоторым нейронным сетям мозга, регулирующим гомеостатические и гедонические аспекты питания (Berthoud, 2011; Inui, 1999; Murphy и Bloom, 2006). Главными среди них являются анорексигенные и орексигенные пути, идущие от дугообразного ядра гипоталамуса (ARC), которое содержит нейроны, продуцирующие проопиомеланокортин (РОМС) и нейропептид Y (NPY)/агути-связанный пептид (AgRP) соответственно, передающие сигнал в паравентрикулярное ядро (PVN) (Atasoy et al., 2012; Cowley et al., 1999; Garfield et al, 2015; Shi et al., 2013). Пути ARC и PVN сходятся в латеральном парабрахиальном ядре, из которого отростки анорексигенных нейронов идут к центральному миндалевидному телу (СеА), экспрессирующему связанный с геном кальцитонина пептид (CGRP) (Carter et al., 2013; Garfield et al., 2015).
Предположительные механизмы воздействия кишечной микробиоты на контроль аппетита хозяина могут включать ее активность в накоплении энергии (Turnbaugh et al., 2006) и продукцию нейроактивных трансмиттеров и метаболитов (Dinan et al., 2015; Forsythe и Kunze, 2013; Sharon et al., 2014). Авторы настоящего изобретения доказали влияние бактериальных белков, которые оказывают локальное действие в кишечнике или системное действие непосредственно на контролирующие аппетит пути. Фактически, было показано, что некоторые бактериальные белки имеют гомологию по последовательности с пептидными гормонами, регулирующими аппетит (Fetissov et al., 2008), и недавно белок ClpB, который продуцируется кишечной симбиотической бактерией Escherichia coli (E. coli), был идентифицирован как миметик антигена α-меланоцит-стимулирующего гормона (α-MSH) (Tennoune et al., 2014). α-MSH представляет собой происходящий из РОМС нейропептид, играющий ключевую роль в сигналинге, регулирующем насыщение, путем активации рецепторов меланокортина 4 (MC4R) (Cone, 2005). Несмотря на то, что опосредованные MC4R анорексигенные эффекты α-MSH в основном приписываются их областям действия в центральной нервной системе (Mul et al., 2013), недавнее исследование показало, что активация MC4R в аргентаффинных клетках кишечника стимулирует высвобождение гормонов насыщения глюкагоноподобного пептида-1 (GLP-1) и пептида YY (PYY) (Panaro et al., 2014). Таким образом, α-MSH-подобные молекулы, происходящие из кишечных бактерий, могут непосредственно действовать на аргентаффинные клетки, синтезирующие гормоны насыщения.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В настоящем изобретении предложены фармацевтические и пищевые композиции, вызывающие (индуцирующие) чувство насыщения и продлевающие чувство сытости у нуждающихся в этом субъектов. В частности, настоящее изобретение определяется формулой изобретения.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения показали, что белок ClpB или бактерия, экспрессирующая белок ClpB, способны вызывать чувство насыщения, продлевать чувство сытости, снижать потребление пищи, контролировать увеличение массы тела и/или стимулировать уменьшение массы тела у нуждающегося в этом субъекта.
Таким образом, согласно одному аспекту настоящего изобретения, предложен способ индукции насыщения у нуждающегося в этом субъекта, включающий введение указанному субъекту эффективного количества белка ClpB или эффективного количества бактерии, экспрессирующей белок ClpB.
Еще один аспект настоящего изобретения относится к способу продления чувства сытости у нуждающегося в этом субъекта, включающему введение указанному субъекту эффективного количества белка ClpB или эффективного количества бактерии, экспрессирующей белок ClpB.
Соответственно, согласно одному аспекту настоящего изобретения, предложен способ снижения объема порции пищи у нуждающегося в этом субъекта, включающий введение указанному субъекту эффективного количества белка ClpB или эффективного количества бактерии, экспрессирующей белок ClpB.
Еще один аспект настоящего изобретения относится к способу снижения потребления пиши у нуждающегося в этом субъекта, включающему введение указанному субъекту эффективного количества белка ClpB или эффективного количества бактерии, экспрессирующей белок ClpB.
Еще один аспект настоящего изобретения также относится к способу контроля увеличения массы тела у нуждающегося в этом субъекта, включающему введение указанному субъекту эффективного количества белка ClpB или эффективного количества бактерии, экспрессирующей белок ClpB.
Еще один аспект настоящего изобретения также относится к способу стимуляции уменьшения массы тела у нуждающегося в этом субъекта, включающему введение указанному субъекту эффективного количества белка ClpB или эффективного количества бактерии, экспрессирующей белок ClpB.
Способы согласно настоящему изобретению предназначены для людей, домашних животных или сельскохозяйственных животных, при этом домашние животные или сельскохозяйственные животные могут быть выбраны из группы, состоящей из собак, кошек, морских свинок, кроликов, свиней, крупного рогатого скота, овец, коз, лошадей и/или домашних птиц. Согласно некоторым вариантам реализации изобретения, субъект представляет собой особь мужского пола или женского пола.
Согласно некоторым вариантам реализации изобретения, субъект не страдает ожирением. Согласно некоторым вариантам реализации изобретения, субъект страдает ожирением. Термин «ожирение» при употреблении в настоящей заявке относится к медицинскому состоянию, при котором ИМТ субъекта предпочтительно составляет >30. «ИМТ», или «индекс массы тела», определяется как значение массы тела субъекта, деленное на значение его роста в квадрате. Повсеместно используемая в медицине формула приводит к получению единицы измерения кг/м2. Как правило, субъект, не страдающий ожирением, имеет нормальную массу тела. «Нормальная масса тела» и «избыточная масса тела без осложнений» относится в настоящей заявке к массе тела, обеспечивающей ИМТ между 18,5 и 30.
Подразумевается, что при употреблении в настоящей заявке термин «чувство сытости» относится по существу к гомеостатическому состоянию, когда субъект ощущает, что его пищевые потребности удовлетворены или минимизированы. Считается, что многие физиологические факторы влияют на чувство сытости субъекта. Например, вкусовые ощущения, или вкус, обонятельные ощущения, или запах, а также чувство полноты желудка могут вносить вклад в чувство «сытости» субъекта. Более конкретно, «чувство сытости» представляет собой состояние, при котором дальнейшее потребление пищи подавляется, и определяет время между приемами пищи и количество еды, потребленной за следующий прием пищи. «Усиление чувства сытости» или т.п. означает чувство сытости, более выраженное и/или более пролонгированное по сравнению с контрольной ситуацией. Термин «насыщение» при употреблении в настоящей заявке относится к состоянию, которое приводит к остановке потребления пищи во время приема пищи, как правило, наблюдаемое в пределах некоторого времени (например, 20-30 мин) после начала потребления пищи. Таким образом, в любом месте в настоящем документе ссылка на «индукцию насыщения» или т.п. означает пробуждение стремления прекратить потребление еды во время приема пищи. Влияние на насыщение можно определить по оценке времени прекращения приема пищи. Эффект насыщения заметен, если количество потребленных калорий по окончании приема пищи значительно меньше по сравнению с контролями, например, по меньшей мере на 1%, 2%, 3%, 4%, 5%, 10% 20% или более. В течение более длительного периода времени (такого как 1, 2, 3, 4, 5 недель или более), можно также оценить снижение массы тела или изменение массы тела по сравнению с контрольной диетой. Масса тела субъекта, которому регулярно вводят некоторое количество исследуемой композиции (например, один раз в день, два раза в день или более), предпочтительно достоверно контролируется (снижается или повышается в меньшей степени) по сравнению с контрольными субъектами. При употреблении в настоящей заявке термин «контрольный субъект» относится к субъектам, которым не вводят пробиотический штамм бактерий согласно настоящему изобретению.
При употреблении в настоящей заявке термин «ClpB» имеет его общепринятое значение в данной области техники и также известен как белок теплового шока F84.1, который является членом семейства Hsp100/ClpB гексамерных ААА+-АТРаз. ClpB был описан как важный фактор для приобретенной термоустойчивости и вирулентности и инфицирующей способности некоторых грамотрицательных и грамположительных патогенных бактерий, таких как Staphylococcus aureus, Francisella turalensis, Listeria monocytogenes, Yersinia enterocolitica и Salmonella thyphimurium. В E. coli K12 шаперонный белок ClpB, также известный как белок теплового шока F84.1 или htpM, представляет собой белок из 857 аминокислот. Как правило, шаперонный белок ClpB содержит или состоит из аминокислотной последовательности шаперонного белка ClpB из Е. Coli K12 с последовательностью SEQ ID NO: 1 (референсный номер в NCBI: NP_417083,1, по наличию на 6 ноября 2013 года и/или номер в UniProtKB/Swiss-Prot: Р63284, по наличию на 6 ноября 2013 г.). Как правило, аминокислотная последовательность шаперонного белок ClpB содержит или состоит из аминокислотной последовательности, на 96-100% идентичной аминокислотной последовательности SEQ ID NO: 1. Предпочтительно, аминокислотная последовательность ClpB на 96, 97, 98, 99 или 100% идентична аминокислотной последовательности 540-550 (ARWTGIPVSR) последовательности SEQ ID NO: 1. В контексте настоящей заявки процент идентичности рассчитывают с помощью полного выравнивания (global alignment, т.е. две последовательности сравнивают по всей длине). Способы определения идентичности двух или более последовательностей хорошо известны в данной области техники. Можно применять, например, программу «Needle», которая использует алгоритм полного выравнивания Нидлмана-Вунша (Needleman and Wunsch, 1970 J. Mol. Biol. 48:443-453) для поиска оптимального выравнивания (включая пропуски) двух последовательностей с учетом полной длины. Программа «Needle» доступна, например, на интернет-сайте ebi.ac.uk. Процент идентичности в соответствии с изобретением предпочтительно рассчитывают с использованием EMBOSS: программа «Needle» (глобальная) с параметром «Gap Open», равным 10.0, параметром «Gap Extend», равным 0,5, и матрицей Blosum62. Согласно изобретению, белок ClpB имитирует белок альфа-MSH для индукции насыщения. Таким образом, согласно некоторым вариантам реализации изобретения, белок ClpB согласно настоящему изобретению распознается антителом против альфа-MSH. Как правило, антитело представляет собой моноклональное антитело. Согласно некоторым вариантам реализации изобретения, антитело представляет собой поликлональное антитело, такое как поликлональные IgG кролика против α-MSH (разведение 1:1000, Peninsula Laboratories, Сан-Карлос, Калифорния, США). Аминокислотная последовательность α-MSH предпочтительно содержит или состоит из аминокислотной последовательности SYSMEHFRWGKPV (SEQ ID NO: 2) (идентификационный номер последовательности в базе данных Gen Pept, PRF: 223274, как доступно на 2 декабря 2013 г.).
SEQ ID NO: 1:

Согласно некоторым вариантам реализации изобретения, белок ClpB вводят указанному субъекту в виде фармацевтической композиции. Согласно некоторым вариантам реализации изобретения, белок ClpB объединен с фармацевтически приемлемыми вспомогательными веществами и необязательно матрицей замедленного высвобождения, такой как биодеградируемые полимеры, для образования фармацевтической композиции. Термин «фармацевтический» или «фармацевтически приемлемый» относится к химическим соединениям и композициям, которые не производят неблагоприятных, аллергических или других нежелательных реакций при введении млекопитающему, в частности, человеку, в зависимости от конкретного случая. Фармацевтически приемлемый носитель или вспомогательное вещество относится к нетоксичному твердому, полутвердому или жидкому наполнителю, разбавителю, инкапсулирующему материалу или вспомогательной форме для введения любого типа. В фармацевтических композициях согласно настоящему изобретению активный компонент отдельно или в комбинации с другим активным компонентом можно вводить животным или человеку в виде стандартной лекарственной формы, в виде смеси со стандартными фармацевтическими добавками. Подходящие единичные формы для введения включают формы для перорального введения, такие как таблетки, желатиновые капсулы, порошки, гранулы и суспензии или растворы для перорального введения, формы для сублингвального или буккального введения, аэрозоли, импланты, формы для подкожного, трансдермального, местного, внутрибрюшинного, внутримышечного, внутривенного, субдермального, трансдермального, интратекального и интраназального введения и формы для ректального введения. Предпочтительно, фармацевтические композиции содержат наполнители, которые являются фармацевтически приемлемыми для лекарственной формы для инъекций. Они могут, в частности, представлять собой изотонический раствор, стерильные, солевые растворы (моно- или дифосфат натрия, хлорид натрия, калия, кальция или магния и т.д. или смеси таких солей), или сухие, в частности, лиофилизированные композиции, которые при добавлении стерильной воды или физиологического солевого раствора (в зависимости от конкретного случая) обеспечивают состояние раствора для инъекций. Фармацевтические формы, подходящие для применения в виде инъекций, включают стерильные водные растворы или дисперсии; лекарственные формы, содержащие кунжутное масло, арахисовое масло или водный пропиленгликоль; и стерильные порошки для приготовления стерильных растворов для инъекций или дисперсий для немедленного приема. Во всех случаях форма должна быть стерильной и должна быть текучей до такой степени, чтобы она могла легко использоваться для наполнения шприца. Она должна являться стабильной при условиях изготовления и хранения и должна быть защищена от контаминирующего действия микроорганизмов, таких как бактерии и грибы. Растворы, содержащие соединения согласно изобретению в виде свободных оснований или фармакологически приемлемой соли, можно получить в воде, подходящим образом смешанной с поверхностно-активным веществом, таким как гидроксипропилцеллюлоза. Дисперсии также можно получить в глицерине, жидких полиэтиленгликолях и их смеси и в маслах. При обычных условиях хранения и использования указанные препараты содержат консервант для предотвращения роста микроорганизмов. Активный ингредиент может быть включен в композицию в нейтральной форме или в форме соли. Фармацевтически приемлемые соли включают соли присоединения кислоты (образованные со свободными белковыми аминогруппами), которые образованы с неорганическими кислотами, такими как, например, соляная кислота или фосфорные кислоты, или такими органическими кислотами, как уксусная, оксалиновая, винная, миндальная и т.д. Соли, образованные со свободными карбоксильными группами, также могут быть получены из неорганических оснований, таких как, например, гидроксиды натрия, калия, аммония, кальция или железа, и такие органические основания, как изопропиламин, триметиламин, гистидин, прокаин и т.д. Носитель также может представлять собой растворитель или дисперсионную среду, содержащую, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и т.д.), их подходящую смесь и растительные масла. Подходящую текучесть можно поддерживать, например, путем использования покрытия, такого как лецитин, путем поддержания требуемого размера частиц в случае дисперсии и путем использования поверхностно-активных веществ. Предотвращение действия микроорганизмов можно осуществлять с помощью различных антибактериальных и противогрибковых агентов, например, парабенов, хлорбутанола, фенола, сорбиновой кислоты, тимерозала и т.д. Во многих случаях предпочтительно включать изотонические агенты, например, сахара или хлорид натрия. Пролонгированное всасывание композиций для инъекции можно осуществлять путем использования в композиции агентов, замедляющих всасывание, например, моностеарата алюминия и желатина. Стерильные растворы для инъекций получают путем включения активных полипептидов в требуемом количестве в подходящем растворителе с различными другими ингредиентами, перечисленными выше, в случае необходимости с последующей стерилизацией фильтрованием. В целом, дисперсии получают путем включения различных стерилизованных активных ингредиентов в стерильный наполнитель, который содержит основную дисперсионную среду и необходимые другие ингредиенты из перечисленных выше. В случае стерильных порошков для получения стерильных растворов для инъекций предпочтительные способы получения представляют собой методы вакуумной сушки и лиофилизирования, которые приводят к получению порошка активного ингредиента вместе с любым дополнительным желаемым ингредиентом из предварительно стерилизованного фильтрованием раствора. При получении лекарственной формы растворы вводят с помощью способа, совместимого с лекарственной формой, и в таком количестве, которое является терапевтически эффективным. Лекарственные формы легко вводят в различных формах дозирования, таких как типы растворов для инъекций, описанные выше, но также можно использовать капсулы с высвобождением лекарственного средства и т.д. Для парентерального введения в водном растворе, например, указанный раствор должен быть при необходимости забуферен подходящим образом, и жидкий разбавитель сначала должен быть доведен до состояния изотонического раствора с помощью достаточного количества солевого раствора или глюкозы. Эти особые водные растворы являются, в частности, подходящими для внутривенного, внутримышечного, подкожного и внутрибрюшинного введения. В связи с этим стерильные водные среды, которые могут использоваться, будут очевидны специалисту в данной области техники в свете настоящего изобретения. Например, одна доза может быть растворена в 1 мл изотонического раствора NaCl и либо добавлена к 1000 мл гиподермаклизисной жидкости, либо инъецирована в предполагаемое место инфузии. Безусловно, доза будет варьировать в зависимости от состояния субъекта, подлежащего лечению. Человек, ответственный за введение, в любом случае будет определять подходящую дозу для конкретного субъекта.
При употреблении в настоящей заявке выражение «бактерия, экспрессирующая С1рВ» относится к бактерии, которая экспрессирует или экспрессирует на увеличенном уровне шаперонный белок ClpB, как определено выше, или полипептид, содержащий или состоящий из аминокислотной последовательности, на от 96 до 100% идентичную аминокислотной последовательности SEQ ID NO: 1, более предпочтительно, на 96, 97, 98, 99 или 100% идентичную аминокислотной последовательности SEQ ID NO: 1.
Согласно некоторым вариантам реализации изобретения, бактерия, экспрессирующая белок ClpB, представляет собой пробиотический штамм бактерий.
Предполагается, что при употреблении в настоящей заявке термин «пробиотический» означает живые микроорганизмы, которые при включении в достаточном количестве оказывают положительный эффект на здоровье, комфорт и самочувствие субъекта помимо стандартных питательных эффектов. Пробиотические микроорганизмы были определены как живые микроорганизмы, которые при введении в подходящем количестве «обеспечивают благоприятный эффект на здоровье хозяина» (FAO/WHO 2001). При употреблении в настоящей заявке выражение «пробиотический штамм бактерий» означает штамм бактерий, которые оказывают благоприятный эффект на здоровье и самочувствие хозяина.
Согласно некоторым вариантам реализации изобретения, пробиотический штамм бактерий согласно настоящему изобретению представляет собой жизнеспособный пробиотический штамм бактерий. Выражение «жизнеспособный пробиотический штамм бактерий» означает микроорганизм, который является метаболически активным и который способен заселять желудочно-кишечный тракт субъекта.
Согласно некоторым вариантам реализации изобретения, пробиотический штамм бактерий согласно настоящему изобретению представляет собой не жизнеспособный пробиотический штамм бактерий, состоящий из смеси бактериальных фрагментов. Согласно некоторым вариантам реализации изобретения, смесь бактериальных фрагментов согласно настоящему изобретению состоит из белков бактериального штамма.
Согласно некоторым вариантам реализации изобретения, пробиотический штамм бактерий согласно настоящему изобретению выбран из пищевых бактерий. Термин «пищевые бактерии» означает бактерии, которые используются и в целом считаются безопасными для употребления в пищу.
Штамм бактерий может представлять собой природный штамм бактерий или может представлять собой штамм бактерий, полученный с помощью генной инженерии.
Согласно некоторым вариантам реализации изобретения, пробиотический штамм бактерий согласно настоящему изобретению представляет собой бактерию, которая конститутивно экспрессирует белок ClpB.
Согласно некоторым вариантам реализации изобретения, белок ClpB экспрессируется на увеличенном уровне в бактерии. В целом экспрессия белка «стимулируется», или белок «экспрессируется на увеличенном уровне», когда он экспрессируется или продуцируется в количестве или с выходом, превышающим конкретный базовый выход, который встречается в природе при стандартных условиях. Увеличенную экспрессию белка можно достигнуть, например, путем изменения любого одного или более из следующих факторов: (a) условия роста или жизни клеток хозяина; (b) полинуклеотида, кодирующий белок; (c) промотора, используемого для контроля экспрессии полинуклеотида и числа его копий в клетке; и (d) самих клеток хозяина.
Согласно некоторым вариантам реализации изобретения, бактерию подвергают условиям стресса таким образом, что экспрессия белка ClpB активируется в указанной бактерии. Стресс может быть выбран из группы, состоящей из нагревания, температурным изменениям, механическому стрессу или длительному хранению, хранению при низкой влажности и/или лиофилизированию или высушиванию распылением.
Согласно некоторым вариантам реализации изобретения, бактерии снабжали питательными веществами по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 раз. Как правило, питательные вещества доставляли с помощью культуральной среды, подходящей для роста указанных бактерий, например, среды Мюллера-Хинтона, как описано в Примере. Согласно некоторым вариантам реализации изобретения, бактерии выделяют на стадии стационарной фазы роста, следующей за указанным повторяющимся снабжением питательными веществами, так как во время этой фазы концентрация белка ClpB является максимальной.
Согласно некоторым вариантам реализации изобретения, бактерии содержат по меньшей мере одну точечную мутацию, приводящую к повышению экспрессии белка ClpB. Термин «точечная мутация» при употреблении в настоящей заявке означает замену и/или делецию нуклеиновой кислоты. Альтернативно (или одновременно), по меньшей мере одна мутация находится в регуляторных последовательностях ДНК гена ClpB, например, в последовательностях контроля транскрипции и трансляции. Предпочтительно, указанная мутация модулирует экспрессию белка. Мутации в регуляторных последовательностях ДНК могут служить для повышения экспрессии белка.
Согласно некоторым вариантам реализации изобретения, пробиотический штамм бактерий согласно настоящему изобретению представляет собой бактерию, созданную с помощью генной инженерии таким образом, чтобы она экспрессировала белок ClpB. Как правило, штамм бактерий трансформируют нуклеиновой кислотой, кодирующей белок ClpB. Термин «трансформация» означает введение «чужеродного» (т.е. постороннего или внеклеточного) гена, последовательности ДНК или РНК в клетку хозяина таким образом, чтобы указанная клетка хозяина экспрессировала введенный ген или последовательность с продукцией желаемого вещества, как правило, белка или фермента, кодируемого введенным геном или последовательностью. Клетка хозяина, которая получает и экспрессирует введенную ДНК или РНК, была «трансформирована». Нуклеиновая кислота может оставаться внехромосомной при трансформации исходного микроорганизма или может быть адаптирована для интеграции в геном микроорганизма. Соответственно, нуклеиновая кислота может включать дополнительные нуклеотидные последовательности, адаптированные таким образом, чтобы способствовать интеграции (например, участок, который обеспечивает гомологичную рекомбинацию и направленную интеграцию в геном хозяина) или стабильной экспрессия и репликации внехромосомной конструкции (например, точку начала репликации, промотор и другие регуляторные последовательности). Согласно некоторым вариантам реализации изобретения, нуклеиновая кислота представляет собой конструкцию или вектор на основе нуклеиновой кислоты. Согласно некоторым вариантам реализации изобретения, конструкция или вектор на основе нуклеиновой кислоты представляет собой экспрессионную конструкцию или вектор, однако другие конструкции и векторы, такие как конструкции и векторы, используемые для клонирования, включены в настоящее изобретение. Согласно некоторым вариантам реализации изобретения, экспрессионная конструкция или вектор представляет собой плазмиду. Как правило, экспрессионная конструкция/вектор также содержит промотор, как описано выше в настоящей заявке. Согласно некоторым вариантам реализации изобретения, промотор обеспечивает конститутивную экспрессию генов под их контролем. Однако также могут использоваться индуцируемые промоторы. Следует иметь в виду, что экспрессионная конструкция/вектор согласно настоящему изобретению может при желании содержать любое количество регуляторных элементов помимо промотора, а также дополнительные гены, подходящие для экспрессии белка ClpB. Способы трансформации бактериальной клетки внеклеточными нуклеиновыми кислотами хорошо известны в данной области техники.
Согласно некоторым вариантам реализации изобретения, пробиотический штамм бактерий представляет собой грамотрицательный штамм.
Согласно некоторым вариантам реализации изобретения, пробиотический штамм бактерий является членом семейства Enterobacteriaceaae.
Согласно некоторым вариантам реализации изобретения, пробиотический штамм бактерий представляет собой штамм Е. coli. Согласно некоторым вариантам реализации изобретения, пробиотические штаммы Е. Coli для применения в соответствии с принципами настоящего изобретения включают непатогенные штаммы Е. Coli, которые проявляют пробиотическую активность. Пример пробиотического штамма Е. Coli представляет собой пробиотический штамм Escherichia coli BU-230-98 (номер депозита в Американской коллекции типовых культур (АТСС) 202226 (DSM 12799)), являющийся изолятом известного коммерчески доступного пробиотического штамма Escherichia coli М-17. Примером непатогенного штамма является E.Coli Nissle 1917. Примером штамма E.coli, который ранее не был известен как пробиотический, является лабораторный штамм E.coli K12.
Как правило, пробиотический штамм бактерий согласно настоящему изобретению поставляется с любой подходящей культуральной средой, хорошо известной в данной области техники. Различные ферментационные среды являются подходящими согласно изобретению, включая, но не ограничиваясь указанными, в первую очередь, например, промышленную среду, в которой культивируют указанный штамм (штаммы) и которая используется в исходном виде или после концентрирования (например, высушивания) или после добавления к другой пищевой основе или продукту. Альтернативно, можно применять бактериальные клетки или бактериальные клетки со средой (например, ферментативным бульоном) или фракции такой среды, содержащей клетки, (т.е. среды с указанным штаммом/штаммами бактерий можно применять). Содержащая клетку или клетки среда содержит живые или жизнеспособные бактериальные клетки и/или мертвые или нежизнеспособные бактериальные клетки штамма (штаммов). Среду, таким образом, можно обрабатывать, путем нагрева и обработки ультразвуком, не ограничиваясь указанными методами. Кроме того, лиофилизированные, или замороженные, бактерии и/или бесклеточные среды (которые могут быть концентрированными) включены в способы получения пробиотического штамма бактерий согласно настоящему изобретению.
Как правило, пробиотический штамм бактерий согласно настоящему изобретению вводят указанному субъекту в результате приема внутрь (т.е. пероральным путем).
Согласно некоторым вариантам реализации изобретения, пробиотический штамм бактерий согласно настоящему изобретению инкапсулируют для защиты от желудочной среды. Соответственно, согласно некоторым вариантам реализации изобретения, пробиотический штамм бактерий согласно настоящему изобретению представлен в виде композиции в инкапсулированной форме таким образом, чтобы срок выживания был значительно улучшен. В таком случае, наличие капсулы может, в частности, задерживать или предотвращать разрушение микроорганизма в желудочно-кишечном тракте. Следует иметь в виду, что композиции согласно вариантам реализации настоящего изобретения могут быть заключены в капсулы или таблетки с контролируемым по времени высвобождением, покрытые кишечнорастворимой оболочкой. Кишечнорастворимое покрытие позволяет капсуле/таблетке сохраняться в неизменном виде (т.е. в нерастворенном виде) по мере ее прохождения через желудочно-кишечный тракт до тех пор, пока она не достигнет тонкого кишечного. Способы инкапсулирования живых бактериальных клеток хорошо известны в данной области техники (см., например, патенты США General Mills Inc., такие как патент США №6723358). Например, микроинкапсулирование с альгинатом и крахмалом Hi-Maize™ с последующей лиофилизацией было признано эффективным для пролонгирования срока хранения бактериальных клеток в молочных продуктах [см., например, Kailasapathy et al. Curr Issues Intest Microbiol. 2002 September; 3(2):39-48]. Альтернативно, инкапсулирование можно осуществлять с помощью глюкоманнановых волокон, таких как экстрагированные из Amorphophallus konjac. Альтернативно, также можно использовать удержание жизнеспособных пробиотиков в эмульсиях на основе кунжутного масла [см., например, Hou et al. J. Dairy Sci. 86:424-428]. Согласно некоторым вариантам реализации изобретения, агенты кишечнорастворимых покрытий предпочтительно представляют собой сополимеры метилакриловой кислоты и алкилакрилата, такие как полимеры Eudragit®. Было показано, что поли(мет)акрилаты, в частности, являются подходящими в качестве материалов покрытий. EUDRAGIT® представляет собой торговое название для сополимеров, происходящих из сложных эфиров акриловой и метилакриловой кислоты, свойства которых определяются функциональными группами. Отдельные виды EUDRAGIT® различаются по соотношению нейтральных, основных или кислотных групп и, таким образом, по физико-химическим свойствам. Профессиональное применение и комбинация различных полимеров EUDRAGIT® дате возможность получения идеальных растворов для контролируемого высвобождения лекарственного средства для различных фармацевтических и технических применений. EUDRAGIT® обеспечивает функциональные пленки для покрытий таблеток и пеллетов с замедленным высвобождением. Полимеры описаны в международных фармакопеях, таких как Ph.Eur., USP/NF, DMF и JPE. Полимеры EUDRAGIT® могут обеспечивать следующие возможности для лекарственного средства с контролируемым высвобождением: нацеливание на область желудочно-кишечного тракта (желудочная устойчивость, высвобождение в толстом кишечнике), защитные покрытия (маскирование вкуса и запаха, защита от влажности) и задержание высвобождение лекарственного средства (лекарственные формы с замедленным высвобождением). Полимеры EUDRAGIT® доступны в широком диапазоне различных концентрации и физических форм, включая водные растворы, водную дисперсию, органические растворы и твердые вещества. Фармацевтические свойства полимеров EUDRAGIT® определяются по химическим свойствам их функциональных групп. Различают:
- поли(мет)акрилаты, растворимые в улучшающих пищеварение жидкостях (путем образования соли): полимеры EUDRAGIT® L (сополимер метакриловой кислоты), EUDRAGIT® S (сополимер метакриловой кислоты), полимеры EUDRAGIT® FS и Е (основный бутилированный сополимер метакрилата) с кислотными или основными группами обеспечивают pH-зависимое высвобождение активного ингредиента. Применение: от простого маскирования вкуса за счет устойчивости к воздействию желудочного сока до контролируемого высвобождения лекарственного средства во всех отделах кишечника.
- поли(мет)акрилаты, нерастворимые в улучшающих пищеварение жидкостях: полимеры EUDRAGIT® RL и RS (сополимеры аммония и метакрилата) с основными группами и полимеры EUDRAGIT® NE с нейтральными группами обеспечивают контролируемое по времени высвобождение активного вещества за счет независимого от pH набухания.
Кишечнорастворимые покрытия EUDRAGIT® предотвращают высвобождение лекарственного средства в желудке и обеспечивают контролируемое высвобождение в кишечнике. Основным критерием для высвобождения является pH-зависимое растворение покрытия, которое происходит в определенном отделе кишечника (pH от 5 до более 7) в отличие от желудка (pH 1-5). Для указанного применения анионные типы EUDRAGIT®, содержащие карбоксильные группы, можно смешивать друг с другом, что обеспечивает возможность тонкой регуляции pH растворения и, таким образом, определения сайта высвобождения лекарственного средства в кишечнике. EUDRAGIT® L и S типа подходят для кишечнорастворимых покрытий. EUDRAGIT® FS 30 D (водная дисперсия анионного сополимера на основе метилакрилата, метилметилакрилата и метилакриловой кислоты) используется для специфичного контролируемого высвобождения в толстом кишечнике.
Как правило, пробиотический штамм бактерий согласно настоящему изобретению вводят указанному субъекту в виде пищевой композиции. Соответственно, еще один аспект настоящего изобретения относится к пищевой композиции, содержащей количество пробиотического штамма бактерий согласно настоящему изобретению.
Согласно некоторым вариантам реализации изобретения, пищевая композиция, содержащая пробиотический штамм бактерий согласно настоящему изобретению, выбрана из полноценной пищевой композиции, пищевых добавок, нутрицевтических композиций и т.д. Композицию согласно настоящему изобретению можно применять в качестве пищевого ингредиента и/или кормового ингредиента.
Пищевой ингредиент может быть представлен в виде раствора или в виде твердого вещества в зависимости от применения и/или способа применения и/или способа введения.
Пробиотический штамм бактерий согласно настоящему изобретению, как правило, добавляют в любое время в процессе получения композиции, например, добавляют к пищевой основе в начале процесса получения или к конечному пищевому продукту.
Термин «пищевой продукта» относится к жидкой (т.е. напитку), твердой или полутвердой питательной композиции, в частности, полноценной пищевой композиции (заместителю пищевого продукта), которая не требует дополнительного употребления питательных веществ или композиций пищевых добавок. Композиции пищевых добавок не полностью заменяют прием питательных веществ другими способами. Пищевые продукты и композиции пищевых добавок представляют собой, например, кисломолочные продукты или молочные продукты, которые предпочтительно вводят или принимают перорально один или более раз в день. Кисломолочные продукты могут быть получены непосредственно с использованием бактерий согласно изобретению в процессе изготовления, например, путем добавления к пищевой основе, с использованием хорошо известных способов. В таких способах штамм (штаммы) согласно изобретению можно применять для добавления к обычно используемым и/или для замещения одного или более или части обычно используемых микроорганизмов. Например, при приготовлении кисломолочных продуктов, таких как йогурт или йогуртные напитки, бактерию согласно изобретению можно добавлять или использовать как часть заквасочной культуры или подходящим образом добавлять в процессе такой ферментации. Необязательно бактерии могут быть инактивирвоанными или убитыми позже в процессе получения. Кисломолочные продукты включают молочные продукты, такие как десерты, йогурты, йогуртные напитки, творог, кефир, ферментированные молочные напитки, молочные коктейли, сыры, заправки, пасты с низким содержанием жира, молодой сыр, соевые напитки, мороженое и т.д., но не ограничиваются указанными. Альтернативно, пищевые продукты и/или композиции пищевых добавок могут представлять собой не молочные или молочными не ферментированные продукты (например, штаммы или бесклеточная среда в неферментированном молоке или в другой пищевой среде). Согласно некоторым вариантам реализации изобретения, пробиотический штамм бактерий согласно настоящему изобретению инкапсулируют и диспергируют в пищевом продукте (например, в молоке) или непищевой среде. Не кисломолочные продукты могут включать мороженое, питательные батончики и заправки и т.д. Не молочные продукты могут включать порошкообразные смеси для приготовления напитков и питательные батончики и т.д. Продукты могут быть получены с использованием известных способов, таких как добавление эффективного количества штамма (штаммов) и/или бесклеточной культуральной среды к пищевой основе, такой как обезжиренное молоко или молоко или молочный состав и ферментация, как известно. Другие пищевые основы, к которым можно добавлять композиции, содержащие бактериальные клетки и/или бесклеточную культуральную среду, представляют собой мясо, заместители мяса или растительные основы.
Композиция, которая содержит пробиотический штамм бактерий согласно настоящему изобретению, может быть твердой, полутвердой или жидкой. Она может быть представлена в виде пищевого продукта или пищевой добавки, например, в виде таблеток, гелей, порошков, капсул, напитков, батончиков и т.д. Например, композиция может быть представлена в виде порошка, упакованного в сашет, который можно растворять в воде, фруктовом соке, молоке или другом напитке.
При употреблении в настоящей заявке термин «пищевой ингредиент» или «кормовой ингредиент» включает лекарственную форму, которая добавлена или может быть добавлена к функциональным пищевым продуктам или продовольственным продуктам в качестве питательной добавки.
Термин «обогащенный пищевой продукт» или «нутрицевтик» или «функциональный пищевой продукт» означает продовольственный продукт, который содержит ингредиенты, оказывающие благоприятные эффекты на здоровье или способные улучшать физиологические функции.
Термин «пищевая добавка» означает продукт питания, направленный на восполнение нормальной пищевого рациона. Пищевая добавка представляет собой концентрированный источник питательных веществ или других веществ, оказывающая питательный или физиологический эффект при ее отдельном приеме или в виде комбинации в малых количествах.
Согласно изобретению, термин «функциональные пищевые продукты» объединяет продукты питания и соответствующие недавно разработанные продукты, важность которых обусловлена не только их питательной и вкусовой ценностью, но и особыми ингредиентными веществами. Согласно изобретению, важное значение имеют среднее и длительное поддержание и обеспечение здоровья. В этом контексте, не терапевтическое применение является предпочтительным. Термины «нутрицевтики», «обогащенные пищевые продукт» и «оригинальные пищевые продукты», которые также представляют собой варианты реализации изобретения, отчасти являются синонимами, однако употребляются также избирательно. Однако профилактический аспект и обеспечение здоровья, а также пищевые характеристики продуктов дают наиболее ясное определение термина «функциональный пищевой продукт». Во многих случаях этот термин связан с продуктами, которые получают путем сортинга и селекции (как и в случае настоящего изобретения), очистки, концентрации, а также в большой степени путем добавления. Выделенные эффективные вещества, в частности, в форме таблеток или пилюлей, не включены. Несмотря на отсутствие официального определения функциональных пищевых продуктов, большинство групп, имеющих отношение к данной области, соглашается, что они представляют собой продукты, реализуемые на рынке как оказывающие специфические эффекты на здоровье субъекта помимо основных питательных свойств. Соответственно, функциональные пищевые продукты представляют собой обычные пищевые продукты, в которые включены компоненты или ингредиенты (такие как описанные в настоящей заявке), придающие указанному пищевому продукту специфическую функциональность, например, медицинский или физиологический благоприятный эффект, отличный от чисто питательного эффекта.
Согласно некоторым вариантам реализации изобретения, напиток представляет собой функциональный напиток или терапевтический напиток, утолитель жажды или обычный напиток. Композицию согласно настоящему изобретению можно использовать, например, в качестве ингредиента для прохладительных напитков, фруктового сока или напитка, содержащего белок молочной сыворотки, оздоровительного чая, какао-напитков, молочных напитков и молочной кислоты бактерий напитки, йогурта и питьевого йогурта, сыра, мороженого, фруктового льда и десертов, кондитерских изделий, бисквитных пирожных и брикетов для кексов, закусочных продуктов, сбалансированных пищевых продуктов и напитков, фруктовых наполнителей, саге глазури, шоколадного наполнителя для хлебобулочных изделий, вкусового наполнителя для творожного пирога, наполнителя для пирожных со вкусом фруктов, глазури для пирожных и пончиков, крема для начинок быстрого приготовления, наполнителей для печенья, готовых к употреблению наполнителей для хлебобулочных изделий, низкокалорийных наполнителей, питательного напитка для взрослых, подкисленного соевого напитка/напитка на основе сока, асептического/стерилизованного шоколадного напитка, смесей для батончиков, порошков для приготовления напитков, обогащенного кальцием соевого молока/молока без добавок и шоколадного молока, обогащенного кальцием кофейного напитка.
Композиция может также применяться в качестве ингредиента пищевых продуктов, таких как американский сырный соус, антислеживающий агент для тертого сыра и сыра в нарезке, чип-дип, сливочный сыр, сухие смеси обезжиренной сметаны для взбивания, полученные замораживанием/оттаиванием молочные сливки для взбивания, полученные замораживанием/оттаиванием стабильные сливки для взбивания, легкий натуральный сыр чеддер с низким содержанием жира, швейцарский йогурт с низким содержанием жира, взбитые замороженные десерты, мороженое в твердой упаковке, мороженое в твердой упаковке с маркировкой безопасности, увеличенной экономичности и гарантии качества, мороженое низкой жирности: мягкое мороженое, соус для барбекю, соус с приправой из сыра, приправа к домашнему сыру, соус Альфредо в виде сухой смеси, смешанный сырный соус, томатный соус в виде сухой смеси и другие.
Согласно некоторым вариантам реализации изобретения, композиция, которая содержит пробиотический штамм бактерий согласно настоящему изоберетнию, применяется с йогуртной продукцией, такой как ферментированный йогуртный напиток, йогурт, питьевой йогурт, сыр, ферментированный крем, молочные десерты и другие. Подходящим образом, композиция также может использоваться в качестве ингредиента в одном или более из продуктов для употребления сыра, мяса, или продуктов, содержащих защитные культуры.
Согласно некоторым вариантам реализации изобретения, пищевая композиция, которая содержит пробиотический штамм бактерий согласно настоящему изобретению, подходит для получения заменителя пищевого продукта. При употреблении в настоящей заявке термин «заменитель пищевого продукта» включает любой питательный продукт, содержащий белки, углеводы, жиры, витамины и минералы, комбинация которых является подходящей в качестве единственного или главного источника питательных веществ для пищевого продукта, если иное специально не указано. Как правило, заменитель пищевого продукта содержит по меньшей мере один источник углеводов, по меньшей мере один источник липидов и/или по меньшей мере один источник белков. В качестве источника белков может использоваться любой подходящий пищевой белок, например, животные белки (такие как молочные белки, мясные белки и яичные белки); растительные белки (такие как соевый белок, пшеничный белок, рисовый белок и гороховый белок); смеси свободных аминокислот или их комбинации. В частности, молочные белки, такие как казеин и белок молочной сыворотки, а также соевые белки являются предпочтительными. Белки могут быть интактными или гидролизированными или представлять собой смесь интактных и гидролизированных белков. Может быть желательно обеспечивать частично гидролизированные белки (степень гидролиза между 2 и 20%), например, для животных, которые находятся в группе риска развития аллергии на коровье молоко. При необходимости гидролизированных белков можно по желанию проводить процесс гидролиза, как известно в данной области техники. Например, белковый гидролизат молочной сыворотки можно получить путем ферментативного гидролиза фракции молочной сыворотки за один или более этапов. Показано, что если фракция молочной сыворотки, используемая в качестве исходного материала, по существу не содержит лактозу, то белок намного меньше страдает от «лизиновой блокады» во время процесса гидролиза. Это обеспечивает снижение степени «лизиновой блокады», составляющее от приблизительно 15% по массе общего лизина до менее чем приблизительно 10% по массе лизина; например, приблизительно 7% по массе лизина, что существенно улучшает пищевую ценность источника белка. Если композиция содержит источник жира, то он предпочтительно обеспечивает от 5% до 40% энергии композиции; например, от 20% до 30% энергии. Подходящий профиль содержания жира может быть достигнут с использованием смеси канолового масла, кукурузного масла и подсолнечного масла с высоким содержанием олеиновой кислоты. Источник углеводов предпочтительно обеспечивает от 40% до 80% энергии композиции. Можно использовать любой подходящий углевод, например, сахарозу, лактозу, глюкозу, фруктозу, сухую кукурузную патоку, мальтодекстрины и их смеси. Как правило, ежедневное замещение продуктов одного приема пищи на низкокалорийные продукты с заменителем пищи способствует поддержанию массы тела после уменьшения массы тела.
Пищевая композиция, которая содержит пробиотический штамм бактерий согласно настоящему изобретению, как правило, содержит носители или наполнители. Термины «носители» или «наполнители» означают материалы, подходящие для введения, и включают любой такой материал, известный в данной области техники, такой как, например, любая жидкость, гель, растворитель, жидкий разбавитель, солюбилизатор и т.п., который является нетоксичным и не приводит к неблагоприятному взаимодействию с любыми компонентами композиции. Примеры приемлемых носителей для питательных продуктов включают, например, воду, солевые растворы, спирт, силикон, воска, вазелиновое масло, растительные масла, полиэтиленгликоли, пропиленгликоль, липосомы, сахара, желатин, лактозу, амилозу, стеарат магния, тальк, поверхностно-активные вещества, кремниевую кислоту, вязкий парафин, парфюмерное масло, моноглицериды и диглицериды жирной кислоты, петролейные эфиры жирных кислот, гидроксиметилцеллюлозу, поливинилпирролидон и т.д.
Согласно некоторым вариантам реализации изобретения, пищевая композиция, которая содержит пробиотический штамм бактерий согласно настоящему изобретению, содержит некоторое количество пищевых волокон. Пищевые волокна проходят по малому кишечнику в непереваренном ферментами виде и функционируют как природные объемообразующие агенты и слабительное. Пищевое волокно может быть растворимым или нерастворимым. В целом, смесь двух типов является предпочтительной. Подходящие источники пищевых волокон включают сою, горох, овес, пектин, гуаровую камедь, гуммиарабик, фруктоолигосахариды, галактоолигосахариды, сиалиллактозу и олигосахариды, происходящие из животного молока. Согласно некоторым вариантам реализации изобретения, пищевое волокно выбрано из маннанов. Маннаны (напроимер, глюкоманнаны и галактоманнаны), такие как гуаровая камедь, камедь бобов рожкового дерева, конжаковая и ксантановая камедь, присутсвуют в клеточных стенках некоторых растений. Глюкоманнаны в целом состоят из (1-4)-β-связанных глюкозных и маннозных единиц, тогда как галактоманнаны в целом состоят из (1-4)-β-маннанового каркаса, содержащего в качестве заместителя одну (1-6)-α-галактозную единицу. Многие бобовые с эндоспермными семенами, такие как гуар и рожковое дерево, содержат галактоманнаны в эндосперме во время развития семени. Глюкоманнаны также могут встречаться в качестве минорного компонента в зернах злаков.
Согласно некоторым вариантам реализации изобретения, пищевая композиция, которая содержит пробиотический штамм бактерий согласно настоящему изобретению, содержит минералы и микропитательные вещества, такие как следовые элементы и витамины, в соответствии с рекомендациями государственных органов, такими как рекомендованные суточные нормы США (USRDA). Например, композиция может содержать один или более из следующих микропитательных веществ в приведенных диапазонах на суточную дозу: от 300 до 500 мг кальция, от 50 до 100 мг магния, от 150 до 250 мг фосфора, от 5 до 20 мг железа, от 1 до 7 мг цинка, от 0,1 до 0,3 мг меди, от 50 до 200 мкг йода, от 5 до 15 мкг селена, от 1000 до 3000 мкг бета-каротина, от 10 до 80 мг витамина С, от 1 до 2 мг витамина В1, от 0,5 до 1,5 мг витамина В6, от 0,5 до 2 мг витамина В2, от 5 до 18 мг ниацина, от 0,5 до 2,0 мкг витамина В12, от 100 до 800 мкг фолиевой кислоты, от 30 до 70 мкг биотина, от 1 до 5 мкг витамина D, от 3 до 10 мкг витамина Е.
Согласно некоторым вариантам реализации изобретения, композиция, которая содержит пробиотический штамм бактерий согласно настоящему изобретению, содержит эмульгаторы. Примеры пищевых эмульгаторов, как правило, включают сложные диацетиловые эфиры моно- и диглицеридов винной кислоты, лецитин и моно- и диглицериды. Подобным образом, могут быть включены подходящие соли и стабилизаторы.
Согласно некоторым вариантам реализации изобретения, пищевая композиция, которая содержит пробиотический штамм бактерий согласно настоящему изобретению, содержит по меньшей мере один пребиотик. Термин «пребиотик» означает пищевые вещества, предназначенные для обеспечения роста пробиотического штамма бактерий согласно настоящему изобретению в кишечнике. Пребиотик может быть выбран из группы, состоящей из олигосахаридов и необязательно включает фруктозу, галактозу, маннозу, сою и/или инулин и/или пищевые волокна.
Согласно некоторым вариантам реализации настоящего изобретения, композиция, содержащая пробиотический штамм бактерий согласно настоящему изобретению, включает защитные гидроколлоиды (такие как камеди, белки, модифицированные крахмалы), связующие вещества, образующие пленку агенты, инкапсулирующие агенты/материалы, материалы стенки/оболочки, матричные соединения, покрытия, эмульгаторы, поверхностно-активные агенты, солюбилизирующие агенты (масла, жиры, воска, лецитины и т.д.), адсорбенты, носители, наполнители, сопутствующие соединения, диспергирующие агенты, увлажнители, вспомогательные вещества, используемые в производственном процессе, растворители, агенты, обеспечивающие текучесть, маскирующие вкус агенты, утяжеляющие агенты, желирующие агенты, гелеобразующие агенты, антиоксиданты и противомикробные агенты. Композиция также может содержать стандартные фармацевтические добавки и адъюванты, вспомогательные вещества и разбавители, включая, но не ограничиваясь указанными, воду, желатин любой природы, растительные камеди, лигнинсульфонат, тальк, сахара, крахмал, гуммиарабик, растительные масла, полиалкиленгликоли, ароматизаторы, консерванты, стабилизаторы, эмульгаторы, буферы, скользящие вещества, красители, смачивающие агенты, наполнители и т.д. Во всех случаях такие дополнительные компоненты выбирают с учетом их пригодности для предполагаемого реципиента.
Согласно некоторым вариантам реализации изобретения, введение белка ClpB (или пробиотического штамма бактерий, экспрессирующего указанный белок) повторяют, например, от 2 до 3 раз в день в течение одного дня или более и в целом в течение продолжительного периода, составляющего по меньшей мере 4 дня или даже от 4 до 15 недель, с одним или более перерывами, если приемлемо. Согласно некоторым вариантам реализации изобретения, белок ClpB вводят одновременно или последовательно с одним приемом пищи субъектом. Согласно некоторым вариантам реализации изобретения, белок ClpB вводят до принятия пищи субъектом.
При употреблении в настоящей заявке термин «эффективное количество» относится к количеству белка ClpB, достаточному для достижения благоприятного эффекта (например, стимуляции насыщения, продления чувства сытости, снижения потребления пищи, контроля увеличения массы тела и/или стимуляции уменьшения массы тела). В контексте настоящего изобретения, количество белка ClpB, которое вводят субъекту, зависит от характеристик указанного субъекта, таких как общее состояние здоровья, возраст, пол и масса тела. Специалист в данной области техники способен определить подходящие дозы в зависимости от указанных и других факторов. Например, когда белок ClpB вводят указанному субъекту в виде пробиотика, штамм согласно настоящему изобретению, способный создавать колонию, будет является достаточным для обеспечения благоприятного эффекта у субъекта. Если пробиотический штамм бактерий вводят в виде пищевого продукта, то указанный пищевой продукт, как правило, содержит между 103 и 1012 КОЕ пробиотического штамма бактерий согласно настоящему изобретению на грамм сухой массы пищевой композиции.
Изобретение также проиллюстрировано следующими фигурами и примерами. Однако следует понимать, что указанные примеры и фигуры никоим образом не ограничивают объем настоящего изобретения.
ФИГУРЫ:
Фигура 1: Эффекты ежедневной внутрижелудочной доставки E.coli K12 дикого типа (WT) или E.coli с делецией гена ClpB (ΔClpB) нормальным взрослым мышам C57Bl6 на массу тела, 24-часовое потребление пищи и режим питания, исследуемые в начале и в конце 3-недельного принудительного кормления. Контрольные мыши (Ctr) не получали какого-либо принудительного кормления. Режим питания оценивали по объему порции пищи, соответствующему среднему количеству пищи, съеденной во время однократного приема пищи, и по частоте приемов пищи, соответствующей количеству принятия пищи за 24 ч, отстоящих друг от друга по меньшей мере на 5 мин. Снижение объема порции пищи отражает более быстрое насыщение, а снижение частоты приема пищи отражает более продолжительное чувство сытости. А. Двухфакторный анализ ANOVA с повторными измерениями, апостериорный критерий Бонферрони, *p<0,05. B, D, G, H, ANOVA, апостериорные критерии Тьюки *p<0,05, **p<0,01, ***p<0,001. Е. t-критерий Стьюдента, *p<0,05.
ПРИМЕР 1:
Материал и способы
Рост E.coli in vitro после регулярного снабжения питательными веществами
Бактерии E.coli K12 культивировали при 37°С в 40 мл среды МН (Becton, Дикинсон, MD), содержащей 30% мясного бульона, 1,75% гидролизата казеина и 0,15% крахмала с pH 7,3 при 25°С в пробирках Falcon на 50 мл. Для моделирования двух предусмотренных графиком ежедневных приемов пищи у людей бактериям обеспечивали свежую среду МН каждые 12 ч в течение 5 последовательных дней. Рост бактерий измеряли как OD при λ=600 нм с помощью спектрофотометра каждые 2 ч после 1-го предоставления среды МН, каждый 1 час после 3-его и каждые 10 мин после 5-го предоставления среды. В конце каждого 12-часового цикла бактерии центрифугировали в течение 5 мин при 6,000 об./мин при комнатной температуре (RT). Супернатанты выливали и замещали эквивалентным объемом (~40 мл) свежей среды МН. После последнего добавления среды МН получали образцы бактерий для экстракции белка в экспоненциальной фазе роста и в последующей стационарной фазе роста.
Экстракция белка
Бактерии E.coli K12 центрифугировали при 4°С в течение 30 мин при скорости 4,000 g. Осадок, содержащий бактерии, растворяли в 2 мл трисгидроксиметиламинометанового буфера (ТРИС-буфера, pH 7,4) и гомогенизировали посредством обработки ультразвуком в течение 3 мин при комнатной температуре. Для отделения белков от нерастворенных фрагментов клеток гомогенат бактерий центрифугировали при 4°С в течение 30 мин при скорости 10,000 g. Супернатант восстанавливали и затем центрифугировали на ультрацентрифуге при 4°С в течение 45 мин при скорости 60,000 g для дальнейшего разделения белков на цитоплазматическую (супернатант) и мембранную (осадок) фракцию. Мембранные белки растворяли в ТРИС-буфере (pH 7,4). Концентрацию белка измеряли с использованием набора 2-D Quant (GE Healthcare, Пискатавей, Нью-Джерси).
Двумерный электрофорез в полиакриламидном геле
Для проведения двумерного электрофореза в полиакриламидном геле (2D-ПААГ) использовали 300 мкг белкового экстракта E.coli для регидратации полосок с иммобилизированным градиентом pH (IPG) (pH 4-7; 18 см; BIO-RAD, Геркулес, Калифорния). Затем белки разделяли в первом направлении путем изоэлектрического фокусирования в общем в течнеие 85,000 В-ч с использованием системы изоэлектрического фокусирования IPGphor (GE Healthcare). После фокусирования полоски IPG инкубировали в течение 15 мин в уравновешивающем буфере [мочевина, 6 моль/л, 30% (об./об.); глицерин, 2% (масс./об.); додецилсульфат натрия (ДСН); Трис-HCl, 50 ммоль/л, pH 8,8; и 0,25% (масс./об.) раствор бромфенолового синего, содержащий 2% (масс./об.) дитиотреитола] и затем алкилировали в течение 15 мин в уравновешивающем буфере, содержащем 4% (масс./об.) йодацетамида. IPG-полоски затем прикрепляли к 10%-ым полиакриламидным градиентным гелям (20 см ⋅ 18 см ⋅ 1 мм) для проведения ДСН-ПААГ. Разделение во втором направлении проводили в течение ночи в системе для вертикального электрофореза Ettan Daltsix (GE Healthcare) при силе электрического тока 12 мА/гель при температуре 25°С. После 2D-ДСН-ПААГ гели фиксировали в течение 2 ч в 2% (об./об.) ортофосфорной кислоте и в 50% (об./об.) метаноле при комнатной температуре. Затем гели промывали водой и белковые пятна визуализировали путем окрашивания красителем Кумасси бриллиантовым синим G-250 (BIO-RAD) [34% (об./об.) метанола, 17% (масс./об.) сульфата аммония, 2% (об./об.) ортофосфорной кислоты и 0,66 г Кумасси бриллиантового синего G-250 /л].
Анализ дифференциальной экспрессии белка
Изображения окрашенных гелей после двумерного электрофореза сканировали с помощью системы ImageScanner II (GE Healthcare), калибровали с помощью маркера шкалы градаций серого (Kodak, Рочестер, Нью-Йорк) и оцифровывали с помощью программой Labscan 6,0 (GE Healthcare). Анализ различающейся экспрессии белка, включающий выявление пятен, количественную оценку, выравнивание и сравнительный анализ, проводили с использованием программы 2D Platinum 5,0 (GE Healthcare). Каждый белковый образец подвергали 2D-ПААГ-электрофоретическому анализу по меньшей мере 3 раза (для мембранных белков) и 4 раза (для цитоплазматических белков) для минимизации изменений, зависимых от цикла, и каждый набор по 3 (или 4) геля сравнивали с помощью программы ImageMaster для подтверждения отсутствия статистически различных пятен в пределах набора гелей. Наиболее репрезентативный гель (миграция в геле, четкость и количество пятен) из каждого набора использовали для сравнения белков Е. Coli, полученных в экспоненциальной и стационарной фазе роста. Уровень экспрессии определяли по относительному объему каждого пятна в геле и выражали как % объем, рассчитанный как объем пятна/∑объемов всех пятен, разделенных в геле. Этот нормированный объем пятна учитывает вариации из-за нагрузки белка и окраски с учетом общего объема по всем точкам, присутствующим в геле. Различия и количество рассчитывали как отношение средних значений % объема для группы точек между 2 фазами. Только пятна с относительным различием объема >1,5 считались значимыми. Отсутствие пятен в пределах геля указывало на отсутствие выявляемой экспрессии белка при выбранных условиях эксперимента. Соответствующие p-значения определяли с помощью t-критерия Стьюдента (уровень значимости p<0,05) после логарифмического преобразования значения объема пятна.
Идентификация белков с помощью жидкостной хроматографии с ионизацией электрораспылением - МС/МС
Интересующие белковые пятна вырезали из окрашенных Кумасси бриллиантовым синим G-250 гелей после двумерного электрофореза с помощью прибора Ettan Spot Picker (GE Healthcare), и переваривание белков в геле осуществляли на автомате Ettan Digester (GE Healthcare), как было ранее описано (Goichon et al., 2011). Белковые экстракты затем ресуспендировали в 10 мкл смеси 5% (об./об.) ацетонитрил/0,1% (об./об.) муравьиная кислота и затем анализировали с помощью системы nano-LC1200, сопряженной с масс-спектрометрической ионной ловушкой 6340 Ion Trap, снабженной источником нанораспыления, и интерфейса HPLC-chip cube (Agilent Technologies, Courtaboeuf, Франция). Вкратце, пептиды концентрировали и обессаливали на предколонке (40 нл, RP-C18) и разделяли на колонке Zorbax С18 (размер пор 30 нм, размер частиц 5 мкм, 43 мм длина × 75 мкм внутренний диаметр; Agilent Technologies). Использовали 9-мин линейный градиент (3%-80% ацетонитрил в 0,1% муравьиной кислоте) при скорости потока 400 нл/мин использовали и элюент анализировали с помощью масс-спектрометрии по типу ионной ловушки.
Для идентификации белка получали перечни пиков МС/МС и сравнивали с базами данных белков с использованием поисковой системы MASCOT Daemon версии 2.2.2 (Matrix Science). Поиск осуществляли по следующим конкретным параметрам: специфичность фермента: трипсин; один пропущенный сайт расщепления допускается; нет фиксированных модификаций; переменные модификации, окисление метионина, карбамидометилирование цистеина, фосфорилирование серина, тирозин и треонина; моноизотопный режим; заряд пептида 2+ и 3+; допуск по массе для ионов-предшественников 1,5 Да; допуск по массе для ионных осколков 0,6 Да; инструмент: ESI-TRAP (масс-спектрометрия с ионизацией электрораспылением и ионной ловушкой); таксономия: Е. coli; база данных: Национальный центр биотехнологической информации (National Center for Biotechnology Information, NCBI [NCBInr 20120531 (18280215 последовательностей, 6265275233 остатков)] (Бетесда, Мэриленд). Совпадения при поиске белка автоматически подтверждались, если они удовлетворяли одному из следующих критериев: идентификация, при которой по меньшей мере два пептида высокого уровня совпадения (жирный шрифт, красный цвет) имеют балл MASCOT больше 54 (p<0,01), или по меньшей мере два пептида высокого уровня совпадения (жирный шрифт, красный цвет) с баллом MASCOT более 47 (p<0,05). Для оценки ложноположительных баллов, все исходные поиски в базе данных осуществляли с использованием опции «ловушка» MASCOT. Результаты считали надежными, если частота ложноположительных совпадений никогда не превышала 1%.
Анализ АТФ
Продукцию АТФ in vitro измеряли с использованием набора для колориметрического/флуориметрического анализа АТФ в соответствии с инструкциями производителя (BioVision, Калифорния). Вкратце, бактериальные белки, полученные в экспоненциальной или стационарной фазе роста, наносили в ряд лунок в двух повторностях для каждой концентрации (1, 10 и 25 мкг/мл в аналитическом буфере для АТФ) и объем доводили до 50 мкл на лунку с помощью аналитического буфера для АТФ. Затем в соответствующие лунки добавляли по 10 мкл раствора различных питательных веществ, 15% сахарозу или среду МН и объем доводили до 50 мкл на лунку с помощью аналитического буфера для АТФ; в контрольные лунки добавляли только буфера для АТФ в объеме 50 мкл на лунку. Планшет инкубировали в течение 2 ч при 37°С. После инкубации в каждую лунку добавляли по 50 мкл реакционной смеси АТФ (содержащей аналитический буфер для АТФ, пробы АТФ, преобразующую и проявляющую смесь для АТФ). Оптическую плотность (OD) измеряли при длине волны 570 нм через 30 мин инкубирования при комнатной температуре в защищенном от дневного света месте.
Разработка и валидация иммуноанализа ClpB
Дизайн анализа для выявления ClpB был основан на нескольких критериях, таких как специфичность и чувствительность выявления в диапазоне линейных концентраций. Определенным условием было выявление ClpB без перекрестной реактивности с α-MSH, которая может возникать из-за наличия α-MSH-подобного эпитопа (эпитопов) в молекуле ClpB. Для облегчения проведения процедуры и выявления сигнала авторы использовали стандартный 96-луночный планшет для ИФА и возможность считывания OD с помощью спектрофотометра. Подробные протоколы ИФА и анализа с помощью Вестерн-блоттинга (WB) ClpB представлены в отдельных разделах.
Для предотвращения связывания «проявляющих» антител (Ab) с «захватывающими» антителами, авторы изобретения получали захватывающие и выявляющие ClpB антитела в различных видах - кроликах и мышах соответственно. Для наиболее эффективного захвата белка ClpB из сложных биологических образцов авторы изобретения покрывали планшет для ИФА поликлональными антителами кролика, имеющими множество эпитопов против ClpB. Для предотвращения перекрестной реактивности между ClpB и α-MSH авторы изобретения использовали в качестве «выявляющих» антител моноклональные антитела мыши против ClpB, которые характеризовались высокий чувствительностью и специфичностью распознавания ClpB по сравнению с α-MSH и были заранее выбраны с помощью ИФА-скрининга нескольких клонов антител. Конъюгированные со щелочной фосфатазой «проявляющие» антитела против антител мыши использовали в качестве общего подхода для ИФА для обеспечения хромогенной ферментативной реакции, регистрируемой как OD, пропорциональная концентрации анализируемого вещества. Получали линейное изменение OD, являющееся результатом анализа 7 последовательных разведений рекомбинантного белка ClpB E.coli, варьирующих от 2 пкМ до 150 пкМ, без достижения плато и без насыщения сигнала OD.
Для того чтобы подтвердить специфичность разработанного анализа ClpB, авторы изобретения измерили концентрацию ClpB в образцах белков, экстрагированных из 10 различных культур бактерий E.coli K12 WT и из культуры мутантного штамма ΔClpB E.coli. Мутантный штамм ΔClpB и соответствующие штаммы дикого типа (WT) были любезно предоставлены доктором Алексом Могком (Dr. Axel Mogk, ZMBH, Гейдельбергский университет, Германия). Более того, исследовали наличие ClpB в указанных образцах бактериальных белков с помощью WB с использованием поликлональных антител кролика против ClpB и сравнивали значения интенсивности сигнала WB-полос и концентрацию ClpB в ИФА. ClpB был выявлен во всех культурах E.coli дикого типа, при этом в 7 культурах концентрация ClpB была выше 1000 пкМ, тогда как ClpB не выявлялся в образце белков, экстрагированных из штамма ΔClpB E.coli. WB-анализ выявил большую полосу с ожидаемым размером 96 кДа у бактерий дикого типа, но не у бактерий ΔClpB E.coli. Уровень OD указанных полос варьировал между отдельными образцами и имел прямую корреляцию с концентрацией ClpB, измеренной с помощью ИФА в тех же образцах культур E.coli. Таким образом, отсутствие выявления ClpB в препаратах белков ΔClpB E.coli подтвердило специфичность анализов, а хорошее соответствие между результата ИФА и WB-анализа обеспечило перекрестную валидацию обоих методов иммунологического выявления ClpB.
Для того чтобы подтвердить, что анализ ClpB в плазме выявляет ClpB, произошедший из кишечных бактерий, авторы изобретения использовали ИФА ClpB для измерения ClpB в плазме мышей, которым в течение 3 недель ежедневно вводили путем внутрижелудочного кормления WT E.coli или ΔClpB E.coli. Образцы плазмы были доступны от предыдущего опубликованного авторами исследования (Tennoune et al., 2014). Авторы изобретения обнаружили, что ClpB в норме присутствовал в плазме крови мышей, включая как контрольных животных, так и мышей, получавших принудительное кормление культуральной жидкостью без бактерий. Важно, что уровень ClpB в плазме был повышен у мышей, получавших WT E.coli, но не изменялся у мышей, которым вводили E.coli с дефицитом ClpB, что подтверждает происхождение плазматического ClpB от кишечных бактерий.
ИФА ClpB
Поликлональные антитела кролика против ClpB E.coli (производится на заказ компанией Delphi Genetics, Госсели, Бельгия), наносили на 96-луночные планшеты Maxisorp (Nunc, Рочестер, Нью-Йорк) в объеме 100 мкл и концентрации 2 мкг/мл в 100 мМ буфере NaHCO3, pH 9,6 на 12 ч при 4°С. Планшеты промывали (5 мин × 3) в фосфатно-солевом буфере (ФСБ), содержавшем 0,05% Tween 20, pH 7,4. Рекомбинантный белок E.coli ClpB (производится на заказ компанией Delphi Genetics), разводили серийно до 5, 10, 25, 50, 70, 100 и 150 пкМ в буфере для образцов (ФСБ, азид натрия 0,02%, pH 7,4) и добавляли в лунки в двух повторностях для получения стандарта. Анализируемые образцы включали: образцы слизистой оболочки толстого кишечника и образцы плазмы мышей и крыс или белки, экстрагированные из культур E.coli K12. Анализируемые образцы добавляли в оставшиеся лунки в двух повторностях и инкубировали в течение 2 ч при комнатной температуре. Планшеты промывали (5 мин × 3) ФСБ, содержащим 0,05% Tween 20, pH 7,4. В лунки добавляли моноклональные антитела мыши против ClpB E.coli (1:500 в буфере для образца, производится на заказ компанией Delphi Genetics) и инкубировали в течение 90 мин при комнатной температуре. Планшеты промывали (5 мин × 3) в ФСБ, содержавшем 0,05% Tween 20, pH 7,4. В лунки добавляли антитела козы против антитела IgG мыши, конъюгированные со щелочной фосфатазой (1:2000 в буфере для образца) от компании Jackson ImmunoResearch Laboratories, Inc. (Вест Гров, Пенсильвания) и инкубировали в течение 90 мин при комнатной температуре. Планшеты промывали (5 мин × 3) в ФСБ 0,05%, содержавшем Tween 20, pH 7,4 и затем добавляли 100 мкл раствора п-нитрофенилфосфата (Sigma, Сент-Луис, Миссури) в качестве субстрата для щелочной фосфатазы. Через 40 мин инкубации при комнатной температуре реакцию останавливали путем добавления 50 мкл 3 н раствора NaOH. OD определяли при длине волны 405 нм с использованием ридера для микротитрационных планшетов Metertech 960 (Metertech Inc., Тайбэй, Тайвань). Значения OD для пустых лунок, полученные в результате считывания планшетов без добавления образцов плазмы или стандартных разведений белка ClpB, вычитали из значений OD для образцов.
Анализ ClpB с помощью Вестерн-блоттинга
Проводили анализ белков, экстрагированных из E.coli K12, с помощью Вестерн-блоттинга. Образцы белков (10 мкг) разделяли в 20% акриламидном геле, содержавшем ДСН, в Трис-глициновом буфере и переносили на нитроцеллюлозную мембрану (GE Healthcare, Орсей, Франция), которую блокировали в течение по меньшей мере 1 ч при комнатной температуре с помощью 5% (масс./об.) обезжиренного сухого молока в TBS (10 ммоль/л трис, pH 8; 150 ммоль/л NaCl) и 0,05% (масс./об.) Tween 20. Затем мембрану инкубировали в течение ночи при 4°C с поликлональными антителами кролика против ClpB E.coli (1:2000, Delphi Genetics). После трехкратной отмывки в блокирующем растворе, содержавшем 5% (масс./об.) обезжиренное сухое молоко в TBS/0,05% Tween 20, мембраны инкубировали в течение 1 ч с конъюгированными с пероксидазой IgG против антител кролика (1:3000, SantaCruz Biotechnology). После трехкратной отмывки пероксидазную реакцию выявляли с использованием набора для выявления ECL (GE Healthcare). Белковые полосы сравнивали со стандартом молекулярных масс (Precision Plus, BioRad), мембраны сканировали с использованием ImageScanner III (GE Healthcare) и анализировали плотность пикселей полос с использованием программы ImageQuant TL версии 7,0 (GE Healthcare).
Кишечное введение белков Е. coli крысам
Животные
Уход и эксперименты на животных проводили в соответствии с нормами, установленными Национальными институтами здравоохранения США и в соответствии правилами Французского и Европейского сообщества (Официальный журнал Европейского сообщества L 358, 18/12/1986). Самок крыс Sprague-Dawley с массой тела 200-250 г (Janvier, Женест-сент-Иль, Франция) содержали в закрытых клетках (по 3 крысы на клетку) в полностью оборудованном виварии с регулируемыми условиями окружения (температура 22±1°С, 12-часовой светотемновой цикл при включении света в 7:30 утра) в течение 1 недели для их акклиматизации в условиях содержания. Животным предоставляли свободный доступ к стандартному гранулированному корму для грызунов (корм RM1, SDS, Великобритания) и питьевой воде.
Эксперимент #1
Данный эксперимент был разработан для оценки соответствия предложенной авторами модели in vitro роста E.coli картине бактериального роста in vivo в кишечнике. Крысам проводили анестезию раствором кетамин (75 мг/кг, Virbac, Каррос, Франция) / ксилазин (5 мг/кг, Bayer, Леверкузен, Франция) в объеме 3:1, 0,1 мл/100 г массы тела, внутрибрюшинно. После лапаратомии толстый кишечник фиксировали путем наложения 2 лигатур: 1-ая в месте соединения слепой и толстого кишки и 2-ая на 4 см ниже. Инфузию толстого кишечника и взятие образцов содержимого просвета проводили с использованием полипропиленового катетера, помещенного в восходящую ободочную кишку и фиксированного с помощью 1-ой лигатуры. 2 мл среды МН или воды осторожно вливали в толстый кишечник и сразу после этого выпивали для измерения OD. После измерения OD образец содержимого толстого кишечника возвращали обратно. Такое взятие образцов содержимого толстого кишечника без добавления новой среды МН или воды повторяли каждые 5 мин во время первых 20 мин и затем через 30 мин и 60 мин. Плотность бактерий измеряли как OD при λ=600 нм с помощью спектрофотометра. Образцы крови отбирали из воротной вены до начала инфузии и через 30 и 60 мин после 1-ой инфузии. Образцы фекалий отбирали из толстого кишечника в конце эксперимента для экстракции ДНК и ПЦР-анализа ClpB.
Количественная полимеразная цепная реакция в реальном времени
Количественную ПЦР (кПЦР) проводили для анализа бактериальной плотности экспрессирующих ДНК ClpB бактерий с использованием прибора CFX 96 qPCR (BioRad, Калифорния). Тотальную ДНК экстрагировали из крысиного кала с использованием копрологического мининабора QAMP DNA stool (QIAGEN Venlo, Нидерланды). Смесь кПЦР включала 5 мкл SYBR Green Master (QIAgen, Уэст-Сассекс, Великобритания), 0,5 мкМ каждого из прямого и обратного праймеров, ДНК из образцов и воду для достижения общего объема 10 мкл. Праймеры были получены из компании Invitrogen (Сержи-Понтуаз, Франция). Проводили трехстадийную ПЦР с 40 циклами. Денатурацию образцов проводили при 95°С в течение 10 мин, отжиг проводили при 60°С в течение 2 мин и удлинение проводили при 95°С в течение 15 сек.
Эксперимент #2
Цель данного эксперимента заключалась в оценке эффектов белков Е. coli на высвобождение пептидов кишечника (GLP-1 и PYY) в системный кровоток. Крысам проводили анестезию и толстый кишечник фиксировали, как описано выше, проводили вливание в толстый кишечник белков E.coli (0,1 мкг/кг белка в 2 мл ФСБ), экстрагированных в экспоненциальной фазе роста (n=6) или стационарной фазе роста (n=6) один раз в течение 20 мин. Образцы крови отбирали из воротной вены до вливания и через 20 мин после вливания для анализа GLP-1, PYY и ClpB. Образцы слизистой оболочки толстого кишечника отбирали в конце эксперимента для анализа ClpB. Анализы GLP-1 и PYY проводили с использованием набора для флуоресцентного ферментативного иммуноферментного анализа (Phoenix Pharmaceutical inc., Калифорния), в соответствии с инструкциями производителя. Флуоресценцию измеряли при 325 нм (возбуждение) и 420 нм (испускание) с использованием ридера для микротитрационных планшетов Chameleon (HIDEX Inc., Турку, Финляндия).
Введение белков Е. coli крысам, потребление пищи и исследование c-fos в головном мозге
Животные
Самцам крыс Wistar с массой тела 200-250 г (Janvier, Женест-сент-Иль, Франция) позволяли акклиматизироваться в условиях содержания. Животных кормили, как описано выше. За три дня до проведения экспериментов крыс переносили в отдельные метаболические клетки (Tecniplast, Лион, Франция), где им обеспечивали свободный доступ к пище с такой же рационом RM1, но в форме порошка (SDS). Обеспечивали свободный доступ к питьевой воде. Крыс аккуратно брали на руки ежедневно на несколько минут во время периода акклиматизации для того, чтобы приучить их к обращению. В конце периода акклиматизации крыс распределяли на три группы для достижения сходной средней массы тела и использовали в экспериментах 1-3. Два эксперимента, включающие ограничение пищи, проводили на одних и тех же крысах с интервалом в 4 дня. 3-ий эксперимент на крысах со свободным доступом к пище включал новые группы.
Эксперимент #1
Цель 1-го эксперимента заключалась в сравнении эффектов мембранных белков E.coli, экстрагированных в экспоненциальной и стационарной фазе роста. Крыс лишали пищи в течение ночи (между 18,00 ч и 10,00 ч), при этом обеспечивали свободный доступ к воде. На следующий день в 10:00 ч крысам после лишения пищи вводили внутрибрюшинно белки E.coli и животных сразу возвращали в их метаболические клетки, которые содержали заранее взвешенное количество еды. Потребление пищи измеряли через 1,2 и 4 ч. 1-ая группа крыс (n=6) получала 0,1 мг/кг мембранных белков, экстрагированных из E.coli в экспоненциальную фазу роста, в 300 мкл ФСБ; 2-ая группа крыс (n=6) получала 0,1 мг/кг мембранных белков, экстрагированных из E.coli в стационарную фазу росту, и контрольная группа (n=6) получала 300 мкл ФСБ.
Эксперимент #2
Цель 2-го эксперимента заключалась в сравнении эффектов цитоплазматических белков E.coli, экстрагированных в экспоненциальной и стационарной фазе роста. Использовали протокол эксперимента, сходный с протоколом для Эксперимента #1.
Эксперимент #3
Настоящий эксперимент был спланирован для оценки эффектов общего белка Е. coli на потребления пищи у крыс со свободным доступом к пище. Инъекции белков E.coli (0,1 мг/кг белка в 300 мкл ФСБ, внутрибрюшинно), экстрагированых в экспоненциальную фазу роста (n=6) или стационарную фазу роста (n=6), или только ФСБ в качестве контроля (n=6) проводили в 19:30 ч и животных возвращали в их метаболические клетки, которые содержали заранее взвешенное количество пищи. Суммарное потребление пищи измеряли через 2 ч. Сразу после этого крысам проводили анестезию с помощью пентобарбитала натрия (0,2 мг/кг, внутрибрюшинно) и перфузию для иммуногистохимического исследования экспрессии c-fos в головном мозге.
Препараты ткани и иммуногистохимический анализ
Головной мозг фиксировали путем перфузии/иммерсии в 4% параформальдегиде, замораживали, делали срезы (14 мкм) на криостате (Leica Microsystems, Нантер, Франция) и затем обрабатывали для иммуногистохимического анализа с использованием набора тирамидного усиления сигнала (TSA) с флуоресцеином (NEN, Бостон, Массачусетс). Для одноцветного окрашивания использовали поликлональную антисыворотку кролика против c-fos (1:4,000, Ab-5, Calbiochem, Merck KGaA, Дармштадт, Германия). Для двойного окрашивания после TSA применяли метод прямой иммунофлуоресценции с использованием моноклональнох антител кролика против β-эндорфина (β-end) 1:1,000 (Life Technologies, Фредерик, Мериленд), которые выявляли с помощью антител Cyanine-3 против антител кролика 1:200 (Jackson ImmunoResearch, Вест Грув, Пенсильвания), или моноклональных IgG мыши против связанного с геном кальцитонина пептида (CGRP) 1:1,000 (Santa Cruz Biotechnology, inc., TX), которые выявляли с помощью конъюгированных с родамином красным антител против антител мыши в разведении 1:200 (Jackson ImmunoResearch). В дугообразном и вентромедиальном ядре гипоталамуса и в центральном ядре миндалевидного тела положительные клетки считали при увеличении х20 на шести последовательных срезах. Среднее количество положительных клеток на крысу использовали для расчета среднего на группу. С помощью программы Adobe Photoshop 6,0 (Adobe Systems, Сан Джоуз, Калифорния) оптимизировали яркость и контраст цифровых изображений.
Продолжительное введение белков Е. coli у мышей
Самцов мышей линии C57Bl6 возрастом два месяца (n=32) получали из компании Janvier Labs и позволяли животным акклиматизироваться в помещении вивария в течение 1 недели при поддержании 12-часового светотемнового цикла (свет включали в 8:00). Затем мышей помещали по отдельности в клетки для мышей BioDAQ (Research Diets, Inc., Нью-Брансуик, Нью-Джерси), снабженные автоматическим регистратором потребления пищи. Через 3 дня акклиматизации в клетках BioDAQ мышей делили на три группы (n=8), каждая из которых получала разное лечение, включающее две ежедневные (в 9:00 и в 18:30) внутрибрюшинные инъекции какого-либо из следующих вариантов: (i) ФСБ, (ii) бактериальные белки, экстрагированные в экспоненциальной фазе роста (0,1 мг/кг массы тела), (iii) бактериальные белки, экстрагированные в стационарной фазе роста (0,1 мг/кг массы тела). Животные имели свободный доступ к пище (SERLAB, Монтатер, Франция) и питьевой воде, массу тела животных измеряли ежедневно. Данные потребления пищи непрерывных регистрировали в течение одной недели и анализировали с помощью средства просмотра данных BioDAQ 2.3.07 (Research Diets). Для анализа паттерна потребления пищи между приемами пищи устанавливали интервал 300 сек. Показатели чувства сытости рассчитывали как продолжительность (сек) интервала после приема пищи, деленная на количество пищи (г), потребленной за предшествующий прием пищи. После эксперимента мышей умерщвляли путем декапитации; извлекали головной мозг и ткань гипоталамуса иссекали для микроматричного анализа мРНК нейропептидов.
Микроматричный анализ мРНК нейропептидов гипоталамуса
Тотальную РНК экстрагировали из гипоталамуса мышей, которым в течение продолжительного времени вводили белки Е. coli, с использованием набора NucleoSpin® РНК II (Macherey-Nagel, Дюрен, Германия) в соответствии с протоколом производителя. Осуществляли обратную транскрипцию переваренной РНК при 42°С в течение 60 мин с использованием системы для обратной транскрипции ImProm-II™ (Promega, Мэдисон, Висконсин). Полученную кДНК использовали в реакции ПЦР в реальном времени. Панель из 9 пар праймеров была создана с помощью программы Primer express (Life Technologies, Сент-Обен, Франция) и проверена на эффективность и специфичность. Смесь для ПЦР, состоящая из 6 мкл кДНК и 6 мкл реакционной смеси Fast SYBR Green Master Mix (Life Technologies), содержащей специфичные обратные и прямые праймеры в концентрации 100 нМ, распределяли в лунки 96-луночных планшетов с помощью системы для работы с жидкостями Bravo (Agilent) и проводили амплификацию с помощью прибора QuantStudio 12K Flex (Life Technologies). Обеспечивали внутреннюю корректировку различий в количестве входной мРНК для кДНК-сигналов для генов-мишеней по сигналам референсных генов. Уровень экспрессии гена затем сравнивали с соответствующей контрольной группой образцов и изменения определяли с помощью метота 2-ΔΔCq.
Регистрация электрофизиологических показателей
Получали срезы головного мозга (250 мкм) взрослых мышей POMC-eGFP (возрастом 5-7 недель; ссылка: C57BL/6J-Tg(Pomc-EGFP)1Low/J, The Jackson Laboratory), как было описано ранее (Fioramonti et al., 2007). Срезы инкубировали при комнатной температуре в оксигенизированной внеклеточной среде, содержавшей: 118 мМ NaCl, 3 мМ KCl, 1 мМ MgCl2, 25 мМ NaHCO3, 1,2 мМ NaH2PO4, 1,5 мМ CaCl2, 5 мМ Hepes, 2,5 мМ D-глюкозу (осмолярность доведена до 310 мОсм с помощью сахарозы, pH 7,4) в течение периода восстановления, составляющего по меньшей мере 60 мин. После помещения срезов в камеру для регистрации проводили их перфузию со скоростью 2-3 мл/мин той же внеклеточной средой. Срезы анализировали с помощью микроскопа Nikon EF600 (Nikon, Шампиньи-сюр-Марн, Франция), оснащенного флуресцеиновым фильтром для флуоресцентного анализа, и ИК-дифференциально-интерференционной контрастной (ДИК) видеомикроскопии. Жизнеспособные РОМС-нейроны дугообразного ядра визуализировали с использованием объектива Х40 с водной иммерсией (Nikon) с помощью флуоресцентной видеокамеры (Nikon). Боросиликатные пипетки (4-6 MΩ; 1,5 мм OD, Sutter Instruments, Новато, Калифорния) наполняли фильтрованной внеклеточной средой. Регистрацию прикрепленных клеток проводили с использованием усилителя Multiclamp 700В, данные оцифровывали с использованием интерфейса Digidata 1440А и получали при 3 кГц с использованием программы pClamp 10,3 (Axon Instruments, Molecular Devices, Саннивейл, Калифорния). Емкостное сопротивление пипеток и клеток было полностью скомпенсировано. После того как был установлен стабильный базовый уровень 1 нМ раствор ClpB (Delphi Genetics) перфузировали в течение 5-10 минут. Частоту испускания импульсов РОМС-нейронов измеряли в течение последних 3 мин перфузии ClpB, через 7-10 мин после перфузии и сравнивали с частотой испускания импульсов, измеренной за 3 мин до начала перфузии.
Статистический анализ
Данные анализировали и строили графики с использованием программы GraphPad Prism 5,02 (GraphPad Software Inc., Сан Диего, Калифорния). Нормальность оценивали с помощью критерия Колмогорова-Смирнова. Различия между группами анализировали с помощью дисперсионного анализа (ANOVA) или непараметрического критерия Краскела-Уоллиса (K-W) с апостериорными критериями Тьюки или Данна в соответствии с результатами нормальности. Если приемлемо, отдельные группы сравнивали с использованием t-критерия Стьюдента и корреляций Пирсона или критерия Манна-Уитни (M-W) в соответствии с результатами нормальности. Эффекты в продолжительных экспериментах анализировали с использованием анализа ANOVA повторных измерений (RM) и апостериорных тестов Бонферрони. Данные представлены как среднее ± стандартная ошибка среднего (SEM). Для всех критериев p<0,05 считали статистически достоверным.
Результаты
Рост E.coli после регулярного снабжения питательными веществами
После 1-го снабжения питательной средой Мюллера-Хинтона (МН) культуры E.coli наблюдали три фазы роста бактерий: 1) латентная фаза - 2 ч; 2) экспоненциальная фаза роста - 4 ч и 3) стационарная фаза роста с оптической плотностью 0,35 (OD), которая оставалась стабильной в течение 6 ч. После 3-го и 5-го снабжения средой МН были обнаружены только две фазы роста: экспоненциальная и стационарная; 1-ая фаза начиналась сразу после снабжения питательными веществами. Каждый новый цикл обеспечения питательных веществ характеризовался более короткой продолжительностью экспоненциальной фазы роста: 2 ч после 3-го и 20 мин после 5-го снабжения. После 5-го снабжения питательными веществами экстрагировали бактериальные белки и разделяли на мембранную и цитоплазматическую фракцию, которые использовали для протеомного анализа и in vivo экспериментов на крысах, не получавших пищу. В новом эксперименте для проверки способности продолжительного регулярного снабжения питательными веществами дополнительно ускорять динамику роста бактерий E.coli K12 снабжали питательными веществами 9 раз. Было обнаружено, что после 7-го и 9-го снабжения питательными веществами экспоненциальная фаза роста больше не менялась и длилась в течение 20 мин с таким же относительным повышением (Δ0,3) OD, что отражает одинаковый рост бактерий после каждого снабжения питательными веществами. В соответствии со стандартами мутности по МакФарланду повышение OD на 0,3 соответствует проросту бактерий на 108-109. После 9-го снабжения питательными веществами бактериальные белки экстрагировали в экспоненциальной и стационарной фазе роста. Была показана общая концентрация белка 0,088 мг/мл и 0,15 мг/мл соответственно. Экстрагированные белки исследовали на содержание ClpB и использовали в анализе продукции АТФ и для in vivo экспериментов, включающих вливание в толстый кишечник и системное введение мышам и крысам, имеющим свободный доступ к пище, с последующим выявлением c-fos в головном мозге.
Протеомный анализ
Для анализа различий в профилях экспрессии белков для вызванных питательными веществами фаз роста бактерий проводили двумерный электрофорез в полиакриламидном геле (2D-ПААГ) отдельно для мембранной и цитоплазматической фракции белков E.coli K12, экстрагированных через 10 мин и 2,3 ч после 5-го добавления среды МН, что соответствует экспоненциальной и стационарной фазе роста соответственно. Общее количество выявленных белковых пятен составляло 2895 (1367 мембранных и 1528 цитоплазматических).
Сравнение 2D-ПААГ для мембранных белков экспоненциальной и стационарной фазы выявило 20 белков с различающейся экспрессией (по меньшей мере в 1,5 раза). Среди них 17 белков имели увеличенную экспрессию в экспоненциальной фазе роста, 15 белков были идентифицированы с помощью масс-спектрометрии. Сравнение 2D-ПААГ цитоплазматических белков показало 20 белков с различающейся экспрессией (по меньшей мере в 1,5 раза). В отличие от мембранных белков большинство (19) цитоплазматических белков имело увеличенную экспрессию в стационарной фазе. Только для одного белкового пятна, соответствующего флагеллину, была показана более высокая экспрессию в экспоненциальной фазе роста. Большинство идентифицированных белков представляли собой белки, участвующие анаболических или катаболических процессах, что указывает на общий смешанный метаболический профиль в обеих фазах роста.
Продукция АТФ белками E.coli in vitro
Для того чтобы изучить, могут ли изменения бактериального протеома в фазах роста бактерий влиять на их способность выделять энергию и продукцию АТФ из питательных веществ, вызванную белками E.coli K12 экспоненциальной и стационарной фазы, исследовали in vitro. Было обнаружено, что белки обеих фаз роста были способны повышать продукцию АТФ из различных источников энергии. Концентрация АТР была выше при использовании белок-содержащего смешанного источника энергии, такого как среда МН, по сравнению с раствором сахарозы. Продукция АТФ повышалась дозозависимым образом с повышением концентрации бактериальных белков, однако не было обнаружено существенных различий между эффектами на продукцию АТФ белков экспоненциальной и стационарной фазы.
Продукция ClpB E.coli in vitro
Авторы изобретения разработали и валидировали твердофазный иммуноферментный анализ (ИФА) для выявления ClpB E.coli, который был использован в настоящем исследовании. Различия в продукции ClpB между фазами роста бактерий исследовали на 4 отдельных культурах E.coli K12. Результаты Вестерн - блоттинга выявили соответствующие полосы ClpB массой 96 кДа во всех белковых препаратах с увеличенным уровнем во время стационарной фазы. Указанные изменения дополнительно подтвердили с использованием ИФА ClpB на тех же препаратах бактериального белка, показавшего, что концентрация ClpB увеличилась в стационарной фазе почти в два раза.
Кишечное вливание питательных веществ и белков E.coli
Для того чтобы проверить, соответствует ли предложенная авторами in vitro модель вызванного питательными веществами роста E.coli динамике роста кишечных бактерий in vivo, крысам, находящимся под анестезией, проводили вливание в толстый кишечник среды МН или воды. Было обнаружено, что медленное вливание среды МН, но не воды, вызывало пролиферацию бактерий в кишечнике с экспоненциальной фазой роста, длящейся в течение 20 мин, что согласуется с данными in vitro. Уровень ClpB в плазме, полученной из воротной вены, значительно не различался через 30 или 60 мин после инфузии МН. Тем не менее, концентрация ClpB в плазме прямо коррелировала с содержанием ДНК ClpB в фекалиях.
Затем для того, чтобы определить, могут ли зависимые от роста бактерий изменения протеома E.coli влиять на механизмы контроля аппетита хозяина локально в кишечнике, в отдельном эксперименте, крысам, находящимся под анестезией, проводили вливание в толстую кишку белков Е. coli экспоненциальной или стационарной фазы роста в концентрации 0,1 мг/кг в течение 20 мин. Концентрация ClpB в слизистой оболочке толстой кишки, измеренная через 20 мин после вливания, была выше у крыс, получавших белки стационарной фазы роста, однако бактериальные белки как экспоненциальной, так и стационарной фазы роста не оказывали влияния на уровень ClpB в плазме. Было обнаружено, что медленное вливание в толстую кишку белков E.coli экспоненциальной фазы роста, но не стационарной фазы роста, приводило к повышению уровня GLP-1 в плазме; повышение уровня PYY было выявлено, напротив, после инфузии белков стационарной фазы, но не экспоненциальной фазы роста, что согласуется с гипотезой авторов о том, что эффекты белков E.coli на сигнальные пути, контролирующие аппетит, хозяина должны зависеть от фазы роста бактерий.
Потребление пищи и экспрессия c-fos в головном мозге после однократных введений белков E.coli у крыс
Поскольку ClpB присутствовал в плазме всех исследуемых крыс и мышей, авторы предположили, что плазматические белки E.coli могут влиять на аппетит благодаря системному действию и что такие эффекты могут быть различными для белков, связанных с разными фазами роста бактерий. В результате проверки этого предположения на крысах, не получавших еду в течение ночи, было обнаружено, что однократное внутрибрюшинное (ВБ) введение (0,1 мг/кг массы тела) мембранной фракции белков E.coli, экстрагированных в стационарной фазе роста, приводило к снижению потребления пищи в течение 1 и 2 часов при возобновлении кормления по сравнению с контрольной группой. Напротив, введение цитоплазматической фракции белков E.coli (0,1 мг/кг массы тела, внутрибрюшинно), экстрагированной в экспоненциальной фазе роста, повышало потребление пищи в течение 4 часов при возобновлении кормления.
Для того, чтобы также исследовать, могут ли общие белки E.coli влиять на спонтанное потребление пищи в зависимости от фазы роста и активировать участки центральной нервной системы, такие как ARC, крысам, имеющим свободный доступ к пище, вводили до начала темновой фазы бактериальные белки (0,1 мг/кг массы тела, внутрибрюшинно). Потребление пищи измеряли в течение 2 ч после инъекции, затем крыс умерщвляли для анализа экспрессии c-fos в головном мозге. Было обнаружено, что крысы, которым вводили бактериальные белки стационарной фазы, потребляли меньше пищи по сравнению с контрольными животными, тогда как введение бактериальных белков, полученных в экспоненциальную фазу роста, значительно не влияло на потребление пищи.
Через два часа после внутрибрюшинной инъекции белков E.coli крысам, имеющим свободный доступ к пище, проводили иммуногистохимический анализ экспрессии c-fos в дугообразном ядре (ARC), вентромедиальном ядре (VMN) гипоталамуса и в центральном ядре миндалевидного тела (СеА). Увеличенное количество c-fos-положительных клеток было обнаружено в ARC и VMN у мышей, получавших бактериальные белки стационарной фазы. Большинство c-fos-экспрессирующих клеток в ARC содержало β-эндорфин (контроли: 71,31±12,81%, экспоненциальная фаза роста E.coli: 73,56±10,45%, стационарная фаза роста E.coli: 80,50±9,68%, ANOVA, р=0,36), т.е. было идентифицировано как анорексигенные РОМС-нейроны. Соответственно, процент оставшихся c-fos-нейронов в ARC значительно не различался между группами (контроли: 28,69±12,81%, экспоненциальная фаза роста E.coli: 26,44±10,45%, стационарная фаза роста E.coli: 19,5±9,68%, ANOVA, р=0,36). Несмотря на то, что общее количество β-эндорфин-положительных клеток значительно не различалось между группами (контроли, 54,82±10,67 клеток, экспоненциальная фаза роста E.coli: 66,03±11,43 клеток, стационарная фаза роста E.coli: 66,03±5,06 клеток, ANOVA, р=0,09), относительное количество активированных β-эндорфиновых нейронов было повышено у крыс, получавших белки стационарной фазы роста по сравнению с контрольными животными и с крысами, получавшими белки экспоненциальной фазы роста. Более того, количество активированных β-эндорфиновых нейронов обратно коррелировало с потреблением пищи (Пирсона r=-0,57, р=0,018).
В СеА количество c-fos-положительных клеток у крыс, которым инъецировали белки стационарной фазы, было повышено по сравнению с двумя другими группами. Почти все c-fos-положительные клетки в СеА были определены как CGRP-экспрессирующие нейроны (контроли, 100±0,01%, экспоненциальная фаза роста E.coli: 100±0,01%, стационарная фаза роста E.coli: 100±0,01%, ANOVA р=0,92). Несмотря на то что общее количество CGRP-положительных нейронов в СеА было сходным среди групп (контроли, 123,8±13,15 клеток, экспоненциальная фаза роста E.coli: 118,3±25,59 клеток, стационарная фаза роста Е.coli: 126,1±6,64 клеток, ANOVA р=0,85), процент CGRP-нейронов с активированным c-fos был повышен только у крыс, получавших белки стационарной фазы. Активация CGRP-нейронов обратно коррелировала с потреблением пищи (Пирсон: r=-0,89, р=0,001).
Паттерн пищевого поведения и нейропептиды гипоталамуса после продолжительного введения белков E.coli мышам
Для того чтобы определить, могут ли бактериальные белки влиять на паттерн пищевого поведения, мышам со свободным доступом к пище проводили ежедневно две инъекции общего белка E.coli (0,1 мг/кг массы тела, внутрибрюшинно) в течение одной недели. Первый день после инъекции характеризовался значительным снижением массы тела и потребления пищи у мышей, получавших бактериальные белки стационарной фазы, но не экспоненциальной фазы роста, по сравнению с контролями. Несмотря на то что ежедневный объем порций пищи значительно не различался между группами, через неделю у мышей, получавших белки стационарной фазы, наблюдалось его снижение по отношению к дню до введения. Частота приема пищи значительно не различалась между группами, несмотря на то что тенденция в сторону повышения наблюдалась у мышей, получавших бактериальные белки стационарной фазы. Несмотря на то что общее потребление пищи среди 3 групп значительно не различалось в течение 6 дней введения, анализ потребления пищи раздельно для светового и темнового периода показал, что у мышей, которым вводили белки фазы экспоненциального роста, наблюдалось увеличенное потребления пищи во время светового периода, но было снижено во время темнового периода. Напротив, у мышей, получавших белки стационарной фазы, наблюдалось более низкое по сравнению с контрольными животными потребление пищи в темновом периоде без каких-либо отличий в световом периоде. Во время первого дня после инъекции показатель чувства сытости был повышен у мышей, получавших белки стационарной фазы, та же группы имела тенденцию к снижению через одну неделю.
Чтобы получить представление о молекулярных изменениях, лежащих в основе изменения паттерна пищевого поведения, наблюдаемого через 6 дней введения бактериальных белков, авторы изобретения анализировали уровень экспрессии в гипоталамусе мРНК некоторых нейропептидов, вовлеченных в контроль аппетита. Было обнаружено, что у мышей, получавших белки стационарной фазы роста, наблюдался увеличенный уровень мРНК-предшественника нейротрофического фактора головного мозга (BDNF) и орексина по сравнению с контрольными животными, а также увеличенный уровень кортикотропин-высвобождающего гормона (CRH) по сравнению с мышами, которым вводили белки фазы экспоненциального роста, которые также имели увеличенный уровень BDNF, но пониженный уровень QRFP.
Электрофизиологическая активация РОМС-нейронов гипоталамуса ClpB
Для того чтобы определить, может ли ClpB как маркер белков E.coli, повышающихся в стационарной фазе роста и миметик α-MSH активировать РОМС-нейроны ARC, его действие проверяли на срезах головного мозга мышей POMC-eGFP с использованием электрофизиологического анализа patch-clamp на прикрепленных клетках. Промывка раствором ClpB (1 нМ) приводила к повышению частоты потенциала действия у ~50% РОМС-нейронов ARC (n=7/13) на 229±109% (исходная частота: 2,02±0,78 Гц по сравнению с ClpB-вызванной: 3,82±1,36 Гц). В целом, РОМС-нейроны не полностью возвращались к исходным показателям частоты испускания импульсов по меньшей мере вплоть до 10 мин после применения ClpB.
Обсуждение
Проведенное авторами исследование выявило, что бактериальные белки могут обеспечивать физиологическую связь между кишечными бактериями и контролем аппетита хозяина, включая как краткосрочные, так и долгосрочные механизмы, связанные с вызванным питательными веществами ростом бактерий в кишечнике и их системными эффектами соответственно. Следующие далее основные результаты подтверждают данное заключение: 1) регулярное снабжение питательными веществами ускоряло и стабилизировало экспоненциальную фазу роста E.coli, длящуюся 20 мин, что согласуется с данными in vivo; 2) стационарная фаза роста E.coli характеризовалась повышением общего содержания бактериальных белков и различающимся протеомом, включая увеличенный ClpB; 3) белки E.coli от обеих фаз роста дозозависимым образом стимулировали продукцию АТФ in vitro; 4) уровень ClpB в плазме не менялся после вызванного питательными веществами роста бактерий в кишечнике, но коррелировал с содержанием ДНК ClpB в кишечной микробиоте; 5) вливание в кишечник белков E.coli, полученных в экспоненциальную и стационарную фазу роста, вызывало стимуляцию GLP-1 и PYY в плазме соответственно; 6) системное введение белков E.coli приводило к значительному снижению потребления пищи только в случае белков, полученных в стационарную фазу, и сопровождалось активацией c-fos в анорексигенных нейронах ARC и СеА; и, наконец 7) ClpB стимулировал частоту испускания импульсов РОМС-нейронов ARC.
Регулярное снабжение питательными веществами и рост бактерий
Среди широкого разнообразия бактерий, которые заселяют желудочно-кишечный тракт человека, E.coli является наиболее распространенным факультативным анаэробом, что объясняет ее применение в качестве модельного организма для изучения симбиотических кишечных бактерий (Foucault et al., 2009). Согласно настоящему изобретению, было обнаружено, что динамика роста E.coli меняется при регулярном снабжении бактерий питательными веществами, где 5-ый цикл обеспечения питательными веществами приводит сразу к экспоненциальному росту, длящемуся 20 мин после стационарной фазы. Затем следуют одинаковые повторения цикла роста после следующего снабжения питательными веществами, что дает основания полагать, что указанное снабжение может обеспечивать цикличность активности внутренних механизмов бактерий. Сходная динамика роста бактерий в ответ на инфузию питательных веществ наблюдалась в толстой кишке крыс, что подтверждает соответствие данных, полученных авторами in vitro, картине in vivo, например, у людей, употребляющих регулярно пищу. Увеличение числа бактерий на 108-109 оставалось стабильным после каждого нового снабжения питательными веществами, что дает основания предполагать, что соответствующая стабильная продукция бактериальной биомассы, включая увеличенное содержание белков в кишечнике, может играть роль фактора вызванной приемом пищи регуляции для хозяина. Учитывая, что средняя фаза приема пищи у людей сходна с продолжительностью экспоненциальной фазы роста бактерий, получающих регулярное снабжение питательными веществами, можно ожидать, что чувство сытости у хозяина может быть вызвано кишечными бактериями, достигающими стационарной фазы роста через 20 мин после контакта с потребленными питательными веществами. Однако бактериальное содержимое в желудочно-кишечном тракте варьирует от 103 в желудке до 1012 в толстом кишечнике. Более того, приблизительно 2 ч необходимо для продвижения поглощенных питательных веществ через желудок и тонкий кишечник и приблизительно 10 ч через толстый кишечник. Такая задержка доставки питательных веществ к большинству кишечных бактерий, вероятно, указывает на то, что помимо непосредственного контакта с питательным болюсом рост бактерий во время фазы приема пищи может также инициироваться питательными веществами, высвобождающимися в просвет кишечника по условному рефлексу Павлова на прием пищи.
Экспрессия бактериальных белков во время фаз роста и восприятие сигнала в кишечнике
Поскольку динамика роста E.coli, регулярно получающих питательные вещества, может быть связана с фазой приема пищи и фазой, следующей за приемом пищи, авторы изобретения исследовали потенциальную возможность связи кишечных бактерий с контролем аппетита хозяина через экспрессию бактериальных белков. Сначала авторы проводили сравнение протеома E.coli в экспоненциальной и стационарной фазе роста. Для этого анализа белки E.coli экстрагировали в середине фазы экспоненциального роста, т.е. через 10 мин после предоставления питательных веществ, и в стационарной фазе роста через 2 ч - это время в норме характеризуется чувством сытости. Обнаружение по меньшей мере 40 белков с различиями в экспрессии между двумя фазами роста подтвердило, что они различаются не только количественно, но и по белковому составу, который увеличивается почти вдвое после пролиферации бактерий, а также меняется качественно. Затем изучали возможное соответствие модифицированного профиля экспрессии белков контролю аппетита хозяина: i) путем сравнения способности бактериальных белков производить источник энергии и ii) путем определения возможных непосредственных эффектов бактериальных белков на пути регуляции аппетита. Возможность последнего подтверждается тем, что белок ClpB E.coli был недавно идентифицирован как конформационный пептидомиметик α-MSH (Tennoune et al., 2014); эти данные подтвердили более раннюю гипотезу авторов о том, что кишечные бактериальные белки, проявляющие гомологию по эпитопам с анорексигенными или орексигенным пептидам, могут отвечать за перекрестную реактивность аутоантител (Fetissov et al., 2008). Таким образом, возможно, что комбинация таких бактериальных пептидомиметиков может влиять непосредственно на аппетит в зависимости от профиля экспрессии белков, связанных с фазой роста бактерий. В результате анализа культур E.coli и слизистой оболочки кишечника был обнаружен увеличенный уровень ClpB, связанный со стационарной фазой роста бактерий. Увеличенный уровень ClpB, таким образом, может вносить вклад в активацию анорексигенных путей после вызванной питательными веществами пролиферации E.coli и повышения продукции бактериальных белков. Важным вопросом является то, на каком этапе бактериальные белки, такие как ClpB, могут действовать на регулирующие аппетит пути.
Несмотря на то что бактериальные белки присутствуют в слизистой оболочке кишечника (Haange et al., 2012), способность их прохождения через стенку кишечника подробно не изучалась (Lim и Rowley, 1985). Теоретически после спонтанного и вызванного лизиса бактерий в кишечнике (Rice и Bayles, 2008), бактериальные компоненты могут проходить через эпителиальный барьер слизистой оболочки в результате всасывания энтероцитами и путем параклеточной диффузии, регулируемой нервной системой кишечника (Neunlist et al., 2012). Например, липополисахарид (ЛПС), который высвобождается при лизисе грамотрицательных бактерий, присутствуют в естественных условиях в плазме здоровых людей и мышей и имеет высокий исходный уровень после употребления пищи с высоким содержанием жиров (Cani et al., 2007). Уровень ЛПС в плазме также повышается после приема пищи (Harte et al., 2012), однако такие данные о бактериальных белках отсутствуют. Авторы настоящего изобретения показали, что уровень ClpB в плазме остается стабильным у крыс после пролиферации бактерий в кишечнике или после вливания в кишечник бактериальных белков. Это указывает на то, что вызванная питательными веществами пролиферация бактерий не оказывает существенного влияния на плазматический ClpB, и наиболее вероятно, другие бактериальные белки, присутствующие в плазме, и, таким образом, указанные белки не могут быть вовлечены в краткосрочный сигналинг в головном мозге, регулирующий насыщение.
Тем не менее, концентрация ClpB в плазме коррелировала с ДНК ClpB в кишечной микробиоте, что дает основания полагать, что количество ClpB-продуцирующих бактерий, которое через длительный период времени должно было бы являться относительно независимым от колебаний, связанных с вызванным питательными веществами ростом бактерий, может являться ключевым фактором, отвечающим за длительное поддержание плазматического уровня ClpB. Это заключение дополнительно подтверждается данными, полученными авторами для валидации ИФА ClpB, показывающими увеличенную концентрацию ClpB в плазме у мышей, постоянно получающих принудительное кормление E.coli в отличие от мышей, которые получали E.coli с мутацией ClpB. В связи с этим возможно, что белки кишечных бактерий, присутствующие в плазме, включая ClpB, могут оказывать системное действие, обеспечивая связь между составом кишечной микробиоты и долгосрочным контролем энергетического метаболизма.
Эффекты белков E.coli на продукцию АТФ in vitro
Энергетический обмен через пищевые цепи отражает универсальную связь между всеми организмами (Yun et al., 2006). АТФ, которая получается в результате катаболизма питательных веществ как у бактерий, так и у животных, служит основным энергетическим субстратом для анаболических процессов. Животные могут чувствовать изменения в АТФ через активность аденозин-5'-монофосфат-активируемой протеинкиназы (АМРК), приводящую к повышению потребления пищи при низком уровне АТФ, и наоборот (Dzamko и Steinberg, 2009). Таким образом, в настоящей работе проводили сравнение способности белков E.coli экспоненциальной фазы роста и стационарной фазы роста производить АТФ in vitro. В самом деле, многие из идентифицированных белков проявляли анаболическую или катаболическую активность. Было обнаружено, что белки E.coli способны стимулировать продукцию АТФ in vitro, что дает основания предполагать, что они могут продолжать катализировать продукцию АТФ после лизиса бактерий в кишечнике. Несмотря на то что не было обнаружено различий в способности продуцировать АТФ между белками от экспоненциальной фазы роста и стационарной фазы роста, зависимая от концентрации бактериальных белков продукция АТФ указывает на то, что повышение содержания бактериальных белков после вызванной питательными веществами пролиферации бактерий должно приводить к более высокому синтезу АТФ. То что эффективность кишечной микробиоты имеет отношение к поглощению энергии в ходе метаболизма хозяина, было ранее установлено путем сравнения полных и стройных людей, а также мышей (Turnbaugh et al., 2006). Данные настоящего изобретения также подтверждают указанные результаты за счет показанной способности белков E.coli производить АТФ. Эти данные также предполагают, что вызванная приемом пищи пролиферация бактерий может приводить к повышению кишечной АТФ, которое может вносить вклад в восприятие через просвет кишечника доступности энергии и расслабления кишечника (Glasgow et al., 1998).
Кишечные эффекты белков E.coli на гормоны насыщения
Далее авторы изобретения исследовали, могут ли белки E.coli в кишечнике стимулировать системное высвобождение кишечных гормонов насыщения, таких как GLP-1 и PYY (Adrian et al., 1985; Batterham et al., 2002; Beglinger и Degen, 2006; Flint et al., 1998). Фактически, оба гормона продуцируются одинаковым или разными аргентаффинными L-клетками, присутствующими во всех отделах кишечника и встречающимися в большом количестве в толстом кишечнике (Eissele et al., 1992), и таким образом, L-клетки непосредственно подвержены действию бактериальных белков. Авторы изобретения обнаружили различные эффекты белков E.coli на высвобождение GLP-1 и PYY, указывающие на стимуляцию GLP-1 белками экспоненциальной фазы роста и стимуляцию PYY белками стационарной фазы роста. Данные результаты указывают на частичное сходство между вызванным питательными веществами ростом бактерий и известной динамикой вызванного приемом пищи высвобождения GLP-1 и PYY. Фактически, как было показано на людях, резкий пик GLP-1 в плазме наблюдается через 15 мин после внутрижелудочной инфузии жидкой пищи, тогда как продолжительное повышение PYY в плазме начинается через 15-30 мин после приема пищи (Edwards et al., 1999; Gerspach et al., 2011). Более долгое высвобождение GLP-1 связывали с захватом жира (van der Klaauw et al., 2013). Таким образом, динамика роста регулярно получающих питательные вещества кишечных бактерий совпадает по времени с динамикой высвобождения GLP-1 и PYY, что предполагает стимулирующую роль кишечных бактерий, и в частности, белков E.coli, в вызванном приемом пищи кишечном сигналинге, регулирующем чувство сытости. Отличающийся эффект белков E.coli, полученных в экспоненциальную фазу роста, на стимуляцию GLP-1, возможно, отражает роль пептидного гормона GLP-1 из семейства инкретинов в гликемическом контроле (Edwards et al., 1999; Steinert et al., 2014). Недавно показанная экспрессия функционального MC4R L-клетками (Panaro et al., 2014) дает основания предполагать их возможную активацию бактериальными белками, имитирующими α-MSH. Увеличенная продукция ClpB во время стационарной фазы роста E.coli, а также увеличенный уровень ClpB в слизистой оболочке кишечника, связанный с увеличенным уровнем PYY в плазме, предполагает непосредственную роль ClpB в активации PYY-продуцирующих L-клеток в толстом кишечнике. С другой стороны, поскольку не идентифицированные белки E.coli, возможно, активируются во время экспоненциальной фазы роста, стимуляция секреции GLP-1 может являться предпочтительной.
Системные эффекты белков E.coli на потребление пищи и регулирующие аппетит мозговые пути
Согласно настоящему изобретению, показано, что периферические инъекции белков E.coli крысам, не получавшим пищу, или крысам, имеющим свободный доступ к пище, а также мышам, имеющим свободный доступ к пище, меняют их потребление пищи в зависимости от фазы роста E.coli. Учитывая, что кишечное вливание питательных веществ не оказывало влияния на уровень ClpB в плазме, который был стабильна в течение короткого периода времени, системное действие бактериальных белков должно было бы объясняться их долгосрочным эффектом, модулирующим аппетит. Более того, из-за короткой продолжительности фазы экспоненциального роста бактериальные белки, которые активируется во время продолжительной стационарной фазы, должно доминировать в плазме, и таким образом, их системное введение может лучше отражать физиологическую картину. Концентрация белков E.coli 0,1 мг/кг, используемая в указанных экспериментах, была сходна с эффективными дозами вызывающих чувство насыщения пептидных гормонов, таких как лептин или PYY, после периферического введения у людей или грызунов (Batterham et al., 2002; Halaas et al., 1995; Heymsfield et al., 1999).
Наблюдали повышение потребления пищи в световую фазу при возобновлении кормления у крыс, не получавших еду, после введения цитоплазматических белков, полученных в экспоненциальную фазу роста, и снижение потребления пищи после введения мембранных белков, полученных в стационарную фазу роста. Данный эксперимент подтвердил тот факт, что различные белковые смеси, полученные от одной и той же бактерии, способны вызывать повышение или снижение потребления пищи в результате их системного действия. Однако в более физиологическом окружении при исследовании эффекта общих белков E.coli у крыс со свободным доступом к пище в темновую фазу, наблюдали только снижение потребления пищи, которое было вызвано белками стационарной фазы.
Результаты повторных инъекций у мышей со свободным доступом к пище также подтвердили роль белков E.coli в создании отрицательного энергетического баланса. В самом деле, первый день инъекций бактериального белка сопровождался снижением потребления пищи и массы тела, которые были значительными у мышей, получавших белки стационарной фазы, и которые также характеризовались относительным увеличением продолжительности чувства сытости. Несмотря на то что потребление пищи у указанных мышей после этого приходило в норму, прогрессирующее снижение объема порции пищи сопровождалось повышением частоты потребления пищи, наиболее вероятно в качестве компенсаторного механизма для поддержания потребления пищи (Meguid et al., 1998). Кроме того, различающиеся эффекты белков E.coli наблюдались во время световой и темновой фаз. Соответственно, паттерн экспрессии мРНК регулирующих аппетит нейропептидов в гипоталамусе показал смешанный ответ с активацией как анорексигенных, так и орехигенных путей. Следует отметить, что в обеих группах мышей, получавших белки E.coli, наблюдалось одинаковое повышение мРНК BDNF - участника анорексигенного пути, более позднего по отношению к MC4R в VMN (Xu et al., 2003). Указанный путь может лежать в основе снижения потребления пищи во время темновой фазы, наблюдаемой в обеих группах, которое была также выражено у мышей, которым вводили белки фазы экспоненциального роста, что указывает на более низкий уровень орексигенных QRFP (Chartrel et al., 2003) и NPY (Herzog, 2003). Напротив, у мышей, которые получали бактериальные белки стационарной фазы, наблюдался усиленный анорексигенный профиль с увеличенным уровнем мРНК CRH, наиболее вероятно задействующего МС4R-экспрессирующие нейроны PVN (Lu et al., 2003). Указанное изменение вместе с увеличенной экспрессией мРНК-предшественника орексина А, который стимулирует частоту приема пищи (Baird et al., 2009), может вносить вклад в снижение объема порции пищи и показатель чувства сытости соответственно у указанных мышей через 6 дней введения.
Авторы изобретения обнаружили у крыс со свободным доступом к пище увеличенную экспрессию c-fos в анорексигенных РОМС-нейронах ARC и VMN, которое давно известно как центр насыщения, взаимосвязанный с РОМС-нейронами ARC (Sternson et al., 2005), что согласуется с гипотезой авторов о том, что бактериальные белки, которые продуцируются во время стационарной фазы, могут активировать некоторые ключевые анорексигенные пути в центральной нервной системе. Полученный паттерн c-fos напоминает паттерн ответа, приводящего к насыщению, во время потребления пищи (Johnstone et al., 2006) или вызванного гормонами чувства сытости, такими как PYY или полипептид поджелудочной железы (Batterham et al., 2002; Challis et al., 2004; Lin et al., 2009). Относительно маленькое число (~10%) c-fos-активных РОМС-нейронов предполагает, что циркулирующие белки E.coli могут оказывать модулирующий эффект на аппетит и массу тела, действуя через этот гипоталамический путь. Несмотря на то что было невозможно определить активацию c-fos нейронами NPY/AgRP, их вклад в сигналинг бактериальными белками не может быть исключен; эти нейроны также экспрессируют MC3R и MC4R (Mounien et al., 2005). Кроме того, более сильная активациях c-fos в CGRP-нейронах СеА (~40%) по сравнению с РОМС-нейронами ARC может говорить о конвергентном действии на более поздних этапах РОМС и NPY/AgRP-нейронов ARC и, возможно, других регулирующих аппетит областей мозга, которые не были исследованы согласно настоящему изобретению, таких как ядро одиночного пучка.
Наконец, для того чтобы определить, может ли активация бактериальными белками регулирующих аппетит участков мозга, таких как РОМС-нейроны ARC, быть вызвана местным действием указанных белков, авторы исследовали способность ClpB при его нанесении на указанные нейроны активировать их электрическую активность. Результаты показали, что приблизительно у половины исследуемых нейронов повышалась частота потенциала действия, при этом указанные нейроны оставались активированными в течение по меньшей мере 10 мин. Продолжительный эффект ClpB соответствует эффекту α-MSH на РОМС-нейроны, экспрессирующие функционально активные рецепторы MC3R и MC4R (Smith et al., 2007). Указанные данные дают основание полагать, что ClpB может являться физиологическим активатором РОМС-нейронов гипоталамуса, отчасти сходным с вызывающими чувство насыщения пептидом PYY и лептином (Batterham et al., 2002; Cowley et al., 2001). Однако остается неизвестным, мог ли белок ClpB непосредственно активировать РОМС-нейроны, или он вызывал их активацию через локальную нейронную сеть.
В целом описанные данные подтверждают роль присутствующих в организме белков E.coli, экспрессия которых повышается в стационарной фазе роста, таких как ClpB, в обеспечении отрицательного энергетического баланса путем активации анорексигенных путей в головном мозге. Эти данные также дают основания полагать, что изменения в составе микробиоты, приводящие к снижению или повышению содержания E.coli и, возможно, других бактерий из семейства Enterobacteriaceae, могут влиять на энергетический баланс хозяина, делая его положительным или отрицательным соответственно.
ПРИМЕР 2
Самцов мышей линии C57Bl6 (Janvier Laboratories) возрастом один месяц содержали, как описано выше, и позволяли животным акклиматизироваться в условиях вивария в течение 1 недели. Мышей распределяли на 3 группы следующим образом: (i) животные, получающие 108 бактерий Е. coli K12 путем принудительного кормления; (ii) животные, получающие 108 бактерий Е. coli K12, дефицитных по ClpB, путем принудительного кормления; (iii) и контрольные животные, которые не получают какого-либо лечения. Штамм, содержащий мутацию в гене ClpB, был создан в лаборатории Бернда Букау (Bernd Bukau, ZMBH, Гейдельбергский университет, Гейдельберг, Германия) и был любезно предоставлен доктором Алексом Могком (Dr Axel Mogk) вместе с соответствующими бактериями Е. coli дикого типа (WT) 5. Мышей помещали по отдельности в клетки BioDAQ (Research Diets) и ежедневно проводили животным внутрижелудочное введение 0,5 мл среды LB, содержащей бактерии, до начала темновой фазы в течение 21 дня. Первые дни кормления сопровождались снижением массы тела и потребления пищи у мышей, получавших WT E.coli в отличие от животных, получающих бактерии, не экспрессирующие белок ClpB (Фигура 1).
ЦИТИРОВАННАЯ ЛИТЕРАТУРА
На всем протяжении настоящей заявки различные ссылки описывают состояние уровня техники, к которому относится настоящее изобретение. Раскрытие указанных ссылок, таким образом, включено в настоящее изобретение посредством ссылки.
Adrian, Т.Е., Ferri, G.L., Bacarese-Hamilton, A.J., Fuessl, H.S., Polak, J.M., and Bloom, S.R. (1985). Human distribution and release of a putative new gut hormone, peptide YY. Gastroenterology 89, 1070-1077.
Atasoy, D., Betley, J.N., Su, H.H., and Sternson, S.M. (2012). Deconstruction of a neural circuit for hunger. Nature 488, 172-177.
Baird, J.-P., Choe, A., Loveland, J.L., Beck, J., Mahoney, C.E., Lord, J.S., and Grigg, L.A. (2009). Orexin-A hyperphagia: hindbrain participation in consummatory feeding responses. Endocrinology 150, 1202-1216.
Batterham, R.L., Cowley, M.A., Small, C.J., Herzog, H., Cohen, M.A., Dakin, C.L., Wren, A.M., Brynes, A.E., Low, M.J., Ghatei, M.A., et al. (2002). Gut hormone PYY(3-36) physiologically inhibits food intake. Nature 418, 650-654.
Beglinger, C., and Degen, L. (2006). Gastrointestinal satiety signals in humans - Physiologic roles for GLP-1 and PYY? Physiology & Behavior 89, 460-464.
Berthoud, H.-R. (2011). Metabolic and hedonic drives in the neural control of appetite: who is the boss? Current Opinion in Neurobiology 21, 888-896.
Cani, P.D., Amar, J., Iglesias, M.A., Poggi, M., Knauf, C., Bastelica, D., Neyrinck, A.M., Fava, F., Tuohy, K.M., Chabo, C., et al. (2007). Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 56, 1761-1772.
Carter, M.E., Soden, M.E., Zweifel, L.S., and Palmiter, R.D. (2013). Genetic identification of a neural circuit that suppresses appetite. Nature 503, 111-114.
Challis, B.G., Coll, A.P., Yeo, G.S.H., Pinnock, S.B., Dickson, S.L., Thresher, R.R., Dixon, J., Zahn, D., Rochford, J.J., White, A., et al. (2004). Mice lacking pro-opiomelanocortin are sensitive to high-fat feeding but respond normally to the acute anorectic effects of peptide-YY3-36. Proc. Natl. Acad. Sci. USA 101, 4695-4700.
Chartrel, N., Dujardin, C., Anouar, Y., Leprince, J., Decker, A., Clerens, S., Do-Rego, J.C., Vandesande, F., Llorens-Cortes, C., Costentin, J., et al. (2003). Identification of 26RFa, a hypothalamic neuropeptide of the RFamide peptide family with orexigenic activity. Proc Natl Acad Sci USA 100, 15247-15252.
Cone, R.D. (2005). Anatomy and regulation of the central melanocortin system. Nat Neurosci 8, 571-578.
Cowley, M.A., Pronchuk, N., Fan, W., Dinulescu, D.M., Colmers, W.F., and Cone, R.D. (1999). Integration of NPY, AGRP, and melanocortin signals in the hypothalamic paraventricular nucleus: evidence of a cellular basis for the adipostat. Neuron 24, 155-163.
Cowley, M.A., Smart, J.L., Rubinstein, M., Cerdan, M.G., Diano, S., Horvath, T.L., Cone, R.D., and Low, M.J. (2001). Leptin activates anorexigenic POMC neurons through a neural network in the arcuate nucleus. Nature 411, 480-484.
Dinan, T.G., Stilling, R.M., Stanton, C., and Cryan, J.F. (2015). Collective unconscious: How gut microbes shape human behavior. Journal of Psychiatric Research 63, 1-9.
Dzamko, N.L., and Steinberg, G.R. (2009). AMPK-dependent hormonal regulation of whole-body energy metabolism. Acta Physiologica 196, 115-127.
Edwards, С.М., Todd, J.F., Mahmoudi, M., Wang, Z., Wang, R.M., Ghatei, M.A., and Bloom, S.R. (1999). Glucagon-like peptide 1 has a physiological role in the control of postprandial glucose in humans: studies with the antagonist exendin 9-39. Diabetes 48, 86-93.
Eissele, R., Goke, R., Willemer, S., Harthus, H.P., Vermeer, H., Arnold, R., and Goke, B. (1992). Glucagon-like peptide-1 cells in the gastrointestinal tract and pancreas of rat, pig and man. European journal of clinical investigation 22, 283-291.
Fetissov, S.O., and  , P. (2011). The new link between gut-brain axis and neuropsychiatric disorders. Curr Opin Clin Nutr Metab Care 14, 477-482.
, P. (2011). The new link between gut-brain axis and neuropsychiatric disorders. Curr Opin Clin Nutr Metab Care 14, 477-482.
Fetissov, S.O., Hamze Sinno, M.,  , M., Bole-Feysot, C.,
, M., Bole-Feysot, C.,  , P.,
, P.,  , Т., and
, Т., and  , P. (2008). Autoantibodies against appetite-regulating peptide hormones and neuropeptides: putative modulation by gut microflora. Nutrition 24, 348-359.
, P. (2008). Autoantibodies against appetite-regulating peptide hormones and neuropeptides: putative modulation by gut microflora. Nutrition 24, 348-359.
Fioramonti, X., Contie, S., Song, Z., Routh, V.H., Lorsignol, A., and Penicaud, L. (2007). Characterization of glucosensing neuron subpopulations in the arcuate nucleus: integration in neuropeptide Y and pro-opio melanocortin networks? Diabetes 56, 1219-1227.
Flint, A., Raben, A., Astrup, A., and Hoist, J.J. (1998). Glucagon-like peptide 1 promotes satiety and suppresses energy intake in humans. The Journal of Clinical Investigation 707, 515-520.
Forsythe, P., and Kunze, W. (2013). Voices from within: gut microbes and the CNS. Cellular and Molecular Life Sciences 70, 55-69.
Foucault, M.-L., Thomas, L., Goussard, S., Branchini, B.R., and Grillot-Courvalin, C. (2009). In vivo bioluminescence imaging for the study of intestinal colonization by Escherichia coli in mice. Applied and Environmental Microbiology 76, 264-274.
Garfield, A.S., Li, C., Madara, J.C., Shah, B.P., Webber, E., Steger, J.S., Campbell, J.N., Gavrilova, O., Lee, C.E., Olson, D.P., et al. (2015). A neural basis for melanocortin-4 receptor-regulated appetite. Nat Neurosci doi: 10,1038/nn,4011.
Gerspach, A.C., Steinert, R.E.,  , L., Graber-Maier, A., and Beglinger, C. (2011). The role of the gut sweet taste receptor in regulating GLP-1, PYY, and CCK release in humans. Am J Physiol Endocrinol Metab 301, E317-E325.
, L., Graber-Maier, A., and Beglinger, C. (2011). The role of the gut sweet taste receptor in regulating GLP-1, PYY, and CCK release in humans. Am J Physiol Endocrinol Metab 301, E317-E325.
Glasgow, I., Mattar, K., and Krantis, A. (1998). Rat gastroduodenal motility in vivo: involvement of NO and ATP in spontaneous motor activity. Am. J. Physiol. Gastrointest. Liver Physiol 275, G889-G896.
Goichon, A.,  , M., Claeyssens, S., Lecleire, S., Cailleux, A.-F., Bole-Feysot, C., Chan, P., Donnadieu, N., Lerebours, E., Lavoinne, A., et al. (2011). Effects of an enteral glucose supply on protein synthesis, proteolytic pathways, and proteome in human duodenal mucosa. The American Journal of Clinical Nutrition 94, 784-794.
, M., Claeyssens, S., Lecleire, S., Cailleux, A.-F., Bole-Feysot, C., Chan, P., Donnadieu, N., Lerebours, E., Lavoinne, A., et al. (2011). Effects of an enteral glucose supply on protein synthesis, proteolytic pathways, and proteome in human duodenal mucosa. The American Journal of Clinical Nutrition 94, 784-794.
Haange, S.B., Oberbach, A., Schlichting, N., Hugenholtz, F., Smidt, H., von Bergen, M., Till, H., and Seifert, J. (2012). Metaproteome analysis and molecular genetics of rat intestinal microbiota reveals section and localization resolved species distribution and enzymatic functionalities. J Proteome Res 11, 5406-5417.
Halaas, J.L., Gajiwala, K.S., Maffei, M., Cohen, S.L., Chait, B.T., Rabinowitz, D., Lallone, R.L., Burley, S.K., and Friedman, J.M. (1995). Weight-reducing effects of the plasma protein encoded by the obese gene. Science 269, 543-546.
Harte, A.L., Varma, M.C., Tripathi, G., McGee, K.C., Al-Daghri, N.M., Al-Attas, O.S., Sabico, S.,  , J.P., Ceriello, A., Saravanan, P., et al. (2012). High fat intake leads to acute postprandial exposure to circulating endotoxin in type 2 diabetic subjects. Diabetes Care 35, 375-382.
, J.P., Ceriello, A., Saravanan, P., et al. (2012). High fat intake leads to acute postprandial exposure to circulating endotoxin in type 2 diabetic subjects. Diabetes Care 35, 375-382.
Herzog, H. (2003). Neuropeptide Y and energy homeostasis: insights from Y receptor knockout models. Eur J Pharmacol 480, 21-29.
Heymsfield, S.B., Greenberg, A.S., Fujioka, K., Dixon, R.M., Kushner, R., Hunt, Т., Lubina, J.A., Patane, J., Self, В., Hunt, P., et al. (1999). Recombinant leptin for weight loss in obese and lean adults: A randomized, controlled, dose-escalation trial. JAMA 282, 1568-1575.
Inui, A. (1999). Feeding and body-weight regulation by hypothalamic neuropeptides--mediation of the actions of leptin. Trends Neurosci 22, 62-67.
Johnstone, L.E., Fong, T.M., and Leng, G. (2006). Neuronal activation in the hypothalamus and brainstem during feeding in rats. Cell Metabolism 4, 313-321.
Ley, R.E., Turnbaugh, P.J., Klein, S., and Gordon, J.I. (2006). Microbial ecology: Human gut microbes associated with obesity. Nature 444, 1022-1023.
Lim, P.L., and Rowley, D. (1985). Intestinal absorption of bacterial antigens in normal adult mice. II. A comparative study of techniques. Int Arch Allergy Appl Immunol 76, 30-36.
Lin, S., Shi, Y.-C, Yulyaningsih, E., Aljanova, A., Zhang, L., Macia, L., Nguyen, A.D., Lin, E.-J.D., During, M.J., Herzog, H., et al. (2009). Critical role of arcuate Y4 receptors and the melanocortin system in pancreatic polypeptide-induced reduction in food intake in mice. PLoS One 4, e8488.
Lu, X.Y., Barsh, G.S., Akil, H., and Watson, S.J. (2003). Interaction between alpha-melanocyte-stimulating hormone and corticotropin-releasing hormone in the regulation of feeding and hypothalamo-pituitary-adrenal responses. J Neurosci 23, 7863-7872.
Manning, S., and Batterham, Rachel L. (2014). Enteroendocrine MC4R and energy balance: linking the long and the short of it. Cell Metabolism 20, 929-931.
Meguid, M.M., Laviano, A., and Rossi-Fanelli, F. (1998). Food intake equals meal size times mean number. Appetite 31, 404.
Mounien, L., Bizet, P., Boutelet, I., Vaudry, H., and  , S. (2005). Expression of melanocortin MC3 and MC4 receptor mRNAs by neuropeptide Y neurons in the rat arcuate nucleus. Neuroendocrinology 82, 164-170.
, S. (2005). Expression of melanocortin MC3 and MC4 receptor mRNAs by neuropeptide Y neurons in the rat arcuate nucleus. Neuroendocrinology 82, 164-170.
Mul, J.D., Spruijt, B.M., Brakkee, J.H., and Adan, R.A.H. (2013). Melanocortin MC4 receptor-mediated feeding and grooming in rodents. European Journal of Pharmacology 719, 192-201.
Murphy, K.G., and Bloom, S.R. (2006). Gut hormones and the regulation of energy homeostasis. Nature 444, 854-859.
Neunlist, M., Van Landeghem, L., Mahe, M.M., Derkinderen, P., Bruley des Varannes, S.B., and Rolli-Derkinderen, M. (2012). The digestive neuronal-glial-epithelial unit: a new actor in gut health and disease. Nat Rev Gastroenterol Hepatol 10, 90-100.
Panaro, B.L., Tough, I.R., Engelstoft, M.S., Matthews, R.T., Digby, G.J., Moller, C.L., Svendsen, В., Gribble, F., Reimann, F., Hoist, J.J., et al. (2014). The melanocortin-4 receptor is expressed in enteroendocrine L cells and regulates the release of peptide YY and glucagon-like peptide 1 in vivo. Cell Metab 20, 1018-1029.
Parks, B.W., Nam, E., Org, E., Kostem, E., Norheim, F., Hui, S.T., Pan, C., Civelek, M., Rau, C.D., Bennett, B.J., et al. (2013). Genetic control of obesity and gut microbiota composition in response to high-fat, high-sucrose diet in mice. Cell Metabolism 17, 141-152.
Power, M.L., and Schulkin, J. (2008). Anticipatory physiological regulation in feeding biology: Cephalic phase responses. Appetite 50, 194-206.
Rice, K.C, and Bayles, K.W. (2008). Molecular control of bacterial death and lysis. Microbiology and Molecular Biology Reviews 72, 85-109.
Sharon, G., Garg, N., Debelius, J., Knight, R., Dorrestein, Pieter C., and Mazmanian, Sarkis K. (2014). Specialized metabolites from the microbiome in health and disease. Cell Metabolism 20, 719-730.
Shi, Y.-C, Lau, J., Lin, Z., Zhang, H., Zhai, L., Sperk, G., Heilbronn, R., Mietzsch, M., Weger, S., Huang, X.-F., et al. (2013). Arcuate NPY controls sympathetic output and BAT function via a relay of tyrosine hydroxylase neurons in the PVN. Cell Metabolism 17, 236-248.
Smith, M.A., Hisadome, K., Al-Qassab, H., Heffron, H., Withers, D.J., and Ashford, M.L. (2007). Melanocortins and agouti-related protein modulate the excitability of two arcuate nucleus neuron populations by alteration of resting potassium conductances. J Physiol 578, 425-438.
Steinert, R.E., Schirra, J., Meyer-Gerspach, A.C, Kienle, P., Fischer, H., Schulte, F., Goeke, В., and Beglinger, С. (2014). Effect of glucagon-like peptide-1 receptor antagonism on appetite and food intake in healthy men. The American Journal of Clinical Nutrition 100, 514-523.
Sternson, S.M., Shepherd, G.M.G., and Friedman, J.M. (2005). Topographic mapping of VMH - arcuate nucleus microcircuits and their reorganization by fasting. Nat Neurosci 8, 1356-1363.
Takagi, K., Legrand, R., Asakawa, A., Amitani, H.,  , M., Tennoune, N.,
, M., Tennoune, N.,  , M., Claeyssens, S., do Rego, J.-C.,
, M., Claeyssens, S., do Rego, J.-C.,  , P., et al. (2013). Anti-ghrelin immunoglobulins modulate ghrelin stability and its orexigenic effect in obese mice and humans. Nat Commun 4:2685.
, P., et al. (2013). Anti-ghrelin immunoglobulins modulate ghrelin stability and its orexigenic effect in obese mice and humans. Nat Commun 4:2685.
Tennoune, N., Chan, P., Breton, J., Legrand, R., Chabane, Y.N., Akkermann, K., Jarv, A., Ouelaa, W., Takagi, K., Ghouzali, I., et al. (2014). Bacterial ClpB heat-shock protein, an antigen-mimetic of the anorexigenic peptide [alpha]-MSH, at the origin of eating disorders. Transl Psychiatry 4, e458.
Tennoune, N., Legrand, R., Ouelaa, W., Breton, J., Lucas, N., Bole-Feysot, C., Rego, J.-C.d.,  , P., and Fetissov, S.O. (2015). Sex-related effects of nutritional supplementation of Escherichia coli: Relevance to eating disorders. Nutrition 31, 498-507.
, P., and Fetissov, S.O. (2015). Sex-related effects of nutritional supplementation of Escherichia coli: Relevance to eating disorders. Nutrition 31, 498-507.
Turnbaugh, P.J., Ley, R.E., Mahowald, M.A., Magrini, V., Mardis, E.R., and Gordon, J.I. (2006). An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 444, 1027-1131.
van der Klaauw, A.A., Keogh, J.M., Henning, E., Trowse, V.M., Dhillo, W.S., Ghatei, M.A., and Farooqi, I.S. (2013). High protein intake stimulates postprandial GLP1 and PYY release. Obesity 21, 1602-1607.
Vijay-Kumar, M., Aitken, J.D., Carvalho, F.A., Cullender, T.C., Mwangi, S., Srinivasan, S., Sitaraman, S.V., Knight, R., Ley, R.E., and Gewirtz, A.T. (2010). Metabolic syndrome and altered gut microbiota in mice lacking toll-like receptor 5. Science 328, 228-231.
Wick, L.M., Quadroni, M., and Egli, T. (2001). Short- and long-term changes in proteome composition and kinetic properties in a culture of Escherichia coli during transition from glucose-excess to glucose-limited growth conditions in continuous culture and vice versa. Environ Microbiol 3, 588-599.
Xu, В., Goulding, E.H., Zang, K., Cepoi, D., Cone, R.D., Jones, K.R., Tecott, L.H., and Reichardt, L.F. (2003). Brain-derived neurotrophic factor regulates energy balance downstream of melanocortin-4 receptor. Nat Neurosci 6, 736-742.
Yun, A.J., Lee, P. Y., Doux, J.D., and Conley, B.R. (2006). A general theory of evolution based on energy efficiency: its implications for diseases. Medical Hypotheses 66, 664-670.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Inserm
<120> ФАРМАЦЕВТИЧЕСКИЕ И ПИЩЕВЫЕ КОМПОЗИЦИИ ДЛЯ ИНДУЦИКЦИИ НАСЫЩЕНИЯ И ПРОДЛЕНИЯ
ЧУВСТВА СЫТОСТИ У НУЖДАЮЩИХСЯ В ЭТОМ СУБЪЕКТОВ
<130> CDM - 692/PCT
<160> 2
<170> PatentIn версия 3.5
<210> 1
<211> 857
<212> БЕЛОК
<213> Escherichia coli
<400> 1
Met Arg Leu Asp Arg Leu Thr Asn Lys Phe Gln Leu Ala Leu Ala Asp
1 5 10 15
Ala Gln Ser Leu Ala Leu Gly His Asp Asn Gln Phe Ile Glu Pro Leu
20 25 30
His Leu Met Ser Ala Leu Leu Asn Gln Glu Gly Gly Ser Val Ser Pro
35 40 45
Leu Leu Thr Ser Ala Gly Ile Asn Ala Gly Gln Leu Arg Thr Asp Ile
50 55 60
Asn Gln Ala Leu Asn Arg Leu Pro Gln Val Glu Gly Thr Gly Gly Asp
65 70 75 80
Val Gln Pro Ser Gln Asp Leu Val Arg Val Leu Asn Leu Cys Asp Lys
85 90 95
Leu Ala Gln Lys Arg Gly Asp Asn Phe Ile Ser Ser Glu Leu Phe Val
100 105 110
Leu Ala Ala Leu Glu Ser Arg Gly Thr Leu Ala Asp Ile Leu Lys Ala
115 120 125
Ala Gly Ala Thr Thr Ala Asn Ile Thr Gln Ala Ile Glu Gln Met Arg
130 135 140
Gly Gly Glu Ser Val Asn Asp Gln Gly Ala Glu Asp Gln Arg Gln Ala
145 150 155 160
Leu Lys Lys Tyr Thr Ile Asp Leu Thr Glu Arg Ala Glu Gln Gly Lys
165 170 175
Leu Asp Pro Val Ile Gly Arg Asp Glu Glu Ile Arg Arg Thr Ile Gln
180 185 190
Val Leu Gln Arg Arg Thr Lys Asn Asn Pro Val Leu Ile Gly Glu Pro
195 200 205
Gly Val Gly Lys Thr Ala Ile Val Glu Gly Leu Ala Gln Arg Ile Ile
210 215 220
Asn Gly Glu Val Pro Glu Gly Leu Lys Gly Arg Arg Val Leu Ala Leu
225 230 235 240
Asp Met Gly Ala Leu Val Ala Gly Ala Lys Tyr Arg Gly Glu Phe Glu
245 250 255
Glu Arg Leu Lys Gly Val Leu Asn Asp Leu Ala Lys Gln Glu Gly Asn
260 265 270
Val Ile Leu Phe Ile Asp Glu Leu His Thr Met Val Gly Ala Gly Lys
275 280 285
Ala Asp Gly Ala Met Asp Ala Gly Asn Met Leu Lys Pro Ala Leu Ala
290 295 300
Arg Gly Glu Leu His Cys Val Gly Ala Thr Thr Leu Asp Glu Tyr Arg
305 310 315 320
Gln Tyr Ile Glu Lys Asp Ala Ala Leu Glu Arg Arg Phe Gln Lys Val
325 330 335
Phe Val Ala Glu Pro Ser Val Glu Asp Thr Ile Ala Ile Leu Arg Gly
340 345 350
Leu Lys Glu Arg Tyr Glu Leu His His His Val Gln Ile Thr Asp Pro
355 360 365
Ala Ile Val Ala Ala Ala Thr Leu Ser His Arg Tyr Ile Ala Asp Arg
370 375 380
Gln Leu Pro Asp Lys Ala Ile Asp Leu Ile Asp Glu Ala Ala Ser Ser
385 390 395 400
Ile Arg Met Gln Ile Asp Ser Lys Pro Glu Glu Leu Asp Arg Leu Asp
405 410 415
Arg Arg Ile Ile Gln Leu Lys Leu Glu Gln Gln Ala Leu Met Lys Glu
420 425 430
Ser Asp Glu Ala Ser Lys Lys Arg Leu Asp Met Leu Asn Glu Glu Leu
435 440 445
Ser Asp Lys Glu Arg Gln Tyr Ser Glu Leu Glu Glu Glu Trp Lys Ala
450 455 460
Glu Lys Ala Ser Leu Ser Gly Thr Gln Thr Ile Lys Ala Glu Leu Glu
465 470 475 480
Gln Ala Lys Ile Ala Ile Glu Gln Ala Arg Arg Val Gly Asp Leu Ala
485 490 495
Arg Met Ser Glu Leu Gln Tyr Gly Lys Ile Pro Glu Leu Glu Lys Gln
500 505 510
Leu Glu Ala Ala Thr Gln Leu Glu Gly Lys Thr Met Arg Leu Leu Arg
515 520 525
Asn Lys Val Thr Asp Ala Glu Ile Ala Glu Val Leu Ala Arg Trp Thr
530 535 540
Gly Ile Pro Val Ser Arg Met Met Glu Ser Glu Arg Glu Lys Leu Leu
545 550 555 560
Arg Met Glu Gln Glu Leu His His Arg Val Ile Gly Gln Asn Glu Ala
565 570 575
Val Asp Ala Val Ser Asn Ala Ile Arg Arg Ser Arg Ala Gly Leu Ala
580 585 590
Asp Pro Asn Arg Pro Ile Gly Ser Phe Leu Phe Leu Gly Pro Thr Gly
595 600 605
Val Gly Lys Thr Glu Leu Cys Lys Ala Leu Ala Asn Phe Met Phe Asp
610 615 620
Ser Asp Glu Ala Met Val Arg Ile Asp Met Ser Glu Phe Met Glu Lys
625 630 635 640
His Ser Val Ser Arg Leu Val Gly Ala Pro Pro Gly Tyr Val Gly Tyr
645 650 655
Glu Glu Gly Gly Tyr Leu Thr Glu Ala Val Arg Arg Arg Pro Tyr Ser
660 665 670
Val Ile Leu Leu Asp Glu Val Glu Lys Ala His Pro Asp Val Phe Asn
675 680 685
Ile Leu Leu Gln Val Leu Asp Asp Gly Arg Leu Thr Asp Gly Gln Gly
690 695 700
Arg Thr Val Asp Phe Arg Asn Thr Val Val Ile Met Thr Ser Asn Leu
705 710 715 720
Gly Ser Asp Leu Ile Gln Glu Arg Phe Gly Glu Leu Asp Tyr Ala His
725 730 735
Met Lys Glu Leu Val Leu Gly Val Val Ser His Asn Phe Arg Pro Glu
740 745 750
Phe Ile Asn Arg Ile Asp Glu Val Val Val Phe His Pro Leu Gly Glu
755 760 765
Gln His Ile Ala Ser Ile Ala Gln Ile Gln Leu Lys Arg Leu Tyr Lys
770 775 780
Arg Leu Glu Glu Arg Gly Tyr Glu Ile His Ile Ser Asp Glu Ala Leu
785 790 795 800
Lys Leu Leu Ser Glu Asn Gly Tyr Asp Pro Val Tyr Gly Ala Arg Pro
805 810 815
Leu Lys Arg Ala Ile Gln Gln Gln Ile Glu Asn Pro Leu Ala Gln Gln
820 825 830
Ile Leu Ser Gly Glu Leu Val Pro Gly Lys Val Ile Arg Leu Glu Val
835 840 845
Asn Glu Asp Arg Ile Val Ala Val Gln
850 855
<210> 2
<211> 13
<212> БЕЛОК
<213> Homo sapiens
<400> 2
Ser Tyr Ser Met Glu His Phe Arg Trp Gly Lys Pro Val
1 5 10
<---
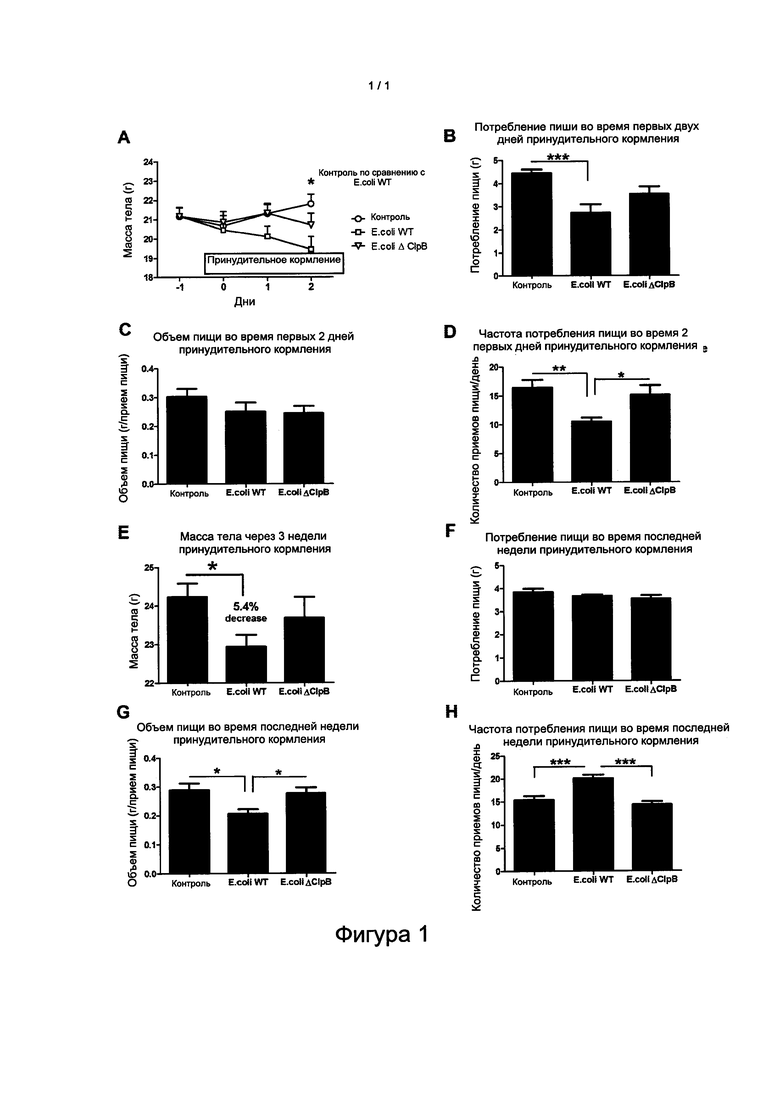
Предложены способ индукции насыщения у нуждающегося в этом субъекта, способ пролонгирования чувства сытости у нуждающегося в этом субъекта, способ снижения потребления пищи у нуждающегося в этом субъекта, способ снижения увеличения массы тела у нуждающегося в этом субъекта, способ стимуляции уменьшения массы тела у нуждающегося в этом субъекта. Изобретения предусматривают пероральное введение указанному субъекту эффективного количества бактерии, которая экспрессирует белок ClpB, который содержит последовательность ARWTGIPVSR. Изобретения позволяют вызывать чувство насыщения, продлевать чувство сытости, снижать потребление пищи, контролировать увеличение массы тела и/или стимулировать уменьшение массы тела у субъекта. 5 н. и 39 з.п. ф-лы, 1 ил., 2 пр.
1. Способ индукции насыщения у нуждающегося в этом субъекта, включающий пероральное введение указанному субъекту эффективного количества бактерии, которая экспрессирует белок ClpB, который содержит последовательность ARWTGIPVSR.
2. Способ пролонгирования чувства сытости у нуждающегося в этом субъекта, включающий пероральное введение указанному субъекту эффективного количества бактерии, которая экспрессирует белок ClpB, который содержит последовательность ARWTGIPVSR.
3. Способ снижения потребления пищи у нуждающегося в этом субъекта, включающий пероральное введение указанному субъекту эффективного количества бактерии, которая экспрессирует белок ClpB, который содержит последовательность ARWTGIPVSR.
4. Способ снижения увеличения массы тела у нуждающегося в этом субъекта, включающий пероральное введение указанному субъекту эффективного количества бактерии, которая экспрессирует белок ClpB, который содержит последовательность ARWTGIPVSR.
5. Способ стимуляции уменьшения массы тела у нуждающегося в этом субъекта, включающий пероральное введение указанному субъекту эффективного количества бактерии, которая экспрессирует белок ClpB, который содержит последовательность ARWTGIPVSR.
6. Способ по пп. 1-4 или 5, отличающийся тем, что указанный субъект представляет собой человека, домашнее животное или сельскохозяйственное животное.
7. Способ по пп. 1-4 или 5, отличающийся тем, что указанный субъект представляет собой особь мужского пола или особь женского пола.
8. Способ по пп. 1-4 или 5, отличающийся тем, что указанный субъект не страдает ожирением.
9. Способ по пп. 1-4 или 5, отличающийся тем, что указанный субъект страдает ожирением.
10. Способ по пп. 1-4 или 5, отличающийся тем, что индекс массы тела (ИМТ) указанного субъекта находится в интервале от 18,5 и до 30.
11. Способ по пп. 1-4 или 5, отличающийся тем, что указанный белок ClpB содержит аминокислотную последовательность, по меньшей мере на 80% идентичную последовательности SEQ ID NO:1.
12. Способ по пп. 1-4 или 5, отличающийся тем, что указанный белок ClpB содержит аминокислотную последовательность, по меньшей мере на 95, 96, 97, 98, 99 или 100 % идентичную последовательности SEQ ID NO:1.
13. Способ по пп. 1-4 или 5, отличающийся тем, что указанную бактерию, которая экспрессирует белок ClpB, вводят указанному субъекту в виде фармацевтической композиции.
14. Способ по пп. 1-4 или 5, отличающийся тем, что указанную бактерию, которая экспрессирует белок ClpB, вводят указанному субъекту в виде пищевой композиции.
15. Способ по п. 13, отличающийся тем, что указанную фармацевтическую композицию вводят указанному субъекту в виде единичной формы для введения, выбранной из группы, состоящей из форм, вводимых пероральным путем, таких как таблетки, желатиновые капсулы, порошки, гранулы и суспензии или растворы для перорального введения, форм для сублингвального или буккального введения.
16. Способ по пп. 1-4 или 5, отличающийся тем, что указанная бактерия, которая экспрессирует белок ClpB, представляет собой пробиотический штамм бактерий.
17. Способ по п. 16, отличающийся тем, что указанный пробиотический штамм бактерий выбран из пищевых бактерий.
18. Способ по п. 16, отличающийся тем, что указанный пробиотический штамм бактерий представляет собой природный штамм бактерий или штамм бактерий, полученный путем генной инженерии.
19. Способ по п. 16, отличающийся тем, что указанный пробиотический штамм бактерий представляет собой бактерию, которую подвергали условиям стресса, приводящим к увеличению экспрессии белка ClpB в указанной бактерии.
20. Способ по п. 16, отличающийся тем, что указанный пробиотический штамм бактерий представляет собой бактерию, которую снабжали питательными веществами по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 раз.
21. Способ по п. 20, отличающийся тем, что указанная бактерия выделена на стадии стационарной фазы роста, которая следует за указанным повторяемым снабжением питательными веществами.
22. Способ по п. 16, отличающийся тем, что указанный пробиотический штамм бактерий содержит по меньшей мере одну точечную мутацию, приводящую к увеличению экспрессии белка ClpB.
23. Способ по п. 16, отличающийся тем, что указанный пробиотический штамм бактерий представляет собой бактерию, сконструированную путем генной инженерии для экспрессии белка ClpB.
24. Способ по п. 16, отличающийся тем, что указанный пробиотический штамм бактерий представляет собой штамм E. coli.
25. Способ по п. 16, отличающийся тем, что указанный пробиотический штамм бактерий инкапсулируют для защиты от среды желудка.
26. Способ по п. 25, отличающийся тем, что указанный пробиотический штамм бактерий инкапсулируют в капсулы или таблетки с кишечнорастворимым покрытием с контролируемым по времени высвобождением.
27. Способ по п. 16, отличающийся тем, что указанный пробиотический штамм бактерий вводят в виде пищевой композиции.
28. Способ по п. 27, отличающийся тем, что указанная пищевая композиция, которая содержит указанный пробиотический штамм бактерий, выбрана из полноценных пищевых композиций, пищевых добавок, нутрицевтических композиций и т.п.
29. Способ по п. 27, отличающийся тем, что указанный пробиотический штамм бактерий используют в качестве пищевого или кормового ингредиента в указанной пищевой композиции.
30. Способ по п. 27, отличающийся тем, что указанная пищевая композиция представляет собой жидкую, твердую или полутвердую пищевую композицию.
31. Способ по п. 27, отличающийся тем, что указанная пищевая композиция представляет собой кисломолочный продукт.
32. Способ по п. 27, отличающийся тем, что указанная пищевая композиция представляет собой функциональный пищевой продукт.
33. Способ по п. 27, отличающийся тем, что указанная пищевая композиция представляет собой напиток.
34. Способ по п. 27, отличающийся тем, что указанная пищевая композиция представляет собой заменитель пищевого продукта.
35. Способ по п. 27, отличающийся тем, что указанная пищевая композиция содержит некоторое количество пищевых волокон.
36. Способ по п. 35, отличающийся тем, что указанное пищевое волокно выбрано из группы, состоящей из сои, гороха, овса, пектина, гуаровой камеди, гуммиарабика, фруктоолигосахаридов, галакто-олигосахаридов, сиалиллактозы и олигосахаридов, полученных из молока животных.
37. Способ по п. 35, отличающийся тем, что указанное пищевое волокно выбрано из маннанов, таких как глюкоманнаны.
38. Способ по п. 37, отличающийся тем, что указанное пищевое волокно выбрано из маннанов, таких как глюкоманнаны Amorphophallus konjac.
39. Способ по п. 27, отличающийся тем, что указанная пищевая композиция содержит по меньшей мере один пребиотик.
40. Способ по п. 39, отличающийся тем, что указанный пребиотик выбран из группы, состоящей из олигосахаридов, и необязательно содержит фруктозу, галактозу, маннозу, сою, и/или инулин, и/или пищевые волокна.
41. Способ по пп. 1-4 или 5, отличающийся тем, что введение бактерии, которая экспрессирует белок ClpB, повторяют от 2 до 3 раз в день, в течение одного дня или более.
42. Способ по п. 41, отличающийся тем, что указанное введение осуществляют в течение продолжительного периода, составляющего по меньшей мере 4 дня, или даже от 4 до 15 недель, или от 1 до 3 месяцев, от 1 до 3 лет, или постоянно.
43. Способ по пп. 1-4 или 5, отличающийся тем, что бактерию, которая экспрессирует белок ClpB, вводят одновременно или последовательно с одним приемом пищи указанным субъектом.
44. Способ по пп. 1-4 или 5, отличающийся тем, что бактерию, которая экспрессирует белок ClpB, вводят до приема пищи указанным субъектом.
| US 6716810 B1, 06.04.2004 | |||
| TENNOUNE N | |||
| Validation du concept du mimetisme moleculaire entre les proteines d'E | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| These de doctorate en Biologie | |||
| Sous la direction de Serguei Fetissov | |||
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| HELLSTROM P.M | |||
| ET AL | |||
| DIGESTIVO Interactions between gastric emptying and | |||

