Область техники, к которой относится изобретение
В настоящем изобретении предложены фармацевтические и пищевые композиции для индуцирования насыщения и продления сытости у субъектов, нуждающихся в этом.
Уровень техники
Состав микрофлоры кишечника связан с метаболическими фенотипами хозяина (Ley et al., 2006), и перенос микрофлоры, характерной для «ожирения», может индуцировать отложение жира (Turnbaugh et al., 2006) и гиперфагию (Vijay-Kumar et al., 2010), указывая на то, что кишечная микрофлора может влиять на пищевое поведение хозяина. Несмотря на то, что механизмы, лежащие в основе воздействия кишечных бактерий на аппетит хозяина, неизвестны, вполне вероятно, что указанные бактерии могут использовать молекулярные сигнальные пути хозяина, контролирующие потребление пищи.
Современная модель контроля потребления пищи включает происходящие из кишечника гормоны голода и насыщения, передающие сигнал в несколько систем межнейронных связей, регулирующих гомеостатические и гедонические аспекты потребления пищи (Berthoud, 2011; Inui, 1999; Murphy and Bloom, 2006). Среди таких систем выделяют анорексигенные и орексигенные пути, берущие начало от дугообразного (аркуатного) ядра (ARC) гипоталамуса, которые включают проопиомеланокортин (РОМС)-содержащие нейроны и нейропептид Y (NPY)/агути- подобный белок (AgRP)-содержащие нейроны, соответственно, связанные с паравентрикулярным ядром (PVN) (Atasoy et al., 2012; Cowley et al., 1999; Garfield et al., 2015; Shi et al., 2013). Пути ARC и PVN сходятся в латеральном парабрахиальном ядре, которое посылает анорексигенные проекции на центральное ядро миндалевидного тела (СеА), экспрессирующее пептид, связанный с геном кальцитонина (CGRP) (Carter et al, 2013; Garfield et al., 2015).
Предполагаемые механизмы воздействия микрофлоры кишечника на контроль аппетита хозяина могут включать активность микрофлоры по накоплению энергии (Turnbaugh et al., 2006) и продукцию нейромедиаторов и метаболитов (Dinan et al., 2015; Forsythe and Kunze, 2013; Sharon et al., 2014). Авторы настоящего изобретения подтвердили наличие бактериальных белков, которые действуют непосредственно на пути, контролирующие аппетит, локально в кишечнике или системно. Было показано, что несколько бактериальных белков демонстрируют гомологию последовательности с пептидными гормонами, регулирующими аппетит (Fetissov et al., 2008), а недавно белок ClpB, продуцируемый симбиотической бактерией кишечника Escherichia coli (E. coli), был идентифицирован как антиген-миметик α-меланоцит-стимулирующего гормона (α-MSH) (Tennoune et al., 2014). α-MSH представляет собой - нейропептид-производное РОМС, играющий ключевую роль в передаче сигнала насыщения путем активации рецепторов меланокортина-4 (MC4R) (Cone, 2005). Несмотря на то, что MC4R- опосредованные анорексигенные эффекты α-MSH в основном относили к его основным областям действия (Mul et al., 2013), недавнее исследование показало, что активация MC4R в энтероэндокринных клетках кишечника стимулирует выработку гормонов насыщения глюкагонподобного пептида-1 (GLP-1) и пептида YY (PYY) (Panaro et al., 2014). Таким образом, α-MSH-подобные молекулы, продуцируемые бактериями кишечника, могут непосредственно воздействовать на энтероэндокринные клетки, синтезирующие гормоны насыщения.
Краткое описание изобретения
В настоящем изобретении предложены фармацевтические и пищевые композиции для индуцирования (индукции) насыщения (чувства насыщения) и продления сытости (чувства сытости) у субъектов, нуждающихся в этом. В частности, настоящее изобретение определено формулой изобретения.
Настоящее изобретение относится к способу индуцирования насыщения у субъектов, нуждающихся в этом, заключающемуся во введении субъекту эффективного количества Hafnia alvei.
Настоящее изобретение, кроме того, относится к способу продления сытости у субъектов, нуждающихся в этом, заключающемуся во введении субъекту эффективного количества Hafnia alvei.
Настоящее изобретение также относится к способу снижения объема пищи у субъектов, нуждающихся в этом, заключающемуся во введении субъекту эффективного количества Hafnia alvei
Другим предметом настоящего изобретения является способ снижения потребления пищи у субъектов, нуждающихся в этом, заключающийся во введении субъекту эффективного количества Hafnia alvei
Настоящее изобретение, помимо того, относится к способу контроля набора веса у субъектов, нуждающихся в этом, заключающемуся во введении субъекту эффективного количества Hafnia alvei
Следующим предметом настоящего изобретения является способ стимулирования снижения веса у субъектов, нуждающихся в этом, заключающийся во введении субъекту эффективного количества Hafnia alvei
В одном из вариантов реализации настоящего изобретения субъект не страдает ожирением. В другом варианте реализации настоящего изобретения субъект страдает ожирением.
В одном из вариантов реализации настоящего изобретения Hafnia alvei вводится субъекту в форме фармацевтической композиции. В другом варианте реализации настоящего изобретения Hafnia alvei вводится субъекту в форме пищевой композиции.
В одном из вариантов реализации настоящего изобретения пищевая композиция выбирается из группы, включающей законченные пищевые композиции, пищевые добавки и нутрицевтические композиции.
В одном из вариантов реализации настоящего изобретения Hafnia alvei представляет собой пробиотик. В другом варианте реализации настоящего изобретения Hafnia alvei инактивирована.
В одном из вариантов реализации настоящего изобретения пищевая композиция включает, по меньшей мере, один пребиотик. В одном из вариантов реализации настоящего изобретения упомянутый один пребиотик представляет собой пищевые волокна.
Подробное описание изобретения
Авторы настоящего изобретения продемонстрировали, что белок ClpB или бактерия, которая экспрессирует белок ClpB, способны индуцировать насыщение, продлевать сытость, уменьшать потребление пищи, контролировать увеличение веса, стимулировать потерю веса и/или уменьшать отношение массы жировой ткани к массе нежировых тканей тела у субъекта, нуждающегося в этом. Авторы настоящего изобретения действительно неожиданно показали, что белок ClpB, который экспрессирует указанная бактерия, оказывает прямое действие на насыщение, сытость и потребление пищи, вероятно, через рецепторы MCR, независимо от любой иммунной реакции, которая могла быть вызвана введением бактерий.
Соответственно, один аспект настоящего изобретения относится к способу индуцирования насыщения у нуждающегося в этом субъекта, включающему введение субъекту эффективного количества белка ClpB или варианта такового, или эффективного количества бактерии, которая экспрессирует белок ClpB или вариант такового.
Другой аспект настоящего изобретения относится к белку ClpB или варианту такового, или к бактерии, которая экспрессирует белок ClpB или вариант такового, для применения в целях индуцирования насыщения у субъекта, нуждающегося в этом, в частности, у субъекта с ожирением.
Еще один аспект настоящего изобретения относится к косметическому нетерапевтическому применению белка ClpB или варианта такового или бактерии, которая экспрессирует белок ClpB или вариант такового, для индуцирования насыщения у субъекта, в частности у субъекта, имеющего нормальную массу тела или избыточную массу тела без осложнений.
Еще один аспект настоящего изобретения относится к способу продления сытости у нуждающегося в этом субъекта, включающему введение субъекту эффективного количества белка ClpB или варианта такового или эффективного количества бактерии, которая экспрессирует белок ClpB или вариант такового.
Другой аспект настоящего изобретения относится к белку ClpB или варианту такового, или к бактерии, которая экспрессирует белок ClpB или вариант такового, для применения для продления сытости у нуждающегося в этом субъекта, в частности, у субъекта с ожирением.
Еще один аспект настоящего изобретения относится к косметическому нетерапевтическому использованию белка ClpB или варианта такового или бактерии, которая экспрессирует белок ClpB или вариант такового, для продления сытости у субъекта, в частности у субъекта, имеющего нормальную массу тела или избыточную массу тела без осложнений.
Соответственно, один аспект настоящего изобретения относится к способу уменьшения количества пищи у нуждающегося в этом субъекта, включающему введение субъекту эффективного количества белка ClpB или варианта такового или эффективного количества бактерии, которая экспрессирует белок ClpB или вариант такового.
Другой аспект настоящего изобретения относится к белку ClpB или варианту такового или к бактерии, которая экспрессирует белок ClpB или вариант такового, для применения для уменьшения количества потребляемой пищи за один прием у нуждающегося в этом субъекта, в частности у субъекта с ожирением.
Еще один аспект настоящего изобретения относится к косметическому нетерапевтическому использованию белка ClpB или варианта такового или бактерии, которая экспрессирует белок ClpB или вариант такового, для уменьшения количества потребляемой пищи за один прием у субъекта, в частности у субъекта, имеющего нормальную массу тела или избыточную массу тела без осложнений.
Еще один аспект настоящего изобретения относится к способу снижения потребления пищи у нуждающегося в этом субъекта, включающему введение субъекту эффективного количества белка ClpB или варианта такового или эффективного количества бактерии, которая экспрессирует белок ClpB или вариант такового.
Другой аспект настоящего изобретения относится к белку ClpB или варианту такового или к бактерии, которая экспрессирует белок ClpB или вариант такового для применения в целях уменьшения потребления пищи у нуждающегося в этом субъекта, в частности, у субъекта с ожирением.
Еще один аспект настоящего изобретения касается косметического нетерапевтического применения белка ClpB или варианта такового или бактерии, которая экспрессирует белок ClpB или вариант такового, для снижения потребления пищи у субъекта, в частности у субъекта, имеющего нормальную массу тела или избыточную массу тела без осложнений.
Еще один аспект настоящего изобретения также относится к способу контролирования, в частности, снижения, увеличения массы тела у субъекта, нуждающегося в этом, включая введение субъекту эффективного количества белка ClpB или варианта такового или эффективного количества бактерии, которая экспрессирует белок ClpB или вариант такового.
Еще один аспект настоящего изобретения относится к белку ClpB или варианту такового или к бактерии, которая экспрессирует белок ClpB или вариант такового, для применения в целях контроля, в частности, снижения, увеличения массы тела, у нуждающегося в этом субъекта, в частности у субъекта с ожирением.
Еще один аспект настоящего изобретения относится к косметическому нетерапевтическому использованию белка ClpB или варианта такового или бактерии, которая экспрессирует белок ClpB или вариант такового, для контроля, в частности, снижения, увеличения массы тела у субъекта, в частности у субъекта, имеющего нормальную массу тела или избыточную массу тела без осложнений.
Еще один аспект настоящего изобретения также относится к способу стимуляции потери массы тела у нуждающегося в этом субъекта, включающему введение субъекту эффективного количества белка ClpB или варианта такового или эффективного количества бактерии, которая экспрессирует белок ClpB или вариант такового.
Другой аспект настоящего изобретения относится к белку ClpB или варианту такового или к бактерии, которая экспрессирует белок ClpB или вариант такового для применения в целях стимулирования потери массы тела у нуждающегося в этом субъекта, в частности, у субъекта с ожирением.
Еще один аспект настоящего изобретения относится к косметическому нетерапевтическому использованию белка ClpB или варианта такового или бактерии, которая экспрессирует белок ClpB или вариант такового, для стимулирования снижения массы тела у субъекта, в частности у субъекта, имеющего нормальную массу тела или избыточную массу тела без осложнений.
Еще один аспект настоящего изобретения относится к способу уменьшения отношения массы жировой ткани к массе нежировых тканей тела у нуждающегося в этом субъекта, включающему введение субъекту эффективного количества белка ClpB или варианта такового или эффективного количества бактерии, которая экспрессирует белок ClpB или вариант такового.
Другой аспект настоящего изобретения относится к белку ClpB или варианту такового или к бактерии, которая экспрессирует белок ClpB или вариант такового для применения в целях уменьшения отношения массы жировой ткани к массе нежировых тканей тела у нуждающегося в этом субъекта, в частности у субъекта с ожирением.
Еще один аспект настоящего изобретения относится к косметическому нетерапевтическому применению белка ClpB или варианта такового или бактерии, которая экспрессирует белок ClpB или вариант такового, для уменьшения отношения массы жировой ткани к массе нежировых тканей тела у субъекта, в частности у субъекта, имеющего нормальную массу тела или избыточную массу тела без осложнений.
В контексте настоящего документа термин «субъект» относится к теплокровным животным, в первую очередь, к людям, домашним животным и сельскохозяйственным животным. В контексте настоящего документа термины «домашнее животное» и «сельскохозяйственное животное» включают, в числе прочего, собак, кошек, морских свинок, кроликов, свиней, крупный рогатый скот, овец, коз, лошадей и птицу. В некоторых вариантах реализации настоящего изобретения субъект является субъектом женского или мужского пола. В некоторых вариантах реализации настоящего изобретения субъект является взрослым субъектом (например, человек в возрасте старше 18 лет). В других вариантах реализации настоящего изобретения субъект является ребенком (например, человек в возрасте младше 18 лет). В некоторых вариантах реализации настоящего изобретения субъект может являться «пациентом», то есть, субъектом, который ожидает получения или получает медицинскую помощь и лечение или был/ является/ будет являться объектом какой-либо медицинской процедуры в соответствии с методами настоящего изобретения, или в отношении которого ведется контроль развития заболевания.
В некоторых вариантах реализации, субъект имеет ожирение. «Ожирение» относится к медицинскому состоянию, в котором субъект предпочтительно имеет ИМТ>30. «ИМТ» или «индекс массы тела» определяют как массу тела субъекта, деленную на квадрат его высоты. Формулы, универсально используемые в медицине, дают единицу измерения кг/м2.
В некоторых вариантах реализации, субъект имеет умеренное ожирение. Субъект с «умеренным ожирением» относится к субъекту, имеющему ИМТ от 30 до 35.
В некоторых вариантах реализации, субъект имеет индекс массы тела от 18,5 до 30.
В некоторых вариантах реализации, субъект не имеет ожирения. Как правило, субъект без ожирения имеет нормальную массу тела. «Нормальная масса тела» относится к массе тела, при которой ИМТ составляет от 18,5 до 25.
В некоторых вариантах реализации, субъект имеет избыточную массу тела. «Избыточная масса тела» относится к массе тела, при которой ИМТ составляет от 25 до 30.
В некоторых вариантах реализации, субъект имеет здоровую избыточную массу тела или избыточную массу тела без осложнений. В настоящем описании, термин «здоровая избыточная масса тела» или «избыточная масса тела без осложнений» относится к субъекту с избыточной массой тела, у которого не обнаружено никаких заболеваний или состояний, непосредственно связанных с его/ее массой.
Согласно некоторым вариантам реализации, субъект находится на диете, способствующей снижению массы тела, и/или хочет снизить массу тела. Согласно другим вариантам реализации, субъект не находится на диете, способствующей снижению массы тела и/или не хочет снизить массу тела.
В настоящем описании, термин «сытость» означает «по существу гомеостатическое состояние», в котором индивидуум чувствует, что его аппетит утолен или снижен. Считается, что многие физиологические факторы влияют на сытость человека. Например, вкусовое ощущение, или вкус, обоняние или запах, а также ощущение наполненности желудка могут способствовать тому, что индивидуум чувствует себя «сытым». Более конкретно, «сытость» - это состояние, в котором дальнейший прием пищи приостановлен и определяет время между приемами пищи и количеством потребляемой пищи, при следующем приеме пищи. «Повышенное чувство сытости» или тому подобное, означает, что чувство сытости является более выраженным и/или более продолжительным по сравнению с ситуацией в контроле.
Используемый в настоящем описании термин «насыщение», относится к состоянию, при которое прекращается прием пищи во время еды, обычно происходящее/наблюдаемое в течение периода времени (например, 20-30 минут) после начала приема пищи. Таким образом, всякий раз, когда в настоящем документе делается ссылка на «индуцирование насыщения» или тому подобное, это означает появление тенденции у субъекта прекратить прием пищи во время еды. Влияние на насыщение может быть определено путем оценивания момента времени окончания приема пищи. Влияние на насыщение наблюдается, если количество потребляемых калорий при прекращении приема пищи достоверно меньше, чем в контроле, например, по меньшей мере на 1%, 2%, 3%, 4%, 5%, 10% 20% и больше. В течение более длительного периода времени (например, 1, 2, 3, 4, 5 недель и больше) можно также подсчитать снижение массы тела или изменение массы тела по сравнению с питанием в контроле. Масса тела субъекта, которому вводят регулярные количества испытываемых композиций (например, один раз в день, два раза в день или более), предпочтительно, значительно контролируется (снижается или уменьшается) по сравнению с контрольными субъектами.
В настоящем описании термин «контрольный субъект / субъект в контроле» относится к субъектам, которым не вводили пробиотические бактериальные штаммы по настоящему изобретению.
В настоящем описании термин «ClpB» имеет общепринятое в данной области значение, и также известен как белок F84.1 теплового шока, который является членом семейства Hsp100/ClpB гексамерных АТФаз ААА+. ClpB был описан как существенный фактор приобретенной термотолерантности и вирулентности и инфекционности у нескольких грамотрицательных и грамположительных патогенных бактерий, таких как Staphylococcus aureus, Francisella turalensis, Listeria monocytogenes, Yersinia enterocolitica и Salmonella thyphimurium. У бактерии E. coli K12, белок-шаперон ClpB также известен как белок теплового шока F84.1 или htpM и представляет собой белок из 857 аминокислот. Как правило, белок-шаперон ClpB включает, или состоит из аминокислотной последовательности белка-шаперона ClpB из Е. coli K12, с последовательностью SEQ ID NO: 1 (номер NCBI: NP_417083.1, как доступно на 6 ноября 2013 года и/или UniProtKB/Swiss-Prot Number: Р63284, как доступно на 6 ноября 2013 года). Как правило, аминокислотная последовательность белка-шаперона ClpB включает или состоит из аминокислотной последовательности на 96-100%, идентичной аминокислотной последовательности SEQ ID NO: 1. Предпочтительно, аминокислотная последовательность ClpB на 96, 97, 98, 99 или 100% идентична аминокислотной последовательности 540-550 (ARWTGIPVSR) SEQ ID NO: 1. В контексте настоящей заявки, процент идентичности вычисляется с использованием глобального выравнивания (т.е. две последовательности сравниваются по всей их длине). Способы сравнения идентичности двух или более последовательностей хорошо известны в данной области. Может, например, быть использована программа «needle», в которой используется глобальный алгоритм выравнивания Needleman-Wunsch (Needleman and Wunsch, 1970 J. Mol. Biol., 48: 443-453), чтобы найти оптимальное выравнивание (включая пробелы) двух последовательностей при рассмотрении всей их длины.
Программа "needle", например, доступна на сайте всемирной сети интернет ebi.ac.uk. Процент идентичности в соответствии с настоящим изобретением, предпочтительно, рассчитывают с использованием программы EMBOSS: программа needle (global) с параметром «Gap Open», равным 10,0, параметром «Gap Extend», равным 0,5, и матрицей Blosum62. Согласно изобретению, белок ClpB имитирует белок α-MSH для индуцирования насыщения. Таким образом, в некоторых вариантах реализации, белок ClpB по настоящему изобретению распознается антителом к альфа-MSH. Как правило, антитело представляет собой моноклональное антитело. В некоторых вариантах реализации, антитело представляет собой поликлональное антитело, например, поликлональное антитело кролика к иммуноглобулину G α-MSH (1:1000, Peninsula Laboratories, San Carlos, С А, США). Аминокислотная последовательность α-MSH, предпочтительно, включает или состоит из аминокислотной последовательности SYSMEHFRWGKPV (SEQ ID NO: 2) (Gen Pept Sequence ID, PRF: 223274, как доступно на 2 декабря 2013 г.).

В контексте настоящего документа «аминокислоты» представлены в виде их полных названий, трехбуквенных кодов или однобуквенных кодов, широко известных в данной области техники. Аминокислотные остатки в составе пептидов имеют следующие сокращения: фенилаланин - Phe или F; лейцин - Leu или L; изолейцин - Не или I; метионин - Met или М; валин - Val или V; серии - Ser или S; пролин - Pro или Р; треонин - Thr или Т; аланин - Ala или А; тирозин - Tyr или Y; гистидин - His или Н; глутамин - Gin или Q; аспарагин - Asn или N; лизин - Lys или K; аспарагиновая кислота - Asp или D; глутаминовая кислота - Glu или Е; цистеин - Cys или С; триптофан - Trp или W; аргинин - Arg или R; и глицин - Gly или G.
В контексте настоящего документа термин «аминокислоты» включает как натуральные, так и синтетические аминокислоты, а также D- и L-аминокислоты. «Стандартная аминокислота» или «природная аминокислота» означает любую из двадцати стандартных L-аминокислот, обычно встречающихся в составе природных пептидов. «Нестандартный аминокислотный остаток» означает любую аминокислоту, за исключением стандартных аминокислот, независимо от того, получена она синтетически или из природного источника. Например, нафтилаланин может использоваться вместо триптофана для содействия синтезу. Другие синтетические аминокислоты, которые могут использоваться в качестве заменителей, включают, в числе прочих, L-гидроксипропил, L-3,4-дигидроксифенилаланин, такие альфа-аминокислоты, как L-альфа-гидроксилизил и D-альфа-метилаланил, L-альфа-метилаланил, бета-аминокислоты, и изохинолил.
В контексте настоящего документа понятие «аминокислота» включает также химически модифицированные аминокислоты, включая, в числе прочего, соли, производные аминокислот (например, амиды) и заменители. Аминокислоты, содержащиеся в полипептидах настоящего изобретения, и, в частности, в составе карбоксильных и амино-концов, могут быть модифицированы путем метилирования, амидирования, ацетилирования или замены на другие химические группы, которые могут изменить период полувыведения полипептида, в то же время не оказывая негативного влияния на его активность. Кроме того, в составе полипептидов настоящего изобретения могут присутствовать или отсутствовать дисульфидные связи.
Белок ClpB или вариант такового по настоящему изобретению могут включать в себя природные стандартные аминокислоты или нестандартные аминокислоты. Полипептиды-миметики включают полипептиды, имеющие следующие модификации:
i) полипептиды, в которых одна или более пептидильных -C(O)NR-связей (групп) замещены на непептидильную связь, такую как -СН2-карбаматная связь (-CH2OC(O)NR-), фосфонатная связь, -СН2-сульфонамидная (-CH2-S(O)2NR-) связь, мочевинная (-NHC(O)NH-) связь, связь -СН2-вторичный амин, или на алкилированную пептидильную связь (-C(O)NR-), где R представляет собой С1-С4 алкил;
ii) полипептиды, в которых N-конец дериватизирован в -NRR1 группу, в -NRC(O)R группу, в -NRC(O)OR группу, в -NRS(O)2R группу, в -NHC(O)NHR группу, где R и R1 представляют собой водород или С1-С4 алкил при условии, что ни R, ни R1 не является водородом;
iii) полипептиды, в которых С-конец дериватизирован в -C(O)R2, где R2 выбирают из группы, состоящей из С1-С4-алкокси, и -NR3R4, где R3 и R4 независимо выбирают из группы, состоящей из водорода и С1-С4 алкила.
В одном варианте реализации настоящего изобретения белок ClpB или вариант такового включает в себя или состоит из аминокислотной последовательности SEQ ID NO: 1 или варианта таковой. В одном варианте реализации настоящего изобретения белок ClpB включает в себя или состоит из аминокислотной последовательности SEQ ID NO: 1.
В одном варианте реализации настоящего изобретения «вариант ClpB» в контексте настоящего документа включает в себя или состоит из последовательности, по крайней мере, на 70% идентичной аминокислотной последовательности SEQ IDNO: 1, предпочтительна идентичность, как минимум, на 80%, еще более предпочтительна идентичность на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более.
В контексте настоящего документа термины «идентичность» или «идентичный» при использовании для описания отношений между двумя и более аминокислотными последовательностями означает степень родства между аминокислотными последовательностями, определяемую числом совпадений между цепями, состоящими из двух или более аминокислотных остатков. «Идентичность» измеряет процент идентичных совпадений в отношении более короткой из двух или более последовательностей с добавлением разрывов (гэпов) (если таковые имеются) с помощью конкретной математической модели или компьютерной программы (то есть, «алгоритмов»). Методы определения степени идентичности двух или более последовательностей хорошо известны в данной области техники. Такие методы включают, в числе прочих, методы, описанные в следующих источниках: Arthur М. Lesk, Computational Molecular Biology: Sources and Methods for Sequence Analysis (New-York: Oxford University Press, 1988); Douglas W. Smith, Biocomputing: Informatics and Genome Projects (New-York: Academic Press, 1993); Hugh G. Griffin and Annette M. Griffin, Computer Analysis of Sequence Data, Part 1 (New Jersey: Humana Press, 1994); Gunnar von Heinje, Sequence Analysis in Molecular Biology: Treasure Trove or Trivial Pursuit (Academic Press, 1987); Michael Gribskov and John DQvereux,Sequence Analysis Primer (New York: M. Stockton Press, 1991); and Carillo etal, 1988. SI AM J. Appl. Math. 48(5): 1073-1082. Предпочтительными методами определения степени идентичности являются те, которые предназначены для максимизации похожести исследуемых последовательностей. Методы определения степени идентичности представлены в виде общедоступных компьютерных программ. Компьютерные программы для определения степени идентичности двух последовательностей включают пакет программ GCG, в том числе, GAP (Devereux et al., 1984. Nucl. Acid. Res. 12(1 Pt l):387-395; Genetics Computer Group, University of Wisconsin Biotechnology Center, Madison, WI), BLASTP, BLASTN, TBLASTN и FASTA (Altschul etal, 1990. J. Mol. Biol. 215(3):403-410). Программа BLASTX находится в общем доступе в Национальном центре биотехнологической информации (National Center for Biotechnology Information (NCBI)) и в других источниках (BLAST Manual, Altschul et al. NCB/NLM/NIH Bethesda, Md. 20894; Altschul et al, 1990. J. Mol. Biol. 215(3):403-410). Предпочтительно использовать программу «needle», которая применяет алгоритм глобального выравнивания Нидлмана-Вунша (Needleman S.B. and Wunsch CD. (1970). J Mol Biol 48, 443-453) для нахождения оптимального выравнивания (включая разрывы) двух последовательностей при рассмотрении их полной длины. Программа «needle», доступна, например, на интернет-сайте ebi.ac.uk. Процент идентичности по настоящему изобретению вычислялся, преимущественно, с помощью пакета EMBOSS: программа needle (глобальное выравнивание) с параметром «Gap Open», равным 10.0, параметром «Gap Extend», равным 0.5, и матрицей Blosum62. Для определения степени идентичности может быть использован также хорошо известный алгоритм Смита-Ватермана.
Термин «вариант» в контексте настоящего документа означает полипептид, который, как правило, отличается от полипептида, особо указанного в настоящем документе, на одну или более замен, делеций, присоединений и/или вставок. Такие варианты могут иметь природное происхождение или могут быть получены синтетически, например, путем модифицирования одной или более вышеупомянутых полипептидных последовательностей и анализа одной или более биологических активностей полипептида, как описано в настоящем документе, и/или используя любой из ряда методов, хорошо известных в данной области техники. Модификации могут быть проведены в отношении структуры полипептидов с получением функциональной молекулы, которая кодирует вариант или производный полипептид с желаемыми свойствами.
Если желательно изменить аминокислотную последовательность полипептида для создания эквивалентного, или даже усовершенствованного, варианта или части лиганда по настоящему изобретению, то специалист в данной области обычно изменяет один или более кодонов в кодирующей последовательности ДНК. Например, конкретные аминокислоты могут быть заменены на другие аминокислоты в структуре белка без существенной потери функции по сравнению с белком ClpB с последовательностью SEQ ID NO: 1.
Определенные замены аминокислотной последовательности могут быть произведены в последовательности белка, и, конечно, соответствующей кодирующей последовательности ДНК, и, тем не менее, привести к получению белка с аналогичными свойствами. Таким образом, предполагается, что в пептидных последовательностях или соответствующих последовательностях ДНК, кодирующих указанные пептиды, могут быть произведены различные изменения без существенной потери их биологической функции или активности. Во многих случаях вариант полипептида будет содержать одну или более консервативных замен. «Консервативная замена» это замена, при которой одна аминокислота замещается другой аминокислотой, имеющей сходные свойства, так что специалист в области пептидной химии может ожидать, что вторичная структура и характер гидропатичности полипептида останутся, по существу, неизменными. Как указано выше, аминокислотные замены, как следствие, основаны, в основном, на относительном сходстве аминокислотных заместителей боковых цепей, например, их гидрофобности, гидрофильности, заряде, размере, и тому подобное. Типовые замены, учитывающие различные вышеизложенные характеристики, хорошо известны специалистам в данной области техники и включают: аргинин и лизин; глутамат и аспартат; серии и треонин; глутамин и аспарагин; а также валин, лейцин и изолейцин. Аминокислотные замены могут быть произведены также на основе сходства полярности, заряда, растворимости, гидрофобности, гидрофильности и/или амфипатической природы остатков. Например, отрицательно заряженные аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту; положительно заряженные аминокислоты включают гистидин, лизин и аргинин; и аминокислоты с полярными незаряженными головками, обладающими аналогичными показателями гидрофильности, включают лейцин, изолейцин и валин; глицин и аланин; аспарагин и глутамин; а также серии, треонин, фенилаланин и тирозин. Другие группы аминокислот, представляющие возможные консервативные замены, включают: (1) Ala, Pro, Gly, Glu, Asp, Gln, Asn, Ser, Thr; (2) Cys, Ser, Tyr, Thr; (3) Val, Ile, Leu, Met, Ala, Phe; (4) Lys, Arg, His; and (5) Phe, Tyr, Trp, His.
В контексте настоящего документа термин «консервативная аминокислотная замена» может быть определен также как аминокислотный обмен внутри одной из следующих пяти групп:
(i) Небольшие алифатические, неполярные или слабо полярные остатки: Ala, Ser, Thr, Pro, Gly;
(ii) Полярные, отрицательно заряженные остатки и их амиды: Asp, Asn, Glu, Gln;
(iii)Полярные, положительно заряженные остатки: His, Arg, Lys;
(iv) Крупные, алифатические, неполярные остатки: Met, Leu, Ile, Val, Cys;
(v) Крупные, ароматические остатки: Phe, Tyr, Trp.
Вариант может также, или в качестве альтернативы, содержать неконсервативные замены. В предпочтительном варианте реализации настоящего изобретения варианты полипептидов отличаются от нативной последовательности заменой, делецией или добавлением 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более аминокислот. Варианты могут также (или в качестве альтернативы) быть модифицированы путем, например, делеции или добавления аминокислот, которые оказывают минимальное влияние на иммуногенность, вторичную структуру и гидропатический характер полипептида.
В одном варианте реализации настоящего изобретения вариант SEQ ID NO: 1 способен индуцировать насыщение, продлевать чувство сытости, снижать потребление пищи, контролировать набор веса, стимулировать снижение веса и/или снижать отношение массы жировой ткани к массе нежировых тканей тела у субъекта, нуждающегося в этом, с эффективностью, по меньшей мере, равной эффективности SEQ ID NO: 1.
В одном варианте реализации настоящего изобретения вариант SEQ ID NO: 1 включает консервативные аминокислотные замены в сравнении с последовательностью SEQ ID NO: 1, а именно, например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более консервативных аминокислотных замен.
В другом варианте реализации настоящего изобретения вариант SEQ ID NO: 1 представляет собой полипептид, в котором 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более аминокислот из последовательности SEQ ID NO: 1 отсутствует (-ют) или заменена (-ны) на любую аминокислоту, или в котором добавлены 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более (смежных или не смежных) аминокислот.
В одном варианте реализации настоящего изобретения белок ClpB или вариант такового, как описано выше в настоящем документе, модифицированы способами, хорошо известными в данной области техники, например, путем добавления одной или более функциональных групп, например, фосфатной, ацетатной, липидной или карбогидратной групп, и/или путем добавления одной или более защитных групп.
Например, белок ClpB или вариант такового могут быть модифицированы путем добавления одной или более функциональных групп, таких как фосфатная, ацетатная или различные липиды и карбогидраты. Белок ClpB или вариант такового также существуют как производные полипептидов.
Термин «производное полипептида» означает соединение, имеющее аминогруппу (--NH--), и более конкретно, пептидную связь. Полипептиды могут рассматриваться как замещенные амиды. Подобно амидной группе, пептидная связь демонстрирует высокую степень резонансной стабилизации. Одинарная С--N связь в пептидном соединении, как правило, на 40 процентов носит характер двойной связи, а двойная связь С=O на 40 процентов носит характер одинарной связи. «Защитные группы» это такие группы, которые предотвращают нежелательные реакции (например, протеолиз) с участием незащищенных функциональных групп. Конкретные примеры аминозащитных групп включают формил, трифторацетил, бензилоксикарбонил, замещенный бензилоксикарбонил, например, (орто- или пара-) хлорбензилоксикарбонил и (орто- или пара-) бромбензилоксикарбонил, и алифатический оксикарбонил, например, трет-бутоксикарбонил и трет-амилоксикарбонил. Карбоксильные группы аминокислот могут быть защищены посредством превращения в сложноэфирные группы. Сложноэфирные группы включают бензиловые эфиры, замещенные бензиловые эфиры, например, метоксибензиловый эфир, алкиловые эфиры, например, циклогексиловый эфир, циклогептиловый эфир или трет-бутиловый эфир. Гуанидиновый компонент может быть защищен при помощи нитрогруппы или арилсульфонила, например тозила, метоксибензенсульфонила или мезитиленсульфонила, даже если он не нуждается в защитной группе. Защитные группы имидазола включают тозил, бензил и динитрофенил. Индольная группа триптофана может быть защищена формилом или может оставаться незащищенной.
Модификация белка ClpB или варианта такового производится, в частности, с целью увеличения продолжительности их жизни in vivo. Один из видов модификации состоит в добавлении к N- или С-концу белка ClpB или варианта такового полиэтиленгликоля (ПЭГ). ПЭГ, как известно специалистам в данной области техники, имеет множество свойств, делающих его идеальным носителем для полипептидов, например, высокую растворимость в воде, высокую подвижность в растворе и низкую иммуногенность. Данная модификация также защищает полипептиды от экзопептидаз и, следовательно, повышает их общую стабильность in vivo.
Другие модификации, используемые для предотвращения разрушения полипептидов эндопептидазами или экзопептидазами, включают модификации N-конца, например, ацетилирование или гликозилирование, модификации С-конца, например, амидирование и использование не встречающихся в природе аминокислот (β-амино и α-трифторметил аминокислоты) в конкретных участках полипептидов.
Другая альтернатива, направленная на повышение размеров полипептидной молекулы, состоит в генетическом слиянии полипептидов с доменом Fc иммуноглобулина человека (включая, например, IgA, IgM и IgG) или в слиянии полипептидов с альбумином.
Белок ClpB или вариант такового, описанные в настоящем документе, могут быть произведены синтетически путем химического синтеза или ферментативного синтеза, как хорошо известно в данной области техники. В качестве альтернативы, нуклеотидные последовательности, кодирующие полипептиды по настоящему изобретению, могут быть включены в состав вектора экспрессии белка и получены в подходящем организме-хозяине (например, бактериях, клетках насекомых, и т.п.), а затем очищены. В одном из вариантов реализации настоящего изобретения белок ClpB или вариант такового получены методом клонирования, например, с использованием любых продуцирующих систем, известных в данной области техники, таких, как Е. coli, дрожжи, бакуловирус - клетка насекомого, или клетки млекопитающих, например, экспрессирующие системы HEK или СНО.
Для целей очистки или идентификации полипептидов может быть добавлен дополнительный полипептид («маркер»). Белковые маркеры делают возможной, например, адсорбцию полипептидов, с высокой аффинностью, на матрице и дальнейшее тщательное промывание матрицы подходящими буферами без существенного вымывания комплекса с последующим селективным элюированием адсорбированного комплекса. Примеры известных специалисту белковых маркеров включают (His)6, Мус, FLAG, гемагглютининовый маркер, глутатионтрансферазный маркер (GST), интеин, несущий хитин-связывающий аффинный маркер, или маркер, представляющий собой мальтозо-связывающий белок (МБР). Данные белковые маркеры могут располагаться на N-конце, на С -конце и/или внутри молекулы.
В некоторых вариантах реализации, белок ClpB вводят субъекту в форме фармацевтической композиции. В некоторых вариантах реализации, белок ClpB комбинируют с фармацевтически приемлемыми вспомогательными веществами (эксципиентами) и, необязательно, с матрицами с замедленным высвобождением, такими как биоразлагаемые полимеры, с целью получения фармацевтических композиций. Термин «фармацевтически» или «фармацевтически приемлемый» относится к молекулярным соединениям и композициям, которые не вызывают неблагоприятную, аллергическую или другую нежелательную реакцию при введении млекопитающему, особенно человеку, соответствующим образом. Фармацевтически приемлемый носитель или вспомогательное вещество относится к нетоксичному твердому, полутвердому или жидкому наполнителю, разбавителю, инкапсулирующему материалу или вспомогательному составу любого типа. В фармацевтических композициях по настоящему изобретению, активное вещество, само по себе, или в комбинации с другим активным веществом, может быть введено в виде индивидуальной формы для введения, в виде смеси с традиционными фармацевтическими носителями, животным и человеку. Подходящие единичные формы для введения включают формы для перорального введения, такие как таблетки, гелевые капсулы, порошки, гранулы и пероральные суспензии или растворы, сублингвальные и буккальные формы для введения, аэрозоли, имплантаты, подкожные, трансдермальные, для местного применения, внутрибрюшинные, внутримышечные, внутривенные, субдермальные, трансдермальные, интратекальные и интраназальные формы введения и формы ректального введения. Предпочтительно, фармацевтические композиции содержат носители, которые фармацевтически приемлемы для состава, пригодного для инъекции. Это могут быть, в частности, изотонические, стерильные, солевые растворы (мононатрий или динатрийфосфат, натрий, калий, кальций или хлорид магния и тому подобное, или смеси таких солей) или сухие, особенно лиофилизированные композиции, которые после добавления, в зависимости от ситуации, стерилизованной воды или физиологического раствора, обеспечивают разбавление инъекционных растворов. Фармацевтические формы, пригодные для инъекционного применения, включают стерильные водные растворы или дисперсии; составы, включающие кунжутное масло, арахисовое масло или водный пропиленгликоль; и стерильные порошки для разового приготовления стерильных растворов или дисперсий для инъекций. Во всех случаях, форма должна быть стерильной и должна быть текучей в той степени, при которой ее легко можно набрать в шприц. Она должна быть стабильной в условиях производства и хранения, и должна быть защищена от загрязняющего действия микроорганизмов, таких как бактерии и грибы. Растворы, содержащие соединения согласно настоящему изобретению в виде свободного основания или фармакологически приемлемых солей, могут быть получены в воде, подходящим образом смешанной с поверхностно-активным веществом, таким как гидроксипропилцеллюлоза. Дисперсии также могут быть получены в глицерине, жидких полиэтиленгликолях и их смесях, и в маслах. В обычных условиях хранения и применения, эти препараты содержат консервант для предотвращения роста микроорганизмов. Активный ингредиент может быть в составе композиции в нейтральной или солевой форме. Фармацевтически приемлемые соли включают кислотно-аддитивные соли (образованные со свободными аминогруппами белка) и которые образованы с неорганическими кислотами, такими как, например, соляная или фосфорная кислоты, или такими органическими кислотами, как уксусная, щавелевая, винная, миндальная, и тому подобное. Соли, образованные с помощью свободных карбоксильных групп, также могут быть получены из неорганических оснований, таких как, например, гидроксиды натрия, калия, аммония, кальция или трехвалентного железа, и таких органических оснований, как изопропиламин, триметиламин, гистидин, про каин и тому подобное. Носитель также может быть растворителем или дисперсионной средой, содержащей, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и т.п.), подходящие их смеси и растительные масла. Необходимую текучесть можно поддерживать, например, путем применения покрытия, такого как лецитин, путем поддержания требуемого размера частиц в случае дисперсии, и путем применения поверхностно-активных веществ. Предотвращение действия микроорганизмов может быть осуществлено различными антибактериальными и противогрибковыми агентами, например, парабенами, хлорбутанолом, фенолом, сорбиновой кислотой, тимеросалом и тому подобным. Во многих случаях, будет предпочтительно включать изотонические агенты, например, сахара или хлорид натрия. Длительное всасывание инъекционных композиций может быть обеспечено использованием в композициях агентов, замедляющих абсорбцию, например, моностеарата алюминия и желатина. Стерильные растворы для инъекций получают путем включения активных полипептидов в требуемом количестве в подходящем растворителе с различными другими ингредиентами, перечисленными выше, с последующей стерилизацией фильтрованием. Как правило, дисперсии получают путем включения различных стерилизованных активных ингредиентов в стерильный носитель, который содержит основную дисперсионную среду и требуемые другие ингредиенты из перечисленных выше. В случае стерильных порошков для получения стерильных растворов для инъекций, предпочтительными способами получения являются способы вакуумной сушки и сушки вымораживанием, которые дают порошок активного ингредиента плюс любой дополнительный желаемый ингредиент из ранее стерилизованного фильтрованием их раствора. После приготовления, растворы вводят способом, совместимым с лекарственным средством, и в таком количестве, которое терапевтически эффективно. Составы легко вводятся в различных лекарственных формах, таких как растворы для инъекций, описанные выше, но также могут быть использованы капсулы для высвобождения лекарственного средства и тому подобное. Для парентерального введения в водном растворе, например, раствор должен быть соответствующим образом забуферен, если необходимо, и жидкий разбавитель сначала доводят до изотонического состояния достаточным количеством солевого раствора или глюкозы. Эти конкретные водные растворы особенно подходят для внутривенного, внутримышечного, подкожного и внутрибрюшинного введения. В связи с этим, стерильные водные среды, которые могут быть использованы, будут известны специалистам в данной области в свете настоящего раскрытия. Например, одну дозу можно растворить в 1 мл изотонического раствора NaCl и, либо добавить в 1000 мл жидкости для гиподермолиза, либо ввести в предполагаемое место инфузии. Некоторое изменение дозировки обязательно произойдет в зависимости от состояния подлежащего лечению субъекта. Лицо, ответственное за введение, в любом случае определяет соответствующую дозу для индивидуального субъекта.
В настоящем описании термин «бактерия, которая экспрессирует ClpB» относится к бактерии, экспрессирующей или сверхэкспрессирующей белок-шаперон ClpB или вариант такового, как определено выше, или полипептид, содержащий или состоящий из аминокислотной последовательности, на 96-100%, идентичной аминокислотной последовательности SEQ ID NO: 1, более предпочтительно на 96, 97, 98, 99 или 100% идентичной аминокислотной последовательности SEQ ID NO: 1.
В некоторых вариантах реализации настоящего изобретения бактерия, которая экспрессирует ClpB или вариант такового, представляет собой пищевую бактерию.
В некоторых вариантах реализации настоящего изобретения бактерия, которая экспрессирует белок ClpB или вариант такового, представляет собой пробиотический бактериальный штамм.
В настоящем описании, термин «пробиотик» предназначен для обозначения живых микроорганизмов, которые, когда они присутствуют в достаточном количестве, оказывают положительное влияние на здоровье, комфорт и хорошее самочувствие, помимо традиционного действия питательных веществ. Пробиотические микроорганизмы определяют как «Живые микроорганизмы, которые при введении в достаточных количествах приносят пользу здоровью хозяина» (FAO/WHO 2001). В настоящем описании, термин «пробиотический бактериальный штамм» обозначает бактериальный штамм, который благотворно влияет на здоровье и физическое самочувствие хозяина.
В некоторых вариантах реализации, бактериальный штамм, в частности пробиотический бактериальный штамм согласно настоящему изобретению, представляет собой жизнеспособный бактериальный штамм, в частности жизнеспособный пробиотический бактериальный штамм. Выражение «жизнеспособный бактериальный штамм» означает микроорганизм, который является метаболически активным и способным колонизировать желудочно-кишечный тракт субъекта.
В некоторых вариантах реализации, бактериальный штамм, в частности, пробиотический бактериальный штамм по настоящему изобретению, представляет собой нежизнеспособный бактериальный штамм, в частности, пробиотический бактериальный штамм, состоящий из смеси бактериальных фрагментов. В некоторых вариантах реализации, смесь бактериальных фрагментов по настоящему изобретению состоит из белков из бактериального штамма.
В некоторых вариантах реализации, пробиотический бактериальный штамм по настоящему изобретению выбран из пищевых бактерий. Термин «пищевые бактерии» обозначает бактерии, которые применяются и обычно считаются безопасными для применения в пищевых продуктах.
Бактериальный штамм может быть природным бактериальным штаммом или он может быть сконструированным путем генной инженерии бактериальным штаммом.
В некоторых вариантах реализации, бактериальный штамм, в частности пробиотический бактериальный штамм по настоящему изобретению, представляет собой бактерию, которая конститутивно экспрессирует белок ClpB или вариант такового.
В некоторых вариантах реализации, белок ClpB или вариант такового сверхэкспрессирован в бактерии. Обычно экспрессия белка «повышена» или белок «сверхэкспрессирован», когда он экспрессируется или продуцируется в количестве или выходе большем, чем ожидаемый основной выход, который имеет место в природе в стандартных условиях. Сверхэкспрессия белка может быть достигнута, например, путем изменения любого одного или нескольких из: (а) условий роста или жизни клеток-хозяев; (b) полинуклеотида, кодирующего белок; (с) промотора, используемого для контроля экспрессии полинуклеотида и числа его копий в клетке; и (d) самих клеток-хозяев.
В некоторых вариантах реализации, бактерию подвергали стрессовым условиям, так что экспрессия белка ClpB или варианта такового в бактерии повышалась. Стресс может быть выбран из группы, состоящей из воздействия тепла, изменения температуры, механического стресса или длительного хранения, хранения при низкой влажности и/или лиофилизации или распылительной сушки.
В некоторых вариантах реализации, бактерии получали питательные вещества по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 раз. Обычно, питательные вещества подают с помощью среды для культивирования, которая подходит для роста бактерий, например, среды Мюллера-Хинтона, как описано в примерах. В некоторых вариантах реализации, бактерии изолируют на стационарной фазе роста, которая следует за указанной многократной подачей питательных веществ, поскольку на этой фазе концентрация белка ClpB или варианта такового является максимальной.
В некоторых вариантах реализации, бактерия содержит по меньшей мере одну точечную мутацию, с тем чтобы экспрессия белка ClpB или варианта такового была повышена. В настоящем описании, термин «точечная мутация» означает замещение и/или делецию нуклеиновой кислоты. В качестве альтернативы или одновременно, по меньшей мере одна мутация расположена внутри регуляторных последовательностях ДНК гена ClpB, например, в последовательностях, контролирующих транскрипцию и трансляцию, предпочтительно, чтобы эта мутация изменяла экспрессию белка. Мутации внутри регуляторных последовательностей ДНК могут служить для регулирования экспрессии белка.
В некоторых вариантах реализации, бактериальный штамм, в частности пробиотический бактериальный штамм по настоящему изобретению, представляет собой бактерию, которая была сконструирована методами генетической инженерии для экспрессии белка ClpB или варианта такового. Как правило, бактериальный штамм трансформировали нуклеиновой кислотой, кодирующей белок ClpB или вариант такового. Термин «трансформация» означает введение в клетку-хозяина «чужеродного» (то есть внешнего или внеклеточного) гена, последовательности ДНК или РНК, для того, чтобы клетка-хозяин стала экспрессировать введенный ген или последовательность для получения желаемого вещества, как правило, белка или фермента, кодируемого введенным геном или последовательностью. Клетка- хозяин, которая принимает и экспрессирует введенную ДНК или РНК, была «трансформирована». Нуклеиновая кислота может оставаться экстрахромосомной после трансформации родительского микроорганизма или может быть предназначена для интеграции в геном микроорганизма. Соответственно, нуклеиновая кислота может включать в себя дополнительные нуклеотидные последовательности, необходимые для интеграции (например, область, которая допускает гомологичную рекомбинацию и целенаправленную интеграцию в геном хозяина) или стабильную экспрессию и репликацию экстрахромосомной конструкции (например, точку начала репликации, промотор и другие регуляторные последовательности). В некоторых вариантах реализации, нуклеиновая кислота представляет собой конструкцию нуклеиновой кислоты, или вектор. В некоторых вариантах реализации, конструкция нуклеиновой кислоты, или вектор, представляет собой экспрессионную конструкцию, или вектор, однако другие конструкции и векторы, такие как те, которые используются для клонирования, охватываются изобретением. В некоторых вариантах реализации, экспрессионная конструкция, или вектор, представляет собой плазмиду. Обычно, экспрессионная конструкция/вектор дополнительно содержит промотор, описанный выше в настоящем документе. В некоторых вариантах реализации, промотор обеспечивает конститутивную экспрессию генов под его контролем. Однако, также могут быть использованы индуцибельные промоторы. Понятно, что экспрессионная конструкция/вектор по настоящему изобретению может содержать любое количество регуляторных элементов в дополнение к промотору, а также дополнительные гены, подходящие для экспрессии белка ClpB или варианта такового, если это необходимо. Способы трансформации бактериальных клеток внеклеточными нуклеиновыми кислотами, хорошо известны в данной области техники.
В некоторых вариантах реализации, бактериальный штамм, в частности, пробиотический бактериальный штамм, представляет собой грамотрицательный штамм.
В некоторых вариантах реализации, бактериальный штамм, в частности пробиотический бактериальный штамм, является членом семейства Entervbacteriaceae. В частности, бактериальный штамм является непатогенным членом семейства Entervbacteriaceae.
Интересно отметить, что авторы настоящего изобретения показали, что белки ClpB всех бактерий Enterobacteriaceae демонстрируют 100% идентичность друг другу.
В некоторых вариантах реализации, бактериальный штамм, в частности, пробиотический бактериальный штамм, является штаммом Е. coli. В некоторых вариантах реализации, штаммы Е. coli, в частности, пробиотические штаммы Е. coli, для применения в соответствии с идеями настоящего изобретения, включают непатогенные штаммы Е. coli, которые проявляют пробиотическую активность. Примером пробиотического штамма E. coli является пробиотический штамм Escherichia coli BU-230-98, номер депозита АТСС №202226 (DSM 12799), который является изолятом известного коммерчески доступного пробиотического штамма Escherichia coli М-17. Примером непатогенного штамма Е. coli является Е. coli Nissle 1917. Примером штамма Е. coli, который не был известен как пробиотик, является лабораторный штамм Е. coli K12.
В других вариантах реализации, бактериальный штамм представляет собой штамм Hafiiia alvei, такой как штамм Hafiiia alvei AF036, коммерческий продукт компании Bioprox. В других вариантах реализации, бактериальный штамм является штаммом Proteus vulgaris.
В других вариантах реализации, используется комбинация бактериальных штаммов, экспрессирующих белок ClpB или вариант такового.
Как правило, бактериальный штамм, в частности пробиотический бактериальный штамм по настоящему изобретению, производят с любой подходящей культуральной средой, хорошо известной в данной области. Согласно настоящему изобретению, пригодны различные ферментационные среды, такие как (но не ограничиваясь ими), например, во-первых, промышленная среда, в которой штамм(ы) выращивают, и которую используют как есть, или после концентрации (например, сушки), или после добавления в другую пищевую основу или продукт. В качестве альтернативы, можно использовать бактериальные клетки или бактериальные клетки со средой (например, ферментационный бульон) или фракции такой среды, содержащей клетки, (то есть среды с указанным бактериальным штаммом/штаммами). Среда, содержащая клетки или клетку, содержит живые или жизнеспособные бактериальные клетки и/или мертвые или нежизнеспособные бактериальные клетки штамма(ов). Таким образом, среда может обрабатываться с помощью, но не ограничиваясь этим, нагревания или ультразвука. Также, лиофилизированные или замороженные, бактерии и/или бесклеточные среды (которые могут быть сконцентрированы) включены в способы получения бактериального штамма, в частности пробиотического бактериального штамма по настоящему изобретению.
В конкретном варианте реализации, бактериальный штамм согласно настоящему изобретению лиофилизируют, а затем, предпочтительно, ресуспендируют перед введением.
Как правило, бактериальный штамм, в частности пробиотический бактериальный штамм по настоящему изобретению, вводят субъекту путем приема внутрь (то есть перорально).
В некоторых вариантах реализации, бактериальный штамм, в частности, пробиотический бактериальный штамм по настоящему изобретению, инкапсулирован для защиты от среды желудка. Соответственно, в некоторых вариантах реализации, бактериальный штамм, в частности пробиотический бактериальный штамм по настоящему изобретению, находится в композициях в капсулированной форме, чтобы значительно улучшить их время выживания. В таком случае, наличие капсулы может, в частности, задерживать или предотвращать деградацию микроорганизма в желудочно-кишечном тракте. Понятно, что композиции по настоящему варианту реализации могут быть инкапсулированы в покрытую кишечнорастворимым покрытием капсулу или таблетку с медленным высвобождением. Кишечнорастворимое покрытие позволяет капсуле/таблетке оставаться в неизмененном состоянии (то есть нерастворенной), когда она проходит через желудочно-кишечный тракт, до тех пор, пока она не достигнет тонкого кишечника. Способы инкапсулирования живых бактериальных клеток хорошо известны в данной области (см., например, патенты США General Mills Inc., такие как патент США №6723358). Например, микроинкапсулирование с альгинатом и крахмалом Hi-Maize™ с последующей сушкой вымораживанием оказалось успешным в продлении срока хранения бактериальных клеток в молочных продуктах (см., например, Kailasapathy K. (2002) Curr Issues Intest Microbiol. 3(2), 39-48]. В качестве альтернативы, инкапсулирование может быть выполнено с помощью глюкоманнановых волокон, таких как волокна, экстрагированные из Amorphophallus konjac. В качестве альтернативы, также можно использовать включение жизнеспособных бактерий в эмульсии из кунжутного масла (см., например, Hou R.C., Lin M.Y., Wang М.М., Tzen J.T. (2003). J. Dairy Sci 86, 424-428). В некоторых вариантах реализации, агенты для кишечнорастворимых покрытий предпочтительно, представляют собой сополимеры метакриловой кислоты и алкилакрилата, такие как полимеры Eudragit®. Поли(мет)акрилаты оказались особенно подходящими в качестве материалов для покрытия. EUDRAGIT® является торговым названием сополимеров, полученных из сложных эфиров акриловой и метакриловой кислот, свойства которых определяются функциональными группами. Индивидуальные марки EUDRAGIT® отличаются пропорциями нейтральных, щелочных или кислотных групп и, следовательно, с точки зрения физико-химических свойств. Эффективное применение и комбинация различных полимеров EUDRAGIT® предлагает идеальные решения для контролируемого высвобождения лекарственного средства в различных фармацевтических и технических областях применения. EUDRAGIT® обеспечивает функциональные пленки для покрытия таблеток и пеллетов с пролонгированным высвобождением. Полимеры описаны в международных фармакопеях, таких как Ph.Eur., USP/NF, DMF и JPE. Полимеры EUDRAGIT® могут обеспечить следующие возможности для контролируемого высвобождения лекарственного средства: направленное действие на желудочно-кишечный тракт (резистентность к действию желудочного сока, высвобождение в толстой кишке), защитные покрытия (маскировка вкуса и запаха, защита от влаги) и отсроченное высвобождение лекарственного средства (препараты с замедленным высвобождением). Полимеры EUDRAGIT® доступны в широком диапазоне различных концентраций и физических форм, включая водные растворы, водную дисперсию, органические растворы и твердые вещества. Фармацевтические свойства полимеров EUDRAGIT® определяются химическими свойствами их функциональных групп. Различают:
- поли(мет)акрилаты, растворимые в пищеварительных соках (путем образования соли): EUDRAGIT® L (сополимер метакриловой кислоты), S (сополимер метакриловой кислоты), FS и Е (основной бутилированный метакрилатный сополимер) полимеры с кислотными или щелочными группами, обеспечивают возможность рН-зависимого высвобождения активного компонента. Применения: от простого маскирования вкуса через резистентность исключительно к желудочному соку, до контролируемого высвобождения лекарственного средства во всех отделах кишечника;
- поли(мет)акрилаты, нерастворимые в пищеварительных соках: полимеры EUDRAGIT® RL и RS (аммонийметакрилатные сополимеры) полимеры с щелочными и EUDRAGIT® NE полимеры с нейтральными группами обеспечивают возможность контролируемого по времени высвобождения активного вещества с помощью не зависящего от рН набухания.
Кишечнорастворимые покрытия EUDRAGIT® обеспечивают защиту от высвобождения лекарственного средства в желудке, и обеспечивают контролируемое высвобождение в кишечнике. Основным критерием для высвобождения является рН-зависимое растворение покрытия, которое происходит в определенной части кишечника (рН от 5 до 7), а не в желудке (рН 1-5). Для этих применений, анионные марки EUDRAGIT®, содержащие карбоксильные группы, можно смешивать друг с другом. Это позволяет точно регулировать рН растворения и, таким образом, определять место высвобождения лекарственного средства в кишечнике. Марки EUDRAGIT® L и S подходят для кишечнорастворимых покрытий. EUDRAGIT® FS 30 D (водная дисперсия анионного сополимера на основе метилакрилата, метилметакрилата и метакриловой кислоты) специально используется для контролируемого высвобождения в толстой кишке.
Как правило, бактериальный штамм, в частности пробиотический бактериальный штамм по настоящему изобретению, вводят субъекту в форме пищевой композиции. Соответственно, еще один аспект настоящего изобретения относится к пищевой композиции, содержащей некоторое количество бактериального штамма, в частности пробиотического бактериального штамма по настоящему изобретению.
В некоторых вариантах реализации, пищевую композицию, которая содержит бактериальный штамм, в частности, пробиотический бактериальный штамм по настоящему изобретению, выбирают из полноценных пищевых композиций, пищевых добавок, нутрицевтических композиций и тому подобного. Композицию по настоящему изобретению можно использовать в качестве пищевого ингредиента и/или кормового ингредиента.
Пищевой ингредиент может быть в форме раствора или в виде твердого вещества в зависимости от использования и/или способа применения и/или способа введения.
Бактериальный штамм, в частности пробиотический бактериальный штамм по настоящему изобретению, обычно добавляют в любое время в процессе получения композиции, например, они могут быть добавлены к пищевой основе в начале производственного процесса или могут быть добавлены в конечный пищевой продукт.
Термин «пища» относится к жидким (т.е. напиткам), твердым или полутвердым пищевым композициям, особенно полноценным пищевым композициям (заменителям пищи), которые не требуют дополнительного приема питательных веществ или пищевых добавок. Композиции пищевых добавок не полностью заменяют потребление питательных веществ другими способами. Пищевые продукты и пищевые добавки представляют собой, например, ферментированные молочные продукты или продукты на основе молочных продуктов, которые, предпочтительно, вводят или принимают внутрь перорально, один или несколько раз в день. Ферментированные молочные продукты могут быть получены непосредственно с использованием бактерий согласно настоящему изобретению в процессе производства, например, путем добавления к пищевой основе, используя методы известные per se. В таких способах, штамм(ы) согласно настоящему изобретению может использоваться в дополнение к обычно используемому микроорганизму и/или может заменить один или несколько, или часть микроорганизма, обычно используемого. Например, при приготовлении ферментированных молочных продуктов, таких как йогурты или напитки на основе йогурта, бактерии согласно настоящему изобретению могут быть добавлены или использованы в качестве части стартовой культуры, или могут быть соответствующим образом добавлены во время такой ферментации. Необязательно, бактерии могут быть инактивированы или убиты позже в процессе производства. Ферментированные молочные продукты включают молочные продукты, такие как (но не ограничиваясь ими) десерты, йогурты, йогуртовые напитки, творог, кефир, кисломолочные напитки, пахта, сыры, соусы, масла с низким содержанием жира, свежий сыр, соевые напитки, мороженое и т.д. В качестве альтернативы, пищевые композиции и/или композиции пищевых добавок могут быть немолочными или молочными неферментированными продуктами (например, штаммами или бесклеточной средой в неферментированном молоке или в другой пищевой среде). В некоторых вариантах реализации, пробиотический бактериальный штамм по настоящему изобретению инкапсулирован и диспергирован в пище (например, в молоке) или в непищевой среде. Неферментированные молочные продукты могут включать мороженое, пищевые батончики и соусы, и тому подобное. Немолочные продукты могут включать в себя порошкообразные напитки и питательные батончики и тому подобное. Продукты могут быть получены с использованием известных способов, таких как добавление эффективного количества штамма(ов) и/или бесклеточной культуральной среды к пищевой основе, такой как обезжиренное молоко, или молоко, или композиция на основе молока и ферментация, как известно. Другими пищевыми основами, к которым могут быть добавлены (композиции, содержащие) бактериальные клетки и/или бесклеточная культуральная среда, являются мясо, заменители мяса или растительные основы.
Композиция, которая содержит бактериальный штамм, в частности пробиотический бактериальный штамм, по настоящему изобретению может быть твердой, полутвердой или жидкой. Она может быть в виде пищевого продукта или пищевой добавки, например, в виде таблеток, гелей, порошков, капсул, напитков, батончиков и т.д. Например, композиция может быть в виде порошка, упакованного в саше, который может быть растворен в воде, фруктовом соке, молоке или другом напитке.
В настоящем описании, термин «пищевой ингредиент» или «кормовой ингредиент» включает композицию, которая добавлена или может быть добавлена к функциональным пищевым продуктам или к пищевым продуктам в качестве пищевой добавки.
Под «обогащенной пищей» или «нутрицевтической» или «функциональной» пищей понимается пищевой продукт, который содержит ингредиенты, имеющие полезные эффекты для здоровья или способные улучшить физиологические функции.
Под «пищевой добавкой» подразумевается пищевой продукт, предназначенный для дополнения нормальной пищи. Пищевая добавка является концентрированным источником питательных веществ или других веществ, обладающих питательным или физиологическим эффектом, когда их принимают отдельно или в виде комбинации в небольших количествах.
Согласно изобретению, термин «функциональные продукты питания» объединяет продукты питания и соответствующие продукты, которые в последнее время разработаны, которым приписывают важность не только потому, что они ценны для питания и вкуса, но и из-за определенных ингредиентов. Согласно изобретению, среднесрочное или долгосрочное поддержание и укрепление здоровья имеют важное значение. В этом контексте, предпочтительными являются нетерапевтические применения. Термины «нутрицевтики», «фудцевтики (foodceuticals)» и «авторские дизайнерские продукты», которые также представляют собой варианты реализации изобретения, используются как синонимы, отчасти, однако, также дифференцированным образом. Тем не менее, превентивный аспект и укрепление здоровья, а также пищевой характер продуктов, лучше всего описываются термином функциональные продукты питания. Во многих случаях, они относятся к продуктам, образованным путем сортировки и отбора (как это имеет место и в настоящем изобретении), очистки, концентрации, все чаще также с помощью добавок. Изолированные эффективные вещества, в частности, в виде таблеток или пилюль, не включены. Несмотря на то, что не существует юридического определения функциональных продуктов питания, большинство сторон, заинтересованных в этой области, согласны с тем, что они являются продуктами питания, которые имеют определенные последствия для здоровья, помимо основных питательных эффектов. Соответственно, функциональные продукты питания представляют собой обычные пищевые продукты, в которые включены компоненты или ингредиенты (такие как описанные здесь), которые придают пищевому продукту определенный функционал, например, медицинскую или физиологическую пользу, кроме чисто питательного эффекта.
В некоторых вариантах реализации, напиток представляет собой функциональный напиток или терапевтический напиток, утолитель жажды или обычный напиток. В качестве примера, композиция по настоящему изобретению может быть использована в качестве ингредиента для безалкогольных напитков, фруктового сока или напитка, содержащего сывороточный белок, чаев для здоровья, какао-напитков, молочных напитков и напитков из молочнокислых бактерий, йогурта и питьевого йогурта, сыра, мороженного, фруктового льда и десертов, кондитерских изделий, бисквитных тортов и смесей для тортов, закусок, сбалансированных продуктов и напитков, фруктовых начинок, глазури для тортов, шоколадной начинки для выпечки, ароматизированной начинки для чизкейка, начинки с фруктовым вкусом для тортов, глазури для тортов и пончиков, кремов для выпечки быстрого приготовления, начинок для печенья, готовых к употреблению пекарских начинок, начинок с низкой калорийностью, питательных напитков для взрослых, подкисленного соевого/сокового напитка, асептического/ретортированного шоколадного напитка, барных смесей, порошков для напитков, обогащенного кальцием соевого/без добавок и шоколадного молока, обогащенного кальцием кофейного напитка.
Композицию можно дополнительно использовать в качестве ингредиента в пищевых продуктах, таких как американский сырный соус, анти-спекающий агент для тертого и измельченного сыра, соус для чипсов, сливочный сыр, сухой взбитый крем, обезжиренная сметана, замороженные/оттаивающие молочные взбитые сливки, замороженный/оттаивающий стабильный взбитый топпинг, маложирный легкий натуральный сыр чеддер, маложирный йогурт в швейцарском стиле, воздушные замороженные десерты, твердое мороженое в упаковке, удобное для этикетки, с улучшенными ценой и вкусовыми качествами твердое мороженое в упаковке, маложирное мороженое: мягкое мороженое, соус для барбекю, сырный соус, творожный соус, сухая смесь для соуса «Альфредо», смесь для сырного соуса, сухая смесь для томатного соуса и другие.
В некоторых вариантах реализации, композиция, которая содержит бактериальный штамм, в частности, пробиотический бактериальный штамм по настоящему изобретению, применяется при производстве йогуртов, таких как ферментированный йогуртовый напиток, йогурт, питьевой йогурт, сыр, ферментированный крем, молочные десерты и другие. Соответственно, композиция может быть дополнительно применена в качестве ингредиента для производства одного или нескольких из сырных, мясных продуктов или продуктов, содержащих защитные культуры.
В некоторых вариантах реализации, пищевая композиция, которая содержит бактериальный штамм, в частности пробиотический бактериальный штамм по настоящему изобретению, подходит для приготовления продуктов-заменителей пищи. Используемый в настоящем описании термин «продукт - заменитель пищи», если не указано иное, включает любой пищевой продукт, содержащий белки, углеводы, липиды, витамины и минералы, комбинация которых пригодна в качестве единственного или основного источника питательных веществ для питания. Как правило, продукт- заменитель пищи содержит по меньшей мере один источник углеводов, по меньшей мере один источник липидов, и/или по меньшей мере один источник белков. В качестве источника белков может быть использован любой подходящий пищевой белок, например, белки животных (такие как молочные белки, мясные белки и яичные белки); растительные белки (такие как соевый белок, белок пшеницы, белок риса и белок гороха); смеси свободных аминокислот; или их комбинации. Белки молока, такие как казеиновый и сывороточный, и соевые белки, являются особенно предпочтительными. Белки могут быть интактными или гидролизованными, или смесью интактных и гидролизованных белков. Может быть желательным поставлять частично гидролизованные белки (степень гидролиза от 2 до 20%), например, для животных, которые, как полагают, имеют риск выработать аллергию на коровье молоко. Если требуются гидролизованные белки, процесс гидролиза может быть осуществлен по желанию и, как известно в данной области. Например, гидролизат сывороточного белка может быть получен ферментативным гидролизом сывороточной фракции в ходе одной или нескольких стадий. Если сывороточная фракция, используемая в качестве исходного материала, по существу не содержит лактозы, установлено, что в процессе гидролиза белок испытывает гораздо меньше блокирования лизина. Это позволяет уменьшить степень блокировки лизина примерно с 15 масс. % от общего лизина до менее чем примерно 10 масс. % лизина; например, до, примерно 7 масс. % лизина, что значительно улучшает питательную ценность источника белка. Если композиция включает источник жира, источник жира предпочтительно обеспечивает от 5% до 40% энергии композиции; например, от 20% до 30% энергии. Подходящий жировой профиль может быть получен с использованием смеси масла канолы, кукурузного масла и высокоолеинового подсолнечного масла. Источник углеводов предпочтительно обеспечивает от 40% до 80% энергии композиции. Может быть использован любой подходящий углевод, например, сахароза, лактоза, глюкоза, фруктоза, твердые вещества кукурузного сиропа, мальтодекстрины и их смеси. Как правило, замена одного ежедневного приема пищи, употреблением низкокалорийного заменителя пищи, способствует поддержанию массы тела после потери массы тела.
Пищевая композиция, которая содержит бактериальный штамм, в частности пробиотический бактериальный штамм, по настоящему изобретению, обычно содержит носители или наполнители. Термины «носители» или «наполнители» означают материалы, подходящие для введения, и включают любой такой материал, известный в данной области, например, любую жидкость, гель, растворитель, жидкий разбавитель, солюбилизатор или тому подобное, который является нетоксичным и который не взаимодействует с какими-либо компонентами, в частности с бактериальным штаммом композиции нежелательным образом. Примеры питательно приемлемых носителей включают, например, воду, солевые растворы, спирт, силикон, воски, вазелин, растительные масла, полиэтиленгликоли, пропиленгликоль, липосомы, сахара, желатин, лактозу, амилозу, стеарат магния, тальк, поверхностно-активные вещества, кремневую кислоту, вязкий парафин, парфюмерное масло, моноглицериды жирных кислот и диглицериды, сложные эфиры петроэтральной жирной кислоты, гидроксиметилцеллюлозу, поливинилпирролидон и тому подобное.
В некоторых вариантах реализации, пищевая композиция, которая содержит бактериальный штамм, в частности пробиотический бактериальный штамм, по настоящему изобретению содержит какое-то количество пищевых волокон. Пищевые волокна проходят через тонкую кишку, непереваренные ферментами, и функционируют как естественный наполнитель и слабительное. Пищевые волокна могут быть растворимыми или нерастворимыми, и, в целом, предпочтительной является смесь двух типов. Подходящими источниками пищевых волокон являются соя, горох, овес, пектин, гуаровая камедь, аравийская камедь, фруктоолигосахариды, галактоолигосахариды, сиалил-лактоза и олигосахариды, полученные из животного молока. В некоторых вариантах осуществления пищевые волокна выбирают из маннанов. Маннаны (такие как глюкоманнаны и галактоманнаны), такие как гуаровая камедь, камедь бобовых, коньяк и ксантановая камедь, присутствуют в некоторых стенках клеток растений. Глюкоманнаны обычно состоят из (1-4)-β-связанных единиц глюкозы и маннозы, в то время как галактоманнаны обычно состоят из (1-4)-β-маннановой основной цепи, замещенной единичными единицами (1-6)-α-галактоза Многие эндоспермальные бобовые, такие как гуар и рожковое дерево, содержат галактоманнаны в эндосперме во время развития семян. Глюкоманнаны также были обнаружены как второстепенный компонент зерна злаков.
В некоторых вариантах реализации, пищевая композиция, которая содержит бактериальный штамм, в частности пробиотический бактериальный штамм, по настоящему изобретению, содержит минералы и микроэлементы, такие как микроэлементы и витамины, в соответствии с рекомендациями государственных органов, таких как USRDA. Например, композиция может содержать на суточную дозу одного или несколько из следующих микроэлементов в указанных диапазонах, от 300 до 500 мг кальция, от 50 до 100 мг магния, от 150 до 250 мг фосфора, от 5 до 20 мг железа, от 1 до 7 мг цинка, от 0,1 до 0,3 мг меди, от 50 до 200 мкг йода, от 5 до 15 мкг селена, от 1000 до 3000 мкг бета-каротина, от 10 до 80 мг витамина С, от 1 до 2 мг витамина В1, от 0,5 до 1,5 мг витамина В6, от 0,5 до 2 мг витамин В2, от 5 до 18 мг ниацина, от 0,5 до 2,0 мкг витамина В12, от 100 до 800 мкг фолиевой кислоты, от 30 до 70 мкг биотина, от 1 до 5 мкг витамина D, от 3 до 10 мкг витамина Е.
В некоторых вариантах реализации, композиция, которая содержит бактериальный штамм, в частности пробиотический бактериальный штамм, по настоящему изобретению содержит эмульгаторы. Примеры эмульгаторов пищевого качества обычно включают сложные эфиры диацетилвинной кислоты моно-и диглицеридов, лецитин и моно-и диглицериды. Аналогичным образом, могут быть включены подходящие соли и стабилизаторы.
В некоторых вариантах реализации, пищевая композиция, которая содержит пробиотический бактериальный штамм по настоящему изобретению, содержит по меньшей мере один пребиотик. «Пребиотик» означает пищевые вещества, предназначенные для стимулирования роста пробиотического бактериального штамма по настоящему изобретению в кишечнике. Пребиотик может быть выбран из группы, состоящей из олигосахаридов и, необязательно, содержит фруктозу, галактозу, маннозу, сою и/или инулин; и/или пищевые волокна.
В некоторых вариантах реализации, композиция, которая содержит бактериальный штамм, в частности пробиотический бактериальный штамм по настоящему изобретению содержит защитные гидроколлоиды (такие как камеди, белки, модифицированные крахмалы), связующие вещества, пленкообразующие агенты, инкапсулирующие агенты/материалы, оболочечные материалы, матричные соединения, покрытия, эмульгаторы, поверхностно-активные вещества, солюбилизирующие агенты (масла, жиры, воски, лецитины и т.д.), адсорбенты, носители, наполнители, ко- соединения, диспергирующие агенты, смачивающие агенты, технологические добавки (растворители), противослеживающие агенты, маскирующие вкус агенты, утяжелители, агенты для затвердевания, гелеобразующие агенты, антиоксиданты и противомикробные средства. Композиция может также содержать обычные фармацевтические добавки и адъюванты, эксципиенты и разбавители, включая, но не ограничиваясь ими, воду, желатин любого происхождения, растительные смолы, лигнинсульфонат, тальк, сахара, крахмал, гуммиарабик, растительные масла, полиалкиленгликоли, ароматизаторы, консерванты, стабилизаторы, эмульгаторы, буферы, лубриканты, красители, смачивающие агенты, наполнители и тому подобное. Во всех случаях, такие дополнительные компоненты будут выбраны с учетом их пригодности для предполагаемого получателя.
В некоторых вариантах реализации, введение белка ClpB или варианта такового, или бактериального штамма, в частности пробиотического бактериального штамма, который экспрессирует белок ClpB или вариант такового, повторяют, например, 2-3 раза в день в течение одного дня или более, и обычно в течение длительного периода продолжительностью по меньшей мере 4 дня или даже от 4 до 15 недель, включая, при необходимости, один или несколько периодов прерывания. В некоторых вариантах реализации, белок ClpB или вариант такового, или бактериальный штамм, который экспрессирует белок ClpB или вариант такового, вводят одновременно или последовательно с одним приемом пищи субъектом. В некоторых вариантах реализации, белок ClpB или вариант такового, или бактериальный штамм, который экспрессирует белок ClpB или вариант такового, вводят до приема пищи субъектом.
В настоящем описании, термин «эффективное количество» относится к достаточному количеству белка ClpB или варианта такового, или бактерий, экспрессирующих белок ClpB или вариант такового, для достижения благоприятного эффекта (например, стимулирования насыщения, продления сытости, снижения потребления пищи, контроля, в частности снижения массы тела, стимулирование потери веса и/или уменьшение отношения массы жировой ткани к массе нежировых тканей тела). В контексте настоящего изобретения, количество белка ClpB или варианта такового, или бактерий, экспрессирующих белок ClpB или вариант такового, вводимых субъекту, будет зависеть от характеристик индивидуума, таких как общее состояние здоровья, возраст, пол, масса тела Квалифицированный специалист сможет определить соответствующие дозы в зависимости от этих и других факторов. Например, когда белок ClpB или вариант такового вводят субъекту в форме пробиотика, штамм по настоящему изобретению может генерировать колонию, которая является достаточной для оказания благоприятного эффекта на субъекта. Если бактериальный штамм, в частности пробиотический бактериальный штамм, вводят в виде пищевого продукта, он обычно может содержать от 103 до 1012 КОЕ бактериального штамма, в частности пробиотического бактериального штамма по настоящему изобретению на г сухой массы пищевой композиции.
Изобретение будет дополнительно проиллюстрировано следующими фигурами и примерами. Однако эти примеры и фигуры никоим образом не должны истолковываться как ограничивающие объем настоящего изобретения.
Краткое описание фигур
Фиг. 1. Эффекты ежедневной внутрижелудочной доставки Е. coli K12 (WT) дикого типа или Е. coli с удаленным геном ClpB (Δ ClpB) у нормальных взрослых мышей С57 В16, на массу тела (А, Е), 24х-часовой прием пищи (В, F) и характер питания (С, D, G, Н), проанализированные в начале и в конце 3-х недель кормления через желудочный зонд.
Контрольные мыши (Ctr) не получали никакого кормления через желудочный зонд. Характер питания оценивали как количество пищи (С, G), соответствующее среднему количеству пищи, съеденной во время одного приема пищи, и как частоту приема пищи (D, Н), соответствующую количеству приемов пищи, разделенных по меньшей мере 5- минутными интервалами в течение 24 часов. Снижение количества потребленной пищи отражает более быстрое насыщение, а снижение частоты приема пищи отражает более продолжительную сытость. А: Двухфакторный дисперсионный анализ с повторными измерениями, апостериорный критерий Бонферрони *р<0,05. B,D,G,H: дисперсионный анализ, апостериорный критерий Тьюки, *р<0,05, **р<0,01, ***р<0,001. Е: t-критерий Стьюдента, *р<0,05.
Фиг. 2. Влияние ClpB на электрическую активность нейронов ARC РОМС. Частота потенциала действия, выраженная в процентах изменения по сравнению с базальным уровнем; среднее значение ± стандартная ошибка среднего. Апостериорные критерии Бонферрони *р<<0,05, **р<<0,01.
Фиг. 3. Динамика массы тела у мышей ob/ob с ожирением до и во время (дни 0-21) кормления через желудочный зонд бактериями Е. coli K12, Е. coli K12 ΔClpB, в обоих случаях содержавшимися в среде Мюллера-Хинтона (МН), или одной только средой МН в качестве контроля (Ктр.,). Двухфакторный дисперсионный анализ, Эффект лечения: р=0,01, апостериорные критерии Бонферрони Ctr. по сравнению с Е. coli K12, *р<0,05 и **р<0,01. Среднее ± стандартная ошибка среднего.
Фиг. 4. Процент изменения средней массы тела (со дня рандомизации = 100%) у мышей ob/ob с ожирением через 3 недели кормления через желудочный зонд бактериями Е. coli К12 (n=8), Е. coli K12 ΔClpB (n=8) в обоих случаях содержащимися в среде Мюллера-Хинтона (МН), или одной только средой МН в качестве контроля (Ктр., N=7). Дисперсионный анализ = 0,01, апостериорный критерий Тьюки *р<0,05. Среднее ± стандартная ошибка среднего.
Фиг. 5. Удельная доля жировой ткани у мышей ob/ob с ожирением, измеренная с помощью ЭХО-ЯМР (EchoMRI) через 3 недели кормления через желудочный зонд бактериями Е. coli K12 (n=8), Е. coli K12 AClpB (п=8), в обоих случаях содержащимися в среде Мюллера-Хинтона (МН), или одной только средой МН в качестве контроля (Ктр., n=7). Дисперсионный анализ р=0,005, апостериорный критерий Тьюки **р<0,01. Среднее ± стандартная ошибка среднего.
Фиг. 6. Отношения массы нежировых тканей к массе жировой ткани у мышей ob/ob с ожирением, измеренные с помощью ЭХО-ЯМР (EchoMRI) через 3 недели кормления через желудочный зонд бактериями Е. coli K12 (n=8), Е. coli K12 ΔClpB (n=8) в обоих случаях содержащимися в среде Мюллера-Хинтона (МН), или одной только средой МН в качестве контроля (Ктр., n=7). Дисперсионный анализ р=0,05, t-критерий Стьюдента *р<0,05. Среднее ± стандартная ошибка среднего.
Фиг. 7. Среднее ежедневное потребление пищи у мышей ob/ob с ожирением в течение 3 недель кормления через желудочный зонд бактериями Е. coli K12 (n=8), Е. coli K12 ΔClpB (n=8), в обоих случаях содержащимися в среде Мюллера-Хинтона (МН), или одной только средой МН в качестве контроля (Ктр., n=7). Дисперсионный анализ <0,0001, апостериорный критерий Тьюки ***р<0,001. Среднее ± стандартная ошибка среднего.
Фиг.8. Среднее количество приемов пищи в сутки у мышей ob/ob с ожирением в течение 3 недель кормления через желудочный зонд бактериями Е. coli K12 (n=8), Е. coli K12 ΔClpB (n=8), в обоих случаях содержащимися в среде Мюллера-Хинтона (МН), или одной только средой МН в качестве контроля (Ктр., n=7). Дисперсионный анализ р<0,0001, апостериорный критерий Тьюки ***р<0,001. Среднее±стандартная ошибка среднего.
Фиг. 9. Ежедневное увеличение массы тела у мышей ob/ob с ожирением во время кормления через желудочный зонд бактериями Е. coli K12, Е. coli Niessle 1917, Е. coli Niessle 1917 лиофилизированными (lyo), во всех случаях содержащимися в среде Мюллера-Хинтона (МН), или одной только средой МН в качестве контроля (Ктр.). Двухфакторный дисперсионный анализ, р=0,02, апостериорный критерий Бонферонни а, Ктр. по сравнению с Е. coli Niessle 1917 lyo, и b, Ктр. по сравнению с Е. coli Niessle 1917, *р<0,05 и **р<0,01. Среднее ± стандартная ошибка среднего.
Фиг. 10. Процент изменения средней массы тела (со дня рандомизации = 100%) у мышей ob/ob с ожирением через 2 недели кормления через желудочный зонд бактериями Е. coli K12, Е. coli Niessle 1917, Е. coli Niessle 1917 лиофилизированными (lyo), во всех случаях содержащимися в среде Мюллера-Хинтона (МН), или одной только средой МН в качестве контроля (Ктр). Критерий Краскела-Уоллиса р<0.01, апостериорный критерий Данна *р<0,05, критерий Манна-Уитни ## р<0.01. Среднее ± стандартная ошибка среднего.
Фиг. 11. Удельная доля жировой ткани у мышей ob/ob с ожирением, измеренная с помощью ЭХО-ЯМР (EchoMRI) через 2 недели кормления через желудочный зонд бактериями Е. coli K12, Е. coli Niessle 1917, Е. coli Niessle 1917 лиофилизированными (lyo), во всех случаях содержащимися в среде Мюллера-Хинтона (МН), или одной только средой МН в качестве контроля (Ктр). t-критерий Стьюдента
**р<0.01. Среднее ± стандартная ошибка среднего.
Фиг. 12. Влияние прямой интрацеребровентрикулярной инъекции различных доз ClpB на потребление пищи у крыс.
Фиг. 13. Увеличение массы тела (в граммах) у мышей ob/ob с ожирением после 17 дней кормления через желудочный зонд бактериями Е. coli Niessle 1917 (Nissle) (n=15), Е. coli K12 (K12 Δ-ClpB) (n=15) или Hajhia alvei AF036 (Hafnia) (n=15), во всех случаях содержавшимися в среде Мюллера-Хинтона (МН), или одной только средой МН в качестве контроля (Контроль) (n=12).
Фиг. 14. Кумулятивное потребление пищи (в граммах) у мышей ob/ob с ожирением после 17 дней кормления через желудочный зонд бактериями Е. coli Niessle 1917 (Nissle) (n=15), Е. coli K12 (K12 Δ-ClpB) (n=15) или Hafnia alvei AF036 (Hafnia) (n=15), во всех случаях содержавшимися в среде Мюллера-Хинтона (МН), или одной только средой МН в качестве контроля (Контроль) (n=12).
Фиг. 15. Увеличение массы жировой и нежировых тканей у мышей ob/ob с ожирением, измеренное методом ЭХО-ЯМР после 17 дней кормления через желудочный зонд бактериями Е. coli Niessle 1917 (Nissle) (n=15), Е. coli K12 (K12 ΔClpB) (n=15) или Hafnia alvei AF036 (H. alvei) (n=15), во всех случаях содержавшимися в среде Мюллера-Хинтона (МН), или одной только средой МН в качестве контроля (Контроль) (n=12). А. Увеличение массы жировой ткани (в граммах). В. Увеличение массы нежировых тканей (в граммах). С.Отношение массы нежировых тканей к массе жировой ткани.
Фиг. 16. Относительная скорость экспрессии протеина гормон-чувствительной липазы (pHSL) (в сравнении со скоростью экспрессии актина в качестве стандарта) у мышей ob/ob с ожирением после 17 дней кормления через желудочный зонд бактериями Е. coli Niessle 1917 (Е. coli Nissle) (n=15), Е. coli K12 (Е. coli K12) (n=15) или Hafnia alvei AF036 (Hafnia alvei) (n=15), во всех случаях содержавшимися в среде Мюллера-Хинтона (МН), или одной только средой МН в качестве контроля (Контроль) (n=12). Скорости экспрессии HSL и актина измерены методом вестерн-блоттинга.
Фиг. 17. Подтверждение эффективности диеты с высоким содержанием жиров. А. Масса тела (в граммах) у мышей на диете с высоким содержанием жиров/ углеводов (ВЖД) (n=48) и у мышей на контрольной диете (Контроль) (n=8). В. Масса жировой ткани (в граммах). С. Масса нежировых тканей (в граммах).
Фиг. 18. Снижение прироста массы тела, начиная с дня 0 (Д0), у мышей с ожирением, вызванным диетой с высоким содержанием жиров (ВЖД) после 14 дней кормления через желудочный зонд бактериями Е. coli K12 (K12 Δ-ClpB) или Hafnia alvei AF036 (Н. alvei), во всех случаях содержавшимися в среде Мюллера-Хинтона (МН), или одной только средой МН в качестве контроля (Контроль).
Фиг. 19. Увеличение массы жировой и нежировых тканей, начиная с дня 0 (Д0), у мышей с ожирением, вызванным диетой с высоким содержанием жиров (ВЖД) после 14 дней кормления через желудочный зонд бактериями Е. coli K12 (K12 Δ-ClpB) или Hafnia alvei AF036 (Н. alvei), во всех случаях содержавшимися в среде Мюллера-Хинтона (МН), или одной только средой МН в качестве контроля (Контроль). А. Увеличение массы жировой ткани (в граммах). В. Увеличение массы нежировых тканей (в граммах).
Фиг. 20. Увеличение массы тела (в ГГПК - Площадь под кривой) у мышей fa/fa с ожирением после 21 дня кормления через желудочный зонд бактериями Е. coli Niessle 1917 (Nissle), Е. coli K12 (K12 ΔClpB) или Hafnia alvei AF036 (H. alvei), во всех случаях содержавшимися в среде Мюллера-Хинтона (МН), или одной только средой МН в качестве контроля (Контроль).
Фиг. 21. Кумулятивное потребление пищи (в ППК) у мышей fa/fa с ожирением после 21 дня кормления через желудочный зонд бактериями Е. coli Niessle 1917 (Nissle), Е. coli K12 (K12 ΔClpB) или Hafnia alvei AF036 (H. alvei), во всех случаях содержавшимися в среде Мюллера-Хинтона (МН), или одной только средой МН в качестве контроля (Контроль).
Фиг. 22. Увеличение массы жировой ткани (в граммах) у мышей fa/fa с ожирением после 21 дня кормления через желудочный зонд бактериями Е. coli Niessle 1917 (Nissle), Е. coli K12 (K12 ΔClpB) или Hafnia alvei AF036 (H. alvei), во всех случаях содержавшимися в среде Мюллера-Хинтона (МН), или одной только средой МН в качестве контроля (Контроль).
Примеры
Пример 1
Материал и методы
In vitro рост E.coli после регулярного снабжения питательными веществами.
Бактерии E.coli K12 культивировали при 37°С в 40 мл среды МН (Becton, Dickinson, MD), содержащей 30% вытяжки говядины, 1,75% гидролизата казеина и 0,15% крахмала рН 7,3 при 25°С в 50 мл флаконах Falcon. Для моделирования двух ожидаемых (в нормальных условиях) ежедневных приемов пищи у людей, бактерии получали новую среду МН каждые 12 ч в течение 5 последующих дней. Бактериальный рост измеряли как оптическую плотность OD при λ=600 нм на спектрофотометре каждые 2 часа после 1-го получения среды МН, каждый 1 ч после 3-го и каждые 10 мин после 5-го получения среды. В конце каждого 12-часового цикла, бактерии центрифугировали в течение 5 мин при 6000 об/мин при комнатной температуре (RT). Супернатанты отбрасывали и заменяли эквивалентным объемом (~40 мл) новой среды МН. После последнего добавления среды МН, бактерии отбирали для экстракции белка в экспоненциальной фазе роста и в последующей стационарной фазе роста.
Экстракция белков
Бактерии E.coli K12 центрифугировали при 4°С в течение 30 мин при 4000 g. Остатки бактерий растворяли в 2 мл буфера трисгидроксиметиламинометана (ТРИС) (рН 7,4) и гомогенизировали ультразвуком в течение 3 мин при комнатной температуре. Для отделения белков от нерастворенных клеточных фрагментов, бактериальный гомогенат центрифугировали при 4°С в течение 30 мин при 10000 g. Супернатант отбирали, и затем ультрацентрифугировали при 4°С в течение 45 мин при 60000 g для дальнейшего разделения белков на цитоплазматическую фракцию (супернатант) и мембранную (остаточную) фракцию. Мембранные белки растворяли в буфере TRIS (рН 7,4). Концентрации белков измеряли с использованием набора 2-D Quant (GE Healthcare, Piscataway, NJ).
Двухмерный полиакриламидный гель-электрофорез
Для 2D-PAGE, использовали 300 мкг белкового экстракта E.coli для регидратации полосок иммобилизованного градиента рН (IPG, immobilized рН gradient) (рН 4-7, 18 см, BIO-RAD, Hercules, СА). Затем белки разделяли в первом направлении с помощью изоэлектрического фокусирования до достижения значения суммарных 85000 вольт-часов с использованием изоэлектрической фокусирующей системы IPGphor (GE Healthcare). После фокусирования, полоски IPG инкубировали в течение 15 мин в уравновешивающем буфере [мочевина 6 моль/л, 30% (об./об.) глицерин, 2% (масс/об.) додецилсульфат натрия (SDS), трис-HCl 50 ммоль/L рН 8,8 и 0,25% (масс/об.) бромфенолового синего, содержащего 2% (масс/об.) дитиотреитола, и затем алкилировали в течение 15 мин в уравновешивающем буфере, содержащем 4% (масс/об.) иодацетамида. Затем полоски IPG помещали на 10% полиакриламидные градиентные гели (20 см ⋅ 18 см ⋅ 1 мм) для SDS-PAGE. Разделение во втором направлении проводили в течение ночи в системе вертикального электрофореза Ettan Daltsix (GE Healthcare) с 12 мА/гель при 25°С. После SDS-PAGE, двумерные гели фиксировали в течение 2 ч в 2% (об./об.) ортофосфорной кислоте и в 50% (об./об.) метаноле при комнатной температуре. Затем гели промывали водой, а белковые пятна визуализировали окрашиванием СВВ G-250 (BIO-RAD) [34% (об./об.) метанол, 17% (масс/об.) аммоний сульфат, 2% (об./об.) ортофосфорной кислоты и 0,66 г СВВ G-250/л].
Анализ дифференциальной экспрессии белка
Изображения окрашенных 2D-гелей сканировали с помощью ImageScanner II (GE Healthcare), калиброванного с помощью нейтрально-серого маркера (Kodak, Rochester, NY) и оцифрованного с помощью программного обеспечения Labscan 6.0 (GE Healthcare). Анализ дифференциальной экспрессии белка, включая определение пятна, количественную оценку, сопоставление и сравнительный анализ, осуществляли с использованием программного обеспечения ImageMaster 2D Platinum 5.0 (GE Healthcare). Каждый образец белка подвергали 2D-PAGE по меньшей мере 3 раза (мембранные белки) и 4 раза (цитоплазматические белки) для минимизации различий от геля к гелю, и каждый набор из 3 (или 4) гелей сравнивали с использованием ImageMaster для подтверждения отсутствия статистически разных пятен в наборе гелей. Наиболее характерный гель (миграция геля, определение пятна и число пятен) каждого набора использовали для сравнения белков E.coli из экспоненциальной и стационарной фаз. Уровень экспрессии определяли по относительному объему каждого пятна в геле и выражали в виде % объема, рассчитанного как объем пятна/Σ объема всех пятен, разрешенных в геле. Этот нормализованный объем пятна учитывает изменения, вызванные нанесением на гель и окрашиванием белка, учитывая общий объем по всем пятнам, присутствующим в геле. Вариации численности были рассчитаны как отношение средних значений % объема для группы пятен между двумя фазами. Учитывали только пятна с коэффициентом вариации объема >1,5. Отсутствие пятна в геле указывало на то, что не было обнаружено никакой детектируемой экспрессии белка в выбранных экспериментальных условиях. Соответствующие значения p определяли с помощью t-критерия Стьюдента (уровень значимости p<0,05) после логарифмического преобразования объема пятна.
Идентификация белка с помощью жидкостной хроматографии-ионизация методом электрораспыления MS/MS, тандемная масс-спектрометрия)
Искомые белковые пятна вырезали из 2D-гелей, окрашенных СВВ G-250 с использованием Ettan Spot Picker (GE Healthcare), а автоматизированный ферментативный гидролиз в геле выполняли на Ettan Digester (GE Healthcare), как описано ранее (Goichon et al., 2011). Затем белковые экстракты ресуспендировали в 10 мкл 5%-ной (об./об.) ацетонитрил/0,1% (об./об.) муравьиной кислоты и затем анализировали с помощью системы nano-LC1200, связанной с масс-спектрометром Ion Trap 6340, снабженным источником наноспрея и приставкой для наномасштабной ВЭЖХ на чипе HPLC-chip cube (Agilent Technologies, Courtaboeuf, Франция). Вкратце, пептиды обогащали и обессоливали на ловушке-колонке с 40 нл RP-C18 и разделяли на Zorbax колонке С18 (с размером пор 30 нм, размером частиц 5 мкм, (длина 43 мм, внутренний диаметр 75 мм, Agilent Technologies). Использовали 9-минутный линейный градиент (3%-80% ацетонитрила в 0,1% муравьиной кислоте) при скорости потока 400 нл/мин и элюент анализировали с помощью масс-спектрометра с ионной ловушкой. Для идентификации белка, списки пиков MS/MS были извлечены и сопоставлены с базами данных о белках с использованием поисковой системы MASCOT Daemon версии 2.2.2 (Matrix Science). Поиски выполнялись со следующими конкретными параметрами: специфичность фермента, трипсин; разрешено одно пропущенное отщепление; никаких фиксированных изменений; вариабельные модификации, окисление метионина, карбамидометилирование цистеина, фосфорилирование серина, тирозина и треонина; моноизотопный; пептидный заряд, 2+ и 3+; допуск по массе для ионов-предшественников, 1,5 Да; допуск по массе для фрагментных ионов, 0,6 Да; ESI-TRAP как инструмент; таксономия, Е. coli; База данных Национального центра биотехнологической информации (NCBI) [NCBInr 20120531 (18280215 последовательностей, 6265275233 остатков)] (Bethesda, MD). Белки были автоматически проверены, на предмет удовлетворения одному из следующих критериев: идентификация с по меньшей мере двумя лидирующими пептидами (жирным и красным), каждый из которых имеет значение MASCOT больше чем 54 (p<<0,01), или по меньшей мере двумя лидирующими пептидами (жирный и красный), каждый из которых имеет значение MASCOT больше 47 (р<<0,05). Чтобы оценить долю ложноположительные показателей, все первоначальные запросы к базе данных были выполнены с использованием опции «decoy» MASCOT. Результаты считались актуальными, если доля ложноположительных показателей никогда не превышала 1%.
Анализ АТФ
Производство АТФ in vitro измеряли с использованием набора для колориметрических/флуорометрических анализов АТФ в соответствии с инструкциями производителя (BioVision, СА). Бактериальные белки из экспоненциальной или стационарной фаз помещали в серию лунок в двух повторностях для каждой концентрации (1, 10 и 25 мкг/мл в буфере для анализа АТФ) и доводили объем до 50 мкл/лунку с помощью буфера для анализа АТФ. Затем к соответствующим лункам добавляли по 10 мкл раствора различных питательных веществ, 15% сахарозы или среды МН и доводили объем до 50 мкл/лунку буфером для анализа АТФ; к контрольным лункам добавляли только 50 мкл/лунку буфера АТФ. Планшет инкубировали 2 часа при 37°С. После инкубации, в каждую лунку добавляли 50 мкл реакционной смеси АТФ (содержащей буфер для анализа АТФ, АТФ-пробы, преобразователь АТФ и проявляющую смесь). OD измеряли при 570 нм после 30 минут инкубации при комнатной температуре, в защищенном от света месте.
Разработка и подтверждение правильности иммуноанализа ClpB
Метод анализа по обнаружению ClpB основывался на нескольких критериях, таких как специфическое и чувствительное обнаружение в линейном диапазоне концентраций. Особым условием было обнаружение ClpB без перекрестной реактивности с α-MSH, что может происходить из-за присутствия α-MSH-подобного эпитопа(ов) в молекуле ClpB. Для простоты процедуры и обнаружения сигнала, использовали стандартную 96-луночную планшету для ELISA (Enzyme-Linked Immunosorbent Assay) и возможность считывания оптической плотности OD с помощью спектрофотометра. Подробные протоколы ELISA ClpB и Вестерн-блоттинга (WB) представлены в отдельных разделах.
Чтобы предотвратить связывание антител для обнаружения захватывающими антителами, производили ClpB-захватывающие антитела (Ab) и ClpB-детекторные антитела в разных видах, кроликах и мышах, соответственно. Для наиболее эффективного захвата белка ClpB из сложных биологических образцов, покрывали планшет ELISA поликлональными антителами кролика, имеющим несколько анти-ClpB эпитопов. Чтобы избежать перекрестной реактивности между ClpB и α-MSH, использовали, в качестве детекторных антител, мышиные моноклональные к ClpB, которые характеризовались высокой чувствительностью и специфичностью для узнавания ClpB, но не α-MSH, предварительно отобранные с помощью ELISA-скрининга нескольких клонов антител. Конъюгированные с щелочной фосфатазой антитела для обнаружения против антител мыши, использовали в качестве общего инструмента ELISA для получения хромогенной ферментативной реакции, считываемой как OD, пропорциональная концентрации анализируемого материала. Линейное изменение OD, возникающее в результате обнаружения 7 последовательных разведений рекомбинантного белка E.coli ClpB в диапазоне от 2 пМ до 150 пМ, было получено без достижения плато и без насыщения OD-сигнала.
Чтобы проверить специфичность разработанного анализа ClpB, измерили концентрации ClpB в образцах белка, экстрагированных из 10 различных культур E.coli K12WT, и из культуры мутантных ΔClpB бактерий E.coli. ΔClpB-мутант и соответствующие штаммы дикого типа (WT) были любезно предоставлены доктором Axel Mogk (ZMBH, Heidelberg University, Germany). Кроме того, проанализировали присутствие ClpB в этих образцах бактериальных белков с помощью WB (Вестерн блоттинга) с использованием поликлональных кроличьих антител к ClpB и сравнили значения интенсивности сигнала между полосами WB и концентрациями ClpB в ELISA. ClpB был обнаружен во всех культурах E.coli WT, причем 7 культур имели концентрации ClpB выше 1000 пМ, тогда как ClpB не обнаруживали в образце белка, экстрагированном из ΔClpB E.coli. Вестерн блоттинг выявил доминантную полосу ожидаемого размера 96 кДа в диком типе WT, но не в ΔClpB E.coli. Интенсивность OD этих полос варьировала между отдельными образцами и положительно коррелировала с концентрациями ClpB, измеренными с помощью ELISA в тех же образцах культур E.coli. Таким образом, отсутствие детекции ClpB в белковых препаратах E.coli ΔClpB подтвердило специфичность анализов, и хорошее соответствие между ELISA и Вестерн блоттингом, и обеспечило перекрестную проверку обоих методов иммунодетекции ClpB.
Чтобы проверить, что ClpB-анализ плазмы обнаруживает ClpB, полученный из бактерий кишечника, использовали ELISA ClpB для измерения ClpB в плазме мышей, которые ежедневно получали питание через желудочный зонд в течение 3 недель, содержащее E.coli WT или E.coli ΔClpB. Образцы плазмы были доступны из нашего ранее опубликованного исследования (Tennoune et al., 2014). Обнаружили, что ClpB обычно присутствует в плазме мышей, включая как контрольных мышей, так и мышей, получавших питание через желудочный зонд, содержащее культуральную среду без бактерий. Важно отметить, что уровни ClpB в плазме повышались у мышей, получавших E.coli WT, но не изменялись у мышей, получавших ClpB-дефицитные E.coli, что подтверждает происхождение плазматического ClpB из бактерий кишечника.
ELISA-анализ ClpB
Кроличьи поликлональные антитела к E.coli ClpB (изготовленные по заказу Delphi Genetics, Gosselies, Belgium) наносили на 96-луночные планшеты Maxisorp (Nunc, Rochester, NY), используя объем 100 мкл и концентрацию 2 мкг/мл в 100 мМ буфере NaHCO3, рН 9,6 в течение 12 ч при 4°С. Планшеты промывали (5 мин × 3) в фосфатно-солевом буферном растворе (PBS) с 0,05% Твин 20, рН 7,4. Рекомбинантный белок ClcB E.coli (изготовленный по заказу Delphi Genetics) последовательно разводили до концентрации 5, 10, 25, 50, 70, 100 и 150 пМ в буфере для образцов (PBS, азид натрия 0,02%, рН 7,4) и добавляли в лунки в двух повторностях, чтобы изготовить стандарт. Образцы анализируемого материала включали: слизистую оболочку толстой кишки и образцы плазмы от мышей и крыс или белки, экстрагированные из культур E.coli K12. Образцы анализируемого материала добавляли в оставшиеся лунки в двух повторностях и инкубировали 2 часа при комнатной температуре. Планшеты промывали (5 мин × 3) в PBS с 0,05% Tween 20, рН 7,4. В лунки добавляли мышиные моноклональные антитела к E.coli ClpB (1: 500 в буфере для образцов, изготовленном Delphi Genetics по заказу) и инкубировали 90 мин при комнатной температуре. Планшеты промывали (5 мин × 3) в PBS с 0,05% Tween 20, рН 7,4. Антитела козы к иммуноглобулину G мыши, конъюгированные с щелочной фосфатазой (1: 2000 в буфере для образцов) от Jackson ImmunoResearch Laboratories, Inc. (West Grove, PA), добавляли в лунки и инкубировали в течение 90 минут при комнатной температуре. Планшеты промывали (5 мин × 3) в PBS с 0,05% Tween 20, рН 7,4 и затем добавляли 100 мкл раствора п-нитрофенилфосфата (Sigma, St. Louis, МО) в качестве субстрата для щелочной фосфатазы. Через 40 мин инкубации при комнатной температуре, реакцию останавливали добавлением 50 мкл 3н NaOH. Оптическую плотность OD определяли при 405 нм с использованием считывающего устройства для микропланшетов Metertech 960 (Metertech Inc., Taipei, Taiwan). Значения OD пустых лунок (blank), полученные в результате считывания планшетов без добавления образцов плазмы или стандартных разведений белка ClpB, вычитали из значений OD образца.
Вестерн-блоттинг анализ ClpB
Вестерн-блоттинг проводили с использованием белков, экстрагированных из E.coli K12. Образцы белка (10 мкг) разделяли на 20%-ном акриламидном SDS-геле в трис-глициновом буфере и переносили на нитроцеллюлозную мембрану (GE Healthcare, Orsay, France), которую блокировали в течение по меньшей мере 1 ч при комнатной температуре с 5% (масс/об.) обезжиренного сухого молока в TBS (10 ммоль/л Tris, рН 8, 150 ммоль/л NaCl) плюс 0,05% (масс/об.) Твин 20. Затем мембрану инкубировали в течение ночи при 4°C с поликлональными антителами кролика к E.coli ClpB (1: 2000, Delphi Genetics). После трех промывок в блокирующем растворе с 5% (масс/об.) обезжиренным сухим молоком в TBS/0,05% Твин 20, мембраны инкубировали в течение 1 часа с антителами к иммуноглобулину G кролика, конъюгированными с пероксидазой (1: 3000, SantaCruz Biotechnology), После трех промывок, пероксидазную реакцию выявляли с использованием набора для детекции ECL (GE Healthcare). Белковые полосы сравнивали с стандартом молекулярных масс (Precision Plus, BioRad), а пленки сканировали с использованием ImageScanner III (GE Healthcare) и анализировали на плотность пикселей полосы с использованием программного обеспечения ImageQuant TL 7.0 (GE Healthcare).
Внутрикишечные введения белков Е. coli крысам.
Животные
Уход за животными и эксперименты проводили в соответствии с рекомендациями, установленными Национальными институтами здравоохранения США (National Institutes of Health, USA), и в соответствии с нормами французского и европейского сообщества (Official Journal of the European Community L 358, 18/12/1986). Самок крыс Sprague-Dawley, с массой тела 200-250 г (Janvier, Genest-Saint-Isle, France), содержали в клетках (3 крысы на клетку) в полностью оборудованном виварии с регулируемыми условиями окружающей среды (22±1°С, 12-часовой цикл чередования света и темноты с включением света в 7:30 утра) в течение 1 недели, чтобы адаптировать их к условиям содержания. Стандартный гранулированный корм для грызунов (RM1 диета, SDS, Великобритания) и питьевая вода были доступны в неограниченном количестве.
Эксперимент № 1
Данный эксперимент был разработан для оценки применимости нашей in vitro модели роста E.coli для случаев in vivo роста бактерий в кишечнике. Крыс анестезировали раствором кетамина (75 мг/кг, Virbac, Carros, France)/ксилазина (5 мг/кг, Bayer, Leverkusen, France), 3: 1 об., в концентрации 0,1 мл/100 г массы тела внутрибрюшинно (I.P.). После лапаротомии, толстая кишка была иммобилизована путем размещения 2 лигатур: 1-ой в месте соединения слепой и ободочной кишки и 2-ой, на 4 см ниже. Инфузию и отбор проб из полости толстой кишки проводили с использованием полипропиленового катетера, вставленного в восходящую ободочную кишку, и зафиксированного 1-й лигатурой. 2 мл среды МН или воды осторожно вводили в толстую кишку и сразу же удаляли для измерения OD. После измерения OD, весь образец содержимого толстой кишки возвращали в толстую кишку. Такой отбор проб содержимого толстой кишки без добавления новой среды МН или воды повторяли каждые 5 мин в течение первых 20 минут, а затем через 30 мин и 60 мин. Плотность бактерий измеряли как OD при λ=600 нм на спектрофотометре. Образцы крови брали из портальной вены до и через 30 и 60 мин после 1-й инфузии. Образцы фекалий брали из толстой кишки в конце эксперимента для экстракции ДНК и ПЦР ClpB.
Количественная полимеразная цепная реакция в реальном времени
Количественную ПЦР (qPCR) проводили для анализа плотности бактерий, экспрессирующих ДНК белка ClpB, с использованием прибора CFX 96 qPCR (BioRad, СА). Тотальную ДНК экстрагировали из фекалий крысы с использованием набора QAMP DNA stool mini kit (QIAGEN Venlo, Netherlands). Реакционная смесь для qPCR включала 5 мкл SYBR Green Master (QIAgen, West Sussex, UK), 0,5 мкМ каждого из прямых и обратных праймеров, ДНК из образцов, и воду до общего объема смеси 10 мкл. Праймеры были приобретены у Invitrogen (Cergy-Pontoise, France). Трехступенчатую ПНР проводили в течение 40 циклов. Образцы денатурировали при 95°С в течение 10 мин, отжигали при 60°С в течение 2 мин и проводили элонгацию при 95°С в течение 15 с.
Эксперимент № 2
Целью эксперимента была оценка влияния белков Е. coli на высвобождение кишечных пептидов (GLP-1 и PYY) в большой круг кровообращения. Крыс подвергали анестезии, и толстую кишку иммобилизовали, как описано выше, инфузии (вливания) в толстую кишку белков E.coli (0,1 мкг/кг белка в 2 мл PBS), экстрагированных в экспоненциальной (n=6) или в стационарной фазе роста (n=6) проводили один раз в течение 20 мин. Образцы крови брали из портальной вены до и после 20 минут инфузий толстой кишки для анализов GLP-1, PYY и ClpB. Образцы слизистой оболочки толстой кишки брали в конце эксперимента для анализа ClpB. Анализы GLP-1 и PYY проводили с использованием набора для флуоресцентного иммуноферментного анализа (Phoenix Pharmaceutical Inc., СА) в соответствии с инструкциями производителя. Флуоресценцию измеряли при 325 нм для возбуждения и 420 нм для излучения с использованием устройства для считывания микропланшетов Chameleon (HIDEX Inc., Турку, Финляндия).
Введение белков E. coli крысам, прием пищи и исследования c-fos мозга
Животные
Самцов крыс Wistar, с массой тела 200-250 г (Janvier, Genest-Saint-Isle, Франция) адаптировали к условиям содержания и кормили, как описано выше. За три дня до экспериментов, крыс переносили в индивидуальные камеры для исследования метаболизма (Tecniplast, Лион, Франция), где их неограниченно кормили таким же кормом RM1, но в порошкообразной форме (SDS). Питьевая вода была доступна всегда. Крыс осторожно трогали/брали руками ежедневно в течение нескольких минут в течение адаптационного периода, чтобы приучить их к манипуляциям. В конце адаптации, крыс распределяли по трем группам для достижения сходной средней массы тела и использовали в экспериментах 1-3. Два эксперимента, включающих ограничение питания, проводили на тех же крысах с интервалом в 4 дня. 3-й эксперимент у крыс с неограниченным кормлением включал их новые партии.
Эксперимент № 1
Первый эксперимент был направлен на сравнение эффектов мембранных белков E.coli, экстрагированных в экспоненциальной и стационарной фазах роста. Крыс лишали пищи в течение ночи (между 18.00 ч. и 10.00 ч.), тогда как вода была доступна без ограничений. На следующий день после лишения пищи, белки E.coli инъецировали внутрибрюшинно, I.P. в 10.00 ч. и крыс немедленно возвращали в их камеры для исследования метаболизма, которые содержали предварительно взвешенное количество пищи. Потребление пищи измеряли через 1, 2 и 4 часа. 1-я группа крыс (n=6) получала 0,1 мг/кг мембранных белков, экстрагированных из E.coli в экспоненциальной фазе в 300 мкл PBS; 2-я группа крыс (n=6) получала 0,1 мг/кг мембранных белков, экстрагированных из E.coli в стационарной фазе, а контрольная группа (n=6) получала 300 мкл PBS.
Эксперимент № 2
Второй эксперимент был направлен на сравнение эффектов цитоплазматических белков E.coli, экстрагированных в экспоненциальной и стационарной фазах. Использовали экспериментальный протокол, аналогичный протоколу эксперимента №1.
Эксперимент № 3
Этот эксперимент был разработан для оценки влияния суммарных белков Е. coli на потребление пищи у крыс с неограниченным кормлением. Инъекции белков E.coli (0,1 мг/кг белка в 300 мкл PBS, внутрибрюшинно I.P.), экстрагированных в экспоненциальной (n=6) или в стационарной фазе (n=6), или только PBS в качестве контроля (n=6), проводили в 19.30 ч. и животных возвращали в камеры для исследования метаболизма, которые содержали предварительно взвешенное количество пищи. Суммарное потребление пищи измеряли через 2 часа. Сразу же после этого крыс анестезировали пентобарбиталом натрия (0,2 мг/кг, I.P.) и перфузировали для иммуногистохимического исследования экспрессии c-fos в головном мозге.
Подготовка тканей и иммуногистохимия
Головной мозг фиксировали перфузией/иммерсией в 4% параформальдегиде, замораживали и делали срезы (14 мкм) на криостате (Leica Microsystems, Nanterre, France), а затем обрабатывали для иммуногистохимии с использованием системы амплификации тирамидного сигнала (TSA, tyramide signal amplification) плюс набор флуоресцеина (NEN, Boston, MA). Для однократного окрашивания, использовали кроличью поликлональную антисыворотку к c-fos (1: 4000, Ab-5, Calbiochem, Merck KGaA, Darmstadt, Germany). Для двойного окрашивания, вслед за TSA, применяли прямой метод иммунофлуоресценции с использованием или моноклональных антител кролика к β-эндорфину (β-конец) 1: 1000 (Life Technologies, Frederick, MD), которые детектировали антителами Cyanine-3 к кролику 1: 200 (Jackson ImmunoResearch, West Grove, PA) или мышиных моноклональных IgG к пептиду, связанному с геном кальцитонина (CGRP) 1: 1000 (Santa Cruz Biotechnology, Inc., TX), которые детектировали коньюгированными с родамином красным антителами к мыши 1: 200 (Jackson ImmunoResearch). В гипоталамических дугообразных и вентромедиальных ядрах и в центральном ядре миндалевидного тела, положительные клетки подсчитывали при увеличении ×20 на шести последовательных срезах. Среднее число положительных клеток на крысу использовали для вычисления среднего значения группы. Цифровые изображения были оптимизированы для яркости и контрастности в программном обеспечении Adobe Photoshop 6.0 (Adobe Systems, San Jose, CA).
Хронические введения белков E. coli мышам
Двухмесячные самцы C57B16 (n=32) были приобретены в Janvier Labs и адаптированы к условиям вивария в течение 1 недели с 12-часовым циклом чередования света и темноты, свет включали в 8:00 ч. утра. Затем мышей помещали индивидуально в клетки для мышей BioDAQ (Research Diets, Inc., New Brunswick, NJ), каждая из которых была оснащена монитором автоматического питания. Через 3 дня адаптации к клеткам для мышей BioDAQ, мышей делили на три группы (n=8), каждая из которых получала различные обработки, включающие две ежедневные внутрибрюшинные I.P. инъекции в 9:00 и 18:30 одного из: (i) PBS, (ii) бактериальных белков, экстрагированных в экспоненциальной фазе (0,1 мг/кг массы тела), (iii) бактериальных белков, экстрагированных в стационарной фазе (0,1 мг/кг массы тела).
Пища (SERLAB, Montataire, Франция) и питьевая вода были доступны без ограничений, и массу тела измеряли ежедневно. Данные о питании непрерывно контролировали в течение одной недели и анализировали с использованием средства просмотра данных BioDAQ 2.3.07 (Research Diets). Для анализа характера питания, интервал между приемами пищи устанавливали на уровне 300 с. Показатель насыщения рассчитывали как время (с) интервала после приема пищи, деленное на количество пищи (г), потребленной в ходе предыдущего приема пищи. После эксперимента, мышей убивали обезглавливанием; мозг удаляли и гипоталамус рассекали для микрочипового анализа нейропептидной мРНК.
Микрочиповый анализ гипоталамической нейропептидной мРНК
Тотальную РНК экстрагировали из гипоталамуса мышей, получавших хроническое введение белков Е. coli, с использованием набора NucleoSpin® РНК II (Macherey-Nagel,  , Germany) в соответствии с протоколом производителей. Обработанную РНК подвергали обратной транскрипции при 42°С в течение 60 мин с использованием набора для обратной транскрипции ImProm-II™ (Promega, Madison, WI). Полученную кДНК использовали для ПЦР в реальном времени. Панель из 9 пар праймеров была разработана с помощью программного обеспечения Primer express (Life Technologies, Saint-Aubin, France) и проверена на эффективность и специфичность. Реакционную смесь для ПЦР, состоящую из 6 мкл кДНК и 6 мкл Fast SYBR Green Master Mix (Life Technologies), содержащую специфические обратные и прямые праймеры в концентрации 100 нМ, распределяли в 96-луночные планшеты с помощью автоматизированной системы для работы с жидкостями Bravo (Agilent) и амплифицировали с помощью QuantStudio 12K Flex (Life Technologies).
, Germany) в соответствии с протоколом производителей. Обработанную РНК подвергали обратной транскрипции при 42°С в течение 60 мин с использованием набора для обратной транскрипции ImProm-II™ (Promega, Madison, WI). Полученную кДНК использовали для ПЦР в реальном времени. Панель из 9 пар праймеров была разработана с помощью программного обеспечения Primer express (Life Technologies, Saint-Aubin, France) и проверена на эффективность и специфичность. Реакционную смесь для ПЦР, состоящую из 6 мкл кДНК и 6 мкл Fast SYBR Green Master Mix (Life Technologies), содержащую специфические обратные и прямые праймеры в концентрации 100 нМ, распределяли в 96-луночные планшеты с помощью автоматизированной системы для работы с жидкостями Bravo (Agilent) и амплифицировали с помощью QuantStudio 12K Flex (Life Technologies).
Сигналы, генерируемые кДНК для генов-мишеней, были внутренне скорректированы с сигналами референсных генов для вариаций в количествах стартовой мРНК. Уровень экспрессии генов затем сравнивали с соответствующей группой контрольных образцов, и регуляции определяли с помощью метода 2-ΔΔCq.
Электрофизиологические записи
Срезы мозга (250 мкм) были получены от взрослых мышей РОМС-eGFP (5-7 недель, реф: C57BL/6J-Tg (Pomc-EGFP) 1Low/J, The Jackson Laboratory), как описано выше (Fioramonti et al., 2007). Срезы инкубировали при RT, в оксигенированной внеклеточной среде, содержащей (в мМ): 118 NaCl, 3 KCl, 1 MgCl2, 25 NaHCO3, 1,2 NaH2PO4, 1,5 CaCl2, 5 HEPES, 2,5 D-глюкозу (осмолярность доводили до 310 мОсм сахарозой, рН 7,4) в течение периода восстановления по меньшей мере 60 мин.
Один раз в записывающей камере срезы перфузировали со скоростью 2-3 мл/мин той же внеклеточной средой. Срезы просматривали с помощью микроскопа Nikon EF600 (Nikon France, Champigny sur Marne), оснащенного для флуоресценции (флуоресцеиновый фильтр) и видеомикроскопии IR-DIC. Жизнеспособные нейроны ARC РОМС визуализировали с использованием водно-иммерсионного объектива Х40 (Nikon) с флуоресцентной видеокамерой (Nikon). Боросиликатные пипетки (4-6 MΩ, 1,5 мм OD, Sutter Instruments, Novato, СА) заполняли фильтрованной внеклеточной средой. Cell-attached запись производилась с использованием усилителя Multiclamp 700В, оцифровывалась с помощью интерфейса Digidata 1440А и собиралась при 3 кГц с использованием программного обеспечения pClamp 10.3 (Axon Instruments, Molecular Devices, Sunnyvale, СА). Емкости пипетки и клетки полностью компенсировались. После установления стабильной базовой линии, 1 нМ ClpB (Delphi Genetics) перфузировали в течение 5-10 минут. Уровень частоты пульсации нейронов РОМС измеряли в течение последних 3 мин перфузии ClpB, через 7-10 мин после перфузии и сравнивали с уровнем частоты пульсации, измеренной за 3 минуты до перфузии.
Статистический анализ
Данные были проанализированы и графики были построены с использованием программного обеспечения GraphPad Prism 5.02 (GraphPad Software Inc., Сан-Диего, Калифорния). Нормальность оценивали с помощью теста Колмогорова-Смирнова. Групповые различия анализировали с помощью дисперсионного анализа или непараметрического критерия Крускал-Уоллиса (K-W) с апостериорными критериями Тьюки или Данна в соответствии с результатами нормальности. Там, где это необходимо, сравнивали отдельные группы, используя t-тест Стьюдента и корреляции Пирсона или тест Манна-Уитни (M-W) в соответствии с результатами нормальности. Эффекты непрерывных экспериментов анализировали с помощью дисперсионного анализа с повторными измерениями (RM) ANOVA и апостериорного критерия Бонферрони. Данные показаны как средние ± стандартная ошибка среднего, и для всех испытаний, p<0,05 считалось статистически значимым.
Результаты
Рост E.coli после регулярного снабжения питательными веществами.
После 1-го добавления питательной среды Мюллера-Хинтона (МН) к культуре Е.coli, наблюдали три фазы бактериального роста:
1) лаг-фазу 2 часа;
2) экспоненциальную фазу 4 часа; и
3) стационарную фазу с оптической плотностью (OD) 0,35, остающейся стабильной в течение 6 ч. После 3-го и 5-го добавления среды МН, наблюдали только две фазы роста: экспоненциальную и стационарную, первая фаза, начиналась сразу же после поступления питательных веществ. Каждый новый цикл кормления характеризовался более короткой продолжительностью фазы экспоненциального роста: 2 часа после 3-го, и 20 минут после 5-го добавления питательных веществ. После 5-го добавления питательных веществ бактериальные белки экстрагировали и разделяли на мембранную и цитоплазматическую фракции, используемые для протеомного анализа и экспериментов in vivo на голодных крысах. В новом эксперименте, чтобы проверить, может ли регулярное добавление питательных веществ дополнительно ускорить динамику роста бактерий, к бактериям Е. coli K12 добавляли питательные вещества 9 раз. Обнаружили, что после 7-го и 9-го добавления питательных веществ, фаза экспоненциального роста впоследствии не изменилась, продолжаясь в течение 20 мин с тем же (Δ0,3) относительным увеличением OD, что отражает одинаковый рост бактерий после каждого поступления питательных веществ. Согласно стандартам McFarland, увеличение 0,3 OD соответствует приросту 108-109 бактерий. После 9-го добавления питательных веществ, бактериальные белки экстрагировали в экспоненциальной и стационарной фазах, показывая концентрацию суммарных белков 0,088 мг/мл и 0,15 мг/мл, соответственно. Выделенные белки проверяли на уровни ClpB и использовали в анализе продуцирования АТФ и для экспериментов in vivo, включая инфузии в толстую кишку и системные инъекции мышам и крысам на свободном кормлении с последующей детекцией c-fos в головном мозге.
Протеомный анализ
Чтобы проанализировать, изменяются ли профили экспрессии белка в зависимости от фаз роста бактерий, индуцированого питательными веществами, двумерный электрофорез в полиакриламидном геле (2D-PAGE) проводили отдельно на мембранной и цитоплазматической фракциях белка K12 E.coli, экстрагированных через 10 мин и 2,3 ч. после 5-го добавления среды МН, в соответствии с экспоненциальной и стационарной фазами, соответственно. Суммарное количество обнаруженных белковых пятен составило 2895 (1367 мембранных и 1528 цитоплазматических).
Сравнение 2D-PAGE мембранных белков из экспоненциальной и стационарной фаз показало 20 дифференциально (по меньшей мере в 1,5 раза) экспрессированных белков. Среди них, 17 белков показали повышенную экспрессию в экспоненциальной фазе и 15 были идентифицированы с помощью масс-спектрометрии. Сравнение 2D-PAGE цитоплазматических белков показало 20 дифференциально (по крайней мере в 1,5 раза) экспрессированных белков. В отличие от мембранных белков, большинство (19) цитоплазматических белков проявляли повышенную экспрессию в течение стационарной фазы. Только одно белковое пятно, соответствующее флагеллину, показывало повышенную экспрессию в экспоненциальной фазе. Большинство идентифицированных белков были вовлечены либо в анаболические, либо в катаболические процессы, показывая метаболический профиль общей смеси в обеих фазах роста.
Производство АТФ белками E.coli in vitro
Чтобы исследовать, может ли влиять изменение бактериального протеома (белковой карты) в фазах роста на их возможности экстракции энергии, было проверено in vitro производство АТФ из питательных веществ белками K12 Е. coli из экспоненциальной и стационарной фаз. Обнаружили, что белки обеих фаз роста способствовали увеличению производства АТФ из разных источников энергии. Концентрации АТФ были выше, когда использовали белок-содержащий смешанный источник энергии, такой как среда МН, по сравнению с раствором сахарозы. Производство АТФ увеличивалось дозозависимо с увеличением концентраций бактериальных белков, однако, не было обнаружено существенных различий между АТФ-продуцирующими эффектами белков из экспоненциальной или стационарной фаз.
Производство ClpB бактериями Е. coli in vitro
Разработали и проверили на применимость иммуноферментный анализ (ELISA) для обнаружения ClpB Е. coli, который был использован в этом исследовании. Различается ли производство белка ClpB на разных фазах роста бактерий, изучали на 4 отдельных культурах Е. coli K12. Вестерн-блоттинг обнаружил соответствующие ClpB полосы размером 96 кДа во всех белковых препаратах, с повышенными уровнями в течение стационарной фазы. Эти изменения были дополнительно подтверждены с использованием ELISA ClpB в тех же бактериальных белковых препаратах, что показывает, что концентрации ClpB почти удваиваются в стационарной фазе.
Внутрикишечные инфузии питательных веществ и белков E.coli
Чтобы проверить, соответствует ли наша in vitro модель роста Е. coli, индуцированного питательными веществами, динамике роста бактерий кишечника in vivo, среду МН или воду вводили в толстую кишку крыс под анестезией. Обнаружили, что инстилляции среды МН, но не воды, индуцировали бактериальную пролиферацию в кишечнике, причем экспоненциальная фаза роста, продолжалась в течение 20 мин, в соответствии с данными in vitro. Уровни ClpB в плазме, измеренные в воротной вене, не отличались существенно через 30 или 60 мин после инфузии МН. Тем не менее концентрации ClpB в плазме коррелировали положительно с содержанием ДНК ClpB в фекалиях.
Затем, чтобы определить, могут ли влиять ли зависящие от роста изменения протеомов E.coli, на механизмы контроля аппетита хозяина локально в кишечнике, в отдельном эксперименте анестезированные крысы получали 20 мин инфузии в толстую кишку белков Е. coli из экспоненциальной или стационарной фаз, в обоих случаях в количестве 0,1 мг/кг. Концентрации ClpB в слизистой оболочке толстой кишки, измеренные через 20 мин после инфузии, были выше у крыс, получающих белки стационарной фазы, однако, на уровни ClpB в плазме не влияли бактериальные белки как из экспоненциальной, так и из стационарной фаз. В соответствии с нашей гипотезой о том, что влияния белков Е. coli на передачу сигналов аппетита хозяина, могут зависеть от фазы роста бактерий, обнаружили, что инстилляции в толстую кишку белков Е. coli из экспоненциальной, но не из стационарной фазы, стимулировали уровни GLP-1 в плазме и, напротив, повышенные уровни PYY в плазме были обнаружены после инфузии белков из стационарной, но не из экспоненциальной фазы.
Потребление пищи и c-fos мозга после однократного введения белков Е. coli крысам
Поскольку ClpB присутствовал в плазме у всех тестируемых крыс и мышей, возможно, что плазматические белки Е. coli могут влиять на аппетит посредством их системного действия, и что такие эффекты могут быть разными для белков, связанных с фазами бактериального роста. Исследуя эту возможность у крыс, голодавших в течение ночи, обнаружили, что одно внутрибрюшинное (IP) введение (0,1 мг/кг массы тела) мембранной фракции белков Е. coli, экстрагированных в стационарной фазе, уменьшало 1 ч.- и 2 ч.- потребление пищи при повторном кормлении по сравнению с контрольной группой. Напротив, введение цитоплазматической фракции белков E.coli (0,1 мг/кг массы тела, внутрибрюшинно I.P.), экстрагированных в экспоненциальной фазе, увеличивало 4 ч.-потребление пищи при повторном кормлении.
Чтобы дополнительно исследовать, могут ли суммарные белки Е. coli влиять на спонтанное потребление пищи, в зависимости от фазы роста, и активировать центральные места, такие как ARC, крысам с неограниченным кормлением инъецировали до наступления фазы темноты бактериальные белки (0,1 мг/кг массы тела, IP). Потребление пищи оценивали в течение 2 ч после инъекций, а затем крыс убивали для анализа экспрессии c-fos в мозге. Обнаружили, что крысы, инъецированные бактериальными белками из стационарной фазы, ели меньше, чем контрольные, тогда как на потребление пищи не оказывали существенного влияния инъекции бактериальными белками из экспоненциальной фазы.
Через два часа после внутрибрюшинной I.P. инъекции белков E.coli крысам на свободном кормлении, экспрессия c-fos была иммуногистохимически проанализирована в ARC и вентромедиальном ядре (VMN) гипоталамуса и в центральном ядре миндалевидного тела (СеА). Увеличенное количество c-fos-позитивных клеток было обнаружено в ARC и VMN у мышей, получавших бактериальные белки из стационарной фазы. Большинство клеток, экспрессирующих c-fos в ARC, содержали β-эндорфин (контрольные образцы, 71,31±12,81%, E.coli экспон. фазы, 73,56±10,45%, Е. coli стационар. фазы, 80,50±9,68%, ANOVA р=0,36), т.е. были идентифицированы как анорексигенные нейроны РОМС. Соответственно, процент оставшихся c-fos-нейронов в ARC существенно не различался среди групп (контрольные образцы, 28,69±12,81%, E.coli экспоненц. фазы, 26,44±10,45%, Е. coli стационарн. фазы, 19,5±9,68%, дисперсионный анализ р=0,36). Несмотря на то, что общее количество β-эндорфин- позитивных клеток существенно не различалось среди групп (контрольные группы, 54,82±10,67 клеток, Е. coli экспоненц. фазы, 66,03±11,43 клетки, Е. coli стационарной фазы, 66,03±5,06 клеток, ANOVA р=0,09), относительное число активированных нейронов β-эндорфинов было увеличено у крыс, получавших белки из стационарной фазы, по сравнению с контрольными крысами и крысами, получавшими белки из экспоненциальной фазы. Кроме того, число активированных β-эндорфиновых нейронов обратно коррелировало с потреблением пищи (Пирсон r=-0,57, р=0,018).
В СеА, количество c-fos-позитивных клеток было увеличено у крыс, которым вводили белки из стационарной фазы по сравнению с двумя другими группами. Почти все c-fos-позитивные клетки в СеА были фенотипированы как CGRP-экспрессирующие нейроны (контрольные, 100±0,01%, E.coli экспоненц. фазы, 100±0,01%, E.coli стационарн. фазы, 100±0,01% дисперсионный анализ р=0,92). Несмотря на то, что суммарное число CGRP-позитивных нейронов в СеА было одинаковым среди групп (контрольные группы, 123,8±13,15 клеток, E.coli экспоненц. фазы, 118,3±25,59 клеток, E. coli стационарн. фазы, 126,1±6,64 клетки, дисперсионный анализ р=0,85), процент c-fos-активированных нейронов CGRP был увеличен только у крыс, получающих белки стационарной фазы. Активация нейронов CGRP обратно коррелировала с потреблением пищи (Пирсон, r=-0,89, р=0,001).
Характер питания и гипоталамические нейропептиды после хронических инъекций белка Е. coli мышам
Чтобы определить, могут ли бактериальные белки влиять на характер питания, две ежедневные инъекции суммарных белков Е. coli (0,1 мг/кг массы тела, внутрибрюшинно, I.P.) делали в течение одной недели мышам на свободном кормлении. Первый день после инъекций характеризовался значительно меньшей массой тела и потреблением пищи у мышей, получавших бактериальные белки из стационарной, но не из экспоненциальной фазы по сравнению с контрольными мышами. Несмотря на то, что ежедневное количество пищи не различалось существенно среди групп, его снижение по сравнению с днем перед инъекциями наблюдалось через неделю у мышей, получавших белки из стационарной фазы. Частота приема пищи не была существенно различной среди групп, несмотря на это тенденция к увеличению наблюдалась у мышей, получавших бактериальные белки из стационарной фазы. Несмотря на то, что суммарный прием пищи среди 3 групп не был существенно различен в течение 6 дней инъекций, при анализе его отдельно для периодов света и темноты, было показано, что мыши, которым вводили белки из экспоненциальной фазы, показывали увеличение потребления пищи в течение светлого периода, но уменьшение во время темного периода. Напротив, мыши, получающие белки из стационарной фазы, показывали уменьшение, по сравнению с контролем потребления пищи в темный период, без какого-либо эффекта в светлый период. В течение первого дня после инъекций, показатель сытости был выше у мышей, получавших белки из стационарной фазы, и эта же группа демонстрировала тенденцию к снижению, через одну неделю.
Чтобы получить представление о молекулярных изменениях, лежащих в основе изменения характера питания, наблюдаемого через 6 дней инъекций бактериальных белков, проанализировали уровни экспрессии гипоталамической мРНК нескольких нейропептидов, участвующих в контроле аппетита. Обнаружили, что мыши, получающие белки стационарной фазы, демонстрировали повышение уровней предшественника мРНК, нейротрофического фактора мозга (BDNF, brain-derived neurotrophic factor) и орэксина, по сравнению с контрольной группой, и кортикотропин- рилизинг-гормона (CRH) по сравнению с мышами, инъецированными белками из экспоненциальной фазы, которые также показывали повышение уровней BDNF, но уменьшение QRFP.
Электрофизиологическая активация гипоталамических нейронов РОМС белками ClpB
Чтобы определить, активирует ли ClpB, как маркер белков E.coli, активированных в стационарной фазе роста, и миметик α-MSH, нейроны ARC РОМС, эффект ClpB исследовали на срезах мозга у мышей POMC-eGFP, используя электрофизиологическую методику patch-clamp с прикрепленной клеткой.
Введение ClpB (1 нМ) в раствор, в котором содержится нейрон, увеличило частоту потенциала действия ~50% нейронов ARC РОМС (n=7/13) на 229±109% (исходная: 2,02±0,78 Гц по сравнению с ClpB: 3,82±1,36 Гц), как показано на Фиг. 2. В целом, нейроны РОМС полностью не возвращались к исходным частотам пульсации нейрона, по меньшей мере, в течение 10 мин после применения ClpB.
Эти результаты, таким образом, указывают на непосредственное влияние белка ClpB на насыщение и сытость.
Обсуждение
Наше исследование показывает, что бактериальные белки могут физиологически связывать бактерии кишечника с контролем аппетита хозяина, включая его краткосрочные и долгосрочные механизмы, связанные с индуцированным питательными веществами ростом бактерий в кишечнике и их системными эффектами, соответственно. Следующие основные результаты подтверждают этот вывод: 1) регулярное предоставление питательных веществ ускоряет и стабилизирует экспоненциальный рост Е. coli, продолжающийся в течение 20 мин, в соответствии с данными in vivo; 2) Стационарная фаза роста E.coli характеризовалась увеличением суммарного содержания бактериальных белков и другим профилем протеома, включая увеличение ClpB; 3) белки Е. coli из обеих фаз роста, стимулировали дозозависимым способом, производство АТФ in vitro; 4) Уровни ClpB в плазме не изменялись после индуцированного питательными веществами роста бактерий в кишечнике, но коррелировали с ДНК ClpB в кишечной микрофлоре; 5) Кишечная инфузия белков E.coli из экспоненциальной и стационарной фаз роста стимулировала плазматические GLP-1 и PYY, соответственно. 6) Системные инъекции белков E.coli значительно снижали потребление пищи только в случае белков из стационарной фазы, сопровождающееся активацией c-fos в анорексигенных нейронах ARC и СеА, и, наконец, 7) белки ClpB стимулировали частоту пульсации нейронов ARC РОМС.
Регулярное предоставление питательных веществ и рост бактерий
Среди широкого спектра бактерий, которые колонизируют желудочно-кишечные тракты людей, Е. coli является наиболее распространенным факультативным анаэробом, обосновывая его использование в качестве модельного организма для симбиотических кишечных бактерий (Foucault et al., 2009). В настоящем исследовании обнаружили, что Е. coli меняет свою динамику роста при регулярной поставке питательных веществ, приводя, после 5-го цикла кормления, к немедленному экспоненциальному росту, продолжающемуся в течение 20 минут, за которым следует стационарная фаза. Цикл роста затем идентично воспроизводился после следующего поступления питательных веществ, указывая на то, что, что он может играть роль ритмоводителя, установленного с помощью механизмов присущих бактериям. Аналогичная динамика роста бактерий в ответ на инфузию питательных веществ наблюдалась в толстой кишке у крыс, подтверждая то, что наши данные in vitro, могут быть актуальными к ситуациям in vivo, например, у людей, регулярно принимающих пищу. Прирост 108-109 числа бактерий оставался стабильным после каждого нового поступления питательных веществ, указывая на то, что соответствующее стабильное производство бактериальной биомассы, включая увеличение содержания белков в кишечнике, может сыграть роль индуцированного пищей регуляторного фактора у хозяина. Учитывая, что средняя прандиальная фаза у людей сходна с продолжительностью экспоненциального роста бактерий, регулярно получающих питательные вещества, возникает соблазн предположить, что сытость хозяина может быть вызвана достижением стационарной фазы кишечными бактериями, через 20 мин после контакта с поглощенными питательными веществами. Однако, содержание бактерий в желудочно-кишечном тракте колеблется от 103 в желудке до 1012 в толстой кишке. Более того, примерно 2 ч. необходимо для продвижения поглощенных питательных веществ через желудок и тонкую кишку и примерно 10 ч. через толстую кишку. Из-за такой задержки поступления питательных веществ к большинству кишечных бактерий, вероятно, что помимо прямого контакта с пищевым комком, рост бактерий во время прандиальной фазы может также инициироваться питательными веществами, высвобождаемыми в просвет кишечника с помощью Павловского рефлекса головного мозга на прием пищи.
Экспрессия бактериальных белков во время фаз роста и чувствительность кишечника
Поскольку динамика роста бактерий Е. coli, регулярно получающих питательные вещества, может быть связана с прандиальной и постпрандиальной фазами хозяина, изучили, может ли потенциально экспрессия бактериальных белков связывать бактерии кишечника с контролем аппетита хозяина. Во-первых, сравнили протеомы Е. coli в экспоненциальной и стационарной фазах роста. Для этого анализа, белки Е. coli были экстрагированы в середине экспоненциальной фазы, то есть через 10 минут после подачи питательных веществ, и в стационарной фазе через 2 часа - время, обычно характеризующееся ощущением сытости. Обнаружение по меньшей мере 40 дифференциально экспрессированных белков между двумя фазами роста подтвердило, что они различаются не только количественно, по содержанию белка, которое почти удваивается после бактериальной пролиферации, но и качественно. Затем была изучена возможная релевантность модифицированного профиля экспрессии белка, контролю аппетита хозяина: i) путем сравнения способности бактериальных белков генерировать энергетический субстрат и ii) путем определения возможных прямых эффектов бактериальных белков на сигнальные пути, регулирующие аппетит. Последняя возможность подтверждается недавней протеомной идентификацией ClpB Е. coli как конформационного белкового миметика a-MSH (Tennoune et al., 2014); данные, которые подтвердили нашу более раннюю гипотезу о том, что бактериальные белки кишечника, показывающие эпитопы, гомологичные анорексигенным или орексигенным пептидам, могут быть ответственны за производство кросс-реактивных аутоантител (Fetissov et al., 2008). Таким образом, можно предположить, что комбинация таких бактериальных белковых миметиков может напрямую влиять на аппетит, в зависимости от профиля экспрессии белка, связанного с фазой роста бактерий. Анализируя культуры Е. coli и слизистую оболочку кишечника, обнаружили повышенные уровни ClpB, связанные со стадией стационарного роста. Таким образом, повышение уровня ClpB может способствовать активации анорексигенных сигнальных путей после индуцированной питательными веществами пролиферации E.coli и увеличения продуцирования бактериальных белков. Важным вопросом является то, что бактериальные белки, такие как ClpB, могут действовать на сигнальные пути, регулирующие аппетит.
Несмотря на то что бактериальные белки присутствуют в слизистой оболочке кишечника (Haange et al., 2012), их прохождение через кишечный барьер не изучалось широко (Lim and Rowley, 1985). Теоретически, после спонтанного и индуцированного бактериального лизиса в кишечнике (Rice and Bayles, 2008), бактериальные компоненты могут проходить через эпителиальный барьер слизистой оболочки путем абсорбции в энтероцитах и путем парацеллюлярной диффузии, регулируемой энтеральной нервной системой (Neunlist et al., 2012). Например, липополисахарид (LPS), который высвобождается при лизисе грамотрицательных бактерий, естественно присутствует в плазме здоровых людей и мышей, с повышением базальных уровней после потребления пищи с высоким содержанием жиров (Cani et al., 2007). Уровни LPS в плазме также увеличиваются после приема пищи (Harte et al., 2012), но таких данных о бактериальных белках нет. В настоящем изобретении показано, что уровни ClpB в плазме у крыс остаются стабильными после бактериальной пролиферации в кишечнике или после кишечной инфузии бактериальных белков. Это указывает на то, что плазматические ClpB, и, скорее всего, другие бактериальные белки, присутствующие в плазме, не испытывают сильного влияния бактериологической пролиферации, вызванной питательными веществами, и, следовательно, они не могут быть вовлечены в сигнализацию сытости, посылаемую в мозг.
Тем не менее концентрации ClpB в плазме коррелировали с ДНК ClpB в кишечной микрофлоре, указывая на то, что число бактерий, продуцирующих ClpB, которое должно быть в долгосрочной перспективе относительно независимым от его колебаний, связанных с бактериальным ростом, обусловленным питательными веществами, может быть основным фактором, для долгосрочного поддержания уровней ClpB в плазме. Этот вывод дополнительно подтвержден и нашими данными, полученными для проверки корректности ELISA ClpB, демонстрирующими повышенные концентрации ClpB в плазме у мышей, хронически получавших через желудочный зонд Е. coli, но не у мышей, получавших ClpB-мутантную E.coli. Следовательно, возможно, что кишечные бактериальные белки, присутствующие в плазме, включая ClpB, могут действовать системно, связывая композицию кишечной микрофлоры с долгосрочным контролем энергетического метаболизма.
Влияние белков Е. coli на продукцию АТФ in vitro
Обмен энергией через пищевые цепи представляет собой универсальную связь между всеми организмами (Yun et al., 2006). АТФ, полученный из катаболизма питательных веществ как у бактерий, так и у животных, служит основным энергетическим субстратом для анаболических процессов. Животные могут ощущать изменения в АТФ посредством активности аденозин-5'-монофосфат-активированной протеинкиназы (АМРК), что приводит к увеличению потребления пищи, когда уровни АТФ низки, и наоборот (Dzamko and Steinberg, 2009). Следовательно, в настоящей работе сравнили способность белков Е. coli в экспоненциальной и стационарной фазах генерировать АТФ in vitro. Действительно, многие из идентифицированных белков проявляли анаболические или катаболические свойства. Обнаружили, что белки Е. coli способны стимулировать выработку АТФ in vitro, указывая на то, что они могут продолжать катализировать выработку АТФ после бактериального лизиса в кишечнике. Несмотря на то, что между белками из экспоненциальной и стационарной фаз не было обнаружено различий в способности продуцировать АТФ, концентрация АТФ, зависящая от концентрации бактериальных белков, указывает на то, что увеличение содержания бактериальных белков после бактериальной пролиферации, индуцированной питательными веществами, должно приводить к более высокому уровню синтеза АТФ. Актуальность эффективности микрофлоры кишечника для сбора энергии для метаболизма хозяина, была ранее установлена путем сравнения тучных и худых людей и мышей (Turnbaugh et al., 2006). Наши данные дополнительно подтверждают эти результаты, демонстрируя способность белков E.coli генерировать АТФ. Это также указывает на то, что бактериальная пролиферация, индуцированная пищей, может привести к увеличению кишечного АТФ, что должно способствовать чувствительности просвета кишечника к доступности энергии и расслаблению кишечника (Glasgow et al., 1998).
Кишечные эффекты белков Е. coli на гормоны насыщения
Затем исследовали, могут ли белки Е. coli в кишечнике стимулировать системное высвобождение гормонов насыщения кишечника, таких как GLP-1 и PYY (Adrian et al., 1985; Batterham et al., 2002; Beglinger and Degen, 2006; Flint et al., 1998). Фактически, оба гормона вырабатываются одними и теми же или различными энтероэндокринными L-клетками, присутствующими во всем кишечнике и многочисленными в толстой кишке (Eissele et al., 1992), и, следовательно, L-клетки непосредственно подвергаются воздействию бактериальных белков. Обнаружили дифференциальные эффекты белков Е. coli на выработку GLP-1 и PYY, демонстрируя стимуляцию GLP-1 белками из экспоненциальной и стимуляцию PYY белками из стационарной фазы роста. Эти результаты указывают на некоторое сходство между индуцированным питательными веществами бактериальным ростом и известной динамикой индуцированной пищей выработки GLP-1 и PYY. Фактически, как было показано у людей, резкий пик GLP-1 в плазме наступает через 15 мин после внутрижелудочной инфузии жидкой пищи, тогда как более продолжительный повышенный уровень PYY в плазме начинается между 15 и 30 мин после приема пищи (Edwards et al., 1999; Gerspach et al., 2011). Более длительная выработка GLP-1 была связана с потреблением жира (van der Klaauw et al., 2013). Таким образом, динамика роста регулярно питающихся кишечных бактерий вписывается в динамику выработки GLP-1 и PYY, что указывает на индуктивную роль кишечных бактерий и, в частности, белков Е. coli, в индуцированной пищей сигнализации кишечной сытости. Дифференциальный эффект белков Е. coli из экспоненциальной фазы для стимуляции GLP-1 может, возможно, отражать роль инкретирования GLP-1 в гликемическом контроле (Edwards et al., 1999; Steinert et al., 2014). Недавняя демонстрация функционального MC4R, экспрессируемого L-клетками (Panaro et al., 2014), дает предпосылки для их возможной активации бактериальными белками-миметиками α-MSH. Увеличение производства ClpB во время стационарной фазы Е. coli, а также повышение уровней ClpB в слизистой оболочке кишечника, связанные с увеличением уровней PYY в плазме, указывают на непосредственную роль ClpB в активации продуцирующих PYY L-клеток в толстой кишке. С другой стороны, до сих пор не идентифицированные белки E.coli, которые могут быть активированы во время фазы экспоненциального роста, могут предпочтительно стимулировать секрецию GLP-1.
Системные эффекты белков E. coli на потребление пищи и сигнальные пути мозга, регулирующие аппетит
Показали в настоящем изобретении, что периферические инъекции белков Е. coli крысам, голодным или с неограниченным кормлением, и мышам с неограниченным кормлением, изменяли потребление пищи в зависимости от фазы роста Е. coli. Учитывая, что на ClpB плазмы не влияла кишечная инфузия питательных веществ, но уровень ClpB плазмы был стабильным в течение короткого времени, системное действие бактериальных белков следует интерпретировать как имеющее значение для их длительных модулирующих эффектов на аппетит. Более того, из-за короткой продолжительности экспоненциальной фазы, бактериальные белки, активированные в течение продолжительной стационарной фазы, должны доминировать в плазме, и, следовательно, их системные введения могут лучше представлять физиологические ситуации. Концентрации белков E.coli 0,1 мг/кг, использованные в этих экспериментах, были сходны с эффективными, способными вызывать чувство насыщения дозами пептидных гормонов, таких как лептин или PYY после периферических введений у людей или грызунов (Batterham et al., 2002; Halaas et al., 1995; Heymsfield et al., 1999).
Увеличение потребления пищи у голодных крыс во время повторного питания в светлой фазе наблюдалось после введения цитоплазматических белков из экспоненциальной фазы, и уменьшение потребления пищи наблюдалось после введения мембранных белков из стационарной фазы. Этот эксперимент подтвердил, что различные белковые смеси от одних и тех же бактерий, способны увеличивать или уменьшать потребление пищи за счет их системного действия. Однако в более физиологической обстановке, путем проверки эффекта суммарных белков Е. coli у крыс с неограниченным кормлением в темной фазе, наблюдалось только снижение потребления пищи, которое было индуцировано белками из стационарной фазы.
Результаты повторных инъекций у мышей с неограниченным кормлением также подтверждают роль протеинов Е. coli в способствовании отрицательному энергетическому балансу. Действительно, первый день инъекций бактериальных белков сопровождался уменьшением потребления пищи и массы тела, значительным у мышей, получавших белки стационарной фазы, и которые также характеризовались повышением показателя сытости. Несмотря на то, что потребление пищи у этих мышей нормализовалось впоследствии, постепенное уменьшение количества пищи сопровождалось увеличением частоты приема пищи, скорее всего, в качестве компенсационного механизма для поддержания потребления пищи (Meguid et al., 1998). Кроме того, дифференциальные эффекты белков Е. coli наблюдали во время светлой и темной фаз. Соответственно, картина экспрессии мРНК регулирующих аппетит нейропептидов в гипоталамусе, показала смешанный ответ с активацией как анорексигенного, так и орексигенного сигнальных путей. Следует отметить, что обе группы мышей, получавших белки Е. coli, показали сходное увеличение мРНК BDNF, анорексигенный сигнальный путь ниже по течению до MC4R в VMN (Xu et al., 2003). Этот сигнальный путь может лежать в основе снижения потребления пищи во время темной фазы, наблюдаемой в обеих группах, и который был дополнительно акцентирован у мышей, которым вводили белки экспоненциальной фазы, демонстрирующие более низкие уровни орексигенного QRFP (Chartrel et al., 2003) и NPY (Herzog, 2003). Напротив, мыши, которые получали бактериальные белки из стационарной фазы, демонстрировали повышенный анорексигенный профиль с повышенными уровнями мРНК CRH, наиболее вероятно, подразумевая нейроны PVN, экспрессирующие MC4R (Lu et al., 2003). Эти изменения в сочетании с повышенной экспрессией предшественника мРНК орексина А, который стимулирует частоту приема пищи (Baird et al., 2009), могут способствовать уменьшению соотношения количества пищи и сытости у этих мышей после 6 дней инъекций.
В соответствии с нашей гипотезой о том, что бактериальные белки, образующиеся в течение стационарной фазы, могут активировать некоторые ключевые центральные анорексигенные сигнальные пути, обнаружили у крыс с неорганиченным кормлением увеличение экспрессии c-fos в анорексигенных нейронах ARC РОМС, а также в VMN, который уже давно известен как центр насыщения и связан с нейронами ARC РОМС (Sternson et al., 2005). Полученная картина c-fos напоминает картину c-fos при возникновении чувства насыщения во время приема пищи (Johnstone et al., 2006) или индуцированую гормонами насыщения, такими как PYY или панкреатический полипептид (Batterham et al., 2002; Challis et al., 2004; Lin et al., 2009). Относительно небольшое количество (~10%) c-fos-активированных нейронов РОМС указывает на то, что циркулирующие белки Е. coli могут оказывать модулирующее действие на аппетит и массу тела, действуя через эти гипоталамические сигнальные пути. Несмотря на то, что было невозможно определить активацию c-fos нейронами NPY/AgRP, их вклад в сигнализацию бактериальными белками нельзя исключить; эти нейроны также экспрессируют MC3R и MC4R (Mounien et al., 2005). Более того, более сильные, чем в ARC РОМС активации c-fos в нейронах СеА CGRP (~40%) могут означать конвергентное нисходящее действие от нейронов ARC РОМС и NPY/AgRP и, возможно, от других зон, регулирующих аппетит, которые здесь не анализировались, например, ядра одиночного пути.
Наконец, чтобы определить, может ли активация регулирующих аппетит участков мозга, например, нейронов ARC РОМС бактериальными белками, быть вызвана их локальным действием, изучали, может ли приложение ClpB к этим нейронам активировать их электрическую активность. Наши результаты показали, что примерно половина исследованных нейронов увеличили частоту их потенциала действия, оставаясь активированными в течение по меньшей мере 10 мин. Продолжительный эффект ClpB согласуется с действием α-MSH на нейроны РОМС, экспрессирующие функциональные MC3R и MC4R (Smith et al., 2007), что указывает на то, что ClpB может быть физиологическим активатором гипоталамических нейронов РОМС, несколько похожим на способные вызывать чувство насыщения PYY и лептин (Batterham et al., 2002; Cowley et al., 2001). Однако пока что неизвестно, мог ли ClpB активировать нейроны РОМС напрямую или через локальную сеть.
В совокупности, эти данные подтверждают роль системно присутствующих белков E.coli, экспрессия которых увеличивается в фазе стационарного роста, например, ClpB, в способствовании отрицательному энергетическому балансу посредством активации анорексигенных путей мозга. Это также указывает на то, что изменения состава микрофлоры, приводящие к низкой или высокой плотности Е. coli и, возможно, других бактерий из семейства Enterobacteriaceae, могут влиять на энергетический баланс хозяина положительным или отрицательным образом, соответственно.
Пример 2
Этот пример демонстрирует влияние бактерий, экспрессирующих ClpB, на потребление пищи.
Одномесячные самцы мышей C57B16 (Janvier Laboratories) были адаптированы к условиям вивария в течение 1 недели и содержались, как описано выше. Мышей распределяли по трем группам следующим образом: (i) получившие через желудочный зонд 108 бактерий Е. coli K12 (экспрессирующих ClpB); (ii) получившие через желудочный зонд 108 Е. coli K12, дефицитных по ClpB; (iii) и контрольные, которые не получали никаких обработок. Мутантный штамм ClpB был получен в Bernd Bukau's Laboratory (ZMBH, Гейдельбергский университет, Гейдельберг, Германия) и любезно предоставлен вместе с соответствующими бактериями E.coli дикого типа (WT) доктором Axel Mogk. Мышей помещали индивидуально в клетки BioDAQ (Research Diets) и вводили им внутрижелудочно с помощью желудочного зонда ежедневно до наступления темной фазы в течение 21 дня 0,5 мл среды LB с бактериями. Первые дни кормления с помощью желудочного зонда сопровождались снижением массы тела и приема пищи у мышей, получавших бактерии Е. coli WT, но не бактерии, которые не экспрессируют белок ClpB (Фиг. 1).
Пример 3
Этот пример демонстрирует влияние бактерий, экспрессирующих ClpB, на мышей ob/ob с ожирением.
Генетически тучных мышей ob/ob адаптировали к условиям вивария в течение 1 недели и содержали, как описано выше. Мышам вводили внутрижелудочно с помощью желудочного зонда (i) 108 бактерий Е. coli K12 (экспрессирующих ClpB); (ii) 108 бактерий Е. coli K12, дефицитных по ClpB; в обоих случаях содержащихся в среде Мюллера-Хинтона (МН), или (iii) только среду МН, в качестве контроля.
Мутантный штамм ClpB был получен в Bernd Bukau's Laboratory (ZMBH, Гейдельбергский университет, Гейдельберг, Германия) и любезно предоставлен вместе с соответствующими бактериями E.coli дикого типа (WT) доктором Axel Mogk. Мышей помещали индивидуально в клетки BioDAQ (Research Diets) и кормили внутрижелудочно с помощью желудочного зонда ежедневно в течение 21 дня, как указано.
Авторы настоящего изобретения показали, что введение с помощью желудочного зонда бактерий E.coli K12 WT индуцировало снижение на 56% прироста массы тела (Фиг. 3 и 4), уменьшение отношения массы жировой ткани/к массе нежировых тканей (Фиг. 5 и 6) и уменьшение на 20% суммарного количества потребления пищи (Фиг. 7 и 8), которое не наблюдалось при введении бактерий Е. coli K12, дефицитных по ClpB.
Пример 4
Этот пример демонстрирует влияние других штаммов бактерий, экспрессирующих ClpB, на мышей ob/ob с ожирением.
Генетически тучных мышей ob/ob адаптировали к условиям вивария в течение 1 недели и содержали, как описано выше. Мышам вводили внутрижелудочно с помощью желудочного зонда (i) 108 бактерий Е. coli K12 (экспрессирующих ClpB); (ii) 108 бактерий Е. coli Niessle 1917 (экспрессирующих ClpB) (iii) 108 бактерий Е. coli Niessle 1917 (экспрессирующих ClpB) в лиофилизированной форме; во всех случаях содержащихся в среде Мюллера-Хинтона (МН) или (iv) одну только среду МН, в качестве контроля. Мышей кормили с помощью желудочного зонда ежедневно в течение 14 дней, как указано.
Авторы настоящего изобретения показали, что введение с помощью желудочного зонда любого штамма бактерий E.coli, экспрессирующих ClpB, индуцировало снижение прироста массы тела (Фиг. 9 и 10) и снижение содержания жира (Фиг. 11).
Пример 5
Этот пример подтверждает прямое действие ClpB, вызывающее чувство насыщения.
Возможное влияние ClpB на пищевое поведение через семейство гипоталамических рецепторов MCR было показано в исследовании на животных на крысах, которые получали интрацеребровентрикулярные (ICV) инъекции ClpB.
Самцов крыс Sprague-Dawley 200-250 г (Janvier, L'Arbresle, France) содержали при 24°С с 12:12-часовым циклом чередования света и темноты (светлый период составлял 7-19 ч) в специализированном виварии с кондиционером. Крыс кормили стандартным гранулированным кормом (RM1 diet, SDS, Essex, UK). Питьевая вода всегда была доступна без ограничений.
Для имплантации канюлей из нержавеющей стали (С311 GA, внешний диаметр 0,9 мм, внутренний 0,58 мм, Plastics One, Roanoke, VA) крыс анестезировали внутрибрюшинной инъекцией кетамина (75 мг/кг)/ксилазина (5 мг/кг) (3: 1 об. - 0,1 мл/100 г массы тела) и помещали в новый стандартный стереотаксический прибор для крыс и мышей New Standard Stereotaxic Instrument for Rats and Mice (Stoelting Europe, Дублин, Ирландия). Канюли были имплантированы под операционным микроскопом (Carl Zeiss, Йена, Германия) в гипоталамическое паравентрикулярное ядро (Брегма: +2,8 мм, латеральное: -0,4 мм от средней линии и вентральное: 8,2 мм от твердой мозговой оболочки), с режущая планка установлена на -3,3 мм. Канюли были прикреплены к черепу, с помощью зубного цемента, поддерживаемого анкерными винтами. После пробуждения, крыс держали отдельно в камерах для исследования метаболизма (Techniplast, Лион, Франция) в течение 1 недели и кормили без ограничений тем же стандартным кормом для грызунов (RM1, SDS), вода была доступна всегда. Физическое состояние и прирост массы тела ежедневно контролировали в послеоперационном периоде для обеспечения хорошего выздоровления.
Затем крыс помещали индивидуально в клетки для крыс BioDAQ (Research Diets, Inc., Нью-Брансуик, Нью-Джерси), каждая из которых была оснащена монитором автоматического кормления. Через 3 дня адаптации к клеткам BioDAQ, крысы были лишены пищи на 12 часов перед инъекцией и разделены на 3 группы (n=3), каждая из которых получала разные дозы однократной инъекции ClpB: 10 нг, 100 нг и 1 мг, разбавленные в 2 мл стерильного искусственного цереброспинального раствора.
Инъекции проводили за 15 минут до начала темной фазы и подачи питания. Потребление пищи измеряли во время темной фазы. У животных наблюдали снижение потребления пищи в зависимости от дозы (Фиг. 12).
Пример 6
Данный пример демонстрирует влияние Hafnia alvei, экспрессирующей ClpB, на мышей ob/ob с ожирением.
Генетически тучных мышей ob/ob адаптировали к условиям вивария в течение 1 недели и содержали, как описано выше. Мышам вводили внутрижелудочно с помощью желудочного зонда (i) 108 бактерий Hafnia alvei AF036 (экспрессирующих ClpB); (ii) 108 бактерий E. coli K12, дефицитных по ClpB; (iii) 108 бактерий E. coli Niessle 1917 (экспрессирующих ClpB); во всех случаях содержавшихся в среде Мюллера-Хинтона (МН) или (iv) одну только среду МН, в качестве контроля.
Авторы настоящего изобретения показали, что введение Hafnia alvei вызвало существенное снижение прироста массы тела в сравнении с тучными мышами контрольной группы (Фиг. 13). Кроме того, снижение прироста массы тела сопровождалось снижением кумулятивного потребления пищи (Фиг. 14), а также уменьшением как массы жировой ткани, так и массы нежировых тканей, без изменения соотношения массы жировой ткани и массы нежировых тканей тела (Фиг. 15).
Гормон-чувствительная липаза (HSL) - это внутриклеточная нейтральная липаза, способная гидролизовать триацилглицериды, диацилглицериды, моноацилглицериды и сложные эфиры холестерина, а также другие жиро- и водорастворимые субстраты. В частности, активация HSL вызывает липолиз. Гормональная активация HSL происходит при посредничестве цАМФ-зависимой протеинкиназы (РКА), которая фосфорилирует HSL. Рост экспрессии фосфорилированной формы протеина HSL (pHSL) означает, таким образом, активацию липолиза. Экспрессия pHSL измерена методом вестерн-блоттинга.
Авторы настоящего изобретения показали, что введение через желудочный зонд бактерий Е. coli Niessle 1917 и Hafnia alvei увеличивало экспрессию pHSL (Фиг. 16) по сравнению с контролем и введением через желудочный зонд бактерии Е. coli K12. Следовательно, бактерии Е. coli Niessle 1917 и Hafnia alvei активируют механизм липолиза.
Пример 7
Данный пример демонстрирует влияние Hafnia alvei, экспрессирующей ClpB, на мышей с ожирением, вызванным диетой с высоким содержанием жиров.
Одномесячных мышей-самцов C57B16 (Janvier Laboratories) в течение 2 недель выдерживали на диете с высоким содержанием жиров/углеводов. Затем мышам вводили внутрижелудочно с помощью желудочного зонда (i) 108 бактерий Hafnia alvei AF036 (экспрессирующих ClpB); (ii) 108 бактерий E. coli K12, дефицитных по ClpB; и те и другие - в среде Мюллера-Хинтона (МН) или (iii) одну только среду МН, в качестве контроля. Каждую мышь помещали в отдельную клетку BioDAQ (Research Diets) и кормили ежедневно внутрижелудочно с помощью зонда в течение 21 дня, как указано.
Возникновение ожирения в результате диеты с высоким содержанием жиров было подтверждено измерением среднего значения массы тела (Фиг. 17А), массы жировой ткани (Фиг. 17 В) и массы нежировых тканей (Фиг. 17С) в группе с индуцированным ожирением, и в группе, где ожирение не индуцировалось.
Авторы настоящего изобретения показали, что введение посредством желудочного зонда любого штамма бактерии Hafnia alvei, экспрессирующей ClpB, вызывало замедление набора веса (Фиг. 18) и снижение содержания жира (Фиг. 19).
Пример 8
Данный пример демонстрирует влияние Hafnia alvei, экспрессирующей ClpB, на крыс Нукера (fa/fa крыс), страдающих ожирением.
Генетически тучных fa/fa крыс Цукера (Janvier Laboratories) адаптировали к условиям вивария в течение 1 недели и содержали, как описано выше. Крысам вводили внутрижелудочно с помощью желудочного зонда (i) 108 бактерий Hafnia alvei AF036 (экспрессирующих ClpB); (ii) 108 бактерий E. coli K12, дефицитных по ClpB; (iii) 108 бактерий E. coli Niessle 1917 (экспрессирующих ClpB); во всех случаях содержавшихся в среде Мюллера-Хинтона (МН) или (iv) одну только среду МН, в качестве контроля. Каждую крысу помещали в отдельную клетку BioDAQ (Research Diets) и кормили ежедневно внутрижелудочно с помощью зонда в течение 21 дня, как указано.
Результаты показывают, что прирост веса тела, в большинстве случаев, снизился в группе, получавшей Н. alvei, по сравнению с крысами, получавшими бактерии Е. coli K12 Δ-ClpB или Е. coli Niessle 1917, или по сравнению с контролем (Фиг. 20). Кроме того, в группе, получавшей Н. alvei, наблюдался самый низкий уровень кумулятивного потребления пищи по сравнению с тремя другими группами (Фиг. 21).
Авторы настоящего изобретения показали также, что прирост массы жировой ткани снизился только в группе, получавшей Н. alvei (Фиг. 22).
В тексте настоящей заявки различные процитированные источники описывают уровень техники, к которому относится настоящее изобретение. Содержание указанных источников полностью включено в описание настоящей заявки посредством ссылки.
Источники информации
Adrian, Т.Е., Ferri, G.L., Bacarese-Hamilton, A.J., Fuessl, H.S., Polak, J.M., and Bloom, S.R. (1985). Human distribution and release of a putative new gut hormone, peptide YY. Gastroenterology 89, 1070-1077.
Atasoy, D., Betley, J.N., Su, H.H., and Sternson, S.M. (2012). Deconstruction of a neural circuit for hunger. Nature 488, 172-177.
Baird, J.-P., Choe, A., Loveland, J.L., Beck, J., Mahoney, C.E., Lord, J.S., and Grigg, L.A. (2009). Orexin-A hyperphagia: hindbrain participation in consummatory feeding responses. Endocrinology 150, 1202-1216.
Batterham, R.L., Cowley, M.A., Small, C.J., Herzog, H., Cohen, M.A., Dakin, C.L.,
Wren, A.M., Brynes, A.E., Low, M.J., Ghatei, M.A., et al., (2002). Gut hormone PYY(3-36) physiologically inhibits food intake. Nature 418, 650-654.
Beglinger, С, and Degen, L. (2006). Gastrointestinal satiety signals in humans - Physiologic roles for GLP-1 and PYY ? Physiology & Behavior 89, 460-464.
Berthoud, H.-R. (2011). Metabolic and hedonic drives in the neural control of appetite: who is the boss? Current Opinion in Neurobiology 21, 888-896.
Cani, P.D., Amar, J., Iglesias, M.A., Poggi, M., Knauf, C, Bastelica, D., Neyrinck, A.M., Fava, F., Tuohy, K.M., Chabo, C, et al, (2007). Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 56, 1761-1772.
Carter, M.E., Soden, M.E., Zweifel, L.S., and Palmiter, R.D. (2013). Genetic identification of a neural circuit that suppresses appetite. Nature 503, 111-114.
Challis, B.G., Coll, A.P., Yeo, G.S.H., Pinnock, S.B., Dickson, S.L., Thresher, R.R., Dixon, J., Zahn, D., Rochford, J.J., White, A., et al., (2004). Mice lacking pro-opiomelanocortin are sensitive to high-fat feeding but respond normally to the acute anorectic effects of peptide- YY3-36. Proc. Natl. Acad. Sci. USA 101, 4695-4700.
Chartrel, N., Dujardin, C, Anouar, Y., Leprince, J., Decker, A., Clerens, S., Do-Rego, J.C., Vandesande, F., Llorens-Cortes, C, Costentin, J., et al., (2003). Identification of 26RFa, a hypothalamic neuropeptide of the RFamide peptide family with orexigenic activity. Proc Natl Acad Sci U S A 100, 15247-15252.
Cone, R.D. (2005). Anatomy and regulation of the central melanocortin system. Nat Neurosci 8, 571-578.
Cowley, M.A., Pronchuk, N., Fan, W., Dinulescu, D.M., Colmers, W.F., and Cone, R.D. (1999). Integration of NPY, AGRP, and melanocortin signals in the hypothalamic paraventricular nucleus: evidence of a cellular basis for the adipostat. Neuron 24, 155- 163.
Cowley, M.A., Smart, J.L., Rubinstein, M., Cerdan, M.G., Diano, S., Horvath, T.L., Cone, R.D., and Low, M.J. (2001). Leptin activates anorexigenic POMC neurons through a neural network in the arcuate nucleus. Nature 411, 480-484.
Dinan, T.G., Stilling, R.M., Stanton, C, and Cryan, J.F. (2015). Collective unconscious: How gut microbes shape human behavior. Journal of Psychiatric Research 63, 1-9.
Dzamko, N.L., and Steinberg, G.R. (2009). AMPK-dependent hormonal regulation of whole-body energy metabolism. Acta Physiologica 196, 115-127.
Edwards, СМ., Todd, J.F., Mahmoudi, M., Wang, Z., Wang, R.M., Ghatei, M.A., and Bloom, S.R. (1999). Glucagon-like peptide 1 has a physiological role in the control of postprandial glucose in humans: studies with the antagonist exendin 9-39. Diabetes 48, 86-93.
Eissele, R., Goke, R, Willemer, S., Harthus, H.P., Vermeer, H., Arnold, R, and Goke, B. (1992). Glucagon-like peptide-1 cells in the gastrointestinal tract and pancreas of rat, pig and man. European journal of clinical investigation 22, 283-291.
Fetissov, S.O., and  P. (2011). The new link between gut-brain axis and neuropsychiatric disorders. Curr Opin Clin Nutr Metab Care 14, 477-482.
P. (2011). The new link between gut-brain axis and neuropsychiatric disorders. Curr Opin Clin Nutr Metab Care 14, 477-482.
Fetissov, S.O., Hamze Sinno, M.,  M., Bole-Feysot, C,
M., Bole-Feysot, C,  P.,
P.,  Т., and Dechelotte, P. (2008). Autoantibodies against appetite-regulating peptide hormones and neuropeptides: putative modulation by gut microflora. Nutrition 24, 348-359.
Т., and Dechelotte, P. (2008). Autoantibodies against appetite-regulating peptide hormones and neuropeptides: putative modulation by gut microflora. Nutrition 24, 348-359.
Fioramonti, X., Contie, S., Song, Z., Routh, V.H., Lorsignol, A., and Penicaud, L. (2007). Characterization of glucosensing neuron subpopulations in the arcuate nucleus: integration in neuropeptide Y and pro-opio melanocortin networks? Diabetes 56, 1219- 1227.
Flint, A., Raben, A., Astrup, A., and Hoist, J.J. (1998). Glucagon-like peptide 1 promotes satiety and suppresses energy intake in humans. The Journal of Clinical Investigation 101, 515-520.
Forsythe, P., and Kunze, W. (2013). Voices from within: gut microbes and the CNS. Cellular and Molecular Life Sciences 70, 55-69.
Foucault, M.-L., Thomas, L., Goussard, S., Branching B.R., and Grillot-Courvalin, C. (2009). In vivo bioluminescence imaging for the study of intestinal colonization by Escherichia coli in mice. Applied and Environmental Microbiology 76, 264-274.
Garfield, A.S., Li, C, Madara, J.C., Shah, B.P., Webber, E., Steger, J.S., Campbell, J.N., Gavrilova, O., Lee, C.E., Olson, D.P., et al., (2015). A neural basis for melanocortin-4 receptor- regulated appetite. Nat Neurosci doi:10.1038/nn.4011.
Gerspach, A.C., Steinert, R.E., Schonenberger, L., Graber-Maier, A., and Beglinger, C. (2011). The role of the gut sweet taste receptor in regulating GLP-1, PYY, and CCK release in humans. Am J Physiol Endocrinol Metab 301, Е317-E325.
Glasgow, I., Mattar, K., and Krantis, A. (1998). Rat gastroduodenal motility in vivo: involvement of NO and ATP in spontaneous motor activity. Am. J. Physiol. Gastrointest. Liver Physiol 275, G889-G896.
Goichon, A.,  M., Claeyssens, S., Lecleire, S., Cailleux, A.-F., Bole-Feysot, C, Chan, P., Donnadieu, N., Lerebours, E., Lavoinne, A., et al., (2011). Effects of an enteral glucose supply on protein synthesis, proteolytic pathways, and proteome in human duodenal mucosa. The American Journal of Clinical Nutrition 94, 784-794.
M., Claeyssens, S., Lecleire, S., Cailleux, A.-F., Bole-Feysot, C, Chan, P., Donnadieu, N., Lerebours, E., Lavoinne, A., et al., (2011). Effects of an enteral glucose supply on protein synthesis, proteolytic pathways, and proteome in human duodenal mucosa. The American Journal of Clinical Nutrition 94, 784-794.
Haange, S.B., Oberbach, A., Schlichting, N., Hugenholtz, F., Smidt, H., von Bergen, M., Till, H., and Seifert, J. (2012). Metaproteome analysis and molecular genetics of rat intestinal microbiota reveals section and localization resolved species distribution and enzymatic functionalities. J Proteome Res 11, 5406-5417.
Halaas, J.L., Gajiwala, K.S., Maffei, M., Cohen, S.L., Chait, B.T., Rabinowitz, D., Lallone, R.L., Burley, S.K., and Friedman, J.M. (1995). Weight-reducing effects of theplasma protein encoded by the obese gene. Science 269, 543-546.
Harte, A.L., Varma, M.C., Tripathi, G., McGee, K.C., Al-Daghri, N.M., Al-Attas, O.S., Sabico, S., O'Hare, J.P., Ceriello, A., Saravanan, P., et al., (2012). High fat intake leads to acute postprandial exposure to circulating endotoxin in type 2 diabetic subjects. Diabetes Care 35, 375-382.
Herzog, H. (2003). Neuropeptide Y and energy homeostasis: insights from Y receptor knockout models. Eur J Pharmacol 480, 21-29.
Heymsfield, S.B., Greenberg, A.S., Fujioka, K., Dixon, R.M., Kushner, R., Hunt, Т., Lubina, J.A., Patane, J., Self, В., Hunt, P., et al., (1999). Recombinant leptin for weight loss in obese and lean adults: A randomized, controlled, dose-escalation trial. JAMA 282, 1568-1575. Inui, A. (1999). Feeding and body-weight regulation by hypothalamic neuropeptides-mediation of the actions of leptin. Trends Neurosci 22, 62-67.
Johnstone, L.E., Fong, T.M., and Leng, G. (2006). Neuronal activation in the hypothalamus and brainstem during feeding in rats. Cell Metabolism 4, 313-321. Ley, R.E., Turnbaugh, P.J., Klein, S., and Gordon, J.I. (2006). Microbial ecology: Human gut microbes associated with obesity. Nature 444, 1022-1023.
Lim, P.L., and Rowley, D. (1985). Intestinal absorption of bacterial antigens in normal adult mice. II. A comparative study of techniques. Int Arch Allergy Appl Immunol 76, 30-36.
Lin, S., Shi, Y.-C, Yulyaningsih, E., Aljanova, A., Zhang, L., Macia, L., Nguyen, A.D., Lin, E.-J.D., During, M.J., Herzog, H., et al., (2009). Critical role of arcuate Y4 receptors and the melanocortin system in pancreatic polypeptide-induced reduction in food intake in mice. PLoS One 4, e8488.
Lu, X.Y., Barsh, G.S., Akil, H., and Watson, S.J. (2003). Interaction between alpha- melanocyte-stimulating hormone and corticotropin-releasing hormone in the regulation of feeding and hypothalamo-pituitary-adrenal responses. J Neurosci 23, 7863-7872.
Manning, S., and Batterham, Rachel L. (2014). Enteroendocrine MC4R and energy balance: linking the long and the short of it. Cell Metabolism 20, 929-931.
Meguid, M.M., Laviano, A., and Rossi-Fanelli, F. (1998). Food intake equals meal size times mean number. Appetite 31, 404.
Mounien, L., Bizet, P., Boutelet, I., Vaudry, H., and Jegou, S. (2005). Expression of melanocortin MC3 and MC4 receptor mRNAs by neuropeptide Y neurons in the rat arcuate nucleus. Neuroendocrinology 82, 164-170.
Mul, J.D., Spruijt, B.M., Brakkee, J.H., and Adan, R.A.H. (2013). Melanocortin MC4 receptor-mediated feeding and grooming in rodents. European Journal of Pharmacology 719, 192-201.
Murphy, K.G., and Bloom, S.R. (2006). Gut hormones and the regulation of energy homeostasis. Nature 444, 854-859.
Neunlist, M., Van Landeghem, L., Mahe, M.M., Derkinderen, P., Bruley des Varannes, S.B., and Rolli-Derkinderen, M. (2012). The digestive neuronal-glial- epithelial unit: a new actor in gut health and disease. Nat Rev Gastroenterol Hepatol 10, 90-100.
Panaro, B.L., Tough, I.R., Engelstoft, M.S., Matthews, R.T., Digby, G.J., Moller, C.L., Svendsen, В., Gribble, F., Reimann, F., Hoist, J.J., et al., (2014). The melanocortin-4 receptor is expressed in enteroendocrine L cells and regulates the release of peptide YY and glucagon- like peptide 1 in vivo. Cell Metab 20, 1018-1029.
Parks, B.W., Nam, E., Org, E., Kostem, E., Norheim, F., Hui, S.T., Pan, C, Civelek, M., Rau, CD., Bennett, B.J., et al., (2013). Genetic control of obesity and gut microbiota composition in response to high-fat, high-sucrose diet in mice. Cell Metabolism 17, 141-152.
Power, M.L., and Schulkin, J. (2008). Anticipatory physiological regulation in feeding biology: Cephalic phase responses. Appetite 50, 194-206.
Rice, K.C., and Bayles, K.W. (2008). Molecular control of bacterial death and lysis. Microbiology and Molecular Biology Reviews 72, 85-109.
Sharon, G., Garg, N., Debelius, J., Knight, R., Dorrestein, Pieter C, and Mazmanian, Sarkis K. (2014). Specialized metabolites from the microbiome in health and disease. Cell Metabolism 20, 719-730.
Shi, Y.-C., Lau, J., Lin, Z., Zhang, H., Zhai, L., Sperk, G., Heilbronn, R., Mietzsch, M., Weger, S., Huang, X.-F., et al., (2013). Arcuate NPY controls sympathetic output and BAT function via a relay of tyrosine hydroxylase neurons in the PVN. Cell Metabolism 17, 236-248.
Smith, M.A., Hisadome, K., Al-Qassab, H., Heffron, H., Withers, D.J., and Ashford, M.L. (2007). Melanocortins and agouti-related protein modulate the excitability of two arcuate nucleus neuron populations by alteration of resting potassium conductances. J Physiol 578, 425- 438.
Steinert, R.E., Schirra, J., Meyer-Gerspach, A.C., Kienle, P., Fischer, H., Schulte, F., Goeke, В., and Beglinger, C. (2014). Effect of glucagon-like peptide-1 receptor antagonism on appetite and food intake in healthy men. The American Journal of Clinical Nutrition 100, 514- 523.
Sternson, S.M., Shepherd, G.M.G., and Friedman, J.M. (2005). Topographic mapping of VMH - arcuate nucleus microcircuits and their reorganization by fasting. Nat Neurosci 8, 1356-1363.
Takagi, K., Legrand, R., Asakawa, A., Amitani, H.,  , M., Tennoune, N.,
, M., Tennoune, N.,  , M., Claeyssens, S., do Rego, J.-C,
, M., Claeyssens, S., do Rego, J.-C,  , P., et al., (2013). Anti-ghrelin immunoglobulins modulate ghrelin stability and its orexigenic effect in obese mice and humans. Nat Commun 4:2685.
, P., et al., (2013). Anti-ghrelin immunoglobulins modulate ghrelin stability and its orexigenic effect in obese mice and humans. Nat Commun 4:2685.
Tennoune, N., Chan, P., Breton, J., Legrand, R., Chabane, Y.N., Akkermann, K., Jarv, A., Ouelaa, W., Takagi, K., Ghouzali, I., et al., (2014). Bacterial ClpB heat-shock protein, an antigen-mimetic of the anorexigenic peptide [alpha]-MSH, at the origin of eating disorders. Transl Psychiatry 4, e458.
Tennoune, N., Legrand, R., Ouelaa, W., Breton, J., Lucas, N., Bole-Feysot, C, Rego, J.-C.d., , P., and Fetissov, S.O. (2015). Sex-related effects of nutritional supplementation of Escherichia coli: Relevance to eating disorders. Nutrition 31, 498-507.
Turnbaugh, P.J., Ley, R.E., Mahowald, M.A., Magrini, V., Mardis, E.R., and Gordon, J.I. (2006). An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 444, 1027-1131.
van der Klaauw, A.A., Keogh, J.M., Henning, E., Trowse, V.M., Dhillo, W.S., Ghatei, M.A., and Farooqi, I.S. (2013). High protein intake stimulates postprandial GLP1 and PYY release. Obesity 21, 1602-1607.
Vijay-Kumar, M., Aitken, J.D., Carvalho, F.A., Cullender, T.C., Mwangi, S., Srinivasan, S., Sitaraman, S.V., Knight, R., Ley, R.E., and Gewirtz, A.T. (2010). Metabolic syndrome and altered gut microbiota in mice lacking toll-like receptor 5. Science 328, 228-231.
Wick, L.M., Quadroni, M., and Egli, T. (2001). Short- and long-term changes in proteome composition and kinetic properties in a culture of Escherichia coli during transition from glucose-excess to glucose-limited growth conditions in continuous culture and vice versa. Environ Microbiol 3, 588-599.
Xu, В., Goulding, E.H., Zang, K., Cepoi, D., Cone, R.D., Jones, K.R., Tecott, L.H., and Reichardt, L.F. (2003). Brain-derived neurotrophic factor regulates energy balance downstream of melanocortin-4 receptor. Nat Neurosci 6, 736-742.
Yun, A.J., Lee, P.Y., Doux, J.D., and Conley, B.R. (2006). A general theory of evolution based on energy efficiency: its implications for diseases. Medical Hypotheses 66, 664-670.
--->
Перечень последовательностей
<110> ИНСЕРМ, СЕНТРЕ ОСПИТАЛЬ УНИВЕРСИТЕР ДЕ РУАН, ТАГЕДИС,
УНИВЕРСИТЕ ДЕ РУАН (FR)
<120> Способы индуцирования и пролонгирования насыщения, уменьшения объёма
и поглощения пищи, контроля веса и стимулирования снижения веса,
пищевая композиция
<150> PCT/EP2017/058116 RU 2018138995
<160> 2
<210> 1
<211> 857
<212> PRT (искусственная последовательность)
<213> Escherichia coli
<400> 1
Met Arg Leu Asp Arg Leu Thr Asn Lys Phe Gln Leu Ala Leu Ala Asp
1 5 10 15
Ala Gln Ser Leu Ala Leu Gly His Asp Asn Gln Phe Ile Glu Pro Leu
20 25 30
His Leu Met Ser Ala Leu Leu Asn Gln Glu Gly Gly Ser Val Ser Pro
35 40 45
Leu Leu Thr Ser Ala Gly Ile Asn Ala Gly Gln Leu Arg Thr Asp Ile
50 55 60
Asn Gln Ala Leu Asn Arg Leu Pro Gln Val Glu Gly Thr Gly Gly Asp
65 70 75 80
Val Gln Pro Ser Gln Asp Leu Val Arg Val Leu Asn Leu Cys Asp Lys
85 90 95
Leu Ala Gln Lys Arg Gly Asp Asn Phe Ile Ser Ser Glu Leu Phe Val
100 105 110
Leu Ala Ala Leu Glu Ser Arg Gly Thr Leu Ala Asp Ile Leu Lys Ala
115 120 125
Ala Gly Ala Thr Thr Ala Asn Ile Thr Gln Ala Ile Glu Gln Met Arg
130 135 140
Gly Gly Glu Ser Val Asn Asp Gln Gly Ala Glu Asp Gln Arg Gln Ala
145 150 155 160
Leu Lys Lys Tyr Thr Ile Asp Leu Thr Glu Arg Ala Glu Gln Gly Lys
165 170 175
Leu Asp Pro Val Ile Gly Arg Asp Glu Glu Ile Arg Arg Thr Ile Gln
180 185 190
Val Leu Gln Arg Arg Thr Lys Asn Asn Pro Val Leu Ile Gly Glu Pro
195 200 205
Gly Val Gly Lys Thr Ala Ile Val Glu Gly Leu Ala Gln Arg Ile Ile
210 215 220
Asn Gly Glu Val Pro Glu Gly Leu Lys Gly Arg Arg Val Leu Ala Leu
225 230 235 240
Asp Met Gly Ala Leu Val Ala Gly Ala Lys Tyr Arg Gly Glu Phe Glu
245 250 255
Glu Arg Leu Lys Gly Val Leu Asn Asp Leu Ala Lys Gln Glu Gly Asn
260 265 270
Val Ile Leu Phe Ile Asp Glu Leu His Thr Met Val Gly Ala Gly Lys
275 280 285
Ala Asp Gly Ala Met Asp Ala Gly Asn Met Leu Lys Pro Ala Leu Ala
290 295 300
Arg Gly Glu Leu His Cys Val Gly Ala Thr Thr Leu Asp Glu Tyr Arg
305 310 315 320
Gln Tyr Ile Glu Lys Asp Ala Ala Leu Glu Arg Arg Phe Gln Lys Val
325 330 335
Phe Val Ala Glu Pro Ser Val Glu Asp Thr Ile Ala Ile Leu Arg Gly
340 345 350
Leu Lys Glu Arg Tyr Glu Leu His His His Val Gln Ile Thr Asp Pro
355 360 365
Ala Ile Val Ala Ala Ala Thr Leu Ser His Arg Tyr Ile Ala Asp Arg
370 375 380
Gln Leu Pro Asp Lys Ala Ile Asp Leu Ile Asp Glu Ala Ala Ser Ser
385 390 395 400
Ile Arg Met Gln Ile Asp Ser Lys Pro Glu Glu Leu Asp Arg Leu Asp
405 410 415
Arg Arg Ile Ile Gln Leu Lys Leu Glu Gln Gln Ala Leu Met Lys Glu
420 425 430
Ser Asp Glu Ala Ser Lys Lys Arg Leu Asp Met Leu Asn Glu Glu Leu
435 440 445
Ser Asp Lys Glu Arg Gln Tyr Ser Glu Leu Glu Glu Glu Trp Lys Ala
450 455 460
Glu Lys Ala Ser Leu Ser Gly Thr Gln Thr Ile Lys Ala Glu Leu Glu
465 470 475 480
Gln Ala Lys Ile Ala Ile Glu Gln Ala Arg Arg Val Gly Asp Leu Ala
485 490 495
Arg Met Ser Glu Leu Gln Tyr Gly Lys Ile Pro Glu Leu Glu Lys Gln
500 505 510
Leu Glu Ala Ala Thr Gln Leu Glu Gly Lys Thr Met Arg Leu Leu Arg
515 520 525
Asn Lys Val Thr Asp Ala Glu Ile Ala Glu Val Leu Ala Arg Trp Thr
530 535 540
Gly Ile Pro Val Ser Arg Met Met Glu Ser Glu Arg Glu Lys Leu Leu
545 550 555 560
Arg Met Glu Gln Glu Leu His His Arg Val Ile Gly Gln Asn Glu Ala
565 570 575
Val Asp Ala Val Ser Asn Ala Ile Arg Arg Ser Arg Ala Gly Leu Ala
580 585 590
Asp Pro Asn Arg Pro Ile Gly Ser Phe Leu Phe Leu Gly Pro Thr Gly
595 600 605
Val Gly Lys Thr Glu Leu Cys Lys Ala Leu Ala Asn Phe Met Phe Asp
610 615 620
Ser Asp Glu Ala Met Val Arg Ile Asp Met Ser Glu Phe Met Glu Lys
625 630 635 640
His Ser Val Ser Arg Leu Val Gly Ala Pro Pro Gly Tyr Val Gly Tyr
645 650 655
Glu Glu Gly Gly Tyr Leu Thr Glu Ala Val Arg Arg Arg Pro Tyr Ser
660 665 670
Val Ile Leu Leu Asp Glu Val Glu Lys Ala His Pro Asp Val Phe Asn
675 680 685
Ile Leu Leu Gln Val Leu Asp Asp Gly Arg Leu Thr Asp Gly Gln Gly
690 695 700
Arg Thr Val Asp Phe Arg Asn Thr Val Val Ile Met Thr Ser Asn Leu
705 710 715 720
Gly Ser Asp Leu Ile Gln Glu Arg Phe Gly Glu Leu Asp Tyr Ala His
725 730 735
Met Lys Glu Leu Val Leu Gly Val Val Ser His Asn Phe Arg Pro Glu
740 745 750
Phe Ile Asn Arg Ile Asp Glu Val Val Val Phe His Pro Leu Gly Glu
755 760 765
Gln His Ile Ala Ser Ile Ala Gln Ile Gln Leu Lys Arg Leu Tyr Lys
770 775 780
Arg Leu Glu Glu Arg Gly Tyr Glu Ile His Ile Ser Asp Glu Ala Leu
785 790 795 800
Lys Leu Leu Ser Glu Asn Gly Tyr Asp Pro Val Tyr Gly Ala Arg Pro
805 810 815
Leu Lys Arg Ala Ile Gln Gln Gln Ile Glu Asn Pro Leu Ala Gln Gln
820 825 830
Ile Leu Ser Gly Glu Leu Val Pro Gly Lys Val Ile Arg Leu Glu Val
835 840 845
Asn Glu Asp Arg Ile Val Ala Val Gln
850 855
<210> 2
<211> 13
<212> PRT
<213> Homo sapiens
<400> 2
Ser Tyr Ser Met Glu His Phe Arg Trp Gly Lys Pro Val
1 5 10
<---
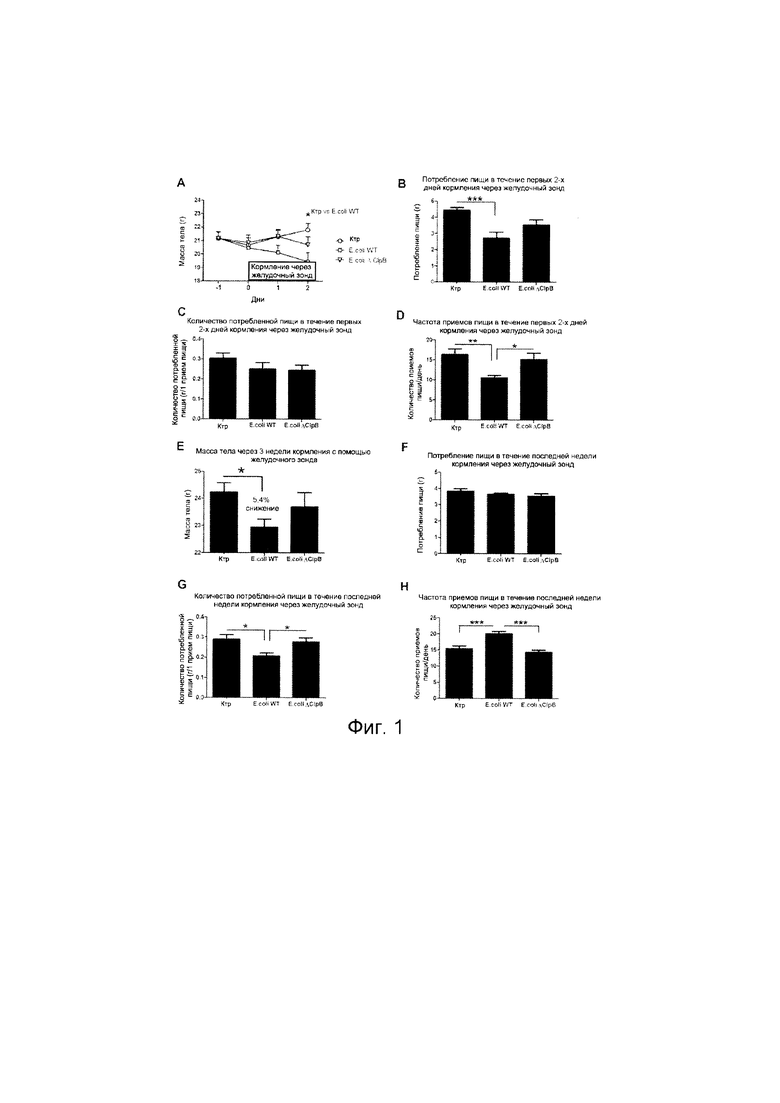
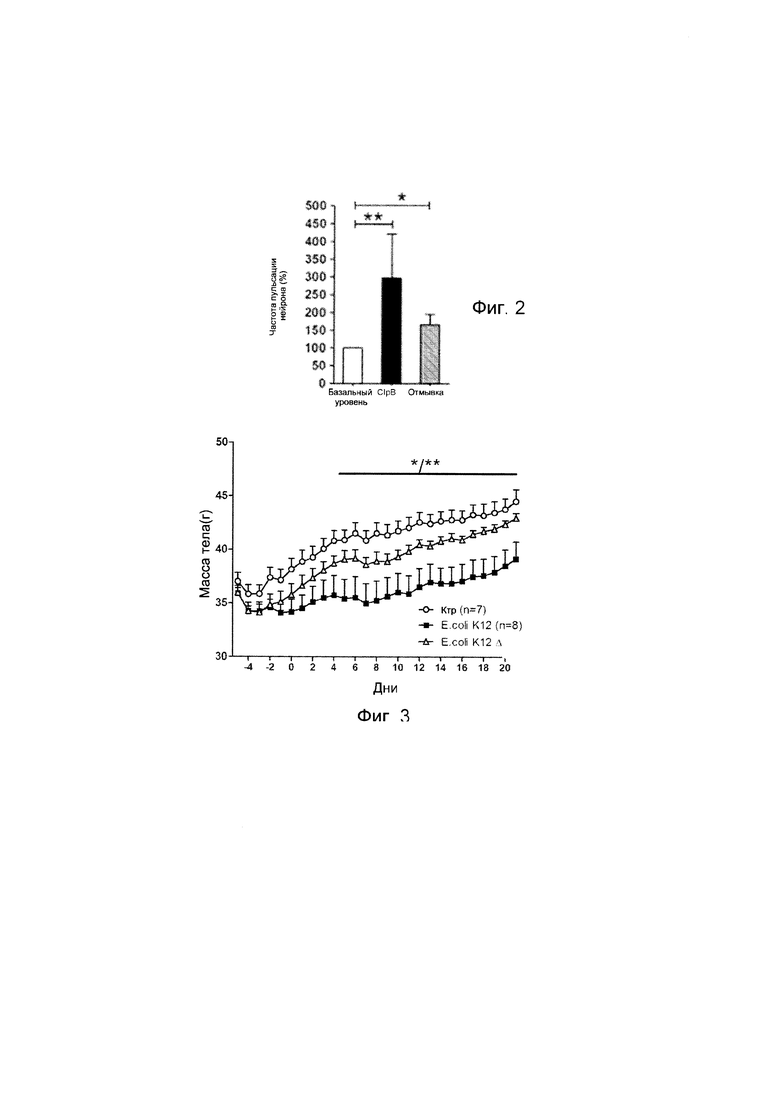
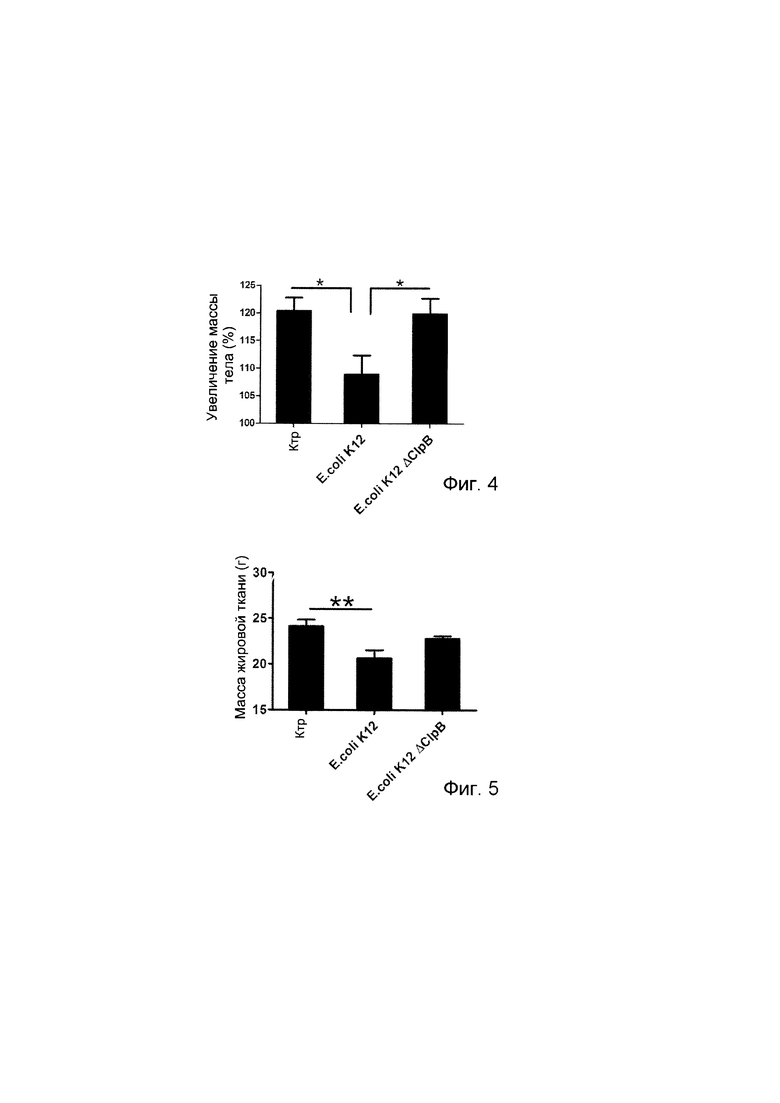
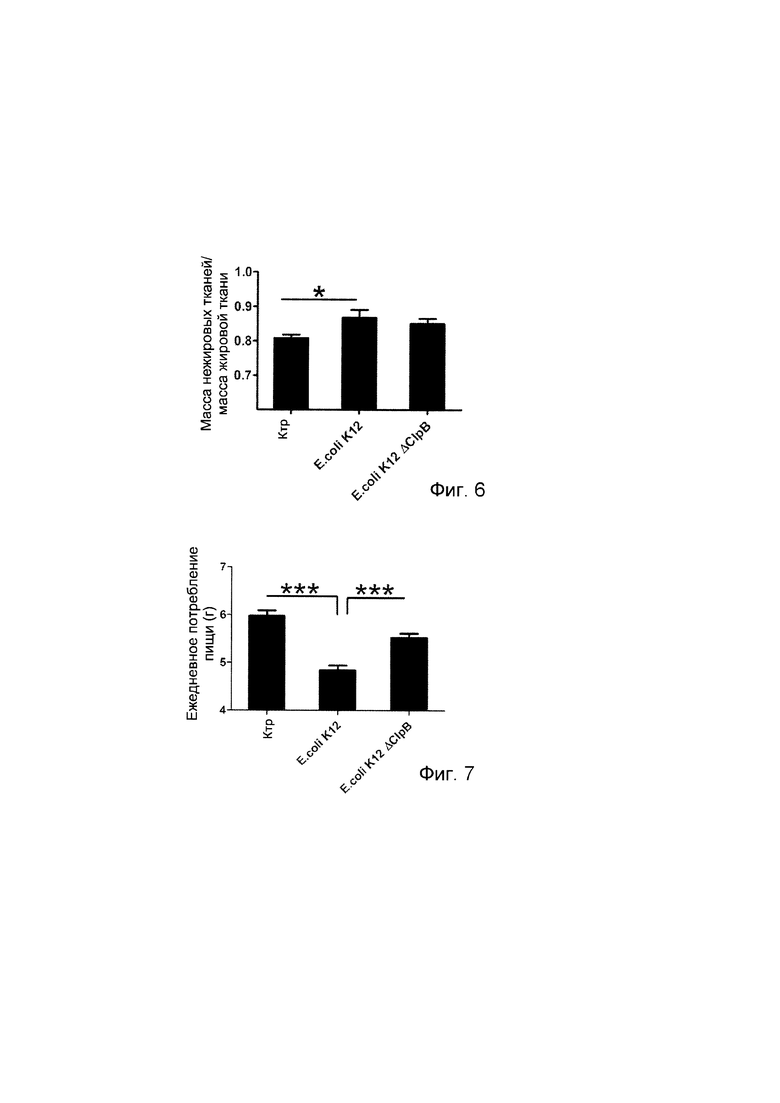
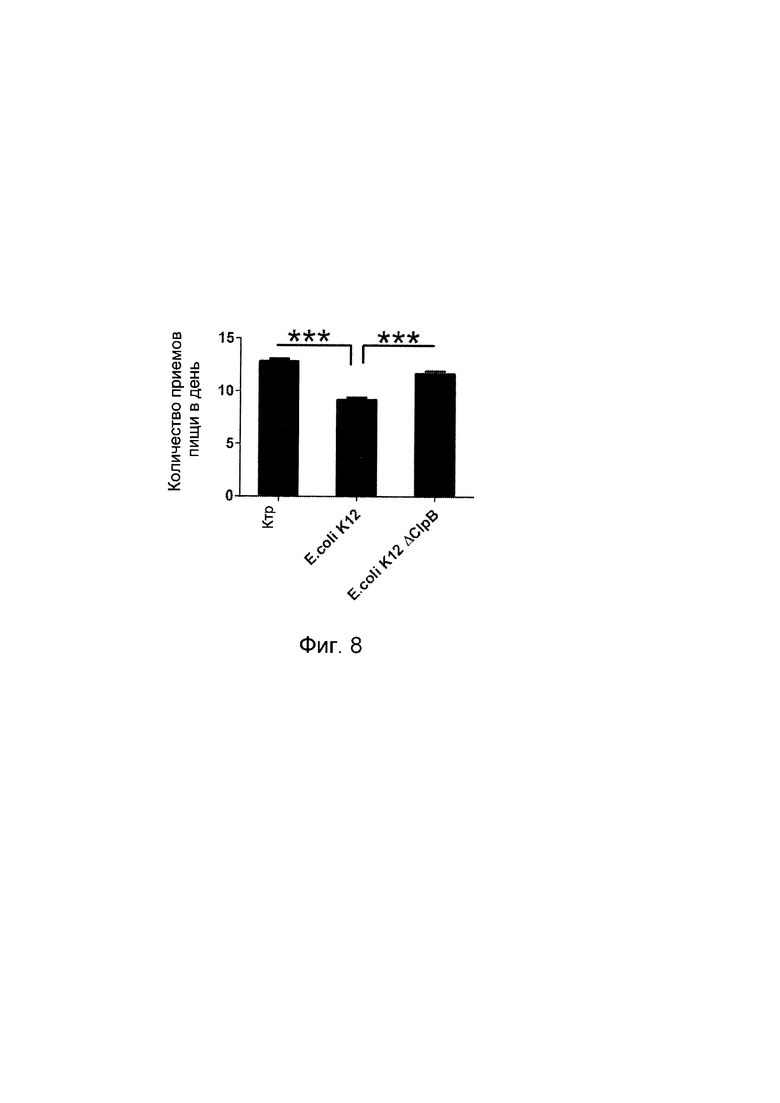
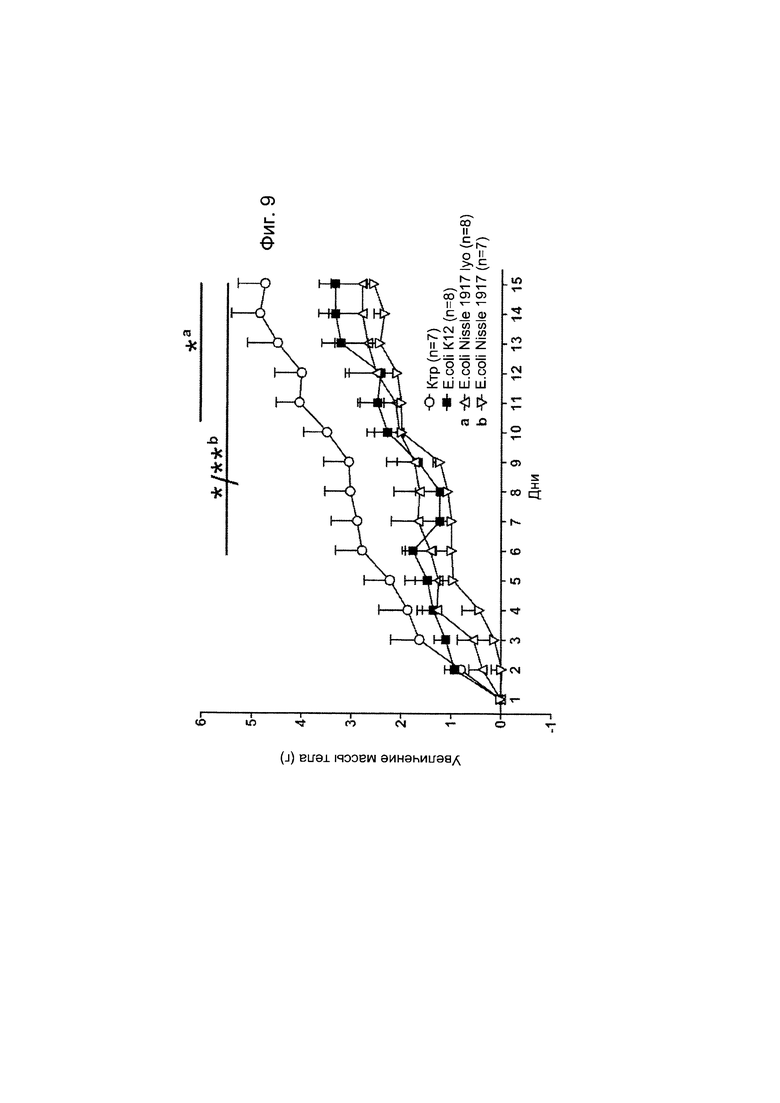
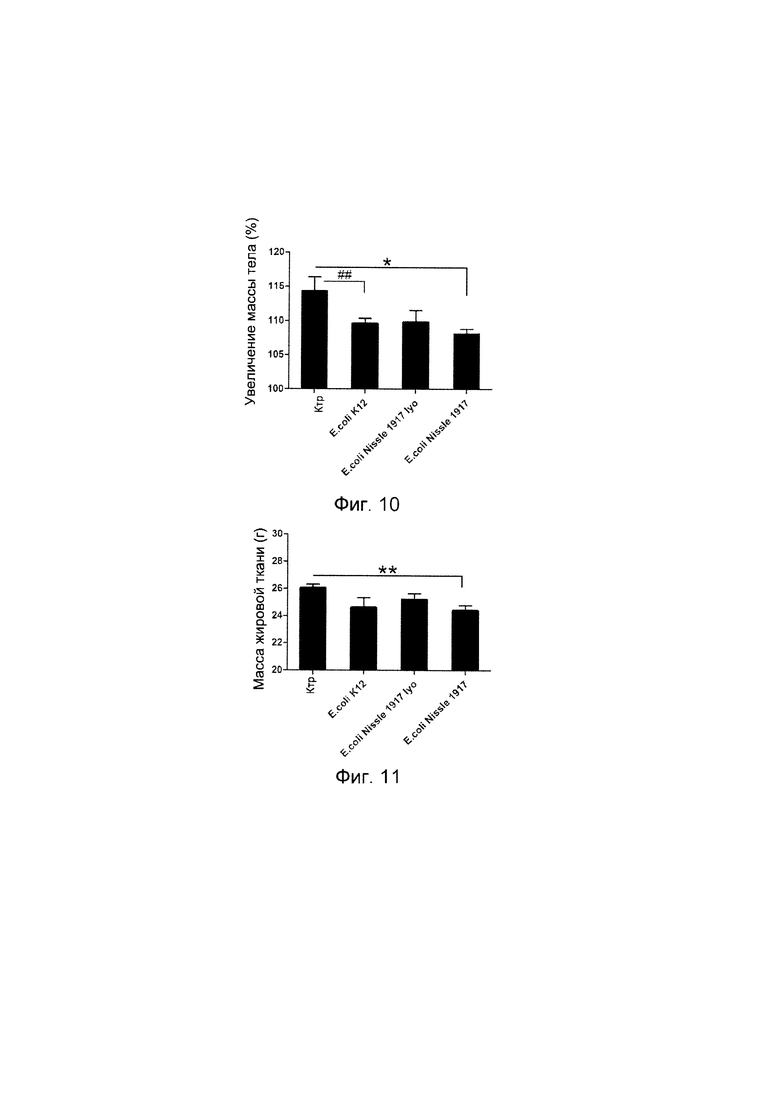
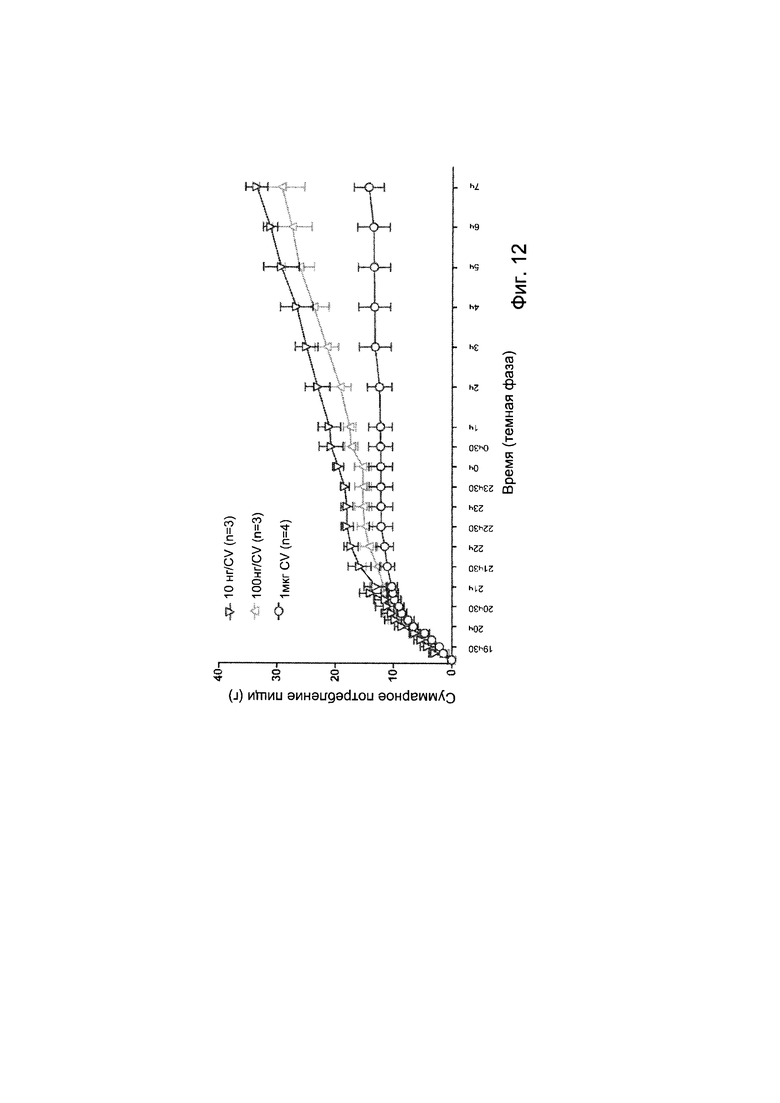
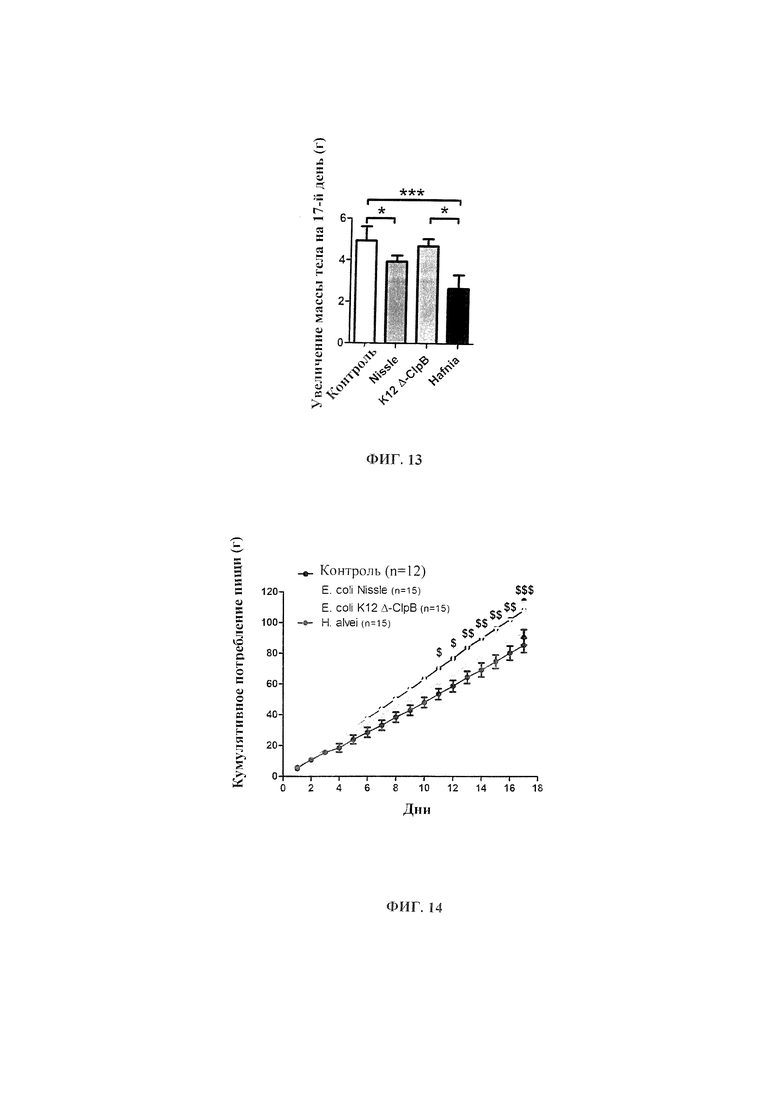
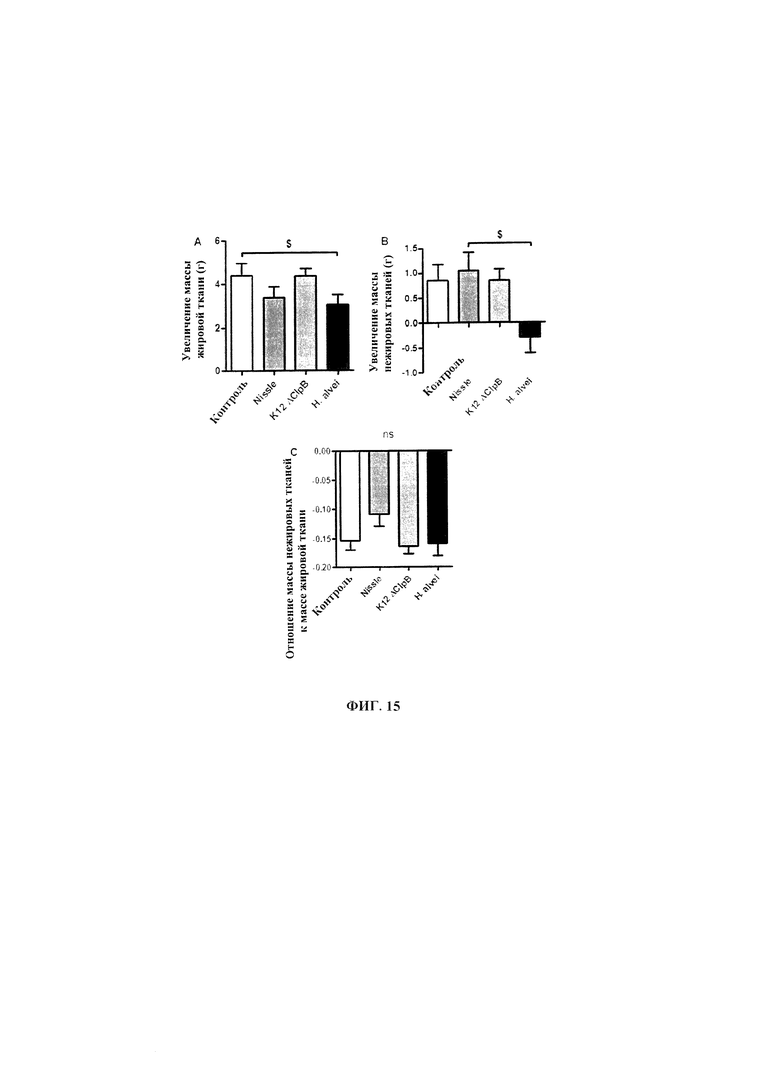
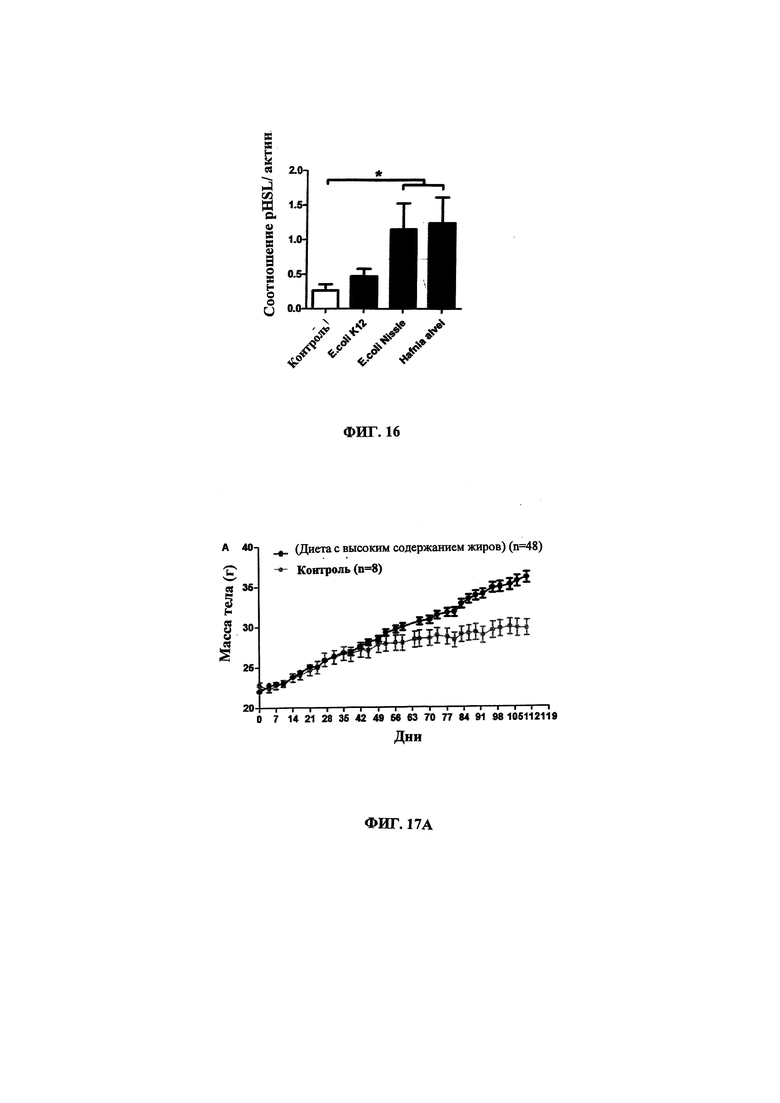
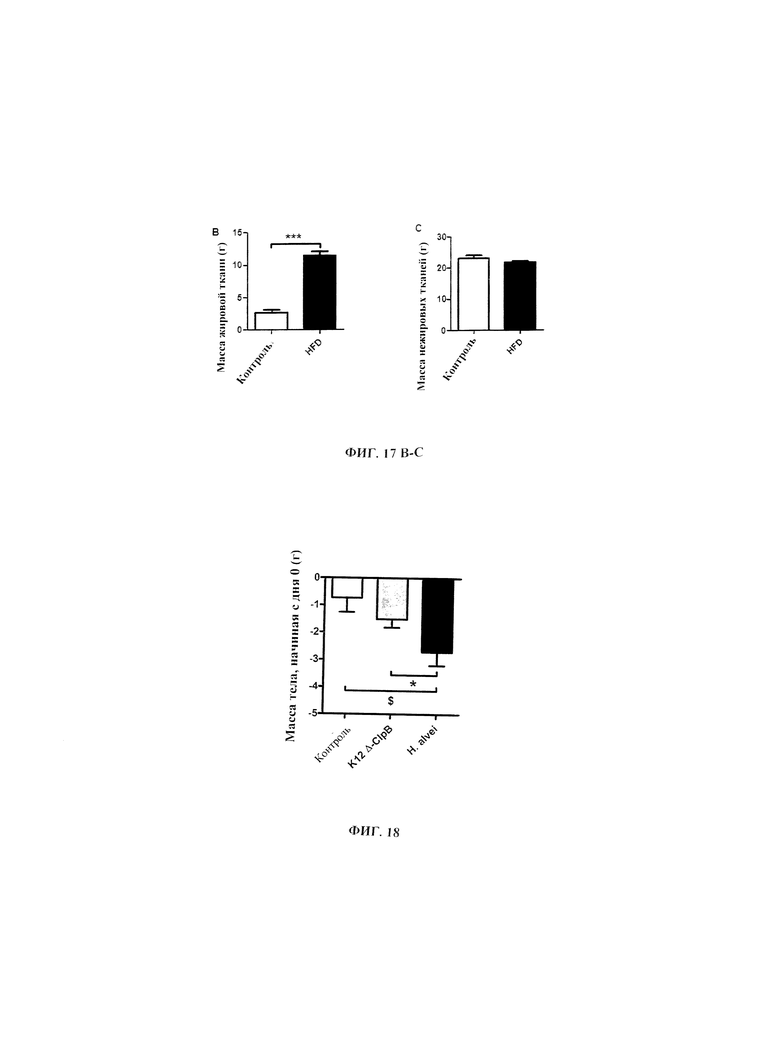
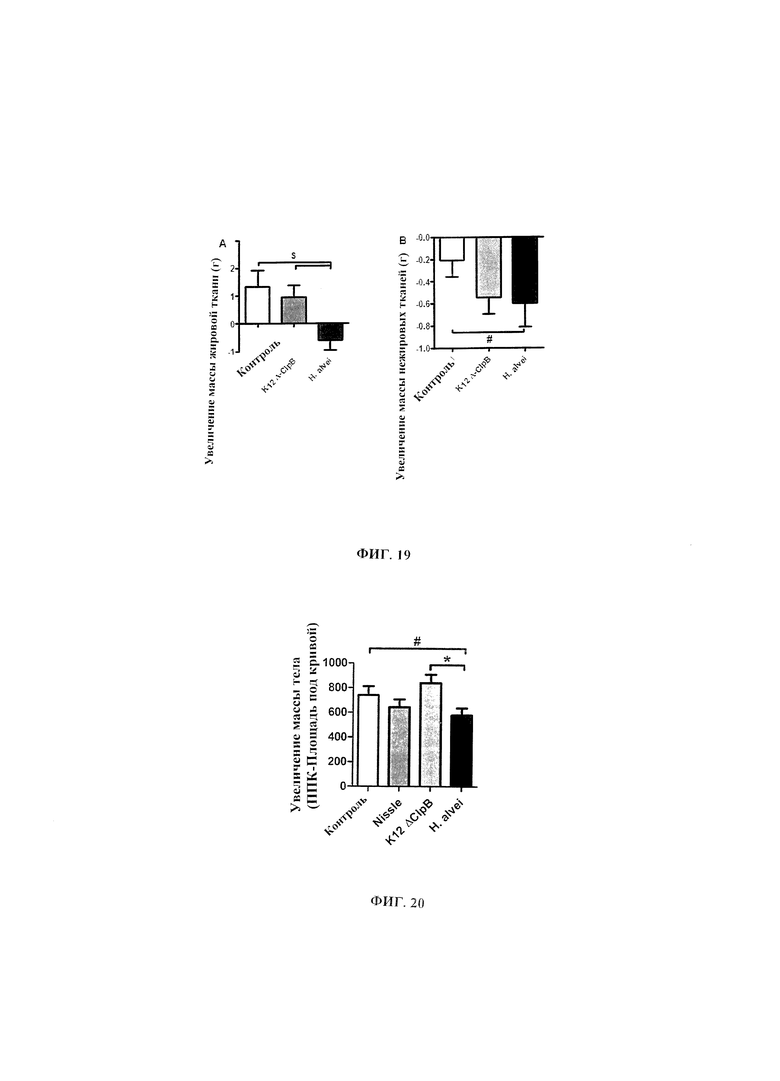
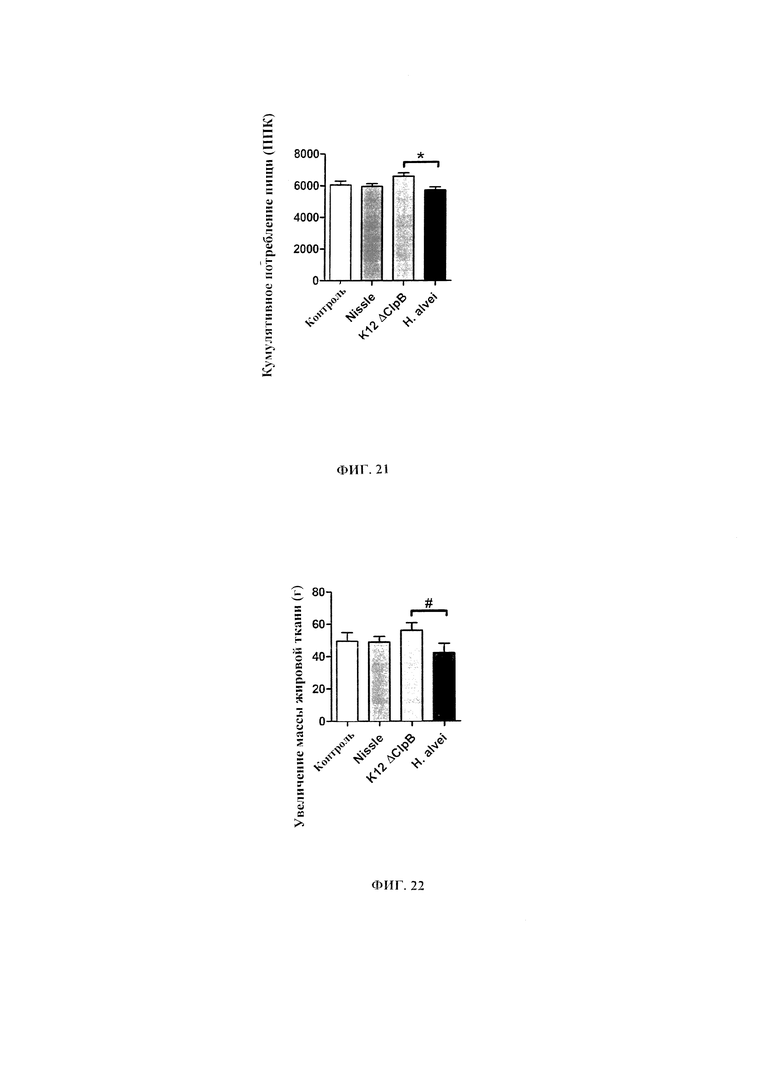
Предложены способ индуцирования насыщения у нуждающегося в этом субъекта, способ продления сытости у нуждающегося в этом субъекта, способ уменьшения количества потребляемой пищи за прием у нуждающегося в этом субъекта, способ уменьшения потребления пищи у нуждающегося в этом субъекта, стимулирования уменьшения массы тела у нуждающегося в этом субъекта. Изобретения предусматривают пероральное введение указанному субъекту эффективного количества Hafnia alvei, экспрессирующей белок ClpB. Изобретения позволяют индуцировать насыщение, продлевать чувство сытости, снижать потребление пищи, уменьшения количества потребляемой пищи за прием и/или стимулировать уменьшение массы тела у субъекта. 5 н. и 9 з.п. ф-лы, 22 ил., 8 пр.
1. Способ индуцирования насыщения у нуждающегося в этом субъекта, включающий пероральное введение указанному субъекту эффективного количества Hafnia alvei, экспрессирующей белок ClpB.
2. Способ продления сытости у нуждающегося в этом субъекта, включающий пероральное введение указанному субъекту эффективного количества Hafnia alvei, экспрессирующей белок ClpB.
3. Способ уменьшения количества потребляемой пищи за прием у нуждающегося в этом субъекта, включающий пероральное введение указанному субъекту эффективного количества Hafnia alvei, экспрессирующей белок ClpB.
4. Способ уменьшения потребления пищи у нуждающегося в этом субъекта, включающий пероральное введение указанному субъекту эффективного количества Hafnia alvei, экспрессирующей белок ClpB.
5. Способ стимулирования уменьшения массы тела у нуждающегося в этом субъекта, включающий пероральное введение указанному субъекту эффективного количества Hafnia alvei, экспрессирующей белок ClpB.
6. Способ по любому из пп. 1-5, отличающийся тем, что указанный субъект не страдает ожирением.
7. Способ по любому из пп. 1-5, отличающийся тем, что указанный субъект страдает ожирением.
8. Способ по любому из пп. 1-7, отличающийся тем, что Hafnia alvei вводят указанному субъекту в форме фармацевтической композиции.
9. Способ по любому из пп. 1-7, отличающийся тем, что Hafnia alvei вводят указанному субъекту в виде пищевой композиции.
10. Способ по п. 9, отличающийся тем, что указанная пищевая композиция выбрана из группы, состоящей из полноценных пищевых композиций, пищевых добавок и нутрицевтических композиций.
11. Способ по любому из пп. 1-10, отличающийся тем, что Hafnia alvei является пробиотиком.
12. Способ по любому из пп. 1-10, отличающийся тем, что Hafnia alvei является инактивированной.
13. Способ по п. 9, отличающийся тем, что указанная пищевая композиция содержит, по меньшей мере, один пребиотик.
14. Способ по п. 13, отличающийся тем, что указанный, по меньшей мере, один пребиотик представляет собой пищевые волокна.
| TENNOUNE N | |||
| ET AL | |||
| Bacterial ClpB heat-shock protein, an antigen-mimetic of the anorexigenic peptide [alpha]-MSH, at the origin of eating disorders | |||
| TRANSLATION PSYCHIATRY, vol | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| база данных UniProtKB-A0A097QZ90, 07.01.2015 | |||
| СПОСОБ ЛЕЧЕНИЯ РОЖИ | 1999 |
|
RU2190438C2 |
| FETISSOV S.O | |||
| ET AL | |||
| Autoantibodies against | |||

