Область техники
Изобретение относится к области биоорганической и биомедицинской химии, биохимии, энзимологии, цитологии и касается новых субстратов для определения ферментативной активности пептидаз, их получения, применения в качестве высокоспецифичных и селективных хромогенных и флуорогенных субстратов для цистеиновых пептидаз семейства папаина (в присутствии ферментов других кланов), применения для специфического тестирования катепсина L в природных смесях различных катепсинов, применения для селективного тестирования активности катепсина L с использованием флуорогенных соединений в системах in vivo, применения при создании тест-систем и тестировании активности цистеиновых пептидаз семейства С1 непосредственно в геле, применения для поиска глутаминспецифичных пептидаз, обладающих способностью расщеплять глиадины и их токсические пептид..
Область техники
Пептидазы (протеазы) играют ключевую роль во всех метаболических процессах. Ферментативная активность пептидаз определяется с помощью различных субстратов. Наиболее удобными и эффективными среди них являются пептидные субстраты, высокоспецифичные и селективные к определенной группе ферментов.
Цистеиновые пептидазы составляют один из основных классов протеолитических ферментов и характеризуются присутствием остатка Cys в активном центре. Большинство цистеиновых пептидаз, известных на данный момент, принадлежат к семейству С1, также называемому семейством папаина (N.D. Rawlings, A.J. Barrett, Introduction: The Clans and Families of Cysteine Peptidases, in: N.D. Rawlings, G.S. Salvesen (Eds.), Handbook of Proteolytic Enzymes, Third edition, Academic Press, London, 2013, v. 2, pp.1743-1776). Цистеиновые пептидазы семейства C1 были найдены в вирусах (G.N. Rudenskaya, D.V. Pupov, Cysteine proteinases of microorganisms and viruses, Biochemistry (Mosc). 73 (2008) 1-13), бактериях (J. Potempa, R. Pike, J. Travis, The multiple forms of trypsin-like activity present in various strains of Porphyromonas gingivalis are due to the presence of either Arg-gingipain or Lys-gingipain, J. Infect. Immunol. 63 (1995) 1176-1182), простейших (С.Serveau, G. Lalmanach, M.A. Juliano, J. Scharfstein, L. Juliano, F. Gauthier, Investigation of the substrate specificity of cruzipain, the major cysteine proteinase of Trypanosoma cruzi, through the use of cystatin-derived substrates and inhibitors, Biochem. J. 313 (1996) 951-956; J. Cazzulo, V Stoka, V. Turk, The major cysteine proteinase of Trypanosoma cruzi: a valid target for chemotherapy of Chagas disease, Curr Pharm Des. 7 (2001) 1143-1156; D.C. Greenbaum, A. Baruch, M. Grainger, Z. Bozdech, K.F. Medziharadszky, J. Engel, J. DeRisi, A.A. Holder, M. Bogyo, A role for the protease falcipain 1 in host cell invasion by the human malaria parasite, Science 298 (2002) 2002-2006; M. Sajid, J.H. McKerrow, Cysteine proteases of parasitic organisms, Mol. Biochem. Parasitol. 120 (2002) 1-21; Y.A. Sabnis, P.V. Desai, P.J. Rosenthal, M.A. Avery, Probing the structure of falcipain-3, a cysteine protease from Plasmodium falciparum: Comparative protein modeling and docking studies, Protein Science 12 (2003) 501-509; C.R. Caffrey, A.P. Lima, D. Steverding, Cysteine peptidases of kinetoplastid parasites, Adv Exp Med Biol. 712 (2011) 84-99; Z. Z. Dou, V.B. Carruthers, Cathepsin proteases in Toxoplasma gondii, Adv. Exp.Med. Biol. 712 (2011) 49-61), растениях (H H.-H. Otto, T. Schirmeister, Cysteine proteases and their inhibitors, Chem. Rev. 97 (1997) 133-171; D.F. Gruis, D.A. Selinger, J.M. Curran, R. Jung, Redundant proteolytic mechanisms process seed storage proteins in the absence of seed-type members of the vacuolar processing enzyme family of cysteine proteases, Plant Cell. 14 (2002) 2863-2882; V.K. Dubey, M.V. Jagannadham, Procerain, a stable cysteine protease from the latex of Calotropis procera, Phytochemistry 62 (2003) 1057-1071) и позвоночных животных (M. Yamashita & S. Konagaya, High activities of cathepsins B, D, H and L in the white muscle of chum salmon in spawning migration, Comp. Biochem. Physiol. 95B, (1990) 149-152, H.-H. Otto, T. Schirmeister, Cysteine proteases and their inhibitors, Chem. Rev. 97 (1997) 133-171; T. S. Uinuk-Ool, N. Takezaki, N. Kuroda, F. Figueroa, A. Sato, I. E. Samonte, W. E. Mayer, J. Klein, Phylogeny of Antigen-Processing Enzymes: Cathepsins of a Cephalochordate, an Agnathan and a Bony Fish, Scand. J. Immunol. 58 (2003) 436-448; M. Fabra &  Ovarian cysteine proteinases in the teleost Fundulus heteroclitus: Molecular cloning and gene expression during vitellogenesis and oocyte maturation, Mol. Reproduct. Development, 67 (2004) 282-294;
Ovarian cysteine proteinases in the teleost Fundulus heteroclitus: Molecular cloning and gene expression during vitellogenesis and oocyte maturation, Mol. Reproduct. Development, 67 (2004) 282-294;  C. Delbarre-Ladrat, M. Lamballerie-Anton, V. Verrez-Bagnis, Calpain and cathepsin activities in post mortem fish and meat muscles, Food Chemistry 101 (2007) 1474-1479; S. Subramanian, S. MacKinnona, N. Rossa, A comparative study on innate immune parameters in the epidermal mucus of various fish species, Comp. Biochem. Physiol. 148B (2007) 256-263; O. Vasiljeva, T. Reinheckel, C. Peters, D. Turk, V. Turk, B. Turk. Emerging roles of cysteine cathepsins in disease and their potential as drug targets. Curr. Pharm.Design, 13 (2007) 385-401; V. Turk, V. Stroka, O. Vasiljeva. M. Renko, T. Sun, B. Turk, D. Turk, Cysteine cathepsins: from structure, function and regulation to new frontiers, Biochim. Biophys. Acta (BBA)-Proteins and Proteomics 1824 (2012) 68-88). В настоящее время серьезная роль отводится пищеварительным цистеиновым пептидазам насекомых, в том числе и таких опасных вредителей, как колорадский жук, большой и малый мучные хрущаки, тараканы и т.д. (Р.Т. Cristofoletti, A.F. Ribeiro, W.R. Terra, The cathepsin L-like proteinases from the midgut of Tenebrio molitor larvae: sequence, properties, immunocytochemical localization and function. Insect Biochem. Mol. Biol. 35 (2005) 883-901; K. Gruden, A.G. Kuipers, G. Guncar, N. Slapar, B. Strukelj, M.A. Jongsma, Molecular basis of Colorado potato beetle adaptation to potato plant defence at the level of digestive cysteine proteinases, Insect Biochem. Mol. Biol. 34 (2004) 365-375; K.S. Vinokurov, E.N. Elpidina, B. Oppert, S. Prabhakar, D.P. Zhuzhikov, Y.E. Dunaevsky, M.A. Belozersky, Fractionation of digestive proteinases from Tenebrio molitor (Coleoptera: Tenebrionidae) larvae and role in protein digestion, Comp. Biochem. Physiol. 145B (2006) 138-146; K.S. Vinokurov, E.N. Elpidina, D.P. Zhuzhikov, B. Oppert, D. Kodrik, F. Sehnal, Digestive proteolysis organization in two closely related Tenebrionid beetles: red flour beetle (Tribolium castaneum) and confused flour beetle (Tribolium confusum), Arch. Insect Biochem. Physiol. 70 (2009) 254-279).
C. Delbarre-Ladrat, M. Lamballerie-Anton, V. Verrez-Bagnis, Calpain and cathepsin activities in post mortem fish and meat muscles, Food Chemistry 101 (2007) 1474-1479; S. Subramanian, S. MacKinnona, N. Rossa, A comparative study on innate immune parameters in the epidermal mucus of various fish species, Comp. Biochem. Physiol. 148B (2007) 256-263; O. Vasiljeva, T. Reinheckel, C. Peters, D. Turk, V. Turk, B. Turk. Emerging roles of cysteine cathepsins in disease and their potential as drug targets. Curr. Pharm.Design, 13 (2007) 385-401; V. Turk, V. Stroka, O. Vasiljeva. M. Renko, T. Sun, B. Turk, D. Turk, Cysteine cathepsins: from structure, function and regulation to new frontiers, Biochim. Biophys. Acta (BBA)-Proteins and Proteomics 1824 (2012) 68-88). В настоящее время серьезная роль отводится пищеварительным цистеиновым пептидазам насекомых, в том числе и таких опасных вредителей, как колорадский жук, большой и малый мучные хрущаки, тараканы и т.д. (Р.Т. Cristofoletti, A.F. Ribeiro, W.R. Terra, The cathepsin L-like proteinases from the midgut of Tenebrio molitor larvae: sequence, properties, immunocytochemical localization and function. Insect Biochem. Mol. Biol. 35 (2005) 883-901; K. Gruden, A.G. Kuipers, G. Guncar, N. Slapar, B. Strukelj, M.A. Jongsma, Molecular basis of Colorado potato beetle adaptation to potato plant defence at the level of digestive cysteine proteinases, Insect Biochem. Mol. Biol. 34 (2004) 365-375; K.S. Vinokurov, E.N. Elpidina, B. Oppert, S. Prabhakar, D.P. Zhuzhikov, Y.E. Dunaevsky, M.A. Belozersky, Fractionation of digestive proteinases from Tenebrio molitor (Coleoptera: Tenebrionidae) larvae and role in protein digestion, Comp. Biochem. Physiol. 145B (2006) 138-146; K.S. Vinokurov, E.N. Elpidina, D.P. Zhuzhikov, B. Oppert, D. Kodrik, F. Sehnal, Digestive proteolysis organization in two closely related Tenebrionid beetles: red flour beetle (Tribolium castaneum) and confused flour beetle (Tribolium confusum), Arch. Insect Biochem. Physiol. 70 (2009) 254-279).
Наиболее обширную группу семейства С1 составляют цистеиновые пептидазы млекопитающих и человека, так называемые катепсины (D.P. Dickinson, Cysteine peptidases of mammals: their biological roles and potential effects in the oral cavity and other tissues in health and disease, Crit. Rev. Oral Biol. Med. 13 (2002) 238-275). По данным биоинформатического анализа генома, только у человека на сегодня известно 11 различных цистеиновых катепсинов В, С, F, Н, K, L, О, S, V(L2), X, W. Главной функцией цистеиновых катепсинов является неспецифическая лизосомальная деградация белков (N.D. Rawlings, A.J. Barrett, Introduction: The Clans and Families of Cysteine Peptidases, in: N.D. Rawlings, G.S. Salvesen (Eds.), Handbook of Proteolytic Enzymes, Third edition, Academic Press, London, 2013, v. 2, pp.1743-1776). Однако показано также их участие в более специфических процессах: регуляции клеточного цикла (V. Turk, V. Stroka, О. Vasiljeva. М. Renko, Т. Sun, В. Turk, D. Turk, Cysteine cathepsins: from structure, function and regulation to new frontiers, Biochim. Biophys. Acta (BBA)-Proteins and Proteomics 1824 (2012) 68-88), процессинге некоторых белков, ферментов и гормонов (Ueno Т., Linder S., Rice W.R., Johansson J., Weaver Т.Е. Processing of pulmonary surfactant protein В by napsin and cathepsin H, J. Biol. Chem. 279 (2004) 16178-16184), презентации антигенов (L.C. Hsing, A.Y. Rudensky. The lysosomal cysteine proteases in MHC class II antigen presentation, Immunol. Rev. 207 (2005) 229-241).
Цистеиновым катепсинам принадлежит особая роль при развитии различных патологических процессов. Так, катепсины В и L участвуют в росте и распространении опухолей (N.D. Rawlings, A.J. Barrett, Introduction: The Clans and Families of Cysteine Peptidases, in: N.D. Rawlings, G.S. Salvesen (Eds.), Handbook of Proteolytic Enzymes, Third edition, Academic Press, London, 2013, v. 2, pp. 1743-1776). Отмечен повышенный уровень катепсина В при множественных травмах и септическом шоке, а также обнаружена секреция зрелых активных катепсинов В и L макрофагами в межклеточную среду при хронических воспалениях (S. Sever, М. Altintas, S. Nankoe,  D. Ko, С. Wei, J. Henderson, Е. del Re, L. Hsing, A. Erickson, C. Cohen, M. Kretzler, D. Kerjaschki, A. Rudensky, B. Nikolic, J. Reiser, Proteolytic processing of dynamin by cytoplasmic cathepsin L is a mechanism for proteinuric kidney disease. J. Clin. Invest. (2007) 117, 2095-104). Катепсины участвуют также в таких воспалительных заболеваниях как панкреатит (катепсин В), ревматоидный артрит и остеоартрит (катепсины В, K, L) (О. Vasiljeva, Т. Reinheckel, С. Peters, D. Turk, V. Turk, В. Turk. Emerging roles of cysteine cathepsins in disease and their potential as drug targets. Curr. Pharm.Design, 13 (2007) 385-401). Катепсины В и H являются возможными маркерами внутритканевых болезней легких (М. Kimura, K. Tani, J. Miyata. The significance of cathepsins, thrombin and aminopeptidase in diffuse interstitial lung diseases. J. Med. Invest. 52 (2005) 93-100). Показано, что катепсин L участвует в возникновении диабета (R. Maehr, J.D. Mintern, А.Е. Herman,
D. Ko, С. Wei, J. Henderson, Е. del Re, L. Hsing, A. Erickson, C. Cohen, M. Kretzler, D. Kerjaschki, A. Rudensky, B. Nikolic, J. Reiser, Proteolytic processing of dynamin by cytoplasmic cathepsin L is a mechanism for proteinuric kidney disease. J. Clin. Invest. (2007) 117, 2095-104). Катепсины участвуют также в таких воспалительных заболеваниях как панкреатит (катепсин В), ревматоидный артрит и остеоартрит (катепсины В, K, L) (О. Vasiljeva, Т. Reinheckel, С. Peters, D. Turk, V. Turk, В. Turk. Emerging roles of cysteine cathepsins in disease and their potential as drug targets. Curr. Pharm.Design, 13 (2007) 385-401). Катепсины В и H являются возможными маркерами внутритканевых болезней легких (М. Kimura, K. Tani, J. Miyata. The significance of cathepsins, thrombin and aminopeptidase in diffuse interstitial lung diseases. J. Med. Invest. 52 (2005) 93-100). Показано, что катепсин L участвует в возникновении диабета (R. Maehr, J.D. Mintern, А.Е. Herman,  D. Mathis, С. Benoist, H.L. Ploegh, Cathepsin L is essential for onset of autoimmune diabetes in NOD mice. J. Clin. Invest. 115 (2005) 2934-2943). Есть данные, что катепсин В участвует в возникновении болезни Альцгеймера, вызывая апоптоз нейронов (Gan L., S. Ye, A. Chu, K. Anton S. Yi, V. Vincent, D. von Schack, D. Chin, J. Murray, S. Lohr, L. Patthy, M. Gonzalez-Zulueta, K. Nikolich, R. Urfer. Identification of cathepsin В as a mediator of neuronal death induced by Abeta-activated microglial cells using a functional genomics approach. J Biol Chem. 279 (2004) 565-572).
D. Mathis, С. Benoist, H.L. Ploegh, Cathepsin L is essential for onset of autoimmune diabetes in NOD mice. J. Clin. Invest. 115 (2005) 2934-2943). Есть данные, что катепсин В участвует в возникновении болезни Альцгеймера, вызывая апоптоз нейронов (Gan L., S. Ye, A. Chu, K. Anton S. Yi, V. Vincent, D. von Schack, D. Chin, J. Murray, S. Lohr, L. Patthy, M. Gonzalez-Zulueta, K. Nikolich, R. Urfer. Identification of cathepsin В as a mediator of neuronal death induced by Abeta-activated microglial cells using a functional genomics approach. J Biol Chem. 279 (2004) 565-572).
Таким образом, учитывая разнообразие и сложность задач, «решаемых» этими ферментами в ходе клеточного протеолиза, очевидно, что имеется серьезная необходимость в создании комплекса высокоспецифичных и селективных пептидных субстратов для тестирования указанного класса ферментов. Между тем, в настоящее время обнаружение цистеиновых пептидаз затруднено вследствие того, что синтетические субстраты, обычно используемые для определения активности этой группы протеиназ, не селективны.
В настоящее время практически все используемые субстраты цистеиновых протеиназ семейства С1 (пептиды, содержащие Arg- и Lys- в P1 положении) не являются селективными для этого класса ферментов, поскольку являются хорошими субстратами трипсиноподобных протеиназ. Известные на сегодняшний день хромогенные субстраты цистеиновых протеиназ семейства С1 можно разделить на несколько групп. Эфиры и амиды бензоиларгинина обладают низкой чувствительностью и плохо растворимы в воде (для папаина KM=14 mM и 33 mM соответственно) (L.J. Brubacher, M.R. Zaher, A kinetic study of hydrophobic interactions at the S1 and S2 sites of papain, Can. J. Biochem. 57 (1979) 1064-1072). Хромофорные n-нитрофениловые субстраты (G. Lowe, Y. Yuthavong, Kinetic specificity in papain-catalysed hydrolyses, Biochem. J. 124 (1971) 107-115) склонны к спонтанному гидролизу, что затрудняет корректное определение активности цистеиновых протеиназ. Наиболее широко применяемыми хромогенными субстратами являются n-нитроанилиды аргинин-содержащих пептидов. Эти соединения обладают хорошей растворимостью и высокой стабильностью в водных смесях, удовлетворительными кинетическими характеристиками (для папаина KM = 3 mM, kcat=0,8 s-1) (J.E. Mole, H.R. Horton, Kinetics of papain-catalyzed hydrolysis of -N-benzoyl-L-arginine-p-nitroanilide, Biochemistry 12 (1973) 816-822). Используемые флуорогенные субстраты цистеиновых протеиназ также обладают двойной специфичностью и ограниченной применимостью при работе с неочищенными ферментными препаратами (Barrett, An improved color reagent for use in Barrett's assay of Cathepsin B, Anal. Biochem. 76 (1976) 374-379; S. Zucker, D. Buttle, J. Nicklin, A. Barrett, The proteolytic activities of chymopapain, papain, and papaya proteinase III, Biochem. Biophys. Res. Commun. 828 (1985) 196-204; J. Tchoupe, T. Moreau, F. Gauthier, J. Bieth, Photometric or fluorometric assay of cathepsin B, L and H and papain using substrates with an aminotrifluoromethylcoumarin leaving group, Biochim. Biophys. Acta, 1076 (1991) 149-151; M. Cezari, L. Puzer, M. Juliano, A. Carmona and A. Juliano, Cathepsin В carboxydipeptidase specificity analysis using internally quenched fluorescent peptides. Biochem J. 368 (2002) 365-369). Главными недостатками всех упомянутых субстратов является необходимость проведения для их получения сложного многостадийного синтеза, и, как результат, низкий выход оптически чистого продукта. Все перечисленные субстраты выпускаются ведущими мировыми компаниями, специализирующимися на производстве реагентов для исследований в области биохимии, молекулярной биологии, биотехнологии: БАХЕМ (Германия), СИГМА (США), ФЛУКА (Швейцария). Такой список влиятельных фирм - производителей субстратов цистеиновых протеиназ, свидетельствует о важности этих продуктов и рыночной потребности в их производстве.
Известен хромогенный пептидный субстрат формулы Glp-Phe-Leu-pNA, селективный для определения активности цистеиновых протеиназ. Синтез субстрата и его использование для тестирования препаратов цистеиновых протеиназ различной степени чистоты был описан в журнале Analytical Biochemistry (I.Yu. Filippova, E.N. Lysogorskaya, E.S. Oksenoit, G.N. Rudenskaya, V.M. Stepanov, L-Pyroglutamyl-L-phenylalanyl-L-leucine-p-nitroanilide - a chromogenic substrate for thiol proteinase assay, Anal Biochem. 143 (1984) 293-297). Этот субстрат производится фирмами БАХЕМ (Германия) и ПЕПТАЙДС ИНСТИТУТС Инк. (Япония) и используется для исследований цистеиновых протеиназ из различных источников, в том числе, катепсинов В из мозга человека и быка (Azaryan А., Agatyan G. and Galoyan A, Cathepsin В from bovine, Biochemistry (Moscow) 52 (1987) 2033-2037), пищеварительных протеиназ одного из опаснейших сельскохозяйственных вредителей - колорадского жука (C.J. Bolter, M.A. Jongsma, Colorado potato beetles (Leptinotarsa decemlineata) adapt to proteinase inhbitors induced in potato leaves by methyl jasmonate, J. Insect Physiol. 41 (1995) 1071-1078). Однако использование этого препарата ограничено его низкой растворимостью в воде и необходимостью использовать высокие концентрации органических растворителей (20% диметилформамид или диметилсульфоксид).
Чтобы избежать этих проблем, был предложен хромогенный субстрат Glp-Phe-Ala-pNA и флуорогенные аналоги этого субстрата, Glp-Phe-Ala-AMC и Glp-Phe-Ala-AFC (Патент №1198082 (СССР), опубл. в Б.И. 1985, N 46; Семашко Т.А. Новые селективные пептидные субстраты цистеиновых пептидаз семейства папаина М., 2011 г. Автореферат дисс. (http://www.chem.msu.su/rus/theses/avtoreferat/2011/2011-06-14-semachko.pdf; Semashko Т.А., Vorotnikova Е.А., Sharikova V.F., Vinokurov K.S., Smirnova Y.A., Dunaevsky Y.E., Belozersky M.A., Oppert В., Elpidina E.N., Filippova I.Y. Selective chromogenic and fluorogenic peptide substrates for the assay of cysteine peptidases in complex mixtures, Anal. Biochem. 49 (2014) 179-187). Эти аланин-содержащие пептидомиметики являются селективными для цистеиновых пептидаз, но проигрывают коммерческим аргинин-содержащим субстратам по эффективности гидролиза. Эти субстраты были успешно использованы как для тестирования очищенных ферментов, так и для характеристики цистеиновых пептидаз в составе сложных многокомпонентных природных смесей (экстрактов пищеварительных пептидаз насекомых.
Известен также хромогенный глутамин-содержащий пептидный субстрат Z-Ala-Ala-Gln-pNA, раскрытый в работе (I.A. Goptar, Т.А. Semashko, S.A. Danilenko, E.N. Lysogorskaya, E.S. Oksenoit, D.P. Zhuzhikov, M.A. Belozersky, Y.E. Dunaevsky, B. Oppert, I.Yu. Filippova, E.N. Elpidina, Cysteine digestive peptidases function as post-glutamine cleaving enzymes in tenebrionid stored product pests, Comparative Biochemistry and Physiology - В Biochemistry and Molecular Biology, 161 (2012) 148-154), однако этот субстрат обладает низкими кинетическими характеристиками.
Из уровня техники также известен субстрат формулы Glp-Phe-Leu-pNA (I.Yu. Filippova, E.N. Lysogorskaya, E.S. Oksenoit, G.N. Rudenskaya, V.M. Stepanov, L-Pyroglutamyl-L-phenylalanyl-L-leucine-p-nitroanilide - a chromogenic substrate for thiol proteinase assay, Anal Biochem. 143 (1984) 293-297).
Наиболее близким аналогом (прототипом) является пептидный субстрат общей формулы A-XaaYaaB, где А=Glp (пироглутамил), Abz (о-аминобензоил); В=pNA (п-нитроанилид), АМС (7амидо-4-метилкумарин) и AFC (7-амидо-4-трифторметилкумарин); Хаа = Phe, Val; Yaa = Ala (Семашко T.A., Новые селективные пептидные субстраты цистеиновых пептидаз семейства папаина, М., 2011 г. (http://www.chem.msu.su/rus/theses/avtoreferat/2011/2011-06-14-semachko.pdf). Недостатком прототипа являются невысокие каталитические (кинетические) параметры, что, несмотря на селективность, снижает эффективность их использования.
Раскрытие изобретения
Задачей настоящего изобретения является создание новых синтетических субстратов для тестирования цистеиновых пептидаз семейства папаина, устойчивых к гидролизу трипсиноподобными пептидазами.
Задача решается новыми пептидными субстратами цистеиновых пептидаз семейства папаина общей формулы (1):

А - N-защитная группа уретанового или ацильного типа;
Хаа - протеиногенная или непротеиногенная аминокислота или дипептид, отвечающие требованиям субстратной специфичности цистеиновых пептидаз семейства папаина (семейства С1);
В - маркерная хромогенная или флуорогенная группировка, для количественного контроля расщепления субстратов спектрофотометрическими или флуорометрическими методами.
Предпочтительно N-защитные группы уретанового типа выбирать из ряда Z (бензилоксикарбонил), Boc (трет-бутилоксикарбонил) или ацильного типа, например, Glp (пироглутамил), Suc (сукцинил), Bz (бензоил), Ас (ацетил), For (формил).
Предпочтительно протеиногенную или непротеиногенную аминокислоты или дипептид, отвечающие требованиям субстратной специфичности цистеиновых пептидаз семейства папаина (семейства С1), выбирать из группы, включающей Phe, Phe-4-Cl (4-хлор-фенилаланин), Phe-3,4-Cl2 (3,4-дихлор-фенилаланин), homoPhe (гомофенилаланин), Phg (фенил-глицин), Tyr, Leu, Val, Ile, Pro, His, Arg, Lys, Thr(OBzl) (О-бензилтреонин), Thr(tBu) (O-трет-бутилтреонин), Ser(OBzl) (О-бензилсерин), Ser(tBu) (O-трет-бутилсерин), Cys(SBzl) (S-бензилцистеин), Ala-Ala, Ala-Gly, Gly-Ala, Gly-Gly.
Предпочтительно хромогенные или флуорогенные группировки выбирать из ряда, включающего АМС (7-амидо-4-метилкумарид), βNA (бета-нафтиламид), AFC (7-амидо-4-трифторметилкумарид), pNA (n-нитроанилид) - для определения ферментативной активности пептидаз. Место гидролиза субстратов показано стрелкой.
Поставленная задача также решается применением заявляемого пептидного субстрата для создания тест-систем детекции пептидаз, при этом детекцию можно проводить в растворе, в геле после проведения электрофореза в нативных условиях, в системах in vivo.
Заявляемый субстрат общей формулы (1) может применяться для определения ферментативной активности пептидаз в качестве высокоспецифичных и селективных хромогенных и флуорогенных субстратов для цистеиновых пептидаз семейства папаина (в присутствии ферментов других кланов); для определения ферментативной активности пептидаз для селективной детекции катепсина L в природных смесях различных катепсинов (например, В и L); для определения ферментативной активности пептидаз для селективного тестирования активности катепсина L с использованием флуорогенных соединений в системах in vivo: клетках и их органеллах, межклеточном пространстве, тканях, а также для определения ферментативной активности пептидаз для поиска глутаминспецифичных пептидаз, обладающих способностью расщеплять глиадины и их токсические пептиды.
Техническим результатом заявляемого пептидного субстрата является его высокая эффективность (кинетические характеристики) и селективность по отношению к цистеиновым пептидазам семейства С1.
Заявляемый субстрат позволяет добиться эффективности гидролиза по сравнению с прототипом и аналогами в 3-5 раз, а в некоторых случаях, например, для катепсина L - даже на порядок. Предлагаемые нами глутамин-содержащие субстраты обладают лучшими кинетическими характеристиками, чем описанные выше аланин-содержащие субстраты и Z-Ala-Ala-Gln-pNA. Все предлагаемые хромогенные и флуорогенные субстраты селективны для цистеиновых пептидаз семейства С1 и не гидролизуются пептидазами других кланов. Это является важным преимуществом по отношению к широко распространенным коммерчески доступным аргинин-содержащим субстратам: Z-Phe-Arg-pNA, Z-Phe-Arg-AMC, Z-Arg-Arg-pNA, Z-Arg-Arg-AMC, которые активно расщепляются также трипсиноподобными сериновыми пептидазами. Это позволяет использовать предложенные глутамин-содержащие субстраты для тестирования цистеиновых пептидаз в присутствии ферментов других кланов.
Также заявляемый пептидный субстрат отличается высокой активностью цистеиновых пептидаз семейства папаина, таких как растительные пептидазы папаин и фицин, а также пептидазы животных - лизосомальный и пищеварительный катепсины L, по предлагаемым селективным субстратам, которые являются высокоспецифичными. В частности, среди катепсинов животного происхождения, учитывая более низкую активность катепсина В по отношению к этим субстратам, их использование позволяет специфически тестировать катепсин L в природных смесях этих двух катепсинов (например, в пищеварительной системе жуков-вредителей запасов зерновых, в лизосомах).
Заявляемый пептидный субстрат отличается селективностью по отношению к катепсину L, являющемуся маркером многих заболеваний, позволяет использовать субстраты для селективного тестирования фермента в системах in vivo. Наличие в субстратах маркерной хромогенной или флуорогенной группировок позволяет использовать их при создании тест-систем и тестирования активности непосредственно в геле после проведения электрофореза в нативных условиях.
Также заявляемый пептидный субстрат характеризуется высокой активностью в отношении постглутаминрасщепляющих пептидаз, гидролизующих глиадины и их токсические пептиды. Это создает возможность использования субстратов для скрининга пептидаз - потенциальных кандидатов для создания препаратов от целиакии (аутоиммунного заболевания ЖКТ, вызываемого недорасщепленными иммуногенными Gln-содержащими пептидами), а также для получения безглютеновых продуктов.
Краткое описание чертежей
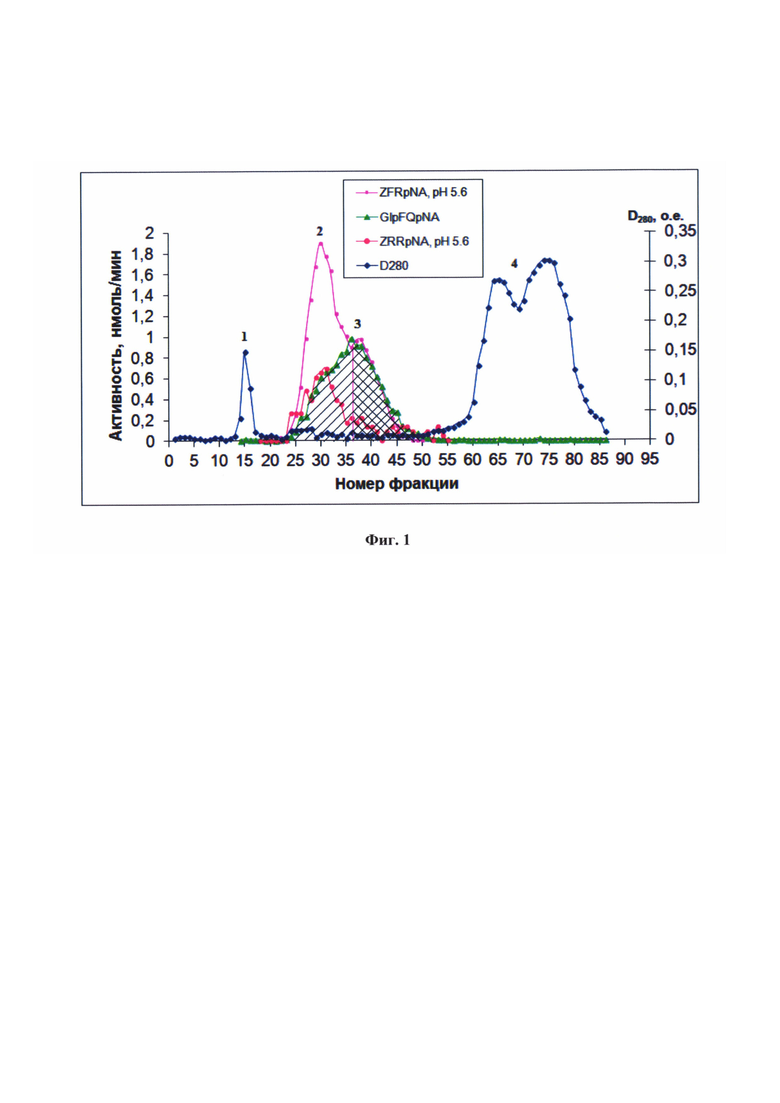
На фиг. 1 показаны профили элюции цистеиновых пептидаз из экстракта кишечника личинок Т. castaneum с сефадекса G-75 по субстратам Glp-Phe-Gln-pNA, Z-Phe-Arg-pNA и Z-Arg-Arg-pNA при рН 5,6 в присутствии 6 мМ Cys. Пики 1 и 4 относятся к неферментативным белковым фракциям. В пиках 2 и 3 содержится активность примесных сериновых трипсиноподобных пептидаз и цистеиновых катепсинов В и L, тестируемая по неселективному субстрату Z-Phe-Arg-pNA. В пике 2 преимущественно содержится активность катепсина В, селективно тестируемая по субстрату Z-Arg-Arg-pNA. В пике 3 элюируется активность преимущественно катепсина L, селективно тестируемая по субстрату Glp-Phe-Gln-pNA.
Подробное описание изобретения
Структура субстратов соответствует требованиям специфичности цистеиновых пепитдаз семейства С1: ключевую позицию P1 (Schechter I., Berger A. On the size of the active site in proteases. I. Papain. Biochem Biophys Res Commun., 1967, 27(2), 157-162) занимает небольшой по размеру остаток глутамина (Gln). В положении Р2, которое является определяющим для специфичности указанных пептидаз, находятся гидрофобные остатки Хаа = Phe, Phe-4-Cl, Phe-3,4-Cl2, homoPhe, Phg, Tyr, Leu, Val, Ile, Pro, His, Thr(OBzl), Thr(tBu), Ser(OBzl), Ser(tBu), Cys(SBzl) или дипептиды Ala-Ala, Ala-Gly, Gly-Ala, Gly-Gly.
Все эти производные удовлетворяют требованиям субстратной специфичности цистеиновых пептидаз семейства С1 (Chang A., Scheer М., Grote A., Schomburg I., Schomburg D. BRENDA, AMENDA and FRENDA the enzyme information system: new content and tools in 2009. Nucleic Acids Res., 2009, 37, 588-592. http://www.brenda-enzymes.org: N.D. Rawlings, G.S. Salvesen (Eds.), Handbook of Proteolytic Enzymes, Third edition, Academic Press, London, 2013, v. 2, pp. 1743-1776).
N-концевая группировка субстратов (А) представлена стандартными N-защитными группировками, используемыми в пептидном синтезе - бензилоксикарбонильной (Z), трет-бутилоксикарбонильной (Boc), бензоильной (Bz), ацетильной (Ас), формильной (For), сукцинильной (Suc) (Гершкович А.А., Кибирев В.К. Химический синтез пептидов. Наукова Думка, Киев, 1992). Кроме того, предлагается использовать в качестве N-концевой группы остаток пироглутаминовой кислоты (Glp). Glp является природной иминокислотой и может выполнять функцию естественной защиты аминогрупп пептидов и белков. Введение остатка Glp способствует также улучшению растворимости субстратов в водно-органических смесях, поскольку пироглутаминовая кислота более гидрофильна чем большинство стандартных N-защитных группировок. Эффективность использования остатка Glp в качестве N-концевой группы была продемонстрирована нами ранее (I.Yu. Filippova, E.N. Lysogorskaya, E.S. Oksenoit, G.N. Rudenskaya, V.M. Stepanov, L-Pyroglutamyl-L-phenylalanyl-L-leucine-p-nitroanilide - a chromogenic substrate for thiol proteinase assay, Anal Biochem. 143 (1984) 293-297). В качестве С-концевых остатков (В) использованы известные маркерные группы: хромогенная n-нитроанилидная (pNA) и флуорогенные 4-амино-7-метилкумарил (АМС), 4-амино-7-трифторметилкумарил (AFC) и бета-нафтиламидная (βNA) группировки, присутствие которых обеспечивает удобство и точность контроля ферментативной активности по изменению спектральных и флуоресцентных характеристик субстратов в ходе гидролиза (Гершкович А.А., Кибирев В.К. Хромогенные и флуорогенные пептидные субстраты протеолитических ферментов. Биоорг. Химия, 1988,14(11), 1461-1488).
В качестве маркерной хромогенной или флуорогенной группировок (В), выбрани из ряда АМС (7-амидо-4-метилкумарид), βNA (бета-нафтиламид), AFC (7-амидо-4-трифторметилкумарид), pNA (n-нитроанилид) - для определения ферментативной активности пептидаз.
Пептидные субстраты могут быть получены любым известным из уровня техники способом (Гершкович А.А., Кибирев В.К. Химический синтез пептидов. Наукова Думка, Киев, 1992; Houben-Weyl Methods of Organic Chemistry: Synthesis of Peptides and Peptidomimetics, 4th edition, 2004. Edited by Goodman M., Toniolo C, Moroder L., Felix A. V. E22a, p. 785. Thieme, Stuttgart, New York). Субстраты указанной выше общей формулы (1) могут быть получены пептидной конденсацией под действием дициклогексилкарбодиимида (ДЦГК) и/или с использованием активированных эфиров в присутствии, например, пентафторфенола. (примеры 1-4).
Карбоксильными компонентами являются: (а) пироглутамил-аминокислоты (см. пример 1 и 2), (б) N-защищенные аминокислоты (Z, Suc, Boc, Bz, Ac, For) или дипептиды (см. пример 3). Аминокомпонент представлен остатком Gln, содержащим С-концевую флуорогенную или хромогенную группу: АМС, βNA, AFC, pNA.
Для синтеза Glp-содержащих карбоксильных компонентов использован подход, основанный на получении соответствующих метиловых эфиров Glp-аминокислот (по методу активированных эфиров) с последующим их ферментативным (пример 1) или щелочным (пример 2) омылением. Для получения N-защищенных аминокислот или дипептидов применяли стандартные приемы введения защитных группировок (пример 3) (Гершкович А. А., Кибирев В.К. Химический синтез пептидов. Наукова Думка, Киев, 1992) или использовали коммерчески доступные N-защищенные производные аминокислот и дипептидов (например, фирмы БАХЕМ, Германия). Для получения пироглутамилпроизводных (аминокислот и пептидов) во всех случаях использовали подход, базирующийся на применении пентафторфенилового эфира пироглутаминовой кислоты GlpOPfp (Семашко и др., 2008)
Пептидную конденсацию проводили в среде безводных органических растворителей (обычно диметилформамида (ДМФ), дихлорметана (ДХМ), ацетонитрила (АН), вводя в реакцию эквимолярные количества карбоксильного и аминокомпонента, пентафторфенола и 5-10% избыток ДЦГК (Гершкович А.А., Кибирев В.К. Химический синтез пептидов. Наукова Думка, Киев, 1992).
Гомогенность веществ определяли на пластинах Silufol (Чехия).
Хроматографию выполняли в следующих системах:
хлороформ - этанол (9:1) (А)
этилацетат-пиридин-уксусная кислота-вода (75:27:17:8) (В);
н-бутанол-пиридин-вода-уксусная кислота (15:1012:3) (С);
н-бутанол-уксусная кислота-вода (3:1:1) (D);
н-бутанол-уксусная кислота-вода (40:7:4) (Е)
бензол-ацетон-уксусная кислота (100:25:4) (F).
Анализ пептидных субстратов проводили на жидкостных хроматографах Милихром А-02 (ЭкоНова, Россия) и Altex Model 110 А (США) на колонке ProntoSil 120-5C18AQ (2,0×75 мм) при элюции раствором 0,1% CF3COOH в линейном градиенте ацетонитрила (скорость потока 1 мл/мин) от 0 до 80% за 20 минут (А) и на колонке Nucleosil С18 (4,6×250 мм) (Биохиммак, Россия) при элюции раствором 1% CF3COOH в линейном градиенте ацетонитрила (скорость потока 1 мл/мин) от 30 до 70% за 30 минут (Б). Детекцию осуществляли при 214 нм, 280 нм и 350 нм.
Спектрофотометрические измерения проводили на спектрофотометрах Genesys 10 S UV (Spectronics, США), а также на микропланшетном фотометре ELx808 (BioTek, США) в 96-луночных планшетах.
Флуоресцентные измерения проводили на микропланшетных флуориметрах Fluoroskan Ascent (Thermo Scientific, США) и Flx808 (BioTek, США) в 96-луночных планшетах, а также на спектрофлуориметре Cary Eclipse (Varian, США).
Структуры полученных соединений подтверждали с помощью 1Н-ЯМР-спектроскопии. Спектры ЯМР регистрировали в дейтеродиметилсульфоксиде на спектрометре "Brucker АС-300" (Германия) при частоте 300 МГц. Химические сдвиги приведены в миллионных долях (δ, м.д.) относительно внутреннего стандарта тетраметилсилана (δ 0,000 м.д.).
Аминокислотный анализ проводили на автоматическом аминокислотном анализаторе Hitachi 835 (Япония) после кислотного гидролиза проб 6 М HCl при 105°С в вакуумированных ампулах в течение 48 ч.
Гидролиз полученных глутаминсодержащих субстратов проводили различными цистеиновыми пептидазами семейства С1 - растительными ферментами: папаином, бромелаином, фицином, а также лизосомальными катепсинами В и L млекопитающих и человека соответственно и пищеварительным катепсином L насекомого вредителя зерновых запасов Tribolium castaneum. Все исследуемые субстраты расщеплялись цистеиновыми пептидазами семейства С1. Предлагаемые нами глутамин-содержащие субстраты обладают лучшими кинетическими характеристиками, чем прототип и его аналоги Glp-Phe-Ala-AMC, Glp-Phe-Ala-AFC, Glp-Phe-Ala-pNA и Z-Ala-Ala-Gln-pNA (пример 5).
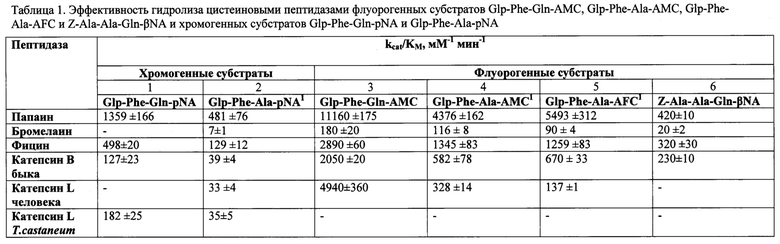
В таблице 1 содержатся данные по эффективности гидролиза цистеиновыми пептидазами различного происхождения хромогенного субстрата Glp-Phe-Gln-pNA и флуорогенных субстратов Glp-Phe-Gln-AMC и Z-Ala-Ala-Gln-βNA. Предложенные глутаминсодержащие субстраты сравнены с прототипами Glp-Phe-Ala-AMC, Glp-Phe-Ala-AFC и Glp-Phe-Ala-pNA. Эффективность гидролиза оценивалась коэффициентом специфичности ферментативной реакции kcat/KM, которая характеризует как скорость ферментативной реакции (величина kcat), так и сродство данного фермента к данному субстрату (величина KM). Чем выше значение kcat/KM, тем субстрат лучше. Флуорогенные субстраты с АМС- и AFC-группировками (столбцы 3-5) примерно на порядок более эффективно расщеплялись всеми изученными ферментами, чем их хромогенные аналоги (столбцы 1 и 2) и флуорогенный субстрат с β-нафтиламидной группой. Сравнение кинетических параметров гидролиза Glp-Phe-Gln-pNA (столбец 1) и Glp-Phe-Gln-AMC (столбец 3) с их аланин-содержащими хромогенным прототипом Glp-Phe-Ala-pNA (столбец 2) и флуорогенными прототипами Glp-Phe-Ala-AMC и Glp-Phe-Ala-AFC (столбцы 4, 5) убедительно показывает преимущество предлагаемых нами глутамин-содержащих субстратов. Наиболее эффективен в отношении предложенных субстратов папаин, но и в случае катепсинов животных значения коэффициента специфичности kcat/KM достаточно высоки.
Для практических целей более удобной оценкой гидролитической эффективности пептидаз является характеристика их удельной активности (примеры 6,7). Величина удельной активности показывает, какое количество субстрата (в молях) расщепляется определенным количеством фермента (в данном случае - молем) в единицу времени (в данном случае - в минуту) (см. примеры 6 и 7). Чем приведенное значение удельной активности выше, тем эффективнее субстрат гидролизуется данным ферментом.
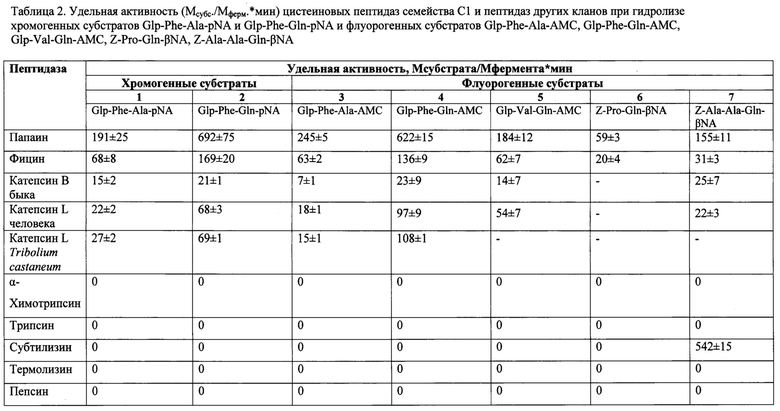
В таблице 2 в соответствующих строках приведены данные по удельной активности ряда исследованных цистеиновых пептидаз семейства С1 по отношению к синтезированному хромогенному субстрату Glp-Phe-Gln-pNA (столбец 2) и флуорогенным субстратам Glp-Phe-Gln-AMC, Glp-Val-Gln-AMC, Z-Pro-Gln-βNA, Z-Ala-Ala-Gln-βNA (столбцы 4-7). В столбцах 1 и 3 приведены значения удельной активности гидролиза указанными цистеиновыми пептидазами аланин-содержащих прототипов: хромогенного субстрата Glp-Phe-Ala-pNA (столбец 1) и флуорогенного субстрата Glp-Phe-Ala-AMC (столбец 3). Среди всех исследованных пептидаз наибольшую активность по всем субстратам проявлял папаин, являющийся родоначальником семейства С1. Наиболее перспективными для всех ферментов среди приведенных субстратов представляются Glp-Phe-Gln-pNA и Glp-Phe-Gln-AMC. Существенным преимуществом этих глутамин-содержащих субстратов (столбцы 2 и 4) по сравнению с их прототипами (столбцы 1 и 3 соответственно) является то, что с их использованием наблюдается более высокая удельная активность всех тестируемых пептидаз, т.е. гидролиз достигается за более короткое время и с меньшим количеством фермента. Особенно заметно преимущество Glp-Phe-Gln-pNA и Glp-Phe-Gln-AMC в случае цистеиновых катепсинов различного происхождения, причем наибольшую активность по указанному субстрату проявляет катепсин L.
Синтезированные хромогенный и флуорогенные субстраты селективны для цистеиновых пептидаз семейства С1 и не гидролизуются пептидазами других кланов - сериновых пептидаз химотрипсина, трипсина и субтилизина, металлопептидазы термолизина и аспартильной пептидазы пепсина (таблица 2). Это позволяет использовать предложенные глутамин-содержащие субстраты для тестирования цистеиновых пептидаз в присутствии ферментов других кланов, и является важным преимуществом по отношению к широко распространенным коммерчески доступным аргинин-содержащим субстратам: Z-Phe-Arg-pNA, Z-Phe-Arg-AMC Z-Arg-Arg-pNA, Z-Arg-Arg-AMC, которые активно расщепляются также трипсиноподобными сериновыми пептидазами.
Учитывая более низкую активность катепсина В (21±1, 23±9 (Мсубстрата/Мфермента*мин) по субстратам Glp-Phe-Gln-pNA и Glp-Phe-Gln-AMC соответственно) по сравнению с катепсином L (68±3, 97±9 (Мсубстрата/Мфермента*мин) по субстратам Glp-Phe-Gln-pNA и Glp-Phe-Gln-AMC соответственно) (таблица 1), их использование позволяет специфически и селективно тестировать катепсин L в природных смесях этих двух катепсинов (например, в пищеварительной системе жуков-вредителей запасов зерновых, в лизосомах).
Селективность предложенных субстратов может быть продемонстрирована в опыте по фракционированию сложной многокомпонентной ферментной смеси из пищеварительного комплекса личинок малого мучного хрущака Tribolium castaneum, в которой среди цистеиновых пептидаз доминировали катепсины L и В (A. Martynov, Е. Elpidina, L. Perkin, В. Oppert, Functional analysis of C1 family cysteine peptidases in the larval gut of Tenebrio molitor and Tribolium castaneum. BMC genomics. 16 (2015) 75-89)) (пример 9). На фиг. 1 показаны профили элюции цистеиновых пептидаз из экстракта кишечника личинок Т. castaneum с сефадекса G-75 по субстратам Glp-Phe-Gln-pNA, Z-Phe-Arg-pNA и Z-Arg-Arg-pNA при рН 5,6 в присутствии 6 мМ Cys. Пики 1 и 4 относятся к неферментативным белковым фракциям. В пиках 2 и 3 содержится активность примесных сериновых трипсиноподобных пептидаз и цистеиновых катепсинов В и L, тестируемая по неселективному субстрату Z-Phe-Arg-pNA. В пике 2 преимущественно содержится активность катепсина В, селективно тестируемая по субстрату Z-Arg-Arg-pNA. В пике 3 элюируется активность преимущественно катепсина L, селективно тестируемая по субстрату Glp-Phe-Gln-pNA. Следует отметить, что только использование Glp-Phe-Gln-pNA позволило селективно тестировать катепсин L и дифференциально выявить катепсины L и В.
Возможно также селективное тестирование активности катепсина L с использованием флуорогенных соединений в системах in vivo: клетках и их органеллах, межклеточном пространстве, тканях.
Для определения ферментативной активности изучаемого фермента в гомогенном растворе используется водный раствор заявляемого субстрата с добавлением полярного органического растворителя, необходимого для полного растворения субстрата. Обычно это водный раствор с добавлением 2-5% органического растворителя, в качестве которого могут быть использованы, например, диметилформамид, диметилсульфоксид, этанол, метанол, пропанол, изопропанол, ацетонитрил, диоксан. Критерием для выбора того или иного растворителя является как растворимость субстрата, (чем растворимость выше, тем лучше), так и влияние данного растворителя на активность определяемого фермента (выбирается такой растворитель, в присутствии которого активность фермента теряется в наименьшей степени).
В случае определения активности в геле, в качестве полимерной матрицы могут быть использованы гели из полиакриламида (ПААГ), крахмала, агарозы, желатина, целлюлозы.
Наличие в субстратах маркерной хромогенной или флуорогенной группировок позволяет использовать их при создании тест-систем в растворе (примеры 6 и 7) и детекции активности непосредственно в геле после проведения электрофореза в нативных условиях (пример 8).
Тест-система для определения ферментативной активности в растворе включает в себя препарат цистеиновой пептидазы, буферный раствор с рН от 5 до 7, содержащий этилендиамнтетрауксусную кислоту (ЭДТА) (конечная концентрация 1 мМ), раствор восстанавливающего SH-реагента (например, цистеин, меркаптоэтанол, дитиотреитол) (конечная концентрация от 2 до 6 мМ), раствор субстрата в 2,5% органическом растворителе (конечная концентрация хромогенных субстратов от 1 до 5 мМ, флуорогенных - от 10 до 100 мкМ) (примеры 6 и 7).
Тест-система для постэлектрофоретического тестирования активности цистеиновых пептидаз семейства С1 непосредственно в геле включает в себя гелевую пластину (полиакриламидную или аналогичную), в которой был проведен электрофорез (ЭФ) исследуемого препарата пептидазы в нативных условиях (в отсутствие додецилсульфата натрия) при рН 6,8-7,2 по методу, описанному в работе (K.S. Vinokurov, В. Oppert, E.N. Elpidina, Anal. Biochem. (2005)) и проявляющий раствор, состоящий из буфера с рН 5-7, содержащего ЭДТА (конечная концентрация 1 мМ), восстанавливающий SH-реагент (например, цистеин, меркаптоэтанол, дитиотреитол) (конечная концентрация от 2 до 6 мМ) с добавлением 50-100 мкМ раствора субстрата в 2,5% органическом растворителе. После инкубации 5-15 мин места локализации протеолитической активности выявляются по образовавшимся флуоресцентным полосам при 350-370 нм (пример 8).
Предлагаемые глутамин-содержащие субстраты могут быть эффективны для поиска глутаминспецифичных пептидаз, обладающих способностью расщеплять глиадины и их токсические пептиды. Это создает перспективу использования субстратов для скрининга пептидаз - потенциальных кандидатов для создания препаратов от целиакии (аутоиммунного заболевания ЖКТ, вызываемого недорасщепленными иммуногенными Gln-содержащими пептидами), а также для получения безглютеновых продуктов.
Таким образом преимуществом заявляемого пептидного субстрата являются:
1. Увеличение эффективности по сравнению с прототипом и аналогами. Эффективность гидролиза оценивалась коэффициентом специфичности ферментативной реакции kcat/KM, которая характеризует как скорость ферментативной реакции (величина kcat), так и сродство данного фермента к данному субстрату (величина KM). Чем выше значение kcat/KM, тем субстрат лучше. Флуорогенные субстраты с АМС- и AFC-группировками (столбцы 3-5) примерно на порядок более эффективно расщеплялись всеми изученными ферментами, чем их хромогенные аналоги (столбцы 1 и 2) и флуорогенный субстрат с β-нафтиламидной группой. Сравнение кинетических параметров гидролиза Glp-Phe-Gln-pNA (столбец 1) и Glp-Phe-Gln-AMC (столбец 3) с их аланин-содержащими хромогенным прототипом Glp-Phe-Ala-pNA (столбец 2) и флуорогенными прототипами Glp-Phe-Ala-AMC и Glp-Phe-Ala-AFC (столбцы 4, 5) убедительно показывает преимущество предлагаемых нами глутамин-содержащих субстратов. (Таблица 1; пример 5).
2. Высокая активность цистеиновых пептидаз семейства папаина, таких как растительные пептидазы папаин и фицин, а также пептидазы животных - лизосомальный и пищеварительный катепсины L, по предлагаемым субстратам, которые являются высокоселективными (таблица 2, примеры 6,7). Существенным преимуществом глутамин-содержащих субстратов по сравнению с их прототипом является то, что с их использованием наблюдается более высокая удельная активность всех тестируемых пептидаз (таблица 2, столбцы 1 и 2, 3 и 4). Это позволяет существенно повысить чувствительность анализа без потери селективности.
3. Селективность подавляющего большинства глутамин-содержащих субстратов по отношению к цистеиновым пептидазам семейства С1. Синтезированные субстраты расщепляются только цистеиновыми пептидазами семейства С1 и не гидролизуются пептидазами других кланов (таблица 2). Это позволяет использовать предложенные глутамин-содержащие субстраты для тестирования цистеиновых пептидаз в присутствии ферментов других кланов, и является важным преимуществом по отношению к широко распространенным коммерчески доступным аргинин-содержащим продуктам: Z-Phe-Arg-pNA, Z-Phe-Arg-AMC Z-Arg-Arg-pNA, Z-Arg-Arg-AMC, которые активно расщепляются также трипсиноподобными сериновыми пептидазами.
4. Возможность селективной детекции различных катепсинов животного происхождения. Учитывая более низкую активность катепсина В (таблица 2) по отношению к глутамин-содержащим субстратам, их использование позволяет специфически тестировать катепсин L в природных смесях этих двух катепсинов (например, в пищеварительной системе жуков-вредителей запасов зерновых, в лизосомах). Селективность предложенных субстратов продемонстрирована в опыте по фракционированию сложной многокомпонентной ферментной смеси из пищеварительного комплекса личинок малого мучного хрущака Tribolium castaneum, в которой среди цистеиновых пептидаз доминировали катепсины L и В (A. Martynov et al, 2015). Тестирование активности катепсинов L и В после хроматографического разделения проводили с помощью предложенного субстрата Glp-Phe-Gln-pNA, обладающего частичной селективностью по отношению к катепсину В коммерческого субстрата Z-Arg-Arg-pNA и неселективного коммерческого субстрата Z-Ala-Phe-Arg-pNA. Только использование Glp-Phe-Gln-pNA позволило селективно тестировать катепсин L и дифференциально выявить катепсины L и В (фиг. 1).
5. Селективное тестирование активности катепсина L с использованием флуорогенных соединений в системах in vivo: клетках и их органеллах, межклеточном пространстве, тканях.
6. Возможности использования предлагаемых субстратов; медико-биологический аспект.
6.1. Наличие в субстратах маркерной хромогенной или флуорогенной группировок позволяет использовать их при создании тест-систем и тестирования активности непосредственно в геле после проведения электрофореза в нативных условиях.
6.2. Селективность предложенных субстратов по отношению к катепсину L, являющемуся маркером многих заболеваний, позволяет использовать их для селективного тестирования фермента в системах in vivo.
6.3. Предлагаемые глутамин-содержащие субстраты могут быть эффективны для поиска глутаминспецифичных пептидаз, обладающих способностью расщеплять глиадины и их токсические пептиды. Это создает перспективу использования субстратов для скрининга пептидаз - потенциальных кандидатов для создания препаратов от целиакии (аутоиммунного заболевания ЖКТ, вызываемого недорасщепленными иммуногенными Gln-содержащими пептидами), а также для получения безглютеновых продуктов.
Настоящее изобретение поясняется конкретными примерами выполнения, которые не являются единственно возможным, но наглядно демонстрирует возможность достижения требуемого технического результата.
Пример 1
Синтез пироглутамил-L-фенилаланил-L-глутаминил-7-амино-4-метилкумарида (Glp-Phe-Gln-AMC)
Пентафторфениловый эфир пироглутаминовой кислоты GlpOPfp
К суспензии 2,58 г (20 ммоль) пироглутаминовой кислоты и 3,70 г (20 ммоль) пентафторфенола в 90 мл абс. этилацетата при 0°С добавляли 4,53 г (22 ммоль) ДЦГК в 10 мл абс. этилацетата. Перемешивали 1 ч при 0°С и 3 ч при 20°С. Осадок дициклогексилмочевины (ДЦГМ) отфильтровывали и промывали этилацетатом. Фильтрат и промывные воды упаривали в вакууме. Сухой остаток промывали гексаном. Сушили над парафином в вакууме. Выход 5,3 г (90%). Rƒ=0,76 (А); 0,5 (В).
Метиловый эфир фенилаланина PheOMe
К суспензии 3,9 г хлоргидрата метилового эфира фенилаланина в 100 мл абс. этилацетата по каплям добавляли 2,5 мл абс. триэтиламина и оставляли при перемешивании на 1 ч. Осадок отфильтровывали, раствор упарили в вакууме. В остатке масло. Выход 3,2 г (98%).
Метиловый эфир пироглутамил-фенилаланина Glp-PheOMe
К 3,2 г (18 ммоль) метилового эфира фенилаланина прибавляли 5,3 г (18 ммоль) пентафторфенилового эфира пироглутаминовой кислоты в 20 мл тетрагидрофурана, перемешивали 20 мин., упаривали в вакууме, остаток растворяли в 80 мл этилацетата и промывали 5% лимонной кислотой, водой. Из промывных вод дополнительно экстрагировали пептид смесью этилацетата и н-бутанола, 8:1. Объединенные экстракты промывали лимонной кислотой и водой. Этилацетатный раствор и экстракты объединяли, сушили безводным Na2SO4 и упаривали в вакууме. Масло кристаллизовали на холоду под абс. эфиром. Выход 3,3 г (63%). Т.пл. 28-30°С, Rf 0,79 (В), 0,83 (С); 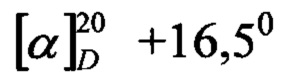 (с 1; ДМФ). Аминокислотный анализ: Glu 1,06; Phe 1,0.
(с 1; ДМФ). Аминокислотный анализ: Glu 1,06; Phe 1,0.
Пироглутамил-L-фенилаланин Glp-Phe
К раствору 1 г (3,38 ммоль) Glp-PheOMe в 27 мл 0,1 М бикарбоната аммония (рН=8), прибавляли 10 мл раствора α-химотрипсина (1 мг/мл в воде). Реакционную смесь перемешивали 5 ч при 37°С, затем упаривали в вакууме. Образовавшееся масло кристаллизовали обработкой эфиром и этиловым спиртом с последующей отгонкой растворителей в вакууме. Сушили над Р2О5. Выход 0,88 г (88%). Т.пл. 159-161°С; Rf 0,48 (С), 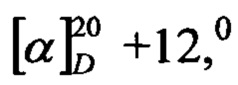 (c 0,5; DMF-вода 1:1).
(c 0,5; DMF-вода 1:1).
Пироглутамил-L-фенилаланил-L-глутаминил-7-амино-4-метилкумарид Glp-Phe-Gln-AMC
К раствору 0,059 г (0,21 ммоль) Glp-Phe и 0,081 г (0,21 ммоль) HBr×Gln-AMC в смеси 1,5 мл ацетонитрила и 5 мл ДМФ, содержащем 30 мкл (0,21 ммоль) триэтиламина при охлаждении до 0°С и перемешивании добавили 0,054 г (0,26 ммоль) ДЦГК. Реакционную смесь оставляли при перемешивании при комнатной температуре на ночь, образовавшийся осадок отфильтровывали, фильтрат упаривали. Полученное маслообразное вещество растворили в смеси этилацетат : бутанол (80:20), Органический слой промывали 3% NaHCO3, водой, 5% лимонной кислотой, сушили над безводным Na2SO4 и упаривали досуха. Продукт кристаллизовали из абс. диэтилового эфира. Выход - 0,060 г (52%). Аминокислотный анализ: Glu:Phe 2,1:1; Rf 0,77 (С), 0,62 (D); ВЭЖХ 13,5 мин (градиент А), 8,9 мин (градиент Б); Mr (рассч./обнар.) 561,5/561,4; ЯМР 1Н, DMSO-D6, δ, м.д. - 1,2 с (3Н, СН3 в АМС), 1,5-1,9 м (2Н, -CH2-CH2-CO-NH2 в Gln и 1Н -NH-CO-СН2-СН2-СН- в Glp), 2-2,4 м (4Н: 2Н, -CH2-CH2-CO-NH2 Gln и 2Н -NH-CO-CH2-CH2-CH- в Glp), 2,5 с (2Н, CH2-СО-О, АМС), 2,7 м (1Н, α-СН Phe), 3,35 д (2Н, CH2- в Phe), 4,5 д (1H, -NH-CO-CH2-CH2-CH- в Glp), 7,2-8,2 м (5Н, С6Н5 в Phe).
Аналогично получали соединения общей формулы Glp-Xaa-Gln-B, где Хаа = Phe-4-Cl, Phe-3,4-Cl2, homoPhe, Phg, Leu, Tyr; В = АМС, βNA, AFC.
Пример 2
Синтез пироглутамил-L-валил-L-глутаминил-7амино-4-метилкумарида (Glp-Val-Gln-AMC)
Хлоргидрат метилового эфира валика HCl ValOMe
5 г (0,043 моль) валина растворяли в 50 мл абс. метанола. К охлажденному до 0°С раствору прибавляли при перемешивании 6 мл (0,043 моль) хлористого тионила. Затем раствор перемешивали в течение 24 ч, упаривали в вакууме досуха и масло вторично обрабатывали метанолом и хлористым тионилом в описанных выше условиях. Полученное кристаллическое вещество промывали абс. эфиром, сушили в вакууме над щелочью. Выход 6,2 г (86%). Rƒ=0,54 (D).
Метиловый эфир пироглутамил-валина Glp-ValOMe
К 3,3 г (20 ммоль) HClxValOMe, растворенному в 30 мл DMF, приливали 3 мл Et3N, перемешивали. Затем к реакционной смеси приливали 5,8 г (20 ммоль) GlpOPfp в 30 мл ДМФ. После окончания реакции осадок отфильтровывали, ДМФ упаривали досуха. Полученное вещество растворяли в 150 мл EtOAc, выпавший осадок отфильтровывали и органическую фазу промывали лимонной кислотой и водой. Сушили над безводным Na2SO4, фильтровали и упаривали в вакууме. Кристаллизовали под петролейным эфиром, сушили над щелочью в вакууме. Выход 2,20 г (48%). Rƒ=0,12 (F).
Пироглутамил-L-валин Glp-Val
0,59 г (2,54 ммоль) Glp-ValOMe растворили в 5 мл метилового спирта, добавили 1,5 мл 2н. NaOH и перемешивали при комнатной температуре 1,5 часа. Растворитель упарили, добавили холодной воды и довели рН до 3. Экстрагировали из воды смесью этилацетат : бутанол (60:40, раствор промывали водой, растворитель упарили и остаток высадили петролейным эфиром. Выход 0,360 г (62%). Rf=0,39 (В) и 0,62 (D).
Пироглутамил-L-валил-L-глутаминил-7-амино-4-метилкумарид Glp-Val-Gln-АМС
К раствору 0,050 г (0,21 ммоль) GlpVal и 0,081 г (0,21 ммоль) HBr×Gln-AMC в смеси 1,5 мл АН и 5 мл ДМФ, содержащем 30 мкл (0,21 ммоль) ТЭА при охлаждении до 0°С и перемешивании добавили 0,054 г (0,26 ммоль) ДЦГК. Реакционную смесь оставляли при перемешивании при комнатной температуре на ночь, образовавшийся осадок отфильтровывали, фильтрат упаривали. Полученное маслообразное вещество растворили в 3,2 мл смеси этилацетат : бутанол (80:20), отфильтровали образовавшийся осадок и довели объем фильтрата до 10 мл все той же смесью этилацетата и бутанола. Органический слой промывали 3% NaHCO3, водой, 5% лимонной кислотой, сушили над безводным Na2SO4 и упаривали досуха. Продукт кристаллизовали из абс. диэтилового эфира. Выход - 0,052 г (48%). Аминокислотный анализ: Glu:Val = 2,2:1;Rf 0,51 (С), 0,41 (D); ВЭЖХ 12,0 мин (градиент А), 7,9 мин (градиент Б); Mr (рассч./ обнар.) 513,2/513,1; ЯМР 1H, DMSO-D6, δ, м.д. -1,2 д (3Н, СН3 в Val), 1,65 д (3Н, CH3 в Val), 2,5 с (3Н, СН3 в АМС), 2,6-2,75 м (2Н, -CH2-CH2-CO-NH2 Gln), 3,3 с (2Н, CH2 АМС) 7,91 с (1Н, -C(O)-NH-СН- в Val, 8,34 д (1H, -C(O)-NH-CH- в Glp).
Аналогично получали соединения общей формулы Glp-Xaa-Gln-B, где Хаа = Val, Pro, Ile, His, Thr(OBzl), Thr(tBu), Ser(OBzl), Ser(tBu), Cys (SBzl); В = АМС, βNA, AFC.
Пример 3
Синтез карбобензокси-L-аланил-L-аланил-L-глутаминил-β-нафтиламида Z-Ala-Ala-Gln-βNA
Бензилоксикарбонил-L-аланил-L-аланин Z-Ala-Ala
При интенсивном перемешивании к раствору 5 г (0,031 моль) аланил-аланина и 7,8 г бикарбоната натрия (0,09 моль) в 60 мл воды прикапывали 9 мл (0,062 моль) карбобензоксихлорида. Реакционную смесь перемешивали 1 ч при 0°С и 2 ч при 20°С. Не вошедший в реакцию карбобензоксихлорид экстрагировали эфиром. Водную фракцию подкисляли 5 н HCl (по конго красному), выпавший осадок фильтровали, промывали 0,2 н соляной кислотой и водой. Сушили в вакууме над щелочью. Выход 8 г (80%). Т.пл. 150-151°C. Rf=0,75 (С), 0,72 (Е).
Карбобензокси-L-аланил-L-аланил-L-глутаминил-β-нафтиламид Z-Ala-Ala-Gln-βNA
К раствору 0,154 г (0,5 ммоль) HCl×Gln-βNA и 0,147 г (0,5 ммоль) Z-Ala-Ala в 4 мл ДМФ, содержащему 70 мкл (0,5 ммоль) ТЭА, при охлаждении и перемешивании добавили 0,134 г (0,65 ммоль) ДЦГК. Реакционную смесь перемешивали 1 ч при охлаждении и оставляли на ночь при комнатной температуре. Образовавшийся осадок ДЦГМ отделяли и упаривали фильтрат досуха. Получившийся твердый остаток растворяли в 15 мл смеси этилацетат : бутанол (80:20). Органический слой промывали 3% NaHCO3 (2×1 мл), водой (2×1 мл), 5% лимонной кислотой (2×1 мл), сушили над безводным Na2SO4 и упаривали досуха. Продукт кристаллизовали под абс. диэтиловым эфиром. Выход 0,134 г (49%). Аминокислотный анализ: Ala:Glu = 1,8:1; Rf 0,81 (С), 0,79 (D); ВЭЖХ 16,8 мин (градиент A); Mr (рассч./ обнар.) 547,6/547,6; ЯМР 1H, DMSO-D6, δ, м.д. -1,1 д (3Н, СН3 в Ala), 1,25 д (3Н, СН3 в Ala), 1,4-1,7 м (2Н, -CH2-CH2-CO-NH2 Gln), 1,75 м (2Н, -СН2-CH2-СО-NH2 Gln), 3,35 м (1H, α-СН Gln), 4,1 м (1Н, α-СН Ala), 4,5 м (1Н, α-СН Ala), 5,3 д (2Н, -О-СН-С6Н5), 6,75 с (1H, ароматич. СН в А-цикле βNA), 7,2-7,7 м (5Н, С6Н5 в Cbz-), 7,8-8,4 м (4Н, ароматические СН в В-цикле βNA).
Аналогично получали соединения общей формулы A-Xaa-Gln-B, где A =Z, Boc, Bz, Ac, For, Suc; Хаа =Arg, Lys, Pro, Ala-Ala, Ala-Gly, Gly-Ala, Gly-Gly; В = АМС, βNA, AFC, pNA.
Пример 4
Синтез пироглутамил-L-фенилаланил-L-глутаминил-n-нитроанилида (Glp-Phe-Gln-pNA)
трет-Бутилоксикарбонил-L-фенилаланил-L-глутаминил-n-нитроанилид Boc-Phe-Gln-pNA
Раствор 4 г (9,3 ммоль) Boc-Phe-OPfp и 3,0 г (7,9 ммоль) H-Gln-pNA в ДМФ перемешивали при комнатной температуре 12 ч. После упаривания на роторном испарителе полученный маслообразный остаток растворили в смеси этилацетата и изопропанола. Органический слой промывали 5% раствором H2SO4, 5% раствором NaHCO3, 20% раствором NaCl, сушили над безводным Na2SO4 и упаривали досуха. Выход: 3,6 г (90%).
Фенилаланил-L-глутаминил n-нитроанилид Glp-Phe-Gln-pNA (трифторацетат) TFA х H-Phe-Gln-pNA
К Boc-Phe-Gln-pNA (3,60 г; 7,0 ммоль) добавили смесь трифторуксусной кислоты (30 мл) и ДХМ (20 мл) и перемешивали 30 мин. Реакционную смесь упарили на роторном испарителе до состояния вязкого масла. К остатку прибавляли диэтиловый эфир, растирали, образовавшийся осадок отфильтровывали и промывали эфиром и петролейным эфиром. Сушили в вакууме. Выход: 3,60 г (97%).
Пироглутамил-L-фенилаланил-L-глутаминил-n-нитроанилида (Glp-Phe-Gln-pNA)
К раствору Glp-OPfp (2,14 г; 7,25 ммоль) в ДМФ добавляли 3,4 г (6,50 ммоль) TFAxPhe-Gln-pNA, 0, 84 мл (6,5 ммоль) триэтиламина и оставляли при перемешивании на ночь. Реакционную смесь упаривали в вакууме, полученный маслообразный остаток растворяли в смеси этилацетата и пропанола и промывали 10% раствором NaHCO3 и насыщенным раствором NaCl. Высушивали раствор над Na2SO4 и упаривали в вакууме. Твердый остаток растирали с сухим этилацетатом и петролейным эфиром. Кристаллический осадок отфильтровывали, промывали этилацетатом и сушили в вакууме.
Выход: 1,763 г (52%). Аминокислотный анализ: Glu:Phe 1,98:1,1; Rf 0,75 (С), 0,60 (D); ВЭЖХ 13,0 мин (градиент А), 8,2 мин (градиент Б).
Пример 5.
Определение кинетических параметров гидролиза Gln-содержащих флуорогенных субстратов цистеиновыми пептидазами семейства С1
В ячейку микропланшета вносили 5 мкл препаратов ферментов известной концентрации. Добавляли 5 мкл 40 мМ ЭДТА и 10 мкл 120 мМ Cys, после чего содержимое ячеек доводили до конечного объема 195 мкл 0,1 М универсальным буфером с рН 5,6. Полученную реакционную смесь инкубировали 20 мин при комнатной температуре. Далее вносили 5 мкл 10-100 мкМ растворов субстратов в ДМФ и измеряли интенсивность флуоресценции в нулевой момент. Затем смесь инкубировали при 37°С, определяя изменение интенсивности флуоресценции через различные промежутки времени. Измерения проводились при λex=360 нм и λem=440 нм для АМС-группировки; λex=355 нм и λem=460 нм для флуорофора βNA (kfl=0,081 мкМ) и при λex=400 нм и λem=505 нм для флуорофора AFC (kfl=0,08 мкМ).
Расчеты кинетических параметров проводили методом нелинейной регрессии при помощи программы OriginPro 7.5 по стандартному уравнению Михаэлиса-Ментен:
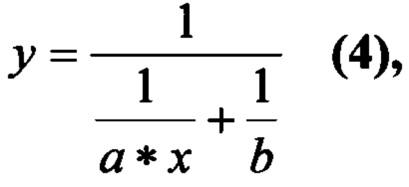
где y - начальная скорость реакции, которая определялась как производная в нулевой момент времени функции зависимости количества выделившегося продукта от времени, М/мин; х - концентрация субстрата в пробе, М; а=kcat*[E]/KM, b=kcat*[Е], где [Е] - концентрация фермента в пробе.
Пример 6
Гидролиз Gln-содержащих флуорогенных субстратов цистеиновыми пептидазами семейства С1 и пептидазами других кланов
В ячейку микропланшета вносили 5 мкл препарата фермента известной концентрации. Содержимое ячеек доводили до конечного объема 195 мкл 0,1 М универсальным буфером рН 5,6, содержащим 0, 0002 ммоль ЭДТА и 0,0012 ммоль Cys для цистеиновыхъ пептидаз, буфером Tris-HCl, рН 8,0, для химотрипсина, трипсина, субтилизина и термолизина и ацетатным буфером, рН 3,5, для пепсина. Реакционную смесь инкубировали 20 мин при комнатной температуре, вносили 5 мкл 1 мМ раствора субстрата в ДМФ и измеряли интенсивность флуоресценции в нулевой момент. Затем смесь инкубировали при 37°С, определяя изменение интенсивности флуоресценции через различные промежутки времени. Измерения проводились при λex=360 нм и λem=440 нм. Активность ферментов рассчитывали по формуле:
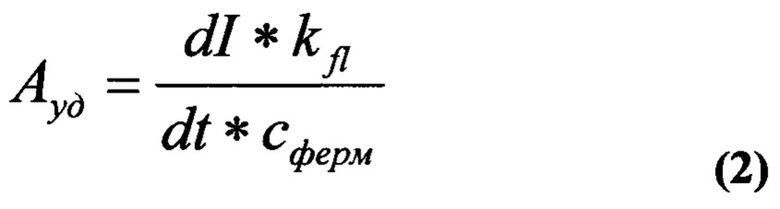
где Ауд - активность препарата, Мсубстр/Мферм мин;
dI/dt - начальная скорость отщепления флуорофора, которая определялась как производная в нулевой момент времени функции зависимости количества выделившегося флуорофора от времени, у.е./мин;
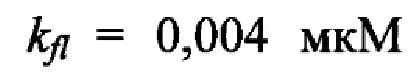 для АМС-группировки, при которой интенсивность флуоресценции раствора равно 1 условной единице (определяется в специальном опыте путем построения калибровочной прямой);
для АМС-группировки, при которой интенсивность флуоресценции раствора равно 1 условной единице (определяется в специальном опыте путем построения калибровочной прямой);
cферм - концентрация фермента в пробе, М
Полученные данные по удельной активности ряда исследованных цистеиновых пептидаз семейства С1 по отношению к флуорогенным Gln-содержащим субстратам Glp-Phe-Gln-AMC, Glp-Val-Gln-AMC, Z-Pro-Gln-βNA, Z-Ala-Ala-Gln-βNA приведены в табл. 2 (столбцы 4-7). В столбце 3 приведены ззначения удельной активности гидролиза указанными цистеиновыми пептидазами аланин-содержащего прототипа Glp-Phe-Ala-AMC.
Пример 7
Гидролиз Gln-содержащих хромогенных субстратов цистеиновыми пептидазами семейства С1
В ячейку микропланшета вносили 5 мкл препарата фермента известной концентрации. Добавляли 5 мкл 40 мМ ЭДТА и 10 мкл 120 мМ Cys, после чего содержимое ячеек доводили до конечного объема 195 мкл 0,1 М универсальным буфером с рН 5,6. Реакционную смесь инкубировали 15 мин при комнатной температуре и 5 мин в термостате при 37°С. Далее вносили 5 мкл 10 мМ, раствора субстрата в ДМФ и измеряли поглощение раствора при 405 нм в начальный момент времени. Затем смесь инкубировали при 37°С, определяя изменение поглощения при 405 нм через каждые 3 минуты.
Для расчета удельной активности Ауд (Мсубстр/Мферм*мин) использовали формулу:
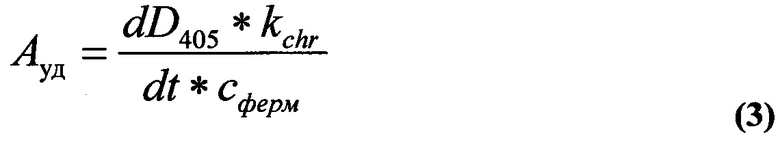
где dD405/dt - начальная скорость отщепления n-нитроанилина в нулевой момент времени (определяется по наклону линейного участка кинетической кривой, о.е./мин;
kchr=157 мкМ для n-нитроанилидной группировки, при которой поглощение раствора равно 1 условной единице (определяется в специальном опыте путем построения калибровочной прямой)
cферм - конечная концентрация фермента в реакционной смеси, М.
Полученные данные по удельной активности ряда исследованных цистеиновых пептидаз семейства С1 по отношению к хромогенным субстратам: Ala-содержащему прототипу Glp-Phe-Ala-pNA и Gln-содержащему субстрату Glp-Phe-Gln-pNA приведены в табл. 2 (столбцы 1 и 2 соответственно).
Пример 8.
Нативный электрофорез и постэлектрофоретическая детекция активности
Электрофорез препаратов проводили в нативных условиях в 12% разделяющем и 4% концентрирующем полиакриламидном геле (ПААГ) в буфере, содержащем 35 мМ HEPES и 43 мМ имидазол, при рН 7,4. На дорожку геля наносили 20 мкл папаина в том же буфере с D280=0,05, 0,1, 0,25, 0,5, 1,0 о.е. Электрофорез проводили обычно в направлении от катода к аноду при силе тока 10 мА в течение 40 мин.
После завершения электрофореза для визуализации белковых полос блок геля помещали в 0,1% раствор кумасси G-250 в смеси этанол/уксусная кислота/вода (30:10:60 объемных %), отмывали тем же раствором без красителя и фотографировали гели.
Постэлектрофоретическое тестирование активности по n-нитроанилидным субстратам проводили методом наложения нитроцеллюлозных мембран, насыщенных специфическими субстратами на блок ПААГ, в котором предварительно проводили электрофоретическое разделение белков. Для этого гель помещали на 20 минут в 10 мл 0,1 М универсального буфера, рН 5,6, содержащий 1 мМ DTT. Параллельно этому в растворе, содержащем 250 мкл 20 мМ субстрата в DMF в 10 мл 0,1 М универсального буфера, рН 5,6, содержащего 1 мМ DTT, замачивали нитроцеллюлозную мембрану и инкубировали в термостате при 37°С 30 минут. Затем нитроцеллюлозную мембрану накладывали на гель после электрофореза и инкубировали при 37°С 60 минут в закрытой чашке Петри. После инкубации мембрану переносили в новую чашку Петри и проводили реакцию диазотирования выделившегося в результате гидролиза n-нитроанилина последовательно тремя свежеприготовленными реагентами:
1. 10 мл водного раствора NaNO2 (0,01 г в 10 мл 1 М HCl), инкубировали 5 минут, затем сливали раствор.
2. 10 мл водного раствора аммоний амидосульфата (0,05 г в 10 мл 1 М HCl), оставляли на 5 минут, отбирали раствор
3. 10 мл раствора N-(1-нафтил)этилендиамин дигидрохлорида (0,005 г в 10 мл) в 50% этиловом спирте, оставляли на 5 минут, отбирали раствор.
Места локализации протеолитической активности определяли по образовавшимся розовым полосам на белом фоне мембраны.
Постэлектрофоретическое тестирование активности по флуорогенным субстратам проводили после инкубации блока геля в растворе флуорогенного субстрата. Гель после электрофореза помещали на 10 минут в 10 мл 0,1 М универсального буфера, рН 5,6, содержащий 1 мМ DTT, 2,5% DMF и 125 мкМ флуорогенный субстрат и инкубировали при 37°С. Места локализации протеолитической активности определяли по визуализации флуоресцентных полос под УФ-лампой при 366 нм.
Полученные результаты свидетельствуют о возможности использования предлагаемых субстратов для прямой детекции и безошибочной локализации в геле белковой полосы, принадлежащей изучаемому ферменту для дальнейшего анализа.
Пример 9
Фракционирование пищеварительных пептидаз из личинок Tribolium castaneum
2,1 мл экстракта (D280=5,9), полученного из кишечника личинок Т. castaneum, после центрифугирования при 16000 об./мин в течение 1 ч наносили при 4°С на колонку с Sephadex G-75 (2,5 × 120 см) в 0,01 М фосфатном буфере, рН 5,6, содержащем 10-4 М TLCK (N- α-тозил-L-лизин-хлорметилкетон) и 10-4 М химостатин. Собирали фракции объемом 9 мл. Концентрацию белка оценивали по величине оптической плотности при 280 нм. Ферментативную активность во фракциях элюата, отбирая аликвоты по 20-80 мкл, определяли с использованием субстратов Glp-Phe-Gln-pNA, Z-Phe-Arg-pNA, Z-Arg-Arg-pNA при рН 5,6 в присутствии 6 мМ цистеина и 1 мМ ЭДТА.
Таким образом, использование предложенных селективных субстратов позволяет однозначно выявить активность цистеиновых пептидаз в столь сложной природной смеси, как пищеварительный комплекс личинок Т. castaneum.
1Данные работы [Semashko Т.А., Vorotnikova Е.А., Sharikova V.F., Vinokurov K.S., Smirnova Y.A., Dunaevsky Y.E., Belozersky M.A., Oppert В., Elpidina E.N., Filippova I.Y. Selective chromogenic and fluorogenic peptide substrates for the assay of cysteine peptidases in complex mixtures, Anal. Biochem. 49 (2014) 179-187].
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ ОПРЕДЕЛЕНИЯ DPP3 И ТЕРАПЕВТИЧЕСКИЕ СПОСОБЫ | 2017 |
|
RU2771824C2 |
| СПЕЦИФИЧЕСКИЕ ПЕПТИДНЫЕ ИНГИБИТОРЫ ЦИСТЕИНОВЫХ КАТЕПСИНОВ | 2020 |
|
RU2752531C1 |
| ИНГИБИРУЮЩЕЕ ДИПЕПТИДИЛПЕПТИДАЗУ IV СРЕДСТВО И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ЕГО ОСНОВЕ | 2011 |
|
RU2485952C2 |
| DPP3 ДЛЯ ОСУЩЕСТВЛЕНИЯ, МОНИТОРИНГА И СТРАТИФИКАЦИИ ТЕРАПИИ NT-ADM АНТИТЕЛАМИ У ПАЦИЕНТОВ С ШОКОМ | 2021 |
|
RU2838370C1 |
| РУКОВОДСТВО ПО ТЕРАПИИ И/ИЛИ МОНИТОРИНГ ТЕРАПИИ ДЛЯ ЛЕЧЕНИЯ ШОКА | 2020 |
|
RU2835616C1 |
| ПРОИЗВОДНЫЕ ОПИОРФИННОГО ПЕПТИДА КАК ПОТЕНЦИАЛЬНЫЕ ИНГИБИТОРЫ ЭКТОПЕПТИДАЗ, УЧАСТВУЮЩИХ В ДЕГРАДАЦИИ ЭНКЕФАЛИНОВ | 2009 |
|
RU2542365C2 |
| ПРИМЕНЕНИЕ ПЕПТИДНОГО СОЕДИНЕНИЯ ДЛЯ ИНДУКЦИИ АПОПТОЗА В ОПУХОЛЕВОЙ КЛЕТКЕ | 2022 |
|
RU2796104C1 |
| ПЕПТИДНЫЕ АГОНИСТЫ GLP-2 | 2010 |
|
RU2551977C2 |
| АКТИВАТОР СИГНАЛЬНОГО ПУТИ ЖАСМОНОВОЙ КИСЛОТЫ | 2016 |
|
RU2728472C2 |
| ПОЛУЧЕННЫЕ С ПОМОЩЬЮ ГЕННОЙ ИНЖЕНЕРИИ АНТИТЕЛА С ЦИСТЕИНОВЫМИ ЗАМЕНАМИ И ИХ КОНЪЮГАТЫ | 2011 |
|
RU2755066C2 |
Изобретение относится к области биоорганической химии и относится к новым субстратам для определения ферментативной активности пептидаз. Предложен пептидный субстрат цистеиновых пептидаз семейства папаина, представляющий собой соединение общей формулы (I) A-Xaa-Gln-B, где А - N-защитная группа, представляющая собой Glp, Хаа - аминокислота Phe, В - маркерная флуорогенная группировка, которая представляет собой 7-амидо-4-метилкумарид (АМС) или 4-нитроанилид (pNA). Пептидный субстрат применяют для создания тест-систем детекции пептидаз, которую проводят в растворе, в геле после проведения электрофореза в нативных условиях, в системах in vivo. 2 н.п. ф-лы, 1 ил., 2 табл., 9 пр.
1. Пептидный субстрат цистеиновых пептидаз семейства папаина общей формулы (I):
A-Xaa-Gln-B (I), где
А - N-защитная группа, представляющая собой Glp,
Хаа - аминокислота Phe,
В - маркерная флуорогенная группировка, представляет собой 7-амидо-4-метилкумарид (АМС) или 4-нитроанилид (pNA).
2. Применение пептидного субстрата по п. 1 для создания тест-систем детекции пептидаз, которую проводят в растворе, в геле после проведения электрофореза в нативных условиях, в системах in vivo.
| Ramjee, Manoj K., et | |||
| al., "Substrate mapping and inhibitor profiling of falcipain-2, falcipain-3 and berghepain-2: Implications for peptidase anti-malarial drug discovery.", Biochemical Journal, 2006, 399(1), pages 47-57 | |||
| Nguyen, Kiet T., et | |||
| al., "Slow-binding inhibition of peptide deformylase by cyclic peptidomimetics as revealed by a new |

