РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Настоящая заявка испрашивает приоритет согласно предварительной заявке США № 62/796417, поданной 24 января 2019 г., предварительной заявке США № 62/800303, поданной 1 февраля 2019 г., предварительной заявке США № 62/796450, поданной 24 января 2019 г., предварительной заявке США № 62/800285, поданной 1 февраля 2019 г., предварительной заявке США № 62/814414, поданной 6 марта 2019 г., предварительной заявке США № 62/814424, поданной 6 марта 2019 г., и предварительной заявке США № 62/857542, поданной 5 июня 2019 г., содержание каждой из которых полностью включено в настоящий документ посредством ссылки.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Настоящая заявка включает перечень последовательностей, поданный в электронном виде в формате ASCII и включенный тем самым в настоящий документ в полном объеме посредством ссылки. Указанная копия в формате ASCII, созданная 24 января 2020 г., названа 131698-03320-Sequence_Listing-FINAL.txt и имеет размер 117124 байтов.
ОБЛАСТЬ ТЕХНИКИ
[0003] Варианты реализации настоящего изобретения относятся к области генной терапии, включая доставку экзогенных последовательностей ДНК в целевую клетку, ткань, орган или организм, а также к модификациям и способам для ингибирования иммунных ответов (например, врожденных иммунных ответов) на указанные последовательности.
УРОВЕНЬ ТЕХНИКИ
[0004] Целью генной терапии является улучшение клинических исходов у пациентов, страдающих либо генетическими мутациями, либо приобретенными заболеваниями, вызванными аберрацией в профиле экспрессии генов. Генная терапия включает лечение или предотвращение медицинских состояний, вызванных дефектными генами или аномальной регуляцией или экспрессией, например, недостаточной экспрессией или избыточной экспрессией, которые могут привести к нарушению, заболеванию, злокачественному новообразованию и т.д. Например, заболевание или нарушение, вызванное дефектным геном, можно лечить, предотвращать или облегчать путем доставки пациенту корректирующего генетического материала, или можно лечить, предотвращать или облегчать путем изменения или подавления у пациента дефектного гена, например, с помощью корректирующего генетического материала, приводящего к терапевтической экспрессии генетического материала в организме пациента.
[0005] Основой генной терапии является обеспечение транскрипционной кассеты с активным генным продуктом (иногда называемым трансгеном), например, который может приводить к положительному эффекту усиления функции, отрицательному эффекту потери функции или другому исходу. Такие исходы могут быть отнесены к экспрессии активирующего антитела или слитого белка либо ингибирующего (нейтрализующего) антитела или слитого белка. Генную терапию также можно применять для лечения заболевания или злокачественного новообразования, вызванного другими факторами. Моногенные нарушения у человека может лечить путем доставки нормального гена в целевые клетки и его экспрессии в них. Доставку и экспрессию корректирующего гена в клетках-мишенях пациента можно осуществлять с применением многочисленных способов, включая применение модифицированных вирусов и вирусных векторов для доставки генов. Среди многочисленных доступных векторов вирусного происхождения (например, рекомбинантный ретровирус, рекомбинантный лентивирус, рекомбинантный аденовирус и т.п.) рекомбинантный аденоассоциированный вирус (рААВ) приобретает все большую популярность в генной терапии в качестве многоцелевого вектора.
[0006] Аденоассоциированные вирусы (ААВ) принадлежат к семейству Parvoviridae и, более конкретно, составляют род депендопарвовирусов. Векторы, происходящие из ААВ (т.е. рекомбинантные ААВ (рААВ) или ААВ-векторы), являются привлекательными для доставки генетического материала, поскольку (i) они способны инфицировать (трансдуцировать) широкий спектр неделящихся и делящихся типов клеток, включая миоциты и нейроны; (ii) они лишены вирусных структурных генов, что уменьшает ответы клеток хозяина на вирусную инфекцию, например, опосредуемые интерфероном ответы; (iii) вирусы дикого типа считаются непатологичными у человека; (iv) в отличие от ААВ дикого типа, которые способны интегрироваться в геном клетки хозяина, дефектные по репликации ААВ-векторы лишены гена rep и обычно персистируют в виде эписом, что ограничивает риск инсерционного мутагенеза или генотоксичности; и (v) по сравнению с другими векторными системами ААВ-векторы обычно считаются относительно слабыми иммуногенами и, следовательно, не запускают значительного иммунного ответа (см. ii), обеспечивая тем самым устойчивость векторной ДНК и, потенциально, долгосрочную экспрессию терапевтических трансгенов.
[0007] Однако применение частиц ААВ в качестве вектора для доставки генов имеет несколько серьезных недостатков. Одним из основных недостатков, ассоциированных с рААВ, является ограниченная емкость вирусной упаковки, составляющая примерно 4,5 т.п.о. гетерологичной ДНК (Dong et al., 1996; Athanasopoulos et al., 2004; Lai et al., 2010), вследствие этого применение ААВ-векторов ограничено их способностью кодировать белки размером менее 150000 Да. Вторым недостатком является то, что в результате распространенности инфекции ААВ дикого типа среди населения кандидаты на генную терапию с применением рААВ должны проходить скрининг на наличие нейтрализующих антител, устраняющих вектор из организма пациента. Третий недостаток связан с иммуногенностью капсида, которая препятствует повторному введению пациентам, которые не были отстранены от начального лечения. Иммунная система пациента может отвечать на вектор, который фактически выступает в роли «бустерной» прививки, стимулирующей выработку иммунной системой высоких титров антител к ААВ, которые исключают возможность лечения в будущем. В некоторых недавних сообщениях выражены опасения, касающиеся иммуногенности в ситуациях, подразумевающих применение высоких доз. Другой весомый недостаток заключается в относительно медленном начале ААВ-опосредуемой экспрессии генов, учитывая, что одноцепочечная ДНК ААВ должна быть преобразована в двухцепочечную ДНК до экспрессии гетерологичного гена.
[0008] Кроме того, обычные вирионы ААВ с капсидами получают путем введения плазмиды или плазмид, содержащих геном, гены rep и гены cap ААВ (Grimm et al., 1998). Однако было обнаружено, что такие вирусные векторы на основе заключенного в капсид ААВ неэффективно трансдуцируют определенные типы клеток и тканей, и что капсиды также индуцируют иммунный ответ. Соответственно, применение векторов на основе аденоассоциированного вируса (ААВ) для генной терапии ограничено из-за однократного введения пациентам (вследствие иммунного ответа пациента), ограниченного диапазона трансгенного генетического материала, подходящего для доставки в ААВ-векторах ввиду минимальной емкости вирусной упаковки (около 4,5 т.п.о.), и медленной ААВ-опосредуемой экспрессии генов.
[0009] Более того, иммунные системы млекопитающих включают ряд механизмов детектирования и устранения вторгающихся патогенов и разных видов аберрантной клеточной активности и процессов, которые могут быть вызваны при введении субъекту вирусного вектора или нуклеиновой кислоты. Например, рецепторы распознавания структур (PRR) представляют собой класс молекул, которые эволюционно развились как сенсоры для детектирования консервативных молекул, ассоциированных с патогенами, таких как чужеродные нуклеиновые кислоты, например, вирусная ДНК и вирусная РНК, и запуска врожденного иммунного ответа. Toll-подобные рецепторы (TLR) представляют собой группу PRR, которые детектируют нуклеиновые кислоты, заключенные в эндосому, и включают TLR9 (детектирует дцДНК, преимущественно неметилированные повторы CpG), TLR3 (детектирует дцРНК) и TLR7 (детектирует оцРНК). Вторая система PRR расположена в цитозоле и служит для детектирования чужеродной нуклеиновой кислоты, в частности, двухцепочечной РНК, внутри инфицированных клеток.1 Эти PRR, называемые «RIG-I-подобными рецепторами» или RLR, включают RIG-I и MDA5. Упомянутые PRR представляют собой геликазы, которые детектируют структурные особенности РНК, такие как 5´-трифосфаты и дифосфаты, промежуточные продукты репликации РНК и/или продукты транскрипции, и инициируют активацию ответа интерферона типа I.1,2 Третий класс PRR запускается цитозольной ДНК, при этом основным внутриклеточным сенсором ДНК является cGAS (цикло-ГМФ-АМФ-синтаза), которая связывается с ДНК и активирует связанный с эндоплазматическим ретикулумом (ЭР) стимулятор генов интерферона (STING), что приводит к активации ответа интерферона типа I и, в некоторых случаях, активации1,4,5 других предложенных сенсоров цитозольной ДНК, включая отсутствующий при меланоме белок (AIM2), индуцируемый IFN-γ белок 16 (IFI16), индуцируемый интерфероном белок X (IFIX), LRRFIP1, DHX9, DHX36, DDX41, Ku70, ДНК-PKc, комплекс MRN (включая MRE11, Rad50 и Nbs1)2,7 и РНК-полимеразу III10. AIM2, IFI16 и IFIX представляют собой белки, содержащие пириновый домен и домен HIN200 (PYHIN).2,6 Кроме того, было показано, что неспаренные нуклеотиды ДНК, фланкирующие короткие участки ДНК со спаренными основаниями, как в структурах «стебель-петля» одноцепочечной ДНК (оцДНК), происходящей из вируса иммунодефицита человека типа 1 (ВИЧ-1), активировали ДНК-сенсор cGAS, индуцирующий интерферон I типа, зависимым от последовательности образом.8,9 Структуры ДНК, содержащие неспаренные гуанозины, фланкирующие короткую (от 12 до 20 п.о.) дцДНК (Y-форма ДНК), обладали сильным стимулирующим действием и специфично усиливали ферментативную активность cGAS.8,9
[0010] Совсем недавно были идентифицированы другие внутриклеточные микробные сенсоры, включая NOD-подобные рецепторы (NLR). Некоторые из NLR также воспринимают немикробные сигналы опасности и образуют большие цитоплазматические белковые комплексы, называемые инфламмасомами, которые являются основным регулятором врожденного иммунитета и воспаления (Martinon et al., Annu. Rev. Immunol. 2009 27: 229-65).
[0011] Инфламмасома состоит из рецепторов семейства NLR или AIM2 и прокаспазы-1. Ассоциированный с апоптозом пятнышкоподобный белок, содержащий домен привлечения каспазы (ASC), представляет собой адаптерный белок и связывает члена семейства NLR с прокаспазой-1. Члены семейства NLR собирают комплекс инфламмасомы с ASC, который, в свою очередь, привлекает и активирует каспазу-1. Несколько членов семейства белков NLR участвуют в образовании различных инфламмасом, включая содержащий пириновый домен член 3 семейства NLR (NLRP3; также известный как циропирин или NALP3), содержащий домен CARD член 4 семейства NLR (NLRC4; также известный как IPAF) и NLRP1. Различные инфламмасомы активируются разными стимулами. Например, NLRP1 активируется летальным токсином, продуцируемым Bacillus anthracis, в то время как NLRC4 отвечает на цитозольный флагеллин в клетках, инфицированных Salmonella, Legionella и Pseudomonas spp. Инфламмасома NLRP3 активируется большим количеством различных стимулов, включая микробные продукты и эндогенные сигналы, такие как кристалл урата, оксид кремния, амилоидные фибриллы и АТФ.
[0012] Сенсорный компонент NOD-подобного рецептора (NLR) (например, криопирин (NLRP3 или NALP3)) распознает сигналы опасности, такие как молекулы молекулярной структуры, ассоциированной с повреждением (DAMP), высвобождаемые во время повреждения ткани или стресса (например, внеклеточный АТФ, кристалл урата, β-амилоид, фрагменты клеток) и молекулярные структуры, ассоциированные с патогенами (PAMP). Инфламмасома собирается в ответ на такую инфекцию патогеном или сигналы «опасности», это требует взаимодействия пириновых доменов криопирина и адаптерного компонента ASC, что приводит к привлечению и активации каспазы-1 (из прокаспазы-1), а затем к созреванию и высвобождению нескольких провоспалительных цитокинов, включая интерлейкин-1β (IL-1β), IL-18 и IL-33.
[0013] Помимо NLR, инфламмасомы могут активироваться членами семейства AIM2. AIM2 характеризуется наличием пиринового домена и ДНК-связывающего домена HIN и активирует каспазу-1 путем детектирования цитозольной ДНК (Fernandes-Alnemri T, et al. 2009. Nature 458:509-513). Сборка инфламмасомы требует предшествующего праймирующего сигнала за счет TLR, который необходим для положительной регуляции экспрессии инфламмасомных рецепторов и субстрата про-IL-1β, прежде чем второй сигнал сможет инициировать образование инфламмасомного комплекса (Bauernfeind FG, et al., 2009, J. Immunol. 183:787-791).
[0014] Несмотря на концептуальную элегантность, перспектива применения молекул нуклеиновых кислот для генной терапии с целью лечения заболеваний человека остается неопределенной. Основная причина этой неопределенности заключается в очевидных нежелательных явлениях, связанных с врожденным иммунным ответом хозяина на терапевтические средства на основе нуклеиновых кислот и, таким образом, путь, с помощью которого эти материалы модулируют экспрессию их предусмотренных мишеней применительно к иммунному ответу. Существующий на сегодняшний день уровень техники, относящийся к созданию, функционированию, поведению и оптимизации молекул нуклеиновых кислот, которые могут быть использованы для клинических приложений, в основном сосредоточен на: (1) антисмысловых олигонуклеотидах и дуплексных РНК, которые непосредственно регулируют трансляцию и экспрессию гена; (2) РНК, подавляющих транскрипцию гена, которые приводят к долгосрочным эпигенетическим модификациям; (3) антисмысловых олигонуклеотидах, которые взаимодействуют со структурами сплайсинга генов и изменяют их; (4) создании синтетических или вирусных векторов, имитирующих физиологические функциональные возможности природного генома ААВ или лентивирусного генома; и (5) доставке терапевтических олигонуклеотидов in vivo. Однако, несмотря на успехи, достигнутые в разработке терапевтических средств на основе нуклеиновых кислот, которые очевидны в недавних клинических достижениях, область генной терапии все еще сильно ограничена непредусмотренными нежелательными явлениями у реципиентов, запускаемыми самими терапевтическими нуклеиновыми кислотами.
[0015] Соответственно, в данной области существует потребность в новой технологии, которая ингибирует (например, уменьшает, облегчает, смягчает, предотвращает) иммунный ответ на введение субъекту векторов или нуклеиновой кислоты, что обеспечивает экспрессию терапевтического белка в клетке, ткани или в организме субъекта для лечения широкого спектра заболеваний.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0016] Согласно настоящему раскрытию предложены способы и фармацевтические композиции для ингибирования (т.е. уменьшения или подавления) иммунного ответа у субъекта, страдающего генетическим нарушением и получающего генную терапию или терапию нуклеиновой кислотой («терапевтические средства на основе нуклеиновых кислот» или «терапевтическую нуклеиновую кислоту» (ТНК)). В настоящем документе предложены невирусные бескапсидные ДНК-векторы с ковалентно замкнутыми концами (зкДНК-векторы (ceDNA vectors)) и ингибиторы для ингибирования иммунного ответа (например, врожденного иммунного ответа). В соответствии с некоторыми вариантами реализации фармацевтические композиции и составы могут включать один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа), таких как рапамицин и аналоги рапамицина, антагонисты TLR (например, антагонисты TLR9), антагонисты cGAS и антагонисты инфламмасомы (например, любой один или более из: ингибитора пути инфламмасомы NLRP3 или ингибитора пути инфламмасомы AIM2, или ингибитора каспазы 1, или любой их комбинации).
[0017] В соответствии с некоторыми аспектами настоящего раскрытия предложены композиции и способы для ингибирования (т.е. уменьшения или подавления) иммунного ответа (например, врожденного иммунного ответа) с применением невирусных бескапсидных ДНК-векторов с ковалентно замкнутыми концами (зкДНК-векторов) для экспрессии ингибитора врожденного иммунного ответа с бескапсидного (например, невирусного) ДНК-вектора с ковалентно замкнутыми концами (называемого в настоящем документе «ДНК-вектором с замкнутыми концами» или «зкДНК-вектором»), причем указанный зкДНК-вектор содержит последовательность нуклеиновой кислоты или ее варианты, оптимизированные по кодонам, ингибитора иммунного ответа (например, врожденного иммунного ответа).
[0018] В соответствии с некоторыми аспектами настоящего раскрытия предложены композиции и способы для ингибирования (т.е. уменьшения или подавления) иммунного ответа (например, врожденного иммунного ответа) с применением невирусных бескапсидных ДНК-векторов с ковалентно замкнутыми концами (зкДНК-векторов) для экспрессии рапамицина и аналогов рапамицина с бескапсидного (например, невирусного) ДНК-вектора с ковалентно замкнутыми концами (называемого в настоящем документе «ДНК-вектором с замкнутыми концами» или «зкДНК-вектором»), причем указанный зкДНК-вектор содержит последовательность нуклеиновой кислоты или ее варианты, оптимизированные по кодонам, рапамицина и аналогов рапамицина. Соответственно, эти зкДНК-векторы можно применять для получения рапамицина и аналогов рапамицина для ингибирования иммунной системы (например, врожденной иммунной системы).
[0019] В соответствии с некоторыми аспектами настоящего раскрытия предложены композиции и способы для ингибирования (т.е. уменьшения или подавления) иммунного ответа (например, врожденного иммунного ответа) с применением невирусных бескапсидных ДНК-векторов с ковалентно замкнутыми концами (зкДНК-векторов) для экспрессии антагониста TLR с бескапсидного (например, невирусного) ДНК-вектора с ковалентно замкнутыми концами (называемого в настоящем документе «ДНК-вектором с замкнутыми концами» или «зкДНК-вектором»), причем указанный зкДНК-вектор содержит последовательность нуклеиновой кислоты или ее варианты, оптимизированные по кодонам, антагониста TLR. Соответственно, эти зкДНК-векторы можно применять для получения антагониста TLR для ингибирования иммунной системы (например, врожденной иммунной системы).
[0020] В соответствии с некоторыми аспектами настоящего раскрытия предложены композиции и способы для ингибирования (т.е. уменьшения или подавления) иммунного ответа (например, врожденного иммунного ответа) с применением невирусных бескапсидных ДНК-векторов с ковалентно замкнутыми концами (зкДНК-векторов) для экспрессии антагониста cGAS с бескапсидного (например, невирусного) ДНК-вектора с ковалентно замкнутыми концами (называемого в настоящем документе «ДНК-вектором с замкнутыми концами» или «зкДНК-вектором»), причем указанный зкДНК-вектор содержит последовательность нуклеиновой кислоты или ее варианты, оптимизированные по кодонам, антагониста cGAS. Соответственно, эти зкДНК-векторы можно применять для получения антагониста cGAS для ингибирования иммунной системы (например, врожденной иммунной системы).
[0021] В соответствии с некоторыми аспектами настоящего раскрытия предложены композиции и способы для ингибирования (т.е. уменьшения или подавления) иммунного ответа (например, врожденного иммунного ответа) с применением невирусных бескапсидных ДНК-векторов с ковалентно замкнутыми концами (зкДНК-векторов) для экспрессии ингибитора пути инфламмасомы NLRP3 или ингибитора пути инфламмасомы AIM2, или ингибитора каспазы 1, или любой их комбинации с бескапсидного (например, невирусного) ДНК-вектора с ковалентно замкнутыми концами (называемого в настоящем документе «ДНК-вектором с замкнутыми концами» или «зкДНК-вектором»), причем указанный зкДНК-вектор содержит последовательность нуклеиновой кислоты или ее варианты, оптимизированные по кодонам, ингибитора пути инфламмасомы NLRP3 или ингибитора пути инфламмасомы AIM2, или ингибитора каспазы 1, или любой их комбинации. Соответственно, эти зкДНК-векторы можно применять для получения ингибитора пути инфламмасомы NLRP3 или ингибитора пути инфламмасомы AIM2, или ингибитора каспазы 1, или любой их комбинации, для ингибирования иммунной системы (например, врожденной иммунной системы).
[0022] В соответствии с некоторыми вариантами реализации фармацевтические композиции и составы могут включать один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа), описанных в настоящем документе, в сочетании с различными типами терапевтических нуклеиновых кислот (ТНК) и носителей (например, липидная наночастица). В соответствии с некоторыми вариантами реализации композиция дополнительно содержит вспомогательное вещество или носитель. В соответствии с некоторыми вариантами реализации фармацевтическая композиция содержит липидную наночастицу (ЛНЧ). Согласно одному варианту реализации ЛНЧ содержит катионный липид. В соответствии с некоторыми вариантами реализации ЛНЧ содержит полиэтиленгликоль (ПЭГ). В соответствии с некоторыми вариантами реализации ЛНЧ содержит холестерин.
[0023] Способы, описанные в настоящем документе, обычно включают применение одного или более ингибиторов иммунного ответа (например, врожденного иммунного ответа) (например, рапамицина и его аналогов, антагонистов TLR, антагонистов cGAS) для предотвращения, уменьшения, ослабления или даже устранения иммунных ответов, ассоциированных с введением трансгена (например, терапевтической нуклеиновой кислоты (ТНК)). В настоящем документе описаны способы, включающие введение указанных ингибиторов.
[0024] Согласно одному варианту реализации терапевтическая нуклеиновая кислота представляет собой молекулу РНК или ее производное. Согласно одному варианту реализации молекула РНК представляет собой антисмысловой олигонуклеотид. Согласно одному варианту реализации антисмысловой олигонуклеотид представляет собой антисмысловую РНК. Согласно одному варианту реализации РНК представляет собой РНК-интерференцию (РНКи).
[0025] Согласно одному варианту реализации терапевтическая нуклеиновая кислота представляет собой молекулу мРНК.
[0026] Согласно одному варианту реализации терапевтическая нуклеиновая кислота представляет собой молекулу ДНК или ее производное.
[0027] Согласно одному варианту реализации терапевтическая нуклеиновая кислота представляет собой антисмысловой олигонуклеотид ДНК. Согласно одному варианту реализации антисмысловой олигонуклеотид ДНК представляет собой нуклеиновую кислоту на основе морфолина. Согласно одному варианту реализации нуклеиновая кислота на основе морфолина представляет собой фосфодиамидат-морфолиновый олигомер (PMO).
[0028] Согласно одному варианту реализации терапевтическая нуклеиновая кислота представляет собой ДНК с замкнутыми концами (зкДНК). Согласно одному варианту реализации зкДНК содержит экспрессионную кассету, содержащую последовательность промотора и трансген. Согласно одному варианту реализации зкДНК содержит экспрессионную кассету, содержащую последовательность полиаденилирования. Согласно одному варианту реализации зкДНК содержит по меньшей мере один инвертированный концевой повтор (ITR), фланкирующий либо 5´-, либо 3´-конец экспрессионной кассеты. Согласно одному варианту реализации экспрессионная кассета фланкирована двумя ITR, причем указанные два ITR содержат один 5´-ITR и один 3´-ITR. Согласно одному варианту реализации экспрессионная кассета соединена с ITR на 3´-конце (3´-ITR). Согласно одному варианту реализации экспрессионная кассета соединена с ITR на 5´-конце (5´-ITR). Согласно одному варианту реализации зкДНК дополнительно содержит спейсерную последовательность между 5´-ITR и экспрессионной кассетой.
[0029] Согласно одному варианту реализации зкДНК дополнительно содержит спейсерную последовательность между 3´-ITR и экспрессионной кассетой. Согласно одному варианту реализации спейсерная последовательность представляет собой по меньшей мере 5 пар оснований в длину. Согласно одному варианту реализации спейсерная последовательность представляет собой от 5 до 200 пар оснований в длину. Согласно одному варианту реализации спейсерная последовательность представляет собой от 5 до 500 пар оснований в длину.
[0030] Согласно одному варианту реализации ITR представляет собой ITR, происходящий из серотипа ААВ. Согласно одному варианту реализации ААВ выбран из группы, состоящей из ААВ1, ААВ2, ААВ3, ААВ4, ААВ5, ААВ6, ААВ7, ААВ8, ААВ9, ААВ10, ААВ11 и ААВ12. Согласно одному варианту реализации ITR происходит из ITR гусиного вируса. Согласно одному варианту реализации ITR происходит из ITR вируса B19. Согласно одному варианту реализации ITR представляет собой ITR дикого типа из парвовируса. Согласно одному варианту реализации ITR представляет собой мутантный ITR. Согласно одному варианту реализации зкДНК содержит два мутантных ITR как на 5´-конце, так и на 3´-конце экспрессионной кассеты.
[0031] Согласно одному варианту реализации зкДНК имеет одноцепочечный разрыв или пропуск.
[0032] Согласно одному варианту реализации зкДНК получают синтетически в бесклеточной среде.
[0033] Согласно одному варианту реализации зкДНК получают в клетке. Согласно одному варианту реализации зкДНК получают в клетках насекомых. Согласно одному варианту реализации клетка насекомого представляет собой Sf9. Согласно одному варианту реализации зкДНК получают в клетке млекопитающего. Согласно одному варианту реализации клетка млекопитающего представляет собой линию клеток человека.
[0034] Согласно одному варианту реализации терапевтическая нуклеиновая кислота представляет собой ДНК с замкнутыми концами, содержащую по меньшей мере одну последовательность-мишень протеломеразы на ее 5´- и 3´-концах экспрессионной кассеты.
[0035] Согласно одному варианту реализации терапевтическая нуклеиновая кислота представляет собой гантелеобразную линейную дуплексную ДНК с замкнутыми концами, содержащую две шпилечные структуры ITR на 5´- и 3´-концах экспрессионной кассеты.
[0036] Согласно одному варианту реализации терапевтическая нуклеиновая кислота представляет собой миникольцо на основе ДНК или MIDGE.
[0037] Согласно одному варианту реализации терапевтическая нуклеиновая кислота представляет собой линейный ДНК-вектор с ковалентно замкнутыми концами. Согласно одному варианту реализации линейный ДНК-вектор с ковалентно замкнутыми концами представляет собой ДНК с минимальной цепью («ministring DNA»).
[0038] Согласно одному варианту реализации терапевтическая нуклеиновая кислота представляет собой doggybone ДНК (dbDNA™).
[0039] Согласно одному варианту реализации терапевтическая нуклеиновая кислота представляет собой миниген.
[0040] Согласно одному варианту реализации терапевтическая нуклеиновая кислота представляет собой плазмиду.
[0041] Соответственно, согласно некоторым аспектам настоящего изобретения предложены способы ингибирования или подавления иммунных ответов при экспрессии трансгена в клетке, включающие: совместное введение в клетку (1) композиции, содержащей невирусный бескапсидный ДНК-вектор с ковалентно замкнутыми концами (зкДНК-вектор), и (2) ингибитора иммунного ответа (например, врожденного иммунного ответа), описанных в настоящем документе. ЗкДНК-вектор содержит гетерологичную последовательность нуклеиновой кислоты, кодирующую трансген, функционально расположенную между двумя разными последовательностями инвертированных концевых повторов (ITR) ААВ, причем один из указанных ITR содержит функциональный сайт концевого разрешения ААВ и сайт связывания Rep, один из указанных ITR содержит делецию, вставку или замену относительно другого ITR так, что зкДНК-вектор, при расщеплении рестрикционным ферментом, имеющим один сайт распознавания на зкДНК-векторе, имеет характерные полосы линейной и непрерывной ДНК, по сравнению с линейными и прерывистыми контрольными ДНК, при анализе на неденатурирующем геле. Как показано в настоящем документе, согласно некоторым вариантам реализации, ингибитор иммунного ответа (например, врожденного иммунного ответа) совместно вводят с применением синтетического наноносителя, описанного в WO 2016/073799, содержание которого полностью включено в настоящий документ посредством ссылки. Согласно некоторым вариантам реализации зкДНК-вектор также присутствует в наноносителе. В соответствии с некоторыми вариантами реализации один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа) выбраны из рапамицина и аналогов рапамицина, антагонистов TLR (например, антагонистов TLR9), антагонистов cGAS и антагонистов инфламмасомы (например, любого одного или более из: ингибитора пути инфламмасомы NLRP3 или ингибитора пути инфламмасомы AIM2, или ингибитора каспазы 1, или любой их комбинации). В соответствии с некоторыми вариантами реализации олигонуклеотид, ингибирующий TLR9, присутствует по меньшей мере на одном из ITR. В соответствии с некоторыми вариантами реализации ингибитор cGAS кодируется зкДНК и функционально связан с промотором, таким как индуцируемый промотор. Согласно другим вариантам реализации ингибитор cGAS не кодируется зкДНК.
[0042] Кроме того, согласно одному аспекту настоящего изобретения предложена композиция, содержащая (i) невирусный бескапсидный ДНК-вектор с ковалентно замкнутыми концами (зкДНК-вектор), причем указанный зкДНК-вектор содержит гетерологичную последовательность нуклеиновой кислоты, кодирующую трансген, функционально расположенную между двумя разными последовательностями инвертированных концевых повторов (ITR) ААВ, причем один из указанных ITR содержит функциональный сайт концевого разрешения ААВ и сайт связывания Rep, один из указанных ITR содержит делецию, вставку или замену относительно другого ITR, при этом указанный зкДНК-вектор, при расщеплении рестрикционным ферментом, имеющим один сайт распознавания на зкДНК-векторе, имеет характерные полосы линейной и непрерывной ДНК, по сравнению с линейными и прерывистыми контрольными ДНК, при анализе на неденатурирующем геле, и (ii) ингибитор иммунного ответа (например, врожденного иммунного ответа). Как показано в настоящем документе, согласно некоторым вариантам реализации, компоненты композиции изготовлены в отдельных синтетических наноносителях. Согласно одному варианту реализации компоненты композиции изготовлены в одном и том же синтетическом наноносителе. В соответствии с некоторыми вариантами реализации один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа) выбраны из рапамицина и аналогов рапамицина, антагонистов TLR (например, антагонистов TLR9), антагонистов cGAS и антагонистов инфламмасомы (например, любого одного или более из: ингибитора пути инфламмасомы NLRP3 или ингибитора пути инфламмасомы AIM2, или ингибитора каспазы 1, или любой их комбинации).
[0043] Невирусные бескапсидные ДНК-векторы, описанные в настоящем документе, могут быть получены в восприимчивых клетках-хозяевах с экспрессионной конструкции (например, плазмиды, бакмиды, бакуловируса или интегрированной линии клеток), например, см. Примеры, раскрытые в международной заявке на патент PCT/US18/49996, поданной 7 сентября 2018 г., или с использованием синтетического способа получения, например, см. Примеры, раскрытые в международной заявке на патент PCT/US19/14122, поданной 6 декабря 2018 г., каждая из которых полностью включена в настоящий документ посредством ссылки. Согласно некоторым вариантам реализации зкДНК-векторы, которые можно применять в способах и композициях, раскрытых в настоящем документе, содержат гетерологичную нуклеиновую кислоту, например, трансген, расположенную между двумя последовательностями инвертированных концевых повторов (ITR). Согласно некоторым вариантам реализации по меньшей мере один из ITR модифицирован путем делеции, вставки и/или замены по сравнению с последовательностью ITR дикого типа (например, ITR ААВ); и по меньшей мере один из ITR содержит функциональный сайт концевого разрешения (TRS) и сайт связывания Rep.
[0044] В соответствии с другим аспектом настоящего раскрытия предложен способ лечения генетического нарушения у субъекта, причем указанный способ включает введение указанному субъекту эффективного количества фармацевтической композиции, раскрытой в настоящем документе.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0045] ФИГ. 1 представляет собой схему, иллюстрирующую один вариант реализации подготовительного способа получения бакуло-инфицированных клеток насекомых (BIIC), которые можно применять для получения зкДНК-вектора в способе, описанном на схеме на ФИГ. 2. i) Две популяции наивных клеток насекомых трансфицируют либо плазмидой, кодирующей белок Rep, либо плазмидой, продуцирующей ДНК-вектор; ii) вирусный супернатант собирают и используют для инфицирования двух новых наивных популяций клеток насекомых с получением BIIC-1 конструкции ДНК-вектора и BIIC-2 (REP). BIIC относится к клеткам насекомых, инфицированным бакуловирусом. Необязательно этап ii) можно повторять один или более раз для получения рекомбинантного бакуловируса в бóльших количествах.
[0046] ФИГ. 2 представляет собой схему, иллюстрирующую один вариант реализации для получения зкДНК-вектора, описанного в настоящем документе.
[0047] ФИГ. 3 представляет собой схему, иллюстрирующую один вариант реализации для характеристики ДНК-вектора (последующий способ), описанного в настоящем документе.
[0048] ФИГ. 4А-ФИГ. 4D представляют собой схематические диаграммы, иллюстрирующие примерные плазмиды и компоненты плазмиды, которые можно применять для получения зкДНК-вектора, раскрытого в настоящем документе. ФИГ. 4A показывает примерную плазмиду Rep, и ФИГ. 4B показывает примерную плазмиду ТТХ-вектора, которая содержит матрицу зкДНК-вектора. ФИГ. 4C и ФИГ. 4D представляют собой схемы примерных функциональных компонентов матрицы ДНК-вектора, которые можно применять для получения зкДНК-векторов, предложенных в настоящем документе. Трансген, который также упоминается как представляющая интерес нуклеиновая кислота (например, репортерная нуклеиновая кислота, такая как люцифераза, или, например, терапевтическая нуклеиновая кислота), расположен между двумя разными ITR. Модифицированный ITR может быть расположен в матрице либо с левой стороны (ФИГ. 4C), либо с правой стороны (ФИГ. 4D). Кроме того, представляющая интерес нуклеиновая кислота может быть функционально связана с промотором, энхансером и элементами терминации. Согласно альтернативным вариантам реализации ITR слева (5´-ITR) или справа (3´-ITR) может быть любого типа. В иллюстративных целях ITR в конструкциях зкДНК на ФИГ. 4C и ФИГ. 4D и в Примерах в настоящем документе показывают модифицированный ITR (ΔITR) и ITR дикого типа (WT-ITR) и представляют собой пример пары асимметричных ITR. Однако в настоящем документе предусмотрены зкДНК-векторы, которые содержат гетерологичную последовательность нуклеиновой кислоты (трансген), расположенную между любыми двумя последовательностями инвертированных концевых повторов (ITR), причем указанные последовательности ITR могут представлять собой пару асимметричных ITR или пару симметричных или по существу симметричных ITR, в соответствии с определением этих терминов в настоящем документе. ЗкДНК-вектор, содержащий NLP, раскрытый в настоящем документе, может содержать последовательности ITR, которые выбраны из любого из: (i) по меньшей мере одного WT-ITR и по меньшей мере одного модифицированного инвертированного концевого повтора (mod-ITR) ААВ (например, асимметричные модифицированные ITR); (ii) двух модифицированных ITR, причем указанная пара mod-ITR имеет различную трехмерную пространственную организацию друг относительно друга (например, асимметричные модифицированные ITR), или (iii) пары симметричных или по существу симметричных WT-WT-ITR, в которой каждый WT-ITR имеет одинаковую трехмерную пространственную организацию, или (iv) пары симметричных или по существу симметричных модифицированных ITR, в которой каждый mod-ITR имеет одинаковую трехмерную пространственную организацию, при этом указанные способы согласно настоящему раскрытию могут дополнительно включать систему доставки, такую как, но не ограничиваясь этим, система доставки на основе липосомных наночастиц.
[0049] ФИГ. 5A и ФИГ. 5B представляют собой чертежи, которые иллюстрируют один вариант реализации для идентификации наличия ДНК-векторов, описанных в настоящем документе. ФИГ. 5А иллюстрирует ДНК, имеющую прерывистую структуру (незамкнутая ДНК, например, контрольная ДНК-кассета, выделенная из матрицы ТТХ-вектора, имеющая открытые концы), и примерные характерные полосы, полученные при разрезании рестрикционной эндонуклеазой, имеющей один сайт распознавания на прерывистой ДНК, например, обнаружение двух фрагментов ДНК разного ожидаемого размера (например, 1 тыс.п.о. и 2 тыс.п.о.) в денатурирующих условиях. ФИГ. 5B иллюстрирует ДНК, имеющую линейную и непрерывную структуру с замкнутыми концами, и примерные характерные полосы, полученные при разрезании рестрикционной эндонуклеазой, имеющей один сайт распознавания на линейной дуплексной непрерывной ДНК, например, обнаружение двух фрагментов ДНК разных размеров (например, 2 тыс.п.о. и 4 тыс.п.о.) в денатурирующих условиях, что в 2 раза больше, чем можно было бы ожидать, если бы ДНК была прерывистой. Несмотря на то, что ДНК денатурирована, комплементарные цепи связаны ковалентно, и полученные денатурированные продукты представляют собой одноцепочечную ДНК, длина которой в два раза превышает длину соответствующих прерывистых продуктов.
[0050] ФИГ. 6 представляет собой примерный неденатурирующий гель, показывающий наличие высокостабильных ДНК-векторов и характерных полос, подтверждающих наличие высокостабильной ДНК с замкнутыми концами (зкДНК-вектора).
[0051] ФИГ. 7 представляет собой гель и стандартную кривую для количественного определения для оценки материала ДНК, полученного с помощью способов, раскрытых в настоящем документе.
[0052] ФИГ. 8 представляет собой анализ методом Вестерн-блоттинга белка FIX, экспрессированного из клеток HEK293, содержащих различные конструкции, и визуализированного с использованием антитела к фактору IX.
[0053] ФИГ. 9 представляет графическое изображение результатов Примера 24. Образцы, введенные гидродинамически, показывают значительное увеличение полного потока (например, экспрессии люциферазы) по сравнению с образцами, введенными негидродинамически, в течение трехдневного периода исследования.
[0054] ФИГ. 10A и 10B представляют данные экспериментов с культивированными клетками THP-1, описанных в Примерах, в которых оценивали ответ интерферона в клетках, обработанных зкДНК-вектором и иммунными ингибиторами. ФИГ. 10А показывает активацию пути интерферона в ответ на зкДНК в клетках THP-1 с интактными путями cGAS/STING и TLR9, но отсутствие активации в этих же клетках, в которых нарушен любой путь. Следует отметить, что включение либо ингибитора A151, либо BX795 аналогичным образом уменьшает активацию пути интерферона. ФИГ. 10В представляет собой сходный эксперимент, показывающий дозозависимое ингибирование индукции интерферона с помощью A151 и AS1411. В каждой группе столбиков доза 2,5 мкМ находится слева, доза 1,25 мкМ находится в середине и доза 0,625 мкМ находится справа.
[0055] ФИГ. 11A и 11B представляет графики данных, полученных в Примере 26. ФИГ. 11А показывает уменьшение индукции NF-κB при введении зкДНК, когда присутствующие в зкДНК CpG метилированы перед введением в клетки. ФИГ. 11B также показывает, что включение иммунного ингибитора A151 уменьшало зкДНК-стимулированную индукцию NF-κB в той же степени, что и метилирование CpG в этом анализе.
[0056] ФИГ. 12A-ФИГ. 12C представляет результаты экспериментов, описанных в Примере 26. ФИГ. 12A и ФИГ. 12B представляют собой графики данных каждого из анализов индукции цитокинов, выполненных на образцах крови, взятых у мышей, обработанных зкДНК-вектором, или мышей, обработанных контрольными ЛНЧ-поли(C), при этом конкретный исследуемый цитокин отражен в верхней части каждого графика. ФИГ. 12С представляет данные анализа экспрессии люциферазы, контролируемой зкДНК, у обработанных мышей, при этом показан полный поток в каждой группе мышей на протяжении исследования. Высокие уровни неметилированного CpG коррелировали с более низким полным потоком, наблюдаемым у мышей.
[0057] ФИГ. 13 представляет данные по полному потоку, полученные в экспериментах, описанных в Примере 27, на мышах на 8-й день неонатального развития. В ходе исследования зкДНК с высоким содержанием CpG вызывала снижение потока в ходе анализа, в то время как зкДНК с уменьшенным содержанием или без неметилированного CpG поддерживала экспрессию люциферазы. Однократное повторное введение дозы умеренно увеличивало наблюдаемые уровни экспрессии в образцах с минимальным содержанием CpG или с отсутствующим CpG, но это устойчивое увеличение после повторного введения дозы не наблюдалось в образцах из группы с высоким содержанием CpG.
[0058] ФИГ. 14A-ФИГ. 14C предоставляет результаты экспериментов, описанных в Примере 28. ФИГ. 14A и ФИГ. 14B представляют собой графики данных каждого из анализов индукции цитокинов, выполненных на образцах крови, взятых у обработанных зкДНК-вектором мышей из генетической линии с мутантным STING, или образцах после обработки контрольным поли(C), при этом конкретный исследуемый цитокин отражен в верхней части каждого графика. За исключением IL-18, в случае зкДНК с низким содержанием CpG и без метилированного CpG наблюдали значительно меньшую индукцию цитокинов. ФИГ. 14С представляет данные анализа экспрессии люциферазы, контролируемой зкДНК, у обработанных мышей, мутантных по STING, при этом показан полный поток в каждой группе мышей на протяжении исследования. Полученные результаты вновь показали корреляцию между высокими уровнями неметилированного CpG в зкДНК и наблюдаемым более низким полным потоком.
[0059] ФИГ. 15А и ФИГ. 15В показывают экспрессию трансгенов Padua FIX и FIX с высокостабильных ДНК-векторов, раскрытых в настоящем документе. Количественный анализ уровней белка FIX, экспрессированного с плазмид или векторов, также оценивали с использованием набора для ИФА VisuLize Factor IX (Affinity Biologicals, #FIX-AG), следуя протоколам, предоставленным поставщиком.
[0060] ФИГ. 16A и 16В показывают результаты исследования устойчивости зкДНК и повторного введения дозы у мышей Rag2, описанного в Примере 10. ФИГ. 16А показывает график зависимости полного потока от времени, наблюдаемой у получавших ЛНЧ-зкДНК-Luc мышей c57bl/6 дикого типа или мышей Rag2. ФИГ. 16B представляет график, показывающий влияние повторной дозы на уровни экспрессии трансгена люциферазы у мышей Rag2, при этом после введения повторной дозы наблюдали повышенную стабильную экспрессию (стрелка указывает время введения повторной дозы).
[0061] ФИГ. 17 представляет данные исследования экспрессии люциферазы с зкДНК у обработанных мышей, описанного в Примере 29, показывающие полный поток в каждой группе мышей на протяжении исследования. Высокие уровни неметилированного CpG коррелировали с более низким полным потоком, наблюдаемым у мышей с течением времени, в то время как использование специфического для печени промотора коррелировало с продолжительной стабильной экспрессией трансгена с зкДНК-вектора на протяжении по меньшей мере 77 дней.
[0062] ФИГ. 18A-18H показывают уровни цитокинов после введения зкДНК-вектора с фармакологическим истощением макрофагов с помощью ингибитора NLRP3 (MCC950) или ингибитора каспазы 1 (VX765). ФИГ. 18A показывает уровни IFN-α, ФИГ. 18B показывает уровни IFN-γ, показывающие значительное снижение IFN-γ при применении ингибитора NLRP3 MCC950 (см. стрелку), ФИГ. 18C показывает уровни IL-β, ФИГ. 18D показывает уровни IL-18, показывающие значительное снижение IFN-γ при применении ингибитора NLRP3 MCC950 (см. стрелку), ФИГ. 18E показывает уровни IL-6, ФИГ. 18F показывает уровни IP-10, ФИГ. 18G показывает уровни MCP-1, ФИГ. 18H показывает уровни TNFα.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0063] Векторы для переноса нуклеиновых кислот и терапевтические агенты на основе нуклеиновых кислот являются перспективными терапевтическими средствами для ряда приложений, таких как экспрессия гена и его модуляция. Вирусные векторы для переноса могут содержать трансгены, кодирующие белки или нуклеиновые кислоты. Примеры таких векторов включают ААВ-векторы, микроРНК (миРНК), короткую интерферирующую РНК (киРНК), а также антисмысловые олигонуклеотиды, которые связывают сайты мутаций в информационной РНК (такие как малая ядерная РНК (мяРНК)). К сожалению, перспективы этих терапевтических средств еще не реализованы, в значительной степени из-за клеточных и гуморальных иммунных ответов, направленных против вирусного вектора для переноса. Эти иммунные ответы включают ответы антител, В-клеток и Т-клеток и часто специфичны в отношении вирусных антигенов вирусного вектора для переноса, таких как вирусные белки капсида или оболочки или их пептиды.
[0064] В настоящее время многие потенциальные пациенты обладают некоторым уровнем уже имеющегося иммунитета против вирусов, на которых основаны вирусные векторы для переноса. Фактически, антитела против вирусных нуклеиновых кислот (как ДНК, так и РНК) или белков широко распространены в человеческой популяции. Кроме того, даже если уровень уже имеющегося иммунитета низок, например, из-за низкой иммуногенности вирусного вектора для переноса, такие низкие уровни все же могут препятствовать успешной трансдукции (например, Jeune, et al., Human Gene Therapy Methods, 24:59-67 (2013)). Таким образом, даже низкие уровни уже имеющегося иммунитета могут препятствовать применению конкретного вирусного вектора для переноса у пациента и могут потребовать от клинициста выбора вирусного вектора для переноса на основе вируса другого серотипа, который может быть не таким эффективным, или даже вообще отказаться в пользу другого типа терапии, если терапия другим вирусным вектором для переноса недоступна.
[0065] Кроме того, вирусные векторы, такие как векторы на основе аденоассоциированного вируса, могут быть высокоиммуногенными и могут вызывать гуморальный и опосредуемый клетками иммунитет, который может снизить эффективность, в частности, в отношении повторного введения. Фактически, клеточные и гуморальные иммунные ответы против вирусного вектора для переноса могут развиться после однократного введения вирусного вектора для переноса. После введения вирусного вектора для переноса титры нейтрализующих антител могут повышаться и оставаться высокими в течение нескольких лет и могут уменьшить эффективность повторного введения вирусного вектора для переноса. Действительно, повторное введение вирусного вектора для переноса обычно приводит к усиленным нежелательным иммунным ответам. Кроме того, могут появляться CD8+ Т-клетки, специфичные в отношении вирусного вектора для переноса, и устранять трансдуцированные клетки, экспрессирующие целевой трансгенный продукт, например, при повторном воздействии вирусной нуклеиновой кислоты или капсидного белка, подобных вирусным антигенам. Например, было показано, что нуклеиновые кислоты или антигены капсида ААВ могут запускать иммуноопосредуемое разрушение гепатоцитов, трансдуцированных вирусным ААВ-вектором для переноса. Полагают, что для многих терапевтических приложений необходимо несколько раундов введения вирусных векторов для переноса, чтобы получить преимущества в долгосрочной перспективе. Однако возможность осуществления этого будет сильно ограничена, в частности, если необходимо повторное введение, без способов и композиций, предложенных в настоящем документе.
[0066] Предложены способы и композиции, которые обеспечивают решения вышеупомянутых препятствий для эффективного применения различных терапевтических средств на основе нуклеиновых кислот, включая вирусные или невирусные (синтетические) векторы для переноса, и другие терапевтические средства на основе нуклеиновых кислот для лечения. Настоящее раскрытие относится к доставке экзогенных последовательностей ДНК в целевую клетку, ткань, орган или организм, а также к модификациям и способам для ингибирования (т.е. уменьшения или подавления) иммунного ответа (например, врожденного иммунного ответа) на них. Такие модификации и способы для ингибирования (т.е. уменьшения или подавления) иммунного ответа (например, врожденного иммунного ответа) можно применять, например, для увеличения продолжительности экспрессии трансгена.
[0067] Неожиданно было обнаружено, что иммунный ответ (например, врожденный иммунный ответ) на ДНК-вектор для переноса может быть ослаблен с помощью способов и соответствующих композиций, предложенных в настоящем документе. Следовательно, способы и композиции потенциально могут повышать эффективность лечения с применением вирусных векторов для переноса и других молекул терапевтических нуклеиновых кислот и обеспечивать терапевтические преимущества в долгосрочной перспективе, даже при повторном введении вирусного вектора для переноса или других терапевтических средств на основе нуклеиновых кислот.
I. Определения
[0068] Если в настоящем документе не указано иное, научные и технические термины, используемые применительно к настоящей заявке, должны иметь стандартные значения, известные специалистами в области техники, к которой относится данное изобретение. Следует понимать, что настоящее изобретение не ограничивается конкретными методиками, протоколами и реагентами, и т.д., описанными в настоящем документе, и, таким образом, может изменяться. Терминология, используемая в настоящем документе, предназначена исключительно для описания конкретных вариантов реализации и не предназначена для ограничения объема настоящего изобретения, который определен исключительно формулой изобретения. Определения распространенных терминов в иммунологии и молекулярной биологии можно найти в следующих источниках: The Merck Manual of Diagnosis and Therapy, 19 изд., опубликовано Merck Sharp & Dohme Corp., 2011 (ISBN 978-0-911910-19-3); Robert S. Porter et al. (под ред.), Fields Virology, 6 изд., опубликовано Lippincott Williams & Wilkins, Philadelphia, PA, USA (2013), Knipe, D.M. and Howley, P.M. (ред.), The Encyclopedia of Molecular Cell Biology and Molecular Medicine, опубликовано Blackwell Science Ltd., 1999-2012 (ISBN 9783527600908); и Robert A. Meyers (ред.), Molecular Biology and Biotechnology: a Comprehensive Desk Reference, опубликовано VCH Publishers, Inc., 1995 (ISBN 1-56081-569-8); Immunology, Werner Luttmann, опубликовано Elsevier, 2006; Janeway's Immunobiology, Kenneth Murphy, Allan Mowat, Casey Weaver (под ред.), Taylor & Francis Limited, 2014 (ISBN 0815345305, 9780815345305); Lewin's Genes XI, опубликовано Jones & Bartlett Publishers, 2014 (ISBN-1449659055); Michael Richard Green and Joseph Sambrook, Molecular Cloning: A Laboratory Manual, 4 изд., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., USA (2012) (ISBN 1936113414); Davis et al., Basic Methods in Molecular Biology, Elsevier Science Publishing, Inc., New York, USA (2012) (ISBN 044460149X); Laboratory Methods in Enzymology: DNA, Jon Lorsch (ред.) Elsevier, 2013 (ISBN 0124199542); Current Protocols in Molecular Biology (CPMB), Frederick M. Ausubel (ред.), John Wiley and Sons, 2014 (ISBN047150338X, 9780471503385), Current Protocols in Protein Science (CPPS), John E. Coligan (ред.), John Wiley and Sons, Inc., 2005; и Current Protocols in Immunology (CPI) (John E. Coligan, Ada M. Kruisbeek, David H. Margulies, Ethan M. Shevach, Warren Strobe (под ред.), John Wiley and Sons, Inc., 2003 (ISBN 0471142735, 9780471142737), содержание которых полностью включено в настоящий документ посредством ссылки.
[0069] В настоящем документе термины «введение», «введенный» и их варианты относятся к введению субъекту композиции или агента (например, терапевтической нуклеиновой кислоты или иммуносупрессора, описанных в настоящем документе) и включают одновременное и последовательное введение одной или более композиций или агентов. «Введение» может относиться, например, к терапевтическим, фармакокинетическим, диагностическим, исследовательским способам, к плацебо и к экспериментальным способам. «Введение» также включает способы лечения in vitro и ex vivo. Введение композиции или агента субъекту осуществляют любым подходящим путем, включая перорально, через легкие, интраназально, парентерально (внутривенно, внутримышечно, внутрибрюшинно или подкожно), ректально, введение в лимфатическую систему, внутрь опухоли или местно. Введение субъекту композиции или агента осуществляют с помощью электропорации. Введение включает самостоятельное введение и введение другим лицом. Введение может быть осуществлено любым подходящим путем. Подходящий путь введения позволяет композиции или агенту выполнять их предусмотренную функцию. Например, если подходящим путем является внутривенный, композицию вводят путем введения указанной композиции или агента в вену субъекта.
[0070] В настоящем документе выражения «терапевтическое средство на основе нуклеиновых кислот», «терапевтическая нуклеиновая кислота» и «ТНК» используются взаимозаменяемо и относятся к любому варианту терапевтического средства с использованием нуклеиновых кислот в качестве активного компонента терапевтического агента для лечения заболевания или нарушения. В настоящем документе эти выражения относятся к терапевтическим средствам на основе РНК и терапевтическим средствам на основе ДНК. Неограничивающие примеры терапевтических средств на основе РНК включают мРНК, антисмысловую РНК и олигонуклеотиды, рибозимы, аптамеры, интерферирующие РНК (РНКи), дцРНК-субстрат Dicer, короткую шпилечную РНК (кшРНК), асимметричную интерферирующую РНК (аиРНК), микроРНК (миРНК). Неограничивающие примеры терапевтических средств на основе ДНК включают миникольцо ДНК, миниген, вирусную ДНК (например, геном лентивируса или ААВ) или невирусные синтетические ДНК-векторы, линейную дуплексную ДНК с замкнутыми концами (зкДНК/CELiD), плазмиды, бакмиды, ДНК-векторы doggybone (dbDNA™), минималистичный вектор для иммунологически определенной экспрессии гена (MIDGE), невирусный ДНК-вектор с минимальной цепью (линейный ковалентно замкнутый ДНК-вектор) или гантелеобразный минимальный ДНК-вектор («гантелеобразную ДНК»).
[0071] В настоящем документе «эффективное количество» или «терапевтически эффективное количество» активного агента или терапевтического агента, такого как иммуносупрессор и/или терапевтическая нуклеиновая кислота, представляет собой количество, достаточное для получения целевого эффекта, например, нормализации или уменьшения иммунного ответа (например, врожденного иммунного ответа) и экспрессии или ингибирования экспрессии целевой последовательности по сравнению с уровнем экспрессии, детектированным в отсутствие терапевтической нуклеиновой кислоты и/или иммуносупрессора. Подходящие анализы для измерения экспрессии целевого гена или целевой последовательности включают, например, исследование уровней белка или РНК с применением методик, известных специалистам в данной области техники, таких как дот-блоттинг, Нозерн-блоттинг, гибридизация in situ, ИФА, иммунопреципитация, функциональный ферментный анализ, а также фенотипические анализы, известные специалистам в данной области техники. Однако уровни дозировки основаны на различных факторах, включая тип повреждения, возраст, массу, пол, медицинское состояние пациента, тяжесть состояния, путь введения и конкретный применяемый активный агент. Соответственно, схема дозирования может значительно изменяться, однако может быть определена врачом обычным путем с применением стандартных способов. Кроме того, термины «терапевтическое количество», «терапевтически эффективные количества» и «фармацевтически эффективные количества» включают профилактические или превентивные количества композиций согласно описанному изобретению. В профилактических или превентивных приложениях описанного изобретения фармацевтические композиции или лекарственные средства вводят пациенту, подверженному или по иным причинам имеющему риск развития заболевания, нарушения или состояния, в количестве, достаточном для того чтобы устранить или уменьшить риск, уменьшить степень тяжести или отсрочить начало указанного заболевания, нарушения или состояния, включая биохимические, гистологические и/или поведенческие симптомы указанного заболевания, нарушения или состояния, его осложнения, и промежуточные патологические фенотипы, возникающие в ходе развития указанного заболевания, нарушения или состояния. Обычно предпочтительным является применение максимальной дозы, т.е. самой высокой безопасной дозы в соответствии с некоторым медицинским заключением. Термины «доза» и «дозировка» используются в настоящем документе взаимозаменяемо.
[0072] В настоящем документе термин «терапевтический эффект» относится к последствиям лечения, результаты которого оценивают как целевые и благоприятные. Терапевтический эффект может включать, прямо или опосредованно, остановку, уменьшение или устранение проявления заболевания. Терапевтический эффект также может включать, прямо или опосредованно, остановку, уменьшение или устранение прогрессирования проявления заболевания.
[0073] Для любого терапевтического агента, описанного в настоящем документе, терапевтически эффективное количество может быть изначально определено на основании предварительных исследований in vitro и/или в моделях на животных. Терапевтически эффективная доза также может быть определена на основании данных у человека. Применяемая доза может быть скорректирована на основании относительных биодоступности и эффективности вводимого соединения. Коррекция дозы для достижения максимальной эффективности на основе описанных выше способов и других хорошо известных способов находится в пределах возможностей обычного специалиста в данной области техники. Общие принципы определения терапевтической эффективности, с которыми можно ознакомиться в главе 1 руководства Goodman and Gilman, The Pharmacological Basis of Therapeutics, 10 изд., McGraw-Hill (New York) (2001), включенного в настоящий документ посредством ссылки, обобщены ниже.
[0074] Фармакокинетические принципы обеспечивают основу для модификации схемы дозирования для получения целевой степени терапевтической эффективности с минимумом неприемлемых нежелательных явлений. Дополнительные рекомендации по модификации дозировки могут быть получены в ситуациях, когда концентрация лекарственного средства в плазме может быть измерена и соотнесена с терапевтическим окном.
[0075] В настоящем документе термины «гетерологичная нуклеотидная последовательность» и «трансген» используются взаимозаменяемо и относятся к представляющей интерес нуклеиновой кислоте (отличной от нуклеиновой кислоты, кодирующей полипептид капсида), которая встроена в зкДНК-вектор и может быть доставлена и экспрессирована зкДНК-вектором, раскрытым в настоящем документе.
[0076] В настоящем документе термины «экспрессионная кассета» и «транскрипционная кассета» используются взаимозаменяемо и относятся к линейному участку нуклеиновых кислот, который включает трансген, функционально связанный с одним или более промоторами или другими регуляторными последовательностями, достаточными для прямой транскрипции трансгена, но который не содержит кодирующих капсид последовательностей, других векторных последовательностей или областей инвертированных концевых повторов. Экспрессионная кассета может дополнительно содержать одну или более цис-действующих последовательностей (например, промоторов, энхансеров или репрессоров), один или более интронов и один или более посттранскрипционных регуляторных элементов.
[0077] Термины «полинуклеотид» и «нуклеиновая кислота», используемые в настоящем документе взаимозаменяемо, относятся к полимерной форме нуклеотидов любой длины, будь то рибонуклеотиды или дезоксирибонуклеотиды. Таким образом, этот термин включает одно-, двух- или многоцепочечные ДНК или РНК, геномную ДНК, кДНК, гибриды ДНК-РНК или полимер, включающий пуриновые и пиримидиновые основания или другие природные, химически или биохимически модифицированные, неприродные или дериватизированные нуклеотидные основания. «Олигонуклеотид» обычно относится к полинуклеотидам, содержащим от примерно 5 до примерно 100 нуклеотидов одно- или двухцепочечной ДНК. Однако для целей настоящего раскрытия верхнего предела длины олигонуклеотида не существует. Олигонуклеотиды также известны как «олигомеры» или «олиго» (oligos) и могут быть выделены из генов или химически синтезированы с помощью способов, известных в данной области техники. Термины «полинуклеотид» и «нуклеиновая кислота» следует понимать как включающие, применительно к описываемым вариантам реализации, одноцепочечные (такие как смысловые или антисмысловые) и двухцепочечные полинуклеотиды. ДНК может быть в форме, например, антисмысловых молекул, плазмидной ДНК, дуплексов ДНК-ДНК, предварительно конденсированной ДНК, продуктов ПЦР, векторов (P1, PAC, BAC, YAC, искусственные хромосомы), экспрессионных кассет, химерных последовательностей, хромосомной ДНК или производных и комбинаций указанных групп. ДНК может быть в форме миникольца, плазмиды, бакмиды, минигена, ДНК с минимальной цепью (линейного ковалентно замкнутого ДНК-вектора), линейной дуплексной ДНК с замкнутыми концами (CELiD или зкДНК), ДНК doggybone (dbDNA™), гантелеобразной ДНК, минималистичного вектора для иммунологически определенной экспрессии гена (MIDGE), вирусного вектора или невирусных векторов. РНК может быть в форме короткой интерферирующей РНК (киРНК), дцРНК-субстрата Dicer, короткой шпилечной РНК (кшРНК), асимметричной интерферирующей РНК (аиРНК), микроРНК (миРНК), мРНК, рРНК, тРНК, вирусной РНК (вРНК) и их комбинаций. Нуклеиновые кислоты включают нуклеиновые кислоты, содержащие известные аналоги нуклеотидов или модифицированные остатки или связи остова, которые являются синтетическими, природными и неприродными, и которые имеют связывающие свойства, аналогичные свойствам референсной нуклеиновой кислоты. Примеры таких аналогов и/или модифицированных остатков включают, без ограничения, фосфотиоаты, фосфодиамидат-морфолиновый олигомер (морфолино), фосфоамидаты, метилфосфонаты, хиральные метилфосфонаты, 2'-O-метилрибонуклеотиды, замкнутую нуклеиновую кислоту (LNA™) и пептидные нуклеиновые кислоты (ПНК). За исключением конкретно указанных ограничений термин включает нуклеиновые кислоты, содержащие известные аналоги природных нуклеотидов, которые имеют связывающие свойства, аналогичные свойствам референсной нуклеиновой кислоты. Если не указано иное, также предусмотрено, что конкретная последовательность нуклеиновой кислоты также включает ее консервативно модифицированные варианты (например, вырожденные замены кодонов), аллели, ортологи, однонуклеотидные полиморфизмы (SNP) и комплементарные последовательности, а также последовательность, указанную явным образом.
[0078] «Нуклеотиды» содержат сахар дезоксирибозу (ДНК) или рибозу (РНК), основание и фосфатную группу. Нуклеотиды соединены между собой посредством фосфатных групп.
[0079] «Основания» включают пурины и пиримидины, которые, в частности, включают природные соединения аденин, тимин, гуанин, цитозин, урацил, инозин и природные аналоги, а также синтетические производные пуринов и пиримидинов, которые включают, но не ограничиваются перечисленными, модификации, которые вводят новые реакционноспособные группы, такие как, но не ограничиваясь перечисленными, амины, спирты, тиолы, карбоксилаты и алкилгалогениды.
[0080] В настоящем документе термин «интерферирующая РНК» или «РНКи», или «последовательность интерферирующей РНК» включает одноцепочечную РНК (например, зрелую миРНК, олигонуклеотиды оцРНКи, олигонуклеотиды оцДНКи), двухцепочечную РНК (т.е. дуплексную РНК, такую как киРНК, дцРНК-субстрат Dicer, кшРНК, аиРНК или пре-миРНК), гибрид ДНК-РНК (см., например, публикацию РСТ № WO 2004/078941) или гибрид ДНК-ДНК (см., например, публикацию РСТ № WO 2004/104199), который способен уменьшать или ингибировать экспрессию целевого гена или последовательности (например, опосредуя разрушение или ингибируя трансляцию мРНК, которые комплементарны последовательности интерферирующей РНК), когда интерферирующая РНК находится в той же клетке, что и целевой ген или последовательность. Таким образом, интерферирующая РНК относится к одноцепочечной РНК, которая комплементарна последовательности мРНК-мишени, или к двухцепочечной РНК, образованной двумя комплементарными цепями или одной самокомплементарной цепью. Интерферирующая РНК может иметь существенную или полную идентичность с целевым геном или последовательностью или может содержать область несовпадения (т.е. мотив несовпадения). Последовательность интерферирующей РНК может соответствовать полноразмерному целевому гену или его подпоследовательности. Предпочтительно молекулы интерферирующих РНК синтезируют химическим путем. Раскрытия каждого из упомянутых выше патентных документов полностью включены в настоящий документ посредством ссылки для всех целей.
[0081] Интерферирующая РНК включает «короткую интерферирующую РНК» или «киРНК», например, интерферирующую РНК из примерно 15-60, 15-50 или 15-40 (дуплекс) нуклеотидов в длину, более типично примерно 15-30, 15-25 или 19-25 (дуплекс) нуклеотидов в длину и предпочтительно составляет примерно 20-24, 21-22 или 21-23 (дуплекс) нуклеотида в длину (например, каждая комплементарная последовательность двухцепочечной киРНК составляет 15-60, 15-50, 15-40, 15-30, 15-25 или 19-25 нуклеотидов в длину, предпочтительно примерно 20-24, 21-22 или 21-23 нуклеотида в длину, и двухцепочечная киРНК составляет примерно 15-60, 15-50, 15-40, 15-30, 15-25 или 19-25 пар оснований в длину, предпочтительно примерно 18-22, 19-20 или 19-21 пару оснований в длину). Дуплексы киРНК могут содержать 3´-липкие концы, имеющие от примерно 1 до примерно 4 нуклеотидов или от примерно 2 до примерно 3 нуклеотидов, и 5´-фосфатные концы. Примеры киРНК включают, без ограничения, двухцепочечную полинуклеотидную молекулу, собранную из двух одноцепочечных молекул, причем одна цепь является смысловой цепью, а другая представляет собой комплементарную антисмысловую цепь; двухцепочечную полинуклеотидную молекулу, собранную из одноцепочечной молекулы, в которой смысловая и антисмысловая области соединены линкером на основе нуклеиновой кислоты или на основе, отличной от нуклеиновой кислоты; двухцепочечную полинуклеотидную молекулу со шпилечной вторичной структурой, имеющей самокомплементарные смысловые и антисмысловые области; и кольцевую одноцепочечную полинуклеотидную молекулу с двумя или более петлевыми структурами и стеблем, имеющим самокомплементарные смысловые и антисмысловые области, причем кольцевой полинуклеотид может подвергаться процессингу in vivo или in vitro с получением активной двухцепочечной молекулы киРНК. В настоящем документе термин «киРНК» включает дуплексы РНК-РНК, а также гибриды ДНК-РНК (см., например, публикацию РСТ № WO 2004/078941).
[0082] В настоящем документе термин «конструкция нуклеиновой кислоты» относится к молекуле нуклеиновой кислоты, будь то одноцепочечная или двухцепочечная, которая выделена из природного гена или которая модифицирована так, чтобы она содержала сегменты нуклеиновых кислот, с помощью способа, который в прочих случаях не существовал бы в природе, или является синтетической. Термин «конструкция нуклеиновой кислоты» является синонимом термина «экспрессионная кассета», когда конструкция нуклеиновой кислоты содержит контрольные последовательности, необходимые для экспрессии кодирующей последовательности согласно настоящему раскрытию. «Экспрессионная кассета» включает кодирующую последовательность ДНК, функционально связанную с промотором.
[0083] Под «гибридизуемая» или «комплементарная», или «по существу комплементарная» подразумевается, что нуклеиновая кислота (например, РНК) включает последовательность нуклеотидов, которая позволяет ей нековалентно связываться, т.е. формировать пары оснований по Уотсону-Крику и/или пары оснований G/U, «ренатурировать» или «гибридизироваться» с другой нуклеиновой кислотой специфичным для последовательности, антипараллельным образом (т.е. нуклеиновая кислота специфично связывается с комплементарной нуклеиновой кислотой) в соответствующих условиях температуры и ионной силы раствора in vitro и/или in vivo. Как известно в данной области техники, стандартное спаривание оснований по Уотсону-Крику включает: спаривание аденина (A) с тимидином (T), спаривание аденина (A) с урацилом (U) и спаривание гуанина (G) с цитозином (C). Кроме того, в данной области техники также известно, что при гибридизации двух молекул РНК (например, дцРНК) гуаниновое (G) основание спаривается с урацилом (U). Например, спаривание оснований G/U частично отвечает за вырожденность (т.е. избыточность) генетического кода в случае спаривания оснований анти-кодонов тРНК с кодонами в мРНК. Применительно к настоящему изобретению гуанин (G) связывающего белок сегмента (дуплекса дцРНК) нацеливающей на ДНК молекулы РНК согласно настоящему изобретению считается комплементарным урацилу (U), и наоборот. Таким образом, когда пара оснований G/U может быть получена в определенном нуклеотидном положении связывающего белок сегмента (дуплекса дцРНК) нацеливающей на ДНК молекулы РНК согласно настоящему изобретению, это положение не рассматривается как некомплементарное, а вместо этого считается комплементарным.
[0084] Термины «пептид», «полипептид» и «белок» используются в настоящем документе взаимозаменяемо и относятся к полимерной форме аминокислот любой длины, которая может включать кодируемые и некодируемые аминокислоты, химически или биохимически модифицированные или дериватизированные аминокислоты, и к полипептидам, имеющим модифицированные пептидные остовы.
[0085] Последовательность ДНК, которая «кодирует» конкретный антагонист инфламмасомы (например, любой один или более из: ингибитора пути инфламмасомы NLRP3 или ингибитора пути инфламмасомы AIM2, или ингибитора каспазы 1, или любой их комбинации), представляет собой последовательность нуклеиновой кислоты ДНК, которая транскрибируется в конкретную РНК и/или белок. Полинуклеотид ДНК может кодировать РНК (мРНК), которая транслируется в белок, или полинуклеотид ДНК может кодировать РНК, которая не транслируется в белок (например, тРНК, рРНК или нацеливающую на ДНК РНК; также называемые «некодирующими» РНК или «нкРНК»).
[0086] В настоящем документе термин «слитый белок», в используемом в настоящем документе значении, относится к полипептиду, который содержит белковые домены по меньшей мере из двух разных белков. Например, слитый белок может содержать (i) один антагонист инфламмасомы (например, любой один или более из: ингибитора пути инфламмасомы NLRP3 или ингибитора пути инфламмасомы AIM2, или ингибитора каспазы 1, или любой их комбинации) или его фрагмент, и (ii) по меньшей мере один белок, не относящийся к представляющему интерес гену (GOI), или, альтернативно, белок другого антагониста инфламмасомы. Слитые белки, предусмотренные в настоящем документе, включают, но не ограничиваются перечисленными, антитело или Fc, или антигенсвязывающий фрагмент антитела, слитый с антагонистом инфламмасомы (например, любым одним или более из: ингибитора пути инфламмасомы NLRP3 или ингибитора пути инфламмасомы AIM2, или ингибитора каспазы 1, или любой их комбинации), например, с внеклеточным доменом рецептора, лигандом, ферментом или пептидом. Антагонист инфламмасомы (например, любой один или более из: ингибитора пути инфламмасомы NLRP3 или ингибитора пути инфламмасомы AIM2, или ингибитора каспазы 1, или любой их комбинации) или его фрагмент, который является частью слитого белка, может представлять собой моноспецифичное антитело или биспецифичное или мультиспецифичное антитело.
[0087] В настоящем документе термин «ген безопасной гавани генома» или «ген безопасной гавани» относится к гену или локусам, в которые последовательность нуклеиновой кислоты может быть вставлена так, чтобы последовательность могла интегрироваться и функционировать предсказуемым образом (например, экспрессировать белок, представляющий интерес) без существенных отрицательных последствий для активности эндогенного гена или стимуляции рака. Согласно некоторым вариантам реализации ген безопасной гавани также представляет собой локусы или ген, в которых вставленная последовательность нуклеиновой кислоты может быть экспрессирована эффективнее и на более высоких уровнях, чем в сайте, не являющемся «безопасной гаванью».
[0088] В настоящем документе термин «доставка гена» означает способ, с помощью которого чужеродную ДНК переносят в клетки-хозяева для приложений генной терапии.
[0089] В настоящем документе термин «концевой повтор» или «TR» включает любой вирусный концевой повтор или любую синтетическую последовательность, которая содержит по меньшей мере одну минимальную требуемую точку начала репликации и область, содержащую палиндромную шпилечную структуру. Связывающая Rep последовательность («RBS») (также называемая RBE (связывающий Rep элемент)) и сайт концевого разрешения («TRS») вместе составляют «минимальную требуемую точку начала репликации» и, соответственно, TR содержит по меньшей мере одну RBS и по меньшей мере один TRS. Каждый из TR, обратно комплементарных друг относительно друга в пределах определенного участка полинуклеотидной последовательности, как правило, называют «инвертированным концевым повтором» или «ITR». Применительно к вирусу ITR опосредуют репликацию, упаковку вируса, интеграцию и освобождение провируса. Как было неожиданно обнаружено, согласно настоящему изобретению TR, не являющиеся обратно комплементарными по всей их длине, все еще могут выполнять обычные функции ITR, и, таким образом, в настоящем документе термин ITR относится к TR в геноме зкДНК или зкДНК-векторе, который способен опосредовать репликацию зкДНК-вектора. Обычный специалист в данной области техники поймет, что в зкДНК-векторах сложной конфигурации может присутствовать более двух пар ITR или асимметричных ITR. ITR может представлять собой ITR ААВ или ITR, не принадлежащий ААВ, или может происходить из ITR ААВ или ITR, не принадлежащего ААВ. Например, ITR может происходить из вируса семейства Parvoviridae, которое включает парвовирусы и депендовирусы (например, парвовирус собак, парвовирус крупного рогатого скота, парвовирус мышей, парвовирус свиней, парвовирус человека B-19), или шпилька SV40, которая служит точкой начала репликации SV40, может применяться в качестве ITR, который может быть дополнительно модифицирован путем усечения, замены, делеции, вставки и/или добавления. Вирусы семейства Parvoviridae состоят из двух подсемейств: Parvovirinae, инфицирующих позвоночных животных, и Densovirinae, инфицирующих беспозвоночных животных. Депендопарвовирусы включают вирусное семейство аденоассоциированных вирусов (ААВ), которые способны к репликации у позвоночных животных-хозяев, включая, но не ограничиваясь перечисленными, человека, виды приматов, бычьих, собачьих, лошадиных и овечьих. Для удобства в настоящем документе ITR, расположенный в 5'-направлении (выше) относительно экспрессионной кассеты в зкДНК-векторе, называется «5'-ITR» или «левым ITR», а ITR, расположенный в 3'-направлении (ниже) относительно экспрессионной кассеты в зкДНК-векторе, называется «3'-ITR» или «правым ITR».
[0090] «ITR дикого типа» или «WT-ITR» относится к последовательности природной последовательности ITR в ААВ или другом депендовирусе, которая сохраняет, например, активность связывания Rep и никирующую способность Rep. Нуклеотидная последовательность WT-ITR из любого серотипа ААВ может незначительно отличаться от канонической природной последовательности из-за вырожденности генетического кода или дрейфа, и, таким образом, последовательности WT-ITR, предусмотренные для применения в настоящем документе, включают последовательности WT-ITR, образующиеся в результате природных изменений, происходящих в процессе продуцирования (например, ошибки репликации).
[0091] В настоящем документе термин «по существу симметричные WT-ITR» или «пара по существу симметричных WT-ITR» относится к паре WT-ITR в одном геноме зкДНК или зкДНК-векторе, которые оба представляют собой ITR дикого типа, которые имеют обратно комплементарные последовательности по всей их длине. Например, ITR может считаться последовательностью дикого типа, даже если он содержит один или более нуклеотидов, отличающихся от нуклеотидов в канонической природной последовательности, при условии, что указанные изменения не влияют на свойства и общую трехмерную структуру последовательности. Согласно некоторым аспектам отличающиеся нуклеотиды представляют собой консервативные изменения последовательности. В качестве одного неограничивающего примера, последовательность, имеющая по меньшей мере 95%, 96%, 97%, 98% или 99% идентичность последовательности с канонической последовательностью (измеренную, например, с использованием BLAST при настройках по умолчанию), также имеет симметричную трехмерную пространственную организацию с другой последовательностью WT-ITR так, что их трехмерные структуры имеют одинаковую форму в геометрическом пространстве. По существу симметричный WT-ITR имеет такие же петли A, C-C' и B-B' в трехмерном пространстве. То, что по существу симметричный WT-ITR представляет собой WT может быть функционально подтверждено путем определения наличия в нем функционального сайта связывания Rep (RBE или RBE´) и сайта концевого разрешения (TRS), который спаривается с соответствующим белком Rep. Могут быть протестированы другие функции, включая экспрессию трансгена в пермиссивных условиях, но не необязательно.
[0092] В настоящем документе выражения «модифицированный ITR» или «mod-ITR», или «мутантный ITR» используются в настоящем документе взаимозаменяемо и относятся к ITR, который имеет мутацию по меньшей мере в одном или более нуклеотидах по сравнению с WT-ITR из того же серотипа. Указанная мутация может приводить к изменению одной или более из областей A, C, C', B, B' в ITR и может приводить к изменению трехмерной пространственной организации (т.е. его трехмерной структуры в геометрическом пространстве) по сравнению с трехмерной пространственной организацией WT-ITR из того же серотипа.
[0093] В настоящем документе термин «асимметричные ITR», также называемые «парой асимметричных ITR», относится к паре ITR в одном геноме зкДНК или зкДНК-векторе, которые не являются обратно комплементарными по их полной длине. В качестве одного неограничивающего примера, асимметричный ITR в паре не имеет симметричной трехмерной пространственной организации с его когнатными ITR так, что их трехмерные структуры имеют разные формы в геометрическом пространстве. Иными словами, асимметричные ITR в паре имеют разную общую геометрическую структуру, т.е. они имеют разную организацию своих петель A, C-C'- и B-B' в трехмерном пространстве (например, один ITR может иметь короткое плечо C-C' и/или короткое плечо B-B' по сравнению с когнатным ITR). Различие по последовательности между двумя ITR может быть связано с добавлением, делецией, усечением или точечной мутацией одного или более нуклеотидов. Согласно одному варианту реализации один ITR из пары асимметричных ITR может представлять собой последовательность ITR ААВ дикого типа, а другой ITR представляет собой модифицированный ITR, как определено в настоящем документе (например, последовательность ITR, не относящуюся к дикому типу, или синтетическую последовательность). Согласно другому варианту реализации ни один ITR из пары асимметричных ITR не является последовательностью ААВ дикого типа, и указанные два ITR представляют собой модифицированные ITR, которые имеют разные формы в геометрическом пространстве (т.е. разную общую геометрическую структуру). Согласно некоторым вариантам реализации один mod-ITR из пары асимметричных ITR может иметь короткое C-C'-плечо, а другой ITR может иметь другую модификацию (например, одно плечо или короткое плечо B-B' и т.д.) так, что они имеют различную трехмерную пространственную организацию по сравнению с когнатным асимметричным mod-ITR.
[0094] В настоящем документе термин «симметричные ITR» относится к паре ITR в одном геноме зкДНК или зкДНК-векторе, которые мутированы или модифицированы относительно последовательностей депендовирусных ITR дикого типа и обратно комплементарны по всей их длине. Ни один из ITR не является последовательностью ITR ААВ2 дикого типа (т.е. они представляют собой модифицированные ITR, также называемые мутантными ITR) и может отличаться по последовательности от ITR дикого типа вследствие добавления, делеции, замены, усечения или точечной мутации нуклеотида. Для удобства в настоящем документе ITR, расположенный в 5'-направлении (выше) относительно экспрессионной кассеты в зкДНК-векторе, называется «5'-ITR» или «левым ITR», а ITR, расположенный в 3'-направлении (ниже) относительно экспрессионной кассеты в зкДНК-векторе, называется «3'-ITR» или «правым ITR».
[0095] В настоящем документе термин «по существу симметричные модифицированные ITR» или «пара по существу симметричных mod-ITR» относится к паре модифицированных ITR в одном геноме зкДНК или зкДНК-векторе, которые оба имеют обратно комплементарные последовательности по всей их длине. Например, модифицированный ITR может считаться по существу симметричным, даже если он имеет некоторые нуклеотидные последовательности, отличающиеся от обратно комплементарной последовательности, при условии, что указанные изменения не влияют на свойства и общую форму. В качестве одного неограничивающего примера приведена последовательность, которая имеет по меньшей мере 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичность последовательности с канонической последовательностью (измеренную с помощью BLAST при настройках по умолчанию), а также имеет симметричную трехмерную пространственную организацию со своим когнатным модифицированным ITR так, что их трехмерные структуры имеют одинаковую форму в геометрическом пространстве. Иными словами, пара по существу симметричных модифицированных ITR имеет одинаково организованные в трехмерном пространстве петли A, C-C' и B-B'. Согласно некоторым вариантам реализации ITR из пары mod-ITR могут иметь разные обратно комплементарные нуклеотидные последовательности, но все же имеют одинаковую симметричную трехмерную пространственную организацию, т.е. оба ITR имеют мутации, которые приводят к одинаковой общей трехмерной форме. Например, один ITR (например, 5'-ITR) в паре mod-ITR может происходить из одного серотипа, а другой ITR (например, 3'-ITR) может происходить из другого серотипа, однако оба могут иметь одинаковую соответствующую мутацию (например, если 5'-ITR имеет делецию в области C, то когнатный модифицированный 3'-ITR из другого серотипа имеет делецию в соответствующем положении в области C') так, что пара модифицированных ITR имеет одинаковую симметричную трехмерную пространственную организацию. В таких вариантах реализации каждый ITR в паре модифицированных ITR может происходить из разных серотипов (например, ААВ1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 и 12), таких как комбинация ААВ2 и ААВ6, при этом модификация в одном ITR отображается в соответствующем положении в когнатном ITR из другого серотипа. Согласно одному варианту реализации пара по существу симметричных модифицированных ITR относится к паре модифицированных ITR (mod-ITR) при условии, что различие в нуклеотидных последовательностях между указанными ITR не влияет на свойства или общую форму, и они имеют по существу одинаковую форму в трехмерном пространстве. В качестве неограничивающего примера приведен mod-ITR, который имеет по меньшей мере 95%, 96%, 97%, 98% или 99% идентичность последовательности с каноническим mod-ITR, определенную стандартными средствами, хорошо известными специалистам в данной области техники, такими как BLAST (Basic Local Alignment Search Tool (средство поиска основного локального выравнивания)) или BLASTN при настройках по умолчанию, а также имеет симметричную трехмерную пространственную организацию так, что их трехмерные структуры имеют одинаковую форму в геометрическом пространстве. Пара по существу симметричных mod-ITR имеет одинаковые петли A, C-C' и B-B' в трехмерном пространстве, например, если модифицированный ITR в паре по существу симметричных mod-ITR имеет делецию плеча C-C', то когнатный mod-ITR имеет соответствующую делецию петли C-C', а также имеет аналогичную трехмерную структуру остальных петель A и B-B', принимающую такую же форму в геометрическом пространстве, что и у когнатного mod-ITR. Термин «фланкирование» относится к относительному положению одной последовательности нуклеиновой кислоты по отношению к другой последовательности нуклеиновой кислоты. Обычно в последовательности ABC A и C фланкируют B. То же самое верно для расположения AxBxC. Соответственно, фланкирующая последовательность предшествует фланкируемой последовательности или следует за ней, но не обязательно должна быть смежной с фланкируемой последовательностью или непосредственно прилегать к ней. Согласно одному варианту реализации термин «фланкирование» относится к концевым повторам на каждом конце линейного дуплексного зкДНК-вектора. В настоящем документе термины «лечить», «лечащий» и/или «лечение» включают подавление, по существу ингибирование, замедление или обращение прогрессирования состояния, по существу облегчение клинических симптомов состояния или по существу предотвращение появления клинических симптомов состояния, получение благоприятных или целевых клинических результатов. Лечение также относится к достижению одного или более из: (a) уменьшения степени тяжести нарушения; (b) ограничения развития симптомов, характерных для нарушения (ий), которое лечат; (c) ограничения ухудшения симптомов, характерных для нарушения (ий), которое лечат; (d) ограничения рецидивирования нарушения (ий) у пациентов, ранее имевших указанное нарушение (ия); и (e) ограничения рецидивирования симптомов у пациентов, ранее не имевших симптомов указанного нарушения (ий). Благоприятные или целевые клинические результаты, такие как фармакологические и/или физиологические эффекты, включают, но не ограничиваются перечисленными, предотвращение возникновения заболевания, нарушения или состояния у субъекта, который может быть предрасположен к указанному заболеванию, нарушению или состоянию, но еще не испытывает симптомов или у него еще не проявляются симптомы указанного заболевания (профилактическое лечение), облегчение симптомов указанного заболевания, нарушения или состояния, уменьшение интенсивности указанного заболевания, нарушения или состояния, стабилизацию (т.е. отсутствие ухудшения) указанного заболевания, нарушения или состояния, предотвращение распространения указанного заболевания, нарушения или состояния, задержку или замедление прогрессирования указанного заболевания, нарушения или состояния, облегчение или временное облегчение указанного заболевания, нарушения или состояния, и комбинации перечисленных, а также увеличение продолжительности выживания по сравнению с ожидаемым выживанием без лечения.
[0096] В настоящем документе термин «увеличение», «усиление», «повышение» (и подобные термины) обычно относится к акту увеличения, прямо или опосредовано, концентрации, уровня, функции, активности или поведения относительно природного, ожидаемого или среднего, или относительно контрольного состояния.
[0097] В настоящем документе термин «подавлять», «снижать», «препятствовать», «ингибировать» и/или «уменьшать» (и другие подобные термины) обычно относится к акту уменьшения, прямо или опосредовано, концентрации, уровня, функции, активности, поведения относительно природного, ожидаемого, среднего, или относительно контрольного состояния. Под «снижение», «снижающий», «уменьшение» или «уменьшающий», применительно к иммунному ответу (например, иммунному ответу (например, врожденному иммунному ответу)), с помощью иммуносупрессора подразумевается детектируемое снижение иммунного ответа на данный иммуносупрессор. Величина снижения иммунного ответа с помощью иммуносупрессора может быть определена относительно уровня иммунного ответа в присутствии иммуносупрессора. Детектируемое снижение может составлять примерно 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 100% или еще ниже, чем иммунный ответ, детектируемый в присутствии иммуносупрессора.
[0098] В настоящем документе термин «липид» относится к группе органических соединений, которые включают, но не ограничиваются перечисленными, сложные эфиры алифатических кислот и характеризуются тем, что они нерастворимы в воде, но растворимы во многих органических растворителях. Обычно они подразделяются по меньшей мере на три класса: (1) «простые липиды», которые включают жиры и масла, а также воски; (2) «сложные липиды», которые включают фосфолипиды и гликолипиды; и (3) «производные липидов», такие как стероиды.
[0099] В настоящем документе термин «липидная частица» включает липидный состав, который можно применять для доставки терапевтического агента, такого как терапевтические средства на основе нуклеиновых кислот и/или иммуносупрессор, к целевому участку, представляющему интерес (например, клетке, ткани, органу и т.п.). Согласно предпочтительным вариантам реализации липидная частица согласно настоящему изобретению представляет собой липидную частицу, содержащую нуклеиновую кислоту, которая, как правило, образована из катионного липида, некатионного липида и необязательно конъюгированного липида, который предотвращает агрегацию частицы. Согласно другим предпочтительным вариантам реализации терапевтический агент, такой как терапевтическая нуклеиновая кислота, может быть инкапсулирован в липидной части частицы, что защищает его от ферментативного разрушения. Согласно другим предпочтительным вариантам реализации иммуносупрессор необязательно может быть включен в липидные частицы, содержащие нуклеиновую кислоту.
[00100] В настоящем документе термин «инкапсулированный в липид» может относиться к липидной частице, которая обеспечивает активный агент или терапевтический агент, такой как нуклеиновая кислота (например, зкДНК), с полной инкапсуляцией, частичной инкапсуляцией или и тем, и другим. Согласно предпочтительному варианту реализации нуклеиновая кислота полностью инкапсулирована в липидной частице (например, с образованием липидной частицы, содержащей нуклеиновую кислоту).
[00101] В настоящем документе термин «липидный конъюгат» относится к конъюгированному липиду, который ингибирует агрегацию липидных частиц. Такие липидные конъюгаты включают, но не ограничиваются перечисленными, конъюгаты ПЭГ-липид, такие как, например, ПЭГ, связанный с диалкилоксипропилами (например, конъюгаты ПЭГ-ДАА), ПЭГ, связанный с диацилглицеринами (например, конъюгаты ПЭГ-ДАГ), ПЭГ, связанный с холестерином, ПЭГ, связанный с фосфатидилэтаноламинами, и ПЭГ, конъюгированный с церамидами (см., например, патент США № 5885613), катионные липиды-ПЭГ, конъюгаты полиоксазолин (POZ)-липид (например, конъюгаты POZ-ДАА; см., например, предварительную заявку США № 61/294828, поданную 13 января 2010 г., и предварительную заявку США № 61/295140, поданную 14 января 2010 г.), полиамидные олигомеры (например, конъюгаты АТТА-липид) и их смеси. Дополнительные примеры конъюгатов POZ-липид описаны в публикации РСТ № WO 2010/006282. ПЭГ или POZ могут быть конъюгированы непосредственно с липидом или могут быть соединены с липидом за счет линкерной группы. Может быть использована любая линкерная группа, подходящая для связывания ПЭГ или POZ с липидом, включая, например, линкерные группы, не содержащие сложного эфира, и линкерные группы, содержащие сложный эфир. Согласно определенным предпочтительным вариантам реализации применяют линкерные группы, не содержащие сложного эфира, такие как амиды или карбаматы. Раскрытия каждого из упомянутых выше патентных документов полностью включены в настоящий документ посредством ссылки для всех целей.
[00102] Типичные примеры фосфолипидов включают, но не ограничиваются перечисленными, фосфатидилхолин, фосфатидилэтаноламин, фосфатидилсерин, фосфатидилинозит, фосфатидную кислоту, пальмитоилолеоилфосфатидилхолин, лизофосфатидилхолин, лизофосфатидилэтаноламин, дипальмитоилфосфатидилхолин, диолеоилфосфатидилхолин, дистеароилфосфатидилхолин и дилинолеоилфосфатидилхолин. Другие соединения, в которых отсутствует фосфор, такие как семейства сфинголипидов и гликосфинголипидов, диацилглицерины и β-ацилоксикислоты, также включены в группу, обозначенную как амфипатические липиды. Кроме того, амфипатические липиды, описанные выше, можно смешивать с другими липидами, включая триглицериды и стерины.
[00103] В настоящем документе термин «нейтральный липид» относится к любой из ряда липидных молекул, которые существуют либо в незаряженной, либо в нейтральной цвиттерионной форме при выбранном pH. При физиологическом значении pH такие липиды включают, например, диацилфосфатидилхолин, диацилфосфатидилэтаноламин, церамид, сфингомиелин, цефалин, холестерин, цереброзиды и диацилглицерины.
[00104] В настоящем документе термин «некатионный липид» относится к любому амфипатическому липиду, а также к любому другому нейтральному липиду или анионному липиду.
[00105] В настоящем документе термин «анионный липид» относится к любому липиду, который отрицательно заряжен при физиологическом значении pH. Эти липиды включают, но не ограничиваются перечисленными, фосфатидилглицерины, кардиолипины, диацилфосфатидилсерины, диацилфосфатидные кислоты, N-додеканоилфосфатидилэтаноламины, N-сукцинилфосфатидилэтаноламины, N-глутарилфосфатидилэтаноламины, лизилфосфатидилглицерины, пальмитоилолеоилфосфатидилглицерин (POPG) и другие анионные модифицирующие группы, присоединенные к нейтральным липидам.
[00106] В настоящем документе термин «гидрофобный липид» относится к соединениям, имеющим неполярные группы, которые включают, но не ограничиваются перечисленными, насыщенные и ненасыщенные алифатические углеводородные группы с длинной цепью и такие группы, необязательно замещенные одной или более ароматическими, циклоалифатическими или гетероциклическими группами. Подходящие примеры включают, но не ограничиваются перечисленными, диацилглицерин, диалкилглицерин, N-N-диалкиламино, 1,2-диацилокси-3-аминопропан и 1,2-диалкил-3-аминопропан.
[00107] В настоящем документе термин «водный раствор» относится к композиции, содержащей в полном объеме, или частично, воду.
[00108] В настоящем документе термин «органический раствор липида» относится к композиции, содержащей в полном объеме, или частично, органический растворитель, содержащий липид.
[00109] В настоящем документе термин «системная доставка» относится к доставке липидных частиц, которая приводит к широкому биораспределению активного агента, такого как интерферирующая РНК (например, киРНК), в организме. Некоторые методики введения могут привести к системной доставке определенных агентов, но не других. Системная доставка означает, что полезное, предпочтительно терапевтическое, количество агента воздействует на большинство частей тела. Для получения широкого биораспределения обычно требуется такое время жизни в крови, при котором агент не подвергается быстрому разложению или выведению (например, органами первого прохождения (печень, легкие и т.д.) или путем быстрого неспецифичного связывания клетками) до достижения очага заболевания, удаленного от места введения. Системную доставку липидных частиц можно осуществлять любыми способами, известными в данной области техники, включая, например, внутривенный, подкожный и внутрибрюшинный. Согласно предпочтительному варианту реализации системную доставку липидных частиц осуществляют путем внутривенной доставки.
[00110] В настоящем документе термин «локальная доставка» относится к доставке активного агента, такого как интерферирующая РНК (например, киРНК), непосредственно в целевой сайт внутри организма. Например, агент может быть локально доставлен путем прямой инъекции в очаг заболевания, такой как опухоль или другой целевой очаг, такой как очаг воспаления или целевой орган, такой как печень, сердце, поджелудочная железа, почка и т.п.
[00111] В настоящем документе термин «концевой повтор» или «TR» включает любой вирусный концевой повтор или любую синтетическую последовательность, которая содержит по меньшей мере одну минимальную требуемую точку начала репликации и область, содержащую палиндромную шпилечную структуру. Связывающая Rep последовательность («RBS») (также называемая RBE (связывающий Rep элемент)) и сайт концевого разрешения («TRS») вместе составляют «минимальную требуемую точку начала репликации», и, соответственно, TR содержит по меньшей мере одну RBS и по меньшей мере один TRS. Каждый из TR, которые обратно комплементарны друг относительно друга в пределах определенного участка полинуклеотидной последовательности, как правило, называют «инвертированным концевым повтором» или «ITR». Применительно к вирусу ITR опосредуют репликацию, упаковку вируса, интеграцию и освобождение провируса. Как было неожиданно обнаружено, согласно настоящему изобретению TR, которые не являются обратно комплементарными по всей их длине, все еще могут выполнять обычные функции ITR, и, таким образом, в настоящем документе термин ITR относится к TR в геноме зкДНК или зкДНК-векторе, который способен опосредовать репликацию зкДНК-вектора. Обычный специалист в данной области техники поймет, что в зкДНК-векторах сложной конфигурации может присутствовать более двух пар ITR или асимметричных ITR. ITR может представлять собой ITR ААВ или ITR, не принадлежащий ААВ, или может происходить из ITR ААВ или из ITR, не принадлежащего ААВ. Например, ITR может происходить из семейства Parvoviridae, которое включает парвовирусы и депендовирусы (например, парвовирус собак, парвовирус крупного рогатого скота, парвовирус мышей, парвовирус свиней, парвовирус человека B-19), или шпилька SV40, которая служит в качестве точки начала репликации SV40, может быть использована как ITR, который дополнительно может быть модифицирован путем усечения, замены, делеции, инсерции и/или добавления. Семейство вирусов Parvoviridae состоит из двух подсемейств: Parvovirinae, инфицирующих позвоночных животных, и Densovirinae, инфицирующих беспозвоночных животных. Депендопарвовирусы включают вирусное семейство аденоассоциированных вирусов (ААВ), которые способны к репликации у позвоночных животных-хозяев, включая, но не ограничиваясь перечисленными, человека, виды приматов, бычьих, собачьих, лошадиных и овечьих. Для удобства в настоящем документе ITR, расположенный в 5' направлении (выше) относительно экспрессионной кассеты в зкДНК-векторе, называется «5'-ITR» или «левым ITR», а ITR, расположенный в 3' направлении (ниже) относительно экспрессионной кассеты в зкДНК-векторе, называется «3'-ITR» или «правым ITR».
[00112] «ITR дикого типа» или «WT-ITR» относится к последовательности природной последовательности ITR ААВ или другого депендовируса, которая сохраняет, например, активность связывания Rep и никирующую способность Rep. Нуклеотидная последовательность WT-ITR из любого серотипа ААВ может незначительно отличаться от канонической природной последовательности из-за вырожденности генетического кода или дрейфа, и, соответственно, последовательности WT-ITR, предусмотренные для применения в настоящем документе, включают последовательности WT-ITR, образованные в результате природных изменений, происходящих в ходе процесса продуцирования (например, ошибки репликации).
[00113] В настоящем документе термин «по существу симметричные WT-ITR» или «пара по существу симметричных WT-ITR» относится к паре WT-ITR в одном зкДНК-геноме или зкДНК-векторе, в которой оба ITR представляют собой ITR дикого типа, которые имеют обратно комплементарную последовательность по всей их длине. Например, ITR может считаться последовательностью дикого типа, даже если он имеет один или более нуклеотидов, отличающихся от канонической природной последовательности, при условии, что указанные изменения не влияют на свойства и общую трехмерную структуру последовательности. Согласно некоторым аспектам отличающиеся нуклеотиды представляют собой консервативные изменения последовательности. В качестве одного неограничивающего примера представлена последовательность, которая имеет по меньшей мере 95%, 96%, 97%, 98% или 99% идентичность последовательности с канонической последовательностью (измеренную, например, с помощью BLAST при настройках по умолчанию), а также имеет симметричную трехмерную пространственную организацию относительно другой последовательности WT-ITR так, что их трехмерные структуры имеют одинаковую форму в геометрическом пространстве. По существу симметричный WT-ITR имеет такие же петли A, C-C' и B-B' в трехмерном пространстве. То, что по существу симметричный WT-ITR представляет собой WT может быть функционально подтверждено путем определения наличия в нем функционального сайта связывания Rep (RBE или RBE´) и сайта концевого разрешения (TRS), который спаривается с соответствующим белком Rep. Могут быть протестированы другие функции, включая экспрессию трансгена в пермиссивных условиях, но необязательно.
[00114] В настоящем документе выражения «модифицированный ITR» или «mod-ITR», или «мутантный ITR» используются в настоящем документе взаимозаменяемо и относятся к ITR, который имеет мутацию по меньшей мере в одном или более нуклеотидах по сравнению с WT-ITR из того же серотипа. Мутация может приводить к изменению одной или более из областей A, C, C', B, B' в ITR и может приводить к изменению трехмерной пространственной организации (т.е. его трехмерной структуры в геометрическом пространстве) по сравнению с трехмерной пространственной организацией WT-ITR из того же серотипа.
[00115] В настоящем документе термин «асимметричные ITR», также называемые «парами асимметричных ITR», относится к паре ITR в одном геноме зкДНК или зкДНК-векторе, которые не являются обратно комплементарными по всей их длине. В качестве одного неограничивающего примера, асимметричный ITR в паре не имеет симметричной трехмерной пространственной организации со своим когнатным ITR так, что их трехмерные структуры имеют разные формы в геометрическом пространстве. Другими словами, асимметричные ITR в паре имеют разные общие геометрические структуры, т.е. они имеют разную организацию их петель A, C-C´ и B-B´ в трехмерном пространстве (например, один ITR может иметь короткое плечо C-C´ и/или короткое плечо B-B´ по сравнению с когнатным ITR). Различие по последовательности между двумя ITR может быть обусловлено добавлением, делецией, усечением или точечной мутацией одного или более нуклеотидов. Согласно одному варианту реализации один ITR из пары асимметричных ITR может представлять собой последовательность ITR ААВ дикого типа, а другой ITR может представлять собой модифицированный ITR, как определено в настоящем документе (например, последовательность ITR, не относящуюся к дикому типу, или синтетическую последовательность ITR). Согласно другому варианту реализации ни один ITR из пары асимметричных ITR не является последовательностью ААВ дикого типа, и указанные два ITR представляют собой модифицированные ITR, которые имеют разные формы в геометрическом пространстве (т.е. разную общую геометрическую структуру). Согласно некоторым вариантам реализации один mod-ITR из пары асимметричных ITR может иметь короткое плечо C-C´, а другой ITR может иметь другую модификацию (например, одно плечо или короткое плечо B-B´ и т.д.) так, что они имеют различную трехмерную пространственную организацию по сравнению с когнатным асимметричным mod-ITR.
[00116] В настоящем документе термин «симметричные ITR» относится к паре ITR в одном зкДНК-геноме или зкДНК-векторе, которые представляют собой депендовирусные последовательности ITR дикого типа или мутированные последовательности (например, модифицированные относительно дикого типа) и обратно комплементарны по всей их длине. В одном неограничивающем примере оба ITR представляют собой последовательности ITR дикого типа из ААВ2. В другом примере ни один из ITR не является последовательностью ITR ААВ2 дикого типа (т.е. они представляют собой модифицированный ITR, также называемый мутантным ITR) и может отличаться по последовательности от ITR дикого типа вследствие добавления, делеции, замены, усечения или точечной мутации нуклеотида. Для удобства в настоящем документе ITR, расположенный в 5'-направлении (выше) относительно экспрессионной кассеты в зкДНК-векторе, называется «5'-ITR» или «левым ITR», а ITR, расположенный в 3'-направлении (ниже) относительно экспрессионной кассеты в зкДНК-векторе, называется «3'-ITR» или «правым ITR».
[00117] В настоящем документе термин «по существу симметричные модифицированные ITR» или «пара по существу симметричных mod-ITR» относится к паре модифицированных ITR в одном геноме зкДНК или зкДНК-векторе, которые оба имеют обратно комплементарные последовательности по всей их длине. Например, модифицированный ITR может считаться по существу симметричным, даже если он имеет некоторые нуклеотидные последовательности, отличающиеся от обратно комплементарной последовательности, при условии, что изменения не влияют на свойства и общую форму. В качестве одного неограничивающего примера представлена последовательность, которая имеет по меньшей мере 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичность последовательности с канонической последовательностью (измеренную с помощью BLAST при настойках по умолчанию), а также имеет симметричную трехмерную пространственную организацию со своим когнатным модифицированным ITR так, что их трехмерные структуры имеют одинаковую форму в геометрическом пространстве. Иными словами, пара по существу симметричных модифицированных ITR имеет петли A, C-C' и B-B', одинаково организованные в трехмерном пространстве. Согласно некоторым вариантам реализации ITR из пары mod-ITR могут иметь разные обратно комплементарные нуклеотидные последовательности, но все же имеют одинаковую симметричную трехмерную пространственную организацию, т.е. оба ITR имеют мутации, которые приводят к одинаковой общей трехмерной форме. Например, один ITR (например, 5'-ITR) в паре mod-ITR может происходить из одного серотипа, а другой ITR (например, 3'-ITR) может происходить из другого серотипа, однако оба они могут иметь одинаковую соответствующую мутацию (например, если 5'-ITR имеет делецию в области C, когнатный модифицированный 3'-ITR из другого серотипа имеет делецию в соответствующем положении в области C') так, что пара модифицированных ITR имеет одинаковую симметричную трехмерную пространственную организацию. Согласно таким вариантам реализации каждый ITR в паре модифицированных ITR может происходить из разных серотипов (например, ААВ1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 и 12), таких как комбинация ААВ2 и ААВ6, при этом модификация в одном ITR отображается в соответствующем положении в когнатном ITR из другого серотипа. Согласно одному варианту реализации пара по существу симметричных модифицированных ITR относится к паре модифицированных ITR (mod-ITR) при условии, что различие в нуклеотидных последовательностях между ITR не влияет на свойства или общую форму, и они имеют по существу одинаковую форму в трехмерном пространстве. В качестве неограничивающего примера представлен mod-ITR, который имеет по меньшей мере 95%, 96%, 97%, 98% или 99% идентичность последовательности с каноническим mod-ITR, как определено с помощью стандартных способов, хорошо известных в данной области техники, таких как BLAST (Basic Local Alignment Search Tool) или BLASTN при настройках по умолчанию, а также имеет симметричную трехмерную пространственную организацию так, что их трехмерная структура имеет одинаковую форму в геометрическом пространстве. Пара по существу симметричных mod-ITR имеет одинаковые петли A, C-C 'и B-B' в трехмерном пространстве, например, если модифицированный ITR в паре по существу симметричных mod-ITR имеет делецию в плече C-C', то когнатный mod-ITR имеет соответствующую делецию в петле C-C', а также имеет сходную трехмерную структуру остальных петель A и B-B', принимающую такую же форму в геометрическом пространстве, что и у его когнатного mod-ITR.
[00118] Термин «фланкирование» относится к относительному положению одной последовательности нуклеиновой кислоты по отношению к другой последовательности нуклеиновой кислоты. Обычно в последовательности ABC A и C фланкируют B. То же самое верно для расположения AxBxC. Таким образом, фланкирующая последовательность предшествует фланкируемой последовательностью или следует за ней, но не обязательно должна быть смежной с фланкируемой последовательностью или непосредственно прилегающей к ней. Согласно одному варианту реализации термин «фланкирование» относится к концевым повторам на каждом конце линейного дуплексного зкДНК-вектора.
[00119] В настоящем документе термин «зкДНК-геном» относится к экспрессионной кассете, которая дополнительно включает по меньшей мере одну область инвертированного концевого повтора. ЗкДНК-геном может дополнительно содержать одну или более спейсерных областей. Согласно некоторым вариантам реализации зкДНК-геном включен в виде межмолекулярного дуплексного полинуклеотида ДНК в плазмиду или вирусный геном.
[00120] В настоящем документе термин «спейсерная область зкДНК» относится к промежуточной последовательности, которая разделяет функциональные элементы в зкДНК-векторе или зкДНК-геноме. Согласно некоторым вариантам реализации спейсерные области зкДНК удерживают два функциональных элемента на целевом расстоянии для оптимальной функциональности. Согласно некоторым вариантам реализации спейсерные области зкДНК обеспечивают или увеличивают генетическую стабильность зкДНК-генома, например, в плазмиде или бакуловирусе. Согласно некоторым вариантам реализации спейсерные области зкДНК облегчают подготовленные генетические манипуляции с зкДНК-геномом, обеспечивая удобное расположение для сайтов клонирования и т.п. Например, согласно определенным аспектам, олигонуклеотидный «полилинкер», содержащий несколько сайтов рестрикционных эндонуклеаз, или последовательность не из открытой рамки считывания, сконструированная так, чтобы в ней отсутствовали известные сайты связывания белка (например, транскрипционного фактора), могут быть размещены в зкДНК-геноме для разделения цис-действующих факторов, например, с помощью 6-членной, 12-членной, 18-членной, 24-членной, 48-членной, 86-членной, 176-членной вставки и т.п. между сайтом концевого разрешения и расположенным в 5´-направлении транскрипционным регуляторным элементом. Аналогичным образом, спейсер может быть встроен между последовательностью сигнала полиаденилирования и 3'-сайтом концевого разрешения.
[00121] В настоящем документе термин «зкДНК-плазмида» относится к плазмиде, которая содержит зкДНК-геном в виде межмолекулярного дуплекса.
[00122] В настоящем документе термин «зкДНК-бакмида» относится к геному инфекционного бакуловируса, содержащему зкДНК-геном в виде межмолекулярного дуплекса, который способен размножаться в E. coli в виде плазмиды и, соответственно, может выполнять роль челночного вектора для бакуловируса.
[00123] В настоящем документе термин «зкДНК-бакуловирус» относится к бакуловирусу, который содержит зкДНК-геном в виде межмолекулярного дуплекса в геноме бакуловируса.
[00124] В настоящем документе термины «инфицированная зкДНК-бакуловирусом клетка насекомого» и «зкДНК-BIIC» используются взаимозаменяемо и относятся к клетке-хозяину беспозвоночного животного (включая, но не ограничиваясь этим, клетку насекомого (например, клетку Sf9)), инфицированной зкДНК-бакуловирусом.
[00125] В настоящем документе термин «ДНК-вектор с замкнутыми концами» относится к бескапсидному ДНК-вектору по меньшей мере с одним ковалентно замкнутым концом, причем по меньшей мере часть вектора имеет структуру внутримолекулярного дуплекса.
[00126] В настоящем документе термин «зкДНК» относится к бескапсидной линейной двухцепочечной (дц) дуплексной ДНК с замкнутыми концами для невирусного переноса генов, синтетической или другой. Подробное описание зкДНК приведено в международной заявке PCT/US2017/020828, поданной 3 марта 2017 г., содержание которой полностью явным образом включено в настоящий документ посредством ссылки. Некоторые способы получения зкДНК, содержащей различные последовательности и конфигурации инвертированных концевых повторов (ITR), с использованием клеточных способов описаны в Примере 1 международной заявки PCT/US18/49996, поданной 7 сентября 2018 г., и PCT/US2018/064242, поданной 6 декабря 2018 г., каждая из которых полностью включена в настоящий документ посредством ссылки. Некоторые способы получения синтетических зкДНК-векторов, содержащих различные последовательности и конфигурации ITR, описаны, например, в международной заявке PCT/US2019/14122, поданной 18 января 2019 г., содержание которой полностью включено в настоящий документ посредством ссылки.
[00127] В настоящем документе термины «зкДНК-вектор» и «зкДНК» используются взаимозаменяемо и относятся к ДНК-вектору с замкнутыми концами, содержащему по меньшей мере один концевой палиндром. Согласно некоторым вариантам реализации зкДНК содержит два ковалентно замкнутых конца.
[00128] В настоящем документе термин «нзДНК» или «никированная зкДНК» относится к ДНК с замкнутыми концами, имеющей одноцепочечный разрыв или пропуск размером 1-100 пар оснований в области «стебля» или спейсерной области, расположенной в 5´-направлении относительно открытой рамки считывания (например, промотора и трансгена, подлежащего экспрессии).
[00129] В настоящем документе термины «пропуск» и «одноцепочечный разрыв» используются взаимозаменяемо и относятся к прерванной части синтетического ДНК-вектора согласно настоящему изобретению, создавая участок части одноцепочечной ДНК в двухцепочечной в других местах зкДНК. Пропуск может иметь от одной 1 пары оснований до 100 пар оснований в длину в одной цепи дуплексной ДНК. Типичные пропуски, разработанные и созданные с применением способов, описанных в настоящем документе, и синтетические векторы, полученные с применением указанных способов, могут иметь, например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59 или 60 пар оснований в длину. Примерные пропуски в настоящем раскрытии могут иметь от 1 пары оснований до 10 пар оснований в длину, от 1 до 20 пар оснований в длину, от 1 до 30 пар оснований в длину.
[00130] В настоящем документе термины «сайт связывания Rep», «элемент связывания Rep», «RBE» и «RBS» используются взаимозаменяемо и относятся к сайту связывания белка Rep (например, Rep 78 ААВ или Rep 68 ААВ), который после связывания белком Rep позволяет белку Rep выполнять его сайт-специфичную эндонуклеазную активность в отношении последовательности, включающей RBS. Последовательность RBS и ее обратный комплемент вместе образуют один RBS. Последовательности RBS известны в данной области техники и включают, например, 5´-GCGCGCTCGCTCGCTC-3′ (SEQ ID NO: 39), представляющую собой последовательность RBS, идентифицированную в ААВ2. В вариантах реализации настоящего изобретения можно применять любую известную последовательность RBS, включая другие известные последовательности RBS ААВ и другие известные природные или синтетические последовательности RBS. Без ограничения какой-либо теорией, считается, что нуклеазный домен белка Rep связывается с дуплексной нуклеотидной последовательностью GCTC и, соответственно, происходит прямое связывание и стабильная сборка двух известных белков Rep ААВ на дуплексном олигонуклеотиде, 5´-(GCGC)(GCTC)(GCTC)(GCTC)-3´ (SEQ ID NO: 39). Кроме того, растворимые агрегированные конформеры (т.е. неопределенное число взаимосвязанных белков Rep) диссоциируют и связываются с олигонуклеотидами, которые содержат сайты связывания Rep. Каждый белок Rep взаимодействует как с азотистыми основаниями, так и с фосфодиэфирным остовом на каждой цепи. Взаимодействия с азотистыми основаниями обеспечивают специфичность в отношении последовательности, в то время как взаимодействия с фосфодиэфирным остовом неспецифичны или менее специфичны в отношении последовательностей и стабилизируют комплекс белка с ДНК.
[00131] В настоящем документе термины «сайт концевого разрешения» и «TRS» используются в настоящем документе взаимозаменяемо и относятся к области, в которой Rep образует тирозин-фосфодиэфирную связь с 5'-тимидином с образованием 3'-ОН, который служит субстратом для удлинения ДНК с помощью клеточной ДНК-полимеразы, например, ДНК-полимеразы дельта или ДНК-полимеразы эпсилон. В качестве альтернативы, комплекс Rep-тимидин может принимать участие в скоординированной реакции лигирования. Согласно некоторым вариантам реализации TRS включает, как минимум, неспаренный тимидин. Согласно некоторым вариантам реализации никирующую эффективность TRS по меньшей мере отчасти можно контролировать за счет расстояния между ним и RBS в пределах одной молекулы. Если акцепторный субстрат представляет собой комплементарный ITR, тогда полученный продукт представляет собой внутримолекулярный дуплекс. Последовательности TRS известны в данной области техники и включают, например, 5´-GGTTGA-3´ (SEQ ID NO: 804), представляющую собой гексануклеотидную последовательность, идентифицированную в ААВ2. В вариантах реализации настоящего изобретения можно применять любую известную последовательность TRS, включая другие известные последовательности TRS ААВ и другие известные природные или синтетические последовательности TRS, такие как AGTT (SEQ ID NO: 085), GGTTGG (SEQ ID NO: 806), AGTTGG (SEQ ID NO: 807), AGTTGA (SEQ ID NO: 808), и другие мотивы, такие как RRTTRR (SEQ ID NO: 809).
[00132] В настоящем документе термины «смысловой» и «антисмысловой» относятся к ориентации структурного элемента на полинуклеотиде. Смысловой и антисмысловой варианты элемента обратно комплементарны друг другу.
[00133] В настоящем документе термин «синтетический ААВ-вектор» и «получение ААВ-вектора синтетическим путем» относится к ААВ-вектору и синтетическим способам его получения в полностью бесклеточной среде.
[00134] В настоящем документе «репортеры» относятся к белкам, которые можно применять для обеспечения детектируемых показателей. Репортеры обычно генерируют измеряемый сигнал, такой как флуоресценция, цвет или люминесценция. Последовательности, кодирующие репортерные белки, кодируют белки, присутствие которых в клетке или в организме легко наблюдать. Например, флуоресцентные белки заставляют клетку флуоресцировать при возбуждении светом с конкретной длиной волны, люциферазы заставляют клетку катализировать реакцию, которая генерирует свет, а ферменты, такие как β-галактозидаза, преобразуют субстрат в окрашенный продукт. Примерные репортерные полипептиды, которые можно применять для экспериментальных или диагностических целей, включают, но не ограничиваются перечисленными, β-лактамазу, β-галактозидазу (LacZ), щелочную фосфатазу (AP), тимидинкиназу (TK), зеленый флуоресцентный белок (GFP) и другие флуоресцентные белки, хлорамфениколацетилтрансферазу (CAT), люциферазу и другие, хорошо известные в данной области техники.
[00135] В настоящем документе термин «эффекторный белок» относится к полипептиду, который обеспечивает детектируемый показатель, либо, например, как репортерный полипептид, либо, более предпочтительно, как полипептид, уничтожающий клетку, например, токсин, или агент, придающий клетке чувствительность к уничтожению выбранным агентом или при его отсутствии. Эффекторные белки включают любой белок или пептид, который непосредственно нацелен на ДНК и/или РНК клетки-хозяина или повреждает их. Например, эффекторные белки могут включать, но не ограничиваются перечисленными, рестрикционную эндонуклеазу, которая нацелена на последовательность ДНК клетки-хозяина (будь то геномный или внехромосомный элемент), протеазу, которая расщепляет полипептид-мишень, необходимый для выживания клетки, ингибитор ДНК-гиразы и токсин рибонуклеазного типа. Согласно некоторым вариантам реализации экспрессия эффекторного белка, контролируемая синтетическим биологическим контуром, описанным в настоящем документе, может принимать участие в качестве фактора в другом синтетическом биологическом контуре, что расширяет диапазон и сложность чувствительности системы биологического контура.
[00136] Транскрипционные регуляторы относятся к транскрипционным активаторам и репрессорам, которые либо активируют, либо репрессируют транскрипцию представляющего интерес гена, такого как антагонист инфламмасомы (например, ингибитор одного или более из пути инфламмасомы NLRP3 и/или AIM2 или ингибитора каспазы 1). Промоторы представляют собой области нуклеиновой кислоты, которые инициируют транскрипцию конкретного гена. Транскрипционные активаторы, как правило, связываются поблизости от транскрипционных промоторов и привлекают РНК-полимеразу для непосредственной инициации транскрипции. Репрессоры связываются с транскрипционными промоторами и стерически затрудняют инициацию транскрипции РНК-полимеразой. Другие транскрипционные регуляторы могут служить либо активаторами, либо репрессорами в зависимости от места их связывания и условий в клетке и в окружающей среде. Неограничивающие примеры классов транскрипционных регуляторов включают, но не ограничиваются перечисленными, белки гомеодомена, белки с цинковыми пальцами, белки с мотивом «крылатая спираль» (белки Forkhead) и белки с лейциновой молнией.
[00137] В настоящем документе «репрессорный белок» или «индукторный белок» представляет собой белок, который связывается с элементом регуляторной последовательности и репрессирует или активирует, соответственно, транскрипцию последовательностей, функционально связанных с указанным элементом регуляторной последовательности. Предпочтительные репрессорные и индукторные белки, описанные в настоящем документе, чувствительны к присутствию или отсутствию по меньшей мере одного вводимого агента или фактора внешней среды. Предпочтительные белки, описанные в настоящем документе, являются модульными по форме и содержат, например, отделяемые ДНК-связывающие и связывающие вводимые агенты, или реагирующие на них, элементы или домены.
[00138] В настоящем документе термин «носитель» включает любые и все возможные растворители, дисперсионные среды, основы, покрытия, разбавители, антибактериальные и противогрибковые агенты, изотонические и замедляющие абсорбцию агенты, буферы, растворы-носители, суспензии, коллоиды и т.п. Применение таких сред и агентов для фармацевтически активных веществ хорошо известно в данной области техники. В композиции также могут быть включены вспомогательные активные ингредиенты. Выражение «фармацевтически приемлемый» относится к молекулярным частицам и композициям, которые не приводят к токсической, аллергической или схожей нежелательной реакции при введении хозяину.
[00139] В настоящем документе термин «домен, реагирующий на вводимый агент» представляет собой домен транскрипционного фактора, который связывает или иначе отвечает на условие или вводимый агент таким способом, который придает восприимчивость присоединенного к нему ДНК-связывающего слитого домена к наличию указанного условия или вводимого агента. Согласно одному варианту реализации наличие указанного условия или вводимого агента приводит к конформационному изменению в отвечающем на вводимый агент домене, или в белке, с которым он слит, которое модифицирует модулирующую транскрипцию активность транскрипционного фактора.
[00140] Термин «in vivo» относится к анализам или способам, которые реализуют у организма или внутри организма, такого как многоклеточное животное. Согласно некоторым аспектам, описанным в настоящем документе, считается, что способ или применение реализуют «in vivo», если используется одноклеточный организм, такой как бактерия. Термин «ex vivo» относится к способам и вариантам применения, которые выполняют с использованием живой клетки с интактной мембраной, находящейся вне организма многоклеточного животного или растения, например, эксплантатов, культивируемых клеток, включая первичные клетки и линии клеток, трансформированных линий клеток, а также экстрагированных тканей или клеток, включая клетки крови, помимо прочего. Термин «in vitro» относится к анализам и способам, которые не требуют наличия клетки с интактной мембраной, например, клеточных экстрактов, и могут относиться к введению программируемого синтетического биологического контура в бесклеточной системе, такой как среда, не содержащая клеток или клеточных систем, таких как клеточные экстракты.
[00141] В настоящем документе термин «промотор» относится к любой последовательности нуклеиновой кислоты, которая регулирует экспрессию другой последовательности нуклеиновой кислоты за счет управления транскрипцией указанной последовательности нуклеиновой кислоты, которая может представлять собой гетерологичный целевой ген, кодирующий белок или РНК. Промоторы могут быть конститутивными, индуцируемыми, репрессируемыми, тканеспецифическими или могут представлять собой любую комбинацию перечисленных. Промотор представляет собой контрольную область последовательности нуклеиновой кислоты, в которой осуществляется контроль инициации и скорости транскрипции остальной части последовательности нуклеиновой кислоты. Промотор также может содержать генетические элементы, с которыми могут связываться регуляторные белки и молекулы, такие как РНК-полимераза и другие транскрипционные факторы. Согласно некоторым вариантам реализации аспектов, описанных в настоящем документе, промотор может управлять экспрессией транскрипционного фактора, который регулирует экспрессию самого промотора. В последовательности промотора может быть расположен сайт инициации транскрипции, а также связывающие белки домены, отвечающие за связывание РНК-полимеразы. Эукариотические промоторы часто, однако не всегда, будут содержать «TATA»-боксы и «CAT»-боксы. Различные промоторы, включая индуцируемые промоторы, можно применять для управления экспрессией трансгенов в зкДНК-векторах, раскрытых в настоящем документе. Последовательность промотора может быть ограничена на ее 3´-конце сайтом инициации транскрипции и простираться в восходящем направлении (в 5´-направлении), чтобы включить минимальное количество оснований или элементов, необходимых для инициации транскрипции на уровнях, детектируемых выше фона.
[00142] В настоящем документе термин «энхансер» относится к цис-действующей регуляторной последовательности (например, 50-1500 пар оснований), которая связывает один или более белков (например, белки-активаторы или транскрипционный фактор) для повышения транскрипционной активации последовательности нуклеиновой кислоты. Энхансеры могут быть расположены на расстоянии до 1000000 пар оснований выше сайта начала гена или ниже сайта начала гена, который они регулируют. Энхансер может быть расположен в пределах интронной области или в экзонной области неродственного гена.
[00143] Можно сказать, что промотор управляет экспрессией или управляет транскрипцией последовательности нуклеиновой кислоты, которую он регулирует. Выражения «функционально связанный», «функционально расположенный», «с функциональной связью», «под контролем» и «под транскрипционным контролем» указывают, что промотор находится в корректном функциональном положении и/или ориентации относительно последовательности нуклеиновой кислоты, которую он регулирует, для контроля инициации транскрипции и/или экспрессии указанной последовательности. В настоящем документе «инвертированный промотор» относится к промотору, в котором последовательность нуклеиновой кислоты располагается в обратной ориентации так, что цепь, которая была смысловой, становится некодирующей цепью, и наоборот. Последовательности инвертированных промоторов можно применять в различных вариантах реализации для регуляции состояния переключателя. Кроме того, согласно различным вариантам реализации, промотор можно применять в сочетании с энхансером.
[00144] Промотор может представлять собой промотор, который в естественных условиях ассоциирован с геном или последовательностью, а также может быть получен путем выделения некодирующих 5´-последовательностей, расположенных в 5´-направлении относительно кодирующего сегмента и/или экзона определенного гена или последовательности. Такой промотор может называться «эндогенным». Аналогичным образом, согласно некоторым вариантам реализации, энхансер может быть представлен энхансером, который в естественных условиях ассоциирован с последовательностью нуклеиновой кислоты и расположен либо в 3', либо в 5'-направлении относительно указанной последовательности.
[00145] Согласно некоторым вариантам реализации кодирующий сегмент нуклеиновой кислоты расположен под контролем «рекомбинантного промотора» или «гетерологичного промотора», оба указанных термина относятся к промотору, который обычно не ассоциирован с кодируемой последовательностью нуклеиновой кислоты, с которой промотор функционально связан в ее естественной среде. Рекомбинантный или гетерологичный энхансер относится к энхансеру, обычно не ассоциированному с конкретной последовательностью нуклеиновой кислоты в ее естественной среде. Такие промоторы или энхансеры могут включать промоторы или энхансеры других генов; промоторы или энхансеры, выделенные из любой другой прокариотической, вирусной или эукариотической клетки; и синтетические промоторы или энхансеры, которые не являются «природными», т.е. содержат различные элементы разных транскрипционных регуляторных областей и/или изменяющие экспрессию мутации, введенные с применением способов генетического конструирования, известных в данной области техники. Наряду с получением последовательностей нуклеиновых кислот промоторов и энхансеров синтетическим путем, последовательности промоторов могут быть получены с использованием рекомбинантного клонирования и/или технологии амплификации нуклеиновых кислот, включая ПЦР, применительно к синтетическим биологическим контурам и модулям, раскрытым в настоящем документе (см., например, патент США № 4683202, патент США № 5928906, каждый из которых включен в настоящий документ посредством ссылки). Кроме того, также предусмотрена возможность применения контрольных последовательностей, которые управляют транскрипцией и/или экспрессией последовательностей в неядерных органеллах, таких как митохондрии, хлоропласты и т.п.
[00146] Как описано в настоящем документе, «индуцируемый промотор» представляет собой промотор, который характеризуется инициацией или усилением транскрипционной активности в присутствии, под влиянием или при контакте с индуктором или индуцирующим агентом. «Индуктор» или «индуцирующий агент», как определено в настоящем документе, может быть эндогенным или обычно является экзогенным соединением или белком, которые вводят таким образом, чтобы обеспечить их активность по индукции транскрипционной активности индуцируемого промотора. Согласно некоторым вариантам реализации индуктор или индуцирующий агент, т.е. химическое вещество, соединение или белок, сам может быть получен в результате транскрипции или экспрессии последовательности нуклеиновой кислоты (т.е. индуктор может представлять собой индукторный белок, экспрессируемый другим компонентом или модулем), который сам может находиться под контролем индуцируемого промотора. Согласно некоторым вариантам реализации индуцируемый промотор индуцируется в отсутствие определенных агентов, таких как репрессор. Примеры индуцируемых промоторов включают, но не ограничиваются перечисленными, чувствительные к тетрациклину, металлотионину, экдизону промоторы, промоторы вирусов млекопитающих (например, поздний промотор аденовируса и промотор длинного концевого повтора вируса опухоли молочной железы мышей (MMTV-LTR)) и другие чувствительные к стероидам промоторы, чувствительные к рапамицину промоторы и т.п.
[00147] Термины «регуляторные последовательности ДНК», «контрольные элементы» и «регуляторные элементы», используемые в настоящем документе взаимозаменяемо, относятся к транскрипционным и трансляционным контрольным последовательностям, таким как промоторы, энхансеры, сигналы полиаденилирования, терминаторы, сигналы разрушения белка и т.п., которые обеспечивают и/или регулируют транскрипцию некодирующей последовательности (например, нацеливающей на ДНК РНК) или кодирующей последовательности (например, сайт-направленного модифицирующего полипептида или полипептида Cas9/Csn1) и/или регулируют трансляцию кодируемого полипептида.
[00148] Выражение «функционально связанный» относится к размещению в непосредственной близости, при котором описываемые так компоненты находятся во взаимосвязи, позволяющей им функционировать предусмотренным для них образом. Например, промотор функционально связан с кодирующей последовательностью, если указанный промотор влияет на ее транскрипцию или экспрессию. «Экспрессионная кассета» включает гетерологичную последовательность ДНК, которая функционально связана с промотором или другой регуляторной последовательностью, достаточными для управления транскрипцией трансгена в зкДНК-векторе. Подходящие промоторы включают, например, тканеспецифические промоторы. Промоторы также могут происходить из ААВ.
[00149] В настоящем документе термин «субъект» относится к человеку или животному, которому предоставляют лечение, включая профилактическое лечение, с применением зкДНК-вектора в соответствии с настоящим изобретением. Обычно животное представляет собой позвоночное животное, такое как, но не ограничиваясь перечисленными, примат, грызун, домашнее животное или промысловое животное. Приматы включают, но не ограничиваются перечисленными, шимпанзе, яванских макак, коат и макак, например, макак-резусов. Грызуны включают мышей, крыс, сурков, хорьков, кроликов и хомяков. Домашние и промысловые животные включают, но не ограничиваются перечисленными, коров, лошадей, свиней, оленей, бизона, буйвола, виды кошачьих, например, домашнюю кошку, виды собачьих, например, собаку, лису, волка, виды птиц, например, курицу, эму, страуса, и рыб, например, форель, сома и лосося. В определенных вариантах реализации аспектов, описанных в настоящем документе, субъект представляет собой млекопитающего, например, примата или человека. Субъект может быть мужского или женского пола. Кроме того, субъект может представлять собой младенца или ребенка. Согласно некоторым вариантам реализации субъект может представлять собой новорожденного или нерожденного субъекта, например, субъект находится in utero. Предпочтительно субъект представляет собой млекопитающего. Млекопитающее может представлять собой человека, примата, отличного от человека, мышь, крысу, собаку, кошку, лошадь или корову, однако указанные примеры не являются ограничивающими. Млекопитающие, отличные от человека, могут быть предпочтительно использованы в качестве субъектов в моделях заболеваний и нарушений на животных. Кроме того, способы и композиции, описанные в настоящем документе, можно применять для одомашненных животных и/или животных-компаньонов. Человек-субъект может быть любого возраста, пола, расы или этнической группы, например, может быть европеоидом (белым), азиатом, африканцем, черным, афроамериканцем, афроевропейцем, испаноамериканцем, представителем ближневосточного этноса и т.д. Согласно некоторым вариантам реализации субъект может представлять собой пациента или другого субъекта в клинических условиях. Согласно некоторым вариантам реализации субъект уже проходит курс лечения. Согласно некоторым вариантам реализации субъект представляет собой эмбрион, плод, новорожденного, младенца, ребенка, подростка или взрослую особь. Согласно некоторым вариантам реализации субъект представляет собой плод человека, новорожденного человека, младенца-человека, ребенка-человека, подростка-человека или взрослого человека. Согласно некоторым вариантам реализации субъект представляет собой эмбрион животного или эмбрион, не принадлежащий человеку, или эмбрион примата, отличного от человека. Согласно некоторым вариантам реализации субъект представляет собой эмбрион человека.
[00150] В настоящем документе термин «клетка-хозяин» включает любой тип клеток, которые являются восприимчивыми к трансформации, трансфекции, трансдукции и т.п. с применением конструкции нуклеиновой кислоты или экспрессионного зкДНК-вектора согласно настоящему раскрытию. В качестве неограничивающих примеров, клетка-хозяин может представлять собой выделенную первичную клетку, плюрипотентные стволовые клетки, клетки CD34+, индуцированные плюрипотентные стволовые клетки или любую из ряда иммортализованных линий клеток (например, клетки HepG2). В качестве альтернативы, клетка-хозяин может представлять собой клетку in situ или in vivo в ткани, органе или организме.
[00151] Термин « экзогенный» относится к веществу, присутствующему в клетке, отличной от его природного источника. В настоящем документе термин «экзогенный» может относиться к нуклеиновой кислоте (например, нуклеиновой кислоте, кодирующей полипептид) или к полипептиду, которые были введены с помощью способа, включающего действия человека, в биологическую систему, такую как клетка или организм, в которых они обычно не обнаруживаются, и введение нуклеиновой кислоты или полипептида в такую клетку или организм является желательным. В качестве альтернативы, «экзогенный» может относиться к нуклеиновой кислоте или полипептиду, которые были введены с помощью способа, включающего действия человека, в биологическую систему, такую как клетка или организм, в которой они обнаруживаются в относительно низких количествах, и увеличение количества нуклеиновой кислоты или полипептида в клетке или организме является желательным, например, для получения эктопической экспрессии или уровней. Напротив, термин «эндогенный» относится к веществу, которое является нативным для биологической системы или клетки.
[00152] Термин «идентичность последовательности» относится к сродству между двумя нуклеотидными последовательностями. Для целей настоящего раскрытия степень идентичности последовательностей между двумя дезоксирибонуклеотидными последовательностями определяется с использованием алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, выше), реализованного в программе Needle пакета EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, выше), предпочтительно версии 3.0.0 или более поздней. Используемые необязательные параметры представляют собой штраф за открытие пропуска 10, штраф за продление пропуска 0,5 и матрицу замены EDNAFULL (версия EMBOSS NCBI NUC4.4). Выходной показатель Needle, называемый «наибольшей длиной идентичного фрагмента» (полученный с применением опции «-nobrief»), используют в качестве процента идентичности и вычисляют следующим образом: (Идентичные дезоксирибонуклеотиды×100)/(Длина выравнивания - Общее число пропусков в выравнивании). Длина выравнивания составляет предпочтительно по меньшей мере 10 нуклеотидов, предпочтительно по меньшей мере 25 нуклеотидов, более предпочтительно по меньшей мере 50 нуклеотидов и наиболее предпочтительно по меньшей мере 100 нуклеотидов.
[00153] В настоящем документе термин «гомология» или «гомологичный» определяется как процент нуклеотидных остатков, которые идентичны нуклеотидным остаткам в соответствующей последовательности на хромосоме-мишени после выравнивания последовательностей и введения пробелов, при необходимости, для достижения максимального процента идентичности последовательностей. Выравнивание для целей определения процента гомологии нуклеотидной последовательности может быть достигнуто различными путями, известными в данной области техники, например, с применением общедоступного компьютерного программного обеспечения, такого как программное обеспечение BLAST, BLAST-2, ALIGN, ClustalW2 или Megalign (DNASTAR). Специалисты в данной области техники могут определить соответствующие параметры для выравнивания последовательностей, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. Согласно некоторым вариантам реализации последовательность нуклеиновой кислоты (например, последовательность ДНК), например, гомологичное плечо, считается «гомологичной», когда последовательность по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или более идентична соответствующей нативной или неотредактированной последовательности нуклеиновой кислоты (например, геномной последовательности) клетки-хозяина.
[00154] В настоящем документе термин «гетерологичный» означает нуклеотидную или полипептидную последовательность, которая не обнаружена в нативной нуклеиновой кислоте или белке, соответственно. Гетерологичная последовательность нуклеиновой кислоты может быть связана с природной последовательностью нуклеиновой кислоты (или ее вариантом) (например, путем генетического конструирования) с получением химерной нуклеотидной последовательности, кодирующей химерный полипептид. Гетерологичная последовательность нуклеиновой кислоты может быть соединена с вариантом полипептида (например, путем генетического конструирования) с получением нуклеотидной последовательности, кодирующей слитый вариант полипептида.
[00155] «Вектор» или «экспрессионный вектор» представляет собой репликон, такой как плазмида, бакмида, фаг, вирус, вирион или космида, к которому может быть присоединен другой сегмент ДНК, т.е. «вставка», чтобы вызвать репликацию присоединенного сегмента в клетке. Вектор может представлять собой конструкцию нуклеиновой кислоты, разработанную для доставки в клетку-хозяина или для переноса между разными клетками-хозяевами. В настоящем документе вектор может быть вирусным или невирусным по происхождению и/или в конечной форме, однако для цели настоящего раскрытия «вектор» обычно относится к зкДНК-вектору в соответствии с использованием этого термин в настоящем документе. Термин «вектор» включает любой генетический элемент, который способен к репликации, когда он ассоциирован с надлежащими контрольными элементами, и который может переносить генные последовательности в клетки. Согласно некоторым вариантам реализации вектор может представлять собой экспрессионный вектор или рекомбинантный вектор.
[00156] В настоящем документе термин «экспрессионный вектор» относится к вектору, который управляет экспрессией РНК или полипептида с последовательностей, связанных с транскрипционными регуляторными последовательностями на векторе. Экспрессируемые последовательности часто, но не обязательно, будут гетерологичными по отношению к клетке. Экспрессионный вектор может содержать дополнительные элементы, например, экспрессионный вектор может иметь две системы репликации, что позволяет поддерживать его в двух организмах, например, в клетках человека для экспрессии и в прокариотическом хозяине для клонирования и амплификации. Термин «экспрессия» относится к клеточным процессам, участвующим в продуцировании РНК и белков и, в соответствующих случаях, в секретировании белков, включая, где это применимо, но не ограничиваясь этим, например, транскрипцию, процессинг транскрипта, трансляцию и укладку, модификацию и процессинг белка. «Продукты экспрессии» включают РНК, транскрибированную с гена, и полипептиды, полученные путем трансляции мРНК, транскрибированной с гена. Термин «ген» означает последовательность нуклеиновой кислоты, которая транскрибируется (ДНК) в РНК in vitro или in vivo, когда она функционально связана с соответствующими регуляторными последовательностями. Ген может включать, но не обязательно, области, предшествующие кодирующей области и следующие за ней, например, 5´-нетранслируемые (5´-UTR) или «лидерные» последовательности и 3´-UTR или «трейлерные» последовательности, а также промежуточные последовательности (интроны) между отдельными кодирующими сегментами (экзонами).
[00157] Под «рекомбинантным вектором» подразумевается вектор, который включает гетерологичную последовательность нуклеиновой кислоты или «трансген», который способен к экспрессии in vivo. Следует понимать, что векторы, описанные в настоящем документе, в некоторых вариантах реализации, можно комбинировать с другими подходящими композициями и видами терапии. Согласно некоторым вариантам реализации вектор является эписомальным. Применение подходящего эписомального вектора обеспечивает средство поддержания представляющего интерес нуклеотида у субъекта в многокопийной внехромосомной ДНК, что устраняет потенциальные эффекты интеграции в хромосому.
[00158] В настоящем документе выражение «генетическое заболевание» относится к заболеванию, частично или полностью, прямо или опосредованно, вызванному одной или более аномалиями в геноме, в частности, к состоянию, присутствующему с момента рождения. Аномалия может представлять собой мутацию, вставку или делецию. Аномалия может влиять на кодирующую последовательность гена или его регуляторную последовательность. Генетическое заболевание может представлять собой, но не ограничивается перечисленными, МДД (мышечную дистрофию Дюшенна), гемофилию, муковисцидоз, хорею Хантингтона, семейную гиперхолестеринемию (дефект рецептора ЛПНП), гепатобластому, болезнь Вильсона, врожденную печеночную порфирию, наследственные нарушения метаболизма печени, синдром Леша-Нихана, серповидноклеточную анемию, талассемии, пигментную ксеродерму, анемию Фанкони, пигментный ретинит, атаксию-телеангиэктазию, синдром Блума, ретинобластому и болезнь Тея-Сакса.
[00159] В настоящем документе термин «ингибирующий полинуклеотид» относится к молекуле ДНК или РНК, которая уменьшает или предотвращает экспрессию (транскрипцию или трансляцию) второго (целевого) полинуклеотида. Ингибирующие полинуклеотиды включают антисмысловые полинуклеотиды, рибозимы и внешние направляющие последовательности. Термин «ингибирующий полинуклеотид» дополнительно включает молекулы ДНК и РНК, например, РНКи, которые кодируют существующие ингибирующие молекулы, такие как молекулы ДНК, которые кодируют рибозимы.
[00160] В настоящем документе «подавление гена» или «подавленный ген» в отношении активности молекулы РНКи, например, киРНК или миРНК, относится к снижению в клетке уровня мРНК гена-мишени (например, мРНК NLRP3, AIM2 или каспазы-1) по меньшей мере примерно на 5%, примерно на 10%, примерно на 20%, примерно на 30%, примерно на 40%, примерно на 50%, примерно на 60%, примерно на 70%, примерно на 80%, примерно на 90%, примерно на 95%, примерно на 99%, примерно на 100% от уровня мРНК, обнаруженного в клетке без присутствия миРНК или молекулы РНК-интерференции. Согласно одному предпочтительному варианту реализации уровни мРНК снижены по меньшей мере примерно на 70%, примерно на 80%, примерно на 90%, примерно на 95%, примерно на 99%, примерно на 100%.
[00161] В настоящем документе термин «РНКи» относится к любому типу интерферирующей РНК, включая, но не ограничиваясь перечисленными, киРНКи, кшРНКи, эндогенную микроРНК и искусственную микроРНК. Например, он включает последовательности, ранее идентифицированные как киРНК, независимо от механизма последующего процессинга РНК (т.е. несмотря на то, что киРНК, как полагают, имеют специфичный способ процессинга in vivo, приводящий к расщеплению мРНК, такие последовательности могут быть включены в векторы между фланкирующими последовательностями, описанными в настоящем документе). Термин «РНКи» может включать как молекулы РНКи, подавляющие ген, так и эффекторные молекулы РНКи, которые активируют экспрессию гена. Только в качестве примера, согласно некоторым вариантам реализации агенты РНКи, которые служат для ингибирования или подавления гена, можно применять в способах, наборах и композициях, раскрытых в настоящем документе, например, для ингибирования иммунного ответа (например, врожденного иммунного ответа).
[00162] В настоящем документе термин «содержащий» или «содержит» используется в отношении композиций, способов и их соответствующего компонента (компонентов), существенных для указанного способа или композиции, но допускающих включение не указанных элементов, будь то существенных или нет.
[00163] В настоящем документе термин «состоящий по существу из» относится к элементам, необходимым для конкретного варианта реализации. Термин допускает наличие элементов, которые не влияют существенным образом на основную и новую или функциональную характеристику (характеристики) указанного варианта реализации. Использование слова «содержащий» указывает на включение, а не на ограничение.
[00164] Термин «состоящий из» относится к композициям, способам и их соответствующим компонентам, описанным в настоящем документе, которые не включают никаких элементов, не перечисленных в указанном описании варианта реализации.
[00165] В настоящем документе термин «состоящий по существу из» относится к элементам, которые необходимы для конкретного варианта реализации. Термин допускает наличие дополнительных элементов, которые не оказывают существенного влияния на основную и новую или функциональную характеристику (характеристики) указанного варианта реализации настоящего изобретения.
[00166] В настоящем описании и прилагаемой формуле изобретения термины в единственном числе (соотв. «a», «an» и «the» в исходном тексте на английском языке) включают их эквиваленты во множественном числе, если иное явным образом не следует из контекста. Соответственно, например, ссылка на «способ» включает один или более способов и/или этапов типа, описанного в настоящем документе, и/или таких, которые будут понятны специалистам в данной области техники после прочтения настоящего раскрытия, и т.п. Аналогичным образом, предусмотрено, что термин «или» включает «и», если из контекста явным образом не следует иное. Несмотря на то, что при практической реализации или тестировании настоящего изобретения могут применяться способы и материалы, аналогичные или эквивалентные тем, которые описаны в настоящем документе, ниже описаны подходящие способы и материалы. В настоящем документе «например» (сокращение «e.g.» в исходном тексте на английском языке, произошедшее от латинского выражения «exempli gratia») указывает на неограничивающий пример. Соответственно, «например» синонимично термину «к примеру».
[00167] За исключением рабочих примеров или если указано иное, все числа, выражающие количества ингредиентов или условий реакции, используемые в настоящем документе, следует понимать как модифицированные термином «примерно» во всех случаях. Термин «примерно» при использовании применительно к процентам может означать ±1%. Настоящее изобретение дополнительно более подробно объяснено в приведенных ниже примерах, однако объем настоящего изобретения ими не ограничен.
[00168] Группы альтернативных элементов или вариантов реализации настоящего изобретения, раскрытых в настоящем документе, не должны рассматриваться как ограничения. На каждый член группы можно ссылаться и заявлять его индивидуально или в любой комбинации с другими членами группы или другими элементами, представленными в настоящем документе. Один или более членов группы могут быть включены или исключены из группы по соображениям удобства и/или патентоспособности. При любом таком включении или исключении в настоящем документе считается, что описание изобретения содержит группу, измененную таким образом, чтобы она удовлетворяла условию письменного описания всех групп Маркуша, используемых в прилагаемой формуле изобретения.
[00169] Согласно некоторым вариантам реализации любого из аспектов раскрытие, описанное в настоящем документе, не касается способа клонирования людей, способов модификации генетической идентичности зародышевой линии человека, вариантов применения эмбрионов человека в промышленных или коммерческих целях, или способов модификации генетической идентичности животных, которые могут причинить им страдания без какой-либо существенной медицинской пользы для человека или животного, а также животных, полученных с помощью таких способов.
[00170] Определения других терминов приведены в настоящем документе в описании различных аспектов настоящего изобретения.
II. Нуклеиновые кислоты
[00171] Нуклеиновые кислоты представляют собой большие, сильно заряженные молекулы, которые быстро разлагаются и выводятся из организма и обычно обладают плохими фармакологическими свойствами, так как они распознаются как чужеродное вещество для организма и становятся мишенью иммунного ответа (например, врожденного иммунного ответа). Следовательно, определенные нуклеиновые кислоты, такие как терапевтические нуклеиновые кислоты или нуклеиновые кислоты, используемые в исследовательских целях (например, антисмысловой олигонуклеотид или вирусные векторы), часто могут запускать иммунные ответы in vivo. Согласно настоящему раскрытию предложены фармацевтические композиции и способы, которые могут облегчать, уменьшать или устранять такие иммунные ответы и повышать эффективность нуклеиновых кислот за счет повышения уровней экспрессии посредством максимального увеличения устойчивости нуклеиновой кислоты при состоянии пониженной иммунологической реактивности у субъекта-реципиента. Это также может минимизировать любые потенциальные нежелательные явления, которые могут привести к повреждению органа или другой токсичности в ходе генной терапии. Многие из композиций и способов, предложенных в настоящем документе, относятся к введению специфичного ингибитора иммунного ответа (например, врожденного иммунного ответа) в сочетании с нуклеиновой кислотой (например, терапевтической нуклеиновой кислотой или нуклеиновой кислотой, используемой в исследовательских целях), что уменьшает иммунный ответ (например, врожденный иммунный ответ), запущенный присутствием нуклеиновой кислоты.
[00172] Иммуногенные/иммуностимулирующие нуклеиновые кислоты могут включать как дезоксирибонуклеиновые кислоты, так и рибонуклеиновые кислоты. Для дезоксирибонуклеиновых кислот (ДНК) было показано, что конкретная последовательность или мотив индуцирует иммунную стимуляцию у млекопитающих. Эти последовательности или мотивы включают, но не ограничиваются перечисленными, мотивы CpG, богатые пиримидином последовательности и палиндромные последовательности. Мотивы CpG в дезоксирибонуклеиновой кислоте часто распознаются эндосомальным Toll-подобным рецептором 9 (TLR-9), который, в свою очередь, запускает как путь стимуляции врожденного иммунитета, так и путь стимуляции приобретенного иммунитета. Определенные иммуностимулирующие последовательности рибонуклеиновой кислоты (РНК) связываются с Toll-подобным рецептором 6 и 7 (TLR-6 и TLR-7) и, как полагают, активируют провоспалительный ответ посредством иммунного ответа (например, врожденного иммунного ответа). Кроме того, двухцепочечная РНК часто может быть иммуностимулирующей из-за ее связывания с TLR-3. Таким образом, чужеродные молекулы нуклеиновых кислот, происходящие из патогенов или терапевтические по своему происхождению, могут быть высокоиммуногенными in vivo.
[00173] В настоящем документе предложена характеристика и разработка молекул нуклеиновых кислот для потенциального терапевтического применения в сочетании с антагонистами иммунного ответа (например, врожденного иммунного ответа). Согласно некоторым вариантам реализации в соответствующих случаях описана химическая модификация олигонуклеотидов с целью изменения и улучшения свойств in vivo (доставки, стабильности, времени жизни, укладки, специфичности в отношении мишени), а также их биологической функции и механизма, которые напрямую коррелируют с терапевтическим приложением.
[00174] Иллюстративные терапевтические нуклеиновые кислоты согласно настоящему изобретению, которые могут быть иммуностимулирующими и требуют применения иммуносупрессоров, раскрытых в настоящем документе, могут включать, но не ограничиваются перечисленными, минигены, плазмиды, миникольца, короткую интерферирующую РНК (киРНК), микроРНК (миРНК), антисмысловые олигонуклеотиды (ASO), рибозимы, двухцепочечную ДНК с замкнутыми концами (например, зкДНК, CELiD, линейную ковалентно замкнутую ДНК («с минимальной цепью»), doggybone (dbDNA™), протеломерную ДНК с замкнутыми концами или гантелеобразную линейную ДНК), дцРНК-субстрат Dicer, короткую шпилечную РНК (кшРНК), асимметричную интерферирующую РНК (аиРНК), микроРНК (миРНК), мРНК, тРНК, рРНК и вирусные ДНК-векторы, вирусный РНК-вектор; и любую их комбинацию.
[00175] киРНК или миРНК, которые могут отрицательно регулировать внутриклеточные уровни конкретных белков посредством процесса, называемого РНК-интерференцией (РНКи), также предусмотрены настоящим изобретением в качестве терапевтических средств на основе нуклеиновых кислот. После введения киРНК или миРНК в цитоплазму клетки-хозяина указанные конструкции двухцепочечной РНК связываются с белком, называемым RISC. Комплекс RISC удаляет смысловую цепь киРНК или миРНК. Комплекс RISC, при объединении с комплементарной мРНК, расщепляет мРНК и высвобождает разрезанные цепи. РНКи осуществляется путем индукции специфичного разрушения мРНК, что приводит к отрицательной регуляции соответствующего белка.
[00176] Антисмысловые олигонуклеотиды (ASO) и рибозимы, которые ингибируют трансляцию мРНК в белок, могут быть терапевтическими средствами на основе нуклеиновых кислот. В случае антисмысловых конструкций эти одноцепочечные дезоксинуклеиновые кислоты имеют последовательность, комплементарную последовательности мРНК целевого белка, и способны связываться с мРНК посредством спаривания оснований по Уотсону-Крику. Такое связывание предотвращает трансляцию целевой мРНК и/или запускает разрушение транскрипта мРНК РНКазой H. В результате антисмысловой олигонуклеотид имеет повышенную специфичность действия (т.е. отрицательную регуляцию специфического белка, связанного с заболеванием).
[00177] Согласно любому из способов, предложенных в настоящем документе, терапевтическая нуклеиновая кислота может представлять собой терапевтическую РНК. Терапевтическая РНК может представлять собой ингибитор трансляции мРНК, агент РНК-интерференции (РНКи), каталитически активную молекулу РНК (рибозим), транспортную РНК (тРНК) или РНК, которая связывает транскрипт мРНК (ASO), белок или другой молекулярный лиганд (аптамер). Согласно любому из способов, предложенных в настоящем документе, агент РНКи может представлять собой двухцепочечную РНК, одноцепочечную РНК, микроРНК, короткую интерферирующую РНК, короткую шпилечную РНК или образующий триплекс олигонуклеотид.
[00178] В соответствии с некоторыми вариантами реализации терапевтическая нуклеиновая кислота представляет собой двухцепочечную ДНК с замкнутыми концами, например, зкДНК. В соответствии с некоторыми вариантами реализации экспрессия и/или продукция терапевтического белка в клетке происходит с невирусного ДНК-вектора, например, зкДНК-вектора. Отличительным преимуществом зкДНК-векторов для экспрессии терапевтического белка по сравнению с обычными ААВ-векторами и даже лентивирусными векторами является отсутствие ограничения по размеру для гетерологичных последовательностей нуклеиновых кислот, кодирующих целевой белок. Таким образом, даже большой терапевтический белок может быть экспрессирован с одного зкДНК-вектора. Таким образом, зкДНК-векторы можно применять для экспрессии терапевтического белка у субъекта, нуждающегося в этом.
[00179] В общем случае зкДНК-вектор для экспрессии терапевтического белка, раскрытый в настоящем документе, содержит, в направлении от 5´ к 3´: первый инвертированный концевой повтор (ITR) аденоассоциированного вируса (ААВ), нуклеотидную последовательность, представляющую интерес (например, экспрессионную кассету, описанную в настоящем документе), и второй ITR ААВ. Последовательности ITR выбирают из любых из: (i) по меньшей мере одного WT-ITR и по меньшей мере одного модифицированного инвертированного концевого повтора ААВ (mod-ITR) (например, асимметричные модифицированные ITR); (ii) двух модифицированных ITR, причем пара mod-ITR имеет различную трехмерную пространственную организацию относительно друг друга (например, асимметричные модифицированные ITR), или (iii) пары симметричных или по существу симметричных WT-WT-ITR, в которой каждый из WT-ITR имеет одинаковую трехмерную пространственную организацию, или (iv) пары симметричных или по существу симметричных модифицированных ITR, в которой каждый из mod-ITR имеет одинаковую трехмерную пространственную организацию.
III. зкДНК-векторы
[00180] В соответствии с некоторыми аспектами настоящего раскрытия предложены невирусные бескапсидные ДНК-векторы с ковалентно замкнутыми концами (зкДНК-вектор), вводимые в сочетании с рапамицином или аналогами рапамицина. Согласно некоторым вариантам реализации рапамицин или аналог рапамицина присутствует в перенасыщенном количестве в синтетическом наноносителе, как описано в WO 2016/073799. Согласно некоторым вариантам реализации зкДНК-вектор также присутствует в том же наноносителе.
[00181] В соответствии с некоторыми аспектами настоящего раскрытия предложены невирусные бескапсидные ДНК-векторы с ковалентно замкнутыми концами (зкДНК), вводимые в сочетании с одним или более антагонистами TLR9. В настоящем документе также предложены конструкции зкДНК, содержащие последовательности, кодирующие, помимо прочего, один или более олигонуклеотидов, ингибирующих TLR9.
[00182] В соответствии с некоторыми аспектами настоящего раскрытия предложены невирусные бескапсидные ДНК-векторы с ковалентно замкнутыми концами (зкДНК), вводимые в сочетании с одним или более антагонистами cGAS. В настоящем документе также предложены конструкции зкДНК, содержащие последовательности, кодирующие, помимо прочего, одну или более РНК или белков, ингибирующих cGAS.
[00183] В соответствии с некоторыми аспектами настоящего раскрытия предложены невирусные бескапсидные ДНК-векторы с ковалентно замкнутыми концами (зкДНК), вводимые в сочетании с одним или более антагонистами инфламмасомы (например, любым одним или более из: ингибитора пути инфламмасомы NLRP3 или ингибитора пути инфламмасомы AIM2, или ингибитора каспазы 1, или любой их комбинации). В настоящем документе также предложены конструкции зкДНК, содержащие последовательности, кодирующие, помимо прочего, один или более антагонистов инфламмасомы (например, любой один или более из: ингибитора пути инфламмасомы NLRP3 или ингибитора пути инфламмасомы AIM2, или ингибитора каспазы 1 или любой их комбинации).
[00184] Как будет понятно, технологии зкДНК-векторов, описанные в настоящем документе, могут быть адаптированы к любому уровню сложности или могут применяться модульно, причем экспрессия различных компонентов ингибитора иммунного ответа (например, врожденного иммунного ответа), таких как те, которые описаны в настоящем документе, может контролироваться независимо. Например, намеренно предусмотрено, что технологии зкДНК-векторов, разработанные в настоящем документе, могут быть простыми, как применение одного зкДНК-вектора для экспрессии одной гетерологичной генной последовательности (например, одного ингибитора иммунного ответа (например, врожденного иммунного ответа), таких как те, которые описаны в настоящем документе, например, в), или могут быть сложными, как применение нескольких зкДНК-векторов, когда каждый вектор экспрессирует несколько ингибиторов иммунного ответа (например, врожденного иммунного ответа), таких как те, которые описаны в настоящем документе, например, или последовательности нуклеиновой кислоты, кодирующей один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа), таких как те, которые описаны в настоящем документе, и, например, ассоциированных кофакторов или вспомогательных белков, каждый из которых независимо контролируется разными промоторами.
[00185] Согласно одному варианту реализации один зкДНК-вектор можно применять для экспрессии одного компонента антагониста инфламмасомы (например, любого одного или более из: ингибитора пути инфламмасомы NLRP3 или ингибитора пути инфламмасомы AIM2, или ингибитора каспазы 1 или любой их комбинации). В качестве альтернативы, один зкДНК-вектор можно применять для экспрессии нескольких компонентов (например, по меньшей мере 2), например, он может экспрессировать два или более ингибиторов пути инфламмасомы NLRP3 и/или два или более ингибиторов пути инфламмасомы AIM2, и/или два или более ингибиторов каспазы 1 или любую их комбинацию под контролем одного промотора (например, сильного промотора), необязательно с использованием последовательности (ей) IRES, чтобы гарантировать надлежащую экспрессию каждого из компонентов, например, кофакторов или вспомогательных белков.
[00186] В настоящем документе также предусмотрен, в другом варианте реализации, один зкДНК-вектор, содержащий по меньшей мере две вставки, причем экспрессия каждой вставки находится под контролем ее собственного промотора. Промоторы могут включать несколько копий одного и того же промотора, несколько разных промоторов или любую их комбинацию. Специалист в данной области техники поймет, что желательной часто является экспрессия несколько антагонистов инфламмасомы на разных уровнях экспрессии, что позволяет контролировать стехиометрию отдельных экспрессируемых компонентов, чтобы гарантировать эффективную экспрессию и, в случае белка, укладывание и комбинирование белка в клетке.
[00187] В соответствии с некоторыми вариантами реализации синтетическую зкДНК получают за счет вырезания из молекулы двухцепочечной ДНК. Синтетическое получение зкДНК-векторов описано в Примерах 2-6 международной заявки PCT/US19/14122, поданной 18 января 2019 г., которая полностью включена в настоящий документ посредством ссылки. Один примерный способ получения зкДНК-вектора с использованием синтетического способа включает вырезание молекулы двухцепочечной ДНК. В общих чертах, зкДНК-вектор может быть получен с применением конструкции двухцепочечной ДНК, например, см. ФИГ. 7A-8E из PCT/US19/14122. Согласно некоторым вариантам реализации конструкция двухцепочечной ДНК представляет собой зкДНК-плазмиду, например, см. ФИГ. 6 в международной заявке на патент PCT/US2018/064242, поданной 6 декабря 2018 г.
[00188] Согласно некоторым вариантам реализации конструкция для получения зкДНК-вектора содержит регуляторный переключатель, описанный в настоящем документе.
[00189] Другой примерный способ получения зкДНК-вектора с использованием синтетического способа, который включает сборку различных олигонуклеотидов, представлен в Примере 3 из PCT/US19/14122, в котором зкДНК-вектор получают путем синтеза 5'-олигонуклеотида и 3'-олигонуклеотида ITR и лигирования указанных олигонуклеотидов ITR в двухцепочечный полинуклеотид, содержащий экспрессионную кассету. На ФИГ. 11B из PCT/US19/14122 представлен примерный способ лигирования олигонуклеотида 5'-ITR и олигонуклеотида 3'-ITR в двухцепочечный полинуклеотид, содержащий экспрессионную кассету.
[00190] В примерном способе получения зкДНК-вектора с использованием синтетического способа, представленном в Примере 4 из PCT/US19/14122, используют одноцепочечную линейную ДНК, содержащую два смысловых ITR, которые фланкируют смысловую последовательность экспрессионной кассеты и ковалентно присоединены к двум антисмысловым ITR, которые фланкируют антисмысловую последовательность экспрессионной кассеты, а концы указанной одноцепочечной линейной ДНК затем лигируют с получением одноцепочечной молекулы с замкнутыми концами. Один неограничивающий пример включает синтез и/или продуцирование молекулы одноцепочечной ДНК, ренатурацию частей указанной молекулы с получением одной линейной молекулы ДНК, которая имеет одну или более областей вторичной структуры со спаренными основаниями, с последующим лигированием свободных 5'- и 3'-концов между собой с получением замкнутой одноцепочечной молекулы.
[00191] Специалист в данной области техники может предусмотреть дополнительные варианты технологий зкДНК-векторов или адаптировать их из методов получения белка с использованием стандартных векторов.
[00192] Невирусные бескапсидные ДНК-векторы могут быть получены в восприимчивых клетках-хозяевах с экспрессионной конструкции (например, плазмиды, бакмиды, бакуловируса или интегрированной линии клеток), например, см. Примеры, раскрытые в международной заявке на патент PCT/US18/49996, поданной 7 сентября 2018 г., или с использованием синтетического способа получения, например, см. Примеры, раскрытые в международной заявке на патент PCT/US19/14122, поданной 6 декабря 2018 г., каждая из которых полностью включена в настоящий документ посредством ссылки. Согласно некоторым вариантам реализации зкДНК-векторы, которые можно применять в способах и композициях, раскрытых в настоящем документе, содержат гетерологичную нуклеиновую кислоту, например, трансген, расположенный между двумя последовательностями инвертированных концевых повторов (ITR). Согласно некоторым вариантам реализации по меньшей мере один из ITR модифицирован путем делеции, вставки и/или замены по сравнению с последовательностью ITR дикого типа (например, ITR ААВ); и по меньшей мере один из ITR содержит функциональный сайт концевого разрешения (TRS) и сайт связывания Rep. Согласно одному варианту реализации по меньшей мере один из ITR имеет по меньшей мере одну делецию, вставку или замену полинуклеотида по отношению к соответствующему ITR ААВ (например, SEQ ID NO: 1 или SEQ ID NO: 51 для ААВ2 дикого типа), чтобы индуцировать репликацию ДНК-вектора в клетке-хозяине в присутствии белка Rep. Как обсуждалось выше, можно использовать любой ITR. В иллюстративных целях ITR в конструкциях зкДНК в Таблице 1A представляют собой модифицированный ITR и WT-ITR. Однако в настоящем документе предусмотрены зкДНК-векторы, которые содержат гетерологичную последовательность нуклеиновой кислоты (трансген), расположенную между двумя последовательностями инвертированных концевых повторов (ITR), причем указанные последовательности ITR могут представлять собой пару асимметричных ITR или пару симметричных или по существу симметричных ITR, в соответствии с определением этих терминов в настоящем документе. ЗкДНК-вектор, содержащий сигнал ядерной локализации (NLS), раскрытый в настоящем документе, может содержать последовательности ITR, которые выбраны из любых из: (i) по меньшей мере одного WT-ITR и по меньшей мере одного модифицированного инвертированного концевого повтора (mod-ITR) ААВ (например, асимметричные модифицированные ITR); (ii) двух модифицированных ITR, причем указанная пара mod-ITR имеет различную трехмерную пространственную организацию друг относительно друга (например, асимметричные модифицированные ITR), или (iii) пары симметричных или по существу симметричных WT-WT-ITR, в которой каждый WT-ITR имеет одинаковую трехмерную пространственную организацию, или (iv) пары симметричных или по существу симметричных модифицированных ITR, в которой каждый mod-ITR имеет одинаковую трехмерную пространственную организацию, при этом указанные способы согласно настоящему раскрытию могут дополнительно включать систему доставки, такую как, но не ограничиваясь этим, система доставки на основе липосомных наночастиц.
[00193] Согласно некоторым вариантам реализации способы и композиции, описанные в настоящем документе, относятся к применению ингибитора иммунного ответа (например, врожденного иммунного ответа), раскрытого в настоящем документе, для совместного введения с любым зкДНК-вектором, включая, но не ограничиваясь перечисленными, зкДНК-вектор, содержащий асимметричные ITR, раскрытый в международной заявке на патент PCT/US18/49996, поданной 7 сентября 2018 г. (см., например, Примеры 1-4); зкДНК-вектор для редактирования генов, раскрытый в международной заявке на патент PCT/US18/64242, поданной 6 декабря 2018 г. (см., например, Примеры 1-7), или зкДНК-вектор для получения антител или слитых белков, раскрытый в международной заявке на патент PCT/US19/18016, поданной 14 февраля 2019 г. (например, см. Примеры 1-4), или зкДНК-вектор для контролируемой экспрессии трансгена, раскрытый в международной заявке на патент PCT/US19/18927, поданной в 22 февраля 2019 г., каждая из которых полностью включена в настоящий документ посредством ссылки. Согласно некоторым вариантам реализации также предусмотрено, что способы и композиции, описанные в настоящем документе, с использованием ингибитора иммунного ответа (например, врожденного иммунного ответа), раскрытого в настоящем документе, можно применять с зкДНК-вектором, полученным синтетическим способом, например, зкДНК-вектором, полученным в бесклеточной системе или системе без использования насекомых для получения зкДНК, как раскрыто в международной заявке PCT/US19/14122, поданной 18 января 2019 г., которая полностью включена в настоящий документ посредством ссылки.
[00194] ЗкДНК-вектор предпочтительно является дуплексным или самокомплементарным, по меньшей мере на протяжении части молекулы, например, трансгена. ЗкДНК-вектор имеет ковалентно замкнутые концы и, соответственно, предпочтительно устойчив к расщеплению экзонуклеазами (например, Exo I или Exo III) более часа при 37°C. Присутствие белка Rep в клетках-хозяевах (например, клетках насекомых или клетках млекопитающих) способствует репликации полинуклеотидной матрицы зкДНК-вектора, которая имеет модифицированный ITR, индуцирующий продуцирование невирусного бескапсидного ДНК-вектора с ковалентно замкнутыми концами. Молекула с ковалентно замкнутыми концами продолжает накапливаться в восприимчивых клетках за счет репликации и предпочтительно является достаточно стабильной с течением времени в присутствии белка Rep в стандартных условиях репликации, например, для накопления со значениями выхода по меньшей мере 1 пг/клетку, предпочтительно по меньшей мере 2 пг/клетку, предпочтительно по меньшей мере 3 пг/клетку, более предпочтительно по меньшей мере 4 пг/клетку, еще более предпочтительно по меньшей мере 5 пг/клетку.
[00195] В частности, в одном варианте реализации ДНК-векторы получают путем обеспечения клеток (например, клеток насекомых или клеток млекопитающих, например, клеток 293 и т.д.), несущих полинуклеотидную матрицу вектора (например, экспрессионную конструкцию), которая содержит два разных ITR (например, ITR ААВ) и представляющую интерес нуклеотидную последовательность (гетерологичную нуклеиновую кислоту, экспрессионную кассету), расположенную между ITR, причем по меньшей мере один из указанных ITR представляет собой модифицированный ITR, содержащий вставку, замену или делецию относительно другого ITR. Полинуклеотидная матрица вектора, описанная в настоящем документе, содержит по меньшей мере один функциональный ITR, который содержит сайт связывания Rep (RBS; например, 5´-GCGCGCTCGCTCGCTC-3´ для ААВ2) и функциональный сайт концевого разрешения (TRS; например, 5´-AGTT). Клетки не экспрессируют вирусные капсидные белки и полинуклеотидная матрица вектора лишена последовательностей, кодирующих вирусный капсид.
[00196] В присутствии Rep полинуклеотидная матрица вектора, имеющая по меньшей мере один модифицированный ITR, реплицируется с получением зкДНК-вектора. Получение зкДНК-вектора проходит в два этапа: во-первых, вырезание («извлечение») матрицы из остова вектора (например, плазмиды, бакмиды, генома и т.д.) с помощью белков Rep, и, во-вторых, Rep-опосредуемая репликация вырезанного векторного генома. Белки Rep и сайты связывания Rep различных серотипов ААВ хорошо известны специалистам в данной области техники. Специалист в данной области техники понимает необходимость выбора белка Rep из серотипа, который связывается с функциональным ITR и реплицирует его.
[00197] Клетки, несущие векторный полинуклеотид, либо уже содержат Rep (например, линия клеток с индуцируемым rep), либо их трансдуцируют вектором, который содержит Rep, а затем выращивают в условиях, обеспечивающих репликацию и высвобождение зкДНК-вектора. Затем ДНК зкДНК-вектора собирают и выделяют из клеток. Наличие ДНК бескапсидного невирусного зкДНК-вектора может быть подтверждено путем расщепления векторной ДНК, выделенной из клеток, рестрикционным ферментом, имеющим один сайт распознавания на ДНК-векторе, и анализа расщепленного ДНК-материала на неденатурирующем геле для подтверждения наличия характерных полос линейной и непрерывной ДНК, по сравнению с линейной и прерывистой ДНК. Например, на ФИГ. 6 представлен гель, подтверждающий продуцирование зкДНК-вектора с нескольких конструкций TTX-плазмиды с использованием одного варианта реализации для получения таких векторов, описанного в Примерах. ЗкДНК-вектор подтверждается характерным профилем полос в геле, как обсуждается в отношении Фигуры 4D. ФИГ. 5A и ФИГ. 5B представляют собой чертежи, иллюстрирующие один вариант реализации для идентификации наличия зкДНК-векторов с замкнутыми концами, продуцируемых с помощью способов согласно настоящему изобретению.
[00198] Матрица для экспрессии векторного полинуклеотида (например, ТТХ-плазмида, бакмида и т.д.) и/или ii) полинуклеотид, который кодирует Rep, могут быть введены в клетки с использованием любых средств, хорошо известных специалистам в данной области техники, включая, но не ограничиваясь перечисленными, трансфекцию (например, фосфат кальция, наночастицу или липосому) или введение с помощью вирусных векторов, например, HSV или бакуловируса. Например, матрица для экспрессии конструкции векторного полинуклеотида, используемая для получения зкДНК-векторов согласно настоящему изобретению, может представлять собой плазмиду (например, ТТХ-плазмиды, например, см. ФИГ. 4В), бакмиду (например, ТТХ-бакмиду) и/или бакуловирус (например, ТТХ-бакуловирус). Согласно одному варианту реализации ТТХ-плазмида содержит сайт рестрикционного клонирования (например, SEQ ID NO: 7), функционально расположенный между ITR, в который может быть вставлена гетерологичная нуклеиновая кислота (например, экспрессионная кассета, содержащая репортерный ген или терапевтическую нуклеиновую кислоту).
[00199] Согласно одному предпочтительному варианту реализации клетки-хозяева, используемые для получения зкДНК-векторов, описанных в настоящем документе, представляют собой клетки насекомых. Согласно другому предпочтительному варианту реализации бакуловирус используется для доставки как полинуклеотида, который кодирует белок Rep, так и полинуклеотидной матрицы для экспрессии конструкции невирусного ДНК-вектора для зкДНК-вектора. Примеры таких способов получения и выделения зкДНК-векторов описаны на ФИГ. 1-33.
[00200] Согласно еще одному аспекту настоящего изобретения предложены линии клеток-хозяев, имеющих стабильно интегрированную в собственный геном матрицу для экспрессии полинуклеотида ДНК-вектора (матрицу зкДНК-вектора), описанную в настоящем документе, для применения в получении невирусного ДНК-вектора. Способы получения таких линий клеток описаны в Lee, L. et al. (2013) Plos One 8(8): e69879, которая полностью включена в настоящий документ посредством ссылки. Предпочтительно белок Rep (например, как описано в Примере 1) добавляют к клеткам-хозяевам при множественности заражения (MOI) равной 3. Согласно одному варианту реализации линия клеток-хозяев представляет собой линию клеток беспозвоночного, предпочтительно клетки насекомых Sf9. Если линия клеток-хозяев представляет собой линию клеток млекопитающего, предпочтительно клетки 293, указанные линии клеток могут содержать стабильно интегрированную полинуклеотидную матрицу вектора, а второй вектор, такой как вирус герпеса, можно применять для введения в клетки белка Rep, что обеспечивает вырезание и амплификацию зкДНК-вектора в присутствии Rep.
[00201] Предпочтительно зкДНК содержит одну или более функциональных полинуклеотидных последовательностей ITR, которые включают сайт связывания Rep (RBS; 5´-GCGCGCTCGCTCGCTC-3´ для ААВ2, SEQ ID NO: 39) и сайт концевого разрешения (TRS; 5´-AGTT).
[00202] Бескапсидные зкДНК-векторы могут быть получены с экспрессионных конструкций (например, ТТХ-плазмид, ТТХ-бакмид, ТТХ-бакуловируса), которые дополнительно включают определенную комбинацию цис-регуляторных элементов, таких как посттранскрипционный регуляторный элемент WHP (WPRE) и поли(A) бычьего гормона роста (BGH). Подходящие экспрессионные кассеты для применения в экспрессионных конструкциях не имеют налагаемых вирусным капсидом ограничений упаковки. Экспрессионные кассеты согласно настоящему раскрытию включают промотор, который может влиять на общие уровни экспрессии, а также на клеточную специфичность. В случае экспрессии трансгена они могут включать высокоактивный немедленный ранний промотор вирусного происхождения. Экспрессионные кассеты могут содержать тканеспецифический эукариотический промотор для ограничения экспрессии трансгена конкретными типами клеток и уменьшения токсических эффектов и иммунных ответов, обусловленных нерегулируемой эктопической экспрессией. Согласно некоторым вариантам реализации экспрессионная кассета может содержать синтетический регуляторный элемент, такой как промотор CAG (SEQ ID NO: 3). Промотор CAG включает (i) ранний энхансерный элемент цитомегаловируса (CMV) (например, SEQ ID NO: 309), (ii) промотор, первый экзон и первый интрон гена бета-актина курицы, и (iii) акцептор сплайсинга гена бета-глобина кролика. В качестве альтернативы, например, экспрессионная кассета может содержать промотор альфа-1-антитрипсина (AAT) (например, SEQ ID NO: 4), специфический для печени (LP1) промотор (например, SEQ ID NO: 5) или промотор HAAT (например, SEQ ID NO: 135) или промотор фактора удлинения-1-альфа (EF1-α) человека (SEQ ID NO: 6) или фрагмент EF1-α (SEQ ID NO: 66) или промотор MND (SEQ ID NO: 70). Согласно некоторым вариантам реализации экспрессионная кассета включает один или более конститутивных промоторов, например, ретровирусный промотор из LTR вируса саркомы Рауса (RSV) (необязательно с энхансером RSV), немедленный ранний промотор цитомегаловируса (CMV) (необязательно с энхансером CMV) и т.п. В качестве альтернативы, можно применять индуцируемый промотор, нативный для трансгена промотор, тканеспецифический промотор или различные промоторы, известные в данной области техники. Согласно одному варианту реализации в экспрессионной кассете для кодирующей ген последовательности используют эндогенный или нативный промотор.
[00203] Индуцируемое редактирование генов с использованием зкДНК-векторов может быть выполнено с использованием методов, описанных, например, в работах Dow et al. Nat Biotechnol 33:390-394 (2015); Zetsche et al. Nat Biotechnol 33:139-42 (2015); Davis et al. Nat Chem Biol 11:316-318 (2015); Polstein et al. Nat Chem Biol 11:198-200 (2015); и/или Kawano et al. Nat Commun 6:6256 (2015), содержание каждой из которых полностью включено в настоящий документ посредством ссылки. Экспрессионные кассеты также могут включать посттранскрипционный элемент, в частности, посттранскрипционный регуляторный элемент вируса гепатита североамериканского сурка (WHP) (WPRE) (SEQ ID NO: 72), для повышения экспрессии трансгена. Можно применять другие посттранскрипционные элементы процессинга, такие как посттранскрипционный элемент гена тимидинкиназы вируса простого герпеса или вируса гепатита B (HBV). Экспрессионные кассеты могут включать последовательность полиаденилирования, известную в данной области техники, или ее вариант, например, природную последовательность, выделенную из бычьей последовательности BGHpA, или вирусную последовательность SV40pA (например, SEQ ID NO: 10), или синтетическую последовательность. Некоторые экспрессионные кассеты также могут включать последовательность 5'-энхансера позднего поли(А)-сигнала (USE) SV40. USE может применяться в комбинации с SV40pA или гетерологичным поли(A)-сигналом.
Время для сбора и накопления ДНК-векторов, описанных в настоящем документе, из клеток может быть выбрано и оптимизировано, чтобы достичь получения ДНК-векторов с высоким выходом. Например, время сбора может быть выбрано с учетом жизнеспособности клеток, морфологии клеток, размножения клеток и т.п. Обычно клетки могут быть собраны по прошествии времени после бакуловирусной инфекции, достаточного для получения ДНК-векторов (например, ТТХ-векторов), но до начала гибели большинства клеток из-за вирусной токсичности. ДНК-векторы могут быть выделены, например, с использованием наборов для очистки плазмид, таких как не содержащие эндотоксинов наборы для очистки плазмид (Endo-Free™ Plasmid kits) от Qiagen. Другие способы, разработанные для выделения плазмид, также могут быть адаптированы для ДНК-векторов. Обычно можно использовать любой способ очистки нуклеиновой кислоты, известный в данной области техники.
Регуляторные последовательности и эффекторы
[00204] Согласно вариантам реализации зкДНК-вектор содержит вторую нуклеотидную последовательность (например, регуляторную последовательность) в дополнение к одной или более нуклеотидным последовательностям, кодирующим терапевтический белок. Согласно вариантам реализации регуляторная последовательность гена функционально связана с нуклеотидной последовательностью, кодирующей терапевтический белок. Согласно вариантам реализации регуляторная последовательность подходит для контроля экспрессии терапевтического белка в клетке-хозяине. Согласно вариантам реализации регуляторная последовательность включает подходящую последовательность промотора, способную управлять транскрипцией гена, функционально связанного с последовательностью промотора, такого как нуклеотидная последовательность, кодирующая терапевтический белок согласно настоящему раскрытию. Согласно вариантам реализации вторая нуклеотидная последовательность включает интронную последовательность, связанную с 5´-концом нуклеотидной последовательности, кодирующей терапевтический белок. Согласно вариантам реализации последовательность энхансера обеспечена в 5´-направлении относительно промотора для повышения эффективности промотора. Согласно вариантам реализации регуляторная последовательность включает энхансер и промотор, причем указанная вторая нуклеотидная последовательность включает интронную последовательность в 5´-направлении относительно нуклеотидной последовательности, кодирующей терапевтический белок, при этом указанный интрон включает один или более сайтов расщепления нуклеазой, и при этом указанный промотор функционально связан с нуклеотидной последовательностью, кодирующей нуклеазу. Согласно вариантам реализации применяемая регуляторная последовательность является нативной для кодирующей последовательности в векторе.
[00205] Промоторы: Подходящие промоторы, включая те, которые описаны выше, могут происходить из вирусов и, соответственно, могут называться вирусными промоторами, или они могут происходить из любого организма, включая прокариотические или эукариотические организмы. Для управления экспрессией с помощью любой РНК-полимеразы (например, pol I, pol II, pol III) можно применять подходящие промоторы. Примерные промоторы включают, но не ограничиваются перечисленными, ранний промотор SV40, промотор из длинного концевого повтора (LTR) вируса опухоли молочной железы мышей; большой поздний промотор аденовируса (Ad MLP); промотор вируса простого герпеса (HSV), промотор цитомегаловируса (CMV), такой как область немедленного раннего промотора CMV (CMVIE), промотор вируса саркомы Рауса (RSV), промотор U6 малой ядерной РНК человека (U6, например, SEQ ID NO: 18) (Miyagishi et al., Nature Biotechnology 20, 497-500 (2002)), улучшенный промотор U6 (например, Xia et al., Nucleic Acids Res. 2003 Sep. 1; 31(17)), промотор H1 человека (H1) (например, SEQ ID NO: 19), промотор CAG, промотор альфа-1-антитрипсина человека (HAAT) (например, SEQ ID NO: 135) и т.п. Согласно вариантам реализации указанные промоторы изменяют на их содержащем интрон 3′-конце так, чтобы они включали один или более сайтов расщепления нуклеазами. Согласно некоторым вариантам реализации ДНК, содержащая сайт(ы) расщепления нуклеазой, является чужеродной для ДНК промотора.
[00206] Промотор может содержать одну или более специфических транскрипционных регуляторных последовательностей для дополнительного усиления экспрессии и/или для пространственного и/или временнóго изменения экспрессии указанного промотора. Промотор также может содержать дистальные энхансерные или репрессорные элементы, которые могут быть локализованы на расстоянии до нескольких тысяч пар оснований от сайта начала транскрипции. Промотор может происходить из источников, включающих вирусы, бактерии, грибы, растения, насекомых и животных. Промотор может регулировать экспрессию генного компонента конститутивно или дифференциально в отношении клетки, ткани или органа, где происходит экспрессия, или в отношении стадии развития, на которой происходит экспрессия, или в ответ на внешние стимулы, такие как физиологические стрессы, патогены, ионы металлов или индуцирующие агенты. Репрезентативные примеры промоторов включают промотор бактериофага T7, промотор бактериофага T3, промотор SP6, оператор/промотор lac, промотор tac, поздний промотор SV40, ранний промотор SV40, промотор RSV-LTR, немедленный ранний промотор CMV, ранний промотор SV40 или поздний промотор SV40 и немедленный ранний промотор CMV, а также промоторы, перечисленные ниже. Такие промоторы и/или энхансеры можно применять для экспрессии любого представляющего интерес гена, например, молекул для редактирования генов, донорной последовательности, терапевтических белков и т.д. Например, вектор может содержать промотор, который функционально связан с последовательностью нуклеиновой кислоты, кодирующей терапевтический белок. Промотор, функционально связанный с последовательностью, кодирующей терапевтический белок, может представлять собой промотор из вируса обезьян 40 (SV40), промотор вируса опухоли молочной железы мышей (MMTV), промотор вируса иммунодефицита человека (ВИЧ), такой как промотор длинного концевого повтора (LTR) вируса иммунодефицита крупного рогатого скота (BIV), промотор вируса Молони, промотор вируса лейкоза птиц (ALV), промотор цитомегаловируса (CMV), такой как немедленный ранний промотор CMV, промотор вируса Эпштейна-Барр (EBV) или промотор вируса саркомы Рауса (RSV). Промотор также может представлять собой промотор из гена человека, такого как убиквитин С человека (hUbC), актин человека, миозин человека, гемоглобин человека, мышечный креатин человека или металлотионеин человека. Промотор также может представлять собой тканеспецифический промотор, такой как специфический для печени промотор, например, альфа-1-антитрипсина человека (HAAT), природный или синтетический. Согласно одному варианту реализации доставку в печень можно обеспечить с использованием специфичного нацеливания на эндогенный ApoE композиции, содержащей зкДНК-вектор, которую направляют в гепатоциты за счет рецептора липопротеина низкой плотности (ЛНП), присутствующего на поверхности гепатоцита.
[00207] Согласно одному варианту реализации используемый промотор представляет собой нативный промотор гена, кодирующего терапевтический белок. Промоторы и другие регуляторные последовательности соответствующих генов, кодирующих терапевтические белки, известны и были охарактеризованы. Используемая промоторная область может дополнительно включать одну или более дополнительных регуляторных последовательностей (например, нативных), например, энхансеров.
[00208] Неограничивающие примеры подходящих промоторов для применения в соответствии с настоящим изобретением включают промотор CAG, например, (SEQ ID NO: 3), промотор HAAT (SEQ ID NO: 135), промотор EF1-α человека (SEQ ID NO: 6) или фрагмент промотора EF1-α (SEQ ID NO: 66) и промотор EF1-α крысы (SEQ ID NO: 310).
[00209] Энхансеры: согласно некоторым вариантам реализации зкДНК, экспрессирующая антагонист инфламмасомы (например, ингибитор одного или более из пути инфламмасомы NLRP3 и/или AIM2, или ингибитор каспазы 1), содержит один или более энхансеров. Согласно некоторым вариантам реализации последовательность энхансера расположена в 5´-направлении относительно последовательности промотора. Согласно некоторым вариантам реализации последовательность энхансера расположена в 3´-направлении относительно последовательности промотора. Примерные энхансеры перечислены в Таблице 1 в настоящем документе.
Таблица 1. Примерные последовательности энхансеров
[00210] Последовательности 5´-UTR и интронные последовательности: согласно некоторым вариантам реализации зкДНК-вектор содержит последовательность 5´-UTR и/или интронную последовательность, которая расположена в 3´-направлении относительно последовательности 5´-ITR. Согласно некоторым вариантам реализации 5´-UTR расположена в 5´-направлении относительно трансгена, например, последовательности, кодирующей антагонист инфламмасомы (например, ингибитор одного или более из пути инфламмасомы NLRP3 и/или AIM2, или ингибитор каспазы 1). Примерные последовательности 5´-UTR перечислены в Таблице 2А.
Таблица 2A: Примерные последовательности 5´-UTR и интронные последовательности
[00211] Последовательности 3´-UTR: согласно некоторым вариантам реализации зкДНК-вектор содержит последовательность 3´-UTR, которая расположена в 5´-направлении относительно последовательности 3´-ITR. Согласно некоторым вариантам реализации 3´-UTR расположена в 3´-направлении от трансгена, например, последовательности, кодирующей антагонист инфламмасомы (например, ингибитор одного или более из пути инфламмасомы NLRP3 и/или AIM2, или ингибитор каспазы 1). Примерные последовательности 3´-UTR перечислены в Таблице 2В.
Таблица 2B: Примерные последовательности 3´-UTR и интронные последовательности
[00212] Последовательности полиаденилирования: последовательность, кодирующая последовательность полиаденилирования, может быть включена в зкДНК-вектор для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), как описано в настоящем документе, для стабилизации мРНК, экспрессируемой с зкДНК-вектора, а также для облечения экспорта из ядра и трансляции. Согласно одному варианту реализации зкДНК-вектор не включает последовательность полиаденилирования. Согласно другим вариантам реализации зкДНК-вектор для экспрессии антагониста инфламмасомы включает по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 10, по меньшей мере 15, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50 или более адениновых динуклеотидов. Согласно некоторым вариантам реализации последовательность полиаденилирования содержит примерно 43 нуклеотида, примерно 40-50 нуклеотидов, примерно 40-55 нуклеотидов, примерно 45-50 нуклеотидов, примерно 35-50 нуклеотидов или любой диапазон между указанными значениями.
[00213] Экспрессионные кассеты могут включать любую последовательность полиаденилирования, известную в данной области техники, или ее вариант. Согласно некоторым вариантам реализации последовательность полиаденилирования (поли(A)) выбрана из любой из последовательностей, перечисленных в Таблице 3. Также можно использовать другие последовательности поли(A), широко известные в данной области техники, например, включая, но не ограничиваясь перечисленными, природную последовательность, выделенную из бычьего BGHpA (например, SEQ ID NO: 9), или поли(A) вируса SV40 (SV40pA) (например, SEQ ID NO: 10), или синтетическую последовательность. Некоторые экспрессионные кассеты также могут включать последовательность 5'-энхансера позднего поли(А)-сигнала (USE) SV40. Согласно некоторым вариантам реализации последовательность USE можно применять в комбинации с SV40pA или гетерологичным сигналом поли(A). Последовательности поли(A) расположены в 3´-направлении относительно трансгена, кодирующего антагонист инфламмасомы.
[00214] Экспрессионные кассеты также могут включать посттранскрипционный элемент для повышения экспрессии трансгена. Согласно некоторым вариантам реализации для повышения экспрессии трансгена применяют посттранскрипционный регуляторный элемент вируса гепатита североамериканского сурка (WPRE) (например, SEQ ID NO: 72). Можно применять другие посттранскрипционные элементы процессинга, такие как посттранскрипционный элемент из гена тимидинкиназы вируса простого герпеса или вируса гепатита B (HBV). С трансгенами могут быть связаны секреторные последовательности, например, последовательности VH-02 и VK-A26, например, SEQ ID NO: 950 и SEQ ID NO: 951.
Таблица 3: Примерные последовательности поли(A)
[00215] Согласно одному варианту реализации векторный полинуклеотид (зкДНК-вектор) содержит пару двух разных ITR, выбранных из группы, состоящей из: SEQ ID NO: 1 и SEQ ID NO: 52; и SEQ ID NO: 2 и SEQ ID NO: 51. Согласно одному варианту реализации каждого из этих аспектов векторный полинуклеотид или невирусные бескапсидные ДНК-векторы с ковалентно замкнутыми концами содержат пару ITR, выбранных из группы состоящий из: SEQ ID NO:101 и SEQ ID NO:102; SEQ ID NO:103 и SEQ ID NO:104, SEQ ID NO:105 и SEQ ID NO:106; SEQ ID NO:107 и SEQ ID NO:108; SEQ ID NO:109 и SEQ ID NO:110; SEQ ID NO:111 и SEQ ID NO:112; SEQ ID NO:113 и SEQ ID NO:114; и SEQ ID NO:115 и SEQ ID NO:116. Согласно некоторым вариантам реализации зкДНК-векторы не имеют ITR, который содержит какую-либо последовательность, выбранную из SEQ ID NO: 500-529.
[00216] Время для сбора и накопления ДНК-векторов, описанных в настоящем документе, из клеток может быть выбрано и оптимизировано, чтобы достичь получения зкДНК-векторов с высоким выходом. Например, время сбора может быть выбрано с учетом жизнеспособности клеток, морфологии клеток, размножения клеток и т.д. Согласно одному варианту реализации клетки культивируют в достаточных условиях и собирают через достаточное время после инфекции бакуловирусом, чтобы получить ДНК-векторы (например, TTX-векторы), но до начала гибели большинства клеток из-за вирусной токсичности. ДНК-векторы могут быть выделены с использованием наборов для очистки плазмид, таких как не содержащие эндотоксинов наборы для выделения плазмид от Qiagen. Другие методы, разработанные для выделения плазмид, также могут быть адаптированы для ДНК-векторов. Обычно могут быть использованы любые способы очистки нуклеиновых кислот.
[00217] ДНК-векторы могут быть очищены с помощью любых способов очистки ДНК, известных специалистам в данной области техники. Согласно одному варианту реализации зкДНК-векторы очищают в виде молекул ДНК. Согласно другому варианту реализации зкДНК-векторы очищают в виде экзосом или микрочастиц.
[00218] Согласно одному варианту реализации бескапсидный невирусный ДНК-вектор содержит или получен из плазмиды, содержащей полинуклеотидную матрицу, содержащую в указанном порядке: первый инвертированный концевой повтор (ITR) аденоассоциированного вируса (ААВ), представляющую интерес нуклеотидную последовательность (например, экспрессионную кассету экзогенной ДНК) и модифицированный ITR ААВ, причем указанная матричная молекула нуклеиновой кислоты лишена способности кодировать капсидный белок ААВ. Согласно дополнительному варианту реализации матричная нуклеиновая кислота согласно настоящему изобретению лишена последовательностей, кодирующих вирусный капсидный белок (т.е. лишена генов капсида ААВ, а также генов капсида других вирусов). Кроме того, согласно конкретному варианту реализации матричная молекула нуклеиновой кислоты также лишена последовательностей, кодирующих белок Rep ААВ. Соответственно, согласно предпочтительному варианту реализации молекула нуклеиновой кислоты согласно настоящему изобретению лишена как функциональных генов cap ААВ, так и rep ААВ.
[00219] Согласно одному варианту реализации зкДНК-вектор может включать структуру ITR, которая мутирована относительно ITR ААВ2 дикого типа, раскрытого в настоящем документе, но все еще сохраняет функциональную часть RBE, trs и RBE´. Согласно некоторым вариантам реализации зкДНК-векторы не имеют ITR, который содержит какую-либо последовательность, выбранную из SEQ ID NO: 500-529.
[00220] Согласно некоторым вариантам реализации трансген, кодирующий антагонист инфламмасомы (например, любой один или более из: ингибитора пути инфламмасомы NLRP3 или ингибитора пути инфламмасомы AIM2, или ингибитора каспазы 1, или любой их комбинации), также может кодировать секреторную последовательность так, что антагонист инфламмасомы направлен к аппарату Гольджи и эндоплазматическому ретикулуму (ЭР), в которых антагонист инфламмасомы (например, любой один или более из: ингибитора пути инфламмасомы NLRP3 или ингибитора пути инфламмасомы AIM2, или ингибитора каспазы 1 или любой их комбинации) будет уложен в правильную конформацию молекулами шаперонов при его прохождении через ЭР и высвобождении из клетки. Примерные секреторные последовательности включают, но не ограничиваются перечисленными, VH-02 (SEQ ID NO: 950) и VK-A26 (SEQ ID NO: 951) и сигнальную последовательность Igκ, а также секреторный сигнал Gluc, обеспечивающий секрецию меченого белка из цитозоля, секреторную последовательность TMD-ST, которая направляет меченый белок в аппарат Гольджи.
[00221] Последовательности ядерной локализации: Согласно некоторым вариантам реализации зкДНК-вектор для экспрессии, например, ингибитора иммунного ответа (например, врожденного иммунного ответа) содержит одну или более последовательностей ядерной локализации (NLS), например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более NLS. Согласно некоторым вариантам реализации одна или более NLS расположены на амино-конце или рядом с ним, на карбокси-конце или рядом с ним, или в их комбинации (например, одна или более NLS на амино-конце и/или одна или более NLS на карбокси-конце). В случае присутствия более чем одной NLS, каждая из них может быть выбрана независимо от других так, что одна NLS будет присутствовать более чем в одной копии и/или в комбинации с одной или более другими NLS, присутствующими в одной или более копиях. Неограничивающие примеры NLS показаны в Таблице 4.
Таблица 4: Сигналы ядерной локализации
[00222] Регуляторные переключатели: Молекулярный регуляторный переключатель представляет собой переключатель, который генерирует измеряемое изменение состояния в ответ на сигнал. Регуляторные переключатели также можно использовать для точной настройки экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), описанного в настоящем документе, так, что ингибитор иммунного ответа экспрессируется предусмотренным образом, включая, но не ограничиваясь этим, экспрессию ингибитора иммунного ответа на целевом уровне экспрессии или в целевом количестве, или, в качестве альтернативы, в присутствии или в отсутствие конкретного сигнала, включая событие клеточной передачи сигнала. Например, как описано в настоящем документе, экспрессия ингибитора иммунного ответа с зкДНК-вектора может быть включена или выключена при возникновении конкретного состояния. Согласно некоторым вариантам реализации переключатель представляет собой двухпозиционный («ON/OFF») переключатель, который разработан для запуска или остановки (т.е. прекращения) экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа) в зкДНК-векторе контролируемым и регулируемым образом. Согласно некоторым вариантам реализации указанный переключатель может включать «аварийный выключатель» (kill switch), который может давать клетке, содержащей указанный зкДНК-вектор, команду для запуска механизма запрограммированной клеточной смерти после активации переключателя. Примерные регуляторные переключатели, предусмотренные для применения в зкДНК-векторе для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), могут применяться для регуляции экспрессии трансгена и более подробно описаны в международной заявке PCT/US18/49996, которая полностью включена в настоящий документ посредством ссылки
(i) Бинарные регуляторные переключатели
[00223] Согласно некоторым вариантам реализации зкДНК-вектор для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа) содержит регуляторный переключатель, который может служить для контролируемой модуляции экспрессии антагониста инфламмасомы. Например, экспрессионная кассета, расположенная между ITR зкДНК-вектора, может дополнительно содержать регуляторную область, например, промотор, цис-элемент, репрессор, энхансер и т.д., которая функционально связана с последовательностью нуклеиновой кислоты, кодирующей ингибитор иммунного ответа (например, врожденного иммунного ответа), причем указанная регуляторная область регулируется одним или более кофакторами или экзогенными агентами. В качестве неограничивающего примера, регуляторные области могут модулироваться низкомолекулярными переключателями или индуцируемыми или репрессируемыми промоторами. Неограничивающие примеры индуцируемых промоторов представляют собой индуцируемые гормонами или индуцируемые металлами промоторы. Другие примерные индуцируемые промоторные/энхансерные элементы включают, но не ограничиваются перечисленными, индуцируемый RU486 промотор, индуцируемый экдизоном промотор, индуцируемый рапамицином промотор и промотор металлотионеина.
(ii) Низкомолекулярные регуляторные переключатели
[00224] Различные известные из уровня техники низкомолекулярные регуляторные переключатели известны в данной области техники и могут быть скомбинированы с ингибитором иммунного ответа (например, врожденного иммунного ответа), раскрытым в настоящем документе, с получением контролируемого регуляторным переключателем зкДНК-вектора. Согласно некоторым вариантам реализации регуляторный переключатель может быть выбран из любого одного или комбинации следующего: пара ортогональный лиганд/ядерный рецептор, например, вариант ретиноидного рецептора/LG335 и GRQCIMFI, вместе с искусственным промотором, контролирующим экспрессию функционально связанного трансгена, как раскрыто в Taylor et al. BMC Biotechnology 10 (2010): 15; сконструированные рецепторы стероидов, например, модифицированный рецептор прогестерона с усеченным С-концом, который не может связывать прогестерон, но связывает RU486 (мифепристон) (патент США № 5364791); рецептор экдизона дрозофилы (Drosophila) и его экдистероидные лиганды (Saez, et al., PNAS, 97(26)(2000), 14512-14517); или переключатель, контролируемый антибиотиком триметопримом (TMP), как раскрыто в Sando R 3rd; Nat Methods. 2013, 10(11):1085-8. Согласно некоторым вариантам реализации регуляторный переключатель для контроля трансгена или экспрессируемый зкДНК-вектором представляет собой переключатель для активации пролекарств, такой как раскрытый в патентах США 8771679 и 6339070.
(iii) Регуляторные переключатели «с кодом доступа»
[00225] Согласно некоторым вариантам реализации регуляторный переключатель может представлять собой «переключатель с кодом доступа» или «контур с кодом доступа». Переключатели с кодом доступа позволяют точно настроить контроль экспрессии трансгена с зкДНК-вектора при возникновении конкретных условий, т.е. для осуществления экспрессии и/или репрессии трансгена должна присутствовать комбинация условий. Например, для осуществления экспрессии трансгена должны возникнуть по меньшей мере условия A и B. Регуляторный переключатель с кодом доступа может представлять собой любое число условий, например, по меньшей мере 2 или по меньшей мере 3, или по меньшей мере 4, или по меньшей мере 5, или по меньшей мере 6, или по меньшей мере 7 или более условий, наличие которых необходимо для осуществления экспрессии трансгена. Согласно некоторым вариантам реализации должны возникнуть по меньшей мере 2 условия (например, условия A, B), а в некоторых вариантах реализации должны возникнуть по меньшей мере 3 условия (например, A, B и C или A, B и D). Только в качестве примера, для осуществления экспрессии гена с зкДНК, содержащей регуляторный переключатель с кодом доступа «ABC», требуется наличие условия A, B и C. Условия A, B и C могут быть следующими; условие A представляет собой наличие состояния или заболевания, условие B представляет собой гормональный ответ, и условие C представляет собой ответ на экспрессию трансгена. Например, если трансген редактирует дефектный ген EPO, то условием A является наличие хронической болезни почек (ХБП), условие B осуществляется при наличии гипоксических условий в почках субъекта, условие C заключается в нарушении привлечения эритропоэтин-продуцирующих клеток (ЭПК) в почках; или, альтернативно, в нарушении активации HIF-2. При повышении уровней кислорода или достижении целевого уровня EPO трансген снова отключается до момента возникновения 3 условий, которые его опять включат.
[00226] Согласно некоторым вариантам реализации регуляторный переключатель с кодом доступа или «контур с кодом доступа», предусмотренный для применения в зкДНК-векторе, содержит гибридные транскрипционные факторы (TF) для расширения диапазона и уровня сложности сигналов окружающей среды, используемых для определения условий биоизоляции. В отличие от блокирующего (deadman) выключателя, который запускает гибель клетки при наличии заранее определенного условия, «контур с кодом доступа» позволяет обеспечить выживание клеток или экспрессию трансгена при наличии конкретного «кода доступа», и может быть легко перепрограммирован, что позволяет обеспечить экспрессию трансгена и/или выживание клеток только при наличии заранее определенного условия окружающей среды или кода доступа.
[00227] Любые и все комбинации регуляторных переключателей, раскрытых в настоящем документе, например, низкомолекулярные переключатели, переключатели на основе нуклеиновых кислот, гибридные переключатели на основе малых молекул и нуклеиновых кислот, посттранскрипционные регуляторные переключатели трансгенов, посттрансляционная регуляция, контролируемые излучением переключатели, опосредуемые гипоксией переключатели и другие регуляторные переключатели, известные специалистам в данной области техники, раскрытые в настоящем документе, могут применяться в регуляторном переключателе с кодом доступа, как раскрыто в настоящем документе. Регуляторные переключатели, предусмотренные для применения, также обсуждаются в обзорной статье Kis et al., JR Soc Interface. 12: 20141000 (2015) и обобщены в Таблице 1 статьи Kis. Согласно некоторым вариантам реализации регуляторный переключатель для применения в системе с кодом доступа может быть выбран из любых переключателей или их комбинации, раскрытых в Таблице 11 международной заявки на патент PCT/US18/49996, которая полностью включена в настоящий документ посредством ссылки.
(iv). Регуляторные переключатели на основе нуклеиновых кислот для контроля экспрессии трансгена
[00228] Согласно некоторым вариантам реализации регуляторный переключатель для контроля экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа) с помощью зкДНК основан на механизме контроля на основе нуклеиновых кислот. Примеры механизмов контроля на основе нуклеиновых кислот известны в данной области техники и предусмотрены для применения. Например, такие механизмы включают рибопереключатели, такие как раскрытые, например, в US 2009/0305253, US2008/0269258, US2017/0204477, WO2018026762A1, патенте США 9222093 и заявке на европейский патент EP288071, а также раскрыты в обзорной статье Villa JK et al. Microbiol Spectr. 2018 May;6(3). Также включены чувствительные к метаболитам транскрипционные биосенсоры, такие как раскрытые в WO2018/075486 и WO2017/147585. Другие известные в данной области техники механизмы, предусмотренные для применения, включают подавление трансгена с применением киРНК или молекулы РНКи (например, miR, кшРНК). Например, зкДНК-вектор может содержать регуляторный переключатель, кодирующий молекулу РНКи, которая комплементарна части трансгена, экспрессируемого зкДНК-вектором. При экспрессии такой РНКи, даже если зкДНК-вектор экспрессирует трансген (например, антагонист инфламмасомы (например, ингибитор одного или более из пути инфламмасомы NLRP3 и/или AIM2, или ингибитор каспазы 1), будет происходить его подавление под действием комплементарной молекулы РНКи, а если РНКи не экспрессируется при экспрессии трансгена (например, антагониста инфламмасомы) зкДНК-вектором, подавление трансгена под действием РНКи не происходит.
[00229] Согласно некоторым вариантам реализации регуляторный переключатель представляет собой тканеспецифический самоинактивирующийся регуляторный переключатель, например, раскрытый в US2002/0022018, благодаря чему регуляторный переключатель преднамеренно выключает трансген (например, антагонист инфламмасомы) в сайте, где в противном случае экспрессия трансгена могла бы быть неблагоприятной. Согласно некоторым вариантам реализации регуляторный переключатель представляет собой обратимую с помощью рекомбиназы систему экспрессии гена, например, раскрытую в US2014/0127162 и патенте США 8324436.
(v). Посттранскрипционные и посттрансляционные регуляторные переключатели.
[00230] Согласно некоторым вариантам реализации регуляторный переключатель для контроля экспрессии зкДНК-вектором ингибитора иммунного ответа (например, врожденного иммунного ответа) представляет собой систему посттранскрипционной модификации. Например, такой регуляторный переключатель может представлять собой рибопереключатель-аптазим, чувствительный к тетрациклину или теофиллину, раскрытый в US2018/0119156, GB201107768, WO2001/064956A3, патенте EP 2707487 и Beilstein et al., ACS Synth. Biol., 2015, 4 (5), pp.526-534; Zhong et al., Elife. 2016 Nov 2;5. pii: e18858. Согласно некоторым вариантам реализации предусмотрено, что обычный специалист в данной области техники может закодировать как трансген, так и ингибиторную киРНК, которая содержит чувствительный к лиганду аптамер (отрицательный переключатель), в результате чего будет получен положительный переключатель, чувствительный к лиганду.
(vi). Другие примерные регуляторные переключатели
[00231] Любой известный регуляторный переключатель может быть использован в зкДНК-векторе для контроля экспрессии зкДНК-вектором ингибитора иммунного ответа (например, врожденного иммунного ответа), включая переключатели, запускаемые изменениями окружающей среды. Дополнительные примеры включают, но не ограничиваются перечисленными; метод BOC согласно Suzuki et al., Scientific Reports 8; 10051 (2018); расширение генетического кода и нефизиологическую аминокислоту; контролируемые излучением или контролируемые ультразвуком положительные/отрицательные (on/off) переключатели (см., например, Scott S. et al., Gene Ther. 2000 Jul; 7 (13): 1121-5; патенты США 5612318; 5571797; 5770581; 5817636; WO1999/025385A1. Согласно некоторым вариантам реализации регуляторный переключатель контролируется имплантируемой системой, например, раскрытой в патенте США 7840263; US2007/0190028A1, в которой экспрессия гена контролируется одной или более формами энергии, включая электромагнитную энергию, что активирует промоторы, функционально связанные с трансгеном в зкДНК-векторе.
[00232] Согласно некоторым вариантам реализации регуляторный переключатель, предусмотренный для применения в зкДНК-векторе, представляет собой опосредуемый гипоксией или активируемый стрессом переключатель, например, такой как те, которые раскрыты в WO1999060142A2, патентах США 5834306; 6218179; 6709858; US2015/0322410; Greco et al., (2004) Targeted Cancer Therapies 9, S368, а также элементы FROG, TOAD и NRSE и условно индуцируемые подавляющие элементы, включая элементы ответа на гипоксию (HRE), элементы ответа на воспаление (IRE) и активируемые сдвиговым напряжением элементы (SSAE), например, как раскрыто в патенте США 9394526. Такой вариант реализации можно применять для «включения» экспрессии трансгена с зкДНК-вектора после ишемии или в ишемических тканях и/или в опухолях.
(vii). «Аварийные выключатели»
[00233] Другие варианты реализации, описанные в настоящем документе, относятся к зкДНК-вектору для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), описанному в настоящем документе, содержащему аварийный выключатель. «Аварийный выключатель», раскрытый в настоящем документе, позволяет уничтожить клетку, содержащую указанный зкДНК-вектор, или подвергнуть ее программируемой клеточной гибели в качестве способа окончательного удаления введенного зкДНК-вектора из системы субъекта. Специалист в данной области техники поймет, что применение «аварийных выключателей» в зкДНК-векторах для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), как правило, будет сопряжено с нацеливанием зкДНК-вектора на ограниченное число клеток, потеря которого является приемлемой для субъекта, или на тип клеток, апоптоз которых желателен (например, на раковые клетки). Во всех аспектах «аварийный выключатель», раскрытый в настоящем документе, разработан таким образом, чтобы обеспечить быстрое и надежное уничтожение клетки, содержащей зкДНК-вектор, в отсутствие вводимого сигнала выживания или другого заданного условия. Другими словами, «аварийный выключатель», кодируемый зкДНК-вектором для экспрессии антагониста инфламмасомы, описанным в настоящем документе, может ограничивать выживание клетки, содержащей зкДНК-вектор, окружающей средой, задаваемой специфическими вводимыми сигналами. Такие «аварийные выключатели» служат для обеспечения биологической функции биоизоляции, если потребуется удалить зкДНК-вектор для экспрессии антагониста инфламмасомы у субъекта или гарантировать отсутствие экспрессии с него кодируемого антагониста инфламмасомы.
[00234] Другие «аварийные выключатели», известные обычному специалисту в данной области техники, предусмотрены для применения в зкДНК-векторе для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), раскрытом в настоящем документе, например, как раскрыто в US2010/0175141; US2013/0009799; US2011/0172826; US2013/0109568, а также «аварийные выключатели», раскрытые в Jusiak et al., Reviews in Cell Biology and Molecular Medicine; 2014; 1-56; Kobayashi et al., PNAS, 2004; 101; 8419-9; Marchisio et al., Int. Journal of Biochem and Cell Biol., 2011; 43; 310-319; и Reinshagen et al., Science Translational Medicine, 2018, 11.
[00235] Соответственно, согласно некоторым вариантам реализации, зкДНК-вектор для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа) может содержать конструкцию нуклеиновой кислоты «аварийного выключателя», которая содержит нуклеиновую кислоту, кодирующую эффекторный токсин или репортерный белок, причем экспрессия эффекторного токсина (например, белка смерти) или репортерного белка контролируется с помощью заранее определенного условия. Например, предварительно определенным условием может быть наличие агента окружающей среды, такого как, например, экзогенный агент, без которого клетка по умолчанию будет экспрессировать эффекторный токсин (например, белок смерти) и будет уничтожена. Согласно альтернативным вариантам реализации предварительно определенным условием является наличие двух или более агентов окружающей среды, например, клетка будет выживать только при подаче двух или более необходимых экзогенных агентов, и без какого-либо из них клетка, содержащая зкДНК-вектор, уничтожается.
[00236] Согласно некоторым вариантам реализации зкДНК-вектор для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа) модифицируют для включения «аварийного выключателя» для разрушения клеток, содержащих указанный зкДНК-вектор, чтобы эффективно остановить экспрессию трансгена, экспрессируемого указанным зкДНК-вектором (например, экспрессию антагониста инфламмасомы) in vivo. В частности, зкДНК-вектор дополнительно генетически модифицирован для экспрессии белка-переключателя, который не функционирует в клетках млекопитающих в нормальных физиологических условиях. Клетки, экспрессирующие белок-переключатель, будут разрушены только после введения лекарственного средства или воздействия условия окружающей среды, специфически нацеленных на указанный белок-переключатель, что остановит экспрессию терапевтического белка или пептида. Например, сообщалось, что клетки, экспрессирующие тимидинкиназу HSV, могут быть уничтожены при введении таких лекарственных средств как ганцикловир и цитозиндезаминаза. См., например, Dey and Evans, Suicide Gene Therapy by Herpes Simplex Virus-1 Thymidine Kinase (HSV-TK), в источнике: Targets in Gene Therapy, под редакцией You (2011); и Beltinger et al., Proc. Natl. Acad. Sci. USA 96(15):8699-8704 (1999). Согласно некоторым вариантам реализации зкДНК-вектор может содержать «аварийный выключатель» на основе киРНК, называемый DISE (гибель, индуцируемая устранением гена выживания, «Death Induced by Survival gene Elimination») (Murmann et al., Oncotarget. 2017; 8:84643-84658. Induction of DISE in ovarian cancer cells in vivo).
[00237] Согласно другому варианту реализации ингибитор иммунного ответа (например, врожденного иммунного ответа), экспрессируемый с зкДНК-векторов, дополнительно содержит дополнительную функциональную возможность, такую как флуоресценция, ферментативная активность, сигнал секреции или активатор иммунных клеток.
[00238] Согласно некоторым вариантам реализации зкДНК, кодирующая ингибитор иммунного ответа (например, врожденного иммунного ответа), может дополнительно содержать линкерный домен, например. В настоящем документе термин «линкерный домен» относится к олиго- или полипептидной области от примерно 2 до 100 аминокислот в длину, которая связывает между собой любые домены/области антагониста инфламмасомы, описанного в настоящем документе. Согласно некоторым вариантам реализации линкеры могут включать или могут состоять из гибких остатков, таких как глицин и серин, так, что смежные белковые домены могут свободно перемещаться друг относительно друга. Более длинные линкеры можно использовать, когда желательно гарантировать, что два смежных домена не будут стерически мешать друг другу. Линкеры могут быть расщепляемыми или нерасщепляемыми. Примеры расщепляемых линкеров включают линкеры 2А (например, Т2А), 2А-подобные линкеры или их функциональные эквиваленты и их комбинации. Линкер может представлять собой линкерную область T2A, происходящую из вируса Thosea asigna.
IV. Способ получения зкДНК-вектора
А. Получение в общем случае
[00239] Определенные способы получения зкДНК-вектора для экспрессии, например, ингибитора иммунного ответа (например, врожденного иммунного ответа), содержащего пару асимметричных ITR или пару симметричных ITR, как определено в настоящем документе, описаны в разделе IV международной заявки PCT/US18/49996, поданной 7 сентября 2018 г., которая полностью включена в настоящий документ посредством ссылки. Согласно некоторым вариантам реализации зкДНК-вектор для экспрессии антагониста инфламмасомы, раскрытый в настоящем документе, может быть получен с использованием клеток насекомых, как описано в настоящем документе. Согласно альтернативным вариантам реализации зкДНК-вектор для экспрессии антагониста инфламмасомы, раскрытый в настоящем документе, может быть получен синтетическим способом и, в некоторых вариантах реализации, бесклеточным способом, как раскрыто в международной заявке PCT/US19/14122, поданной 18 января 2019 г., которая полностью включена в настоящий документ посредством ссылки.
[00240] Как описано в настоящем документе, согласно одному варианту реализации, зкДНК-вектор для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), например, может быть получен, например, с помощью способа, включающего этапы: a) инкубации популяции клеток-хозяев (например, клеток насекомых), несущих полинуклеотидную матрицу экспрессионной конструкции (например, зкДНК-плазмиду, зкДНК-бакмиду и/или зкДНК-бакуловирус), лишенную последовательностей, кодирующих вирусный капсид, в присутствии белка Rep в эффективных условиях и в течение периода времени, достаточного для того чтобы индуцировать продукцию указанного зкДНК-вектора в клетках-хозяевах, и при этом указанные клетки-хозяева не содержат последовательностей, кодирующих вирусный капсид; и b) сбора и выделения зкДНК-вектора из клеток-хозяев. Присутствие белка Rep индуцирует репликацию векторного полинуклеотида с модифицированным ITR для продуцирования зкДНК-вектора в клетке-хозяине. Однако вирусные частицы (например, вирионы ААВ) не экспрессируются. Таким образом, отсутствует ограничение по размеру, такое как присутствующее в естественных условиях в ААВ-векторах или других вирусных векторах.
[00241] Присутствие зкДНК-вектора, выделенного из клеток-хозяев, может быть подтверждено путем расщепления ДНК, выделенной из клетки-хозяина, рестрикционным ферментом, имеющим один сайт распознавания на зкДНК-векторе, и анализа расщепленного ДНК-материала на неденатурирующем геле для подтверждения присутствия характерных полос линейной и непрерывной ДНК по сравнению с линейной и прерывистой ДНК.
[00242] Согласно еще одному аспекту настоящего изобретения предложено применение линий клеток-хозяев, имеющих стабильно интегрированную в собственный геном полинуклеотидную матрицу для экспрессии ДНК-вектора (зкДНК-матрицу), в получении невирусного ДНК-вектора, например, как описано в Lee, L. et al. (2013) Plos One 8(8): e69879. Предпочтительно Rep добавляют к клеткам-хозяевам при MOI, равной примерно 3. Когда линия клеток-хозяев представляет собой линию клеток млекопитающего, например, клетки HEK293, линии клеток могут иметь стабильно интегрированную полинуклеотидную матрицу вектора, и второй вектор, такой как вирус герпеса, может использоваться для введения в клетки белка Rep, что позволяет вырезать и амплифицировать зкДНК в присутствии Rep и вспомогательного вируса.
[00243] Согласно одному варианту реализации клетки-хозяева, используемые для получения зкДНК-векторов для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), например, как описано в настоящем документе, представляют собой клетки насекомых, а для доставки как полинуклеотида, который кодирует белок Rep, так и полинуклеотидной матрицы экспрессионной конструкции невирусного ДНК-вектора для зкДНК используют бакуловирус, например, как описано на ФИГ. 4A-4D и в Примере 1. Согласно некоторым вариантам реализации клетку-хозяина модифицируют таким образом, чтобы она экспрессировала белок Rep.
[00244] Затем зкДНК-вектор собирают и выделяют из клеток-хозяев. Время сбора и накопления из клеток зкДНК-векторов, описанных в настоящем документе, может быть выбрано и оптимизировано так, чтобы достичь получения зкДНК-векторов с высоким выходом. Например, время сбора может быть выбрано с учетом жизнеспособности клеток, морфологии клеток, размножения клеток и т.д. Согласно одному варианту реализации клетки культивируют в достаточных условиях и собирают через достаточное время после инфекции бакуловирусом, чтобы получить зкДНК-векторы, но до начала гибели большинства клеток из-за токсичности бакуловируса. ДНК-векторы могут быть выделены с использованием наборов для очистки плазмид, таких как не содержащие эндотоксинов наборы для очистки плазмид (Endo-Free Plasmid Kits) от Qiagen. Другие способы, разработанные для выделения плазмид, также могут быть адаптированы для ДНК-векторов. Обычно могут быть использованы любые способы очистки нуклеиновых кислот.
[00245] ДНК-векторы могут быть очищены любыми способами очистки ДНК, известными специалистам в данной области техники. Согласно одному варианту реализации зкДНК-векторы очищают в виде молекул ДНК. Согласно другому варианту реализации зкДНК-векторы очищают в виде экзосом или микрочастиц.
Присутствие зкДНК-вектора для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа) может быть подтверждено путем расщепления векторной ДНК, выделенной из клеток, рестрикционным ферментом, имеющим один сайт распознавания на ДНК-векторе, и анализа как расщепленного, так и нерасщепленного ДНК-материала с использованием гель-электрофореза, чтобы подтвердить присутствие характерных полос линейной и непрерывной ДНК по сравнению с линейной и прерывистой ДНК. ФИГ. 4C и ФИГ. 4D иллюстрируют один вариант реализации для идентификации наличия зкДНК-векторов с замкнутыми концами, продуцируемых с помощью способов, описанных в настоящем документе.
B. зкДНК-плазмида
[00246] ЗкДНК-плазмида представляет собой плазмиду, используемую для последующего получения зкДНК-вектора для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), например. Согласно некоторым вариантам реализации зкДНК-плазмида может быть сконструирована с использованием известных методик, чтобы обеспечить по меньшей мере следующие элементы в виде функционально связанных компонентов в направлении транскрипции: (1) модифицированную последовательность 5´-ITR; (2) экспрессионную кассету, содержащую цис-регуляторный элемент, например, промотор, индуцируемый промотор, регуляторный переключатель, энхансеры и т.п.; и (3) модифицированную последовательность 3´-ITR, причем указанная последовательность 3´-ITR симметрична относительно последовательности 5´-ITR. Согласно некоторым вариантам реализации экспрессионная кассета, фланкированная ITR, содержит сайт клонирования для введения экзогенной последовательности. Экспрессионная кассета заменяет кодирующие rep и cap области геномов ААВ.
[00247] Согласно одному аспекту зкДНК-вектор для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), например, получают из плазмиды, называемой в настоящем документе «зкДНК-плазмидой», кодирующей, в указанном порядке: первый инвертированный концевой повтор (ITR) аденоассоциированного вируса (ААВ), экспрессионную кассету, содержащую трансген, и мутированный или модифицированный ITR ААВ, причем указанная зкДНК-плазмида лишена последовательностей, кодирующих капсидный белок ААВ. Согласно альтернативным вариантам реализации зкДНК-плазмида кодирует, в указанном порядке: первый (или 5') модифицированный или мутированный ITR ААВ, экспрессионную кассету, содержащую трансген, и второй (или 3') модифицированный ITR ААВ, причем указанная зкДНК-плазмида лишена последовательностей, кодирующих капсидный белок ААВ, и при этом указанные 5'- и 3'-ITR являются симметричными друг относительно друга. Согласно альтернативным вариантам реализации зкДНК-плазмида кодирует, в указанном порядке: первый (или 5') модифицированный или мутированный ITR ААВ, экспрессионную кассету, содержащую трансген, и второй (или 3') мутированный или модифицированный ITR ААВ, причем указанная зкДНК-плазмида лишена последовательностей, кодирующих капсидный белок ААВ, и при этом указанные модифицированные 5'- и 3'-ITR имеют одинаковые модификации (т.е. они являются обратно комплементарными или симметричными друг относительно друга).
[00248] Согласно дополнительному варианту реализации система зкДНК-плазмиды лишена последовательностей, кодирующих вирусный капсидный белок (т.е. лишена генов капсида AAV, а также генов капсидов других вирусов). Кроме того, согласно конкретному варианту реализации, зкДНК-плазмида также лишена последовательностей, кодирующих белок Rep ААВ. Соответственно, в предпочтительном варианте реализации, зкДНК-плазмида лишена функциональных генов cap ААВ и rep ААВ, GG-3′ для ААВ2, а также вариабельной палиндромной последовательности, обеспечивающей образование шпильки.
[00249] ЗкДНК-плазмида согласно настоящему изобретению может быть получена с использованием природных нуклеотидных последовательностей из геномов любых серотипов ААВ, хорошо известных в данной области техники. Согласно одному варианту реализации остов зкДНК-плазмиды происходит из генома ААВ1, ААВ2, ААВ3, ААВ4, ААВ5, ААВ5, ААВ7, ААВ8, ААВ9, ААВ10, ААВ11, ААВ12, ААВrh8, ААВrh10, ААВ-DJ и ААВ-DJ8. Например, NCBI: NC002077; NC001401; NC001729; NC001829; NC006152; NC006260; NC006261; из указателя вирусов Kotin and Smith («The Springer Index of Viruses»), доступного по ссылке, поддерживаемой Springer (веб-адрес: oesys.springer.de/viruses/database/mkchapter.asp?virID=42.04.) (примечание: ссылки на указатель URL или базу данных подразумевают содержание доступного по ссылке ресурса URL или базы данных на фактическую дату подачи данной заявки). В конкретном варианте реализации остов зкДНК-плазмиды происходит из генома ААВ2. В другом конкретном варианте реализации остов зкДНК-плазмиды представляет собой синтетический остов, сконструированный методами генной инженерии с включением на его 5'- и 3'-концах ITR, происходящих из одного из указанных геномов ААВ.
[00250] ЗкДНК-плазмида необязательно может включать селектируемый или селективный маркер для использования при создании линии клеток, продуцирующей зкДНК-вектор. Согласно одному варианту реализации селективный маркер может быть вставлен после (т.е. в 3´-направлении) последовательности 3´-ITR. Согласно другому варианту реализации селективный маркер может быть вставлен перед (т.е. в 5´-направлении) последовательностью 5´-ITR. Подходящие селективные маркеры включают, например, маркеры, придающие устойчивость к лекарственному средству. Селективные маркеры могут представлять собой, например, гены устойчивости к бластицидину S, канамицину, генетицину и т.п. Согласно предпочтительному варианту реализации селективный маркер для отбора по чувствительности к лекарственному средству представляет собой ген устойчивости к бластицидину S.
[00251] Примерный зкДНК-вектор (например, рААВ0) для экспрессии антагониста инфламмасомы (например, ингибитора одного или более из пути инфламмасомы NLRP3 и/или AIM2, или ингибитора каспазы 1) получают из плазмиды рААВ. Способ получения рААВ-вектора может включать: (а) доставку в клетку-хозяина плазмиды рААВ, как описано выше, при этом как клетка-хозяин, так и плазмида лишены генов, кодирующих капсидный белок, (b) культивирование клетки-хозяина в условиях, обеспечивающих продуцирование генома зкДНК; и (c) сбор клеток и выделение генома ААВ, продуцированного указанными клетками.
C. Примерный способ получения зкДНК-векторов из зкДНК-плазмид
[00252] В настоящем документе также предложены способы получения бескапсидных зкДНК-векторов для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), в частности, способ с достаточно высоким выходом для обеспечения достаточного количества вектора для экспериментов in vivo.
[00253] Согласно некоторым вариантам реализации способ получения зкДНК-вектора для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), например, включает следующие этапы: (1) введение конструкции нуклеиновой кислоты, содержащей экспрессионную кассету и последовательности двух симметричных ITR, в клетку-хозяина (например, клетки Sf9), (2) необязательно создание клональной линии клеток, например, с использованием селективного маркера, присутствующего на плазмиде, (3) введение кодирующего Rep гена (либо путем трансфекции, либо инфекции бакуловирусом, несущим указанный ген) в указанную клетку насекомого; и (4) сбор клеток и очистку зкДНК-вектора. Конструкция нуклеиновой кислоты, содержащая экспрессионную кассету и две последовательности ITR, описанная выше для получения зкДНК-вектора, может иметь форму зкДНК-плазмиды или бакмиды, или бакуловируса, полученных с использованием зкДНК-плазмиды, как описано ниже. Конструкция нуклеиновой кислоты может быть введена в клетку-хозяина путем трансфекции, вирусной трансдукции, стабильной интеграции или других способов, известных в данной области техники.
D. Линии клеток:
[00254] Линии клеток-хозяев, используемые в получении зкДНК-вектора для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), например, могут включать линии клеток насекомых, происходящие из Spodoptera frugiperda, такие как клетки Sf9, Sf21, или клетки Trichoplusia ni, или линии клеток других беспозвоночных животных, позвоночных животных или других эукариот, включая клетки млекопитающих. Также могут быть использованы другие линии клеток, известные обычному специалисту в данной области техники, такие как HEK293, Huh-7, HeLa, HepG2, HeplA, 911, CHO, COS, MeWo, NIH3T3, A549, HT1 180, моноциты, а также зрелые и незрелые дендритные клетки. Линии клеток-хозяев могут быть трансфицированы для стабильной экспрессии зкДНК-плазмиды для получения зкДНК-вектора с высоким выходом.
[00255] ЗкДНК-плазмиды могут быть введены в клетки Sf9 путем кратковременной трансфекции с использованием реагентов (например, липосомальных, фосфата кальция) или физических средств (например, электропорации), известных в данной области техники. В качестве альтернативы, могут быть созданы стабильные линии клеток Sf9 с зкДНК-плазмидой, стабильно интегрированной в их геномы. Такие стабильные линии клеток могут быть созданы путем включения селективного маркера в зкДНК-плазмиду согласно описанию выше. Если зкДНК-плазмида, используемая для трансфекции линии клеток, включает селективный маркер, например, для антибиотика, клетки, которые были трансфицированы зкДНК-плазмидой и интегрировали ДНК зкДНК-плазмиды в их геном, могут быть отобраны путем добавления антибиотика в среды для культивирования клеток. Затем устойчивые клоны клеток могут быть выделены с помощью методик разведения до получения отдельных клеток или переноса колоний и размножены.
E. Выделение и очистка зкДНК-векторов:
[00256] Примеры способа получения и выделения зкДНК-векторов описаны на ФИГ. 1-7 и в конкретных примерах ниже. зкДНК-векторы для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), например, раскрытые в настоящем документе, могут быть получены из клеток-продуцентов, экспрессирующих белок (белки) Rep ААВ, дополнительно трансформированных зкДНК-плазмидой, зкДНК-бакмидой или зкДНК-бакуловирусом. Плазмиды, которые можно применять для получения зкДНК-векторов, включают плазмиды, которые кодируют антагонист инфламмасомы, или плазмиды, кодирующие один или более белков REP.
[00257] Согласно одному аспекту полинуклеотид кодирует белок Rep ААВ (Rep 78 или 68), доставляемый в продуцирующую клетку в плазмиде (Rep-плазмиде), бакмиде (Rep-бакмиде) или бакуловирусе (Rep-бакуловирусе). Rep-плазмида, Rep-бакмида и Rep-бакуловирус могут быть получены с помощью способов, описанных выше.
[00258] В настоящем документе описаны способы получения зкДНК-вектора для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), например. Экспрессионные конструкции, используемые для получения зкДНК-вектора для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), описанного в настоящем документе, могут представлять собой плазмиду (например, зкДНК-плазмиды), бакмиду (например, зкДНК-бакмиду) и/или бакуловирус (например, зкДНК-бакуловирус). Только в качестве примера, зкДНК-вектор может быть получен из клеток, коинфицированных зкДНК-бакуловирусом и Rep-бакуловирусом. Белки Rep, продуцированные с Rep-бакуловируса, могут реплицировать зкДНК-бакуловирус с получением зкДНК-векторов. В качестве альтернативы, зкДНК-векторы для экспрессии антагониста инфламмасомы могут быть получены из клеток, стабильно трансфицированных конструкцией, содержащей последовательность, кодирующую белок Rep ААВ (Rep78/52), доставленной в Rep-плазмидах, Rep-бакмидах или Rep-бакуловирусе. зкДНК-бакуловирус может быть кратковременно трансфицирован в клетки, реплицирован с помощью белка Rep и может продуцировать зкДНК-векторы.
[00259] Бакмида (например, зкДНК-бакмида) может быть трансфицирована в восприимчивые клетки насекомых, такие как клетки Sf9, Sf21, клетки Tni (Trichoplusia ni), клетки High Five, и продуцировать зкДНК-бакуловирус, который представляет собой рекомбинантный бакуловирус, включающий последовательности, содержащие симметричные ITR и экспрессионную кассету. Клетки насекомых могут быть снова инфицированы зкДНК-бакуловирусом для получения следующего поколения рекомбинантного бакуловируса. Необязательно этап может быть повторен один или более раз для получения большего количества рекомбинантного бакуловируса.
[00260] Время для сбора и накопления из клеток зкДНК-векторов для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), описанных в настоящем документе, может быть выбрано и оптимизировано, чтобы достичь получения зкДНК-векторов с высоким выходом. Например, время сбора может быть выбрано с учетом жизнеспособности клеток, морфологии клеток, размножения клеток и т.д. Обычно клетки могут быть собраны через период времени после инфекции бакуловирусом, достаточный для получения зкДНК-векторов (например, зкДНК-векторов), но до начала гибели большинства клеток из-за вирусной токсичности. ЗкДНК-векторы могут быть выделены из клеток Sf9 с использованием наборов для очистки плазмид, таких как наборы Qiagen ENDO-FREE PLASMID®. Другие способы, разработанные для выделения плазмид, также могут быть адаптированы для зкДНК-векторов. Обычно можно использовать любые известные в данной области техники способы очистки нуклеиновых кислот, а также коммерчески доступные наборы для экстракции ДНК.
[00261] В качестве альтернативы, очистку можно осуществлять путем щелочного лизиса клеточного осадка, центрифугирования полученного лизата и выполнения хроматографического разделения. В качестве одного неограничивающего примера, указанный способ можно выполнять путем загрузки супернатанта на ионообменную колонку (например, SARTOBIND Q®), которая удерживает нуклеиновые кислоты, с последующим элюированием (например, 1,2 М раствором NaCl) и выполнением дальнейшей хроматографической очистки на гель-фильтрационной колонке (например, на 6 Fast Flow GE). Бескапсидный ААВ-вектор затем выделяют, например, путем осаждения.
[00262] Согласно некоторым вариантам реализации зкДНК-векторы для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа) также могут быть очищены в форме экзосом или микрочастиц. В данной области техники известно, что многие типы клеток высвобождают не только растворимые белки, но и сложные грузы белок/нуклеиновая кислота за счет отделения мембранных микровезикул (Cocucci et al., 2009; EP 10306226.1). Такие везикулы включают микровезикулы (также называемые микрочастицами) и экзосомы (также называемые нановезикулами); и первые, и вторые содержат в качестве груза белки и РНК. Микровезикулы образуются в результате прямого отпочковывания плазматической мембраны, а экзосомы высвобождаются во внеклеточную среду при слиянии мультивезикулярных эндосом с плазматической мембраной. Соответственно, содержащие зкДНК-вектор микровезикулы и/или экзосомы могут быть выделены из клеток, трансдуцированных зкДНК-плазмидой либо бакмидой или бакуловирусом, полученными с использованием зкДНК-плазмиды.
[00263] Микровезикулы могут быть выделены путем фильтрации или ультрацентрифугирования культуральной среды при 20000×g, а экзосомы - при 100000×g. Оптимальная продолжительность ультрацентрифугирования может быть определена экспериментально и зависит от конкретного типа клеток, из которых выделяют везикулы. Предпочтительно культуральную среду сначала очищают с помощью низкоскоростного центрифугирования (например, при 2000×g в течение 5-20 минут) и подвергают центробежному концентрированию с использованием, например, центрифужной колонки AMICON® (Millipore®, Уотфорд, Великобритания). Микровезикулы и экзосомы могут быть дополнительно очищены с помощью FACS или MACS с использованием специфичных антител, распознающих конкретные поверхностные антигены, присутствующие на микровезикулах и экзосомах. Другие способы очистки микровезикул и экзосом включают, но не ограничиваются перечисленными, иммунопреципитацию, аффинную хроматографию, фильтрацию и магнитные гранулы, покрытые специфичными антителами или аптамерами. После очистки везикулы промывают, например, фосфатно-солевым буфером (ФСБ). Одним из преимуществ использования микровезикул или экзосом для доставки содержащих зкДНК везикул является то, что такие везикулы могут быть нацелены на клетки различных типов путем включения в их мембраны белков, распознаваемых специфическими рецепторами на соответствующих типах клеток. (см. также EP 10306226)
[00264] Другой аспект настоящего изобретения относится к способам очистки зкДНК-векторов из линий клеток-хозяев, которые стабильно интегрировали конструкцию зкДНК в их собственный геном. Согласно одному варианту реализации зкДНК-векторы очищают в виде молекул ДНК. Согласно другому варианту реализации зкДНК-векторы очищают в виде экзосом или микрочастиц.
[00265] На ФИГ. 5 международной заявки PCT/US18/49996 показан гель, подтверждающий продуцирование зкДНК из нескольких конструкций зкДНК-плазмид с использованием способа, описанного в Примерах. ЗкДНК подтверждается на основании характерного профиля полос на геле (см. ФИГ. 5A).
V. Фармацевтические композиции и составы
[00266] Настоящее изобретение предусматривает фармацевтические композиции и составы, содержащие терапевтическую нуклеиновую кислоту и один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа), описанных в настоящем документе. Согласно некоторым вариантам реализации фармацевтическая композиция, содержащая терапевтическую нуклеиновую кислоту и один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа), может включать фармацевтически приемлемое вспомогательное вещество или носитель. В соответствии с некоторыми вариантами реализации фармацевтическая композиция содержит ДНК-вектор с замкнутыми концами, например, зкДНК-вектор, описанный в настоящем документе, и рапамицин или аналог рапамицина, а также фармацевтически приемлемый носитель или разбавитель. В соответствии с некоторыми вариантами реализации фармацевтическая композиция содержит ДНК-вектор с замкнутыми концами, например, зкДНК-вектор, описанный в настоящем документе, и ингибитор TLR (например, ингибитор TLR9), а также фармацевтически приемлемый носитель или разбавитель. В соответствии с некоторыми вариантами реализации фармацевтическая композиция содержит ДНК-вектор с замкнутыми концами, например, зкДНК-вектор, описанный в настоящем документе, и ингибитор cGAS, а также фармацевтически приемлемый носитель или разбавитель. В соответствии с некоторыми вариантами реализации фармацевтическая композиция содержит ДНК-вектор с замкнутыми концами, например, зкДНК-вектор, описанный в настоящем документе, и антагонист инфламмасомы (например, любой один или более из: ингибитора пути инфламмасомы NLRP3 или ингибитора пути инфламмасомы AIM2, или ингибитора каспазы 1, или любой их комбинации), а также фармацевтически приемлемый носитель или разбавитель.
[00267] ДНК-векторы, раскрытые в настоящем документе, могут быть включены в фармацевтические композиции, подходящие для введения субъекту для доставки in vivo в клетки, ткани или органы субъекта, включая, в некоторых вариантах реализации, фармацевтические композиции, содержащие ингибиторы иммунного ответа (например, врожденного иммунного ответа), описанные в настоящем документе. Как правило, фармацевтическая композиция содержит зкДНК-векторы, раскрытые в настоящем документе, и фармацевтически приемлемый носитель. Например, TTX-векторы согласно настоящему изобретению могут быть включены в фармацевтическую композицию, подходящую для целевого пути терапевтического введения (например, парентерального введения). Также предусмотрена пассивная трансдукция ткани путем внутривенной или внутриартериальной инфузии под высоким давлением, а также внутриклеточной инъекции, такой как внутриядерная микроинъекция или внутрицитоплазматическая инъекция. Фармацевтические композиции для терапевтических целей могут быть изготовлены в виде раствора, микроэмульсии, дисперсии, липосом или другой упорядоченной структуры, подходящей для высокой концентрации TTX-вектора. Стерильные растворы для инъекций могут быть приготовлены путем включения соединения TTX-вектора в необходимом количестве в соответствующем буфере с одним ингредиентом или комбинацией ингредиентов, перечисленных выше, при необходимости, с последующей стерилизацией путем фильтрации.
[00268] Фармацевтически активные композиции, содержащие TTX-вектор, могут быть изготовлены для доставки трансгена в нуклеиновой кислоте в клетки реципиента, что приводит к терапевтической экспрессии в них указанного трансгена. Композиция также может включать фармацевтически приемлемый носитель.
[00269] Композиции и векторы, предложенные в настоящем документе, можно применять для доставки трансгена с различными целями. Согласно некоторым вариантам реализации трансген кодирует белок или функциональную РНК, которая предназначена для применения в исследовательских целях, например, для создания соматической трансгенной модели на животных, несущей трансген, например, для исследования функции продукта трансгена. В другом примере трансген кодирует белок или функциональную РНК, который(ая) предназначен(а) для применения с целью создания модели заболевания на животных. Согласно некоторым вариантам реализации трансген кодирует один или более пептидов, полипептидов или белков, которые можно применять для лечения или предотвращения патологических состояний у млекопитающего-субъекта. Трансген может быть перенесен в организм пациента (например, экспрессирован у пациента) в количестве, достаточном для лечения заболевания, ассоциированного с уменьшенной экспрессией, отсутствием экспрессии или дисфункцией указанного гена. Согласно некоторым вариантам реализации трансген представляет собой молекулу для редактирования генов (например, нуклеазу). Согласно определенным вариантам реализации нуклеаза представляет собой CRISPR-ассоциированную нуклеазу (Cas-нуклеазу).
[00270] В соответствии с некоторыми вариантами реализации фармацевтически активные композиции, описанные в настоящем документе, могут быть введены в комбинации с антигистаминным агентом или стероидом. В соответствии с некоторыми вариантами реализации антигистаминный агент или стероид вводят в той же композиции, что и фармацевтически активные композиции, описанные в настоящем документе. В соответствии с некоторыми вариантами реализации антигистаминный агент или стероид вводят в отдельной композиции, чем фармацевтически активные композиции, описанные в настоящем документе. В соответствии с некоторыми вариантами реализации антигистаминный агент или стероид вводят одновременно с фармацевтически активной композицией. В соответствии с некоторыми вариантами реализации антигистаминный агент или стероид вводят последовательно с фармацевтически активной композицией. Любой антигистаминный агент, известный в данной области техники, можно применять в вариантах реализации в настоящем документе. В соответствии с некоторыми вариантами реализации антигистаминный агент представляет собой одно или более из омфенирамина, буклизина, хлорфенирамина, циннаризина, клемастина, циклизина, ципрогептадина, дифенгидрамина, дифенилпиралина, доксиламина, меклозина, фенирамина, прометазина, трипролидина, акривастина, астемизола, цетиризина, дезлоратадина, фексофенадина, левоцетиризина, лоратадина, мизоластина, терфенадина, их фармацевтически приемлемой соли или их комбинации. Любой стероид, известный в данной области техники, можно применять в вариантах реализации в настоящем документе. В соответствии с некоторыми вариантами реализации стероид представляет собой один или более из по меньшей мере одного из флуоксиместерона, местеролона, метандростенолона, нандролон-ундеканоата, нандролон-циплоната, оксандролона, оксиметолона, нандролон-гексилоксифенилпропионата, тестостерона, преднизона, кортизола, кортизона, преднизолона, дексаметазона, бетаметазона, триамцинолона, беклометазона, флудрокортизона, дезоксикортикостерона, альдостерона и станозолола.
[00271] Фармацевтические композиции для терапевтических целей, как правило, должны быть стерильными и стабильными в условиях изготовления и хранения. Стерильные растворы для инъекций могут быть приготовлены путем включения соединения зкДНК-вектора в необходимом количестве в соответствующем буфере с одним ингредиентом или комбинацией ингредиентов, перечисленных выше, при необходимости, с последующей стерилизацией путем фильтрации.
Единичная лекарственная форма
[00272] В соответствии с некоторыми вариантами реализации фармацевтические композиции могут быть представлены в виде единичной лекарственной формы. Единичная лекарственная форма, как правило, адаптирована к одному или более конкретным путям введения фармацевтической композиции. Согласно некоторым вариантам реализации единичная лекарственная форма адаптирована для введения путем ингаляции. Согласно некоторым вариантам реализации единичная лекарственная форма адаптирована для введения с помощью испарителя. Согласно некоторым вариантам реализации единичная лекарственная форма адаптирована для введения с помощью небулайзера. Согласно некоторым вариантам реализации единичная лекарственная форма адаптирована для введения с помощью генератора аэрозоля. Согласно некоторым вариантам реализации единичная лекарственная форма адаптирована для перорального введения, для буккального введения или для сублингвального введения. Согласно некоторым вариантам реализации единичная лекарственная форма адаптирована для внутривенного, внутримышечного или подкожного введения. Согласно некоторым вариантам реализации единичная лекарственная форма адаптирована для интратекального или интрацеребровентрикулярного введения. Согласно некоторым вариантам реализации фармацевтическая композиция изготовлена для местного введения. Количество активного ингредиента, которое можно комбинировать с материалом носителя для получения единичной лекарственной формы, обычно будет представлять собой количество соединения, которое вызывает терапевтический эффект.
IV. Введение и дозирование
[00273] Раскрытие, предложенное в настоящем документе, описывает способы предотвращения, уменьшения или устранения нежелательного иммунного ответа (например, врожденного иммунного ответа) у субъекта (например, субъекта-человека) путем введения указанному субъекту по меньшей мере одного ингибитора иммунного ответа (например, врожденного иммунного ответа), описанного в настоящем документе, и нуклеиновой кислоты (например, терапевтической нуклеиновой кислоты, нуклеиновой кислоты, используемой для исследовательских целей), причем указанные введения ингибитора иммунного ответа (например, врожденного иммунного ответа) и введение нуклеиновой кислоты соотнесены во времени так, чтобы обеспечить модуляцию иммунного ответа (например, врожденного иммунного ответа), когда введение двух агентов обеспечено в виде комбинации. Эти два агента могут быть введены одновременно в одном составе, в одно и то же время в разных составах, или они могут быть введены по отдельности в разные моменты времени.
[00274] Согласно одному варианту реализации экспрессированный ингибитор иммунного ответа (например, врожденного иммунного ответа), раскрытый в настоящем документе, не вызывает реакции иммунной системы, а скорее подавляет врожденную иммунную систему у субъекта по меньшей мере на 10% или 20%, или 30%, или 40%, или 50%, или 60%, или 70%, или 80%, или 90%, или 95%, или 98%, или 99%, или 100% по сравнению с отсутствием введения зкДНК-вектора, экспрессирующего ингибитор.
[00275] Технология, описанная в настоящем документе, в целом направлена на способы введения субъекту ДНК-векторов с замкнутыми концами совместно с одним или более ингибиторами иммунного ответа (например, врожденного иммунного ответа), выбранными из одного или более или комбинации рапамицина или аналогов рапамицина, ингибиторов TLR (например, TLR9), ингибиторов cGAS и одного или более антагонистов инфламмасомы (например, любого одного или более из: ингибитора пути инфламмасомы NLRP3 или ингибитора пути инфламмасомы AIM2, или ингибитора каспазы 1, или любой их комбинации), описанных в настоящем документе. Согласно некоторым вариантам реализации ДНК-вектор с замкнутыми концами включает, но не ограничивается перечисленными, зкДНК-векторы, раскрытые в настоящем документе, и мРНК, антисмысловую РНК и олигонуклеотид, рибозимы, аптамер, интерферирующие РНК (РНКи), дцРНК-субстрат Dicer, короткую шпилечную РНК (кшРНК), асимметричную интерферирующую РНК (аиРНК), микроРНК (миРНК), миникольцо ДНК, миниген, вирусную ДНК (например, геном лентивируса или ААВ) или невирусные синтетические ДНК-векторы, линейную дуплексную ДНК с замкнутыми концами (зкДНК/CELiD), плазмиды, бакмиды, ДНК-векторы doggybone (dbDNA™), минималистичный вектор для иммунологически определенной экспрессии гена (MIDGE), невирусный ДНК-вектор с минимальной цепью (линейный ковалентно замкнутый ДНК-вектор) или гантелеобразный минимальный ДНК-вектор («гантелеобразную ДНК»). (см., например, WO2010/0086626, содержание которого полностью включено в настоящий документ посредством ссылки). В соответствии с некоторыми вариантами реализации ингибиторы врожденного иммунного ответа и нуклеиновые кислоты могут быть введены субъекту или пациенту в любой комбинации. Например, может быть введен один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа). В соответствии с некоторыми вариантами реализации субъекту или пациенту вводят ингибитор иммунного ответа (например, врожденного иммунного ответа), описанный в настоящем документе, и нуклеиновые кислоты (например, миникольцо, миниген, минимальную цепь ковалентно замкнутой ДНК, ДНК doggybone (dbDNA™), гантелеобразную ДНК, линейную дуплексную ДНК с замкнутыми концами (зкДНК и CELiD), кольцевой вектор на основе плазмиды, антисмысловой олигонуклеотид (ASO), РНКи, киРНК, мРНК и т.д.). В соответствии с некоторыми вариантами реализации субъекту или пациенту вводят рапамицин или аналоги рапамицина, один или более ингибиторов TLR9 и нуклеиновые кислоты. В соответствии с некоторыми вариантами реализации субъекту или пациенту вводят рапамицин или аналоги рапамицина, один или более ингибиторов cGAS и нуклеиновые кислоты. В соответствии с некоторыми вариантами реализации субъекту или пациенту вводят рапамицин или аналоги рапамицина, один или более антагонистов инфламмасомы и нуклеиновые кислоты. В соответствии с некоторыми вариантами реализации субъекту или пациенту вводят рапамицин или аналоги рапамицина, один или более ингибиторов TLR9, один или более ингибиторов cGAS и нуклеиновые кислоты. В соответствии с некоторыми вариантами реализации субъекту или пациенту вводят рапамицин или аналоги рапамицина, один или более ингибиторов TLR9, один или более антагонистов инфламмасомы и нуклеиновые кислоты. В соответствии с некоторыми вариантами реализации субъекту или пациенту вводят один или более ингибиторов TLR9, один или более ингибиторов cGAS и зкДНК-вектор, содержащий нуклеиновые кислоты. В соответствии с некоторыми вариантами реализации субъекту или пациенту вводят один или более ингибиторов TLR9, один или более ингибиторов cGAS, один или более антагонистов инфламмасомы и нуклеиновые кислоты. В соответствии с некоторыми вариантами реализации субъекту или пациенту вводят рапамицин или аналоги рапамицина, один или более ингибиторов TLR9, один или более ингибиторов cGAS, один или более антагонистов инфламмасомы и нуклеиновые кислоты.
[00276] Согласно некоторым вариантам реализации субъекту можно одновременно вводить один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа) и одну или более нуклеиновых кислот (например, миникольцо, миниген, минимальную цепь ковалентно замкнутой ДНК, ДНК doggybone (dbDNA™), гантелеобразную ДНК, линейную дуплексную ДНК с замкнутыми концами (зкДНК и CELiD), кольцевой вектор на основе плазмиды, антисмысловой олигонуклеотид (ASO), РНКи, киРНК, мРНК и т.д.). Например, способ может включать введение субъекту ингибитора иммунного ответа (например, врожденного иммунного ответа) и терапевтического средства на основе нуклеиновой кислоты в виде двух отдельных составов, но одновременно. В другом примере способ может включать одновременное введение субъекту ингибитора иммунного ответа (например, врожденного иммунного ответа) и терапевтической нуклеиновой кислоты в одном составе в одно и то же время.
[00277] Согласно некоторым вариантам реализации субъекту можно последовательно вводить один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа) и одну или более нуклеиновых кислот (например, миникольцо, миниген, минимальную цепь ковалентно замкнутой ДНК, ДНК doggybone (dbDNA™), гантелеобразную ДНК, линейную дуплексную ДНК с замкнутыми концами (зкДНК и CELiD), кольцевой вектор на основе плазмиды, антисмысловой олигонуклеотид (ASO), РНКи, киРНК, мРНК и т.д.). Например, ингибитор иммунного ответа (например, врожденного иммунного ответа) может быть введен до введения терапевтической нуклеиновой кислоты.
[00278] В случаях последовательного введения может быть период задержки между введением одного или более ингибиторов иммунного ответа (например, врожденного иммунного ответа) и ТНК. Например, ингибитор иммунного ответа (например, врожденного иммунного ответа) может быть введен за часы, дни или недели до введения ТНК (например, по меньшей мере за 30 минут, по меньшей мере за 1 час, по меньшей мере за 2 часа, по меньшей мере за 3 часа, по меньшей мере за 4 часа, по меньшей мере за 5 часов, по меньшей мере за 6 часов, по меньшей мере за 7 часов, по меньшей мере за 8 часов, по меньшей мере за 9 часов, по меньшей мере за 10 часов, по меньшей мере за 11 часов, по меньшей мере за 12 часов, по меньшей мере за 13 часов, по меньшей мере за 14 часов, по меньшей мере за 15 часов, по меньшей мере за 16 часов, по меньшей мере за 17 часов, по меньшей мере за 18 часов, по меньшей мере за 19 часов, по меньшей мере за 20 часов, по меньшей мере за 21 час, по меньшей мере за 22 часа, по меньшей мере за 23 часа, по меньшей мере за 24 часа, по меньшей мере примерно за 2 дня, по меньшей мере примерно за 3 дня, по меньшей мере примерно за 4 дня, по меньшей мере примерно за 5 дней, по меньшей мере примерно за 6 дней, по меньшей мере примерно за 1 неделю, по меньшей мере примерно за 2 недели, по меньшей мере примерно за 3 недели и по меньшей мере примерно за 4 недели до введения нуклеиновой кислоты). Согласно некоторым вариантам реализации ингибитор иммунного ответа (например, врожденного иммунного ответа) может быть введен примерно за тридцать (30) минут до введения ТНК. Согласно некоторым вариантам реализации ингибитор иммунного ответа (например, врожденного иммунного ответа) может быть введен примерно за один (1) час до введения нуклеиновой кислоты. Согласно некоторым вариантам реализации ингибитор иммунного ответа (например, врожденного иммунного ответа) может быть введен примерно за два (2) часа до введения нуклеиновой кислоты. Согласно некоторым вариантам реализации ингибитор иммунного ответа (например, врожденного иммунного ответа) может быть введен примерно за три (3) часа до введения нуклеиновой кислоты. Согласно некоторым вариантам реализации ингибитор иммунного ответа (например, врожденного иммунного ответа) может быть введен примерно за четыре (4) часа до введения нуклеиновой кислоты. Согласно некоторым вариантам реализации ингибитор иммунного ответа (например, врожденного иммунного ответа) может быть введен примерно за пять (5) часов до введения нуклеиновой кислоты. Согласно некоторым вариантам реализации ингибитор иммунного ответа (например, врожденного иммунного ответа) может быть введен примерно за шесть (6) часов до введения нуклеиновой кислоты. Согласно некоторым вариантам реализации ингибитор иммунного ответа (например, врожденного иммунного ответа) может быть введен примерно за семь (7) часов до введения нуклеиновой кислоты. Согласно некоторым вариантам реализации ингибитор иммунного ответа (например, врожденного иммунного ответа) может быть введен примерно за восемь (8) часов до введения нуклеиновой кислоты. Согласно некоторым вариантам реализации ингибитор иммунного ответа (например, врожденного иммунного ответа) может быть введен примерно за девять (9) часов до введения нуклеиновой кислоты. Согласно некоторым вариантам реализации ингибитор иммунного ответа (например, врожденного иммунного ответа) может быть введен примерно за десять (10) часов до введения нуклеиновой кислоты.
[00279] Согласно одному варианту реализации ингибитор иммунного ответа (например, врожденного иммунного ответа) вводят не более чем примерно за 1 час, примерно за 2 часа, примерно за 3 часа, примерно за 4 часа, примерно за 5 часов, примерно за 6 часов, примерно за 7 часов, примерно за 8 часов, примерно за 9 часов, примерно за 10 часов, примерно за 11 часов, примерно за 12 часов, примерно за 13 часов, примерно за 14 часов, примерно за 15 часов, примерно за 16 часов, примерно за 17 часов, примерно за 18 часов, примерно за 19 часов, примерно за 20 часов, примерно за 21 час, примерно за 22 часа, примерно за 23 часа или 24 часа до введения нуклеиновой кислоты. Согласно некоторым вариантам реализации ингибитор иммунного ответа (например, врожденного иммунного ответа) может быть введен не более чем примерно за 1 день, примерно за 2 дня, примерно за 3 дня, примерно за 4 дня, примерно за 6 дней или примерно за 7 дней до введения нуклеиновой кислоты.
[00280] Согласно некоторым вариантам реализации ингибитор иммунного ответа (например, врожденного иммунного ответа) может быть введен примерно через 30 минут, примерно через 1 час, примерно через 2 часа, примерно через 3 часа, примерно через 4 часа, примерно через 5 часов, примерно через 6 часов, примерно через 7 часов, примерно через 8 часов, примерно через 9 часов, примерно через 10 часов, примерно через 11 часов, примерно через 12 часов, примерно через 13 часов, примерно через 14 часов, примерно через 15 часов, примерно через 16 часов, примерно через 17 часов, примерно через 18 часов, примерно через 19 часов, примерно через 20 часов, примерно через 21 час, примерно через 22 часа, примерно через 23 часа или через 24 часа после введения нуклеиновой кислоты. Согласно некоторым вариантам реализации ингибитор иммунного ответа (например, врожденного иммунного ответа) может быть введен примерно через 1 день, примерно через 2 дня, примерно через 3 дня, примерно через 4 дня, примерно через 6 дней или примерно через 7 дней после введения нуклеиновой кислоты.
[00281] Согласно одному варианту реализации ингибитор иммунного ответа (например, врожденного иммунного ответа) вводят не более чем примерно через 1 час, примерно через 2 часа, примерно через 3 часа, примерно через 4 часа, примерно через 5 часов, примерно через 6 часов, примерно через 7 часов, примерно через 8 часов, примерно через 9 часов, примерно через 10 часов, примерно через 11 часов, примерно через 12 часов, примерно через 13 часов, примерно через 14 часов, примерно через 15 часов, примерно через 16 часов, примерно через 17 часов, примерно через 18 часов, примерно через 19 часов, примерно через 20 часов, примерно через 21 час, примерно через 22 часа, примерно через 23 часа или через 24 часа после введения нуклеиновой кислоты. Согласно некоторым вариантам реализации ингибитор иммунного ответа (например, врожденного иммунного ответа) может быть введен не более чем примерно через 1 день, примерно через 2 дня, примерно через 3 дня, примерно через 4 дня, примерно через 6 дней или примерно через 7 дней после введения нуклеиновой кислоты.
[00282] Согласно некоторым вариантам реализации один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа) могут быть введены несколько раз до, одновременно и/или после введения нуклеиновой кислоты.
[00283] Согласно некоторым вариантам реализации нуклеиновая кислота (например, зкДНК-вектор) может быть введена в виде однократной дозы или в виде многократных доз. В соответствии с некоторыми вариантами реализации субъекту может быть введено более одной дозы. Многократные дозы могут быть введены по мере необходимости, поскольку зкДНК-вектор не вызывает иммунного ответа хозяина против капсида ввиду отсутствия вирусного капсида. В соответствии с некоторыми вариантами реализации количество вводимых доз может, например, составлять от 2 до 10 или более доз, например, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более.
[00284] Согласно некоторым вариантам реализации нуклеиновая кислота может быть введена и повторно дозирована несколько раз в сочетании с одним или более ингибиторами иммунного ответа (например, врожденного иммунного ответа), раскрытыми в настоящем документе. Например, терапевтическая нуклеиновая кислота может быть введена в день 0 с одним или более ингибиторами иммунного ответа, которые вводят до, после или одновременно с введением нуклеиновой кислоты в первой схеме дозирования. После начального лечения в день 0 второе дозирование (повторная доза) может быть выполнено примерно через 1 неделю, примерно через 2 недели, примерно через 3 недели, примерно через 4 недели, примерно через 5 недель, примерно через 6 недель, примерно через 7 недель, примерно через 8 недель или примерно через 3 месяца, примерно через 4 месяца, примерно через 5 месяцев, примерно через 6 месяцев, примерно через 7 месяцев, примерно через 8 месяцев, примерно через 9 месяцев, примерно через 10 месяцев, примерно через 11 месяцев или примерно через 1 год, примерно через 2 года, примерно через 3 года, примерно через 4 года, примерно через 5 лет, примерно через 6 лет, примерно через 7 лет, примерно через 8 лет, примерно через 9 лет, примерно через 10 лет, примерно через 11 лет, примерно через 12 лет, примерно через 13 лет, примерно через 14 лет, примерно через 15 лет, примерно через 16 лет, примерно через 17 лет, примерно через 18 лет, примерно через 19 лет, примерно через 20 лет, примерно через 21 год, примерно через 22 года, примерно через 23 года, примерно через 24 года, примерно через 25 лет, примерно через 26 лет, примерно через 27 лет, примерно через 28 лет, примерно через 29 лет, примерно через 30 лет, примерно через 31 год, примерно через 32 года, примерно через 33 года, примерно через 34 года, примерно через 35 лет, примерно через 36 лет, примерно через 37 лет, примерно через 38 лет, примерно через 39 лет, примерно через 40 лет, примерно через 41 год, примерно через 42 года, примерно через 43 года, примерно через 44 года, примерно через 45 лет, примерно через 46 лет, примерно через 47 лет, примерно через 48 лет, примерно через 49 лет или примерно через 50 лет после начального лечения нуклеиновой кислотой, предпочтительно с одним или более ингибиторами иммунного ответа (например, врожденного иммунного ответа), раскрытыми в настоящем документе.
[00285] В соответствии с некоторыми вариантами реализации повторное дозирование нуклеиновой кислоты приводит к увеличению экспрессии нуклеиновой кислоты. В соответствии с некоторыми вариантами реализации увеличение экспрессии нуклеиновой кислоты после повторного дозирования, по сравнению с экспрессией нуклеиновой кислоты после первой дозы, от примерно 0,5 раза до примерно 10 раз, от примерно 1 раза до примерно 5 раз, от примерно 1 раза до примерно 2 раз или примерно в 0,5 раза, примерно в 1 раз, примерно в 2 раза, примерно в 3 раза, примерно в 4 раза, примерно в 5 раз, примерно в 6 раз, примерно в 7 раз, примерно в 8 раз, примерно в 9 раз или примерно в 10 раз выше после повторного дозирования нуклеиновой кислоты.
[00286] Обычно дозировка будет варьироваться в зависимости от конкретных характеристик зкДНК-вектора, эффективности экспрессии, а также в зависимости от возраста, состояния и пола пациента. Дозировка может быть определена специалистом в данной области техники и, в отличие от обычных ААВ-векторов, также может быть скорректирована индивидуальным врачом в случае любого осложнения, поскольку зкДНК-векторы не содержат активирующие иммунную систему капсидные белки, которые препятствуют повторному дозированию.
[00287] В соответствии с некоторыми вариантами реализации для достижения целевого уровня экспрессии гена на протяжении различных периодов времени, например, ежедневно, еженедельно, ежемесячно, ежегодно и т.д., можно применять более одного введения (например, два, три, четыре или более введений) нуклеиновой кислоты (например, зкДНК-вектора) для экспрессии белка, как раскрыто в настоящем документе.
[00288] В соответствии с любым из вариантов реализации, раскрытых в настоящем документе, нуклеиновая кислота может представлять собой терапевтическую нуклеиновую кислоту.
Терапевтический эффект
[00289] Эффективность зкДНК-вектора, экспрессирующего ингибитор иммунного ответа (например, врожденного иммунного ответа), раскрытого в настоящем документе, для подавления или ослабления врожденной иммунной системы, может быть определена квалифицированным клиницистом. Однако лечение считается «эффективным лечением», в соответствии с использованием этого термина в настоящем документе, если любой или все признаки или симптомы врожденной иммунной системы уменьшаются и/или изменяются благоприятным образом, или если другие клинически принятые симптомы или маркеры заболевания улучшаются или облегчаются, например, по меньшей мере на 10% после лечения зкДНК-вектором, кодирующим ингибитор иммунного ответа (например, врожденного иммунного ответа), раскрытым в настоящем документе. Примерные маркеры и симптомы обсуждаются в Примерах в настоящем документе. Эффективность также может быть измерена по отсутствию у индивидуума ухудшения, которое оценивается по стабилизации заболевания или по потребности в медицинских вмешательствах (т.е. прогрессирование заболевания прекращается или по меньшей мере замедляется). Способы измерения этих показателей известны специалистам в данной области техники и/или описаны в настоящем документе. Лечение включает любое лечение заболевания у индивидуума или животного (некоторые неограничивающие примеры включают человека или млекопитающее) и включает: (1) ингибирование заболевания, например, прекращение или замедление прогрессирования заболевания или нарушения; или (2) облегчение заболевания, например, вызывающее регресс симптомов; и (3) предотвращение или уменьшение вероятности развития заболевания, или предотвращение вторичных заболеваний/нарушений, ассоциированных с указанным заболеванием, таких как печеночная или почечная недостаточность. Эффективное количество для лечения заболевания означает такое количество, которое, при введении нуждающемуся в этом млекопитающему, является достаточным для обеспечения эффективного лечения, в соответствии с определением этого термина в настоящем документе, этого заболевания.
[00290] Эффективность агента может быть определена путем оценки физических показателей, которые являются специфическими для данного заболевания. Стандартные методы анализа показателей заболевания известны в данной области техники. Например, физические показатели для врожденной иммунной системы включают, например, но не ограничиваются перечисленными, растворимый CD14 (sCD14) и IL-18, IL-22 в плазме или крови, белки инфламмасомы, такие как AIM2, NLRP3, NLRP1, ASC и каспазу-1 в спинномозговой жидкости или крови, активация цитокиновых путей может быть использована как функциональный показатель активации пути инфламмасомы NLRP3 и/или AIM2 или активации каспазы 1, и включает биомаркеры, такие как, но не ограничиваясь перечисленными: интерлейкин (IL)-1β, IL-6, IL-8, IL-18, интерферон (IFN)-γ, интерферон (IFN)-α, хемоаттрактантный белок моноцитов (MCP)-1 и/или фактор некроза опухоли (TNF)-α.
[00291] Согласно одному варианту реализации зкДНК-вектор содержит последовательность нуклеиновой кислоты для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), раскрытого в настоящем документе, например, который является функциональным для подавления врожденной иммунной системы. Согласно предпочтительному варианту реализации ингибитор иммунного ответа (например, врожденного иммунного ответа), раскрытый в настоящем документе, например, как раскрыто в настоящем документе, не вызывает реакции иммунной системы, а скорее подавляет или ослабляет иммунную систему у субъекта.
[00292] Фармацевтические композиции для терапевтических целей, как правило, должны быть стерильными и стабильными в условиях изготовления и хранения. Композиция может быть изготовлена в виде раствора, микроэмульсии, дисперсии, липосом или другой упорядоченной структуры, подходящей для высокой концентрации ДНК-вектора с замкнутыми концами, например, зкДНК-вектора. Стерильные растворы для инъекций могут быть приготовлены путем включения зкДНК-вектора в необходимом количестве в соответствующем буфере с одним ингредиентом или комбинацией ингредиентов, перечисленных выше, при необходимости, с последующей стерилизацией путем фильтрации.
[00293] ДНК-вектор с замкнутыми концами, включая зкДНК-вектор, и ингибитор иммунного ответа (например, врожденного иммунного ответа), раскрытые в настоящем документе, могут быть включены в фармацевтическую композицию, подходящую для местного, системного, внутриамниотического, интратекального, внутричерепного, внутриартериального, внутривенного, внутрилимфатического, внутрибрюшинного, подкожного, трахеального, внутритканевого (например, внутримышечного, внутрисердечного, внутрипеченочного, внутрипочечного, интрацеребрального), интратекального, интравезикального, конъюнктивального (например, экстраорбитального, интраорбитального, ретроорбитального, интраретинального, субретинального, хориоидального, субхориоидального, интрастромального, интракамерального и интравитреального), интракохлеарного и мукозального (например, перорального, ректального, назального) введения. Также предусмотрена пассивная трансдукция ткани путем внутривенной или внутриартериальной инфузии под высоким давлением, а также внутриклеточной инъекции, такой как внутриядерная микроинъекция или внутрицитоплазматическая инъекция.
[00294] В некоторых аспектах способы, предложенные в настоящем документе, включают доставку в клетку-хозяина одного или более ДНК-векторов с замкнутыми концами, включая зкДНК-вектор, и ингибитора иммунного ответа (например, врожденного иммунного ответа), описанных в настоящем документе. В настоящем документе также предложены клетки, полученные с помощью таких способов, и организмы (такие как животные, растения или грибы), содержащие такие клетки или полученные из них. Способы доставки нуклеиновых кислот могут включать липофекцию, нуклеофекцию, микроинъекцию, биолистику, липосомы, иммунолипосомы, конъюгаты поликатион или липид:нуклеиновая кислота, «незащищенную» (naked) ДНК и усиленное агентом поглощение ДНК. Липофекция описана, например, в патентах США №№ 5049386, 4946787 и 4897355, и реагенты для липофекции продаются коммерчески (например, трансфектам (Transfectam™) и липофектин (Lipofectin™)). Доставка может быть осуществлена в клетки (например, введение in vitro или ex vivo) или в ткани-мишени (например, введение in vivo).
[00295] В данной области техники известны различные методики и способы доставки нуклеиновых кислот в клетки. Например, ДНК-вектор с замкнутыми концами, включая зкДНК-вектор, и рапамицин или аналог рапамицина, описанные в настоящем документе, могут быть изготовлены в виде состава липидных наночастиц (ЛНЧ), липидоидов, липосом, липидных наночастиц, липоплексов или наночастиц типа ядро-оболочка. Как правило, ЛНЧ состоят из молекул нуклеиновой кислоты (например, зкДНК), одного или более ионизируемых или катионных липидов (или их солей), одного или более неионных или нейтральных липидов (например, фосфолипида), молекулы, которая предотвращает агрегацию (например, ПЭГ или конъюгата ПЭГ-липид), и необязательно стерина (например, холестерина).
[00296] Другой способ доставки в клетку ДНК-вектора с замкнутыми концами, включая зкДНК-вектор, и ингибитора иммунного ответа (например, врожденного иммунного ответа), описанных в настоящем документе, заключается в конъюгации нуклеиновой кислоты с лигандом, который интернализуется клеткой. Например, лиганд может связывать рецептор на поверхности клетки и интернализоваться за счет эндоцитоза. Лиганд может быть ковалентно связан с нуклеотидом в нуклеиновой кислоте. Примерные конъюгаты для доставки нуклеиновых кислот в клетку описаны, например, в WO2015/006740, WO2014/025805, WO2012/037254, WO2009/082606, WO2009/073809, WO2009/018332, WO2006/112872, WO2004/090108, WO2004/091515 и WO2017/177326.
[00297] Нуклеиновые кислоты и ДНК-вектор с замкнутыми концами, включая зкДНК-вектор, описанные в настоящем документе, также могут быть доставлены в клетку путем трансфекции. Подходящие способы трансфекции включают, но не ограничиваются перечисленными, опосредуемую липидами трансфекцию, опосредуемую катионным полимером трансфекцию или осаждение фосфатом кальция. Реагенты для трансфекции хорошо известны в данной области техники и включают, но не ограничиваются перечисленными, реагент для трансфекции TurboFect (Thermo Fisher Scientific®), реагент Pro-Ject (Thermo Fisher Scientific®), реагент для трансфекции белков TRANSPASS™ P (New England Biolabs®), реагент для доставки белков CHARIOT™ (Active Motif), реагент для трансфекции белков PROTEOJUICE™ (EMD Millipore®), 293fectin, LIPOFECTAMINE™ 2000, LIPOFECTAMINE™ 3000 (Thermo Fisher Scientific®), LIPOFECTAMINE™ (Thermo Fisher Scientific®), LIPOFECTIN™ (Thermo Fisher Scientific®), DMRIE-C, CELLFECTIN™ (Thermo Fisher Scientific®), OLIGOFECTAMINE™ (Thermo Fisher Scientific®), LIPOFECTACE™, FUGENE™ (Roche®, Базель, Швейцария), FUGENE™ HD (Roche®), TRANSFECTAM™ (трансфектам, Promega®, Мэдисон, Висконсин), TFX-10™ (Promega®), TFX-20™ (Promega®), TFX-50™ (Promega®), TRANSFECTIN™ (BioRad®, Геркулес, Калифорния), SILENTFECT™ (Bio-Rad®), Effectene™ (Qiagen®, Валенсия, Калифорния), DC-chol (Avanti Polar Lipids), GENEPORTER™ (Gene Therapy Systems®, Сан-Диего, Калифорния), DHARMAFECT 1™ (Dharmacon®, Лафайет, Колорадо), DHARMAFECT 2™ (Dharmacon®), DHARMAFECT 3™ (Dharmacon®), DHARMAFECT 4™ (Dharmacon®), ESCORT™ III (Sigma®, Сент-Луис, Миссури) и ESCORT™ IV (Sigma Chemical Co.). Нуклеиновые кислоты, такие как зкДНК, также могут быть доставлены в клетку с помощью микроструйных способов, известных специалистам в данной области техники.
[00298] ДНК-вектор с замкнутыми концами, включая зкДНК-вектор, и ингибитор иммунного ответа (например, врожденного иммунного ответа), описанные в настоящем документе, также могут быть введены непосредственно в организм для трансдукции клеток in vivo. Введение осуществляют любым из путей, обычно используемых для приведения молекулы в максимальный контакт с клетками крови или тканей, включая, но не ограничиваясь перечисленным, инъекцию, инфузию, местное нанесение и электропорацию. Подходящие способы введения таких нуклеиновых кислот доступны и хорошо известны специалистам в данной области техники, и хотя для введения конкретной композиции можно использовать более одного пути, конкретный путь часто может обеспечить более быструю и более эффективную реакцию по сравнению с другим путем введения.
[00299] Способы введения ДНК-вектора с замкнутыми концами, включая зкДНК-вектор, и ингибитора врожденного иммунного ответа, описанных в настоящем документе, могут включать доставку в гематопоэтические стволовые клетки, например, с помощью способов, описанных, например, в патенте США №5928638.
[00300] ДНК-вектор с замкнутыми концами, включая зкДНК-вектор, и ингибитор иммунного ответа (например, врожденного иммунного ответа), описанные в настоящем документе, могут быть добавлены к липосомам для доставки в клетку или орган-мишень у субъекта. Липосомы представляют собой везикулы, которые имеют по меньшей мере один липидный бислой. Липосомы, как правило, используются в качестве носителей для доставки лекарственных/терапевтических средств в контексте фармацевтической разработки. Они работают за счет слияния с клеточной мембраной и перегруппировки ее липидной структуры для доставки лекарственного средства или активного фармацевтического ингредиента (АФИ). Липосомальные композиции для такой доставки состоят из фосфолипидов, в частности, соединений, содержащих фосфатидилхолиновую группу, однако указанные композиции также могут включать другие липиды. Примерные липосомы и липосомальные составы раскрыты в международной заявке PCT/US2018/050042, поданной 7 сентября 2018 г., и в международной заявке PCT/US2018/064242, поданной 6 декабря 2018 г., например, см. раздел, озаглавленный «Фармацевтические составы».
[00301] Различные способы доставки, известные в данной области техники, или их модификации могут быть использованы для доставки ДНК-вектора с замкнутыми концами, включая зкДНК-вектор, и ингибитора иммунного ответа (например, врожденного иммунного ответа), описанных в настоящем документе, in vitro или in vivo. Например, согласно некоторым вариантам реализации, зкДНК-векторы доставляют, образуя кратковременное отверстие в клеточной мембране с использованием механической, электрической, ультразвуковой, гидродинамической или лазерной энергии так, чтобы облегчить вход ДНК в целевые клетки. Например, зкДНК-вектор может быть доставлен путем кратковременного разрыва клеточной мембраны путем продавливания клетки через канал ограниченного размера или другими способами, известными в данной области техники. В некоторых случаях зкДНК-вектор по отдельности напрямую вводят путем инъекции незащищенной ДНК в клетки кожи, вилочковой железы, сердечной мышцы, скелетных мышц или печени. В некоторых случаях зкДНК-вектор доставляют с помощью генной пушки. Сферические частицы золота или вольфрама (диаметром 1-3 мкм), покрытые бескапсидными AAV-векторами, могут быть разогнаны до высокой скорости с помощью сжатого газа для проникновения в клетки целевой ткани.
[00302] Композиции, содержащие ДНК-вектор с замкнутыми концами, включая зкДНК-вектор, и рапамицин или аналог рапамицина, описанные в настоящем документе, и фармацевтически приемлемый носитель, конкретно предусмотрены в настоящем документе. Согласно некоторым вариантам реализации зкДНК-вектор изготовлен с липидной системой доставки, например, липосомами, как описано в настоящем документе. Согласно некоторым вариантам реализации такие композиции вводят любым путем, желательным для квалифицированного специалиста. Композиции могут быть введены субъекту различными путями, включая пероральный, парентеральный, сублингвальный, трансдермальный, ректальный, трансмукозальный, местный, путем ингаляции, путем буккального введения, внутриплевральный, внутривенный, внутриартериальный, внутрибрюшинный, подкожный, внутримышечный, интраназальный, интратекальный и внутрисуставный или их комбинации. Для ветеринарного применения композиция может быть введена в виде достаточно приемлемого состава в соответствии с обычной ветеринарной практикой. Ветеринар может легко определить схему дозирования и путь введения, которые наиболее подходят для конкретного животного. Композиции могут быть ведены с помощью обычных шприцев, безыгольных инъекционных устройств, «генных пушек с бомбардировкой микрочастицами» или других физических методов, таких как электропорация («ЭП»), гидродинамические методы или применение ультразвука.
[00303] В некоторых случаях ДНК-вектор с замкнутыми концами, включая зкДНК-вектор, и один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа), описанных в настоящем документе, доставляют с помощью гидродинамической инъекции, которая является простым и высокоэффективным методом прямой внутриклеточной доставки любых водорастворимых соединений и частиц во внутренние органы и скелетные мышцы по всей конечности.
[00304] В некоторых случаях ДНК-вектор с замкнутыми концами, включая зкДНК-вектор, и один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа), описанных в настоящем документе, доставляют с помощью ультразвука путем создания наноскопических пор в мембране для облегчения внутриклеточной доставки частиц ДНК в клетки внутренних органов или опухолей, поэтому размер и концентрация ДНК-вектора с замкнутыми концами имеют большое значение для эффективности системы. В некоторых случаях ДНК-векторы с замкнутыми концами, включая зкДНК-вектор, и один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа), описанных в настоящем документе, доставляют с помощью магнитофекции с использованием магнитных полей для концентрирования частиц, содержащих нуклеиновую кислоту, в клетках-мишенях.
[00305] В некоторых случаях могут быть использованы химические системы доставки, например, с использованием наномерных комплексов, которые включают уплотнение отрицательно заряженной нуклеиновой кислоты поликатионными наномерными частицами, в составе катионной липосомы/мицеллы или катионных полимеров. Катионные липиды, используемые в указанном способе доставки, включают, но не ограничиваются перечисленными, моновалентные катионные липиды, поливалентные катионные липиды, гуанидин-содержащие соединения, соединения производных холестерина, катионные полимеры (например, полиэтиленимин, поли-L-лизин, протамин, другие катионные полимеры) и гибриды липид-полимер.
А. Экзосомы:
[00306] Согласно некоторым вариантам реализации ДНК-вектор с замкнутыми концами, включая зкДНК-вектор, и один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа), описанных в настоящем документе, доставляют упакованными в экзосоме. Экзосомы представляют собой небольшие мембранные везикулы эндоцитарного происхождения, которые высвобождаются во внеклеточную среду после слияния мультивезикулярных телец с плазматической мембраной. Их поверхность состоит из липидного бислоя клеточной мембраны донорной клетки, они содержат цитозоль из клетки, которая продуцировала экзосому, и экспонируют на поверхности мембранные белки родительской клетки. Экзосомы продуцируются различными типами клеток, в том числе эпителиальными клетками, В- и Т-лимфоцитами, тучными клетками (MC), а также дендритными клетками (DC). Согласно некоторым вариантам реализации для применения предусмотрены экзосомы с диаметром от 10 нм до 1 мкм, от 20 нм до 500 нм, от 30 нм до 250 нм, от 50 до 100 нм. Экзосомы могут быть выделены для доставки в целевые клетки либо с использованием их донорных клеток, либо путем введения в них конкретных нуклеиновых кислот. Для получения экзосом, содержащих бескапсидные ААВ-векторы согласно настоящему изобретению, могут быть использованы различные подходы, известные в данной области техники.
B. Микрочастицы/наночастицы:
[00307] Согласно некоторым вариантам реализации ДНК-вектор с замкнутыми концами, включая зкДНК-вектор, и рапамицин или аналог рапамицина, описанные в настоящем документе, доставляют с помощью липидной наночастицы. Обычно липидные наночастицы содержат ионизируемый аминолипид (например, гептатриаконта-6,9,28,31-тетраен-19-ил-4-(диметиламино)бутаноат, DLin-MC3-DMA, фосфатидилхолин(1,2-дистеароил-sn-глицеро-3-фосфохолин, DSPC), холестерин и липид оболочки (полиэтиленгликоль-димиристоилглицерин, ПЭГ-ДМГ), например, как описано Tam et al. (2013). Advances in Lipid Nanoparticles for siRNA delivery. Pharmaceuticals 5(3): 498-507. Согласно некоторым вариантам реализации липидная наночастица имеет средний диаметр от примерно 10 до примерно 1000 нм. Согласно некоторым вариантам реализации липидная наночастица имеет диаметр менее 300 нм. Согласно некоторым вариантам реализации липидная наночастица имеет диаметр от примерно 10 до примерно 300 нм. Согласно некоторым вариантам реализации липидная наночастица имеет диаметр менее 200 нм. Согласно некоторым вариантам реализации липидная наночастица имеет диаметр от примерно 25 до примерно 200 нм. Согласно некоторым другим вариантам реализации липидные частицы, содержащие терапевтическую нуклеиновую кислоту и/или иммуносупрессор, как правило, имеют средний диаметр от примерно 20 нм до примерно 100 нм, от 30 нм до примерно 150 нм, от примерно 40 нм до примерно 150 нм, от примерно 50 нм до примерно 150 нм, от примерно 60 нм до примерно 130 нм, от примерно 70 нм до примерно 110 нм, от примерно 70 нм до примерно 100 нм, от примерно 80 нм до примерно 100 нм, от примерно 90 нм до примерно 100 нм, от примерно 70 до примерно 90 нм, от примерно 80 нм до примерно 90 нм, от примерно 70 нм до примерно 80 нм или примерно 30 нм, 35 нм, 40 нм, 45 нм, 50 нм, 55 нм, 60 нм, 65 нм, 70 нм, 75 нм, 80 нм, 85 нм, 90 нм, 95 нм, 100 нм, 105 нм, 110 нм, 115 нм, 120 нм, 125 нм, 130 нм, 135 нм, 140 нм, 145 нм или 150 нм, чтобы обеспечить эффективную доставку. Липидные частицы, содержащие нуклеиновую кислоту, и способ их получения раскрыты, например, в PCT/US18/50042, публикациях патентов США №№ 20040142025 и 20070042031, раскрытия которых полностью включены в настоящий документ посредством ссылки для всех целей. Согласно некоторым вариантам реализации препарат липидных наночастиц (например, композиция, содержащая множество липидных наночастиц) имеет распределение по размерам, в котором средний размер (например, диаметр) составляет от примерно 70 нм до примерно 200 нм, и более типично средний размер составляет примерно 100 нм или менее.
[00308] В соответствии с некоторыми вариантами реализации жидкая фармацевтическая композиция, содержащая нуклеиновую кислоту (например, терапевтическую нуклеиновую кислоту, нуклеиновую кислоту, используемую для исследовательских целей) и/или ингибитор иммунного ответа (например, врожденного иммунного ответа) согласно настоящему изобретению, может быть изготовлена в виде состава липидных частиц. Согласно некоторым вариантам реализации липидная частица, содержащая нуклеиновую кислоту, может быть образована из катионного липида. Согласно некоторым другим вариантам реализации липидная частица, содержащая нуклеиновую кислоту, может быть образована из некатионного липида. Согласно предпочтительному варианту реализации липидная частица согласно настоящему изобретению представляет собой липидную частицу, содержащую нуклеиновую кислоту, которая образована из катионного липида, содержащего нуклеиновую кислоту, выбранную из группы, состоящей из мРНК, антисмысловой РНК и олигонуклеотида, рибозимов, аптамера, интерферирующих РНК (РНКи), дцРНК-субстрата Dicer, короткой шпилечной РНК (кшРНК), асимметричной интерферирующей РНК (аиРНК), микроРНК (миРНК), миникольца ДНК, минигена, вирусной ДНК (например, генома лентивируса или ААВ) или невирусных синтетических ДНК-векторов, линейной дуплексной ДНК с замкнутыми концами (зкДНК/CELiD), плазмид, бакмид, ДНК-векторов doggybone (dbDNA™), минималистичного вектора для иммунологически определенной экспрессии гена (MIDGE), невирусного ДНК-вектора с минимальной цепью (линейный, ковалентно замкнутый ДНК-вектор) или гантелеобразного минимального ДНК-вектора («гантелеобразной ДНК»).
[00309] Для доставки ДНК-вектора с замкнутыми концами, включая зкДНК-вектор, описанный в настоящем документе, могут быть использованы различные липидные наночастицы, известные в данной области техники. Например, различные способы доставки с использованием липидных наночастиц описаны в патентах США №№ 9404127, 9006417 и 9518272.
[00310] Согласно некоторым вариантам реализации ДНК-вектор с замкнутыми концами, включая зкДНК-вектор, и один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа), описанных в настоящем документе, доставляют с помощью наночастицы золота. Обычно нуклеиновая кислота может быть ковалентно связана с наночастицей золота или нековалентно связана с наночастицей золота (например, связана заряд-зарядным взаимодействием), например, как описано Ding et al. (2014). Gold Nanoparticles for Nucleic Acid Delivery. Mol. Ther. 22(6); 1075-1083. Согласно некоторым вариантам реализации конъюгаты наночастицы золота с нуклеиновой кислотой получают с использованием способов, описанных, например, в патенте США № 6812334.
C. Конъюгаты
[00311] Согласно некоторым вариантам реализации ДНК-вектор с замкнутыми концами, включая зкДНК-вектор, и один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа), описанных в настоящем документе, как раскрыто в настоящем документе, конъюгированы (например, ковалентно связаны) с агентом, который увеличивает поглощение клетками. «Агент, который увеличивает поглощение клетками» представляет собой молекулу, которая облегчает транспорт нуклеиновой кислоты через липидную мембрану. Например, нуклеиновая кислота может быть конъюгирована с липофильным соединением (например, холестерином, токоферолом и т.д.), проникающим в клетки пептидом (СРР) (например, пенетратином, ТАТ, Syn1B и т.д.) и полиаминами (например, спермином). Дополнительные примеры агентов, которые увеличивают поглощение клетками, раскрыты, например, в Winkler (2013). Oligonucleotide conjugates for therapeutic applications. Ther. Deliv. 4(7); 791-809.
[00312] Согласно некоторым вариантам реализации ДНК-вектор с замкнутыми концами, включая зкДНК-вектор, и один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа), описанных в настоящем документе, как раскрыто в настоящем документе, конъюгируют с полимером (например, полимерной молекулой) или молекулой фолата (например, молекулой фолиевой кислоты). В целом, доставка нуклеиновых кислот, конъюгированных с полимерами, известна в данной области техники, например, описана в WO 2000/34343 и WO 2008/022309. Согласно некоторым вариантам реализации зкДНК-вектор, раскрытый в настоящем документе, конъюгирован с полиамидным полимером, например, согласно описанию в патенте США №8987377. Согласно некоторым вариантам реализации нуклеиновая кислота, описанная в настоящем раскрытии, конъюгирована с молекулой фолиевой кислоты согласно описанию в патенте США №8507455.
[00313] Согласно некоторым вариантам реализации ДНК-вектор с замкнутыми концами, включая зкДНК-вектор, и рапамицин или аналог рапамицина, описанные в настоящем документе, как раскрыто в настоящем документе, конъюгируют с углеводом, например, как описано в патенте США №8450467.
[00314] Согласно некоторым вариантам реализации липидные наночастицы могут быть конъюгированы с другими группами для предотвращения агрегации. Такие липидные конъюгаты включают, но не ограничиваются перечисленными, конъюгаты ПЭГ-липид, такие как, например, ПЭГ, связанный с диалкилоксипропилами (например, конъюгаты ПЭГ-ДАА), ПЭГ, связанный с диацилглицеринами (например, конъюгаты ПЭГ-ДАГ), ПЭГ, связанный с холестерином, ПЭГ, связанный с фосфатидилэтаноламинами, и ПЭГ, конъюгированный с церамидами (см., например, патент США № 5885613), катионные липиды-ПЭГ, конъюгаты полиоксазолин (POZ)-липид (например, конъюгаты POZ-ДАА; см., например, предварительную заявку США № 61/294828, поданную 13 января 2010 г., и предварительную заявку США № 61/295140, поданную 14 января 2010 г.), полиамидные олигомеры (например, конъюгаты АТТА-липид) и их смеси. Дополнительные примеры конъюгатов POZ-липид описаны в публикации РСТ № WO 2010/006282. ПЭГ или POZ могут быть конъюгированы непосредственно с липидом или могут быть связаны с липидом за счет линкерной группы. Может быть использована любая линкерная группа, подходящая для связывания ПЭГ или POZ с липидом, включая, например, не содержащие сложного эфира линкерные группы и содержащие сложный эфир линкерные группы. Согласно определенным предпочтительным вариантам реализации применяют линкерные группы, не содержащие сложного эфира, такие как амиды или карбаматы. Раскрытия каждого из упомянутых выше патентных документов полностью включены в настоящий документ посредством ссылки для всех целей.
D. Нанокапсула
[00315] В качестве альтернативы, могут быть использованы нанокапсульные составы ДНК-вектора с замкнутыми концами, включая зкДНК-вектор, и рапамицина или аналога рапамицина, описанных в настоящем документе, как раскрыто в настоящем документе. Обычно нанокапсулы могут захватывать вещества стабильным и воспроизводимым образом. Чтобы избежать побочных эффектов, обусловленных внутриклеточной перегрузкой полимерами, такие ультрадисперсные частицы (размером около 0,1 мкм) следует конструировать с использованием полимеров, способных разлагаться in vivo. Предусмотрено применение биоразлагаемых полиалкил-цианоакрилатных наночастиц, которые удовлетворяют таким требованиям.
Е. Липосомы
[00316] ДНК-вектор с замкнутыми концами, включая зкДНК-вектор, и один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа), описанные в настоящем документе, могут быть добавлены к липосомам для доставки в клетку или орган-мишень у субъекта. Липосомы представляют собой везикулы, которые имеют по меньшей мере один липидный бислой. Липосомы, как правило, используются в качестве носителей для доставки лекарственных/терапевтических средств в контексте фармацевтической разработки. Они работают за счет слияния с клеточной мембраной и перегруппировки ее липидной структуры для доставки лекарственного средства или активного фармацевтического ингредиента (АФИ). Липосомальные композиции для такой доставки состоят из фосфолипидов, в частности, соединений, содержащих фосфатидилхолиновую группу, однако указанные композиции также могут включать другие липиды.
[00317] Изготовление и применение липосом в целом известно специалистам в данной области техники. Были разработаны липосомы с улучшенной стабильностью в сыворотке и временем полужизни в кровотоке (патент США № 5741516). Кроме того, были описаны различные способы получения липосом и липосомоподобных препаратов в качестве потенциальных носителей лекарственных средств (патенты США №5567434; 5552157; 5565213; 5738868 и 5795587).
F. Примерные композиции липосом и липидных наночастиц (ЛНЧ)
[00318] ДНК-вектор с замкнутыми концами, включая зкДНК-вектор, и один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа), описанные в настоящем документе, могут быть добавлены к липосомам для доставки в клетку, например, клетку, в которой требуется экспрессия трансгена. Липосомы представляют собой везикулы, которые имеют по меньшей мере один липидный бислой. Липосомы, как правило, используют в качестве носителей для доставки лекарственных/терапевтических средств в контексте фармацевтической разработки. Они работают за счет слияния с клеточной мембраной и перегруппировки ее липидной структуры для доставки лекарственного средства или активного фармацевтического ингредиента (АФИ). Липосомальные композиции для такой доставки состоят из фосфолипидов, в частности, соединений, содержащих фосфатидилхолиновую группу, однако указанные композиции также могут включать другие липиды.
[00319] Липидные наночастицы (ЛНЧ), содержащие зкДНК, раскрыты в международной заявке PCT/US2018/050042, поданной 7 сентября 2018 г., и международной заявке PCT/US2018/064242, поданной 6 декабря 2018 г., каждая из которых полностью включена в настоящий документ посредством ссылки, и предусмотрены для применения в способах и композициях, раскрытых в настоящем документе.
[00320] Согласно некоторым аспектам настоящего раскрытия предложен липосомальный состав, который включает одно или более соединений с функциональной группой полиэтиленгликоля (ПЭГ) (так называемых «пегилированных соединений»), которые могут уменьшать иммуногенность/антигенность соединения (соединений), обеспечивать их гидрофильность и гидрофобность, а также уменьшать частоту дозирования. В других случаях, липосомальный состав просто включает полимер полиэтиленгликоля (ПЭГ) в качестве дополнительного компонента. В таких аспектах молекулярная масса ПЭГ или функциональной группы ПЭГ может составлять от 62 Да до примерно 5000 Да.
[00321] Согласно некоторым аспектам настоящего раскрытия предложен липосомальный состав, который доставит АФИ с профилем пролонгированного или контролируемого высвобождения на протяжении периода времени от нескольких часов до недель. Согласно некоторым родственным аспектам липосомальный состав может содержать водные камеры, которые ограничены липидными бислоями. Согласно другим родственным аспектам липосомальный состав инкапсулирует АФИ с компонентами, которые претерпевают физический переход при повышенной температуре, который высвобождает АФИ в течение периода от нескольких часов до недель.
[00322] Согласно некоторым аспектам липосомальный состав содержит сфингомиелин и один или более липидов, раскрытых в настоящем документе. Согласно некоторым аспектам липосомальный состав содержит оптисомы.
[00323] Согласно некоторым аспектам настоящего раскрытия предложен липосомальный состав, который включает один или более липидов, выбранных из: натриевой соли N-(карбонилметоксиполиэтиленгликоль 2000)-1,2-дистеароил-sn-глицеро-3-фосфоэтаноламина, (дистеароил-sn-глицерофосфоэтаноламина), MPEG (метоксиполиэтиленгликоль)-конъюгированного липида, HSPC (гидрогенизированного соевого фосфатидилхолина); ПЭГ (полиэтиленгликоля); DSPE (дистеароил-sn-глицерофосфоэтаноламина); DSPC (дистеароилфосфатидилхолина); DOPC (диолеоилфосфатидилхолина); DPPG (дипальмитоилфосфатидилглицерина); EPC (яичного фосфатидилхолина); DOPS (диолеоилфосфатидилсерина); POPC (пальмитоилолеоилфосфатидилхолина); SM (сфингомиелина); MPEG (метоксиполиэтиленгликоля); DMPC (димиристоилфосфатидилхолина); DMPG (димиристоилфосфатидилглицерина); DSPG (дистеароилфосфатидилглицерина); DEPC (диэрукоилфосфатидилхолина); DOPE (диолеоил-sn-глицерофосфоэтаноламина), сульфата холестерина (CS), дипальмитоилфосфатидилглицерина (DPPG), DOPC (диолеоил-sn-глицерофосфатидилхолина) или любой их комбинации.
[00324] Согласно некоторым аспектам настоящего раскрытия предложен липосомальный состав, содержащий фосфолипид, холестерин и пегилированный липид в молярном соотношении 56:38:5. Согласно некоторым аспектам общее содержание липидов в липосомальном составе составляет 2-16 мг/мл. Согласно некоторым аспектам настоящего раскрытия предложен липосомальный состав, содержащий липид, содержащий фосфатидилхолиновую функциональную группу, липид, содержащий этаноламиновую функциональную группу, и пегилированный липид. Согласно некоторым аспектам настоящего раскрытия предложен липосомальный состав, содержащий липид, содержащий фосфатидилхолиновую функциональную группу, липид, содержащий этаноламиновую функциональную группу, и пегилированный липид, в молярном соотношении 3:0,015:2, соответственно. Согласно некоторым аспектам настоящего раскрытия предложен липосомальный состав, содержащий липид, содержащий фосфатидилхолиновую функциональную группу, холестерин и пегилированный липид. Согласно некоторым аспектам настоящего раскрытия предложен липосомальный состав, содержащий липид, содержащий фосфатидилхолиновую функциональную группу, и холестерин. Согласно некоторым аспектам пегилированный липид представляет собой ПЭГ-2000-DSPE. Согласно некоторым аспектам настоящего раскрытия предложен липосомальный состав, содержащий DPPG, соевый ФХ, липидный конъюгат MPEG-DSPE и холестерин.
[00325] Согласно некоторым аспектам настоящего раскрытия предложен липосомальный состав, содержащий один или более липидов, содержащих фосфатидилхолиновую функциональную группу, и один или более липидов, содержащих этаноламиновую функциональную группу. Согласно некоторым аспектам настоящего раскрытия предложен липосомальный состав, содержащий одно или более из: липидов, содержащих фосфатидилхолиновую функциональную группу, липидов, содержащих этаноламиновую функциональную группу, и стеринов, например, холестерина. Согласно некоторым аспектам липосомальный состав содержит DOPC/DEPC; и DOPE.
[00326] Согласно некоторым аспектам настоящего раскрытия предложен липосомальный состав, дополнительно содержащий одно или более фармацевтических вспомогательных веществ, например, сахарозу и/или глицин.
[00327] Согласно некоторым аспектам настоящего раскрытия предложен липосомальный состав с униламеллярной или мультиламеллярной структурой. Согласно некоторым аспектам настоящего раскрытия предложен липосомальный состав, который содержит мультивезикулярные частицы и/или частицы на основе пены. Согласно некоторым аспектам настоящего раскрытия предложен липосомальный состав, относительный размер которого больше, чем у обычных наночастиц, и составляет примерно от 150 до 250 нм. Согласно некоторым аспектам липосомальный состав представляет собой лиофилизированный порошок.
[00328] Согласно некоторым аспектам настоящего раскрытия предложен липосомальный состав, который получают и нагружают зкДНК-векторами, раскрытыми или описанными в настоящем документе, путем добавления слабого основания к смеси, содержащей выделенную зкДНК вне липосомы. Такое добавление повышает значение pH вне липосом приблизительно до 7,3 и стимулирует вход АФИ в липосому. Согласно некоторым аспектам настоящего раскрытия предложен липосомальный состав, имеющий кислый pH внутри липосомы. В таких случаях значение pH внутри липосом может составлять 4-6,9 и более предпочтительно pH 6,5. Согласно другим аспектам настоящего раскрытия предложен липосомальный состав, полученный с применением технологии внутрилипосомальной стабилизации лекарственных средств. В таких случаях используют полимерные или неполимерные высокозаряженные анионы и внутрилипосомальные захватывающие агенты, например, полифосфат или октасульфат сахарозы.
[00329] Согласно некоторым аспектам настоящего раскрытия предложена липидная наночастица, содержащая ДНК-вектор, включая зкДНК-вектор, описанный в настоящем документе, и ионизируемый липид. Например, состав липидных наночастиц, который получают и нагружают зкДНК, полученной с помощью способа, раскрытого в международной заявке PCT/US2018/050042, поданной 7 сентября 2018 г., которая включена в настоящий документ. Это можно осуществлять путем интенсивного смешивания этанольных растворов липидов с водным раствором зкДНК при низком pH, при котором происходит протонирование ионизируемого липида и обеспечиваются благоприятные энергетические характеристики для ассоциации зкДНК/липида и нуклеации частиц. Частицы могут быть дополнительно стабилизированы путем разбавления водой и удаления органического растворителя. Частицы могут быть сконцентрированы до целевого уровня.
[00330] Обычно липидные частицы получают при соотношении общего количества липида и зкДНК (массовом или весовом) от примерно 10:1 до 30:1. Согласно некоторым вариантам реализации соотношение липида и зкДНК (массовое соотношение; отношение масс./масс.) может находиться в диапазоне от примерно 1:1 до примерно 25:1, от примерно 10:1 до примерно 14:1, от примерно 3:1 до примерно 15:1, от примерно 4:1 до примерно 10:1, от примерно 5:1 до примерно 9:1 или от примерно 6:1 до примерно 9:1. В соответствии с некоторыми вариантами реализации любого из аспектов или вариантами реализации в настоящем документе композиция имеет соотношение общего количества липида и зкДНК примерно 15:1. В соответствии с некоторыми вариантами реализации любого из аспектов или вариантами реализации в настоящем документе композиция имеет соотношение общего количества липида и зкДНК примерно 30:1. В соответствии с некоторыми вариантами реализации любого из аспектов или вариантами реализации в настоящем документе композиция имеет соотношение общего количества липида и зкДНК примерно 40:1. В соответствии с некоторыми вариантами реализации любого из аспектов или вариантами реализации в настоящем документе композиция имеет соотношение общего количества липида и зкДНК примерно 50:1. Количества липидов и зкДНК могут быть скорректированы, чтобы обеспечить целевое соотношение N/P, например, соотношение N/P, равное 3, 4, 5, 6, 7, 8, 9, 10 или выше. Обычно общее содержание липидов в составе липидных частиц может быть в диапазоне от примерно 5 мг/мл до примерно 30 мг/мл.
[00331] Ионизируемый липид, как правило, применяют для конденсации груза нуклеиновой кислоты, например, зкДНК, при низком pH, и для стимуляции ассоциации мембран и их фузогенности. Обычно ионизируемые липиды представляют собой липиды, содержащие по меньшей мере одну аминогруппу, которая положительно заряжена или становится протонированной в кислых условиях, например, при рН 6,5 или ниже. В настоящем документе ионизируемые липиды также называют катионными липидами.
[00332] Примерные ионизируемые липиды описаны в международных РСТ публикациях патентов WO2015/095340, WO2015/199952, WO2018/011633, WO2017/049245, WO2015/061467, WO2012/040184, WO2012/000104, WO2015/074085, WO2016/081029, WO2017/004143, WO2017/075531, WO2017/117528, WO2011/022460, WO2013/148541, WO2013/116126, WO2011/153120, WO2012/044638, WO2012/054365, WO2011/090965, WO2013/016058, WO2012/162210, WO2008/042973, WO2010/129709, WO2010/144740, WO2012/099755, WO2013/049328, WO2013/086322, WO2013/086373, WO2011/071860, WO2009/132131, WO2010/048536, WO2010/088537, WO2010/054401, WO2010/054406, WO2010/054405, WO2010/054384, WO2012/016184, WO2009/086558, WO2010/042877, WO2011/000106, WO2011/000107, WO2005/120152, WO2011/141705, WO2013/126803, WO2006/007712, WO2011/038160, WO2005/121348, WO2011/066651, WO2009/127060, WO2011/141704, WO2006/069782, WO2012/031043, WO2013/006825, WO2013/033563, WO2013/089151, WO2017/099823, WO2015/095346 и WO2013/086354, и публикациях патентов США US2016/0311759, US2015/0376115, US2016/0151284, US2017/0210697, US2015/0140070, US2013/0178541, US2013/0303587, US2015/0141678, US2015/0239926, US2016/0376224, US2017/0119904, US2012/0149894, US2015/0057373, US2013/0090372, US2013/0274523, US2013/0274504, US2013/0274504, US2009/0023673, US2012/0128760, US2010/0324120, US2014/0200257, US2015/0203446, US2018/0005363, US2014/0308304, US2013/0338210, US2012/0101148, US2012/0027796, US2012/0058144, US2013/0323269, US2011/0117125, US2011/0256175, US2012/0202871, US2011/0076335, US2006/0083780, US2013/0123338, US2015/0064242, US2006/0051405, US2013/0065939, US2006/0008910, US2003/0022649, US2010/0130588, US2013/0116307, US2010/0062967, US2013/0202684, US2014/0141070, US2014/0255472, US2014/0039032, US2018/0028664, US2016/0317458 и US2013/0195920, содержание которых полностью включено в настоящий документ посредством ссылки.
[00333] Согласно некоторым вариантам реализации ионизируемый липид представляет собой MC3 (6Z,9Z,28Z,31Z)-гептатриаконта-6,9,28,31-тетраен-19-ил-4-(диметиламино)бутаноат (DLin-MC3-DMA или MC3), имеющий следующую структуру:
 .
.
[00334] Липид DLin-MC3-DMA описан в работе Jayaraman et al., Angew. Chem. Int. Ed Engl. (2012), 51(34): 8529-8533, содержание которой полностью включено в настоящий документ посредством ссылки.
[00335] Согласно некоторым вариантам реализации ионизируемый липид представляет собой липид ATX-002, описанный в WO 2015/074085, содержание которого полностью включено в настоящий документ посредством ссылки.
[00336] Согласно некоторым вариантам реализации ионизируемый липид представляет собой (13Z,16Z)-N,N-диметил-3-нонилдокоза-13,16-диен-1-амин (Соединение 32), описанный в WO 2012/040184, содержание которого полностью включено в настоящий документ посредством ссылки.
[00337] Согласно некоторым вариантам реализации ионизируемый липид представляет собой Соединение 6 или Соединение 22, описанные в WO 2015/199952, содержание которого полностью включено в настоящий документ посредством ссылки.
[00338] Без ограничений, ионизируемый липид может составлять 20-90% (мол.) от общего количества липидов, присутствующих в липидной наночастице. Например, молярное содержание ионизируемого липида может составлять 20-70% (мол.), 30-60% (мол.) или 40-50% (мол.) от общего количества липидов, присутствующих в липидной наночастице. Согласно некоторым вариантам реализации ионизируемый липид составляет от примерно 50 мол. % до примерно 90 мол. %, от общего количества липидов, присутствующих в липидной наночастице.
[00339] Согласно некоторым аспектам липидная наночастица может дополнительно содержать некатионный липид. Неионные липиды включают амфипатические липиды, нейтральные липиды и анионные липиды. Соответственно, некатионный липид может представлять собой нейтральный незаряженный, цвиттерионный или анионный липид. Некатионные липиды, как правило, применяют для усиления фузогенности.
[00340] Примерные некатионные липиды, предусмотренные для применения в способах и композициях, содержащих ДНК-вектор, включая зкДНК-вектор, описанный в настоящем документе, описаны в международной заявке PCT/US 2018/050042, поданной 7 сентября 2018 г., и PCT/US 2018/064242, поданной 6 декабря 2018 г., которые полностью включены в настоящий документ.
[00341] Примерные некатионные липиды описаны в публикации международной заявки WO 2017/099823 и публикации патента США US 2018/0028664, содержание которых полностью включено в настоящий документ посредством ссылки.
[00342] Некатионный липид может составлять 0-30% (мол.) от общего количества липидов, присутствующих в липидной наночастице. Например, содержание некатионного липида составляет 5-20% (мол.) или 10-15% (мол.) от общего количества липидов, присутствующих в липидной наночастице. Согласно различным вариантам реализации молярное соотношение ионизируемого липида и нейтрального липида колеблется от примерно 2:1 до примерно 8:1.
[00343] Согласно некоторым вариантам реализации липидные наночастицы не содержат каких-либо фосфолипидов. Согласно некоторым аспектам липидная наночастица может дополнительно содержать компонент, такой как стерин, для обеспечения целостности мембраны.
[00344] Один примерный стерин, который подходит для применения в липидной наночастице, представляет собой холестерин и его производные. Примерные производные холестерина описаны в международной заявке WO 2009/127060 и публикации патента США US 2010/0130588, содержание которых полностью включено в настоящий документ посредством ссылки.
[00345] Компонент, обеспечивающий целостность мембраны, такой как стерин, может составлять 0-50% (мол.) от общего количества липидов, присутствующих в липидной наночастице. Согласно некоторым вариантам реализации такой компонент составляет 20-50% (мол.) 30-40% (мол.) от общего содержания липидов в липидной наночастице.
[00346] Согласно некоторым аспектам липидная наночастица может дополнительно содержать полиэтиленгликоль (ПЭГ) или молекулу конъюгированного липида. Обычно их используют для ингибирования агрегации липидных наночастиц и/или для обеспечения стерической стабилизации. Примерные конъюгированные липиды включают, но не ограничиваются перечисленными, конъюгаты ПЭГ-липид, конъюгаты полиоксазолин (POZ)-липид, конъюгаты полиамид-липид (такие как конъюгаты ATTA-липид), конъюгаты катионная группа-полимер-липид (CPL) и их смеси. Согласно некоторым вариантам реализации молекула конъюгированного липида представляет собой конъюгат ПЭГ-липид, например, метоксиполиэтиленгликоль-конъюгированный липид. Примерные конъюгаты ПЭГ-липид включают, но не ограничиваются перечисленными, ПЭГ-диацилглицерин (ДАГ) (такой как 1-(монометоксиполиэтиленгликоль)-2,3-димиристоилглицерин (ПЭГ-ДМГ)), ПЭГ-диалкилоксипропил (ДАА), ПЭГ-фосфолипид, ПЭГ-церамид (Cer), пегилированный фосфатидилэтаноламин (ПЭГ-ФЭ), ПЭГ-сукцинатдиацилглицерин (PEGS-DAG) (такой как 4-O-(2',3'-ди(тетрадеканоилокси)пропил-1-O)-(w-метокси(полиэтокси)этил)бутандиоат (PEG-S-DMG)), ПЭГ-диалкоксипропилкарбам, натриевая соль N-(карбонилметоксиполиэтиленгликоль 2000)-1,2-дистеароил-sn-глицеро-3-фосфоэтаноламина или их смесь. Дополнительные примерные конъюгаты ПЭГ-липид описаны, например, в US 5885613, US 6287591, US 2003/0077829, US 2003/0077829, US 2005/0175682, US 2008/0020058, US 2011/0117125, US 2010/0130588, US 2016/0376224 и US 2017/0119904, содержание которых полностью включено в настоящий документ посредством ссылки.
[00347] Согласно некоторым вариантам реализации ПЭГ-липид представляет собой соединение, раскрытое в US 2018/0028664, содержание которого полностью включено в настоящий документ посредством ссылки.
[00348] Согласно некоторым вариантам реализации ПЭГ-липид раскрыт в US20150376115 или в US 2016/0376224, содержание которых полностью включено в настоящий документ посредством ссылки.
[00349] Конъюгат ПЭГ-ДАА может представлять собой, например, ПЭГ-дилаурилоксипропил, ПЭГ-димиристилоксипропил, ПЭГ-дипальмитилоксипропил или ПЭГ-дистеарилоксипропил. ПЭГ-липид может представлять собой одно или более из ПЭГ-ДМГ, ПЭГ-дилаурилглицерина, ПЭГ-дипальмитоилглицерина, ПЭГ-дистерилглицерина, ПЭГ-дилаурилгликамида, ПЭГ-димиристилгликамида, ПЭГ-дипальмитоилгликамида, ПЭГ-дистерилгликамида, ПЭГ-холестерина (1-[8'-(холест-5-ен-3[бета]-окси)карбоксамидо-3',6'-диоксаоктанил]карбамоил-[омега]-метилполиэтиленгликоль, ПЭГ-DMB (простой эфир 3,4-дитетрадекоксилбензил-[омега]-метилполиэтиленгликоля) и 1,2-димиристоил-sn-глицеро-3-фосфоэтаноламин-N-[метоксиполиэтиленгликоль-2000]. В некоторых примерах ПЭГ-липид может быть выбран из группы, состоящей из ПЭГ-ДМГ, 1,2-димиристоил-sn-глицеро-3-фосфоэтаноламин-N-[метоксиполиэтиленгликоля-2000].
[00350] Липиды, конъюгированные с молекулой, отличной от ПЭГ, также могут быть использованы вместо ПЭГ-липида. Например, конъюгаты полиоксазолин (POZ)-липид, конъюгаты полиамид-липид (такие как конъюгаты ATTA-липид) и конъюгаты катионная группа-полимер-липид (CPL) могут использоваться вместо ПЭГ-липида или вместе с ним. Примерные конъюгированные липиды, т.е. ПЭГ-липиды, конъюгаты (POZ)-липид, конъюгаты АТТА-липид и катионный полимер-липиды, описаны в публикациях международных заявок на патент WO 1996/010392, WO 1998/051278, WO 2002/087541, WO 2005/026372, WO 2008/147438, WO 2009/086558, WO 2012/000104, WO 2017/117528, WO 2017/099823, WO 2015/199952, WO 2017/004143, WO 2015/095346, WO 2012/000104, WO 2012/000104 и WO 2010/006282, публикациях заявок на патент США US 2003/0077829, US 2005/0175682, US 2008/0020058, US 2011/0117125, US 2013/0303587, US 2018/0028664, US 2015/0376115, US 2016/0376224, US 2016/0317458, US 2013/0303587, US 2013/0303587 и US 20110123453, и патентах США US 5885613, US 6287591, US 6320017 и US 6586559, содержание которых полностью включено в настоящий документ посредством ссылки.
[00351] Согласно некоторым вариантам реализации одно или более дополнительных соединений могут представлять собой терапевтический агент. Терапевтический агент может быть выбран из любого класса, подходящего для применения в терапевтических целях. Другими словами, терапевтический агент может быть выбран из любого класса, подходящего для применения в терапевтических целях. Другими словами, терапевтический агент может быть выбран в соответствии с терапевтической целью и целевым биологическим действием. Например, если зкДНК в ЛНЧ можно применять для лечения рака, дополнительное соединение может представлять собой противораковый агент (например, химиотерапевтический агент, нацеленную противораковую терапию (включая, но не ограничиваясь перечисленными, малую молекулу, антитело или конъюгат антитела с лекарственным средством). Согласно другому примеру, если ЛНЧ, содержащую зкДНК, можно применять для лечения инфекции, дополнительное соединение может представлять собой антимикробный агент (например, антибиотик или противовирусное соединение). Согласно еще одному примеру, если ЛНЧ, содержащую зкДНК, можно применять для лечения иммунного заболевания или нарушения, дополнительное соединение может представлять собой соединение, которое модулирует иммунный ответ (например, иммуносупрессор, иммуностимулирующее соединение или соединение, модулирующее один или более конкретных иммунных путей). Согласно некоторым вариантам реализации в композициях и способах согласно настоящему изобретению можно применять другие смеси различных липидных наночастиц, содержащих другие соединения, такие как зкДНК, кодирующая другой белок или другое соединение, такое как терапевтическое средство.
[00352] Согласно некоторым вариантам реализации дополнительное соединение представляет собой иммуномодулирующий агент. Например, дополнительное соединение представляет собой иммуносупрессор. Согласно некоторым вариантам реализации дополнительное соединение представляет собой иммуностимулирующий агент.
[00353] В настоящем документе также предложена фармацевтическая композиция, содержащая инкапсулированный в липидную наночастицу зкДНК-вектор и рапамицин или аналог рапамицина, описанные в настоящем документе, и фармацевтически приемлемый носитель или вспомогательное вещество. В настоящем документе также предложена фармацевтическая композиция, содержащая инкапсулированный в липидную наночастицу зкДНК-вектор и фармацевтически приемлемый носитель или вспомогательное вещество, причем указанный рапамицин или аналог рапамицина совместно вводят субъекту в другой композиции, как описано в настоящем документе.
[00354] Согласно некоторым аспектам настоящего изобретения предложен состав липидных наночастиц, дополнительно содержащий одно или более фармацевтических вспомогательных веществ. Согласно некоторым вариантам реализации состав липидных наночастиц дополнительно содержит сахарозу, Tris, трегалозу и/или глицин.
[00355] ДНК-вектор с замкнутыми концами, включая зкДНК-вектор, и необязательно один или более ингибиторов иммунного ответа (например, врожденного иммунного ответа), описанные в настоящем документе, могут образовывать комплексы с липидной частью (portion) частицы или могут быть инкапсулированы в липидной части (position) липидной наночастицы. Согласно некоторым вариантам реализации ДНК-вектор, включая зкДНК-вектор, описанный в настоящем документе, может быть полностью инкапсулирован в липидной части липидной наночастицы, что защищает его от разрушения нуклеазой, например, в водном растворе. Согласно некоторым вариантам реализации ДНК-вектор в липидной наночастице, включая зкДНК-вектор, описанный в настоящем документе, не разрушается в значительной степени после воздействия на липидную наночастицу нуклеазы при 37°С в течение по меньшей мере примерно 20, 30, 45 или 60 минут. Согласно некоторым вариантам реализации зкДНК в липидной наночастице существенно не разрушается после инкубации указанной частицы в сыворотке при 37°С в течение по меньшей мере примерно 30, 45 или 60 минут или по меньшей мере примерно 2, 3, 4, 5, 6, 7, 8, 9, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34 или 36 часов.
[00356] Согласно определенным вариантам реализации липидные наночастицы по существу нетоксичны для субъекта, например, млекопитающего, такого как человек. Согласно некоторым аспектам состав липидных наночастиц представляет собой лиофилизированный порошок.
[00357] Согласно некоторым вариантам реализации липидные наночастицы представляют собой частицы с твердой сердцевиной, которые имеют по меньшей мере один липидный бислой. Согласно другим вариантам реализации липидные наночастицы имеют небислойную структуру, т.е. имеют неламеллярную (т.е. небислойную) морфологию. Без ограничений, небислойная морфология может включать, например, трехмерные трубки, стержни, частицы кубической симметрии и т.д. Например, морфологию липидных наночастиц (ламеллярных в сравнении с неламеллярными) можно легко оценить и охарактеризовать с использованием, например, анализа методом «крио-ТЭМ» (криотрансмиссионная электронная микроскопия), как описано в US2010/0130588, содержание которого полностью включено в настоящий документ посредством ссылки.
[00358] Согласно некоторым другим вариантам реализации липидные наночастицы, имеющие неламеллярную морфологию, являются электронно-плотными. Согласно некоторым аспектам настоящего раскрытия предложена липидная наночастица, которая является либо униламеллярной, либо мультиламеллярной по структуре. Согласно некоторым аспектам настоящего раскрытия предложен состав липидных наночастиц, который содержит мультивезикулярные частицы и/или частицы на основе пены.
Контролируя композицию и концентрацию липидных компонентов, можно контролировать скорость, с которой липидный конъюгат высвобождается из липидной частицы, и, в свою очередь, скорость, с которой липидная наночастица становится фузогенной. Кроме того, другие переменные, включая, например, pH, температуру или ионную силу, можно применять для изменения и/или контроля скорости, с которой липидная наночастица становится фузогенной. Другие способы, которые можно применять для контроля скорости, с которой указанная липидная наночастица становится фузогенной, будут понятны обычным специалистам в данной области техники после ознакомления с указанным описанием. Также будет понятно, что размер липидных частиц можно контролировать путем контроля композиции и концентрации липидного конъюгата. pKa катионных липидов в составах может быть сопоставлена с эффективностью ЛНЧ для доставки нуклеиновых кислот (см. Jayaraman et al, Angewandte Chemie, International Edition (2012), 51(34), 8529-8533; Semple et al., Nature Biotechnology 28, 172-176 (2010), обе работы полностью включены в настоящий документ посредством ссылки). Предпочтительный диапазон pKa составляет от ~5 до ~7. pKa каждого катионного липида в липидных наночастицах может быть определена с использованием анализа, основанного на флуоресценции 2-(п-толуидино)-6-нафталинсульфоновой кислоты (TNS).
VI. Ингибиторы иммунного ответа
[00359] В настоящем документе предложены ингибиторы иммунного ответа. В соответствии с вариантами реализации ингибиторы иммунного ответа представляют собой ингибиторы врожденного иммунного ответа.
Рапамицин или аналоги рапамицина
[00360] В соответствии с некоторыми аспектами настоящего раскрытия предложены невирусные бескапсидные ДНК-векторы с ковалентно замкнутыми концами (зкДНК-вектор), вводимые в сочетании с рапамицином или аналогами рапамицина. Согласно некоторым вариантам реализации рапамицин или аналог рапамицина присутствует в перенасыщенном количестве в синтетическом наноносителе, как описано в WO 2016/073799. Согласно некоторым вариантам реализации зкДНК-вектор также присутствует в том же наноносителе.
[00361] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, рапамицин или аналог рапамицина вводят субъекту совместно с зкДНК-вектором. Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, зкДНК-вектор и рапамицин или аналог рапамицина вводят совместно в одном составе. Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, рапамицин или аналог рапамицина присутствует в перенасыщенной концентрации в синтетическом наноносителе, как описано в WO 2016/073799. Согласно некоторым вариантам реализации зкДНК-вектор также присутствует в том же наноносителе. Согласно некоторым вариантам реализации зкДНК-вектор, изготовленный в виде состава с липидной наночастицей, также присутствует в том же наноносителе.
[00362] Согласно некоторым вариантам реализации аналог рапамицина представляет собой любой из аналогов рапамицина, известных в данной области техники, такой как любой из аналогов рапамицина, описанных в патенте США 5138051 или WO 2017/040341, содержание каждого из которых полностью включено в настоящий документ посредством ссылки.
[00363] Согласно некоторым вариантам реализации аналог рапамицина представляет собой соединение Формулы I, показанное ниже:
[00364] Согласно некоторым вариантам реализации аналог рапамицина представляет собой соединение Формулы II, в котором конфигурация заместителей на C-33 Формулы I представляет собой R-конфигурацию, как показано ниже:
[00365] Согласно некоторым вариантам реализации аналог рапамицина представляет собой соединение Формулы III, как показано ниже:
или его фармацевтически приемлемая соль, где: R1 представляет собой ОН или ОСН3 R2 представляет собой Н или F R3 представляет собой Н, ОН или ОСН3; и R4 представляет собой ОН или ОСН3.
[00366] Согласно некоторым вариантам реализации аналог рапамицина представляет собой соединение Формулы III в чистой хиральной форме в виде одного диастереомера Формулы IV, как показано ниже:
[00367] Согласно некоторым вариантам реализации аналог рапамицина представляет собой соединение Формулы III в чистой хиральной форме в виде одного диастереомера Формулы V, как показано ниже:
[00368] Согласно некоторым вариантам реализации аналог рапамицина представляет собой соединение Формулы III в чистой хиральной форме в виде одного диастереомера Формулы VI, как показано ниже:
[00369] Согласно некоторым вариантам реализации аналог рапамицина представляет собой соединение Формулы III в чистой хиральной форме в виде одного диастереомера Формулы VII, как показано ниже:
[00370] Согласно некоторым вариантам реализации аналог рапамицина представляет собой соединение Формулы III в чистой хиральной форме в виде одного диастереомера Формулы VIII, как показано ниже:
[00371] Согласно некоторым вариантам реализации аналог рапамицина представляет собой соединение Формулы IX, как показано ниже:
или его фармацевтически приемлемая соль, где R2 представляет собой H или F, R3 представляет собой OH или OCH3; и R4 представляет собой ОСН3 или ОН. Согласно определенным вариантам реализации R4 представляет собой OCH3. Согласно определенным вариантам реализации R4 представляет собой OCH3, R2 представляет собой F и R3 представляет собой OCH3. Согласно определенным вариантам реализации R4 представляет собой OCH3, R2 представляет собой H и R3 представляет собой OH. Согласно определенным вариантам реализации R2 представляет собой H, R3 представляет собой H и R4 представляет собой OH. Согласно различным вариантам реализации соединения Формулы IX представлены в виде рацемической смеси.
[00372] Соответственно, в некоторых вариантах реализации аналог рапамицина выбран из любой из Формул I-IX или их производного.
[00373] Согласно некоторым вариантам реализации рапамицин или аналог рапамицина доставляют или вводят с использованием синтетического наноносителя, как описано в WO 2016/073799, полностью включенном в настоящий документ посредством ссылки.
[00374] Как описано в WO 2016/073799, концентрация рапамицина в составе во время получения синтетического наноносителя, относительно предела растворимости рапамицина в указанном составе, может значительно влиять на способность полученных синтетических наноносителей индуцировать иммунную толерантность. Кроме того, характер диспергирования рапамицина в синтетических наноносителях может влиять на возможность осуществления первоначальной стерилизации путем фильтрации полученных синтетических наноносителей. Соответственно, в некоторых вариантах реализации, синтетические наноносители, созданные в условиях, которые приводят к концентрации рапамицина, превышающей его растворимость в полученной суспензии наноносителей, используют в композициях и способах, описанных в настоящем документе. Такие синтетические наноносители могут обеспечивать более продолжительную иммунную толерантность и могут быть первоначально стерилизованы путем фильтрации.
[00375] Согласно некоторым вариантам реализации зкДНК-вектор вводят совместно с композицией, содержащей синтетические наноносители, содержащие материал гидрофобного полиэфирного носителя и рапамицин или аналог рапамицина, причем указанный рапамицин или аналог рапамицина присутствует в синтетических наноносителях в стабильном, перенасыщенном количестве, которое составляет менее 50% (масс./масс.) в расчете на массу рапамицина или аналога рапамицина по отношению к массе материала гидрофобного полиэфирного носителя.
[00376] Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, значения массы представляют собой рецептурные значения массы материалов, которые комбинируют во время изготовления синтетических наноносителей. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, значения массы представляют собой значения массы материалов в полученной композиции синтетического наноносителя.
[00377] Согласно некоторым вариантам реализации любых из композиций или способов, предложенных в настоящем документе, рапамицин или аналог рапамицина присутствует в стабильном, перенасыщенном количестве, которое составляет менее 45% (масс./масс.). Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, рапамицин или аналог рапамицина присутствует в стабильном, перенасыщенном количестве, которое составляет менее 40% (масс./масс.). Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, рапамицин или аналог рапамицина присутствует в стабильном, перенасыщенном количестве, которое составляет менее 35% (масс./масс.). Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, рапамицин или аналог рапамицина присутствует в стабильном, перенасыщенном количестве, которое составляет менее 30% (масс./масс.). Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, рапамицин или аналог рапамицина присутствует в стабильном, перенасыщенном количестве, которое составляет менее 25% (масс./масс.). Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, рапамицин или аналог рапамицина присутствует в стабильном, перенасыщенном количестве, которое составляет менее 20% (масс./масс.). Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, рапамицин или аналог рапамицина присутствует в стабильном, перенасыщенном количестве, которое составляет менее 15% (масс./масс.). Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, рапамицин или аналог рапамицина присутствует в стабильном, перенасыщенном количестве, которое составляет менее 10% (масс./масс.). Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, рапамицин или аналог рапамицина присутствует в стабильном, перенасыщенном количестве, которое превышает 7% (масс./масс.).
[00378] Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, материал гидрофобного полиэфирного носителя содержит PLA, PLG, PLGA или поликапролактон. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, материал гидрофобного полиэфирного носителя дополнительно содержит PLA-ПЭГ, PLGA-ПЭГ или PCL-ПЭГ.
[00379] Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, количество материала гидрофобного полиэфирного носителя в синтетических наноносителях составляет 5-95% (масс./масс.) материала гидрофобного полиэфирного носителя/общее количество твердых веществ. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, количество материала гидрофобного полиэфирного носителя в синтетических наноносителях составляет 60-95% (масс./масс.) материала гидрофобного полиэфирного носителя/общее количество твердых веществ.
[00380] Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, синтетические наноносители дополнительно содержат неионное поверхностно-активное вещество со значением HLB менее или равным 10. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, неионное поверхностно-активное вещество со значением HLB менее или равным 10 содержит сложный эфир сорбитана, алифатический спирт, сложный эфир алифатической кислоты, этоксилированный алифатический спирт, полоксамер, алифатическую кислоту, холестерин, производное холестерина или желчную кислоту или соль. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, неионное поверхностно-активное вещество со значением HLB менее или равным 10 содержит SPAN 40, SPAN 20, олеиловый спирт, стеариловый спирт, изопропилпальмитат, моностеарат глицерина, BRIJ 52, BRIJ 93, плюроновую кислоту P-123, плюроновую кислоту L-31, пальмитиновую кислоту, додекановую кислоту, трипальмитат глицерина или трилинолеат глицерина. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, неионное поверхностно-активное вещество со значением HLB менее или равным 10 представляет собой SPAN 40.
[00381] Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, неионное поверхностно-активное вещество со значением HLB менее или равным 10 инкапсулировано в синтетических наноносителях, присутствует на поверхности синтетических наноносителей или и то, и другое. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, количество неионного поверхностно-активного вещества со значением HLB менее или равным 10 составляет >0,1, но <15% (масс./масс.) неионного поверхностно-активного вещества со значением HLB менее или равным 10/материал гидрофобного полиэфирного носителя. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, количество неионного поверхностно-активного вещества со значением HLB менее или равным 10 составляет >1, но <13% (масс./масс.) неионного поверхностно-активного вещества со значением HLB менее или равным 10/материал гидрофобного полиэфирного носителя. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, количество неионного поверхностно-активного вещества со значением HLB менее или равным 10 составляет >1, но <9% (масс./масс.) неионного поверхностно-активного вещества со значением HLB менее или равным 10/материал гидрофобного полиэфирного носителя.
[00382] Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, композиция первоначально может быть стерилизована путем фильтрации через фильтр с размером пор 0,22 мкм.
[00383] Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, среднее значение распределения частиц по размеру, полученное с использованием динамического светорассеяния синтетических наноносителей, представляет собой диаметр более 120 нм. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, диаметр составляет более 150 нм. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, диаметр составляет более 200 нм. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, диаметр составляет более 250 нм. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, диаметр составляет менее 300 нм. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, диаметр составляет менее 250 нм. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, диаметр составляет менее 200 нм.
[00384] Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, рапамицин или аналог рапамицина инкапсулирован в синтетических наноносителях.
[00385] Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, композиция дополнительно содержит фармацевтически приемлемый носитель.
[00386] Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, рапамицин или аналог рапамицина присутствует в перенасыщенном количестве, которое по меньшей мере на 1% превышает предел насыщения рапамицина или аналога рапамицина в материале гидрофобного полиэфирного носителя. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, рапамицин или аналог рапамицина присутствует в перенасыщенном количестве, которое по меньшей мере на 5% превышает предел насыщения рапамицина или аналога рапамицина в материале гидрофобного полиэфирного носителя. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, рапамицин или аналог рапамицина присутствует в перенасыщенном количестве, которое по меньшей мере на 10% превышает предел насыщения рапамицина или аналога рапамицина в материале гидрофобного полиэфирного носителя. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, рапамицин или аналог рапамицина присутствует в перенасыщенном количестве, которое по меньшей мере на 15% превышает предел насыщения рапамицина или аналога рапамицина в материале гидрофобного полиэфирного носителя. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, рапамицин или аналог рапамицина присутствует в перенасыщенном количестве, которое по меньшей мере на 20% превышает предел насыщения рапамицина или аналога рапамицина в материале гидрофобного полиэфирного носителя. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, рапамицин или аналог рапамицина присутствует в перенасыщенном количестве, которое по меньшей мере на 25% превышает предел насыщения рапамицина или аналога рапамицина в материале гидрофобного полиэфирного носителя. Согласно одному варианту реализации любых из композиций или способов, предложенных в настоящем документе, рапамицин или аналог рапамицина присутствует в перенасыщенном количестве, которое по меньшей мере на 30% превышает предел насыщения рапамицина или аналога рапамицина в материале гидрофобного полиэфирного носителя.
[00387] Согласно другому варианту реализации любых из композиций или способов, предложенных в настоящем документе, количество рапамицина или аналога рапамицина превышает предел насыщения по меньшей мере на 1%. Согласно другому варианту реализации количество рапамицина или аналога рапамицина превышает предел насыщения по меньшей мере на 5%. Согласно другому варианту реализации количество рапамицина или аналога рапамицина превышает предел насыщения по меньшей мере на 10%. Согласно другому варианту реализации количество рапамицина или аналога рапамицина превышает предел насыщения по меньшей мере на 15%. Согласно другому варианту реализации количество рапамицина или аналога рапамицина превышает предел насыщения по меньшей мере на 20%. Согласно другому варианту реализации количество рапамицина или аналога рапамицина превышает предел насыщения по меньшей мере на 25%. Согласно другому варианту реализации количество рапамицина или аналога рапамицина превышает предел насыщения по меньшей мере на 30%.
Ингибиторы cGAS
[00388] В соответствии с некоторыми аспектами настоящего раскрытия предложены невирусные бескапсидные ДНК-векторы с ковалентно замкнутыми концами (зкДНК), вводимые в сочетании с одним или более антагонистами cGAS. В настоящем документе также предложены конструкции зкДНК, содержащие последовательности, кодирующие, помимо прочего, одну или более РНК или белков, ингибирующих cGAS.
[00389] cGAS представляет собой другой класс PRR, запускаемых цитозольной ДНК, который связывается с ДНК и активирует связанный с ЭР стимулятор генов интерферонов (STING). Это приводит к активации ответа интерферона типа I и, в некоторых случаях, к активации других предполагаемых сенсоров цитозольной ДНК, включая отсутствующий при меланоме белок (AIM2), IFN-γ-индуцируемый белок 16 (IFI16), интерферон-индуцируемый белок X (IFIX), LRRFIP1, DHX9, DHX36, DDX41, Ku70, DNA-PKc, комплекс MRN (включая MRE11, Rad50 и Nbs1) и РНК-полимеразу III. AIM2, IFI16 и IFIX представляют собой белки, содержащие пириновый домен и домен HIN200 (PYHIN). Кроме того, было показано, что неспаренные нуклеотиды ДНК, фланкирующие короткие участки ДНК со спаренными основаниями, как в структурах «стебель-петля» одноцепочечной ДНК (оцДНК), происходящей из вируса иммунодефицита человека типа 1 (ВИЧ-1), активировали индуцируемый интерфероном типа I ДНК-сенсор cGAS зависимым от последовательности образом. Структуры ДНК, содержащие неспаренные гуанозины, фланкирующие короткую (от 12 до 20 п.о.) дцДНК (Y-форма ДНК), обладали сильным стимулирующим действием и специфично усиливали ферментативную активность cGAS
[00390] cGAS непосредственно связывает ДНК за счет взаимодействий с сахаро-фосфатным остовом обеих цепей ДНК (S.R. Paluden. Microbiology and Molecular Biology Reviews. 2015. 79(2): 225). Это вызывает конформационное изменение фермента, позволяющее нуклеотидным субстратам АТФ и ГТФ получать доступ к активному сайту, что приводит к синтезу цГАМФ (A. Dempsey and A.G. Bowie, Virology 2015 May, 0: 146-152). цГАМФ затем связывает STING, что приводит к продуцированию интерферона I типа (A. Dempsey and A.G. Bowie, Virology 2015 May, 0: 146-152). Важно отметить, что cGAS контактирует с дцДНК только за счет фосфатного остова ДНК, что приводит к независимой от нуклеотидной последовательности чувствительности (A. Dempsey and A.G. Bowie, Virology 2015 May, 0: 146-152). Также было показано, что cGAS может активироваться неспаренными нуклеотидами ДНК, в частности, гуанозинами, фланкирующими короткие спаренные по основаниям участки ДНК из 12-20 п.о., как в структурах «стебель-петля» одноцепочечной ДНК (оцДНК), происходящей из вируса иммунодефицита человека типа 1 (ВИЧ-1) (M.H. Christnesen and S.R. Paluden. Cellular and Molecular Immunology. 2017. 14:4-13; A-M Herzner et al., 2015. Nature Immunology).
[00391] Соответственно, структурные особенности зкДНК, важные для активации врожденного иммунитета под действием PRR, включают, но не ограничиваются перечисленными, модифицированные последовательности инвертированных концевых повторов (ITR) ААВ, включая сайт связывания Rep (RBS) и сайт концевого разрешения (TRS); шпилечные последовательности в ITR; богатую CG последовательность RBS; отсутствие метилирования ДНК; и линейную дуплексную структуру ДНК с фланкирующими ITR, которые могут содержать, например, одноцепочечную ДНК в виде петли.
[00392] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор cGAS вводят субъекту совместно с зкДНК. Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, в которых ингибитор cGAS представляет собой последовательность РНК или белка, зкДНК кодирует РНК или белковый ингибитор cGAS.
[00393] Согласно некоторым вариантам реализации ингибитор cGAS представляет собой противомалярийный препарат (J. An et al., J. Immunol. March 27, 2015). Согласно некоторым вариантам реализации противомалярийный препарат представляет собой противомалярийный препарат на основе аминохинолина или аминоакридина (J. An et al., J. Immunol. March 27, 2015). Согласно некоторым вариантам реализации противомалярийный препарат выбран из хинакрина (QC), 9-амино-6-хлор-2-метоксиакридина (AMCA), гидроксихлорохина (HCQ) и хлорохина (CQ) (J. An et al., J. Immunol. March 27, 2015).
[00394] Согласно некоторым вариантам реализации ингибитор cGAS представляет собой низкомолекулярное соединение, которое связывается с каталитическим карманом cGAS (J. Vincent et al., Nature Communications, 8:750). Согласно некоторым вариантам реализации низкомолекулярное соединение, которое связывается с каталитическим карманом cGAS, выбрано из RU166365, RU281332, RU320521, RU320519, RU320461, RU320462, RU320520, RU320467 и RU320582 (J. Vincent et al., Nature Communications, 8:750). Согласно некоторым вариантам реализации низкомолекулярное соединение, которое связывается с каталитическим карманом cGAS, представляет собой RU320521 (J. Vincent et al., Nature Communications, 8:750). Согласно некоторым вариантам реализации низкомолекулярное соединение, которое связывается с каталитическим карманом cGAS, выбрано из соединения 15, соединения 16, соединения 17, соединения 18, соединения 19 и PF-06928215 (J. Vincent et al., Nature Communications, 8:750; PLOS ONE. September 21, 2017). Согласно некоторым вариантам реализации низкомолекулярное соединение, которое связывается с каталитическим карманом cGAS, представляет собой PF-06928215 (PLOS ONE. September 21, 2017).
[00395] Согласно некоторым вариантам реализации ингибитор cGAS представляет собой любое из низкомолекулярных соединений, описанных в US 20160068560, содержание которого полностью включено в настоящий документ посредством ссылки.
[00396] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор cGAS кодируется зкДНК, вводимой субъекту (включая, например, последующую доставку зкДНК). Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор cGAS, кодируемый зкДНК, вводимой субъекту, представляет собой белок ORF52 герпесвируса, ассоциированного с саркомой Капоши, имеющий аминокислотную последовательность MAAPRGRPKKDLTMEDLTAKISQLTVENRELRKALGSTADPRDRPLTATEKEAQLTATVGALSAAAAKKIEARVRTIFSKVVTQKQVDDALKGLSLRIDVCMSDGGTAKPPPGANNRRRRGASTTRAGVDD (SEQ ID NO: 882), или его вариант, который ингибирует cGAS (M.H. Christnesen and S.R. Paluden. Cellular and Molecular Immunology. 2017. 14:4-13). Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор cGAS, кодируемый зкДНК, вводимой субъекту, представляет собой ортолог ORF52 гаммагерпесвируса.
[00397] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор cGAS, кодируемый зкДНК, вводимой субъекту, представляет собой цитоплазматическую изоформу LANA (ассоциированный с латентностью ядерный антиген) герпесвируса, ассоциированного с саркомой Капоши, также называемую в настоящем документе «цитоплазматической изоформой LANA», или ее вариант, который ингибирует cGAS (Zhang G. et al., Proc Natl Acad Sci U S A. 2016 Feb 23;113(8):E1034-43). LANA или ORF73 имеет последовательность из следующих 1129 аминокислот: MAPPGMRLRSGRSTGAPLTRGSCRKRNRSPERCDLGDDLHLQPRRKHVADSVDGRECGPHTLPIPGSPTVFTSGLPAFVSSPTLPVAPIPSPAPATPLPPPALLPPVTTSSSPIPPSHPVSPGTTDTHSPSPALPPTQSPESSQRPPLSSPTGRPDSSTPMRPPPSQQTTPPHSPTTPPPEPPSKSSPDSLAPSTLRSLRKRRLSSPQGPSTLNPICQSPPVSPPRCDFANRSVYPPWATESPIYVGSSSDGDTPPRQPPTSPISIGSSSPSEGSWGDDTAMLVLLAEIAEEASKNEKECSENNQAGEDNGDNEISKESQVDKDDNDNKDDEEEQETDEEDEEDDEEDDEEDDEEDDEEDDEEDDEEDDEEEDEEEDEEEDEEEDEEEEEDEEDDDDEDNEDEEDDEEEDKKEDEEDGGDGNKTLSIQSSQQQQEPQQQEPQQQEPQQQEPQQQEPQQQEPQQQEPQQQEPQQREPQQREPQQREPQQREPQQREPQQREPQQREPQQREPQQREPQQREPQQREPQQREPQQQEPQQQEPQQQEPQQQEPQQQEPQQQEPQQQEPQQQEPQQQEPQQQEPQQQEPQQQEPQQQDEQQQDEQQQDEQQQDEQQQDEQQQDEQQQDEQQQDEQEQQDEQQQDEQQQQDEQEQQEEQEQQEEQQQDEQQQDEQQQDEQQQDEQEQQDEQQQDEQQQQDEQEQQEEQEQQEEQEQQEEQEQQEEQEQELEEQEQELEEQEQELEEQEQELEEQEQELEEQEQELEEQEQELEEQEQELEEQEQELEEQEQELEEQEQELEEQEQELEEQEQELEEQEQELEEQEQEQELEEVEEQEQEQEEQELEEVEEQEQEQEEQEEQELEEVEEQEEQELEEVEEQEEQELEEVEEQEQQGVEQQEQETVEEPIILHGSSSEDEMEVDYPVVSTHEQIASSPPGDNTPDDDPQPGPSREYRYVLRTSPPHRPGVRMRRVPVTHPKKPHPRYQQPPVPYRQIDDCPAKARPQHIFYRRFLGKDGRRDPKCQWKFAVIFWGNDPYGLKKLSQAFQFGGVKAGPVSCLPHPGPDQSPITYCVYVYCQNKDTSKKVQMARLAWEASHPLAGNLQSSIVKFKKPLPLTQPGENQGPGDSPQEMT (SEQ ID NO: 883).
[00398] Неограничивающий пример усеченной цитоплазматической изоформы LANA для применения с зкДНК, описанными в настоящем документе, представляет собой LANAΔ161 или SEQ ID NO: 532 (в которой отсутствуют аминокислоты 161-1162 из SEQ ID NO: 884).
[00399] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор cGAS представляет собой антитело или антигенсвязывающий фрагмент, который связывает cGAS. Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, антитело или антигенсвязывающий фрагмент, который связывает cGAS, кодируется зкДНК.
[00400] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор cGAS представляет собой ингибитор cGAS на основе РНК, такой как киРНК, специфичная в отношении cGAS. Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор cGAS на основе РНК кодируется зкДНК.
[00401] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор cGAS представляет собой ингибитор cGAS на основе миРНК, такой как miR-25 (GGCCAGTGTTGAGAGGCGGAGACTTGGGCAATTGCTGGACGCTGCCCTGGGCATTGCACTTGTCTCGGTCTGACAGTGCCGGCC; SEQ ID NO: 885) и miR-93 (CTGGGGGCTCCAAAGTGCTGTTCGTGCAGGTAGTGTGATTACCCAACCTACTGCTGAGCTAGCACTTCCCGAGCCCCCGG; SEQ ID NO: 886)11. Как полагают, miR-25 и miR-93 нацелены на коактиватор ядерного рецептора 3 (NCOA3), эпигенетический фактор, который поддерживает начальные уровни экспрессии cGAS, что ведет к репрессии cGAS (Wu et al. 2017. Nat. Cell Biol. 19(10):1286-1296). Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор cGAS на основе миРНК кодируется зкДНК.
Ингибиторы TLR
[00402] В соответствии с некоторыми аспектами настоящего раскрытия предложены невирусные бескапсидные ДНК-векторы с ковалентно замкнутыми концами (зкДНК), вводимые в сочетании с одним или более антагонистами TLR. В настоящем документе также предложены конструкции зкДНК, содержащие последовательности, кодирующие, помимо прочего, один или более олигонуклеотидов, ингибирующих TLR. В соответствии с некоторыми аспектами настоящего раскрытия предложены невирусные бескапсидные ДНК-векторы с ковалентно замкнутыми концами (зкДНК), вводимые в сочетании с одним или более антагонистами TLR9. В настоящем документе также предложены конструкции зкДНК, содержащие последовательности, кодирующие, помимо прочего, один или более олигонуклеотидов, ингибирующих TLR9.
[00403] В соответствии с некоторыми вариантами реализации ингибитор TLR9 представляет собой низкомолекулярный антагонист. Согласно другому варианту реализации ингибитор TLR9 представляет собой антитело к TLR9. В соответствии с некоторыми вариантами реализации антитело к TLR9 представляет собой моноклональное антитело. Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, один или более концевых структурных элементов зкДНК, таких как последовательности ITR, содержат последовательность олигонуклеотида, ингибирующего TLR9.
[00404] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, олигонуклеотид, ингибирующий TLR9, имеет один или более из следующих признаков (i) три последовательных нуклеотида G на 3´-конце; (ii) триплет CC(T) на 5´-конце; и (iii) расстояние между 5´-CC(T) и последующим триплетом GGG оптимально составляет 3-5 нуклеотидов в длину. Согласно некоторым вариантам реализации олигонуклеотид, ингибирующий TLR9, имеет последовательность 5´CCTN(3-5)G(3-5)RR3´ (SEQ ID NO: 887). Согласно некоторым вариантам реализации олигонуклеотид, ингибирующий TLR9, не имеет внутрицепочечной и/или межцепочечной водородной связи по Хугстину между смежными G.
[00405] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, олигонуклеотид, ингибирующий TLR9, представляет собой ингибирующий TLR9 олигонуклеотид класса G, имеющий характеристики стэкинга G4, и содержит несколько триплетов G3 или тетрад G4, такой как ингибиторный олигонуклеотид, содержащий TTAGGGn (SEQ ID NO: 888). Неограничивающие примеры такого ингибирующего TLR9 олигонуклеотида класса G, включают ODN-2088 (TCCTGGCGGGGAAGT, SEQ ID NO: 889), ODN-2114 (TCCTGGAGGGGAAGT, SEQ ID NO: 890), поли(G) (GGGGGGGGGGGGGGGGGGGG, SEQ ID NO: 891), ODN-A151 (TTAGGGTTAGGGTTAGGGTTAGGG, SEQ ID NO: 892), G-ODN (CTCCTATTGGGGGTTTCCTAT, SEQ ID NO: 893), а также IRS-869 (TCCTGGAGGGGTTGT, SEQ ID NO: 894) и AS1411 (GGTGGTGGTGGTTGTGGTGGTGGTGG, SEQ ID NO: 903).
[00406] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибирующий TLR9 олигонуклеотид представляет собой ингибирующий TLR9 олигонуклеотид класса R, обладающий характеристиками, включающими палиндромность и/или наличие коротких 5´ или 3´-липких концов, такой как ингибиторный олигонуклеотид INH-1. Неограничивающие примеры такого ингибирующего TLR9 олигонуклеотида класса R включают INH-1 (CCTGGATGGGAATTCCCATCCAGG, SEQ ID NO: 895), INH-4 (TTCCCATCCAGGCCTGGATGGGAA, SEQ ID NO: 896) и IRS-661 (TGCTTGCAAGCTTGCAAGCA, SEQ ID NO: 897).
[00407] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибирующий TLR9 олигонуклеотид представляет собой ингибирующий TLR9 олигонуклеотид класса B, имеющий линейные характеристики и мотив 5´-CC(T)→GGG-3´, такой как ингибиторный олигонуклеотид INH-18. Неограничивающие примеры такого ингибирующего TLR9 олигонуклеотида класса B включают ODN-2088 (TCCTGGCGGGGAAGT, SEQ ID NO: 889), ODN-2114 (TCCTGGAGGGGAAGT, SEQ ID NO: 890), 4024 (TCCTGGATGGGAAGT, SEQ ID NO: 898), 4084F (CCTGGATGGGAA, SEQ ID NO: 899), INH-13 (CTTACCGCTGCACCTGGATGGGAA, SEQ ID NO: 900), INH-18 (CCTGGATGGGAACTTACCGCTGCA, SEQ ID NO: 901), G-ODN (CTCCTATTGGGGGTTTCCTAT, SEQ ID NO: 893), IRS-869 (TCCTGGAGGGGTTGT, SEQ ID NO: 864), IRS-954 TGCTCCTGGAGGGGTTGT, SEQ ID NO: 902) и AS1411 (GGTGGTGGTGGTTGTGGTGGTGGTGG, SEQ ID NO: 903).
[00408] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, кодирующая последовательность, кодируемая зкДНК, такая как последовательность трансгена, модифицирована так, что динуклеотиды CpG, локализованные в триплете кодона для выбранной аминокислоты, заменяют триплетом кодона для той же аминокислоты, лишенным динуклеотида CpG.
[00409] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, в которых ингибитор TLR9 представляет собой последовательность РНК или белка, зкДНК кодирует РНК или белковый ингибитор TLR9. Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор TLR9 представляет собой антитело или антигенсвязывающий фрагмент, который связывает TLR9. Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, антитело или антигенсвязывающий фрагмент, который связывает TLR9, кодируется зкДНК.
[00410] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор TLR9 вводят субъекту совместно с зкДНК. Неограничивающие примеры ингибиторов TLR9 можно найти в «Classification, Mechanisms of Action, and Therapeutic Applications of Inhibitory Oligonucleotides for Toll-Like Receptors (TLR) 7 and 9», P.S. Lenert, Mediators of Inflammation, Vol. 2010, 986596; US20150203850; и US2017026800, содержание каждого из которых полностью включено в настоящий документ посредством ссылки.
[00411] Соответственно, в некоторых вариантах реализации композиций и способов, описанных в настоящем документе, ингибитор TLR9 вводят субъекту совместно с зкДНК.
[00412] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор TLR9 кодируется в цис-форме с помощью зкДНК, вводимой субъекту (включая, например, последующую доставку зкДНК). Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор TLR9 вводят в транс-форме с помощью зкДНК, вводимой субъекту.
[00413] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор TLR9 представляет собой олигонуклеотид, ингибирующий TLR9.
[00414] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, олигонуклеотид, ингибирующий TLR9, имеет один или более из следующих признаков (i) три последовательных нуклеотида G на 3´-конце; (ii) триплет CC(T) на 5´-конце; и (iii) расстояние между 5´CC(T) и последующим триплетом GGG оптимально составляет 3-5 нуклеотидов в длину. Согласно некоторым вариантам реализации олигонуклеотид, ингибирующий TLR9, имеет последовательность 5´CCTN(3-5)G(3-5)RR3´ (SEQ ID NO: 887). Согласно некоторым вариантам реализации олигонуклеотид, ингибирующий TLR9, не имеет внутрицепочечной и/или межцепочечной водородной связи по Хугстину между смежными G.
[00415] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор TLR9 представляет собой антитело или антигенсвязывающий фрагмент, который связывает TLR9. Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, антитело или антигенсвязывающий фрагмент, который связывает TLR9, кодируется зкДНК.
[00416] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор TLR9 представляет собой ингибитор эндосомального подкисления, например, хлорохин.
Антагонисты инфламмасом
Ингибиторы пути инфламмасомы NLRP3:
[00417] Согласно некоторым вариантам реализации антагонист инфламмасомы ингибирует NLRP3. Термин «NLRP3», также упоминаемый как криопирин, относится к содержащей пириновый домен семейства NOD-подобных рецепторов инфламмасоме 3 или к содержащему домены NACHT, LRR и PYD белку 3 (NALP3), также известному как криопирин, индуцируемый холодом аутовоспалительный синдром 1 (CIAS1), гусеничноподобный рецептор 1.1 (CLR1.1) или содержащий пириновый домен Apafl-подобный белок 1 (PYPAF1). NALP3 также известен под другими названиями: NLRP3 PYD-NACHT-NAD-LRR NALP3 Cias1, Pypaf1, Mmig1 PYD-NACHT-NAD-LRR. NLRP3 является компонентом полибелкового олигомера, состоящего из белка NLRP3, ASC (ассоциированного с апоптозом пятнышкоподобного белка, содержащего CARD) и прокаспазы 1.
[00418] Ингибиторы NLRP3, предусмотренные для применения в способах и композициях, описанных в настоящем документе, раскрыты в Shao, Bo-Zong, et al. «NLRP3 inflammasome and its inhibitors: a review.» Frontiers in pharmacology 6 (2015): 262., и Wang et al., Lab investigation, 2017, 97; 922-934, которые полностью включены в настоящий документ посредством ссылки.
[00419] Согласно некоторым вариантам реализации ингибитор инфламмасомы NLRP3 представляет собой MCC950 или его функциональное производное. MCC950 имеет формулу:
и является сильным и селективным ингибитором NLRP3. MCC950 блокирует высвобождение IL-1β, индуцированное активаторами NLRP3, такими как АТФ, MSU и нигерицин, предотвращая олигомеризацию адаптерного белка инфламмасомы ASC (ассоциированного с апоптозом пятнышкоподобного белка, содержащего CARD) (Coll RC. et al., 2015. A small-molecule inhibitor of the NLRP3 inflammasome for the treatment of inflammatory diseases. Nature Med 21(3), 248-255.; Guo H. et al., 2015. Inflammasomes: mechanism of action, role in disease, and therapeutics. Nat Med. 21(7):677-87; Ren, Honglei, et al. «Selective NLRP3 (Pyrin Domain-Containing Protein 3) Inflammasome Inhibitor Reduces Brain Injury After Intracerebral Hemorrhage.» Stroke (2017): STROKEAHA-117.).
[00420] Согласно некоторым вариантам реализации ингибитор инфламмасомы NLRP3 представляет собой Bay11-7082, который имеет следующую структуру:
и, как сообщалось, селективно ингибирует активность инфламмасомы NLRP3 в макрофагах, независимо от их ингибирующего действия на активность NF-κB (Juliana C. et al, 2010. Anti-inflammatory Compounds Parthenolide and Bay11-7082 Are Direct Inhibitors of the Inflammasome. J. Biol Chem. 285(13): 9792-9802].
[00421] Согласно некоторым вариантам реализации ингибитор инфламмасомы NLRP3 представляет собой глибенкламид (также известный как глибурид), который имеет следующую структуру:
который блокирует созревание каспазы-1 и про-IL-1β путем ингибирования оттока K+ (Laliberte RE. et al., 1999. ATP treatment of human monocytes promotes caspase-1 maturation and externalization. J Biol Chem. 274(52):36944-51). Глибенкламид также сильно блокирует активацию инфламмасомы NRLP3, индуцированную PAMP, DAMP и кристаллическими веществами (Lamkanfi M. et al., 2009. Glyburide inhibits the Cryopyrin/Nalp3 inflammasome. J. Cell Biol., 187: 61 - 70; Dostert C. et al., 2009. Malarial hemozoin is a Nalp3 inflammasome activating danger signal. PLoS One. 4(8): e6510).
[00422] Согласно некоторым вариантам реализации ингибитор инфламмасомы NLRP3 представляет собой изоликвиритигенин (также известный как ILG), который имеет следующую структуру:
который представляет собой флавоноид халконового типа, выделенный из корня солодки (Glycyrrhiza uralensis) и, как сообщается, ингибирует NLRP3-активированную олигомеризацию ASC (Honda H. et al., 2014. Isoliquiritigenin is a potent inhibitor of NLRP3 inflammasome activation and diet-induced adipose tissue inflammation. J Leukoc Biol. 96(6):1087-100.). NLRP3-зависимая продукция IL-1β ингибировалась низкими концентрациями изоликвиритигенина (от 1 до 10 мкМ), это свидетельствует о том, что изоликвиритигенин может блокировать инфламмасому NLRP3 как на стадии праймирования, так и на стадии активации.
[00423] Согласно некоторым вариантам реализации ингибитор инфламмасомы NLRP3 представляет собой 6673-34-0; (5-хлор-2-метокси-N-[2-(4-сульфамоилфенил)этил]бензамид)), который раскрыт в заявке США US20160052876, которая полностью включена в настоящий документ посредством ссылки. Согласно некоторым вариантам реализации ингибитор инфламмасомы NLRP3 представляет собой любое из низкомолекулярных соединений, описанных в US20160052876, содержание которого полностью включено в настоящий документ посредством ссылки.
[00424] Согласно некоторым вариантам реализации ингибитор инфламмасомы NLRP3 представляет собой антагонист рецептора цистеинил-лейкотриена, раскрытый в Ozaki et al., 2015; Coll et al., 2011; Haerter et al., 2009 и патенте US 7498460, которые полностью включены в настоящий документ посредством ссылки. Сообщалось, что антагонист рецептора цистеинил-лейкотриена ингибирует процессинг IL-1, индуцированный как инфламмасомой NLRP3, так и AIM2, путем предотвращения олигомеризации ASC, а также, по-видимому, имеет другие функции во врожденных иммунных ответах, отличные от его роли адаптера для образования инфламмасомы (Ozaki et al., 2015).
[00425] Согласно некоторым вариантам реализации низкомолекулярные ингибиторы, нацеленные на NLRP3 и AIM2, были охарактеризованы и широко описаны в (Ozaki et al., 2015). Подавляющее большинство из них являются фармакологическими ингибиторами, которые были перепрофилированы для нацеливания на инфламмасому (Guo et al., 2015) и включают: партенолид (Juliana et al., 2010), Bay 11-708 (Juliana et al., 2010), CRID3 (Coll et al., 2011), ауранофин (Isakov et al., 2014), изоликвиритигенин (Honda et al., 2014), 3,4-метилендиокси-*-нитростирен (He et al., 2014), циклопентеноновый простагландин 15d-PJ2 (Maier et al., 2015) и 25-гидроксихолестерин (25-HC) (Reboldi et al., 2014). Более того, было показано, что интерферон типа I также подавляет активацию инфламмасомы посредством слабо изученного механизма (Guarda et al., 2011). Однако недавно было продемонстрировано, что IFN-стимулируемый генный продукт, холестерин-25-гидроксилаза (Ch25h), оказывает антагонистическое действие как на транскрипцию IL1b, так и активацию инфламмасом NLRP3, NLRC4 и AIM2, это свидетельствует о том, что Ch25h обладает широкой ингибирующей активностью на несколько инфламмасом (Reboldi et al., 2014).
[00426] Кодирующая последовательность NLRP3 представлена под номерами доступа NCBI NM-004895.1 (SEQ ID NO: 530), NM-183395 (SEQ ID NO: 531), NM-001079821 (SEQ ID NO: 532), NM-001127461 (SEQ ID NO: 533) и NM-001127462 (SEQ ID NO: 534). В данном случае кодон инициации трансляции в NLRP3 предпочтительно представляет собой кодон, расположенный через 6 нуклеотидов после кодона инициации трансляции, описанного в каждом из этих номеров доступа NCBI. Примеры мутантного гена NLRP3 включают ген NLRP3, в котором аденин в положении 1709, считая от кодона инициации трансляции (в случае кодирующей области, показанной под номерами доступа NCBI, положение 1715, считая от кодона инициации трансляции), представляет собой гуанин, цитозин в положении 1043 (положение 1049 в кодирующей области, показанной под номерами доступа NCBI), считая от кодона инициации трансляции, представляет собой тимин или гуанин в положении 587 (положение 593 в кодирующей области, показанной под номерами доступа NCBI), считая от кодона инициации трансляции, представляет собой аденин. NLRP3 предпочтительно представляет собой вариант, в котором нуклеотид в положении 1079 мутирован на гуанин. Как поймет специалист в данной области техники, могут существовать варианты гена NLRP3, которые кодируют функционально эквивалентный NLRP3, который сохраняет функцию, по меньшей мере частично, активации каспазы-1 и/или стимуляции созревания воспалительных цитокинов, таких как интерлейкин 1β и интерлейкин 18. Такой функционально эквивалентный NLRP3 может, соответственно, включать аминокислотные замены, делеции или добавления, которые не устраняют активность.
[00427] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор инфламмасомы NLRP3 представляет собой ингибитор NLRP3 на основе РНК (РНКи), такой как киРНК, специфичная в отношении NLRP3. Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор NLRP3 на основе РНК кодируется зкДНК. КиРНК NLRP3 может быть коммерчески доступной, например, SI03060323 (Qiagen®).
[00428] Согласно некоторым вариантам реализации ингибитор NLRP3 представляет собой РНКи, кодируемую в зкДНК. Во избежание каких-либо сомнений, аминокислотная последовательность белка NLRP3 человека соответствует NM-004895.1 (SEQ ID NO: 539) и представлена ниже:
MKMASTRCKLARYLEDLEDVDLKKFKMHLEDYPPQKGCIPLPRG QTEKADHVDLATLMIDFNGEEKAWAMAVWIFAAINRRDLYEKAKRDEPKWGSDNARVS NPTVICQEDSIEEEWMGLLEYLSRISICKMKKDYRKKYRKYVRSRFQCIEDRNARLGE SVSLNKRYTRLRLIKEHRSQQEREQELLAIGKTKTCESPVSPIKMELLFDPDDEHSEP VHTVVFQGAAGIGKTILARKMMLDWASGTLYQDRFDYLFYIHCREVSLVTQRSLGDLI MSCCPDPNPPIHKIVRKPSRILFLMDGFDELQGAFDEHIGPLCTDWQKAERGDILLSS LIRKKLLPEASLLITTRPVALEKLQHLLDHPRHVEILGFSEAKRKEYFFKYFSDEAQA RAAFSLIQENEVLFTMCFIPLVCWIVCTGLKQQMESGKSLAQTSKTTTAVYVFFLSSL LQPRGGSQEHGLCAHLWGLCSLAADGIWNQKILFEESDLRNHGLQKADVSAFLRMNLF QKEVDCEKFYSFIHMTFQEFFAAMYYLLEEEKEGRTNVPGSRLKLPSRDVTVLLENYG KFEKGYLIFVVRFLFGLVNQERTSYLEKKLSCKISQQIRLELLKWIEVKAKAKKLQIQ PSQLELFYCLYEMQEEDFVQRAMDYFPKIEINLSTRMDHMVSSFCIENCHRVESLSLG FLHNMPKEEEEEEKEGRHLDMVQCVLPSSSHAACSHGLVNSHLTSSFCRGLFSVLSTS QSLTELDLSDNSLGDPGMRVLCETLQHPGCNIRRLWLGRCGLSHECCFDISLVLSSNQ KLVELDLSDNALGDFGIRLLCVGLKHLLCNLKKLWLVSCCLTSACCQDLASVLSTSHS LTRLYVGENALGDSGVAILCEKAKNPQCNLQKLGLVNSGLTSVCCSALSSVLSTNQNL THLYLRGNTLGDKGIKLLCEGLLHPDCKLQVLELDNCNLTSHCCWDLSTLLTSSQSLR KLSLGNNDLGDLGVMMFCEVLKQQSCLLQNLGLSEMYFNYETKSALETLQEEKPELTV
VFEPSW (SEQ ID NO: 539)
[00429] Белок NLRP3 человека кодируется геном NLRP3, содержащим последовательности нуклеиновой кислоты NM-004895.1 (SEQ ID NO: 530), NM-183395 (SEQ ID NO: 531), NM-001079821 (SEQ ID NO: 532), NM-001127461 (SEQ ID NO: 533) и NM-001127462 (SEQ ID NO: 534), и белок NLRP3 человека имеет аминокислотную последовательность NM-004895 (SEQ ID NO: 539).
[00430] Ингибиторы NLRP3 дополнительно включают антисмысловые полинуклеотиды, которые можно применять для ингибирования транскрипции гена NLRP3 и, следовательно, активации инфламмасомы NLRP3. Полинуклеотиды, комплементарные сегменту полинуклеотида, кодирующего NLRP3 (например, полинуклеотида, представленного в SEQ ID NO: 530-534), разработаны для связывания с мРНК, кодирующей NLRP3, и для ингибирования трансляции такой мРНК. Антисмысловые полинуклеотиды могут кодироваться зкДНК-вектором, раскрытым в настоящем документе, и необязательно могут быть функционально связаны с тканеспецифическим или индуцируемым промотором, раскрытым в настоящем документе.
[00431] Ингибирование мРНК NLRP3 можно выполнять с помощью молекул подавляющих ген РНКи в соответствии со способами, обычно известными специалистам в данной области техники. Например, олигонуклеотидные дуплексы подавляющих ген киРНК, специфично нацеленных на NLRP3 человека (NM-004895.1), можно легко применять для подавления экспрессии NLRP3. Нацеливание на мРНК NLRP3 может быть успешно выполнено с использованием киРНК; и другие молекулы киРНК могут быть легко получены специалистами в данной области техники на основе известной последовательности мРНК-мишени. Соответственно, во избежание каких-либо сомнений, обычный специалист в данной области техники может сконструировать ингибиторы нуклеиновых кислот, такие как агенты РНКи (подавляющие РНК), для последовательности нуклеиновой кислоты NM-004895.1, которая представлена ниже:
1 gtagatgagg aaactgaagt tgaggaatag tgaagagttt gtccaatgtc atagccccgt
61 aatcaacggg acaaaaattt tcttgctgat gggtcaagat ggcatcgtga agtggttgtt
121 caccgtaaac tgtaatacaa tcctgtttat ggatttgttt gcatattttt ccctccatag
181 ggaaaccttt cttccatggc tcaggacaca ctcctggatc gagccaacag gagaactttc
241 tggtaagcat ttggctaact tttttttttt tgagatggag tcttgctgtg tcgcctaggc
301 tggagtgcag tggcgtgatc ttggctcact gcagcctcca cttcccgggt tcaatcaatt
361 ctcctacctc aacttcctga gtagctggga ttacaggcgc ccgccaccac acccggctca
421 tttttgtact tttagtagag acacagtttt gccatgttgg ccaggctggt cttgaattcc
481 tcagctcagg tgatctgcct gccttggcct ctcaaagtgc tgggattaca ggcgtgagcc
541 actgtgcccg gccttggcta acttttcaaa attaaagatt ttgacttgtt acagtcatgt
601 gacatttttt tctttctgtt tgctgagttt ttgataattt atatctctca aagtggagac
661 tttaaaaaag actcatccgt gtgccgtgtt cactgcctgg tatcttagtg tggaccgaag
721 cctaaggacc ctgaaaacag ctgcagatga agatggcaag cacccgctgc aagctggcca
781 ggtacctgga ggacctggag gatgtggact tgaagaaatt taagatgcac ttagaggact
841 atcctcccca gaagggctgc atccccctcc cgaggggtca gacagagaag gcagaccatg
901 tggatctagc cacgctaatg atcgacttca atggggagga gaaggcgtgg gccatggccg
961 tgtggatctt cgctgcgatc aacaggagag acctttatga gaaagcaaaa agagatgagc
1021 cgaagtgggg ttcagataat gcacgtgttt cgaatcccac tgtgatatgc caggaagaca
1081 gcattgaaga ggagtggatg ggtttactgg agtacctttc gagaatctct atttgtaaaa
1141 tgaagaaaga ttaccgtaag aagtacagaa agtacgtgag aagcagattc cagtgcattg
1201 aagacaggaa tgcccgtctg ggtgagagtg tgagcctcaa caaacgctac acacgactgc
1261 gtctcatcaa ggagcaccgg agccagcagg agagggagca ggagcttctg gccatcggca
1321 agaccaagac gtgtgagagc cccgtgagtc ccattaagat ggagttgctg tttgaccccg
1381 atgatgagca ttctgagcct gtgcacaccg tggtgttcca gggggcggca gggattggga
1441 aaacaatcct ggccaggaag atgatgttgg actgggcgtc ggggacactc taccaagaca
1501 ggtttgacta tctgttctat atccactgtc gggaggtgag ccttgtgaca cagaggagcc
1561 tgggggacct gatcatgagc tgctgccccg acccaaaccc acccatccac aagatcgtga
1621 gaaaaccctc cagaatcctc ttcctcatgg acggcttcga tgagctgcaa ggtgcctttg
1681 acgagcacat aggaccgctc tgcactgact ggcagaaggc cgagcgggga gacattctcc
1741 tgagcagcct catcagaaag aagctgcttc ccgaggcctc tctgctcatc accacgagac
1801 ctgtggccct ggagaaactg cagcacttgc tggaccatcc tcggcatgtg gagatcctgg
1861 gtttctccga ggccaaaagg aaagagtact tcttcaagta cttctctgat gaggcccaag
1921 ccagggcagc cttcagtctg attcaggaga acgaggtcct cttcaccatg tgcttcatcc
1981 ccctggtctg ctggatcgtg tgcactggac tgaaacagca gatggagagt ggcaagagcc
2041 ttgcccagac atccaagacc accaccgcgg tgtacgtctt cttcctttcc agtttgctgc
2101 agccccgggg agggagccag gagcacggcc tctgcgccca cctctggggg ctctgctctt
2161 tggctgcaga tggaatctgg aaccagaaaa tcctgtttga ggagtccgac ctcaggaatc
2221 atggactgca gaaggcggat gtgtctgctt tcctgaggat gaacctgttc caaaaggaag
2281 tggactgcga gaagttctac agcttcatcc acatgacttt ccaggagttc tttgccgcca
2341 tgtactacct gctggaagag gaaaaggaag gaaggacgaa cgttccaggg agtcgtttga
2401 agcttcccag ccgagacgtg acagtccttc tggaaaacta tggcaaattc gaaaaggggt
2461 atttgatttt tgttgtacgt ttcctctttg gcctggtaaa ccaggagagg acctcctact
2521 tggagaagaa attaagttgc aagatctctc agcaaatcag gctggagctg ctgaaatgga
2581 ttgaagtgaa agccaaagct aaaaagctgc agatccagcc cagccagctg gaattgttct
2641 actgtttgta cgagatgcag gaggaggact tcgtgcaaag ggccatggac tatttcccca
2701 agattgagat caatctctcc accagaatgg accacatggt ttcttccttt tgcattgaga
2761 actgtcatcg ggtggagtca ctgtccctgg ggtttctcca taacatgccc aaggaggaag
2821 aggaggagga aaaggaaggc cgacaccttg atatggtgca gtgtgtcctc ccaagctcct
2881 ctcatgctgc ctgttctcat ggattggtga acagccacct cacttccagt ttttgccggg
2941 gcctcttttc agttctgagc accagccaga gtctaactga attggacctc agtgacaatt
3001 ctctggggga cccagggatg agagtgttgt gtgaaacgct ccagcatcct ggctgtaaca
3061 ttcggagatt gtggttgggg cgctgtggcc tctcgcatga gtgctgcttc gacatctcct
3121 tggtcctcag cagcaaccag aagctggtgg agctggacct gagtgacaac gccctcggtg
3181 acttcggaat cagacttctg tgtgtgggac tgaagcacct gttgtgcaat ctgaagaagc
3241 tctggttggt cagctgctgc ctcacatcag catgttgtca ggatcttgca tcagtattga
3301 gcaccagcca ttccctgacc agactctatg tgggggagaa tgccttggga gactcaggag
3361 tcgcaatttt atgtgaaaaa gccaagaatc cacagtgtaa cctgcagaaa ctggggttgg
3421 tgaattctgg ccttacgtca gtctgttgtt cagctttgtc ctcggtactc agcactaatc
3481 agaatctcac gcacctttac ctgcgaggca acactctcgg agacaagggg atcaaactac
3541 tctgtgaggg actcttgcac cccgactgca agcttcaggt gttggaatta gacaactgca
3601 acctcacgtc acactgctgc tgggatcttt ccacacttct gacctccagc cagagcctgc
3661 gaaagctgag cctgggcaac aatgacctgg gcgacctggg ggtcatgatg ttctgtgaag
3721 tgctgaaaca gcagagctgc ctcctgcaga acctggggtt gtctgaaatg tatttcaatt
3781 atgagacaaa aagtgcgtta gaaacacttc aagaagaaaa gcctgagctg accgtcgtct
3841 ttgagccttc ttggtaggag tggaaacggg gctgccagac gccagtgttc tccggtccct
3901 ccagctgggg gccctcaggt ggagagagct gcgatccatc caggccaaga ccacagctct
3961 gtgatccttc cggtggagtg tcggagaaga gagcttgccg acgatgcctt cctgtgcaga
4021 gcttgggcat ctcctttacg ccagggtgag gaagacacca ggacaatgac agcatcgggt
4081 gttgttgtca tcacagcgcc tcagttagag gatgttcctc ttggtgacct catgtaatta
4141 gctcattcaa taaagcactt tctttatttt tctcttctct gtctaacttt ctttttccta
4201 tcttttttct tctttgttct gtttactttt gctcatatca tcattcccgc tatctttcta
4261 ttaactgacc ataacacaga actagttgac tatatattat gttgaaattt tatggcagct
4321 atttatttat ttaaattttt tgtaacagtt ttgttttcta ataagaaaaa tccatgcttt
4381 ttgtagctgg ttgaaaattc aggaatatgt aaaacttttt ggtatttaat taaattgatt
ccttttctta attttaaaaa aaaaaaaaaa (SEQ ID NO: 530)
[00432] Согласно некоторым вариантам реализации ингибитор инфламмасомы NLRP3 представляет собой киРНК, ингибируя посредством этого мРНК инфламмасомы NLRP3. Согласно некоторым вариантам реализации ингибитор инфламмасомы NLRP3 представляет собой GUGCAUUGAAGACAGGAAUTT (SEQ ID NO: 540) (Wang et al., Laboratory Invest. (2017) 97: 922-934, которая полностью включена в настоящий документ посредством ссылки), который ингибирует экспрессию NLRP3 человека или его фрагмента или гомолога, который по меньшей мере на 50% или по меньшей мере на 60%, или по меньшей мере на 70%, или по меньшей мере на 80% или по меньшей мере на 90% идентичен ему. Согласно некоторым вариантам реализации ингибитор инфламмасомы NLRP3 представляет собой коммерчески доступную киРНК, такую как доступная от Santa Cruz® (номер по каталогу sc-40327).
[00433] Согласно некоторым вариантам реализации ингибитор инфламмасомы NLRP3 представляет собой РНКи, которая комплементарна последовательности-мишени РНКи в NM_001079821.2 человека, ген NCBI 114548 (NLRP3). Агент РНКи, который ингибирует NLRP3, может представлять собой нуклеиновую кислоту, которая комплементарна 17-21 последовательному основанию SEQ ID NO: 541-551, показанным в Таблице 5A.
[00434] Таблица 5A: Последовательности-мишени РНКи для ингибирования NLRP3:
[00435] Согласно некоторым вариантам реализации ингибитор инфламмасомы NLRP3 представляет собой агент киРНК. Примерные последовательности киРНК, которые ингибируют NLRP3, показаны в Таблице 5B.
[00436] Таблица 5B: Примерные киРНК, которые ингибируют NLRP3
[00437] Согласно некоторым вариантам реализации ингибитор инфламмасомы NLRP3 представляет собой миРНК (miR), которая ингибирует экспрессию NLRP3, или агонист miR, которая ингибирует экспрессию NLRP3. Примерные miR, которые ингибируют NLRP3, представляют собой miR-9 и miR-223.
[00438] miR-9 ингибирует активацию инфламмасомы NLRP3 (Wang, Yue, et al. «MicroRNA-9 inhibits NLRP3 inflammasome activation in human atherosclerosis inflammation cell models through the JAK1/STAT signaling pathway.» Cellular Physiology and Biochemistry 41.4 (2017): 1555-1571.). Соответственно, пре-miR-9 (предшественник MiR-9) или miR-9 можно применять для ингибирования NLRP3. Последовательность зрелой miR-9 (MIMAT0000441) представляет собой 5´-UCU UUG GUU AUC U AG CUG UAU GA-3´ (SEQ ID NO: 587). hsa-miR-9-5p (UCUUUGGUUAUCUAGCUGUAUGA) (SEQ ID NO: 588). Согласно некоторым вариантам реализации ингибитор инфламмасомы NLRP3 представляет собой агонист miR-9 SQ22538 (SQ; 9-(тетрагидро-2-фуранил)-9H-пурин-6-амин), который, как сообщалось, повышает экспрессию miR-9 (Ham, Onju, et al. «Small molecule-mediated induction of miR-9 suppressed vascular smooth muscle cell proliferation and neointima formation after balloon injury.» Oncotarget 8.55 (2017): 93360.). Формула SQ22538 показана ниже:
[00439] miR-223 ингибирует активность инфламмасомы NLRP3. (Bauernfeind, Franz, et al. «NLRP3 inflammasome activity is negatively controlled by miR-223.» The Journal of Immunology 189.8 (2012): 4175-4181; Feng, Zunyong, et al. «Ly6G+ neutrophil-derived miR-223 inhibits the NLRP3 inflammasome in mitochondrial DAMP-induced acute lung injury.» Cell death & disease 8.11 (2017): e3170). miR-223 может быть синтезирована как mmu-miR-223. По меньшей мере один или 2- или 3, или 4 блока последовательности, комплементарной miR-223 (5´-TGGGGTATTTGACAAACTGACA-3´ (SEQ ID NO:589), можно применять для ингибирования NLRP3. cbn-mir-233 MI0024890 имеет последовательность: UCGCCCAUCCCGUUGUUCCAAUAUUCCAACAACAAGUGAUUAUUGAGCAAUGCGCAUGUGCGG (SEQ ID NO: 590); cbr-mir-233 MI0000530 имеет последовательность: AAGCAUUUUUCUGUCCCGCGCAUCCCUUUGUUCCAAUAUUCAAACCAGUAGAAAGAUUAUUGAGCAAUGCGCAUGUGCGGGACAGAUUGAAUAGCUG (SEQ ID NO: 591); cel-mir-233 MI0000308 имеет последовательность: AUAUAGCAUCUUUCUGUCUCGCCCAUCCCGUUGCUCCAAUAUUCUAACAACAAGUGAUUAUUGAGCAAUGCGCAUGUGCGGGAUAGACUGAUGGCUGC (SEQ ID NO: 592); crm-mir-233 MI0011059 имеет последовательность: UGAAGCGUCUCUCUGUCCCGCUCAUCCUGUUGUUCCAAUAUUCCAACAGCCCAGUGAUUAUUGAGCAAUGCGCAUGUGCGGGACAGAUUGUAUGCUGCCAU (SEQ ID NO: 593).
[00440] Согласно некоторым вариантам реализации ингибитор инфламмасомы NLRP3 представляет собой анти-миРНК (анти-miR), которая ингибирует экспрессию miR, подавляющей экспрессию или функцию NLRP3. Примерные анти-miR представляют собой анти-miR-22 и анти-miR-33. Было продемонстрировано, что miR22 поддерживает экспрессию NLRP3 (Li, S., et al., «MiR-22 sustains NLRP3 expression and attenuates H. pylori-induced gastric carcinogenesis.» Oncogene 37.7 (2018): 884.). Зрелая последовательность miR-22 представляет собой hsa-miR-22 (hsa-miR-22-5p MIMAT000449) и имеет следующий вид: AGUUCUUCAGUGGCAAGCUUUA (SEQ ID NO: 594) с последовательностью «стебель-петля», которая представлена далее: hsa-mir-22 MI0000078 имеет последовательность: GGCUGAGCCGCAGUAGUUCUUCAGUGGCAAGCUUUAUGUCCUGACCCAGCUAAAGCUGCCAGUUGAAGAACUGUUGCCCUCUGCC (SEQ ID NO: 595).
[00441] Сообщалось, что miR-33 положительно регулирует экспрессию мРНК и белка NLRP3, а также активность каспазы-1 в первичных макрофагах (Xie, Qingyun, et al. «MicroRNA - 33 regulates the NLRP3 inflammasome signaling pathway in macrophages.» Molecular medicine reports 17.2 (2018): 3318-3327). Зрелая последовательность miR-33 представляет собой mmu-miR-33-5p или MIMAT0000667; и представляет собой: GUGCAUUGUAGUUGCAUUGCA (SEQ ID NO: 596); при этом последовательность «стебель-петля» имеет следующий вид: mmu-mir-33 MI0000707: CUGUGGUGCAUUGUAGUUGCAUUGCAUGUUCUGGCAAUACCUGUGCAAUGUUUCCACAGUGCAUCACGG (SEQ ID NO: 597).
[00442] Соответственно, в некоторых вариантах реализации, ингибитор NLRP3 представляет собой анти-miR-22, которая комплементарна по меньшей мере части, например, 15-25 звеньям SEQ ID NO: 594 или SEQ ID NO: 595, или анти-miR-33, которая комплементарна по меньшей мере части, например, 15-21 звену SEQ ID NO: 596 или SEQ ID NO: 597.
[00443] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор инфламмасомы NLRP3 представляет собой антитело к NLRP3 человека (номер по каталогу AF6789) от R&D Systems (Миннеаполис, Миннесота). Согласно некоторым вариантам реализации антитело-ингибитор NLRP3 кодируется зкДНК.
Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор NLRP3 представляет собой антитело или антигенсвязывающий фрагмент, который связывает NLRP3. Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, антитело или антигенсвязывающий фрагмент, который связывает NLRP3, кодируется зкДНК.
Ингибитор инфламмасомы NLRP3 относится к соединениям, которые ингибируют или по меньшей мере уменьшают активность инфламмасомы, включая глибурид и его функционально эквивалентные предшественники или производные, ингибиторы каспазы-1, активаторы аденозинмонофосфат-активируемой протеинкиназы (AMPK) и ингибиторы P2X7. Ингибирование инфламмасомы NLRP3 может быть достигнуто с помощью одного соединения или комбинации соединений, которые ингибируют инфламмасому или каспазу-1, но не приводят к изменениям ферментативной активности цитохрома P450 (cyp), включая изоформы cyp, 3A4, 2C9 и 2C19, что может отрицательно повлиять на метаболизм статинов и тем самым уменьшить биодоступность статинов.
Ингибиторы пути инфламмасомы AIM2
[00444] Согласно некоторым вариантам реализации антагонист инфламмасомы ингибирует AIM2. AIM2, альтернативно известный как PISA, представляет собой полипептид из 343 аминокислот (см. номер доступа Genbank AF024714.1; номер доступа RefSeq NP_004824.1) (SEQ ID NO: 598). AIM2 является членом семейства IFI20X/IF116 и, как известно, экспрессируется в селезенке, тонком кишечнике, лейкоцитах периферической крови и семенниках. AIM2 содержит домен PYD, который участвует во взаимодействии с ASC, а также домен HIN200, который участвует во взаимодействии с дцДНК. AIM2, как предполагают, играет роль в онкогенной реверсии и может контролировать пролиферацию клеток. Экспрессия AIM2 индуцируется интерфероном-гамма.
[00445] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор AIM2 представляет собой антитело или антигенсвязывающий фрагмент, который связывает AIM2. Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, антитело или антигенсвязывающий фрагмент, который связывает NLRP3, кодируется зкДНК. Ингибиторы AIM2 раскрыты в Farshchian et al., Oncotarget 2017; 8(28); 45825-45836, которая полностью включена в настоящий документ посредством ссылки.
[00446] Согласно некоторым вариантам реализации ингибитор инфламмасомы AIM2 представляет собой моноклональное антитело к ASC человека (клон 23-4, MBL, Нагоя, Япония), которое, как сообщается, создает помехи для PYD из ASC. Согласно некоторым вариантам реализации ингибитор инфламмасомы AIM2 представляет собой антитело к AIM2 человека (номер по каталогу 8055) (Cell Signaling Technology® (Беверли, Массачусетс). Согласно некоторым вариантам реализации ингибитор инфламмасомы AIM2 представляет собой эндогенный ингибитор AIM2, такой как пиринсодержащие белки, недавно описанные (Khare et al., 2014; de Almeida et al., 2015), или антимикробные пептиды кателицидины, о которых сообщили Schauber и коллеги (Dombrowski et al., 2011). Согласно некоторым вариантам реализации ингибитор инфламмасомы AIM2 представляет собой любое соединение, раскрытое в небольшом обзоре Miriam Canavase «the duality of AIM2 inflammasome: A focus on its role in autoimmunity and Skin diseases. Am. J. Pharm & Toxicology; 2016).
[00447] Согласно некоторым вариантам реализации ингибитор инфламмасомы AIM2 представляет собой P202, который представляет собой тетрамер p202 и, как сообщается, уменьшает активацию AIM2 и предотвращает зависимую от дцДНК кластеризацию ASC и активацию инфламмасомы AIM2 (Fernandes-Alnemri, Teresa, et al. «The AIM2 inflammasome is critical for innate immunity to Francisella tularensis.» Nature immunology 11.5 (2010): 385; Yin, Qian, et al. «Molecular mechanism for p202-mediated specific inhibition of AIM2 inflammasome activation.» Cell reports 4.2 (2013): 327-339). Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, P202 кодируется зкДНК.
[00448] Согласно некоторым вариантам реализации ингибитор инфламмасомы AIM2 представляет собой любое из низкомолекулярных соединений, описанных в WO 2017138586A или US 2013/0158100 A1, содержание каждого из которых полностью включено в настоящий документ посредством ссылки.
[00449] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор AIM2 представляет собой ингибитор AIM2 на основе РНК, такой как киРНК, специфичная в отношении AIM2. Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор AIM2 на основе РНК кодируется зкДНК. Белок AIM2 человека кодируется геном AIM2, содержащим последовательность нуклеиновой кислоты NM_004833.2 (SEQ ID NO: 600), и белок AIM2 человека имеет аминокислотную последовательность NP_004824.1 (SEQ ID NO: 598). Ингибиторы AIM2 дополнительно включают антисмысловые полинуклеотиды, которые можно применять для ингибирования транскрипции гена AIM2 и, соответственно, активации инфламмасомы AIM2. Полинуклеотиды, комплементарные сегменту полинуклеотида, кодирующего AIM2 (например, полинуклеотида, представленного в SEQ ID NO: 600), разработаны для связывания с мРНК, кодирующей AIM2, и ингибирования трансляции такой мРНК. Антисмысловые полинуклеотиды могут кодироваться зкДНК-вектором, раскрытым в настоящем документе, и необязательно могут быть функционально связаны с тканеспецифическим или индуцируемым промотором, раскрытым в настоящем документе. Ингибирование мРНК AIM2 можно осуществлять с помощью молекул подавляющих ген РНКи в соответствии со способами, обычно известными специалистам в данной области техники. Например, олигонуклеотидные дуплексы подавляющих ген киРНК, специфично нацеленных на AIM2 человека (NM_004833.2), можно легко применять для подавления экспрессии AIM2. Нацеливание на мРНК AIM2 может быть успешно выполнено с использованием киРНК; и другие молекулы киРНК могут быть легко получены специалистами в данной области техники на основе известной последовательности мРНК-мишени. Соответственно, во избежание каких-либо сомнений, обычный специалист в данной области техники может разработать ингибиторы на основе нуклеиновых кислот, такие как агенты РНКи (подавляющие РНК), для последовательности нуклеиновой кислоты NM_004833.2, которая представлена ниже:
1 atagacattt tcttctgtgg ctgctagtga gaacccaaac cagctcagcc aattagagct
61 ccagttgtca ctcctaccca cactgggcct gggggtgaag ggaagtgttt attaggggta
121 catgtgaagc cgtccagaag tgtcagagtc tttgtagctt tgaaagtcac ctaggttatt
181 tgggcatgct ctcctgagtc ctctgctagt taagctctct gaaaagaagg tggcagaccc
241 ggtttgctga tcgccccagg gatcaggagg ctgatcccaa agttgtcaga tggagagtaa
301 atacaaggag atactcttgc taacaggcct ggataacatc actgatgagg aactggatag
361 gtttaagttc tttctttcag acgagtttaa tattgccaca ggcaaactac atactgcaaa
421 cagaatacaa gtagctacct tgatgattca aaatgctggg gcggtgtctg cagtgatgaa
481 gaccattcgt atttttcaga agttgaatta tatgcttttg gcaaaacgtc ttcaggagga
541 gaaggagaaa gttgataagc aatacaaatc ggtaacaaaa ccaaagccac taagtcaagc
601 tgaaatgagt cctgctgcat ctgcagccat cagaaatgat gtcgcaaagc aacgtgctgc
661 accaaaagtc tctcctcatg ttaagcctga acagaaacag atggtggccc agcaggaatc
721 tatcagagaa gggtttcaga agcgctgttt gccagttatg gtactgaaag caaagaagcc
781 cttcacgttt gagacccaag aaggcaagca ggagatgttt catgctacag tggctacaga
841 aaaggaattc ttctttgtaa aagtttttaa tacactgctg aaagataaat tcattccaaa
901 gagaataatt ataatagcaa gatattatcg gcacagtggt ttcttagagg taaatagcgc
961 ctcacgtgtg ttagatgctg aatctgacca aaaggttaat gtcccgctga acattatcag
1021 aaaagctggt gaaaccccga agatcaacac gcttcaaact cagccccttg gaacaattgt
1081 gaatggtttg tttgtagtcc agaaggtaac agaaaagaag aaaaacatat tatttgacct
1141 aagtgacaac actgggaaaa tggaagtact gggggttaga aacgaggaca caatgaaatg
1201 taaggaagga gataaggttc gacttacatt cttcacactg tcaaaaaatg gagaaaaact
1261 acagctgaca tctggagttc atagcaccat aaaggttatt aaggccaaaa aaaaaacata
1321 gagaagtaaa aaggaccaat tcaagccaac tggtctaagc agcatttaat tgaagaatat
1381 gtgatacagc ctcttcaatc agattgtaag ttacctgaaa gctgcagttc acaggctcct
1441 ctctccacca aattaggata gaataattgc tggataaaca aattcagaat atcaacagat
gatcacaata aacatctgtt tctcattcaa aaaaaaaaaa aaaaaaaaaa aaaaaaaa
(SEQ ID NO: 600)
Согласно некоторым вариантам реализации ингибитор инфламмасомы AIM2 представляет собой киРНК, ингибируя посредством этого мРНК инфламмасомы AIM2. Согласно некоторым вариантам реализации ингибитор инфламмасомы AIM2 представляет собой 5´-CCCGAAGATCAACACGCTTCA-3´ (SEQ ID NO: 601) или 5´-AAAGGTTAATGTCCCGCTGAA-3´ (SEQ ID NO: 665) (обе из Farshchian et al. Oncotarget (2017) 8: 45825-45836), которая ингибирует экспрессию AIM2 человека или его фрагмента или гомолога, который по меньшей мере на 50% или по меньшей мере на 60%, или по меньшей мере на 70%, или по меньшей мере на 80%, или по меньшей мере на 90% идентичен ему.
[00450] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор инфламмасомы AIM2 представляет собой ингибитор AIM2 на основе РНК, такой как киРНК, специфичная в отношении AIM2. Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор AIM2 на основе РНК кодируется зкДНК. КиРНК AIM2 может быть коммерчески доступна, например, SI04261432 (Qiagen®); или RCN0000096104 (#1), TRCN0000096105 (#2), TRCN0000096106 (#3) от OpenBiosystems® (Хантсвилл, Алабама).
[00451] Согласно некоторым вариантам реализации ингибитор инфламмасомы AIM2 представляет собой A151 (5´-TTAGGGTTAGGGTTAGGGTTAGGG-3´ (SEQ ID NO: 602) или C151 (5´-TTCAAATTCAAATTCAAATTCAAA-3´ (SEQ ID NO: 603), который синтезирован с фосфотиоатным (PO) остовом. A151 (также называемый ODN TTAGGG) представляет собой синтетический олигонуклеотид (ODN), содержащий 4 повтора иммуносупрессорного мотива TTAGGG (SEQ ID NO: 604), обычно обнаруживаемого в теломерной ДНК млекопитающих (Steinhagen F. et al., 2017. Suppressive oligodeoxynucleotides containing TTAGGG motifs inhibit cGAS activation in human monocytes. Eur J Immunol). A151 блокирует активацию инфламмасомы AIM2 в ответ на цитозольную дцДНК, но требует фосфотиоатного (PO) остова (Kaminsji et al., J Immunol 2013; 191:3876-3883, Synthetic Oligodeoxynucleotides Containing Suppressive TTAGGG Motifs Inhibit AIM2 Inflammasome Activation; Eichholz K. et al., 2016. Immune-Complexed Adenovirus Induce AIM2-Mediated Pyroptosis in Human Dendritic Cells. PLoS Pathog. 12(9): e1005871). Согласно некоторым вариантам реализации ингибитор инфламмасомы AIM2 представляет собой A151 (5′-TTAGGGTTAGGGTTAGGGTTAGGG-3′ (SEQ ID NO: 602) или по меньшей мере один повтор TTAGGG (SEQ ID NO: 604), каждый с фосфотиоатным (PO) остовом. Согласно некоторым вариантам реализации ингибитор инфламмасомы AIM2 представляет собой A151 (5´-TTAGGGTTAGGGTTAGGGTTAGGG-3´ (SEQ ID NO: 602) или по меньшей мере один повтор TTAGGG (SEQ ID NO: 604), который не имеет фосфодиэфирного (PE) остова. Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор инфламмасомы AIM2 кодируется зкДНК, вводимой субъекту (включая, например, последующую доставку зкДНК). Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор инфламмасомы AIM2, кодируемый зкДНК, вводимой субъекту, представляет собой A151 (SEQ ID NO: 602).
[00452] Согласно некоторым вариантам реализации ингибитор инфламмасомы AIM2 представляет собой РНКи, которая комплементарна последовательности-мишени РНКи в NM_001348247.1 человека (SEQ ID NO: 566), ген 9447 NCBI (AIM2). Агент РНКи, который ингибирует AIM2, может представлять собой нуклеиновую кислоту, комплементарную 17-21 последовательному основанию SEQ ID NO: 605-610, показанным в Таблице 5C.
[00453] Таблица 5C: Последовательности-мишени РНКи для ингибирования AIM2:
[00454] Согласно некоторым вариантам реализации ингибитор инфламмасомы AIM2 представляет собой агент киРНК. Примерные последовательности киРНК, которые ингибируют AIM2, показаны в Таблице 5D.
[00455] Таблица 5D: Примерные киРНК, которые ингибируют AIM2
[00456] Согласно некоторым вариантам реализации ингибитор инфламмасомы AIM2 представляет собой миРНК (miR), которая ингибирует экспрессию AIM2, или агонист miR, которая ингибирует экспрессию AIM2. Примерные miR, которые ингибируют AIM2, представляют собой miR-223 (Yang, Fan, et al. «MicroRNA-223 acts as an important regulator to Kupffer cells activation at the early stage of Con A-induced acute liver failure via AIM2 signaling pathway.» Cellular Physiology and Biochemistry 34.6 (2014): 2137-2152). Соответственно, ингибитор AIM2 для применения в настоящем изобретении представляет собой miR-223, соответствующую любой из SEQ ID NO: 589-593.
[00457] Инфламмасому AIM2, восстановленную в бесклеточной системе in vitro, можно применять в качестве инструмента для скрининга ингибиторов инфламмасомы AIM2 в соответствии с методами, раскрытыми в Kaneko et al., 2015, или методами, раскрытыми в заявке США US2013/0158100A1, которая полностью включена в настоящий документ посредством ссылки.
Ингибиторы каспазы-1
[00458] Согласно некоторым вариантам реализации антагонист инфламмасомы ингибирует каспазу-1. Согласно некоторым вариантам реализации ингибитор каспазы-1 для применения в способах и композициях представляет собой белнаказан (VX-765). VX-765 представляет собой всасываемое перорально пролекарство VRT-043198, сильного и селективного ингибитора каспаз, принадлежащих к подсемейству ICE/каспазы-1, и имеет следующую формулу:
(см. Wannamaker W. et al., 2007. (S)-1-((S)-2-{[1-(4-amino-3-chloro-phenyl)-methanoyl]-amino}-3,3-dimethyl-butanoyl)-pyrrolidine-2-carboxylic acid ((2R,3S)-2-ethoxy-5-oxo-tetrahydro-furan-3-yl)-amide (VX-765), an orally available selective interleukin (IL)-converting enzyme/caspase-1 inhibitor, exhibits potent anti-inflammatory activities by inhibiting the release of IL-1beta and IL-18. J Pharmacol Exp Ther. 321(2):509-16).
[00459] Согласно некоторым вариантам реализации ингибитор каспазы-1 представляет собой Z-VAD-FMK, который имеет следующую структуру:
и представляет собой проникающий в клетки ингибитор одной или более каспаз и сильный ингибитор активации каспазы-1 в NLRP3-индуцированных клетках (Dostert C. et al., 2009. Malarial hemozoin is a Nalp3 inflammasome activating danger signal. PLoS One. 4(8):e6510.). Z-VAD-FMK необратимо связывается с каталитическим сайтом протеаз каспаз (Slee EA. et al., 1996. Benzyloxycarbonyl-Val-Ala-Asp (OMe) fluoromethylketone (Z-VAD.FMK) inhibits apoptosis by blocking the processing of CPP32. Biochem J. 315 (Pt 1):21-4.).
[00460] Согласно некоторым вариантам реализации ингибитор каспазы-1 представляет собой Ac-YVAD-cmk, который имеет следующую структуру:
и представляет собой ингибитор каспазы-1 и хлорметилкетон тетрапептид на основе последовательности-мишени в proIL-1β (YVHD). Сообщалось, что Ac-YVAD cmk блокирует активацию инфламмасомы и, следовательно, проявляет противовоспалительный, антиапоптотический и антипироптотический эффекты.
[00461] Согласно некоторым вариантам реализации ингибитор каспазы-1 представляет собой Ac-YVAD-CHO, который имеет следующую структуру:
(Brenner, B., et al. 1998. Cell Death Differ. 5: 29-37. PMID: 10200443) Субстрат каспазы-1 (CAS 143305-11-7)
[00462] Согласно некоторым вариантам реализации ингибитор каспазы-1 представляет собой партенолид, который имеет следующую структуру:
[00463] Партенолид, сесквитерпеновый лактон, полученный из пиретрума, является известным ингибитором активации NF-κB, а также непосредственным ингибитором каспазы-1 и множества инфламмасом, включая инфламмасомы NLRP3 и NLRP1 (Juliana C. et al., 2010. Anti-inflammatory compounds parthenolide and Bay 11-7082 are direct inhibitors of the inflammasome. J Biol Chem. 285(13):9792-802.). Партенолид непосредственно ингибирует инфламмасому NLRP3, нарушая активность АТФазы NLRP3.
[00464] Согласно некоторым вариантам реализации ингибитор каспазы-1 представляет собой один или комбинацию: пралнаказана (VX-740), который имеет следующую структуру:
[00465] Z-WEHD-FMK (также известного как бензилоксикарбонил-V-A-D-O-метилфторметилкетон).
[00466] Согласно некоторым вариантам реализации ингибитор каспазы-1 представляет собой шиконин или ацетилшиконин, причем шиконин представляет собой:
и ацетилшиконин представляет собой:
Шиконин представляет собой высоколипофильный нафтохинон, обнаруженный в корнях Lithospermum erythrorhizon, который используется в традиционной китайской медицине из-за его плейотропных эффектов, и подавляет активацию инфламмасомы NLRP3 (Zorman et al., PLOS One, 2016; 11(7); e0159826.).
[00467] Согласно некоторым вариантам реализации ингибитор каспазы-1 может представлять собой низкомолекулярный ингибитор, как будет понятно специалисту в данной области техники. Неограничивающие примеры включают содержащие цианопропанат молекулы, такие как (S)-3-((S)-1-((S)-2-(4-амино-3-хлорбензамидо)-3,3-диметилбутаноил)пирролидин-2-карбоксамидо)-3-цианопропановая кислота, а также другие низкомолекулярные ингибиторы каспазы-1, такие как ((2R,3S)-2-этокси-5-оксотетрагидрофуран-3-ил)амид (S)-1-((S)-2-{[1-(4-амино-3-хлорфенил)метаноил]амино}-3,3-диметилбутаноил)пирролидин-2-карбоновой кислоты. Такие ингибиторы могут быть синтезированы химическим путем.
[00468] Согласно некоторым вариантам реализации ингибитор каспазы-1 может представлять собой прямой ингибитор ферментативной активности каспазы-1 или может представлять собой непрямой ингибитор, который ингибирует инициацию сборки инфламмасомы или распространение сигнала инфламмасомы. Ингибиторы каспазы-1 для применения в настоящем изобретении могут представлять собой антиоксиданты, включая ингибиторы активных форм кислорода (АФК). Примеры таких ингибиторов каспазы-1 включают, но не ограничиваются перечисленными, флавоноиды, включая флавоны, такие как апигенин, лютеолин и диосмин; флавонолы, такие как мирицетин, физетин и кверцетин; флаванолы и их полимеры, такие как катехин, галлокатехин, эпикатехин, эпигаллокатехин, эпигаллокатехин-3-галлат и теафлавин; изофлавоновые фитоэстрогены; и стильбеноиды, такие как ресвератрол. Также включены фенольные кислоты и их сложные эфиры, такие как галловая кислота и салициловая кислота; терпеноиды или изопреноиды, такие как андрографолид и партенолид; витамины, такие как витамины А, С и Е; кофакторы витаминов, такие как кофермент Q10, марганец и йодид, другие органические антиоксиданты, такие как лимонная кислота, щавелевая кислота, фитиновая кислота и альфа-липоевая кислота, и экстракт лакового дерева Rhus verniciflua Stokes. Ингибитор каспазы-1 может представлять собой комбинацию этих соединений, например, комбинацию α-липоевой кислоты, кофермента Q10 и витамина E, или комбинацию ингибитора (ов) каспазы 1 с другим ингибитором инфламмасомы, таким как глибурид или его функционально эквивалентный предшественник или производное.
[00469] Примеры дозировок некоторых ингибиторов инфламмасом перечислены далее: апигенин (примерно 0,1-10 мг/кг), лютеолин (примерно 1-100 мг), диосмин (примерно 100-900 мг), мирицетин (примерно 10-300 мг), кверцетин (примерно 10-1000 мг), физетин (1-200 мг/кг), экстракт Rhus verniciflua Stokes (1-100 мг/кг), катехин (примерно 50-500 мг), галлокатехин (примерно 100-1000 мг), эпикатехин (примерно 0,1-10 мг/кг), эпигаллокатехин (примерно 100-1000 мг), эпигаллокатехин-3-галлат (примерно 100-1000 мг), теафлавин (примерно 75-750 мг), изофлавоновые фитоэстрогены (примерно 25-250 мг), ресвератрол (примерно 100-1000 мг), андрографолид (примерно 100-500 мг), партенолид (примерно 0,1-50 мг), витамин A (примерно 5000-20000 МЕ), витамин C (примерно 100-2000 мг), кофермент Q10 (примерно 30-500 мг), витамин Е (примерно 10-1000 МЕ), α-липоевая кислота (примерно 10-1000 мг), кофермент Q10 (30-500 мг), марганец (примерно 1-100 мг), α-липоевая кислота, кофермент Q10 и витамин E (примерно 10-1000 мг, 30-500 мг, 10-1000 МЕ, соответственно), глибурид (примерно 1-20 мг) и производное глибурида, в котором отсутствует группа циклогексилмочевины (примерно 1-200 мг).
[00470] Согласно некоторым вариантам реализации ингибитор каспазы-1 представляет собой любое из низкомолекулярных соединений, описанных в патентах США 6355618; 6632962, 5756466 или международных заявках: WO2001/042216; WO2004/064713, WO98/16502, WO 97/24339, EP623592 и Dolle et al., J. Med. Chem. 39, 2438 (1996); Dolle et al., J. Med. Chem. 40, 1941 (1997), содержание каждого из которых полностью включено в настоящий документ посредством ссылки. Согласно некоторым вариантам реализации, как сообщалось, ингибитор каспазы-1 представляет собой непептидные ингибиторы каспазы-1. Патент США № (Bemis et al.);
[00471] Согласно некоторым вариантам реализации ингибитор каспазы-1 представляет собой ингибиторы ICE (каспазы-1), имеющие структуру:
где R1 представляет собой, помимо прочего, R3CO-, R3 представляет собой, помимо прочего, C1-C6 алкил, арил, гетероарил, -(CHR)n-арил и -(CHR)n-гетероарил, и R2 выбран из различных групп. Согласно некоторым вариантам реализации ингибитор каспазы-1 представляет собой ингибитор ICE (каспазы-1), имеющий структуру:
где R1 включает арил и гетероарил; А представляет собой аминокислоту; n равно 0-4; m равно 0 или 1; и R2 представляет собой арил. Согласно некоторым вариантам реализации ингибитор каспазы-1 представляет собой ингибиторы ICE (каспазы-1), имеющие структуру:
где R1 включает арил и гетероарил; AA1 и AA2 представляют собой одинарные связи или аминокислотные остатки; Tet представляет собой тетразольное кольцо; Z представляет собой алкилен, алкенилен, O, S и т.д.; и E представляет собой H, алкил и т.д.
[00472] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор каспазы-1 представляет собой ингибитор каспазы-1 на основе РНК, такой как киРНК, специфичная в отношении каспазы-1. Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор AIM2 на основе РНК кодируется зкДНК.
[00473] Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор каспазы-1 представляет собой ингибитор каспазы-1 на основе РНК, такой как киРНК, специфичная в отношении каспазы-1. Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор каспазы-1 на основе РНК кодируется зкДНК. Примеры последовательностей киРНК для каспазы-1, предусмотренных для применения в наборах и композициях, описанных в настоящем документе, раскрыты в WO2008/033285; Keller, M., et al. Cell. 2008; 132(5): 818-831; Artlett, C.M., et al. Arthritis and Rheumatology. 2011 Jul; 63 (11): 3563-3574; Burdette, D., et al. J Gen Virology. 2012, 93: 235-246, которые полностью включены в настоящий документ посредством ссылки. Последовательности киРНК для каспазы-1 также коммерчески доступны и известны обычным специалистам.
[00474] Белок каспазы-1 человека кодируется геном CASP1, содержащим последовательность нуклеиновой кислоты NM_033292.3 (SEQ ID NO: 611), и белок каспазы-1 человека имеет аминокислотную последовательность NP_150634.1 (SEQ ID NO: 612). Ингибиторы каспазы-1 дополнительно включают антисмысловые полинуклеотиды, которые можно применять для ингибирования транскрипции гена каспазы-1 и ингибирования посредством этого каспазы-1 и последующих путей инфламмасомы NLRP3 и инфламмасомы AIM2. Полинуклеотиды, которые комплементарные сегменту полинуклеотида, кодирующего каспазу-1 (например, полинуклеотида, представленного в SEQ ID NO: 611), разработаны для связывания с мРНК, кодирующей каспазу-1, и для ингибирования трансляции такой мРНК. Антисмысловые полинуклеотиды могут кодироваться зкДНК-вектором, раскрытым в настоящем документе, и необязательно могут быть функционально связаны с тканеспецифическим или индуцируемым промотором, раскрытым в настоящем документе.
[00475] Ингибирование мРНК каспазы-1 или прокаспазы-1 может быть выполнено с помощью молекул подавляющих ген РНКи в соответствии со способами, обычно известными специалистам в данной области техники. Например, олигонуклеотидные дуплексы подавляющих ген киРНК, специфично нацеленных на каспазу-1 человека (NM_033292.3), можно легко применять для подавления экспрессии прокаспазы-1. Нацеливание на мРНК каспазы-1 может быть успешно выполнено с использованием киРНК; и другие молекулы киРНК могут быть легко получены специалистами в данной области техники на основе известной последовательности мРНК-мишени. Соответственно, во избежание каких-либо сомнений, обычный специалист в данной области техники может разработать ингибиторы нуклеиновых кислот, такие как агенты РНКи (подавляющие РНК), для последовательности нуклеиновой кислоты NM_033292.3, которая представлена ниже:
1 atactttcag tttcagtcac acaagaaggg aggagagaaa agccatggcc gacaaggtcc
61 tgaaggagaa gagaaagctg tttatccgtt ccatgggtga aggtacaata aatggcttac
121 tggatgaatt attacagaca agggtgctga acaaggaaga gatggagaaa gtaaaacgtg
181 aaaatgctac agttatggat aagacccgag ctttgattga ctccgttatt ccgaaagggg
241 cacaggcatg ccaaatttgc atcacataca tttgtgaaga agacagttac ctggcaggga
301 cgctgggact ctcagcagat caaacatctg gaaattacct taatatgcaa gactctcaag
361 gagtactttc ttcctttcca gctcctcagg cagtgcagga caacccagct atgcccacat
421 cctcaggctc agaagggaat gtcaagcttt gctccctaga agaagctcaa aggatatgga
481 aacaaaagtc ggcagagatt tatccaataa tggacaagtc aagccgcaca cgtcttgctc
541 tcattatctg caatgaagaa tttgacagta ttcctagaag aactggagct gaggttgaca
601 tcacaggcat gacaatgctg ctacaaaatc tggggtacag cgtagatgtg aaaaaaaatc
661 tcactgcttc ggacatgact acagagctgg aggcatttgc acaccgccca gagcacaaga
721 cctctgacag cacgttcctg gtgttcatgt ctcatggtat tcgggaaggc atttgtggga
781 agaaacactc tgagcaagtc ccagatatac tacaactcaa tgcaatcttt aacatgttga
841 ataccaagaa ctgcccaagt ttgaaggaca aaccgaaggt gatcatcatc caggcctgcc
901 gtggtgacag ccctggtgtg gtgtggttta aagattcagt aggagtttct ggaaacctat
961 ctttaccaac tacagaagag tttgaggatg atgctattaa gaaagcccac atagagaagg
1021 attttatcgc tttctgctct tccacaccag ataatgtttc ttggagacat cccacaatgg
1081 gctctgtttt tattggaaga ctcattgaac atatgcaaga atatgcctgt tcctgtgatg
1141 tggaggaaat tttccgcaag gttcgatttt catttgagca gccagatggt agagcgcaga
1201 tgcccaccac tgaaagagtg actttgacaa gatgtttcta cctcttccca ggacattaaa
1261 ataaggaaac tgtatgaatg tctgtgggca ggaagtgaag agatccttct gtaaaggttt
1321 ttggaattat gtctgctgaa taataaactt ttttgaaata ataaatctgg tagaaaaatg
1381 aaaaaaaaaa aaa (SEQ ID NO: 611)
[00476] Согласно некоторым вариантам реализации ингибитор каспазы-1 представляет собой РНКи, которая комплементарна последовательности-мишени РНКи в NM_033292.3 (SEQ ID NO: 611); также обозначается как ген 834 NCBI (CASP1). В настоящее время транскрипты каспазы-1 дикого типа включают: NM_001223.4, NM_001257118.2, NM_001257119.2, NM_033292.3 (SEQ ID NO: 611), NM_033293.3, NM_033294.3, NM_033295.3, XM_017018393.1, XM_017018394.1, XM_017018395.1, XM_017018396.1. Агент РНКи, который ингибирует каспазу-1, может представлять собой нуклеиновую кислоту, комплементарную 17-21 последовательному основанию SEQ ID NO: 613-619, которые показаны в Таблице 5E.
[00477] Таблица 5E: Последовательности-мишени РНКи для ингибирования каспазы-1:
[00478] Согласно некоторым вариантам реализации ингибитор каспазы-1 представляет собой агент киРНК. Примерные последовательности киРНК, которые ингибируют каспазу-1, показаны в Таблице 5F.
[00479] Таблица 5F: Примерные киРНК, которые ингибируют каспазу-1
[00480] Согласно некоторым вариантам реализации ингибитор каспазы-1 представляет собой киРНК, ингибируя посредством этого мРНК каспазы-1 (или пробелка прокаспазы-1), что приводит к ингибированию последующих путей инфламмасомы NLRP3 и/или инфламмасомы AIM2. Согласно некоторым вариантам реализации ингибитор каспазы-1 представляет собой GAA GGC CCA UAU AGA GAA A (SEQ ID NO: 904; показана последовательность смысловой цепи), который ингибирует экспрессию каспазы-1 человека или ее фрагмента или гомолога, который по меньшей мере на 50% или по меньшей мере на 60%, или по меньшей мере на 70%, или по меньшей мере на 80%, или по меньшей мере на 90% идентичен ей. Примеры последовательностей киРНК для каспазы-1, предусмотренных для применения в наборах и композициях, описанных в настоящем документе, раскрыты в WO 2008/033285 или заявке США US20090280058, Keller, M., et al. Cell. 2008; 132(5): 818-831; Artlett, C.M., et al. Arthritis and Rheumatology. 2011 Jul; 63 (11): 3563-3574; Burdette, D., et al. J Gen Virology. 2012, 93: 235-246; которые полностью включены в настоящий документ посредством ссылки.
[00481] Изготовленные на заказ киРНК для NLRP3, AIM2 и каспазы-1 могут быть созданы по заказу в Dharmacon Research, Inc., Лафайет, Колорадо. Другие источники для изготовления на заказ киРНК включают Xeragon Oligonucleotides, Хантсвилл, Алабама, и Ambion из Остина, Техас. В качестве альтернативы, киРНК могут быть синтезированы химическим путем с использованием фосфоамидитов рибонуклеозидов и синтезатора ДНК/РНК. Согласно некоторым вариантам реализации РНКи или киРНК для NLRP3, AIM2 и каспазы-1 могут кодироваться в зкДНК, раскрытых в настоящем документе.
[00482] Согласно некоторым вариантам реализации ингибитор каспазы-1 представляет собой субстрат каспазы-1 (CAS 143305-11-7), имеющий структуру:
и который имеет следующую последовательность: Asn-Glu-Ala-Tyr-Val-His-Asp-Ala-Pro-Val-Arg-Ser-Leu-Asn (SEQ ID NO: 538). Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор каспазы-1 кодируется зкДНК, вводимой субъекту (включая, например, последующую доставку зкДНК). Согласно некоторым вариантам реализации композиций и способов, описанных в настоящем документе, ингибитор каспазы-1, кодируемый зкДНК, вводимой субъекту, представляет собой субстрат каспазы-1 (SEQ ID NO: 538).
[00483] РНКи может быть разработана для нацеливания на различные мРНК. Общая стратегия разработки РНКи, например, киРНК, включает начало со стоп-кодона AUG и затем сканирование длины целевой кДНК-мишени для поиска динуклеотидных последовательностей АА. 19 3´-концевых нуклеотидов, примыкающих к последовательностям АА, как сообщалось, являются потенциальными сайтами-мишенями киРНК. Затем потенциальные сайты-мишени сравнивали с соответствующей базой данных генома так, что могли быть отброшены любые последовательности-мишени, которые имеют значительную гомологию с нецелевыми генами. Было локализовано множество последовательностей-мишеней по длине гена, поэтому последовательности-мишени происходили из 3´, 5´ и медиальной частей мРНК. КиРНК для отрицательного контроля получали с использованием той же нуклеотидной композиции, что и киРНК согласно настоящему изобретению, но скремблировали и проверяли на отсутствие гомологии последовательностей с какими-либо генами трансфицируемых клеток. (Elbashir, S. M., et al., 2001, Nature, 411, 494-498; Ambion siRNA Design Protocol, at www.ambion.com).
[00484] Последовательности-мишени могут иметь 17-25 оснований в длину, и оптимально 21 основание в длину, начиная с AA. РНКи или киРНК, которые связываются с последовательностями-мишенями, были модифицированы тиоловой группой у 5 атома углерода С6 на одной цепи.
VII. Способы применения
[00485] ЗкДНК-вектор для экспрессии, например, ингибитора иммунного ответа (например, врожденного иммунного ответа), раскрытый в настоящем документе, также можно применять в способе доставки представляющей интерес нуклеотидной последовательности (например, кодирующей ингибитор врожденного иммунного ответа) в клетку-мишень (например, клетку-хозяина). Способ может представлять собой, в частности, способ доставки ингибитора иммунного ответа (например, врожденного иммунного ответа) в клетку субъекта, который нуждается в этом, и лечения иммунного нарушения, или ослабления или подавления врожденной иммунной системы. Настоящее изобретение обеспечивает экспрессию in vivo ингибитора иммунного ответа (например, врожденного иммунного ответа), закодированного в зкДНК-векторе, в клетке у субъекта так, чтобы возник терапевтический эффект экспрессии антагониста инфламмасомы. Эти результаты наблюдаются как в in vivo, так и в in vitro режимах доставки зкДНК-вектора.
[00486] Кроме того, согласно настоящему изобретению предложен способ доставки ингибитора иммунного ответа (например, врожденного иммунного ответа), например, в клетку субъекта, нуждающегося в этом, включающий многократные введения зкДНК-вектора согласно настоящему изобретению, кодирующего указанный антагонист инфламмасомы. Поскольку зкДНК-вектор согласно настоящему изобретению не индуцирует иммунный ответ, подобный тому, который обычно наблюдается против инкапсидированных вирусных векторов, такая стратегия многократного введения, вероятно, будет иметь больший успех в системе на основе зкДНК. ЗкДНК-вектор вводят в количествах, достаточных для трансфекции клеток целевой ткани и обеспечения достаточных уровней переноса генов и экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), например, без неоправданных нежелательных эффектов. Обычные и фармацевтически приемлемые пути введения включают, но не ограничиваются перечисленными, введение в сетчатку (например, субретинальную инъекцию, супрахориоидальную инъекцию или интравитреальную инъекцию), внутривенное введение (например, в липосомальном составе), прямую доставку в выбранный орган (например, в любую одну или более тканей, выбранных из: печени, почек, желчного пузыря, предстательной железы, надпочечников, сердца, кишечника, легких и желудка), внутримышечный и другие парентеральные пути введения. Пути введения могут быть скомбинированы, если требуется.
[00487] Доставка зкДНК-вектора для экспрессии, например, ингибитора иммунного ответа (например, врожденного иммунного ответа), описанного в настоящем документе, не ограничивается доставкой экспрессируемого ингибитора. Например, зкДНК-векторы, полученные с помощью обычного способа (например, с использованием способа продуцирования на основе клеток (например, способов продуцирования в клетках насекомых)) или синтетического способа, как описано в настоящем документе, можно применять с другими предложенными системами доставки, чтобы обеспечить часть генной терапии. Один неограничивающий пример системы, которую можно комбинировать с зкДНК-векторами в соответствии с настоящим раскрытием, включает системы, которые доставляют по отдельности один или более кофакторов или иммунных супрессоров для эффективной генной экспрессии зкДНК-вектора, экспрессирующего ингибитор.
[00488] Согласно настоящему изобретению также предложен способ подавления у субъекта иммунного ответа, например, врожденного иммунного ответа, включающий введение в нуждающуюся в этом целевую клетку (в частности, в мышечную клетку или ткань) указанного субъекта терапевтически эффективного количества зкДНК-вектора, необязательно с фармацевтически приемлемым носителем. Несмотря на то, что указанный зкДНК-вектор может быть введен в присутствии носителя, такой носитель не является необходимым. Выбранный зкДНК-вектор содержит нуклеотидную последовательность, кодирующую ингибитор иммунного ответа (например, врожденного иммунного ответа), например, который можно применять для лечения или подавления иммунной системы. В частности, указанный зкДНК-вектор может содержать целевую последовательность антагониста инфламмасомы, функционально связанную с контрольными элементами, способными управлять транскрипцией целевого антагониста инфламмасомы, кодируемого указанной экзогенной последовательностью ДНК, при введении субъекту. Указанный зкДНК-вектор может быть введен любым подходящим путем, как описано выше и в других разделах настоящего документа.
[00489] Композиции и векторы, предложенные в настоящем документе, можно применять для доставки ингибитора иммунного ответа (например, врожденного иммунного ответа), например, для различных целей. Согласно некоторым вариантам реализации трансген кодирует ингибитор иммунного ответа (например, врожденного иммунного ответа), который предназначен для применения в исследовательских целях, например, для создания соматической трансгенной модели на животных, несущей трансген, например, для исследования функции ингибитора иммунного ответа (например, врожденного иммунного ответа). В другом примере трансген кодирует ингибитор иммунного ответа (например, врожденного иммунного ответа), который предназначен для применения в создании модели подавленной иммунной системы на животных или у субъекта с ослабленным иммунитетом. Согласно некоторым вариантам реализации кодируемый ингибитор иммунного ответа (например, врожденного иммунного ответа) можно применять для лечения или предотвращения повышенных иммунных ответов или состояния повышенной реактивности врожденного иммунитета у субъекта, например, в ответ на генную терапию или подобное, у субъекта-млекопитающего. Ингибитор иммунного ответа (например, врожденного иммунного ответа) может быть перенесен пациенту (например, экспрессирован у пациента) в количестве, достаточном для уменьшения или предотвращения повышенных иммунных ответов у субъекта.
[00490] ЗкДНК-вектор не ограничивается одной молекулой зкДНК-вектора. Таким образом, в другом аспекте, несколько зкДНК-векторов, экспрессирующих разные белки или одни и те же ингибиторы иммунного ответа (например, врожденного иммунного ответа), например, но функционально связанные с различными промоторами или цис-регуляторными элементами, могут быть доставлены одновременно или последовательно в целевую клетку, ткань, орган или в организм субъекта. Таким образом, эта стратегия может обеспечить одновременную генную терапию несколькими антагонистами инфламмасом или доставку их генов. Также можно разместить разные части ингибитора в отдельных зкДНК-векторах (например, разные домены и/или кофакторы, необходимые для функциональности ингибитора иммунного ответа (например, врожденного иммунного ответа)), например, которые могут быть введены одновременно или в разные моменты времени, и которые можно регулировать по отдельности, добавляя посредством этого дополнительный уровень контроля экспрессии одного и более ингибиторов. Доставку также можно выполнять несколько раз и, что важно для генной терапии в клинических условиях, с последующим увеличением или снижением доз, учитывая отсутствие иммунного ответа хозяина против капсида благодаря отсутствию вирусного капсида. Ожидается, что ответ против капсида не возникнет из-за отсутствия капсида.
[00491] Согласно настоящему изобретению также предложен способ подавления у субъекта иммунного ответа, например, врожденного иммунного ответа, включающий введение в нуждающуюся в этом целевую клетку (в частности, в мышечную клетку или ткань) указанного субъекта терапевтически эффективного количества зкДНК-вектора, раскрытого в настоящем документе, необязательно с фармацевтически приемлемым носителем. Несмотря на то, что указанный зкДНК-вектор может быть введен в присутствии носителя, такой носитель не является необходимым. Реализованный зкДНК-вектор содержит представляющую интерес нуклеотидную последовательность, например, ингибитор иммунного ответа, который можно применять для подавления врожденной иммунной системы или ослабления состояния повышенной иммунореактивности у субъекта. В частности, зкДНК-вектор может содержать целевую экзогенную последовательность ДНК, функционально связанную с контрольными элементами, способными управлять транскрипцией целевого полипептида, белка или олигонуклеотида, кодируемого указанной экзогенной последовательностью ДНК, при введении субъекту. ЗкДНК-вектор может быть введен любым подходящим путем, как описано выше и в других разделах настоящего документа.
Лечение ex vivo
[00492] Согласно некоторым вариантам реализации клетки извлекают из организма субъекта, вводят в них зкДНК-вектор для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), например, раскрытый в настоящем документе, и затем клетки возвращают в организм субъекта. Способы извлечения клеток из организма субъекта для лечения ex vivo с последующим введением обратно в организм субъекта известны в данной области техники (см., например, патент США № 5399346; раскрытие которого полностью включено в настоящий документ). В качестве альтернативы, зкДНК-вектор вводят в клетки, полученные от другого субъекта, в культивируемые клетки или в клетки из любого другого подходящего источника, и указанные клетки вводят нуждающемуся в этом субъекту.
[00493] Клетки, трансдуцированные зкДНК-вектором для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), например, раскрытым в настоящем документе, предпочтительно вводят субъекту в «терапевтически эффективном количестве» в комбинации с фармацевтическим носителем. Специалисты в данной области техники поймут, что терапевтические эффекты не обязательно должны быть полными или обеспечивать исцеление при условии, что для субъекта они в какой-то мере благоприятны.
[00494] Согласно некоторым вариантам реализации зкДНК-вектор для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), например, раскрытый в настоящем документе, может кодировать антагонист инфламмасомы, описанный в настоящем документе (иногда называемый трансгеном или гетерологичной нуклеотидной последовательностью), который должен продуцироваться в клетке in vitro, ex vivo или in vivo. Например, в отличие от применения зкДНК-векторов, описанных в настоящем документе, в способе лечения, который обсуждается в настоящем документе, в некоторых вариантах реализации зкДНК-вектор для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа) может быть введен в культивируемые клетки, и экспрессированный антагонист инфламмасомы может быть выделен из клеток, например, для получения антител и слитых белков. Согласно некоторым вариантам реализации культивируемые клетки, содержащие зкДНК-вектор для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), раскрытый в настоящем документе, можно применять для коммерческого получения антител или слитых белков, например, в качестве источника клеток для мелкомасштабного или крупномасштабного биоизготовления антител или слитых белков. Согласно альтернативным вариантам реализации зкДНК-вектор для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), раскрытый в настоящем документе, вводят в клетки у субъекта-хозяина, не являющегося человеком, для продуцирования антител или слитых белков in vivo, включая мелкомасштабное получение, а также коммерческое крупномасштабное получение антагониста инфламмасомы.
[00495] ЗкДНК-векторы для экспрессии ингибитора иммунного ответа (например, врожденного иммунного ответа), раскрытые в настоящем документе, можно применять как в ветеринарных, так и в медицинских приложениях. Подходящие субъекты для способов доставки генов ex vivo, описанных выше, включают как птиц (например, кур, уток, гусей, перепелов, индеек и фазанов), так и млекопитающих (например, человека, крупный рогатый скот, овечьих, козьих, лошадиных, кошачьих, собачьих и зайцеобразных), при этом предпочтительными являются млекопитающие. Субъекты-люди являются наиболее предпочтительными. Субъекты-люди включают новорожденных, младенцев, детей и подростков, и взрослых людей.
[00496] Все патенты и другие публикации; включая литературные ссылки, выданные патенты, опубликованные заявки на патент и заявки на патент, находящиеся на рассмотрении одновременно с настоящей заявкой; упомянутые в настоящем описании, явным образом включены в настоящий документ посредством ссылки с целью описания и раскрытия, например, методологий, описанных в таких публикациях, которые могут быть использованы в связи с технологией, описанной в настоящем документе. Эти публикации представлены исключительно по причине их раскрытия до даты подачи настоящей заявки. Ничто в этом отношении не должно истолковываться как признание того, что авторы изобретения не обладают правами на более раннее изобретение по отношению к такому раскрытию в силу предшествующего изобретения или по любой другой причине. Все утверждения, касающиеся дат, или представления относительно содержания этих документов основаны на информации, доступной заявителям, и не являются допущением каким-либо образом признания правильности дат или содержания этих документов.
[00497] Описание вариантов реализации настоящего раскрытия не должно рассматриваться как исчерпывающее или ограничивающее настоящее изобретение точной раскрытой формой. Несмотря на то, что в настоящем документе описаны в иллюстративных целях конкретные варианты реализации и примеры раскрытия, возможны различные эквивалентные модификации в пределах объема раскрытия, как будет понятно специалистам в соответствующей области техники. Например, хотя стадии или функции способа представлены в указанном порядке, согласно альтернативным вариантам реализации функции могут выполняться в другом порядке, или функции могут выполняться по существу одновременно. Принципиальные положения настоящего раскрытия, предоставленные в настоящем документе, могут быть применены к другим процедурам или способам в зависимости от ситуации. Различные варианты реализации, описанные в настоящем документе, могут быть объединены для обеспечения дополнительных вариантов реализации. Аспекты настоящего раскрытия могут быть модифицированы, при необходимости, для применения составов, функций и концепций вышеупомянутых ссылок и применения для обеспечения дополнительных вариантов реализации настоящего раскрытия. Кроме того, из соображений биологической функциональной эквивалентности могут быть сделаны некоторые изменения в структуре белка, не влияющие на биологическое или химическое действие в качественном или количественном отношении. Эти и другие изменения могут быть выполнены в раскрытии в свете подробного описания. Все такие модификации предусмотрены, как включенные в объем прилагаемой формулы изобретения.
[00498] Конкретные элементы любого из вышеприведенных вариантов реализации могут быть объединены или заменены элементами других вариантов реализации. Кроме того, хотя преимущества, связанные с некоторыми вариантами реализации настоящего раскрытия, были описаны в контексте этих вариантов реализации, другие варианты реализации также могут демонстрировать такие преимущества, и не все варианты реализации должны обязательно демонстрировать такие преимущества, чтобы попадать в объем раскрытия.
[00499] Описанная в настоящем документе технология дополнительно иллюстрируется следующими примерами, которые никоим образом не следует истолковывать как дополнительные ограничения. Следует понимать, что настоящее изобретение не ограничено конкретной методологией, протоколами и реагентами, и т.п., описанными в настоящем документе, и потому допускает варианты. Терминология, используемая в настоящем документе, предназначена исключительно для описания конкретных вариантов реализации, и не предназначена для ограничения объема настоящего изобретения, который определен исключительно формулой изобретения.
ПРИМЕРЫ
ПРИМЕР 1: Конструирование ТТХ-плазмид
[00500] Получали плазмиды в формате ТТХ, структурная схема которых показана на ФИГ. 4C (TTX-R) или ФИГ. 4D (TTX-L). Примеры плазмид TTX-R и TTX-L описаны в Таблице 6A ниже. Плазмиды TTX-R и TTX-L различаются положением мутированной последовательности ITR ААВ2, как показано на ФИГ. 4C и ФИГ. 4D, соответственно. Плазмиды ТТХ-R (ТТХ-плазмида 1, 3, 5 и 7) получали путем молекулярного клонирования, раскрытого в настоящем документе, с получением ТТХ-векторов. Плазмиды ТТХ-L (ТТХ-плазмида 2, 4, 6 и 8) для применения в получении ТТХ-векторов (ТТХ-вектор 2, 4, 6, 8). Каждая из плазмид TTX-R содержит (а) инвертированный концевой повтор дикого типа (ITR) ААВ2; (b) экспрессионную кассету и (c) модифицированный инвертированный концевой повтор (ITR) ААВ2, как проиллюстрировано на ФИГ. 4D.
[00501] Плазмиды зкДНК (т.е. плазмиды, содержащие матрицу зкДНК-вектора, используемую для последующего получения зкДНК-вектора) могут быть сконструированы с использованием известных методик, чтобы предпочтительно обеспечить по меньшей мере следующие элементы в виде функционально связанных компонентов в направлении транскрипции: 5´-ITR (мутантный или дикого типа ААВ); контрольные элементы, включая промотор, представляющую интерес экзогенную последовательность ДНК; область терминации транскрипции; и 3´-ITR (мутантный или дикий тип соответствующего ITR ААВ). Примечательно, что нуклеотидные последовательности в ITR по существу заменяют области, кодирующие rep и cap. Несмотря на то, что последовательности rep идеально кодируются вспомогательной плазмидой или вектором, альтернативно они могут быть перенесены самой векторной плазмидой. В таких случаях последовательности rep предпочтительно расположены вне области, помещенной между ITR, но также могут быть расположены внутри области, помещенной между ITR. Целевая экзогенная последовательность ДНК функционально связана с контрольными элементами, которые управляют транскрипцией или экспрессией кодируемого полипептида, белка или их олигонуклеотида в клетке, ткани, органе или в организме субъекта (т.е. in vitro, ex vivo или in vivo). Такие контрольные элементы могут содержать контрольные последовательности, обычно ассоциированные с выбранным геном. В качестве альтернативы, можно применять гетерологичные контрольные последовательности. Подходящие гетерологичные контрольные последовательности обычно включают те, которые происходят из последовательностей, кодирующих гены млекопитающих или вирусов.
[00502] Целевая экзогенная последовательность ДНК в зкДНК-векторе может быть функционально связана с контрольными элементами, которые управляют транскрипцией или экспрессией кодируемого полипептида, белка или их олигонуклеотида в клетке, ткани, органе или в организме субъекта (т.е. in vitro, ex vivo или in vivo). Такие контрольные элементы могут содержать контрольные последовательности, обычно ассоциированные с выбранным геном. В качестве альтернативы, можно применять гетерологичные контрольные последовательности. Подходящие гетерологичные контрольные последовательности обычно включают те, которые происходят из последовательностей, кодирующих гены млекопитающих или вирусов. Примеры включают, но не ограничиваются перечисленными, промоторы, такие как ранний промотор SV40; промотор из длинного концевого повтора (LTR) вируса опухоли молочной железы мышей; большой поздний промотор аденовируса (Ad MLP); промоторы вируса простого герпеса (HSV); промотор цитомегаловируса (CMV), такой как область немедленного раннего промотора CMV (CMVIE); промотор вируса саркомы Рауса (RSV); синтетические промоторы; гибридные промоторы; и тому подобное. Кроме того, в настоящем изобретении также можно применять последовательности, происходящие из невирусных генов, таких как ген металлотионеина мыши. Известны последовательности ITR многих серотипов ААВ.
[00503] Экспрессионная кассета каждой из TTX-плазмид (как TTX-R, так и TTX-L) включает следующие элементы, расположенные между последовательностями ITR: (i) энхансер/промотор; (ii) сайт клонирования для трансгена; (iii) посттранскрипционный элемент ответа WHP (WPRE); и (iv) сигнал полиаденилирования из гена бычьего гормона роста (BGHpA). Между каждым компонентом также были введены уникальные сайты распознавания рестрикционных эндонуклеаз (R1-6) (например, см. ФИГ. 4C и ФИГ. 4D), чтобы облегчить введение новых генетических компонентов в конкретные сайты в конструкции. Сайты ферментов R3 и R4 конструируют в сайте клонирования для введения открытой рамки считывания трансгена. Эти последовательности клонировали в плазмиду pFastBac HT B, полученную от ThermoFisher Scientific.
[00504] Все TTX-плазмиды дополнительно содержат экзогенную последовательность, открытая рамка считывания которой для трансгена (люциферазы светлячка или «Luc» либо фактора IX человека или «FIX») также была получена путем вставки экзогенной последовательности в сайт клонирования. Каждую из структур нескольких примеров ТТХ-плазмид, представленных в Таблице 6А, конструировали по схеме, показанной на ФИГ. 4D (правый мутированный ITR ААВ) или ФИГ. 4C (левый мутированный ITR). Каждая TTX-плазмида включала энхансер/промотор и трансген (например, люциферазу с различными промоторами или FIX с промотором CAG), посттрансляционный регуляторный элемент (WPRE) и терминационный сигнал полиаденилирования (поли(А) BGH), фланкированные: (a) полинуклеотидной последовательностью мутированного инвертированного концевого повтора (ITR) ААВ2, кодируемой в плазмиде либо на левой (L), либо на правой (R) стороне экспрессионной кассеты, и (b) последовательностью ITR ААВ2 дикого типа (немутированного) на противоположном конце экспрессионной кассеты.
[00505] ТТХ-плазмиды в Таблице 6А конструировали с WPRE, содержащим SEQ ID NO: 8, и BGHpA, содержащим SEQ ID NO: 9, в качестве компонентов между трансгеном люциферазы и правым ITR. Кроме того, каждая из TTX-плазмид (от TTX-1 до TTX-10) также содержала сайт клонирования R3/R4 (SEQ ID NO: 7) по обе стороны от ORF репортерной последовательности люциферазы или фактора IX (Padua FIX из SEQ ID NO: 12 или FIX из SEQ ID NO: 11).
[00506] Касательно Таблицы 6A:
• «wt-L» относится к ITR ААВ2 дикого типа, кодируемому в плазмиде на левой стороне экспрессионной кассеты (содержащему полинуклеотидную последовательность SEQ ID NO: 51);
• «wt-R» относится к ITR ААВ2 дикого типа, кодируемому в плазмиде на правой стороне экспрессионной кассеты (содержащему полинуклеотидную последовательность SEQ ID NO: 1);
• «mut-L» относится к мутированной последовательности ITR ААВ2, представленной в SEQ ID NO: 52;
• «mut-R» относится к мутированной последовательности ITR ААВ2, представленной в SEQ ID NO: 2;
• «CAG» относится к синтетическому промотору, сконструированному из (C) элементов немедленного раннего энхансера и промотора цитомегаловируса, (A) первого экзона и первого интрона гена бета-актина курицы, (G) акцептора сплайсинга гена бета-глобина кролика, из SEQ ID NO: 3;
• «AAT w/SV40 intr» относится к AAT (альфа-1-антитрипсину человека) с интроном большого Т-антигена SV40 из SEQ ID NO: 4; и
• «hEF1-α» относится к фактору удлинения-1 альфа (EF-1 альфа) человека из SEQ ID NO: 6.
Таблица 6A
Таблица 6B
Каждая конструкция в Таблице 6B содержит модифицированную последовательность поли(A) SV40 (SEQ ID NO: 10), расположенную в 3´-нетранслируемой области (UTR) между трансгеном и mut-R ITR.
«LP-1 β» относится к промотору LP-1 β (SEQ ID NO: 16), который аналогичен промотору LP-1 (SEQ ID NO: 5) с 2 дополнительными сайтами рестрикционного фермента.
[00507] Согласно одному варианту реализации векторный полинуклеотид (зкДНК-вектор) содержит пару двух разных ITR, выбранных из группы, состоящей из: SEQ ID NO: 1 и SEQ ID NO: 52; и SEQ ID NO: 2 и SEQ ID NO: 51. Согласно одному варианту реализации каждого из этих аспектов векторный полинуклеотид или невирусные бескапсидные ДНК-векторы с ковалентно замкнутыми концами содержит (ат) пару ITR, выбранных из группы, состоящей из: SEQ ID NO: 101 и SEQ ID NO: 102; SEQ ID NO: 103 и SEQ ID NO: 104, SEQ ID NO: 105 и SEQ ID NO: 106; SEQ ID NO: 107 и SEQ ID NO: 108; SEQ ID NO: 109 и SEQ ID NO: 110; SEQ ID NO: 111 и SEQ ID NO: 112; SEQ ID NO: 113 и SEQ ID NO: 114; и SEQ ID NO: 115 и SEQ ID NO: 116. Согласно некоторым вариантам реализации зкДНК-векторы не имеют ITR, который содержит какую-либо последовательность, выбранную из SEQ ID NO: 500-529.
ПРИМЕР 2: Бакмида и бакуловирус для получения линейных, непрерывных и бескапсидных ДНК-векторов
[00508] Компетентные клетки DH10Bac (компетентные клетки MAX Efficiency® DH10Bac™, Thermo Fisher, номер по каталогу 10361012) трансформировали либо ТТХ, либо контрольными плазмидами в соответствии с протоколом, предоставленным поставщиком, доступным на их веб-сайте (Thermo Fisher, можно найти в Интернете по адресу: https://www.thermofisher.com/order/catalog/product/10361012). Чтобы получить рекомбинантные бакмиды («TTX-бакмиды»), в клетках DH10Bac индуцировали рекомбинацию между плазмидой и бакуловирусным челночным вектором. Рекомбинантные бакмиды отбирали путем положительного отбора на основании скрининга по сине-белому окрашиванию E. coli (маркер Φ80dlacZΔM15 обеспечивает α-комплементацию гена β-галактозидазы из бакмидного вектора) на чашке с бактериальным агаром, содержащим X-gal и IPTG. Белые колонии собирали и культивировали в 10 мл сред.
[00509] Рекомбинантные бакмиды («TTX-бакмиды») выделяли из E. coli и трансфицировали в клетки насекомых Sf9 или Sf21 с использованием FugeneHD™ с получением инфекционного бакуловируса. Адгезивные клетки насекомых Sf9 или Sf21 культивировали в 50 мл сред в колбах T25 при 25°С. Через четыре дня культуральную среду (содержащую вирус P0) удаляли от клеток, фильтровали через фильтр с размером пор 0,45 мкм и отделяли инфекционные рекомбинантные бакуловирусные частицы («TTX-бакуловирус» или «сравнительный бакуловирус») от клеток в культуре.
[00510] Необязательно первое поколение бакуловируса (P0) амплифицировали путем инфицирования наивных клеток насекомых Sf9 или Sf21 в 50-500 мл сред. Клетки культивировали при 130 об/мин и 25°С, отслеживая диаметр и жизнеспособность клеток до тех пор, пока клетки не достигали диаметра 18-19 нм (в отличие от диаметра наивных клеток, равного 14-15 нм) и плотности ~4,0×106 клеток/мл. Через 3-8 дней после инфицирования бакуловирусные частицы P1 в среде собирали после центрифугирования для удаления клеток и клеточных фрагментов с последующей фильтрацией через фильтр с размером пор 0,45 мкм.
[00511] Собирали TTX-бакуловирус и определяли инфекционную активность бакуловируса. В частности, 4×20 мл культур клеток Sf9 при 2,5×106 клеток/мл обрабатывали бакуловирусом P1 в следующих разведениях 1/1000, 1/10000, 1/50000, 1/100000 и инкубировали. Инфективность определяли ежедневно на протяжении 4-5 дней по показателю увеличения диаметра клеток и остановке клеточного цикла, а также изменению жизнеспособности клеток.
[00512] Последовательность Rep 78 (SEQ ID NO: 13) функционально связывали с фрагментом промотора IE1 (SEQ ID NO: 15), а затем вставляли в сайт рестрикции BamHI/KpnI вектора для двойной экспрессии pFASTBAC™ (ThermoFisher, номер по каталогу 10712024) так, что последовательность Rep 78 связана с последовательностью поли(A) TK HSV на 3´-конце. Последовательность Rep 52 (SEQ ID NO: 14) затем клонировали в сайт SalI-HindIII вектора, чтобы функционально связать последовательность Rep52 с промотором pPH на 5´ и последовательностью поли(A) SV40 на 3´. Полученная конструкция называется в настоящем документе «Rep-плазмидой».
[00513] Rep-плазмиду трансформировали в компетентные клетки DH10Bac (компетентные клетки MAX Efficiency® DH10Bac™, Thermo Fisher, номер по каталогу 10361012) в соответствии с протоколом, предоставленным поставщиком, доступным на их веб-сайте (Thermo Fisher®, https://www.thermofisher.com/order/catalog/product/10361012). Для получения рекомбинантных бакмид («Rep-бакмид») в клетках DH10Bac индуцировали рекомбинацию между Rep-плазмидой и бакуловирусным челночным вектором. Рекомбинантные бакмиды отбирали путем положительного отбора на основании скрининга по сине-белому окрашиванию E. coli (маркер Φ80dlacZΔM15 обеспечивает α-комплементацию гена β-галактозидазы из бакмидного вектора) на чашке с бактериальным агаром, содержащим X-gal и IPTG. Одиночные белые колонии собирали и инокулировали в 10 мл селективных сред (канамицин, гентамицин, тетрациклин в бульоне LB). Рекомбинантные бакмиды (Rep-бакмиды) выделяли из E.coli, и трансфицировали клетки насекомых Sf9 или Sf21 указанными Rep-бакмидами, чтобы получить инфекционный бакуловирус.
[00514] Клетки насекомых Sf9 или Sf21 культивировали в 50 мл сред в течение 4 дней и выделяли из культуры инфекционный рекомбинантный бакуловирус («Rep-бакуловирус»). Необязательно Rep-бакуловирус первого поколения (P0) амплифицировали путем инфицирования наивных клеток насекомых Sf9 или Sf21 и культивировали в 50-500 мл сред. Через 3-8 дней после инфицирования бакуловирусные частицы P1 в среде собирали, отделяя клетки путем центрифугирования или фильтрации, или используя другой способ фракционирования. Собирали Rep-бакуловирус и определяли инфекционную активность бакуловируса. В частности, 4×20 мл культур клеток Sf9 при 2,5×106 клеток/мл обрабатывали бакуловирусом P1 в следующих разведениях: 1/1000, 1/10000, 1/50000, 1/100000 и инкубировали. Инфективность определяли ежедневно на протяжении 4-5 дней по показателю увеличения диаметра клеток и остановке клеточного цикла, а также изменению жизнеспособности клеток.
[00515] Среды для культивирования клеток Sf, содержащие либо (1) ТТХ или α (альфа)-бакуловирус, либо (2) Rep-бакуловирус, описанный выше, затем добавляли к свежей культуре клеток Sf9 (2,5×106 клеток/мл, 20 мл) при соотношении 1:1000 и 1:10000, соответственно. Затем клетки культивировали при 130 об/мин при 25°С. Через 4-5 дней после коинфицирования детектировали диаметр и жизнеспособность клеток. Когда диаметр клеток достигал 18-20 нм при жизнеспособности ~70-80%, клеточные культуры центрифугировали, среду удаляли и собирали клеточные осадки. Клеточные осадки сначала ресуспендировали в достаточном объеме водной среды, либо воды, либо буфера. ТТХ или α (альфа)-векторы выделяли и очищали из клеток с использованием протокола очистки Qiagen Midi Plus (Qiagen, номер по каталогу 12945, 0,2 мг массы осадка клеток, обработанной на колонку).
[00516] Значения выхода ДНК-векторов (например, TTX-векторов), полученных и очищенных из клеток насекомых Sf9, первоначально определяли на основании поглощения в УФ-диапазоне при 260 нм. Значения выхода различных TTX-ДНК векторов, определенные на основании поглощения в УФ-диапазоне, представлены в Таблице 7 ниже.
Таблица 7
(Диаметр в микрометрах)
Жизнеспособность: 53,3%
Диаметр: 18,4 мкм
Жизнеспособность: 81,8%
Диаметр: 18,7 мкм
Жизнеспособность: 74,5%
Диаметр: 18,5 мкм
Жизнеспособность: 84,4%
Диаметр: 19,7 мкм
Жизнеспособность: 77,4%
Диаметр: 18,6 мкм
ПРИМЕР 3: Денатурирующий гель-электрофорез для идентификации продукции зкДНК-вектора
[00517] Чтобы качественным образом продемонстрировать, что выделенный материал ДНК-векторов имеет ковалентно замкнутые концы, как требует их определение, образцы расщепляют рестрикционной эндонуклеазой, идентифицированной на основании последовательности ДНК-вектора как эндонуклеаза с одним сайтом рестрикции, что предпочтительно приводит к получению двух продуктов расщепления неравного размера (пример: 1000 п.о. и 2000 п.о.). После расщепления и электрофореза на денатурирующем геле (который разделяет две комплементарные цепи ДНК) линейная, незамкнутая ковалентно ДНК будет разделяться на фрагменты размером 1000 п.о. и 2000 п.о., в то время как ковалентно замкнутая ДНК будет разделяться на фрагменты в 2 раза большего размера (2000 п.о. и 4000 п.о.), так как две цепи ДНК связаны, уже развернуты и имеют вдвое большую длину (но одноцепочечные). Кроме того, при расщеплении мономерных, димерных и n-мерных форм ДНК-вектора все они будут разделяться на фрагменты одинакового размера из-за связанных между собой концов мультимерного ДНК-вектора (см. ФИГ. 5B).
[00518] В настоящем документе выражение «Анализ для идентификации ДНК-вектора с помощью электрофореза в агарозном геле в условиях нативного и денатурирующего геля» относится к следующему анализу. В качестве рестрикционной эндонуклеазы выбирают фермент, выполняющий единственный разрез, чтобы получить продукты, длина которых составляет приблизительно 1/3× и 2/3× от длины ДНК-вектора. Это обеспечивает разделение полос как на нативном, так и на денатурирующем гелях. Перед денатурацией важно удалить буфер из образца. Набор для очистки ПЦР Qiagen (номер по каталогу Qiagen 28104) или «центрифужные колонки» для обессоливания, например, колонки GE HealthCare Ilustra™ MicroSpin™ G-25 (GE Healthcare, номер по каталогу 27532501), хорошо работают с расщеплением эндонуклеазами.
1. Расщепите ДНК соответствующей рестрикционной эндонуклеазой (ами)
2. Внесите в набор для очистки ПЦР Qiagen, элюируйте dH2O (30 мкл)
3. Добавьте 4 мкл 10-кратного денатурирующего раствора (10× = 0,5 М NaOH, 10 мМ ЭДТА)
4. Добавьте 6 мкл 10-кратного раствора для загрузки в гель (краситель плюс глицерин или фиколл, НЕ забуференный)
5. Маркеры молекулярной массы ДНК могут быть приготовлены без набора Qiagen путем добавления 10-кратного денатурирующего раствора до конечной концентрации 4×.
6. Подготовьте 0,8-1,0% гель в H2O в микроволновой печи до кипения, оставьте при температуре окружающей среды на несколько минут.
7. Вылейте в лоток для геля с гребенкой и поместите в холодную комнату для ускорения полимеризации (2 часа)
8. Поместите лоток в бокс для электрофореза и уравновесьте 1 мМ ЭДТА и 200 мМ NaOH в течение 2 часов, периодически встряхивая, чтобы обеспечить одинаковую концентрацию NaOH в геле и в боксе для геля.
9. Приготовьте 1 л 1-кратного денатурирующего раствора (50 мМ NaOH, 1 мМ ЭДТА)
10. Налейте в бокс для геля объем, достаточный для погружения геля на глубину более 0,5 см.
11. Большие гели (15-20 см) анализируйте в течение ночи при 25 В. Средние гели (8-11 см) анализируйте в течение ночи при 20 В.
Последующий анализ геля
12. Перенесите гель в лоток и промойте dH2O
13. Слейте воду и нейтрализуйте гель в 1-кратном TBE или TAE (20 минут при осторожном перемешивании)
14. Перенесите гель в dH2O (или 1-кратный TBE/TAE) с 1-кратным SYBR Gold (20 мин при осторожном встряхивании)
Thermo Fisher, краситель для окрашивания нуклеиновых кислот в геле SYBR® Gold (10000-кратный концентрат в ДМСО), номер по каталогу: S11494
15. Визуализируйте гель, используя эпифлуоресцентный свет (синий) или УФ (312 нм)
[00519] Выделенные ДНК-векторы/вектор идентифицируют с помощью электрофореза в агарозном геле в нативных или денатурирующих условиях, как проиллюстрировано на ФИГ. 5 и ФИГ. 6. ДНК-вектор дает несколько полос на нативных гелях, как представлено на ФИГ. 5А. Каждая полоса может представлять векторы, имеющие различную конформацию в нативном состоянии, например, непрерывные, прерывистые, мономерные, димерные и т.д.
[00520] Структуры выделенного зкДНК-вектора анализировали далее путем расщепления ДНК, полученной из коинфицированных клеток Sf9 (как описано в настоящем документе), рестрикционными эндонуклеазами, выбранными на основании a) наличия только одного сайта разрезания в ДНК-векторе, и b) полученных фрагментов достаточно большого размера, чтобы они были отчетливо видны при фракционировании на 0,8% денатурирующем агарозном геле (>800 п.о.).
[00521] В частности, равные количества (2 мкг на основании OD260) ТТХ-плазмиды и ТТХ-вектора расщепляли рестрикционными эндонуклеазами при 37°C в течение 1 часа. После расщепления материал ДНК-вектора выделяли с использованием колонки QIAquick и элюировали водой. Образцы денатурировали в денатурирующем растворе (0,05 М NaOH, 1 мМ ЭДТА), в то время как 0,8% агарозный гель, приготовленный на воде, предварительно уравновешивали в течение 2 часов в буфере для уравновешивания (1 мМ ЭДТА, 200 мМ NaOH). Затем образцы, погруженные в 1-кратный денатурирующий раствор (50 мМ NaOH, 1 мМ ЭДТА), анализировали на геле в течение ночи при 4°C. На следующий день гель промывали, нейтрализовали в TBE в течение 20 минут, вымачивали в 1-кратном водном растворе SYBR Gold в течение 1 часа и визуализировали в УФ/синем свете.
[00522] Первоначально наличие ДНК-вектора идентифицируют на основании характерных многополосных профилей (первичная и вторичная полосы находятся на некотором расстоянии, чтобы показать, что вторичная полоса представляет собой материал, масса которого примерно в два раза превышает массу первичной полосы) на нативном геле, а затем подтверждают на денатурированном геле на основании характерного многополосного профиля, проиллюстрированного на правой стороне ФИГ. 5А. Как проиллюстрировано на ФИГ. 5B, линейные ДНК-векторы с прерывистой структурой и TTX-вектор с линейной и непрерывной структурой можно различить по размеру их продуктов реакции - например, ожидается, что ДНК-вектор с прерывистой структурой будет давать фрагменты размером 1 т.п.о. и 2 т.п.о., в то время как бескапсидный вектор с непрерывной структурой, как ожидается, даст фрагменты размером 2 т.п.о. и 4 т.п.о.
[00523] На ФИГ. 6 представлено иллюстративное изображение реального денатурирующего геля с ТТХ-векторами 1 и 2, 3 и 4, 5 и 6 и 7 и 8 (все они описаны в Таблице 1A выше) с (+) расщеплением эндонуклеазой или без него (-). После эндонуклеазной реакции каждый ТТХ-вектор давал две полосы (*). Размеры двух полос для указанных векторов, определенные на основании маркера размеров, приведены в нижней части изображения. Размеры полос подтверждают, что каждый из TTX-векторов имеет непрерывную структуру.
[00524] Вклад ТТХ-плазмиды в поглощение в УФ-диапазоне оценивали путем сравнения интенсивности флуоресценции ТТХ-вектора со стандартом. Например, если на основании поглощения в УФ-диапазоне определено, что на гель загружено 4 мкг TTX-вектора, а интенсивность флуоресценции TTX-вектора эквивалентна полосе 2 т.п.о. при известной массе 1 мкг, то масса TTX-вектора равна 1 мкг. Таким образом, TTX-вектор составляет 25% от общего количества поглощающего УФ материала. Затем строят график зависимости интенсивности полосы на геле от вычисленного введенного количества, которому соответствует полоса - например, если общее количество TTX-вектора соответствует 8 т.п.о., а вырезанная сравнительная полоса соответствует 2 т.п.о., то интенсивность указанной полосы на графике будет соответствовать 25% от общего введенного количества, что в данном случае составит 0,25 мкг при введении 1,0 мкг. Используя титрование TTX-плазмиды для построения стандартной кривой, рассчитывают количество для полосы TTX-вектора, используя уравнение линии регрессии, которое затем можно использовать для определения процента от общего введенного количества, представленного TTX-вектором, или процента чистоты (ФИГ. 7).
ПРИМЕР 4: ДНК-векторы экспрессируют кодируемый трансгеном белок in vitro
[00525] Последовательность SA кДНК дикого типа для мРНК фактора IX человека («wtFIX», SEQ ID NO: 11) или вариант Padua последовательности кДНК («PaduaFIX», SEQ ID NO: 12) вводили в сайт клонирования ТТХ-плазмиды 1, чтобы получить ТТХ-плазмиду 1-wtFIX и ТТХ-плазмиду 1-PaduaFIX, соответственно. Эти плазмиды вводили в клетки насекомых Sf9 и использовали для получения TTX-бакмиды 1-wtFIX и TTX-бакмиды 1-PaduaFIX, а также TTX-бакуловируса 1-wtFIX и TTX-бакуловируса 1-PaduaFIX, соответственно, с использованием способов, описанных в настоящем документе. Экспрессию белка in vitro с ТТХ-плазмид и ТТХ-векторов тестировали путем трансфекции клеток HEK293 (2×105 клеток/лунку, 96-луночный планшет) с использованием 250 нг/лунку (1) TTX-плазмиды 1-wtFIX, (2) ТТХ-плазмиды 1-PaduaFIX, (3) ТТХ-вектора 1-wtFIX, (4) ТТХ-вектора 1-PaduaFIX, (5) β (бета)-плазмиды 1-wtFIX или (6) β (бета)-вектора 1-wtFIX, используя реагент для трансфекции Fugene6 (3:1 Fugene6:ДНК). Результат анализа методом Вестерн-блоттинга представлен на ФИГ. 8. Реакция FIX-антитело выявила полосы массой 55 кДа, которые соответствуют массе продуцируемых белков FIX. Лизаты для отрицательного контроля, трансфицированные β (бета)-плазмидой 1-wtFIX или β (бета)-вектором 1-wtFIX, не продуцировали детектируемого количества белка FIX. Этот результат подтверждает, что ТТХ-вектор 1 можно применять для эффективного переноса и экспрессии терапевтического гена, такого как ген, кодирующий фактор IX человека.
[00526] ИФА: В общих чертах, культуральные среды от трансфицированных клеток добавляли с двумя повторами в лунки, обработанные антителом к FIX, и инкубировали в течение 1 часа с последующей промывкой и инкубацией с детектирующим антителом в течение 1 часа при комнатной температуре. Образцы снова промывали, добавляли субстрат TMB и проявляли окрашивание в течение 10 минут, останавливали реакцию и немедленно считывали поглощение образцов при 450 нм. Пример образцов после реакций с субстратом TMB представлен на ФИГ. 15А, а концентрация FIX в каждом образце, определенная на основании поглощения образца при 450 нм, представлена на ФИГ. 15А. Был детектирован высокий уровень экспрессии белка FIX с ТТХ-плазмиды 1 и ТТХ-вектора 1, при этом не была детектирована значительная экспрессия FIX с β (сравнительной)-плазмиды или β (сравнительного)-вектора.
[00527] Это еще раз подтверждает, что ТТХ-вектор 1, полученный из ТТХ-плазмиды 1, содержащий, в направлении от 5´- к 3´-концу, репликативную полинуклеотидную последовательность WT (SEQ ID NO: 51), промотор CAG (SEQ ID NO: 3), сайт клонирования R3/R4 (SEQ ID NO: 7), WPRE (SEQ ID NO: 8), BGHpA (SEQ ID NO: 9) и модифицированную репликативную полинуклеотидную последовательность (SEQ ID NO: 2), значительно более эффективно индуцирует экспрессию трансгена по сравнению с α (альфа)-вектором 1, полученным из α (альфа)-плазмиды 1, которая не включает WPRE (SEQ ID NO: 8) и BGHpA (SEQ ID NO: 9).
ПРИМЕР 5: Получение зкДНК, коэкспрессирующей фактор IX и ингибитор cGAS
[00528] Белок ORF52 герпесвируса, ассоциированного с саркомой Капоши (SEQ ID NO: 882), или его вариант, который ингибирует cGAS, или усеченную цитоплазматическую изоформу LANA (LANAΔ161 или SEQ ID NO: 884), в которой отсутствуют аминокислоты 161-1162 из SEQ ID NO: 882), функционально связывают с промотором и вставляют в сайт рестрикционного клонирования R5 плазмиды TTX 9 или TTX 10, кодирующей трансген фактора IX, как описано в Примере 1 и Примере 4. Таким образом, получают зкДНК, которая кодирует как фактор IX, так и ингибитор cGAS, как описано в Примерах 2-3.
ПРИМЕР 6: Подтверждение экспрессии ингибитора cGAS, экспрессируемого зкДНК
[00529] Экспрессия целевого ингибитора cGAS, коэкспрессируемого зкДНК, такого как белок ORF52 герпесвируса, ассоциированного с саркомой Капоши (SEQ ID NO: 882), или его вариант, который ингибирует cGAS, или усеченная цитоплазматическая изоформа LANA (SEQ ID NO: 884), может быть подтверждена с использованием клеток HeLa и антител, специфичных в отношении ингибитора cGAS, таких как антитело к ORF52, описанное в Li et al. («Kaposi´s sarcoma-associated herpesvirus inhibitor of cGAS (KicGAS) Encoded by ORF52, is an Abundant Tegument protein and Is Required for Production of Infectious Progeny Viruses», J. Virol. 2016, 90(11): 5329). Например, культивируют клетки HeLa и выполняют кратковременные трансфекции конструкций, коэкспрессирующих фактор IX и целевой ингибитор cGAS, используя, например, реагент для трансфекции Fusegene6 (3:1; fusgene6:ДНК). Для детектирования экспрессии ингибитора cGAS используют методики Вестерн-блоттинга и/или проточной цитометрии, как известно специалистам в данной области техники. Экспрессию фактора IX подтверждают, как описано в Примере 4.
ПРИМЕР 7: Получение зкДНК, коэкспрессирующей фактор IX и ингибитор TLR-9
[00530] Олигонуклеотиды, которые могут образовывать шпилечную структуру, содержащие следующие последовательности, такие как (TCCTGGCGGGGAAGT, SEQ ID NO: 889), ODN-2114 (TCCTGGAGGGGAAGT, SEQ ID NO: 890), поли(G) (GGGGGGGGGGGGGGGGGGGG, SEQ ID NO: 891), ODN-A151 (TTAGGGTTAGGGTTAGGGTTAGGG, SEQ ID NO: 892), G-ODN (CTCC-TATTGGGGGTTTCCTAT, SEQ ID NO: 893), IRS-869 (TCCTGGAGGGGTTGT, SEQ ID NO: 894), INH-1 (CCTGGATGGGAATTCCCATCCAGG, SEQ ID NO: 895), INH-4 (TTCCCATCCAGGCCTGGATGGGAA, SEQ ID NO: 896), (IRS-661 TGCTTGCAAGCTT-GCAAGCA, SEQ ID NO: 897), 4024 (TCCTGGATGGGAAGT, SEQ ID NO: 898), 4084F (CCTGGATGGGAA, SEQ ID NO: 899), INH-13 (CTTACCGCTGCACCTGGATGGGAA, SEQ ID NO: 900), INH-18 (CCTGGATGGGAACTTACCGCTGCA, SEQ ID NO: 901) и IRS-954 TGCTCCTGGAGGGGTTGT, SEQ ID NO: 902), конструируют так, чтобы получить липкие концы после ренатурации комплементарных цепей 5´-3´ и 3´-5´ так, что они могут быть вставлены путем лигирования в предварительно выбранный сайт рестрикционного клонирования, например, R5 или другой сайт плазмиды TTX 9 или TTX 10, кодирующей трансген фактора IX, как описано в Примере 1 и Примере 4.
[00531] Например, олигонуклеотиды с подходящим сайтом рестрикции ренатурируют путем смешивания каждой цепи в равных молярных количествах в подходящем буфере: например, 100 мМ ацетата калия; 30 мМ HEPES, pH 7,5, нагревания до 94°C в течение 2 минут и постепенного охлаждения. Таким образом, для олигонуклеотидов с прогнозируемой значительной вторичной структурой благоприятна более плавная стадия охлаждения/ренатурации. Это можно осуществить, поместив раствор олигонуклеотидов на водяную баню или в термостат и отключив/выключив устройство. Ренатурированные олигонуклеотиды можно разбавить в буфере, не содержащем нуклеаз, и хранить в двухцепочечной ренатурированной форме при 4°С. Затем зкДНК-плазмиду с олигонуклеотидной последовательностью, ингибирующей TLR-9, очищают (например, с помощью гель-электрофореза или колонки) и используют для получения зкДНК-вектора. Может быть получена зкДНК, которая кодирует фактор IX и которая содержит антагонист TLR-9.
ПРИМЕР 8: Контролируемая экспрессия трансгена с зкДНК: экспрессия трансгена с зкДНК-вектора in vivo может поддерживаться и/или повышаться путем повторного введения дозы.
[00532] ЗкДНК-вектор получали в соответствии со способами, описанными в Примере 1 выше, используя зкДНК-плазмиду, содержащую промотор CAG (SEQ ID NO: 3), при этом трансген люциферазы (SEQ ID NO: 71) используют как примерный антагонист инфламмасомы, фланкированный асимметричными ITR (например, 5´-WT-ITR (SEQ ID NO: 1) и 3´ mod-ITR (SEQ ID NO: 2)), и оценивали в различных программах лечения in vivo. Этот зкДНК-вектор использовали во всех последующих экспериментах, описанных в Примерах 6-10. В Примере 6 зкДНК-вектор очищали, изготавливали в виде состава с липидной наночастицей (ЛНЧ-зкДНК) и вводили путем инъекции в хвостовую вену каждой мыши CD-1® IGS. Липосомы изготавливали с использованием подходящей смеси липидов, содержащей четыре компонента для образования липосом на основе липидных наночастиц (ЛНЧ), включая катионные липиды, вспомогательные липиды, холестерин и ПЭГ-липиды.
[00533] Для оценки устойчивой экспрессии трансгена in vivo с зкДНК-вектора на протяжении длительного периода времени ЛНЧ-зкДНК вводили в стерильном ФСБ путем внутривенной инъекции в хвостовую вену мышам CD-1® IGS в возрасте приблизительно 5-7 недель. Была проведена оценка трех групп с разными дозировками: 0,1 мг/кг, 0,5 мг/кг и 1,0 мг/кг, по десять мышей на группу (за исключением группы 1,0 мг/кг, в которой было 15 мышей на группу). Инъекции вводили в день 0. Пять мышей из каждой группы получали инъекцию дополнительной идентичной дозы в день 28. Экспрессию люциферазы измеряли путем визуализации IVIS после внутривенного введения мышам CD-1® IGS (Charles River Laboratories; мыши дикого типа (WT)). Экспрессию люциферазы оценивали с помощью визуализации IVIS после введения путем внутрибрюшинной инъекции 150 мг/кг субстрата люциферина в дни 3, 4, 7, 14, 21, 28, 31, 35 и 42 и регулярно (например, еженедельно, два раза в неделю или через каждые 10 суток, или через каждые 2 недели) в период между днями 42-110. Экспрессия трансгена люциферазы, в качестве примерного антагониста инфламмасомы, измеренная с помощью визуализации IVIS в течение по меньшей мере 132 дней после 3 разных протоколов введения (данные не показаны).
[00534] Расширенное исследование выполняли для изучения эффекта повторной дозы, например, повторного введения ЛНЧ-зкДНК, экспрессирующей люциферазу, субъектам, получавшим ЛНЧ-зкДНК. В частности, оценивали возможность повышения уровней экспрессии путем одного или более дополнительных введений зкДНК-вектора.
[00535] В этом исследовании биораспределение экспрессии люциферазы с зкДНК-вектора оценивали с помощью IVIS у мышей CD-1® IGS после первоначального внутривенного введения 1,0 мг/кг (т.е. праймирующей дозы) в дни 0 и 28 (Группа A). Второе введение зкДНК-вектора осуществляли путем инъекции в хвостовую вену 3 мг/кг (Группа B) или 10 мг/кг (Группа C) в 1,2 мл в хвостовую вену в день 84. В этом исследовании использовали по пять (5) мышей CD-1® в каждой из Групп A, B и C. Визуализацию IVIS экспрессии люциферазы у мышей выполняли перед введением дополнительных доз в дни 49, 56, 63 и 70, как описано выше, а также после введения повторных доз в день 84 и в дни 91, 98, 105, 112 и 132. Экспрессию люциферазы оценивали и детектировали во всех трех Группах A, B и C по меньшей мере до дня 110 (самый продолжительный оцененный период).
[00536] Было показано, что уровень экспрессии люциферазы повышается с помощью повторной дозы (т.е. повторного введения композиции зкДНК) ЛНЧ-зкДНК-Luc, согласно определению путем оценки активности люциферазы в присутствии люциферина. Экспрессия трансгена люциферазы, в качестве примерного антагониста инфламмасомы, измеренная с помощью визуализации IVIS в течение по меньшей мере 110 дней после 3 разных протоколов введения (Группы A, B и C). У мышей, не получавших никакой дополнительной повторной дозы (праймирующая доза 1 мг/кг (т.е. Группа А)), наблюдали стабильную экспрессию люциферазы на протяжении всего исследования. Мыши в группе B, которым вводили повторную дозу 3 мг/кг зкДНК-вектора, показали приблизительно семикратное увеличение наблюдаемой светимости по сравнению с мышами в Группе C. Неожиданно, мыши, получившие повторную дозу 10 мг/кг зкДНК-вектора, имели 17-кратное увеличение наблюдаемой светимости люциферазы по сравнению с мышами, не получавшими какой-либо повторной дозы (Группа А).
[00537] Группа A мышей CD-1® IGS показывает экспрессию люциферазы после внутривенного введения 1 мг/кг зкДНК-вектора в хвостовую вену в дни 0 и 28. Группы B и C мышей CD-1® IGS показывают экспрессию люциферазы после введения 1 мг/кг зкДНК-вектора в первый момент времени (день 0) и введения повторной дозы зкДНК-вектора во второй момент времени в день 84. Второе введение (т.е. повторная доза) зкДНК-вектора повысило экспрессию по меньшей мере в 7 раз и даже до 17 раз.
[00538] 3-кратное увеличение дозы (т.е. количества) зкДНК-вектора при повторном введении дозы в Группе В (т.е. при повторном введении дозы 3 мг/кг) привело к 7-кратному увеличению экспрессии люциферазы. Также неожиданно 10-кратное увеличение количества зкДНК-вектора при повторном введении дозы (т.е. при повторном введении дозы 10 мг/кг) в Группе С привело к 17-кратному увеличению экспрессии люциферазы. Таким образом, второе введение (т.е. повторная доза) зкДНК повысило экспрессию по меньшей мере в 7 раз и даже до 17 раз. Это показывает, что повышение экспрессии трансгена в результате повторной дозы больше, чем ожидалось, и зависит от дозы или количества зкДНК-вектора при введении повторной дозы и, по-видимому, является синергическим по отношению к начальной экспрессии трансгена после первоначального праймирующего введения в день 0. Таким образом, дозозависимое повышение экспрессии трансгена не является аддитивным, скорее, уровень экспрессии трансгена является дозозависимым и сверхпропорционален суммарному количеству зкДНК-вектора, введенному в каждый момент времени.
[00539] Обе группы B и C показали значительное дозозависимое повышение экспрессии люциферазы по сравнению с контрольными мышами (Группа A), которые не получали повторной дозы зкДНК-вектора во второй момент времени. В совокупности эти данные показывают, что экспрессия трансгена с зкДНК-вектора может быть повышена дозозависимым образом с помощью повторной дозы (т.е. повторного введения) зкДНК-вектора по меньшей мере во второй момент времени.
[00540] В совокупности эти данные демонстрируют, что уровень экспрессии трансгена, например, антагониста инфламмасомы, с зкДНК-векторов может поддерживаться на устойчивом уровне на протяжении по меньшей мере 84 дней и может быть повышен in vivo после повторной дозы зкДНК-вектора, введенной по меньшей мере во второй момент времени.
ПРИМЕР 9: Синтетические наноносители с перенасыщенными количествами рапамицина
[00541] Композиции наноносителей, содержащие полимеры PLGA (лактид:гликолид 3:1, характеристическая вязкость 0,39 дл/г) и PLA-ПЭГ (блок ПЭГ 5 кДа, характеристическая вязкость 0,36 дл/г), а также агент рапамицин (RAPA), могут быть синтезированы с использованием метода испарения эмульсии типа «масло-в-воде». Органическая фаза образуется путем растворения полимеров и RAPA в дихлорметане. Эмульсия образуется путем гомогенизации органической фазы в водной фазе, содержащей поверхностно-активное вещество поливиниловый спирт (ПВС). Затем эмульсию объединяют с большим количеством водного буфера и перемешивают, чтобы обеспечить испарение растворителя. Содержание RAPA в различных композициях варьируют так, что при увеличении содержания RAPA в композиции превышается предел насыщения RAPA системы. Содержание RAPA на пределе насыщения для композиции рассчитывают с использованием растворимости RAPA в водной фазе и в дисперсной фазе наноносителя. Для композиций, содержащих ПВС в качестве основного растворенного вещества в водной фазе, обнаружено, что растворимость RAPA в водной фазе пропорциональна концентрации ПВС так, что RAPA растворим при массовом соотношении 1:125 к растворенному ПВС. Для композиций, содержащих описанные PLGA и PLA-ПЭГ в качестве полимеров-наноносителей, обнаружено, что растворимость RAPA в дисперсной фазе наноносителя составляет 7,2% масс./масс. Для расчета содержания RAPA на пределе насыщения для композиции можно использовать следующую формулу:
Содержание RAPA= V(0,008cПВС + 0,072cpol)
[00542] где cПВС представляет собой массовую концентрацию ПВС, cpol представляет собой общую массовую концентрацию полимеров, а V представляет собой объем суспензии наноносителя по завершении испарения.
Таблица 8
[00543] Для 1, 2 и 3 не поддается выделению постоянная доля в 60% RAPA, это указывает на равновесное состояние субнасыщения между водной и органической фазами. Для остальных наноносителей, содержащих большие количества RAPA, не поддается выделению постоянная масса в 6,8 мг RAPA. Эта постоянная абсолютная потеря массы указывает на то, что система находится в состоянии перенасыщения (т.е. перенасыщена в одной или более фазах).
ПРИМЕР 10: Синтетические наноносители с перенасыщенным количеством рапамицина устраняют или задерживают выработку антител
[00544] Композиции наноносителей, содержащие полимеры PLGA (лактид:гликолид 3:1, характеристическая вязкость 0,39 дл/г) и PLA-ПЭГ (блок ПЭГ 5 кДа, характеристическая вязкость 0,36 дл/г), а также агент RAPA, могут быть синтезированы с использованием метода испарения эмульсии типа «масло-в-воде», описанного в Примере 5. Содержание RAPA в различных композициях варьируют так, что при увеличении содержания RAPA в композиции превышается предел насыщения RAPA системы.
Таблица 9
[00545] Чтобы оценить способность композиций индуцировать иммунную толерантность, мышам совместно вводят наноноситель и гемоцианин лимфы улитки (KLH) путем внутривенной инъекции три раза в неделю, а затем еженедельно стимулируют только KLH. Затем после стимуляции KLH сыворотки мышей анализируют на наличие антител к KLH. Композиции, полученные в перенасыщенном состоянии и имеющие конечную нагрузку RAPA 8% или выше, приводили к отсутствию или задержке выработки антител к KLH в большей степени, чем композиции, созданные при насыщении или не достигая его и имеющие конечную нагрузку RAPA 5% или ниже.
ПРИМЕР 11: Синтетические наноносители с перенасыщенными количествами рапамицина
[00546] Композиции наноносителей, содержащие полимеры PLA (характеристическая вязкость 0,41 дл/г) и PLA-ПЭГ (блок ПЭГ 5 кДа, характеристическая вязкость 0,50 дл/г), а также агент RAPA, синтезировали с использованием метода испарения эмульсии типа «масло-в-воде», описанного в Примере 9. Содержание RAPA в различных композициях варьировали так, что при увеличении содержания RAPA в композициях превышался предел насыщения RAPA системы. Содержание RAPA на пределе насыщения для композиции рассчитывали с использованием метода, описанного в Примере 9. Для композиций, содержащих описанные PLA и PLA-ПЭГ в качестве полимеров-наноносителей, было обнаружено, что растворимость RAPA в дисперсионной фазе наноносителя составляет 8,4% масс./масс. Для расчета содержания RAPA на пределе насыщения для композиции можно использовать следующую формулу:
Содержание RAPA = V(0,008cПВС + 0,084cpol)
[00547] где cПВС представляет собой массовую концентрацию ПВС, cpol представляет собой общую массовую концентрацию полимеров, а V представляет собой объем суспензии наноносителя по завершении испарения. Все партии наноносителей фильтруют через фильтры с размером пор 0,22 мкм по завершении изготовления.
Таблица 10
(%)
Диаметр (нм)
Пропускная способность (г/м2)
Несмотря на добавление повышающегося количества RAPA к наноносителям 12-15, конечное содержание RAPA в наноносителях не увеличивалось, в то время как пропускная способность фильтра уменьшалась. Это указывает на то, что композиции были перенасыщены RAPA, и избыток RAPA удаляется во время промывки и/или фильтрации.
ПРИМЕР 12: Фактор IX или VIII для лечения гемофилии B с применением зкДНК, кодирующей фактор IX или фактор VIII, вводимой совместно с рапамицином
[00548] Эксперимент проводят на мышах с дефицитом фактора IX или фактора VIII, которые содержат введенную (knock-in) последовательность hFIX или hFVIII с вредной мутацией (например, R333Q для hFIX). Самцы мышей с подавленным (knockout) фактором IX или FVIII получают однократные или повторные дозы ЛНЧ-зкДНК (липидные наночастицы с зкДНК), вводимые совместно с рапамицином или аналогом рапамицина, причем указанные ЛНЧ-зкДНК и рапамицин или аналог рапамицина содержатся в отдельных композициях. ЛНЧ с зкДНК-векторами вводят соответствующим мышам в дозах от 0,3 до 5 мг/кг в объеме 1,2 мл совместно с наноносителем с рапамицином (например, перенасыщенным рапамицином (например, SVP-рапамицином), как описано в Примерах 9-11), или его аналогом, вводимым при, например, 0,05 мг/кг, 0,1 мг/кг и до 5 мг/кг. Терапевтически эффективные дозы определяют путем мониторинга эффективности ингибирования иммунного ответа (например, при однократном и повторном дозировании) и измерения целевой величины экспрессии трансгена. Каждая доза может быть введена путем в/в введения. SVP-Rap может быть совместно введен, например, в день 0 и день 14.
[00549] Экспрессию фактора IX или фактора VIII в плазме оценивают с помощью ИФА, как описано в Примере 4, в различные моменты времени, например, через 10, 20, 30, 40, 50, 1000 и 200 дней или более и т.д. Активированное частичное тромбопластиновое время и время кровотечения также можно измерить как определение эффективности и эффекта совместного введения рапамицина, или аналога, на экспрессию фактора VIII или фактора IX. Ожидается, что у мышей, которые получают зкДНК-вектор, вводимый совместно с рапамицином, повышенная и/или устойчивая экспрессия фактора IX или фактора VIII будет наблюдаться в течение более длительного периода времени по сравнению с мышами, которые получают только зкДНК-вектор, но не рапамицин, или его аналог. Кроме того, ожидается, что при повторном введении дозы у мышей, которые получают повторную дозу зкДНК-вектора и рапамицина, будет наблюдаться меньшая активация секреции цитокинов и увеличенная продолжительность экспрессии трансгена и терапевтическая эффективность по сравнению с мышами, получившими повторную дозу зкДНК-вектора, которым не вводили рапамицин. Время совместного введения может быть разнесено на 0, 1, 2, 3, 4, 5, 6, 7, 8 часов.
ПРИМЕР 13: Фактор IX для лечения гемофилии B с применением зкДНК, кодирующей фактор IX и антагонист cGAS
[00550] Эксперимент проводят на мышах с дефицитом фактора IX, которые имеют вставленную последовательность hFIX с вредной мутацией (R333Q). Самцы мышей с подавленным фактором IX получают однократные или повторные дозы ЛНЧ-зкДНК (липидные наночастицы с зкДНК). Используют два варианта ЛНЧ с зкДНК-вектором; 1) ЛНЧ с зкДНК-вектором, кодирующим фактор IX человека (либо нативную последовательность человека, либо варианты Padua FIX) и кодирующим белок ORF52 вируса герпеса, ассоциированного с саркомой Капоши; ЛНЧ с зкДНК-вектором, кодирующим только фактор IX, но не ингибитор cGAS, в качестве сравнительного зкДНК-вектора. ЛНЧ с зкДНК-векторами вводят соответствующим мышам в дозах от 0,3 до 5 мг/кг в объеме 1,2 мл. Каждая доза должна быть введена путем в/в гидродинамического введения. Экспрессию фактора IX в плазме оценивают с помощью ИФА, как описано в настоящем документе, в различные моменты времени, например, через 10, 20, 30, 40, 50, 1000 и 200 дней или более и т.д. Активированное частичное тромбопластиновое время и время кровотечения также измеряют как определение эффективности. Ожидается, что у мышей, которые получают зкДНК-вектор, экспрессирующий как hFIX, так и ORF52, повышенная и/или устойчивая экспрессия фактора IX будет наблюдаться в течение более длительного периода времени по сравнению с мышами, которые получают зкДНК-вектор, экспрессирующий только фактор IX, но не ORF52 или другой ингибитор cGAS. Кроме того, ожидается, что при повторном введении дозы у мышей, которые получают повторную дозу зкДНК-вектора, содержащего как ORF52, так и фактор IX, будет наблюдаться меньшая активация секреции цитокинов и увеличенная продолжительность экспрессии трансгена и терапевтическая эффективность по сравнению с мышами, получившими повторную дозу зкДНК-вектора, кодирующего только фактор IX. Ингибитор cGAS и фактор IX могут быть доставлены на разных зкДНК-векторах, но предпочтительно они кодируются одним и тем же вектором, и, соответственно, ингибирование cGAS происходит в той же клетке, что и клетка, которая получает зкДНК-вектор, кодирующий трансген, такой как фактор IX.
ПРИМЕР 14: Определение эффектов совместного введения зкДНК и антагонистов cGAS на врожденные иммунные ответы и продолжительность экспрессии фактора IX
[00551] Чтобы исследовать эффекты совместного введения представляющей интерес зкДНК и ингибитора cGAS или антагониста cGAS на иммунный ответ (например, врожденный иммунный ответ) in vitro, можно использовать репортерные линии для функциональных анализов, в которых исследуется активация cGAS. Репортерная линия клеток cGAS, которую можно применять для таких анализов in vitro, может представлять собой стабильно котрансфицированную линию клеток, которая экспрессирует полноразмерный cGAS человека и репортерный ген, такой как репортерный ген секретируемой щелочной фосфатазы (SEAP), под транскрипционным контролем элемента ответа транскрипционного фактора, такого как сайт связывания NF-kB, сайт связывания AP-1 или их комбинация. Например, репортерные клетки высевают в 96-луночные планшеты. По истечении заранее определенного периода времени, например, 16 часов, клетки стимулируют различными количествами композиций, содержащих зкДНК, экспрессирующую фактор IX, с ингибитором cGAS или без него. Активность репортерного гена, такого как SEAP, может быть проанализирована с использованием любого метода или анализа, известного специалисту в данной области техники, для сравнения уровня активации cGAS в присутствии представляющей интерес зкДНК с ингибитором cGAS или без него. Ожидается, что в присутствии ингибитора cGAS будет наблюдаться меньшая активация репортерной молекулы.
[00552] Кроме того, можно использовать репортерные линии с подавленным cGAS, такие как линии, происходящие из моноцитов THP-1 человека, которые представляют собой линию клеток, часто используемую для исследования путей, чувствительных к ДНК, поскольку они экспрессируют все сенсоры цитозольной ДНК, идентифицированные к настоящему времени (за исключением DAI). Такие репортерные линии с подавленным cGAS могут экспрессировать один или более индуцируемых генов секретируемых репортеров, таких как люцифераза Lucia и SEAP (секретируемая эмбриональная щелочная фосфатаза). Репортерный ген может находиться под контролем минимального промотора ISG54 (гена, стимулируемого интерфероном) в сочетании с одним или более, например, пятью, элементами ответа, стимулируемыми IFN. Репортерный ген также может находиться под контролем минимального промотора IFN-β, слитого с одной или более, например, пятью, копиями элемента ответа, такого как элемент ответа NF-kB. Активность cGAS в присутствии ингибиторов cGAS в комбинации с зкДНК, описанными в настоящем документе, можно сравнить в линии клеток с подавленным геном в сопоставлении с исходной линией клеток.
[00553] Чтобы исследовать эффекты совместного введения представляющей интерес зкДНК и ингибитора cGAS или антагониста cGAS на активацию cGAS и STING иммунного ответа (например, врожденного иммунного ответа) ex vivo, моноциты человека могут быть выделены, например, с помощью центрифугирования периферической крови в градиенте плотности и магнитного разделения. Эти моноциты можно исследовать до и после контакта и/или активации представляющей интерес зкДНК с ингибитором cGAS или без него, с подходящими контролями. После обработки сыворотку и клеточные супернатанты используют для измерения одного или более цитокиновых путей в качестве функционального показателя активации пути cGAS/STING, таких как интерлейкин (IL)-1β, IL-6, IL-8, интерферон (IFN)-γ, моноцитарный хемоаттрактантный белок (MCP)-1 и/или фактор некроза опухоли (TNF)-α, с использованием любого анализа или метода, известного специалисту в данной области техники. Кроме того, ядерные экстракты можно использовать для проверки активации NF-κB с использованием любого анализа или метода, известного специалисту в данной области техники. Ожидается, что при введении зкДНК в присутствии ингибитора cGAS будет наблюдаться меньшая активация цитокиновых путей и секреция цитокинов, что приводит к увеличению продолжительности экспрессии трансгена и терапевтической эффективности.
[00554] Чтобы исследовать эффекты совместного введения представляющей интерес зкДНК и ингибитора cGAS или антагониста cGAS на активацию cGAS и STING иммунного ответа (например, врожденного иммунного ответа) in vivo, можно использовать модель на мышах. Образцы сыворотки или лимфоцитов мыши исследуют до и после контакта и/или активации зкДНК, экспрессирующей представляющий интерес трансген, такой как фактор IX, с ингибитором cGAS или без него, с подходящими контролями. После обработки сыворотку и клеточные супернатанты используют для измерения одного или более цитокиновых путей в качестве функционального показателя активации пути cGAS/STING, таких как интерлейкин (IL)-1β, IL-6, IL-8, интерферон (IFN)-γ, моноцитарный хемоаттрактантный белок (MCP)-1 и/или фактор некроза опухоли (TNF)-α, с использованием любого анализа или метода, известного специалисту в данной области техники. Кроме того, ядерные экстракты можно использовать для проверки активации NF-κB с использованием любого анализа или метода, известного специалисту в данной области техники. Ожидается, что при введении зкДНК в присутствии ингибитора cGAS будет наблюдаться меньшая активация и секреция цитокинов, что приводит к увеличению продолжительности экспрессии трансгена и терапевтической эффективности.
ПРИМЕР 15: Фактор IX для лечения гемофилии B с применением зкДНК, кодирующей фактор IX и антагонист TLR-9
[00555] Эксперимент проводят на мышах с дефицитом фактора IX, которые имеют вставленную последовательность hFIX с вредной мутацией (R333Q). Самцы мышей с подавленным фактором IX получают однократные или повторные дозы ЛНЧ-зкДНК (липидные наночастицы с зкДНК). Используют два варианта ЛНЧ с зкДНК-вектором; 1) ЛНЧ-зкДНК, кодирующей фактор IX человека (либо нативную последовательность человека, либо варианты Padua FIX) и кодирующей белок ORF52 вируса герпеса, ассоциированного с саркомой Капоши; ЛНЧ-зкДНК, кодирующей только фактор IX, но не ингибитор cGAS, в качестве сравнительного зкДНК-вектора. ЛНЧ с зкДНК-векторами вводят соответствующим мышам в дозах от 0,3 до 5 мг/кг в объеме 1,2 мл. Каждая доза должна быть введена путем в/в гидродинамического введения. Экспрессию фактора IX в плазме оценивают с помощью ИФА, как описано в Примере 4, в различные моменты времени, например, через 10, 20, 30, 40, 50, 1000 и 200 дней или более и т.д. Активированное частичное тромбопластиновое время и время кровотечения также измеряют как определение эффективности. Ожидается, что у мышей, которые получают зкДНК-вектор, содержащий антагонист TLR9 и экспрессирующий фактор hFIX, повышенная и/или устойчивая экспрессия фактора IX будет наблюдаться в течение более длительного периода времени по сравнению с мышами, которые получают зкДНК-вектор, экспрессирующий только фактор IX, но не ингибитор TLR-9. Кроме того, ожидается, что при повторном введении дозы у мышей, которые получают повторную дозу зкДНК-вектора, содержащего ингибитор TLR9, например, шпилечную олигонуклеотидную последовательность, и фактор IX, будет наблюдаться меньшая активация секреции цитокинов и увеличенная продолжительность экспрессии трансгена и терапевтическая эффективность по сравнению с мышами, получившими повторную дозу зкДНК-вектора, кодирующего только фактор IX. Ингибитор TLR-9 и фактор IX могут быть доставлены на разных зкДНК-векторах, в транс-конфигурации, но предпочтительно они кодируются одним и тем же вектором, и, соответственно, ингибирование TLR-9 происходит в той же клетке, что и клетка, которая получает зкДНК-вектор, кодирующий трансген, такой как фактор IX.
ПРИМЕР 16: Определение эффектов зкДНК и антагонистов TLR на врожденные иммунные ответы и продолжительность экспрессии трансгена
[00556] Чтобы исследовать эффекты совместного введения представляющей интерес зкДНК и ингибитора TLR9 или антагониста TLR9 на врожденные иммунные ответы in vitro, можно использовать репортерные линии для TLR9-зависимых функциональных анализов, в которых исследуются последующие эффекты активации TLR9. Репортерная линия клеток TLR9 может представлять собой стабильно котрансфицированную линию клеток, которая экспрессирует полноразмерный Toll-подобный рецептор 9 (TLR9) человека и репортерный ген, такой как репортерный ген секретируемой щелочной фосфатазы (SEAP), под транскрипционным контролем элемента ответа фактора транскрипции, такого как сайт связывания NF-kB, сайт связывания AP-1 или их комбинация. Например, репортерные клетки высевают в 96-луночные планшеты. По истечении заранее определенного периода времени, например, 16 часов, клетки стимулируют различными количествами композиций, содержащих зкДНК, экспрессирующую представляющий интерес трансген, с антагонистом TLR9 или без него. Такой антагонист может представлять собой олигонуклеотид, ингибирующий TLR. Активность репортерного гена, такого как SEAP, может быть проанализирована с использованием любого метода или анализа, известного специалисту в данной области техники, для определения уровня активации TLR9 в присутствии представляющей интерес зкДНК с антагонистом TLR9 или без него. Ожидается, что в присутствии ингибитора TLR9 будет наблюдаться меньшая активация репортерной молекулы.
[00557] Чтобы исследовать эффекты совместного введения представляющей интерес зкДНК и ингибитора TLR9 или антагониста TLR9 на TLR9-опосредуемую активацию врожденных иммунных ответов ex vivo, моноциты человека могут быть выделены, например, с помощью центрифугирования периферической крови в градиенте плотности и магнитного разделения. Эти моноциты можно исследовать до и после контакта и/или активации представляющей интерес зкДНК с антагонистом TLR9 или без него, с подходящими контролями. После обработки сыворотку и клеточные супернатанты используют для измерения одного или более цитокиновых путей в качестве функционального показателя активации TLR9, таких как интерлейкин (IL)-1β, IL-6, IL-8, интерферон (IFN)-γ, моноцитарный хемоаттрактантный белок (MCP)-1 и/или фактор некроза опухоли (TNF)-α, с использованием любого анализа или метода, известного специалисту в данной области техники. Кроме того 0914800, ядерные экстракты можно использовать для проверки активации NF-κB с использованием любого анализа или метода, известного специалисту в данной области техники. Ожидается, что при введении зкДНК в присутствии ингибитора TLR9 будет наблюдаться меньшая активация цитокиновых путей и секреция цитокинов, что приводит к увеличению продолжительности экспрессии трансгена и терапевтической эффективности.
[00558] Чтобы исследовать эффекты совместного введения представляющей интерес зкДНК и ингибитора TLR9 или антагониста TLR9 на TLR9-опосредуемую активацию врожденных иммунных ответов in vivo, можно использовать модель на мышах. Образцы сыворотки или лимфоцитов мыши исследуют до и после контакта и/или активации зкДНК, экспрессирующей представляющий интерес трансген, такой как фактор IX, с ингибитором TLR9 или без него, с подходящими контролями. После обработки сыворотку и клеточные супернатанты используют для измерения одного или более цитокиновых путей в качестве функционального показателя активации пути cGAS/STING, таких как интерлейкин (IL)-1β, IL-6, IL-8, интерферон (IFN)-γ, моноцитарный хемоаттрактантный белок (MCP)-1 и/или фактор некроза опухоли (TNF)-α, с использованием любого анализа или метода, известного специалисту в данной области техники. Кроме того, ядерные экстракты можно использовать для проверки активации NF-κB с использованием любого анализа или метода, известного специалисту в данной области техники. Ожидается, что при введении зкДНК в присутствии ингибитора TLR9 будет наблюдаться меньшая активация и секреция цитокинов, что приводит к увеличению продолжительности экспрессии трансгена и терапевтической эффективности.
ПРИМЕР 17: Изготовление состава зкДНК с RAPA в виде ЛНЧ-векторов
Согласно некоторым вариантам реализации желательной может быть упаковка рапамицина непосредственно в зкДНК-вектор. Далее описан один неограничивающий пример такого непосредственного изготовления состава зкДНК и RAPA.
Комбинации зкДНК с рапамицином в липидных наночастицах (ЛНЧ) могут быть приготовлены путем смешивания спиртового раствора липидов, содержащего рапамицин, с водным раствором зкДНК с использованием микроструйного устройства (например, NanoAssemblr™) в соотношении 1:3 (об./об.) при общей скорости потока 12 мл/мин. Массовое соотношение общего количества липидов и зкДНК может составлять приблизительно от 10:1 до 30:1. В общих чертах, ионизируемый липид (например, MC3), некатионный липид (например, дистеароилфосфатидилхолин (DSPC)), компонент для обеспечения целостности мембраны (такой как стерин, например, холестерин) и молекулу конъюгированного липида (например, ПЭГ-липид, например, 1-(монометоксиполиэтиленгликоль)-2,3-димиристоилглицерин, со средней молекулярной массой ПЭГ 2000 («ПЭГ-DMG»)) солюбилизируют в спирте (например, этаноле) в молярном соотношении 50:10:38,5:1,5. Затем рапамицин растворяют в растворе липидов до целевой концентрации. ЗкДНК разбавляют до 0,2 мг/мл в 25 мМ буфере на основе ацетата натрия, pH 4. После образования ЛНЧ (с использованием, например, NanoAssemblr™) спирт удаляют, а буфер на основе ацетата натрия заменяют ФСБ путем диализа. Удаление спирта и одновременная замена буфера могут быть выполнены, например, путем диализа или фильтрации с тангенциальным потоком. Полученные липидные наночастицы фильтруют через стерильный фильтр с размером пор 0,2 мкм и хранят так же, как и ЛНЧ с зкДНК-векторами, описанные выше.
ПРИМЕР 18: Определение эффектов совместного введения зкДНК-вектора и рапамицина или аналога рапамицина на врожденные иммунные ответы и продолжительность экспрессии фактора IX
[00559] Чтобы исследовать эффекты совместного введения представляющей интерес зкДНК и рапамицина или аналога на врожденные иммунные ответы in vitro, можно использовать репортерные линии для функциональных анализов, в которых исследуются последующие эффекты активации TLR и mTORC1. Репортерная линия клеток TLR9 может представлять собой стабильно котрансфицированную линию клеток, которая экспрессирует полноразмерный Toll-подобный рецептор 9 (TLR9) человека и репортерный ген, такой как репортерный ген секретируемой щелочной фосфатазы (SEAP), под транскрипционным контролем элемент ответа фактора транскрипции, такого как сайт связывания NF-kB, сайт связывания AP-1 или их комбинация. Например, репортерные клетки высевают в 96-луночные планшеты. По истечении заранее определенного периода времени, например, 16 часов, клетки стимулируют различными количествами композиций, содержащих зкДНК, экспрессирующую представляющий интерес трансген, с рапамицином или его аналогом, или без него. Активность репортерного гена, такого как SEAP, может быть проанализирована с использованием любого метода или анализа, известного специалисту в данной области техники, для определения уровня активации mTORC1 в присутствии представляющей интерес зкДНК с рапамицином, или его аналогом, или без него. Ожидается, что в присутствии рапамицина будет наблюдаться более значительная активация репортерной молекулы, и что индукция STAT3 цитокина IL-10 и других цитокинов будет уменьшена.
[00560] Чтобы исследовать эффекты совместного введения представляющей интерес зкДНК и рапамицина на активацию врожденных иммунных ответов ex vivo, моноциты человека могут быть выделены, например, с помощью центрифугирования периферической крови в градиенте плотности и магнитного разделения. Эти моноциты можно исследовать до и после контакта и/или активации представляющей интерес зкДНК с рапамицином или его аналогом, или без него, с подходящими контролями. После обработки сыворотку и клеточные супернатанты используют для измерения одного или более цитокиновых путей в качестве функционального показателя, такого как активация mTORC1 и/или IL-10, с использованием любого анализа или метода, известного специалисту в данной области техники. Кроме того, ядерные экстракты можно использовать для проверки активации NF-κB с использованием любого анализа или метода, известного специалисту в данной области техники. Ожидается, что при введении зкДНК в присутствии рапамицина или его аналога будет наблюдаться меньшая активация цитокиновых путей и секреция цитокинов, например, IL-10 и IFN типа I, что приводит к увеличению продолжительности экспрессии трансгена и терапевтической эффективности.
ПРИМЕР 19: Получение зкДНК-вектора, коэкспрессирующего фактор IX и ингибитор TLR-9
[00561] Олигонуклеотиды, которые могут образовывать шпилечную структуру, содержащие следующие последовательности, такие как (TCCTGGCGGGGAAGT, SEQ ID NO: 889), ODN-2114 (TCCTGGAGGGGAAGT, SEQ ID NO: 890), поли(G) (GGGGGGGGGGGGGGGGGGGG, SEQ ID NO: 891), ODN-A151 (TTAGGGTTAGGGTTAGGGTTAGGG, SEQ ID NO: 892), G-ODN (CTCC-TATTGGGGGTTTCCTAT, SEQ ID NO: 893), IRS-869 (TCCTGGAGGGGTTGT, SEQ ID NO: 894), INH-1 (CCTGGATGGGAATTCCCATCCAGG, SEQ ID NO: 895), INH-4 (TTCCCATCCAGGCCTGGATGGGAA, SEQ ID NO: 896), (IRS-661 TGCTTGCAAGCTT-GCAAGCA, SEQ ID NO: 897), 4024 (TCCTGGATGGGAAGT, SEQ ID NO: 898), 4084F (CCTGGATGGGAA, SEQ ID NO: 899), INH-13 (CTTACCGCTGCACCTGGATGGGAA, SEQ ID NO: 900), INH-18 (CCTGGATGGGAACTTACCGCTGCA, SEQ ID NO: 901) и IRS-954 TGCTCCTGGAGGGGTTGT, SEQ ID NO: 902), конструируют так, чтобы получить липкие концы после ренатурации комплементарных цепей 5´-3´ и 3´-5´ так, что они могут быть вставлены путем лигирования в предварительно выбранный сайт рестрикционного клонирования, например, R5 или другой сайт плазмиды TTX 9 или TTX 10, кодирующей трансген фактора IX, как описано в Примере 1 и Примере 4.
[00562] Например, олигонуклеотиды с подходящим сайтом рестрикции ренатурируют путем смешивания каждой цепи в равных молярных количествах в подходящем буфере: например, 100 мМ ацетата калия; 30 мМ HEPES, pH 7,5, нагревания до 94°C в течение 2 минут и постепенного охлаждения. Таким образом, для олигонуклеотидов с прогнозируемой значительной вторичной структурой благоприятна более плавная стадия охлаждения/ренатурации. Это можно осуществить, поместив раствор олигонуклеотида на водяную баню или в термостат и отключив/выключив устройство. Ренатурированные олигонуклеотиды можно разбавить не содержащим нуклеазы буфером и хранить в двухцепочечной ренатурированной форме при 4°С. Затем зкДНК-вектор с олигонуклеотидной последовательностью, ингибирующей TLR-9, очищают (например, с помощью гель-электрофореза или колонки) и используют для получения зкДНК-вектора. Может быть получен зкДНК-вектор, который кодирует фактор IX и содержит антагонист TLR-9, как описано в Примерах 2-3. В настоящем документе описаны способы определения эффектов совместного введения зкДНК-вектора, экспрессирующего ингибитор TLR-9, и рапамицина или аналога рапамицина.
ПРИМЕР 20: Получение зкДНК-вектора, коэкспрессирующего фактор IX и ингибитор cGAS
[00563] Белок ORF52 герпесвируса, ассоциированного с саркомой Капоши (SEQ ID NO: 882), или его вариант, который ингибирует cGAS, или усеченную цитоплазматическую изоформу LANA (LANAΔ161 или SEQ ID NO: 884), в которой отсутствуют аминокислоты 161-1162 из SEQ ID NO: 883), функционально связывают с промотором и вставляют в сайт рестрикционного клонирования R5 плазмиды TTX 9 или TTX 10, кодирующей трансген фактора IX, как описано в Примере 1 и Примере 4. Таким образом, получают зкДНК-вектор, который кодирует как фактор IX, так и ингибитор cGAS, как описано в Примерах 2-3. В настоящем документе описаны способы определения эффектов совместного введения зкДНК-вектора, экспрессирующего ингибитор cGAS, и рапамицина или аналога рапамицина.
ПРИМЕР 21: Устойчивая экспрессия трансгена in vivo зкДНК-векторами, изготовленными в виде составов с ЛНЧ
[00564] Воспроизводимость результатов, полученных в Примере 7, с другой липидной наночастицей оценивали in vivo на мышах. В день 0 мышам вводили дозу либо зкДНК-вектора, содержащего трансген люциферазы, управляемый промотором CAG, который был инкапсулирован в ЛНЧ, отличающиеся от использованных в Примере 6, либо тех же ЛНЧ, содержащих поли(C), но без зкДНК или гена люциферазы. В частности, самцам мышей CD-1® в возрасте приблизительно 4 недель вводили путем однократной инъекции 0,5 мг/кг ЛНЧ-TTX-люциферазы или контрольных ЛНЧ-поли(C), вводимых внутривенно через боковую хвостовую вену в день 0. В день 14 животным вводили системно люциферин в дозе 150 мг/кг путем внутрибрюшинной инъекции при 2,5 мл/кг. Приблизительно через 15 минут после введения люциферина каждое животное визуализировали с использованием системы визуализации in vivo («IVIS»).
[00565] Значительная флуоресценция в печени наблюдалась у всех четырех мышей, получивших зкДНК, при этом, за исключением места инъекции, у животных наблюдалась очень незначительная флуоресценция, это указывает на то, что ЛНЧ опосредовали специфичную для печени доставку конструкции зкДНК, и что доставленный зкДНК-вектор был способен к контролируемой устойчивой экспрессии своего трансгена в течение по меньшей мере двух недель после введения.
ПРИМЕР 22: Устойчивая экспрессия трансгена в печени in vivo после введения зкДНК-вектора
[00566] В отдельном эксперименте оценивали локализацию зкДНК, доставленной с помощью ЛНЧ, в печени обработанных животных. ЗкДНК-вектор, содержащий представляющий интерес функциональный трансген, инкапсулировали в те же ЛНЧ, что и использованные в Примере 17, и вводили мышам in vivo при уровне дозы 0,5 мг/кг путем внутривенной инъекции. Через 6 часов мышей умерщвляли и отбирали образцы печени, фиксировали формалином и заливали парафином с использованием стандартных протоколов. Анализы методом гибридизации in situ RNAscope® выполняли для визуализации зкДНК-векторов в ткани с использованием зонда, специфичного в отношении трансгена зкДНК, и детектирования с использованием хромогенной реакции и окрашивания гематоксилином (Advanced Cell Diagnostics). Визуализирующий анализ подтвердил, что зкДНК присутствовала в образцах гепатоцитов, взятых у обработанных мышей. Специалист поймет, что люциферазу в зкДНК-векторе можно заменить любой последовательностью нуклеиновой кислоты, выбранной из Таблицы 5.
ПРИМЕР 23: Устойчивая экспрессия трансгена зкДНК в глазах in vivo
[00567] Устойчивость экспрессии трансгена зкДНК-вектора в тканях, отличных от печени, оценивали для определения переносимости и экспрессии зкДНК-вектора после введения в глаза in vivo. Несмотря на то, что люциферазу использовали в качестве примерного трансгена, обычный специалист может легко заменить трансген люциферазы последовательностью антагониста инфламмасомы из любой из последовательностей, перечисленных в Таблице 5A-5F.
[00568] В день 0 самцам крыс Sprague Dawley в возрасте приблизительно 9 недель вводили путем субретинальной инъекции 5 мкл либо зкДНК-вектора, содержащего трансген люциферазы, изготовленного в виде состава с реагентом для трансфекции jetPEI® (Polyplus), либо плазмидной ДНК, кодирующей люциферазу, изготовленной в виде состава с jetPEI®, в обоих случаях в концентрации 0,25 мкг/мкл. В каждой группе тестировали по четыре крысы. Животным давали седативное средство и вводили исследуемый препарат в правый глаз путем субретинальной инъекции с помощью иглы калибра 33. Левый глаз каждого животного оставляли интактным. Сразу после инъекции глаза проверяли методом оптической когерентной томографии или визуализации глазного дна, чтобы подтвердить наличие субретинального пузырька. Крысы получали бупренорфин и местную мазь с антибиотиком в соответствии со стандартными процедурами.
[00569] В дни 7, 14, 21, 28 и 35 животным в обеих группах вводили системно свежеприготовленный люциферин в дозе 150 мг/кг путем внутрибрюшинной инъекции при 2,5 мл/кг. Через 5-15 минут после введения люциферина всех животных визуализировали с использованием IVIS под анестезией изофлураном. Полный поток [фотон/с] и средний поток (фотон/с/ср/см2) в представляющей интерес области, включающей глаз, получали при экспозиции 5 минут. Результаты представлены графически в виде средней светимости в обработанном глазу («инъецированный») для каждой группы лечения относительно средней светимости в необработанном глазу («неинъецированный») для каждой группы лечения. Значительная флуоресценция легко детектировалась в глазах, в которые был введен зкДНК-вектор, но была намного слабее в глазах, в которые была введена плазмида. Через 35 дней крыс, которым инъецировали плазмиду, умерщвляли, продолжая исследование для крыс, получивших зкДНК, с инъекцией люциферина и визуализацией IVIS в дни 42, 49, 56, 63, 70 и 99. Результаты демонстрируют, что зкДНК-вектор, введенный в глаз крысы путем однократной инъекции, опосредует экспрессию трансгена in vivo и что такая экспрессия поддерживалась на высоком уровне на протяжении по меньшей мере 99 дней после инъекции.
ПРИМЕР 24: Гидродинамическая доставка зкДНК
[00570] Гидродинамическая инъекция в хвостовую вену является хорошо известным методом введения нуклеиновой кислоты в печень грызунов. В этой системе инъекция под давлением большого объема неинкапсулированной нуклеиновой кислоты приводит к кратковременному увеличению проницаемости клеток и доставке непосредственно в ткани и клетки. Это обеспечивает экспериментальный механизм обхода многих иммунных систем хозяина, таких как привлечение макрофагов. Соответственно, экспрессию люциферазы, наблюдаемую после гидродинамической инъекции незащищенного (naked) зкДНК-вектора, сравнивали с экспрессией, наблюдаемой после более традиционной внутривенной инъекции зкДНК, инкапсулированной в ЛНЧ. Для этого эксперимента в зкДНК-векторах использовали левый ITR ААВ2 дикого типа и мутированный правый ITR.
[00571] В общих чертах, готовили зкДНК-вектор, кодирующий люциферазу под контролем промотора CAG, который либо инкапсулировали в ЛНЧ, либо оставили неинкапсулированным. Взрослым самцам мышей CD-1 вводили путем инъекции в хвостовую вену либо (i) инкапсулированный в ЛНЧ зкДНК-вектор в дозе 0,5 мг/кг в общем объеме 5 мл/кг, либо (ii) тот же вектор, но неинкапсулированный, в дозе 0,01 мг/кг в общем объеме 1,2 мл. В каждой группе лечения было по три мыши. Значения массы тела регистрировали в дни 1, 2 и 3. Прижизненную визуализацию выполняли в дни 1 и 3 с использованием системы визуализации in vivo (IVIS). Для визуализации каждой мыши вводили путем внутрибрюшинной инъекции 150 мг/кг люциферина при 2,5 мл/кг. Через 15 минут каждую мышь анестезировали и визуализировали.
[00572] Даже при введении в 50-кратно более низкой дозе экспрессия люциферазы, наблюдаемая у мышей, получивших гидродинамическую инъекцию, была намного выше (~1010 максимальный полный поток), чем у мышей, не получивших гидродинамическую инъекцию (~107 максимальный полный поток) (ФИГ. 9). В предыдущих исследованиях было обнаружено, что введение ЛНЧ по отдельности без груза зкДНК-вектора не вызывало иммунного ответа (данные не показаны), и, таким образом, разница между двумя группами дозирования может быть обусловлена связыванием ЛНЧ-инкапсулированного зкДНК-вектора одной или более иммунными системами хозяина и избеганием этих систем с помощью гидродинамического введения.
ПРИМЕР 25: Модуляция иммунных путей в культивируемых клетках и влияние на экспрессию зкДНК-вектора
[00573] Был разработан клеточный анализ, чтобы облегчить исследование вклада различных иммунных путей в ответ хозяина на введение зкДНК. В анализе используются клетки THP-1 (линия клеток острого моноцитарного лейкоза) в нескольких вариантах: клетки THP-1 Dual™ (Invitrogen) со стабильной интеграцией репортерных конструкций для детектирования как активации NF-κB (путь TLR9, за счет детектирования SEAP с использованием Quanti-Blue™), так и активации пути IRF (за счет секретируемой люциферазы с использованием Quanti-Luc™), клетки THP-1 с конститутивным подавлением иммунного пути cGAS и клетки THP-1 с конститутивным подавлением иммунного пути STING. Используя известные ингибиторы определенных путей, можно лучше понять относительный вклад эндогенных иммунных путей в наблюдаемый иммунный ответ на конкретный стимул.
[00574] В общих чертах, клетки THP-1 в культуре разбавляли до 0,5×106/мл в средах Opti-MEM™ (ThermoFisher) и добавляли по 150 мкл в каждую лунку 96-луночного планшета. Клетки предварительно обрабатывали ингибиторами: целевые ингибиторы разбавляли в Opti-MEM™ и добавляли в предусмотренные лунки с образцами. Для этого эксперимента в каждой лунке с образцом использовали A151 (олигонуклеотид TTAGGGTTAGGGTTAGGGTTAGGG (SEQ ID NO:892) и BX795 (N-[3-[[5-йод-4-[[3-[(2-тиенилкарбонил)амино]пропил]амино]-2-пиримидинил]амино]фенил]-1-пирролидинкарбоксамид, CAS 702675-74-9) в конечных концентрациях 0 мкМ, 0,625 мкМ, 1,25 мкМ или 2,5 мкМ. Планшеты инкубировали при 37°C в течение 2 часов. 200 нг целевого зкДНК-вектора разводили 1:3 в липофектамине™ 3000 и инкубировали в течение 5-10 мин при комнатной температуре. Затем в лунки с образцами добавляли комплекс зкДНК вектор-липофектамин. Планшеты инкубировали в течение 24 часов при 37°C. Величину активации NF-κB и активации IRF2 количественно оценивали с использованием наборов Quanti-Blue™ и Quanti-Luc™, соответственно, в соответствии с инструкциями производителя.
[00575] Введение двух разных препаратов зкДНК-вектора в репортерные клетки THP-1 Dual™ привело к значительной индукции интерферона для обоих препаратов, это указывает на активацию по меньшей мере одного иммунного пути (ФИГ. 10A). Примечательно, что индукцию интерферона не наблюдали, когда любая из двух линий THP-1 с подавленными генами была обработана зкДНК в одинаковой концентрации (ФИГ. 10А), это указывает на то, что путь cGAS/STING участвует в индукции цитокинов в ответ на введение зкДНК. Аналогичный результат был обнаружен при обработке репортерных клеток THP-1 Dual™ как зкДНК, так и BX795; BX795 является специфичным ингибитором пути STING, и то, что он устраняет зкДНК-индуцированную индукцию интерферона, снова указывает на вовлечение пути STING (ФИГ. 10A). Известно, что A151 ингибирует путь cGAS/STING, путь TLR9, а также опосредуемые инфламмасомами иммунные пути. Его эффект был аналогичен тому, который наблюдали при обработке BX795 (ФИГ. 10A).
[00576] Во втором эксперименте анализировали концентрации ингибитора, необходимые для наблюдения защитного эффекта при введении зкДНК (ФИГ. 10B). Как для A151, так и для AS1411 наблюдаемое ингибирование индукции интерферона зависело от концентрации, при этом максимальное ингибирование наблюдали при концентрации 2,5 мкМ (ФИГ. 10B).
ПРИМЕР 26: Влияние модуляции содержания неметилированного CpG в зкДНК на иммунный ответ
[00577] Известно, что мотивы CpG в последовательности гена стимулируют путь TLR9, чувствительный к ДНК. Соответственно, изучали влияние уменьшения количества мотивов CpG в последовательности конструкции зкДНК на активацию пути врожденного иммунитета при введении этой последовательности in vivo.
A. Клеточные анализы для тестирования влияния минимизации содержания неметилированного CpG в зкДНК
[00578] Выполняли исследования для оценки (i) активации пути TLR9 в ответ на введение зкДНК, и (ii) эффекта модуляции присутствия/статуса метилирования CpG на такую активацию. Для этого конкретного исследования использовали зкДНК-вектор, который экспрессировал зеленый флуоресцентный белок и содержал левый ITR дикого типа и мутантный правый ITR.
[00579] Клетки HEK-293, экспрессирующие TLR9 человека (клетки HEK-BLUE.hTLR9, InvivoGen), высевали в 96-луночный планшет при 50000 клеток на лунку. Планшеты инкубировали в течение ночи при 37°C. Для образцов зкДНК, подвергнутых предварительной обработке метилированием, зкДНК-вектор, буфер, S-аденозилметионин, CpG-метилтрансферазу и воду смешивали до достижения общего реакционного объема 50 мкл в соответствии с методами, известными в данной области техники. Реакционную смесь инкубировали при 37°C в течение 1 часа, затем останавливали реакцию нагреванием до 65°C в течение 20 минут. ЗкДНК очищали из реакционной смеси с использованием коммерчески доступного набора для очистки (набор для очистки ПЦР, Qiagen®) и измеряли концентрацию полученной ДНК.
[00580] Клетки предварительно обрабатывали в течение 3 часов любыми целевыми ингибиторами, в этом эксперименте A151 использовали в конечной концентрации 10 мкМ на лунку. После предварительной обработки клетки трансфицировали 300 нг зкДНК в соотношении 1:3 с липофектамином 3000, разведенным в Opti-MEM™, или положительным контролем ODN2006, который, как известно, стимулирует путь TLR9. Клетки инкубировали в течение 24 часов при 37°C и 5% CO2. Затем измеряли экспрессию SEAP (компонент пути TLR9) с использованием Quanti-BLUE™ (InvivoGen).
[00581] Как показано на ФИГ. 11A, ODN2006 индуцирует устойчивый ответ NF-κB; конструкция зкДНК индуцировала менее значительный ответ, а при предварительном метилировании ответ снижался до фонового уровня. При комбинировании с A151 (известно, что он ингибирует путь TLR9) образцы, обработанные зкДНК, также показали минимальные уровни индукции NF-κB (ФИГ. 11B). Во-первых, это демонстрирует, что путь TLR9 вносит вклад в иммунный ответ хозяина на введение зкДНК. Кроме того, минимизация содержания CpG путем метилирования устраняла большую часть активации TLR9 под действием зкДНК, и этот эффект мог быть имитирован путем предварительной обработки клеток A151 без изменения содержания или статуса метилирования CpG.
B. Исследования на мышах, в которых оценивают влияние минимизации содержания неметилированного CpG в зкДНК
[00582] Влияние минимизации содержания CpG в зкДНК-векторах также оценивали на мышах. Измеряли цитокиновый ответ и экспрессию генов, кодируемых зкДНК, при введении зкДНК-векторов мышам.
[00583] Использовали три разных зкДНК-вектора, каждый из которых кодировал люциферазу в качестве трансгена. Первый зкДНК-вектор имел большое количество неметилированного CpG (~350) («зкДНК с высоким содержанием CpG») и содержал конститутивный промотор CAG; второй имел умеренное количество неметилированного CpG (~60) («зкДНК с низким содержанием CpG») и содержал специфический для печени промотор hAAT; и третий представлял собой метилированную форму второго варианта так, что он не содержал неметилированного CpG (««зкДНК без CpG») и также содержал промотор hAAT. В остальном зкДНК-векторы были идентичны. Векторы готовили, как описано выше.
[00584] Четыре группы по четыре самца мышей CD-1, в возрасте приблизительно 4 недель, получали один из зкДНК-векторов, инкапсулированных в ЛНЧ, или контроль поли(C). В день 0 каждой мыши водили путем однократной внутривенной инъекции в хвостовую вену 0,5 мг/кг зкДНК-вектора в объеме 5 мл/кг. Значения массы тела регистрировали в дни -1, -, 1, 2, 3, 7 и еженедельно после этого, до умерщвления мышей. Образцы цельной крови и сыворотки брали в дни 0, 1 и 35. Прижизненную визуализацию проводили в дни 7, 14, 21, 28 и 35, и затем еженедельно с использованием системы визуализации in vivo (IVIS). Для визуализации каждой мыши вводили путем внутрибрюшинной инъекции 150 мг/кг люциферина при 2,5 мл/кг. Через 15 минут каждую мышь анестезировали и визуализировали. Мышей умерщвляли в день 93 и собирали терминальные ткани, включая печень и селезенку. Измерения цитокинов проводили через 6 часов после введения дозы в день 0.
[00585] Сходную потерю массы тела наблюдали в каждой из групп мышей, обработанных зкДНК (5-7%), с последующим быстрым восстановлением ко дню 7. Анализы цитокинов из образцов со дня 0 показали, что, хотя многие из оцененных цитокинов были одинаково повышены во всех группах лечения, интерферон альфа, фактор некроза опухоли альфа и MIP-1 альфа были все снижены в образцах с низким содержанием CpG или без CpG по сравнению с образцами с высоким содержанием CpG (ФИГ. 12A и ФИГ. 12B).
[00586] В то время как у мышей, обработанных либо зкДНК с низким содержанием CpG, либо зкДНК с высоким содержанием CpG, наблюдали значительную флуоресценцию в дни 7 и 14, у мышей, обработанных зкДНК с высоким содержанием CpG, флуоресценция быстро снижалась после 14 дня и непрерывно снижалась до конца исследования. Напротив, полный поток для мышей, обработанных зкДНК с низким содержанием CpG, и мышей, обработанных зкДНК без CpG, оставался на постоянном высоком уровне (ФИГ. 12C), это указывает на то, что сохранение наличия неметилированного CpG в векторе зкДНК ниже некоторого порога и, соответственно, отсутствие запуска пути TLR9, помогает избежать в этом исследовании более быстрого снижения экспрессии белка, кодируемого зкДНК (и, следовательно, флуоресценции), наблюдаемого в других случаях.
ПРИМЕР 27: Экспрессия и ответ хозяина у новорожденных мышей
[00587] Предыдущие эксперименты показали, что путь cGAS/STING по меньшей мере частично участвует в ответе индукции цитокинов, наблюдаемом при введении зкДНК-вектора в клетки. Известно, что этот путь становится активным позже в процессе развития, поэтому у новорожденных мышей с незрелой иммунной системой отсутствует активный путь cGAS/STING. Соответственно, проводили эксперимент на новорожденных мышах, чтобы исследовать эффект отсутствия данного пути на экспрессию и устойчивость зкДНК-вектора.
[00588] Использовали зкДНК-вектор, кодирующий люциферазу в качестве трансгена, с левым ITR ААВ2 дикого типа и правым мутантным ITR и промотором CAG. ЗкДНК-вектор готовили, как описано выше. Образцы зкДНК-вектора или контроль поли(C) вводили путем внутривенной инъекции в хвостовую вену новорожденных (8-дневных) самцов мышей CD-1 при уровне дозы 0,1 или 0,5 мг/кг в объеме до 5 мл/кг. В каждую группу образцов включали пять повторов. Значения массы тела регистрировали в первый день и в последующие три дня. Прижизненную визуализацию выполняли в дни 7, 14 и 21 с использованием системы визуализации in vivo (IVIS). Для визуализации каждой мыши вводили путем внутрибрюшинной инъекции 150 мг/кг люциферина при 2,5 мл/кг. Через 15 минут каждую мышь анестезировали и визуализировали.
[00589] Примечательно, что после инъекции нулевого дня потерю массы не наблюдали ни в одной из групп лечения. Высокие уровни полного потока (репрезентативные для экспрессии люциферазы с введенных зкДНК-векторов) наблюдали у всех животных, которым вводили зкДНК, при этом доза 0,5 мг/кг приводила к уровню экспрессии приблизительно на 1 log выше, чем доза 0,1 мг/кг, в первые 14 дней (ФИГ. 13). После этого уровень экспрессии стабилизировался и сохранялся на том же уровне в обеих группах дозирования. По сравнению с аналогичными исследованиями на взрослых мышах CD-1 уровень экспрессии зкДНК-вектора у новорожденных мышей даже через 14 дней был по меньшей мере на два log выше (данные не показаны). Этот результат свидетельствует о том, что избегание активации пути cGAS/STING является благоприятным для стимулирования экспрессии и устойчивости зкДНК-вектора.
ПРИМЕР 28: Влияние модуляции нескольких иммунных путей на устойчивость и экспрессию зкДНК, а также индукцию цитокинов зкДНК
[00590] В предыдущих исследованиях оценивали эффекты модуляции пути TLR9 как в культивируемых клеточных системах, так и в системах на мышах. Однако известно, что в ответ хозяина на чужеродную ДНК вовлечено несколько молекулярных путей, и влияние избегания запуска пути TLR9 может быть нелегко обнаружить, если один или более других путей все еще задействованы введением зкДНК. Чтобы проверить это, CpG-минимизированные зкДНК-векторы тестировали на линии мышей goldenticket, имеющих мутацию, которая устраняет функцию STING. Таким образом, эксперимент позволил исследовать путь TLR9, без искажающего эффекта активности пути cGAS/STING.
[00591] Использовали три разных зкДНК-вектора, каждый из которых кодировал люциферазу в качестве трансгена. Первый зкДНК-вектор имел большое количество неметилированного CpG (~350) («зкДНК с высоким содержанием CpG») и содержал конститутивный промотор (cET), второй имел умеренное количество неметилированного CpG (~60) («зкДНК с низким содержанием CpG»), а третий имел небольшое количество CpG (~36), но был метилирован таким образом, что не содержал неметилированного CpG («зкДНК без CpG»). Вторая и третья конструкции содержали промоторы hAAT, специфические для печени. В остальном зкДНК-векторы были идентичны. Векторы готовили, как описано выше.
[00592] Каждый из образцов зкДНК-вектора или контроль поли(C) вводили путем внутривенной инъекции в хвостовую вену взрослым самцам мышей goldenticket (Tmem173gt) при уровне дозы 0,5 мг/кг в объеме 5 мл/кг. В некоторых случаях мышам вводили вторую дозу образца зкДНК-вектора в день 22. В каждую группу образцов включали четыре повтора. Значения массы тела регистрировали в дни введения дозы и в последующие три дня. Образцы цельной крови и сыворотки брали в день 0 (через 6 часов после введения дозы) и день 22 (через 6 часов после введения дозы). Прижизненную визуализацию выполняли в дни 7, 14, 22, 29, 36 и 43 с использованием системы визуализации in vivo (IVIS). Для визуализации каждой мыши вводили путем внутрибрюшинной инъекции 150 мг/кг люциферина при 2,5 мл/кг. Через 15 минут каждую мышь анестезировали и визуализировали. Мышей умерщвляли в день 43 и собирали терминальные ткани, включая печень и селезенку. Измерения цитокинов осуществляли в крови, взятой в День 0 и 22.
[00593] Потеря массы тела после введения зкДНК составляла менее 5% и по существу восстанавливалась во всех случаях ко дню 3. После повторного введения в день 22 обработанные мыши снова потеряли <5% массы тела и быстро восстановили ее в следующие дни. Индукцию цитокинов оценивали в образцах крови со дня 0 (ФИГ. 14A). За исключением IL-18, уровни каждого из анализируемых цитокинов коррелировали со степенью присутствия CpG в конструкции зкДНК (ФИГ. 14A), при этом у мышей, обработанных зкДНК без CpG, индукция IFN-альфа, IFN-гамма, IL-6, IP-10, MCP-1, MIP-1 альфа, MIP-1 бета и RANTES была незначительной или отсутствовала. У мышей, получивших повторную дозу, во всех образцах наблюдали увеличение уровней всех цитокинов относительно показателей в день 0 (ФИГ. 14B), но снова во всех случаях, кроме IL-18, степень активации коррелировала с количеством CpG, присутствующего во введенном зкДНК-векторе.
[00594] Экспрессия люциферазы в различных группах обработки была сходной до дня 22 (ФИГ. 14C). После этой временной точки в образцах мышей, получивших либо однократную дозу, либо повторную дозу зкДНК с высоким содержанием CpG, наблюдали резкое снижение полного потока, в то время как в группах, получивших зкДНК с низким содержанием CpG и зкДНК без CpG, либо сохранялись одинаковые измерения полного потока, либо потеря сигнала была слабее относительно группы, получившей зкДНК с высоким содержанием CpG. Объединенные результаты демонстрируют, что минимизация содержания CpG во введенных зкДНК-векторах и, как следствие, избегание задействования пути TLR9 врожденного иммунитета, внесла вклад в выраженное падение индукции цитокинов и более надежную устойчивость экспрессии генов с зкДНК у обработанных мышей goldenticket.
[00595] ПРИМЕР 29: Длительное введение доз и введение повторных доз зкДНК-вектора у мышей Rag2.
[00596] В ситуациях, когда один или более трансгенов, кодируемых в экспрессионной генной кассете зкДНК-вектора, экспрессируются в среде хозяина (например, в клетке или в организме субъекта), где экспрессированный белок распознается как чужеродный, существует вероятность того, что у хозяина выработается адаптивный иммунный ответ, который может привести к нежелательному истощению продукта экспрессии, которое может быть ошибочно принято за отсутствие экспрессии. В некоторых случаях это может происходить с репортерной молекулой, которая является гетерологичной для нормальной среды хозяина. Соответственно, экспрессию трансгена зкДНК-вектора оценивали in vivo в модели на мышах Rag2, в которой отсутствуют B- и T-клетки и, следовательно, не вырабатывается адаптивный иммунный ответ на ненативные для мышей белки, такие как люцифераза. В общих чертах, в день 0 мышам линий c57bl/6 и Rag2 с подавленными генами вводили путем внутривенной инъекции в хвостовую вену 0,5 мг/кг инкапсулированного в ЛНЧ зкДНК-вектора, экспрессирующего люциферазу, или контроля поли(C), а в день 21 некоторым мышам вводили повторную дозу того же самого ЛНЧ-инкапсулированного зкДНК-вектора при том же уровне дозы. Все тестируемые группы состояли из 4 мышей в каждой. Визуализацию IVIS выполняли после инъекции люциферина с недельными интервалами.
[00597] При сравнении полного потока, наблюдаемого в анализах IVIS, флуоресценция, наблюдаемая у мышей дикого типа (косвенная мера присутствия экспрессированной люциферазы), которым вводили ЛНЧ-зкДНК-вектор-Luc, постепенно снижалась после дня 21, в то время как мыши Rag2, которым вводили тот же самый препарат, проявили относительно постоянную устойчивую экспрессию люциферазы на протяжении 42 дней эксперимента (ФИГ. 16А). Наблюдаемое приблизительно в день 21 снижение у мышей дикого типа соответствует временному интервалу, в котором можно ожидать появления адаптивного иммунного ответа. Повторное введение ЛНЧ с зкДНК-вектором у мышей Rag2 приводило к заметному увеличению экспрессии, которая была устойчивой в течение по меньшей мере 21 дня, на протяжении которых ее отслеживали в этом исследовании (ФИГ. 16B). Результаты позволяют предположить, что адаптивный иммунитет может играть определенную роль при экспрессии ненативного белка с зкДНК-вектора у хозяина, и что наблюдаемое снижение экспрессии в период времени 20+ дней с момента первоначального введения может указывать на мешающий адаптивный иммунный ответ на экспрессируемую молекулу, а не на снижение экспрессии (или в дополнение к ней). Следует отметить, что этот ответ, как ожидается, будет низким в случае экспрессии у хозяина нативных белков, когда, предположительно, хозяин будет правильно распознавать экспрессированные молекулы как собственные, и у него не будет вырабатываться такой иммунный ответ.
ПРИМЕР 30: Влияние специфической для печени экспрессии и модуляции CpG на устойчивую экспрессию
[00598] Как описано в Примере 29, нежелательный иммунный ответ хозяина в некоторых случаях может искусственно ослаблять то, что иначе могло бы быть устойчивой экспрессией одного или более целевых трансгенов с введенного зкДНК-вектора. Чтобы оценить влияние избегания и/или ослабления потенциального иммунного ответа хозяина на устойчивую экспрессию с зкДНК-вектора, использовали два подхода. Во-первых, поскольку вектор зкДНК-Luc, использованный в предшествующих примерах, находился под контролем конститутивного промотора CAG, была получена аналогичная конструкция с использованием специфического для печени промотора (hAAT) или другого конститутивного промотора (hEF-1), чтобы определить, будет ли избегание длительного воздействия миелоидных клеток или тканей, не относящихся к печени, уменьшать любые наблюдаемые иммунные эффекты. Во-вторых, некоторые конструкции зкДНК-люцифераза модифицировали так, чтобы иметь уменьшенное содержание CpG, известного триггера иммунной реакции хозяина. Измеряли экспрессию гена люциферазы, кодируемой зкДНК, при введении мышам таких модифицированных зкДНК-векторов с замененным промотором.
[00599] Использовали три разных зкДНК-вектора, каждый из которых кодировал люциферазу в качестве трансгена. Первый зкДНК-вектор имел большое количество неметилированного CpG (~350) и содержал конститутивный промотор CAG («зкДНК CAG»); второй имел умеренное количество неметилированного CpG (~60) и содержал специфический для печени промотор hAAT («зкДНК hAAT с низким содержанием CpG»); и третий представлял собой метилированную форму второго, не содержащую неметилированного CpG, и также содержал промотор hAAT («зкДНК hAAT без CpG»). В остальном зкДНК-векторы были идентичны. Векторы готовили, как описано выше.
[00600] Четыре группы по четыре самца мышей CD-1®, в возрасте приблизительно 4 недель, получали один из зкДНК-векторов, инкапсулированных в ЛНЧ, или контроль поли(C). В день 0 каждой мыши вводили путем однократной внутривенной инъекции в хвостовую вену 0,5 мг/кг зкДНК-вектора в объеме 5 мл/кг. Значения массы тела регистрировали в дни -1, -, 1, 2, 3, 7 и еженедельно после этого, до умерщвления мышей. Образцы цельной крови и сыворотки брали в дни 0, 1 и 35. Прижизненную визуализацию выполняли в дни 7, 14, 21, 28 и 35, и затем еженедельно с использованием системы визуализации in vivo (IVIS). Для визуализации каждой мыши вводили путем внутрибрюшинной инъекции 150 мг/кг люциферина при 2,5 мл/кг. Через 15 минут каждую мышь анестезировали и визуализировали. Мышей умерщвляли в день 93 и собирали терминальные ткани, включая печень и селезенку. Измерения цитокинов проводили через 6 часов после введения дозы в день 0.
[00601] Несмотря на то, что у всех получивших зкДНК мышей наблюдали значительную флуоресценцию в дни 7 и 14, у мышей, получивших зкДНК CAG, флуоресценция быстро снижалась после дня 14 и более постепенно снижалась на протяжении оставшейся части исследования. Напротив, у мышей, получивших зкДНК hAAT с низким содержанием CpG и без CpG, полный поток оставался на постоянном высоком уровне (ФИГ. 17). Это позволяет предположить, что направление доставки зкДНК-вектора специфически в печень приводило к устойчивой и длительной экспрессии трансгена с вектора в течение по меньшей мере 77 дней после однократной инъекции. Конструкции с минимизированным содержанием CpG или с полным отсутствием CpG имели сходные профили длительной устойчивой экспрессии, в то время как конструкция с конститутивным промотором и высоким содержанием CpG проявила снижение экспрессии с течением времени, это позволяет предположить, что иммунная активация хозяина при введении зкДНК-вектора может играть определенную роль в любом снижении экспрессии, наблюдаемом при введении такого вектора субъекту. Эти результаты обеспечивают альтернативные методы адаптации продолжительности ответа к желаемому уровню путем выбора ограниченного конкретной тканью промотора и/или изменения содержания CpG в зкДНК-векторе в том случае, если наблюдается иммунный ответ хозяина - потенциально, специфичный в отношении трансгена ответ.
ПРИМЕР 31: Экспрессия антагониста инфламмасомы in vivo
[00602] После подтверждения надлежащей экспрессии и функции белка в клетках-реципиентах in vitro, зкДНК-вектор с последовательностями, кодирующими антагонист инфламмасомы, может быть изготовлен с липидными наночастицами и введен мышам с дефицитом функциональной экспрессии продукта соответствующего белка в различные моменты времени (in utero, новорожденный, в возрасте 4 недель и 8 недель) для проверки экспрессии и функции белка in vivo.
[00603] ЛНЧ с зкДНК-векторами вводят соответствующим мышам в дозах от 0,3 до 5 мг/кг в объеме 1,2 мл. Каждая доза должна быть введена путем внутривенного гидродинамического введения или будет введена, например, с помощью внутрибрюшинной инъекции. Введение нормальным мышам является контролем, а также может быть использовано для детектирования наличия и количества терапевтического белка.
[00604] После однократного введения, например, однократной дозы ЛНЧ-зкДНК, экспрессия в ткани печени у мыши-реципиента будет определяться в различные моменты времени, например, через 10, 20, 30, 40, 50, 1000 и 200 дней или более. и т.д. В частности, образцы печени и желчного протока мыши будут получены и проанализированы на наличие белка с использованием иммуноокрашивания срезов ткани. Наличие белка будет оцениваться количественно, а также в отношении надлежащей локализации в ткани и клетках в ней. Клетки в печени (например, печеночные и эпителиальные) и желчного протока (например, холангиоциты) будут оцениваться для определения экспрессии белка.
ПРИМЕР 32: Получение зкДНК, коэкспрессирующей терапевтический ген (например, фактор IX) и ингибитор пути инфламмасомы NLRP3.
[00605] A151 (SEQ ID NO: 892) или его вариант, который ингибирует AIM2, функционально связывают с промотором и вставляют в сайт рестрикционного клонирования R5 зкДНК-вектора, как описано в Примере 1. Таким образом, получают зкДНК, которая кодирует как фактор IX, так и ингибитор AIM2.
ПРИМЕР 33: Подтверждение экспрессии ингибитора инфламмасомы NLRP3, экспрессируемого зкДНК
[00606] Экспрессию целевого ингибитора NLRP3 или AIM2, или каспазы-1, коэкспрессируемого зкДНК, такого как A151 (SEQ ID NO: 892), можно подтвердить с использованием клеток HeLa и антител, специфичных в отношении ингибитора. Например, культивируют клетки HeLa и выполняют кратковременные трансфекции конструкций, коэкспрессирующих фактор IX и целевой ингибитор NLRP3 или AIM2, или каспазы-1, используя, например, реагент для трансфекции Fusegene6 (3:1; fusgene6:ДНК). Для детектирования экспрессии ингибитора NLRP3 или AIM2, или каспазы-1 используют методики Вестерн-блоттинга и/или проточной цитометрии, как известно специалистам в данной области техники.
ПРИМЕР 34: Фактор IX для лечения гемофилии B с применением зкДНК, кодирующей фактор IX и ингибитор пути инфламмасомы NLRP3.
[00607] Эксперимент проводят на мышах с дефицитом фактора IX, которые имеют вставленную последовательность hFIX с вредной мутацией (R333Q). Самцы мышей с подавленным фактором IX получают однократные или повторные дозы ЛНЧ-зкДНК (липидные наночастицы с зкДНК). Используют два варианта ЛНЧ с зкДНК-вектором; 1) ЛНЧ-зкДНК, кодирующей фактор IX человека (либо нативную последовательность человека, либо варианты Padua FIX) и кодирующей A151 (SEQ ID NO: 892); ЛНЧ-зкДНК, кодирующей только фактор IX, но не ингибитор cGAS, в качестве сравнительного зкДНК-вектора. ЛНЧ с зкДНК-векторами вводят соответствующим мышам в дозах от 0,3 до 5 мг/кг в объеме 1,2 мл. Каждая доза должна быть введена путем в/в гидродинамического введения. Экспрессию фактора IX в плазме оценивают с помощью ИФА в различные моменты времени, например, через 7, 14 и 21 день или более и т.д. Активированное частичное тромбопластиновое время и время кровотечения также измеряют как определение эффективности. Ожидается, что у мышей, которые получают зкДНК-вектор, экспрессирующий как hFIX, так и A151, повышенная и/или устойчивая экспрессия фактора IX будет наблюдаться в течение более длительного периода времени по сравнению с мышами, которые получают зкДНК-вектор, экспрессирующий только фактор IX, но не A151 или другой ингибитор NLRP3 или AIM2, или каспазы-1. Кроме того, ожидается, что после повторного введения дозы у мышей, которые получают повторную дозу зкДНК-вектора, содержащего как A151, так и фактор IX, будет наблюдаться меньшая активация секреции цитокинов и увеличенная продолжительность экспрессии трансгена и терапевтическая эффективность по сравнению с мышами, получившими повторную дозу зкДНК-вектора, кодирующего только фактор IX. Ингибитор пути инфламмасомы NLRP3 и фактор IX могут быть доставлены на разных зкДНК-векторах, но предпочтительно они кодируются одним и тем же вектором, и, соответственно, ингибирование ингибитором пути инфламмасомы NLRP3 происходит в той же клетке, что и клетка, которая получает зкДНК-вектор, кодирующий трансген, такой как фактор IX.
ПРИМЕР 35: Определение эффектов совместного введения зкДНК и ингибитора инфламмасомы NLRP3 на врожденные иммунные ответы и продолжительность экспрессии фактора IX
[00608] Чтобы исследовать эффекты совместного введения представляющей интерес зкДНК и ингибитора пути инфламмасомы NLRP3 на врожденные иммунные ответы in vitro, можно использовать репортерные линии для функциональных анализов, в которых исследуется активация инфламмасомы NLRP3 или каспазы-1. Репортерная линия клеток инфламмасомы NLRP3, которую можно применять для таких анализов in vitro, может представлять собой стабильно котрансфицированную линию клеток, которая экспрессирует полноразмерный NLRP3 и репортерный ген, такой как репортерный ген секретируемой щелочной фосфатазы (SEAP), под транскрипционным контролем элемента ответа транскрипционного фактора, такого как сайт связывания NF-kB, сайт связывания AP-1 или их комбинация. Например, репортерные клетки высевают в 96-луночные планшеты. По истечении заранее определенного периода времени, например, 16 часов, клетки стимулируют различными количествами композиций, содержащих зкДНК, экспрессирующую фактор IX, с ингибитором инфламмасомы NLRP3 или без него. Активность репортерного гена, такого как SEAP, может быть проанализирована с использованием любого метода или анализа, известного специалисту в данной области техники, для сравнения уровня активации каспазы-1 или активации инфламмасомы NLRP3 в присутствии представляющей интерес зкДНК с ингибитором пути инфламмасомы NLRP3 или без него. Ожидается, что в присутствии ингибитора инфламмасомы NLRP3 будет наблюдаться меньшая активация репортерной молекулы. Аналогичный репортерный анализ можно использовать для оценки ингибиторов каспазы-1.
[00609] Аналогичным образом, чтобы исследовать эффекты совместного введения представляющей интерес зкДНК и ингибитора пути инфламмасомы AIM2 на врожденные иммунные ответы in vitro, можно использовать репортерные линии для функциональных анализов, в которых исследуется активация инфламмасомы AIM2 или каспазы-1. Репортерная линия клеток инфламмасомы AIM2, которую можно применять для таких анализов in vitro, может представлять собой стабильно котрансфицированную линию клеток, которая экспрессирует полноразмерный AIM2 человека и репортерный ген, такой как репортерный ген секретируемой щелочной фосфатазы (SEAP), под транскрипционным контролем элемента ответа транскрипционного фактора, такого как сайт связывания NF-kB, сайт связывания AP-1 или их комбинация. Анализ может быть проведен так же, как и репортерный анализ инфламмасомы NLRP3, в котором репортерные клетки, например, высеянные в 96-луночные планшеты, после заранее определенного периода времени, стимулируют различными количествами композиций, содержащих зкДНК, экспрессирующую фактор IX, с ингибитором инфламмасомы AIM2 или без него. Активность репортерного гена, такого как SEAP, может быть проанализирована с использованием любого метода или анализа, известного специалисту в данной области техники, для сравнения уровня активации каспазы-1 или активации инфламмасомы AIM2 в присутствии представляющей интерес зкДНК с ингибитором пути инфламмасомы AIM2 или без него. Ожидается, что в присутствии ингибитора инфламмасомы AIM2 будет наблюдаться меньшая активация репортерной молекулы. Аналогичный репортерный анализ можно использовать для оценки ингибиторов каспазы-1.
[00610] Кроме того, могут быть использованы репортерные линии с подавлением инфламмасомы NLRP3 или инфламмасомы AIM2, такие как клетки THP1-defNLRP3 (InvivoGen) или клетки THP-1, сверхэкспрессирующие TRIM11, подавляющий инфламмасому AIM2 (Liu et al., Cell Reports (2016) 16: 1988-2002), и другие линии клеток, известные в данной области техники. Такие репортерные линии с подавленным AIM2 или NLRP3 могут экспрессировать один или более индуцируемых генов секретируемых репортеров, таких как люцифераза Lucia и SEAP (секретируемая эмбриональная щелочная фосфатаза). Репортерный ген может находиться под контролем минимального промотора ISG54 (гена, стимулируемого интерфероном) в сочетании с одним или более, например, пятью, элементами ответа, стимулируемыми IFN. Репортерный ген также может находиться под контролем минимального промотора IFN-β, слитого с одной или более, например, пятью, копиями элемента ответа, такого как элемент ответа NF-kB. Активность NLRP3 или AIM2, или каспазы-1 в присутствии по меньшей мере одного ингибитора NLRP3 или AIM2, или каспазы-1 в комбинации с зкДНК, описанными в настоящем документе, можно сравнить в линии клеток с подавленным геном в сопоставлении с исходной линией клеток.
[00611] Чтобы исследовать эффекты совместного введения представляющей интерес зкДНК и ингибитора инфламмасомы NLRP3 и/или ингибитора инфламмасомы AIM2, или антагониста NLRP3, или антагониста AIM2 на путь активации инфламмасомы NLRP3 и/или AIM2 ex vivo, моноциты человека могут быть выделены, например, с помощью центрифугирования периферической крови в градиенте плотности и магнитного разделения. Эти моноциты можно исследовать до и после контакта и/или активации представляющей интерес зкДНК с ингибитором инфламмасомы NLRP3 и/или ингибитором инфламмасомы AIM2, или антагонистом NLRP3, или антагонистом AIM2, или ингибитором каспазы-1, или без них, с подходящими контролями. После обработки сыворотку и клеточные супернатанты используют для измерения одного или более цитокиновых путей в качестве функционального показателя активации пути инфламмасомы NLRP3 и/или ингибитора пути инфламмасомы AIM2, такого как интерлейкин (IL)-1β, IL-6, IL-8, IL-18, интерферон (IFN)-γ, интерферон (IFN)-α, моноцитарный хемоаттрактантный белок (MCP)-1, IP-10 и/или фактор некроза опухоли (TNF)-α, используя любой анализ или метод, известный специалисту в данной области техники. Кроме того, ядерные экстракты можно использовать для проверки активации NF-κB с использованием любого анализа или метода, известного специалисту в данной области техники. Ожидается, что при введении зкДНК в присутствии ингибитора пути инфламмасомы NLRP3 и/или AIM2 или ингибитора каспазы 1 будет наблюдаться меньшая активация цитокиновых путей и секреция цитокинов, что способствует увеличению продолжительности экспрессии трансгена и терапевтической эффективности.
[00612] Чтобы исследовать эффекты совместного введения представляющей интерес зкДНК и ингибитора пути инфламмасомы NLRP3 и/или AIM2, или ингибитора каспазы 1 на активацию пути инфламмасомы NLRP3 и/или AIM2 или активацию каспазы 1 in vivo, можно использовать модель на мышах. Образцы сыворотки или лимфоцитов мыши исследуют до и после контакта и/или активации зкДНК, экспрессирующей представляющий интерес трансген, такой как фактор IX, с ингибитором пути инфламмасомы NLRP3 и/или AIM2 или ингибитором каспазы-1, или без них, с подходящими контролями. После обработки сыворотку и клеточные супернатанты используют для измерения одного или более цитокиновых путей в качестве функционального показателя активации пути инфламмасомы NLRP3 и/или AIM2 или активации каспазы 1, такого как интерлейкин (IL)-1β, IL-6, IL-8, IL-18, интерферон (IFN)-γ, интерферон (IFN)-α, моноцитарный хемоаттрактантный белок (MCP)-1 и/или фактор некроза опухоли (TNF)-α, используя любой анализ или метод, известный специалисту в данной области техники. Кроме того, ядерные экстракты можно использовать для проверки активации NF-κB с использованием любого анализа или метода, известного специалисту в данной области техники. Ожидается, что при введении зкДНК в присутствии ингибитора пути инфламмасомы NLRP3 и/или AIM2 или ингибитора каспазы 1 будет наблюдаться менее значительная иммунная активация и секреция цитокинов, что способствует увеличению продолжительности экспрессии трансгена и терапевтической эффективности.
[00613] Оценивали совместное введение представляющей интерес зкДНК, экспрессирующей фактор IX человека, продуцируемый с плазмиды ТТХ-9, и ингибитора инфламмасомы NLRP3 и каспазы-1. Группы мышей C57bL (n = 8) оценивали, как показано в Таблице 11.
[00614] Таблица 11:
[00615] В общих чертах, животных предварительно обрабатывали ингибитором активации макрофагов или контролем в соответствии с группами, как показано в Таблице 12. Животным вводили MCC950 (ингибитор NLRP2) (Группа 5) или VX765 (белнаказан; селективный ингибитор каспазы-1) (Группа 4) в/б за 12-16 часов, а затем также за 1 час до введения 0,5 мг/кг зкДНК (TTX9-ЛНЧ) (Группа 1) или ЛНЧ-киРНК (отрицательный контроль) (Группа 1) путем в/в введения через боковую хвостовую вену. Контрольной группе предварительной обработки вводили только клондронат (контроль растворителя) (Группа 3). Цельную кровь собирали в каждой группе в дни 0, 1, 7 и 21 путем отбора образцов крови через хвостовую вену или лицевую вену, или из орбитальной области.
Таблица 12: Введение предварительной обработки
п/о
(Ингибитор NLRP3)
[00616] aПервое п/о введение будет выполнено за 12-16 часов до обработки зкДНК в День 0.
[00617] bВторое пероральное введение будет выполнено за 1 час до обработки зкДНК в День 0.
[00618] № = номер; ROA = путь введения; п/о = желудочный зонд; в/б = внутрибрюшинно; в/в = внутривенно; TA = исследуемый препарат; NA = неприменимо.
[00619] Уровни цитокинов определяли количественно и оценивали с использованием мультиплексного иммуноанализа ProcartaPlex (Invitrogen) в соответствии с инструкциями производителя, который представляет собой количественный мультиплексный иммуноанализ на основе гранул для измерения уровней различных цитокинов и хемокинов с использованием технологической платформы Luminex. Образцы, полученные от исследуемых мышей, смешивали с предварительно смешанным изготовленным на заказ 8-плексным набором для определения цитокинов мыши, магнитными гранулами и анализировали уровни IFN-α, IFN-γ, IL-6, IP-10, IL-18, IL-1β, MCP-1 и TNF-альфа. На ФИГ. 18A-18H показана оценка уровней цитокинов после введения TTX-9 с фармакологическим истощением макрофагов с помощью ингибитора NLRP3 (MCC950) или ингибитора каспазы 1 (VX765). Уровни IFNγ и IL-18 были значительно уменьшены при обработке MCC950 (ингибитор NLRP3) (ФИГ. 18B и ФИГ. 18D), а уровни IP-10 уменьшались при обработке MCC950 (ФИГ. 18F). Уровни IL-18 также были уменьшены при обработке VX765 (ингибитор каспазы-1) (ФИГ. 18D).
Перечень последовательностей без соблюдения формальностей
Примечание. Последовательность впоследствии была оптимизирована по кодонам в компании GenScript.
Примечание. Последовательность впоследствии была оптимизирована по кодонам в компании GenScript.
(промотор U6 малой ядерной РНК человека)
(Промотор H1 человека)
ATGGAGTTGGTGGGCTGGCTCGTGGACAAAGGCATTACTTCGGAAAAGCAGTGGATTCAGGAGGATCAGGCATCTTACATCTCATTCAACGCTGCCAGTAACTCGAGGTCCCAGATCAAGGCAGCGCTGGACAACGCGGGAAAGATTATGAGTCTGACCAAAACTGCTCCAGACTACCTCGTTGGTCAGCAACCGGTGGAAGATATCTCCAGCAACAGGATCTACAAGATTCTGGAGCTCAACGGCTACGACCCTCAATACGCTGCCTCAGTGTTCTTGGGTTGGGCCACCAAGAAATTCGGCAAGAGAAACACTATCTGGCTGTTCGGCCCCGCTACCACTGGAAAGACAAACATCGCAGAAGCGATTGCTCACACGGTGCCATTCTACGGCTGCGTCAACTGGACAAACGAGAACTTCCCGTTCAACGACTGTGTCGATAAGATGGTTATCTGGTGGGAGGAAGGAAAGATGACGGCCAAAGTGGTCGAAAGCGCCAAGGCAATTCTGGGTGGCTCTAAAGTGCGCGTCGACCAGAAGTGCAAATCTTCAGCTCAAATCGATCCTACCCCCGTTATTGTGACATCAAACACGAACATGTGTGCCGTGATCGACGGAAACAGTACAACGTTCGAACACCAGCAACCTCTCCAGGATCGTATGTTCAAGTTCGAGCTCACCCGCCGTTTGGACCATGATTTCGGCAAGGTCACTAAACAAGAGGTTAAGGACTTCTTCCGCTGGGCTAAAGATCACGTTGTGGAGGTTGAACATGAGTTCTACGTCAAGAAAGGAGGTGCTAAGAAACGTCCAGCCCCGTCGGACGCAGATATCTCCGAACCTAAGAGGGTGAGAGAGTCGGTCGCACAGCCAAGCACTTCTGACGCAGAAGCTTCCATTAACTACGCAGATAGGTACCAAAACAAGTGCAGCAGACACGTGGGTATGAACTTGATGCTGTTCCCATGCCGCCAGTGTGAGCGTATGAACCAAAACTCTAACATCTGTTTCACACATGGCCAGAAGGACTGCCTCGAATGTTTCCCTGTGTCAGAGAGTCAGCCCGTCTCAGTCGTTAAGAAAGCTTACCAAAAGTTGTGCTACATCCACCATATTATGGGTAAAGTCCCTGATGCCTGTACCGCTTGTGATCTGGTCAACGTGGATTTGGACGACTGTATTTTCGAGCAATAA
(CR456383.1 Клон кДНК полноразмерной открытой рамки считывания (ORF) ARSA Homo sapiens (клон кДНК C22ORF pGEM.ARSA.V2))
(Идуронат-2-сульфатаза (IDS) Genbank Homo sapiens, RefSeqGene на хромосоме X)
(Rattus norvegicus BAC CH230-35L12 (Исследовательский институт Детской больницы Окленда), полная последовательность. Идентификатор последовательности: gi|49615137|AC097023.6)
(SEQ ID: 509)
Источники
1. Intracellular Nucleic Acid Detection in Autoimmunity. J.T. Crowl и др., Annu. Rev. Immunol. 2017, 35: 313-336
2. Innate Immune Recognition of DNA: a recent history. A. Dempsey и A.G. Bowie, Virology, Май 2015 г., 0: 146-152
3. Cutting Edge: Antimalarial Drugs Inhibit IFN-β Production through Blockade of Cyclic GMP-AMP Synthase-DNA Interaction. J. An и др., J. Immunol. 27 марта 2015 г.
4. Small molecule inhibition of cGAS reduces interferon expression in primary macrophages from autoimmune mice. J. Vincent и др., Nature Communications, 8:750
5. Discovery of PF-06928215 as a high affinity inhibitor of cGAS enabled by a novel fluorescence polarization assay. PLOS ONE. 21 сентября 2017 г.
6. The functional interactome of PYHIN immune regulators reveals IFIX is a sensor of viral DNA. B.A. Diner и др., Molecular Systems Biology, 2015, 11:787
7. Activation and Regulation of DNA-Driven Immune Responses. S.R. Paluden. Microbiology and Molecular Biology Reviews. 2015. 79(2): 225
8. Viral Evasion of DNA-stimulated innate immune responses. M.H. Christnesen и S.R. Paluden. Cellular and Molecular Immunology. 2017. 14:4-13
9. Sequence-specific activation of the DNA sensor cGAS by Y-form DNA structures as found in primary HIV-1 cDNA. A-M Herzner и др., 2015. Nature Immunology
10. RNA Polymerase III Detects Cytosolic DNA and Induces Type-I Interferons Through the RIG-I Pathway. Y-H Chiu и др. 2009. Cell. 138(3): 576-591
11. Mi-R-25/93 mediates hypoxia-induced immunosuppression by repressing cGAS. M.Z. Wu и др. 2017. Nat. Cell Biol. 19(10):1286-1296
12. Cytoplasmic isoforms of Kaposi sarcoma herpesvirus LANA recruit and antagonize the innate immune DNA sensor cGAS. Zhang G. и др., Proc Natl Acad Sci U S A., 23 февраля 2016 г.;113(8):E1034-43.
13. Kaposi´s sarcoma-associated herpesvirus inhibitor of cGAS (KicGAS) Encoded by ORF52, is an Abundant Tegument protein and Is Required for Production of Infectious Progeny Viruses. W. Li, и др., J. Virol. 2016, 90(11): 5329.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ДЖЕНЕРЕЙШЕН БИО КО.
<120> ДНК с замкнутыми концами (зкДНК) и применение в способах уменьшения
иммунного ответа, связанного с генной терапией или терапией
нуклеиновой кислотой
<130> 131698-03320
<140> PCT/US2020/015026
<141> 2020-01-24
<150> 62/857,542
<151> 2019-06-05
<150> 62/814,424
<151> 2019-03-06
<150> 62/814,414
<151> 2019-03-06
<150> 62/800,303
<151> 2019-02-01
<150> 62/800,285
<151> 2019-02-01
<150> 62/796,450
<151> 2019-01-24
<150> 62/796,417
<151> 2019-01-24
<160> 1220
<170> PatentIn version 3.5
<210> 1
<211> 141
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg gctttgcccg ggcggcctca gtgagcgagc 120
gagcgcgcag ctgcctgcag g 141
<210> 2
<211> 130
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 2
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg gcggcctcag tgagcgagcg agcgcgcagc 120
tgcctgcagg 130
<210> 3
<211> 1923
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 3
tcaatattgg ccattagcca tattattcat tggttatata gcataaatca atattggcta 60
ttggccattg catacgttgt atctatatca taatatgtac atttatattg gctcatgtcc 120
aatatgaccg ccatgttggc attgattatt gactagttat taatagtaat caattacggg 180
gtcattagtt catagcccat atatggagtt ccgcgttaca taacttacgg taaatggccc 240
gcctggctga ccgcccaacg acccccgccc attgacgtca ataatgacgt atgttcccat 300
agtaacgcca atagggactt tccattgacg tcaatgggtg gagtatttac ggtaaactgc 360
ccacttggca gtacatcaag tgtatcatat gccaagtccg ccccctattg acgtcaatga 420
cggtaaatgg cccgcctggc attatgccca gtacatgacc ttacgggact ttcctacttg 480
gcagtacatc tacgtattag tcatcgctat taccatggtc gaggtgagcc ccacgttctg 540
cttcactctc cccatctccc ccccctcccc acccccaatt ttgtatttat ttatttttta 600
attattttgt gcagcgatgg gggcgggggg gggggggggg cgcgcgccag gcggggcggg 660
gcggggcgag gggcggggcg gggcgaggcg gagaggtgcg gcggcagcca atcagagcgg 720
cgcgctccga aagtttcctt ttatggcgag gcggcggcgg cggcggccct ataaaaagcg 780
aagcgcgcgg cgggcgggag tcgctgcgac gctgccttcg ccccgtgccc cgctccgccg 840
ccgcctcgcg ccgcccgccc cggctctgac tgaccgcgtt actcccacag gtgagcgggc 900
gggacggccc ttctcctccg ggctgtaatt agcgcttggt ttaatgacgg cttgtttctt 960
ttctgtggct gcgtgaaagc cttgaggggc tccgggaggg ccctttgtgc gggggggagc 1020
ggctcggggg gtgcgtgcgt gtgtgtgtgc gtggggagcg ccgcgtgcgg cccgcgctgc 1080
ccggcggctg tgagcgctgc gggcgcggcg cggggctttg tgcgctccgc agtgtgcgcg 1140
aggggagcgc ggccgggggc ggtgccccgc ggtgcggggg gggctgcgag gggaacaaag 1200
gctgcgtgcg gggtgtgtgc gtgggggggt gagcaggggg tgtgggcgcg gcggtcgggc 1260
tgtaaccccc ccctgcaccc ccctccccga gttgctgagc acggcccggc ttcgggtgcg 1320
gggctccgta cggggcgtgg cgcggggctc gccgtgccgg gcggggggtg gcggcaggtg 1380
ggggtgccgg gcggggcggg gccgcctcgg gccggggagg gctcggggga ggggcgcggc 1440
ggcccccgga gcgccggcgg ctgtcgaggc gcggcgagcc gcagccattg ccttttatgg 1500
taatcgtgcg agagggcgca gggacttcct ttgtcccaaa tctgtgcgga gccgaaatct 1560
gggaggcgcc gccgcacccc ctctagcggg cgcggggcga agcggtgcgg cgccggcagg 1620
aaggaaatgg gcggggaggg ccttcgtgcg tcgccgcgcc gccgtcccct tctccctctc 1680
cagcctcggg gctgtccgcg gggggacggc tgccttcggg ggggacgggg cagggcgggg 1740
ttcggcttct ggcgtgtgac cggcggctct agagcctctg ctaaccatgt tttagccttc 1800
ttctttttcc tacagctcct gggcaacgtg ctggttattg tgctgtctca tcatttgtcg 1860
acagaattcc tcgaagatcc gaaggggttc aagcttggca ttccggtact gttggtaaag 1920
cca 1923
<210> 4
<211> 1272
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 4
aggctcagag gcacacagga gtttctgggc tcaccctgcc cccttccaac ccctcagttc 60
ccatcctcca gcagctgttt gtgtgctgcc tctgaagtcc acactgaaca aacttcagcc 120
tactcatgtc cctaaaatgg gcaaacattg caagcagcaa acagcaaaca cacagccctc 180
cctgcctgct gaccttggag ctggggcaga ggtcagagac ctctctgggc ccatgccacc 240
tccaacatcc actcgacccc ttggaatttc ggtggagagg agcagaggtt gtcctggcgt 300
ggtttaggta gtgtgagagg gtccgggttc aaaaccactt gctgggtggg gagtcgtcag 360
taagtggcta tgccccgacc ccgaagcctg tttccccatc tgtacaatgg aaatgataaa 420
gacgcccatc tgatagggtt tttgtggcaa ataaacattt ggtttttttg ttttgttttg 480
ttttgttttt tgagatggag gtttgctctg tcgcccaggc tggagtgcag tgacacaatc 540
tcatctcacc acaaccttcc cctgcctcag cctcccaagt agctgggatt acaagcatgt 600
gccaccacac ctggctaatt ttctattttt agtagagacg ggtttctcca tgttggtcag 660
cctcagcctc ccaagtaact gggattacag gcctgtgcca ccacacccgg ctaatttttt 720
ctatttttga cagggacggg gtttcaccat gttggtcagg ctggtctaga ggtaccggat 780
cttgctacca gtggaacagc cactaaggat tctgcagtga gagcagaggg ccagctaagt 840
ggtactctcc cagagactgt ctgactcacg ccaccccctc caccttggac acaggacgct 900
gtggtttctg agccaggtac aatgactcct ttcggtaagt gcagtggaag ctgtacactg 960
cccaggcaaa gcgtccgggc agcgtaggcg ggcgactcag atcccagcca gtggacttag 1020
cccctgtttg ctcctccgat aactggggtg accttggtta atattcacca gcagcctccc 1080
ccgttgcccc tctggatcca ctgcttaaat acggacgagg acagggccct gtctcctcag 1140
cttcaggcac caccactgac ctgggacagt gaatccggac tctaaggtaa atataaaatt 1200
tttaagtgta taatgtgtta aactactgat tctaattgtt tctctctttt agattccaac 1260
ctttggaact ga 1272
<210> 5
<211> 547
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 5
ccctaaaatg ggcaaacatt gcaagcagca aacagcaaac acacagccct ccctgcctgc 60
tgaccttgga gctggggcag aggtcagaga cctctctggg cccatgccac ctccaacatc 120
cactcgaccc cttggaattt ttcggtggag aggagcagag gttgtcctgg cgtggtttag 180
gtagtgtgag aggggaatga ctcctttcgg taagtgcagt ggaagctgta cactgcccag 240
gcaaagcgtc cgggcagcgt aggcgggcga ctcagatccc agccagtgga cttagcccct 300
gtttgctcct ccgataactg gggtgacctt ggttaatatt caccagcagc ctcccccgtt 360
gcccctctgg atccactgct taaatacgga cgaggacagg gccctgtctc ctcagcttca 420
ggcaccacca ctgacctggg acagtgaatc cggactctaa ggtaaatata aaatttttaa 480
gtgtataatg tgttaaacta ctgattctaa ttgtttctct cttttagatt ccaacctttg 540
gaactga 547
<210> 6
<211> 1179
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 6
ggctccggtg cccgtcagtg ggcagagcgc acatcgccca cagtccccga gaagttgggg 60
ggaggggtcg gcaattgaac cggtgcctag agaaggtggc gcggggtaaa ctgggaaagt 120
gatgtcgtgt actggctccg cctttttccc gagggtgggg gagaaccgta tataagtgca 180
gtagtcgccg tgaacgttct ttttcgcaac gggtttgccg ccagaacaca ggtaagtgcc 240
gtgtgtggtt cccgcgggcc tggcctcttt acgggttatg gcccttgcgt gccttgaatt 300
acttccacct ggctgcagta cgtgattctt gatcccgagc ttcgggttgg aagtgggtgg 360
gagagttcga ggccttgcgc ttaaggagcc ccttcgcctc gtgcttgagt tgaggcctgg 420
cctgggcgct ggggccgccg cgtgcgaatc tggtggcacc ttcgcgcctg tctcgctgct 480
ttcgataagt ctctagccat ttaaaatttt tgatgacctg ctgcgacgct ttttttctgg 540
caagatagtc ttgtaaatgc gggccaagat ctgcacactg gtatttcggt ttttggggcc 600
gcgggcggcg acggggcccg tgcgtcccag cgcacatgtt cggcgaggcg gggcctgcga 660
gcgcggccac cgagaatcgg acgggggtag tctcaagctg gccggcctgc tctggtgcct 720
ggtctcgcgc cgccgtgtat cgccccgccc tgggcggcaa ggctggcccg gtcggcacca 780
gttgcgtgag cggaaagatg gccgcttccc ggccctgctg cagggagctc aaaatggagg 840
acgcggcgct cgggagagcg ggcgggtgag tcacccacac aaaggaaaag ggcctttccg 900
tcctcagccg tcgcttcatg tgactccacg gagtaccggg cgccgtccag gcacctcgat 960
tagttctcga gcttttggag tacgtcgtct ttaggttggg gggaggggtt ttatgcgatg 1020
gagtttcccc acactgagtg ggtggagact gaagttaggc cagcttggca cttgatgtaa 1080
ttctccttgg aatttgccct ttttgagttt ggatcttggt tcattctcaa gcctcagaca 1140
gtggttcaaa gtttttttct tccatttcag gtgtcgtga 1179
<210> 7
<211> 8
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 7
gtttaaac 8
<210> 8
<211> 581
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 8
gagcatctta ccgccattta ttcccatatt tgttctgttt ttcttgattt gggtatacat 60
ttaaatgtta ataaaacaaa atggtggggc aatcatttac atttttaggg atatgtaatt 120
actagttcag gtgtattgcc acaagacaaa catgttaaga aactttcccg ttatttacgc 180
tctgttcctg ttaatcaacc tctggattac aaaatttgtg aaagattgac tgatattctt 240
aactatgttg ctccttttac gctgtgtgga tatgctgctt tatagcctct gtatctagct 300
attgcttccc gtacggcttt cgttttctcc tccttgtata aatcctggtt gctgtctctt 360
ttagaggagt tgtggcccgt tgtccgtcaa cgtggcgtgg tgtgctctgt gtttgctgac 420
gcaaccccca ctggctgggg cattgccacc acctgtcaac tcctttctgg gactttcgct 480
ttccccctcc cgatcgccac ggcagaactc atcgccgcct gccttgcccg ctgctggaca 540
ggggctaggt tgctgggcac tgataattcc gtggtgttgt c 581
<210> 9
<211> 225
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 9
tgtgccttct agttgccagc catctgttgt ttgcccctcc cccgtgcctt ccttgaccct 60
ggaaggtgcc actcccactg tcctttccta ataaaatgag gaaattgcat cgcattgtct 120
gagtaggtgt cattctattc tggggggtgg ggtggggcag gacagcaagg gggaggattg 180
ggaagacaat agcaggcatg ctggggatgc ggtgggctct atggc 225
<210> 10
<211> 213
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 10
taagatacat tgatgagttt ggacaaacca caactagaat gcagtgaaaa aaatgcttta 60
tttgtgaaat ttgtgatgct attgctttat ttgtaaccat tataagctgc aataaacaag 120
ttaacaacaa caattgcatt cattttatgt ttcaggttca gggggaggtg tgggaggttt 180
tttaaagcaa gtaaaacctc tacaaatgtg gta 213
<210> 11
<211> 1386
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 11
atgcagcgcg tgaacatgat catggccgag agccccggcc tgatcaccat ctgcctgctg 60
ggctacctgc tgagcgccga gtgcaccgtg ttcctggacc acgagaacgc caacaagatc 120
ctgaaccgcc ccaagcgcta caacagcggc aagctggagg agttcgtgca gggcaacctg 180
gagcgcgagt gcatggagga gaagtgcagc ttcgaggagg cccgcgaggt gttcgagaac 240
accgagcgca ccaccgagtt ctggaagcag tacgtggacg gcgaccagtg cgagagcaac 300
ccctgcctga acggcggcag ctgcaaggac gacatcaaca gctacgagtg ctggtgcccc 360
ttcggcttcg agggcaagaa ctgcgagctg gacgtgacct gcaacatcaa gaacggccgc 420
tgcgagcagt tctgcaagaa cagcgccgac aacaaggtgg tgtgcagctg caccgagggc 480
taccgcctgg ccgagaacca gaagagctgc gagcccgccg tgcccttccc ctgcggcagg 540
gtgagcgtga gccagaccag caagctgacc cgcgccgagg ccgtgttccc cgacgtggac 600
tacgtgaaca gcaccgaggc cgagaccatc ctggacaaca tcacccagag cacccagagc 660
ttcaacgact tcacccgcgt ggtgggcggc gaggacgcca agcccggcca gttcccctgg 720
caggtggtgc tgaacggcaa ggtggacgcc ttctgcggcg gcagcatcgt gaacgagaag 780
tggatcgtga ccgccgccca ctgcgtggag accggcgtga agatcaccgt ggtggccggc 840
gagcacaaca tcgaggagac cgagcacacc gagcagaagc gcaacgtgat ccgcatcatc 900
ccccaccaca actacaacgc cgccatcaac aagtacaacc acgacatcgc cctgctggag 960
ctggacgagc ccctggtgct gaacagctac gtgaccccca tctgcatcgc cgacaaggag 1020
tacaccaaca tcttcctgaa gttcggcagc ggctacgtga gcggctgggg ccgcgtgttc 1080
cacaagggcc gcagcgccct ggtgctgcag tacctgcgcg tgcccctggt ggaccgcgcc 1140
acctgcctgc gcagcaccaa gttcaccatc tacaacaaca tgttctgcgc cggcttccac 1200
gagggcggca gggacagctg ccagggcgac agcggcggcc cccacgtgac cgaggtggag 1260
ggcaccagct tcctgaccgg catcatcagc tggggcgagg agtgcgccat gaagggcaag 1320
tacggcatct acaccaaggt gagccgctac gtgaactgga tcaaggagaa gaccaagctg 1380
acctaa 1386
<210> 12
<211> 1386
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 12
atgcagcgcg tgaacatgat catggccgag agccccggcc tgatcaccat ctgcctgctg 60
ggctacctgc tgagcgccga gtgcaccgtg ttcctggacc acgagaacgc caacaagatc 120
ctgaaccgcc ccaagcgcta caacagcggc aagctggagg agttcgtgca gggcaacctg 180
gagcgcgagt gcatggagga gaagtgcagc ttcgaggagg cccgcgaggt gttcgagaac 240
accgagcgca ccaccgagtt ctggaagcag tacgtggacg gcgaccagtg cgagagcaac 300
ccctgcctga acggcggcag ctgcaaggac gacatcaaca gctacgagtg ctggtgcccc 360
ttcggcttcg agggcaagaa ctgcgagctg gacgtgacct gcaacatcaa gaacggccgc 420
tgcgagcagt tctgcaagaa cagcgccgac aacaaggtgg tgtgcagctg caccgagggc 480
taccgcctgg ccgagaacca gaagagctgc gagcccgccg tgcccttccc ctgcggcagg 540
gtgagcgtga gccagaccag caagctgacc cgcgccgagg ccgtgttccc cgacgtggac 600
tacgtgaaca gcaccgaggc cgagaccatc ctggacaaca tcacccagag cacccagagc 660
ttcaacgact tcacccgcgt ggtgggcggc gaggacgcca agcccggcca gttcccctgg 720
caggtggtgc tgaacggcaa ggtggacgcc ttctgcggcg gcagcatcgt gaacgagaag 780
tggatcgtga ccgccgccca ctgcgtggag accggcgtga agatcaccgt ggtggccggc 840
gagcacaaca tcgaggagac cgagcacacc gagcagaagc gcaacgtgat ccgcatcatc 900
ccccaccaca actacaacgc cgccatcaac aagtacaacc acgacatcgc cctgctggag 960
ctggacgagc ccctggtgct gaacagctac gtgaccccca tctgcatcgc cgacaaggag 1020
tacaccaaca tcttcctgaa gttcggcagc ggctacgtga gcggctgggg ccgcgtgttc 1080
cacaagggcc gcagcgccct ggtgctgcag tacctgcgcg tgcccctggt ggaccgcgcc 1140
acctgcctgc tgagcaccaa gttcaccatc tacaacaaca tgttctgcgc cggcttccac 1200
gagggcggca gggacagctg ccagggcgac agcggcggcc cccacgtgac cgaggtggag 1260
ggcaccagct tcctgaccgg catcatcagc tggggcgagg agtgcgccat gaagggcaag 1320
tacggcatct acaccaaggt gagccgctac gtgaactgga tcaaggagaa gaccaagctg 1380
acctaa 1386
<210> 13
<211> 1876
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 13
cgcagccacc atggcggggt tttacgagat tgtgattaag gtccccagcg accttgacgg 60
gcatctgccc ggcatttctg acagctttgt gaactgggtg gccgagaagg aatgggagtt 120
gccgccagat tctgacatgg atctgaatct gattgagcag gcacccctga ccgtggccga 180
gaagctgcag cgcgactttc tgacggaatg gcgccgtgtg agtaaggccc cggaggccct 240
tttctttgtg caatttgaga agggagagag ctacttccac atgcacgtgc tcgtggaaac 300
caccggggtg aaatccatgg ttttgggacg tttcctgagt cagattcgcg aaaaactgat 360
tcagagaatt taccgcggga tcgagccgac tttgccaaac tggttcgcgg tcacaaagac 420
cagaaatggc gccggaggcg ggaacaaggt ggtggatgag tgctacatcc ccaattactt 480
gctccccaaa acccagcctg agctccagtg ggcgtggact aatatggaac agtatttaag 540
cgcctgtttg aatctcacgg agcgtaaacg gttggtggcg cagcatctga cgcacgtgtc 600
gcagacgcag gagcagaaca aagagaatca gaatcccaat tctgatgcgc cggtgatcag 660
atcaaaaact tcagccaggt acatggagct ggtcgggtgg ctcgtggaca aggggattac 720
ctcggagaag cagtggatcc aggaggacca ggcctcatac atctccttca atgcggcctc 780
caactcgcgg tcccaaatca aggctgcctt ggacaatgcg ggaaagatta tgagcctgac 840
taaaaccgcc cccgactacc tggtgggcca gcagcccgtg gaggacattt ccagcaatcg 900
gatttataaa attttggaac taaacgggta cgatccccaa tatgcggctt ccgtctttct 960
gggatgggcc acgaaaaagt tcggcaagag gaacaccatc tggctgtttg ggcctgcaac 1020
taccgggaag accaacatcg cggaggccat agcccacact gtgcccttct acgggtgcgt 1080
aaactggacc aatgagaact ttcccttcaa cgactgtgtc gacaagatgg tgatctggtg 1140
ggaggagggg aagatgaccg ccaaggtcgt ggagtcggcc aaagccattc tcggaggaag 1200
caaggtgcgc gtggaccaga aatgcaagtc ctcggcccag atagacccga ctcccgtgat 1260
cgtcacctcc aacaccaaca tgtgcgccgt gattgacggg aactcaacga ccttcgaaca 1320
ccagcagccg ttgcaagacc ggatgttcaa atttgaactc acccgccgtc tggatcatga 1380
ctttgggaag gtcaccaagc aggaagtcaa agactttttc cggtgggcaa aggatcacgt 1440
ggttgaggtg gagcatgaat tctacgtcaa aaagggtgga gccaagaaaa gacccgcccc 1500
cagtgacgca gatataagtg agcccaaacg ggtgcgcgag tcagttgcgc agccatcgac 1560
gtcagacgcg gaagcttcga tcaactacgc agacaggtac caaaacaaat gttctcgtca 1620
cgtgggcatg aatctgatgc tgtttccctg cagacaatgc gagagaatga atcagaattc 1680
aaatatctgc ttcactcacg gacagaaaga ctgtttagag tgctttcccg tgtcagaatc 1740
tcaacccgtt tctgtcgtca aaaaggcgta tcagaaactg tgctacattc atcatatcat 1800
gggaaaggtg ccagacgctt gcactgcctg cgatctggtc aatgtggatt tggatgactg 1860
catctttgaa caataa 1876
<210> 14
<211> 1194
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 14
atggagctgg tcgggtggct cgtggacaag gggattacct cggagaagca gtggatccag 60
gaggaccagg cctcatacat ctccttcaat gcggcctcca actcgcggtc ccaaatcaag 120
gctgccttgg acaatgcggg aaagattatg agcctgacta aaaccgcccc cgactacctg 180
gtgggccagc agcccgtgga ggacatttcc agcaatcgga tttataaaat tttggaacta 240
aacgggtacg atccccaata tgcggcttcc gtctttctgg gatgggccac gaaaaagttc 300
ggcaagagga acaccatctg gctgtttggg cctgcaacta ccgggaagac caacatcgcg 360
gaggccatag cccacactgt gcccttctac gggtgcgtaa actggaccaa tgagaacttt 420
cccttcaacg actgtgtcga caagatggtg atctggtggg aggaggggaa gatgaccgcc 480
aaggtcgtgg agtcggccaa agccattctc ggaggaagca aggtgcgcgt ggaccagaaa 540
tgcaagtcct cggcccagat agacccgact cccgtgatcg tcacctccaa caccaacatg 600
tgcgccgtga ttgacgggaa ctcaacgacc ttcgaacacc agcagccgtt gcaagaccgg 660
atgttcaaat ttgaactcac ccgccgtctg gatcatgact ttgggaaggt caccaagcag 720
gaagtcaaag actttttccg gtgggcaaag gatcacgtgg ttgaggtgga gcatgaattc 780
tacgtcaaaa agggtggagc caagaaaaga cccgccccca gtgacgcaga tataagtgag 840
cccaaacggg tgcgcgagtc agttgcgcag ccatcgacgt cagacgcgga agcttcgatc 900
aactacgcag accgctacca aaacaaatgt tctcgtcacg tgggcatgaa tctgatgctg 960
tttccctgca gacaatgcga gagaatgaat cagaattcaa atatctgctt cactcacgga 1020
cagaaagact gtttagagtg ctttcccgtg tcagaatctc aacccgtttc tgtcgtcaaa 1080
aaggcgtatc agaaactgtg ctacattcat catatcatgg gaaaggtgcc agacgcttgc 1140
actgcctgcg atctggtcaa tgtggatttg gatgactgca tctttgaaca ataa 1194
<210> 15
<211> 141
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 15
aataaacgat aacgccgttg gtggcgtgag gcatgtaaaa ggttacatca ttatcttgtt 60
cgccatccgg ttggtataaa tagacgttca tgttggtttt tgtttcagtt gcaagttggc 120
tgcggcgcgc gcagcacctt t 141
<210> 16
<211> 556
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 16
ccctaaaatg ggcaaacatt gcaagcagca aacagcaaac acacagccct ccctgcctgc 60
tgaccttgga gctggggcag aggtcagaga cctctctggg cccatgccac ctccaacatc 120
cactcgaccc cttggaattt cggtggagag gagcagaggt tgtcctggcg tggtttaggt 180
agtgtgagag gggaatgact cctttcggta agtgcagtgg aagctgtaca ctgcccaggc 240
aaagcgtccg ggcagcgtag gcgggcgact cagatcccag ccagtggact tagcccctgt 300
ttgctcctcc gataactggg gtgaccttgg ttaatattca ccagcagcct cccccgttgc 360
ccctctggat ccactgctta aatacggacg aggacactcg agggccctgt ctcctcagct 420
tcaggcacca ccactgacct gggacagtga atccggacat cgattctaag gtaaatataa 480
aatttttaag tgtataattt gttaaactac tgattctaat tgtttctctc ttttagattc 540
caacctttgg aactga 556
<210> 17
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 17
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 18
<211> 241
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 18
gagggcctat ttcccatgat tccttcatat ttgcatatac gatacaaggc tgttagagag 60
ataattggaa ttaatttgac tgtaaacaca aagatattag tacaaaatac gtgacgtaga 120
aagtaataat ttcttgggta gtttgcagtt ttaaaattat gttttaaaat ggactatcat 180
atgcttaccg taacttgaaa gtatttcgat ttcttggctt tatatatctt gtggaaagga 240
c 241
<210> 19
<211> 215
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 19
gaacgctgac gtcatcaacc cgctccaagg aatcgcgggc ccagtgtcac taggcgggaa 60
cacccagcgc gcgtgcgccc tggcaggaag atggctgtga gggacagggg agtggcgccc 120
tgcaatattt gcatgtcgct atgtgttctg ggaaatcacc ataaacgtga aatgtctttg 180
gatttgggaa tcgtataaga actgtatgag accac 215
<210> 20
<211> 150
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 20
ataaacgata acgccgttgg tggcgtgagg catgtaaaag gttacatcat tatcttgttc 60
gccatccggt tggtataaat agacgttcat gttggttttt gtttcagttg caagttggct 120
gcggcgcgcg cagcaccttt gcggccatct 150
<210> 21
<400> 21
000
<210> 22
<400> 22
000
<210> 23
<400> 23
000
<210> 24
<400> 24
000
<210> 25
<400> 25
000
<210> 26
<400> 26
000
<210> 27
<400> 27
000
<210> 28
<400> 28
000
<210> 29
<400> 29
000
<210> 30
<400> 30
000
<210> 31
<400> 31
000
<210> 32
<400> 32
000
<210> 33
<400> 33
000
<210> 34
<400> 34
000
<210> 35
<400> 35
000
<210> 36
<400> 36
000
<210> 37
<400> 37
000
<210> 38
<400> 38
000
<210> 39
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 39
gcgcgctcgc tcgctc 16
<210> 40
<400> 40
000
<210> 41
<400> 41
000
<210> 42
<400> 42
000
<210> 43
<400> 43
000
<210> 44
<400> 44
000
<210> 45
<400> 45
000
<210> 46
<400> 46
000
<210> 47
<400> 47
000
<210> 48
<400> 48
000
<210> 49
<400> 49
000
<210> 50
<400> 50
000
<210> 51
<211> 141
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 51
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc t 141
<210> 52
<211> 130
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 52
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcgtcg ggcgaccttt 60
ggtcgcccgg cctcagtgag cgagcgagcg cgcagagagg gagtggccaa ctccatcact 120
aggggttcct 130
<210> 53
<211> 3123
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 53
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggaaacc cgggcgtgcc 60
cgggcgcctc agtgagcgag cgagcgcgca gagagggagt ggccaactcc atcactaggg 120
gttcctgaac agagaaacag gagaatatgg gccaaacagg atatctgtgg taagcagttc 180
ctgccccggc tcagggccaa gaacagttgg aacagcagaa tatgggccaa acaggatatc 240
tgtggtaagc agttcctgcc ccggctcagg gccaagaaca gatggtcccc agatgcggtc 300
ccgccctcag cagtttctag agaaccatca gatgtttcca gggtgcccca aggacctgaa 360
atgaccctgt gccttatttg aactaaccaa tcagttcgct tctcgcttct gttcgcgcgc 420
ttctgctccc cgagctctat ataagcagag ctcgtttagt gaaccgtcag atcgcctgga 480
gacgccatcc acgctgtttt gacttccata gaaggccgcc accatggaag acgccaaaaa 540
cataaagaaa ggcccggcgc cattctatcc gctggaagat ggaaccgctg gagagcaact 600
gcataaggct atgaagagat acgccctggt tcctggaaca attgctttta cagatgcaca 660
tatcgaggtg gacatcactt acgctgagta cttcgaaatg tccgttcggt tggcagaagc 720
tatgaaacga tatgggctga atacaaatca cagaatcgtc gtatgcagtg aaaactctct 780
tcaattcttt atgccggtgt tgggcgcgtt atttatcgga gttgcagttg cgcccgcgaa 840
cgacatttat aatgaacgtg aattgctcaa cagtatgggc atttcgcagc ctaccgtggt 900
gttcgtttcc aaaaaggggt tgcaaaaaat tttgaacgtg caaaaaaagc tcccaatcat 960
ccaaaaaatt attatcatgg attctaaaac ggattaccag ggatttcagt cgatgtacac 1020
gttcgtcaca tctcatctac ctcccggttt taatgaatac gattttgtgc cagagtcctt 1080
cgatagggac aagacaattg cactgatcat gaactcctct ggatctactg gtctgcctaa 1140
aggtgtcgct ctgcctcata gaactgcctg cgtgagattc tcgcatgcca gagatcctat 1200
ttttggcaat caaatcattc cggatactgc gattttaagt gttgttccat tccatcacgg 1260
ttttggaatg tttactacac tcggatattt gatatgtgga tttcgagtcg tcttaatgta 1320
tagatttgaa gaagagctgt ttctgaggag ccttcaggat tacaagattc aaagtgcgct 1380
gctggtgcca accctattct ccttcttcgc caaaagcact ctgattgaca aatacgattt 1440
atctaattta cacgaaattg cttctggtgg cgctcccctc tctaaggaag tcggggaagc 1500
ggttgccaag aggttccatc tgccaggtat caggcaagga tatgggctca ctgagactac 1560
atcagctatt ctgattacac ccgaggggga tgataaaccg ggcgcggtcg gtaaagttgt 1620
tccatttttt gaagcgaagg ttgtggatct ggataccggg aaaacgctgg gcgttaatca 1680
aagaggcgaa ctgtgtgtga gaggtcctat gattatgtcc ggttatgtaa acaatccgga 1740
agcgaccaac gccttgattg acaaggatgg atggctacat tctggagaca tagcttactg 1800
ggacgaagac gaacacttct tcatcgttga ccgcctgaag tctctgatta agtacaaagg 1860
ctatcaggtg gctcccgctg aattggaatc catcttgctc caacacccca acatcttcga 1920
cgcaggtgtc gcaggtcttc ccgacgatga cgccggtgaa cttcccgccg ccgttgttgt 1980
tttggagcac ggaaagacga tgacggaaaa agagatcgtg gattacgtcg ccagtcaagt 2040
aacaaccgcg aaaaagttgc gcggaggagt tgtgtttgtg gacgaagtac cgaaaggtct 2100
taccggaaaa ctcgacgcaa gaaaaatcag agagatcctc ataaaggcca agaagggcgg 2160
aaagatcgcc gtgtaagagc atcttaccgc catttattcc catatttgtt ctgtttttct 2220
tgatttgggt atacatttaa atgttaataa aacaaaatgg tggggcaatc atttacattt 2280
ttagggatat gtaattacta gttcaggtgt attgccacaa gacaaacatg ttaagaaact 2340
ttcccgttat ttacgctctg ttcctgttaa tcaacctctg gattacaaaa tttgtgaaag 2400
attgactgat attcttaact atgttgctcc ttttacgctg tgtggatatg ctgctttata 2460
gcctctgtat ctagctattg cttcccgtac ggctttcgtt ttctcctcct tgtataaatc 2520
ctggttgctg tctcttttag aggagttgtg gcccgttgtc cgtcaacgtg gcgtggtgtg 2580
ctctgtgttt gctgacgcaa cccccactgg ctggggcatt gccaccacct gtcaactcct 2640
ttctgggact ttcgctttcc ccctcccgat cgccacggca gaactcatcg ccgcctgcct 2700
tgcccgctgc tggacagggg ctaggttgct gggcactgat aattccgtgg tgttgtctgt 2760
gccttctagt tgccagccat ctgttgtttg cccctccccc gtgccttcct tgaccctgga 2820
aggtgccact cccactgtcc tttcctaata aaatgaggaa attgcatcgc attgtctgag 2880
taggtgtcat tctattctgg ggggtggggt ggggcaggac agcaaggggg aggattggga 2940
agacaatagc aggcatgctg gggatgcggt gggctctatg gcaggaaccc ctagtgatgg 3000
agttggccac tccctctctg cgcgctcgct cgctcactga ggccgggcga ccaaaggtcg 3060
cccgacgccc gggctttgcc cgggcggcct cagtgagcga gcgagcgcgc agctgcctgc 3120
agg 3123
<210> 54
<211> 3117
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 54
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc tgaacagaga aacaggagaa tatgggccaa acaggatatc 180
tgtggtaagc agttcctgcc ccggctcagg gccaagaaca gttggaacag cagaatatgg 240
gccaaacagg atatctgtgg taagcagttc ctgccccggc tcagggccaa gaacagatgg 300
tccccagatg cggtcccgcc ctcagcagtt tctagagaac catcagatgt ttccagggtg 360
ccccaaggac ctgaaatgac cctgtgcctt atttgaacta accaatcagt tcgcttctcg 420
cttctgttcg cgcgcttctg ctccccgagc tctatataag cagagctcgt ttagtgaacc 480
gtcagatcgc ctggagacgc catccacgct gttttgactt ccatagaagg ccgccaccat 540
ggaagacgcc aaaaacataa agaaaggccc ggcgccattc tatccgctgg aagatggaac 600
cgctggagag caactgcata aggctatgaa gagatacgcc ctggttcctg gaacaattgc 660
ttttacagat gcacatatcg aggtggacat cacttacgct gagtacttcg aaatgtccgt 720
tcggttggca gaagctatga aacgatatgg gctgaataca aatcacagaa tcgtcgtatg 780
cagtgaaaac tctcttcaat tctttatgcc ggtgttgggc gcgttattta tcggagttgc 840
agttgcgccc gcgaacgaca tttataatga acgtgaattg ctcaacagta tgggcatttc 900
gcagcctacc gtggtgttcg tttccaaaaa ggggttgcaa aaaattttga acgtgcaaaa 960
aaagctccca atcatccaaa aaattattat catggattct aaaacggatt accagggatt 1020
tcagtcgatg tacacgttcg tcacatctca tctacctccc ggttttaatg aatacgattt 1080
tgtgccagag tccttcgata gggacaagac aattgcactg atcatgaact cctctggatc 1140
tactggtctg cctaaaggtg tcgctctgcc tcatagaact gcctgcgtga gattctcgca 1200
tgccagagat cctatttttg gcaatcaaat cattccggat actgcgattt taagtgttgt 1260
tccattccat cacggttttg gaatgtttac tacactcgga tatttgatat gtggatttcg 1320
agtcgtctta atgtatagat ttgaagaaga gctgtttctg aggagccttc aggattacaa 1380
gattcaaagt gcgctgctgg tgccaaccct attctccttc ttcgccaaaa gcactctgat 1440
tgacaaatac gatttatcta atttacacga aattgcttct ggtggcgctc ccctctctaa 1500
ggaagtcggg gaagcggttg ccaagaggtt ccatctgcca ggtatcaggc aaggatatgg 1560
gctcactgag actacatcag ctattctgat tacacccgag ggggatgata aaccgggcgc 1620
ggtcggtaaa gttgttccat tttttgaagc gaaggttgtg gatctggata ccgggaaaac 1680
gctgggcgtt aatcaaagag gcgaactgtg tgtgagaggt cctatgatta tgtccggtta 1740
tgtaaacaat ccggaagcga ccaacgcctt gattgacaag gatggatggc tacattctgg 1800
agacatagct tactgggacg aagacgaaca cttcttcatc gttgaccgcc tgaagtctct 1860
gattaagtac aaaggctatc aggtggctcc cgctgaattg gaatccatct tgctccaaca 1920
ccccaacatc ttcgacgcag gtgtcgcagg tcttcccgac gatgacgccg gtgaacttcc 1980
cgccgccgtt gttgttttgg agcacggaaa gacgatgacg gaaaaagaga tcgtggatta 2040
cgtcgccagt caagtaacaa ccgcgaaaaa gttgcgcgga ggagttgtgt ttgtggacga 2100
agtaccgaaa ggtcttaccg gaaaactcga cgcaagaaaa atcagagaga tcctcataaa 2160
ggccaagaag ggcggaaaga tcgccgtgta agagcatctt accgccattt attcccatat 2220
ttgttctgtt tttcttgatt tgggtataca tttaaatgtt aataaaacaa aatggtgggg 2280
caatcattta catttttagg gatatgtaat tactagttca ggtgtattgc cacaagacaa 2340
acatgttaag aaactttccc gttatttacg ctctgttcct gttaatcaac ctctggatta 2400
caaaatttgt gaaagattga ctgatattct taactatgtt gctcctttta cgctgtgtgg 2460
atatgctgct ttatagcctc tgtatctagc tattgcttcc cgtacggctt tcgttttctc 2520
ctccttgtat aaatcctggt tgctgtctct tttagaggag ttgtggcccg ttgtccgtca 2580
acgtggcgtg gtgtgctctg tgtttgctga cgcaaccccc actggctggg gcattgccac 2640
cacctgtcaa ctcctttctg ggactttcgc tttccccctc ccgatcgcca cggcagaact 2700
catcgccgcc tgccttgccc gctgctggac aggggctagg ttgctgggca ctgataattc 2760
cgtggtgttg tctgtgcctt ctagttgcca gccatctgtt gtttgcccct cccccgtgcc 2820
ttccttgacc ctggaaggtg ccactcccac tgtcctttcc taataaaatg aggaaattgc 2880
atcgcattgt ctgagtaggt gtcattctat tctggggggt ggggtggggc aggacagcaa 2940
gggggaggat tgggaagaca atagcaggca tgctggggat gcggtgggct ctatggcagg 3000
aacccctagt gatggagttg gccactccct ctctgcgcgc tcgctcgctc actgaggccg 3060
cccgggaaac ccgggcgtgc gcctcagtga gcgagcgagc gcgcagctgc ctgcagg 3117
<210> 55
<211> 2841
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 55
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggaaacc cgggcgtgcc 60
cgggcgcctc agtgagcgag cgagcgcgca gagagggagt ggccaactcc atcactaggg 120
gttcctgaac agagaaacag gagaatatgg gccaaacagg atatctgtgg taagcagttc 180
ctgccccggc tcagggccaa gaacagttgg aacagcagaa tatgggccaa acaggatatc 240
tgtggtaagc agttcctgcc ccggctcagg gccaagaaca gatggtcccc agatgcggtc 300
ccgccctcag cagtttctag agaaccatca gatgtttcca gggtgcccca aggacctgaa 360
atgaccctgt gccttatttg aactaaccaa tcagttcgct tctcgcttct gttcgcgcgc 420
ttctgctccc cgagctctat ataagcagag ctcgtttagt gaaccgtcag atcgcctgga 480
gacgccatcc acgctgtttt gacttccata gaaggccgcc accatgatca tggccgagag 540
ccctggcctg atcaccatct gcctgctggg ctacctgctg agcgccgagt gcaccgtgtt 600
cctggaccac gagaacgcca acaagatcct gaaccggccc aagagataca acagcggcaa 660
gctggaggag ttcgtgcagg gcaacctgga gagggagtgc atggaggaga agtgcagctt 720
cgaggaggcc agggaagtgt tcgagaacac cgagcggacc accgagttct ggaagcagta 780
cgtggacggc gaccagtgcg agagcaaccc ttgcctgaac ggcggcagct gcaaggacga 840
catcaacagc tacgagtgct ggtgcccttt cggcttcgag ggcaagaact gcgagctgga 900
cgtgacctgc aacatcaaga acggccgctg cgagcagttc tgcaagaaca gcgccgacaa 960
caaagtggtg tgtagctgca ccgagggcta cagactggcc gagaaccaga agagctgcga 1020
gcccgccgtg cccttcccct gcggcagagt gagcgtgtcc cagaccagca agctgaccag 1080
agccgagacc gtgttccccg acgtggacta cgtgaatagc accgaggccg agaccatcct 1140
ggacaacatc acccagagca cccagtcctt caacgacttc accagagttg tgggcggcga 1200
ggacgccaag cccggccagt tcccctggca ggtggtgctg aacggcaaag tggatgcctt 1260
ctgcggcggc agcatcgtga acgagaagtg gatcgtgaca gccgcccact gcgtggagac 1320
cggcgtgaag atcaccgtgg tggccggcga acacaatatc gaggagaccg agcacaccga 1380
gcagaagcgg aacgtcatcc ggattatccc ccaccacaac tacaacgccg ccatcaacaa 1440
gtacaaccac gacatcgccc tgctggagct ggacgagcct ctggtgctga atagctacgt 1500
gacccccatc tgcatcgccg acaaggagta caccaacatc ttcctgaagt tcggcagcgg 1560
ctacgtgtcc ggctggggca gagtgttcca caagggcaga agcgccctgg tgctgcagta 1620
cctgagagtg cccctggtgg acagagccac ctgcctgttg agcaccaagt tcaccatcta 1680
caacaacatg ttctgcgccg gcttccacga gggcggcaga gacagctgcc agggcgacag 1740
cggcggaccc cacgtgaccg aagtggaggg caccagcttc ctgaccggca tcatcagctg 1800
gggcgaggag tgcgccatga agggcaagta cggcatctac accaaagtga gccggtacgt 1860
gaactggatc aaggagaaaa ccaagctgac ctgagagcat cttaccgcca tttattccca 1920
tatttgttct gtttttcttg atttgggtat acatttaaat gttaataaaa caaaatggtg 1980
gggcaatcat ttacattttt agggatatgt aattactagt tcaggtgtat tgccacaaga 2040
caaacatgtt aagaaacttt cccgttattt acgctctgtt cctgttaatc aacctctgga 2100
ttacaaaatt tgtgaaagat tgactgatat tcttaactat gttgctcctt ttacgctgtg 2160
tggatatgct gctttatagc ctctgtatct agctattgct tcccgtacgg ctttcgtttt 2220
ctcctccttg tataaatcct ggttgctgtc tcttttagag gagttgtggc ccgttgtccg 2280
tcaacgtggc gtggtgtgct ctgtgtttgc tgacgcaacc cccactggct ggggcattgc 2340
caccacctgt caactccttt ctgggacttt cgctttcccc ctcccgatcg ccacggcaga 2400
actcatcgcc gcctgccttg cccgctgctg gacaggggct aggttgctgg gcactgataa 2460
ttccgtggtg ttgtctgtgc cttctagttg ccagccatct gttgtttgcc cctcccccgt 2520
gccttccttg accctggaag gtgccactcc cactgtcctt tcctaataaa atgaggaaat 2580
tgcatcgcat tgtctgagta ggtgtcattc tattctgggg ggtggggtgg ggcaggacag 2640
caagggggag gattgggaag acaatagcag gcatgctggg gatgcggtgg gctctatggc 2700
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 2760
ccgggcgacc aaaggtcgcc cgacgcccgg gctttgcccg ggcggcctca gtgagcgagc 2820
gagcgcgcag ctgcctgcag g 2841
<210> 56
<211> 2835
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 56
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc tgaacagaga aacaggagaa tatgggccaa acaggatatc 180
tgtggtaagc agttcctgcc ccggctcagg gccaagaaca gttggaacag cagaatatgg 240
gccaaacagg atatctgtgg taagcagttc ctgccccggc tcagggccaa gaacagatgg 300
tccccagatg cggtcccgcc ctcagcagtt tctagagaac catcagatgt ttccagggtg 360
ccccaaggac ctgaaatgac cctgtgcctt atttgaacta accaatcagt tcgcttctcg 420
cttctgttcg cgcgcttctg ctccccgagc tctatataag cagagctcgt ttagtgaacc 480
gtcagatcgc ctggagacgc catccacgct gttttgactt ccatagaagg ccgccaccat 540
gatcatggcc gagagccctg gcctgatcac catctgcctg ctgggctacc tgctgagcgc 600
cgagtgcacc gtgttcctgg accacgagaa cgccaacaag atcctgaacc ggcccaagag 660
atacaacagc ggcaagctgg aggagttcgt gcagggcaac ctggagaggg agtgcatgga 720
ggagaagtgc agcttcgagg aggccaggga agtgttcgag aacaccgagc ggaccaccga 780
gttctggaag cagtacgtgg acggcgacca gtgcgagagc aacccttgcc tgaacggcgg 840
cagctgcaag gacgacatca acagctacga gtgctggtgc cctttcggct tcgagggcaa 900
gaactgcgag ctggacgtga cctgcaacat caagaacggc cgctgcgagc agttctgcaa 960
gaacagcgcc gacaacaaag tggtgtgtag ctgcaccgag ggctacagac tggccgagaa 1020
ccagaagagc tgcgagcccg ccgtgccctt cccctgcggc agagtgagcg tgtcccagac 1080
cagcaagctg accagagccg agaccgtgtt ccccgacgtg gactacgtga atagcaccga 1140
ggccgagacc atcctggaca acatcaccca gagcacccag tccttcaacg acttcaccag 1200
agttgtgggc ggcgaggacg ccaagcccgg ccagttcccc tggcaggtgg tgctgaacgg 1260
caaagtggat gccttctgcg gcggcagcat cgtgaacgag aagtggatcg tgacagccgc 1320
ccactgcgtg gagaccggcg tgaagatcac cgtggtggcc ggcgaacaca atatcgagga 1380
gaccgagcac accgagcaga agcggaacgt catccggatt atcccccacc acaactacaa 1440
cgccgccatc aacaagtaca accacgacat cgccctgctg gagctggacg agcctctggt 1500
gctgaatagc tacgtgaccc ccatctgcat cgccgacaag gagtacacca acatcttcct 1560
gaagttcggc agcggctacg tgtccggctg gggcagagtg ttccacaagg gcagaagcgc 1620
cctggtgctg cagtacctga gagtgcccct ggtggacaga gccacctgcc tgttgagcac 1680
caagttcacc atctacaaca acatgttctg cgccggcttc cacgagggcg gcagagacag 1740
ctgccagggc gacagcggcg gaccccacgt gaccgaagtg gagggcacca gcttcctgac 1800
cggcatcatc agctggggcg aggagtgcgc catgaagggc aagtacggca tctacaccaa 1860
agtgagccgg tacgtgaact ggatcaagga gaaaaccaag ctgacctgag agcatcttac 1920
cgccatttat tcccatattt gttctgtttt tcttgatttg ggtatacatt taaatgttaa 1980
taaaacaaaa tggtggggca atcatttaca tttttaggga tatgtaatta ctagttcagg 2040
tgtattgcca caagacaaac atgttaagaa actttcccgt tatttacgct ctgttcctgt 2100
taatcaacct ctggattaca aaatttgtga aagattgact gatattctta actatgttgc 2160
tccttttacg ctgtgtggat atgctgcttt atagcctctg tatctagcta ttgcttcccg 2220
tacggctttc gttttctcct ccttgtataa atcctggttg ctgtctcttt tagaggagtt 2280
gtggcccgtt gtccgtcaac gtggcgtggt gtgctctgtg tttgctgacg caacccccac 2340
tggctggggc attgccacca cctgtcaact cctttctggg actttcgctt tccccctccc 2400
gatcgccacg gcagaactca tcgccgcctg ccttgcccgc tgctggacag gggctaggtt 2460
gctgggcact gataattccg tggtgttgtc tgtgccttct agttgccagc catctgttgt 2520
ttgcccctcc cccgtgcctt ccttgaccct ggaaggtgcc actcccactg tcctttccta 2580
ataaaatgag gaaattgcat cgcattgtct gagtaggtgt cattctattc tggggggtgg 2640
ggtggggcag gacagcaagg gggaggattg ggaagacaat agcaggcatg ctggggatgc 2700
ggtgggctct atggcaggaa cccctagtga tggagttggc cactccctct ctgcgcgctc 2760
gctcgctcac tgaggccgcc cgggaaaccc gggcgtgcgc ctcagtgagc gagcgagcgc 2820
gcagctgcct gcagg 2835
<210> 57
<211> 3912
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 57
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggaaacc cgggcgtgcc 60
cgggcgcctc agtgagcgag cgagcgcgca gagagggagt ggccaactcc atcactaggg 120
gttcctggct cagaggctca gaggcacaca ggagtttctg ggctcaccct gcccccttcc 180
aacccctcag ttcccatcct ccagcagctg tttgtgtgct gcctctgaag tccacactga 240
acaaacttca gcctactcat gtccctaaaa tgggcaaaca ttgcaagcag caaacagcaa 300
acacacagcc ctccctgcct gctgaccttg gagctggggc agaggtcaga gacctctctg 360
ggcccatgcc acctccaaca tccactcgac cccttggaat ttcggtggag aggagcagag 420
gttgtcctgg cgtggtttag gtagtgtgag agggtccggg ttcaaaacca cttgctgggt 480
ggggagtcgt cagtaagtgg ctatgccccg accccgaagc ctgtttcccc atctgtacaa 540
tggaaatgat aaagacgccc atctgatagg gtttttgtgg caaataaaca tttggttttt 600
ttgttttgtt ttgttttgtt ttttgagatg gaggtttgct ctgtcgccca ggctggagtg 660
cagtgacaca atctcatctc accacaacct tcccctgcct cagcctccca agtagctggg 720
attacaagca tgtgccacca cacctggcta attttctatt tttagtagag acgggtttct 780
ccatgttggt cagcctcagc ctcccaagta actgggatta caggcctgtg ccaccacacc 840
cggctaattt tttctatttt tgacagggac ggggtttcac catgttggtc aggctggtct 900
agaggtaccg gatcttgcta ccagtggaac agccactaag gattctgcag tgagagcaga 960
gggccagcta agtggtactc tcccagagac tgtctgactc acgccacccc ctccaccttg 1020
gacacaggac gctgtggttt ctgagccagg tacaatgact cctttcggta agtgcagtgg 1080
aagctgtaca ctgcccaggc aaagcgtccg ggcagcgtag gcgggcgact cagatcccag 1140
ccagtggact tagcccctgt ttgctcctcc gataactggg gtgaccttgg ttaatattca 1200
ccagcagcct cccccgttgc ccctctggat ccactgctta aatacggacg aggacagggc 1260
cctgtctcct cagcttcagg caccaccact gacctgggac agtgccgcca ccatggaaga 1320
cgccaaaaac ataaagaaag gcccggcgcc attctatccg ctggaagatg gaaccgctgg 1380
agagcaactg cataaggcta tgaagagata cgccctggtt cctggaacaa ttgcttttac 1440
agatgcacat atcgaggtgg acatcactta cgctgagtac ttcgaaatgt ccgttcggtt 1500
ggcagaagct atgaaacgat atgggctgaa tacaaatcac agaatcgtcg tatgcagtga 1560
aaactctctt caattcttta tgccggtgtt gggcgcgtta tttatcggag ttgcagttgc 1620
gcccgcgaac gacatttata atgaacgtga attgctcaac agtatgggca tttcgcagcc 1680
taccgtggtg ttcgtttcca aaaaggggtt gcaaaaaatt ttgaacgtgc aaaaaaagct 1740
cccaatcatc caaaaaatta ttatcatgga ttctaaaacg gattaccagg gatttcagtc 1800
gatgtacacg ttcgtcacat ctcatctacc tcccggtttt aatgaatacg attttgtgcc 1860
agagtccttc gatagggaca agacaattgc actgatcatg aactcctctg gatctactgg 1920
tctgcctaaa ggtgtcgctc tgcctcatag aactgcctgc gtgagattct cgcatgccag 1980
agatcctatt tttggcaatc aaatcattcc ggatactgcg attttaagtg ttgttccatt 2040
ccatcacggt tttggaatgt ttactacact cggatatttg atatgtggat ttcgagtcgt 2100
cttaatgtat agatttgaag aagagctgtt tctgaggagc cttcaggatt acaagattca 2160
aagtgcgctg ctggtgccaa ccctattctc cttcttcgcc aaaagcactc tgattgacaa 2220
atacgattta tctaatttac acgaaattgc ttctggtggc gctcccctct ctaaggaagt 2280
cggggaagcg gttgccaaga ggttccatct gccaggtatc aggcaaggat atgggctcac 2340
tgagactaca tcagctattc tgattacacc cgagggggat gataaaccgg gcgcggtcgg 2400
taaagttgtt ccattttttg aagcgaaggt tgtggatctg gataccggga aaacgctggg 2460
cgttaatcaa agaggcgaac tgtgtgtgag aggtcctatg attatgtccg gttatgtaaa 2520
caatccggaa gcgaccaacg ccttgattga caaggatgga tggctacatt ctggagacat 2580
agcttactgg gacgaagacg aacacttctt catcgttgac cgcctgaagt ctctgattaa 2640
gtacaaaggc tatcaggtgg ctcccgctga attggaatcc atcttgctcc aacaccccaa 2700
catcttcgac gcaggtgtcg caggtcttcc cgacgatgac gccggtgaac ttcccgccgc 2760
cgttgttgtt ttggagcacg gaaagacgat gacggaaaaa gagatcgtgg attacgtcgc 2820
cagtcaagta acaaccgcga aaaagttgcg cggaggagtt gtgtttgtgg acgaagtacc 2880
gaaaggtctt accggaaaac tcgacgcaag aaaaatcaga gagatcctca taaaggccaa 2940
gaagggcgga aagatcgccg tgtaagagca tcttaccgcc atttattccc atatttgttc 3000
tgtttttctt gatttgggta tacatttaaa tgttaataaa acaaaatggt ggggcaatca 3060
tttacatttt tagggatatg taattactag ttcaggtgta ttgccacaag acaaacatgt 3120
taagaaactt tcccgttatt tacgctctgt tcctgttaat caacctctgg attacaaaat 3180
ttgtgaaaga ttgactgata ttcttaacta tgttgctcct tttacgctgt gtggatatgc 3240
tgctttatag cctctgtatc tagctattgc ttcccgtacg gctttcgttt tctcctcctt 3300
gtataaatcc tggttgctgt ctcttttaga ggagttgtgg cccgttgtcc gtcaacgtgg 3360
cgtggtgtgc tctgtgtttg ctgacgcaac ccccactggc tggggcattg ccaccacctg 3420
tcaactcctt tctgggactt tcgctttccc cctcccgatc gccacggcag aactcatcgc 3480
cgcctgcctt gcccgctgct ggacaggggc taggttgctg ggcactgata attccgtggt 3540
gttgtctgtg ccttctagtt gccagccatc tgttgtttgc ccctcccccg tgccttcctt 3600
gaccctggaa ggtgccactc ccactgtcct ttcctaataa aatgaggaaa ttgcatcgca 3660
ttgtctgagt aggtgtcatt ctattctggg gggtggggtg gggcaggaca gcaaggggga 3720
ggattgggaa gacaatagca ggcatgctgg ggatgcggtg ggctctatgg caggaacccc 3780
tagtgatgga gttggccact ccctctctgc gcgctcgctc gctcactgag gccgggcgac 3840
caaaggtcgc ccgacgcccg ggctttgccc gggcggcctc agtgagcgag cgagcgcgca 3900
gctgcctgca gg 3912
<210> 58
<211> 3906
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 58
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc tggctcagag gctcagaggc acacaggagt ttctgggctc 180
accctgcccc cttccaaccc ctcagttccc atcctccagc agctgtttgt gtgctgcctc 240
tgaagtccac actgaacaaa cttcagccta ctcatgtccc taaaatgggc aaacattgca 300
agcagcaaac agcaaacaca cagccctccc tgcctgctga ccttggagct ggggcagagg 360
tcagagacct ctctgggccc atgccacctc caacatccac tcgacccctt ggaatttcgg 420
tggagaggag cagaggttgt cctggcgtgg tttaggtagt gtgagagggt ccgggttcaa 480
aaccacttgc tgggtgggga gtcgtcagta agtggctatg ccccgacccc gaagcctgtt 540
tccccatctg tacaatggaa atgataaaga cgcccatctg atagggtttt tgtggcaaat 600
aaacatttgg tttttttgtt ttgttttgtt ttgttttttg agatggaggt ttgctctgtc 660
gcccaggctg gagtgcagtg acacaatctc atctcaccac aaccttcccc tgcctcagcc 720
tcccaagtag ctgggattac aagcatgtgc caccacacct ggctaatttt ctatttttag 780
tagagacggg tttctccatg ttggtcagcc tcagcctccc aagtaactgg gattacaggc 840
ctgtgccacc acacccggct aattttttct atttttgaca gggacggggt ttcaccatgt 900
tggtcaggct ggtctagagg taccggatct tgctaccagt ggaacagcca ctaaggattc 960
tgcagtgaga gcagagggcc agctaagtgg tactctccca gagactgtct gactcacgcc 1020
accccctcca ccttggacac aggacgctgt ggtttctgag ccaggtacaa tgactccttt 1080
cggtaagtgc agtggaagct gtacactgcc caggcaaagc gtccgggcag cgtaggcggg 1140
cgactcagat cccagccagt ggacttagcc cctgtttgct cctccgataa ctggggtgac 1200
cttggttaat attcaccagc agcctccccc gttgcccctc tggatccact gcttaaatac 1260
ggacgaggac agggccctgt ctcctcagct tcaggcacca ccactgacct gggacagtgc 1320
cgccaccatg gaagacgcca aaaacataaa gaaaggcccg gcgccattct atccgctgga 1380
agatggaacc gctggagagc aactgcataa ggctatgaag agatacgccc tggttcctgg 1440
aacaattgct tttacagatg cacatatcga ggtggacatc acttacgctg agtacttcga 1500
aatgtccgtt cggttggcag aagctatgaa acgatatggg ctgaatacaa atcacagaat 1560
cgtcgtatgc agtgaaaact ctcttcaatt ctttatgccg gtgttgggcg cgttatttat 1620
cggagttgca gttgcgcccg cgaacgacat ttataatgaa cgtgaattgc tcaacagtat 1680
gggcatttcg cagcctaccg tggtgttcgt ttccaaaaag gggttgcaaa aaattttgaa 1740
cgtgcaaaaa aagctcccaa tcatccaaaa aattattatc atggattcta aaacggatta 1800
ccagggattt cagtcgatgt acacgttcgt cacatctcat ctacctcccg gttttaatga 1860
atacgatttt gtgccagagt ccttcgatag ggacaagaca attgcactga tcatgaactc 1920
ctctggatct actggtctgc ctaaaggtgt cgctctgcct catagaactg cctgcgtgag 1980
attctcgcat gccagagatc ctatttttgg caatcaaatc attccggata ctgcgatttt 2040
aagtgttgtt ccattccatc acggttttgg aatgtttact acactcggat atttgatatg 2100
tggatttcga gtcgtcttaa tgtatagatt tgaagaagag ctgtttctga ggagccttca 2160
ggattacaag attcaaagtg cgctgctggt gccaacccta ttctccttct tcgccaaaag 2220
cactctgatt gacaaatacg atttatctaa tttacacgaa attgcttctg gtggcgctcc 2280
cctctctaag gaagtcgggg aagcggttgc caagaggttc catctgccag gtatcaggca 2340
aggatatggg ctcactgaga ctacatcagc tattctgatt acacccgagg gggatgataa 2400
accgggcgcg gtcggtaaag ttgttccatt ttttgaagcg aaggttgtgg atctggatac 2460
cgggaaaacg ctgggcgtta atcaaagagg cgaactgtgt gtgagaggtc ctatgattat 2520
gtccggttat gtaaacaatc cggaagcgac caacgccttg attgacaagg atggatggct 2580
acattctgga gacatagctt actgggacga agacgaacac ttcttcatcg ttgaccgcct 2640
gaagtctctg attaagtaca aaggctatca ggtggctccc gctgaattgg aatccatctt 2700
gctccaacac cccaacatct tcgacgcagg tgtcgcaggt cttcccgacg atgacgccgg 2760
tgaacttccc gccgccgttg ttgttttgga gcacggaaag acgatgacgg aaaaagagat 2820
cgtggattac gtcgccagtc aagtaacaac cgcgaaaaag ttgcgcggag gagttgtgtt 2880
tgtggacgaa gtaccgaaag gtcttaccgg aaaactcgac gcaagaaaaa tcagagagat 2940
cctcataaag gccaagaagg gcggaaagat cgccgtgtaa gagcatctta ccgccattta 3000
ttcccatatt tgttctgttt ttcttgattt gggtatacat ttaaatgtta ataaaacaaa 3060
atggtggggc aatcatttac atttttaggg atatgtaatt actagttcag gtgtattgcc 3120
acaagacaaa catgttaaga aactttcccg ttatttacgc tctgttcctg ttaatcaacc 3180
tctggattac aaaatttgtg aaagattgac tgatattctt aactatgttg ctccttttac 3240
gctgtgtgga tatgctgctt tatagcctct gtatctagct attgcttccc gtacggcttt 3300
cgttttctcc tccttgtata aatcctggtt gctgtctctt ttagaggagt tgtggcccgt 3360
tgtccgtcaa cgtggcgtgg tgtgctctgt gtttgctgac gcaaccccca ctggctgggg 3420
cattgccacc acctgtcaac tcctttctgg gactttcgct ttccccctcc cgatcgccac 3480
ggcagaactc atcgccgcct gccttgcccg ctgctggaca ggggctaggt tgctgggcac 3540
tgataattcc gtggtgttgt ctgtgccttc tagttgccag ccatctgttg tttgcccctc 3600
ccccgtgcct tccttgaccc tggaaggtgc cactcccact gtcctttcct aataaaatga 3660
ggaaattgca tcgcattgtc tgagtaggtg tcattctatt ctggggggtg gggtggggca 3720
ggacagcaag ggggaggatt gggaagacaa tagcaggcat gctggggatg cggtgggctc 3780
tatggcagga acccctagtg atggagttgg ccactccctc tctgcgcgct cgctcgctca 3840
ctgaggccgc ccgggaaacc cgggcgtgcg cctcagtgag cgagcgagcg cgcagctgcc 3900
tgcagg 3906
<210> 59
<211> 3630
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 59
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggaaacc cgggcgtgcc 60
cgggcgcctc agtgagcgag cgagcgcgca gagagggagt ggccaactcc atcactaggg 120
gttcctggct cagaggctca gaggcacaca ggagtttctg ggctcaccct gcccccttcc 180
aacccctcag ttcccatcct ccagcagctg tttgtgtgct gcctctgaag tccacactga 240
acaaacttca gcctactcat gtccctaaaa tgggcaaaca ttgcaagcag caaacagcaa 300
acacacagcc ctccctgcct gctgaccttg gagctggggc agaggtcaga gacctctctg 360
ggcccatgcc acctccaaca tccactcgac cccttggaat ttcggtggag aggagcagag 420
gttgtcctgg cgtggtttag gtagtgtgag agggtccggg ttcaaaacca cttgctgggt 480
ggggagtcgt cagtaagtgg ctatgccccg accccgaagc ctgtttcccc atctgtacaa 540
tggaaatgat aaagacgccc atctgatagg gtttttgtgg caaataaaca tttggttttt 600
ttgttttgtt ttgttttgtt ttttgagatg gaggtttgct ctgtcgccca ggctggagtg 660
cagtgacaca atctcatctc accacaacct tcccctgcct cagcctccca agtagctggg 720
attacaagca tgtgccacca cacctggcta attttctatt tttagtagag acgggtttct 780
ccatgttggt cagcctcagc ctcccaagta actgggatta caggcctgtg ccaccacacc 840
cggctaattt tttctatttt tgacagggac ggggtttcac catgttggtc aggctggtct 900
agaggtaccg gatcttgcta ccagtggaac agccactaag gattctgcag tgagagcaga 960
gggccagcta agtggtactc tcccagagac tgtctgactc acgccacccc ctccaccttg 1020
gacacaggac gctgtggttt ctgagccagg tacaatgact cctttcggta agtgcagtgg 1080
aagctgtaca ctgcccaggc aaagcgtccg ggcagcgtag gcgggcgact cagatcccag 1140
ccagtggact tagcccctgt ttgctcctcc gataactggg gtgaccttgg ttaatattca 1200
ccagcagcct cccccgttgc ccctctggat ccactgctta aatacggacg aggacagggc 1260
cctgtctcct cagcttcagg caccaccact gacctgggac agtgccgcca ccatgatcat 1320
ggccgagagc cctggcctga tcaccatctg cctgctgggc tacctgctga gcgccgagtg 1380
caccgtgttc ctggaccacg agaacgccaa caagatcctg aaccggccca agagatacaa 1440
cagcggcaag ctggaggagt tcgtgcaggg caacctggag agggagtgca tggaggagaa 1500
gtgcagcttc gaggaggcca gggaagtgtt cgagaacacc gagcggacca ccgagttctg 1560
gaagcagtac gtggacggcg accagtgcga gagcaaccct tgcctgaacg gcggcagctg 1620
caaggacgac atcaacagct acgagtgctg gtgccctttc ggcttcgagg gcaagaactg 1680
cgagctggac gtgacctgca acatcaagaa cggccgctgc gagcagttct gcaagaacag 1740
cgccgacaac aaagtggtgt gtagctgcac cgagggctac agactggccg agaaccagaa 1800
gagctgcgag cccgccgtgc ccttcccctg cggcagagtg agcgtgtccc agaccagcaa 1860
gctgaccaga gccgagaccg tgttccccga cgtggactac gtgaatagca ccgaggccga 1920
gaccatcctg gacaacatca cccagagcac ccagtccttc aacgacttca ccagagttgt 1980
gggcggcgag gacgccaagc ccggccagtt cccctggcag gtggtgctga acggcaaagt 2040
ggatgccttc tgcggcggca gcatcgtgaa cgagaagtgg atcgtgacag ccgcccactg 2100
cgtggagacc ggcgtgaaga tcaccgtggt ggccggcgaa cacaatatcg aggagaccga 2160
gcacaccgag cagaagcgga acgtcatccg gattatcccc caccacaact acaacgccgc 2220
catcaacaag tacaaccacg acatcgccct gctggagctg gacgagcctc tggtgctgaa 2280
tagctacgtg acccccatct gcatcgccga caaggagtac accaacatct tcctgaagtt 2340
cggcagcggc tacgtgtccg gctggggcag agtgttccac aagggcagaa gcgccctggt 2400
gctgcagtac ctgagagtgc ccctggtgga cagagccacc tgcctgttga gcaccaagtt 2460
caccatctac aacaacatgt tctgcgccgg cttccacgag ggcggcagag acagctgcca 2520
gggcgacagc ggcggacccc acgtgaccga agtggagggc accagcttcc tgaccggcat 2580
catcagctgg ggcgaggagt gcgccatgaa gggcaagtac ggcatctaca ccaaagtgag 2640
ccggtacgtg aactggatca aggagaaaac caagctgacc tgagagcatc ttaccgccat 2700
ttattcccat atttgttctg tttttcttga tttgggtata catttaaatg ttaataaaac 2760
aaaatggtgg ggcaatcatt tacattttta gggatatgta attactagtt caggtgtatt 2820
gccacaagac aaacatgtta agaaactttc ccgttattta cgctctgttc ctgttaatca 2880
acctctggat tacaaaattt gtgaaagatt gactgatatt cttaactatg ttgctccttt 2940
tacgctgtgt ggatatgctg ctttatagcc tctgtatcta gctattgctt cccgtacggc 3000
tttcgttttc tcctccttgt ataaatcctg gttgctgtct cttttagagg agttgtggcc 3060
cgttgtccgt caacgtggcg tggtgtgctc tgtgtttgct gacgcaaccc ccactggctg 3120
gggcattgcc accacctgtc aactcctttc tgggactttc gctttccccc tcccgatcgc 3180
cacggcagaa ctcatcgccg cctgccttgc ccgctgctgg acaggggcta ggttgctggg 3240
cactgataat tccgtggtgt tgtctgtgcc ttctagttgc cagccatctg ttgtttgccc 3300
ctcccccgtg ccttccttga ccctggaagg tgccactccc actgtccttt cctaataaaa 3360
tgaggaaatt gcatcgcatt gtctgagtag gtgtcattct attctggggg gtggggtggg 3420
gcaggacagc aagggggagg attgggaaga caatagcagg catgctgggg atgcggtggg 3480
ctctatggca ggaaccccta gtgatggagt tggccactcc ctctctgcgc gctcgctcgc 3540
tcactgaggc cgggcgacca aaggtcgccc gacgcccggg ctttgcccgg gcggcctcag 3600
tgagcgagcg agcgcgcagc tgcctgcagg 3630
<210> 60
<211> 3624
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 60
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc tggctcagag gctcagaggc acacaggagt ttctgggctc 180
accctgcccc cttccaaccc ctcagttccc atcctccagc agctgtttgt gtgctgcctc 240
tgaagtccac actgaacaaa cttcagccta ctcatgtccc taaaatgggc aaacattgca 300
agcagcaaac agcaaacaca cagccctccc tgcctgctga ccttggagct ggggcagagg 360
tcagagacct ctctgggccc atgccacctc caacatccac tcgacccctt ggaatttcgg 420
tggagaggag cagaggttgt cctggcgtgg tttaggtagt gtgagagggt ccgggttcaa 480
aaccacttgc tgggtgggga gtcgtcagta agtggctatg ccccgacccc gaagcctgtt 540
tccccatctg tacaatggaa atgataaaga cgcccatctg atagggtttt tgtggcaaat 600
aaacatttgg tttttttgtt ttgttttgtt ttgttttttg agatggaggt ttgctctgtc 660
gcccaggctg gagtgcagtg acacaatctc atctcaccac aaccttcccc tgcctcagcc 720
tcccaagtag ctgggattac aagcatgtgc caccacacct ggctaatttt ctatttttag 780
tagagacggg tttctccatg ttggtcagcc tcagcctccc aagtaactgg gattacaggc 840
ctgtgccacc acacccggct aattttttct atttttgaca gggacggggt ttcaccatgt 900
tggtcaggct ggtctagagg taccggatct tgctaccagt ggaacagcca ctaaggattc 960
tgcagtgaga gcagagggcc agctaagtgg tactctccca gagactgtct gactcacgcc 1020
accccctcca ccttggacac aggacgctgt ggtttctgag ccaggtacaa tgactccttt 1080
cggtaagtgc agtggaagct gtacactgcc caggcaaagc gtccgggcag cgtaggcggg 1140
cgactcagat cccagccagt ggacttagcc cctgtttgct cctccgataa ctggggtgac 1200
cttggttaat attcaccagc agcctccccc gttgcccctc tggatccact gcttaaatac 1260
ggacgaggac agggccctgt ctcctcagct tcaggcacca ccactgacct gggacagtgc 1320
cgccaccatg atcatggccg agagccctgg cctgatcacc atctgcctgc tgggctacct 1380
gctgagcgcc gagtgcaccg tgttcctgga ccacgagaac gccaacaaga tcctgaaccg 1440
gcccaagaga tacaacagcg gcaagctgga ggagttcgtg cagggcaacc tggagaggga 1500
gtgcatggag gagaagtgca gcttcgagga ggccagggaa gtgttcgaga acaccgagcg 1560
gaccaccgag ttctggaagc agtacgtgga cggcgaccag tgcgagagca acccttgcct 1620
gaacggcggc agctgcaagg acgacatcaa cagctacgag tgctggtgcc ctttcggctt 1680
cgagggcaag aactgcgagc tggacgtgac ctgcaacatc aagaacggcc gctgcgagca 1740
gttctgcaag aacagcgccg acaacaaagt ggtgtgtagc tgcaccgagg gctacagact 1800
ggccgagaac cagaagagct gcgagcccgc cgtgcccttc ccctgcggca gagtgagcgt 1860
gtcccagacc agcaagctga ccagagccga gaccgtgttc cccgacgtgg actacgtgaa 1920
tagcaccgag gccgagacca tcctggacaa catcacccag agcacccagt ccttcaacga 1980
cttcaccaga gttgtgggcg gcgaggacgc caagcccggc cagttcccct ggcaggtggt 2040
gctgaacggc aaagtggatg ccttctgcgg cggcagcatc gtgaacgaga agtggatcgt 2100
gacagccgcc cactgcgtgg agaccggcgt gaagatcacc gtggtggccg gcgaacacaa 2160
tatcgaggag accgagcaca ccgagcagaa gcggaacgtc atccggatta tcccccacca 2220
caactacaac gccgccatca acaagtacaa ccacgacatc gccctgctgg agctggacga 2280
gcctctggtg ctgaatagct acgtgacccc catctgcatc gccgacaagg agtacaccaa 2340
catcttcctg aagttcggca gcggctacgt gtccggctgg ggcagagtgt tccacaaggg 2400
cagaagcgcc ctggtgctgc agtacctgag agtgcccctg gtggacagag ccacctgcct 2460
gttgagcacc aagttcacca tctacaacaa catgttctgc gccggcttcc acgagggcgg 2520
cagagacagc tgccagggcg acagcggcgg accccacgtg accgaagtgg agggcaccag 2580
cttcctgacc ggcatcatca gctggggcga ggagtgcgcc atgaagggca agtacggcat 2640
ctacaccaaa gtgagccggt acgtgaactg gatcaaggag aaaaccaagc tgacctgaga 2700
gcatcttacc gccatttatt cccatatttg ttctgttttt cttgatttgg gtatacattt 2760
aaatgttaat aaaacaaaat ggtggggcaa tcatttacat ttttagggat atgtaattac 2820
tagttcaggt gtattgccac aagacaaaca tgttaagaaa ctttcccgtt atttacgctc 2880
tgttcctgtt aatcaacctc tggattacaa aatttgtgaa agattgactg atattcttaa 2940
ctatgttgct ccttttacgc tgtgtggata tgctgcttta tagcctctgt atctagctat 3000
tgcttcccgt acggctttcg ttttctcctc cttgtataaa tcctggttgc tgtctctttt 3060
agaggagttg tggcccgttg tccgtcaacg tggcgtggtg tgctctgtgt ttgctgacgc 3120
aacccccact ggctggggca ttgccaccac ctgtcaactc ctttctggga ctttcgcttt 3180
ccccctcccg atcgccacgg cagaactcat cgccgcctgc cttgcccgct gctggacagg 3240
ggctaggttg ctgggcactg ataattccgt ggtgttgtct gtgccttcta gttgccagcc 3300
atctgttgtt tgcccctccc ccgtgccttc cttgaccctg gaaggtgcca ctcccactgt 3360
cctttcctaa taaaatgagg aaattgcatc gcattgtctg agtaggtgtc attctattct 3420
ggggggtggg gtggggcagg acagcaaggg ggaggattgg gaagacaata gcaggcatgc 3480
tggggatgcg gtgggctcta tggcaggaac ccctagtgat ggagttggcc actccctctc 3540
tgcgcgctcg ctcgctcact gaggccgccc gggaaacccg ggcgtgcgcc tcagtgagcg 3600
agcgagcgcg cagctgcctg cagg 3624
<210> 61
<211> 4257
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 61
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc ttgtagttaa tgattaaccc gccatgctac ttatctacgt 180
agccatgctc tagagcggcc gcacgcgtag atcttcaata ttggccatta gccatattat 240
tcattggtta tatagcataa atcaatattg gctattggcc attgcatacg ttgtatctat 300
atcataatat gtacatttat attggctcat gtccaatatg accgccatgt tggcattgat 360
tattgactag ttattaatag taatcaatta cggggtcatt agttcatagc ccatatatgg 420
agttccgcgt tacataactt acggtaaatg gcccgcctgg ctgaccgccc aacgaccccc 480
gcccattgac gtcaataatg acgtatgttc ccatagtaac gccaataggg actttccatt 540
gacgtcaatg ggtggagtat ttacggtaaa ctgcccactt ggcagtacat caagtgtatc 600
atatgccaag tccgccccct attgacgtca atgacggtaa atggcccgcc tggcattatg 660
cccagtacat gaccttacgg gactttccta cttggcagta catctacgta ttagtcatcg 720
ctattaccat ggtcgaggtg agccccacgt tctgcttcac tctccccatc tcccccccct 780
ccccaccccc aattttgtat ttatttattt tttaattatt ttgtgcagcg atgggggcgg 840
gggggggggg ggggcgcgcg ccaggcgggg cggggcgggg cgaggggcgg ggcggggcga 900
ggcggagagg tgcggcggca gccaatcaga gcggcgcgct ccgaaagttt ccttttatgg 960
cgaggcggcg gcggcggcgg ccctataaaa agcgaagcgc gcggcgggcg ggagtcgctg 1020
cgacgctgcc ttcgccccgt gccccgctcc gccgccgcct cgcgccgccc gccccggctc 1080
tgactgaccg cgttactccc acaggtgagc gggcgggacg gcccttctcc tccgggctgt 1140
aattagcgct tggtttaatg acggcttgtt tcttttctgt ggctgcgtga aagccttgag 1200
gggctccggg agggcccttt gtgcgggggg gagcggctcg gggggtgcgt gcgtgtgtgt 1260
gtgcgtgggg agcgccgcgt gcggcccgcg ctgcccggcg gctgtgagcg ctgcgggcgc 1320
ggcgcggggc tttgtgcgct ccgcagtgtg cgcgagggga gcgcggccgg gggcggtgcc 1380
ccgcggtgcg gggggggctg cgaggggaac aaaggctgcg tgcggggtgt gtgcgtgggg 1440
gggtgagcag ggggtgtggg cgcggcggtc gggctgtaac ccccccctgc acccccctcc 1500
ccgagttgct gagcacggcc cggcttcggg tgcggggctc cgtacggggc gtggcgcggg 1560
gctcgccgtg ccgggcgggg ggtggcggca ggtgggggtg ccgggcgggg cggggccgcc 1620
tcgggccggg gagggctcgg gggaggggcg cggcggcccc cggagcgccg gcggctgtcg 1680
aggcgcggcg agccgcagcc attgcctttt atggtaatcg tgcgagaggg cgcagggact 1740
tcctttgtcc caaatctgtg cggagccgaa atctgggagg cgccgccgca ccccctctag 1800
cgggcgcggg gcgaagcggt gcggcgccgg caggaaggaa atgggcgggg agggccttcg 1860
tgcgtcgccg cgccgccgtc cccttctccc tctccagcct cggggctgtc cgcgggggga 1920
cggctgcctt cgggggggac ggggcagggc ggggttcggc ttctggcgtg tgaccggcgg 1980
ctctagagcc tctgctaacc atgttttagc cttcttcttt ttcctacagc tcctgggcaa 2040
cgtgctggtt attgtgctgt ctcatcattt gtcgacagaa ttcctcgaag atccgaaggg 2100
gttcaagctt ggcattccgg tactgttggt aaagccacca tggaagacgc caaaaacata 2160
aagaaaggcc cggcgccatt ctatccgctg gaagatggaa ccgctggaga gcaactgcat 2220
aaggctatga agagatacgc cctggttcct ggaacaattg cttttacaga tgcacatatc 2280
gaggtggaca tcacttacgc tgagtacttc gaaatgtccg ttcggttggc agaagctatg 2340
aaacgatatg ggctgaatac aaatcacaga atcgtcgtat gcagtgaaaa ctctcttcaa 2400
ttctttatgc cggtgttggg cgcgttattt atcggagttg cagttgcgcc cgcgaacgac 2460
atttataatg aacgtgaatt gctcaacagt atgggcattt cgcagcctac cgtggtgttc 2520
gtttccaaaa aggggttgca aaaaattttg aacgtgcaaa aaaagctccc aatcatccaa 2580
aaaattatta tcatggattc taaaacggat taccagggat ttcagtcgat gtacacgttc 2640
gtcacatctc atctacctcc cggttttaat gaatacgatt ttgtgccaga gtccttcgat 2700
agggacaaga caattgcact gatcatgaac tcctctggat ctactggtct gcctaaaggt 2760
gtcgctctgc ctcatagaac tgcctgcgtg agattctcgc atgccagaga tcctattttt 2820
ggcaatcaaa tcattccgga tactgcgatt ttaagtgttg ttccattcca tcacggtttt 2880
ggaatgttta ctacactcgg atatttgata tgtggatttc gagtcgtctt aatgtataga 2940
tttgaagaag agctgtttct gaggagcctt caggattaca agattcaaag tgcgctgctg 3000
gtgccaaccc tattctcctt cttcgccaaa agcactctga ttgacaaata cgatttatct 3060
aatttacacg aaattgcttc tggtggcgct cccctctcta aggaagtcgg ggaagcggtt 3120
gccaagaggt tccatctgcc aggtatcagg caaggatatg ggctcactga gactacatca 3180
gctattctga ttacacccga gggggatgat aaaccgggcg cggtcggtaa agttgttcca 3240
ttttttgaag cgaaggttgt ggatctggat accgggaaaa cgctgggcgt taatcaaaga 3300
ggcgaactgt gtgtgagagg tcctatgatt atgtccggtt atgtaaacaa tccggaagcg 3360
accaacgcct tgattgacaa ggatggatgg ctacattctg gagacatagc ttactgggac 3420
gaagacgaac acttcttcat cgttgaccgc ctgaagtctc tgattaagta caaaggctat 3480
caggtggctc ccgctgaatt ggaatccatc ttgctccaac accccaacat cttcgacgca 3540
ggtgtcgcag gtcttcccga cgatgacgcc ggtgaacttc ccgccgccgt tgttgttttg 3600
gagcacggaa agacgatgac ggaaaaagag atcgtggatt acgtcgccag tcaagtaaca 3660
accgcgaaaa agttgcgcgg aggagttgtg tttgtggacg aagtaccgaa aggtcttacc 3720
ggaaaactcg acgcaagaaa aatcagagag atcctcataa aggccaagaa gggcggaaag 3780
atcgccgtgt aattctagag tcggggcggc cggccgcttc gagcagacat gataagatac 3840
attgatgagt ttggacaaac cacaactaga atgcagtgaa aaaaatgctt tatttgtgaa 3900
atttgtgatg ctattgcttt atttgtaacc attataagct gcaataaaca agttaacaac 3960
aacaattgca ttcattttat gtttcaggtt cagggggagg tgtgggaggt tttttaaagc 4020
aagtaaaacc tctacaaatg tggtaaaatc gataagtgcg gaccgagcgg ccgctctaga 4080
gcatggctac gtagataagt agcatggcgg gttaatcatt aactacacct gcaggcagct 4140
gcgcgctcgc tcgctcactg aggccgcccg ggcgtcgggc gacctttggt cgcccggcct 4200
cagtgagcga gcgagcgcgc agagagggag tggccaactc catcactagg ggttcct 4257
<210> 62
<211> 2510
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 62
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc ttgtagttaa tgattaaccc gccatgctac ttatctacgt 180
agccatgctc tagagcggcc gcgctagccc ctaaaatggg caaacattgc aagcagcaaa 240
cagcaaacac acagccctcc ctgcctgctg accttggagc tggggcagag gtcagagacc 300
tctctgggcc catgccacct ccaacatcca ctcgacccct tggaatttcg gtggagagga 360
gcagaggttg tcctggcgtg gtttaggtag tgtgagaggg gaatgactcc tttcggtaag 420
tgcagtggaa gctgtacact gcccaggcaa agcgtccggg cagcgtaggc gggcgactca 480
gatcccagcc agtggactta gcccctgttt gctcctccga taactggggt gaccttggtt 540
aatattcacc agcagcctcc cccgttgccc ctctggatcc actgcttaaa tacggacgag 600
gacactcgag ggccctgtct cctcagcttc aggcaccacc actgacctgg gacagtgaat 660
ccggacatcg attctaaggt aaatataaaa tttttaagtg tataatttgt taaactactg 720
attctaattg tttctctctt ttagattcca acctttggaa ctgaattcta gaccaccatg 780
cagagggtga acatgatcat ggctgagagc cctggcctga tcaccatctg cctgctgggc 840
tacctgctgt ctgctgagtg cactgtgttc ctggaccatg agaatgccaa caagatcctg 900
aacaggccca agagatacaa ctctggcaag ctggaggagt ttgtgcaggg caacctggag 960
agggagtgca tggaggagaa gtgcagcttt gaggaggcca gggaggtgtt tgagaacact 1020
gagaggacca ctgagttctg gaagcagtat gtggatgggg accagtgtga gagcaacccc 1080
tgcctgaatg ggggcagctg caaggatgac atcaacagct atgagtgctg gtgccccttt 1140
ggctttgagg gcaagaactg tgagctggat gtgacctgca acatcaagaa tggcagatgt 1200
gagcagttct gcaagaactc tgctgacaac aaggtggtgt gcagctgcac tgagggctac 1260
aggctggctg agaaccagaa gagctgtgag cctgctgtgc cattcccatg tggcagagtg 1320
tctgtgagcc agaccagcaa gctgaccagg gctgaggctg tgttccctga tgtggactat 1380
gtgaacagca ctgaggctga aaccatcctg gacaacatca cccagagcac ccagagcttc 1440
aatgacttca ccagggtggt ggggggggag gatgccaagc ctggccagtt cccctggcaa 1500
gtggtgctga atggcaaggt ggatgccttc tgtgggggca gcattgtgaa tgagaagtgg 1560
attgtgactg ctgcccactg tgtggagact ggggtgaaga tcactgtggt ggctggggag 1620
cacaacattg aggagactga gcacactgag cagaagagga atgtgatcag gatcatcccc 1680
caccacaact acaatgctgc catcaacaag tacaaccatg acattgccct gctggagctg 1740
gatgagcccc tggtgctgaa cagctatgtg acccccatct gcattgctga caaggagtac 1800
accaacatct tcctgaagtt tggctctggc tatgtgtctg gctggggcag ggtgttccac 1860
aagggcaggt ctgccctggt gctgcagtac ctgagggtgc ccctggtgga cagggccacc 1920
tgcctgagga gcaccaagtt caccatctac aacaacatgt tctgtgctgg cttccatgag 1980
gggggcaggg acagctgcca gggggactct gggggccccc atgtgactga ggtggagggc 2040
accagcttcc tgactggcat catcagctgg ggggaggagt gtgccatgaa gggcaagtat 2100
ggcatctaca ccaaagtctc cagatatgtg aactggatca aggagaagac caagctgacc 2160
taatgactcc atggttcgaa tgctttattt gtgaaatttg tgatgctatt gctttatttg 2220
taaccattat aagctgcaat aaacaagtta acaacaacaa ttgcattcat tttatgtttc 2280
aggttcaggg ggaggtgtgg gaggtttttt aaaactagtg cggccgctct agagcatggc 2340
tacgtagata agtagcatgg cgggttaatc attaactaca aggaacccct agtgatggag 2400
ttggccactc cctctctgcg cgctcgctcg ctcactgagg ccgggcgacc aaaggtcgcc 2460
cgacgcccgg gcggcctcag tgagcgagcg agcgcgcagc tgcctgcagg 2510
<210> 63
<211> 126
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 63
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggaaacc cgggcgtgcc 60
cgggcgcctc agtgagcgag cgagcgcgca gagagggagt ggccaactcc atcactaggg 120
gttcct 126
<210> 64
<211> 120
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 64
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgcccggga aacccgggcg tgcgcctcag tgagcgagcg agcgcgcagc tgcctgcagg 120
<210> 65
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 65
gcgcgctcgc tcgctcactg aggccgcccg ggcgtcgggc gacctttggt cgcccggcct 60
cagtgagcga gcgagcgcgc 80
<210> 66
<211> 141
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 66
aataaacgat aacgccgttg gtggcgtgag gcatgtaaaa ggttacatca ttatcttgtt 60
cgccatccgg ttggtataaa tagacgttca tgttggtttt tgtttcagtt gcaagttggc 120
tgcggcgcgc gcagcacctt t 141
<210> 67
<211> 1876
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 67
cgcagccacc atggcggggt tttacgagat tgtgattaag gtccccagcg accttgacga 60
gcatctgccc ggcatttctg acagctttgt gaactgggtg gccgagaagg aatgggagtt 120
gccgccagat tctgacatgg atctgaatct gattgagcag gcacccctga ccgtggccga 180
gaagctgcag cgcgactttc tgacggaatg gcgccgtgtg agtaaggccc cggaggccct 240
tttctttgtg caatttgaga agggagagag ctacttccac atgcacgtgc tcgtggaaac 300
caccggggtg aaatccatgg ttttgggacg tttcctgagt cagattcgcg aaaaactgat 360
tcagagaatt taccgcggga tcgagccgac tttgccaaac tggttcgcgg tcacaaagac 420
cagaaatggc gccggaggcg ggaacaaggt ggtggatgag tgctacatcc ccaattactt 480
gctccccaaa acccagcctg agctccagtg ggcgtggact aatatggaac agtatttaag 540
cgcctgtttg aatctcacgg agcgtaaacg gttggtggcg cagcatctga cgcacgtgtc 600
gcagacgcag gagcagaaca aagagaatca gaatcccaat tctgatgcgc cggtgatcag 660
atcaaaaact tcagccaggt acatggagct ggtcgggtgg ctcgtggaca aggggattac 720
ctcggagaag cagtggatcc aggaggacca ggcctcatac atctccttca atgcggcctc 780
caactcgcgg tcccaaatca aggctgcctt ggacaatgcg ggaaagatta tgagcctgac 840
taaaaccgcc cccgactacc tggtgggcca gcagcccgtg gaggacattt ccagcaatcg 900
gatttataaa attttggaac taaacgggta cgatccccaa tatgcggctt ccgtctttct 960
gggatgggcc acgaaaaagt tcggcaagag gaacaccatc tggctgtttg ggcctgcaac 1020
taccgggaag accaacatcg cggaggccat agcccacact gtgcccttct acgggtgcgt 1080
aaactggacc aatgagaact ttcccttcaa cgactgtgtc gacaagatgg tgatctggtg 1140
ggaggagggg aagatgaccg ccaaggtcgt ggagtcggcc aaagccattc tcggaggaag 1200
caaggtgcgc gtggaccaga aatgcaagtc ctcggcccag atagacccga ctcccgtgat 1260
cgtcacctcc aacaccaaca tgtgcgccgt gattgacggg aactcaacga ccttcgaaca 1320
ccagcagccg ttgcaagacc ggatgttcaa atttgaactc acccgccgtc tggatcatga 1380
ctttgggaag gtcaccaagc aggaagtcaa agactttttc cggtgggcaa aggatcacgt 1440
ggttgaggtg gagcatgaat tctacgtcaa aaagggtgga gccaagaaaa gacccgcccc 1500
cagtgacgca gatataagtg agcccaaacg ggtgcgcgag tcagttgcgc agccatcgac 1560
gtcagacgcg gaagcttcga tcaactacgc agacaggtac caaaacaaat gttctcgtca 1620
cgtgggcatg aatctgatgc tgtttccctg cagacaatgc gagagaatga atcagaattc 1680
aaatatctgc ttcactcacg gacagaaaga ctgtttagag tgctttcccg tgtcagaatc 1740
tcaacccgtt tctgtcgtca aaaaggcgta tcagaaactg tgctacattc atcatatcat 1800
gggaaaggtg ccagacgctt gcactgcctg cgatctggtc aatgtggatt tggatgactg 1860
catctttgaa caataa 1876
<210> 68
<211> 129
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 68
atcatggaga taattaaaat gataaccatc tcgcaaataa ataagtattt tactgttttc 60
gtaacagttt tgtaataaaa aaacctataa atattccgga ttattcatac cgtcccacca 120
tcgggcgcg 129
<210> 69
<211> 1203
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 69
gccgccacca tggagttggt gggctggctc gtggacaaag gcattacttc ggaaaagcag 60
tggattcagg aggatcaggc atcttacatc tcattcaacg ctgccagtaa ctcgaggtcc 120
cagatcaagg cagcgctgga caacgcggga aagattatga gtctgaccaa aactgctcca 180
gactacctcg ttggtcagca accggtggaa gatatctcca gcaacaggat ctacaagatt 240
ctggagctca acggctacga ccctcaatac gctgcctcag tgttcttggg ttgggccacc 300
aagaaattcg gcaagagaaa cactatctgg ctgttcggcc ccgctaccac tggaaagaca 360
aacatcgcag aagcgattgc tcacacggtg ccattctacg gctgcgtcaa ctggacaaac 420
gagaacttcc cgttcaacga ctgtgtcgat aagatggtta tctggtggga ggaaggaaag 480
atgacggcca aagtggtcga aagcgccaag gcaattctgg gtggctctaa agtgcgcgtc 540
gaccagaagt gcaaatcttc agctcaaatc gatcctaccc ccgttattgt gacatcaaac 600
acgaacatgt gtgccgtgat cgacggaaac agtacaacgt tcgaacacca gcaacctctc 660
caggatcgta tgttcaagtt cgagctcacc cgccgtttgg accatgattt cggcaaggtc 720
actaaacaag aggttaagga cttcttccgc tgggctaaag atcacgttgt ggaggttgaa 780
catgagttct acgtcaagaa aggaggtgct aagaaacgtc cagccccgtc ggacgcagat 840
atctccgaac ctaagagggt gagagagtcg gtcgcacagc caagcacttc tgacgcagaa 900
gcttccatta actacgcaga taggtaccaa aacaagtgca gcagacacgt gggtatgaac 960
ttgatgctgt tcccatgccg ccagtgtgag cgtatgaacc aaaactctaa catctgtttc 1020
acacatggcc agaaggactg cctcgaatgt ttccctgtgt cagagagtca gcccgtctca 1080
gtcgttaaga aagcttacca aaagttgtgc tacatccacc atattatggg taaagtccct 1140
gatgcctgta ccgcttgtga tctggtcaac gtggatttgg acgactgtat tttcgagcaa 1200
taa 1203
<210> 70
<211> 388
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 70
gaacagagaa acaggagaat atgggccaaa caggatatct gtggtaagca gttcctgccc 60
cggctcaggg ccaagaacag ttggaacagc agaatatggg ccaaacagga tatctgtggt 120
aagcagttcc tgccccggct cagggccaag aacagatggt ccccagatgc ggtcccgccc 180
tcagcagttt ctagagaacc atcagatgtt tccagggtgc cccaaggacc tgaaatgacc 240
ctgtgcctta tttgaactaa ccaatcagtt cgcttctcgc ttctgttcgc gcgcttctgc 300
tccccgagct ctatataagc agagctcgtt tagtgaaccg tcagatcgcc tggagacgcc 360
atccacgctg ttttgacttc catagaag 388
<210> 71
<211> 1662
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 71
gccgccacca tggaagacgc caaaaacata aagaaaggcc cggcgccatt ctatccgctg 60
gaagatggaa ccgctggaga gcaactgcat aaggctatga agagatacgc cctggttcct 120
ggaacaattg cttttacaga tgcacatatc gaggtggaca tcacttacgc tgagtacttc 180
gaaatgtccg ttcggttggc agaagctatg aaacgatatg ggctgaatac aaatcacaga 240
atcgtcgtat gcagtgaaaa ctctcttcaa ttctttatgc cggtgttggg cgcgttattt 300
atcggagttg cagttgcgcc cgcgaacgac atttataatg aacgtgaatt gctcaacagt 360
atgggcattt cgcagcctac cgtggtgttc gtttccaaaa aggggttgca aaaaattttg 420
aacgtgcaaa aaaagctccc aatcatccaa aaaattatta tcatggattc taaaacggat 480
taccagggat ttcagtcgat gtacacgttc gtcacatctc atctacctcc cggttttaat 540
gaatacgatt ttgtgccaga gtccttcgat agggacaaga caattgcact gatcatgaac 600
tcctctggat ctactggtct gcctaaaggt gtcgctctgc ctcatagaac tgcctgcgtg 660
agattctcgc atgccagaga tcctattttt ggcaatcaaa tcattccgga tactgcgatt 720
ttaagtgttg ttccattcca tcacggtttt ggaatgttta ctacactcgg atatttgata 780
tgtggatttc gagtcgtctt aatgtataga tttgaagaag agctgtttct gaggagcctt 840
caggattaca agattcaaag tgcgctgctg gtgccaaccc tattctcctt cttcgccaaa 900
agcactctga ttgacaaata cgatttatct aatttacacg aaattgcttc tggtggcgct 960
cccctctcta aggaagtcgg ggaagcggtt gccaagaggt tccatctgcc aggtatcagg 1020
caaggatatg ggctcactga gactacatca gctattctga ttacacccga gggggatgat 1080
aaaccgggcg cggtcggtaa agttgttcca ttttttgaag cgaaggttgt ggatctggat 1140
accgggaaaa cgctgggcgt taatcaaaga ggcgaactgt gtgtgagagg tcctatgatt 1200
atgtccggtt atgtaaacaa tccggaagcg accaacgcct tgattgacaa ggatggatgg 1260
ctacattctg gagacatagc ttactgggac gaagacgaac acttcttcat cgttgaccgc 1320
ctgaagtctc tgattaagta caaaggctat caggtggctc ccgctgaatt ggaatccatc 1380
ttgctccaac accccaacat cttcgacgca ggtgtcgcag gtcttcccga cgatgacgcc 1440
ggtgaacttc ccgccgccgt tgttgttttg gagcacggaa agacgatgac ggaaaaagag 1500
atcgtggatt acgtcgccag tcaagtaaca accgcgaaaa agttgcgcgg aggagttgtg 1560
tttgtggacg aagtaccgaa aggtcttacc ggaaaactcg acgcaagaaa aatcagagag 1620
atcctcataa aggccaagaa gggcggaaag atcgccgtgt aa 1662
<210> 72
<211> 581
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 72
gagcatctta ccgccattta ttcccatatt tgttctgttt ttcttgattt gggtatacat 60
ttaaatgtta ataaaacaaa atggtggggc aatcatttac atttttaggg atatgtaatt 120
actagttcag gtgtattgcc acaagacaaa catgttaaga aactttcccg ttatttacgc 180
tctgttcctg ttaatcaacc tctggattac aaaatttgtg aaagattgac tgatattctt 240
aactatgttg ctccttttac gctgtgtgga tatgctgctt tatagcctct gtatctagct 300
attgcttccc gtacggcttt cgttttctcc tccttgtata aatcctggtt gctgtctctt 360
ttagaggagt tgtggcccgt tgtccgtcaa cgtggcgtgg tgtgctctgt gtttgctgac 420
gcaaccccca ctggctgggg cattgccacc acctgtcaac tcctttctgg gactttcgct 480
ttccccctcc cgatcgccac ggcagaactc atcgccgcct gccttgcccg ctgctggaca 540
ggggctaggt tgctgggcac tgataattcc gtggtgttgt c 581
<210> 73
<211> 225
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 73
tgtgccttct agttgccagc catctgttgt ttgcccctcc cccgtgcctt ccttgaccct 60
ggaaggtgcc actcccactg tcctttccta ataaaatgag gaaattgcat cgcattgtct 120
gagtaggtgt cattctattc tggggggtgg ggtggggcag gacagcaagg gggaggattg 180
ggaagacaat agcaggcatg ctggggatgc ggtgggctct atggc 225
<210> 74
<211> 1177
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 74
ggctcagagg ctcagaggca cacaggagtt tctgggctca ccctgccccc ttccaacccc 60
tcagttccca tcctccagca gctgtttgtg tgctgcctct gaagtccaca ctgaacaaac 120
ttcagcctac tcatgtccct aaaatgggca aacattgcaa gcagcaaaca gcaaacacac 180
agccctccct gcctgctgac cttggagctg gggcagaggt cagagacctc tctgggccca 240
tgccacctcc aacatccact cgaccccttg gaatttcggt ggagaggagc agaggttgtc 300
ctggcgtggt ttaggtagtg tgagagggtc cgggttcaaa accacttgct gggtggggag 360
tcgtcagtaa gtggctatgc cccgaccccg aagcctgttt ccccatctgt acaatggaaa 420
tgataaagac gcccatctga tagggttttt gtggcaaata aacatttggt ttttttgttt 480
tgttttgttt tgttttttga gatggaggtt tgctctgtcg cccaggctgg agtgcagtga 540
cacaatctca tctcaccaca accttcccct gcctcagcct cccaagtagc tgggattaca 600
agcatgtgcc accacacctg gctaattttc tatttttagt agagacgggt ttctccatgt 660
tggtcagcct cagcctccca agtaactggg attacaggcc tgtgccacca cacccggcta 720
attttttcta tttttgacag ggacggggtt tcaccatgtt ggtcaggctg gtctagaggt 780
accggatctt gctaccagtg gaacagccac taaggattct gcagtgagag cagagggcca 840
gctaagtggt actctcccag agactgtctg actcacgcca ccccctccac cttggacaca 900
ggacgctgtg gtttctgagc caggtacaat gactcctttc ggtaagtgca gtggaagctg 960
tacactgccc aggcaaagcg tccgggcagc gtaggcgggc gactcagatc ccagccagtg 1020
gacttagccc ctgtttgctc ctccgataac tggggtgacc ttggttaata ttcaccagca 1080
gcctcccccg ttgcccctct ggatccactg cttaaatacg gacgaggaca gggccctgtc 1140
tcctcagctt caggcaccac cactgacctg ggacagt 1177
<210> 75
<400> 75
000
<210> 76
<400> 76
000
<210> 77
<400> 77
000
<210> 78
<400> 78
000
<210> 79
<400> 79
000
<210> 80
<400> 80
000
<210> 81
<400> 81
000
<210> 82
<400> 82
000
<210> 83
<400> 83
000
<210> 84
<400> 84
000
<210> 85
<400> 85
000
<210> 86
<400> 86
000
<210> 87
<400> 87
000
<210> 88
<400> 88
000
<210> 89
<400> 89
000
<210> 90
<400> 90
000
<210> 91
<400> 91
000
<210> 92
<400> 92
000
<210> 93
<400> 93
000
<210> 94
<400> 94
000
<210> 95
<400> 95
000
<210> 96
<400> 96
000
<210> 97
<400> 97
000
<210> 98
<400> 98
000
<210> 99
<400> 99
000
<210> 100
<400> 100
000
<210> 101
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 101
gcgcgctcgc tcgctcactg aggccgcccg ggaaacccgg gcgtgcgcct cagtgagcga 60
gcgagcgcgc 70
<210> 102
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 102
gcgcgctcgc tcgctcactg aggcgcacgc ccgggtttcc cgggcggcct cagtgagcga 60
gcgagcgcgc 70
<210> 103
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 103
gcgcgctcgc tcgctcactg aggccgtcgg gcgacctttg gtcgcccggc ctcagtgagc 60
gagcgagcgc gc 72
<210> 104
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 104
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacggc ctcagtgagc 60
gagcgagcgc gc 72
<210> 105
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 105
gcgcgctcgc tcgctcactg aggccgcccg ggcaaagccc gggcgtcggc ctcagtgagc 60
gagcgagcgc gc 72
<210> 106
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 106
gcgcgctcgc tcgctcactg aggccgacgc ccgggctttg cccgggcggc ctcagtgagc 60
gagcgagcgc gc 72
<210> 107
<211> 83
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 107
gcgcgctcgc tcgctcactg aggccgcccg ggcaaagccc gggcgtcggg ctttgcccgg 60
cctcagtgag cgagcgagcg cgc 83
<210> 108
<211> 83
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 108
gcgcgctcgc tcgctcactg aggccgggca aagcccgacg cccgggcttt gcccgggcgg 60
cctcagtgag cgagcgagcg cgc 83
<210> 109
<211> 77
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 109
gcgcgctcgc tcgctcactg aggccgaaac gtcgggcgac ctttggtcgc ccggcctcag 60
tgagcgagcg agcgcgc 77
<210> 110
<211> 77
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 110
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgtt tcggcctcag 60
tgagcgagcg agcgcgc 77
<210> 111
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 111
gcgcgctcgc tcgctcactg aggcaaagcc tcagtgagcg agcgagcgcg c 51
<210> 112
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 112
gcgcgctcgc tcgctcactg aggctttgcc tcagtgagcg agcgagcgcg c 51
<210> 113
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 113
gcgcgctcgc tcgctcactg aggccgcccg ggcgtcgggc gacctttggt cgcccggcct 60
cagtgagcga gcgagcgcgc 80
<210> 114
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 114
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 115
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 115
gcgcgctcgc tcgctcactg aggcgcccgg gcgtcgggcg acctttggtc gcccggcctc 60
agtgagcgag cgagcgcgc 79
<210> 116
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 116
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cgggcgcctc 60
agtgagcgag cgagcgcgc 79
<210> 117
<211> 89
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 117
gcgcgctcgc tcgctcactg aggccgcccg ggcaaagccc gggcgtcggg cgactttgtc 60
gcccggcctc agtgagcgag cgagcgcgc 89
<210> 118
<211> 89
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 118
gcgcgctcgc tcgctcactg aggccgggcg acaaagtcgc ccgacgcccg ggctttgccc 60
gggcggcctc agtgagcgag cgagcgcgc 89
<210> 119
<211> 87
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 119
gcgcgctcgc tcgctcactg aggccgcccg ggcaaagccc gggcgtcggg cgattttcgc 60
ccggcctcag tgagcgagcg agcgcgc 87
<210> 120
<211> 87
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 120
gcgcgctcgc tcgctcactg aggccgggcg aaaatcgccc gacgcccggg ctttgcccgg 60
gcggcctcag tgagcgagcg agcgcgc 87
<210> 121
<211> 85
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 121
gcgcgctcgc tcgctcactg aggccgcccg ggcaaagccc gggcgtcggg cgtttcgccc 60
ggcctcagtg agcgagcgag cgcgc 85
<210> 122
<211> 85
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 122
gcgcgctcgc tcgctcactg aggccgggcg aaacgcccga cgcccgggct ttgcccgggc 60
ggcctcagtg agcgagcgag cgcgc 85
<210> 123
<211> 89
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 123
gcgcgctcgc tcgctcactg aggccgcccg ggaaacccgg gcgtcgggcg acctttggtc 60
gcccggcctc agtgagcgag cgagcgcgc 89
<210> 124
<211> 89
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 124
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cgggtttccc 60
gggcggcctc agtgagcgag cgagcgcgc 89
<210> 125
<211> 87
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 125
gcgcgctcgc tcgctcactg aggccgcccg gaaaccgggc gtcgggcgac ctttggtcgc 60
ccggcctcag tgagcgagcg agcgcgc 87
<210> 126
<211> 87
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 126
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cggtttccgg 60
gcggcctcag tgagcgagcg agcgcgc 87
<210> 127
<211> 85
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 127
gcgcgctcgc tcgctcactg aggccgcccg aaacgggcgt cgggcgacct ttggtcgccc 60
ggcctcagtg agcgagcgag cgcgc 85
<210> 128
<211> 85
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 128
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cgtttcgggc 60
ggcctcagtg agcgagcgag cgcgc 85
<210> 129
<211> 83
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 129
gcgcgctcgc tcgctcactg aggccgccca aagggcgtcg ggcgaccttt ggtcgcccgg 60
cctcagtgag cgagcgagcg cgc 83
<210> 130
<211> 83
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 130
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc ctttgggcgg 60
cctcagtgag cgagcgagcg cgc 83
<210> 131
<211> 81
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 131
gcgcgctcgc tcgctcactg aggccgccaa aggcgtcggg cgacctttgg tcgcccggcc 60
tcagtgagcg agcgagcgcg c 81
<210> 132
<211> 81
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 132
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc tttggcggcc 60
tcagtgagcg agcgagcgcg c 81
<210> 133
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 133
gcgcgctcgc tcgctcactg aggccgcaaa gcgtcgggcg acctttggtc gcccggcctc 60
agtgagcgag cgagcgcgc 79
<210> 134
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 134
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgct ttgcggcctc 60
agtgagcgag cgagcgcgc 79
<210> 135
<211> 546
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 135
ccctaaaatg ggcaaacatt gcaagcagca aacagcaaac acacagccct ccctgcctgc 60
tgaccttgga gctggggcag aggtcagaga cctctctggg cccatgccac ctccaacatc 120
cactcgaccc cttggaattt ttcggtggag aggagcagag gttgtcctgg cgtggtttag 180
gtagtgtgag aggggaatga ctcctttcgg taagtgcagt ggaagctgta cactgcccag 240
gcaaagcgtc cgggcagcgt aggcgggcga ctcagatccc agccagtgga cttagcccct 300
gtttgctcct ccgataactg gggtgacctt ggttaatatt caccagcagc ctcccccgtt 360
gcccctctgg atccactgct taaatacgga cgaggacagg gccctgtctc ctcagcttca 420
ggcaccacca ctgacctggg acagtgaatc cggactctaa ggtaaatata aaatttttaa 480
gtgtataatg tgttaaacta ctgattctaa ttgtttctct cttttagatt ccaacctttg 540
gaactg 546
<210> 136
<400> 136
000
<210> 137
<400> 137
000
<210> 138
<400> 138
000
<210> 139
<400> 139
000
<210> 140
<400> 140
000
<210> 141
<400> 141
000
<210> 142
<400> 142
000
<210> 143
<400> 143
000
<210> 144
<400> 144
000
<210> 145
<400> 145
000
<210> 146
<400> 146
000
<210> 147
<400> 147
000
<210> 148
<400> 148
000
<210> 149
<400> 149
000
<210> 150
<400> 150
000
<210> 151
<211> 1530
<212> ДНК
<213> Homo sapiens
<400> 151
atgtccatgg gggcaccgcg gtccctcctc ctggccctgg ctgctggcct ggccgttgcc 60
cgtccgccca acatcgtgct gatctttgcc gacgacctcg gctatgggga cctgggctgc 120
tatgggcacc ccagctctac cactcccaac ctggaccagc tggcggcggg agggctgcgg 180
ttcacagact tctacgtgcc tgtgtctctg tgcacaccct ctagggccgc cctcctgacc 240
ggccggctcc cggttcggat gggcatgtac cctggcgtcc tggtgcccag ctcccggggg 300
ggcctgcccc tggaggaggt gaccgtggcc gaagtcctgg ctgcccgagg ctacctcaca 360
ggaatggccg gcaagtggca ccttggggtg gggcctgagg gggccttcct gcccccccat 420
cagggcttcc atcgatttct aggcatcccg tactcccacg accagggccc ctgccagaac 480
ctgacctgct tcccgccggc cactccttgc gacggtggct gtgaccaggg cctggtcccc 540
atcccactgt tggccaacct gtccgtggag gcgcagcccc cctggctgcc cggactagag 600
gcccgctaca tggctttcgc ccatgacctc atggccgacg cccagcgcca ggatcgcccc 660
ttcttcctgt actatgcctc tcaccacacc cactaccctc agttcagtgg gcagagcttt 720
gcagagcgtt caggccgcgg gccatttggg gactccctga tggagctgga tgcagctgtg 780
gggaccctga tgacagccat aggggacctg gggctgcttg aagagacgct ggtcatcttc 840
actgcagaca atggacctga gaccatgcgt atgtcccgag gcggctgctc cggtctcttg 900
cggtgtggaa agggaacgac ctacgagggc ggtgtccgag agcctgcctt ggccttctgg 960
ccaggtcata tcgctcccgg cgtgacccac gagctggcca gctccctgga cctgctgcct 1020
accctggcag ccctggctgg ggccccactg cccaatgtca ccttggatgg ctttgacctc 1080
agccccctgc tgctgggcac aggcaagagc cctcggcagt ctctcttctt ctacccgtcc 1140
tacccagacg aggtccgtgg ggtttttgct gtgcggactg gaaagtacaa ggctcacttc 1200
ttcacccagg gctctgccca cagtgatacc actgcagacc ctgcctgcca cgcctccagc 1260
tctctgactg ctcatgagcc cccgctgctc tatgacctgt ccaaggaccc tggtgagaac 1320
tacaacctgc tggggggtgt ggccggggcc accccagagg tgctgcaagc cctgaaacag 1380
cttcagctgc tcaaggccca gttagacgca gctgtgacct tcggccccag ccaggtggcc 1440
cggggcgagg accccgccct gcagatctgc tgtcatcctg gctgcacccc ccgcccagct 1500
tgctgccatt gcccagatcc ccatgcctga 1530
<210> 152
<211> 1653
<212> ДНК
<213> Homo sapiens
<400> 152
atgccgccac cccggaccgg ccgaggcctt ctctggctgg gtctggttct gagctccgtc 60
tgcgtcgccc tcggatccga aacgcaggcc aactcgacca cagatgctct gaacgttctt 120
ctcatcatcg tggatgacct gcgcccctcc ctgggctgtt atggggataa gctggtgagg 180
tccccaaata ttgaccaact ggcatcccac agcctcctct tccagaatgc ctttgcgcag 240
caagcagtgt gcgccccgag ccgcgtttct ttcctcactg gcaggagacc tgacaccacc 300
cgcctgtacg acttcaactc ctactggagg gtgcacgctg gaaacttctc caccatcccc 360
cagtacttca aggagaatgg ctatgtgacc atgtcggtgg gaaaagtctt tcaccctggg 420
atatcttcta accataccga tgattctccg tatagctggt cttttccacc ttatcatcct 480
tcctctgaga agtatgaaaa cactaagaca tgtcgagggc cagatggaga actccatgcc 540
aacctgcttt gccctgtgga tgtgctggat gttcccgagg gcaccttgcc tgacaaacag 600
agcactgagc aagccataca gttgttggaa aagatgaaaa cgtcagccag tcctttcttc 660
ctggccgttg ggtatcataa gccacacatc cccttcagat accccaagga atttcagaag 720
ttgtatccct tggagaacat caccctggcc cccgatcccg aggtccctga tggcctaccc 780
cctgtggcct acaacccctg gatggacatc aggcaacggg aagacgtcca agccttaaac 840
atcagtgtgc cgtatggtcc aattcctgtg gactttcagc ggaaaatccg ccagagctac 900
tttgcctctg tgtcatattt ggatacacag gtcggccgcc tcttgagtgc tttggacgat 960
cttcagctgg ccaacagcac catcattgca tttacctcgg atcatgggtg ggctctaggt 1020
gaacatggag aatgggccaa atacagcaat tttgatgttg ctacccatgt tcccctgata 1080
ttctatgttc ctggaaggac ggcttcactt ccggaggcag gcgagaagct tttcccttac 1140
ctcgaccctt ttgattccgc ctcacagttg atggagccag gcaggcaatc catggacctt 1200
gtggaacttg tgtctctttt tcccacgctg gctggacttg caggactgca ggttccacct 1260
cgctgccccg ttccttcatt tcacgttgag ctgtgcagag aaggcaagaa ccttctgaag 1320
cattttcgat tccgtgactt ggaagaggat ccgtacctcc ctggtaatcc ccgtgaactg 1380
attgcctata gccagtatcc ccggccttca gacatccctc agtggaattc tgacaagccg 1440
agtttaaaag atataaagat catgggctat tccatacgca ccatagacta taggtatact 1500
gtgtgggttg gcttcaatcc tgatgaattt ctagctaact tttctgacat ccatgcaggg 1560
gaactgtatt ttgtggattc tgacccattg caggatcaca atatgtataa tgattcccaa 1620
ggtggagatc ttttccagtt gttgatgcct tga 1653
<210> 153
<400> 153
000
<210> 154
<400> 154
000
<210> 155
<400> 155
000
<210> 156
<400> 156
000
<210> 157
<400> 157
000
<210> 158
<400> 158
000
<210> 159
<400> 159
000
<210> 160
<400> 160
000
<210> 161
<400> 161
000
<210> 162
<400> 162
000
<210> 163
<400> 163
000
<210> 164
<400> 164
000
<210> 165
<400> 165
000
<210> 166
<400> 166
000
<210> 167
<400> 167
000
<210> 168
<400> 168
000
<210> 169
<400> 169
000
<210> 170
<400> 170
000
<210> 171
<400> 171
000
<210> 172
<400> 172
000
<210> 173
<400> 173
000
<210> 174
<400> 174
000
<210> 175
<400> 175
000
<210> 176
<400> 176
000
<210> 177
<400> 177
000
<210> 178
<400> 178
000
<210> 179
<400> 179
000
<210> 180
<400> 180
000
<210> 181
<400> 181
000
<210> 182
<400> 182
000
<210> 183
<400> 183
000
<210> 184
<400> 184
000
<210> 185
<400> 185
000
<210> 186
<400> 186
000
<210> 187
<400> 187
000
<210> 188
<400> 188
000
<210> 189
<400> 189
000
<210> 190
<400> 190
000
<210> 191
<400> 191
000
<210> 192
<400> 192
000
<210> 193
<400> 193
000
<210> 194
<400> 194
000
<210> 195
<400> 195
000
<210> 196
<400> 196
000
<210> 197
<400> 197
000
<210> 198
<400> 198
000
<210> 199
<400> 199
000
<210> 200
<400> 200
000
<210> 201
<400> 201
000
<210> 202
<400> 202
000
<210> 203
<400> 203
000
<210> 204
<400> 204
000
<210> 205
<400> 205
000
<210> 206
<400> 206
000
<210> 207
<400> 207
000
<210> 208
<400> 208
000
<210> 209
<400> 209
000
<210> 210
<400> 210
000
<210> 211
<400> 211
000
<210> 212
<400> 212
000
<210> 213
<400> 213
000
<210> 214
<400> 214
000
<210> 215
<400> 215
000
<210> 216
<400> 216
000
<210> 217
<400> 217
000
<210> 218
<400> 218
000
<210> 219
<400> 219
000
<210> 220
<400> 220
000
<210> 221
<400> 221
000
<210> 222
<400> 222
000
<210> 223
<400> 223
000
<210> 224
<400> 224
000
<210> 225
<400> 225
000
<210> 226
<400> 226
000
<210> 227
<400> 227
000
<210> 228
<400> 228
000
<210> 229
<400> 229
000
<210> 230
<400> 230
000
<210> 231
<400> 231
000
<210> 232
<400> 232
000
<210> 233
<400> 233
000
<210> 234
<400> 234
000
<210> 235
<400> 235
000
<210> 236
<400> 236
000
<210> 237
<400> 237
000
<210> 238
<400> 238
000
<210> 239
<400> 239
000
<210> 240
<400> 240
000
<210> 241
<400> 241
000
<210> 242
<400> 242
000
<210> 243
<400> 243
000
<210> 244
<400> 244
000
<210> 245
<400> 245
000
<210> 246
<400> 246
000
<210> 247
<400> 247
000
<210> 248
<400> 248
000
<210> 249
<400> 249
000
<210> 250
<400> 250
000
<210> 251
<400> 251
000
<210> 252
<400> 252
000
<210> 253
<400> 253
000
<210> 254
<400> 254
000
<210> 255
<400> 255
000
<210> 256
<400> 256
000
<210> 257
<400> 257
000
<210> 258
<400> 258
000
<210> 259
<400> 259
000
<210> 260
<400> 260
000
<210> 261
<400> 261
000
<210> 262
<400> 262
000
<210> 263
<400> 263
000
<210> 264
<400> 264
000
<210> 265
<400> 265
000
<210> 266
<400> 266
000
<210> 267
<400> 267
000
<210> 268
<400> 268
000
<210> 269
<400> 269
000
<210> 270
<400> 270
000
<210> 271
<400> 271
000
<210> 272
<400> 272
000
<210> 273
<400> 273
000
<210> 274
<400> 274
000
<210> 275
<400> 275
000
<210> 276
<400> 276
000
<210> 277
<400> 277
000
<210> 278
<400> 278
000
<210> 279
<400> 279
000
<210> 280
<400> 280
000
<210> 281
<400> 281
000
<210> 282
<400> 282
000
<210> 283
<400> 283
000
<210> 284
<400> 284
000
<210> 285
<400> 285
000
<210> 286
<400> 286
000
<210> 287
<400> 287
000
<210> 288
<400> 288
000
<210> 289
<400> 289
000
<210> 290
<400> 290
000
<210> 291
<400> 291
000
<210> 292
<400> 292
000
<210> 293
<400> 293
000
<210> 294
<400> 294
000
<210> 295
<400> 295
000
<210> 296
<400> 296
000
<210> 297
<400> 297
000
<210> 298
<400> 298
000
<210> 299
<400> 299
000
<210> 300
<400> 300
000
<210> 301
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 301
gcgcgctcgc tcgctc 16
<210> 302
<211> 8
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 302
actgaggc 8
<210> 303
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 303
cgggcgacca aaggtcgccc ga 22
<210> 304
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 304
cgcccgggcg 10
<210> 305
<211> 8
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 305
gcctcagt 8
<210> 306
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 306
gagcgagcga gcgcgc 16
<210> 307
<400> 307
000
<210> 308
<211> 317
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 308
ggtgtggaaa gtccccaggc tccccagcag gcagaagtat gcaaagcatg catctcaatt 60
agtcagcaac caggtgtgga aagtccccag gctccccagc aggcagaagt atgcaaagca 120
tgcatctcaa ttagtcagca accatagtcc cgcccctaac tccgcccatc ccgcccctaa 180
ctccgcccag ttccgcccat tctccgcccc atggctgact aatttttttt atttatgcag 240
aggccgaggc cgcctcggcc tctgagctat tccagaagta gtgaggaggc ttttttggag 300
gcctaggctt ttgcaaa 317
<210> 309
<211> 576
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 309
tagtaatcaa ttacggggtc attagttcat agcccatata tggagttccg cgttacataa 60
cttacggtaa atggcccgcc tggctgaccg cccaacgacc cccgcccatt gacgtcaata 120
atgacgtatg ttcccatagt aacgccaata gggactttcc attgacgtca atgggtggag 180
tatttacggt aaactgccca cttggcagta catcaagtgt atcatatgcc aagtacgccc 240
cctattgacg tcaatgacgg taaatggccc gcctggcatt atgcccagta catgacctta 300
tgggactttc ctacttggca gtacatctac gtattagtca tcgctattac catggtgatg 360
cggttttggc agtacatcaa tgggcgtgga tagcggtttg actcacgggg atttccaagt 420
ctccacccca ttgacgtcaa tgggagtttg ttttggcacc aaaatcaacg ggactttcca 480
aaatgtcgta acaactccgc cccattgacg caaatgggcg gtaggcgtgt acggtgggag 540
gtctatataa gcagagctgg tttagtgaac cgtcag 576
<210> 310
<211> 1313
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 310
ggagccgaga gtaattcata caaaaggagg gatcgccttc gcaaggggag agcccaggga 60
ccgtccctaa attctcacag acccaaatcc ctgtagccgc cccacgacag cgcgaggagc 120
atgcgcccag ggctgagcgc gggtagatca gagcacacaa gctcacagtc cccggcggtg 180
gggggagggg cgcgctgagc gggggccagg gagctggcgc ggggcaaact gggaaagtgg 240
tgtcgtgtgc tggctccgcc ctcttcccga gggtggggga gaacggtata taagtgcggt 300
agtcgccttg gacgttcttt ttcgcaacgg gtttgccgtc agaacgcagg tgagtggcgg 360
gtgtggcttc cgcgggcccc ggagctggag ccctgctctg agcgggccgg gctgatatgc 420
gagtgtcgtc cgcagggttt agctgtgagc attcccactt cgagtggcgg gcggtgcggg 480
ggtgagagtg cgaggcctag cggcaacccc gtagcctcgc ctcgtgtccg gcttgaggcc 540
tagcgtggtg tccgccgccg cgtgccactc cggccgcact atgcgttttt tgtccttgct 600
gccctcgatt gccttccagc agcatgggct aacaaaggga gggtgtgggg ctcactctta 660
aggagcccat gaagcttacg ttggatagga atggaagggc aggaggggcg actggggccc 720
gcccgccttc ggagcacatg tccgacgcca cctggatggg gcgaggcctg tggctttccg 780
aagcaatcgg gcgtgagttt agcctacctg ggccatgtgg ccctagcact gggcacggtc 840
tggcctggcg gtgccgcgtt cccttgcctc ccaacaaggg tgaggccgtc ccgcccggca 900
ccagttgctt gcgcggaaag atggccgctc ccggggccct gttgcaagga gctcaaaatg 960
gaggacgcgg cagcccggtg gagcgggcgg gtgagtcacc cacacaaagg aagagggcct 1020
tgcccctcgc cggccgctgc ttcctgtgac cccgtggtct atcggccgca tagtcacctc 1080
gggcttctct tgagcaccgc tcgtcgcggc ggggggaggg gatctaatgg cgttggagtt 1140
tgttcacatt tggtgggtgg agactagtca ggccagcctg gcgctggaag tcattcttgg 1200
aatttgcccc tttgagtttg gagcgaggct aattctcaag cctcttagcg gttcaaaggt 1260
attttctaaa cccgtttcca ggtgttgtga aagccaccgc taattcaaag caa 1313
<210> 311
<400> 311
000
<210> 312
<400> 312
000
<210> 313
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 313
Met Asp Trp Thr Trp Arg Ile Leu Phe Leu Val Ala Ala Ala Thr Gly
1 5 10 15
Ala His Ser
<210> 314
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 314
Met Leu Pro Ser Gln Leu Ile Gly Phe Leu Leu Leu Trp Val Pro Ala
1 5 10 15
Ser Arg Gly
<210> 315
<211> 7
<212> БЕЛОК
<213> Обезьяний вирус 40
<400> 315
Pro Lys Lys Lys Arg Lys Val
1 5
<210> 316
<211> 16
<212> БЕЛОК
<213> Неизвестный
<220>
<223> Описание неизвестного:
Последовательность двойного NLS нуклеоплазмина
<400> 316
Lys Arg Pro Ala Ala Thr Lys Lys Ala Gly Gln Ala Lys Lys Lys Lys
1 5 10 15
<210> 317
<211> 9
<212> БЕЛОК
<213> Неизвестный
<220>
<223> Описание неизвестного:
Последовательность C-myc NLS
<400> 317
Pro Ala Ala Lys Arg Val Lys Leu Asp
1 5
<210> 318
<211> 11
<212> БЕЛОК
<213> Неизвестный
<220>
<223> Описание неизвестного:
Последовательность C-myc NLS
<400> 318
Arg Gln Arg Arg Asn Glu Leu Lys Arg Ser Pro
1 5 10
<210> 319
<211> 38
<212> БЕЛОК
<213> Homo sapiens
<400> 319
Asn Gln Ser Ser Asn Phe Gly Pro Met Lys Gly Gly Asn Phe Gly Gly
1 5 10 15
Arg Ser Ser Gly Pro Tyr Gly Gly Gly Gly Gln Tyr Phe Ala Lys Pro
20 25 30
Arg Asn Gln Gly Gly Tyr
35
<210> 320
<211> 42
<212> БЕЛОК
<213> Неизвестный
<220>
<223> Описание неизвестного:
Домен IBB из последовательности импортина-альфа
<400> 320
Arg Met Arg Ile Glx Phe Lys Asn Lys Gly Lys Asp Thr Ala Glu Leu
1 5 10 15
Arg Arg Arg Arg Val Glu Val Ser Val Glu Leu Arg Lys Ala Lys Lys
20 25 30
Asp Glu Gln Ile Leu Lys Arg Arg Asn Val
35 40
<210> 321
<211> 8
<212> БЕЛОК
<213> Неизвестный
<220>
<223> Описание неизвестного:
Последовательность белка Т миомы
<400> 321
Val Ser Arg Lys Arg Pro Arg Pro
1 5
<210> 322
<211> 8
<212> БЕЛОК
<213> Неизвестный
<220>
<223> Описание неизвестного:
Последовательность белка Т миомы
<400> 322
Pro Pro Lys Lys Ala Arg Glu Asp
1 5
<210> 323
<211> 8
<212> БЕЛОК
<213> Homo sapiens
<400> 323
Pro Gln Pro Lys Lys Lys Pro Leu
1 5
<210> 324
<211> 12
<212> БЕЛОК
<213> Mus musculus
<400> 324
Ser Ala Leu Ile Lys Lys Lys Lys Lys Met Ala Pro
1 5 10
<210> 325
<211> 5
<212> БЕЛОК
<213> Вирус гриппа
<400> 325
Asp Arg Leu Arg Arg
1 5
<210> 326
<211> 7
<212> БЕЛОК
<213> Вирус гриппа
<400> 326
Pro Lys Gln Lys Lys Arg Lys
1 5
<210> 327
<211> 10
<212> БЕЛОК
<213> Вирус гепатита дельта
<400> 327
Arg Lys Leu Lys Lys Lys Ile Lys Lys Leu
1 5 10
<210> 328
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 328
Arg Glu Lys Lys Lys Phe Leu Lys Arg Arg
1 5 10
<210> 329
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 329
Lys Arg Lys Gly Asp Glu Val Asp Gly Val Asp Glu Val Ala Lys Lys
1 5 10 15
Lys Ser Lys Lys
20
<210> 330
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 330
Arg Lys Cys Leu Gln Ala Gly Met Asn Leu Glu Ala Arg Lys Thr Lys
1 5 10 15
Lys
<210> 331
<400> 331
000
<210> 332
<400> 332
000
<210> 333
<400> 333
000
<210> 334
<400> 334
000
<210> 335
<400> 335
000
<210> 336
<400> 336
000
<210> 337
<400> 337
000
<210> 338
<400> 338
000
<210> 339
<400> 339
000
<210> 340
<400> 340
000
<210> 341
<400> 341
000
<210> 342
<400> 342
000
<210> 343
<400> 343
000
<210> 344
<400> 344
000
<210> 345
<400> 345
000
<210> 346
<400> 346
000
<210> 347
<400> 347
000
<210> 348
<400> 348
000
<210> 349
<400> 349
000
<210> 350
<400> 350
000
<210> 351
<400> 351
000
<210> 352
<400> 352
000
<210> 353
<400> 353
000
<210> 354
<400> 354
000
<210> 355
<400> 355
000
<210> 356
<400> 356
000
<210> 357
<400> 357
000
<210> 358
<400> 358
000
<210> 359
<400> 359
000
<210> 360
<400> 360
000
<210> 361
<400> 361
000
<210> 362
<400> 362
000
<210> 363
<400> 363
000
<210> 364
<400> 364
000
<210> 365
<400> 365
000
<210> 366
<400> 366
000
<210> 367
<400> 367
000
<210> 368
<400> 368
000
<210> 369
<400> 369
000
<210> 370
<400> 370
000
<210> 371
<400> 371
000
<210> 372
<400> 372
000
<210> 373
<400> 373
000
<210> 374
<400> 374
000
<210> 375
<400> 375
000
<210> 376
<400> 376
000
<210> 377
<400> 377
000
<210> 378
<400> 378
000
<210> 379
<400> 379
000
<210> 380
<400> 380
000
<210> 381
<400> 381
000
<210> 382
<400> 382
000
<210> 383
<400> 383
000
<210> 384
<400> 384
000
<210> 385
<400> 385
000
<210> 386
<400> 386
000
<210> 387
<400> 387
000
<210> 388
<400> 388
000
<210> 389
<400> 389
000
<210> 390
<400> 390
000
<210> 391
<400> 391
000
<210> 392
<400> 392
000
<210> 393
<400> 393
000
<210> 394
<400> 394
000
<210> 395
<400> 395
000
<210> 396
<400> 396
000
<210> 397
<400> 397
000
<210> 398
<400> 398
000
<210> 399
<400> 399
000
<210> 400
<400> 400
000
<210> 401
<400> 401
000
<210> 402
<400> 402
000
<210> 403
<400> 403
000
<210> 404
<400> 404
000
<210> 405
<400> 405
000
<210> 406
<400> 406
000
<210> 407
<400> 407
000
<210> 408
<400> 408
000
<210> 409
<400> 409
000
<210> 410
<400> 410
000
<210> 411
<400> 411
000
<210> 412
<400> 412
000
<210> 413
<400> 413
000
<210> 414
<400> 414
000
<210> 415
<400> 415
000
<210> 416
<400> 416
000
<210> 417
<400> 417
000
<210> 418
<400> 418
000
<210> 419
<400> 419
000
<210> 420
<400> 420
000
<210> 421
<400> 421
000
<210> 422
<400> 422
000
<210> 423
<400> 423
000
<210> 424
<400> 424
000
<210> 425
<400> 425
000
<210> 426
<400> 426
000
<210> 427
<400> 427
000
<210> 428
<400> 428
000
<210> 429
<400> 429
000
<210> 430
<400> 430
000
<210> 431
<400> 431
000
<210> 432
<400> 432
000
<210> 433
<400> 433
000
<210> 434
<400> 434
000
<210> 435
<400> 435
000
<210> 436
<400> 436
000
<210> 437
<400> 437
000
<210> 438
<400> 438
000
<210> 439
<400> 439
000
<210> 440
<400> 440
000
<210> 441
<400> 441
000
<210> 442
<400> 442
000
<210> 443
<400> 443
000
<210> 444
<400> 444
000
<210> 445
<400> 445
000
<210> 446
<400> 446
000
<210> 447
<400> 447
000
<210> 448
<400> 448
000
<210> 449
<400> 449
000
<210> 450
<400> 450
000
<210> 451
<400> 451
000
<210> 452
<400> 452
000
<210> 453
<400> 453
000
<210> 454
<400> 454
000
<210> 455
<400> 455
000
<210> 456
<400> 456
000
<210> 457
<400> 457
000
<210> 458
<400> 458
000
<210> 459
<400> 459
000
<210> 460
<400> 460
000
<210> 461
<400> 461
000
<210> 462
<400> 462
000
<210> 463
<400> 463
000
<210> 464
<400> 464
000
<210> 465
<400> 465
000
<210> 466
<400> 466
000
<210> 467
<400> 467
000
<210> 468
<400> 468
000
<210> 469
<400> 469
000
<210> 470
<400> 470
000
<210> 471
<400> 471
000
<210> 472
<400> 472
000
<210> 473
<400> 473
000
<210> 474
<400> 474
000
<210> 475
<400> 475
000
<210> 476
<400> 476
000
<210> 477
<400> 477
000
<210> 478
<400> 478
000
<210> 479
<400> 479
000
<210> 480
<400> 480
000
<210> 481
<400> 481
000
<210> 482
<400> 482
000
<210> 483
<400> 483
000
<210> 484
<400> 484
000
<210> 485
<400> 485
000
<210> 486
<400> 486
000
<210> 487
<400> 487
000
<210> 488
<400> 488
000
<210> 489
<400> 489
000
<210> 490
<400> 490
000
<210> 491
<400> 491
000
<210> 492
<400> 492
000
<210> 493
<400> 493
000
<210> 494
<400> 494
000
<210> 495
<400> 495
000
<210> 496
<400> 496
000
<210> 497
<400> 497
000
<210> 498
<400> 498
000
<210> 499
<400> 499
000
<210> 500
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 500
gcccgctggt ttccagcggg ctgcgggccc gaaacgggcc cgc 43
<210> 501
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 501
cgggcccgtg cgggcccaaa gggcccgc 28
<210> 502
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 502
gcccgggcac gcccgggttt cccgggcg 28
<210> 503
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 503
cgtgcgggcc caaagggccc gc 22
<210> 504
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 504
cgggcgacca aaggtcgccc g 21
<210> 505
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 505
cgcccgggct ttgcccgggc 20
<210> 506
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 506
cgggcgacca aaggtcgccc gacgcccggg ctttgcccgg gc 42
<210> 507
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 507
cgggcgacca aaggtcgccc g 21
<210> 508
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 508
cgcccgggct ttgcccgggc 20
<210> 509
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 509
cgggcgacca aaggtcgccc gacgcccggg cggc 34
<210> 510
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 510
cgggcgacca aaggtcgccc g 21
<210> 511
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 511
cgcccgggct ttgcccgggc 20
<210> 512
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 512
cggggcccga cgcccgggct ttgcccgggc 30
<210> 513
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 513
cgggcgacca aaggtcgccc g 21
<210> 514
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 514
cgcccgggct ttgcccgggc 20
<210> 515
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 515
cgggcccgac gcccgggctt tgcccgggc 29
<210> 516
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 516
cgggcgacca aaggtcgccc g 21
<210> 517
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 517
cgcccgggct ttgcccgggc 20
<210> 518
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 518
gcccgggcaa agcccgggcg 20
<210> 519
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 519
cgggcgacct ttggtcgccc g 21
<210> 520
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 520
gcccgggcaa agcccgggcg tcgggcgacc tttggtcgcc cg 42
<210> 521
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 521
gcccgggcaa agcccgggcg 20
<210> 522
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 522
cgggcgacct ttggtcgccc g 21
<210> 523
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 523
gcccgggcaa agcccgggcg 20
<210> 524
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 524
cgggcgacct ttggtcgccc g 21
<210> 525
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 525
gccgcccggg cgacgggcga cctttggtcg cccg 34
<210> 526
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 526
gcccgggcaa agcccgggcg 20
<210> 527
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 527
cgggcgacct ttggtcgccc g 21
<210> 528
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 528
gcccgggcaa agcccgggcg 20
<210> 529
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 529
cgggcgacct ttggtcgccc g 21
<210> 530
<211> 4470
<212> ДНК
<213> Homo sapiens
<400> 530
gtagatgagg aaactgaagt tgaggaatag tgaagagttt gtccaatgtc atagccccgt 60
aatcaacggg acaaaaattt tcttgctgat gggtcaagat ggcatcgtga agtggttgtt 120
caccgtaaac tgtaatacaa tcctgtttat ggatttgttt gcatattttt ccctccatag 180
ggaaaccttt cttccatggc tcaggacaca ctcctggatc gagccaacag gagaactttc 240
tggtaagcat ttggctaact tttttttttt tgagatggag tcttgctgtg tcgcctaggc 300
tggagtgcag tggcgtgatc ttggctcact gcagcctcca cttcccgggt tcaatcaatt 360
ctcctacctc aacttcctga gtagctggga ttacaggcgc ccgccaccac acccggctca 420
tttttgtact tttagtagag acacagtttt gccatgttgg ccaggctggt cttgaattcc 480
tcagctcagg tgatctgcct gccttggcct ctcaaagtgc tgggattaca ggcgtgagcc 540
actgtgcccg gccttggcta acttttcaaa attaaagatt ttgacttgtt acagtcatgt 600
gacatttttt tctttctgtt tgctgagttt ttgataattt atatctctca aagtggagac 660
tttaaaaaag actcatccgt gtgccgtgtt cactgcctgg tatcttagtg tggaccgaag 720
cctaaggacc ctgaaaacag ctgcagatga agatggcaag cacccgctgc aagctggcca 780
ggtacctgga ggacctggag gatgtggact tgaagaaatt taagatgcac ttagaggact 840
atcctcccca gaagggctgc atccccctcc cgaggggtca gacagagaag gcagaccatg 900
tggatctagc cacgctaatg atcgacttca atggggagga gaaggcgtgg gccatggccg 960
tgtggatctt cgctgcgatc aacaggagag acctttatga gaaagcaaaa agagatgagc 1020
cgaagtgggg ttcagataat gcacgtgttt cgaatcccac tgtgatatgc caggaagaca 1080
gcattgaaga ggagtggatg ggtttactgg agtacctttc gagaatctct atttgtaaaa 1140
tgaagaaaga ttaccgtaag aagtacagaa agtacgtgag aagcagattc cagtgcattg 1200
aagacaggaa tgcccgtctg ggtgagagtg tgagcctcaa caaacgctac acacgactgc 1260
gtctcatcaa ggagcaccgg agccagcagg agagggagca ggagcttctg gccatcggca 1320
agaccaagac gtgtgagagc cccgtgagtc ccattaagat ggagttgctg tttgaccccg 1380
atgatgagca ttctgagcct gtgcacaccg tggtgttcca gggggcggca gggattggga 1440
aaacaatcct ggccaggaag atgatgttgg actgggcgtc ggggacactc taccaagaca 1500
ggtttgacta tctgttctat atccactgtc gggaggtgag ccttgtgaca cagaggagcc 1560
tgggggacct gatcatgagc tgctgccccg acccaaaccc acccatccac aagatcgtga 1620
gaaaaccctc cagaatcctc ttcctcatgg acggcttcga tgagctgcaa ggtgcctttg 1680
acgagcacat aggaccgctc tgcactgact ggcagaaggc cgagcgggga gacattctcc 1740
tgagcagcct catcagaaag aagctgcttc ccgaggcctc tctgctcatc accacgagac 1800
ctgtggccct ggagaaactg cagcacttgc tggaccatcc tcggcatgtg gagatcctgg 1860
gtttctccga ggccaaaagg aaagagtact tcttcaagta cttctctgat gaggcccaag 1920
ccagggcagc cttcagtctg attcaggaga acgaggtcct cttcaccatg tgcttcatcc 1980
ccctggtctg ctggatcgtg tgcactggac tgaaacagca gatggagagt ggcaagagcc 2040
ttgcccagac atccaagacc accaccgcgg tgtacgtctt cttcctttcc agtttgctgc 2100
agccccgggg agggagccag gagcacggcc tctgcgccca cctctggggg ctctgctctt 2160
tggctgcaga tggaatctgg aaccagaaaa tcctgtttga ggagtccgac ctcaggaatc 2220
atggactgca gaaggcggat gtgtctgctt tcctgaggat gaacctgttc caaaaggaag 2280
tggactgcga gaagttctac agcttcatcc acatgacttt ccaggagttc tttgccgcca 2340
tgtactacct gctggaagag gaaaaggaag gaaggacgaa cgttccaggg agtcgtttga 2400
agcttcccag ccgagacgtg acagtccttc tggaaaacta tggcaaattc gaaaaggggt 2460
atttgatttt tgttgtacgt ttcctctttg gcctggtaaa ccaggagagg acctcctact 2520
tggagaagaa attaagttgc aagatctctc agcaaatcag gctggagctg ctgaaatgga 2580
ttgaagtgaa agccaaagct aaaaagctgc agatccagcc cagccagctg gaattgttct 2640
actgtttgta cgagatgcag gaggaggact tcgtgcaaag ggccatggac tatttcccca 2700
agattgagat caatctctcc accagaatgg accacatggt ttcttccttt tgcattgaga 2760
actgtcatcg ggtggagtca ctgtccctgg ggtttctcca taacatgccc aaggaggaag 2820
aggaggagga aaaggaaggc cgacaccttg atatggtgca gtgtgtcctc ccaagctcct 2880
ctcatgctgc ctgttctcat ggattggtga acagccacct cacttccagt ttttgccggg 2940
gcctcttttc agttctgagc accagccaga gtctaactga attggacctc agtgacaatt 3000
ctctggggga cccagggatg agagtgttgt gtgaaacgct ccagcatcct ggctgtaaca 3060
ttcggagatt gtggttgggg cgctgtggcc tctcgcatga gtgctgcttc gacatctcct 3120
tggtcctcag cagcaaccag aagctggtgg agctggacct gagtgacaac gccctcggtg 3180
acttcggaat cagacttctg tgtgtgggac tgaagcacct gttgtgcaat ctgaagaagc 3240
tctggttggt cagctgctgc ctcacatcag catgttgtca ggatcttgca tcagtattga 3300
gcaccagcca ttccctgacc agactctatg tgggggagaa tgccttggga gactcaggag 3360
tcgcaatttt atgtgaaaaa gccaagaatc cacagtgtaa cctgcagaaa ctggggttgg 3420
tgaattctgg ccttacgtca gtctgttgtt cagctttgtc ctcggtactc agcactaatc 3480
agaatctcac gcacctttac ctgcgaggca acactctcgg agacaagggg atcaaactac 3540
tctgtgaggg actcttgcac cccgactgca agcttcaggt gttggaatta gacaactgca 3600
acctcacgtc acactgctgc tgggatcttt ccacacttct gacctccagc cagagcctgc 3660
gaaagctgag cctgggcaac aatgacctgg gcgacctggg ggtcatgatg ttctgtgaag 3720
tgctgaaaca gcagagctgc ctcctgcaga acctggggtt gtctgaaatg tatttcaatt 3780
atgagacaaa aagtgcgtta gaaacacttc aagaagaaaa gcctgagctg accgtcgtct 3840
ttgagccttc ttggtaggag tggaaacggg gctgccagac gccagtgttc tccggtccct 3900
ccagctgggg gccctcaggt ggagagagct gcgatccatc caggccaaga ccacagctct 3960
gtgatccttc cggtggagtg tcggagaaga gagcttgccg acgatgcctt cctgtgcaga 4020
gcttgggcat ctcctttacg ccagggtgag gaagacacca ggacaatgac agcatcgggt 4080
gttgttgtca tcacagcgcc tcagttagag gatgttcctc ttggtgacct catgtaatta 4140
gctcattcaa taaagcactt tctttatttt tctcttctct gtctaacttt ctttttccta 4200
tcttttttct tctttgttct gtttactttt gctcatatca tcattcccgc tatctttcta 4260
ttaactgacc ataacacaga actagttgac tatatattat gttgaaattt tatggcagct 4320
atttatttat ttaaattttt tgtaacagtt ttgttttcta ataagaaaaa tccatgcttt 4380
ttgtagctgg ttgaaaattc aggaatatgt aaaacttttt ggtatttaat taaattgatt 4440
ccttttctta attttaaaaa aaaaaaaaaa 4470
<210> 531
<211> 3845
<212> ДНК
<213> Homo sapiens
<400> 531
gttcctgagg ctggcatctg gatgaggaaa ctgaagttga ggaatagtga agagtttgtc 60
caatgtcata gccccgtaat caacgggaca aaaattttct tgctgatggg tcaagatggc 120
atcgtgaagt ggttgttcac cgtaaactgt aatacaatcc tgtttatgga tttgtttgca 180
tatttttccc tccataggga aacctttctt ccatggctca ggacacactc ctggatcgag 240
ccaacaggag aactttctgg taagcatttg gctaactttt ttttttttga gatggagtct 300
tgctgtgtcg cctaggctgg agtgcagtgg cgtgatcttg gctcactgca gcctccactt 360
cccgggttca atcaattctc ctacctcaac ttcctgagta gctgggatta caggcgcccg 420
ccaccacacc cggctcattt ttgtactttt agtagagaca cagttttgcc atgttggcca 480
ggctggtctt gaattcctca gctcaggtga tctgcctgcc ttggcctctc aaagtgctgg 540
gattacaggc gtgagccact gtgcccggcc ttggctaact tttcaaaatt aaagattttg 600
acttgttaca gtcatgtgac atttttttct ttctgtttgc tgagtttttg ataatttata 660
tctctcaaag tggagacttt aaaaaagact catccgtgtg ccgtgttcac tgcctggtat 720
cttagtgtgg accgaagcct aaggaccctg aaaacagctg cagatgaaga tggcaagcac 780
ccgctgcaag ctggccaggt acctggagga cctggaggat gtggacttga agaaatttaa 840
gatgcactta gaggactatc ctccccagaa gggctgcatc cccctcccga ggggtcagac 900
agagaaggca gaccatgtgg atctagccac gctaatgatc gacttcaatg gggaggagaa 960
ggcgtgggcc atggccgtgt ggatcttcgc tgcgatcaac aggagagacc tttatgagaa 1020
agcaaaaaga gatgagccga agtggggttc agataatgca cgtgtttcga atcccactgt 1080
gatatgccag gaagacagca ttgaagagga gtggatgggt ttactggagt acctttcgag 1140
aatctctatt tgtaaaatga agaaagatta ccgtaagaag tacagaaagt acgtgagaag 1200
cagattccag tgcattgaag acaggaatgc ccgtctgggt gagagtgtga gcctcaacaa 1260
acgctacaca cgactgcgtc tcatcaagga gcaccggagc cagcaggaga gggagcagga 1320
gcttctggcc atcggcaaga ccaagacgtg tgagagcccc gtgagtccca ttaagatgga 1380
gttgctgttt gaccccgatg atgagcattc tgagcctgtg cacaccgtgg tgttccaggg 1440
ggcggcaggg attgggaaaa caatcctggc caggaagatg atgttggact gggcgtcggg 1500
gacactctac caagacaggt ttgactatct gttctatatc cactgtcgag aggtgagcct 1560
tgtgacacag aggagcctgg gggacctgat catgagctgc tgccccgacc caaacccacc 1620
catccacaag atcgtgagaa aaccctccag aatcctcttc ctcatggacg gcttcgatga 1680
gctgcaaggt gcctttgacg agcacatagg accgctctgc actgactggc agaaggccga 1740
gcggggagac attctcctga gcagcctcat cagaaagaag ctgcttcccg aggcctctct 1800
gctcatcacc acgagacctg tggccctgga gaaactgcag cacttgctgg accatcctcg 1860
gcatgtggag atcctgggtt tctccgaggc caaaaggaaa gagtacttct tcaagtactt 1920
ctctgatgag gcccaagcca gggcagcctt cagtctgatt caggagaacg aggtcctctt 1980
caccatgtgc ttcatccccc tggtctgctg gatcgtgtgc actggactga aacagcagat 2040
ggagagtggc aagagccttg cccagacatc caagaccacc accgcggtgt acgtcttctt 2100
cctttccagt ttgctgcagc cccggggagg gagccaggag cacggcctct gcgcccacct 2160
ctgggggctc tgctctttgg ctgcagatgg aatctggaac cagaaaatcc tgtttgagga 2220
gtccgacctc aggaatcatg gactgcagaa ggcggatgtg tctgctttcc tgaggatgaa 2280
cctgttccaa aaggaagtgg actgcgagaa gttctacagc ttcatccaca tgactttcca 2340
ggagttcttt gccgccatgt actacctgct ggaagaggaa aaggaaggaa ggacgaacgt 2400
tccagggagt cgtttgaagc ttcccagccg agacgtgaca gtccttctgg aaaactatgg 2460
caaattcgaa aaggggtatt tgatttttgt tgtacgtttc ctctttggcc tggtaaacca 2520
ggagaggacc tcctacttgg agaagaaatt aagttgcaag atctctcagc aaatcaggct 2580
ggagctgctg aaatggattg aagtgaaagc caaagctaaa aagctgcaga tccagcccag 2640
ccagctggaa ttgttctact gtttgtacga gatgcaggag gaggacttcg tgcaaagggc 2700
catggactat ttccccaaga ttgagatcaa tctctccacc agaatggacc acatggtttc 2760
ttccttttgc attgagaact gtcatcgggt ggagtcactg tccctggggt ttctccataa 2820
catgcccaag gaggaagagg aggaggaaaa ggaaggccga caccttgata tggtgcagtg 2880
tgtcctccca agctcctctc atgctgcctg ttctcatggg ttggggcgct gtggcctctc 2940
gcatgagtgc tgcttcgaca tctccttggt cctcagcagc aaccagaagc tggtggagct 3000
ggacctgagt gacaacgccc tcggtgactt cggaatcaga cttctgtgtg tgggactgaa 3060
gcacctgttg tgcaatctga agaagctctg gttggtgaat tctggcctta cgtcagtctg 3120
ttgttcagct ttgtcctcgg tactcagcac taatcagaat ctcacgcacc tttacctgcg 3180
aggcaacact ctcggagaca aggggatcaa actactctgt gagggactct tgcaccccga 3240
ctgcaagctt caggtgttgg aattagacaa ctgcaacctc acgtcacact gctgctggga 3300
tctttccaca cttctgacct ccagccagag cctgcgaaag ctgagcctgg gcaacaatga 3360
cctgggcgac ctgggggtca tgatgttctg tgaagtgctg aaacagcaga gctgcctcct 3420
gcagaacctg gggttgtctg aaatgtattt caattatgag acaaaaagtg cgttagaaac 3480
acttcaagaa gaaaagcctg agctgaccgt cgtctttgag ccttcttggt aggagtggaa 3540
acggggctgc cagacgccag tgttctccgg tccctccagc tgggggccct caggtggaga 3600
gagctgcgat ccatccaggc caagaccaca gctctgtgat ccttccggtg gagtgtcgga 3660
gaagagagct tgccgacgat gccttcctgt gcagagcttg ggcatctcct ttacgccagg 3720
gtgaggaaga caccaggaca atgacagcat cgggtgttgt tgtcatcaca gcgcctcagt 3780
tagaggatgt tcctcttggt gacctcatgt aattagctca ttcaataaag cactttcttt 3840
atttt 3845
<210> 532
<211> 3545
<212> ДНК
<213> Homo sapiens
<400> 532
gttcctgagg ctggcatctg gggaaacctt tcttccatgg ctcaggacac actcctggat 60
cgagccaaca ggagaacttt ctgtgtggac cgaagcctaa ggaccctgaa aacagctgca 120
gatgaagatg gcaagcaccc gctgcaagct ggccaggtac ctggaggacc tggaggatgt 180
ggacttgaag aaatttaaga tgcacttaga ggactatcct ccccagaagg gctgcatccc 240
cctcccgagg ggtcagacag agaaggcaga ccatgtggat ctagccacgc taatgatcga 300
cttcaatggg gaggagaagg cgtgggccat ggccgtgtgg atcttcgctg cgatcaacag 360
gagagacctt tatgagaaag caaaaagaga tgagccgaag tggggttcag ataatgcacg 420
tgtttcgaat cccactgtga tatgccagga agacagcatt gaagaggagt ggatgggttt 480
actggagtac ctttcgagaa tctctatttg taaaatgaag aaagattacc gtaagaagta 540
cagaaagtac gtgagaagca gattccagtg cattgaagac aggaatgccc gtctgggtga 600
gagtgtgagc ctcaacaaac gctacacacg actgcgtctc atcaaggagc accggagcca 660
gcaggagagg gagcaggagc ttctggccat cggcaagacc aagacgtgtg agagccccgt 720
gagtcccatt aagatggagt tgctgtttga ccccgatgat gagcattctg agcctgtgca 780
caccgtggtg ttccaggggg cggcagggat tgggaaaaca atcctggcca ggaagatgat 840
gttggactgg gcgtcgggga cactctacca agacaggttt gactatctgt tctatatcca 900
ctgtcgagag gtgagccttg tgacacagag gagcctgggg gacctgatca tgagctgctg 960
ccccgaccca aacccaccca tccacaagat cgtgagaaaa ccctccagaa tcctcttcct 1020
catggacggc ttcgatgagc tgcaaggtgc ctttgacgag cacataggac cgctctgcac 1080
tgactggcag aaggccgagc ggggagacat tctcctgagc agcctcatca gaaagaagct 1140
gcttcccgag gcctctctgc tcatcaccac gagacctgtg gccctggaga aactgcagca 1200
cttgctggac catcctcggc atgtggagat cctgggtttc tccgaggcca aaaggaaaga 1260
gtacttcttc aagtacttct ctgatgaggc ccaagccagg gcagccttca gtctgattca 1320
ggagaacgag gtcctcttca ccatgtgctt catccccctg gtctgctgga tcgtgtgcac 1380
tggactgaaa cagcagatgg agagtggcaa gagccttgcc cagacatcca agaccaccac 1440
cgcggtgtac gtcttcttcc tttccagttt gctgcagccc cggggaggga gccaggagca 1500
cggcctctgc gcccacctct gggggctctg ctctttggct gcagatggaa tctggaacca 1560
gaaaatcctg tttgaggagt ccgacctcag gaatcatgga ctgcagaagg cggatgtgtc 1620
tgctttcctg aggatgaacc tgttccaaaa ggaagtggac tgcgagaagt tctacagctt 1680
catccacatg actttccagg agttctttgc cgccatgtac tacctgctgg aagaggaaaa 1740
ggaaggaagg acgaacgttc cagggagtcg tttgaagctt cccagccgag acgtgacagt 1800
ccttctggaa aactatggca aattcgaaaa ggggtatttg atttttgttg tacgtttcct 1860
ctttggcctg gtaaaccagg agaggacctc ctacttggag aagaaattaa gttgcaagat 1920
ctctcagcaa atcaggctgg agctgctgaa atggattgaa gtgaaagcca aagctaaaaa 1980
gctgcagatc cagcccagcc agctggaatt gttctactgt ttgtacgaga tgcaggagga 2040
ggacttcgtg caaagggcca tggactattt ccccaagatt gagatcaatc tctccaccag 2100
aatggaccac atggtttctt ccttttgcat tgagaactgt catcgggtgg agtcactgtc 2160
cctggggttt ctccataaca tgcccaagga ggaagaggag gaggaaaagg aaggccgaca 2220
ccttgatatg gtgcagtgtg tcctcccaag ctcctctcat gctgcctgtt ctcatggatt 2280
ggtgaacagc cacctcactt ccagtttttg ccggggcctc ttttcagttc tgagcaccag 2340
ccagagtcta actgaattgg acctcagtga caattctctg ggggacccag ggatgagagt 2400
gttgtgtgaa acgctccagc atcctggctg taacattcgg agattgtggt tggggcgctg 2460
tggcctctcg catgagtgct gcttcgacat ctccttggtc ctcagcagca accagaagct 2520
ggtggagctg gacctgagtg acaacgccct cggtgacttc ggaatcagac ttctgtgtgt 2580
gggactgaag cacctgttgt gcaatctgaa gaagctctgg ttggtcagct gctgcctcac 2640
atcagcatgt tgtcaggatc ttgcatcagt attgagcacc agccattccc tgaccagact 2700
ctatgtgggg gagaatgcct tgggagactc aggagtcgca attttatgtg aaaaagccaa 2760
gaatccacag tgtaacctgc agaaactggg gttggtgaat tctggcctta cgtcagtctg 2820
ttgttcagct ttgtcctcgg tactcagcac taatcagaat ctcacgcacc tttacctgcg 2880
aggcaacact ctcggagaca aggggatcaa actactctgt gagggactct tgcaccccga 2940
ctgcaagctt caggtgttgg aattagacaa ctgcaacctc acgtcacact gctgctggga 3000
tctttccaca cttctgacct ccagccagag cctgcgaaag ctgagcctgg gcaacaatga 3060
cctgggcgac ctgggggtca tgatgttctg tgaagtgctg aaacagcaga gctgcctcct 3120
gcagaacctg gggttgtctg aaatgtattt caattatgag acaaaaagtg cgttagaaac 3180
acttcaagaa gaaaagcctg agctgaccgt cgtctttgag ccttcttggt aggagtggaa 3240
acggggctgc cagacgccag tgttctccgg tccctccagc tgggggccct caggtggaga 3300
gagctgcgat ccatccaggc caagaccaca gctctgtgat ccttccggtg gagtgtcgga 3360
gaagagagct tgccgacgat gccttcctgt gcagagcttg ggcatctcct ttacgccagg 3420
gtgaggaaga caccaggaca atgacagcat cgggtgttgt tgtcatcaca gcgcctcagt 3480
tagaggatgt tcctcttggt gacctcatgt aattagctca ttcaataaag cactttcttt 3540
atttt 3545
<210> 533
<211> 4015
<212> ДНК
<213> Homo sapiens
<400> 533
gttcctgagg ctggcatctg gatgaggaaa ctgaagttga ggaatagtga agagtttgtc 60
caatgtcata gccccgtaat caacgggaca aaattttctt gctgatgggt caagatggca 120
tcgtgaagtg gttgttcacc gtaaactgta atacaatcct gtttatggat ttgtttgcat 180
atttttccct ccatagggaa acctttcttc catggctcag gacacactcc tggatcgagc 240
caacaggaga actttctggt aagcatttgg ctaacttttt tttttttgag atggagtctt 300
gctgtgtcgc ctaggctgga gtgcagtggc gtgatcttgg ctcactgcag cctccacttc 360
ccgggttcaa tcaattctcc tacctcaact tcctgagtag ctgggattac aggcgcccgc 420
caccacaccc ggctcatttt tgtactttta gtagagacac agttttgcca tgttggccag 480
gctggtcttg aattcctcag ctcaggtgat ctgcctgcct tggcctctca aagtgctggg 540
attacaggcg tgagccactg tgcccggcct tggctaactt ttcaaaatta aagattttga 600
cttgttacag tcatgtgaca tttttttctt tctgtttgct gagtttttga taatttatat 660
ctctcaaagt ggagacttta aaaaagactc atccgtgtgc cgtgttcact gcctggtatc 720
ttagtgtgga ccgaagccta aggaccctga aaacagctgc agatgaagat ggcaagcacc 780
cgctgcaagc tggccaggta cctggaggac ctggaggatg tggacttgaa gaaatttaag 840
atgcacttag aggactatcc tccccagaag ggctgcatcc ccctcccgag gggtcagaca 900
gagaaggcag accatgtgga tctagccacg ctaatgatcg acttcaatgg ggaggagaag 960
gcgtgggcca tggccgtgtg gatcttcgct gcgatcaaca ggagagacct ttatgagaaa 1020
gcaaaaagag atgagccgaa gtggggttca gataatgcac gtgtttcgaa tcccactgtg 1080
atatgccagg aagacagcat tgaagaggag tggatgggtt tactggagta cctttcgaga 1140
atctctattt gtaaaatgaa gaaagattac cgtaagaagt acagaaagta cgtgagaagc 1200
agattccagt gcattgaaga caggaatgcc cgtctgggtg agagtgtgag cctcaacaaa 1260
cgctacacac gactgcgtct catcaaggag caccggagcc agcaggagag ggagcaggag 1320
cttctggcca tcggcaagac caagacgtgt gagagccccg tgagtcccat taagatggag 1380
ttgctgtttg accccgatga tgagcattct gagcctgtgc acaccgtggt gttccagggg 1440
gcggcaggga ttgggaaaac aatcctggcc aggaagatga tgttggactg ggcgtcgggg 1500
acactctacc aagacaggtt tgactatctg ttctatatcc actgtcgaga ggtgagcctt 1560
gtgacacaga ggagcctggg ggacctgatc atgagctgct gccccgaccc aaacccaccc 1620
atccacaaga tcgtgagaaa accctccaga atcctcttcc tcatggacgg cttcgatgag 1680
ctgcaaggtg cctttgacga gcacatagga ccgctctgca ctgactggca gaaggccgag 1740
cggggagaca ttctcctgag cagcctcatc agaaagaagc tgcttcccga ggcctctctg 1800
ctcatcacca cgagacctgt ggccctggag aaactgcagc acttgctgga ccatcctcgg 1860
catgtggaga tcctgggttt ctccgaggcc aaaaggaaag agtacttctt caagtacttc 1920
tctgatgagg cccaagccag ggcagccttc agtctgattc aggagaacga ggtcctcttc 1980
accatgtgct tcatccccct ggtctgctgg atcgtgtgca ctggactgaa acagcagatg 2040
gagagtggca agagccttgc ccagacatcc aagaccacca ccgcggtgta cgtcttcttc 2100
ctttccagtt tgctgcagcc ccggggaggg agccaggagc acggcctctg cgcccacctc 2160
tgggggctct gctctttggc tgcagatgga atctggaacc agaaaatcct gtttgaggag 2220
tccgacctca ggaatcatgg actgcagaag gcggatgtgt ctgctttcct gaggatgaac 2280
ctgttccaaa aggaagtgga ctgcgagaag ttctacagct tcatccacat gactttccag 2340
gagttctttg ccgccatgta ctacctgctg gaagaggaaa aggaaggaag gacgaacgtt 2400
ccagggagtc gtttgaagct tcccagccga gacgtgacag tccttctgga aaactatggc 2460
aaattcgaaa aggggtattt gatttttgtt gtacgtttcc tctttggcct ggtaaaccag 2520
gagaggacct cctacttgga gaagaaatta agttgcaaga tctctcagca aatcaggctg 2580
gagctgctga aatggattga agtgaaagcc aaagctaaaa agctgcagat ccagcccagc 2640
cagctggaat tgttctactg tttgtacgag atgcaggagg aggacttcgt gcaaagggcc 2700
atggactatt tccccaagat tgagatcaat ctctccacca gaatggacca catggtttct 2760
tccttttgca ttgagaactg tcatcgggtg gagtcactgt ccctggggtt tctccataac 2820
atgcccaagg aggaagagga ggaggaaaag gaaggccgac accttgatat ggtgcagtgt 2880
gtcctcccaa gctcctctca tgctgcctgt tctcatggat tggtgaacag ccacctcact 2940
tccagttttt gccggggcct cttttcagtt ctgagcacca gccagagtct aactgaattg 3000
gacctcagtg acaattctct gggggaccca gggatgagag tgttgtgtga aacgctccag 3060
catcctggct gtaacattcg gagattgtgg ttggggcgct gtggcctctc gcatgagtgc 3120
tgcttcgaca tctccttggt cctcagcagc aaccagaagc tggtggagct ggacctgagt 3180
gacaacgccc tcggtgactt cggaatcaga cttctgtgtg tgggactgaa gcacctgttg 3240
tgcaatctga agaagctctg gttggtgaat tctggcctta cgtcagtctg ttgttcagct 3300
ttgtcctcgg tactcagcac taatcagaat ctcacgcacc tttacctgcg aggcaacact 3360
ctcggagaca aggggatcaa actactctgt gagggactct tgcaccccga ctgcaagctt 3420
caggtgttgg aattagacaa ctgcaacctc acgtcacact gctgctggga tctttccaca 3480
cttctgacct ccagccagag cctgcgaaag ctgagcctgg gcaacaatga cctgggcgac 3540
ctgggggtca tgatgttctg tgaagtgctg aaacagcaga gctgcctcct gcagaacctg 3600
gggttgtctg aaatgtattt caattatgag acaaaaagtg cgttagaaac acttcaagaa 3660
gaaaagcctg agctgaccgt cgtctttgag ccttcttggt aggagtggaa acggggctgc 3720
cagacgccag tgttctccgg tccctccagc tgggggccct caggtggaga gagctgcgat 3780
ccatccaggc caagaccaca gctctgtgat ccttccggtg gagtgtcgga gaagagagct 3840
tgccgacgat gccttcctgt gcagagcttg ggcatctcct ttacgccagg gtgaggaaga 3900
caccaggaca atgacagcat cgggtgttgt tgtcatcaca gcgcctcagt tagaggatgt 3960
tcctcttggt gacctcatgt aattagctca ttcaataaag cactttcttt atttt 4015
<210> 534
<211> 4016
<212> ДНК
<213> Homo sapiens
<400> 534
gttcctgagg ctggcatctg gatgaggaaa ctgaagttga ggaatagtga agagtttgtc 60
caatgtcata gccccgtaat caacgggaca aaaattttct tgctgatggg tcaagatggc 120
atcgtgaagt ggttgttcac cgtaaactgt aatacaatcc tgtttatgga tttgtttgca 180
tatttttccc tccataggga aacctttctt ccatggctca ggacacactc ctggatcgag 240
ccaacaggag aactttctgg taagcatttg gctaactttt ttttttttga gatggagtct 300
tgctgtgtcg cctaggctgg agtgcagtgg cgtgatcttg gctcactgca gcctccactt 360
cccgggttca atcaattctc ctacctcaac ttcctgagta gctgggatta caggcgcccg 420
ccaccacacc cggctcattt ttgtactttt agtagagaca cagttttgcc atgttggcca 480
ggctggtctt gaattcctca gctcaggtga tctgcctgcc ttggcctctc aaagtgctgg 540
gattacaggc gtgagccact gtgcccggcc ttggctaact tttcaaaatt aaagattttg 600
acttgttaca gtcatgtgac atttttttct ttctgtttgc tgagtttttg ataatttata 660
tctctcaaag tggagacttt aaaaaagact catccgtgtg ccgtgttcac tgcctggtat 720
cttagtgtgg accgaagcct aaggaccctg aaaacagctg cagatgaaga tggcaagcac 780
ccgctgcaag ctggccaggt acctggagga cctggaggat gtggacttga agaaatttaa 840
gatgcactta gaggactatc ctccccagaa gggctgcatc cccctcccga ggggtcagac 900
agagaaggca gaccatgtgg atctagccac gctaatgatc gacttcaatg gggaggagaa 960
ggcgtgggcc atggccgtgt ggatcttcgc tgcgatcaac aggagagacc tttatgagaa 1020
agcaaaaaga gatgagccga agtggggttc agataatgca cgtgtttcga atcccactgt 1080
gatatgccag gaagacagca ttgaagagga gtggatgggt ttactggagt acctttcgag 1140
aatctctatt tgtaaaatga agaaagatta ccgtaagaag tacagaaagt acgtgagaag 1200
cagattccag tgcattgaag acaggaatgc ccgtctgggt gagagtgtga gcctcaacaa 1260
acgctacaca cgactgcgtc tcatcaagga gcaccggagc cagcaggaga gggagcagga 1320
gcttctggcc atcggcaaga ccaagacgtg tgagagcccc gtgagtccca ttaagatgga 1380
gttgctgttt gaccccgatg atgagcattc tgagcctgtg cacaccgtgg tgttccaggg 1440
ggcggcaggg attgggaaaa caatcctggc caggaagatg atgttggact gggcgtcggg 1500
gacactctac caagacaggt ttgactatct gttctatatc cactgtcgag aggtgagcct 1560
tgtgacacag aggagcctgg gggacctgat catgagctgc tgccccgacc caaacccacc 1620
catccacaag atcgtgagaa aaccctccag aatcctcttc ctcatggacg gcttcgatga 1680
gctgcaaggt gcctttgacg agcacatagg accgctctgc actgactggc agaaggccga 1740
gcggggagac attctcctga gcagcctcat cagaaagaag ctgcttcccg aggcctctct 1800
gctcatcacc acgagacctg tggccctgga gaaactgcag cacttgctgg accatcctcg 1860
gcatgtggag atcctgggtt tctccgaggc caaaaggaaa gagtacttct tcaagtactt 1920
ctctgatgag gcccaagcca gggcagcctt cagtctgatt caggagaacg aggtcctctt 1980
caccatgtgc ttcatccccc tggtctgctg gatcgtgtgc actggactga aacagcagat 2040
ggagagtggc aagagccttg cccagacatc caagaccacc accgcggtgt acgtcttctt 2100
cctttccagt ttgctgcagc cccggggagg gagccaggag cacggcctct gcgcccacct 2160
ctgggggctc tgctctttgg ctgcagatgg aatctggaac cagaaaatcc tgtttgagga 2220
gtccgacctc aggaatcatg gactgcagaa ggcggatgtg tctgctttcc tgaggatgaa 2280
cctgttccaa aaggaagtgg actgcgagaa gttctacagc ttcatccaca tgactttcca 2340
ggagttcttt gccgccatgt actacctgct ggaagaggaa aaggaaggaa ggacgaacgt 2400
tccagggagt cgtttgaagc ttcccagccg agacgtgaca gtccttctgg aaaactatgg 2460
caaattcgaa aaggggtatt tgatttttgt tgtacgtttc ctctttggcc tggtaaacca 2520
ggagaggacc tcctacttgg agaagaaatt aagttgcaag atctctcagc aaatcaggct 2580
ggagctgctg aaatggattg aagtgaaagc caaagctaaa aagctgcaga tccagcccag 2640
ccagctggaa ttgttctact gtttgtacga gatgcaggag gaggacttcg tgcaaagggc 2700
catggactat ttccccaaga ttgagatcaa tctctccacc agaatggacc acatggtttc 2760
ttccttttgc attgagaact gtcatcgggt ggagtcactg tccctggggt ttctccataa 2820
catgcccaag gaggaagagg aggaggaaaa ggaaggccga caccttgata tggtgcagtg 2880
tgtcctccca agctcctctc atgctgcctg ttctcatggg ttggggcgct gtggcctctc 2940
gcatgagtgc tgcttcgaca tctccttggt cctcagcagc aaccagaagc tggtggagct 3000
ggacctgagt gacaacgccc tcggtgactt cggaatcaga cttctgtgtg tgggactgaa 3060
gcacctgttg tgcaatctga agaagctctg gttggtcagc tgctgcctca catcagcatg 3120
ttgtcaggat cttgcatcag tattgagcac cagccattcc ctgaccagac tctatgtggg 3180
ggagaatgcc ttgggagact caggagtcgc aattttatgt gaaaaagcca agaatccaca 3240
gtgtaacctg cagaaactgg ggttggtgaa ttctggcctt acgtcagtct gttgttcagc 3300
tttgtcctcg gtactcagca ctaatcagaa tctcacgcac ctttacctgc gaggcaacac 3360
tctcggagac aaggggatca aactactctg tgagggactc ttgcaccccg actgcaagct 3420
tcaggtgttg gaattagaca actgcaacct cacgtcacac tgctgctggg atctttccac 3480
acttctgacc tccagccaga gcctgcgaaa gctgagcctg ggcaacaatg acctgggcga 3540
cctgggggtc atgatgttct gtgaagtgct gaaacagcag agctgcctcc tgcagaacct 3600
ggggttgtct gaaatgtatt tcaattatga gacaaaaagt gcgttagaaa cacttcaaga 3660
agaaaagcct gagctgaccg tcgtctttga gccttcttgg taggagtgga aacggggctg 3720
ccagacgcca gtgttctccg gtccctccag ctgggggccc tcaggtggag agagctgcga 3780
tccatccagg ccaagaccac agctctgtga tccttccggt ggagtgtcgg agaagagagc 3840
ttgccgacga tgccttcctg tgcagagctt gggcatctcc tttacgccag ggtgaggaag 3900
acaccaggac aatgacagca tcgggtgttg ttgtcatcac agcgcctcag ttagaggatg 3960
ttcctcttgg tgacctcatg taattagctc attcaataaa gcactttctt tatttt 4016
<210> 535
<400> 535
000
<210> 536
<400> 536
000
<210> 537
<400> 537
000
<210> 538
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 538
Asn Glu Ala Tyr Val His Asp Ala Pro Val Arg Ser Leu Asn
1 5 10
<210> 539
<211> 1036
<212> БЕЛОК
<213> Homo sapiens
<400> 539
Met Lys Met Ala Ser Thr Arg Cys Lys Leu Ala Arg Tyr Leu Glu Asp
1 5 10 15
Leu Glu Asp Val Asp Leu Lys Lys Phe Lys Met His Leu Glu Asp Tyr
20 25 30
Pro Pro Gln Lys Gly Cys Ile Pro Leu Pro Arg Gly Gln Thr Glu Lys
35 40 45
Ala Asp His Val Asp Leu Ala Thr Leu Met Ile Asp Phe Asn Gly Glu
50 55 60
Glu Lys Ala Trp Ala Met Ala Val Trp Ile Phe Ala Ala Ile Asn Arg
65 70 75 80
Arg Asp Leu Tyr Glu Lys Ala Lys Arg Asp Glu Pro Lys Trp Gly Ser
85 90 95
Asp Asn Ala Arg Val Ser Asn Pro Thr Val Ile Cys Gln Glu Asp Ser
100 105 110
Ile Glu Glu Glu Trp Met Gly Leu Leu Glu Tyr Leu Ser Arg Ile Ser
115 120 125
Ile Cys Lys Met Lys Lys Asp Tyr Arg Lys Lys Tyr Arg Lys Tyr Val
130 135 140
Arg Ser Arg Phe Gln Cys Ile Glu Asp Arg Asn Ala Arg Leu Gly Glu
145 150 155 160
Ser Val Ser Leu Asn Lys Arg Tyr Thr Arg Leu Arg Leu Ile Lys Glu
165 170 175
His Arg Ser Gln Gln Glu Arg Glu Gln Glu Leu Leu Ala Ile Gly Lys
180 185 190
Thr Lys Thr Cys Glu Ser Pro Val Ser Pro Ile Lys Met Glu Leu Leu
195 200 205
Phe Asp Pro Asp Asp Glu His Ser Glu Pro Val His Thr Val Val Phe
210 215 220
Gln Gly Ala Ala Gly Ile Gly Lys Thr Ile Leu Ala Arg Lys Met Met
225 230 235 240
Leu Asp Trp Ala Ser Gly Thr Leu Tyr Gln Asp Arg Phe Asp Tyr Leu
245 250 255
Phe Tyr Ile His Cys Arg Glu Val Ser Leu Val Thr Gln Arg Ser Leu
260 265 270
Gly Asp Leu Ile Met Ser Cys Cys Pro Asp Pro Asn Pro Pro Ile His
275 280 285
Lys Ile Val Arg Lys Pro Ser Arg Ile Leu Phe Leu Met Asp Gly Phe
290 295 300
Asp Glu Leu Gln Gly Ala Phe Asp Glu His Ile Gly Pro Leu Cys Thr
305 310 315 320
Asp Trp Gln Lys Ala Glu Arg Gly Asp Ile Leu Leu Ser Ser Leu Ile
325 330 335
Arg Lys Lys Leu Leu Pro Glu Ala Ser Leu Leu Ile Thr Thr Arg Pro
340 345 350
Val Ala Leu Glu Lys Leu Gln His Leu Leu Asp His Pro Arg His Val
355 360 365
Glu Ile Leu Gly Phe Ser Glu Ala Lys Arg Lys Glu Tyr Phe Phe Lys
370 375 380
Tyr Phe Ser Asp Glu Ala Gln Ala Arg Ala Ala Phe Ser Leu Ile Gln
385 390 395 400
Glu Asn Glu Val Leu Phe Thr Met Cys Phe Ile Pro Leu Val Cys Trp
405 410 415
Ile Val Cys Thr Gly Leu Lys Gln Gln Met Glu Ser Gly Lys Ser Leu
420 425 430
Ala Gln Thr Ser Lys Thr Thr Thr Ala Val Tyr Val Phe Phe Leu Ser
435 440 445
Ser Leu Leu Gln Pro Arg Gly Gly Ser Gln Glu His Gly Leu Cys Ala
450 455 460
His Leu Trp Gly Leu Cys Ser Leu Ala Ala Asp Gly Ile Trp Asn Gln
465 470 475 480
Lys Ile Leu Phe Glu Glu Ser Asp Leu Arg Asn His Gly Leu Gln Lys
485 490 495
Ala Asp Val Ser Ala Phe Leu Arg Met Asn Leu Phe Gln Lys Glu Val
500 505 510
Asp Cys Glu Lys Phe Tyr Ser Phe Ile His Met Thr Phe Gln Glu Phe
515 520 525
Phe Ala Ala Met Tyr Tyr Leu Leu Glu Glu Glu Lys Glu Gly Arg Thr
530 535 540
Asn Val Pro Gly Ser Arg Leu Lys Leu Pro Ser Arg Asp Val Thr Val
545 550 555 560
Leu Leu Glu Asn Tyr Gly Lys Phe Glu Lys Gly Tyr Leu Ile Phe Val
565 570 575
Val Arg Phe Leu Phe Gly Leu Val Asn Gln Glu Arg Thr Ser Tyr Leu
580 585 590
Glu Lys Lys Leu Ser Cys Lys Ile Ser Gln Gln Ile Arg Leu Glu Leu
595 600 605
Leu Lys Trp Ile Glu Val Lys Ala Lys Ala Lys Lys Leu Gln Ile Gln
610 615 620
Pro Ser Gln Leu Glu Leu Phe Tyr Cys Leu Tyr Glu Met Gln Glu Glu
625 630 635 640
Asp Phe Val Gln Arg Ala Met Asp Tyr Phe Pro Lys Ile Glu Ile Asn
645 650 655
Leu Ser Thr Arg Met Asp His Met Val Ser Ser Phe Cys Ile Glu Asn
660 665 670
Cys His Arg Val Glu Ser Leu Ser Leu Gly Phe Leu His Asn Met Pro
675 680 685
Lys Glu Glu Glu Glu Glu Glu Lys Glu Gly Arg His Leu Asp Met Val
690 695 700
Gln Cys Val Leu Pro Ser Ser Ser His Ala Ala Cys Ser His Gly Leu
705 710 715 720
Val Asn Ser His Leu Thr Ser Ser Phe Cys Arg Gly Leu Phe Ser Val
725 730 735
Leu Ser Thr Ser Gln Ser Leu Thr Glu Leu Asp Leu Ser Asp Asn Ser
740 745 750
Leu Gly Asp Pro Gly Met Arg Val Leu Cys Glu Thr Leu Gln His Pro
755 760 765
Gly Cys Asn Ile Arg Arg Leu Trp Leu Gly Arg Cys Gly Leu Ser His
770 775 780
Glu Cys Cys Phe Asp Ile Ser Leu Val Leu Ser Ser Asn Gln Lys Leu
785 790 795 800
Val Glu Leu Asp Leu Ser Asp Asn Ala Leu Gly Asp Phe Gly Ile Arg
805 810 815
Leu Leu Cys Val Gly Leu Lys His Leu Leu Cys Asn Leu Lys Lys Leu
820 825 830
Trp Leu Val Ser Cys Cys Leu Thr Ser Ala Cys Cys Gln Asp Leu Ala
835 840 845
Ser Val Leu Ser Thr Ser His Ser Leu Thr Arg Leu Tyr Val Gly Glu
850 855 860
Asn Ala Leu Gly Asp Ser Gly Val Ala Ile Leu Cys Glu Lys Ala Lys
865 870 875 880
Asn Pro Gln Cys Asn Leu Gln Lys Leu Gly Leu Val Asn Ser Gly Leu
885 890 895
Thr Ser Val Cys Cys Ser Ala Leu Ser Ser Val Leu Ser Thr Asn Gln
900 905 910
Asn Leu Thr His Leu Tyr Leu Arg Gly Asn Thr Leu Gly Asp Lys Gly
915 920 925
Ile Lys Leu Leu Cys Glu Gly Leu Leu His Pro Asp Cys Lys Leu Gln
930 935 940
Val Leu Glu Leu Asp Asn Cys Asn Leu Thr Ser His Cys Cys Trp Asp
945 950 955 960
Leu Ser Thr Leu Leu Thr Ser Ser Gln Ser Leu Arg Lys Leu Ser Leu
965 970 975
Gly Asn Asn Asp Leu Gly Asp Leu Gly Val Met Met Phe Cys Glu Val
980 985 990
Leu Lys Gln Gln Ser Cys Leu Leu Gln Asn Leu Gly Leu Ser Glu Met
995 1000 1005
Tyr Phe Asn Tyr Glu Thr Lys Ser Ala Leu Glu Thr Leu Gln Glu
1010 1015 1020
Glu Lys Pro Glu Leu Thr Val Val Phe Glu Pro Ser Trp
1025 1030 1035
<210> 540
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<220>
<223> Описание комбинированной молекулы ДНК/РНК: синтетический
олигонуклеотид
<400> 540
gugcauugaa gacaggaaut t 21
<210> 541
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 541
ggctgtaaca ttcggagatt g 21
<210> 542
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 542
tcatcattcc cgctatcttt c 21
<210> 543
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 543
ccgtaagaag tacagaaagt a 21
<210> 544
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 544
gagactcagg agtcgcaatt t 21
<210> 545
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 545
cctcatgtaa ttagctcatt c 21
<210> 546
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 546
gtggatctag ccacgctaat g 21
<210> 547
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 547
ccacagtgta acctgcagaa a 21
<210> 548
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 548
ccagccagag tctaactgaa t 21
<210> 549
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 549
gcgttagaaa cacttcaaga a 21
<210> 550
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 550
gctggaattg ttctactgtt t 21
<210> 551
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 551
ccacatgact ttccaggagt t 21
<210> 552
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 552
ggctgtaaca ttcggagatt g 21
<210> 553
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 553
ccggggctgt aacattcgga gattgctcga gcaatctccg aatgttacag cctttttg 58
<210> 554
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 554
aattcaaaaa ggctgtaaca ttcggagatt gctcgagcaa tctccgaatg ttacagcc 58
<210> 555
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 555
tcatcattcc cgctatcttt c 21
<210> 556
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 556
ccggtcatca ttcccgctat ctttcctcga ggaaagatag cgggaatgat gatttttg 58
<210> 557
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 557
aattcaaaaa tcatcattcc cgctatcttt cctcgaggaa agatagcggg aatgatga 58
<210> 558
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 558
ccgtaagaag tacagaaagt a 21
<210> 559
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 559
ccggccgtaa gaagtacaga aagtactcga gtactttctg tacttcttac ggtttttg 58
<210> 560
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 560
aattcaaaaa ccgtaagaag tacagaaagt actcgagtac tttctgtact tcttacgg 58
<210> 561
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 561
gagactcagg agtcgcaatt t 21
<210> 562
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 562
ccgggagact caggagtcgc aatttctcga gaaattgcga ctcctgagtc tctttttg 58
<210> 563
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 563
aattcaaaaa gagactcagg agtcgcaatt tctcgagaaa ttgcgactcc tgagtctc 58
<210> 564
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 564
cctcatgtaa ttagctcatt c 21
<210> 565
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 565
ccggcctcat gtaattagct cattcctcga ggaatgagct aattacatga ggtttttg 58
<210> 566
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 566
aattcaaaaa cctcatgtaa ttagctcatt cctcgaggaa tgagctaatt acatgagg 58
<210> 567
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 567
gtggatctag ccacgctaat g 21
<210> 568
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 568
ccgggtggat ctagccacgc taatgctcga gcattagcgt ggctagatcc actttttg 58
<210> 569
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 569
aattcaaaaa gtggatctag ccacgctaat gctcgagcat tagcgtggct agatccac 58
<210> 570
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 570
ccacagtgta acctgcagaa a 21
<210> 571
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 571
aattcaaaaa ccacagtgta acctgcagaa actcgagttt ctgcaggtta cactgtgg 58
<210> 572
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 572
ccagccagag tctaactgaa t 21
<210> 573
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 573
ccggccagcc agagtctaac tgaatctcga gattcagtta gactctggct ggtttttg 58
<210> 574
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 574
aattcaaaaa ccagccagag tctaactgaa tctcgagatt cagttagact ctggctgg 58
<210> 575
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 575
gcgttagaaa cacttcaaga a 21
<210> 576
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 576
ccgggcgtta gaaacacttc aagaactcga gttcttgaag tgtttctaac gctttttg 58
<210> 577
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 577
aattcaaaaa gcgttagaaa cacttcaaga actcgagttc ttgaagtgtt tctaacgc 58
<210> 578
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 578
gctggaattg ttctactgtt t 21
<210> 579
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 579
ccgggctgga attgttctac tgtttctcga gaaacagtag aacaattcca gctttttg 58
<210> 580
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 580
aattcaaaaa gctggaattg ttctactgtt tctcgagaaa cagtagaaca attccagc 58
<210> 581
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 581
ccacatgact ttccaggagt t 21
<210> 582
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 582
ccggccacat gactttccag gagttctcga gaactcctgg aaagtcatgt ggtttttg 58
<210> 583
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 583
aattcaaaaa ccacatgact ttccaggagt tctcgagaac tcctggaaag tcatgtgg 58
<210> 584
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 584
gaaagccaaa gctaagaagt a 21
<210> 585
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 585
ccgggaaagc caaagctaag aagtactcga gtacttctta gctttggctt tctttttg 58
<210> 586
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 586
aattcaaaaa gaaagccaaa gctaagaagt actcgagtac ttcttagctt tggctttc 58
<210> 587
<211> 23
<212> РНК
<213> Homo sapiens
<400> 587
ucuuugguua ucuagcugua uga 23
<210> 588
<211> 23
<212> РНК
<213> Homo sapiens
<400> 588
ucuuugguua ucuagcugua uga 23
<210> 589
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 589
tggggtattt gacaaactga ca 22
<210> 590
<211> 63
<212> РНК
<213> Caenorhabditis brenneri
<400> 590
ucgcccaucc cguuguucca auauuccaac aacaagugau uauugagcaa ugcgcaugug 60
cgg 63
<210> 591
<211> 97
<212> РНК
<213> Caenorhabditis briggsae
<400> 591
aagcauuuuu cugucccgcg caucccuuug uuccaauauu caaaccagua gaaagauuau 60
ugagcaaugc gcaugugcgg gacagauuga auagcug 97
<210> 592
<211> 98
<212> РНК
<213> Caenorhabditis elegans
<400> 592
auauagcauc uuucugucuc gcccaucccg uugcuccaau auucuaacaa caagugauua 60
uugagcaaug cgcaugugcg ggauagacug auggcugc 98
<210> 593
<211> 101
<212> РНК
<213> Caenorhabditis remanei
<400> 593
ugaagcgucu cucugucccg cucauccugu uguuccaaua uuccaacagc ccagugauua 60
uugagcaaug cgcaugugcg ggacagauug uaugcugcca u 101
<210> 594
<211> 22
<212> РНК
<213> Homo sapiens
<400> 594
aguucuucag uggcaagcuu ua 22
<210> 595
<211> 85
<212> РНК
<213> Homo sapiens
<400> 595
ggcugagccg caguaguucu ucaguggcaa gcuuuauguc cugacccagc uaaagcugcc 60
aguugaagaa cuguugcccu cugcc 85
<210> 596
<211> 21
<212> РНК
<213> Mus musculus
<400> 596
gugcauugua guugcauugc a 21
<210> 597
<211> 69
<212> РНК
<213> Mus musculus
<400> 597
cuguggugca uuguaguugc auugcauguu cuggcaauac cugugcaaug uuuccacagu 60
gcaucacgg 69
<210> 598
<211> 343
<212> БЕЛОК
<213> Homo sapiens
<400> 598
Met Glu Ser Lys Tyr Lys Glu Ile Leu Leu Leu Thr Gly Leu Asp Asn
1 5 10 15
Ile Thr Asp Glu Glu Leu Asp Arg Phe Lys Phe Phe Leu Ser Asp Glu
20 25 30
Phe Asn Ile Ala Thr Gly Lys Leu His Thr Ala Asn Arg Ile Gln Val
35 40 45
Ala Thr Leu Met Ile Gln Asn Ala Gly Ala Val Ser Ala Val Met Lys
50 55 60
Thr Ile Arg Ile Phe Gln Lys Leu Asn Tyr Met Leu Leu Ala Lys Arg
65 70 75 80
Leu Gln Glu Glu Lys Glu Lys Val Asp Lys Gln Tyr Lys Ser Val Thr
85 90 95
Lys Pro Lys Pro Leu Ser Gln Ala Glu Met Ser Pro Ala Ala Ser Ala
100 105 110
Ala Ile Arg Asn Asp Val Ala Lys Gln Arg Ala Ala Pro Lys Val Ser
115 120 125
Pro His Val Lys Pro Glu Gln Lys Gln Met Val Ala Gln Gln Glu Ser
130 135 140
Ile Arg Glu Gly Phe Gln Lys Arg Cys Leu Pro Val Met Val Leu Lys
145 150 155 160
Ala Lys Lys Pro Phe Thr Phe Glu Thr Gln Glu Gly Lys Gln Glu Met
165 170 175
Phe His Ala Thr Val Ala Thr Glu Lys Glu Phe Phe Phe Val Lys Val
180 185 190
Phe Asn Thr Leu Leu Lys Asp Lys Phe Ile Pro Lys Arg Ile Ile Ile
195 200 205
Ile Ala Arg Tyr Tyr Arg His Ser Gly Phe Leu Glu Val Asn Ser Ala
210 215 220
Ser Arg Val Leu Asp Ala Glu Ser Asp Gln Lys Val Asn Val Pro Leu
225 230 235 240
Asn Ile Ile Arg Lys Ala Gly Glu Thr Pro Lys Ile Asn Thr Leu Gln
245 250 255
Thr Gln Pro Leu Gly Thr Ile Val Asn Gly Leu Phe Val Val Gln Lys
260 265 270
Val Thr Glu Lys Lys Lys Asn Ile Leu Phe Asp Leu Ser Asp Asn Thr
275 280 285
Gly Lys Met Glu Val Leu Gly Val Arg Asn Glu Asp Thr Met Lys Cys
290 295 300
Lys Glu Gly Asp Lys Val Arg Leu Thr Phe Phe Thr Leu Ser Lys Asn
305 310 315 320
Gly Glu Lys Leu Gln Leu Thr Ser Gly Val His Ser Thr Ile Lys Val
325 330 335
Ile Lys Ala Lys Lys Lys Thr
340
<210> 599
<400> 599
000
<210> 600
<211> 1558
<212> ДНК
<213> Homo sapiens
<400> 600
atagacattt tcttctgtgg ctgctagtga gaacccaaac cagctcagcc aattagagct 60
ccagttgtca ctcctaccca cactgggcct gggggtgaag ggaagtgttt attaggggta 120
catgtgaagc cgtccagaag tgtcagagtc tttgtagctt tgaaagtcac ctaggttatt 180
tgggcatgct ctcctgagtc ctctgctagt taagctctct gaaaagaagg tggcagaccc 240
ggtttgctga tcgccccagg gatcaggagg ctgatcccaa agttgtcaga tggagagtaa 300
atacaaggag atactcttgc taacaggcct ggataacatc actgatgagg aactggatag 360
gtttaagttc tttctttcag acgagtttaa tattgccaca ggcaaactac atactgcaaa 420
cagaatacaa gtagctacct tgatgattca aaatgctggg gcggtgtctg cagtgatgaa 480
gaccattcgt atttttcaga agttgaatta tatgcttttg gcaaaacgtc ttcaggagga 540
gaaggagaaa gttgataagc aatacaaatc ggtaacaaaa ccaaagccac taagtcaagc 600
tgaaatgagt cctgctgcat ctgcagccat cagaaatgat gtcgcaaagc aacgtgctgc 660
accaaaagtc tctcctcatg ttaagcctga acagaaacag atggtggccc agcaggaatc 720
tatcagagaa gggtttcaga agcgctgttt gccagttatg gtactgaaag caaagaagcc 780
cttcacgttt gagacccaag aaggcaagca ggagatgttt catgctacag tggctacaga 840
aaaggaattc ttctttgtaa aagtttttaa tacactgctg aaagataaat tcattccaaa 900
gagaataatt ataatagcaa gatattatcg gcacagtggt ttcttagagg taaatagcgc 960
ctcacgtgtg ttagatgctg aatctgacca aaaggttaat gtcccgctga acattatcag 1020
aaaagctggt gaaaccccga agatcaacac gcttcaaact cagccccttg gaacaattgt 1080
gaatggtttg tttgtagtcc agaaggtaac agaaaagaag aaaaacatat tatttgacct 1140
aagtgacaac actgggaaaa tggaagtact gggggttaga aacgaggaca caatgaaatg 1200
taaggaagga gataaggttc gacttacatt cttcacactg tcaaaaaatg gagaaaaact 1260
acagctgaca tctggagttc atagcaccat aaaggttatt aaggccaaaa aaaaaacata 1320
gagaagtaaa aaggaccaat tcaagccaac tggtctaagc agcatttaat tgaagaatat 1380
gtgatacagc ctcttcaatc agattgtaag ttacctgaaa gctgcagttc acaggctcct 1440
ctctccacca aattaggata gaataattgc tggataaaca aattcagaat atcaacagat 1500
gatcacaata aacatctgtt tctcattcaa aaaaaaaaaa aaaaaaaaaa aaaaaaaa 1558
<210> 601
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 601
cccgaagatc aacacgcttc a 21
<210> 602
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 602
ttagggttag ggttagggtt aggg 24
<210> 603
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 603
ttcaaattca aattcaaatt caaa 24
<210> 604
<211> 6
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 604
ttaggg 6
<210> 605
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 605
agccactaag tcaagctgaa a 21
<210> 606
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 606
ccaactggtc taagcagcat t 21
<210> 607
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 607
gaaacgagga cacaatgaaa t 21
<210> 608
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 608
gccactaagt caagctgaaa t 21
<210> 609
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 609
ctggagttca tagcaccata a 21
<210> 610
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 610
cccgctgaac attatcagaa a 21
<210> 611
<211> 1393
<212> ДНК
<213> Homo sapiens
<400> 611
atactttcag tttcagtcac acaagaaggg aggagagaaa agccatggcc gacaaggtcc 60
tgaaggagaa gagaaagctg tttatccgtt ccatgggtga aggtacaata aatggcttac 120
tggatgaatt attacagaca agggtgctga acaaggaaga gatggagaaa gtaaaacgtg 180
aaaatgctac agttatggat aagacccgag ctttgattga ctccgttatt ccgaaagggg 240
cacaggcatg ccaaatttgc atcacataca tttgtgaaga agacagttac ctggcaggga 300
cgctgggact ctcagcagat caaacatctg gaaattacct taatatgcaa gactctcaag 360
gagtactttc ttcctttcca gctcctcagg cagtgcagga caacccagct atgcccacat 420
cctcaggctc agaagggaat gtcaagcttt gctccctaga agaagctcaa aggatatgga 480
aacaaaagtc ggcagagatt tatccaataa tggacaagtc aagccgcaca cgtcttgctc 540
tcattatctg caatgaagaa tttgacagta ttcctagaag aactggagct gaggttgaca 600
tcacaggcat gacaatgctg ctacaaaatc tggggtacag cgtagatgtg aaaaaaaatc 660
tcactgcttc ggacatgact acagagctgg aggcatttgc acaccgccca gagcacaaga 720
cctctgacag cacgttcctg gtgttcatgt ctcatggtat tcgggaaggc atttgtggga 780
agaaacactc tgagcaagtc ccagatatac tacaactcaa tgcaatcttt aacatgttga 840
ataccaagaa ctgcccaagt ttgaaggaca aaccgaaggt gatcatcatc caggcctgcc 900
gtggtgacag ccctggtgtg gtgtggttta aagattcagt aggagtttct ggaaacctat 960
ctttaccaac tacagaagag tttgaggatg atgctattaa gaaagcccac atagagaagg 1020
attttatcgc tttctgctct tccacaccag ataatgtttc ttggagacat cccacaatgg 1080
gctctgtttt tattggaaga ctcattgaac atatgcaaga atatgcctgt tcctgtgatg 1140
tggaggaaat tttccgcaag gttcgatttt catttgagca gccagatggt agagcgcaga 1200
tgcccaccac tgaaagagtg actttgacaa gatgtttcta cctcttccca ggacattaaa 1260
ataaggaaac tgtatgaatg tctgtgggca ggaagtgaag agatccttct gtaaaggttt 1320
ttggaattat gtctgctgaa taataaactt ttttgaaata ataaatctgg tagaaaaatg 1380
aaaaaaaaaa aaa 1393
<210> 612
<211> 404
<212> БЕЛОК
<213> Homo sapiens
<400> 612
Met Ala Asp Lys Val Leu Lys Glu Lys Arg Lys Leu Phe Ile Arg Ser
1 5 10 15
Met Gly Glu Gly Thr Ile Asn Gly Leu Leu Asp Glu Leu Leu Gln Thr
20 25 30
Arg Val Leu Asn Lys Glu Glu Met Glu Lys Val Lys Arg Glu Asn Ala
35 40 45
Thr Val Met Asp Lys Thr Arg Ala Leu Ile Asp Ser Val Ile Pro Lys
50 55 60
Gly Ala Gln Ala Cys Gln Ile Cys Ile Thr Tyr Ile Cys Glu Glu Asp
65 70 75 80
Ser Tyr Leu Ala Gly Thr Leu Gly Leu Ser Ala Asp Gln Thr Ser Gly
85 90 95
Asn Tyr Leu Asn Met Gln Asp Ser Gln Gly Val Leu Ser Ser Phe Pro
100 105 110
Ala Pro Gln Ala Val Gln Asp Asn Pro Ala Met Pro Thr Ser Ser Gly
115 120 125
Ser Glu Gly Asn Val Lys Leu Cys Ser Leu Glu Glu Ala Gln Arg Ile
130 135 140
Trp Lys Gln Lys Ser Ala Glu Ile Tyr Pro Ile Met Asp Lys Ser Ser
145 150 155 160
Arg Thr Arg Leu Ala Leu Ile Ile Cys Asn Glu Glu Phe Asp Ser Ile
165 170 175
Pro Arg Arg Thr Gly Ala Glu Val Asp Ile Thr Gly Met Thr Met Leu
180 185 190
Leu Gln Asn Leu Gly Tyr Ser Val Asp Val Lys Lys Asn Leu Thr Ala
195 200 205
Ser Asp Met Thr Thr Glu Leu Glu Ala Phe Ala His Arg Pro Glu His
210 215 220
Lys Thr Ser Asp Ser Thr Phe Leu Val Phe Met Ser His Gly Ile Arg
225 230 235 240
Glu Gly Ile Cys Gly Lys Lys His Ser Glu Gln Val Pro Asp Ile Leu
245 250 255
Gln Leu Asn Ala Ile Phe Asn Met Leu Asn Thr Lys Asn Cys Pro Ser
260 265 270
Leu Lys Asp Lys Pro Lys Val Ile Ile Ile Gln Ala Cys Arg Gly Asp
275 280 285
Ser Pro Gly Val Val Trp Phe Lys Asp Ser Val Gly Val Ser Gly Asn
290 295 300
Leu Ser Leu Pro Thr Thr Glu Glu Phe Glu Asp Asp Ala Ile Lys Lys
305 310 315 320
Ala His Ile Glu Lys Asp Phe Ile Ala Phe Cys Ser Ser Thr Pro Asp
325 330 335
Asn Val Ser Trp Arg His Pro Thr Met Gly Ser Val Phe Ile Gly Arg
340 345 350
Leu Ile Glu His Met Gln Glu Tyr Ala Cys Ser Cys Asp Val Glu Glu
355 360 365
Ile Phe Arg Lys Val Arg Phe Ser Phe Glu Gln Pro Asp Gly Arg Ala
370 375 380
Gln Met Pro Thr Thr Glu Arg Val Thr Leu Thr Arg Cys Phe Tyr Leu
385 390 395 400
Phe Pro Gly His
<210> 613
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 613
cacacgtctt gctctcatta t 21
<210> 614
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 614
ctacaactca atgcaatctt t 21
<210> 615
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 615
ccagatatac tacaactcaa t 21
<210> 616
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 616
gaagagtttg aggatgatgc t 21
<210> 617
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 617
ccatgggtga aggtacaata a 21
<210> 618
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 618
gctttgattg actccgttat t 21
<210> 619
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 619
gaaggtacaa taaatggctt a 21
<210> 620
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 620
cacacgtctt gctctcatta t 21
<210> 621
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 621
ccggcacacg tcttgctctc attatctcga gataatgaga gcaagacgtg tgtttttg 58
<210> 622
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 622
aattcaaaaa cacacgtctt gctctcatta tctcgagata atgagagcaa gacgtgtg 58
<210> 623
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 623
ctacaactca atgcaatctt t 21
<210> 624
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 624
ccggctacaa ctcaatgcaa tctttctcga gaaagattgc attgagttgt agtttttg 58
<210> 625
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 625
aattcaaaaa ctacaactca atgcaatctt tctcgagaaa gattgcattg agttgtag 58
<210> 626
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 626
ccagatatac tacaactcaa t 21
<210> 627
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 627
ccggccagat atactacaac tcaatctcga gattgagttg tagtatatct ggtttttg 58
<210> 628
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 628
aattcaaaaa ccagatatac tacaactcaa tctcgagatt gagttgtagt atatctgg 58
<210> 629
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 629
tgtatgaatg tctgctgggc a 21
<210> 630
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 630
ccggtgtatg aatgtctgct gggcactcga gtgcccagca gacattcata catttttg 58
<210> 631
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 631
aattcaaaaa tgtatgaatg tctgctgggc actcgagtgc ccagcagaca ttcataca 58
<210> 632
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 632
caaggacctg aaggagaaga a 21
<210> 633
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 633
ccggcaagga cctgaaggag aagaactcga gttcttctcc ttcaggtcct tgtttttg 58
<210> 634
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 634
aattcaaaaa caaggacctg aaggagaaga actcgagttc ttctccttca ggtccttg 58
<210> 635
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 635
caatgtctgt gggaggaaga a 21
<210> 636
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 636
ccggcaatgt ctgtgggagg aagaactcga gttcttcctc ccacagacat tgtttttg 58
<210> 637
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 637
aattcaaaaa caatgtctgt gggaggaaga actcgagttc ttcctcccac agacattg 58
<210> 638
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 638
caaggtcctg tagggagaag a 21
<210> 639
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 639
ccggcaaggt cctgtaggga gaagactcga gtcttctccc tacaggacct tgtttttg 58
<210> 640
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 640
aattcaaaaa caaggtcctg tagggagaag actcgagtct tctccctaca ggaccttg 58
<210> 641
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 641
caaggtcctg tagggagaag a 21
<210> 642
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 642
ccggcaaggt cctgtaggga gaagactcga gtcttctccc tacaggacct tgtttttg 58
<210> 643
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 643
aattcaaaaa caaggtcctg tagggagaag actcgagtct tctccctaca ggaccttg 58
<210> 644
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 644
acaagcccaa ggtgatcatt a 21
<210> 645
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 645
ccggacaagc ccaaggtgat cattactcga gtaatgatca ccttgggctt gttttttg 58
<210> 646
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 646
aattcaaaaa acaagcccaa ggtgatcatt actcgagtaa tgatcacctt gggcttgt 58
<210> 647
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 647
caaggacttg aaggagaaga a 21
<210> 648
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 648
ccggcaagga cttgaaggag aagaactcga gttcttctcc ttcaagtcct tgtttttg 58
<210> 649
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 649
aattcaaaaa caaggacttg aaggagaaga actcgagttc ttctccttca agtccttg 58
<210> 650
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 650
cccaagtttg aagtacaagt a 21
<210> 651
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 651
ccggcccaag tttgaagtac aagtactcga gtacttgtac ttcaaacttg ggtttttg 58
<210> 652
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 652
aattcaaaaa cccaagtttg aagtacaagt actcgagtac ttgtacttca aacttggg 58
<210> 653
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 653
cccaggacat gataataaga t 21
<210> 654
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 654
ccggcccagg acatgataat aagatctcga gatcttatta tcatgtcctg ggtttttg 58
<210> 655
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 655
aattcaaaaa cccaggacat gataataaga tctcgagatc ttattatcat gtcctggg 58
<210> 656
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 656
gaatttgaca gtttcctgcc a 21
<210> 657
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 657
ccgggaattt gacagtttcc tgccactcga gtggcaggaa actgtcaaat tctttttg 58
<210> 658
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 658
aattcaaaaa gaatttgaca gtttcctgcc actcgagtgg caggaaactg tcaaattc 58
<210> 659
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 659
cccaagtttg aggtcaaagt t 21
<210> 660
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 660
ccggcccaag tttgaggtca aagttctcga gaactttgac ctcaaacttg ggtttttg 58
<210> 661
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 661
aattcaaaaa cccaagtttg aggtcaaagt tctcgagaac tttgacctca aacttggg 58
<210> 662
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 662
cgacaagatg ttctccctca a 21
<210> 663
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 663
ccggcgacaa gatgttctcc ctcaactcga gttgagggag aacatcttgt cgtttttg 58
<210> 664
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 664
aattcaaaaa cgacaagatg ttctccctca actcgagttg agggagaaca tcttgtcg 58
<210> 665
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 665
aaaggttaat gtcccgctga a 21
<210> 666
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 666
agccactaag tcaagctgaa a 21
<210> 667
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 667
ccggagccac taagtcaagc tgaaactcga gtttcagctt gacttagtgg cttttttg 58
<210> 668
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 668
aattcaaaaa agccactaag tcaagctgaa actcgagttt cagcttgact tagtggct 58
<210> 669
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 669
ccaactggtc taagcagcat t 21
<210> 670
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 670
ccggccaact ggtctaagca gcattctcga gaatgctgct tagaccagtt ggtttttg 58
<210> 671
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 671
aattcaaaaa ccaactggtc taagcagcat tctcgagaat gctgcttaga ccagttgg 58
<210> 672
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 672
gaaacgagga cacaatgaaa t 21
<210> 673
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 673
ccgggaaacg aggacacaat gaaatctcga gatttcattg tgtcctcgtt tctttttg 58
<210> 674
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 674
aattcaaaaa gaaacgagga cacaatgaaa tctcgagatt tcattgtgtc ctcgtttc 58
<210> 675
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 675
gccactaagt caagctgaaa t 21
<210> 676
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 676
ccgggccact aagtcaagct gaaatctcga gatttcagct tgacttagtg gctttttg 58
<210> 677
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 677
aattcaaaaa gccactaagt caagctgaaa tctcgagatt tcagcttgac ttagtggc 58
<210> 678
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 678
ctggagttca tagcaccata a 21
<210> 679
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 679
ccggctggag ttcatagcac cataactcga gttatggtgc tatgaactcc agtttttg 58
<210> 680
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 680
aattcaaaaa ctggagttca tagcaccata actcgagtta tggtgctatg aactccag 58
<210> 681
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 681
cccgctgaac attatcagaa a 21
<210> 682
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 682
ccggcccgct gaacattatc agaaactcga gtttctgata atgttcagcg ggtttttg 58
<210> 683
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 683
aattcaaaaa cccgctgaac attatcagaa actcgagttt ctgataatgt tcagcggg 58
<210> 684
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 684
agccactaag tcaagctgaa a 21
<210> 685
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 685
ccggagccac taagtcaagc tgaaactcga gtttcagctt gacttagtgg cttttttg 58
<210> 686
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 686
aattcaaaaa agccactaag tcaagctgaa actcgagttt cagcttgact tagtggct 58
<210> 687
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 687
ccaactggtc taagcagcat t 21
<210> 688
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 688
ccggccaact ggtctaagca gcattctcga gaatgctgct tagaccagtt ggtttttg 58
<210> 689
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 689
aattcaaaaa ccaactggtc taagcagcat tctcgagaat gctgcttaga ccagttgg 58
<210> 690
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 690
gaaacgagga cacaatgaaa t 21
<210> 691
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 691
ccgggaaacg aggacacaat gaaatctcga gatttcattg tgtcctcgtt tctttttg 58
<210> 692
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 692
aattcaaaaa gaaacgagga cacaatgaaa tctcgagatt tcattgtgtc ctcgtttc 58
<210> 693
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 693
gccactaagt caagctgaaa t 21
<210> 694
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 694
ccgggccact aagtcaagct gaaatctcga gatttcagct tgacttagtg gctttttg 58
<210> 695
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 695
aattcaaaaa gccactaagt caagctgaaa tctcgagatt tcagcttgac ttagtggc 58
<210> 696
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 696
ctggagttca tagcaccata a 21
<210> 697
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 697
ccggctggag ttcatagcac cataactcga gttatggtgc tatgaactcc agtttttg 58
<210> 698
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 698
aattcaaaaa ctggagttca tagcaccata actcgagtta tggtgctatg aactccag 58
<210> 699
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 699
cccgctgaac attatcagaa a 21
<210> 700
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 700
ccggcccgct gaacattatc agaaactcga gtttctgata atgttcagcg ggtttttg 58
<210> 701
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 701
aattcaaaaa cccgctgaac attatcagaa actcgagttt ctgataatgt tcagcggg 58
<210> 702
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 702
agccactaag tcaagctgaa a 21
<210> 703
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 703
ccggagccac taagtcaagc tgaaactcga gtttcagctt gacttagtgg cttttttg 58
<210> 704
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 704
aattcaaaaa agccactaag tcaagctgaa actcgagttt cagcttgact tagtggct 58
<210> 705
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 705
ccaactggtc taagcagcat t 21
<210> 706
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 706
ccggccaact ggtctaagca gcattctcga gaatgctgct tagaccagtt ggtttttg 58
<210> 707
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 707
aattcaaaaa ccaactggtc taagcagcat tctcgagaat gctgcttaga ccagttgg 58
<210> 708
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 708
gaaacgagga cacaatgaaa t 21
<210> 709
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 709
ccgggaaacg aggacacaat gaaatctcga gatttcattg tgtcctcgtt tctttttg 58
<210> 710
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 710
aattcaaaaa gaaacgagga cacaatgaaa tctcgagatt tcattgtgtc ctcgtttc 58
<210> 711
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 711
gccactaagt caagctgaaa t 21
<210> 712
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 712
ccgggccact aagtcaagct gaaatctcga gatttcagct tgacttagtg gctttttg 58
<210> 713
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 713
aattcaaaaa gccactaagt caagctgaaa tctcgagatt tcagcttgac ttagtggc 58
<210> 714
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 714
ctggagttca tagcaccata a 21
<210> 715
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 715
ccggctggag ttcatagcac cataactcga gttatggtgc tatgaactcc agtttttg 58
<210> 716
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 716
aattcaaaaa ctggagttca tagcaccata actcgagtta tggtgctatg aactccag 58
<210> 717
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 717
cccgctgaac attatcagaa a 21
<210> 718
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 718
ccggcccgct gaacattatc agaaactcga gtttctgata atgttcagcg ggtttttg 58
<210> 719
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 719
aattcaaaaa cccgctgaac attatcagaa actcgagttt ctgataatgt tcagcggg 58
<210> 720
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 720
agccactaag tcaagctgaa a 21
<210> 721
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 721
ccggagccac taagtcaagc tgaaactcga gtttcagctt gacttagtgg cttttttg 58
<210> 722
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 722
aattcaaaaa agccactaag tcaagctgaa actcgagttt cagcttgact tagtggct 58
<210> 723
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 723
ccaactggtc taagcagcat t 21
<210> 724
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 724
ccggccaact ggtctaagca gcattctcga gaatgctgct tagaccagtt ggtttttg 58
<210> 725
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 725
aattcaaaaa ccaactggtc taagcagcat tctcgagaat gctgcttaga ccagttgg 58
<210> 726
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 726
gaaacgagga cacaatgaaa t 21
<210> 727
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 727
ccgggaaacg aggacacaat gaaatctcga gatttcattg tgtcctcgtt tctttttg 58
<210> 728
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 728
aattcaaaaa gaaacgagga cacaatgaaa tctcgagatt tcattgtgtc ctcgtttc 58
<210> 729
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 729
gccactaagt caagctgaaa t 21
<210> 730
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 730
ccgggccact aagtcaagct gaaatctcga gatttcagct tgacttagtg gctttttg 58
<210> 731
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 731
aattcaaaaa gccactaagt caagctgaaa tctcgagatt tcagcttgac ttagtggc 58
<210> 732
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 732
ctggagttca tagcaccata a 21
<210> 733
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 733
ccggctggag ttcatagcac cataactcga gttatggtgc tatgaactcc agtttttg 58
<210> 734
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 734
aattcaaaaa ctggagttca tagcaccata actcgagtta tggtgctatg aactccag 58
<210> 735
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 735
cccgctgaac attatcagaa a 21
<210> 736
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 736
ccggcccgct gaacattatc agaaactcga gtttctgata atgttcagcg ggtttttg 58
<210> 737
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 737
aattcaaaaa cccgctgaac attatcagaa actcgagttt ctgataatgt tcagcggg 58
<210> 738
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 738
agccactaag tcaagctgaa a 21
<210> 739
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 739
ccggagccac taagtcaagc tgaaactcga gtttcagctt gacttagtgg cttttttg 58
<210> 740
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 740
aattcaaaaa agccactaag tcaagctgaa actcgagttt cagcttgact tagtggct 58
<210> 741
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 741
ccaactggtc taagcagcat t 21
<210> 742
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 742
ccggccaact ggtctaagca gcattctcga gaatgctgct tagaccagtt ggtttttg 58
<210> 743
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 743
gaaacgagga cacaatgaaa t 21
<210> 744
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 744
ccgggaaacg aggacacaat gaaatctcga gatttcattg tgtcctcgtt tctttttg 58
<210> 745
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 745
aattcaaaaa gaaacgagga cacaatgaaa tctcgagatt tcattgtgtc ctcgtttc 58
<210> 746
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 746
gccactaagt caagctgaaa t 21
<210> 747
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 747
ccgggccact aagtcaagct gaaatctcga gatttcagct tgacttagtg gctttttg 58
<210> 748
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 748
aattcaaaaa gccactaagt caagctgaaa tctcgagatt tcagcttgac ttagtggc 58
<210> 749
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 749
ctggagttca tagcaccata a 21
<210> 750
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 750
ccggctggag ttcatagcac cataactcga gttatggtgc tatgaactcc agtttttg 58
<210> 751
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 751
aattcaaaaa ctggagttca tagcaccata actcgagtta tggtgctatg aactccag 58
<210> 752
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 752
cccgctgaac attatcagaa a 21
<210> 753
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 753
ccggcccgct gaacattatc agaaactcga gtttctgata atgttcagcg ggtttttg 58
<210> 754
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 754
aattcaaaaa cccgctgaac attatcagaa actcgagttt ctgataatgt tcagcggg 58
<210> 755
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 755
gattgtttca acacaagagt a 21
<210> 756
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 756
ccgggattgt ttcaacacaa gagtactcga gtactcttgt gttgaaacaa tctttttg 58
<210> 757
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 757
aattcaaaaa gattgtttca acacaagagt actcgagtac tcttgtgttg aaacaatc 58
<210> 758
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 758
agccactaag tcaagctgaa a 21
<210> 759
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 759
ccggagccac taagtcaagc tgaaactcga gtttcagctt gacttagtgg cttttttg 58
<210> 760
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 760
aattcaaaaa agccactaag tcaagctgaa actcgagttt cagcttgact tagtggct 58
<210> 761
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 761
ccaactggtc taagcagcat t 21
<210> 762
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 762
ccggccaact ggtctaagca gcattctcga gaatgctgct tagaccagtt ggtttttg 58
<210> 763
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 763
aattcaaaaa ccaactggtc taagcagcat tctcgagaat gctgcttaga ccagttgg 58
<210> 764
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 764
gaaacgagga cacaatgaaa t 21
<210> 765
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 765
ccgggaaacg aggacacaat gaaatctcga gatttcattg tgtcctcgtt tctttttg 58
<210> 766
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 766
aattcaaaaa gaaacgagga cacaatgaaa tctcgagatt tcattgtgtc ctcgtttc 58
<210> 767
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 767
gccactaagt caagctgaaa t 21
<210> 768
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 768
ccgggccact aagtcaagct gaaatctcga gatttcagct tgacttagtg gctttttg 58
<210> 769
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 769
aattcaaaaa gccactaagt caagctgaaa tctcgagatt tcagcttgac ttagtggc 58
<210> 770
<400> 770
000
<210> 771
<400> 771
000
<210> 772
<400> 772
000
<210> 773
<400> 773
000
<210> 774
<400> 774
000
<210> 775
<400> 775
000
<210> 776
<400> 776
000
<210> 777
<400> 777
000
<210> 778
<400> 778
000
<210> 779
<400> 779
000
<210> 780
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 780
ctggagttca tagcaccata a 21
<210> 781
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 781
ccggctggag ttcatagcac cataactcga gttatggtgc tatgaactcc agtttttg 58
<210> 782
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 782
aattcaaaaa ctggagttca tagcaccata actcgagtta tggtgctatg aactccag 58
<210> 783
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 783
cccgctgaac attatcagaa a 21
<210> 784
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 784
ccggcccgct gaacattatc agaaactcga gtttctgata atgttcagcg ggtttttg 58
<210> 785
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 785
aattcaaaaa cccgctgaac attatcagaa actcgagttt ctgataatgt tcagcggg 58
<210> 786
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 786
agccactaag tcaagctgaa a 21
<210> 787
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 787
ccggagccac taagtcaagc tgaaactcga gtttcagctt gacttagtgg cttttttg 58
<210> 788
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 788
aattcaaaaa agccactaag tcaagctgaa actcgagttt cagcttgact tagtggct 58
<210> 789
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 789
ccaactggtc taagcagcat t 21
<210> 790
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 790
ccggccaact ggtctaagca gcattctcga gaatgctgct tagaccagtt ggtttttg 58
<210> 791
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 791
aattcaaaaa ccaactggtc taagcagcat tctcgagaat gctgcttaga ccagttgg 58
<210> 792
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 792
gaaacgagga cacaatgaaa t 21
<210> 793
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 793
ccgggaaacg aggacacaat gaaatctcga gatttcattg tgtcctcgtt tctttttg 58
<210> 794
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 794
aattcaaaaa gaaacgagga cacaatgaaa tctcgagatt tcattgtgtc ctcgtttc 58
<210> 795
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 795
gccactaagt caagctgaaa t 21
<210> 796
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 796
ccgggccact aagtcaagct gaaatctcga gatttcagct tgacttagtg gctttttg 58
<210> 797
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 797
aattcaaaaa gccactaagt caagctgaaa tctcgagatt tcagcttgac ttagtggc 58
<210> 798
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 798
ctggagttca tagcaccata a 21
<210> 799
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 799
ccggctggag ttcatagcac cataactcga gttatggtgc tatgaactcc agtttttg 58
<210> 800
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 800
aattcaaaaa ctggagttca tagcaccata actcgagtta tggtgctatg aactccag 58
<210> 801
<211> 21
<212> ДНК
<213> Homo sapiens
<400> 801
cccgctgaac attatcagaa a 21
<210> 802
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 802
ccggcccgct gaacattatc agaaactcga gtttctgata atgttcagcg ggtttttg 58
<210> 803
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 803
aattcaaaaa cccgctgaac attatcagaa actcgagttt ctgataatgt tcagcggg 58
<210> 804
<211> 6
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 804
ggttga 6
<210> 805
<211> 4
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 805
agtt 4
<210> 806
<211> 6
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 806
ggttgg 6
<210> 807
<211> 6
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 807
agttgg 6
<210> 808
<211> 6
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 808
agttga 6
<210> 809
<211> 6
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 809
rrttrr 6
<210> 810
<400> 810
000
<210> 811
<400> 811
000
<210> 812
<400> 812
000
<210> 813
<400> 813
000
<210> 814
<400> 814
000
<210> 815
<400> 815
000
<210> 816
<400> 816
000
<210> 817
<400> 817
000
<210> 818
<400> 818
000
<210> 819
<400> 819
000
<210> 820
<400> 820
000
<210> 821
<400> 821
000
<210> 822
<400> 822
000
<210> 823
<400> 823
000
<210> 824
<400> 824
000
<210> 825
<400> 825
000
<210> 826
<400> 826
000
<210> 827
<400> 827
000
<210> 828
<400> 828
000
<210> 829
<400> 829
000
<210> 830
<400> 830
000
<210> 831
<400> 831
000
<210> 832
<400> 832
000
<210> 833
<400> 833
000
<210> 834
<400> 834
000
<210> 835
<400> 835
000
<210> 836
<400> 836
000
<210> 837
<400> 837
000
<210> 838
<400> 838
000
<210> 839
<400> 839
000
<210> 840
<400> 840
000
<210> 841
<400> 841
000
<210> 842
<400> 842
000
<210> 843
<400> 843
000
<210> 844
<400> 844
000
<210> 845
<400> 845
000
<210> 846
<400> 846
000
<210> 847
<400> 847
000
<210> 848
<400> 848
000
<210> 849
<400> 849
000
<210> 850
<400> 850
000
<210> 851
<400> 851
000
<210> 852
<400> 852
000
<210> 853
<400> 853
000
<210> 854
<400> 854
000
<210> 855
<400> 855
000
<210> 856
<400> 856
000
<210> 857
<400> 857
000
<210> 858
<400> 858
000
<210> 859
<400> 859
000
<210> 860
<400> 860
000
<210> 861
<400> 861
000
<210> 862
<400> 862
000
<210> 863
<400> 863
000
<210> 864
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 864
tcctggaggg gttgt 15
<210> 865
<400> 865
000
<210> 866
<400> 866
000
<210> 867
<400> 867
000
<210> 868
<400> 868
000
<210> 869
<400> 869
000
<210> 870
<400> 870
000
<210> 871
<400> 871
000
<210> 872
<400> 872
000
<210> 873
<400> 873
000
<210> 874
<400> 874
000
<210> 875
<400> 875
000
<210> 876
<400> 876
000
<210> 877
<400> 877
000
<210> 878
<400> 878
000
<210> 879
<400> 879
000
<210> 880
<400> 880
000
<210> 881
<400> 881
000
<210> 882
<211> 131
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 882
Met Ala Ala Pro Arg Gly Arg Pro Lys Lys Asp Leu Thr Met Glu Asp
1 5 10 15
Leu Thr Ala Lys Ile Ser Gln Leu Thr Val Glu Asn Arg Glu Leu Arg
20 25 30
Lys Ala Leu Gly Ser Thr Ala Asp Pro Arg Asp Arg Pro Leu Thr Ala
35 40 45
Thr Glu Lys Glu Ala Gln Leu Thr Ala Thr Val Gly Ala Leu Ser Ala
50 55 60
Ala Ala Ala Lys Lys Ile Glu Ala Arg Val Arg Thr Ile Phe Ser Lys
65 70 75 80
Val Val Thr Gln Lys Gln Val Asp Asp Ala Leu Lys Gly Leu Ser Leu
85 90 95
Arg Ile Asp Val Cys Met Ser Asp Gly Gly Thr Ala Lys Pro Pro Pro
100 105 110
Gly Ala Asn Asn Arg Arg Arg Arg Gly Ala Ser Thr Thr Arg Ala Gly
115 120 125
Val Asp Asp
130
<210> 883
<211> 1129
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 883
Met Ala Pro Pro Gly Met Arg Leu Arg Ser Gly Arg Ser Thr Gly Ala
1 5 10 15
Pro Leu Thr Arg Gly Ser Cys Arg Lys Arg Asn Arg Ser Pro Glu Arg
20 25 30
Cys Asp Leu Gly Asp Asp Leu His Leu Gln Pro Arg Arg Lys His Val
35 40 45
Ala Asp Ser Val Asp Gly Arg Glu Cys Gly Pro His Thr Leu Pro Ile
50 55 60
Pro Gly Ser Pro Thr Val Phe Thr Ser Gly Leu Pro Ala Phe Val Ser
65 70 75 80
Ser Pro Thr Leu Pro Val Ala Pro Ile Pro Ser Pro Ala Pro Ala Thr
85 90 95
Pro Leu Pro Pro Pro Ala Leu Leu Pro Pro Val Thr Thr Ser Ser Ser
100 105 110
Pro Ile Pro Pro Ser His Pro Val Ser Pro Gly Thr Thr Asp Thr His
115 120 125
Ser Pro Ser Pro Ala Leu Pro Pro Thr Gln Ser Pro Glu Ser Ser Gln
130 135 140
Arg Pro Pro Leu Ser Ser Pro Thr Gly Arg Pro Asp Ser Ser Thr Pro
145 150 155 160
Met Arg Pro Pro Pro Ser Gln Gln Thr Thr Pro Pro His Ser Pro Thr
165 170 175
Thr Pro Pro Pro Glu Pro Pro Ser Lys Ser Ser Pro Asp Ser Leu Ala
180 185 190
Pro Ser Thr Leu Arg Ser Leu Arg Lys Arg Arg Leu Ser Ser Pro Gln
195 200 205
Gly Pro Ser Thr Leu Asn Pro Ile Cys Gln Ser Pro Pro Val Ser Pro
210 215 220
Pro Arg Cys Asp Phe Ala Asn Arg Ser Val Tyr Pro Pro Trp Ala Thr
225 230 235 240
Glu Ser Pro Ile Tyr Val Gly Ser Ser Ser Asp Gly Asp Thr Pro Pro
245 250 255
Arg Gln Pro Pro Thr Ser Pro Ile Ser Ile Gly Ser Ser Ser Pro Ser
260 265 270
Glu Gly Ser Trp Gly Asp Asp Thr Ala Met Leu Val Leu Leu Ala Glu
275 280 285
Ile Ala Glu Glu Ala Ser Lys Asn Glu Lys Glu Cys Ser Glu Asn Asn
290 295 300
Gln Ala Gly Glu Asp Asn Gly Asp Asn Glu Ile Ser Lys Glu Ser Gln
305 310 315 320
Val Asp Lys Asp Asp Asn Asp Asn Lys Asp Asp Glu Glu Glu Gln Glu
325 330 335
Thr Asp Glu Glu Asp Glu Glu Asp Asp Glu Glu Asp Asp Glu Glu Asp
340 345 350
Asp Glu Glu Asp Asp Glu Glu Asp Asp Glu Glu Asp Asp Glu Glu Asp
355 360 365
Asp Glu Glu Glu Asp Glu Glu Glu Asp Glu Glu Glu Asp Glu Glu Glu
370 375 380
Asp Glu Glu Glu Glu Glu Asp Glu Glu Asp Asp Asp Asp Glu Asp Asn
385 390 395 400
Glu Asp Glu Glu Asp Asp Glu Glu Glu Asp Lys Lys Glu Asp Glu Glu
405 410 415
Asp Gly Gly Asp Gly Asn Lys Thr Leu Ser Ile Gln Ser Ser Gln Gln
420 425 430
Gln Gln Glu Pro Gln Gln Gln Glu Pro Gln Gln Gln Glu Pro Gln Gln
435 440 445
Gln Glu Pro Gln Gln Gln Glu Pro Gln Gln Gln Glu Pro Gln Gln Gln
450 455 460
Glu Pro Gln Gln Gln Glu Pro Gln Gln Arg Glu Pro Gln Gln Arg Glu
465 470 475 480
Pro Gln Gln Arg Glu Pro Gln Gln Arg Glu Pro Gln Gln Arg Glu Pro
485 490 495
Gln Gln Arg Glu Pro Gln Gln Arg Glu Pro Gln Gln Arg Glu Pro Gln
500 505 510
Gln Arg Glu Pro Gln Gln Arg Glu Pro Gln Gln Arg Glu Pro Gln Gln
515 520 525
Arg Glu Pro Gln Gln Gln Glu Pro Gln Gln Gln Glu Pro Gln Gln Gln
530 535 540
Glu Pro Gln Gln Gln Glu Pro Gln Gln Gln Glu Pro Gln Gln Gln Glu
545 550 555 560
Pro Gln Gln Gln Glu Pro Gln Gln Gln Glu Pro Gln Gln Gln Glu Pro
565 570 575
Gln Gln Gln Glu Pro Gln Gln Gln Glu Pro Gln Gln Gln Glu Pro Gln
580 585 590
Gln Gln Asp Glu Gln Gln Gln Asp Glu Gln Gln Gln Asp Glu Gln Gln
595 600 605
Gln Asp Glu Gln Gln Gln Asp Glu Gln Gln Gln Asp Glu Gln Gln Gln
610 615 620
Asp Glu Gln Gln Gln Asp Glu Gln Glu Gln Gln Asp Glu Gln Gln Gln
625 630 635 640
Asp Glu Gln Gln Gln Gln Asp Glu Gln Glu Gln Gln Glu Glu Gln Glu
645 650 655
Gln Gln Glu Glu Gln Gln Gln Asp Glu Gln Gln Gln Asp Glu Gln Gln
660 665 670
Gln Asp Glu Gln Gln Gln Asp Glu Gln Glu Gln Gln Asp Glu Gln Gln
675 680 685
Gln Asp Glu Gln Gln Gln Gln Asp Glu Gln Glu Gln Gln Glu Glu Gln
690 695 700
Glu Gln Gln Glu Glu Gln Glu Gln Gln Glu Glu Gln Glu Gln Gln Glu
705 710 715 720
Glu Gln Glu Gln Glu Leu Glu Glu Gln Glu Gln Glu Leu Glu Glu Gln
725 730 735
Glu Gln Glu Leu Glu Glu Gln Glu Gln Glu Leu Glu Glu Gln Glu Gln
740 745 750
Glu Leu Glu Glu Gln Glu Gln Glu Leu Glu Glu Gln Glu Gln Glu Leu
755 760 765
Glu Glu Gln Glu Gln Glu Leu Glu Glu Gln Glu Gln Glu Leu Glu Glu
770 775 780
Gln Glu Gln Glu Leu Glu Glu Gln Glu Gln Glu Leu Glu Glu Gln Glu
785 790 795 800
Gln Glu Leu Glu Glu Gln Glu Gln Glu Leu Glu Glu Gln Glu Gln Glu
805 810 815
Leu Glu Glu Gln Glu Gln Glu Gln Glu Leu Glu Glu Val Glu Glu Gln
820 825 830
Glu Gln Glu Gln Glu Glu Gln Glu Leu Glu Glu Val Glu Glu Gln Glu
835 840 845
Gln Glu Gln Glu Glu Gln Glu Glu Gln Glu Leu Glu Glu Val Glu Glu
850 855 860
Gln Glu Glu Gln Glu Leu Glu Glu Val Glu Glu Gln Glu Glu Gln Glu
865 870 875 880
Leu Glu Glu Val Glu Glu Gln Glu Gln Gln Gly Val Glu Gln Gln Glu
885 890 895
Gln Glu Thr Val Glu Glu Pro Ile Ile Leu His Gly Ser Ser Ser Glu
900 905 910
Asp Glu Met Glu Val Asp Tyr Pro Val Val Ser Thr His Glu Gln Ile
915 920 925
Ala Ser Ser Pro Pro Gly Asp Asn Thr Pro Asp Asp Asp Pro Gln Pro
930 935 940
Gly Pro Ser Arg Glu Tyr Arg Tyr Val Leu Arg Thr Ser Pro Pro His
945 950 955 960
Arg Pro Gly Val Arg Met Arg Arg Val Pro Val Thr His Pro Lys Lys
965 970 975
Pro His Pro Arg Tyr Gln Gln Pro Pro Val Pro Tyr Arg Gln Ile Asp
980 985 990
Asp Cys Pro Ala Lys Ala Arg Pro Gln His Ile Phe Tyr Arg Arg Phe
995 1000 1005
Leu Gly Lys Asp Gly Arg Arg Asp Pro Lys Cys Gln Trp Lys Phe
1010 1015 1020
Ala Val Ile Phe Trp Gly Asn Asp Pro Tyr Gly Leu Lys Lys Leu
1025 1030 1035
Ser Gln Ala Phe Gln Phe Gly Gly Val Lys Ala Gly Pro Val Ser
1040 1045 1050
Cys Leu Pro His Pro Gly Pro Asp Gln Ser Pro Ile Thr Tyr Cys
1055 1060 1065
Val Tyr Val Tyr Cys Gln Asn Lys Asp Thr Ser Lys Lys Val Gln
1070 1075 1080
Met Ala Arg Leu Ala Trp Glu Ala Ser His Pro Leu Ala Gly Asn
1085 1090 1095
Leu Gln Ser Ser Ile Val Lys Phe Lys Lys Pro Leu Pro Leu Thr
1100 1105 1110
Gln Pro Gly Glu Asn Gln Gly Pro Gly Asp Ser Pro Gln Glu Met
1115 1120 1125
Thr
<210> 884
<211> 160
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 884
Met Ala Pro Pro Gly Met Arg Leu Arg Ser Gly Arg Ser Thr Gly Ala
1 5 10 15
Pro Leu Thr Arg Gly Ser Cys Arg Lys Arg Asn Arg Ser Pro Glu Arg
20 25 30
Cys Asp Leu Gly Asp Asp Leu His Leu Gln Pro Arg Arg Lys His Val
35 40 45
Ala Asp Ser Val Asp Gly Arg Glu Cys Gly Pro His Thr Leu Pro Ile
50 55 60
Pro Gly Ser Pro Thr Val Phe Thr Ser Gly Leu Pro Ala Phe Val Ser
65 70 75 80
Ser Pro Thr Leu Pro Val Ala Pro Ile Pro Ser Pro Ala Pro Ala Thr
85 90 95
Pro Leu Pro Pro Pro Ala Leu Leu Pro Pro Val Thr Thr Ser Ser Ser
100 105 110
Pro Ile Pro Pro Ser His Pro Val Ser Pro Gly Thr Thr Asp Thr His
115 120 125
Ser Pro Ser Pro Ala Leu Pro Pro Thr Gln Ser Pro Glu Ser Ser Gln
130 135 140
Arg Pro Pro Leu Ser Ser Pro Thr Gly Arg Pro Asp Ser Ser Thr Pro
145 150 155 160
<210> 885
<211> 84
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 885
ggccagtgtt gagaggcgga gacttgggca attgctggac gctgccctgg gcattgcact 60
tgtctcggtc tgacagtgcc ggcc 84
<210> 886
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 886
ctgggggctc caaagtgctg ttcgtgcagg tagtgtgatt acccaaccta ctgctgagct 60
agcacttccc gagcccccgg 80
<210> 887
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<220>
<221> модифицированное_основание
<222> (4)..(8)
<223> a, c, t, g, неизвестно или другое
<220>
<221> прочий_признак
<222> (4)..(8)
<223> Эта область может включать 3-5 нуклеотидов
<220>
<221> прочий_признак
<222> (9)..(13)
<223> Эта область может включать 3-5 нуклеотидов
<400> 887
cctnnnnngg gggrr 15
<210> 888
<211> 6
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 888
ttaggg 6
<210> 889
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 889
tcctggcggg gaagt 15
<210> 890
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 890
tcctggaggg gaagt 15
<210> 891
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 891
gggggggggg gggggggggg 20
<210> 892
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 892
ttagggttag ggttagggtt aggg 24
<210> 893
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 893
ctcctattgg gggtttccta t 21
<210> 894
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 894
tcctggaggg gttgt 15
<210> 895
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 895
cctggatggg aattcccatc cagg 24
<210> 896
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 896
ttcccatcca ggcctggatg ggaa 24
<210> 897
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 897
tgcttgcaag cttgcaagca 20
<210> 898
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 898
tcctggatgg gaagt 15
<210> 899
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 899
cctggatggg aa 12
<210> 900
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 900
cttaccgctg cacctggatg ggaa 24
<210> 901
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 901
cctggatggg aacttaccgc tgca 24
<210> 902
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 902
tgctcctgga ggggttgt 18
<210> 903
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 903
ggtggtggtg gttgtggtgg tggtgg 26
<210> 904
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 904
gaaggcccau auagagaaa 19
<210> 905
<211> 143
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 905
ttgcccactc cctctctgcg cgctcgctcg ctcggtgggg cctgcggacc aaaggtccgc 60
agacggcaga ggtctcctct gccggcccca ccgagcgagc gacgcgcgca gagagggagt 120
gggcaactcc atcactaggg taa 143
<210> 906
<211> 143
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 906
ttaccctagt gatggagttg cccactccct ctctgcgcgc gtcgctcgct cggtggggcc 60
ggcagaggag acctctgccg tctgcggacc tttggtccgc aggccccacc gagcgagcga 120
gcgcgcagag agggagtggg caa 143
<210> 907
<211> 141
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 907
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc t 141
<210> 908
<211> 141
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 908
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg gctttgcccg ggcggcctca gtgagcgagc 120
gagcgcgcag ctgcctgcag g 141
<210> 909
<211> 144
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 909
ttggccactc cctctatgcg cactcgctcg ctcggtgggg cctggcgacc aaaggtcgcc 60
agacggacgt gggtttccac gtccggcccc accgagcgag cgagtgcgca tagagggagt 120
ggccaactcc atcactagag gtat 144
<210> 910
<211> 144
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 910
atacctctag tgatggagtt ggccactccc tctatgcgca ctcgctcgct cggtggggcc 60
ggacgtggaa acccacgtcc gtctggcgac ctttggtcgc caggccccac cgagcgagcg 120
agtgcgcata gagggagtgg ccaa 144
<210> 911
<211> 127
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 911
ttggccactc cctctatgcg cgctcgctca ctcactcggc cctggagacc aaaggtctcc 60
agactgccgg cctctggccg gcagggccga gtgagtgagc gagcgcgcat agagggagtg 120
gccaact 127
<210> 912
<211> 127
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 912
agttggccac attagctatg cgcgctcgct cactcactcg gccctggaga ccaaaggtct 60
ccagactgcc ggcctctggc cggcagggcc gagtgagtga gcgagcgcgc atagagggag 120
tggccaa 127
<210> 913
<211> 166
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 913
tcccccctgt cgcgttcgct cgctcgctgg ctcgtttggg ggggcgacgg ccagagggcc 60
gtcgtctggc agctctttga gctgccaccc ccccaaacga gccagcgagc gagcgaacgc 120
gacagggggg agagtgccac actctcaagc aagggggttt tgtaag 166
<210> 914
<211> 166
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 914
cttacaaaac ccccttgctt gagagtgtgg cactctcccc cctgtcgcgt tcgctcgctc 60
gctggctcgt ttgggggggt ggcagctcaa agagctgcca gacgacggcc ctctggccgt 120
cgccccccca aacgagccag cgagcgagcg aacgcgacag ggggga 166
<210> 915
<211> 144
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 915
ttgcccactc cctctaatgc gcgctcgctc gctcggtggg gcctgcggac caaaggtccg 60
cagacggcag aggtctcctc tgccggcccc accgagcgag cgagcgcgca tagagggagt 120
gggcaactcc atcactaggg gtat 144
<210> 916
<211> 144
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 916
atacccctag tgatggagtt gcccactccc tctatgcgcg ctcgctcgct cggtggggcc 60
ggcagaggag acctctgccg tctgcggacc tttggtccgc aggccccacc gagcgagcga 120
gcgcgcatta gagggagtgg gcaa 144
<210> 917
<211> 120
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 917
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
cgcacgcccg ggtttcccgg gcggcctcag tgagcgagcg agcgcgcagc tgcctgcagg 120
<210> 918
<211> 122
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 918
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgacgcccg ggctttgccc gggcggcctc agtgagcgag cgagcgcgca gctgcctgca 120
gg 122
<210> 919
<211> 129
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 919
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg gcgcctcagt gagcgagcga gcgcgcagct 120
gcctgcagg 129
<210> 920
<211> 101
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 920
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ctttgcctca gtgagcgagc gagcgcgcag ctgcctgcag g 101
<210> 921
<211> 139
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 921
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgaca aagtcgcccg acgcccgggc tttgcccggg cggcctcagt gagcgagcga 120
gcgcgcagct gcctgcagg 139
<210> 922
<211> 137
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 922
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgaaa atcgcccgac gcccgggctt tgcccgggcg gcctcagtga gcgagcgagc 120
gcgcagctgc ctgcagg 137
<210> 923
<211> 135
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 923
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgaaa cgcccgacgc ccgggctttg cccgggcggc ctcagtgagc gagcgagcgc 120
gcagctgcct gcagg 135
<210> 924
<211> 133
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 924
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcaaag cccgacgccc gggctttgcc cgggcggcct cagtgagcga gcgagcgcgc 120
agctgcctgc agg 133
<210> 925
<211> 139
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 925
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg gtttcccggg cggcctcagt gagcgagcga 120
gcgcgcagct gcctgcagg 139
<210> 926
<211> 137
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 926
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg tttccgggcg gcctcagtga gcgagcgagc 120
gcgcagctgc ctgcagg 137
<210> 927
<211> 135
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 927
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgt ttcgggcggc ctcagtgagc gagcgagcgc 120
gcagctgcct gcagg 135
<210> 928
<211> 133
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 928
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgccctt tgggcggcct cagtgagcga gcgagcgcgc 120
agctgcctgc agg 133
<210> 929
<211> 131
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 929
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgccttt ggcggcctca gtgagcgagc gagcgcgcag 120
ctgcctgcag g 131
<210> 930
<211> 129
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 930
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgctttg cggcctcagt gagcgagcga gcgcgcagct 120
gcctgcagg 129
<210> 931
<211> 127
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 931
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgtttcg gcctcagtga gcgagcgagc gcgcagctgc 120
ctgcagg 127
<210> 932
<211> 122
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 932
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacggcctc agtgagcgag cgagcgcgca gctgcctgca 120
gg 122
<210> 933
<211> 130
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 933
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg gcggcctcag tgagcgagcg agcgcgcagc 120
tgcctgcagg 130
<210> 934
<211> 120
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 934
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggaaacc cgggcgtgcg 60
cctcagtgag cgagcgagcg cgcagagagg gagtggccaa ctccatcact aggggttcct 120
<210> 935
<211> 122
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 935
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgt cgggcgacct ttggtcgccc 60
ggcctcagtg agcgagcgag cgcgcagaga gggagtggcc aactccatca ctaggggttc 120
ct 122
<210> 936
<211> 122
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 936
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
ggcctcagtg agcgagcgag cgcgcagaga gggagtggcc aactccatca ctaggggttc 120
ct 122
<210> 937
<211> 129
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 937
cctgcaggca gctgcgcgct cgctcgctca ctgaggcgcc cgggcgtcgg gcgacctttg 60
gtcgcccggc ctcagtgagc gagcgagcgc gcagagaggg agtggccaac tccatcacta 120
ggggttcct 129
<210> 938
<211> 101
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 938
cctgcaggca gctgcgcgct cgctcgctca ctgaggcaaa gcctcagtga gcgagcgagc 60
gcgcagagag ggagtggcca actccatcac taggggttcc t 101
<210> 939
<211> 139
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 939
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacttt gtcgcccggc ctcagtgagc gagcgagcgc gcagagaggg agtggccaac 120
tccatcacta ggggttcct 139
<210> 940
<211> 137
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 940
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgatttt cgcccggcct cagtgagcga gcgagcgcgc agagagggag tggccaactc 120
catcactagg ggttcct 137
<210> 941
<211> 135
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 941
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgtttcg cccggcctca gtgagcgagc gagcgcgcag agagggagtg gccaactcca 120
tcactagggg ttcct 135
<210> 942
<211> 133
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 942
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggctttgcc cggcctcagt gagcgagcga gcgcgcagag agggagtggc caactccatc 120
actaggggtt cct 133
<210> 943
<211> 139
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 943
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggaaacc cgggcgtcgg 60
gcgacctttg gtcgcccggc ctcagtgagc gagcgagcgc gcagagaggg agtggccaac 120
tccatcacta ggggttcct 139
<210> 944
<211> 137
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 944
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccggaaaccg ggcgtcgggc 60
gacctttggt cgcccggcct cagtgagcga gcgagcgcgc agagagggag tggccaactc 120
catcactagg ggttcct 137
<210> 945
<211> 135
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 945
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgaaacggg cgtcgggcga 60
cctttggtcg cccggcctca gtgagcgagc gagcgcgcag agagggagtg gccaactcca 120
tcactagggg ttcct 135
<210> 946
<211> 133
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 946
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccaaagggcg tcgggcgacc 60
tttggtcgcc cggcctcagt gagcgagcga gcgcgcagag agggagtggc caactccatc 120
actaggggtt cct 133
<210> 947
<211> 131
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 947
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc caaaggcgtc gggcgacctt 60
tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca actccatcac 120
taggggttcc t 131
<210> 948
<211> 129
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 948
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc aaagcgtcgg gcgacctttg 60
gtcgcccggc ctcagtgagc gagcgagcgc gcagagaggg agtggccaac tccatcacta 120
ggggttcct 129
<210> 949
<211> 127
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 949
cctgcaggca gctgcgcgct cgctcgctca ctgaggccga aacgtcgggc gacctttggt 60
cgcccggcct cagtgagcga gcgagcgcgc agagagggag tggccaactc catcactagg 120
ggttcct 127
<210> 950
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 950
Met Asp Trp Thr Trp Arg Ile Leu Phe Leu Val Ala Ala Ala Thr Gly
1 5 10 15
Ala His Ser
<210> 951
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 951
Met Leu Pro Ser Gln Leu Ile Gly Phe Leu Leu Leu Trp Val Pro Ala
1 5 10 15
Ser Arg Gly
<210> 952
<211> 141
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 952
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg gctttgcccg ggcggcctca gtgagcgagc 120
gagcgcgcag ctgcctgcag g 141
<210> 953
<211> 141
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 953
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc t 141
<210> 954
<211> 130
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 954
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg gcggcctcag tgagcgagcg agcgcgcagc 120
tgcctgcagg 130
<210> 955
<211> 130
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 955
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcgtcg ggcgaccttt 60
ggtcgcccgg cctcagtgag cgagcgagcg cgcagagagg gagtggccaa ctccatcact 120
aggggttcct 130
<210> 956
<211> 143
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 956
ttgcccactc cctctctgcg cgctcgctcg ctcggtgggg cctgcggacc aaaggtccgc 60
agacggcaga ggtctcctct gccggcccca ccgagcgagc gacgcgcgca gagagggagt 120
gggcaactcc atcactaggg taa 143
<210> 957
<211> 144
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 957
ttggccactc cctctatgcg cactcgctcg ctcggtgggg cctggcgacc aaaggtcgcc 60
agacggacgt gggtttccac gtccggcccc accgagcgag cgagtgcgca tagagggagt 120
ggccaactcc atcactagag gtat 144
<210> 958
<211> 127
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 958
ttggccactc cctctatgcg cgctcgctca ctcactcggc cctggagacc aaaggtctcc 60
agactgccgg cctctggccg gcagggccga gtgagtgagc gagcgcgcat agagggagtg 120
gccaact 127
<210> 959
<211> 166
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 959
tcccccctgt cgcgttcgct cgctcgctgg ctcgtttggg ggggcgacgg ccagagggcc 60
gtcgtctggc agctctttga gctgccaccc ccccaaacga gccagcgagc gagcgaacgc 120
gacagggggg agagtgccac actctcaagc aagggggttt tgtaag 166
<210> 960
<211> 144
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 960
ttgcccactc cctctaatgc gcgctcgctc gctcggtggg gcctgcggac caaaggtccg 60
cagacggcag aggtctcctc tgccggcccc accgagcgag cgagcgcgca tagagggagt 120
gggcaactcc atcactaggg gtat 144
<210> 961
<211> 143
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 961
ttaccctagt gatggagttg cccactccct ctctgcgcgc gtcgctcgct cggtggggcc 60
ggcagaggag acctctgccg tctgcggacc tttggtccgc aggccccacc gagcgagcga 120
gcgcgcagag agggagtggg caa 143
<210> 962
<211> 144
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 962
atacctctag tgatggagtt ggccactccc tctatgcgca ctcgctcgct cggtggggcc 60
ggacgtggaa acccacgtcc gtctggcgac ctttggtcgc caggccccac cgagcgagcg 120
agtgcgcata gagggagtgg ccaa 144
<210> 963
<211> 127
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 963
agttggccac attagctatg cgcgctcgct cactcactcg gccctggaga ccaaaggtct 60
ccagactgcc ggcctctggc cggcagggcc gagtgagtga gcgagcgcgc atagagggag 120
tggccaa 127
<210> 964
<211> 166
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 964
cttacaaaac ccccttgctt gagagtgtgg cactctcccc cctgtcgcgt tcgctcgctc 60
gctggctcgt ttgggggggt ggcagctcaa agagctgcca gacgacggcc ctctggccgt 120
cgccccccca aacgagccag cgagcgagcg aacgcgacag ggggga 166
<210> 965
<211> 144
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 965
atacccctag tgatggagtt gcccactccc tctatgcgcg ctcgctcgct cggtggggcc 60
ggcagaggag acctctgccg tctgcggacc tttggtccgc aggccccacc gagcgagcga 120
gcgcgcatta gagggagtgg gcaa 144
<210> 966
<211> 120
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 966
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
cgcacgcccg ggtttcccgg gcggcctcag tgagcgagcg agcgcgcagc tgcctgcagg 120
<210> 967
<211> 122
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 967
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgacgcccg ggctttgccc gggcggcctc agtgagcgag cgagcgcgca gctgcctgca 120
gg 122
<210> 968
<211> 129
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 968
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg gcgcctcagt gagcgagcga gcgcgcagct 120
gcctgcagg 129
<210> 969
<211> 101
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 969
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ctttgcctca gtgagcgagc gagcgcgcag ctgcctgcag g 101
<210> 970
<211> 139
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 970
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgaca aagtcgcccg acgcccgggc tttgcccggg cggcctcagt gagcgagcga 120
gcgcgcagct gcctgcagg 139
<210> 971
<211> 137
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 971
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgaaa atcgcccgac gcccgggctt tgcccgggcg gcctcagtga gcgagcgagc 120
gcgcagctgc ctgcagg 137
<210> 972
<211> 135
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 972
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgaaa cgcccgacgc ccgggctttg cccgggcggc ctcagtgagc gagcgagcgc 120
gcagctgcct gcagg 135
<210> 973
<211> 133
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 973
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcaaag cccgacgccc gggctttgcc cgggcggcct cagtgagcga gcgagcgcgc 120
agctgcctgc agg 133
<210> 974
<211> 139
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 974
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg gtttcccggg cggcctcagt gagcgagcga 120
gcgcgcagct gcctgcagg 139
<210> 975
<211> 137
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 975
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg tttccgggcg gcctcagtga gcgagcgagc 120
gcgcagctgc ctgcagg 137
<210> 976
<211> 135
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 976
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgt ttcgggcggc ctcagtgagc gagcgagcgc 120
gcagctgcct gcagg 135
<210> 977
<211> 133
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 977
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgccctt tgggcggcct cagtgagcga gcgagcgcgc 120
agctgcctgc agg 133
<210> 978
<211> 131
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 978
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgccttt ggcggcctca gtgagcgagc gagcgcgcag 120
ctgcctgcag g 131
<210> 979
<211> 129
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 979
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgctttg cggcctcagt gagcgagcga gcgcgcagct 120
gcctgcagg 129
<210> 980
<211> 127
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 980
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgtttcg gcctcagtga gcgagcgagc gcgcagctgc 120
ctgcagg 127
<210> 981
<211> 122
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 981
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacggcctc agtgagcgag cgagcgcgca gctgcctgca 120
gg 122
<210> 982
<211> 130
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 982
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg gcggcctcag tgagcgagcg agcgcgcagc 120
tgcctgcagg 130
<210> 983
<211> 120
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 983
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggaaacc cgggcgtgcg 60
cctcagtgag cgagcgagcg cgcagagagg gagtggccaa ctccatcact aggggttcct 120
<210> 984
<211> 122
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 984
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgt cgggcgacct ttggtcgccc 60
ggcctcagtg agcgagcgag cgcgcagaga gggagtggcc aactccatca ctaggggttc 120
ct 122
<210> 985
<211> 122
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 985
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
ggcctcagtg agcgagcgag cgcgcagaga gggagtggcc aactccatca ctaggggttc 120
ct 122
<210> 986
<211> 129
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 986
cctgcaggca gctgcgcgct cgctcgctca ctgaggcgcc cgggcgtcgg gcgacctttg 60
gtcgcccggc ctcagtgagc gagcgagcgc gcagagaggg agtggccaac tccatcacta 120
ggggttcct 129
<210> 987
<211> 101
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 987
cctgcaggca gctgcgcgct cgctcgctca ctgaggcaaa gcctcagtga gcgagcgagc 60
gcgcagagag ggagtggcca actccatcac taggggttcc t 101
<210> 988
<211> 139
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 988
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacttt gtcgcccggc ctcagtgagc gagcgagcgc gcagagaggg agtggccaac 120
tccatcacta ggggttcct 139
<210> 989
<211> 137
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 989
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgatttt cgcccggcct cagtgagcga gcgagcgcgc agagagggag tggccaactc 120
catcactagg ggttcct 137
<210> 990
<211> 135
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 990
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgtttcg cccggcctca gtgagcgagc gagcgcgcag agagggagtg gccaactcca 120
tcactagggg ttcct 135
<210> 991
<211> 133
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 991
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggctttgcc cggcctcagt gagcgagcga gcgcgcagag agggagtggc caactccatc 120
actaggggtt cct 133
<210> 992
<211> 139
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 992
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggaaacc cgggcgtcgg 60
gcgacctttg gtcgcccggc ctcagtgagc gagcgagcgc gcagagaggg agtggccaac 120
tccatcacta ggggttcct 139
<210> 993
<211> 137
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 993
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccggaaaccg ggcgtcgggc 60
gacctttggt cgcccggcct cagtgagcga gcgagcgcgc agagagggag tggccaactc 120
catcactagg ggttcct 137
<210> 994
<211> 135
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 994
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgaaacggg cgtcgggcga 60
cctttggtcg cccggcctca gtgagcgagc gagcgcgcag agagggagtg gccaactcca 120
tcactagggg ttcct 135
<210> 995
<211> 133
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 995
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccaaagggcg tcgggcgacc 60
tttggtcgcc cggcctcagt gagcgagcga gcgcgcagag agggagtggc caactccatc 120
actaggggtt cct 133
<210> 996
<211> 131
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 996
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc caaaggcgtc gggcgacctt 60
tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca actccatcac 120
taggggttcc t 131
<210> 997
<211> 129
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 997
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc aaagcgtcgg gcgacctttg 60
gtcgcccggc ctcagtgagc gagcgagcgc gcagagaggg agtggccaac tccatcacta 120
ggggttcct 129
<210> 998
<211> 127
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 998
cctgcaggca gctgcgcgct cgctcgctca ctgaggccga aacgtcgggc gacctttggt 60
cgcccggcct cagtgagcga gcgagcgcgc agagagggag tggccaactc catcactagg 120
ggttcct 127
<210> 999
<211> 122
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 999
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacggcctc agtgagcgag cgagcgcgca gctgcctgca 120
gg 122
<210> 1000
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический олигонуклеотид
<400> 1000
cgatcgttcg at 12
<210> 1001
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический олигонуклеотид
<400> 1001
atcgaaccat cg 12
<210> 1002
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1002
atcgaacgat cg 12
<210> 1003
<211> 165
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1003
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgcccgggc aaagcccggg cgtcgggcga cctttggtcg cccggcctca gtgagcgagc 120
gagcgcgcag agagggagtg gccaactcca tcactagggg ttcct 165
<210> 1004
<211> 140
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1004
cccctagtga tggagttggc cactccctct ctgcgcgctc gctcgctcac tgaggccgcc 60
cgggcaaagc ccgggcgtcg ggcgaccttt ggtcgcccgg cctcagtgag cgagcgagcg 120
cgcagagaga tcactagggg 140
<210> 1005
<211> 91
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1005
gcgcgctcgc tcgctcactg aggccgcccg ggcaaagccc gggcgtcggg cgacctttgg 60
tcgcccggcc tcagtgagcg agcgagcgcg c 91
<210> 1006
<211> 91
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1006
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cgggctttgc 60
ccgggcggcc tcagtgagcg agcgagcgcg c 91
<210> 1007
<211> 1662
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1007
gccgccacca tggaagacgc caaaaacata aagaaaggcc cggcgccatt ctatccgctg 60
gaagatggaa ccgctggaga gcaactgcat aaggctatga agagatacgc cctggttcct 120
ggaacaattg cttttacaga tgcacatatc gaggtggaca tcacttacgc tgagtacttc 180
gaaatgtccg ttcggttggc agaagctatg aaacgatatg ggctgaatac aaatcacaga 240
atcgtcgtat gcagtgaaaa ctctcttcaa ttctttatgc cggtgttggg cgcgttattt 300
atcggagttg cagttgcgcc cgcgaacgac atttataatg aacgtgaatt gctcaacagt 360
atgggcattt cgcagcctac cgtggtgttc gtttccaaaa aggggttgca aaaaattttg 420
aacgtgcaaa aaaagctccc aatcatccaa aaaattatta tcatggattc taaaacggat 480
taccagggat ttcagtcgat gtacacgttc gtcacatctc atctacctcc cggttttaat 540
gaatacgatt ttgtgccaga gtccttcgat agggacaaga caattgcact gatcatgaac 600
tcctctggat ctactggtct gcctaaaggt gtcgctctgc ctcatagaac tgcctgcgtg 660
agattctcgc atgccagaga tcctattttt ggcaatcaaa tcattccgga tactgcgatt 720
ttaagtgttg ttccattcca tcacggtttt ggaatgttta ctacactcgg atatttgata 780
tgtggatttc gagtcgtctt aatgtataga tttgaagaag agctgtttct gaggagcctt 840
caggattaca agattcaaag tgcgctgctg gtgccaaccc tattctcctt cttcgccaaa 900
agcactctga ttgacaaata cgatttatct aatttacacg aaattgcttc tggtggcgct 960
cccctctcta aggaagtcgg ggaagcggtt gccaagaggt tccatctgcc aggtatcagg 1020
caaggatatg ggctcactga gactacatca gctattctga ttacacccga gggggatgat 1080
aaaccgggcg cggtcggtaa agttgttcca ttttttgaag cgaaggttgt ggatctggat 1140
accgggaaaa cgctgggcgt taatcaaaga ggcgaactgt gtgtgagagg tcctatgatt 1200
atgtccggtt atgtaaacaa tccggaagcg accaacgcct tgattgacaa ggatggatgg 1260
ctacattctg gagacatagc ttactgggac gaagacgaac acttcttcat cgttgaccgc 1320
ctgaagtctc tgattaagta caaaggctat caggtggctc ccgctgaatt ggaatccatc 1380
ttgctccaac accccaacat cttcgacgca ggtgtcgcag gtcttcccga cgatgacgcc 1440
ggtgaacttc ccgccgccgt tgttgttttg gagcacggaa agacgatgac ggaaaaagag 1500
atcgtggatt acgtcgccag tcaagtaaca accgcgaaaa agttgcgcgg aggagttgtg 1560
tttgtggacg aagtaccgaa aggtcttacc ggaaaactcg acgcaagaaa aatcagagag 1620
atcctcataa aggccaagaa gggcggaaag atcgccgtgt aa 1662
<210> 1008
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<220>
<221> MOD_RES
<222> (1)..(1)
<223> Любая аминокислота
<400> 1008
Xaa Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ala Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Phe Asp Gly Thr Lys Lys Tyr Tyr Thr Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Thr Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Arg Gly Ile Gly Ala Arg Arg Gly Pro Tyr Tyr Met Asp
100 105 110
Val Trp Gly Lys Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly
450
<210> 1009
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полипептид
<400> 1009
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 1010
<211> 1310
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1010
ggagccgaga gtaattcata caaaaggagg gatcgccttc gcaaggggag agcccaggga 60
ccgtccctaa attctcacag acccaaatcc ctgtagccgc cccacgacag cgcgaggagc 120
atgcgctcag ggctgagcgc ggggagagca gagcacacaa gctcatagac cctggtcgtg 180
ggggggagga ccggggagct ggcgcggggc aaactgggaa agcggtgtcg tgtgctggct 240
ccgccctctt cccgagggtg ggggagaacg gtatataagt gcggcagtcg ccttggacgt 300
tctttttcgc aacgggtttg ccgtcagaac gcaggtgagg ggcgggtgtg gcttccgcgg 360
gccgccgagc tggaggtcct gctccgagcg ggccgggccc cgctgtcgtc ggcggggatt 420
agctgcgagc attcccgctt cgagttgcgg gcggcgcggg aggcagagtg cgaggcctag 480
cggcaacccc gtagcctcgc ctcgtgtccg gcttgaggcc tagcgtggtg tccgcgccgc 540
cgccgcgtgc tactccggcc gcactctggt cttttttttt tttgttgttg ttgccctgct 600
gccttcgatt gccgttcagc aataggggct aacaaaggga gggtgcgggg cttgctcgcc 660
cggagcccgg agaggtcatg gttggggagg aatggaggga caggagtggc ggctggggcc 720
cgcccgcctt cggagcacat gtccgacgcc acctggatgg ggcgaggcct ggggtttttc 780
ccgaagcaac caggctgggg ttagcgtgcc gaggccatgt ggccccagca cccggcacga 840
tctggcttgg cggcgccgcg ttgccctgcc tccctaacta gggtgaggcc atcccgtccg 900
gcaccagttg cgtgcgtgga aagatggccg ctcccgggcc ctgttgcaag gagctcaaaa 960
tggaggacgc ggcagcccgg tggagcgggc gggtgagtca cccacacaaa ggaagagggc 1020
ctggtccctc accggctgct gcttcctgtg accccgtggt cctatcggcc gcaatagtca 1080
cctcgggctt ttgagcacgg ctagtcgcgg cggggggagg ggatgtaatg gcgttggagt 1140
ttgttcacat ttggtgggtg gagactagtc aggccagcct ggcgctggaa gtcatttttg 1200
gaatttgtcc ccttgagttt tgagcggagc taattctcgg gcttcttagc ggttcaaagg 1260
tatcttttaa accctttttt aggtgttgtg aaaaccaccg ctaattcaaa 1310
<210> 1011
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1011
gcgcgctcgc tcgctc 16
<210> 1012
<211> 6
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1012
ggttga 6
<210> 1013
<211> 4
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1013
agtt 4
<210> 1014
<211> 6
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1014
ggttgg 6
<210> 1015
<211> 6
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1015
agttgg 6
<210> 1016
<211> 6
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1016
agttga 6
<210> 1017
<211> 6
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1017
rrttrr 6
<210> 1018
<211> 581
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1018
gagcatctta ccgccattta ttcccatatt tgttctgttt ttcttgattt gggtatacat 60
ttaaatgtta ataaaacaaa atggtggggc aatcatttac atttttaggg atatgtaatt 120
actagttcag gtgtattgcc acaagacaaa catgttaaga aactttcccg ttatttacgc 180
tctgttcctg ttaatcaacc tctggattac aaaatttgtg aaagattgac tgatattctt 240
aactatgttg ctccttttac gctgtgtgga tatgctgctt tatagcctct gtatctagct 300
attgcttccc gtacggcttt cgttttctcc tccttgtata aatcctggtt gctgtctctt 360
ttagaggagt tgtggcccgt tgtccgtcaa cgtggcgtgg tgtgctctgt gtttgctgac 420
gcaaccccca ctggctgggg cattgccacc acctgtcaac tcctttctgg gactttcgct 480
ttccccctcc cgatcgccac ggcagaactc atcgccgcct gccttgcccg ctgctggaca 540
ggggctaggt tgctgggcac tgataattcc gtggtgttgt c 581
<210> 1019
<211> 225
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1019
tgtgccttct agttgccagc catctgttgt ttgcccctcc cccgtgcctt ccttgaccct 60
ggaaggtgcc actcccactg tcctttccta ataaaatgag gaaattgcat cgcattgtct 120
gagtaggtgt cattctattc tggggggtgg ggtggggcag gacagcaagg gggaggattg 180
ggaagacaat agcaggcatg ctggggatgc ggtgggctct atggc 225
<210> 1020
<211> 8
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1020
actgaggc 8
<210> 1021
<211> 8
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1021
gcctcagt 8
<210> 1022
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1022
gagcgagcga gcgcgc 16
<210> 1023
<211> 1923
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1023
tcaatattgg ccattagcca tattattcat tggttatata gcataaatca atattggcta 60
ttggccattg catacgttgt atctatatca taatatgtac atttatattg gctcatgtcc 120
aatatgaccg ccatgttggc attgattatt gactagttat taatagtaat caattacggg 180
gtcattagtt catagcccat atatggagtt ccgcgttaca taacttacgg taaatggccc 240
gcctggctga ccgcccaacg acccccgccc attgacgtca ataatgacgt atgttcccat 300
agtaacgcca atagggactt tccattgacg tcaatgggtg gagtatttac ggtaaactgc 360
ccacttggca gtacatcaag tgtatcatat gccaagtccg ccccctattg acgtcaatga 420
cggtaaatgg cccgcctggc attatgccca gtacatgacc ttacgggact ttcctacttg 480
gcagtacatc tacgtattag tcatcgctat taccatggtc gaggtgagcc ccacgttctg 540
cttcactctc cccatctccc ccccctcccc acccccaatt ttgtatttat ttatttttta 600
attattttgt gcagcgatgg gggcgggggg gggggggggg cgcgcgccag gcggggcggg 660
gcggggcgag gggcggggcg gggcgaggcg gagaggtgcg gcggcagcca atcagagcgg 720
cgcgctccga aagtttcctt ttatggcgag gcggcggcgg cggcggccct ataaaaagcg 780
aagcgcgcgg cgggcgggag tcgctgcgac gctgccttcg ccccgtgccc cgctccgccg 840
ccgcctcgcg ccgcccgccc cggctctgac tgaccgcgtt actcccacag gtgagcgggc 900
gggacggccc ttctcctccg ggctgtaatt agcgcttggt ttaatgacgg cttgtttctt 960
ttctgtggct gcgtgaaagc cttgaggggc tccgggaggg ccctttgtgc gggggggagc 1020
ggctcggggg gtgcgtgcgt gtgtgtgtgc gtggggagcg ccgcgtgcgg cccgcgctgc 1080
ccggcggctg tgagcgctgc gggcgcggcg cggggctttg tgcgctccgc agtgtgcgcg 1140
aggggagcgc ggccgggggc ggtgccccgc ggtgcggggg gggctgcgag gggaacaaag 1200
gctgcgtgcg gggtgtgtgc gtgggggggt gagcaggggg tgtgggcgcg gcggtcgggc 1260
tgtaaccccc ccctgcaccc ccctccccga gttgctgagc acggcccggc ttcgggtgcg 1320
gggctccgta cggggcgtgg cgcggggctc gccgtgccgg gcggggggtg gcggcaggtg 1380
ggggtgccgg gcggggcggg gccgcctcgg gccggggagg gctcggggga ggggcgcggc 1440
ggcccccgga gcgccggcgg ctgtcgaggc gcggcgagcc gcagccattg ccttttatgg 1500
taatcgtgcg agagggcgca gggacttcct ttgtcccaaa tctgtgcgga gccgaaatct 1560
gggaggcgcc gccgcacccc ctctagcggg cgcggggcga agcggtgcgg cgccggcagg 1620
aaggaaatgg gcggggaggg ccttcgtgcg tcgccgcgcc gccgtcccct tctccctctc 1680
cagcctcggg gctgtccgcg gggggacggc tgccttcggg ggggacgggg cagggcgggg 1740
ttcggcttct ggcgtgtgac cggcggctct agagcctctg ctaaccatgt tttagccttc 1800
ttctttttcc tacagctcct gggcaacgtg ctggttattg tgctgtctca tcatttgtcg 1860
acagaattcc tcgaagatcc gaaggggttc aagcttggca ttccggtact gttggtaaag 1920
cca 1923
<210> 1024
<211> 1272
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1024
aggctcagag gcacacagga gtttctgggc tcaccctgcc cccttccaac ccctcagttc 60
ccatcctcca gcagctgttt gtgtgctgcc tctgaagtcc acactgaaca aacttcagcc 120
tactcatgtc cctaaaatgg gcaaacattg caagcagcaa acagcaaaca cacagccctc 180
cctgcctgct gaccttggag ctggggcaga ggtcagagac ctctctgggc ccatgccacc 240
tccaacatcc actcgacccc ttggaatttc ggtggagagg agcagaggtt gtcctggcgt 300
ggtttaggta gtgtgagagg gtccgggttc aaaaccactt gctgggtggg gagtcgtcag 360
taagtggcta tgccccgacc ccgaagcctg tttccccatc tgtacaatgg aaatgataaa 420
gacgcccatc tgatagggtt tttgtggcaa ataaacattt ggtttttttg ttttgttttg 480
ttttgttttt tgagatggag gtttgctctg tcgcccaggc tggagtgcag tgacacaatc 540
tcatctcacc acaaccttcc cctgcctcag cctcccaagt agctgggatt acaagcatgt 600
gccaccacac ctggctaatt ttctattttt agtagagacg ggtttctcca tgttggtcag 660
cctcagcctc ccaagtaact gggattacag gcctgtgcca ccacacccgg ctaatttttt 720
ctatttttga cagggacggg gtttcaccat gttggtcagg ctggtctaga ggtaccggat 780
cttgctacca gtggaacagc cactaaggat tctgcagtga gagcagaggg ccagctaagt 840
ggtactctcc cagagactgt ctgactcacg ccaccccctc caccttggac acaggacgct 900
gtggtttctg agccaggtac aatgactcct ttcggtaagt gcagtggaag ctgtacactg 960
cccaggcaaa gcgtccgggc agcgtaggcg ggcgactcag atcccagcca gtggacttag 1020
cccctgtttg ctcctccgat aactggggtg accttggtta atattcacca gcagcctccc 1080
ccgttgcccc tctggatcca ctgcttaaat acggacgagg acagggccct gtctcctcag 1140
cttcaggcac caccactgac ctgggacagt gaatccggac tctaaggtaa atataaaatt 1200
tttaagtgta taatgtgtta aactactgat tctaattgtt tctctctttt agattccaac 1260
ctttggaact ga 1272
<210> 1025
<211> 1177
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1025
ggctcagagg ctcagaggca cacaggagtt tctgggctca ccctgccccc ttccaacccc 60
tcagttccca tcctccagca gctgtttgtg tgctgcctct gaagtccaca ctgaacaaac 120
ttcagcctac tcatgtccct aaaatgggca aacattgcaa gcagcaaaca gcaaacacac 180
agccctccct gcctgctgac cttggagctg gggcagaggt cagagacctc tctgggccca 240
tgccacctcc aacatccact cgaccccttg gaatttcggt ggagaggagc agaggttgtc 300
ctggcgtggt ttaggtagtg tgagagggtc cgggttcaaa accacttgct gggtggggag 360
tcgtcagtaa gtggctatgc cccgaccccg aagcctgttt ccccatctgt acaatggaaa 420
tgataaagac gcccatctga tagggttttt gtggcaaata aacatttggt ttttttgttt 480
tgttttgttt tgttttttga gatggaggtt tgctctgtcg cccaggctgg agtgcagtga 540
cacaatctca tctcaccaca accttcccct gcctcagcct cccaagtagc tgggattaca 600
agcatgtgcc accacacctg gctaattttc tatttttagt agagacgggt ttctccatgt 660
tggtcagcct cagcctccca agtaactggg attacaggcc tgtgccacca cacccggcta 720
attttttcta tttttgacag ggacggggtt tcaccatgtt ggtcaggctg gtctagaggt 780
accggatctt gctaccagtg gaacagccac taaggattct gcagtgagag cagagggcca 840
gctaagtggt actctcccag agactgtctg actcacgcca ccccctccac cttggacaca 900
ggacgctgtg gtttctgagc caggtacaat gactcctttc ggtaagtgca gtggaagctg 960
tacactgccc aggcaaagcg tccgggcagc gtaggcgggc gactcagatc ccagccagtg 1020
gacttagccc ctgtttgctc ctccgataac tggggtgacc ttggttaata ttcaccagca 1080
gcctcccccg ttgcccctct ggatccactg cttaaatacg gacgaggaca gggccctgtc 1140
tcctcagctt caggcaccac cactgacctg ggacagt 1177
<210> 1026
<211> 547
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1026
ccctaaaatg ggcaaacatt gcaagcagca aacagcaaac acacagccct ccctgcctgc 60
tgaccttgga gctggggcag aggtcagaga cctctctggg cccatgccac ctccaacatc 120
cactcgaccc cttggaattt ttcggtggag aggagcagag gttgtcctgg cgtggtttag 180
gtagtgtgag aggggaatga ctcctttcgg taagtgcagt ggaagctgta cactgcccag 240
gcaaagcgtc cgggcagcgt aggcgggcga ctcagatccc agccagtgga cttagcccct 300
gtttgctcct ccgataactg gggtgacctt ggttaatatt caccagcagc ctcccccgtt 360
gcccctctgg atccactgct taaatacgga cgaggacagg gccctgtctc ctcagcttca 420
ggcaccacca ctgacctggg acagtgaatc cggactctaa ggtaaatata aaatttttaa 480
gtgtataatg tgttaaacta ctgattctaa ttgtttctct cttttagatt ccaacctttg 540
gaactga 547
<210> 1027
<211> 556
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1027
ccctaaaatg ggcaaacatt gcaagcagca aacagcaaac acacagccct ccctgcctgc 60
tgaccttgga gctggggcag aggtcagaga cctctctggg cccatgccac ctccaacatc 120
cactcgaccc cttggaattt cggtggagag gagcagaggt tgtcctggcg tggtttaggt 180
agtgtgagag gggaatgact cctttcggta agtgcagtgg aagctgtaca ctgcccaggc 240
aaagcgtccg ggcagcgtag gcgggcgact cagatcccag ccagtggact tagcccctgt 300
ttgctcctcc gataactggg gtgaccttgg ttaatattca ccagcagcct cccccgttgc 360
ccctctggat ccactgctta aatacggacg aggacactcg agggccctgt ctcctcagct 420
tcaggcacca ccactgacct gggacagtga atccggacat cgattctaag gtaaatataa 480
aatttttaag tgtataattt gttaaactac tgattctaat tgtttctctc ttttagattc 540
caacctttgg aactga 556
<210> 1028
<211> 1179
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1028
ggctccggtg cccgtcagtg ggcagagcgc acatcgccca cagtccccga gaagttgggg 60
ggaggggtcg gcaattgaac cggtgcctag agaaggtggc gcggggtaaa ctgggaaagt 120
gatgtcgtgt actggctccg cctttttccc gagggtgggg gagaaccgta tataagtgca 180
gtagtcgccg tgaacgttct ttttcgcaac gggtttgccg ccagaacaca ggtaagtgcc 240
gtgtgtggtt cccgcgggcc tggcctcttt acgggttatg gcccttgcgt gccttgaatt 300
acttccacct ggctgcagta cgtgattctt gatcccgagc ttcgggttgg aagtgggtgg 360
gagagttcga ggccttgcgc ttaaggagcc ccttcgcctc gtgcttgagt tgaggcctgg 420
cctgggcgct ggggccgccg cgtgcgaatc tggtggcacc ttcgcgcctg tctcgctgct 480
ttcgataagt ctctagccat ttaaaatttt tgatgacctg ctgcgacgct ttttttctgg 540
caagatagtc ttgtaaatgc gggccaagat ctgcacactg gtatttcggt ttttggggcc 600
gcgggcggcg acggggcccg tgcgtcccag cgcacatgtt cggcgaggcg gggcctgcga 660
gcgcggccac cgagaatcgg acgggggtag tctcaagctg gccggcctgc tctggtgcct 720
ggtctcgcgc cgccgtgtat cgccccgccc tgggcggcaa ggctggcccg gtcggcacca 780
gttgcgtgag cggaaagatg gccgcttccc ggccctgctg cagggagctc aaaatggagg 840
acgcggcgct cgggagagcg ggcgggtgag tcacccacac aaaggaaaag ggcctttccg 900
tcctcagccg tcgcttcatg tgactccacg gagtaccggg cgccgtccag gcacctcgat 960
tagttctcga gcttttggag tacgtcgtct ttaggttggg gggaggggtt ttatgcgatg 1020
gagtttcccc acactgagtg ggtggagact gaagttaggc cagcttggca cttgatgtaa 1080
ttctccttgg aatttgccct ttttgagttt ggatcttggt tcattctcaa gcctcagaca 1140
gtggttcaaa gtttttttct tccatttcag gtgtcgtga 1179
<210> 1029
<211> 141
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1029
aataaacgat aacgccgttg gtggcgtgag gcatgtaaaa ggttacatca ttatcttgtt 60
cgccatccgg ttggtataaa tagacgttca tgttggtttt tgtttcagtt gcaagttggc 120
tgcggcgcgc gcagcacctt t 141
<210> 1030
<211> 317
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1030
ggtgtggaaa gtccccaggc tccccagcag gcagaagtat gcaaagcatg catctcaatt 60
agtcagcaac caggtgtgga aagtccccag gctccccagc aggcagaagt atgcaaagca 120
tgcatctcaa ttagtcagca accatagtcc cgcccctaac tccgcccatc ccgcccctaa 180
ctccgcccag ttccgcccat tctccgcccc atggctgact aatttttttt atttatgcag 240
aggccgaggc cgcctcggcc tctgagctat tccagaagta gtgaggaggc ttttttggag 300
gcctaggctt ttgcaaa 317
<210> 1031
<211> 241
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1031
gagggcctat ttcccatgat tccttcatat ttgcatatac gatacaaggc tgttagagag 60
ataattggaa ttaatttgac tgtaaacaca aagatattag tacaaaatac gtgacgtaga 120
aagtaataat ttcttgggta gtttgcagtt ttaaaattat gttttaaaat ggactatcat 180
atgcttaccg taacttgaaa gtatttcgat ttcttggctt tatatatctt gtggaaagga 240
c 241
<210> 1032
<211> 215
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1032
gaacgctgac gtcatcaacc cgctccaagg aatcgcgggc ccagtgtcac taggcgggaa 60
cacccagcgc gcgtgcgccc tggcaggaag atggctgtga gggacagggg agtggcgccc 120
tgcaatattt gcatgtcgct atgtgttctg ggaaatcacc ataaacgtga aatgtctttg 180
gatttgggaa tcgtataaga actgtatgag accac 215
<210> 1033
<211> 546
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1033
ccctaaaatg ggcaaacatt gcaagcagca aacagcaaac acacagccct ccctgcctgc 60
tgaccttgga gctggggcag aggtcagaga cctctctggg cccatgccac ctccaacatc 120
cactcgaccc cttggaattt ttcggtggag aggagcagag gttgtcctgg cgtggtttag 180
gtagtgtgag aggggaatga ctcctttcgg taagtgcagt ggaagctgta cactgcccag 240
gcaaagcgtc cgggcagcgt aggcgggcga ctcagatccc agccagtgga cttagcccct 300
gtttgctcct ccgataactg gggtgacctt ggttaatatt caccagcagc ctcccccgtt 360
gcccctctgg atccactgct taaatacgga cgaggacagg gccctgtctc ctcagcttca 420
ggcaccacca ctgacctggg acagtgaatc cggactctaa ggtaaatata aaatttttaa 480
gtgtataatg tgttaaacta ctgattctaa ttgtttctct cttttagatt ccaacctttg 540
gaactg 546
<210> 1034
<211> 576
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1034
tagtaatcaa ttacggggtc attagttcat agcccatata tggagttccg cgttacataa 60
cttacggtaa atggcccgcc tggctgaccg cccaacgacc cccgcccatt gacgtcaata 120
atgacgtatg ttcccatagt aacgccaata gggactttcc attgacgtca atgggtggag 180
tatttacggt aaactgccca cttggcagta catcaagtgt atcatatgcc aagtacgccc 240
cctattgacg tcaatgacgg taaatggccc gcctggcatt atgcccagta catgacctta 300
tgggactttc ctacttggca gtacatctac gtattagtca tcgctattac catggtgatg 360
cggttttggc agtacatcaa tgggcgtgga tagcggtttg actcacgggg atttccaagt 420
ctccacccca ttgacgtcaa tgggagtttg ttttggcacc aaaatcaacg ggactttcca 480
aaatgtcgta acaactccgc cccattgacg caaatgggcg gtaggcgtgt acggtgggag 540
gtctatataa gcagagctgg tttagtgaac cgtcag 576
<210> 1035
<211> 150
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1035
ataaacgata acgccgttgg tggcgtgagg catgtaaaag gttacatcat tatcttgttc 60
gccatccggt tggtataaat agacgttcat gttggttttt gtttcagttg caagttggct 120
gcggcgcgcg cagcaccttt gcggccatct 150
<210> 1036
<211> 1313
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1036
ggagccgaga gtaattcata caaaaggagg gatcgccttc gcaaggggag agcccaggga 60
ccgtccctaa attctcacag acccaaatcc ctgtagccgc cccacgacag cgcgaggagc 120
atgcgcccag ggctgagcgc gggtagatca gagcacacaa gctcacagtc cccggcggtg 180
gggggagggg cgcgctgagc gggggccagg gagctggcgc ggggcaaact gggaaagtgg 240
tgtcgtgtgc tggctccgcc ctcttcccga gggtggggga gaacggtata taagtgcggt 300
agtcgccttg gacgttcttt ttcgcaacgg gtttgccgtc agaacgcagg tgagtggcgg 360
gtgtggcttc cgcgggcccc ggagctggag ccctgctctg agcgggccgg gctgatatgc 420
gagtgtcgtc cgcagggttt agctgtgagc attcccactt cgagtggcgg gcggtgcggg 480
ggtgagagtg cgaggcctag cggcaacccc gtagcctcgc ctcgtgtccg gcttgaggcc 540
tagcgtggtg tccgccgccg cgtgccactc cggccgcact atgcgttttt tgtccttgct 600
gccctcgatt gccttccagc agcatgggct aacaaaggga gggtgtgggg ctcactctta 660
aggagcccat gaagcttacg ttggatagga atggaagggc aggaggggcg actggggccc 720
gcccgccttc ggagcacatg tccgacgcca cctggatggg gcgaggcctg tggctttccg 780
aagcaatcgg gcgtgagttt agcctacctg ggccatgtgg ccctagcact gggcacggtc 840
tggcctggcg gtgccgcgtt cccttgcctc ccaacaaggg tgaggccgtc ccgcccggca 900
ccagttgctt gcgcggaaag atggccgctc ccggggccct gttgcaagga gctcaaaatg 960
gaggacgcgg cagcccggtg gagcgggcgg gtgagtcacc cacacaaagg aagagggcct 1020
tgcccctcgc cggccgctgc ttcctgtgac cccgtggtct atcggccgca tagtcacctc 1080
gggcttctct tgagcaccgc tcgtcgcggc ggggggaggg gatctaatgg cgttggagtt 1140
tgttcacatt tggtgggtgg agactagtca ggccagcctg gcgctggaag tcattcttgg 1200
aatttgcccc tttgagtttg gagcgaggct aattctcaag cctcttagcg gttcaaaggt 1260
attttctaaa cccgtttcca ggtgttgtga aagccaccgc taattcaaag caa 1313
<210> 1037
<211> 213
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1037
taagatacat tgatgagttt ggacaaacca caactagaat gcagtgaaaa aaatgcttta 60
tttgtgaaat ttgtgatgct attgctttat ttgtaaccat tataagctgc aataaacaag 120
ttaacaacaa caattgcatt cattttatgt ttcaggttca gggggaggtg tgggaggttt 180
tttaaagcaa gtaaaacctc tacaaatgtg gta 213
<210> 1038
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 1038
Pro Lys Lys Lys Arg Lys Val
1 5
<210> 1039
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 1039
Met Asp Trp Thr Trp Arg Ile Leu Phe Leu Val Ala Ala Ala Thr Gly
1 5 10 15
Ala His Ser
<210> 1040
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 1040
Met Leu Pro Ser Gln Leu Ile Gly Phe Leu Leu Leu Trp Val Pro Ala
1 5 10 15
Ser Arg Gly
<210> 1041
<211> 7
<212> БЕЛОК
<213> Обезьяний вирус 40
<400> 1041
Pro Lys Lys Lys Arg Lys Val
1 5
<210> 1042
<211> 21
<212> ДНК
<213> Обезьяний вирус 40
<400> 1042
cccaagaaga agaggaaggt g 21
<210> 1043
<211> 16
<212> БЕЛОК
<213> Неизвестный
<220>
<223> Описание неизвестного:
Последовательность двойного NLS нуклеоплазмина
<400> 1043
Lys Arg Pro Ala Ala Thr Lys Lys Ala Gly Gln Ala Lys Lys Lys Lys
1 5 10 15
<210> 1044
<211> 9
<212> БЕЛОК
<213> Неизвестный
<220>
<223> Описание неизвестного:
Последовательность C-myc NLS
<400> 1044
Pro Ala Ala Lys Arg Val Lys Leu Asp
1 5
<210> 1045
<211> 11
<212> БЕЛОК
<213> Неизвестный
<220>
<223> Описание неизвестного:
Последовательность C-myc NLS
<400> 1045
Arg Gln Arg Arg Asn Glu Leu Lys Arg Ser Pro
1 5 10
<210> 1046
<211> 38
<212> БЕЛОК
<213> Homo sapiens
<400> 1046
Asn Gln Ser Ser Asn Phe Gly Pro Met Lys Gly Gly Asn Phe Gly Gly
1 5 10 15
Arg Ser Ser Gly Pro Tyr Gly Gly Gly Gly Gln Tyr Phe Ala Lys Pro
20 25 30
Arg Asn Gln Gly Gly Tyr
35
<210> 1047
<211> 42
<212> БЕЛОК
<213> Неизвестный
<220>
<223> Описание неизвестного:
Домен IBB из последовательности импортина-альфа
<400> 1047
Arg Met Arg Ile Glx Phe Lys Asn Lys Gly Lys Asp Thr Ala Glu Leu
1 5 10 15
Arg Arg Arg Arg Val Glu Val Ser Val Glu Leu Arg Lys Ala Lys Lys
20 25 30
Asp Glu Gln Ile Leu Lys Arg Arg Asn Val
35 40
<210> 1048
<211> 8
<212> БЕЛОК
<213> Неизвестный
<220>
<223> Описание неизвестного:
Последовательность белка Т миомы
<400> 1048
Val Ser Arg Lys Arg Pro Arg Pro
1 5
<210> 1049
<211> 8
<212> БЕЛОК
<213> Неизвестный
<220>
<223> Описание неизвестного:
Последовательность белка Т миомы
<400> 1049
Pro Pro Lys Lys Ala Arg Glu Asp
1 5
<210> 1050
<211> 8
<212> БЕЛОК
<213> Homo sapiens
<400> 1050
Pro Gln Pro Lys Lys Lys Pro Leu
1 5
<210> 1051
<211> 12
<212> БЕЛОК
<213> Mus musculus
<400> 1051
Ser Ala Leu Ile Lys Lys Lys Lys Lys Met Ala Pro
1 5 10
<210> 1052
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1052
gcgcgctcgc tcgctcactg aggccgcccg ggaaacccgg gcgtgcgcct cagtgagcga 60
gcgagcgcgc 70
<210> 1053
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1053
gcgcgctcgc tcgctcactg aggcgcacgc ccgggtttcc cgggcggcct cagtgagcga 60
gcgagcgcgc 70
<210> 1054
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1054
gcgcgctcgc tcgctcactg aggccgtcgg gcgacctttg gtcgcccggc ctcagtgagc 60
gagcgagcgc gc 72
<210> 1055
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1055
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacggc ctcagtgagc 60
gagcgagcgc gc 72
<210> 1056
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1056
gcgcgctcgc tcgctcactg aggccgcccg ggcaaagccc gggcgtcggc ctcagtgagc 60
gagcgagcgc gc 72
<210> 1057
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1057
gcgcgctcgc tcgctcactg aggccgacgc ccgggctttg cccgggcggc ctcagtgagc 60
gagcgagcgc gc 72
<210> 1058
<211> 83
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1058
gcgcgctcgc tcgctcactg aggccgcccg ggcaaagccc gggcgtcggg ctttgcccgg 60
cctcagtgag cgagcgagcg cgc 83
<210> 1059
<211> 83
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1059
gcgcgctcgc tcgctcactg aggccgggca aagcccgacg cccgggcttt gcccgggcgg 60
cctcagtgag cgagcgagcg cgc 83
<210> 1060
<211> 77
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1060
gcgcgctcgc tcgctcactg aggccgaaac gtcgggcgac ctttggtcgc ccggcctcag 60
tgagcgagcg agcgcgc 77
<210> 1061
<211> 77
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1061
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgtt tcggcctcag 60
tgagcgagcg agcgcgc 77
<210> 1062
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1062
gcgcgctcgc tcgctcactg aggcaaagcc tcagtgagcg agcgagcgcg c 51
<210> 1063
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1063
gcgcgctcgc tcgctcactg aggctttgcc tcagtgagcg agcgagcgcg c 51
<210> 1064
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1064
gcgcgctcgc tcgctcactg aggccgcccg ggcgtcgggc gacctttggt cgcccggcct 60
cagtgagcga gcgagcgcgc 80
<210> 1065
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1065
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 1066
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1066
gcgcgctcgc tcgctcactg aggcgcccgg gcgtcgggcg acctttggtc gcccggcctc 60
agtgagcgag cgagcgcgc 79
<210> 1067
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1067
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cgggcgcctc 60
agtgagcgag cgagcgcgc 79
<210> 1068
<211> 5
<212> БЕЛОК
<213> Вирус гриппа
<400> 1068
Asp Arg Leu Arg Arg
1 5
<210> 1069
<211> 7
<212> БЕЛОК
<213> Вирус гриппа
<400> 1069
Pro Lys Gln Lys Lys Arg Lys
1 5
<210> 1070
<211> 10
<212> БЕЛОК
<213> Вирус гепатита дельта
<400> 1070
Arg Lys Leu Lys Lys Lys Ile Lys Lys Leu
1 5 10
<210> 1071
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 1071
Arg Glu Lys Lys Lys Phe Leu Lys Arg Arg
1 5 10
<210> 1072
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 1072
Lys Arg Lys Gly Asp Glu Val Asp Gly Val Asp Glu Val Ala Lys Lys
1 5 10 15
Lys Ser Lys Lys
20
<210> 1073
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 1073
Arg Lys Cys Leu Gln Ala Gly Met Asn Leu Glu Ala Arg Lys Thr Lys
1 5 10 15
Lys
<210> 1074
<211> 8
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1074
gtttaaac 8
<210> 1075
<211> 8
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1075
ttaattaa 8
<210> 1076
<211> 141
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1076
aataaacgat aacgccgttg gtggcgtgag gcatgtaaaa ggttacatca ttatcttgtt 60
cgccatccgg ttggtataaa tagacgttca tgttggtttt tgtttcagtt gcaagttggc 120
tgcggcgcgc gcagcacctt t 141
<210> 1077
<211> 317
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1077
ggtgtggaaa gtccccaggc tccccagcag gcagaagtat gcaaagcatg catctcaatt 60
agtcagcaac caggtgtgga aagtccccag gctccccagc aggcagaagt atgcaaagca 120
tgcatctcaa ttagtcagca accatagtcc cgcccctaac tccgcccatc ccgcccctaa 180
ctccgcccag ttccgcccat tctccgcccc atggctgact aatttttttt atttatgcag 240
aggccgaggc cgcctcggcc tctgagctat tccagaagta gtgaggaggc ttttttggag 300
gcctaggctt ttgcaaa 317
<210> 1078
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1078
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacggc ctcagtgagc 60
gagcgagcgc gc 72
<210> 1079
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1079
gagacagaca cactcctgct atgggtactg ctgctctggg ttccaggttc cactggtgac 60
<210> 1080
<211> 1260
<212> ДНК
<213> Аденоассоциированный вирус - 2
<400> 1080
atggagctgg tcgggtggct cgtggacaag gggattacct cggagaagca gtggatccag 60
gaggaccagg cctcatacat ctccttcaat gcggcctcca actcgcggtc ccaaatcaag 120
gctgccttgg acaatgcggg aaagattatg agcctgacta aaaccgcccc cgactacctg 180
gtgggccagc agcccgtgga ggacatttcc agcaatcgga tttataaaat tttggaacta 240
aacgggtacg atccccaata tgcggcttcc gtctttctgg gatgggccac gaaaaagttc 300
ggcaagagga acaccatctg gctgtttggg cctgcaacta ccgggaagac caacatcgcg 360
gaggccatag cccacactgt gcccttctac gggtgcgtaa actggaccaa tgagaacttt 420
cccttcaacg actgtgtcga caagatggtg atctggtggg aggaggggaa gatgaccgcc 480
aaggtcgtgg agtcggccaa agccattctc ggaggaagca aggtgcgcgt ggaccagaaa 540
tgcaagtcct cggcccagat agacccgact cccgtgatcg tcacctccaa caccaacatg 600
tgcgccgtga ttgacgggaa ctcaacgacc ttcgaacacc agcagccgtt gcaagaccgg 660
atgttcaaat ttgaactcac ccgccgtctg gatcatgact ttgggaaggt caccaagcag 720
gaagtcaaag actttttccg gtgggcaaag gatcacgtgg ttgaggtgga gcatgaattc 780
tacgtcaaaa agggtggagc caagaaaaga cccgccccca gtgacgcaga tataagtgag 840
cccaaacggg tgcgcgagtc agttgcgcag ccatcgacgt cagacgcgga agcttcgatc 900
aactacgcag acaggtacca aaacaaatgt tctcgtcacg tgggcatgaa tctgatgctg 960
tttccctgca gacaatgcga gagaatgaat cagaattcaa atatctgctt cactcacgga 1020
cagaaagact gtttagagtg ctttcccgtg tcagaatctc aacccgtttc tgtcgtcaaa 1080
aaggcgtatc agaaactgtg ctacattcat catatcatgg gaaaggtgcc agacgcttgc 1140
actgcctgcg atctggtcaa tgtggatttg gatgactgca tctttgaaca ataaatgatt 1200
taaatcaggt atggctgccg atggttatct tccagattgg ctcgaggaca ctctctctga 1260
<210> 1081
<211> 1932
<212> ДНК
<213> Аденоассоциированный вирус - 2
<400> 1081
atgccggggt tttacgagat tgtgattaag gtccccagcg accttgacga gcatctgccc 60
ggcatttctg acagctttgt gaactgggtg gccgagaagg aatgggagtt gccgccagat 120
tctgacatgg atctgaatct gattgagcag gcacccctga ccgtggccga gaagctgcag 180
cgcgactttc tgacggaatg gcgccgtgtg agtaaggccc cggaggccct tttctttgtg 240
caatttgaga agggagagag ctacttccac atgcacgtgc tcgtggaaac caccggggtg 300
aaatccatgg ttttgggacg tttcctgagt cagattcgcg aaaaactgat tcagagaatt 360
taccgcggga tcgagccgac tttgccaaac tggttcgcgg tcacaaagac cagaaatggc 420
gccggaggcg ggaacaaggt ggtggatgag tgctacatcc ccaattactt gctccccaaa 480
acccagcctg agctccagtg ggcgtggact aatatggaac agtatttaag cgcctgtttg 540
aatctcacgg agcgtaaacg gttggtggcg cagcatctga cgcacgtgtc gcagacgcag 600
gagcagaaca aagagaatca gaatcccaat tctgatgcgc cggtgatcag atcaaaaact 660
tcagccaggt acatggagct ggtcgggtgg ctcgtggaca aggggattac ctcggagaag 720
cagtggatcc aggaggacca ggcctcatac atctccttca atgcggcctc caactcgcgg 780
tcccaaatca aggctgcctt ggacaatgcg ggaaagatta tgagcctgac taaaaccgcc 840
cccgactacc tggtgggcca gcagcccgtg gaggacattt ccagcaatcg gatttataaa 900
attttggaac taaacgggta cgatccccaa tatgcggctt ccgtctttct gggatgggcc 960
acgaaaaagt tcggcaagag gaacaccatc tggctgtttg ggcctgcaac taccgggaag 1020
accaacatcg cggaggccat agcccacact gtgcccttct acgggtgcgt aaactggacc 1080
aatgagaact ttcccttcaa cgactgtgtc gacaagatgg tgatctggtg ggaggagggg 1140
aagatgaccg ccaaggtcgt ggagtcggcc aaagccattc tcggaggaag caaggtgcgc 1200
gtggaccaga aatgcaagtc ctcggcccag atagacccga ctcccgtgat cgtcacctcc 1260
aacaccaaca tgtgcgccgt gattgacggg aactcaacga ccttcgaaca ccagcagccg 1320
ttgcaagacc ggatgttcaa atttgaactc acccgccgtc tggatcatga ctttgggaag 1380
gtcaccaagc aggaagtcaa agactttttc cggtgggcaa aggatcacgt ggttgaggtg 1440
gagcatgaat tctacgtcaa aaagggtgga gccaagaaaa gacccgcccc cagtgacgca 1500
gatataagtg agcccaaacg ggtgcgcgag tcagttgcgc agccatcgac gtcagacgcg 1560
gaagcttcga tcaactacgc agacaggtac caaaacaaat gttctcgtca cgtgggcatg 1620
aatctgatgc tgtttccctg cagacaatgc gagagaatga atcagaattc aaatatctgc 1680
ttcactcacg gacagaaaga ctgtttagag tgctttcccg tgtcagaatc tcaacccgtt 1740
tctgtcgtca aaaaggcgta tcagaaactg tgctacattc atcatatcat gggaaaggtg 1800
ccagacgctt gcactgcctg cgatctggtc aatgtggatt tggatgactg catctttgaa 1860
caataaatga tttaaatcag gtatggctgc cgatggttat cttccagatt ggctcgagga 1920
cactctctct ga 1932
<210> 1082
<211> 1876
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1082
cgcagccacc atggcggggt tttacgagat tgtgattaag gtccccagcg accttgacgg 60
gcatctgccc ggcatttctg acagctttgt gaactgggtg gccgagaagg aatgggagtt 120
gccgccagat tctgacatgg atctgaatct gattgagcag gcacccctga ccgtggccga 180
gaagctgcag cgcgactttc tgacggaatg gcgccgtgtg agtaaggccc cggaggccct 240
tttctttgtg caatttgaga agggagagag ctacttccac atgcacgtgc tcgtggaaac 300
caccggggtg aaatccatgg ttttgggacg tttcctgagt cagattcgcg aaaaactgat 360
tcagagaatt taccgcggga tcgagccgac tttgccaaac tggttcgcgg tcacaaagac 420
cagaaatggc gccggaggcg ggaacaaggt ggtggatgag tgctacatcc ccaattactt 480
gctccccaaa acccagcctg agctccagtg ggcgtggact aatatggaac agtatttaag 540
cgcctgtttg aatctcacgg agcgtaaacg gttggtggcg cagcatctga cgcacgtgtc 600
gcagacgcag gagcagaaca aagagaatca gaatcccaat tctgatgcgc cggtgatcag 660
atcaaaaact tcagccaggt acatggagct ggtcgggtgg ctcgtggaca aggggattac 720
ctcggagaag cagtggatcc aggaggacca ggcctcatac atctccttca atgcggcctc 780
caactcgcgg tcccaaatca aggctgcctt ggacaatgcg ggaaagatta tgagcctgac 840
taaaaccgcc cccgactacc tggtgggcca gcagcccgtg gaggacattt ccagcaatcg 900
gatttataaa attttggaac taaacgggta cgatccccaa tatgcggctt ccgtctttct 960
gggatgggcc acgaaaaagt tcggcaagag gaacaccatc tggctgtttg ggcctgcaac 1020
taccgggaag accaacatcg cggaggccat agcccacact gtgcccttct acgggtgcgt 1080
aaactggacc aatgagaact ttcccttcaa cgactgtgtc gacaagatgg tgatctggtg 1140
ggaggagggg aagatgaccg ccaaggtcgt ggagtcggcc aaagccattc tcggaggaag 1200
caaggtgcgc gtggaccaga aatgcaagtc ctcggcccag atagacccga ctcccgtgat 1260
cgtcacctcc aacaccaaca tgtgcgccgt gattgacggg aactcaacga ccttcgaaca 1320
ccagcagccg ttgcaagacc ggatgttcaa atttgaactc acccgccgtc tggatcatga 1380
ctttgggaag gtcaccaagc aggaagtcaa agactttttc cggtgggcaa aggatcacgt 1440
ggttgaggtg gagcatgaat tctacgtcaa aaagggtgga gccaagaaaa gacccgcccc 1500
cagtgacgca gatataagtg agcccaaacg ggtgcgcgag tcagttgcgc agccatcgac 1560
gtcagacgcg gaagcttcga tcaactacgc agacaggtac caaaacaaat gttctcgtca 1620
cgtgggcatg aatctgatgc tgtttccctg cagacaatgc gagagaatga atcagaattc 1680
aaatatctgc ttcactcacg gacagaaaga ctgtttagag tgctttcccg tgtcagaatc 1740
tcaacccgtt tctgtcgtca aaaaggcgta tcagaaactg tgctacattc atcatatcat 1800
gggaaaggtg ccagacgctt gcactgcctg cgatctggtc aatgtggatt tggatgactg 1860
catctttgaa caataa 1876
<210> 1083
<211> 1194
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1083
atggagctgg tcgggtggct cgtggacaag gggattacct cggagaagca gtggatccag 60
gaggaccagg cctcatacat ctccttcaat gcggcctcca actcgcggtc ccaaatcaag 120
gctgccttgg acaatgcggg aaagattatg agcctgacta aaaccgcccc cgactacctg 180
gtgggccagc agcccgtgga ggacatttcc agcaatcgga tttataaaat tttggaacta 240
aacgggtacg atccccaata tgcggcttcc gtctttctgg gatgggccac gaaaaagttc 300
ggcaagagga acaccatctg gctgtttggg cctgcaacta ccgggaagac caacatcgcg 360
gaggccatag cccacactgt gcccttctac gggtgcgtaa actggaccaa tgagaacttt 420
cccttcaacg actgtgtcga caagatggtg atctggtggg aggaggggaa gatgaccgcc 480
aaggtcgtgg agtcggccaa agccattctc ggaggaagca aggtgcgcgt ggaccagaaa 540
tgcaagtcct cggcccagat agacccgact cccgtgatcg tcacctccaa caccaacatg 600
tgcgccgtga ttgacgggaa ctcaacgacc ttcgaacacc agcagccgtt gcaagaccgg 660
atgttcaaat ttgaactcac ccgccgtctg gatcatgact ttgggaaggt caccaagcag 720
gaagtcaaag actttttccg gtgggcaaag gatcacgtgg ttgaggtgga gcatgaattc 780
tacgtcaaaa agggtggagc caagaaaaga cccgccccca gtgacgcaga tataagtgag 840
cccaaacggg tgcgcgagtc agttgcgcag ccatcgacgt cagacgcgga agcttcgatc 900
aactacgcag accgctacca aaacaaatgt tctcgtcacg tgggcatgaa tctgatgctg 960
tttccctgca gacaatgcga gagaatgaat cagaattcaa atatctgctt cactcacgga 1020
cagaaagact gtttagagtg ctttcccgtg tcagaatctc aacccgtttc tgtcgtcaaa 1080
aaggcgtatc agaaactgtg ctacattcat catatcatgg gaaaggtgcc agacgcttgc 1140
actgcctgcg atctggtcaa tgtggatttg gatgactgca tctttgaaca ataa 1194
<210> 1084
<211> 1876
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1084
cgcagccacc atggcggggt tttacgagat tgtgattaag gtccccagcg accttgacgg 60
gcatctgccc ggcatttctg acagctttgt gaactgggtg gccgagaagg aatgggagtt 120
gccgccagat tctgacatgg atctgaatct gattgagcag gcacccctga ccgtggccga 180
gaagctgcag cgcgactttc tgacggaatg gcgccgtgtg agtaaggccc cggaggccct 240
tttctttgtg caatttgaga agggagagag ctacttccac atgcacgtgc tcgtggaaac 300
caccggggtg aaatccatgg ttttgggacg tttcctgagt cagattcgcg aaaaactgat 360
tcagagaatt taccgcggga tcgagccgac tttgccaaac tggttcgcgg tcacaaagac 420
cagaaatggc gccggaggcg ggaacaaggt ggtggatgag tgctacatcc ccaattactt 480
gctccccaaa acccagcctg agctccagtg ggcgtggact aatatggaac agtatttaag 540
cgcctgtttg aatctcacgg agcgtaaacg gttggtggcg cagcatctga cgcacgtgtc 600
gcagacgcag gagcagaaca aagagaatca gaatcccaat tctgatgcgc cggtgatcag 660
atcaaaaact tcagccaggt acatggagct ggtcgggtgg ctcgtggaca aggggattac 720
ctcggagaag cagtggatcc aggaggacca ggcctcatac atctccttca atgcggcctc 780
caactcgcgg tcccaaatca aggctgcctt ggacaatgcg ggaaagatta tgagcctgac 840
taaaaccgcc cccgactacc tggtgggcca gcagcccgtg gaggacattt ccagcaatcg 900
gatttataaa attttggaac taaacgggta cgatccccaa tatgcggctt ccgtctttct 960
gggatgggcc acgaaaaagt tcggcaagag gaacaccatc tggctgtttg ggcctgcaac 1020
taccgggaag accaacatcg cggaggccat agcccacact gtgcccttct acgggtgcgt 1080
aaactggacc aatgagaact ttcccttcaa cgactgtgtc gacaagatgg tgatctggtg 1140
ggaggagggg aagatgaccg ccaaggtcgt ggagtcggcc aaagccattc tcggaggaag 1200
caaggtgcgc gtggaccaga aatgcaagtc ctcggcccag atagacccga ctcccgtgat 1260
cgtcacctcc aacaccaaca tgtgcgccgt gattgacggg aactcaacga ccttcgaaca 1320
ccagcagccg ttgcaagacc ggatgttcaa atttgaactc acccgccgtc tggatcatga 1380
ctttgggaag gtcaccaagc aggaagtcaa agactttttc cggtgggcaa aggatcacgt 1440
ggttgaggtg gagcatgaat tctacgtcaa aaagggtgga gccaagaaaa gacccgcccc 1500
cagtgacgca gatataagtg agcccaaacg ggtgcgcgag tcagttgcgc agccatcgac 1560
gtcagacgcg gaagcttcga tcaactacgc agacaggtac caaaacaaat gttctcgtca 1620
cgtgggcatg aatctgatgc tgtttccctg cagacaatgc gagagaatga atcagaattc 1680
aaatatctgc ttcactcacg gacagaaaga ctgtttagag tgctttcccg tgtcagaatc 1740
tcaacccgtt tctgtcgtca aaaaggcgta tcagaaactg tgctacattc atcatatcat 1800
gggaaaggtg ccagacgctt gcactgcctg cgatctggtc aatgtggatt tggatgactg 1860
catctttgaa caataa 1876
<210> 1085
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1085
ctaggccgcc cgggcaaagc ccgggcgtcg ggcgaccttt ggtcgcccgg c 51
<210> 1086
<211> 65
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1086
ctaggactga ggccgcccgg gcaaagcccg ggcgtcgggc gacctttggt cgcccggcct 60
cagtc 65
<210> 1087
<211> 67
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1087
ggactgaggc cgcccgggca aagcccgggc gtcgggcgac ctttggtcgc ccggcctcag 60
tcctgca 67
<210> 1088
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1088
gtgcgggcga ccaaaggtcg cccgacgccc gggcgcactc a 41
<210> 1089
<211> 56
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1089
ggactgaggc cgggcgacca aaggtcgccc gacgcccggg cggcctcagt cctgca 56
<210> 1090
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1090
ctaggactga ggccgcccgg gcgtcgggcg acctttggtc gcccggcctc agtc 54
<210> 1091
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1091
ggactgaggc cgggcgacca aaggtcgccc gacggcctca gtcctgca 48
<210> 1092
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1092
ctaggactga ggccgtcggg cgacctttgg tcgcccggcc tcagtc 46
<210> 1093
<211> 67
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1093
ggactgaggc ccgggcgacc aaaggtcgcc cgacgcccgg gctttgcccg ggcgcctcag 60
tcctgca 67
<210> 1094
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1094
atacctaggc acgcgtgtta ctagttatta atagtaatca attacgg 47
<210> 1095
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1095
atacctaggg gccgcacgcg tgttactag 29
<210> 1096
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1096
atacactcag tgcctgcagg cacgtggtcc ggagatccag ac 42
<210> 1097
<211> 3754
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1097
cctaggtgag cgagcgagcg cgcagagagg gagtggccaa ctccatcact aggggttcct 60
tgtagttaat gattaacccg ccatgctact tatcgcggcc gctcaatatt ggccattagc 120
catattattc attggttata tagcataaat caatattggc tattggccat tgcatacgtt 180
gtatctatat cataatatgt acatttatat tggctcatgt ccaatatgac cgccatgttg 240
gcattgatta ttgactagtt attaatagta atcaattacg gggtcattag ttcatagccc 300
atatatggag ttccgcgtta cataacttac ggtaaatggc ccgcctggct gaccgcccaa 360
cgacccccgc ccattgacgt caataatgac gtatgttccc atagtaacgc caatagggac 420
tttccattga cgtcaatggg tggagtattt acggtaaact gcccacttgg cagtacatca 480
agtgtatcat atgccaagtc cgccccctat tgacgtcaat gacggtaaat ggcccgcctg 540
gcattatgcc cagtacatga ccttacggga ctttcctact tggcagtaca tctacgtatt 600
agtcatcgct attaccatgg tcgaggtgag ccccacgttc tgcttcactc tccccatctc 660
ccccccctcc ccacccccaa ttttgtattt atttattttt taattatttt gtgcagcgat 720
gggggcgggg gggggggggg ggcgcgcgcc aggcggggcg gggcggggcg aggggcgggg 780
cggggcgagg cggagaggtg cggcggcagc caatcagagc ggcgcgctcc gaaagtttcc 840
ttttatggcg aggcggcggc ggcggcggcc ctataaaaag cgaagcgcgc ggcgggcggg 900
agtcgctgcg acgctgcctt cgccccgtgc cccgctccgc cgccgcctcg cgccgcccgc 960
cccggctctg actgaccgcg ttactcccac aggtgagcgg gcgggacggc ccttctcctc 1020
cgggctgtaa ttagcgcttg gtttaatgac ggcttgtttc ttttctgtgg ctgcgtgaaa 1080
gccttgaggg gctccgggag ggccctttgt gcggggggga gcggctcggg gggtgcgtgc 1140
gtgtgtgtgt gcgtggggag cgccgcgtgc ggcccgcgct gcccggcggc tgtgagcgct 1200
gcgggcgcgg cgcggggctt tgtgcgctcc gcagtgtgcg cgaggggagc gcggccgggg 1260
gcggtgcccc gcggtgcggg gggggctgcg aggggaacaa aggctgcgtg cggggtgtgt 1320
gcgtgggggg gtgagcaggg ggtgtgggcg cggcggtcgg gctgtaaccc ccccctgcac 1380
ccccctcccc gagttgctga gcacggcccg gcttcgggtg cggggctccg tacggggcgt 1440
ggcgcggggc tcgccgtgcc gggcgggggg tggcggcagg tgggggtgcc gggcggggcg 1500
gggccgcctc gggccgggga gggctcgggg gaggggcgcg gcggcccccg gagcgccggc 1560
ggctgtcgag gcgcggcgag ccgcagccat tgccttttat ggtaatcgtg cgagagggcg 1620
cagggacttc ctttgtccca aatctgtgcg gagccgaaat ctgggaggcg ccgccgcacc 1680
ccctctagcg ggcgcggggc gaagcggtgc ggcgccggca ggaaggaaat gggcggggag 1740
ggccttcgtg cgtcgccgcg ccgccgtccc cttctccctc tccagcctcg gggctgtccg 1800
cggggggacg gctgccttcg ggggggacgg ggcagggcgg ggttcggctt ctggcgtgtg 1860
accggcggct ctagagcctc tgctaaccat gttttagcct tcttcttttt cctacagctc 1920
ctgggcaacg tgctggttat tgtgctgtct catcatttgt cgacagaatt cctcgaagat 1980
ccgaaggggt tcaagcttgg cattccggta ctgttggtaa agccagttta aacgccgcca 2040
ccatggtgag caagggcgag gagctgttca ccggggtggt gcccatcctg gtcgagctgg 2100
acggcgacgt aaacggccac aagttcagcg tgtccggcga gggcgagggc gatgccacct 2160
acggcaagct gaccctgaag ttcatctgca ccaccggcaa gctgcccgtg ccctggccca 2220
ccctcgtgac caccctgacc tacggcgtgc agtgcttcag ccgctacccc gaccacatga 2280
agcagcacga cttcttcaag tccgccatgc ccgaaggcta cgtccaggag cgcaccatct 2340
tcttcaagga cgacggcaac tacaagaccc gcgccgaggt gaagttcgag ggcgacaccc 2400
tggtgaaccg catcgagctg aagggcatcg acttcaagga ggacggcaac atcctggggc 2460
acaagctgga gtacaactac aacagccaca acgtctatat catggccgac aagcagaaga 2520
acggcatcaa ggtgaacttc aagatccgcc acaacatcga ggacggcagc gtgcagctcg 2580
ccgaccacta ccagcagaac acccccatcg gcgacggccc cgtgctgctg cccgacaacc 2640
actacctgag cacccagtcc gccctgagca aagaccccaa cgagaagcgc gatcacatgg 2700
tcctgctgga gttcgtgacc gccgccggga tcactctcgg catggacgag ctgtacaagt 2760
aattaattaa gagcatctta ccgccattta ttcccatatt tgttctgttt ttcttgattt 2820
gggtatacat ttaaatgtta ataaaacaaa atggtggggc aatcatttac atttttaggg 2880
atatgtaatt actagttcag gtgtattgcc acaagacaaa catgttaaga aactttcccg 2940
ttatttacgc tctgttcctg ttaatcaacc tctggattac aaaatttgtg aaagattgac 3000
tgatattctt aactatgttg ctccttttac gctgtgtgga tatgctgctt tatagcctct 3060
gtatctagct attgcttccc gtacggcttt cgttttctcc tccttgtata aatcctggtt 3120
gctgtctctt ttagaggagt tgtggcccgt tgtccgtcaa cgtggcgtgg tgtgctctgt 3180
gtttgctgac gcaaccccca ctggctgggg cattgccacc acctgtcaac tcctttctgg 3240
gactttcgct ttccccctcc cgatcgccac ggcagaactc atcgccgcct gccttgcccg 3300
ctgctggaca ggggctaggt tgctgggcac tgataattcc gtggtgttgt ctgtgccttc 3360
tagttgccag ccatctgttg tttgcccctc ccccgtgcct tccttgaccc tggaaggtgc 3420
cactcccact gtcctttcct aataaaatga ggaaattgca tcgcattgtc tgagtaggtg 3480
tcattctatt ctggggggtg gggtggggca ggacagcaag ggggaggatt gggaagacaa 3540
tagcaggcat gctggggatg cggtgggctc tatggctcta gagcatggct acgtagataa 3600
gtagcatggc gggttaatca ttaactacac ctgcagcagg aacccctagt gatggagttg 3660
gccactccct ctctgcgcgc tcgctcgctc cctgcaggac tgaggccggg cgaccaaagg 3720
tcgcccgacg cccgggcggc ctcagtcctg cagg 3754
<210> 1098
<211> 8418
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1098
ggcagctgcg cgctcgctcg ctcacctagg ccgcccgggc aaagcccggg cgtcgggcga 60
cctttggtcg cccggcctag gtgagcgagc gagcgcgcag agagggagtg gccaactcca 120
tcactagggg ttccttgtag ttaatgatta acccgccatg ctacttatcg cggccgctca 180
atattggcca ttagccatat tattcattgg ttatatagca taaatcaata ttggctattg 240
gccattgcat acgttgtatc tatatcataa tatgtacatt tatattggct catgtccaat 300
atgaccgcca tgttggcatt gattattgac tagttattaa tagtaatcaa ttacggggtc 360
attagttcat agcccatata tggagttccg cgttacataa cttacggtaa atggcccgcc 420
tggctgaccg cccaacgacc cccgcccatt gacgtcaata atgacgtatg ttcccatagt 480
aacgccaata gggactttcc attgacgtca atgggtggag tatttacggt aaactgccca 540
cttggcagta catcaagtgt atcatatgcc aagtccgccc cctattgacg tcaatgacgg 600
taaatggccc gcctggcatt atgcccagta catgacctta cgggactttc ctacttggca 660
gtacatctac gtattagtca tcgctattac catggtcgag gtgagcccca cgttctgctt 720
cactctcccc atctcccccc cctccccacc cccaattttg tatttattta ttttttaatt 780
attttgtgca gcgatggggg cggggggggg gggggggcgc gcgccaggcg gggcggggcg 840
gggcgagggg cggggcgggg cgaggcggag aggtgcggcg gcagccaatc agagcggcgc 900
gctccgaaag tttcctttta tggcgaggcg gcggcggcgg cggccctata aaaagcgaag 960
cgcgcggcgg gcgggagtcg ctgcgacgct gccttcgccc cgtgccccgc tccgccgccg 1020
cctcgcgccg cccgccccgg ctctgactga ccgcgttact cccacaggtg agcgggcggg 1080
acggcccttc tcctccgggc tgtaattagc gcttggttta atgacggctt gtttcttttc 1140
tgtggctgcg tgaaagcctt gaggggctcc gggagggccc tttgtgcggg ggggagcggc 1200
tcggggggtg cgtgcgtgtg tgtgtgcgtg gggagcgccg cgtgcggccc gcgctgcccg 1260
gcggctgtga gcgctgcggg cgcggcgcgg ggctttgtgc gctccgcagt gtgcgcgagg 1320
ggagcgcggc cgggggcggt gccccgcggt gcgggggggg ctgcgagggg aacaaaggct 1380
gcgtgcgggg tgtgtgcgtg ggggggtgag cagggggtgt gggcgcggcg gtcgggctgt 1440
aacccccccc tgcacccccc tccccgagtt gctgagcacg gcccggcttc gggtgcgggg 1500
ctccgtacgg ggcgtggcgc ggggctcgcc gtgccgggcg gggggtggcg gcaggtgggg 1560
gtgccgggcg gggcggggcc gcctcgggcc ggggagggct cgggggaggg gcgcggcggc 1620
ccccggagcg ccggcggctg tcgaggcgcg gcgagccgca gccattgcct tttatggtaa 1680
tcgtgcgaga gggcgcaggg acttcctttg tcccaaatct gtgcggagcc gaaatctggg 1740
aggcgccgcc gcaccccctc tagcgggcgc ggggcgaagc ggtgcggcgc cggcaggaag 1800
gaaatgggcg gggagggcct tcgtgcgtcg ccgcgccgcc gtccccttct ccctctccag 1860
cctcggggct gtccgcgggg ggacggctgc cttcgggggg gacggggcag ggcggggttc 1920
ggcttctggc gtgtgaccgg cggctctaga gcctctgcta accatgtttt agccttcttc 1980
tttttcctac agctcctggg caacgtgctg gttattgtgc tgtctcatca tttgtcgaca 2040
gaattcctcg aagatccgaa ggggttcaag cttggcattc cggtactgtt ggtaaagcca 2100
gtttaaacgc cgccaccatg gtgagcaagg gcgaggagct gttcaccggg gtggtgccca 2160
tcctggtcga gctggacggc gacgtaaacg gccacaagtt cagcgtgtcc ggcgagggcg 2220
agggcgatgc cacctacggc aagctgaccc tgaagttcat ctgcaccacc ggcaagctgc 2280
ccgtgccctg gcccaccctc gtgaccaccc tgacctacgg cgtgcagtgc ttcagccgct 2340
accccgacca catgaagcag cacgacttct tcaagtccgc catgcccgaa ggctacgtcc 2400
aggagcgcac catcttcttc aaggacgacg gcaactacaa gacccgcgcc gaggtgaagt 2460
tcgagggcga caccctggtg aaccgcatcg agctgaaggg catcgacttc aaggaggacg 2520
gcaacatcct ggggcacaag ctggagtaca actacaacag ccacaacgtc tatatcatgg 2580
ccgacaagca gaagaacggc atcaaggtga acttcaagat ccgccacaac atcgaggacg 2640
gcagcgtgca gctcgccgac cactaccagc agaacacccc catcggcgac ggccccgtgc 2700
tgctgcccga caaccactac ctgagcaccc agtccgccct gagcaaagac cccaacgaga 2760
agcgcgatca catggtcctg ctggagttcg tgaccgccgc cgggatcact ctcggcatgg 2820
acgagctgta caagtaatta attaagagca tcttaccgcc atttattccc atatttgttc 2880
tgtttttctt gatttgggta tacatttaaa tgttaataaa acaaaatggt ggggcaatca 2940
tttacatttt tagggatatg taattactag ttcaggtgta ttgccacaag acaaacatgt 3000
taagaaactt tcccgttatt tacgctctgt tcctgttaat caacctctgg attacaaaat 3060
ttgtgaaaga ttgactgata ttcttaacta tgttgctcct tttacgctgt gtggatatgc 3120
tgctttatag cctctgtatc tagctattgc ttcccgtacg gctttcgttt tctcctcctt 3180
gtataaatcc tggttgctgt ctcttttaga ggagttgtgg cccgttgtcc gtcaacgtgg 3240
cgtggtgtgc tctgtgtttg ctgacgcaac ccccactggc tggggcattg ccaccacctg 3300
tcaactcctt tctgggactt tcgctttccc cctcccgatc gccacggcag aactcatcgc 3360
cgcctgcctt gcccgctgct ggacaggggc taggttgctg ggcactgata attccgtggt 3420
gttgtctgtg ccttctagtt gccagccatc tgttgtttgc ccctcccccg tgccttcctt 3480
gaccctggaa ggtgccactc ccactgtcct ttcctaataa aatgaggaaa ttgcatcgca 3540
ttgtctgagt aggtgtcatt ctattctggg gggtggggtg gggcaggaca gcaaggggga 3600
ggattgggaa gacaatagca ggcatgctgg ggatgcggtg ggctctatgg ctctagagca 3660
tggctacgta gataagtagc atggcgggtt aatcattaac tacacctgca gcaggaaccc 3720
ctagtgatgg agttggccac tccctctctg cgcgctcgct cgctccctgc aggactgagg 3780
ccgggcgacc aaaggtcgcc cgacgcccgg gcggcctcag tcctgcaggg agcgagcgag 3840
cgcgcagctg cctgcacggg cgcgccggta ccgggagatg ggggaggcta actgaaacac 3900
ggaaggagac aataccggaa ggaacccgcg ctatgacggc aataaaaaga cagaataaaa 3960
cgcacgggtg ttgggtcgtt tgttcataaa cgcggggttc ggtcccaggg ctggcactct 4020
gtcgataccc caccgagacc ccattgggac caatacgccc gcgtttcttc cttttcccca 4080
ccccaacccc caagttcggg tgaaggccca gggctcgcag ccaacgtcgg ggcggcaagc 4140
cctgccatag ccactacggg tacgtaggcc aaccactaga actatagcta gagtcctggg 4200
cgaacaaacg atgctcgcct tccagaaaac cgaggatgcg aaccacttca tccggggtca 4260
gcaccaccgg caagcgccgc gacggccgag gtctaccgat ctcctgaagc cagggcagat 4320
ccgtgcacag caccttgccg tagaagaaca gcaaggccgc caatgcctga cgatgcgtgg 4380
agaccgaaac cttgcgctcg ttcgccagcc aggacagaaa tgcctcgact tcgctgctgc 4440
ccaaggttgc cgggtgacgc acaccgtgga aacggatgaa ggcacgaacc cagttgacat 4500
aagcctgttc ggttcgtaaa ctgtaatgca agtagcgtat gcgctcacgc aactggtcca 4560
gaaccttgac cgaacgcagc ggtggtaacg gcgcagtggc ggttttcatg gcttgttatg 4620
actgtttttt tgtacagtct atgcctcggg catccaagca gcaagcgcgt tacgccgtgg 4680
gtcgatgttt gatgttatgg agcagcaacg atgttacgca gcagcaacga tgttacgcag 4740
cagggcagtc gccctaaaac aaagttaggt ggctcaagta tgggcatcat tcgcacatgt 4800
aggctcggcc ctgaccaagt caaatccatg cgggctgctc ttgatctttt cggtcgtgag 4860
ttcggagacg tagccaccta ctcccaacat cagccggact ccgattacct cgggaacttg 4920
ctccgtagta agacattcat cgcgcttgct gccttcgacc aagaagcggt tgttggcgct 4980
ctcgcggctt acgttctgcc caggtttgag cagccgcgta gtgagatcta tatctatgat 5040
ctcgcagtct ccggcgagca ccggaggcag ggcattgcca ccgcgctcat caatctcctc 5100
aagcatgagg ccaacgcgct tggtgcttat gtgatctacg tgcaagcaga ttacggtgac 5160
gatcccgcag tggctctcta tacaaagttg ggcatacggg aagaagtgat gcactttgat 5220
atcgacccaa gtaccgccac ctaacaattc gttcaagccg agatcggctt cccggccgcg 5280
gagttgttcg gtaaattgtc acaacgccgc gaatatagtc tttaccatgc ccttggccac 5340
gcccctcttt aatacgacgg gcaatttgca cttcagaaaa tgaagagttt gctttagcca 5400
taacaaaagt ccagtatgct ttttcacagc ataactggac tgatttcagt ttacaactat 5460
tctgtctagt ttaagacttt attgtcatag tttagatcta ttttgttcag tttaagactt 5520
tattgtccgc ccacacccgc ttacgcaggg catccattta ttactcaacc gtaaccgatt 5580
ttgccaggtt acgcggctgg tctgcggtgt gaaataccgc acagatgcgt aaggagaaaa 5640
taccgcatca ggcgctcttc cgcttcctcg ctcactgact cgctgcgctc ggtcgttcgg 5700
ctgcggcgag cggtatcagc tcactcaaag gcggtaatac ggttatccac agaatcaggg 5760
gataacgcag gaaagaacat gtgagcaaaa ggccagcaaa aggccaggaa ccgtaaaaag 5820
gccgcgttgc tggcgttttt ccataggctc cgcccccctg acgagcatca caaaaatcga 5880
cgctcaagtc agaggtggcg aaacccgaca ggactataaa gataccaggc gtttccccct 5940
ggaagctccc tcgtgcgctc tcctgttccg accctgccgc ttaccggata cctgtccgcc 6000
tttctccctt cgggaagcgt ggcgctttct caatgctcac gctgtaggta tctcagttcg 6060
gtgtaggtcg ttcgctccaa gctgggctgt gtgcacgaac cccccgttca gcccgaccgc 6120
tgcgccttat ccggtaacta tcgtcttgag tccaacccgg taagacacga cttatcgcca 6180
ctggcagcag ccactggtaa caggattagc agagcgaggt atgtaggcgg tgctacagag 6240
ttcttgaagt ggtggcctaa ctacggctac actagaagga cagtatttgg tatctgcgct 6300
ctgctgaagc cagttacctt cggaaaaaga gttggtagct cttgatccgg caaacaaacc 6360
accgctggta gcggtggttt ttttgtttgc aagcagcaga ttacgcgcag aaaaaaagga 6420
tctcaagaag atcctttgat cttttctacg gggtctgacg ctcagtggaa cgaaaactca 6480
cgttaaggga ttttggtcat gagattatca aaaaggatct tcacctagat ccttttaaat 6540
taaaaatgaa gttttaaatc aatctaaagt atatatgagt aaacttggtc tgacagttac 6600
caatgcttaa tcagtgaggc acctatctca gcgatctgtc tatttcgttc atccatagtt 6660
gcctgactcc ccgtcgtgta gataactacg atacgggagg gcttaccatc tggccccagt 6720
gctgcaatga taccgcgaga cccacgctca ccggctccag atttatcagc aataaaccag 6780
ccagccggaa gggccgagcg cagaagtggt cctgcaactt tatccgcctc catccagtct 6840
attaattgtt gccgggaagc tagagtaagt agttcgccag ttaatagttt gcgcaacgtt 6900
gttgccattg ctacaggcat cgtggtgtca cgctcgtcgt ttggtatggc ttcattcagc 6960
tccggttccc aacgatcaag gcgagttaca tgatccccca tgttgtgcaa aaaagcggtt 7020
agctccttcg gtcctccgat cgttgtcaga agtaagttgg ccgcagtgtt atcactcatg 7080
gttatggcag cactgcataa ttctcttact gtcatgccat ccgtaagatg cttttctgtg 7140
actggtgagt actcaaccaa gtcattctga gaatagtgta tgcggcgacc gagttgctct 7200
tgcccggcgt caatacggga taataccgcg ccacatagca gaactttaaa agtgctcatc 7260
attggaaaac gttcttcggg gcgaaaactc tcaaggatct taccgctgtt gagatccagt 7320
tcgatgtaac ccactcgtgc acccaactga tcttcagcat cttttacttt caccagcgtt 7380
tctgggtgag caaaaacagg aaggcaaaat gccgcaaaaa agggaataag ggcgacacgg 7440
aaatgttgaa tactcatact cttccttttt caatattatt gaagcattta tcagggttat 7500
tgtctcatga gcggatacat atttgaatgt atttagaaaa ataaacaaat aggggttccg 7560
cgcacatttc cccgaaaagt gccacctgaa attgtaaacg ttaatatttt gttaaaattc 7620
gcgttaaatt tttgttaaat cagctcattt tttaaccaat aggccgaaat cggcaaaatc 7680
ccttataaat caaaagaata gaccgagata gggttgagtg ttgttccagt ttggaacaag 7740
agtccactat taaagaacgt ggactccaac gtcaaagggc gaaaaaccgt ctatcagggc 7800
gatggcccac tacgtgaacc atcaccctaa tcaagttttt tggggtcgag gtgccgtaaa 7860
gcactaaatc ggaaccctaa agggagcccc cgatttagag cttgacgggg aaagccggcg 7920
aacgtggcga gaaaggaagg gaagaaagcg aaaggagcgg gcgctagggc gctggcaagt 7980
gtagcggtca cgctgcgcgt aaccaccaca cccgccgcgc ttaatgcgcc gctacagggc 8040
gcgtcccatt cgccattcag gctgcaaata agcgttgata ttcagtcaat tacaaacatt 8100
aataacgaag agatgacaga aaaattttca ttctgtgaca gagaaaaagt agccgaagat 8160
gacggtttgt cacatggagt tggcaggatg tttgattaaa aacataacag gaagaaaaat 8220
gccccgctgt gggcggacaa aatagttggg aactgggagg ggtggaaatg gagtttttaa 8280
ggattattta gggaagagtg acaaaataga tgggaactgg gtgtagcgtc gtaagctaat 8340
acgaaaatta aaaatgacaa aatagtttgg aactagattt cacttatctg gttcggatct 8400
cctagtgagc tccctgca 8418
<210> 1099
<211> 225
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1099
tgtgccttct agttgccagc catctgttgt ttgcccctcc cccgtgcctt ccttgaccct 60
ggaaggtgcc actcccactg tcctttccta ataaaatgag gaaattgcat cgcattgtct 120
gagtaggtgt cattctattc tggggggtgg ggtggggcag gacagcaagg gggaggattg 180
ggaagacaat agcaggcatg ctggggatgc ggtgggctct atggc 225
<210> 1100
<211> 1177
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1100
ggctcagagg ctcagaggca cacaggagtt tctgggctca ccctgccccc ttccaacccc 60
tcagttccca tcctccagca gctgtttgtg tgctgcctct gaagtccaca ctgaacaaac 120
ttcagcctac tcatgtccct aaaatgggca aacattgcaa gcagcaaaca gcaaacacac 180
agccctccct gcctgctgac cttggagctg gggcagaggt cagagacctc tctgggccca 240
tgccacctcc aacatccact cgaccccttg gaatttcggt ggagaggagc agaggttgtc 300
ctggcgtggt ttaggtagtg tgagagggtc cgggttcaaa accacttgct gggtggggag 360
tcgtcagtaa gtggctatgc cccgaccccg aagcctgttt ccccatctgt acaatggaaa 420
tgataaagac gcccatctga tagggttttt gtggcaaata aacatttggt ttttttgttt 480
tgttttgttt tgttttttga gatggaggtt tgctctgtcg cccaggctgg agtgcagtga 540
cacaatctca tctcaccaca accttcccct gcctcagcct cccaagtagc tgggattaca 600
agcatgtgcc accacacctg gctaattttc tatttttagt agagacgggt ttctccatgt 660
tggtcagcct cagcctccca agtaactggg attacaggcc tgtgccacca cacccggcta 720
attttttcta tttttgacag ggacggggtt tcaccatgtt ggtcaggctg gtctagaggt 780
accggatctt gctaccagtg gaacagccac taaggattct gcagtgagag cagagggcca 840
gctaagtggt actctcccag agactgtctg actcacgcca ccccctccac cttggacaca 900
ggacgctgtg gtttctgagc caggtacaat gactcctttc ggtaagtgca gtggaagctg 960
tacactgccc aggcaaagcg tccgggcagc gtaggcgggc gactcagatc ccagccagtg 1020
gacttagccc ctgtttgctc ctccgataac tggggtgacc ttggttaata ttcaccagca 1080
gcctcccccg ttgcccctct ggatccactg cttaaatacg gacgaggaca gggccctgtc 1140
tcctcagctt caggcaccac cactgacctg ggacagt 1177
<210> 1101
<211> 1326
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1101
ctgcagggcc cactagtgga gccgagagta attcatacaa aaggagggat cgccttcgca 60
aggggagagc ccagggaccg tccctaaatt ctcacagacc caaatccctg tagccgcccc 120
acgacagcgc gaggagcatg cgcccagggc tgagcgcggg tagatcagag cacacaagct 180
cacagtcccc ggcggtgggg ggaggggcgc gctgagcggg ggccagggag ctggcgcggg 240
gcaaactggg aaagtggtgt cgtgtgctgg ctccgccctc ttcccgaggg tgggggagaa 300
cggtatataa gtgcggtagt cgccttggac gttctttttc gcaacgggtt tgccgtcaga 360
acgcaggtga gtggcgggtg tggcttccgc gggccccgga gctggagccc tgctctgagc 420
gggccgggct gatatgcgag tgtcgtccgc agggtttagc tgtgagcatt cccacttcga 480
gtggcgggcg gtgcgggggt gagagtgcga ggcctagcgg caaccccgta gcctcgcctc 540
gtgtccggct tgaggcctag cgtggtgtcc gccgccgcgt gccactccgg ccgcactatg 600
cgttttttgt ccttgctgcc ctcgattgcc ttccagcagc atgggctaac aaagggaggg 660
tgtggggctc actcttaagg agcccatgaa gcttacgttg gataggaatg gaagggcagg 720
aggggcgact ggggcccgcc cgccttcgga gcacatgtcc gacgccacct ggatggggcg 780
aggcctgtgg ctttccgaag caatcgggcg tgagtttagc ctacctgggc catgtggccc 840
tagcactggg cacggtctgg cctggcggtg ccgcgttccc ttgcctccca acaagggtga 900
ggccgtcccg cccggcacca gttgcttgcg cggaaagatg gccgctcccg gggccctgtt 960
gcaaggagct caaaatggag gacgcggcag cccggtggag cgggcgggtg agtcacccac 1020
acaaaggaag agggccttgc ccctcgccgg ccgctgcttc ctgtgacccc gtggtctatc 1080
ggccgcatag tcacctcggg cttctcttga gcaccgctcg tcgcggcggg gggaggggat 1140
ctaatggcgt tggagtttgt tcacatttgg tgggtggaga ctagtcaggc cagcctggcg 1200
ctggaagtca ttcttggaat ttgccccttt gagtttggag cgaggctaat tctcaagcct 1260
cttagcggtt caaaggtatt ttctaaaccc gtttccaggt gttgtgaaag ccaccgctaa 1320
ttcaaa 1326
<210> 1102
<211> 573
<212> ДНК
<213> Mus musculus
<400> 1102
gtaagagttt tatgtttttt catctctgct tgtatttttc tagtaatgga agcctggtat 60
tttaaaatag ttaaattttc ctttagtgct gatttctaga ttattattac tgttgttgtt 120
gttattattg tcattatttg catctgagaa cccttaggtg gttatattat tgatatattt 180
ttggtatctt tgatgacaat aatgggggat tttgaaagct tagctttaaa tttcttttaa 240
ttaaaaaaaa atgctaggca gaatgactca aattacgttg gatacagttg aatttattac 300
ggtctcatag ggcctgcctg ctcgaccatg ctatactaaa aattaaaagt gtgtgttact 360
aattttataa atggagtttc catttatatt tacctttatt tcttatttac cattgtctta 420
gtagatattt acaaacatga cagaaacact aaatcttgag tttgaatgca cagatataaa 480
cacttaacgg gttttaaaaa taataatgtt ggtgaaaaaa tataactttg agtgtagcag 540
agaggaacca ttgccacctt cagattttcc tgt 573
<210> 1103
<211> 1993
<212> ДНК
<213> Mus musculus
<400> 1103
acgatcggga actggcatct tcagggagta gcttaggtca gtgaagagaa gaacaaaaag 60
cagcatatta cagttagttg tcttcatcaa tctttaaata tgttgtgtgg tttttctctc 120
cctgtttcca cagacaagag tgagatcgcc catcggtata atgatttggg agaacaacat 180
ttcaaaggcc tgtaagttat aatgctgaaa gcccacttaa tatttctggt agtattagtt 240
aaagttttaa aacacctttt tccaccttga gtgtgagaat tgtagagcag tgctgtccag 300
tagaaatgtg tgcattgaca gaaagactgt ggatctgtgc tgagcaatgt ggcagccaga 360
gatcacaagg ctatcaagca ctttgcacat ggcaagtgta actgagaagc acacattcaa 420
ataatagtta attttaattg aatgtatcta gccatgtgtg gctagtagct cctttcctgg 480
agagagaatc tggagcccac atctaacttg ttaagtctgg aatcttattt tttatttctg 540
gaaaggtcta tgaactatag ttttgggggc agctcactta ctaactttta atgcaataag 600
atctcatggt atcttgagaa cattattttg tctctttgta gtactgaaac cttatacatg 660
tgaagtaagg ggtctatact taagtcacat ctccaacctt agtaatgttt taatgtagta 720
aaaaaatgag taattaattt atttttagaa ggtcaatagt atcatgtatt ccaaataaca 780
gaggtatatg gttagaaaag aaacaattca aaggacttat ataatatcta gccttgacaa 840
tgaataaatt tagagagtag tttgcctgtt tgcctcatgt tcataaatct attgacacat 900
atgtgcatct gcacttcagc atggtagaag tccatattcc tttgcttgga aaggcaggtg 960
ttcccattac gcctcagaga atagctgacg ggaagaggct ttctagatag ttgtatgaaa 1020
gatatacaaa atctcgcagg tatacacagg catgatttgc tggttgggag agccacttgc 1080
ctcatactga ggtttttgtg tctgcttttc agagtcctga ttgccttttc ccagtatctc 1140
cagaaatgct catacgatga gcatgccaaa ttagtgcagg aagtaacaga ctttgcaaag 1200
acgtgtgttg ccgatgagtc tgccgccaac tgtgacaaat cccttgtgag taccttctga 1260
ttttgtggat ctactttcct gctttctgga actctgtttc aaagccaatc atgactccat 1320
cacttaaggc cccgggaaca ctgtggcaga gggcagcaga gagattgata aagccagggt 1380
gatgggaatt ttctgtggga ctccatttca tagtaattgc agaagctaca atacactcaa 1440
aaagtctcac cacatgactg cccaaatggg agcttgacag tgacagtgac agtagatatg 1500
ccaaagtgga tgagggaaag accacaagag ctaaaccctg taaaaagaac tgtaggcaac 1560
taaggaatgc agagagaaga agttgccttg gaagagcata ccaactgcct ctccaatacc 1620
aatggtcatc cctaaaacat acgtatgaat aacatgcaga ctaagcaggc tacatttagg 1680
aatatacatg tatttacata aatgtatatg catgtaacaa caatgaatga aaactgaggt 1740
catggatctg aaagagagca agggggctta catgagaggg tttggaggga ggggttggag 1800
ggagggaggt attattcttt agttttacag ggaacgtagt aaaaacatag gcttctccca 1860
aaggagcaga gcccatgagg agctgtgcaa ggttccccag cttgatttta cctgctcctc 1920
aaattccctt gatttgtttt tattataatg actttactcc tagcttttag tgtcagatag 1980
aaaacatgga agg 1993
<210> 1104
<211> 1350
<212> ДНК
<213> Homo sapiens
<400> 1104
taggaggctg aggcaggagg atcgcttgag cccaggagtt cgagaccagc ctgggcaaca 60
tagtgtgatc ttgtatctat aaaaataaac aaaattagct tggtgtggtg gcgcctgtag 120
tccccagcca cttggagggg tgaggtgaga ggattgcttg agcccgggat ggtccaggct 180
gcagtgagcc atgatcgtgc cactgcactc cagcctgggc gacagagtga gaccctgtct 240
cacaacaaca acaacaacaa caaaaaggct gagctgcacc atgcttgacc cagtttctta 300
aaattgttgt caaagcttca ttcactccat ggtgctatag agcacaagat tttatttggt 360
gagatggtgc tttcatgaat tcccccaaca gagccaagct ctccatctag tggacaggga 420
agctagcagc aaaccttccc ttcactacaa aacttcattg cttggccaaa aagagagtta 480
attcaatgta gacatctatg taggcaatta aaaacctatt gatgtataaa acagtttgca 540
ttcatggagg gcaactaaat acattctagg actttataaa agatcacttt ttatttatgc 600
acagggtgga acaagatgga ttatcaagtg tcaagtccaa tctatgacat caattattat 660
acatcggagc cctgccaaaa aatcaatgtg aagcaaatcg cagcccgcct cctgcctccg 720
ctctactcac tggtgttcat ctttggtttt gtgggcaaca tgctggtcat cctcatcctg 780
ataaactgca aaaggctgaa gagcatgact gacatctacc tgctcaacct ggccatctct 840
gacctgtttt tccttcttac tgtccccttc tgggctcact atgctgccgc ccagtgggac 900
tttggaaata caatgtgtca actcttgaca gggctctatt ttataggctt cttctctgga 960
atcttcttca tcatcctcct gacaatcgat aggtacctgg ctgtcgtcca tgctgtgttt 1020
gctttaaaag ccaggacggt cacctttggg gtggtgacaa gtgtgatcac ttgggtggtg 1080
gctgtgtttg cgtctctccc aggaatcatc tttaccagat ctcaaaaaga aggtcttcat 1140
tacacctgca gctctcattt tccatacagt cagtatcaat tctggaagaa tttccagaca 1200
ttaaagatag tcatcttggg gctggtcctg ccgctgcttg tcatggtcat ctgctactcg 1260
ggaatcctaa aaactctgct tcggtgtcga aatgagaaga agaggcacag ggctgtgagg 1320
cttatcttca ccatcatgat tgtttatttt 1350
<210> 1105
<211> 1223
<212> ДНК
<213> Homo sapiens
<400> 1105
tgacagagac tcttgggatg acgcactgct gcatcaaccc catcatctat gcctttgtcg 60
gggagaagtt cagaaactac ctcttagtct tcttccaaaa gcacattgcc aaacgcttct 120
gcaaatgctg ttctattttc cagcaagagg ctcccgagcg agcaagctca gtttacaccc 180
gatccactgg ggagcaggaa atatctgtgg gcttgtgaca cggactcaag tgggctggtg 240
acccagtcag agttgtgcac atggcttagt tttcatacac agcctgggct gggggtgggg 300
tgggagaggt cttttttaaa aggaagttac tgttatagag ggtctaagat tcatccattt 360
atttggcatc tgtttaaagt agattagatc ttttaagccc atcaattata gaaagccaaa 420
tcaaaatatg ttgatgaaaa atagcaacct ttttatctcc ccttcacatg catcaagtta 480
ttgacaaact ctcccttcac tccgaaagtt ccttatgtat atttaaaaga aagcctcaga 540
gaattgctga ttcttgagtt tagtgatctg aacagaaata ccaaaattat ttcagaaatg 600
tacaactttt tacctagtac aaggcaacat ataggttgta aatgtgttta aaacaggtct 660
ttgtcttgct atggggagaa aagacatgaa tatgattagt aaagaaatga cacttttcat 720
gtgtgatttc ccctccaagg tatggttaat aagtttcact gacttagaac caggcgagag 780
acttgtggcc tgggagagct ggggaagctt cttaaatgag aaggaatttg agttggatca 840
tctattgctg gcaaagacag aagcctcact gcaagcactg catgggcaag cttggctgta 900
gaaggagaca gagctggttg ggaagacatg gggaggaagg acaaggctag atcatgaaga 960
accttgacgg cattgctccg tctaagtcat gagctgagca gggagatcct ggttggtgtt 1020
gcagaaggtt tactctgtgg ccaaaggagg gtcaggaagg atgagcattt agggcaagga 1080
gaccaccaac agccctcagg tcagggtgag gatggcctct gctaagctca aggcgtgagg 1140
atgggaagga gggaggtatt cgtaaggatg ggaaggaggg aggtattcgt gcagcatatg 1200
aggatgcaga gtcagcagaa ctg 1223
<210> 1106
<211> 215
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1106
gaacgctgac gtcatcaacc cgctccaagg aatcgcgggc ccagtgtcac taggcgggaa 60
cacccagcgc gcgtgcgccc tggcaggaag atggctgtga gggacagggg agtggcgccc 120
tgcaatattt gcatgtcgct atgtgttctg ggaaatcacc ataaacgtga aatgtctttg 180
gatttgggaa tcttataagt tctgtatgag accac 215
<210> 1107
<211> 141
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1107
cctgcaggca gctgcgcgct cgctcgctca cctaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctaggtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc t 141
<210> 1108
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1108
gcgcgctcgc tcgctcacc 19
<210> 1109
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1109
ctaggtgagc gagcgagcgc gc 22
<210> 1110
<211> 75
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1110
cctgcaggac tgaggccgcc cgggcaaagc ccgggcgtcg ggcgaccttt ggtcgcccgg 60
cctcagtcct gcagg 75
<210> 1111
<211> 130
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1111
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagt 60
gcgggcgacc aaaggtcgcc cgacgcccgg gcgcactcag tgagcgagcg agcgcgcagc 120
tgcctgcagg 130
<210> 1112
<211> 142
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1112
cctgcaggca gctgcgcgct cgctcgctcc ctaggactga ggccgcccgg gcgtcgggcg 60
acctttggtc gcccggcctc agtcctaggg agcgagcgag cgcgcagaga gggagtggcc 120
aactccatca ctaggggttc ct 142
<210> 1113
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1113
gcgcgctcgc tcgctcactg agtgcgggcg accaaaggtc gcccgacgcc cgggcgcact 60
cagtgagcga gcgagcgcgc 80
<210> 1114
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1114
gcgcgctcgc tcgctcactg a 21
<210> 1115
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1115
gtgagcgagc gagcgcgc 18
<210> 1116
<211> 89
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1116
gcgcgctcgc tcgctcactg aggccgcccg ggcaaagccc gggcgtcggg cgactttgtc 60
gcccggcctc agtgagcgag cgagcgcgc 89
<210> 1117
<211> 89
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1117
gcgcgctcgc tcgctcactg aggccgggcg acaaagtcgc ccgacgcccg ggctttgccc 60
gggcggcctc agtgagcgag cgagcgcgc 89
<210> 1118
<211> 87
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1118
gcgcgctcgc tcgctcactg aggccgcccg ggcaaagccc gggcgtcggg cgattttcgc 60
ccggcctcag tgagcgagcg agcgcgc 87
<210> 1119
<211> 87
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1119
gcgcgctcgc tcgctcactg aggccgggcg aaaatcgccc gacgcccggg ctttgcccgg 60
gcggcctcag tgagcgagcg agcgcgc 87
<210> 1120
<211> 85
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1120
gcgcgctcgc tcgctcactg aggccgcccg ggcaaagccc gggcgtcggg cgtttcgccc 60
ggcctcagtg agcgagcgag cgcgc 85
<210> 1121
<211> 85
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1121
gcgcgctcgc tcgctcactg aggccgggcg aaacgcccga cgcccgggct ttgcccgggc 60
ggcctcagtg agcgagcgag cgcgc 85
<210> 1122
<211> 89
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1122
gcgcgctcgc tcgctcactg aggccgcccg ggaaacccgg gcgtcgggcg acctttggtc 60
gcccggcctc agtgagcgag cgagcgcgc 89
<210> 1123
<211> 89
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1123
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cgggtttccc 60
gggcggcctc agtgagcgag cgagcgcgc 89
<210> 1124
<211> 87
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1124
gcgcgctcgc tcgctcactg aggccgcccg gaaaccgggc gtcgggcgac ctttggtcgc 60
ccggcctcag tgagcgagcg agcgcgc 87
<210> 1125
<211> 87
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1125
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cggtttccgg 60
gcggcctcag tgagcgagcg agcgcgc 87
<210> 1126
<211> 85
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1126
gcgcgctcgc tcgctcactg aggccgcccg aaacgggcgt cgggcgacct ttggtcgccc 60
ggcctcagtg agcgagcgag cgcgc 85
<210> 1127
<211> 85
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1127
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cgtttcgggc 60
ggcctcagtg agcgagcgag cgcgc 85
<210> 1128
<211> 83
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1128
gcgcgctcgc tcgctcactg aggccgccca aagggcgtcg ggcgaccttt ggtcgcccgg 60
cctcagtgag cgagcgagcg cgc 83
<210> 1129
<211> 83
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1129
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc ctttgggcgg 60
cctcagtgag cgagcgagcg cgc 83
<210> 1130
<211> 81
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1130
gcgcgctcgc tcgctcactg aggccgccaa aggcgtcggg cgacctttgg tcgcccggcc 60
tcagtgagcg agcgagcgcg c 81
<210> 1131
<211> 81
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1131
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc tttggcggcc 60
tcagtgagcg agcgagcgcg c 81
<210> 1132
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1132
gcgcgctcgc tcgctcactg aggccgcaaa gcgtcgggcg acctttggtc gcccggcctc 60
agtgagcgag cgagcgcgc 79
<210> 1133
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1133
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgct ttgcggcctc 60
agtgagcgag cgagcgcgc 79
<210> 1134
<211> 81
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1134
ctgcgcgctc gctcgctcac tgaggccgaa acgtcgggcg acctttggtc gcccggcctc 60
agtgagcgag cgagcgcgca g 81
<210> 1135
<211> 81
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1135
ctgcgcgctc gctcgctcac tgaggccggg cgaccaaagg tcgcccgacg tttcggcctc 60
agtgagcgag cgagcgcgca g 81
<210> 1136
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1136
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacggc ctcagtgagc 60
gagcgagcgc gc 72
<210> 1137
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1137
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 1138
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1138
gcgcgctcgc tcgctcactg aggcgcccgg gcgtcgggcg acctttggtc gcccggcctc 60
agtgagcgag cgagcgcgc 79
<210> 1139
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1139
ggagtcaaag ttctgtttgc cctgatctgc atcgctgtgg ccgaggcc 48
<210> 1140
<211> 99
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1140
attcatacca acttgaagaa aaagttcagc ctcttcatcc tggtctttct cctgttcgca 60
gtcatctgtg tttggaagaa agggagcgac tatgaggcc 99
<210> 1141
<211> 588
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1141
gcccctctcc ctcccccccc cctaacgtta ctggccgaag ccgcttggaa taaggccggt 60
gtgcgtttgt ctatatgtta ttttccacca tattgccgtc ttttggcaat gtgagggccc 120
ggaaacctgg ccctgtcttc ttgacgagca ttcctagggg tctttcccct ctcgccaaag 180
gaatgcaagg tctgttgaat gtcgtgaagg aagcagttcc tctggaagct tcttgaagac 240
aaacaacgtc tgtagcgacc ctttgcaggc agcggaaccc cccacctggc gacaggtgcc 300
tctgcggcca aaagccacgt gtataagata cacctgcaaa ggcggcacaa ccccagtgcc 360
acgttgtgag ttggatagtt gtggaaagag tcaaatggct ctcctcaagc gtattcaaca 420
aggggctgaa ggatgcccag aaggtacccc attgtatggg atctgatctg gggcctcggt 480
gcacatgctt tacatgtgtt tagtcgaggt taaaaaaacg tctaggcccc ccgaaccacg 540
gggacgtggt tttcctttga aaaacacgat gataatatgg ccacaacc 588
<210> 1142
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1142
gcccgggcgt cgggcgacct ttggtcgccc g 31
<210> 1143
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1143
gcccgggcgt cgggcgacct ttggtcgccc g 31
<210> 1144
<211> 2218
<212> ДНК
<213> Homo sapiens
<400> 1144
cagggcagcc ttcagtctga ttcaggagaa cgaggtcctc ttcaccatgt gcttcatccc 60
cctggtctgc tggatcgtgt gcactggact gaaacagcag atggagagtg gcaagagcct 120
tgcccagaca tccaagacct ccaccgcggt gtacgtcttc ttcctttcca gtttgctgca 180
gccccgggga gggagccagg agcacggcct ctgcgcccac ctctgggggc tctgctcttt 240
ggctgcagat ggaatctgga accagaaaat cctgtttgaa gagtccgacc tcaggaatca 300
tggactgcag aaggcggatg tgtctgcttt cctgaggatg aacctgttcc aaaaggaagt 360
ggactgcgag aagttctaca gcttcatcca catgactttc caggagttct ttgccgccat 420
gtactacctg ctggaagagg aaaaggaagg aaggacgaac gttccaggga gtcgtttgaa 480
gcttcccagc cgagacgtga cagtccttct ggaaaactat ggcaaattcg aaaaggggta 540
tttgattttt gttgtacgtt tcctctttgg cctggtaaac caggagagga cctcctactt 600
ggagaagaaa ttaagttgca tgatctctca gcaaatcagg ctggagctgc tgaaatggat 660
tgaagtgaaa gccaaagcta aaaagctgca tgatcagccc agccagctgg aattgttcta 720
ctgtttgtac gagatgcagg aggaggactt cgtgcaaagg gccatggact atttccccaa 780
gattgagatc aatctctcca ccagaatgga ccacatggtt tcctcctttt gcattgagaa 840
ctgtcatcgg gtggagtcac tgtccctggg gtttctccat aacatgccca aggaggaaga 900
ggaggaggaa aaggaaggcc gacaccttga tatggtgcag tgtgtcctcc caagctcctc 960
tcatgctgcc tgttctcatg ggttggggcg ctgtggcctc tcccatgagt gctgcttcga 1020
catctccttg gtcctcagca gcaaccagaa gctggtggag ctggacctga gtgacaacgc 1080
cctcggtgac ttcggaatca gacttctgtg tgtgggactg aagcacctgt tgtgcaatct 1140
gaagaagctc tggttggtga attctgcctt acgtcagtct gttgttcagc tttgtcctcg 1200
gtactcagca ctaatcagaa tctcacgcac ctttactgcg aggcaacact ctcggagaca 1260
agggatcaaa ctactctgtg agggactctt gcaccccgac tgcaagcttc aggtgttgga 1320
attagacaac tgcaacctca cgtcacactg ctgctgggat ctttccacac ttctgacctc 1380
cagccagagc ctgcgaaagc tgagcctggg caacaatgac ctgggcgacc tgggggtcat 1440
gatgttctgt gaagtgctga aacagcagag ctgcctcctg cagaacctgg ggttgtctga 1500
aatgtatttc aattatgaga caaaaagtgc gttagaaaca cttcaagaag aaaagcctga 1560
gctgaccgtc gtctttgagc cttcttggta ggagtggaaa cggggctgcc agacgccagt 1620
gttctccggt ccctccagct gggggccctc aggtggagag agctgcgatc catccaggcc 1680
aagaccacag ctctgtgatc cttccggtgg agtgtcggag aagagagctt gccgacgatg 1740
ccttcctgtg cagagcttgg gcatctcctt tacgccaggg tgaggaagac accaggacaa 1800
tgacagcatc gggtgttgtt gtcatcacag cgcctcagtt agaggatgtt cctctggtga 1860
cctcatgtaa ttagctcatt caataaagca ctttctttat ttttctcttc tctgtctaac 1920
cttctttttc ctatcttttt ttcttctttg ttctgtttac ttttgctcat atcatcattc 1980
ccgctatctt tctattaact gaccataaca cagaactagt tgactatata ttatgttgaa 2040
attttatggc agctatttat ttatttaaat tttttgtaat agttttgttt tctaataaga 2100
aaaatccatg ctttttgtag ctggttgaaa attcaggaat atgtaaaact ttttggtatt 2160
taattaaatt gattcctttt cttaatttta aaaaaaaaaa aaaaaaaaaa aaaaaaaa 2218
<210> 1145
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1145
ccggccacag tgtaacctgc agaaactcga gtttctgcag gttacactgt ggtttttg 58
<210> 1146
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1146
gcgcgctcgc tcgctc 16
<210> 1147
<211> 518
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1147
tcaatattgg ccattagcca tattattcat tggttatata gcataaatca atattggcta 60
ttggccattg catacgttgt atctatatca taatatgtac atttatattg gctcatgtcc 120
aatatgaccg ccatgttggc attgattatt gactagttat taatagtaat caattacggg 180
gtcattagtt catagcccat atatggagtt ccgcgttaca taacttacgg taaatggccc 240
gcctggctga ccgcccaacg acccccgccc attgacgtca ataatgacgt atgttcccat 300
agtaacgcca atagggactt tccattgacg tcaatgggtg gagtatttac ggtaaactgc 360
ccacttggca gtacatcaag tgtatcatat gccaagtccg ccccctattg acgtcaatga 420
cggtaaatgg cccgcctggc attatgccca gtacatgacc ttacgggact ttcctacttg 480
gcagtacatc tacgtattag tcatcgctat taccatgg 518
<210> 1148
<211> 777
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1148
aggctcagag gcacacagga gtttctgggc tcaccctgcc cccttccaac ccctcagttc 60
ccatcctcca gcagctgttt gtgtgctgcc tctgaagtcc acactgaaca aacttcagcc 120
tactcatgtc cctaaaatgg gcaaacattg caagcagcaa acagcaaaca cacagccctc 180
cctgcctgct gaccttggag ctggggcaga ggtcagagac ctctctgggc ccatgccacc 240
tccaacatcc actcgacccc ttggaatttc ggtggagagg agcagaggtt gtcctggcgt 300
ggtttaggta gtgtgagagg gtccgggttc aaaaccactt gctgggtggg gagtcgtcag 360
taagtggcta tgccccgacc ccgaagcctg tttccccatc tgtacaatgg aaatgataaa 420
gacgcccatc tgatagggtt tttgtggcaa ataaacattt ggtttttttg ttttgttttg 480
ttttgttttt tgagatggag gtttgctctg tcgcccaggc tggagtgcag tgacacaatc 540
tcatctcacc acaaccttcc cctgcctcag cctcccaagt agctgggatt acaagcatgt 600
gccaccacac ctggctaatt ttctattttt agtagagacg ggtttctcca tgttggtcag 660
cctcagcctc ccaagtaact gggattacag gcctgtgcca ccacacccgg ctaatttttt 720
ctatttttga cagggacggg gtttcaccat gttggtcagg ctggtctaga ggtaccg 777
<210> 1149
<211> 427
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1149
gagtcaatgg gaaaaaccca ttggagccaa gtacactgac tcaataggga ctttccattg 60
ggttttgccc agtacataag gtcaataggg ggtgagtcaa caggaaagtc ccattggagc 120
caagtacatt gagtcaatag ggactttcca atgggttttg cccagtacat aaggtcaatg 180
ggaggtaagc caatgggttt ttcccattac tgacatgtat actgagtcat tagggacttt 240
ccaatgggtt ttgcccagta cataaggtca ataggggtga atcaacagga aagtcccatt 300
ggagccaagt acactgagtc aatagggact ttccattggg ttttgcccag tacaaaaggt 360
caataggggg tgagtcaatg ggtttttccc attattggca catacataag gtcaataggg 420
gtgacta 427
<210> 1150
<211> 83
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1150
cgggggaggc tgctggtgaa tattaaccaa ggtcacccca gttatcggag gagcaaacag 60
gggctaagtc cacacgcgtg gta 83
<210> 1151
<211> 777
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1151
aggctcagag gcacacagga gtttctgggc tcaccctgcc cccttccaac ccctcagttc 60
ccatcctcca gcagctgttt gtgtgctgcc tctgaagtcc acactgaaca aacttcagcc 120
tactcatgtc cctaaaatgg gcaaacattg caagcagcaa acagcaaaca cacagccctc 180
cctgcctgct gaccttggag ctggggcaga ggtcagagac ctctctgggc ccatgccacc 240
tccaacatcc actcgacccc ttggaatttc ggtggagagg agcagaggtt gtcctggcgt 300
ggtttaggta gtgtgagagg gtccgggttc aaaaccactt gctgggtggg gagtcgtcag 360
taagtggcta tgccccgacc ccgaagcctg tttccccatc tgtacaatgg aaatgataaa 420
gacgcccatc tgatagggtt tttgtggcaa ataaacattt ggtttttttg ttttgttttg 480
ttttgttttt tgagatggag gtttgctctg tcgcccaggc tggagtgcag tgacacaatc 540
tcatctcacc acaaccttcc cctgcctcag cctcccaagt agctgggatt acaagcatgt 600
gccaccacac ctggctaatt ttctattttt agtagagacg ggtttctcca tgttggtcag 660
cctcagcctc ccaagtaact gggattacag gcctgtgcca ccacacccgg ctaatttttt 720
ctatttttga cagggacggg gtttcaccat gttggtcagg ctggtctaga ggtactg 777
<210> 1152
<211> 66
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1152
gtttgctgct tgcaatgttt gcccatttta gggtggacac aggacgctgt ggtttctgag 60
ccaggg 66
<210> 1153
<211> 212
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1153
ggaggggtgg agtcgtgacc cctaaaatgg gcaaacattg caagcagcaa acagcaaaca 60
cacagccctc cctgcctgct gaccttggag ctggggcaga ggtcagagac ctctctgggc 120
ccatgccacc tccaacatcc actcgacccc ttggaatttc ggtggagagg agcagaggtt 180
gtcctggcgt ggtttaggta gtgtgagagg gg 212
<210> 1154
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1154
aggctcagag gcacacagga gtttctgggc tcaccctgcc cccttccaac ccctcagttc 60
ccatcctcca gcagctgttt gtgtgctgcc tctgaagtcc acactgaaca aacttcagcc 120
tactcatgtc cctaaaatgg gcaaacattg caagcagcaa acagcaaaca cacagccctc 180
cctgcctgct gaccttggag ctggggcaga ggtcagagac ctctctgggc ccatgccacc 240
tccaacatcc actcgacccc ttggaatttc ggtggagagg agcagaggtt gtcctggcgt 300
ggtttaggta gtgtgagagg ggtacccggg 330
<210> 1155
<211> 194
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1155
ccctaaaatg ggcaaacatt gcaagcagca aacagcaaac acacagccct ccctgcctgc 60
tgaccttgga gctggggcag aggtcagaga cctctctggg cccatgccac ctccaacatc 120
cactcgaccc cttggaattt ttcggtggag aggagcagag gttgtcctgg cgtggtttag 180
gtagtgtgag aggg 194
<210> 1156
<211> 240
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1156
gggcctgaaa taacctctga aagaggaact tggttaggta ccttctgagg ctgaaagaac 60
cagctgtgga atgtgtgtca gttagggtgt ggaaagtccc caggctcccc agcaggcaga 120
agtatgcaaa gcatgcatct caattagtca gcaaccaggt gtggaaagtc cccaggctcc 180
ccagcaggca gaagtatgca aagcatgcat ctcaattagt cagcaaccat agtcccacta 240
<210> 1157
<211> 73
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1157
cgggggaggc tgctggtgaa tattaaccaa ggtcacccca gttatcggag gagcaaacag 60
gggctaagtc cac 73
<210> 1158
<211> 100
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1158
aggttaattt ttaaaaagca gtcaaaagtc caagtggccc ttggcagcat ttactctctc 60
tgtttgctct ggttaataat ctcaggagca caaacattcc 100
<210> 1159
<211> 296
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1159
gttacataac ttatggtaaa tggcctgcct ggctgactgc ccaatgaccc ctgcccaatg 60
atgtcaataa tgatgtatgt tcccatgtaa tgccaatagg gactttccat tgatgtcaat 120
gggtggagta tttatggtaa ctgcccactt ggcagtacat caagtgtatc atatgccaag 180
tatgccccct attgatgtca atgatggtaa atggcctgcc tggcattatg cccagtacat 240
gaccttatgg gactttccta cttggcagta catctatgta ttagtcattg ctatta 296
<210> 1160
<211> 235
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1160
ggcctgaaat aacctctgaa agaggaactt ggttaggtac cttctgaggc ggaaagaacc 60
agctgtggaa tgtgtgtcag ttagggtgtg gaaagtcccc aggctcccca gcaggcagaa 120
gtatgcaaag catgcatctc aattagtcag caaccaggtg tggaaagtcc ccaggctccc 180
cagcaggcag aagtatgcaa agcatgcatc tcaattagtc agcaaccata gtccc 235
<210> 1161
<211> 1127
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1161
ggagtcgctg cgacgctgcc ttcgccccgt gccccgctcc gccgccgcct cgcgccgccc 60
gccccggctc tgactgaccg cgttactccc acaggtgagc gggcgggacg gcccttctcc 120
tccgggctgt aattagcgct tggtttaatg acggcttgtt tcttttctgt ggctgcgtga 180
aagccttgag gggctccggg agggcccttt gtgcgggggg gagcggctcg gggggtgcgt 240
gcgtgtgtgt gtgcgtgggg agcgccgcgt gcggcccgcg ctgcccggcg gctgtgagcg 300
ctgcgggcgc ggcgcggggc tttgtgcgct ccgcagtgtg cgcgagggga gcgcggccgg 360
gggcggtgcc ccgcggtgcg gggggggctg cgaggggaac aaaggctgcg tgcggggtgt 420
gtgcgtgggg gggtgagcag ggggtgtggg cgcggcggtc gggctgtaac ccccccctgc 480
acccccctcc ccgagttgct gagcacggcc cggcttcggg tgcggggctc cgtacggggc 540
gtggcgcggg gctcgccgtg ccgggcgggg ggtggcggca ggtgggggtg ccgggcgggg 600
cggggccgcc tcgggccggg gagggctcgg gggaggggcg cggcggcccc cggagcgccg 660
gcggctgtcg aggcgcggcg agccgcagcc attgcctttt atggtaatcg tgcgagaggg 720
cgcagggact tcctttgtcc caaatctgtg cggagccgaa atctgggagg cgccgccgca 780
ccccctctag cgggcgcggg gcgaagcggt gcggcgccgg caggaaggaa atgggcgggg 840
agggccttcg tgcgtcgccg cgccgccgtc cccttctccc tctccagcct cggggctgtc 900
cgcgggggga cggctgcctt cgggggggac ggggcagggc ggggttcggc ttctggcgtg 960
tgaccggcgg ctctagagcc tctgctaacc atgttttagc cttcttcttt ttcctacagc 1020
tcctgggcaa cgtgctggtt attgtgctgt ctcatcattt gtcgacagaa ttcctcgaag 1080
atccgaaggg gttcaagctt ggcattccgg tactgttggt aaagcca 1127
<210> 1162
<211> 93
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1162
ctctaaggta aatataaaat ttttaagtgt ataatgtgtt aaactactga ttctaattgt 60
ttctctcttt tagattccaa cctttggaac tga 93
<210> 1163
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1163
gccctgtctc ctcagcttca ggcaccacca ctgacctggg acagtgaatc cgga 54
<210> 1164
<211> 173
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1164
ctgccttctc cctcctgtga gtttggtaag tcactgactg tctatgcctg ggaaagggtg 60
ggcaggagat ggggcagtgc aggaaaagtg gcactatgaa ccctgcagcc ctagacaatt 120
gtactaacct tcttctcttt cctctcctga caggttggtg tacagtagct tcc 173
<210> 1165
<211> 91
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1165
aagaggtaag ggtttaaggg atggttggtt ggtggggtat taatgtttaa ttacctggag 60
cacctgcctg aaatcacttt ttttcaggtt g 91
<210> 1166
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1166
gccctgtctc ctcagcttca ggcaccacca ctgacctggg acagtgaata atta 54
<210> 1167
<211> 147
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1167
gccctgtctc ctcagcttca ggcaccacca ctgacctggg acagtgaatc cggactctaa 60
ggtaaatata aaatttttaa gtgtataatg tgttaaacta ctgattctaa ttgtttctct 120
cttttagatt ccaacctttg gaactga 147
<210> 1168
<211> 147
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1168
gccctgtctc ctcagcttca ggcaccacca ctgacctggg acagtgaata attactctaa 60
ggtaaatata aaatttttaa gtgtataatg tgttaaacta ctgattctaa ttgtttctct 120
cttttagatt ccaacctttg gaactga 147
<210> 1169
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1169
tcctcagctt caggcaccac cactgacctg ggacagtgaa tcgccacc 48
<210> 1170
<211> 128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1170
gctagcaggt aagtgccgtg tgtggttccc gcgggcctgg cctctttacg ggttatggcc 60
cttgcgtgcc ttgaattact gacactgaca tccacttttt ctttttctcc acaggtttaa 120
acgccacc 128
<210> 1171
<211> 98
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1171
aagaggtaag ggtttaagtt atcgttagtt cgtgcaccat taatgtttaa ttacctggag 60
cacctgcctg aaatcatttt tttttcaggt tggctagt 98
<210> 1172
<211> 172
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1172
gcttagtgct gagcacatcc agtgggtaaa gttccttaaa atgctctgca aagaaattgg 60
gacttttcat taaatcagaa attttacttt tttcccctcc tgggagctaa agatatttta 120
gagaagaatt aaccttttgc ttctccagtt gaacatttgt agcaataagt ca 172
<210> 1173
<211> 160
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1173
gccctgtctc ctcagcttca ggcaccacca ctgacctggg acagtgaatc cggactctaa 60
ggtaaatata aaatttttaa gtgtataatg tgttaaacta ctgattctaa ttgtttctct 120
cttttagatt ccaacctttg gaactgaatt ctagaccacc 160
<210> 1174
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1174
accactttca caatctgcta gcaaaggtt 29
<210> 1175
<211> 133
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1175
gtaagtatca aggttacaag acaggtttaa ggagaccaat agaaactggg cttgtcgaga 60
cagagaagac tcttgcgttt ctgataggca cctattggtc ttactgacat ccactttgcc 120
tttctctcca cag 133
<210> 1176
<211> 341
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1176
tgggcaggaa ctgggcactg tgcccagggc atgcactgcc tccacgcagc aaccctcaga 60
gtcctgagct gaaccaagaa ggaggagggg gtcgggcctc cgaggaaggc ctagccgctg 120
ctgctgccag gaattccagg ttggaggggc ggcaacctcc tgccagcctt caggccactc 180
tcctgtgcct gccagaagag acagagcttg aggagagctt gaggagagca ggaaagcctc 240
ccccgttgcc cctctggatc cactgcttaa atacggacga ggacagggcc ctgtctcctc 300
agcttcaggc accaccactg acctgggaca gtgaatcgac a 341
<210> 1177
<211> 316
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1177
tctagagaag ctttattgcg gtagtttatc acagttaaat tgctaacgca gtcagtgctt 60
ctgacacaac agtctcgaac ttaagctgca gtgactctct taaggtagcc ttgcagaagt 120
tggtcgtgag gcactgggca ggtaagtatc aaggttacaa gacaggttta aggagaccaa 180
tagaaactgg gcttgtcgag acagagaaga ctcttgcgtt tctgataggc acctattggt 240
cttactgaca tccactttgc ctttctctcc acaggtgtcc actcccagtt caattacagc 300
tcttaaggcc ctgcag 316
<210> 1178
<211> 76
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1178
caaagtccag gcccctctgc tgcagcgccc gcgcgtccag aggccctgcc agacacgcgc 60
gaggttcgag gctgag 76
<210> 1179
<211> 127
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1179
agaatgatga aaaccgaggt tggaaaaggt tgtgaaacct tttaactctc cacagtggag 60
tccattattt cctctggctt cctcaaattc atattcacag ggtcgttggc tgtgggttgc 120
aattacc 127
<210> 1180
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1180
atagcagagc aatcaccacc aagcctggaa taactgcaag ggctctgctg acatcttcct 60
gaggtgccaa ggaaatgagg 80
<210> 1181
<211> 208
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1181
gggtcaccac cacctccaca gcacagacag acactcagga gccagccagc caggtaagtt 60
tagtcttttt gtcttttatt tcaggtcccg gatccggtgg tggtgcaaat caaagaactg 120
ctcctcagtg gatgttgcct ttacttctag gcctgtacgg aagtgttact tctgctctaa 180
aagctgcgga attgtacccg cggccgcg 208
<210> 1182
<211> 159
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1182
aagcttctgc cttctccctc ctgtgagttt ggtaagtcac tgactgtcta tgcctgggaa 60
agggtgggca ggagatgggg cagtgcagga aaagtggcac tatgaaccct gcagccctag 120
acaattgtac taaccttctt ctctttcctc tcctgacag 159
<210> 1183
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1183
cgcgcctagc agtgtcccag ccgggttcgt gtcgcc 36
<210> 1184
<211> 141
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1184
acgccgcctg ggtcccagtc cccgtcccat cccccggcgg cctaggcagc gtttccagcc 60
ccgagaactt tgttcttttt gtcccgcccc ctgcgcccaa ccgcctgcgc cgccttccgg 120
cccgagttct ggagactcaa c 141
<210> 1185
<211> 110
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1185
gttggatgaa accttcctcc tactgcacag cccgcccccc tacagccccg gtccccacgc 60
ctagaagaca gcggaactaa gaaaagaaga ggcctgtgga cagaacaatc 110
<210> 1186
<211> 164
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1186
ggtggggcgg ggttgagtcg gaaccacaat agccaggcga agaaactaca actcccaggg 60
cgtcccggag caggccaacg ggactacggg aagcagcggg cagcggcccg cgggaggcac 120
ctcggagatc tgggtgcaaa agcccagggt taggaaccgt aggc 164
<210> 1187
<211> 127
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1187
ggccaccgga attaaccctt cagggctggg ggccgcgcta tgccccgccc cctccccagc 60
cccagacacg gaccccgcag gagatgggtg cccccatccg cacactgtcc tttggccacc 120
ggacatc 127
<210> 1188
<211> 341
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1188
tgggcaggaa ctgggcactg tgcccagggc atgcactgcc tccacgcagc aaccctcaga 60
gtcctgagct gaaccaagaa ggaggagggg gtcgggcctc cgaggaaggc ctagccgctg 120
ctgctgccag gaattccagg ttggaggggc ggcaacctcc tgccagcctt caggccactc 180
tcctgtgcct gccagaagag acagagcttg aggagagctt gaggagagca ggaaagcctc 240
ccccgttgcc cctctggatt cactgcttaa atacggacga ggacagggcc ctgtctcctc 300
agcttcaggc accaccactg acctgggaca gtgaatcgac a 341
<210> 1189
<211> 77
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1189
tccataaagt aggaaacact acacgattcc ataaagtagg aaacactaca tcactccata 60
aagtaggaaa cactaca 77
<210> 1190
<211> 88
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1190
tgaaagatgg atttccaagg ttaattcatt ggaattgaaa attaacagag atctagagct 60
gaattcctgc agccaggggg atcagcct 88
<210> 1191
<211> 395
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1191
taaaatacag catagcaaaa ctttaacctc caaatcaagc ctctacttga atccttttct 60
gagggatgaa taaggcatag gcatcagggg ctgttgccaa tgtgcattag ctgtttgcag 120
cctcaccttc tttcatggag tttaagatat agtgtatttt cccaaggttt gaactagctc 180
ttcatttctt tatgttttaa atgcactgac ctcccacatt ccctttttag taaaatattc 240
agaaataatt taaatacatc attgcaatga aaataaatgt tttttattag gcagaatcca 300
gatgctcaag gcccttcata atatccccca gtttagtagt tggacttagg gaacaaagga 360
acctttaata gaaattggac agcaagaaag cgagc 395
<210> 1192
<211> 800
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1192
agtcaatatg ttcaccccaa aaaagctgtt tgttaacttg ccaacctcat tctaaaatgt 60
atatagaagc ccaaaagaca ataacaaaaa tattcttgta gaacaaaatg ggaaagaatg 120
ttccactaaa tatcaagatt tagagcaaag catgagatgt gtggggatag acagtgaggc 180
tgataaaata gagtagagct cagaaacaga cccattgata tatgtaagtg acctatgaaa 240
aaaatatggc attttacaat gggaaaatga tggtcttttt cttttttaga aaaacaggga 300
aatatattta tatgtaaaaa ataaaaggga acccatatgt cataccatac acacaaaaaa 360
attccagtga attataagtc taaatggaga aggcaaaact ttaaatcttt tagaaaataa 420
tatagaagca tgccatcaag acttcagtgt agagaaaaat ttcttatgac tcaaagtcct 480
aaccacaaag aaaagattgt taattagatt gcatgaatat taagacttat ttttaaaatt 540
aaaaaaccat taagaaaagt caggccatag aatgacagaa aatatttgca acaccccagt 600
aaagagaatt gtaatatgca gattataaaa agaagtctta caaatcagta aaaaataaaa 660
ctagacaaaa atttgaacag atgaaagaga aactctaaat aatcattaca catgagaaac 720
tcaatctcag aaatcagaga actatcattg catatacact aaattagaga aatattaaaa 780
ggctaagtaa catctgtggc 800
<210> 1193
<211> 407
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1193
aattatctct aaggcatgtg aactggctgt cttggttttc atctgtactt catctgctac 60
ctctgtgacc tgaaacatat ttataattcc attaagctgt gcatatgata gatttatcat 120
atgtattttc cttaaaggat ttttgtaaga actaattgaa ttgatacctg taaagtcttt 180
atcacactac ccaataaata ataaatctct ttgttcagct ctctgtttct ataaatatgt 240
accagtttta ttgtttttag tggtagtgat tttattctct ttctatatat atacacacac 300
atgtgtgcat tcataaatat atacaatttt tatgaataaa aaattattag caatcaatat 360
tgaaaaccac tgatttttgt ttatgtgagc aaacagcaga ttaaaag 407
<210> 1194
<211> 186
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1194
catcacattt aaaagcatct cagcctacca tgagaataag agaaagaaaa tgaagatcaa 60
aagcttattc atctgttttt ctttttcgtt ggtgtaaagc caacaccctg tctaaaaaac 120
ataaatttct ttaatcattt tgcctctttt ctctgtgctt caattaataa aaaatggaaa 180
gaatct 186
<210> 1195
<211> 395
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1195
taaaatacag catagcaaaa ctttaacctc caaatcaagc ctctacttga atccttttct 60
gagggatgaa taaggcatag gcatcagggg ctgttgccaa tgtgcattag ctgtttgcag 120
cctcaccttc tttcatggag tttaagatat agtgtatttt cccaaggttt gaactagctc 180
ttcatttctt tatgttttaa atgcactgac ctcccacatt ccctttttag taaaatattc 240
agaaataatt taaatacatc attgcaatga aaataaatgt tttttattag gcagaatcca 300
gatgctcaag gcccttcata atatccccca gtttagtagt tggacttagg gaacaaagga 360
acctttaata gaaattggac agcaagaaag ccagc 395
<210> 1196
<211> 580
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1196
gagcatctta ccgccattta ttcccatatt tgttctgttt ttcttgattt gggtatacat 60
ttaaatgtta ataaaacaaa atggtggggc aatcatttac atttttaggg atatgtaatt 120
actagttcag gtgtattgcc acaagacaaa catgttaaga aactttcccg ttatttacgc 180
tctgttcctg ttaatcaacc tctggattac aaaatttgtg aaagattgac tgatattctt 240
aactatgttg ctccttttac gctgtgtgga tatgctgctt tatagcctct gtatctagct 300
attgcttccc gtacggcttt cgttttctcc tccttgtata aatcctggtt gctgtctctt 360
ttagaggagt tgtggcccgt tgtccgtcaa cgtggcgtgg tgtgctctgt gtttgctgac 420
gcaaccccca ctggctgggg cattgccacc acctgtcaac tcctttctgg gactttcgct 480
ttccccctcc cgatcgccac ggcagaactc atcgccgcct gccttgcccg ctgctggaca 540
ggggctaggt tgctgggcac tgataattcc gtggtgttgt 580
<210> 1197
<211> 64
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1197
cctcgccccg gacctgccct cccgccaggt gcacccacct gcaataaatg cagcgaagcc 60
ggga 64
<210> 1198
<211> 247
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1198
gataatcaac ctctggatta caaaatttgt gaaagattga ctggtattct taactatgtt 60
gctcctttta cgctatgtgg atacgctgct ttaatgcctt tgtatcatgc tattgcttcc 120
cgtatggctt tcattttctc ctccttgtat aaatcctggt tagttcttgc cacggcggaa 180
ctcatcgccg cctgccttgc ccgctgctgg acaggggctc ggctgttggg cactgacaat 240
tccgtgg 247
<210> 1199
<211> 144
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1199
aaatacatca ttgcaatgaa aataaatgtt ttttattagg cagaatccag atgctcaagg 60
cccttcataa tatcccccag tttagtagtt ggacttaggg aacaaaggaa cctttaatag 120
aaattggaca gcaagaaagc gagc 144
<210> 1200
<211> 62
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1200
gagcatctta ccgccattta ttcccatatt tgttctgttt ttcttgattt gggtatacat 60
tt 62
<210> 1201
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1201
aataaaagat ctttattttc attagatctg tgtgttggtt ttttgtgtg 49
<210> 1202
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1202
gcggccgcaa taaaagatca gagctctaga gatctgtgtg ttggtttttt gtgt 54
<210> 1203
<211> 74
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1203
ggatccaata aaatatcttt attttcatta catctgtgtg ttggtttttt gtgtgttttc 60
ctgtaacgat cggg 74
<210> 1204
<211> 143
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1204
ctcgatgctt tatttgtgaa atttgtgatg ctattgcttt atttgtaacc attataagct 60
gcaataaaca agttaacaac aacaattgca ttcattttat gtttcaggtt cagggggagg 120
tgtgggaggt tttttaaact agt 143
<210> 1205
<211> 228
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1205
ctactgtgcc ttctagttgc cagccatctg ttgtttgccc ctcccccttg ccttccttga 60
ccctggaagg tgccactccc actgtccttt cctaataaaa tgaggaaatt gcatcacatt 120
gtctgagtag gtgtcattct attctggggg gtggggtggg gcaggacagc aagggggagg 180
attgggaaga caatagcagg catgctgggg atgcagtggg ctctatgg 228
<210> 1206
<211> 222
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1206
cagacatgat aagatacatt gatgagtttg gacaaaccac aactagaatg cagtgaaaaa 60
aatgctttat ttgtgaaatt tgtgatgcta ttgctttatt tgtaaccatt ataagctgca 120
ataaacaagt taacaacaac aattgcattc attttatgtt tcaggttcag ggggagatgt 180
gggaggtttt ttaaagcaag taaaacctct acaaatgtgg ta 222
<210> 1207
<211> 226
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1207
ccagacatga taagatacat tgatgagttt ggacaaacca caactagaat gcagtgaaaa 60
aaatgcttta tttgtgaaat ttgtgatgct attgctttat ttgtaaccat tataagctgc 120
aataaacaag ttaacaacaa caattgcatt cattttatgt ttcaggttca gggggaggtg 180
tgggaggttt tttaaagcaa gtaaaacctc tacaaatgtg gtatgg 226
<210> 1208
<211> 129
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1208
gttaacaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 60
aaaaaaaaaa tgcatccccc cccccccccc cccccccccc ccccccaaag gctcttttca 120
gagccacca 129
<210> 1209
<211> 232
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1209
gcggccgcgg ggatccagac atgataagat acattgatga gtttggacaa accacaacta 60
gaatgcagtg aaaaaaatgc tttatttgtg aaatttgtga tgctattgct ttatttgtaa 120
ccattataag ctgcaataaa caagttaaca acaacaattg cattcatttt atgtttcagg 180
ttcaggggga ggtgtgggag gttttttagt cgaccatgct ggggagagat ct 232
<210> 1210
<211> 135
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1210
gatccagaca tgataagata cattgatgag tttggacaaa ccacaactag aatgcagtga 60
aaaaaatgct ttatttgtga aatttgtgat gctattgctt tatttgtaac cattataagc 120
tgcaataaac aagtt 135
<210> 1211
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1211
cggcaataaa aagacagaat aaaacgcacg ggtgttgggt cgtttgttc 49
<210> 1212
<211> 226
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1212
ccataccaca tttgtagagg ttttacttgc tttaaaaaac ctcccacacc tccccctgaa 60
cctgaaacat aaaatgaatg caattgttgt tgttaacttg tttattgcag cttataatgg 120
ttacaaataa agcaatagca tcacaaattt cacaaataaa gcattttttt cactgcattc 180
tagttgtggt ttgtccaaac tcatcaatgt atcttatcat gtctgg 226
<210> 1213
<211> 416
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1213
catcacattt aaaagcatct cagcctacca tgagaataag agaaagaaaa tgaagatcaa 60
aagcttattc atctgttttt ctttttcgtt ggtgtaaagc caacaccctg tctaaaaaac 120
ataaatttct ttaatcattt tgcctctttt ctctgtgctt caattaataa aaaatggaaa 180
gaatctaata gagtggtaca gcactgttat ttttcaaaga tgtgttgcta tcctgaaaat 240
tctgtaggtt ctgtggaagt tccagtgttc tctcttattc cacttcggta gaggatttct 300
agtttcttgt gggctaatta aataaatcat taatactctt ctaagttatg gattataaac 360
attcaaaata atattttgac attatgataa ttctgaataa aagaacaaaa accatg 416
<210> 1214
<211> 415
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1214
atcacattta aaagcatctc agcctaccat gagaataaga gaaagaaaat gaagatcaaa 60
agcttattca tctgtttttc tttttcgttg gtgtaaagcc aacaccctgt ctaaaaaaca 120
taaatttctt taatcatttt gcctcttttc tctgtgcttc aattaataaa aaatggaaag 180
aatctaatag agtggtacag cactgttatt tttcaaagat gtgttgctat cctgaaaatt 240
ctgtaggttc tgtggaagtt ccagtgttct ctcttattcc acttcggtag aggatttcta 300
gtttcttgtg ggctaattaa ataaatcatt aatactcttc taagttatgg attataaaca 360
ttcaaaataa tattttgaca ttatgataat tctgaataaa agaacaaaaa ccatg 415
<210> 1215
<211> 122
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1215
taagatacat tgatgagttt ggacaaacca caactagaat gcagtgaaaa aaatgcttta 60
tttgtgaaat ttgtgatgct attgctttat ttgtaaccat tataagctgc aataaacaag 120
tt 122
<210> 1216
<211> 133
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический полинуклеотид
<400> 1216
tgctttattt gtgaaatttg tgatgctatt gctttatttg taaccattat aagctgcaat 60
aaacaagtta acaacaacaa ttgcattcat tttatgtttc aggttcaggg ggaggtgtgg 120
gaggtttttt aaa 133
<210> 1217
<211> 64
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 1217
aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa aaaaaaaaaa 60
aaaa 64
<210> 1218
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 1218
Tyr Val Ala Asp
1
<210> 1219
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 1219
Tyr Val His Asp
1
<210> 1220
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 1220
Trp Glu His Asp
1
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МОДУЛЯЦИЯ АКТИВНОСТИ REP БЕЛКА ПРИ ПОЛУЧЕНИИ ДНК С ЗАМКНУТЫМИ КОНЦАМИ (ЗКДНК) | 2020 |
|
RU2812850C2 |
| НЕВИРУСНЫЕ ДНК-ВЕКТОРЫ И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ ДЛЯ ЭКСПРЕССИИ ТЕРАПЕВТИЧЕСКОГО СРЕДСТВА НА ОСНОВЕ ФАКТОРА VIII (FVIII) | 2020 |
|
RU2812852C2 |
| НЕВИРУСНЫЕ ДНК-ВЕКТОРЫ И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ ДЛЯ ЭКСПРЕССИИ ТЕРАПЕВТИЧЕСКОГО СРЕДСТВА НА ОСНОВЕ ФЕНИЛАЛАНИНГИДРОКСИЛАЗЫ (PAH) | 2020 |
|
RU2814137C2 |
| ДНК-ВЕКТОРЫ С ЗАМКНУТЫМИ КОНЦАМИ, ПОЛУЧАЕМЫЕ ПУТЕМ БЕСКЛЕТОЧНОГО СИНТЕЗА, И СПОСОБ ПОЛУЧЕНИЯ зкДНК-ВЕКТОРОВ | 2019 |
|
RU2820586C2 |
| КОНТРОЛИРУЕМАЯ ЭКСПРЕССИЯ ТРАНСГЕНОВ С ИСПОЛЬЗОВАНИЕМ ДНК-ВЕКТОРОВ С ЗАМКНУТЫМИ КОНЦАМИ (зкДНК) | 2019 |
|
RU2816871C2 |
| НЕВИРУСНЫЕ ДНК-ВЕКТОРЫ И ИХ ПРИМЕНЕНИЕ ДЛЯ ПРОДУЦИРОВАНИЯ АНТИТЕЛ И СЛИТЫХ БЕЛКОВ | 2019 |
|
RU2800914C2 |
| МОДИФИЦИРОВАННАЯ ДНК С ЗАМКНУТЫМИ КОНЦАМИ (зкДНК) | 2018 |
|
RU2800026C2 |
| СПОСОБЫ ТЕРАПИИ НА ОСНОВЕ CAR Т-КЛЕТОК С ПОВЫШЕННОЙ ЭФФЕКТИВНОСТЬЮ | 2016 |
|
RU2788131C2 |
| АНТИТЕЛА К ENTPD2, ВИДЫ КОМБИНИРОВАННОЙ ТЕРАПИИ И СПОСОБЫ ПРИМЕНЕНИЯ АНТИТЕЛ И ВИДОВ КОМБИНИРОВАННОЙ ТЕРАПИИ | 2019 |
|
RU2790991C2 |
| CD20 ТЕРАПИЯ, CD22 ТЕРАПИЯ И КОМБИНИРОВАННАЯ ТЕРАПИЯ КЛЕТКАМИ, ЭКСПРЕССИРУЮЩИМИ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) K CD19 | 2016 |
|
RU2752918C2 |
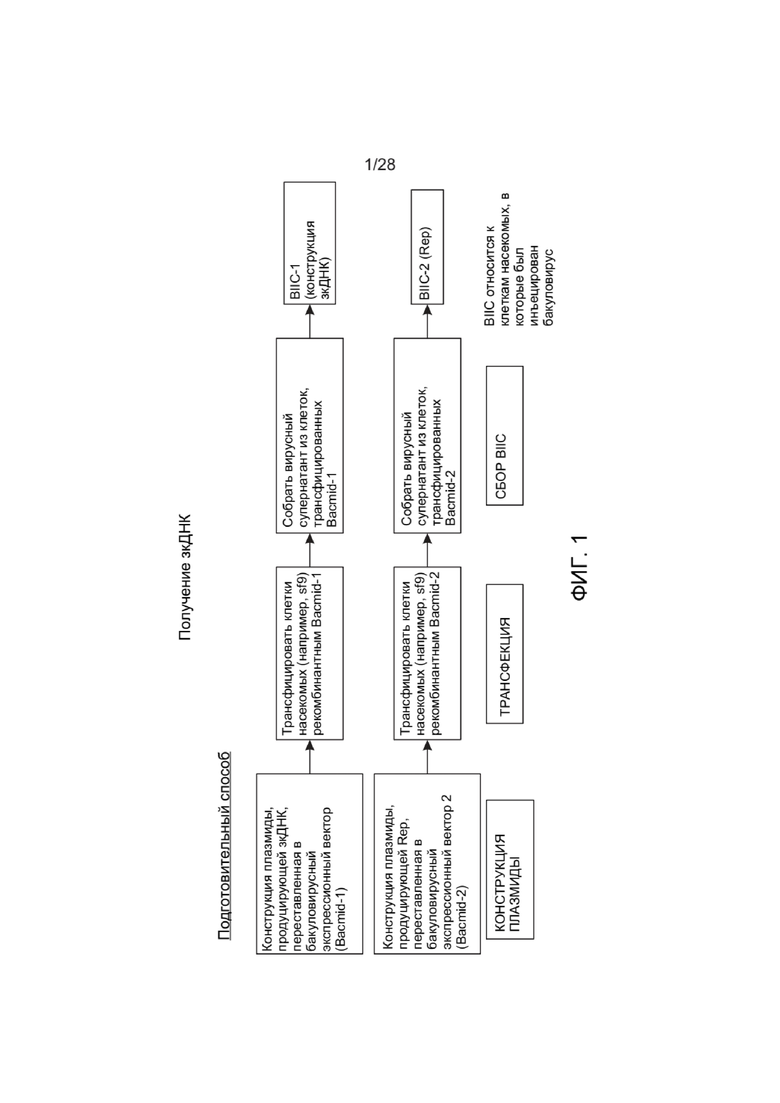
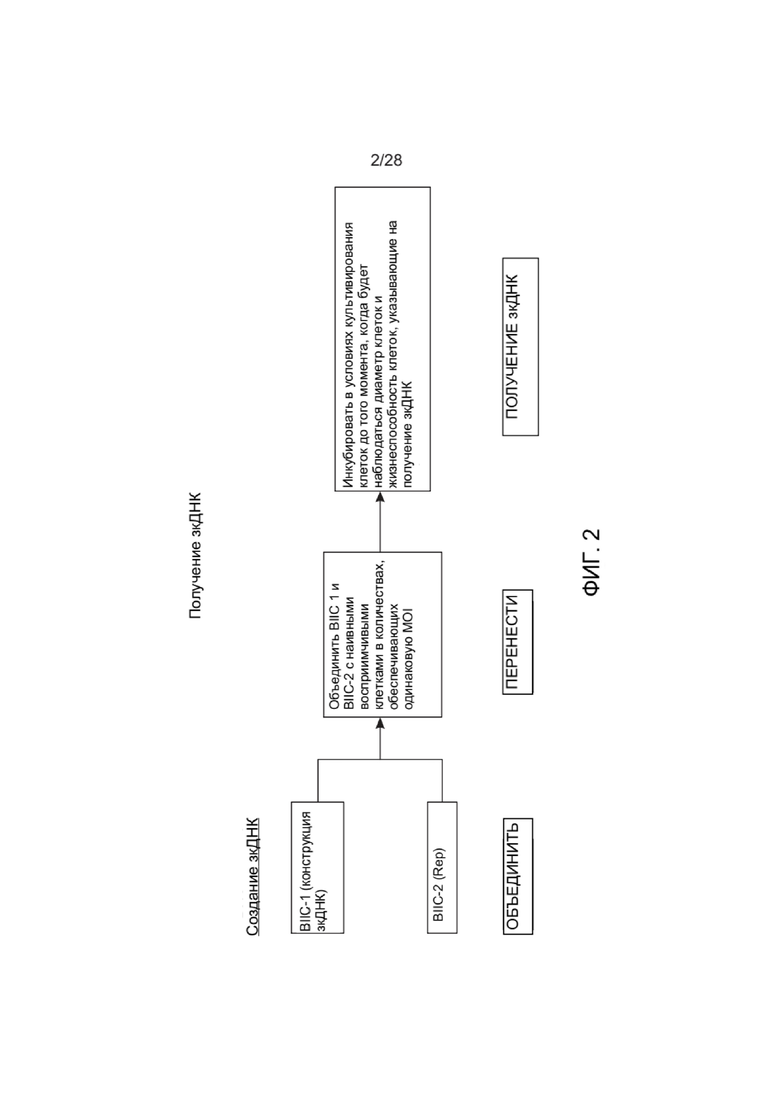
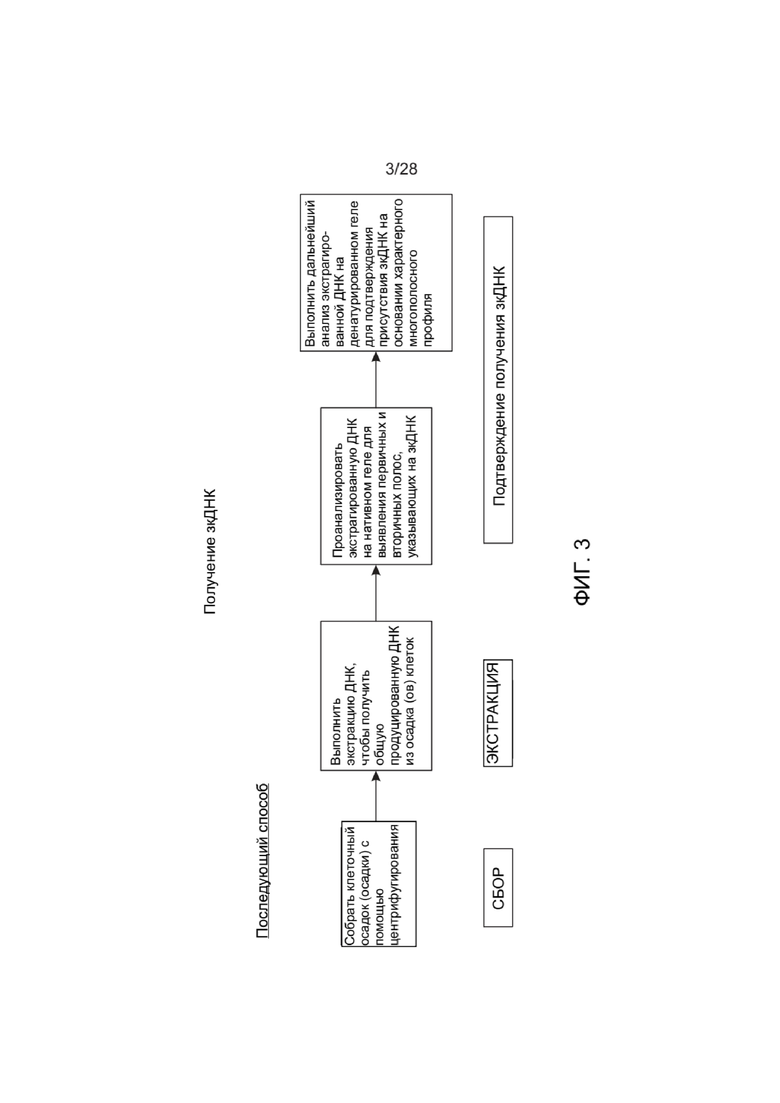
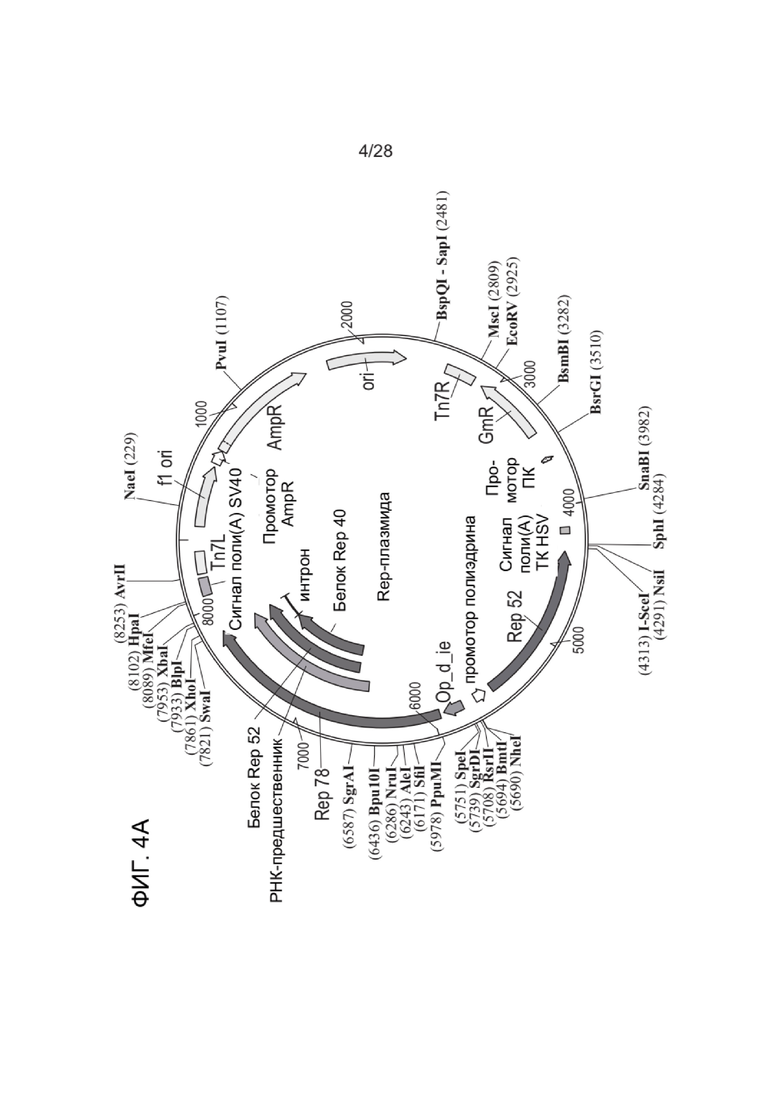
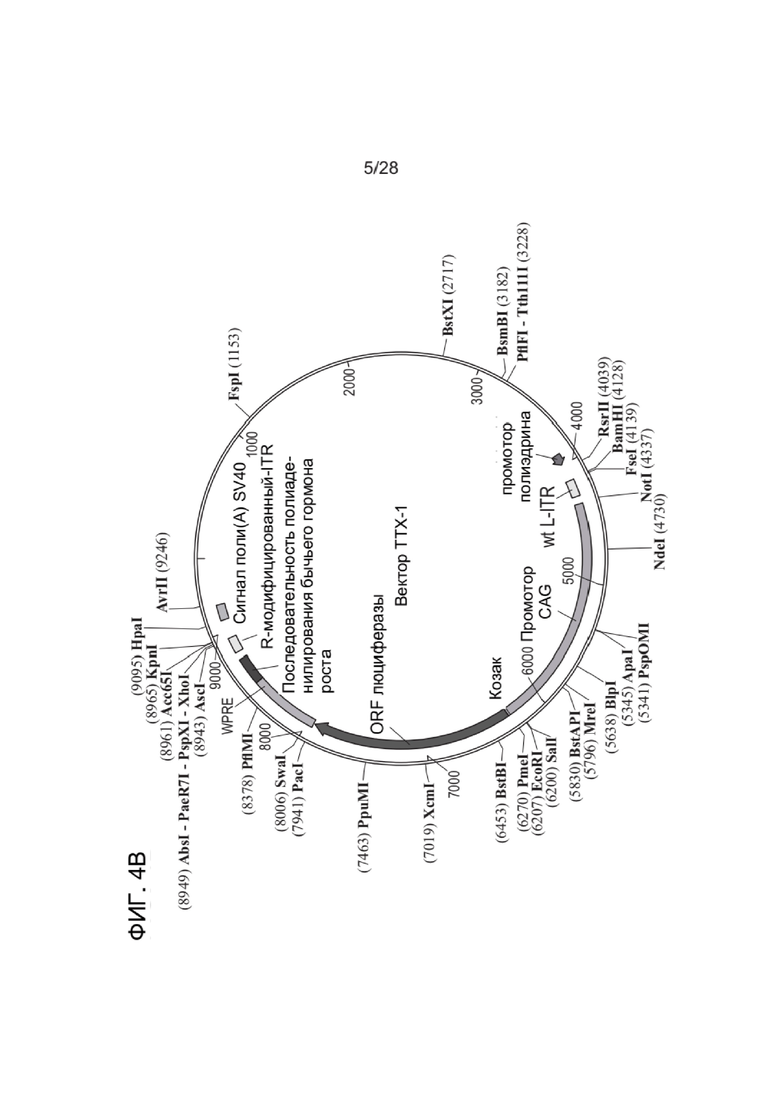
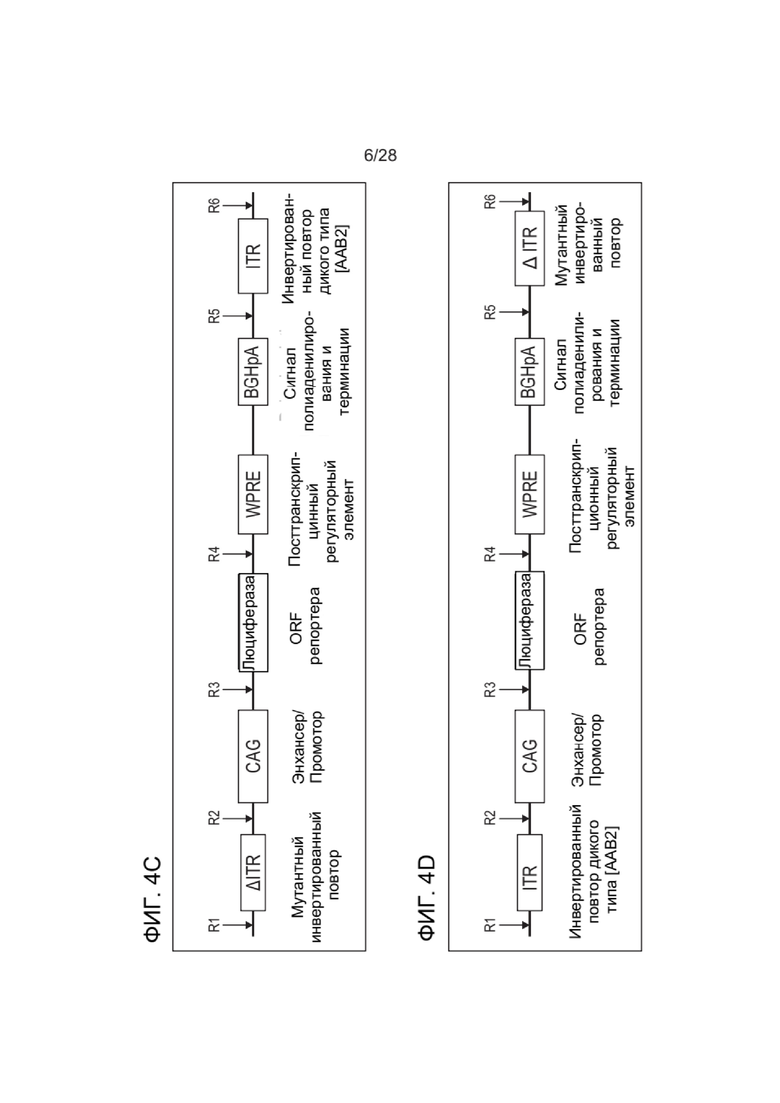
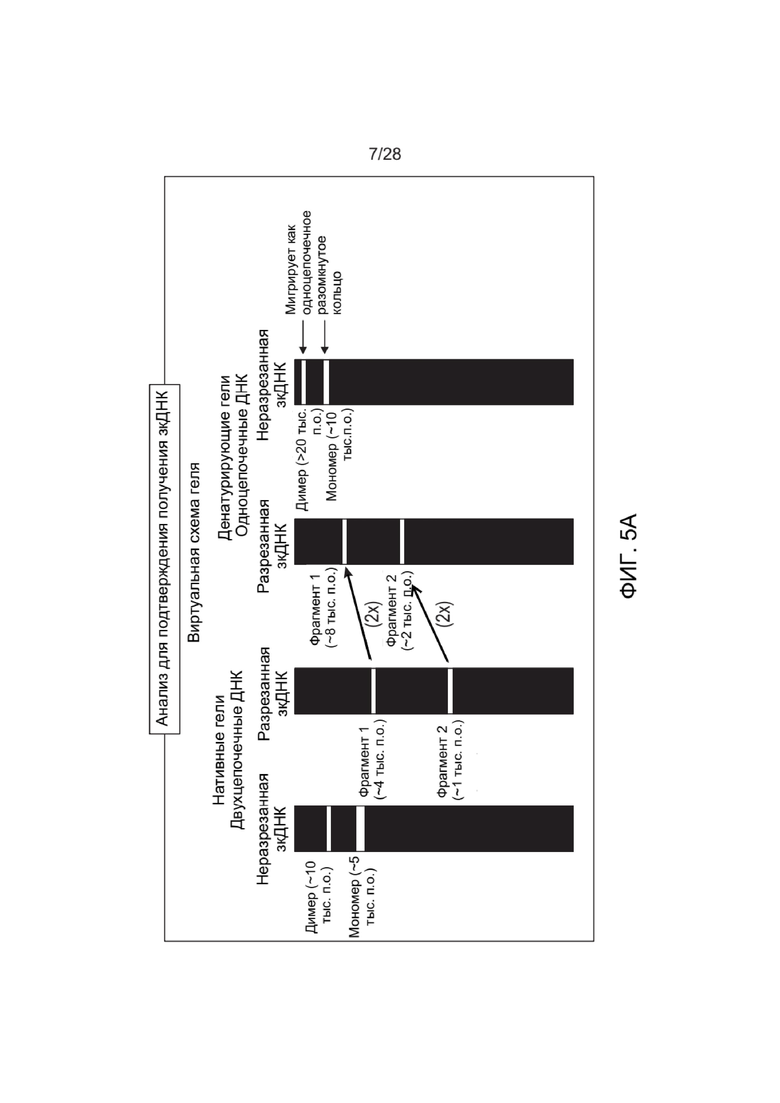

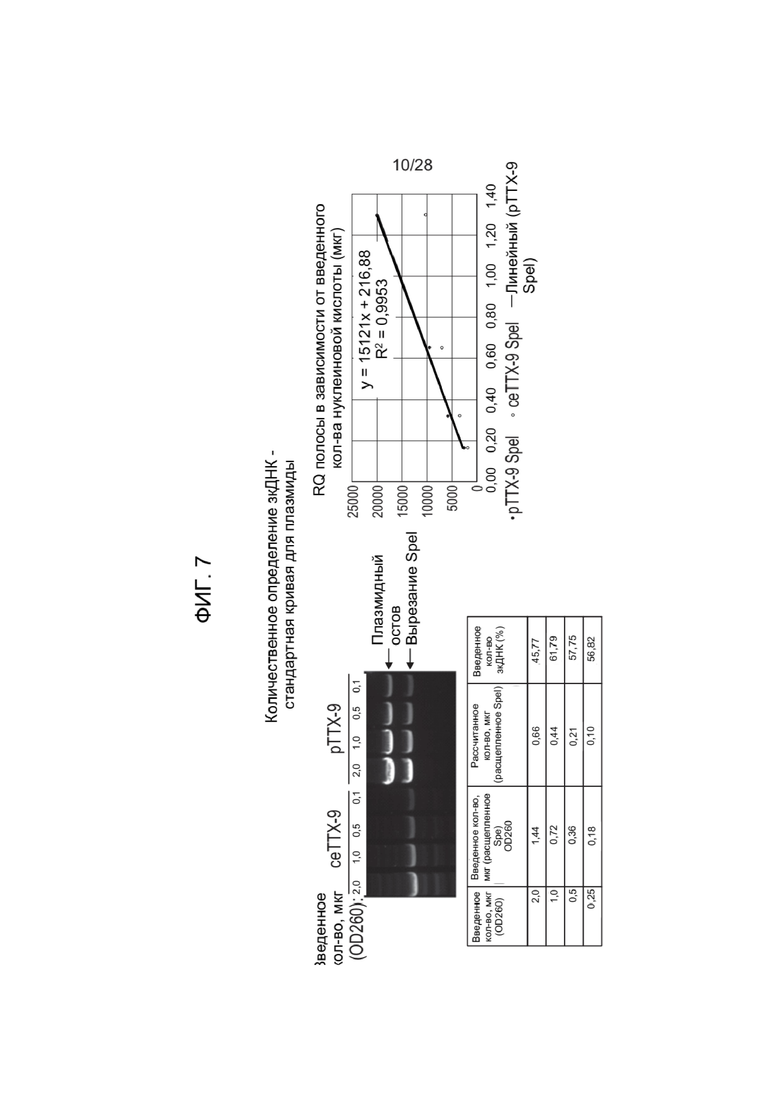
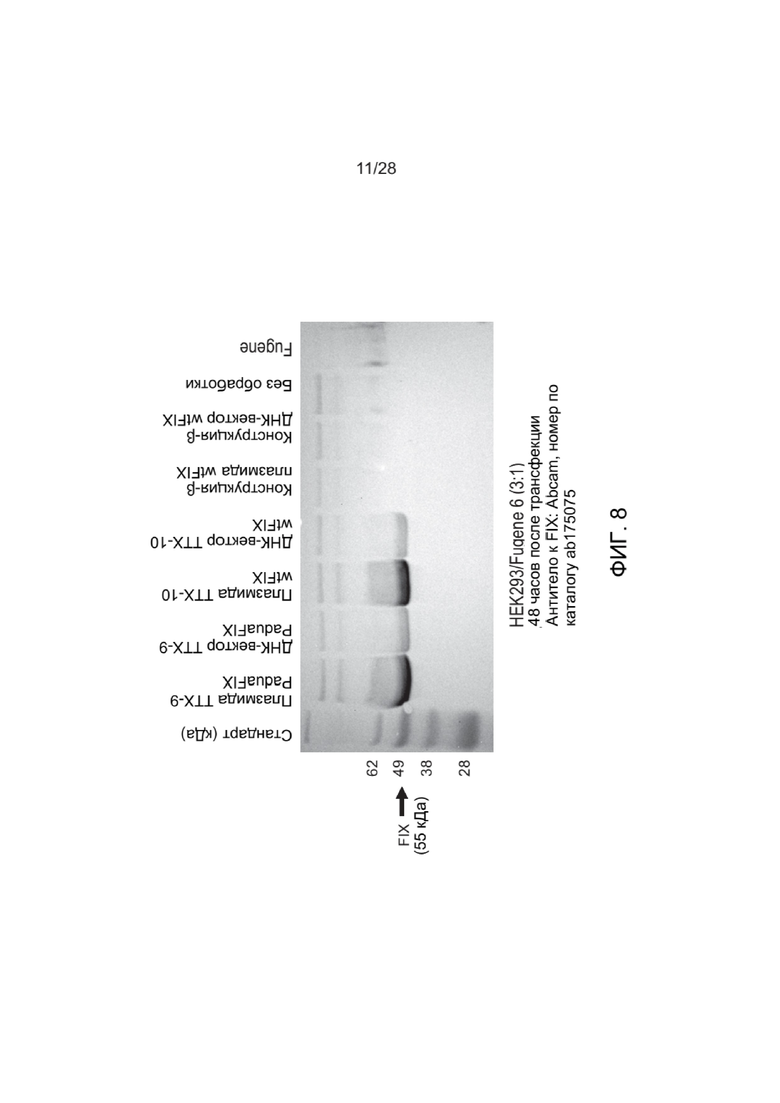
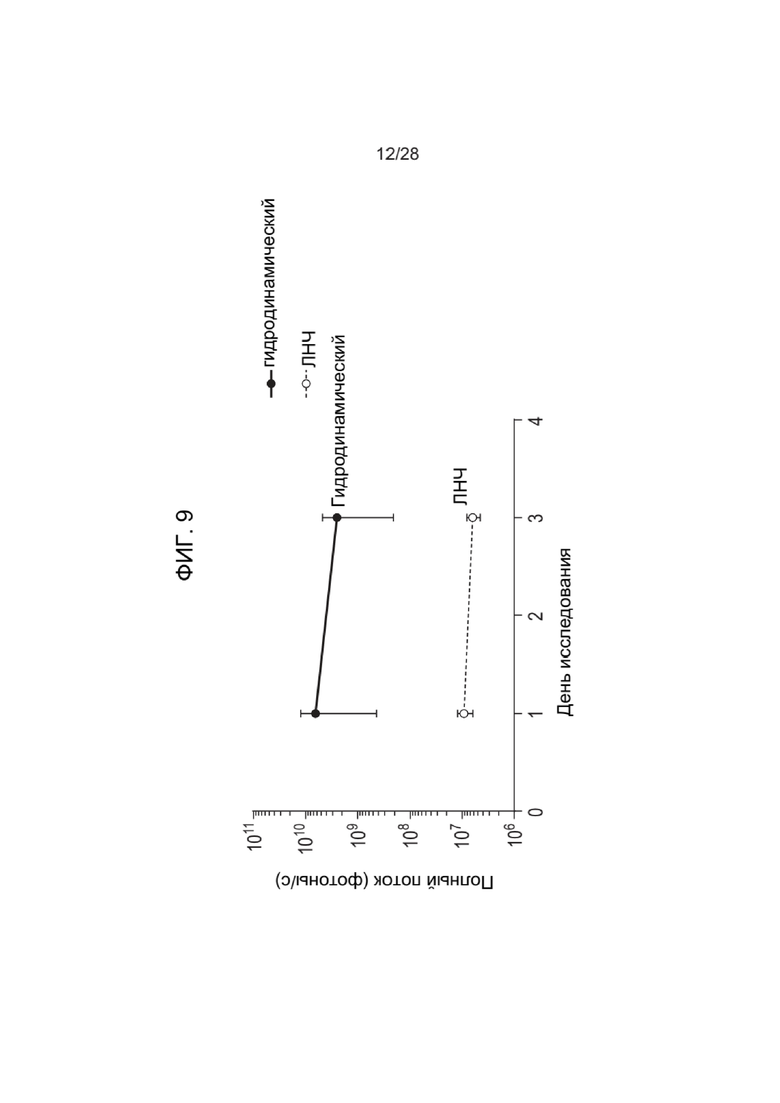
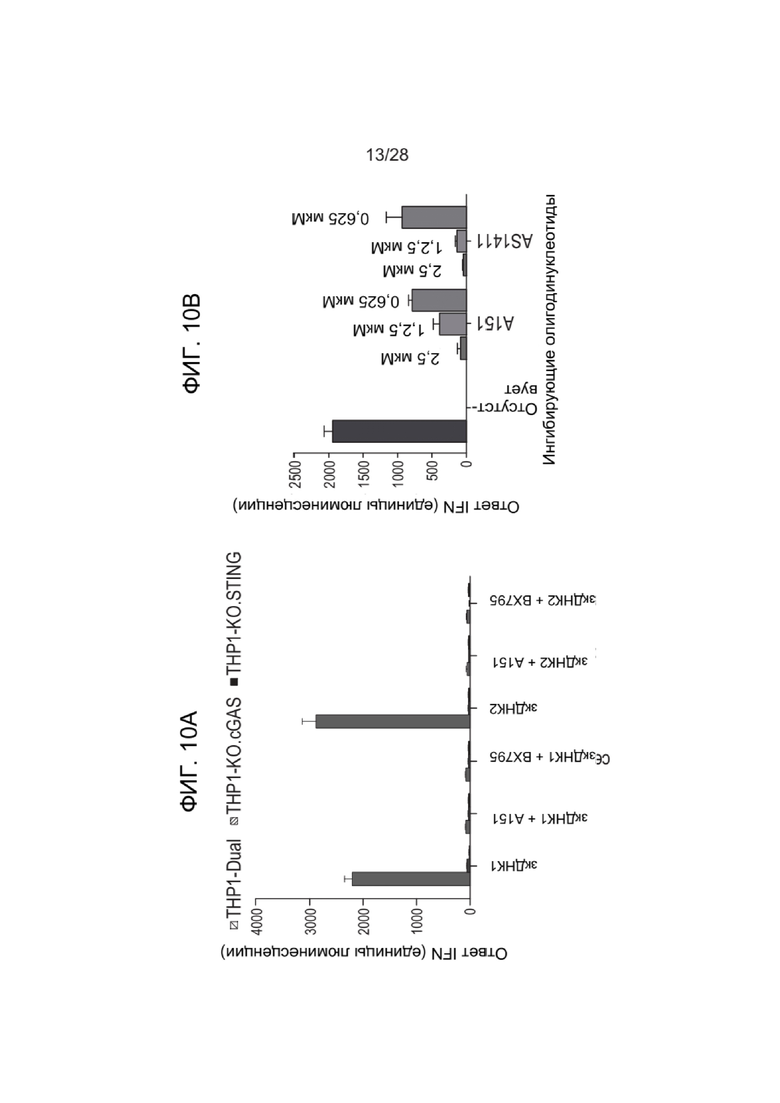


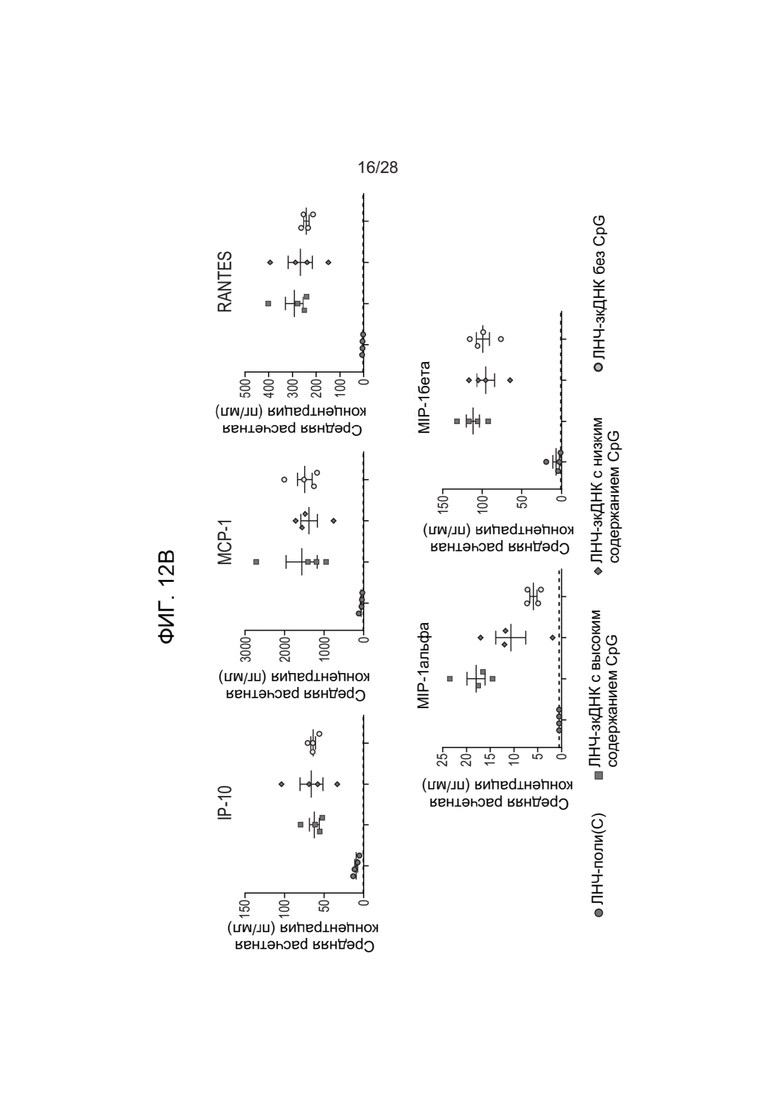
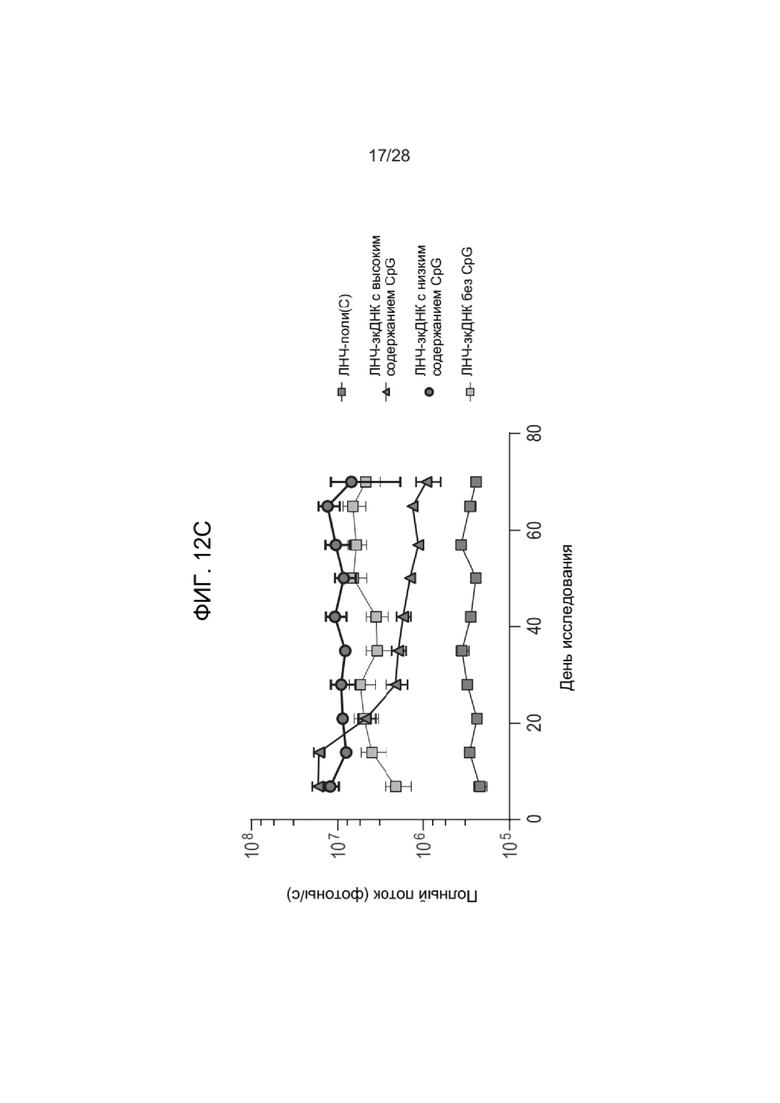
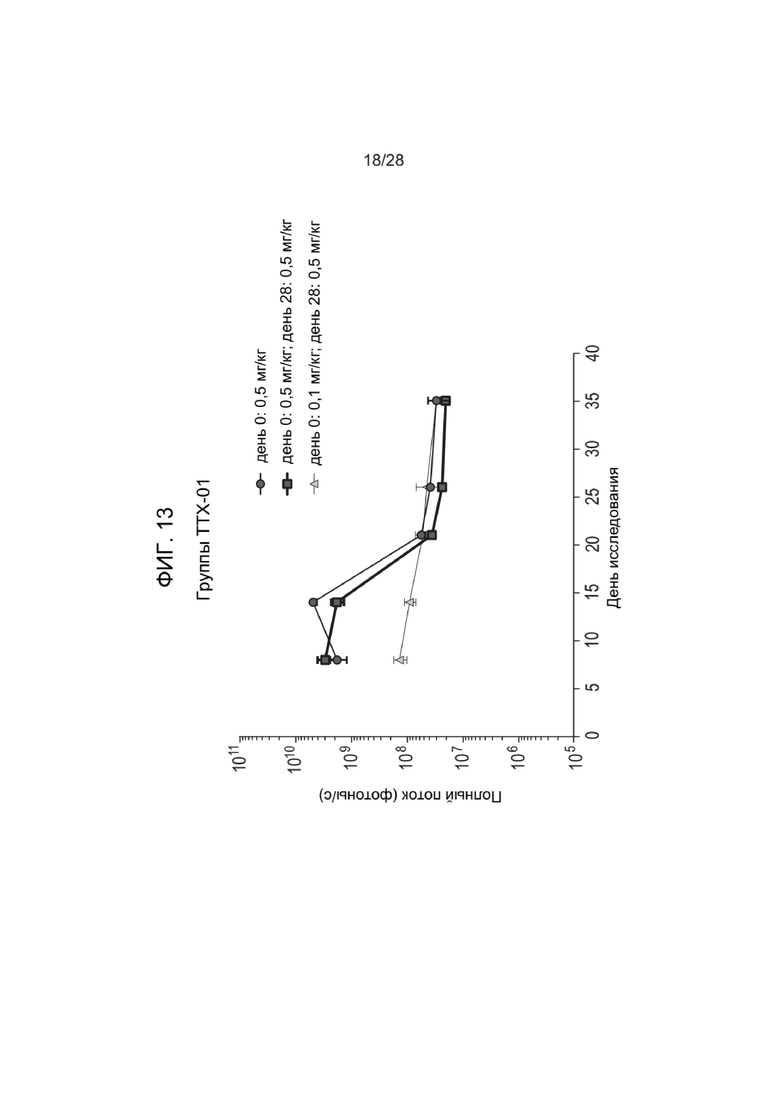

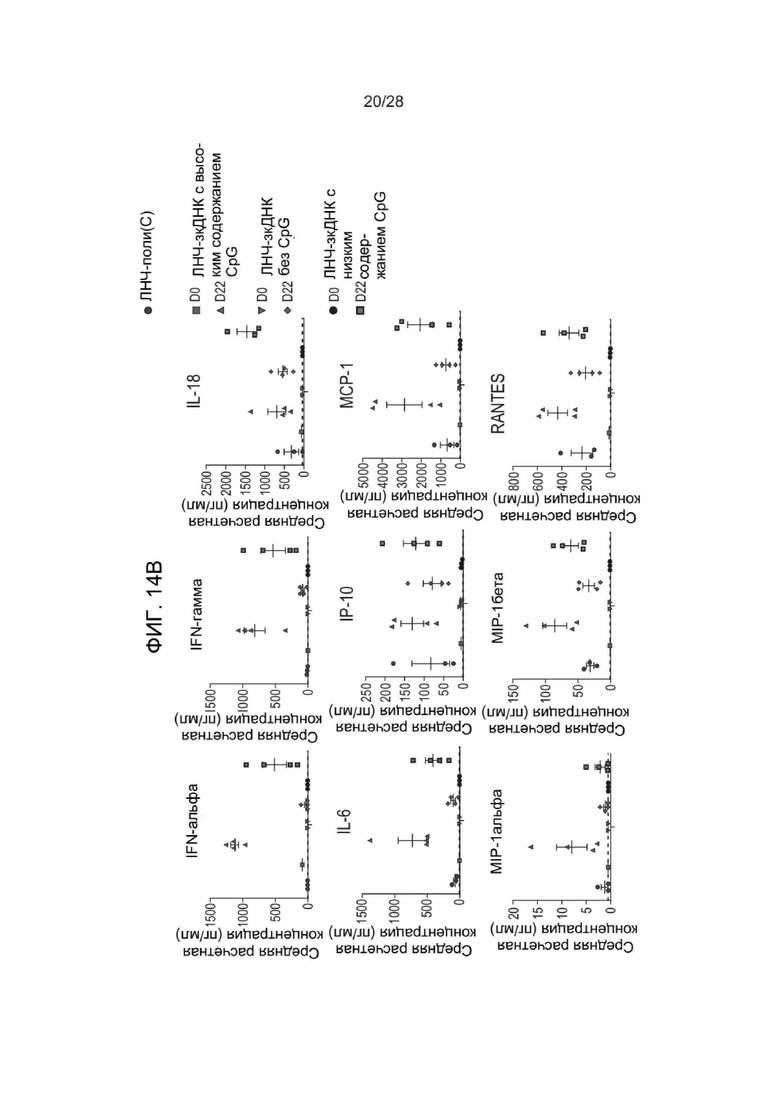
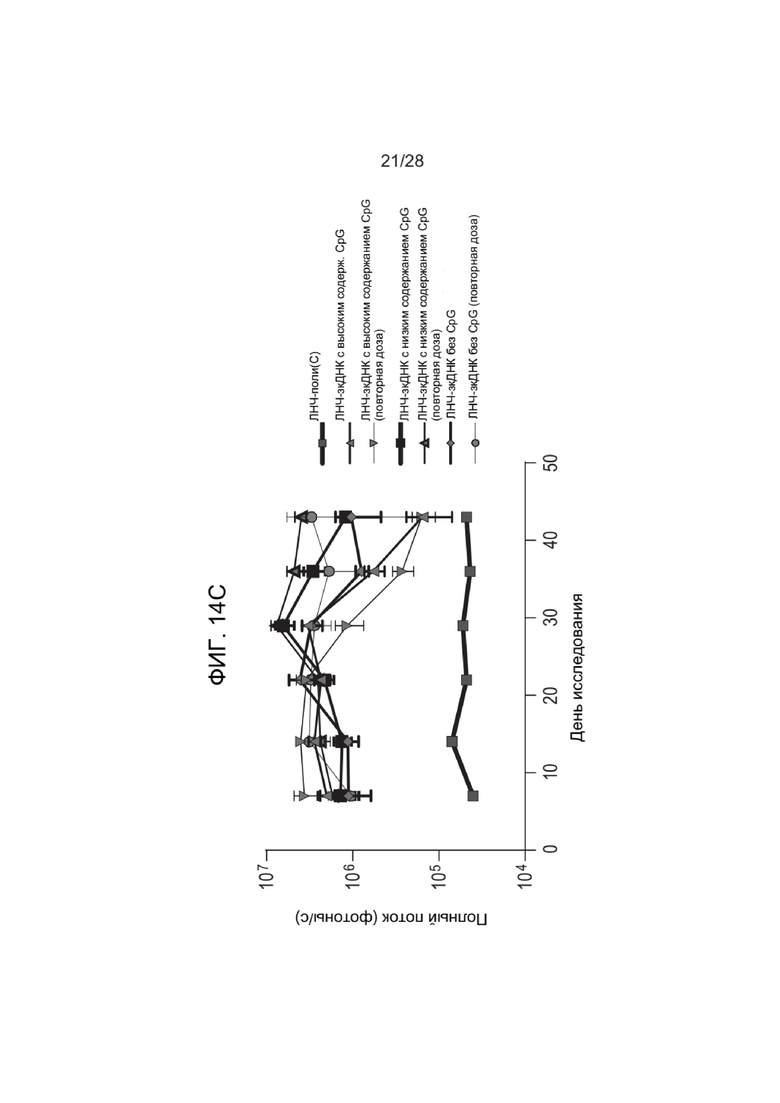
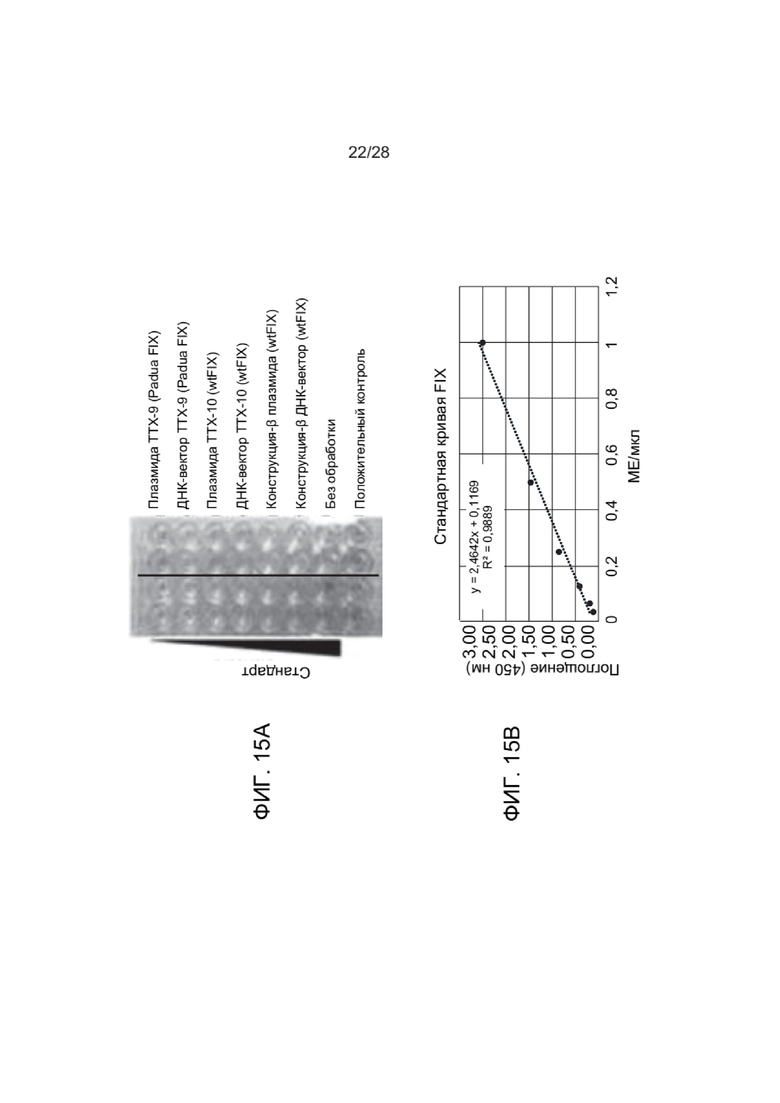
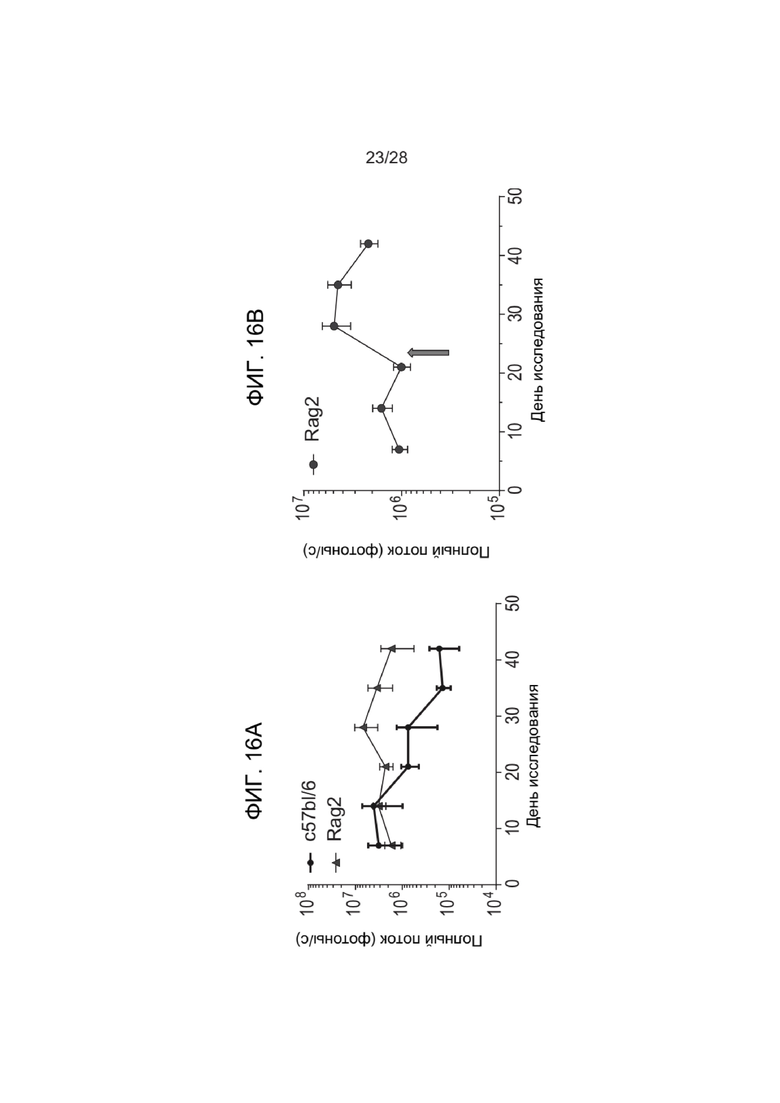
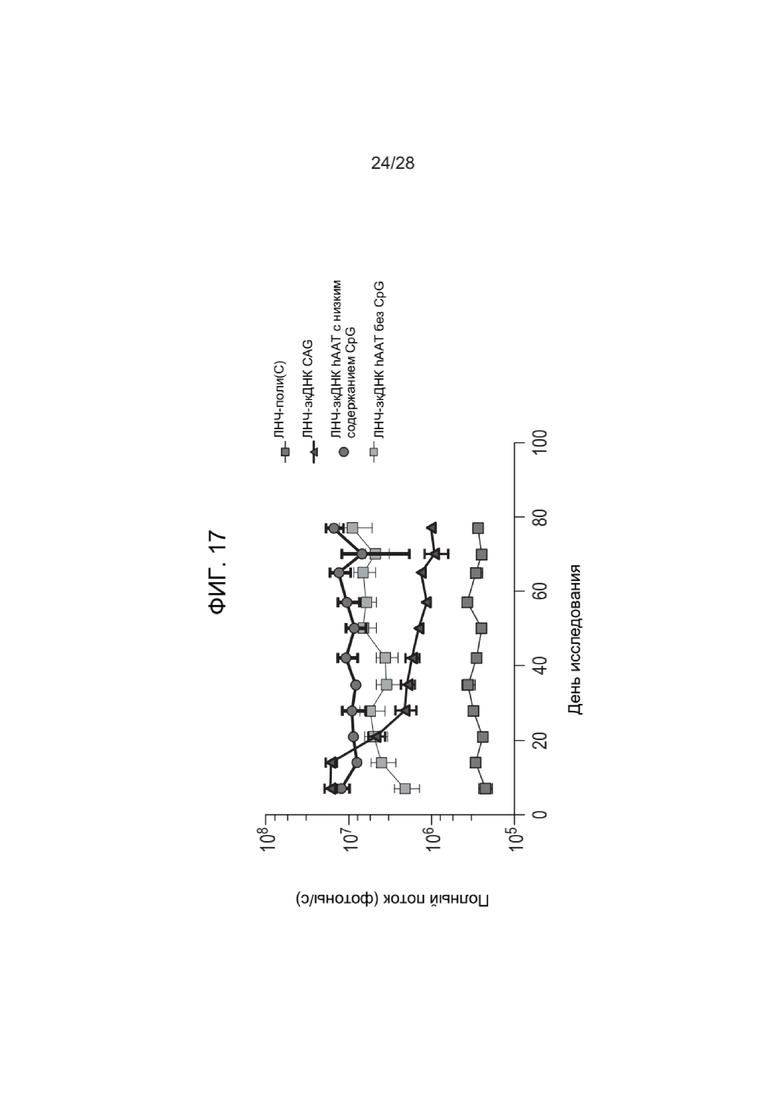
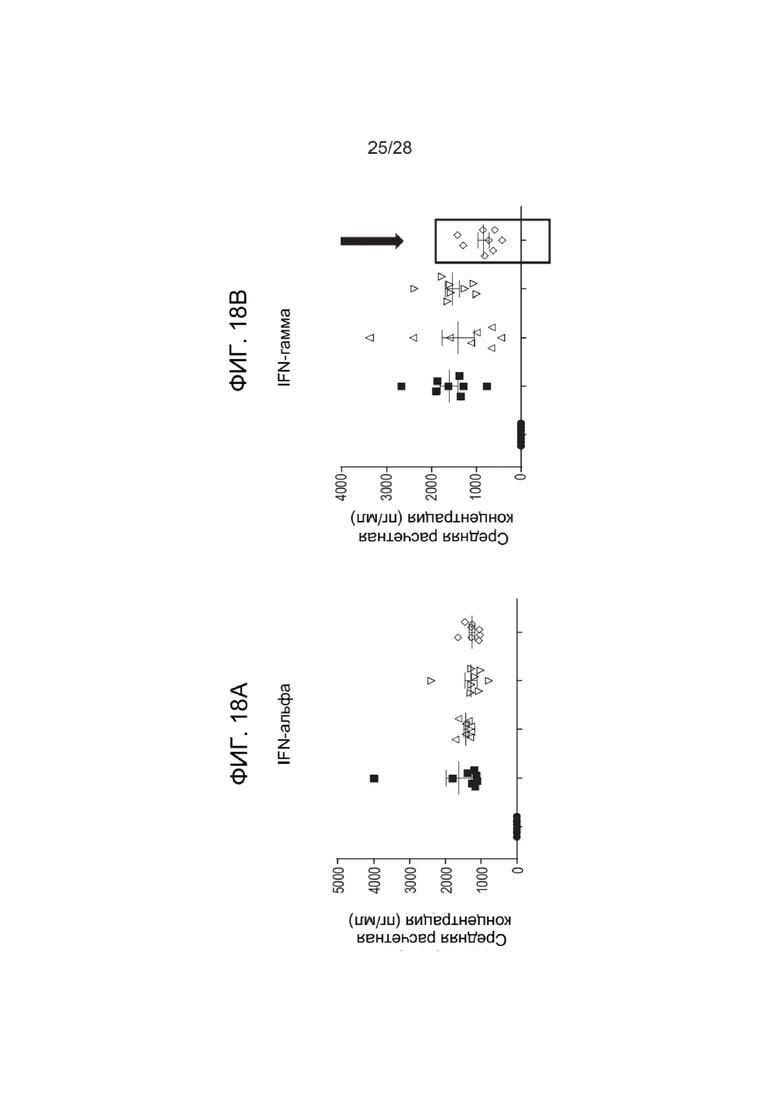
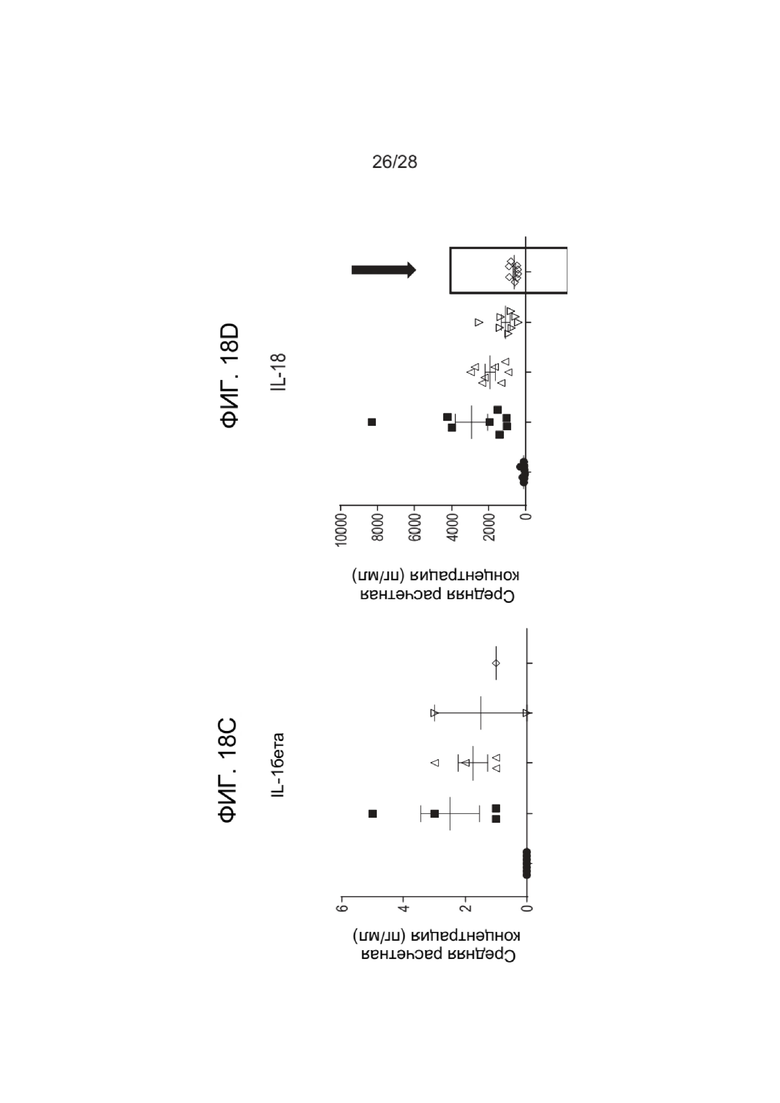
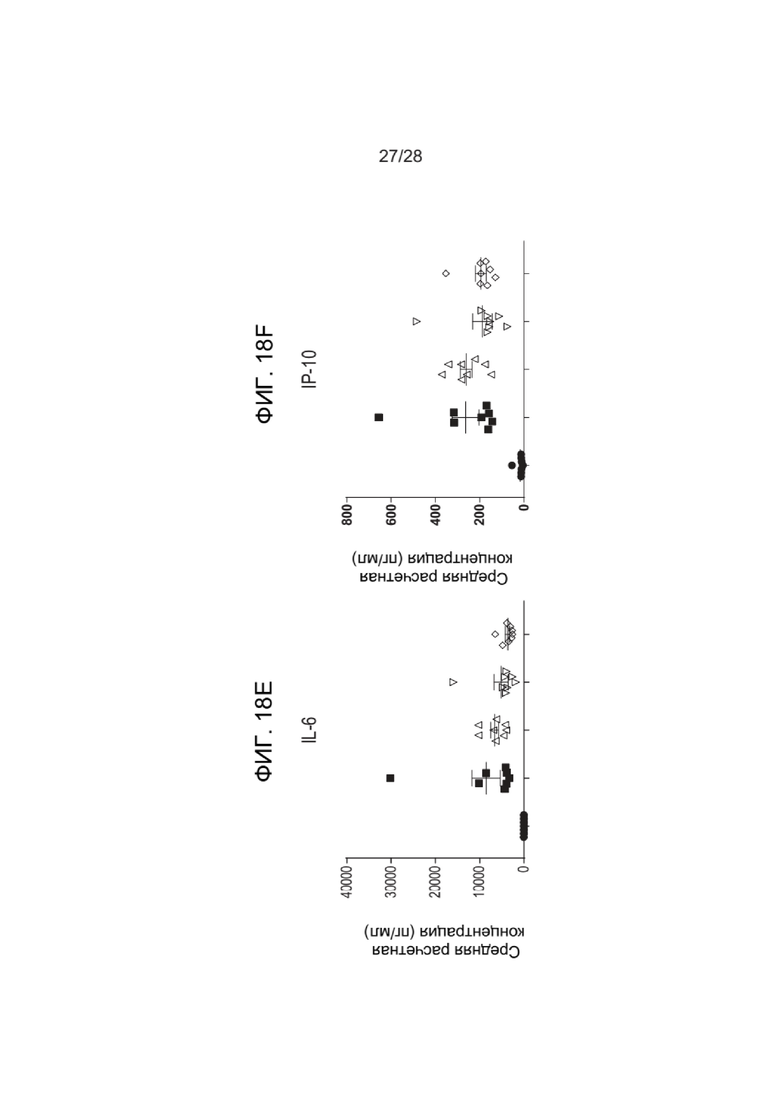
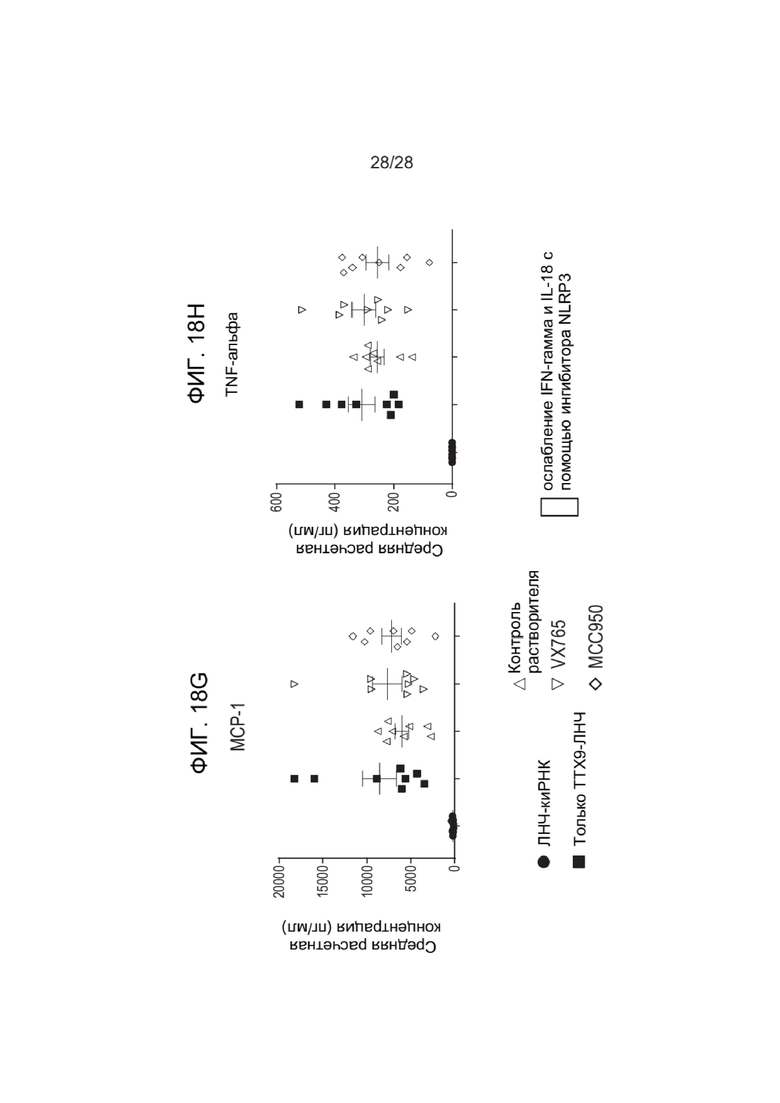
Изобретение относится к области биотехнологии, в частности к способу ингибирования иммунного ответа, включающему введение ингибитора иммунного ответа и невирусного бескапсидного ДНК-вектора с ковалентно замкнутыми концами, содержащего по меньшей мере один трансген, в клетку, в которой экспрессируется по меньшей мере один трансген. Также раскрыты композиция, фармацевтическая композиция, клетка для экспрессии ингибитора иммунного ответа и набор для ингибирования иммунного ответа. Изобретение эффективно для ингибирования иммунного ответа, при котором по меньшей мере один трансген экспрессируется в клетке. 6 н. и 130 з.п. ф-лы, 18 ил., 12 табл., 35 пр.
1. Способ ингибирования иммунного ответа, при котором по меньшей мере один трансген экспрессируется в клетке, отличающийся тем, что указанный способ включает:
введение в указанную клетку по меньшей мере одного ингибитора иммунного ответа, причем ингибитор иммунного ответа представляет собой ингибитор Toll-подобного рецептора (TLR), ингибитор инфламмасомы NLRP3, ингибитор инфламмасомы AIM2, ингибитор каспазы-1, или ингибитор цикло-ГМФ-АМФ-синтазы (cGAS); и
невирусного бескапсидного ДНК-вектора с ковалентно замкнутыми концами (зкДНК-вектор), причем указанный зкДНК-вектор содержит указанный по меньшей мере один трансген, расположенный между двумя фланкирующими инвертированными концевыми повторами (ITR).
2. Способ по п. 1, отличающийся тем, что указанный иммунный ответ представляет собой врожденный иммунный ответ.
3. Способ по п. 1 или 2, отличающийся тем, что указанный ингибитор иммунного ответа представляет собой ингибитор врожденного иммунного ответа.
4. Способ по любому из пп. 1-3, отличающийся тем, что указанный зкДНК-вектор дополнительно кодирует указанный по меньшей мере один ингибитор иммунного ответа.
5. Способ по любому из пп. 1-4, отличающийся тем, что указанный ингибитор иммунного ответа вводят отдельно, в разных составах, от указанного зкДНК-вектора.
6. Способ по п. 5, отличающийся тем, что указанный ингибитор иммунного ответа вводят в одно и то же время, что и указанный зкДНК-вектор.
7. Способ по п. 5, отличающийся тем, что указанный ингибитор иммунного ответа вводят до введения указанного зкДНК-вектора.
8. Способ по п. 5, отличающийся тем, что указанный ингибитор иммунного ответа вводят после введения указанного зкДНК-вектора.
9. Способ по любому из пп. 1-4, отличающийся тем, что указанный ингибитор иммунного ответа и указанный зкДНК-вектор вводят в одно и то же время в одном составе.
10. Способ по п. 1, отличающийся тем, что указанный ингибитор TLR представляет собой ингибитор TLR9.
11. Способ по п. 1, отличающийся тем, что указанный ингибитор TLR представляет собой рапамицин или аналог рапамицина.
12. Способ по п. 1, отличающийся тем, что указанный ингибитор инфламмасомы AIM2, представляет собой A151 из SEQ ID NO: 602 или его вариант, который ингибирует инфламмасому AIM2 или каспазу-1.
13. Способ по п. 1, отличающийся тем, что указанный ингибитор инфламмасомы NLRP3, ингибитор инфламмасомы AIM2, или ингибитор каспазы-1, представляет собой миРНК, специфичную в отношении инфламмасомы NLRP3, инфламмасомы AIM2, или каспазы-1, соответственно.
14. Способ по п. 1, отличающийся тем, что указанный ингибитор инфламмасомы NLRP3, ингибитор инфламмасомы AIM2, или ингибитор каспазы-1, представляет собой киРНК, специфичную в отношении инфламмасомы NLRP3, инфламмасомы AIM2 или каспазы-1, соответственно.
15. Способ по п. 1, отличающийся тем, что указанный ингибитор инфламмасомы NLRP3, ингибитор инфламмасомы AIM2, или ингибитор каспазы-1, представляет собой антитело или антигенсвязывающий фрагмент, который связывает инфламмасому NLRP3, инфламмасому AIM2 или каспазу-1, соответственно.
16. Способ по п. 1, отличающийся тем, что указанный ингибитор инфламмасомы NLRP3 выбран из группы, состоящей из: MCC950, 16673-34-0, Bay11-7082, глибенкламида и изоликвиритигенина.
17. Способ по п. 1, отличающийся тем, что указанный ингибитор инфламмасомы AIM2 выбран из группы, состоящей из: A151 (SEQ ID NO: 602), miR-223 и моноклонального антитела к ASC человека.
18. Способ по п. 1, отличающийся тем, что указанный ингибитор каспазы-1 выбран из группы, состоящей из: белнаказана, шиконина, ацетилшиконина, пралнаказана, Z-VAD-FMK, Z-WEHD-FMK; Ac-YVAD-cmk, Ac-YVAD-CHO и партенолида.
19. Способ по п. 1, отличающийся тем, что указанный ингибитор cGAS представляет собой белок ORF52 герпесвируса, ассоциированного с саркомой Капоши, из SEQ ID NO: 882 или его вариант, который ингибирует cGAS, или ортолог ORF52 гаммагерпесвируса.
20. Способ по п. 1, отличающийся тем, что указанный ингибитор cGAS представляет собой цитоплазматическую изоформу связанного с латентностью ядерного антигена (LANA) герпесвируса, ассоциированного с саркомой Капоши, из SEQ ID NO: 883.
21. Способ по п. 20, отличающийся тем, что указанная цитоплазматическая изоформа LANA герпесвируса, ассоциированного с саркомой Капоши, представляет собой LANAΔ161 SEQ ID NO: 884.
22. Способ по п. 1, отличающийся тем, что указанный ингибитор cGAS представляет собой миРНК, специфичную в отношении cGAS.
23. Способ по п. 22, отличающийся тем, что указанная миРНК, специфичная в отношении cGAS, выбрана из группы, состоящей из miR-25 SEQ ID NO: 885 и miR-93 SEQ ID NO: 886.
24. Способ по п. 1, отличающийся тем, что указанный ингибитор cGAS представляет собой киРНК, специфичную в отношении cGAS.
25. Способ по п. 1, отличающийся тем, что указанный ингибитор cGAS представляет собой антитело или антигенсвязывающий фрагмент, который связывает cGAS.
26. Способ по п. 1, отличающийся тем, что указанный ингибитор cGAS представляет собой противомалярийный препарат.
27. Способ по п. 26, отличающийся тем, что указанный противомалярийный препарат представляет собой противомалярийный препарат на основе аминохинолина или аминоакридина.
28. Способ по п. 27, отличающийся тем, что указанный противомалярийный препарат выбран из группы, состоящей из хинакрина (QC), 9-амино-6-хлор-2-метоксиакридина (AMCA), гидроксихлорохина (HCQ) и хлорохина (CQ).
29. Способ по п. 1, отличающийся тем, что указанный ингибитор cGAS представляет собой низкомолекулярное соединение, которое связывается с каталитическим карманом cGAS.
30. Способ по п. 29, отличающийся тем, что указанное низкомолекулярное соединение, которое связывается с каталитическим карманом cGAS, выбрано из группы, состоящей из RU166365, RU281332, RU320521, RU320519, RU320461, RU320462, RU320520, RU320467, и RU320582.
31. Способ по п. 30, отличающийся тем, что указанное низкомолекулярное соединение, которое связывается с каталитическим карманом cGAS, представляет собой RU320521.
32. Способ по п. 29, отличающийся тем, что указанное низкомолекулярное соединение, которое связывается с каталитическим карманом cGAS, представляет собой PF-06928215.
33. Способ по п. 10, отличающийся тем, что указанный ингибитор TLR9 представляет собой олигонуклеотид, ингибирующий TLR9.
34. Способ по п. 33, отличающийся тем, что указанная олигонуклеотидная последовательность, ингибирующая TLR9, имеет один или более из следующих признаков (i) три последовательных нуклеотида G на 3'-конце; (ii) триплет CC(T) на 5'-конце; и (iii) расстояние между тремя последовательными нуклеотидами G на 3'-конце и триплетом CC(T) на 5'-конце составляет 3-5 нуклеотидов в длину.
35. Способ по п. 33, отличающийся тем, что указанная олигонуклеотидная последовательность, ингибирующая TLR9, содержит последовательность TTAGGGn SEQ ID NO: 888.
36. Способ по п. 33, отличающийся тем, что указанная олигонуклеотидная последовательность, ингибирующая TLR9, выбрана из группы, состоящей из ODN-2088 TCCTGGCGGGGAAGT, SEQ ID NO: 889, ODN-2114 TCCTGGAGGGGAAGT, SEQ ID NO: 890, poly-G GGGGGGGGGGGGGGGGGGGG, SEQ ID NO: 891, ODN-A151 TTAGGGTTAGGGTTAGGGTTAGGG, SEQ ID NO: 892, G-ODN CTCC-TATTGGGGGTTTCCTAT, SEQ ID NO: 893, IRS-869 TCCTGGAGGGGTTGT, SEQ ID NO: 894, INH-1 CCTGGATGGGAATTCCCATCCAGG, SEQ ID NO: 895, INH-4 TTCCCATCCAGGCCTGGATGGGAA, SEQ ID NO: 896, IRS-661 TGCTTGCAAGCTT-GCAAGCA, SEQ ID NO: 897, 4024 TCCTGGATGGGAAGT, SEQ ID NO: 898, 4084F CCTGGATGGGAA, SEQ ID NO: 899, INH-13 CTTACCGCTGCACCTGGATGGGAA, SEQ ID NO: 900, INH-18 CCTGGATGGGAACTTACCGCTGCA, SEQ ID NO: 901, и IRS-954 TGCTCCTGGAGGGGTTGT, SEQ ID NO: 902, и AS1411 GGTGGTGGTGGTTGTGGTGGTGGTGG, SEQ ID NO: 903.
37. Способ по п. 10, отличающийся тем, что указанный ингибитор TLR9 представляет собой миРНК, специфичную в отношении TLR9.
38. Способ по п. 10, отличающийся тем, что указанный ингибитор TLR9 представляет собой киРНК, специфичную в отношении TLR9.
39. Способ по п. 10, отличающийся тем, что указанный ингибитор TLR9 представляет собой антитело или антигенсвязывающий фрагмент, который связывает TLR9.
40. Способ по любому из пп. 1-39, отличающийся тем, что указанный по меньшей мере один трансген функционально расположен между двумя фланкирующими инвертированными концевыми повторами дикого типа (WT-ITR).
41. Способ по любому из пп. 1-39, отличающийся тем, что указанный по меньшей мере один трансген расположен между двумя фланкирующими мутантными инвертированными концевыми повторами (мутантные ITR).
42. Способ по любому из пп. 1-39, отличающийся тем, что указанный по меньшей мере один трансген расположен между двумя фланкирующими инвертированными концевыми повторами, причем один фланкирующий ITR представляет собой WT-ITR, и один фланкирующий ITR представляет собой мутантный ITR.
43. Способ по любому из пп. 1-42, отличающийся тем, что один или оба из указанных фланкирующих ITR происходят из вируса, выбранного из группы, состоящей из парвовируса, депендовируса и аденоассоциированного вируса (ААВ).
44. Способ по любому из пп. 1-43, отличающийся тем, что указанные фланкирующие ITR являются симметричными или асимметричными ITR.
45. Способ по п. 44, отличающийся тем, что указанные фланкирующие ITR являются симметричными или по существу симметричными ITR.
46. Способ по п. 44, отличающийся тем, что указанные фланкирующие ITR являются асимметричными ITR.
47. Способ по любому из пп. 1-39, отличающийся тем, что один из указанных фланкирующих ITR представляет собой ITR дикого типа, или оба из указанных фланкирующих ITR представляют собой ITR дикого типа.
48. Способ по любому из пп. 1-39, отличающийся тем, что указанные фланкирующие ITR происходят из разных вирусных серотипов.
49. Способ по любому из пп. 1-39, отличающийся тем, что по меньшей мере один из указанных фланкирующих ITR изменен относительно последовательности ITR ААВ дикого типа путем делеции, добавления и/или замены, что влияет на общую трехмерную конформацию указанного ITR.
50. Способ по п. 49, отличающийся тем, что один или оба из указанных фланкирующих ITR происходят из серотипа ААВ, выбранного из группы, состоящей из ААВ1, ААВ2, ААВ3, ААВ4, ААВ5, ААВ6, ААВ7, ААВ8, ААВ9, ААВ10, ААВ11 и ААВ12.
51. Способ по любому из пп. 1-50, отличающийся тем, что один или оба из указанных ITR являются синтетическими.
52. Способ по п. 50 или 51, отличающийся тем, что один из указанных фланкирующих ITR не относится к ITR дикого типа, или оба из указанных фланкирующих ITR не относятся к ITR дикого типа.
53. Способ по любому из пп. 1-50, отличающийся тем, что один или оба из указанных фланкирующих ITR модифицированы путем делеции, вставки и/или замены по меньшей мере в одной из областей ITR, выбранных из группы, состоящей из A, A', B, B', С, С', D и D' областей.
54. Способ по п. 53, отличающийся тем, что указанная делеция, вставка и/или замена приводит к удалению всей или части структуры «стебель-петля», образуемой указанными областями A, A', B, B', C и/или C'.
55. Способ по п. 53, отличающийся тем, что один или оба из указанных фланкирующих ITR модифицированы путем делеции, вставки и/или замены, что приводит к удалению всей или части структуры «стебель-петля», образуемой указанными областями B и B'.
56. Способ по п. 53, отличающийся тем, что один или оба из указанных фланкирующих ITR модифицированы путем делеции, вставки и/или замены, что приводит к удалению всей или части структуры «стебель-петля», образуемой указанными областями C и C'.
57. Способ по п. 53, отличающийся тем, что один или оба из указанных фланкирующих ITR модифицированы путем делеции, вставки и/или замены, что приводит к удалению всей или части структуры «стебель-петля», образуемой указанными областями B и B', и/или всей или части структуры «стебель-петля», образуемой указанными областями C и C'.
58. Способ по любому из пп. 1-50, отличающийся тем, что один или оба из указанных фланкирующих ITR содержат одну структуру «стебель-петля» в области, которая, в ITR дикого типа, содержит первую структуру «стебель-петля», образуемую указанными областями B и B', и вторую структуру «стебель-петля», образуемую указанными областями C и C'.
59. Способ по любому из пп. 1-50, отличающийся тем, что один или оба из указанных фланкирующих ITR содержат один стебель и две петли в области, которая, в ITR дикого типа, содержит первую структуру «стебель-петля», образуемую указанными областями B и B', и вторую структуру «стебель-петля», образуемую указанными областями C и C'.
60. Способ по любому из пп. 1-50, отличающийся тем, что один или оба из указанных фланкирующих ITR содержат один стебель и одну петлю в области, которая, в ITR дикого типа, содержит первую структуру «стебель-петля», образуемую указанными областями B и B', и вторую структуру «стебель-петля», образуемую указанными областями C и C'.
61. Способ по любому из пп. 1-60, отличающийся тем, что оба фланкирующих ITR изменяют так, что это приводит к общей трехмерной симметрии, когда указанные ITR инвертированы друг относительно друга.
62. Способ по любому из пп. 1-61, отличающийся тем, что указанный зкДНК-вектор содержит по меньшей мере один регуляторный переключатель.
63. Способ по п. 62, отличающийся тем, что указанный по меньшей мере один регуляторный переключатель выбран из группы, состоящей из бинарного регуляторного переключателя, низкомолекулярного регуляторного переключателя, регуляторного переключателя «с кодом доступа», регуляторного переключателя на основе нуклеиновой кислоты, посттранскрипционного регуляторного переключателя, контролируемого излучением или контролируемого ультразвуком регуляторного переключателя, опосредуемого гипоксией регуляторного переключателя, регуляторного переключателя воспалительного ответа, активируемого сдвигом регуляторного переключателя и аварийного выключателя («kill switch»).
64. Способ по любому из пп. 1-63, отличающийся тем, что указанный зкДНК-вектор и/или ингибитор иммунного ответа инкапсулированы.
65. Способ по любому из пп. 1-63, отличающийся тем, что указанный зкДНК-вектор и/или ингибитор иммунного ответа находятся в наноносителе.
66. Способ по п. 65, отличающийся тем, что указанный наноноситель содержит липидную наночастицу (ЛНЧ).
67. Способ по п. 1, отличающийся тем, что указанная зкДНК при расщеплении рестрикционным ферментом, имеющим один сайт распознавания на указанном зкДНК-векторе, и анализе на неденатурирующем геле, демонстрирует присутствие характерных полос линейной и непрерывной ДНК по сравнению с контрольной линейной и прерывистой ДНК.
68. Способ по любому из предшествующих пунктов, отличающийся тем, что указанный зкДНК-вектор содержит по меньшей мере один промотор, связанный с указанным по меньшей мере одним трансгеном.
69. Способ по п. 68, отличающийся тем, что указанный промотор выбран из группы, состоящей из промотора CAG, промотора HAAT, промотора EF1-α человека или его фрагмента и промотора EF1-α крысы.
70. Способ по любому из предшествующих пунктов, отличающийся тем, что указанный зкДНК-вектор дополнительно содержит энхансер, содержащий последовательность нуклеиновой кислоты, выбранную из группы, состоящей из SEQ ID NO: 1147-1160.
71. Способ по любому из предшествующих пунктов, отличающийся тем, что указанный зкДНК-вектор содержит последовательность 5'-нетранслируемой области (UTR) и/или интрона, содержащую последовательность нуклеиновой кислоты, выбранную из группы, состоящей из SEQ ID NO: 1161-1188.
72. Способ по любому из предшествующих пунктов, отличающийся тем, что указанный зкДНК-вектор содержит 3'-UTR, содержащую последовательность нуклеиновой кислоты, выбранную из группы, состоящей из SEQ ID NO: 8 и 1189-1200.
73. Способ по любому из предшествующих пунктов, отличающийся тем, что указанный зкДНК-вектор содержит по меньшей мере одну последовательность поли(A), содержащую последовательность нуклеиновой кислоты, выбранную из группы, состоящей из SEQ ID NO: 9 и 1201-1216.
74. Способ по любому из предшествующих пунктов, отличающийся тем, что увеличение количества указанного зкДНК-вектора, который вводят в клетку, повышает экспрессию указанного по меньшей мере одного трансгена в указанной клетке.
75. Способ по любому из предшествующих пунктов, отличающийся тем, что указанный по меньшей мере один трансген кодирует терапевтический трансген, и при этом целевой уровень экспрессии указанного трансгена представляет собой терапевтически эффективное количество.
76. Способ по любому из предшествующих пунктов, отличающийся тем, что указанный по меньшей мере один трансген, при его транскрипции или трансляции, модифицирует аномальное количество эндогенного белка у субъекта.
77. Способ по любому из предшествующих пунктов, отличающийся тем, что указанный по меньшей мере один трансген, при его транскрипции или трансляции, модифицирует аномальную функцию или активность эндогенного белка или пути у субъекта.
78. Способ по п. 76 или 77, отличающийся тем, что указанный субъект представляет собой пациента-человека, нуждающегося в лечении.
79. Способ по любому из предшествующих пунктов, отличающийся тем, что указанный по меньшей мере один трансген кодирует или содержит нуклеотидную молекулу, выбранную из группы, состоящей из РНКи, киРНК, миРНК, днкРНК и антисмыслового олиго- или полинуклеотида.
80. Способ по любому из пп. 1-79, отличающийся тем, что указанный по меньшей мере один трансген кодирует белок.
81. Способ по п. 80, отличающийся тем, что указанный белок представляет собой маркерный белок.
82. Способ по любому из пп. 1-79, отличающийся тем, что указанный по меньшей мере один трансген кодирует агонист или антагонист эндогенного белка или пути, ассоциированного с указанным заболеванием или нарушением.
83. Способ по любому из пп. 1-79, отличающийся тем, что указанный по меньшей мере один трансген кодирует антитело.
84. Способ по любому из предшествующих пунктов, дополнительно включающий введение дополнительного ингибитора иммунного ответа.
85. Способ по любому из пп. 1-84, отличающийся тем, что указанный зкДНК-вектор получен способом, включающим этапы: (a) инкубации популяции клеток насекомых, несущих экспрессионную конструкцию зкДНК, в присутствии по меньшей мере одного белка Rep, при этом указанная экспрессионная конструкция зкДНК кодирует указанный зкДНК-вектор в эффективных условиях и на протяжении времени, достаточного для того чтобы индуцировать продуцирование указанного зкДНК-вектора в указанных клетках насекомых; и (b) выделения указанного зкДНК-вектора из указанных клеток насекомых.
86. Способ по п. 85, отличающийся тем, что указанная экспрессионная конструкция зкДНК выбрана из группы, состоящей из зкДНК-плазмиды, зкДНК-бакмиды и зкДНК-бакуловируса.
87. Способ п. 85, отличающийся тем, что указанные клетки насекомых экспрессируют по меньшей мере один белок Rep.
88. Способ по п. 87, отличающийся тем, что указанный по меньшей мере один белок Rep происходит из вируса, выбранного из группы, состоящей из парвовируса, депендовируса и аденоассоциированного вируса (ААВ).
89. Способ по п. 88, отличающийся тем, что указанный по меньшей мере один белок Rep происходит из серотипа ААВ, выбранного из группы, состоящей из ААВ1, ААВ2, ААВ3, ААВ4, ААВ5, ААВ6, ААВ7, ААВ8, ААВ9, ААВ10, ААВ11 и ААВ12.
90. Композиция для ингибирования иммунного ответа, содержащая по меньшей мере один ингибитор иммунного ответа, причем ингибитор иммунного ответа представляет собой ингибитор Toll-подобного рецептора (TLR), ингибитор инфламмасомы NLRP3, ингибитор инфламмасомы AIM2, ингибитор каспазы-1, или ингибитор цикло-ГМФ-АМФ-синтазы (cGAS); и
невирусный бескапсидный ДНК-вектор с ковалентно замкнутыми концами (зкДНК-вектор), где указанный зкДНК-вектор содержит по меньшей мере один трансген, расположенный между двумя фланкирующими инвертированными концевыми повторами (ITR).
91. Композиция по п. 90, отличающаяся тем, что указанный зкДНК-вектор дополнительно кодирует указанный по меньшей мере один ингибитор иммунного ответа.
92. Композиция по п. 90 или 91, отличающаяся тем, что указанный иммунный ответ представляет собой врожденный иммунный ответ.
93. Композиция по п. 90 или 92, отличающаяся тем, что указанный по меньшей мере один трансген расположен между двумя фланкирующими инвертированными концевыми повторами дикого типа (WT-ITR).
94. Композиция по п. 90 или 92, отличающаяся тем, что указанный по меньшей мере один трансген расположен между двумя фланкирующими мутантными инвертированными концевыми повторами (мутантными ITR).
95. Композиция по п.90 или 92, отличающаяся тем, что указанный по меньшей мере один трансген расположен между двумя фланкирующими инвертированными концевыми повторами (ITR), причем один их фланкирующих ITR представляет собой WT-ITR и другой фланкирующий ITR представляет собой мутантный ITR.
96. Композиция по п. 90 или 92, отличающаяся тем, что один или оба из указанных фланкирующих ITR происходят из вируса, выбранного из группы, состоящей из парвовируса, депендовируса и аденоассоциированного вируса (ААВ).
97. Композиция по п. 96, отличающаяся тем, что указанные фланкирующие ITR являются симметричными или асимметричными ITR.
98. Композиция по п. 90 или 92, отличающаяся тем, что оба из указанных ITR являются ITR дикого типа.
99. Композиция по п. 90 или 92, отличающаяся тем, что указанные фланкирующие ITR происходят из разных вирусных серотипов.
100. Композиция по п. 90 или 92, отличающаяся тем, что по меньшей мере один из указанных фланкирующих ITR изменен относительно последовательности ITR ААВ дикого типа путем делеции, добавления и/или замены, что влияет на общую трехмерную конформацию указанного ITR.
101. Композиция по п. 90 или 92, отличающаяся тем, что один или оба из указанных ITR происходят из серотипа ААВ, выбранного из группы, состоящей из ААВ1, ААВ2, ААВ3, ААВ4, ААВ5, ААВ6, ААВ7, ААВ8, ААВ9, ААВ10, ААВ11 и ААВ12.
102. Композиция по любому из пп. 90 или 92-101, отличающаяся тем, что один или более из указанных фланкирующих ITR являются синтетическими.
103. Композиция по п. 90 или 92, отличающаяся тем, что оба из указанных фланкирующих ITR не являются ITR дикого типа.
104. Композиция по п. 90 или 92, отличающаяся тем, что один или оба из указанных фланкирующих ITR модифицируют путем делеции, вставки и/или замены по меньшей мере в одной из областей ITR, выбранных из группы, состоящей из областей A, A', B, B', С, С', D и D'.
105. Композиция по п. 104, отличающаяся тем, что указанная делеция, вставка и/или замена приводит к удалению всей или части структуры «стебель-петля», образуемой указанными областями A, A', B, B', C и/или C'.
106. Композиция по п. 90 или 92, отличающаяся тем, что один или оба из указанных фланкирующих ITR модифицированы путем делеции, вставки и/или замены, что приводит к удалению всей или части структуры «стебель-петля», образуемой указанными областями B и B'.
107. Композиция по п. 90 или 92, отличающаяся тем, что один или оба из указанных фланкирующих ITR модифицированы путем делеции, вставки и/или замены, что приводит к удалению всей или части структуры «стебель-петля», образуемой указанными областями C и C'.
108. Композиция по п. 90 или 92, отличающаяся тем, что один или оба из указанных фланкирующих ITR модифицированы путем делеции, вставки и/или замены, что приводит к удалению всей или части структуры «стебель-петля», образуемой указанными областями B и B', и/или всей или части структуры «стебель-петля», образуемой указанными областями C и C'.
109. Композиция по п. 90 или 92, отличающаяся тем, что один или оба из указанных фланкирующих ITR содержат одну структуру «стебель-петля» в области, которая, в ITR дикого типа, содержит первую структуру «стебель-петля», образуемую указанными областями B и B', и вторую структуру «стебель-петля», образуемую указанными областями C и C'.
110. Композиция по п. 90 или 92, отличающаяся тем, что один или оба из указанных фланкирующих ITR содержат один стебель и две петли в области, которая, в ITR дикого типа, содержит первую структуру «стебель-петля», образуемую указанными областями B и B', и вторую структуру «стебель-петля», образуемую указанными областями C и C'.
111. Композиция по п. 90 или 92, отличающаяся тем, что один или оба из указанных фланкирующих ITR содержат один стебель и одну петлю в области, которая, в ITR дикого типа, содержит первую структуру «стебель-петля», образуемую указанными областями B и B', и вторую структуру «стебель-петля», образуемую указанными областями C и C'.
112. Композиция по п. 90 или 92, отличающаяся тем, что оба фланкирующих ITR изменяют так, что это приводит к общей трехмерной симметрии, когда указанные ITR инвертированы друг относительно друга.
113. Композиция по любому из пп. 90 и 92-112, отличающаяся тем, что указанные фланкирующие ITR основаны на последовательностях из вируса, выбранного из группы, состоящей из парвовируса, депендовируса и аденоассоциированного вируса (ААВ).
114. Композиция по любому из пп. 90 и 92-113, отличающаяся тем, что указанный зкДНК-вектор включает по меньшей мере один регуляторный переключатель.
115. Композиция по п. 114, отличающаяся тем, что по меньшей мере один регуляторный переключатель выбран из группы, состоящей из бинарного регуляторного переключателя, низкомолекулярного регуляторного переключателя, регуляторного переключателя «с кодом доступа», регуляторного переключателя на основе нуклеиновой кислоты, посттранскрипционного регуляторного переключателя, контролируемого излучением или контролируемого ультразвуком регуляторного переключателя, опосредуемого гипоксией регуляторного переключателя, регуляторного переключателя воспалительного ответа, активируемого сдвигом регуляторного переключателя и аварийного выключателя.
116. Композиция по любому из пп. 90 и 92-115, отличающаяся тем, что указанный зкДНК-вектор инкапсулирован.
117. Композиция по любому из пп. 90 и 92-116, отличающаяся тем, что указанный зкДНК-вектор находится в наноносителе.
118. Композиция по п. 117, отличающаяся тем, что указанный наноноситель содержит липидную наночастицу (ЛНЧ).
119. Композиция по любому из пп. 90 и 92-118, отличающаяся тем, что указанная зкДНК при расщеплении рестрикционным ферментом, имеющим один сайт распознавания на указанном зкДНК-векторе, и при анализе на неденатурирующем геле, демонстрирует присутствие характерных полос линейной и непрерывной ДНК по сравнению с контрольной линейной и прерывистой ДНК.
120. Композиция по любому из пп. 90 и 92-119, отличающаяся тем, что указанный зкДНК-вектор содержит энхансер, содержащий последовательность нуклеиновой кислоты, выбранную из группы, состоящей из SEQ ID NO: 1147-1160.
121. Композиция по любому из пп. 90 и 92-120, отличающаяся тем, что указанный зкДНК-вектор содержит последовательность 5'-UTR и/или интрона, содержащую последовательность нуклеиновой кислоты, выбранную из группы, состоящей из SEQ ID NO: 1161-1188.
122. Композиция по любому из пп. 90 и 92-121, отличающаяся тем, что указанный зкДНК-вектор содержит 3'-UTR, содержащую последовательность нуклеиновой кислоты, выбранную из группы, состоящей из SEQ ID NO: 8 и 1189- 1200.
123. Композиция по любому из пп. 90 и 92-122, отличающаяся тем, что указанный зкДНК-вектор содержит по меньшей мере одну последовательность поли(A), содержащую последовательность нуклеиновой кислоты, выбранную из группы, состоящей из SEQ ID NO: 9 и 1201-1216.
124. Композиция по любому из пп. 90 и 92-123, отличающаяся тем, что указанный зкДНК-вектор содержит по меньшей мере один промотор, связанный с указанным по меньшей мере одним трансгеном.
125. Композиция по любому из пп. 90 и 92-124, отличающаяся тем, что по меньшей мере один из фланкирующих ITR содержит функциональный сайт концевого разрешения и сайт связывания Rep, и при этом один из указанных фланкирующих ITR содержит делецию, вставку и/или замену относительно другого фланкирующего ITR.
126. Композиция по любому из пп. 90 и 92-125, отличающаяся тем, что указанный по меньшей мере один трансген оптимизирован по кодонам для экспрессии в эукариотической клетке.
127. Композиция по любому из пп. 90 и 92-126, дополнительно содержащая фармацевтически приемлемый носитель.
128. Композиция по п. 90, отличающаяся тем, что указанный ингибитор TLR представляет собой ингибитор TLR9.
129. Композиция по п. 90, отличающаяся тем, что указанный ингибитор TLR представляет собой рапамицин или аналог рапамицина.
130. Способ экспрессии ингибитора иммунного ответа в клетке, отличающийся тем, что указанный способ включает приведение указанной клетки в контакт с композицией по любому из пп. 90 и 92-129.
131. Способ по п. 130, отличающийся тем, что указанная клетка находится в условиях in vitro или in vivo.
132. Фармацевтическая композиция для ингибирования иммунного ответа, при котором трансген экспрессируется в клетке, содержащая композицию по любому из пп. 90 и 92-129.
133. Фармацевтическая композиция по п. 132, отличающаяся тем, что указанный ингибитор иммунного ответа представляет собой ингибитор врожденного иммунного ответа.
134. Фармацевтическая композиция по п. 133, отличающаяся тем, что указанный ингибитор врожденного иммунного ответа изготовлен отдельно от указанного зкДНК-вектора.
135. Клетка для экспрессии ингибитора иммунного ответа, содержащая композицию по любому из пп. 90 и 92-129.
136. Набор для ингибирования иммунного ответа, при котором по меньшей мере один трансген экспрессируется в клетке, содержащий композицию по любому из пп. 90 и 92-129, фармацевтическую композицию по любому из пп. 132-134 или клетку по п. 135, и фармацевтически приемлемый носитель.
| WO 2009108235 A2, 03.09.2009 | |||
| LINA LI et al., Production and Characterization of Novel Recombinant Adeno-Associated Virus Replicative-Form Genomes: A Eukaryotic Source of DNA for Gene Transfer, PLoS One., 2013, v | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| СПОСОБ ОБЖИГА ПОДИНЫ АЛЮМИНИЕВОГО ЭЛЕКТРОЛИЗЕРА | 0 |
|
SU209770A1 |
| WO 2019094548 А1, 16.05.2019 | |||
| ВОЛКОВ М.Ю., Роль Toll-подобных рецепторов и их эндогенных | |||

