ОБЛАСТЬ ТЕХНИКИ
В данном документе представлен слитый белок на основе антитела к ЕТА и TGF-β Trap. В данном документе также представлена фармацевтическая композиция, содержащая слитый белок на основе антитела к ЕТА и TGF-β Trap. Кроме того, в данном документе представлен способ лечения, предупреждения возникновения или улучшения течения одного или более симптомов легочной артериальной гипертензии, легочной гипертензии или легочного фиброза посредством применения слитого белка на основе антитела к ЕТА и TGF-β Trap.
УРОВЕНЬ ТЕХНИКИ
Легочная артериальная гипертензия (PAH) представляет собой редкое и прогрессирующее патологическое состояние, характеризующееся значительным повышением давления крови в легочной артерии. Легочная артериальная гипертензия стала патологическим состоянием, угрожающим здоровью человека. Данные показывают, что ежегодная заболеваемость различными типами легочной артериальной гипертензии во всем мире составляет от 2,4 до 7,6 на миллион, а ее распространенность составляет от 15 до 26 на миллион населения. Она стала третьим по частоте сердечно-сосудистым патологическим состоянием после ишемической болезни сердца и гипертензии. Причина легочной артериальной гипертензии еще полностью не выяснена. В связи с бессимптомным началом у большинства пациентов нарушение функции сердца при легочной артериальной гипертензии уже характеризуется III-IV степенью тяжести. Сопутствующие симптомы легочной артериальной гипертензии обычно включают одышку (особенно во время нагрузки), боль в груди, периодические обмороки и т.д. Кроме того, по мере развития патологического состояния постоянное высокое давление в легочных артериях приведет к тому, что правый желудочек не сможет беспрерывно снабжать кровью легкие, что в конечном итоге приведет к правожелудочковой недостаточности. Сердечная недостаточность представляет собой наиболее частую причину смерти пациентов с легочной артериальной гипертензией.
В настоящее время лечение легочной артериальной гипертензии отсутствует, и медикаментозная терапия представляет собой поддерживающую терапию первого выбора при легочной артериальной гипертензии. Лекарственные средства, одобренные FDA для лечения легочной артериальной гипертензии, представляют собой вазодилататоры, которые в зависимости от механизма действия можно разделить на блокаторы кальциевых каналов, агонисты рецепторов простациклина, ингибиторы фосфодиэстеразы 5 типа (PDE5), ингибиторы эндотелиновых рецепторов и т.п.
Легочная артериальная гипертензия вызывается компенсаторным повышением кровяного давления в легких со стороны сердца после того, как кровоснабжение в легких со стороны сердца становится недостаточным из-за вазоконстрикции сосудов легких или сосудов, связанных с легкими. Ее микроскопические проявления включают утолщение интимы легочных артериол и сужение, ремоделирование, ригидность или локальную окклюзию сосудов, вызванную тромбозом, за счет чего повышается сопротивление кровеносных сосудов легочному кровотоку (Simonneau et al., 2004, J. Am. Coll. Cardiol. 43:5S-12S; Barst et al., 2004, J. Am. Coll. Cardiol. 43:40S-47S).
Ингибиторы эндотелиновых рецепторов (например, эндотелинового рецептора A (ETA или ETAR)) могут эффективно блокировать повышение сосудистого давления, вызванное эндотелином, с облегчением симптомов легочной артериальной гипертензии и улучшением способности переносить нагрузку и гемодинамики у пациента (Serasli et al., 2010, Recent Pat. Cardiovasc. Drug Discov. 5:184-95).
Ряд исследований показал, что трансформирующий фактор роста β1 (TGF-β1) играет важную роль в развитии различных патологических состояний, особенно в развитии кардиоваскулярных патологических состояний. TGF-β1 участвует в таких процессах, как пролиферация, дифференцировка, миграция и апоптоз клеток, и активирует такие процессы, как пролиферация гладкомышечных клеток, отложение внеклеточного матрикса и эндотелиально-мезенхимальный переход (EndMT) посредством регуляции множества сигнальных путей, за счет чего индуцируется возникновение и развитие легочной артериальной гипертензии (Yan et al., 2016, Int. J. Cardiol., 222:368-374; Graham et al., 2013, Circulation, 128:1354-1364; Liu et al., 2016, Int. J. Biochem. Cell. Biol., 77:141-154). Пирфенидон в качестве одобренного лекарственного средства для лечения легочного фиброза может ингибировать экспрессию TGF-β, уменьшать пролиферацию фибробластов и снижать уровни мРНК и белка α-гладкомышечного актина (α-SMA), индуцированного TGF-β.
TGF-β Trap состоит из внеклеточного конца рецептора TGF-β2 типа и может связываться с TGF-β1 или TGF-β3, за счет чего эффективно блокируются процессы, опосредованные TGF-β, такие как пролиферация и дифференцировка гладкомышечных клеток сосудов, отложение внеклеточного матрикса и эндотелиально-мезенхимальный переход при легочной артериальной гипертензии. Он может способствовать уменьшению фиброза и ремоделированию легочных артериол, за счет чего снижается сопротивление легочного кровотока и улучшается функция правых отделов сердца (Goumans et al., 2018, Cold Spring Harb. Perspect. Biol., 10: a022210; Yung et al., 2016, Am. J. Respir. Crit. Care Med., 194:1140-1151).
В данном документе представлен слитый белок на основе антитела, ингибирующего ЕТА, и TGF-β Trap. С одной стороны, слитый белок на основе антитела, ингибирующего ЕТА, и TGF-β Trap может блокировать сигнальный путь ЕТА и снижать сопротивление кровеносных сосудов легочному кровотоку; с другой стороны, он может блокировать сигнальный путь TGF-β и облегчать течение фиброза и ремоделирование легочных артериол. Цели лечения легочной артериальной гипертензии и улучшения функции правых отделов сердца достигаются за счет двойного механизма действия.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В данном документе представлен слитый белок на основе антитела к ЕТА и TGF-β Trap. Также в данном документе представлен способ лечения, предупреждения возникновения или улучшения одного или более симптомов легочной артериальной гипертензии, легочной гипертензии, легочного фиброза и кардиоваскулярного фиброза посредством применения слитого белка.
В данном документе представлен слитый белок на основе антитела к ETA и TGF-β Trap, структурно характеризующийся тем, что слитый белок содержит антитело к ETA и фрагмент TGF-β Trap.
В данном документе представлен слитый белок на основе антитела к ETA и TGF-β Trap, структурно характеризующийся тем, что слитый белок содержит антитело к ETA, один, два, три, четыре, пять, шесть, семь или восемь TGF-β Trap и соответствующее количество пептидных линкеров (линкеров); при этом в слитом белке аминоконец TGF-β Trap связан с карбоксильным концом легкой цепи или тяжелой цепи антитела к ЕТА посредством пептидной линкерной (линкерной) последовательности или в слитом белок карбоксильный конец TGF-β Trap связан с аминоконцом легкой цепи или тяжелой цепи антитела к ЕТА посредством пептидной линкерной (линкерной) последовательности.
В данном документе представлен слитый белок на основе антитела к ETA и TGF-β Trap, структурно характеризующийся тем, что слитый белок содержит антитело к ETA, один, два, три или четыре TGF-β Trap и соответствующее количество пептидных линкеров (линкеров); при этом в слитом белке аминоконец TGF-β Trap связан с карбоксильным концом легкой цепи или тяжелой цепи антитела к ЕТА посредством пептидной линкерной (линкерной) последовательности или в слитом белок карбоксильный конец TGF-β Trap связан с аминоконцом легкой цепи или тяжелой цепи антитела к ЕТА посредством пептидной линкерной (линкерной) последовательности.
В данном документе представлен слитый белок на основе антитела к ETA и TGF-β Trap, структурно характеризующийся тем, что слитый белок содержит антитело к ETA, два TGF-β Trap и два пептидных линкера (линкера); при этом в слитом белке аминоконец TGF-β Trap связан с карбоксильным концом легкой цепи или тяжелой цепи антитела к ЕТА посредством пептидной линкерной (линкерной) последовательности или в слитом белок карбоксильный конец TGF-β Trap связан с аминоконцом легкой цепи или тяжелой цепи антитела к ЕТА посредством пептидной линкерной (линкерной) последовательности.
В данном документе представлен слитый белок на основе антитела к ETA и TGF-β Trap, структурно характеризующийся тем, что слитый белок содержит антитело к ETA, TGF-β Trap и пептидный линкер (линкер); при этом в слитом белке аминоконец TGF-β Trap связан с карбоксильным концом легкой цепи или тяжелой цепи антитела к ЕТА посредством пептидной линкерной (линкерной) последовательности или в слитом белок карбоксильный конец TGF-β Trap связан с аминоконцом легкой цепи или тяжелой цепи антитела к ЕТА посредством пептидной линкерной (линкерной) последовательности.
В данном документе представлен слитый белок на основе антитела к ETA и TGF-β Trap, структурно характеризующийся тем, что антитело к ETA, TGF-β Trap и пептидная линкерная последовательность слиты с образованием слитого белка с помощью одного из следующих средств:
(1) посредством пептидной линкерной (линкера) последовательности аминоконец TGF-β Trap связан с карбоксиконцом тяжелой/легкой цепи антитела к ЕТА: N'-R-линкер-TGF-β Trap-C'; и
(2) посредством пептидной линкерной (линкера) последовательности карбоксиконец TGF-β Trap связан с аминоконцом легкой цепи или тяжелой цепи антитела к ETA: N'-TGF-β Trap-линкер-R-C';
при этом N' представляет собой аминоконец полипептидной цепи, C' представляет собой карбоксиконец полипептидной цепи, TGF-β Trap представляет собой фрагмент TGF-β Trap, R представляет собой аминокислотную последовательность легкой цепи или тяжелой цепи антитела к ЕТА, и линкер представляет собой пептидный линкер.
В данном документе представлен полинуклеотид, кодирующий слитый белок на основе антитела к ЕТА и TGF-β Trap, описанных в данном документе.
В данном документе представлен вектор, содержащий полинуклеотид, кодирующий слитый белок на основе антитела к ЕТА и TGF-β Trap, описанных в данном документе.
В данном документе представлена клетка-хозяин, содержащая вектор, описанный в данном документе.
В данном документе представлена фармацевтическая композиция, содержащая слитый белок на основе антитела к ETA и TGF-β Trap, описанные в данном документе, и фармацевтически приемлемый носитель.
В данном документе представлено применение слитого белка на основе антитела к ЕТА и TGF-β Trap, описанных в данном документе, в получении лекарственного препарата, предназначенного для лечения, предупреждения возникновения или улучшения одного или более симптомов легочной артериальной гипертензии и заболеваний, связанных с легочной артериальной гипертензией.
В данном документе представлено применение слитого белка на основе антитела к ЕТА и TGF-β Trap, описанных в данном документе, в получении лекарственного препарата, предназначенного для лечения, предупреждения возникновения или улучшения одного или более симптомов легочной гипертензии и заболеваний, связанных с легочной гипертензией.
В данном документе представлено применение слитого белка на основе антитела к ЕТА и TGF-β Trap, описанных в данном документе, в получении лекарственного препарата, предназначенного для лечения, предупреждения возникновения или улучшения одного или более симптомов легочного фиброза и заболеваний, связанных с легочным фиброзом.
В данном документе представлено применение слитого белка на основе антитела к ЕТА и TGF-β Trap, описанных в данном документе, в получении лекарственного препарата, предназначенного для лечения, предупреждения возникновения или улучшения одного или более симптомов кардиоваскулярного фиброза и заболеваний, связанных с кардиоваскулярным фиброзом.
В данном документе представлено применение слитого белка на основе антитела к ЕТА и TGF-β Trap, описанных в данном документе, в получении лекарственного препарата, предназначенного для одновременного лечения, предупреждения возникновения или улучшения одного или более симптомов двух или более заболеваний из легочной артериальной гипертензии, легочной гипертензии, легочного фиброза или кардиоваскулярного фиброза.
В данном документе представлен способ лечения, предупреждения возникновения или улучшения одного или более симптомов легочной артериальной гипертензии и заболеваний, связанных с легочной артериальной гипертензией, предусматривающий введение субъектам терапевтически эффективной дозы слитого белка на основе антитела к ЕТА и TGF-β Trap, описанных в данном документе.
В данном документе представлен способ лечения, предупреждения возникновения или улучшения одного или более симптомов легочной гипертензии и заболеваний, связанных с легочной гипертензией, предусматривающий введение субъектам терапевтически эффективной дозы слитого белка на основе антитела к ЕТА и TGF-β Trap, описанных в данном документе.
В данном документе представлен способ лечения, предупреждения возникновения или улучшения одного или более симптомов легочного фиброза и заболеваний, связанных с легочным фиброзом, предусматривающий введение субъектам терапевтически эффективной дозы слитого белка на основе антитела к ЕТА и TGF-β Trap, описанных в данном документе.
В данном документе представлен способ лечения, предупреждения возникновения или улучшения одного или более симптомов кардиоваскулярного фиброза и заболеваний, связанных с кардиоваскулярным фиброзом, предусматривающий введение субъектам терапевтически эффективной дозы слитого белка на основе антитела к ЕТА и TGF-β Trap, описанных в данном документе.
В данном документе представлен способ лечения, предупреждения возникновения или улучшения одного или более симптомов двух или более заболеваний из легочной артериальной гипертензии, легочной гипертензии, легочного фиброза или кардиоваскулярного фиброза, предусматривающий введение субъектам терапевтически эффективной дозы слитого белка на основе антитела к ЕТА и TGF-β Trap, описанных в данном документе.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На фигуре 1 показаны результаты ингибирования слитым белком h15F3-(G4S)4-TGF-β Trap (содержащим SEQ ID NO: 162, SEQ ID NO: 190, SEQ ID NO: 207 и SEQ ID NO: 210) на основе антитела к ETA и TGF-β Trap изменений в Ca2+, опосредованных ETA человека.
На фигуре 2 показаны результаты блокирования TGF-β1, обусловленного активацией рецептора TGF-β1, посредством слитого белка h15F3-(G4S)4-TGF-β Trap на основе антитела к ETA и TGF-β Trap.
На фигуре 3 показано, что слитый белок h15F3-(G4S)4-TGF-β Trap на основе антитела к ETA и TGF-β Trap характеризуется значительно улучшенным эффектом в отношении легочного фиброза.
На фигуре 4 показано, что слитый белок h15F3-(G4S)4-TGF-β Trap на основе антитела к ETA и TGF-β Trap может значительно улучшать выживаемость/период выживаемости мышей с легочным фиброзом.
ПОДРОБНОЕ ОПИСАНИЕ
Определения
Если в данном документе не указано иное, научные и технические термины имеют значения, понятные специалисту в данной области техники. Как правило, номенклатура и методики, относящиеся к фармакологии, биологии, биохимии, культивированию клеток и тканей, биологии, молекулярной биологии, иммунологии, микробиологии, генетике и химии белков и нуклеиновых кислот, а также гибридизации, хорошо известны и широко используются в данной области техники.
Стандартные однобуквенные или трехбуквенные сокращения используются в данном документе для обозначения полинуклеотидных и полипептидных последовательностей. Если не указано иное, аминоконец полипептидной последовательности расположен слева, а ее карбоксиконец расположен справа, при этом 5'-концы расположенных выше цепей однонитевых и двухнитевых последовательностей нуклеиновых кислот расположены слева, а их 3'-концы расположены справа. Конкретная часть полипептида может быть представлена количеством аминокислотных остатков, как, например, аминокислотами от 80 до 130, или представлена действительным остатком сайта, как, например, от Lys80 до Lys130. Конкретная полипептидная или полинуклеотидная последовательность также может быть описана с объяснением ее отличия от референтной последовательности.
Термины "пептид", "полипептид" и "белок" относятся к молекуле, содержащей две или более аминокислот, которые связаны пептидной связью. Эти термины охватывают, например, природные и искусственные белки, полипептидные аналоги белковых фрагментов и белковых последовательностей (таких как мутантные белки, варианты и слитые белки) и белки, которые посттранскрипционно или иным образом ковалентно или нековалентно модифицированы. Пептид, полипептид или белок может быть мономером или полимером.
Термин "полипептидный фрагмент" относится к полипептиду, который содержит аминоконец и/или карбоксиконец, отсутствующий у соответствующего полноразмерного белка. Длина фрагмента может составлять, например, по меньшей мере 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 20, 50, 70, 80, 90, 100, 150 или 200 аминокислот. Длина фрагмента может составлять, например, до 1000, 750, 500, 250, 200, 175, 150, 125, 100, 90, 80, 70, 60, 50, 40, 30, 20, 15, 14, 13, 12, 11 или 10 аминокислот. Фрагмент может дополнительно содержать одну или более дополнительных аминокислот на одном конце или на обоих, как, например, аминокислотные последовательности из различных природных белков (например, Fc-доменов или доменов лейциновая застежка) или искусственные аминокислотные последовательности (например, искусственные линкерные последовательности).
Полипептиды в данном документе включают полипептиды, модифицированные по любой причине и любыми способами, например, для: (1) снижения чувствительности к протеолизу, (2) снижения чувствительности к окислению, (3) изменения аффинности в отношении образованных белковых комплексов, (4) изменения аффинности связывания и (4) придания или модификации других физико-химических или функциональных свойств. Аналоги предусматривают мутантный белок полипептида. Например, одна или более аминокислотных замен (например, консервативные аминокислотные замены) могут быть выполнены в природных последовательностях (например, вне домена полипептида, который образует внутримолекулярный контакт). "Консервативная аминокислотная замена" представляет собой замену, которая существенно не изменяет структурные характеристики исходной последовательности (например, замена аминокислот не должна разрушать спирали, присутствующие в исходной последовательности, или мешать другим типам вторичной структуры, которые придают характеристики родительской последовательности или необходимы для ее функции).
"Вариант" полипептида включает аминокислотную последовательность, в которой один или более аминокислотных остатков вставлены, удалены и/или заменены в аминокислотной последовательности по сравнению с другой полипептидной последовательности. Варианты в данном документе включают слитые белки.
"Производное" полипептида представляет собой химически модифицированный полипептид, например путем связывания с другими химическими компонентами, такими как полиэтиленгликоль и альбумин (например, сывороточный альбумин человека), фосфорилирования и гликозилирования.
Если не указано иное, термин "антитело" включает антитела с двумя полноразмерными тяжелыми цепями и двумя полноразмерными легкими цепями, а также их производные, варианты, фрагменты и мутантные белки, примеры которых перечислены ниже.
Термин "антитело" представляет собой белок, который содержит антигенсвязывающую часть и необязательно остов или каркасную часть, которые позволяют антигенсвязывающей части принимать конформацию, способствующую связыванию антитела с антигеном. Примеры антител включают интактные антитела, фрагменты антител (такие как антигенсвязывающая часть антитела), производные антител и аналоги антител. Например, антитело может содержать альтернативные белковые каркасы или искусственные каркасы с трансплантированными CDR или производными CDR. Каркас содержит без ограничения каркас на основе антитела, который вводится, такой как каркас, который стабилизирует трехмерную структуру антитела, и полностью синтетический каркас, который содержит, например, биологически совместимый полимер. См., например, Korndorfer et al., 2003, Proteins: Structure, Function and Bioinformatics 53:121-129; Roque et al., 2004, Biotechnol. Prog. 20:639-654. Кроме того, можно использовать пептидоподобные антитела ("PAM") и каркасы на основе имитаторов антител, в которых используются фибриновые лиганды в качестве каркасов.
Антитела могут иметь структуру, например, природного иммуноглобулина. "Иммуноглобулин" представляет собой тетрамерную молекулу. В природном иммуноглобулине каждый тетрамер состоит из двух идентичных пар полипептидных цепей, при этом каждая пара содержит "легкую" цепь (приблизительно 25 кДа) и "тяжелую" цепь (приблизительно 50-70 кДа). Аминоконец каждой цепи содержит вариабельный домен из приблизительно 100-110 или больше аминокислот, который главным образом связан с распознаванием антигена. Карбоксильный конец каждой цепи определяет константную область, главным образом связанную с действием эффекторов. Легкие цепи человеческого антитела делятся на легкие цепи κ и λ. Тяжелые цепи делятся на тяжелые цепи μ, δ, α или ε, и они определяют изотипы антигена, такие как IgM, IgD, IgG, IgA и IgE. В легкой и тяжелой цепях вариабельные и константные области связаны областью "J" из приблизительно 12 или больше аминокислот, а тяжелая цепь также содержит область "D" из приблизительно 10 или больше аминокислот. См. Fundamental Immunology Ch.7 (edited by Paul, 2nd edition, Raven Press, 1989) (раскрытие которого включено в данный документ посредством ссылки в полном объеме для любой цели). Вариабельные области каждой пары легкой/тяжелой цепи образуют сайты связывания антител, поэтому интактный иммуноглобулин содержит два сайта связывания.
Цепи природного иммуноглобулина демонстрируют ту же основную структуру, что и относительно консервативная каркасная область (FR), соединенная тремя высоковариабельными областями, также известными как определяющие комплементарность области или CDR. От N-конца до C-конца легкая и тяжелая цепи содержат домены FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Распределение аминокислот в каждом домене согласуется с определением в Kabat et al. Sequences of Proteins of Immunological Interest, 5th edition, US Dept. of Health and Human Services, PHS, NIH, NIH Publication No. 91-3242, 1991.
Если не указано иное, "антитело" означает либо интактный иммуноглобулин, либо его антигенсвязывающую часть, которая может конкурировать с интактным антителом за специфическое связывание. Антигенсвязывающая часть может быть получена посредством методик рекомбинантной ДНК или ферментативного или химического расщепления интактных антител. Антигенсвязывающая часть содержит, в частности, Fab, Fab', F(ab')2, Fv, доменные антитела (dAb), фрагменты, содержащие определяющие комплементарность области (CDR), одноцепочечные антитела (scFv), химерные антитела, диантитела, триантитела, тетраантитела и полипептиды, содержащие по меньшей мере часть иммуноглобулина, достаточную для придания полипептиду специфического связывания с антигеном.
Fab-фрагмент представляет собой одновалентный фрагмент с доменами VL, VH, CL и CH1; F(ab')2-фрагмент представляет собой двухвалентный фрагмент с двумя Fab-фрагментами, соединенными дисульфидной связью в шарнирной области; Fd-фрагмент содержит VH- или VL-домен; и dAb-фрагмент содержит VH-домен, VL-домен или антигенсвязывающий фрагмент VH- или VL-доменов (патенты США №№ US 6846634 и US 6696245; публикации патентных заявок США №№ US 2005/0202512, US 2004/0202995, US 2004/0038291, US 2004/0009507 и US 2003/0039958; Ward et al., 1989, Nature 341:544-546).
Одноцепочечное антитело (scFv) представляет собой антитело, в котором области VL и VH соединены линкером (например, синтетической последовательностью из аминокислотных остатков) с образованием непрерывного белка, при этом линкер достаточно длинный, чтобы позволить белкой цепи укладываться обратно и образовывать одновалентный антигенсвязывающий сайт (см., например, Bird et al., 1988, Science 242:423-26 и Huston et al., 1988, Proc. Natl. Acad. Sci. USA 85:5879-83).
Диантитело представляет собой бивалентное антитело, содержащее две полипептидные цепи, каждая из которых содержит VH- и VL-домены, связанные линкером, который является настолько коротким, что не позволяет спариваться двум доменам одной и той же цепи. Следовательно, обеспечивается спаривание каждого домена с комплементарным доменом другой полипептидной цепи (см., например, Holliger et al., 1993, Proc. Natl. Acad. Sci. USA 90:6444-48 и Poljak et al., 1994, Structure 2:1121-23). Если диантитело содержит две идентичные полипептидные цепи, то диантитело, полученное в результате спаривания двух идентичных полипептидных цепей, будет содержать один и тот же антигенсвязывающий сайт. Полипептидные цепи с разными последовательностями можно использовать для получения диантител с разными антигенсвязывающими сайтами. Аналогично триантитела и тетраантитела представляют собой антитела, которые содержат три и четыре полипептидные цепи соответственно и образуют три и четыре антигенсвязывающих сайта соответственно, которые могут быть одинаковыми или разными.
Способ, описанный в Kabat et al. Sequences of Proteins of Immunological Interest, 5th edition, US Dept. of Health and Human Services, PHS, NIH, NIH Publication No. 91-3242, 1991, может использоваться для идентификации определяющих комплементарность областей (CDR) и каркасных областей (FR) данного антитела. Одна или более CDR могут быть встроены в молекулу либо ковалентно, либо нековалентно для получения антитела. В антителе можно встраивать более крупную полипептидную цепь в CDR. CDR могут быть ковалентно связаны с другой полипептидной цепью или встроены в нее нековалентно. CDR позволяют антителам специфически связываться со специфическими ассоциированными антигенами.
Антитела могут иметь один или более сайтов связывания. Если существует более одного сайта связывания, то сайт связывания может быть таким же или отличаться от другого. Например, природный человеческий иммуноглобулин обычно содержит два идентичных сайта связывания, тогда как "биспецифическое" или "бифункциональное" антитело содержит два разных сайта связывания.
Термин "мышиное антитело" включает все антитела, содержащие одну или более вариабельных и константных областей, полученных из последовательностей мышиного иммуноглобулина.
Термин "гуманизированное антитело" представляет собой антитело, полученное путем трансплантации последовательности определяющих комплементарность областей молекул мышиных антител в каркас вариабельных областей человеческих антител.
Термин "антигенсвязывающий домен", "антигенсвязывающая область" или "антигенсвязывающий сайт" представляет собой часть антитела, которая содержит аминокислотные остатки (или другие части), которое взаимодействует с антигеном и вносят вклад в его специфичность и аффинность антитела в отношении антигена. В случае антитела, которое специфически связывается со своим антигеном, они будут включать по меньшей мере часть по меньшей мере одного из доменов CDR.
Термин "эпитоп" обозначает молекулы, которая связывается с антителом (например, связывается антителом). Эпитоп может содержать прерывистую часть молекулы (например, в полипептиде, аминокислотные остатки, прерывистые в первичной последовательности полипептида, достаточно близки друг к другу в третичной и четвертичной структурах полипептида для связывания антителом).
"Процент идентичности" двух полинуклеотидных или двух полипептидных последовательностей определяют с помощью компьютерной программы GAP (GCG Wisconsin Package; часть версии 10.3 (Accelrys, Сан-Диего, Калифорния)) с использованием стандартных параметров для сравнения последовательностей.
Термины "полинуклеотид", "олигонуклеотид" и "нуклеиновая кислота" могут взаимозаменяемо использоваться по всему тексту и включают молекулы ДНК (например, кДНК или геномной ДНК), молекулы РНК (например, мРНК), аналоги ДНК или РНК и их гибриды, полученные с использованием нуклеотидных аналогов (например, пептидных нуклеиновых кислот и неприродных нуклеотидных аналогов). Молекулы нуклеиновой кислоты могут быть одно- или двухнитевыми. В одном варианте осуществления молекулы нуклеиновой кислоты в данном документе содержат непрерывную открытую рамку считывания, кодирующую антитело или его фрагменты, производные, мутантные белки или их варианты, представленные в данном документе.
Если последовательности двух одноцепочечных полинуклеотидов могут быть расположены в обратном порядке и параллельно, два одноцепочечных полинуклеотида "комплементарны" друг другу, поэтому каждый нуклеотид в одном полинуклеотиде противоположен комплементарному нуклеотиду в другом полинуклеотиде, гэпы не вводят и не обнаруживают неспаренные нуклеотиды на 5'- или 3'-концах каждой последовательности. Если два полинуклеотида могут гибридизоваться друг с другом в умеренно жестких условиях, то один полинуклеотид "комплементарен" другому полинуклеотиду. Таким образом, один полинуклеотид может быть комплементарен другому полинуклеотиду, но не его комплементарной последовательности.
Термин "вектор" представляет собой нуклеиновую кислоту, которую можно использовать для введения другой связанной с ней нуклеиновой кислоты в клетку. Одним типом вектора является "плазмида", которая относится к линейной или кольцевой молекуле двухцепочечной ДНК, с которой могут быть связаны дополнительные сегменты нуклеиновой кислоты. Другим типом вектора является вирусный вектор (например, дефективные по репликации ретровирусы, аденовирусы и аденовирус-ассоциированные вирусы), в котором дополнительные сегменты ДНК могут быть введены в вирусный геном. Некоторые векторы могут автономно реплицироваться в клетке-хозяине, в которую они введены (например, бактериальные векторы, содержащие точку начала бактериальный репликации, и эписомальные векторы млекопитающих). Другие векторы (например, неэписомальные векторы млекопитающих) интегрируются в геном клетки-хозяина при введении в клетку-хозяина и, таким образом, реплицируются с геномом хозяина.
"Вектор экспрессии" представляет собой тип вектора, который может управлять экспрессией выбранного полинуклеотида.
Если регуляторная последовательность влияет на экспрессию нуклеотидной последовательности (например, уровень экспрессии, время или сайт), то нуклеотидная последовательность "функционально связана" с регуляторной последовательностью. "Регуляторная последовательность" представляет собой нуклеиновую кислоту, которая влияет на экспрессию (например, уровень экспрессии, время или сайт) нуклеиновой кислоты, с которой она функционально связана. Например, регуляторные гены действуют непосредственно на регулируемые нуклеиновые кислоты или через одну или более других молекул (например, полинуклеотидов, которые связываются с регуляторными последовательностями и/или нуклеиновыми кислотами). Примеры регуляторных последовательностей включают промоторы, энхансеры и другие элементы контроля экспрессии (например, сигналы полиаденилирования). Дополнительные примеры регуляторных последовательностей могут быть описаны, например, в Goeddel, 1990, Gene Expression Technology: Methods in Enzymology, Volume 185, Academic Press, San Diego, CA и Baron et al., 1995, Nucleic Acids Res. 23:3605-06.
Термин "клетка-хозяин" относится к клетке, используемой для экспрессии нуклеиновой кислоты, такой как нуклеиновая кислота, представленная в данном документе. Клетка-хозяин может представлять собой прокариотов, таких как E. coli, или эукариотов, таких как одноклеточные эукариоты (например, дрожжи или другие грибы), растительные клетки (например, клетки растений табака или томата), клетки животных (например, клетки человека, клетки обезьяны, клетки хомяка, клетки крысы, клетки мыши или клетки насекомых), или гибридомы. Обычно клетка-хозяин представляет собой культуральную клетку, которую можно трансформировать или трансфицировать нуклеиновой кислотой, кодирующей полипептид, которая затем может быть экспрессирована в клетке-хозяине. Фраза "рекомбинантная клетка-хозяин" может быть использована для описания клетки-хозяина, трансформированной или трансфицированной нуклеиновой кислотой с ожидаемой экспрессией. Клетка-хозяин также может быть клеткой, которая содержит нуклеиновую кислоту, но не экспрессирует ее на требуемом уровне, если только регуляторные последовательности не введены в клетку-хозяина, так что она функционально связана с нуклеиновой кислотой. Следует учитывать, что термин "клетка-хозяин" относится не только к конкретной рассматриваемой клетке, но также к потомству или возможному потомству этой клетки. Вследствие определенных модификаций, происходящих в последующих поколениях, вызванных, например, мутациями или влиянием факторов окружающей среды, потомство может фактически отличаться от родительской клетки, но все же находится в пределах объема терминологии, используемой в данном документе.
Эндотелиновый рецептор
Эндотелиновый рецептор (ETA) относится к подсемейству A семейства 7-трансмембранных рецепторов, которое связано с одним или более внутриклеточными сигнальными путями посредством гетеротримерного гуанин-нуклеотид-связывающего белка (G-белка) (Jelinek et al., 1993, Science 259:1614-1616 и Segre et al., 1993, Trends Endocrinol. Metab. 4:309-314). Используемые в данном документе термины "эндотелиновый рецептор" и "ETA" или "ETAR" могут использоваться взаимозаменяемо.
В одном варианте осуществления антитело, описанное в данном документе, может быть выбрано для связывания с мембрано-связанными эндотелиновыми рецепторами, экспрессируемыми на клетках, и ингибирования или блокирования эндотелинового сигнального пути, опосредованного эндотелиновыми рецепторами. В одном варианте осуществления антитело, описанное в данном документе, специфически связывается с человеческими эндотелиновыми рецепторами. В другом варианте осуществления антитело, которое связывается с человеческими эндотелиновыми рецепторами, также может связываться с эндотелиновыми рецепторами других видов, таких как крысы. В следующих примерах представлено получение мышиных антител, которые связываются с человеческими мембранно-связанными эндотелиновыми рецепторами, и в дополнительном варианте осуществления мышиные антитела также связываются с эндотелиновыми рецепторами других видов.
Известны полинуклеотидные и полипептидные последовательности эндотелиновых рецепторов нескольких видов. Под SEQ ID NO: 1-SEQ ID NO: 6 показаны человеческие, обезьяньи и крысиные последовательности. Данные о последовательности взяты из базы данных GenBank Национального центра биотехнологической информации США.
Информация о последовательности эндотелинового рецептора A (ETA) представлена ниже:
полинуклеотидная последовательность человека (Homo sapiens) (SEQ ID NO: 1); номер доступа: S63938;
аминокислотная последовательность человека (Homo sapiens) (SEQ ID NO: 2); номер доступа: AAB20278;
полинуклеотидная последовательность обезьяны (Cynomolgus monkey) (SEQ ID NO: 3); номер доступа: JV635771;
аминокислотная последовательность обезьяны (Cynomolgus monkey) (SEQ ID NO: 4); номер доступа: AFJ71111;
полинуклеотидная последовательность крысы (Rattus norvegicus) (SEQ ID NO: 5); номер доступа: M60786;
аминокислотная последовательность крысы (Rattus norvegicus) (SEQ ID NO: 6); номер доступа: AAA41114.
Антитело к эндотелиновому рецептору А (ETA)
В одном варианте осуществления антитело к ETA, описанное в данном документе, содержит одну, две, три, четыре, пять или шесть аминокислотных последовательностей, где каждая аминокислотная последовательность независимо выбрана из аминокислотных последовательностей, перечисленных ниже:
a. аминокислотных последовательностей CDR1 легкой цепи под SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14, SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20, SEQ ID NO: 22, SEQ ID NO: 24, SEQ ID NO: 26, SEQ ID NO: 28 и SEQ ID NO: 30;
b. аминокислотных последовательностей CDR2 легкой цепи под SEQ ID NO: 32, SEQ ID NO: 34, SEQ ID NO: 36, SEQ ID NO: 38, SEQ ID NO: 40, SEQ ID NO: 42, SEQ ID NO: 44, SEQ ID NO: 46, и SEQ ID NO: 48;
c. аминокислотных последовательностей CDR3 легкой цепи под SEQ ID NO: 50, SEQ ID NO: 52, SEQ ID NO: 54, SEQ ID NO: 56, SEQ ID NO: 58, SEQ ID NO: 60, SEQ ID NO: 62, SEQ ID NO: 64, SEQ ID NO: 66, SEQ ID NO: 68 и SEQ ID NO: 205;
d. аминокислотных последовательностей CDR1 тяжелой цепи под SEQ ID NO: 70, SEQ ID NO: 72, SEQ ID NO: 74, SEQ ID NO: 76, SEQ ID NO: 78, SEQ ID NO: 80, SEQ ID NO: 82, SEQ ID NO: 84, SEQ ID NO: 86, SEQ ID NO: 88 и SEQ ID NO: 90;
e. аминокислотных последовательностей CDR2 тяжелой цепи под SEQ ID NO: 92, SEQ ID NO: 94, SEQ ID NO: 96, SEQ ID NO: 98, SEQ ID NO: 100, SEQ ID NO: 102, SEQ ID NO: 104, SEQ ID NO: 106, SEQ ID NO: 108, SEQ ID NO: 110, SEQ ID NO: 112 и SEQ ID NO: 114 и
f. аминокислотных последовательностей CDR3 тяжелой цепи под SEQ ID NO: 116, SEQ ID NO: 118, SEQ ID NO: 120, SEQ ID NO: 122, SEQ ID NO: 124, SEQ ID NO: 126, SEQ ID NO: 128, SEQ ID NO: 130, SEQ ID NO: 132, SEQ ID NO: 134 и SEQ ID NO: 136.
В таблице 1 перечислены аминокислотные последовательности CDR легкой цепи антитела к ЕТА, описанного в данном документе, и их соответствующие полинуклеотидные кодирующие последовательности. В таблице 2 перечислены аминокислотные последовательности CDR тяжелой цепи антитела к ЕТА, описанного в данном документе, и их соответствующие полинуклеотидные кодирующие последовательности.
Таблица 1. Аминокислотные последовательности CDR легкой цепи и их полинуклеотидные кодирующие последовательности
нуклеотидная последовательность
(SEQ ID NO: 7)
(SEQ ID NO: 31)
(SEQ ID NO: 49)
аминокислотная последовательность
(SEQ ID NO: 8)
(SEQ ID NO: 32)
(SEQ ID NO: 50) или QHSYSWPWT
(SEQ ID NO: 205)
нуклеотидная последовательность
(SEQ ID NO: 9)
(SEQ ID NO: 33)
(SEQ ID NO: 51)
аминокислотная последовательность
(SEQ ID NO: 10)
(SEQ ID NO: 34)
(SEQ ID NO: 52)
нуклеотидная последовательность
(SEQ ID NO: 11)
(SEQ ID NO: 35)
(SEQ ID NO: 53)
аминокислотная последовательность
(SEQ ID NO: 12)
(SEQ ID NO: 36)
(SEQ ID NO: 54)
нуклеотидная последовательность
(SEQ ID NO: 13)
(SEQ ID NO: 37)
(SEQ ID NO: 55)
аминокислотная последовательность
(SEQ ID NO: 14)
(SEQ ID NO: 38)
(SEQ ID NO: 56)
нуклеотидная последовательность
(SEQ ID NO: 39)
(SEQ ID NO: 57)
аминокислотная последовательность
(SEQ ID NO: 40)
(SEQ ID NO: 58)
нуклеотидная последовательность
acacactcaattagcc
(SEQ ID NO: 17)
(SEQ ID NO: 41)
(SEQ ID NO: 59)
аминокислотная последовательность
(SEQ ID NO: 18)
(SEQ ID NO: 42)
(SEQ ID NO: 60)
нуклеотидная последовательность
(SEQ ID NO: 19)
(SEQ ID NO: 43)
(SEQ ID NO: 61)
аминокислотная последовательность
(SEQ ID NO: 20)
(SEQ ID NO: 44)
(SEQ ID NO: 62)
нуклеотидная последовательность
(SEQ ID NO: 21)
(SEQ ID NO: 43)
(SEQ ID NO: 63)
аминокислотная последовательность
(SEQ ID NO: 22)
(SEQ ID NO: 44)
(SEQ ID NO: 62)
нуклеотидная последовательность
(SEQ ID NO: 23)
(SEQ ID NO: 43)
(SEQ ID NO: 61)
аминокислотная последовательность
(SEQ ID NO: 24)
(SEQ ID NO: 44)
(SEQ ID NO: 62)
нуклеотидная последовательность
(SEQ ID NO: 25)
(SEQ ID NO: 45)
(SEQ ID NO: 63)
аминокислотная последовательность
(SEQ ID NO: 26)
(SEQ ID NO: 46)
(SEQ ID NO: 64)
нуклеотидная последовательность
(SEQ ID NO: 27)
(SEQ ID NO: 45)
(SEQ ID NO: 65)
аминокислотная последовательность
(SEQ ID NO: 28)
(SEQ ID NO: 46)
(SEQ ID NO: 66)
нуклеотидная последовательность
(SEQ ID NO: 29)
(SEQ ID NO: 47)
(SEQ ID NO: 67)
аминокислотная последовательность
(SEQ ID NO: 30)
(SEQ ID NO: 48)
(SEQ ID NO: 68)
Таблица 2. Аминокислотные последовательности CDR тяжелой цепи и их полинуклеотидные кодирующие последовательности
нуклеотидная последовательность
(SEQ ID NO: 69)
(SEQ ID NO: 91)
(SEQ ID NO: 115)
аминокислотная последовательность
(SEQ ID NO: 70)
(SEQ ID NO: 92)
нуклеотидная последовательность
(SEQ ID NO: 117)
аминокислотная последовательность
нуклеотидная последовательность
аминокислотная последовательность
(SEQ ID NO: 74)
(SEQ ID NO: 96)
(SEQ ID NO: 120)
нуклеотидная последовательность
(SEQ ID NO: 75)
(SEQ ID NO: 97)
(SEQ ID NO: 121)
аминокислотная последовательность
(SEQ ID NO: 76)
(SEQ ID NO: 122)
нуклеотидная последовательность
(SEQ ID NO: 77)
(SEQ ID NO: 99)
(SEQ ID NO: 123)
аминокислотная последовательность
(SEQ ID NO: 100)
(SEQ ID NO: 124)
нуклеотидная последовательность
(SEQ ID NO: 79)
(SEQ ID NO: 101)
аминокислотная последовательность
(SEQ ID NO: 102)
(SEQ ID NO: 126)
нуклеотидная последовательность
(SEQ ID NO: 81)
(SEQ ID NO: 103)
(SEQ ID NO: 127)
аминокислотная последовательность
(SEQ ID NO: 104)
(SEQ ID NO: 128)
нуклеотидная последовательность
(SEQ ID NO: 81)
(SEQ ID NO: 105)
(SEQ ID NO: 127)
аминокислотная последовательность
(SEQ ID NO: 106)
(SEQ ID NO: 128)
нуклеотидная последовательность
(SEQ ID NO: 129)
аминокислотная последовательность
(SEQ ID NO: 108)
(SEQ ID NO: 130)
нуклеотидная последовательность
(SEQ ID NO: 85)
(SEQ ID NO: 109)
аминокислотная последовательность
(SEQ ID NO: 86)
(SEQ ID NO: 110)
(SEQ ID NO: 132)
нуклеотидная последовательность
(SEQ ID NO: 87)
(SEQ ID NO: 111)
(SEQ ID NO: 133)
аминокислотная последовательность
(SEQ ID NO: 88)
(SEQ ID NO: 112)
(SEQ ID NO: 134)
нуклеотидная последовательность
(SEQ ID NO: 89)
(SEQ ID NO: 113)
(SEQ ID NO: 135)
аминокислотная последовательность
(SEQ ID NO: 90)
(SEQ ID NO: 114)
(SEQ ID NO: 136)
В одном варианте осуществления антитело, описанное в данном документе, содержит последовательность, отличающуюся от такой из аминокислотных последовательностей CDR, перечисленных в таблицах 1 и 2, пятью, четырьмя, тремя, двумя или одним (одной) добавлением, заменой и/или делецией одной аминокислоты. В другом варианте осуществления антитело, описанное в данном документе, содержит последовательность, отличающуюся от такой из аминокислотных последовательностей CDR, перечисленных в таблицах 1 и 2, четырьмя, тремя, двумя или одним (одной) добавлением, заменой и/или делецией одной аминокислоты. В другом варианте осуществления антитело, описанное в данном документе, содержит последовательность, отличающуюся от такой из аминокислотных последовательностей CDR, перечисленных в таблицах 1 и 2, тремя, двумя или одним (одной) добавлением, заменой и/или делецией одной аминокислоты. В другом варианте осуществления антитело, описанное в данном документе, содержит последовательность, отличающуюся от такой из аминокислотных последовательностей CDR, перечисленных в таблицах 1 и 2, двумя или одним (одной) добавлением, заменой и/или делецией одной аминокислоты. В другом варианте осуществления антитело, описанное в данном документе, содержит последовательность, отличающуюся от такой из аминокислотных последовательностей CDR, перечисленных в таблицах 1 и 2, одним (одной) добавлением, заменой и/или делецией одной аминокислоты.
В другом варианте осуществления антитело к ETA (антитело к ETA-1), описанное в данном документе, содержит одну или две аминокислотные последовательности, где каждая аминокислотная последовательность независимо выбрана из аминокислотных последовательностей, перечисленных ниже:
a. аминокислотных последовательностей CDR1 легкой цепи под SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14, SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20, SEQ ID NO: 22, SEQ ID NO: 24, SEQ ID NO: 26, SEQ ID NO: 28 и SEQ ID NO: 30 и
b. аминокислотных последовательностей CDR1 тяжелой цепи под SEQ ID NO: 70, SEQ ID NO: 72, SEQ ID NO: 74, SEQ ID NO: 76, SEQ ID NO: 78, SEQ ID NO: 80, SEQ ID NO: 82, SEQ ID NO: 84, SEQ ID NO: 86, SEQ ID NO: 88, и SEQ ID NO: 90.
В одном аспекте антитело к ETA-1 дополнительно содержит одну или две аминокислотные последовательности, где каждая аминокислотная последовательность независимо выбрана из аминокислотных последовательностей, перечисленных ниже:
a. аминокислотных последовательностей CDR2 легкой цепи под SEQ ID NO: 32, SEQ ID NO: 34, SEQ ID NO: 36, SEQ ID NO: 38, SEQ ID NO: 40, SEQ ID NO: 42, SEQ ID NO: 44, SEQ ID NO: 46, и SEQ ID NO: 48 и
b. аминокислотных последовательностей CDR2 тяжелой цепи под SEQ ID NO: 92, SEQ ID NO: 94, SEQ ID NO: 96, SEQ ID NO: 98, SEQ ID NO: 100, SEQ ID NO: 102, SEQ ID NO: 104, SEQ ID NO: 106, SEQ ID NO: 108, SEQ ID NO: 110, SEQ ID NO: 112 и SEQ ID NO: 114.
В другом аспекте антитело к ETA-1 дополнительно содержит одну или две аминокислотные последовательности, где каждая аминокислотная последовательность независимо выбрана из аминокислотных последовательностей, перечисленных ниже:
a. аминокислотных последовательностей CDR3 легкой цепи под SEQ ID NO: 50, SEQ ID NO: 52, SEQ ID NO: 54, SEQ ID NO: 56, SEQ ID NO: 58, SEQ ID NO: 60, SEQ ID NO: 62, SEQ ID NO: 64, SEQ ID NO: 66, SEQ ID NO: 68, и SEQ ID NO: 205 и
b. аминокислотных последовательностей CDR3 тяжелой цепи под SEQ ID NO: 116, SEQ ID NO: 118, SEQ ID NO: 120, SEQ ID NO: 122, SEQ ID NO: 124, SEQ ID NO: 126, SEQ ID NO: 128, SEQ ID NO: 130, SEQ ID NO: 132, SEQ ID NO: 134 и SEQ ID NO: 136.
В другом варианте осуществления антитело к ETA (антитело к ETA-2), описанное в данном документе, содержит одну или две аминокислотные последовательности, где каждая аминокислотная последовательность независимо выбрана из аминокислотных последовательностей, перечисленных ниже:
a. аминокислотных последовательностей CDR2 легкой цепи под SEQ ID NO: 32, SEQ ID NO: 34, SEQ ID NO: 36, SEQ ID NO: 38, SEQ ID NO: 40, SEQ ID NO: 42, SEQ ID NO: 44, SEQ ID NO: 46 и SEQ ID NO: 48 и
b. аминокислотных последовательностей CDR2 тяжелой цепи под SEQ ID NO: 92, SEQ ID NO: 94, SEQ ID NO: 96, SEQ ID NO: 98, SEQ ID NO: 100, SEQ ID NO: 102, SEQ ID NO: 104, SEQ ID NO: 106, SEQ ID NO: 108, SEQ ID NO: 110, SEQ ID NO: 112 и SEQ ID NO: 114.
В одном аспекте антитело к ETA-2 дополнительно содержит одну или две аминокислотные последовательности, где каждая аминокислотная последовательность независимо выбрана из аминокислотных последовательностей, перечисленных ниже:
a. аминокислотных последовательностей CDR1 легкой цепи под SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14, SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20, SEQ ID NO: 22, SEQ ID NO: 24, SEQ ID NO: 26, SEQ ID NO: 28 и SEQ ID NO: 30 и
b. аминокислотных последовательностей CDR1 тяжелой цепи под SEQ ID NO: 70, SEQ ID NO: 72, SEQ ID NO: 74, SEQ ID NO: 76, SEQ ID NO: 78, SEQ ID NO: 80, SEQ ID NO: 82, SEQ ID NO: 84, SEQ ID NO: 86, SEQ ID NO: 88 и SEQ ID NO: 90.
В другом аспекте антитело к ETA-2 дополнительно содержит одну или две аминокислотные последовательности, где каждая аминокислотная последовательность независимо выбрана из аминокислотных последовательностей, перечисленных ниже:
a. аминокислотных последовательностей CDR3 легкой цепи под SEQ ID NO: 50, SEQ ID NO: 52, SEQ ID NO: 54, SEQ ID NO: 56, SEQ ID NO: 58, SEQ ID NO: 60, SEQ ID NO: 62, SEQ ID NO: 64, SEQ ID NO: 66, SEQ ID NO: 68 и SEQ ID NO: 205 и
b. аминокислотных последовательностей CDR3 тяжелой цепи под SEQ ID NO: 116, SEQ ID NO: 118, SEQ ID NO: 120, SEQ ID NO: 122, SEQ ID NO: 124, SEQ ID NO: 126, SEQ ID NO: 128, SEQ ID NO: 130, SEQ ID NO: 132, SEQ ID NO: 134 и SEQ ID NO: 136.
В другом варианте осуществления антитело к ETA (антитело к ETA-3), описанное в данном документе, содержит одну или две аминокислотные последовательности, где каждая аминокислотная последовательность независимо выбрана из аминокислотных последовательностей, перечисленных ниже:
a. аминокислотных последовательностей CDR3 легкой цепи под SEQ ID NO: 50, SEQ ID NO: 52, SEQ ID NO: 54, SEQ ID NO: 56, SEQ ID NO: 58, SEQ ID NO: 60, SEQ ID NO: 62, SEQ ID NO: 64, SEQ ID NO: 66, SEQ ID NO: 68 и SEQ ID NO: 205 и
b. аминокислотных последовательностей CDR3 тяжелой цепи под SEQ ID NO: 116, SEQ ID NO: 118, SEQ ID NO: 120, SEQ ID NO: 122, SEQ ID NO: 124, SEQ ID NO: 126, SEQ ID NO: 128, SEQ ID NO: 130, SEQ ID NO: 132, SEQ ID NO: 134 и SEQ ID NO: 136.
В одном аспекте антитело к ETA-3 дополнительно содержит одну или две аминокислотные последовательности, где каждая аминокислотная последовательность независимо выбрана из аминокислотных последовательностей, перечисленных ниже:
a. аминокислотных последовательностей CDR1 легкой цепи под SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14, SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20, SEQ ID NO: 22, SEQ ID NO: 24, SEQ ID NO: 26, SEQ ID NO: 28 и SEQ ID NO: 30 и
b. аминокислотных последовательностей CDR1 тяжелой цепи под SEQ ID NO: 70, SEQ ID NO: 72, SEQ ID NO: 74, SEQ ID NO: 76, SEQ ID NO: 78, SEQ ID NO: 80, SEQ ID NO: 82, SEQ ID NO: 84, SEQ ID NO: 86, SEQ ID NO: 88 и SEQ ID NO: 90.
В другом аспекте антитело к ETA-3 дополнительно содержит одну или две аминокислотные последовательности, где каждая аминокислотная последовательность независимо выбрана из аминокислотных последовательностей, перечисленных ниже:
a. аминокислотных последовательностей CDR2 легкой цепи под SEQ ID NO: 32, SEQ ID NO: 34, SEQ ID NO: 36, SEQ ID NO: 38, SEQ ID NO: 40, SEQ ID NO: 42, SEQ ID NO: 44, SEQ ID NO: 46 и SEQ ID NO: 48 и
b. аминокислотных последовательностей CDR2 тяжелой цепи под SEQ ID NO: 92, SEQ ID NO: 94, SEQ ID NO: 96, SEQ ID NO: 98, SEQ ID NO: 100, SEQ ID NO: 102, SEQ ID NO: 104, SEQ ID NO: 106, SEQ ID NO: 108, SEQ ID NO: 110, SEQ ID NO: 112 и SEQ ID NO: 114.
В одном варианте осуществления антитело к ETA, описанное в данном документе, содержит а. аминокислотную последовательность CDR1 легкой цепи, независимо выбранную из перечня, приведенного ниже: SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 14, SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20, SEQ ID NO: 22, SEQ ID NO: 24, SEQ ID NO: 26, SEQ ID NO: 28 и SEQ ID NO: 30;
b. аминокислотную последовательность CDR2 легкой цепи, независимо выбранную из перечня, приведенного ниже: SEQ ID NO: 32, SEQ ID NO: 34, SEQ ID NO: 36, SEQ ID NO: 38, SEQ ID NO: 40, SEQ ID NO: 42, SEQ ID NO: 44, SEQ ID NO: 46 и SEQ ID NO: 48;
с. аминокислотную последовательность CDR3 легкой цепи, независимо выбранную из перечня, приведенного ниже: SEQ ID NO: 50, SEQ ID NO: 52, SEQ ID NO: 54, SEQ ID NO: 56, SEQ ID NO: 58, SEQ ID NO: 60, SEQ ID NO: 62, SEQ ID NO: 64, SEQ ID NO: 66 и SEQ ID NO: 68;
d. аминокислотную последовательность CDR1 тяжелой цепи, независимо выбранную из перечня, приведенного ниже: SEQ ID NO: 70, SEQ ID NO: 72, SEQ ID NO: 74, SEQ ID NO: 76, SEQ ID NO: 78, SEQ ID NO: 80, SEQ ID NO: 82, SEQ ID NO: 84, SEQ ID NO: 86, SEQ ID NO: 88 и SEQ ID NO: 90;
е. аминокислотную последовательность CDR2 тяжелой цепи, независимо выбранную из перечня, приведенного ниже: SEQ ID NO: 92, SEQ ID NO: 94, SEQ ID NO: 96, SEQ ID NO: 98, SEQ ID NO: 100, SEQ ID NO: 102, SEQ ID NO: 104, SEQ ID NO: 106, SEQ ID NO: 108, SEQ ID NO: 110, SEQ ID NO: 112 и SEQ ID NO: 114 и
f. аминокислотную последовательность CDR3 тяжелой цепи, независимо выбранную из перечня, приведенного ниже: SEQ ID NO: 116, SEQ ID NO: 118, SEQ ID NO: 120, SEQ ID NO: 122, SEQ ID NO: 124, SEQ ID NO: 126, SEQ ID NO: 128, SEQ ID NO: 130, SEQ ID NO: 132, SEQ ID NO: 134, и SEQ ID NO: 136.
В одном варианте осуществления антитело к ETA, описанное в данном документе, содержит аминокислотную последовательность CDR3 легкой цепи, независимо выбранную из перечня, приведенного ниже: SEQ ID NO: 50, SEQ ID NO: 52, SEQ ID NO: 54, SEQ ID NO: 56, SEQ ID NO: 58, SEQ ID NO: 60, SEQ ID NO: 62, SEQ ID NO: 64, SEQ ID NO: 66, SEQ ID NO: 68, и SEQ ID NO: 205. В другом варианте осуществления антитело к ETA, описанное в данном документе, содержит аминокислотную последовательность CDR3 тяжелой цепи, независимо выбранную из перечня, приведенного ниже: SEQ ID NO: 116, SEQ ID NO: 118, SEQ ID NO: 120, SEQ ID NO: 122, SEQ ID NO: 124, SEQ ID NO: 126, SEQ ID NO: 128, SEQ ID NO: 130, SEQ ID NO: 132, SEQ ID NO: 134 и SEQ ID NO: 136.
В другом варианте осуществления антитело к ETA, описанное в данном документе, содержит комбинацию аминокислотных последовательностей CDR3 легкой цепи и тяжелой цепи, независимо выбранных из перечня, приведенного ниже: SEQ ID NO: 50 и SEQ ID NO: 116, SEQ ID NO: 50 и SEQ ID NO: 205, SEQ ID NO: 62 и SEQ ID NO: 128, SEQ ID NO: 62 и SEQ ID NO: 130, SEQ ID NO: 64 и SEQ ID NO: 132, SEQ ID NO: 66 и SEQ ID NO: 134 и SEQ ID NO: 68 и SEQ ID NO: 136.
В одном варианте осуществления антитело к ETA, описанное в данном документе, содержит:
(a) аминокислотную последовательность CDR1 легкой цепи под SEQ ID NO: 8;
аминокислотную последовательность CDR2 легкой цепи под SEQ ID NO: 32;
аминокислотную последовательность CDR3 легкой цепи под SEQ ID NO: 50 или SEQ ID NO: 205;
аминокислотную последовательность CDR1 тяжелой цепи под SEQ ID NO: 70;
аминокислотную последовательность CDR2 тяжелой цепи под SEQ ID NO: 92 и
аминокислотную последовательность CDR3 тяжелой цепи под SEQ ID NO: 116;
(b) аминокислотную последовательность CDR1 легкой цепи под SEQ ID NO: 10;
аминокислотную последовательность CDR2 легкой цепи под SEQ ID NO: 34;
аминокислотную последовательность CDR3 легкой цепи под SEQ ID NO: 52;
аминокислотную последовательность CDR1 тяжелой цепи под SEQ ID NO: 72;
аминокислотную последовательность CDR2 тяжелой цепи под SEQ ID NO: 94 и
аминокислотную последовательность CDR3 тяжелой цепи под SEQ ID NO: 118;
(c) аминокислотную последовательность CDR1 легкой цепи под SEQ ID NO: 12;
аминокислотную последовательность CDR2 легкой цепи под SEQ ID NO: 36;
аминокислотную последовательность CDR3 легкой цепи под SEQ ID NO: 54;
аминокислотную последовательность CDR1 тяжелой цепи под SEQ ID NO: 74;
аминокислотную последовательность CDR2 тяжелой цепи под SEQ ID NO: 96 и
аминокислотную последовательность CDR3 тяжелой цепи под SEQ ID NO: 120;
(d) аминокислотную последовательность CDR1 легкой цепи под SEQ ID NO: 14;
аминокислотную последовательность CDR2 легкой цепи под SEQ ID NO: 38;
аминокислотную последовательность CDR3 легкой цепи под SEQ ID NO: 56;
аминокислотную последовательность CDR1 тяжелой цепи под SEQ ID NO: 76;
аминокислотную последовательность CDR2 тяжелой цепи под SEQ ID NO: 98 и
аминокислотную последовательность CDR3 тяжелой цепи под SEQ ID NO: 122;
(e) аминокислотную последовательность CDR1 легкой цепи под SEQ ID NO: 16;
аминокислотную последовательность CDR2 легкой цепи под SEQ ID NO: 40;
аминокислотную последовательность CDR3 легкой цепи под SEQ ID NO: 58;
аминокислотную последовательность CDR1 тяжелой цепи под SEQ ID NO: 78;
аминокислотную последовательность CDR2 тяжелой цепи под SEQ ID NO: 100 и
аминокислотную последовательность CDR3 тяжелой цепи под SEQ ID NO: 124;
(f) аминокислотную последовательность CDR1 легкой цепи под SEQ ID NO: 18;
аминокислотную последовательность CDR2 легкой цепи под SEQ ID NO: 42;
аминокислотную последовательность CDR3 легкой цепи под SEQ ID NO: 60;
аминокислотную последовательность CDR1 тяжелой цепи под SEQ ID NO: 80;
аминокислотную последовательность CDR2 тяжелой цепи под SEQ ID NO: 102 и
аминокислотную последовательность CDR3 тяжелой цепи под SEQ ID NO: 126;
(g) аминокислотную последовательность CDR1 легкой цепи под SEQ ID NO: 20 или SEQ ID NO: 22;
аминокислотную последовательность CDR2 легкой цепи под SEQ ID NO: 44;
аминокислотную последовательность CDR3 легкой цепи под SEQ ID NO: 62;
аминокислотную последовательность CDR1 тяжелой цепи под SEQ ID NO: 82;
аминокислотную последовательность CDR2 тяжелой цепи под SEQ ID NO: 104 или 106 и
аминокислотную последовательность CDR3 тяжелой цепи под SEQ ID NO: 128;
(h) аминокислотную последовательность CDR1 легкой цепи под SEQ ID NO: 24;
аминокислотную последовательность CDR2 легкой цепи под SEQ ID NO: 44;
аминокислотную последовательность CDR3 легкой цепи под SEQ ID NO: 62;
аминокислотную последовательность CDR1 тяжелой цепи под SEQ ID NO: 84;
аминокислотную последовательность CDR2 тяжелой цепи под SEQ ID NO: 108 и
аминокислотную последовательность CDR3 тяжелой цепи под SEQ ID NO: 130;
(i) аминокислотную последовательность CDR1 легкой цепи под SEQ ID NO: 26;
аминокислотную последовательность CDR2 легкой цепи под SEQ ID NO: 46;
аминокислотную последовательность CDR3 легкой цепи под SEQ ID NO: 64;
аминокислотную последовательность CDR1 тяжелой цепи под SEQ ID NO: 86;
аминокислотную последовательность CDR2 тяжелой цепи под SEQ ID NO: 110 и
аминокислотную последовательность CDR3 тяжелой цепи под SEQ ID NO: 132;
(j) аминокислотную последовательность CDR1 легкой цепи под SEQ ID NO: 28;
аминокислотную последовательность CDR2 легкой цепи под SEQ ID NO: 46;
аминокислотную последовательность CDR3 легкой цепи под SEQ ID NO: 66;
аминокислотную последовательность CDR1 тяжелой цепи под SEQ ID NO: 88;
аминокислотную последовательность CDR2 тяжелой цепи под SEQ ID NO: 112 и
аминокислотную последовательность CDR3 тяжелой цепи под SEQ ID NO: 134 или
(k) аминокислотную последовательность CDR1 легкой цепи под SEQ ID NO: 30;
аминокислотную последовательность CDR2 легкой цепи под SEQ ID NO: 48;
аминокислотную последовательность CDR3 легкой цепи под SEQ ID NO: 68;
аминокислотную последовательность CDR1 тяжелой цепи под SEQ ID NO: 90;
аминокислотную последовательность CDR2 тяжелой цепи под SEQ ID NO: 114 и
аминокислотную последовательность CDR3 тяжелой цепи под SEQ ID NO: 136.
В другом варианте осуществления антитело к ETA, описанное в данном документе, содержит
аминокислотную последовательность CDR1 легкой цепи под SEQ ID NO: 28;
аминокислотную последовательность CDR2 легкой цепи под SEQ ID NO: 46;
аминокислотную последовательность CDR3 легкой цепи под SEQ ID NO: 66;
аминокислотную последовательность CDR1 тяжелой цепи под SEQ ID NO: 88;
аминокислотную последовательность CDR2 тяжелой цепи под SEQ ID NO: 112 и
аминокислотную последовательность CDR3 тяжелой цепи под SEQ ID NO: 134.
В другом варианте осуществления антитело к ETA, описанное в данном документе, содержит одну или две аминокислотные последовательности, где каждая аминокислотная последовательность независимо выбрана из аминокислотных последовательностей, перечисленных ниже:
a. аминокислотных последовательностей вариабельного домена легкой цепи под SEQ ID NO: 138 (L1), SEQ ID NO: 140 (L2), SEQ ID NO: 142 (L3), SEQ ID NO: 144 (L4), SEQ ID NO: 146 (L5), SEQ ID NO: 148 (L6), SEQ ID NO: 150 (L7), SEQ ID NO: 152 (L8), SEQ ID NO: 154 (L9), SEQ ID NO: 156 (L10), SEQ ID NO: 158 (L11), SEQ ID NO: 160 (L12), SEQ ID NO: 162 (L13) и SEQ ID NO: 164 (L14) и аминокислотной последовательности, на по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90% или по меньшей мере 95% идентичной им; и
b. аминокислотных последовательностей вариабельного домена тяжелой цепи под SEQ ID NO: 166 (H1), SEQ ID NO: 168 (H2), SEQ ID NO: 170 (H3), SEQ ID NO: 172 (H4), SEQ ID NO: 174 (H5), SEQ ID NO: 176 (H6), SEQ ID NO: 178 (H7), SEQ ID NO: 180 (H8), SEQ ID NO: 182 (H9), SEQ ID NO: 184 (H10), SEQ ID NO: 186 (H11), SEQ ID NO: 188 (H12), SEQ ID NO: 190 (H13) и SEQ ID NO: 192 (H14) и аминокислотной последовательности, на по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90% или по меньшей мере 95% идентичной им.
В другом варианте осуществления полинуклеотидная кодирующая последовательность антитела к ETA, описанного в данном документе, содержит одну или две полинуклеотидные последовательности, где каждая полинуклеотидная последовательность независимо выбрана из полинуклеотидных последовательностей, перечисленных ниже:
a. полинуклеотидных кодирующих последовательностей вариабельного домена легкой цепи под SEQ ID NO: 137, SEQ ID NO: 139, SEQ ID NO: 141, SEQ ID NO: 143, SEQ ID NO: 145, SEQ ID NO: 147, SEQ ID NO: 149, SEQ ID NO: 151, SEQ ID NO: 153, SEQ ID NO: 155, SEQ ID NO: 157, SEQ ID NO: 159, SEQ ID NO: 161 и SEQ ID NO: 163 и полинуклеотидной последовательности, на по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90% или по меньшей мере 95% идентичной им; и
b. полинуклеотидных кодирующих последовательностей вариабельного домена тяжелой цепи под SEQ ID NO: 165, SEQ ID NO: 167, SEQ ID NO: 169, SEQ ID NO: 171, SEQ ID NO: 173, SEQ ID NO: 175, SEQ ID NO: 177, SEQ ID NO: 179, SEQ ID NO: 181, SEQ ID NO: 183, SEQ ID NO: 185, SEQ ID NO: 187, SEQ ID NO: 189 и SEQ ID NO: 191 и полинуклеотидной последовательности, на по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90% или по меньшей мере 95% идентичной им.
В другом варианте осуществления антитело к ETA, описанное в данном документе, содержит
а. аминокислотную последовательность вариабельного домена легкой цепи, независимо выбранную из перечня, приведенного ниже: SEQ ID NO: 138 (L1), SEQ ID NO: 140 (L2), SEQ ID NO: 142 (L3), SEQ ID NO: 144 (L4), SEQ ID NO: 146 (L5), SEQ ID NO: 148 (L6), SEQ ID NO: 150 (L7), SEQ ID NO: 152 (L8), SEQ ID NO: 154 (L9), SEQ ID NO: 156 (L10), SEQ ID NO: 158 (L11), SEQ ID NO: 160 (L12), SEQ ID NO: 162 (L13), и SEQ ID NO: 164 (L14) и аминокислотную последовательность, на по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90% или по меньшей мере 95% идентичную им; и
b. аминокислотную последовательность вариабельного домена тяжелой цепи, независимо выбранную из перечня, приведенного ниже: SEQ ID NO: 166 (H1), SEQ ID NO: 168 (H2), SEQ ID NO: 170 (H3), SEQ ID NO: 172 (H4), SEQ ID NO: 174 (H5), SEQ ID NO: 176 (H6), SEQ ID NO: 178 (H7), SEQ ID NO: 180 (H8), SEQ ID NO: 182 (H9), SEQ ID NO: 184 (H10), SEQ ID NO: 186 (H11), SEQ ID NO: 188 (H12), SEQ ID NO: 190 (H13) и SEQ ID NO: 192 (H14) и аминокислотную последовательность, на по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90% или по меньшей мере 95% идентичную им.
В другом варианте осуществления антитело к ETA, описанное в данном документе, содержит
а. аминокислотную последовательность вариабельного домена легкой цепи, независимо выбранную из перечня, приведенного ниже: SEQ ID NO: 138 (L1), SEQ ID NO: 140 (L2), SEQ ID NO: 142 (L3), SEQ ID NO: 144 (L4), SEQ ID NO: 146 (L5), SEQ ID NO: 148 (L6), SEQ ID NO: 150 (L7), SEQ ID NO: 152 (L8), SEQ ID NO: 154 (L9), SEQ ID NO: 156 (L10), SEQ ID NO: 158 (L11), SEQ ID NO: 160 (L12), SEQ ID NO: 162 (L13), и SEQ ID NO: 164 (L14); и
b. аминокислотную последовательность вариабельного домена тяжелой цепи, независимо выбранную из перечня, приведенного ниже: SEQ ID NO: 166 (H1), SEQ ID NO: 168 (H2), SEQ ID NO: 170 (H3), SEQ ID NO: 172 (H4), SEQ ID NO: 174 (H5), SEQ ID NO: 176 (H6), SEQ ID NO: 178 (H7), SEQ ID NO: 180 (H8), SEQ ID NO: 182 (H9), SEQ ID NO: 184 (H10), SEQ ID NO: 186 (H11), SEQ ID NO: 188 (H12), SEQ ID NO: 190 (H13) и SEQ ID NO: 192 (H14).
В другом варианте осуществления антитело к ETA, описанное в данном документе, содержит комбинацию аминокислотных последовательностей вариабельного домена легкой цепи и тяжелой цепи, независимо выбранных из перечня, приведенного ниже: SEQ ID NO: 138 и SEQ ID NO: 166 ( L1H1), SEQ ID NO: 140 и SEQ ID NO: 168 (L2H2), SEQ ID NO: 142 и SEQ ID NO: 170 (L3H3), SEQ ID NO: 144 и SEQ ID NO: 172 (L4H4), SEQ ID NO: 146 и SEQ ID NO: 174 (L5H5), SEQ ID NO: 148 и SEQ ID NO: 176 (L6H6), SEQ ID NO: 150 и SEQ ID NO: 178 (L7H7), SEQ ID NO: 152 и SEQ ID NO: 180 (L8H8), SEQ ID NO: 154 и SEQ ID NO: 182 (L9H9), SEQ ID NO: 156 и SEQ ID NO: 184 (L10H10), SEQ ID NO: 158 и SEQ ID NO: 186 (L11H11), SEQ ID NO: 160 и SEQ ID NO: 188 (L12H12), SEQ ID NO: 162 и SEQ ID NO: 190 (L13H13) и SEQ ID NO: 164 и SEQ ID NO: 192 (L14H14). В другом варианте осуществления антитело к ETA, описанное в данном документе, содержит комбинацию аминокислотных последовательностей вариабельного домена легкой цепи и тяжелой цепи: SEQ ID NO: 162 и SEQ ID NO: 190 (L13H13).
Символ "LxHy" также может использоваться в данном документе в отношении антитела к ETA, описанного в данном документе, где "x" соответствует вариабельной области легкой цепи, а "y" соответствует вариабельной области тяжелой цепи. Например, L2H1 относится к антителу с вариабельной областью легкой цепи, содержащей аминокислотную последовательность под SEQ ID NO: 140 (L2), и с вариабельной областью легкой цепи, содержащей аминокислотную последовательность под SEQ ID NO: 166 (H1).
В другом варианте осуществления антитело к ЕТА, описанное в данном документе, содержит вариабельную область легкой цепи, выбранную из L1-L14, или вариабельную область тяжелой цепи, выбранную из H1-H14, и их фрагменты, производные, мутантные белки или их варианты.
В другом варианте осуществления антитело к ETA, описанное в данном документе, содержит комбинацию аминокислотных последовательностей CDR3 легкой цепи и тяжелой цепи, независимо выбранных из перечня, приведенного ниже: SEQ ID NO: 138 и SEQ ID NO: 166, SEQ ID NO: 150 и SEQ ID NO: 178, SEQ ID NO: 152 и SEQ ID NO: 180, SEQ ID NO: 154 и SEQ ID NO: 182, SEQ ID NO: 156 и SEQ ID NO: 184, SEQ ID NO: 158 и SEQ ID NO: 186, SEQ ID NO: 160 и SEQ ID NO: 188, SEQ ID NO: 162 и SEQ ID NO: 190 и SEQ ID NO: 164 и SEQ ID NO: 192.
В одном варианте осуществления антитело к ETA, описанное в данном документе, содержит аминокислотную последовательность вариабельного домена легкой цепи под SEQ ID NO: 138 или аминокислотную последовательность вариабельного домена тяжелой цепи под SEQ ID NO: 166. В другом варианте осуществления антитело к ETA, описанное в данном документе, содержит комбинацию аминокислотной последовательности вариабельного домена легкой цепи под SEQ ID NO: 138 и аминокислотной последовательности вариабельного домена тяжелой цепи под SEQ ID NO: 166. В другом варианте осуществления антитело к ЕТА, описанное в данном документе, дополнительно содержит аминокислотные последовательности константного домена, где каждая аминокислотная последовательность константного домена независимо выбрана из аминокислотных последовательностей, перечисленных ниже: a. аминокислотных последовательностей константного домена легкой цепи под SEQ ID NO: 194 и SEQ ID NO: 196 и b. аминокислотных последовательностей константного домена тяжелой цепи под SEQ ID NO: 198 и SEQ ID NO: 206.
В другом варианте осуществления антитело к ETA, описанное в данном документе, дополнительно содержит одну или две аминокислотные последовательности константного домена, где каждая аминокислотная последовательность константного домена независимо выбрана из комбинации аминокислотных последовательностей константного домена легкой цепи и тяжелой цепи, перечисленных ниже:
а. комбинации аминокислотной последовательности константного домена легкой цепи под SEQ ID NO: 194 и аминокислотной последовательности константного домена тяжелой цепи под SEQ ID NO: 198;
b. комбинации аминокислотной последовательности константного домена легкой цепи под SEQ ID NO: 194 и аминокислотной последовательности константного домена тяжелой цепи под SEQ ID NO: 206;
с. комбинации аминокислотной последовательности константного домена легкой цепи под SEQ ID NO: 196 и аминокислотной последовательности константного домена тяжелой цепи под SEQ ID NO: 198;
d. комбинации аминокислотной последовательности константного домена легкой цепи под SEQ ID NO: 196 и аминокислотной последовательности константного домена тяжелой цепи под SEQ ID NO: 206.
В одном варианте осуществления антитело, описанное в данном документе, содержит аминокислотные последовательности CDR и FR (каркасной области) легкой цепи и тяжелой цепи, приведенные в данном документе. В одном варианте осуществления антитело содержит последовательность CDR1 легкой цепи, приведенную в данном документе. В другом варианте осуществления антитело содержит последовательность CDR2 легкой цепи, приведенную в данном документе. В другом варианте осуществления антитело содержит последовательность CDR3 легкой цепи, приведенную в данном документе. В другом варианте осуществления антитело содержит последовательность CDR1 тяжелой цепи, приведенную в данном документе. В другом варианте осуществления антитело содержит последовательность CDR2 тяжелой цепи, приведенную в данном документе. В другом варианте осуществления антитело содержит последовательность CDR3 тяжелой цепи, приведенную в данном документе. В другом варианте осуществления антитело содержит последовательность FR1 легкой цепи, приведенную в данном документе. В другом варианте осуществления антитело содержит последовательность FR2 легкой цепи, приведенную в данном документе. В другом варианте осуществления антитело содержит последовательность FR3 легкой цепи, приведенную в данном документе. В другом варианте осуществления антитело содержит последовательность FR4 легкой цепи, приведенную в данном документе. В другом варианте осуществления антитело содержит последовательность FR1 тяжелой цепи, приведенную в данном документе. В другом варианте осуществления антитело содержит последовательность FR2 тяжелой цепи, приведенную в данном документе. В другом варианте осуществления антитело содержит последовательность FR3 тяжелой цепи, приведенную в данном документе. В другом варианте осуществления антитело содержит последовательность FR4 тяжелой цепи, приведенную в данном документе.
В одном варианте осуществления последовательность CDR3 антитела отличается от комбинации аминокислотных последовательностей CDR3 легкой и тяжелой цепей под SEQ ID NO: 50 и SEQ ID NO: 116, приведенных в данном документе, не более чем шестью, пятью, четырьмя, тремя, двумя или одним (одной) добавлением, заменой и/или делецией одной аминокислоты. В другом варианте осуществления последовательность CDR3 легкой цепи антитела отличается от аминокислотной последовательности CDR3 легкой цепи под SEQ ID NO: 50, приведенной в данном документе, не более чем шестью, пятью, четырьмя, тремя, двумя или одним (одной) добавлением, заменой и/или делецией одной аминокислоты. В другом варианте осуществления последовательность CDR3 легкой цепи антитела отличается от аминокислотной последовательности CDR3 легкой цепи под SEQ ID NO: 50, приведенной в данном документе, не более чем шестью, пятью, четырьмя, тремя, двумя или одним (одной) добавлением, заменой и/или делецией одной аминокислоты, и последовательность CDR3 тяжелой цепи антитела отличается от аминокислотной последовательности CDR3 тяжелой цепи под SEQ ID NO: 116 или SEQ ID NO: 118, приведенной в данном документе, не более чем шестью, пятью, четырьмя, тремя, двумя или одним (одной) добавлением, заменой и/или делецией одной аминокислоты. В другом варианте осуществления антитело дополнительно содержит одну, две, три, четыре, пять или шесть комбинаций последовательностей CDR легкой и тяжелой цепей, приведенных в данном документе. В другом варианте осуществления антитело дополнительно содержит одну, две, три, четыре, пять или шесть комбинаций последовательностей CDR легкой и тяжелой цепей, при этом каждая последовательность в отдельности отличается от комбинации аминокислотных последовательностей CDR3 легкой и тяжелой цепей под SEQ ID NO: 50 и SEQ ID NO: 116, приведенных в данном документе, не более чем шестью, пятью, четырьмя, тремя, двумя или одной отдельной аминокислотой. В другом варианте осуществления антитело содержит CDR вариабельной области легкой цепи и CDR вариабельной области тяжелой цепи, приведенные в данном документе. В другом варианте осуществления антитело содержит одну, две, три, четыре, пять и/или шесть комбинаций последовательностей CDR легкой и тяжелой цепей, приведенных в данном документе.
В одном варианте осуществления антитело (например, антитело или фрагмент антитела) содержит последовательность вариабельного домена легкой цепи L1, приведенную в данном документе. В одном варианте осуществления вариабельный домен легкой цепи содержит аминокислотную последовательность, которая отличается пятнадцатью, четырнадцатью, тринадцатью, двенадцатью, одиннадцатью, десятью, девятью, восемью, семью, шестью, пятью, четырьмя, тремя, двумя или одной аминокислотой от последовательности вариабельного домена легкой цепи L1, где отличие в каждой последовательности независимо представляет собой делецию, вставку или замену аминокислотного остатка. В другом варианте осуществления вариабельный домен легкой цепи содержит аминокислотную последовательность, на по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97% или по меньшей мере 99% идентичную последовательности вариабельного домена легкой цепи L1. В другом варианте осуществления полинуклеотидная кодирующая последовательность вариабельного домена легкой цепи содержит нуклеотидную кодирующую последовательность, на по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере на 97% или по меньшей мере 99% идентичную полинуклеотидной кодирующей последовательности L1. В другом варианте осуществления полинуклеотидная кодирующая последовательность вариабельного домена легкой цепи содержит полинуклеотидную последовательность, которая гибридизуется с комплементарной последовательностью полинуклеотидной кодирующей последовательности вариабельного домена легкой цепи L1 в умеренно жестких условиях. В другом варианте осуществления полинуклеотидная кодирующая последовательность вариабельного домена легкой цепи содержит полинуклеотидную последовательность, которая гибридизуется с комплементарной последовательностью полинуклеотидной кодирующей последовательности вариабельного домена легкой цепи L1 в жестких условиях.
В одном варианте осуществления антитело (например, антитело или фрагмент антитела) содержит последовательность вариабельного домена тяжелой цепи H1, приведенную в данном документе. В другом варианте осуществления вариабельный домен содержит аминокислотную последовательность, которая отличается пятнадцатью, четырнадцатью, тринадцатью, двенадцатью, одиннадцатью, десятью, девятью, восемью, семью, шестью, пятью, четырьмя, тремя, двумя или одной аминокислотой от последовательности вариабельного домена тяжелой цепи H1, где отличие в каждой последовательности независимо представляет собой делецию, вставку или замену аминокислотного остатка. В другом варианте осуществления вариабельный домен тяжелой цепи содержит аминокислотную последовательность, на по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97% или по меньшей мере 99% идентичную последовательности вариабельного домена тяжелой цепи H1. В другом варианте осуществления полинуклеотидная кодирующая последовательность вариабельного домена тяжелой цепи содержит нуклеотидную кодирующую последовательность, на по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере на 97% или по меньшей мере 99% идентичную полинуклеотидной кодирующей последовательности H1. В другом варианте осуществления полинуклеотидная кодирующая последовательность вариабельного домена тяжелой цепи содержит полинуклеотидную последовательность, которая гибридизуется с комплементарной последовательностью полинуклеотидной кодирующей последовательности вариабельного домена тяжелой цепи H1 в умеренно жестких условиях. В одном варианте осуществления полинуклеотидная кодирующая последовательность вариабельного домена тяжелой цепи содержит полинуклеотидную последовательность, которая гибридизуется с комплементарной последовательностью полинуклеотидной кодирующей последовательности вариабельного домена тяжелой цепи H1 в жестких условиях.
В одном варианте осуществления антитело, описанное в данном документе, включает антитело, содержащее комбинацию L1H1, L2H2, L3H3, L4H4, L5H5, L6H6, L7H7, L8H8, L9H9, L10H10, L11H11, L12H12, L13H13 или L14H14, или их требуемый фенотип (например, IgA, IgG1, IgG2a, IgG2b, IgG3, IgM, IgE и IgD), или их Fab- или F(ab')2-фрагмент.
В одном варианте осуществления антитело, описанное в данном документе, включает антитело, содержащее комбинацию L1H1, или антитело с переключением его класса (например, IgA, IgG1, IgG2a, IgG2b, IgG3, IgM, IgE и IgD) или его Fab- или F(ab')2-фрагмент.
Антитело, описанное в данном документе (например, антитело, фрагмент антитела и производное антитела), может содержать любую из константных областей, известных из уровня техники. Константная область легкой цепи может, например, представлять собой константную область легкой цепи κ или λ, такую как мышиная константная область легкой цепи κ или λ. Константная область тяжелой цепи может, например, представлять собой константную область тяжелой цепи α, δ, ε, γ или μ, такую как мышиная константная область тяжелой цепи α, δ, ε, γ или μ. В одном варианте осуществления константная область легкой или тяжелой цепи представляет собой фрагмент, производное, вариант или мутантный белок природной константной области.
В одном варианте осуществления антитело, описанное в данном документе, дополнительно содержит константный домен легкой цепи κ или λ или его фрагмент. Последовательности константной области легкой цепи и их полинуклеотидные кодирующие последовательности представлены ниже: полинуклеотидная последовательность (κ), (SEQ ID NO: 193); аминокислотная последовательность (κ), (SEQ ID NO: 194); полинуклеотидная последовательность (λ), (SEQ ID NO: 195); аминокислотная последовательность (λ), (SEQ ID NO: 196).
В другом варианте осуществления антитело, описанное в данном документе, дополнительно содержит константный домен тяжелой цепи или его фрагмент. Последовательности константной области тяжелой цепи и их полинуклеотидные кодирующие последовательности представлены ниже: полинуклеотидная последовательность (IgG4), (SEQ ID NO: 197); аминокислотная последовательность (IgG4), (SEQ ID NO: 198).
В одном варианте осуществления антитело к ЕТА, описанное в данном документе, выбрано из мышиного антитела, гуманизированного антитела, химерного антитела, моноклонального антитела, поликлонального антитела, рекомбинантного антитела, фрагмента антигенсвязывающего антитела, одноцепочечного антитела, диантитела, триантитела, тетраантитела, Fab-фрагмента, F(ab')x-фрагмента, доменного антитела, антитела IgD, антитела IgE, антитела IgM, антитела IgGl, антитела IgG2, антитела IgG3 и антитела IgG4. В другом варианте осуществления антитело к ETA, описанное в данном документе, представляет собой моноклональное антитело к ETA. В другом варианте осуществления антитело к ETA, описанное в данном документе, представляет собой мышиное антитело к ETA. Антитело к ЕТА, описанное в данном документе, представляет собой гуманизированное антитело к ЕТА.
В одном варианте осуществления антитело к ETA, описанное в данном документе, представляет собой моноклональное антитело A-1 (содержащее SEQ ID NO: 138 и SEQ ID NO: 166), A-7 (содержащее SEQ ID NO: 150 и SEQ ID NO: 178), A-8 (содержащее SEQ ID NO: 152 и SEQ ID NO: 180), A-9 (содержащее SEQ ID NO: 154 и SEQ ID NO: 182), A-10 (содержащее SEQ ID NO: 156 и SEQ ID NO: 184), A-11 (содержащее SEQ ID NO: 158 и SEQ ID NO: 186), A-12 (содержащее SEQ ID NO: 160 и SEQ ID NO: 188), A-13 (содержащее SEQ ID NO: 162 и SEQ ID NO: 190) или A-14 (содержащее SEQ ID NO: 164 и SEQ ID NO: 192).
Антитела и фрагменты антител
В одном варианте осуществления антитело, описанное в данном документе, представляет собой интактное антитело (в том числе поликлональные, моноклональные, химерные, гуманизированные или человеческие антитела с полноразмерными тяжелыми и/или легкими цепями). В другом варианте осуществления антитело, описанное в данном документе, представляет собой фрагмент антитела, например, F(ab')2-, Fab-, Fab'-, Fv-, Fc- или Fd-фрагмент, и он может быть включен в однодоменные антитела, одноцепочечные антитела, максиантитела, миниантитела, интраантитела, диантитела, триантитела, тетраантитела, v-NAR и бис-scFv (см., например, Hollinger and Hudson, 2005, Nature Biotechnology, 23:1126-1136). В другом варианте осуществления антитело, описанное в данном документе, также включает полипептидные антитела, такие как раскрытые в патенте США № US 6703199, в том числе моноклональные антитела с полипептидом фибронектина. В другом варианте осуществления антитело, описанное в данном документе, также содержит полипептиды других антител, такие как полипептиды, раскрытые в публикации заявки на патент США № US 2005/0238646, которые представляют собой одноцепочечные полипептиды.
В одном варианте осуществления вариабельные области генов, экспрессирующих родственные моноклональные антитела в гибридомах, амплифицируют с использованием нуклеотидных праймеров. Эти праймеры могут быть синтезированы специалистом в данной области техники или могут быть приобретены из коммерческих источников (см., например, Stratagene, La Jolla и California), при этом указанные поставщики продают праймеры для мышиных и человеческих вариабельных областей, в том числе праймеры для областей VHa, VHb, VHc, VHd, CH1, VL и CL. Эти праймеры могут быть использованы для амплификации вариабельных областей тяжелой или легкой цепей, которые затем могут быть встроены в векторы, такие как IMMUNOZAPTMH или ILLLFFLUNOZAPTML (Stratagene) соответственно. Затем эти векторы могут быть введены в экспрессионные системы на основе E. coli, дрожжей или клеток млекопитающих. С применением этих способов можно получить большие количества слитых одноцепочечных белков, содержащих домены VH и VL (см. Bird et al., 1988, Science 242: 423-426).
После получения антитело-продуцирующих клеток в соответствии с настоящей заявкой с применением любой из вышеупомянутых иммунизации и других методик гены специфических антител могут быть клонированы посредством выделения генов и амплификации из них ДНК или мРНК в соответствии со стандартными способами, описанными в данном документе. Антитело, продуцируемое из них, секвенируют и идентифицируют CDR. ДНК, кодирующую CDR, можно подвергать манипуляции, как описано ранее, с получением других антител, описанных в данном документе.
Предпочтительно антитело, описанное в данном документе, может в клеточных анализах, описанных в данном документе, и/или анализах in vivo, описанных в данном документе, модулировать передачу сигнала, опосредованную эндотелином, и/или перекрестно блокировать связывание одного из антител, описанных в настоящей заявке, и/или перекрестно блокироваться связыванием с ETA посредством одного из антител, описанных в настоящей заявке. Следовательно, анализы, описанные в данном документе, могут быть использованы для идентификации таких связывающих средств.
В некоторых вариантах осуществления антитело получают посредством первого идентифицирования в клеточных анализах и/или анализах in vivo, описанных в данном документе, при этом антитело, которое связывается с клетками, сверхэкспрессирующими ETA, и/или нейтрализует и/или перекрестно блокирует антитела, описанные в настоящей заявке, и/или перекрестно блокируется связыванием с ETA посредством одного из антител, описанных в настоящей заявке.
Специалисту в данной области техники будет понятно, что некоторые белки, такие как антитела, можно подвергать ряду посттрансляционных модификаций. Типы и степени этих модификаций зависят от линий клеток-хозяев, используемых для экспрессии белка, а также от условий культивирования. Такие модификации включают вариации в гликозилировании, окислении метионина, образовании дикетопиперизина, изомеризации аспартата и дезамидировании аспарагина. Частые модификации вследствие действия карбоксипептидаз приводят к потере основных карбоксиконцевых остатков (таких как лизин или аргинин) (что описано в Harris, 1995, Journal of Chromatography 705:129-134).
Альтернативный способ получения мышиных моноклональных антител заключается в инъекции клеток гибридом в брюшную полость сингенным мышам, например, мышам, которых обрабатывали (например, первичной иммунизацией пристаном), чтобы способствовать образованию асцитной жидкости, содержащей моноклональные антитела. Моноклональные антитела могут быть выделены и очищены посредством многих хорошо известных методик. Такие методики выделения включают аффинную хроматографию с белком А-сефарозой, эксклюзионную хроматографию и ионообменную хроматографию (см., например, Coligan, pages 2.7.1-2.7.12 and pages 2.9.1-2.9.3 и Baines et al., "Purification of Immunoglobulin G (IgG)", Methods in Molecular Biology, vol. 10, pages 79-104 (The Humana Press, Inc. 1992)). Моноклональное антитело можно очистить с помощью аффинной хроматографии с использованием подходящего лиганда, выбранного исходя из конкретных свойств антитела (например, изотипа тяжелой или легкой цепи, специфичности связывания и т.д.). Примеры подходящих лигандов, иммобилизованных на твердой подложке, включают белок A, белок G, антитело к константной области (легкой цепи или тяжелой цепи), антиидиотипическое антитело и белок, связывающий TGF-β, или его фрагмент или вариант.
Молекулярная эволюция определяющих комплементарность областей (CDR) в центре сайта связывания антитела была использована для выделения антител с повышенными значениями аффинности, например антител, характеризующихся повышенной аффинностью в отношении c-erbB-2, что описано в Schier et al., 1996, J. Mol. Biol. 263:551-567. Соответственно, такие методики применимы для получения антител к человеческим эндотелиновым рецепторам. Антитела к человеческим эндотелиновым рецепторам можно использовать, например, в анализах для выявления присутствия эндотелинового рецептора либо in vitro, либо in vivo.
Антитела также могут быть получены посредством любой из традиционных методик. Например, такие антитела могут быть очищены от клеток, которые естественным образом экспрессируют их (например, антитело может быть очищено от гибридомы, которая продуцирует его), или получены в рекомбинантных системах экспрессии с использованием любой методики, известной из уровня техники. См., например, Monoclonal Antibodies, Hybridomas: A New Dimension in Biological Analyses, Kennet et al. (eds.), Plenum Press, (1980) и Antibodies: A Laboratory Manual, Harlow and Land (eds.), Cold Spring Harbor Laboratory Press (1988). Это обсуждается в приведенном ниже разделе "Нуклеиновая кислота".
Антитела могут быть получены и подвергнуты скринингу в отношении необходимых свойств посредством любых известных методик. Некоторые методики относятся к выделению нуклеиновых кислот, кодирующих полипептидные цепи (или их части) родственных антител (например, антител к эндотелиновому рецептору), и манипуляции с нуклеиновыми кислотами посредством методик рекомбинантной ДНК. Нуклеиновые кислоты могут быть слиты с другой соответствующей нуклеиновой кислотой или модифицированы (например, посредством мутагенеза или других традиционных методик) с добавлением, делецией или заменой одного или более аминокислотных остатков.
Если требуется улучшение аффинности антител по настоящему изобретению, содержащих одну или более из вышеупомянутых CDR, можно использовать ряд протоколов созревания аффинности, в том числе поддержание CDR (Yang et al., 1995, J. Mol. Biol., 254:392-403), шаффлинг цепей (Marks et al., 1992, Bio/Technology, 10:779-783), применение мутантных штаммов E. coli. (Low et al., 1996, J. Mol. Biol., 250:350-368), реаранжировку ДНК (Patten et al., 1997, Curr. Opin. Biotechnol., 8:724-733), фаговый дисплей (Thompson et al., 1996, J. Mol. Biol., 256:7-88) и дополнительные методики ПЦР (Crameri et al., 1998, Nature, 391:288-291). Все эти методики созревания аффинности обсуждаются в Vaughan et al., 1998, Nature Biotechnology, 16:535-539.
В одном варианте осуществления антитело, описанное в данном документе, представляет собой фрагмент антитела к эндотелиновому рецептору. Фрагмент может полностью состоять из последовательностей, полученных из антитела, или может содержать дополнительные последовательности. Примеры антигенсвязывающих фрагментов включают Fab, F(ab')2, одноцепочечные антитела, диантитела, триантитела, тетраантитела и доменные антитела. Другие примеры представлены в Lunde et al., 2002, Biochem. Soc. Trans. 30:500-06.
Одноцепочечные антитела могут быть образованы путем связывания фрагментов вариабельного домена тяжелой и легкой цепей (Fv-области) посредством аминокислотного мостика (короткого пептидного линкера), в результате чего образуется одна полипептидная цепь. Такие одноцепочечные Fv (scFv) были получены путем слияния ДНК, кодирующей пептидный линкер между ДНК, кодирующими полипептиды двух вариабельных доменов (VL и VH). Образованные полипептиды могут обратно укладываться сами по себе с образованием антигенсвязывающих мономеров или они могут образовывать полимеры (например, димеры, тримеры или тетрамеры) в зависимости от длины гибкого линкера между двумя вариабельными доменами (Kortt et al., 1997, Prot. Eng. 10:423 и Kortt et al., 2001, Biomol. Eng. 18:95-108). Посредством комбинирования различных полипептидов, содержащих VL и VH, могут быть образованы полимерные scFv, которые связываются с разными фенотипами (Kriangkum et al., 2001, Biomol. Eng. 18:31-40). Методики, разработанные для получения одноцепочечных антител, включают методики, описанные в патенте США № US 4946778; Bird, 1988, Science 242:423; Huston et al., 1988, PNAS USA 85:5879-5883; Ward et al., 1989, Nature 334:544-546; deGraaf et al., 2002, Methods Mol. Biol. 178:379-87. Одноцепочечные антитела, полученные из описанных в данном документе антител, включающих без ограничения scFv, содержащие комбинацию вариабельных доменов L1H1, охватываются объемом, описанным в данном документе.
Антигенсвязывающие фрагменты, полученные из антител, также могут быть получены посредством протеолитического гидролиза антител, например, путем расщепления интактного антитела пепсином или папаином в соответствии с традиционными способами. В качестве примера, фрагменты антител можно получить путем ферментативного расщепления антител пепсином с получением SS-фрагмента, называемого F(ab')2. Данный фрагмент может быть дополнительно расщеплен с использованием сульфгидрильного восстанавливающего средства с получением одновалентных 3,5S Fab'-фрагментов. Факультативная схема заключается в проведении реакции расщепления сульфгидрильной защитной группой с получением расщепления дисульфидной связи; кроме того, ферментативное расщепление папаином непосредственно приводит к получению двух одновалентных Fab-фрагментов и Fc-фрагмента. Эти способы описаны, например, в патенте США № US 4331647; Nisonoffet et al., 1960, Arch. Biochem. Biophys. 89:230; Porter, 1959, Biochem. J. 73:119; Edelman et al., Methods in Enzymology 1:422 (Academic Press, 1967) и Andrews and Titus, J. A. Current Protocols in Immunology (Coligan et al., (eds)), John Wiley & Sons, 2003), pages 2.8.1-2.8.10 and 2.10A.1-2.10A.5. Другие способы расщепления антител, такие как получение тяжелых цепей с образованием одновалентных фрагментов тяжелых и легких цепей (Fd), дополнительное расщепление фрагментов или другие ферментативные, химические или генетические методики, также могут быть использованы, при условии, что фрагменты связываются с антигеном, который распознается интактным антителом.
Другой формой фрагмента антитела является пептид, содержащий одну или более определяющих комплементарность областей (CDR) антитела. CDR могут быть получены путем конструирования полинуклеотидов, которые кодируют соответствующие CDR. Такие полинуклеотиды могут быть получены, например, посредством полимеразной цепной реакции для синтеза вариабельной области с использованием мРНК или антитело-продуцирующих клеток в качестве матрицы (см., например, Larrick et al., 1991, Methods: A Companion to Methods in Enzymology 2:106; Courtenay-Luck, "Genetic Manipulation of Monoclonal Antibodies", Monoclonal Antibodies: Production, Engineering and Clinical Application, Ritter et al. (eds.), page 166 (Cambridge University Press, 1995) и Ward et al., "Genetic Manipulation and Expression of Antibodies", Monoclonal Antibodies: Principles and Applications, Birch et al., (eds.), p. 137 (Wiley-Liss, Inc. 1995). Фрагмент антитела может дополнительно содержать по меньшей мере один домен вариабельной области антитела, описанного в данном документе. Таким образом, например, домен V-области может быть мономерным и представлять собой домен VH или VL, который может независимо связываться с эндотелиновым рецептором с аффинностью, составляющей по меньшей мере 1×10-7 M или меньше, как описано ниже.
Домен вариабельной области может быть любым встречающимся в природе вариабельным доменом или его генетически сконструированным вариантом. Под генетически сконструированным вариантом подразумевается домен вариабельной области, полученный с применением методик конструирования рекомбинантной ДНК. Такие генетически сконструированные варианты включают те, которые получены, например, из специфической вариабельной области антитела путем вставок, делеций или изменений в или относительно аминокислотных последовательностей конкретного антитела. Конкретные примеры включают генетически сконструированные домены вариабельной области, содержащие только одну CDR и необязательно одну или более аминокислот каркаса из одного антитела и оставшуюся часть домена вариабельной области из другого антитела.
Домен вариабельной области может быть ковалентно связан по С-концевой аминокислоте с по меньшей мере одним другим доменом антитела или его фрагментом. Таким образом, в качестве примера, домен VH, который присутствует в домене вариабельной области, может быть связан с доменом CH1 иммуноглобулина или его фрагмента. Аналогичным образом домен VL может быть связан с доменом CK или его фрагментом. Таким образом, например, антитело может представлять собой Fab-фрагмент, в котором антигенсвязывающий домен содержит объединенные домены VH и VL, ковалентно связанные на своих C-концах с доменами CH1 и CK соответственно. Домен CH1 может быть расширен дополнительными аминокислотами, например, для обеспечения шарнирной области или части шарнирной области домена, выявленной в Fab'-фрагменте, или для обеспечения дополнительных доменов, таких как домены CH2 и CH3 антитела.
Производные и варианты антител
Нуклеотидные последовательности L1 и H1, кодирующие аминокислотную последовательность А-1, могут быть изменены, например, посредством рандомного мутагенеза или сайт-направленного мутагенеза (например, олигонуклеотид-индуцированного сайт-направленного мутагенеза) для создания измененного полинуклеотида, содержащего одну или более конкретных нуклеотидных замен, делеций или вставок по сравнению с немутантным полинуклеотидом. Примеры методик осуществления таких изменений описаны в Walder et al., 1986, Gene 42:133; Bauer et al., 1985, Gene 37:73; Craik, 1985, BioTechniques, 3:12-19; Smith et al., 1981, Genetic Engineering: Principles and Methods, Plenum Press и в патентах США №№ US 4518584 и US 4737462. Эти и другие способы можно использовать для получения, например, производных антител к эндотелиновым рецепторам с требуемыми свойствами, такими как повышенная аффинность, авидность или специфичность в отношении эндотелиновых рецепторов, повышенная активность или стабильность in vivo или in vitro или сниженные побочные эффекты in vivo по сравнению с недериватизированным антителом.
Другие производные антител к эндотелиновым рецепторам из уровня техники включают ковалентные или агрегированные конъюгаты антител к эндотелиновым рецепторам или их фрагментов с другими белками или полипептидами, например, полученные посредством экспрессии рекомбинантных слитых белков, содержащих гетерологичные полипептиды, слитые с N-концом или С-концом полипептида антитела к эндотелиновым рецепторам. Например, конъюгированный пептид может представлять собой гетерологичный сигнальный (или лидерный) полипептид, например, лидерный пептид альфа-фактора дрожжей или пептид, меченный эпитопом. Антитело, содержащее слитые белки, может содержать пептиды, добавленные для облегчения очистки или идентификации антитела (например, поли-His). Антитело также может быть связано с FLAG-пептидом, что описано в Hopp et al., 1988, Bio/Technology 6:1204 и патенте США № US 5011912. FLAG-пептид является высокоантигенным и обеспечивает эпитоп, обратимо связываемый специфическим моноклональным антителом (mAb), что позволяет проводить экспресс-анализ и легкую очистку экспрессированного рекомбинантного белка. Реагенты, используемые для получения слитых белков, в которых FLAG-пептид слит с данным полипептидом, коммерчески доступны (Sigma-Aldrich, Сент-Луис, Миссури). В другом варианте осуществления олигомеры, которые содержат одно или более антител, можно использовать в качестве антагонистов эндотелиновых рецепторов или высших олигомеров. Олигомеры могут быть в форме ковалентно связанных или нековалентно связанных димеров, тримеров или высших олигомеров. Предполагается применение олигомеров, содержащих два или более антител, одним из примеров которых является гомодимер. Другие олигомеры включают гетеродимеры, гомотримеры, гетеротримеры, гомотетрамеры, гетеротетрамеры и т.д.
Один вариант осуществления относится к олигомерам, содержащим несколько антител, которые связаны посредством ковалентных или нековалентных взаимодействий между пептидными фрагментами, слитыми с антителами. Такие пептиды могут быть пептидными линкерами (спейсерами) или пептидами, которые обладают свойством обеспечения олигомеризации. Лейциновые застежки и определенные полипептиды, полученные из антител, представляют собой пептиды, которые могут обеспечивать олигомеризацию антител, что более подробно описано ниже.
В конкретных вариантах осуществления олигомеры содержат от двух до четырех антител. Антитела в составе олигомеров могут иметь любую форму, такую как любая из форм, описанных выше, например, варианты или фрагменты. Предпочтительно олигомеры содержат антитела, характеризующиеся связывающей активностью в отношении эндотелиновых рецепторов.
В одном варианте осуществления олигомеры получают с использованием полипептидов, полученных из иммуноглобулинов. Получение слитых белков, содержащих определенные гетерологичные полипептиды, слитые с различными частями полипептидов, полученных из антител (в том числе Fc-домена), было описано, например, в Ashkenazi et al., 1991, PNAS USA 88:10535; Byrn et al., 1990, Nature 344:677 и Hollenbaugh et al., Construction of Immunoglobulin Fusion Proteins, Current Protocols in Immunology, Suppl. 4, pp.ages 10.19.1-10.19.11. Один вариант осуществления в данном документе относится к димеру, содержащему два слитых белка, получаемых посредством слияния эндотелин-связывающего фрагмента антитела к эндотелиновым рецепторам с Fc-областью антитела. Димер может быть получен посредством следующих способов: например, посредством вставки слитого гена, кодирующего слитый белок, в соответствующий вектор экспрессии, экспрессии слитого гена в клетках-хозяевах, трансформированных рекомбинантным вектором экспрессии, и обеспечения возможности сборки экспрессированного слитого белка, равно как и молекуле антитела, после чего межцепочечные дисульфидные связи между Fc-фрагментами образуют димер.
Термин "полипептид Fc", используемый в данном документе, включает полипептиды в форме природных белков и мутантных белков, полученных из Fc-области антител. Также включены усеченные формы таких полипептидов, содержащие шарнирную область, которая способствует димеризации. Слитые белки, содержащие Fc фрагменты (и образованные из них олигомеры), обладают преимуществом простой очистки посредством аффинной хроматографии на колонках с белком A или белком G.
Подходящий Fc-полипептид, описанный в заявке РСТ WO 93/10151 (включенной в данный документ посредством ссылки), представляет собой одноцепочечный полипептид, расположенный в направлении от N-концевой шарнирной области до природного С-конца Fc-области антитела IgG1 человека. Другим применимым Fc-полипептидом является мутантный белок Fc, описанный в патенте США № US 5457035 и Baum et al., 1994, EMBO J. 13:3992-4001. Аминокислотная последовательность данного мутантного белка идентична последовательности природного Fc, показанной в WO 93/10151, за исключением того, что аминокислота 19 была заменена с Leu на Ala, аминокислота 20 была заменена с Leu на Glu, а аминокислота 22 была заменена с Gly на Ala. Мутантный белок характеризуется сниженной аффинностью в отношении Fc-рецепторов. В других вариантах осуществления тяжелая цепь и/или легкая цепь антитела к эндотелиновому рецептору может быть заменена вариабельной частью его тяжелой цепи и/или легкой цепи.
В качестве альтернативы олигомер представляет собой слитый белок, содержащий несколько антител с пептидными линкерами (спейсерными пептидами) или без них. Подходящие пептидные линкеры описаны в патентах США №№ US 4751180 и US 4935233.
Другой способ получения олигомерных антител предусматривает применение лейциновой застежки. Домены лейциновая застежка представляют собой пептиды, которые способствуют олигомеризации белков, в которых они обнаружены. Лейциновые застежки были первоначально идентифицированы в нескольких ДНК-связывающих белках (Landschulz et al., 1988, Science 240:1759), а позже обнаружены в ряде различных белков. Среди известных лейциновых застежек имеются встречающиеся в природе пептиды или их производные, которые могут быть димеризованы или тримеризованы. Примеры доменов лейциновая застежка, подходящих для получения растворимых олигомерных белков, описаны в заявке РСТ WO 94/10308, а лейциновые застежки, полученные из легочного сурфактантного белка D (SPD), описаны в Hoppe et al., 1994, FEBS Letters 344:191, включенной в данный документ посредством ссылки. Применение модифицированной лейциновой застежки, которая способствует стабильной тримеризации слитого с ней гетерологичного белка, описано в Fanslow et al., 1994, Semin. Immunol. 6:267-78. В одном способе рекомбинантные слитые белки, содержащие фрагмент антитела к эндотелиновым рецепторам или его производное, слитый с пептидом с лейциновой застежкой, экспрессируют в подходящих клетках-хозяевах, а растворимый олигомерный фрагмент антитела к эндотелиновым рецепторам или его производное выделяют из культурального супернатанта.
В другом варианте осуществления производные антитела могут содержать по меньшей мере одну из CDR, раскрытых в данном документе. В качестве примера одна или более CDR могут быть интегрированы в известные каркасные области антитела (IgG1, IgG2 и т.д.) или конъюгированы с подходящим носителем для увеличения его периода полужизни. Подходящие носители включают без ограничения Fc, альбумин, трансферрин и т.п. Эти и другие подходящие носители известны из уровня техники. Такие конъюгированные пептиды CDR могут быть в мономерной, димерной, тетрамерной или других формах. В одном варианте осуществления один или более водорастворимых полимеров связываются в одном или более конкретных сайтах, например, на аминоконце связывающего средства. В одном примере производное антитела включает одно или более водорастворимых полимерных звеньев, в том числе без ограничения, полиэтиленгликоль, полиоксиэтиленгликоль или полипропиленгликоль. См., например, патенты США №№ US 4640835, US 4496689, US 4301144, US 4670417, US 4791192 и US 4179337. В некоторых вариантах осуществления производное предусматривает одно или более из монометоксиполиэтиленгликоля, декстрана, целлюлозы или других полимеров на основе углеводов, поли(N-винилпирролидон)-полиэтиленгликоля, полиоксиэтилированных полиолов (например, глицерина) и поливинилового спирта, а также смеси таких полимеров. В некоторых вариантах осуществления один или более водорастворимых полимеров рандомно присоединяются к одной или более боковым цепям. В некоторых вариантах осуществления PEG может действовать с улучшением терапевтического эффекта связывающего средства, такого как антитело. Некоторые такие способы описаны, например, в патенте США № US 6133426, который включен в данный документ посредством ссылки для любой цели.
Следует учитывать, что антитело, описанное в данном документе, может содержать по меньшей мере одну аминокислотную замену, при условии, что антитело сохраняет специфичность связывания. Следовательно, модификации структур антитела охватываются объемом, описанным в данном документе. Данные модификации могут включать аминокислотные замены, которые могут быть консервативными или неконсервативными, которые не нарушают способность антитела связывать эндотелиновые рецепторы. Консервативные аминокислотные замены могут охватывать не встречающиеся в природе аминокислотные остатки, которые обычно интегрируются путем химического синтеза пептидов, но не путем синтеза в биологических системах. Данные аминокислотные остатки включают пептидомиметики и другие обращенные или инвертированные формы аминокислотных фрагментов. Консервативные аминокислотные замены также могут включать замену встречающегося в природе аминокислотного остатка на не встречающийся в природе остаток, что несущественно или вообще не влияет на полярность или заряд аминокислотного остатка в этом сайте. Неконсервативные замены могут включать замену представителя одного класса аминокислот или аналогов аминокислот на представителя другого класса с другими физическими свойствами (например, размером, полярностью, гидрофобностью и зарядом).
Более того, специалист в данной области техники может создать варианты для тестирования, которые содержат аминокислотную замену в каждом требуемом аминокислотном остатке. Варианты можно подвергнуть скринингу с применением анализов активности, известных специалисту в данной области техники. Такие варианты можно использовать для сбора информации о подходящих вариантах. В качестве примера, если было обнаружено, что изменение определенного аминокислотного остатка приводит к нарушению, нежелательному снижению или неприемлемой активности, то следует избегать использования вариантов с таким изменением. Другими словами на основе информации, собранной в ходе таких стандартных экспериментов, специалист в данной области техники может легко определить аминокислоты, дальнейших замен в которых следует избегать, либо по отдельности, либо в комбинации с другими мутациями.
Специалист в данной области техники сможет определить подходящие варианты полипептида, изложенные в данном документе, с применением известных методик. В некоторых вариантах осуществления специалист в данной области техники может идентифицировать подходящие области молекулы, которые можно изменить без нарушения активности посредством нацеливания на области, которые не важны для проявления активности. В некоторых вариантах осуществления могут быть идентифицированы остатки или части молекул, которые являются консервативными среди подобных полипептидов. В некоторых вариантах осуществления даже консервативные замены областей, которые важны для биологической активности или структуры, не нарушают биологическую активность и не оказывают неблагоприятного воздействия на структуру полипептида. Кроме того, специалист в данной области техники может проанализировать исследования структуры-функции для идентификации остатков в подобных полипептидах, которые важны для проявления активности или структуры. С учетом такого сравнения можно предсказать важность аминокислотных остатков в белке, которые соответствуют аминокислотным остаткам, важным для активности или структуры в подобных белках. Специалист в данной области техник может сделать выбор в пользу химически подобных аминокислотных замен для таких предсказанных важных аминокислотных остатков.
Специалист в данной области техники также может проанализировать трехмерную структуру и аминокислотную последовательность по отношению к этой структуре в подобных полипептидах. Принимая во внимание такую информацию, специалист в данной области техники может предсказать выравнивание аминокислотных остатков антитела относительно его трехмерной структуры. В некоторых вариантах осуществления специалист в данной области техники может решить не вносить радикальных изменений в аминокислотные остатки, которые, как предполагается, находятся на поверхности белка, поскольку такие остатки могут участвовать в важных взаимодействиях с другими молекулами. Ряд научных публикаций посвящен прогнозированию вторичной структуры. См. Moult, 1996, Curr. Op. Biotech. 7:422-427; Chou et al., 1974, Biochemistry 13:222-245; Chou et al., 1974, Biochemistry 113:211-222; Chou et al., 1978, Adv. Enzymol. Relat. Areas Mol. Biol. 47:45-148; Chou et al., 1979, Ann. Rev. Biochem. 47:251-276 и Chou et al., Biophys. J. 26:367-384. Более того, в настоящее время доступны компьютерные программы для помощи в прогнозировании вторичной структуры. В качестве примера два полипептида или белка, которые характеризуются идентичностью последовательности, превышающей 30%, или подобием последовательности, превышающим 40%, часто характеризуются сходными высокоуровневыми структурами. Недавнее расширение базы данных белков (PDB) обеспечило повышенную прогнозируемость вторичной структуры, включая потенциальное число складок в структуре полипептида или белка. См. Holm et al., 1999, Nucl. Acid. Res. 27:244-247. Было высказано предположение (Brenner et al., 1997, Curr. Op. Struct. Biol. 7:369-376), что существует ограниченное количество складок в данном полипептиде или белке, и что как только критическое число структур будет определено, прогнозирование структуры станет значительно точнее. Дополнительные способы прогнозирования вторичной структуры включают "протягивание" (Jones, 1997, Curr. Opin. Struct. Biol. 7:377-87 и Sippl et al., 1996, Structure 4:15-19); "анализ профиля" (Bowie et al., 1991, Science 253:164-170; Gribskov et al., 1990, Meth. Enzym. 183:146-159 и Gribskov et al., 1987, PNAS USA 84:4355-4358), и "эволюционную связь" (см. Holm выше (1999) и Brenner выше (1997). В некоторых вариантах осуществления варианты антител включают варианты гликозилирования, в которых число и/или тип сайтов гликозилирования были изменены по сравнению с аминокислотными последовательностями исходного полипептида. В некоторых вариантах осуществления варианты содержат большее или меньшее количество сайтов N-связанного гликозилирования, чем нативные белки. В качестве альтернативы исключение такой последовательности путем замен удаляет существующую N-связанную углеводную цепь. Также представлена перестройка N-связанных углеводных цепей, при которой один или более сайтов N-связанных углеводных цепей (обычно те, которые встречаются в природе) исключаются, и создается один или более новых N-связанных сайтов. Дополнительные предпочтительные варианты антител включают цистеиновые варианты, в которых один или более цистеиновых остатков удалены или заменены другой аминокислотой (например, серином) по сравнению с исходной аминокислотной последовательностью. Цистеиновые варианты могут быть применимы, если антитела должны быть уложены в биологически активную конформацию (например, после выделения растворимых телец включения). Цистеиновые варианты обычно имеют меньше цистеиновых остатков, чем природные белки, и обычно имеют четное количество цистеинов, чтобы минимизировать взаимодействия, возникающие вследствие неспаренных цистеиновых остатков.
Необходимые аминокислотные замены (либо консервативные, либо неконсервативные) могут быть определены специалистом в данной области техники в то время, когда такие замены необходимы. В некоторых вариантах осуществления аминокислотные замены могут быть использованы для идентификации важных остатков антител к человеческим эндотелиновым рецепторам или для повышения или снижения аффинности антител к человеческим эндотелиновым рецепторам, описанных в данном документе. В соответствии с некоторыми вариантами осуществления предпочтительными аминокислотными заменами являются те, которые (1) снижают чувствительность к протеолизу, (2) снижают чувствительность к окислению, (3) изменяют аффинность связывания в отношении образованных белковых комплексов, (4) изменяют аффинность связывания и/или (4) придают или модифицируют другие физико-химические или функциональные свойства в таких полипептидах. В соответствии с некоторыми вариантами осуществления одиночные или множественные аминокислотные замены (в некоторых вариантах осуществления консервативные аминокислотные замены) могут быть выполнены во встречающихся в природе последовательностях (в некоторых вариантах осуществления в части полипептида вне домен (доменов), образующих внутримолекулярные контакты). В некоторых вариантах осуществления консервативные аминокислотные замены обычно не могут приводить к существенному изменению структурных характеристик исходной последовательности (например, замена аминокислот не должна приводить к нарушению спиралей, присутствующих в исходной последовательности, или приводить к нарушению других типов вторичной структуры, которые характеризуют исходную последовательность). Примеры известных в данной области техники вторичных и третичных структур полипептидов описаны в Proteins, Structures and Molecular Principles, Creighton, ed., W.H. Freeman and Company, (1984); Introduction to Protein Structure, Branden and Tooze, eds., Garl and Publishing, (1991) и Thornton et al., 1991, Nature 354:105, каждая из которых включена в данный документ посредством ссылки.
В некоторых вариантах осуществления антитело, описанное в данном документе, может быть химически связано с полимерами, липидами или другими фрагментами. Антигенсвязывающие средства могут содержать по меньшей мере одну из CDR, описанную в данном документе, включенную в биологически совместимую каркасную структуру. В одном примере биологически совместимая каркасная структура содержит полипептид или его часть, достаточную для образования конформационно стабильной структурной основы, или каркаса, или остова, которые способны представлять одну или более аминокислотных последовательностей, которые связываются с антигеном (например, CDR, вариабельные области и т.д.) в локализованной области поверхности. Такие структуры могут представлять собой встречающийся в природе полипептид или полипептидную "складку" (структурный мотив) или могут иметь одну или более модификаций, таких как добавления, делеции или замены аминокислот, по сравнению с встречающимися в природе полипептидом или складкой. Данные остовы могут быть получены из полипептида любого вида (или более чем одного вида), такого как человек, другие млекопитающие, другие позвоночные, беспозвоночные, бактерии или вирусы. Обычно биологически совместимая каркасная структура основана на белковом остове или каркасе, но не на домене иммуноглобулина. В качестве примера можно использовать эти белковые остовы на основе фибронектина, анкирина, липокалина, неокарзиностаина, цитохрома b, белка с цинковыми пальцами CP1, PST1, белка двойной спирали, LACI-D1, домена Z и домена тендамистата (см., например, Nygren and Uhlen, 1997, Current Opinion in Structural Biology 7:463-469).
Кроме того, специалисту в данной области техники будет понятно, что подходящие связывающие средства включают в себя части этих антител, такие как одну или более из CDR1, CDR2 и CDR3 тяжелой цепи, и CDR1, CDR2 и CDR3 легкой цепи, которые конкретно раскрыты в данном документе. По меньшей мере одна из областей CDR1, CDR2, CDR3 тяжелой цепи, CDR1, CDR2 и CDR3 легкой цепи может содержать по меньшей мере одну аминокислотную замену, при условии, что антитело сохраняет специфичность связывания незамещенной CDR. Отличная от CDR часть антитела может представлять собой небелковую молекулу, в которой связывающее средство перекрестно блокирует связывание антитела, раскрытого в данном документе, с человеческим ETA и/или ингибирует опосредованную эндотелином передачу сигнала посредством рецептора. Отличная от CDR часть антитела может представлять собой небелковую молекулу, в которой антитело характеризуется типом связывания, аналогичным типу связывания по меньшей мере одного из антител A-1/A-2 с пептидом человеческого ETA, и/или нейтрализует активность эндотелина в анализе конкурентного связывания. Отличная от CDR часть антитела может состоять из аминокислот, где антитело представляет собой рекомбинантный связывающий белок или синтетический пептид, при этом рекомбинантный связывающий белок перекрестно блокирует связывание антитела, раскрытого в данном документе, с человеческим ЕТА и/или нейтрализует активность эндотелина in vivo или in vitrо. Отличная от CDR часть антитела может состоять из аминокислот, при этом антитело представляет собой рекомбинантное антитело, которое характеризуется типом связывания, аналогичным типу связывания по меньшей мере одного из антител A-1/A-2 с пептидом человеческого ETA, и/или нейтрализует опосредованную эндотелином передачу сигнала в анализе конкурентного связывания.
Нуклеиновые кислоты
В одном аспекте в данном документе представлена выделенная молекула нуклеиновой кислоты. Молекула нуклеиновой кислоты предусматривает, например, полинуклеотиды, которые кодируют все антитело или его часть, такие как одна или обе цепи антитела, описанного в данном документе, или его фрагмент, производное, мутантный белок или его вариант; полинуклеотиды, достаточные для применения в качестве зондов для гибридизации; праймеры для ПЦР или праймеры для секвенирования для идентификации, анализа, мутирования или амплификации полинуклеотида, кодирующего полипептид; антисмысловые нуклеиновые кислоты для ингибирования экспрессии полинуклеотида и их комплементарные последовательности. Нуклеиновая кислота может быть любой длины. Длина нуклеиновой кислоты может составлять, например, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 75, 100, 125, 150, 175, 200, 250, 300, 350, 400, 450, 500, 750, 1000, 1500, 3000, 5000 или больше нуклеотидов и/или она может содержать одну или более дополнительных последовательностей, например, регуляторные последовательности, и/или может быть частью более крупной нуклеиновой кислоты, например вектора. Нуклеиновая кислота может быть однонитевой или двухнитевой и может содержать нуклеотиды РНК и/или ДНК, а также их искусственные варианты (например, пептидные нуклеиновые кислоты).
Нуклеиновая кислота, кодирующая полипептид антитела (например, тяжелую или легкую цепь, только вариабельный домен или полноразмерные), может быть выделена из B-клеток мышей, которые были иммунизированы антигеном ЕТА. Нуклеиновая кислота может быть выделена традиционными способами, такими как полимеразная цепная реакция (ПЦР).
Последовательности нуклеиновой кислоты, кодирующие вариабельные области тяжелой и легкой цепей, показаны выше. Специалисту в данной области техники будет понятно, что вследствие вырожденности генетического кода каждая из полипептидных последовательностей, раскрытых в данном документе, может быть закодирована большим количеством других последовательностей нуклеиновых кислот. В данном документе представлена каждая вырожденная нуклеотидная последовательность, кодирующая антитело, описанное в данном документе.
Кроме того, в данном документе представлены нуклеиновые кислоты, которые гибридизуются с другими нуклеиновыми кислотами (например, нуклеиновыми кислотами, содержащими нуклеотидную последовательность любого из A-1/A-2) при определенных условиях гибридизации. Способы гибридизации нуклеиновых кислот хорошо известны из уровня техники. См., например, Current Protocols in Molecular Biology, John Wiley & Sons, N.Y. (1989), 6.3.1-6.3.6. Как определено в данном документе, например, в умеренно жестких условиях гибридизации используют раствор для предварительной промывки, содержащий 5× хлорид натрия/цитрат натрия (SSC), 0,5% SDS, 1,0 мМ EDTA (pH 8,0), гибридизационный буфер из приблизительно 50% формамида, 6× SSC, температуру гибридизации 55°C (или другие подобные растворы для гибридизации, такие как раствор, содержащий приблизительно 50% формамида, температуру гибридизации 42°C) и условия элюирования 60°C в 0,5× SSC и 0,1% SDS. В жестких условиях гибридизации гибридизацию осуществляют в 6× SSC при 45°C, после чего следует одна или более промывок в 0,1× SSC и 0,2% SDS при 68°C. Кроме того, специалист в данной области техники может изменять условия гибридизации и/или промывки для повышения или снижения жесткости гибридизации, поэтому нуклеиновые кислоты, содержащие нуклеотидные последовательности, на по меньшей мере 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98% или 99% гомологичные друг другу, обычно остаются гибридизированными друг с другом. Основные параметры, влияющие на выбор условий гибридизации, и руководство по разработке подходящих условий изложены, например, в Sambrook, Fritsch and Maniatis, 1989, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, chapters 9 and 11 и Current Protocols in Molecular Biology, 1995, Ausubel et al., eds., John Wiley & Sons, Inc., sections 2.10 and 6.3-6.4, и они могут быть легко определены специалистом в данной области техники на основе, например, длины и/или состава оснований ДНК. Изменения могут быть внесены посредством введения мутаций в нуклеиновую кислоту, за счет чего происходят изменения аминокислотной последовательности полипептида (например, антигенсвязывающего белка), кодируемого ею. Мутации можно вводить с использованием любой методики, известной из уровня техники. В одном варианте осуществления один или более конкретных аминокислотных остатков изменяют с применением, например, протокола сайт-направленного мутагенеза. В другом варианте осуществления один или более случайно выбранных остатков изменяют с применением, например, протокола рандомного мутагенеза. Независимо от того, как он получен, мутантный полипептид можно экспрессировать и подвергать скринингу в отношении требуемых свойств.
Мутации можно вводить в нуклеиновую кислоту без существенного изменения биологической активности полипептида, кодируемого ею. Например, могут быть выполнены замены нуклеотидов, приводящие к аминокислотным заменам в остатках заменимых аминокислот. В одном варианте осуществления нуклеотидные последовательности, представленные в данном документе для L-1-L-2 и H-1-H-2 или их фрагментов, вариантов или производных, мутированы таким образом, что аминокислотная последовательность, кодируемая ими, содержит по сравнению с последовательностью, кодируемой L-1-L-2 и H-1-H-2, как показано в данном документе, одну или более делеций или замен аминокислотных остатков, что приводит к образованию последовательностей, содержащих два или более отличающихся остатков. В другом варианте осуществления мутагенез приводит к вставке аминокислоты, смежной с одним или более аминокислотными остатками L-1-L-2 и H-1-H-2, как показано в данном документе, что приводит к образованию последовательностей, содержащих два или более отличающихся остатков. В качестве альтернативы одна или более мутаций могут быть введены в нуклеиновую кислоту для селективного изменения биологической активности (например, связывания с ETA) полипептида, кодируемого ею. Например, мутация может количественно или качественно изменять биологическую активность. Примеры количественных изменений предусматривают увеличение, уменьшение или устранение активности. Примеры качественных изменений предусматривают изменение антигенной специфичности антитела.
В другом аспекте в данном документе представлены молекулы нуклеиновой кислоты, которые являются подходящими для применения в качестве праймеров или гибридизационных зондов для выявления последовательностей нуклеиновых кислот, представленных в данном документе. Молекула нуклеиновой кислоты в данном документе может содержать только часть последовательности нуклеиновой кислоты, кодирующей полноразмерный полипептид по данному документу, например фрагмент, который можно использовать в качестве зонда или праймера, или фрагмент, кодирующий активную часть (например, часть, связывающую ЕТА) полипептида по данному документу. Зонды на основе последовательности нуклеиновой кислоты по данному документу, можно использовать для выявления нуклеиновой кислоты или подобных нуклеиновых кислот, например транскриптов, кодирующих полипептид по данному документу. Зонды могут содержать метящую группу, например, радиоизотоп, флуоресцентное соединение, фермент или ферментативный кофактор. Такие зонды можно использовать для идентификации клетки, экспрессирующей полипептид.
В другом аспекте в данном документе представлен вектор, содержащий нуклеиновую кислоту, кодирующую полипептид по данному документу или его часть. Примеры векторов включают без ограничения плазмиды, вирусные векторы, неэписомные векторы млекопитающих и векторы экспрессии, например рекомбинантные векторы экспрессии. Рекомбинантные векторы экспрессии в данном документе могут содержать нуклеиновую кислоту по данному документу в форме, подходящей для экспрессии нуклеиновой кислоты в клетке-хозяине. Рекомбинантные векторы экспрессии содержат одну или более регуляторных последовательностей, которые подвергаются скринингу с учетом клеток-хозяев, используемых для экспрессии, и они функционально связаны с предварительно экспрессируемой последовательностью нуклеиновой кислоты. Регуляторные последовательности включают те последовательности, которые управляют конститутивной экспрессией нуклеотидной последовательности во многих типах клеток-хозяев (например, энхансер раннего гена SV40, промотор вируса саркомы Рауса и промотор цитомегаловируса), те, которые управляют экспрессией нуклеотидной последовательности только в определенных клетках-хозяевах (например, тканеспецифическая регуляторная последовательность, см. Voss et al., 1986, Trends Biochem. Sci. 11:287 и Maniatis et al., 1987, Science 236:1237, раскрытие каждой из которых включено в данный документ посредством ссылки в полном объеме), и те, которые управляют индуцируемой экспрессией нуклеотидной последовательности в ответ на конкретную обработку или конкретное условие (например, промотор металлотионина в клетках млекопитающих и отвечающий на тетрациклин и/или отвечающий на стрептомицин промотор как в прокариотической, так и в эукариотической системах (выше)). Специалисту в данной области техники будет понятно, что конструирование вектора экспрессии может зависеть от таких факторов, как выбор клетки-хозяина, подлежащей трансформации, и требуемый уровень экспрессии белка. Вектор экспрессии в данном документе может быть введен в клетку-хозяина, за счет чего продуцируется белок или пептид, в том числе слитый белок или пептид, кодируемый нуклеиновой кислотой, описанной в данном документе.
В другом аспекте в данном документе представлена клетка-хозяин, в которую может быть введен вектор экспрессии. Клетка-хозяин может представлять собой любую прокариотическую или эукариотическую клетку. Прокариотические клетки-хозяева включают грамотрицательные или грамположительные организмы, например, E. coli или бациллы. Высшие эукариотические клетки включают клетки насекомых, дрожжевые клетки и сформированные линии клеток млекопитающих. Примеры подходящих линий клеток-хозяев млекопитающих включают клетки яичника китайского хомячка (CHO) или их производные, такие как Veggie CHO и родственные клеточные линии, которые растут в бессывороточной среде (см. Rasmussen et al., 1998, Cytotechnology 28:31), или линию CHO DXB-11, дефектную по DHFR (см. Urlaub et al., 1980, PNAS USA 77:4216-20). Дополнительные клеточные линии CHO включают CHO-K1 (№ CCL-61 в ATCC), EM9 (№ CRL-1861 в ATCC) и UV20 (№ CRL-1862 в ATCC). Дополнительные клетки-хозяева включают линию COS-7 клеток почки обезьяны (№ CRL-1651 в ATCC) (см. Gluzman et al., 1981, Cell 23:175), клетки L, клетки C127, клетки 3T3 (№ CCL-163 в ATCC), клетки AM-1/D (описанные в патенте США № US 6210924), клетки HeLa, линии клеток BHK (№ CRL-10 в ATCC), линию клеток CV1/EBNA, полученную из линии клеток CV1 почки африканской зеленой мартышки (№ CCL-70 в ATCC) (см. McMahan et al., 1991, EMBO J. 10:2821), клетки почки человеческого эмбриона, такие как 293, 293 EBNA или MSR 293, человеческие эпидермальные клетки A431, человеческие клетки C010220, другие трансформированные линии клеток приматов, нормальные диплоидные клетки, клеточные линии, полученные из in vitro культуры первичной ткани, первичные эксплантаты, клетки HL-60, U937, HaK или Jurkat. Подходящие векторы клонирования и экспрессии для применения с бактериальными, грибными, дрожжевыми клетками-хозяевами и клетками-хозяевами млекопитающих описаны Pouwels et al. (Cloning Vectors: A Laboratory Manual, Elsevier, 1985).
ДНК-вектор может быть введен в прокариотические или эукариотические клетки посредством традиционных методик трансформации или трансфекции. Что касается стабильной трансфекции клеток млекопитающих, известно, что в зависимости от используемых вектора экспрессии и методики трансфекции лишь небольшая часть клеток может интегрировать чужеродную ДНК в свой геном. Для идентификации и скрининга данных интегрантов ген, который кодирует селективный маркер (например, антибиотикорезистентности), обычно вводят в клетку-хозяина вместе с представляющим интерес геном. Предпочтительные селективные маркеры предусматривают маркеры, которые придают резистентность к лекарственным средствам, таким как G418, гигромицин и метотрексат. Среди других методик клетки, стабильно трансфицированные введенной нуклеиновой кислотой, можно идентифицировать путем скрининга с использованием лекарственного средства (например, клетки, в которые интегрирован селектируемый ген, выживут, в то время как другие клетки погибнут).
Трансформированные клетки можно культивировать в условиях, которые способствуют экспрессии полипептида, и при этом полипептид может быть выделен с применением стандартных способов очистки белка. Один такой способ очистки описан в примерах ниже. Полипептиды, предназначенные для применения в данном документе, включают по сути гомогенные рекомбинантные полипептиды антител к эндотелиновым рецепторам млекопитающих, которые по сути не содержат контаминирующих эндогенных материалов.
Активность антител
В одном варианте осуществления антитело, описанное в данном документе, специфически связывается с эндотелиновыми рецепторами, ингибирует передачу сигналов и проявляет терапевтические биологические эффекты, такие как снижение легочной артериальной гипертензии, в животных моделях. В другом варианте осуществления антитело, описанное в данном документе, представляет собой мышиное антитело или гуманизированное антитело, которые могут специфически связываться с человеческими эндотелиновыми рецепторами. Такие антитела включают антагонистические или нейтрализующие антитела, которые могут приводить к снижению или нейтрализации опосредованной эндотелином передачи сигнала.
В одном варианте осуществления антитело, описанное в данном документе, характеризуется значением Kd, составляющим от примерно 0,01 до 1000 нМ, от 0,1 до 500 нМ, от 0,5 до 200 нМ, от 1 до 200 нМ или от 10 до 100 нМ, при связывании с человеческим эндотелиновым рецептором ЕТА. В другом варианте осуществления антитело, описанное в данном документе, характеризуется значением Kd, составляющим от примерно 1 до 200 нМ, при связывании с человеческим эндотелиновым рецептором ETA. В другом варианте осуществления антитело, описанное в данном документе, характеризуется значением Kd, составляющим от примерно 10 до 100 нМ, при связывании с человеческим эндотелиновым рецептором ETA. В другом варианте осуществления антитело, описанное в данном документе, характеризуется значением Kd, составляющим примерно 1 нМ, 2 нМ, 5 нМ, 10 нМ, 20 нМ, 30 нМ, 40 нМ, 50 нМ, 60 нМ, 70 нМ, 80 нМ, 90 нМ или 100 нМ, при связывании с человеческим эндотелиновым рецептором ЕТА.
В одном варианте осуществления антитело, описанное в данном документе, характеризуется значением IC50, составляющим от примерно 0,01 до 500 нМ, от 0,1 до 200 нМ, от 0,5 до 200 нМ, от 1 до 200 нМ или от 10 до 100 нМ, при снижении опосредованной человеческим эндотелином передачи сигнала. В другом варианте осуществления антитело, описанное в данном документе, характеризуется значением IC50, составляющим от примерно 1 до 200 нМ, при снижении опосредованной человеческим эндотелином передачи сигнала. В другом варианте осуществления антитело, описанное в данном документе, характеризуется значением IC50, составляющим от примерно 10 до 100 нМ, при снижении опосредованной человеческим эндотелином передачи сигнала. В другом варианте осуществления антитело, описанное в данном документе, характеризуется значением IC50, составляющим примерно 1 нМ, 2 нМ, 5 нМ, 10 нМ, 20 нМ, 30 нМ, 40 нМ, 50 нМ, 60 нМ, 70 нМ, 80 нМ, 90 нМ или 100 нМ, при снижении опосредованной человеческим эндотелином передачи сигнала.
В одном варианте осуществления антитело к ЕТА характеризуется одним или более из следующих свойств:
a. характеризуется значением Kd, которое равняется такому значению у референтного антитела или превосходит его, при связывании с человеческим эндотелиновым рецептором ETA;
b. характеризуется значением IC50, которое равняется такому значению у референтного антитела или превосходит его, при ингибировании активации человеческого эндотелинового рецептора ЕТА эндотелином и
c. проявляет перекрестную конкуренцию с референтным антителом за связывание с человеческим эндотелиновым рецептором ЕТА.
В другом варианте осуществления антитело к ЕТА, описанное в данном документе, характеризуется одним или более из следующих свойств:
a. характеризуется значением Kd, которое равняется такому значению у референтного антитела к ETA или превосходит его, при связывании с человеческим эндотелиновым рецептором ETA;
b. характеризуется значением IC50, которое равняется такому значению у референтного антитела к ЕТА или превосходит его, при ингибировании активации человеческого эндотелинового рецептора ЕТА эндотелином и
с. проявляет перекрестную конкуренцию с референтным антителом к ЕТА за связывание с человеческим эндотелиновым рецептором ЕТА.
В одном аспекте референтное антитело содержит комбинацию аминокислотной последовательности вариабельного домена легкой цепи под SEQ ID NO: 138 и аминокислотной последовательности вариабельного домена тяжелой цепи под SEQ ID NO: 166. В другом аспекте референтное антитело представляет собой моноклональное антитело A-1, A-2, A-7, A-9 или A-12. В данном документе выражение "по сути подобный" означает, что значение IC50 или Kd для антитела, описанного в данном документе, сопоставимо со значением для референтного антитела или составляет примерно 200%, 180%, 160%, 150%, 140%, 120%, 110%, 100%, 99%, 98%, 97%, 95%, 90%, 85%, 80%, 75%, 70%, 65% или 50% от значения для референтного антитела. В одном варианте осуществления референтное антитело включает, например, антитело, содержащее комбинацию тяжелой цепи и легкой цепи L1H1 или L2H2. В другом варианте осуществления референтное антитело включает антитело к ETA A-1. В одном варианте осуществления антитело к ЕТА, описанное в данном документе, может специфически связываться с человеческими эндотелиновыми рецепторами и может приводить к снижению легочной артериальной гипертензии в животных моделях. В одном варианте осуществления легочная артериальная гипертензия снижается на 2% по сравнению с необработанными животными. В другом варианте осуществления легочная артериальная гипертензия снижается на приблизительно 5% по сравнению с необработанными животными. В другом варианте осуществления легочная артериальная гипертензия снижается на приблизительно 10% по сравнению с необработанными животными. В другом варианте осуществления легочная артериальная гипертензия снижается на приблизительно 15% по сравнению с необработанными животными. В другом варианте осуществления легочная артериальная гипертензия снижается на приблизительно 20% по сравнению с необработанными животными. В другом варианте осуществления легочная артериальная гипертензия снижается на приблизительно 25% по сравнению с необработанными животными. Степень снижения легочной артериальной гипертензии контролируется дозой. Для пациентов-животных или пациентов-людей терапевтически эффективная доза представляет собой дозу, которая приводит к снижению легочной артериальной гипертензии до диапазона значений нормы.
TGF-β Trap
В одном варианте осуществления TGF-β Trap, описанный в данном документе, представляет собой белок, который может блокировать функцию TGF-β. В другом варианте осуществления TGF-β Trap, описанный в данном документе, содержит аминокислотную последовательность под SEQ ID NO: 207. В другом варианте осуществления аминокислотная последовательность TGF-β Trap, описанная в данном документе, представляет собой SEQ ID NO: 207.
Пептидный линкер (линкер)
В одном варианте осуществления пептидный линкер (линкер), описанный в данном документе, содержит аминокислотную последовательность, независимо выбранную из одного из следующего: SEQ ID NO: 208, SEQ ID NO: 209 и SEQ ID NO: 210.
В одном варианте осуществления аминокислотная последовательность пептидного линкера, описанная в данном документе, представляет собой SEQ ID NO: 208. В другом варианте осуществления аминокислотная последовательность пептидного линкера, описанная в данном документе, представляет собой SEQ ID NO: 209. В другом варианте осуществления аминокислотная последовательность пептидного линкера, описанная в данном документе, представляет собой SEQ ID NO: 210.
Слитый белок на основе антитела к ЕТА и TGF-β Trap
В одном варианте осуществления слитый белок на основе антитела к ETA и TGF-β Trap, представленных в данном документе, содержит антитело к ETA, описанное в данном документе, и фрагмент TGF-β Trap.
В другом варианте осуществления слитый белок на основе антитела к ETA и TGF-β Trap, представленных в данном документе, содержит антитело к ETA, описанное в данном документе, один, два, три, четыре, пять, шесть, семь или восемь TGF-β Trap и такое же количество пептидных линкеров (линкеров); при этом в слитом белке аминоконец TGF-β Trap соединен с карбоксиконцом легкой цепи или тяжелой цепи антитела к ЕТА посредством пептидной линкерной последовательности или в слитом белке карбоксильный конец TGF-β Trap соединен с аминоконцом легкой цепи или тяжелой цепи антитела к ЕТА посредством пептидной линкерной последовательности.
В другом варианте осуществления слитый белок на основе антитела к ETA и TGF-β Trap, представленных в данном документе, содержит антитело к ETA, описанное в данном документе, один, два, три или четыре TGF-β Trap и такое же количество пептидных линкеров (линкеров); при этом в слитом белке аминоконец TGF-β Trap соединен с карбоксиконцом легкой цепи или тяжелой цепи антитела к ЕТА посредством пептидной линкерной последовательности или в слитом белке карбоксильный конец TGF-β Trap соединен с аминоконцом легкой цепи или тяжелой цепи антитела к ЕТА посредством пептидной линкерной последовательности.
В другом варианте осуществления слитый белок на основе антитела к ETA и TGF-β Trap, представленных в данном документе, содержит антитело к ETA, описанное в данном документе, один или два TGF-β Trap и такое же количество пептидных линкеров (линкеров); при этом в слитом белке аминоконец TGF-β Trap соединен с карбоксиконцом легкой цепи или тяжелой цепи антитела к ЕТА посредством пептидной линкерной последовательности или в слитом белке карбоксильный конец TGF-β Trap соединен с аминоконцом легкой цепи или тяжелой цепи антитела к ЕТА посредством пептидной линкерной последовательности.
В другом варианте осуществления слитый белок на основе антитела к ETA и TGF-β Trap, представленных в данном документе, содержит антитело к ETA, описанное в данном документе, два TGF-β Trap и два пептидных линкера (линкера); при этом в слитом белке аминоконец TGF-β Trap соединен с карбоксиконцом легкой цепи или тяжелой цепи антитела к ЕТА посредством пептидной линкерной последовательности или в слитом белке карбоксильный конец TGF-β Trap соединен с аминоконцом легкой цепи или тяжелой цепи антитела к ЕТА посредством пептидной линкерной последовательности.
В одном варианте осуществления в слитом белке на основе антитела к ETA и TGF-β Trap, представленных в данном документе, аминоконец TGF-β Trap соединен с карбоксиконцом легкой цепи или тяжелой цепи антитела к ETA посредством пептидной линкерной последовательности. В другом варианте осуществления в слитом белке на основе антитела к ETA и TGF-β Trap, представленных в данном документе, аминоконец TGF-β Trap соединен с карбоксиконцом легкой цепи антитела к ETA посредством пептидной линкерной последовательности. В другом варианте осуществления в слитом белке на основе антитела к ETA и TGF-β Trap, представленных в данном документе, аминоконец TGF-β Trap соединен с карбоксиконцом тяжелой цепи антитела к ETA посредством пептидной линкерной последовательности.
В одном варианте осуществления слитый белок на основе антитела к ETA и TGF-β Trap, представленных в данном документе, содержит аминокислотные последовательности SEQ ID NO: 162, SEQ ID NO: 190, SEQ ID NO: 207 и SEQ ID NO: 210.
В одном варианте осуществления в данном документе представлен слитый белок на основе антитела к ETA и TGF-β Trap, где антитело к ETA, TGF-β Trap и пептидная линкерная последовательность слиты с образованием слитого белка с помощью одного из следующих средств:
(3) посредством пептидной линкерной последовательности аминоконец TGF-β Trap связан с карбоксиконцом тяжелой/легкой цепи антитела к ЕТА: N'-R-линкер-TGF-β Trap-C'; и
(4) посредством пептидной линкерной последовательности карбоксиконец TGF-β Trap связан с аминоконцом легкой цепи или тяжелой цепи антитела к ETA: N'-TGF-β Trap-линкер-R-C';
при этом N' представляет собой аминоконец полипептидной цепи, C' представляет собой карбоксиконец полипептидной цепи, TGF-β Trap представляет собой фрагмент TGF-β Trap, R представляет собой аминокислотную последовательность легкой цепи или тяжелой цепи антитела к ЕТА, и линкер представляет собой пептидный линкер.
В одном варианте осуществления слитый белок на основе антитела к ETA и TGF-β Trap, представленных в данном документе, содержит аминокислотную последовательность SEQ ID NO: 211 или SEQ ID NO: 212. В другом варианте осуществления слитый белок на основе антитела к ETA и TGF-β Trap, представленных в данном документе, содержит аминокислотные последовательности SEQ ID NO: 211 и SEQ ID NO: 212.
Биологическая активность слитого белка на основе антитела к ЕТА и TGF-β Trap
Биологическая активность слитого белка на основе антитела к ETA и TGF-β Trap предусматривает биологическую активность TGF-β Trap и биологическую активность антитела к ЕТА. Ингибиторы ЕТА на основе антител могут эффективно блокировать повышение сосудистого давления, вызванное эндотелином, с облегчением тяжести симптомов легочной артериальной гипертензии и улучшением способности переносить нагрузку и гемодинамики у пациента. "Биологическая активность TGF-β Trap" означает, что слитый белок на основе антитела к ETA и TGF-β Trap связывается с TGF-β in vivo и блокирует вызванную этим клеточную реакцию на стресс, а также проявляет биологическую активность терапевтических эффектов в отношении, например, других связанных симптомов легочной артериальной гипертензии, легочной гипертензии и легочного фиброза. Вышеупомянутая клеточная реакция на стресс включает без ограничения снижение легочного артериального давления, снижение давления в аорте и связанные с этим изменения в ремоделировании сердечных и легочных сосудов. Благодаря комбинации биологической активности TGF-β Trap и антитела к ETA, слитый белок на основе TGF-β Trap, описанный в данном документе, можно использовать для лечения различных патологических состояний и заболеваний, ассоциированных с TGF-β и ETA. Слитый белок проявляет свои биологические эффекты за счет воздействия на TGF-β и/или ETA, и, следовательно, слитый белок на основе TGF-β Trap, описанный в данном документе, можно использовать для лечения субъектов с патологическими состояниями и заболеваниями, которые благоприятно отвечают на "сниженную стимуляцию TGF-β" или "сниженную стимуляцию ETA". Эти субъекты упоминаются в качестве субъектов, которым "требуется сниженная стимуляция TGF-β" или "требуется сниженная стимуляция ETA". Включены другие сопутствующие симптомы легочной артериальной гипертензии, легочной гипертензии, легочного фиброза и кардиоваскулярного фиброза.
В одном варианте осуществления изменение биологической активности слитого белка на основе антитела к ETA или TGF-β Trap выявляют с помощью анализа потока кальция и методики выявления репортерного гена для количественной оценки функции слитого белка на основе антитела к ETA или TGF-β Trap для ингибирования ETA и блокирования TGF-β in vitro.
Фармацевтическая композиция
В одном варианте осуществления в данном документе представлена фармацевтическая композиция, содержащая слитый белок на основе антитела к ЕТА и TGF-β Trap, представленных в данном документе, и один или более фармацевтически приемлемых носителей.
В одном варианте осуществления фармацевтическая композиция, описанная в данном документе, предназначена для внутривенной или подкожной инъекции.
Способы лечения
В одном варианте осуществления в данном документе представлено применение слитого белка на основе антитела к ЕТА и TGF-β Trap, описанных в данном документе, в получении лекарственного препарата, предназначенного для лечения, предупреждения возникновения или улучшения течения легочной артериальной гипертензии и заболеваний, связанных с легочной артериальной гипертензией.
В другом варианте осуществления в данном документе представлено применение слитого белка на основе антитела к ЕТА и TGF-β Trap, описанных в данном документе, в получении лекарственного препарата, предназначенного для лечения, предупреждения возникновения или улучшения течения легочной гипертензии и заболеваний, связанных с легочной гипертензией.
В другом варианте осуществления в данном документе представлено применение слитого белка на основе антитела к ЕТА и TGF-β Trap, описанных в данном документе, в получении лекарственного препарата, предназначенного для лечения, предупреждения возникновения или улучшения течения легочного фиброза и заболеваний, связанных с легочным фиброзом.
В другом варианте осуществления в данном документе представлено применение слитого белка на основе антитела к ЕТА и TGF-β Trap, описанных в данном документе, в получении лекарственного препарата, предназначенного для лечения, предупреждения возникновения или улучшения кардиоваскулярного фиброза и заболеваний, связанных с кардиоваскулярным фиброзом.
В дополнительном варианте осуществления в данном документе представлено применение слитого белка на основе антитела к ЕТА и TGF-β Trap, описанных в данном документе, в получении лекарственного препарата, предназначенного для одновременного лечения, предупреждения возникновения или улучшения течения двух или более заболеваний из легочной артериальной гипертензии, легочной гипертензии, легочного фиброза или кардиоваскулярного фиброза.
В данном документе термин "субъект" относится к млекопитающим, в том числе к людям, и он используется взаимозаменяемо с термином "пациент".
Выражение "лечение" включает облегчение тяжести по меньшей мере одного симптома или других аспектов патологического состояния или облегчение тяжести состояния. Слитый белок на основе антитела к ETA и TGF-β Trap, представленных в данном документе, может представлять собой эффективное терапевтическое средство без необходимости достижения полного эффекта излечения или устранения всех симптомов или проявлений патологического состояния. Как известно из уровня техники, лекарственное средство в качестве терапевтического средства можно использовать для снижения тяжести данного патологического состояния, но необязательно для устранения всех проявлений патологического состояния, чтобы его можно было рассматривать в качестве эффективного терапевтического средства. Аналогично профилактическое лечение необязательно должно быть полностью эффективным для предупреждения возникновения симптомов, чтобы оно составляло эффективное профилактическое средство. Достаточным является уменьшение влияния патологического состояния (например, посредством уменьшения количества или тяжести его симптомов, или посредством усиления другого лечебного эффекта, или посредством создания другого эффективного воздействия) или уменьшение вероятности возникновения или обострения состояния у субъектов. Один вариант осуществления в данном документе относится к способу, предусматривающему введение пациенту слитого белка на основе антитела к ETA и TGF-β Trap в количестве и в течение времени, достаточных для индуцирования непрерывного улучшения показателя, который отражает тяжесть конкретного заболевания, выше исходного уровня.
Фармацевтическая композиция, содержащая слитый белок на основе антитела к ETA и TGF-β Trap, может вводиться с применением любой подходящей методики, в том числе без ограничения парентерального, местного или ингаляционного введения. В случае инъекции фармацевтическая композиция может быть введена, например, внутрисуставным, внутривенным, внутримышечным, внутриочаговым, внутрибрюшинным или подкожным путем, в виде быстрой инъекции или непрерывной инфузии. Например, может рассматриваться местное введение в месте патологического состояния или повреждения, такое как трансдермальное введение и введение имплантатов с замедленным высвобождением. Ингаляционное введение включает, например, назальную или пероральную ингаляцию, спреи, ингаляцию антител в форме аэрозолей и т.п. Другие варианты включают составы для перорального введения, в том числе таблетки, сиропы или пастилки.
Предпочтительным является введение слитого белка на основе антитела к ЕТА и TGF-β Trap, представленных в данном документе, в форме композиции, содержащей один или более других компонентов, таких как физиологически приемлемые носитель, наполнитель или разбавитель. Композиция может необязательно дополнительно содержать один или более физиологически активных средств, описанных ниже. В различных конкретных вариантах осуществления композиция содержит один, два, три, четыре, пять или шесть физиологически активных средств, кроме одного или более слитых белков на основе антитела (такого как мышиное антитело или гуманизированное антитело) и TGF-β, представленных в данном документе.
В одном варианте осуществления фармацевтическая композиция содержит слитый белок на основе мышиного антитела или гуманизированного антитела и TGF-β Trap, представленных в данном документе, и одно или более веществ, выбранных из буферов с показателем pH, подходящим для слитого белка на основе антитела и TGF-β Trap, антиоксидантов, таких как аскорбиновая кислота, низкомолекулярные полипептиды (например, полипептиды, содержащие менее 10 аминокислот), белки, аминокислоты, углеводы, такие как декстрин, комплексообразователи, такие как EDTA, глутатион, стабилизаторы и наполнители. Согласно соответствующим отраслевым стандартам также могут добавляться консерванты. Композиция может быть составлена в виде лиофилизированного порошка с использованием в качестве разбавителя подходящего раствора наполнителя. Соответствующие компоненты нетоксичны для пациентов в используемых дозе и концентрации. Дополнительные примеры компонентов, которые могут быть использованы в фармацевтических составах, описаны в Remington's Pharmaceutical Sciences, 16th edition (1980) and 20th edition (2000). Mack Publishing Company предоставляет набор для применения практикующими врачами, который содержит один или более слитых белков на основе антитела и TGF-β Trap, представленных в данном документе, а также ярлыки или другие инструкции для лечения любого из заболеваний, обсуждаемых в данном документе. В одном варианте осуществления набор содержит стерильный препарат одного или более слитых белков на основе антитела и TGF-β Trap в одном или более флаконах пробирочного типа в форме вышеупомянутой композиции.
Доза и частота введения могут варьироваться в зависимости от следующих факторов: способа введения, используемого слитого белка на основе специфического антитела и TGF-β Trap, природы и тяжести заболевания, подлежащего лечению, наличия острых или хронических симптомов, а также размера тела и общих симптомов у пациента. Подходящую дозу можно определить с применением способов, хорошо известных в данной области техники, например, исследованиями по расчету дозы в клинических испытаниях.
Слитые белки на основе антитела и TGF-β Trap, представленные в данном документе, можно вводить, например, один или более раз с равными интервалами в течение определенного периода времени. В конкретных вариантах осуществления слитый белок на основе мышиного антитела или гуманизированного антитела и TGF-β Trap вводят один раз в по меньшей мере месяц или более длительно, например, в течение одного, двух или трех месяцев или даже неопределенно долго. Для лечения хронических симптомов долгосрочное лечение обычно является наиболее эффективным. Однако для лечения острых симптомов достаточно кратковременного введения, например, в течение от одной до шести недель. Как правило, человеческие антитела вводят до тех пор, пока пациент не продемонстрирует медицински значимое улучшение выбранного признака или показателя выше исходного уровня.
Пример схемы лечения, представленной в данном документе, включает подкожную инъекцию слитого белка на основе антитела и TGF-β Trap в соответствующей дозе один раз в неделю или более длительно для лечения симптомов легочной артериальной гипертензии, легочной гипертензии, легочного фиброза и кардиоваскулярного фиброза. Слитый белок на основе антитела и TGF-β Trap можно вводить еженедельно или ежемесячно до тех пор, пока не будет достигнут необходимый результат, например, пока у пациента не исчезнут симптомы. При необходимости лечение можно возобновить или в качестве альтернативы можно вводить поддерживающую дозу.
Давление в легочной артерии пациента можно контролировать до, в ходе и/или после лечения слитым белком на основе антитела и TGF-β Trap для выявления любых изменений давления. При некоторых заболеваниях изменения легочного артериального давления могут зависеть от таких факторов, как прогрессирование заболевания. Легочное артериальное давление может быть измерено посредством известных методик.
Конкретные варианты осуществления способов и композиций, представленных в данном документе, включают применение, например, слитого белка на основе антитела и TGF-β Trap и одного или более антагонистов эндотелина, двух или более слитых белков на основе антитела и TGF-β Trap, представленных в данном документе, или слитого белка на основе антитела и TGF-β Trap, представленных в данном документе, и одного или более других антагонистов эндотелина. В другом варианте осуществления слитый белок на основе антитела и TGF-β Trap, представленных в данном документе, вводят по отдельности или в комбинации с другими средствами для лечения тяжелых симптомов у пациента. Примеры данных средств включают белковые и небелковые лекарственные средства. Если несколько лекарственных средств вводят в комбинации, их дозу следует соответственно корректировать, что хорошо известно в данной области техники. Комбинированная терапия посредством "комбинированного введения" не ограничивается одновременным введением, а также включает режим лечения, при котором антиген и белок вводят по меньшей мере один раз в течение курса лечения, включающего введение пациенту по меньшей мере одного другого терапевтического средства.
В другом аспекте в данном документе представлен способ лечения симптомов легочной артериальной гипертензии, легочной гипертензии и легочного фиброза, предусматривающий применение смеси, содержащей слитый белок на основе антитела и TGF-β Trap, представленных в данном документе, и фармацевтически приемлемые наполнители с целью лечения заболеваний, связанных с вышеупомянутыми патологическими состояниями. Способ получения средств описан выше.
Кроме того, в данном документе представлены композиции, наборы и способы, относящиеся к слитому белку на основе антитела, которое может специфически связываться с человеческим ETA, и TGF-β Trap. Представлены последовательности нуклеиновых кислот, их производные и фрагменты, содержащие полинуклеотидные последовательности, кодирующие весь слитый белок или его часть на основе антитела, связывающего человеческий ETA, и TGF-β Trap, такие как последовательности нуклеиновой кислоты, кодирующие весь слитый белок или его часть на основе антитела, связывающего человеческий ETA, фрагмента антитела, производного антитела и TGF-β Trap. Кроме того, в данном документе представлены вектор, содержащий последовательность нуклеиновой кислоты, и клетка, содержащая нуклеиновую кислоту и/или вектор. Представленные способы включают, например, способы получения, идентифицирования или выделения слитого белка на основе антитела, связывающего человеческий ETA, и TGF-β Trap, способы определения того, способен ли слитый белок на основе антитела, связывающего человеческий ETA, и TGF-β Trap по-прежнему связываться с ETA, способы определения передачи сигнала, опосредованной слитым белком на основе антитела, связывающего человеческий ETA, и TGF-β, блокирующего TGF-β, и способы введения слитого белка на основе антитела, связывающего человеческий ETA, и TGF-β Trap модельным животным для измерения биологической активности in vivo.
Следующие ниже конкретные примеры используются для дополнительной иллюстрации технических решений в данном документе.
Если не указано иное, в данном документе используемые материалы, оборудование и т.п. могут быть приобретены на рынке или обычно используются в данной области техники. Если не указано иное, способы в следующих примерах являются общепринятыми в данной области техники.
1. Клонирование и субклонирование генов антител
Собирали клетки гибридомы, секретирующие антитела, и в соответствии с протоколом производителя набора для экстракции мРНК QIAGEN экстрагировали мРНК клеток гибридомы. Затем экстрагированную мРНК обратно транскрибировали в кДНК. Праймеры для обратной транскрипции представляют собой специфические праймеры для мышиных константных областей легкой и тяжелой цепей, при этом праймер для обратной транскрипции тяжелой цепи представляет собой (5'-TTTGGRGGGAAGATGAAGAC-3') (SEQ ID NO: 199), а праймеры для обратной транскрипции легкой цепи представляют собой (5'-TTAACACTCTCCCCTGTTGAA-3') (SEQ ID NO: 200) и (5'-TTAACACTCATTCCTGTTGAA-3') (SEQ ID NO: 201). Условия реакции для RT-PCR: 25°C, 5 мин (минут); 50°C, 60 мин; 70°C, 15 мин. Обратно транскрибируемую кДНК разбавляли до 500 мкл с использованием 0,1 мМ ТЕ, добавляли в пробирку для ультрафильтрации (Amicon Ultra-0.5) и центрифугировали при 2000 g в течение 10 мин; фильтрат удаляли, и добавляли 500 мкл 0,1 мМ ТЕ, и центрифугировали при 2000 g в течение 10 мин; фильтрат удаляли, а пробирку для приготовления опрокидывали в новую центрифужную пробирку и центрифугировали при 2000 g в течение 10 мин с получением очищенной кДНК; 10 мкл очищенной кДНК отбирали в качестве матрицы и добавляли 4 мкл 5x буфера для наращивания (Promega, коммерчески доступный), 4 мкл дАТФ (1 мМ) и 10 ед. концевой трансферазы (Promega, коммерчески доступная), тщательно перемешивали и инкубировали при 37°C в течение 5 мин, затем при 65°C в течение 5 мин, а затем кДНК с поли-А-хвостом использовали в качестве матрицы для амплификации генов вариабельных областей легкой и тяжелой цепей антитела с применением ПЦР. Все прямые праймеры представляют собой OligodT, обратные праймеры для тяжелой цепи представляют собой (5'-TGGACAGGGATCCAGAGTTCC-3') (SEQ ID NO: 202) и (5'-TGGACAGGGCTCCATAGTTCC-3') (SEQ ID NO: 203), а обратный праймер для легкой цепи представляет собой (5'-ACTCGTCCTTGGTCAACGTG-3') (SEQ ID NO: 204). Условия для реакции ПЦР: 95°C, 5 мин; 95°C, 30 с (секунд), 56°C, 30 с, 72 °C, 1 мин, 40 циклов; 72°C, 7 мин. ПЦР-продукт лигировали с вектором PMD 18-T (Takara Bio, коммерчески доступный) и затем секвенировали. Последовательности CDR клонированного антитела показаны в таблице 1 и таблице 2.
Праймеры для ПЦР конструировали на основе секвенированных последовательностей ДНК антитела, и, таким образом, полные сигнальные пептиды легкой цепи и тяжелой цепи, а также вариабельные домены и константная область мышиного IgG1 связаны с вектором экспрессии pTM5.
2. Получение плазмиды экспрессии для гена слитого белка на основе антитела
Последовательности гена человеческого TGF-β Trap (hTGF-β Trap) сливали с С-концом тяжелой цепи антитела к ETA посредством способа перекрывающейся ПЦР. Сайт для рестриктазы Nhe1 добавляли на 5'-конце вариабельной области тяжелой цепи слитого белка, а сайт для рестриктазы Not1 добавляли на 3'-конце слитого белка с использованием праймеров для ПЦР, так что полную тяжелую цепь и ген слитого белка на основе hTGF-β Trap загружали в вектор экспрессии pTM5. Аналогично сайт для рестриктазы Nhe1 добавляли на 5'-конце вариабельной области легкой цепи, а сайт для рестриктазы Bsiw1 добавляли на ее 3'-конце, так что полная последовательность вариабельной области легкой цепи была связана с вектором экспрессии pTM5, который был загружен константной областью легкой цепи.
3. Временная экспрессия слитого белка на основе антитела
5×105/мл суспендированных экспрессирующих клеточных линий HEK293 или CHO инокулировали в колбу с мешалкой и культивировали с вращением в течение 24 ч (часов) при 37°C и 5% CO2 для достижения плотности 1×106/мл, а затем использовали для трансфекции. В процессе трансфекции полиэтиленимин (PEI) использовали в качестве среды для трансфекции и смешивали с ДНК (количество ДНК составляло 0,5 мкг на 1×106 клеток, при этом соотношение легкой цепи и тяжелой цепи антитела составляло 3:2), где предпочтительное соотношение PEI и ДНК составляло 3:1. Смесь из обоих добавляли к культуре клеток после 15 минут инкубации в стоячем положении. После получения смеси PEI и ДНК клетки культивировали с перемешиванием в течение 24 ч при 37°C и 5% CO2, а затем к раствору клеточной культуры добавляли 0,5% триптон в качестве добавки, требуемой для экспрессии, и в конечном итоге клеточный супернатант собирали после завершения экспрессии (через 96 ч) для очистки и разделения антител.
4. Очистка и разделение слитого белка на основе антитела
Собранный клеточный супернатант центрифугировали с высокой скоростью (8000 об/мин, 15 мин) с удалением клеток и клеточного дебриса, а затем фильтровали и осветляли посредством фильтровальной мембраны с размером пор 0,45 мкм. Осветленную надосадочную жидкость использовали для очистки. Процесс очистки завершали хроматографированием. Супернатант вначале пропускали через колонку для аффинной хроматографии с белком G, в ходе чего антитело, содержащееся в супернатанте, связывалось с лигандом колонки для аффинной хроматографии с белком G и оставалось в колонке. Затем буфер для элюирования с низким значением pH (менее или равным 3,0) использовали для промывки хроматографической колонки для диссоциации связанного с ней антитела, и собранный элюат антитела с низким значением pH быстро нейтрализовали 1 М трис-HCl для защиты антитела от инактивации. Полученный элюат, содержащий слитый белок на основе антитела, заменяли буферной системой PBS через 16 ч после диализа.
5. С помощью анализа потока кальция выявляли биологическую активность слитого белка на основе антитела к ETA и TGF-β Trap, блокирующего ETA in vitro
Клетки CHO-DHFR, коэкспрессирующие hETA-Aequorin, инокулировали по 25000 клеток на лунку в черный 96-луночный планшет для культивирования клеток и культивировали в течение ночи при 37°C. На следующий день клеточный супернатант удаляли, и добавляли 50 мкл субстрата целентеразина h (Promega, коммерчески доступный), и инкубировали в течение 2 ч в темноте, а затем добавляли 50 мкл очищенного антитела или слитого белка и инкубировали в течение 30 мин. Автоматический пробоотборник использовали для перфузии эндотелина 1 в микропланшетном ридере SpectraMax L Molecular Devices. Мгновенное изменение потока кальция выявляли в течение 40 с, и фиксировали время достижения пика и пиковое значение. На фигуре 1 показано, что антитело к ETA и слитый белок на основе антитела ETA и TGF-β Trap могли эффективно ингибировать изменения внутриклеточной концентрации Ca2+, опосредованные человеческим ETA.
6. В эксперименте с репортерным геном выявляли слитый белок на основе антитела к ETA и TGF-β Trap, блокирующий путь передачи сигнала in vitro, опосредованный TGF-β.
Клетки 4T1, стабильно экспрессирующие SBE-люциферазу, инокулировали в количестве 34000 клеток на лунку в 96-луночном планшете для культивирования клеток и культивировали в течение ночи при 37°C. На второй день предварительно инкубировали 3 нг/мл TGF-β1 и градиентно разбавленный слитый белок на основе антитела к ETA и TGF-β Trap в равном объеме в течение 30 мин при комнатной температуре. В течение предварительной инкубации супернатант среды удаляли из 96-луночного планшета для культивирования клеток, поверхность клеток дважды промывали бессывороточной средой, а оставшуюся жидкость аспирировали. 100 мкл ранее вступившего в реакцию реакционного раствора добавляли в 96-луночный планшет для культивирования клеток, и смесь инкубировали при 37°C в течение 6 часов. После инкубации добавляли 100 мкл хемилюминесцентного субстрата Bright Glo (Promega) и затем клеточный лизат переносили в белый 96-луночный планшет, а относительную интенсивность флуоресценции считывали на микропланшет-ридере SpectraMax L (Molecular Devices). На фигуре 2 показана ингибирующая кривая, полученная по результатам эксперимента с репортерным геном, в котором выявляли слитый белок на основе антитела к ETA и TGF-β Trap, блокирующий TGF-β1 от активации рецептора TGF-β1.
7. Активность слитого белка антитела in vivo в отношении блеомицин-индуцированной модели легочного фиброза у мышей C57BL/6
Блеомицин (BLM) вводили интратрахеально для индуцирования модели легочного фиброза (BPF) у здоровых мышей C57BL/6, которую использовали для оценки эффекта внутрибрюшинной инъекции h15F3-(G4S)4-TGF-β Trap в лечении легочного фиброза в данной модели. Мышей анестезировали с помощью 2% раствора изофлурана и фиксировали на экспериментальном столе в положении лежа на спине, а в их трахею быстро имплантировали иглу. Микрошприцом объемом 100 мкл набирали определенное количество физиологического солевого раствора (контрольная группа здоровых животных) или раствора блеомицина (2,5 мг/кг) в соответствии с массой тела и вводили их через имплантированную иглу в пространство между кольцами трахеи с целью медленного поступления медицинского раствора в трахею в дозе 50 мкл/20 г. После инъекции мышей быстро открепляли и вращали для того, чтобы лекарственный раствор равномерно распределялся в легких. В произвольном порядке животных группировали в соответствии с массой тела, 12 - в группе здоровых животных, 20 - в модельной группе и по 14 в каждой из групп с низкой и высокой дозами h15F3-(G4S)4-TGF-β Trap. h15F3-(G4S)4-TGF-β Trap или контроль вводили в соответствии с протоколом через один день после создания модели, а затем непрерывно в течение 3-х недель. Эффективность препарата оценивали по таким показателям, как выживаемость животных и поражения легких. На фигуре 3 показано, что в модельной группе согласно данным окрашивания легочной ткани по Массону большая часть альвеолярной структуры исчезала, альвеолярная перегородка расширялась, и было выявлено отложение большого количества коллагеновых волокон, что свидетельствовало о том, что введение блеомицина в трахею эффективно индуцировало легочный фиброз у здоровых мышей C57BL/6; при этом у животных в группах с низкой и высокой дозами h15F3-(G4S)4-TGF-β Trap выявили значительное улучшение вышеупомянутых маркеров легочного фиброза, а взаимосвязь между дозой и эффектом была очевидной. Кроме того, на фигуре 4 показано, что обработка с использованием h15F3-(G4S)4-TGF-β Trap может увеличивать время выживаемости мышей, что указывало на то, что данное лекарственное средство способно ингибировать процесс легочного фиброза.
Вышеупомянутые варианты осуществления представляют собой только предпочтительные решения настоящего изобретения, которые никоим образом не следует рассматривать как ограничивающие настоящее изобретение, и при этом существуют другие варианты и модификации, если они не выходят за рамки технических решений, определенных в формуле изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Gmax Biopharm LLC.
<120> СЛИТЫЙ БЕЛОК НА ОСНОВЕ АНТИТЕЛА К ETA И TGF-β TRAP, СОДЕРЖАЩАЯ ЕГО
ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ И ИХ ПРИМЕНЕНИЕ
<130> 2019
<160> 212
<170> PatentIn версия 3.5
<210> 1
<211> 1868
<212> ДНК
<213> Homo sapiens
<400> 1
gaattcggga aaaagtgaag gtgtaaaagc agcacaagtg caataagaga tatttcctca 60
aatttgcctc aagatggaaa ccctttgcct cagggcatcc ttttggctgg cactggttgg 120
atgtgtaatc agtgataatc ctgagagata cagcacaaat ctaagcaatc atgtggatga 180
tttcaccact tttcgtggca cagagctcag cttcctggtt accactcatc aacccactaa 240
tttggtccta cccagcaatg gctcaatgca caactattgc ccacagcaga ctaaaattac 300
ttcagctttc aaatacatta acactgtgat atcttgtact attttcatcg tgggaatggt 360
ggggaatgca actctgctca ggatcattta ccagaacaaa tgtatgagga atggccccaa 420
cgcgctgata gccagtcttg cccttggaga ccttatctat gtggtcattg atctccctat 480
caatgtattt aagctgctgg ctgggcgctg gccttttgat cacaatgact ttggcgtatt 540
tctttgcaag ctgttcccct ttttgcagaa gtcctcggtg gggatcaccg tcctcaacct 600
ctgcgctctt agtgttgaca ggtacagagc agttgcctcc tggagtcgtg ttcagggaat 660
tgggattcct ttggtaactg ccattgaaat tgtctccatc tggatcctgt cctttatcct 720
ggccattcct gaagcgattg gcttcgtcat ggtacccttt gaatataggg gtgaacagca 780
taaaacctgt atgctcaatg ccacatcaaa attcatggag ttctaccaag atgtaaagga 840
ctggtggctc ttcgggttct atttctgtat gcccttggtg tgcactgcga tcttctacac 900
cctcatgact tgtgagatgt tgaacagaag gaatggcagc ttgagaattg ccctcagtga 960
acatcttaag cagcgtcgag aagtggcaaa aacagttttc tgcttggttg taatttttgc 1020
tctttgctgg ttccctcttc atttaagccg tatattgaag aaaactgtgt ataacgagat 1080
ggacaagaac cgatgtgaat tacttagttt cttactgctc atggattaca tcggtattaa 1140
cttggcaacc atgaattcat gtataaaccc catagctctg tattttgtga gcaagaaatt 1200
taaaaattgt ttccagtcat gcctctgctg ctgctgttac cagtccaaaa gtctgatgac 1260
ctcggtcccc atgaacggaa caagcatcca gtggaagaac cacgatcaaa acaaccacaa 1320
cacagaccgg agcagccata aggacagcat gaactgacca cccttagaag cactcctcgg 1380
tactcccata atcctctcgg agaaaaaaat cacaaggcaa ctgtgagtcc gggaatctct 1440
tctctgatcc ttcttcctta attcactccc acacccaaga agaaatgctt tccaaaaccg 1500
caagggtaga ctggtttatc cacccacaac atctacgaat cgtacttctt taattgatct 1560
aatttacata ttctgcgtgt tgtattcagc actaaaaaat ggtgggagct gggggagaat 1620
gaagactgtt aaatgaaacc agaaggatat ttactacttt tgcatgaaaa tagagctttc 1680
aagtacatgg ctagctttta tggcagttct ggtgaatgtt caatgggaac tggtcaccat 1740
gaaactttag agattaacga caagattttc tacttttttt aagtgatttt tttgtccttc 1800
agccaaacac aatatgggct caagtcactt ttatttgaaa tgtcatttgg tgccagtatc 1860
ccgaattc 1868
<210> 2
<211> 427
<212> БЕЛОК
<213> Homo sapiens
<400> 2
Met Glu Thr Leu Cys Leu Arg Ala Ser Phe Trp Leu Ala Leu Val Gly
1 5 10 15
Cys Val Ile Ser Asp Asn Pro Glu Arg Tyr Ser Thr Asn Leu Ser Asn
20 25 30
His Val Asp Asp Phe Thr Thr Phe Arg Gly Thr Glu Leu Ser Phe Leu
35 40 45
Val Thr Thr His Gln Pro Thr Asn Leu Val Leu Pro Ser Asn Gly Ser
50 55 60
Met His Asn Tyr Cys Pro Gln Gln Thr Lys Ile Thr Ser Ala Phe Lys
65 70 75 80
Tyr Ile Asn Thr Val Ile Ser Cys Thr Ile Phe Ile Val Gly Met Val
85 90 95
Gly Asn Ala Thr Leu Leu Arg Ile Ile Tyr Gln Asn Lys Cys Met Arg
100 105 110
Asn Gly Pro Asn Ala Leu Ile Ala Ser Leu Ala Leu Gly Asp Leu Ile
115 120 125
Tyr Val Val Ile Asp Leu Pro Ile Asn Val Phe Lys Leu Leu Ala Gly
130 135 140
Arg Trp Pro Phe Asp His Asn Asp Phe Gly Val Phe Leu Cys Lys Leu
145 150 155 160
Phe Pro Phe Leu Gln Lys Ser Ser Val Gly Ile Thr Val Leu Asn Leu
165 170 175
Cys Ala Leu Ser Val Asp Arg Tyr Arg Ala Val Ala Ser Trp Ser Arg
180 185 190
Val Gln Gly Ile Gly Ile Pro Leu Val Thr Ala Ile Glu Ile Val Ser
195 200 205
Ile Trp Ile Leu Ser Phe Ile Leu Ala Ile Pro Glu Ala Ile Gly Phe
210 215 220
Val Met Val Pro Phe Glu Tyr Arg Gly Glu Gln His Lys Thr Cys Met
225 230 235 240
Leu Asn Ala Thr Ser Lys Phe Met Glu Phe Tyr Gln Asp Val Lys Asp
245 250 255
Trp Trp Leu Phe Gly Phe Tyr Phe Cys Met Pro Leu Val Cys Thr Ala
260 265 270
Ile Phe Tyr Thr Leu Met Thr Cys Glu Met Leu Asn Arg Arg Asn Gly
275 280 285
Ser Leu Arg Ile Ala Leu Ser Glu His Leu Lys Gln Arg Arg Glu Val
290 295 300
Ala Lys Thr Val Phe Cys Leu Val Val Ile Phe Ala Leu Cys Trp Phe
305 310 315 320
Pro Leu His Leu Ser Arg Ile Leu Lys Lys Thr Val Tyr Asn Glu Met
325 330 335
Asp Lys Asn Arg Cys Glu Leu Leu Ser Phe Leu Leu Leu Met Asp Tyr
340 345 350
Ile Gly Ile Asn Leu Ala Thr Met Asn Ser Cys Ile Asn Pro Ile Ala
355 360 365
Leu Tyr Phe Val Ser Lys Lys Phe Lys Asn Cys Phe Gln Ser Cys Leu
370 375 380
Cys Cys Cys Cys Tyr Gln Ser Lys Ser Leu Met Thr Ser Val Pro Met
385 390 395 400
Asn Gly Thr Ser Ile Gln Trp Lys Asn His Asp Gln Asn Asn His Asn
405 410 415
Thr Asp Arg Ser Ser His Lys Asp Ser Met Asn
420 425
<210> 3
<211> 2493
<212> ДНК
<213> Яванский макак
<400> 3
cgggggtggc tgtgtcccag gatagctgga aggttaggac gctcttgcgg tcccagagtg 60
gagtggaagt tctggagctt tgggaggaga cggggaggac agactggagg cgtgttcctc 120
cggagttttc ttttccgtgc gagccctcgc gcgcgcgtac agtcatcccg ctggtctgac 180
gattgtggag aggaggtgga gaggcttcat ccatcccacc cggtcgtcgc ggggaattgg 240
ggtcccagcg agacctcccc cggagaagca gtgcccagga agttttctga agccggggta 300
gctgtgcagc cggagccgcc gccgcgccgg agcccgggac accggccacc ctccgcgcca 360
cccacccttg ccggctccgg cttcctctgg cccaggcgcc tcgcggaccc ggcagctgtc 420
tgcgcccgcc gagctccacg gtgaaaaaaa tagtgaaggt gtaaaagcag cacaagtgca 480
ataagagata tttcctcaaa tttgcctcaa gatggaaacc gtttgcctca gggcatcctt 540
ttggctggca ctggttggat gtgtaatcag tgataatgct gagagataca gcacaaatct 600
aagcaatcat gtggatgatt tcaccacttt tcatggcaca gagctcagcc tcctggttac 660
cactcatcaa cccactaact tggtcctacc cagcaatggc tcaatgcaca actattgccc 720
acagcagact aaaattactt cagcttttaa atacattaac actgtgatat cttgtactat 780
tttcatcgtg ggaatggtgg ggaatgcaac tctgctcagg atcatttacc agaacaaatg 840
tatgaggaat ggccccaacg cgctgatagc cagccttgcc cttggagacc ttatctatgt 900
ggtcattgat ctccctatca atgtatttaa gctgctggct gggcgctggc cttttgatca 960
caatgacttt ggcgtatttc tttgcaagct gttccccttt ttgcagaagt cctcagtggg 1020
gatcaccgtc ctcaacctct gcgctcttag tgttgacagg tacagagcag ttgcctcctg 1080
gagtcgtgtt cagggaattg ggattccttt ggtaactgcc attgaaattg tctccatctg 1140
gatcctgtcc ttcatcctgg ccattcctga agcgattggc ttcgtcatgg taccctttga 1200
atataggggt gaacagcata aaacctgtat gctcaatgct acgtcaaaat tcatggagtt 1260
ctaccaagat gtaaaggact ggtggctctt tgggttctat ttctgtatgc ccttggtgtg 1320
cactgcgatc ttctacaccc tcatgacttg tgagatgttg aacagaagga atggcagctt 1380
gagaattgcc ctcagtgaac atcttaagca gcgtcgagaa gtggcaaaaa cagttttctg 1440
cttggtcgta atttttgctc tgtgctggtt ccctcttcat ttaagccgta tattgaagaa 1500
aaccgtgtat aatgagatgg acaagaaccg atgtgaatta cttagtttct tgctgctcat 1560
ggattacatc ggtattaact tggcaaccat gaattcatgt ataaacccca tagctctgta 1620
ttttgtgagc aagaaattta aaaattgttt ccagtcatgc ctctgctgct gctgttacca 1680
gtccaaaagt ctgatgacct cggtccccat gaacggaaca agcatccagt ggaagaacca 1740
cgaacaaaac aaccacaaca cagaccggag cagccacaag gacagcatga actgaccacc 1800
ctgcgaagca ctcctgggta ctcccataat cctctgggag aaaaaaatca caaggcaact 1860
gtgactccgg aaatctcttc tctgatcctt cttccttaac tcactcccac acccaagaag 1920
aaatgctttc caaaaccgca agggtagacc ggtttagcca cccacgacat ctaccaatcg 1980
tacttcttta attaatctga tttacatatt ccgcgtgttg tattcagcac taaaaaatgg 2040
tgggagctgg gagagaatga agactgttca atgaaaccag aaggatattt actacttttg 2100
catgaaaata gagctttcaa gtacatgggt agcttttatg gcagttctgg tgaatgttca 2160
gtgggaactg gtcaccatga aactttagag attatgacaa gattttctac tttttttaac 2220
tgattttttg tccttcagcc aaacacaata tgggctcaag ttacttttat ttgaaatgtc 2280
atttggtgcc agtatttttt aactgcataa tagcctaaca tgactatttg aacttattta 2340
cacatagttt gcaaaaaaaa aaagacaaaa atagtattca ggtgagcaat taggttagta 2400
ttttctatgt cactgtttat ttttttaaaa cacaaattct aaagctacaa caaatactac 2460
aggcccttaa aacacagtct gatgatacat ttg 2493
<210> 4
<211> 427
<212> БЕЛОК
<213> Яванский макак
<400> 4
Met Glu Thr Val Cys Leu Arg Ala Ser Phe Trp Leu Ala Leu Val Gly
1 5 10 15
Cys Val Ile Ser Asp Asn Ala Glu Arg Tyr Ser Thr Asn Leu Ser Asn
20 25 30
His Val Asp Asp Phe Thr Thr Phe His Gly Thr Glu Leu Ser Leu Leu
35 40 45
Val Thr Thr His Gln Pro Thr Asn Leu Val Leu Pro Ser Asn Gly Ser
50 55 60
Met His Asn Tyr Cys Pro Gln Gln Thr Lys Ile Thr Ser Ala Phe Lys
65 70 75 80
Tyr Ile Asn Thr Val Ile Ser Cys Thr Ile Phe Ile Val Gly Met Val
85 90 95
Gly Asn Ala Thr Leu Leu Arg Ile Ile Tyr Gln Asn Lys Cys Met Arg
100 105 110
Asn Gly Pro Asn Ala Leu Ile Ala Ser Leu Ala Leu Gly Asp Leu Ile
115 120 125
Tyr Val Val Ile Asp Leu Pro Ile Asn Val Phe Lys Leu Leu Ala Gly
130 135 140
Arg Trp Pro Phe Asp His Asn Asp Phe Gly Val Phe Leu Cys Lys Leu
145 150 155 160
Phe Pro Phe Leu Gln Lys Ser Ser Val Gly Ile Thr Val Leu Asn Leu
165 170 175
Cys Ala Leu Ser Val Asp Arg Tyr Arg Ala Val Ala Ser Trp Ser Arg
180 185 190
Val Gln Gly Ile Gly Ile Pro Leu Val Thr Ala Ile Glu Ile Val Ser
195 200 205
Ile Trp Ile Leu Ser Phe Ile Leu Ala Ile Pro Glu Ala Ile Gly Phe
210 215 220
Val Met Val Pro Phe Glu Tyr Arg Gly Glu Gln His Lys Thr Cys Met
225 230 235 240
Leu Asn Ala Thr Ser Lys Phe Met Glu Phe Tyr Gln Asp Val Lys Asp
245 250 255
Trp Trp Leu Phe Gly Phe Tyr Phe Cys Met Pro Leu Val Cys Thr Ala
260 265 270
Ile Phe Tyr Thr Leu Met Thr Cys Glu Met Leu Asn Arg Arg Asn Gly
275 280 285
Ser Leu Arg Ile Ala Leu Ser Glu His Leu Lys Gln Arg Arg Glu Val
290 295 300
Ala Lys Thr Val Phe Cys Leu Val Val Ile Phe Ala Leu Cys Trp Phe
305 310 315 320
Pro Leu His Leu Ser Arg Ile Leu Lys Lys Thr Val Tyr Asn Glu Met
325 330 335
Asp Lys Asn Arg Cys Glu Leu Leu Ser Phe Leu Leu Leu Met Asp Tyr
340 345 350
Ile Gly Ile Asn Leu Ala Thr Met Asn Ser Cys Ile Asn Pro Ile Ala
355 360 365
Leu Tyr Phe Val Ser Lys Lys Phe Lys Asn Cys Phe Gln Ser Cys Leu
370 375 380
Cys Cys Cys Cys Tyr Gln Ser Lys Ser Leu Met Thr Ser Val Pro Met
385 390 395 400
Asn Gly Thr Ser Ile Gln Trp Lys Asn His Glu Gln Asn Asn His Asn
405 410 415
Thr Asp Arg Ser Ser His Lys Asp Ser Met Asn
420 425
<210> 5
<211> 1436
<212> ДНК
<213> Rattus norvegicus
<400> 5
gtgagaccaa cataacagga cgtttcttca gatccacatt aagatgggtg tcctttgctt 60
tctggcgtcc ttttggctgg ccctggtggg aggcgcaatc gctgacaatg ctgagagata 120
cagtgctaat ctaagcagcc acgtggagga cttcacccct tttccaggga cagagttcga 180
ctttctgggc accacccttc gaccccctaa tttggccctg cctagcaatg gctcaatgca 240
tggctattgc ccacagcaga caaaaatcac gacggctttc aaatatatca acactgtgat 300
atcctgtacc attttcatcg tgggaatggt ggggaacgcc actctcctaa gaatcattta 360
ccaaaacaag tgtatgagga acggccccaa tgcgctcata gccagcctgg cccttggaga 420
ccttatctac gtggtcattg atctccccat caatgtgttt aagctgttgg cggggcgctg 480
gccttttgac cacaatgatt ttggagtgtt tctctgcaag ctgttcccct ttttgcagaa 540
gtcgtccgtg ggcatcactg tcctgaatct ctgcgctctc agtgtggaca ggtacagagc 600
agtggcttcc tggagccggg ttcaaggaat cgggatcccc ttgattaccg ccattgaaat 660
tgtctccatc tggatccttt cctttatctt ggccatccca gaagcaatcg gcttcgtcat 720
ggtacccttc gaatacaagg gcgagcagca caggacctgc atgctcaacg ccacgaccaa 780
gttcatggag ttttaccaag acgtgaagga ctggtggctc tttggattct acttctgcat 840
gcccttggtg tgcacagcaa tcttctatac cctcatgacc tgtgagatgc tcaacagaag 900
gaatgggagc ttgcggattg ccctcagcga acacctcaag cagcgtcgag aggtggcaaa 960
gaccgtcttc tgcttggttg tcatcttcgc cctgtgctgg ttccctcttc acttaagccg 1020
aattttgaag aaaaccgtct atgatgagat ggataagaac cggtgtgaac tgctcagctt 1080
cttgctgctc atggattaca ttggcattaa cctggcaacc atgaactctt gcataaaccc 1140
aatagctctg tattttgtga gcaagaaatt caaaaattgt tttcagtcat gcctctgttg 1200
ctgttgtcac cagtccaaaa gcctcatgac ctcggtcccc atgaatggaa cgagtatcca 1260
gtggaagaac caggagcaga accacaacac agaacggagc agccacaagg acagcatgaa 1320
ctaaccctgt gcagaagcac cgagcagtgt gccttcgagt cccaggatga aacggtcacg 1380
cagcagctgc gctcccaaaa cctcccaggt ctctcccctg ctttttgtct aagctt 1436
<210> 6
<211> 426
<212> БЕЛОК
<213> Rattus norvegicus
<400> 6
Met Gly Val Leu Cys Phe Leu Ala Ser Phe Trp Leu Ala Leu Val Gly
1 5 10 15
Gly Ala Ile Ala Asp Asn Ala Glu Arg Tyr Ser Ala Asn Leu Ser Ser
20 25 30
His Val Glu Asp Phe Thr Pro Phe Pro Gly Thr Glu Phe Asp Phe Leu
35 40 45
Gly Thr Thr Leu Arg Pro Pro Asn Leu Ala Leu Pro Ser Asn Gly Ser
50 55 60
Met His Gly Tyr Cys Pro Gln Gln Thr Lys Ile Thr Thr Ala Phe Lys
65 70 75 80
Tyr Ile Asn Thr Val Ile Ser Cys Thr Ile Phe Ile Val Gly Met Val
85 90 95
Gly Asn Ala Thr Leu Leu Arg Ile Ile Tyr Gln Asn Lys Cys Met Arg
100 105 110
Asn Gly Pro Asn Ala Leu Ile Ala Ser Leu Ala Leu Gly Asp Leu Ile
115 120 125
Tyr Val Val Ile Asp Leu Pro Ile Asn Val Phe Lys Leu Leu Ala Gly
130 135 140
Arg Trp Pro Phe Asp His Asn Asp Phe Gly Val Phe Leu Cys Lys Leu
145 150 155 160
Phe Pro Phe Leu Gln Lys Ser Ser Val Gly Ile Thr Val Leu Asn Leu
165 170 175
Cys Ala Leu Ser Val Asp Arg Tyr Arg Ala Val Ala Ser Trp Ser Arg
180 185 190
Val Gln Gly Ile Gly Ile Pro Leu Ile Thr Ala Ile Glu Ile Val Ser
195 200 205
Ile Trp Ile Leu Ser Phe Ile Leu Ala Ile Pro Glu Ala Ile Gly Phe
210 215 220
Val Met Val Pro Phe Glu Tyr Lys Gly Glu Gln His Arg Thr Cys Met
225 230 235 240
Leu Asn Ala Thr Thr Lys Phe Met Glu Phe Tyr Gln Asp Val Lys Asp
245 250 255
Trp Trp Leu Phe Gly Phe Tyr Phe Cys Met Pro Leu Val Cys Thr Ala
260 265 270
Ile Phe Tyr Thr Leu Met Thr Cys Glu Met Leu Asn Arg Arg Asn Gly
275 280 285
Ser Leu Arg Ile Ala Leu Ser Glu His Leu Lys Gln Arg Arg Glu Val
290 295 300
Ala Lys Thr Val Phe Cys Leu Val Val Ile Phe Ala Leu Cys Trp Phe
305 310 315 320
Pro Leu His Leu Ser Arg Ile Leu Lys Lys Thr Val Tyr Asp Glu Met
325 330 335
Asp Lys Asn Arg Cys Glu Leu Leu Ser Phe Leu Leu Leu Met Asp Tyr
340 345 350
Ile Gly Ile Asn Leu Ala Thr Met Asn Ser Cys Ile Asn Pro Ile Ala
355 360 365
Leu Tyr Phe Val Ser Lys Lys Phe Lys Asn Cys Phe Gln Ser Cys Leu
370 375 380
Cys Cys Cys Cys His Gln Ser Lys Ser Leu Met Thr Ser Val Pro Met
385 390 395 400
Asn Gly Thr Ser Ile Gln Trp Lys Asn Gln Glu Gln Asn His Asn Thr
405 410 415
Glu Arg Ser Ser His Lys Asp Ser Met Asn
420 425
<210> 7
<211> 33
<212> ДНК
<213> Mus musculus
<400> 7
agggccagtc agaacattgg cacaagcata cac 33
<210> 8
<211> 11
<212> БЕЛОК
<213> Mus musculus
<400> 8
Arg Ala Ser Gln Asn Ile Gly Thr Ser Ile His
1 5 10
<210> 9
<211> 33
<212> ДНК
<213> Mus musculus
<400> 9
cgagcaagtg aaaatattta cagttattta gca 33
<210> 10
<211> 11
<212> БЕЛОК
<213> Mus musculus
<400> 10
Arg Ala Ser Glu Asn Ile Tyr Ser Tyr Leu Ala
1 5 10
<210> 11
<211> 39
<212> ДНК
<213> Mus musculus
<400> 11
cagagcctct ttgatattga tggaaagaca tatttgaat 39
<210> 12
<211> 13
<212> БЕЛОК
<213> Mus musculus
<400> 12
Gln Ser Leu Phe Asp Ile Asp Gly Lys Thr Tyr Leu Asn
1 5 10
<210> 13
<211> 33
<212> ДНК
<213> Mus musculus
<400> 13
cgggcaagtc aggacattgg tggtagctta aac 33
<210> 14
<211> 11
<212> БЕЛОК
<213> Mus musculus
<400> 14
Arg Ala Ser Gln Asp Ile Gly Gly Ser Leu Asn
1 5 10
<210> 15
<211> 33
<212> ДНК
<213> Mus musculus
<400> 15
agggccagcc agactattag cgacttctta cac 33
<210> 16
<211> 11
<212> БЕЛОК
<213> Mus musculus
<400> 16
Arg Ala Ser Gln Thr Ile Ser Asp Phe Leu His
1 5 10
<210> 17
<211> 33
<212> ДНК
<213> Mus musculus
<400> 17
agggcaagtg aggacataca cactcaatta gcc 33
<210> 18
<211> 11
<212> БЕЛОК
<213> Mus musculus
<400> 18
Arg Ala Ser Glu Asp Ile His Thr Gln Leu Ala
1 5 10
<210> 19
<211> 48
<212> ДНК
<213> Mus musculus
<400> 19
agatctagtc agtacattgt tcatagtact ggaaccacct atttagaa 48
<210> 20
<211> 16
<212> БЕЛОК
<213> Mus musculus
<400> 20
Arg Ser Ser Gln Tyr Ile Val His Ser Thr Gly Thr Thr Tyr Leu Glu
1 5 10 15
<210> 21
<211> 48
<212> ДНК
<213> Mus musculus
<400> 21
agatctagtc attaccttgt tcatgataac ggaaacacct atgttgaa 48
<210> 22
<211> 16
<212> БЕЛОК
<213> Mus musculus
<400> 22
Arg Ser Ser His Tyr Leu Val His Asp Asn Gly Asn Thr Tyr Val Glu
1 5 10 15
<210> 23
<211> 48
<212> ДНК
<213> Mus musculus
<400> 23
agatctagtc agaacattgt ccatagtact ggaaacacct atttagaa 48
<210> 24
<211> 16
<212> БЕЛОК
<213> Mus musculus
<400> 24
Arg Ser Ser Gln Asn Ile Val His Ser Thr Gly Asn Thr Tyr Leu Glu
1 5 10 15
<210> 25
<211> 30
<212> ДНК
<213> Mus musculus
<400> 25
agtgtcagct caagtgtaag ttacatacac 30
<210> 26
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 26
Ser Val Ser Ser Ser Val Ser Tyr Ile His
1 5 10
<210> 27
<211> 30
<212> ДНК
<213> Mus musculus
<400> 27
agtgccagct caagtgtaag ttacatgtgc 30
<210> 28
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 28
Ser Ala Ser Ser Ser Val Ser Tyr Met Cys
1 5 10
<210> 29
<211> 18
<212> ДНК
<213> Mus musculus
<400> 29
cagggcatta acaattat 18
<210> 30
<211> 6
<212> БЕЛОК
<213> Mus musculus
<400> 30
Gln Gly Ile Asn Asn Tyr
1 5
<210> 31
<211> 21
<212> ДНК
<213> Mus musculus
<400> 31
tatgcttcta agtctatatc t 21
<210> 32
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 32
Tyr Ala Ser Lys Ser Ile Ser
1 5
<210> 33
<211> 21
<212> ДНК
<213> Mus musculus
<400> 33
aatgcaaaaa ccttagcaga a 21
<210> 34
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 34
Asn Ala Lys Thr Leu Ala Glu
1 5
<210> 35
<211> 21
<212> ДНК
<213> Mus musculus
<400> 35
ctggtgtctg aattggactc t 21
<210> 36
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 36
Leu Val Ser Glu Leu Asp Ser
1 5
<210> 37
<211> 21
<212> ДНК
<213> Mus musculus
<400> 37
gccacatcca gcttagattc t 21
<210> 38
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 38
Ala Thr Ser Ser Leu Asp Ser
1 5
<210> 39
<211> 21
<212> ДНК
<213> Mus musculus
<400> 39
tatgcttccc aatccatctc t 21
<210> 40
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 40
Tyr Ala Ser Gln Ser Ile Ser
1 5
<210> 41
<211> 21
<212> ДНК
<213> Mus musculus
<400> 41
ggtgcagcca gtttgaaaag t 21
<210> 42
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 42
Gly Ala Ala Ser Leu Lys Ser
1 5
<210> 43
<211> 21
<212> ДНК
<213> Mus musculus
<400> 43
aaagtttcca accgattttc t 21
<210> 44
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 44
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 45
<211> 21
<212> ДНК
<213> Mus musculus
<400> 45
gacacatcca aactggcttc t 21
<210> 46
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 46
Asp Thr Ser Lys Leu Ala Ser
1 5
<210> 47
<211> 21
<212> ДНК
<213> Mus musculus
<400> 47
tatacatcaa ctttacagtc a 21
<210> 48
<211> 7
<212> БЕЛОК
<213> Mus musculus
<400> 48
Tyr Thr Ser Thr Leu Gln Ser
1 5
<210> 49
<211> 27
<212> ДНК
<213> Mus musculus
<400> 49
caacatagtt atagcttccc gtggacg 27
<210> 50
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 50
Gln His Ser Tyr Ser Phe Pro Trp Thr
1 5
<210> 51
<211> 27
<212> ДНК
<213> Mus musculus
<400> 51
cagcatcatt atggtattcc gttcacg 27
<210> 52
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 52
Gln His His Tyr Gly Ile Pro Phe Thr
1 5
<210> 53
<211> 27
<212> ДНК
<213> Mus musculus
<400> 53
tggcaaggta cacattttcc gctcacg 27
<210> 54
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 54
Trp Gln Gly Thr His Phe Pro Leu Thr
1 5
<210> 55
<211> 27
<212> ДНК
<213> Mus musculus
<400> 55
ctacaatatg ctagttctcc gtatacg 27
<210> 56
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 56
Leu Gln Tyr Ala Ser Ser Pro Tyr Thr
1 5
<210> 57
<211> 27
<212> ДНК
<213> Mus musculus
<400> 57
caaagtggta acacctttcc gtggacg 27
<210> 58
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 58
Gln Ser Gly Asn Thr Phe Pro Trp Thr
1 5
<210> 59
<211> 27
<212> ДНК
<213> Mus musculus
<400> 59
caacagtata ggagtattcc gtggacg 27
<210> 60
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 60
Gln Gln Tyr Arg Ser Ile Pro Trp Thr
1 5
<210> 61
<211> 27
<212> ДНК
<213> Mus musculus
<400> 61
tttcaaggtt cacattttcc attcacg 27
<210> 62
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 62
Phe Gln Gly Ser His Phe Pro Phe Thr
1 5
<210> 63
<211> 27
<212> ДНК
<213> Mus musculus
<400> 63
tttcaaggtt cacatttccc attcacg 27
<210> 64
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 64
His Gln Trp Ser Thr Asn Pro Pro Thr
1 5
<210> 65
<211> 27
<212> ДНК
<213> Mus musculus
<400> 65
cagcagtgga gtagtaaccc acccacg 27
<210> 66
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 66
Gln Gln Trp Ser Ser Asn Pro Pro Thr
1 5
<210> 67
<211> 24
<212> ДНК
<213> Mus musculus
<400> 67
cagcagttta gtaaacttcg gaca 24
<210> 68
<211> 8
<212> БЕЛОК
<213> Mus musculus
<400> 68
Gln Gln Phe Ser Lys Leu Arg Thr
1 5
<210> 69
<211> 36
<212> ДНК
<213> Mus musculus
<400> 69
gggttctcac tgaccacttc tggcttgggt gttgcc 36
<210> 70
<211> 12
<212> БЕЛОК
<213> Mus musculus
<400> 70
Gly Phe Ser Leu Thr Thr Ser Gly Leu Gly Val Ala
1 5 10
<210> 71
<211> 30
<212> ДНК
<213> Mus musculus
<400> 71
ggctacacct ttactagcta ctggatacac 30
<210> 72
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 72
Gly Tyr Thr Phe Thr Ser Tyr Trp Ile His
1 5 10
<210> 73
<211> 30
<212> ДНК
<213> Mus musculus
<400> 73
ggcctcaaca ttaaagacat ctatattcac 30
<210> 74
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 74
Gly Leu Asn Ile Lys Asp Ile Tyr Ile His
1 5 10
<210> 75
<211> 30
<212> ДНК
<213> Mus musculus
<400> 75
ggttactcat tcaccaacta ctggatacac 30
<210> 76
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 76
Gly Tyr Ser Phe Thr Asn Tyr Trp Ile His
1 5 10
<210> 77
<211> 30
<212> ДНК
<213> Mus musculus
<400> 77
ggattcactt tcagtgacta tcccatgtct 30
<210> 78
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 78
Gly Phe Thr Phe Ser Asp Tyr Pro Met Ser
1 5 10
<210> 79
<211> 30
<212> ДНК
<213> Mus musculus
<400> 79
ggattcactt tcagtagctt tggcatgtct 30
<210> 80
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 80
Gly Phe Thr Phe Ser Ser Phe Gly Met Ser
1 5 10
<210> 81
<211> 30
<212> ДНК
<213> Mus musculus
<400> 81
ggattcactt tcagtaccta tggcatgtct 30
<210> 82
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 82
Gly Phe Thr Phe Ser Thr Tyr Gly Met Ser
1 5 10
<210> 83
<211> 30
<212> ДНК
<213> Mus musculus
<400> 83
ggattcactt tcagtagtta tggcatgtct 30
<210> 84
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 84
Gly Phe Thr Phe Ser Ser Tyr Gly Met Ser
1 5 10
<210> 85
<211> 36
<212> ДНК
<213> Mus musculus
<400> 85
gggttttcac tgaccacttc tggtatgggt gtaggc 36
<210> 86
<211> 12
<212> БЕЛОК
<213> Mus musculus
<400> 86
Gly Phe Ser Leu Thr Thr Ser Gly Met Gly Val Gly
1 5 10
<210> 87
<211> 36
<212> ДНК
<213> Mus musculus
<400> 87
ggattttcac tgagcacttc tggtttgggt gtaggc 36
<210> 88
<211> 12
<212> БЕЛОК
<213> Mus musculus
<400> 88
Gly Phe Ser Leu Ser Thr Ser Gly Leu Gly Val Gly
1 5 10
<210> 89
<211> 24
<212> ДНК
<213> Mus musculus
<400> 89
ggattcacct tcagtgatta ttac 24
<210> 90
<211> 8
<212> БЕЛОК
<213> Mus musculus
<400> 90
Gly Phe Thr Phe Ser Asp Tyr Tyr
1 5
<210> 91
<211> 48
<212> ДНК
<213> Mus musculus
<400> 91
cacatttggt cggatggtga cacgcgctat tacccagccc tgaagaac 48
<210> 92
<211> 16
<212> БЕЛОК
<213> Mus musculus
<400> 92
His Ile Trp Ser Asp Gly Asp Thr Arg Tyr Tyr Pro Ala Leu Lys Asn
1 5 10 15
<210> 93
<211> 36
<212> ДНК
<213> Mus musculus
<400> 93
tacattaatc ctgacactga ttatagtgag tacaat 36
<210> 94
<211> 12
<212> БЕЛОК
<213> Mus musculus
<400> 94
Tyr Ile Asn Pro Asp Thr Asp Tyr Ser Glu Tyr Asn
1 5 10
<210> 95
<211> 36
<212> ДНК
<213> Mus musculus
<400> 95
aggattgatc ctgcgaacgg taagactgca tatgac 36
<210> 96
<211> 12
<212> БЕЛОК
<213> Mus musculus
<400> 96
Arg Ile Asp Pro Ala Asn Gly Lys Thr Ala Tyr Asp
1 5 10
<210> 97
<211> 36
<212> ДНК
<213> Mus musculus
<400> 97
atgattgatc cttccgatgc tgaaactggg ttaaat 36
<210> 98
<211> 12
<212> БЕЛОК
<213> Mus musculus
<400> 98
Met Ile Asp Pro Ser Asp Ala Glu Thr Gly Leu Asn
1 5 10
<210> 99
<211> 24
<212> ДНК
<213> Mus musculus
<400> 99
gttagtgatg gtggtggttc cacc 24
<210> 100
<211> 8
<212> БЕЛОК
<213> Mus musculus
<400> 100
Val Ser Asp Gly Gly Gly Ser Thr
1 5
<210> 101
<211> 24
<212> ДНК
<213> Mus musculus
<400> 101
attagtagtg ctggtagttt cacc 24
<210> 102
<211> 8
<212> БЕЛОК
<213> Mus musculus
<400> 102
Ile Ser Ser Ala Gly Ser Phe Thr
1 5
<210> 103
<211> 51
<212> ДНК
<213> Mus musculus
<400> 103
accattaata ctaatggtgg taccacctat tatcgagaca gtgtgaaggg c 51
<210> 104
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 104
Thr Ile Asn Thr Asn Gly Gly Thr Thr Tyr Tyr Arg Asp Ser Val Lys
1 5 10 15
Gly
<210> 105
<211> 51
<212> ДНК
<213> Mus musculus
<400> 105
accataaata ctaatggtgg taacacctat tattcagaca atgtgaaggg c 51
<210> 106
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 106
Thr Ile Asn Thr Asn Gly Gly Asn Thr Tyr Tyr Ser Asp Asn Val Lys
1 5 10 15
Gly
<210> 107
<211> 51
<212> ДНК
<213> Mus musculus
<400> 107
accattagta ctaatggtgc caccgccaat tatccagaca gtgtgaaggg c 51
<210> 108
<211> 17
<212> БЕЛОК
<213> Mus musculus
<400> 108
Thr Ile Ser Thr Asn Gly Ala Thr Ala Asn Tyr Pro Asp Ser Val Lys
1 5 10 15
Gly
<210> 109
<211> 48
<212> ДНК
<213> Mus musculus
<400> 109
cacatttggt gggatgatga taagtactat aatccatccc tgaagagc 48
<210> 110
<211> 16
<212> БЕЛОК
<213> Mus musculus
<400> 110
His Ile Trp Trp Asp Asp Asp Lys Tyr Tyr Asn Pro Ser Leu Lys Ser
1 5 10 15
<210> 111
<211> 48
<212> ДНК
<213> Mus musculus
<400> 111
cacatttggt gggatgatga taagtactat aatccatccc ttaagaga 48
<210> 112
<211> 16
<212> БЕЛОК
<213> Mus musculus
<400> 112
His Ile Trp Trp Asp Asp Asp Lys Tyr Tyr Asn Pro Ser Leu Lys Arg
1 5 10 15
<210> 113
<211> 30
<212> ДНК
<213> Mus musculus
<400> 113
attagaaatc gggctaatgg ttacacaaca 30
<210> 114
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 114
Ile Arg Asn Arg Ala Asn Gly Tyr Thr Thr
1 5 10
<210> 115
<211> 30
<212> ДНК
<213> Mus musculus
<400> 115
atgaaggatg atagtcttta ctttgacaac 30
<210> 116
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 116
Met Lys Asp Asp Ser Leu Tyr Phe Asp Asn
1 5 10
<210> 117
<211> 30
<212> ДНК
<213> Mus musculus
<400> 117
gcaagtgctg gttattattt ttttgacttc 30
<210> 118
<211> 10
<212> БЕЛОК
<213> Mus musculus
<400> 118
Ala Ser Ala Gly Tyr Tyr Phe Phe Asp Phe
1 5 10
<210> 119
<211> 15
<212> ДНК
<213> Mus musculus
<400> 119
ggtagggggg cccac 15
<210> 120
<211> 5
<212> БЕЛОК
<213> Mus musculus
<400> 120
Gly Arg Gly Ala His
1 5
<210> 121
<211> 33
<212> ДНК
<213> Mus musculus
<400> 121
gcaagaattg gcgattacta taatatggac tac 33
<210> 122
<211> 11
<212> БЕЛОК
<213> Mus musculus
<400> 122
Ala Arg Ile Gly Asp Tyr Tyr Asn Met Asp Tyr
1 5 10
<210> 123
<211> 45
<212> ДНК
<213> Mus musculus
<400> 123
acaagacatg cttcctacta tagctacgac cattctatgg actac 45
<210> 124
<211> 15
<212> БЕЛОК
<213> Mus musculus
<400> 124
Thr Arg His Ala Ser Tyr Tyr Ser Tyr Asp His Ser Met Asp Tyr
1 5 10 15
<210> 125
<211> 36
<212> ДНК
<213> Mus musculus
<400> 125
gcaagacggg ggtacgacgt tgggtgcttt gaccac 36
<210> 126
<211> 12
<212> БЕЛОК
<213> Mus musculus
<400> 126
Ala Arg Arg Gly Tyr Asp Val Gly Cys Phe Asp His
1 5 10
<210> 127
<211> 27
<212> ДНК
<213> Mus musculus
<400> 127
gcaagagact acggggctat ggactac 27
<210> 128
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 128
Ala Arg Asp Tyr Gly Ala Met Asp Tyr
1 5
<210> 129
<211> 27
<212> ДНК
<213> Mus musculus
<400> 129
gcaactgaaa agggagctat gggctac 27
<210> 130
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 130
Ala Thr Glu Lys Gly Ala Met Gly Tyr
1 5
<210> 131
<211> 57
<212> ДНК
<213> Mus musculus
<400> 131
gctcgaagaa ctgagactat gattacgaca gtgctatatt actatgctat ggactac 57
<210> 132
<211> 19
<212> БЕЛОК
<213> Mus musculus
<400> 132
Ala Arg Arg Thr Glu Thr Met Ile Thr Thr Val Leu Tyr Tyr Tyr Ala
1 5 10 15
Met Asp Tyr
<210> 133
<211> 54
<212> ДНК
<213> Mus musculus
<400> 133
gctcgaagga gggaagttaa cttcggtatt aactattact attctatgga ctac 54
<210> 134
<211> 18
<212> БЕЛОК
<213> Mus musculus
<400> 134
Ala Arg Arg Arg Glu Val Asn Phe Gly Ile Asn Tyr Tyr Tyr Ser Met
1 5 10 15
Asp Tyr
<210> 135
<211> 36
<212> ДНК
<213> Mus musculus
<400> 135
gtaagagatt cctatcacta cgggtacttc gatgtc 36
<210> 136
<211> 12
<212> БЕЛОК
<213> Mus musculus
<400> 136
Val Arg Asp Ser Tyr His Tyr Gly Tyr Phe Asp Val
1 5 10
<210> 137
<211> 324
<212> ДНК
<213> Mus musculus
<400> 137
gacatcttgc tgactcagtc tccagccatc ctgtctgtga gtccaggaaa aagagtcagt 60
ttctcctgca gggccagtca gaacattggc acaagcatac actggtatca gcaaagaaca 120
aatggttctc caaggcttct cataaagtat gcttctaagt ctatatctgg gatttcttcc 180
aggtttagtg gcagtggctc agggacagat tttactctta gtatcaacag tgtggagtct 240
gaagatattg cagcttatta ctgtcaacat agttatagct tcccgtggac gttcggtgga 300
ggcaccaagc tggaaatcaa acgg 324
<210> 138
<211> 108
<212> БЕЛОК
<213> Mus musculus
<400> 138
Asp Ile Leu Leu Thr Gln Ser Pro Ala Ile Leu Ser Val Ser Pro Gly
1 5 10 15
Lys Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Asn Ile Gly Thr Ser
20 25 30
Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Lys Ser Ile Ser Gly Ile Ser Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser
65 70 75 80
Glu Asp Ile Ala Ala Tyr Tyr Cys Gln His Ser Tyr Ser Phe Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
100 105
<210> 139
<211> 324
<212> ДНК
<213> Mus musculus
<400> 139
gacatccaga tgactcagtc tccagcctcc ctatctacat ctgtgggaga aactgtcacc 60
atcacatgtc gagcaagtga aaatatttac agttatttag catggtatca gcagagacag 120
ggaaaatctc ctcacctcct ggtcaataat gcaaaaacct tagcagaagg tgtgccatca 180
aggttcagtg gcagtggatc aggcacacat ttttctctga ggatcagcgg cctgcagcct 240
gaagattttg ggagttatta ctgtcagcat cattatggta ttccgttcac gttcggaggg 300
gggaccaagt tgtcaataaa acgg 324
<210> 140
<211> 108
<212> БЕЛОК
<213> Mus musculus
<400> 140
Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ser Thr Ser Val Gly
1 5 10 15
Glu Thr Val Thr Ile Thr Cys Arg Ala Ser Glu Asn Ile Tyr Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Arg Gln Gly Lys Ser Pro His Leu Leu Val
35 40 45
Asn Asn Ala Lys Thr Leu Ala Glu Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr His Phe Ser Leu Arg Ile Ser Gly Leu Gln Pro
65 70 75 80
Glu Asp Phe Gly Ser Tyr Tyr Cys Gln His His Tyr Gly Ile Pro Phe
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Ser Ile Lys Arg
100 105
<210> 141
<211> 339
<212> ДНК
<213> Mus musculus
<400> 141
gatgttgtga tgacccagat tccactcact ttgtcggtta ccattggaca accagcctcc 60
atctcttgca agtcaagtca gagcctcttt gatattgatg gaaagacata tttgaattgg 120
ttgttacaga ggccaggcca gtctccaaag cgcctaatct atctggtgtc tgaattggac 180
tctggagtcc ctgacaggtt cactggcagt ggatcaggga cagatttcac actgaaaatc 240
agcagagtgg aggctgagga tttgggagtt tactattgtt ggcaaggtac acattttccg 300
ctcacgttcg gtgctgggac caagctggag ctgaaacgg 339
<210> 142
<211> 113
<212> БЕЛОК
<213> Mus musculus
<400> 142
Asp Val Val Met Thr Gln Ile Pro Leu Thr Leu Ser Val Thr Ile Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Phe Asp Ile
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Leu Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Lys Arg Leu Ile Tyr Leu Val Ser Glu Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Trp Gln Gly
85 90 95
Thr His Phe Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105 110
Arg
<210> 143
<211> 324
<212> ДНК
<213> Mus musculus
<400> 143
gacatccaga tgacccagtc tccatcctcc ttatctgcct ctctgggaga aagagtcagt 60
ctcacttgtc gggcaagtca ggacattggt ggtagcttaa actggcttca gcagaaacca 120
gatggaacta ttaaacgcct gatctacgcc acatccagct tagattctgg tgtccccaaa 180
aggttcagtg gcagtaggtc tgggtcagtt ttttctctca ccatcaccag ccttgagtct 240
gaagattttg tagactattt ctgtctacaa tatgctagtt ctccgtatac gttcggaggg 300
gggaccaagc tggaaataaa acgg 324
<210> 144
<211> 108
<212> БЕЛОК
<213> Mus musculus
<400> 144
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Ser Leu Thr Cys Arg Ala Ser Gln Asp Ile Gly Gly Ser
20 25 30
Leu Asn Trp Leu Gln Gln Lys Pro Asp Gly Thr Ile Lys Arg Leu Ile
35 40 45
Tyr Ala Thr Ser Ser Leu Asp Ser Gly Val Pro Lys Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Ser Val Phe Ser Leu Thr Ile Thr Ser Leu Glu Ser
65 70 75 80
Glu Asp Phe Val Asp Tyr Phe Cys Leu Gln Tyr Ala Ser Ser Pro Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
100 105
<210> 145
<211> 324
<212> ДНК
<213> Mus musculus
<400> 145
gacattgtga tgactcagtc tccagccacc ctgtctgtga ctccaggaga tagagtctct 60
ctttcctgca gggccagcca gactattagc gacttcttac actggtatca acaaaaatca 120
catgagtctc caaggcttct catcaaatat gcttcccaat ccatctctgg gatcccctcc 180
aggttcagtg gcactggatc agggtcagat ttcactctca ctatcaacag tgtggaacct 240
gaagatgttg gagtgtatta ctgtcaaagt ggtaacacct ttccgtggac gttcggtgga 300
ggcaccaagc tggaaatcaa acgg 324
<210> 146
<211> 108
<212> БЕЛОК
<213> Mus musculus
<400> 146
Asp Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Val Thr Pro Gly
1 5 10 15
Asp Arg Val Ser Leu Ser Cys Arg Ala Ser Gln Thr Ile Ser Asp Phe
20 25 30
Leu His Trp Tyr Gln Gln Lys Ser His Glu Ser Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Thr Gly Ser Gly Ser Asp Phe Thr Leu Thr Ile Asn Ser Val Glu Pro
65 70 75 80
Glu Asp Val Gly Val Tyr Tyr Cys Gln Ser Gly Asn Thr Phe Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
100 105
<210> 147
<211> 324
<212> ДНК
<213> Mus musculus
<400> 147
gacatccaga tgacacaatc ttcatcctcc ttttctggat ttctaggaga cagagtcacc 60
attacttgca gggcaagtga ggacatacac actcaattag cctggtatca gcagaaacca 120
ggaaatgctc ctaggctctt aatatctggt gcagccagtt tgaaaagtgg ggttccttca 180
agattcagtg gcactggatc tggaaaggat tacactctca gcattaccag tcttcagact 240
gaagatgttg ctacatatta ctgtcaacag tataggagta ttccgtggac gttcggtgga 300
ggcaccaagc tggaaatcaa acgg 324
<210> 148
<211> 108
<212> БЕЛОК
<213> Mus musculus
<400> 148
Asp Ile Gln Met Thr Gln Ser Ser Ser Ser Phe Ser Gly Phe Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Asp Ile His Thr Gln
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Asn Ala Pro Arg Leu Leu Ile
35 40 45
Ser Gly Ala Ala Ser Leu Lys Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Thr Gly Ser Gly Lys Asp Tyr Thr Leu Ser Ile Thr Ser Leu Gln Thr
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Gln Tyr Arg Ser Ile Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
100 105
<210> 149
<211> 339
<212> ДНК
<213> Mus musculus
<400> 149
gatgttttga tgacccaaac tccgctctcc ctgcctgtca gtcttggaga tcacgcctcc 60
atctcttgca gatctagtca gtacattgtt catagtactg gaaccaccta tttagaatgg 120
tacctacaga aaccaggcca gtctccacag ctcctgatct acaaagtttc caaccgattt 180
tctggggtcc cagacaggtt cactggcagt ggatcaggga cagatttcac actcaggatc 240
agcagagtgg aggctgagga tctgggagtt tatttctgct ttcaaggttc acattttcca 300
ttcacgttcg gctcggggac aaagttggaa ataaaacgg 339
<210> 150
<211> 113
<212> БЕЛОК
<213> Mus musculus
<400> 150
Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp His Ala Ser Ile Ser Cys Arg Ser Ser Gln Tyr Ile Val His Ser
20 25 30
Thr Gly Thr Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Arg Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Phe Cys Phe Gln Gly
85 90 95
Ser His Phe Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg
<210> 151
<211> 339
<212> ДНК
<213> Mus musculus
<400> 151
gaagttgtga tgacccaaac tccactctcc ttgcctgtca gtcttggaga tcaagcctcc 60
atctcttgca gatctagtca ttaccttgtt catgataacg gaaacaccta tgttgaatgg 120
tacctgcaga agccaggcca gtctccaaag ctcctgatct acaaggtttc caaccgattt 180
tctggagtcc cagacaggtt tactggcagt ggttcaggga cagatttcac actcaagatc 240
agcagagtgg agtctgagga tctgggaatt tattactgct ttcaaggttc acatttccca 300
ttcacgttcg gctcggggac agagttggaa ataaaacgg 339
<210> 152
<211> 113
<212> БЕЛОК
<213> Mus musculus
<400> 152
Glu Val Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser His Tyr Leu Val His Asp
20 25 30
Asn Gly Asn Thr Tyr Val Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ser Glu Asp Leu Gly Ile Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Phe Pro Phe Thr Phe Gly Ser Gly Thr Glu Leu Glu Ile Lys
100 105 110
Arg
<210> 153
<211> 339
<212> ДНК
<213> Mus musculus
<400> 153
gatgttttga tgacccaaac tccactctcc ctgcctgtca gtcttggaga tcaagcctcc 60
atctcttgca gatctagtca gaacattgtc catagtactg gaaacaccta tttagaatgg 120
tacctgcaga aaccaggcca gtctccaaag ctcctgattt ataaagtttc caaccgattt 180
tctggggtcc caaacaggtt ccgtggcagt ggatcaggga cagatttcac actcaagatc 240
accagagtgg aggctgagga tctgggaatt tattactgct ttcaaggttc acattttcca 300
ttcacgttcg gctcggggac aaagttggaa ataaaacgg 339
<210> 154
<211> 113
<212> БЕЛОК
<213> Mus musculus
<400> 154
Asp Val Leu Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Asn Ile Val His Ser
20 25 30
Thr Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asn Arg Phe Arg Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Thr Arg Val Glu Ala Glu Asp Leu Gly Ile Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Phe Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg
<210> 155
<211> 321
<212> ДНК
<213> Mus musculus
<400> 155
caaattgttc tcacccagtc tccagcaatc atgtctgcat ctccagggga gaaggtcacc 60
atgacctgca gtgtcagctc aagtgtaagt tacatacact ggtaccaaca gaagtcaggc 120
acctccccca aaagatggat ttatgacaca tccaaactgg cttctggagt ccctgctcgc 180
ttcagtggca gtgggtctgg gacctcttac tctctcacaa tcagcagcat ggaggctgaa 240
gatgctgcca cttattactg ccaccagtgg agtactaacc cacccacgtt cggagggggg 300
accaagctgg aaataagacg g 321
<210> 156
<211> 107
<212> БЕЛОК
<213> Mus musculus
<400> 156
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Val Ser Ser Ser Val Ser Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys His Gln Trp Ser Thr Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Arg Arg
100 105
<210> 157
<211> 321
<212> ДНК
<213> Mus musculus
<400> 157
caaattgttc tcacccagtc tccagcactc atgtctgcat ctccagggga gaaggtcacc 60
atgacctgca gtgccagctc aagtgtaagt tacatgtgct ggtaccagca gaagccaaga 120
tcctccccca aaccctggat ttatctcaca tccaacctgg cttctggagt ccctgctcgc 180
ttcagtggca gtgggtctgg gacctcttac tctctcacaa tcagtagcat ggaggctgaa 240
gatgctgcca cttattactg ccagcagtgg agtagtaacc cacccacgtt cggtgctggg 300
accaagctgg agctgaaacg g 321
<210> 158
<211> 107
<212> БЕЛОК
<213> Mus musculus
<400> 158
Gln Ile Val Leu Thr Gln Ser Pro Ala Leu Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Cys Trp Tyr Gln Gln Lys Pro Arg Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Leu Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Pro Thr
85 90 95
Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg
100 105
<210> 159
<211> 321
<212> ДНК
<213> Mus musculus
<400> 159
gaaatccaga tgacacagac tccatcctcc ctgtctgcct ctctgggaga cagagtcacc 60
atcacttgca gtgcaagtca gggcattaac aattatttga actggtatca gcagaaacca 120
ggtggaaaga ctagactcct catctattat acatcaactt tacagtcagg agtcccatca 180
aggttcagtg gcagtgggtc tgggacacat tattctctca ccatcagcaa tctggaacct 240
gaagatattg ccacttacta ttgtcagcag tttagtaaac ttcggacatt cggtggaggc 300
accaggctgg aaatcaaacg g 321
<210> 160
<211> 107
<212> БЕЛОК
<213> Mus musculus
<400> 160
Glu Ile Gln Met Thr Gln Thr Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Gln Gly Ile Asn Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gly Lys Thr Arg Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr His Tyr Ser Leu Thr Ile Ser Asn Leu Glu Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Phe Ser Lys Leu Arg Thr
85 90 95
Phe Gly Gly Gly Thr Arg Leu Glu Ile Lys Arg
100 105
<210> 161
<211> 324
<212> ДНК
<213> Homo sapiens
<400> 161
gagattgtgc tgactcagag tccagacttc cagtcagtga cccccaagga gaaagtcacc 60
atcacatgcc gggcaagcca gaacatcggc acaagcattc actggtacca gcagaagccc 120
gatcagtccc ctaagctgct gatcaaatat gcctctaaga gtatttcagg ggtgccctct 180
agattcagcg gctccgggtc tggaacagac tttactctga ccattaactc cctggaggct 240
gaagatgccg ctacttacta ttgtcagcat agctactcat tcccttggac attcgggcag 300
gggaccaaag tggaaatcaa acgt 324
<210> 162
<211> 108
<212> БЕЛОК
<213> Homo sapiens
<400> 162
Glu Ile Val Leu Thr Gln Ser Pro Asp Phe Gln Ser Val Thr Pro Lys
1 5 10 15
Glu Lys Val Thr Ile Thr Cys Arg Ala Ser Gln Asn Ile Gly Thr Ser
20 25 30
Ile His Trp Tyr Gln Gln Lys Pro Asp Gln Ser Pro Lys Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Lys Ser Ile Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln His Ser Tyr Ser Phe Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
100 105
<210> 163
<211> 339
<212> ДНК
<213> Homo sapiens
<400> 163
gatattgtga tgacccaaac tccgctctcc ctgtccgtca cccctggaca gccggcctcc 60
atctcttgca gatctagtca gaacattgtt catagtactg gaaacaccta tttagaatgg 120
tacctacaga aaccaggcca gtctccacag ctcctgatct acaaagtttc caaccgattt 180
tctggggtcc cagacaggtt cagtggcagt ggatcaggga cagatttcac actcaaaatc 240
agcagagtgg aggctgagga tgttggagtt tattactgct ttcaaggttc acattttcca 300
ttcacgttcg gccaagggac caaggtggaa atcaaacgt 339
<210> 164
<211> 113
<212> БЕЛОК
<213> Homo sapiens
<400> 164
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Asn Ile Val His Ser
20 25 30
Thr Gly Asn Thr Tyr Leu Glu Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Phe Gln Gly
85 90 95
Ser His Phe Pro Phe Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg
<210> 165
<211> 360
<212> ДНК
<213> Mus musculus
<400> 165
caggttactc tgaaagagtc tggccctggg atattgcagc cctcccagac cctcagtctg 60
acttgttctt tctctgggtt ctcactgacc acttctggct tgggtgttgc ctggattcgt 120
cagccttcag ggaagggtct ggagtggctg gcacacattt ggtcggatgg tgacacgcgc 180
tattacccag ccctgaagaa ccgactgaca atctccaagg attcctccag caaccaggtc 240
ttcctcaaga tcgcccgtgt ggacactgca gatactgcca catactactg tgctcgaatg 300
aaggatgata gtctttactt tgacaactgg ggccaaggca ctattttcac agtctcctca 360
<210> 166
<211> 120
<212> БЕЛОК
<213> Mus musculus
<400> 166
Gln Val Thr Leu Lys Glu Ser Gly Pro Gly Ile Leu Gln Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Phe Ser Gly Phe Ser Leu Thr Thr Ser
20 25 30
Gly Leu Gly Val Ala Trp Ile Arg Gln Pro Ser Gly Lys Gly Leu Glu
35 40 45
Trp Leu Ala His Ile Trp Ser Asp Gly Asp Thr Arg Tyr Tyr Pro Ala
50 55 60
Leu Lys Asn Arg Leu Thr Ile Ser Lys Asp Ser Ser Ser Asn Gln Val
65 70 75 80
Phe Leu Lys Ile Ala Arg Val Asp Thr Ala Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Arg Met Lys Asp Asp Ser Leu Tyr Phe Asp Asn Trp Gly Gln
100 105 110
Gly Thr Ile Phe Thr Val Ser Ser
115 120
<210> 167
<211> 351
<212> ДНК
<213> Mus musculus
<400> 167
cagatccagt tggtgcagtc tggggctgaa ctggcaaaac ctggggcctc agtgaggatg 60
tcctgcgaga cttctggcta cacctttact agctactgga tacactggat aaaagagagg 120
cctggacagg gtctggaatg gattggatac attaatcctg acactgatta tagtgagtac 180
aatcagaaat tcaaggacaa ggccagattg actgcagaca aatcctccac cacagcctac 240
atggagctga acagcctgac atttgatgat tctgcagtct attactgtgc aagtgctggt 300
tattattttt ttgacttctg gggccaaggc accactctca cagtctcctc a 351
<210> 168
<211> 117
<212> БЕЛОК
<213> Mus musculus
<400> 168
Gln Ile Gln Leu Val Gln Ser Gly Ala Glu Leu Ala Lys Pro Gly Ala
1 5 10 15
Ser Val Arg Met Ser Cys Glu Thr Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Ile His Trp Ile Lys Glu Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Asp Thr Asp Tyr Ser Glu Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Arg Leu Thr Ala Asp Lys Ser Ser Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Asn Ser Leu Thr Phe Asp Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Ala Gly Tyr Tyr Phe Phe Asp Phe Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser
115
<210> 169
<211> 336
<212> ДНК
<213> Mus musculus
<400> 169
gaggttcagc tgcagcagtc tggggcagaa cttgtgaaac caggggcctc agtcaagttg 60
tcctgtacaa cttctggcct caacattaaa gacatctata ttcactgggt gaagcagagg 120
cctgaacagg gcctggagtg gattgggagg attgatcctg cgaacggtaa gactgcatat 180
gacctgaagt tccaggccaa ggccactata acagcagaca catcttccaa aacagcctac 240
ctgcacctca gcagcctgac atctgaggac actgccgtct attactgtgg taggggggcc 300
cactggggcc aaggcaccac tctcacagtc tcctca 336
<210> 170
<211> 112
<212> БЕЛОК
<213> Mus musculus
<400> 170
Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Thr Thr Ser Gly Leu Asn Ile Lys Asp Ile
20 25 30
Tyr Ile His Trp Val Lys Gln Arg Pro Glu Gln Gly Leu Glu Trp Ile
35 40 45
Gly Arg Ile Asp Pro Ala Asn Gly Lys Thr Ala Tyr Asp Leu Lys Phe
50 55 60
Gln Ala Lys Ala Thr Ile Thr Ala Asp Thr Ser Ser Lys Thr Ala Tyr
65 70 75 80
Leu His Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Gly Arg Gly Ala His Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
100 105 110
<210> 171
<211> 354
<212> ДНК
<213> Mus musculus
<400> 171
cagatccagt tggtgcagtc tgggcctcag ctggttaggc ctggggcttc agtgaagata 60
tcctgcgagg cttctggtta ctcattcacc aactactgga tacactgggt gaagcagagg 120
cctggacagg gtcttgagtg gattggcatg attgatcctt ccgatgctga aactgggtta 180
aatcagaagt tcaaggacaa ggccacattg actgtagaca aatcctccag cacagcctac 240
atgcaactca gcagcccgac atctgaagac tctgcggtct attactgtgc aagaattggc 300
gattactata atatggacta ctggggtcaa ggaacctcag tcaccgtctc ctca 354
<210> 172
<211> 118
<212> БЕЛОК
<213> Mus musculus
<400> 172
Gln Ile Gln Leu Val Gln Ser Gly Pro Gln Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Glu Ala Ser Gly Tyr Ser Phe Thr Asn Tyr
20 25 30
Trp Ile His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Met Ile Asp Pro Ser Asp Ala Glu Thr Gly Leu Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Pro Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ile Gly Asp Tyr Tyr Asn Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser
115
<210> 173
<211> 366
<212> ДНК
<213> Mus musculus
<400> 173
gaagtgaagg tggtggagtc tgggggaggt ttagtgcagc ctggagggtc cctgaaactc 60
tcctgtgcag cctctggatt cactttcagt gactatccca tgtcttgggt tcgccagact 120
ccagagaaga gactggagtg ggtcgcatac gttagtgatg gtggtggttc cacctactat 180
ccagacattg taaagggccg attcaccatc tcccgagaca atgccaagaa caccctgtac 240
cttcaaatga gcagtctgaa gtctgaggac acggccatgt atttctgtac aagacatgct 300
tcctactata gctacgacca ttctatggac tactggggtc agggaacctc agtcaccgtc 360
tcatca 366
<210> 174
<211> 122
<212> БЕЛОК
<213> Mus musculus
<400> 174
Glu Val Lys Val Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Pro Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Tyr Val Ser Asp Gly Gly Gly Ser Thr Tyr Tyr Pro Asp Ile Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Lys Ser Glu Asp Thr Ala Met Tyr Phe Cys
85 90 95
Thr Arg His Ala Ser Tyr Tyr Ser Tyr Asp His Ser Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 175
<211> 357
<212> ДНК
<213> Mus musculus
<400> 175
cagatccagt tggtgcagtc tgggggagac ttagtgaggc ctggagggtc cctgaaactc 60
tcctgtgcag cctctggatt cactttcagt agctttggca tgtcttggat tcgccagact 120
ccagacaaga ggctggagtg ggtcgcaacc attagtagtg ctggtagttt cacctactat 180
ccagacagtg tgaagggccg attcaccatc tccagagaca atgccaggaa caccctgtat 240
ctacaaatga acagtctgaa gtctgaggac tcagccatgt attactgtgc aagacggggg 300
tacgacgttg ggtgctttga ccactggggc cgaggcacca ctctcacagt ctcctca 357
<210> 176
<211> 119
<212> БЕЛОК
<213> Mus musculus
<400> 176
Gln Ile Gln Leu Val Gln Ser Gly Gly Asp Leu Val Arg Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Gly Met Ser Trp Ile Arg Gln Thr Pro Asp Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Ser Ala Gly Ser Phe Thr Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Arg Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Ser Glu Asp Ser Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Tyr Asp Val Gly Cys Phe Asp His Trp Gly Arg Gly
100 105 110
Thr Thr Leu Thr Val Ser Ser
115
<210> 177
<211> 348
<212> ДНК
<213> Mus musculus
<400> 177
gaggtgcacc tggtggagtc tgggggaggc ttagtgcagc ctggagggtc cctgaaactc 60
tcctgtgcag cctctggatt cactttcagt acctatggca tgtcttgggt tcgccagact 120
ccagacaaga ggctggagtt ggtcgcgacc attaatacta atggtggtac cacctattat 180
cgagacagtg tgaagggccg attcaccatc tccagagaca atgccaagaa caccctgtac 240
ctgcaaatga gcagtctgaa gtctgatgac acagccatgt attactgtgc aagagactac 300
ggggctatgg actactgggg tcaaggaacc tcagtcaccg tctcctca 348
<210> 178
<211> 116
<212> БЕЛОК
<213> Mus musculus
<400> 178
Glu Val His Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Thr Pro Asp Lys Arg Leu Glu Leu Val
35 40 45
Ala Thr Ile Asn Thr Asn Gly Gly Thr Thr Tyr Tyr Arg Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Lys Ser Asp Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Asp Tyr Gly Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser Val
100 105 110
Thr Val Ser Ser
115
<210> 179
<211> 345
<212> ДНК
<213> Mus musculus
<400> 179
gatgtgcacc tggtggagtc tgggggaggc ttagtgcagc ctggagggtc cctgacagtc 60
tcctgcgcag cctctggatt cactttcagt acctatggca tgtcttgggt tcgccagact 120
cgagacaaga ggctggagtt ggtcgcaacc ataaatacta atggtggtaa cacctattat 180
tcagacaatg tgaagggccg attcaccatt tccagagaca atgccaagaa caccctgtat 240
ttggaaatga gaggtctgag gtctggggac acagccatgt attactgtgc aagagactac 300
ggggctatgg actactgggg tcaaggaacc tcagtcaccg tctct 345
<210> 180
<211> 115
<212> БЕЛОК
<213> Mus musculus
<400> 180
Asp Val His Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Thr Val Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Thr Arg Asp Lys Arg Leu Glu Leu Val
35 40 45
Ala Thr Ile Asn Thr Asn Gly Gly Asn Thr Tyr Tyr Ser Asp Asn Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Glu Met Arg Gly Leu Arg Ser Gly Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Asp Tyr Gly Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser Val
100 105 110
Thr Val Ser
115
<210> 181
<211> 348
<212> ДНК
<213> Mus musculus
<400> 181
gaggtgcagc tgcagcagcc tgggggaggc ttagtacagc ctggagggtc cctgacactc 60
tcctgtgcaa cctctggatt cactttcagt agttatggca tgtcttgggt tcgccagact 120
ccagccaaga ggctggagtt ggtcgcaacc attagtacta atggtgccac cgccaattat 180
ccagacagtg tgaagggccg attcaccatc tccagagaca atgccaagag caccctgtac 240
ctacaaatgc gcagtctgaa gtctgaggac acagccatgt attactgtgc aactgaaaag 300
ggagctatgg gctactgggg tcaaggaacc tcagtcaccg tctcctca 348
<210> 182
<211> 116
<212> БЕЛОК
<213> Mus musculus
<400> 182
Glu Val Gln Leu Gln Gln Pro Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Thr Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Thr Pro Ala Lys Arg Leu Glu Leu Val
35 40 45
Ala Thr Ile Ser Thr Asn Gly Ala Thr Ala Asn Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Arg Ser Leu Lys Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Thr Glu Lys Gly Ala Met Gly Tyr Trp Gly Gln Gly Thr Ser Val
100 105 110
Thr Val Ser Ser
115
<210> 183
<211> 381
<212> ДНК
<213> Mus musculus
<400> 183
caagttactc taaaagagtc tggccctggg atattgaagc cctcacagac cctcagtctg 60
acttgttctt tctctgggtt ttcactgacc acttctggta tgggtgtagg ctggattcgt 120
cagccttcag ggaagggtct ggagtggctg gcacacattt ggtgggatga tgataagtac 180
tataatccat ccctgaagag ccaggtcaca atctccaagg acacctccag aaaccaggta 240
ttcctcaaga tcaccagtgt ggacactgca gatactgcca cttactactg tgctcgaaga 300
actgagacta tgattacgac agtgctatat tactatgcta tggactactg gggtcaagga 360
acctcagtca ccgtctcctc a 381
<210> 184
<211> 127
<212> БЕЛОК
<213> Mus musculus
<400> 184
Gln Val Thr Leu Lys Glu Ser Gly Pro Gly Ile Leu Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Phe Ser Gly Phe Ser Leu Thr Thr Ser
20 25 30
Gly Met Gly Val Gly Trp Ile Arg Gln Pro Ser Gly Lys Gly Leu Glu
35 40 45
Trp Leu Ala His Ile Trp Trp Asp Asp Asp Lys Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Gln Val Thr Ile Ser Lys Asp Thr Ser Arg Asn Gln Val
65 70 75 80
Phe Leu Lys Ile Thr Ser Val Asp Thr Ala Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Arg Arg Thr Glu Thr Met Ile Thr Thr Val Leu Tyr Tyr Tyr
100 105 110
Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser
115 120 125
<210> 185
<211> 378
<212> ДНК
<213> Mus musculus
<400> 185
caagttactc taaaagagtc tggccctggg atattgaagc cctcacagac cctcagtctg 60
acttgttctt tctctggatt ttcactgagc acttctggtt tgggtgtagg ctggattcgt 120
cagccttcag ggaagggtct ggagtggctg gcacacattt ggtgggatga tgataagtac 180
tataatccat cccttaagag acagatcaca atctccaagg attcctccag aaaccaggta 240
ttcctcaaga tcaccaatgt ggacactgca gatactgcca cttactactg tgctcgaagg 300
agggaagtta acttcggtat taactattac tattctatgg actactgggg tcaaggaacc 360
tcagtcaccg tctcctca 378
<210> 186
<211> 126
<212> БЕЛОК
<213> Mus musculus
<400> 186
Gln Val Thr Leu Lys Glu Ser Gly Pro Gly Ile Leu Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Phe Ser Gly Phe Ser Leu Ser Thr Ser
20 25 30
Gly Leu Gly Val Gly Trp Ile Arg Gln Pro Ser Gly Lys Gly Leu Glu
35 40 45
Trp Leu Ala His Ile Trp Trp Asp Asp Asp Lys Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Arg Gln Ile Thr Ile Ser Lys Asp Ser Ser Arg Asn Gln Val
65 70 75 80
Phe Leu Lys Ile Thr Asn Val Asp Thr Ala Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Arg Arg Arg Glu Val Asn Phe Gly Ile Asn Tyr Tyr Tyr Ser
100 105 110
Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser
115 120 125
<210> 187
<211> 363
<212> ДНК
<213> Mus musculus
<400> 187
gaggtgaagc tggtggagtc tggaggacgc ttggtacagc ctgggaattc tctgagactc 60
tcctgtgcaa cttctggatt caccttcagt gattattaca tgagttgggt ccgccagact 120
ccaggaaggg cacttgagtg gttgagtttt attagaaatc gggctaatgg ttacacaaca 180
gagtacagtg catctgtgaa gggtcgattc accatctcca gagataattc ccaaagcatc 240
ctctatcttc acatgagcac cctgagacct gaggacagtg ccacttatta ctgtgtaaga 300
gattcctatc actacgggta cttcgatgtc tggggcgcag ggaccacggt caccgtctcc 360
tca 363
<210> 188
<211> 121
<212> БЕЛОК
<213> Mus musculus
<400> 188
Glu Val Lys Leu Val Glu Ser Gly Gly Arg Leu Val Gln Pro Gly Asn
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Thr Pro Gly Arg Ala Leu Glu Trp Leu
35 40 45
Ser Phe Ile Arg Asn Arg Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gln Ser Ile
65 70 75 80
Leu Tyr Leu His Met Ser Thr Leu Arg Pro Glu Asp Ser Ala Thr Tyr
85 90 95
Tyr Cys Val Arg Asp Ser Tyr His Tyr Gly Tyr Phe Asp Val Trp Gly
100 105 110
Ala Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 189
<211> 360
<212> ДНК
<213> Homo sapiens
<400> 189
caggtgaccc tgaaggaatc cgggcctact ctggtgaaac ctacccagac tctgactctg 60
acttgtactt ttagcggctt ctcactgacc acatctggac tgggagtggc ttggatcaga 120
cagcctcctg gaaaggccct ggagtggctg gctcacattt ggagcgacgg cgatactcgg 180
tactatccag ccctgaaaaa cagactgact atcaccaagg acacatccaa aaaccaggtg 240
gtcctgacaa tgactaatat ggaccccgtc gataccgcaa catactattg cgcccatatg 300
aaggatgact ctctgtactt tgataactgg gggcagggaa ctctggtgac cgtgagcagc 360
<210> 190
<211> 120
<212> БЕЛОК
<213> Homo sapiens
<400> 190
Gln Val Thr Leu Lys Glu Ser Gly Pro Thr Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu Thr Thr Ser
20 25 30
Gly Leu Gly Val Ala Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu
35 40 45
Trp Leu Ala His Ile Trp Ser Asp Gly Asp Thr Arg Tyr Tyr Pro Ala
50 55 60
Leu Lys Asn Arg Leu Thr Ile Thr Lys Asp Thr Ser Lys Asn Gln Val
65 70 75 80
Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala His Met Lys Asp Asp Ser Leu Tyr Phe Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 191
<211> 348
<212> ДНК
<213> Homo sapiens
<400> 191
gaggtgcagc tgctggaatc tgggggggga ctggtgcagc ctggaggaag cctgagactg 60
agttgtgccg caagtgggtt tacatttagc tcctacggaa tgagctgggt gaggcaggct 120
ccaggcaagg gactggagtg ggtctctgca atcagtacca acggagccac agcttactat 180
gccgactccg tgaagggccg gttcactatc tcaagagata acagcaagaa caccctgtat 240
ctgcagatga attctctgcg ggcagaagac acagccgtct actattgcgc tactgagaaa 300
ggggcaatga gccactgggg acagggcaca ctggtgaccg tgagttcc 348
<210> 192
<211> 116
<212> БЕЛОК
<213> Homo sapiens
<400> 192
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Thr Asn Gly Ala Thr Ala Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Glu Lys Gly Ala Met Ser His Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 193
<211> 318
<212> ДНК
<213> Homo sapiens
<400> 193
accgtggccg ccccctccgt gttcatcttc cccccctccg acgagcagct gaagtccggc 60
accgcctccg tggtgtgcct gctgaacaac ttctacccca gggaggccaa ggtgcagtgg 120
aaggtggaca acgccctgca gtccggcaac tcccaggagt ccgtgaccga gcaggactcc 180
aaggactcca cctactccct gtcctccacc ctgaccctgt ccaaggccga ctacgagaag 240
cacaaggtgt acgcctgcga ggtgacccac cagggcctgt cctcccccgt gaccaagtcc 300
ttcaacaggg gcgagtgc 318
<210> 194
<211> 106
<212> БЕЛОК
<213> Homo sapiens
<400> 194
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
1 5 10 15
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
20 25 30
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
35 40 45
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
50 55 60
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
65 70 75 80
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
85 90 95
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 195
<211> 318
<212> ДНК
<213> Homo sapiens
<400> 195
ggacagccaa aggcagcacc atctgtgacc ctgttcccac ctagctccga ggagctgcag 60
gccaacaagg ccaccctggt gtgcctgatc tccgactttt acccaggagc agtgacagtg 120
gcatggaagg ccgattctag ccctgtgaag gccggcgtgg agaccacaac cccatctaag 180
cagagcaaca ataagtacgc cgcctcctct tatctgtccc tgacccccga gcagtggaag 240
tctcaccgga gctattcctg ccaggtgaca cacgagggca gcacagtgga gaagaccgtg 300
gcccctacag agtgttcc 318
<210> 196
<211> 106
<212> БЕЛОК
<213> Homo sapiens
<400> 196
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 197
<211> 978
<212> ДНК
<213> Homo sapiens
<400> 197
gcctccacca agggcccctc cgtgttcccc ctggccccct gctccaggtc cacctccgag 60
tccaccgccg ccctgggctg cctggtgaag gactacttcc ccgagcccgt gaccgtgtcc 120
tggaactccg gcgccctgac ctccggcgtg cacaccttcc ccgccgtgct gcagtcctcc 180
ggcctgtact ccctgtcctc cgtggtgacc gtgccctcct cctccctggg caccaagacc 240
tacacctgca acgtggacca caagccctcc aacaccaagg tggacaagag ggtggagtcc 300
aagtacggcc ccccctgccc cccctgcccc gcccccgagg ccgccggcgg cccctccgtg 360
ttcctgttcc cccccaagcc caaggacacc ctgatgatct ccaggacccc cgaggtgacc 420
tgcgtggtgg tggacgtgtc ccaggaggac cccgaggtgc agttcaactg gtacgtggac 480
ggcgtggagg tgcacaacgc caagaccaag cccagggagg agcagttcaa ctccacctac 540
agggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa ggagtacaag 600
tgcaaggtgt ccaacaaggg cctgccctcc tccatcgaga agaccatctc caaggccaag 660
ggccagccca gggagcccca ggtgtacacc ctgcccccct cccaggagga gatgaccaag 720
aaccaggtgt ccctgacctg cctggtgaag ggcttctacc cctccgacat cgccgtggag 780
tgggagtcca acggccagcc cgagaacaac tacaagacca ccccccccgt gctggactcc 840
gacggctcct tcttcctgta ctccaggctg accgtggaca agtccaggtg gcaggagggc 900
aacgtgttct cctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 960
ctgtccctgt ccctgggc 978
<210> 198
<211> 326
<212> БЕЛОК
<213> Homo sapiens
<400> 198
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly
325
<210> 199
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 199
tttggrggga agatgaagac 20
<210> 200
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 200
ttaacactct cccctgttga a 21
<210> 201
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 201
ttaacactca ttcctgttga a 21
<210> 202
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 202
tggacaggga tccagagttc c 21
<210> 203
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 203
tggacagggc tccatagttc c 21
<210> 204
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 204
actcgtcctt ggtcaacgtg 20
<210> 205
<211> 9
<212> БЕЛОК
<213> Mus musculus
<400> 205
Gln His Ser Tyr Ser Trp Pro Trp Thr
1 5
<210> 206
<211> 326
<212> БЕЛОК
<213> Homo sapiens
<400> 206
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ser Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 207
<211> 137
<212> БЕЛОК
<213> Homo sapiens
<400> 207
Gly Ile Pro Pro His Val Gln Lys Ser Val Asn Asn Asp Met Ile Val
1 5 10 15
Thr Asp Asn Asn Gly Ala Val Lys Phe Pro Gln Leu Cys Lys Phe Cys
20 25 30
Asp Val Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met Ser Asn
35 40 45
Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys Val Ala
50 55 60
Val Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val Cys His
65 70 75 80
Asp Pro Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala Ala Ser
85 90 95
Pro Lys Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr Phe Phe
100 105 110
Met Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile Phe Ser
115 120 125
Glu Glu Tyr Asn Thr Ser Asn Pro Asp
130 135
<210> 208
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 208
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 209
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 209
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 210
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 210
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 211
<211> 214
<212> БЕЛОК
<213> Homo sapiens
<400> 211
Glu Ile Val Leu Thr Gln Ser Pro Asp Phe Gln Ser Val Thr Pro Lys
1 5 10 15
Glu Lys Val Thr Ile Thr Cys Arg Ala Ser Gln Asn Ile Gly Thr Ser
20 25 30
Ile His Trp Tyr Gln Gln Lys Pro Asp Gln Tyr Pro Lys Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Lys Ser Ile Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln His Ser Tyr Ser Phe Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 212
<211> 603
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 212
Gln Val Thr Leu Lys Glu Ser Gly Pro Thr Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu Thr Thr Ser
20 25 30
Gly Leu Gly Val Ala Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu
35 40 45
Trp Leu Ala His Ile Trp Ser Asp Gly Asp Thr Arg Tyr Tyr Pro Ala
50 55 60
Leu Lys Asn Arg Leu Thr Ile Thr Lys Asp Thr Ser Lys Asn Gln Val
65 70 75 80
Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala His Met Lys Asp Asp Ser Leu Tyr Phe Asp Asn Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
210 215 220
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
260 265 270
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly
435 440 445
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
450 455 460
Gly Ser Gly Ile Pro Pro His Val Gln Lys Ser Val Asn Asn Asp Met
465 470 475 480
Ile Val Thr Asp Asn Asn Gly Ala Val Lys Phe Pro Gln Leu Cys Lys
485 490 495
Phe Cys Asp Val Arg Phe Ser Thr Cys Asp Asn Gln Lys Ser Cys Met
500 505 510
Ser Asn Cys Ser Ile Thr Ser Ile Cys Glu Lys Pro Gln Glu Val Cys
515 520 525
Val Ala Val Trp Arg Lys Asn Asp Glu Asn Ile Thr Leu Glu Thr Val
530 535 540
Cys His Asp Pro Lys Leu Pro Tyr His Asp Phe Ile Leu Glu Asp Ala
545 550 555 560
Ala Ser Pro Lys Cys Ile Met Lys Glu Lys Lys Lys Pro Gly Glu Thr
565 570 575
Phe Phe Met Cys Ser Cys Ser Ser Asp Glu Cys Asn Asp Asn Ile Ile
580 585 590
Phe Ser Glu Glu Tyr Asn Thr Ser Asn Pro Asp
595 600
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА К PD-1 СОБАК | 2014 |
|
RU2732604C2 |
| АНТИТЕЛО К АЛЬФА-РЕЦЕПТОРУ ИНТЕРЛЕЙКИНА-4 ЧЕЛОВЕКА, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2807060C1 |
| АНТИТЕЛА ПРОТИВ СОБАЧЬЕГО CTLA-4 | 2020 |
|
RU2818586C2 |
| АНТИГЕНСВЯЗЫВАЮЩИЙ ПОЛИПЕПТИД, СВЯЗЫВАЮЩИЙСЯ С CD47, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2833294C2 |
| АНТИТЕЛО К GIPR И ЕГО СЛИТЫЙ С GLP-1 БЕЛОК, А ТАКЖЕ ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ЕГО ОСНОВЕ И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2800370C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРЕДУПРЕЖДЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2017 |
|
RU2766586C2 |
| АНТИТЕЛО К CD40, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2779128C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2019 |
|
RU2781542C2 |
| Антитело против PCSK9 и его применение | 2016 |
|
RU2756012C2 |
| Антитела к TSLP человека и их применение | 2021 |
|
RU2825460C1 |
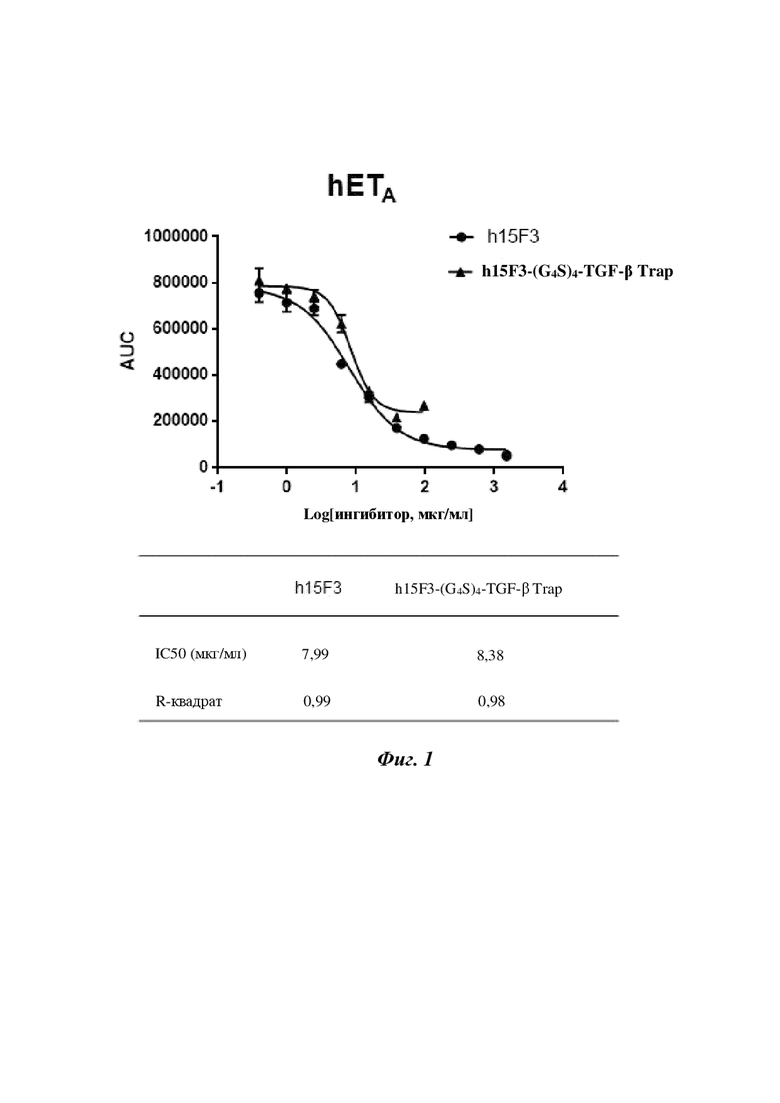
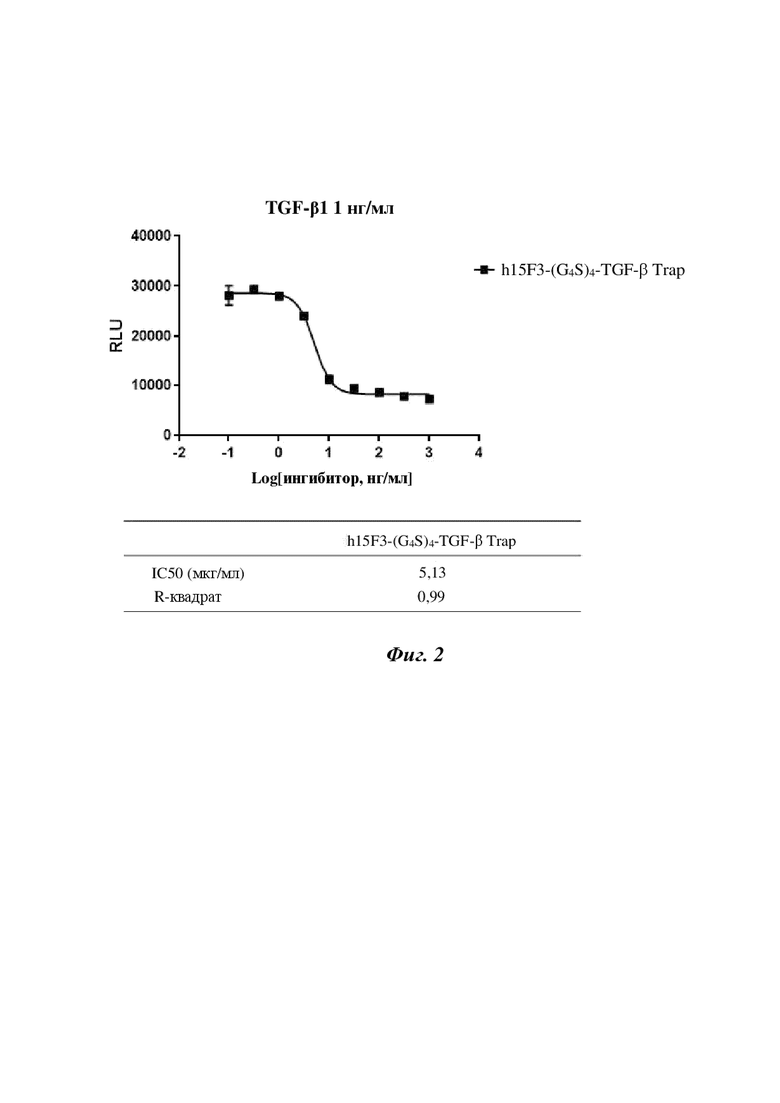

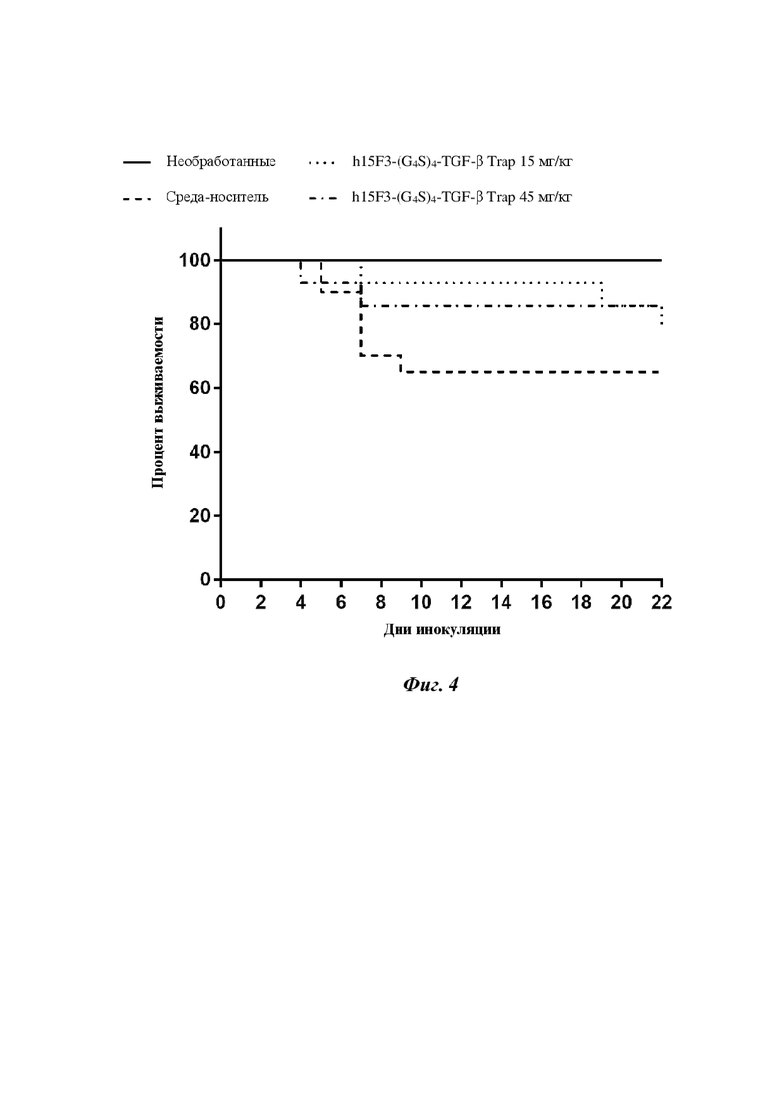
Изобретение относится к области биотехнологии, конкретно к слитым белкам на основе антитела к эндотелиновому рецептору A (ETA)и TGF-β Trap, и может быть использовано в медицине в терапии легочной артериальной гипертензии, легочного фиброза или кардиоваскулярного фиброза. Предложен слитый белок, содержащий антитело к ETA и TGF-β Trap или его фрагмент, которые способны блокировать функцию TGF-β. При этом указанный TGF-β Trap состоит из внеклеточного конца рецептора TGF-β 2 типа, а указанное антитело к ETA характеризуется наличием трех CDR в легкой цепи с SEQ ID NO: 8, 32 и 50 или 205 и трех CDR в тяжелой цепи с SEQ ID NO: 70, 92 и 116. Изобретение обеспечивает получение слитого белка с двойным механизмом действия: с одной стороны, он может блокировать сигнальный путь ЕТАR и снижать сопротивление кровеносных сосудов легочному кровотоку, с другой стороны, он может блокировать сигнальный путь TGF-β и облегчать течение фиброза и ремоделирование легочных артериол. 9 н. и 21 з.п. ф-лы, 4 ил., 2 табл., 7 пр.
1. Слитый белок антитела к эндотелиновому рецептору A (ETA) и TGF-β Trap или его фрагмента для блокирования сигнального пути ETA и сигнального пути TGF-β, структурно характеризующийся тем, что слитый белок содержит антитело к ETA и TGF-β Trap или его фрагмент, где указанный TGF-β Trap состоит из внеклеточного конца рецептора TGF-β 2 типа, и указанные TGF-β Trap или его фрагмент могут блокировать функцию TGF-β, и указанное антитело к ETA содержит:
аминокислотную последовательность CDR1 легкой цепи под SEQ ID NO: 8;
аминокислотную последовательность CDR2 легкой цепи под SEQ ID NO: 32;
аминокислотную последовательность CDR3 легкой цепи под SEQ ID NO: 50 или SEQ ID NO: 205;
аминокислотную последовательность CDR1 тяжелой цепи под SEQ ID NO: 70;
аминокислотную последовательность CDR2 тяжелой цепи под SEQ ID NO: 92 и
аминокислотную последовательность CDR3 тяжелой цепи под SEQ ID NO: 116.
2. Слитый белок по п. 1, где антитело к ETA содержит одну или две аминокислотные последовательности, где каждая аминокислотная последовательность независимо выбрана из аминокислотных последовательностей, перечисленных ниже:
a. аминокислотных последовательностей вариабельного домена легкой цепи под SEQ ID NO: 138 и SEQ ID NO: 162; и
b. аминокислотных последовательностей вариабельного домена тяжелой цепи под SEQ ID NO: 166 и SEQ ID NO: 190.
3. Слитый белок по п. 1, где полинуклеотидная кодирующая последовательность антитела к ЕТА содержит одну или две полинуклеотидные последовательности, при этом каждая полинуклеотидная последовательность независимо выбрана из полинуклеотидных последовательностей, приведенных ниже:
a. полинуклеотидных кодирующих последовательностей вариабельного домена легкой цепи под SEQ ID NO: 137 и SEQ ID NO: 161; и
b. полинуклеотидных кодирующих последовательностей вариабельного домена тяжелой цепи под SEQ ID NO: 165 и SEQ ID NO: 189.
4. Слитый белок по п. 1, где антитело к ЕТА содержит комбинацию аминокислотных последовательностей вариабельного домена легкой цепи и тяжелой цепи, независимо выбранных из перечня, приведенного ниже: SEQ ID NO: 138 и SEQ ID NO: 166, и SEQ ID NO: 162 и SEQ ID NO: 190.
5. Слитый белок по п. 4, где антитело к ЕТА содержит комбинацию аминокислотных последовательностей под SEQ ID NO: 138 и SEQ ID NO: 166.
6. Слитый белок по п. 4, где антитело к ЕТА содержит комбинацию аминокислотных последовательностей под SEQ ID NO: 162 и SEQ ID NO: 190.
7. Слитый белок по п. 1, где антитело к ETA содержит одну или две аминокислотные последовательности, где каждая аминокислотная последовательность независимо выбрана из аминокислотных последовательностей, перечисленных ниже:
a. аминокислотных последовательностей константной области легкой цепи под SEQ ID NO: 194 и SEQ ID NO: 196;
b. аминокислотных последовательностей константной области тяжелой цепи под SEQ ID NO: 198 и SEQ ID NO: 206.
8. Слитый белок по п. 1, где антитело к ЕТА предусматривает мышиное антитело к ЕТА или гуманизированное антитело к ЕТА.
9. Слитый белок по п. 1, где антитело к ЕТА предусматривает моноклональное антитело к ЕТА.
10. Слитый белок по п. 1, где антитело к ЕТА характеризуется значением IC50, составляющим от примерно 1 нМ до 200 нМ или от 10 нМ до 100 нМ, при снижении опосредованной человеческим эндотелином передачи сигнала.
11. Слитый белок по п. 1, где слитый белок содержит антитело к ЕТА, один, два, три, четыре, пять, шесть, семь или восемь TGF-β Trap и то же число пептидных линкеров; при этом в слитом белке аминоконец TGF-β Trap связан с карбоксиконцом легкой цепи или тяжелой цепи антитела к ETA посредством пептидной линкерной последовательности или в слитом белке карбоксиконец TGF-β Trap связан с аминоконцом легкой цепи или тяжелой цепи антитела к ЕТА посредством пептидной линкерной последовательности.
12. Слитый белок по п. 11, где слитый белок содержит антитело к ЕТА, один, два, три или четыре TGF-β Trap и то же число пептидных линкеров.
13. Слитый белок по п. 11, где слитый белок содержит антитело к ЕТА, два TGF-β Trap и два пептидных линкера.
14. Слитый белок по п. 11, где слитый белок содержит антитело к ЕТА, TGF-β Trap и пептидный линкер.
15. Слитый белок по п. 11, где в слитом белке аминоконец TGF-β Trap связан с карбоксиконцом легкой цепи или тяжелой цепи антитела к ЕТА посредством пептидной линкерной последовательности.
16. Слитый белок по п. 11, где в слитом белке аминоконец TGF-β Trap связан с карбоксиконцом тяжелой цепи антитела к ЕТА посредством пептидной линкерной последовательности.
17. Слитый белок по п. 11, где слитый белок содержит следующие аминокислотные последовательности: SEQ ID NO: 162, SEQ ID NO: 190, SEQ ID NO: 207 и SEQ ID NO: 210.
18. Слитый белок по п. 11, где антитело к ЕТА, TGF-β Trap и пептидная линкерная последовательность слиты с образованием слитого белка с помощью одного из следующих средств:
(1) посредством пептидной линкерной последовательности аминоконец TGF-β Trap связан с карбоксиконцом тяжелой/легкой цепи антитела к ЕТА: N'-R-линкер-TGF-β Trap-C'; и
(2) посредством пептидной линкерной последовательности карбоксиконец TGF-β Trap связан с аминоконцом легкой цепи или тяжелой цепи антитела к ETA: N'-TGF-β Trap-линкер-R-C';
при этом N' представляет собой аминоконец полипептидной цепи, C' представляет собой карбоксиконец полипептидной цепи, TGF-β Trap представляет собой фрагмент TGF-β Trap, R представляет собой аминокислотную последовательность легкой цепи или тяжелой цепи антитела к ЕТА, и линкер представляет собой пептидный линкер.
19. Слитый белок по п. 1, где TGF-β Trap содержит аминокислотную последовательность под SEQ ID NO: 207.
20. Слитый белок по п. 11, где каждая последовательность пептидного линкера независимо содержит одну из следующих аминокислотных последовательностей: SEQ ID NO: 208, SEQ ID NO: 209 и SEQ ID NO: 210.
21. Слитый белок по п. 20, где последовательность пептидного линкера содержит аминокислотную последовательность под SEQ ID NO: 210.
22. Полинуклеотид, кодирующий слитый белок по любому из пп. 1-21.
23. Экспрессионный вектор, содержащий полинуклеотид по п. 22.
24. Клетка-хозяин для экспрессии слитого белка по любому из пп. 1-21, содержащая экспрессионный вектор по п. 23, где указанная клетка не является эмбриональной клеткой человека.
25. Фармацевтическая композиция для предупреждения возникновения, улучшения течения или лечения легочной артериальной гипертензии, легочного фиброза или кардиоваскулярного фиброза, содержащая терапевтически эффективную дозу слитого белка по любому из пп. 1-21 и фармацевтически приемлемый носитель.
26. Применение фармацевтической композиции по п. 25 в получении лекарственного препарата, предназначенного для предупреждения возникновения, улучшения течения или лечения легочной артериальной гипертензии.
27. Применение фармацевтической композиции по п. 25 в получении лекарственного препарата, предназначенного для предупреждения возникновения, улучшения течения или лечения легочной гипертензии.
28. Применение фармацевтической композиции по п. 25 в получении лекарственного препарата, предназначенного для предупреждения возникновения, улучшения течения или лечения легочного фиброза.
29. Применение фармацевтической композиции по п. 25 в получении лекарственного препарата, предназначенного для предупреждения возникновения, улучшения течения или лечения кардиоваскулярного фиброза.
30. Применение по любому из пп. 26-29, где лекарственный препарат подлежит внутривенному или подкожному введению.
| ROMAN B.L | |||
| et al., Catching a Disease: A Molecular Trap as a Therapy for Pulmonary Arterial Hypertension, American Journal of Respiratory and Critical Care Medicine, 2014, v.194, n.9, p.1047-1049 | |||
| YUNG LAI-MING et al., A Selective Transforming Growth Factor-β Ligand Trap Attenuates Pulmonary Hypertension, Am J Respir Crit Care Med., 2016, v.194, |

