Область техники
Изобретение относится к области биотехнологии и фармацевтики, и может быть использовано для стимуляции восстановления периферической иннервации после травматического повреждения. Изобретение представляет собой плазмидную бицистронную конструкцию, одновременно несущую по одной копии нуклеотидной последовательности, кодирующей: мозговой нейротрофический фактор (BDNF), стимулирующий рост аксонов, формирование сосудов, кровоснабжающих нерв, выживание мотонейронов, лишенных травмой иннервируемых ими мишеней; и активатор плазминогена урокиназного типа или урокиназу (uPA), как активирующий факторы роста и нейротрофины, так и самостоятельно стимулирующий необходимые для регенерации процессы - клеточную миграцию и клеточную пролиферацию. Последовательность расположения участков, кодирующих BDNF и uPA - BDNF перед uPA и разделяющая их последовательность IRES-HCV обеспечивают необходимую преимущественную экспрессию BDNF. uPA экспрессируется в количестве примерно в 10 раз меньшем, чем BDNF.
Изобретение может быть использовано для приготовления лекарственного средства для стимуляции восстановления иннервации после травматического повреждения периферических нервов.
Уровень техники
Разработка способов и средств регенерация тканей и органов является актуальной биомедицинской задачей. Различные травмы, заболевания, хирургические вмешательства часто сопровождаются повреждением нервов. Данная проблема затрагивает миллионы людей по всему миру, в том числе трудоспособное население и не редко приводит к инвалидности когнитивного, моторного или психотического характера, что ведет к снижению качества жизни, а также вызывает социальные и экономические проблемы [Afsoun Seddighi, Amir Nikouei, Amir Saied Seddighi, Ali Reza Zali, Seyed Mahmood Tabatabaei, Ali Reza Sheykhi, Fatemeh Yourdkhani, Shoayb Naeimian. Peripheral Nerve Injury: A Review Article. International Clinical Neuroscience Journal Vol 3, No 1, p. 1-6. JTS Pettikiriarachchi, CL Parish, MS Shoichet, JS Forsythe, DR Nisbet. Biomaterials for brain tissue engineering. Australian journal of chemistry 63 (8), 1143-1154.]. По данным 2015 года повреждения периферических нервов встречаются у 13-23 пациентов из 100 000 и представляют одну из основных проблем современной травматологии и неврологии.
[Karp, J., Dalton, P., & Shoichet, M. (2003). Scaffolds for Tissue Engineering. MRS Bulletin, 28(4), 301-306. doi:10.1557/mrs2003.85]. Этиология травмам периферических нервов включает в себя проникающие травмы, сдавливание, вытягивание, ишемию и менее распространенные механизмы, такие как тепловой, электрический шок, излучение, перкуссия и вибрация [Afsoun Seddighi, Amir Nikouei, Amir Saied Seddighi, Ali Reza Zali, Seyed Mahmood Tabatabaei, Ali Reza Sheykhi, Fatemeh Yourdkhani, Shoayb Naeimian. Peripheral Nerve Injury: A Review Article. International Clinical Neuroscience Journal Vol 3, No 1, p. 1-6.]. Поврежденные нервы периферической нервной системы обладают способностью к регенерации. Однако спонтанная регенерация нерва при отсутствии какого-либо медицинского вмешательства чаще всего не приводит к регенерации поврежденного или утраченного участка нерва и восстановлению иннервации травмированных тканей. Хирургическое вмешательство, при котором проводится пространственное совмещение концов разорванного/пересеченного нерва посредством сшивания его концов с (или без) использованием трансплантатов, также часто не приводит к восстановлению функций. Нейриты перестают расти, не достигая иннервируемых мишеней и в местах остановки роста формируются невриномы. Дополнительными ограничениями являются: длина необходимого трансплантата, недостаток донорского материала, возможность развития различных заболеваний, образованием невриномы, рубцевание [Karp, J., Dalton, Р., & Shoichet, М. (2003). Scaffolds for Tissue Engineering. MRS Bulletin, 28(4), 301-306. doi:10.1557/mrs2003.85]. Использование аллогенных трансплантатов, может вызывать иммунный ответ [J.M. Karp, P.D. Dalton, M.S. Shoichet, F. Cahn. Scaffolds for Tissue Engineering. MRS BULLETIN. April 2003, pp. 301-306]. Активно разрабатываются подходы стимуляции и восстановления периферической иннервации за счет обеспечения определенных органов, тканей и их отдельных зон необходимыми полипептидными продуктами (гормонами, ферментами, ростовыми факторами и т.д.), при этом наиболее перспективным подходом считается применение векторных конструкций, включающих гены, которые кодируют нужные белки, являющиеся ключевыми факторами, стимулирующими рост нервов и восстановление периферической иннервации.
Из уровня техники известен способ стимуляции регенеративных процессов в ишемизированных тканях, включающий введение в организм больного среды культивирования стромальных клеток из жировой ткани человека, содержащей факторы роста VEGF, HGF, ангиопоэтин и ангиогенин для эффективного стимулирования регенеративных процессов в ишемизированных тканях [RU 2497529, 10.11.2013].
Также из уровня техники известен способ получения зрелых человеческих нейротрофических белков семейства NGF/BDNF, которые полностью биологически активны. Кроме того, раскрыт ген, кодирующий человеческий BDNF.
Доказано, что нейротрофические факторы могут быть полезными при лечении дегенерации нервных клеток и потери дифференцированной функции, которая происходит при различных нейродегенеративных заболеваний, таких как болезнь Альцгеймера или Паркинсона, либо после таких травматических повреждений, как инсульт или физическая травма спинного мозга [US 5235043, 10.08.1993].
Из уровня техники известен способ стимуляции восстановления иннервации поврежденной ткани с помощью введения в область повреждения векторной конструкции, обеспечивающей высокий уровень экспрессии фактора, участвующего в процессе регенерации нерва, который заключается в том, что в поврежденную мышцу вводят терапевтически эффективное количество рекомбинантной плазмиды с нуклеотидной последовательностью SEQ ID NO: l, которая содержит модифицированную нуклеотидную последовательность, кодирующую BDNF человека, и последовательность Козак, в качестве плазмидного вектора использован pVax1 Invitrogen [RU 2538621, 10.01.2015]. Данный способ позволяет ускорить восстановление структуры и проводимости периферических нервов после травм, путем трансфекции мышц, иннервируемых поврежденным нервом, плазмидной конструкцией, кодирующей мозговой нейротрофический фактор (BDNF). Недостатком указанного способа можно отметить то, что данная плазмида вызывает повышение локальной продукции только одного из нейротрофических факторов - BDNF. Белок с данной плазмиды считывается в виде предшественника, что необходимо для его правильного созревания. Для реализации нейротрофической активности предшественник должен подвергнуться протеолитической активации, без которой он вместо поддержки выживания нейронов стимулирует их гибель.
Из уровня техники известен способ стимуляции восстановления иннервации поврежденной ткани у млекопитающих с помощью введения в область повреждения векторной конструкции, обеспечивающей высокий уровень экспрессии фактора, участвующего в процессе регенерации нерва, заключающийся в том, что в поврежденную мышцу вводят терапевтически эффективное количество рекомбинантной плазмиды, содержащей плазмидный вектор pVax1 со встроенным по сайтам рестрикции EcoRI и EcoRV фрагментом ДНК, кодирующим оптимизированную для экспрессии в клетках млекопитающих урокиназу, содержащий последовательность Козак, слитую с модифицированным геном урокиназы, характеризующуюся нуклеотидной последовательностью, приведенной в SEQ ID NO: 1, рекомбинантная плазмида, предназначенная для экспрессии гена урокиназы в клетках млекопитающих, содержащая плазмидный вектор pVaxl со встроенным по сайтам рестрикции EcoRI и EcoRV фрагментом ДНК, кодирующим оптимизированную для экспрессии в клетках млекопитающих урокиназу, содержащим последовательность Козак, слитую с модифицированным геном урокиназы, характеризующаяся нуклеотидной последовательностью, приведенной в SEQ ID N0:1, а также фрагмент ДНК, кодирующий оптимизированную для экспрессии в клетках млекопитающих урокиназу, содержащий последовательность Козак, слитую с модифицированным геном урокиназы, и характеризующийся нуклеотидной последовательностью, приведенной в SEQ ID NO: 2 [RU 2563541, 20.09.2015]. Известный способ позволяет ускорить восстановление структуры и проводимости периферических нервов после травм и ишемии путем трансфекции мышц, иннервируемых поврежденным нервом. Недостатком данного способа можно указать то, что данная плазмида кодирует только урокиназу, которая, несмотря на то что обладает способностью стимулировать процессы регенерации не обладает собственными нейротрофическими эффектами и может лишь активировать эндогенные нейротрофины, экспрессия которых при травматическом повреждении недостаточна для обеспечения эффективной реиннервации.
Наиболее близким аналогом (прототипом) предлагаемого изобретения является способ стимуляции восстановления периферической иннервации ткани после травмы, заключающийся во введении терапевтически эффективного количества (1) плазмидного вектора, содержащего нуклеотидную последовательность, кодирующую BDNF, или (2) плазмидного вектора, содержащего нуклеотидную последовательность, кодирующую человеческую uPA дикого типа (NM 002658), или их комбинации (3), или (4) комбинации плазмидного вектора, содержащего нуклеотидную последовательность, кодирующую BDNF с плазмидным вектором, содержащим оптимизированную нуклеотидную последовательность, кодирующую хемокин, продуцируемый стромальными клетками SDF-1 [RU 2486918, 10.07.2013]. Известный способ позволяет ускорить восстановление структуры и проводимости периферических нервов после травм за счет локального увеличения продукции нейротрофических факторов. При этом указывается, что комбинация (1:1 по весу кДНК) плазмид pVax1-BDNFopt + pVax1-uPA обеспечивает дополнительное 1,5-2-кратное (в сравнении с экспрессией только BDNF или uPA) ускорение восстановления проводимости и структуры нерва, а, следовательно, и иннервации соответствующих скелетных мышц.
Однако в случае применения смеси плазмид не гарантирована их скоординированная работа. Проникновение плазмид имеющих разную массу и разную последовательность вставки в ткани происходит независимо, их активность регулируется и реализуется в разных клетках также независимо и поэтому невозможно достигнуть оптимального соотношения экспрессирующихся в тканях белков и поэтому невозможно достигнуть максимальной эффективности структуры и функциональной активности поврежденных периферических нервов.
Таким образом, с учетом накопленных данных, существует потребность в разработке генно-инженерных конструкций нового поколения для создания эффективных генотерапевтических препаратов для стимуляции восстановления структуры и функциональной активности периферических нервов после травматических повреждений.
Раскрытие изобретения
Техническим результатом заявляемого изобретения является скоординированная локализованная экспрессия двух белков с достижением оптимального соотношения продукции рекомбинантного human-BDNF и продукции human-uPA. BDNF экспрессируется в количестве, на порядок большем, чем uPA, при этом достигается продукция рекомбинантного human-BDNF не менее 70 нг/мл и продукция human-uPA не менее 9 нг/мл кондиционированной среды при минимально возможном увеличении размера плазмиды (не более 5.5 тыс. п.н.). Данное соотношение human-BDNF и human-uPA является более предпочтительным, чем соотношение, возникающее при трансфекции отдельными плазмидами, поскольку избыток human-uPA может привести к избыточной активации ММП и повреждению/аутолизу ткани. Оптимальное соотношение продукции human-BDNF и human-uPA обеспечивается благодаря наличию в составе генетической конструкции IRES (internal ribosome entry site) вируса гепатита С, который обеспечивает продукцию первого белка на порядок более высокую, чем второго белка. К преимуществам данной генетической конструкции, помимо наличия IRES можно отнести:
1. Наличие SV40 энхансера, встроенного между промотором и энхансером цитомегаловируса. Добавление этой последовательности было призвано увеличить активный транспорт плазмиды в ядро целевой клетки, за счет распознавания SV40 энхансера множеством транскрипционных факторов.
2. Короткого искусственного интрона (КИИ), который призван усилить транспорт бицистронной мРНК в цитоплазму клетки. Добавление этой последовательности (~100 п.н.) в плазмиду перед старт-кодоном human-BDNF увеличило продукцию белков human-BDNF и human-uPA. Это было подтверждено экспериментально: 70 нг/мл human-BDNF в кондиционированной среде для плазмиды с КИИ и 55 нг/мл human-BDNF в кондиционированной среде для плазмиды без КИИ.
3. В качестве вставки в сконструированной плазмиде pNCure были использованы природные кДНК генов мозгового нейротрофического фактора и активатора плазминогена урокиназного типа человека, полученные из культуры мезенхимных стромальных клеток подкожного жира здорового донора методом обратной транскрипции (RT-PCR).
 Технический результат достигается генно-инженерной конструкцией, для стимуляции посттравматической реиннервации периферических нервов, представляющей собой экспрессионный вектор - плазмиду, в которую клонированы вставки кДНК, кодирующие мозговой нейротрофический фактор человека (hBDNF), активатор плазминогена урокиназного типа (урокиназу, huPA) и последовательность IRES (сайт внутренней посадки рибосомы вируса гепатита С или др. функционально родственные вирусные последовательности IRES). В качестве регуляторных элементов экспрессии и трансляции конструкции использованы энхансер, внедренный в последовательность цитомегаловирусного промотера, искусственный интрон с последовательностью SEQ ID NO: 1. При этом последовательность кДНК мозгового нейротрофического фактора человека (hBDNF) встроена в вектор с 853 н.п. по 1599 н.п.; активатор плазминогена урокиназного типа человека (huPA)- с 1983 н.п. по 3275 н.п.; регуляторная последовательность содержит: энхансер SV40 или CMV с 417 н.п. по 478 н.п., внедренный в последовательность цитомегаловирусного промотера с 86 н.п. по 691 н.п., искусственный интрон - с 761 н.п. по 839 н.п., и сайт внутренней посадки рибосомы вируса гепатита С (IREShcv) с 1608 н.п. по 1982 н.п.
Технический результат достигается генно-инженерной конструкцией, для стимуляции посттравматической реиннервации периферических нервов, представляющей собой экспрессионный вектор - плазмиду, в которую клонированы вставки кДНК, кодирующие мозговой нейротрофический фактор человека (hBDNF), активатор плазминогена урокиназного типа (урокиназу, huPA) и последовательность IRES (сайт внутренней посадки рибосомы вируса гепатита С или др. функционально родственные вирусные последовательности IRES). В качестве регуляторных элементов экспрессии и трансляции конструкции использованы энхансер, внедренный в последовательность цитомегаловирусного промотера, искусственный интрон с последовательностью SEQ ID NO: 1. При этом последовательность кДНК мозгового нейротрофического фактора человека (hBDNF) встроена в вектор с 853 н.п. по 1599 н.п.; активатор плазминогена урокиназного типа человека (huPA)- с 1983 н.п. по 3275 н.п.; регуляторная последовательность содержит: энхансер SV40 или CMV с 417 н.п. по 478 н.п., внедренный в последовательность цитомегаловирусного промотера с 86 н.п. по 691 н.п., искусственный интрон - с 761 н.п. по 839 н.п., и сайт внутренней посадки рибосомы вируса гепатита С (IREShcv) с 1608 н.п. по 1982 н.п.
Технический результат также достигается лекарственной формой, содержащей в качестве активного компонента заявляемую генотерапевтическую конструкцию и фармацевтически приемлемые растворители для парентерального введения, стабилизаторы и консерванты.
Также технический результат достигается способом стимуляции локальной экспрессии мозгового нейротрофического фактора и активатора плазминогена урокиназного типа в периневральных тканях в области травмы, включающий введение лекарственной формы, содержащей генотерапевтическую конструкцию, в терапевтически-эффективном количестве. При этом введение лекарственной формы осуществляют путем единомоментной (разовой) многоточечной периневральной внутримышечной инъекции в области травмы нерва, повторяемой от 3 до 5 раз с периодичностью 1 раз в 2-4 недели до достижения терапевтического эффекта.
Технический результат также достигается применением заявляемой генотерапевтической конструкции в качестве генотерапевтического средства для лечения травм периферических нервов.
При этом кДНК hBDNF встроена в модифицированную, оптимизированную плазмиду сконструированную на основе pVax1 сразу после промотора CMV и консенсусной последовательности Козак (GCCACC) с использованием сайтов рестрикции NheI и HindIII. кДНК huPA (клонированная с использованием сайтов рестрикции Асс651 и EagI) отделена от hBDNF с помощью IRES HCV [Lukavsky PJ. Structure and function of HCV IRES domains. Virus Res. 2009; 139(2): 166-71], собранного из синтетических олигонуклеотидов (амплифицированных с помощью ПЦР и клонированных с использованием сайтов рестрикции HindIII и Acc65I). Каждая кДНК несет свои собственные стартовый и стоп-кодоны.
Основные элементы конструкции:
 последовательность гена hBDNF
последовательность гена hBDNF
 последовательность гена huPA
последовательность гена huPA
 последовательность IRES (IRES вируса гепатита С или др. функционально родственные вирусные последовательности IRES). Регуляторные элементы экспрессии и трансляции конструкции:
последовательность IRES (IRES вируса гепатита С или др. функционально родственные вирусные последовательности IRES). Регуляторные элементы экспрессии и трансляции конструкции:
 Промотор - pCMV промотор цитомегаловируса (могут быть использованы и др. функционально родственные последовательности)
Промотор - pCMV промотор цитомегаловируса (могут быть использованы и др. функционально родственные последовательности)
 Энхансер 1 - (SV40 энхансер или CMV экхансер, или др. функционально родственные последовательности)
Энхансер 1 - (SV40 энхансер или CMV экхансер, или др. функционально родственные последовательности)
 Короткий искусственный интрон, повышающий эффективность трансляции
Короткий искусственный интрон, повышающий эффективность трансляции
 Терминатор - bGH poly А (участок полиаденилирования гена бычьего гормона роста)
Терминатор - bGH poly А (участок полиаденилирования гена бычьего гормона роста)
Регуляторные элементы экспрессии и трансляции конструкции в бактерии Е. coli:
 Точка начала репликации (ori)
Точка начала репликации (ori)
 Селективный маркер для отбора трансформированных бактерий: - ген устойчивости к канамицину
Селективный маркер для отбора трансформированных бактерий: - ген устойчивости к канамицину
В использованный вектор сконструированный на основе pVaxl были внесены следующие изменения:
- исходно присутствующий в векторе pVax1 ген устойчивости к канамицину заменен на гомологичный,
- в промоторную область вектора pVax1 добавлена ранее отсутствовавшая там последовательность энхансера SV40, призванная улучшить транспорт плазмиды в ядро,
перед вставкой, кодирующей терапевтические белки, был добавлен искусственный интрон (который способствовал увеличению продукции целевых белков более, чем на 30%).
- между целевыми вставками был помещен сайт внутренней посадки рибосомы вируса гепатита С (IREShcv), который обеспечивает эффективное считывание обоих лекарственных белков с преобладанием экспрессии первого белка по сравнению со вторым примерно на порядок. Возможно использование функционально аналогичных последовательностей IRES (вирусных, прокариотических и эукариотических сайтов внутренней посадки рибосомы)
Комбинацию использованных в генетической конструкции энхансера SV40, с коротким искусственным интроном и КИИ, и с сайтом внутренней посадки рибосомы вируса гепатита С (IREShcv) можно рассматривать как неотъемлемые характеристические свойства созданной плазмиды.
Для реализации данного изобретения потенциально могут быть использованы любые терапевтические вектора и др. плазмиды, отвечающие следующим требованиям: небольшой размер, наличие последовательности, обеспечивающей селекцию, высокая копийность на этапе наработки, пригодность для экспрессии белков в эукариотических клеточных системах.
Любой из функциональных элементов может быть заменен на существующий (из других генетических конструкций) или заново синтезированный.
Краткое описание чертежей
Изобретение поясняется следующими чертежами.
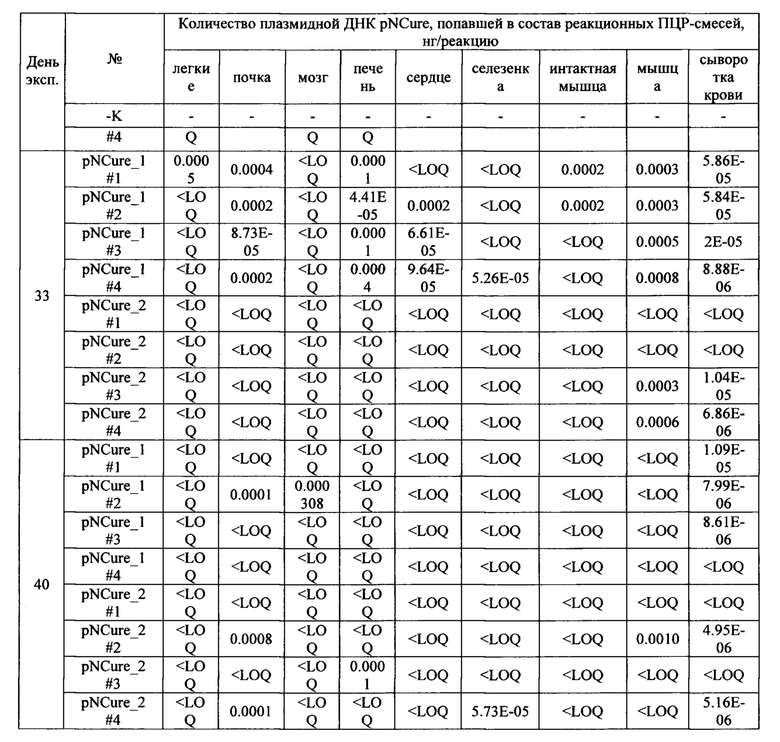
На фиг. 1 показано содержание hBDNF и huPA в кондиционированной среде после трансфекции HEK293 вариантами плазмиды pNCure.
На фиг. 2 представлен обобщенный механизм действия плазмиды pNCure.
На фиг. 3 представлена структура плазмидной ДНК pNCure, кодирующей пробелки мозгового нейротрофического фактора человека (human-proBDNF) и активатора плазминогена урокиназного типа человека (human-uPA).
На фиг. 4 представлено обоснование преимущества многоточечного режима введения лекарственного средства. Введение в несколько точек обеспечивает более высокий уровень экспрессии трансгенов по сравнению с введением препарата в одну точку (окрашивание b-галактозидазой - верхние панели и левая нижняя панель, 100х, n=3; иммуноферментный анализ экспрессии трансгенов - нижняя средняя и правая панели).
На фиг. 5 показано стимулирующее влияние BDNF и uPA на дифференцировку клеток Neuro2a в нейральном направлении, выражающееся в изменениях морфологии клеток. pVax - среда культивирования НЕК293 клеток, трансфицированных контрольной плазмидой; pNCure - среда культивирования НЕК293 клеток, трансфицированных плазмидой, содержащей BDNF и uPA; 1% ФБС - среда культивирования клеток, содержащая 1% ФБС, положительный контроль; 10% ФБС - среда культивирования клеток, содержащая 10% ФБС, отрицательный контроль.
На фиг. 6 представлены результаты статистического анализа влияния продуцируемых клетками НЕК293, трансфицированных плазмидой pNCure нейротрофических факторов (BDNF и uPA) на число клеток, формирующих нейриты на in vitro модели нейральной дифференцировки клеток Neuro2a. pVax - среда культивирования НЕК293 клеток, трансфицированных контрольной плазмидой; pNCure -среда культивирования НЕК293 клеток, трансфицированных плазмидой, содержащей BDNF и uPA; 1% ФБС - среда культивирования клеток, содержащая 1% ФБС, положительный контроль; 10% ФБС - среда культивирования клеток, содержащая 10% ФБС, отрицательный контроль. Статистический обсчет нейритов в одиннадцати случайно выбранных полях зрения трех лунок. Представлены результаты 4-х независимых экспериментов, * и **р<0,05 по сравнению с группой pVax, N=11.
На фиг. 7 представлены результаты анализа влияния продуцируемых клетками НЕК293, трансфицированных плазмидой pNCure нейротрофических факторов (BDNF и uPA) на формирование нейритов на модели эксплантов спинальных ганглиев мыши в Матригеле, сделанные на 4-е сутки. pVax - среда культивирования HEK293 клеток, трансфицированных контрольной плазмидой. Стрелками обозначены мигрирующие клетки (панель pVax); или отрастающие аксоны (на панели pNCure). На нижней панели представлены увеличенные области, демонстрирующие мигрирующие клетки (панель pVax) и регенерирующие аксоны (панель pNCure).
На фиг. 8 показано влияние продуцируемых клетками НЕК293, трансфицированных плазмидой pNCure нейротрофических факторов (BDNF и uPA), на миграцию клеток из эксплантов спинальных ганглиев мыши в Матригель. Ядра клеток окрашены DAPI. Белой прерывистой линией обозначена граница экспланта; стрелками указан путь миграции клеток из эксплантов в Матригель. На верхней панели результаты использования кондиционированной среды клеток НЕК293, трансфецированных контрольной плазмидой pVax. На нижней панели результаты использования кондиционированной среды клеток НЕК293, трансфицированных плазмидой pNCure, продуцирующей нейротрофические факторы (BDNF и uPA).
На фиг. 9 показано влияние генетической конструкции pNCure в дозах 60 мкг (pNCure_1) и 120 мкг (pNCure_2) мкг на восстановление функции поврежденного общего малоберцового нерва. Амплитуда суммарного потенциала действия нерва отражает число активных нейритов в нерве. Электрофизиологическое исследование амплитуды СПДН на 24 сутки эксперимента. Значения представлены в виде медианы и процентилей 25% и 75% (* - п>5; р<0.05 по сравнению с группой "-К").
На фиг. 10 показано влияние генетической конструкции pNCure в дозах 60 мкг (pNCure_1) и 120 мкг (pNCure_2) мкг на восстановление функции поврежденного общего малоберцового нерва. Латентный период СПДН обратно пропорционален скорости проведения импульса по нерву. Электрофизиологическое исследование латентного периода СПДН через 24 суток после начала эксперимента. Значения представлены в виде медианы и процентилей 25% и 75% (n>5).
На фиг. 11 представлены данные подтверждающие достоверное увеличение количества аксонов на 24-е сутки от начала эксперимента при введении генетической конструкции pNCure. На верхних панелях показаны поперечные срезы нерва с окрашенными нейритами. Данные представлены виде медианы и процентилей 25% и 75%. Пунктиром отмечено соответствующее значение для интактного нерва (n>5, p<0.05 по сравнению с группой "-К").
На фиг. 12 представлены результаты, подтверждающие локальный характер экспрессии лекарственного белка в ткани экспериментального животного. Иммуноферментный анализ образцов тканей мыши, подвергшейся инъекции генетической конструкции, на содержание рекомбинантного мозгового нейротрофического фактора человека.
Осуществление изобретения
Все используемые реагенты являются коммерчески доступными, все процедуры, если не оговорено особо, осуществляли при комнатной температуре или температуре окружающей среды, то есть в диапазоне от 18 до 25°С.
Ниже представлено более подробное описание заявляемого изобретения. Настоящее изобретение может подвергаться различным изменениям и модификациям, понятным специалисту на основе прочтения данного описания. Такие изменения не ограничивают объем притязаний. Например, могут изменяться стратегия сборки генетической конструкции (используемые праймеры, сайты клонирования и т.д.). Методы наращивания, выделения и очистки плазмид, способ трансфекции, методы оценки концентраций АФР и др.
Термин «фармацевтически приемлемый» относится к нетоксическому материалу, который не взаимодействует с действием активного компонента фармацевтической композиции. "Фармацевтически приемлемый носитель" относится к биосовместимому раствору, который в достаточной степени имеет такие характеристики, как стерильность, p[Eta], изотоничность, стабильность и подобные, и может включать любые растворители, разбавители, включая стерильный физиологический раствор, раствор хлорида натрия для инъекций, раствор Рингера для инъекций, раствор декстрозы для инъекций, раствор декстрозы и хлорида натрия для инъекций, содержащий лактат раствор Рингера для инъекций и другие водные буферные растворы, дисперсионные среды, покрытия, антибактериальные и противогрибковые средства, изотонические вещества и подобные. Фармацевтически приемлемый носитель также может содержать стабилизаторы, консерванты, антиоксиданты или другие добавки, которые хорошо известны специалистам в данной области, или другой наполнитель, известный из уровня техники.
Действующей основой препарата является плазмидная ДНК pNCure (фиг. 3), кодирующая одновременно два лекарственных белка: пробелок мозговой нейротрофический фактор человека (human-proBDNF) и активатор плазминогена урокиназного типа человека (human-uPA).
В качестве основы для конструирования плазмидного вектора был выбран одобренный для клинического использования вектор pVax1, соответствующий требованиям Food and Drug Administration (FDA, 1996), изначально предназначенный для иммунизации животных с помощью ДНК, кодирующих белки-мишени, минуя стадии экспрессии белка в клетках продуцентах и их очистки.
Для улучшения эффективности, уменьшения размера и повышения безопасности исходный pVax1 был значительно модифицирован:
- был заменен ген устойчивости к канамицину,
- в промоторную область добавлена последовательность энхансера SV40, призванная улучшить транспорт плазмиды в ядро,
- перед вставкой, кодирующей терапевтические белки, был добавлен искусственный интрон (который способствовал увеличению продукции целевых белков более, чем на 30%).
- между целевыми вставками был помещен сайт внутренней посадки рибосомы вируса гепатита С (IRESHCV), который обеспечивает эффективное считывание обоих лекарственных белков с преобладанием экспрессии первого белка (human-proBDNF) по сравнению со вторым (human-uPA) не менее, чем в 10 раз.
В качестве вставки были использованы природные кДНК генов мозгового нейротрофического фактора и активатора плазминогена урокиназного типа человека, полученные из культуры мезенхимных стромальных клеток подкожного жира здорового донора методом обратной транскрипции (RT-PCR).
Обобщенный механизм действия pNCure представлен на фиг. 2 и описан подробнее ниже.
Мозговой нейротрофический фактор секретируется клетками в виде пробелка и активируется во внеклеточной среде. В зрелой форме через связывание с рецептором trkB он обеспечивает выживание и регенерацию, а также стимулирует рост аксонов различных типов нейронов. Однако, в виде пробелка он обладает большим сродством к альтернативному рецептору р75 на поверхности нейронов и Шванновских клеток, который запускает апоптоз. Одной из основных эндопротеаз, осуществляющих внеклеточную активацию BDNF, является эндопротеаза плазмин, которая в свою очередь нуждается в активации активатором плазминогена урокиназного типа (или урокиназой).
Кроме того, большинство нейротрофинов и факторов роста, так же как и мозговой нейротрофический фактор синтезируются в виде пробелков и для обеспечения их активности нуждаются в протеолитической активации. Локально синтезируемый после введения препарата активатор плазминогена, кроме активации синтезируемого после введения препарата мозгового нейротрофического фактора, активирует эндогенные нейротрофические факторы и факторы роста, также способствуя процессу регенерации.
Это важно в связи с тем, что ранее многократно показано нарушение процесса активации факторов роста в очагах ишемии, формирующихся при травматическом повреждении тканей.
Помимо активации BDNF, других нейротрофинов и факторов роста плазмин и урокиназный активатор плазминогена способствуют разрушению фибрина (интраневральные кровоизлияния с последующим образованием фибринового тромба неизбежно сопровождают травматические повреждения нервных стволов), который обладает нейротоксическим действием и является ингибитором роста аксонов, а также активируют резидентные (эндогенные) факторы роста.
Помимо этого, урокиназный активатор плазминогена регулирует направление роста нервных волокон через связывание с рецептором uPAR на поверхности конуса роста нервных клеток. Однако для запуска необходимых протеолитических каскадов и реализации навигационной функции количество урокиназного активатора плазминогена не должна быть слишком высоким, так как его излишняя активность может быть причиной нежелательных явлений, например, излишний протеолиз матриксных белков.
Использованный дизайн плазмидного вектора, с одной стороны, позволил вместить в плазмиду кДНК двух факторов роста при минимально возможном увеличении размера плазмиды, а с другой позволил обеспечить близкое к оптимальному соотношение продуцируемых белков, т.е. высокую продукцию рекомбинантного human-BDNF и умеренную продукцию human-uPA, различающуюся в среднем на порядок.
Бицистронная генетическая конструкция была собрана следующим образом: кДНК hBDNF была встроена в модифицированный вектор pVax1 сразу после промотора CMV и консенсусной последовательности Козака (GCCACC) с использованием сайтов рестрикции NheI и HindIII [Kozak М, 1984, "Point mutations close to the AUG initiator codon affect the efficiency of translation of rat preproinsulin in vivo". Nature. 308 (5956): 241-246. doi:10.1038/308241a0]. кДНК huPA (клонированная с использованием сайтов рестрикции Асс651 и EagI) была отделена от hBDNF с помощью IRES HCV [Lukavsky PJ. Structure and function of HCV IRES domains. Virus Res. 2009; 139(2): 166-71], собранного из синтетических олигонуклеотидов (амплифицированных с помощью ПЦР и клонированных с использованием сайтов рестрикции HindIII и Асс651). Согласно исследованию R. Jubin [Jubin R, Vantuno NE, Kieft JS, et al. Hepatitis С virus internal ribosome entry site (IRES) stem loop Hid contains a phylogenetically conserved GGG triplet essential for translation and IRES folding. J Virol. 2000; 74(22): 10430-7] IRES HCV обеспечивает продукцию второго белка (huPA) в 10 раз меньше в молярном отношении, чем первого (hBDNF). Каждая кДНК несет свои собственные стартовый и стоп-кодоны (старт-кодон huPA является встроенным в IRES HCV). Для повышения продуктивности плазмиды были сконструированы несколько вариантов плазмиды: с синтетическим интроном и последовательностью WPRE (фиг. 1). Синтетический интрон с последовательностью SEQ ID NO: 1 был сконструирован de novo, собран из синтетических олигонуклеотидов и введен между промотором CMV и консенсусной последовательностью Kozak (сайт рестрикции NheI) (жирным шрифтом выделены ключевые последовательности интрона). Было проведено исследование каждого варианта плазмиды с прямой и обратной ориентацией интрона (f/r-Intr) с оценкой влияния ориентации интрона (прямая или обратная) на продукцию белка. Последовательность WPRE была клонирована из pAAV-EFla-tdTomato-WPRE-pGHpA (Botond Roska, Addgene плазмида # 67527) [Wertz A, Trenholm S, Yonehara K, Hillier D, Raics Z, Leinweber M, Szalay G, Ghanem A, Keller G, Rozsa B, Conzelmann KK, Roska B. Science. 2015 Jul 3; 349(6243):70-4. doi: 10.1126/science.aab1687] и введена в варианты плазмидного вектора, содержащего f/r-Intr между стоп-кодоном huPA и сигналом полиаденилирования бычьего гормона роста (сайт рестрикции XbaI).
Анализ кондиционированной среды HEK293, трансфицированных вариантами бицистронного вектора, на содержание hBDNF и huPA и анализ эффективности трансфекции показали, что оптимальным соотношением продуктивности и размера обладает вариант вектора с прямой ориентацией синтетического интрона и без WPRE. Полученная плазмида pVaxlm-f-Intr-hBDNF-IRES-huPA была названа «плазмидой для лечения нервов» (для краткости pNCure).
После травмы нерва у человека экспрессия нейротрофинов и мозгового нейротрофического фактора, в частности, повышается примерно на 1 месяц, а экспрессия урокиназы повышается примерно на 1 неделю, чего, с учетом низкой скорости роста нейритов у человека, обычно недостаточно для полноценной реиннервации, в особенности, при высокоуровневой травме нервного ствола.
pNCure относится к фармакологической группе биологических препаратов для генной терапии, стимуляторов восстановления иннервации после травм периферического нерва. Последовательность pNCure (pVax2-fIntron-hBDNF-IRESHCV-huPA) SEQ ID NO: 2 с обозначением функциональных фрагментов: 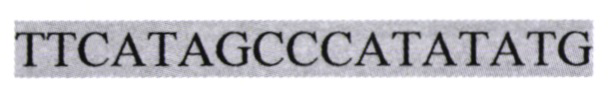 - промотор цитомегаловируса;
- промотор цитомегаловируса;  - SV40 энхансер;
- SV40 энхансер;  - искусственный интрон;
- искусственный интрон;  - Козак-последовательность;
- Козак-последовательность;  или
или  - старт-кодон;
- старт-кодон;  или
или  - стоп-кодон; GCGACCATCCTTTTCCTT - кДНК hBDNF;
- стоп-кодон; GCGACCATCCTTTTCCTT - кДНК hBDNF; 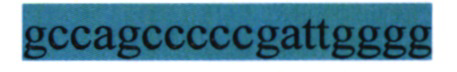 - IRESHCV;
- IRESHCV;  - начало кДНК huPA в составе IRESHCV; AGAGCCCTGCTGGCGCGC - кДНК huPA; CTGTGCCTTCXAGTTGCC - сигнал полиаденилирования бычьего гормона роста; AGCCATATXCAACGGGAA - ген устойчивости к канамицину; CCCGTAGAAAAGATCAAA - сайт начала репликации плазмиды.
- начало кДНК huPA в составе IRESHCV; AGAGCCCTGCTGGCGCGC - кДНК huPA; CTGTGCCTTCXAGTTGCC - сигнал полиаденилирования бычьего гормона роста; AGCCATATXCAACGGGAA - ген устойчивости к канамицину; CCCGTAGAAAAGATCAAA - сайт начала репликации плазмиды.
Способ стимуляции локальной экспрессии мозгового нейротрофического фактора и активатора плазминогена урокиназного типа в тканях животных моделей за счет введения генотерапевтической конструкции.
Внутримышечное введение pNCure в область травмы периферического нерва, вызывает длительную локальную продукцию мозгового нейротрофического фактора и активатора плазминогена урокиназного типа и, тем самым, стимулирует выживание нейронов, утративших связи с мишенями, восстановление проведения нервного импульса по сохранившимся нейритам, восстановление иннервации тканей за счет стимуляции роста нейритов, активацию нейротрофинов и факторов роста, необходимых для обеспечения всех компонентов механизмов регенерации, важных для восстановления иннервации.
Для эффективного запуска и поддержания процессов регенерации необходимо обеспечить продукцию трансгенов длительное время и на достаточном уровне, что требует адекватной доставки генетической конструкции в клетки-продуценты (мышечные волокна). Однократное введение плазмиды в мышечную ткань обеспечивает экспрессию трансгенов в течении не более чем одного месяца. Оптимальным способом доставки генетической конструкции является многократное внутримышечное введение плазмиды. Причем разовая доза лекарственного препарата должна вводиться малыми объемами в несколько точек мышцы в периневральной области. Это нужно с одной стороны для того чтобы увеличить число мышечных волокон, которые будут локально производить лекарственные белки, и для того чтобы уменьшить потери лекарственного средства за счет минимизации возможности обратного вытекания раствора при введении большого объема.
Для стимуляции восстановления нерва переднюю большеберцовую мышцу (иннервируется поврежденным нервом) обкалывали раствором плазмиды pNCure (60 мкл, 1 мкг/мкл) - 6 инъекций в разные точки мышцы.
Сравнение эффективности введения дозы лекарственного средства в несколько точек и в одну точку мышцы проводили с помощью гистологического окрашивания криосрезов мышцы после введения маркерной плазмиды, кодирующей бета-галактозидазу и с помощью иммуноферментного анализом среды культивирования мышечных эксплантов на hBDNF и huPA (Фиг. 4).
Применение заявляемой генотерапевтической конструкции в качестве генотерапевтического средства для лечения травм периферических нервов.
Примеры специфической фармакологической активности pNCure in vitro. В качестве тест-системы использовали линейные клетки НЕК293 и Neuro2a.
1. Содержание урокиназы (uPA) и BDNF в кондиционной среде культивирования клеток НЕК293, трансфицированных плазмидой pNCure и контрольной плазмидой pVax
Клетки НЕК293 представляют собой линию эмбрионального почечного эпителия человека (АТСС® CRL-1573™), которые легко трансфицируются (эффективность трансфекции до 95%), выдерживают большое количество пассажей, и поэтому являются общепринятым объектом для оценки эффективности экспрессии трансгенов после трансфекции исследуемыми плазмидами.
Клетки линии НЕК293 культивировали в стандартных асептических условиях СО2 инкубатора с использованием среды DMEM (Gibco или HyClone), содержащей 10% фетальной бычьей сыворотки (ФБС) (Gibco или HyClone) и 1% антибиотика-антимикотика (Gibco или HyClone). Пассирование клеток осуществляли раствором трипсина/ЭДТА.
Для оценки функциональной активности нейротрофных факторов BDNF и uPA использовали кондиционированную среду культивирования, полученную от НЕК293 (АТСС® CRL-1573™), трансфицированных плазмидой pNCure. Для трансфекции клетки высевали в ч. Петри из расчета 104 клеток/см2 и культивировали в СО2 инкубаторе при +37°С, используя укомплектованную среду роста. Для проведения трансфекции использовали клетки HEK293, достигшие плотности 70% монослоя. Перед трансфекцией клеткам меняли укомплектованную среду роста на DMEM, не содержащий сыворотку и антибиотик. Плазмиду pNCure и контрольную pVax в количестве 5 мкг смешивали с 250 мкл бессывороточной среды Opti-MEM (GIBCO). Через 5 мин в каждый раствор добавляли по 40 мкл реагента Lipofectamine2000™ (Invirogen), разведенного предварительно также в 250 мкл среды Opti-MEM. Полученную смесь плазмид и трансфекционного реагента инкубировали 30 мин при комнатной температуре, далее растворы добавляли в среду культивирования клеток HEK293 и инкубировали 6 часов. Через 6 ч. клеткам меняли среду на укомплектованную среду роста DMEM.
Для увеличения концентрации факторов BDNF и uPA в кондиционированной среде роста НЕК после трансфекции их pNCure клеточную массу наращивали до количества 5×107, собирали среду и концентрировали с помощью мембранной фильтрации в условиях центрифугирования. Диаметр пор фильтров должен быль таким, чтобы в конечном растворе содержались белки, молекулярная масса которых больше 20 кДа. Те же манипуляции осуществляли с контрольной средой. Для этого через 72 ч среды культивирования собирали, центрифугировали при 1000 g 10 мин для очищения среды от клеток и полученный супернатант переносили в чистые пробирки. Далее образцы концентрировали в 1000 раз с использованием фильтров (Centrycon YM-10, Millipore), задерживающих белки с молекулярной массой более 20 кДа.
Далее методом ИФА определяли концентрацию BDNF и uPA с использованием коммерческих наборов: для BDNF человека использовали коммерческий набор Human Free BDNF Quantikine ELISA Kit (R&D Systems); для uPA человека использовали Human uPA ELISA Kit (Abeam). Процедуру осуществляли в строгом соответствии с протоколом изготовителя. Используя стандартные образцы BDNF и uPA, готовили калибровочные пробы известной концентрации так, чтобы пробы №1 содержали 500 пг/мл BDNF и uPA, каждая последующая содержала BDNF и uPA в 2 раза меньше. Пробы №0 содержали только контрольный раствор Sample Diluent. Собранные образцы кондиционированной среды в количестве 800 мкл, а также стандартные образцы BDNF и uPA наносили в лунки 96-ти луночных планшет в трех повторах и инкубировали на шейкере в течение 12 ч. при +4°С. Далее планшеты промывали раствором Wash Buffer и в каждую лунку добавляли по 100 мкл биотинилированных моноклональных антител против BDNF и uPA человека. Через 3 ч инкубации планшеты отмывали и в лунки вносили по 100 мкл раствора комплекса стрептавидин-пероксидаза хрена. Через 1 ч планшеты промывали и вносили в лунки по 100 мкл прилагаемого к набору раствора субстрата ТМВ/Е. Планшеты инкубировали при комнатной температуре в течение 15 мин, затем в каждую лунку добавляли по 100 мкл буфера Stop Buffer, и оценивали интенсивность реакции на спектрофотометре Zenith 3200 Anthos при длине волны 450 нм.
Данные, полученные в результате измерения концентрации BDNF и uPA методом ИФА в кондиционированных средах клеток НЕК 293 после трансфекции pNCure или контрольной плазмидой pVax, представлены в Таблицах 1 и 2.
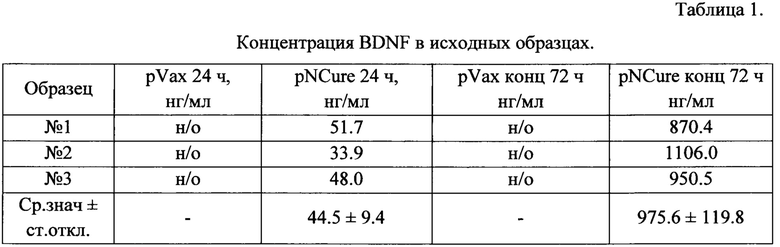
н/о - не обнаружено
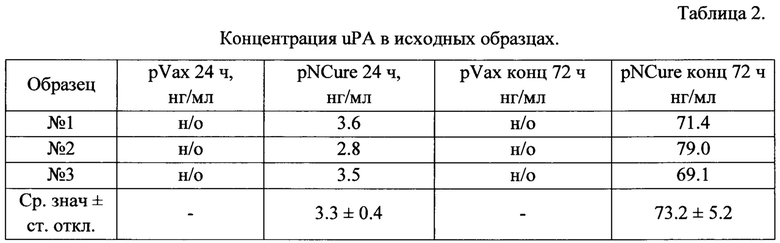
н/о - не обнаружено
Суммированные данные по определению концентраций BDNF и uPA в кондиционированной среде клеток НЕК293, трансфицированных pNCure или контрольной плазмидой pVax, приведены в Таблице 3.
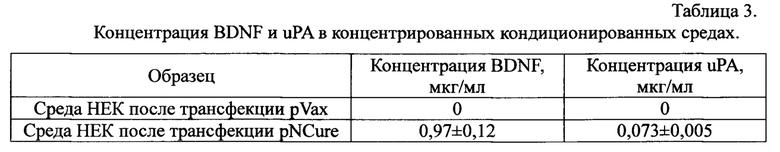
Коммерческие наборы Human Free BDNF Quantikine ELISA Kit (R&D Systems) и Human uPA ELISA Kit (Abeam) для измерения концентрации BDNF и uPA в среде культивирования определяют пороговую концентрацию BDNF от 62,5 пг/мл и uPA от 62,5 пг/мл. В результате наших измерений было обнаружено, что экспрессия белков в контрольной кондиционированной среде ниже порогового уровня определения.
Таким образом, при трансфекции клеток НЕК293 контрольной плазмидой pVax содержание BDNF и uPA в кондиционированной среде не обнаруживается. При трансфекции клеток НК293 pNCure происходит длительное и многократное увеличение экспрессии нейротрофных факторов BDNF и uPA в концентрациях, достаточных для проведения углубленного изучения биологической активности in vitro. Для проведения дальнейших экспериментов использовали контрольный образец кондиционированной среды и образец кондиционированной среды, содержащий нейротрофные факторы.
Таким образом, кондиционированная среда клеток НЕК293, трансфицированных pNCure содержит факторы BDNF и uPA в концентрациях, достаточных для оценки их биологической активности in vitro.
2. Влияние кондиционной среды культивирования клеток НЕК293, трансфицированных плазмидой pNCure и контрольной плазмидой pVax, на скорость роста нейритов нейробластомы мыши Neuro2a (модель нейрогенеза)
Линия Neuro2a (АТСС® CCL-131™) представляет собой линию клеток нейробластомы мыши, выдерживает большое количество пассажей, и способна дифференцироваться в нейральном направлении при снижении содержания сыворотки в среде культивирования до 1%. Данная линия также является хорошо описанной и воспроизводимой моделью для оценки влияния тестируемых веществ на скорость роста нейритов (модель нейритогенеза in vitro).
Для изучения биологического влияния продуцируемых плазмидой нейротрофных факторов BDNF и uPA, была проведена оценка их способности стимулировать дифференцировку клеток нейробластомы Neuro2a в нейральном направлении. При дифференцировке в нейрональном направлении клетки формируют отростки - нейриты. Для оценки влияния на нейральную дифференцировку Neuro2a клетки культивировали примерно до 40% монослоя в полной среде роста, содержащей DMEM, 10% раствор ФБС, 1% раствор антибиотика-антимикотика, и 1% раствор незаменимых аминокислот (NEAA, Non Essential Amino Acids, Invitogen). В день эксперимента клеткам меняли среду культивирования на кондиционированную среду клеток НЕК293, трансфицированных pNCure или контрольной плазмидой pVax. Среду разводили таким образом, что конечная концентрация BDNF составила 100 нг/мл. Контрольную среду разводили в той же пропорции. В качестве отрицательного контроля клеткам меняли среду на полную среду роста клеток, содержащую 10% раствор ФБС; в качестве положительного контроля для дифференцировки использовали среду роста с 1% пониженным содержанием ФБС. После этого клетки инкубировали 72 ч. Далее были проанализированы отростки, формирующиеся от тел клеток Neuro2a. Визуализацию осуществляли с использованием светового микроскопа, оборудованного CDD камерой. Подсчет числа отростков (зон ветвления) осуществляли с использованием программы ClickCounter2.
Было обнаружено, что через 72 ч в группе pNCure происходит значительное усиление формирования отростков, чего не наблюдается в группе отрицательного контроля (10% ФБС), где визуализируются лишь единичные клетки, имеющие отростки. На фиг. 5 показаны клетки, приобретающие морфологию нейронов (группы pNCure, 1% ФБС и группа pVax). Примечательно, что в группе pVax также наблюдается незначительное стимулирующее влияние на формирование нейритов по сравнению с отрицательным контролем 10% ФБС. Это может объяснять тем, что кондиционированная среда клеток НЕК, хоть и не содержит в высоких концентрациях исследуемые факторы BDNF и uPA, тем не менее, содержит другие факторы, которые также способны стимулировать нейрогенез. Тем не менее, при сравнении результатов, полученных в группах pNCure и pVax, было подтверждено статистически значимое влияние продуктов экспрессии pNCure (BDNF и uPA) на формирование нейритов. На фиг. 6 представлены результаты статистической обработки, подтверждающие стимулирующий эффект, оказываемый факторами продуцируемыми при трансфекции плазмидой pNCure, на клетки Neuro2a к дифференцировке в нейральном направлении.
Таким образом, показано, что трансфекция клеток НЕК293 плазмидой pNCure, содержащей кДНК BDNF человека и uPA человека вызывает значительное увеличение содержания этих факторов в среде культивирования по сравнению с контролем pVax. Этот эффект длительный, что свидетельствует о том, что введение трансгена в клетки обеспечивает стабильную его экспрессию на протяжении 72 ч. в культуре клеток. Кроме того, нейротрофные факторы BDNF и uPA, продуцируемые клетками, вызывают дифференцировку клеток Neuro2a в нейральном направлении, о чем свидетельствует увеличение формирование отростков (нейритов) от тел клеток.
3. Рост аксонов и миграция нейральных клеток из эксплантов в Матригель. содержащий среду культивирования клеток НЕК293, трансфицированных плазмидой pNCure и контрольной плазмидой pVax на трехмерной эксплантной модели спинального ганглия мыши в Матригеле
Для выделения ганглиев использовали постнатальных мышей линии C57BL в возрасте 3 дней. Культивировали спинальные ганглии ex vivo в геле (Матригеле). Метод позволяет изучать процессы, происходящие в тканевом экспланте спинального ганглия в трехмерном пространстве. В Матригель вводили кондиционированную среду культивирования клеток НЕК293, трансфицированных плазмидой pNCure (или контрольной плазмидой pVax), анализировали их влияние на экспланты в динамике в течение 7 д от момента помещения эксплантов в Матригель. На каждый эксплант наносили 60 мкл Матригеля, в который предварительно добавляли кондиционированную среду клеток НЕК293, трансфицированных pNCure (концентрация по BDNF составляла 100 нг/мл) или контрольной плазмидой (разведенной в одинаковой пропорции с опытной средой). Планшет помещали на 10 мин. в СО2 инкубатор (+37°С) для полимеризации Матригеля. Далее в лунки вносили по 400 мкл теплой среды RPMI1640, в которую также добавляли кондиционированную среду клеток НЕК293, трансфицированных препаратом pNCure (концентрация по BDNF 100 нг/мл) или контрольной плазмидой и помещали планшет в инкубатор. Визуализацию эксплантов осуществляли на 4 сутки при помощи инвертированного флуоресцентного микроскопа Axiovert 200М (Zeiss).
На фиг. 7 показаны микрофотографии трехмерных эксплантов СГ, сделанные на 4-е сутки культивирования эксплантов в Матригеле. Наличие нейротрофных факторов: BDNF и uPA (белковые продукты pNCure) стимулирует рост нейритов из эксплантов СГ в Матригель (стрелки на вкладке pNCure) по сравнению с контрольным образцом (pVax). При этом в контроле, как и в опыте, происходит миграция нейральных клеток из СГ в Матригель (стрелки на вкладке pVax), однако роста нейритов практически не наблюдается. Дополнительно на нижней панели фигуры приведены увеличенные области, в которых показаны мигрирующие клетки в контрольной группе pVax и растущие нейриты в опытной группе pNCure. Исходя из этих данных видно, что присутствие нейротрофных факторов BDNF и uPA обладает мощным стимулом для регенерации поврежденных путем денервации аксонов спинальных ганглиев.
Визуализацию ядер нейральных клеток, мигрировавших из эксплантов СГ в Матригель оценивать с использованием конфокальной микроскопии (микроскоп Leica TCS SP5; программное обеспечение LAS AF). Флуоресцентный краситель DAPI визуализировали с помощью лазера длиной волны 405 нм. Изображения со всех лунок получали при одинаковых настройках микроскопа.
При анализе изображений оказалось, что присутствие в Матригеле нейротрофных факторов BDNF и uPA, содержащихся в кондиционированной среде клеток, трансфицированных pNCure, стимулирует как скорость миграции нейральных клеток (стрелки на Фиг. 8), так и их число (синяя флуоресценция) по сравнению с контролем. Обнаруженная разница между группой pNCure и контроль pVax была очевидна при визуальном анализе.
Таким образом, при изучении биологической активности препарата in vitro было обнаружено, что трансфекция клеток НЕК293 плазмидой pNCure, содержащей кДНК BDNF человека и uPA человека вызывает значительное увеличение содержания этих факторов в среде культивирования. Этот эффект является длительным и наблюдается на протяжении как минимум 72 ч. Нейротрофные факторы BDNF и uPA усиливают дифференцировку клеток Neuro2a в нейральном направлении, усиливая формирование отростков (нейритов) от тел клеток. Более того, присутствие нейротрофных факторов BDNF и uPA обладает мощным стимулом для регенерации нейритов и миграции клеток спинального ганглия на трехмерной эксплантной модели СГ мыши в Матригеле.
4. Восстановление проводимости поврежденного нерва при действии плазмиды pNCure
Восстановление проводимости нерва оценивали с помощью регистрации вызванных потенциалов действия, регистрируемых с поверхностной ветви общего малоберцового нерва. Мышей на 24-е сутки после повреждения нерва и введения лекарственного препарата эксперимента подвергали эвтаназии. Стимуляцию нерва проводили в электрофизиологической камере с раствором Лайли на 7-8 мм проксимальнее места повреждения с использованием пары серебряных электродов. Параметры стимуляции: частота импульса 1 Гц, продолжительность импульса 0.05 мс и супрамаксимальная амплитуда 10 В. Промежуток времени от забоя животного до момента регистрации вызванных потенциалов действия составлял около 35-45 минут для каждого животного. Суммарные потенциалы действия нерва (СПДН) регистрировали 3 раза для каждого нерва с использованием монополярного аспирирующего электрода, который располагался на 10 мм дистальнее места повреждения. Регистрирующий электрод был связан через усилитель сигнала и аналого-цифровой преобразователь (Е14-140 L-card) с компьютером. Регистрацию и анализ данных осуществляли с помощью программы PowerGraph Professional 3.3. Регистрируемый СПДН характеризуется двумя параметрами - латентным периодом и амплитудой. Латентный период - время от момента стимуляции нерва до момента регистрации вызванного потенциала действия характеризует скорость проведения возбуждения по нерву, причем его величина обратно пропорциональна скорости проведения возбуждения по нерву. Таким образом, увеличение латентного периода после повреждения свидетельствует о снижении скорости проведения вызванных потенциалов по поврежденному нерву. Поскольку скорость проведения вызванных потенциалов действия по нерву определяется качеством миелиновой оболочки нервных волокон, то укорочение латентного периода свидетельствует о лучшем ее восстановлении.
Амплитуда характеризует количество нервных волокон, участвующих в проведении возбуждения (Fugleholm К, 2000; Vleggeert-Lankamp CLAM, 2007). Чем меньше амплитуда СПДН, тем меньшее количество активных нервных волокон содержит поврежденный нерв.
Повреждение общего малоберцового нерва методом раздавливания вызывало нарушение функции нерва, о чем свидетельствовало уменьшение амплитуды и увеличение латентного периода СПДН. Результаты демонстрируют, что на 24 сутки после начала эксперимента при измерении СПДН с поверхностной ветви общего малоберцового нерва с помощью электрофизиологической методики у животных, которым делали инъекцию pNCure в дозе 60 мкг (1 мг/мл), амплитуда СПДН была достоверно выше по сравнению с животными, которым инъецировали контрольный вектор pVax. Так, амплитуда СПДН поверхностной ветви общего малоберцового нерва у мышей, получивших 60 мкг pNCure, на 24 день проведения эксперимента была в 1.7 раза больше, чем амплитуда СПДН у мышей из группы "-К": 0.475 (0.338; 0.612) и 0.285 (0.245; 0.353) мВ (n=10; р<0.001), соответственно. Сравнимый рост амплитуды СПДН наблюдали и в группе положительного контроля (+К) - 0.457 (0.329; 0.573) мВ (n=10; p<0.05). В группе экспериментальных животных, которым делали инъекции 120 мкг pNCure ("pNCure_2"), на 24 день проведения эксперимента амплитуда СПДН была в 2.2 раза больше, чем амплитуда СПДН у мышей из группы "-К" и составила 0.627 (0.506; 0.832) мВ (n=5; p<0.001). Хотя достоверно показатели амплитуды СПДН между группами "pNCure_1" и "pNCure_2" не отличались (р=0.174), видно, что инъекция 120 мкг плазмиды pNCure способствует лучшему восстановлению проводимости нервных волокон, чем инъекция 60 мкг pNCure (фиг. 9).
Латентный период СПДН поврежденного нерва мышей, которых подвергали лечению pNCure, как в дозе 60 мкг, так и в дозе 120 мкг, на 24 день проведения эксперимента достоверно не отличался от аналогичного показателя групп контроля (фиг. 10). Так в группе "pNCure_1" латентный период составил 0.715 (0.622; 0.893) мс, а в группе "pNCure_2" - 0.507 (0.45; 0.887) мс. В группах контроля латентный период составил 0.861 (0.68; 0.983) мс в группе "-К" и 0.771 (0.7; 0.94) мс в группе "+К". Хотя статистически достоверной разницы по величине латентного периода СПДН между группами обнаружено не было, наблюдается тенденция к уменьшению латентного периода СПДН (увеличению скорости проведения СПДН) после введения pNCure, причем наблюдается дозовая зависимость. Поскольку данная постановка эксперимента предполагала многократные инъекции плазмидного препарата (по причине ориентации исследования на последующее применение препарата у человека) и не могла продолжаться менее 24 сут, то отсутствие статистически достоверной разницы в величине латентного периода между экспериментальными и контрольными группами, вероятно, связано, как с относительно щадящим методом повреждения нерва, так и с более высоким эндогенным регенераторным потенциалом мышей. Благодаря этому существующая в начальные моменты разница в продолжительности латентного периода СПДН между экспериментальными и контрольными группами на 24 сут уменьшается.
Полученные данные свидетельствуют в пользу того, что нервы животных, которым делали инъекцию плазмиды pNCure в дозе 60 и 120 мкг, содержат большее количество функциональных нервных волокон, чем у животных из группы контроля (показатель амплитуда СПДН), причем более выраженный эффект наблюдается при инъекции двукратной дозы - 120 мкг.
5. Оценка восстановления структуры нерва при действии плазмиды pNCure Оценку влияния генотерапевтического лекарственного средства на восстановление структуры поврежденного нерва осуществляли с помощью иммунофлуоресцентного анализа срезов поврежденных нервов. Для этого нервы после измерения проводимости на 24-й день эксперимента фиксировали 4% раствором формальдегида в фосфатном буфере в течение 3 часов. Фиксированные препараты отмывали от формалина в фосфатно-солевом буфере в течение 3 часов и замораживали с помощью погружения в жидкий азот в среде для замораживания Tissue-Tek (Sakura Finetek). Из участка нерва, расположенного на 3 мм дистальнее места повреждения, готовили поперечные срезы толщиной 6 мкм. Полученные срезы отмывали в фосфатно-солевом буфере (рН 7.0) 3 раза по 5 минут. Первичные и вторичные антитела разводили в фосфатно-солевом буфере, содержащем 0,1% Triton X-100 и 5% нормальной козьей сыворотки. На срезы наносили разведенные в 80 раз первичные кроличьи антитела к белку цитоскелета аксонов NF200 (Abeam, ab8135). Часть срезов окрашивают кроличьими неспецифическими IgG в качестве отрицательного контроля (Thermo scientific, #NC-100-P0). Срезы с нанесенными на них первичными антителами инкубировали при 4°С в течение ночи. Затем срезы отмывали в фосфатно-солевом буфере (рН 7.0) 3 раза по 5 минут и наносили на них вторичные антитела: козьи антитела против иммуноглобулинов кролика, конъюгированные с Alexa 488 (Invitrogen). Срезы инкубировали в темноте во влажной камере в течение 1 часа. Затем срезы отмывали в фосфатно-солевом буфере (рН 7.0) 3 раза по 5 минут и наносили на них 300 мМ раствор 4-,6-диамидино-2-фенилиндола (DAPI) (Invitrogen) в фосфатно-солевом буфере. Срезы инкубировали в течение 2-х минут, а затем снова отмывали в фосфатно-солевом буфере (рН 7.0) 3 раза по 5 минут. Со срезов удаляли избыточную жидкость и заключали в нефлюоресцирующую среду.
Съемку полученных срезов проводили с помощью микроскопа Leica DM6000, оборудованного CCD DFC490 камерой (Leica Microsystems). Возбуждение красителя Alexa 488 проводят при длине волны 488 нм, а регистрацию вызванного возбуждения - при максимуме длин волн 546 нм. Количество окрашенных аксонов на полученных снимках оценивают при помощи программы Metamorph 7.1.
При анализе окрашенных криосрезов поврежденных нервов к маркеру зрелых аксонов NF200 оказалось, что через 24 суток после начала эксперимента у животных, которым вводили плазмиду pNCure в дозе 60 мкг, количество и размер структур, положительно окрашенных на аксоны, было больше по сравнению с животными, которым вводили контрольный вектор pVax. В нервах, полученных от животных из группы "pNCure_1" наблюдали в 2.1 раза больше аксонов по сравнению с нервами животных из группы контроля: 573 (489; 619) и 276 (234; 311) (n>5, p<0.05), соответственно (фиг. 11). В нервах, полученных от мышей из групп "pNCure_2" и "+К", количество нервных волокон на срез составило 554 (518; 633) и 461 (407; 545), что составляет статистически достоверную разницу с аналогичным показателем мышей из группы "-К", но достоверно не отличается от группы "pNCure_1".
Полученные данные свидетельствуют о более эффективном восстановлении структуры нерва (числа нейритов) в поврежденных нервах у животных, которым вводили генетическую конструкцию pNCure.
Таким образом, при изучении биологической активности препарата in vivo, установлено, что плазмида pNCure достоверно стимулирует восстановление структуры нерва. Помимо восстановления структуры поврежденного нерва плазмида pNCure стимулирует восстановление его функции, характеризующееся увеличением числа нервных волокон, проводящих потенциалы действия (увеличение амплитуды вызванных потенциалов действия по сравнению с контролем).
Исследуемый препарат показал себя практически нетоксичным и может быть использован для терапии повреждений нервной ткани у человека. За минимальную терапевтически эффективную дозу рекомендуется принимать дозу, эквивалентную исследуемой дозе препарата 20 мкг/кг для животных моделей. При пересчете дозы на человека использовано соотношение масс животного и человека с использованием коэффициента безопасности, рекомендованного различными руководствами (Миронов А.Н., Бунатян Н.Д. и др. Руководство по проведению доклинических исследований лекарственных средств, Москва, 2012. - 944 с.). Таким образом, начальная доза для человека составляет 2 мкг/кг веса (140 мкл на пациента весом 70 кг). Учитывая небольшой объем, в котором возможно периневральное многоточечное введение препарата, максимальный объем введения принят равным 2 мл (2 мг) (данный объем распределен на многоточечное введение - обкалывание зоны повреждения). В любом случае введение препарата в соответствии с настоящим изобретением определяет лечащий врач на основании специфических параметров пациента, таких как возраст, масса тела, пол, степень повреждения нервной ткани и т.п.
Примеры, демонстрирующие безопасность.
Исследование диссеминации плазмиды pNCure в органы и ткани мыши после многократного внутримышечного введения.
Для изучения способности исследуемой плазмиды pNCure диссеминировать из места введения (передняя большеберцовая мышца) в органы и ткани использовали самцов мышей линии С57/В16 в возрасте 10-12 недель и массой 23-27 г. Все манипуляции с животными выполняли в соответствии с «Правилами проведения работ с использованием экспериментальных животных», утвержденными этическим комитетом при ФФМ МГУ. На протяжении всего эксперимента мыши находились на стандартном режиме в виварии и получали стандартный гранулированный корм (ООО «Аллер Петфуд», Россия) ad libitum при свободном доступе к воде. С 1-го по 7-ой день мышей выдерживали при описанных выше стандартных условиях без осуществления манипуляций. На 8-ой день эксперимента мышам делали инъекцию генетической конструкции pNCure с целью исследования ее диссеминации из места введения. Для этого каждую из мышей анестезировали инъекцией 400 мкл 2.5% раствора 2,2,2-трибромэтанола («Sigma»), фиксировали ее на операционном столике животом вверх. Голень левой задней конечности очищали от шерсти и дезинфицировали 70% спиртом. Инъекцию генетической конструкции pNCure осуществляли инсулиновым шприцем (BD) с толщиной иглы 29G путем обкалывания передней большеберцовой мышцы (6 инъекций, суммарный объем - 60 мкл). Обкалывание осуществляли чрескожно от нижней трети голени вдоль оси мышцы к верхней трети голени. Каждый раз иглу вводили срезом вверх, а по ходу продвижения иглы понемногу вводили раствор генетической конструкции. После введения раствора плазмиды шприц с иглой каждый раз очень быстро извлекали. Мышь отвязывали от столика, присваивали ей номер экспериментальной группы и помещали в клетку.
В исследовании было 2 группы экспериментальных животных.
В первой группе ("pNCure_1") мышам делали внутримышечную инъекцию плазмиды pNCure в концентрации 1 мг/мл.
Во второй группе ("pNCure_2") мышам делали внутримышечную инъекцию плазмиды pNCure, в концентрации 2 мг/мл.
Мышам обоих групп ("pNCure_1" и "pNCure_2") за период проведения всего исследования делали по 4 инъекции: на 8-й, 11-й, 15-й, 19-й дни эксперимента. Всего в эксперименте использовали 26 мышей: по 12 мышей в каждой группе "pNCure_1" и "pNCure_2". Еще двум мышам не делали инъекций генетических конструкций и использовали их в качестве доноров органов и тканей для приготовления контрольных и калибровочных образцов.
Через 1, 2 и 4 недели после окончания курса внутримышечных инъекций pNCure, т.е. на 26-й, 33-й и 40-й день эксперимента из каждой группы забивали по 4 мыши методом цервикальной дислокации. У них забирали образцы крови (300-400 мкл), а также легких, печени, почек, селезенки, сердца, головного мозга, мышечной ткани из места введения и из контралатеральной конечности.
Образцы органов размером приблизительно 2×2×3 мм (масса около 20-30 мг) взвешивали и помещали в полипропиленовые криопробирки, замораживали в жидком азоте и хранили при -80°С до момента проведения исследования.
Образцам крови давали свернуться (30 минут при комнатной температуре) и позволяли сыворотке крови отделиться от тромба (18 часов при +4°С). Затем пробирки со свернувшейся кровью центрифугировали при 10000g в течение 15 минут, отделившуюся сыворотку крови переносили в криопробирки, замораживали в жидком азоте и хранили при -80°С до момента анализа.
По завершении эксперимента полученные в разные временные точки эксперимента образцы органов и тканей лизировали при 56°С в течение 18-24 часов (до полного растворения) в 200 мкл раствора, содержащего 50 мМ Tris (рН 8.0), 100 мМ ЭДТА, 0.5% SDS и 0.7 мг протеиназы К на 1 мл раствора. Полученные лизаты разводили деионизованной водой в 10 раз и использовали в качестве матрицы для ПЦР в реальном времени. Лизаты органов и тканей, содержащих ДНК, до момента исследования хранили при -20°С. Из образцов сыворотки крови ДНК выделяли фенол-хлороформным методом с последующим переосаждением изопропанолом и восстановлением деионизованной водой до исходного объема сыворотки крови.
Оценку диссеминации плазмидной ДНК из места введения в органы и ткани оценивали методом ПЦР в реальном времени. Для этого использовали пары праймеров, приведенные в таблице 4.

Амплификацию контрольных и экспериментальных образцов производили в термоциклере iCycler с системой детекции iQ5 (Bio-Rad) при помощи набора для ПЦР в реальном времени qPCRmix-HS SYBR (#PK147L) согласно прописи реакционной смеси и программе амплификации рекомендуемой производителем. Количество раствора вносимой ДНК составило 1 мкл на 25 мкл тотального объема реакционной смеси. Отжиг праймеров на матрицу осуществляли при 57.5°С, а элонгацию осуществляли при 72°С в течение 20 секунд. Регистрацию флуоресцентного сигнала (SYBR) в канале FAM и анализ полученных результатов осуществляли при помощи программы CFX Manager Software (Bio-Rad). В качестве анализируемого параметра использовали параметр ΔCt, обратно пропорциональный количеству исходной ДНК в реакционной смеси и характеризующий цикл, когда уровень сигнала превысил значение фона. Параллельно с обнаружением плазмидной ДНК в органах и тканях мыши определяли количество геномной ДНК (ген мышечного актина альфа-1 - Acta1). Данный тест использовали в качестве внутреннего контроля для сравнения качества выделения ДНК из образцов и органов мыши от образца к образцу и контроля на деградацию ДНК. Нормирование определенной в образце плазмидной ДНК осуществляли на массу лизированного образца органа или ткани.
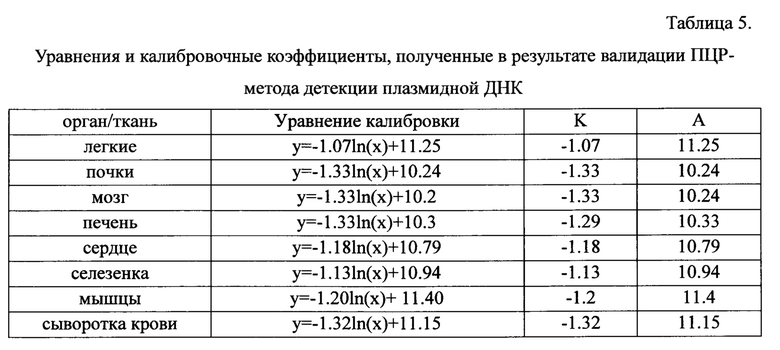
Методика анализа содержания плазмидной и геномной ДНК в образцах сыворотки крови, образцов органов и тканей была валидирована. Для валидации использовались растворы плазмиды pNCure на деионизованной воде концентрацией 0.5 нг/мкл, 5 пг/мкл, 50 фг/мкл, 0.5 фг/мкл, а также растворы плазмиды pNCure на экстракте сыворотки крови и лизатах образцов органов и тканей с концентрацией pNCure 0.5 нг/мкл, 5 пг/мкл, 50 фг/мкл, 0.5 фг/мкл. Калибровочные образцы подвергали анализу на содержание плазмидной ДНК методом ПЦР в реальном времени с использованием праймеров pNCure-f и pNCure-r (в 3 независимых повторах). Полученные данные использовали для построения калибровочных графиков, выведения уравнений с целью последующего вычисления концентрации плазмиды pNCure в образцах сыворотки крови, образцах органов и тканей. Для получения калибровочных графиков использовали логарифмическую интерполяцию  зависимости порогового цикла (ΔCt) от количества внесенной плазмидной ДНК.
зависимости порогового цикла (ΔCt) от количества внесенной плазмидной ДНК.
Выполненная валидация подтвердила, что плазмидная ДНК одинаково хорошо и воспроизводимо определяется во всех типах матриц: от деионизованной воды до экстрактов сыворотки крови, образцов органов и тканей. Поскольку как таковой экстракции ДНК из образцов органов и тканей (за исключением сыворотки крови) не проводилось (лишь лизис и разбавление в 10 раз), то и эффективность экстракции плазмидной ДНК из образцов органов и тканей не оценивали. В эксперименте также анализу подвергали образцы, содержащие плазмидную ДНК в количестве 5 аг на ПЦР-реакцию. Однако полученные для них параметры ΔCt оказались ниже предела чувствительности метода.
Определение количества плазмидной ДНК в образцах органов и тканей С использованием калибровочных кривых плазмидной ДНК в образцах крови, а также лизатах органов и тканей были получены уравнения, позволяющие определять количество плазмидной ДНК в экспериментальных образцах органов и тканей. Общий вид полученного уравнения:
 где
где
Р - искомое количество плазмидной ДНК в экспериментальном образце,
ΔCt - значение порогового цикла для соответствующего образца
А и k - коэффициенты из уравнения , описывающего калибровочную кривую. Уравнения, а также калибровочные коэффициенты, полученные в результате валидации отражены в таблице 5.
Определение пределов чувствительности (LOD) и количественного определения (LOQ) ПЦР-метода детекции плазмидной ДНК
Для определения пределов чувствительности (Limit of Detection) и пределов количественного определения (LOQ) плазмидной ДНК в образцах органов и тканей, а также в сыворотке крови использовали данные измерений, полученные в ходе валидации методики при анализе «холостых» проб лизатов и экстрактов, не содержащих плазмидную ДНК. LOD оценивается как количество плазмидной ДНК в реакционной смеси, соответствующее утроенному среднему значению ΔCt (из 3), полученному при анализе «холостых» проб лизатов или экстрактов, a LOQ - количество плазмидной ДНК, соответствующее десяти средним значениям ΔCt, полученных при анализе «холостых» проб [3]. Формулы для вычисления LOD и LOQ приведены ниже:
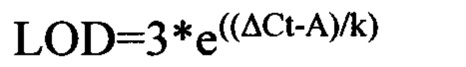
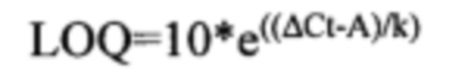 где
где
LOD - предел обнаружения,
LOQ - предел измерения,
Р - искомое количество плазмидной ДНК в экспериментальном образце,
ΔCt - значение порогового цикла для соответствующего образца
А и k - коэффициенты из уравнения 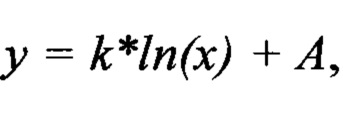 описывающего калибровочную кривую.
описывающего калибровочную кривую.
Результаты определения пределов чувствительности (LOD) и количественного определения (LOQ) ПЦР-метода детекции плазмидной ДНК приведены в таблице 6.

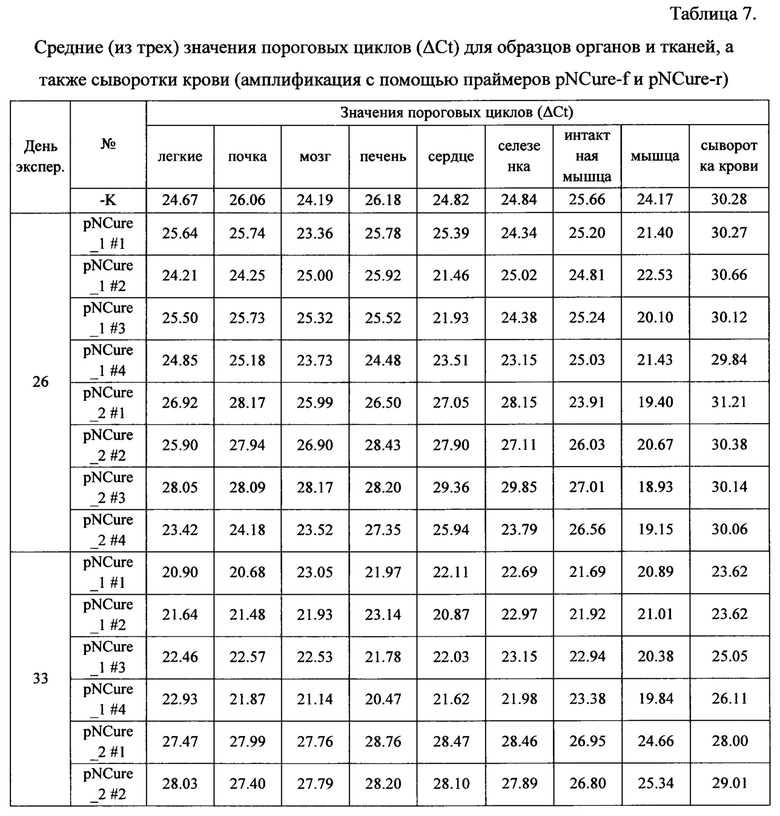
Результаты исследования диссеминации плазмиды pNCure в органы и ткани мышей из области введения приведены в таблицах.

Расчет количества плазмидной ДНК, попавшей в состав реакционной ПЦР-смеси, осуществляли при помощи приведенной выше формулы 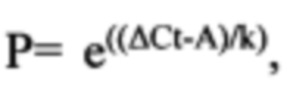 с использованием калибровочных коэффициентов.
с использованием калибровочных коэффициентов.
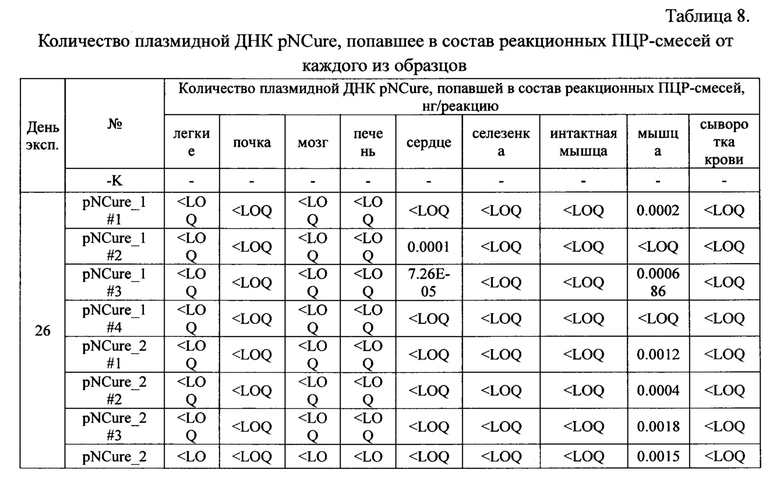
Расчет количества плазмидной ДНК в исходных образцах сыворотки крови, а также образцах органов и тканей осуществляли путем нормирования данных на массу образца с учетом всех примененных разбавлений. Полученные данные приведены в Таблице 9.
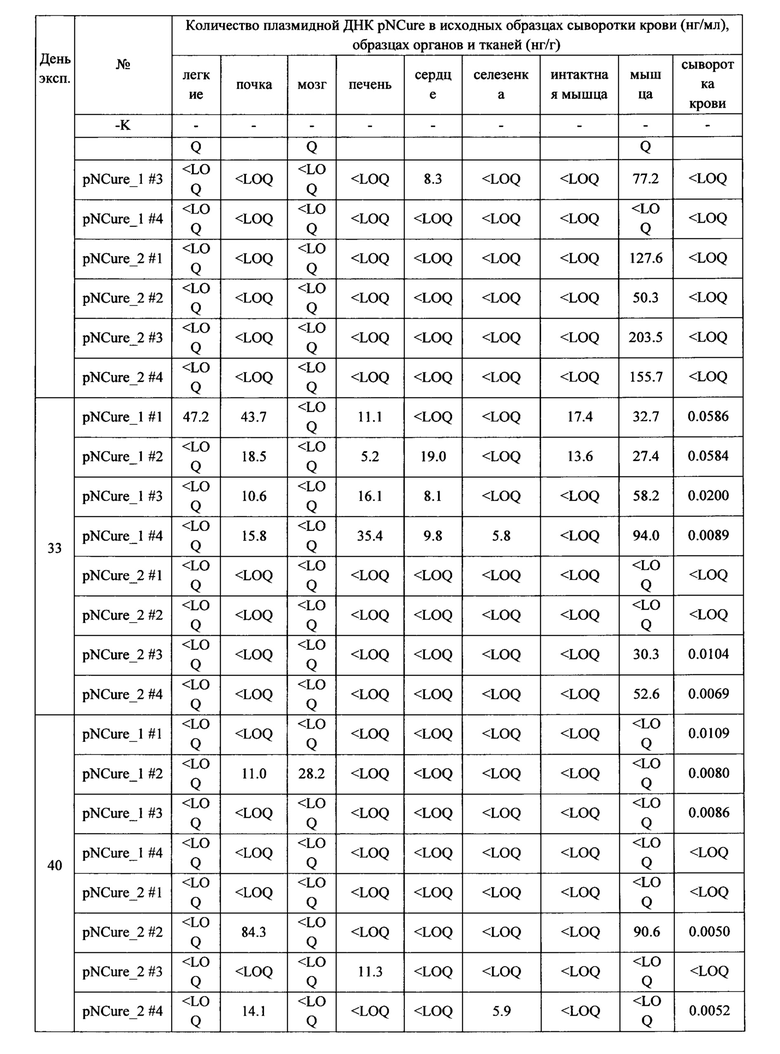
Полученные данные демонстрируют, что в абсолютном большинстве образцов плазмидная ДНК отсутствует или находится ниже предела чувствительности метода обнаружения. В некоторых образцах обнаруживаются следовые количества выбранной специфической последовательности плазмидной ДНК.
Для проверки экспрессии данной генетической конструкции вне сайта введения был проведен иммуноферментный анализ на мозговой нейротрофический фактор человека в гомогенатах образцов органов и тканей (Human Free BDNF Immunoassay, R&D, #DBD00). Результаты исследования приведены на фиг. 12. Видно, что достоверного повышения уровня экспрессии белков, кодируемых данной плазмидой, в органах и тканях животных (за исключением мышцы, куда делали инъекцию) не наблюдается.
На основании полученных результатов можно сделать следующие выводы о том, что диссеминация плазмиды pNCure после ее многократного внутримышечного введения из места введения в органы и ткани мыши не значительна. Увеличение количества вводимой плазмиды pNCure в 2 раза в целом отражается на большем количестве плазмиды в месте инъекции, однако, согласно полученным данным, не способствует усилению ее диссеминации из места введения. С течением времени количество плазмиды pNCure в органах и тканях животного прогрессивно убывает, хотя следы выбранной специфической последовательности плазмидной ДНК можно обнаружить в незначительных количествах в органах и тканях части животных через 4 недели после последней инъекции. При этом максимально-детектируемые количества выбранной специфической последовательности плазмидной ДНК составляют не более 0.2% от стандартной введенной дозы (60 мкг) и не сопровождаются эктопической экспрессией трансгена. По-видимому, обнаруживаемые фрагменты плазмидной ДНК не являются функционально-активными.
Генотоксичность
Исследование выполнено на половозрелых самцах мышей Fi (СВАхС57В1/6), массой 20-22 г., в возрасте 8-9 недель, полученных из питомника лабораторных животных «Столбовая». Условия содержания животных соответствовали стандартам, указанным в: Guide for the care and use of laboratory animals/ National Academy press; Washington, D.C., 2011; Приказе Минздрава России от 01.04.2016 №199н "Об утверждении Правил надлежащей лабораторной практики"; СП 2.2.1.3218-14 «Санитарно-эпидемиологические требования к устройству, оборудованию и содержанию экспериментально-биологических клиник (вивариев)» от 29 августа 2014 г. До начала эксперимента мыши содержались в лаборатории в течение не менее 5 дней. Во время этого периода осуществлялся ежедневный осмотр внешнего состояния животных. Мыши с обнаруженными в ходе осмотра отклонениями от нормы исключались из эксперимента.
Оценку ДНК-повреждающей активности препарата проводили методом ДНК-комет в щелочной версии. Препарат из расчета 20 и 200 мг/кг вводили животным в бедренную мышцу, однократно, на сроки 3 и 24 часа. В качестве отрицательного контроля использовали животных, которым внутримышечно вводили физиологический раствор в эквивалентных количествах. В качестве положительного контроля использовали животных, которым вводили внутрибрюшинно метилметансульфонат в дозе 40 мг/кг на срок 3 часа. Умерщвление животных осуществляли смещением шейных позвонков. Выделяли бедренную мышцу, в которую проводили инъекцию препарата, обе бедренные кости, печень и головной мозг. Эпифизы бедренных костей срезали и вымывали клетки костного мозга 2 мл предварительно охлажденного до 4°С фосфатно-солевого буфера (ФСБ), содержащего 20 mM EDTA-Na2 и 10% ДМСО (рН 7.5). Печень и головной мозг измельчали в стеклянной пробирке в 2 мл того же буфера и раздавливали стеклянной палочкой. Бедренную мышцу гомогенизировали в 2 мл ФСБ с использованием гомогенизатора IKA Т25 при 8000 об/мин в течение 15 секунд. Пробирки с измельченными тканями выдерживали 5 мин при комнатной температуре для осаждения крупных фрагментов. Полученные суспензии клеток в объеме 60 мкл вносили в пробирки, содержащие 240 мкл 1% раствора легкоплавкой агарозы в буфере ФСБ, подогретого до 36°С (микротермостат «Термит», Россия) и ресуспендировали. Затем 60 мкл раствора агарозы с клетками наносили на предварительно покрытые 1% универсальной агарозой предметные стекла, покрывали покровным стеклом и помещали на лед. Далее все операции проводили в затемненном помещении при желтом свете. После затвердевания агарозы (около 5-10 минут) покровные стекла осторожно удаляли, микропрепараты помещали в стеклянную кювету (тип Шиффендекер), заливали предварительно охлажденным до 4°С лизирующим буфером (10 mM Tris-HCl [рН 10], 2.5М NaCl, 100 mM EDTA-Na2, 1% Triton Х-100, 10% ДМСО) и инкубировали не менее 1 часа. После окончания лизиса микропрепараты переносили в охлажденный до 4°С буфер для электрофореза - 300 mM NaOH, 1 mM EDTA-Na2 [рН>13]. Инкубировали в течение 20 мин для реализации щелочно-лабильных сайтов и щелочной денатурации ДНК. После микропрепараты переносили в камеру для электрофореза (SubCell GT, "Bio-Rad"), заполненную свежим охлажденным буфером и проводили электрофорез в течение 20 минут при напряженности поля 1 V/cm и силе тока ~300 mA. По окончанию электрофореза микропрепараты переносили в стеклянную кювету, отмывали в течение 10 мин в фосфатно-солевом буфере и фиксировали в 70% растворе этилового спирта в течение 15 минут. После фиксации микропрепараты высушивали и хранили до анализа при комнатной температуре.
Непосредственно перед микроскопированием препараты окрашивали флуоресцирующим красителем SYBR Green I (1:10000 в ТЕ-буфере [рН 8.5] с 50% глицерином,) в течение 30 мин в темноте. Анализ проводили на эпифлуоресцентном микроскопе Микмед-2 12Т («Ломо», Россия), совмещенном с цифровой камерой высокого разрешения (VEC-335, «ЭВС», Россия), при увеличении х200. Полученные с микропрепаратов изображения ДНК-комет анализировали с использованием программного обеспечения CASP 1.2.2. В качестве показателя поврежденности ДНК использовали процентное содержание ДНК в хвосте ДНК-комет (%ДНК в хвосте). С каждого микропрепарата снимали и анализировали не менее 100 ДНК-комет.
Перед статистической обработкой выборки каждой экспериментальной группы проверялись на нормальность и гомогенность дисперсий показателей с использованием критерия Шапиро-Уилка и критерия Барлетта, соответственно. При соответствии указанным условиям проводили статистическую оценку с использованием непараметрического критерия Даннета. Если распределение отличалось от нормального, и/или выявлялись отличия в дисперсиях, в соответствии с рекомендуемыми подходами к статистической обработке данных метода ДНК-комет проводили логарифмическое преобразование исходных показателей с последующим анализом с использованием непараметрического критерия Даннета.
Поврежденность ДНК, оцениваемая по показателю %ДНК в хвосте, в органах/тканях животных негативного контроля находилась в пределах исторических контролей лаборатории. У животных группы позитивного контроля введение метилметансульфоната в дозе 40 мг/кг привело к статистически значимому увеличению ДНК-повреждений в исследованных органах/тканях животных.
В клетках костного мозга животных группы негативного контроля уровень ДНК-повреждений зарегистрирован на уровне 3.7±1.6% ДНК в хвосте. У животных, обработанных препаратом в дозах 20 и 200 мг/кг, данный показатель составил 3.4±0.9% и 3.2±0.9% ДНК в хвосте, соответственно. Сопоставление полученных данных с показателем соответствующего негативного контроля не выявило статистически значимых отличий.
Влияние препарата pNCure на уровень ДНК-повреждений в органах и тканях мышей в условиях 3-часовой экспозиции
В клетках печени животных, обработанных растворителем, уровень ДНК-повреждений зарегистрирован на уровне 3.5±0.3% ДНК в хвосте. Не выявлено значимых отличий по сравнению с контролем при введении животным препарата в дозах 20 и 200 мг/кг: 3.2±0.7 и 3.0±0.5% ДНК в хвосте, соответственно.
Уровень ДНК-повреждений в клетках головного мозга контрольных животных зарегистрирован на уровне 1.9±0.6% ДНК в хвосте. После введения препарата в дозах 20 и 200 мг/кг показатель составил, соответственно, 1.8±0.2 и 1.8±0.5 %ДНК в хвосте, что статистически значимо не отличалось от показателя соответствующего негативного контроля.
В клетках мышечной ткани животных группы негативного контроля уровень ДНК-повреждений зарегистрирован на уровне 5.4±0.9% ДНК в хвосте. У животных, обработанных препаратом в дозах 20 и 200 мг/кг, данный показатель составил 5.8±0.8 и 5.3±0.7% ДНК в хвосте. Сопоставление полученных данных с показателем соответствующего негативного контроля не выявило статистически значимых отличий.
Влияние препарата pNCure на уровень ДНК-повреждений в органах и тканях мышей в условиях 24-часовой экспозиции
В клетках костного мозга животных, обработанных растворителем, уровень ДНК-повреждений зарегистрирован на уровне 2.4±1.0% ДНК в хвосте. После введения препарата в дозах 20 и 200 мг/кг данный показатель оказался сходным: соответственно, 2.4±0.8 и 2.4±0.5% ДНК в хвосте.
Аналогично, в клетках печени не выявлено значимых отличий по сравнению с контролем при введении препарата в дозах 20 и 200 мг/кг: 2.0±0.3 и 1.9±0.5% ДНК в хвосте против 2.3±0.3% для негативного контроля.
Уровень ДНК-повреждений в клетках головного мозга животных негативного контроля зарегистрирован на уровне 1.1±0.3% ДНК в хвосте. После введения препарата в дозах 20 и 200 мг/кг показатель составил, соответственно, 1.2±0.3 и 1.3±0.7% ДНК в хвосте, что статистически значимо не отличалось от показателя соответствующего негативного контроля.
В клетках мышечной ткани мышей группы негативного контроля уровень ДНК-повреждений зарегистрирован на уровне 5.3±1.5% ДНК в хвосте. У животных, обработанных препаратом в дозах 20 и 200 мг/кг, данный показатель составил 4.9±1.0 и 5.5±0.9% ДНК в хвосте, соответственно. Попарное сопоставление полученных данных с показателями соответствующего негативного контроля не выявило статистически значимых отличий.
Таким образом, установлено, что препарат pNCure при внутримышечном введении мышам в дозах 20 и 200 мг/кг в условиях 3-й 24-часовой экспозиции не индуцирует ДНК-повреждений в клетках костного мозга, печени, головного мозга и мышечной ткани.
Результаты проведенных экспериментов позволяют сделать следующие выводы: Препарат pNCure при однократном внутримышечном введении в дозах 20 и 200 мг/кг на срок 3 часа не индуцирует повреждения ДНК в клетках костного мозга, печени, головного мозга и мышечной ткани мышей. Препарат pNCure при однократном внутримышечном введении в дозах 20 и 200 мг/кг на срок 24 часа не индуцирует повреждения ДНК в клетках костного мозга, печени, головного мозга и мышечной ткани мышей.
Полученные результаты позволяют заключить, что препарат pNCure в диапазоне исследованных дозировок не проявляет эффектов, свидетельствующих о его генотоксической активности.
Исследование субхронической токсичности плазмидной ДНК pNCure
С целью исследования возможных нежелательных явлений при многократном введении исследуемого препарата выполнены токсикологические исследования, направленные на выявление возможного повреждающего действия лекарственного средства при его длительном введении, на выявление наиболее чувствительных органов и систем организма.
В ходе исследования учету подлежали случаи гибели животных, оценивалась неврологическая симптоматика, регистрировался вес животных, учитывалось потребление животными пищи и воды, макроскопическая и гистологическая картина состояния внутренних органов (у каждого погибшего или эвтаназированного животного), клинический анализ крови при помощи автоматического анализатора клеток крови, биохимический анализ сыворотки крови при помощи автоматического анализатора, ЭКГ во втором стандартном отведении, поведение животных в тесте «Открытое поле. Исследования выполнялись в соответствии с требованиями нормативного документа «Правила лабораторной практики».
Для проведения исследования хронической токсичности использовали крыс линии Wistar обоего пола массой 180-200 г и кроликов породы «Шиншилла» обоего пола массой 2,2-2,5 кг (на момент поступления в виварий). Использованные виды лабораторных животных являются стандартными тест-системами в опытах для изучения хронического токсического действия фармакологических веществ. Крысы были получены из питомника лабораторных животных «Пущино», кролики - из ФГУП «Питомник лабораторных животных «Рапполово». Состояние здоровья животных при поступлении подтверждено ветеринарными сертификатами.
До начала исследования животные содержались 14 д в стандартных условиях под наблюдением без проведения исследовательских манипуляций.
При изучении хронического токсического действия ФС использован внутримышечный путь введения, запланированный для клинического применения. Курс введения препарата в эксперименте составил 30 сут. Выбор доз и шага доз осуществляли с учетом результатов исследования острой токсичности ФС (в выбранных дозах падеж животных отсутствовал, признаки токсического действия ФС обнаружены не были). В качестве минимальной дозы была выбрана терапевтическая доза, которая, при пересчете предполагаемой терапевтической дозы для человека (1,4 мг на 70 кг массы тела или 20 мкг/кг), составила для крыс - 0,118 мг/кг, для кроликов - 0,064 мг/кг. В качестве максимальной дозы была выбрана доза 2 мг/кг (доза, использованная в исследовании острого токсического действия фармацевтической субстанции, в сто раз превышающая предполагаемую терапевтическую дозу для человека) для обоих видов животных. Введение промежуточной дозы при отсутствии признаков острого токсического действия фармацевтической субстанции показалось нецелесообразным. Таким образом: Дозы для в/м введения крысам: 0 мг/кг, 0,118 мг/кг, 2 мг/кг.Дозы для в/м введения кроликам: 0 мг/кг, 0,064 мг/кг, 2 мг/кг.
Животные были случайным образом распределены по группам: 1 группа (контрольная) - 20 крыс обоего пола, внутримышечно ежедневно в течение 30 дней вводили физиологический раствор; 2 группа (опытная) - 20 крыс обоего пола, внутримышечно ежедневно в течение 30 дней вводили ФС в дозе 0,118 мг/кг; 3 группа (опытная) - 20 крыс обоего пола, внутримышечно ежедневно в течение 30 дней вводили ФС в дозе 2 мг/кг; 4 группа (опытная) - 12 кроликов обоего пола, внутримышечно ежедневно в течение 30 д вводили физиологический раствор; 5 группа (контрольная) - 12 кроликов обоего пола, внутримышечно ежедневно в течение 30 дней вводили ФС в дозе 0,064 мг/кг; 6 группа (опытная) - 12 кроликов обоего пола, внутримышечно ежедневно в течение 30 дней вводили ФС в дозе 2 мг/кг. Общее количество экспериментальных крыс -60 голов (30 самок и 30 самцов), кроликов - 36 голов (18 самок и 18 самцов).
С первого по 30 день эксперимента животным вводили физиологический раствор или ФС в дозах в соответствии с номером группы. На 30 день эксперимента эвтаназировали для последующей диагностической аутопсии и забора органов на гистологический анализ половину животных в каждой группе (5 самцов и 5 самок в каждой группе для крыс, 3 самца и 3 самки в каждой группе для кроликов). Вторая половина животных оставлена для наблюдения обратимости действия ФС. Последующие наблюдения длились 30 сут (с 31 до 60 сут эксперимента), затем животных эвтаназировали аналогичным образом. Таким образом, общая продолжительность введения ФС составит 30 дней, общая продолжительность наблюдений за животными - 60 дней.
В результате проведенных исследований определена зависимость проявлений токсического эффекта ФС на фиксируемые в вышеописанных тестах физиологические параметры от величины примененной дозы препарата при хроническом введении и обратимость токсических изменений органов и тканей.
Результаты и обсуждение результатов исследования
Гибель животных. Внутримышечное тридцатикратное ежедневное введение ФС в рассмотренных дозах не привело к гибели ни одного животного в ходе всего времени эксперимента и в течение периода последующего наблюдения в 30 дней.
Осмотр с элементами неврологического тестирования. Параметры общего состояния животных в течение срока наблюдения (осмотры проводили на 1, 8, 15, 22, 29, 38, 45, 52, 59 дни исследования) соответствовали норме и оставались стабильными. Животные проявляли нормальную активность в клетке, отсутствовал тремор, внешний вид животных (состояние шерсти, кожи, доступных для наблюдения слизистых оболочек) соответствовал таковому у крыс и кроликов, проживающих в удовлетворительных для них условиях. Параметры индивидуального эмоционального статуса (реакция на перемещение в новую обстановку, прикосновение, вокализации при осмотре) соответствовали характерному для популяции распределению, признаки эмоциональной напряженности в клетке (барберинг) отсутствовали. На все стимулы простых рефлексов (вздрагивание, переворачивание, роговичный, ушной) животные реагировали нормально, что позволяет охарактеризовать отсутствие токсического действия ФС при тридцатикратном ежедневном применении на глубинные механизмы нервной деятельности.
Макроскопическое и гистологическое исследование внутренних органов. На 30 и 60 сутки эксперимента проводили диагностическую аутопсию животных. Нижеприведенное описание в полной мере относится к каждому животному, независимо от группы.
При наружном осмотре выделений из естественных отверстий не обнаружено. Шерсть блестящая, опрятного вида, очагов облысения нет, у некоторых кроликов - признаки линьки. Зубы сохранены. Видимые слизистые оболочки бледной окраски, блестящие. Молочные железы самок без уплотнений на ощупь, выделения из сосков отсутствовали. Половые органы самцов развиты правильно, без крипторхизма. Деформации или отека конечностей нет.
Грудная и брюшная полости выпота не содержали. Положение внутренних органов грудной и брюшной полостей не нарушено. Париетальный и висцеральный листки плевры и брюшина тонкие, блестящие, гладкие.
Подчелюстные лимфатические узлы и слюнные железы овальной формы, бледно-желтого или розоватого цвета, с гладкой поверхностью, тонкой капсулой, не спаяны между собой и подлежащими тканями. Поверхность разреза однородной окраски.
Тимус треугольной формы, беловатого цвета, умеренно плотной консистенции, обычных размеров.
Интима аорты гладкая, блестящая, беловатого цвета. Диаметр аорты не изменен. Листки перикарда тонкие, прозрачные, гладкие. Величина и форма сердца без изменений. Мышца сердца на разрезе однородной коричневатой окраски, умеренно плотная.
Просвет трахеи и крупных бронхов не изменен, слизистая оболочка блестящая, гладкая, бледного цвета. Легкие воздушные, без уплотнений на ощупь, бледно-розовой окраски.
Слизистая пищевода блестящая, гладкая, бледного цвета. Желудок обычной величины и формы, заполнен пищевым содержимым. Слизистая оболочка безжелезистой части желудка складчатая, розоватая, блестящая, без изъязвлений. Слизистая тела желудка складчатая, розоватая, блестящая, без изъязвлений. Слизистая оболочка тонкого кишечника бледно-розового цвета, блестящая, гладкая, заполнена химусом. Слизистая оболочка толстой кишки сероватого цвета, блестящая, гладкая, заполнена каловыми массами нормальной консистенции.
Форма и величина печени без изменений. Поверхность печени гладкая, однородной темно-красной окраски, капсула тонкая, прозрачная. Ткань печени на разрезе полнокровная, умеренно плотная. У кроликов - желчный пузырь нормальный, без признаков патологических изменений.
Поджелудочная железа кружевной формы, бледно-розового цвета, дольчатая, умеренно плотной консистенции.
Селезенка обычной формы, светло-вишневого цвета, умеренно плотной консистенции. Поверхность органа гладкая, капсула тонкая. На разрезе на темно-красном фоне селезенки видны мелкие сероватого цвета фолликулы.
Величина и форма почек без изменений. Поверхность почек коричневатого цвета, гладкая, капсула тонкая, прозрачная, легко снимаемая. На разрезе органа хорошо различимы корковое и мозговое вещество.
Надпочечники округлой формы, бледно-коричневого цвета, с гладкой поверхностью, умеренно плотные. На разрезе четко выделяется темно окрашенное мозговое вещество.
Мочевой пузырь опорожнен или заполнен прозрачной мочой. Слизистая оболочка пузыря гладкая, блестящая, бледной окраски.
Тело матки самок обычной плотности, величины и формы. Рога матки тонкие, слизистая - блестящая, бледная. Яичники темно-красного цвета, с неровной поверхностью, умеренно плотные.
Семенники самцов беловатого цвета, обычных размеров и плотности.
Оболочки головного мозга тонкие, прозрачные. Вещество мозга обычной плотности, поверхность мозга гладкая. На фронтальных разрезах мозга отчетливо видны серое и белое вещество. Желудочки мозга обычной величины, расширения нет.
Раздражения кожи, отеков или инфильтратов в месте внутрибрюшинного введения не отмечалось.
Нижеприведенное описание гистологических препаратов изученных органов в полной мере относится к каждому животному, независимо от группы.
Клетки эндотелия внутренней оболочки аорты с четкими ядрами. Деструкций эластических волокон средней оболочки нет. Поперечная исчерченность миофибрилл отчетливая, ядра кардиомиоцитов содержат достаточное количество хроматина, ядерная оболочка тонкая. Очагов нарушений тинкториальных свойств цитоплазмы нет. Кардиофиброза нет.
Эпителий гортани, трахеи, крупных бронхов не изменен, ядра четкие. Альвеолы всех долей легких содержат воздух. Ядра альвеолярного эпителия четкие, цитоплазма оксифильная. Эпителий внутрилегочных бронхов без изменений. Острые воспалительные изменения легочной ткани отсутствовали.
Трабекулярное строение печени на срезах - без видимых изменений. Границы гепатоцитов отчетливые, цитоплазма зернистая, слабо оксифильная. Синусоиды печени полнокровные.
Капилляры клубочков и интерстициальной ткани почек полнокровные, цитоплазма эпителия проксимальных канальцев почек оксифильная, клеточные границы различимы, ядра нефроэпителия светлые, четкие.
Сосуды коры и мозгового вещества надпочечников полнокровные. Все зоны коры надпочечников отчетливо выражены. Цитоплазма клеток пучковой зоны вакуолизирована. Клетки мозгового вещества крупные, овальной формы, объединенные в гроздья и тяжи.
Лимфатические узлы состояли из коркового и мозгового вещества. В корковом веществе видны фолликулы, состоящие из лимфоидных элементов. Лимфоциты имели четкие ядра, тонкую мембрану и достаточное количество хроматина. Наряду с лимфоидными элементами в узлах присутствовали плазматические клетки, ретикулярные клетки и макрофаги.
Атрофии или деструкции фолликулов не наблюдалось.
Слизистая оболочка безжелезистой части желудка - без видимых изменений. Покровный эпителий слизистой тела желудка - без дефектов эпителиальной выстилки. Главные и обкладочные клетки желез тела желудка не изменены. Дефектов многослойного плоского неороговевающего эпителия слизистой пищевода нет.
Дольчатое строение поджелудочной железы сохранено. Сосуды стромы поджелудочной железы умеренно полнокровные.
Фолликулы щитовидной железы заполнены небольшим количеством оксифильного, слабовакуолизированного коллоида. Эпителий фолликул обычной высоты, ядра четкие. Сосуды стромы умеренно полнокровные.
Тимус сохранил выраженное дольчатое строение. Корковое вещество долек заполнено лимфоцитами/тимоцитами. Мозговое вещество тимуса содержало меньшее количество лимфоцитов, а также светлые эпителиальные клетки, местами образующие концентрические фигуры и тельца Гассаля. Строма тимуса умеренно полнокровная.
В корковом веществе яичников самок видны фолликулы разной величины и степени созревания. Фолликулярный эпителий не изменен, ядра светлые, четкие, мозговое вещество яичников полнокровное. Клетки семенных канальцев яичек самцов находились на разных стадиях сперматогенеза. Эпителий семенных канальцев и интерстициальные клетки не изменены. Ядра четкие.
Таким образом, по результатам макроскопического и гистологического исследования тридцатикратное ежедневное внутримышечное введение ФС в указанных дозах крысам и кроликам обоего пола не сопровождается развитием дистрофических, деструктивных, очаговых склеротических изменений в паренхиматозных клетках и строме внутренних органов.
Взвешивание внутренних органов животных при проведении диагностической аутопсии на 30 и 60 сутки исследования не показало статистически значимых различий как между кроликами в исследованных группах, так и между кроликами после курса введения исследуемого препарата и после периода наблюдения, последовавшего за курсом введения препарата. Аналогичным образом, не обнаружено статистически значимых различий по указанным показателям для самцов и самок крыс.
Клинический анализ крови. Кровь для проведения клинического анализа крови забирали у животных на 30 и 60 сутки эксперимента.
Результаты анализа у кроликов показали отсутствие отклонений показателей от физиологической нормы, например, гематокрит (норма от 35% до 48%) у кроликов на 30 день в контрольной группе составляет 39,51% при стандартном отклонении 3,03, в группе 0,064 - 39,5% при стандартном отклонении 3,15, в группе 2 - 38,73% при стандартном отклонении 3,12; гематокрит на 60 день у групп в том же порядке составляет: 43,42±3,04%, 42,95±3,24%, 42,12±4,13%. Т.е. статистически значимые различия отсутствуют и между пробами разных групп и между пробами, взятыми на 30 и 60 сутки исследования (Таблица 2.16).
Результаты анализа у самцов крыс показали отсутствие отклонений показателей от физиологической нормы. Гемоглобин (норма от 12 до 18 г/дл) у самцов крыс на 30 день в контрольной группе составляет 14,19 г/дл при стандартном отклонении 0,41, в группе 0,118 - 14,46 г/дл при стандартном отклонении 0,35, в группе 2 - 14,9 г/дл при стандартном отклонении 0,16; гемоглобин на 60 день у групп в том же порядке составляет: 14,82±0,29 г/дл, 14,21±0,3 г/дл, 14,4±0,24 г/дл. То есть, статистически значимые различия отсутствуют и между пробами разных групп и между пробами, взятыми на 30 и 60 сутки исследования.
Результаты анализа у самок крыс показали отсутствие отклонений показателей от физиологической нормы. Содержание эритроцитов (норма от 7,2 до 9,6*1012/л) у самок крыс на 30 день в контрольной группе составляет 8,11*1012/л при стандартном отклонении 0,2, в группе 0,118-8,13*1012/л при стандартном отклонении 0,13, в группе 2-8,11*1012/л при стандартном отклонении 0,19; содержание эритроцитов на 60 день у групп в том же порядке составляет: 7,95±0,18*1012/л, 8,08±0,15*1012/л, 7,92±0,18*1012/л. То есть, статистически значимые различия отсутствуют, и между пробами разных групп и между пробами, взятыми на 30 и 60 сутки исследования.
Таким образом, при выбранном режиме введения исследуемого препарата по показателям клинического анализа крови не наблюдалось различий между контрольными и опытными животными на 30 и 60 дни исследования.
Биохимическое исследование сыворотки крови. Кровь для проведения биохимического анализа плазмы забирали у животных на 30 и 60 сутки эксперимента.
Результаты анализа у кроликов показали отсутствие отклонений показателей от физиологической нормы. Уровень мочевины (норма от 14 до 40 мг/дл) у кроликов на 30 день в контрольной группе составляет 25,9 мг/дл при стандартном отклонении 10,78, в группе 0,064-27,7 мг/дл при стандартном отклонении 5,86, в группе 2-30,85 мг/дл при стандартном отклонении 6,42; уровень мочевины на 60 день у групп в том же порядке составляет: 19,38±7,25 мг/дл, 24,92±8,2, 29,3±8,23 мг/дл. То есть, статистически значимые различия отсутствуют и между пробами разных групп и между пробами, взятыми на 30 и 60 сутки исследования (Таблица 2.18).
Результаты анализа у самцов крыс показали отсутствие отклонений показателей от физиологической нормы. Уровень общего белка (норма от 62 до 72 г/дл) у самцов крыс на 30 день в контрольной группе составляет 66,35 г/дл при стандартном отклонении 3,08, в группе 0,118-63,45 г/дл при стандартном отклонении 3,8, в группе 2-63,25 г/дл при стандартном отклонении 3,0; уровень общего белка на 60 день у групп в том же порядке составляет: 63,4±3,81 г/дл, 65,0±5,6 г/дл, 65,2±3,76 г/дл. То есть, статистически значимые различия отсутствуют и между пробами разных групп и между пробами, взятыми на 30 и 60 сутки исследования.
Результаты анализа у самок крыс показали отсутствие отклонений показателей от физиологической нормы. Активность лактатдегидрогеназы (норма от 200 до 500 МЕ/л) у самок крыс на 30 день в контрольной группе составляет 336,7 МЕ/л при стандартном отклонении 31,61, в группе 0,118-346,32 МЕ/л при стандартном отклонении 13,61, в группе 2-301,92 МЕ/л при стандартном отклонении 28,59; активность лактатдегидрогеназы на 60 день у групп в том же порядке составляет: 310,8±29,06 МЕ/л, 303,03±24,46 МЕ/л, 315,24±36,39 МЕ/л. То есть, статистически значимые различия отсутствуют и между пробами разных групп и между пробами, взятыми на 30 и 60 сутки исследования.
Таким образом, при выбранном режиме введения исследуемого препарата по показателям клинического анализа крови не наблюдалось различий между контрольными и опытными животными на 30 и 60 дни исследования.
ЭКГ во втором стандартном отведении. Электрокардиограмму у животных регистрировали на 29 и 59 сутки эксперимента.
Результаты ЭКГ у кроликов показали отсутствие отклонений показателей от физиологической нормы. Частота сердечных сокращений (норма от 190 до 354 уд/мин) у кроликов на 29 день в контрольной группе составляет 291 уд/мин при стандартном отклонении 2,88, в группе 0,064-287 уд/мин при стандартном отклонении 8,68, в группе 2-276,5 уд/мин при стандартном отклонении 21,4; частота сердечных сокращений на 59 день у групп в том же порядке составляет: 277±13,19 уд/мин, 267,5±15,33 уд/мин, 271,5±14,58 уд/мин. То есть, статистически значимые различия отсутствуют и между параметрами разных групп и между параметрами тестирования на 29 и 59 сутки исследования.
Результаты ЭКГ у самцов крыс показали отсутствие отклонений показателей от физиологической нормы. Интервал QT у самцов крыс на 29 день в контрольной группе составляет 160,6 мс при стандартном отклонении 12,34, в группе 0,118-160,4 мс при стандартном отклонении 14,54, в группе 2-153,8 мс при стандартном отклонении 11,82. Итервал QT на 59 день у групп в том же порядке составляет: 155,4±10,14 мс, 155,0±17,03 мс, 159,6±14,57 мс. То есть, статистически значимые различия отсутствуют и между параметрами разных групп, и между параметрами тестирования на 29 и 59 сутки исследования.
Результаты ЭКГ у самок крыс показали отсутствие отклонений показателей от физиологической нормы. Величина зубца R (норма от 0,07 до 0,25 мВ) у самок крыс на 29 день в контрольной группе составляет 0,16 мВ при стандартном отклонении 0,01, в группе 0,118-0,16 мВ при стандартном отклонении 0,02, в группе 2-0,15 мВ при стандартном отклонении 0,01. Величина зубца R на 59 день у групп в том же порядке составляет: 0,15±0,01 мВ, 0,16±0,01 мВ, 0,16±0,02 мВ. То есть, статистически значимые различия отсутствуют и между параметрами разных групп, и между параметрами тестирования на 29 и 59 сутки исследования.
Таким образом, при выбранном режиме введения исследуемого препарата по показателям электрокардиограммы не наблюдалось различий между контрольными и опытными животными на 29 и 59 дни исследования.
Исследование поведения в тесте «Открытое поле». На 29 и 59 сутки эксперимента исследовали поведение крыс в тесте «Открытое поле» по основным показателям, отражающим локомоторную активность (пройденный путь, время в движении), ориентировочно-исследовательское поведение (количество обследованных норок, количество стоек, количество выходов в центр арены) и тревожность (время груминга, количество дефекаций, время, проведенное в центре арены, латентный период выхода в центр арены).
Результаты анализа параметров теста у самцов крыс показали отсутствие отклонений показателей от физиологической нормы. Количество стоек у самцов крыс на 29 день в контрольной группе составляет 9,4 шт при стандартном отклонении 5,55, в группе 0,118-10,6 шт при стандартном отклонении 4,28, в группе 2 - 7,6 шт при стандартном отклонении 2,97; количество стоек на 59 день у групп в том же порядке составляет: 10,8±3,56 шт, 8,2±4,6 шт, 8,0±5,15 шт. То есть, статистически значимые различия отсутствуют и между параметрами разных групп и между параметрами тестирования на 29 и 59 сутки исследования (Таблица 2.24).
Результаты анализа параметров теста у самок крыс показали отсутствие отклонений показателей от физиологической нормы. Латентный период выхода в центр у самок крыс на 29 день в контрольной группе составляет 85,43 с при стандартном отклонении 41,02, в группе 0,118-87,12 с при стандартном отклонении 36,89, в группе 2-63,4 с при стандартном отклонении 44,74; латентный период выхода в центр на 59 день у групп в том же порядке составляет: 77,72±39,71 с, 75,2±37,14 с, 58,48±38,76 с. Т.е. статистически значимые различия отсутствуют и между параметрами разных групп и между параметрами тестирования на 29 и 59 сутки исследования.
Таким образом, при выбранном режиме введения исследуемого препарата по показателям локомоторной активности, ориентировочно-исследовательского поведения и тревожности (психо-эмоционального статуса) в тесте «Открытое поле» не наблюдалось различий между контрольными и опытными животными на 29 и 59 дни исследования.
Общие выводы из полученных результатов.
В целом, применение исследуемой ФС в дозах 0,064 для кроликов, 0,118 для крыс и 2 мг/кг для обоих видов животных в течение 30 дней при внутримышечном пути введения не влияло на внешний вид, общее состояние и поведение животных обоего пола, не оказывало негативного влияния на параметры крови и основные физиологические функции организма, не вызывало патоморфологических изменений исследованных органов и основных систем органов, что свидетельствует о хорошей переносимости и безвредности препарата в условиях проведенного исследования. Препарат не оказывал токсического действия при ежедневном тридцатикратном введении экспериментальным животным. Таким образом, оказалось невозможным продемонстрировать обратимость токсического действия исследованной ФС: в период наблюдения, как и в период введения препарата, исследованные параметры во всех группах находились в пределах физиологической нормы. Последнее, в свою очередь, может свидетельствовать об отсутствии отставленных токсических эффектов в указанный период наблюдения.
Проведенные исследования показывают отсутствие токсического эффекта ФС в терапевтической дозе (пересчитанной для использованных видов животных) и стократной терапевтической дозе (для человека) и свидетельствуют об отсутствии противопоказаний по показателям хронической токсичности для клинических испытаний фармакологической субстанции лекарственного средства на основе генно-инженерной конструкции, содержащей последовательности кДНК мозгового нейротрофического фактора и активатора плазминогена урокиназного типа, для лечения травм периферических нервов pNCure. Применение заявляемой генотерапевтической конструкции обеспечивает увеличение скорости восстановления параметров проведения нервного импульса через поврежденный участок нерва, восстановления функциональной активности мышц, иннервируемых поврежденным нервом и прорастания нейритов через поврежденный участок нерва. Применение лекарственного средства, включающего заявляемую генотерапевтическую конструкцию, позволит сократить сроки лечения травм периферических нервов и снизить инвалидизации при травмах периферических нервов.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ СТИМУЛИРОВАНИЯ ВОССТАНОВЛЕНИЯ ПЕРИФЕРИЧЕСКОЙ ИННЕРВАЦИИ ТКАНЕЙ С ПОМОЩЬЮ ВЕКТОРНЫХ КОНСТРУКЦИЙ | 2011 |
|
RU2486918C1 |
| СПОСОБ СТИМУЛЯЦИИ ВОССТАНОВЛЕНИЯ ИННЕРВАЦИИ ПОВРЕЖДЕННОЙ ТКАНИ | 2014 |
|
RU2563541C1 |
| СПОСОБ СТИМУЛИРОВАНИЯ ВОССТАНОВЛЕНИЯ ИННЕРВАЦИИ ТКАНЕЙ ПОСЛЕ ТРАВМ И ИШЕМИИ С ПОМОЩЬЮ ВЕКТОРНОЙ КОНСТРУКЦИИ | 2013 |
|
RU2538621C2 |
| Лекарственное средство для лечения фиброза печени, способ его получения и способ лечения фиброза печени | 2015 |
|
RU2615445C2 |
| Препарат для стимулирования регенерации и нейропротекции нервной ткани и способ его использования | 2023 |
|
RU2828544C1 |
| Ген H5, кодирующий стабилизированную форму белка гемагглютинина вируса гриппа A (H5N8), рекомбинантная плазмидная ДНК pVAX-H5, содержащая ген H5 и обеспечивающая его экспрессию, и стабилизированная форма белка гемагглютинина вируса гриппа А (H5/N8), продуцируемая рекомбинантной плазмидной ДНК pVAX-H5 в организме животных и индуцирующая специфический иммунный ответ против высокопатогенного вируса гриппа A (H5N8) | 2024 |
|
RU2832515C1 |
| ЛЕЧЕНИЕ ЗАБОЛЕВАНИЙ, СВЯЗАННЫХ С НЕЙТРОФИЧЕСКИМ ФАКТОРОМ ГОЛОВНОГО МОЗГА (BDNF), ПУТЕМ ИНГИБИРОВАНИЯ ПРИРОДНОГО АНТИСМЫСЛОВОГО ТРАНСКРИПТА BDNF | 2013 |
|
RU2661104C2 |
| ДНК-ФРАГМЕНТ, КОДИРУЮЩИЙ НЕЙРОТРОПНЫЙ ФАКТОР МОЗГА (BDNF), БЕЛОК BDNF И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 1990 |
|
RU2131926C1 |
| СПОСОБ ПОЛУЧЕНИЯ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК МЛЕКОПИТАЮЩИХ, МОДИФИЦИРОВАННЫХ ГЕНОМ ФАКТОРА РОСТА НЕРВОВ ЧЕЛОВЕКА | 2011 |
|
RU2458984C1 |
| КОМПОЗИЦИЯ ДЛЯ НЕЙРОПРОТЕКЦИИ И СТИМУЛЯЦИИ НЕЙРОРЕГЕНЕРАЦИИ ГОЛОВНОГО МОЗГА ПОСЛЕ ПОВРЕЖДЕНИЯ, СРЕДСТВО НА ЕЕ ОСНОВЕ, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2022 |
|
RU2803286C1 |



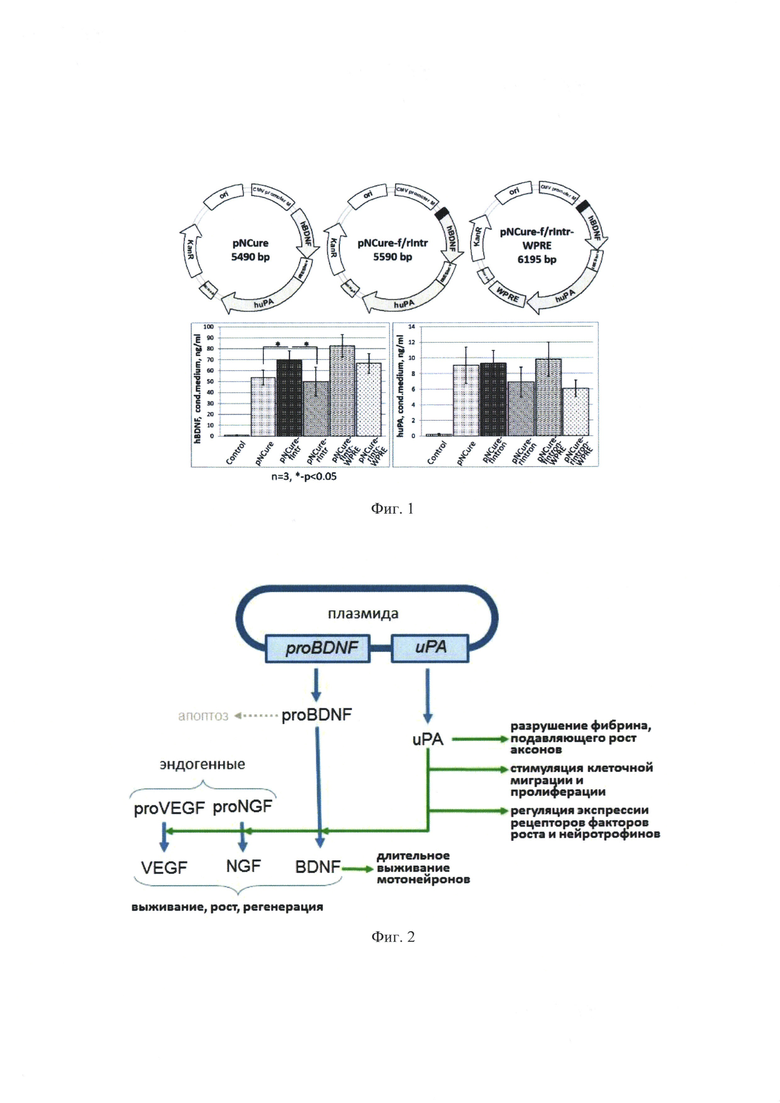
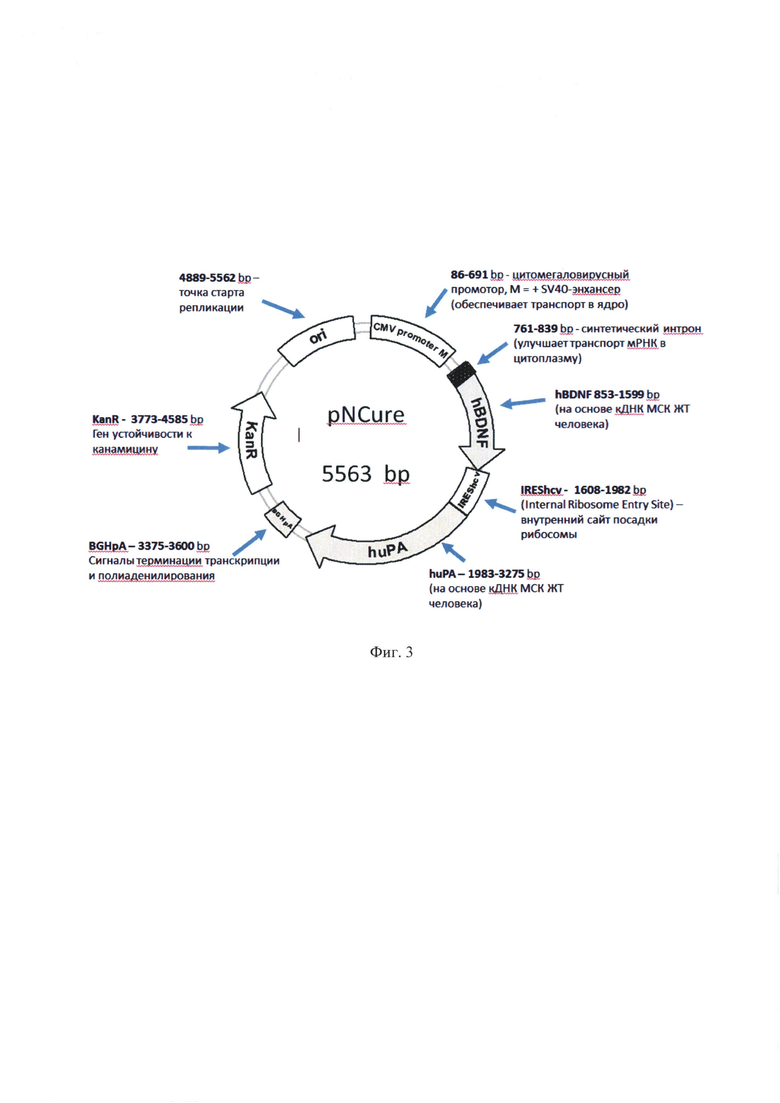
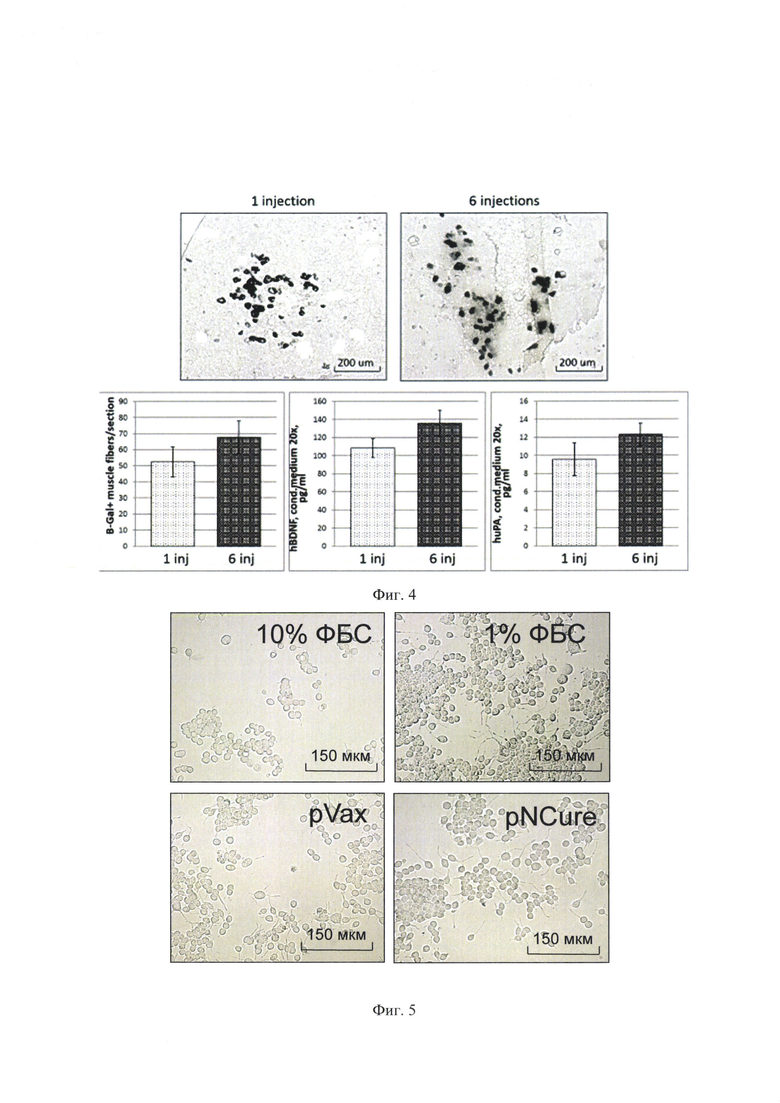
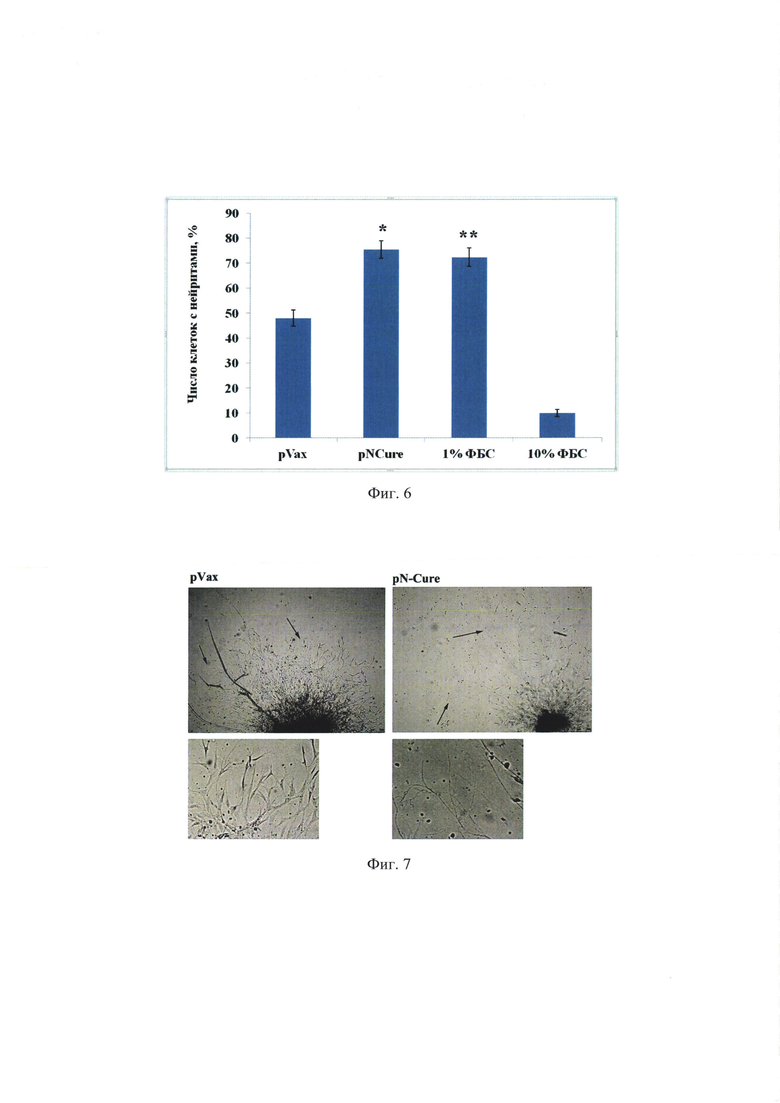
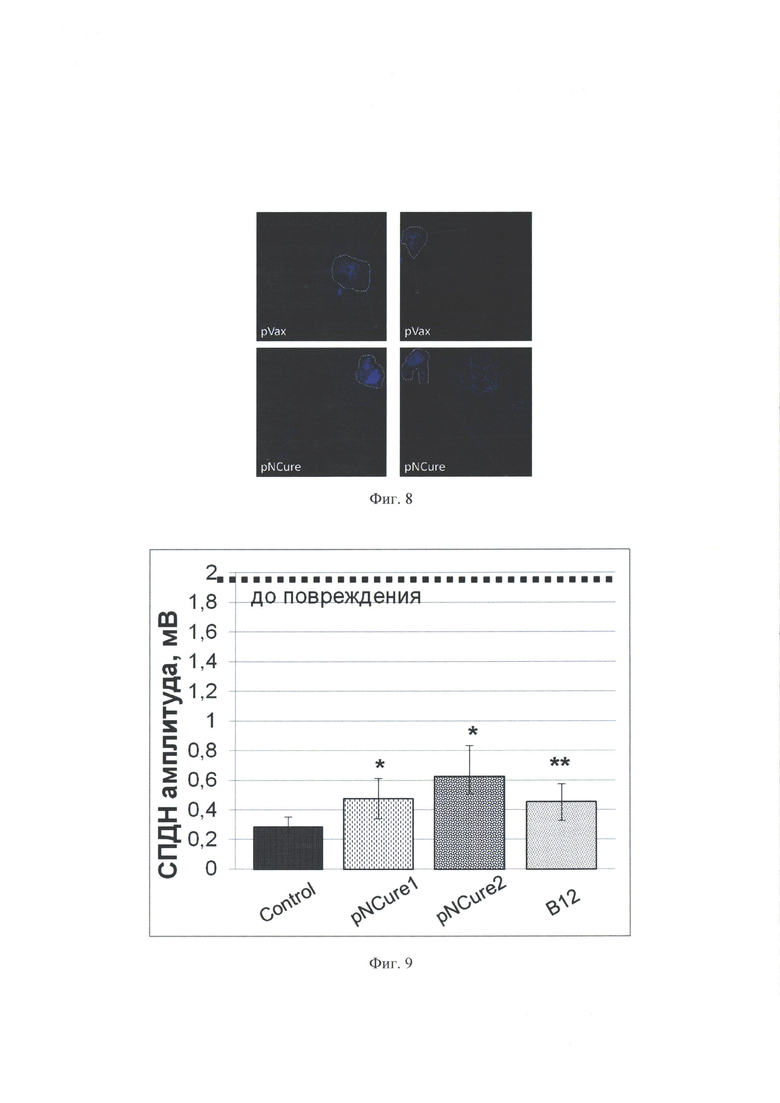


Предложенная группа изобретений относится к области медицины, в частности к биотехнологии и фармацевтике. Предложена генно-инженерная конструкция для стимуляции посттравматической регенерации периферических нервов, представляющая собой вектор со встроенными последовательностями кДНК hBDNF и huPA и регуляторную последовательность, включающую энхансер, внедренный в последовательность цитомегаловирусного промотера, искусственный интрон и сайт внутренней посадки рибосомы вируса гепатита С. Предложено применение вышеуказанной генно-инженерной конструкции в качестве генотерапевтического средства для лечения травм периферических нервов. Предложены также лекарственная форма, содержащая в качестве активного компонента вышеуказанную генно-инженерную конструкцию, и способ стимуляции локальной экспрессии hBDNF и huPA в периневральных тканях в области травмы, включающий введение лекарственной формы. Предложенная группа изобретений обеспечивает скоординированную локализованную экспрессию двух белков с достижением оптимального соотношения продукции рекомбинантного hBDNF и продукции huPA. 4 н. и 2 з.п. ф-лы, 12 ил., 9 табл., 7 пр.
1. Генно-инженерная конструкция для стимуляции посттравматической регенерации периферических нервов, представляющая собой вектор со встроенными последовательностями кДНК мозгового нейротрофического фактора человека (hBDNF) и активатора плазминогена урокиназного типа человека (huPA) и регуляторную последовательность, включающую энхансер, внедренный в последовательность цитомегаловирусного промотера, искусственный интрон с последовательностью SEQ ID NO: 1 и сайт внутренней посадки рибосомы вируса гепатита С (IRESHCV).
2. Генно-инженерная конструкция по п. 1, характеризующаяся тем, что последовательность кДНК мозгового нейротрофического фактора человека (hBDNF) встроена в вектор с 853 н.п. по 1599 н.п.; активатор плазминогена урокиназного типа человека (huPA) - с 1983 н.п. по 3275 н.п.; регуляторная последовательность содержит: энхансер SV40 или CMV с 417 н.п. по 478 н.п., внедренный в последовательность цитомегаловирусного промотера с 86 н.п. по 691 н.п., искусственный интрон - с 761 н.п. по 839 н.п., и сайт внутренней посадки рибосомы вируса гепатита С (IRESHCV) с 1608 н.п. по 1982 н.п.
3. Лекарственная форма, содержащая в качестве активного компонента генно-инженерную конструкцию по п. 1 и фармацевтически приемлемые растворители для парентерального введения, стабилизаторы и консерванты.
4. Способ стимуляции локальной экспрессии мозгового нейротрофического фактора и активатора плазминогена урокиназного типа в периневральных тканях в области травмы, включающий введение лекарственной формы по п. 3 в терапевтически эффективном количестве.
5. Способ по п. 4, характеризующийся тем, что введение лекарственной формы осуществляют путем единомоментной многоточечной периневральной внутримышечной инъекции в области травмы с периодичностью введения 1 раз в 2-4 недели, повторяемой от 3 до 5 раз до достижения терапевтического эффекта.
6. Применение генно-инженерной конструкции по п. 1 в качестве генотерапевтического средства для лечения травм периферических нервов.
| KARAGYAUR M | |||
| et al | |||
| Non-viral transfer of BDNF and uPA stimulates peripheral nerve regeneration | |||
| Biomed Pharmacother | |||
| Устройство для закрепления лыж на раме мотоциклов и велосипедов взамен переднего колеса | 1924 |
|
SU2015A1 |
| СПОСОБ СТИМУЛИРОВАНИЯ ВОССТАНОВЛЕНИЯ ПЕРИФЕРИЧЕСКОЙ ИННЕРВАЦИИ ТКАНЕЙ С ПОМОЩЬЮ ВЕКТОРНЫХ КОНСТРУКЦИЙ | 2011 |
|
RU2486918C1 |
| WO 2017072498 A1, 04.05.2017. | |||

