Область техники
Изобретение относится к иммунологии, в частности, к способу определения иммуногенности антигенов, входящих в состав векторных и субъединичных вакцин.
Предшествующий уровень техники
На сегодняшний день инфекционные заболевания являются одной из ведущих причин смертности в мире. По данным Всемирной организации здравоохранения в 2015 г около 30% всех смертей в мире были связаны с инфекционными заболеваниями. Наиболее эффективным способом защиты от инфекционных заболеваний является вакцинация. Использование вакцин не только позволило существенно снизить заболеваемость многими опасными инфекциями, но и полностью ликвидировать некоторые из них (например, натуральную оспу). Однако, остается еще более 300 инфекционных агентов, против которых пока не разработано профилактических средств защиты.
Разработка вакцин является длительным и дорогостоящим процессом, требующим больших трудозатрат. Так, опираясь на опубликованные данные, разработка вакцинного препарата в среднем занимает около 10,7 лет (PLoS One. 2013; 8(3): е57755. doi: 10.1371/journal.pone.0057755. Epub 2013 Mar 20. Risk in vaccine research and development quantified. Pronker ES1, Weenen TC, Commandeur H, Claassen EH, Osterhaus AD.), а стоимость разработки находится в районе 200-500 миллионов долларов США. (Dev Biol (Basel). 2002; 110:25-9). Одним из важнейших факторов, которые определяют успешность данного процесса, является правильный выбор вакцинного антигена. Главным критерием при подборе вакцинного антигена, помимо протективности (способности обеспечить защиту животных от летальных доз инфекционного агента после вакцинации), является его иммуногенность (способность антигена вызывать Т или В клеточный антиген-специфический иммунный ответ). Обобщая, можно заключить, что уровень иммуногенности антигена прямо пропорционально влияет на его итоговую протективность. В связи с этим, перед проведением полномасштабных доклинических исследований эффективности кандидатной вакцины, в рамках которых не допускается внесение каких либо изменений в качественные и количественные характеристики используемого вакцинного антигена, целесообразным является применение предварительного теста, позволяющего надежно оценивать иммуногенные свойства большого количества вариантов антигена и выбрать для последующей работы его наиболее иммуногенный вариант.
Так, известен способ оценки иммуногенности вакцинных антигенов путем иммунизации антигенами лабораторных животных с последующим выделением лимфоцитов селезенки, рестимуляции их антигеном и определением процента пролиферирующих лимфоцитов (J Exp Med. 1969 Aug 1; 130(2):327-43. Cellular immunity in vitro. Clonal proliferation of antigen-stimulated lymphocytes. Marshall WH, Valentine FT, Lawrence HS.). Недостатками указанного способа является продолжительность процедуры иммунизации лабораторных животных (для формирования антиген-специфичных клонов Т и В лимфоцитов требуется не менее 2 недель); необходимость траты большого количества животных, позволяющих получить статистически достоверные значения об иммуногенности исследуемых антигенов; а также отсутствие информации о типе клеток пролиферирующих в ответ на добавление антигена (В, Т лимфоциты, CD4+ или CD8+ Т лимфоциты и др.).
Другим вариантом является оценка иммуногенности антигенов in silico на основании специальных биоинформатических программ (Tools for the Prediction of T-Cell Epitopes, 2018), позволяющих предсказывать наличие T- и В- клеточных эпитопов в исследуемых антигенах. Применение данных методов не гарантирует, что полученные in silico результаты полностью воспроизведутся в экспериментальной работе, однако могут быть применены в начале работы для уменьшения количества вариантов антигена подлежащих последующему изучению in vitro и in vivo.
В. Maillere et al. (EP 2376925 A1) был предложен способ определения иммуногенности вакцинных антигенов, предполагающий моделирование реакций иммунной системы in vitro. Для этого автор предлагает, используя периферическую кровь человека, проводить совместное культивирование дендритных клеток, праймированных антигеном, с Т-лимфоцитами, с дальнейший оценкой пролиферации лимфоцитов. Разработанный метод позволяет анализировать иммуногенность вакцин, адъювантов, кандидатных иммунотерапевтических препаратов и других химических соединений. Данный способ как наиболее близкий по техническому решению был выбран авторами заявляемого изобретения за прототип.
Описанный выше способ позволяет проверять иммуногенность широкого спектра антигенов, за счет близкого моделирования in vitro тех иммунных процессов, которые происходят в организме человека. Однако недостатком данного способа является то, что он не учитывает вклад иммуносупрессорных доминант (эпитопов для Т регуляторных лимфоцитов) антигена в развитие иммунного ответа.
Таким образом, в науке существует острая потребность в разработке легко масштабируемого и надежного метода, позволяющего быстро оценивать иммунный ответ на кандидатный вакцинный антиген в лабораторных условиях, лишенного указанного недостатка.
Раскрытие настоящего изобретения
Задачей настоящего изобретения является разработка легко масштабируемого надежного метода первичной оценки иммуногенности вакцинного антигена в условиях in vitro, с одновременным определением наличия иммунносупрессорных эпитопов для Т регуляторных клеток.
Указанная задача решается за счет того, что создан способ проверки иммуногенности антигенов, включающий приготовление клеточной культуры, содержащей дендритные клетки, включающий праймирование антигеном и совместное культивирование с Т эффекторными клетками, отличающийся тем, что в культуру клеток добавляют Т регуляторные клетки и оценивают пролиферацию Т эффекторных клеток.
Также создан способ выбора антигена, заключающийся в том, что выбирают из исследуемых антигенов по способу, описанному в способе проверки иммуногенности антигенов, тот антиген, который не угнетает пролиферацию Т эффекторных клеток в присутствии Т регуляторных клеток.
Заявлен также способ повышения иммуногенности, заключающийся в том, что антиген вначале проверяют иммуногенность антигенов, затем модифицируют выбранный антиген путем уменьшения количества эпитопов к Т регуляторным клеткам, после чего синтезируют модифицированный антиген и второй раз проверяют иммуногенность антигена.
Как известно, эффективность вакцинации связана с индукцией протективного гуморального и/или клеточного иммунного ответа, который зависит от многих факторов и в том числе от особенностей структуры вакцинного антигена. Было показано, что важную роль в формировании иммунного ответа играет наличие Т- и В- клеточных эпитопов в структуре антигена, которые являются иммунодоминантами и приводят к стимуляции специфических иммунных ответов.
При попадании в организм, вакцинный антиген распознается и поглощается антиген-презентирующими клетками (АПК). Попадая внутрь клетки, антиген расщепляется на фрагменты, которые взаимодействуют с молекулами МНС и в комплексе с ними представляются Т лимфоцитам. Т лимфоциты распознают антигенные пептиды на поверхности АПК с помощью Т-клеточных рецепторов (TCR) и с участием корецепторов CD4 (у Т-хелперов) или CD8 (у цитотоксических Т клеток). После распознавания антигена Т-клетки получают дополнительные активационные стимулы и начинают экспрессировать IL2, в результате аутокринного действия которого начинают активно делиться. На следующем этапе CD4+ клетки дифференцируются в Th1- и Th2-клетки, которые отвечают за развитие клеточного и гуморального иммунного ответа, соответственно.
Таким образом, очевидно, что чем больше количество Т-клеточных эпитопов содержится в структуре вакцинного антигена, тем большее количество лимфоцитов будет активировано и тем мощнее будет иммунный ответ.
Основываясь на этих данных, В. Maillere et al. (ЕР 2376925 А1) предложили метод оценки иммуногенности веществ, при котором in vitro моделируются иммунные реакции, которые возникают в организме при введении антигена. Для этого авторы предлагают использовать дендритные клетки, которые являются антиген-презентирующими клетками и Т-лимфоциты, активация и пролиферация которых является маркерным событием, показывающим насколько иммуногенным является антиген. Однако недостатком данного метода является то, что он не учитывает работу еще одного важного звена иммунитета: Т регуляторных клеток (Т рег). Основной функцией Т рег клеток является регулирование силы и продолжительности иммунного ответа. Как и другие Т лимфоциты данные клетки активируются после распознавания антигенных пептидов, представленных на поверхности АПК. Однако, после активации Т рег клетки вырабатывают ряд факторов, участвующих в угнетении иммунного ответа. Мишенями действия Treg-клеток являются как Т-эффекторные клетки, так и дендритные клетки, ответственные за презентацию антигена и активацию Т-клеток. Соответственно, чем больше Т рег эпитопов содержит антиген, тем меньший иммунный ответ будет развиваться после его введения. Таким образом, выбор вакцинного антигена без учета вклада Т регуляторных эпитопов в его иммуногенность, может привести к значительному снижению эффективности вакцинации.
Разработанный авторами патента метод предполагает моделирование in vitro взаимодействий, происходящих в организме человека при иммунизации, а именно презентацию антигена дендритными клетками, активацию и пролиферацию Т лимфоцитов, при этом дополнительно оценивается способность антигена активировать Т регуляторные клетки. Данный способ не только позволяет сделать вывод об иммуногенности кандидатного вакцинного антигена, но также, в случае низкой иммуногенности, понять возможную причину этого, например, недостаточное количество Т клеточных эпитопов, либо наличие большого количества эпитопов к Т регуляторным клеткам.
Краткое описание фигур
На фиг. 1 представлена схема эксперимента по оценке иммуногенности вакцинного антигена.
А экспериментальная группа, учитывающая вклад Т регуляторных клеток в общую иммуногенность антигена
Б экспериментальная группа, не учитывающая активацию Т регуляторных клеток
1 - Дендритные клетки
2 - Добавление анализируемого антигена
3 - Т эффекторные лимфоциты
4 - Т регуляторные лимфоциты
5 - Совместное культивирование клеток
На фиг. 2 представлена стратегия выделения группы пролиферирующих CD3+CD4+ и CD3+CD8+ лимфоцитов.
А. ось ординат - боковое светорассеяние (SSC)
Ось абсцисс - малоугольное рассеяние света (FSC)
1 Лимфоциты
Б. ось ординат - флуоресценция красителя, который конъюгирован с антителами к CD3
Ось абсцисс - малоугольное рассеяние света (FSC)
2 CD3+ лимфоциты
В. ось ординат - флуоресценция красителя, который конъюгирован с антителами к CD8
Ось абсцисс - флуоресценция красителя, который конъюгирован с антителами к CD4
3 CD3+CD8+ лимфоциты
4 CD3+CD4+ лимфоциты
Г. ось ординат - флуоресценция красителя, который конъюгирован с антителами к CD8
Ось абсцисс - флуоресценция CFSE
5 Пролиферирующие CD3+CD8+ лимфоциты
6 Пролиферирующие CD3+CD4+ лимфоциты
Пример 1
Получение культуры дендритных клеток.
Дедритные клетки были получены из моноцитов человека согласно методике, описанной Porsch et al [Posch, W.,  C, Wilflingseder, D. Generation of Human Monocyte-derived Dendritic Cells from Whole Blood. J. Vis. Exp. (118), e54968, doi:10.3791/54968 (2016)]. На первом этапе производили отбор крови здоровых добровольцев. Далее кровь помещали в стерильные пробирки для центрифугирования и доводили объем жидкости до 50 мл фосфатно-солевым буферным раствором в модификации Дульбеко (D-PBS). Пробирки центрифугировали 400xg в течение 15 мин при комнатной температуре. Верхний слой, содержащий плазму и тромбоциты, удаляли используя стерильную 10 мл серологическую пипетку, а пограничный слой, содержащий мононуклеарные клетки, переносили в новые 50 мл пробирки, объединяя образцы из двух пробирок. Объем жидкости в пробирках доводили до 50 мл D-PBS. Затем производили центрифугирование на скорости 400xg в течение 15 мин при комнатной температуре. Верхний слой, содержащий плазму и тромбоциты, удаляли используя стерильную 10 мл серологическую пипетку, а пограничный слой, содержащий мононуклеарные клетки переносили в новую пробирку. Объем жидкости в пробирке доводили до 50 мл D-PBS. Далее производили разделение клеток центрифугированием с градиентом плотности. Для этого в 50 мл пробирку помещали 15 мл Ficoll и 25 мл клеточной суспензии в D-PBS. Пробирки центрифугировали на скорости 700xg в течение 30 минут при комнатной температуре. Далее отбирали интерфазу, содержащую мононуклеарные клетки. Далее клетки дважды промывали D-PBS. Для этого выделенные клетки переносили в новую пробирку и доводили до 50 мл D-PBS. Затем производили центрифугирование на скорости 400xg в течение 15 мин при комнатной температуре. Выделенные мононуклеарные клетки разводили D-PBS до концентрации 8*107 клеток/мл.
C, Wilflingseder, D. Generation of Human Monocyte-derived Dendritic Cells from Whole Blood. J. Vis. Exp. (118), e54968, doi:10.3791/54968 (2016)]. На первом этапе производили отбор крови здоровых добровольцев. Далее кровь помещали в стерильные пробирки для центрифугирования и доводили объем жидкости до 50 мл фосфатно-солевым буферным раствором в модификации Дульбеко (D-PBS). Пробирки центрифугировали 400xg в течение 15 мин при комнатной температуре. Верхний слой, содержащий плазму и тромбоциты, удаляли используя стерильную 10 мл серологическую пипетку, а пограничный слой, содержащий мононуклеарные клетки, переносили в новые 50 мл пробирки, объединяя образцы из двух пробирок. Объем жидкости в пробирках доводили до 50 мл D-PBS. Затем производили центрифугирование на скорости 400xg в течение 15 мин при комнатной температуре. Верхний слой, содержащий плазму и тромбоциты, удаляли используя стерильную 10 мл серологическую пипетку, а пограничный слой, содержащий мононуклеарные клетки переносили в новую пробирку. Объем жидкости в пробирке доводили до 50 мл D-PBS. Далее производили разделение клеток центрифугированием с градиентом плотности. Для этого в 50 мл пробирку помещали 15 мл Ficoll и 25 мл клеточной суспензии в D-PBS. Пробирки центрифугировали на скорости 700xg в течение 30 минут при комнатной температуре. Далее отбирали интерфазу, содержащую мононуклеарные клетки. Далее клетки дважды промывали D-PBS. Для этого выделенные клетки переносили в новую пробирку и доводили до 50 мл D-PBS. Затем производили центрифугирование на скорости 400xg в течение 15 мин при комнатной температуре. Выделенные мононуклеарные клетки разводили D-PBS до концентрации 8*107 клеток/мл.
Моноциты выделяли из полученной клеточной суспензии с помощью набора Anti-Human CD14 Magnetic Particles - DM (BD, cat#557769). Полученную клеточную суспензию моноцитов центрифугировали на скорости 400xg в течение 15 мин при комнатной температуре. Осадок разводили предварительно нагретой до 37°С культуральной средой (среда RPMI 1640, дополненной 10% инактивированной нагреванием фетальной бычьей сывороткой, 1% раствором L-глутамина, 100 нг/мл ГМ-КСФ и 50 нг/мл ИЛ-4) до концентрации 1*106 клеток/мл. Далее клеточную суспензию разливали по 3 мл/лунку 6-луночного планшета. На пятый день незрелые дендритные клетки собирали путем пипетирования культивируемых в 6-луночном планшете клеток. Полученные клетки подсчитывали и окрашивали антителами против CD11c, MHCII (I-A/Е), вместе красителем, позволяющим оценить жизнеспособность клеток. Анализ образцов проводили методом проточной цитометрии.
Пример 2.
Выделение лимфоцитов
На первом этапе производили выделение мононуклеарных клеток из крови здоровых добровольцев, как описано в примере 1. Выделенные клетки разводили до концентрации 1×106 кл/мл и помещали в культуральные флаконы 75 см2. Далее проводили инкубацию клеток при температуре 37°С, 5% СO2. Во время культивирования моноциты прикрепляются к культуральному флакону, тогда как лимфоциты остаются неприкрепленными. Через 1 час осторожно собирали культуральную среду вместе с лимфоцитами.
Далее из полученных лимфоцитов с помощью сортера BD FACS AriaIII выделяли следующие популяции клеток.
Т эфф CD45+ CD3+CD25-
Т реги: CD45+ CD3+, CD25+, CD127-,
Пример 3.
Оценка пролиферации лимфоцитов.
Для определения количества пролиферирующих лимфоцитов использовали метод окраски клеток красителем CFSE. Принцип этого метода заключается в том, что флуоресцентный не токсичный краситель CFSE способен беспрепятственно проникать в клетки. После стимуляции клеток антигеном лимфоциты начинают пролиферировать, при этом краситель, находящийся в родительской клетке, распределяется поровну между дочерними. Таким образом, используя проточный цитофлюориметр через определенное время после повторной стимуляции клеток антигеном возможно определить процент клеток с низкой флуоресценцией, которая будет только у тех клеток, которые делились за время эксперимента.
Окраску CFSE осуществляли следующим образом. В пустую стерильную центрифужную пробирку объемом 15 мл вносили выделенные лимфоциты (пример 2) в количестве 5×106 клеток. Объем жидкости доводили до 1 мл путем добавления D-PBS. Далее в пробирку добавляли 1 мкл 5 мМ раствора красителя CFSE) и быстро перемешивали. Затем проводили инкубацию клеток в течение 5 мин в темноте. Далее в пробирку добавляли 10 мл полной среды RPMI (РАА, США) для клеток включающей 10% фетальной сыворотки крупного рогатого скота (Thermo Scientific, США), 100 мкг/мл стрептомицина (ПанЭко, Россия), 100Ед./мл пенициллина (ПанЭко, Россия), 0,18% бикарбоната натрия (ПанЭко, Россия), 292 мг L-глутамина (ПанЭко, Россия). Затем пробирки центрифугировали на скорости 200xg в течение 10 мин. С помощью пипетки Пастера удаляли надосадочную жидкость. Осадок ресуспендировали в 10 мл стерильного изотоничного фосфатного буфера, содержащего 1% фетальной сыворотки крупного рогатого скота (Thermo Scientific, США). Затем пробирки центрифугировали на скорости 200xg в течение 10 мин. С помощью пипетки Пастера удаляли надосадочную жидкость. Осадок ресуспендировали в 5 мл полной среды RPMI.
Пример 4.
Оценка иммуногенности вакцинного антигена GP
На первом этапе дендритные клетки (ДК), получение которых описано в примере 1, были разделены на две чашки. В одну чашку добавляли 200 мг/мл антигена GP, поверхностного белка вируса Эбола (ДК+АГ), а в другую пробирку добавили D-PBS, который использовался в качестве отрицательного контроля (ДК-АГ). Все пробирки инкубировали в течение ночи. На следующий день все клетки центрифугировали, надосадочную жидкость удаляли, а осадок ресуспендировали в свежей среде RPMI. Процедуру промывки повторяли два раза. Далее дендритные клетки, и Тэфф клетки, окрашенные CFSE (пример 3) и Т регуляторные клетки смешивались в лунках планшета в различных сочетаниях, по 3 повтора:
Группа 1. (ДК+АГ) + Тэфф клетки (соотношение 1:10)
Группа 2. (ДК-АГ) + Тэфф клетки (соотношение 1:10)
Группа 3. (ДК+АГ) + Тэфф клетки + Т рег клетки (соотношение 1:10:1)
Группа 4. (ДК-АГ) - Тэфф клетки + Т рег клетки (соотношение 1:10:1)
Через 72 часа клетки ресуспендировали в лунках планшета и перенесли в центрифужные пробирки объемом 1,5 мл, содержащие 1 мл D-PBS. Далее пробирки центрифугировали при 200 g/мин в течение 10 мин. Затем надосадочную жидкость удалили, а осадок ресуспендировали в 1 мл D-PBS. Далее пробирки центрифугировали при 200 g/мин в течение 10 мин. Затем надосадочную жидкость удалили, а осадок ресуспендировали в 100 мкл буфера для окраски Stain Buffer (BD Pharmingen, США). В каждую пробирку добавили по 5 мкл антител против CD3, CD4, CD8:
anti-CD3 АРС (BD Biosciences, кат.номер: 553066),
anti-CD4 АРС-Су7 (BD Biosciences, кат.номер: 552051),
anti-CD8 PE-CF594 (BD Biosciences, кат.номер: 562283).
Пробирки инкубировали в темноте в течение 30 минут при +4°С.
Далее с помощью проточного цитофлюориметра BD FACS AriaIII определяли пролиферирующие (с меньшим количеством красителя CFSE клеток) Т лимфоциты. Стратегия гейтирования представлена на рис. 2. На первом этапе выделяли лимфоциты (1) в шкале FSC (малоугольное рассеяние света) и SSC (боковое светорассеяние). Далее, ориентируясь на флуоресценцию аллофикоцианина (АРС), выделяли популяцию CD3-позитивных клеток. Затем, выделяли популяции CD4+ и CD8+ лимфоцитов. Далее анализировали интенсивность флуоресценции CFSE среди этих групп клеток.
Данные были получены в трех повторах и для каждой группы было посчитано среднее значение и стандартное отклонения. Результаты представлены в таблице 1
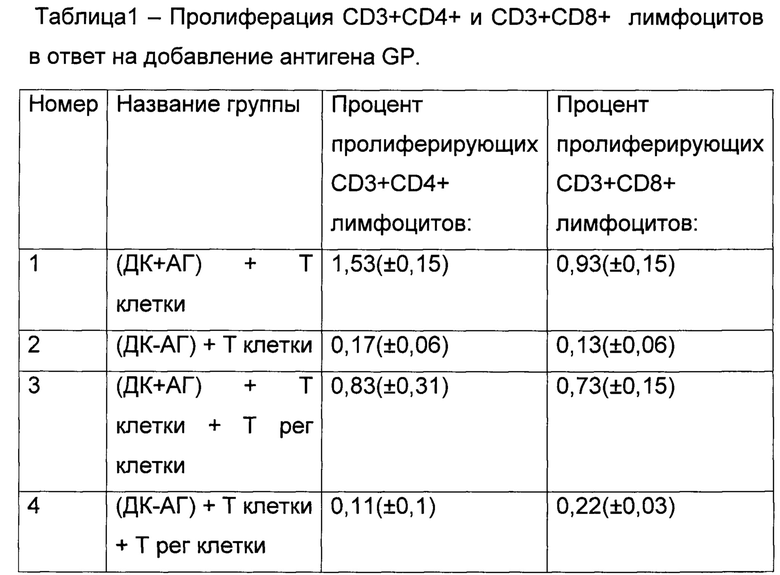
Результирующий процент пролиферирующих клеток в каждом образце определяли путем вычитания результата, полученного при анализе контрольной группы клеток нестимулированных антигеном (например, (ДК-АГ) + Т клетки), из результата, полученного в группе клеток стимулированных антигеном (например, (ДК+АГ) + Т клетки).
Таким образом, результирующий процент пролиферирующих CD4+T лимфоцитов в отсутствии Т регуляторных клеток составил 1,36%, тогда как присутствие Т регуляторных клеток снижало процент пролиферирующих CD4+T лимфоцитов до 0,72%. Результирующий процент пролиферирующих CD8+T лимфоцитов без Т регуляторных клеток составил 0,80%, добавление же Т регуляторных клеток приводило к снижению пролиферирующих CD3+CD8+ лимфоцитов до 0,51%. Как видно из представленных результатов, добавление Т регуляторных клеток достоверно (p<0,05) приводит к ингибированию пролиферации Т клеток, а следовательно исследуемый антиген GP вируса Эбола содержит эпитопы к Т регуляторным клеткам и активирует их.
Пример 5
Увеличение иммуногенности вакцинного антигена
Сиквенс антигена GP был проанализирован с целью поиска эпитопов к Т регуляторным клеткам, на основе имеющихся баз данных. На первом этапе с помощью программного обеспечения с порогом 8,5 (что является разницей в медианном процентильном ранге от 26 опорных аллелей, установленных для МНС класса II) были предсказаны области или пептиды, активирующие Т регуляторные клетки. Эти пептиды образовывались из белка с 15-мерным шагом и 10-кратным перекрытием. На втором этапе рассматривался каждый пептид, активирующий Т регуляторные клетки и подбиралась аминокислотная замена для выбранных пептидов. Замена подбиралась таким образом, чтобы не создавать новый эпитоп для Т регуляторных клеток в соседних пептидах и не нарушать эпитопы для Т эфф. клеток. На основе разработанного модифицированного сиквенса был получен рекомбинантный антиген GPмод
Иммуногенность GPмод была проверена с помощью метода, описанного в примере 4.
Результаты эксперимента представлены в таблице 2.
Согласно результатам эксперимента, результирующий процент пролиферирующих CD4+T лимфоцитов в отсутствие Т регуляторных клеток составил 1,49%, а в присутствие Т регуляторных клеток - 1,45%. Результирующий процент пролиферирующих CD8+T лимфоцитов в отсутствие Т регуляторных клеток составил 0,91%, а в присутствие Т регуляторных клеток - 0,88%. Как видно из представленных результатов, добавление Т регуляторных клеток не приводило к достоверным отличиям в пролиферации Т клеток, а следовательно, исследуемый антиген GPмод вируса Эбола не активировал Т регуляторные клетки
Таким образом, специалисту среднего уровня должно быть очевидно, что использование модифицированного антигена GPмод, с уменьшенным количеством Т эпитопов, приведет к увеличению иммуногенности вакцины против вируса Эбола. Данный подход, заключающийся в оценке способности вакцинного антигена ингибировать пролиферацию Т клеток через активацию Т регуляторных клеток, может быть использован при разработке любых вакцин и других препаратов, действие которых реализуется через индукцию иммунного ответа (например, противоопухолевых препаратов).
Все приведенные примеры подтверждают выполнение технической задачи, а также промышленную. Применимость данного изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| Полиэпитопная противоопухолевая вакцинная конструкция, содержащая эпитопы опухоль-ассоциированных антигенов, фармацевтическая композиция и ее применение для стимуляции специфического противоопухолевого иммунного ответа | 2016 |
|
RU2684235C2 |
| ПРОФИЛАКТИЧЕСКАЯ ИЛИ ТЕРАПЕВТИЧЕСКАЯ ПОЛИЭПИТОПНАЯ ПРОТИВОТУБЕРКУЛЕЗНАЯ ВАКЦИННАЯ КОНСТРУКЦИЯ, ОБЕСПЕЧИВАЮЩАЯ ИНДУКЦИЮ КЛЕТОЧНОГО ИММУННОГО ОТВЕТА CD4+ ИЛИ CD8+ Т-ЛИМФОЦИТОВ | 2011 |
|
RU2539035C2 |
| Способ получения in vitro популяций активированных антигенспецифических противоопухолевых цитотоксических Т-лимфоцитов, специфичных к эпитопам опухоль-ассоциированного антигена | 2016 |
|
RU2619186C1 |
| ВИЧ ВАКЦИНА, ОСНОВАННАЯ НА НАПРАВЛЕННОСТИ МАКСИМИЗИРОВАННЫХ Gag И Nef НА ДЕНДРИТНЫЕ КЛЕТКИ | 2009 |
|
RU2539765C2 |
| ПОЛИЭПИТОПНЫЕ ИММУНОГЕННЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2598265C2 |
| ВАКЦИНА НА ОСНОВЕ В-КЛЕТОК, НАГРУЖЕННЫХ ЛИГАНДОМ Т-КЛЕТОК-ПРИРОДНЫХ КИЛЛЕРОВ И АНТИГЕНОМ | 2006 |
|
RU2420311C2 |
| СПОСОБ ГЕНЕРАЦИИ СПЕЦИФИЧЕСКОГО ИММУННОГО ОТВЕТА ПРОТИВ АНТИГЕНОВ ВИРУСА ИММУНОДЕФИЦИТА ЧЕЛОВЕКА | 2011 |
|
RU2465324C1 |
| СПОСОБ ПОЛУЧЕНИЯ АНТИГЕН-СПЕЦИФИЧЕСКИХ ЦИТОТОКСИЧЕСКИХ КЛЕТОК, ОБЛАДАЮЩИХ АКТИВНОСТЬЮ ПРОТИВ КЛЕТОК РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ | 2013 |
|
RU2596920C2 |
| КОМБИНИРОВАННЫЙ ПРЕПАРАТ ДЛЯ УВЕЛИЧЕНИЯ ИММУНОГЕННОСТИ ВАКЦИНЫ (ВАРИАНТЫ) | 2008 |
|
RU2491090C2 |
| ПРОФИЛАКТИЧЕСКАЯ ПРОТИВОРАКОВАЯ ВАКЦИНА | 2004 |
|
RU2556128C2 |
Предложенная группа изобретений относится к области медицины, в частности к иммунологии. Предложен способ проверки иммуногенности вакцинных антигенов in vitro для получения высокоэффективных вакцин против опасных инфекций, включающий приготовление группы клеточных культур. В каждую пробирку добавляют антитела против CD3, CD4, CD8, далее определяют пролиферирующие CD3+CD4+ и CD3+CD8+ Т лимфоциты, затем анализируют интенсивность пролиферации лимфоцитов среди этих групп клеток. Из исследуемых антигенов выбирают тот, который не угнетает пролиферацию Т эффекторных клеток в присутствии Т регуляторных клеток. Предложен способ повышения иммуногенности вакцины, заключающийся в том, что вакцинный антиген вначале проверяют вышеописанным способом, затем его модифицируют путем уменьшения количества эпитопов к Т регуляторным клеткам, синтезируют модифицированный вакцинный антиген и второй раз проверяют. Предложенная группа изобретений обеспечивает повышение иммуногенности вакцины и антигена. 2 н. и 1 з.п. ф-лы, 2 ил., 2 табл., 5 пр.
1. Способ проверки иммуногенности вакцинных антигенов in vitro для получения высокоэффективных вакцин против опасных инфекций, включающий приготовление группы клеточных культур, содержащих:
группа 1 - дендритные клетки (ДК), вакцинный антиген (АГ+), Т эффекторные клетки (Тэфф) (в соотношении ДК к Тэфф 1:10);
группа 2 - дендритные клетки (ДК), фосфатно-солевой буфер в качестве отрицательного контроля (АГ-), Т эффекторные клетки (Тэфф) (в соотношении ДК к Тэфф 1:10);
группа 3 - дендритные клетки (ДК), вакцинный антиген (АГ+), Т эффекторные клетки (Тэфф), Т регуляторные клетки (Трег) (в соотношении ДК к Тэфф к Трег 1:10:1);
группа 4 - дендритные клетки (ДК), фосфатно-солевой буфер в качестве отрицательного контроля (АГ-), Т эффекторные клетки (Тэфф), Т регуляторные клетки (Трег) (в соотношении ДК к Тэфф к Трег 1:10:1),
после чего в каждую пробирку добавляют антитела против CD3, CD4, CD8, далее определяют пролиферирующие CD3+CD4+ и CD3+CD8+ Т лимфоциты, затем анализируют интенсивность пролиферации лимфоцитов среди этих групп клеток, после чего из исследуемых антигенов выбирают тот, который не угнетает пролиферацию Т эффекторных клеток в присутствии Т регуляторных клеток.
2. Способ повышения иммуногенности вакцины, заключающийся в том, что вакцинный антиген вначале проверяют способом по п. 1, затем вакцинный антиген модифицируют путем уменьшения количества эпитопов к Т регуляторным клеткам, затем синтезируют модифицированный вакцинный антиген и второй раз проверяют способом по п. 1.
3. Способ проверки иммуногенности вакцинных антигенов по п. 1, в котором проверяют антигены вируса Эбола.
| ПЫЛЕСОС С КОНСТРУКТИВНЫМИ ЭЛЕМЕНТАМИ ИЗ ВСПУЧЕННОГО ПОЛИПРОПИЛЕНА | 2005 |
|
RU2376925C2 |
| АНАЛИЗЫ Т-КЛЕТОК | 2007 |
|
RU2476884C2 |
| WO 03038057 A2, 08.05.2003 | |||
| MENORET S | |||
| et al | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| Methods Mol Biol | |||
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| ЖЕЛЕЗНИКОВА Г.Ф | |||
| Регуляторные Т-лимфоциты в иммунном ответе на инфекцию | |||
| Журнал инфектологии | |||
| Способ приготовления лака | 1924 |
|
SU2011A1 |

