Область техники
Настоящее изобретение относится к:
способам, облегчающим опосредованное антигенсвязывающими молекулами поглощение антигена клетками;
способам увеличения количества антигенов, с которыми может связываться одна антигенсвязывающая молекула;
способам, ускоряющим снижение концентрации антигена в плазме посредством введения антигенсвязывающих молекул;
способам, улучшающим фармакокинетику антигенсвязывающих молекул;
способам снижения общей концентрации антигена или концентрации свободного антигена в плазме;
антигенсвязывающим молекулам, которые увеличивают поглощение антигена клетками;
антигенсвязывающим молекулам, которые обладают способностью связываться с увеличенным количеством антигенов;
антигенсвязывающим молекулам, способным ускорять снижение концентрации антигена в плазме посредством введения молекул;
антигенсвязывающим молекулам с улучшенной фармакокинетикой;
фармацевтическим композициям, содержащим антигенсвязывающие молекулы;
способам получения таковых, описанных выше; и т.п.
Приоритет
По настоящему изобретению испрашивается приоритет патентной заявки Японии № 2010-079667, поданной 30 марта 2010 года, и патентной заявки Японии № 2010-250830, поданной 9 ноября 2010 года, содержание которых в полном объеме включено в настоящий документ в качестве ссылки.
Предшествующий уровень техники
Антитела привлекают внимание в качестве лекарственных средств, так как они обладают высокой стабильностью в плазме и обладают малым количеством побочных эффектов. В настоящее время, ряд подобных IgG лекарственных средств являются коммерчески доступными и на сегодняшний день разрабатываются многие лекарственные средства на основе антител (NPL 1 и 2). Между тем, описаны различные способы, применимые ко второму поколению лекарственных средств на основе антител, включающие те, которые усовершенствуют эффекторную функцию, антигенсвязывающую способность, фармакокинетику и стабильность, и те, которые снижают риск развития иммуногенности (NPL 3). В основном, требуемая доза лекарственного средства на основе антитела является очень высокой. Это, в свою очередь, привело к возникновению проблем, таким как высокая стоимость производства, а также к трудностям при получении подкожных составов. В теории, доза фармацевтического средства на основе антитела может быть снижена посредством улучшения фармакокинетики антитела или улучшения аффинности между антителами и антигенами.
В литературе описаны способы улучшения фармакокинетики антител с использованием искусственных замен аминокислот в константных областях (NPL 4 и 5). Аналогично, описано созревание аффинности как способ улучшения антигенсвязывающей способности или антигеннейтрализующей активности (NPL 6). Этот способ дает возможность увеличения антигенсвязывающей способности посредством введения аминокислотной мутации в область CDR вариабельной области или подобной. Повышение антигенсвязывающей способности обеспечивает улучшение биологической активности in vitro или обеспечивает возможность снижения дозы и дополнительно обеспечивает повышенную эффективность in vivo (NPL 7).
Антигеннейтрализующая способность одной молекулы антитела зависит от ее аффинности. При увеличении аффинности антиген можно нейтрализовать меньшим количеством антитела. Для увеличения аффинности антитела можно использовать различные способы (NPL 6). Кроме того, если аффинность можно сделать очень большой посредством ковалентного связывания антитела с антигеном, одна молекула антитела может нейтрализовать одну молекулу антигена (двухвалентное антитело может нейтрализовать две молекулы антигена). Однако, стехиометрия нейтрализации одного антитела против одного антигена (одного двухвалентного антитела против двух антигенов) является лимитирующим фактором для предшествующих способов, и таким образом невозможно полностью нейтрализовать антиген количеством антитела, меньшим, чем количество антигена. Другими словами, эффект, усиливающий аффинность, имеет предельное значение (NPL 9). Для увеличения периода действия нейтрализующего эффекта нейтрализующего антитела в течение определенного периода, антитело должно быть введено в дозе, более высокой, чем количество антигена, продуцируемое в организме в течение такого же периода. При улучшении фармакокинетики антител или только технологии созревания аффинности, описанной выше, при снижении требуемой дозы антитела существует, таким образом, ограничивающий фактор. Таким образом, с целью поддержания антигеннейтрализующего эффекта антител в течение целевого периода времени с помощью антител в количестве, меньшем, чем количество антигена, одно антитело должно нейтрализовать множество антигенов. Недавно описано антитело, которое связывается с антигеном способом, зависящим от значения рН, в качестве нового способа для достижения описанной выше задачи (PTL 1). Зависимые от значения рН антигенсвязывающие антитела, которые прочно связываются с антигеном при нейтральных условиях в плазме и диссоциируют от антигена при кислых условиях в энодосоме, могут диссоциировать от антигена в эндосоме. Если зависимое от значения рН антигенсвязывающие антитело диссоциирует от антигена, которое рециркулирует в плазме посредством FcRn, то оно может повторно связываться с другим антителом. Таким образом, одно зависимое от значения рН антигенсвязывающее антитело может многократно связываться с рядом антигенов.
Кроме того, время удержания антигена в плазме является очень коротким по сравнению со временем удержания антител, рециркулирующими в плазме посредством связывания с FcRn. Если антитело с таким продолжительным временем удержания в плазме связывает антиген, время удержания в плазме комплекса антиген-антитело увеличивается на тот же период, что и время удержания в плазме антитела. Таким образом, посредством связывания с антителом повышается время удержания антигена, и таким образом концентрация антигена в плазме увеличивается.
Антитело IgG имеет более длительный период времени удержания в плазме в результате связывания с FcRn. Связывание между IgG и FcRn наблюдают только при кислых условиях (pH 6,0). В отличие от этого, наличие связывания практически не наблюдается в нейтральных условиях (pH 7,4). Антитело IgG поглощается клеткой неспецифическим способом. Антитело возвращается к клеточной поверхности посредством связывания с эндосомальным FcRn в кислых условиях эндосомы, а затем диссоциирует от FcRn при нейтральных условиях в плазме. Если связывание с FcRn при кислых условиях является невозможным вследствие введения мутаций в Fc-домен IgG, отсутствие рециркуляции антитела в плазму из эндосомы заметно снижает период времени удержания антитела в плазме. Способ, описывающий увеличение времени удержания антитела IgG, представляет собой способ, повышающий связывание с FcRn при кислых условиях. Для улучшения связывания при кислых условиях в Fc-домен антитела IgG вводят аминокислотные мутации. Это увеличивает эффективность рециркуляции в плазму из эндосомы, что приводит в результате к увеличению времени удержания в плазме. Важным требованием при замене аминокислот является отсутствие увеличения эффективности связывания с FcRn в нейтральных условиях. Если антитело IgG связывается с FcRn в нейтральных условиях, антитело, которое возвращается к клеточной поверхности посредством связывания с FcRn в кислых условиях эндосомы, не диссоциирует от FcRn в нейтральных условиях плазмы. В этом случае, время удержания скорее уменьшится, так как антитело IgG не рециркулирует в плазму. Например, как описано в J Immunol. (2002) 169(9): 5171-80, антитело IgG1, модифицированное посредством введения аминокислотных замен таким образом, что в результате получают антитело, способное связываться с FcRn мыши в нейтральных условиях (pH 7,4), как сообщают, демонстрирует очень низкое значение времени удержания при введении мышам. Кроме того, как описано в J Immunol. (2009) 182(12): 7663-71; J Biol Chem. 2007 Jan 19; 282(3): 1709-17; и J Immunol. 2002 Nov 1; 169(9): 5171-80, антитело IgG1 модифицировали посредством введения замен аминокислот таким образом, что полученное в результате антитело демонстрирует повышенную способность к связыванию с FcRn человека в кислых условиях (pH 6,0), и в тоже время становиться способным к связыванию с FcRn человека в нейтральных условиях (pH 7,4). Описано, что полученное в результате антитело демонстрирует отсутствие увеличения времени удержания или изменений во времени удержания при введении яванским макакам. Таким образом, технология конструирования антител для усовершенствования функции антител сосредоточена только на времени удержания антител в плазме посредством увеличения связывания FcRn человека в кислых условиях, без увеличения времени удержания в нейтральных условиях (pH 7,4). На сегодняшний день не существует сообщений, описывающих преимущества, получаемые при увеличении связывания с FcRn человека в нейтральных условиях (pH 7,4) посредством введения замены аминокислот в Fc-домен антитела IgG. Даже если увеличивается аффинность антигена к антителу, не может быть увеличена элиминация плазменного антигена. Опубликовано, что описанные выше зависимые от pH антигенсвязывающие антитела более эффективны как способ увеличения элиминации плазменного антигена, по сравнению с типичными антителами (PTL 1).
Таким образом, одно зависимое от значения рН антигенсвязывающие антитело связывает ряд антигенов и способно, по сравнению с типичными антителами, облегчать элиминацию антигена в плазме. Таким образом, зависящие от рН антигенсвязывающие антитела имеют действие, которое нельзя осуществить посредством типичных антител. Однако на сегодняшний день не существует сообщений о способах конструирования антител для дополнительного увеличения способности зависящих от рН антигенсвязывающих антител к повторному связыванию с антигенами и для получения эффекта увеличения элиминации антигена в плазме. Документы, касающиеся известного уровня техники, относящиеся к настоящему изобретению, представлены ниже:
СПИСОК ССЫЛОК
Патентная литература
[PTL 1] WO 2009/125825, ANTIGEN-BINDING MOLECULE CAPABLE OF BINDING TO TWO OR MORE ANTIGEN MOLECULES REPEATEDLY
Непатентная литература
[NPL 1] Monoclonal antibody successes in the clinic, Janice M Reichert, Clark J Rosensweig, Laura B Faden & Matthew C Dewitz, Nature Biotechnology 23, 1073-1078 (2005)
[NPL 2] Pavlou AK, Belsey MJ., The therapeutic antibodies market to 2008., Eur J Pharm Biopharm. 2005 Apr; 59(3): 389-96
[NPL 3] Kim SJ, Park Y, Hong HJ., Antibody engineering for the development of therapeutic antibodies., Mol Cells. 2005 Aug 31; 20(1): 17-29. Review
[NPL 4] Hinton PR, Xiong JM, Johlfs MG, Tang MT, Keller S, Tsurushita N., An engineered human IgG1 antibody with longer serum half-life., J Immunol. 2006 Jan 1; 176(1): 346-56
[NPL 5] Ghetie V, Popov S, Borvak J, Radu C, Matesoi D, Medesan C, Ober RJ, Ward ES., Increasing the serum persistence of an IgG fragment by random mutagenesis., Nat Biotechnol. 1997 Jul; 15(7): 637-40
[NPL 6] Proc Natl Acad Sci U S A. 2005 Jun 14; 102(24): 8466-71. Epub 2005 Jun 6. A general method for greatly improving the affinity of antibodies by using combinatorial libraries. Rajpal A, Beyaz N, Haber L, Cappuccilli G, Yee H, Bhatt RR, Takeuchi T, Lerner RA, Crea R
[NPL 7] Wu H, Pfarr DS, Johnson S, Brewah YA, Woods RM, Patel NK, White WI, Young JF, Kiener PA. Development of Motavizumab, an Ultra-potent Antibody for the Prevention of Respiratory Syncytial Virus Infection in the Upper and Lower Respiratory Tract. J Mol Biol. (2007) 368: 652-665
[NPL 8] Hanson CV, Nishiyama Y, Paul S. Catalytic antibodies and their applications. Curr Opin Biotechnol. 2005 Dec; 16(6): 631-6
[NPL 9] Rathanaswami P, Roalstad S, Roskos L, Su QJ, Lackie S, Babcook J. Demonstration of an in vivo generated sub-picomolar affinity fully human monoclonal antibody to interleukin-8. Biochem Biophys Res Commun. 2005 Sep 9; 334(4): 1004-13
Сущность изобретения
Техническая проблема
Настоящее изобретение осуществляют, учитывая обстоятельства, описанные выше. Задачей по настоящему изобретению является разработка способов, облегчающих поглощение антигена клетками посредством использования антигенсвязывающих молекул, способов увеличения количества антигенов, с которыми может связываться одна антигенсвязывающая молекула, способов, ускоряющих снижение концентрации антигена в плазме посредством введения антигенсвязывающих молекул, способов улучшения фармакокинетики антигенсвязывающих молекул, антигенсвязывающих молекул, которые облегчают поглощение антигена клетками, антигенсвязывающих молекул, которые обладают способностью связываться с повышенным количеством связываемых антигенов, антигенсвязывающих молекул, способных ускорять снижение концентрации антигена в плазме посредством введения, антигенсвязывающих молекул с улучшенной фармакокинетикой, фармацевтических композиций, содержащих антигенсвязывающие молекулы, и способов получения таковых, описанных выше.
Решение проблемы
Авторы настоящего изобретения провели целенаправленные исследования способов, облегчающих поглощение антигена клетками посредством антигенсвязывающих молекул (молекулы, такие как полипептиды, которые обладают антигенсвязывающей способностью), способов, позволяющих антигенсвязывающим молекулам неоднократно связываться с антигенами, способов, ускоряющих снижение концентрации антигена в плазме посредством введения антигенсвязывающих молекул, и способов увеличения времени удержания антигенсвязывающих молекул. В результате авторы настоящего изобретения открыли, что антигенсвязывающие молекулы, которые обладают способностью связывать FcRn человека при низких эндосомальных pH и которые обладают более высокой активностью связывания с FcRn человека, чем активность связывания интактного иммуноглобулина человека типа IgG при рН плазмы, могли способствовать поглощению антигена клетками. Авторы настоящего изобретения также обнаружили, что опосредованное антигенсвязывающими молекулами поглощение антигена клетками может быть дополнительно улучшено, и количество антигенов, с которыми может связываться одна молекула, может быть увеличено посредством использования антигенсвязывающей молекулы, которая обладает более слабой антигенсвязывающей активностью при низких значениях рН в эндосоме, чем при рН плазмы; посредством введения такой антигенсвязывающей молекулы можно ускорить снижение концентрации антигена в плазме; и можно улучшить фармакокинетику антигенсвязывающей молекулы.
Конкретно, настоящее изобретение относится к:
способам, облегчающим опосредованное антигенсвязывающими молекулами поглощение антигена клетками;
способам увеличения количества антигенов, с которыми может связываться одна антигенсвязывающая молекула;
способам, ускоряющим снижение концентрации антигена в плазме посредством введения антигенсвязывающих молекул;
способам, улучшающим фармакокинетику антигенсвязывающих молекул;
способам снижения общей концентрации антигена или концентрации свободного антигена в плазме;
антигенсвязывающим молекулам, которые увеличивают поглощение антигена клетками;
антигенсвязывающим молекулам, которые обладают способностью связываться с увеличенным количеством антигенов;
антигенсвязывающим молекулам, способным ускорять снижение концентрации антигена в плазме посредством введения молекул;
антигенсвязывающим молекулам с улучшенной фармакокинетикой;
фармацевтическим композициям, содержащим антигенсвязывающие молекулы;
способам получения таковых, описанных выше; и т.п. Более конкретно, настоящее изобретение относится к:
[1] антигенсвязывающей молекуле, содержащей антигенсвязывающий домен и FcRn-связывающий домен человека, обладающей активностью связывания с FcRn человека в кислом и нейтральном диапазонах pH, где активность связывания с FcRn человека в диапазоне нейтральных значений рН представлена микромолярностью со значением более чем 3,2;
[2] антигенсвязывающей молекуле, содержащей антигенсвязывающий домен и FcRn-связывающий домен человека, которая обладает активностью связывания с FcRn человека в диапазоне нейтральных значений pH, где активность связывания с FcRn человека в диапазоне нейтральных значений рН представлена значением, в 28 раз большим, чем для интактного IgG человека;
[3] антигенсвязывающей молекуле, содержащей антигенсвязывающий домен и FcRn-связывающий домен человека, которая обладает активностью связывания с FcRn человека в диапазоне нейтральных значений pH, где активность связывания с FcRn человека в диапазоне нейтральных значений рН представлена микромолярностью со значением более чем 2,3;
[4] антигенсвязывающей молекуле, содержащей антигенсвязывающий домен и FcRn-связывающий домен человека, которая обладает активностью связывания с FcRn человека в диапазоне нейтральных значений pH, где активность связывания с FcRn человека в диапазоне нейтральных значений рН представлена значением, в 38 раз большим, чем для интактного IgG человека;
[5] антигенсвязывающей молекуле по любому из пп.[1]-[4], где диапазон нейтральных значений рН представлен значениями pH от 7,0 до 8,0;
[6] антигенсвязывающей молекуле, содержащей антигенсвязывающий домен и FcRn-связывающий домен человека, для которой общая концентрация антигена в плазме после введения антигенсвязывающей молекулы не являющемуся человеком животному представлена значением, меньшим, чем значение концентрации антигена в плазме после введения контрольной антигенсвязывающей молекулы не являющемуся человеком животному, содержащей такой же антигенсвязывающий домен и интактный Fc-домен IgG человека в качестве FcRn-связывающего домена человека;
[7] антигенсвязывающей молекуле, для которой концентрация антигена в плазме после введения антигенсвязывающей молекулы не являющемуся человеком животному представлена значением, меньшим, чем значение общей концентрации антигена в плазме, полученное для не являющегося человеком животного, которому не вводили антигенсвязывающую молекулу;
[8] антигенсвязывающей молекуле, содержащей антигенсвязывающий домен и FcRn-связывающий домен человека, где молярное соотношение "антиген/антигенсвязывающая молекула" (C) для антигенсвязывающей молекулы. рассчитанное, как указано далее:
C=A/B,
представляет собой значение, меньшее, чем молярное соотношение "антиген/антигенсвязывающая молекула" (C’) для антигенсвязывающей молекулы, содержащей тот же антигенсвязывающий домен и интактный Fc-домен IgG человека в качестве FcRn-связывающего домена человека, рассчитанное следующим образом:
C’=A’/B’,
где:
A представляет собой значение общей концентрации антигена в плазме после введения антигенсвязывающей молекулы не являющемуся человеком животному,
B представляет собой значение концентрации антигенсвязывающей молекулы в плазме после введения антигенсвязывающие молекулы не являющемуся человеком животному,
A’ представляет собой значение общей концентрации антигена в плазме после введения контрольной антигенсвязывающей молекулы не являющемуся человеком животному,
B’ представляет собой значение концентрации антигенсвязывающей молекулы в плазме после введения контрольной антигенсвязывающей молекулы не являющемуся человеком животному;
[9] антигенсвязывающей молекуле по любому из пп.[6]-[8], где не являющееся человеком животное представляет собой трансгенную мышь с FcRn человека;
[10] антигенсвязывающей молекулы по любому из пп.[6]-[9], где концентрация антигена в плазме представляет собой постоянную общую концентрацию антигена в плазме;
[11] антигенсвязывающей молекуле по любому из пп.[6]-[9], где концентрация антигена в плазме представляет собой временную общую концентрацию антигена в плазме;
[12] антигенсвязывающей молекуле, содержащей антигенсвязывающий домен и FcRn-связывающий домен человека, которая обладает активностью связывания с FcRn человека в диапазонах кислых и нейтральных значений рН, где активность связывания с FcRn человека в диапазоне нейтральных значений pH представлена значением, более высоким, чем для интактного IgG человека;
[13] антигенсвязывающей молекуле по любому из пп.[1]-[11], где антигенсвязывающая активность антигенсвязывающего домена в диапазоне кислых значений рН представлена значением, меньшим, чем антигенсвязывающая активность антигенсвязывающего домена в диапазоне нейтральных значений рН;
[14] антигенсвязывающей молекуле по п.[12] или [13], где отношение антигенсвязывающей активности в диапазоне кислых и нейтральных значений рН представляет собой по меньшей мере 2 для значения KD (в кислом диапазоне рН)/KD (в нейтральном диапазоне рН);
[15] антигенсвязывающей молекуле по любому из пп.[12]-[14], которая содержит аминокислотную мутацию антигенсвязывающего домена, который содержит замену гистидина на по меньшей мере одну аминокислоту для антигенсвязывающего домена или содержит вставку по меньшей мере из одного гистидина;
[16] антигенсвязывающей молекулы по любому из пп.[12]-[14], где антигенсвязывающий домен получен из библиотеки антигенсвязывающих доменов;
[17] антигенсвязывающей молекуле по любому из пп.[1]-[16], которая содержит, в качестве FcRn-связывающего домена человека, Fc-домен, полученный в результате замещения различных аминокислот на по меньшей мере одну аминокислоту в Fc-домене родительского IgG;
[18] антигенсвязывающей молекуле по любому из пп.[1]-[17], где FcRn-связывающий домен человека представляет собой FcRn-связывающий домен человека, содержащий аминокислотную последовательность с заменой различных аминокислот на по меньшей мере одну аминокислоту, выбранную из аминокислот, расположенных в позициях 237, 238, 239, 248, 250, 252, 254, 255, 256, 257, 258, 265, 270, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 325, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434 и 436 (нумерация EU) в Fc-домене родительского IgG;
[19] антигенсвязывающей молекуле по любому из пп.[1]-[18], которая содержит FcRn-связывающий домен человека, содержащий аминокислотную замену в Fc-домене родительского IgG, который включает по меньшей мере одну аминокислотную замену, выбранную из:
замена аминокислоты Met на Gly в положении 237;
замена аминокислоты Ala на Pro в положении 238;
замена аминокислоты Lys на Ser в положении 239;
замена аминокислоты Ile на Lys в положении 248;
замена аминокислоты Ala, Phe, Ile, Met, Gln, Ser, Val, Trp или Tyr на Thr в положении 250;
замена аминокислоты Phe, Trp или Tyr на Met в положении 252;
замена аминокислоты Thr на Ser в положении 254;
замена аминокислоты Glu на Arg в положении 255;
замена аминокислоты Asp, Glu или Gln на Thr в положении 256;
замена аминокислот Ala, Gly, Ile, Leu, Met, Asn, Ser, Thr, или Val на Pro в положении 257;
замена аминокислоты His на Glu в положении 258;
замена аминокислоты Ala на Asp в положении 265;
замена аминокислоты Phe на Asp в положении 270;
замена аминокислоты Ala или Glu на Asn в положении 286;
замена аминокислоты His на Thr в положении 289;
замена аминокислоты Ala на Asn в положении 297;
замена аминокислоты Gly на Ser в положении 298;
замена аминокислоты Ala на Val в положении 303;
замена аминокислоты Ala на Val в положении 305;
замена аминокислот Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr на Thr в положении 307;
замена аминокислот Ala, Phe, Ile, Leu, Met, Pro, Gln или Thr на Val в положении 308;
замена аминокислот Ala, Asp, Glu, Pro или Arg на Leu или Val в положении 309;
замена аминокислот Ala, His или Ile на Gln в положении 311;
замена аминокислот Ala или His на Asp в положении 312;
замена аминокислот Lys или Arg на Leu в положении 314;
замена аминокислот Ala или His на Asn в положении 315;
замена аминокислоты Ala на Lys в положении 317;
замена аминокислоты Gly на Asn в положении 325;
замена аминокислоты Val на Ile в положении 332;
замена аминокислоты Leu на Lys в положении 334;
замена аминокислоты His на Lys в положении 360;
замена аминокислоты Ala на Asp в положении 376;
замена аминокислоты Ala на Glu в положении 380;
замена аминокислоты Ala на Glu в положении 382;
замена аминокислоты Ala на Asn или Ser в положении 384;
замена аминокислоты Asp или His на Gly в положении 385;
замена аминокислоты Pro на Gln в положении 386;
замена аминокислоты Glu на Pro в положении 387;
замена аминокислоты Ala или Ser на Asn в положении 389;
замена аминокислоты Ala на Ser в положении 424;
замена аминокислот Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr на Met в положении 428;
замена аминокислоты Lys на His в положении 433;
замена аминокислот Ala, Phe, His, Ser, Trp или Tyr на Asn в положении 434;
и замена аминокислоты His или Phe на Tyr в положении 436 в нумерации EU;
[20] антигенсвязывающей молекуле по любому из пп.[1]-[18], в которой FcRn-связывающий домен человека содержит по меньшей мере одну аминокислоту, выбранную из:
Met в положении аминокислоты 237;
Ala в положении аминокислоты 238;
Lys в положении аминокислоты 239;
Ile в положении аминокислоты 248;
Ala, Phe, Ile, Met, Gln, Ser, Val, Trp или Tyr в положении аминокислоты 250;
Phe, Trp или Tyr в положении аминокислоты 252;
Thr в положении аминокислоты 254;
Glu в положении аминокислоты 255;
Asp, Glu или Gln в положении аминокислоты 256;
Ala, Gly, Ile, Leu, Met, Asn, Ser, Thr или Val в положении аминокислоты 257;
His в положении аминокислоты 258;
Ala в положении аминокислоты 265;
Phe в положении аминокислоты 270;
Ala или Glu в положении аминокислоты 286;
His в положении аминокислоты 289;
Ala в положении аминокислоты 297;
Gly в положении аминокислоты 298;
Ala в положении аминокислоты 303;
Ala в положении аминокислоты 305;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr в положении аминокислоты 307;
Ala, Phe, Ile, Leu, Met, Pro, Gln или Thr в положении аминокислоты 308;
Ala, Asp, Glu, Pro или Arg в положении аминокислоты 309;
Ala, His или Ile в положении аминокислоты 311;
Ala или His в положении аминокислоты 312;
Lys или Arg в положении аминокислоты 314;
Ala или His в положении аминокислоты 315;
Ala в положении аминокислоты 317;
Gly в положении аминокислоты 325;
Val в положении аминокислоты 332;
Leu в положении аминокислоты 334;
His в положении аминокислоты 360;
Ala в положении аминокислоты 376;
Ala в положении аминокислоты 380;
Ala в положении аминокислоты 382;
Ala в положении аминокислоты 384;
Asp или His в положении аминокислоты 385;
Pro в положении аминокислоты 386;
Glu в положении аминокислоты 387;
Ala или Ser в положении аминокислоты 389;
Ala в положении аминокислоты 424;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr в положении аминокислоты 428;
Lys в положении аминокислоты 433;
Ala, Phe, His, Ser, Trp или Tyr в положении аминокислоты 434;
и His или Phe в положении аминокислоты 436 (нумерация EU) в Fc-домене родительского IgG;
[21] антигенсвязывающей молекуле по любому из пп.[18]-[20], где родительский IgG выбран из IgG, полученного от не являющегося человеком животного;
[22] антигенсвязывающей молекуле по любому из пп.[18]-[20], где родительский IgG представляет собой IgG человека;
[23] антигенсвязывающей молекуле по любому из пп.[1]-[22], которая обладает антагонистической активностью;
[24] антигенсвязывающей молекуле по пп.[1]-[23], которая связывается с мембранным антигеном или растворимым антигеном;
[25] антигенсвязывающей молекуле по любому из пп.[1]-[24], где антигенсвязывающий домен содержит искусственный лиганд, который связывается с рецептором;
[26] антигенсвязывающей молекуле по любому из пп.[1]-[24], где антигенсвязывающий домен содержит искусственный рецептор, который связывается с лигандом;
[27] антигенсвязывающей молекуле по любому из пп.[1]-[24], которая представляет собой антитело;
[28] антигенсвязывающей молекуле [27], где антитело выбрано из химерного антитела, гуманизированного антитела или антитела человека;
[29] фармацевтическим композициям, содержащим любую из антигенсвязывающих молекул по пп.[1]-[28];
[30] способу облегчения опосредованного антигенсвязывающей молекулой поглощения антигена клетками посредством увеличения ее активности связывания с FcRn человека в диапазоне нейтральных значений рН, где антигенсвязывающая молекула содержит антигенсвязывающий домен и FcRn-связывающий домен человека и обладает активностью связывания с FcRn человека в диапазоне кислых значений рН;
[31] способу облегчения опосредованного антигенсвязывающей молекулой поглощения антигена клеткой посредством увеличения активности связывания антигенсвязывающей молекулы с FcRn человека в диапазоне нейтральных значений рН и снижения ее антигенсвязывающей активности в диапазоне кислых значений рН до значения, меньшего, чем в диапазоне нейтральных значений рН, где антигенсвязывающая молекула содержит антигенсвязывающий домен и FcRn-связывающий домен человека и имеет активность связывания с FcRn человека в диапазоне кислых значений рН;
[32] способу увеличения количества антигенов, с которыми может связываться одна антигенсвязывающая молекула, посредством увеличения ее активности связывания с FcRn человека в диапазоне нейтральных значений рН, где антигенсвязывающая молекула содержит антигенсвязывающий домен и FcRn-связывающий домен человека и обладает активностью связывания с FcRn человека в диапазоне кислых значений рН;
[33] способу увеличения количества антигенов, с которыми может связываться одна антигенсвязывающая молекула, посредством увеличения ее активности связывания с FcRn человека в диапазоне нейтральных значений рН и снижения ее антигенсвязывающей активности в диапазоне кислых значений рН до значения, меньшего, чем диапазоне нейтральных значений рН, где антигенсвязывающая молекула содержит антигенсвязывающий домен и FcRn-связывающий домен человека и обладает активностью связывания с FcRn человека в диапазоне кислых значений рН;
[34] способу увеличения способности антигенсвязывающей молекулы к элиминации антигена из плазмы посредством увеличения ее активности связывания с FcRn человека в диапазоне нейтральных значений рН, где антигенсвязывающая молекула содержит антигенсвязывающий домен и FcRn-связывающий домен человека и обладает активностью связывания с FcRn человека в диапазоне кислых значений рН;
[35] способу увеличения способности антигенсвязывающей молекулы к элиминации антигена из плазмы посредством увеличения ее активности связывания с FcRn человека в диапазоне нейтральных значений рН и снижения ее антигенсвязывающей активности в диапазоне кислых значений рН до значения, меньшего, чем в диапазоне нейтральных значений рН, где антигенсвязывающая молекула содержит антигенсвязывающий домен и FcRn-связывающий домен человека и обладает активностью связывания с FcRn человека в диапазоне кислых значений рН;
[36] способу улучшения фармакокинетики антигенсвязывающей молекулы посредством увеличения ее активности связывания с FcRn человека в диапазоне нейтральных значений рН, где антигенсвязывающая молекула содержит антигенсвязывающий домен и FcRn-связывающий домен человека и обладает активностью связывания с FcRn человека в диапазоне кислых значений рН;
[37] способу улучшения фармакокинетики антигенсвязывающей молекулы посредством увеличения ее активности связывания с FcRn человека в диапазоне нейтральных значений рН и снижения ее антигенсвязывающей активности в диапазоне кислых значений рН до значения, меньшего, чем в диапазоне нейтральных значений рН, где антигенсвязывающая молекула содержит антигенсвязывающий домен и FcRn-связывающий домен человека и обладает активностью связывания с FcRn человека в диапазоне кислых значений рН;
[38] способу, облегчающему внутриклеточную диссоциацию антигена, связанного с антигенсвязывающей молекулой за пределами клетки, от антигенсвязывающей молекулы, посредством увеличения ее активности связывания с FcRn человека в диапазоне нейтральных значений рН и снижения ее антигенсвязывающей активности в диапазоне кислых значений рН до значения, меньшего, чем в диапазоне нейтральных значений рН, где антигенсвязывающая молекула содержит антигенсвязывающий домен и FcRn-связывающий домен человека и обладает активностью связывания с FcRn человека в диапазоне кислых значений рН;
[39] способу, облегчающему внеклеточное высвобождение антигенсвязывающей молекулы в форме, свободной от антигена, поглощенной клеткой в форме, связанной с антигеном, посредствам повышения ее активности связывания с FcRn человека в диапазоне нейтральных значений рН и снижения ее антигенсвязывающей активности в диапазоне кислых значений рН до значения, меньшего, чем в диапазоне нейтральных значений рН, где антигенсвязывающая молекула содержит антигенсвязывающий домен и FcRn-связывающий домен человека и обладает активностью связывания с FcRn человека в диапазоне кислых значений рН;
[40] способу снижения в плазме общей концентрации антигена или концентрации свободного антигена посредством увеличения его активности связывания с FcRn человека в диапазоне нейтральных значений рН, где антигенсвязывающая молекула содержит антигенсвязывающий домен и FcRn-связывающий домен человека и обладает активностью связывания с FcRn человека в диапазоне кислых значений рН;
[41] способу снижения общей концентрации антигена или концентрации свободного антигена в плазме посредством увеличения его активности связывания с FcRn человека в диапазоне нейтральных значений рН и снижением его антигенсвязывающей активности в диапазоне кислых значений рН до значения, меньшего, чем в диапазоне нейтральных значений рН, где антигенсвязывающая молекула содержит антигенсвязывающий домен и FcRn-связывающий домен человека и обладает активностью связывания с FcRn человека в диапазоне кислых значений рН;
[42] способу по любому из пп.[30]-[41], где диапазон кислых значений pH представляет собой значения pH от 5,5 до 6,5, и диапазон нейтральных значений рН представляет собой значения pH от 7,0 до 8,0;
[43] способу по любому из пп.[30]-[41], где увеличение активности связывания с FcRn человека в диапазоне нейтральных значений рН представляет собой увеличение посредством замены различных аминокислот на по меньшей мере одну аминокислоту в родительском Fc-домене IgG FcRn-связывающего домена человека;
[44] способу по любому из пп.[30]-[41], где увеличение активности связывания с FcRn человека в диапазоне нейтральных значений рН представляет собой увеличение полсредством замены различных аминокислот на по меньшей мере одну аминокислоту, выбранную из аминокислот, представленных в положениях 237, 238, 239, 248, 250, 252, 254, 255, 256, 257, 258, 265, 270, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 325, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434 и 436 (нумерация EU) в родительском Fc-домене IgG FcRn-связывающего домена человека;
[45] способу по любому из пп.[31], [33], [35], [37]-[39] и [41], где антигенсвязывающую активность антигенсвязывающей молекулы в диапазоне кислых значений рН снижают до значения, меньшего, чем в диапазоне нейтральных значений рН, посредством замены гистидина на по меньшей мере одну аминокислоту в антигенсвязывающей молекуле или посредством вставки по меньшей мере одного гистидина;
[46] способу по любому из пп.[31], [33], [35], [37]-[39] и [41], где антигенсвязывающий домен получают из библиотеки антигенсвязывающих доменов;
[47] способу по любому из пп.[31], [33], [35], [37]-[39] и [41], где снижение антигенсвязывающей активности представлено увеличением значения KD (в диапазоне кислых значений рН)/KD (в диапазоне нейтральных значений рН), которое представляет собой отношение антигенсвязывающей активности в диапазоне кислых значений рН и в диапазоне нейтральных значений рН по отношению к этому значению до введения гистидиновой замены или вставки гистидина;
[48] способу получения антигенсвязывающей молекулы, который включает стадии:
(a) отбора антигенсвязывающей молекулы, которая имеет активность связывания с FcRn человека в диапазоне нейтральных значений рН, более высокую, чем 3,2 микромоля, полученную посредством замены по меньшей мере одной аминокислоты в FcRn-связывающем домене человека антигенсвязывающей молекулы;
(b) получения гена, кодирующего антигенсвязывающую молекулу, в которой FcRn-связывающий домен человека и антигенсвязывающий домен, полученные на стадии (a), сцеплены; и
(c) получения антигенсвязывающей молекулы с использованием гена, полученного на стадии (b);
[49] способу получения антигенсвязывающей молекулы, который включает стадии:
(a) отбора антигенсвязывающей молекулы, которая имеет активность связывания с FcRn человека в диапазоне нейтральных значений рН, более высокую, чем до внесения изменений по меньшей мере в одну аминокислоту FcRn-связывающего домена человека в антигенсвязывающей молекуле, имеющей активность связывания с FcRn человека в диапазоне кислых значений рН;
(b) изменения по меньшей мере одной аминокислоты в антигенсвязывающем домене антигенсвязывающей молекулы и отбора антигенсвязывающей молекулы, которая в диапазоне нейтральных значений рН обладает более высокой антигенсвязывающей активностью, чем в диапазоне кислых значений рН;
(c) получения гена, кодирующего антигенсвязывающую молекулу, в которой FcRn-связывающий домен человека и антигенсвязывающий домен, полученные на стадиях (a) и (b), сцеплены; и
(d) получения антигенсвязывающей молекулы с использованием гена, полученного на стадии(c); и
[50] способу получения антигенсвязывающей молекулы, который включает стадии:
(a) отбора антигенсвязывающей молекулы, которая имеет более высокую активность связывания с FcRn человека в диапазоне нейтральных значений рН, чем активность связывания до изменения по меньшей мере одной аминокислоты в FcRn-связывающем домене антигенсвязывающей молекулы человека, обладающей активностью связывания с FcRn человека в диапазоне кислых значений рН;
(b) отбора антигенсвязывающей молекулы, которая в диапазоне нейтральных значений рН обладает более высокой антигенсвязывающей активностью, чем в диапазоне кислых значений рН;
(c) получения гена, кодирующего антигенсвязывающую молекулу, в которой FcRn-связывающий домен человека и антигенсвязывающий домен, полученные на стадиях (a) и (b), сцеплены; и
(d) получение антигенсвязывающей молекулы с использованием гена, полученного на стадии (c);
[51] антигенсвязывающей молекуле, полученной посредством способа получения по любому из пп.[48]-[50];
[52] способу скрининга антигенсвязывающей молекулы, который включает стадии:
(a) отбора антигенсвязывающей молекулы, которая обладает активностью связывания с FcRn человека в диапазоне нейтральных значений рН, более высокой, чем 3,2 микромоля, где антигенсвязывающая молекула получена посредством изменения по меньшей мере одной аминокислоты в FcRn-связывающем домене человека антигенсвязывающей молекулы;
(b) получения гена, кодирующего антигенсвязывающую молекулу, в которой FcRn-связывающий домен человека и антигенсвязывающий домен, полученные на стадии (a), сцеплены; и
(c) получения антигенсвязывающей молекулы с использованием гена, полученного на стадии (b);
[53] способу скрининга антигенсвязывающей молекулы, который содержит стадии:
(a) отбора антигенсвязывающей молекулы, которая обладает активностью связывания с FcRn человека в диапазоне нейтральных значений рН, более высокой, чем до изменения по меньшей мере одной аминокислоты в FcRn-связывающем домене антигенсвязывающей молекулы человека, обладающей активностью связывания с FcRn человека в диапазоне кислых значений рН;
(b) изменения по меньшей мере одной аминокислоты в антигенсвязывающем домене антигенсвязывающей молекулы и отбора антигенсвязывающей молекулы, которая обладает антигенсвязывающей активностью в диапазоне нейтральных значений рН, более высокой, чем в диапазоне кислых значений рН;
(c) получения гена, кодирующего антигенсвязывающую молекулу, в которой FcRn-связывающий домен человека и антигенсвязывающий домен, полученные на стадиях (a) и (b), сцеплены; и
(d) получения антигенсвязывающие молекулы с использованием гена, полученного на стадии (c);
[54] способу скрининга антигенсвязывающей молекулы, который включает стадии:
(a) отбора антигенсвязывающей молекулы, которая обладает активностью связывания с FcRn человека в диапазоне нейтральных значений рН, более высокой, чем до изменения по меньшей мере одной аминокислоты в FcRn-связывающем домене человека антигенсвязывающей молекулы, обладающей активностью связывания с FcRn человека в диапазоне кислых значений рН;
(b) отбора антигенсвязывающей молекулы, которая обладает антигенсвязывающей активностью в диапазоне нейтральных значений рН, более высокой, чем в диапазоне кислых значений рН;
(c) получения гена, кодирующего антигенсвязывающую молекулу, в которой FcRn-связывающий домен человека и антигенсвязывающий домен, полученные на стадиях (a) и (b), сцеплены; и
(d) получения антигенсвязывающей молекулы с использованием гена, полученного на стадии (c);
[55] способу по любому из пп.[30]-[54], где антигенсвязывающий домен содержит искусственный лиганд, который связывается с рецептором;
[56] способу по любому из пп.[30]-[54], где антигенсвязывающий домен содержит искусственный рецептор, который связывается с лигандом; и
[57] способу по любому из пп.[30]-[54], где антигенсвязывающая молекула представляет собой антитело.
Преимущества изобретения
Настоящее изобретение относится к:
способам, облегчающим опосредованное антигенсвязывающими молекулами поглощение антигена клетками; способам увеличения количества антигенов, с которыми может связываться одна антигенсвязывающая молекула; и способам, ускоряющим снижение концентрации антигена в плазме посредством введения антигенсвязывающих молекул. При облегчении опосредованного антигенсвязывающими молекулами поглощения антигена клетками, снижение концентрации антигена в плазме может быть ускорено посредством введения таких антигенсвязывающих молекул, и фармакокинетика антигенсвязывающей молекулы может быть усовершенствована для увеличения количества антигенов, с которыми может связываться одна антигенсвязывающая молекула. Таким образом, антигенсвязывающие молекулы могут создавать более лучшие эффекты in vivo, чем общепринятые антигенсвязывающие молекулы.
Краткое описание чертежей
[фиг.1] На фиг.1 в графической форме представлена, в зависимости от времени, концентрация в плазме растворимой формы рецептора IL-6 человека после введения антитела против рецептора IL-6 человека трансгенным мышам с FcRn человека (линия 276), где концентрация в плазме растворимой формы рецептора IL-6 человека является постоянной (стационарная модель инфузии).
[фиг.2] На фиг.2 представлено схематическое изображение, которое показывает, что диссоциация молекулы антитела IgG от растворимого антигена в эндосоме приводит в результате к увеличению элиминации антигена, что приводит к новому раунду связывания с другим антигеном.
[фиг.3] На фиг.3 в графической форме представлена, в зависимости от времени, концентрация антитела в плазме у трансгенных мышей с FcRn человека.
[фиг.4] На фиг.4 в графической форме представлена, в зависимости от времени, концентрация в плазме растворимой формы рецептора IL-6 человека у трансгенных мышей с FcRn человека.
[фиг.5] На фиг.5 в графической форме представлена, в зависимости от времени, концентрация антитела в плазме у нормальной мыши.
[фиг.6] На фиг.6 в графической форме представлена, в зависимости от времени, концентрация в плазме растворимой формы рецептора IL-6 человека у нормальной мыши.
[фиг.7] На фиг.7 в графической форме представлена, в зависимости от времени, концентрация в плазме несвязанной растворимой формы рецептора IL-6 человека у нормальной мыши.
[фиг.8] На фиг.8 в графической форме представлена, в зависимости от времени, концентрация в плазме растворимой формы рецептора IL-6 человека у трансгенных мышей с FcRn человека.
[фиг.9] На фиг.9 в графической форме представлена, в зависимости от времени, концентрация в плазме растворимой формы рецептора IL-6 человека после введения Fv4-IgG1-F14 в низкой дозе (0,01 мг/кг) или 1 мг/кг.
[фиг.10] На фиг.10 в графической форме представлена, в зависимости от времени, концентрация антитела в плазме после введения Fv4-IgG1-F14 в низкой дозе (0,01 мг/кг) или 1 мг/кг.
[фиг.11] На фиг.11 в графической форме представлена, в зависимости от времени, концентрация в плазме растворимой формы рецептора IL-6 человека после введения антитела против рецептора IL-6 человека нормальной мыши, где концентрация в плазме растворимой формы рецептора IL-6 человека является постоянной.
[фиг.12] На фиг.12 в графической форме представлена, в зависимости от времени, концентрация антитела в плазме после совместной инъекции hsIL-6R и антитела против рецептора IL-6 человека трансгенным мышам с FcRn человека (линия 276).
[фиг.13] На фиг.13 в графической форме представлена, в зависимости от времени, концентрация в плазме растворимой формы рецептора IL-6 человека после совместной инъекции hsIL-6R и антитела против рецептора IL-6 человека трансгенным мышам с FcRn человека (линия 276).
[фиг.14] На фиг.14 представлена зависимость между аффинностью связывания Fc-вариантов с FcRn человека при pH 7,0 и концентрацией в плазме hsIL-6R в сутки 1 после совместной инъекции hsIL-6R и антитела против рецептора IL-6 человека трансгенным мышам с FcRn человека (линия 276).
[фиг.15] На фиг.15 представлена зависимость между аффинностью связывания Fc-вариантов с FcRn человека при pH 7,0 и концентрация антитела в плазме в сутки 1 после совместной инъекции hsIL-6R и антитела против рецептора IL-6 человека трансгенным мышам с FcRn человека (линия 276).
[фиг.16] На фиг.16 представлено в зависимости от времени молярное отношение антиген/антитело (значение C) после совместной инъекции hsIL-6R и антитела против рецептора IL-6 человека трансгенным мышам с FcRn человека (линия 276).
[фиг.17] На фиг.17 представлена зависимость между аффинностью связывания Fc-вариантов с FcRn человека при pH 7,0 и молярное отношение антиген/антитело (значение C) в сутки 1 после совместной инъекции hsIL-6R и антитела против рецептора IL-6 человека трансгенным мышам с FcRn человека (линия 276).
[фиг.18] На фиг.18 в графической форме представлена, в зависимости от времени, концентрация в плазме hsIL-6R после введения Fv4-IgG1-F14 в низких дозах (0,01 или 0,2 мг/кг) или 1 мг/кг трансгенным мышам с FcRn человека (линия 276), где концентрация в плазме hsIL-6R является постоянной (стационарная модель инфузии).
[фиг.19] На фиг.19 представлена, в зависимости от времени, концентрация hsIL-6R в плазме трансгенных мышей с FcRn человека линии 276 и линии 32 после совместной инъекции трансгенным мышам с FcRn человека (линия 276 и 32) hsIL-6R и антитела против рецептора IL-6 человека.
[фиг.20] На фиг.20 представлена, в зависимости от времени, концентрация антитела в плазме трансгенной мыши с FcRn человека линии 276 и линии 32 после совместной инъекции hsIL-6R и антитела против рецептора IL-6 человека трансгенным мышам с FcRn человека (линия 276 и 32).
[фиг.21] На фиг.21 в графической форме представлена, в зависимости от времени, концентрация в плазме hsIL-6R после введения антитела против рецептора IL-6 человека трансгенным мышам с FcRn человека (линия 32), где концентрация в плазме hsIL-6R является постоянной (стационарная модель инфузии).
[фиг.22] На фиг.22 в графической форме представлена, в зависимости от времени, концентрация антитела в плазме после введения антитела против рецептора IL-6 человека трансгенным мышам с FcRn человека (линия 32), где концентрация в плазме hsIL-6R является постоянной (стационарная модель инфузии).
[фиг.23] На фиг.23 представлено, в зависимости от времени, молярное отношение антиген/антитело (значение C) после введения антитела против рецептора IL-6 человека трансгенным мышам с FcRn человека (линия 32), где концентрация в плазме hsIL-6R является постоянной (стационарная модель инфузии).
[фиг.24] На фиг.24 представлена зависимость между аффинностью связывания Fc-вариантов с FcRn человека при pH 7,0 и молярное отношение антиген/антитело (значение C) в сутки 1 после введения антитела против рецептора IL-6 человека трансгенным мышам с FcRn человека (линия 32), где концентрация в плазме hsIL-6R является постоянной (стационарная модель инфузии).
[фиг.25] На фиг.25 в графической форме представлена, в зависимости от времени, концентрация антитела в плазме после введения антитела против рецептора IL-6 человека, обладающего Fc-вариантом F11, F39, F48 и F264, трансгенным мышам с FcRn человека (линия 32), где концентрация в плазме hsIL-6R является постоянной (стационарная модель инфузии).
[фиг.26] на фиг.26 в графической форме представлена, в зависимости от времени, концентрация в плазме hsIL-6R после введения антител против рецептора IL-6 человека, обладающих Fc-вариантом F11, F39, F48 и F264, трансгенным мышам с FcRn человека (линия 32), где концентрация в плазме hsIL-6R является постоянной (стационарная модель инфузии).
[фиг.27] На фиг.27 в графической форме представлена, в зависимости от времени, концентрация антитела в плазме после введения антитела против рецептора IL-6 человека, обладающего Fc-вариантом F157, F196 и F262, трансгенным мышам с FcRn человека (линия 32), где концентрация в плазме hsIL-6R является постоянной (стационарная модель инфузии).
[фиг.28] На фиг.28 в графической форме представлена, в зависимости от времени, концентрация в плазме hsIL-6R после введения антитела против рецептора IL-6 человека, обладающего Fc-вариантом F157, F196 и F262, трансгенным мышам с FcRn человека (линия 32), где концентрация в плазме hsIL-6R является постоянной (стационарная модель инфузии).
[фиг.29] На фиг.29 представлена фармакокинетическая модель, используемая для исследований in silico традиционных антител и антител, элиминирующих антигены.
[фиг.30] На фиг.30 в графической форме представлена, в зависимости от времени, концентрация в плазме IL-6 человека после совместного инъецирования нормальной мыши IL-6 человека и антитела против IL-6 человека.
[фиг.31] На фиг.31 в графической форме представлена, в зависимости от времени, концентрация антитела в плазме после совместной инъекции нормальной мыши IL-6 человека и антитела против IL-6.
[фиг.32] На фиг.32 представлены сенсограммы связывания IgA человека со слитым белком CD89-Fc при pH 7,4 и pH 6,0 с использованием Biacore.
[фиг.33] На фиг.33 в графической форме представлена, в зависимости от времени, концентрация в плазме IgA человека после совместной инъекции нормальной мыши IgA человека и слитого белка CD89-Fc.
[фиг.34] На фиг.34 в графической форме представлена, в зависимости от времени, концентрация антитела в плазме после совместной инъекции IgA человека и слитого белка CD89-Fc нормальной мыши.
[фиг.35] На фиг.35 представлено графическое изображение концентрации в плазме растворимого плексина A1 человека в момент времени, соответствующий 7 часам, после совместной инъекции нормальной мыши растворимого плексина A1 человека и антитела против плексина A1.
Описание вариантов осуществления
Настоящее изобретение относится к способам, облегчающим опосредованное антигенсвязывающей молекулой поглощение антигена клетками. Более конкретно, настоящее изобретение относится к способам, облегчающим поглощение антигена клетками посредством антигенсвязывающей молекулы, обладающей активностью связывания с FcRn человека в диапазоне кислых значений рН, где способы основаны на повышении активности связывания с FcRn человека антигенсвязывающей молекулы в диапазоне нейтральных значений рН. Настоящее изобретение также относится к способам увеличения поглощения антигена клетками посредством антигенсвязывающей молекулы, обладающей активностью связывания с FcRn человека в диапазоне кислых значений рН, где способы основаны на изменении по меньшей мере одной аминокислоты в FcRn-связывающем домене антигенсвязывающей молекулы человека.
Настоящее изобретение также относится к способам, облегчающим поглощение антигена клетками посредством антигенсвязывающей молекулы, обладающей активностью связывания с FcRn человека в диапазоне кислых значений рН, где способы основаны на использовании FcRn-связывающего домена человека, содержащего аминокислотную последовательность с заменой различных аминокислот на по меньшей мере одну аминокислоту, выбранную из таковых аминокислот в положениях 237, 238, 239, 248, 250, 252, 254, 255, 256, 257, 258, 265, 270, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 325, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434 и 436 (нумерация EU) в Fc-домене родительского IgG FcRn-связывающего домена человека, включающего Fc-домен родительского IgG.
Настоящее изобретение также относится к способам, облегчающим опосредованное антигенсвязывающими молекулами поглощение антигена клетками, посредством снижения антигенсвязывающей активности (связывающей активности) описанной выше антигенсвязывающей молекулы в диапазоне кислых значений рН до ее антигенсвязывающей активности, меньшей, чем в диапазоне нейтральных значений рН; и это облегчает поглощение антигена клетками. Настоящее изобретение также относится к способам, увеличивающим опосредованное антигенсвязывающей молекулой поглощение антигена клетками, где способы основаны на изменении по меньшей мере одной аминокислоты в антигенсвязывающем домене описанной выше антигенсвязывающей молекулы, которая облегчает поглощение антигена клетками. Настоящее изобретение также относится к способам, облегчающим опосредованное антигенсвязывающей молекулой поглощение антигена клетками, где способы основаны на замене гистидина на по меньшей мере одну аминокислоту или основаны на вставке по меньшей мере одного гистидина в антигенсвязывающий домен описанной выше антигенсвязывающей молекулы, которая увеличивает поглощение антигена клетками.
В настоящем документе, «поглощение антигена клетками», опосредованное антигенсвязывающей молекулой, обозначает, что антигены поглощаются клетками посредством эндоцитоза. При этом в настоящем документе, выражение «увеличивает поглощение клетками» обозначает, что скорость внутриклеточного поглощения антигенсвязывающей молекулы, связанной с антигеном в плазме, увеличена, и/или количество повторного использования поглощенного антигена в плазме снижено. Это обозначает, что скорость поглощения антигена клетками увеличивают по сравнению со скоростью поглощения антигена антигенсвязывающей молекулой до увеличения активности связывания с FcRn человека антигенсвязывающей молекулы в диапазоне нейтральных значений рН, или до увеличения активности связывания с FcRn человека и снижении антигенсвязывающей активности (связывающей активности) антигенсвязывающей молекулы в диапазоне кислых значений рН до значения, меньшего, чем ее антигенсвязывающая активность в диапазоне нейтральных значений рН. Предпочтительно, скорость увеличивают по сравнению с интактным IgG человека, и более предпочтительно по сравнению с интактным IgG человека. Таким образом, в настоящем изобретении, можно оценивать, произошло ли ускорение поглощения антигена клетками посредством антигенсвязывающей молекулы, на основе увеличения скорости поглощения антигена клетками. Скорость поглощения антигена клетками можно рассчитать, например, посредством мониторинга во времени снижения концентрации антигена в среде для культивирования, содержащей клетки, экспрессирующие FcRn, после добавления антигена и антигенсвязывающей молекулы в среду, или мониторингом во времени количества антигена, поглощенного клетками, экспрессирующими FcRn человека. Используя способы по настоящему изобретению для увеличения скорости опосредованного антигенсвязывающими молекулами поглощения антигена клетками, можно увеличить, например скорость элиминации антигена из плазмы посредством введения антигенсвязывающей молекулы. Таким образом, можно оценивать, произошло ли увеличение опосредованного антигенсвязывающими молекулами поглощения антигена клетками, например, посредством тестирования того, увеличивается ли скорость элиминации антигена из плазмы или снижается ли общая концентрация антигена в плазме посредством введения антигенсвязывающей молекулы.
В настоящем документе, «общая концентрация антигена в плазме» обозначает сумму концентраций антигена, связанного с антигенсвязывающей молекулой, и несвязанного антигена, или «концентрация свободного антигена в плазме», которая представляет собой концентрацию антигена, не связанного с антигенсвязывающей молекулой. Различные способы измерения «общей концентрации антигена в плазме» или «концентрации свободного антигена в плазме» хорошо известно в данной области, как описано в настоящем документе далее.
«Интактный IgG человека», как применяют в настоящем документе, обозначает немодифицированный IgG человека и не ограниченный конкретным классом IgG. Это обозначает, что IgG1, IgG2, IgG3 или IgG4 человека можно использовать в качестве «интактного IgG человека» при условии, что он может связываться с FcRn человека в диапазоне кислых значений рН. Предпочтительно, «интактный IgG человека» может представлять собой IgG1 человека.
Настоящее изобретение также относится к способам увеличения количества антигенов, с которыми может связываться одна антигенсвязывающая молекула. Более конкретно, настоящее изобретение относится к способам увеличения количества антигенов, с которыми может связываться одна антигенсвязывающая молекула, обладающая активностью связывания с FcRn человека в диапазоне кислых значений рН, посредством повышения активности связывания с FcRn человека антигенсвязывающей молекулы в диапазоне нейтральных значений рН. Настоящее изобретение также относится к способам увеличения количества антигенов, с которыми может связываться одна антигенсвязывающая молекула, обладающая активностью связывания с FcRn человека в диапазоне кислых значений рН, посредством изменения по меньшей мере одной аминокислоты в FcRn-связывающем домене человека антигенсвязывающей молекулы.
Настоящее изобретение также относится к способам увеличения количества антигенов, с которыми может связываться одна антигенсвязывающая молекула, обладающая активностью связывания с FcRn человека в диапазоне кислых значений рН, посредством использования FcRn-связывающего домена человека, включающего аминокислотную последовательность, в которой по меньшей мере одну аминокислоту, выбранную из аминокислот в положениях 237, 238, 239, 248, 250, 252, 254, 255, 256, 257, 258, 265, 270, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 325, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434 и 436 (нумерация EU) в родительском Fc-домене IgG FcRn-связывающего домена человека, содержащего родительский Fc-домен IgG, заменяют другой аминокислотой.
«Родительский IgG», как применяют в настоящем документе, обозначает немодифицированный IgG, который затем модифицируют для получения варианта при условии, что модифицированный вариант родительского IgG может связываться с FcRn человека в диапазоне кислых значений рН (таким образом, для родительского IgG не требуется наличие связывающей активности с FcRn человека в кислых условиях). Родительский IgG может представлять собой природный IgG, или вариант или сконструированную версию природного IgG. Родительский IgG может относиться к самому полипептиду, композициям, которые содержат родительский IgG, или аминокислотной последовательности, которая кодирует родительский IgG. Следует отметить, что «родительский IgG» включает известный, коммерчески доступный, полученный рекомбинантным способом IgG, как указано выше. Оригинальный «родительский IgG» не ограничен и может быть получен из любых организмов не являющихся человеком животных или от человека. Предпочтительно, организм выбран из мыши, крысы, морской свинки, хомяка, песчанки, кошки, кролика, собаки, козы, овцы, коровы, лошади, верблюда и не являющегося человеком примата. В другом варианте осуществления «родительский IgG» можно также получать от яванского макака, мартышки, макаки резус, шимпанзе или человека. Предпочтительно, «родительский IgG» получают от IgG1 человека без ограничения конкретным классом IgG. Это обозначает, что IgG1, IgG2, IgG3 или IgG4 человека можно соответствующим образом использовать в качестве «родительского IgG». Подобным способом, любой класс или подкласс IgG из любого организма, указанного выше, можно предпочтительно использовать в качестве «родительского IgG». Пример варианта или сконструированной версии природного IgG описан в Curr Opin Biotechnol. 2009 Dec; 20(6): 685-91, Curr Opin Immunol. 2008 Aug; 20(4): 460-70, Protein Eng Des Sel. 2010 Apr; 23(4): 195-202, WO 2009/086320, WO 2008/092117, WO 2007/041635 и WO 2006/105338, но не ограничивается ими.
Кроме того, настоящее изобретение относится к способам увеличения количества антигенов, с которыми может связываться одна антигенсвязывающая молекула, посредством снижения антигенсвязывающей активности (связывающей активности) в диапазоне кислых значений рН описанной выше антигенсвязывающей молекулы, которая обладает повышенным количеством событий связывания антигена, меньшим, чем ее антигенсвязывающая активность в диапазоне нейтральных значений рН. Настоящее изобретение также относится к способам увеличения количества антигенов, с которыми может связываться одна антигенсвязывающая молекула, посредством изменения по меньшей мере одной аминокислоты в антигенсвязывающем домене описанной выше антигенсвязывающей молекулы, которая обладает повышенным количеством событий связывания антигена. Настоящее изобретение также относится к способам увеличения количества антигенов, с которыми может связываться одна молекула антигена, посредством замены гистидина на по меньшей мере одну аминокислоту или посредством вставки по меньшей мере одного гистидина в антигенсвязывающий домен описанной выше антигенсвязывающей молекулы, которая обладает повышенным количеством событий связывания антигена.
В настоящем документе, «количество антигенов, с которыми может связываться одна антигенсвязывающая молекула» обозначает количество антигенов, с которыми может связываться одна антигенсвязывающая молекула, прежде чем молекулу элиминируют вследствие деградации. В настоящем документе, «увеличение количества антигенов, с которыми может связываться одна антигенсвязывающая молекула» обозначает увеличение количества циклов, достигаемое, прежде чем антигенсвязывающую молекулу элиминируют вследствие деградации, где каждый цикл состоит из связывания антигена с антигенсвязывающей молекулой в плазме, внутриклеточного поглощения антигенсвязывающей молекулы, связанной с антигеном, и диссоциации от антигена в эндосоме, с последующим возвращением антигенсвязывающей молекулы в плазму. Это обозначает, что количество циклов увеличено по сравнению с антигенсвязывающей молекулой до повышения активности связывания с FcRn человека антигенсвязывающей молекулы в диапазоне нейтральных значений рН, или до повышения активности связывания с FcRn человека и снижения антигенсвязывающей активности (связывающей активности) антигенсвязывающей молекулы в диапазоне кислых значений рН до меньшей, чем ее антигенсвязывающая активность в диапазоне нейтральных значений рН. Таким образом, можно оценивать, увеличено ли количество циклов, тестируя, вышеописанное «облегчено ли внутриклеточное поглощение» или «улучшена ли фармакокинетика», как описано ниже.
Настоящее изобретение также относится к способам облегчения внутриклеточной диссоциации антигена из антигенсвязывающей молекулы, которая связывается с антигеном за пределами клеток. Более конкретно, настоящее изобретение относится к способам облегчения внутриклеточной диссоциации антигена из антигенсвязывающей молекулы, которая связывается с антигеном за пределами клеток, посредством увеличения активности связывания с FcRn человека в диапазоне нейтральных значений рН антигенсвязывающей молекулы, которая обладает активностью связывания с FcRn человека в диапазоне кислых значений рН, и снижением ее антигенсвязывающей активности в диапазоне кислых значений рН до значения, меньшего, чем диапазоне нейтральных значений рН. Настоящее изобретение также относится к способам облегчения внутриклеточной диссоциации антигена из антигенсвязывающей молекулы, которая связывается с антигеном за пределами клеток, где способы основаны на изменении по меньшей мере одной аминокислоты в антигенсвязывающем домене антигенсвязывающей молекулы и одновременно изменении по меньшей мере одной аминокислоты в FcRn-связывающем домене антигенсвязывающей молекулы человека, обладающей активностью связывания с FcRn человека в диапазоне кислых значений рН. Настоящее изобретение также относится к способам облегчения внутриклеточной диссоциации антигена из антигенсвязывающей молекулы, которая связывается с антигеном за пределами клетки, посредством замены гистидина на по меньшей мере одну аминокислоту или посредством вставки по меньшей мере одного гистидина в антигенсвязывающий домен антигенсвязывающей молекулы и одновременно замещением по меньшей мере одной аминокислоты, выбранной из аминокислот в положениях 237, 238, 239, 248, 250, 252, 254, 255, 256, 257, 258, 265, 270, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 325, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434 и 436 (нумерация EU) в родительском Fc-домене IgG FcRn-связывающего домена человека, другой аминокислотой.
В настоящем изобретении антигены могут диссоциировать из антигенсвязывающей молекулы в любом месте внутри клетки; однако, предпочтительный участок диссоциации представляет собой раннюю эндосому. В настоящем документе, «внутриклеточная диссоциация связанного с антигенсвязывающей молекулой антигена из антигенсвязывающей молекулы за пределами клеток» не обязательно означает, что все антигены, поглощенные клетками посредством связывания с антигенсвязывающей молекулой, диссоциируют от антигенсвязывающей молекулы в пределах клетки. Таким образом, допустимо, что доля антигенов, диссоциировавших от антигенсвязывающей молекулы в пределах клетки, увеличивается по сравнению с долей молекул до снижения антигенсвязывающей активности антигенсвязывающей молекулы в диапазоне кислых значений рН до значения, меньшего, чем в диапазоне нейтральных значений рН, и одновременно увеличение активности связывания с FcRn человека в диапазоне нейтральных значений рН. Такой способ облегчения внутриклеточной диссоциации антигена из антигенсвязывающей молекулы, связанной с антигеном за пределами клетки, является тождественным способу, который наделяет антигенсвязывающую молекулу способностью облегчать внутриклеточную диссоциацию антигена из антигенсвязывающей молекулы посредством облегчения поглощения антигенсвязывающей молекулы, связанной с антигеном.
Настоящее изобретение также относится к способам облегчения внеклеточного высвобождения свободной от антигена антигенсвязывающей молекулы, поглощенной клетками в форме, связанной с антигеном. Более конкретно, настоящее изобретение относится к способам облегчения внеклеточного высвобождения свободной от антигена антигенсвязывающей молекулы, поглощенной клетками в форме, связанной с антигеном, посредством увеличения активности связывания с FcRn человека в диапазоне нейтральных значений рН антигенсвязывающей молекулы, которая обладает активностью связывания с FcRn человека в диапазоне кислых значений рН, и снижением ее антигенсвязывающей активности в диапазоне кислых значений рН до значения, меньшего, чем в диапазоне нейтральных значений рН. Настоящее изобретение также относится к способам облегчения внеклеточного высвобождения свободной от антигена антигенсвязывающей молекулы, поглощенной клетками в форме, связанной с антигеном, где способы основаны на изменении по меньшей мере одной аминокислоты в антигенсвязывающей молекуле и одновременно изменением по меньшей мере одной аминокислоты в FcRn-связывающем домене человека. Настоящее изобретение также относится к способам облегчения внеклеточного высвобождения свободной от антигена антигенсвязывающей молекулы, поглощенной клетками в форме, связанной с антигеном, посредством замены гистидина на по меньшей мере одну аминокислоту или посредством вставки по меньшей мере одного гистидина в антигенсвязывающую молекулу, и одновременно замещением по меньшей мере одной аминокислоты, выбранной из аминокислот в положениях 237, 238, 239, 248, 250, 252, 254, 255, 256, 257, 258, 265, 270, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 325, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434 и 436 (нумерация EU) в родительском Fc-домене IgG человека FcRn-связывающего домена, другой аминокислотой.
В настоящем документе, «внеклеточное высвобождение свободной от антигена антигенсвязывающей молекулы, поглощенной клетками в форме, связанной с антигенами» не обязательно обозначает, что каждая из поглощенных клетками антигенсвязывающих молекул, связанных с антигеном, высвобождается в свободной от антигена форме за пределами клетки. Допустимо, что доля антигенсвязывающих молекул, высвобожденных в свободной от антигена форме за пределами клеток, увеличивается по сравнению с долей молекул до снижения антигенсвязывающей активности антигенсвязывающей молекулы в диапазоне кислых значений рН до значения, меньшего, чем в диапазоне нейтральных значений рН, и увеличение активности связывания с FcRn человека в диапазоне нейтральных значений рН. Антигенсвязывающая молекула, высвобожденная за пределами клеток, предпочтительно сохраняет активность связывания с антигеном. Такой способ облегчения внеклеточного высвобождения свободной от антигена антигенсвязывающей молекулы, поглощенной клетками в свободной от антигена форме, является тождественным способу, который наделяет антигенсвязывающую молекулу способностью облегчать внеклеточное высвобождение свободной от антигена антигенсвязывающей молекулы, поглощенной клетками в связанной с антигеном форме, посредством облегчения поглощения клетками антигенсвязывающих молекул, связанных с антигеном в клетках.
Настоящее изобретение также относится к способам увеличения способности к элиминации плазменного антигена посредством введения антигенсвязывающей молекулы. В настоящем изобретении «способы увеличения способности к элиминации плазменного антигена» являются тождественными «способам усиления способности антигенсвязывающей молекулы к элиминации антигена из плазмы». Более конкретно, настоящее изобретение относится к способам увеличения способности к элиминации плазменного антигена посредством антигенсвязывающей молекулы, обладающей активностью связывания с FcRn человека в диапазоне кислых значений рН, посредством увеличения активности связывания антигенсвязывающей молекулы с FcRn человека в диапазоне нейтральных значений рН. Настоящее изобретение также относится к способам увеличения способности к элиминации плазменного антигена посредством антигенсвязывающей молекулы, обладающей активностью связывания с FcRn человека в диапазоне кислых значений рН, где способы основаны на изменении по меньшей мере одной аминокислоты в FcRn-связывающем домене человека антигенсвязывающей молекулы.
Настоящее изобретение также относится к способам увеличения способности к элиминации плазменного антигена посредством антигенсвязывающей молекулы, обладающей активностью связывания с FcRn человека в диапазоне кислых значений рН, посредством использования FcRn-связывающего домена человека, содержащего аминокислотную последовательность с заменой по меньшей мере одной аминокислоты, выбранной из аминокислот в положениях 237, 238, 239, 248, 250, 252, 254, 255, 256, 257, 258, 265, 270, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 325, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434 и 436 (нумерация EU) в родительском Fc-домене IgG FcRn-связывающего домена человека, содержащего Fc-домен родительского IgG, на другую аминокислоту.
Настоящее изобретение также относится к способам увеличения способности к элиминации плазменного антигена посредством антигенсвязывающей молекулы, посредством снижения антигенсвязывающей активности в диапазоне кислых значений рН описанной выше антигенсвязывающей молекулы с повышенной способностью к элиминации плазменного антигена по сравнению с антигенсвязывающей активностью в диапазоне нейтральных значений рН. Настоящее изобретение также относится к способам увеличения способности к элиминации плазменного антигена посредством антигенсвязывающей молекулы, посредством изменения по меньшей мере одной аминокислоты в антигенсвязывающем домене описанной выше антигенсвязывающей молекулы с повышенной способностью к элиминации плазменного антигена. Настоящее изобретение также относится к способам увеличения способности к элиминации плазменного антигена посредством введения антигенсвязывающей молекулы, посредством замены гистидина на по меньшей мере одну аминокислоту или посредством вставки по меньшей мере одного гистидина в антигенсвязывающий домен описанной выше антигенсвязывающей молекулы с повышенной способностью к элиминации плазменного антигена.
В настоящем документе, «способность к элиминации плазменного антигена» обозначает способность к элиминации антигена из плазмы при введении антигенсвязывающей молекулы или секреции in vivo. Таким образом, «повышение способности антигенсвязывающей молекулы к элиминации плазменного антигена» в настоящем документе обозначает, что скорость элиминации антигена из плазмы ускоряется при введении антигенсвязывающей молекулы по сравнению с элиминацией антигена до повышения активности связывания с FcRn человека антигенсвязывающей молекулы в диапазоне нейтральных значений рН или до повышения активности связывания с FcRn человека и одновременно снижением ее антигенсвязывающей активности в диапазоне кислых значений рН до значения, меньшего, чем диапазоне нейтральных значений рН. Повышение активности антигенсвязывающей молекулы для элиминации антигена из плазмы можно оценивать, например, посредством введения растворимого антигена и антигенсвязывающей молекулы in vivo, и измерением концентрации растворимого антигена в плазме после введения. Если концентрация растворимого антигена в плазме после введения растворимого антигена и антигенсвязывающей молекулы снижают посредством увеличения активности связывания антигенсвязывающей молекулы с FcRn человека в диапазоне нейтральных значений рН, или посредством повышения ее активности связывания с FcRn человека и одновременно снижением ее антигенсвязывающей активности в диапазоне кислых значений рН до значения, меньшего, чем диапазоне нейтральных значений рН, то способность антигенсвязывающей молекулы к элиминации плазменного антигена может считаться увеличенной. Форма растворимого антигена может представлять собой антигенсвязывающую молекулу, связанную с антигеном, или антигенсвязывающую молекулу, не связанную с антигеном, чью концентрацию можно определять как «концентрация в плазме антигенсвязывающей молекулы, связанной с антигеном» и «концентрация в плазме антигенсвязывающей молекулы, несвязанной с антигеном», соответственно (последнее является синонимом «концентрации свободного антигена в плазме»). Так как «общая концентрация антигена в плазме» обозначает сумму концентраций антигенсвязывающей молекулы, связанной с антигеном, и не связанной с антигеном, или «концентрацию свободного антигена в плазме», которая представляет собой концентрацию антигенсвязывающей молекулы, не связанной с антигеном, концентрацию растворимого антигена можно определять как «общую концентрацию антигена в плазме». Различные способы измерения «общей концентрации антигена в плазме» или «концентрации свободного антигена в плазме» хорошо известны в данной области, как описано в настоящем документе далее.
Настоящее изобретение также относится к способам улучшения фармакокинетики антигенсвязывающих молекул. Более конкретно, настоящее изобретение относится к способам улучшения фармакокинетики антигенсвязывающей молекулы, обладающей активностью связывания с FcRn человека в диапазоне кислых значений рН, посредством повышения активности связывания с FcRn человека антигенсвязывающей молекулы в диапазоне нейтральных значений рН. Кроме того, настоящее изобретение относится к способам улучшения фармакокинетики антигенсвязывающей молекулы, обладающей активностью связывания с FcRn человека в диапазоне кислых значений рН, посредством изменения по меньшей мере одной аминокислоты в FcRn-связывающем домене человека антигенсвязывающей молекулы.
Настоящее изобретение также относится к способам улучшения фармакокинетики антигенсвязывающей молекулы, обладающей активностью связывания с FcRn человека в диапазоне кислых значений рН, посредством использования FcRn-связывающего домена человека, содержащего аминокислотную последовательность с заменой различных аминокислот по меньшей мере на одну аминокислоту, выбранную из аминокислот в положениях 237, 238, 239, 248, 250, 252, 254, 255, 256, 257, 258, 265, 270, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 325, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434 и 436 (нумерация EU) в родительском Fc-домене IgG FcRn-связывающего домена человека, содержащего Fc-домен IgG.
Кроме того, настоящее изобретение относится к способам улучшения фармакокинетики антигенсвязывающей молекулы посредством снижения антигенсвязывающей активности в диапазоне кислых значений рН описанной выше антигенсвязывающей молекулы с улучшенной фармакокинетикой до значения, меньшего, чем ее антигенсвязывающая активность в диапазоне нейтральных значений рН. Настоящее изобретение также относится к способам улучшения фармакокинетики антигенсвязывающей молекулы, обладающей активностью связывания с FcRn человека в диапазоне кислых значений рН, посредством изменения по меньшей мере одной аминокислоты в антигенсвязывающем домене описанной выше антигенсвязывающей молекулы с улучшенной фармакокинетикой. Настоящее изобретение также относится к способам улучшения фармакокинетики посредством замены гистидина на по меньшей мере одну аминокислоту или посредством вставки по меньшей мере одного гистидина в антигенсвязывающий домен описанной выше антигенсвязывающей молекулы с улучшенной фармакокинетикой.
В настоящем документе, «улучшение фармакокинетики», «усовершенствование фармакокинетики», и «фармакокинетика лучшего качества» можно иначе формулировать как «улучшение удержания в плазме (крови)», «усовершенствование удержания в плазме (крови)», «лучшего качества удержание в плазме (крови). Эти термины являются синонимами.
В настоящем документе, «усовершенствование фармакокинетики» обозначает не только увеличение периода до элиминации из плазмы (например, до деградации антигенсвязывающей молекулы внутриклеточно или т.п. и невозможности ее возвращения в плазму) после введения антигенсвязывающей молекулы людям или не являющимся человеком животным, таким как мыши, крысы, обезьяны, кролики и собаки, но также увеличением времени удержания антигенсвязывающей молекулы в форме, которая обеспечивает связывание антигена (например, в свободной от антигена форме антигенсвязывающей молекулы) в течение периода введения для элиминации вследствие деградации. Интактный IgG человека может связываться с FcRn не являющегося человеком животного. Например, предпочтительно можно использовать мышь для введения с целью подтверждения свойства антигенсвязывающей молекулы по изобретению, так как интактный IgG человека может связываться с FcRn мыши сильнее, чем с FcRn человека (Int Immunol. 2001 Dec; 13(12): 1551-9). В качестве другого примера, мышь, у которой ее природные гены FcRn нарушены и которая содержит для экспрессии трансген в виде гена FcRn человека (Methods Mol Biol. 2010; 602: 93-104), предпочтительно также можно использовать для введения с целью подтверждения свойства антигенсвязывающей молекулы по изобретению, описываемого ниже в настоящем документе. Конкретно, «улучшение фармакокинетики» также включает увеличение периода до элиминации вследствие деградации антигенсвязывающей молекулы, не связанной с антигенами (свободная от антигена форма антигенсвязывающей молекулы). Антигенсвязывающая молекула в плазме не связывается с новым антигеном, если антигенсвязывающая молекула уже связана с антигеном. Таким образом, чем более длительным является период, за который антигенсвязывающая молекула не связывается с антигеном, тем более длительным является период, за который она может связаться с новым антигеном (более высокий шанс связывания с другим антигеном). Это позволяет уменьшить период времени, в течение которого антиген освобождают от антигенсвязывающей молекулы in vivo, и увеличить период, в течение которого антиген связывают с антигенсвязывающей молекулой. Концентрацию в плазме свободной от антигена формы антигенсвязывающей молекулы можно увеличивать, и можно увеличивать период, за который антиген связывают с антигенсвязывающей молекулой, посредством ускорения элиминации антигена из плазмы за счет введения антигенсвязывающей молекулы. Конкретно, в настоящем документе «улучшение фармакокинетики антигенсвязывающей молекулы» включает улучшение фармакокинетического параметра свободной от антигена формы антигенсвязывающей молекулы (любое увеличение времени полувыведение из плазмы, увеличение среднего времени удержания в плазме и снижение плазменного клиренса), увеличение периода, за который антиген связывают с антигенсвязывающей молекулой после введения антигенсвязывающей молекулы, и ускорение элиминации антигена из плазмы, опосредованной антигенсвязывающей молекулой. Улучшение фармакокинетики антигенсвязывающей молекулы можно оценивать посредством определения любого из параметров: полувыведение из плазмы, среднее время удержания в плазме, и плазменный клиренс антигенсвязывающей молекулы или ее свободной от антигена формы («Pharmacokinetics: Enshu-niyoru Rikai (Understanding through practice)» Nanzando). Например, концентрацию в плазме антигенсвязывающей молекулы или ее свободной от антигена формы определяют после введения антигенсвязывающие молекулы мышам, крысам, обезьянам, кроликам, собакам или людям. Затем определяют каждый параметр. Если период полувыведения из плазмы или среднее время удержания в плазме увеличивают, то фармакокинетику антигенсвязывающей молекулы можно считать улучшенной. Параметры можно определять способами, известными специалистам в данной области. Параметры можно оценивать, соответствующим образом, например, посредством бескомпартментного анализа с использованием программного обеспечения для фармакокинетического анализа WinNonlin (Pharsight) согласно прилагаемой инструкции по эксплуатации. Концентрация в плазме свободной от антигена антигенсвязывающей молекулы можно определять способами, известными специалистам в данной области, например, используя способ анализа, описанный в Clin Pharmacol. 2008 Apr; 48(4): 406-17.
В настоящем документе, «улучшение фармакокинетики» также включает увеличение периода, за который антиген связывают с антигенсвязывающей молекулой после введения антигенсвязывающей молекулы. Увеличен ли период, за который антиген связывают с антигенсвязывающей молекулой после введения антигенсвязывающей молекулы, можно оценивать, определяя концентрацию свободного антигена в плазме. О наличии увеличения можно судить, основываясь на определенную концентрацию свободного антигена в плазме или период времени, требуемый для увеличения значения отношения концентрации свободного антигена к общей концентрации антигена.
Концентрацию в плазме свободного антигена, не связанного с антигенсвязывающей молекулой, или отношение концентрации свободного антигена к общей концентрации антигена можно определять способами, известными специалистам в данной области, например, способом, описанным в Pharm Res. 2006 Jan; 23 (1): 95-103. Альтернативно, если антиген демонстрирует конкретную функцию in vivo, достигается ли связывание антиген с антигенсвязывающей молекулой, которая нейтрализует функцию антигена (антагонистическая молекула), можно оценивать посредством тестирования того, нейтрализована ли функция антигена. Произошла ли нейтрализация функции антигена, можно оценивать посредством исследования маркера in vivo, который отражает функцию антигена. Связывается ли антиген с антигенсвязывающей молекулой, которая активирует функцию антигена (агонистическая молекула), можно оценивать, исследуя маркер in vivo, который отражает функцию антигена.
Определение концентрации свободного антигена в плазме и отношение количества свободного антигена в плазме к количеству общего антигена в плазме, анализ маркера in vivo, и подобные измерения конкретно не ограничены; однако, анализы предпочтительно осуществляют после определенного периода времени, прошедшего после введения антигенсвязывающей молекулы. В настоящем изобретении период времени после введения антигенсвязывающей молекулы конкретно не ограничен; специалисты в данной области могут определять подходящий период в зависимости от свойств и т.п. введенной антигенсвязывающей молекулы. Такие периоды включают, например, одни сутки после введения антигенсвязывающей молекулы, трое суток после введения антигенсвязывающей молекулы, семь суток после введения антигенсвязывающей молекулы, 14 суток после введения антигенсвязывающей молекулы и 28 суток после введения антигенсвязывающей молекулы. В настоящем документе «концентрация антигена в плазме» обозначает или «общую концентрацию антигена в плазме», которая представляет собой сумму концентрации связанного с антигенсвязывающей молекулой антигена и концентрации несвязанного антигена, или «концентрацию свободного антигена в плазме», которая представляет собой концентрацию антигена, не связанного с антигенсвязывающей молекулой.
Общая концентрация антигена в плазме может быть снижена посредством антигенсвязывающей молекулы по настоящему изобретению, в 2 раза, 5 раз, 10 раз, 20 раз, 50 раз, 100 раз, 200 раз, 500 раз, 1000 раз или даже больше по сравнению с введением контрольной антигенсвязывающей молекулы, содержащей интактный Fc-домен IgG человека в качестве FcRn-связывающего домена человека или по сравнению с ситуацией, когда молекулу антигенсвязывающего домена не вводят.
Молярное отношение "антиген/антигенсвязывающая молекула" можно рассчитать, как представлено ниже:
значение A: молярная концентрация антигена в каждый момент времени
значение B: молярная концентрация антигенсвязывающей молекулы в каждый момент времени
значение C: молярная концентрация антигена в расчете на молярную концентрацию антигенсвязывающей молекулы (молярное отношение антиген/антигенсвязывающая молекула) в каждый момент времени
C=A/B.
Низкое значение C показывает более высокую эффективность элиминации антигена в расчете на антигенсвязывающую молекулу, в то время как повышенное значение C показывает низкую эффективность элиминации антигена из расчета на антигенсвязывающую молекулу.
Молярное отношение антиген/антигенсвязывающая молекула можно рассчитать, как описано выше.
Молярное отношение антиген/антигенсвязывающая молекула можно снижать посредством введения антигенсвязывающей молекулы по настоящему изобретению в 2 раза, 5 раз, 10 раз, 20 раз, 50 раз, 100 раз, 200 раз, 500 раз, 1000 раз, или еще больше по сравнению с введением контрольной антигенсвязывающей молекулы, содержащей интактный Fc-домен IgG человека в качестве FcRn-связывающего домена человека.
В настоящем документе интактный IgG1, IgG2, IgG3 или IgG4 человека предпочтительно используют в качестве интактного IgG человека с целью использования контрольного интактного IgG человека, сравниваемого с антигенсвязывающими молекулами по их активности связывания с FcRn человека или активности in vivo. Предпочтительно, можно соответствующим образом использовать контрольную антигенсвязывающую молекулу, содержащую тот же антигенсвязывающий домен в качестве представляющей интерес антигенсвязывающей молекулы и интактный Fc-домен IgG человека в качестве FcRn-связывающего домена человека. Более предпочтительно, интактный IgG1 человека применяют с целью контрольного интактного IgG человека, сравниваемого с антигенсвязывающими молекулами по их активности связывания с FcRn человека или активности in vivo.
Снижение общей концентрации антигена в плазме или молярного отношения антиген/антитело можно оценивать, как описано в примерах 6, 8 и 13. Более конкретно, при использовании трансгенной линии 32 мышей с FcRn человека или линии 276 с FcRn человека (Jackson Laboratories, Methods Mol Biol. 2010; 602: 93-104), их можно оценивать либо посредством модели совместной инъекции антиген-антитела, либо посредством модели стационарной инфузии антигена, если антигенсвязывающая молекула не дает перекрестную реакцию с соответствующим антигеном мыши. Если антигенсвязывающая молекула дает перекрестную реакцию с соответствующим антигеном мыши, их можно оценивать простым инъецированием антигенсвязывающей молекулы трансгенной линии 32 мышей с FcRn человека или линии 276 с FcRn человека (Jackson Laboratories). В модели совместной инъекции смесь антигенсвязывающей молекулы и антигена вводят мыши. В модели стационарной инфузии антигена, инфузионный насос, содержащий раствор антигена, имплантируют мыши для достижения постоянной концентрации антигена в плазме, а затем мыши инъецируют антигенсвязывающую молекулу. Тестируемую антигенсвязывающую молекулу вводят в той же дозе. Общую концентрацию антигена в плазме, концентрацию свободного антигена в плазме и концентрацию антигенсвязывающей молекулы в плазме измеряют в подходящий момент времени с использованием способа, известного специалистам в данной области.
Общую концентрацию антигена или концентрацию свободного антигена в плазме и молярное отношение антиген/антигенсвязывающая молекула можно измерять через 2, 4, 7, 14, 28, 56 или 84 суток после введения для оценки длительности эффекта по настоящему изобретению. Другими словами, для оценки свойств антигенсвязывающей молекулы по настоящему изобретению определяют длительную концентрацию антигена в плазме, измеряя общую концентрацию общего антигена или концентрацию свободного антигена в плазме и молярное отношение антиген/антигенсвязывающая молекула через 2, 4, 7, 14, 28, 56 или 84 суток после введения антигенсвязывающей молекулы. Достигли ли снижения концентрации антигена в плазме или молярного отношения антиген/антигенсвязывающая молекула посредством антигенсвязывающей молекулы, описанной в настоящем изобретении, можно определять, оценивая снижение в каждый из моментов времени или в нескольких моментах времени, описанных выше.
Общую концентрацию антигена или концентрацию свободного антигена в плазме и молярное отношение антиген/антигенсвязывающая молекула можно измерять через 15 мин, 1, 2, 4, 8, 12 или 24 часа после введения для оценки кратковременного эффекта по настоящему изобретению. Другими словами, кратковременную концентрацию антигена в плазме определяют, измеряя общую концентрацию антигена или концентрацию свободного антигена в плазме и молярное отношение антиген/антигенсвязывающая молекула через 15 мин, 1, 2, 4, 8, 12 или 24 часа после введения антигенсвязывающей молекулы для оценки свойств антигенсвязывающей молекулы по настоящему изобретению.
Путь введения антигенсвязывающей молекулы по настоящему изобретению можно выбирать из интрадермального, внутривенного, интравитреального, подкожного, интраперитонеального, парентерального и внутримышечной инъекции.
В настоящем изобретении предпочтительным является усовершенствование фармакокинетики у человека.
Если время удержания у человека является трудным для определения, оно может быть рассчитано на основе времени удержания у мыши (например, нормальных мышах, трансгенных мышах, экспрессирующих антиген человека, трансгенных мышах, экспрессирующих FcRn человека) или обезьянах (например, яванских макаках).
В настоящем документе диапазон кислых значений рН, как правило, относится к значениям pH от 4,0 до 6,5. Диапазон кислых значений рН предпочтительно представляет собой диапазон, указанный посредством любого значения рН в пределах pH от 5,5 до 6,5, предпочтительно выбранного из 5,5, 5,6, 5,7, 5,8, 5,9, 6,0, 6,1, 6,2, 6,3, 6,4 и 6,5, более предпочтительно pH от 5,8 до 6,0, которые представляют собой значения, близкие к значению рН в эндосоме in vivo. Между тем, в настоящем документе диапазон нейтральных значений рН, как правило, относится к значению pH от 6,7 до 10,0. Диапазон нейтральных значений рН предпочтительно представляет собой диапазон, указанный посредством любого значения pH в пределах значений pH от 7,0 до 8,0, предпочтительно выбранного из pH 7,0, 7,1, 7,2, 7,3, 7,4, 7,5, 7,6, 7,7, 7,8, 7,9 и 8,0, более предпочтительно pH 7,4, которые представляют собой значения, близкие к значению рН в плазме (крови) in vivo. pH 7,0 можно использовать в качестве альтернативы pH 7,4, если является затруднительным оценить аффинность связывания между FcRn-связывающим доменом человека и FcRn человека вследствие низкой аффинности последнего при pH 7,4. Так как температура используется в условиях анализа, аффинность связывания между FcRn-связывающим доменом человека и FcRn человека можно оценивать при любой температуре от 10 градусов C до 50 градусов C. Предпочтительно, температуру от 15 градусов C до 40 градусов C используют для определения аффинности связывания между FcRn-связывающим доменом человека и FcRn человека. Более предпочтительно, любую температуру от 20 градусов C до 35 градусов C, как и любую из 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 и 35 градусов C также используют для определения аффинности связывания между FcRn-связывающим доменом человека и FcRn человека. Температура 25 градусов C, описанная в примере 5, представляет собой один из примеров для варианта осуществления настоящего изобретения.
Таким образом, в настоящем документе «снижение антигенсвязывающей активности антигенсвязывающей молекулы в диапазоне кислых значений рН до значения, меньшего, чем в диапазоне нейтральных значений рН» означает, что антигенсвязывающая активность антигенсвязывающей молекулы при pH от 4,0 до pH 6,5 хуже по сравнению с ее антигенсвязывающей активностью при pH от 6,7 до 10,0. Предпочтительно, выражение выше обозначает, что антигенсвязывающая активность антигенсвязывающей молекулы при pH от 5,5 до 6,5 хуже по сравнению с антигенсвязывающей активностью при pH от 7,0 до 8,0, более предпочтительно обозначает, что ее антигенсвязывающая активность при значениях рН, близких к значению рН в ранней эндосоме, хуже, по сравнению с ее антигенсвязывающей активностью при рН в плазме in vivo. Конкретно, антигенсвязывающая активность антигенсвязывающей молекулы при pH от 5,8 до 6,0 хуже по сравнению с антигенсвязывающей активностью антигенсвязывающей молекулы при pH 7,4.
Между тем, в настоящем документе выражение «снижение антигенсвязывающей активности антигенсвязывающей молекулы в диапазоне кислых значений рН до значения, меньшего, чем в диапазоне нейтральных значений рН» также обозначают как «увеличение антигенсвязывающей активности антигенсвязывающей молекулы в диапазоне нейтральных значений рН до значения большего, чем в диапазоне кислых значений рН». Конкретно, в настоящем изобретении является возможным увеличение отношения антигенсвязывающей активности антигенсвязывающей молекулы между диапазонами кислых и нейтральных значений рН. Например, значение KD (pH 5,8)/KD (pH 7,4) увеличивают в варианте осуществления, описанном ниже. Отношение антигенсвязывающей активности антигенсвязывающей молекулы между диапазонами кислых и нейтральных значений рН можно увеличивать, например, посредством снижения ее антигенсвязывающей активности в диапазоне кислых значений рН, увеличением ее антигенсвязывающей активности в диапазоне нейтральных значений рН или в обоих случаях.
В настоящем документе выражение «ухудшение антигенсвязывающей активности в диапазоне кислых значений рН по сравнению с антигенсвязывающей активностью в диапазоне нейтральных значений рН» иногда используют вместо выражения «снижение антигенсвязывающей активности в диапазоне кислых значений рН до значения, меньшего, чем диапазоне нейтральных значений рН».
В настоящем документе активность связывания с FcRn человека в диапазоне кислых значений рН обозначает активность связывания с FcRn человека при pH от 4,0 до 6,5, предпочтительно активность связывания с FcRn человека при pH от 5,5 до 6,5, и более предпочтительно активность связывания с FcRn человека при pH от 5,8 до pH 6,0, что является сравнимым с рН ранней эндосомы in vivo. Между тем, в настоящем документе активность связывания с FcRn человека в диапазоне нейтральных значений рН обозначает активность связывания с FcRn человека при pH от 6,7 до 10,0, предпочтительно активность связывания с FcRn человека при pH от 7,0 до 8,0 и более предпочтительно активность связывания с FcRn человека при pH 7,4, что является сравнимым со значением рН в плазме in vivo.
Антигенсвязывающие молекулы по настоящему изобретению обладают FcRn-связывающим доменом человека. FcRn-связывающий домен человека конкретно не ограничен, при условии, что антигенсвязывающие молекулы демонстрируют активность связывания с FcRn человека в диапазоне кислых и нейтральных значений pH. Альтернативно, домен может обладать прямой или непрямой активностью связывания с FcRn человека. Такие домены включают, например, Fc-домен иммуноглобулина типа IgG, альбумин, домен 3 альбумина, антитела против FcRn человека, пептиды против FcRn человека и каркасные молекулы против FcRn человека, каждый из которых обладает активностью прямого связывания с FcRn человека; и молекулы, которые связываются с IgG или альбумином, которые обладают активностью непрямого связывания с FcRn человека. Такие предпочтительные домены по настоящему изобретению обладают активностью связывания с FcRn человека в диапазонах кислых и нейтральных значений рН. Возможно использование доменов без каких-либо изменений при условии, что они уже обладают активностью связывания с FcRn человека в диапазоне кислых и нейтральных значений рН. Если домены обладают только слабой активностью связывания с FcRn человека или не обладают активностью связывания с FcRn человека в диапазоне кислых и/или нейтральных зачений pH, активность связывания с FcRn человека можно обеспечить посредством изменения аминокислот в антигенсвязывающих молекулах. Однако, предпочтительно, чтобы активность связывания с FcRn человека в диапазоне кислых и/или нейтральных значений pH обеспечивали посредством изменения аминокислот в FcRn-связывающем домене человека. Альтернативно, аминокислоты в доменах, которые уже обладают активностью связывания с FcRn человека в диапазоне кислых и/или нейтральных значений pH, можно изменять для увеличения активности связывания с FcRn человека. Желаемые аминокислотные изменения в FcRn-связывающем домене человека можно выбирать, сравнивая активность связывания с FcRn человека в диапазоне кислых и/или нейтральных значений pH до и после аминокислотной замены.
Предпочтительный FcRn-связывающий домен человека представляет собой область, которая напрямую связывается с FcRn человека. Такие предпочтительные FcRn-связывающие области включают, например, Fc-домены антитела. Между тем, области, способные к связыванию с полипептидом, таким как альбумин или IgG, которые обладают активностью связывания с FcRn человека, могут непрямым образом связываться с FcRn человека посредством альбумина, IgG или подобных. Таким образом, такая FcRn-связывающая область человека по настоящему изобретению может представлять собой область, которая связывается с полипептидом, обладающим активностью связывания с FcRn человека.
Антигенсвязывающие молекулы по настоящему изобретению конкретно не ограничены, при условии, что они включают антигенсвязывающий домен, обладающий активностью связывания, специфичной для целевого антигена. Такие предпочтительные антигенсвязывающие домены содержат, например, домены, обладающие антигенсвязывающей областью антитела. Антигенсвязывающая область антитела содержит, например, CDR и вариабельные области. Если антигенсвязывающая область антитела представляет собой CDR, она может содержать все шесть CDR из целого антитела, или одну, две, или более CDR. Если CDR включают в качестве связывающей области антитела, они могут включать делеции, замены, добавления и/или вставки аминокислот, или могут представлять собой часть CDR.
С другой стороны, антигенсвязывающие молекулы, которые используют в способах по настоящему изобретению, содержат антигенсвязывающие молекулы, которые обладают антагонистической активностью (антагонистические антигенсвязывающие молекулы), антигенсвязывающие молекулы, которые обладают агонистической активностью (агонистическая антигенсвязывающая молекула), и молекулы, обладающие цитотоксичностью. В предпочтительном варианте осуществления антигенсвязывающие молекулы содержат антагонистические антигенсвязывающие молекулы, в частности, антагонистические антигенсвязывающие молекулы, которые распознают антиген, такие как рецептор или цитокин.
В настоящем изобретении представляющая интерес антигенсвязывающая молекула конкретно не ограничена и может представлять собой любые антигенсвязывающие молекулы. Антигенсвязывающая молекула по настоящему изобретению предпочтительно включает и антигенсвязывающую активность (антигенсвязывающий домен), и FcRn-связывающий домен человека. В частности, предпочтительная антигенсвязывающая молекула по настоящему изобретению относится к домену, который связывается с FcRn человека. Антигенсвязывающая молекула, содержащая и антигенсвязывающий домен, и FcRn-связывающий домен человека, включает, например, антитела. Предпочтительные антитела в контексте настоящего изобретения включают, например, антитела IgG. Если используемое антитело представляет собой антитело IgG, тип IgG не ограничен; можно использовать IgG, относящийся к любому изотипу (подклассу), такому как IgG1, IgG2, IgG3 или IgG4. Кроме того, антигенсвязывающие молекулы по настоящему изобретению могут включать константную область антитела, и в константную область можно вводить аминокислотные мутации. Вводимые аминокислотные мутации включают, например, те, которые делают возможным или нарушают связывание с рецептором Fcгамма (Proc Natl Acad Sci USA. 2006 Mar 14; 103(11): 4005-10), но не ограничены этими примерами. Альтернативно, также возможно изменять зависимое от рН связывание посредством отбора соответствующей константной области, такой как из IgG2.
Если представляющая интерес антигенсвязывающая молекула по настоящему изобретению представляет собой антитело, это может быть антитело, полученное от любого животного, такого как антитело мыши, антитело человека, антитело крысы, антитело кролика, антитело козы или антитело верблюда. Кроме того, антитело может представлять собой измененное антитело, например, химерное антитело, и в частности, измененное антитело, включающее замену аминокислоты в последовательности гуманизированного антитела и т.д. Антитела также включают биспецифические антитела, продукты модификации антител, продукты, сцепленные с различными молекулами, и полипептиды, включающие фрагменты антител.
«Химерные антитела» представляют собой антитела, полученные посредством комбинирования последовательностей, полученных от различных животных. Конкретно, химерное антитело включает, например, антитела, обладающие вариабельными областями (V) тяжелой и легкой цепи из антитела мыши и константными областями (C) тяжелой и легкой цепи из антитела человека.
«Гуманизированные антитела», также обозначаемые как реконструированные антитела человека, представляют собой антитела, в которых определяющие комплементарность области (CDR) антитела, полученные от не принадлежащему человеку млекопитающего, например, мыши, трансплантируют в CDR антитела человека. Способы идентификации CDR известны (Kabat et al., Sequence of Proteins of Immunological Interest (1987), National Institute of Health, Bethesda, Md.; Chothia et al., Nature (1989) 342: 877). Основные технологии генетической рекомбинации, подходящие для этой цели, также хорошо известны (см. европейскую заявку на патент EP 125023; и WO 96/02576).
Биспецифическое антитело относится к антителу, которое обладает, в случае некоторых молекул антител, вариабельными областями, которые распознают различные эпитопы. Биспецифическое антитело может представлять собой антитело, которое распознает два или более различных антигенов, или антитело, которое распознает два или более различных эпитопов на одном и том же антигене.
Кроме того, полипептиды, включающие фрагменты антител включают, например, Fab-фрагменты, F(ab’)2 фрагменты, scFvs (Nat Biotechnol. 2005 Sep; 23(9): 1126-36), домены антител (dAb) (WO 2004/058821, WO 2003/002609), scFv-Fc (WO 2005/037989), dAb-Fc, и слитые с Fc белки. Fc-домен можно использовать в качестве FcRn-связывающего домена человека, если молекула включает Fc-домен. Альтернативно, FcRn-связывающий домен может быть слит с этими молекулами.
Дополнительно, антигенсвязывающие молекулы, которые применимы к настоящему изобретению, могут представлять собой подобные антителам молекулы. Подобные антителам молекулы (каркасная молекула, пептидная молекула) представляют собой молекулу, которая может проявлять свои функции посредством связывания с целевой молекулой (Current Opinion in Biotechnology (2006) 17: 653-658; Current Opinion in Biotechnology (2007) 18: 1-10; Current Opinion in Structural Biology (1997) 7: 463-469; Protein Science (2006) 15: 14-27), и включает, например, DARPin (WO 2002/020565), аффитело (WO 1995/001937), Авимер (WO 2004/044011; WO 2005/040229) и Аднектин (WO 2002/032925). Если эти подобные антителам молекулы могут связываться с молекулами зависимым от рН способом и/или обладать активностью связывания с FcRn человека в диапазоне нейтральных значений рН, то возможно облегчать поглощение антигена клетками посредством антигенсвязывающих молекул, способствуя снижению концентрации антигена в плазме посредством введения антигенсвязывающих молекул, и улучшению фармакокинетики антигенсвязывающих молекул, и увеличению количества антигенов, с которыми может связываться одна антигенсвязывающая молекула.
Кроме того, антигенсвязывающая молекула может представлять собой белок, полученный в результате слияния между FcRn-связывающим доменом человека и рецепторным белком, который связывается с мишенью, включающей лиганд, и включает, например, слитые белки TNFR-Fc, слитые белки IL1R-Fc, слитые белки VEGFR-Fc и слитые белки CTLA4-Fc (Nat Med. 2003, Jan; 9(1): 47-52; BioDrugs. (2006) 20(3): 151-60). Если такие слитые белки рецептор-FcRn-связывающий домен человека связываются зависимым от рН способом с целевой молекулой, включающей лиганд, и/или обладают активностью связывания с FcRn человека в диапазоне нейтральных значений рН, возможно облегчать поглощение антигена клетками посредством антигенсвязывающих молекул, способствуя снижению концентрации антигена в плазме посредством введения антигенсвязывающих молекул, и улучшать фармакокинетики антигенсвязывающих молекул, и увеличивать количество антигенов, с которыми может связываться одна антигенсвязывающая молекула. Рецепторный белок конструируют соответствующим образом и модифицируют таким образом, чтобы включить связывающий домен рецепторного белка для мишени, включающей лиганд. Как изложено в примере выше, включающем слитые белки TNFR-Fc, слитые белки IL1R-Fc, слитые белки VEGFR-Fc и слитые белки CTLA4-Fc, растворимую рецепторную молекулу, содержащую внеклеточный домен такого рецепторного белка, который необходим для связывания с такими мишенями, включающими лиганды, преимущественно используют в настоящем изобретении. Такие сконструированные и модифицированные рецепторные молекулы обозначают в настоящем изобретении как искусственный рецептор. Способ, используемый для конструирования и модификации рецепторной молекулы для конструирования искусственной рецепторной молекулы, известен в данной области.
Кроме того, антигенсвязывающая молекула может представлять собой слитый белок, в котором искусственный белок-лиганд, который связывается с мишенью и обладает нейтрализующим эффектом, подвергают слиянию с FcRn-связывающим доменом человека, и искусственный белок-лиганд включает, например, мутант IL-6 (EMBO J. 1994 Dec 15; 13(24): 5863-70). Если такие искусственные слитые белки-лиганды могут связываться с целевыми молекулами зависимым от рН способом и/или обладать активностью связывания с FcRn человека в диапазоне нейтральных значений рН, возможно облегчать поглощение антигена клетками посредством антигенсвязывающих молекул, облегчать снижение концентрации антигена в плазме посредством введения антигенсвязывающих молекул и улучшать фармакокинетику антигенсвязывающих молекул, и увеличивать количество антигенов, с которыми может связываться одна антигенсвязывающая молекула.
Кроме того, антитела по настоящему изобретению могут включать модифицированные цепи сахаров. Антитела с модифицированными цепями сахаров включают, например, антитела с модифицированным гликозилированием (WO 99/54342), антитела, которые являются дефектными по фукозе, которую добавляют к цепи сахара (WO 00/61739; WO 02/31140; WO 2006/067847; WO2 006/067913), и антитела, обладающие цепями сахаров, разделенные GlcNAc (WO 02/79255).
Условия, используемые в анализе антигенсвязывающей активности или активности связывания с FcRn человека, отличные от рН, могут быть соответствующим образом выбраны специалистами в данной области, и условия конкретно не ограничены. Например, условия с использованием буфера MES при 37 градусах C, как описано в WO 2009/125825, можно использовать для определения активности. В другом варианте осуществления Na-фосфатный буфер при 25 градусах C, как описано в примерах 4 или 5, можно использовать для определения активности. Между тем, антигенсвязывающую активность и активность связывания с FcRn человека антигенсвязывающей молекулы можно определять способами, известными специалистам в данной области, например, с использованием Biacore (GE Healthcare) или подобных. Если антиген представляет собой растворимый антиген, активность антигенсвязывающей молекулы для связывания с растворимым антигеном можно определять посредством помещения антигена в качестве анализируемого вещества на чип с иммобилизованной антигенсвязывающей молекулой. Альтернативно, если антиген представляет собой антиген мембранного типа, активность антигенсвязывающей молекулы для связывания с антигеном мембранного типа можно определять посредством помещения антигенсвязывающей молекулы в качестве анализируемого вещества на чип с иммобилизованным антигеном. Активность связывания с FcRn человека антигенсвязывающей молекулы можно определять посредством помещения FcRn человека или антигенсвязывающей молекулы в качестве анализируемого вещества на чип с иммобилизованной антигенсвязывающей молекулой или FcRn человека, соответственно.
В настоящем изобретении отношение между антигенсвязывающей активностью в диапазоне кислых значений рН и антигенсвязывающей активностью в диапазоне нейтральных значений pH конкретно не ограничено при условии, что антигенсвязывающая активность в диапазоне кислых значений рН является ниже, чем таковая в диапазоне нейтральных значений рН. Однако, значение KD (pH 5,8)/KD (pH 7,4), которое представляет собой отношение константы диссоциации (KD) по отношению к антигену при pH 5,8 и pH 7,4, составляет предпочтительно 2 или выше, более предпочтительно 10 или выше, и еще более предпочтительно 40 или выше. Верхний предел значения KD (pH 5,8)/KD(pH 7,4) конкретно не ограничен, и может представлять собой любое значение, например, 400, 1000, или 10000, при условии, что производство возможно с помощью технологий, известных специалистам в данной области.
Если антиген представляет собой растворимый антиген, значение антигенсвязывающей активности может быть представлено в отношении константы диссоциации (KD). С другой стороны, если антиген представляет собой антиген мембранного типа, активность может быть представлена в отношении кажущейся константы диссоциации (кажущаяся KD). Константу диссоциации (KD) и кажущуюся константу диссоциации (кажущаяся KD) можно определять способами, известными специалистам в данной области, например, с использованием Biacore (GE Healthcare), графика Скетчарда, проточного цитометра или подобных.
В настоящем изобретении другие параметры, которые характерны для отношения антигенсвязывающей активности между значениями в диапазоне кислых и нейтральных значений рН включают, например, константу скорости диссоциации kd. Если константу скорости диссоциации (kd) применяют вместо константы диссоциации (KD) в качестве параметра, характерного для отношения активности связывания, значение kd (в диапазоне кислых значений рН)/kd(в диапазоне нейтральных значений рН), которое представляет собой отношение kd (константа скорость диссоциации) в отношении антигена в диапазоне кислых значений рН и нейтральных значений рН, предпочтительно составляет 2 или выше, более предпочтительно 5 или выше, даже более предпочтительно 10 или выше и еще более предпочтительно 30 или выше. Верхний предел значения kd (в диапазоне кислых значений рН)/kd (в диапазоне нейтральных значений рН) конкретно не ограничен и может представлять собой любое значение, например, 50, 100 или 200, при условии, что производство возможно с помощью технологий, известных специалистам в данной области.
Если антиген представляет собой растворимый антиген, значение антигенсвязывающей активности можно представлять с использованием константы скорости диссоциации (kd). Альтернативно, если антиген представляет собой антиген мембранного типа, значение может быть представлено в отношении кажущейся kd (кажущаяся константа скорости диссоциации). Константу скорости диссоциации (kd) и кажущуюся константу скорости диссоциации (кажущаяся kd) можно определять способами, известными специалистам в данной области, например, с использованием Biacore (GE Healthcare), проточного цитометра или т.п.
В настоящем изобретении, если антигенсвязывающую активность антигенсвязывающей молекулы определяют при различных значениях pH, предпочтительно, чтобы условия при измерении являлись постоянными, за исключением рН.
Способы снижения (ухудшения) антигенсвязывающей активности антигенсвязывающей молекулы в диапазоне кислых значений рН до значения, меньшего, чем диапазоне нейтральных значений рН (способы для придания зависимой от рН связывающей активности), конкретно не ограничены и могут достигаться любыми способами. Конкретно, как описано в WO 2009/125825, способы включают, например, способы снижения (ухудшения) антигенсвязывающей активности в диапазоне кислых значений рН до значения, меньшего, чем в диапазоне нейтральных значений рН, посредством замены гистидина на аминокислоту в антигенсвязывающей молекуле или встраиванием гистидина в антигенсвязывающую молекулу. Уже известно, что антителу можно придать зависимую от рН антигенсвязывающую активность посредством замены гистидина на аминокислоту в антителе (FEBS Letter (1992) 309(1): 85-88). Такая гистидиновая мутация (замена) или участки встраивания конкретно не ограничены; и гистидин можно заменять на аминокислоту в любом участке или встраивать в любой участок. Предпочтительные участки для гистидиновой мутации (замены) или вставки включают, например, области, где мутация или вставка имеет влияние на антигенсвязывающую активность антигенсвязывающей молекулы. Такие области включают участки, где мутация или вставка снижает (ухудшает) антигенсвязывающую активность в диапазоне кислых значений рН до значения, меньшего, чем в диапазоне нейтральных значений рН (значение KD (в диапазоне кислых значений рН)/KD (в диапазоне нейтральных значений рН) увеличивается) по сравнению со значением до введения мутации или вставки. Если антигенсвязывающая молекула представляет собой антитело, такие области включают, например, вариабельные области антитела и CDR. Количество гистидиновых мутаций или вставок, подлежащих введению (достигаемых), может соответсвующим образом определить специалист в данной области. Гистидиновую замену можно вводить только в одном участке или в двух или более. Альтернативно, гистидин можно встраивать только в одном участке или в двух или более. Кроме того, мутации, отличные от гистидиновой мутации (мутация (делеция, добавление, вставка и/или замена) с аминокислотой, отличной от гистидина), можно вводить в дополнение к гистидиновой мутации. Альтернативно, гистидиновую мутацию можно комбинировать с гистидиновой вставкой. Такую гистидиновую замену или вставку можно проводить посредством вероятностного способа, такого как гистидиновое сканирование, которое проводят с использованием гистидина вместо аланина в аланиновом сканировании, известном специалистам в данной области. Далее, антигенсвязывающие молекулы, обладающие более высоким значением KD (в диапазоне кислых значений рН)/KD(в диапазоне нейтральных значений рН), чем до введения мутации, можно выбирать из библиотеки антигенсвязывающих молекул с введенными случайными гистидиновыми мутациями или вставками.
Если гистидин заменяют аминокислотой в антигенсвязывающей молекуле или гистидин встраивают в антигенсвязывающую молекулу, антигенсвязывающая активность антигенсвязывающей молекулы в диапазоне нейтральных значений рН после гистидиновой замены или вставки предпочтительно является эквивалентной антигенсвязывающей активности антигенсвязывающей молекулы в диапазоне нейтральных значений рН до гистидиновой замены или вставки, но конкретно этим не ограничена. В настоящем документе «антигенсвязывающая активность антигенсвязывающей молекулы в диапазоне нейтральных значений рН после гистидиновой замены или вставки является эквивалентной антигенсвязывающей активности антигенсвязывающей молекулы в диапазоне нейтральных значений рН до гистидиновой замены или вставки» обозначает, что антигенсвязывающая молекула после гистидиновой замены или вставки сохраняет 10% или более, предпочтительно 50% или более, более предпочтительно 80% или более, и еще более предпочтительно 90% или более антигенсвязывающей активности антигенсвязывающей молекулы до гистидиновой замены или вставки. Если антигенсвязывающая активность антигенсвязывающей молекулы снижена вследствие гистидиновой замены или вставки, антигенсвязывающую активность можно регулировать посредством замены, делеции, добавления и/или вставки одной или нескольких аминокислот в антигенсвязывающую молекулу таким образом, что антигенсвязывающая активность становится эквивалентной антигенсвязывающей активности до гистидиновой замены или вставки. Настоящее изобретение также содержит такие антигенсвязывающие молекулы, чью активность связывания сделали эквивалентной в результате замены, делеции, добавления и/или вставки одной или нескольких аминокислот в антигенсвязывающую молекулу после гистидиновой замены или вставки.
Другие способы снижения (ухудшения) антигенсвязывающей активности антигенсвязывающей молекулы в диапазоне кислых значений рН до значения, меньшего, чем в диапазоне нейтральных значений рН, включают способы замены аминокислот на неприродные аминокислоты в антигенсвязывающей молекуле или вставки неприродных аминокислот в антигенсвязывающую молекулу. Известно, что pKa можно искусственно регулировать, используя неприродные аминокислоты (Angew. Chem. Int. Ed. 2005, 44, 34; Chem Soc Rev. 2004 Sep 10, 33 (7): 422-30; Amino Acids. (1999) 16(3-4): 345-79). Таким образом, в настоящем изобретении, возможно использование неприродных аминокислот вместо гистидина, описанного выше. Участки, куда вводят неприродные аминокислоты, конкретно не ограничены, и можно заменять неприродными аминокислотами или встраивать их в любом участке. Предпочтительные участки замены на неприродные аминокислоты или вставка включают, например, области, где замены или вставки обладают влиянием на антигенсвязывающую активность связывания антигенсвязывающей молекулы. Например, если антигенсвязывающая молекула представляет собой антитело, такие области включают вариабельные области антитела и определяющие комплементарность области (CDR). Между тем, количество неприродных аминокислот, подлежащих введению, конкретно не ограничено; и возможна замена неприродными аминокислотами только в одном участке или двух или более участках. Альтернативно, неприродные аминокислоты можно встраивать только в одном участке или двух или более участках. Кроме того, другие аминокислоты можно делетировать, добавлять, вставлять и/или заменять в дополнение к замене или вставке неприродных аминокислот. Кроме того, неприродными аминокислотами можно заменять и/или встраивать их в комбинации с описанными выше гистидиновыми заменами и/или вставками. Любая неприродная аминокислота может быть использована в настоящем изобретении. Возможно использование неприродных аминокислот, известных специалистам в данной области.
В настоящем изобретении, если антигенсвязывающая молекула представляет собой антитело, возможные участки замены на гистидин или неприродные аминокислоты включают, например, последовательности CDR и последовательности, отвечающие за структуру CDR антитела, включая, например, участки, описанные в WO 2009/125825. Положения аминокислот указаны согласно нумерации по Kabat (Kabat EA et al. (1991) Sequences of Proteins of Immunological Interest, NIH).
Систему нумерации Kabat в основном используют, когда речь идет об остатках в вариабельном домене (приблизительно остатки 1-107 легкой цепи и остатки 1-113 тяжелой цепи) (например, Kabat et al., Sequences of Immunological Interest. 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991)). «Система нумерации EU» или «индекс EU» в основном используют, когда речь идет об остатках в константной области тяжелой цепи иммуноглобулина (например, индекс EU, опубликованный в Kabat et al., выше). «Индекс EU, как в Kabat» относится к системе нумерации EU остатков антитела IgG1 человека. Если не указано иное в настоящем документе, ссылка на номера остатков в вариабельном домене антител обозначает нумерацию остатков посредством системы нумерации Kabat. Если не указано иначе в настоящем документе, ссылка на номера остатков в константном домене антител обозначает нумерацию остатков посредством системы нумерации EU (см. например, WO 2006073941).
Тяжелая цепь: H27, H31, H32, H33, H35, H50, H58, H59, H61, H62, H63, H64, H65, H99, H100b и H102.
Легкая цепь: L24, L27, L28, L32, L53, L54, L56, L90, L92 и L94.
Из этих участков изменений, H32, H61, L53, L90 и L94, как предполагается, являются высоко распространенными участками изменений.
Если антиген представляет собой рецептор IL-6 (например, рецептор IL-6 человека), предпочтительные участки изменения включают следующие. Однако участки изменения конкретно не ограничены ими.
Тяжелая цепь: H27, H31, H32, H35, H50, H58, H61, H62, H63, H64, H65, H100b и H102.
Легкая цепь: L24, L27, L28, L32, L53, L56, L90, L92 и L94.
Конкретно, предпочтительные комбинации участков замены на гистидин или неприродные аминокислоты включают, например, комбинацию из H27, H31 и H35; комбинацию из H27, H31, H32, H35, H58, H62 и H102; комбинацию из L32 и L53; и комбинацию из L28, L32 и L53. Кроме того, предпочтительные комбинации участков замены в тяжелых и легких цепях включают, например, комбинацию из H27, H31, L32 и L53.
Их этих участков, гистидин или неприродные аминокислоты замешены только в одном участке или во многих участках.
Между тем, если антигенсвязывающая молекула представляет собой средство, обладающее константной областью антитела, способы снижения (ухудшения) антигенсвязывающей активности антигенсвязывающей молекулы в диапазоне кислых значений рН до значения, меньшего, чем в диапазоне нейтральных значений рН включают, например, способы изменения аминокислот в константной области антитела. Конкретно, такие способы включают, например, способы замещения константной области, описанные в WO 2009/125825 (SEQ ID NO: 11, 12, 13 и 14). Между тем, способы изменения константной области антитела включают, например, способы оценки константных областей различных изотипов (IgG1, IgG2, IgG3 и IgG4) и отбор изотипов, которые снижают антигенсвязывающую активность в диапазоне кислых значений рН (увеличивают скорость диссоциации в диапазоне кислых значений рН). Такие способы также включают способы снижения антигенсвязывающей активности в диапазоне кислых значений рН (увеличение скорости диссоциации в диапазоне кислых значений рН) посредством введения замены аминокислот в аминокислотные последовательности изотипов дикого типа (аминокислотные последовательности дикого типа IgG1, IgG2, IgG3 или IgG4). Последовательность шарнирной области в константной области антитела существенно отличается среди изотипов (IgG1, IgG2, IgG3 и IgG4), и различия в аминокислотной последовательности шарнирной области имеют большое влияние на антигенсвязывающую активность. Таким образом, возможен отбор подходящего изотипа для снижения антигенсвязывающей активности в диапазоне кислых значений рН (увеличение скорости диссоциации в диапазоне кислых значений рН) в зависимости от типа антигена или эпитопа. Кроме того, так как различия в аминокислотной последовательности шарнирной области имеют большое влияние на антигенсвязывающую активность, предпочтительные участки замен аминокислот в аминокислотных последовательностях изотипов дикого типа допускают в пределах шарнирной области.
Если антигенсвязывающую активность антигенсвязывающей молекулы в диапазоне кислых значений рН снижают (уменьшают) до значения, меньшего, чем в диапазоне нейтральных значений рН (когда значение KD (в диапазоне кислых значений рН)/KD (в диапазоне нейтральных значений рН) увеличено) посредством описанных выше способов и т.п., в основном важно, чтобы значение KD (в диапазоне кислых значений рН)/KD(в диапазоне нейтральных значений рН) представляло собой значение, в два раза или более высокое, предпочтительно в пять раз или более высокое, и более предпочтительно в десять раз или более высокое по сравнению с оригинальным антителом, но конкретно не ограничено этими значениями.
Описанные выше способы можно использовать для получения антигенсвязывающих молекул, чья антигенсвязывающая активность в диапазоне кислых значений рН снижена (уменьшена) до значения, меньшего, чем в диапазоне нейтральных значений рН (антигенсвязывающие молекулы, которые связываются зависимым от рН способом), посредством замены аминокислот или вставки из антигенсвязывающих молекул, которые не обладают такими свойствами. Другие способы включают способы прямого получения антигенсвязывающих молекул, обладающих описанными выше свойствами. Например антитела, обладающие представляющим интерес желаемым свойством, можно непосредственно отбирать посредством скрининга с использованием зависимого от рН связывания с антигеном в качестве индикатора из антител, полученных посредством иммунизирования антигеном животных (мышей, крыс, хомяков, кроликов, трансгенных мышей с иммуноглобулином человека, трансгенных крыс с иммуноглобулином человека, трансгенных кроликов с иммуноглобулином человека, лам, верблюдов и т.д.). Антитела можно получать посредством гибридомной технологии или технологией B-клеточного клонирования (Proc Natl Acad Sci USA. 1996 Jul 23; 93(15): 7843-8; J Immunol Methods. 2006 Oct 20; 316(1-2): 133-43; Journal of the Association for Laboratory Automation; WO 2004/106377; WO 2008/045140; и WO 2009/113742), которые представляют собой способы, известные специалистам в данной области, но не ограничены этими способами. Альтернативно, антитела, которые обладают представляющими интерес свойствами, можно непосредственно отбирать посредством скрининга с использованием зависимого от рН связывания антигена в качестве индикатора, из библиотеки, предоставляющей антигенсвязывающий домен in vitro. Такая библиотека включает природную библиотеку человека, библиотеку иммунизированных не являющихся человеком животных и человека, полусинтетическую библиотеку и синтетическую библиотеку, которые представляют собой библиотеки, хорошо известные специалистам в данной области (Methods Mol Biol. 2002; 178: 87-100; J Immunol Methods. 2004 Jun; 289(1-2): 65-80; и Expert Opin Biol Ther. 2007 May; 7(5): 763-79), но не ограничены ими. Однако способы конкретно не ограничены этими примерами.
В настоящем изобретении использовали различие в pH в качестве различий в условиях окружающей среды между плазмой и эндосомой по отношению к различной аффинности связывания антигенсвязывающей молекулы с антигеном в плазме и эндосоме (сильное связывание в плазме и слабое связывание в эндосоме). Так как различие в условиях окружающей среды между плазмой и эндосомой не ограничены различием в рН, свойство рН-зависимого связывания антигенсвязывающей молекулы с антигеном можно заменять, используя другие факторы, чья концентрация является различной в пределах плазмы и эндосомы. Такой фактор также можно использовать для получения антитела, которое связывается с антигеном в пределах плазмы, но диссоциирует от антигена в пределах эндосомы. Таким образом, настоящее изобретение также включает антигенсвязывающую молекулу, содержащую антигенсвязывающий домен и FcRn-связывающий домен человека, который обладает активностью связывания с FcRn человека в диапазоне кислых и нейтральных значений рН, и более низкую антигенсвязывающую активность в эндосоме, чем в плазме, где активность связывания с FcRn человека в плазме является более высокой, чем активность связывания интактного IgG человека.
Способы увеличения активности связывания с FcRn человека FcRn-связывающего домена человека в антигенсвязывающей молекуле по настоящему изобретению в диапазоне нейтральных значений рН конкретно не ограничены, и активность связывания может быть увеличена посредством любых способов. Конкретно, если Fc-домен иммуноглобулина типа IgG применяют в качестве FcRn-связывающего домена человека, активность связывания с FcRn человека в диапазоне нейтральных значений рН можно увеличивать посредством изменения его аминокислот. Такой предпочтительный Fc-домен иммуноглобулина типа IgG, подлежащий изменению, включает, например, Fc-домен родительского IgG человека (IgG1, IgG2, IgG3 или IgG4 и их сконструированные варианты). Аминокислоты в любых участках можно заменять на другие аминокислоты при условии, что они придают активность связывания с FcRn человека или увеличивают ее в диапазоне нейтральных значений рН. Если антигенсвязывающая молекула IgG1 обладает Fc-доменом человека в качестве FcRn-связывающего домена человека, предпочтительно, чтобы молекула имела замены, которые делают возможным связывание с FcRn человека в диапазоне нейтральных значений рН по сравнению с родительским IgG1 человека. Аминокислоты, для которых можно совершать такие изменения, включают, например, аминокислоты в положениях от 221 до 225, 227, 228, 230, 232, 233 до 241, от 243 до 252, от 254 до 260, от 262 до 272, 274, 276, 278 до 289, от 291 до 312, от 315 до 320, 324, 325, 327 до 339, 341, 343, 345, 360, 362, 370, от 375 до 378, 380, 382, от 385 до 387, 389, 396, 414, 416, 423, 424, от 426 до 438, 440 и 442 (нумерация EU). Более конкретно, такие изменения аминокислот включают, например, те, которые перечислены в таблице 1. Связывание с FcRn человека Fc-домена иммуноглобулина типа IgG в диапазоне нейтральных значений рН можно сделать возможным посредством использования изменений, описанных выше.
Кроме того, изменения, которые делают возможным связывание с FcRn человека в диапазоне кислых значений рН, по сравнению с родительским IgG человека, представлены в качестве примера в таблице 2. Если подходящие изменения, которые также делают возможным связывание с FcRn человека в диапазоне нейтральных значений рН, выбраны из перечисленных выше изменений, их можно применять в настоящем изобретении. Между тем, комбинации изменений, которые делают возможным связывание Fv4-IgG1 с FcRn человека в кислых условиях, представлены в таблицах 6-1 и 6-2. Особенно предпочтительные аминокислоты, подлежащие изменению в Fc-домене человека родительского IgG, включают, например, аминокислоты в положениях 237, 238, 239, 248, 250, 252, 254, 255, 256, 257, 258, 265, 270, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 325, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434 и 436 (нумерация EU). Активность связывания антигенсвязывающей молекулы с FcRn человека можно увеличивать в диапазоне нейтральных значений рН посредством замены различных аминокислот на по меньшей мере одну аминокислоту, выбранную из перечисленных выше аминокислот.
Особенно предпочтительные изменения включают, например
замену аминокислоты Met на Gly в положении 237;
замену аминокислоты Ala на Pro в положении 238;
замену аминокислоты Lys на Ser в положении 239;
замену аминокислоты Ile на Lys в положении 248;
замену аминокислоты Ala, Phe, Ile, Met, Gln, Ser, Val, Trp, или Tyr на Thr в положении 250;
замену аминокислоты Phe, Trp или Tyr на Met в положении 252;
замену аминокислоты Thr на Ser в положении 254;
замену аминокислоты Glu на Arg в положении 255;
замену аминокислот Asp, Glu или Gln на Thr в положении 256;
замену аминокислот Ala, Gly, Ile, Leu, Met, Asn, Ser, Thr или Val на Pro в положении 257;
замену аминокислоты His на Glu в положении 258;
замену аминокислоты Ala на Asp в положении 265;
замену аминокислоты Phe на Asp в положении 270;
замену аминокислот Ala или Glu на Asn в положении 286;
замену аминокислоты His на Thr в положении 289;
замену аминокислоты Ala на Asn в положении 297;
замену аминокислоты Gly на Ser в положении 298;
замену аминокислоты Ala на Val в положении 303;
замену аминокислот Ala на Val в положении 305;
замену аминокислот Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr на Thr в положении 307;
замену аминокислот Ala, Phe, Ile, Leu, Met, Pro, Gln или Thr на Val в положении 308;
замену аминокислот Ala, Asp, Glu, Pro или Arg на Leu или Val в положении 309;
замену аминокислот Ala, His или Ile на Gln в положении 311;
замену аминокислот Ala или His на Asp в положении 312;
замену аминокислот Lys или Arg на Leu в положении 314;
замену аминокислоты Ala или His на Asn в положении 315;
замену аминокислоты Ala на Lys в положении 317;
замену аминокислоты Gly на Asn в положении 325;
замену аминокислоты Val на Ile в положении 332;
замену аминокислоты Leu на Lys в положении 334;
замену аминокислоты His на Lys в положении 360;
замену аминокислоты Ala на Asp в положении 376;
замену аминокислоты Ala на Glu в положении 380;
замену аминокислоты Ala на Glu в положении 382;
замену аминокислоты Ala на Asn или Ser в положении 384;
замену аминокислот Asp или His на Gly в положении 385;
замену аминокислоты Pro на Gln в положении 386;
замену аминокислоты Glu на Pro в положении 387;
замену аминокислот Ala или Ser на Asn в положении 389;
замену аминокислоты Ala на Ser в положении 424;
замену аминокислоты Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr на Met в положении 428;
замену аминокислоты Lys на His в положении 433;
замену аминокислот Ala, Phe, His, Ser, Trp или Tyr на Asn в положении 434;
и замену аминокислот His или Phe на Tyr в положении 436 (нумерация EU) в Fc-домене родительского IgG. Между тем, количество аминокислот, подлежащих изменению, конкретно не ограничено; и возможно изменение аминокислот только в одном участке или двух или более участках. Комбинации двух или более изменений аминокислот включают, например, те, которые представлены в таблице 3. Между тем комбинации изменений, которые предоставляют возможность связывания с FcRn человека в диапазоне кислых значений рН, по сравнению с родительским IgG человека, представлены в таблицах от 4-1 до 4-5. Если подходящие комбинации изменений, которые могут предоставлять возможность связывания с FcRn человека в диапазоне нейтральных значений рН, выбраны из описанных выше изменений, их можно использовать в настоящем изобретении. Кроме того, комбинации изменений, которые предоставляют возможность связывания Fv4-IgG1 с FcRn человека в нейтральных условиях, представлены в таблицах 6-1 и 6-2.
Символ «^» в таблицах указывает на вставку аминокислоты после указанного номера в нумерации EU. Например, ^281S обозначает, что S встраивают между положениями 281 и 282 в нумерации EU.

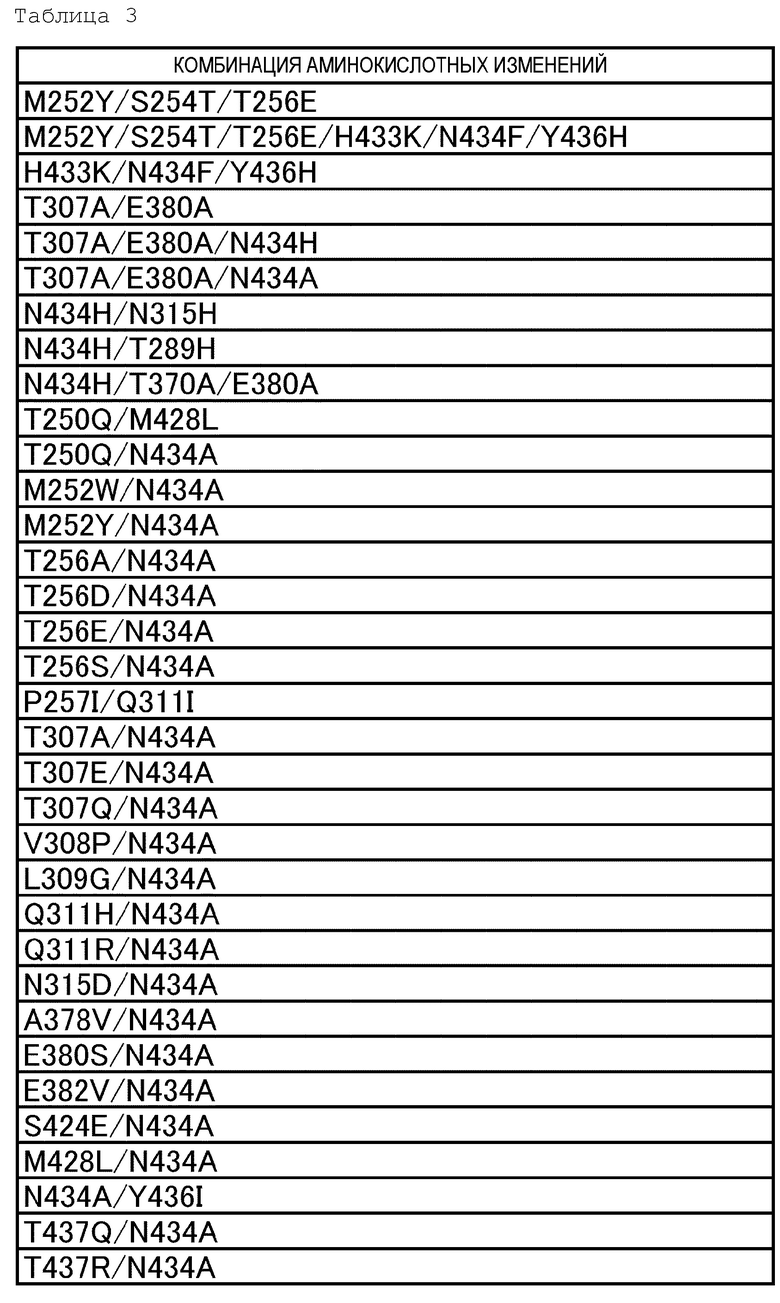

Таблица 4-2 является продолжением таблицы 4-1.
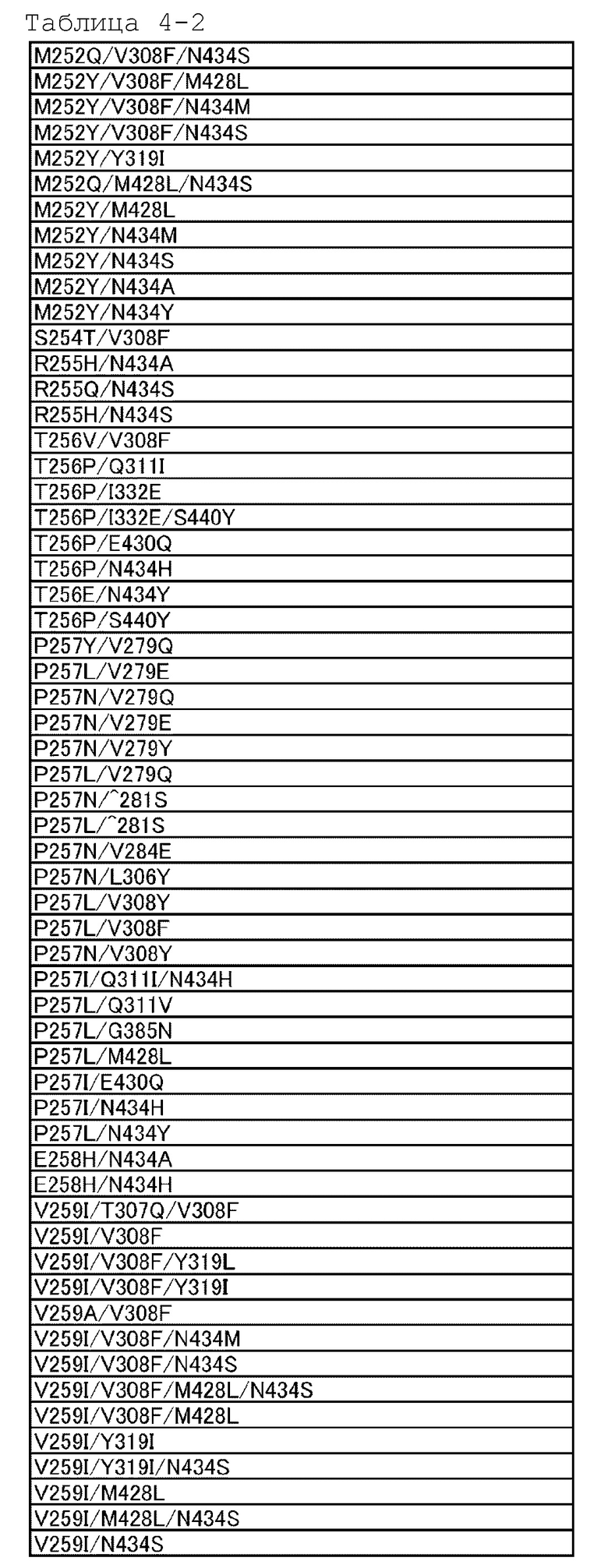
Таблица 4-3 является продолжением таблицы 4-2.
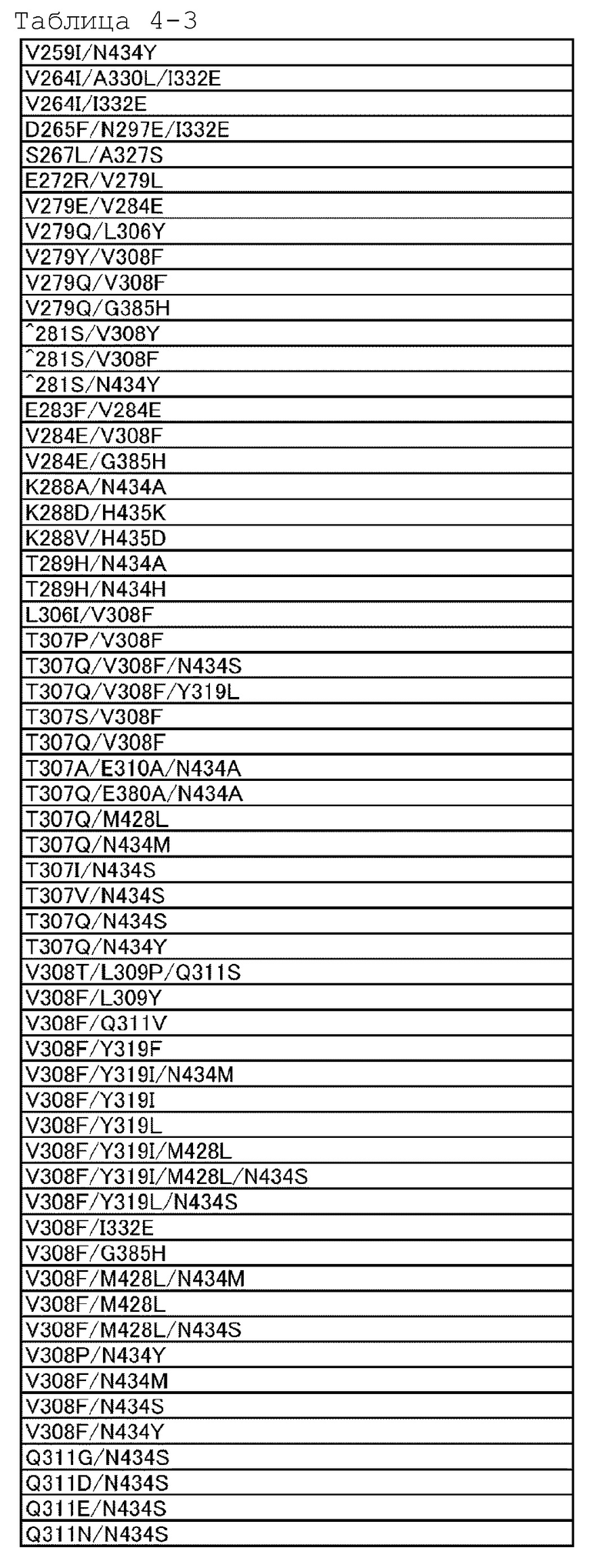
Таблица 4-4 является продолжением таблицы 4-3.
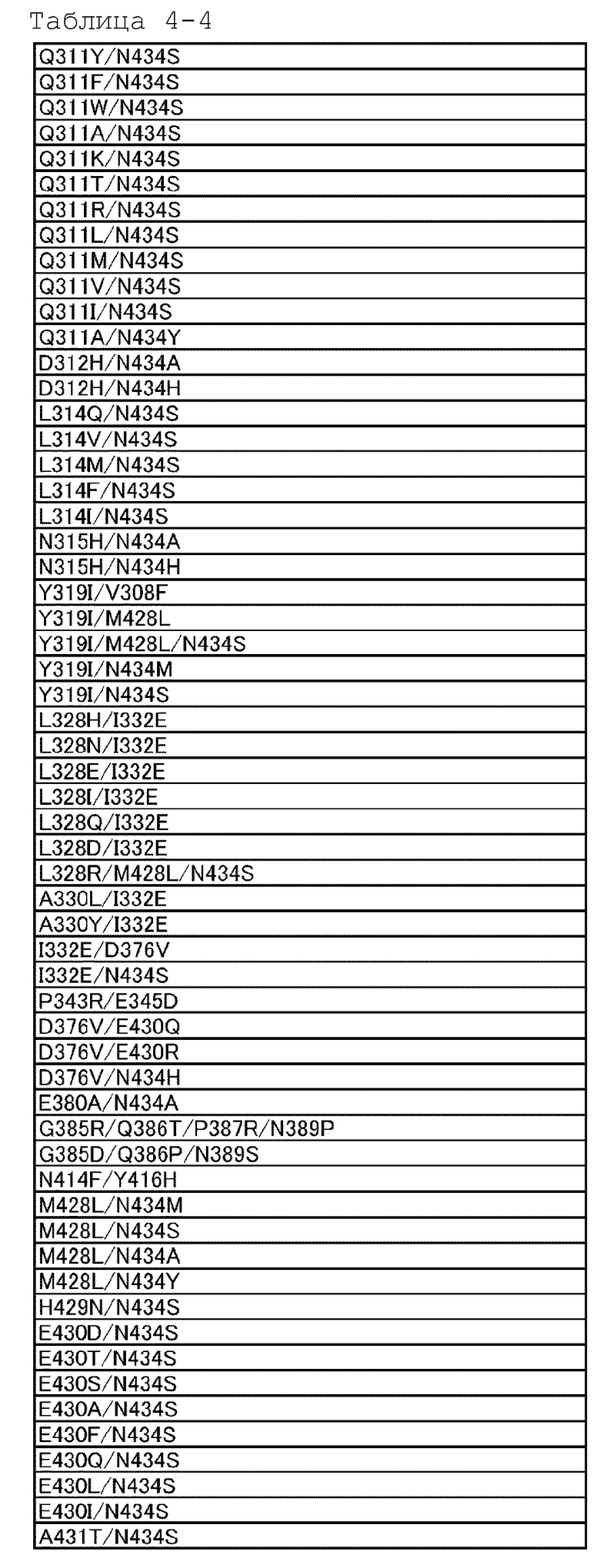
Таблица 4-5 является продолжением таблицы 4-4.
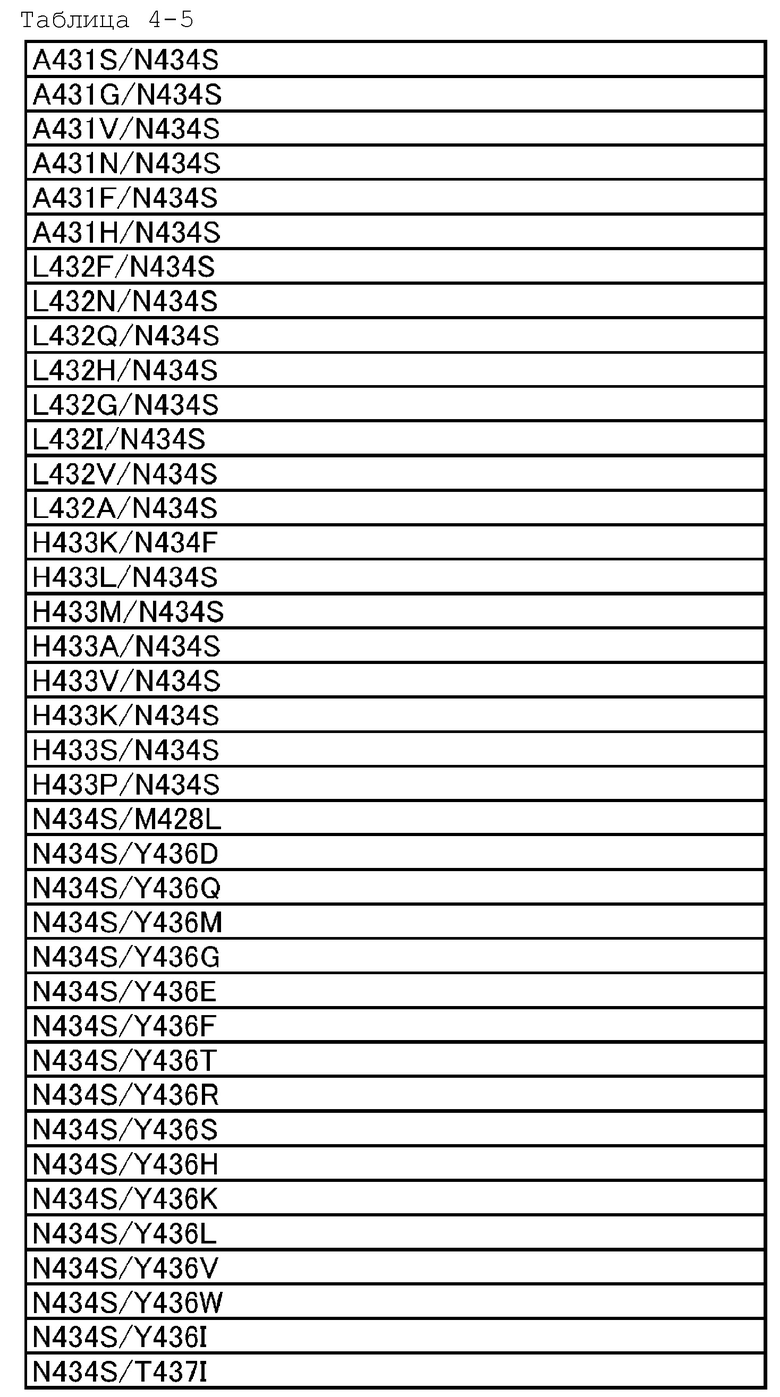
Такие изменения аминокислот соответствующим образом можно вводить, используя известные способы. Например, изменения в Fc-домене интактного IgG1 человека описаны в Drug Metab Dispos. 2007 Jan 35(1): 86-94; Int Immunol. 2006 Dec 18, (12): 1759-69; J Biol Chem. 2001 Mar. 2, 276(9): 6591-604; J Biol Chem. (2007) 282(3): 1709-17; J Immunol. (2002) 169(9): 5171-80; J Immunol. (2009) 182(12): 7663-71; Molecular Cell, Vol. 7, 867-877, April, 2001; Nat Biotechnol. 1997 July 15, (7): 637-40; Nat Biotechnol. 2005 Oct. 23, (10): 1283-8; Proc Natl Acad Sci USA. 2006 Dec. 5, 103(49): 18709-14; EP 2154157; US 20070141052; WO 2000/042072; WO 2002/060919; WO 2006/020114; WO 2006/031370; WO 2010/033279; WO 2006/053301; и WO 2009/086320.
Согласно Journal of Immunology (2009) 182: 7663-7671, активность связывания с FcRn человека интактного IgG1 человека в диапазоне кислых значений рН (pH 6,0) составляет KD, равную 1,7 микромолей (мкМ), в то время как в диапазоне нейтральных значений рН активность является почти неопределяемой. Таким образом, в предпочтительном варианте осуществления антигенсвязывающие молекулы, подлежащие использованию в способах по настоящему изобретению, относятся к антигенсвязывающим молекулам, чья активность связывания с FcRn человека в диапазоне кислых значений рН составляет KD, равную 20 микромолям или больше, и является идентичной или большей в диапазоне нейтральных значений рН, чем таковая интактного IgG человека. В более предпочтительном варианте осуществления антигенсвязывающая молекула включает антигенсвязывающие молекулы, чья активность связывания с FcRn человека представляет собой значение KD, равное 2,0 микромолям или больше в диапазоне кислых значений рН, и KD, равную 40 микромолям или больше в диапазоне нейтральных значений рН. В еще более предпочтительном варианте осуществления антигенсвязывающая молекула включает антигенсвязывающие молекулы, чья активность связывания с FcRn человека представляет собой значение KD, равное 0,5 микромолей или более в диапазоне кислых значений рН, и KD, равную 15 микромолям или больше в диапазоне нейтральных значений рН. Описанные выше KD определяют способом, описанным в Journal of Immunology (2009) 182: 7663-7671 (посредством иммобилизации антигенсвязывающей молекулы на чипе и нанесения FcRn человека в качестве анализируемого вещества).
Константу диссоциации (KD) можно использовать в качестве значения активности связывания с FcRn человека. Однако активность связывания с FcRn человека интактного IgG человека имеет небольшую активность связывания с FcRn человека в диапазоне нейтральных значений рН (pH 7,4), и таким образом трудно рассчитать активность в виде KD. Способы оценки того, является ли активность связывания с FcRn человека более высокой, чем у интактного IgG человека при pH 7,4, включает способы оценки посредством сравнения интенсивности ответа в Biacore после нанесения анализируемого вещества в одинаковой концентрации. Конкретно, если ответ после нанесения на чип FcRn человека, иммобилизованного с антигенсвязывающей молекулой при pH 7,4, является более сильным, чем ответ после нанесения FcRn человека на чип, иммобилизованный с интактным IgG человека при pH 7,4, активность связывания с FcRn человека антигенсвязывающих молекул оценивается как более высокая, чем таковая для интактного IgG человека при pH 7,4.
pH 7,0 также можно использовать в качестве рН нейтрального диапазона. Использование pH 7,0 в качестве нейтрального pH может способствовать слабому взаимодействию между FcRn человека и FcRn-связывающим доменом. Так как температуру используют в условиях анализа, аффинность связывания можно оценивать при любой температуре от 10 градусов C до 50 градусов C. Предпочтительно, температуру от 15 градусов C до 40 градусов C используют для определения аффинности связывания между FcRn-связывающим доменом человека и FcRn человек. Более предпочтительно, для определения аффинности связывания между FcRn-связывающим доменом человека и FcRn человека также используют любую температуру от 20 градусов C до 35 градусов C, как любое значение из 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 и 35 градусов C. Температура в 25 градусов C, описанная в примере 5, является одним из примеров варианта осуществления по настоящему изобретению. В предпочтительном варианте осуществления взаимодействие между FcRn человека и FcRn-связывающим доменом можно измерять при pH 7,0 и при 25 градусов C, как описано в примере 5. Аффинность связывания антигенсвязывающей молекулы с FcRn человека можно измерять посредством Biacore, как описано в примере 5.
В более предпочтительном варианте осуществления антигенсвязывающие молекулы по настоящему изобретению обладают активностью связывания с FcRn человека при pH 7,0 и при 25 градусов C, которая представляет собой значение, более высокое, чем для интактного IgG человека. В более предпочтительном варианте осуществления активность связывания с FcRn человека при pH 7,0 и при 25 градусов C в 28 раз больше, чем для интактного IgG человека, или больше, чем KD, равная 3,2 микромолям. В более предпочтительном варианте осуществления активность связывания с FcRn человека при pH 7,0 и при 25 градусов C в 38 раз больше, чем для интактного IgG человека, или больше, чем KD, равная 2,3 микромолям.
Интактный IgG1, IgG2, IgG3 или IgG4 человека предпочтительно используют в качестве интактного IgG человека с целью контрольного интактного IgG человека, подлежащего сравнению с антигенсвязывающими молекулами по их активности связывания с FcRn человека или активности in vivo. Предпочтительно, можно соответствующим образом использовать контрольную антигенсвязывающую молекулу, содержащую тот же антигенсвязывающий домен в качестве представляющей интерес антигенсвязывающей молекулы и интактный Fc-домен IgG человека в качестве FcRn-связывающего домена человека. Более предпочтительно, интактный IgG1 человека применяют с целью контрольного интактного IgG человека, подлежащего сравнению с антигенсвязывающими молекулами по их активности связывания с FcRn человека или активности in vivo.
Более конкретно, антигенсвязывающие молекулы с долговременным эффектом активности по отношению к элиминации антигена в плазме, описанные в настоящем изобретении, обладают активностью связывания с FcRn человека при pH 7,0 и при 25 градусов C в пределах диапазона значений, от 28 раз до 440 раз больших, чем для интактного IgG1 человека, или KD в пределах диапазона от 3,0 до 0,2 микромолей. Долговременную концентрацию антигена в плазме определяют, измеряя концентрацию общего или свободного антигена в плазме и молярное отношение антиген/антигенсвязывающая молекула через 2, 4, 7, 14, 28, 56 или 84 суток после введения антигенсвязывающей молекулы для оценки влияния долговременного эффекта антигенсвязывающей молекулы по настоящему изобретению на активность элиминации антигена в плазме. Достигают ли снижения концентрации антигена в плазме или молярного отношения антиген/антигенсвязывающая молекула посредством антигенсвязывающей молекулы, описанной в настоящем изобретении можно определить, оценивая снижение в любом одном или нескольких моментах времени, описанных выше.
Еще более конкретно, антигенсвязывающие молекулы с кратковременным эффектом по отношению к активности элиминации антигена в плазме, описанной в настоящем изобретении, обладают активностью связывания с FcRn человека при pH 7,0 и при 25 градусов C, в 440 раз большей, чем для интактного IgG человека, или KD, большей, чем 0,2 микромоля. Кратковременную концентрацию антигена в плазме определяют, измеряя общую концентрацию антигена или концентрацию свободного антигена в плазме и молярное отношение антиген/антигенсвязывающая молекула через 15 мин, 1, 2, 4, 8, 12 или 24 часа после введения антигенсвязывающей молекулы для оценки кратковременного эффекта антигенсвязывающей молекулы по настоящему изобретению на активность элиминации антигена в плазме.
Способы по настоящему изобретению применимы к любым антигенсвязывающим молекулам независимо от типа целевого антигена.
Антигены, которые распознаются антигенсвязывающими молекулами, такими как представляющие интерес антитела, в способах по настоящему изобретению конкретно не ограничены. Такие представляющие интерес антитела могут распознавать любой антиген. Антитела, чью фармакокинетику увеличивают посредством способов по настоящему изобретению, включают, например, рецепторные белки (мембраносвязанные рецепторы и растворимые рецепторы), антитела, которые распознают мембранный антиген, такой как поверхностные клеточные маркеры, и антитела, которые распознают растворимый антиген, такой как цитокины. Конкретные примеры антигена, который распознается антителом, чья фармакокинетика улучшена способами по настоящему изобретению, включают, например: 17-IA, 4-1 BB, 4Dc, 6-кето-PGF1a, 8-изо-PGF2a, 8-оксо-dG, аденозиновый рецептор A1, A33, ACE, ACE-2, Активин, Активин A, Активин AB, Активин B, Активин C, Активин RIA, Активин RIA ALK-2, Активин RIB ALK-4, Активин RIIA, Активин RIIB, ADAM, ADAM10, ADAM12, ADAM15, ADAM17/TACE, ADAM8, ADAM9, ADAMTS, ADAMTS4, ADAMTS5, Адрессин, адипонектин, АДФ-рибозилциклаза-1, aFGF, AGE, ALCAM, ALK, ALK-1, ALK-7, аллерген, альфа1-антихемотрипсин, альфа1-антитрипсин, альфа-синуклеин, альфа-V/бета-1 антагонист, аминин, амилин, амилоид бета, амилоидные иммуноглобулиновые вариабельные области тяжелой цепи, амилоидные иммуноглобулиновые вариабельные области легкой цепи, Андроген, ANG, ангиотензиноген, лиганд ангиопоэтин-2, анти-Id, антитромбин III, антракс, APAF-1, APE, APJ, апо A1, апо сывороточный амилоид A Apo-SAA, APP, APRIL, AR, ARC, ART, артемин, аспарагиновую кислоту, атриальный натрийуретический фактор, атриальный натрийуретический пептид, атриальные натрийуретические пептиды A, атриальные натрийуретические пептиды B, атриальные натрийуретические пептиды C, интегрин av/b3, Ax1, B7-1, B7-2, B7-H, BACE, BACE-1, протективный антиген Bacillus anthracis, Bad, BAFF, BAFF-R, Bag-1, BAK, Bax, BCA-1, BCAM, Bel, BCMA, BDNF, b-ECGF, бета-2-микроглобулин, беталактамазу, bFGF, BID, Bik, BIM, BLC, BL-CAM, BLK, стимулятор B-лимфоцитов (BIyS), BMP, BMP-2 (BMP-2a), BMP-3 (остеогенин), BMP-4 (BMP-2b), BMP-5, BMP-6 (Vgr-1), BMP-7 (OP-1), BMP-8 (BMP-8a), BMPR, BMPR-IA (ALK-3), BMPR-IB (ALK-6), BMPR-II (BRK-3), BMP, BOK, бомбезин, нейротрофический фактор костного происхождения, бычий гормон роста, BPDE, BPDE-ДНК, BRK-2, BTC, молекулы клеточной адгезии B-лимфоцитов, C10, C1-ингибитор, C1q, C3, C3a, C4, C5, C5a (комплемент 5a), CA125, CAD-8, Кадгерин-3, Кальцитонин, цАМФ, Карбоангидраза-IX, карциноэмбриональный антиген (CEA), ассоциированный с карциномой антиген, кардиотрофин-1, Катепсин A, Катепсин B, Катепсин C/DPPI, Катепсин D, Катепсин E, Катепсин H, Катепсин L, Катепсин O, Катепсин S, Катепсин V, Катепсин X/Z/P, CBL, CCI, CCK2, CCL, CCL1/I-309, CCLll/эотаксин, CCL12/MCP-5, CCL13/MCP-4, CCL14/HCC-1, CCL15/HCC-2, CCL16/HCC-4, CCL17/TARC, CCL18/PARC, CCL19/ELC, CCL2/MCP-1, CCL20/MIP-3-альфа, CCL21/SLC, CCL22/MDC, CCL23/MPIF-1, CCL24/Эотаксин-2, CCL25/TECK, CCL26/Эотаксин-3, CCL27/CTACK, CCL28/MEC, CCL3/M1P-1-альфа, CCL3L1/LD-78-бета, CCL4/MIP-1-бета, CCL5/RANTES, CCL6/C10, CCL7/MCP-3, CCL8/MCP-2, CCL9/10/MTP-1-гамма, CCR, CCR1, CCR10, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CD1, CD10, CD105, CD11a, CD11b, CD11c, CD123, CD13, CD137, CD138, CD14, CD140a, CD146, CD147, CD148, CD15, CD152, CD16, CD164, CD18, CD19, CD2, CD20, CD21, CD22, CD23, CD25, CD26, CD27L, CD28, CD29, CD3, CD30, CD30L, CD32, CD33 (белки p67), CD34, CD37, CD38, CD3E, CD4, CD40, CD40L, CD44, CD45, CD46, CD49a, CD49b, CD5, CD51, CD52, CD54, CD55, CD56, CD6, CD61, CD64, CD66e, CD7, CD70, CD74, CD8, CD80 (B7-1), CD89, CD95, CD105, CD158a, CEA, CEACAM5, CFTR, cGMP, рецептор CGRP, CINC, CKb8-1, Клаудин18, CLC, токсин Clostridium botulinum, токсин Clostridium difficile, токсин Clostridium perfringens, c-Met, CMV, CMV UL, CNTF, CNTN-1, фактор комплемента 3 (C3), фактор комплемента D, глобулин, связывающий кортикостероид, рецептор колониестимулирующиго фактора-1, COX, C-Ret, CRG-2, CRTH2, CT-1, CTACK, CTGF, CTLA-4, CX3CL1/фракталкин, CX3CR1, CXCL, CXCLl/Gro-альфа, CXCL10, CXCL11/I-TAC, CXCL12/SDF-1-альфа/бета, CXCL13/BCA-1, CXCL14/BRAK, CXCL15/Lungkine, CXCL16, CXCL16, CXCL2/Gro-бета CXCL3/Gro-гамма, CXCL3, CXCL4/PF4, CXCL5/ENA-78, CXCL6/GCP-2, CXCL7/NAP-2, CXCL8/IL-8, CXCL9/Mig, CXCLIO/IP-10, CXCR, CXCR1, CXCR2, CXCR3, CXCR4, CXCR5, CXCR6, цистатин C, опухолеассоциированный антиген цитокератин, DAN, DCC, DcR3, DC-SIGN, комплементзависимый стимулятор гемолиза, белок дельта-подобный лиганд 4, des(1-3)-IGF-1 (IGF-1 головного мозга), Dhh, оксидаза DHICA, Dickkopf-1, дигоксин, Дипептидилпептидаза IV, DK1, DNAM-1, ДНКазу, Dpp, DPPIV/CD26, Dtk, ECAD, EDA, EDA-A1, EDA-A2, EDAR, EGF, EGFR (ErbB-1), домен подобный EGF содержащий белок 7, Эластаза, эластин, EMA, EMMPRIN, ENA, ENA-78, эндосиалин, эндотелиновый рецептор, эндотоксин, энкефалиназа, eNOS, Eot, Эотаксин, Эотаксин-2, эотаксины, EpCAM, эфрин B2/EphB4, тирозинкиназный рецептор Epha2, рецептор эпидермального фактора роста (EGFR), рецептор ErbB2, тирозинкиназный рецептор ErbB3, ERCC, эритропоэтин (EPO), эритропоэтиновый рецептор, E-селектин, ET-1, Exodus-2, F белок RSV, F10, F11, F12, F13, F5, F9, Фактор Ia, Фактор IX, Фактор Xa, Фактор VII, Фактор VIII, Фактор VIIIc, Fas, FcальфаR, FcэпсилонRI, FcгаммаIIb, FcгаммаRI, FcгаммаRIIa, FcгаммаRIIIa, FcгаммаRIIIb, FcRn, FEN-1, ферритин, FGF, FGF-19, FGF-2, рецептор FGF-2, FGF-3, FGF-8, кислый FGF, основной FGF, FGFR, FGFR-3, фибрин, белок активации фибробластов (FAP), фактор роста фибробластов, фактор роста фибробластов-10, фибронектин, FL, FLIP, Flt-3, лиганд FLT3, фолатный рецептор, фолликулостимулирующий гормон (FSH), фракталкин (CX3C), свободная тяжелая цепь, свободная легкая цепь, FZD1, FZD10, FZD2, FZD3, FZD4, FZD5, FZD6, FZD7, FZD8, FZD9, G250, Gas 6, GCP-2, GCSF, G-CSF, рецептор G-CSF, GD2, GD3, GDF, GDF-1, GDF-15 (MIC-1), GDF-3 (Vgr-2), GDF-5 (BMP-14/CDMP-1), GDF-6 (BMP-13/CDMP-2), GDF-7 (BMP-12/CDMP-3), GDF-8 (миостатин), GDF-9, GDNF, гельсолин, GFAP, GF-CSF, GFR-альфа1, GFR-альфа2, GFR-альфа3, GF-бета1, гликопротеин оболочки gH, GITR, глюкагон, глюкагоновый рецептор, рецептор глюкагоноподобного пептида 1, Glut 4, глутаминаткарбоксипептидаза II, рецепторы гликопротеинового гормона, гликопротеин IIb/IIIa (GP IIb/IIIa), глипикан-3, GM-CSF, рецептор GM-CSF, gp130, gp140, gp72, гранулоцит-CSF (G-CSF), GRO/MGSA, фактор, высвобождающий гормон роста, GRO-бета, GRO-гамма, H. pylori, Гаптен (NP-cap или NIP-cap), HB-EGF, HCC, HCC 1, оболочечный гликопротеин HCMV gB, HCMV UL, гематопоэтический фактор роста (HGF), Hep B gp120, гепараназа, гепариновый кофактор II, фактор роста печени, протектиновый антиген Bacillus anthracis, гликопротеин E2 вируса гепатита C, гепатит E, гепсидин, Her1, Her2/neu (ErbB-2), Her3 (ErbB-3), Her4 (ErbB-4), гликопротеин gB вируса простого герпеса (HSV), HGF, HGFA, высокомолекулярный антиген, ассоциированный с меланомой (HMW-MAA), оболочечные белки ВИЧ, такие как GP120, MIB gp 120 с петлей V3 ВИЧ, HLA, HLA-DR, HM1.24, HMFG PEM, HMGB-1, HRG, Hrk, HSP47, Hsp90, гликопротеин gD HSV, сердечный миозин человека, цитомегаловирус человека (HCMV), гормон роста человека (hGH), сывороточный альбумин человека, активатор плазминогена тканевого типа человека (t-PA), Хантингтин, HVEM, IAP, ICAM, ICAM-1, ICAM-3, ICE, ICOS, IFN-альфа, IFN-бета, IFN-гамма, IgA, рецептор IgA, IgE, IGF, белки, связывающие IGF, IGF-1, IGF-1 R, IGF-2, IGFBP, IGFR, IL, IL-1, IL-10, рецепторы IL-10, IL-11, рецепторы IL-11, IL-12, рецепторы IL-12, IL-13, рецепторы IL-13, IL-15, рецепторы IL-15, IL-16, рецепторы IL-16, IL-17, рецепторы IL-17, IL-18 (IGIF), рецепторы IL-18, IL-1альфа, IL-1бета, рецепторы IL-1, IL-2, рецепторы IL-2, IL-20, рецепторы IL-20, IL-21, рецепторы IL-21, IL-23, рецепторы IL-23, рецепторы IL-2, IL-3, рецепторы IL-3, IL-31, рецепторы IL-31, рецепторы IL-3, IL-4, рецепторы IL-4, IL-5, рецепторы IL-5, IL-6, рецепторы IL-6, IL-7, рецепторы IL-7, IL-8, рецепторы IL-8, IL-9, рецепторы IL-9, иммуноглобулиновый иммунный комплекс, иммуноглобулины, INF-альфа, рецепторы INF-альфа, INF-бета, рецепторы INF-бета, INF-гамма, рецепторы INF-гамма, IFN типа I, рецептор IFN типа I, грипп, ингибин, ингибин альфа, ингибин бета, iNOS, инсулин, A-цепь инсулина, B-цепь инсулина, инсулиноподобный фактор роста 1, инсулиноподобный фактор роста 2, белки, связывающие инсулиноподобный фактор роста, интегрин, интегрин алфа2, интегрин алфа3, интегрин алфа4, интегрин альфа4/бета1, интегрин альфа-V/бета-3, интегрин альфа-V/бета-6, интегрин альфа4/альфа7, интегрин альфа5/альфа1, интегрин альфа5/бета3, интегрин альфа5/бета6, интегрин альфа-дельта (альфаV), интегрин альфа-тета, интегрин бета1, интегрин бета2, интегрин бета3(GPIIb-IIIa), IP-10, I-TAC, JE, калликлеин, калликреин 11, калликреин 12, калликреин 14, калликреин 15, калликреин 2, калликреин 5, калликреин 6, калликреин L1, калликреин L2, калликреин L3, калликреин L4, каллистатин, KC, KDR, фактор роста кератиноцитов (KGF), фактор роста кератиноцитов-2 (KGF-2), KGF, иммуноглобулиноподобный рецептор киллерных клеток, лиганд kit (KL), тирозинкиназа kit, ламинин 5, LAMP, LAPP (амилин, островковый амилоидный полипептид), LAP (TGF-1), латентно-ассоциированный пептид, латентный TGF-1, латентный TGF-1 bp1, LBP, LDGF, LDL, рецептор LDL, LECT2, Lefty, лептин, лютеинезирующий гормон (LH), антиген Lewis-Y, Lewis-Y связанный антиген, LFA-1, LFA-3, рецепторы LFA-3, Lfo, LIF, LIGHT, липопротеины, LIX, LKN, Lptn, L-Селектин, LT-a, LT-b, LTB4, LTBP-1, поверхностно-активное вещество легких, лютеинизирующий гормон, лимфотактин, рецептор лимфотоксин бета, рецептор лизосфинголипид, Mac-1, макрофаг-CSF (M-CSF), MAdCAM, MAG, MAP2, MARC, маспин, MCAM, MCK-2, MCP, MCP-1, MCP-2, MCP-3, MCP-4, MCP-I (MCAF), M-CSF, MDC, MDC (67 a.к.), MDC (69 a.к.), мегсин, Mer, рецептор семейства тирозинкиназ MET, металлопротеазы, мембранный гликопротеин OX2, мезотелин, рецептор MGDF, MGMT, MHC (HLA-DR), микробный белок, MIF, MIG, MIP, MIP-1 альфа, MIP-1 бета, MIP-3 альфа, MIP-3 бета, MIP-4, MK, MMAC1, MMP, MMP-1, MMP-10, MMP-11, MMP-12, MMP-13, MMP-14, MMP-15, MMP-2, MMP-24, MMP-3, MMP-7, MMP-8, MMP-9, моноцитарный белок-аттрактант, моноцитарный колониеингибирующий фактор, мышиный ассоциированный с гонадотропином пептид, MPIF, Mpo, MSK, MSP, MUC-16, MUC18, муцин (Mud), ингибирующее вещество Мюллера, Mug, MuSK, ассоциированный с миелином гликопротеин, миелоидный предшественник ингибирующего фактора-1 (MPIF-1), NAIP, нанотело, NAP, NAP-2, NCA 90, NCAD, N-кадгерин, NCAM, неприлизин, молекула адгезии нервных клеток, неросерпин, нейрональный фактор роста (NGF), нейротрофин-3, нейротрофин-4, нейротрофин-6, нейропилин 1, нейротурин, NGF-бета, NGFR, NKG20, N-метионильный гормон роста человека, nNOS, NO, Nogo-A, рецептор Nogo, неструктурный белок типа 3 (NS3) из вируса гепатита C, NOS, Npn, NRG-3, NT, NT-3, NT-4, NTN, OB, OGG1, онкостатин M, OP-2, OPG, OPN, OSM, рецепторы OSM, остеоиндуктивные факторы, остеопонтин, OX40L, OX40R, окисленный LDL, p150, p95, PADPr, паратиреоидный гормон, PARC, PARP, PBR, PBSF, PCAD, P-Кадгерин, PCNA, PCSK9, PDGF, PDGF рецептор, PDGF-AA, PDGF-AB, PDGF-BB, PDGF-D, PDK-1, PEC AM, PEDF, PEM, PF-4, PGE, PGF, PGI2, PGJ2, PIGF, PIN, PLA2, плацентарный фактор роста, плацентарная щелочная фосфатаза (PLAP), плацентарный лактоген, ингибитор активатора плазминогена-1, тромбоцитарный фактор роста, plgR, PLP, полигликолевые цепи различного размера (например, PEG-20, PEG-30, PEG40), PP14, прекалликреин, прионный белок, прокальцитонин, белок 1 программируемой гибели клеток, проинсулин, пролактин, пропротеинконвертаза PC9, прорелаксин, специфический мембранный антиген предстательной железы (PSMA), белок A, белок C, белок D, белок S, белок Z, PS, PSA, PSCA, PsmAr, PTEN, PTHrp, Ptk, PTN, гликопротеиновый лиганд P-селектина 1, R51, RAGE, RANK, RANKL, RANTES, релаксин, A-цепь релаксина, B-цепь релаксина, ренин, респираторно-синцитиальный вирус (RSV) F, Ret, ретикулон 4, ревматоидные факторы, RLI P76, RPA2, RPK-1, RSK, RSV Fgp, S100, RON-8, SCF/KL, SCGF, склеростин, SDF-1, SDF1 альфа, SDF1 бета, СЕРИН, сывороточный амилоид P, сывороточный альбумин, sFRP-3, Shh, шига-подобный токсин II, SIGIRR, SK-1, SLAM, SLPI, SMAC, SMDF, SMOH, SOD, SPARC, рецептор сфингозин-1-фосфата 1, стафилоккоковая липотейхоевая кислота, Stat, STEAP, STEAP-II, фактор стволовых клеток (SCF), стрептокиназа, супероксиддисмутаза, синдекан-1, TACE, TACI, TAG-72 (ассоциированный с опухолью гликопротеин 72), TARC, TB, TCA-3, альфа/бета T-клеточный рецептор, TdT, TECK, TEM1, TEM5, TEM7, TEM8, тенасцин, TERT, тестикулярная PLAP подобная щелочная фосфатаза, TfR, TGF, TGF-альфа, TGF-бета, пан-специфический TGF-бета, TGF-бета RII, TGF-бета RIIb, TGF-бета RIII, TGF-бета R1 (ALK-5), TGF-бета1, TGF-бета2, TGF-бета3, TGF-бета4, TGF-бета5, TGF-I, тромбин, тромбопоэтин (TPO), рецептор тимического стромального лимфопротеина, Тимус Ck-1, тиреотропный гормон (TSH), тироксин, тироксинсвязывающий глобулин, Tie, TIMP, TIQ, тканевой фактор, протеазный ингибитор тканевого фактора, белок тканевого фактора, TMEFF2, Tmpo, TMPRSS2, рецептор TNF I, рецептор TNF II, TNF-альфа, TNF-бета, TNF-бета2, TNFc, TNF-RI, TNF-RII, TNFRSF10A (TRAIL R1 Apo-2/DR4), TNFRSF10B (TRAIL R2 DR5/KILLER/TRICK-2A/TRICK-B), TNFRSF10C (TRAIL R3 DcR1/LIT/TRID), TNFRSF10D (TRAIL R4 DcR2/TRUNDD), TNFRSF11A (RANK ODF R/TRANCE R), TNFRSF11B (OPG OCIF/TR1), TNFRSF12 (TWEAK R FN14), TNFRSF12A, TNFRSF13B (TACI), TNFRSF13C (BAFF R), TNFRSF14 (HVEM ATAR/HveA/LIGHT R/TR2), TNFRSF16 (NGFR p75NTR), TNFRSF17 (BCMA), TNFRSF18 (GITR AITR), TNFRSF19 (TROY TAJ/TRADE), TNFRSF19L (RELT), TNFRSF1A (TNF R1CD120a/p55-60), TNFRSF1B (TNF RII CD120b/p75-80), TNFRSF21 (DR6), TNFRSF22 (DcTRAIL R2 TNFRH2), TNFRSF25 (DR3 Apo-3/LARD/TR-3/TRAMP/WSL-1), TNFRSF26 (TNFRH3), TNFRSF3 (LTbR TNF RIII/TNFC R), TNFRSF4 (OX40 ACT35/TXGP1 R), TNFRSF5 (CD40 p50), TNFRSF6 (Fas Apo-1/APT1/CD95), TNFRSF6B (DcR3 M68/TR6), TNFRSF7 (CD27), TNFRSF8 (CD30), TNFRSF9 (4-1 BB CD137/ILA), TNFRST23 (DcTRAIL R1 TNFRH1), TNFSF10 (TRAIL Apo-2 лиганд/TL2), TNFSF11 (TRANCE/RANK лиганд ODF/OPG лиганд), TNFSF12 (TWEAK Apo-3 лиганд/DR3 лиганд), TNFSF13 (APRIL TALL2), TNFSF13B (BAFF BLYS/TALL1/THANK/TNFSF20), TNFSF14 (LIGHT HVEM лиганд/LTg), TNFSF15 (TL1A/VEGI), TNFSF18 (GITR лиганд AITR лиганд/TL6), TNFSF1A (TNF-a конектин/DIF/TNFSF2), TNFSF1B (TNF-b LTa/TNFSF1), TNFSF3 (LTb TNFC/p33), TNFSF4 (OX40 лиганд gp34/TXGP1), TNFSF5 (CD40 лиганд CD154/gp39/HIGM1/IMD3/TRAP), TNFSF6 (Fas лиганд Apo-1 лиганд/APT1 лиганд), TNFSF7 (CD27 лиганд CD70), TNFSF8 (CD30 лиганд CD153), TNFSF9 (4-1 BB лиганд CD137 лиганд), TNF-альфа, TNF-бета, TNIL-I, токсический метаболит, TP-1, t-PA, Tpo, TRAIL, TRAIL R, TRAIL-R1, TRAIL-R2, TRANCE, рецептор трансферрина, трансформирующие факторы роста (TGF), такие как TGF-альфа и TGF-бета, трансмембранный гликопротеин NMB, транстиретин, TRF, Trk, TROP-2, трофобластный гликопротеин, TSG, TSLP, фактор некроза опухоли (TNF), опухолеассоциированный антиген CA 125, опухолеассоциированный антиген, экспрессирующий родственные Lewis Y углеводы, TWEAK, TXB2, Ung, uPAR, uPAR-1, урокиназа, VAP-1, фактор роста эндотелия сосудов (VEGF), васпин, VCAM, VCAM-1, VECAD, VE-кадгерин, VE-кадгерин-2, VEFGR-1 (flt-1), VEFGR-2, рецептор VEGF (VEGFR), VEGFR-3 (flt-4), VEGI, VIM, вирусные антигены, рецептор VitB12, рецептор витронектина, VLA, VLA-1, VLA-4, интегрин VNR, фактор фон Виллебранда (vWF), WIF-1, WNT1, WNT10A, WNT10B, WNT11, WNT16, WNT2, WNT2B/13, WNT3, WNT3A, WNT4, WNT5A, WNT5B, WNT6, WNT7A, WNT7B, WNT8A, WNT8B, WNT9A, WNT9B, XCL1, XCL2/SCM-1-бета, XCL1/лимфотактин, XCR1, XEDAR, XIAP и XPD.
Антигенсвязывающие молекулы, описанные в настоящем изобретении, способны к снижению общей концентрации перечисленных выше антигенов в плазме. Антигенсвязывающие молекулы, описанные в настоящем изобретении, также способны к элиминации из плазмы вируса, бактерий и грибов посредством связывания со структурными компонентами вируса, бактерий и грибов. Конкретно, в качестве структурных компонентов вируса, бактерий и грибов можно использовать F белок RSV, липотейхоевую кислоту стафилококка, токсин Clostridium difficile, шига-подобный токсин II, протективный антиген Bacillus anthracis и E2 гликопротеин вируса гепатита C.
Хотя способы по настоящему изобретению не ограничены какой-либо конкретной теорией, взаимосвязь между снижением (ухудшением) антигенсвязывающей способности антигенсвязывающей молекулы в диапазоне кислых значений рН до значения, меньшего, чем в диапазоне нейтральных значений рН, и/или увеличением (усилением) активности связывания с FcRn человека в диапазоне нейтральных значений рН и увеличением количества антигенов, с которыми может связываться одна антигенсвязывающая молекула, вследствие облегчения поглощения антигенсвязывающих молекул клетками, и усилением элиминации антигена из плазмы можно объяснить следующим образом.
Например, если антигенсвязывающая молекула представляет собой антитело, которое связывается с мембранным антигеном, антитело, введенное в организм связанным с антигеном, затем поглощается посредством интернализации внутри эндосом в клетки вместе с антигеном, так как антитело остается связанным с антигеном. Затем антитело перемещается в лизосомы, так как антитело является связанным с антигеном, и антитело подвергается деградации лизосомой вместе с антигеном. Опосредованную интернализацией элиминацию из плазмы называют антигензависимой элиминацией, и такой тип элиминации уже описан для ряда молекул антител (Drug Discov Today. 2006 Jan; 11(1-2): 81-8). Если одна молекула антитела IgG связывается с антигенами двухвалентным способом, интернализуется одна молекула антитела, так как антитело является связанным с двумя молекулами антигена, и подвергается деградации в лизосоме. Таким образом, в случае типичных антител, одна молекула антитела IgG не может связывать три или более молекул антигена. Например, одна молекула антитела IgG, обладающая нейтрализующей активностью, не может нейтрализовать три или более молекулы антигена.
Относительно более удлиненное время удержания (медленная элиминация) молекул IgG в плазме является следствием функции FcRn человека, который известен как рецептор спасения молекул IgG. При поглощении эндосомами посредством пиноцитоза, молекулы IgG связываются с FcRn человека, экспрессируемым в эндосомах в кислых условиях эндосомы. Так как молекулы IgG, которые не связываются с FcRn человека, перемещаются в лизосомы, где деградируют, молекулы IgG, которые связаны с FcRn человека, перемещаются на клеточную поверхность и снова возвращаются в плазму посредством диссоциирования от FcRn человека в нейтральных условиях в плазме.
Альтернативно, если антигенсвязывающая молекула представляет собой антитело, которое связывается с растворимым антигеном, введенное в организм антитело связывается с антигеном, а затем поглощается клетками, так как антитело является связанным с антигеном. Много антител, поглощенных клетками, высвобождаются за пределы клетки посредством FcRn. Однако так как антитела высвобождаются за пределы клеток, с антителами остаются связаны антигены, и антитела не могут связаться с антигенами снова. Таким образом, подобно антителам, которые связываются с мембранными антигенами, в случае типичных антител одна молекула антитела IgG не может связываться с тремя или более молекулами антигенов.
pH-зависимые антигенсвязывающие антитела, которые сильно связаны с антигеном в нейтральных условиях в плазме, но диссоциируют от антигена в кислых условиях в эндосоме (антитела, которые связываются в нейтральных условиях, но диссоциируют в кислых условиях), могут диссоциировать от антигена в эндосоме. Такие pH-зависимые антигенсвязывающие антитела могут связываться с антигенами снова, если они рециркулируют в плазму посредством FcRn после диссоциации антигена; таким образом, каждое антитело может повторно связываться с рядом антигенов. Кроме того, антиген, связанный с антигенсвязывающей молекулой, диссоциирует в эндосоме и не возвращается в плазму. Это облегчает опосредованное антигенсвязывающей молекулой поглощение антигена клетками. Таким образом, введение антигенсвязывающей молекулы может усиливать элиминацию антигена и снижать, тем самым, концентрацию антигенов в плазме.
Опосредованное антигенсвязывающими молекулами поглощение антигена клетками можно дополнительно увеличивать посредством обеспечения активности связывания FcRn человека в нейтральных условиях (pH 7,4) с антителом, которое связывает антиген зависимым от рН способом (связывает в нейтральных условиях, но диссоциирует в кислых условиях). Таким образом, введение антигенсвязывающей молекулы может усиливать элиминацию антигена и снижать, тем самым, концентрацию антигена в плазме. Обычно, как антитело, так и комплекс антиген-антитело поглощаются клетками посредством неспецифического эндоцитоза, а затем транспортируются на клеточную поверхность посредством связывания с FcRn в кислых условиях в эндосоме. Антитело и комплекс антиген-антитело рециркулируют в плазму посредством диссоциации от FcRn в нейтральных условиях на клеточную поверхность. Таким образом, если антитело, которое демонстрирует надлежащую зависимость от рН при связывании антигена (связывается в нейтральных условиях, но диссоциирует в кислых условиях) связывается с антигеном в плазме, а затем диссоциирует от связанного антигена в эндосоме, скорость элиминации антигена оценивают как равную скорости поглощения антигена клетками посредством неспецифического эндоцитоза. С другой стороны, если зависимость от рН является ненадлежащей, антиген, который не диссоциирует в эндосоме, также рециркулирует в плазму. Между тем, если зависимость от рН является pH надлежащей, скорость определяющей стадией при элиминации антигена является поглощение клетками посредством неспецифического эндоцитоза. Некоторые из FcRn считают локализованными на клеточной поверхности, так как FcRn переносят антитела от эндосомы на клеточную поверхность.
Авторы настоящего изобретения предположили, что иммуноглобулины типа IgG, которые представляют собой одну из антигенсвязывающих молекул, обладают, как правило, небольшой FcRn-связывающей активностью в диапазоне нейтральных значений рН, но те, которые демонстрируют наличие FcRn-связывающей активности в диапазоне нейтральных значений рН, могут связываться с FcRn на клеточной поверхности и таким образом поглощаться клетками зависимым от FcRn способом посредством связывания FcRn, представленным на клеточной поверхности. Скорость опосредованного FcRn поглощения клетками является более высокой, чем скорость поглощения клетками посредством неспецифического эндоцитоза. Таким образом, скорость элиминации антигена можно дополнительно увеличить посредством обеспечения наличия FcRn-связывающей активности в диапазоне нейтральных значений рН. Конкретно, антигенсвязывающая молекула с FcRn-связывающей активностью в диапазоне нейтральных значений рН перемещает антиген в клетки более быстро, чем типичный иммуноглобулин типа IgG (интактный человека), а затем антигенсвязывающая молекула диссоциирует из антигена в эндосоме. Антигенсвязывающая молекула рециркулирует на клеточную поверхность или в плазму и снова связывается с другим антигеном или поглощается клетками посредством FcRn. Скорость этого циклического процесса можно увеличить посредством повышения FcRn-связывающей активности в диапазоне нейтральных значений рН, увеличивая, тем самым, скорость элиминации антигена из плазмы. Кроме того, эффективность можно дополнительно увеличить посредством снижения антигенсвязывающей активности антигенсвязывающей молекулы в диапазоне кислых значений рН до значения, меньшего, чем диапазоне нейтральных значений рН. Кроме того, как предполагают, количество антигенов, с которыми может связываться одна антигенсвязывающая молекула, увеличивается с увеличением количества циклов, совершаемых одной антигенсвязывающей молекулой. Антигенсвязывающая молекула по настоящему изобретению содержит антигенсвязывающий домен и FcRn-связывающий домен. Так как FcRn-связывающий домен не оказывает влияния на связывание с антигеном, или с точки зрения механизма, описанного выше, увеличение опосредованного антигенсвязывающими молекулами поглощения антигена клетками можно ожидать независимо от типа антигена, и в результате увеличивать скорость элиминации антигена посредством снижения антигенсвязывающей активности антигенсвязывающих молекул в диапазоне кислых значений рН (связывающей активности) до значения, меньшего, чем диапазоне нейтральных значений рН, и/или увеличивать ее активность связывания с FcRn при pH плазмы.
<Средства, которые служат в качестве антигенсвязывающей молекулы>
Кроме того, настоящее изобретение относится к антигенсвязывающим молекулам, которые обладают активностью связывания с FcRn человека в диапазонах кислых и нейтральных значений рН и тем, чья антигенсвязывающая активность в диапазоне кислых значений рН является более низкой, чем в диапазоне нейтральных значений рН. Конкретные примеры антигенсвязывающих молекул включают такие молекулы, которые обладают активностью связывания с FcRn человека при от pH 5,8 до pH 6,0 и pH 7,4, которые, как предполагают, составляют значения pH in vivo ранней эндосомы и плазмы, соответственно, и к тем, чья антигенсвязывающая активность является более низкой при pH 5,8, чем при pH 7,4. Антигенсвязывающую молекулу, чья антигенсвязывающая активность является более низкой при pH 5,8, чем при pH 7,4, можно также обозначать как антигенсвязывающую молекулу, чья антигенсвязывающая активность является более сильной при pH 7,4, чем при pH 5,8.
Антигенсвязывающие молекулы по настоящему изобретению, обладающие активностью связывания с FcRn человека в диапазоне кислых и нейтральных значений pH, предпочтительно представляют собой антигенсвязывающие молекулы, которые также обладают активностью связывания с FcRn человека в диапазоне кислых значений рН и более сильной активностью связывания с FcRn человека, чем для интактного IgG человека, в диапазоне нейтральных значений рН. Отношение активности связывания не ограничено, при условии, что их активность связывания с FcRn человека является даже ненамного более сильной при pH 7,4.
Согласно Journal of Immunology (2009) 182: 7663-7671, активность связывания с FcRn человека и интактного IgG1 человека является KD 1,7 микромолей в диапазоне кислых значений рН (pH 6,0), тогда как активность является почти недетектируемой в диапазоне нейтральных значений рН. Таким образом, в предпочтительном варианте осуществления антигенсвязывающие молекулы по настоящему изобретению с активностью связывания с FcRn человека в диапазоне кислых и нейтральных значений pH включают антигенсвязывающие молекулы, которые обладают активностью связывания с FcRn человека с KD, составляющей 20 микромолей или более высокой в диапазоне кислых значений рН, что является равной или более сильной, чем таковая в диапазоне нейтральных значений рН интактного IgG человека. В более предпочтительном варианте осуществления антигенсвязывающие молекулы по настоящему изобретению включают антигенсвязывающие молекулы, чья активность связывания с FcRn человека представляет собой KD, равную 2,0 микромолям или более высокую в диапазоне кислых значений рН, и KD, равную 40 микромолям или более высокую в диапазоне нейтральных значений рН. В еще более предпочтительном варианте осуществления, антигенсвязывающие молекулы по настоящему изобретению включают антигенсвязывающие молекулы, чья активность связывания с FcRn человека представляет собой KD, равную 0,5 микромолей или более высокой в диапазоне кислых значений рН, и KD, составляющую 15 микромолей или более высокой в диапазоне нейтральных значений рН. Указанные выше значения KD определяют способом, описанным в Journal of Immunology (2009) 182: 7663-7671 (посредством иммобилизирования антигенсвязывающей молекулы на чипе и нанесения FcRn человека в качестве анализируемого средства).
Настоящее изобретение относится к антигенсвязывающей молекуле, содержащей антигенсвязывающий домен и FcRn-связывающий домен человека, где антигенсвязывающая молекула обладает активностью связывания с FcRn человека в диапазоне кислых и нейтральных значений pH, в которой FcRn человека и более низкая антигенсвязывающая активность в диапазоне кислых значений рН, чем в диапазоне нейтральных значений рН, представляет собой более высокое значение, чем KD, равная 3,2 микромолям. Настоящее изобретение также относится к антигенсвязывающей молекуле, содержащей антигенсвязывающий домен и FcRn-связывающий домен человека, которая обладает активностью связывания с FcRn человека в диапазоне нейтральных значений pH, где активность связывания с FcRn человека в диапазоне нейтральных значений рН представляет собой значение, в 28 раз более высокое, чем таковое для интактного IgG человека. Антигенсвязывающие молекулы по настоящему изобретению обладают активностью связывания с FcRn человека при pH 7,0 и при 25 градусов C, которая представляет собой более высокое значение, чем таковая для интактного IgG человека. В более предпочтительном варианте осуществления активность связывания с FcRn человека при pH 7,0 и при 25 градусов C представляет собой значение, в 28 раз более высокое, чем для интактного IgG человека, или с KD, равной более чем 3,2 микромолей.
Настоящее изобретение относится к антигенсвязывающей молекуле, содержащей антигенсвязывающий домен и FcRn-связывающий домен человека, где антигенсвязывающая молекула обладает активностью связывания с FcRn человека в диапазоне нейтральных значений рН, где активность связывания с FcRn человека в диапазоне нейтральных значений рН представляет собой KD, равную более чем 2,3 микромолям. Настоящее изобретение также относится к антигенсвязывающей молекуле, содержащей антигенсвязывающий домен и FcRn-связывающий домен человека, где антигенсвязывающая молекула обладает активностью связывания с FcRn человека в диапазоне нейтральных значений рН, где активность связывания с FcRn человека в диапазоне нейтральных значений рН является в 38 раз более высокой, чем таковая для интактного IgG человека.
В настоящем документе, диапазон кислых значений рН, как правило, относится к значению pH от 4,0 до pH 6,5. Диапазон кислых значений рН предпочтительно представляет собой диапазон, указанный посредством любого значения pH в пределах pH от 5,5 до pH 6,5, предпочтительно выбранного из 5,5, 5,6, 5,7, 5,8, 5,9, 6,0, 6,1, 6,2, 6,3, 6,4 и 6,5, более предпочтительно pH от 5,8 до pH 6,0, что представляет собой значения, близкие к значению pH в ранней эндосоме in vivo. Между тем, в настоящем документе диапазон нейтральных значений рН, как правило, относится к pH от 6,7 до pH 10,0. Диапазон нейтральных значений рН предпочтительно представляет собой диапазон, указанный посредством любого значения pH в пределах pH от 7,0 до pH 8,0, предпочтительно выбранного из pH 7,0, 7,1, 7,2, 7,3, 7,4, 7,5, 7,6, 7,7, 7,8, 7,9 и 8,0, более предпочтительно pH 7,4, что представляет собой значение, близкое к значению рН в плазме (крови) in vivo. Значение pH 7,0 можно использовать в качестве альтернативного для значения pH 7,4, если затруднительно определить аффинность связывания между FcRn-связывающим доменом человека и FcRn человека вследствие низкой аффинности связывания последнего при pH 7,4. Так как температура используется в условиях анализа, аффинность связывания между FcRn-связывающим доменом человека и FcRn человека можно оценивать при любой температуре от 10 градусов C до 50 градусов C. Предпочтительно, температуру от 15 градусов C до 40 градусов C используют для определения аффинности связывания между FcRn-связывающим доменом человека и FcRn человека. Более предпочтительно, любую температуру от 20 градусов C до 35 градусов C, как и любую из 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 и 35 градусов C также используют для определения аффинности связывания между FcRn-связывающим доменом человека и FcRn-человека. Температура 25 градусов C, описанная в примере 5, представляет собой один из примеров для варианта осуществления настоящего изобретения.
В более предпочтительном варианте осуществления активность связывания с FcRn человека при pH 7,0 и при 25 градусов C является в 38 раз более высокой, чем для интактного IgG человека, или более высокой, чем KD, равная 2,3 микромолям. Интактный IgG1 человека, IgG2, IgG3 или IgG4 применяют в качестве интактного IgG человека с целью контрольного интактного IgG человека, сравниваемого с антигенсвязывающими молекулами по их активности связывания с FcRn человека. Более предпочтительно, интактный IgG1 человека применяют с целью контрольного интактного IgG человека, сравниваемого с антигенсвязывающими молекулами по их активности связывания с FcRn человека.
Настоящее изобретение относится к антигенсвязывающей молекуле, содержащей антигенсвязывающий домен и FcRn-связывающий домен человека, где общая концентрация антигена в плазме после введения антигенсвязывающей молекулы не относящемуся к человеку животному является более низкой, чем общая концентрация антигена в плазме после введения контрольной антигенсвязывающей молекулы не являющемуся человеком животному.
Настоящее изобретение также относится к антигенсвязывающей молекуле, для которой концентрация антигена в плазме после введения антигенсвязывающей молекулы не относящемуся к человеку животному является более низкой, чем общая концентрация антигена в плазме, полученная для не являющегося человеком животного, которому не вводили антигенсвязывающую молекулу.
Общую концентрацию антигена в плазме можно снижать посредством введения антигенсвязывающей молекулы по настоящему изобретению в 2 раза, 5 раз, 10 раз, 20 раз, 50 раз, 100 раз, 200 раз, 500 раз и 1000 раз или даже более по сравнению с введением контрольной антигенсвязывающей молекулы, содержащей Fc-домен интактного IgG человека в качестве FcRn-связывающего домена человека, или по сравнению со случаем, когда не вводят молекулу антигенсвязывающего домена по настоящему изобретению.
В другом варианте осуществления настоящее изобретение относится к антигенсвязывающей молекуле, содержащей антигенсвязывающий домен и FcRn-связывающий домен человека, для которой молярное отношение антиген/антигенсвязывающая молекула (C) антигенсвязывающей молекулы рассчитывается следующим способом:
C=A/B,
является более низким, чем молярное отношение антиген/антигенсвязывающая молекула (C’) антигенсвязывающей молекулы, содержащей тот же антигенсвязывающий домен и интактный Fc-домен IgG человека в качестве FcRn-связывающего домена человека, рассчитанного по формуле:
C’=A’/B’,
где:
A представляет собой общую концентрацию антигена в плазме после введения антигенсвязывающей молекулы не являющемуся человеком животному,
B представляет собой концентрацию в плазме антигенсвязывающей молекулы после введения антигенсвязывающей молекулы не являющемуся человеком животному,
A’ представляет собой общую концентрацию антигена в плазме после введения контрольной антигенсвязывающей молекулы не являющемуся человеком животному,
B’ представляет собой концентрацию в плазме антигенсвязывающей молекулы после введения контрольной антигенсвязывающей молекулы не являющемуся человеком животному.
Молярное отношение антиген/антигенсвязывающая молекула можно снижать посредством введения антигенсвязывающей молекулы по настоящее изобретение в 2 раза, 5 раза, 10 раз, 20 раз, 50 раз, 100 раз, 200 раз, 500 раз и 1000 раз или даже больше по сравнению с введением антигенсвязывающей молекулы, содержащей интактный Fc-домен IgG человека в качестве FcRn-связывающего домена человека.
Снижение общей концентрации антигена в плазме или молярное отношение антиген/антитело можно оценивать, как описано в примерах 6, 8 и 13. Более конкретно, используя трансгенную линию 32 или линию 276 мышей с FcRn человека (Jackson Laboratories, Methods Mol Biol. (2010) 602: 93-104.), их можно оценивать либо посредством модели совместной инъекции антиген-антитело, либо стационарной модели инфузии антигена, если антигенсвязывающая молекула не дает перекрестную реакцию с соответствующим антигеном мыши. Если антигенсвязывающая молекула вступает в перекрестную реакцию с соответствующим антигеном мыши, их можно оценивать посредством простой инъекции антигенсвязывающей молекулы трансгенной линии 32 или 276 мышей с FcRn человека (Jackson Laboratories). В модели совместной инъекции, смесь антигенсвязывающей молекулы и антигена вводят мыши. В модели стационарной инфузии антигена, инфузионный насос, содержащий раствор антигена, имплантируют мыши для достижения постоянной концентрации антигена в плазме, а затем мыши инъецируют антигенсвязывающую молекулу. Тестируемую антигенсвязывающую молекулу вводят в той же дозе. Общую концентрацию антигена в плазме, концентрацию свободного антигена в плазме и концентрацию антигенсвязывающей молекулы в плазме измеряют в соответствующий момент времени, используя способы, известные специалистам в данной области.
Путь введения антигенсвязывающей молекулы по настоящему изобретению можно выбирать из интрадермального, внутривенного, интравитреального, подкожного, интраперитонеального, парентерального и внутримышечной инъекции.
Более конкретно, антигенсвязывающие молекулы с долгосрочным эффектом на активность элиминации антигена в плазме, описанные в настоящем изобретении, обладают активностью связывания с FcRn человека при pH 7,0 и 25 градусов C в пределах диапазона, от 28 раз до 440 раз более высокого, чем для интактного IgG1 человека, или с KD в пределах диапазона 3,0 микромолей до 0,2 микромолей. Краткосрочную концентрацию антигена в плазме определяют посредством измерения общей концентрации антигена или концентрации свободного антигена в плазме и молярное отношение антиген/антигенсвязывающая молекула через 2, 4, 7, 14, 28, 56 или 84 суток после введения антигенсвязывающей молекулы с целью оценки долгосрочного эффекта антигенсвязывающей молекулы по настоящему изобретению на активность элиминации антигена в плазме. Достигают ли снижения концентрации антигена в плазме или молярного отношения антиген/антигенсвязывающая молекула посредством антигенсвязывающей молекулы, описанной в настоящем изобретении, можно определять посредством оценки снижения в любой один момент времени или несколько моментов времени, описанных выше.
Еще более конкретно, антигенсвязывающие молекулы с кратковременным эффектом на элиминацию антигена в плазме, описанные в настоящем изобретении, обладают активностью связывания с FcRn человека при pH 7,0 и при 25 градусов C, в 440 раз более высокой, чем интактный IgG человека, или с KD, более высокой, чем 0,2 микромоля. Кратковременную концентрацию антигена в плазме определяют посредством измерения общей концентрации антигена или концентрации свободного антигена в плазме и молярного отношения антиген/антигенсвязывающая молекула через 15 мин, 1, 2, 4, 8, 12 или 24 часов после введения антигенсвязывающих молекул с целью оценки кратковременного эффекта антигенсвязывающих молекул по настоящему изобретению на активность элиминации антигена в плазме.
Кроме того, в антигенсвязывающей молекуле по настоящему изобретению, которая обладает более низкой антигенсвязывающей активностью в диапазоне кислых значений рН, чем в диапазоне нейтральных значений рН, отношение активности связывания не ограничено, при условии, что антигенсвязывающая активность является более низкой в диапазоне кислых значений рН, чем в диапазоне нейтральных значений рН. При условии, что антигенсвязывающая активность в диапазоне кислых значений рН является даже немного более низкой, антигенсвязывающая молекула является подходящей. В предпочтительном варианте осуществления антигенсвязывающие молекулы по настоящему изобретению включают антигенсвязывающие молекулы, чья антигенсвязывающая активность при pH 7,4 является удвоенной или более, чем таковая при pH 5,8. В более предпочтительном варианте осуществления антигенсвязывающие молекулы по настоящему изобретению включают антигенсвязывающие молекулы, чья антигенсвязывающая активность при pH 7,4 является в 10 раз или более высокой, чем таковая при pH 5,8. В еще более предпочтительном варианте осуществления, антигенсвязывающие молекулы по настоящему изобретению включают антигенсвязывающие молекулы, чья антигенсвязывающая активность при pH 7,4 является в 40 раз или более высокой, чем таковая при pH 5,8.
Конкретно, антигенсвязывающие молекулы по настоящему изобретению включают, например, варианты осуществления, описанные в WO 2009/125825. Более конкретно, в предпочтительном варианте осуществления, антигенсвязывающая молекула по настоящему изобретению обладает антигенсвязывающей активностью при pH 5,8, которая является более низкой, чем таковая при pH 7,4, где значение KD (pH 5,8)/KD (pH 7,4), которое представляет собой отношение KD для антигена при pH 5,8 и KD при pH 7,4, предпочтительно представляет собой 2 или более, более предпочтительно 10 или более и еще более предпочтительно 40 или более. Верхний предел значения KD (pH 5,8)/KD (pH7,4) конкретно не ограничен и может представлять собой любое значение, например, 400, 1000 или 10000, при условии, что получение является возможным с использованием технологий, известных специалистам в данной области.
В другом предпочтительном варианте осуществления антигенсвязывающая молекула по настоящему изобретению, чья антигенсвязывающая активность при pH 5,8 является более низкой, чем таковая при pH 7,4, имеет значение kd(pH 5,8)/kd(pH 7,4), которое представляет собой отношение kd для антигена при pH 5,8 и kd для антигена при pH 7,4, которое представляет собой 2 или более, более предпочтительно 5 или более, даже более предпочтительно 10 или более и еще более предпочтительно 30 или более. Верхний предел значения kd (pH 5,8)/kd (pH 7,4) конкретно не ограничен и может представлять собой любое значение, например, 50, 100 или 200, при условии, что получение является возможным с использованием технологий, известных специалистам в данной области.
Условия, отличные от pH, при которых измеряют антигенсвязывающую активность и активность связывания с FcRn человека может быть соответствующим образом выбрана специалистами в данной области, и условия конкретно не ограничены; однако, измерения можно проводить, например, в условиях с MES буфером и при 37 градусов C, как описано в примерах. Кроме того, антигенсвязывающую активность антигенсвязывающей молекулы можно определять способами, известными специалистам в данной области, например, с использованием Biacore T100 (GE Healthcare) или т.п., как описано в примерах.
Антигенсвязывающие молекулы по настоящему изобретению усиливают поглощение антигена клетками. Молекулы легко диссоциируют от антигена в эндосоме, а затем высвобождаются за пределы клетки посредством связывания с FcRn человека. Антигенсвязывающие молекулы по настоящему изобретению, как предполагают, легко связываются с антигеном в плазме снова. Таким образом, например, если антигенсвязывающая молекула по настоящему изобретению представляет собой нейтрализующую антигенсвязывающую молекулу, снижение концентрации антигена в плазме можно усиливать посредством введенных молекул. Таким образом, антигенсвязывающая молекула, которая обладает активностью связывания с FcRn человека в диапазоне кислых значений рН, обладает более низкой антигенсвязывающей активностью в диапазоне кислых значений рН, чем в диапазоне нейтральных значений рН; и антигенсвязывающая молекула, которая обладает активностью связывания с FcRn человека в диапазоне нейтральных значений рН, вероятно, представляет собой антигенсвязывающую молекулу, которая обладает лучшей фармакокинетикой и может связывать больше антигенов на молекулу.
В предпочтительном варианте осуществления такие антигенсвязывающие молекулы, обладающие активностью связывания с FcRn человека в диапазоне кислых и нейтральных значений pH, включают те, которые содержат FcRn-связывающий домен человека, обладающий способностью прямо или опосредованно связываться с FcRn человека. Если домен уже обладает связывающей активностью с FcRn человека в диапазоне кислых и нейтральных значений pH, его можно использовать в низменном виде. Альтернативно, даже если домен обладает активностью связывания с FcRn человека в диапазоне кислых значений рН, но демонстрирует только слабую или отсутствие активности связывания с FcRn человека в диапазоне нейтральных значений рН, его можно использовать после изменения аминокислот в домене для придания активность связывания с FcRn человека в диапазоне нейтральных значений рН. Альтернативно, возможно увеличить активность связывания с FcRn человека посредством изменения аминокислот в домене, который уже обладает активностью связывания с FcRn человека в диапазонах кислых и нейтральных значений рН. Такие антигенсвязывающие молекулы включают, например, те, которые обладают аминокислотной последовательностью домена Fc IgG, которая содержит изменение по меньшей мере одной аминокислоты. Изменение аминокислот конкретно не ограничено; и изменение можно осуществлять в любом участке при условии, что активность связывания с FcRn человека в диапазоне нейтральных значений рН является более высокой, чем до изменения.
Конкретно, аминокислотные изменения, которые обеспечивают активность связывания с FcRn человека в диапазоне кислых и нейтральных значений pH, включают, например, изменение аминокислоты в положениях от 221 до 225, 227, 228, 230, 232, от 233 до 241, от 243 до 252, от 254 до 260, от 262 до 272, 274, 276, от 278 до 289, от 291 до 312, от 315 до 320, 324, 325, от 327 до 339, 341, 343, 345, 360, 362, 370, от 375 до 378, 380, 382, от 385 до 387, 389, 396, 414, 416, 423, 424, от 426 до 438, 440 и 442 (нумерация EU) в родительском Fc-домене IgG, описанном выше. Более конкретно, изменения аминокислот включают изменения в участках (в нумерации EU), представленные в таблицах 1, 2, 6-1, 6-2 и 9. Предпочтительные антигенсвязывающие молекулы включают антигенсвязывающие молекулы, содержащие аминокислотную последовательность, которая получена в результате изменения по меньшей мере одной аминокислоты, выбранной из аминокислот в положениях 237, 238, 239, 248, 250, 252, 254, 255, 256, 257, 258, 265, 270, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 325, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434 и 436 в нумерации EU. В предпочтительном варианте осуществления такие аминокислотные изменения включают:
замену аминокислоты Met на Gly в положении 237;
замену аминокислоты Ala на Pro в положении 238;
замену аминокислоты Lys на Ser в положении 239;
замену аминокислоты Ile на Lys в положении 248;
замену аминокислоты Ala, Phe, Ile, Met, Gln, Ser, Val, Trp, или Tyr на Thr в положении 250;
замену аминокислоты Phe, Trp или Tyr на Met в положении 252;
замену аминокислоты Thr на Ser в положении 254;
замену аминокислоты Glu на Arg в положении 255;
замену аминокислот Asp, Glu или Gln на Thr в положении 256;
замену аминокислот Ala, Gly, Ile, Leu, Met, Asn, Ser, Thr или Val на Pro в положении 257;
замену аминокислоты His на Glu в положении 258;
замену аминокислоты Ala на Asp в положении 265;
замену аминокислоты Phe на Asp в положении 270;
замену аминокислот Ala или Glu на Asn в положении 286;
замену аминокислоты His на Thr в положении 289;
замену аминокислоты Ala на Asn в положении 297;
замену аминокислоты Gly на Ser в положении 298;
замену аминокислоты Ala на Val в положении 303;
замену аминокислот Ala на Val в положении 305;
замену аминокислот Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr на Thr в положении 307;
замену аминокислот Ala, Phe, Ile, Leu, Met, Pro, Gln или Thr на Val в положении 308;
замену аминокислот Ala, Asp, Glu, Pro или Arg на Leu или Val в положении 309;
замену аминокислот Ala, His или Ile на Gln в положении 311;
замену аминокислот Ala или His на Asp в положении 312;
замену аминокислот Lys или Arg на Leu в положении 314;
замену аминокислоты Ala или His на Asn в положении 315;
замену аминокислоты Ala на Lys в положении 317;
замену аминокислоты Gly на Asn в положении 325;
замену аминокислоты Val на Ile в положении 332;
замену аминокислоты Leu на Lys в положении 334;
замену аминокислоты His на Lys в положении 360;
замену аминокислоты Ala на Asp в положении 376;
замену аминокислоты Ala на Glu в положении 380;
замену аминокислоты Ala на Glu в положении 382;
замену аминокислоты Ala на Asn или Ser в положении 384;
замену аминокислот Asp или His на Gly в положении 385;
замену аминокислоты Pro на Gln в положении 386;
замену аминокислоты Glu на Pro в положении 387;
замену аминокислот Ala или Ser на Asn в положении 389;
замену аминокислоты Ala на Ser в положении 424;
замену аминокислоты Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr на Met в положении 428;
замену аминокислоты Lys на His в положении 433;
замену аминокислот Ala, Phe, His, Ser, Trp или Tyr на Asn в положении 434;
и замену аминокислот His или Phe на Tyr в положении 436 (нумерация EU).
Количество аминокислот, подлежащих изменению, конкретно не ограничено; возможно изменение аминокислоты только в одном участке или двух или более участках. Комбинации двух или более аминокислотных изменений включают, например, те, которые представлены в таблицах 3, 4-1 до 4-5, 6-1, 6-2 и 9.
Между тем, домены, которые уже обладают связывающей активностью с FcRn человека в диапазонах кислых и нейтральных значений pH, включают, например, FcRn-связывающие домены человека, содержащие по меньшей мере одну аминокислоту, выбранную из:
Met в положении аминокислоты 237;
Ala в положении аминокислоты 238;
Lys в в положении аминокислоты 239;
Ile в положении аминокислоты 248;
Ala, Phe, Ile, Met, Gln, Ser, Val, Trp или Tyr в положении аминокислоты 250;
Phe, Trp или Tyr в положении аминокислоты 252;
Thr в положении аминокислоты 254;
Glu в положении аминокислоты 255;
Asp, Glu, или Gln в положении аминокислоты 256;
Ala, Gly, Ile, Leu, Met, Asn, Ser, Thr или Val в положении аминокислоты 257;
His в положении аминокислоты 258;
Ala в положении аминокислоты 265;
Phe в положении аминокислоты 270;
Ala или Glu в положении аминокислоты 286;
His в положении аминокислоты 289;
Ala в положении аминокислоты 297;
Gly в положении аминокислоты 298;
Ala в положении аминокислоты 303;
Ala в положении аминокислоты 305;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr в положении аминокислоты 307;
Ala, Phe, Ile, Leu, Met, Pro, Gln или Thr в положении аминокислоты 308;
Ala, Asp, Glu, Pro или Arg в положении аминокислоты 309;
Ala, His или Ile в положении аминокислоты 311;
Ala или His в положении аминокислоты 312;
Lys или Arg в положении аминокислоты 314;
Ala или His в положении аминокислоты 315;
Ala в положении аминокислоты 317;
Gly в положении аминокислоты 325;
Val в положении аминокислоты 332;
Leu в положении аминокислоты 334;
His в положении аминокислоты 360;
Ala в положении аминокислоты 376;
Ala в положении аминокислоты 380;
Ala в положении аминокислоты 382;
Ala в положении аминокислоты 384;
Asp или His в положении аминокислоты 385;
Pro в положении аминокислоты 386;
Glu в положении аминокислоты 387;
Ala или Ser в положении аминокислоты 389;
Ala в положении аминокислоты 424;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr в положении аминокислоты 428;
Lys в положении аминокислоты 433;
Ala, Phe, His, Ser, Trp или Tyr в положении аминокислоты 434;
и His или Phe в положении аминокислоты 436 (нумерация EU) в родительском Fc-домене IgG.
Аминокислота в участке или аминокислоты в двух или более участках могут иметь эти аминокислоты. Комбинации аминокислот в двух или более положениях включают, например, те, которые представлены в таблице 3, 4-1 до 4-5, 6-1, 6-2 и 9.
Альтернативно, в предпочтительном варианте осуществления антигенсвязывающая молекула, чья антигенсвязывающая активность в диапазоне кислых значений рН является более низкой, чем таковая в диапазоне нейтральных значений рН, включает антигенсвязывающие молекулы, в которых по меньшей мере одну аминокислоту в антигенсвязывающей молекуле заменяют на гистидин или неприродную аминокислоту, или в которой по меньшей мере один гистидин или неприродная аминокислота уже встроены. Участок, в который вводят мутацию гистидина или неприродной аминокислоты, конкретно не ограничен и может представлять собой любой участок, при условии, что антигенсвязывающая активность в диапазоне кислых значений рН является более слабой, чем таковая в диапазоне нейтральных значений рН, и значение KD (в диапазоне кислых значений рН)/KD (в диапазоне нейтральных значений рН) является большим, или kd (в диапазоне кислых значений рН)/kd (в диапазоне нейтральных значений рН) является большим) по сравнению с таковым до замены. Примеры включают вариабельные области и CDR антитела в случае, если антигенсвязывающая молекула представляет собой антитело. Количество аминокислот, подлежащих замене на гистидин или неприродную аминокислоту, и количество встраиваемых аминокислот, подлежащих встраиванию, могут соответствующим образом определять специалисты в данной области. Одну аминокислоту можно заменить гистидином или неприродной аминокислотой, или можно встраивать одну аминокислоту, или две или более аминокислот можно заменять гистидином или неприродными аминокислотами, или две или более аминокислоты можно встраивать. Кроме того, за исключением замен на гистидин или неприродную аминокислоту или вставку гистидина или неприродной аминокислоты, делецию, добавление, вставку, и/или замену и других аминокислот можно также осуществлять одновременно. Замены гистидина или неприродной аминокислоты или вставку гистидина или неприродной аминокислоты можно проводить случайным образом, используя способ, такой как гистидиновое сканирование, в котором используют гистидин вместо аланина в аланиновом сканировании, которое известно специалистам в данной области. Антигенсвязывающие молекулы, чья KD (pH 5,8)/KD (pH 7,4) или kd (pH 5,8)/kd (pH 7,4) увеличена по сравнению с таковой до мутации, можно выбирать из антигенсвязывающих молекул, в которой мутацию с гистидином или неприродной аминокислотой вводят случайным образом.
Предпочтительные антигенсвязывающие молекулы с мутацией на гистидин или на неприродную аминокислоту, и чья антигенсвязывающая активность в диапазоне кислых значений рН является более низкой, чем таковая в диапазоне нейтральных значений рН, включают, например, антигенсвязывающие молекулы, чья антигенсвязывающая активность при pH 7,4 после мутации на гистидин или на неприродную аминокислоту является эквивалентной антигенсвязывающей активности при pH 7,4 до мутации на гистидин или неприродную аминокислоту. В настоящем изобретении «антигенсвязывающая молекула после мутации на гистидин или неприродную аминокислоту обладает антигенсвязывающей активностью, которая является эквивалентной антигенсвязывающей активности антигенсвязывающей молекулы до мутации на гистидин или неприродную аминокислоту» обозначает, что если антигенсвязывающую активность антигенсвязывающей молекулы до мутации на гистидин или неприродную аминокислоту определяют как 100%, антигенсвязывающая активность антигенсвязывающей молекулы после мутации на гистидин или неприродную аминокислоту составляет по меньшей мере 10% или более, предпочтительно 50% или более, более предпочтительно 80% или более и еще более предпочтительно 90% или более. Антигенсвязывающая активность при pH 7,4 после мутации на гистидин или неприродную аминокислоту может быть более высокой, чем антигенсвязывающая активность при pH 7,4 до мутации на гистидин или неприродную аминокислоту. Если антигенсвязывающая активность антигенсвязывающей молекулы снижается вследствие замены или вставки гистидина или неприродной аминокислоты, антигенсвязывающую активность можно регулировать посредством введения замены, делеции, добавления и/или вставки таких одной или нескольких аминокислот в антигенсвязывающую молекулу таким образом, что антигенсвязывающая активность становиться эквивалентной антигенсвязывающей активности до гистидиновой замены или вставки. Настоящее изобретение также относится к таким антигенсвязывающим молекулам, чья активность связывания стала эквивалентной в результате замены, делеции, добавления и/или вставки одной или нескольких аминокислот после гистидиновой замены или вставки.
Дополнительно, если антигенсвязывающая молекула представляет собой средство, включающее константную область антитела, в другом предпочтительном варианте осуществления - антигенсвязывающие молекулы, чья антигенсвязывающая активность при pH 5,8 является более низкой, чем антигенсвязывающая активность при pH 7,4, настоящее изобретение относится к способам изменения константных областей антител, содержащихся в антигенсвязывающих молекулах. Конкретные примеры константных областей антитела после изменения включают константные области, описанные в примерах в WO 2009/125825 (SEQ ID NO: 11, 12, 13 и 14).
Если антигенсвязывающую активность антигенсвязывающего средства при pH 5,8 снижают по сравнению с антигенсвязывающей активностью при pH 7,4 (если значение KD (pH 5,8)/KD (pH7,4) увеличено) посредством описанных выше способов, то таким образом, в основном является предпочтительным значение KD (pH 5,8)/KD (pH 7,4), которое представляет собой значение, в два раза или более, более предпочтительно в пять раз или более, и даже более предпочтительно в десять раз или более высокое по сравнению с таковым исходного антитела, но конкретно не ограничено этими значениями.
Кроме того, настоящее изобретение относится к антигенсвязывающим молекулам, с заменой гистидина или неприродной аминокислотой по меньшей мере на одну аминокислоту в одном из участков, описанных выше. Положения аминокислот представлены согласно нумерации по Kabat (Kabat EA et al. (1991) Sequences of Proteins of Immunological Interest, NIH).
Тяжелая цепь: H27, H31, H32, H33, H35, H50, H58, H59, H61, H62, H63, H64, H65, H99, H100b и H102.
Легкая цепь: L24, L27, L28, L32, L53, L54, L56, L90, L92 и L94.
Из всех участков изменений, как предполагают, H32, H61, L53, L90 и L94 представляют собой наиболее важные участки для изменений.
Конкретно, предпочтительные комбинации участков для замены на гистидин или неприродную аминокислоту включают, например, комбинацию из H27, H31 и H35; комбинацию из H27, H31, H32, H35, H58, H62 и H102; комбинацию из L32 и L53; и комбинацию из L28, L32 и L53. Кроме того, предпочтительные комбинации участков для замен в тяжелых и легких цепях включают, например, комбинацию из H27, H31, L32 и L53.
Антигенсвязывающая молекула по настоящему изобретению может обладать другими свойствами, и например, может представлять собой агонистическую или антагонистическую антигенсвязывающую молекулу, при условии, что ее антигенсвязывающая активность является более низкой в диапазоне кислых значений рН, чем в диапазоне нейтральных значений рН, и она обладает активностью связывания с FcRn человека в диапазоне кислых и нейтральных значений рН. Предпочтительные антигенсвязывающие молекулы по настоящему изобретению включают, например, антагонистические антигенсвязывающие молекулы. Такая антагонистическая антигенсвязывающая молекула, как правило, представляет собой антигенсвязывающую молекулу, которая ингибирует рецептор-опосредованный внутриклеточный путь передачи сигнала посредством блокирования связывания между лигандом (агонистом) и рецептором.
Между тем, антигенсвязывающая молекула по настоящему изобретению может распознавать любой антиген. Конкретно, антигены, распознаваемые антигенсвязывающей молекулой по настоящему изобретению, включают, например, описанные выше рецепторные белки (мембраносвязанные рецепторы и растворимые рецепторы), мембранные антигены, такие как маркеры клеточной поверхности и растворимые антигены, такие как цитокины. Такие антигены включают, например, антигены, описанные выше.
В предпочтительном варианте осуществления антигенсвязывающие молекулы по настоящему изобретению включают иммуноглобулины типа IgG (антитела IgG) с антигенсвязывающим доменом и FcRn-связывающим доменом человека. Если антитело IgG применяют в качестве антигенсвязывающей молекулы, тип не ограничен; и возможно использовать IgG1, IgG2, IgG3, IgG4 и подобные.
Происхождение антигенсвязывающих молекул по настоящему изобретению конкретно не ограничено, и они могут быть любого происхождения. Возможно использование, например, антител мыши, антител человека, антител крысы, антител кролика, антител козы, антител верблюда и других. Кроме того, антитела, например, могут представлять собой описанные выше химерные антитела, и в частности, измененные антитела с заменами аминокислотной последовательности, такие как гуманизированные антитела. Антитела могут представлять собой описанные выше биспецифические антитела, продукты модификации антител, с которыми сцеплены различные молекулы, полипептиды, включающие фрагменты антител, и антитела с модифицированными цепями сахаров.
Биспецифическое антитело относится к антителу, которое имеет в той же молекуле антитела вариабельные области, которые распознают различные эпитопы. Биспецифическое или полиспецифическое антитело может представлять собой антитело, которое распознает два или более различных антигенов, или антитело, которое распознает два или более различных эпитопов на одном и том же антигене.
Кроме того, полипептиды, включающие фрагменты антител включают, например, фрагменты Fab, фрагменты F(ab’)2, scFvs (Nat Biotechnol. 2005 Sep; 23(9): 1126-36), доменные антитела (dAb) (WO 2004/058821, WO 2003/002609), scFv-Fc (WO 2005/037989), dAb-Fc и слитые белки Fc. Fc-домен можно использовать в качестве FcRn-связывающего домена человека, если молекула включает Fc-домен. Альтернативно, FcRn-связывающий домен может быть слит с этими молекулами.
Дополнительно, антигенсвязывающие молекулы, которые применимы в настоящем изобретении, могут представлять собой антителоподобные молекулы. Антителоподобная молекула (каркасная молекула, пептидная молекула) представляет собой молекулу, которая может осуществлять функции посредством связывания с целевой молекулой (Current Opinion in Biotechnology (2006) 17: 653-658; Current Opinion in Biotechnology (2007) 18: 1-10; Current Opinion in Structural Biology (1997) 7: 463-469; Protein Science (2006) 15: 14-27) и включает, например, DARPins (WO 2002/020565), Аффитело (WO 1995/001937), Авимер (WO 2004/044011; WO 2005/040229) и Аднектин (WO 2002/032925). Если эти антителоподобные молекулы могут связывать целевую молекулу зависимым от pH способом и/или обладать активностью связывания с FcRn человека в диапазоне нейтральных значений рН, возможно облегчить поглощение антигена клетками посредством антигенсвязывающих молекул, способствовать снижению концентрация антигена в плазме посредством введения антигенсвязывающих молекул, улучшать фармакокинетику антигенсвязывающей молекулы и увеличивать количество антигенов, с которыми может связываться одна антигенсвязывающая молекула.
Кроме того, антигенсвязывающая молекула может представлять собой белок, полученный в результате слияния между FcRn-связывающим доменом человека и рецепторным белком, который связывается с мишенью, включающей лиганд и включает, например, слитые белки TNFR-Fc, слитые белки ILIR-Fc, слитые белки VEGFR-Fc и слитые белки CTLA4-Fc (Nat Med. 2003, Jan; 9(1): 47-52; BioDrugs. (2006) 20(3): 151-60). Если такие слитые белки рецептор-FcRn-связывающий домен человека связывают целевую молекулу, включающую лиганд, зависимым от pH способом, и/или обладают активностью связывания с FcRn человека в диапазоне нейтральных значений рН, возможно облегчить поглощение антигена клетками посредством антигенсвязывающих молекул, способствовать снижению концентрации антигена в плазме посредством введения антигенсвязывающей молекулы и улучшить фармакокинетику антигенсвязывающих молекул, и увеличить количество антигенов, с которыми может связываться одна антигенсвязывающая молекула. Рецепторный белок соответствующим образом конструируют и модифицируют для включения связывающего домена рецепторого белка с мишенью, включающей лиганд. Как изложено в примере выше, включающем слитые белки TNFR-Fc, слитые белки ILIR-Fc, слитые белки VEGFR-Fc и слитые белки CTLA4-Fc, молекула растворимого рецептора, содержащая внеклеточный домен таких рецепторных белков, которые необходимы для связывания с такими мишенями, включающими лиганды, являются предпочтительно используемыми в настоящем изобретении. Такую сконструированную и модифицированную рецепторную молекулу именуют в этой заявке как искусственный рецептор. Способ, используемый для конструирования и модифицирования рецепторной молекулы для конструирования искусственной рецепторной молекулы, известен в данной области.
Кроме того, антигенсвязывающая молекула может представлять собой слитый белок, в котором искусственный белок-лиганд, который связывается с мишенью и обладает нейтрализующим эффектом, подвергают слиянию с FcRn-связывающим доменом человека, и искусственные белки-лиганды включают, например, мутантный IL-6 (EMBO J. 1994 Dec 15; 13(24): 5863-70). Если такие искусственные слитые белки-лиганды могут связываться с целевыми молекулами зависимым от pH способом и/или обладают активностью связывания с FcRn человека в диапазоне нейтральных значений рН, то возможно облегчить поглощения антигена клетками посредством антигенсвязывающих молекул, способствовать снижению концентрации антигена в плазме посредством введения антигенсвязывающих молекул, улучшить фармакокинетику антигенсвязывающих молекул и увеличить количества антигенов, с которыми может связываться одна антигенсвязывающая молекула.
Кроме того, антитела по настоящему изобретению могут включать модифицированные цепи сахаров. Антитела с модифицированными цепями сахаров включают, например, антитела с модифицированным гликозилированием (WO 99/54342), антитела, которые лишены фукозы, которую присоединяют к цепи сахаров (WO 00/61739; WO 02/31140; WO 2006/067847; W02 006/067913), и антитела, разделенные GlcNAc (WO 02/79255).
Условия, используемые в анализе антигенсвязывающей активности или активности связывания с FcRn человека, отличные от pH, соответствующим образом могут выбрать специалисты в данной области, и условия конкретно не ограничены. Например, для определения активности можно использовать условия использования MES буфера при 37 градусов C, как описано в WO 2009/125825. Между тем, антигенсвязывающую активность и активность связывания с FcRn человека антигенсвязывающей молекулы можно определять способами, известными специалистам в данной области, например, с использованием Biacore (GE Healthcare) или подобных. Если антиген представляет собой растворимый антиген, активность антигенсвязывающей молекулы к связыванию растворимого антигена можно определять посредством нанесения антигена в качестве анализируемого вещества на чип с иммобилизованной антигенсвязывающей молекулой. Альтернативно, если антиген представляет собой антиген мембранного типа, активность антигенсвязывающей молекулы для связывания с антигеном мембранного типа можно определять посредством нанесения антигенсвязывающей молекулы в качестве анализируемого вещества на чип с иммобилизованным антигеном. Активность связывания с FcRn человека антигенсвязывающей молекулы можно определять посредством нанесения FcRn человека или антигенсвязывающей молекулы в качестве анализируемого вещества на чип с иммобилизованной антигенсвязывающей молекулой или FcRn человека, соответственно.
Генерация химерных антител известна. В случае химерного антитела человек-мышь, например, ДНК, кодирующую область V антитела, может быть сцеплена с ДНК, кодирующей область C антитела человека; эту конструкцию можно встраивать в экспрессирующий вектор и вводить в хозяина для получения химерного антитела.
«Гуманизированные антитела» также обозначают как преобразованные антитела человека, и они представляют собой антитела, в которых определяющая комплементарность область (CDR) от не принадлежащего человеку млекопитающего, например мыши, трансплантируют в CDR антитела человека. Способы идентификации CDR известны (Kabat et al., Sequence of proteins of Im munological Interest (1987), National Institute of Health, Bethesda, Md.; Chothia et al., Nature (1989) 342: 877). Общие технологии генетической рекомбинации, подходящие для этой цели, также известны (см. европейскую заявку на патент EP 125023; и WO 96/02576). Гуманизированные антитела можно получать известными способами, например, можно определять CDR антитела мыши и получают ДНК, кодирующую антитело, в котором CDR сцеплена с каркасной областью (FR) антитела человека. Гуманизированные антитела затем можно получать с использованием системы, которая использует традиционные экспрессирующие векторы. Такую ДНК можно синтезировать путем ПЦР, с использованием в качестве праймеров некоторые олигонуклеотиды, полученные таким образом, что они обладают областями, которые перекрываются с концевыми областями и CDR, и FR (см. способ, описанный в WO 98/13388). FR антитела человека, сцепленного с CDR, выбирают таким образом, что CDR формирует подходящий антигенсвязывающий участок. Если необходимо, аминокислоты в FR вариабельной области антитела можно изменять таким образом, что CDR преобразованного антитела человека могут формировать подходящий антигенсвязывающий участок (Sato et al., Cancer Res. (1993) 53: 10.01-6). Аминокислотные остатки в FR, которые можно изменять, включают участки, которые непосредственно связываются с антигеном посредством нековалентных связей (Amit et al., Science (1986) 233: 747-53), участки, которые оказывают влияние или имеют эффект на структуру CDR (Chothia et al., J. Mol. Biol. (1987) 196: 901-17), и участки, вовлеченные во взаимодействие VH-VL (EP 239400).
Если антигенсвязывающие молекулы по настоящему изобретению представляют собой химерные антитела или гуманизированные антитела, C-области этих антител предпочтительно получают от антитела человека. Например, C-гамма1, C-гамма2, C-гамма3 и C-гамма4 можно использовать для цепи H, тогда как C-каппа и C-лямбда можно использовать для цепи L. Кроме того, если необходимо, аминокислотные мутации можно вводить в область С антитела человека для усиления или уменьшения связывания с рецептором Fc-гамма или для увеличения стабильности антитела или продуктивности. Химерное антитело по настоящему изобретению предпочтительно включает вариабельную область антитела, полученного от не принадлежащего человеку млекопитающего, и константную область, полученного из антитела человека. Между тем, гуманизированное антитело предпочтительно включает CDR антитела, полученного из не принадлежащего человеку млекопитающего, и FR, и области C, полученные из антитело человека. Константные области, полученные из антитела человека, предпочтительно включают FcRn-связывающую область человека. Такие антитела включают, например, IgG (IgG1, IgG2, IgG3 и IgG4). Константные области, используемые для гуманизированных антител по настоящему изобретению, могут представлять собой константные области антител лобого изотипа. Константную область, полученную из IgG1 человека, предпочтительно используют, хотя константные области не ограничены ею. FR, полученные из антитела человека, которые применяют для гуманизированных антител, конкретно не ограничены и могут быть получены из антитела любого изотипа.
Вариабельные и константные области химерных и гуманизированных антител по настоящему изобретению можно изменять посредством делеции, замены, вставки и/или добавления и подобных, при условии, что исходные антитела демонстрируют специфичность связывания.
Так как иммуногенность в организме человека снижают, химерные и гуманизированные антитела с использованием происходящих от человека последовательностей, как предполагают, являются полезными при введении людям с терапевтическими и подобными целями.
Такие антигенсвязывающие молекулы по настоящему изобретению можно получать любым способом. Например, антигенсвязывающую молекулу, которая исходно не обладает активностью связывания с FcRn человека в диапазонах кислых и нейтральных значений рН, антигенсвязывающую молекулу, которая обладает более сильной антигенсвязывающей активностью в диапазоне кислых значений рН, чем в диапазоне нейтральных значений рН, или антигенсвязывающую молекулу, которая обладает сравнимой антигенсвязывающей активностью в диапазонах кислых или нейтральных значений рН, можно искусственным образом преобразовывать в антигенсвязывающую молекулу, обладающую желаемой активностью, посредством указанных выше аминокислотных изменений или подобных. Альтернативно, антитело, обладающее желаемой активностью, можно выбирать посредством скрининга из ряда антител, полученных из библиотеки антител или гибридом, описанных ниже.
Если изменяют аминокислоты в антигенсвязывающей молекуле, возможно использование известной последовательности для аминокислотной последовательности антигенсвязывающей молекулы до изменения или аминокислотной последовательности антигенсвязывающей молекулы, вновь индентифицированной посредством способов, известных специалистам в данной области. Например, если антигенсвязывающая молекула представляет собой антитело, ее можно получать из библиотеки антител или клонированием гена, кодирующего антитело, из гибридом, продуцирующих антитела.
Относительно библиотек антител, много библиотек антител уже известны, и способы получения библиотек антител также известны; таким образом, специалисты в данной области могут соответствующим образом получать библиотеки антител. Например, относительно фаговых библиотек, можно сослаться на литературу, такую как Clackson et al., Nature (1991) 352: 624-8; Marks et al., J. Mol. Biol. (1991) 222: 581-97; Waterhouses et al., Nucleic Acids Res. (1993) 21: 2265-6; Griffiths et al., EMBO J. (1994) 13: 324,0-60; Vaughan et al., Nature Biotechnology (1996) 14: 309-14; и публикация патента Японии Kohyo № (JP-A) H20-504970 (нерассмотренная публикация национальной фазы Японии, соответствующая международной публикации не в Японии). Кроме того, возможно использование известных способов, таких как способы с использованием эукариотический клеток в качестве библиотеки (WO 95/15393), и способы на основе рибосомного дисплея. Кроме того, технологии получения антител человека посредством пэннинга с использованием библиотек антител человека также известны. Например, вариабельные области антител человека можно экспрессировать на поверхности фагов в виде одноцепочечного антитела (scFvs) с использованием способов фагового дисплея, и можно выбирать фаги, которые связываются с антигенами. Генетическим анализом отобранных фагов можно определять последовательности ДНК, кодирующие вариабельные области антител человека, которые связываются с антигенами. При выявлении последовательностей ДНК scFvs, которые связываются с антигенами, на основе этих последовательностей можно получать подходящие экспрессирующие векторы для получения антител человека. Эти способы уже хорошо известны, и можно сослаться на WO 92/01047, WO 92/20791, WO 93/06213, WO 93/11236, WO 93/19172, WO 95/01438, и WO 95/15388.
Что касается способов получения генов, кодирующих антитела из гибридом, по существу можно использовать известные технологии, которые включают использование желаемых антигенов или клеток, экспрессирующих желаемые антигены в качестве сенсибилизирующих антигенов, используя их для иммунизаций согласно общепринятым способам иммунизации, слияние полученных в результате иммунных клеток с известными родительскими клетками посредством общепринятых способов слияния клеток, скрининга клеток, продуцирующих моноклональные антитела (гибридомы), общепринятыми способами скрининга, синтезированием кДНК вариабельных областей антитела (области V) из мРНК полученных гибридом с использованием обратной транскриптазы, и связывания их с ДНК, кодирующей требуемые константные области антител (C области).
Более конкретно, сенсибилизирующие антигены для получения описанных выше генов антигенсвязывающей молекулы, кодирующей H цепи и L цепи могут включать, например, и полные антигены с иммуногенностью и неполные антигены, включающие гаптены и т.п. без иммуногенности; однако антигены не ограничены этими примерами. Например, возможно использование целых белков и неполных пептидов представляющих интерес белков. Кроме того, известно, что средства, содержащие полисахариды, нуклеиновые кислоты, липиды и подобные, могут представлять собой антигены. Таким образом, антигены антигенсвязывающих молекул по настоящему изобретению конкретно не ограничены. Антигены можно получать способами, известными специалистам в данной области, например, способами на основе бакуловируса (например, WO 98/46777) и подобные. Гибридомы можно получать, например, способом Milstein et al. (G. Kohler and C. Milstein, Methods Enzymol. (1981) 73: 3-46) и подобным. Если иммуногенность антигена является низкой, иммунизацию можно проводить после сцепления антигена с макромолекулой, обладающей иммуногенностью, такой как альбумин. Альтернативно, если необходимо, антигены можно преобразовать в растворимые антигены посредством сцепления с другими молекулами. Если трансмембранные молекулы, такие как мембранные антигены (например, рецепторы), используют в качестве антигенов, части внеклеточных областей мембранных антигенов можно использовать в качестве фрагмента, или в качестве иммуногенов можно использовать клетки, экспрессирующие трансмембранные молекулы на своей клеточной поверхности.
Клетки, продуцирующие антигенсвязывающие молекулы, можно получать посредством иммунизации животных с использованием подходящих сенсибилизирующих антигенов, описанных выше. Альтернативно, клетки, продуцирующие антигенсвязывающие молекулы, можно получать иммунизацией лимфоцитов in vitro, которые продуцируют антигенсвязывающие молекулы. Для иммунизации можно использовать различных млекопитающих; так широко используемые животные включают грызунов, зайцеобразных и приматов. Такие животные включают, например, грызунов, таких как мыши, крысы и хомяки; зайцеобразных, таких как кролики; и приматов, включая обезьян, таких как яванские макаки, макаки-резус, бабуины и шимпанзе. Кроме того, иммунный репертуар трансгенных животных, несущих ген антитела человека, также известен, и антитела человека можно получать, используя таких животных (см. WO 96/34096; Mendez et al., Nat. Genet. (1997) 15: 146-56). Вместо использования таких трансгенных животных, например, желаемые антитела человека с активностью связывания против антигенов можно получать сенсибилизацией лимфоцитов человека in vitro требуемыми антигенами или клетками, экспрессирующими требуемые антигены, а затем слиянием сенсибилизированных лимфоцитов с миеломными клетками человека, такими как U266 (см. публикацию патентной заявкт Японии Kokoku № (JP-B) H01-59878 (изученная, одобренная патентная заявка Японии, опубликованная для опротестования)). Кроме того, требуемые антитела человека можно получать иммунизацией трансгенных животных, несущих полный репертуар генов антитела человека, желаемыми антигенами (см. WO 93/12227, WO 92/03918, WO 94/02602, WO 96/34096 и WO 96/33735).
Иммунизацию животных можно проводить, соответствующим образом разводя и суспендируя сенсибилизирующий антиген в фосфатно-солевом буфере (PBS), физиологическом растворе, или подобных, и смешиванием их с адъювантом для эмульсификации, если необходимо. Затем его интраперитонеально или подкожно инъецируют животным. Затем, сенсибилизирующий антиген, смешанный с неполным адъювантом Фрейнда, вводят предпочтительно несколько раз в течение 21 суток. Продукцию антител можно подтверждать посредством измерения титра представляющих интерес антител в сыворотке животных, используя общепринятые способы.
Клетки, продуцирующие антигенсвязывающие молекулы, полученные из лимфоцитов или животных, иммунизированных требуемым антигеном, можно подвергать слиянию с миеломными клетками для получения гибридом, используя общепринятые средства для слияния (например, полиэтиленгликоль) (Goding, Monoclonal antibody: Principles and Practice, Academic Press, (1986) 59-103). Если необходимо, гибридомные клетки можно культивировать и выращивать, и специфичность связывания антигенсвязывающей молекулы, полученной из таких гибридом, можно измерять с использованием известных способов анализа, таких как иммунопреципитация, радиоимунный анализ (RIA) и твердофазный иммуноферментный анализ (ELISA). Впоследствии, если необходимо, гибридомы, продуцирующие представляющие интерес антигенсвязывающие молекулы, чья специфичность, аффинность или активность определена, могут быть субклонированы способами, такими как предельное разведение.
Затем гены, кодирующие отобранные антигенсвязывающие молекулы, можно клонировать из гибридом или клеток, продуцирующих антигенсвязывающие молекулы (сенсибилизированные лимфоциты, и подобные), с использованием зондов, которые специфично связываются с антигенсвязывающими молекулами (например, олигонуклеотиды, комплиментарные последовательностям, кодирующим константные области антитела). Также возможно клонировать гены из мРНК, используя ОТ-ПЦР. Иммуноглобулины классифицируются на пять различных классов, IgA, IgD, IgE, IgG и IgM. Эти классы дополнительно подразделяются на несколько подклассов (изотипов) (например, IgG-1, IgG-2, IgG-3 и IgG-4; IgA-1 и IgA-2; и подобные). Цепи H и L, используемые в настоящем изобретении для получения антигенсвязывающих молекул, конкретно не ограничены и могут происходить из антител, принадлежащих к любому из этих классов или подклассов; однако, особенно предпочтительным является IgG.
В настоящем документе, возможно изменение генов, кодирующих Н цепь, и генов, кодирующих L цепь, с использованием методов генной инженерии. Генетически измененные антитела, такие как химерные антитела и гуманизированные антитела, которые искуственно изменены с целью снижения гетерологичной иммуногенности и подобной в отношении людей, можно соответствующим образом продуцировать для антител, таких как антитела мыши, крысы, кролика, хомяка, овцы и верблюда. Химерные антитела представляют собой антитела, включающие вариабельные области H и L цепей антитела не принадлежащего человеку млекопитающего, такого как антитело мыши, и константные области H и L цепей антитела человека. Химерные антитела можно получать, лигируя ДНК, кодирующую вариабельную область антитела мыши, с ДНК, кодирующей константную область антитела человека, встраивая указанное в экспрессирующий вектор, и вводя вектор в хозяина для получения антител. Гуманизированное антитело, которое также называют "преобразованное антитело человека", можно синтезировать посредством ПЦР с использованием некоторых олигонуклеотидов, полученных таким образом, что они имеют перекрывающиеся участки на концах последовательностей ДНК, предназначенных для сцепления с определяющими комплементарность областями (CDR) антитела не принадлежащего человеку млекопитающего, такого как мышь. Полученную в результате ДНК можно лигировать с ДНК, кодирующей константную область антитела человека. Лигированную ДНК можно встраивать в экспрессирующий вектор, и вектор можно вводить в хозяина, продуцирующего антитело (см. EP 239400 и WO 96/02576). FR антитела человека, которые лигируют с CDR, выбирают, если CDR образуют подходящий антигенсвязывающий участок. Если необходимо, аминокислоты в каркасной области вариабельной области антитела можно замещать таким образом, что CDR перестроенного антитела человека образует подходящий антигенсвязывающий участок (K. Sato et al., Cancer Res. (1993) 53: 10.01-10.06).
В дополнение к гуманизации, описанной выше, антитела можно изменять для усиления их биологических свойств, например, связывания с антигенами. В настоящем изобретении такие изменения могут быть достигнуты посредством способов, таких как сайт-специфический мутагенез (см. например, Kunkel (1910,0) Proc. Natl. Acad. Sci. USA 82: 488), мутагенез ПЦР и кассетный мутагенез. В основном, мутантные антитела, чьи биологические свойства улучшены, демонстрируют гомологию аминокислотной последовательности и/или сходство на 70% или более, более предпочтительно 80% или более, и даже более предпочтительно 90% или более (например, 95% или более, 97%, 98% или 99%), по сравнению с аминокислотной последовательностью вариабельной области исходной антитела. В настоящем документе, гомология последовательности и/или сходство определяют как отношение аминокислотных остатков, которые являются гомологичными (один и тот же остаток) или схожими (аминокислотные остатки, классифицируемые в одну и ту же группу, на основе основных свойств боковых цепей аминокислот) с остатками исходного антитела, затем для значения гомологии последовательности достигали максимума посредством выравнивания последовательностей и введением пропуска, если необходимо. В основном, природные аминокислотные остатки классифицируют в группы, основываясь на характеристиках их боковых цепей следующим образом:
(1) гидрофобные: аланин, изолейцин, валин, метионин и лейцин;
(2) нейтральные гидрофильные: аспарагин, глутамин, цистеин, треонин и серин;
(3) кислые: аспарагиновая кислота и глутаминовая кислота;
(4) основные: аргинин, гистидин и лизин;
(5) остатки, которые оказывают действие на ориентацию цепи: глицин и пролин; и
(6) ароматические: тирозин, триптофан и фенилаланин.
В основном, всего шесть определяющих комплементарность областей (CDR; гипервариабельные области), представленные в вариабельных областях H и L цепей, взаимодействуют друг с другом с образованием антигенсвязывающего участка антитела. Как известно, вариабельная область сама по себе также является способной распознавать и связывать антиген, хотя ее аффинность является более низкой, чем аффинность целого участка связывания. Таким образом, гены, кодирующие H и L цепи по настоящему изобретению, могут кодировать каждый фрагмент, включая антигенсвязывающий участок H или L цепи, при условии, что полипептид, кодируемый геном, сохраняет активность связывания с желаемым антигеном.
Как описано выше, вариабельная область тяжелой цепи в основном состоит из трех CDR и четырех FR. В предпочтительном варианте осуществления настоящего изобретения, аминокислотные остатки, подлежащие «изменению», могут быть соответствующим образом выбраны из аминокислотных остатков, например, в CDR или FR. В основном, изменения аминокислотных остатков в CDR могут снижать антигенсвязывающую способность. Таким образом, подходящие аминокислотные остатки подлежащие «изменению» в настоящем изобретении предпочтительно выбирают из аминокислотных остатков в FR, но конкретно ими не ограничены. Возможно выбирать аминокислоты в CDR при условии, что подтверждено, что изменения не снижают связывающую активность. Альтернативно, при использовании общедоступных баз данных или подобных, специалисты в данной области могут получать подходящие последовательности, которые можно использовать в качестве FR вариабельной области антитела организма, такого как человек или мышь.
Кроме того, настоящее изобретение относится к генам, кодирующим антигенсвязывающие молекулы по настоящему изобретению. Гены, кодирующие антигенсвязывающие молекулы по настоящему изобретению могут представлять собой любые гены и могут представлять собой ДНК, РНК, аналоги нуклеиновых кислот или т.п.
Кроме того, настоящее изобретение также относится к клеткам-хозяевам, несущим гены, описанные выше. Клетки-хозяева конкретно не ограничены и включают, например, E. coli и клетки различных животных. Клетки-хозяева можно использовать, например, в качестве системы продукции для получения и экспрессии антител по настоящему изобретению. Системы продукции in vitro и in vivo являются доступными для систем продукции полипептидов. Такие системы продукции in vitro включают, например, системы продукции с использованием эукариотических или прокариотических клеток.
Эукариотические клетки, которые можно использовать в качестве клеток-хозяев включают, например, клетки животных, растительные клетки и клетки грибов. Клетки животных включают: клетки млекопитающих, например, CHO (J. Exp. Med. (1995) 108: 94,0), COS, HEK293, 3T3, миеломы, BHK (почки детенышей хомячка), HeLa и Vero; клетки амфибий, такие как ооциты Xenopus laevis (VaIle et al., Nature (1981) 291: 338-340); и клетки насекомых, такие как Sf9, Sf21 и Tn5. CHO-DG44, CHO-DX11B, клетки COS7, клетки HEK293 и клетки BHK предпочтительно используют для экспрессии антител по настоящему изобретению. Среди клеток животных, конкретно клетки CHO являются предпочтительными для экспрессии в крупном масштабе. Векторы можно вводить в клетки-хозяева, например, посредством способов с фосфатом кальция, способов с DEAE-декстраном, способов с использованием катионной липосомы DOTAP (Boehringer-Mannheim), способов электропорации и способов липофекции.
Относительно растительных клеток, например, клетки, полученные из Nicotiana tabacum и ряски (Lemna minor), известны в качестве системы продукции белка. Для получения антигенсвязывающих молекул по настоящему изобретению из этих клеток можно культивировать каллюсы. Относительно клеток грибов, известные белковые экспрессирующие системы представляют собой те, в которых используют дрожжевые клетки, например, клетки вида Saccharomyces (такие как Saccharomyces cerevisiae и Saccharomyces pombe); и клетки нитевидных грибов, например, вида Aspergillus (такие как Aspergillus niger). Эти клетки можно использовать в качестве хозяина для получения антигенсвязывающих молекул по настоящему изобретению.
Бактериальные клетки можно использовать в прокариотических системах продукции. Относительно бактериальных клеток, системы продукции с использованием Bacillus subtilis известны в дополнение к системам продукции с использованием E. Coli, описанных выше. Такие системы можно использовать в получении антигенсвязывающих молекул по настоящему изобретению.
<Способы скрининга>
Настоящее изобретение относится к способам скрининга антигенсвязывающих молекул, обладающих активностью связывания с FcRn человека в диапазонах кислых и нейтральных значений рН. Настоящее изобретение также относится к способам скрининга антигенсвязывающих молекул, которые обладают активностью связывания с FcRn человека в диапазонах кислых и нейтральных значений рН и более низкой антигенсвязывающей активностью в диапазоне кислых значений рН, чем в диапазоне нейтральных значений рН. Настоящее изобретение также относится к способам скрининга антигенсвязывающих молекул, способных облегчать поглощение антигена клетками. Настоящее изобретение также относится к способам скрининга антигенсвязывающих молекул, которые модифицированы таким образом, что являются способными к связыванию с большим количеством антигенов на молекулу. Настоящее изобретение также относится к способам скрининга антигенсвязывающих молекул, способных облегчать элиминацию антигена. Настоящее изобретение дополнительно относится к способам скрининга антигенсвязывающих молекул с улучшенной фармакокинетикой. Настоящее изобретение также относится к способам скрининга антигенсвязывающих молекул с увеличенной внутриклеточной диссоциацией от связанного с ними антигена за пределами клеток. Настоящее изобретение также относится к способам скрининга антигенсвязывающих молекул с увеличенным внеклеточным высвобождением в форме, свободной от антигена, после поглощения клетками в форме, связанной с антигеном. Настоящее изобретение дополнительно относится к способам скрининга антигенсвязывающих молекул, которые являются особенно пригодными в качестве фармацевтических композиций. Описанные выше способы пригодны для скрининга антигенсвязывающих молекул, которые являются особенно превосходными по времени удержания и обладают превосходной способностью к элиминации антигенов из плазмы.
Конкретно, настоящее изобретение относится к способам скрининга антигенсвязывающих молекул, где способы включают следующие стадии:
(a) отбора антигенсвязывающей молекулы, которая обладает более высокой активностью связывания с FcRn человека в диапазоне нейтральных значений рН, чем таковая до изменения по меньшей мере одной аминокислоты в FcRn-связывающем домене человека антигенсвязывающей молекулы, обладающей активностью связывания с FcRn человека в диапазоне кислых значений рН; и
(b) изменения по меньшей мере одной аминокислоты в антигенсвязывающем домене антигенсвязывающей молекулы и отбора антигенсвязывающей молекулы, которая обладает более высокой антигенсвязывающей активностью в диапазоне нейтральных значений рН, чем в диапазоне кислых значений рН.
Стадии (a) и (b) можно проводить в любом порядке. Кроме того, каждую стадию можно повторять дважды или более раз. Число повторов для стадий (a) и (b) конкретно не ограничено; однако, число, как правило, составляет десять раз или менее.
В способах скрининга по настоящему изобретению антигенсвязывающая активность антигенсвязывающей молекулы в диапазоне нейтральных значений рН конкретно не ограничена при условии, что она представляет собой антигенсвязывающую активность в диапазоне pH от 6,7 до 10,0. Например, включают варианты осуществления, описанные в WO 2009/125825. Предпочтительные активности связывания с антигеном включают антигенсвязывающую активность в диапазоне pH от 7,0 до 8,0. Более предпочтительная активность связывания с антигеном включает антигенсвязывающую активность при pH 7,4. Между тем, антигенсвязывающая активность антигенсвязывающих молекул в диапазоне кислых значений рН конкретно не ограничена, при условии, что она представляет собой антигенсвязывающую активность в диапазоне pH от 4,0 до 6,5. Предпочтительная активность связывания с антигеном включает антигенсвязывающую активность в диапазоне pH 5,5 и 6,5. Более предпочтительная активность связывания с антигеном включает антигенсвязывающую активность при pH 5,8 или pH 5,5.
Активность связывания с FcRn человека антигенсвязывающей молекулы в диапазоне нейтральных значений рН конкретно не ограничена, при условии, что она представляет собой активность связывания с FcRn человека в диапазоне pH от 6,7 до 10,0. Предпочтительная активность связывания с FcRn человека включает активность связывания с FcRn человека в диапазоне pH от 7,0 до 8,0. Более предпочтительная активность связывания с FcRn человека включает активность связывания с FcRn человека при pH 7,4.
Активность связывания с FcRn человека антигенсвязывающих молекул в диапазоне кислых значений рН конкретно не ограничена, при условии, что она представляет собой активность связывания с FcRn человека в диапазоне pH от 4,0 до 6,5. Предпочтительная активность связывания с FcRn человека включает активность связывания с FcRn человека в диапазоне pH от 5,5 до 6,5. Более предпочтительная активность связывания с FcRn человека включает активность связывания с FcRn человека в диапазоне pH 5,8 до 6,0.
В настоящем документе, диапазон кислых значений рН, как правило, относится к pH от 4,0 до pH 6,5. Предпочтительно, диапазон кислых значений рН представляет собой диапазон, представленный любым значением pH в пределах pH от 5,5 до pH 6,5, предпочтительно выбранным из 5,5, 5,6, 5,7, 5,8, 5,9, 6,0, 6,1, 6,2, 6,3, 6,4 или 6,5, особенно предпочтительно значение pH от 5,8 до pH 6,0, которое представляет собой значение, близкое к рН в ранней эндосоме in vivo. Между тем, в настоящем документе диапазон нейтральных значений рН, как правило, относится к pH от 6,7 до pH 10,0. Диапазон нейтральных значений рН предпочтительно представляет собой диапазон, представленный любым значением pH в пределах pH от 7,0 до pH 8,0, предпочтительно выбранным из pH 7,0, 7,1, 7,2, 7,3, 7,4, 7,5, 7,6, 7,7, 7,8, 7,9 и 8,0, более предпочтительно pH 7,4, которое представляет собой значение, близкое к рН плазмы (крови) in vivo. pH 7,0 можно использовать в качестве альтернативы pH 7,4, если является затруднительным оценить аффинность связывания между FcRn-связывающим доменом человека и FcRn человека вследствие низкой аффинности последнего при pH 7,4. Так как температуру использовали в условиях анализа, аффинность связывания между FcRn-связывающим доменом человека и FcRn человека можно оценивать при любой температуре от 10 градусов C до 50 градусов C. Предпочтительно, температуру от 15 градусов C до 40 градусов C используют для определения аффинности связывания между FcRn-связывающим доменом человека и FcRn человека. Более предпочтительно, любую температуру от 20 градусов C до 35 градусов C, такую как любое значение из 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 и 35 градусов C также используют для определения аффинности связывания между FcRn-связывающим доменом человека и FcRn человека. Температура 25 градусов C, описанная в примере 5, представляет собой один из примеров варианта осуществления настоящего изобретения.
Антигенсвязывающую активность и активность связывания с FcRn человека антигенсвязывающих молекул можно определять способами, известными специалистам в данной области. Подходящие условия, наряду с pH, могут выбрать специалисты в данной области. Антигенсвязывающую активность и активность связывания с FcRn человека антигенсвязывающих молекул можно оценивать, используя KD (константу диссоциации), кажущуюся KD (кажущуюся константу диссоциации), скорость диссоциации kd (скорость диссоциации), кажущуюся kd (кажущаяся диссоциация: кажущаяся скорость диссоциации) или т.п. Их можно определять способами, известными специалистам в данной области, например, с использованием Biacore (GE Healthcare), графика Скетчарда, проточного цитометра или подобных.
Согласно Journal of Immunology (2009) 182: 7663-7671, активность связывания с FcRn человека интактного IgG1 человека представляет собой KD 1,7 микромолей в диапазоне кислых значений рН (pH 6,0), в то время как активность является почти недетектируемой в диапазоне нейтральных значений рН. Таким образом, в предпочтительном варианте осуществления можно подвергать скринингу антигенсвязывающие молекулы по настоящему изобретению, которые обладают активностью связывания с FcRn человека в диапазонах кислых и нейтральных значений рН, включая антигенсвязывающие молекулы, которые обладают активностью связывания с FcRn человека с KD, равной 20 микромолям или больше в диапазоне кислых значений рН, которая является равной или большей, чем таковая интактного IgG человека в диапазоне нейтральных значений рН. В более предпочтительном варианте осуществления можно подвергать скринингу антигенсвязывающие молекулы по настоящему изобретению, включающие антигенсвязывающие молекулы, чья активность связывания с FcRn человека представляет собой KD, равную 2,0 микромолям или больше в диапазоне кислых значений рН, и KD, равную 40 микромолям или больше в диапазоне нейтральных значений рН. В еще более предпочтительном варианте осуществления можно подвергать скринингу антигенсвязывающие молекулы по настоящему изобретению, включающие антигенсвязывающие молекулы, чья активность связывания с FcRn человека представляет собой значение KD, равное 0,5 микромолям или больше в диапазоне кислых значений рН, и KD, равную 15 микромолям или больше в диапазоне нейтральных значений рН. Значения вышеуказанной KD определяют способом, описанным в Journal of Immunology (2009) 182: 7663-7671 (посредством иммобилизации антигенсвязывающей молекулы на чипе и нанесении FcRn человека в качестве анализируемого вещества).
Настоящее изобретение относится к способу скрининга антигенсвязывающей молекулы, где способ включает стадии:
(a) отбора антигенсвязывающей молекулы, которая обладает более высокой активностью связывания с FcRn человека в диапазоне нейтральных значений рН, чем KD, равная 3,2 микромолей, полученной посредством изменения по меньшей мере одной аминокислоты в FcRn-связывающем домене человека антигенсвязывающей молекулы,
(b) получения гена, кодирующего антигенсвязывающую молекулу, в которой FcRn-связывающий домен человека и антигенсвязывающий домен, полученные на стадии (a), сцеплены; и
(c) получения антигенсвязывающей молекулы с использованием гена, полученного в (b).
В одном из вариантов осуществления можно подвергать скринингу антигенсвязывающую молекулу, содержащую антигенсвязывающий домен и FcRn-связывающий домен человека, который обладает активностью связывания с FcRn человека в диапазонах кислых и нейтральных значений рН, где активность связывания с FcRn человека и более низкая антигенсвязывающая активность в диапазоне кислых значений рН, чем в диапазоне нейтральных значений рН, является большей, чем KD, равная 3,2 микромолям, согласно способам, используемым специалистами в данной области, как описано в настоящем документе ранее. В более предпочтительном варианте осуществления активность связывания с FcRn человека при pH 7,0 и при 25 градусов C является более высокой, чем KD, являющаяся 3,2 микромолей.
Настоящее изобретение относится к способу скрининга антигенсвязывающей молекулы, содержащей антигенсвязывающий домен и FcRn-связывающий домен человека, которая обладает активностью связывания с FcRn человека в диапазоне нейтральных значений pH, где активность связывания с FcRn человека в диапазоне нейтральных значений рН является более высокой, чем KD, равная 2,3 микромолям. Настоящее изобретение также относится к способу скринига антигенсвязывающей молекулы, содержащей антигенсвязывающий домен и FcRn-связывающий домен человека, которая обладает активностью связывания с FcRn человека в диапазоне нейтральных значений pH, где активность связывания с FcRn человека в диапазоне нейтральных значений рН является в 38 раз более высокой, чем для интактного IgG человека.
Антигенсвязывающие молекулы по настоящему изобретению, обладающие активностью связывания с FcRn человека в диапазоне нейтральных значений рН, конкретно не ограничены, при условии, что они обладают активностью связывания с FcRn человека при pH от 6,7 до 10,0. Однако, активность связывания с FcRn человека при pH от 6,7 до 10,0 предпочтительных антигенсвязывающих молекул является более высокой, чем таковая интактного IgG человека.
Антигенсвязывающие молекулы по настоящему изобретению, обладающие активностью связывания с FcRn человека в диапазоне кислых значений рН, конкретно не ограничены, при условии, что они обладают активностью связывания с FcRn человека при pH от 4,0 до 6,5. Однако, активность связывания с FcRn человека при pH от 5,5 до 6,5 предпочтительных антигенсвязывающих молекул является сравнимой или более высокой, чем таковая интактного IgG человека.
В настоящем документе, стадия отбора антигенсвязывающей молекулы, которая обладает более высокой антигенсвязывающей активностью в диапазоне нейтральных значений рН, чем в диапазоне кислых значений рН является аналогичной стадии отбора антигенсвязывающей молекулы, которая обладает более низкой антигенсвязывающей активностью в диапазоне кислых значений рН, чем в диапазоне нейтральных значений рН.
Отношение антигенсвязывающей активности между диапазонами нейтральных и кислых значений pH конкретно не ограничено при условии, что антигенсвязывающая активность в диапазоне нейтральных значений рН, является более высокой, чем в диапазоне кислых значений рН. Однако, антигенсвязывающая активность при pH от 6,7 до 10,0 предпочтительно в два или более раза, более предпочтительно в десять или более раз, и еще более предпочтительно в 40 или более раз превышает антигенсвязывающую активность при pH от 4,0 до 6,5.
В способах скрининга по настоящему изобретению возможно использовать библиотеки, такие как фаговые библиотеки.
В способах по настоящему изобретению, антиген и антигенсвязывающая молекула могут связываться вместе в любом состоянии, и таким образом состояние конкретно не ограничено. Например, антиген может взаимодействовать с иммобилизованной антигенсвязывающей молекулой для достижения их связывания. Альтернативно, антигенсвязывающая молекула может взаимодействовать с иммобилизованным антигеном для достижения их связывания. Альтернативно, антигенсвязывающая молекула и антиген могут взаимодействовать друг с другом в растворе для достижения их связывания.
Антигенсвязывающие молекулы, подлежащие скринингу посредством способов скрининга по настоящему изобретению, можно получать любым способом. Например, возможно использовать библиотеки предсуществующих антител, предсуществующих антигенсвязывающих доменов (фаговые библиотеки, и т.д.), библиотеки антител или антигенсвязывающих доменов, полученные из B-клеток иммунизированных животных, или гибридом, полученных посредством иммунизирования животных, библиотеки антител или антигенсвязывающих доменов, полученные посредством введения случайных аминокислотных изменений в описанные выше библиотеки антител или антигенсвязывающих доменов, библиотеки антител или антигенсвязывающих доменов, с введенными мутациями гистидина или неприродной аминокислоты (библиотеки с высоким содержанием гистидина или неприродной аминокислоты, библиотеки антигенсвязывающих доменов, с введенной мутацией гистидина или неприродной аминокислоты мутации в конкретные участки и т.д.), или т.п.
Антигенсвязывающие молекулы, которые связываются с антигеном много раз, которые, таким образом, являются более усовершенствованными по значению времени удержания в плазме, можно получать посредством способа скринига по настоящему изобретению. Таким образом, способы скрининга по настоящему изобретению можно использовать в качестве способов скрининга для получения антигенсвязывающих молекул, которые являются более усовершенствованными по значению времени удержания в плазме.
Кроме того, антигенсвязывающие молекулы, которые могут связываться с антигеном два или более раз, при введении животным, таким как люди, мыши или обезьяны, можно получать способами скрининга по настоящему изобретению. Таким образом, способы скрининга по настоящему изобретению можно использовать в качестве способов скрининга для получения антигенсвязывающих молекул, которые могут связываться с антигеном два или более раз.
Кроме того, антигенсвязывающие молекулы, которые способны к связыванию с большим количеством антигенов по сравнению с числом из антигенсвязывающих участков, при введении животным, таким как люди, мыши или обезьяны, можно получать способами скрининга по настоящему изобретению. Таким образом, способы скрининга по настоящему изобретению можно использовать в качестве способов скрининга для получения антигенсвязывающих молекул, которые способны к связыванию большего количества антигенов по сравнению с количеством их антигенсвязывающих участков. Например, если антитело представляет собой нейтрализующее антитело, способы скрининга по настоящему изобретению можно использовать в качестве способов скрининга для получения антигенсвязывающих молекул, которые могут нейтрализовать большее количество антигенов по сравнению с количеством антигенсвязывающих участков антигенсвязывающих молекул.
Кроме того, антигенсвязывающие молекулы, которые способны к диссоциации в пределах клетки от внеклеточно связанного антигена при введении животным, таким как люди, мыши или обезьяны, можно получать способами скрининга по настоящему изобретению.
Таким образом, способы скрининга по настоящему изобретению можно использовать в качестве способов скрининга для получения антигенсвязывающих молекул, которые способны к диссоциации в пределах клетки от внеклеточно связанного антигена.
Кроме того, антигенсвязывающие молекулы, которые связываются с антигеном и поглощаются клетками и высвобождаются за пределами клеток в форме, свободной от антигена, при введении животным, таким как люди, мыши или обезьяны можно получать посредством способов скрининга по настоящему изобретению. Таким образом, способы скрининга по настоящему изобретению можно использовать в качестве способов скрининга для получения антигенсвязывающих молекул, которые связываются с антигеном и поглощаются клетками и высвобождаются за пределами клеток в свободной от антигена форме.
Кроме того, антигенсвязывающие молекулы, которые могут быстро элиминировать антигены в плазме при введении животным, таким как люди, мыши или обезьяны, можно получать способами скрининга по настоящему изобретению. Таким образом, способы скрининга по настоящему изобретению можно использовать в качестве способов скрининга для получения антигенсвязывающих молекул с повышенной (высокой) способностью к элиминации антигенов в плазме.
Кроме того, такие антигенсвязывающие молекулы, как ожидают, являются особенно превосходными в качестве лекарственных средств, так как можно снижать дозу и частоту введения пациентам, и как результат, можно снижать общую дозу. Таким образом, способы скрининга по настоящему изобретению можно использовать в качестве способов скрининга антигенсвязывающих молекул для применения в качестве фармацевтических композиций.
<Способы получения антигенсвязывающих молекул>
Настоящее изобретение относится к способам получения антигенсвязывающих молекул, которые обладают активностью связывания с FcRn человека при эндосомальном pH и pH плазмы, и более низкой антигенсвязывающей активностью при эндосомальном pH, чем при pH плазмы. Настоящее изобретение также относится к способам получения антигенсвязывающих молекул, которые являются более совершенными в фармакокинетике и в ускорении снижения концентрации антигена в плазме при введении. Настоящее изобретение также относится к способам получения антигенсвязывающих молекул, которые являются особенно пригодными при использовании в качестве фармацевтических композиций.
Конкретно, настоящее изобретение относится к способам получения антигенсвязывающих молекул, где способ включает стадии:
(a) отбора антигенсвязывающей молекулы, которая обладает более высокой активностью связывания с FcRn человека в диапазоне нейтральных значений рН, чем таковая до изменения по меньшей мере одной аминокислоты в FcRn-связывающем домене человека антигенсвязывающей молекулы, обладающей активностью связывания с FcRn человека в диапазоне кислых значений рН;
(b) изменения по меньшей мере одной аминокислоты в антигенсвязывающем домене антигенсвязывающей молекулы и отбора антигенсвязывающей молекулы, которая обладает более высокой антигенсвязывающей активностью в диапазоне нейтральных значений рН, чем в диапазоне кислых значений рН;
(c) получения гена, кодирующего антигенсвязывающую молекулу, в которой FcRn-связывающий домен человека и антигенсвязывающий домен, полученные на стадиях (a) и (b), сцеплены; и
(d) получения антигенсвязывающей молекулы с использованием гена, полученного на стадии (c).
Стадии (a) и (b) можно осуществлять в любом порядке. Кроме того, каждую стадию можно повторять дважды или более. Число повторов для стадий (a) и (b) конкретно не ограничено; однако, число, как правило, составляет десять раз или менее.
Линкер, функционально связывающий FcRn-связывающий домен человека и антигенсвязывающий домен, полученные на стадиях (a) и (b), не ограничен какой-либо формой. FcRn-связывающий домен человека и антигенсвязывающий домен могут быть связаны посредством либо ковалентных, либо нековалентных связей. В частности, линкер может представлять собой пептидный линкер или химический линкер или связывающие пары, подобные комбинации биотина и стрептавидина. Модификация полипептида, включающая FcRn-связывающий домен человека и антигенсвязывающий домен, известна в данной области. В другом варианте осуществления FcRn-связывающий домен человека и антигенсвязывающий домен по настоящему изобретению можно связывать посредством образования слитого белка между FcRn-связывающим доменом человека и антигенсвязывающим доменом. Для конструирования слитого белка между FcRn-связывающим доменом человека и антигенсвязывающим доменом, гены, кодирующие FcRn-связывающий домен человека и антигенсвязывающий домен, могут быть функционально связаны таким образом, чтобы сформировать слитый полипептид в рамке. Соответствующим образом, линкер, содержащий пептид, состоящий из нескольких аминокислот, можно встраивать между FcRn-связывающим доменом человека и антигенсвязывающим доменом. Различные гибкие линкеры, такие как линкер, чья последовательность состоит из (GGGGS)n,известны в данной области.
Антигенсвязывающие молекулы, которые применяют в способах получения по настоящему изобретению, можно получать посредством любого способа. Например, возможно использование предсуществующих антител, предсуществующих библиотек (фаговые библиотеки и т.п.), антител и библиотек, которые получают из гибридом, полученных посредством иммунизации животных или из B-клеток иммунизированных животных, антител и библиотек, полученных посредством введения случайных аминокислотных изменений в описанные выше антитела и библиотеки, антител и библиотек, полученных посредством введения гистидина или неприродных аминокислотных мутаций в описанные выше антитела, и библиотеки (библиотеки с высоким содержанием гистидина или неприродной аминокислоты, библиотеки с введенным гистидином или неприродной аминокислотой в конкретные участки и т.п.) и подобных.
В описанных выше способах получения, активность связывания с FcRn человека антигенсвязывающих молекул в диапазоне нейтральных значений рН конкретно не ограничена при условии, что она представляет собой активность связывания с FcRn человека в диапазоне pH от 6,7 до 10,0. Предпочтительная активность связывания с FcRn человека включает активность связывания с FcRn человека в диапазоне pH от 7,0 до 8,0. Более предпочтительная активность связывания с FcRn человека включает активность связывания с FcRn человека при pH 7,4.
Активность связывания с FcRn человека антигенсвязывающей молекулы в диапазоне кислых значений рН конкретно не ограничена, при условии, что она представляет собой активность связывания с FcRn человека в диапазоне pH от 4,0 до 6,5. Предпочтительная активность связывания с FcRn человека включает активность связывания с FcRn человека в диапазоне pH от 5,5 до 6,5. Более предпочтительная активность связывания с FcRn человека включает активность связывания с FcRn человека при pH 6,0.
В настоящем документе, диапазон кислых значений рН, как правило, относится к pH от 4,0 до pH 6,5. Диапазон кислых значений рН предпочтительно представляет собой диапазон, представленный любым значением pH в пределах pH от 5,5 до pH 6,5, предпочтительно выбранный из 5,5, 5,6, 5,7, 5,8, 5,9, 6,0, 6,1, 6,2, 6,3, 6,4 и 6,5, более предпочтительно pH от 5,8 до pH 6,0, что представляет собой значение, близкое к значению pH в ранней эндосоме in vivo. Между тем, в настоящем документе диапазон нейтральных значений рН, как правило, относится к pH от 6,7 до pH 10,0. Диапазон нейтральных значений рН предпочтительно представляет собой диапазон, представленный любым значением pH в пределах pH от 7,0 до pH 8,0, предпочтительно выбранный из pH 7,0, 7,1, 7,2, 7,3, 7,4, 7,5, 7,6, 7,7, 7,8, 7,9 и 8,0, более предпочтительно pH 7,4, что представляет собой значение, близкое к значению рН плазмы (крови) in vivo. pH 7,0 можно использовать в качестве альтернативы pH 7,4, если является затруднительным оценить аффинность связывания между FcRn-связывающим доменом человека и FcRn человека вследствие низкой аффинности последнего при pH 7,4. Так как температуру используют в условиях анализа, аффинность связывания между FcRn-связывающим доменом человека и FcRn человека можно оценивать при любой температуре от 10 градусов C до 50 градусов C. Предпочтительно, температура от 15 градусов C до 40 градусов C используют для определения аффинности связывания между FcRn-связывающим доменом человека и FcRn человека. Более предпочтительно, любую температуру от 20 градусов C до 35 градусов C, такую как одна из 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 и 35 градусов C, так же используют для определения аффинности связывания между FcRn-связывающим доменом человека и FcRn человека. Температура 25 градусов C, описанная в примере 5, представляет собой один из примеров для варианта осуществления настоящего изобретения.
Настоящее изобретение относится к способу получения антигенсвязывающей молекулы, где способ содержит стадии:
(a) отбора антигенсвязывающей молекулы, которая обладает более высокой активностью связывания с FcRn человека в диапазоне нейтральных значений рН, чем KD, равная 3,2 микромолей, где молекула получена посредством изменения по меньшей мере одной аминокислоты в FcRn-связывающем домене человека антигенсвязывающей молекулы;
(b) получения гена, кодирующего антигенсвязывающую молекулу, в которой FcRn-связывающий домен человека и антигенсвязывающий домен, полученные на стадии (a), сцеплены; и
(c) получения антигенсвязывающей молекулы с использованием гена, полученного на стадии (b).
В предпочтительном варианте осуществления можно получать антигенсвязывающие молекулы по настоящему изобретению, обладающие активностью связывания с FcRn человека в диапазонах кислых и нейтральных значений рН, включающие антигенсвязывающие молекулы, которые обладают активностью связывания с FcRn человека со значением KD, равным 20 микромолей или более высоким в диапазоне кислых значений рН, которое равно или более высокое, чем таковое интактного IgG человека в диапазоне нейтральных значений рН. В более предпочтительном варианте осуществления можно также получать антигенсвязывающие молекулы по настоящему изобретению, включающие антигенсвязывающие молекулы, чья активность связывания с FcRn человека представляет собой KD, равную 2,0 микромолей или более высокую в диапазоне кислых значений рН, и KD, равную 40 микромолей или более высокой в диапазоне нейтральных значений рН. В еще более предпочтительном варианте осуществления можно предпочтительно получать антигенсвязывающие молекулы по настоящему изобретению, включающие антигенсвязывающие молекулы, чья активность связывания с FcRn человека представляет собой KD, равную 0,5 микромолям или более высокую в диапазоне кислых значений рН, и KD, равную 15 микромолям или более высокую в диапазоне нейтральных значений рН. Значения KD, указанные выше, определяют способом, описанным в Journal of Immunology (2009) 182: 7663-7671 (посредством иммобилизации антигенсвязывающей молекулы на чипе и нанесения FcRn человека в качестве анализируемого вещества). В одном из вариантов осуществления антигенсвязывающая молекула, содержащая антигенсвязывающий домен и FcRn-связывающий домен человека, где антигенсвязывающая молекула обладает активностью связывания с FcRn человека в диапазонах кислых и нейтральных значений рН, где активность связывания с FcRn человека и более низкая антигенсвязывающая активность в диапазоне кислых значений рН, чем в диапазоне нейтральных значений рН, является более высокой, чем KD, равная 3,2 микромолям, можно получать согласно способам, используемых специалистами в данной области, как описано в настоящем документе выше. В более предпочтительном варианте осуществления активность связывания с FcRn человека полученных таким образом антигенсвязывающих молекул при pH 7,0 и 25 градусов C является более высокой, чем KD, равная 3,2 микромолям.
Настоящее изобретение относится к способу получения антигенсвязывающей молекулы, содержащей антигенсвязывающий домен и FcRn-связывающий домен человека, которая обладает активностью связывания с FcRn человека в диапазоне нейтральных значений pH, где активность связывания с FcRn человека в диапазоне нейтральных значений рН является более высокой, чем KD, равная 2,3 микромолям. Настоящее изобретение также относится к способу получения антигенсвязывающей молекулы, содержащей антигенсвязывающий домен и FcRn-связывающий домен человека, которая обладает активностью связывания с FcRn человека в диапазоне нейтральных значений pH, где активность связывания с FcRn человека в диапазоне нейтральных значений рН является в 38 раз более высокой, чем у интактного IgG человека.
В описанных выше способах получения, антигенсвязывающая активность антигенсвязывающей молекулы в диапазоне нейтральных значений рН конкретно не ограничена, при условии, что антигенсвязывающая активность представляет собой антигенсвязывающую активность при pH между pH 6,7 и pH 10,0 и включает, например, вариант осуществления, описанный в WO 2009/125825. Предпочтительная антигенсвязывающая активность представляет собой антигенсвязывающую активность при pH между значениями pH 7,0 и pH 8,0, и более предпочтительная антигенсвязывающая активность представляет собой антигенсвязывающую активность при pH 7,4. Альтернативно, антигенсвязывающая активность антигенсвязывающей молекулы в диапазоне кислых значений рН конкретно не ограничена, при условии, что антигенсвязывающая активность представляет собой антигенсвязывающеую активность при pH между значениями pH 4,0 и pH 6,5. Предпочтительная антигенсвязывающая активность представляет собой антигенсвязывающую активность при pH между значениями pH от 5,5 до pH 6,5, и более предпочтительная антигенсвязывающая активность представляет собой антигенсвязывающую активность при pH 5,8 или pH 5,5.
Антигенсвязывающую активность и активность связывания с FcRn человека антигенсвязывающей молекулы можно определять способами, известными специалистам в данной области. Условия, помимо pH, могут соответствующим образом определить специалисты в данной области.
В способах получения по настоящему изобретению, антигенсвязывающие молекулы, обладающие активностью связывания с FcRn человека в диапазоне нейтральных значений рН, конкретно не ограничены при условии, что они обладают активностью связывания с FcRn человека при pH от 6,7 до 10,0. Однако активность связывания с FcRn человека антигенсвязывающих молекул при pH от 6,7 до 10,0 предпочтительно является более высокой, чем таковая интактного IgG человека. Более предпочтительны антигенсвязывающие молекулы, обладающие более высокой активностью связывания с FcRn человека, чем KD, равная 40 микромолям, еще более предпочтительно KD, равная более чем 15 микромолям.
В способах получения по настоящему изобретению, антигенсвязывающие молекулы, обладающие активностью связывания с FcRn человека в диапазоне кислых значений рН, конкретно не ограничены при условии, что они обладают активностью связывания с FcRn человека при pH от 4,0 до 6,5. Однако, при pH от 5,5 до 6,5, антигенсвязывающие молекулы предпочтительно обладают активностью связывания с FcRn человека, более высокой, чем KD, равная 20 микромолям. Активность связывания с FcRn человека более предпочтительно является сравнимой или более высокой, чем таковая интактного IgG1 человека (более высокая, чем KD, равная 1,7 микромолей), более предпочтительно более высокой, чем KD, равная 0,5 микромолям.
Значения KD, описанные выше, определяют способом, описанным в «The Journal of Immunology, (2009) 182: 7663-7671» (посредством иммобилизации антигенсвязывающей молекулы на чипе и нанесении FcRn человека в качестве анализируемого средства).
В способах получения по настоящему изобретению стадия отбора антигенсвязывающих молекул, чья антигенсвязывающая активность при pH от 6,7 до pH 10,0 является более высокой, чем таковая при pH от 4,0 до pH 6,5 является аналогичной стадии отбора антигенсвязывающих молекул, чья антигенсвязывающая активность при pH от 4,0 до pH 6,5 является более низкой, чем таковая при pH от 6,7 до pH 10,0.
Отношение между антигенсвязывающей активностью в диапазоне нейтральных значений рН и в диапазоне кислых значений рН конкретно не ограничено при условии, что антигенсвязывающая активность в диапазоне нейтральных значений рН является более высокой, чем таковая в диапазоне кислых значений рН. Антигенсвязывающая активность при pH от 6,7 до pH 10,0 предпочтительно является в два раза или больше, более предпочтительно в десять раз или больше, и еще более предпочтительно в 40 раз или больше, чем антигенсвязывающая активность при pH от 4,0 до pH 6,5.
В описанных выше способах получения антиген и антигенсвязывающая молекула могут связываться друг с другом в любом состоянии, и FcRn человека и антигенсвязывающая молекула могут связываться друг с другом в любом состоянии. Состояние конкретно не ограничено; например, антиген или FcRn человека может взаимодействовать с иммобилизированной антигенсвязывающей молекулой для связывания с антигенсвязывающей молекулой. Альтернативно, антигенсвязывающая молекула может взаимодействовать с иммобилизованным антигеном или FcRn человека для связывания с антигенсвязывающей молекулой. Альтернативно, антигенсвязывающая молекула может взаимодействовать с антигеном или FcRn человека в растворе для связывания с антигенсвязывающей молекулой.
Антигенсвязывающие молекулы, полученные посредством описанных выше способов, могут представлять собой антигенсвязывающую молекулу; и предпочтительные антигенсвязывающие молекулы включают, например, те, которые обладают антигенсвязывающим доменом и FcRn-связывающим доменом человека, который содержит изменение по меньшей мере одной аминокислоты в FcRn-связывающем домене человека, и гистидиновую замену на аминокислоту(ы) или вставку по меньшей мере одного гистидина.
Изменения аминокислот в FcRn-связывающем домене человека конкретно не ограничены, при условии, что они увеличивают активность связывания с FcRn человека в диапазоне нейтральных значений рН. Изменения включают, например те аминокислоты, которые располагаются в положении от 221 до 225, 227, 228, 230, 232, от 233 до 241, от 243 до 252, от 254 до 260, от 262 до 272, 274, 276, от 278 до 289, от 291 до 312, от 315 до 320, 324, 325, от 327 до 339, 341, 343, 345, 360, 362, 370, от 375 до 378, 380, 382, от 385 до 387, 389, 396, 414, 416, 423, 424, от 426 до 438, 440 и 442 (нумерация EU) в описанном выше Fc-домене IgG. Более конкретно, изменения аминокислот включают те аминокислоты, которые располагаются в положениях аминокислот, представленных в таблицах 1, 2, 6-1 и 6-2 (в нумерации EU). Предпочтительно, активность связывания с FcRn человека можно увеличивать в диапазоне нейтральных значений рН посредством изменения по меньшей мере одной аминокислоты, выбранной из аминокислот в положениях 237, 238, 239, 248, 250, 252, 254, 255, 256, 257, 258, 265, 270, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 325, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434 и 436 (нумерация EU). Число аминокислот, подлежащих изменению, конкретно не ограничено; и возможно изменять аминокислоты только в одном участке или двух или более участках. Комбинация из двух или более изменений аминокислот включают, например, те, которые представлены в таблицах 3, от 4-1 до 4-5, 6-1 и 6-2.8
Между тем, участок, в который вводят гистидиновую мутацию, конкретно не ограничен, и таким образом гистидиновую мутацию можно вводить в любой участок при условии, что гистидиновая мутация снижает антигенсвязывающую активность в диапазоне кислых значений рН до значения, меньшего, чем в диапазоне нейтральных значений рН. Такие гистидиновые мутации можно вводить в один или более участков.
Таким образом, способы получения по настоящему изобретению могут дополнительно содержать стадии изменения описанных выше аминокислот и гистидиновую замену или гистидиновую вставку. В способах получения по настоящему изобретению неприродные аминокислоты можно использовать вместо гистидина. Таким образом, настоящее изобретение может быть также понято посредством замещения указанного выше гистидина неприродными аминокислотами.
Кроме того, в другом варианте осуществления антигенсвязывающие молекулы, которые получают способами получения, описанными выше, включают, например, антигенсвязывающие молекулы, содержащие измененные константные области антитела. Таким образом, способы получения по настоящему изобретению могут дополнительно содержать стадию изменения аминокислот константных областей антитела.
Антигенсвязывающие молекулы, полученные способами получения по настоящему изобретению, вводят для ускорения снижения концентрации антигена в плазме. Таким образом, способы получения по настоящему изобретению можно использовать в качестве способа получения антигенсвязывающих молекул для ускорения снижения концентрации антигена в плазме при введении.
Альтернативно, антигенсвязывающие молекулы, полученные посредством способов получения по настоящему изобретению, обладают улучшенной фармакокинетикой. Таким образом, способы получения по настоящему изобретению можно использовать в качестве способа получения антигенсвязывающих молекул с усовершенствованной фармакокинетикой.
Альтернативно, антигенсвязывающие молекулы, полученные посредством способов получения по настоящему изобретению, могут увеличивать количество антигенов, с которыми может связываться одна антигенсвязывающая молекула при введении животному, такому как люди, мыши и обезьяны. Таким образом, способы получения по настоящему изобретению можно использовать в качестве способа получения антигенсвязывающих молекул, которые обладают увеличенным количеством антигенов, с которыми может связываться одна антигенсвязывающая молекула.
Кроме того, антигенсвязывающие молекулы, полученные посредством способов получения по настоящему изобретению, как ожидают, являются способными в пределах клетки к диссоциации из внеклеточно связанного антигена, при введении животным, таким как люди, мыши или обезьяны. Таким образом, способы получения по настоящему изобретению можно использовать в качестве способов получения антигенсвязывающих молекул, которые способны к диссоциации в пределах клетки из внеклеточносвязанного антигена.
Кроме того, антигенсвязывающие молекулы, полученные посредством способов получения по настоящему изобретению, как ожидают, являются способными к связыванию с антигенами и поглощаются клетками, а также высвобождаются наружу клеток в форме, свободной от антигена, при введении животным, таким как люди, мыши или обезьяны. Таким образом, способы получения по настоящему изобретению можно использовать в качестве способов получения антигенсвязывающих молекул, которые способны связывать антиген и поглощаться клетками и высвобождаться наружу клеток в форме, свободной от антигена.
Кроме того, так как при введении такие антигенсвязывающие молекулы обладают более высокой активностью для снижения концентрации антигена в плазме, по сравнению с типичными антигенсвязывающими молекулами, они, как ожидают, особенно подходящи в качестве лекарственных средств. Таким образом, способы получения по настоящему изобретению можно использовать в качестве способов получения антигенсвязывающих молекул для применения в качестве фармацевтических композиций.
Гены, полученные посредством способов получения по настоящему изобретению, как правило, несут (вставлены) в соответствующие векторы, а затем вводятся в клетки-хозяева. Векторы конкретно не ограничены при условии, что они стабильно сохраняют встроенные нуклеиновые кислоты. Например, если E. coli применяют в качестве хозяина, предпочтительные векторы для клонирования включают вектор pBluescript (Stratagene); однако, можно использовать различные коммерчески доступные векторы. При использовании векторов для получения антигенсвязывающих молекул по настоящему изобретению, особенно пригодны экспрессирующие векторы. Экспрессирующие векторы конкретно не ограничены при условии, что векторы экспрессируют антигенсвязывающие молекулы in vitro, в E. coli, в культуре клеток или в организме. Например, вектор pBEST (Promega) является предпочтительным для экспрессии in vitro; вектор pET (Invitrogen) является предпочтительным для E. coli; вектор pME18S-FL3 (номер доступа GenBank AB009864) является предпочтительным для культуры клеток; и вектор pME18S (Mol Cell Biol. (1988) 8: 466-472) является предпочтительным для организмов. ДНК по настоящему изобретению можно встраивать в векторы общепринятыми способами, например, посредством лигирования с использованием участков распознавания рестрикционных ферментов (Current Protocols in Molecular Biology, edit. Ausubel et al., (1987) Publish. John Wiley & Sons, Section 11.4-11.11).
Клетки-хозяева, указанные выше, конкретно не ограничены, и различные клетки-хозяева можно использовать в зависимости от цели. Примеры клеток для экспрессии антигенсвязывающих молекул включают бактериальные клетки (такие как Streptococcus, Staphylococcus, E. coli, Streptomyces и Bacillus subtilis), эукариотические клетки (такие как дрожжи и Aspergillus), клетки насекомых (таких как Drosophila S2 и Spodoptera SF9), клетки животных (такие как CHO, COS, HeLa, C127, 3T3, BHK, HEK293 и клетки меланомы Bowes), и растительные клетки. Векторы можно встраивать в клетку-хозяина известными способами, например, способами осаждения с фосфатом кальция, способами электропорации (Current Protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons, Section 9.1-9.9), способами липофекции и микроинъекции.
Клетки-хозяева можно культивировать известными способами. Например, при использовании клеток животных в качестве хозяина, DMEM, MEM, RPMI1640 или IMDM можно использовать в качестве среды для культивирования. Среды можно использовать с добавками сыворотки, такими как FBS или эмбриональная телячья сыворотка (FCS). Клетки можно культивировать в культурах без сыворотки. Предпочтительный pH приблизительно составляет от 6 до 8 в течение курса культивирования. Инкубацию проводят, как правило, от 30 до 40 градусов C приблизительно в течение от 15 до 200 часов. По мере необходимости среду заменяют, аэрируют или перемешивают.
Подходящие сигналы секреции можно включать в представляющие интерес полипептиды, при условии, что антигенсвязывающие молекулы, экспрессируемые клеткой-хозяином, секретируются в просвет эндоплазматического ретикулума, в периплазматическое пространство или во внеклеточное окружение. Эти сигналы могут являться эндогенными для представляющих интерес антигенсвязывающих молекул или могут представлять собой гетерологичные сигналы.
С другой стороны, например, системы продукции, использующие животных или растения, можно использовать в качестве систем получения полипептидов in vivo. Представляющий интерес полинуклеотид вводят животному или в растение, и полипептид продуцируют в организме животного или растения, а затем собирают. «Хозяева» по настоящему изобретению включают таких животных и растения.
Система продукции с использованием животных включают системы продукции с использованием млекопитающих или насекомых. Возможно использовать млекопитающих, таких как козы, свиньи, овцы, мыши и быки (Vicki Glaser SPECTRUM Biotechnology Applications (1993)). Млекопитающие могут представлять собой трансгенных животных.
Например, полинуклеотид, кодирующий антигенсвязывающую молекулу по настоящему изобретению, получают в виде гена, слитого с геном, кодирующим полипептид, продуцируемый, в частности, в молоке, такой как бета-казеин козы. Далее, в эмбрионы козы инъецируют полинуклеотидые фрагменты, содержащие слитый ген, а затем трансплантируют самкам коз. Требуемые антигенсвязывающие молекулы можно получать из молока, полученного от трансгенных коз, которые рождены от коз с введенными эмбрионами или от их потомства. При необходимости можно вводить гормоны для увеличения объема молока, содержащего антигенсвязывающую молекулу, продуцируемую трансгенной козой (Ebert et al., Bio/Technology (1994) 12: 699-702).
Насекомых, таких как тутовый шелкопряд, можно использовать для получения антигенсвязывающих молекул по настоящему изобретению. При использовании тутового шелкопряда можно использовать бакуловирусы, несущие полинуклеотид, кодирующий представляющую интерес антигенсвязывающую молекулу, для инфицирования тутового шелкопряда, и из жидкостей его организма можно получать представляющую интерес антигенсвязывающую молекулу.
Кроме того, если растения применяют для получения антигенсвязывающих молекул по настоящему изобретению, например, можно использовать табак. Если применяют табак, полинуклеотид, кодирующий представляющую интерес антигенсвязывающую молекулу, встраивают в экспрессионный вектор растения, например, pMON 530, а затем вектор встраивают в бактерии, такие как Agrobacterium tumefaciens. Бактериям затем позволяют инфицировать табак, такой как Nicotiana tabacum, и требуемые антигенсвязывающие молекулы можно собирать из его листьев (Ma et al., Eur. J. Immunol. (1994) 24: 131-138). Альтернативно, возможно инфицировать ряску (Lemna minor) похожими бактериями. После клонирования, требуемые антигенсвязывающие молекулы можно получать из клеток ряски (Cox KM et al., Nat. Biotechnol. 2006 Dec; 24(12): 1591-1597).
Полученные таким образом антигенсвязывающие молекулы можно выделять из внутреннего или внешнего пространства клеток-хозяев (такого как среда и молоко) и очищать по существу в виде чистых и гомогенных антигенсвязывающих молекул. Способы выделения и очистки антигенсвязывающих молекул конкретно не ограничены, и можно использовать способы выделения и очистки, используемые, как правило, для очистки полипептидов. Антигенсвязывающие молекулы можно выделять и очищать посредством подходящего отбора и комбинирования, например, хроматографией на колонках, фильтрацией, ультрафильтрацией, высаливанием, осаждением с растворителем, экстракцией растворителем, дистилляцией, иммунопреципитацией, SDS-электрофорезом в полиакриламидном геле, изоэлектрофокусированием, диализом и перекристаллизацией.
Хроматография включает, например, аффинную хроматографию, ионообменную хроматографию, гидрофобную хроматографию, гель-фильтрацию, хроматографию с обращенной фазой и адсорбционную хроматографию (Strategies for Protein Purification and Characterization: A Laboratory Course Manual. Ed Daniel R. Marshak et al., (1996) Cold Spring Harbor Laboratory Press). Такие хроматографические способы можно проводить, используя жидкофазную хроматографию, такую как ВЭЖХ и FPLC. Колонки, используемые для аффинной хроматографии, включают колонки с белком A и белком G. Колонки, использующие белок A, включают, например, Hyper D, POROS и Sepharose F. F. (Pharmacia).
Если необходимо, антигенсвязывающую молекулу можно модифицировать произвольно, и пептиды могут быть частично делетированы, обеспечивая подходящую белковую модификацию фермента для действия до или после очистки антигенсвязывающей молекулы. Такие белковые модификации ферментов включают, например, трипсин, химотрипсин, лизилэндопептидазы, протеинкиназы и глюкозидазы.
<Фармацевтические композиции>
Настоящее изобретение также относится к фармацевтическим композициям, которые включают антигенсвязывающие молекулы по настоящему изобретению, антигенсвязывающие молекулы, выделенные способами скрининга по настоящему изобретению, или антигенсвязывающие молекулы, полученные посредством способов получения по настоящему изобретению. Антигенсвязывающие молекулы по настоящему изобретению и антигенсвязывающие молекулы, полученные посредством способов получения по настоящему изобретению, обладают более высокой активностью снижения концентрации антигена в плазме при введении по сравнению с типичными антигенсвязывающими молекулами, и являются, таким образом, пригодными в качестве фармацевтических композиций. Фармацевтическая композиция по настоящему изобретению может включать фармацевтически приемлемые носители.
В настоящем изобретении, фармацевтические композиции как правило, относятся к средствам для лечения или предупреждения или тестирования и диагностики заболевания.
Фармацевтические композиции по настоящему изобретению можно составлять способами, известными специалистам в данной области. Например, фармацевтические композиции можно использовать парентерально, в форме инъекций стерильных растворов или суспензий, включающих воду или другую фармацевтически приемлемую жидкость. Например, такие композиции можно составлять посредством смешивания в форме стандартной дозы, в основном требуемой в утвержденной медицинской технологии производства посредством комбинирования, соответствующим образом, с фармацевтически приемлемыми носителями или средой, конкретно со стерильной водой, физиологическим раствором, растительным маслом, эмульгатором, суспензией, поверхностно-активным веществом, стабилизатором, ароматизатором, эксципиентом, носителем, консервантом, связывающим средством или подобным. В таких составах регулируют количество активного ингредиента для получения подходящего количества в заранее определенном диапазоне.
Стерильные композиции для инъекций можно составлять, используя носители, такие как дистиллированная вода для инъекций, согласно стандартной практике составления. Водные растворы для инъекций включают, например, физиологический раствор и изотонические растворы, содержащие декстрозу или другие адъюванты (например, D-сорбит, D-манноза, D-маннит и хлорид натрия). Также возможно использование в комбинации подходящих солюбилизаторов, например, спиртов (этанола и подобных), полиспиртов (пропиленгликоля, полиэтиленгликоля и подобных), неионных поверхностно-активных веществ (полисорбата 80(TM), HCO-50 и подобных).
Масла включают кунжутное и соевое масла. Бензилбензоат и/или бензиловый спирт можно использовать в комбинации в качестве солюбилизаторов. Также возможно комбинировать буферы (например, фосфатный буфер и буфер с ацетатом натрия), успокаивающие средства (например, прокаин-гидрохлорид), стабилизаторы (например, бензиловый спирт и фенол) и/или антиоксиданты. Подходящие ампулы заполняют полученными растворами для инъекций.
Фармацевтические композиции по настоящему изобретению предпочтительно вводят парентерально. Например, композиции могут быть в лекарственной форме для инъекций, трансназального введения, транспульмонарного введения или трансдермального введения. Например, фармацевтические композиции можно вводить системно или местно посредством внутривенной инъекции, внутримышечной инъекции, интраперитонеальной инъекции, подкожной инъекции или подобных.
Способы введения можно соответствующим образом выбирать, учитывая возраст и симптомы пациента. Доза фармацевтической композиции, содержащей антигенсвязывающую молекулу, может представлять собой дозу, например, от 0,0001 до 1000 мг/кг для каждого введения. Альтернативно, доза может представлять собой, например, дозу от 0,001 до 100000 мг на пациента. Однако, настоящее изобретение не ограничено числовыми значениями, описанными выше. Дозы и способы введения варьируют в зависимости от массы, возраста, симптомов пациента и т.п.. Специалисты в данной области могут установить подходящие дозы и способы введения, учитывая факторы, описанные выше.
Аминокислоты, содержащиеся в аминокислотных последовательностях по настоящему изобретению, могут быть модифицированы после трансляции. Например, модификация N-концевого глутамина в пироглутаминовую кислоту посредством пироглутамилирования хорошо известна специалистам в данной области. Как и следовало ожидать, такие модификации аминокислот после трансляции включают в аминокислотные последовательности в настоящем изобретении.
Все известные документы о уровне техники, процитированные в описании, включены в настоящий документ путем ссылки.
ПРИМЕРЫ
Ниже в настоящем документе настоящее изобретение будет конкретно описано со ссылкой на примеры, но их не следует рассматривать как ограничивающие.
[Пример 1] Исследование усиления ускоряющего эффекта антитела на элиминацию антигена
Антитело против рецептора IL-6
Получение антитела против рецептора IL-6 человека, обладающего активностью связывания с FcRn в нейтральных условиях
H54/L28-IgG1, содержащий H54 (SEQ ID NO: 1) и L28 (SEQ ID NO: 2), описанный в WO 2009/125825, представляет собой гуманизированное антитело против рецептора IL-6. Мутацию вводят в H54 (SEQ ID NO: 1) для увеличения связывания с FcRn в условиях нейтральных значений рН (pH 7,4). Конкретно, H54-IgG1-F14 (SEQ ID NO: 3) получали из константной области тяжелой цепи IgG1 посредством замены Trp на Met в положении 252 и Trp на Asn в положении 434 в нумерации EU. Замены аминокислот вводили способами, известными специалистам в данной области, описанными в ссылочном примере 1.
H54/L28-IgG1, содержащий H54 (SEQ ID NO: 1) и L28 (SEQ ID NO: 2), и H54/L28-IgG1-F14, содержащий H54-IgG1-F14 (SEQ ID NO: 3) и L28 (SEQ ID NO: 2), экспрессировали и очищали способом, известным специалистам в данной области, описанным в ссылочном примера 2.
In vivo исследование антител посредством стационарной модели инфузии с использованием трансгенной линии мышей 276 с FcRn человека
Используя H54/L28-IgG1 и H54/L28-IgG1-F14, полученный, как описано выше, проводили тестирование in vivo посредством стационарой модели инфузии с использованием трансгенной линии 276 мышей с FcRn человека. Инфузионный насос (MINI-OSMOTIC PUMP MODEL 2004; alzet), содержащий растворимый рецептор IL-6 человека, имплантировали под кожу спинки мышей трансгенной линии 276 с FcRn человека (B6.mFcRn-/-.hFcRn Tg линия 27 6+/+ мышь (B6.mFcRn-/-hFCRnTg276B6.Cg-Fcgrt<tm1Dcr>Tg(FCGRT)276Dcr(Jackson#4919)), Jackson Laboratories; Methods Mol Biol. (2010) 602: 93-104) для получения модели животных, у которых концентрацию в плазме растворимого рецептора IL-6 человека сохраняли постоянной. Антитела против рецептора IL-6 человека вводили модельным животным для оценки динамики in vivo после введения растворимого рецептора IL-6 человека. Моноклональное антитело против CD4 мыши (R&D) вводили из расчета 20 мг/кг до имплантирования инфузионного насоса и через 14 суток после введения антитела в хвостовую вену для супрессии продукции нейтрализующего антитела против растворимого рецептора IL-6 человека. Затем, инфузионный насос, содержащий 92,8 микрограм/мл растворимого рецептора IL-6 человека, имплантировали под кожу спинки мышей. Через трое суток после имплантации инфузионного насоса, антитела против рецептора IL-6 человека (H54/L28-IgG1 и H54/L28-IgG1-F14) вводили из растчета 1 мг/кг единожды в хвостовую вену. Кровь собирали через 15 минут, семь часов, одни сутки, двое суток, трое суток, четверо суток, семь суток, 14 суток, 21 сутки и 28 суток после введения антитела против рецептора IL-6 человека. Собранную кровь немедленно центрифугировали при 15000 об/мин и 4 градусов C в течение 15 минут для отделения плазмы. Отделенную плазму хранили в холодильнике при -20 градусов C или ниже до проведения анализа.
Определение концентрации hsIL-6R в плазме посредством электрохемилюминесцентного анализа
Концентрацию hsIL-6R в плазме мыши измеряли посредством электрохемилюминесценции. Образцы калибровочной кривой hsIL-6R, доводили до концентрации 2000, 1000, 500, 250, 125, 62,5 и 31,25 пг/мл, и получали образцы плазмы мыши, разведенные в 50 раз или более раз. Образцы смешивали с раствором моноклонального антитела против IL-6R человека (R&D), меченного рутением, со сложным эфиром сульфо-Tag-NHS (Meso Scale Discovery), биотинилированным антителом против IL-6R человека (R&D) и WT-IgG1, а затем оставляли для реакции в течение ночи при 37 градусов C. Конечная концентрация WT-IgG1 в качестве антитела против рецептора IL-6 человека, содержащего H (WT) (SEQ ID NO: 4) и L (WT) (SEQ ID NO: 5), составляла 333 микрограмм/мл, что превышает концентрацию антитела против рецептора IL-6 человека, содержащегося в образцах, с целью связывания практически всех молекул hsIL-6R в образцах с WT-IgG1. Затем образцы переносили на планшет MA400 PR со стрептавидином (Meso Scale Discovery), и оставляли для реакции в течение одного часа при комнатной температуре, и осуществляли промывание. Немедленно затем вносили буфер Read Buffer T (×4) (Meso Scale Discovery), измерения проводили посредством Sector PR 400 Reader (Meso Scale Discovery). Концентрацию hsIL-6R рассчитывали, основываясь на данных калибровочной кривой, используя аналитическое программное обеспечение SOFTmax PRO (Molecular Devices). Динамика концентрации hsIL-6R в плазме после внутривенного введения H54/L28-IgG1 и H54/L28-IgG1-F14, как измерено в этом способе, представлена на фиг.1.
Как показано на фиг.1, по сравнению с базовой линией концентрации hsIL-6R без антитела, введение H54/L28-IgG1 приводило в результате к значимому повышению концентрации в плазме hsIL-6R. С другой стороны, введение H54/L28-IgG1-F14 приводило в результате к снижению повышения концентрации hsIL-6R в плазме по сравнению с H54/L28-IgG1. Это снижение в повышении получают в результате повышенного связывания с FcRn человека при нейтральных pH в H54/L28-IgG1-F14 по сравнению с H54/L28-IgG1. Это демонстрирует, что повышение аффинности связывания антитела с FcRn при нейтральных pH может увеличить клиренс антигена, хотя степень усиления клиренса антигена была небольшой для H54/L28-IgG1-F14 по сравнению с H54/L28-IgG1.
[Пример 2] Исследование усиления ускоряющего эффекта элиминации антигена антигенсвязывающих антител, зависимых от рН (получение антител)
Рассмотрение зависимых от рН антител, связывающих рецептор IL-6 человека
H54/L28-IgG1, содержащий H54 (SEQ ID NO: 1) и L28 (SEQ ID NO: 2), описанный в WO 2009/125825, представляет собой гуманизированное антитело против рецептора IL-6. Fv4-IgG1, содержащий VH3-IgG1 (SEQ ID NO: 6) и VL3-CK (SEQ ID NO: 7), представляет собой гуманизированное антитело против рецептора IL-6, которое получается в результате придания H54/L28-IgG1 свойства связывать растворимый рецептор IL-6 человека зависимым от рН способом (которое связывает при pH 7,4, но диссоциирует при pH 5,8). Тест in vivo, описанный в WO 2009/125825, с использованием мышей продемонстрировал, что элиминация растворимого рецептора IL-6 человека была значительно увеличена в группе, которой вводили смесь Fv4-IgG1 и растворимого рецептора IL-6 человека в качестве антигена по сравнению с группой, которой вводили смесь H54/L28-IgG1 и растворимого рецептора IL-6 человека в качестве антигена.
Растворимый рецептор IL-6 человека связывается с рядовым антителом, которое связывается с растворимым рецептором IL-6 человека и рециркулирует в плазму вместе с антителом посредством FcRn. Между тем, антитело, которое связывается с растворимым рецептором IL-6 человека зависимым от рН способом, диссоциирует от растворимого рецептора IL-6 человека, который связан с антителом в кислых условиях в эндосоме. Диссоциировавший растворимый рецептор IL-6 человека деградирует в лизосоме. Это может сильно усиливать элиминацию растворимого рецептора IL-6 человека. Затем антитело, которое связывается с растворимым рецептором IL-6 человека зависимым от pH способом, рециркулирует в плазму посредством FcRn. Рециркулированное антитело может связываться с другим растворимым рецептором IL-6 человека снова. Посредством повторения этого цикла, одна молекула антитела может неоднократно связываться с растворимым рецептором IL-6 человека много раз (фиг.2).
Антитела, которые связываются с антигенами зависимым от рН способом, увеличивают элиминацию растворимого антигена. Антитела осуществляют действие посредством неоднократного связывания с растворимым антигеном множество раз. Таким образом, такие антитела являются очень полезными. Способ усиления связывания FcRn в нейтральных условиях (pH 7,4) тестировали для дополнительного увеличения ускоряющего эффекта элиминации антигена.
Получение зависимых от рН антител, связывающих рецептор IL-6 человека, обладающих активностью связывания с FcRn в нейтральных условиях
Мутации вводят в Fv4-IgG1, содержащий VH3-IgG1 (SEQ ID NO: 6) и VL3-CK (SEQ ID NO: 7), для увеличения связывания с FcRn в нейтральных условиях (pH 7,4). Конкретно, VH3-IgG1-v1 (SEQ ID NO: 8) получали из константной области тяжелой цепи IgG1 посредством замены Tyr на Met в положении 252, Thr на Ser в положении 254, и Glu на Thr в положении 256 в нумерации EU, тогда как VH3-IgG1-v2 (SEQ ID NO: 9) конструировали из константной области тяжелой цепи IgG1 посредством замены Trp на Asn в положении 434 в нумерации EU. Замены аминокислот вводят посредством способа, известного специалистам в данной области, описанным в ссылочном примере 1.
H54/L28-IgG1, содержащий H54 (SEQ ID NO: 1) и L28 (SEQ ID NO: 2), Fv4-IgG1, содержащий VH3-IgG1 (SEQ ID NO: 6) и VL3-CK (SEQ ID NO: 7), Fv4-IgG1-v1, содержащий VH3-IgG1-v1 (SEQ ID NO: 8) и VL3-CK (SEQ ID NO: 7), и Fv4-IgG1-v2, содержащий VH3-IgG1-v2 (SEQ ID NO: 9) и VL3-CK (SEQ ID NO: 7), экспрессировали и очищали способом, известным специалистам в данной области, описанным в ссылочном примере 2.
[Пример 3] Исследование увеличения ускоряющего элиминацию антигена эффекта зависимого от рН антигенсвязывающего антитела (тест in vivo)
Тест in vivo с использованием трансгенных мышей с FcRn человека и нормальных мышей
Кинетики in vivo hsIL-6R (растворимый рецептор IL-6 человека: получено, как описано в справочном примере 3) и антитела против рецептора IL-6 человека оценивали после введения трансгенным мышам с FcRn человека (B6.mFcRn-/-.hFcRn Tg линия 276 +/+ мышь, Jackson Laboratories; Methods Mol Biol. (2010) 602: 93-104) и нормальным мышам (мыши C57BL/6J; Charles River Japan) hsIL-6R по отдельности или hsIL-6R и антитела против рецептора IL-6 человека в комбинации. Раствор hsIL-6R (5 микрограмм/мл) или раствор смеси, содержащей hsIL-6R и антитела против рецептора IL-6 человека (5 микрограмм/мл и 0,1 мг/мл, соответственно), вводили единожды в дозе 10 мл/кг в хвостовую вену. В этом случае, антитела против рецептора IL-6 человека представлены в избытке над hsIL-6R, и таким образом почти каждый hsIL-6R, как предполагают, связываются с антителом. Кровь собирали через 15 минут, семь часов, одни сутки, двое суток, трое суток, четверо суток, семь суток, 14 суток, 21 сутки и 28 суток после введения. Собранную кровь немедленно центрифугировали при 15000 об/мин и 4 градусов C в течение 15 минут для отделения плазмы. Отделенную плазму хранили в холодильнике при -20 градусов или ниже перед выполнением анализа. Используемые антитела против рецептора IL-6 человека представляют собой: описанные выше H54/L28-IgG1, Fv4-IgG1 и Fv4-IgG1-v2 для трансгенных мышей с FcRn человека и описанные выше H54/L28-IgG1, Fv4-IgG1, Fv4-IgG1-v1 и Fv4-IgG1-v2 для нормальных мышей.
Измерение концентрации антител против рецептора IL-6 человека в плазме посредством ELISA
Концентрацию антитела против рецептора IL-6 человека в плазме мыши измеряли посредством ELISA. (Fab’)2 фрагмент антитела против IgG человека (специфичного к гамма-цепи) (Sigma) переносили на Nunc-ImmunoPlate MaxiSorp (Nalge Nunc International) и оставляли в течение ночи при 4 градусов C для получения планшетов с иммобилизованными антителами против IgG человека. Получали образцы калибровочной кривой, имеющие концентрацию в плазме 0,8, 0,4, 0,2, 0,1, 0,05, 0,025 и 0,0125 микрограмм/мл, и образцы плазмы мыши, разведенные в 100 раз и более. 200 микролитров (мкл) 20 нг/мл hsIL-6R добавляли к 100 микролитрам образцов калибровочной кривой и образцам плазмы, а затем образцы оставляли в течение одного часа при комнатной температуре. Затем образцы переносили на планшеты с иммобилизованными антителами против IgG человека и оставляли в течение одного часа при комнатной температуре. Затем биотинилированное антитело против IL-6R человека (R&D) добавляли для реакции в течение одного часа при комнатной температуре. Затем стрептавидин-ПолиHRP80 (Stereospecific Detection Technologies) добавляли для осуществлении реакции в течение одного часа при комнатной температуре, и хромогенную реакцию проводили с использованием TMP One Component HRP Microwell Substrate (BioFX Laboratories) в качестве субстрата. После остановки реакции 1 н. серной кислотой (Showa Chemical), поглощение при 450 нм измеряли посредством ридера для микропланшетов. Концентрацию в плазме мыши рассчитывали из оптической плотности калибровочной кривой, используя аналитическое программное обеспечение SOFTmax PRO (Molecular Devices). Динамика концентрации в плазме после внутривенного введения, как измерено этим способом, представлена на фиг.3 для трансгенных мышей с FcRn человека и на фиг.5 для нормальных мышей.
Измерение концентрации hsIL-6R в плазме посредством электрохемилюминесцентного анализа
Концентрацию hsIL-6R в плазме мыши измеряли посредством электрохемилюминисценции. Получали образцы калибровочной кривой hsIL-6R, доведенные до концентрации 2000, 1000, 500, 250, 125, 62,5 и 31,25 пг/мл, и образцы плазмы мыши, разведенные в 50 раз или более. Образцы смешивали с раствором моноклонального антитела против IL-6R (R&D), меченного рутением, со сложным эфиром сульфо-Tag NHS (Meso Scale Discovery), биотинилированного антитела против IL-6R человека (R&D) и WT-IgG1, а затем оставляли для реакции в течение ночи при 37 градусов C. Конечная концентрация WT-IgG1 в качестве антитела против рецептора IL-6 человека, содержащего H (WT) (SEQ ID NO: 4) и L (WT) (SEQ ID NO: 5), составила 333 микрограмм/мл, что является больше концентрации антитела против рецептора IL-6 человека, содержащегося в образцах, с целью связывания практически всех молекул hsIL-6R в образцах с WT-IgG1. Затем образцы переносили на планшет со стрептавидином MA400 PR (Meso Scale Discovery) и оставляли для реакции в течение одного часа при комнатной температуре, и проводили промывание. Немедленно затем вносили буфер Read Buffer T (×4) (Meso Scale Discovery), измерения проводили посредством Sector PR 400 Reader (Meso Scale Discovery). Концентрацию hsIL-6R рассчитывали, основываясь на данных калибровочной кривой, используя аналитическое программное обеспечение SOFTmax PRO (Molecular Devices). Динамика концентрации hsIL-6R в плазме после внутривенного введения H54/L28-IgG1 и H54/L28-IgG1-F14, как измерено в этом способе, представлена на фиг.4 для трансгенных мышей с FcRn человека и на фиг.6 для нормальных мышей.
Определение концентрации свободного hsIL-6R в плазме посредством электрохемилюминесцентного анализа
Для оценки степени нейтрализации растворимого рецептора IL-6 человека в плазме, концентрацию растворимого рецептора IL-6 человека, свободного от (ненейтрализованного посредством) антитела против рецептора IL-6 человека (концентрация свободного hsIL-6R) в плазме мыши, определяли электрохемилюминесцентным анализом. Все антитела типа IgG (IgG мыши, антитела против рецептора IL-6 человека и комплекс антитело против рецептора IL-6 человека-растворимый рецептор IL-6 человека) в плазме адсорбировали на белке A посредством добавления 12 микролитров каждого стандартного образца hsIL-6R, полученного из расчета 10000, 5000, 2500, 1250, 625, 312,5, или 156,25 пг/мл, и образцов плазмы мыши на соответствующее количество смолы rProtein A Sepharose Fast Flow (GE Healthcare), высушенной на 0,22-микрометровой чашке фильтра (Millipore). Затем, раствор в чашке осаждали с использованием высокоскоростной центрифуги для сбора раствора, который пропускали через смолу. Пропущенный через смолу раствор не содержал связанный с белком A комплекс антитело против рецептора IL-6 человека-растворимый рецептор IL-6 человека. Таким образом, концентрацию свободного hsIL-6R в плазме можно определять, измеряя концентрацию hsIL-6R в пропущенном через смолу растворе.
Затем пропущенный через смолу раствор смешивали с моноклональным антителом против IL-6R человека (R&D), меченным рутением, со сложным эфиром сульфо-Tag NHS (Meso Scale Discovery) и биотинилированного антитела против IL-6R человека (R&D). Полученную в результате смесь инкубировали при комнатной температуре в течение одного часа, а затем добавляли на планшет со стрептавидином MA400 PR, разделяя на аликвоты (Meso Scale Discovery). После одного часа инкубации при комнатной температуре планшет отмывали и также разделяли на аликвоты Read Bufer T (×4) (Meso Scale Discovery). Немедленно результаты реакции на планшете измеряли в SECTOR PR 400 Reader (Meso Scale Discovery). Концентрацию hsIL-6R рассчитывали, основываясь на данных в стандартной кривой с использованием анализа на программном обеспечении SOFTmax PRO (Molecular Devices). Динамику изменения концентрации свободного hsIL-6R в плазме нормальных мышей после внутривенного введения определяли описанным выше способом, как показано на фиг.7.
Эффект зависимого от рН связывания с рецептором IL-6 человека
H54/L28-IgG1 и Fv4-IgG1, которые связываются с рецептором IL-6 человека зависимым от рН способом тестировали in vivo, и результаты сравнивали друг с другом. Как показано на фиг.3 и 5, удержание антитела в плазме было сравнимым. Между тем, как показано на фиг.4 и 6, hsIL-6R, введенный одновременно с Fv4-IgG1, который связывается с рецептором IL-6 человека зависимым от рН способом, как обнаружили, увеличивает элиминацию hsIL-6R по сравнению с hsIL-6R, введенным одновременно с H54/L28-IgG1. Тенденцию, описанную выше, наблюдали и для трансгенных мышей с FcRn человека, и нормальных мышей; таким образом, показано, что посредством придания зависимой от рН способности связывания с рецептором IL-6 человека, концентрация hsIL-6R в плазме через четверо суток после введения может быть снижена приблизительно в 17 и 34 раза, соответственно.
Эффект связывания с FcRn в нейтральных условиях (pH 7,4)
Сообщалось, что интактный IgG1 человека плохо связывался с FcRn человека (обладает очень низкой аффиностью связывания) в нейтральных условиях (pH 7,4). Связывание с FcRn человека в нейтральных условиях (pH 7,4), как сообщают, увеличивается при замене Trp на Asn в положении 434 (нумерация EU) в интактном IgG1 человека (J Immunol. (2009) 182 (12): 7663-71). Fv4-IgG1-v2, который получают в результате введения указанной выше аминокислотной замены в Fv4-IgG1, тестировали посредством теста in vivo с использованием трансгенных мышей с FcRn человека. Результаты теста сравнивали с результатами, полученными для Fv4-IgG1. Как показано на фиг.3, время удержания антител являлось сравнимым между двумя антителами. Между тем, как показано на фиг.4, hsIL-6R вводят одновременно с Fv4-IgG1-v2, который демонстрирует повышенное связывание с FcRn человека в нейтральных условиях (pH 7,4), как обнаружено, элиминируется быстрее по сравнению с hsIL-6R введенным одновременно с Fv4-IgG1. Таким образом, показано, что посредством придания способности связываться с FcRn человека в нейтральных условиях (pH 7,4), концентрация в плазме hsIL-6R через четверо суток после введения может быть снижена приблизительно в четыре раза.
Основываясь на гомологии между FcRn человека и FcRn мыши, замена Trp на Asn в положении 434 в нумерации EU, как предполагают, увеличивает связывание с FcRn мыши в нейтральных условиях (pH 7,4). Между тем, связывание с FcRn мыши в нейтральных условиях (pH 7,4), как сообщалось, увеличивается посредством замещения Tyr на Met в положении 252, Thr на Ser в положении 254 и Glu на Thr в положении 256 в нумерации EU (J Immunol. (2002) 169(9): 5171-80). Fv4-IgG1-v1 и Fv4-IgG1-v2, которые получены в результате введения описанных выше аминокислотных замен в Fv4-IgG1, тестировали in vivo, используя нормальных мышей. Результаты тестирования сравнивали с результатами, полученными для Fv4-IgG1. Как показано на фиг.5, время удержания в плазме Fv4-IgG1-v1 и Fv4-IgG1-v2, которых усовершенстовали для увеличения связывания с FcRn мыши в нейтральных условиях (pH 7,4), являлось немного укороченным (концентрации в плазме нейтрализующего антитела через одни сутки после введения снижалась приблизительно в 1,5 и 1,9 раза, соответственно) по сравнению с Fv4-IgG1.
Как показано на фиг.6, hsIL-6R, введенный одновременно с Fv4-IgG1-v1 или Fv4-IgG1-v2, которых усовершенствовали для увеличения связывания с FcRn мыши в нейтральных условиях (pH 7,4), как показали, элиминируется явно быстрее по сравнению с hsIL-6R, введенным одновременно с Fv4-IgG1. Fv4-IgG1-v1 и Fv4-IgG1-v2 снижали концентрацию hsIL-6R в плазме через одни сутки после введения приблизительно в 32 и 80 раз, соответственно. Таким образом, выявили, что концентрацию в плазме можно снижать посредством придания способности связывания с FcRn мыши в нейтральных условиях (pH 7,4). Как описано выше, посредством придания способности связывания с FcRn мыши в нейтральных условиях (pH 7,4), концентрация антитела в плазме немного снижалась; однако, получили эффект снижения концентрации hsIL-6R в плазме, который в значительной степени превосходит снижение концентрации антитела. Кроме того, hsIL-6R, введенный одновременно с Fv4-IgG1-v1 или Fv4-IgG1-v2, как обнаружили, элиминируется даже быстрее по сравнению с группой, которой hsIL-6R вводили отдельно. Как показано на фиг.6, показано, что hsIL-6R, введенный одновременно с Fv4-IgG1-v1 или Fv4-IgG1-v2, может снижать концентрацию hsIL-6R в плазме через одни сутки после введения приблизительно в 4 или 11 раз, соответственно, по сравнению с hsIL-6R отдельно. Конкретно, это обозначает, что элиминация растворимого рецептора IL-6 может быть ускорена посредством введения антитела, которое связывается с растворимым рецептором IL-6 зависимым от рН способом и которому придают способность связывания с FcRn мыши в нейтральных условиях (pH 7,4). Конкретно, концентрацию антигена в плазме можно снижать in vivo посредством введения такого антитела в организм.
Как показано на фиг.7, свободный hsIL-6R был представлен в диапазоне детектируемой концентрации в течение семи суток после введения H54/L28-IgG1, в то время как hsIL-6R являлся неопределяемым через одни сутки после введения Fv4-IgG1. С другой стороны, свободный hsIL-6R являлся неопределяемым через семь часов после введения Fv4-IgG1-v1 или Fv4-IgG1-v2. Конкретно, концентрация свободного hsIL-6R была более низкой в присутствии Fv4-IgG1, который связывается с hsIL-6R зависимым от рН способом, по сравнению с H54/L28-IgG1, что предполагает более сильный нейтрализующий эффект hsIL-6R, который получают посредством обеспечения зависимой от рН способности связывания с hsIL-6R. Кроме того, свободная концентрация hsIL-6R была более низкой в присутствии Fv4-IgG1-v1 или Fv4-IgG1-v2, и тот и другой модифицируют из Fv4-IgG1 для повышения способности связывания с FcRn при pH 7,4. Это демонстрирует, что более сильный нейтрализующий эффект hsIL-6R можно получать посредством увеличения активности связывания с FcRn при pH 7,4.
При введении, обыкновенное нейтрализующее антитело, такое как H54/L28-IgG1, снижает клиренс связанного антигена, что приводит к удлиненному времени удержания антигена в плазме. Не является предпочтительным, что введенные антитела пролонгируют время удержания антигена, чье действие должно быть нейтрализовано антителами. Время удержания антигена можно укорачивать посредством обеспечения процессу связывания антигена зависимости от pH (антитело связывается в нейтральных условиях, но подвергается диссоциации в кислых условиях). В настоящем изобретении, время удержания антигена в плазме можно было бы дополнительно укорачивать посредством дополнительного обеспечения способности связывания с FcRn человека в нейтральных условиях (pH 7,4). Кроме того, показано, что по сравнению с клиренсом антигена отдельно, клиренс антигена можно было бы дополнительно увеличивать посредством введения антитела, которое связывается с антигеном зависимым от рН способом, и который обеспечивают способностью связывания с FcRn в нейтральных условиях (pH 7,4). В настоящее время, не существует способа, пригодного для увеличения клиренса антигена посредством введения антитела, относительно клиренса антигена отдельно. Таким образом, способы, осуществленные, как описано в этом ПРИМЕРЕ, являются очень полезными в качестве способа элиминации антигенов из плазмы посредством введения антител. Кроме того, авторы настоящего изобретения первыми открыли преимущество увеличения способности связывания с FcRn в нейтральных условиях (pH 7,4). Кроме того, и v4-IgG1-v1 и Fv4-IgG1-v2, которые имеют различные аминокислотные замены, увеличивающие способность связывания с FcRn в нейтральных условиях (pH 7,4), производили сравнимый эффект. Это позволяет предположить, что несмотря на тип аминокислотных замен, каждая аминокислотная замена, которая увеличивает способность связывания с FcRn человека в нейтральных условиях (pH 7,4), потенциально обладает эффектом, увеличивающим элиминацию антигена. Конкретно, молекулы антитител, которые при введении элиминируют антигены из плазмы, можно получать, используя следующие аминокислотные замены как в отдельности, так и в комбинации:
замена аминокислоты Ile на Pro в положении 257 и замену аминокислоты Ile на Gln в положении 311 в нумерация EU, каждая из которых описана в J Biol Chem. 2007, 282(3): 1709-17; замена аминокислоты Ala, Tyr или Trp на Asn в положении 434, замена аминокислоты Tyr на Met в положении 252, замена аминокислоты Gln на Thr в положении 307, замена аминокислоты Pro на Val в положении 308, замена аминокислоты Gln на Thr в положении 250, замена аминокислоты Leu на Met в положении 428, замена аминокислоты Ala на Glu в положении 380, замена аминокислоты Val на Ala в положении 378, замена аминокислоты Ile на Tyr в положении 436 в нумерации EU, каждая из которых описана в J Immunol. (2009) 182(12): 7663-71; замена аминокислоты Tyr на Met в положении 252, замена аминокислоты Thr на Ser в положении 254, замена аминокислоты Glu на Thr в положении 256 в нумерации EU, каждая из которых описана в J Biol Chem. 2006 Aug. 18, 281(33): 23514-24; замена аминокислоты Lys на His в положении 433, замена аминокислоты Phe на Asn в положении 434, и замена аминокислоты His на Tyr в положении 436 в нумерации EU, каждая из которых описана в Nat Biotechnol. 2005 октября 23(10): 1283-8; и т.п.
[Пример 4] Оценка активности связывания с FcRn человека
Для системы анализа, основанной на Biacore для тестирования взаимодействия между антителом и FcRn, систему, в которой иммобилизируют антитело на сенсорный чип и используют FcRn человека в качестве анализируемого средства, описывают в J Immunol. (2009) 182(12): 7663-71. Для этой цели, FcRn человека получали, как описано в ссылочном примере 4. Fv4-IgG1, Fv4-IgG1-v1 и Fv4-IgG1-v2 оценивали по активности связывания с FcRn человека (константа диссоциации (KD)) при pH 6,0 и pH 7,4 посредством использования описанной выше системы. Антитела тестировали в качестве тестируемого вещества после непосредственной иммобилизации на сенсорный чип Series S CM5. С использованием связанного с аминами набора по инструкции производителя, антитела иммобилизовали на сенсорном чипе так, чтобы гарантировать количество иммобилизации 500 RU (резонансных единиц). Используемый подвижный буфер представлял собой 50 ммоль/1 Na-фосфат/150 ммоль/1 NaCl, содержащий 0,05% (об./об.%) поверхностно-активного вещества P20 (pH 6,0).
С получением сенсорных чипов, анализ проводили с использованием в качестве подвижного буфера, 50 ммоль/1 Na-фосфат/150 ммоль/1 NaCl, содержащий 0,05% поверхностно-активного вещества P20 (pH 6,0) или 50 ммоль/1 Na-фосфат/150 ммоль/1 NaCl, содержащий 0,05% поверхностно-активного вещества P20 (pH 7,4). Анализы проводили исключительно при 25 градусов C. Разведенные растворы FcRn человека и подвижный буфер в качестве контрольного раствора инъецировали при скорости потока 5 микролитров/в мин в течение десяти минут для обеспечения FcRn человеку возможности взаимодействовать с антителом на чипе. Затем подвижный буфер инъецировали при скорости потока 5 микролитров/мин в течение одной минуты для контроля за диссоциацией FcRn. Затем сенсорный чип регенерировали посредством двух раундов инъецирования инъекцией 20 ммоль/1 Tris-HCl/150 ммоль/1 NaCl (pH 8,1) при скорости потока 30 микролитров/мин в течение 15 секунд.
Результаты анализировали с использованием программного обеспечения Biacore T100 Evaluation (Ver. 2.0.1). Посредством стационарного аффинного способа рассчитывали константу диссоциации (KD) из анализа результатов при шести различных концентрациях FcRn. Результаты активности связывания с FcRn человека (константы диссоциации (KD)) Fv4-IgG1, Fv4-IgG1-v1 и Fv4-IgG1-v2 при pH 6,0 и pH 7,4 представлены в таблице 5 ниже.
При pH 7,4, связывание FcRn человека с Fv4-IgG1 было слишком слабым для определения значения KD (NA). Между тем наблюдали, что Fv4-IgG1-v1 и Fv4-IgG1-v2 связывается с FcRn человека при pH 7,4, и значение KD определяли как 36,55 и 11,03 микромолей, соответственно. Значения KD для FcRn человека при pH 6,0 определяли как 1,99, 0,32 и 0,11 микромолей. Как показано на фиг.3, при сравнении с Fv4-IgG1, Fv4-IgG1-v2 усиливалась элиминация hsIL-6R у трасгенных мышей с FcRn человека. Таким образом, можно прогнозировать, что элиминация антигена ускоряется посредством увеличения связывания с FcRn человека при pH 7,4, по меньшей мере , более высокой, чем 11,03 микромолей посредством изменения IgG1 человека. Между тем, как описано в J Immunol. (2002) 169(9): 5171-80, IgG1 человека связывается приблизительно в десять раз более сильно с FcRn мыши, чем с FcRn человека. По этой причине, Fv4-IgG1-v1 и Fv4-IgG1-v2, как прогнозируют, тоже связывается приблизительно в десять раз более сильно с FcRn мыши, чем FcR человека при pH 7,4. Усиление элиминации hsIL-6R посредством Fv4-IgG1-v1 или Fv4-IgG1-v2 у нормальной мыши, показанное на фиг.6, является более значимым, чем ускорение элиминации посредством Fv4-IgG1-v2 у трасгенных мышей с FcRn человека, показанных на фиг.4. Это позволяют предположить, что увеличение элиминации hsIL-6R усиливается согласно силе связывания с FcRn при pH 7,4.
[Пример 5] Получение зависимых от рН антител, связывающих рецептор IL-6 человека с увеличенным связыванием FcRn человека в нейтральных условиях
Различные изменения для увеличения связывания с FcRn человека в нейтральных условиях вводили в Fv4-IgG1 для дополнительного увеличения эффекта элиминации антигена зависимых от рН антител, связывающих рецептор IL-6 человека у трасгенных мышей с FcRn человека. Конкретно, аминокислотные изменения, представленные в таблицах 6-1 и 6-2, вводили в константную область тяжелой цепи Fv4-IgG1 для получения различных мутантов (количество участков мутаций аминокислот представлено согласно нумерации EU). Замены аминокислот вводили способами, известными специалистам в данной области, как описано в ссылочном примере 1.


Таблица 6-2 является продолжением таблицы 6-1.

Каждый вариант, содержащий полученную тяжелую цепь и L (WT) (SEQ ID NO: 5) экспрессировали и очищали способами, известными known специалистам в данной области, как описано в ссылочном примере 2.
Оценка связывания с FcRn человека
Связывание между антителом и FcRn человека анализировали кинетически с использованием Biacore T100 (GE Healthcare). Для этой цели, FcRn человека получали, как описано в ссылочном примере 4. Подходящее количество белка L (ACTIGEN) иммобилизовали на сенсорном чипе CM4 (GE Healthcare) посредством способа на основе связанных аминов, и обеспечивали удержание на чипе представляющих интерес антител. Затем, разведенные растворы FcRn и подвижный буфер (в качестве контрольного раствора) инъецировали для обеспечения FcRn человека возможности взаимодействовать с антителом, удержанным на сенсорным чипе. Используемый подвижный буфер содержал 50 ммоль/1 фосфат натрия, 150 ммоль/1 NaCl и 0,05% (масс./об.) Tween20 (pH 7,0). FcRn разбавляли с использованием каждого буфера. Чип регенерировали с использованием 10 ммоль/1 глицин-HCl (pH 1,5). Анализы проводили исключительно при 25 градусов C. Константу скорости ассоциации ka (1/Мс) и константу скорости диссоциации kd (1/с), каждая из которых представляет собой кинетические параметры, рассчитывали, основываясь на сенсограммах, полученных в анализах, и KD (M) каждого антитела для FcRn человека определяли из этих значений. Каждый параметр рассчитывали с использованием программного обеспечения Biacore T100 Evaluation (GE Healthcare).
Результат оценки связывания FcRn человека в нейтральных условиях (pH 7,0) посредством Biacore представлен в таблицах 6-1 и 6-2. KD интактного IgG1 не возможно было рассчитать, так как он продемонстрировал только очень слабое связывание. Таким образом, KD представлено как ND в таблице 6-1.
[Пример 6] Тестирование in vivo зависимых от рН антител, связывающих рецептор IL-6 человека, с увеличенным связыванием FcRn человека в нейтральных условиях
Зависимые от рН антитела, связывающие рецептор IL-6 человека со способностью связывания FcRn человека в нейтральных условиях, получали с использованием тяжелых цепей, полученных, как описано в пример 4, для придания способности связывания с FcRn человека в нейтральных условиях. Антитела оценивали на их эффект элиминации антигена in vivo. Конкретно, антитела, перечисленные ниже, экспрессировали и очищали способами, известными специалистам в данной области, как описано в ссылочном примере 2:
Fv4-IgG1, содержащий VH3-IgG1 и VL3-CK;
Fv4-IgG1-v2, содержащий VH3-IgG1-v2 и VL3-CK;
Fv4-IgG1-F14, содержащий VH3-IgG1-F14 и VL3-CK;
Fv4-IgG1-F20, содержащий VH3-IgG1-F20 и VL3-CK;
Fv4-IgG1-F21, содержащий VH3-IgG1-F21 и VL3-CK;
Fv4-IgG1-F25, содержащий VH3-IgG1-F25 и VL3-CK;
Fv4-IgG1-F29, содержащий VH3-IgG1-F29 и VL3-CK;
Fv4-IgG1-F35, содержащий VH3-IgG1-F35 и VL3-CK;
Fv4-IgG1-F48, содержащий VH3-IgG1-F48 и VL3-CK;
Fv4-IgG1-F93, содержащий VH3-IgG1-F93 и VL3-CK; и
Fv4-IgG1-F94, содержащий VH3-IgG1-F94 и VL3-CK.
Посредством таких способов, описанных в примере 3, полученные зависимые от рН антитела, связывающие рецептор IL-6 человека, тестировали in vivo с использованием трансгенных мышей с FcRn человека (B6.mFcRn-/-.hFcRn Tg линия 276 +/+ мышь, Jackson Laboratories; Methods Mol Biol. (2010) 602: 93-104).
Динамика концентрации в плазме растворимого рецептора IL-6 человека после внутривенного введения трансгенным мышам с FcRn человека представлена на фиг.8. Результаты тестирования показали, что концентрация в плазме растворимого рецептора IL-6 человека сохранялась на низком уровне в течение времени в присутствии любого из зависимых от рН антител, связывающих рецептор IL-6 человека, с увеличенным связыванием FcRn человека в нейтральных условиях, по сравнению со связыванием в присутствии Fv4-IgG1, который почти не обладает способностью связывания с FcRn человека в нейтральных условиях. Наряду с другими, антитела, для которых получали заметный эффект, включают, например, Fv4-IgG1-F14. Концентрация в плазме растворимого рецептора IL-6 человека, введенного одновременно с Fv4-IgG1-F14, продемонстрировала снижение приблизительно в 54 раза через одни сутки после введения по сравнению с таковой растворимого рецептора IL-6 человека, введенного одновременно с Fv4-IgG1. Кроме того, концентрация в плазме растворимого рецептора IL-6 человека, введенного одновременно с Fv4-IgG1-F21, продемонстрировала снижение приблизительно в 24 раза через семь часов после введения по сравнению с таковой растворимого рецептора IL-6 человека, введенного одновременно с Fv4-IgG1. Кроме того, концентрация в плазме растворимого рецептора IL-6 человека, введенного одновременно с Fv4-IgG1-F25 через семь часов после введения, была ниже предела детекции (1,56 нг/мл). Таким образом, Fv4-IgG1-F25, как и ожидалось, способствует поразительному снижению в 200 или более раз в концентрации растворимого рецептора IL-6 человека относительно концентрации растворимого рецептора IL-6 человека, введенного одновременно с Fv4-IgG1. Полученные результаты, описанные выше, продемонстрировали, что увеличение связывания с FcRn человека зависимых от pH антигенсвязывающих антител в нейтральных условиях является высокоэффективным для усиления эффекта элиминации антигена. Между тем, тип аминокислотных изменений для усиления связывания FcRn человека в нейтральных условиях, которые вводят для усиления эффекта элиминации антигена, конкретно не ограничен; и такие изменения включают те, которые представлены в таблицах 6-1 и 6-2. Эффект элиминации антигена можно предсказывать как увеличенный посредством любого введенного изменения in vivo.
Кроме того, концентрацию в плазме растворимого рецептора IL-6 человека вводили одновременно с одним из четырех типов зависимых от рН антител, связывающих рецептор IL-6 человека, Fv4-IgG1-F14, Fv4-IgG1-F21, Fv4-IgG1-F25 и Fv4-IgG1-F48, сохраняющийся на более низком уровне в течение времени, чем таковая растворимого рецептора IL-6 человека, введенного отдельно. Такие зависимые от pH антитела, связывающие рецептор IL-6 человека, можно вводить в организм, где концентрацию в плазме растворимого рецептора IL-6 человека сохраняют постоянной (стационарной) для сохранения в плазме концентрации растворимого рецептора IL-6 человека на более низком уровне, чем стационарная концентрация в плазме. Конкретно, концентрацию антигена в плазме in vivo можно снижать посредством введения такого антитела в организм.
[Пример 7] Оценка эффективности низких доз (0,01 мг/кг) Fv4-IgG1-F14
Fv4-IgG1-F14, полученный, как описано в примере 6, тестировали в низких дозах (0,01 мг/кг) посредством такого же способа тестирования in vivo, как описано в примере 6. Результаты (показаны на фиг.9) сравнивали с результатами, описанными в примере 6, которые получали посредством введения Fv4-IgG1 и Fv4-IgG1-F14 при 1 мг/кг.
Результаты продемонстрировали, что хотя концентрация антитела в плазме в группе, которой вводили Fv4-IgG1-F14 при 0,01 мг/кг, являлась приблизительно в 100 раз более низкой по сравнению с группой, которой вводили 1 мг/кг (фиг.10), показатели динамики концентрации в плазме растворимого рецептора IL-6 человека являлись сравнимыми друг с другом. Кроме того, показано, что концентрация в плазме растворимого рецептора IL-6 человека через семь часов после введения в группе, которой вводили Fv4-IgG1-F14 из расчета 0,01 мг/кг, снижали приблизительно в три раза по сравнению с таковой в группе, которой вводили Fv4-IgG1 из расчета 1 мг/кг. Кроме того, в присутствии Fv4-IgG1-F14, концентрация в плазме растворимого рецептора IL-6 человека, который вводили в различных дозах, являлась пониженной в течение времени в обеих группах, по сравнению с группой, которой вводили растворимый рецептор IL-6 человека по отдельности.
Результаты демонстрируют, что даже при введении в дозе, составляющей сотую, Fv4-IgG1, Fv4-IgG1-F14, которые получены в результате модификации Fv4-IgG1 для усиления связывания с FcRn человека в нейтральных условиях, эффективно снижают концентрацию в плазме растворимого рецептора IL-6 человека. Конкретно, предполагают, что антигены могут быть эффективно элиминированы даже в более низкой дозе, если зависимые от pH антигенсвязывающие антитела модифицируют для усиления их способности связывания с FcRn в нейтральных условиях.
[Пример 8] Тест in vivo, основанный на стационарной модели с использованием нормальных мышей
Оценка связывания с FcRn мыши в нейтральных условиях
VH3/L (WT)-IgG1, содержащий VH3-IgG1 (SEQ ID NO: 6) и L (WT) (SEQ ID NO: 5), VH3/L (WT)-IgG1-v2, содержащий VH3-IgG1-v2 (SEQ ID NO: 9) и L (WT) (SEQ ID NO: 5), и VH3/L (WT)-IgG1-F20, содержащий VH3-IgG1-F20 (SEQ ID NO: 10) и L (WT) (SEQ ID NO: 5), каждый из которых получали, как описано в примере 5, оценивали на связывание с FcRn мыши в нейтральных условиях (pH 7,4) посредством способа, описанного ниже.
Связывание между антителом и FcRn мыши анализировали кинетически с использованием Biacore T100 (GE Healthcare). Подходящее количество белка L (ACTIGEN) иммобилизовали на сенсорном чипе CM4 (GE Healthcare) посредством способа на основе связанных аминов, и обеспечивали фиксацию представляющего интерес антигена на чипе. Затем разведенные растворы FcRn и подвижный буфер (в качестве контрольного раствора) инъецировали для обеспечения возможности взаимодействия FcRn мыши с антителом, зафиксированном на сенсорном чипе. Используемый подвижный буфер содержал 50 ммоль/1 фосфата натрия, 150 ммоль/1 NaCl и 0,05% (масс./об.) Tween 20 (pH 7,4). FcRn разбавляли с использованием каждого буфера. Чип подвергали регенерации с использованием 10 ммоль/1 глицин-HCl (pH 1,5). Анализы проводили исключительно при 25 градусов C. Константу скорости ассоциации ka (1/Мс) и константу скорости диссоциации kd (1/с), каждая из которых представляет собой кинетические параметры, рассчитывали, основываясь на сенсограммах, полученных в анализах, и KD (M) каждого антитела для FcRn мыши определяли из этих значений. Каждый параметр рассчитывали с использованием программного обеспечения Biacore T100 Evaluation (GE Healthcare).
Результаты представлены в таблице 7 (аффинность для FcRn мыши при pH 7,4). VH3/L (WT)-IgG1 (IgG1 в таблице 7), чья константная область представляет собой интактный IgG1, который демонстрирует только очень слабое связывание с FcRn мыши. Таким образом, KD невозможно рассчитать, и ее обозначают как ND в таблице 7. Анализ результатов показал, что измененные антитела с повышенным связыванием с FcRn человека в нейтральных условиях также продемонстрировали повышенное связывание с FcRn мыши в нейтральных условиях.
Тестирование in vivo с использованием нормальных мышей с постоянной концентрацией в плазме растворимого рецептора IL-6 человека
С использованием H54/L28-IgG1, Fv4-IgG1, Fv4-IgG1-v2 и Fv4-IgG1-F20, полученных, как описано в примерах 1 и 5, тестирование in vivo проводили способом, описанным ниже.
Инфузионный тест in vivo с использованием нормальных мышей
Инфузионный насос (MINI-OSMOTIC PUMP MODEL 2004; alzet), содержащий растворимый рецептор IL-6 человека, имплантировали под кожу спины нормальных мышей (мыши C57BL/6J; Charles River Japan) для получения модели животных, где концентрацию в плазме растворимого рецептора IL-6 человека сохраняют постоянной. Антитела против рецептора IL-6 человека вводили модели животных для оценки динамики in vivo после введения растворимого рецептора IL-6 человека. Моноклональное антитело против CD4 мыши (R&D) вводили из расчета 20 мг/кг единожды в хвостовую вену для супрессии продукции нейтрализующего антитела против растворимого рецептора IL-6 человека. Затем, инфузионный насос, содержащий 92,8 микрограмм/мл растворимого рецептора IL-6 человека, имплантировали под кожу спины мыши. Через трое суток после имплантации инфузионного насоса, антитела против рецептора IL-6 человека вводили из расчета 1 мг/кг единожды в хвостовую вену. Кровь собирали через 15 минут, семь часов, одни сутки, двое суток, трое суток, четверо суток, семь суток, 14 суток, 21 сутки и через 28 суток после введения антитела против рецептора IL-6 человека. Собранную кровь немедленно центрифугировали при 15000 об/мин и 4 градусов C в течение 15 минут для отделения плазмы. Перед проведением анализа отделенную плазму хранили в холодильнике при -20 градусов C или ниже.
Определение концентрации в плазме антитела против рецептора IL-6 человека посредством ELISA
Используемый способ был таким же, как описано в примере 3.
Определение концентарции в плазме hsIL-6R посредством электрохемилюминесцентного анализа
Используемый способ был таким же, как описано в примере 1.
Как показано на фиг.11, концентрацию в плазме растворимого рецептора IL-6 человека повышали до 650 нг/мл (в 15 раз до введения), когда H54/L28-IgG1, нейтрализующее антитело против растворимого рецептора IL-6 человека, вводили нормальным мышам (группа hsIL-6R), у которых концентрация в плазме растворимого рецептора IL-6 человека сохранялась постоянной приблизительно в количестве 40 нг/мл. С другой стороны, концентрацию в плазме растворимого рецептора IL-6 человека поддерживали в количестве приблизительно 70 нг/мл в группе, которой вводили Fv4-IgG1, который получен в результате обеспечения H54/L28-IgG1 зависимой от pH спосбности связывания с антигеном. Это позволяет предположить, что увеличение концентрации в плазме растворимого рецептора IL-6 человека, вызванное введением H54/L28-IgG1, типичного нейтрализующего антитела, можно снижать до приблизительно одной десятой посредством обеспечения способности связывания, зависящей от рН.
Кроме того, концентрация в плазме растворимого рецептора IL-6 человека, как показано, сохраняется постоянной при одной десятой или ниже одной десятой от стационарной концентрацией посредством введение Fv-IgG1-v2 или Fv-IgG1-F20, каждый из которых получен в результате введения изменения в зависимые от pH антитела, связывающие рецептор IL-6, для снижения связывания FcRn в нейтральных условиях. Если вводили Fv-IgG1-v2, концентрация в плазме растворимого рецептора IL-6 человека через 14 суток после введения составляла приблизительно 2 нг/мл. Таким образом, Fv-IgG1-v2 мог снижать концентрацию до 1/20 от уровня до введения. Между тем, если вводили Fv-IgG1-F20, концентрация в плазме растворимого рецептора IL-6 человека через семь часов, одни сутки, двое суток и четверо суток после введения была ниже предела детекции (1,56 нг/мл). Это позволяет предположить, что Fv-IgG1-F20 снижал концентрацию до 1/25 или ниже 1/25 от уровня до введения.
Результаты, описанные выше, демонстрируют, что концентрацию антигена в плазме можно значительно снизить посредством увеличения скорости элиминации антигена в плазме, посредством введения антитела, обладающего и зависимой от рН антигенсвязывающей способностью, и активностью связывания с FcRn в нейтральных условиях на модели животных, для которой концентрацию антигена в плазме сохраняют постоянной.
Типичные антитела, такие как H54/L28-IgG1, могут только нейтрализовать действие целевого антигена посредством связывания с целевым антигеном, и даже более того, они увеличивают концентрацию антигена в плазме. В отличие от этого, антитела, обладающие и зависимой от рН антигенсвязывающей способностью, и активностью связывания с FcRn в нейтральных условиях, как обнаружено, способны не только нейтрализовать целевой антиген, но также и снижать концентрацию в плазме целевого антигена. Можно ожидать, что эффект удаления антигена из плазмы является более существенным, чем нейтрализация. Кроме того, удаление антигена может также содействовать целевым антигенам, для которых удаление является недостаточно эффективными посредством одной нейтрализации.
[Пример 9] Идентификация порогового уровня аффинности связывания с FcRn человека в нейтральном pH, требуемого для увеличения элиминации антигена, и зависимость между элиминацией антигена и аффинностью связывания с FcRn человека при нейтральном pH
Получение антитела для исследования in vivo
Получали варианты Fc Fv4-IgG1, содержащего VH3-IgG1 (SEQ ID NO: 6) и VL3-CK (SEQ ID NO: 7), с увеличенным связыванием с FcRn в нейтральных условиях pH. Конкретно, получали VH3-M73 (SEQ ID NO: 15) и VH3-IgG1-v1 (SEQ ID NO: 8). Замены аминокислот вводили способами, известными специалистам в данной области, как описано в ссылочном примере 1.
H54/L28-IgG1, содержащий H54 (SEQ ID NO: 1) и L28 (SEQ ID NO: 2), Fv4-IgG1, содержащий VH3-IgG1 (SEQ ID NO: 6) и VL3-CK (SEQ ID NO: 7), Fv4-M73, содержащий VH3-M73 (SEQ ID NO: 15) и VL3-CK (SEQ ID NO: 7), Fv4-IgG1-v1, содержащий VH3-IgG1-v1 (SEQ ID NO: 8) и VL3-CK (SEQ ID NO: 7), и Fv4-IgG1-v2, содержащий VH3-IgG1-v2 (SEQ ID NO: 9) и VL3-CK (SEQ ID NO: 7), экспрессировали и очищали способом, известным специалистам в данной области, описанным в ссылочном примере 2.
Оценка аффинности связывания антитела с FcRn человека в условиях нейтральных значений pH
VH3/L (WT)-IgG1, содержащий VH3-IgG1 (SEQ ID NO: 6) и L (WT) (SEQ ID NO: 5), VH3/L (WT)-M73, содержащий VH3-M73 (SEQ ID NO: 15) и L (WT) (SEQ ID NO: 5), VH3/L (WT)-IgG1-v1, содержащий VH3-IgG1-v1 (SEQ ID NO: 8) и L (WT) (SEQ ID NO: 5), и VH3/L (WT)-IgG1-v2, содержащий VH3-IgG1-v2 (SEQ ID NO: 9) и L (WT) (SEQ ID NO: 5), каждый из которых получали, как описано в примере 2, оценивали на связывание с FcRn человека в условиях нейтральных pH (pH 7,0).
Активность связывания VH3/L (WT)-IgG1-v1 и VH3/L (WT)-IgG1-v2 с FcRn человека измеряли с использованием способа, описанного в примере 5. Вследствие низкой активности связывания VH3/L (WT)-IgG1 и VH3/L (WT)-M73 c FcRn человека, активность связывания c FcRn человека не может быть измерена с использованием способа, описанного в примере 5, таким образом, эти антитела оценивали посредством способа, описанного ниже. Связывание между антителом и FcRn человека анализировали кинетически с использованием Biacore T100 (GE Healthcare). Подходящее количество белка L (ACTIGEN) иммобилизовали на сенсорном чипе CM4 (GE Healthcare) посредством способа на основе связанных аминов, и обеспечивали захват представляющего интерес антитела на чипе. Затем, разведенные растворы FcRn и подвижный буфер в качестве контрольного раствора инъецировали для обеспечения возможности взаимодействия FcRn человека с антителом, зафиксированном на сенсорном чипе. Используемый подвижный буфер содержал 50 ммоль/1 фосфата натрия, 150 ммоль/1 NaCl и 0,05% (масс./об.) Tween 20 (pH 7,0). FcRn разбавляли с использованием каждого буфера. Чип регенерировали с использованием 10 ммоль/1 глицин-HCl (pH 1,5). Анализы проводили при 25 градусов C.
KD (M) каждого антитела получали из данных сенсограммы с исользованием програмного обеспечения Biacore T100 Evaluation (GE Healthcare), которая одновременно соответствует фазам ассоциации и диссоциации сенсограмм и в целом соответствует всем кривым в рабочем наборе. Сенсограммы соответствовали модели связывания 1:1, модели «связывание Ленгмюра», задаваемой программным обеспечением Biacore T100 Evaluation. Для некоторых из связывающих взаимодействий, KD получали посредством нелинейного регрессионного анализа графиков из Req, реакции равновесного связывания, в зависимости от log концентрации анализируемого вещества с использованием равновесного подхода.
Результаты связывания с FcRn человека в нейтральных условиях (pH 7,0) посредством Biacore представлены в таблице 8.
Исследование in vivo эффекта антитела на элиминацию антигена в модели совместной инъекции с использованием трансгенной линии 276 мышей с FcRn человека
In vivo исследование антител с использованием модели совместной инъекции проводили, как описано в примере 3. Антитела против рецептора IL-6 человека, используемые в этом исследовании, представляют собой описанные выше H54/L28-IgG1, Fv4-IgG1, Fv4-M73, Fv4-IgG1-v1 и Fv4-IgG1-v2. Мыши, используемые в этом исследовании, представляют собой трансгенных мышей с FcRn человека (B6.mFcRn-/-.hFcRn Tg линия 276 +/+ мышь, Jackson Laboratories; Methods Mol Biol. (2010) 602: 93-104).
Как показано на фиг.12, фармакокинетики H54/L28-IgG1, Fv4-IgG1, Fv4-M73, Fv4-IgG1-v1 и Fv4-IgG1-v2 являлись сравнимыми, и эти антитела сохранялись в одинаковой концентрации в плазме в течение исследования.
Динамика концентрации hsIL-6R в плазме представлена на фиг.13. По сравнению с hsIl-6R, введенным с Fv4-IgG1, hsIL-6R, введенный с Fv4-IgG1-v2, продемонстрировал повышенный клиренс, в то время как hsIL-6R, введенный с Fv4-M73 и Fv4-IgG1-v1, продемонстрировал сниженный клиренс. Хотя все варианты Fc, M73, v1 и v2 обладают повышенной аффинностью связывания с FcRn человека в нейтральных условиях pH (pH 7,0), показано, что только Fv4-IgG1-v2, но не Fv4-M73 и Fv4-IgG1-v1, продемонстрировал повышенный клиренс hsIL-6R. Это показывает, что для увеличения клиренса антигена аффинность связывания антитела с FcRn человека при pH 7,0 должна быть по меньшей мере большей, чем для IgG1-v1, чья аффинность связывания с FcRn человека при pH 7,0 составляет KD 3,2 микромоля или в 28 раз более высокая, чем у интактного IgG1 человека (аффинность связывания с FcRn человека представлена KD, равной 88 микромолям).
На фиг.14 описано взаимосвязь между аффинностью связывания вариантов Fc с FcRn человека при pH 7,0 и концентрацией в плазме hsIL-6R через сутки 1 после совместной инъекции hsIL-6R и вариантов Fc. Варианты Fc, описанные в этом примере и примере 6 (Fv4-IgG1, Fv4-M73, Fv4-IgG1-v1, Fv4-IgG1-v2, Fv4-IgG1-F14, Fv4-IgG1-F20, Fv4-IgG1-F21, Fv4-IgG1-F25, Fv4-IgG1-F29, Fv4-IgG1-F35, Fv4-IgG1-F48, Fv4-IgG1-F93 и Fv4-IgG1-F94), представлены на графике. Посредством повышения аффинности связывания антитела с FcRn человека при pH 7,0, концентрация в плазме hsIL-6R, которая отражает клиренс антигена, вначале увеличивается, но затем стремительно снижается. Это показывает, что для увеличения клиренса антигена, по сравнению с интактным IgG1 человека, аффинность связывания антитела с FcRn человека при pH 7,0 предпочтительно должна быть более высокой, чем KD, равная 2,3 микромолям (значение, полученное из кривой аппроксимации, фиг.14). Аффинность связывания антитела с FcRn человека между KD, равной 88 микромолям, и KD, равной 2,3 микромолей, будет скорее снижать клиренс антигена (более высокая концентрация hsIL-6R). Другими словами, аффинность связывания антитела с FcRn человека при pH 7,0 предпочтительно должна быть в 38 раз более высокой, чем для интактного IgG1 человека для увеличения элиминации антигена или снижать клиренс антигена другим способом.
На фиг.15 представлена зависимость между аффинностью связывания вариантов Fc с FcRn человека при pH 7,0 и концентрация антитела в плазме через сутки 1 после совместной инъекции hsIL-6R и вариантов Fc. Варианты Fc, описанные в этом примере и примере 6 (Fv4-IgG1, Fv4-M73, Fv4-IgG1-v1, Fv4-IgG1-v2, Fv4-IgG1-F14, Fv4-IgG1-F20, Fv4-IgG1-F21, Fv4-IgG1-F25, Fv4-IgG1-F29, Fv4-IgG1-F35, Fv4-IgG1-F48, Fv4-IgG1-F93 и Fv4-IgG1-F94), изображают на графике. Посредством увеличения аффинности связывания антитела с FcRn человека при pH 7,0, концентрация в плазме антитела, которая отражает фармакокинетики антител (клиренс), вначале увеличивается, но затем стремительно снижается. Это демонстрирует, что для поддержания фармакокинетики антитела, аналогичной интактному IgG1 человека (аффинность связывания с FcRn человека представляет собой KD, равную 88 микромолям), аффинность антитела по отношению к FcRn человека при pH 7,0 должна быть более слабой, чем KD, равная 0,2 микромолям (значение, полученное из аппроксимирующей кривой на фиг.15). Аффинность связывания антитела для FcRn человека более высокая, чем KD, равная 0,2 микромолям, увеличивала клиренс антитела (т.е. более быстрая элиминация антитела из плазмы). Другими словами, аффинность связывания антитела с FcRn человека, которая при pH 7,0 должна быть в 440 раз более высокой, чем для интактного IgG1 человека, чтобы продемонстрировать аналогичную фармакокинетику антител в качестве интактного IgG1 человека, или другим способом, будет приводить к быстрой элиминации антитела из плазмы.
Принимая во внимание и фиг.14 и 15, для увеличения клиренса антигена (т.е. снижения концентрации антигена в плазме) по сранению с IgG1, в то время как сохраняется фармакокинетика антител, аналогичная интактному IgG1 человека, аффинность связывания антитела с FcRn человека при pH 7,0 должна быть между 2,3 микромолей и 0,2 микромоле, или другими словами, аффинность связывания антитела с FcRn человека при pH 7,0 должна быть в пределах диапазона от 38 раз до 440 раз более высокой, чем для интактного IgG1 человека. Такое антитело с аналогичной фармакокинетикой как у IgG1 с длительной активностью элиминации антигена может быть полезным для терапевтических средств на основе антитела, которым требуется более длительные интервалы дозирования, как в случае хронического заболевания вследствие его пролонгированного действия.
С другой стороны, посредством увеличения аффинности связывания антитела с FcRn человека при pH 7,0 более чем KD, равная 0,2 микромоля, или другими словами, посредством увеличения аффинности связывания антитела с FcRn человека при pH 7,0 более чем в 440 раз по сравнению с интактным IgG1 человека, увеличивается клиренс антигена до большей степени в пределах кратковременного действия, хотя антитело элимируют из плазмы быстрее, чем для интактного IgG1 человека. Такое антитело со способностью индуцировать быстрое и сильное снижение концентрации антигена будет являться полезным для терапевтического средства на основе антитела, такого как для острого заболевания, где связанный с заболеванием антиген необходимо удалять из плазмы вследствие его быстродействующего свойства.
Количество антигена, элиминируемое из плазмы посредством антитела, представляет собой важный фактор для оценки эффективности элиминации антигена посредством введения вариантов Fc антитела, обладающих повышенной аффинностью связывания с FcRn человека при pH 7,0. Для оценки эффективности элиминации антигена посредством антитела проводили следующий расчет в каждый момент времени исследования in vivo, описанный в этом примере и примере 6.
Значение A: молярная концентрация антигена в каждый момент времени;
Значение B: молярная концентрация антигена в каждый момент времени;
Значение C: молярная концентрация антигена по отношению к молярной концентрации антитела (молярное отношение антигена/антитело) в каждый момент времени
C=A/B
Зависимость от времени значения C (молярное отношение антиген/антитело) для каждого антитела описана на фиг.16. Низкие значения C показывают высокую элиминацию антигена посредством антитела, тогда как высокое значение C показывает низкую эффективность элиминации антигена по отношению к антителу. Низкое значение C по сравнению с IgG1 показывает, что более высокая эффективность элиминации антигена достигается посредством вариантов Fc, в то время как высокие значения C по сравнению с IgG1 показывают, что варианты Fc обладают отрицательным эффектом на эффективность элиминации антигена. Все варианты Fc за исключением Fv4-M73 и Fv4-IgG1-v1 продемонстрировал повышенную эффективность элиминации антигена по сравнению с Fv4-IgG1. Fv4-M73 и Fv4-IgG1-v1 продемонстрировали негативное последствие на эффективность элиминации антигена, который согласуется с фиг.14.
На фиг.17 представлена зависимость между аффинностью связывания Fc-вариантов с FcRn человека при pH 7,0 и значение C (молярное отношение антиген/антитело) через сутки 1 после совместной инъекции hsIL-6R и вариантов Fc. Варианты Fc, описанные в этом примере и примере 6 (Fv4-IgG1, Fv4-M73, Fv4-IgG1-v1, Fv4-IgG1-v2, Fv4-IgG1-F14, Fv4-IgG1-F20, Fv4-IgG1-F21, Fv4-IgG1-F25, Fv4-IgG1-F29, Fv4-IgG1-F35, Fv4-IgG1-F48, Fv4-IgG1-F93, и Fv4-IgG1-F94), представлены графически. Это показывает, что для достижения более высокой эффективности элиминации антигена по сравнению с интактным IgG1 человека, аффинность антитела к FcRn человека при pH 7,0 должна быть более высокой, чем KD, равная 3,0 микромолям (значение, полученное из аппроксимирующей кривой на фиг.17). Другими словами, аффинность связывания антитела с FcRn человека при pH 7,0 должна быть по меньшей мере в 29 раз более высокой, чем для интактного IgG1 человека для достижения более высокой эффективности элиминации антигена по сравнению с интактным IgG1 человека.
В заключение, группа вариантов антител, обладающая аффинностью связывания с FcRn при pH 7,0 между KD, равной 3,0 микромоля и 0,2 микромоля, или другими словами, группа вариантов антител, обладающих аффинностью связывания с FcRn при pH 7,0 в пределах диапазона от 29 раз до 440 раз более высокого, чем для интактного IgG1 человека, обладает фармакокинетикой, аналогичной фармакокинетикам антител для IgG1, но имеют повышенную способность к элиминации антитела из плазмы. Таким образом, такое антитело демонстрирует повышенную эффективность элиминации антигена по сравнению с IgG1. Фармакокинетика, аналогичная IgG1, будет давать возможность длительной элиминации антигена из плазмы (элиминация антигена длительного действия), и таким образом дает возможность создания более длительных интервалов дозирования, что может являться преимуществом для терапевтических средством на основе антител, предназначенных для лечения хронических заболеваний. Группа вариантов антител, обладающих аффинностью связывания с FcRn при pH 7,0 более высокой, чем KD, равная 0,2 микромоля, или другими словами, группа вариантов антител, обладающая аффинностью связывания с FcRn при pH 7,0, в 440 раз более высокой, чем для интактного IgG1 человека, обладает ускоренным клиренсом антигена (кратковременная элиминация антитела). Однако, так как такое антитело , дает возможность даже более ускоренного клиренса антитела (быстродействующая элиминация антигена), таким образом, такое антитело также демонстрирует повышенную эффективность элиминации антигена по сравнению с IgG1. Как показано в примере 8, Fv4-IgG1-F20 у нормальных мышей будет индуцировать обширную элиминацию антигена из плазмы в очень короткий период, но эффект элиминации антигена не является продолжительным. Такой профиль будет предпочтительным для воздействия на острое заболевание, где антиген, связанный с заболеванием, необходимо быстро и интенсивно выводить из плазмы в очень короткий период.
[Пример 10] Исследование in vivo Fv4-IgG1-F14 посредством стационарной модели инфузии с использованием трансгенной линии 276 мышей с FcRn человека
Исследование in vivo Fv4-IgG1-F14 посредством стационарной модели инфузии с использованием трансгенных мышей линии 276 с FcRn человека проводили, как описано в примере 1. Группа исследования состоит из контрольной группы (без антитела), Fv4-IgG1 в дозе 1 мг/кг и Fv4-IgG1-F14 в дозе 1 мг/кг, 0,2 мг/кг и 0,01 мг/кг.
На фиг.18 описывают временную зависимость концентрации в плазме hsIL-6R после введения антитела. По сравнению с базовым уровнем hsIL-6R без антитела, введение 1 мг/кг Fv4-IgG1 приводило в результате к повышению в несколько раз концентрации hsIL-6R в плазме. С другой стороны, введение 1 мг/кг Fv4-IgG1-F14 привело в результате к значительному снижению концентрации в плазме по сравнению с группой Fv4-IgG1 и контрольной группой. Через сутки 2, концентрация в плазме hsIL-6R не определялась (нижний предел определения концентрации в плазме hsIL-6R представляет собой 1,56 нг/мл в этой системе измерения), и это длилось вплоть до 14 суток.
Как показано в примере 1, H54/L28-IgG1-F14 продемонстрировал снижение концентрации в плазме hsIL-6R по сравнению с H54/L28-IgG1, но степень снижения была небольшой. Степень снижения была более высокой для вариабельной области Fv4, которая обладала зависимым от рН свойством связывания с hsIL-6R. Это демонстрирует, что хотя увеличивание аффинности связывания с FcRn человека при pH 7,0 является эффективным для снижения концентрации антигена в плазме, комбинация зависимого от рН связывания антигена и повышенная аффинность связывания с FcRn человека при нейтральном pH значимо увеличивает элиминацию антигена.
Исследование с использованием более низких доз Fv4-IgG1-F14 продемонстрировало, что даже при 0,01 мг/кг, 1/100 от 1 мг/кг снижается концентрация антигена в плазме ниже базовой линии, демонстрируя значимую эффективность молекулы для удаления антигена из плазмы.
[Пример 11] Сравнение трансгенной линии 276 и линии 32 мышей с FcRn человека в модели совместной инъекции
Предшествующие исследование in vivo провели с использованием трансгенной линии 276 мышей с FcRn человека (Jackson Laboratories). Для сравнения различий между трансгенной линии 276 мышей с FcRn человека и другой трансгенной линией, линия 32, предприняли исследование совместной инъекцией H54/L28-IgG1, Fv4-IgG1 и Fv4-IgG1-v2 с использованием трансгенной линии 32 мыши FcRn человека (B6.mFcRn-/-.hFcRn Tg линия 32 +/+ мышь (B6.mFcRn-/- hFCRN Tg32; B6.Cg-Fcgrt<tmlDcr> Tg(FCGRT)32Dcr) (Jackson #4915)), Jackson Laboratories; Mehtods Mol Biol. (2010) 602: 93-104). Способ исследования был аналогичен способу примера 3, но трансгенную линию 32 мышей с FcRn человека применяли вместо трансгенной линии 276 мышей с FcRn человека.
На фиг.19 описана зависимость от времени концентрации в плазме hsIL-6R и в трансгенной линии 276 мышей с FcRn человека и в трансгенной линии 32 мышей с FcRn человека. H54/L28-IgG1, Fv4-IgG1 и Fv4-IgG1-v2 продемонстрировали аналогичные временные зависимости концентраций в плазме hsIL-6R. В обеих линиях мышей увеличение аффинности связывания с FcRn человека при pH 7,0 увеличивало элиминацию антигена из плазмы (сравнение Fv4-IgG1 и Fv4-IgG1-v2) в одинаковой степени.
На фиг.20 описывают временную зависимость концентрации антитела в плазме и в трансгенной линии 276 мышей с FcRn человека и в трансгенной линии 32 мышей с FcRn человека. H54/L28-IgG1, Fv4-IgG1 и Fv4-IgG1-v2 продемонстрировало аналогичные временные зависимости концентрации антител в плазме.
В заключение, незначимое отличие наблюдали между линиями 276 и 32, демонстрируя, что вариант Fc для увеличения аффинности связывания с FcRn человека при pH 7,0 был более эффективным в двух различных трансгенных линиях мышей, экспрессирующих FcRn человека для увеличения элиминации концентрации антигена в плазме.
[Пример 12] Создание различных вариантов антитела Fc, обладающих повышенной аффинностью связывания с FcRn человека при нейтральном pH
Создание вариантов Fc
Различные мутации для увеличения аффинности связывания с FcRn человека в нейтральном диапазоне pH вводили в Fv4-IgG1 для дополнительного увеличения профиля элиминации антигена. Конкретно, аминокислотные мутации, представленные в таблицах от 9-1 до 9-14, вводили в константную область тяжелой цепи Fv4-IgG1 для создания вариантов Fc (число участков аминокислотных мутаций описаны согласно нумерация EU). Замены аминокислот вводили способом, известным специалистам в данной области, описанным в ссылочном примере 1.
Дополнительные варианты (от IgG1-F100 до IgG1-F599), каждый содержащий полученную тяжелую цепь и L (WT) (SEQ ID NO: 5), экспрессируют и очищают способами, известными специалистам в данной области, как описано в ссылочном примере 2.
Оценка связывания FcRn человека
Связывание между антителом и FcRn человека кинетически анализировали, как описано в примере 5 для IgG1-v1, IgG1-v2 и от IgG1-F2 до IgG1-F599 или примере 9 для IgG1 и M73. Результаты связывания с FcRn человека в нейтральных условиях (pH 7,0) представлены посредством Biacore в таблицах от 9-1 до 9-14.
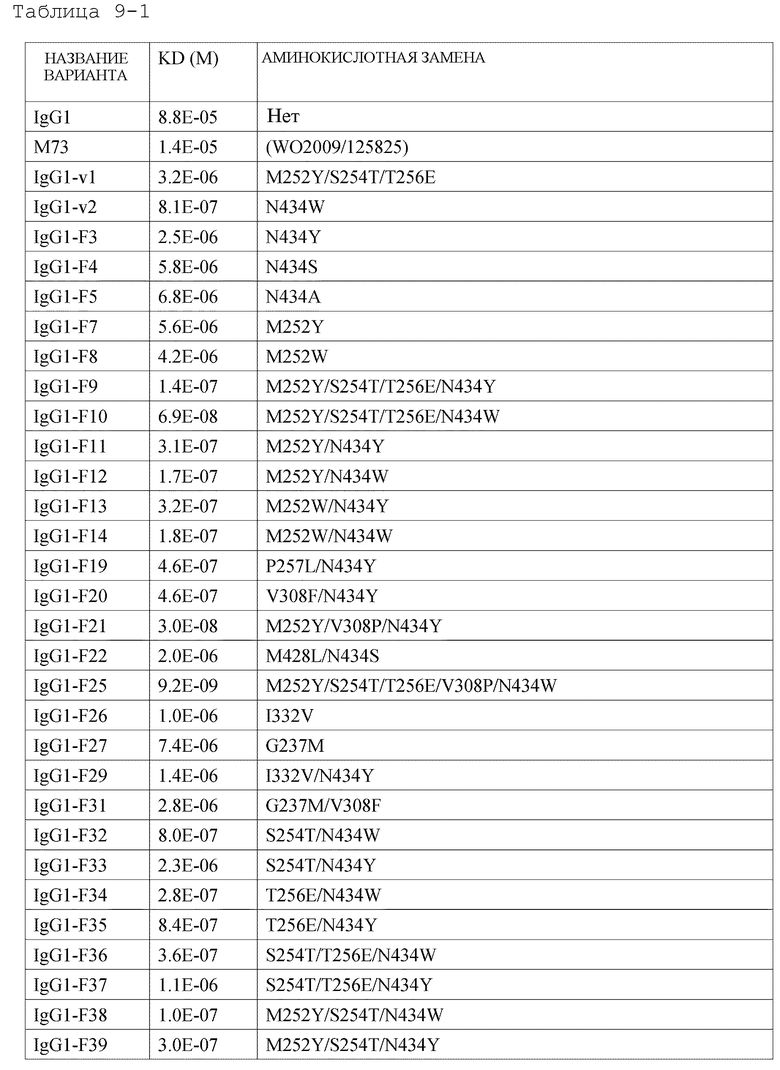
Таблица 9-2 представляет собой продолжение таблицы 9-1.
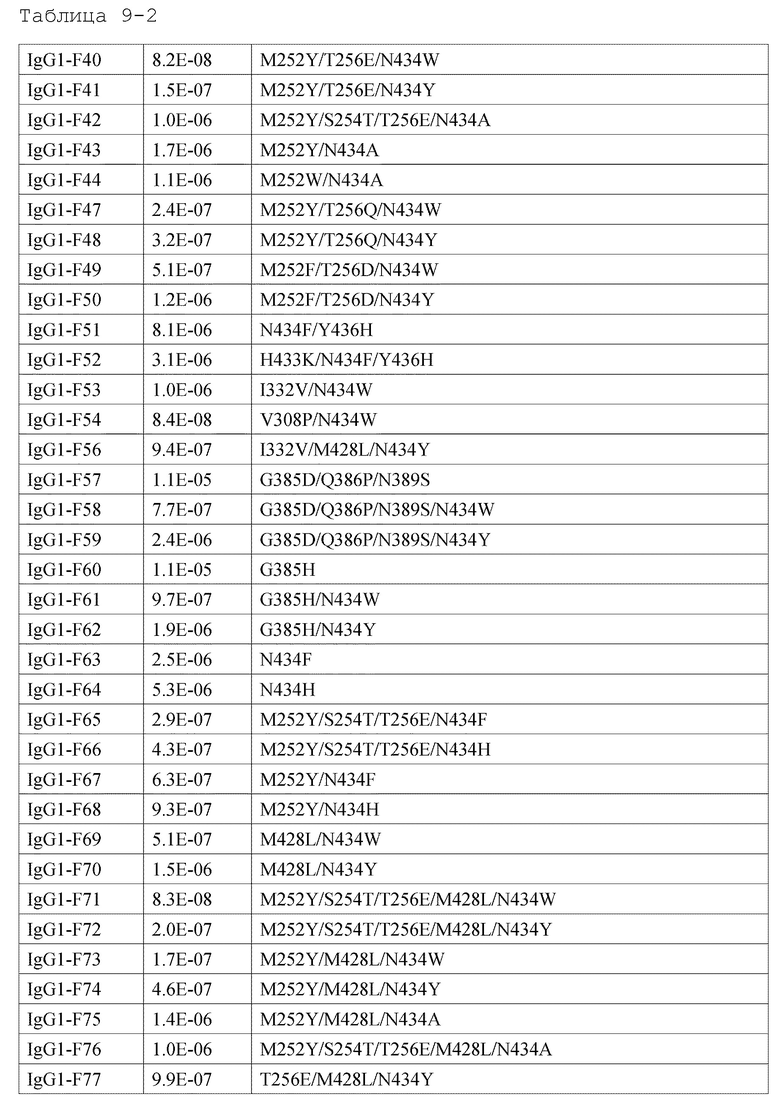
Таблица 9-3 представляет собой продолжение таблицы 9-2.
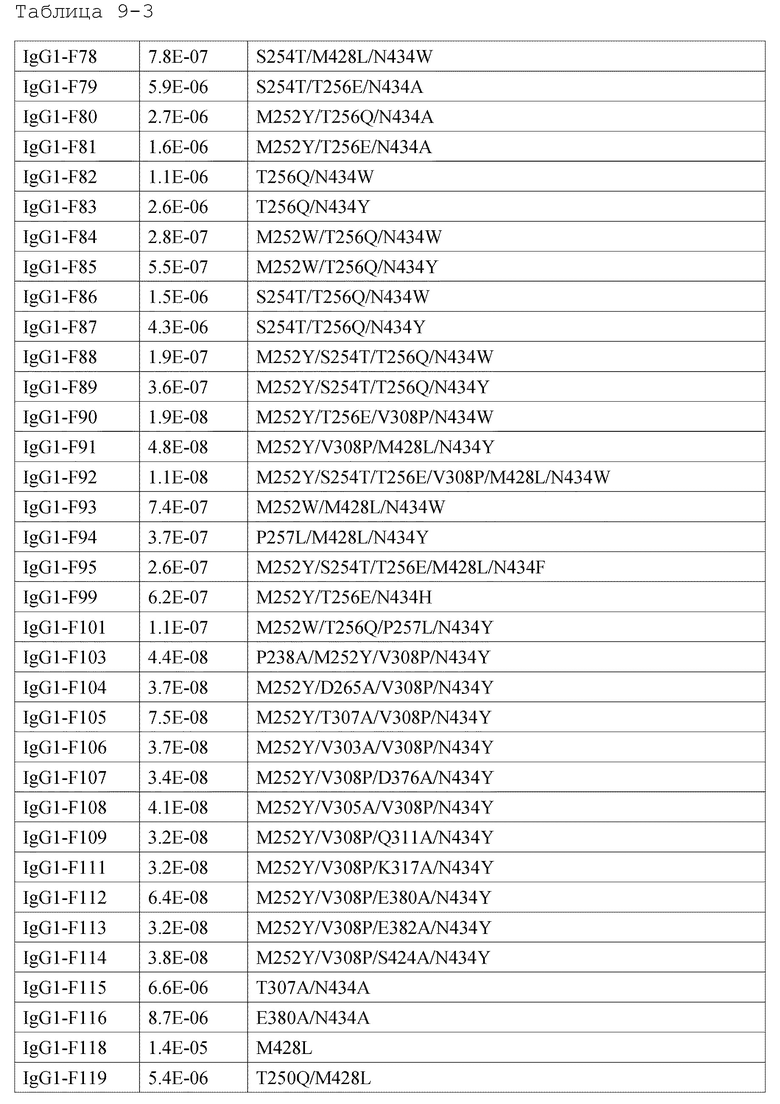
Таблица 9-4 представляет собой продолжение таблицы 9-3.
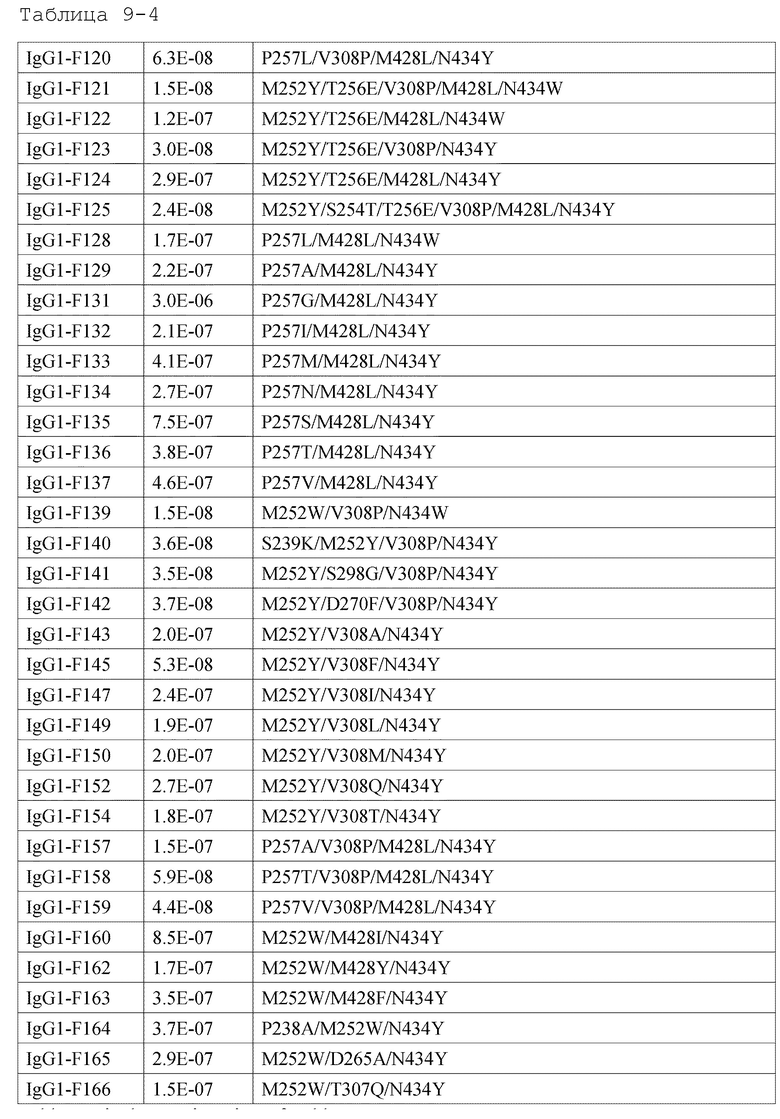
Таблица 9-5 представляет собой продолжение таблицы 9-4.
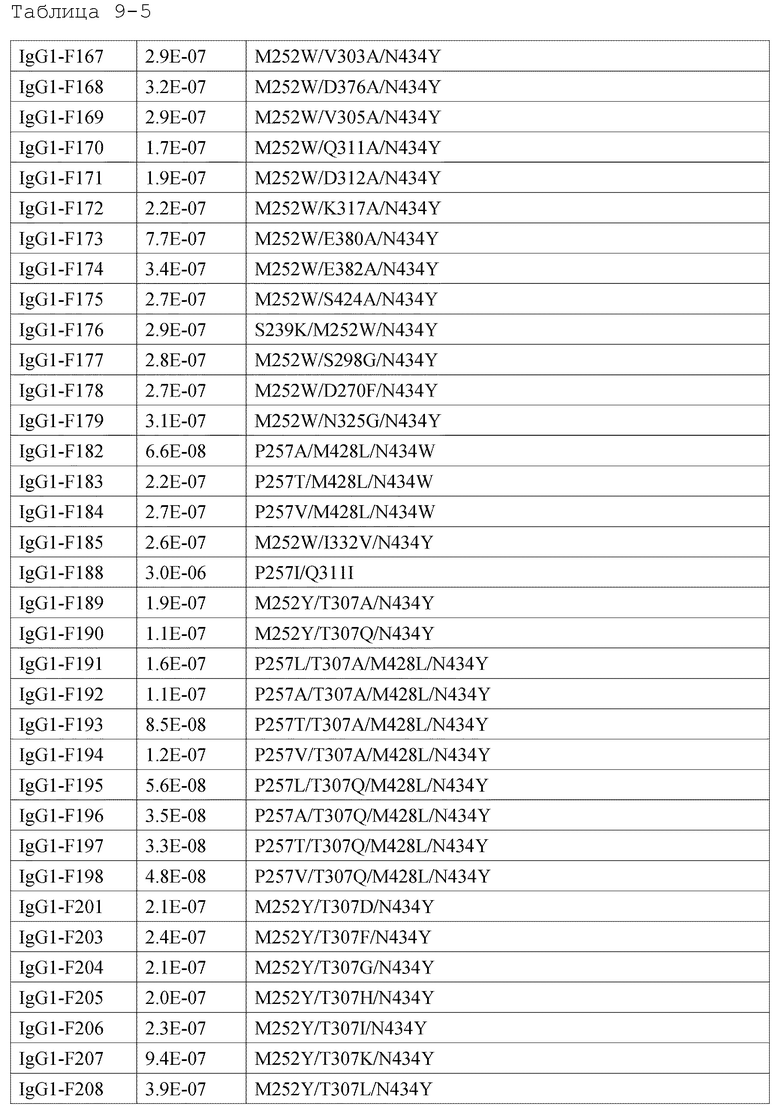
Таблица 9-6 представляет собой продолжение таблицы 9-5.
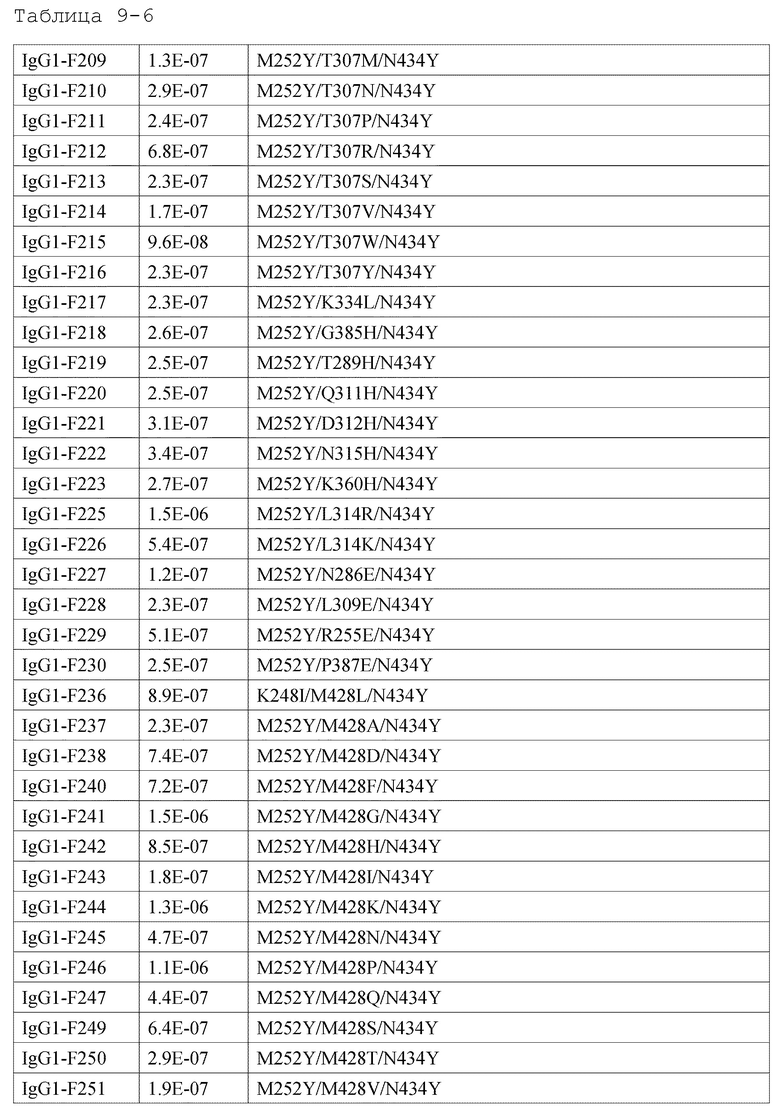
Таблица 9-7 представляет собой продолжение таблицы 9-6.

Таблица 9-8 представляет собой продолжение таблицы 9-7.
Таблица 9-9 представляет собой продолжение таблицы 9-8.

Таблица 9-10 представляет собой продолжение таблицы 9-9.
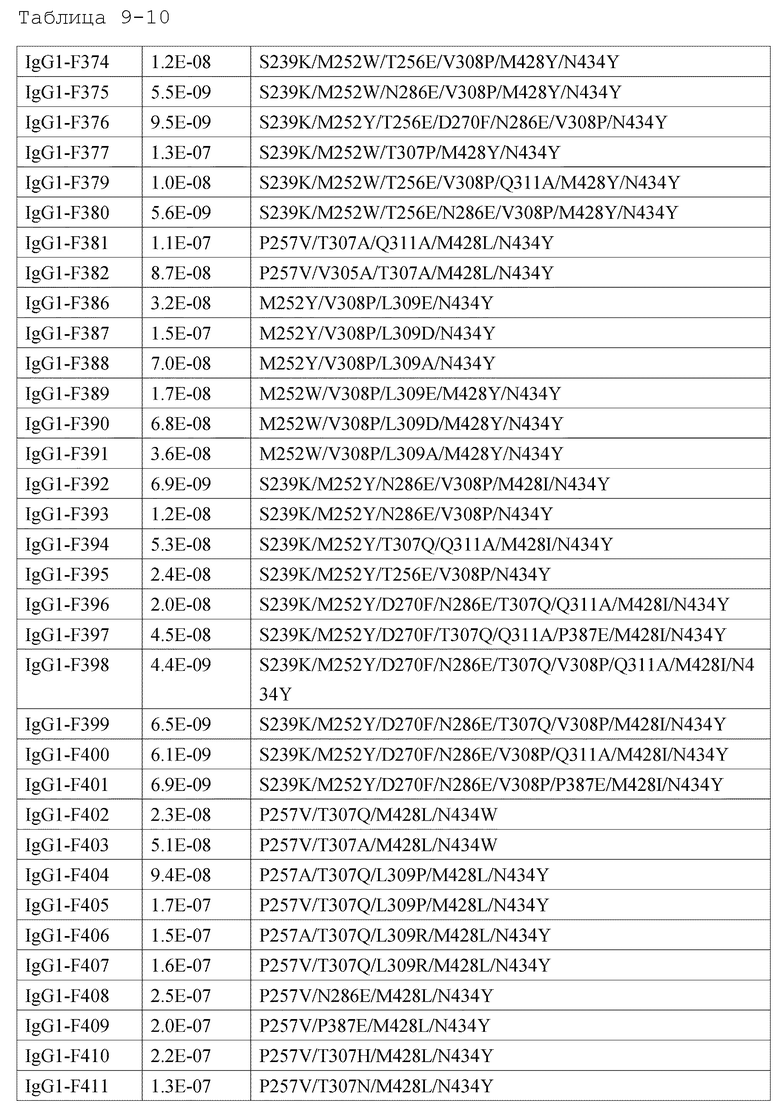
Таблица 9-11 представляет собой продолжение таблицы 9-10.
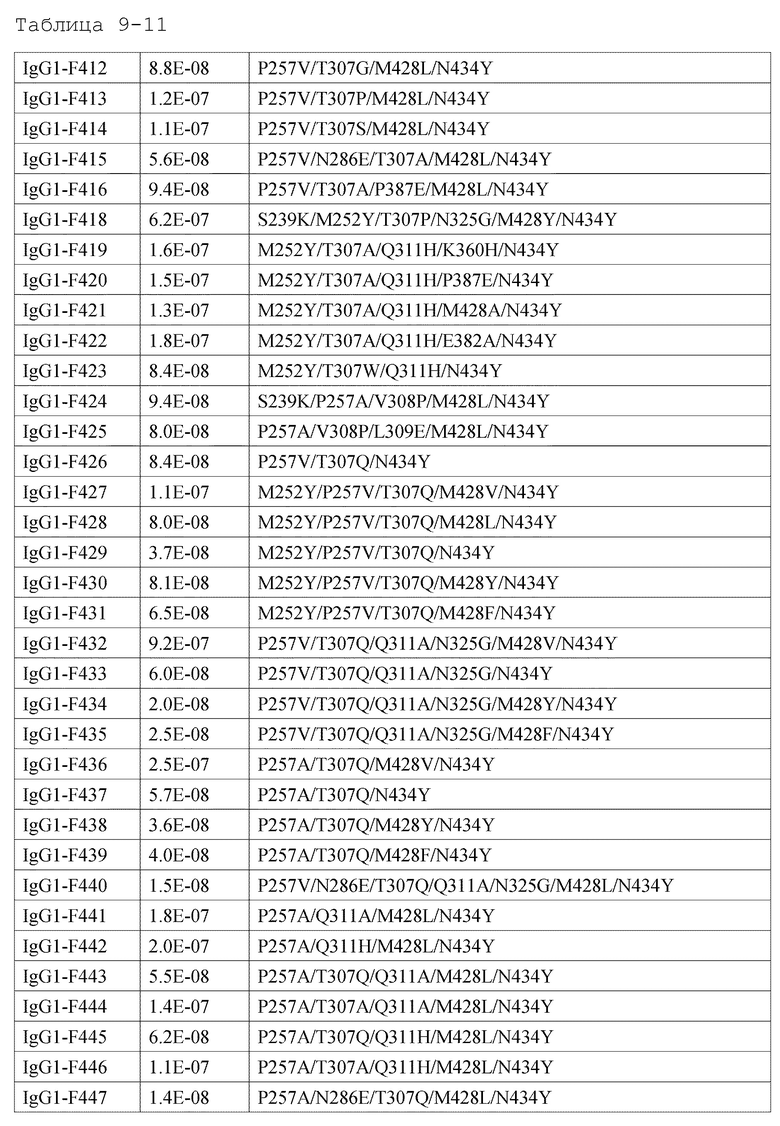
Таблица 9-12 представляет собой продолжение таблицы 9-11.

Таблица 9-13 представляет собой продолжение таблицы 9-12.
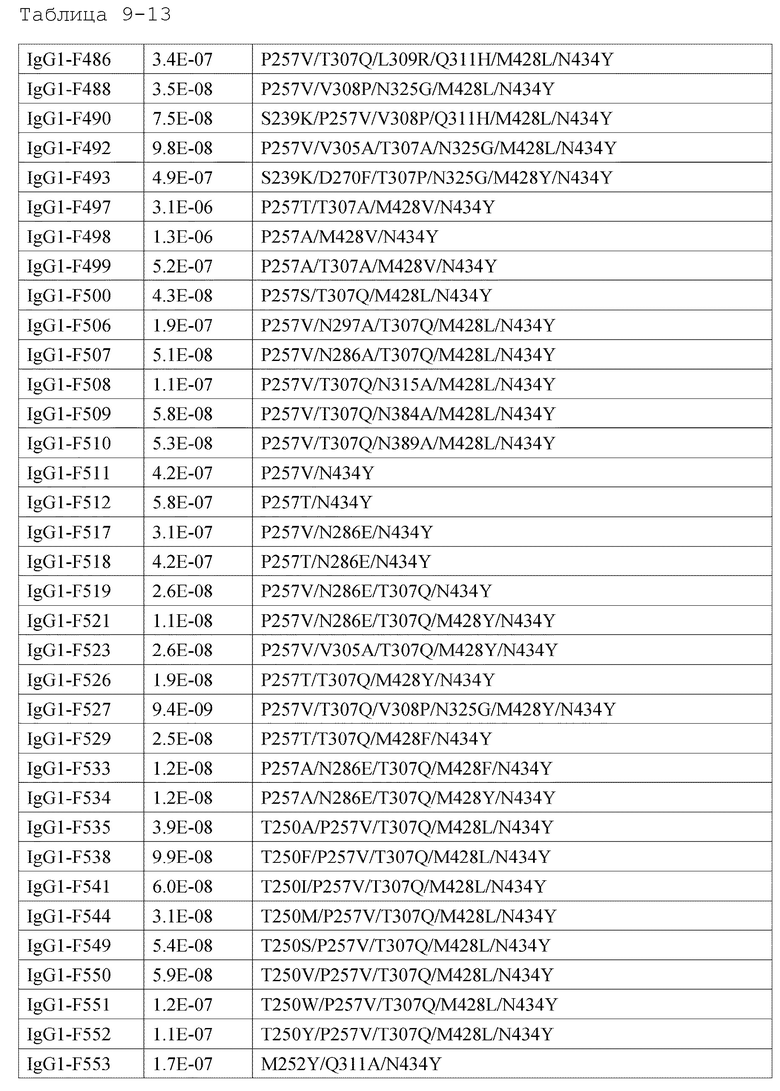
Таблица 9-14 представляет собой продолжение таблицы 9-13.
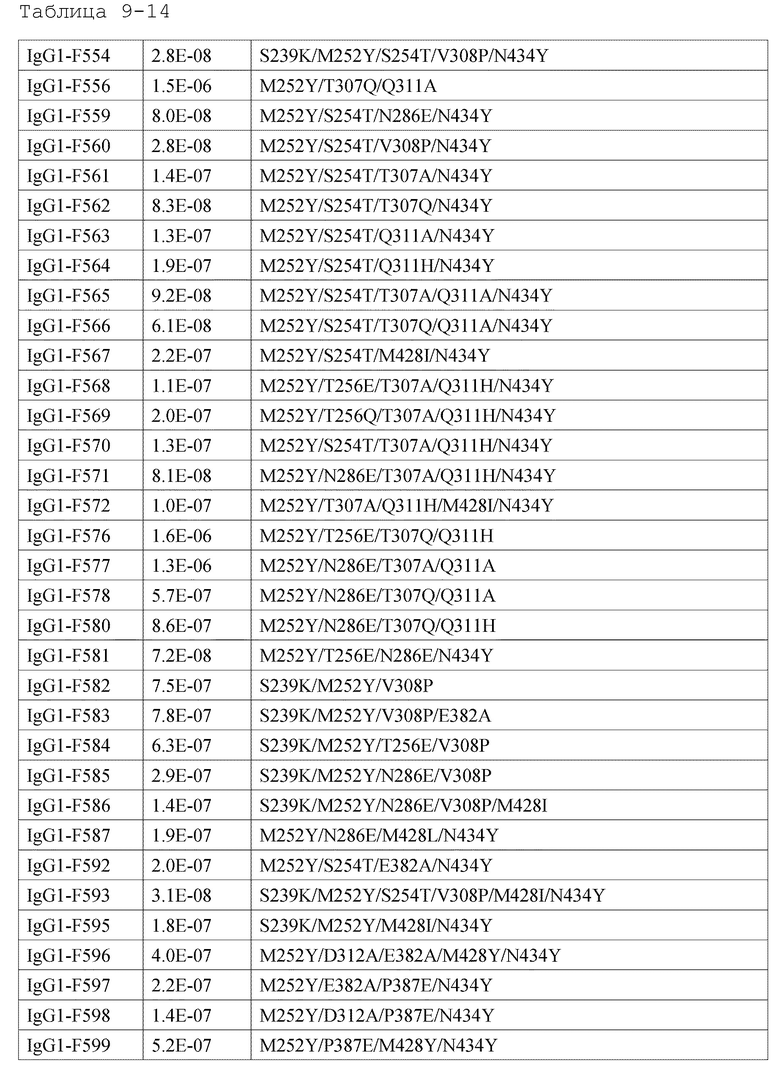
[Пример 13] Исследование in vivo различных вариантов Fc антител посредством модели стационарной инфузии с использованием трансгенной линии 32 мышей с FcRn человека
Варианты Fc, полученные в примере 12, протестировали на их способность к элиминации антигена из плазмы в модели стационарной инфузии с использованием трансгенной линии 32 мышей с FcRn человека. Исследование стационарной модели инфузии in vivo осуществляли, как описано в примере 1, но применяли трансгенную линию 32 мышей с FcRn человека вместо линии 276, и дважды инъецировали моноклональные антитела против CD4 мыши (до имплантации инфузионного насоса и через 14 суток после инъекции антитела) или три раза (до имплантации инфузионного насоса и через 10 и через 20 суток после инъекции антитела).
Из вариантов Fc, описанных в таблицах от 9-1 до 9-14, выбранные Fc варианты антитела, перечисленные ниже, экспрессировали и очищали способами, известными специалистам в данной области, как описано в ссылочном примере 2:
Fv4-IgG1, содержащий VH3-IgG1 и VL3-CK;
Fv4-IgG1-F11, содержащий VH3-IgG1-F11 и VL3-CK;
Fv4-IgG1-F14, содержащий VH3-IgG1-F14 и VL3-CK;
Fv4-IgG1-F39, содержащий VH3-IgG1-F39 и VL3-CK;
Fv4-IgG1-F48, содержащий VH3-IgG1-F48 и VL3-CK;
Fv4-IgG1-F140, содержащий VH3-IgG1-F140 и VL3-CK;
Fv4-IgG1-F157, содержащий VH3-IgG1-F157 и VL3-CK;
Fv4-IgG1-F194, содержащий VH3-IgG1-F194 и VL3-CK;
Fv4-IgG1-F196, содержащий VH3-IgG1-F196 и VL3-CK;
Fv4-IgG1-F198, содержащий VH3-IgG1-F198 и VL3-CK;
Fv4-IgG1-F262, содержащий VH3-IgG1-F262 и VL3-CK;
Fv4-IgG1-F264, содержащий VH3-IgG1-F264 и VL3-CK;
Fv4-IgG1-F393, содержащий VH3-IgG1-F393 и VL3-CK;
Fv4-IgG1-F424, содержащий VH3-IgG1-F434 и VL3-CK; и
Fv4-IgG1-F447, содержащий VH3-IgG1-F447 и VL3-CK.
Эти антитела вводили трансгенной линии 32 мышей с FcRn человека в дозе 1 мг/кг.
На фиг.21 описывается временная зависимость концентрации в плазме hsIL-6R у мышей. По сравнению с Fv4-IgG1, все варианты Fc, обладающие повышенной аффинностью связывания с FcRn человека при pH 7,0, продемонстрировали снижение концентрации hsIL-6R в плазме, таким образом увеличили элиминацию антигена из плазмы. Хотя степень и продолжительность снижения концентрации антигена была различной среди вариантов Fc, все варианты неуклонно снижали концентрацию hsIL-6R в плазме по сравнению с IgG1, демонстрируя, что повышенная аффинность связывания с FcRn человека при pH 7,0 будет универсально увеличивать элиминацию антигена из плазмы. На фиг.22 описывают временную зависимость концентрации антитела в плазме у мышей. Фармакокинетики антител были различными среди вариантов Fc.
Как описано в примере 9, количество антигена, элиминированного из плазмы посредством антитела, является важным фактором для оценки эффективности элиминации антигена посредством введения вариантов Fc антител, обладающих повышенной аффинностью связывания с FcRn человека при pH 7,0. Таким образом, временная зависимость значения C (молярное отношение антиген/антитело) для каждого антитела описана в фиг.23. На фиг.24 представлена зависимость между аффинностью связывания вариантов Fc с FcRn человека при pH 7,0 и значение C (молярное отношение антиген/антитело) через сутки 1 после введения антитела. Это показывает, что все варианты Fc антител, протестированные в этом иследовании, обладают более низким значением C по сравнению с Fv4-IgG1. Так как все варианты Fc, протестированные в этом исследовании, обладают аффиностью связывания с FcRn человека при pH 7,0 более высокой, чем KD, равная 3,0 микромолям, они достигают более высокой эффективностью элиминации антигена по сравнению с интактным IgG1 человека. Это согласуется с результатами, полученным в примере 9 (фиг.17).
На фиг.25 описано, что среди вариантов Fc, протестированных в этом исследовании, антитела, обладающие вариантами Fc F11, F39, F48 и F264, продемонстрировали аналогичную с IgG1 фармакокинетику. Так как это исследование проводят с использованием трансгенных мышей с FcRn человека, эти варианты Fc, как ожидают, обладают длительным периодом полувыведения, аналогичным IgG1 и у человека. На фиг.26 описана временная зависимость концентрации в плазме hsIL-6R у мыши, которой инъецировали антитела, обладающие фармакокинетикой, аналогичной интактному IgG1 человека (F11, F39, F48 и F264). Эти варианты снижали концентрацию hsIL-6R в плазме по сравнению с IgG1 приблизительно в 10 раз. Кроме того, эти антитела снижали концентрацию hsIL-6R ниже базовой линии концентрации hsIL-6R (концентрация без антитела). Таким образом, эти антитела будут давать возможность длительной элиминации антигена из плазмы, и таким образом обеспечивать длительные интервалы между дозами, которые будут предпочтительными для терапевтических средств на основе антител, необходимых для лечения хронических заболеваний.
На фиг.27 и 28 описана временная зависимость концентрации антитела в плазме и концентрации в плазме hsIL-6R для IgG1 и варианта Fc F157, F196 и F262, соответственно. Неожиданно, хотя фармакокинетики антител F157 и F262 показали значимо ускоренный клиренс из плазмы по сравнению с интактным IgG1 человека, F157 и F262 продемонстрировали очень интенсивную и длительную элиминацию hsIL-6R из плазмы. Конкретно, концентрация в плазме hsIL-6R для F157 была ниже предела детекции (1,56 нг/мл), от 1 до 28 суток (за исключением суток 14), и концентрация в плазме hsIL-6R для F262 была ниже предела детекции (1,56 нг/мл) от 14 до 28 суток. С другой стороны, для F196 с более медленным клиренсом антитела по сравнению с F157, концентрация антигена начала повышаться на 14 сутки и вернулась обратно к базовой линии на 28 сутки. Среди вариантов Fc, протестированных в этом исследовании, F157 и F262 были только варианты Fc, которые являлись способными к снижению концентрации hsIL-6R в плазме ниже 1,56 нг/мл на 28 сутки.
Такой длительный эффект F157 и F262 является неожиданным из фармакокинетики антитела, так как антитела элиминируются из плазмы очень быстро по сравнению с интактным IgG1 человека. В частности, концентрация антител в плазме F157 не определялась на 21 сутки. Однако, концентрация hsIL-6R в плазме продолжала снижаться до уровня, более низкого, чем предел детекции 1,56 нг/мл в сутки 21 и 28. Этот неожиданный эффект, как предполагают, существует в результате наличия антитела на поверхности эндотелиальных клеток сосудов в виде связанной с FcRn формой. Хотя эти антитела продемонстрировали низкую концентрацию в плазме, эти антитела все еще присутствуют в сосудистом компартменте в виде связанной с FcRn формы (которую нельзя измерить как концентрацию антитела в плазме). Это связанное с FcRn антитело может все еще быть связанным с антигеном в плазме, и затем FcRn опосредовал поглощение комплекса антиген/антитело, антиген высвобождается в пределах эндосомы и деградируется лизосомой, тогда как антитело рециркулирует обратно к клеточной поверхности в виде связанной с FcRn формой. Таким образом эти связанные с FcRn антитела вносят вклад в элиминацию антигена. Это объясняет факт того, что эти антитела сохраняют способность к элиминации антигена даже после того, как концентрация антитела в плазме становится низкой.
[Пример 14] Сравнительное исследование in silico общепринятого антитела и антитела, элиминирующего антиген
В примере 13 показано, что антитело, которое связывает антиген зависимым от рН способом и увеличивает аффинность связывания с FcRn человека при нейтральном pH, способно к элиминации антигена из плазмы. Таким образом, такие элиминирующие антиген антитела пригодны для антиген-антительного нацеливания, в котором простое связывание и нейтрализация является недостаточным для лечения заболевания, и требуется удаление антигена из плазмы.
Антитела, элиминирующие антигены, являются также пригодным для антиген-антительного нацеливания, для которых является достаточным простое связывание и нейтрализация. Антитела, связывающие и нейтрализующие антиген, требуют по меньшей мере молярного количества антитела, аналогичного количеству антигена в плазме (если антитело обладает бесконечно большой аффинностью к антигену, антиген можно нейтрализовать посредством аналогичного антигену молярного количества антитела). В отличие от общепринятого антитела (антитело без зависимого от pH связывания антигена и конструирования Fc), антитела, элиминирующие антиген, могут снижать концентрацию антигена в плазме. Это обозначает, что концентрацию антитела, требуемую для нейтрализации антигена, можно снижать. Если антитело, элиминирующее антиген, снижало концентрацию антигена в плазме в 10 раз по сравнению с общепринятым антителом, концентрацию антитела, требуемого для нейтрализации антигена, можно также снижать в 10 раз. Таким образом, в терапевтическом плане, антитело, элиминирующее антиген, может снижать дозу антигена или увеличивать интервал между дозами по сравнению с общепринятым антителом.
Варианты Fc, такие как F11, F39, F48 и F264, способны к снижению концентрация антигена в плазме по сравнению с IgG1 приблизительно в 10 раз. Для оценки эффекта таких антител, элиминирующих антиген, по сравнению с общепринятым антителом, осуществляли оценку in silico дозы антитела, требуемой для сохранения нейтрализации антигена в терапевтическом плане и для общепринятого антитела, и для антитела, элиминирующего антиген. Определили дозу, требуемую для поддержания нейтрализации, посредством интервала дозирования через 3 месяца. (т.е доза, требуемая для Q3M).
Конструирование фармакокинетической модели
Конструировали фармакокинетическую модель (PK) с использованием программного обеспечения SAAM II для анализа PK (The SAAM Institute, Inc.). Модель PK конструируют, как описано в Pharmacokinet Pharmacodyn. 2001 Dec; 28(6): 507-32 и Br J Clin Pharmacol. 2007 May; 63(5): 548-61. Концепция модели PK представлена на фиг.29. Количество каждого компартмента описывали посредством следующих дифференциальных уравнений.
[Math.1]
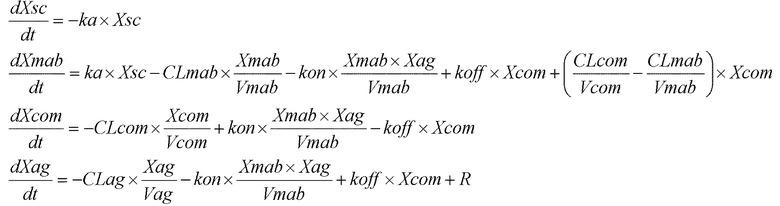
Xsc: количество антитела в подкожной ткани
Xmab: количество свободного антитела в сыворотке
Xcom: количество иммунного комплекса антитела и антигена (=комплекс)
Xag: количество свободного антигена в сыворотке
ka: константа скорости всасывания
В этой модели, биодоступность (F) оценена как 1 для всех антител, и скорость биосинтеза антигена (R) оценивают посредством следующего уравнения.
[Math.2]
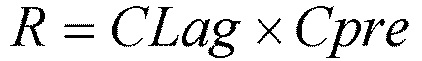
Cpre: стационарная концентрация антигена в сыворотке.
Фармакокинетические параметры и кинетические параметры связывания антигена, используемые в этом исследовании in silico, описаны в таблице 10.
Моделирование для рассчета эффекта антитела, элиминирующего антиген, и созревания аффинности
Стационарная концентрация (Cpre) до введения антитела была установлена на уровне 2400 нг/мл. С конструированием модели PK, оценивали минимальную дозу антитела для поддержания концентрации свободного антигена ниже 35 нг/мл через 84 суток после одного подкожного введения. Молекулярную массу антигена устанавливали как 190 кДа и молекулярную массу терапевтического антитела для всех устанавливали как 150 кДа.
В качестве антитела, в этом исследовании in silico применяли общепринятое антитело и антитело, элиминирующее антиген с различной аффинностью связывания (различная степень созревания аффинности от родительского антитела с KD 1 нМ). Эффект антитела, элиминирующего антиген, отражается в виде более быстрого клиренса комплекса антиген-антитело, чем для общепринятого антитела. Параметры клиренса комплекса антиген-антитело (CLcom) описаны а таблице 11.
Эффект созревания аффинности родительского антитела с KD 1 нМ также рассматривали (аффинность варьировала в 100 кратном диапазоне). KD 1 нМ, 300 пМ, 100 пМ, 30 пМ и 10 пМ применяют в этом исследовании in silico. Эффект созревания аффинности отображают как уменьшение koff. Значения koff варьирует в 100 кратном диапазоне (koff = 53,05, 17,68, 5,30, 1,77, 0,53 [1/сутки]).
Для общепринятого антитела и антитела, элиминирующего антиген, с аффинностью связывания (KD) 1 нМ, 300 пМ, 100 пМ, 30 пМ и 10 пМ получали дозу антитела для введения в организм с целью поддержания концентрации свободного антигена ниже 35 нг/мл через 84 суток после однократного подкожного введения Результаты описаны в таблице 12.
Для родительского общепринятого антитела с аффинностью связывания 1 нМ требуется 2,868 мг для достижения дозы Q3M. Хотя дозу антитела можно снижать посредством увеличения аффинности связывания с антигеном, уменьшение дозы достигает максимального значения. Это максимальное значение получают исходя из того факта, что для антитела, связывающего и нейтрализующего антиген, требуется по меньшей мере одинаковое молярное количество антитела относительно антигена в плазме. Даже с аффинностью связывания 10 пМ, для общепринятого антитела требуется 475 мг для достижения дозы Q3M, которая представляет собой дозу, которая не может быть введена подкожно посредством инъекции, вследствие ограничения концентрации составленного антитела и объемом, вводимым подкожно.
С другой стороны, посредством конструирования общепринятого антитела в антитело, элиминирующее антиген, посредством конструирования зависимости от pH при связывании антигена (или посредством прямого создания антитела с зависимым от pH связыванием) и конструированием области Fc для получения повышенной аффинности связывания с FcRn при нейтральном pH, дозу антитела можно значительно снижать. Для антитела, элиминирующего антиген с аффинностью связывания 1 нМ, требуется только 180 мг для достижения дозы Q3M. Этот уровень дозы не может быть достигнут посредством общепринятого антитела даже с безграничной аффинностью. Посредством увеличения аффинности связывания антитело, элиминирующего антиген, до 10 пМ, дозу можно снизить до 33 мг, которая представляет собой дозу, которая может быть легко введена подкожно.
Таким образом, это исследование in silico продемонстрировало, что антитело, элиминирующее антиген, обладает значительным преимуществом над общепринятым антителом. Дозу антитела можно снижать до уровня, который является недостижимым для общепринятого антитела даже с безграничной аффинностью. В отношении интервала дозирования, если антитело, элиминирующее антиген, вводят в той же дозе, что и доза общепринятого антитела, антитело, элиминирующее антиген будет иметь более продолжительный эффект, что таким образом дает возможность значимо более удлиненного интервала дозирования. И снижение дозы и увеличение длительности интервала дозирования посредством антитела, элиминирующего антиген, будет обеспечивать значительное преимуществом над общепринятым антителом.
Следует отметить, что, как описано в примере 1, антителу, элиминирующему антиген, не обязательно требуется зависимое от pH связывание с антигеном. Зависимое от pH связывание с антигеном может значимо увеличивать активность элиминации антигена для антитела. Кроме того, свойство зависимого от pH связывания можно заменять посредством использования других факторов, концентрация которых является различной в пределах плазмы и эндосомы. Такой фактор также можно использовать для получения антитела, которое связывается с антигеном в пределах плазмы, но диссоциирует от антигена в пределах эндосомы.
[Пример 15] Исследование увеличения эффекта усиления элиминации IL-6 человека зависимых от рН антител против IL-6 человека
Создание зависимых от pH антител, связывающих IL-6 человека
CLB8-IgG1, содержащий CLB8H-IgG1 (SEQ ID NO: 16) и CLB8L-CK (SEQ ID NO: 17), описанный в WO 2009/125825, представляет собой химерное антитело против IL-6. H16/L13-IgG1, содержащий H16-IgG1 (SEQ ID NO: 18) и L13-CK (SEQ ID NO: 19), представляет собой химерное антитело против IL-6, которое получают в результате предоставления CLB8-IgG1 свойства связываться с IL-6 человека зависимым от рН способом (которое связывается при pH 7,4, но диссоциирует при pH 5,8).
Оценка зависимой от рН активности связывания химерного антитела против IL-6 с IL-6 человека
CLB8-IgG1 и H16/L13-IgG1 оценивали по активности связывания с IL-6 человека (константа диссоциации (KD)) при pH 5,5 и pH 7,4 с использованием Biacore T100 (GE Healthcare). Анализ проводили, используя 10 ммоль/1 ACES/150 ммоль/1 NaCl, содержащий 0,05% поверхностно-активного вещества P20 (pH 7,4 и pH 6,0) в качестве подвижного буфера. Затем антитела связывали с рекомбинантным белком A/G (Thermo Scientific), иммобилизованном на сенсорных чипах с использованием способа на основе связанных аминов, инъецировали подходящие концентрации IL-6 человека (TORAY) в качестве анализируемого средства. Анализы проводили при 37 градусов C. Результаты анализировали с использованием программного обеспечения Biacore T100 Evaluation (GE Healthcare), и константу скорости ассоциации, ka (1/Мс), и константу скорости диссоциации, kd (1/с), рассчитывали из результатов анализа. Затем KD (M) рассчитывали из ka и kd (Таблица 13). Кроме того, зависимое от pH связывание оценивали для рассчета отношения KD между pH 7,4 и pH 6,0 для каждого антитела.
Получение зависимых от pH антител против IL-6 человека, обладающих активностью связывания с FcRn в нейтральных условиях
Мутации вводили в H16/L13-IgG1, содержащий H16-IgG1 (SEQ ID NO: 18) и L13-CK (SEQ ID NO: 19), для повышения связывания FcRn в нейтральных условиях (pH 7,4). Конкретно, H16-IgG1-v2 (SEQ ID NO: 20) получали из константной области тяжелой цепи IgG1 посредством замены Trp на Asn в положении 434 в нумерации EU, тогда как H16-F14 (SEQ ID NO: 21) конструировали из константной области тяжелой цепи IgG1 посредством замены Tyr на Met в положении 252 и Trp на Asn в положении 434 в нумерации EU. Замены аминокислот вводили способом, известным специалистам в данной области, описанным в ссылочном примере 1.
CLB8-IgG1, содержащий CLB8H-IgG1 (SEQ ID NO: 16) и CLB8L-CK (SEQ ID NO: 17), H16/L13-IgG1, содержащий H16-IgG1 (SEQ ID NO: 18) и L13-CK (SEQ ID NO: 19), H16/L13-IgG1-v2, содержащий H16-IgG1-v2 (SEQ ID NO: 20) и L13-CK (SEQ ID NO: 19), и H16/L13-F14, содержащий H16-F14 (SEQ ID NO: 21) и L13-CK (SEQ ID NO: 19), экспрессировали и очищали способом, известным специалистам в данной области, описанным в ссылочном примере 2.
Оценка связывающей активности с FcRn мыши вариантов Fc при нейтральном pH
VH3/L(WT)-IgG1, содержащий VH3-IgG1 и L (WT), VH3/L(WT)-IgG1-v2, содержащий VH3-IgG1-v2 и L (WT), и VH3/L (WT)-IgG1-F14, содержащий VH3-IgG1-F14 и L (WT), каждый из которых получали, как описано в примере 5, оценивали на связывание с FcRn мыши в нейтральных условиях (pH 7,4) посредством способа, описанного в примере 8.
Результаты представлены в таблице 14. IgG1 продемонстрировал очень слабую активность связывания, тогда как IgG1-v2 и IgG1-F14 показали более высокую аффинность связывания с FcRn мыши при pH 7,4.
Тест in vivo с использованием нормальных мышей
Кинетики in vivo для IL-6 человека (hIL-6; TORAY) и антитела против IL-6 человека оценивали после введения нормальной мыши (мыши C57BL/6J; Charles River Japan) hIL-6 отдельно или hIL-6 и антитела против IL-6 человека. Раствор hIL-6 (5 микрограмм/мл) или раствор смеси, содержащей hIL-6 и антитело против IL-6 человека (группа CLB8-IgG1; 5 микрограмм/мл hIL-6 и 0,025 мг/мл CLB8-IgG1, H16/L13-IgG1, H16/L13-IgG1-v2 и H16/L13-IgG1-F14 группа; 5 микрограмм/мл hIL-6 и 0,14 мг/мл H16/L13-IgG1, H16/L13-IgG1-v2 и H16/L13-IgG1-F14, соответственно) вводили однократно в дозе 10 мл/кг в хвостовую вену. Дозу антитела устанавливали такой, чтобы больше, чем 99,8% IL-6 человека связывалось с антителом в растворе для введения. Кровь собирали через 5 минут, 30 минут, два часа, четыре часа, семь часов, одни сутки после введения отдельно hIL-6 и через 5 минут, семь часов, одни сутки, двое суток, трое суток, четверо суток, семеро суток, 14 суток, 21 сутки и 30 суток после введения hIL-6 и раствора смеси с антителом против IL-6 человека. Собранную кровь немедленно центрифугировали при 15000 об/мин и 4 градусов C в течение 15 минут для отделения плазмы. Отделенную плазму хранили в холодильнике при -20 градусов C или ниже до проведения анализа.
Измерение концентрации IL-6 человека в плазме посредством ELISA
Концентрацию IL-6 человека в плазме мыши измеряли посредством использования набора Quantikine HS ELISA (R&D). Получали образцы калибровочной кривой, обладающие концентрацией в плазме, равной 20, 10, 5, 2,5, 1,25, 0,625 и 0,3125 нг/мл, и образцы плазмы мыши, разведенные в 100 раз или более. Для получения IL-6 человека, связанного полностью в образце с CLB8-IgG1, 150 микролитров 5 микрограмм/мл CLB8-IgG1 добавляли к 150 микролитрам образцов калибровочной кривой и к образцам плазмы, а затем образцы оставляли в течение одного часа при комнатной температуре. Затем, образцы переносили на планшеты, предоставленные в наборе для ELISA (R&D), и оставляли в течение одного часа при комнатной температуре. Затем конъюгат IL-6, предоставленный в наборе ELISA (R&D), добавляли для осуществления реакции в течение одного часа при комнатной температуре, и раствор субстрата, предоставленный в наборе ELISA (R&D), добавляли для осуществления реакции в течение одного часа при комнатной температуре. Далее, проводили хромогенную реакцию для осуществления реакции в течение получаса при комнатной температуре с использованием раствора Amplifier, предоставленного в наборе для ELISA (R&D) в качестве субстрата. После остановки реакции останавливающим раствором, предоставленным в наборе для ELISA (R&D), измеряли оптическую плотность при 490 нм посредством ридера для микропланшетов. Концентрацию в плазме мыши рассчитывали из оптической плотности калибровочной кривой, используя аналитическое программное обеспечение SOFTmax PRO (Molecular Devices). Зависимость от времени концентрации hIL-6 в плазме после внутривенного введения, как измерено этим способом, представлена на фиг.30 для нормальных мышей.
Измерение концентрации в плазме антитела против IL-6 посредством ELISA
Концентрацию антитела против IL-6 человека в плазме мыши измеряли посредством ELISA. F(ab’)2 фрагмент антитела против IgG человека (специфичная гамма-цепь) (Sigma) переносили на Nunc-ImmunoPlate MaxiSorp (Nalge Nunc International) и оставляли в течение ночи при 4 градусов C для получения планшетов с иммобилизованным антителом против IgG. Получали образцы калибровочной кривой, обладающие концентрацией в плазме 1,6, 0,8, 0,4, 0,2, 0,1, 0,05 и 0,025 микрограмм/мл, и образцы мышиной плазмы, разведенные в 100 раз и более. Для получения антитела против IL-6 человека, полностью связанного в образце с IL-6 человека, 200 микролитров IL-6 человека в количестве 1 микрограмм/мл добавляли к 100 микролитрам образцов калибровочной кривой и образцам плазмы, а затем образцы оставляли в течение одного часа при комнатной температуре. Затем, образцы переносили на планшеты с иммобилизованным антителом против IgG человека, и оставляли в течение одного часа при комнатной температуре. Затем, антитело козы против IgG человека (специфичная гамма-цепь), конъюгированное с биотином (BIOT) (Southern Biotech Association) добавляли для осуществления реакции в течение одного часа при комнатной температуре. Затем, стрептавидин-полиHRP80 (Stereospecific Detection Technologies) добавляли для осуществления реакции в течение одного часа при комнатной температуре, и проводили хромогенную реакцию с использованием TMB One Component HRP Microwell Substrate (BioFX Laboratories) в качестве субстрата. После остановки реакции 1 н. серной кислотой (Showa Chemical), оптическую плотность при 450 нм измеряли посредством ридера для микропланшетов. Концентрацию в плазме мыши рассчитывали из оптической плотности калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (Molecular Devices). Зависимость от времени концентрации антитела в плазме после внутривенного введения, как измерено этим способом, представлена для нормальных мышей на фиг.31.
Эффект зависимого от рН связывания с IL-6 человека
CLB8-IgG1 и H16/L13-IgG1, которые связываются с IL-6 человека зависимым от pH способом, тестировали in vivo, и результаты сравнивали между собой. Как показано на фиг.31, фармакокинетика антитела продемонстрировала линейный клиренс. Между тем, как показано на фиг.30, hIL-6, введенный одновременно с H16/L13-IgG1, который связывается с IL-6 человека зависимым от рН способом, как обнаружили, ускоряет элиминацию hIL-6 по сравнению с hIL-6, введенным одновременно с CLB8-IgG1. Таким образом, показано, что посредством придания зависимой от pH способности связывания с IL-6 человека, концентрация hIL-6 в плазме через четверо суток после введения могла быть снижена приблизительно в 76 раз.
Эффект связывания с FcRn в нейтральных условиях (pH 7,4)
В дополнение к H16/L13-IgG1, H16/L13-IgG1-v2 и H16/L13-F14, которые получены в результате введения описанных выше аминокислотных замен в H16/L13-IgG1, тестировали in vivo с использованием нормальных мышей. Результаты теста сравнивали с результатами, полученными для H16/L13-IgG1. Как показано на фиг.31, концентрация в плазме антитела H16/L13-IgG1-v2, которое увеличило связывание с FcRn мыши в нейтральных условиях (pH 7,4), была в 2,9 раз ниже по сравнению с H16/L13-IgG1 в сутки один после введения. Альтернативно, концентрация в плазме антитела H16/L13-F14, которое дополнительно увеличило связывание с FcRn мыши в нейтральных условиях (pH 7,4), была в 21 раз более низкой, чем для H16/L13-IgG1 через 7 часов после введения.
Как показано на фиг.30, для hIL-6, введенного одновременно с H16/L13-IgG1-v2 или H16/L13-F14, что увеличивало связывание с FcRn мыши в нейтральных условиях (pH 7,4), показана более быстрая элиминация по сравнению с hIL-6, введенным одновременно с H16/L13-IgG1. H16/L13-IgG1-v2 снижал концентрацию в плазме hIL-6 приблизительно в 10 раз по сравнению с H16/L13-IgG1 в сутки 1. H16/L13-F14 снижал концентрацию в плазме hIL-6 приблизительно в 38 раз по сравнению с H16/L13-IgG1 через семь часов. Таким образом, выявлено, что концентрация в плазме IL-6 человека может быть снижена посредством придания способности связывания с FcRn мыши в нейтральных условиях (pH 7,4). Как описано выше, посредством обеспечения способности связывания с FcRn мыши в нейтральных условиях (pH 7,4), концентрацию антитела в плазме снижают; однако, получали эффект снижения концентрации hIL-6 в плазме, который в значительной степени превысил снижение в концентрации антитела. Конкретно, это обозначает, что элиминацию IL-6 человека можно увеличивать посредством введения антитела, которое связывается с IL-6 человека зависимым от рН способом и которому придают способность связывания с FcRn мыши в нейтральных условиях (pH 7,4).
Полученные результаты, описанные выше, продемонстрировали, что концентрацию антигена в плазме, не только растворимого рецептора IL-6 человека, но также антигена, такого как IL-6 человека, можно значительно снижать посредством введения антитела, обладающего и зависимой от pH способностью связывания с антигеном и способностью связывания с FcRn в нейтральных условиях.
[Пример 16] Исследование увеличения эффекта ускорения элиминиции IgA человека слитого белка рецептора Fc, который связывается с IgA человека зависимым от рН способом
Получение слитого белка рецептора Fc, который связывается с IgA человека зависимым от рН способом
A0-IgG1, содержащий димер A0H-IgG1 (SEQ ID NO: 22) представляет собой слитый белок CD89-Fc человека. Как описано в J. Mol. Biol. (2003) 324: 645-657, CD89 человека, также известный как Fc альфа рецептор I человека, связывается с IgA человека зависимым от рН способом (т.е. сильно связывается с IgA человека при нейтральном значении pH, но слабо связывается с IgA человека при кислых значениях pH).
Оценка зависимой от рН активности связывания слитого белка CD89-Fc с IgA человека
A0-IgG1 оценивали по отношению к активности связывания с IgA человека (константа диссоциации (KD)) при pH 6,0 и pH 7,4 с использованием Biacore T100 (GE Healthcare). Анализ проводили с использованием 10 ммоль/1 ACES/150 ммоль/1 NaCl, содержащего 0,05% поверхностно-активного вещества P20 (pH 7,4 и pH 6,0) в качестве подвижного буфера. Затем слитый белок CD89-Fc связывали с рекомбинантным белком A/G (Thermo Scientific), иммобилизованном на сенсорных чипах с использованием способа на основе связанных аминов, в качестве анализируемого вещества инъецировали подходящие концентрации hIgA (IgA человека: получали, как описано в ссылочном примере 5). Анализы проводили при 37 градусов C. Результаты анализировали с использованием программного обеспечения Biacore T100 Evaluation (GE Healthcare), и полученные сенсограммы представлены на фиг.32. Четко показано, что слитый белок CD89-Fc обладает зависимой от рН активностью связывания с IgA человека, где слитый белок сильно связывается с IgA человека при нейтральном значении pH, но слабо связывается с IgA человека при кислых значениях pH.
Получение зависимого от рН слитого белка рецептора Fc, обладающего активностью связывания с FcRn в нейтральных условиях
Мутации вводили в A0-IgG1, содержащий димер A0H-IgG1 (SEQ ID NO: 22), для увеличения связывания с FcRn в нейтральных условиях (pH 7,4). Конкретно, A0-IgG1-v2 получали из константной области тяжелой цепи IgG1 посредством замены Trp на Asn в положении 426 в A0-IgG1. Замены аминокислот вводили способом, известным специалистам в данной области, описанным в ссылочном примере 1.
A0-IgG1, содержащий димер A0H-IgG1 (SEQ ID NO: 22), и A0-IgG1-v2, содержащий димер A0H-IgG1-v2 (SEQ ID NO: 23), экспрессировали и очищали способом, известным специалистам в данной области, описанным в ссылочном примере 2.
Тест in vivo с использованием нормальных мышей
Кинетики in vivo IgA человека (hIgA) и слитого белка CD89-Fc оценивали после введения отдельно hIgA или hIgA и слитого белка CD89-Fc (A0H-IgG1 или A0H-IgG1-v2) у нормальной мыши (мыши C57BL/6J; Charles River Japan). Раствор hIgA (80 микрограмм/мл) или раствор смеси, содержащей hIgA и слитый белок CD89-Fc (80 микрограмм/мл и 1,5 мг/мл, соответственно, в котором большая часть hIgA связана со слитым белком CD89-Fc), вводили однократно в дозе 10 мл/кг в хвостовую вену. Кровь собирали через 15 минут, семь часов, одни сутки, двое суток, четверо суток и семь суток после введения. Собранную кровь немедленно центрифугировали при 15000 об/мин и 4 градусов C в течение 15 минут для отделения плазмы. Отделенную плазму хранили в холодильнике при -20 градусов C или ниже до проведения анализа.
Измерение концентрации в плазме IgA человека посредством ELISA
Концентрацию IgA человека в плазме мыши измеряли посредством ELISA с использованием hsIL-6R, так как рекомбинантный IgA человека обладает вариабельной областью против hsIL-6R. Антитело козы против IgA человека (Bethyl Laboratories) переносили на Nunc-ImmunoPlate MaxiSorp (Nalge Nunc International) и оставляли в течение ночи при 4 градусов C для получения планшетов с иммобилизованными антителами против IgA человека. Получали образцы калибровочной кривой, имеющие концентрацию в плазме 0,4, 0,2, 0,1, 0,05, 0,025, 0,0125 или 0,00625 микрограмм/мл, и образцы мышиной плазмы, разведенные в 100 раз и более. Для получения IgA человека, полностью связанного в образце с hsIL-6R, 200 микролитров 10 микрограмм/мл hsIL-6R добавляли к 100 микролитрам образцов калибровочной кривой и образцам плазмы, а затем образцы оставляли в течение одного часа при комнатной температуре. Затем, образцы преносили на планшеты с иммобилизованным антителом против IgA человека и оставляли в течение одного часа при комнатной температуре. Затем биотинилированное антитело против IL-6R человека (R&D) добавляли для осуществления реакции в течение одного часа при комнатной температуре. Затем стрептавидин-полиHRP80 (Stereospecific Detection Technologies) добавляли для осуществления реакции в течение одного часа при комнатной температуре, и проводили хромогенную реакцию, используя TMB One Component HRP Microwell Substrate (BioFX Laboratories) в качестве субстрата. После остановки реакции 1 н. серной кислотой (Showa Chemical), измеряли оптическую плотность при 450 нм посредством ридера для микропланшетов. Концентрацию в плазме мыши рассчитывали из оптической плотности калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (Molecular Devices). Зависимость от времени концентрации hIgA в пазме после внутривенного введения, измеренная этим способом, представлена на фиг.33 для нормальных мышей.
Измерение концентрации в плазме слитого белка CD89-Fc посредством ELISA
Концентрацию слитого белка CD89-Fc в плазме мыши измеряли посредством ELISA. F(ab’)2 фрагмент антитела против IgG человека (специфичная гамма-цепь) (Sigma) переносили на Nunc-ImmunoPlate MaxiSorp (Nalge Nunc International) и оставляли в течение ночи при 4 градусов C для получения планшетов с иммобилизованным антителом против IgG человека. Получали образцы калибровочной кривой, обладающие концентрацией в плазме 25,6, 12,8, 6,4, 3,2, 1,6, 0,8, и 0,4 микрограмм/мл, и образцы мышиной плазмы, разведенные в 100 раз и более. Для получения слитого белка CD89-Fc в образце, полностью связанного с IgA человека, 200 микролитров 5 микрограмм/мл IgA человека добавляли к 100 микролитрам образцов калибровочной кривой и образцам плазмы, а затем образцы оставляли в течение одного часа при комнатной температуре. Затем образцы переносили на планшеты с иммобилизованным антителом против IgG человека, и оставляли в течение одного часа при комнатной температуре. Затем конъюгат антитела козы против IgG человека (Fc специфичный)-щелочная фосфатаза (SIGMA) добавляли для осуществления реакции в течение одного часа при комнатной температуре. Далее проводили хромогенную реакцию с использованием системы BluePhos Microwell Phosphatase Substrates System (Kirkegaard & Perry Laboratories) в качестве субстрата, и оптическую плотность при 650 нм измеряли посредством ридера для микропланшетов. Концентрацию в плазме мыши рассчитывали из оптической плотности калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (Molecular Devices). Зависимость от времени концентрации в плазме слитого белка CD89-Fc после внутривенного введения, как измерено этим способом, представлена на фиг.34 для нормальных мышей.
Эффект связывания с FcRn в нейтральных условиях (pH 7,4)
В дополнение к A0-IgG1, A0-IgG1-v2, который получен в результате введения указанных выше замен аминокислот в A0-IgG1, тестировали in vivo с использованием нормальных мышей. Результаты тестов сравнивали с результатами A0-IgG1. Как показано на фиг.34, концентрация в плазме A0-IgG1-v2, который увеличил связывание с FcRn мыши в нейтральных условиях (pH 7,4), была в 1,8 раз меньшей, чем для A0-IgG1 через двое суток после введения.
Как показано на фиг.33, hIgA, введенный одновременно с A0-IgG1-v2, который увеличил связывание с FcRn мыши в нейтральных условиях (pH 7,4), показал более быструю элиминацию по сравнению с hIgA, введенным одновременно с A0-IgG1. A0-IgG1-v2 снизил концентрацию в плазме hIgA приблизительно в 5,7 раз по сравнению с A0-IgG1 через двое суток. Как описано выше, посредством придания способности связывания с FcRn мыши в нейтральных условиях (pH 7,4), концентрацию антитела в плазме снижали; однако, получали эффект снижение концентрации hIgA в плазме, который в значительной степени превысил снижение в концентрации антитела. Конкретно, это обозначает, что элиминацию IgA человека можно ускорить посредством введения слитого белка рецептора Fc, который связывается с IgA человека зависимым от рН способом и которому придают способность связывания с FcRn мыши в нейтральных условиях (pH 7,4).
Результаты, описанные выше, показывают, что концентрацию антигена в плазме, такого как концентрация IgA человека, можно также значительно снижать посредством введения слитого белка рецептора Fc, обладающего и зависимой от рН антигенсвязывающей способностью и способностью связывания с FcRn в нейтральных условиях. Таким образом, слитый белок рецептора Fc можно также конструировать для придания способности к элиминации концентрации антигена (или лиганда) в плазме из плазмы.
[Пример 17] Исследование увеличения эффекта ускорения элиминации плексина A1 зависимых от рН антител против плексина A1 (получение антител)
Рассмотрение зависимого от рН антитела, связывающего плексин A1 человека
PX268-IgG1, содержащий PX268H-IgG1 (SEQ ID NO: 24) и PX268L-CK (SEQ ID NO: 25), представляет собой химерное антитело против плексина A1. PX141-IgG1, содержащий PX141H-IgG1 (SEQ ID NO: 26) и PX141L-CK (SEQ ID NO: 27), представляет собой химерное антитело против плексина A1, которое связывается с растворимым плексином A1 человека зависимым от рН способом (т.е. сильно связывается с растворимым плексином A1 человека при нейтральном значении pH, но слабо связывается с растворимым плексином A1 человека при кислом значении pH).
Оценка зависимой от pH активности связывания антитела против плексина A1 человека
PX268-IgG1 и PX141-IgG1 оценивали относительно активности связывания с плексином А1 человека (константа диссоциации (KD)) при pH 6,0 и pH 7,4 с Biacore T100 (GE Healthcare). Анализ проводили с использованием 10 ммоль/1 ACES/150 ммоль/1 NaCl, содержащего 0,05% поверхностно-активного вещества P20 (pH 7,4 и pH 6,0) в качестве подвижного буфера. Затем антитела связывали с рекомбинантным белком A/G (Thermo Scientific), иммобилизованным на сенсорных чипах с использованием способа на основе связанных аминов, подходящие концентрации hsPlexin A1 (растворимый плексин A1 человека: получен, как описано в ссылочном примере 5) инъецировали в качестве анализируемого средства. Анализы проводили при 37 градусов C. Результаты анализировали с использованием программного обеспечения Biacore T100 Evaluation (GE Healthcare), и константу скорости ассоциации, ka (1/Мс), и константу скорости диссоциации, kd (1/с), рассчитывали из результатов анализа. Затем KD (M) рассчитывали из ka и kd (таблица 15). Кроме того, зависимое от рН связывание оценивали для рассчета отношения KD между pH 7,4 и pH 6,0 для каждого антитела.
Получение зависимых от pH антител против плексина А1 человека, обладающих активностью связывания с FcRn в нейтральных условиях
Мутации вводили в PX141-IgG1, содержащий PX141H-IgG1 (SEQ ID NO: 26) и PX141L-CK (SEQ ID NO: 27), для увеличения связывания с FcRn в нейтральных условиях (pH 7,4). Конкретно, PX141H-IgG1-v2 (SEQ ID NO: 28) получали из константной области тяжелой цепи IgG1 посредством замены Trp на Asn в положении 434 в нумерации EU. Замены аминокислот вводили способом, известным специалистам в данной области, описанным в ссылочном примере 1.
PX268-IgG1, содержащий PX268H-IgG1 (SEQ ID NO: 24) и PX268L-CK (SEQ ID NO: 25), PX141-IgG1, содержащий PX141H-IgG1 (SEQ ID NO: 26) и PX141L-CK (SEQ ID NO: 27), и PX141-IgG1-v2, содержащий PX141H-IgG1-v2 (SEQ ID NO: 28) и PX141L-CK (SEQ ID NO: 27), экспрессировали и очищали способом, известным специалистам в данной области, описанным в ссылочном примере 2.
Тестирование in vivo с использованием нормальных мышей
Кинетики in vivo растворимого плексина A1 человека (hsPlexin A1) и антитело против плексина A1 оценивали после введения hsPlexin A1 отдельно или hsPlexin A1 и антитела против плексина A1 нормальной мыши (мыши C57BL/6J; Charles River Japan). Раствор hsPlexin A1 (100 микрограмм/мл) или раствор смеси, содержащей hsPlexin A1 и антитело против плексина A1 (группа PX268-IgG1; 100 микрограмм/мл hsPlexin A1 и 1,2 мг/мл PX268-IgG1, группа PX141-IgG1 и PX141-IgG1-v2; 100 микрограмм/мл hsPlexin A1 и 1,0 мг/мл PX141-IgG1 и PX141-IgG1-v2, соответственно) вводили однократно в дозе 10 мл/кг в хвостовую вену.
Дозу антитела устанавливали такой, чтобы более чем 99,9% растворимого плексина A1 человека было связано с антителом в растворе введения. Кровь собирали через 15 минут, семь часов, одни сутки, двое суток, четверо суток, семеро суток после введения hsPlexin A1 и смеси раствора с антителом против плексина A1. Собранную кровь немедленно центрифугировали при 15000 об/мин и 4 градусов C в течение 15 минут для отделения плазмы. Отделенную плазму хранили в холодильнике при -20 градусов C или ниже, до проведения анализа.
Измерение концентрации в плазме плексина A1 человека посредством ELISA после введения hsPlexin A1 отдельно
Концентрацию плексина A1 человека в плазме мыши измеряли посредством ELISA с использованием биотинилированного антитела против FLAG M2 (Sigma), так как рекомбинантый плексин A1 человека обладает концевой последовательностью FLAG-метки на С конце. Поликлональное антитело кролика против плексина A1 человека, полученное посредством иммунизирования плексином A1 кролика, переносили на Nunc-ImmunoPlate MaxiSorp (Nalge Nunc International) и оставляли в течение ночи при 4 градусов C для получения планшетов с иммобилизированным антителом против плексина A1. Получали образцы калибровочной кривой, имеющие концентрацию в плазме 25,6, 12,8, 6,4, 3,2, 1,6, и 0,8 микрограмм/мл, и образцы мышиной плазмы, разведенные в 100 раз и более. Затем образцы переносили на планшеты с иммобилизованным антителом против плексина A1 человека и оставляли в течение одного часа при комнатной температуре. Затем биотинилированное антитело против FLAG M2 (Sigma) добавляли для осуществления реакции в течение одного часа при комнатной температуре. Затем стрептавидин-полиHRP80 (Stereospecific Detection Technologies) добавляли для осуществления реакции в течение одного часа при комнатной температуре, и проводили хромогенную реакцию с использованием TMB One Component HRP Microwell Substrate (BioFX Laboratories) в качестве субстрата. После остановки реакции 1 н. серной кислотой (Showa Chemical) измеряли оптическую плотность при 450 нм посредством ридера для микропланшетов. Концентрацию в плазме мыши рассчитывали из оптической плотности калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (Molecular Devices). Зависимость от времени концентрации в плазме hsPlexin A1 после внутривенного введения, измеренная этим способом, представлена на фиг.35.
Измерение концентрации в плазме плексина A1 человека в группе PX268-IgG1 посредством ELISA
Концентрацию плексина A1 человека в плазме мыши измеряли посредством ELISA с использованием биотинилированного антитела против FLAG M2 (Sigma), так как рекомбинантый плексин A1 человека обладает концевой последовательностью FLAG-метки на С конце. Поликлональное антитело кролика против плексина A1 человека, полученное посредством иммунизирования плексином A1 кролика, переносили на Nunc-ImmunoPlate MaxiSorp (Nalge Nunc International) и оставляли в течение ночи при 4 градусов C для получения планшетов с иммобилизированным антителом против плексина A1 человека. Получали образцы калибровочной кривой, имеющие концентрацию в плазме 25,6, 12,8, 6,4, 3,2, 1,6 и 0,8 микрограмм/мл, и образцы мышиной плазмы, разведенные в 50 раз и более. Для того, чтобы плексин A1 человека полностью связался в образце с PX268-IgG1, 150 микролитров PX268-IgG1 в концентрации 40 микрограмм/мл добавляли к 150 микролитрам образцов калибровочной кривой и образцам плазмы, а затем образцы оставляли в течение ночи при 37 градусов C. Затем образцы переносили на планшеты с иммобилизованным антителом к плексину A1 человека и оставляли в течение одного часа при комнатной температуре (или 4 градусов C). Затем биотинилированное антитело против FLAG M2 (Sigma) добавляли для осуществления реакции в течение одного часа при комнатной температуре (или 4 градусов C). Затем стрептавидин-полиHRP80 (Stereospecific Detection Technologies) добавляли для осуществления реакции в течение одного часа при комнатной температуре (или 4 градусов C), и проводили хромогенную реакцию с использованием TMB One Component HRP Microwell Subsrate (BioFX Laboratories) в качестве субстрата. После остановки реакции 1 н. серной кислотой (Showa Chemical) измеряли оптическую плотность при 450 нм посредством ридера для микропланшетов. Концентрацию в плазме мыши рассчитывали из оптической плотности калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (Molecular Devices). Зависимость от времени концентрации в плазме hsPlexin A1 после внутривенного введения, измеренного этим способом, представлена на фиг.35.
Измерение концентрации в плазме плексина A1 человека в группе PX141-IgG1 и PX141-IgG1-v2 посредством ELISA
Концентрацию плексина A1 человека в плазме мыши измеряли ELISA с использованием биотинилированного антитела против FLAG M2 (Sigma), так как рекомбинантый плексин A1 человека обладает концевой последовательностью FLAG-метки на С конце. PX268-IgG1 переносили на Nunc-ImmunoPlate MaxiSorp (Nalge Nunc International) и оставляли в течение ночи при 4 градусов C для получения планшетов с иммобилизированным антителом против плексина A1. Получали образцы калибровочной кривой, имеющие концентрацию в плазме 25,6, 12,8, 6,4, 3,2, 1,6 и 0,8 микрограмм/мл, и образцы мышиной плазмы, разведенные в 50 раз и более. Для того чтобы плексин A1 человека полностью связался в образце с PX141-IgG1 или PX141-IgG1-v2, 150 микролитров PX141-IgG1 или PX141-IgG1-v2 в концентрации 40 микрограмм/мл добавляли к 150 микролитрам образцов калибровочной кривой и образцам плазмы, а затем образцы оставляли в течение ночи при 37 градусов C. Затем, образцы переносили на планшеты с иммобилизованным антителом к плексину A1 человека и оставляли в течение одного часа при комнатной температуре (или 4 градусов C). Затем биотинилированное антитело против FLAG M2 (Sigma) добавляли для осуществления реакции в течение одного часа при комнатной температуре (или 4 градусов C). Затем стрептавидин-полиHRP80 (Stereospecific Detection Technologies) добавляли для осуществления реакции в течение одного часа при комнатной температуре (или 4 градусов C), и проводили хромогенную реакцию с использованием TMB One Component HRP Microwell Subsrate (BioFX Laboratories) в качестве субстрата. После остановки реакции 1 Н серной кислотой (Showa Chemical) измеряли оптическую плотность при 450 нм посредством ридера для микропланшетов. Концентрацию в плазме мыши рассчитывали из оптической плотности калибровочной кривой с использованием аналитического программного обеспечения SOFTmax PRO (Molecular Devices). Концентрация hsPlexin A1 в плазме через 7 часов после внутривенного введения, измеренная этим способом, представлена на фиг.35.
Эффект зависимого от рН связывания с растворимым плексином A1 человека
PX268-IgG1 и PX141-IgG1, которые связываются с IL-6 человека зависимым от рН способом, тестировали in vivo, и результаты сравнивали друг с другом. Между тем, как показано на фиг.35, для hsPlexin A1, введенного одновременно с PX141-IgG1, который связывается с растворимым плексином A1 человека зависимым от рН способом, обнаружено снижение общей концентрации в плазме hsPlexin A1 по сравнению с hsPlexin A1, введенным одновременно с PX268-IgG1.
Эффект связывания с FcRn в нейтральных условиях (pH 7,4)
В дополнение к PX141-IgG1, PX141-IgG1-v2, который получен в результате введения описанных выше аминокислотных замен в PX141-IgG1, тестировали in vivo с использованием нормальных мышей. Результаты тестирования сравнивали с результатами, полученными для PX 141-IgG1.
Как показано на фиг.35, hsPlexin A1, введенный одновременно с PX141-IgG1-v2, который увеличивал связывание с FcRn мыши в нейтральных условиях (pH 7,4), показал снижение общей концентрации в плазме hsPlexin A1 до недетектируемого уровня (предел детекции представляет собой 0,8 микрограмм/мл). Таким образом, показано, что концентрация растворимого плексина A1 человека может быть снижена посредством придания способности связывания с FcRn мыши в нейтральных условиях (pH 7,4). Конкретно, это обозначает, что элиминацию растворимого плексина A1 человека можно увеличивать посредством введения антитела, которое связывается с плексином A1 человека зависимым от рН способом и которому придают способность связывания с FcRn мыши в нейтральных условиях (pH 7,4).
Результаты, описанные выше, демонстрируют, что концентрацию антигена в плазме, не только растворимого рецептора IL-6 человека, но также антигена, такого как IL-6 человека, IgA человека и растворимого плексина A1 человека можно также значительно снижать посредством введение антитела, обладающего и pH-зависимой антигенсвязывающей способностью и способностью связывания с FcRn в нейтральных условиях.
[Ссылочный пример 1] Конструирование экспрессирующих векторов для антител IgG, с введенными заменами аминокислот
Мутанты были получены с использовнием набора для сайт-специфического мутагенеза QuikChange (Stratagene) или набора для клонирования In-Fusion HD (Clontech) согласно способу, описанному в предоставленной инструкции, и полученные в результате фрагменты плазмиды встраивали в экспрессирующий вектор клетки млекопитающего для получения желаемых векторов, экспрессирующих H цепь, и векторов, экспрессирующих L цепь. Нуклеотидные последовательности полученных экспрессирующих векторов определяли с использованием общепринятых способов, известных специалистам в данной области.
[Ссылочный пример 2] Экспрессия и очистка антитела IgG
Антитела экспрессировали способом, описанным ниже. Антитела экспрессировали в FreestyleHEK293 (Invitrogen), как описано в протоколе, предоставленном производителем, или в клеточной линии HEK293H (Invitrogen). Клеточную линию HEK293H полученную из клеток эмбриональной почечной карциномы человека (Invitrogen), суспендировали в DMEM (Invitrogen), обогащенной 10%, эмбриональной телячьей сывороткой (Invitrogen). Клетки высевали из расчета 10 мл на чашку в чашки для прикрепления клеток (10 см в диаметре; CORNING) из расчета клеточной плотности 5-6×105 клеток/мл и культивировали в CO2 инкубаторе (37 градусов C, 5% CO2) в течение суток и ночь. Затем среду удаляли посредством аспирации и добавляли 6,9 мл среды CHO-S-SFM-II (Invitrogen). Полученную плазмиду вводили в клетки способом липофекции. Полученные в результате культуры супернатантов собирали, центрифугировали (приблизительно 2000×g, 5 мин, комнатная температура) для удаления клеток, и подвергали стерилизации посредством фильтрования через 0,22-микрометровый фильтр MIILEX (зарегистрированный товарный знак)-GV (Millipore) для получения супернатантов. Антитела очищали от полученных супернатантов культуры способом, известным специалистам в данной областис использованием rProtein A Sepharose™ Fast Flow (Amersham Biosciences). Для определения концентрации очищенного антитела измеряли оптическую плотность при 280 нм с использованием спектрофотометра. Концентрации антитела рассчитывали из определенных значений с использованием коэффициента оптической палотности, рассчитанного способом, описанным в Protein Science (1995) 4: 2411-2423.
[Ссылочный пример 3] Получение растворимого рецептора IL-6 человека (hsIL-6R)
Рекомбинантный рецептор IL-6 человека в качестве антигена получали следующим способом. Клеточную линию, конститутивно экспрессирующую растворимый рецептор IL-6 человека (далее в настоящем документе, обозначаемый как hsIL-6R), обладающую аминокислотной последовательностью в положениях от 1 до 357 от N-конца, как описано в J. Immunol. 152: 4958-4968 (1994), создавали способом, известным специалистам в данной области. Клетки культивировали для экспрессии hsIL-6R. hsIL-6R очищали от супернатанта культуры двумя стадиями: хроматографией на колонке с Blue Sepharose 6 FF и гель-фильтрацией. Фракции, элюированные в качестве основного пика на конечной стадии, применяли как конечный продукт очистки.
[Ссылочный пример 4] Получение FcRn человека
FcRn представляет собой комплекс FcRn и beta2-микроглобулина. Олиго-ДНК праймеры получали, основываясь на опубликованную последовательность гена FcRn человека (J Exp Med. 1994 Dec 1; 180(6): 2377-81). Фрагмент ДНК, кодирующий полный ген, получали посредством ПЦР с использованием кДНК человека (Human Placenta Marathon-Ready cDNA, Clontech) в качестве матрицы и полученных праймеров. Используя полученный фрагмент ДНК в качестве матрицы, фрагмент ДНК, кодирующий внеклеточный домен, содержащий сигнальную последовательность (Metl-Leu290), амплифицировали ПЦР, и встраивали в экспрессирующий вектор клетки млекопитающего. Кроме того, олиго-ДНК праймеры получали, основываясь на опубликованной последовательности гена бета2-микроглобулина человека (Proc. Natl. Acad. Sci. U.S.A. 99 (26): 16899-16903 (2002)). Фрагмент ДНК, кодирующий полный ген, получали посредством ПЦР, используя кДНК человека (Human Placenta Marathon-Ready кДНК, Clontech) в качестве матрицы и полученные праймеры. Используя полученный фрагмент ДНК в качестве матрицы, фрагмент ДНК, кодирующий полный белок, содержащий сигнальную последовательность (Met1-Met119), амплифицировали посредством ПЦР и встраивали в экспрессирующий вектор клеток млекопитающего.
Растворимый FcRn человека экспрессировали посредством следующего способа. Плазмиды, сконструированные для экспрессии FcRn человека (SEQ ID NO: 30) и бета2-микроглобулина (SEQ ID NO: 31), встраивали в клеточную линию HEK293H, полученную из клеток эмбриональной почечной карциномы человека (Invitrogen), посредством способа липофекции, используя PEI (Polyscience). Полученные в результате супернатанты культуры собирали, и FcRn очищали, используя IgG Sepharose 6 Fast Flow (Amersham Biosciences), с последующей дополнительной очисткой с использованием HiTrap Q HP (GE Healthcare) (J Immunol. 2002 Nov 1; 169(9): 5171-80).
[Ссылочный пример 5] Получение IgA человека (hIgA)
hIgA, содержащий H (WT)-IgA1 (SEQ ID NO: 29) и L (WT) (SEQ ID NO: 5), экспрессировали и очищали способом, известным специалистам в данной области с использованием rProtein L-агарозы (ACTIgen) с последующей гель-фильтрацией.
[Ссылочный пример 6] Получение растворимого плексина A1 человека (hsPlexin A1)
Рекомбинантный растворимый плексин A1 человека в качестве антигена (далее в настоящем документе, обозначаемый как hsPlexin A1) получали, как указано далее. hsPlexin A1 конструировали посредством обращения к NCBI Reference Sequence (NP_115618). Конкретно, hsPlexin A1 содержал аминокислотную последовательность в положениях 27-1243 из указанной выше NCBI Reference FLAG-tag (DYKDDDDK), которую соединяли с ее C концом. hsPlexin A1 временно экспрессировали с использованием FreeStyle293 (Invitrogen) и очищали от супернатанта культуры посредством двух стадий: хроматографией на колонке с антителом против FLAG и гель-фильтрацией. Фракции, элюированные в качестве основного пика на конечной стадии, применяли как конечный продукт очистки.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CHUGAI SEIYAKU KABUSHIKI KAISHA
<120> МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ АНТИГЕНЫ, КОТОРЫЕ УВЕЛИЧИВАЮТ КЛИРЕНС
АНТИГЕНОВ
<130> C1-A1004Y1P
<150> JP 2010-079667
<151> 2010-03-30
<150> JP 2010-250830
<151> 2010-11-09
<160> 31
<170> PatentIn version 3.4
<210> 1
<211> 447
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 1
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser Asp Asp
20 25 30
Gln Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Gly Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Ala Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 2
<211> 214
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 2
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Tyr Gly Ser Glu Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Ser Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 3
<211> 447
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 3
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser Asp Asp
20 25 30
Gln Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Gly Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Ala Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Trp Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Trp His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 4
<211> 449
<212> БЕЛОК
<213> Homo sapiens
<400> 4
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Arg Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Ser Ile Thr Ser Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Arg Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Thr Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Met Leu Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Arg Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Ser Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 5
<211> 214
<212> БЕЛОК
<213> Homo sapiens
<400> 5
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 6
<211> 447
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 6
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 7
<211> 214
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 7
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Gln Ala Ser Thr Asp Ile Ser Ser His
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Tyr Gly Ser His Leu Leu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gly Gln Gly Asn Arg Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 8
<211> 447
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 8
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Tyr Ile Thr
245 250 255
Arg Glu Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 9
<211> 447
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 9
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Trp His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 10
<211> 447
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 10
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Phe Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Tyr His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 11
<211> 324
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 11
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro
<210> 12
<211> 324
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 12
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Ser Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro
<210> 13
<211> 326
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 13
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Ala His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 14
<211> 324
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 14
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Ser Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Ala His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro
<210> 15
<211> 443
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 15
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Ser Cys Val Glu
210 215 220
Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu
290 295 300
Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440
<210> 16
<211> 447
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 16
Glu Val Gln Leu Val Glu Ser Gly Gly Lys Leu Leu Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Ala Met Ser Trp Phe Arg Gln Ser Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Glu Ile Ser Ser Gly Gly Ser Tyr Thr Tyr Tyr Pro Asp Thr Val
50 55 60
Thr Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Glu Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Leu Trp Gly Tyr Tyr Ala Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 17
<211> 213
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 17
Gln Ile Val Leu Ile Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Arg Leu Leu Ile Tyr
35 40 45
Asp Thr Ser Asn Leu Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 18
<211> 447
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 18
Glu Val Gln Leu Val Glu Ser Gly Gly Lys Leu Leu Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Ala Met Ser Trp Phe Arg Gln Ser Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Glu Ile Ser Ser Gly Gly Ser Tyr Thr Tyr His Pro His Thr Val
50 55 60
Thr Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Glu Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Leu Trp Gly His Tyr Ala Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 19
<211> 213
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 19
Gln Ile Val Leu Ile Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Arg Leu Leu Ile Tyr
35 40 45
Asp Thr Ser His His Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys His Gln Trp Ser Gly His Pro Tyr Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 20
<211> 447
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 20
Glu Val Gln Leu Val Glu Ser Gly Gly Lys Leu Leu Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Ala Met Ser Trp Phe Arg Gln Ser Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Glu Ile Ser Ser Gly Gly Ser Tyr Thr Tyr His Pro His Thr Val
50 55 60
Thr Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Glu Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Leu Trp Gly His Tyr Ala Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Trp His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 21
<211> 447
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 21
Glu Val Gln Leu Val Glu Ser Gly Gly Lys Leu Leu Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Ala Met Ser Trp Phe Arg Gln Ser Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Glu Ile Ser Ser Gly Gly Ser Tyr Thr Tyr His Pro His Thr Val
50 55 60
Thr Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Glu Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Leu Trp Gly His Tyr Ala Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Trp Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Trp His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 22
<211> 436
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 22
Gln Glu Gly Asp Phe Pro Met Pro Phe Ile Ser Ala Lys Ser Ser Pro
1 5 10 15
Val Ile Pro Leu Asp Gly Ser Val Lys Ile Gln Cys Gln Ala Ile Arg
20 25 30
Glu Ala Tyr Leu Thr Gln Leu Met Ile Ile Lys Asn Ser Thr Tyr Arg
35 40 45
Glu Ile Gly Arg Arg Leu Lys Phe Trp Asn Glu Thr Asp Pro Glu Phe
50 55 60
Val Ile Asp His Met Asp Ala Asn Lys Ala Gly Arg Tyr Gln Cys Gln
65 70 75 80
Tyr Arg Ile Gly His Tyr Arg Phe Arg Tyr Ser Asp Thr Leu Glu Leu
85 90 95
Val Val Thr Gly Leu Tyr Gly Lys Pro Phe Leu Ser Ala Asp Arg Gly
100 105 110
Leu Val Leu Met Pro Gly Glu Asn Ile Ser Leu Thr Cys Ser Ser Ala
115 120 125
His Ile Pro Phe Asp Arg Phe Ser Leu Ala Lys Glu Gly Glu Leu Ser
130 135 140
Leu Pro Gln His Gln Ser Gly Glu His Pro Ala Asn Phe Ser Leu Gly
145 150 155 160
Pro Val Asp Leu Asn Val Ser Gly Ile Tyr Arg Cys Tyr Gly Trp Tyr
165 170 175
Asn Arg Ser Pro Tyr Leu Trp Ser Phe Pro Ser Asn Ala Leu Glu Leu
180 185 190
Val Val Thr Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser His Thr Ser Pro Pro Ser Pro Ala Pro
210 215 220
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
225 230 235 240
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
245 250 255
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
260 265 270
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
275 280 285
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
290 295 300
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
305 310 315 320
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
325 330 335
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys
340 345 350
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
355 360 365
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
370 375 380
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
385 390 395 400
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
405 410 415
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
420 425 430
Leu Ser Leu Ser
435
<210> 23
<211> 436
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 23
Gln Glu Gly Asp Phe Pro Met Pro Phe Ile Ser Ala Lys Ser Ser Pro
1 5 10 15
Val Ile Pro Leu Asp Gly Ser Val Lys Ile Gln Cys Gln Ala Ile Arg
20 25 30
Glu Ala Tyr Leu Thr Gln Leu Met Ile Ile Lys Asn Ser Thr Tyr Arg
35 40 45
Glu Ile Gly Arg Arg Leu Lys Phe Trp Asn Glu Thr Asp Pro Glu Phe
50 55 60
Val Ile Asp His Met Asp Ala Asn Lys Ala Gly Arg Tyr Gln Cys Gln
65 70 75 80
Tyr Arg Ile Gly His Tyr Arg Phe Arg Tyr Ser Asp Thr Leu Glu Leu
85 90 95
Val Val Thr Gly Leu Tyr Gly Lys Pro Phe Leu Ser Ala Asp Arg Gly
100 105 110
Leu Val Leu Met Pro Gly Glu Asn Ile Ser Leu Thr Cys Ser Ser Ala
115 120 125
His Ile Pro Phe Asp Arg Phe Ser Leu Ala Lys Glu Gly Glu Leu Ser
130 135 140
Leu Pro Gln His Gln Ser Gly Glu His Pro Ala Asn Phe Ser Leu Gly
145 150 155 160
Pro Val Asp Leu Asn Val Ser Gly Ile Tyr Arg Cys Tyr Gly Trp Tyr
165 170 175
Asn Arg Ser Pro Tyr Leu Trp Ser Phe Pro Ser Asn Ala Leu Glu Leu
180 185 190
Val Val Thr Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
195 200 205
Gly Ser Gly Gly Gly Gly Ser His Thr Ser Pro Pro Ser Pro Ala Pro
210 215 220
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
225 230 235 240
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
245 250 255
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
260 265 270
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
275 280 285
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
290 295 300
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
305 310 315 320
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
325 330 335
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys
340 345 350
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
355 360 365
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
370 375 380
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
385 390 395 400
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
405 410 415
Cys Ser Val Met His Glu Ala Leu His Trp His Tyr Thr Gln Lys Ser
420 425 430
Leu Ser Leu Ser
435
<210> 24
<211> 448
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 24
Gln Ser Leu Glu Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Ala Ser
1 5 10 15
Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Ser Phe Ser Ser Asn Tyr
20 25 30
Trp Ile Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ala Cys Ile Tyr Ile Gly Ser Asp Ser Ala Tyr Tyr Ala Ser Trp Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Lys Thr Ser Ser Thr Thr Val Thr Leu
65 70 75 80
Gln Met Thr Ser Leu Thr Ala Ala Asp Thr Ala Thr Tyr Phe Cys Ala
85 90 95
Arg Trp Asp Asn Ser Gly Arg Ala Leu Lys Leu Trp Gly Pro Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 25
<211> 219
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 25
Gln Met Leu Thr Gln Thr Ala Ser Pro Val Ser Ala Ala Val Gly Gly
1 5 10 15
Thr Val Thr Ile Lys Cys Gln Ser Ser Gln Ser Val Ala Asp Asn Asn
20 25 30
His Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu
35 40 45
Ile Tyr Phe Ala Ser Thr Leu Ala Ser Gly Val Ser Ser Arg Phe Lys
50 55 60
Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Asp Val Gln
65 70 75 80
Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Leu Gly Asn Tyr Asp Cys Gly
85 90 95
Ser Ala Asp Cys His Ala Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 26
<211> 454
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 26
Gln Glu Gln Leu Glu Glu Ser Gly Gly Gly Leu Val Lys Pro Glu Gly
1 5 10 15
Ser Leu Thr Leu Thr Cys Lys Ala Ser Gly Phe Asp Phe Ser Ser Tyr
20 25 30
Tyr Tyr Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Cys Ile Val Thr Gly Ser Tyr Gly Arg Ser Trp Tyr Ala Ser
50 55 60
Trp Ala Lys Gly Arg Phe Thr Ile Thr Arg Ser Thr Ser Leu Asn Thr
65 70 75 80
Val Thr Leu Gln Leu Asn Ser Leu Thr Ala Ala Asp Thr Ala Thr Tyr
85 90 95
Phe Cys Ala Arg Asp Pro Phe Val Ile Ala Ser Ser His Tyr Gln Asn
100 105 110
Leu Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys
450
<210> 27
<211> 218
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 27
Ala Leu Val Met Thr Gln Thr Pro Ser Pro Val Ser Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Ser Cys Gln Ser Ser Pro Asn Ile Tyr Asn Asn
20 25 30
Tyr Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Ala Ser Asn Leu Glu Thr Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Asp Val Gln
65 70 75 80
Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Tyr Lys Ser Tyr Asp
85 90 95
Asn Asp Asp Asn Ala Phe Gly Gly Gly Thr Glu Val Val Val Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 28
<211> 454
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 28
Gln Glu Gln Leu Glu Glu Ser Gly Gly Gly Leu Val Lys Pro Glu Gly
1 5 10 15
Ser Leu Thr Leu Thr Cys Lys Ala Ser Gly Phe Asp Phe Ser Ser Tyr
20 25 30
Tyr Tyr Met Cys Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Cys Ile Val Thr Gly Ser Tyr Gly Arg Ser Trp Tyr Ala Ser
50 55 60
Trp Ala Lys Gly Arg Phe Thr Ile Thr Arg Ser Thr Ser Leu Asn Thr
65 70 75 80
Val Thr Leu Gln Leu Asn Ser Leu Thr Ala Ala Asp Thr Ala Thr Tyr
85 90 95
Phe Cys Ala Arg Asp Pro Phe Val Ile Ala Ser Ser His Tyr Gln Asn
100 105 110
Leu Trp Gly Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Trp His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys
450
<210> 29
<211> 472
<212> БЕЛОК
<213> Искусственная
<220>
<223> Искусственно синтезированная последовательность
<400> 29
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Arg Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Ser Ile Thr Ser Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Arg Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Thr Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Met Leu Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Arg Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Ser Leu Val Thr Val Ser Ser Ala Ser Pro Thr Ser Pro Lys Val Phe
115 120 125
Pro Leu Ser Leu Cys Ser Thr Gln Pro Asp Gly Asn Val Val Ile Ala
130 135 140
Cys Leu Val Gln Gly Phe Phe Pro Gln Glu Pro Leu Ser Val Thr Trp
145 150 155 160
Ser Glu Ser Gly Gln Gly Val Thr Ala Arg Asn Phe Pro Pro Ser Gln
165 170 175
Asp Ala Ser Gly Asp Leu Tyr Thr Thr Ser Ser Gln Leu Thr Leu Pro
180 185 190
Ala Thr Gln Cys Leu Ala Gly Lys Ser Val Thr Cys His Val Lys His
195 200 205
Tyr Thr Asn Pro Ser Gln Asp Val Thr Val Pro Cys Pro Val Pro Ser
210 215 220
Thr Pro Pro Thr Pro Ser Pro Ser Thr Pro Pro Thr Pro Ser Pro Ser
225 230 235 240
Cys Cys His Pro Arg Leu Ser Leu His Arg Pro Ala Leu Glu Asp Leu
245 250 255
Leu Leu Gly Ser Glu Ala Asn Leu Thr Cys Thr Leu Thr Gly Leu Arg
260 265 270
Asp Ala Ser Gly Val Thr Phe Thr Trp Thr Pro Ser Ser Gly Lys Ser
275 280 285
Ala Val Gln Gly Pro Pro Glu Arg Asp Leu Cys Gly Cys Tyr Ser Val
290 295 300
Ser Ser Val Leu Pro Gly Cys Ala Glu Pro Trp Asn His Gly Lys Thr
305 310 315 320
Phe Thr Cys Thr Ala Ala Tyr Pro Glu Ser Lys Thr Pro Leu Thr Ala
325 330 335
Thr Leu Ser Lys Ser Gly Asn Thr Phe Arg Pro Glu Val His Leu Leu
340 345 350
Pro Pro Pro Ser Glu Glu Leu Ala Leu Asn Glu Leu Val Thr Leu Thr
355 360 365
Cys Leu Ala Arg Gly Phe Ser Pro Lys Asp Val Leu Val Arg Trp Leu
370 375 380
Gln Gly Ser Gln Glu Leu Pro Arg Glu Lys Tyr Leu Thr Trp Ala Ser
385 390 395 400
Arg Gln Glu Pro Ser Gln Gly Thr Thr Thr Phe Ala Val Thr Ser Ile
405 410 415
Leu Arg Val Ala Ala Glu Asp Trp Lys Lys Gly Asp Thr Phe Ser Cys
420 425 430
Met Val Gly His Glu Ala Leu Pro Leu Ala Phe Thr Gln Lys Thr Ile
435 440 445
Asp Arg Leu Ala Gly Lys Pro Thr His Val Asn Val Ser Val Val Met
450 455 460
Ala Glu Val Asp Gly Thr Cys Tyr
465 470
<210> 30
<211> 365
<212> БЕЛОК
<213> Homo sapiens
<400> 30
Met Gly Val Pro Arg Pro Gln Pro Trp Ala Leu Gly Leu Leu Leu Phe
1 5 10 15
Leu Leu Pro Gly Ser Leu Gly Ala Glu Ser His Leu Ser Leu Leu Tyr
20 25 30
His Leu Thr Ala Val Ser Ser Pro Ala Pro Gly Thr Pro Ala Phe Trp
35 40 45
Val Ser Gly Trp Leu Gly Pro Gln Gln Tyr Leu Ser Tyr Asn Ser Leu
50 55 60
Arg Gly Glu Ala Glu Pro Cys Gly Ala Trp Val Trp Glu Asn Gln Val
65 70 75 80
Ser Trp Tyr Trp Glu Lys Glu Thr Thr Asp Leu Arg Ile Lys Glu Lys
85 90 95
Leu Phe Leu Glu Ala Phe Lys Ala Leu Gly Gly Lys Gly Pro Tyr Thr
100 105 110
Leu Gln Gly Leu Leu Gly Cys Glu Leu Gly Pro Asp Asn Thr Ser Val
115 120 125
Pro Thr Ala Lys Phe Ala Leu Asn Gly Glu Glu Phe Met Asn Phe Asp
130 135 140
Leu Lys Gln Gly Thr Trp Gly Gly Asp Trp Pro Glu Ala Leu Ala Ile
145 150 155 160
Ser Gln Arg Trp Gln Gln Gln Asp Lys Ala Ala Asn Lys Glu Leu Thr
165 170 175
Phe Leu Leu Phe Ser Cys Pro His Arg Leu Arg Glu His Leu Glu Arg
180 185 190
Gly Arg Gly Asn Leu Glu Trp Lys Glu Pro Pro Ser Met Arg Leu Lys
195 200 205
Ala Arg Pro Ser Ser Pro Gly Phe Ser Val Leu Thr Cys Ser Ala Phe
210 215 220
Ser Phe Tyr Pro Pro Glu Leu Gln Leu Arg Phe Leu Arg Asn Gly Leu
225 230 235 240
Ala Ala Gly Thr Gly Gln Gly Asp Phe Gly Pro Asn Ser Asp Gly Ser
245 250 255
Phe His Ala Ser Ser Ser Leu Thr Val Lys Ser Gly Asp Glu His His
260 265 270
Tyr Cys Cys Ile Val Gln His Ala Gly Leu Ala Gln Pro Leu Arg Val
275 280 285
Glu Leu Glu Ser Pro Ala Lys Ser Ser Val Leu Val Val Gly Ile Val
290 295 300
Ile Gly Val Leu Leu Leu Thr Ala Ala Ala Val Gly Gly Ala Leu Leu
305 310 315 320
Trp Arg Arg Met Arg Ser Gly Leu Pro Ala Pro Trp Ile Ser Leu Arg
325 330 335
Gly Asp Asp Thr Gly Val Leu Leu Pro Thr Pro Gly Glu Ala Gln Asp
340 345 350
Ala Asp Leu Lys Asp Val Asn Val Ile Pro Ala Thr Ala
355 360 365
<210> 31
<211> 119
<212> БЕЛОК
<213> Homo sapiens
<400> 31
Met Ser Arg Ser Val Ala Leu Ala Val Leu Ala Leu Leu Ser Leu Ser
1 5 10 15
Gly Leu Glu Ala Ile Gln Arg Thr Pro Lys Ile Gln Val Tyr Ser Arg
20 25 30
His Pro Ala Glu Asn Gly Lys Ser Asn Phe Leu Asn Cys Tyr Val Ser
35 40 45
Gly Phe His Pro Ser Asp Ile Glu Val Asp Leu Leu Lys Asn Gly Glu
50 55 60
Arg Ile Glu Lys Val Glu His Ser Asp Leu Ser Phe Ser Lys Asp Trp
65 70 75 80
Ser Phe Tyr Leu Leu Tyr Tyr Thr Glu Phe Thr Pro Thr Glu Lys Asp
85 90 95
Glu Tyr Ala Cys Arg Val Asn His Val Thr Leu Ser Gln Pro Lys Ile
100 105 110
Val Lys Trp Asp Arg Asp Met
115
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ИЗМЕНЕНИЯ УДЕРЖАНИЯ В ПЛАЗМЕ И ИММУНОГЕННОСТИ АНТИГЕНСВЯЗЫВАЮЩЕЙ МОЛЕКУЛЫ | 2012 |
|
RU2799423C1 |
| ВАРИАНТЫ Fc-ОБЛАСТИ С МОДИФИЦИРОВАННЫМ СВЯЗЫВАНИЕМ FcRn И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2714116C2 |
| ВАРИАНТЫ Fc-ОБЛАСТИ С МОДИФИЦИРОВАННЫМИ СВОЙСТВАМИ СВЯЗЫВАНИЯ FcRn И БЕЛКА А | 2015 |
|
RU2713131C1 |
| ВАРИАНТ Fc И ЕГО ПОЛУЧЕНИЕ | 2021 |
|
RU2828852C1 |
| ВАРИАНТЫ С FC-ФРАГМЕНТОМ, ОБЛАДАЮЩИЕ ПОВЫШЕННОЙ АФФИННОСТЬЮ К FCRN И ПОВЫШЕННОЙ АФФИННОСТЬЮ ПО МЕНЬШЕЙ МЕРЕ К ОДНОМУ РЕЦЕПТОРУ FC-ФРАГМЕНТА | 2018 |
|
RU2820162C2 |
| ГЕННО-ИНЖЕНЕРНЫЕ ПОЛИПЕПТИДЫ И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2822498C2 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, СПОСОБНАЯ МНОГОКРАТНО СВЯЗЫВАТЬСЯ С МНОЖЕСТВОМ АНТИГЕННЫХ МОЛЕКУЛ | 2018 |
|
RU2757786C2 |
| Биспецифическое антитело и его применение | 2019 |
|
RU2785292C2 |
| АНТИТЕЛА ПРОТИВ SARS-COV-2 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2021 |
|
RU2832025C1 |
| КОМПОЗИЦИИ ДЛЯ УВЕЛИЧЕНИЯ ПЕРИОДА ПОЛУЖИЗНИ ТЕРАПЕВТИЧЕСКОГО СРЕДСТВА У СОБАК И СПОСОБЫ ПРИМЕНЕНИЯ | 2020 |
|
RU2828219C2 |
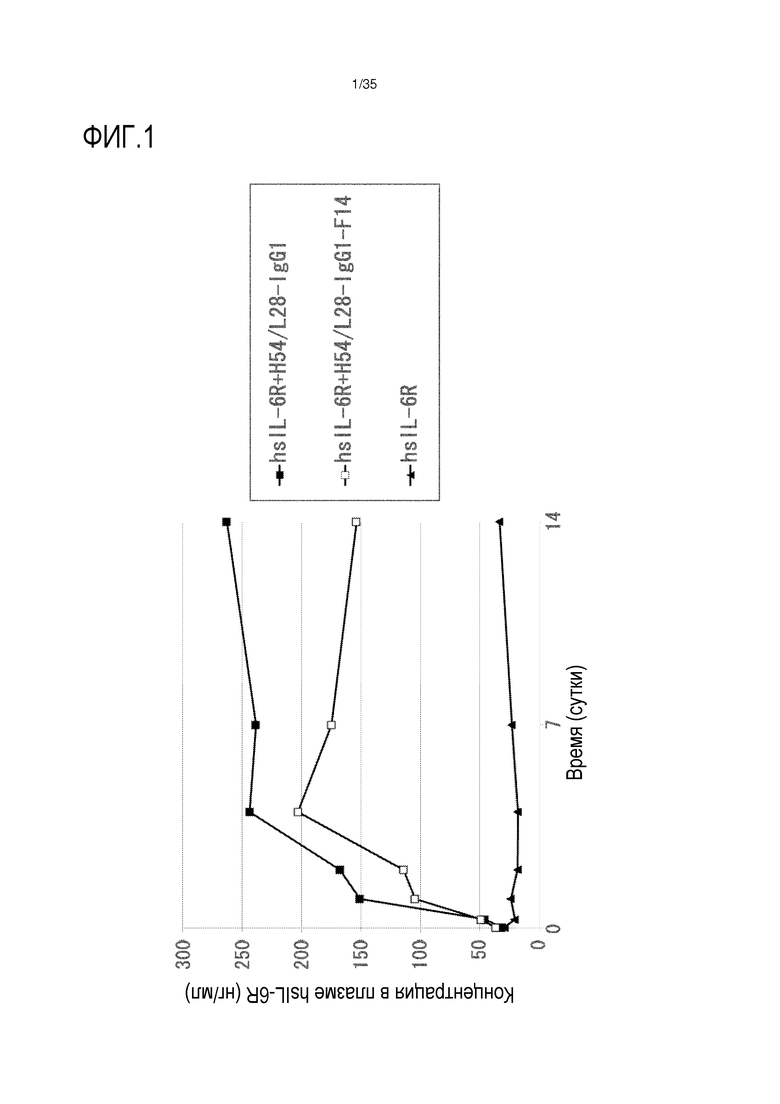
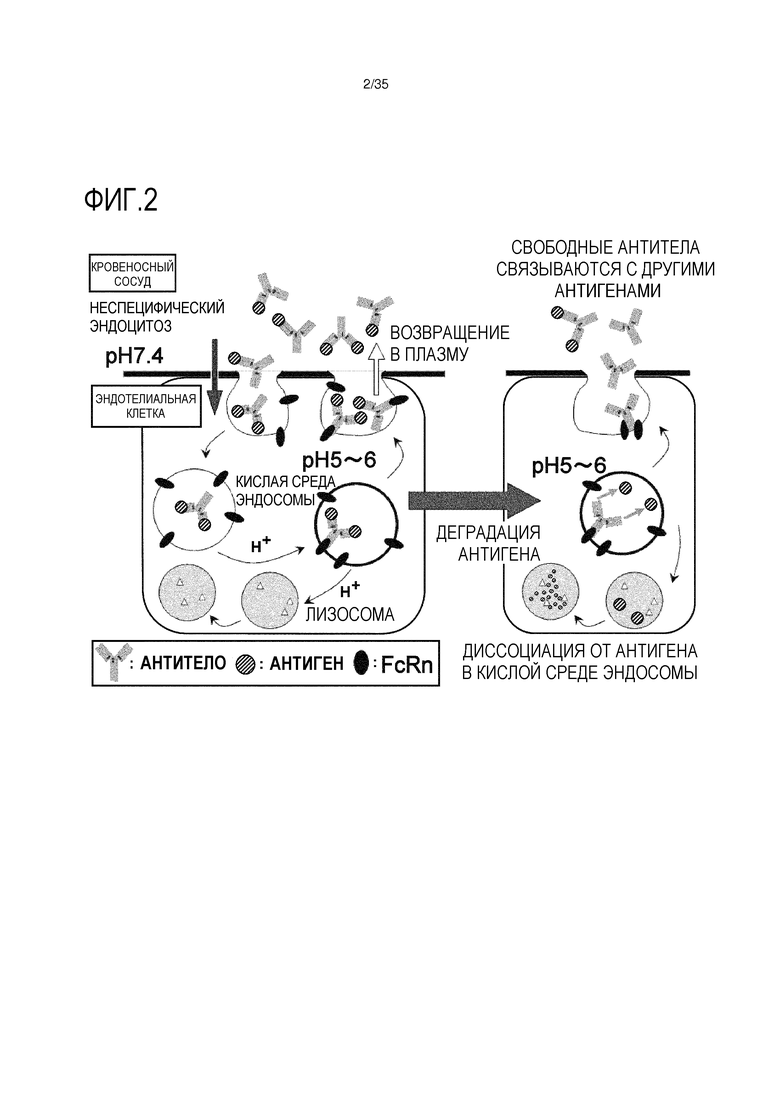
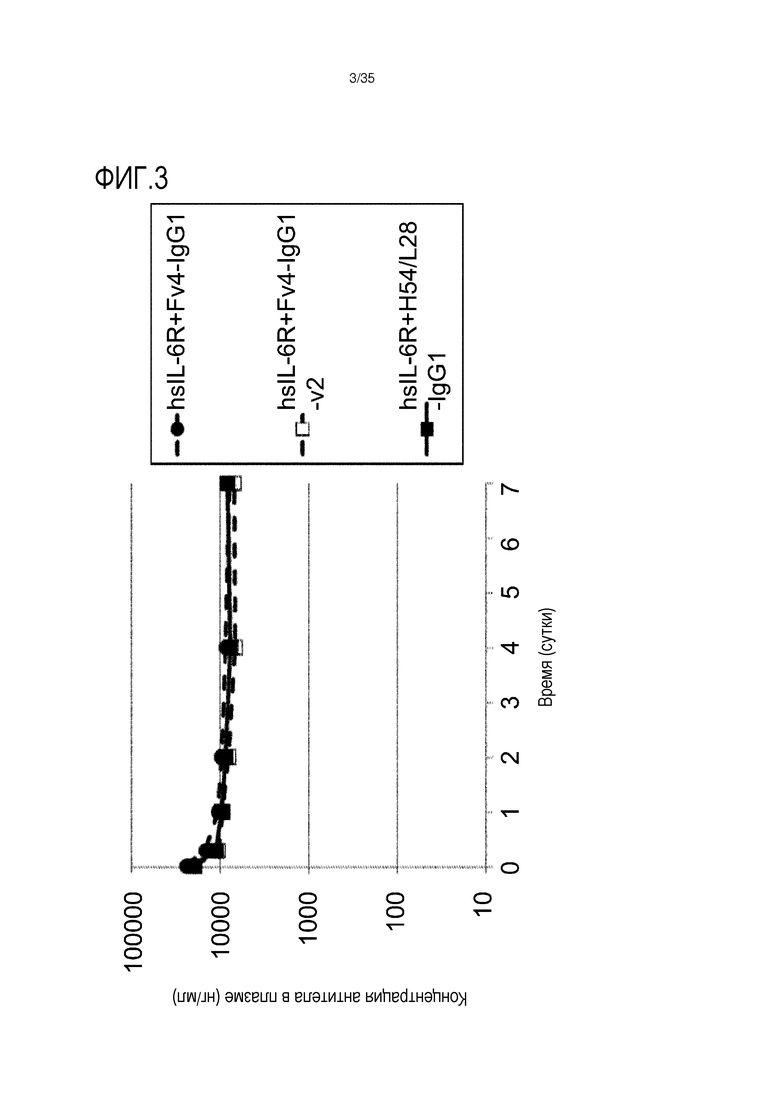
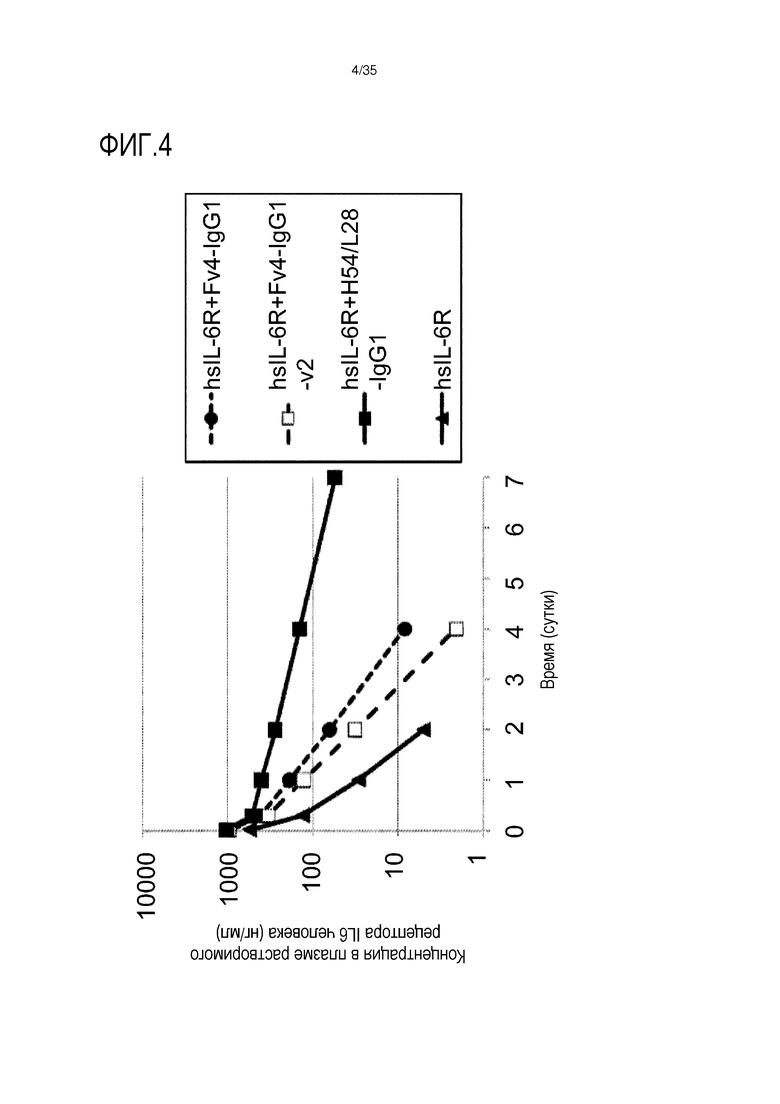

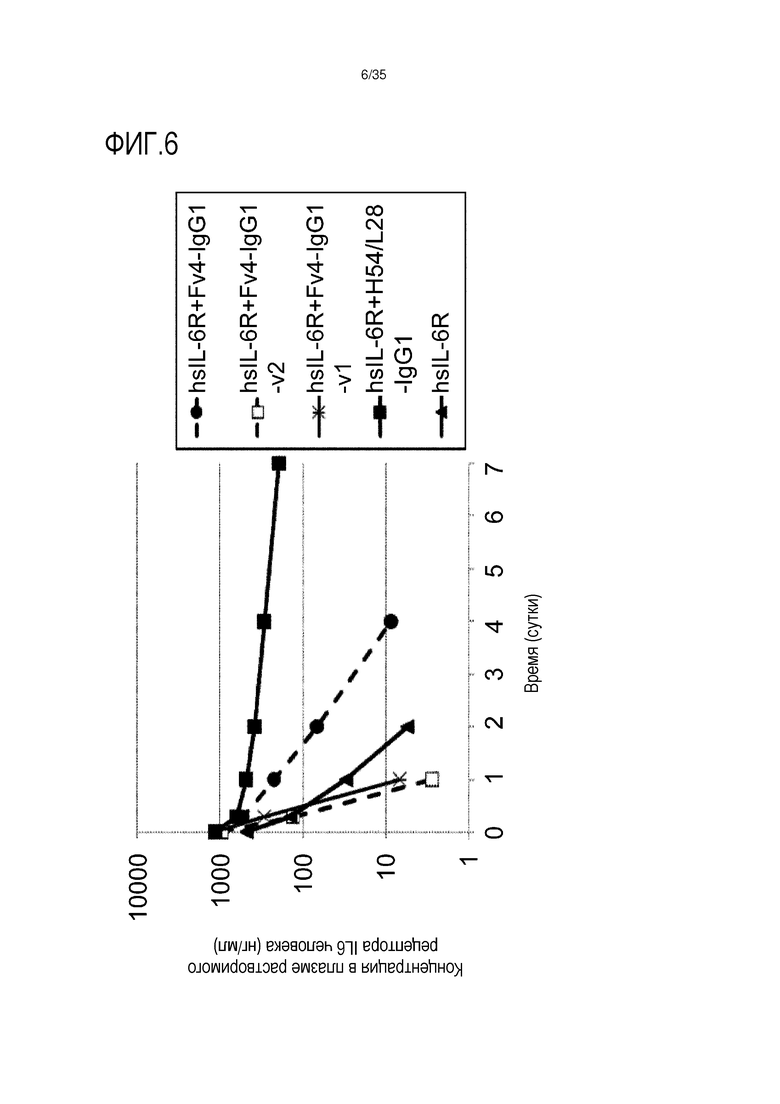
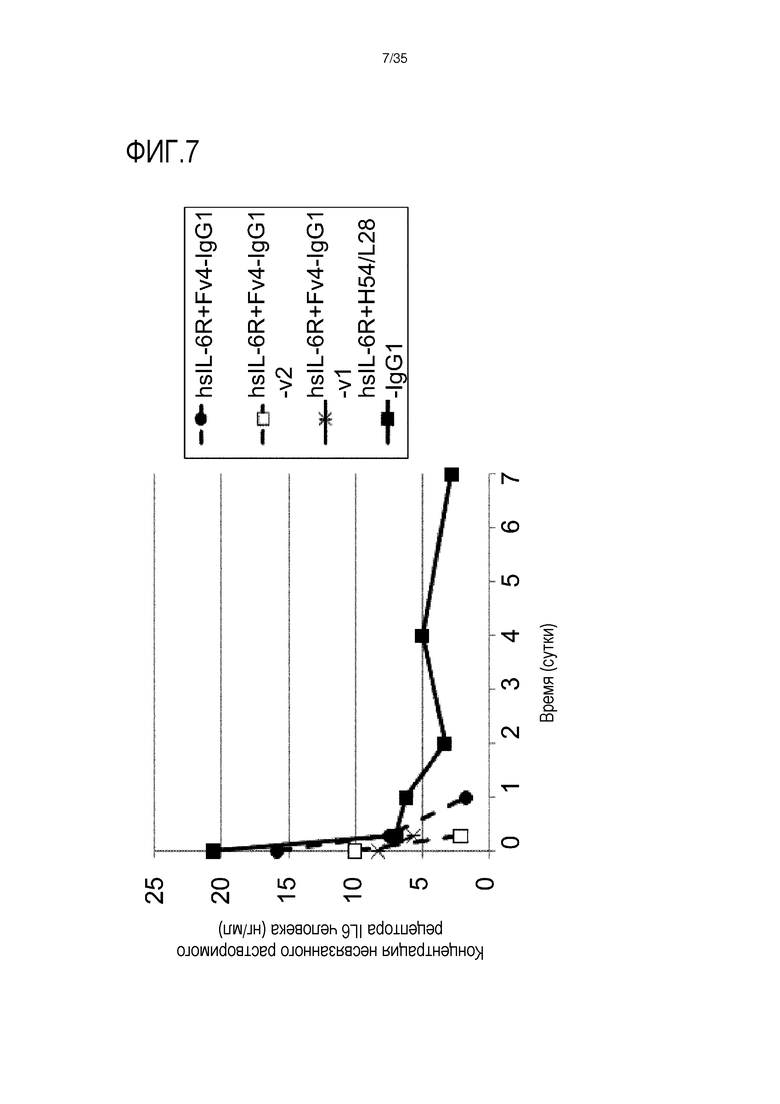
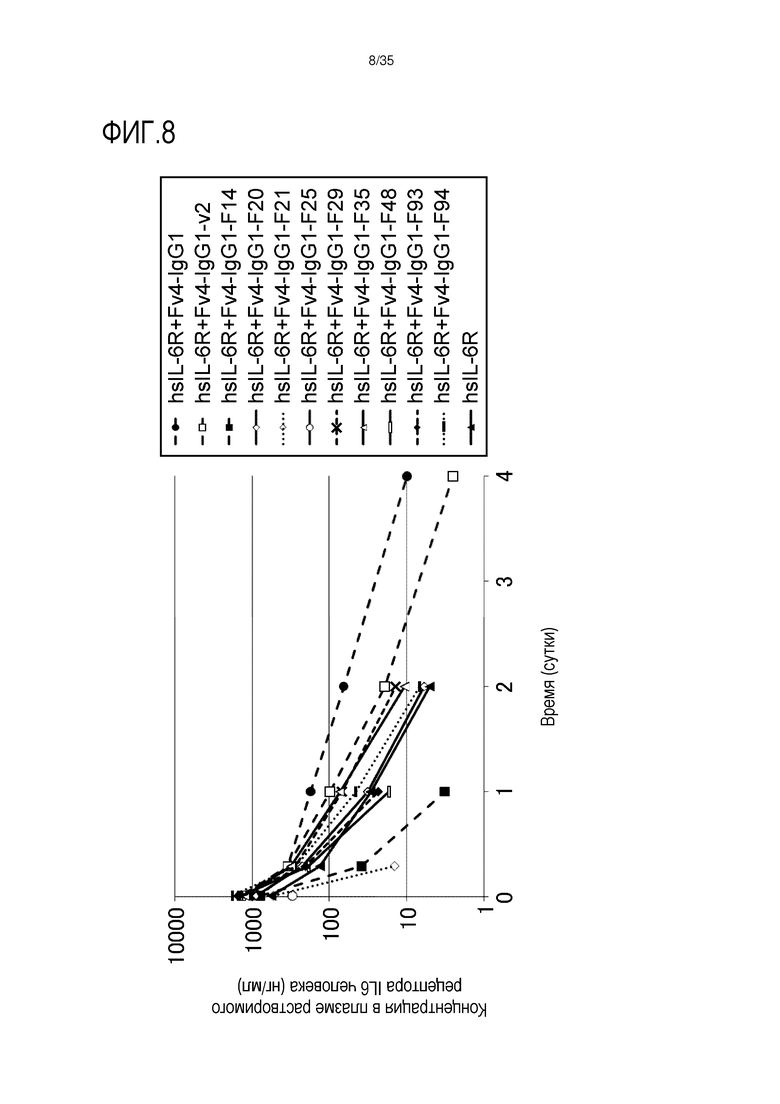

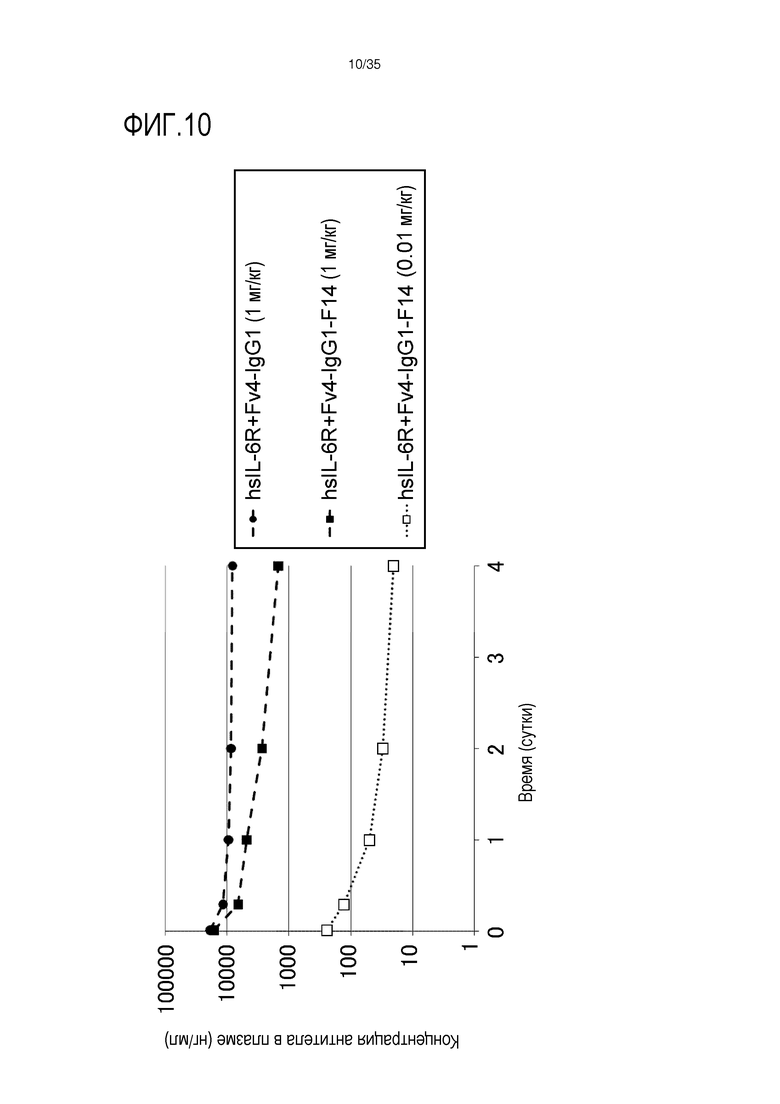
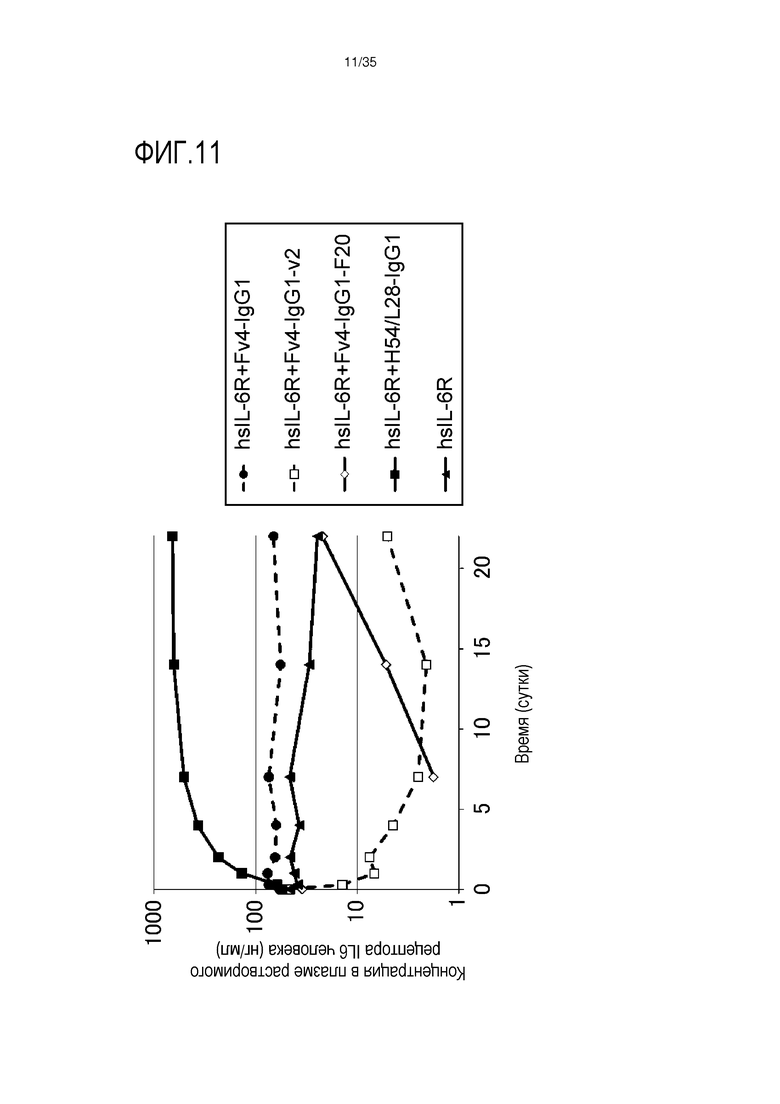


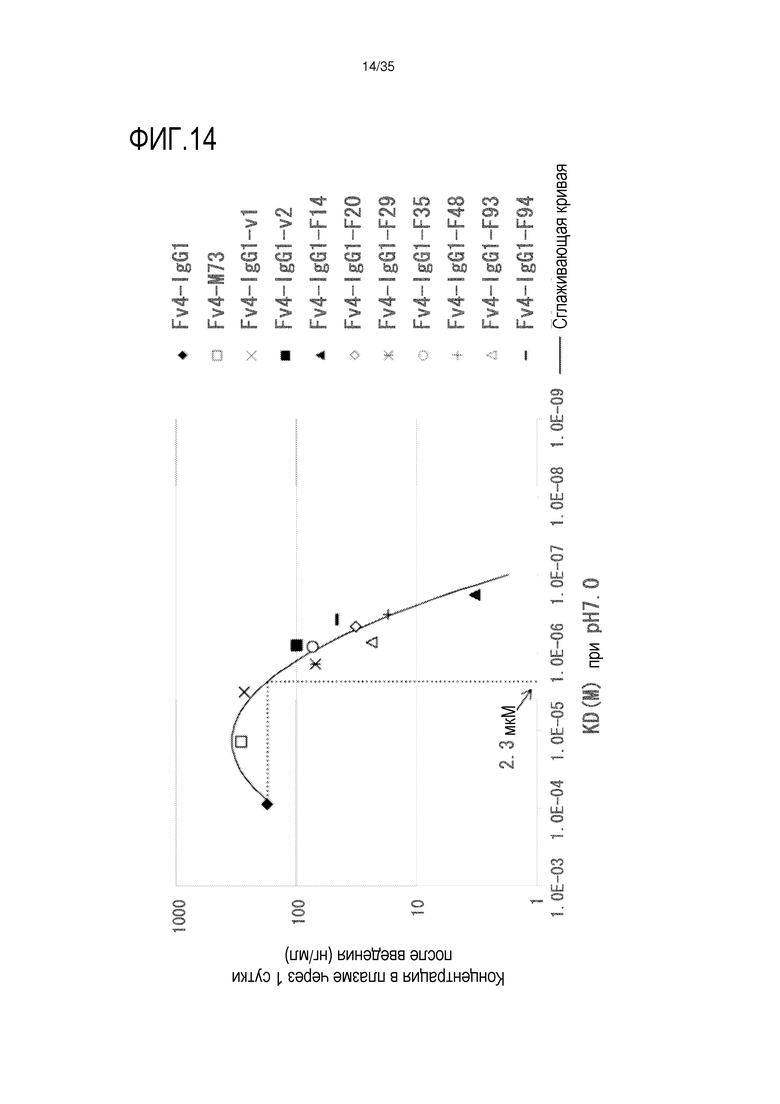
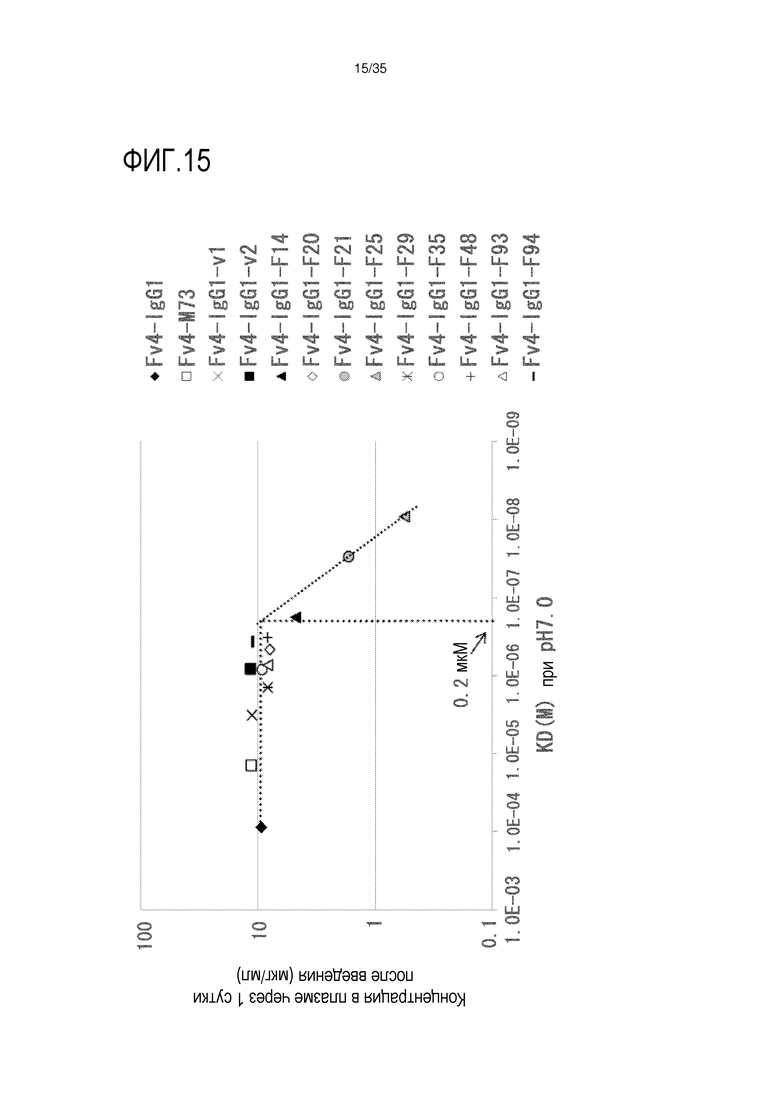

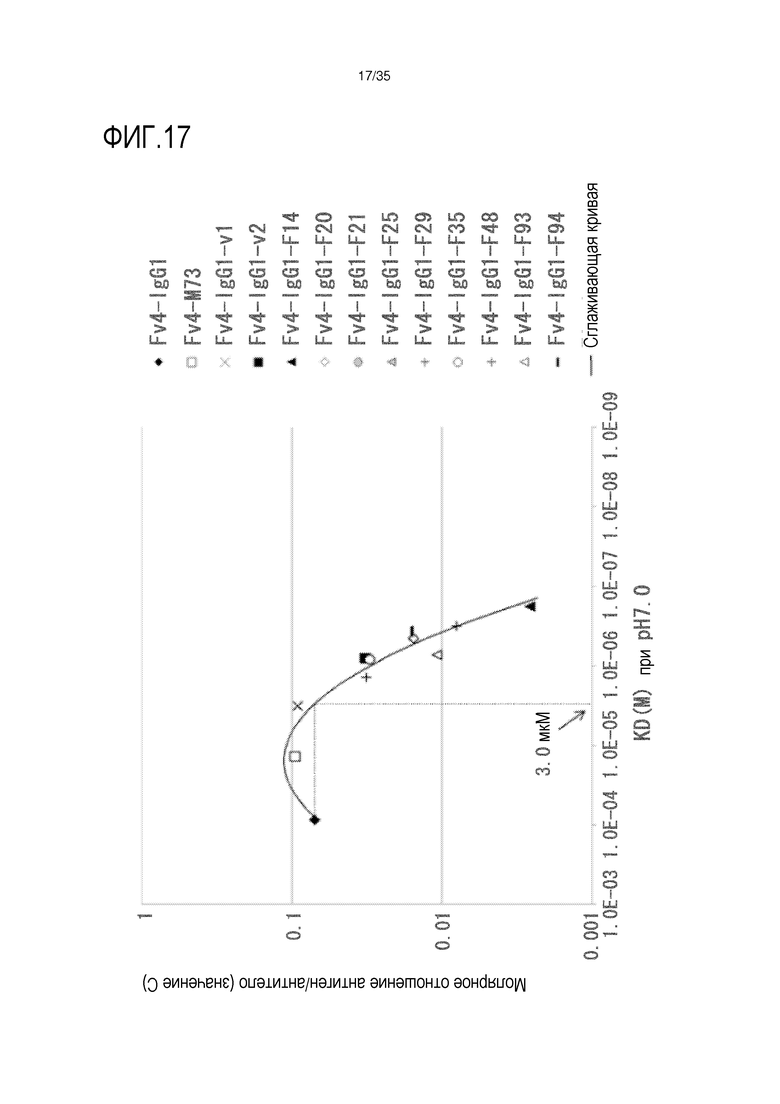
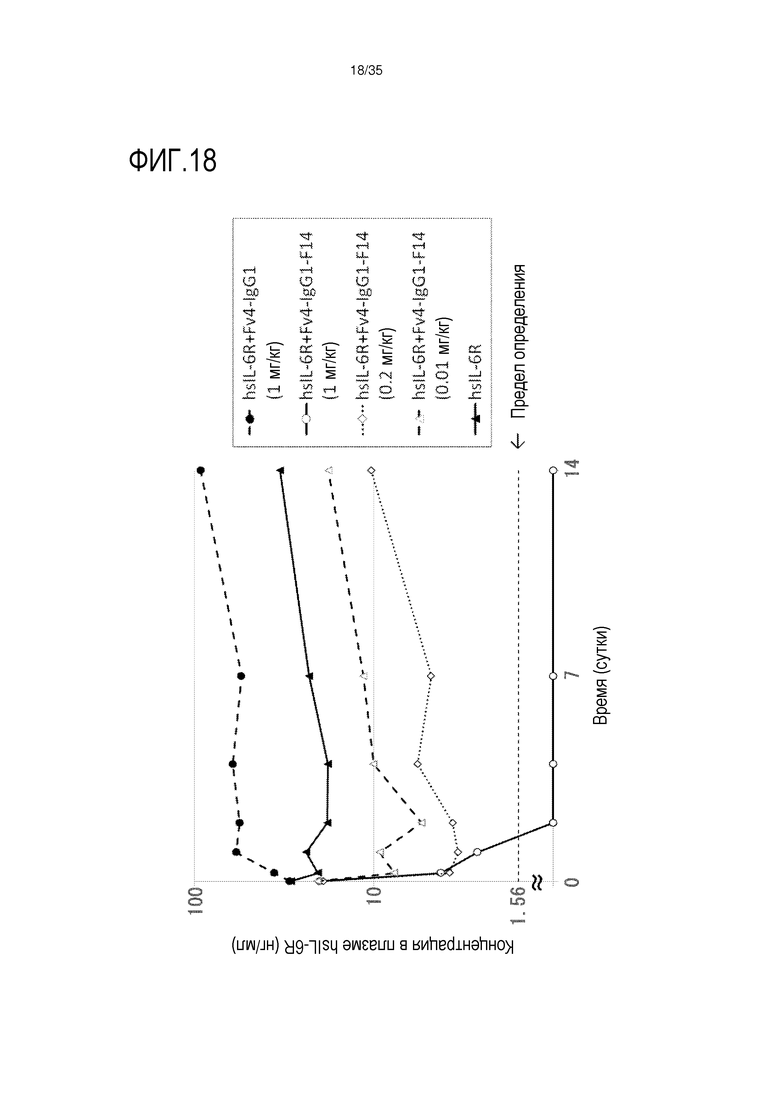

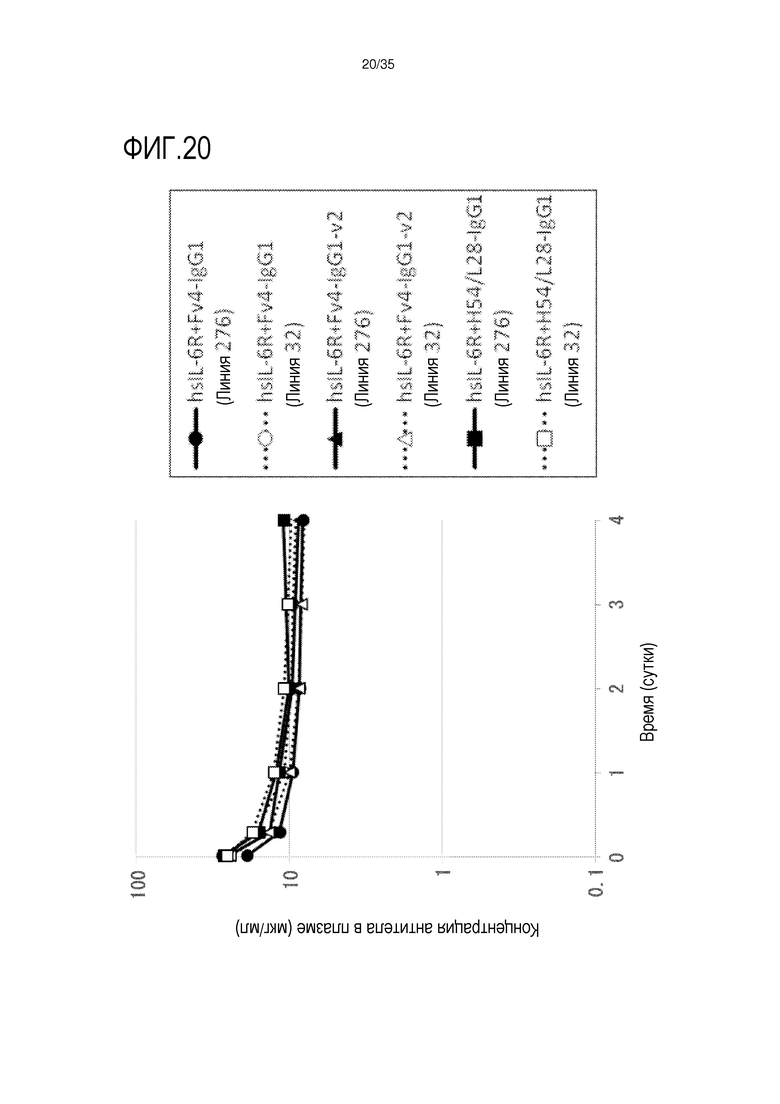
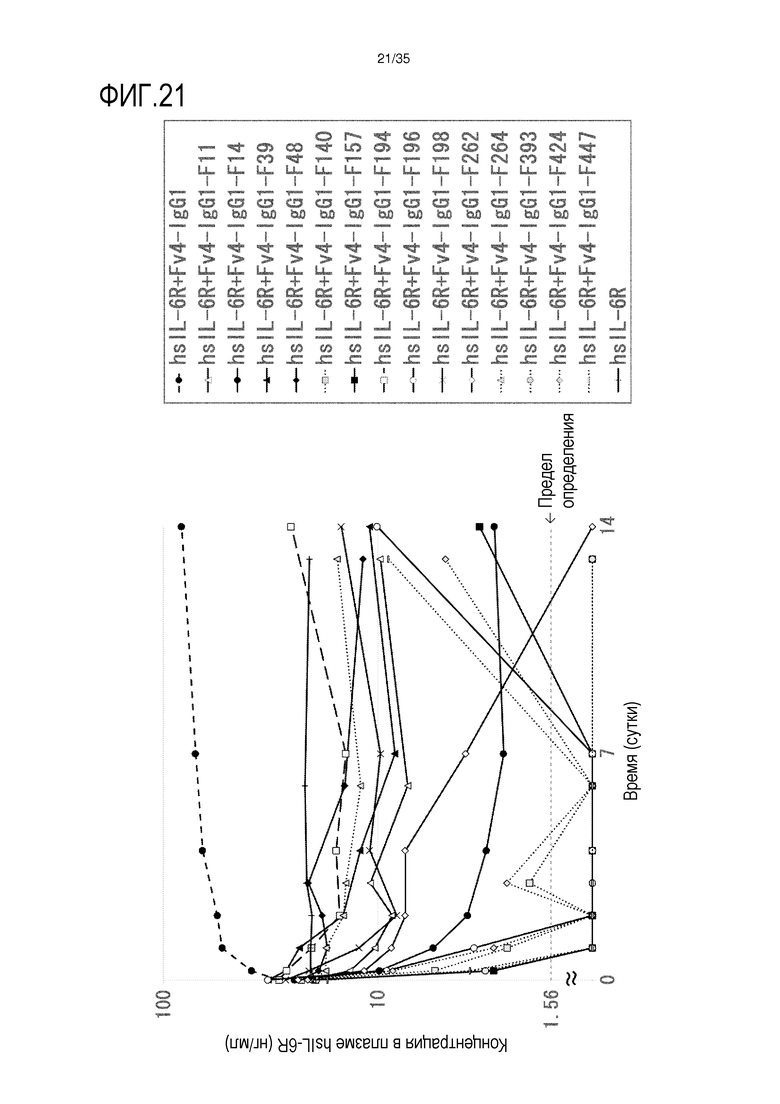
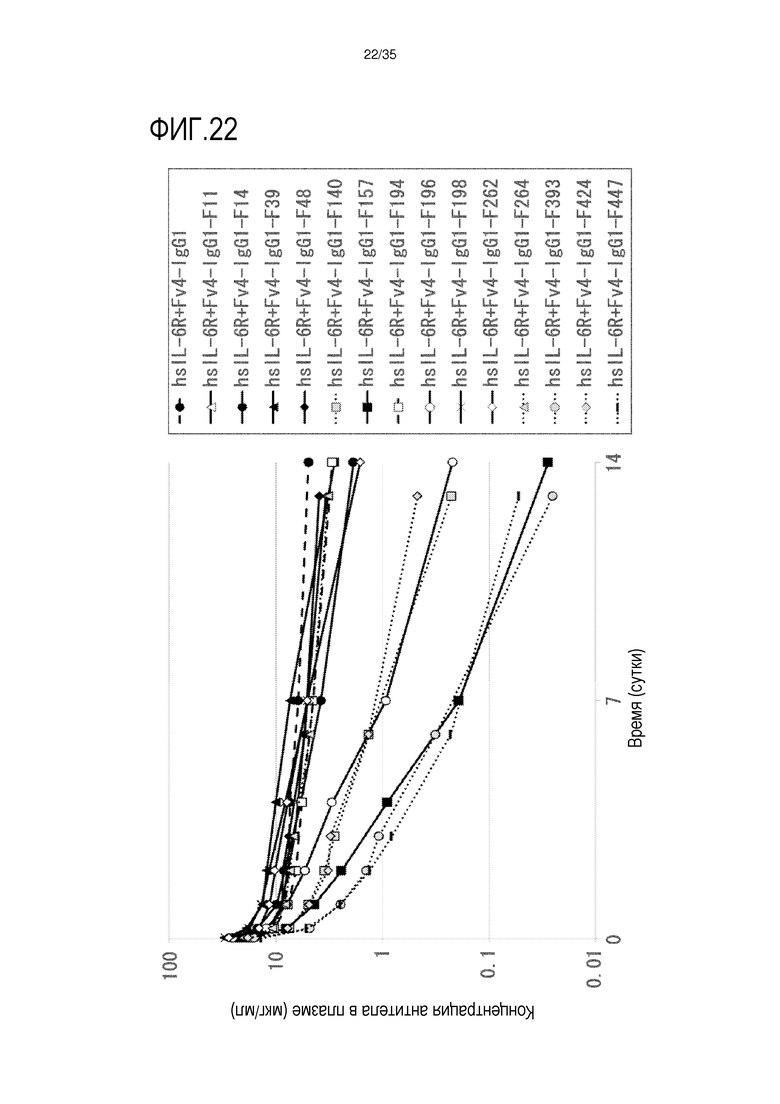
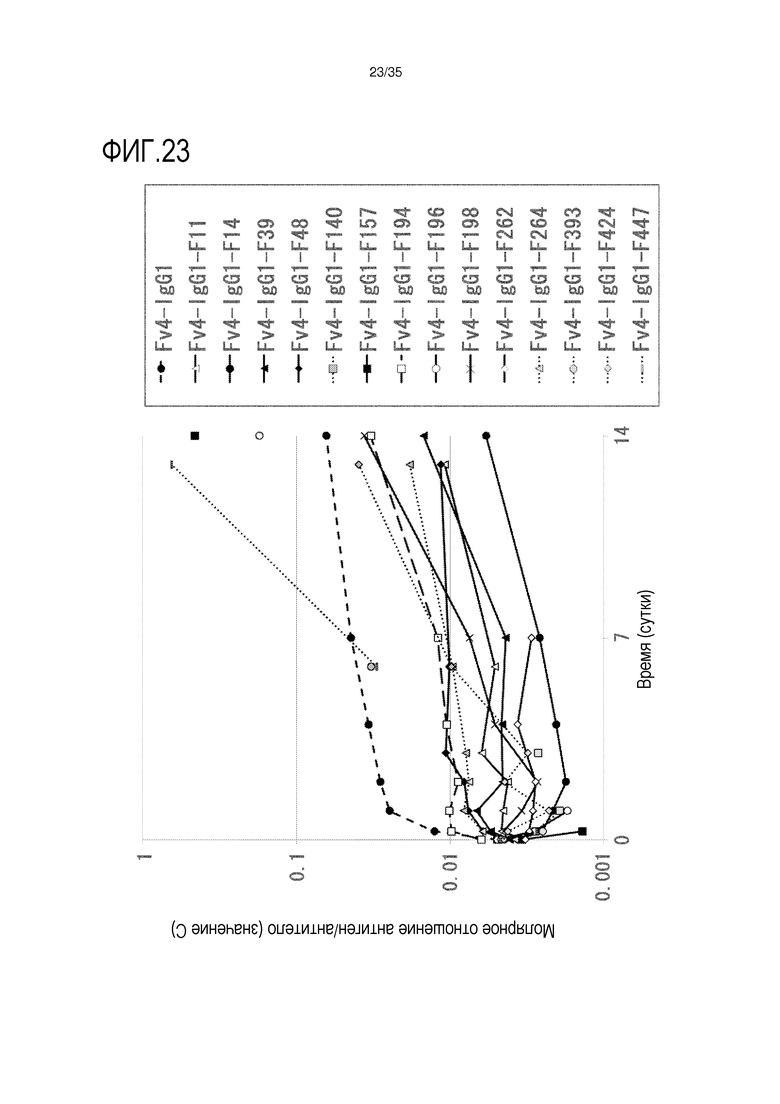
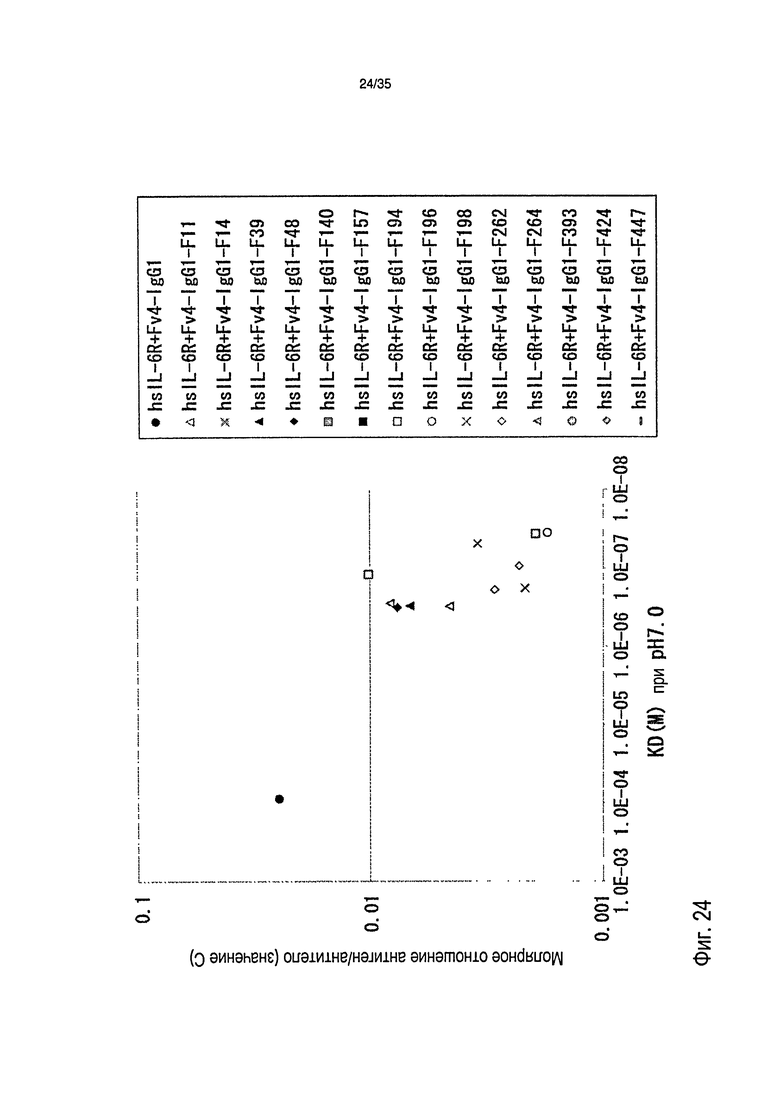
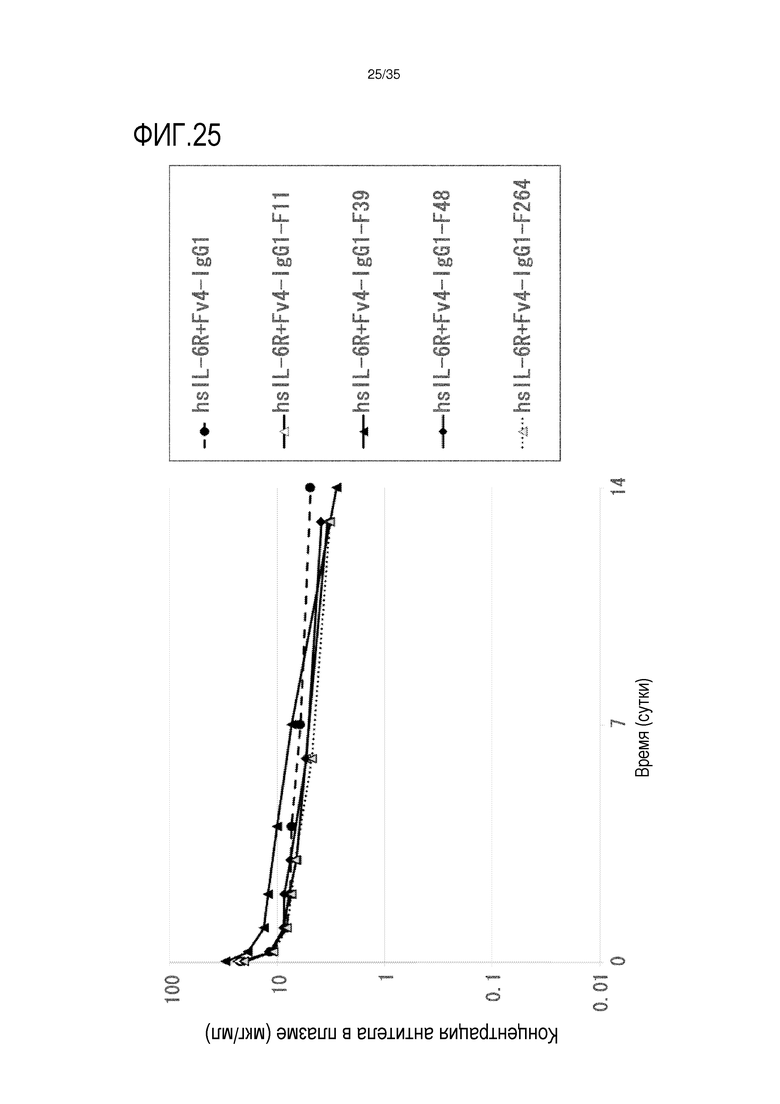
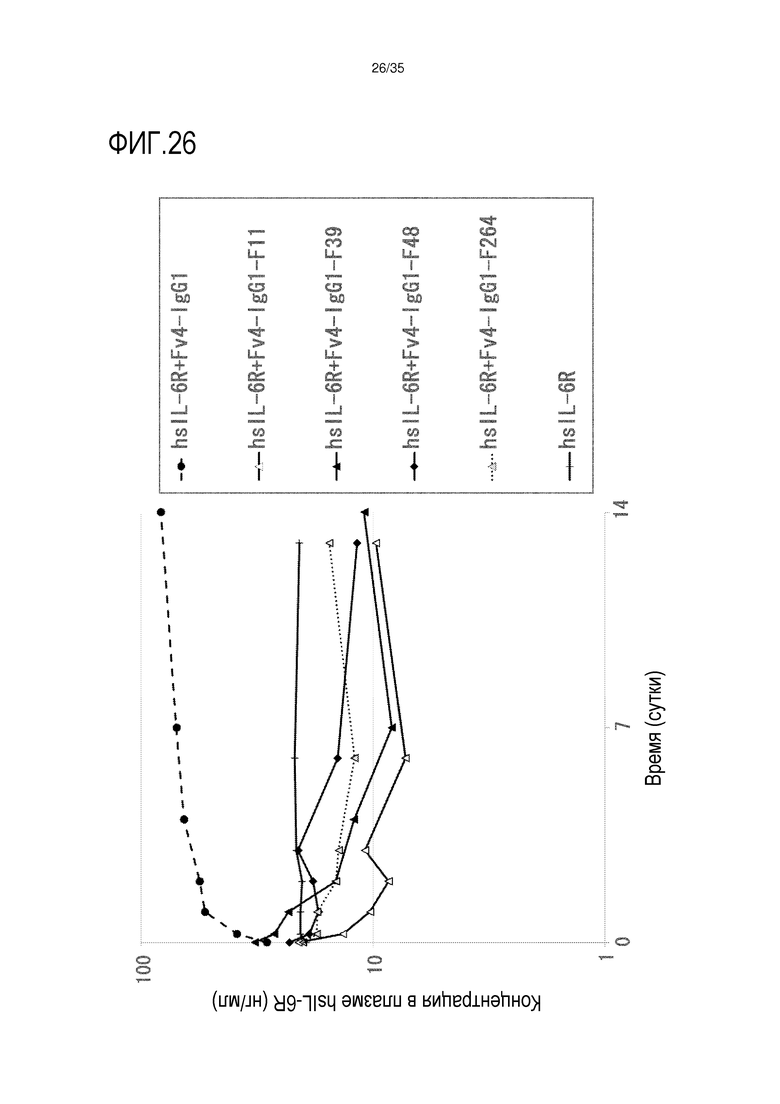
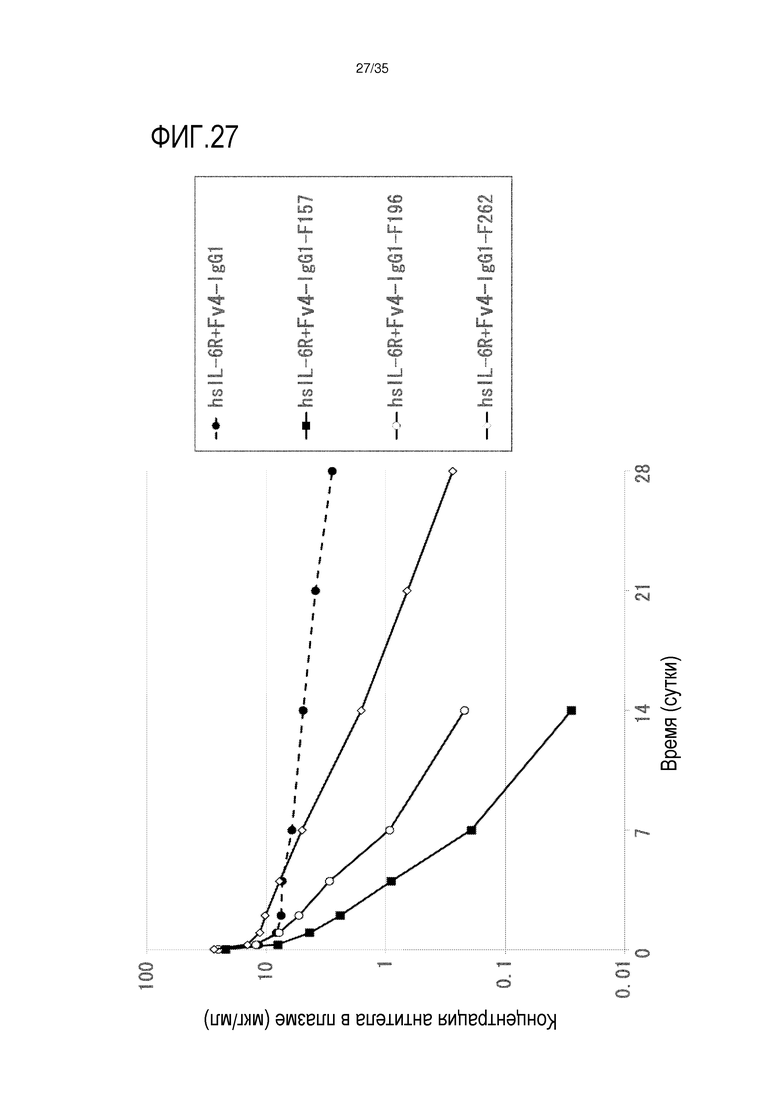
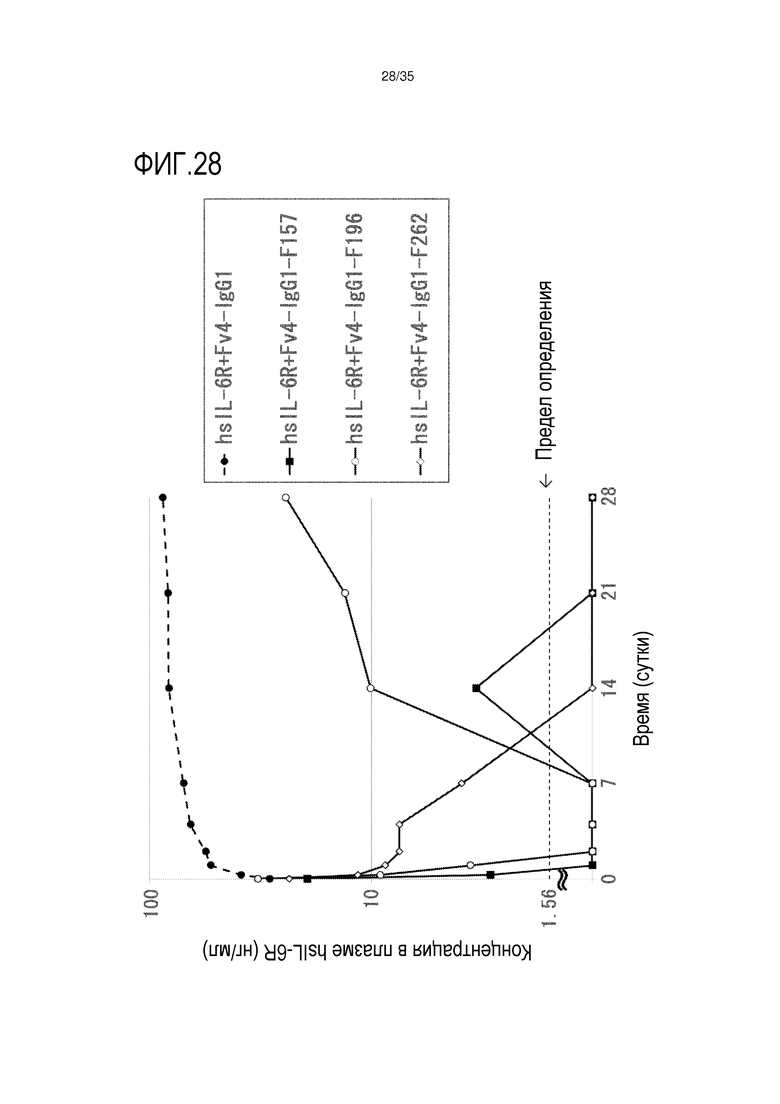
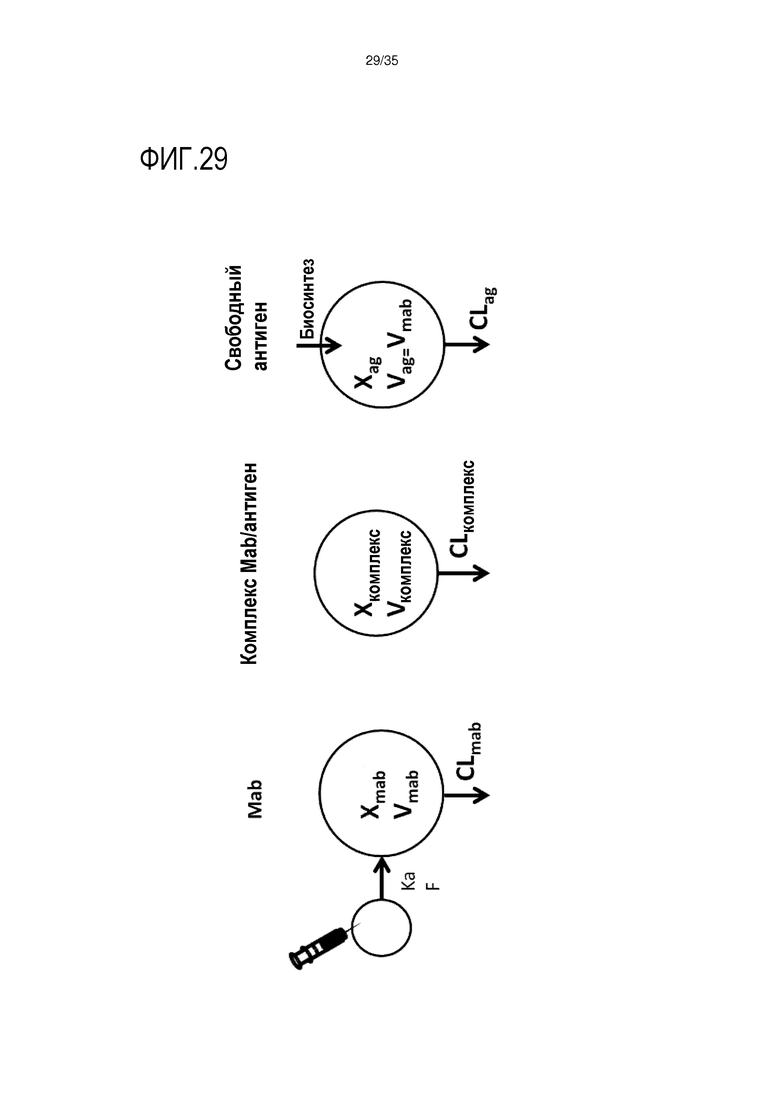

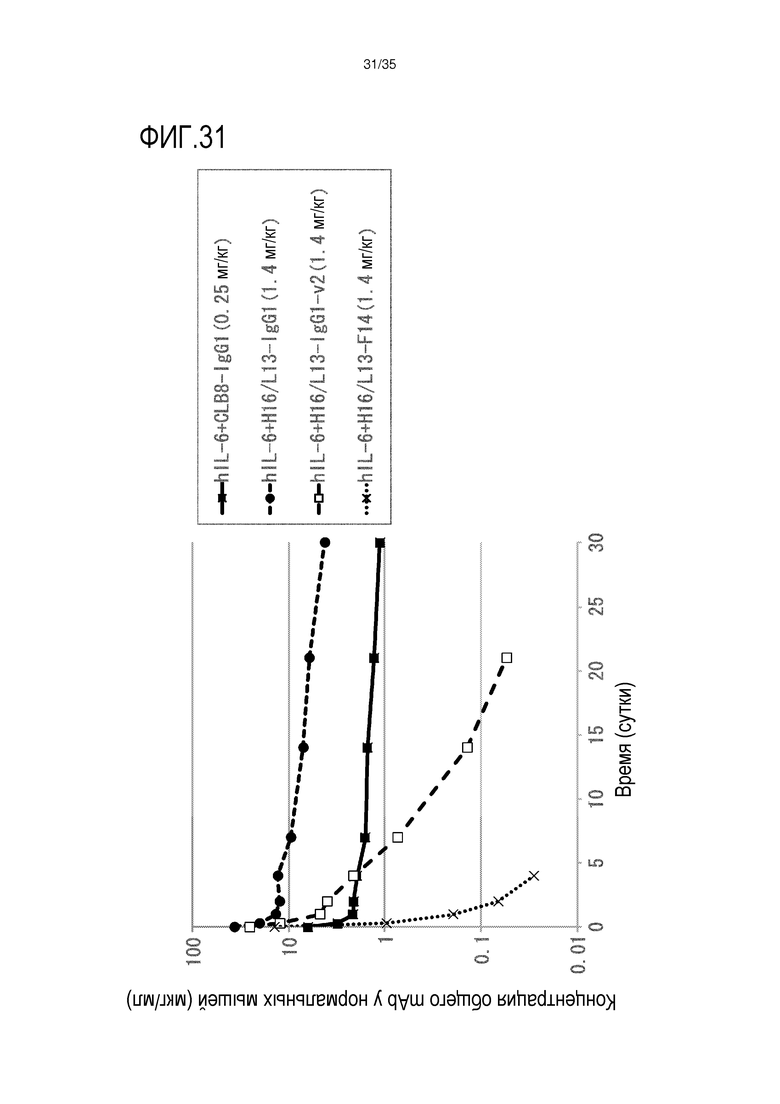
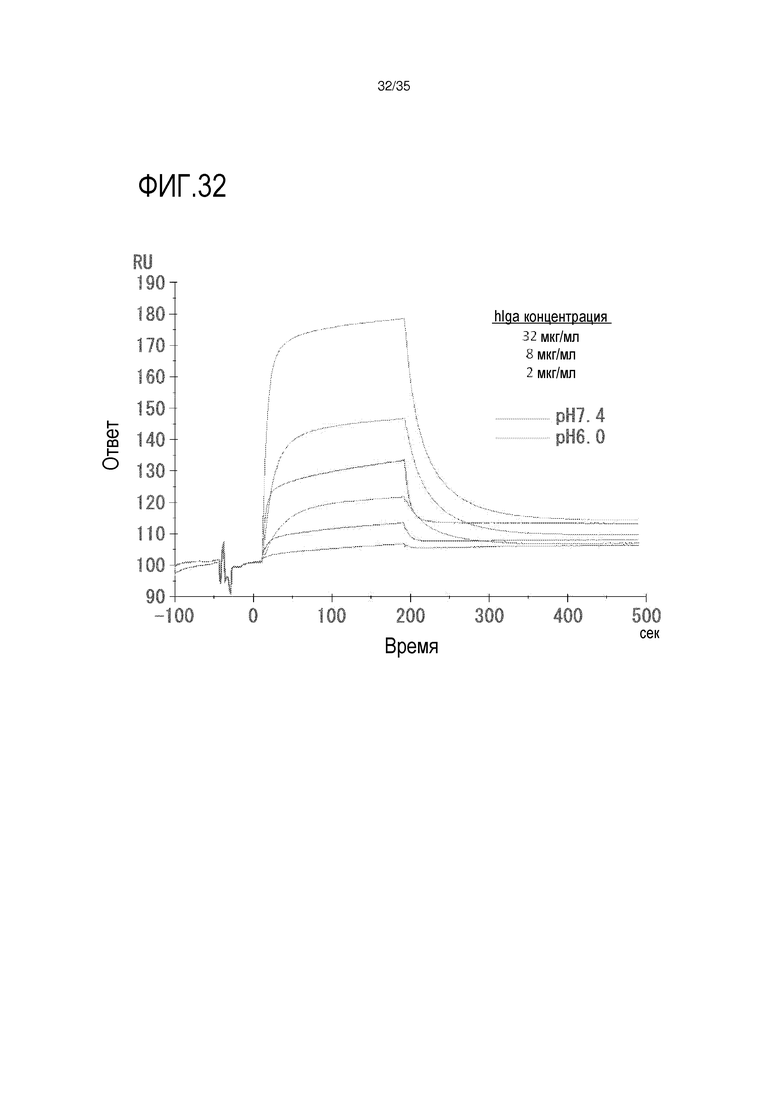
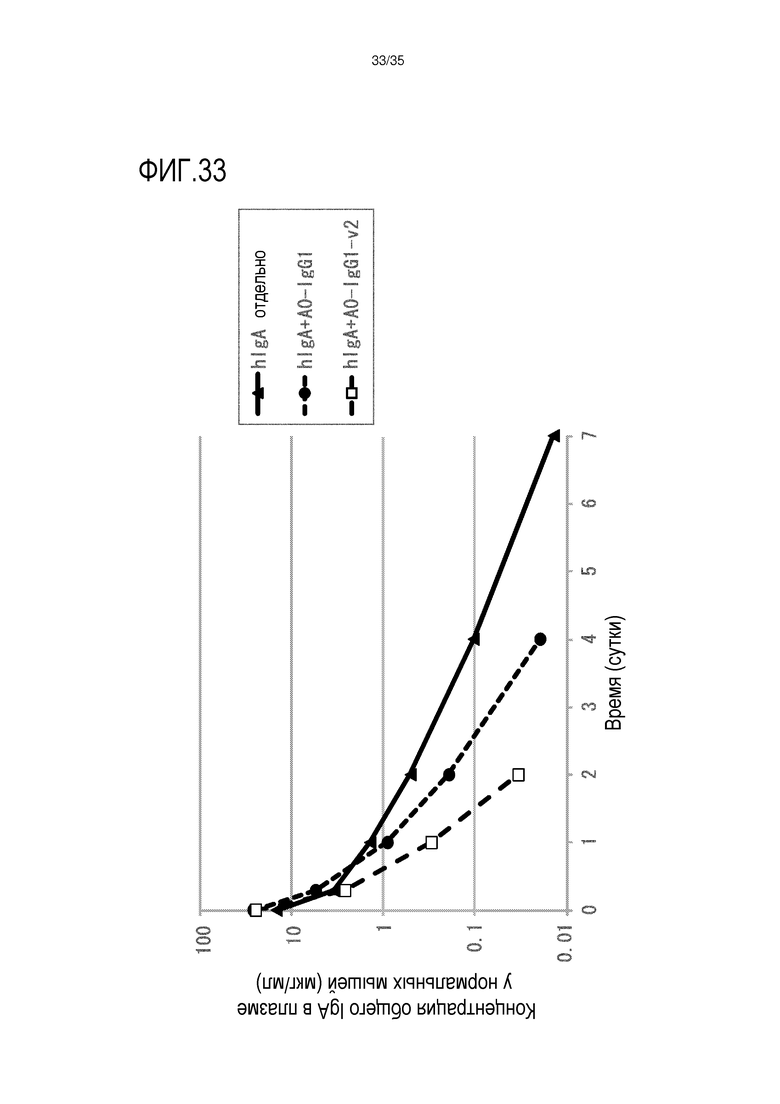


Изобретение относится к области биотехнологии, в частности к способу облегчения опосредованного антителом поглощения антигена клетками, включающему введение в указанное антитело аминокислотных мутаций. При этом вышеуказанное антитело обладает активностью связывания человеческого FcRn при рН 5.8 и рН 7. Также раскрыты способ увеличения количества антигенов, с которыми может связываться одно антитело, способ увеличения способности антитела к элиминации антигена из плазмы, способ усиления элиминации антигена из плазмы, способ облегчения внеклеточного высвобождения свободной от антигена формы антитела, поглощенного клеткой в связанной с антигеном форме, способ снижения общей концентрации антигена или концентрации свободного антигена в плазме, а также способ получения антитела, предусматривающие ввод в антитело вышеуказанных мутаций. Изобретение также относится к способу скрининга антитела, предусматривающему отбор антитела, которое обладает активностью связывания человеческого FcRn при рН 5.8 и рН 7.0 и более высокой активностью связывания человеческого FcRn при рН 7.0. Изобретение позволяет эффективно получать антитело с улучшенной фармакокинетикой. 10 н. и 1 з.п. ф-лы, 35 ил., 15 табл., 17 пр.
1. Способ облегчения опосредованного антителом поглощения антигена клетками, который включает стадии:
(а) введения в указанное антитело IgG1, IgG2, IgG3 или IgG4, которое содержит антигенсвязывающий домен и FcRn-связывающий домен человека, включающий Fc домен IgG1 человека, IgG2 человека, IgG3 человека или IgG4 человека, по меньшей мере одной из следующих аминокислотных замен в положениях, соответствующих нумерации EU в Fc-домене:
N434W;
N434Y;
M252Y/S254T/T256E/N434Y;
M252Y/S254T/T256E/N434W;
M252Y/N434Y;
M252Y/N434W;
M252W/N434Y;
M252W/N434W;
P257L/N434Y;
V308F/N434Y;
M252Y/V308P/N434Y;
M428L/N434S;
M252Y/S254T/T256E/V308P/N434W;
I332V;
I332V/N434Y;
G237M/V308F;
S254T/N434W;
S254T/N434Y;
T256E/N434W;
T256E/N434Y;
S254T/T256E/N434W;
S254T/T256E/N434Y;
M252Y/S254T/N434W;
M252Y/S254T/N434Y;
M252Y/T256E/N434W;
M252Y/T256E/N434Y;
M252Y/S254T/T256E/N434A;
M252Y/N434A;
M252W/N434A;
M252Y/T256Q/N434W;
M252Y/T256Q/N434Y;
M252F/T256D/N434W;
M252F/T256D/N434Y;
H433K/N434F/Y436H;
I332V/N434W;
V308P/N434W;
I332V/M428L/N434Y;
G385D/Q386P/N389S/N434W;
G385D/Q386P/N389S/N434Y;
G385H/N434W;
G385H/N434Y;
N434F;
M252Y/S254T/T256E/N434F;
M252Y/S254T/T256E/N434H;
M252Y/N434F;
M252Y/N434H;
M428L/N434W;
M428L/N434Y;
M252Y/S254T/T256E/M428L/N434W;
M252Y/S254T/T256E/M428L/N434Y;
M252Y/M428L/N434W;
M252Y/M428L/N434Y;
M252Y/M428L/N434A;
M252Y/S254T/T256E/M428L/N434A;
T256E/M428L/N434Y;
S254T/M428L/N434W;
M252Y/T256Q/N434A;
M252Y/T256E/N434A;
T256Q/N434W;
T256Q/N434Y;
M252W/T256Q/N434W;
M252W/T256Q/N434Y;
S254T/T256Q/N434W;
M252Y/S254T/T256Q/N434W;
M252Y/S254T/T256Q/N434Y;
M252Y/T256E/V308P/N434W;
M252Y/V308P/M428L/N434Y;
M252Y/S254T/T256E/V308P/M428L/N434W;
M252W/M428L/N434W;
P257L/M428L/N434Y;
M252Y/S254T/T256E/M428L/N434F;
M252Y/T256E/N434H;
M252W/T256Q/P257L/N434Y;
P238A/M252Y/V308P/N434Y;
M252Y/D265A/V308P/N434Y;
M252Y/T307A/V308P/N434Y;
M252Y/V303A/V308P/N434Y;
M252Y/V308P/D376A/N434Y;
M252Y/V305A/V308P/N434Y;
M252Y/V308P/Q311A/N434Y;
M252Y/V308P/K317A/N434Y;
M252Y/V308P/E380A/N434Y;
M252Y/V308P/E382A/N434Y;
M252Y/V308P/S424A/N434Y;
P257L/V308P/M428L/N434Y;
M252Y/T256E/V308P/M428L/N434W;
M252Y/T256E/M428L/N434W;
M252Y/T256E/V308P/N434Y;
M252Y/T256E/M428L/N434Y;
M252Y/S254T/T256E/V308P/M428L/N434Y;
P257L/M428L/N434W;
P257A/M428L/N434Y;
P257G/M428L/N434Y;
P257I/M428L/N434Y;
P257M/M428L/N434Y;
P257N/M428L/N434Y;
P257S/M428L/N434Y;
P257T/M428L/N434Y;
P257V/M428L/N434Y;
M252W/V308P/N434W;
S239K/M252Y/V308P/N434Y;
M252Y/S298G/V308P/N434Y;
M252Y/D270F/V308P/N434Y;
M252Y/V308A/N434Y;
M252Y/V308F/N434Y;
M252Y/V308I/N434Y;
M252Y/V308L/N434Y;
M252Y/V308M/N434Y;
M252Y/V308Q/N434Y;
M252Y/V308T/N434Y;
P257A/V308P/M428L/N434Y;
P257T/V308P/M428L/N434Y;
P257V/V308P/M428L/N434Y;
M252W/M428I/N434Y;
M252W/M428Y/N434Y;
M252W/M428F/N434Y;
P238A/M252W/N434Y;
M252W/D265A/N434Y;
M252W/T307Q/N434Y;
M252W/V303A/N434Y;
M252W/D376A/N434Y;
M252W/V305A/N434Y;
M252W/Q311A/N434Y;
M252W/D312A/N434Y;
M252W/K317A/N434Y;
M252W/E380A/N434Y;
M252W/E382A/N434Y;
M252W/S424A/N434Y;
S239K/M252W/N434Y;
M252W/S298G/N434Y;
M252W/D270F/N434Y;
M252W/N325G/N434Y;
P257A/M428L/N434W;
P257T/M428L/N434W;
P257V/M428L/N434W;
M252W/I332V/N434Y;
P257I/Q311I;
M252Y/T307A/N434Y;
M252Y/T307Q/N434Y;
P257L/T307A/M428L/N434Y;
P257A/T307A/M428L/N434Y;
P257T/T307A/M428L/N434Y;
P257V/T307A/M428L/N434Y;
P257L/T307Q/M428L/N434Y;
P257A/T307Q/M428L/N434Y;
P257T/T307Q/M428L/N434Y;
P257V/T307Q/M428L/N434Y;
M252Y/T307D/N434Y;
M252Y/T307F/N434Y;
M252Y/T307G/N434Y;
M252Y/T307H/N434Y;
M252Y/T307I/N434Y;
M252Y/T307K/N434Y;
M252Y/T307L/N434Y;
M252Y/T307M/N434Y;
M252Y/T307N/N434Y;
M252Y/T307P/N434Y;
M252Y/T307R/N434Y;
M252Y/T307S/N434Y;
M252Y/T307V/N434Y;
M252Y/T307W/N434Y;
M252Y/T307Y/N434Y;
M252Y/K334L/N434Y;
M252Y/G385H/N434Y;
M252Y/T289H/N434Y;
M252Y/Q311H/N434Y;
M252Y/D312H/N434Y;
M252Y/N315H/N434Y;
M252Y/K360H/N434Y;
M252Y/L314R/N434Y;
M252Y/L314K/N434Y;
M252Y/N286E/N434Y;
M252Y/L309E/N434Y;
M252Y/R255E/N434Y;
M252Y/P387E/N434Y;
K248I/M428L/N434Y;
M252Y/M428A/N434Y;
M252Y/M428D/N434Y;
M252Y/M428F/N434Y;
M252Y/M428G/N434Y;
M252Y/M428H/N434Y;
M252Y/M428I/N434Y;
M252Y/M428K/N434Y;
M252Y/M428N/N434Y;
M252Y/M428P/N434Y;
M252Y/M428Q/N434Y;
M252Y/M428S/N434Y;
M252Y/M428T/N434Y;
M252Y/M428V/N434Y;
M252Y/M428W/N434Y;
M252Y/M428Y/N434Y;
M252W/T307Q/M428Y/N434Y;
M252W/Q311A/M428Y/N434Y;
M252W/T307Q/Q311A/M428Y/N434Y;
M252Y/T307A/M428Y/N434Y;
M252Y/T307Q/M428Y/N434Y;
M252Y/D270F/N434Y;
M252Y/T307A/Q311A/N434Y;
M252Y/T307Q/Q311A/N434Y;
M252Y/T307A/Q311H/N434Y;
M252Y/T307Q/Q311H/N434Y;
M252Y/E382A/N434Y;
M252Y/E382A/M428Y/N434Y;
M252Y/T307A/E382A/M428Y/N434Y;
M252Y/T307Q/E382A/M428Y/N434Y;
P238A/M252Y/M428F/N434Y;
M252Y/V305A/M428F/N434Y;
M252Y/N325G/M428F/N434Y;
M252Y/D376A/M428F/N434Y;
M252Y/E380A/M428F/N434Y;
M252Y/E382A/M428F/N434Y;
M252Y/E380A/E382A/M428F/N434Y;
S239K/M252Y/V308P/E382A/N434Y;
M252Y/D270F/V308P/E382A/N434Y;
S239K/M252Y/V308P/M428Y/N434Y;
M252Y/T307Q/V308P/E382A/N434Y;
M252Y/V308P/Q311H/E382A/N434Y;
S239K/M252Y/D270F/V308P/N434Y;
M252Y/V308P/E382A/M428F/N434Y;
M252Y/V308P/E382A/M428L/N434Y;
M252Y/V308P/E382A/M428Y/N434Y;
M252Y/V308P/M428Y/N434Y;
M252Y/V308P/M428F/N434Y;
S239K/M252Y/V308P/E382A/M428Y/N434Y;
M252Y/V308P/E380A/E382A/M428F/N434Y;
M252Y/T256E/E382A/N434Y;
M252Y/T256E/M428Y/N434Y;
M252Y/T256E/E382A/M428Y/N434Y;
S239K/M252Y/V308P/E382A/M428I/N434Y;
M252Y/V308P/E380A/E382A/M428I/N434Y;
S239K/M252Y/V308P/M428F/N434Y;
S239K/M252Y/E380A/E382A/M428F/N434Y;
M252Y/Q311A/M428Y/N434Y;
M252Y/D312A/M428Y/N434Y;
M252Y/Q311A/D312A/M428Y/N434Y;
S239K/M252Y/V308P/Q311A/M428Y/N434Y;
S239K/M252Y/V308P/D312A/M428Y/N434Y;
S239K/M252Y/V308P/Q311A/D312A/M428Y/N434Y;
M252Y/T256E/T307P/N434Y;
M252Y/T307P/M428Y/N434Y;
M252W/V308P/M428Y/N434Y;
M252Y/T256E/V308P/E382A/N434Y;
M252W/V308P/E382A/N434Y;
S239K/M252W/V308P/E382A/N434Y;
S239K/M252W/V308P/E382A/M428Y/N434Y;
S239K/M252W/V308P/E382A/M428I/N434Y;
S239K/M252W/V308P/M428F/N434Y;
S239K/M252W/E380A/E382A/M428F/N434Y;
S239K/M252Y/T307P/M428Y/N434Y;
M252Y/T256E/Q311A/D312A/M428Y/N434Y;
S239K/M252Y/M428Y/N434Y;
S239K/M252Y/D270F/M428Y/N434Y;
S239K/M252Y/D270F/V308P/M428Y/N434Y;
S239K/M252Y/V308P/M428I/N434Y;
S239K/M252Y/V308P/N325G/E382A/M428I/N434Y;
S239K/M252Y/D270F/V308P/N325G/N434Y;
S239K/M252Y/D270F/T307P/V308P/N434Y;
S239K/M252Y/T256E/D270F/V308P/N434Y;
S239K/M252Y/D270F/T307Q/V308P/N434Y;
S239K/M252Y/D270F/T307Q/V308P/Q311A/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/N434Y;
S239K/M252Y/T256E/D270F/T307Q/Q311A/N434Y;
S239K/M252Y/D270F/V308P/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/N434Y;
S239K/M252Y/D270F/V308P/L309E/N434Y;
S239K/M252Y/D270F/V308P/P387E/N434Y;
S239K/M252Y/D270F/T307Q/L309E/Q311A/N434Y;
S239K/M252Y/D270F/V308P/N325G/M428I/N434Y;
S239K/M252Y/T256E/D270F/V308P/M428I/N434Y;
S239K/M252Y/D270F/T307Q/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/M428I/N434Y;
S239K/M252Y/D270F/V308P/L309E/M428I/N434Y;
S239K/M252Y/D270F/V308P/M428L/N434Y;
S239K/M252Y/V308P/M428L/N434Y;
S239K/M252Y/V308P/L309P/M428L/N434Y;
S239K/M252Y/V308P/L309R/M428L/N434Y;
S239K/M252Y/L309P/M428L/N434Y;
S239K/M252Y/L309R/M428L/N434Y;
S239K/M252Y/T307Q/V308P/M428L/N434Y;
S239K/M252Y/D270F/V308P/L309E/P387E/N434Y;
S239K/M252Y/T256E/D270F/V308P/N325G/M428I/N434Y;
S239K/M252Y/T307Q/N434Y;
P257V/T307Q/M428I/N434Y;
P257V/T307Q/M428V/N434Y;
P257V/T307Q/N325G/M428L/N434Y;
P257V/T307Q/Q311A/M428L/N434Y;
P257V/T307Q/Q311A/N325G/M428L/N434Y;
P257V/V305A/T307Q/M428L/N434Y;
S239K/M252Y/E258H/D270F/T307Q/V308P/Q311A/N434Y;
S239K/M252Y/D270F/V308P/N325G/E382A/M428I/N434Y;
M252Y/P257V/T307Q/M428I/N434Y;
S239K/M252W/V308P/M428Y/N434Y;
S239K/M252W/V308P/Q311A/M428Y/N434Y;
S239K/M252W/T256E/V308P/M428Y/N434Y;
S239K/M252W/N286E/V308P/M428Y/N434Y;
S239K/M252Y/T256E/D270F/N286E/V308P/N434Y;
S239K/M252W/T307P/M428Y/N434Y;
S239K/M252W/T256E/V308P/Q311A/M428Y/N434Y;
S239K/M252W/T256E/N286E/V308P/M428Y/N434Y;
P257V/T307A/Q311A/M428L/N434Y;
P257V/V305A/T307A/M428L/N434Y;
M252Y/V308P/L309E/N434Y;
M252Y/V308P/L309D/N434Y;
M252Y/V308P/L309A/N434Y;
M252W/V308P/L309E/M428Y/N434Y;
M252W/V308P/L309D/M428Y/N434Y;
M252W/V308P/L309A/M428Y/N434Y;
S239K/M252Y/N286E/V308P/M428I/N434Y;
S239K/M252Y/N286E/V308P/N434Y;
S239K/M252Y/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/T256E/V308P/N434Y;
S239K/M252Y/D270F/N286E/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/P387E/M428I/N434Y;
S239K/M252Y/D270F/N286E/T307Q/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/T307Q/V308P/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/P387E/M428I/N434Y;
P257V/T307Q/M428L/N434W;
P257V/T307A/M428L/N434W;
P257A/T307Q/L309P/M428L/N434Y;
P257V/T307Q/L309P/M428L/N434Y;
P257A/T307Q/L309R/M428L/N434Y;
P257V/T307Q/L309R/M428L/N434Y;
P257V/N286E/M428L/N434Y;
P257V/P387E/M428L/N434Y;
P257V/T307H/M428L/N434Y;
P257V/T307N/M428L/N434Y;
P257V/T307G/M428L/N434Y;
P257V/T307P/M428L/N434Y;
P257V/T307S/M428L/N434Y;
P257V/N286E/T307A/M428L/N434Y;
P257V/T307A/P387E/M428L/N434Y;
S239K/M252Y/T307P/N325G/M428Y/N434Y;
M252Y/T307A/Q311H/K360H/N434Y;
M252Y/T307A/Q311H/P387E/N434Y;
M252Y/T307A/Q311H/M428A/N434Y;
M252Y/T307A/Q311H/E382A/N434Y;
M252Y/T307W/Q311H/N434Y;
S239K/P257A/V308P/M428L/N434Y;
P257A/V308P/L309E/M428L/N434Y;
P257V/T307Q/N434Y;
M252Y/P257V/T307Q/M428V/N434Y;
M252Y/P257V/T307Q/M428L/N434Y;
M252Y/P257V/T307Q/N434Y;
M252Y/P257V/T307Q/M428Y/N434Y;
M252Y/P257V/T307Q/M428F/N434Y;
P257V/T307Q/Q311A/N325G/M428V/N434Y;
P257V/T307Q/Q311A/N325G/N434Y;
P257V/T307Q/Q311A/N325G/M428Y/N434Y;
P257V/T307Q/Q311A/N325G/M428F/N434Y;
P257A/T307Q/M428V/N434Y;
P257A/T307Q/N434Y;
P257A/T307Q/M428Y/N434Y;
P257A/T307Q/M428F/N434Y;
P257V/N286E/T307Q/Q311A/N325G/M428L/N434Y;
P257A/Q311A/M428L/N434Y;
P257A/Q311H/M428L/N434Y;
P257A/T307Q/Q311A/M428L/N434Y;
P257A/T307A/Q311A/M428L/N434Y;
P257A/T307Q/Q311H/M428L/N434Y;
P257A/T307A/Q311H/M428L/N434Y;
P257A/N286E/T307Q/M428L/N434Y;
P257A/N286E/T307A/M428L/N434Y;
S239K/M252Y/D270F/T307P/N325G/M428Y/N434Y;
S239K/M252Y/T307P/L309E/N325G/M428Y/N434Y;
P257S/T307A/M428L/N434Y;
P257M/T307A/M428L/N434Y;
P257N/T307A/M428L/N434Y;
P257I/T307A/M428L/N434Y;
P257V/T307Q/M428Y/N434Y;
P257V/T307Q/M428F/N434Y;
S239K/P257V/V308P/M428L/N434Y;
P257V/T307Q/V308P/N325G/M428L/N434Y;
P257V/T307Q/V308P/Q311A/N325G/M428L/N434Y;
P257V/T307A/V308P/N325G/M428L/N434Y;
P257A/V308P/N325G/M428L/N434Y;
P257A/T307A/V308P/M428L/N434Y;
P257A/T307Q/V308P/M428L/N434Y;
P257V/N286E/T307Q/N325G/M428L/N434Y;
T256E/P257V/N434Y;
T256E/P257T/N434Y;
S239K/P257T/V308P/M428L/N434Y;
P257T/V308P/N325G/M428L/N434Y;
T256E/P257T/V308P/N325G/M428L/N434Y;
P257T/V308P/N325G/E382A/M428L/N434Y;
P257T/V308P/N325G/P387E/M428L/N434Y;
P257T/V308P/L309P/N325G/M428L/N434Y;
P257T/V308P/L309R/N325G/M428L/N434Y;
T256E/P257V/T307Q/M428L/N434Y;
P257V/T307Q/E382A/M428L/N434Y;
P257V/T307Q/P387E/M428L/N434Y;
P257L/V308P/N434Y;
P257T/T307Q/N434Y;
P257V/T307Q/N325G/N434Y;
P257V/T307Q/Q311A/N434Y;
P257V/V305A/T307Q/N434Y;
P257V/N286E/T307A/N434Y;
P257V/T307Q/L309R/Q311H/M428L/N434Y;
P257V/V308P/N325G/M428L/N434Y;
S239K/P257V/V308P/Q311H/M428L/N434Y;
P257V/V305A/T307A/N325G/M428L/N434Y;
S239K/D270F/T307P/N325G/M428Y/N434Y;
P257T/T307A/M428V/N434Y;
P257A/M428V/N434Y;
P257A/T307A/M428V/N434Y;
P257S/T307Q/M428L/N434Y;
P257V/N297A/T307Q/M428L/N434Y;
P257V/N286A/T307Q/M428L/N434Y;
P257V/T307Q/N315A/M428L/N434Y;
P257V/T307Q/N384A/M428L/N434Y;
P257V/T307Q/N389A/M428L/N434Y;
P257V/N434Y;
P257T/N434Y;
P257V/N286E/N434Y;
P257T/N286E/N434Y;
P257V/N286E/T307Q/N434Y;
P257V/N286E/T307Q/M428Y/N434Y;
P257V/V305A/T307Q/M428Y/N434Y;
P257T/T307Q/M428Y/N434Y;
P257V/T307Q/V308P/N325G/M428Y/N434Y;
P257T/T307Q/M428F/N434Y;
P257A/N286E/T307Q/M428F/N434Y;
P257A/N286E/T307Q/M428Y/N434Y;
T250A/P257V/T307Q/M428L/N434Y;
T250F/P257V/T307Q/M428L/N434Y;
T250I/P257V/T307Q/M428L/N434Y;
T250M/P257V/T307Q/M428L/N434Y;
T250S/P257V/T307Q/M428L/N434Y;
T250V/P257V/T307Q/M428L/N434Y;
T250W/P257V/T307Q/M428L/N434Y;
T250Y/P257V/T307Q/M428L/N434Y;
M252Y/Q311A/N434Y;
S239K/M252Y/S254T/V308P/N434Y;
M252Y/T307Q/Q311A;
M252Y/S254T/N286E/N434Y;
M252Y/S254T/V308P/N434Y;
M252Y/S254T/T307A/N434Y;
M252Y/S254T/T307Q/N434Y;
M252Y/S254T/Q311A/N434Y;
M252Y/S254T/Q311H/N434Y;
M252Y/S254T/T307A/Q311A/N434Y;
M252Y/S254T/T307Q/Q311A/N434Y;
M252Y/S254T/M428I/N434Y;
M252Y/T256E/T307A/Q311H/N434Y;
M252Y/T256Q/T307A/Q311H/N434Y;
M252Y/S254T/T307A/Q311H/N434Y;
M252Y/N286E/T307A/Q311H/N434Y;
M252Y/T307A/Q311H/M428I/N434Y;
M252Y/T256E/T307Q/Q311H;
M252Y/N286E/T307A/Q311A;
M252Y/N286E/T307Q/Q311A;
M252Y/N286E/T307Q/Q311H;
M252Y/T256E/N286E/N434Y;
S239K/M252Y/V308P;
S239K/M252Y/V308P/E382A;
S239K/M252Y/T256E/V308P;
S239K/M252Y/N286E/V308P;
S239K/M252Y/N286E/V308P/M428I;
M252Y/N286E/M428L/N434Y;
M252Y/S254T/E382A/N434Y;
S239K/M252Y/S254T/V308P/M428I/N434Y;
S239K/M252Y/M428I/N434Y;
M252Y/D312A/E382A/M428Y/N434Y;
M252Y/E382A/P387E/N434Y;
M252Y/D312A/P387E/N434Y или
M252Y/P387E/M428Y/N434Y; и
(b) введения в указанное антитело по меньшей мере одной из гистидиновых замен или инсерций в антигенсвязывающий домен тяжелой цепи в положение, выбранное из группы, состоящей из Н27, Н31, Н32, Н33, Н35, Н50, Н58, Н59, Н61, Н62, Н63, Н64, Н65, Н99, H100b и Н102 в соответствии с нумерацией Kabat, и/или антигенсвязывающий домен легкой цепи в положение, выбранное из группы, состоящей из L24, L27, L28, L32, L53, L54, L56, L90, L92 и L94 в соответствии с нумерацией Kabat;
где указанное антитело, содержащее антигенсвязывающий домен и человеческий FcRn-связывающий домен, полученные в (а) и (b), связывается с растворимым антигеном, обладает активностью связывания человеческого FcRn при рН 5.8 и рН 7.0, где его активность связывания FcRn человека при рН 7.0 сильнее, чем интактного IgG человека, и обладает более низкой антигенсвязывающей активностью при рН 5.8, чем при рН 7.4, где отношение антигенсвязывающей активности при рН 5.8 и рН 7.4 равно по меньшей мере 2 при величине KD при рН 5.8 / KD при рН 7.4.
2. Способ увеличения количества антигенов, с которыми может связываться одно антитело, который включает стадии:
(а) введения в указанное антитело IgG1, IgG2, IgG3 или IgG4, которое содержит антигенсвязывающий домен и FcRn-связывающий домен человека, включающий Fc домен IgG1 человека, IgG2 человека, IgG3 человека или IgG4 человека, по меньшей мере одной из следующих аминокислотных замен в положениях, соответствующих нумерации EU в Fc-домене:
N434W;
N434Y;
M252Y/S254T/T256E/N434Y;
M252Y/S254T/T256E/N434W;
M252Y/N434Y;
M252Y/N434W;
M252W/N434Y;
M252W/N434W;
P257L/N434Y;
V308F/N434Y;
M252Y/V308P/N434Y;
M428L/N434S;
M252Y/S254T/T256E/V308P/N434W;
I332V;
I332V/N434Y;
G237M/V308F;
S254T/N434W;
S254T/N434Y;
T256E/N434W;
T256E/N434Y;
S254T/T256E/N434W;
S254T/T256E/N434Y;
M252Y/S254T/N434W;
M252Y/S254T/N434Y;
M252Y/T256E/N434W;
M252Y/T256E/N434Y;
M252Y/S254T/T256E/N434A;
M252Y/N434A;
M252W/N434A;
M252Y/T256Q/N434W;
M252Y/T256Q/N434Y;
M252F/T256D/N434W;
M252F/T256D/N434Y;
H433K/N434F/Y436H;
I332V/N434W;
V308P/N434W;
I332V/M428L/N434Y;
G385D/Q386P/N389S/N434W;
G385D/Q386P/N389S/N434Y;
G385H/N434W;
G385H/N434Y;
N434F;
M252Y/S254T/T256E/N434F;
M252Y/S254T/T256E/N434H;
M252Y/N434F;
M252Y/N434H;
M428L/N434W;
M428L/N434Y;
M252Y/S254T/T256E/M428L/N434W;
M252Y/S254T/T256E/M428L/N434Y;
M252Y/M428L/N434W;
M252Y/M428L/N434Y;
M252Y/M428L/N434A;
M252Y/S254T/T256E/M428L/N434A;
T256E/M428L/N434Y;
S254T/M428L/N434W;
M252Y/T256Q/N434A;
M252Y/T256E/N434A;
T256Q/N434W;
T256Q/N434Y;
M252W/T256Q/N434W;
M252W/T256Q/N434Y;
S254T/T256Q/N434W;
M252Y/S254T/T256Q/N434W;
M252Y/S254T/T256Q/N434Y;
M252Y/T256E/V308P/N434W;
M252Y/V308P/M428L/N434Y;
M252Y/S254T/T256E/V308P/M428L/N434W;
M252W/M428L/N434W;
P257L/M428L/N434Y;
M252Y/S254T/T256E/M428L/N434F;
M252Y/T256E/N434H;
M252W/T256Q/P257L/N434Y;
P238A/M252Y/V308P/N434Y;
M252Y/D265A/V308P/N434Y;
M252Y/T307A/V308P/N434Y;
M252Y/V303A/V308P/N434Y;
M252Y/V308P/D376A/N434Y;
M252Y/V305A/V308P/N434Y;
M252Y/V308P/Q311A/N434Y;
M252Y/V308P/K317A/N434Y;
M252Y/V308P/E380A/N434Y;
M252Y/V308P/E382A/N434Y;
M252Y/V308P/S424A/N434Y;
P257L/V308P/M428L/N434Y;
M252Y/T256E/V308P/M428L/N434W;
M252Y/T256E/M428L/N434W;
M252Y/T256E/V308P/N434Y;
M252Y/T256E/M428L/N434Y;
M252Y/S254T/T256E/V308P/M428L/N434Y;
P257L/M428L/N434W;
P257A/M428L/N434Y;
P257G/M428L/N434Y;
P257I/M428L/N434Y;
P257M/M428L/N434Y;
P257N/M428L/N434Y;
P257S/M428L/N434Y;
P257T/M428L/N434Y;
P257V/M428L/N434Y;
M252W/V308P/N434W;
S239K/M252Y/V308P/N434Y;
M252Y/S298G/V308P/N434Y;
M252Y/D270F/V308P/N434Y;
M252Y/V308A/N434Y;
M252Y/V308F/N434Y;
M252Y/V308I/N434Y;
M252Y/V308L/N434Y;
M252Y/V308M/N434Y;
M252Y/V308Q/N434Y;
M252Y/V308T/N434Y;
P257A/V308P/M428L/N434Y;
P257T/V308P/M428L/N434Y;
P257V/V308P/M428L/N434Y;
M252W/M428I/N434Y;
M252W/M428Y/N434Y;
M252W/M428F/N434Y;
P238A/M252W/N434Y;
M252W/D265A/N434Y;
M252W/T307Q/N434Y;
M252W/V303A/N434Y;
M252W/D376A/N434Y;
M252W/V305A/N434Y;
M252W/Q311A/N434Y;
M252W/D312A/N434Y;
M252W/K317A/N434Y;
M252W/E380A/N434Y;
M252W/E382A/N434Y;
M252W/S424A/N434Y;
S239K/M252W/N434Y;
M252W/S298G/N434Y;
M252W/D270F/N434Y;
M252W/N325G/N434Y;
P257A/M428L/N434W;
P257T/M428L/N434W;
P257V/M428L/N434W;
M252W/I332V/N434Y;
P257I/Q311I;
M252Y/T307A/N434Y;
M252Y/T307Q/N434Y;
P257L/T307A/M428L/N434Y;
P257A/T307A/M428L/N434Y;
P257T/T307A/M428L/N434Y;
P257V/T307A/M428L/N434Y;
P257L/T307Q/M428L/N434Y;
P257A/T307Q/M428L/N434Y;
P257T/T307Q/M428L/N434Y;
P257V/T307Q/M428L/N434Y;
M252Y/T307D/N434Y;
M252Y/T307F/N434Y;
M252Y/T307G/N434Y;
M252Y/T307H/N434Y;
M252Y/T307I/N434Y;
M252Y/T307K/N434Y;
M252Y/T307L/N434Y;
M252Y/T307M/N434Y;
M252Y/T307N/N434Y;
M252Y/T307P/N434Y;
M252Y/T307R/N434Y;
M252Y/T307S/N434Y;
M252Y/T307V/N434Y;
M252Y/T307W/N434Y;
M252Y/T307Y/N434Y;
M252Y/K334L/N434Y;
M252Y/G385H/N434Y;
M252Y/T289H/N434Y;
M252Y/Q311H/N434Y;
M252Y/D312H/N434Y;
M252Y/N315H/N434Y;
M252Y/K360H/N434Y;
M252Y/L314R/N434Y;
M252Y/L314K/N434Y;
M252Y/N286E/N434Y;
M252Y/L309E/N434Y;
M252Y/R255E/N434Y;
M252Y/P387E/N434Y;
K248I/M428L/N434Y;
M252Y/M428A/N434Y;
M252Y/M428D/N434Y;
M252Y/M428F/N434Y;
M252Y/M428G/N434Y;
M252Y/M428H/N434Y;
M252Y/M428I/N434Y;
M252Y/M428K/N434Y;
M252Y/M428N/N434Y;
M252Y/M428P/N434Y;
M252Y/M428Q/N434Y;
M252Y/M428S/N434Y;
M252Y/M428T/N434Y;
M252Y/M428V/N434Y;
M252Y/M428W/N434Y;
M252Y/M428Y/N434Y;
M252W/T307Q/M428Y/N434Y;
M252W/Q311A/M428Y/N434Y;
M252W/T307Q/Q311A/M428Y/N434Y;
M252Y/T307A/M428Y/N434Y;
M252Y/T307Q/M428Y/N434Y;
M252Y/D270F/N434Y;
M252Y/T307A/Q311A/N434Y;
M252Y/T307Q/Q311A/N434Y;
M252Y/T307A/Q311H/N434Y;
M252Y/T307Q/Q311H/N434Y;
M252Y/E382A/N434Y;
M252Y/E382A/M428Y/N434Y;
M252Y/T307A/E382A/M428Y/N434Y;
M252Y/T307Q/E382A/M428Y/N434Y;
P238A/M252Y/M428F/N434Y;
M252Y/V305A/M428F/N434Y;
M252Y/N325G/M428F/N434Y;
M252Y/D376A/M428F/N434Y;
M252Y/E380A/M428F/N434Y;
M252Y/E382A/M428F/N434Y;
M252Y/E380A/E382A/M428F/N434Y;
S239K/M252Y/V308P/E382A/N434Y;
M252Y/D270F/V308P/E382A/N434Y;
S239K/M252Y/V308P/M428Y/N434Y;
M252Y/T307Q/V308P/E382A/N434Y;
M252Y/V308P/Q311H/E382A/N434Y;
S239K/M252Y/D270F/V308P/N434Y;
M252Y/V308P/E382A/M428F/N434Y;
M252Y/V308P/E382A/M428L/N434Y;
M252Y/V308P/E382A/M428Y/N434Y;
M252Y/V308P/M428Y/N434Y;
M252Y/V308P/M428F/N434Y;
S239K/M252Y/V308P/E382A/M428Y/N434Y;
M252Y/V308P/E380A/E382A/M428F/N434Y;
M252Y/T256E/E382A/N434Y;
M252Y/T256E/M428Y/N434Y;
M252Y/T256E/E382A/M428Y/N434Y;
S239K/M252Y/V308P/E382A/M428I/N434Y;
M252Y/V308P/E380A/E382A/M428I/N434Y;
S239K/M252Y/V308P/M428F/N434Y;
S239K/M252Y/E380A/E382A/M428F/N434Y;
M252Y/Q311A/M428Y/N434Y;
M252Y/D312A/M428Y/N434Y;
M252Y/Q311A/D312A/M428Y/N434Y;
S239K/M252Y/V308P/Q311A/M428Y/N434Y;
S239K/M252Y/V308P/D312A/M428Y/N434Y;
S239K/M252Y/V308P/Q311A/D312A/M428Y/N434Y;
M252Y/T256E/T307P/N434Y;
M252Y/T307P/M428Y/N434Y;
M252W/V308P/M428Y/N434Y;
M252Y/T256E/V308P/E382A/N434Y;
M252W/V308P/E382A/N434Y;
S239K/M252W/V308P/E382A/N434Y;
S239K/M252W/V308P/E382A/M428Y/N434Y;
S239K/M252W/V308P/E382A/M428I/N434Y;
S239K/M252W/V308P/M428F/N434Y;
S239K/M252W/E380A/E382A/M428F/N434Y;
S239K/M252Y/T307P/M428Y/N434Y;
M252Y/T256E/Q311A/D312A/M428Y/N434Y;
S239K/M252Y/M428Y/N434Y;
S239K/M252Y/D270F/M428Y/N434Y;
S239K/M252Y/D270F/V308P/M428Y/N434Y;
S239K/M252Y/V308P/M428I/N434Y;
S239K/M252Y/V308P/N325G/E382A/M428I/N434Y;
S239K/M252Y/D270F/V308P/N325G/N434Y;
S239K/M252Y/D270F/T307P/V308P/N434Y;
S239K/M252Y/T256E/D270F/V308P/N434Y;
S239K/M252Y/D270F/T307Q/V308P/N434Y;
S239K/M252Y/D270F/T307Q/V308P/Q311A/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/N434Y;
S239K/M252Y/T256E/D270F/T307Q/Q311A/N434Y;
S239K/M252Y/D270F/V308P/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/N434Y;
S239K/M252Y/D270F/V308P/L309E/N434Y;
S239K/M252Y/D270F/V308P/P387E/N434Y;
S239K/M252Y/D270F/T307Q/L309E/Q311A/N434Y;
S239K/M252Y/D270F/V308P/N325G/M428I/N434Y;
S239K/M252Y/T256E/D270F/V308P/M428I/N434Y;
S239K/M252Y/D270F/T307Q/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/M428I/N434Y;
S239K/M252Y/D270F/V308P/L309E/M428I/N434Y;
S239K/M252Y/D270F/V308P/M428L/N434Y;
S239K/M252Y/V308P/M428L/N434Y;
S239K/M252Y/V308P/L309P/M428L/N434Y;
S239K/M252Y/V308P/L309R/M428L/N434Y;
S239K/M252Y/L309P/M428L/N434Y;
S239K/M252Y/L309R/M428L/N434Y;
S239K/M252Y/T307Q/V308P/M428L/N434Y;
S239K/M252Y/D270F/V308P/L309E/P387E/N434Y;
S239K/M252Y/T256E/D270F/V308P/N325G/M428I/N434Y;
S239K/M252Y/T307Q/N434Y;
P257V/T307Q/M428I/N434Y;
P257V/T307Q/M428V/N434Y;
P257V/T307Q/N325G/M428L/N434Y;
P257V/T307Q/Q311A/M428L/N434Y;
P257V/T307Q/Q311A/N325G/M428L/N434Y;
P257V/V305A/T307Q/M428L/N434Y;
S239K/M252Y/E258H/D270F/T307Q/V308P/Q311A/N434Y;
S239K/M252Y/D270F/V308P/N325G/E382A/M428I/N434Y;
M252Y/P257V/T307Q/M428I/N434Y;
S239K/M252W/V308P/M428Y/N434Y;
S239K/M252W/V308P/Q311A/M428Y/N434Y;
S239K/M252W/T256E/V308P/M428Y/N434Y;
S239K/M252W/N286E/V308P/M428Y/N434Y;
S239K/M252Y/T256E/D270F/N286E/V308P/N434Y;
S239K/M252W/T307P/M428Y/N434Y;
S239K/M252W/T256E/V308P/Q311A/M428Y/N434Y;
S239K/M252W/T256E/N286E/V308P/M428Y/N434Y;
P257V/T307A/Q311A/M428L/N434Y;
P257V/V305A/T307A/M428L/N434Y;
M252Y/V308P/L309E/N434Y;
M252Y/V308P/L309D/N434Y;
M252Y/V308P/L309A/N434Y;
M252W/V308P/L309E/M428Y/N434Y;
M252W/V308P/L309D/M428Y/N434Y;
M252W/V308P/L309A/M428Y/N434Y;
S239K/M252Y/N286E/V308P/M428I/N434Y;
S239K/M252Y/N286E/V308P/N434Y;
S239K/M252Y/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/T256E/V308P/N434Y;
S239K/M252Y/D270F/N286E/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/P387E/M428I/N434Y;
S239K/M252Y/D270F/N286E/T307Q/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/T307Q/V308P/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/P387E/M428I/N434Y;
P257V/T307Q/M428L/N434W;
P257V/T307A/M428L/N434W;
P257A/T307Q/L309P/M428L/N434Y;
P257V/T307Q/L309P/M428L/N434Y;
P257A/T307Q/L309R/M428L/N434Y;
P257V/T307Q/L309R/M428L/N434Y;
P257V/N286E/M428L/N434Y;
P257V/P387E/M428L/N434Y;
P257V/T307H/M428L/N434Y;
P257V/T307N/M428L/N434Y;
P257V/T307G/M428L/N434Y;
P257V/T307P/M428L/N434Y;
P257V/T307S/M428L/N434Y;
P257V/N286E/T307A/M428L/N434Y;
P257V/T307A/P387E/M428L/N434Y;
S239K/M252Y/T307P/N325G/M428Y/N434Y;
M252Y/T307A/Q311H/K360H/N434Y;
M252Y/T307A/Q311H/P387E/N434Y;
M252Y/T307A/Q311H/M428A/N434Y;
M252Y/T307A/Q311H/E382A/N434Y;
M252Y/T307W/Q311H/N434Y;
S239K/P257A/V308P/M428L/N434Y;
P257A/V308P/L309E/M428L/N434Y;
P257V/T307Q/N434Y;
M252Y/P257V/T307Q/M428V/N434Y;
M252Y/P257V/T307Q/M428L/N434Y;
M252Y/P257V/T307Q/N434Y;
M252Y/P257V/T307Q/M428Y/N434Y;
M252Y/P257V/T307Q/M428F/N434Y;
P257V/T307Q/Q311A/N325G/M428V/N434Y;
P257V/T307Q/Q311A/N325G/N434Y;
P257V/T307Q/Q311A/N325G/M428Y/N434Y;
P257V/T307Q/Q311A/N325G/M428F/N434Y;
P257A/T307Q/M428V/N434Y;
P257A/T307Q/N434Y;
P257A/T307Q/M428Y/N434Y;
P257A/T307Q/M428F/N434Y;
P257V/N286E/T307Q/Q311A/N325G/M428L/N434Y;
P257A/Q311A/M428L/N434Y;
P257A/Q311H/M428L/N434Y;
P257A/T307Q/Q311A/M428L/N434Y;
P257A/T307A/Q311A/M428L/N434Y;
P257A/T307Q/Q311H/M428L/N434Y;
P257A/T307A/Q311H/M428L/N434Y;
P257A/N286E/T307Q/M428L/N434Y;
P257A/N286E/T307A/M428L/N434Y;
S239K/M252Y/D270F/T307P/N325G/M428Y/N434Y;
S239K/M252Y/T307P/L309E/N325G/M428Y/N434Y;
P257S/T307A/M428L/N434Y;
P257M/T307A/M428L/N434Y;
P257N/T307A/M428L/N434Y;
P257I/T307A/M428L/N434Y;
P257V/T307Q/M428Y/N434Y;
P257V/T307Q/M428F/N434Y;
S239K/P257V/V308P/M428L/N434Y;
P257V/T307Q/V308P/N325G/M428L/N434Y;
P257V/T307Q/V308P/Q311A/N325G/M428L/N434Y;
P257V/T307A/V308P/N325G/M428L/N434Y;
P257A/V308P/N325G/M428L/N434Y;
P257A/T307A/V308P/M428L/N434Y;
P257A/T307Q/V308P/M428L/N434Y;
P257V/N286E/T307Q/N325G/M428L/N434Y;
T256E/P257V/N434Y;
T256E/P257T/N434Y;
S239K/P257T/V308P/M428L/N434Y;
P257T/V308P/N325G/M428L/N434Y;
T256E/P257T/V308P/N325G/M428L/N434Y;
P257T/V308P/N325G/E382A/M428L/N434Y;
P257T/V308P/N325G/P387E/M428L/N434Y;
P257T/V308P/L309P/N325G/M428L/N434Y;
P257T/V308P/L309R/N325G/M428L/N434Y;
T256E/P257V/T307Q/M428L/N434Y;
P257V/T307Q/E382A/M428L/N434Y;
P257V/T307Q/P387E/M428L/N434Y;
P257L/V308P/N434Y;
P257T/T307Q/N434Y;
P257V/T307Q/N325G/N434Y;
P257V/T307Q/Q311A/N434Y;
P257V/V305A/T307Q/N434Y;
P257V/N286E/T307A/N434Y;
P257V/T307Q/L309R/Q311H/M428L/N434Y;
P257V/V308P/N325G/M428L/N434Y;
S239K/P257V/V308P/Q311H/M428L/N434Y;
P257V/V305A/T307A/N325G/M428L/N434Y;
S239K/D270F/T307P/N325G/M428Y/N434Y;
P257T/T307A/M428V/N434Y;
P257A/M428V/N434Y;
P257A/T307A/M428V/N434Y;
P257S/T307Q/M428L/N434Y;
P257V/N297A/T307Q/M428L/N434Y;
P257V/N286A/T307Q/M428L/N434Y;
P257V/T307Q/N315A/M428L/N434Y;
P257V/T307Q/N384A/M428L/N434Y;
P257V/T307Q/N389A/M428L/N434Y;
P257V/N434Y;
P257T/N434Y;
P257V/N286E/N434Y;
P257T/N286E/N434Y;
P257V/N286E/T307Q/N434Y;
P257V/N286E/T307Q/M428Y/N434Y;
P257V/V305A/T307Q/M428Y/N434Y;
P257T/T307Q/M428Y/N434Y;
P257V/T307Q/V308P/N325G/M428Y/N434Y;
P257T/T307Q/M428F/N434Y;
P257A/N286E/T307Q/M428F/N434Y;
P257A/N286E/T307Q/M428Y/N434Y;
T250A/P257V/T307Q/M428L/N434Y;
T250F/P257V/T307Q/M428L/N434Y;
T250I/P257V/T307Q/M428L/N434Y;
T250M/P257V/T307Q/M428L/N434Y;
T250S/P257V/T307Q/M428L/N434Y;
T250V/P257V/T307Q/M428L/N434Y;
T250W/P257V/T307Q/M428L/N434Y;
T250Y/P257V/T307Q/M428L/N434Y;
M252Y/Q311A/N434Y;
S239K/M252Y/S254T/V308P/N434Y;
M252Y/T307Q/Q311A;
M252Y/S254T/N286E/N434Y;
M252Y/S254T/V308P/N434Y;
M252Y/S254T/T307A/N434Y;
M252Y/S254T/T307Q/N434Y;
M252Y/S254T/Q311A/N434Y;
M252Y/S254T/Q311H/N434Y;
M252Y/S254T/T307A/Q311A/N434Y;
M252Y/S254T/T307Q/Q311A/N434Y;
M252Y/S254T/M428I/N434Y;
M252Y/T256E/T307A/Q311H/N434Y;
M252Y/T256Q/T307A/Q311H/N434Y;
M252Y/S254T/T307A/Q311H/N434Y;
M252Y/N286E/T307A/Q311H/N434Y;
M252Y/T307A/Q311H/M428I/N434Y;
M252Y/T256E/T307Q/Q311H;
M252Y/N286E/T307A/Q311A;
M252Y/N286E/T307Q/Q311A;
M252Y/N286E/T307Q/Q311H;
M252Y/T256E/N286E/N434Y;
S239K/M252Y/V308P;
S239K/M252Y/V308P/E382A;
S239K/M252Y/T256E/V308P;
S239K/M252Y/N286E/V308P;
S239K/M252Y/N286E/V308P/M428I;
M252Y/N286E/M428L/N434Y;
M252Y/S254T/E382A/N434Y;
S239K/M252Y/S254T/V308P/M428I/N434Y;
S239K/M252Y/M428I/N434Y;
M252Y/D312A/E382A/M428Y/N434Y;
M252Y/E3 82А/РЗ 87E/N434Y;
M252Y/D312A/P387E/N434Y или
M252Y/P387E/M428Y/N434Y; и
(b) введения в указанное антитело по меньшей мере одной из гистидиновых замен или инсерций в антигенсвязывающий домен тяжелой цепи в положение, выбранное из группы, состоящей из Н27, Н31, Н32, Н33, Н35, Н50, Н58, Н59, Н61, Н62, Н63, Н64, Н65, Н99, H100b и H102 в соответствии с нумерацией по Kabat, и/или антигенсвязывающий домен легкой цепи в положение, выбранное из группы, состоящей из L24, L27, L28, L32, L53, L54, L56, L90, L92 и L94 в соответствии с нумерацией по Kabat;
где указанное антитело, содержащее антигенсвязывающий домен и человеческий FcRn-связывающий домен, полученные в (а) и (b), связывается с растворимым антигеном, обладает активностью связывания человеческого FcRn при рН 5.8 и рН 7.0, где активность связывания FcRn человека при рН 7.0 сильнее, чем интактного IgG человека, и обладает более низкой антигенсвязывающей активностью при рН 5.8, чем при рН 7.4, где отношение антигенсвязывающей активности при рН 5.8 и рН 7.4 равно по меньшей мере 2 при величине KD при рН 5.8 / KD при рН 7.4.
3. Способ увеличения способности антитела к элиминации антигена из плазмы, который включает стадии:
(а) введения в указанное антитело IgG1, IgG2, IgG3 или IgG4, которое содержит антигенсвязывающий домен и FcRn-связывающий домен человека, включающий Fc домен IgG1 человека, IgG2 человека, IgG3 человека или IgG4 человека, по меньшей мере одной из следующих аминокислотных замен в положениях, соответствующих нумерации EU в Fc-домене:
N434W;
N434Y;
M252Y/S254T/T256E/N434Y;
M252Y/S254T/T256E/N434W;
M252Y/N434Y;
M252Y/N434W;
M252W/N434Y;
M252W/N434W;
P257L/N434Y;
V308F/N434Y;
M252Y/V308P/N434Y;
M428L/N434S;
M252Y/S254T/T256E/V308P/N434W;
I332V;
I332V/N434Y;
G237M/V308F;
S254T/N434W;
S254T/N434Y;
T256E/N434W;
T256E/N434Y;
S254T/T256E/N434W;
S254T/T256E/N434Y;
M252Y/S254T/N434W;
M252Y/S254T/N434Y;
M252Y/T256E/N434W;
M252Y/T256E/N434Y;
M252Y/S254T/T256E/N434A;
M252Y/N434A;
M252W/N434A;
M252Y/T256Q/N434W;
M252Y/T256Q/N434Y;
M252F/T256D/N434W;
M252F/T256D/N434Y;
H433K/N434F/Y436H;
I332V/N434W;
V308P/N434W;
I332V/M428L/N434Y;
G385D/Q386P/N389S/N434W;
G385D/Q386P/N389S/N434Y;
G385H/N434W;
G385H/N434Y;
N434F;
M252Y/S254T/T256E/N434F;
M252Y/S254T/T256E/N434H;
M252Y/N434F;
M252Y/N434H;
M428L/N434W;
M428L/N434Y;
M252Y/S254T/T256E/M428L/N434W;
M252Y/S254T/T256E/M428L/N434Y;
M252Y/M428L/N434W;
M252Y/M428L/N434Y;
M252Y/M428L/N434A;
M252Y/S254T/T256E/M428L/N434A;
T256E/M428L/N434Y;
S254T/M428L/N434W;
M252Y/T256Q/N434A;
M252Y/T256E/N434A;
T256Q/N434W;
T256Q/N434Y;
M252W/T256Q/N434W;
M252W/T256Q/N434Y;
S254T/T256Q/N434W;
M252Y/S254T/T256Q/N434W;
M252Y/S254T/T256Q/N434Y;
M252Y/T256E/V308P/N434W;
M252Y/V308P/M428L/N434Y;
M252Y/S254T/T256E/V308P/M428L/N434W;
M252W/M428L/N434W;
P257L/M428L/N434Y;
M252Y/S254T/T256E/M428L/N434F;
M252Y/T256E/N434H;
M252W/T256Q/P257L/N434Y;
P238A/M252Y/V308P/N434Y;
M252Y/D265A/V308P/N434Y;
M252Y/T307A/V308P/N434Y;
M252Y/V303A/V308P/N434Y;
M252Y/V308P/D376A/N434Y;
M252Y/V305A/V308P/N434Y;
M252Y/V308P/Q311A/N434Y;
M252Y/V308P/K317A/N434Y;
M252Y/V308P/E380A/N434Y;
M252Y/V308P/E382A/N434Y;
M252Y/V308P/S424A/N434Y;
P257L/V308P/M428L/N434Y;
M252Y/T256E/V308P/M428L/N434W;
M252Y/T256E/M428L/N434W;
M252Y/T256E/V308P/N434Y;
M252Y/T256E/M428L/N434Y;
M252Y/S254T/T256E/V308P/M428L/N434Y;
P257L/M428L/N434W;
P257A/M428L/N434Y;
P257G/M428L/N434Y;
P257I/M428L/N434Y;
P257M/M428L/N434Y;
P257N/M428L/N434Y;
P257S/M428L/N434Y;
P257T/M428L/N434Y;
P257V/M428L/N434Y;
M252W/V308P/N434W;
S239K/M252Y/V308P/N434Y;
M252Y/S298G/V308P/N434Y;
M252Y/D270F/V308P/N434Y;
M252Y/V308A/N434Y;
M252Y/V308F/N434Y;
M252Y/V308I/N434Y;
M252Y/V308L/N434Y;
M252Y/V308M/N434Y;
M252Y/V308Q/N434Y;
M252Y/V308T/N434Y;
P257A/V308P/M428L/N434Y;
P257T/V308P/M428L/N434Y;
P257V/V308P/M428L/N434Y;
M252W/M428I/N434Y;
M252W/M428Y/N434Y;
M252W/M428F/N434Y;
P238A/M252W/N434Y;
M252W/D265A/N434Y;
M252W/T307Q/N434Y;
M252W/V303A/N434Y;
M252W/D376A/N434Y;
M252W/V305A/N434Y;
M252W/Q311A/N434Y;
M252W/D312A/N434Y;
M252W/K317A/N434Y;
M252W/E380A/N434Y;
M252W/E382A/N434Y;
M252W/S424A/N434Y;
S239K/M252W/N434Y;
M252W/S298G/N434Y;
M252W/D270F/N434Y;
M252W/N325G/N434Y;
P257A/M428L/N434W;
P257T/M428L/N434W;
P257V/M428L/N434W;
M252W/I332V/N434Y;
P257I/Q311I;
M252Y/T307A/N434Y;
M252Y/T307Q/N434Y;
P257L/T307A/M428L/N434Y;
P257A/T307A/M428L/N434Y;
P257T/T307A/M428L/N434Y;
P257V/T307A/M428L/N434Y;
P257L/T307Q/M428L/N434Y;
P257A/T307Q/M428L/N434Y;
P257T/T307Q/M428L/N434Y;
P257V/T307Q/M428L/N434Y;
M252Y/T307D/N434Y;
M252Y/T307F/N434Y;
M252Y/T307G/N434Y;
M252Y/T307H/N434Y;
M252Y/T307I/N434Y;
M252Y/T307K/N434Y;
M252Y/T307L/N434Y;
M252Y/T307M/N434Y;
M252Y/T307N/N434Y;
M252Y/T307P/N434Y;
M252Y/T307R/N434Y;
M252Y/T307S/N434Y;
M252Y/T307V/N434Y;
M252Y/T307W/N434Y;
M252Y/T307Y/N434Y;
M252Y/K334L/N434Y;
M252Y/G385H/N434Y;
M252Y/T289H/N434Y;
M252Y/Q311H/N434Y;
M252Y/D312H/N434Y;
M252Y/N315H/N434Y;
M252Y/K360H/N434Y;
M252Y/L314R/N434Y;
M252Y/L314K/N434Y;
M252Y/N286E/N434Y;
M252Y/L309E/N434Y;
M252Y/R255E/N434Y;
M252Y/P387E/N434Y;
K248I/M428L/N434Y;
M252Y/M428A/N434Y;
M252Y/M428D/N434Y;
M252Y/M428F/N434Y;
M252Y/M428G/N434Y;
M252Y/M428H/N434Y;
M252Y/M428I/N434Y;
M252Y/M428K/N434Y;
M252Y/M428N/N434Y;
M252Y/M428P/N434Y;
M252Y/M428Q/N434Y;
M252Y/M428S/N434Y;
M252Y/M428T/N434Y;
M252Y/M428V/N434Y;
M252Y/M428W/N434Y;
M252Y/M428Y/N434Y;
M252W/T307Q/M428Y/N434Y;
M252W/Q311A/M428Y/N434Y;
M252W/T307Q/Q311A/M428Y/N434Y;
M252Y/T307A/M428Y/N434Y;
M252Y/T307Q/M428Y/N434Y;
M252Y/D270F/N434Y;
M252Y/T307A/Q311A/N434Y;
M252Y/T307Q/Q311A/N434Y;
M252Y/T307A/Q311H/N434Y;
M252Y/T307Q/Q311H/N434Y;
M252Y/E382A/N434Y;
M252Y/E382A/M428Y/N434Y;
M252Y/T307A/E382A/M428Y/N434Y;
M252Y/T307Q/E382A/M428Y/N434Y;
P238A/M252Y/M428F/N434Y;
M252Y/V305A/M428F/N434Y;
M252Y/N325G/M428F/N434Y;
M252Y/D376A/M428F/N434Y;
M252Y/E380A/M428F/N434Y;
M252Y/E382A/M428F/N434Y;
M252Y/E380A/E382A/M428F/N434Y;
S239K/M252Y/V308P/E382A/N434Y;
M252Y/D270F/V308P/E382A/N434Y;
S239K/M252Y/V308P/M428Y/N434Y;
M252Y/T307Q/V308P/E382A/N434Y;
M252Y/V308P/Q311H/E382A/N434Y;
S239K/M252Y/D270F/V308P/N434Y;
M252Y/V308P/E382A/M428F/N434Y;
M252Y/V308P/E382A/M428L/N434Y;
M252Y/V308P/E382A/M428Y/N434Y;
M252Y/V308P/M428Y/N434Y;
M252Y/V308P/M428F/N434Y;
S239K/M252Y/V308P/E382A/M428Y/N434Y;
M252Y/V308P/E380A/E382A/M428F/N434Y;
M252Y/T256E/E382A/N434Y;
M252Y/T256E/M428Y/N434Y;
M252Y/T256E/E382A/M428Y/N434Y;
S239K/M252Y/V308P/E382A/M428I/N434Y;
M252Y/V308P/E380A/E382A/M428I/N434Y;
S239K/M252Y/V308P/M428F/N434Y;
S239K/M252Y/E380A/E382A/M428F/N434Y;
M252Y/Q311A/M428Y/N434Y;
M252Y/D312A/M428Y/N434Y;
M252Y/Q311A/D312A/M428Y/N434Y;
S239K/M252Y/V308P/Q311A/M428Y/N434Y;
S239K/M252Y/V308P/D312A/M428Y/N434Y;
S239K/M252Y/V308P/Q311A/D312A/M428Y/N434Y;
M252Y/T256E/T307P/N434Y;
M252Y/T307P/M428Y/N434Y;
M252W/V308P/M428Y/N434Y;
M252Y/T256E/V308P/E382A/N434Y;
M252W/V308P/E382A/N434Y;
S239K/M252W/V308P/E382A/N434Y;
S239K/M252W/V308P/E382A/M428Y/N434Y;
S239K/M252W/V308P/E382A/M428I/N434Y;
S239K/M252W/V308P/M428F/N434Y;
S239K/M252W/E380A/E382A/M428F/N434Y;
S239K/M252Y/T307P/M428Y/N434Y;
M252Y/T256E/Q311A/D312A/M428Y/N434Y;
S239K/M252Y/M428Y/N434Y;
S239K/M252Y/D270F/M428Y/N434Y;
S239K/M252Y/D270F/V308P/M428Y/N434Y;
S239K/M252Y/V308P/M428I/N434Y;
S239K/M252Y/V308P/N325G/E382A/M428I/N434Y;
S239K/M252Y/D270F/V308P/N325G/N434Y;
S239K/M252Y/D270F/T307P/V308P/N434Y;
S239K/M252Y/T256E/D270F/V308P/N434Y;
S239K/M252Y/D270F/T307Q/V308P/N434Y;
S239K/M252Y/D270F/T307Q/V308P/Q311A/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/N434Y;
S239K/M252Y/T256E/D270F/T307Q/Q311A/N434Y;
S239K/M252Y/D270F/V308P/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/N434Y;
S239K/M252Y/D270F/V308P/L309E/N434Y;
S239K/M252Y/D270F/V308P/P387E/N434Y;
S239K/M252Y/D270F/T307Q/L309E/Q311A/N434Y;
S239K/M252Y/D270F/V308P/N325G/M428I/N434Y;
S239K/M252Y/T256E/D270F/V308P/M428I/N434Y;
S239K/M252Y/D270F/T307Q/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/M428I/N434Y;
S239K/M252Y/D270F/V308P/L309E/M428I/N434Y;
S239K/M252Y/D270F/V308P/M428L/N434Y;
S239K/M252Y/V308P/M428L/N434Y;
S239K/M252Y/V308P/L309P/M428L/N434Y;
S239K/M252Y/V308P/L309R/M428L/N434Y;
S239K/M252Y/L309P/M428L/N434Y;
S239K/M252Y/L309R/M428L/N434Y;
S239K/M252Y/T307Q/V308P/M428L/N434Y;
S239K/M252Y/D270F/V308P/L309E/P387E/N434Y;
S239K/M252Y/T256E/D270F/V308P/N325G/M428I/N434Y;
S239K/M252Y/T307Q/N434Y;
P257V/T307Q/M428I/N434Y;
P257V/T307Q/M428V/N434Y;
P257V/T307Q/N325G/M428L/N434Y;
P257V/T307Q/Q311A/M428L/N434Y;
P257V/T307Q/Q311A/N325G/M428L/N434Y;
P257V/V305A/T307Q/M428L/N434Y;
S239K/M252Y/E258H/D270F/T307Q/V308P/Q311A/N434Y;
S239K/M252Y/D270F/V308P/N325G/E382A/M428I/N434Y;
M252Y/P257V/T307Q/M428I/N434Y;
S239K/M252W/V308P/M428Y/N434Y;
S239K/M252W/V308P/Q311A/M428Y/N434Y;
S239K/M252W/T256E/V308P/M428Y/N434Y;
S239K/M252W/N286E/V308P/M428Y/N434Y;
S239K/M252Y/T256E/D270F/N286E/V308P/N434Y;
S239K/M252W/T307P/M428Y/N434Y;
S239K/M252W/T256E/V308P/Q311A/M428Y/N434Y;
S239K/M252W/T256E/N286E/V308P/M428Y/N434Y;
P257V/T307A/Q311A/M428L/N434Y;
P257V/V305A/T307A/M428L/N434Y;
M252Y/V308P/L309E/N434Y;
M252Y/V308P/L309D/N434Y;
M252Y/V308P/L309A/N434Y;
M252W/V308P/L309E/M428Y/N434Y;
M252W/V308P/L309D/M428Y/N434Y;
M252W/V308P/L309A/M428Y/N434Y;
S239K/M252Y/N286E/V308P/M428I/N434Y;
S239K/M252Y/N286E/V308P/N434Y;
S239K/M252Y/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/T256E/V308P/N434Y;
S239K/M252Y/D270F/N286E/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/P387E/M428I/N434Y;
S239K/M252Y/D270F/N286E/T307Q/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/T307Q/V308P/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/P387E/M428I/N434Y;
P257V/T307Q/M428L/N434W;
P257V/T307A/M428L/N434W;
P257A/T307Q/L309P/M428L/N434Y;
P257V/T307Q/L309P/M428L/N434Y;
P257A/T307Q/L309R/M428L/N434Y;
P257V/T307Q/L309R/M428L/N434Y;
P257V/N286E/M428L/N434Y;
P257V/P387E/M428L/N434Y;
P257V/T307H/M428L/N434Y;
P257V/T307N/M428L/N434Y;
P257V/T307G/M428L/N434Y;
P257V/T307P/M428L/N434Y;
P257V/T307S/M428L/N434Y;
P257V/N286E/T307A/M428L/N434Y;
P257V/T307A/P387E/M428L/N434Y;
S239K/M252Y/T307P/N325G/M428Y/N434Y;
M252Y/T307A/Q311H/K360H/N434Y;
M252Y/T307A/Q311H/P387E/N434Y;
M252Y/T307A/Q311H/M428A/N434Y;
M252Y/T307A/Q311H/E382A/N434Y;
M252Y/T307W/Q311H/N434Y;
S239K/P257A/V308P/M428L/N434Y;
P257A/V308P/L309E/M428L/N434Y;
P257V/T307Q/N434Y;
M252Y/P257V/T307Q/M428V/N434Y;
M252Y/P257V/T307Q/M428L/N434Y;
M252Y/P257V/T307Q/N434Y;
M252Y/P257V/T307Q/M428Y/N434Y;
M252Y/P257V/T307Q/M428F/N434Y;
P257V/T307Q/Q311A/N325G/M428V/N434Y;
P257V/T307Q/Q311A/N325G/N434Y;
P257V/T307Q/Q311A/N325G/M428Y/N434Y;
P257V/T307Q/Q311A/N325G/M428F/N434Y;
P257A/T307Q/M428V/N434Y;
P257A/T307Q/N434Y;
P257A/T307Q/M428Y/N434Y;
P257A/T307Q/M428F/N434Y;
P257V/N286E/T307Q/Q311A/N325G/M428L/N434Y;
P257A/Q311A/M428L/N434Y;
P257A/Q311H/M428L/N434Y;
P257A/T307Q/Q311A/M428L/N434Y;
P257A/T307A/Q311A/M428L/N434Y;
P257A/T307Q/Q311H/M428L/N434Y;
P257A/T307A/Q311H/M428L/N434Y;
P257A/N286E/T307Q/M428L/N434Y;
P257A/N286E/T307A/M428L/N434Y;
S239K/M252Y/D270F/T307P/N325G/M428Y/N434Y;
S239K/M252Y/T307P/L309E/N325G/M428Y/N434Y;
P257S/T307A/M428L/N434Y;
P257M/T307A/M428L/N434Y;
P257N/T307A/M428L/N434Y;
P257I/T307A/M428L/N434Y;
P257V/T307Q/M428Y/N434Y;
P257V/T307Q/M428F/N434Y;
S239K/P257V/V308P/M428L/N434Y;
P257V/T307Q/V308P/N325G/M428L/N434Y;
P257V/T307Q/V308P/Q311A/N325G/M428L/N434Y;
P257V/T307A/V308P/N325G/M428L/N434Y;
P257A/V308P/N325G/M428L/N434Y;
P257A/T307A/V308P/M428L/N434Y;
P257A/T307Q/V308P/M428L/N434Y;
P257V/N286E/T307Q/N325G/M428L/N434Y;
T256E/P257V/N434Y;
T256E/P257T/N434Y;
S239K/P257T/V308P/M428L/N434Y;
P257T/V308P/N325G/M428L/N434Y;
T256E/P257T/V308P/N325G/M428L/N434Y;
P257T/V308P/N325G/E382A/M428L/N434Y;
P257T/V308P/N325G/P387E/M428L/N434Y;
P257T/V308P/L309P/N325G/M428L/N434Y;
P257T/V308P/L309R/N325G/M428L/N434Y;
T256E/P257V/T307Q/M428L/N434Y;
P257V/T307Q/E382A/M428L/N434Y;
P257V/T307Q/P387E/M428L/N434Y;
P257L/V308P/N434Y;
P257T/T307Q/N434Y;
P257V/T307Q/N325G/N434Y;
P257V/T307Q/Q311A/N434Y;
P257V/V305A/T307Q/N434Y;
P257V/N286E/T307A/N434Y;
P257V/T307Q/L309R/Q311H/M428L/N434Y;
P257V/V308P/N325G/M428L/N434Y;
S239K/P257V/V308P/Q311H/M428L/N434Y;
P257V/V305A/T307A/N325G/M428L/N434Y;
S239K/D270F/T307P/N325G/M428Y/N434Y;
P257T/T307A/M428V/N434Y;
P257A/M428V/N434Y;
P257A/T307A/M428V/N434Y;
P257S/T307Q/M428L/N434Y;
P257V/N297A/T307Q/M428L/N434Y;
P257V/N286A/T307Q/M428L/N434Y;
P257V/T307Q/N315A/M428L/N434Y;
P257V/T307Q/N384A/M428L/N434Y;
P257V/T307Q/N389A/M428L/N434Y;
P257V/N434Y;
P257T/N434Y;
P257V/N286E/N434Y;
P257T/N286E/N434Y;
P257V/N286E/T307Q/N434Y;
P257V/N286E/T307Q/M428Y/N434Y;
P257V/V305A/T307Q/M428Y/N434Y;
P257T/T307Q/M428Y/N434Y;
P257V/T307Q/V308P/N325G/M428Y/N434Y;
P257T/T307Q/M428F/N434Y;
P257A/N286E/T307Q/M428F/N434Y;
P257A/N286E/T307Q/M428Y/N434Y;
T250A/P257V/T307Q/M428L/N434Y;
T250F/P257V/T307Q/M428L/N434Y;
T250I/P257V/T307Q/M428L/N434Y;
T250M/P257V/T307Q/M428L/N434Y;
T250S/P257V/T307Q/M428L/N434Y;
T250V/P257V/T307Q/M428L/N434Y;
T250W/P257V/T307Q/M428L/N434Y;
T250Y/P257V/T307Q/M428L/N434Y;
M252Y/Q311A/N434Y;
S239K/M252Y/S254T/V308P/N434Y;
M252Y/T307Q/Q311A;
M252Y/S254T/N286E/N434Y;
M252Y/S254T/V308P/N434Y;
M252Y/S254T/T307A/N434Y;
M252Y/S254T/T307Q/N434Y;
M252Y/S254T/Q311A/N434Y;
M252Y/S254T/Q311H/N434Y;
M252Y/S254T/T307A/Q311A/N434Y;
M252Y/S254T/T307Q/Q311A/N434Y;
M252Y/S254T/M428I/N434Y;
M252Y/T256E/T307A/Q311H/N434Y;
M252Y/T256Q/T307A/Q311H/N434Y;
M252Y/S254T/T307A/Q311H/N434Y;
M252Y/N286E/T307A/Q311H/N434Y;
M252Y/T307A/Q311H/M428I/N434Y;
M252Y/T256E/T307Q/Q311H;
M252Y/N286E/T307A/Q311A;
M252Y/N286E/T307Q/Q311A;
M252Y/N286E/T307Q/Q311H;
M252Y/T256E/N286E/N434Y;
S239K/M252Y/V308P;
S239K/M252Y/V308P/E382A;
S239K/M252Y/T256E/V308P;
S239K/M252Y/N286E/V308P;
S239K/M252Y/N286E/V308P/M428I;
M252Y/N286E/M428L/N434Y;
M252Y/S254T/E382A/N434Y;
S239K/M252Y/S254T/V308P/M428I/N434Y;
S239K/M252Y/M428I/N434Y;
M252Y/D312A/E382A/M428Y/N434Y;
M252Y/E3 82А/РЗ 87E/N434Y;
M252Y/D312A/P387E/N434Y или
M252Y/P387E/M428Y/N434Y; и
(b) введения в указанное антитело по меньшей мере одной из гистидиновых замен или инсерций в антигенсвязывающий домен тяжелой цепи в положение, выбранное из группы, состоящей из Н27, Н31, Н32, Н33, Н35, Н50, Н58, Н59, Н61, Н62, Н63, Н64, Н65, Н99, H100b и H102 в соответствии с нумерацией по Kabat, и/или антигенсвязывающий домен легкой цепи в положение, выбранное из группы, состоящей из L24, L27, L28, L32, L53, L54, L56, L90, L92 и L94 в соответствии с нумерацией по Kabat;
где указанное антитело, содержащее антигенсвязывающий домен и человеческий FcRn-связывающий домен, полученные в (а) и (b), связывается с растворимым антигеном, обладает активностью связывания человеческого FcRn при рН 5.8 и рН 7.0, где активность связывания FcRn человека при рН 7.0 сильнее, чем интактного IgG человека, и обладает более низкой антигенсвязывающей активностью при рН 5.8, чем при рН 7.4, где отношение антигенсвязывающей активности при рН 5.8 и рН 7.4 равно по меньшей мере 2 при величине KD при рН 5.8 / KD при рН 7.4.
4. Способ усиления элиминации антигена из плазмы посредством введения антитела пациенту, который включает стадии:
(а) введения в указанное антитело IgG1, IgG2, IgG3 или IgG4, которое содержит антигенсвязывающий домен и FcRn-связывающий домен человека, включающий Fc домен IgG1 человека, IgG2 человека, IgG3 человека или IgG4 человека, по меньшей мере одной из следующих аминокислотных замен в положениях, соответствующих нумерации EU в Fc-домене:
N434W;
N434Y;
M252Y/S254T/T256E/N434Y;
M252Y/S254T/T256E/N434W;
M252Y/N434Y;
M252Y/N434W;
M252W/N434Y;
M252W/N434W;
P257L/N434Y;
V308F/N434Y;
M252Y/V308P/N434Y;
M428L/N434S;
M252Y/S254T/T256E/V308P/N434W;
I332V;
I332V/N434Y;
G237M/V308F;
S254T/N434W;
S254T/N434Y;
T256E/N434W;
T256E/N434Y;
S254T/T256E/N434W;
S254T/T256E/N434Y;
M252Y/S254T/N434W;
M252Y/S254T/N434Y;
M252Y/T256E/N434W;
M252Y/T256E/N434Y;
M252Y/S254T/T256E/N434A;
M252Y/N434A;
M252W/N434A;
M252Y/T256Q/N434W;
M252Y/T256Q/N434Y;
M252F/T256D/N434W;
M252F/T256D/N434Y;
H433K/N434F/Y436H;
I332V/N434W;
V308P/N434W;
I332V/M428L/N434Y;
G385D/Q386P/N389S/N434W;
G385D/Q386P/N389S/N434Y;
G385H/N434W;
G385H/N434Y;
N434F;
M252Y/S254T/T256E/N434F;
M252Y/S254T/T256E/N434H;
M252Y/N434F;
M252Y/N434H;
M428L/N434W;
M428L/N434Y;
M252Y/S254T/T256E/M428L/N434W;
M252Y/S254T/T256E/M428L/N434Y;
M252Y/M428L/N434W;
M252Y/M428L/N434Y;
M252Y/M428L/N434A;
M252Y/S254T/T256E/M428L/N434A;
T256E/M428L/N434Y;
S254T/M428L/N434W;
M252Y/T256Q/N434A;
M252Y/T256E/N434A;
T256Q/N434W;
T256Q/N434Y;
M252W/T256Q/N434W;
M252W/T256Q/N434Y;
S254T/T256Q/N434W;
M252Y/S254T/T256Q/N434W;
M252Y/S254T/T256Q/N434Y;
M252Y/T256E/V308P/N434W;
M252Y/V308P/M428L/N434Y;
M252Y/S254T/T256E/V308P/M428L/N434W;
M252W/M428L/N434W;
P257L/M428L/N434Y;
M252Y/S254T/T256E/M428L/N434F;
M252Y/T256E/N434H;
M252W/T256Q/P257L/N434Y;
P238A/M252Y/V308P/N434Y;
M252Y/D265A/V308P/N434Y;
M252Y/T307A/V308P/N434Y;
M252Y/V303A/V308P/N434Y;
M252Y/V308P/D376A/N434Y;
M252Y/V305A/V308P/N434Y;
M252Y/V308P/Q311A/N434Y;
M252Y/V308P/K317A/N434Y;
M252Y/V308P/E380A/N434Y;
M252Y/V308P/E382A/N434Y;
M252Y/V308P/S424A/N434Y;
P257L/V308P/M428L/N434Y;
M252Y/T256E/V308P/M428L/N434W;
M252Y/T256E/M428L/N434W;
M252Y/T256E/V308P/N434Y;
M252Y/T256E/M428L/N434Y;
M252Y/S254T/T256E/V308P/M428L/N434Y;
P257L/M428L/N434W;
P257A/M428L/N434Y;
P257G/M428L/N434Y;
P257I/M428L/N434Y;
P257M/M428L/N434Y;
P257N/M428L/N434Y;
P257S/M428L/N434Y;
P257T/M428L/N434Y;
P257V/M428L/N434Y;
M252W/V308P/N434W;
S239K/M252Y/V308P/N434Y;
M252Y/S298G/V308P/N434Y;
M252Y/D270F/V308P/N434Y;
M252Y/V308A/N434Y;
M252Y/V308F/N434Y;
M252Y/V308I/N434Y;
M252Y/V308L/N434Y;
M252Y/V308M/N434Y;
M252Y/V308Q/N434Y;
M252Y/V308T/N434Y;
P257A/V308P/M428L/N434Y;
P257T/V308P/M428L/N434Y;
P257V/V308P/M428L/N434Y;
M252W/M428I/N434Y;
M252W/M428Y/N434Y;
M252W/M428F/N434Y;
P238A/M252W/N434Y;
M252W/D265A/N434Y;
M252W/T307Q/N434Y;
M252W/V303A/N434Y;
M252W/D376A/N434Y;
M252W/V305A/N434Y;
M252W/Q311A/N434Y;
M252W/D312A/N434Y;
M252W/K317A/N434Y;
M252W/E380A/N434Y;
M252W/E382A/N434Y;
M252W/S424A/N434Y;
S239K/M252W/N434Y;
M252W/S298G/N434Y;
M252W/D270F/N434Y;
M252W/N325G/N434Y;
P257A/M428L/N434W;
P257T/M428L/N434W;
P257V/M428L/N434W;
M252W/I332V/N434Y;
P257I/Q311I;
M252Y/T307A/N434Y;
M252Y/T307Q/N434Y;
P257L/T307A/M428L/N434Y;
P257A/T307A/M428L/N434Y;
P257T/T307A/M428L/N434Y;
P257V/T307A/M428L/N434Y;
P257L/T307Q/M428L/N434Y;
P257A/T307Q/M428L/N434Y;
P257T/T307Q/M428L/N434Y;
P257V/T307Q/M428L/N434Y;
M252Y/T307D/N434Y;
M252Y/T307F/N434Y;
M252Y/T307G/N434Y;
M252Y/T307H/N434Y;
M252Y/T307I/N434Y;
M252Y/T307K/N434Y;
M252Y/T307L/N434Y;
M252Y/T307M/N434Y;
M252Y/T307N/N434Y;
M252Y/T307P/N434Y;
M252Y/T307R/N434Y;
M252Y/T307S/N434Y;
M252Y/T307V/N434Y;
M252Y/T307W/N434Y;
M252Y/T307Y/N434Y;
M252Y/K334L/N434Y;
M252Y/G385H/N434Y;
M252Y/T289H/N434Y;
M252Y/Q311/N434Y;
M252Y/D312H/N434Y;
M252Y/N315H/N434Y;
M252Y/K360H/N434Y;
M252Y/L314R/N434Y;
M252Y/L314K/N434Y;
M252Y/N286E/N434Y;
M252Y/L309E/N434Y;
M252Y/R255E/N434Y;
M252Y/P387E/N434Y;
K248I/M428L/N434Y;
M252Y/M428A/N434Y;
M252Y/M428D/N434Y;
M252Y/M428F/N434Y;
M252Y/M428G/N434Y;
M252Y/M428H/N434Y;
M252Y/M428I/N434Y;
M252Y/M428K/N434Y;
M252Y/M428N/N434Y;
M252Y/M428P/N434Y;
M252Y/M428Q/N434Y;
M252Y/M428S/N434Y;
M252Y/M428T/N434Y;
M252Y/M428V/N434Y;
M252Y/M428W/N434Y;
M252Y/M428Y/N434Y;
M252W/T307Q/M428Y/N434Y;
M252W/Q311A/M428Y/N434Y;
M252W/T307Q/Q311A/M428Y/N434Y;
M252Y/T307A/M428Y/N434Y;
M252Y/T307Q/M428Y/N434Y;
M252Y/D270F/N434Y;
M252Y/T307A/Q311A/N434Y;
M252Y/T307Q/Q311A/N434Y;
M252Y/T307A/Q311H/N434Y;
M252Y/T307Q/Q311H/N434Y;
M252Y/E382A/N434Y;
M252Y/E382A/M428Y/N434Y;
M252Y/T307A/E382A/M428Y/N434Y;
M252Y/T307Q/E382A/M428Y/N434Y;
P238A/M252Y/M428F/N434Y;
M252Y/V305A/M428F/N434Y;
M252Y/N325G/M428F/N434Y;
M252Y/D376A/M428F/N434Y;
M252Y/E380A/M428F/N434Y;
M252Y/E382A/M428F/N434Y;
M252Y/E380A/E382A/M428F/N434Y;
S239K/M252Y/V308P/E382A/N434Y;
M252Y/D270F/V308P/E382A/N434Y;
S239K/M252Y/V308P/M428Y/N434Y;
M252Y/T307Q/V308P/E382A/N434Y;
M252Y/V308P/Q311H/E382A/N434Y;
S239K/M252Y/D270F/V308P/N434Y;
M252Y/V308P/E382A/M428F/N434Y;
M252Y/V308P/E382A/M428L/N434Y;
M252Y/V308P/E382A/M428Y/N434Y;
M252Y/V308P/M428Y/N434Y;
M252Y/V308P/M428F/N434Y;
S239K/M252Y/V308P/E382A/M428Y/N434Y;
M252Y/V308P/E380A/E382A/M428F/N434Y;
M252Y/T256E/E382A/N434Y;
M252Y/T256E/M428Y/N434Y;
M252Y/T256E/E382A/M428Y/N434Y;
S239K/M252Y/V308P/E382A/M428I/N434Y;
M252Y/V308P/E380A/E382A/M428I/N434Y;
S239K/M252Y/V308P/M428F/N434Y;
S239K/M252Y/E380A/E382A/M428F/N434Y;
M252Y/Q311A/M428Y/N434Y;
M252Y/D312A/M428Y/N434Y;
M252Y/Q311A/D312A/M428Y/N434Y;
S239K/M252Y/V308P/Q311A/M428Y/N434Y;
S239K/M252Y/V308P/D312A/M428Y/N434Y;
S239K/M252Y/V308P/Q311A/D312A/M428Y/N434Y;
M252Y/T256E/T307P/N434Y;
M252Y/T307P/M428Y/N434Y;
M252W/V308P/M428Y/N434Y;
M252Y/T256E/V308P/E382A/N434Y;
M252W/V308P/E382A/N434Y;
S239K/M252W/V308P/E382A/N434Y;
S239K/M252W/V308P/E382A/M428Y/N434Y;
S239K/M252W/V308P/E382A/M428I/N434Y;
S239K/M252W/V308P/M428F/N434Y;
S239K/M252W/E380A/E382A/M428F/N434Y;
S239K/M252Y/T307P/M428Y/N434Y;
M252Y/T256E/Q311A/D312A/M428Y/N434Y;
S239K/M252Y/M428Y/N434Y;
S239K/M252Y/D270F/M428Y/N434Y;
S239K/M252Y/D270F/V308P/M428Y/N434Y;
S239K/M252Y/V308P/M428I/N434Y;
S239K/M252Y/V308P/N325G/E382A/M428I/N434Y;
S239K/M252Y/D270F/V308P/N325G/N434Y;
S239K/M252Y/D270F/T307P/V308P/N434Y;
S239K/M252Y/T256E/D270F/V308P/N434Y;
S239K/M252Y/D270F/T307Q/V308P/N434Y;
S239K/M252Y/D270F/T307Q/V308P/Q311A/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/N434Y;
S239K/M252Y/T256E/D270F/T307Q/Q311A/N434Y;
S239K/M252Y/D270F/V308P/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/N434Y;
S239K/M252Y/D270F/V308P/L309E/N434Y;
S239K/M252Y/D270F/V308P/P387E/N434Y;
S239K/M252Y/D270F/T307Q/L309E/Q311A/N434Y;
S239K/M252Y/D270F/V308P/N325G/M428I/N434Y;
S239K/M252Y/T256E/D270F/V308P/M428I/N434Y;
S239K/M252Y/D270F/T307Q/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/M428I/N434Y;
S239K/M252Y/D270F/V308P/L309E/M428I/N434Y;
S239K/M252Y/D270F/V308P/M428L/N434Y;
S239K/M252Y/V308P/M428L/N434Y;
S239K/M252Y/V308P/L309P/M428L/N434Y;
S239K/M252Y/V308P/L309R/M428L/N434Y;
S239K/M252Y/L309P/M428L/N434Y;
S239K/M252Y/L309R/M428L/N434Y;
S239K/M252Y/T307Q/V308P/M428L/N434Y;
S239K/M252Y/D270F/V308P/L309E/P387E/N434Y;
S239K/M252Y/T256E/D270F/V308P/N325G/M428I/N434Y;
S239K/M252Y/T307Q/N434Y;
P257V/T307Q/M428I/N434Y;
P257V/T307Q/M428V/N434Y;
P257V/T307Q/N325G/M428L/N434Y;
P257V/T307Q/Q311A/M428L/N434Y;
P257V/T307Q/Q311A/N325G/M428L/N434Y;
P257V/V305A/T307Q/M428L/N434Y;
S239K/M252Y/E258H/D270F/T307Q/V308P/Q311A/N434Y;
S239K/M252Y/D270F/V308P/N325G/E382A/M428I/N434Y;
M252Y/P257V/T307Q/M428I/N434Y;
S239K/M252W/V308P/M428Y/N434Y;
S239K/M252W/V308P/Q311A/M428Y/N434Y;
S239K/M252W/T256E/V308P/M428Y/N434Y;
S239K/M252W/N286E/V308P/M428Y/N434Y;
S239K/M252Y/T256E/D270F/N286E/V308P/N434Y;
S239K/M252W/T307P/M428Y/N434Y;
S239K/M252W/T256E/V308P/Q311A/M428Y/N434Y;
S239K/M252W/T256E/N286E/V308P/M428Y/N434Y;
P257V/T307A/Q311A/M428L/N434Y;
P257V/V305A/T307A/M428L/N434Y;
M252Y/V308P/L309E/N434Y;
M252Y/V308P/L309D/N434Y;
M252Y/V308P/L309A/N434Y;
M252W/V308P/L309E/M428Y/N434Y;
M252W/V308P/L309D/M428Y/N434Y;
M252W/V308P/L309A/M428Y/N434Y;
S239K/M252Y/N286E/V308P/M428I/N434Y;
S239K/M252Y/N286E/V308P/N434Y;
S239K/M252Y/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/T256E/V308P/N434Y;
S239K/M252Y/D270F/N286E/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/P387E/M428I/N434Y;
S239K/M252Y/D270F/N286E/T307Q/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/T307Q/V308P/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/P387E/M428I/N434Y;
P257V/T307Q/M428L/N434W;
P257V/T307A/M428L/N434W;
P257A/T307Q/L309P/M428L/N434Y;
P257V/T307Q/L309P/M428L/N434Y;
P257A/T307Q/L309R/M428L/N434Y;
P257V/T307Q/L309R/M428L/N434Y;
P257V/N286E/M428L/N434Y;
P257V/P387E/M428L/N434Y;
P257V/T307H/M428L/N434Y;
P257V/T307N/M428L/N434Y;
P257V/T307G/M428L/N434Y;
P257V/T307P/M428L/N434Y;
P257V/T307S/M428L/N434Y;
P257V/N286E/T307A/M428L/N434Y;
P257V/T307AT387E/M428L/N434Y;
S239K/M252Y/T307P/N325G/M428Y/N434Y;
M252Y/T307A/Q311H/K360H/N434Y;
M252Y/T307A/Q311H/P387E/N434Y;
M252Y/T307A/Q311H/M428A/N434Y;
M252Y/T307A/Q311H/E382A/N434Y;
M252Y/T307W/Q311H/N434Y;
S239K/P257A/V308P/M428UN434Y;
P257A/V308P/L309E/M428L/N434Y;
P257V/T307Q/N434Y;
M252Y/P257V/T307Q/M428V/N434Y;
M252Y/P257V/T307Q/M428L/N434Y;
M252Y/P257V/T307Q/N434Y;
M252Y/P257V/T307Q/M428Y/N434Y;
M252Y/P257V/T307Q/M428F/N434Y;
P257V/T307Q/Q311A/N325G/M428V/N434Y;
P257V/T307Q/Q311A/N325G/N434Y;
P257V/T307Q/Q311A/N325G/M428Y/N434Y;
P257V/T307Q/Q311A/N325G/M428F/N434Y;
P257A/T307Q/M428V/N434Y;
P257A/T307Q/N434Y;
P257A/T307Q/M428Y/N434Y;
P257A/T307Q/M428F/N434Y;
P257V/N286E/T307Q/Q311A/N325G/M428L/N434Y;
P257A/Q311A/M428L/N434Y;
P257A/Q311H/M428L/N434Y;
P257A/T307Q/Q311A/M428L/N434Y;
P257A/T307A/Q311A/M428L/N434Y;
P257A/T307Q/Q311H/M428L/N434Y;
P257A/T307A/Q311H/M428L/N434Y;
P257A/N286E/T307Q/M428L/N434Y;
P257A/N286E/T307A/M428L/N434Y;
S239K/M252Y/D270F/T307P/N325G/M428Y/N434Y;
S239K/M252Y/T307P/L309E/N325G/M428Y/N434Y;
P257S/T307A/M428L/N434Y;
P257M/T307A/M428L/N434Y;
P257N/T307A/M428L/N434Y;
P257I/T307A/M428L/N434Y;
P257V/T307Q/M428Y/N434Y;
P257V/T307Q/M428F/N434Y;
S239K/P257V/V308P/M428L/N434Y;
P257V/T307Q/V308P/N325G/M428L/N434Y;
P257V/T307Q/V308P/Q311A/N325G/M428L/N434Y;
P257V/T307A/V308P/N325G/M428L/N434Y;
P257A/V308P/N325G/M428L/N434Y;
P257A/T307A/V308P/M428L/N434Y;
P257A/T307Q/V308P/M428L/N434Y;
P257V/N286E/T307Q/N325G/M428L/N434Y;
T256E/P257V/N434Y;
T256E/P257T/N434Y;
S239K/P257T/V308P/M428L/N434Y;
P257T/V308P/N325G/M428L/N434Y;
T256E/P257T/V308P/N325G/M428L/N434Y;
P257T/V308P/N325G/E382A/M428L/N434Y;
P257T/V308P/N325G/P387E/M428L/N434Y;
P257T/V308P/L309P/N325G/M428L/N434Y;
P257T/V308P/L309R/N325G/M428L/N434Y;
Т256Е/P257V/T307Q/M428L/N434Y;
P257V/T307Q/E382A/M428L/N434Y;
P257V/T307Q/P387E/M428L/N434Y;
P257L/V308P/N434Y;
P257T/T307Q/N434Y;
P257V/T307Q/N325G/N434Y;
P257V/T307Q/Q31L4/N434Y;
P257V/V305A/T307Q/N434Y;
P257V/N286E/T307A/N434Y;
P257V/T307Q/L309R/Q311H/M428L/N434Y;
P257V/V308P/N325G/M428L/N434Y;
S239K/P257V/V308P/Q311H/M428L/N434Y;
P257V/V305A/T307A/N325G/M428L/N434Y;
S239K/D270F/T307P/N325G/M428Y/N434Y;
P257T/T307A/M428V/N434Y;
P257A/M428V/N434Y;
P257A/T307A/M428V/N434Y;
P257S/T307Q/M428L/N434Y;
P257V/N297A/T307Q/M428L/N434Y;
P257V/N286A/T307Q/M428L/N434Y;
P257V/T307Q/N315A/M428L/N434Y;
P257V/T307Q/N384A/M428L/N434Y;
P257V/T307Q/N389A/M428L/N434Y;
P257V/N434Y;
P257T/N434Y;
P257V/N286E/N434Y;
P257T/N286E/N434Y;
P257V/N286E/T307Q/N434Y;
P257V/N286E/T307Q/M428Y/N434Y;
P257V/V305A/T307Q/M428Y/N434Y;
P257T/T307Q/M428Y/N434Y;
P257V/T307Q/V308P/N325G/M428Y/N434Y;
P257T/T307Q/M428F/N434Y;
P257A/N286E/T307Q/M428F/N434Y;
P257A/N286E/T307Q/M428Y/N434Y;
T250A/P257V/T307Q/M428L/N434Y;
T250F/P257V/T307Q/M428L/N434Y;
T250I/P257V/T307Q/M428L/N434Y;
T250M/P257V/T307Q/M428L/N434Y;
T250S/P257V/T307Q/M428L/N434Y;
T250V/P257V/T307Q/M428L/N434Y;
T250W/P257V/T307Q/M428L/N434Y;
T250Y/P257V/T307Q/M428L/N434Y;
M252Y/Q311A/N434Y;
S239K/M252Y/S254T/V308P/N434Y;
M252Y/T307Q/Q311A M252Y/S254T/N286E/N434Y;
M252Y/S254T/V308P/N434Y;
M252Y/S254T/T307A/N434Y;
M252Y/S254T/T307Q/N434Y;
M252Y/S254T/Q311A/N434Y;
M252Y/S254T/Q311H/N434Y;
M252Y/S254T/T307A/Q311A/N434Y;
M252Y/S254T/T307Q/Q311A/N434Y;
M252Y/S254T/M428I/N434Y;
M252Y/T256E/T307A/Q311H/N434Y;
M252Y/T256Q/T307A/Q311H/N434Y;
M252Y/S254T/T307A/Q311H/N434Y;
M252Y/N286E/T307A/Q311H/N434Y;
M252Y/T307A/Q311H/M428I/N434Y;
M252Y/T256E/T307Q/Q311H;
M252Y/N286E/T307A/Q311A;
M252Y/N286E/T307Q/Q311A;
M252Y/N286E/T307Q/Q311H;
M252Y/T256E/N286E/N434Y;
S239K/M252Y/V308P;
S239K/M252Y/V308P/E382A;
S239K/M252Y/T256E/V308P;
S239K/M252Y/N286E/V308P;
S239K/M252Y/N286E/V308P/M428I;
M252Y/N286E/M428L/N434Y;
M252Y/S254T/E382A/N434Y;
S239K/M252Y/S254T/V308P/M428I/N434Y;
S239K/M252Y/M428I/N434Y;
M252Y/D312A/E382A/M428Y/N434Y;
M252Y/E382A/P387E/N434Y;
M252Y/D312A/P387E/N434Y или
M252Y/P387E/M428Y/N434Y; и
(b) введения в указанное антитело по меньшей мере одной из гистидиновых замен или инсерций в антигенсвязывающий домен тяжелой цепи в положение, выбранное из группы, состоящей из Н27, Н31, Н32, Н33, Н35, Н50, Н58, Н59, Н61, Н62, Н63, Н64, Н65, Н99, H100b и H102 в соответствии с нумерацией по Kabat, и/или антигенсвязывающий домен легкой цепи в положение, выбранное из группы, состоящей из L24, L27, L28, L32, L53, L54, L56, L90, L92 и L94 в соответствии с нумерацией по Kabat;
где указанное антитело, содержащее антигенсвязывающий домен и человеческий FcRn-связывающий домен, полученные в (а) и (b), связывается с растворимым антигеном, обладает активностью связывания человеческого FcRn при рН 5.8 и обладает более низкой антигенсвязывающей активностью при рН 5.8, чем при рН 7.4, где отношение антигенсвязывающей активности при рН 5.8 и рН 7.4 равно по меньшей мере 2 при величине KD при рН 5.8 / KD при рН 7.4.
5. Способ облегчения внеклеточного высвобождения свободной от антигена формы антитела, поглощенного клеткой в связанной с антигеном форме, который включает стадии:
(а) введения в антитело IgG1, IgG2, IgG3 или IgG4, которое содержит антигенсвязывающий домен и FcRn-связывающий домен человека, включающий Fc домен IgG1 человека, IgG2 человека, IgG3 человека или IgG4 человека, по меньшей мере одной из следующих аминокислотных замен в положениях, соответствующих нумерации EU в Fc-домене:
N434W;
N434Y;
M252Y/S254T/T256E/N434Y;
M252Y/S254T/T256E/N434W;
M252Y/N434Y;
M252Y/N434W;
M252W/N434Y;
M252W/N434W;
P257L/N434Y;
V308F/N434Y;
M252Y/V308P/N434Y;
M428L/N434S;
M252Y/S254T/T256E/V308P/N434W;
I332V;
I332V/N434Y;
G237M/V308F;
S254T/N434W;
S254T/N434Y;
T256E/N434W;
T256E/N434Y;
S254T/T256E/N434W;
S254T/T256E/N434Y;
M252Y/S254T/N434W;
M252Y/S254T/N434Y;
M252Y/T256E/N434W;
M252Y/T256E/N434Y;
M252Y/S254T/T256E/N434A;
M252Y/N434A;
M252W/N434A;
M252Y/T256Q/N434W;
M252Y/T256Q/N434Y;
M252F/T256D/N434W;
M252F/T256D/N434Y;
H433K/N434F/Y436H;
I332V/N434W;
V308P/N434W;
I332V/M428L/N434Y;
G385D/Q386P/N389S/N434W;
G385D/Q386P/N389S/N434Y;
G385H/N434W;
G385H/N434Y;
N434F;
M252Y/S254T/T256E/N434F;
M252Y/S254T/T256E/N434H;
M252Y/N434F;
M252Y/N434H;
M428L/N434W;
M428L/N434Y;
M252Y/S254T/T256E/M428L/N434W;
M252Y/S254T/T256E/M428L/N434Y;
M252Y/M428L/N434W;
M252Y/M428L/N434Y;
M252Y/M428L/N434A;
M252Y/S254T/T256E/M428L/N434A;
T256E/M428L/N434Y;
S254T/M428L/N434W;
M252Y/T256Q/N434A;
M252Y/T256E/N434A;
T256Q/N434W;
T256Q/N434Y;
M252W/T256Q/N434W;
M252W/T256Q/N434Y;
S254T/T256Q/N434W;
M252Y/S254T/T256Q/N434W;
M252Y/S254T/T256Q/N434Y;
M252Y/T256E/V308P/N434W;
M252Y/V308P/M428L/N434Y;
M252Y/S254T/T256E/V308P/M428L/N434W;
M252W/M428L/N434W;
P257L/M428L/N434Y;
M252Y/S254T/T256E/M428L/N434F;
M252Y/T256E/N434H;
M252W/T256Q/P257L/N434Y;
P238A/M252Y/V308P/N434Y;
M252Y/D265A/V308P/N434Y;
M252Y/T307A/V308P/N434Y;
M252Y/V303A/V308P/N434Y;
M252Y/V308P/D376A/N434Y;
M252Y/V305A/V308P/N434Y;
M252Y/V308P/Q311A/N434Y;
M252Y/V308P/K317A/N434Y;
M252Y/V308P/E380A/N434Y;
M252Y/V308P/E382A/N434Y;
M252Y/V308P/S424A/N434Y;
P257L/V308P/M428L/N434Y;
M252Y/T256E/V308P/M428L/N434W;
M252Y/T256E/M428L/N434W;
M252Y/T256E/V308P/N434Y;
M252Y/T256E/M428L/N434Y;
M252Y/S254T/T256E/V308P/M428L/N434Y;
P257L/M428L/N434W;
P257A/M428L/N434Y;
P257G/M428L/N434Y;
P257I/M428L/N434Y;
P257M/M428L/N434Y;
P257N/M428L/N434Y;
P257S/M428L/N434Y;
P257T/M428L/N434Y;
P257V/M428L/N434Y;
M252W/V308P/N434W;
S239K/M252Y/V308P/N434Y;
M252Y/S298G/V308P/N434Y;
M252Y/D270F/V308P/N434Y;
M252Y/V308A/N434Y;
M252Y/V308F/N434Y;
M252Y/V308I/N434Y;
M252Y/V308L/N434Y;
M252Y/V308M/N434Y;
M252Y/V308Q/N434Y;
M252Y/V308T/N434Y;
P257A/V308P/M428L/N434Y;
P257T/V308P/M428L/N434Y;
P257V/V308P/M428L/N434Y;
M252W/M428I/N434Y;
M252W/M428Y/N434Y;
M252W/M428F/N434Y;
P238A/M252W/N434Y;
M252W/D265A/N434Y;
M252W/T307Q/N434Y;
M252W/V303A/N434Y;
M252W/D376A/N434Y;
M252W/V305A/N434Y;
M252W/Q311A/N434Y;
M252W/D312A/N434Y;
M252W/K317A/N434Y;
M252W/E380A/N434Y;
M252W/E382A/N434Y;
M252W/S424A/N434Y;
S239K/M252W/N434Y;
M252W/S298G/N434Y;
M252W/D270F/N434Y;
M252W/N325G/N434Y;
P257A/M428L/N434W;
P257T/M428L/N434W;
P257V/M428L/N434W;
M252W/I332V/N434Y;
P257I/Q311I;
M252Y/T307A/N434Y;
M252Y/T307Q/N434Y;
P257L/T307A/M428L/N434Y;
P257A/T307A/M428L/N434Y;
P257T/T307A/M428L/N434Y;
P257V/T307A/M428L/N434Y;
P257L/T307Q/M428L/N434Y;
P257A/T307Q/M428L/N434Y;
P257T/T307Q/M428L/N434Y;
P257V/T307Q/M428L/N434Y;
M252Y/T307D/N434Y;
M252Y/T307F/N434Y;
M252Y/T307G/N434Y;
M252Y/T307H/N434Y;
M252Y/T307I/N434Y;
M252Y/T307K/N434Y;
M252Y/T307L/N434Y;
M252Y/T307M/N434Y;
M252Y/T307N/N434Y;
M252Y/T307P/N434Y;
M252Y/T307R/N434Y;
M252Y/T307S/N434Y;
M252Y/T307V/N434Y;
M252Y/T307W/N434Y;
M252Y/T307Y/N434Y;
M252Y/K334L/N434Y;
M252Y/G385H/N434Y;
M252Y/T289H/N434Y;
M252Y/Q311H/N434Y;
M252Y/D312H/N434Y;
M252Y/N315H/N434Y;
M252Y/K360H/N434Y;
M252Y/L314R/N434Y;
M252Y/L314K/N434Y;
M252Y/N286E/N434Y;
M252Y/L309E/N434Y;
M252Y/R255E/N434Y;
M252Y/P387E/N434Y;
K248I/M428L/N434Y;
M252Y/M428A/N434Y;
M252Y/M428D/N434Y;
M252Y/M428F/N434Y;
M252Y/M428G/N434Y;
M252Y/M428H/N434Y;
M252Y/M428I/N434Y;
M252Y/M428K/N434Y;
M252Y/M428N/N434Y;
M252Y/M428P/N434Y;
M252Y/M428Q/N434Y;
M252Y/M428S/N434Y;
M252Y/M428T/N434Y;
M252Y/M428V/N434Y;
M252Y/M428W/N434Y;
M252Y/M428Y/N434Y;
M252W/T307Q/M428Y/N434Y;
M252W/Q311A/M428Y/N434Y;
M252W/T307Q/Q311A/M428Y/N434Y;
M252Y/T307A/M428Y/N434Y;
M252Y/T307Q/M428Y/N434Y;
M252Y/D270F/N434Y;
M252Y/T307A/Q311A/N434Y;
M252Y/T307Q/Q311A/N434Y;
M252Y/T307A/Q311H/N434Y;
M252Y/T307Q/Q311H/N434Y;
M252Y/E382A/N434Y;
M252Y/E382A/M428Y/N434Y;
M252Y/T307A/E382A/M428Y/N434Y;
M252Y/T307Q/E382A/M428Y/N434Y;
P238A/M252Y/M428F/N434Y;
M252Y/V305A/M428F/N434Y;
M252Y/N325G/M428F/N434Y;
M252Y/D376A/M428F/N434Y;
M252Y/E380A/M428F/N434Y;
M252Y/E382A/M428F/N434Y;
M252Y/E380A/E382A/M428F/N434Y;
S239K/M252Y/V308P/E382A/N434Y;
M252Y/D270F/V308P/E382A/N434Y;
S239K/M252Y/V308P/M428Y/N434Y;
M252Y/T307Q/V308P/E382A/N434Y;
M252Y/V308P/Q311H/E382A/N434Y;
S239K/M252Y/D270F/V308P/N434Y;
M252Y/V308P/E382A/M428F/N434Y;
M252Y/V308P/E382A/M428L/N434Y;
M252Y/V308P/E382A/M428Y/N434Y;
M252Y/V308P/M428Y/N434Y;
M252Y/V308P/M428F/N434Y;
S239K/M252Y/V308P/E382A/M428Y/N434Y;
M252Y/V308P/E380A/E382A/M428F/N434Y;
M252Y/T256E/E382A/N434Y;
M252Y/T256E/M428Y/N434Y;
M252Y/T256E/E382A/M428Y/N434Y;
S239K/M252Y/V308P/E382A/M428I/N434Y;
M252Y/V308P/E380A/E382A/M428I/N434Y;
S239K/M252Y/V308P/M428F/N434Y;
S239K/M252Y/E380A/E382A/M428F/N434Y;
M252Y/Q311A/M428Y/N434Y;
M252Y/D312A/M428Y/N434Y;
M252Y/Q311A/D312A/M428Y/N434Y;
S239K/M252Y/V308P/Q311A/M428Y/N434Y;
S239K/M252Y/V308P/D312A/M428Y/N434Y;
S239K/M252Y/V308P/Q311A/D312A/M428Y/N434Y;
M252Y/T256E/T307P/N434Y;
M252Y/T307P/M428Y/N434Y;
M252W/V308P/M428Y/N434Y;
M252Y/T256E/V308P/E382A/N434Y;
M252W/V308P/E382A/N434Y;
S239K/M252W/V308P/E382A/N434Y;
S239K/M252W/V308P/E382A/M428Y/N434Y;
S239K/M252W/V308P/E382A/M428I/N434Y;
S239K/M252W/V308P/M428F/N434Y;
S239K/M252W/E380A/E382A/M428F/N434Y;
S239K/M252Y/T307P/M428Y/N434Y;
M252Y/T256E/Q311A/D312A/M428Y/N434Y;
S239K/M252Y/M428Y/N434Y;
S239K/M252Y/D270F/M428Y/N434Y;
S239K/M252Y/D270F/V308P/M428Y/N434Y;
S239K/M252Y/V308P/M428I/N434Y;
S239K/M252Y/V308P/N325G/E382A/M428I/N434Y;
S239K/M252Y/D270F/V308P/N325G/N434Y;
S239K/M252Y/D270F/T307P/V308P/N434Y;
S239K/M252Y/T256E/D270F/V308P/N434Y;
S239K/M252Y/D270F/T307Q/V308P/N434Y;
S239K/M252Y/D270F/T307Q/V308P/Q311A/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/N434Y;
S239K/M252Y/T256E/D270F/T307Q/Q311A/N434Y;
S239K/M252Y/D270F/V308P/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/N434Y;
S239K/M252Y/D270F/V308P/L309E/N434Y;
S239K/M252Y/D270F/V308P/P387E/N434Y;
S239K/M252Y/D270F/T307Q/L309E/Q311A/N434Y;
S239K/M252Y/D270F/V308P/N325G/M428I/N434Y;
S239K/M252Y/T256E/D270F/V308P/M428I/N434Y;
S239K/M252Y/D270F/T307Q/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/M428I/N434Y;
S239K/M252Y/D270F/V308P/L309E/M428I/N434Y;
S239K/M252Y/D270F/V308P/M428L/N434Y;
S239K/M252Y/V308P/M428L/N434Y;
S239K/M252Y/V308P/L309P/M428L/N434Y;
S239K/M252Y/V308P/L309R/M428L/N434Y;
S239K/M252Y/L309P/M428L/N434Y;
S239K/M252Y/L309R/M428L/N434Y;
S239K/M252Y/T307Q/V308P/M428L/N434Y;
S239K/M252Y/D270F/V308P/L309E/P387E/N434Y;
S239K/M252Y/T256E/D270F/V308P/N325G/M428I/N434Y;
S239K/M252Y/T307Q/N434Y;
P257V/T307Q/M428I/N434Y;
P257V/T307Q/M428V/N434Y;
P257V/T307Q/N325G/M428L/N434Y;
P257V/T307Q/Q311A/M428L/N434Y;
P257V/T307Q/Q311A/N325G/M428L/N434Y;
P257V/V305A/T307Q/M428L/N434Y;
S239K/M252Y/E258H/D270F/T307Q/V308P/Q311A/N434Y;
S239K/M252Y/D270F/V308P/N325G/E382A/M428I/N434Y;
M252Y/P257V/T307Q/M428I/N434Y;
S239K/M252W/V308P/M428Y/N434Y;
S239K/M252W/V308P/Q311A/M428Y/N434Y;
S239K/M252W/T256E/V308P/M428Y/N434Y;
S239K/M252W/N286E/V308P/M428Y/N434Y;
S239K/M252Y/T256E/D270F/N286E/V308P/N434Y;
S239K/M252W/T307P/M428Y/N434Y;
S239K/M252W/T256E/V308P/Q311A/M428Y/N434Y;
S239K/M252W/T256E/N286E/V308P/M428Y/N434Y;
P257V/T307A/Q311A/M428L/N434Y;
P257V/V305A/T307A/M428L/N434Y;
M252Y/V308P/L309E/N434Y;
M252Y/V308P/L309D/N434Y;
M252Y/V308P/L309A/N434Y;
M252W/V308P/L309E/M428Y/N434Y;
M252W/V308P/L309D/M428Y/N434Y;
M252W/V308P/L309A/M428Y/N434Y;
S239K/M252Y/N286E/V308P/M428I/N434Y;
S239K/M252Y/N286E/V308P/N434Y;
S239K/M252Y/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/T256E/V308P/N434Y;
S239K/M252Y/D270F/N286E/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/P387E/M428I/N434Y;
S239K/M252Y/D270F/N286E/T307Q/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/T307Q/V308P/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/P387E/M428I/N434Y;
P257V/T307Q/M428L/N434W;
P257V/T307A/M428L/N434W;
P257A/T307Q/L309P/M428L/N434Y;
P257V/T307Q/L309P/M428L/N434Y;
P257A/T307Q/L309R/M428L/N434Y;
P257V/T307Q/L309R/M428L/N434Y;
P257V/N286E/M428L/N434Y;
P257V/P387E/M428L/N434Y;
P257V/T307H/M428L/N434Y;
P257V/T307N/M428L/N434Y;
P257V/T307G/M428L/N434Y;
P257V/T307P/M428L/N434Y;
P257V/T307S/M428L/N434Y;
P257V/N286E/T307AM428L/N434Y;
P257V/T307A/P387E/M428L/N434Y;
S239K/M252Y/T307P/N325G/M428Y/N434Y;
M252Y/T307A/Q311H/K360H/N434Y;
M252Y/T307A/Q311H/P387E/N434Y;
M252Y/T307A/Q311H/M428A/N434Y;
M252Y/T307A/Q311H/E382A/N434Y;
M252Y/T307W/Q311H/N434Y;
S239K/P257A/V308P/M428L/N434Y;
P257A/V308P/L309E/M428L/N434Y;
P257V/T307Q/N434Y;
M252Y/P257V/T307Q/M428V/N434Y;
M252Y/P257V/T307Q/M428L/N434Y;
M252Y/P257V/T307Q/N434Y;
M252Y/P257V/T307Q/M428Y/N434Y;
M252Y/P257V/T307Q/M428F/N434Y;
P257V/T307Q/Q311A/N325G/M428V/N434Y;
P257V/T307Q/Q311A/N325G/N434Y;
P257V/T307Q/Q311A/N325G/M428Y/N434Y;
P257V/T307Q/Q311A/N325G/M428F/N434Y;
P257AT307Q/M428V/N434Y;
P257A/T307Q/N434Y;
P257A/T307Q/M428Y/N434Y;
P257A/T307Q/M428F/N434Y;
P257V/N286E/T307Q/Q311A/N325G/M428L/N434Y;
P257A/Q311A/M428L/N434Y;
P257A/Q311H/M428L/N434Y;
P257A/T307Q/Q311A/M428L/N434Y;
P257A/T307A/Q311A/M428L/N434Y;
P257A/T307Q/Q311H/M428L/N434Y;
P257A/T307A/Q311H/M428L/N434Y;
P257A/N286E/T307Q/M428L/N434Y;
P257A/N286E/T307A/M428L/N434Y;
S239K/M252Y/D270F/T307P/N325G/M428Y/N434Y;
S239K/M252Y/T307P/L309E/N325G/M428Y/N434Y;
P257S/T307A/M428L/N434Y;
P257M/T307A/M428L/N434Y;
P257N/T307A/M428L/N434Y;
P257I/T307A/M428L/N434Y;
P257V/T307Q/M428Y/N434Y;
P257V/T307Q/M428F/N434Y;
S239K/P257V/V308P/M428L/N434Y;
P257V/T307Q/V308P/N325G/M428L/N434Y;
P257V/T307Q/V308P/Q311A/N325G/M428L/N434Y;
P257V/T307A/V308P/N325G/M428L/N434Y;
P257A/V308P/N325G/M428L/N434Y;
P257A/T307A/V308P/M428L/N434Y;
P257A/T307Q/V308P/M428L/N434Y;
P257V/N286E/T307Q/N325G/M428L/N434Y;
T256E/P257V/N434Y;
T256E/P257T/N434Y;
S239K/P257T/V308P/M428L/N434Y;
P257T/V308P/N325G/M428L/N434Y;
T256E/P257T/V308P/N325G/M428L/N434Y;
P257T/V308P/N325G/E382A/M428L/N434Y;
P257T/V308P/N325G/P387E/M428L/N434Y;
P257T/V308P/L309P/N325G/M428L/N434Y;
P257T/V308P/L309R/N325G/M428L/N434Y;
T256E/P257V/T307Q/M428L/N434Y;
P257V/T307Q/E382A/M428L/N434Y;
P257V/T307Q/P387E/M428L/N434Y;
P257L/V308P/N434Y;
P257T/T307Q/N434Y;
P257V/T307Q/N325G/N434Y;
P257V/T307Q/Q311A/N434Y;
P257V/V305A/T307Q/N434Y;
P257V/N286E/T307A/N434Y;
P257V/T307Q/L309R/Q311H/M428L/N434Y;
P257V/V308P/N325G/M428L/N434Y;
S239K/P257V/V308P/Q311H/M428L/N434Y;
P257V/V305A/T307A/N325G/M428L/N434Y;
S239K/D270F/T307P/N325G/M428Y/N434Y;
P257T/T307A/M428V/N434Y;
P257A/M428V/N434Y;
P257A/T307A/M428V/N434Y;
P257S/T307Q/M428L/N434Y;
P257V/N297A/T307Q/M428L/N434Y;
P257V/N286A/T307Q/M428L/N434Y;
P257V/T307Q/N315A/M428L/N434Y;
P257V/T307Q/N384A/M428L/N434Y;
P257V/T307Q/N389A/M428L/N434Y;
P257V/N434Y;
P257T/N434Y;
P257V/N286E/N434Y;
P257T/N286E/N434Y;
P257V/N286E/T307Q/N434Y;
P257V/N286E/T307Q/M428Y/N434Y;
P257V/V305A/T307Q/M428Y/N434Y;
P257T/T307Q/M428Y/N434Y;
P257V/T307Q/V308P/N325G/M428Y/N434Y;
P257T/T307Q/M428F/N434Y;
P257A/N286E/T307Q/M428F/N434Y;
P257A/N286E/T307Q/M428Y/N434Y;
T250A/P257V/T307Q/M428L/N434Y;
T250F/P257V/T307Q/M428L/N434Y;
T250EP257\7T307Q/M428L/N434Y;
T250M/P257V/T307Q/M428L/N434Y;
T250S/P257V/T307Q/M428L/N434Y;
T250V/P257V/T307Q/M428L/N434Y;
T250W/P257V/T307Q/M428L/N434Y;
T250Y/P257V/T307Q/M428L/N434Y;
M252Y/Q311A/N434Y;
S239K/M252Y/S254T/V308P/N434Y;
M252Y/T307Q/Q311AM252Y/S254T/N286E/N434Y;
M252Y/S254T/V308P/N434Y;
M252Y/S254T/T307A/N434Y;
M252Y/S254T/T307Q/N434Y;
M252Y/S254T/Q311A/N434Y;
M252Y/S254T/Q311H/N434Y;
M252Y/S254T/T307A/Q311A/N434Y;
M252Y/S254T/T307Q/Q311A/N434Y;
M252Y/S254T/M428I/N434Y;
M252Y/T256E/T307A/Q311H/N434Y;
M252Y/T256Q/T307A/Q311H/N434Y;
M252Y/S254T/T307A/Q311H/N434Y;
M252Y/N286E/T307A/Q311H/N434Y;
M252Y/T307A/Q311H/M428I/N434Y;
M252Y/T256E/T307Q/Q311H;
M252Y/N286E/T307A/Q311A;
M252Y/N286E/T307Q/Q311A;
M252Y/N286E/T307Q/Q311H;
M252Y/T256E/N286E/N434Y;
S239K/M252Y/V308P;
S239K/M252Y/V308P/E382A;
S239K/M252Y/T256E/V308P;
S239K/M252Y/N286E/V308P;
S239K/M252Y/N286E/V308P/M428I;
M252Y/N286E/M428L/N434Y;
M252Y/S254T/E382A/N434Y;
S239K/M252Y/S254T/V308P/M428I/N434Y;
S239K/M252Y/M428I/N434Y;
M252Y/D312A/E382A/M428Y/N434Y;
M252Y/E382A/P387E/N434Y;
M252Y/D312A/P387E/N434Y или
M252Y/P387E/M428Y/N434Y; и
(b) введения в указанное антитело по меньшей мере одной из гистидиновых замен или инсерций в антигенсвязывающий домен тяжелой цепи в положение, выбранное из группы, состоящей из Н27, Н31, Н32, Н33, Н35, Н50, Н58, Н59, Н61, Н62, Н63, Н64, Н65, Н99, H100b и H102 в соответствии с нумерацией по Kabat, и/или антигенсвязывающий домен легкой цепи в положение, выбранное из группы, состоящей из L24, L27, L28, L32, L53, L54, L56, L90, L92 и L94 в соответствии с нумерацией по Kabat;
где указанное антитело, содержащее антигенсвязывающий домен и человеческий FcRn-связывающий домен, полученные в (а) и (b), связывается с растворимым антигеном, обладает активностью связывания человеческого FcRn при рН 5.8 и рН 7.0, где его активность связывания FcRn человека при рН 7.0 сильнее, чем интактного IgG человека, и обладает более низкой антигенсвязывающей активностью при рН 5.8, чем при рН 7.4, где отношение антигенсвязывающей активности при рН 5.8 и рН 7.4 равно по меньшей мере 2 при величине KD при рН 5.8 / KD при рН 7.4.
6. Способ снижения общей концентрации антигена или концентрации свободного антигена в плазме, который включает стадии:
(а) введения в антитело IgG1, IgG2, IgG3 или IgG4, которое содержит антигенсвязывающий домен и FcRn-связывающий домен человека, включающий Fc домен IgG1 человека, IgG2 человека, IgG3 человека или IgG4 человека, по меньшей мере одной из следующих аминокислотных замен в положениях, соответствующих нумерации EU в Fc-домене:
N434W;
N434Y;
M252Y/S254T/T256E/N434Y;
M252Y/S254T/T256E/N434W;
M252Y/N434Y;
M252Y/N434W;
M252W/N434Y;
M252W/N434W;
P257L/N434Y;
V308F/N434Y;
M252Y/V308P/N434Y;
M428L/N434S;
M252Y/S254T/T256E/V308P/N434W;
I332V;
I332V/N434Y;
G237M/V308F;
S254T/N434W;
S254T/N434Y;
T256E/N434W;
T256E/N434Y;
S254T/T256E/N434W;
S254T/T256E/N434Y;
M252Y/S254T/N434W;
M252Y/S254T/N434Y;
M252Y/T256E/N434W;
M252Y/T256E/N434Y;
M252Y/S254T/T256E/N434A;
M252Y/N434A;
M252W/N434A;
M252Y/T256Q/N434W;
M252Y/T256Q/N434Y;
M252F/T256D/N434W;
M252F/T256D/N434Y;
H433K/N434F/Y436H;
I332V/N434W;
V308P/N434W;
I332V/M428L/N434Y;
G385D/Q386P/N389S/N434W;
G385D/Q386P/N389S/N434Y;
G385H/N434W;
G385H/N434Y;
N434F;
M252Y/S254T/T256E/N434F;
M252Y/S254T/T256E/N434H;
M252Y/N434F;
M252Y/N434H;
M428L/N434W;
M428L/N434Y;
M252Y/S254T/T256E/M428L/N434W;
M252Y/S254T/T256E/M428L/N434Y;
M252Y/M428L/N434W;
M252Y/M428L/N434Y;
M252Y/M428L/N434A;
M252Y/S254T/T256E/M428L/N434A;
T256E/M428L/N434Y;
S254T/M428L/N434W;
M252Y/T256Q/N434A;
M252Y/T256E/N434A;
T256Q/N434W;
T256Q/N434Y;
M252W/T256Q/N434W;
M252W/T256Q/N434Y;
S254T/T256Q/N434W;
M252Y/S254T/T256Q/N434W;
M252Y/S254T/T256Q/N434Y;
M252Y/T256E/V308P/N434W;
M252Y/V308P/M428L/N434Y;
M252Y/S254T/T256E/V308P/M428L/N434W;
M252W/M428L/N434W;
P257L/M428L/N434Y;
M252Y/S254T/T256E/M428L/N434F;
M252Y/T256E/N434H;
M252W/T256Q/P257L/N434Y;
P238A/M252Y/V308P/N434Y;
M252Y/D265A/V308P/N434Y;
M252Y/T307A/V308P/N434Y;
M252Y/V303A/V308P/N434Y;
M252Y/V308P/D376A/N434Y;
M252Y/V305A/V308P/N434Y;
M252Y/V308P/Q311A/N434Y;
M252Y/V308P/K317A/N434Y;
M252Y/V308P/E380A/N434Y;
M252Y/V308P/E382A/N434Y;
M252Y/V308P/S424A/N434Y;
P257L/V308P/M428L/N434Y;
M252Y/T256E/V308P/M428L/N434W;
M252Y/T256E/M428L/N434W;
M252Y/T256E/V308P/N434Y;
M252Y/T256E/M428L/N434Y;
M252Y/S254T/T256E/V308P/M428L/N434Y;
P257L/M428L/N434W;
P257A/M428L/N434Y;
P257G/M428L/N434Y;
P257I/M428L/N434Y;
P257M/M428L/N434Y;
P257N/M428L/N434Y;
P257S/M428L/N434Y;
P257T/M428L/N434Y;
P257V/M428L/N434Y;
M252W/V308P/N434W;
S239K/M252Y/V308P/N434Y;
M252Y/S298G/V308P/N434Y;
M252Y/D270F/V308P/N434Y;
M252Y/V308A/N434Y;
M252Y/V308F/N434Y;
M252Y/V308I/N434Y;
M252Y/V308L/N434Y;
M252Y/V308M/N434Y;
M252Y/V308Q/N434Y;
M252Y/V308T/N434Y;
P257A/V308P/M428L/N434Y;
P257T/V308P/M428L/N434Y;
P257V/V308P/M428L/N434Y;
M252W/M428I/N434Y;
M252W/M428Y/N434Y;
M252W/M428F/N434Y;
P238A/M252W/N434Y;
M252W/D265A/N434Y;
M252W/T307Q/N434Y;
M252W/V303A/N434Y;
M252W/D376A/N434Y;
M252W/V305A/N434Y;
M252W/Q311A/N434Y;
M252W/D312A/N434Y;
M252W/K317A/N434Y;
M252W/E380A/N434Y;
M252W/E382A/N434Y;
M252W/S424A/N434Y;
S239K/M252W/N434Y;
M252W/S298G/N434Y;
M252W/D270F/N434Y;
M252W/N325G/N434Y;
P257A/M428L/N434W;
P257T/M428L/N434W;
P257V/M428L/N434W;
M252W/I332V/N434Y;
P257I/Q311I;
M252Y/T307A/N434Y;
M252Y/T307Q/N434Y;
P257L/T307A/M428L/N434Y;
P257A/T307A/M428L/N434Y;
P257T/T307A/M428L/N434Y;
P257V/T307A/M428L/N434Y;
P257L/T307Q/M428L/N434Y;
P257A/T307Q/M428L/N434Y;
P257T/T307Q/M428L/N434Y;
P257V/T307Q/M428L/N434Y;
M252Y/T307D/N434Y;
M252Y/T307F/N434Y;
M252Y/T307G/N434Y;
M252Y/T307H/N434Y;
M252Y/T307I/N434Y;
M252Y/T307K/N434Y;
M252Y/T307L/N434Y;
M252Y/T307M/N434Y;
M252Y/T307N/N434Y;
M252Y/T307P/N434Y;
M252Y/T307R/N434Y;
M252Y/T307S/N434Y;
M252Y/T307V/N434Y;
M252Y/T307W/N434Y;
M252Y/T307Y/N434Y;
M252Y/K334L/N434Y;
M252Y/G385H/N434Y;
M252Y/T289H/N434Y;
M252Y/Q311H/N434Y;
M252Y/D312H/N434Y;
M252Y/N315H/N434Y;
M252Y/K360H/N434Y;
M252Y/L314R/N434Y;
M252Y/L314K/N434Y;
M252Y/N286E/N434Y;
M252Y/L309E/N434Y;
M252Y/R255E/N434Y;
M252Y/P387E/N434Y;
K248I/M428L/N434Y;
M252Y/M428A/N434Y;
M252Y/M428D/N434Y;
M252Y/M428F/N434Y;
M252Y/M428G/N434Y;
M252Y/M428H/N434Y;
M252Y/M428I/N434Y;
M252Y/M428K/N434Y;
M252Y/M428N/N434Y;
M252Y/M428P/N434Y;
M252Y/M428Q/N434Y;
M252Y/M428S/N434Y;
M252Y/M428T/N434Y;
M252Y/M428V/N434Y;
M252Y/M428W/N434Y;
M252Y/M428Y/N434Y;
M252W/T307Q/M428Y/N434Y;
M252W/Q311A/M428Y/N434Y;
M252W/T307Q/Q311A/M428Y/N434Y;
M252Y/T307A/M428Y/N434Y;
M252Y/T307Q/M428Y/N434Y;
M252Y/D270F/N434Y;
M252Y/T307A/Q311A/N434Y;
M252Y/T307Q/Q311A/N434Y;
M252Y/T307A/Q311H/N434Y;
M252Y/T307Q/Q311H/N434Y;
M252Y/E382A/N434Y;
M252Y/E382A/M428Y/N434Y;
M252Y/T307A/E382A/M428Y/N434Y;
M252Y/T307Q/E382A/M428Y/N434Y;
P238A/M252Y/M428F/N434Y;
M252Y/V305A/M428F/N434Y;
M252Y/N325G/M428F/N434Y;
M252Y/D376A/M428F/N434Y;
M252Y/E380A/M428F/N434Y;
M252Y/E382A/M428F/N434Y;
M252Y/E380A/E382A/M428F/N434Y;
S239K/M252Y/V308P/E382A/N434Y;
M252Y/D270F/V308P/E382A/N434Y;
S239K/M252Y/V308P/M428Y/N434Y;
M252Y/T307Q/V308P/E382A/N434Y;
M252Y/V308P/Q311H/E382A/N434Y;
S239K/M252Y/D270F/V308P/N434Y;
M252Y/V308P/E382A/M428F/N434Y;
M252Y/V308P/E382A/M428L/N434Y;
M252Y/V308P/E382A/M428Y/N434Y;
M252Y/V308P/M428Y/N434Y;
M252Y/V308P/M428F/N434Y;
S239K/M252Y/V308P/E382A/M428Y/N434Y;
M252Y/V308P/E380A/E382A/M428F/N434Y;
M252Y/T256E/E382A/N434Y;
M252Y/T256E/M428Y/N434Y;
M252Y/T256E/E382A/M428Y/N434Y;
S239K/M252Y/V308P/E382A/M428I/N434Y;
M252Y/V308P/E380A/E382A/M428I/N434Y;
S239K/M252Y/V308P/M428F/N434Y;
S239K/M252Y/E380A/E382A/M428F/N434Y;
M252Y/Q311A/M428Y/N434Y;
M252Y/D312A/M428Y/N434Y;
M252Y/Q311A/D312A/M428Y/N434Y;
S239K/M252Y/V308P/Q311A/M428Y/N434Y;
S239K/M252Y/V308P/D312A/M428Y/N434Y;
S239K/M252Y/V308P/Q311A/D312A/M428Y/N434Y;
M252Y/T256E/T307P/N434Y;
M252Y/T307P/M428Y/N434Y;
M252W/V308P/M428Y/N434Y;
M252Y/T256E/V308P/E382A/N434Y;
M252W/V308P/E382A/N434Y;
S239K/M252W/V308P/E382A/N434Y;
S239K/M252W/V308P/E382A/M428Y/N434Y;
S239K/M252W/V308P/E382A/M428I/N434Y;
S239K/M252W/V308P/M428F/N434Y;
S239K/M252W/E380A/E382A/M428F/N434Y;
S239K/M252Y/T307P/M428Y/N434Y;
M252Y/T256E/Q311A/D312A/M428Y/N434Y;
S239K/M252Y/M428Y/N434Y;
S239K/M252Y/D270F/M428Y/N434Y;
S239K/M252Y/D270F/V308P/M428Y/N434Y;
S239K/M252Y/V308P/M428I/N434Y;
S239K/M252Y/V308P/N325G/E382A/M428I/N434Y;
S239K/M252Y/D270F/V308P/N325G/N434Y;
S239K/M252Y/D270F/T307P/V308P/N434Y;
S239K/M252Y/T256E/D270F/V308P/N434Y;
S239K/M252Y/D270F/T307Q/V308P/N434Y;
S239K/M252Y/D270F/T307Q/V308P/Q311A/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/N434Y;
S239K/M252Y/T256E/D270F/T307Q/Q311A/N434Y;
S239K/M252Y/D270F/V308P/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/N434Y;
S239K/M252Y/D270F/V308P/L309E/N434Y;
S239K/M252Y/D270F/V308P/P387E/N434Y;
S239K/M252Y/D270F/T307Q/L309E/Q311A/N434Y;
S239K/M252Y/D270F/V308P/N325G/M428I/N434Y;
S239K/M252Y/T256E/D270F/V308P/M428I/N434Y;
S239K/M252Y/D270F/T307Q/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/M428I/N434Y;
S239K/M252Y/D270F/V308P/L309E/M428I/N434Y;
S239K/M252Y/D270F/V308P/M428L/N434Y;
S239K/M252Y/V308P/M428L/N434Y;
S239K/M252Y/V308P/L309P/M428L/N434Y;
S239K/M252Y/V308P/L309R/M428L/N434Y;
S239K/M252Y/L309P/M428L/N434Y;
S239K/M252Y/L309R/M428L/N434Y;
S239K/M252Y/T307Q/V308P/M428L/N434Y;
S239K/M252Y/D270F/V308P/L309E/P387E/N434Y;
S239K/M252Y/T256E/D270F/V308P/N325G/M428I/N434Y;
S239K/M252Y/T307Q/N434Y;
P257V/T307Q/M428I/N434Y;
P257V/T307Q/M428V/N434Y;
P257V/T307Q/N325G/M428L/N434Y;
P257V/T307Q/Q311A/M428L/N434Y;
P257V/T307Q/Q311A/N325G/M428L/N434Y;
P257V/V305A/T307Q/M428L/N434Y;
S239K/M252Y/E258H/D270F/T307Q/V308P/Q311A/N434Y;
S239K/M252Y/D270F/V308P/N325G/E382A/M428I/N434Y;
M252Y/P257V/T307Q/M428I/N434Y;
S239K/M252W/V308P/M428Y/N434Y;
S239K/M252W/V308P/Q311A/M428Y/N434Y;
S239K/M252W/T256E/V308P/M428Y/N434Y;
S239K/M252W/N286E/V308P/M428Y/N434Y;
S239K/M252Y/T256E/D270F/N286E/V308P/N434Y;
S239K/M252W/T307P/M428Y/N434Y;
S239K/M252W/T256E/V308P/Q311A/M428Y/N434Y;
S239K/M252W/T256E/N286E/V308P/M428Y/N434Y;
P257V/T307A/Q311A/M428L/N434Y;
P257V/V305A/T307A/M428L/N434Y;
M252Y/V308P/L309E/N434Y;
M252Y/V308P/L309D/N434Y;
M252Y/V308P/L309A/N434Y;
M252W/V308P/L309E/M428Y/N434Y;
M252W/V308P/L309D/M428Y/N434Y;
M252W/V308P/L309A/M428Y/N434Y;
S239K/M252Y/N286E/V308P/M428I/N434Y;
S239K/M252Y/N286E/V308P/N434Y;
S239K/M252Y/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/T256E/V308P/N434Y;
S239K/M252Y/D270F/N286E/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/P387E/M428I/N434Y;
S239K/M252Y/D270F/N286E/T307Q/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/T307Q/V308P/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/P387E/M428I/N434Y;
P257V/T307Q/M428L/N434W;
P257V/T307A/M428L/N434W;
P257A/T307Q/L309P/M428L/N434Y;
P257V/T307Q/L309P/M428L/N434Y;
P257A/T307Q/L309R/M428L/N434Y;
P257V/T307Q/L309R/M428L/N434Y;
P257V/N286E/M428L/N434Y;
P257V/P387E/M428L/N434Y;
P257V/T307H/M428L/N434Y;
P257V/T307N/M428L/N434Y;
P257V/T307G/M428L/N434Y;
P257V/T307P/M428L/N434Y;
P257V/T307S/M428L/N434Y;
P257V/N286E/T307A/M428L/N434Y;
P257V/T307A/P387E/M428L/N434Y;
S239K/M252Y/T307P/N325G/M428Y/N434Y;
M252Y/T307A/Q311H/K360H/N434Y;
M252Y/T307A/Q311H/P387E/N434Y;
M252Y/T307A/Q311H/M428A/N434Y;
M252Y/T307A/Q311H/E382A/N434Y;
M252Y/T307W/Q311H/N434Y;
S239K/P257A/V308P/M428L/N434Y;
P257A/V308P/L309E/M428L/N434Y;
P257V/T307Q/N434Y;
M252Y/P257V/T307Q/M428V/N434Y;
M252Y/P257V/T307Q/M428L/N434Y;
M252Y/P257V/T307Q/N434Y;
M252Y/P257V/T307Q/M428Y/N434Y;
M252Y/P257V/T307Q/M428F/N434Y;
P257V/T307Q/Q311A/N325G/M428V/N434Y;
P257V/T307Q/Q311A/N325G/N434Y;
P257V/T307Q/Q311A/N325G/M428Y/N434Y;
P257V/T307Q/Q311A/N325G/M428F/N434Y;
P257A/T307Q/M428V/N434Y;
P257A/T307Q/N434Y;
P257A/T307Q/M428Y/N434Y;
P257A/T307Q/M428F/N434Y;
P257V/N286E/T307Q/Q311A/N325G/M428L/N434Y;
P257A/Q311A/M428L/N434Y;
P257A/Q311H/M428L/N434Y;
P257A/T307Q/Q311A/M428L/N434Y;
P257A/T307A/Q311A/M428L/N434Y;
P257A/T307Q/Q311H/M428L/N434Y;
P257AT307A/Q311H/M428L/N434Y;
P257A/N286E/T307Q/M428L/N434Y;
P257A/N286E/T307A/M428L/N434Y;
S239K/M252Y/D270F/T307P/N325G/M428Y/N434Y;
S239K/M252Y/T307P/L309E/N325G/M428Y/N434Y;
P257S/T307A/M428L/N434Y;
P257M/T307A/M428L/N434Y;
P257N/T307A/M428L/N434Y;
P257I/T307A/M428L/N434Y;
P257V/T307Q/M428Y/N434Y;
P257V/T307Q/M428F/N434Y;
S239K/P257V/V308P/M428L/N434Y;
P257V/T307Q/V308P/N325G/M428L/N434Y;
P257V/T307Q/V308P/Q311A/N325G/M428L/N434Y;
P257V/T307A/V308P/N325G/M428L/N434Y;
P257A/V308P/N325G/M428L/N434Y;
P257A/T307A/V308P/M428L/N434Y;
P257A/T307Q/V308P/M428L/N434Y;
P257V/N286E/T307Q/N325G/M428L/N434Y;
T256E/P257V/N434Y;
T256E/P257T/N434Y;
S239K/P257T/V308P/M428L/N434Y;
P257T/V308P/N325G/M428L/N434Y;
T256E/P257T/V308P/N325G/M428L/N434Y;
P257T/V308P/N325G/E382A/M428L/N434Y;
P257T/V308P/N325G/P387E/M428L/N434Y;
P257T/V308P/L309P/N325G/M428L/N434Y;
P257T/V308P/L309R/N325G/M428L/N434Y;
T256E/P257V/T307Q/M428L/N434Y;
P257V/T307Q/E382A/M428L/N434Y;
P257V/T307Q/P387E/M428L/N434Y;
P257L/V308P/N434Y;
P257T/T307Q/N434Y;
P257V/T307Q/N325G/N434Y;
P257V/T307Q/Q311A/N434Y;
P257V/V305A/T307Q/N434Y;
P257V/N286E/T307A/N434Y;
P257V/T307Q/L309R/Q311H/M428L/N434Y;
P257V/V308P/N325G/M428L/N434Y;
S239K/P257V/V308P/Q311H/M428L/N434Y;
P257V/V305A/T307A/N325G/M428L/N434Y;
S239K/D270F/T307P/N325G/M428Y/N434Y;
P257T/T307A/M428V/N434Y;
P257VM428V/N434Y;
P257A/T307A/M428V/N434Y;
P257S/T307Q/M428L/N434Y;
P257V/N297A/T307Q/M428L/N434Y;
P257V/N286A/T307Q/M428L/N434Y;
P257V/T307Q/N315A/M428L/N434Y;
P257V/T307Q/N384A/M428L/N434Y;
P257V/T307Q/N389A/M428L/N434Y;
P257V/N434Y;
P257T/N434Y;
P257V/N286E/N434Y;
P257T/N286E/N434Y;
P257V/N286E/T307Q/N434Y;
P257V/N286E/T307Q/M428Y/N434Y;
P257V/V305A/T307Q/M428Y/N434Y;
P257T/T307Q/M428Y/N434Y;
P257V/T307Q/V308P/N325G/M428Y/N434Y;
P257T/T307Q/M428F/N434Y;
P257AN286E/T307Q/M428F/N434Y;
P257A/N286E/T307Q/M428Y/N434Y;
T250A/P257V/T307Q/M428L/N434Y;
T250F/P257V/T307Q/M428L/N434Y;
T250I/P257V/T307Q/M428L/N434Y;
T250M/P257V/T307Q/M428L/N434Y;
T250S/P257V/T307Q/M428L/N434Y;
T250V/P257V/T307Q/M428L/N434Y;
T250W/P257V/T307Q/M428L/N434Y;
T25OY/P257V/T307Q/M428L/N434Y;
M252Y/Q311A/N434Y;
S239K/M252Y/S254T/V308P/N434Y;
M252Y/T307Q/Q311A;
M252Y/S254T/N286E/N434Y;
M252Y/S254T/V308P/N434Y;
M252Y/S254T/T307A/N434Y;
M252Y/S254T/T307Q/N434Y;
M252Y/S254T/Q311A/N434Y;
M252Y/S254T/Q311H/N434Y;
M252Y/S254T/T307A/Q311A/N434Y;
M252Y/S254T/T307Q/Q311A/N434Y;
M252Y/S254T/M428I/N434Y;
M252Y/T256E/T307A/Q311H/N434Y;
M252Y/T256Q/T307A/Q311H/N434Y;
M252Y/S254T/T307A/Q311H/N434Y;
M252Y/N286E/T307A/Q311H/N434Y;
M252Y/T307A/Q311H/M428I/N434Y;
M252Y/T256E/T307Q/Q311H;
M252Y/N286E/T307A/Q311A;
M252Y/N286E/T307Q/Q311A;
M252Y/N286E/T307Q/Q311H;
M252Y/T256E/N286E/N434Y;
S239K/M252Y/V308P;
S239K/M252Y/V308P/E382A;
S239K/M252Y/T256E/V308P;
S239K/M252Y/N286E/V308P;
S239K/M252Y/N286E/V308P/M428I;
M252Y/N286E/M428L/N434Y;
M252Y/S254T/E382A/N434Y;
S239K/M252Y/S254T/V308P/M428I/N434Y;
S239K/M252Y/M428I/N434Y;
M252Y/D312A/E382A/M428Y/N434Y;
M252Y/E382A/P387E/N434Y;
M252Y/D312A/P387E/N434Y или
M252Y/P387E/M428Y/N434Y; и
(b) введения в указанное антитело по меньшей мере одной из гистидиновых замен или инсерций в антигенсвязывающий домен тяжелой цепи в положение, выбранное из группы, состоящей из Н27, Н31, Н32, Н33, Н35, Н50, Н58, Н59, Н61, Н62, Н63, Н64, Н65, Н99, H100b и H102 в соответствии с нумерацией по Kabat, и/или антигенсвязывающий домен легкой цепи в положение, выбранное из группы, состоящей из L24, L27, L28, L32, L53, L54, L56, L90, L92 и L94 в соответствии с нумерацией по Kabat;
где указанное антитело, содержащее антигенсвязывающий домен и человеческий FcRn-связывающий домен, полученные в (а) и (b), связывается с растворимым антигеном, обладает активностью связывания человеческого FcRn при рН 5.8 и рН 7.0, где его активность связывания FcRn человека при рН 7.0 сильнее, чем интактного IgG человека, и обладает более низкой антигенсвязывающей активностью при рН 5.8, чем при рН 7.4, где отношение антигенсвязывающей активности при рН 5.8 и рН 7.4 равно по меньшей мере 2 при величине KD при рН 5.8 / KD при рН 7.4.
7. Способ по п. 6, где снижение антигенсвязывающей активности представлено увеличением величины KD при рН 5.8 / KD при рН 7.4, которое представляет собой соотношение антигенсвязывающей активности при рН 5.8 и при рН 7.4, относительно этого значения до гистидиновой замены или инсерций.
8. Способ получения антитела, где способ включает этапы:
(а) отбора антитела, которое обладает активностью связывания человеческого FcRn при рН 5.8 и рН 7.0 и более высокой активностью связывания человеческого FcRn при рН 7.0, чем до изменения по меньшей мере одной аминокислоты в человеческом FcRn-связывающем домене антитела IgG1, IgG2, IgG3 или IgG4, включающем Fc домен IgG1 человека, IgG2 человека, IgG3 человека или IgG4 человека и обладающем активностью связывания человеческого FcRn при рН 5.8, где указанный человеческий FcRn-связывающий домен содержит одну из следующих аминокислотных замен в положениях, соответствующих нумерации EU в Fc-домене:
N434W;
N434Y;
M252Y/S254T/T256E/N434Y;
M252Y/S254T/T256E/N434W;
M252Y/N434Y;
M252Y/N434W;
M252W/N434Y;
M252W/N434W;
P257L/N434Y;
V308F/N434Y;
M252Y/V308P/N434Y;
M428L/N434S;
M252Y/S254T/T256E/V308P/N434W;
I332V;
I332V/N434Y;
G237M/V308F;
S254T/N434W;
S254T/N434Y;
T256E/N434W;
T256E/N434Y;
S254T/T256E/N434W;
S254T/T256E/N434Y;
M252Y/S254T/N434W;
M252Y/S254T/N434Y;
M252Y/T256E/N434W;
M252Y/T256E/N434Y;
M252Y/S254T/T256E/N434A;
M252Y/N434A;
M252W/N434A;
M252Y/T256Q/N434W;
M252Y/T256Q/N434Y;
M252F/T256D/N434W;
M252F/T256D/N434Y;
H433K/N434F/Y436H;
I332V/N434W;
V308P/N434W;
I332V/M428L/N434Y;
G385D/Q386P/N389S/N434W;
G385D/Q386P/N389S/N434Y;
G385H/N434W;
G385H/N434Y;
N434F;
M252Y/S254T/T256E/N434F;
M252Y/S254T/T256E/N434H;
M252Y/N434F;
M252Y/N434H;
M428L/N434W;
M428L/N434Y;
M252Y/S254T/T256E/M428L/N434W;
M252Y/S254T/T256E/M428L/N434Y;
M252Y/M428L/N434W;
M252Y/M428L/N434Y;
M252Y/M428L/N434A;
M252Y/S254T/T256E/M428L/N434A;
T256E/M428L/N434Y;
S254T/M428L/N434W;
M252Y/T256Q/N434A;
M252Y/T256E/N434A;
T256Q/N434W;
T256Q/N434Y;
M252W/T256Q/N434W;
M252W/T256Q/N434Y;
S254T/T256Q/N434W;
M252Y/S254T/T256Q/N434W;
M252Y/S254T/T256Q/N434Y;
M252Y/T256E/V308P/N434W;
M252Y/V308P/M428L/N434Y;
M252Y/S254T/T256E/V308P/M428L/N434W;
M252W/M428L/N434W;
P257L/M428L/N434Y;
M252Y/S254T/T256E/M428L/N434F;
M252Y/T256E/N434H;
M252W/T256Q/P257L/N434Y;
P238A/M252Y/V308P/N434Y;
M252Y/D265A/V308P/N434Y;
M252Y/T307A/V308P/N434Y;
M252Y/V303A/V308P/N434Y;
M252Y/V308P/D376A/N434Y;
M252Y/V305A/V308P/N434Y;
M252Y/V308P/Q311A/N434Y;
M252Y/V308P/K317A/N434Y;
M252Y/V308P/E380A/N434Y;
M252Y/V308P/E382A/N434Y;
M252Y/V308P/S424A/N434Y;
P257L/V308P/M428L/N434Y;
M252Y/T256E/V308P/M428L/N434W;
M252Y/T256E/M428L/N434W;
M252Y/T256E/V308P/N434Y;
M252Y/T256E/M428L/N434Y;
M252Y/S254T/T256E/V308P/M428L/N434Y;
P257L/M428L/N434W;
P257A/M428L/N434Y;
P257G/M428L/N434Y;
P257I/M428L/N434Y;
P257M/M428L/N434Y;
P257N/M428L/N434Y;
P257S/M428L/N434Y;
P257T/M428L/N434Y;
P257V/M428L/N434Y;
M252W/V308P/N434W;
S239K/M252Y/V308P/N434Y;
M252Y/S298G/V308P/N434Y;
M252Y/D270F/V308P/N434Y;
M252Y/V308A/N434Y;
M252Y/V308F/N434Y;
M252Y/V308I/N434Y;
M252Y/V308L/N434Y;
M252Y/V308M/N434Y;
M252Y/V308Q/N434Y;
M252Y/V308T/N434Y;
P257A/V308P/M428L/N434Y;
P257T/V308P/M428L/N434Y;
P257V/V308P/M428L/N434Y;
M252W/M428I/N434Y;
M252W/M428Y/N434Y;
M252W/M428F/N434Y;
P238A/M252W/N434Y;
M252W/D265A/N434Y;
M252W/T307Q/N434Y;
M252W/V303A/N434Y;
M252W/D376A/N434Y;
M252W/V305A/N434Y;
M252W/Q311A/N434Y;
M252W/D312A/N434Y;
M252W/K317A/N434Y;
M252W/E380A/N434Y;
M252W/E382A/N434Y;
M252W/S424A/N434Y;
S239K/M252W/N434Y;
M252W/S298G/N434Y;
M252W/D270F/N434Y;
M252W/N325G/N434Y;
P257A/M428L/N434W;
P257T/M428L/N434W;
P257V/M428L/N434W;
M252W/I332V/N434Y;
P257I/Q311I;
M252Y/T307A/N434Y;
M252Y/T307Q/N434Y;
P257L/T307A/M428L/N434Y;
P257A/T307A/M428L/N434Y;
P257T/T307A/M428L/N434Y;
P257V/T307A/M428L/N434Y;
P257L/T307Q/M428L/N434Y;
P257A/T307Q/M428L/N434Y;
P257T/T307Q/M428L/N434Y;
P257V/T307Q/M428L/N434Y;
M252Y/T307D/N434Y;
M252Y/T307F/N434Y;
M252Y/T307G/N434Y;
M252Y/T307H/N434Y;
M252Y/T307I/N434Y;
M252Y/T307K/N434Y;
M252Y/T307L/N434Y;
M252Y/T307M/N434Y;
M252Y/T307N/N434Y;
M252Y/T307P/N434Y;
M252Y/T307R/N434Y;
M252Y/T307S/N434Y;
M252Y/T307V/N434Y;
M252Y/T307W/N434Y;
M252Y/T307Y/N434Y;
M252Y/K334L/N434Y;
M252Y/G385H/N434Y;
M252Y/T289H/N434Y;
M252Y/Q311H/N434Y;
M252Y/D312H/N434Y;
M252Y/N315H/N434Y;
M252Y/K360H/N434Y;
M252Y/L314R/N434Y;
M252Y/L314K/N434Y;
M252Y/N286E/N434Y;
M252Y/L309E/N434Y;
M252Y/R255E/N434Y;
M252Y/P387E/N434Y;
K248I/M428L/N434Y;
M252Y/M428A/N434Y;
M252Y/M428D/N434Y;
M252Y/M428F/N434Y;
M252Y/M428G/N434Y;
M252Y/M428H/N434Y;
M252Y/M428I/N434Y;
M252Y/M428K/N434Y;
M252Y/M428N/N434Y;
M252Y/M428P/N434Y;
M252Y/M428Q/N434Y;
M252Y/M428S/N434Y;
M252Y/M428T/N434Y;
M252Y/M428V/N434Y;
M252Y/M428W/N434Y;
M252Y/M428Y/N434Y;
M252W/T307Q/M428Y/N434Y;
M252W/Q311A/M428Y/N434Y;
M252W/T307Q/Q311A/M428Y/N434Y;
M252Y/T307A/M428Y/N434Y;
M252Y/T307Q/M428Y/N434Y;
M252Y/D270F/N434Y;
M252Y/T307A/Q311A/N434Y;
M252Y/T307Q/Q311A/N434Y;
M252Y/T307A/Q311H/N434Y;
M252Y/T307Q/Q311H/N434Y;
M252Y/E382A/N434Y;
M252Y/E382A/M428Y/N434Y;
M252Y/T307A/E382A/M428Y/N434Y;
M252Y/T307Q/E382A/M428Y/N434Y;
P238A/M252Y/M428F/N434Y;
M252Y/V305A/M428F/N434Y;
M252Y/N325G/M428F/N434Y;
M252Y/D376A/M428F/N434Y;
M252Y/E380A/M428F/N434Y;
M252Y/E382A/M428F/N434Y;
M252Y/E380A/E382A/M428F/N434Y;
S239K/M252Y/V308P/E382A/N434Y;
M252Y/D270F/V308P/E382A/N434Y;
S239K/M252Y/V308P/M428Y/N434Y;
M252Y/T307Q/V308P/E382A/N434Y;
M252Y/V308P/Q311H/E382A/N434Y;
S239K/M252Y/D270F/V308P/N434Y;
M252Y/V308P/E382A/M428F/N434Y;
M252Y/V308P/E382A/M428L/N434Y;
M252Y/V308P/E382A/M428Y/N434Y;
M252Y/V308P/M428Y/N434Y;
M252Y/V308P/M428F/N434Y;
S239K/M252Y/V308P/E382A/M428Y/N434Y;
M252Y/V308P/E380A/E382A/M428F/N434Y;
M252Y/T256E/E382A/N434Y;
M252Y/T256E/M428Y/N434Y;
M252Y/T256E/E382A/M428Y/N434Y;
S239K/M252Y/V308P/E382A/M428I/N434Y;
M252Y/V308P/E380A/E382A/M428I/N434Y;
S239K/M252Y/V308P/M428F/N434Y;
S239K/M252Y/E380A/E382A/M428F/N434Y;
M252Y/Q311A/M428Y/N434Y;
M252Y/D312A/M428Y/N434Y;
M252Y/Q311A/D312A/M428Y/N434Y;
S239K/M252Y/V308P/Q311A/M428Y/N434Y;
S239K/M252Y/V308P/D312A/M428Y/N434Y;
S239K/M252Y/V308P/Q311A/D312A/M428Y/N434Y;
M252Y/T256E/T307P/N434Y;
M252Y/T307P/M428Y/N434Y;
M252W/V308P/M428Y/N434Y;
M252Y/T256E/V308P/E382A/N434Y;
M252W/V308P/E382A/N434Y;
S239K/M252W/V308P/E382A/N434Y;
S239K/M252W/V308P/E382A/M428Y/N434Y;
S239K/M252W/V308P/E382A/M428I/N434Y;
S239K/M252W/V308P/M428F/N434Y;
S239K/M252W/E380A/E382A/M428F/N434Y;
S239K/M252Y/T307P/M428Y/N434Y;
M252Y/T256E/Q311A/D312A/M428Y/N434Y;
S239K/M252Y/M428Y/N434Y;
S239K/M252Y/D270F/M428Y/N434Y;
S239K/M252Y/D270F/V308P/M428Y/N434Y;
S239K/M252Y/V308P/M428I/N434Y;
S239K/M252Y/V308P/N325G/E382A/M428I/N434Y;
S239K/M252Y/D270F/V308P/N325G/N434Y;
S239K/M252Y/D270F/T307P/V308P/N434Y;
S239K/M252Y/T256E/D270F/V308P/N434Y;
S239K/M252Y/D270F/T307Q/V308P/N434Y;
S239K/M252Y/D270F/T307Q/V308P/Q311A/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/N434Y;
S239K/M252Y/T256E/D270F/T307Q/Q311A/N434Y;
S239K/M252Y/D270F/V308P/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/N434Y;
S239K/M252Y/D270F/V308P/L309E/N434Y;
S239K/M252Y/D270F/V308P/P387E/N434Y;
S239K/M252Y/D270F/T307Q/L309E/Q311A/N434Y;
S239K/M252Y/D270F/V308P/N325G/M428I/N434Y;
S239K/M252Y/T256E/D270F/V308P/M428I/N434Y;
S239K/M252Y/D270F/T307Q/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/M428I/N434Y;
S239K/M252Y/D270F/V308P/L309E/M428I/N434Y;
S239K/M252Y/D270F/V308P/M428L/N434Y;
S239K/M252Y/V308P/M428L/N434Y;
S239K/M252Y/V308P/L309P/M428L/N434Y;
S239K/M252Y/V308P/L309R/M428L/N434Y;
S239K/M252Y/L309P/M428L/N434Y;
S239K/M252Y/L309R/M428L/N434Y;
S239K/M252Y/T307Q/V308P/M428L/N434Y;
S239K/M252Y/D270F/V308P/L309E/P387E/N434Y;
S239K/M252Y/T256E/D270F/V308P/N325G/M428I/N434Y;
S239K/M252Y/T307Q/N434Y;
P257V/T307Q/M428I/N434Y;
P257V/T307Q/M428V/N434Y;
P257V/T307Q/N325G/M428L/N434Y;
P257V/T307Q/Q311A/M428L/N434Y;
P257V/T307Q/Q311A/N325G/M428L/N434Y;
P257V/V305A/T307Q/M428L/N434Y;
S239K/M252Y/E258H/D270F/T307Q/V308P/Q311A/N434Y;
S239K/M252Y/D270F/V308P/N325G/E382A/M428I/N434Y;
M252Y/P257V/T307Q/M428I/N434Y;
S239K/M252W/V308P/M428Y/N434Y;
S239K/M252W/V308P/Q311A/M428Y/N434Y;
S239K/M252W/T256E/V308P/M428Y/N434Y;
S239K/M252W/N286E/V308P/M428Y/N434Y;
S239K/M252Y/T256E/D270F/N286E/V308P/N434Y;
S239K/M252W/T307P/M428Y/N434Y;
S239K/M252W/T256E/V308P/Q311A/M428Y/N434Y;
S239K/M252W/T256E/N286E/V308P/M428Y/N434Y;
P257V/T307A/Q311A/M428L/N434Y;
P257V/V305A/T307A/M428L/N434Y;
M252Y/V308P/L309E/N434Y;
M252Y/V308P/L309D/N434Y;
M252Y/V308P/L309A/N434Y;
M252W/V308P/L309E/M428Y/N434Y;
M252W/V308P/L309D/M428Y/N434Y;
M252W/V308P/L309A/M428Y/N434Y;
S239K/M252Y/N286E/V308P/M428I/N434Y;
S239K/M252Y/N286E/V308P/N434Y;
S239K/M252Y/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/T256E/V308P/N434Y;
S239K/M252Y/D270F/N286E/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/P387E/M428I/N434Y;
S239K/M252Y/D270F/N286E/T307Q/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/T307Q/V308P/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/P387E/M428I/N434Y;
P257V/T307Q/M428L/N434W;
P257V/T307A/M428L/N434W;
P257A/T307Q/L309P/M428L/N434Y;
P257V/T307Q/L309P/M428L/N434Y;
P257A/T307Q/L309R/M428L/N434Y;
P257V/T307Q/L309R/M428L/N434Y;
P257V/N286E/M428L/N434Y;
P257V/P387E/M428L/N434Y;
P257V/T307H/M428L/N434Y;
P257V/T307N/M428L/N434Y;
P257V/T307G/M428L/N434Y;
P257V/T307P/M428L/N434Y;
P257V/T307S/M428L/N434Y;
P257V/N286E/T307A/M428L/N434Y;
P257V/T307A/P387E/M428L/N434Y;
S239K/M252Y/T307P/N325G/M428Y/N434Y;
M252Y/T307A/Q311H/K360H/N434Y;
M252Y/T307A/Q311H/P387E/N434Y;
M252Y/T307A/Q311H/M428A/N434Y;
M252Y/T307A/Q311H/E382A/N434Y;
M252Y/T307W/Q311H/N434Y;
S239K/P257A/V308P/M428L/N434Y;
P257A/V308P/L309E/M428L/N434Y;
P257V/T307Q/N434Y;
M252Y/P257V/T307Q/M428V/N434Y;
M252Y/P257V/T307Q/M428L/N434Y;
M252Y/P257V/T307Q/N434Y;
M252Y/P257V/T307Q/M428Y/N434Y;
M252Y/P257V/T307Q/M428F/N434Y;
P257V/T307Q/Q311A/N325G/M428V/N434Y;
P257V/T307Q/Q311A/N325G/N434Y;
P257V/T307Q/Q311A/N325G/M428Y/N434Y;
P257V/T307Q/Q311A/N325G/M428F/N434Y;
P257A/T307Q/M428V/N434Y;
P257A/T307Q/N434Y;
P257A/T307Q/M428Y/N434Y;
P257A/T307Q/M428F/N434Y;
P257V/N286E/T307Q/Q311A/N325G/M428L/N434Y;
P257A/Q311A/M428L/N434Y;
P257A/Q311H/M428L/N434Y;
P257A/T307Q/Q311A/M428L/N434Y;
P257A/T307A/Q311A/M428L/N434Y;
P257A/T307Q/Q311H/M428L/N434Y;
P257A/T307A/Q311H/M428L/N434Y;
P257A/N286E/T307Q/M428L/N434Y;
P257A/N286E/T307A/M428L/N434Y;
S239K/M252Y/D270F/T307P/N325G/M428Y/N434Y;
S239K/M252Y/T307P/L309E/N325G/M428Y/N434Y;
P257S/T307A/M428L/N434Y;
P257M/T307A/M428L/N434Y;
P257N/T307A/M428L/N434Y;
P257I/T307A/M428L/N434Y;
P257V/T307Q/M428Y/N434Y;
P257V/T307Q/M428F/N434Y;
S239K/P257V/V308P/M428L/N434Y;
P257V/T307Q/V308P/N325G/M428L/N434Y;
P257V/T307Q/V308P/Q311A/N325G/M428L/N434Y;
P257V/T307A/V308P/N325G/M428L/N434Y;
P257A/V308P/N325G/M428L/N434Y;
P257A/T307A/V308P/M428L/N434Y;
P257A/T307Q/V308P/M428L/N434Y;
P257V/N286E/T307Q/N325G/M428L/N434Y;
T256E/P257V/N434Y;
T256E/P257T/N434Y;
S239K/P257T/V308P/M428L/N434Y;
P257T/V308P/N325G/M428L/N434Y;
T256E/P257T/V308P/N325G/M428L/N434Y;
P257T/V308P/N325G/E382A/M428L/N434Y;
P257T/V308P/N325G/P387E/M428I/N434Y;
P257T/V308P/L309P/N325G/M428L/N434Y;
P257T/V308P/L309R/N325G/M428L/N434Y;
T256E/P257V/T307Q/M428I/N434Y;
P257V/T307Q/E382A/M428L/N434Y;
P257V/T307Q/P387E/M428L/N434Y;
P257L/V308P/N434Y;
P257T/T307Q/N434Y;
P257V/T307Q/N325G/N434Y;
P257V/T307Q/Q311A/N434Y;
P257V/V305A/T307Q/N434Y;
P257V/N286E/T307A/N434Y;
P257V/T307Q/L309R/Q311H/M428L/N434Y;
P257V/V308P/N325G/M428L/N434Y;
S239K/P257V/V308P/Q311H/M428L/N434Y;
P257V/V305A/T307A/N325G/M428L/N434Y;
S239K/D270F/T307P/N325G/M428Y/N434Y;
P257T/T307A/M428V/N434Y;
P257A/M428V/N434Y;
P257A/T307A/M428V/N434Y;
P257S/T307Q/M428L/N434Y;
P257V/N297A/T307Q/M428L/N434Y;
P257V/N286A/T307Q/M428L/N434Y;
P257V/T307Q/N315A/M428L/N434Y;
P257V/T307Q/N384A/M428L/N434Y;
P257V/T307Q/N389A/M428L/N434Y;
P257V/N434Y;
P257T/N434Y;
P257V/N286E/N434Y;
P257T/N286E/N434Y;
P257V/N286E/T307Q/N434Y;
P257V/N286E/T307Q/M428Y/N434Y;
P257V/V305A/T307Q/M428Y/N434Y;
P257T/T307Q/M428Y/N434Y;
P257V/T307Q/V308P/N325G/M428Y/N434Y;
P257T/T307Q/M428F/N434Y;
P257A/N286E/T307Q/M428F/N434Y;
P257A/N286E/T307Q/M428Y/N434Y;
T250A/P257V/T307Q/M428L/N434Y;
T250F/P257V/T307Q/M428L/N434Y;
T250I/P257V/T307Q/M428L/N434Y;
T250M/P257V/T307Q/M428L/N434Y;
T250S/P257V/T307Q/M428L/N434Y;
T250V/P257V/T307Q/M428L/N434Y;
T250W/P257V/T307Q/M428L/N434Y;
T250Y/P257V/T307Q/M428L/N434Y;
M252Y/Q311A/N434Y;
S239K/M252Y/S254T/V308P/N434Y;
M252Y/T307Q/Q311A;
M252Y/S254T/N286E/N434Y;
M252Y/S254T/V308P/N434Y;
M252Y/S254T/T307A/N434Y;
M252Y/S254T/T307Q/N434Y;
M252Y/S254T/Q311A/N434Y;
M252Y/S254T/Q311H/N434Y;
M252Y/S254T/T307A/Q311A/N434Y;
M252Y/S254T/T307Q/Q311A/N434Y;
M252Y/S254T/M428I/N434Y;
M252Y/T256E/T307A/Q311H/N434Y;
M252Y/T256Q/T307A/Q311H/N434Y;
M252Y/S254T/T307A/Q311H/N434Y;
M252Y/N286E/T307A/Q311H/N434Y;
M252Y/T307A/Q311H/M428I/N434Y;
M252Y/T256E/T307Q/Q311H;
M252Y/N286E/T307A/Q311A;
M252Y/N286E/T307Q/Q311A;
M252Y/N286E/T307Q/Q311H;
M252Y/T256E/N286E/N434Y;
S239K/M252Y/V308P;
S239K/M252Y/V308P/E382A;
S239K/M252Y/T256E/V308P;
S239K/M252Y/N286E/V308P;
S239K/M252Y/N286E/V308P/M428I;
M252Y/N286E/M428L/N434Y;
M252Y/S254T/E382A/N434Y;
S239K/M252Y/S254T/V308P/M428I/N434Y;
S239K/M252Y/M428I/N434Y;
M252Y/D312A/E382A/M428Y/N434Y;
M252Y/E3 82А/РЗ 87E/N434Y;
M252Y/D312A/P387E/N434Y; или
M252Y/P387E/M428Y/N434Y; и
(b) замены гистидином аминокислоты или инсерции гистидина в положении антигенсвязывающего домена тяжелой цепи, которое выбрано из группы, состоящей из Н27, Н31, Н32, Н33, Н35, Н50, Н58, Н59, Н61, Н62, Н63, Н64, Н65, Н99, H100b и Н102 в соответствии с нумерацией по Kabat, и/или в антигенсвязывающем домене легкой цепи, которое выбрано из группы, состоящей из L24, L27, L28, L32, L53, L54, L56, L90, L92 и L94 в соответствии с нумерацией по Kabat, и отбор антитела, которое связывается с растворимым антигеном и обладает более высокой антигенсвязывающей активностью при рН 7.4, чем при рН 5.8, где отношение антигенсвязывающей активности при рН 5.8 и рН 7.4 равно по меньшей мере 2 при величине KD при рН 5.8 / KD при рН 7.4;
(c) получения гена, кодирующего антитело, в котором человеческий FcRn-связывающий домен и антигенсвязывающий домен, отобранные на стадиях (а) и (b), сцеплены; и
(d) получения антитела с использованием гена, полученного на стадии (с).
9. Способ получения антитела, включающий стадии:
(а) отбора антитела, которое содержит человеческий FcRn-связывающий домен, обладающий активностью связывания FcRn человека при рН 5.8 и рН 7.0 и более высокой активностью связывания человеческого FcRn при рН 7.0, чем до изменения по меньшей мере одной аминокислоты в человеческом FcRn-связывающем домене антитела IgG1, IgG2, IgG3 или IgG4, включающем Fc домен IgG1 человека, IgG2 человека, IgG3 человека или IgG4 и обладающем активностью связывания человеческого FcRn при рН 5.8, где указанный человеческий FcRn-связывающий домен после вышеуказанного изменения содержит одну из следующих аминокислотных замен в положениях, соответствующих нумерации EU в Fc-домене:
N434W;
N434Y;
M252Y/S254T/T256E/N434Y;
M252Y/S254T/T256E/N434W;
M252Y/N434Y;
M252Y/N434W;
M252W/N434Y;
M252W/N434W;
P257L/N434Y;
V308F/N434Y;
M252Y/V308P/N434Y;
M428L/N434S;
M252Y/S254T/T256E/V308P/N434W;
I332V;
I332V/N434Y;
G237M/V308F;
S254T/N434W;
S254T/N434Y;
T256E/N434W;
T256E/N434Y;
S254T/T256E/N434W;
S254T/T256E/N434Y;
M252Y/S254T/N434W;
M252Y/S254T/N434Y;
M252Y/T256E/N434W;
M252Y/T256E/N434Y;
M252Y/S254T/T256E/N434A;
M252Y/N434A;
M252W/N434A;
M252Y/T256Q/N434W;
M252Y/T256Q/N434Y;
M252F/T256D/N434W;
M252F/T256D/N434Y;
H433K/N434F/Y436H;
I332V/N434W;
V308P/N434W;
I332V/M428L/N434Y;
G385D/Q386P/N389S/N434W;
G385D/Q386P/N389S/N434Y;
G385H/N434W;
G385H/N434Y;
N434F;
M252Y/S254T/T256E/N434F;
M252Y/S254T/T256E/N434H;
M252Y/N434F;
M252Y/N434H;
M428L/N434W;
M428L/N434Y;
M252Y/S254T/T256E/M428L/N434W;
M252Y/S254T/T256E/M428L/N434Y;
M252Y/M428L/N434W;
M252Y/M428L/N434Y;
M252Y/M428L/N434A;
M252Y/S254T/T256E/M428L/N434A;
T256E/M428L/N434Y;
S254T/M428L/N434W;
M252Y/T256Q/N434A;
M252Y/T256E/N434A;
T256Q/N434W;
T256Q/N434Y;
M252W/T256Q/N434W;
M252W/T256Q/N434Y;
S254T/T256Q/N434W;
M252Y/S254T/T256Q/N434W;
M252Y/S254T/T256Q/N434Y;
M252Y/T256E/V308P/N434W;
M252Y/V308P/M428L/N434Y;
M252Y/S254T/T256E/V308P/M428L/N434W;
M252W/M428L/N434W;
P257L/M428L/N434Y;
M252Y/S254T/T256E/M428L/N434F;
M252Y/T256E/N434H;
M252W/T256Q/P257L/N434Y;
P238A/M252Y/V308P/N434Y;
M252Y/D265A/V308P/N434Y;
M252Y/T307A/V308P/N434Y;
M252Y/V303A/V308P/N434Y;
M252Y/V308P/D376A/N434Y;
M252Y/V305A/V308P/N434Y;
M252Y/V308P/Q311A/N434Y;
M252Y/V308P/K317A/N434Y;
M252Y/V308P/E380A/N434Y;
M252Y/V308P/E382A/N434Y;
M252Y/V308P/S424A/N434Y;
P257L/V308P/M428L/N434Y;
M252Y/T256E/V308P/M428L/N434W;
M252Y/T256E/M428L/N434W;
M252Y/T256E/V308P/N434Y;
M252Y/T256E/M428L/N434Y;
M252Y/S254T/T256E/V308P/M428L/N434Y;
P257L/M428L/N434W;
P257A/M428L/N434Y;
P257G/M428L/N434Y;
P257I/M428L/N434Y;
P257M/M428L/N434Y;
P257N/M428L/N434Y;
P257S/M428L/N434Y;
P257T/M428L/N434Y;
P257V/M428L/N434Y;
M252W/V308P/N434W;
S239K/M252Y/V308P/N434Y;
M252Y/S298G/V308P/N434Y;
M252Y/D270F/V308P/N434Y;
M252Y/V308A/N434Y;
M252Y/V308F/N434Y;
M252Y/V308I/N434Y;
M252Y/V308L/N434Y;
M252Y/V308M/N434Y;
M252Y/V308Q/N434Y;
M252Y/V308T/N434Y;
P257A/V308P/M428IVN434Y;
P257T/V308P/M428L/N434Y;
P257V/V308P/M428IVN434Y;
M252W/M428I/N434Y;
M252W/M428Y/N434Y;
M252W/M428F/N434Y;
P238A/M252W/N434Y;
M252W/D265A/N434Y;
M252W/T307Q/N434Y;
M252W/V303A/N434Y;
M252W/D376A/N434Y;
M252W/V305A/N434Y;
M252W/Q311A/N434Y;
M252W/D312A/N434Y;
M252W/K317A/N434Y;
M252W/E380A/N434Y;
M252W/E382A/N434Y;
M252W/S424A/N434Y;
S239K/M252W/N434Y;
M252W/S298G/N434Y;
M252W/D270F/N434Y;
M252W/N325G/N434Y;
P257A/M428L/N434W;
P257T/M428L/N434W;
P257V/M428L/N434W;
M252W/I332V/N434Y;
P257I/Q311I;
M252Y/T307A/N434Y;
M252Y/T307Q/N434Y;
P257L/T307A/M428L/N434Y;
P257A/T307A/M428L/N434Y;
P257T/T307A/M428L/N434Y;
P257V/T307A/M428L/N434Y;
P257L/T307Q/M428L/N434Y;
P257A/T307Q/M428L/N434Y;
P257T/T307Q/M428L/N434Y;
P257V/T307Q/M428L/N434Y;
M252Y/T307D/N434Y;
M252Y/T307F/N434Y;
M252Y/T307G/N434Y;
M252Y/T307H/N434Y;
M252Y/T307I/N434Y;
M252Y/T307K/N434Y;
M252Y/T307L/N434Y;
M252Y/T307M/N434Y;
M252Y/T307N/N434Y;
M252Y/T307P/N434Y;
M252Y/T307R/N434Y;
M252Y/T307S/N434Y;
M252Y/T307V/N434Y;
M252Y/T307W/N434Y;
M252Y/T307Y/N434Y;
M252Y/K334L/N434Y;
M252Y/G385H/N434Y;
M252Y/T289H/N434Y;
M252Y/Q311H/N434Y;
M252Y/D312H/N434Y;
M252Y/N315H/N434Y;
M252Y/K360H/N434Y;
M252Y/L314R/N434Y;
M252Y/L314K/N434Y;
M252Y/N286E/N434Y;
M252Y/L309E/N434Y;
M252Y/R255E/N434Y;
M252Y/P387E/N434Y;
K248I/M428L/N434Y;
M252Y/M428A/N434Y;
M252Y/M428D/N434Y;
M252Y/M428F/N434Y;
M252Y/M428G/N434Y;
M252Y/M428H/N434Y;
M252Y/M428I/N434Y;
M252Y/M428K/N434Y;
M252Y/M428N/N434Y;
M252Y/M428P/N434Y;
M252Y/M428Q/N434Y;
M252Y/M428S/N434Y;
M252Y/M428T/N434Y;
M252Y/M428V/N434Y;
M252Y/M428W/N434Y;
M252Y/M428Y/N434Y;
M252W/T307Q/M428Y/N434Y;
M252W/Q311A/M428Y/N434Y;
M252W/T307Q/Q311A/M428Y/N434Y;
M252Y/T307A/M428Y/N434Y;
M252Y/T307Q/M428Y/N434Y;
M252Y/D270F/N434Y;
M252Y/T307A/Q311A/N434Y;
M252Y/T307Q/Q311A/N434Y;
M252Y/T307A/Q311H/N434Y;
M252Y/T307Q/Q311H/N434Y;
M252Y/E382A/N434Y;
M252Y/E382A/M428Y/N434Y;
M252Y/T307A/E382A/M428Y/N434Y;
M252Y/T307Q/E382A/M428Y/N434Y;
P238A/M252Y/M428F/N434Y;
M252Y/V305A/M428F/N434Y;
M252Y/N325G/M428F/N434Y;
M252Y/D376A/M428F/N434Y;
M252Y/E380A/M428F/N434Y;
M252Y/E382A/M428F/N434Y;
M252Y/E380A/E382A/M428F/N434Y;
S239K/M252Y/V308P/E382A/N434Y;
M252Y/D270F/V308P/E382A/N434Y;
S239K/M252Y/V308P/M428Y/N434Y;
M252Y/T307Q/V308P/E382A/N434Y;
M252Y/V308P/Q311H/E382A/N434Y;
S239K/M252Y/D270F/V308P/N434Y;
M252Y/V308P/E382A/M428F/N434Y;
M252Y/V308P/E382A/M428L/N434Y;
M252Y/V308P/E382A/M428Y/N434Y;
M252Y/V308P/M428Y/N434Y;
M252Y/V308P/M428F/N434Y;
S239K/M252Y/V308P/E382A/M428Y/N434Y;
M252Y/V308P/E380A/E382A/M428F/N434Y;
M252Y/T256E/E382A/N434Y;
M252Y/T256E/M428Y/N434Y;
M252Y/T256E/E382A/M428Y/N434Y;
S239K/M252Y/V308P/E382A/M428I/N434Y;
M252Y/V308P/E380A/E382A/M428I/N434Y;
S239K/M252Y/V308P/M428F/N434Y;
S239K/M252Y/E380A/E382A/M428F/N434Y;
M252Y/Q311A/M428Y/N434Y;
M252Y/D312A/M428Y/N434Y;
M252Y/Q311A/D312A/M428Y/N434Y;
S239K/M252Y/V308P/Q311A/M428Y/N434Y;
S239K/M252Y/V308P/D312A/M428Y/N434Y;
S239K/M252Y/V308P/Q311A/D312A/M428Y/N434Y;
M252Y/T256E/T307P/N434Y;
M252Y/T307P/M428Y/N434Y;
M252W/V308P/M428Y/N434Y;
M252Y/T256E/V308P/E382A/N434Y;
M252W/V308P/E382A/N434Y;
S239K/M252W/V308P/E382A/N434Y;
S239K/M252W/V308P/E382A/M428Y/N434Y;
S239K/M252W/V308P/E382A/M428I/N434Y;
S239K/M252W/V308P/M428F/N434Y;
S239K/M252W/E380A/E382A/M428F/N434Y;
S239K/M252Y/T307P/M428Y/N434Y;
M252Y/T256E/Q311A/D312A/M428Y/N434Y;
S239K/M252Y/M428Y/N434Y;
S239K/M252Y/D270F/M428Y/N434Y;
S239K/M252Y/D270F/V308P/M428Y/N434Y;
S239K/M252Y/V308P/M428I/N434Y;
S239K/M252Y/V308P/N325G/E382A/M428I/N434Y;
S239K/M252Y/D270F/V308P/N325G/N434Y;
S239K/M252Y/D270F/T307P/V308P/N434Y;
S239K/M252Y/T256E/D270F/V308P/N434Y;
S239K/M252Y/D270F/T307Q/V308P/N434Y;
S239K/M252Y/D270F/T307Q/V308P/Q311A/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/N434Y;
S239K/M252Y/T256E/D270F/T307Q/Q311A/N434Y;
S239K/M252Y/D270F/V308P/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/N434Y;
S239K/M252Y/D270F/V308P/L309E/N434Y;
S239K/M252Y/D270F/V308P/P387E/N434Y;
S239K/M252Y/D270F/T307Q/L309E/Q311A/N434Y;
S239K/M252Y/D270F/V308P/N325G/M428I/N434Y;
S239K/M252Y/T256E/D270F/V308P/M428I/N434Y;
S239K/M252Y/D270F/T307Q/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/M428I/N434Y;
S239K/M252Y/D270F/V308P/L309E/M428I/N434Y;
S239K/M252Y/D270F/V308P/M428L/N434Y;
S239K/M252Y/V308P/M428L/N434Y;
S239K/M252Y/V308P/L309P/M428L/N434Y;
S239K/M252Y/V308P/L309R/M428L/N434Y;
S239K/M252Y/L309P/M428L/N434Y;
S239K/M252Y/L309R/M428L/N434Y;
S239K/M252Y/T307Q/V308P/M428L/N434Y;
S239K/M252Y/D270F/V308P/L309E/P387E/N434Y;
S239K/M252Y/T256E/D270F/V308P/N325G/M428I/N434Y;
S239K/M252Y/T307Q/N434Y;
P257V/T307Q/M428I/N434Y;
P257V/T307Q/M428V/N434Y;
P257V/T307Q/N325G/M428L/N434Y;
P257V/T307Q/Q311A/M428L/N434Y;
P257V/T307Q/Q311A/N325G/M428L/N434Y;
P257V/V305A/T307Q/M428L/N434Y;
S239K/M252Y/E258H/D270F/T307Q/V308P/Q311A/N434Y;
S239K/M252Y/D270F/V308P/N325G/E382A/M428I/N434Y;
M252Y/P257V/T307Q/M428I/N434Y;
S239K/M252W/V308P/M428Y/N434Y;
S239K/M252W/V308P/Q311A/M428Y/N434Y;
S239K/M252W/T256E/V308P/M428Y/N434Y;
S239K/M252W/N286E/V308P/M428Y/N434Y;
S239K/M252Y/T256E/D270F/N286E/V308P/N434Y;
S239K/M252W/T307P/M428Y/N434Y;
S239K/M252W/T256E/V308P/Q311A/M428Y/N434Y;
S239K/M252W/T256E/N286E/V308P/M428Y/N434Y;
P257V/T307A/Q311A/M428L/N434Y;
P257V/V305A/T307A/M428L/N434Y;
M252Y/V308P/L309E/N434Y;
M252Y/V308P/L309D/N434Y;
M252Y/V308P/L309A/N434Y;
M252W/V308P/L309E/M428Y/N434Y;
M252W/V308P/L309D/M428Y/N434Y;
M252W/V308P/L309A/M428Y/N434Y;
S239K/M252Y/N286E/V308P/M428I/N434Y;
S239K/M252Y/N286E/V308P/N434Y;
S239K/M252Y/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/T256E/V308P/N434Y;
S239K/M252Y/D270F/N286E/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/P387E/M428I/N434Y;
S239K/M252Y/D270F/N286E/T307Q/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/T307Q/V308P/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/P387E/M428I/N434Y;
P257V/T307Q/M428IVN434W;
P257V/T307A/M428L/N434W;
P257A/T307Q/L309P/M428L/N434Y;
P257V/T307Q/L309P/M428L/N434Y;
P257A/T307Q/L309R/M428L/N434Y;
P257V/T307Q/L309R/M428L/N434Y;
P257V/N286E/M428L/N434Y;
P257V/P387E/M428L/N434Y;
P257V/T307H/M428IVN434Y;
P257V/T307N/M428L/N434Y;
P257V/T307G/M428L/N434Y;
P257V/T307P/M428L/N434Y;
P257V/T307S/M428L/N434Y;
P257V/N286E/T307A/M428L/N434Y;
P257V/T307A/P387E/M428L/N434Y;
S239K/M252Y/T307P/N325G/M428Y/N434Y;
M252Y/T307A/Q311H/K360H/N434Y;
M252Y/T307A/Q311H/P387E/N434Y;
M252Y/T307A/Q311H/M428A/N434Y;
M252Y/T307A/Q311H/E382A/N434Y;
M252Y/T307W/Q311H/N434Y;
S239K/P257A/V308P/M428L/N434Y;
P257A/V308P/L309E/M428L/N434Y;
P257V/T307Q/N434Y;
M252Y/P257V/T307Q/M428V/N434Y;
M252Y/P257V/T307Q/M428L/N434Y;
M252Y/P257V/T307Q/N434Y;
M252Y/P257V/T307Q/M428Y/N434Y;
M252Y/P257V/T307Q/M428F/N434Y;
P257V/T307Q/Q311A/N325G/M428V/N434Y;
P257V/T307Q/Q311A/N325G/N434Y;
P257V/T307Q/Q311A/N325G/M428Y/N434Y;
P257V/T307Q/Q311A/N325G/M428F/N434Y;
P257A/T307Q/M428V/N434Y;
P257A/T307Q/N434Y;
P257A/T307Q/M428Y/N434Y;
P257A/T307Q/M428F/N434Y;
P257V/N286E/T307Q/Q311A/N325G/M428L/N434Y;
P257A/Q311A/M428L/N434Y;
P257A/Q311H/M428L/N434Y;
P257A/T307Q/Q311A/M428L/N434Y;
P257A/T307A/Q311A/M428L/N434Y;
P257A/T307Q/Q311H/M428L/N434Y;
P257A/T307A/Q311H/M428L/N434Y;
P257A/N286E/T307Q/M428L/N434Y;
P257A/N286E/T307A/M428L/N434Y;
S239K/M252Y/D270F/T307P/N325G/M428Y/N434Y;
S239K/M252Y/T307P/L309E/N325G/M428Y/N434Y;
P257S/T307A/M428L/N434Y;
P257M/T307A/M428L/N434Y;
P257N/T307A/M428L/N434Y;
P257I/T307A/M428L/N434Y;
P257V/T307Q/M428Y/N434Y;
P257V/T307Q/M428F/N434Y;
S239K/P257V/V308P/M428IVN434Y;
P257V/T307Q/V308P/N325G/M428L/N434Y;
P257V/T307Q/V308P/Q311A/N325G/M428L/N434Y;
P257V/T307A/V308P/N325G/M428L/N434Y;
P257A/V308P/N325G/M428L/N434Y;
P257A/T307A/V308P/M428L/N434Y;
P257A/T307Q/V308P/M428L/N434Y;
P257V/N286E/T307Q/N325G/M428L/N434Y;
T256E/P257V/N434Y;
T256E/P257T/N434Y;
S239K/P257T/V308P/M428L/N434Y;
P257T/V308P/N325G/M428L/N434Y;
T256E/P257T/V308P/N325G/M428L/N434Y;
P257T/V308P/N325G/E382A/M428L/N434Y;
P257T/V308P/N325G/P387E/M428L/N434Y;
P257T/V308P/L309P/N325G/M428L/N434Y;
P257T/V308P/L309R/N325G/M428L/N434Y;
T256E/P257V/T307Q/M428IVN434Y;
P257V/T307Q/E382A/M428L/N434Y;
P257V/T307Q/P387E/M428L/N434Y;
P257L/V308P/N434Y;
P257T/T307Q/N434Y;
P257V/T307Q/N325G/N434Y;
P257V/T307Q/Q311A/N434Y;
P257V/V305A/T307Q/N434Y;
P257V/N286E/T307A/N434Y;
P257V/T307Q/L309R/Q311H/M428L/N434Y;
P257V/V308P/N325G/M428L/N434Y;
S239K/P257V/V308P/Q311H/M428L/N434Y;
P257V/V305A/T307A/N325G/M428L/N434Y;
S239K/D270F/T307P/N325G/M428Y/N434Y;
P257T/T307A/M428V/N434Y;
P257A/M428V/N434Y;
P257A/T307A/M428V/N434Y;
P257S/T307Q/M428L/N434Y;
P257V/N297A/T307Q/M428L/N434Y;
P257V/N286A/T307Q/M428L/N434Y;
P257V/T307Q/N315A/M428L/N434Y;
P257V/T307Q/N384A/M428L/N434Y;
P257V/T307Q/N389A/M428L/N434Y;
P257V/N434Y;
P257T/N434Y;
P257V/N286E/N434Y;
P257T/N286E/N434Y;
P257V/N286E/T307Q/N434Y;
P257V/N286E/T307Q/M428Y/N434Y;
P257V/V305A/T307Q/M428Y/N434Y;
P257T/T307Q/M428Y/N434Y;
P257V/T307Q/V308P/N325G/M428Y/N434Y;
P257T/T307Q/M428F/N434Y;
P257A/N286E/T307Q/M428F/N434Y;
P257A/N286E/T307Q/M428Y/N434Y;
T250A/P257V/T307Q/M428L/N434Y;
T250F/P257V/T307Q/M428L/N434Y;
T250I/P257V/T307Q/M428L/N434Y;
T250M/P257V/T307Q/M428L/N434Y;
T250S/P257V/T307Q/M428L/N434Y;
T250V/P257V/T307Q/M428L/N434Y;
T250W/P257V/T307Q/M428L/N434Y;
T250Y/P257V/T307Q/M428L/N434Y;
M252Y/Q311A/N434Y;
S239K/M252Y/S254T/V308P/N434Y;
M252Y/T307Q/Q311A;
M252Y/S254T/N286E/N434Y;
M252Y/S254T/V308P/N434Y;
M252Y/S254T/T307A/N434Y;
M252Y/S254T/T307Q/N434Y;
M252Y/S254T/Q311A/N434Y;
M252Y/S254T/Q311H/N434Y;
M252Y/S254T/T307A/Q311A/N434Y;
M252Y/S254T/T307Q/Q311A/N434Y;
M252Y/S254T/M428I/N434Y;
M252Y/T256E/T307A/Q311H/N434Y;
M252Y/T256Q/T307A/Q311H/N434Y;
M252Y/S254T/T307A/Q311H/N434Y;
M252Y/N286E/T307A/Q311H/N434Y;
M252Y/T307A/Q311H/M428FN434Y;
M252Y/T256E/T307Q/Q311H;
M252Y/N286E/T307A/Q311A;
M252Y/N286E/T307Q/Q311A;
M252Y/N286E/T307Q/Q311H;
M252Y/T256E/N286E/N434Y;
S239K/M252Y/V308P;
S239K/M252Y/V308P/E382A;
S239K/M252Y/T256E/V308P;
S239K/M252Y/N286E/V308P;
S239K/M252Y/N286E/V308P/M428I;
M252Y/N286E/M428L/N434Y;
M252Y/S254T/E382A/N434Y;
S239K/M252Y/S254T/V308P/M428I/N434Y;
S239K/M252Y/M428I/N434Y;
M252Y/D312A/E382A/M428Y/N434Y;
M252Y/E382A/P387E/N434Y;
M252Y/D312A/P387E/N434Y или
M252Y/P387E/M428Y/N434Y;
(b) отбора антитела, которое связывается с растворимым антигеном и обладает более высокой антигенсвязывающей активностью при рН 7.4, чем при рН 5.8, где отношение антигенсвязывающей активности при рН 5.8 и рН 7.4 равно по меньшей мере 2 при величине KD при рН 5.8 / KD при рН 7.4, где указанное антитело содержит антигенсвязывающий домен, который получают из библиотеки антигенсвязывающих доменов, в которые ввели по меньшей мере одну замену гистидином или инсерцию в антигенсвязывающий домен тяжелой цепи в положение, выбранное из группы, состоящей из Н27, Н31, Н32, Н33 Н35, Н50, Н58, Н59, Н61, Н62, Н63, Н64, Н65, Н99, H100b и Н102 в соответствии с нумерацией по Kabat, и/или антигенсвязывающий домен легкой цепи в положение, выбранное из группы, состоящей из L24, L27, L28, L32, L53, L54, L56, L90, L92 и L94 в соответствии с нумерацией по Kabat;
(c) получения гена, кодирующего антитело, в котором человеческий FcRn-связывающий домен и антигенсвязывающий домен, отобранные на стадиях (а) и (b), сцеплены; и
(d) получения антитела с использованием гена, полученного на стадии (с).
10. Способ скрининга антитела, включающий стадии:
(а) отбора антитела, которое обладает активностью связывания человеческого FcRn при рН 5.8 и рН 7.0 и более высокой активностью связывания человеческого FcRn при рН 7.0, чем до изменения по меньшей мере одной аминокислоты в человеческом FcRn-связывающем домене антитела IgG1, IgG2, IgG3 или IgG4, которое содержит Fc домен IgG1 человека, IgG2 человека, IgG3 человека или IgG4 человека, обладающего активностью связывания человеческого FcRn при рН 5.8, где человеческий FcRn-связывающий домен содержит одну из следующих аминокислотных замен в положениях, соответствующих нумерации EU в Fc-домене:
N434W;
N434Y;
M252Y/S254T/T256E/N434Y;
M252Y/S254T/T256E/N434W;
M252Y/N434Y;
M252Y/N434W;
M252W/N434Y;
M252W/N434W;
P257L/N434Y;
V308F/N434Y;
M252Y/V308P/N434Y;
M428L/N434S;
M252Y/S254T/T256E/V308P/N434W;
I332V;
I332V/N434Y;
G237M/V308F;
S254T/N434W;
S254T/N434Y;
T256E/N434W;
T256E/N434Y;
S254T/T256E/N434W;
S254T/T256E/N434Y;
M252Y/S254T/N434W;
M252Y/S254T/N434Y;
M252Y/T256E/N434W;
M252Y/T256E/N434Y;
M252Y/S254T/T256E/N434A;
M252Y/N434A;
M252W/N434A;
M252Y/T256Q/N434W;
M252Y/T256Q/N434Y;
M252F/T256D/N434W;
M252F/T256D/N434Y;
H433K/N434F/Y436H;
I332V/N434W;
V308P/N434W;
I332V/M428L/N434Y;
G385D/Q386P/N389S/N434W;
G385D/Q386P/N389S/N434Y;
G385H/N434W;
G385H/N434Y;
N434F;
M252Y/S254T/T256E/N434F;
M252Y/S254T/T256E/N434H;
M252Y/N434F;
M252Y/N434H;
M428L/N434W;
M428L/N434Y;
M252Y/S254T/T256E/M428L/N434W;
M252Y/S254T/T256E/M428L/N434Y;
M252Y/M428L/N434W;
M252Y/M428L/N434Y;
M252Y/M428L/N434A;
M252Y/S254T/T256E/M428L/N434A;
T256E/M428L/N434Y;
S254T/M428L/N434W;
M252Y/T256Q/N434A;
M252Y/T256E/N434A;
T256Q/N434W;
T256Q/N434Y;
M252W/T256Q/N434W;
M252W/T256Q/N434Y;
S254T/T256Q/N434W;
M252Y/S254T/T256Q/N434W;
M252Y/S254T/T256Q/N434Y;
M252Y/T256E/V308P/N434W;
M252Y/V308P/M428L/N434Y;
M252Y/S254T/T256E/V308P/M428L/N434W;
M252W/M428L/N434W;
P257L/M428L/N434Y;
M252Y/S254T/T256E/M428L/N434F;
M252Y/T256E/N434H;
M252W/T256Q/P257L/N434Y;
P238A/M252Y/V308P/N434Y;
M252Y/D265A/V308P/N434Y;
M252Y/T307A/V308P/N434Y;
M252Y/V303A/V308P/N434Y;
M252Y/V308P/D376A/N434Y;
M252Y/V305A/V308P/N434Y;
M252Y/V308P/Q311A/N434Y;
M252Y/V308P/K317A/N434Y;
M252Y/V308P/E380A/N434Y;
M252Y/V308P/E382A/N434Y;
M252Y/V308P/S424A/N434Y;
P257L/V308P/M428L/N434Y;
M252Y/T256E/V308P/M428L/N434W;
M252Y/T256E/M428L/N434W;
M252Y/T256E/V308P/N434Y;
M252Y/T256E/M428L/N434Y;
M252Y/S254T/T256E/V308P/M428L/N434Y;
P257L/M428L/N434W;
P257A/M428L/N434Y;
P257G/M428L/N434Y;
P257I/M428L/N434Y;
P257M/M428L/N434Y;
P257N/M428L/N434Y;
P257S/M428L/N434Y;
P257T/M428L/N434Y;
P257V/M428L/N434Y;
M252W/V308P/N434W;
S239K/M252Y/V308P/N434Y;
M252Y/S298G/V308P/N434Y;
M252Y/D270F/V308P/N434Y;
M252Y/V308A/N434Y;
M252Y/V308F/N434Y;
M252Y/V308I/N434Y;
M252Y/V308L/N434Y;
M252Y/V308M/N434Y;
M252Y/V308Q/N434Y;
M252Y/V308T/N434Y;
P257A/V308P/M428L/N434Y;
P257T/V308P/M428L/N434Y;
P257V/V308P/M428L/N434Y;
M252W/M428I/N434Y;
M252W/M428Y/N434Y;
M252W/M428F/N434Y;
P238A/M252W/N434Y;
M252W/D265A/N434Y;
M252W/T307Q/N434Y;
M252W/V303A/N434Y;
M252W/D376A/N434Y;
M252W/V305A/N434Y;
M252W/Q311A/N434Y;
M252W/D312A/N434Y;
M252W/K317A/N434Y;
M252W/E380A/N434Y;
M252W/E382A/N434Y;
M252W/S424A/N434Y;
S239K/M252W/N434Y;
M252W/S298G/N434Y;
M252W/D270F/N434Y;
M252W/N325G/N434Y;
P257A/M428L/N434W;
P257T/M428L/N434W;
P257V/M428L/N434W;
M252W/I332V/N434Y;
P257I/Q311I;
M252Y/T307A/N434Y;
M252Y/T307Q/N434Y;
P257L/T307A/M428L/N434Y;
P257A/T307A/M428IVN434Y;
P257T/T307A/M428L/N434Y;
P257V/T307A/M428L/N434Y;
P257L/T307Q/M428IVN434Y;
P257A/T307Q/M428L/N434Y;
P257T/T307Q/M428L/N434Y;
P257V/T307Q/M428IVN434Y;
M252Y/T307D/N434Y;
M252Y/T307F/N434Y;
M252Y/T307G/N434Y;
M252Y/T307H/N434Y;
M252Y/T307I/N434Y;
M252Y/T307K/N434Y;
M252Y/T307L/N434Y;
M252Y/T307M/N434Y;
M252Y/T307N/N434Y;
M252Y/T307P/N434Y;
M252Y/T307R/N434Y;
M252Y/T307S/N434Y;
M252Y/T307V/N434Y;
M252Y/T307W/N434Y;
M252Y/T307Y/N434Y;
M252Y/K334L/N434Y;
M252Y/G385H/N434Y;
M252Y/T289H/N434Y;
M252Y/Q311H/N434Y;
M252Y/D312H/N434Y;
M252Y/N315H/N434Y;
M252Y/K360H/N434Y;
M252Y/L314R/N434Y;
M252Y/L314K/N434Y;
M252Y/N286E/N434Y;
M252Y/L309E/N434Y;
M252Y/R255E/N434Y;
M252Y/P387E/N434Y;
K248I/M428L/N434Y;
M252Y/M428A/N434Y;
M252Y/M428D/N434Y;
M252Y/M428F/N434Y;
M252Y/M428G/N434Y;
M252Y/M428H/N434Y;
M252Y/M428I/N434Y;
M252Y/M428K/N434Y;
M252Y/M428N/N434Y;
M252Y/M428P/N434Y;
M252Y/M428Q/N434Y;
M252Y/M428S/N434Y;
M252Y/M428T/N434Y;
M252Y/M428V/N434Y;
M252Y/M428W/N434Y;
M252Y/M428Y/N434Y;
M252W/T307Q/M428Y/N434Y;
M252W/Q311A/M428Y/N434Y;
M252W/T307Q/Q311A/M428Y/N434Y;
M252Y/T307A/M428Y/N434Y;
M252Y/T307Q/M428Y/N434Y;
M252Y/D270F/N434Y;
M252Y/T307A/Q311A/N434Y;
M252Y/T307Q/Q311A/N434Y;
M252Y/T307A/Q311H/N434Y;
M252Y/T307Q/Q311H/N434Y;
M252Y/E382A/N434Y;
M252Y/E382A/M428Y/N434Y;
M252Y/T307A/E382A/M428Y/N434Y;
M252Y/T307Q/E382A/M428Y/N434Y;
P238A/M252Y/M428F/N434Y;
M252Y/V305A/M428F/N434Y;
M252Y/N325G/M428F/N434Y;
M252Y/D376A/M428F/N434Y;
M252Y/E380A/M428F/N434Y;
M252Y/E382A/M428F/N434Y;
M252Y/E380A/E382A/M428F/N434Y;
S239K/M252Y/V308P/E382A/N434Y;
M252Y/D270F/V308P/E382A/N434Y;
S239K/M252Y/V308P/M428Y/N434Y;
M252Y/T307Q/V308P/E382A/N434Y;
M252Y/V308P/Q311H/E382A/N434Y;
S239K/M252Y/D270F/V308P/N434Y;
M252Y/V308P/E382A/M428F/N434Y;
M252Y/V308P/E382A/M428L/N434Y;
M252Y/V308P/E382A/M428Y/N434Y;
M252Y/V308P/M428Y/N434Y;
M252Y/V308P/M428F/N434Y;
S239K/M252Y/V308P/E382A/M428Y/N434Y;
M252Y/V308P/E380A/E382A/M428F/N434Y;
M252Y/T256E/E382A/N434Y;
M252Y/T256E/M428Y/N434Y;
M252Y/T256E/E382A/M428Y/N434Y;
S239K/M252Y/V308P/E382A/M428I/N434Y;
M252Y/V308P/E380A/E382A/M428I/N434Y;
S239K/M252Y/V308P/M428F/N434Y;
S239K/M252Y/E380A/E382A/M428F/N434Y;
M252Y/Q311A/M428Y/N434Y;
M252Y/D312A/M428Y/N434Y;
M252Y/Q311A/D312A/M428Y/N434Y;
S239K/M252Y/V308P/Q311A/M428Y/N434Y;
S239K/M252Y/V308P/D312A/M428Y/N434Y;
S239K/M252Y/V308P/Q311A/D312A/M428Y/N434Y;
M252Y/T256E/T307P/N434Y;
M252Y/T307P/M428Y/N434Y;
M252W/V308P/M428Y/N434Y;
M252Y/T256E/V308P/E382A/N434Y;
M252W/V308P/E382A/N434Y;
S239K/M252W/V308P/E382A/N434Y;
S239K/M252W/V308P/E382A/M428Y/N434Y;
S239K/M252W/V308P/E382A/M428I/N434Y;
S239K/M252W/V308P/M428F/N434Y;
S239K/M252W/E380A/E382A/M428F/N434Y;
S239K/M252Y/T307P/M428Y/N434Y;
M252Y/T256E/Q311A/D312A/M428Y/N434Y;
S239K/M252Y/M428Y/N434Y;
S239K/M252Y/D270F/M428Y/N434Y;
S239K/M252Y/D270F/V308P/M428Y/N434Y;
S239K/M252Y/V308P/M428I/N434Y;
S239K/M252Y/V308P/N325G/E382A/M428I/N434Y;
S239K/M252Y/D270F/V308P/N325G/N434Y;
S239K/M252Y/D270F/T307P/V308P/N434Y;
S239K/M252Y/T256E/D270F/V308P/N434Y;
S239K/M252Y/D270F/T307Q/V308P/N434Y;
S239K/M252Y/D270F/T307Q/V308P/Q311A/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/N434Y;
S239K/M252Y/T256E/D270F/T307Q/Q311A/N434Y;
S239K/M252Y/D270F/V308P/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/N434Y;
S239K/M252Y/D270F/V308P/L309E/N434Y;
S239K/M252Y/D270F/V308P/P387E/N434Y;
S239K/M252Y/D270F/T307Q/L309E/Q311A/N434Y;
S239K/M252Y/D270F/V308P/N325G/M428I/N434Y;
S239K/M252Y/T256E/D270F/V308P/M428I/N434Y;
S239K/M252Y/D270F/T307Q/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/M428I/N434Y;
S239K/M252Y/D270F/V308P/L309E/M428I/N434Y;
S239K/M252Y/D270F/V308P/M428L/N434Y;
S239K/M252Y/V308P/M428L/N434Y;
S239K/M252Y/V308P/L309P/M428L/N434Y;
S239K/M252Y/V308P/L309R/M428L/N434Y;
S239K/M252Y/L309P/M428L/N434Y;
S239K/M252Y/L309R/M428L/N434Y;
S239K/M252Y/T307Q/V308P/M428L/N434Y;
S239K/M252Y/D270F/V308P/L309E/P387E/N434Y;
S239K/M252Y/T256E/D270F/V308P/N325G/M428I/N434Y;
S239K/M252Y/T307Q/N434Y;
P257V/T307Q/M428I/N434Y;
P257V/T307Q/M428V/N434Y;
P257V/T307Q/N325G/M428L/N434Y;
P257V/T307Q/Q311A/M428L/N434Y;
P257V/T307Q/Q311A/N325G/M428L/N434Y;
P257V/V305A/T307Q/M428L/N434Y;
S239K/M252Y/E258H/D270F/T307Q/V308P/Q311A/N434Y;
S239K/M252Y/D270F/V308P/N325G/E382A/M428I/N434Y;
M252Y/P257V/T307Q/M428I/N434Y;
S239K/M252W/V308P/M428Y/N434Y;
S239K/M252W/V308P/Q311A/M428Y/N434Y;
S239K/M252W/T256E/V308P/M428Y/N434Y;
S239K/M252W/N286E/V308P/M428Y/N434Y;
S239K/M252Y/T256E/D270F/N286E/V308P/N434Y;
S239K/M252W/T307P/M428Y/N434Y;
S239K/M252W/T256E/V308P/Q311A/M428Y/N434Y;
S239K/M252W/T256E/N286E/V308P/M428Y/N434Y;
P257V/T307A/Q311A/M428L/N434Y;
P257V/V305A/T307A/M428L/N434Y;
M252Y/V308P/L309E/N434Y;
M252Y/V308P/L309D/N434Y;
M252Y/V308P/L309A/N434Y;
M252W/V308P/L309E/M428Y/N434Y;
M252W/V308P/L309D/M428Y/N434Y;
M252W/V308P/L309A/M428Y/N434Y;
S239K/M252Y/N286E/V308P/M428I/N434Y;
S239K/M252Y/N286E/V308P/N434Y;
S239K/M252Y/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/T256E/V308P/N434Y;
S239K/M252Y/D270F/N286E/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/P387E/M428I/N434Y;
S239K/M252Y/D270F/N286E/T307Q/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/T307Q/V308P/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/P387E/M428I/N434Y;
P257V/T307Q/M428L/N434W;
P257V/T307A/M428L/N434W;
P257A/T307Q/L309P/M428L/N434Y;
P257V/T307Q/L309P/M428L/N434Y;
P257A/T307Q/L309R/M428L/N434Y;
P257V/T307Q/L309R/M428L/N434Y;
P257V/N286E/M428L/N434Y;
P257V/P387E/M428L/N434Y;
P257V/T307H/M428L/N434Y;
P257V/T307N/M428L/N434Y;
P257V/T307G/M428IVN434Y;
P257V/T307P/M428L/N434Y;
P257V/T307S/M428L/N434Y;
P257V/N286E/T307A/M428L/N434Y;
P257V/T307A/P387E/M428L/N434Y;
S239K/M252Y/T307P/N325G/M428Y/N434Y;
M252Y/T307A/Q311H/K360H/N434Y;
M252Y/T307A/Q311H/P387E/N434Y;
M252Y/T307A/Q311H/M428A/N434Y;
M252Y/T307A/Q311H/E382A/N434Y;
M252Y/T307W/Q311H/N434Y;
S239K/P257A/V308P/M428L/N434Y;
P257A/V308P/L309E/M428L/N434Y;
P257V/T307Q/N434Y;
M252Y/P257V/T307Q/M428V/N434Y;
M252Y/P257V/T307Q/M428L/N434Y;
M252Y/P257V/T307Q/N434Y;
M252Y/P257V/T307Q/M428Y/N434Y;
M252Y/P257V/T307Q/M428F/N434Y;
P257V/T307Q/Q311A/N325G/M428V/N434Y;
P257V/T307Q/Q311A/N325G/N434Y;
P257V/T307Q/Q311A/N325G/M428Y/N434Y;
P257V/T307Q/Q311A/N325G/M428F/N434Y;
P257A/T307Q/M428V/N434Y;
P257A/T307Q/N434Y;
P257A/T307Q/M428Y/N434Y;
P257A/T307Q/M428F/N434Y;
P257V/N286E/T307Q/Q311A/N325G/M428L/N434Y;
P257A/Q311A/M428L/N434Y;
P257A/Q311H/M428L/N434Y;
P257A/T307Q/Q311A/M428L/N434Y;
P257A/T307A/Q311A/M428L/N434Y;
P257A/T307Q/Q311H/M428L/N434Y;
P257A/T307A/Q311H/M428L/N434Y;
P257A/N286E/T307Q/M428L/N434Y;
P257A/N286E/T307A/M428L/N434Y;
S239K/M252Y/D270F/T307P/N325G/M428Y/N434Y;
S239K/M252Y/T307P/L309E/N325G/M428Y/N434Y;
P257S/T307A/M428L/N434Y;
P257M/T307A/M428L/N434Y;
P257N/T307A/M428IVN434Y;
P257I/T307A/M428L/N434Y;
P257V/T307Q/M428Y/N434Y;
P257V/T307Q/M428F/N434Y;
S239K/P257V/V308P/M428IVN434Y;
P257V/T307Q/V308P/N325G/M428L/N434Y;
P257V/T307Q/V308P/Q311A/N325G/M428L/N434Y;
P257V/T307A/V308P/N325G/M428L/N434Y;
P257A/V308P/N325G/M428L/N434Y;
P257A/T307A/V308P/M428L/N434Y;
P257A/T307Q/V308P/M428L/N434Y;
P257V/N286E/T307Q/N325G/M428L/N434Y;
T256E/P257V/N434Y;
T256E/P257T/N434Y;
S239K/P257T/V308P/M428L/N434Y;
P257T/V308P/N325G/M428L/N434Y;
T256E/P257T/V308P/N325G/M428L/N434Y;
P257T/V308P/N325G/E382A/M428L/N434Y;
P257T/V308P/N325G/P387E/M428IVN434Y;
P257T/V308P/L309P/N325G/M428L/N434Y;
P257T/V308P/L309R/N325G/M428L/N434Y;
T256E/P257V/T307Q/M428L/N434Y;
P257V/T307Q/E382A/M428L/N434Y;
P257V/T307Q/P387E/M428IVN434Y;
P257L/V308P/N434Y;
P257T/T307Q/N434Y;
P257V/T307Q/N325G/N434Y;
P257V/T307Q/Q311A/N434Y;
P257V/V305A/T307Q/N434Y;
P257V/N286E/T307A/N434Y;
P257V/T307Q/L309R/Q311H/M428L/N434Y;
P257V/V308P/N325G/M428L/N434Y;
S239K/P257V/V308P/Q311H/M428L/N434Y;
P257V/V305A/T307A/N325G/M428L/N434Y;
S239K/D270F/T307P/N325G/M428Y/N434Y;
P257T/T307A/M428V/N434Y;
P257A/M428V/N434Y;
P257A/T307A/M428V/N434Y;
P257S/T307Q/M428L/N434Y;
P257V/N297A/T307Q/M428L/N434Y;
P257V/N286A/T307Q/M428L/N434Y;
P257V/T307Q/N315A/M428L/N434Y;
P257V/T307Q/N384A/M428L/N434Y;
P257V/T307Q/N389A/M428L/N434Y;
P257V/N434Y;
P257T/N434Y;
P257V/N286E/N434Y;
P257T/N286E/N434Y;
P257V/N286E/T307Q/N434Y;
P257V/N286E/T307Q/M428Y/N434Y;
P257V/V305A/T307Q/M428Y/N434Y;
P257T/T307Q/M428Y/N434Y;
P257V/T307Q/V308P/N325G/M428Y/N434Y;
P257T/T307Q/M428F/N434Y;
P257A/N286E/T307Q/M428F/N434Y;
P257A/N286E/T307Q/M428Y/N434Y;
T250A/P257V/T307Q/M428L/N434Y;
T250F/P257V/T307Q/M428L/N434Y;
T250I/P257V/T307Q/M428L/N434Y;
T250M/P257V/T307Q/M428L/N434Y;
T250S/P257V/T307Q/M428L/N434Y;
T250V/P257V/T307Q/M428L/N434Y;
T250W/P257V/T307Q/M428L/N434Y;
T250Y/P257V/T307Q/M428L/N434Y;
M252Y/Q311A/N434Y;
S239K/M252Y/S254T/V308P/N434Y;
M252Y/T307Q/Q311A;
M252Y/S254T/N286E/N434Y;
M252Y/S254T/V308P/N434Y;
M252Y/S254T/T307A/N434Y;
M252Y/S254T/T307Q/N434Y;
M252Y/S254T/Q311A/N434Y;
M252Y/S254T/Q311H/N434Y;
M252Y/S254T/T307A/Q311A/N434Y;
M252Y/S254T/T307Q/Q311A/N434Y;
M252Y/S254T/M428I/N434Y;
M252Y/T256E/T307A/Q311H/N434Y;
M252Y/T256Q/T307A/Q311H/N434Y;
M252Y/S254T/T307A/Q311H/N434Y;
M252Y/N286E/T307A/Q311H/N434Y;
M252Y/T307A/Q311H/M428I/N434Y;
M252Y/T256E/T307Q/Q311H;
M252Y/N286E/T307A/Q311A;
M252Y/N286E/T307Q/Q311A;
M252Y/N286E/T307Q/Q311H;
M252Y/T256E/N286E/N434Y;
S239K/M252Y/V308P;
S239K/M252Y/V308P/E382A;
S239K/M252Y/T256E/V308P;
S239K/M252Y/N286E/V308P;
S239K/M252Y/N286E/V308P/M428I;
M252Y/N286E/M428L/N434Y;
M252Y/S254T/E382A/N434Y;
S239K/M252Y/S254T/V308P/M428I/N434Y;
S239K/M252Y/M428I/N434Y;
M252Y/D312A/E382A/M428Y/N434Y;
M252Y/E3 82А/Р387E/N434Y;
M252Y/D312A/P387E/N434Y или
M252Y/P387E/M428Y/N434Y;
(b) замены на гистидин аминокислоты или инсерции гистидина в антигенсвязывающем домене тяжелой цепи в положении, выбранном из группы, состоящей из Н27, Н31, Н32, Н33, Н35, Н50, Н58, Н59, Н61, Н62, Н63, Н64, Н65, Н99, H100b и H102 в соответствии с нумерацией по Rabat, и/или антигенсвязывающем домене легкой цепи в положении, выбранном из группы, состоящей из L24, L27, L28, L32, L53, L54, L56, L90, L92 и L94 в соответствии с нумерацией по Kabat,
и отбора антитела, которое связывается с растворимым антигеном и обладает более высокой антигенсвязывающей активностью при рН 5.8, чем при рН 7.4, где отношение антигенсвязывающей активности при рН 5.8 и рН 7.4 равно по меньшей мере 2 при величине KD при рН 5.8 / KD при рН 7.4; и
(c) отбора антитела, в котором человеческий FcRn-связывающий домен и антигенсвязывающий домен, отобранные на стадиях (а) и (b), сцеплены.
11. Способ скрининга антитела, включающий стадии:
(а) отбора антитела, которое содержит человеческий FcRn-связывающий домен, обладающий активностью связывания человеческого FcRn при рН 5.8 и рН 7.0 и более высокой активностью связывания человеческого FcRn при рН 7.0, чем до изменения по меньшей мере одной аминокислоты в человеческом FcRn-связывающем домене антитела IgG1, IgG2, IgG3 или IgG4, включающем Fc домен IgG1 человека, IgG2 человека, IgG3 человека или IgG4 человека и обладающем активностью связывания человеческого FcRn при рН 5.8, где указанный человеческий FcRn-связывающий домен после вышеуказанного изменения содержит одну из следующих аминокислотных замен в положениях, соответствующих нумерации EU в Fc-домене:
N434W;
N434Y;
M252Y/S254T/T256E/N434Y;
M252Y/S254T/T256E/N434W;
M252Y/N434Y;
M252Y/N434W;
M252W/N434Y;
M252W/N434W;
P257L/N434Y;
V308F/N434Y;
M252Y/V308P/N434Y;
M428L/N434S;
M252Y/S254T/T256E/V308P/N434W;
I332V;
I332V/N434Y;
G237M/V308F;
S254T/N434W;
S254T/N434Y;
T256E/N434W;
T256E/N434Y;
S254T/T256E/N434W;
S254T/T256E/N434Y;
M252Y/S254T/N434W;
M252Y/S254T/N434Y;
M252Y/T256E/N434W;
M252Y/T256E/N434Y;
M252Y/S254T/T256E/N434A;
M252Y/N434A;
M252W/N434A;
M252Y/T256Q/N434W;
M252Y/T256Q/N434Y;
M252F/T256D/N434W;
M252F/T256D/N434Y;
H433K/N434F/Y436H;
I332V/N434W;
V308P/N434W;
I332V/M428L/N434Y;
G385D/Q386P/N389S/N434W;
G385D/Q386P/N389S/N434Y;
G385H/N434W;
G385H/N434Y;
N434F;
M252Y/S254T/T256E/N434F;
M252Y/S254T/T256E/N434H;
M252Y/N434F;
M252Y/N434H;
M428L/N434W;
M428L/N434Y;
M252Y/S254T/T256E/M428L/N434W;
M252Y/S254T/T256E/M428L/N434Y;
M252Y/M428L/N434W;
M252Y/M428L/N434Y;
M252Y/M428L/N434A;
M252Y/S254T/T256E/M428L/N434A;
T256E/M428L/N434Y;
S254T/M428L/N434W;
M252Y/T256Q/N434A;
M252Y/T256E/N434A;
T256Q/N434W;
T256Q/N434Y;
M252W/T256Q/N434W;
M252W/T256Q/N434Y;
S254T/T256Q/N434W;
M252Y/S254T/T256Q/N434W;
M252Y/S254T/T256Q/N434Y;
M252Y/T256E/V308P/N434W;
M252Y/V308P/M428L/N434Y;
M252Y/S254T/T256E/V308P/M428L/N434W;
M252W/M428L/N434W;
P257L/M428L/N434Y;
M252Y/S254T/T256E/M428L/N434F;
M252Y/T256E/N434H;
M252W/T256Q/P257L/N434Y;
P238A/M252Y/V308P/N434Y;
M252Y/D265A/V308P/N434Y;
M252Y/T307A/V308P/N434Y;
M252Y/V303A/V308P/N434Y;
M252Y/V308P/D376A/N434Y;
M252Y/V305A/V308P/N434Y;
M252Y/V308P/Q311A/N434Y;
M252Y/V308P/K317A/N434Y;
M252Y/V308P/E380A/N434Y;
M252Y/V308P/E382A/N434Y;
M252Y/V308P/S424A/N434Y;
P257L/V308P/M428L/N434Y;
M252Y/T256E/V308P/M428L/N434W;
M252Y/T256E/M428L/N434W;
M252Y/T256E/V308P/N434Y;
M252Y/T256E/M428L/N434Y;
M252Y/S254T/T256E/V308P/M428L/N434Y;
P257L/M428L/N434W;
P257A/M428L/N434Y;
P257G/M428L/N434Y;
P257I/M428L/N434Y;
P257M/M428L/N434Y;
P257N/M428L/N434Y;
P257S/M428L/N434Y;
P257T/M428L/N434Y;
P257V/M428L/N434Y;
M252W/V308P/N434W;
S239K/M252Y/V308P/N434Y;
M252Y/S298G/V308P/N434Y;
M252Y/D270F/V308P/N434Y;
M252Y/V308A/N434Y;
M252Y/V308F/N434Y;
M252Y/V308I/N434Y;
M252Y/V308L/N434Y;
M252Y/V308M/N434Y;
M252Y/V308Q/N434Y;
M252Y/V308T/N434Y;
P257A/V308P/M428L/N434Y;
P257T/V308P/M428L/N434Y;
P257V/V308P/M428IVN434Y;
M252W/M428I/N434Y;
M252W/M428Y/N434Y;
M252W/M428F/N434Y;
P238A/M252W/N434Y;
M252W/D265A/N434Y;
M252W/T307Q/N434Y;
M252W/V303A/N434Y;
M252W/D376A/N434Y;
M252W/V305A/N434Y;
M252W/Q311A/N434Y;
M252W/D312A/N434Y;
M252W/K317A/N434Y;
M252W/E380A/N434Y;
M252W/E382A/N434Y;
M252W/S424A/N434Y;
S239K/M252W/N434Y;
M252W/S298G/N434Y;
M252W/D270F/N434Y;
M252W/N325G/N434Y;
P257A/M428L/N434W;
P257T/M428L/N434W;
P257V/M428L/N434W;
M252W/I332V/N434Y;
P257I/Q311I;
M252Y/T307A/N434Y;
M252Y/T307Q/N434Y;
P257L/T307A/M428IVN434Y;
P257A/T307A/M428L/N434Y;
P257T/T307A/M428L/N434Y;
P257V/T307A/M428L/N434Y;
P257L/T307Q/M428L/N434Y;
P257A/T307Q/M428L/N434Y;
P257T/T307Q/M428L/N434Y;
P257V/T307Q/M428L/N434Y;
M252Y/T307D/N434Y;
M252Y/T307F/N434Y;
M252Y/T307G/N434Y;
M252Y/T307H/N434Y;
M252Y/T307I/N434Y;
M252Y/T307K/N434Y;
M252Y/T307L/N434Y;
M252Y/T307M/N434Y;
M252Y/T307N/N434Y;
M252Y/T307P/N434Y;
M252Y/T307R/N434Y;
M252Y/T307S/N434Y;
M252Y/T307V/N434Y;
M252Y/T307W/N434Y;
M252Y/T307Y/N434Y;
M252Y/K334L/N434Y;
M252Y/G385H/N434Y;
M252Y/T289H/N434Y;
M252Y/Q311H/N434Y;
M252Y/D312H/N434Y;
M252Y/N315H/N434Y;
M252Y/K360H/N434Y;
M252Y/L314R/N434Y;
M252Y/L314K/N434Y;
M252Y/N286E/N434Y;
M252Y/L309E/N434Y;
M252Y/R255E/N434Y;
M252Y/P387E/N434Y;
K248I/M428L/N434Y;
M252Y/M428A/N434Y;
M252Y/M428D/N434Y;
M252Y/M428F/N434Y;
M252Y/M428G/N434Y;
M252Y/M428H/N434Y;
M252Y/M428I/N434Y;
M252Y/M428K/N434Y;
M252Y/M428N/N434Y;
M252Y/M428P/N434Y;
M252Y/M428Q/N434Y;
M252Y/M428S/N434Y;
M252Y/M428T/N434Y;
M252Y/M428V/N434Y;
M252Y/M428W/N434Y;
M252Y/M428Y/N434Y;
M252W/T307Q/M428Y/N434Y;
M252W/Q311A/M428Y/N434Y;
M252W/T307Q/Q311A/M428Y/N434Y;
M252Y/T307A/M428Y/N434Y;
M252Y/T307Q/M428Y/N434Y;
M252Y/D270F/N434Y;
M252Y/T307A/Q311A/N434Y;
M252Y/T307Q/Q311A/N434Y;
M252Y/T307A/Q311H/N434Y;
M252Y/T307Q/Q311H/N434Y;
M252Y/E382A/N434Y;
M252Y/E382A/M428Y/N434Y;
M252Y/T307A/E382A/M428Y/N434Y;
M252Y/T307Q/E382A/M428Y/N434Y;
P238A/M252Y/M428F/N434Y;
M252Y/V305A/M428F/N434Y;
M252Y/N325G/M428F/N434Y;
M252Y/D376A/M428F/N434Y;
M252Y/E380A/M428F/N434Y;
M252Y/E382A/M428F/N434Y;
M252Y/E380A/E382A/M428F/N434Y;
S239K/M252Y/V308P/E382A/N434Y;
M252Y/D270F/V308P/E382A/N434Y;
S239K/M252Y/V308P/M428Y/N434Y;
M252Y/T307Q/V308P/E382A/N434Y;
M252Y/V308P/Q311H/E382A/N434Y;
S239K/M252Y/D270F/V308P/N434Y;
M252Y/V308P/E382A/M428F/N434Y;
M252Y/V308P/E382A/M428L/N434Y;
M252Y/V308P/E382A/M428Y/N434Y;
M252Y/V308P/M428Y/N434Y;
M252Y/V308P/M428F/N434Y;
S239K/M252Y/V308P/E382A/M428Y/N434Y;
M252Y/V308P/E380A/E382A/M428F/N434Y;
M252Y/T256E/E382A/N434Y;
M252Y/T256E/M428Y/N434Y;
M252Y/T256E/E382A/M428Y/N434Y;
S239K/M252Y/V308P/E382A/M428I/N434Y;
M252Y/V308P/E380A/E382A/M428I/N434Y;
S239K/M252Y/V308P/M428F/N434Y;
S239K/M252Y/E380A/E382A/M428F/N434Y;
M252Y/Q311A/M428Y/N434Y;
M252Y/D312A/M428Y/N434Y;
M252Y/Q311A/D312A/M428Y/N434Y;
S239K/M252Y/V308P/Q311A/M428Y/N434Y;
S239K/M252Y/V308P/D312A/M428Y/N434Y;
S239K/M252Y/V308P/Q311A/D312A/M428Y/N434Y;
M252Y/T256E/T307P/N434Y;
M252Y/T307P/M428Y/N434Y;
M252W/V308P/M428Y/N434Y;
M252Y/T256E/V308P/E382A/N434Y;
M252W/V308P/E382A/N434Y;
S239K/M252W/V308P/E382A/N434Y;
S239K/M252W/V308P/E382A/M428Y/N434Y;
S239K/M252W/V308P/E382A/M428I/N434Y;
S239K/M252W/V308P/M428F/N434Y;
S239K/M252W/E380A/E382A/M428F/N434Y;
S239K/M252Y/T307P/M428Y/N434Y;
M252Y/T256E/Q311A/D312A/M428Y/N434Y;
S239K/M252Y/M428Y/N434Y;
S239K/M252Y/D270F/M428Y/N434Y;
S239K/M252Y/D270F/V308P/M428Y/N434Y;
S239K/M252Y/V308P/M428I/N434Y;
S239K/M252Y/V308P/N325G/E382A/M428I/N434Y;
S239K/M252Y/D270F/V308P/N325G/N434Y;
S239K/M252Y/D270F/T307P/V308P/N434Y;
S239K/M252Y/T256E/D270F/V308P/N434Y;
S239K/M252Y/D270F/T307Q/V308P/N434Y;
S239K/M252Y/D270F/T307Q/V308P/Q311A/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/N434Y;
S239K/M252Y/T256E/D270F/T307Q/Q311A/N434Y;
S239K/M252Y/D270F/V308P/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/N434Y;
S239K/M252Y/D270F/V308P/L309E/N434Y;
S239K/M252Y/D270F/V308P/P387E/N434Y;
S239K/M252Y/D270F/T307Q/L309E/Q311A/N434Y;
S239K/M252Y/D270F/V308P/N325G/M428I/N434Y;
S239K/M252Y/T256E/D270F/V308P/M428I/N434Y;
S239K/M252Y/D270F/T307Q/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/M428I/N434Y;
S239K/M252Y/D270F/V308P/L309E/M428I/N434Y;
S239K/M252Y/D270F/V308P/M428L/N434Y;
S239K/M252Y/V308P/M428L/N434Y;
S239K/M252Y/V308P/L309P/M428L/N434Y;
S239K/M252Y/V308P/L309R/M428L/N434Y;
S239K/M252Y/L309P/M428L/N434Y;
S239K/M252Y/L309R/M428L/N434Y;
S239K/M252Y/T307Q/V308P/M428L/N434Y;
S239K/M252Y/D270F/V308P/L309E/P387E/N434Y;
S239K/M252Y/T256E/D270F/V308P/N325G/M428I/N434Y;
S239K/M252Y/T307Q/N434Y;
P257V/T307Q/M428I/N434Y;
P257V/T307Q/M428V/N434Y;
P257V/T307Q/N325G/M428L/N434Y;
P257V/T307Q/Q311A/M428L/N434Y;
P257V/T307Q/Q311A/N325G/M428L/N434Y;
P257V/V305A/T307Q/M428L/N434Y;
S239K/M252Y/E258H/D270F/T307Q/V308P/Q311A/N434Y;
S239K/M252Y/D270F/V308P/N325G/E382A/M428I/N434Y;
M252Y/P257V/T307Q/M428I/N434Y;
S239K/M252W/V308P/M428Y/N434Y;
S239K/M252W/V308P/Q311A/M428Y/N434Y;
S239K/M252W/T256E/V308P/M428Y/N434Y;
S239K/M252W/N286E/V308P/M428Y/N434Y;
S239K/M252Y/T256E/D270F/N286E/V308P/N434Y;
S239K/M252W/T307P/M428Y/N434Y;
S239K/M252W/T256E/V308P/Q311A/M428Y/N434Y;
S239K/M252W/T256E/N286E/V308P/M428Y/N434Y;
P257V/T307A/Q311A/M428L/N434Y;
P257V/V305A/T307A/M428L/N434Y;
M252Y/V308P/L309E/N434Y;
M252Y/V308P/L309D/N434Y;
M252Y/V308P/L309A/N434Y;
M252W/V308P/L309E/M428Y/N434Y;
M252W/V308P/L309D/M428Y/N434Y;
M252W/V308P/L309A/M428Y/N434Y;
S239K/M252Y/N286E/V308P/M428I/N434Y;
S239K/M252Y/N286E/V308P/N434Y;
S239K/M252Y/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/T256E/V308P/N434Y;
S239K/M252Y/D270F/N286E/T307Q/Q311A/M428I/N434Y;
S239K/M252Y/D270F/T307Q/Q311A/P387E/M428I/N434Y;
S239K/M252Y/D270F/N286E/T307Q/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/T307Q/V308P/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/Q311A/M428I/N434Y;
S239K/M252Y/D270F/N286E/V308P/P387E/M428I/N434Y;
P257V/T307Q/M428L/N434W;
P257V/T307A/M428L/N434W;
P257A/T307Q/L309P/M428L/N434Y;
P257V/T307Q/L309P/M428L/N434Y;
P257A/T307Q/L309R/M428L/N434Y;
P257V/T307Q/L309R/M428L/N434Y;
P257V/N286E/M428IVN434Y;
P257V/P387E/M428L/N434Y;
P257V/T307H/M428L/N434Y;
P257V/T307N/M428IVN434Y;
P257V/T307G/M428L/N434Y;
P257V/T307P/M428L/N434Y;
P257V/T307S/M428L/N434Y;
P257V/N286E/T307A/M428L/N434Y;
P257V/T307A/P387E/M428IVN434Y;
S239K/M252Y/T307P/N325G/M428Y/N434Y;
M252Y/T307A/Q311H/K360H/N434Y;
M252Y/T307A/Q311H/P387E/N434Y;
M252Y/T307A/Q311H/M428A/N434Y;
M252Y/T307A/Q311H/E382A/N434Y;
M252Y/T307W/Q311H/N434Y;
S239K/P257A/V308P/M428L/N434Y;
P257A/V308P/L309E/M428L/N434Y;
P257V/T307Q/N434Y;
M252Y/P257V/T307Q/M428V/N434Y;
M252Y/P257V/T307Q/M428L/N434Y;
M252Y/P257V/T307Q/N434Y;
M252Y/P257V/T307Q/M428Y/N434Y;
M252Y/P257V/T307Q/M428F/N434Y;
P257V/T307Q/Q311A/N325G/M428V/N434Y;
P257V/T307Q/Q311A/N325G/N434Y;
P257V/T307Q/Q311A/N325G/M428Y/N434Y;
P257V/T307Q/Q311A/N325G/M428F/N434Y;
P257A/T307Q/M428V/N434Y;
P257A/T307Q/N434Y;
P257A/T307Q/M428Y/N434Y;
P257A/T307Q/M428F/N434Y;
P257V/N286E/T307Q/Q311A/N325G/M428L/N434Y;
P257A/Q311A/M428L/N434Y;
P257A/Q311H/M428L/N434Y;
P257A/T307Q/Q311A/M428L/N434Y;
P257A/T307A/Q311A/M428L/N434Y;
P257A/T307Q/Q311H/M428L/N434Y;
P257A/T307A/Q311H/M428L/N434Y;
P257A/N286E/T307Q/M428L/N434Y;
P257A/N286E/T307A/M428L/N434Y;
S239K/M252Y/D270F/T307P/N325G/M428Y/N434Y;
S239K/M252Y/T307P/L309E/N325G/M428Y/N434Y;
P257S/T307A/M428L/N434Y;
P257M/T307A/M428L/N434Y;
P257N/T307A/M428L/N434Y;
P257I/T307A/M428L/N434Y;
P257V/T307Q/M428Y/N434Y;
P257V/T307Q/M428F/N434Y;
S239K/P257V/V308P/M428L/N434Y;
P257V/T307Q/V308P/N325G/M428L/N434Y;
P257V/T307Q/V308P/Q311A/N325G/M428L/N434Y;
P257V/T307A/V308P/N325G/M428L/N434Y;
P257A/V308P/N325G/M428L/N434Y;
P257A/T307A/V308P/M428L/N434Y;
P257A/T307Q/V308P/M428L/N434Y;
P257V/N286E/T307Q/N325G/M428L/N434Y;
T256E/P257V/N434Y;
T256E/P257T/N434Y;
S239K/P257T/V308P/M428L/N434Y;
P257T/V308P/N325G/M428L/N434Y;
T256E/P257T/V308P/N325G/M428L/N434Y;
P257T/V308P/N325G/E382A/M428L/N434Y;
P257T/V308P/N325G/P387E/M428L/N434Y;
P257T/V308P/L309P/N325G/M428L/N434Y;
P257T/V308P/L309R/N325G/M428L/N434Y;
T256E/P257V/T307Q/M428L/N434Y;
P257V/T307Q/E382A/M428L/N434Y;
P257V/T307Q/P387E/M428L/N434Y;
P257L/V308P/N434Y;
P257T/T307Q/N434Y;
P257V/T307Q/N325G/N434Y;
P257V/T307Q/Q311A/N434Y;
P257V/V305A/T307Q/N434Y;
P257V/N286E/T307A/N434Y;
P257V/T307Q/L309R/Q311H/M428L/N434Y;
P257V/V308P/N325G/M428L/N434Y;
S239K/P257V/V308P/Q311H/M428L/N434Y;
P257V/V305A/T307A/N325G/M428L/N434Y;
S239K/D270F/T307P/N325G/M428Y/N434Y;
P257T/T307A/M428V/N434Y;
P257A/M428V/N434Y;
P257A/T307A/M428V/N434Y;
P257S/T307Q/M428L/N434Y;
P257V/N297A/T307Q/M428L/N434Y;
P257V/N286A/T307Q/M428L/N434Y;
P257V/T307Q/N315A/M428L/N434Y;
P257V/T307Q/N384A/M428L/N434Y;
P257V/T307Q/N389A/M428L/N434Y;
P257V/N434Y;
P257T/N434Y;
P257V/N286E/N434Y;
P257T/N286E/N434Y;
P257V/N286E/T307Q/N434Y;
P257V/N286E/T307Q/M428Y/N434Y;
P257V/V305A/T307Q/M428Y/N434Y;
P257T/T307Q/M428Y/N434Y;
P257V/T307Q/V308P/N325G/M428Y/N434Y;
P257T/T307Q/M428F/N434Y;
P257A/N286E/T307Q/M428F/N434Y;
P257A/N286E/T307Q/M428Y/N434Y;
T250A/P257V/T307Q/M428L/N434Y;
T250F/P257V/T307Q/M428IVN434Y;
T250I/P257V/T307Q/M428L/N434Y;
T250M/P257V/T307Q/M428L/N434Y;
T250S/P257V/T307Q/M428L/N434Y;
T250V/P257V/T307Q/M428L/N434Y;
T250W/P257V/T307Q/M428L/N434Y;
T250Y/P257V/T307Q/M428L/N434Y;
M252Y/Q311A/N434Y;
S239K/M252Y/S254T/V308P/N434Y;
M252Y/T307Q/Q311A M252Y/S254T/N286E/N434Y;
M252Y/S254T/V308P/N434Y;
M252Y/S254T/T307A/N434Y;
M252Y/S254T/T307Q/N434Y;
M252Y/S254T/Q311A/N434Y;
M252Y/S254T/Q311H/N434Y;
M252Y/S254T/T307A/Q311A/N434Y;
M252Y/S254T/T307Q/Q311A/N434Y;
M252Y/S254T/M428I/N434Y;
M252Y/T256E/T307A/Q311H/N434Y;
M252Y/T256Q/T307A/Q311H/N434Y;
M252Y/S254T/T307A/Q311H/N434Y;
M252Y/N286E/T307A/Q311H/N434Y;
M252Y/T307A/Q311H/M428FN434Y;
M252Y/T256E/T307Q/Q311H;
M252Y/N286E/T307A/Q311A;
M252Y/N286E/T307Q/Q311A;
M252Y/N286E/T307Q/Q311H;
M252Y/T256E/N286E/N434Y;
S239K/M252Y/V308P;
S239K/M252Y/V308P/E382A;
S239K/M252Y/T256E/V308P;
S239K/M252Y/N286E/V308P;
S239K/M252Y/N286E/V308P/M428I;
M252Y/N286E/M428L/N434Y;
M252Y/S254T/E382A/N434Y;
S239K/M252Y/S254T/V308P/M428I/N434Y;
S239K/M252Y/M428I/N434Y;
M252Y/D312A/E382A/M428Y/N434Y;
M252Y/E3 82А/РЗ 87E/N434Y;
M252Y/D312A/P387E/N434Y или
M252Y/P387E/M428Y/N434Y; и
(b) отбора антитела, которое связывается с растворимым антигеном и обладает более высокой антигенсвязывающей активностью при рН 7.4, чем при рН 5.8, где отношение антигенсвязывающей активности при рН 5.8 и рН 7.4 равно по меньшей мере 2 при величине KD при рН 5.8 / KD при рН 7.4, где антитело содержит антигенсвязывающий домен, который получают из библиотеки антигенсвязывающих доменов, в которые введена по меньшей мере одна замена на гистидин или инсерция в антигенсвязывающем домене тяжелой цепи в положение, выбранное из группы, состоящей из Н27, Н31, Н32, Н33, Н35, Н50, Н58, Н59, Н61, Н62, Н63, Н64, Н65, Н99, H100b и H102 в соответствии с нумерацией по Kabat, и/или антигенсвязывающем домене легкой цепи в положение, выбранное из группы, состоящей из L24, L27, L28, L32, L53, L54, L56, L90, L92 и L94 в соответствии с нумерацией по Kabat; и
(с) отбор антитела, в котором человеческий FcRn-связывающий домен и антигенсвязывающий домен, отобранные на стадиях (а) и (b), сцеплены.
| YIK ANDY YEUNG et al., Engineering Human IgG1 Affinity to Human Neonatal Fc Receptor: Impact of Affinity Improvement on Pharmacokinetics in Primates, J Immunol, 2009, Vol.182, pp | |||
| Устройство для укрепления оттяжек антенн | 1927 |
|
SU7663A1 |
| US 20070148164 A1, 28.06.2007 | |||
| AMITA DATTA-MANNAN et al., Monoclonal Antibody Clearance | |||
| IMPACT OF MODULATING THE INTERACTION OF IgG WITH THE NEONATAL Fc | |||

