Область техники
Настоящее изобретение относится к способам улучшения фармакокинетики антигенсвязывающих молекул и способам увеличения числа раз связывания с антигенами антигенсвязывающих молекул, а также к антигенсвязывающим молекулам, обладающим улучшенной фармакокинетикой, антигенсвязывающим молекулам, характеризующимся увеличенным числом раз связывания с антигенами, и способам продуцирования таких молекул.
Предпосылки создания изобретения
Антитела обращают на себя внимание в качестве лекарственных средств, поскольку они являются в высокой степени устойчивыми в плазме и имеют немногочисленные побочные эффекты. В настоящее время на рынке имеются лекарственные средства в виде антител IgG-типа, и намного больше лекарственных средств в виде антител в настоящее время находятся в процессе разработки (непатентные документы 1 и 2). Между тем, были разработаны различные способы, применимые для получения лекарственных средств в виде антител второго поколения, в том числе те, которые усиливают эффекторную функцию, способность к связыванию антигенов, фармакокинетику и устойчивость, и те, которые уменьшают угрозу иммуногенности (непатентный документ 3). Обычно требуемая доза лекарственного средства в виде антитела является очень высокой. Это, в свою очередь, привело к таким проблемам, как высокие затраты на продукцию, а также трудности производства препаратов для подкожного введения. Теоретически дозу лекарственного средства в виде антитела можно снизить путем улучшения фармакокинетики антител или увеличения аффинности между антителами и антигенами.
В литературе были приведены сообщения о способах улучшения фармакокинетики антител, используя искусственную замену аминокислот в константных областях (непатентные документы 4 и 5). Так же сообщалось о созревании аффинности в качестве метода усиления способности к связыванию антигенов или антигеннейтрализующей активности (непатентный документ 6). Этот метод позволяет усилить антигенсвязывающую активность путем введения аминокислотных мутаций в участок CDR вариабельной области или т.п. Усиление способности к связыванию антигенов позволяет увеличить in vitro биологическую активность или снизить дозу и, кроме того, позволяет увеличить in vivo эффективность (непатентный документ 7).
Способность к нейтрализации антигенов одной молекулы антитела зависит от ее аффинности. При увеличении аффинности антиген можно нейтрализовать меньшим количеством антитела. Для увеличения аффинности антитела можно использовать различные способы. Кроме того, если бы аффинность можно было сделать очень большой при ковалентном связывании антитела с антигеном, одна молекула антитела могла бы нейтрализовать одну молекулу антигена (двухвалентное антитело может нейтрализовать две молекулы антигена). Однако нейтрализация в стехиометрическом соотношении: одно антитело против одного антигена (одно двухвалентное антитело против двух антигенов) является пределом уже существующих способов, и, следовательно, невозможно полностью нейтрализовать антиген количеством антитела, меньшим количества антигена. Другими словами, эффект увеличения аффинности имеет предел (непатентный документ 9). Для продления эффекта нейтрализации нейтрализующего антитела на определенный период времени антитело должно вводиться в дозе, превышающей количество антигена, продуцируемого в организме в течение этого же периода. Несмотря на описанное выше улучшение фармакокинетики антител или метод созревания аффинности, существует, таким образом, предел снижения требуемой дозы антитела.
В результате, для поддержания антигеннейтрализующего эффекта в течение целевого периода времени с использованием количества антитела, меньшего количества антигена, одно антитело должно нейтрализовать множество антигенов. Способы нейтрализации множества антигенов одним антителом включают инактивацию антигенов с использованием каталитических антител, которые являются антителами, которым сообщена каталитическая функция. Когда антиген является белком, он может быть инактивирован при гидролизе его пептидных связей. Антитело может многократно нейтрализовать антигены путем катализирования такого гидролиза (непатентный документ 8). Существует множество предшествующих опубликованных сообщений о каталитических антителах и методах их продуцирования. Однако не было сообщений о каталитических антителах, обладающих достаточной каталитической активностью, в качестве фармацевтического средства. В частности, при in vivo исследовании антител в отношении определенного антигена не было публикации, касающейся каталитических антител, которые, по сравнению с обычным некаталитическим нейтрализующим антителом, могут вызывать сравнимый или более сильный эффект даже при низких дозах или вызывать более продолжительный эффект даже при одной и той же дозе.
Как описано выше, не было сообщений об антителах, которые могут вызывать больший in vivo эффект, чем обычные нейтрализующие антитела, благодаря нейтрализации множества молекул антигена одним антителом. Таким образом, с точки зрения снижения дозы и удлинения срока службы, существует потребность в новых методах, которые позволяют продуцировать новые молекулы антител, обладающие более сильным in vivo эффектом, чем обычные нейтрализующие антитела, благодаря нейтрализации в индивидуальном порядке множества молекул антигена.
Имеющие отношение к настоящему изобретению документы, касающиеся известного уровня техники, представлены ниже:
Документы, касающиеся известного уровня техники
Непатентные документы
Непатентный документ 1: Monoclonal antibody successes in the clinic. Janice M Reichert, Clark J Rosensweig, Laura B Faden & Matthew C Dewitz, Nature Biotechnology 23, 1073-1078 (2005).
Непатентный документ 2: Pavlou AK, Belsey MJ. The therapeutic antibodies market to 2008. Eur. J. Pharm. Biopharm. 2005 Apr; 59(3): 389-396.
Непатентный документ 3: Kim SJ, Park Y, Hong HJ. Antibody engineering for the development of therapeutic antibodies. Mol Cells. 2005 Aug 31; 20(1): 17-29. Review.
Непатентный документ 4: Hinton PR, Xiong JM, Johlfs MG, Tang MT, Keller S, Tsurushita N. An engineered human IgG1 antibody with longer serum half-life. J Immunol. 2006 Jan 1; 176(1): 346-356.
Непатентный документ 5: Ghetie V, Popov S, Borvak J, Radu C, Matesoi D, Medesan C, Ober RJ, Ward ES. Increasing the serum persistence of an IgG fragment by random mutagenesis. Nat. Biotechnol. 1997 Jul; 15(7): 637-640.
Непатентный документ 6: Rajpal A, Beyaz N, Haber L, Cappuccilli G, Yee H, Bhatt RR, Takeuchi T, Lerner RA, Crea R. A general method for greatly improving the affinity of antibodies by using combinatorial libraries. Proc. Natl. Acad. Sci USA. 2005 Jun 14; 102(24): 8466-8471, Epub 2005 Jun 6.
Непатентный документ 7: Wu H, Pfarr DS, Johnson S, Brewah YA, Woods RM, Patel NK, White WI, Young JF, Kiener PA. Development of Motavizumab, an Ultra-potent Antibody for the Prevention of Respiratory Syncytial Virus Infection in the Upper and Lower Respiratory Tract. J. Mol. Biol. 2007, 368, 652-665.
Непатентный документ 8: Hanson CV, Nishiyama Y, Paul S. Catalytic antibodies and their applications. Curr. Opin. Biotechnol. 2005 Dec; 16(6): 631-636.
Непатентный документ 9: Rathanaswami P, Roalstad S, Roskos L, Su QJ, Lackie S, Babcook J. Demonstration of an in vivo generated sub-picomolar affinity fully human monoclonal antibody to interleukin-8. Biochem. Biophys. Res. Commun. 2005 Sep 9; 334(4): 1004-1013.
Краткое изложение сущности изобретения
Задачи, решаемые настоящим изобретением
Отмеченные выше обстоятельства привели к открытиям настоящего изобретения. В результате, целью настоящего изобретения является обеспечение способов многократного связывания антигенсвязывающих молекул с антигенами и способов улучшения фармакокинетики антигенсвязывающих молекул, а также антигенсвязывающих молекул, которые способны к многократному связыванию с антигенами, антигенсвязывающих молекул, обладающих улучшенной фармакокинетикой, фармацевтических композиций, содержащих такие антигенсвязывающие молекулы, и способов продуцирования таких молекул и композиций.
Способ решения задач
В настоящем описании были проведены исследования, посвященные способам многократного связывания полипептидов, обладающих способностью к связыванию антигенов, таких как антигенсвязывающие молекулы, с антигенами и способам увеличения времени полужизни таких молекул в плазме (крови) (улучшения их фармакокинетики). В результате было обнаружено, что, если антигенсвязывающая активность антигенсвязывающей молекулы при pH в ранних эндосомах ниже ее антигенсвязывающей активности при pH плазмы (крови), она могла связываться с антигенами многократно и иметь большее время полужизни в плазме.
В результате, настоящее изобретение относится к способам многократного связывания антигенсвязывающих молекул с антигенами, способам улучшения фармакокинетики антигенсвязывающих молекул и способам продуцирования антигенсвязывающих молекул с улучшенной фармакокинетикой; настоящее изобретение также относится к антигенсвязывающим молекулам, которые способны к многократному связыванию с антигенами, и к антигенсвязывающим молекулам с улучшенной фармакокинетикой. Конкретнее, настоящим изобретением обеспечиваются:
[1] антигенсвязывающая молекула, имеющая значение KD (pH 5,8)/KD (pH 7,4), определяемое как соотношение между KD для антигена при pH 5,8 и KD для антигена при pH 7,4, составляющее 2 или больше;
[2] антигенсвязывающая молекула в соответствии с [1], значение KD (pH 5,8)/KD (pH 7,4) которой составляет 10 или больше;
[3] антигенсвязывающая молекула в соответствии с [1], значение KD (pH 5,8)/KD (pH 7,4) которой составляет 40 или больше;
[4] антигенсвязывающая молекула в соответствии с любым из [1]-[3], по крайней мере одна аминокислота которой была замещена гистидином, или в которую был вставлен по крайней мере один гистидин;
[5] антигенсвязывающая молекула в соответствии с любым из [1]-[4], которая обладает антагонистической активностью;
[6] антигенсвязывающая молекула в соответствии с любым из [1]-[5], которая связывается с мембранным антигеном или растворимым антигеном;
[7] антигенсвязывающая молекула в соответствии с любым из [1]-[6], которая является антителом;
[8] фармацевтическая композиция, включающая антигенсвязывающую молекулу в соответствии с любым из [1]-[7];
[9] способ улучшения фармакокинентики антигенсвязывающей молекулы путем ослабления антигенсвязывающей активности антигенсвязывающей молекулы при pH 5,8 по сравнению с указанной активностью при pH 7,4;
[10] способ увеличения числа раз связывания с антигеном для антигенсвязывающей молекулы путем ослабления антигенсвязывающей активности антигенсвязывающей молекулы при pH 5,8 по сравнению с указанной активностью при pH 7,4;
[11] способ увеличения числа антигенов, которое может быть связано антигенсвязывающей молекулой, путем ослабления антигенсвязывающей активности антигенсвязывающей молекулы при pH 5,8 по сравнению с указанной активностью при pH 7,4;
[12] способ диссоциации внутри клетки антигена от связавшейся с ним внеклеточно антигенсвязывающей молекулы путем ослабления антигенсвязывающей активности антигенсвязывающей молекулы при pH 5,8 по сравнению с указанной активностью при pH 7,4;
[13] способ высвобождения антигенсвязывающей молекулы, которая связалась с антигеном и подверглась интернализации в клетку, в свободной от антигена форме наружу из клетки путем ослабления антигенсвязывающей активности антигенсвязывающей молекулы при pH 5,8 по сравнению с указанной активностью при pH 7,4;
[14] способ увеличения способности антигенсвязывающей молекулы к элиминации антигена в плазме путем ослабления антигенсвязывающей активности антигенсвязывающей молекулы при pH 5,8 по сравнению с указанной активностью при pH 7,4;
[15] способ в соответствии с любым из [9]-[14], в котором значение KD (pH 5,8)/KD (pH 7,4), определяемое как соотношение между KD для антигена при pH 5,8 и KD для антигена при pH 7,4, составляет 2 или больше;
[16] способ в соответствии с любым из [9]-[14], в котором значение KD (pH 5,8)/KD (pH 7,4) составляет 10 или больше;
[17] способ в соответствии с любым из [9]-[14], в котором значение KD (pH 5,8)/KD (pH 7,4) составляет 40 или больше;
[18] способ улучшения фармакокинетики антигенсвязывающей молекулы путем замены по крайней мере одной аминокислоты антигенсвязывающей молекулы гистидином или вставки по крайней мере одного гистидина в антигенсвязывающую молекулу;
[19] способ увеличения числа раз связывания с антигеном для антигенсвязывающей молекулы путем замены по крайней мере одной аминокислоты антигенсвязывающей молекулы гистидином или вставки по крайней мере одного гистидина в антигенсвязывающую молекулу;
[20] способ увеличения числа антигенов, которые могут быть связаны антигенсвязывающей молекулой, путем замены по крайней мере одной аминокислоты антигенсвязывающей молекулы гистидином или вставки по крайней мере одного гистидина в антигенсвязывающую молекулу;
[21] способ диссоциации внутри клетки антигена от связавшейся с ним внеклеточно антигенсвязывающей молекулы путем замены по крайней мере одной аминокислоты антигенсвязывающей молекулы гистидином или вставки по крайней мере одного гистидина в антигенсвязывающую молекулу;
[22] способ высвобождения антигенсвязывающей молекулы, которая связалась с антигеном и подверглась интернализации в клетку, в свободной от антигена форме наружу из клетки путем замены по крайней мере одной аминокислоты антигенсвязывающей молекулы гистидином или вставки по крайней мере одного гистидина в антигенсвязывающую молекулу;
[23] способ увеличения способности антигенсвязывающей молекулы к элиминации антигена в плазме путем замены по крайней мере одной аминокислоты антигенсвязывающей молекулы гистидином или вставки по крайней мере одного гистидина в антигенсвязывающую молекулу;
[24] способ в соответствии с любым из [18]-[23], в котором замена гистидином или его вставка увеличивает значение KD (pH 5,8)/KD (pH 7,4), определяемое как соотношение между антигенсвязывающей активностью при pH 5,8 и антигенсвязывающей активностью при pH 7,4, по сравнению со значением KD (pH 5,8)/KD (pH 7,4) до замены гистидином или его вставки;
[25] способ в соответствии с любым из [9]-[24], в котором антигенсвязывающая молекула обладает антагонистической активностью;
[26] способ в соответствии с любым из [9]-[25], в котором антигенсвязывающая молекула связывается с мембранным антигеном или растворимым антигеном;
[27] способ в соответствии с любым из [9]-[26], в котором антигенсвязывающей молекулой является антитело;
[28] способ скрининга антигенсвязывающих молекул, который включает стадии:
(a) определения антигенсвязывающей активности антигенсвязывающей молекулы при pH от 6,7 до 10,0;
(b) определения антигенсвязывающей активности антигенсвязывающей молекулы при pH от 4,0 до 6,5; и
(c) отбора антигенсвязывающей молекулы, антигенсвязывающая активность которой при pH от 6,7 до 10,0 выше антигенсвязывающей активности при pH от 4,0 до 6,5;
[29] способ скрининга в соответствии с [28], который включает стадию отбора антитела, антигенсвязывающая активность которого при pH от 6,7 до 10,0 – дважды или более антигенсвязывающей активности при pH от 4,0 до 6,5;
[30] способ скрининга антигенсвязывающих молекул, который включает стадии:
(a) связывания антигенсвязывающей молекулы с антигеном при параметре pH, составляющем от 6,7 до 10,0;
(b) помещения антигенсвязывающей молекулы, которая связалась с антигеном на стадии (a), при параметре pH, составляющем от 4,0 до 6,5; и
(c) получения антигенсвязывающей молекулы, которая подвергалась диссоциации при параметре pH, составляющем от 4,0 до 6,5;
[31] способ скрининга для получения антигенсвязывающей молекулы, активность связывания которой при первом pH превышает активность связывания при втором pH, который включает стадии:
(a) связывания антигенсвязывающей молекулы с колонкой с иммобилизованным антигеном при параметре первого pH;
(b) элюирования антигенсвязывающей молекулы, которая связалась с колонкой при первом pH, с колонки при параметре второго pH; и
(c) сбора подвергшейся элюции антигенсвязывающей молекулы;
[32] способ скрининга для получения антигенсвязывающей молекулы, активность связывания которой при первом pH превышает активность связывания при втором pH, который включает стадии:
(a) связывания библиотеки антигенсвязывающих молекул с колонкой с иммобилизованным антигеном при параметре первого pH;
(b) элюирования антигенсвязывающей молекулы с колонки при параметре второго pH;
(c) амплификации гена, кодирующего элюируемую антигенсвязывающую молекулу гена; и
(d) получения элюируемой антигенсвязывающей молекулы;
[33] способ скрининга в соответствии с [31] или [32], в котором первый pH составляет от 6,7 до 10,0, а второй pH составляет от 4,0 до 6,5;
[34] способ скрининга в соответствии с любым из [28]-[33], в котором по крайней мере одна или более аминокислот антигенсвязывающей молекулы была замещена гистидином, или по крайней мере один гистидин был вставлен в антигенсвязывающую молекулу;
[35] способ скрининга в соответствии с любым из [28]-[33] для получения антигенсвязывающей молекулы, которая превосходит в сохранении в плазме;
[36] способ скрининга в соответствии с любым из [28]-[33] для получения антигенсвязывающей молекулы, которая способна к связыванию антигена два или более раз;
[37] способ скрининга в соответствии с любым из [28]-[33] для получения антигенсвязывающей молекулы, которая способна к связыванию большего числа антигеном, чем число ее антигенсвязывающих сайтов;
[38] способ скрининга в соответствии с любым из [28]-[33] для получения антигенсвязывающей молекулы, которая подвергает диссоциации связавшийся с ней внеклеточно антиген внутри клетки;
[39] способ скрининга в соответствии с любым из [28]-[33] для получения антигенсвязывающей молекулы, которая связывается с антигеном и подвергается интернализацию в клетку, и высвобождается наружу из клетки в свободной от антигена форме;
[40] способ скрининга в соответствии с любым из [28]-[33] для получения антигенсвязывающей молекулы, которая обладает увеличенной способностью к элиминации антигена в плазме;
[41] способ скрининга в соответствии с любым из [28]-[40], в котором антигенсвязывающая молекула используется в виде фармацевтической композиции;
[42] способ скрининга в соответствии с любым из [28]-[41], в котором антигенсвязывающей молекулой является антитело;
[43] способ продуцирования антигенсвязывающей молекулы, который включает стадии:
(a) определения антигенсвязывающей активности антигенсвязывающей молекулы при pH от 6,7 до 10,0;
(b) определения антигенсвязывающей активности антигенсвязывающей молекулы при pH от 4,0 до 6,5;
(c) отбора антигенсвязывающей молекулы, антигенсвязывающая активность которой при pH от 6,7 до 10,0 выше антигенсвязывающей активности при pH от 4,0 до pH 6,5;
(d) получения гена, кодирующего антигенсвязывающую молекулу, отобранную на стадии (c); и
(e) продуцирования антигенсвязывающей молекулы, используя полученный на стадии (d) ген;
[44] способ продуцирования антигенсвязывающей молекулы, который включает стадии:
(a) связывания антигенсвязывающей молекулы с антигеном при параметре pH, составляющем от 6,7 до 10,0;
(b) допущения нахождения антигенсвязывающей молекулы, связанной с антигеном на стадии (a), при параметре pH, составляющем от 4,0 до 6,5;
(c) сбора антигенсвязывающей молекулы, которая подвергалась диссоциации при параметре pH, составляющем от 4,0 до 6,5;
(d) получения гена, кодирующего антигенсвязывающую молекулу, полученную на стадии (c); и
(e) продуцирования антигенсвязывающей молекулы, используя полученный на стадии (d) ген;
[45] способ продуцирования антигенсвязывающей молекулы, активность связывания которой при первом pH превышает активность связывания при втором pH, который включает стадии:
(a) связывания антигенсвязывающей молекулы с колонкой с иммобилизованным антигеном при параметре первого pH;
(b) элюирования антигенсвязывающей молекулы, которая связалась с колонкой при первом pH, с колонки при параметре второго pH;
(c) сбора подвергшейся элюции антигенсвязывающей молекулы;
(d) получения гена, кодирующего антигенсвязывающую молекулу, полученную на стадии (c); и
(e) продуцирования антигенсвязывающей молекулы, используя полученный на стадии (d) ген;
[46] способ продуцирования антигенсвязывающей молекулы, активность связывания которой при первом pH превышает активность связывания при втором pH, который включает стадии:
(a) связывания библиотеки антигенсвязывающих молекул с колонкой с иммобилизованным антигеном при параметре первого pH;
(b) элюирования антигенсвязывающей молекулы с колонки при параметре второго pH;
(c) амплификации гена, кодирующего элюируемую антигенсвязывающую молекулу;
(d) сбора подвергшейся элюции антигенсвязывающей молекулы;
(e) получения гена, кодирующего антигенсвязывающую молекулу, собранную на стадии (d); и
(f) продуцирования антигенсвязывающей молекулы, используя полученный на стадии (е) ген;
[47] способ продуцирования в соответствии с [45] или [46], в котором первый pH составляет от 6,7 до 10,0, а второй pH составляет от 4,0 до 6,5;
[48] способ продуцирования в соответствии с любым из [43]-[47], который, кроме того, включает стадию замещения по крайней мере одной аминокислоты антигенсвязывающей молекулы гистидином или вставки по крайней мере одного гистидина в антигенсвязывающую молекулу;
[49] способ продуцирования в соответствии с любым из [43]-[48], в котором антигенсвязывающей молекулой является антитело;
[50] фармацевтическая композиция, включающая антигенсвязывающую молекулу, полученную способом продуцирования в соответствии с любым из [43]-[49].
Эффекты настоящего изобретения
Настоящим изобретением обеспечиваются способы изготовления связывающих один антиген молекул для многократного связывания с множеством молекул антигена. Когда антигенсвязывающая молекула связывается с множеством молекул антигена, фармакокинетику антигенсвязывающей молекулы можно улучшить, и такая молекула может вызывать in vivo эффекты, превосходящие эффекты, вызываемые обычными антигенсвязывающими молекулами.
Краткое описание чертежей
Фиг. 1 представляет собой схему, на которой представлен путь деградации антител, связавшихся со связанным с мембраной антигеном.
Фиг. 2 представляет собой схему, на которой представлен механизм возвращения молекул IgG с помощью FcRn.
Фиг. 3 представляет собой схему, представляющую повторное связывание молекул IgG с новым антигеном после диссоциации от связанного с мембраной антигена внутри эндосом.
Фиг. 4 представляет собой схему, представляющую повторное связывание молекул IgG с новым антигеном после диссоциации от растворимого антигена внутри эндосом.
Фиг. 5 представляет собой схему, на которой представлен процесс пэннинга, в котором используется колонка с иммобилизованным антигеном.
Фиг. 6 представляет собой графики, на которых представлены результаты ELISA фагов для клонов, полученных с помощью пэннинга, в котором используется колонка. Верхний график отображает WT, а нижний график отображает CL5.
Фиг. 7 представляет собой график, на котором отображена биологическая нейтрализующая активность связывающихся в зависимости от pH антител против рецептора IL-6.
Фиг. 8 представляет собой графики, на которых представлены результаты Biacore – сенсограммы для связывания связывающихся в зависимости от pH антител против рецептора IL-6 с растворимым рецептором IL-6 при pH 7,4. Верхний график отображает WT; второй график сверху отображает H3pI/L73; третий график сверху отображает H170/L82, и нижний график отображает CLH5/L73.
Фиг. 9 представляет собой графики, на которых представлены результаты Biacore – сенсограммы для связывания связывающихся в зависимости от pH антител против рецептора IL-6 с растворимым рецептором IL-6 при pH 5,8. Верхний график отображает WT; второй график сверху отображает H3pI/L73; третий график сверху отображает H170/L82, и нижний график отображает CLH5/L73.
Фиг. 10 представляет собой графики, на которых представлены результаты Biacore – сенсограммы для ассоциации (при pH 7,4) и диссоциации (при pH 5,8) связывающихся в зависимости от pH антител против рецептора IL-6 с рецептором IL-6 мембранного типа и от него, соответственно. Верхний график отображает WT; второй график сверху отображает H3pI/L73; третий график сверху отображает H170/L82, и нижний график отображает CLH5/L73.
Фиг. 11 представляет собой полученную с помощью Biacore сенсограмму, свидетельствующую о многократном связывании связывающихся в зависимости от pH антител против рецептора IL-6 с SR344.
Фиг. 12 представляет собой график, на котором представлено общее количество связанного антигена в эксперименте повторного связывания связывающихся в зависимости от pH антител против рецептора IL-6 с SR344.
Фиг. 13 представляет собой график, на котором представлены зависимости от времени концентраций в плазме связывающихся в зависимости от pH антител против рецептора IL-6 у трансгенных по рецептору IL-6 человека мышей.
Фиг. 14 представляет собой график, на котором представлены зависимости от времени концентраций в плазме связывающихся в зависимости от pH антител против рецептора IL-6 у яванских макак.
Фиг. 15 представляет собой график, на котором представлены зависимости от времени концентраций CRP у яванских макак, относительно связывающихся в зависимости от pH антител против рецептора IL-6.
Фиг. 16 представляет собой график, на котором представлены зависимости от времени концентраций рецептора IL-6 яванского макака несвязанного типа у яванских макак, относительно связывающихся в зависимости от pH антител против рецептора IL-6.
Фиг. 17 представляет собой графики, на которых представлены результаты Biacore – сенсограммы для ассоциации (при pH 7,4) и диссоциации (при pH 5,8) связывающихся в зависимости от pH антител против рецептора IL-6 с рецептором IL-6 мембранного типа и от него, соответственно. В порядке перемещения сверху вниз представлены результаты для WT, H3pI/L73-IgG1, Fv2-IgG1 и Fv4-IgG1.
Фиг. 18 представляет собой график, на котором представлены зависимости от времени концентраций в плазме связывающихся в зависимости от pH антител против рецептора IL-6 (WT, H3pI/L73-IgG1, Fv2-IgG1 и Fv4-IgG1) у трансгенных по рецептору IL-6 человека мышей.
Фиг. 19 представляет собой графики, на которых представлены результаты Biacore – сенсограммы для ассоциации (при pH 7,4) и диссоциации (при pH 5,8) связывающихся в зависимости от pH антител против рецептора IL-6 с рецептором IL-6 мембранного типа и от него, соответственно. В порядке перемещения сверху вниз представлены результаты для WT, Fv4-IgG1, Fv4-IgG2 и Fv4-M58.
Фиг. 20 представляет собой график, на котором представлены зависимости от времени концентраций в плазме связывающихся в зависимости от pH антител против рецептора IL-6 (WT, Fv4-IgG1, Fv4-IgG2 и Fv4-M58) у трансгенных по рецептору IL-6 человека мышей.
Фиг. 21 представляет собой графики, на которых представлены результаты Biacore – сенсограммы для ассоциации (при pH 7,4) и диссоциации (при pH 5,8) связывающихся в зависимости от pH антител против рецептора IL-6 с рецептором IL-6 мембранного типа и от него, соответственно. Перемещаясь сверху вниз, представлены результаты для Fv1-M71, Fv1-M73, Fv3-M71 и Fv3-M73.
Фиг. 22 представляет собой график, на котором представлены зависимости от времени концентраций в плазме связывающихся в зависимости от pH антител против рецептора IL-6 у яванских макак, во время введения H3pI/L73-IgG1, Fv1-M71, Fv1-M73, Fv2-IgG1, Fv3-M73 и Fv4-M73 в дозе 0,5 мг/кг и во время введения Ат с большой аффинностью в дозе 1,0 мг/кг.
Фиг. 23 представляет собой график, на котором представлены зависимости от времени концентраций CRP у яванских макак, относительно связывающихся в зависимости от pH антител против рецептора IL-6 (групп, которым вводили H3pI/L73-IgG1, Fv1-M71, Fv1-M73, Fv2-IgG1, Fv3-M73, Fv4-M73 и Ат с большой аффинностью).
Фиг. 24 представляет собой график, на котором представлены зависимости от времени концентраций рецептора IL-6 яванского макака несвязанного типа у яванских макак, относительно связывающихся в зависимости от pH антител против рецептора IL-6 (групп, которым вводили H3pI/L73-IgG1, Fv1-M71, Fv1-M73, Fv2-IgG1, Fv3-M73, Fv4-M73 и Ат с большой аффинностью).
Фиг. 25 представляет собой схематическое представление, отображающее FR1, FR2, FR3 и FR4 вместе с CDR1, CDR2 и CDR3 тяжелых цепей (VH1, VH2, VH3, VH4) и легких цепей (VL1, VL2, VL3). Звездочки указывают на положения, в которых существуют аминокислотные мутации в совмещенных последовательностях.
Фиг. 26 представляет собой полученную с помощью Biacore сенсограмму, отображающую зависящее от pH связывание антитела против IL-6, анти-IL6 клона 2, с IL-6 при pH 7,4 и pH 5,5. Кривые в сенсограмме при pH 7,4 соответствуют 100, 50, 25, 12,5 и 6,25 нг/мл IL-6, сверху.
Фиг. 27 представляет собой полученную с помощью Biacore сенсограмму, отображающую зависящее от pH связывание антитела против рецептора IL-31, анти-IL31R клона 1, с рецептором IL-31 при pH 7,4 и pH 5,5. Кривые в сенсограмме при pH 5,5 соответствуют 100, 50, 25 и 12,5 нг/мл рецептора IL-31, сверху.
На фиг. 28 представлена зависимость от времени концентрации антитела в плазме после внутривенного введения мыши раствора смеси, содержащего SR344 и антитело против рецептора IL-6 человека.
На фиг. 29 представлена зависимость от времени концентрации SR344 в плазме после внутривенного введения мыши раствора смеси, содержащего SR344 и антитело против рецептора IL-6 человека.
Вариант осуществления изобретения
Настоящим изобретением обеспечиваются способы увеличения числа раз связывания с антигенами для антигенсвязывающих молекул. Конкретнее, настоящим изобретением обеспечиваются способы увеличения числа раз связывания с антигенами для антигенсвязывающих молекул путем ослабления способности к связыванию антигенов антигенсвязывающих молекул при кислотном pH по сравнению с указанной способностью при нейтральном pH. Кроме того, настоящим изобретением обеспечиваются способы увеличения числа раз связывания с антигенами для антигенсвязывающих молекул путем замещения гистидином по крайней мере одной аминокислоты в антигенсвязывающих молекулах или вставки по крайней мере одного гистидина в антигенсвязывающие молекулы. Кроме того, настоящим изобретением обеспечиваются способы увеличения числа раз связывания с антигенами для антигенсвязывающих молекул путем замены, делеции, добавления и/или вставки аминокислот в характерную для антитела константную область антигенсвязывающих молекул.
Настоящим изобретением также обеспечиваются способы увеличения числа антигенов, которое может быть связано антигенсвязывающей молекулой. Конкретнее, настоящим изобретением обеспечиваются способы увеличения числа антигенов, которое может быть связано антигенсвязывающей молекулой, путем ослабления способности к связыванию антигенов при кислотном pH по сравнению с указанной способностью при нейтральном pH. Кроме того, настоящим изобретением обеспечиваются способы увеличения числа антигенов, которое может быть связано антигенсвязывающей молекулой, путем замещения гистидином по крайней мере одной аминокислоты в антигенсвязывающих молекулах или вставки по крайней мере одного гистидина в антигенсвязывающие молекулы. Кроме того, настоящим изобретением обеспечиваются способы увеличения числа антигенов, которое может быть связано антигенсвязывающей молекулой, благодаря замене, делеции, добавлению и/или вставке аминокислот в характерную для антитела константную область антигенсвязывающих молекул.
Настоящим изобретением также обеспечиваются способы диссоциации внутри клетки антигена от связавшейся с ним внеклеточно антигенсвязывающей молекулы. Конкретнее, настоящим изобретением обеспечиваются способы диссоциации внутри клетки антигена от связавшейся с ним внеклеточно антигенсвязывающей молекулы путем ослабления способности к связыванию антигена при кислотном pH по сравнению с указанной способностью при нейтральном pH. Кроме того, настоящим изобретением обеспечиваются способы диссоциации внутри клетки антигена от связавшейся с ним внеклеточно антигенсвязывающей молекулы путем замещения гистидином по крайней мере одной аминокислоты в антигенсвязывающих молекулах или вставки по крайней мере одного гистидина в антигенсвязывающую молекулу. Кроме того, настоящим изобретением обеспечиваются способы диссоциации внутри клетки антигена от связавшейся с ним внеклеточно антигенсвязывающей молекулы благодаря замене, делеции, добавлению и/или вставке аминокислот в характерную для антитела константную область антигенсвязывающих молекул.
Настоящим изобретением также обеспечиваются способы высвобождения антигенсвязывающей молекулы, которая была связана с антигеном и подвергнута интернализации в клетку, в свободной от антигена форме наружу из клетки. Конкретнее, настоящим изобретением обеспечиваются способы высвобождения антигенсвязывающей молекулы, которая была связана с антигеном и подвергнута интернализации в клетку, в свободной от антигена форме наружу из клетки путем ослабления способности к связыванию антигенов при кислотном pH по сравнению с указанной способностью при нейтральном pH. Кроме того, настоящим изобретением обеспечиваются способы высвобождения антигенсвязывающей молекулы, которая была связана с антигеном и подвергнута интернализации в клетку, в свободной от антигена форме наружу из клетки путем замещения гистидином по крайней мере одной аминокислоты в антигенсвязывающей молекуле или вставки по крайней мере одного гистидина в антигенсвязывающую молекулу. Кроме того, настоящим изобретением обеспечиваются способы высвобождения антигенсвязывающей молекулы, которая была связана с антигеном и подвергнута интернализации в клетку, в свободной от антигена форме наружу из клетки благодаря замене, делеции, добавлению и/или вставке аминокислот в характерную для антитела константную область антигенсвязывающей молекулы.
Настоящим изобретением также обеспечиваются способы увеличения способности антигенсвязывающей молекулы к элиминации антигенов в плазме. Конкретнее, настоящим изобретением обеспечиваются способы увеличения способности антигенсвязывающей молекулы к элиминации антигенов в плазме путем ослабления способности к связыванию антигенов при кислотном pH по сравнению с указанной способностью при нейтральном pH. Кроме того, настоящим изобретением обеспечиваются способы увеличения способности антигенсвязывающей молекулы к элиминации антигенов в плазме путем замещения гистидином по крайней мере одной аминокислоты в антигенсвязывающих молекулах или вставки по крайней мере одного гистидина в антигенсвязывающие молекулы. Кроме того, настоящим изобретением обеспечиваются способы увеличения способности антигенсвязывающей молекулы к элиминации антигенов в плазме благодаря замене, делеции, добавлению и/или вставке аминокислот в характерную для антитела константную область антигенсвязывающей молекулы.
Настоящим изобретением также обеспечиваются способы улучшения фармакокинетики антигенсвязывающих молекул. Конкретнее, настоящим изобретением обеспечиваются способы улучшения фармакокинетики антигенсвязывающих молекул (удлинению сохранения в плазме) путем ослабления способности к связыванию антигенов при кислотном pH по сравнению с указанной способностью при нейтральном pH. Кроме того, настоящим изобретением обеспечиваются способы улучшения фармакокинетики антигенсвязывающих молекул путем замещения гистидином по крайней мере одной аминокислоты в антигенсвязывающих молекулах или вставки по крайней мере одного гистидина в антигенсвязывающие молекулы. Кроме того, настоящим изобретением обеспечиваются способы улучшения фармакокинетики антигенсвязывающих молекул путем замены, делеции, добавления и/или вставки аминокислот в характерную для антитела константную область антигенсвязывающих молекул.
Кроме того, настоящим изобретением обеспечиваются способы увеличения способности антигенсвязывающих молекул к элиминации антигенов в плазме. Конкретнее, настоящим изобретением обеспечиваются способы увеличения способности антигенсвязывающих молекул к элиминации антигенов в плазме путем ослабления способности к связыванию антигенов при кислотном pH по сравнению с указанной способностью при нейтральном pH. Кроме того, настоящим изобретением обеспечиваются способы увеличения способности антигенсвязывающих молекул к элиминации антигенов в плазме путем замещения гистидином по крайней мере одной аминокислоты в антигенсвязывающих молекулах или вставки по крайней мере одного гистидина в антигенсвязывающие молекулы. Кроме того, настоящим изобретением обеспечиваются способы увеличения способности антигенсвязывающих молекул к элиминации антигенов в плазме путем замены, делеции, добавления и/или вставки аминокислот в характерную для антитела константную область антигенсвязывающей молекулы.
Здесь выражения «улучшение фармакокинетики», «лучшая фармакокинетика» равнозначны выражениям «увеличение сохранения в плазме (крови)» и «лучшее сохранение в плазме (крови)», соответственно, и эти выражения являются синонимами.
Здесь ослабление антигенсвязывающей активности при кислотном pH по сравнению с указанной активностью при нейтральном pH означает, что способность к связыванию антигена антигенсвязывающей молекулы при pH, составляющем от 4,0 до 6,5, снижена по сравнению с указанной способностью при pH, составляющем от 6,7 до 10,0, предпочтительно, что антигенсвязывающая активность антигенсвязывающей молекулы при pH, составляющем от 5,5 до 6,5, снижена по сравнению с указанной активностью при pH, составляющем от 7,0 до 8,0, и более предпочтительно, что антигенсвязывающая активность антигенсвязывающей молекулы при pH 5,8 снижена по сравнению с указанной активностью при pH 7,4. В результате, в настоящем изобретении кислотный pH составляет обычно от 4,0 до 6,5, предпочтительно от 5,5 до 6,5 и более предпочтительно 5,8. Альтернативно, в настоящем изобретении нейтральный pH составляет обычно от 6,7 до 10,0, предпочтительно от 7,0 до 8,0 и более предпочтительно 7,4.
Здесь выражение «ослабление способности антигенсвязывающей молекулы к связыванию антигена при кислотном pH по сравнению с указанной способностью при нейтральном pH» равнозначно выражению «увеличение способности антигенсвязывающей молекулы к связыванию антигена при нейтральном pH по сравнению с указанной способностью при кислотном pH». Другими словами, в настоящем изобретении должно быть увеличено различие между кислотным и нейтральным pH в способности антигенсвязывающей молекулы к связыванию антигена. Например, должно быть увеличено, как описано ниже, значение KD (pH 5,8)/KD (pH 7,4). Различие между кислотным и нейтральным pH в способности антигенсвязывающей молекулы к связыванию антигена может быть увеличено, например, путем любого из двух или обоих действий: ослабления способности к связыванию антигена при кислотном pH и увеличения способности к связыванию антигена при нейтральном pH.
Отличные от pH условия для определения антигенсвязывающей активности могут быть выбраны надлежащим образом квалифицированными в данной области техники специалистами, и эти условия особенно не ограничиваются. Антигенсвязывающую активность можно определить, например, в условиях использования буфера MES и 37°C, как описано здесь в примерах. Кроме того, антигенсвязывающую активность антигенсвязывающей молекулы можно определить с помощью известных квалифицированным в данной области техники специалистам способов, например, используя Biacore (GE Healthcare) или т.п., как описано в данном описании в примерах. Когда антигеном является растворимый антиген, активность связывания с растворимым антигеном можно определить путем инъецирования антигена в качестве аналита в чип, на котором иммобилизована антигенсвязывающая молекула. Альтернативно, когда антигеном является мембранный антиген, активность связывания с мембранным антигеном можно определить путем инъецирования антигенсвязывающей молекулы в качестве аналита в чип с иммобилизованным на нем антигеном.
В настоящем изобретении различие между кислотным и нейтральным pH в антигенсвязывающей активности особенно не ограничивается при условии, что антигенсвязывающая активность при кислотном pH ниже антигенсвязывающей активности при нейтральном pH. Однако значение KD (pH 5,8)/KD (pH 7,4), которое является соотношением между константами диссоциации (KD) от антигена при pH 5,8 и pH 7,4, составляет предпочтительно 2 или больше, более предпочтительно 10 или больше, и еще предпочтительнее 40 или больше. Верхний предел значения KD (pH 5,8)/KD (pH 7,4) особенно не ограничивается и может быть любым значением, например, 400, 1000 или 10000, при условии, что молекулу можно продуцировать с помощью методов, известных квалифицированным в данной области техники специалистам. Когда антигеном является растворимый антиген, антигенсвязывающую активность можно представить через константу диссоциации (KD). Альтернативно, когда антигеном является мембранный антиген, антигенсвязывающую активность можно представить через кажущуюся константу диссоциации. Константу диссоциации (KD) и кажущуюся константу диссоциации (кажущуюся KD) можно определить с помощью способов, известных квалифицированным в данной области техники специалистам, например, используя Biacore (GE Healthcare), график Скэтчарда или FACS.
Альтернативно, возможно использование, например, kd, константы скорости диссоциации, в качестве показателя различия между кислотным и нейтральным pH в антигенсвязывающей активности. Когда в качестве показателя различия в активности связывания используется константа скорости диссоциации (kd) вместо константы диссоциации (KD), значение kd (pH 5,8)/kd (pH 7,4), которое является соотношением между константами скорости диссоциации (kd) от антигена при pH 5,8 и pH 7,4, составляет предпочтительно 2 или больше, более предпочтительно 5 или больше, даже предпочтительнее 10 или больше или еще предпочтительнее 30 или больше. Верхний предел значения kd (pH 5,8)/kd (pH 7,4) особенно не ограничивается и может быть любым значением, например, 50, 100 или 200, при условии, что молекулу можно продуцировать с помощью методов, обычных для квалифицированных в данной области техники специалистов.
Когда антигеном является растворимый антиген, антигенсвязывающую активность можно представить через константу скорости диссоциации (kd). Альтернативно, когда антигеном является мембранный антиген, антигенсвязывающую активность можно представить через кажущуюся константу скорости диссоциации. Константу скорости диссоциации (kd) и кажущуюся константу скорости диссоциации (кажущуюся kd) можно определить с помощью способов, известных квалифицированным в данной области техники специалистам, например, используя Biacore (GE Healthcare) или FACS.
В настоящем изобретении, когда антигенсвязывающую активность антигенсвязывающей молекулы определяют при различных pH, предпочтительно, чтобы условия проведения измерений, за исключением pH, были постоянными.
Способы ослабления антигенсвязывающей активности антигенсвязывающей молекулы при pH 5,8 по сравнению с указанной активностью при pH 7,4 (способы сообщения способности к связыванию в зависимости от pH) особенно не ограничиваются, и могут использоваться любые способы. Такие способы включают, например, способы ослабления антигенсвязывающей активности при pH 5,8 по сравнению с указанной активностью при pH 7,4 путем замещения гистидином аминокислот в антигенсвязывающей молекуле или вставки гистидина в антигенсвязывающую молекулу. Уже известно, что антителу можно сообщить зависящую от pH антигенсвязывающую активность путем замещения гистидином аминокислот в антителе (FEBS Letter, 309 (1), 8588 (1992)). Точки такой мутации (замены) на гистидин или вставки гистидина особенно не ограничиваются, и допустима любая точка при условии, что антигенсвязывающая активность при pH 5,8 понижается относительно указанной активности при pH 7,4 (значение KD (pH 5,8)/KD (pH 7,4) становится больше) по сравнению с тем, что наблюдалось до мутации или вставки. Когда антигенсвязывающей молекулой является антитело, такие точки включают, например, точки в пределах вариабельной области антитела. Подходящее число точек мутаций на гистидин или вставок гистидина может быть надлежащим образом определено квалифицированными в данной области техники специалистами. Гистидин может быть заменен или вставлен в одной (одну) точке(у), или двух (две) или более точках(ек). Также возможно введения мутации не на гистидин (мутации на аминокислоты, отличные от гистидина) в то же самое время. Кроме того, мутация на гистидин может быть введена одновременно со вставкой гистидина. Возможна замена гистидином или его вставка случайным образом, используя такой способ, как сканирование гистидином, в котором используется гистидин вместо аланина при сканировании аланином, известном квалифицированным в данной области техники специалистам. Альтернативно, антигенсвязывающие молекулы, значение KD (pH 5,8)/KD (pH 7,4) которых увеличивается по сравнению с указанным значением до мутации, можно отобрать из библиотеки антигенсвязывающих молекул с мутацией на гистидин или вставкой гистидина случайным образом.
Когда гистидином замещают аминокислоты антигенсвязывающей молекулы, или его вставляют между аминокислотами такой молекулы, предпочтительно, но не обязательно, чтобы антигенсвязывающая активность антигенсвязывающей молекулы при pH 7,4 после замены гистидином или его вставки была сравнима с указанной активностью при pH 7,4 до замены гистидином или его вставки. Выражение «антигенсвязывающая активность антигенсвязывающей молекулы при pH 7,4 после замены гистидином или его вставки сравнима с указанной активностью при pH 7,4 до замены гистидином или его вставки» означает, что даже после замены гистидином или его вставки антигенсвязывающая молекула сохраняет 10% или более, предпочтительно 50% или более, более предпочтительно 80% или более, и еще предпочтительнее 90% или более антигенсвязывающей активности до замены гистидином или его вставки. Когда антигенсвязывающая активность антигенсвязывающей молекулы была ослаблена вследствие замены гистидином или его вставки, антигенсвязывающую активность можно скорректировать путем введения замены, делеции, добавления и/или вставки одной или нескольких аминокислот в антигенсвязывающую молекулу, так что антигенсвязывающая активность становится сравнимой с антигенсвязывающей активностью до замены гистидином или его вставки. Настоящее изобретение также включает такие антигенсвязывающие молекулы, которые обладают сравнимой активностью связывания в результате замены, делеции, добавления и/или вставки одной или нескольких аминокислот после замены гистидином или его вставки.
Альтернативные способы ослабления антигенсвязывающей активности антигенсвязывающей молекулы при pH 5,8 по сравнению с указанной активностью при pH 7,4 включают способы замещения не встречающимися в природе аминокислотами аминокислот в антигенсвязывающей молекуле или вставки не встречающихся в природе аминокислот в аминокислоты антигенсвязывающей молекулы. Известно, что pKa можно искусственно контролировать, используя не встречающиеся в природе аминокислоты (Angew. Chem. Int. Ed. 2005, 44, 34; Chem Soc Rev. 2004 Sep 10; 33(7): 422-430; Amino Acids. 1999; 16(3-4): 345-379). Таким образом, в настоящем изобретении не встречающиеся в природе аминокислоты можно использовать вместо гистидина, описываемого выше. Замену такой не встречающейся в природе аминокислотой и/или ее вставку можно вводить одновременно с заменой гистидином и/или его вставкой, описанной выше. В настоящем изобретении можно использовать любые не встречающиеся в природе аминокислоты. Возможно использование не встречающихся в природе аминокислот, известных квалифицированным в данной области техники специалистам.
Кроме того, когда антигенсвязывающей молекулой является вещество, имеющее константную область антитела, альтернативные способы ослабления антигенсвязывающей активности антигенсвязывающей молекулы при pH 5,8 по сравнению с указанной активностью при pH 7,4 включают способы модифицирования константной области антитела, содержащейся в антигенсвязывающей молекуле. Такие способы модифицирования константной области антитела включают, например, способы замещения константной области, описанные здесь в примерах.
Альтернативные способы модифицирования константной области антитела включают, например, способы оценки различных изотипов константных областей (IgG1, IgG2, IgG3 и IgG4) и отбор изотипа, который ослабляет антигенсвязывающую активность при pH 5,8 (увеличивает скорость диссоциации при pH 5,8). Альтернативно, способы включают способы ослабления антигенсвязывающей активности при pH 5,8 (увеличения скорости диссоциации при pH 5,8) путем замещения аминокислот в аминокислотной последовательности изотипа дикого типа (аминокислотной последовательности IgG1, IgG2, IgG3 или IgG4 дикого типа). Последовательность шарнирной области константной области антитела является в значительной степени различной среди изотипов (IgG1, IgG2, IgG3 и IgG4), и различие в аминокислотной последовательности шарнирной области оказывает большое влияние на антигенсвязывающую активность. Следовательно, можно выбрать изотип, подходящий для ослабления антигенсвязывающей активности при pH 5,8 (для увеличения скорости диссоциации при at pH 5,8), с учетом типа антигена или эпитопа. Кроме того, поскольку различие в аминокислотной последовательности шарнирной области оказывает большое влияние на антигенсвязывающую активность, считают, что предпочтительные точки аминокислотных замен в аминокислотной последовательности изотипа дикого типа находятся внутри шарнирной области.
Когда антигенсвязывающую активность антигенсвязывающего вещества при pH 5,8 ослабляют по сравнению с указанной активностью при pH 7,4 (когда увеличивают значение KD (pH 5,8)/KD (pH 7,4)) с помощью описанных выше способов и т.п., обычно предпочтительно, чтобы значение KD (pH 5,8)/KD (pH 7,4) превышало в два раза и более, более предпочтительно в пять раз или более, и даже предпочтительнее в десять раз или более указанное значение исходного антитела, хотя настоящее изобретение особенно не ограничивается ими.
Здесь выражение «улучшение фармакокинетики» означает увеличение времени, требуемого для элиминации антигенсвязывающей молекулы из плазмы (например, достижения состояния, при котором антигенсвязывающая молекула не может возвращаться в плазму вследствие деградации в клетках, или по другим причинам) после введения животному, такому как человек, мышь, крыса, обезьяна, кролик или собака, а также увеличение времени сохранения в плазме антигенсвязывающей молекулы, находящейся в форме, способной к связыванию с антигенами (например, находящейся в свободной от антигена форме), в течение периода времени, пока она не элиминируется из плазмы после введения. Даже если антигенсвязывающая молекула циркулирует в плазме, она не может связываться с антигеном, когда она уже связана с другим антигеном. В результате, период времени, в течение которого антигенсвязывающая молекула может вновь связываться с другим антигеном, увеличивается (увеличивается шанс связать другой антиген) при увеличении периода времени, в течение которого антигенсвязывающая молекула находится в свободной от антигена форме. Это делает возможным укорочение периода времени, в течение которого антиген свободен от антигенсвязывающих молекул in vivo (другими словами, увеличение периода времени, в течение которого антиген связан антигенсвязывающей молекулой). Например, отношение антигенов, связанных с антигенсвязывающими молекулами, к антигенам в организме в плазме (общему количеству молекул антигена, связанных с антигенсвязывающими молекулами и свободных от них) обычно уменьшается в определенный период времени после введения антигенсвязывающих молекул. Однако такое уменьшение можно остановить (например, степень уменьшения можно сделать меньшей) путем увеличения времени сохранения антигенсвязывающих молекул в форме, способной к связыванию с антигенами. Это приводит к увеличению отношения антигенов, связанных с антигенсвязывающими молекулами, к антигенам в организме в определенный период времени после введения антитела.
В частности, в настоящем изобретении «улучшение фармакокинетики» не означает обязательно увеличение (удлинение) периода времени, требуемого для элиминации антигенсвязывающей молекулы после введения. Даже если период времени, требуемый для элиминации антигенсвязывающей молекулы после введения, остается неизменным, можно сказать, что фармакокинетики «улучшена» в настоящем изобретении, если:
увеличено время сохранения в плазме антигенсвязывающей молекулы, находящейся в форме, способной к связыванию с антигеном, (например, антигенсвязывающей молекулы, находящейся в свободной от антигена форме);
укорочен период времени, в течение которого антиген свободен от антигенсвязывающей молекулы в организме (другими словами, увеличен период времени, в течение которого антигенсвязывающая молекула связана с антигеном); и
увеличено отношение антигенов, связанных с антигенсвязывающими молекулами, к антигенам в организме. Таким образом, в настоящем изобретении «улучшение фармакокинетики» включает по крайней мере:
(1) увеличение времени, требуемого для элиминации антигенсвязывающей молекулы из плазмы после введения антигенсвязывающей молекулы;
(2) увеличение времени сохранения в плазме антигенсвязывающей молекулы в форме, способной к связыванию с антигеном, после введения антигенсвязывающей молекулы;
(3) укорочение периода времени, в течение которого антиген свободен от антигенсвязывающей молекулы в организме после введения антигенсвязывающей молекулы, (увеличение периода времени, в течение которого антигенсвязывающая молекула связана с антигеном в организме); и
(4) увеличение отношения антигенов, связанных с антигенсвязывающими молекулами, к антигенам в организме.
Когда антигеном является растворимый антиген, присутствующий в плазме, даже если фармакокинетика антигенсвязывающей молекулы (скорость элиминации из плазмы) является эквивалентной, существуют случаи, когда элиминация антигена, связанного с антигенсвязывающей молекулой, ускоряется. Ухудшение фармакокинетики антигена (ускорение элиминации из плазмы) приводит к относительному улучшению фармакокинетики антигенсвязывающей молекулы, и, следовательно, приводит к увеличению времени, в течение которого антигенсвязывающая молекула присутствует в плазме в форме, способной к связыванию с антигенами. Таким образом, в одном варианте осуществления, «улучшение фармакокинетики» антигенсвязывающей молекулы настоящего изобретения включает увеличение скорости элиминации растворимых антигенов из плазмы после введения антигенсвязывающих молекул (способности антигенсвязывающей молекулы к элиминации антигенов из плазмы).
В настоящем изобретении, когда антигеном является мембранный антиген, можно определить, связывается ли связывающая один антиген молекула с множеством антигенов, с помощью проверки того, улучшена ли фармакокинетика антигенсвязывающей молекулы. «Улучшение фармакокинетики» можно определить с помощью следующего способа. Например, увеличено ли время, требуемое для элиминации антигенсвязывающей молекулы после введения, можно оценить с помощью определения любого одного из параметров для антигенсвязывающей молекулы, таких как время полужизни в плазме, среднее время сохранения в плазме и клиренс в плазме (“Pharmacokinetics: Enshu-niyoru Rikai (Understanding through practice)” Nanzando). Например, когда увеличивается время полужизни в плазме или среднее время сохранения в плазме антигенсвязывающей молекулы, введенной мышам, крысам, обезьянам, кроликам, собакам, людям или животным, считают, что фармакокинетика антигенсвязывающей молекулы улучшена. Эти параметры можно определить с помощью известных квалифицированным в данной области техники специалистам способов. Например, параметры можно надлежащим образом определить с помощью некомпартментного анализа, используя программное обеспечение для анализа фармакокинетики WinNonlin (Pharsight) в соответствии с прилагаемым руководством.
Альтернативно, увеличено ли время сохранения в плазме антигенсвязывающей молекулы в форме, способной к связыванию с антигенами, после введения антигенсвязывающей молекулы, можно определить с помощью измерения концентрации в плазме свободной от антигена антигенсвязывающей молекулы и определения любого одного из параметров для свободной от антигена антигенсвязывающей молекулы, таких как время полужизни в плазме, среднее время сохранения в плазме и клиренс в плазме. Концентрацию свободной от антигена антигенсвязывающей молекулы в плазме можно измерить с помощью известных квалифицированным в данной области техники специалистам способов. Например, такие измерения описываются в Clin Pharmacol. 2008 Apr; 48(4): 406-417.
Кроме того, укорочен ли период времени, в течение которого антиген свободен от антигенсвязывающих молекул в организме после введения антигенсвязывающих молекул (увеличен ли период времени, в течение которого антигенсвязывающая молекула связана с антигеном), можно оценить с помощью определения концентрации в плазме несвязанного антигена, который свободен от антигенсвязывающих молекул, и учета периода времени, в течение которого остается низкой концентрация свободного антигена в плазме или отношение количества свободного антигена к общему количеству антигена. Концентрацию в плазме свободного антигена и отношение количества свободного антигена к общему количеству антигена можно определить с помощью известных квалифицированным в данной области техники специалистам способов. Например, такие измерения описываются в Pharm Res. 2006 Jan; 23(1): 95-103. Альтернативно, когда антиген проявляет некоторое действие in vivo, можно определить, связан ли антиген антигенсвязывающей молекулой, которая нейтрализует действие антигена, (антагонистической молекулой), с помощью проверки того, нейтрализовано ли действие антигена. Нейтрализацию действия антигена можно определить с помощью анализа in vivo маркера, отражающего действие антигена. Связан ли антиген антигенсвязывающей молекулой, которая активирует действие антигена, (агонистической молекулой), можно определить с помощью анализа in vivo маркера, отражающего действие антигена.
Нет особого ограничения в отношении определения концентрации в плазме свободного антигена и отношения количества свободного антигена к общему количеству антигена, и анализа in vivo маркера, но определение предпочтительно проводят после определенного периода времени после введения антигенсвязывающего вещества. В настоящем изобретении такой период времени после введения антигенсвязывающего вещества особенно не ограничивается, и подходящий период времени может быть определен квалифицированными в данной области техники специалистами в зависимости от свойств вводимого антигенсвязывающего вещества и т.п. Примерами такого периода времени являются один день после введения антигенсвязывающего вещества, три дня после введения антигенсвязывающего вещества, семь дней после введения антигенсвязывающего вещества, 14 дней после введения антигенсвязывающего вещества и 28 дней после введения антигенсвязывающего вещества.
В настоящем изобретении предпочтительным является улучшение фармакокинетики у человека. Даже когда трудно определить сохранение в плазме человека, его можно предсказать на основе сохранения в плазме мышей (например, нормальных мышей, экспрессирующих антиген человека трансгенных мышей и экспрессирующих FcRn человека трансгенных мышей) или обезьян (например, яванских макак).
Способы определения сохранения в плазме особенно не ограничиваются. Определение можно провести, например, в соответствии со способами, описанными здесь в примерах.
Способна ли антигенсвязывающая молекула к многократному связыванию с антигенами, можно определить с помощью проверки того, подвергается ли диссоциации в кислотных условиях, одинаковых с условиями в эндосоме, антиген, связанный с антигенсвязывающей молекулой в нейтральных условиях, одинаковых с условиями в плазме, и со сколькими антигенами может вновь связаться антигенсвязывающая молекула в нейтральных условиях. В частности, определение можно провести путем предоставления антигенсвязывающей молекуле и антигену возможности образовывать комплекс в нейтральных условиях, подвергания комплекса воздействию кислотных условий в течение заданного периода времени и затем проверки того, может ли антигенсвязывающая молекула вновь связываться с антигеном в нейтральных условиях, используя устройство для анализа взаимодействий антигенсвязывающих молекул с антигенами, такое как Biacore. Когда способность к связыванию антигенов антигенсвязывающей молекулы, которой сообщена способность к связыванию в зависимости от pH, улучшена с удвоением такой способности антигенсвязывающей молекулы до модификации, можно считать, что число раз связывания антигенсвязывающей молекулы, которой сообщена способность к связыванию в зависимости от pH, увеличено вдвое по сравнению с таким числом для антигенсвязывающей молекулы до модификации. Альтернативно, когда антигеном является мембранный антиген, и, следовательно, антигенсвязывающая молекула элиминируется из плазмы посредством опосредованного антигенами поглощения и деградации в липосоме, увеличено ли число раз связывания антигенсвязывающей молекулы, которой сообщена способность к связыванию в зависимости от pH, по сравнению с таким числом до модификации, можно определить путем сравнения фармакокинетики или длительности связывания с антигеном между антигенсвязывающей молекулой, которой сообщена способность к связыванию в зависимости от pH, и антигенсвязывающей молекулой до модификации. Например, когда длительность связывания с антигеном антигенсвязывающей молекулы, которой сообщена способность к связыванию в зависимости от pH, увеличивается вдвое относительно указанной длительности для антигенсвязывающей молекулы до модификации, считают, что число раз связывания антигенсвязывающей молекулы, которой сообщена способность к связыванию в зависимости от pH, увеличивается с удвоением такого числа для антигенсвязывающей молекулы до модификации. Альтернативно, когда определяют концентрацию в плазме несвязанного антигена, который свободен от антигенсвязывающей молекулы, и увеличивается вдвое период времени, в течение которого остается низкой концентрация в плазме свободного антигена или отношение количества свободного антигена к общему количеству антигена, считают, что число раз связывания антигенсвязывающей молекулы, которой сообщена способность к связыванию в зависимости от pH, увеличивается вдвое по сравнению с таким числом для антигенсвязывающей молекулы до модификации.
Когда антигеном является растворимый антиген, если антиген, связанный с антигенсвязывающей молекулой в нейтральных условиях в плазме, подвергается диссоциации в эндосоме, а антигенсвязывающая молекула возвращается в плазму, антигенсвязывающая молекула может снова связываться с антигеном в нейтральных условиях в плазме. Таким образом, антигенсвязывающая молекула, которая обладает свойствами диссоциироваться от антигена в кислотных условиях в эндосоме, способна к многократному связыванию с антигенами. По сравнению с тем, когда связанный с антигенсвязывающей молекулой антиген не подвергается диссоциации в эндосоме (антиген остается связанным с антигенсвязывающей молекулой после возвращения в плазму), когда связанный с антигенсвязывающей молекулой антиген подвергается диссоциации в эндосомах, антиген доставляется в лизосому и затем подвергается деградации, и, таким образом, скорость элиминации антигена из плазмы увеличивается. Т.е., также можно определить, способна ли антигенсвязывающая молекула к многократному связыванию с антигенами, используя в качестве показателя скорость элиминации антигена из плазмы. Скорость элиминации антигена из плазмы можно определить, например, с путем введения антигенов (например, мембранного антигена) и антигенсвязывающей молекулы in vivo и затем измерения концентрации антигенов в плазме. Когда антиген (например, мембранный антиген) продуцируется или секретируется in vivo, концентрация антигена в плазме уменьшается, если увеличивается скорость элиминации антигена из плазмы. Следовательно, можно также определить, способна ли антигенсвязывающая молекула к многократному связыванию с антигенами, используя в качестве показателя концентрацию антигена в плазме.
Здесь «увеличение числа раз связывания с антигеном антигенсвязывающей молекулы» означает, что увеличивается число циклов при принятии за один цикл процесс, в котором антигенсвязывающая молекула, введенная человеку, мыши, обезьяне или т.п., связывается с антигеном и подвергается интернализации в клетку. В частности, здесь выражение «антигенсвязывающая молекула связывается дважды с антигеном» означает, что связанная с антигеном антигенсвязывающая молекула подвергается интернализации в клетку и высвобождается в свободной от антигена форме наружу из клетки, и выброшенная антигенсвязывающая молекула вновь связывается с другим антигеном и снова подвергается интернализации в клетку.
Когда антигенсвязывающая молекула подвергается интернализации в клетку, она может быть в форме, связанной одним антигеном, или двумя или более антигенами.
Здесь выражение «число раз связывания с антигеном антигенсвязывающей молекулы увеличивается» не означает обязательно, что число раз связывания с антигеном увеличивается для каждой из антигенсвязывающих молекул. Например, среди антигенсвязывающих молекул в композиции антигенсвязывающих молекул может увеличиваться доля антигенсвязывающих молекул, которые связываются с антигенами два и более раз, или может увеличиваться среднее число событий связывания антигенсвязывающих молекул в композиции антигенсвязывающих молекул.
В настоящем изобретении предпочтительно, когда число раз связывания с антигеном антигенсвязывающей молекулы увеличивается при введении молекулы человеку. Однако когда трудно определить число раз связывания с антигеном у человека, такое число у человека можно предсказать на основе результатов, полученных с помощью in vitro анализа или измерения с использованием мышей (например, экспрессирующих антиген человека трансгенных мышей и экспрессирующих FcRn человека трансгенных мышей) или обезьян (например, яванских макак).
В настоящем изобретении предпочтительно, когда антигенсвязывающая молекула связывается с антигенами два или более раз. Например, предпочтительно, когда из антигенсвязывающих молекул в композиции антигенсвязывающих молекул по крайней мере 10% или больше, предпочтительно 30% или больше, более предпочтительно 50% или больше, и еще предпочтительнее 80% или больше (например, 90% или больше, 95% или больше и т.д.) связывается с антигенами два или более раз.
Здесь выражение «увеличение числа антигенов, которое может быть связано антигенсвязывающей молекулой» означает увеличение числа антигенов, которое может быть связано антигенсвязывающей молекулой в течение периода времени, пока антигенсвязывающая молекула не подверглась деградации в лизосоме клетки, после введения антигенсвязывающей молекулы животному, такому как человек, мышь или обезьяна.
Как правило, антитела, такие как IgG, имеют два связывающих домена, и, следовательно, одно антитело связывается максимум с двумя антигенами. Связанное с антигеном(ами) антитело подвергается интернализации в клетку, и антитело и антиген(ы) подвергаются деградации в лизосоме. Как правило, антитела, такие как IgG, могут связываться максимум с двумя антигенами. Когда антигенсвязывающая активность антигенсвязывающей молекулы, такой как антитело, при эндосомальном pH ослабляют по сравнению с такой активностью при pH в плазме с помощью способов настоящего изобретения, интернализованная в клетку антигенсвязывающая молекула, такая как антитело, подвергается диссоциации и высвобождается наружу из клетки, и, следовательно, может снова связываться с другим антигеном. Другими словами, способы настоящего изобретения делают возможным связывание антигенсвязывающей молекулы с числом антигеном, превышающим число ее антигенсвязывающих сайтов. В частности, при использовании способов настоящего изобретения, например, IgG, имеющий два антигенсвязывающих сайта, может связываться с тремя или более антигенами, предпочтительно четырьмя или более антигенами, в течение периода времени, пока антитело не подверглось деградации, после введения. Например, когда антителом является нейтрализующее антитело, «увеличение числа антигенов, которое может быть связано антигенсвязывающей молекулой» равнозначно «увеличению числа антигенов, которое антигенсвязывающая молекула может нейтрализовать». Таким образом, «связывает» может быть заменено словом «нейтрализует», когда антителом является нейтрализующее антитело.
В настоящем изобретении «увеличение числа антигенов, которое может быть связано антигенсвязывающей молекулой» не означает обязательно увеличение числа антигенов, которое может быть связано каждой антигенсвязывающей молекулой. Например, может увеличиваться среднее число антигенов, которое может быть связано антигенсвязывающей молекулой в композиции антигенсвязывающих молекул, или может увеличиваться доля антигенсвязывающих молекул, которые могут связываться с количеством антигенов, превышающим число ее антигенсвязывающих сайтов.
В настоящем изобретении предпочтительно, когда число антигенов, которое может быть связано антигенсвязывающей молекулой, увеличивается при введении молекулы человеку. Однако когда трудно определить такое число у человека, его можно предсказать на основе результатов, полученных с помощью in vitro анализа или измерения с использованием мышей (например, экспрессирующих антиген человека трансгенных мышей и экспрессирующих FcRn человека трансгенных мышей) или обезьян (например, яванских макак). Когда антителом является нейтрализующее антитело, обычно считают, что описываемое выше число раз связывания с антигенами антигенсвязывающей молекулы коррелирует с числом антигенов, которое может нейтрализовано антигенсвязывающей молекулой. Таким образом, число антигенов, которое может быть нейтрализовано антигенсвязывающей молекулой, можно определить с помощью способов, одинаковых с те, которые описаны выше для определения числа раз связывания антигенсвязывающей молекулы.
Кроме того, настоящим изобретением обеспечиваются способы связывания антигенсвязывающей молекулы с антигенами два или более раз в организме при введении антигенсвязывающей молекулы, антигенсвязывающая активность которой при кислотном pH ниже указанной активности при нейтральном pH.
Настоящее изобретением также относится к способам нейтрализации числа антигенов, превышающего число антигенсвязывающих сайтов антигенсвязывающей молекулы, обладающей нейтрализующей активностью, путем введения антигенсвязывающей молекулы, антигенсвязывающая активность которой при кислотном pH ниже указанной активности при нейтральном pH. Предпочтительно настоящее изобретение относится к способам нейтрализации трех или более антигенов, предпочтительно четырех или более антигенов путем введения IgG, антигенсвязывающая активность которого при кислотном pH ниже указанной активности при нейтральном pH.
Настоящее изобретением также относится к способам диссоциации внутри клетки антигена от связавшейся с ним внеклеточно антигенсвязывающей молекулы путем ослабления способности к связыванию с антигенами антигенсвязывающей молекулы при кислотном pH по сравнению с указанной способностью при нейтральном pH. В настоящем изобретении антиген может подвергаться диссоциации от антигенсвязывающей молекулы где-либо внутри клетки; однако предпочтительно, когда антиген подвергается диссоциации внутри ранней эндосомы. В настоящем изобретении выражение «антиген подвергается диссоциации внутри клетки от связавшейся с ним внеклеточно антигенсвязывающей молекулы» не означает обязательно, что каждый антиген, интернализованный в клетку благодаря связыванию с антигенсвязывающей молекулой, подвергается диссоциации от антигенсвязывающей молекулы внутри клетки. Приемлемо, когда доля антигена, которая подвергается диссоциации от антигенсвязывающей молекулы внутри клетки, увеличивается по сравнению с такой долей до ослабления способности к связыванию с антигенами антигенсвязывающей молекулы при кислотном pH по сравнению с такой способностью при нейтральном pH.
Кроме того, настоящее изобретение относится к способам усиления внутриклеточного связывания антигенсвязывающей молекулы, свободной от антигена, с FcRn путем ослабления способности к связыванию с антигенами антигенсвязывающей молекулы при кислотном pH по сравнению с указанной способностью при нейтральном pH. Как правило, FcRn связывается с антигенсвязывающей молекулой внутри эндосомы. Однако полагают, что связанная с мембранным антигеном антигенсвязывающая молекула не связывается с FcRn. Поэтому в предпочтительном варианте осуществления, когда антигеном является связанный с мембраной антиген, настоящее изобретение включает способы усиления эндосомальной диссоциации антигенов от антигенсвязывающих молекул и, следовательно, усиления связывания с FcRn антигенсвязывающих молекул путем ослабления способности к связыванию с антигенами антигенсвязывающей молекулы при эндосомальном pH (кислотном pH) по сравнению с указанной способностью при pH в плазме (нейтральном pH). Когда антигеном является растворимый антиген, антигенсвязывающая молекула может связываться с FcRn в присутствии антигена или в его отсутствие. Если диссоциацию антигена от антигенсвязывающей молекулы внутри эндосом можно ускорить путем ослабления способности к связыванию с антигенами антигенсвязывающей молекулы при pH внутри эндосом (кислотном pH) по сравнению с указанной способностью при pH в плазме (нейтральном pH), связывание с FcRn антигенсвязывающей молекулы, которая «свободна от антигена», можно усилить с помощью способов настоящего изобретения.
Независимо от того, является ли антиген связанным с мембраной или растворимым, если антигенсвязывающая молекула, свободная от антигена, может возвращаться в плазму с помощью FcRn, антигенсвязывающая молекула может снова связывается с антигеном. При повторении этого процесса антигенсвязывающая молекула может связываться с антигеном многократно. В настоящем изобретении выражение «усиления связывания с FcRn антигенсвязывающей молекулы внутри клетки не означает обязательно, что каждая антигенсвязывающая молекула связывается с FcRn. Приемлемо, когда увеличивается доля антигенсвязывающей молекулы, свободной от антигена, которая связывается с FcRn внутри клетки, по сравнению с такой долей до ослабления способности к связыванию с антигенами антигенсвязывающей молекулы при эндосомальном pH по сравнению с указанной способностью при pH в плазме. Антигенсвязывающие молекулы, предпочтительные в способах настоящего изобретения для усиления внутриклеточного связывания между антигенсвязывающей молекулой и FcRn, включают, например, антигенсвязывающие молекулы, которые связываются со связанными с мембранами антигенами (мембранными антигенами), такими как мембранные белки. Другие предпочтительные антигенсвязывающие молекулы включают антигенсвязывающие молекулы, которые связываются с растворимыми антигенами, таким как растворимые белки.
Способы усиления связывания антигенсвязывающей молекулы с FcRn внутри клетки иначе определяются как способы ускорения связывания с FcRn антигенсвязывающей молекулы внутри клетки, например, внутри эндосом.
Кроме того, настоящее изобретение относится к способам высвобождения антигенсвязывающей молекулы, которая связалась с антигеном и подверглась интернализации в клетку, в свободной от антигена форме наружу из клетки путем ослабления способности к связыванию с антигенами антигенсвязывающей молекулы при кислотном pH по сравнению с указанной способностью при нейтральном pH. В настоящем изобретении выражение «высвобождение антигенсвязывающей молекулы, которая связалась с антигеном и подверглась интернализации в клетку, в свободной от антигена форме наружу из клетки» не означает обязательно, что каждая антигенсвязывающая молекула, которая связалась с антигеном и подверглась интернализации в клетку, высвобождается в свободной от антигена форме наружу из клетки. Приемлемо, когда увеличивается доля антигенсвязывающих молекул, которые высвобождаются наружу из клетки, по сравнению с указанной долей до ослабления способности к связыванию с антигенами антигенсвязывающей молекулы при кислотном pH по сравнению с указанной способностью при нейтральном pH. Предпочтительно, когда высвобождаемая наружу из клетки антигенсвязывающая молекула сохраняет способность к связыванию антигенов. Кроме того, способ высвобождения антигенсвязывающей молекулы, которая связалась с антигеном и подверглась интернализации в клетку, в свободной от антигена форме наружу из клетки может также упоминаться как способ сообщения антигенсвязывающей молекуле такого свойства, что антигенсвязывающая молекула становится более легко высвобождаемой наружу из клетки в свободной от антигена форме, когда антигенсвязывающая молекула связывается с антигеном и подвергается интернализации в клетку.
Кроме того, настоящее изобретение относится к способам увеличения способности антигенсвязывающих молекул к элиминации антигенов в плазме путем ослабления способности к связыванию с антигенами антигенсвязывающей молекулы при кислотном pH по сравнению с указанной способностью при нейтральном pH. В настоящем изобретении выражение «способность к элиминации антигенов в плазме» относится е способности к элиминации из плазмы антигенов, которые присутствуют в плазме, когда антигенсвязывающие молекулы вводятся in vivo или секретируются in vivo. Таким образом, в настоящем изобретении выражение «увеличение способности антигенсвязывающей молекулы к элиминации антигена из плазмы» означает, что при введении антигенсвязывающих молекул in vivo увеличивается скорость элиминации антигенов из плазмы по сравнению с указанной скоростью до ослабления способности к связыванию с антигенами антигенсвязывающих молекул при кислотном pH по сравнению с такой способностью при нейтральном pH. Увеличивается ли способность антигенсвязывающей молекулы к элиминации антигенов в плазмы, можно определить, например, путем введения растворимых антигенов и антигенсвязывающих молекулы in vivo и затем измерения концентрации растворимых антигенов в плазме. Когда концентрация растворимых антигенов в плазме после введения растворимых антигенов и антигенсвязывающих молекул понижается при ослаблении способности к связыванию с антигенами антигенсвязывающей молекулы при кислотном pH по сравнению с такой способностью при нейтральном pH, может быть решено, что увеличивается способность антигенсвязывающей молекулы к элиминации антигенов в плазме.
Настоящее изобретение также относится к способам улучшения фармакокинетики антигенсвязывающей молекулы путем замещения гистидином или не встречающейся в природе аминокислотой по крайней мере одной аминокислоты в антигенсвязывающей молекуле или вставки гистидина или не встречающейся в природе аминокислоты в указанную молекулу.
Кроме того, настоящим изобретением обеспечиваются способы увеличения числа раз связывания с антигенами антигенсвязывающей молекулы путем замещения гистидином или не встречающейся в природе аминокислотой по крайней мере одной аминокислоты в антигенсвязывающей молекуле или вставки гистидина или не встречающейся в природе аминокислоты в указанную молекулу.
Кроме того, настоящее изобретение относится к способам увеличения числа антигенов, которое может быть связано антигенсвязывающей молекулой, путем замещения гистидином или не встречающейся в природе аминокислотой по крайней мере одной аминокислоты в антигенсвязывающей молекуле или вставки гистидина или не встречающейся в природе аминокислоты в указанную молекулу.
Настоящим изобретением также обеспечиваются способы диссоциации антигена внутри клетки от связавшейся с ним внеклеточно антигенсвязывающей молекулы путем замещения гистидином или не встречающейся в природе аминокислотой по крайней мере одной аминокислоты в антигенсвязывающей молекуле или вставки гистидина или не встречающейся в природе аминокислоты в указанную молекулу.
Настоящим изобретением также обеспечиваются способы высвобождения антигенсвязывающей молекулы, которая связалась с антигеном и подверглась интернализации в клетку, в свободной от антигена форме наружу из клетки путем замещения гистидином или не встречающейся в природе аминокислотой по крайней мере одной аминокислоты в антигенсвязывающей молекуле или вставки гистидина или не встречающейся в природе аминокислоты в указанную молекулу.
Настоящим изобретением также обеспечиваются способы увеличения способности антигенсвязывающей молекулы к элиминации антигенов в плазме путем замещения гистидином или не встречающейся в природе аминокислотой по крайней мере одной аминокислоты в антигенсвязывающей молекуле или вставки гистидина или не встречающейся в природе аминокислоты в указанную молекулу.
Точка мутации (замены, вставки и т.д.) на гистидин или на не встречающуюся в природе аминокислоту особенно не ограничивается. Гистидином или не встречающейся в природе аминокислотой может быть заменена аминокислота в любой точке или он (она) может быть вставлен(а) в любую точку. Предпочтительные точки замены гистидином или не встречающейся в природе аминокислотой или его (ее) вставки включают, например, точки внутри района, который оказывает влияние на способность к связыванию с антигеном антигенсвязывающий молекулы. Например, когда антигенсвязывающей молекулой является антитело, такие точки включают вариабельную область или CDR антитела. Число мутаций на гистидин или не встречающуюся в природе аминокислоту особенно не ограничивается. Замена гистидином или не встречающейся в природе аминокислотой или его (ее) вставка может быть осуществлена в одной точке, или в двух или более точках. Кроме того, может быть введена делеция, добавление, вставка и/или замена других аминокислот одновременно с заменой гистидином или не встречающейся в природе аминокислотой или с его (ее) вставкой.
В настоящем изобретении, когда антигенсвязывающей молекулой является антитело, возможные точки замены гистидином или не встречающейся в природе аминокислотой включают, например, точки внутри последовательности CDR или последовательности, ответственной за структуру CDR антитела. Такие точки включают, например, перечисленные ниже точки. Положения аминокислот пронумерованы на основе нумерации по Kabat (Kabat EA et al., (1991) Sequences of Proteins of Immunological Interest, NIH).
Тяжелая цепь: H27, H31, H32, H33, H35, H50, H58, H59, H61, H62, H63, H64, H65, H99, H100b и H102
Легкая цепь: L24, L27, L28, L32, L53, L54, L56, L90, L92 и L94
Среди указанных выше точек H32, H61, L53, L90 и L94 могли бы быть универсальными точками модификаций.
Когда антигеном является рецептор для IL-6 (например, рецептор для IL-6 человека), предпочтительные точки модификаций включают следующие точки. Однако точки модификаций особенно не ограничиваются ими.
Тяжелая цепь: H27, H31, H32, H35, H50, H58, H61, H62, H63, H64, H65, H100b и H102
Легкая цепь: L24, L27, L28, L32, L53, L56, L90, L92 и L94
Когда гистидином или не встречающейся в природе аминокислотой заменяют аминокислоты во множестве точек, предпочтительные комбинации точек замен включают, например, комбинацию H27, H31 и H35; комбинацию H27, H31, H32, H35, H58, H62 и H102; комбинацию L32 и L53 и комбинацию L28, L32 и L53. Кроме того, предпочтительные комбинации точек замен в тяжелой и легкой цепи включают комбинацию H27, H31, L32 и L53.
Когда антигеном является IL-6 (например, IL-6 человека), предпочтительные точки модификаций включают следующие точки. Однако точки модификаций особенно не ограничиваются ими.
Тяжелая цепь: H32, H59, H61 и H99
Легкая цепь: L53, L54, L90 и L94
Когда антигеном является рецептор для IL-31 (например, рецептор для IL-31 человека), предпочтительные точки модификаций включают Н33. Однако точки модификаций особенно не ограничиваются им.
Касательно указанных выше точек, аминокислота только в одной точке может быть замещена гистидином или не встречающейся в природе аминокислотой. Альтернативно, аминокислоты во множестве точек могут быть замещены гистидином или не встречающейся в природе аминокислотой.
Способы настоящего изобретения применимы к любым антигенсвязывающим молекулам, независимо от типа антигена-мишени.
Антигенсвязывающие молекулы по настоящему изобретению особенно не ограничиваются при условии, что они обладают специфической активностью связывания представляющего интерес антигена. Предпочтительные антигенсвязывающие молекулы по настоящему изобретению включают, например, вещества, имеющие антигенсвязывающий домен антитела. Антигенсвязывающий домен антитела включает, например, CDR или вариабельную область. Когда антигенсвязывающим доменом антитела является CDR, антигенсвязывающая молекула может включать все шесть CDR целого антитела, или один, или два или более из них. Альтернативно, когда антигенсвязывающая молекула включает CDR в качестве связывающего домена антитела, CDR может включать делецию, замену, добавление и/или вставку аминокислоты или может представлять собой частичный CDR.
Кроме того, когда антигенсвязывающая молекула включает характерную для антитела константную область, настоящее изобретение относится к способам улучшения фармакокинетики антигенсвязывающих молекул путем модификации (например, замены, делеции, добавления и/или вставки аминокислоты) характерной для антитела константной области в антигенсвязывающей молекуле.
Кроме того, когда антигенсвязывающая молекула включает характерную для антитела константную область, настоящим изобретением обеспечиваются способы увеличения числа раз связывания с антигенами антигенсвязывающей молекулы путем модификации (например, замены, делеции, добавления и/или вставки аминокислоты) характерной для антитела константной области в антигенсвязывающей молекуле.
Кроме того, когда антигенсвязывающая молекула включает характерную для антитела константную область, настоящее изобретение относится к способам увеличения числа антигенов, которое может быть связано антигенсвязывающей молекулой, путем модификации (например, замены, делеции, добавления и/или вставки аминокислоты) характерной для антитела константной области в антигенсвязывающей молекуле.
Кроме того, когда антигенсвязывающая молекула включает характерную для антитела константную область, настоящее изобретение относится к способам диссоциации внутри клетки антигена от связавшейся с ним внеклеточно антигенсвязывающей молекулы путем модификации (например, замены, делеции, добавления и/или вставки аминокислоты) характерной для антитела константной области в антигенсвязывающей молекуле.
Кроме того, когда антигенсвязывающая молекула включает характерную для антитела константную область, настоящее изобретение относится к способам высвобождения антигенсвязывающей молекулы, которая связалась с антигеном и подверглась интернализации в клетку, в свободной от антигена форме наружу из клетки путем модификации (например, замены, делеции, добавления и/или вставки аминокислоты) характерной для антитела константной области в антигенсвязывающей молекуле.
Кроме того, когда антигенсвязывающая молекула включает характерную для антитела константную область, настоящее изобретение относится к способам увеличения способности антигенсвязывающей молекулы к элиминации антигена в плазме путем модификации (например, замены, делеции, добавления и/или вставки аминокислоты) характерной для антитела константной области в антигенсвязывающей молекуле.
В предпочтительном варианте осуществления антигенсвязывающее вещество по настоящему изобретению включает антигенсвязывающие вещества, включающие FcRn-связывающий район. После интернализации в клетки антигенсвязывающие вещества, включающие FcRn-связывающий район, могут возвращаться в плазму путем возращения благодаря FcRn. Связывающий FcRn район является предпочтительно доменом, который непосредственно связывается с FcRn. Предпочтительный FcRn-связывающий район включает, например, Fc-области антитела. Однако FcRn-связывающим районом настоящего изобретения может быть район, который может связываться с полипептидом, обладающим способностью к связыванию с FcRn, таким как альбумин или IgG, поскольку такой район, который может связываться с полипептидом, обладающим способностью к связыванию FcRn, может связываться с FcRn опосредованно через альбумин, IgG и т.д.
Антигены, распознаваемые антигенсвязывающими молекулами, такими как антитела, представляющими интерес в способах по настоящему изобретению, особенно не ограничиваются. Такие представляющие интерес антитела могут распознавать любой антиген. Антитела, фармакокинетика которых должна быть улучшена с помощью способов по настоящему изобретению, включают, например, антитела, которые распознают мембранные антигены, такие как рецепторные белки (связанные с мембранами рецепторы и растворимые рецепторы) и маркеры клеточной поверхности, и антитела, которые распознают растворимые антигены, такие как цитокины. Предпочтительные примеры мембранных антигенов по настоящему изобретению включают мембранные белки. Примеры растворимых антигенов по настоящему изобретению включают растворимые белки. Антигены, распознаваемые антителами, фармакокинетика которых должна быть улучшена с помощью способов по настоящему изобретению, включают, например, IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12, IL-15, IL-31, IL-23, рецептор IL-2, рецептор IL-6, рецептор OSM, gp130, рецептор IL-5, CD40, CD4, Fas, остеопонтин, CRTH2, CD26, PDGF-D, CD20, вызывающий хемотаксис моноцитов фактор, CD23, TNF-α, HMGB-1, α4 интегрин, ICAM-1, CCR2, CD11a, CD3, IFNγ, BLyS, HLA-DR, TGF-β, CD52 и рецептор IL-31. Особенно предпочтительные антигены включают рецептор IL-6.
Кроме того, антигенсвязывающая молекула, представляющая интерес в способах по настоящему изобретению, включает антигенсвязывающие молекулы, обладающие антагонистической активностью, (антагонистические антигенсвязывающие молекулы) и антигенсвязывающие молекулы, обладающие агонистической активностью, (агонистические антигенсвязывающие молекулы). В предпочтительном варианте осуществления антигенсвязывающая молекула включает антагонистические антигенсвязывающие молекулами, в частности, антагонистические антигенсвязывающие молекулы, которые распознают мембранные антигены, такие как рецепторы, или растворимые антигены, такие как цитокины. Например, антагонистическая антигенсвязывающая молекула, которая распознает рецептор, ингибирует связывание лиганда с рецептором путем связывания с рецептором и, таким образом, ингибирует передачу сигнала, опосредуемую рецептором.
В настоящем изобретении представляющая интерес антигенсвязывающая молекула особенно не ограничивается и может быть любой антигенсвязывающей молекулой. Антигенсвязывающая молекула по настоящему изобретению предпочтительно включает как антигенсвязывающую активность (антигенсвязывающий район), так и FcRn-связывающий район. В частности, предпочтительная антигенсвязывающая молекула настоящего изобретения включает район, который связывается с FcRn человека. Антигенсвязывающая молекула, включающая как антигенсвязывающую активность, так и FcRn-связывающий район, включает, например, антитела. Антитела, предпочтительные в рамках настоящего изобретения, включают, например, антитела IgG-типа. Когда используемым антителом является антитело IgG-типа, тип IgG особенно не ограничивается; может использоваться IgG, относящийся к любому изотипу (субклассу), такому как IgG1, IgG2, IgG3 или IgG4. Кроме того, в константную область любого из этих изотипов IgG могут быть введены затрагивающие аминокислоты мутации (например, M73). Вводимые, затрагивающие аминокислоты мутации включают, например, мутации, усиливающие или ослабляющие связывание с рецептором Fcγ (Proc. Natl. Acad. Sci. USA. 2006 Mar 14; 103(11): 4005-4010), и мутации, усиливающие или ослабляющие связывание с FcRn (J. Biol. Chem. 2001 Mar 2; 276(9): 6591-604), но не ограничиваются этими примерами. Альтернативно, можно также изменить связывание в зависимости от pH путем отбора подходящей константной области, например, IgG2.
Когда представляющей интерес в настоящем изобретении антигенсвязывающей молекулой является антитело, она может быть антителом, полученным из любого животного, таким как антитело мыши, антитело человека, антитело крысы, антитело кролика, антитело козы или антитело верблюда. Кроме того, антитело может быть модифицированным антителом, например, химерным антителом, и, в частности, модифицированным антителом, включающим аминокислотную замену в последовательности гуманизированного антитела и т.д. Антитела также включают биспецифические антитела, продукты модификации антител, связанные с различными молекулами, и полипептиды, включающие фрагменты антител.
«Химерные антитела» представляют собой антитела, получены объединением последовательностей, происходящих из различных животных. В частности, химерное антитело включает, например, антитела, имеющие вариабельные (V) области тяжелой и легкой цепей из мышиного антитела и константные (С) области тяжелой и легкой цепей из антитела человека.
«Гуманизированные антитела», также называемые реконструированными человеческими антителами, представляют собой антитела, в которых определяющие комплементарность участки (CDR) антитела, происходящего из не являющегося человеком млекопитающего, например, мыши, пересажены в CDR антитела человека. Способы идентификации CDR известны (Kabat et al., Sequence of Proteins of Immunological Interest (1987), National Institute of Health, Bethesda, Md.; Chothia et al., Nature (1989) 342: 877). Также известны общие методы генетической рекомбинации, подходящие для этой цели (см. заявку на Европейский патент EP 125023 и WO 96/02576).
Биспецифическое антитело относится к антителу, которое имеет, в одной и той же молекуле антитела, вариабельные области, которые распознают различные эпитопы. Биспецифическим антителом может быть антитело, которое распознает два или более различных антигенов, или антитело, которое распознает два или более различных эпитопа на одном и том же антигене.
Кроме того, полипептиды, включающие фрагменты антитела, включают, например, Fab-фрагменты, F(abʹ)2-фрагменты, scFv (Nat Biotechnol. 2005 Sep; 23(9): 1126-1136), доменные антитела (dAb) (WO 2004/058821, WO 2003/002609), scFv-Fc (WO 2005/037989), dAb-Fc и слитые с Fc белки. Конечно, молекулы, включающие Fc-домен, обладают активностью связывания с FcRn и, следовательно, подходят для использования в способах, раскрытых в настоящем изобретении.
Кроме того, используемыми в настоящем изобретении антигенсвязывающими молекулами могут быть антителоподобные молекулы. Антителоподобная молекула является молекулой, которая может проявлять действия при связывании с молекулой-мишенью (Current Opinion in Biotechnology 2006, 17: 653-658; Current Opinion in Biotechnology 2007, 18: 1-10; Current Opinion in Structural Biology 1997, 7: 463-469; Protein Science 2006, 15: 14-27), и включает, например, сконструированные белки с анкириновым повтором (DARPin) (WO 2002/020565), аффитело (Affibody) (WO 1995/001937), авимер (Avimer) (WO 2004/044011; WO 2005/040229), и аднектин (Adnectin) (WO 2002/032925). Если эти антителоподобные молекулы могут связываться с молекулами-мишенями pH-зависимым образом, возможно связывание одной молекулы с множеством молекул-мишеней.
Кроме того, антигенсвязывающей молекулой может быть рецепторный белок или слитый белок рецептор-Fc, который связывается с мишенью, включающий, например, слитый белок TNFR-Fc, слитый белок IL1R-Fc, слитый белок VEGFR-Fc и слитый белок CTLA4-Fc (Nat. Med. 2003 Jan; 9(1): 47-52; BioDrugs. 2006; 20(3): 151-60). Если такие рецепторные белки и слитые белки рецептор-Fc могут связываться с молекулами-мишенями pH-зависимым образом, возможно связывание одной молекулы с множеством молекул-мишеней.
Кроме того, антигенсвязывающая молекула может быть искусственным белком-лигандом или слитым с искусственным лигандом белком, который связывается с мишенью и обладает нейтрализующим эффектом, и включает, например, мутантный IL-6 (EMBO J. 1994 Dec. 15; 13(24): 5863-5870). Если такие искусственные белки-лиганды и слитые с искусственным лигандом белки могут связываться с молекулами-мишенями pH-зависимым образом, возможно связывание одной молекулы с множеством молекул-мишеней.
Кроме того, антитела по настоящему изобретению могут включать модифицированные сахарные цепи. Антитела с модифицированными сахарными цепями включают, например, антитела с модифицированным гликозилированием (WO 99/54342), антитела, лишенные фукозы, которую добавляют в сахарную цепь (WO 00/61739; WO 02/31140; WO 2006/067847; WO 2006/067913), и антитела, имеющие сахарные цепи с делящим пополам N-ацетилглюкозамином (GlcNAc) (WO 02/79255).
Хотя способы по настоящему изобретению не ограничиваются какой-либо конкретной теорией, связь между созданием более слабой способности к связыванию антигена при кислотном pH, чем при нейтральном pH, улучшением фармакокинетики и многократным связыванием с антигеном можно объяснить, например, следующим образом.
Например, когда антителом является антитело, которое связывается с мембранным антигеном, введенное в организм антитело связывается с антигеном и затем поступает посредством интернализации в эндосомы в клетках вместе с антигеном, в то время как антитело продолжает быть связанным с антигеном. Затем антитело перемещается в лизосомы, в то время как антитело продолжает быть связанным с антигеном, и антитело подвергается деградации лизосомой вместе с антигеном. Опосредованную интернализацией элиминацию из плазмы называют антигензависимой элиминацией, и о такой элиминации сообщалось для многочисленных молекул антител (Drug Discov. Today. 2006 Jan; 11(1-2): 81-88). Когда одна молекула антитела IgG-типа связывается с антигенами двухвалентно, одна молекула антитела подвергается интернализации, в то время как антитело продолжает быть связанным с двумя молекулами антигена, и подвергается деградации в лизосоме. В результате, в случае обычных антител, одна молекула антитела IgG-типа не может связываться с тремя или более молекулами антигена. Например, одна молекула антитела IgG-типа, обладающая нейтрализующей активностью, не может нейтрализовать три или более молекулы антигена.
Относительно длительное сохранение (медленная элиминация) молекул IgG в плазме обусловлено действием FcRn, который известен как рецептор для возвращения молекул IgG. После поступления в эндосомы посредством пиноцитоза молекулы IgG связываются с FcRn, экспрессируемым в эндосомах, в кислотных условиях в эндосомах. В то время как молекулы IgG, которые не связались с FcRn, переносятся в лизосомы, где они подвергаются деградации, молекулы IgG, связавшиеся в FcRn, перемещаются на клеточную поверхность и снова возвращаются в плазму при диссоциации от FcRn в нейтральных условиях в плазме.
Альтернативно, когда антителом является антитело, которое связывается с растворимым антигеном, введенное в организм антитело связывается с антигеном и затем поступает в клетки, в то время как антитело продолжает быть связанным с антигеном. Многие поступившие в клетки антитела высвобождаются наружу из клеток благодаря FcRn. Однако, поскольку наружу из клеток высвобождаются антитела, которые продолжают быть связанными с антигенами, антитела не могут снова связываться с антигенами. Таким образом, подобно антителам, которые связываются с мембранными антигенами, в случае обычных антител, одна молекула антитела IgG-типа не может связываться с тремя или более молекулами антигена.
Авторы настоящего изобретения рассуждали, что, когда антитела, связанные с антигенами, такими как мембранные антигены, поступают в эндосомы посредством интернализации, в то время как антитела, которые продолжают быть связанными с антигенами, перемещаются в лизосомы и подвергаются деградации, антитела IgG-типа, от которых антигены подверглись диссоциации в эндосомах, могли бы связываться с FcRn, которые экспрессируются в эндосомах. В частности, авторы настоящего изобретения обнаружили, что антитело, которое сильно связывается с антигеном в плазме, но слабо связывается с антигеном внутри эндосомы, может связаться с антигеном в плазме и поступить, все еще образуя комплекс с антигеном, в эндосомы в клетках посредством интернализации, подвергнуться диссоциации от антигена в эндосоме, затем связываться с FcRn и переместиться на клеточную поверхность, и снова возвратиться в плазму в не связанном с антигенами состоянии для нейтрализации множества связанных с мембранными антигенов. Кроме того, авторы настоящего изобретения обнаружили, что антитело, обладающее способностью к сильному связыванию с антигенами в плазме, но к слабому связыванию с антигенами в эндосоме, может подвергаться диссоциации от антигенов в эндосоме, даже когда антитело связалось с такими антигенами, как растворимые антигены; поэтому они снова высвобождаются в плазму в не связанном с антигенами состоянии и могут нейтрализовать множество растворимых антигенов.
В частности, авторы настоящего изобретения обратили внимание на то, что pH в плазме отличен от pH в эндосомах, и таким образом обнаружили, что антитела, которые сильно связываются с антигенами при параметре pH в плазме, но слабо связываются с антигенами при параметре pH в эндосомах, превосходят в сохранении в плазме, поскольку одна молекула антитела может связываться с множеством антигенов.
Эндосомы, которые являются мембранными везикулами, образуют сети в цитоплазме эукариотических клеток и ответственны за метаболизм макромолекул в ходе развития процесса от клеточной мембраны в лизосомы. Сообщалось, что pH в эндосомах является обычно кислотным pH в диапазоне от 5,5 до 6,0 (Nat. Rev. Mol. Cell Biol. 2004 Feb; 5(2): 121-32). Между тем, известно, что pH в плазме является почти нейтральным (обычно pH 7,4).
В результате, антигенсвязывающая молекула, антигенсвязывающая активность которой при кислотном pH слабее антигенсвязывающей активности при нейтральном pH, связывается с антигеном в плазме, которая имеет нейтральный pH, поступает в клетки и затем подвергается диссоциации от антигена в эндосомах, которые имеют кислотный pH. Антигенсвязывающая молекула, которая подверглась диссоциации от антигена, связывается с FcRn, перемещается на клеточную поверхность и снова возвращается в плазму в не связанном с антигенами состоянии. В результате, антигенсвязывающая молекула может связываться с антигенами многократно, и фармакокинетика улучшается.
Вещества в виде антигенсвязывающих молекул
Кроме того, настоящим изобретением обеспечиваются антигенсвязывающие молекулы, антигенсвязывающая активность которых при pH от 4,0 до 6,5 ниже антигенсвязывающей активности при pH от 6,7 до 10,0, предпочтительно антигенсвязывающие молекулы, антигенсвязывающая активность которых при pH от 5,0 до 6,0 ниже антигенсвязывающей активности при pH от 7,0 до 8,0. В частности, антигенсвязывающие молекулы, антигенсвязывающая активность которых при pH от 4,0 до 6,5 ниже антигенсвязывающей активности при pH от 6,7 до 10,0, включают, например, антигенсвязывающие молекулы, антигенсвязывающая активность которых при pH 5,8 ниже антигенсвязывающей активности при pH 7,4. Антигенсвязывающие молекулы, антигенсвязывающая активность которых при pH 5,8 ниже антигенсвязывающей активности при pH от 7,4, можно также определить как антигенсвязывающие молекулы, антигенсвязывающая активность которых при pH 7,4 выше антигенсвязывающей активности при pH от 5,8.
Что касается антигенсвязывающих молекул настоящего изобретения, антигенсвязывающая активность которых при pH 5,8 ниже антигенсвязывающей активности при pH от 7,4, при условии, что антигенсвязывающая активность при pH 5,8 ниже антигенсвязывающей активности при pH от 7,4, нет ограничения в отношении различия в активности связывания, а требуется только, чтобы антигенсвязывающая активность при pH 5,8 была ниже, даже слегка.
Предпочтительный вариант антигенсвязывающей молекулы по настоящему изобретению, антигенсвязывающая активность которой при pH 5,8 ниже антигенсвязывающей активности при pH от 7,4, включает антигенсвязывающие молекулы, антигенсвязывающая активность которых при pH 7,4 – дважды или более антигенсвязывающей активности при pH 5,8. Более предпочтительный вариант антигенсвязывающей молекулы включает антигенсвязывающие молекулы, антигенсвязывающая активность которых при pH 7,4 выше в десять раз или более антигенсвязывающей активности при pH 5,8. Еще более предпочтительный вариант антигенсвязывающей молекулы включает антигенсвязывающие молекулы, антигенсвязывающая активность которых при pH 7,4 выше в 40 раз или более антигенсвязывающей активности при pH 5,8.
В частности, в предпочтительном варианте осуществления антигенсвязывающая молекула по настоящему изобретению обладает антигенсвязывающей активностью при pH 5,8, которая ниже антигенсвязывающей активности при pH 7,4, причем значение KD (pH 5,8)/KD (pH 7,4), которое является соотношением между KD для антигена при pH 5,8 и KD для антигена при pH 7,4, составляет предпочтительно 2 или больше, более предпочтительно 10 или больше, и еще предпочтительнее 40 или больше. Верхний предел для значения KD (pH 5,8)/KD (pH 7,4) особенно не ограничивается и может быть любым значением, например, 400, 1000 или 10000, при условии возможности ее продуцирования с использованием известных квалифицированным в данной области техники специалистам методов.
В другом предпочтительном варианте осуществления антигенсвязывающая молекула по настоящему изобретению, антигенсвязывающая которой при pH 5,8 ниже антигенсвязывающей активности при pH 7,4, имеет значение kd (pH 5,8)/kd (pH 7,4), которое является соотношением между kd для антигена при pH 5,8 и kd для антигена при pH 7,4, которое составляет 2 или больше, более предпочтительно 5 или больше, даже предпочтительнее 10 или больше и еще предпочтительнее 30 или больше. Верхний предел для значения kd (pH 5,8)/kd (pH 7,4) особенно не ограничивается и может быть любым значением, например, 50, 100 или 200, при условии возможности ее продуцирования с использованием известных квалифицированным в данной области техники специалистам методов.
Отличные от pH условия, при которых определяется антигенсвязывающая активность, могут быть выбраны надлежащим образом квалифицированными в данной области техники специалистами, и эти условия особенно не ограничиваются; однако измерения можно выполнить, например, в условиях использования буфера MES и 37°C, как описано в примерах. Кроме того, антигенсвязывающую активность антигенсвязывающей молекулы можно определить с помощью известных квалифицированным в данной области техники специалистам способов, например, используя Biacore Т100 (GE Healthcare) или т.п., как описано в примерах.
Считается, что такая антигенсвязывающая молекула, которая слабо связывается с антигеном при кислотном pH, легко подвергается диссоциации от антигена в кислотных условиях в эндосомах, и что после интернализации в клетки она связывается с FcRn и легко высвобождается наружу из клеток. Антигенсвязывающая молекула, выброшенная наружу из клеток, при этом не будучи деградированной внутри клеток, может снова связываться с другими антигенами. В результате, когда антигенсвязывающей молекулой является, например, антигенсвязывающая нейтрализующая молекула, антигенсвязывающая молекула, которая легко подвергается диссоциации от антигена в кислотных условиях в эндосомах, может связывать и нейтрализовать антигены многократно. В результате, антигенсвязывающие молекулы, антигенсвязывающая активность которых при pH от 4,0 до 6 ниже антигенсвязывающей активности при pH от 6,7 до 10,0, превосходят в сохранении в плазме.
В предпочтительном варианте осуществления антигенсвязывающая молекула, антигенсвязывающая активность которой при pH 5,8 ниже антигенсвязывающей молекулы при pH 7,4, включает антигенсвязывающие молекулы, в которых по крайней мере одна аминокислота в антигенсвязывающей молекуле замещена гистидином или не встречающейся в природе аминокислотой, или в которые вставлен по крайней мере один гистидин или не встречающаяся в природе аминокислота. Точка, в которую вводится мутация на гистидин или не встречающуюся в природе аминокислоту, особенно не ограничивается и может быть любой точкой при условии, что антигенсвязывающая активность при pH 5,8 слабее антигенсвязывающей активности при pH 7,4 (больше значение KD (pH 5,8)/KD (pH 7,4) или больше значение kd (pH 5,8)/kd (pH 7,4)) по сравнению с тем, что наблюдается до замены. Примеры включают вариабельные области и CDR антитела в том случае, когда антигенсвязывающей молекулой является антитело. Число аминокислот, замещаемых гистидином или не встречающейся в природе аминокислотой, и число вставляемых аминокислот может быть надлежащим образом определено квалифицированными в данной области техники специалистами. Может быть замещена гистидином или не встречающейся в природе аминокислотой одна аминокислота, или может быть вставлена одна аминокислота, или могут быть замещены гистидином или не встречающими в природе аминокислотами две или более аминокислот, или могут быть вставлены две или более аминокислот. Кроме того, помимо замен на гистидин или не встречающуюся в природе аминокислоту или вставки гистидина или не встречающейся в природе аминокислоты, может быть также одновременно осуществлена делеция, добавление, вставка и/или замена и т.п. других аминокислот. Замены на гистидин или не встречающуюся в природе аминокислоту или вставка гистидина или не встречающейся в природе аминокислоты могут быть проведены случайным образом, используя такой способ, как сканирование гистидином, в котором используется гистидин вместо аланина при сканировании аланином, известном квалифицированным в данной области техники специалистам. Антигенсвязывающие молекулы, значение KD (pH 5,8)/KD (pH 7,4) или kd (pH 5,8)/kd (pH 7,4) которых увеличивается по сравнению с указанным значением до мутации, можно выбрать из антигенсвязывающих молекул, в которые мутация на гистидин или не встречающуюся в природе аминокислоту была введена случайным образом.
Предпочтительные антигенсвязывающие молекулы с мутацией на гистидин или не встречающуюся в природе аминокислоту, антигенсвязывающая активность которых pH 5,8 ниже антигенсвязывающей активности при pH 7,4, включают, например, антигенсвязывающие молекулы, антигенсвязывающая активность которых при pH 7,4 после мутации на гистидин или не встречающуюся в природе аминокислоту эквивалента антигенсвязывающей активности при pH 7,4 до мутации на гистидин или не встречающуюся в природе аминокислоту. В настоящем изобретении выражение «антигенсвязывающая молекула после мутации на гистидин или не встречающуюся в природе аминокислоту имеет антигенсвязывающую активность, которая эквивалентна антигенсвязывающей активности антигенсвязывающей молекулы до мутации на гистидин или не встречающуюся в природе аминокислоту» означает, что, когда за 100% принимают антигенсвязывающую активность антигенсвязывающей молекулы до мутации на гистидин или не встречающуюся в природе аминокислоту, антигенсвязывающая активность антигенсвязывающей молекулы после мутации на гистидин или не встречающуюся в природе аминокислоту составляет по крайней мере 10% или больше, предпочтительно 50% или больше, более предпочтительно 80% или больше и еще предпочтительнее 90% или больше. Антигенсвязывающая активность при pH 7,4 после мутации на гистидин или не встречающуюся в природе аминокислоту может превышать антигенсвязывающую активность при pH 7,4 до мутации на гистидин или не встречающуюся в природе аминокислоту. Когда антигенсвязывающая активность антигенсвязывающей молекулы уменьшается вследствие замены гистидином или не встречающейся в природе аминокислотой или его (ее) вставки, антигенсвязывающую активность можно скорректировать путем введения замены, делеции, добавления и/или вставки и т.п. одной или нескольких аминокислот в антигенсвязывающую молекулу, так что антигенсвязывающая активность становится эквивалентной антигенсвязывающей активности до замены гистидином или его вставки. Настоящее изобретение также включает такие антигенсвязывающие молекулы, активность связывания которых сделана эквивалентной в результате замены, делеции, добавления и/или вставки одной или нескольких аминокислот после замены гистидином или его вставки.
Кроме того, когда антигенсвязывающей молекулой является вещество, включающее константную область антитела, в другом предпочтительном варианте антигенсвязывающей молекулы, антигенсвязывающая активность которой при pH 5,8 ниже антигенсвязывающей активности при pH 7,4, настоящее изобретение включают способы модифицирования константной области антитела, содержащейся в антигенсвязывающих молекулах. Конкретные примеры константных областей антител после модификации включают константные области, описанные в примерах.
Когда антигенсвязывающую активность антигенсвязывающего вещества при pH 5,8 ослабляют по сравнению с указанной активностью при pH 7,4 (когда увеличивают значение KD (pH 5,8)/KD (pH 7,4)) с помощью описанных выше способов и т.п., обычно предпочтительно, когда значение KD (pH 5,8)/KD (pH 7,4) больше в два раза и более, более предпочтительно в пять раз или более и даже предпочтительнее в десять раз или более указанного значения исходного антитела, но особенно не ограничивается ими.
Антигенсвязывающие молекулы по настоящему изобретению могут, кроме того, обладать любым другим свойством при условии, что их антигенсвязывающая активность при pH от 4,0 до 6,5 ниже указанной активности при pH от 6,7 до 10,0. Например, антигенсвязывающие молекулы могут быть агонистическими или антагонистическими антигенсвязывающими молекулами. Предпочтительные антигенсвязывающие молекулы по настоящему изобретению включают, например, антагонистические антигенсвязывающие молекулы. Как правило, антагонистическая антигенсвязывающая молекула ингибируют опосредованную рецептором внутриклеточную передачу сигналов путем ингибирования связывания между лигандом (агонистом) и рецептором.
Кроме того, настоящим изобретением обеспечиваются антитела, в которых аминокислота в по крайней мере одной точке, указанной ниже, замещена гистидином или не встречающейся в природе аминокислотой. Положения аминокислот указаны на основе нумерации по Kabat (Kabat EA et al., (1991) Sequences of Proteins of Immunological Interest, NIH).
Тяжелая цепь: H27, H31, H32, H33, H35, H50, H58, H59, H61, H62, H63, H64, H65, H99, H100b и H102
Легкая цепь: L24, L27, L28, L32, L53, L54, L56, L90, L92 и L94
Среди указанных выше точек H32, H61, L53, L90 и L94 могли бы быть универсальными точками модификаций.
Когда антигеном является рецептор для IL-6 (например, рецептор для IL-6 человека), предпочтительные точки модификаций включают следующие точки. Однако точки модификаций особенно не ограничиваются ими.
Тяжелая цепь: H27, H31, H32, H35, H50, H58, H61, H62, H63, H64, H65, H100b и H102
Легкая цепь: L24, L27, L28, L32, L53, L56, L90, L92 и L94
Когда гистидином или не встречающейся в природе аминокислотой заменяют аминокислоты во множестве точек, предпочтительные комбинации точек замен включают, например, комбинацию H27, H31 и H35; комбинацию H27, H31, H32, H35, H58, H62 и H102; комбинацию L32 и L53 и комбинацию L28, L32 и L53. Кроме того, предпочтительные комбинации точек замен в тяжелой и легкой цепи включают комбинацию H27, H31, L32 и L53.
Когда антигеном является IL-6 (например, IL-6 человека), предпочтительные точки модификаций включают следующие точки. Однако точки модификаций особенно не ограничиваются ими.
Тяжелая цепь: H32, H59, H61 и H99
Легкая цепь: L53, L54, L90 и L94
Когда антигеном является рецептор для IL-31 (например, рецептор для IL-31 человека), предпочтительные точки модификаций включают Н33. Однако точки модификаций особенно не ограничиваются им.
Антигенсвязывающие молекулы по настоящему изобретению могут распознавать любой антиген. Распознаваемые антителами настоящего изобретения антигены включают, в частности, вышеупомянутые рецепторные белки (связанные с мембранами рецепторы или растворимые рецепторы), мембранные антигены, такие как маркеры клеточной поверхности, и растворимые антигены, такие как цитокины, например, IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12, IL-15, IL-31, IL-23, рецептор IL-2, рецептор IL-6, рецептор OSM, gp130, рецептор IL-5, CD40, CD4, Fas, остеопонтин, CRTH2, CD26, PDGF-D, CD20, вызывающий хемотаксис моноцитов фактор, CD23, TNF-α, HMGB-1, α4 интегрин, ICAM-1, CCR2, CD11a, CD3, IFNγ, BLyS, HLA-DR, TGF-β, CD52 и рецептор IL-31.
Особенно предпочтительные антигены включают рецептор IL-6.
Антигенсвязывающие молекулы настоящего изобретения описаны выше.
В предпочтительном варианте осуществления настоящего изобретения антигенсвязывающие молекулы включают антитела. Антитела, имеющие антигенсвязывающую активность и FcRn-связывающий район, включают, например, антитела IgG-типа. Когда используемым антителом является антитело IgG-типа, его тип не ограничивается. Возможно использование IgG1, IgG2, IgG3, IgG4 и т.п.
Происхождение антитела по настоящему изобретению особенно не ограничивается, и может использоваться антитело любого происхождения. Возможно использование, например, антител мыши, антител человека, антител крысы, антител кролика, антител козы, антител верблюда и других. Кроме того, антитела могут быть, например, описанными выше химерными антителами и, в частности, модифицированными антителами с заменами в аминокислотных последовательностях, такими как гуманизированные антитела. Антитела могут также быть описанными выше биспецифическими антителами, продуктами модификации антител, с которыми связаны различные молекулы, полипептидами, включающими фрагменты антител, и антителами с модифицированными сахарными цепями.
Создание химерных антител известно. В случае химерного человеческого-мышиного антитела, например, ДНК, кодирующую V-область антитела мыши, можно соединить с ДНК, кодирующей С-область антитела человека; соединенную ДНК можно встроить в экспрессионный вектор и ввести в хозяина для продуцирования химерного антитела.
«Гуманизированные антитела», также называемые реконструированными человеческими антителами, представляют собой антитела, в которых определяющий комплементарность участок (CDR) антитела, происходящего из не являющегося человеком млекопитающего, например, мыши, пересажен в CDR антитела человека. Способы идентификации CDR известны (Kabat et al., Sequence of Proteins of Immunological Interest (1987), National Institute of Health, Bethesda, Md.; Chothia et al., Nature (1989) 342: 877). Также известны общие методы генетической рекомбинации, подходящие для этой цели (см. заявку на Европейский патент EP 125023 и WO 96/02576). Гуманизированные антитела можно продуцировать известными способами, например, можно определить CDR антитела мыши, и получить ДНК, кодирующую антитело, в котором эта CDR соединена с каркасной область (FR) антитела человека. Гуманизированные антитела затем можно продуцировать, используя систему, в которой используются общепринятые экспрессионные векторы. Такие ДНК можно синтезировать с помощью ПЦР, используя в качестве праймеров несколько олигонуклеотидов, приготовленных так, чтобы они имели части, которые перекрываются с концевыми районами как CDR, так и FR (см. способ, описанный в WO 98/13388). FR антитела человека, соединенные через CDR, выбирают так, чтобы CDR образовывали подходящий антигенсвязывающий сайт. При необходимости аминокислоты в FR вариабельной области антитела могут быть замещены так, чтобы CDR реконструированного антитела человека могли образовывать подходящий антигенсвязывающий сайт (Sato, K. et al., Cancer Res. (1993) 53: 10.01-6). Аминокислотные остатки в FR, которые можно модифицировать, включают части, которые непосредственно связываются с антигеном посредством нековалентных связей (Amit et al., Science (1986) 233: 747-53), части, оказывающие влияние или эффект на структуру CDR (Chothia et al., J. Mol. Biol. (1987) 196: 901-917), и части, вовлеченные во взаимодействия VH-VL (EP 239400).
Когда антителами по настоящему изобретению являются химерные антитела или гуманизированные антитела, C-области этих антител предпочтительно происходят из антител человека. Например, Сγ1, Сγ2, Сγ3 и Сγ4 можно использовать для Н-цепи, в то время как Сκ и Сλ можно использовать для L-цепи. Кроме того, при необходимости, аминокислотные мутации могут быть введены в С-область антитела человека для усиления или ослабления связывания с рецептором Fcγ или FcRn или увеличения устойчивости антитела или его продуктивности. Химерное антитело по настоящему изобретению предпочтительно включает вариабельную область антитела, происходящего из не являющегося человеком млекопитающего, и константную область, происходящую из антитела человека. Между тем, гуманизированное антитело предпочтительно включает CDR антитела, происходящего из не являющегося человеком млекопитающего, и FR и С-области, происходящие из антитела человека. Константные области, происходящие из антител человека, предпочтительно включают FcRn-связывающий район. Такие антитела включают, например, IgG (IgG1, IgG2, IgG3 и IgG4). Константными областями, используемыми для гуманизированных антител по настоящему изобретению, могут быть константные области антител любого изотипа. Предпочтительно используют константную область IgG человека, хотя она не ограничивается ею. Происходящие из антитела человека FR, которые используются для гуманизированных антител, также особенно не ограничиваются и могут происходить из антитела любого изотипа.
Вариабельные и константные области химерных и гуманизированных антител по настоящему изобретению могут быть модифицированы с помощью делеции, замены, вставки и/или добавления и т.п. при условии, что демонстрируется специфичность связывания исходных антител.
Полагают, что вследствие понижения иммуногенности для человека химерные и гуманизированные антитела, в которых используются последовательности человеческого происхождения, применимы в случае введения людям для терапевтических целей и т.п.
Антитела по настоящему изобретению можно получить любым способом. Например, антитела, антигенсвязывающая активность которых при pH 5,8 первоначально превышает антигенсвязывающую активность при pH 7,4 или сравнима с ней, можно искусственно модифицировать благодаря описанной выше замене гистидином или т.п. так, чтобы их антигенсвязывающая активность при pH 5,8 стала ниже антигенсвязывающей активности при pH 7,4. Альтернативно, антитела, антигенсвязывающая активность которых при pH 5,8 ниже антигенсвязывающей активности при pH 7,4, можно отобрать путем скрининга ряда антител, полученных из библиотеки антител или гибридом, как описано ниже.
При замещении гистидином аминокислот в антителе можно использовать известные аминокислотные последовательности для H-цепи или L-цепи антитела до введения мутаций на гистидин, или можно также использовать аминокислотные последовательности антител, недавно полученных с помощью известных квалифицированным в данной области техники специалистам способов. Например, антитела можно получить из библиотеки антител, или их можно получить путем клонирования генов, кодирующих антитела, из гибридом, продуцирующих моноклональные антитела.
Что касается библиотек антител, уже известно множество библиотек антител, и также известны способы получения библиотек антител; поэтому квалифицированные в данной области техники специалисты могут надлежащим образом получить библиотеки антител. Например, относительно библиотек последовательностей антител в фагах можно обратиться к литературе, такой как Clackson et al., Nature 1991, 352: 624-628; Marks et al., J. Mol. Biol. 1991, 222: 581-597; Waterhouses et al., Nucleic Acids Res. 1993, 21: 2265-2266; Griffiths et al., EMBO J. 1994, 13: 324.0-60; Vaughan et al., Nature Biotechnology 1996, 14: 309-14; и публикация заявки Kohyo на патент Японии № (JP-A) H20-504970 (публикация не рассмотренной заявки на японской национальной фазе рассмотрения, соответствующая публикации не японской международной заявки). Кроме того, можно использовать известные способы, такие как способы, в которых используются эукариотические клетки в качестве библиотек (WO 95/15393), и способы рибосомного дисплея. Кроме того, также известны методы получения антител человека с помощью пэннинга, используя библиотеки антител человека. Например, вариабельные области антител человека можно экспрессировать на поверхности фагов в виде одноцепочечных антител (scFv), используя способы фагового дисплея, и можно отобрать фаги, которые связываться с антигенами. С помощью генетического анализа отобранных фагов можно определить последовательности ДНК, кодирующие вариабельные области антител человека, которые связываются с антигенами. Как только ДНК-последовательности scFvs, которые связываются с антигенами, выявлены, на основе этих последовательностей можно получить подходящие экспрессионные векторы для получения антител человека. Эти способы уже широко известны, и можно обратиться к WO 92/01047, WO 92/20791, WO 93/06213, WO 93/11236, WO 93/19172, WO 95/01438 и WO 95/15388.
Что касается способов получения генов, кодирующих антитела из гибридом, можно использовать в своей основе известные методы, которые включают использование желаемых антигенов или клеток, экспрессирующих желаемые антигены, в качестве сенсибилизирующих антигенов, используя их для выполнения иммунизаций в соответствии с общепринятыми способами иммунизации, сливание результирующих иммунных клеток с известными родительскими клетками с помощью общепринятых способов слияния клеток, скрининг продуцирующих моноклональные антитела клеток (гибридом) с помощью общепринятых способов скрининга, синтез кДНК вариабельных областей (V-областей) антител на основе мРНК из полученных гибридом, используя обратную транскриптазу, и соединение их с ДНК, кодирующими константные области (C-области) желаемых антител.
Конкретнее, сенсибилизирующие антигены для получения описанных выше генов антител, кодирующих H-цепи и L-цепи, включают как полные антигены, обладающие иммуногенностью, так и неполные антигены, включающие гаптены и т.п., не обладающие иммуногенностью; однако они не ограничиваются этими примерами. Например, можно использовать целые белки и частичные пептиды белков, представляющие интерес. Кроме того, известно, что антигенами могут быть вещества, включающие полисахариды, нуклеиновые кислоты, липиды и т.п. Следовательно, антигены для антител по настоящему изобретению особенно не ограничиваются. Антигены можно получить известными квалифицированным в данной области техники специалистам способами, например, способами на основе бакуловирусов (например, WO 98/46777) и т.п. Гибридомы можно получить, например, способами Milstein и др. (G. Kohler и C. Milstein, Methods Enzymol. 1981, 73: 3-46) и т.п. При низкой иммуногенности антигена иммунизацию можно выполнить после соединения антигена с обладающей иммуногенностью макромолекулой, такой как альбумин. Альтернативно, при необходимости, антигены можно преобразовать в растворимые антигены путем соединения их с другими молекулами. Когда в качестве антигенов используются трансмембранные молекулы, такие как мембранные антигены (например, рецепторы), в качестве фрагмента можно использовать части внеклеточных областей мембранных антигенов, или в качестве иммуногенов можно использовать клетки, экспрессирующие трансмембранные молекулы на своей клеточной поверхности.
Продуцирующие антитела клетки можно получить иммунизацией животных, используя подходящие сенсибилизирующие антигены, описанные выше. Альтернативно, продуцирующие антитела клетки можно приготовить с помощью in vitro иммунизации лимфоцитов, которые могут продуцировать антитела. Для иммунизации могут использоваться различные животные; такие обычно используемые животные включают грызунов, зайцеобразных и приматов. Такие животные включают, например, грызунов, таких как мыши, крысы и хомяки; зайцеобразных, таких как кролики, и приматов, включающих обезьян, таких как яванские макаки, макаки-резус, павианы и шимпанзе. Кроме того, также известны трансгенные животные, несущие наборы генов антител человека, и антитела человека можно получить, используя этих животных (см. WO 96/34096; Mendez et al., Nat. Genet. 1997, 15: 146-156). Вместо использования таких трансгенных животных, например, желаемые антитела человека, обладающие антигенсвязывающей активностью, можно получить путем in vitro сенсибилизации лимфоцитов человека с использованием желаемых антигенов или клеток, экспрессирующих желаемые антигены, и затем слияния сенсибилизированных лимфоцитов с клетками миеломы человека, такими как U266 (см. публикацию заявки Kokoku на патент Японии № (JP-B) H01-59878 (рассмотренную, принятую заявку на патент Японии, опубликованную для опротестования)). Кроме того, желаемые антитела человека можно получить путем иммунизации трансгенных животных, несущих полный набор генов антител человека, с использованием желаемых антигенов (см. WO 93/12227, WO 92/03918, WO 94/02602, WO 96/34096 и WO 96/33735).
Иммунизацию животных можно выполнить путем соответствующего разведения и суспендирования сенсибилизирующего антигена в забуференном фосфатом солевом растворе (PBS), физиологическом растворе или т.п., и смешивания его с адъювантом с приготовлением эмульсии, при необходимости. Затем ее внутрибрюшинно или подкожно инъецируют животным. Затем сенсибилизирующий антиген, смешанный с неполным адъювантом Фрейнда, предпочтительно вводят несколько раз через каждые 4-21 дня. Продукцию антител можно подтвердить путем измерения титра представляющего интерес антитела в сыворотках животных, используя общепринятые способы.
Продуцирующие антитела клетки, полученные из лимфоцитов или животных, иммунизированных желаемым антигеном, можно слить с миеломными клетками для создания гибридом, используя общепринятые агенты для слияния (например, полиэтиленгликоль) (Goding, Monoclonal Antibodies: Principles и Practice, Academic Press, 1986, 59-103). При необходимости, гибридомные клетки можно культивировать и выращивать, и специфичность связывания антител, продуцируемых этими гибридомами, можно определить, используя известные способы анализа, такие как иммунопреципитация, радиоиммуноанализ (RIA) и иммуноферментный твердофазный анализ (ELISA). После этого гибридомы, продуцирующие представляющие интерес антитела, специфичность, аффинность или активность которых было определена, можно субклонировать с использованием таких способов, как предельное разведение.
Затем кодирующие отобранные антитела гены можно клонировать из гибридом или продуцирующих антитела клеток (сенсибилизированных лимфоцитов и т.п.), используя зонды, которые специфически связываются с антителами (например, олигонуклеотиды, комплементарные последовательностям, кодирующим константные области антител). Можно также клонировать гены из мРНК, используя ОТ-ПЦР. Иммуноглобулины делят на пять различных классов: IgA, IgD, IgE, IgG и IgM. Эти классы далее делят на несколько субклассов (изотипов) (например, IgG-1, IgG-2, IgG-3 и IgG-4; IgA-1 и IgA-2; и т.п.). H-цепи и L-цепи, используемые в настоящем изобретении для получения антител, особенно не ограничиваются и могут происходить из антител, относящихся к любому из этих классов или субклассов; однако особенно предпочтительным является IgG.
Здесь можно модифицировать кодирующие H-цепи гены и кодирующие L-цепи гены, используя методы генетической инженерии. Генетически модифицированные антитела, такие как химерные антитела и гуманизированные антитела, которые были искусственно модифицированы с целью уменьшения гетерологичной иммуногенности и т.п. в отношении людей, можно надлежащим образом продуцировать для таких антител, как антитела мыши, антитела крысы, антитела кролика, антитела хомячка, антитела овцы и антитела верблюда. Химерными антителами являются антитела, включающие вариабельные области H-цепи и L-цепи антитела млекопитающего, не являющегося человеком, такого как антитело мыши, и константные области H-цепи и L-цепи антитела человека. Химерные антитела можно получить путем лигирования ДНК, кодирующей вариабельную область антитела мыши, с ДНК, кодирующей константную область антитела человека, встраивания лигированной ДНК в экспрессионный вектор и введения вектора в хозяина для продукции антитела. Гуманизированное антитело, также называемое реконструированным человеческим антителом, можно синтезировать с помощью ПЦР, используя несколько олигонуклеотидов, приготовленных так, чтобы они имели перекрывающиеся части на концах ДНК-последовательностей, предназначенных для соединения определяющих комплементарность участков (CDR) антитела не являющегося человеком млекопитающего, такого как мышь. Результирующую ДНК можно лигировать с ДНК, кодирующей константную область антитела человека. Лигированную ДНК можно встроить в экспрессионный вектор, и вектор можно ввести в хозяина для продукции антитела (см. EP 239400 и WO 98/13388). FR антитела человека, которые лигируют через CDR, отбирают, когда CDR образуют подходящий антигенсвязывающий сайт. При необходимости аминокислоты в каркасной области вариабельной области антитела могут быть замещены так, чтобы CDR реконструированного антитела человека образовывали подходящий антигенсвязывающий сайт (K. Sato et al., Cancer Res. (1993) 53: 10.01-6).
Помимо описанной выше гуманизации, антитела можно модифицировать для улучшения их биологических свойств, например, связывания с антигеном. В настоящем изобретению такие модификации можно успешно выполнить с помощью таких способов, как сайт-направленный мутагенез (см., например, Kunkel (1910.0) Proc. Natl. Acad. Sci. USA 82: 488), мутагенез с использованием ПЦР и кассетный мутагенез. Как правило, мутантные антитела, биологические свойства которых были улучшены, демонстрируют гомологичность и/или схожесть аминокислотных последовательностей, составляющую 70% или более, более предпочтительно 80% или более и даже предпочтительнее 90% или более (например, 95% или более, 97%, 98% или 99%), после сравнения с аминокислотной последовательностью вариабельной области исходного антитела. Здесь гомологию и/или схожесть последовательностей определяют как отношение аминокислотных остатков, которые являются гомологичными (одинаковыми) или схожими (аминокислотными остатками, отнесенными в одну и ту же группу на основе общих свойств боковых цепей аминокислот), к остаткам исходного антитела, после того как значение гомологии последовательностей было максимизировано при совмещении последовательностей и введения пробелов, при необходимости. Обычно природные аминокислотные остатки делят на основе свойств их боковых цепей на следующие группы:
(1) гидрофобные: аланин, изолейцин, валин, метионин и лейцин;
(2) нейтральные гидрофильные: аспарагин, глютамин, цистеин, треонин и серин;
(3) кислотные: аспарагиновая кислота и глютаминовая кислота;
(4) основные: аргинин, гистидин и лизин;
(5) остатки, влияющие на ориентацию цепи: глицин и пролин; и
(6) ароматические: тирозин, триптофан и фенилаланин
Обычно всего шесть определяющих комплементарность участков (CDR; гипервариабельных участков), присутствующих в вариабельных областях H-цепи и L-цепи, взаимодействуют друг с другом с образованием антигенсвязывающего сайта антитела. Также известно, что вариабельная область сама по себе способна к распознаванию антигена и связыванию с ним, хотя ее аффинность меньше аффинности целого связывающего сайта. Таким образом, гены, кодирующие H-цепь и L-цепь антитела настоящего изобретения, могут кодировать фрагменты, каждый из которых включает антигенсвязывающий сайт H-цепи или L-цепи, при условии, что полипептид, кодируемый этим геном, сохраняет активность связывания с желаемым антигеном.
Как описывается выше, вариабельная область тяжелой цепи обычно образуется тремя CDR и четырьмя FR. В предпочтительном варианте осуществления настоящего изобретения «модифицируемые» аминокислотные остатки могут быть соответствующим образом выбраны из аминокислотных остатков, например, в CDR или FR. Как правило, модификации аминокислотных остатков в CDR могут уменьшить способность к связыванию антигенов. Поэтому подходящие для «модификации» в настоящем изобретении аминокислотные остатки предпочтительно выбирают из аминокислотных остатков в FR, но не ограничиваются ими. Можно выбрать аминокислотные остатки в CDR при условии, что модификация, как подтверждено, не уменьшает способность к связыванию. Альтернативно, используя общедоступные базы данных или т.п., квалифицированные в данной области техники специалисты могут получить соответствующие последовательности, которые могут использоваться в качестве FR вариабельной области антитела такого организма, как человек или мышь.
Кроме того, настоящим изобретением обеспечиваются гены, кодирующие антитела по настоящему изобретению. Гены, кодирующие антитела по настоящему изобретению, могут быть любыми генами и могут быть ДНК, РНК, аналогами нуклеиновых кислот или т.п.
Кроме того, настоящим изобретением обеспечиваются клетки-хозяева, несущие описанные выше гены. Клетки-хозяева особенно не ограничиваются и включают, например, E.coli и клетки различных животных. Клетки-хозяева могут использоваться, например, в качестве продукционной системы для продуцирования и экспрессии антител настоящего изобретения. Имеются in vitro и in vivo продукционные системы для систем продукции полипептидов. Такие in vitro продукционные системы включают, например, продукционные системы, в которых используются эукариотические клетки или прокариотические клетки.
Эукариотические клетки, которые можно использовать в качестве клеток-хозяев, включают, например, клетки животных, клетки растений и клетки грибов. Клетки животных включают клетки млекопитающих, например, клетки CHO (J. Exp. Med. (1995) 108: 94.0), COS, HEK293, 3T3, клетки миеломы, клетки BHK (почки детеныша хомячка), HeLa и Vero; клетки земноводных, такие как ооциты Xenopus laevis (Valle et al., Nature (1981) 291: 338-340); и клетки насекомых, такие как Sf9, Sf21 и Tn5. Для экспрессии антител по настоящему изобретению предпочтительно используются клетки CHO-DG44, CHO-DX11B, COS7, HEK293 и клетки BHK. Среди клеток животных, клетки CHO cells являются особенно предпочтительными для высокомасштабной экспрессии. Векторы могут вводиться в клетки-хозяева с помощью, например, способов с использованием кальция фосфата, способов с использованием DEAE-декстрана, способов, в которых используются катионная липосома DOTAP (Boehringer-Mannheim), способов электропорации и способов липофекции.
Что касается клеток растений, например, происходящие из Nicotiana tabacum клетки и клетки ряски (Lemna minor) известны в качестве системы для продукции белков. Для продуцирования антител по настоящему изобретению из этих клеток можно культивировать каллюсы. Что касается клеток грибов, известными системами для экспрессии белков являются системы, в которых используются дрожжевые клетки, например, клетки рода Saccharomyces (например, Saccharomyces cerevisiae и Saccharomyces pombe), и клетки нитевидных грибов, например, рода Aspergillus (например, Aspergillus niger). Эти клетки могут использоваться в качестве хозяина для продукции антител по настоящему изобретению.
Бактериальные клетки могут использоваться в прокариотических продукционных системах. Что касается бактериальных клеток, известны продукционные системы, в которых используется Bacillus subtilis, помимо описанных выше продукционных систем, в которых используется E.coli. Такие системы могут использоваться для продукции антител настоящего изобретения.
Способы скрининга
Настоящим изобретением обеспечиваются способы скрининга для получения антигенсвязывающих молекул, антигенсвязывающая активность которых при кислотном pH ниже антигенсвязывающей активности при нейтральном pH. Настоящим изобретением также обеспечиваются способы скрининга для получения антигенсвязывающих молекул, которые могут в индивидуальном порядке связываться с множеством антигенов. Настоящим изобретением также обеспечиваются способы скрининга для получения антигенсвязывающих молекул, которые превосходят в сохранении в плазме. Настоящим изобретением также обеспечиваются способы скрининга для получения антигенсвязывающей молекулы, которая подвергается диссоциации внутри клетки от связавшегося с ней внеклеточно антигена. Настоящим изобретением также обеспечиваются способы скрининга для получения антигенсвязывающей молекулы, которая связывается с антигеном и подвергается интернализации в клетку, и высвобождается наружу из клетки в свободной от антигена форме. Настоящим изобретением также обеспечиваются способы скрининга для получения антигенсвязывающей молекулы, которая обладает увеличенной способностью к элиминации антигенов в плазме. Кроме того, настоящим изобретением также обеспечиваются способы скрининга для получения антигенсвязывающих молекул, которые особенно полезны при применении в виде фармацевтических композиций.
В частности, настоящим изобретением обеспечиваются способы скрининга антигенсвязывающих молекул, которые включают стадии:
(a) определения антигенсвязывающей активности антигенсвязывающей молекулы при pH от 6,7 до 10,0;
(b) определения антигенсвязывающей активности антигенсвязывающей молекулы при pH от 4,0 до 6,5; и
(c) отбора антигенсвязывающей молекулы, антигенсвязывающая активность которой при pH от 6,7 до 10,0 выше антигенсвязывающей активности при pH от 4,0 до 6,5.
В способах скрининга по настоящему изобретению антигенсвязывающая активность антигенсвязывающей молекулы при pH от 6,7 до 10 особенно не ограничивается при условии, что она является антигенсвязывающей активностью при pH между 6,7 и 10,0. Однако, например, предпочтительной антигенсвязывающей активностью является антигенсвязывающая активность при pH между 7,0 и 8,0, а более предпочтительной антигенсвязывающей активностью является антигенсвязывающая активность при pH 7,4. Кроме того, антигенсвязывающая активность антигенсвязывающей молекулы при pH от 4,0 до 6,5 особенно не ограничивается при условии, что она является антигенсвязывающей активностью при pH между 4,0 и 6,5. Однако, например, предпочтительной антигенсвязывающей активностью является антигенсвязывающая активность при pH между 5,5 и 6,5, а более предпочтительной антигенсвязывающей активностью является антигенсвязывающая активность при pH 5,8 или pH 5,5.
Антигенсвязывающую активность антигенсвязывающей молекулы можно определить известными квалифицированным в данной области техники специалистам способами. Отличные от pH условия могут быть надлежащим образом определены квалифицированными в данной области техники специалистами. Антигенсвязывающую активность антигенсвязывающей молекулы можно определить как константу диссоциации (KD), кажущуюся константу диссоциации (кажущуюся KD), скорость диссоциации (kd), кажущуюся скорость диссоциации (кажущуюся kd) или т.п. Эти константы можно определить способами, известными квалифицированным в данной области техники специалистам, например, используя Biacore (GE Healthcare), график Скэтчарда или FACS.
Здесь выражение «стадия отбора антигенсвязывающей молекулы, антигенсвязывающая активность которой при pH от 6,7 до 10,0 выше антигенсвязывающей активности при pH от 4,0 до 6,5» равнозначно выражению «стадия отбора антигенсвязывающей молекулы, антигенсвязывающая активность которой при pH от 4,0 до 6,5 ниже антигенсвязывающей активности при pH от 6,7 до 10,0».
Различие между антигенсвязывающей активностью при pH от 6,7 до 10,0 и антигенсвязывающей активностью при pH от 4,0 до 10,0 особенно не ограничивается при условии, что антигенсвязывающая активность при pH от 6,7 до 10,0 выше антигенсвязывающей активности при pH от 4,0 до 6,5. Однако антигенсвязывающая активность при pH от 6,7 до 10,0 предпочтительно выше в два раза или более, более предпочтительно в десять раз или более и еще предпочтительнее в 40 раз или более антигенсвязывающей активности при pH от 4,0 до 6,5.
Кроме того, настоящим изобретением также обеспечиваются способы скрининга антигенсвязывающих молекул, которые включают стадии:
(a) связывания антигенсвязывающей молекулы с антигеном при параметре pH, составляющем от 6,7 до 10,0;
(b) помещения антигенсвязывающей молекулы, которая связалась с антигеном на стадии (a), при параметре pH, составляющем от 4,0 до 6,5; и
(c) получения антигенсвязывающей молекулы, которая подвергалась диссоциации при параметре pH, составляющем от 4,0 до 6,5.
Кроме того, настоящим изобретением также обеспечиваются способы скрининга антигенсвязывающих молекул, которые включают стадии:
(a) отбора антигенсвязывающей молекулы, которая не связывается с антигеном при параметре pH, составляющем от 4,0 до 6,5;
(b) связывания антигенсвязывающей молекулы, отобранной на стадии (а), с антигеном при параметре pH, составляющем от 6,7 до 10,0; и
(c) получения антигенсвязывающей молекулы, которая связалась с антигеном при параметре pH, составляющем от 6,7 до 10,0.
Кроме того, настоящим изобретением также обеспечиваются способы скрининга антигенсвязывающих молекул, которые включают стадии:
(a) связывания антигенсвязывающей молекулы с антигеном при параметре pH, составляющем от 6,7 до 10,0;
(b) помещения антигенсвязывающей молекулы, которая связалась с антигеном на стадии (a), при параметре pH, составляющем от 4,0 до 6,5;
(c) получения антигенсвязывающей молекулы, которая подвергалась диссоциации при параметре pH, составляющем от 4,0 до 6,5;
(d) амплификации гена, кодирующего антигенсвязывающую молекулу, которая подверглась диссоциации; и
(e) получения элюируемой антигенсвязывающей молекулы.
Стадии (a)-(d) могут быть повторены два или более раз. Таким образом, настоящим изобретением обеспечиваются описанные выше способы, дополнительно включающие стадию повторения стадий (a)-(d) два или более раз. Число повторов стадий (a)-(d) особенно не ограничивается; однако это число обычно составляет десять или меньше.
Кроме того, настоящим изобретением также обеспечиваются способы скрининга антигенсвязывающих молекул, которые включают стадии:
(a) отбора антигенсвязывающей молекулы, которая не связывается с антигеном при параметре pH, составляющем от 4,0 до 6,5;
(b) связывания антигенсвязывающей молекулы, отобранной на стадии (а), с антигеном при параметре pH, составляющем от 6,7 до 10,0;
(c) получения антигенсвязывающей молекулы, которая связалась с антигеном при параметре pH, составляющем от 6,7 до 10,0;
(d) амплификации гена, кодирующего антигенсвязывающую молекулу, которая подверглась диссоциации; и
(e) сбора элюируемой антигенсвязывающей молекулы.
Стадии (a)-(d) могут быть повторены два или более раз. Таким образом, настоящим изобретением обеспечиваются описанные выше способы, дополнительно включающие стадию повторения стадий (a)-(d) два или более раз. Число повторов стадий (a)-(d) особенно не ограничивается; однако это число обычно составляет десять или меньше.
Когда в способах скрининга по настоящему изобретению используется библиотека фагов или т.п., стадия амплификации гена, кодирующего антигенсвязывающую молекулу, может также быть стадией амплификации фагов.
В способах по настоящему изобретению связывание антигенсвязывающей молекулы и антигена может проводиться в любой форме, без особого ограничения. Например, связывание антигенсвязывающей молекулы и антигена может проводиться путем приведения антигена в контакт с иммобилизованной антигенсвязывающей молекулой или путем приведения антигенсвязывающей молекулы в контакт с иммобилизованным антигеном. Альтернативно, связывание антигенсвязывающей молекулы и антигена может проводиться путем контактирования антигена и антигенсвязывающей молекулы в растворе.
Кроме того, настоящим изобретением также обеспечиваются способы скрининга для получения антигенсвязывающих молекул, антигенсвязывающая активность которых при первом pH превышает антигенсвязывающую активность при втором pH, которые включают стадии:
(a) связывания антигенсвязывающей молекулы с колонкой с иммобилизованным антигеном при параметре первого pH;
(b) элюирования антигенсвязывающей молекулы, которая связалась с колонкой при первом pH, с колонки при параметре второго pH; и
(c) сбора подвергшейся элюции антигенсвязывающей молекулы.
Кроме того, настоящим изобретением также обеспечиваются способы скрининга для получения антигенсвязывающих молекул, антигенсвязывающая активность которых при первом pH меньше антигенсвязывающей активности при втором pH, которые включают стадии:
(a) пропускания антигенсвязывающей молекулы через колонку с иммобилизованным антигеном при параметре первого pH;
(b) сбора антигенсвязывающей молекулы, которая вымылась без связывания с колонкой на стадии (a);
(c) связывания антигенсвязывающей молекулы, собранной на стадии (b), с колонной при параметре второго pH; и
(d) получения антигенсвязывающей молекулы, которая связалась с колонкой на стадии (c).
Кроме того, настоящим изобретением также обеспечиваются способы скрининга для получения антигенсвязывающих молекул, антигенсвязывающая активность которых при первом pH превышает антигенсвязывающую активность при втором pH, которые включают стадии:
(a) связывания библиотеки антигенсвязывающих молекул с колонкой с иммобилизованным антигеном при параметре первого pH;
(b) элюирования антигенсвязывающей молекулы с колонки при параметре второго pH;
(c) амплификации гена, кодирующего элюируемую антигенсвязывающую молекулу; и
(d) получения элюируемой антигенсвязывающей молекулы.
Стадии (a)-(с) могут быть повторены два или более раз. Таким образом, настоящим изобретением обеспечиваются описанные выше способы, дополнительно включающие стадию повторения стадий (a)-(с) два или более раз. Число повторов стадий (a)-(с) особенно не ограничивается; однако это число обычно составляет десять или меньше.
В настоящем изобретении каждый из первого и второго pH может быть любым pH при условии, что они не являются одинаковыми. В предпочтительной комбинации первого и второго pH, например, первый pH находится между 6,7 и 10,0, а второй pH находится между 4,0 и 6,5; в более предпочтительной комбинации первый pH находится между 7,0 и 8,0, а второй pH находится между 5,5 и 6,5; и в еще более предпочтительной комбинации первый pH составляет 7,4, а второй pH составляет 5,8 или 5,5.
В другой предпочтительной комбинации первого и второго pH, например, первый pH находится между 4,0 и 6,5, а второй pH находится между 6,7 и 10,0; в более предпочтительной комбинации первый pH находится между 5,5 и 6,5, а второй pH находится между 7,0 и 8,0; и в еще более предпочтительной комбинации первый pH составляет 5,8 или 5,5, а второй pH составляет 7,4.
Антигенсвязывающими молекулами, скринируемыми с помощью способов настоящего изобретения, могут быть любые антигенсвязывающие молекулы. Например, можно использовать описанные выше антигенсвязывающие молекулы в скрининге настоящего изобретения. Например, можно скринировать антигенсвязывающие молекулы, включающие природные последовательности, или антигенсвязывающие молекулы, включающие аминокислотные последовательности с заменами. Предпочтительные антигенсвязывающие молекулы, скринируемые в настоящем изобретении, включают, например, антигенсвязывающие молекулы, в которых по крайней мере одна аминокислота замещена гистидином, или в которые вставлен по крайней мере один гистидин. Точка введения замены на гистидин или его вставки особенно не ограничивается и может быть любой точкой. Кроме того, замена гистидином или его вставка может быть введена в одной точке или может быть введена в двух или более точках. Кроме того, предпочтительные антигенсвязывающие молекулы, скринируемые в настоящем изобретении, включают, например, антигенсвязывающие молекулы, включающие модифицированные константные области антител.
Антигенсвязывающими молекулами, скринируемыми способами по настоящему изобретению, может быть ряд различных антигенсвязывающих молекул, в которые введены замены на гистидин или вставки гистидина в различных точках, например, при сканировании гистидином.
Таким образом, способы скрининга по настоящему изобретению могут, кроме того, включать стадию замещения по крайней мере одной аминокислоты в антигенсвязывающей молекуле гистидином или вставки по крайней мере одного гистидина в антигенсвязывающую молекулу.
В способах скрининга по настоящему изобретению не встречающиеся в природе аминокислоты могут использоваться вместо гистидина. Следовательно, настоящее изобретением может быть также следующим из замены вышеупомянутого гистидина не встречающими в природе аминокислотами.
Кроме того, способы скрининга по настоящему изобретению могут, кроме того, включать стадию модифицирования аминокислот константных областей антител.
Антигенсвязывающие вещества, скринируемые способами скрининга по настоящему изобретению, могут быть получены любыми способами. Например, возможно использование ранее существовавших антител, ранее существовавших библиотек (библиотек фагов и т.п.), антител и библиотек, которые готовят на основе гибридом, полученных путем иммунизации животных, или на основе В-клеток иммунизированных животных, антител и библиотек (библиотек с высоким содержанием гистидина или не встречающейся в природе аминокислоты, библиотек, в которые введен гистидин или не встречающая в природе аминокислота в специфических точках, и т.п.), приготовленных путем введения мутаций на гистидин или мутаций на не встречающиеся в природе аминокислоты в описанные выше антитела и библиотеки, и т.д.
Антигенсвязывающие молекулы, многократно связывающиеся с антигеном, которые, следовательно, превосходят в сохранении в плазме, можно получить способами скрининга по настоящему изобретению. Таким образом, способы скрининга настоящего изобретения могут использоваться в качестве способов скрининга для получения антигенсвязывающих молекул, которые превосходят в сохранении в плазме.
Кроме того, способами скрининга по настоящему изобретению можно получить антигенсвязывающие молекулы, которые могут связываться с антигеном два или более раз после введения животным, таким как люди, мыши или обезьяны. Таким образом, способы скрининга настоящего изобретения могут использоваться в качестве способов скрининга для получения антигенсвязывающих молекул, которые могут связываться с антигеном два или более раз.
Кроме того, способами скрининга по настоящему изобретению можно получить антигенсвязывающие молекулы, которые способны к связыванию большего числа антигенов, чем число их антигенсвязывающих сайтов, после введения животным, таким как люди, мыши или обезьяны. Таким образом, способы скрининга настоящего изобретения могут использоваться в качестве способов скрининга для получения антигенсвязывающих молекул, которые способны к связыванию большего числа антигенов, чем число их антигенсвязывающих сайтов. Например, когда антителом является нейтрализующее антитело, способы скрининга настоящего изобретения могут использоваться в качестве способов скрининга для получения антигенсвязывающих молекул, которые могут нейтрализовать большее число антигенов, чем число антигенсвязывающих сайтов антигенсвязывающих молекул.
Кроме того, способами скрининга по настоящему изобретению можно получить антигенсвязывающие молекулы, которые способны подвергаться диссоциации внутри клетки от связавшегося с ними внеклеточно антигена после введения животным, таким как люди, мыши или обезьяны. Таким образом, способы скрининга настоящего изобретения могут использоваться в качестве способов скрининга для получения антигенсвязывающих молекул, которые способны подвергаться диссоциации внутри клетки от связавшегося с ними внеклеточно антигена.
Кроме того, способами скрининга по настоящему изобретению можно получить антигенсвязывающие молекулы, которые связываются с антигеном и подвергаются интернализации в клетку, и высвобождаются наружу из клетки в свободной от антигена форме после введения животным, таким как люди, мыши или обезьяны. Таким образом, способы скрининга настоящего изобретения могут использоваться в качестве способов скрининга для получения антигенсвязывающих молекул, которые связываются с антигеном и подвергаются интернализации в клетку, и высвобождаются наружу из клетки в свободной от антигена форме.
Кроме того, способами скрининга по настоящему изобретению можно получить антигенсвязывающие молекулы, которые могут быстро элиминировать антигены в плазме после введения животным, таким как люди, мыши или обезьяны. Таким образом, способы скрининга настоящего изобретения могут использоваться в качестве способов скрининга для получения антигенсвязывающих молекул с увеличенной (высокой) способностью к элиминации антигенов в плазме.
Кроме того, предполагается, что антигенсвязывающие молекулы намного лучше в качестве лекарственных средств, поскольку можно снизить дозу и частоту введения пациентам, и в результате можно снизить общую дозу. Таким образом, способы скрининга настоящего изобретения могут использоваться в качестве способов скрининга для получения антигенсвязывающих молекул для применения в виде фармацевтических композиций.
Кроме того, настоящим изобретением обеспечиваются библиотеки, в которых увеличено содержание гистидина по сравнению с исходными библиотеками. Библиотеки, содержащие антигенсвязывающие молекулы с увеличенным содержанием гистидина, могут использоваться в описанных выше способах скрининга и способах продуцирования, описываемых ниже.
Библиотеки с увеличенным содержанием гистидина можно получить известными квалифицированным в данной области техники специалистам способами, которые включают следующий способ. 20 типов триплетных кодонов (тринуклеотидов), кодирующих 20 типов аминокислот, можно включить с равной частотой при синтезе нуклеиновых кислот для получения библиотеки способом с использованием тринуклеотидов (J. Mol. Biol. 2008 Feb 29; 376(4): 1182-1200). В результате, можно сделать так, чтобы в мутируемом для получения библиотеки положении содержалось 20 типов аминокислот с равной вероятностью. Частоту гистидина в мутируемом для получения библиотеки положении можно увеличить путем увеличения доли кодирующего гистидин тринуклеотида по сравнению с тринуклеотидами для остальных аминокислот среди 20 типов при синтезе.
Способы продуцирования антигенсвязывающих молекул
Настоящим изобретением обеспечиваются способы продуцирования антигенсвязывающих молекул, антигенсвязывающая активность которых при pH в эндосоме ниже антигенсвязывающей активности при pH в плазме. Настоящим изобретением также обеспечиваются способы продуцирования антигенсвязывающих молекул, которые превосходят в сохранении в плазме. Настоящим изобретением также обеспечиваются способы продуцирования антигенсвязывающих молекул, которые особенно полезны при применении в виде фармацевтических композиций.
В частности, настоящим изобретением обеспечиваются способы продуцирования антигенсвязывающих молекул, которые включает стадии:
(a) определения антигенсвязывающей активности антигенсвязывающей молекулы при pH от 6,7 до 10,0;
(b) определения антигенсвязывающей активности антигенсвязывающей молекулы при pH от 4,0 до 6,5;
(c) отбора антигенсвязывающей молекулы, антигенсвязывающая активность которой при pH от 6,7 до 10,0 выше антигенсвязывающей активности при pH от 4,0 до pH 6,5;
(d) получения гена, кодирующего антигенсвязывающую молекулу, отобранную на стадии (c); и
(e) продуцирования антигенсвязывающей молекулы, используя полученный на стадии (d) ген.
Настоящим изобретением также обеспечиваются способы продуцирования антигенсвязывающих молекул, которые включают стадии:
(a) связывания антигенсвязывающей молекулы с антигеном при параметре pH, составляющем от 6,7 до 10,0;
(b) допущения нахождения антигенсвязывающей молекулы, связанной с антигеном на стадии (a), при параметре pH, составляющем от 4,0 до 6,5;
(c) сбора антигенсвязывающей молекулы, которая подвергалась диссоциации при параметре pH, составляющем от 4,0 до 6,5;
(d) получения гена, кодирующего антигенсвязывающую молекулу, полученную на стадии (c); и
(e) продуцирования антигенсвязывающей молекулы, используя полученный на стадии (d) ген.
Кроме того, настоящим изобретением обеспечиваются способы продуцирования антигенсвязывающих молекул, которые включают стадии:
(a) отбора антигенсвязывающей молекулы, которая не связывается с антигеном при параметре pH, составляющем от 4,0 до 6,5;
(b) связывания антигена при параметре pH, составляющем от 6,7 до 10,0, с антигенсвязывающей молекулой, отобранной на стадии (а);
(c) сбора антигенсвязывающей молекулы, которая связалась с антигеном при параметре pH, составляющем от 6,7 до 10,0;
(d) получения гена, кодирующего антигенсвязывающую молекулу, собранную на стадии (c); и
(e) продуцирования антигенсвязывающей молекулы, используя полученный на стадии (d) ген.
Кроме того, настоящим изобретением обеспечиваются способы продуцирования антигенсвязывающих молекул, которые включают стадии:
(a) связывания антигенсвязывающей молекулы с антигеном при параметре pH, составляющем от 6,7 до 10,0;
(b) допущения нахождения антигенсвязывающей молекулы, которая связалась с антигеном на стадии (a), при параметре pH, составляющем от 4,0 до 6,5;
(c) сбора антигенсвязывающей молекулы, которая подвергалась диссоциации при параметре pH, составляющем от 4,0 до 6,5;
(d) амплификации гена, кодирующего подвергнувшуюся диссоциации антигенсвязывающую молекулу;
(e) сбора элюируемой антигенсвязывающей молекулы;
(f) получения гена, кодирующего антигенсвязывающую молекулу, собранную на стадии (e); и
(g) продуцирования антигенсвязывающей молекулы, используя ген, полученный на стадии (f).
Стадии (a)-(d) могут быть повторены два или более раз. Таким образом, настоящим изобретением обеспечиваются описанные выше способы, дополнительно включающие стадию повторения стадий (a)-(d) два или более раз. Число повторов стадий (a)-(d) особенно не ограничивается; однако это число обычно составляет десять или меньше.
Кроме того, настоящим изобретением обеспечиваются способы продуцирования антигенсвязывающих молекул, которые включают стадии:
(a) отбора антигенсвязывающей молекулы, которая не связывается с антигеном при параметре pH, составляющем от 4,0 до 6,5;
(b) связывания антигена при параметре pH, составляющем от 6,7 до 10,0, с антигенсвязывающей молекулой, отобранной на стадии (а);
(c) сбора антигенсвязывающей молекулы, которая связалась с антигеном при параметре pH, составляющем от 6,7 до 10,0;
(d) амплификации гена, кодирующего антигенсвязывающую молекулу, которая подверглась диссоциации;
(e) сбора элюируемой антигенсвязывающей молекулы;
(f) получения гена, кодирующего антигенсвязывающую молекулу, собранную на стадии (e); и
(g) продуцирования антигенсвязывающей молекулы, используя полученный на стадии (f) ген.
Стадии (a)-(d) могут быть повторены два или более раз. Таким образом, настоящим изобретением обеспечиваются описанные выше способы, дополнительно включающие стадию повторения стадий (a)-(d) два или более раз. Число повторов стадий (a)-(d) особенно не ограничивается; однако это число обычно составляет десять или меньше.
Кроме того, настоящим изобретением обеспечиваются способы продуцирования антигенсвязывающих молекул, антигенсвязывающая активность которых при первом pH превышает антигенсвязывающую активность при втором pH, которые включают стадии:
(a) связывания антигенсвязывающей молекулы с колонкой с иммобилизованным антигеном при параметре первого pH;
(b) элюирования антигенсвязывающей молекулы, которая связалась с колонкой при первом pH, с колонки при параметре второго pH;
(c) сбора подвергшейся элюции антигенсвязывающей молекулы;
(d) получения гена, кодирующего антигенсвязывающую молекулу, собранную на стадии (c); и
(e) продуцирования антигенсвязывающей молекулы, используя полученный на стадии (d) ген.
Кроме того, настоящим изобретением обеспечиваются способы продуцирования антигенсвязывающих молекул, антигенсвязывающая активность которых при первом pH превышает антигенсвязывающую активность при втором pH, которые включают стадии:
(a) связывания библиотеки антигенсвязывающих молекул с колонкой с иммобилизованным антигеном при параметре первого pH;
(b) элюирования антигенсвязывающей молекулы с колонки при параметре второго pH;
(c) амплификации гена, кодирующего элюируемую антигенсвязывающую молекулу;
(d) сбора подвергшейся элюции антигенсвязывающей молекулы;
(e) получения гена, кодирующего антигенсвязывающую молекулу, собранную на стадии (d); и
(f) продуцирования антигенсвязывающей молекулы, используя полученный в (е) ген.
Стадии (a)-(с) могут быть повторены два или более раз. Таким образом, настоящим изобретением обеспечиваются описанные выше способы, дополнительно включающие стадию повторения стадий (a)-(с) два или более раз. Число повторов стадий (a)-(с) особенно не ограничивается; однако это число обычно составляет десять или меньше.
Когда в способах продуцирования настоящего изобретения используется библиотека фагов или т.п., стадия амплификации гена, кодирующего антигенсвязывающую молекулу, может быть стадией амплификации фагов.
Антигенсвязывающие вещества, используемые в способах продуцирования настоящего изобретения, могут быть получены любым способом. Например, возможно использование ранее существовавших антител, ранее существовавших библиотек (библиотек фагов и т.п.), антител и библиотек, которые готовят на основе гибридом, полученных путем иммунизации животных, или на основе В-клеток иммунизированных животных, антител и библиотек (библиотек с высоким содержанием гистидина или не встречающейся в природе аминокислоты, библиотек, в которые введен гистидин или не встречающая в природе аминокислота в специфических точках, и т.п.), приготовленных путем введения мутаций на гистидин или мутаций на не встречающиеся в природе аминокислоты в описанные выше антитела и библиотеки и т.д.
В описанных выше способах продуцирования антигенсвязывающая активность антигенсвязывающей молекулы при pH от 6,7 до 10, особенно не ограничивается при условии, что она является антигенсвязывающей активностью при pH между 6,7 и 10,0. Предпочтительной антигенсвязывающей активностью является антигенсвязывающая активность при pH между 7,0 и 8,0, а более предпочтительной антигенсвязывающей активностью является антигенсвязывающая активность при pH 7,4. Альтернативно, антигенсвязывающая активность антигенсвязывающей молекулы при pH от 4,0 до 6,5 особенно не ограничивается при условии, что она является антигенсвязывающей активностью при pH между 4,0 и 6,5. Предпочтительной антигенсвязывающей активностью является антигенсвязывающая активность при pH между 5,5 и 6,5, а более предпочтительной антигенсвязывающей активностью является антигенсвязывающая активность при pH 5,8 или pH 5,5.
Антигенсвязывающую активность антигенсвязывающей молекулы можно определить способами, известными квалифицированным в данной области техники специалистам. Условия, за исключением pH, могут быть надлежащим образом определены квалифицированными в данной области техники специалистами.
Выражение «стадия отбора антигенсвязывающих молекул, антигенсвязывающая активность которых при pH от 6,7 до 10,0 выше антигенсвязывающей активности при pH от 4,0 до 6,5» является синонимом выражения «стадия отбора антигенсвязывающих молекул, антигенсвязывающая активность которой при pH от 4,0 до 6,5 ниже антигенсвязывающей активности при pH от 6,7 до 10,0».
Различие в антигенсвязывающих активностях при pH от 6,7 до 10,0 и при pH от 4,0 до 10,0 особенно не ограничивается при условии, что антигенсвязывающая активность при pH от 6,7 до 10,0 выше антигенсвязывающей активности при pH от 4,0 до 6,5. Антигенсвязывающая активность при pH от 6,7 до 10,0 предпочтительно выше в два раза или более, более предпочтительно в десять раз или более и еще предпочтительнее в 40 раз или более антигенсвязывающей активности при pH от 4,0 до 6,5.
В описанных выше способах продуцирования антигенсвязывающую молекулу моно связать с антигеном в любых условиях, и эти условия особенно не ограничиваются. Например, антигенсвязывающую молекулу можно связать с антигеном путем приведения антигена в контакт с иммобилизованной антигенсвязывающей молекулой или путем приведения антигенсвязывающей молекулы в контакт с иммобилизованным антигеном. Альтернативно, антигенсвязывающую молекулу можно связать с антигеном путем контактирования антигена и антигенсвязывающей молекулы в растворе.
В описанных выше способах продуцирования каждый из первого и второго pH может быть любым pH при условии, что они не являются одинаковыми. В предпочтительной комбинации первого и второго pH, например, первый pH находится между 6,7 и 10,0, а второй pH находится между 4,0 и 6,5; в более предпочтительной комбинации первый pH находится между 7,0 и 8,0, а второй pH находится между 5,5 и 6,5; и в еще более предпочтительной комбинации первый pH составляет 7,4, а второй pH составляет 5,8 или 5,5.
В другой предпочтительной комбинации первого и второго pH, например, первый pH находится между 4,0 и 6,5, а второй pH находится между 6,7 и 10,0; в более предпочтительной комбинации первый pH находится между 5,5 и 6,5, а второй pH находится между 7,0 и 8,0; и в еще более предпочтительной комбинации первый pH составляет 5,8 или 5,5, а второй pH составляет 7,4.
Антигенсвязывающими молекулами, продуцируемыми описанными выше способами продуцирования, могут быть любые антигенсвязывающие молекулы. Предпочтительные антигенсвязывающие молекулы включают, например, антигенсвязывающие молекулы, в которых по крайней мере одна аминокислота замещена гистидином, или в которые вставлен по крайней мере один гистидин. Точка введения мутации на гистидин особенно не ограничивается и может быть любой точкой. Кроме того, мутация на гистидин может быть введена в одной точке или в двух или более точках.
Таким образом, способы продуцирования по настоящему изобретению могут, кроме того, включать стадию замещения по крайней мере одной аминокислоты в антигенсвязывающей молекуле гистидином или вставки по крайней мере одного гистидина в антигенсвязывающую молекулу.
В способах продуцирования по настоящему изобретению не встречающиеся в природе аминокислоты могут использоваться вместо гистидина. Следовательно, настоящее изобретение может быть также следующим из замены вышеупомянутого гистидина не встречающими в природе аминокислотами.
Кроме того, в другом варианте осуществления, антигенсвязывающие молекулы, продуцируемые описанными выше способами продуцирования, включают, например, антигенсвязывающие молекулы, включающие модифицированные константные области антител. В результате, способы продуцирования по настоящему изобретению могут, кроме того, включать стадию модифицирования аминокислот константных областей антител.
Антигенсвязывающие молекулы, продуцируемые способами продуцирования по настоящему изобретению, превосходят в сохранении в плазме. Таким образом, способы продуцирования по настоящему изобретению могут использоваться в качестве способов продуцирования антигенсвязывающих молекул, которые превосходят в сохранении в плазме.
Кроме того, предполагается, что антигенсвязывающие молекулы, продуцируемые способами продуцирования по настоящему изобретению, способны к связыванию с антигеном два или более раз после введения животным, таким как люди, мыши или обезьяны. Таким образом, способы продуцирования настоящего изобретения могут использоваться в качестве способов продуцирования антигенсвязывающих молекул, которые способны к связыванию с антигеном два или более раз.
Кроме того, предполагается, что антигенсвязывающие молекулы, продуцируемые способами продуцирования по настоящему изобретению, способны к связыванию большего числа антигенов, чем число их антигенсвязывающих сайтов, после введения животным, таким как люди, мыши или обезьяны. Таким образом, способы продуцирования по настоящему изобретению могут использоваться в качестве способов продуцирования антигенсвязывающих молекул, которые способны к связыванию большего числа антигенов, чем число их антигенсвязывающих сайтов.
Кроме того, предполагается, что антигенсвязывающие молекулы, продуцируемые способами продуцирования по настоящему изобретению, способны подвергаться диссоциации внутри клетки от связавшегося с ними внеклеточно антигена после введения животным, таким как люди, мыши или обезьяны. Таким образом, способы продуцирования по настоящему изобретению могут использоваться в качестве способов продуцирования антигенсвязывающих молекул, которые способны подвергаться диссоциации внутри клетки от связавшегося с ними внеклеточно антигена.
Кроме того, предполагается, что антигенсвязывающие молекулы, продуцируемые способами продуцирования по настоящему изобретению, могут связываться с антигеном и подвергаться интернализации в клетку, а также быть выброшенными наружу из клетки в свободной от антигена форме после введения животным, таким как люди, мыши или обезьяны. Таким образом, способы продуцирования по настоящему изобретению могут использоваться в качестве способов продуцирования антигенсвязывающих молекул, которые могут связываться с антигеном и подвергаться интернализации в клетку, и быть выброшенными наружу из клетки в свободной от антигена форме.
Кроме того, предполагается, что антигенсвязывающие молекулы, продуцируемые способами продуцирования по настоящему изобретению, способны к быстрой элиминации антигенов из плазмы после введения животным, таким как люди, мыши или обезьяны. Таким образом, способы продуцирования по настоящему изобретению могут использоваться в качестве способов продуцирования антигенсвязывающих молекул с увеличенной (высокой) способностью к элиминации антигенов в плазме.
Кроме того, такие антигенсвязывающие молекулы могут снизить число доз, вводимых пациентам, и, как предполагается, намного лучше в качестве лекарственных средств. Таким образом, способы продуцирования по настоящему изобретению могут использоваться в качестве способов продуцирования антигенсвязывающих молекул для применения в виде фармацевтических композиций.
Гены, получаемые способами продуцирования по настоящему изобретению, обычно несут соответствующие векторы (их встраивают в такие векторы) и затем вводят в клетки-хозяева. Векторы особенно не ограничиваются при условии, что они устойчиво удерживают встроенные нуклеиновые кислоты. Например, когда в качестве хозяина используется Escherichia coli (E.coli), предпочтительные векторы для клонирования включают вектор pBluescript (Stratagene); однако можно использовать различные, имеющиеся в продаже векторы. При использовании векторов для продукции антигенсвязывающих молекул по настоящему изобретению особенно применимы экспрессионные векторы. Экспрессионные векторы особенно не ограничиваются при условии, что векторы экспрессируют антигенсвязывающие молекулы in vitro, в E.coli, в клетках культуры или в теле организма. Например, вектор pBEST (Promega) является предпочтительным для in vitro экспрессии; вектор pET (Invitrogen) является предпочтительным для E.coli; вектор pME18S-FL3 (входящий № в GenBank - AB009864) является предпочтительным для клеток культуры; и вектор pME18S (Mol. Cell Biol. 8: 466-472 (1988)) является предпочтительным для тел организмов. ДНК настоящего изобретения можно встроить в векторы общепринятыми способами, например, путем лигирования с использованием сайтов для ферментов рестрикции (Current protocols in Molecular Biology, edit. Ausubel et al., (1987) Publish. John Wiley & Sons, Section 11.4-11.11).
Указанные выше клетки-хозяева особенно не ограничиваются, и в зависимости от цели могут использоваться различные клетки-хозяева. Примеры клеток для экспрессии антигенсвязывающих молекул включают бактериальные клетки (такие как клетки Streptococcus, Staphylococcus, E.coli, Streptomyces и Bacillus subtilis), эукариотические клетки (такие как клетки дрожжей и Aspergillus), клетки насекомых (такие как Drosophila S2 и Spodoptera SF9), клетки животных (такие как CHO, COS, HeLa, C127, 3T3, BHK, HEK293 и клетки меланомы Боуэса) и клетки растений. Векторы могут вводиться в клетку-хозяина с помощью известных способов, например, способов преципитации кальция фосфатом, способов электропорации (Current protocols in Molecular Biology edit. Ausubel et al. (1987) Publish. John Wiley & Sons, Section 9.1-9.9), способов липофекции и способов микроинъкции.
Клетки-хозяева можно культивировать с использованием известных способов. Например, при использовании клеток животных в качестве хозяина DMEM, MEM, RPMI 1640 или IMDM могут использоваться в качестве среды для культивирования. Они могут использоваться с сывороточными добавками, такими как FBS или фетальная телячья сыворотка (FCS). Клетки могут культивироваться в бессывороточных культурах. Предпочтительный pH составляет приблизительно 6-8 в течение хода культивирования. Инкубацию обычно выполняют при 30-40°C в течение приблизительно 15-200 часов. Среду меняют, аэрируют или перемешивают, в соответствии с необходимостью.
В представляющие интерес полипептиды могут быть включены подходящие сигналы секреции, так что антигенсвязывающие молекулы, экспрессируемые в клетке-хозяине, секретируются в просвет эндоплазматического ретикулума, в периплазматическое пространство или в внеклеточную среду. Эти сигналы могут быть эндогенными относительно представляющих интерес антигенсвязывающих молекул или могут быть гетерологичными сигналами.
С другой стороны, например, продукционные системы, в которых используются животные или растения, могут использоваться в качестве систем для продуцирования полипептидов in vivo. Представляющий интерес полинуклеотид вводят в животное или растение, и полипептид продуцируется в организме животного или растения и затем извлекается. «Хозяева» настоящего изобретения включают таких животных и такие растения.
Продукционные системы, в которых используются животные, включают системы, в которых используются млекопитающие или насекомые. Возможно использование таких млекопитающих, как козы, свиньи, овцы, мыши и жвачные животные (Vicki Glaser SPECTRUM Biotechnology Applications (1993)). Млекопитающие могут быть трансгенными животными.
Например, полинуклеотид, кодирующий антигенсвязывающую молекулу по настоящему изобретению, получают в виде гена, слитого с геном, кодирующим полипептид, специфически продуцируемый в молоке, такой как β-казеин козы. Затем в эмбрионы коз инъецируют полинуклеотидные фрагменты, содержащие слитый ген, и затем их трансплантируют козам. Желаемые антигенсвязывающие молекулы можно получить из молока, продуцируемого трансгенными козами, которые родились от коз, получивших эмбрионы, или от их потомства. Для увеличения объема содержащего антигенсвязывающую молекулу молока, продуцируемого трансгенными козами, могут вводиться по обстоятельствам гормоны (Ebert et al., Bio/Technology (1994) 12: 699-702).
Для продуцирования антигенсвязывающих молекул по настоящему изобретению могут использоваться насекомые, такие как тутовые шелкопряды. При использовании тутовых шелкопрядов для их инфицирования могут использоваться бакуловирусы, несущие полинуклеотид, кодирующий представляющую интерес антигенсвязывающую молекулу, и представляющую интерес антигенсвязывающую молекулу можно получить из жидкостей их организмов.
Кроме того, когда растения используются для продуцирования антигенсвязывающих молекул по настоящему изобретению, может использоваться, например, табак. При использовании табака полинуклеотид, кодирующий представляющую интерес антигенсвязывающую молекулу, встраивают в вектор для экспрессии в растениях, например, pMON 530, и затем вектор вводят в бактерии, такие как Agrobacterium tumefaciens. Бактериям затем предоставляют возможность инфицировать табак, например, Nicotiana tabacum, и желаемые антигенсвязывающие молекулы можно извлечь из их листьев (Ma et al., Eur. J. Immunol. (1994) 24: 131-138). Альтернативно, можно инфицировать ряску (Lemna minor) схожими бактериями. После клонирования желаемые антигенсвязывающие молекулы можно получить из клеток ряски (Cox KM et al., Nat. Biotechnol. 2006 Dec; 24(12): 1591-1597).
Полученные таким образом антигенсвязывающие молекулы можно выделить из внутреннего или внешнего (например, среды и молока) пространств клеток-хозяев и очистить до по существу чистых и гомогенных антигенсвязывающих молекул. Способы выделения и очистки антигенсвязывающих молекул особенно не ограничиваются, и могут использоваться способы выделения и очистки, обычно используемые для очистки полипептидов. Антигенсвязывающие молекулы можно выделить и очистить с помощью соответствующего выбора и комбинирования, например, хроматографических колонок, фильтрации, ультрафильтрации, высаливания, преципитации в растворе, экстракции растворителем, дистилляции, иммунопреципитации, электрофореза в SDS-ПААГ, изоэлектрофокусировки, диализа и перекристаллизации.
Хроматография включает, например, аффинную хроматографию, ионообменную хроматографию, хроматографию на основе гидрофобных взаимодействий, гель-фильтрацию, хроматографию с обращенной фазой и адсорбционную хроматографию (Strategies for Protein Purification и Characterization: A Laboratory Course Manual. Ed Daniel R. Marshak et al., (1996) Cold Spring Harbor Laboratory Press). Такие хроматографические способы можно проводить, используя жидкостную хроматографию, такую как HPLC (жидкостная хроматография высокого давления) и FPLC (жидкостная хроматография быстрого разрешения). Колонки, используемые для аффинной хроматографии, включают колонки с белком A и колонки с белком G. Колонки, в которых используется белок A, включают, например, Hyper D, POROS и Сефарозу F.F. (Pharmacia).
При необходимости антигенсвязывающую молекулу можно произвольно модифицировать, и можно частично удалить пептиды путем предоставления подходящему для модификации белка ферменту возможности оказывать действие до или после очистки антигенсвязывающей молекулы. Такие ферменты для модификации белков включают, например, трипсин, химотрипсин, лизилэндопептидазы, протеинкиназы и глюкозидазы.
Антитела против рецептора для IL-6
Кроме того, настоящим изобретением обеспечиваются указанные ниже в (a)-(m) антитела против рецептора для IL-6:
(a) антитело, которое включает вариабельную область тяжелой цепи, включающую аминокислотную последовательность, в которой на His замещена по крайней мере одна из следующих аминокислот: Tyr в положении 27, Asp в положении 31, Asp в положении 32, Trp в положении 35, Tyr в положении 51, Asn в положении 59, Ser в положении 63, Met в положении 106 и Tyr в положении 108 в аминокислотной последовательности SEQ ID NO: 1 (вариабельной области H53);
(b) антитело, которое включает вариабельную область тяжелой цепи (H3pI), имеющую аминокислотную последовательность, в которой Tyr в положении 27, Asp в положении 31 и Trp в положении 35 в аминокислотной последовательности SEQ ID NO: 1 (вариабельной области H53) замещены на His;
(c) антитело, которое включает вариабельную область тяжелой цепи, имеющую аминокислотную последовательность, в которой Tyr в положении 27, Asp в положении 31, Asp в положении 32, Trp в положении 35, Asn в положении 59, Ser в положении 63 и Tyr в положении 108 в аминокислотной последовательности SEQ ID NO: 1 (вариабельной области H53) замещены на His;
(d) антитело, которое включает вариабельную область тяжелой цепи (H170), имеющую аминокислотную последовательность, в которой Tyr в положении 27, Asp в положении 31, Asp в положении 32, Trp в положении 35, Asn в положении 59, Ser в положении 63 и Tyr в положении 108 замещены на His, и в которой Ser в положении 99 замещен на Val, а Thr в положении 103 замещен на Ile в аминокислотной последовательности SEQ ID NO: 1 (вариабельной области H53);
(e) антитело, которое включает вариабельную область тяжелой цепи, имеющую аминокислотную последовательность, в которой Asp в положении 31, Tyr в положении 51, Ser в положении 63, Met в положении 106 и Tyr в положении 108 в аминокислотной последовательности SEQ ID NO: 1 (вариабельной области H53) замещены на His;
(f) антитело, которое включает вариабельную область тяжелой цепи (CLH5), имеющую аминокислотную последовательность, в которой Asp в положении 31, Tyr в положении 51, Ser в положении 63, Met в положении 106 и Tyr в положении 108 замещены на His, и в которой Ser в положении 99 замещен на Phe, а Thr в положении 103 замещен на Ile в аминокислотной последовательности SEQ ID NO: 1 (вариабельной области H53);
(g) антитело, которое включает вариабельную область легкой цепи, имеющую аминокислотную последовательность, в которой на His замещена по крайней мере одна из следующих аминокислот: Asp в положении 28, Tyr в положении 32, Glu в положении 53, Ser в положении 56 и Asn в положении 92 в аминокислотной последовательности SEQ ID NO: 2 (вариабельной области PF1L);
(h) антитело, которое включает вариабельную область легкой цепи (L73), имеющую аминокислотную последовательность, в которой Asp в положении 28, Tyr в положении 32 и Glu в положении 53 в аминокислотной последовательности SEQ ID NO: 2 (вариабельной области PF1L) замещены на His;
(i) антитело, которое включает вариабельную область легкой цепи (L82), имеющую аминокислотную последовательность, в которой Tyr в положении 32 и Glu в положении 53 в аминокислотной последовательности SEQ ID NO: 1 (вариабельной области H53) замещены на His;
(j) антитело, которое включает вариабельную область легкой цепи (CLL5), имеющую аминокислотную последовательность, в которой Tyr в положении 32, Glu в положении 53, Ser в положении 56 и Asn в положении 92 в аминокислотной последовательности SEQ ID NO: 2 (вариабельной области PF1L) замещены на His;
(k) антитело, которое включает вариабельную область тяжелой цепи антитела (b) и вариабельную область легкой цепи антитела (h);
(l) антитело, которое включает вариабельную область тяжелой цепи антитела (d) и вариабельную область легкой цепи антитела (i); и
(m) антитело, которое включает вариабельную область тяжелой цепи антитела (f) и вариабельную область легкой цепи антитела (h).
Конкретные примеры вариабельной области тяжелой цепи, имеющей аминокислотную последовательность, в которой на His замещена по крайней мере одна из следующих аминокислот: Tyr в положении 27, Asp в положении 31, Asp в положении 32, Trp в положении 35, Tyr в положении 51, Asn в положении 59, Ser в положении 63, Met в положении 106 и Tyr в положении 108 в аминокислотной последовательности SEQ ID NO: 1 (вариабельной области H53), включают, например, следующие вариабельные области тяжелой цепи:
вариабельную область тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 3 (H3pI)
вариабельную область тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 4 (H170)
вариабельную область тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 5 (CLH5).
Конкретные примеры вариабельной области легкой цепи, имеющей аминокислотную последовательность, в которой на His замещена по крайней мере одна из следующих аминокислот: Asp в положении 28, Tyr в положении 32, Glu в положении 53, Ser в положении 56 и Asn в положении 92 в аминокислотной последовательности SEQ ID NO: 2 (вариабельной области PF1L), включают, например, следующие вариабельные области легкой цепи:
вариабельную область легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 6 (L73)
вариабельную область легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 7 (L82)
вариабельную область легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 8 (CLL5).
Положения аминокислот и аминокислотные замены в каждом из описанных выше антител H3pI, H170, CLH5, L73, L82 и CLL5 представлены ниже в таблице 1. Положения аминокислот указаны на основе нумерации по Kabat.
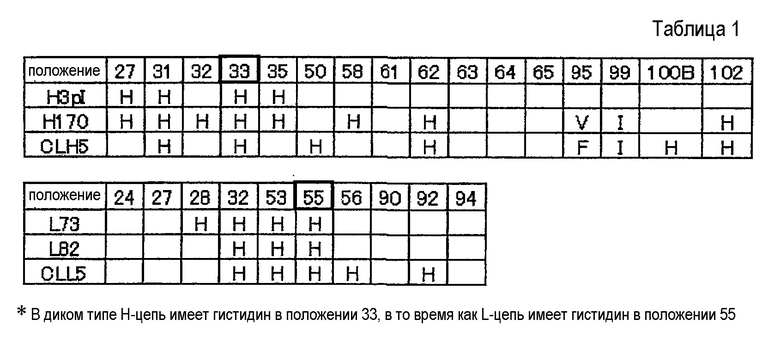
Настоящим изобретением обеспечиваются антитела, включающие по крайней мере одну из аминокислотных замен, описанных выше в (a)-(j), и способы продуцирования таких антител. Таким образом, антитела по настоящему изобретению также включают антитела, включающие не только любую из аминокислотных замен, описанных выше в (a)-(j), но также аминокислотную замену(ы), отличную от аминокислотных замен, описанных выше в (a)-(j). Аминокислотные замены, отличные от аминокислотных замен, описанных выше в (a)-(j), включают, например, замену, делецию, добавление и/или вставку в аминокислотную последовательность CDR и FR.
Кроме того, настоящим изобретением обеспечиваются указанные ниже в (1)-(28) антитела против рецептора для IL-6:
(1) антитело, которое включает вариабельную область тяжелой цепи (вариабельную область VH1-IgG1), имеющую аминокислотную последовательность с положения 1 до положения 119 в SEQ ID NO: 21 (VH1-IgG1);
(2) антитело, которое включает вариабельную область тяжелой цепи (вариабельную область VH2-IgG1), имеющую аминокислотную последовательность с положения 1 до положения 119 в SEQ ID NO: 22 (VH2-IgG1);
(3) антитело, которое включает вариабельную область тяжелой цепи (вариабельную область VH3-IgG1), имеющую аминокислотную последовательность с положения 1 до положения 119 в SEQ ID NO: 23 (VH3-IgG1);
(4) антитело, которое включает вариабельную область тяжелой цепи (вариабельную область VH4-IgG1), имеющую аминокислотную последовательность с положения 1 до положения 119 в SEQ ID NO: 24 (VH4-IgG1);
(5) антитело, которое включает вариабельную область легкой цепи (вариабельную область VL1-CK), имеющую аминокислотную последовательность с положения 1 до положения 107 в SEQ ID NO: 25 (VL1-CK);
(6) антитело, которое включает вариабельную область легкой цепи (вариабельную область VL2-CK), имеющую аминокислотную последовательность с положения 1 до положения 107 в SEQ ID NO: 26 (VL2-CK);
(7) антитело, которое включает вариабельную область легкой цепи (вариабельную область VL3-CK), имеющую аминокислотную последовательность с положения 1 до положения 107 в SEQ ID NO: 27 (VL3-CK);
(8) антитело (Fv1-IgG1), которое включает вариабельную область тяжелой цепи антитела (2) и вариабельную область легкой цепи антитела (6);
(9) антитело (Fv2-IgG1), которое включает вариабельную область тяжелой цепи антитела (1) и вариабельную область легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 7 (L82);
(10) антитело (Fv3-IgG1), которое включает вариабельную область тяжелой цепи антитела (4) и вариабельную область легкой цепи антитела (5);
(11) антитело (Fv4-IgG1), которое включает вариабельную область тяжелой цепи антитела (3) и вариабельную область легкой цепи антитела (7);
(12) антитело (VH3-IgG2ΔGK), которое включает тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 33;
(13) антитело (VH3-M58), которое включает тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 34;
(14) антитело (VH3-M73), которое включает тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 35;
(15) антитело (Fv4-IgG2ΔGK), которое включает тяжелую цепь антитела (12) и легкую цепь, имеющую аминокислотную последовательность SEQ ID NO: 27 (VL3-CK);
(16) антитело (Fv4-M58), которое включает тяжелую цепь антитела (13) и легкую цепь, имеющую аминокислотную последовательность SEQ ID NO: 27 (VL3-CK);
(17) антитело (Fv4-M73), которое включает тяжелую цепь антитела (14) и легкую цепь, имеющую аминокислотную последовательность SEQ ID NO: 27 (VL3-CK);
(18) антитело (VH2-M71), которое включает тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 36 (VH2-M71);
(19) антитело (VH2-M73), которое включает тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 37 (VH2-M73);
(20) антитело (VH4-M71), которое включает тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 38 (VH4-M71);
(21) антитело (VH4-M73), которое включает тяжелую цепь, имеющую аминокислотную последовательность SEQ ID NO: 39 (VH4-M73);
(22) антитело (Fv1-M71), которое включает тяжелую цепь антитела (18) и легкую цепь, имеющую аминокислотную последовательность SEQ ID NO: 26 (VL2-CK);
(23) антитело (Fv1-M73), которое включает тяжелую цепь антитела (19) и легкую цепь, имеющую аминокислотную последовательность SEQ ID NO: 26 (VL2-CK);
(24) антитело (Fv3-M71), которое включает тяжелую цепь антитела (20) и легкую цепь, имеющую аминокислотную последовательность SEQ ID NO: 25 (VL1-CK);
(25) антитело (Fv3-M73), которое включает тяжелую цепь антитела (21) и легкую цепь, имеющую аминокислотную последовательность SEQ ID NO: 25 (VL1-CK);
(26) антитело, которое включает легкую цепь, имеющую аминокислотную последовательность SEQ ID NO: 25 (VL1-CK);
(27) антитело, которое включает легкую цепь, имеющую аминокислотную последовательность SEQ ID NO: 26 (VL2-CK); и
(28) антитело, которое включает легкую цепь, имеющую аминокислотную последовательность SEQ ID NO: 27 (VL3-CK).
Кроме того, настоящим изобретением обеспечиваются FR и CDR, указанные ниже в (a)-(v):
(a) CDR1 тяжелой цепи, имеющий SEQ ID NO: 40 (VH1, 2, 3, 4);
(b) CDR2 тяжелой цепи, имеющий SEQ ID NO: 41 (VH1, 2);
(c) CDR2 тяжелой цепи, имеющий SEQ ID NO: 42 (VH3);
(d) CDR2 тяжелой цепи, имеющий SEQ ID NO: 43 (VH4);
(e) CDR3 тяжелой цепи, имеющий SEQ ID NO: 44 (VH1, 2);
(f) CDR3 тяжелой цепи, имеющий SEQ ID NO: 45 (VH3, 4);
(g) FR1 тяжелой цепи, имеющая SEQ ID NO: 46 (VH1, 2);
(h) FR1 тяжелой цепи, имеющая SEQ ID NO: 47 (VH3, 4);
(i) FR2 тяжелой цепи, имеющая SEQ ID NO: 48 (VH1, 2, 3, 4);
(j) FR3 тяжелой цепи, имеющая SEQ ID NO: 49 (VH1);
(k) FR3 тяжелой цепи, имеющая SEQ ID NO: 50 (VH2);
(l) FR3 тяжелой цепи, имеющая SEQ ID NO: 51 (VH3, 4);
(m) FR4 тяжелой цепи, имеющая SEQ ID NO: 52 (VH1, 2, 3, 4);
(n) CDR1 легкой цепи, имеющий SEQ ID NO: 53 (VL1, 2);
(o) CDR1 легкой цепи, имеющий SEQ ID NO: 54 (VL3);
(p) CDR2 легкой цепи, имеющий SEQ ID NO: 55 (VL1, VL3);
(q) CDR2 легкой цепи, имеющий SEQ ID NO: 56 (VL2);
(r) CDR3 легкой цепи, имеющий SEQ ID NO: 57 (VL1, 2, 3);
(s) FR1 легкой цепи, имеющая of SEQ ID NO: 58 (VL1, 2, 3);
(t) FR2 легкой цепи, имеющая SEQ ID NO: 59 (VL1, 2, 3);
(u) FR3 легкой цепи, имеющая SEQ ID NO: 60 (VL1, 2, 3); и
(v) FR4 легкой цепи, имеющая SEQ ID NO: 61 (VL1, 2, 3).
Соответствующие последовательности FR и CDR, указанных выше в (a)-(v), представлены на фиг. 25. Кроме того, настоящим изобретением обеспечиваются полипептиды, включающие любой(ую) из FR и CDR, указанных выше в (a)-(v).
Антитела против рецептора для IL-6 по настоящему изобретению также включают фрагменты и модифицированные продукты антител, включающие любую из описанных выше аминокислотных замен. Такие фрагменты антител включают, например, Fab, F(abʹ)2, Fv, одноцепочечный Fv (scFv), H- и L-цепи которого соединены вместе через подходящий линкер, однодоменную H-цепь и однодоменную L-цепь (см., например, Nat. Biotechnol. 2005 Sep; 23(9): 1126-36), унитело (WO 2007059782 A1) и SMIP (WO 2007014278 A2). Происхождение антител особенно не ограничивается. Антитела включают антитела человека, мыши, крысы и кролика. Антитела по настоящему изобретению могут также быть химерными, гуманизированными, полностью гуманизированными антителами или т.п.
В частности, такие фрагменты антител получают с помощью обработки антител ферментами, например, папаином или пепсином, или с помощью конструирования генов, кодирующих такие фрагменты антител, встраивания их в экспрессионные векторы и затем их экспрессии в подходящих клетках-хозяевах (см., например, Co, M.S. et al., J. Immunol. (1994) 152, 2968-2976; Better, M. & Horwitz, A.H. Methods in Enzymology (1989) 178, 476-496; Plueckthun, A. & Skerra, A. Methods in Enzymology (1989) 178, 497-515; Lamoyi, E., Methods in Enzymology (1989) 121, 652-663; Rousseaux, J. et al., Methods in Enzymology (1989) 121, 663-66; Bird, R.E. et al., TIBTECH (1991) 9, 132-137).
Настоящим изобретением обеспечиваются способы продуцирования (i) полипептида настоящего изобретения или (ii) полипептида, кодируемого геном, кодирующим полипептид по настоящему изобретению, причем способы включают стадию культивирования клетки-хозяина, включающей вектор, в который встроен полинуклеотид, кодирующий полипептид по настоящему изобретению.
Конкретнее, настоящим изобретением обеспечиваются способы продуцирования полипептида по настоящему изобретению, которые включают стадии:
(a) культивирования клетки-хозяина, включающей вектор, в который встроен ген, кодирующий полипептид по настоящему изобретению; и
(b) получения полипептида, кодируемого этим геном.
scFv получают путем соединения V-областей H- и L-цепей антитела. В таком scFv V-область Н-цепи соединена с V-областью L-цепи через линкер, предпочтительно пептидный линкер (Huston, J.S. et al., Proc. Natl. Acad. Sci. U.S.A. (1988) 10.0, 5879-5883). V-области H-цепи и L-цепи в scFv могут происходить из любого из описанных выше антител. Пептидный линкер для соединения V-областей включает, например, произвольные одноцепочечные пептиды из 12-19 аминокислотных остатков.
Когда антитело против рецептора для IL-6 по настоящему изобретению включает константную область, константная область может быть константной областью любого типа, могут использоваться, например, IgG1, IgG2 или IgG4. Константной областью предпочтительно является константная область антитела человека. Альтернативно, константной областью может быть модифицированная форма, включающая замену, делецию, добавление и/или вставку в аминокислотной последовательности константных областей IgG1 человека, IgG2 человека или IgG4 человека.
Предпочтительным рецептором для IL-6, с которым связывается антитело против рецептора IL-6 по настоящему изобретению, является рецептор IL-6 человека.
Антитела против рецептора для IL-6 по настоящему изобретению превосходят в сохранении в плазме, и они существуют в плазме в течение длительного периода в форме, способной к связыванию с антигеном, т.е., растворимыми или связанными с мембранами рецепторами для IL-6. Таким образом, антитела против рецептора для IL-6 связываются in vivo с растворимыми или связанными с мембранами рецепторами для IL-6 в течение длительного периода времени. Кроме того, антитела против рецептора для IL-6 способны к связыванию с рецепторами для IL-6 два или более раз, и, следовательно, как предполагается, способны нейтрализовать три или более рецепторов для IL-6.
Фармацевтические композиции
Настоящее изобретение также относится к фармацевтическим композициям, включающим антигенсвязывающие молекулы по настоящему изобретению, антигенсвязывающие молекулы, выделенные способами скрининга по настоящему изобретению, или антигенсвязывающие молекулы, продуцированные способами продуцирования по настоящему изобретению. Антигенсвязывающие молекулы по настоящему изобретению и антигенсвязывающие молекулы, продуцированные способами по настоящему изобретению, превосходят в сохранении в плазме, и, следовательно, как предполагается, снижают частоту введения антигенсвязывающих молекул, и, следовательно, применимы в виде фармацевтических композиций. Фармацевтическая композиция по настоящему изобретению может включать фармацевтически приемлемые носители.
В настоящем изобретении фармацевтические композиции обычно относятся к средствам для лечения или предупреждения, или лечения и диагностики заболеваний.
Фармацевтические композиции по настоящему изобретению могут быть составлены с использованием известных квалифицированным в данной области техники специалистам способов. Например, они могут использоваться парентерально, в форме инъекций стерильных растворов или суспензий, включающих воду или другую фармацевтически приемлемую жидкость. Например, такие композиции могут быть составлены путем смешивания в форме стандартной дозы, требуемой в способе производства полностью разрешенных к применению лекарств, при соответствующем объединении с фармацевтически приемлемыми носителями или средами, в частности, со стерильной водой, физиологическим раствором, растительным маслом, эмульгатором, суспензией, поверхностно-активным веществом, стабилизатором, корригентом, наполнителем, носителем, консервантом, связующим веществом или т.п. В таких композициях количество активного ингредиента регулируют для получения соответствующего количества в заранее определенном диапазоне.
Стерильные композиции для инъекции могут быть составлены, используя носители, такие как дистиллированная вода для инъекции, в соответствии с обычным способом составления.
Водные растворы для инъекции включают, например, физиологический раствор или изотонические растворы, содержащие декстрозу или другие вспомогательные вещества (например, D-сорбит, D-маннозу, D-маннит и натрия хлорид). Также возможно использование в комбинации соответствующих солюбилизаторов, например, спиртов (этилового спирта и т.п.), полиспиртов (пропиленгликоля, полиэтиленгликоля и т.п.), неионных поверхностно-активных веществ (полисорбата 80 (TM), HCO-50 и т.п.).
Масла включают кунжутное масло и соевое масло. Бензилбензоат и/или бензиловый спирт могут использоваться в комбинации в качестве солюбилизаторов. Также возможно комбинирование буферов (например, фосфатного буфера и натрийацетатного буфера), успокаивающих средств (например, прокаина гидрохлорида), стабилизаторов (например, бензилового спирта и фенола) и/или антиоксидантов. Подходящие ампулы заполняют приготовленными инъекционными растворами.
Фармацевтические композиции по настоящему изобретению предпочтительно вводят парентерально. Например, композиции могут быть в лекарственной форме для инъецирований, введения через нос, введения через легкие или чрескожного введения. Например, они могут вводиться системно или местно с помощью внутривенной инъекции, внутримышечной инъекции, внутрибрюшинной инъекции, подкожной инъекции или т.п.
Способы введения могут быть надлежащим образом выбраны с учетом возраста и симптомов пациента. Доза фармацевтической композиции, содержащей антигенсвязывающую молекулу, может составлять, например, от 0,0001 до 1000 мг/кг для каждого введения. Альтернативно, доза может составлять, например, от 0,001 до 100000 мг для каждого пациента. Однако настоящее изобретение не ограничивается численными значениями, указанными выше. Дозы и способы введения могут меняться в зависимости от веса, возраста, симптомов пациента и т.п. Квалифицированные в данной области техники специалисты могут установить соответствующие дозы и пути введения с учетом описанных выше факторов.
Аминокислоты, содержащиеся в аминокислотных последовательностях настоящего изобретения, могут подвергаться посттрасляционным модификациях. Квалифицированным в данной области техники специалистам хорошо известна, например, модификация N-концевого остатка глютамина (Gln) в остаток пироглютаминовой кислоты (pGlu) при пироглютамилировании. Конечно, такие посттрансляционно модифицированные аминокислоты включены в аминокислотные последовательности настоящего изобретения.
Все приведенные в описании документы, касающиеся известного уровня техники, включены в данное описание посредством ссылки.
Примеры
Ниже настоящее изобретение будет описано специально со ссылкой на примеры, но не должно быть истолковано, как ограниченное ими.
Пример 1. Продуцирование модифицированного гуманизированого антитела PM1
Приготовление рекомбинантного растворимого рецептора для IL-6 человека (SR344)
Рекомбинантный рецептор IL-6 человека из рецептора IL-6 человека, который служил в качестве антигена, был получен следующим образом. Была создана линия клеток CHO, которая постоянно экспрессирует растворимый рецептор IL-6 человека (упоминаемый ниже как SR344) (Yamasaki, et al., Science 1988; 241: 825-828 (GenBank #X12830)), состоящий из аминокислотной последовательности с 1-ой аминокислоты до 344-ой аминокислоты с N-конца, как сообщалось в J. Biochem., 108, 673-676 (1990).
SR344 очищали из супернатанта культуры, полученного из SR344-экспрессирующих клеток CHO, используя три колоночные хроматографии: хроматографию с использованием колонки с голубой сефарозой 6 FF, аффинную хроматографию с использованием колонки, на которой иммобилизовано специфичное для SR344, и хроматографию с использованием колонки для гель-фильтрации. Элюируемую в виде основного пика фракцию использовали в качестве конечного очищенного продукта.
Приготовление рекомбинантного растворимого рецептора IL-6 яванского макака (cIL-6R)
Олигодезоксинуклеотидные праймеры Rhe6Rf1 (SEQ ID NO: 16) и Rhe6Rr2 (SEQ ID NO: 17) были созданы на основе общедоступной последовательности гена рецептора IL-6 макака-резус (Birney et al., Ensemble 2006, Nucleic Acids Res., 2006, Jan. 1; 34 (Database issue): D556-61). Используя полученную из поджелудочной железы яванского макака кДНК в качестве матрицы, был получен ДНК-фрагмент, кодирующий ген рецептора IL-6 яванского макака полной длины, с помощью ПЦР, используя праймеры Rhe6Rf1 и Rhe6Rr2. Используя результирующий ДНК-фрагмент в качестве матрицы, ДНК-фрагмент размером 1131 п.о. (SEQ ID NO: 20), кодирующий белок, в котором 6xHis добавлен к С-концу растворимого района (Met1-Pro363), содержащего сигнальную область гена рецептора IL-6 яванского макака, был амплифицирован с помощью ПЦР, используя олигодезоксинуклеотидные праймеры CynoIL6R N-EcoRI (SEQ ID NO: 18) и CynoIL6R C-NotI-His (SEQ ID NO: 19). Результирующий ДНК-фрагмент расщепляли ферментами EcoRI и NotI и встраивали в вектор для экспрессии в клетках млекопитающих, и результирующий вектор затем использовали для создания линии клеток CHO со стабильной экспрессией (cyno.sIL-6R-продуцирующих клеток CHO).
Среду после культивирования cyno.sIL-6R-продуцирующих клеток CHO очищали с использованием колонки HisTrap (GE Healthcare Biosciences), концентрировали, используя Amicon Ultra-15 Ultracel-10k (Millipore), и далее очищали с использованием колонки Superdex 200 пг 16/60 для гель-фильтрации (GE Healthcare Biosciences) для получения конечного очищенного продукта растворимого рецептора IL-6 яванского макака (ниже упоминаемого как cIL-6R).
Приготовление рекомбинантного IL-6 яванского макака (cIL-6)
IL-6 яванского макака готовили следующим образом. Нуклеотидную последовательность, кодирующую 212 аминокислот, зарегистрированную под входящим в SWISSPROT № P79341, получали, клонировали в вектор для экспрессии в клетках млекопитающих и вводили в клетки CHO для создания линии клеток со стабильной экспрессией (cyno.IL-6-продуцирующих клеток CHO). Среду от культивирования cyno.IL-6-продуцирующих клеток CHO очищали с использованием колонки с SP-Сефарозой/FF (GE Healthcare Biosciences), концентрировали, используя Ultra-15 Ultracel-5k (Millipore), и затем далее очищали с использованием колонки Superdex 75 пг 26/60 для гель-фильтрации (GE Healthcare Biosciences). Результирующий препарат концентрировали, используя Amicon Ultra-15 Ultracel-5k (Millipore), для получения конечного очищенного продукта IL-6 яванского макака (ниже упоминаемого как cIL-6).
Создание линии клеток BaF3, экспрессирующих gp130 человека.
Линию клеток BaF3, экспрессирующих gp130 человека, создавали, как указано ниже, для того, чтобы получить линию клеток, демонстрирующих IL-6-зависимый рост.
кДНК полноразмерного gp130 человека (Hibi et al., Cell 1990; 63: 1149-1157 (GenBank #NM_002184)) амплифицировали с помощью ПЦР и клонировали в экспрессионный вектор pCOS2Zeo, который был получен путем удаления сайта экспрессии гена DHFR из pCHOI (Hirata, et al., FEBS Letter 1994; 356: 244-248) и вставки сайта экспрессии гена устойчивости к зеоцину, для конструирования pCOS2Zeo/gp130. кДНК полноразмерного IL-6R человека амплифицировали с помощью ПЦР и клонировали в pcDNA3.1(+) (Invitrogen) для конструирования hIL-6R/pcDNA3.1(+). 10 мкг pCOS2Zeo/gp130 вмешивали в клетки BaF3 (0,8×107 клеток), суспендированные в PBS, и на смесь посылали импульсы, используя генератор импульсов для генов (Bio-Rad) при напряжении 0,33 кВ и емкости 950 мкФ Клетки BaF3, подвергшиеся введению гена с помощью обработки - электропорации, культивировали целый день и ночь в среде RPMI1640 (Invitrogen), содержащей 0,2 нг/мл интерлейкина-3 мыши (Peprotech) и 10% фетальной бычьей сыворотки (ниже упоминаемой как FBS, HyClone), и затем скринировали путем добавления среды RPMI1640, содержащей 100 нг/мл интерлейкина-6 человека (R&D Systems), 100 нг/мл растворимого рецептора интерлейкина-6 человека (R&D Systems) и 10% FBS, для создания линии клеток BaF3, экспрессирующих gp130 человека (ниже упоминаемой как BaF3/gp130). Поскольку эта линия BaF3/gp130 подвергается пролиферации в присутствии интерлейкина-6 человека (R&D Systems) и SR344, ее можно использовать для оценки ингибиторной в отношении роста активности (а именно, нейтрализующей рецептор IL-6 активности) антитела против рецептора IL-6.
Продуцирование гуманизированного антитела против рецептора для IL-6
В рамках текущего примера и встречающего в данном описании, WT является сокращением термина «дикий тип», H (WT) является сокращением термина «Н-цепь дикого типа» (с аминокислотной последовательностью: SEQ ID NO: 9), и L (WT) является сокращением термина «L-цепь дикого типа» (с аминокислотной последовательностью: SEQ ID NO: 10). В этой связи были введены мутации в последовательность каркасной области и последовательность CDR гуманизированного антитела мыши PM1, описанного в Cancer Res. 1993, Feb. 15; 53(4): 851-856, для получения модифицированных H-цепей H53 (аминокислотной последовательности: SEQ ID NO: 1) и PF1H (аминокислотной последовательности: SEQ ID NO: 11), и модифицированных L-цепей L28 (аминокислотной последовательности: SEQ ID NO: 12) и PF1L (аминокислотной последовательности: SEQ ID NO: 2). Конкретнее, мутанты были получены, используя набор для сайт-направленного мутагенеза QuikChange (Stratagene) в соответствии со способом, описанным в предоставленных инструкциях, и результирующие плазмидные фрагменты встраивали в вектор для экспрессии в клетках млекопитающих для получения векторов для экспрессии желаемых H-цепей и векторов для экспрессии желаемых L-цепей. Нуклеотидные последовательности полученных экспрессионных векторов определяли, используя общепринятые методики, известные квалифицированным в данной области техники специалистам.
Экспрессия и очистка гуманизированного антитела против рецептора IL-6
Экспрессию антител осуществляли с использованием описанного ниже способа. Линию клеток HEK293H, происходящих из почки человеческого эмбриона (Invitrogen), суспендировали в среде DMEM (Invitrogen), дополненной 10% фетальной бычьей сыворотки (Invitrogen). Клетки засевали в объеме 10 мл на чашку в чашки для прилипающих клеток (диаметром 10 см; CORNING) при плотности клеток, составляющей 5-6×105 клеток/мл, и культивировали в термостате с CO2 (37°C, 5% CO2) в течение одного целого дня и ночи. Затем среду удаляли с помощью аспирации, и добавляли 6,9 мл среды CHO-S-SFM-II (Invitrogen). Приготовленную плазмиду вводили в клетки способом липофекции. Супернатанты результирующей культуры собирали, центрифугировали (при приблизительно 2000×g, в течение 5 мин, при комнатной температуре) для удаления клеток и стерилизовали фильтрованием через 0,22 мкм фильтр MILLEX®-GV (Millipore) для получения супернатантов. Антитела очищали от полученных супернатантов культур с помощью способа, известного квалифицированным в данной области техники специалистам, используя Sepharose™ Fast Flow с рекомбинантным белком А (Amersham Biosciences). Для определения концентрации очищенного антитела измеряли оптическую плотность при 280 нм, используя спектрофотометр. Концентрации антител рассчитывали на основе определенных значений, используя коэффициент поглощения, рассчитанный с помощью способа PACE (Protein Science 1995; 4: 2411-2423).
Пример 2. Продуцирование антитела H3pI/L73, связывающегося в зависимости от pH.
Способ создания антитела, способного к многократной нейтрализации антигенов.
Поскольку молекулы IgG являются двухвалентными, одна молекула IgG может нейтрализовать вплоть до двух молекул антигена, когда два сайта связываются с двумя антигенами; однако она не может нейтрализовать три или более молекул антигена. Поэтому для сохранения нейтрализующего эффекта нейтрализующего антитела в течение определенного периода времени необходимо вводить количество антитела, равное количеству антигена, продуцируемого в течение этого периода времени, или превышающее такое количество. Таким образом, существует предел степени, до которой может быть снижена требуемая доза антитела при улучшении фармакокинетики или аффинности антитела. Следовательно, если бы была возможной нейтрализация двух или более молекул антитела одной молекулой IgG, та же самая доза могла бы увеличить продолжительность нейтрализующего эффекта, или в альтернативном случае могла бы быть снижена доза антитела, требуемая для достижения той же самой продолжительности.
Для нейтрализующих антител существует два типа антигенов-мишеней: антигены растворимого типа, которые присутствуют в плазме, и связанные с мембранами антигены, которые экспрессируются на поверхности клеток.
Когда антигеном является связанный с мембраной антиген, введенное антитело связывается с мембранным антигеном на клеточной поверхности, и антитело впоследствии поступает в эндосомы внутри клетки посредством интернализации вместе с мембранным антигеном, связанным с антителом. Затем антитело, которое продолжает быть связанным с антигеном, перемещается в лизосому, где оно подвергается деградации лизосомой вместе с антигеном. Опосредованную интернализацией элиминацию антитела из плазмы мембранным антигеном называют антигензависимой элиминацией, и о такой элиминации сообщалось для многочисленных молекул антител (Drug Discov. Today. 2006 Jan; 11(1-2): 81-88). Поскольку одна молекула антитела IgG-типа связывается с двумя молекулами антигена, когда она связывается двухвалентно с антигенами, и затем подвергается интернализации, и непосредственно подвергается деградации в лизосоме, одна обычная молекула антитела IgG-типа не может нейтрализовать две или более молекулы антигена (фиг. 1).
Причиной длительного сохранения (медленной элиминации) молекул IgG в плазме является действие FcRn, который известен как рецептор для возвращения молекул IgG (Nat. Rev. Immunol. 2007 Sep; 7(9): 715-25). Молекулы IgG, поступившие в эндосомы посредством пиноцитоза, связываются с FcRn, экспрессируемым в эндосомах, в кислотных условиях внутри эндосом. Молекулы IgG, связавшиеся с FcRn, перемещаются на клеточную поверхность, где они подвергаются диссоциации от FcRn в нейтральных условиях в плазме и возвращаются в плазму, в то время как молекулы IgG, не способные к связыванию с FcRn, отправляются в лизосомы, где они подвергаются деградации (фиг. 2).
Молекулы IgG, связавшиеся с мембранным антигеном, поступают во внутриклеточные эндосомы посредством интернализации, перемещаются, будучи связанными с антигеном, в лизосомы и подвергаются деградации. Когда антитело IgG-типа связывается двухвалентно с антигенами, оно нейтрализует две молекулы антигена и повергается деградации вместе с антигенами. Если антитело IgG-типа, после поступления во внутриклеточные эндосомы посредством интернализации, может подвергаться диссоциации от антигена в кислотных условиях внутри эндосом, подвергшееся диссоциации антитело может быть способным к связыванию с FcRn, экспрессируемым в эндосомах. Молекула IgG, подвергшаяся диссоциации от антигена и связавшаяся с FcRn, перемещается на клеточную поверхность и затем подвергается диссоциации от FcRn в нейтральных условиях в плазме, и, таким образом, снова возвращается в плазму. Молекула IgG, которая возвратилась в плазму, снова способна к связыванию с новым мембранным антигеном. Повторение этого процесса делает возможным многократное связывание одной молекулы IgG с мембранными антигенами, делая, таким образом, возможной нейтрализацию множества антигенов одной молекулой IgG (Fig. 3).
В случае растворимого антигена введенное антитело связывается с антигеном в плазме и остается в плазме в форме комплекса антиген-антитело. Обычно, в то время как сохранение антитела в плазме является очень длительным (скорость элиминация является очень медленной) благодаря действию FcRn, описанному выше, сохранение антигена в плазме является коротким (скорость элиминации является быстрой). Поэтому связанные с антителами антигены демонстрируют сохранение в плазме, соизмеримое с сохранение в плазме антитела (скорость элиминации является очень медленной). Антигены продуцируются в организме с постоянной скоростью и, в отсутствие антитела, наблюдаются в плазме в концентрации, при которой скорость продуцирования антигена и скорость элиминации антигена находятся в равновесии. В присутствии антитела большая часть антигенов связывается с антителами, что приводит к очень медленной элиминации антигенов. Таким образом, концентрация антигена в плазме увеличивается по сравнению с указанной концентрацией в отсутствие антитела (Kidney Int. 2003, 64, 697-703; J. National Cancer Institute 2002, 94(19), 1484-1493; J. Allergy и Clinical Immunology 1997, 100(1), 110-121; Eur. J. Immunol. 1993, 23; 2026-2029). Даже если аффинность антитела в отношении антигена является очень большой, концентрация антигена повышается, поскольку антитело медленно элиминируется из плазмы, и нейтрализующий эффект антитела прекращается после того, как концентрации антитела и антигена становятся равными. Хотя антитела с большей константой диссоциации (KD) могут нейтрализовать растворимые антигены при более низкой концентрации антитела, антитела в концентрации, составляющей половину или меньше от концентрации присутствующего антигена, не способны нейтрализовать антигены независимо от того, насколько большой является аффинность антитела (Biochem. Biophys. Res. Commun. 2005 Sep 9; 334(4): 1004-1013). Как и в случае с молекулами IgG, не связавшимися с антигенами, молекулы IgG, связавшиеся с антигенами в плазме, также поступают в эндосомы посредством пиноцитоза и связываются с FcRn, экспрессируемым в эндосомах, в кислотных условиях внутри эндосом. Молекулы IgG, связавшиеся с FcRn, перемещаются на клеточную поверхность, в то время как антитело продолжает быть связанным с антигеном, и затем подвергаются диссоциации от FcRn в нейтральных условиях в плазме. Поскольку в плазму возвращаются молекулы IgG, будучи связанными с антигенами, они не могут связываться с новыми антигенами в плазме. В том случае, если молекулы IgG могут подвергаться диссоциации от антигена в кислотных условиях внутри эндосом, подвергшийся диссоциации антиген не будет способен к связыванию с FcRn и, таким образом, может подвергаться деградации лизосомами. С другой стороны, молекулы IgG могут возвращаться в плазму снова благодаря связыванию с FcRn. Поскольку молекулы IgG, которые возвратились в плазму, уже подверглись диссоциации от антигена в эндосомах, они снова способны к связыванию с новым антигеном в плазме. Повторение этого процесса делает возможным многократное связывание одной молекулы IgG с растворимыми антигенами. Это делает возможной нейтрализацию множества антигенов одной молекулой IgG (Фиг. 4).
Таким образом, независимо от того, является ли антиген мембранным антигеном или растворимым антигеном, если возможна диссоциация антитела IgG-типа от антигена в кислотных условиях внутри эндосом, одна молекула IgG могла бы быть способной к многократной нейтрализации антигенов. Для того, чтобы антитела IgG-типа подвергались диссоциации от антигенов в кислотных условиях внутри эндосом, необходимо, чтобы связывание антигена с антителом было значительно слабее в кислотных условиях, чем в нейтральных условиях. Поскольку необходимо нейтрализовать мембранные антигены на клеточной поверхности, антитела должны сильно связываться с антигенами при pH на клеточной поверхности, а именно pH 7,4. Поскольку pH внутри эндосом, как сообщалось, обычно находиться между 5,5 и 6,0 (Nat. Rev. Mol. Cell. Biol. 2004 Feb; 5(2): 121-32), считают, что антитело, которое слабо связывается с антигеном при pH от 5,5 до 6,0, подвергается диссоциации от антигена в кислотных условиях внутри эндосом. Конкретнее, одна молекула IgG, которая сильно связывается с антигеном при pH 7,4 на клеточной поверхности и слабо связывается с антигеном при pH от 5,5 до 6,0 внутри эндосом, может быть способна к нейтрализации множества антигенов и, таким образом, улучшению фармакокинетики.
Как правило, белок-белковые взаимодействия состоят из гидрофобного взаимодействия, электростатического взаимодействия и соединения с помощью водородных связей, и силу связывания обычно представляют через константу связывания (аффинность) или кажущуюся константу связывания (авидность). Связывание в зависимости от pH, сила которого меняется при переходе от нейтральных условий (pH 7,4) к кислотным условиям (pH от 5,5 до 6,0), присутствует во встречающихся в природе белок-белковых взаимодействиях. Например, вышеотмеченное связывание между молекулами IgG и FcRn, известным как рецептор для возвращения молекул IgG, является сильным в кислотных условиях (pH от 5,5 до 6,0), но необыкновенно слабым в нейтральных условиях (pH 7,4). Большинство таких изменений в зависимости от pH в белок-белковых взаимодействиях связано с остатками гистидина. Поскольку pKa остатка гистидина находится около pH 6,0-6,5, уровень протонной диссоциации для остатков гистидина меняется при переходе от нейтральных условий (pH 7,4) к кислотным условиям (pH от 5,5 до 6,0). Дело в том, что остатки гистидина являются не заряженными и функционируют в качестве акцепторов атомов водорода в нейтральных условиях (pH 7,4), в то время как в кислотных условиях (pH от 5,5 до 6,0) они становятся положительно заряженными и функционируют в качестве доноров атомов водорода. Сообщалось, что связывание в зависимости от pH при описанном выше IgG-FcRn взаимодействии также связано с остатками гистидина, присутствующими в IgG (Mol. Cell. 2001 Apr; 7(4): 867-877).
Следовательно, pH-зависимость можно сообщить белок-белковым взаимодействиям путем замещения аминокислотного остатка, вовлеченного в белок-белковые взаимодействия, остатком гистидина или путем введения гистидина в сайт взаимодействия. Такие попытки были также предприняты для белок-белковых взаимодействий между антителами и антигенами, и было успешно получено мутантное антитело с уменьшенной в кислотных условиях способностью к связыванию антигена путем введения гистидина в последовательность CDR антитела против лизоцима яичного белка (FEBS Letter (vol. 309, No. 1, 85-88, 1992)). Кроме того, сообщалось (WO 2003-105757) об антителе, которое получают путем введения гистидина в последовательность его CDR и которое специфически связывается с антигеном при низком pH раковых тканей, но слабо связывается в нейтральных условиях.
Хотя способы установления зависимости от pH во взаимодействиях антигена с антителом, описанные выше, были опубликованы, не сообщалось о молекуле IgG, которая нейтрализует множество антигенов путем сильного связывания с антигенами при pH жидкостей организма, составляющем 7,4, но слабого связывания антигенов при pH внутри эндосом, составляющем от 5,5 до 6,0. Другими словами, не было сообщений, относящихся к модификациям, которые значительно уменьшают связывание в кислотных условиях при сохранении связывания в нейтральных условиях, так что, по сравнению с немодифицированным антителом, модифицированное антитело связывается с антигенами многократно in vivo и, таким образом, демонстрирует улучшенную фармакокинетику, а также увеличенную продолжительность нейтрализующего эффекта в той же самой дозе.
Рецептор для IL-6 присутствует в организме в форме либо растворимого рецептора IL-6, либо мембранного рецептора IL-6 (Nat. Clin. Pract. Rheumatol. 2006 Nov; 2(11): 619-626). Антитела против рецептора IL-6 связываются как с растворимым рецептором IL-6, так и с мембранным рецептором IL-6 и нейтрализуют их биологическое действие. Считается, что после связывания с мембранным рецептором IL-6 антитела против рецептора IL-6, будучи связанными с мембранным рецептором IL-6, поступают во внутриклеточные эндосомы посредством интернализации, затем перемещаются в лизосомы, в то время как антитела продолжают быть связанными с мембранным рецептором IL-6, и подвергаются деградации лизосомами вместе с мембранным рецептором IL-6. В действительности, сообщалось, что гуманизированное антитело против рецептора IL-6 демонстрирует нелинейный клиренс, и его антигензависимая элиминация вносит большой вклад в элиминацию гуманизированного антитела против рецептора IL-6 (The Journal of Rheumatology, 2003, 30; 71426-71435). Таким образом, одно гуманизированное антитело против рецептора IL-6 связывается с одним или двумя мембранными рецепторами IL-6 (одновалентно или двухвалентно), и затем подвергается интернализации и деградации в лизосомах. Следовательно, если возможно продуцирование модифицированных антител, которые демонстрируют крайне сниженную способность к связыванию в кислотных условиях, но сохраняют такую же способность к связыванию, как у гуманизированного антитела против рецептора IL-6 дикого типа в нейтральных условиях, (антитела против рецептора IL-6 со связыванием в зависимости от pH), множество рецепторов IL-6 может быть нейтрализовано одним гуманизированным антителом против рецептора IL-6. Таким образом, по сравнению с гуманизированными антителами против рецептора IL-6 дикого типа антитела против рецептора IL-6 со связыванием в зависимости от pH могут увеличить продолжительность нейтрализующего эффекта in vivo в той же самой дозе.
Продуцирование связывающегося в зависимости от pH гуманизированного антитела против рецептора IL-6 H3pI/L73:
Сообщалось о введении гистидина в CDR в качестве способа установления связывания в зависимости от pH при взаимодействии антигена с антителом (FEBS Letter (vol. 309, No. 1, 85-88, 1992)). Для нахождения аминокислотных остатков, экспонированных на поверхности вариабельной области H53/PF1L, полученном в примере 1, и вероятных остатков, взаимодействующих с антигеном, была создана модель Fv-фрагмента H53/PF1L с помощью гомологичного моделирования, используя программное обеспечение MOE (Chemical Computing Group Inc.). Пространственную модель, созданную на основе информации о последовательности H53/PF1L, использовали для выбора Н27, H31, H35, L28, L32 и L53 (нумерация по Kabat, Kabat, E.A. et al., 1991, Sequences of Proteins of Immunological Interest, NIH) в качестве точек мутаций, которые могут установить связывание в зависимости от pH с антигеном при введении гистидина. Продукт, в котором остатки в H27, H31 и H35 в H53, полученной в примере 1, замещены гистидинами, был назван H3pI (с аминокислотной последовательностью: SEQ ID NO: 3), а продукт, в котором остатки в L28, L32 и L53 в PF1L, полученной в примере 1, были замещены гистидином, был назван L73 (с аминокислотной последовательностью: SEQ ID NO: 6).
Получение, экспрессия и очистка вектора для экспрессии H3pI/L73
Выполняли аминокислотную модификацию для получения модифицированных в выбранных точках антител. Мутации были введены в H53 (нуклеотидную последовательность: SEQ ID NO: 13) и PF1L (нуклеотидную последовательность: SEQ ID NO: 14), полученных в примере 1, для получения H3pI (аминокислотной последовательности: SEQ ID NO: 3) и L73 (аминокислотной последовательности: SEQ ID NO: 6). Конкретнее, набор для сайт-направленного мутагенеза QuikChange (Stratagene) использовали в соответствии со способом, описанным в предоставленных инструкциях, и результирующие плазмидные фрагменты встраивали в вектор для экспрессии в клетках млекопитающих для получения вектора для экспрессии желаемой H-цепи и вектора для экспрессии желаемой L-цепи. Нуклеотидные последовательности результирующих экспрессионных векторов определяли, используя известный квалифицированным в данной области техники специалистам способ. H3pI/L73, в котором использованы H3pI вместо H-цепи и L73 вместо L-цепи, экспрессировали и очищали с помощью описанного в примере 1 способа.
Пример 3. Сообщение способности к связыванию в зависимости от pH с антигеном путем модификации на His в CDR, используя технологию фагового дисплея
Продуцирование молекул scFv гуманизированного антитела PM1
Гуманизированное антитело PM1, которое является гуманизированным антителом против рецептора IL-6 (Cancer Res. 1993 Feb 15; 53(4): 851-6), превращали в scFv. Области VH и VL амплифицировали с помощью ПЦР, и получали scFv из HL гуманизированного РМ1, имеющий линкерную последовательность GGGGSGGGGSGGGGS (SEQ ID NO. 15) между VH и VL.
Отбор положений, в которые может быть введен гистидин, путем сканирования гистидином
ПЦР проводили, используя полученную ДНК scFv из HL гуманизированного РМ1 в качестве матрицы для получения гистидиновой библиотеки, в которой любая одна из аминокислот CDR замещена гистидином. Части библиотеки создавали с помощью ПЦР, используя праймеры, в которых кодон для аминокислоты, которую желали мутировать для создания библиотеки, был замещен CAT, соответствующим гистидину кодоном, а другие части создавали с помощью нормальной ПЦР. Эти части затем соединяли с помощью ПЦР с компоновкой. Созданную библиотеку затем расщепляли SfiI, встраивали в фагемидный вектор pELBG lacI, который был также расщеплен SfiI, и затем использовали для трансформации XL1-Blue (Stratagene). Результирующие колонии использовали для оценки связывания с антигеном с помощью ELISA фагов и анализа последовательности scFv из HL. ELISA фагов проводили, используя планшет, покрытый SR344 в концентрации 1 мкг/мл, в соответствии с J. Mol. Biol. 1992; 227: 381-388. Клоны, которые, как установлено, связываются с SR344, подвергали анализу последовательностей, используя специфические праймеры.
Титр фагов определяли с помощью ELISA с использованием антитела против Etag (GE Healthcare) и антитела против M13 GE Healthcare). Это значение затем использовали для отбора положений, в которых замена остатка в CDR гистидином не изменяла значительно способность к связыванию по сравнению с scFv из HL гуманизированного РМ1, на основе результатов ELISA фагов в отношении SR344. Отобранные положения представлены в таблице 2. Нумерация каждого остатка была в соответствии с нумерацией по Kabat (Kabat, et al., 1991, Sequences of Proteins of Immunological Interest, NIH).
Положения замены на гистидин, не оказывающей значительное влияние на способность к связыванию
L24, L26, L27, L28, L30, L31, L32, L52, L53, L54, L56, L90, L92, L93, L94
Конструирование библиотеки гистидин-модифицированных CDR
Была сконструирована библиотека, в которой аминокислоты остатков CDR, которые не изменяли значительно способность к связыванию после замещения гистидином, представленные в таблице 2 (положения, в которые может быть введен гистидин), представляют собой аминокислоты их исходной последовательности (последовательности дикого типа) или гистидин. Библиотеку конструировали на основе последовательностей H-цепи PF1H и L-цепи PF1L, полученных в примере 1, так что мутированные для создания библиотеки положения имеют исходные последовательности или гистидины (либо исходная последовательность, либо гистидины).
Части библиотеки конструировали с помощью ПЦР с использованием праймеров, которые были разработаны так, чтобы в положении, которое желали мутировать для создания библиотеки, находился кодон для исходной аминокислоты или кодон для гистидина, а другие части получали с помощью нормальной ПЦР, или с помощью ПЦР с использованием синтетических праймеров как в частях библиотеки. Эти части затем соединяли с помощью ПЦР с компоновкой (J. Mol. Biol. 1996; 256: 77-88).
Эту библиотеку использовали для конструирования библиотеки рибосомного дисплея в соответствии с J. Immunological Methods 1999; 231: 119-135. Для выполнения in vitro трансляции вне клеток Escherichia coli последовательность SDA (сайт связывания рибосом) и промотор T7 добавляли с 5ʹ стороны, и частичную последовательность гена 3, служащую в качестве линкера для рибосомного дисплея, присоединяли с 3ʹ стороны, используя SfiI.
Получение связывающегося в зависимости от pH scFv из библиотеки с помощью пэннига с использованием гранул
Для обогащения исключительно scFv, обладающим способностью к связыванию с SR344, дважды выполняли пэннинг с использованием метода рибосомного дисплея в соответствии с Nature Biotechnology 2000 Dec; 18: 1287-1292. Для получения антигена приготовленный SR344 биотинилировали, используя NHS-PEO4-биотин (Pierce). Пэннинг выполняли, используя 40 нМ биотинилированный антиген.
Используя результирующий пул ДНК в качестве матрицы, был реконструирован HL scFv с помощью ПЦР с использованием специфических праймеров. После расщепления ферментом SfiI расщепленный HL scFv встраивали в фагемидный вектор pELBG lacI, который был также расщеплен SfiI, и затем использовали для трансформации XL1-Blue (Stratagene).
Клетки Escherichia coli, несущие желаемую плазмиду, выращивали до 0,4-0,6 ОП/мл в среде 2YT, содержащей 100 мкг/мл ампициллина и 2% глюкозы. К ним добавляли фаг-помошник (M13KO7, 4,5×1011 бляшкообразующих единиц), статически культивировали в течение 30 минут при 37°C и затем культивировали со встряхиванием в течение 30 минут 37°C. Культуру переносили в 50 мл пробирку Фэлкона, центрифугировали в течение 10 минут при 3000 оборотов/мин, ресуспендировали в среде 2YT, содержащей 100 мкг/мл ампициллина, 25 мкг/мл канамицина и 0,5 мМ IPTG, и затем инкубировали в течение ночи при 30°C.
Подвергнутую культивированию в течение ночи культуру осаждали с использованием 2,5 M NaCl и 10% PEG и затем разбавляли PBS для получения раствора библиотеки фагов. К раствору библиотеки фагов добавляли 10% M-PBS (PBS, содержащий 10% снятого молока) и 1 M Трис-HCl до конечной концентрации, составляющей 2,5% M-PBS, и pH 7,4. Пэннинг выполняли с помощью обычного способа пэннинга с использованием антигена, иммобилизованного на магнитных гранулах (J. Immunol. Methods 2008 Mar 20; 332(1-2): 2-9; J. Immunol. Methods 2001 Jan 1; 247(1-2): 191-203; Biotechnol. Prog. 2002 Mar-Apr; 18(2): 212-20). Конкретнее, 40 пмоль меченного биотином SR344 добавляли к приготовленной библиотеки фагов, и библиотеку контактировали с антигеном в течение 60 минут при 37°C. Добавляли покрытые стрептавидином гранулы (Dynal M-280), промытые 5% M-PBS (PBS, содержащим 5% снятого молока), и им предоставляли возможность связываться в течение 15 минут при 37°C. Гранулы промывали пять раз как 0,5 мл PBST (PBS, содержащего 0,1% Tween-20, pH 7,4), так и PBS (pH 7,4). Гранулы затем суспендировали в 1 мл PBS (pH 5,5) при 37°C, и сразу же извлекали фаг. Раствор извлеченных фагов добавляли к 10 мл клеток XL1-Blue в фазе логарифмического роста (ОП600, составляющая от 0,4 до 0,5), и смеси позволяли находиться в течение 30 минут при 37°C для инфицирования. Инфицированные E.coli высевали на чашку 225Ч225 мм, содержащую 2 YT, 100 мкг/мл ампициллина и 2% глюкозы. Эти E.coli использовали для начала дополнительного культивирования фагов таким же образом, как описано выше, и повторения пэннинга 8 раз.
Оценка с помощью ELISA фагов
Указанные выше одиночные колонии инокулировали в 100 мкл 2YT, 100 мкг/мл ампициллина, 2% глюкозы и 12,5 мкг/мл тетрациклина и культивировали в течение ночи при 30°C. 2 мкл этой культуры инокулировали в 300 мкл 2YT, 100 мкг/мл ампициллина и 2% глюкозы и затем культивировали в течение 4 часов при 37°C. К культуре добавляли фаг-помошник (M13KO7) в количестве 9Ч108 бляшкообразующих единиц, смеси позволяли стоять в течение 30 минут при 37°C, а затем ее встряхивали в течение 30 минут при 37°C для инфицирования. Впоследствии среду заменяли 300 мкл 2 YT, 100 мкг/мл ампициллина, 25 мкг/мл канамицина и 0,5 мМ IPTG. После культивирования в течение ночи при 30°C получали супернатант после центрифугирования. К 40 мкл супернатанта после центрифугирования добавляли 360 мкл 50 мМ PBS (pH 7,4), и смесь подвергали ELISA. 96-луночный титрационный микропланшет StreptaWell (Roche) покрывали в течение ночи 100 мкл PBS, содержащего 62,5 нг/мл меченного биотином SR344. После удаления антигена промывкой PBST выполняли блокирование с использованием 250 мкл 2% BSA-PBS в течение 1 часа или более. После удаления 2% BSA-PBS добавляли приготовленный супернатант культуры, и планшету позволяли стоять в течение 1 часа при 37°C для связывания антитела. После промывки добавляли 50 мМ PBS (pH 7,4) или 50 мМ PBS (pH 5,5), и проводили инкубацию при нахождении планшета при 37°C в течение 30 минут. После промывки выполняли детектирование с использованием конъюгированного с HRP антитела против M13 (Amersham Pharmacia Biotech), разведенного 2% BSA-PBS, и одного раствора TMB (Zymed), с последующим добавлением серной кислоты для остановки реакции и измерением оптической плотности при 450 нм.
Однако с помощью этого пэннинга, в котором используется иммобилизованный на магнитных гранулах антиген, не были получены клоны, демонстрирующие способность к связыванию в сильной зависимости от pH. Клоны, которые, как установлено, демонстрировали способность к связыванию со слабой pH-зависимостью, были подвергнуты анализу последовательностей, используя специфические праймеры. Положения в этих клонах, в которых гистидин присутствовал с высокой частотой, представлены в таблице 3.
Положения замены на гистидин, выявленные с использованием библиотеки фагов (пэннинга с использованием магнитных гранул)
L24, L27, L28, L32, L53, L56, L90, L92, L94
Получение связывающегося в зависимости от pH scFv из библиотеки с помощью пэннинга с использованием колонки
С помощью обычного пэннинга, в котором используется иммобилизованный на магнитных гранулах антиген, не были получены клоны, обладающие способностью к связыванию в сильной зависимости от pH. Это может быть обусловлено следующими причинами. В пэннинге, в котором используется иммобилизованный на магнитных гранулах или планшете антиген, собирают все фаги, подвергшиеся диссоциации от магнитных гранул или планшета в кислотных условиях. Поэтому вместе полученные фаговые клоны, обладающие слабой pH-зависимостью, уменьшают вероятность включения клонов, обладающих сильной pH-зависимостью, в конечном счете обогащенные клоны.
Поэтому в качестве более строгого способа пэнинга был исследован пэннинг, в котором используется колонка с иммобилизованным антигеном (Фиг. 5). Не было предшествующих сообщений, касающихся получения клонов, обладающих способностью к связыванию в зависимости от pH, путем использования пэннинга с использованием колонки с иммобилизованным антигеном. В пэннинге, в котором используется колонка с иммобилизованным антигеном, когда подвергают элюции в кислотных условиях фаги, связавшиеся в нейтральных условиях, клоны, обладающие слабой pH-зависимостью, повторно связываются с антигеном внутри колонки и, следовательно, в меньшей степени подвергаются элюции, что делает возможной избирательную элюцию с колонки клонов с сильной pH-зависимостью, которые в меньшей степени повторно связываются внутри колонки. Кроме того, хотя в пэннинге, в котором используется иммобилизованный на магнитных гранулах антиген, извлекают «все» фаги, подвергшиеся диссоциации в кислотных условиях, пэннинг, в котором используется колонка с иммобилизованным антигеном, делает возможным избирательное извлечение фагов, обладающих способностью к связыванию с сильной зависимостью от pH, путем допущения протекания через колонку кислотного буфера для вызова начала элюции и получения только «подходящих фракций».
Сначала готовили колонку, на которой иммобилизован антиген SR344. 200 мкл стрептавидин-сефарозы (GE Healthcare) промывали 1 мл PBS, суспендировали в 500 мкл PBS и приводили в контакт с 400 пмолями меченого биотином SR344 в течение 1 часа при комнатной температуре. Впоследствии пустую колонку (Amersham Pharmacia Biotech) заполняли указанной выше сефарозой и промывали приблизительно 3 мл PBS. Вышеупомянутую библиотеку фагов, осажденных с использованием PEG, разводили в соотношении 1/25 раствором 0,5% BSA-PBS (pH 7,4), пропускали через 0,45 нм фильтр и затем добавляли в колонку. После промывки приблизительно 6 мл PBS (pH 7,4) допускали протекание через колонку 50 мМ MES-NaCl (pH 5,5) для элюции антител, которые подвергаются диссоциации при низком pH. Собирали подходящие подвергшиеся элюции фракции, и полученный раствор фагов добавляли к 10 мл клеток XL1-Blue в фазе логарифмического роста (ОП600, составляющая от 0,4 до 0,5), и допускали стояние смеси в течение 30 минут при 37°C.
Инфицированные E.coli высевали на чашку 225×225 мм, содержащую 2 YT, 100 мкг/мл ампициллина и 2% глюкозы. Эти E.coli использовали для начала дополнительного культивирования фагов таким же образом, как описано выше, и повторения пэннинга 8 раз.
Оценка с помощью ELISA фагов
Результирующие фаги оценивали с помощью ELISA фагов. Клоны, которые, как установлено, обладают сильной зависимостью от pH, подвергали анализу последовательностей, используя специфические праймеры. В результате было получено несколько клонов, демонстрирующих связывание с сильной зависимостью от pH по сравнению с WT. Было установлено, как продемонстрировано на фиг. 6, что клон CL5 (H-цепь: CLH5, L-цепь: CLL5) (CLH5: аминокислотная последовательность SEQ ID NO: 5, CLL5: аминокислотная последовательность SEQ ID NO: 8) демонстрируют связывание с особенно сильной зависимостью от pH по сравнению с WT. Таким образом, было подтверждено, что при невозможности получения с помощью обычного пэннинга, в котором используется иммобилизованный на магнитных гранулах антиген, антител, демонстрирующих связывание с сильной зависимостью от pH, их можно получить с помощью пэннинга, в котором используется колонка с иммобилизованным антигеном. Следовательно, было установлено, что пэннинг, в котором используется колонка с иммобилизованным антигеном, является высокоэффективным способом получения связывающихся в зависимости pH антител из библиотеки. Аминокислотные последовательности клонов, демонстрирующих связывание в зависимости от pH, были проанализированы, и положения, в которых гистидин присутствует с высокой вероятностью в обогащенных клонах, представлены в таблице 4.
Положения замены на гистидин, выявленные с использованием библиотеки фагов (пэннинга с использованием колонки)
L24, L27, L28, L32, L53, L56, L90, L92, L94
Пример 4. Экспрессия и очистка гуманизированного антитела против рецептора IL-6 с модификацией на гистидин
Получение, экспрессия и очистка вектора для экспрессии гуманизированного антитела против рецептора IL-6 с модификацией на гистидин
Для превращения клонов, демонстрирующих сильную pH-зависимость в ELISA фагов, в IgG VH и VL были соответственно амплифицированы с помощью ПЦР, расщеплены ферментами XhoI/NheI и EcoRI и встроены в вектор для экспрессии в клетках млекопитающих. Нуклеотидную последовательность каждого фрагмента ДНК определяли с помощью известного квалифицированным в данной области техники специалистам способа. CLH5/L73, в котором были использованы CLH5 вместо H-цепи и L73, полученную в примере 2, вместо L-цепи, экспрессировали и очищали как IgG. Экспрессия и очистка выполнялись описанным в примере 1 способом.
Путем комбинирования точек мутаций получали антитело, обладающее даже большей pH-зависимостью. Основываясь на положениях, в которых был сосредоточен His в библиотеке фагов, а также касающейся структуры информации и т.п., H32, H58, H62 и H102 в H3pI, которая была получена как H-цепь в примере 2, были замещены гистидином, и, кроме того, H95 и H99 были замещены валином и изолейцином, соответственно, для получения H170 (SEQ ID NO: 4). Продуцирование варианта выполняли, используя описанный в примере 1 способ. Кроме того, получали L82 (SEQ ID NO: 7) путем замещения 28-ого гистидина L73, которая была получена как L-цепь в примере 2, аспарагиновой кислотой. Продуцирование варианта выполняли, используя описанный в примере 1 способ. H170/L82, в котором были использованы H170 вместо H-цепи и L82 вместо L-цепи, экспрессировали и очищали как IgG, используя описанный в примере 1 способ.
Пример 5. Оценка IL-6R-нейтрализующей активности связывающегося в зависимости от pH антитела
Оценка нейтрализующей рецептор IL-6 человека активности клонов, превращенных в IgG.
Нейтрализующую рецептор IL-6 активность оценивали для четырех антител: гуманизированного антитела PM1 (WT) и H3pI/L73, CLH5/L73 и H170/L82, полученных в примерах 2 и 4.
Конкретнее, нейтрализующую рецептор IL-6 активность оценивали, используя клетки BaF3/gp130, демонстрирующие зависимый от IL-6/рецептора IL-6 рост. Клетки BaF3/gp130 промывали трижды средой RPMI1640, содержащей 10% FBS, затем суспендировали до концентрации 5×104 клеток/мл в среде RPMI1640, содержащей 60 нг/мл интерлейкина-6 человека (Toray), 60 нг/мл рекомбинантного растворимого рецептора IL-6 человека (SR344) и 10% FBS. В каждую лунку 96-луночного планшета (Corning) вносили 50 мкл суспензии. Затем очищенное антитело разводили средой RMPI1640, содержащей 10% FBS, и в каждую лунку вмешивали 50 мкл антитела. После культивирования в течение 3 дней при 37°C и 5% CO2 добавляли 20 мкл/лунку реагента WST-8 (Cell Counting Kit-8, Dojindo Laboratories), разведенного в два раза PBS, и затем сразу же измеряли оптическую плотность при 450 нм (контрольная длина волны: 620 нм), используя Sunrise Classic (Tecan). После культивирования в течение 2 часов снова измеряли оптическую плотность при 450 нм (контрольная длина волны: 620 нм). Нейтрализующую рецептор IL-6 активность определяли на основе изменения оптической плотности через 2 часа. В результате было установлено, как продемонстрировано на фиг. 7, что H3pI/L73, CLH5/L73 и H170/L82 обладают биологической нейтрализующей активностью, эквивалентной таковой гуманизированного антитела PM1 (WT).
Пример 6. Анализ с использованием Biacore связывающегося в зависимости от pH антитела
Анализ связывания клонов со связыванием в зависимости от pH с растворимым рецептором IL-6
Кинетические анализы взаимодействий антигена с антителом при pH 5,8 и pH 7,4 были выполнены с использованием Biacore T100 (GE Healthcare) на четырех антителах: гуманизированном антителе PM1 (WT) и H3pI/L73, CLH5/L73 и H170/L82, полученных в примерах 2 и 4 (в буфере: 10 мМ MES (pH 7,4 или pH 5,8), 150 мМ NaCl и 0,05% Tween 20). Различные антитела связывали на сенсорном чипе, на котором был иммобилизован рекомбинантный белок A/G (Pierce) путем присоединения к амину. SR344, приведенный к концентрациям, составляющим от 9,8 до 400 нМ, инъецировали в чип в качестве аналита. Ассоциацию и диссоциацию связывающихся в зависимости от pH клонов с SR344 наблюдали в режиме реального времени (фиг. 8 и 9). Все измерения проводили при 37°C. Константы скорости ассоциации ka (1/Мс) и константы скорости диссоциации kd (1/с) рассчитывали, используя программное обеспечение для оценки анализа с использованием Biacore T100 (GE Healthcare), а константы диссоциации KD (M) рассчитывали на основе этих значений (таблица 5). Кроме того, для каждого клона рассчитывали отношение аффинности при pH 5,8 к аффинности при pH 7,4 для оценки связывания в зависимости от pH. Все измерения проводили при 37°C.
В результате расчета отношения аффинности при pH 5,8 к аффинности при pH 7,4 для каждого клона, связывание (аффинность) в зависимости от pH H3pI/L73, H170/L82 и CLH5/L73 с SR344 было 41-кратным, 394-кратным и 66-кратным, соответственно, при этом каждое из этих антител продемонстрировало связывание в зависимости от pH, превышающее более чем в 15 раз такое связывание для антитела дикого типа.
Еще не сообщалось об антителах против рецептора IL-6, которые сильно связываются с антигеном при pH плазмы, составляющем 7,4, но слабо связываются с антигеном при pH внутри эндосом, составляющем от 5,5 до 6,0. В этом исследовании были получены антитела, которые сохраняют биологическую нейтрализующую активность, эквивалентную такой активности гуманизированного антитела против рецептора IL-6 дикого типа, и аффинность при pH 7,4, но проявляют аффинность при pH 5,8, которая было специально уменьшена более чем в 10 раз.

Анализ связывания связывающихся в зависимости от pH клонов с мембранным рецептором IL-6
Для полученных выше связывающихся в зависимости от pH клонов следили за взаимодействиями антитела с антигеном в виде мембранного рецептора IL-6 при pH 5,8 и pH 7,4, используя Biacore T100 (GE Healthcare). Связывание с мембранным рецептором IL-6 оценивали с помощью определения связывания с рецептором IL-6, который был иммобилизован на сенсорном чипе. SR344 биотинилировали в соответствии с известным квалифицированным в данной области техники специалистам способом, и биотинилированный SR344 иммобилизовали на сенсорном чипе через стрептавидин, используя сродство между стрептавидином и биотином. Все измерения проводили при 37°C, и буфер подвижной фазы содержал 10 мМ MES (pH 5,8), 150 мМ NaCl и 0,05% Tween 20. Связывающиеся в зависимости от рН клоны инъецировали в чип при параметре pH 7,4, и им предоставляли возможность связываться с SR344 (буфер для инъекционных образцов: 10 мМ MES (pH 7,4), 150 мМ NaCl, 0,05% Tween 20), и следили за pH-зависимой диссоциацией каждого клона при pH 5,8 подвижной фазы (фиг. 10).
Скорость диссоциации (kd (1/с)) при pH 5,8 рассчитывали, используя программное обеспечение для оценки анализа с использованием Biacore T100 (GE Healthcare), путем подбора по точкам только стадии диссоциации при pH 5,8, при этом 0,5 мкг/мл образца связывали в 10 мМ MES (pH 7,4), 150 мМ NaCl и 0,05% Tween 20 и подвергали диссоциации в 10 мМ MES (pH 5,8), 150 мМ NaCl и 0,05% Tween 20. Аналогично, скорость диссоциации (kd (1/с)) при pH 7,4 рассчитывали, используя программное обеспечение для оценки анализа с использованием Biacore T100 (GE Healthcare), путем подбора по точкам только стадии диссоциации при pH 7,4, при этом 0,5 мкг/мл образца связывали в 10 мМ MES (pH 7,4), 150 мМ NaCl и 0,05% Tween 20 и подвергали диссоциации в 10 мМ MES (pH 7,4), 150 мМ NaCl и 0,05% Tween 20. Константа скорости pH-зависимой диссоциации каждого клона представлена в таблице 6.
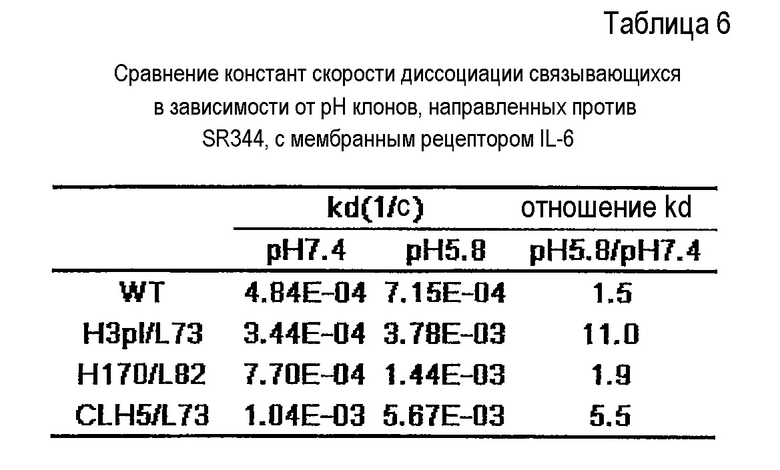
Наибольшая зависимость от pH отношения констант скорости диссоциации наблюдалась для H3pI/L73, а затем для CLH5/L73 и 170/L82 в нисходящем порядке, и каждый клон продемонстрировал диссоциацию с большей зависимости от pH от мембранного рецептора IL-6, чем WT. Однако уровень pH-зависимой ассоциации/диссоциации отличался между растворимым рецептором IL-6 и мембранным рецептором IL-6. Было обнаружено, что H170/L82, которое продемонстрировало связывание с наибольшей зависимостью от pH в анализе связывания с растворимым рецептором IL-6, демонстрировало связывание с наименьшей зависимостью от pH в анализе связывания с мембранным рецептором IL-6. В общем, известно, что, в то время как молекулы IgG связывается с растворимым антигеном одновалентно (проявляют аффинность), они связываются двухвалентно с мембранными антигенами (проявляют авидность). Полагают, что различие в образе связывания между растворимыми антигенами и мембранными антигенами оказало влияние на связывание в зависимости от pH H170/L82.
Пример 7. Подтверждение многократного связывания антигена связывающимся в зависимости от pH антителом
Как описано в примере 2, связывающиеся в зависимости от pH антитела могут быть способны к многократному связыванию с антигенами. Дело в том, что связывающееся в зависимости от pH антитело, которое связалось с антигеном, неспецифическим образом поступает в эндосомы, но подвергается диссоциации от растворимого антигена в кислотных условиях внутри эндосом. Антитело связывается с FcRn и, таким образом, возвращается в плазму. Поскольку возвратившееся в плазму антитело не связано с антигеном, оно способно к связыванию с новым антигеном снова. Повторение этого процесса делает возможным многократное связывание связывающихся в зависимости от pH антител с антигенами. Однако от антител IgG-типа, не обладающих способностью к связыванию в зависимости от рН, диссоциации в кислотных условиях внутри эндосом подвергаются не все антигены. Поэтому такие антитела, возвратившиеся в плазму с помощью FcRn, остаются связанными с антигеном и, следовательно, не могут связываться с новыми антигенами. Следовательно, в почти во всех случаях, каждая одна молекула антител IgG-типа способна к нейтрализации только двух антигенов (в случае двухвалентного связывания).
Поэтому была проведена оценка того, способны ли три связывающихся в зависимости от pH антитела (H3pI/L73, CLH5/L73 и H170/L82), созданных в примерах 2 и 4, к связыванию с антигеном SR344 многократно по сравнению с гуманизированным антителом PM1 (дикого типа, WT).
Для оценки способности к многократному связыванию с антигеном антител, связывающихся при pH 7,4 и подвергающихся диссоциации при pH 5,8, использовали Biacore (GE Healthcare). Оцениваемое антитело связывали с сенсорным чипом с иммобилизованным на нем рекомбинантным белком A/G (Pierce) путем присоединения к амину, и допускали протекание подвижной фазы с pH 7,4 (стадия 1). Затем допускали протекание раствора SR344, доведенного до pH 7,4, в качестве аналита для связывания SR344 с антителом при pH 7,4 (стадия 2). Это связывание при pH 7,4 воспроизводит связывание с антигеном в плазме. Впоследствии добавляли буфер, доведенный до pH 5,8, сам по себе (не содержащий SR344) в качестве аналита для подвергания связанного с антителом антигена воздействию кислотных условий (стадия 3). Эта диссоциация при pH 5,8 воспроизводит уровень связывания комплексов антиген-антитело в эндосомах. Впоследствии повторяли стадию 2. Это воспроизводит связывание снова антитела, возвратившегося в плазму с помощью FcRn, с новым антигеном. Впоследствии повторяли стадию 3 для подвергания комплекса антитело-антиген воздействию кислотных условий. Повторение «стадии 2–стадии 3» множество раз при 37°C, как описано выше, может воспроизводить in vivo ситуацию, при которой антитела повторно поступают из плазмы в эндосомы посредством пиноцитоца и возвращаются в плазму с помощью FcRn (Nat. Rev. Immunol. 2007 Sep; 7(9): 715-25).
Полученные связывающиеся в зависимости от pH клоны, описанные выше, анализировали, используя Biacore T100 (GE Healthcare), в отношении их способности к многократному связыванию с антигеном SR344 при pH 5,8 и pH 7,4. Конкретнее, анализ проводили следующим образом. Все измерения проводили при 37°C. Сначала описанные выше выборочные антитела связывали на сенсорном чипе с иммобилизованным на нем рекомбинатном белком A/G (Pierce) путем присоединения к амину, при этом буфер подвижной фазы представлял собой 10 мМ MES (pH 5,8), 150 мМ NaCl и 0,05% Tween 20 (стадия 1). SR344, приведенный к концентрации, составляющей приблизительно 40 нМ, инъецировали в качестве аналита в течение 3 минут при pH 7,4, и ему предоставляли возможность связываться (буфер для инъецируемого SR344 представлял собой 10 мМ MES (pH 7,4), 150 мМ NaCl и 0,05% Tween 20) (стадия 2). Впоследствии инъецирование SR344 прекращали, и допускали протекание подвижной фазы с pH 5,8 в течение приблизительно 70 секунд для подвергания комплекса антитело/SR344 воздействию кислотных условий (стадия 3). Десять непрерывных повторов циклического процесса из связывания (стадии 2)/воздействия указанной степени кислотности (стадии 3) были осуществлены для наблюдения сенсограммы в режиме реального времени, которая продемонстрирована на фиг. 11. WT продемонстрировал меньшую диссоциацию SR344 во время кислотного воздействия на стадии 3, и, следовательно, доля антитела, способного к связыванию с новыми антигенами в последующей стадии 2 была крайне низкой. Напротив, было установлено, что связывающиеся в зависимости от pH клоны, в частности, H170/L82 и CLH5/L73, продемонстрировали настолько сильную диссоциацию во время кислотного воздействия на стадии 3, что большая часть связанного SR344 подверглась диссоциации, и поэтому почти все антитела были способны к связыванию с новыми антигенами на последующей стадии 2. При 10 повторах циклического процесса, состоящего из связывания (стадии 2) и кислотного воздействия (стадии), почти все антитела H170/L82 и CLH5/L73 были способны к связыванию с новыми антигенами в каждом процессе.
Полученные сенсограммы использовали для расчета количества связывающегося SR344 в каждом цикле для каждого образца, используя программное обеспечение для оценки анализа с использованием Biacore T100 (GE Healthcare). Суммарные значения в зависимости от времени 10 циклов продемонстрированы на фиг. 12. Суммарные значения в RU (относительных единицах), полученные в 10-ом цикле, эквиваленты общему количеству антигенов, связавшихся во время десяти циклов. Связывающиеся в зависимости от pH клоны, в частности, H170/L82 и CLH5/L73, продемонстрировали наибольшие общие количества связавшихся антигенов по сравнению с WT и, как установлено, были способны к повторному связыванию с количеством антигенов, превышающим приблизительно в четыре раза количество, связывающееся диким типом. В результате было выявлено, что путем сообщения дикому типу способности к связыванию в зависимости от pH такие антитела могут многократно связываться с антигенами и, таким образом, нейтрализовать множество антигенов.
Пример 8. Фармакокинетический/фармакодинамический анализ связывающегося в зависимости от pH антитела с использованием трансгенных по рецептору IL-6 человека мышей.
Рецепторы IL-6 присутствуют в организме как в форме растворимого рецептора IL-6, так и в форме рецептора IL-6 мембранного типа (Nat. Clin. Pract. Rheumatol. 2006 Nov; 2(11): 619-626). Антитела против рецептора IL-6 связываются с растворимыми рецепторами IL-6 и рецепторами IL-6 мембранного типа и нейтрализуют их биологическое действие. Полагают, что антитело против рецептора IL-6 связывается с рецептором IL-6 мембранного типа и впоследствии поступает в эндосомы внутри клетки посредством интернализации, в то время как антитело продолжает быть связанным с рецептором IL-6 мембранного типа, и затем перемещается в лизосому, все еще продолжая быть связанным с рецептором IL-6 мембранного типа, где оно подвергается деградации лизосомой вместе с рецептором IL-6 мембранного типа. Если H3pI/L73, CLH5/L73 и H170/L82, которые являются связывающимися в зависимости от pH антителами против рецептора IL-6, оцененными в примере 6, способны к возвращению в плазму благодаря FcRn в результате диссоциации в кислотных условиях внутри эндосом, возвратившиеся в плазму антитела могут снова связываться с антигенами. Это делает возможной нейтрализацию множества рецепторов IL-6 мембранного типа одной молекулой антитела. Достигается ли возвращение в плазму благодаря FcRn в результате диссоциации в кислотных условиях внутри эндосом при использовании созданных, связывающихся в зависимости от pH антител против рецептора IL-6, можно определить с помощью оценки того, улучшается ли фармакокинетика этих антител по сравнению с антителом дикого типа.
Таким образом, фармакокинетику в трансгенных по рецептору IL-6 мышах (мышах hIL-6R tg, Proc. Natl. Acad. Sci. USA 1995 May 23; 92(11): 4862-4866) оценивали для четырех типов антител, т.е. гуманизированного антитела PM1 (дикого типа: WT) и H3pI/L73, CLH5/L73 и H170/L82, созданных в примерах 2 и 4. WT, H3pI/L73, CLH5/L73 или H170/L82 вводили мышам hIL-6R tg с помощью внутривенного введения одной дозы в расчете 25 мг/кг, и образцы крови отбирали до введения и в динамике времени. Отобранные образцы сразу же центрифугировали в течение 15 минут при 15000 оборотов/мин и 4°C для получения плазмы. Отделенную плазму хранили в камере замораживания, установленной на -20°C или ниже, до проведения измерений.
Определение концентрации в плазме мышей проводили с помощью ELISA. Готовили образцы для калибровочной кривой с концентрациями в плазме, составляющими 6,4, 3,2, 1,6, 0,8, 0,4, 0,2 и 0,1 мкг/мл. Образцы калибровочной кривой и образцы для измерений в плазме мышей распределяли по иммунопланшету (Nunc-Immuno Plate, MaxiSorp (Nalge Nunc International)) с иммобилизованным на нем F(abʹ)2 против IgG человека (специфичным для γ-цепи) (Sigma), и им позволяли находиться в спокойном состоянии в течение одного часа при комнатной температуре. Впоследствии допускали вступление во взаимодействие козьего анти-IgG человека-BIOT (Southern Biotechnology Associates) и конъюгата стрептавидин-щелочная фосфатаза (Roche Diagnostics), и проводили хромогенную реакцию путем применения системы субстратов для фосфатазы при использовании в микролунках BluePhos (Kirkegaard & Perry Laboratories) в качестве субстрата. С использованием считывающего устройства для микропланшетов определяли оптическую плотность при 650 нм. Концентрации в плазме мышей рассчитывали, исходя из оптической плотности калибровочной кривой, используя аналитическое программное обеспечение SOFTmax PRO (Molecular Devices). Зависимости от времени концентраций в плазме WT, а также H3pI/L73, CLH5/L73 и H170/L82 продемонстрированы на фиг. 13.
Фармакокинетика была улучшена для всех из H3pI/L73, CLH5/L73 и H170/L82 по сравнению с WT. В частности, была значительно улучшена фармакокинетика H3pI/L73 и CLH5/L73. Антитело против рецептора IL-6 дикого типа (WT), связавшееся с рецептором IL-6 мембранного типа, поступает в эндосому внутри клетки посредством интернализации, перемещается в лизосому, в то время как антитело продолжает быть связанным с антигеном, и затем подвергается деградации; поэтому оно имеет короткое время нахождения в плазме. Напротив, так как фармакокинетика связывающихся в зависимости от pH антител против рецептора IL-6 была значительно улучшена, полагали, что связывающиеся в зависимости от pH антитела против рецептора IL-6 возвращаются снова в плазму благодаря FcRn в результате диссоциации от антигена, рецептора IL-6 мембранного типа, в кислотных условиях внутри эндосом.
Хотя фармакокинетика была улучшена для всех из H3pI/L73, CLH5/L73 и H170/L82 по сравнению с WT, эффект увеличения времени удерживания в плазме H170/L82 был слабее такового H3pI/L73 и CLH5/L73. Поскольку молекулы IgG, как полагают, обычно связываются двухвалентно со связанным с мембраной антигеном, полагают, что антитела против рецептора IL-6 также связываются двухвалентно (проявляют авидность) с рецепторами IL-6 мембранного типа и затем подвергаются интернализации. Как указано в примере 6, с помощью анализа с использованием Biacore установлено, что H170/L82 быстро диссоциировалось от рецептора IL-6 при pH 5,8 после связывания с растворимым рецептором IL-6 (фиг. 9), но скорость его диссоциации от рецептора IL-6 при pH 5,8 после связывания с рецептором IL-6 мембранного типа было крайне низкой (фиг. 10). Исходя из этого результата, полагают, что причиной слабого эффекта увеличения времени удерживания в плазме H170/L82 является то, что антитело не было способно к диссоциации в достаточной мере внутри эндосом после интернализации вследствие его медленной диссоциации при pH 5,8 после связывания с рецептором IL-6 мембранного типа. А именно, как и для случая, относящегося к мембранным антигенам, было установлено, что для нейтрализации множества мембранных антигенов одной молекулой IgG более важна pH-зависимость диссоциации от молекулы с двухвалентным связыванием (проявляющей авидность), чем pH-зависимость диссоциации от молекулы с одновалентным связыванием (проявляющей аффинность).
Пример 9. Фармакокинетический/фармакодинамический анализ связывающегося в зависимости от pH антитела с использованием яванских макак.
Так как фармакокинетика связывающихся в зависимости от pH антител против рецептора IL-6 была значительно улучшена в примере 8, полагали, что связывающиеся в зависимости от pH антитела против рецептора IL-6 возвращаются в плазму благодаря FcRn в результате диссоциации от антигена, рецептора IL-6 мембранного типа, в кислотных условиях внутри эндосом. Если возвратившиеся в плазму антитела могут снова связываться с рецепторами IL-6 мембранного типа, полагают, что нейтрализация антигена, рецептора IL-6 мембранного типа, связывающимися в зависимости от pH антителами против рецептора IL-6 сохраняется дольше, чем нейтрализация антителом против рецептора IL-6 дикого типа, в одной и той же дозе. Кроме того, поскольку среди рецепторов IL-6 также присутствует растворимый рецептор IL-6, полагают, что в одной и той же дозе продолжительность нейтрализации дольше также по отношению к растворимому рецептору IL-6.
Фармакокинетику в яванских макаках оценивали для WT и H3pI/L73. WT или H3pI/L73 вводили яванским макакам с помощью внутривенного введения одной дозы в расчете 1 мг/кг, и образцы крови отбирали до введения и в динамике времени. Отобранные образцы сразу же центрифугировали в течение 15 минут при 15000 оборотов/мин и 4°C для получения плазмы. Отделенную плазму хранили в камере замораживания, установленной на -20°C или ниже, до проведения измерений.
Определение концентрации в плазме яванских макак проводили с помощью ELISA. Сначала F(abʹ)2-фрагмент антитела против IgG человека (специфичного для γ-цепи) (Sigma) распределяли по иммунопланшету Nunc-ImmunoPlate MaxiSorp (Nalge Nunc International) и позволяли находиться в спокойном состоянии в течение ночи при 4°С для приготовления планшетов с иммобилизованным на них фрагментом антитела против IgG человека. Готовили образцы для калибровочной кривой, имеющие концентрации в плазме, составляющие 3,2, 1,6, 0,8, 0,4, 0,2, 0,1 и 0,05 мкг/мл, и разведенные в 100 раз или более образцы для измерений в плазме яванских макак; 200 мкл 20 нг/мл рецептора IL-6 яванского макака добавляли к 100 мкл образцов для калибровочной кривой и образцов для измерений в плазме; и затем смесям позволяли находиться в спокойном состоянии в течение одного часа при комнатной температуре. Впоследствии образцы распределяли по планшету с иммобилизованным на нем фрагментом антитела против IgG человека и позволяли находиться в спокойном состоянии в течение еще одного часа при комнатной температуре. Допускали вступление во взаимодействие биотинилированого антитела против рецептора IL-6 человека (R&D) в течение одного часа при комнатной температуре, а затем допускали вступление во взаимодействие конъюгата стрептавидин-PolyHRP80 (Stereospecific Detection Technologies) в течение одного часа. Проводили хромогенную реакцию путем применения однокомпонентного субстрата для HRP при использовании в микролунках TMP (BioFX Laboratories) в качестве субстрата; затем реакцию останавливали с помощью 1 н серной кислоты (Showa Chemical), и с использованием считывающего устройства для микропланшетов определяли оптическую плотность при 450 нм. Концентрации в плазме яванских макак рассчитывали, исходя из оптической плотности калибровочной кривой, используя аналитическое программное обеспечение SOFTmax PRO (Molecular Devices). Зависимости от времени концентраций в плазме WT и H3pI/L73 после внутривенного введения продемонстрированы на фиг. 14. В результате, фармакокинетика H3pI/L73 была значительно улучшена по сравнению с WT у яванских макак таким же образом, как и у трансгенных по рецептору IL-6 человека трансгенных мышей. Поскольку фармакокинетика связывающегося в зависимости от pH антитела против рецептора IL-6, H3pI/L73, была значительно улучшена, полагали, что H3pI/L73 возвращается в плазму благодаря FcRn в результате диссоциации от антигена, рецептора IL-6 мембранного типа, в кислотных условиях внутри эндосом.
Для оценки степени, в которой рецептор IL-6 мембранного типа яванского макака нейтрализуется при внутривенном введении WT и H3pI/L73, изучали эффекты выборочных антител на C-реактивный белок плазмы (CRP), индуцируемый IL-6 яванского макака. Поскольку CRP секретируется, когда IL-6 связывается с рецепторами IL-6 мембранного типа, CRP служит в качестве индикатора нейтрализации рецепторов IL-6 мембранного типа. IL-6 яванского макака (cyno.IL-6, приготовленный в примере 1), содержащий 1% инактивированной плазмы яванского макака, вводили подкожно в нижние части спины животных ежедневно в дозе 5 мкг/кг с дня 3 до дня 10 после введения WT или H3pI/L73. Брали образцы крови из подкожной вены ноги непосредственно перед началом введения IL-6 яванского макака (день 3) и после введения с 24-часовыми интервалами (со дня 4 до дня 11), затем отделяли плазму. Концентрации CRP у индивидуальных животных определяли с использованием Cias R CRP (Kanto Chemical), используя автоматизированный анализатор (TBA-120FR, Toshiba Medical Systems). Зависимости от времени концентрации CRP после индукции с помощью IL-6 яванского макака относительно WT и H3pI/L73 продемонстрированы на фиг. 15. В результате, продолжительность подавления продукции CRP была значительно увеличена с помощью H3pI/L73 по сравнению с WT. Основываясь на этих полученных данных, полагали, что связывающееся в зависимости от pH антитело против рецептора IL-6, H3pI/L73, возвращается в плазму благодаря FcRn в результате диссоциации от своего антигена, рецептора IL-6 мембранного типа, в кислотных условиях внутри эндосом и нейтрализует рецептор IL-6 мембранного типа при связывании с ним снова, и, таким образом, подавляет продукцию CRP в течение большего периода времени, чем WT. Другими словами, установлено, что H3pI/L73 в виде одной молекулы антитела способен к связыванию с рецептором IL-6 мембранного типа и его нейтрализации более одного раза. Поскольку продолжительность подавления продукции CRP с помощью H3pI/L73 увеличивается по сравнению с таковой с помощью WT, длительность времени, в течение которого антиген, рецептор IL-6 мембранного типа, связывается антителами, как было показано, увеличена для H3pI/L73 по сравнению с WT.
Для оценки степени, в которой растворимый рецептор IL-6 яванского макака нейтрализуется при внутривенном введении WT и H3pI/L73, определяли концентрацию несвязанного растворимого рецептора IL-6 яванского макака в плазме яванских макак. Все антитела IgG-типа (IgG яванского макака, антитело против рецептора IL-6 человека и комплекс антитела против рецептора IL-6 человека с растворимым рецептором IL-6 яванского макака), присутствующие в плазме, адсорбировали на белке A путем добавления 30 мкл плазмы яванского макака к соответствующему количеству смолы Sepharose Fast Flow с рекомбинантным белком А (GE Healthcare), обезвоженной в чашке 0,22 мкм фильтра (Millipore). После центрифугирования с использованием высокоскоростной центрифуги получали прошедший через смолу раствор (ниже упоминаемый как «пропущенный раствор»). Поскольку пропущенный раствор не содержит комплекс антитела против рецептора IL-6 человека с растворимым рецептором IL-6 яванского макака, который связывается с белком A, концентрацию несвязанного растворимого рецептора IL-6 можно определить путем измерения концентрации растворимого рецептора IL-6 яванского макака в пропущенном растворе. Моноклональное антитело против рецептора IL-6 человека (R&D), которое было меченным рутением с использованием N-гидроксисукцимидного эфира Sulfo-Tag (Meso Scale Discovery) и биотинилированным антителом против рецептора IL-6 человека (R&D), смешивали с образцами для калибровочной кривой рецептора IL-6 яванского макака, приведенного к концентрациям, составляющим 4000, 2000, 1000, 500, 250, 125 и 62,5 пг/мл, и образцами плазмы, обработанными белком A, как описано выше. Смесям предоставляли возможность вступать во взаимодействие в течение одного часа при комнатной температуре. Впоследствии смеси распределяли по 96-луночному планшету SA-Coated Standard MA2400 (Meso Scale Discovery). После допущения взаимодействия в течение еще одного часа и промывки распределяли буфер T для считывания (x4) (Meso Scale Discovery). Сразу же после этого проводили измерение с использованием секторного визуализатора 2400 (Meso Scale Discovery). Концентрации рецептора IL-6 яванского макака рассчитывали, исходя из зависимости калибровочной кривой, используя аналитическое программное обеспечение SOFTmax PRO (Molecular Devices). Зависимости от времени концентраций несвязанного растворимого рецептора IL-6 яванского макака для WT и H3pI/L73 продемонстрированы на фиг. 16. В результате, продолжительность нейтрализации растворимого рецептора IL-6 яванского макака с помощью H3pI/L73 была значительно увеличена по сравнению с WT. Основываясь на этих полученных данных, полагали, что связывающееся в зависимости от pH антитело против рецептора IL-6, H3pI/L73, подвергается диссоциации от своего антигена, растворимого рецептора IL-6, в кислотных условиях в эндосомах и возвращается в плазму благодаря FcRn, и связывается с растворимым рецептором IL-6 и нейтрализует его снова. Поскольку продолжительность супрессии несвязанного растворимого рецептора IL-6 яванского макака с помощью H3pI/L73 увеличивается по сравнению с таковой с помощью WT, длительность времени, в течение которого антиген, растворимый рецептор IL-6, связывается антителами, как было показано, увеличена для H3pI/L73 по сравнению с WT.
Исходя из этих данных, было уставлено, что период времени до исчезновения антитела из плазмы, а также период времени, в течение которого рецепторы IL-6, растворимый и мембранного типа, связываются антителом в организме, значительно увеличены для связывающихся в зависимости от pH антител против рецептора IL-6, которые были созданы так, что они сильно связываются с антигеном при pH 7,4, который является pH в плазме, но слабо связываются с антигеном при pH 5,8, который является pH внутри эндосом, по сравнению с антителом против рецептора IL-6 дикого типа. Это делает возможным снижение дозы и частоты введения пациентам и в результате общую вводимую дозу. Поэтому полагают, что связывающееся в зависимости от pH антитело против рецептора IL-6 особенно полезно в качестве лекарственного средства для применения в качестве антагониста IL-6.
Пример 10. Улучшение связывания в зависимости от pH с рецептором IL-6 мембранного типа путем оптимизации вариабельной области.
Оптимизация вариабельных областей H3pI/L73 и CLH5/L82
Было установлено, что антитела, обладающие способностями к связыванию в зависимости от pH, демонстрируют лучшие эффекты в примере 9. Поэтому для дальнейшего улучшения способностей к связыванию в зависимости от pH вводились мутации в последовательность CDR CLH5, полученной в примере 3, для конструирования VH1-IgG1 (SEQ ID NO: 21) и VH2-IgG1 (SEQ ID NO: 22). Кроме того, мутации вводились в последовательность каркасной области и последовательно CDR H3pI для конструирования модифицированных Н-цепей VH3-IgG1 (SEQ ID NO: 23) и VH4-IgG1 (SEQ ID NO: 24). Мутации вводились в последовательности CDR L73 и L82 для конструирования модифицированных L-цепей VL1-CK (SEQ ID NO: 25), VL2-CK (SEQ ID NO: 26) и VL3-CK (SEQ ID NO: 27). Конкретнее, мутанты были созданы, используя набор для сайт-направленного мутагенеза QuikChange (Stratagene) в соответствии со способом, описанным в прилагаемых инструкциях, и результирующие плазмидные фрагменты встраивали в вектор для экспрессии в клетках млекопитающих для конструирования векторов для экспрессии желаемых H-цепей и векторов для экспрессии желаемых L-цепей. Нуклеотидные последовательности результирующих экспрессионных векторов определяли с помощью способов, известных специалистам со средним уровнем компетентности в данной области техники.
Антитело, имеющее VH2-IgG1 (SEQ ID NO: 22) в качестве H-цепи и VL2-CK (SEQ ID NO: 26) в качестве L-цепи, назвали Fv1-IgG1, антитело, имеющее VH1-IgG1 (SEQ ID NO: 21) в качестве H-цепи и L82 в качестве L-цепи, назвали Fv2-IgG1, антитело, имеющее VH4-IgG1 (SEQ ID NO: 24) в качестве H-цепи и VL1-CK (SEQ ID NO: 25) в качестве L-цепи, назвали Fv3-IgG1, и антитело, имеющее VH3-IgG1 (SEQ ID NO: 23) в качестве H-цепи и VL3-CK (SEQ ID NO: 27) в качестве L-цепи, назвали Fv4-IgG1. Из них были экспрессированы и очищены Fv2-IgG1 и Fv4-IgG1. Экспрессию и очистку выполняли с помощью описанного в примере 1 способа.
Анализ связывания связывающихся в зависимости от pH клонов с растворимым рецептором IL-6
Кинетические анализы взаимодействий антигена с антителом при pH 7,4 были выполнены на четырех типах антител, т.е. гуманизированном антителе PM1 (дикого типа: WT) и WT, H3pI/L73-IgG1, Fv2-IgG1, и Fv4-IgG1, созданных в примерах 2 и 10, с использованием Biacore T100 (GE Healthcare) (в буфере: 10 мМ MES (pH 7,4), 150 мМ NaCl и 0,05% Tween 20). Каждое антитело связывали на сенсорном чипе, на котором был иммобилизован специфичный для γ-цепи IgG F(ab)2 (Pierce) путем присоединения к амину, и затем SR344, приведенный к концентрациям, составляющим от 9,8 до 400 нМ, инъецировали в чип в качестве аналита. Ассоциацию с SR344 и диссоциацию от него наблюдали в режиме реального времени для связывающихся в зависимости от pH клонов. Все измерения проводили при 37°C. Константы скорости ассоциации ka (1/Мс) а константы скорости диссоциации kd (1/с) рассчитывали, используя программное обеспечение для оценки анализа с использованием Biacore T100 (GE Healthcare), а константы диссоциации KD (M) рассчитывали на основе этих значений (таблица 7).

В результате расчета аффинности pH 7,4 для каждого клона, константы диссоциации (аффинность, значение KD) WT, H3pI/L73-IgG1, Fv2-IgG1 и Fv4-IgG1 от SR344 составляли, соответственно, 2,7 нМ, 1,4 нМ, 2,0 нМ и 1,4 нМ, и они были почти эквивалентными. Было установлено, что Fv2-IgG1 и Fv4-IgG1 обладают способностью к связыванию с растворимым рецептором IL-6, равной указанной способности WT или превышающей ее.
Анализ связывания связывающихся в зависимости от pH клонов с рецептором IL-6 мембранного типа
Для четырех типов созданных клонов, WT, H3pI/L73-IgG1, Fv2-IgG1 и Fv4-IgG1, следили за взаимодействиями антитела с антигеном в виде рецептора IL-6 мембранного типа при pH 5,8 и pH 7,4, используя Biacore T100 (GE Healthcare). Связывание с рецептором IL-6 мембранного типа оценивали с помощью определения связывания с рецептором IL-6, который был иммобилизован на сенсорном чипе. SR344 биотинилировали в соответствии со способом, известным среди специалистов со средним уровнем компетентности в данной области техники, и биотинилированный SR344 иммобилизовали на сенсорном чипе через стрептавидин, используя сродство между стрептавидином и биотином. Все измерения проводили при 37°C. Буфер подвижной фазы представлял собой 10 мМ MES (pH 5,8), 150 мМ NaCl и 0,05% Tween 20. Связывающиеся в зависимости от pH клоны инъецировали в чип при pH 7,4 для предоставления им возможность связаться с SR344 (буфер для инъекционных образцов: 10 мМ MES (pH 7,4), 150 мМ NaCl, 0,05% Tween 20), затем следили за pH-зависимой диссоциацией каждого клона при pH подвижной фазы, составляющем 5,8 (фиг. 17).
Концентрации в образцах устанавливали на 0,25 мкг/мл. Связывание проводили в 10 мМ MES (pH 7,4), 150 мМ NaCl и 0,05% Tween 20. Диссоциацию проводили в 10 мМ MES (pH 5,8), 150 мМ NaCl и 0,05% Tween 20. Для этого случая константы скорости диссоциации (kd (1/с)) при pH 5,8 рассчитывали путем подбора по точкам только стадии диссоциации при pH 5,8, используя программное обеспечение для оценки анализа с использованием Biacore T100 (GE Healthcare). Аналогично, концентрации в образцах устанавливали на 0,25 мкг/мл, связывание проводили в 10 мМ MES (pH 7,4), 150 мМ NaCl и 0,05% Tween 20, диссоциацию проводили в 10 мМ MES (pH 5,8), 150 мМ NaCl и 0,05% Tween 20, и константы скорости диссоциации (kd (1/с)) при pH 7,4 рассчитывали путем подбора по точкам только стадии диссоциации при pH 7,4, используя программное обеспечение для оценки анализа с использованием Biacore T100 (GE Healthcare). Константа скорости pH-зависимой диссоциации каждого клона представлена в таблице 8.

В результате расчета зависимости от pH для каждого клона зависимость от pH связывания с рецептором IL-6 мембранного типа, SR344, каждого из четырех клонов, WT, H3pI/L73-IgG1, Fv2-IgG1 и Fv4-IgG1 была 1,0-кратной, 2,59-кратной, 7,18-кратной и 5,56-кратной, соответственно. Fv2-IgG1 и Fv4-IgG1 продемонстрировали диссоциацию с большей зависимостью от pH от рецептора IL-6 мембранного типа, чем H3pI/L73-IgG1.
На основе вышеотмеченного установлено, что Fv2-IgG1 и Fv4-IgG1 демонстрируют связывание с большей зависимостью от pH с рецептором IL-6 мембранного типа, чем H3pI/L73-IgG1 при сохранении аффинности в отношении растворимого рецептора IL-6, равной такой аффинности WT или превышающей ее.
Пример 11. Фармакокинетический/фармакодинамический анализ связывающихся в зависимости от pH антител с оптимизированными вариабельными областями, используя трансгенных по рецептору IL-6 человека мышей.
Фармакокинетику Fv2-IgG1 и Fv4-IgG1, а также WT и H3pI/L73-IgG1, приготовленных и оцененных в примере 10, оценивали, используя трансгенных по рецептору IL-6 человека мышей, используемых в примере 8. WT, H3pI/L73-IgG1, Fv2-IgG1 или Fv4-IgG1 вводили мышам hIL-6R tg с помощью внутривенного введения одной дозы в расчете 25 мг/кг, и концентрацию каждого антитела в плазме определяли таким же образом, как описано в примере 8. Зависимости от времени концентраций в плазме WT, H3pI/L73-IgG1, Fv2-IgG1 и Fv4-IgG1 продемонстрированы на фиг. 18.
Фармакокинетика H3pI/L73-IgG1 улучшалась по сравнению с WT таким же образом, как и в примере 8, в то время как фармакокинетика Fv2-IgG1 и Fv4-IgG1 была дополнительно улучшена по сравнению с H3pI/L73-IgG1. Измерение в отношении концентраций несвязанного рецептора IL-6, определяемых у яванских макак в примере 9, выполняли у мышей hIL-6R tg в этом анализе, используя такой же способ. В результате подтвердили увеличение продолжительности нейтрализации растворимого рецептора IL-6 для Fv2-IgG1 и Fv4-IgG1 по сравнению с указанной продолжительностью для H3pI/L73-IgG1 (не представленные данные). Как указано в примере 10, зависящее от pH связывание с рецептором IL-6 мембранного типа было улучшено для Fv2-IgG1 и Fv4-IgG1 по сравнению с H3pI/L73-IgG1. Следовательно, было определено, что возможно дальнейшее улучшение фармакокинетики и продолжительности нейтрализации растворимого рецептора IL-6 в сравнении с таковыми H3pI/L73-IgG1 путем улучшения зависящего от pH связывания с рецептором IL-6 мембранного типа.
Пример 12. Улучшение связывания в зависимости от pH с рецептором IL-6 мембранного типа путем оптимизации константной области.
Оптимизация константной области Fv4-IgG1
Сообщалось, что вообще связывание со связанными с мембранами антигена меняется в зависимости от константной области антитела (J. Immunol. Methods 1997 Jun 23; 205(1): 67-72). Константные области связывающихся в зависимости от pH антител, приготовленных выше, были константными областями изотипа IgG1. Поэтому было проведено исследование в отношении оптимизации константной области для улучшения зависящего от pH связывания с рецептором IL-6 мембранного типа.
Была введена мутация во встречающуюся в природе константную область, т.е., константную область IgG2 (SEQ ID NO: 28), для конструирования константной области IgG2ΔGK (SEQ ID NO: 29). Другая мутация была введена в константную область IgG2ΔGK для конструирования константной области M58 (SEQ ID NO: 30). Мутации были, кроме того, введены в константные области IgG2 и M58 для конструирования константных областей M71 (SEQ ID NO: 31) и M73 (SEQ ID NO: 32).
VH3-IgG2ΔGK (SEQ ID NO: 33) создавали путем замещения константной области VH3-IgG1, приготовленной в примере 10, на IgG2ΔGK, VH3-M58 (SEQ ID NO: 34) создавали путем замещения константной области на M58, и VH3-M73 (SEQ ID NO: 35) создавали путем замещения константной области на M73. Конкретнее, были сконструированы экспрессионные векторы, в которых часть константной области VH3, используемой в примере 10, была замещена желаемой константной областью путем расщепления ферментами NheI/NotI и лигирования. Нуклеотидные последовательности результирующих экспрессионных векторов определяли, используя способ, известный среди специалистов со средним уровнем компетентности в данной области техники.
Выполняли экспрессию и очистку следующих фрагментов антител: Fv4-IgG2, в котором использованы VH3-IgG2ΔGK (SEQ ID NO: 33) вместо H-цепи и VL3-CK (SEQ ID NO: 27) вместо L-цепи; Fv4-M58, в котором использованы VH3-M58 (SEQ ID NO: 34) вместо H-цепи и VL3-CK (SEQ ID NO: 27) вместо L-цепи; и Fv4-M73, в котором использованы VH3-M73 (SEQ ID NO: 35) вместо H-цепи и VL3-CK (SEQ ID NO: 27) вместо L-цепи. Экспрессию и очистку выполняли с использованием описанного в примере 1 способа.
Анализ связывания Fv4, имеющего оптимизированную константную область, с растворимым рецептором IL-6
Ассоциацию с SR344 и диссоциацию от него наблюдали в режиме реального времени, используя такой же способ, как и в примере 10, для приготовленных таким способом Fv4-IgG1, Fv4-IgG2, Fv4-M58 и Fv4-M73, а также WT. После анализа рассчитывали константы скорости ассоциации ka (1/Мс) и константы скорости диссоциации kd (1/с) таким же образом, и затем константы диссоциации KD (M) рассчитывали на основе этих значений (таблица 9).

В результате расчета аффинности pH 7,4 для каждого клона, константы диссоциации (аффинность, значение KD) Fv4-IgG1, Fv4-IgG2, Fv4-M58 и Fv4-M73 от SR344 составляли, соответственно, 1,4 нМ, 1,3 нМ, 1,4 нМ и 1,4 нМ, и они были почти эквивалентными. Это указывает на то, что способность к связыванию связывающихся в зависимости от pH клонов с растворимым рецептором IL-6, SR344, не изменяется даже после модифицирования константной области. Основываясь на этих данных, считали, что способность к связыванию с растворимым рецептором IL-6 не изменяется для Fv1, Fv2 и Fv3, даже если аналогичным образом была изменена константная область.
Анализ связывания Fv4, имеющего оптимизированную константную область, с рецептором IL-6 мембранного типа
Следили за взаимодействиями антитела с антигеном в виде рецептора IL-6 мембранного типа при pH 5,8 и pH 7,4 таким же образом, как описано в примере 10, используя Biacore T100 (GE Healthcare), для приготовленных таким способом Fv4-IgG1, Fv4-IgG2, Fv4-M58 и Fv4-M73, а также WT. Результаты, полученные путем инъецирования связывающихся в зависимости от рН клонов при параметре pH 7,4 для предоставления возможности связывания с SR344 и слежения за pH-зависимой диссоциацией каждого клона в подвижной фазе с pH 5,8, представлены в таблице 10. Дальнейшие анализы выполняли таким же образом, как и в примере 10, и скорости pH-зависимой диссоциации для каждого клона представлены в таблице 10.

В результате расчета зависимости от pH для каждого клона pH-зависимость связывания Fv4-IgG1, Fv4-IgG2, Fv4-M58 и Fv4-M73 с SR344 была 5,6-кратной, 17,0-кратной, 17,6-кратной и 10,1-кратной, соответственно; таким образом все из Fv4-IgG2, Fv4-M58 и Fv4-M73 продемонстрировали диссоциацию с большей зависимостью от pH от рецептора IL-6 мембранного типа, чем Fv4-IgG1.
Основываясь на результатах анализа связывания с растворимым рецептором IL-6 и связывания с рецептором IL-6 мембранного типа, используя вариабельную область Fv4, было установлено, что замена константной области из IgG1 на IgG2, M58 или M73 может улучшить зависящее от pH связывание с рецептором IL-6 мембранного типа, без вызова изменения аффинности в отношении растворимого рецептора IL-6. Считали, что это имеют силу аналогично для Fv1, Fv2 и Fv3.
Пример 13. Фармакокинетический/фармакодинамический анализ связывающихся в зависимости от pH антител, имеющих оптимизированную константную область, используя трансгенных по рецептору IL-6 человека мышей.
Фармакокинетику Fv4-IgG1, Fv4-IgG2 и Fv4-M58, приготовленных в примере 13, оценивали, используя трансгенных по рецептору IL-6 человека мышей (мышей hIL-6R tg), используемых в примере 8, для исследования эффектов константной области на фармакокинетику. WT, Fv4-IgG1, Fv4-IgG2 или Fv4-M58 вводили мышам hIL-6R tg с помощью внутривенного введения одной дозы в расчете 25 мг/кг, и затем измерение концентраций каждого антитела в плазме выполняли таким же образом, как описано в примере 8. Зависимости от времени концентраций в плазме WT, Fv4-IgG1, Fv4-IgG2 и Fv4-M58 продемонстрированы на фиг. 20.
Аналогично примеру 11, фармакокинетика Fv4-IgG1 была улучшена по сравнению с WT, а фармакокинетика Fv4-IgG2 и Fv4-M58 была дополнительно улучшена по сравнению с Fv4-IgG1. Измерение в отношении концентраций несвязанного рецептора IL-6, определяемых у яванских макак в примере 9, выполняли у мышей hIL-6R tg в этом анализе, используя такой же способ. В результате подтвердили увеличение продолжительности нейтрализации растворимого рецептора IL-6 для Fv4-IgG2 и Fv4-M58 по сравнению с указанной продолжительностью для Fv4-IgG1 (не представленные данные). Как указано в примере 10, pH-зависимость связывания с рецептором IL-6 мембранного типа была увеличена для Fv4-IgG2 и Fv4-M58 по сравнению с Fv4-IgG1. Следовательно, было установлено, что возможно улучшение зависящего от pH связывания с рецептором IL-6 мембранного типа и улучшение фармакокинетики и продолжительности нейтрализации растворимого рецептора IL-6 путем замены константной области из IgG1 на IgG2 или M58. Основываясь на этих данных, полагали, что фармакокинетика и продолжительность нейтрализации растворимого рецептора IL-6, не только в случае Fv4, но также в случаях Fv1, Fv2 и Fv3, могут быть улучшены по сравнению с IgG1 путем замены константной области из IgG1 на IgG2 или M58.
Пример 14. Создание связывающихся в зависимости от pH антител, имеющих оптимизированные вариабельные и константные области
Используя такой же способом, как и описанный выше способ, были созданы VH2-M71 (SEQ ID NO: 36) и VH2-M73 (SEQ ID NO: 37), имеющие M71 и M73 вместо константной области VH2-IgG1, и VH4-M71 (SEQ ID NO: 38) и VH4-M73 (SEQ ID NO: 39), имеющие M71 и M73 вместо константной области VH4-IgG1.
Были экспрессированы и очищены Fv1-M71, в котором использованы VH2-M71 вместо H-цепи и VL2-CK вместо L-цепи, Fv1-M73, в котором использованы VH2-M73 вместо H-цепи и VL2-CK вместо L-цепи, Fv3-M71, в котором использованы VH4-M71 вместо H-цепи и VL1-CK вместо L-цепи, и Fv3-M73, в котором использованы VH4-M73 вместо H-цепи и VL1-CK вместо L-цепи. Экспрессию и очистку выполняли с использованием описанного в примере 1 способа.
Анализ связывания связывающихся в зависимости от pH антител, имеющих оптимизированные вариабельные и константные области, с растворимым рецептором IL-6
Ассоциацию с SR344 и диссоциацию от него наблюдали в режиме реального времени, используя такой же способ, как и в примере 10, для одиннадцати типов антител: гуманизированного антитела PM1 (дикого типа: WT) и H3pI/L73-IgG1, Fv1-M71, Fv1-M73, Fv2-IgG1, Fv3-M71, Fv3-M73, Fv4-IgG1, Fv4-IgG2, Fv4-M58 и Fv4-M73, созданных как описано выше. Константы скорости ассоциации ka (1/Мс) и константы скорости диссоциации kd (1/с) рассчитывали при анализе таким же образом, а константы диссоциации KD (M) рассчитывали на основе этих значений (таблица 11).
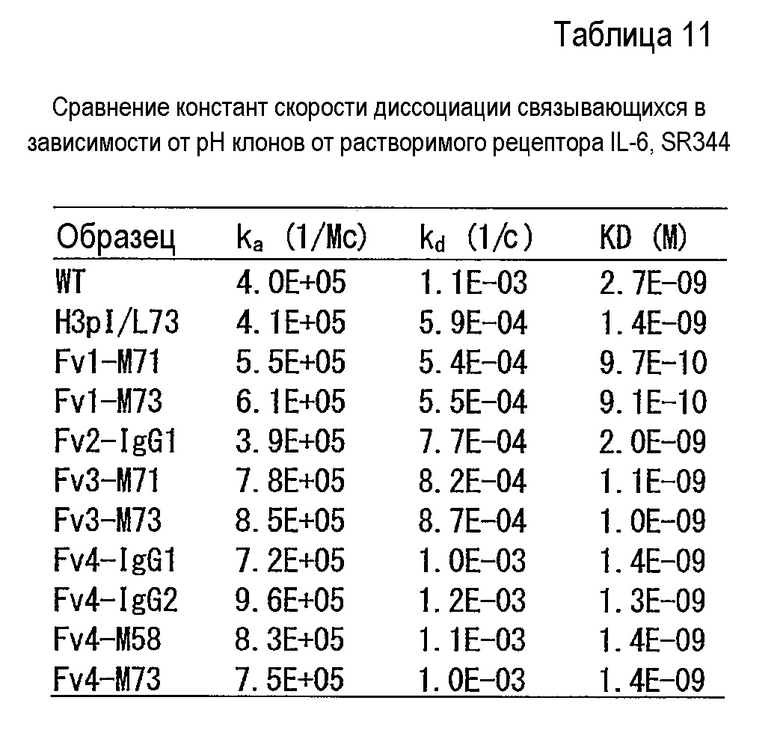
Установлено, что все результирующие десять типов связывающихся в зависимости от pH клонов имеют константы диссоциации (аффинность, значения KD) от растворимого рецептора IL-6, равные константе диссоциации WT или превышающие ее.
Анализ связывания связывающихся в зависимости от pH антител, имеющих оптимизированные вариабельные и константные области, с рецептором IL-6 мембранного типа
Следили за взаимодействиями антитела с антигеном в виде рецептора IL-6 мембранного типа при pH 5,8 и pH 7,4 таким же образом, как описано в примере 10, используя Biacore T100 (GE Healthcare), для одиннадцати типов антител: гуманизированного антитела PM1 (дикого типа: WT) и H3pI/L73-IgG1, Fv1-M71, Fv1-M73, Fv2-IgG1, Fv3-M71, Fv3-M73, Fv4-IgG1, Fv4-IgG2, Fv4-M58 и Fv4-M73, приготовленных как описано выше. Связывающиеся в зависимости от pH клоны инъецировали при параметре pH 7,4 для предоставления возможности связывания с SR344, и затем следили за pH-зависимой диссоциацией каждого клона при pH подвижной фазе, составляющем 5,8. Результаты продемонстрированы на фиг. 21 (результаты для Fv1-M71, Fv1-M73, Fv3-M71 и Fv3-M73 продемонстрированы на фиг. 21, в то время как результаты для других клонов продемонстрированы на фиг. 17 и 19). Анализы выполняли таким же образом, как и в примере 10, а pH-зависимости констант скорости диссоциации всех одиннадцати типов клонов представлены в таблице 12.
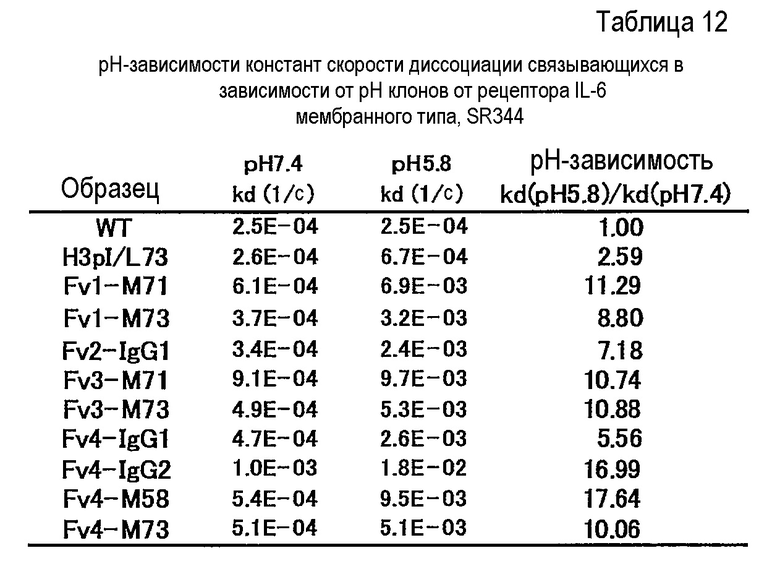
Десять типов полученных связывающихся в зависимости от pH клонов продемонстрировали способность к связыванию в зависимости от pH с рецептором IL-6 мембранного типа. Кроме того, установлено, что все из Fv1-M71, Fv1-M73, Fv2-IgG1, Fv3-M71, Fv3-M73, Fv4-IgG1, Fv4-IgG2, Fv4-M58 и Fv4-M73 демонстрируют связывание с увеличенной зависимостью от pH с рецептором IL-6 мембранного типа, по сравнению с H3pI/L73-IgG1, для которых период времени до исчезновения антитела из плазмы, а также период времени, в течение которого рецепторы IL-6, растворимый и мембранного типа, связываются антителом в организме, как установлено, были значительно увеличены по сравнению с WT, как продемонстрировано у яванских макак в примере 9.
Пример 15. Фармакокинетический/фармакодинамический анализ связывающихся в зависимости от pH антител, имеющих оптимизированные вариабельные и константные области, используя яванских макак.
Создание известного антитела против рецептора IL-6 с большой аффинностью
Для экспрессии антитела против рецептора IL-6 с большой аффинностью VQ8F11-21 hIgG1, описанного в US 2007/0280945 A1 (US 2007/0280945 A1, аминокислотные последовательности 19 и 27), в качестве известного антитела против рецептора IL-6 с большой аффинностью был сконструирован вектор для экспрессии в клетках млекопитающих. Вариабельную область антитела конструировали с использованием объединяющих синтетических олигодезоксинуклеотидов для ПЦР (ПЦР с компоновкой). Константную область амплифицировали с помощью ПЦР с экспрессионного вектора, используемого в примере 1. Вариабельную область антитела и константную область антитела лигировали с помощью ПЦР с компоновкой и встраивали в вектор для экспрессии в млекопитающих. Результирующие фрагменты ДНК H-цепи и L-цепи встраивали в векторы для экспрессии в клетках млекопитающих для конструирования представляющих интерес вектора для экспрессии H-цепи и вектора для экспрессии L-цепи. Нуклеотидные последовательности результирующих экспрессионных векторов определяли способом, известным среди специалистов со средним уровнем компетентности в данной области техники. Экспрессию и очистку выполняли, используя сконструированные экспрессионные векторы. Экспрессию и очистку выполняли, используя описанный в примере 1 способ, для получения антитела против рецептора IL-6 с большой аффинностью (Ат с большой аффинностью).
Фармакокинетический/фармакодинамический анализ в яванских макаках
Фармакокинетику и фармакологическую эффективность оценивали у яванских макаках для связывающихся в зависимости от pH антител H3pI/L73-IgG1 и Fv1-M71, Fv1-M73, Fv2-IgG1, Fv3-M73, и Fv4-M73 и известного антитела против рецептора IL-6 с большой аффинностью (Ат с большой аффинностью). H3pI/L73-IgG1, Fv1-M71, Fv1-M73, Fv2-IgG1, Fv3-M73 или Fv4-M73 вводили яванским макакам с помощью внутривенного введения одной дозы в расчете 0,5 мг/кг, в то время как Ат с большой аффинностью вводили с помощью внутривенного введения одной дозы в расчете 1,0 мг/кг. Образцы крови отбирали до введения и в динамике времени. Концентрацию каждого антитела в плазме определяли таким же образом, как и в примере 9. Зависимости от времени концентраций в плазме H3pI/L73-IgG1, Fv1-M71, Fv1-M73, Fv2-IgG1, Fv3-M73 и Fv4-M73, а также Ат с большой аффинностью продемонстрированы на фиг. 21. Для оценки фармакологической эффективности по показателю степени, в которой нейтрализуется рецептор IL-6 мембранного типа яванского макака, IL-6 яванского макака вводили подкожно в нижние части спины животных ежедневно в дозе 5 мкг/кг с дня 3 до дня 10 (с дня 6 до дня 10, что касается Ат с большой аффинностью) после введения антитела, таким же образом, как и в примере 9. Концентрацию CRP у каждого животного определяли через 24 часа после каждого введения. Зависимости от времени концентраций CRP при введении каждого антитела продемонстрированы на фиг. 22. Для оценки фармакологической эффективности по показателю степени нейтрализации растворимого рецептора IL-6 яванского макака определяли концентрацию несвязанного растворимого рецептора IL-6 яванского макака в плазме яванских макак таким же образом, как и в примере 9. Зависимости от времени концентраций несвязанного растворимого рецептора IL-6 яванского макака при введении каждого антитела продемонстрированы на фиг. 23.
В результате, концентрации антител в плазме сохранялись высокими для каждого из Fv1-M71, Fv1-M73, Fv2-IgG1, Fv3-M73 и Fv4-M73 по сравнению с H3pI/L73-IgG1, в то время как концентрации CRP и несвязанного растворимого рецептора IL-6 яванского макака сохранялись на низких уровнях. А именно, этот результат показал, что период времени, в течение которого рецепторы IL-6, мембранного типа и растворимые, связываются антителом, (или, другими словами, продолжительность нейтрализации) увеличен с помощью указанных антител по сравнению с H3pI/L73-IgG1.
Кроме того, для этих связывающихся в зависимости от pH антител против рецептора IL-6 были подтверждены их нейтрализующие эффекты и устойчивая эффективность, которые были равны или превышали таковые известного антитела против рецептора IL-6 с большой аффинностью (Ат с большой аффинностью), вводимого в дозе 1,0 мг/кг, при введении указанных антител только в половине дозы известного антитела, т.е., в дозе 0,5 мг/кг. Следовательно, было уяснено, что связывающиеся в зависимости от pH антитела имеют нейтрализующие эффекты и устойчивую эффективность, превышающие таковые известного антитела против рецептора IL-6 с большой аффинностью.
Было также подтверждено, что те из представленных в таблице 12 антител, для которых фармакокинетический/фармакодинамический анализ не проводился с использованием яванских макак в этом анализе, демонстрируют связывание с увеличенной pH-зависимостью с рецептором IL-6 мембранного типа по сравнению с H3pI/L73-IgG1. Поэтому также полагали, что период времени, в течение которого рецепторы IL-6, мембранного типа и растворимые, связываются антителами, (или, другими словами, продолжительность нейтрализации и устойчивые нейтрализующие эффекты) увеличен для этих антител по сравнению с H3pI/L73-IgG1.
В примере 9, для H3pI/L73-IgG1 установлено, что период времени до исчезновения антитела из плазмы, а также период времени, в течение которого растворимый рецептор IL-6 и рецептор IL-6 мембранного типа связываются антителом в организме, (устойчивые нейтрализующие эффекты) значительно увеличен по сравнению с WT. Поэтому полагали, что Fv1-M71, Fv1-M73, Fv2-IgG1, Fv3-M71, Fv3-M73, Fv4-IgG1, Fv4-IgG2, Fv4-M58 и Fv4-M73, имеющие устойчивые нейтрализующие эффекты, превосходящие таковые H3pI/L73-IgG1, имеют поразительно увеличенные устойчивые нейтрализующие эффекты по сравнению с WT.
В противоположность антителам против рецептора IL-6, связывающиеся в зависимости от pH антитела против рецептора IL-6, созданные так, что они сильно связываются с антигеном при pH в плазме, составляющем 7,4, но исключительно слабо связываются с антигеном при pH в эндосомах, составляющем 5,8, делают возможным снижение дозы и частоты введения пациентам антитела против рецептора IL-6, и в результате они могут значительно снизить общее вводимое количество. Поэтому полагают, что связывающиеся в зависимости от pH антитела против рецептора IL-6 намного лучше в качестве лекарственного средства для применения в качестве антагониста IL-6.
Пример 16. Создание связывающегося в зависимости от pH антитела против IL-6
Экспрессия и очистка антитела против IL-6
В примерах 1-15 было с успехом создано множество гуманизированных антител против рецептора IL-6, которые связываются в зависимости от pH с рецептором IL-6, путем сообщения pH-зависимости благодаря введению замен на гистидин и т.п. в вариабельную область, в частности, в последовательности CDR гуманизированных антител против рецептора IL-6. Было установлено, что все из этих антител многократно связываются с рецептором IL-6 и демонстрируют значительное улучшение в фармакокинетике/фармакодинамике.
Следовательно, было подтверждено, что способность антитела к связыванию в зависимости от pH с антигеном можно сообщить другому антителу, которое связывается с антигеном, отличным от рецептора IL-6, используя аналогичный способ. В качестве антигена был выбран IL-6 человека, и было создано антитело против IL-6, включающее H-цепь (WT) (аминокислотную последовательность: SEQ ID NO: 62) и L-цепь (WT) (аминокислотную последовательность: SEQ ID NO: 63), которое связывается с IL-6, описанное в WO 2004/039826, («антитело против IL6 дикого типа»). Используя известный квалифицированным в данной области техники специалистам способ, фрагменты генов, кодирующие представляющие интерес аминокислотные последовательности антитела, встраивали в векторы для экспрессии в клетках млекопитающих для конструирования представляющих интерес вектора для экспрессии H-цепи и вектора для экспрессии L-цепи. Нуклеотидные последовательности результирующих экспрессионных векторов определяли, используя известный квалифицированному специалисту способ. Антитело против IL6 дикого типа экспрессировали и очищали с помощью описанного в пример 1 способа.
Создание антитела против IL-6 с pH-зависимостью
Для сообщения способности антитела к связыванию в зависимости от pH с IL-6 были введены замены на гистидин в аминокислоты CDR антитела против IL-6 (анти-IL6 дикого типа), включающего H-цепь (WT) (аминокислотную последовательность: SEQ ID NO: 62) и L-цепь (WT) (аминокислотную последовательность: SEQ ID NO: 63). Путем замещения на гистидин в аминокислотах CDR и последующего скрининга было получено несколько клонов, демонстрирующих связывание в зависимости от pH. Связывание при pH 5,5 было значительно ослаблено по сравнению со связыванием при pH 7,4. Положения замены на гистидин в клонах с pH-зависимостью представлены в таблице 13. Примеры включают «анти-IL6 клон 1», включающий H-цепь (c1) (аминокислотную последовательность: SEQ ID NO: 64) и L-цепь (c1) (аминокислотную последовательность: SEQ ID NO: 65), и «анти-IL-6 клон 2», включающий H-цепь (c1) (аминокислотную последовательность: SEQ ID NO: 64) и L-цепь (c2) (аминокислотную последовательность: SEQ ID NO: 66). Анти-IL6 клон 1 и анти-IL-6 клон 2 экспрессировали и очищали с помощью описанного в примере 1 способа.
Положения замены на гистидин в клонах с pH-зависимостью
L53, L54, L90, L94
Анализ связывания клонов с pH-зависимостью с IL-6 человека
Кинетический анализ взаимодействий антигена с антителом при pH 5,5 и pH 7,4 был выполнен с использованием Biacore T100 (GE Healthcare) на трех типах антител, приготовленных, как отмечено выше: анти-IL6 дикого типа, анти-IL6 клоне 1 и анти-IL-6 клоне 2 (в буфере: DPBS(-) (pH 7,4 или pH 5,5), 150 мМ NaCl). Антитела связывали на сенсорном чипе, на котором был иммобилизован рекомбинантный белок A/G (Pierce) путем присоединения к амину, и затем IL-6 человека (Toray), приведенный к соответствующей концентрации, инъецировали в чип в качестве аналита. Все измерения проводили при 37°C. Константы скорости ассоциации ka (1/Мс) и константы скорости диссоциации kd (1/с) рассчитывали, используя программное обеспечение для оценки анализа с использованием Biacore T100 (GE Healthcare), а константы диссоциации KD (M) рассчитывали на основе этих значений (таблица 14). Кроме того, для каждого клона рассчитывали отношение аффинности при pH 5,5 к аффинности при pH 7,4 для оценки связывания в зависимости от pH.
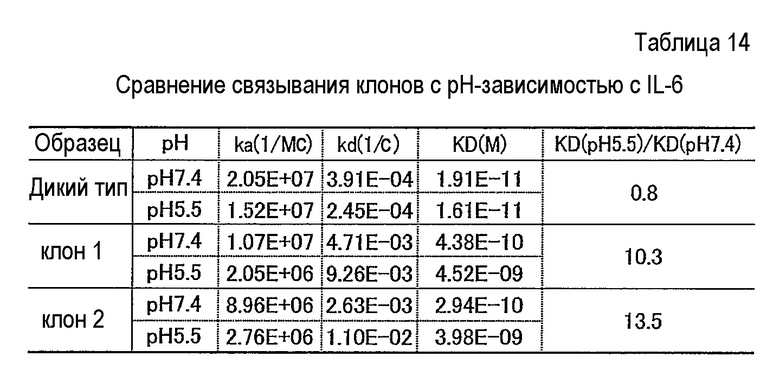
Рассчитанное отношение аффинности при pH 5,5 к аффинности при pH 7,4 ((KD) (pH 5,5)/(KD) (pH 7,4)), которое свидетельствует о зависящем от pH связывании с рецептором IL-6 человека, составляло 0,8, 10,3 и 13,5 для анти-IL6 дикого типа, анти-IL6 клона 1 и анти-IL6 клона 2, соответственно. Т.е. способность к связыванию в зависимости от pH каждого клона превышала более чем в 10 раз такую способность WT. Сенсограммы анти-IL-6 клона 2 при pH 7,4 и pH 5,5 продемонстрированы на фиг. 26.
Таким образом, было установлено, что, как и в случае антител против рецептора IL-6, связывающиеся в зависимости от рН антитела против IL-6, которые сильно связываются с антигеном в нейтральных условиях в плазме, но слабо в кислотных условиях внутри эндосом, можно создать путем введения замен на гистидин и т.п., главным образом, в аминокислотные последовательности CDR. Как указано в примерах 1-15, антитело против рецептора IL-6, которое обладает способностью к связыванию в зависимости от pH, многократно связывается с рецептором IL-6, и его фармакокинетика/фармакодинамика значительно улучшена. Т.е. было высказано предположение, что анти-IL-6 клон 1 и анти-IL-6 клон 2, обладающие способностью к связыванию в зависимости от pH, многократно связываются с большим числом антигенов со значительно улучшенной фармакокинетикой/фармакодинамикой, по сравнению с антителом против IL6 дикого типа.
Пример 17. Создание связывающегося в зависимости от pH антитела против рецептора IL-31
Экспрессия и очистка антитела против рецептора IL-31
В примерах 1-15 было с успехом создано множество гуманизированных антител против рецептора IL-6, которые связываются в зависимости от pH с рецептором IL-6, путем сообщения pH-зависимости благодаря введению замен на гистидин и т.п. в вариабельную область, в частности, в последовательности CDR гуманизированных антител против рецептора IL-6. Было установлено, что все из этих антител многократно связываются с рецептором IL-6 и демонстрируют значительное улучшение в фармакокинетике/фармакодинамике.
Следовательно, было подтверждено, что способность антитела к связыванию в зависимости от pH с антигеном можно сообщить другому антителу, которое связывается с антигеном, отличным от рецептора IL-6, используя аналогичный способ. В качестве антигена был выбран рецептор IL-31 мыши, и было создано антитело против IL-31, включающее H-цепь (WT) (аминокислотную последовательность: SEQ ID NO: 67) и L-цепь (WT) (аминокислотную последовательность: SEQ ID NO: 68), которое связывается с рецептором IL-31 мыши, описанное в WO 2007/142325, («антитело против рецептора IL31 дикого типа»). Используя известный квалифицированным в данной области техники специалистам способ, фрагменты генов, кодирующие представляющие интерес аминокислотные последовательности, встраивали в векторы для экспрессии в клетках млекопитающих для конструирования представляющих интерес вектора для экспрессии H-цепи и вектора для экспрессии L-цепи. Нуклеотидные последовательности результирующих экспрессионных векторов определяли, используя известный квалифицированному специалисту способ. Антитело против рецептора IL31 дикого типа экспрессировали и очищали с помощью описанного в пример 1 способа.
Создание антитела против IL-31 с pH-зависимостью
Для сообщения способности антитела к связыванию в зависимости от pH с рецептором IL-31 были введены замены на гистидин в аминокислоты CDR антитела против рецептора IL-31 (анти-IL31R дикого типа), включающего H-цепь (WT) (аминокислотную последовательность: SEQ ID NO: 67) и L-цепь (WT) (аминокислотную последовательность: SEQ ID NO: 68). Путем замещения на гистидин в аминокислотах CDR и последующего скрининга было получено несколько клонов, демонстрирующих связывание в зависимости от pH. Связывание при pH 5,5 было значительно ослаблено по сравнению со связыванием при pH 7,4. Положение замены на гистидин в клонах с pH-зависимостью представлено в таблице 15. Примером является «анти-IL31R клон 1», включающий H-цепь (c1) (аминокислотную последовательность: SEQ ID NO: 69) и L-цепь (WT). Анти-IL31R клон 1 экспрессировали и очищали с помощью описанного в примере 1 способа.
Положение замены на гистидин в клонах с pH-зависимостью
Анализ связывания клонов с pH-зависимостью с растворимым рецептором IL-31
Кинетический анализ взаимодействий антигена с антителом при pH 5,5 и pH 7,4 был выполнен с использованием Biacore T100 (GE Healthcare) для двух типов антител, приготовленных, как отмечено выше: анти-IL31R дикого типа и анти-IL31R клона 1 (в буфере: DPBS(-) (pH 7,4 или pH 5,5), 150 мМ NaCl, 0,01% Tween 20, 0,02% NaN3). Антитела связывали на сенсорном чипе, на котором был иммобилизован рекомбинантный белок A/G (Pierce) путем присоединения к амину, и затем растворимый рецептор IL-31 мыши (приготовленный в соответствии со способом, описанным в WO 2007/142325), приведенный к соответствующей концентрации, инъецировали в чип в качестве аналита. Все измерения проводили при 25°C. Константы скорости ассоциации ka (1/Мс) и константы скорости диссоциации kd (1/с) рассчитывали, используя программное обеспечение для оценки анализа с использованием Biacore T100 (GE Healthcare), а константы диссоциации KD (M) рассчитывали на основе этих значений (таблица 16). Кроме того, для каждого клона рассчитывали отношение аффинности при pH 5,5 к аффинности при pH 7,4 для оценки связывания в зависимости от pH.
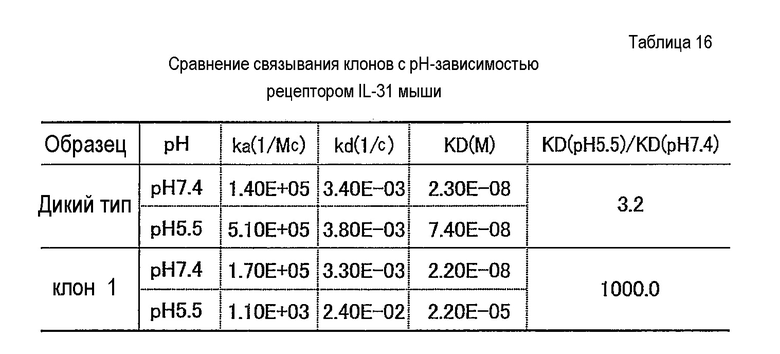
Рассчитанное отношение аффинности при pH 5,5 к аффинности при pH 7,4 ((KD) (pH 5,5)/(KD) (pH 7,4)), которое свидетельствует о зависящем от рН связывании с рецептором IL-31 мыши, составляло 3,2 и 1000 для анти-IL31R дикого типа и анти-IL31R клона 1, соответственно. Т.е. способность к связыванию в зависимости от pH анти-IL31R клона 1 превышала приблизительно в 300 раз такую способность WT. Сенсограммы для анти-IL31R клона 1 при pH 7,4 и pH 5,5 продемонстрированы на фиг. 27.
Таким образом, было установлено, что, как и в случаях антител против рецептора IL-6 и антител против IL-6, связывающиеся в зависимости от рН антитела против рецептора IL-31, которые сильно связываются с антигеном в нейтральных условиях в плазме, но слабо в кислотных условиях внутри эндосом, можно создать путем введения замен на гистидин и т.п., главным образом, в аминокислотные последовательности CDR. Как указано в примерах 1-15, антитело против рецептора IL-6, которое обладает способностью к связыванию в зависимости от pH, многократно связывается с рецептором IL-6, и его фармакокинетика/фармакодинамика поразительно улучшена. Т.е. было высказано предположение, что анти-IL31R клон 1, обладающий способностью к связыванию в зависимости от pH, многократно связывается с большим числом антигенов со значительно улучшенной фармакокинетикой/фармакодинамикой, по сравнению с антителом против рецептора IL-31 дикого типа.
Пример 18. Многократное связывание с антигеном связывающегося в зависимости от pH антитела
Экспрессия и очистка вводимого мышам антитела
Были приготовлены четыре типа гуманизированных антител против рецептора IL-6, описанных ниже. Как и антитела, которые не связываются в зависимости от pH с рецептором IL-6, WT-IgG1, включающий H (WT) (аминокислотную последовательность: SEQ ID NO: 9) и L (WT) (аминокислотную последовательность: SEQ ID NO: 10), и H54/L28-IgG1, включающий H54 (аминокислотную последовательность: SEQ ID NO: 70) и L28 (аминокислотную последовательность: SEQ ID NO: 12), экспрессировали и очищали, используя указанный в примере 1 способ. Как и антитела, которые связываются в зависимости от pH с рецептором IL-6, H170/L82-IgG1 примера 3, включающий H170 (аминокислотную последовательность: SEQ ID NO: 4) и L82 (аминокислотную последовательность: SEQ ID NO: 7), и Fv4-IgG1 примера 10, включающий VH3-IgG1 (SEQ ID NO: 23) и VL3-CK (SEQ ID NO: 27), экспрессировали и очищали, используя указанный в примере 1 способ.
Анализ связывания каждого типа антитела с растворимым рецептором IL-6
Кинетический анализ взаимодействий антигена с антителом при pH 7,4 и pH 5,8 был выполнен с использованием Biacore T100 (GE Healthcare) для четырех типов приготовленных антител: WT-IgG1, H54/L28-IgG1, H170/L82-IgG1 и Fv4-IgG1 (в буфере: 10 мМ MES (pH 7,4 или pH 5,8), 150 мМ NaCl, 0,05% поверхностно-активного вещества-P20). Антитела связывали на сенсорном чипе, на котором был иммобилизован рекомбинантный белок A/G (Pierce) путем присоединения к амину, и SR344, приведенный к соответствующей концентрации, инъецировали в чип в качестве аналита. Ассоциацию с SR344 и диссоциацию от него каждого типа антитела наблюдали в режиме реального времени. Все измерения проводили при 37°C. Константы скорости ассоциации ka (1/Мс) и константы скорости диссоциации kd (1/с) рассчитывали, используя программное обеспечение для оценки анализа с использованием Biacore T100 (GE Healthcare), а константы диссоциации KD (M) рассчитывали на основе этих значений (таблица 17).

Рассчитывали отношение аффинности при pH 5,8 к аффинности при pH 7,4 для каждого антител. Отношение KD, которое свидетельствует о зависящем от pH связывании с SR344, составляло 1,6, 0,7, 61,9 и 27,3 для WT-IgG1, H54/L28-IgG1, H170/L82-IgG1 и Fv4-IgG1, соответственно. Кроме того, рассчитывали отношение скорости диссоциации (kd) при pH 5,8 к kd при pH 7,4 для каждого антитела. Отношение kd, которое свидетельствует о скорости зависящей от pH диссоциации от SR344, составляло 2,9, 2,0, 11,4 и 38,8 для WT-IgG1, H54/L28-IgG1, H170/L82-IgG1 и Fv4-IgG1, соответственно. Таким образом, было подтверждено, что H170/L82-IgG1 и Fv4-IgG1 демонстрируют зависящее от pH связывание, в то время как обычные антитела WT-IgG1 и H54/L28-IgG1 едва проявляют эту способность. Кроме того, поскольку аффинности (KD) этих антител при pH 7,4 почти равны, полагали, что их способности к связыванию с SR344 в плазме эквивалентны.
In Vivo фармакокинетический анализ с использованием мышей
Фармакокинетику SR344 и антитела против рецептора IL-6 человека оценивали после введения SR344 (рецептора IL-6 человека, приготовленного в примере 1) самого по себе или после одновременного введения SR344 и антитела против рецептора IL-6 человека мышам, которые не экспрессируют рецептор IL-6 человека (C57BL/6J; антитела против рецептора IL-6 человека не связываются с рецептором IL-6 мыши). Раствор SR344 (5 мкг/мл) или смешанный раствор, содержащий SR344 и антитело против рецептора IL-6 человека (5 мкг/мл и 0,1 мг/мл, соответственно), вводили в хвостовую вену с помощью введения одной дозы, составляющей 10 мл/кг. Поскольку антитело против рецептора IL-6 человека присутствовало в достаточном избыточном количестве относительно SR344, полагали, что почти все молекулы SR344 были связаны антителом. Образцы крови брали через 15 минут, 2 часа, 8 часов, 1 день, 2 дня, 3 дня, 4 дня, 7 дней, 14 дней, 21 день и 28 дней после введения. Взятые образцы крови сразу же подвергали центрифугированию в течение 15 минут при 15000 оборотов/мин и 4°C для получения плазмы. Отделенную плазму хранили в камере замораживания, установленной на -20°C или ниже, до проведения измерений. В качестве антител против рецептора IL-6 человека использовали описанные выше WT-IgG1, H54/L28-IgG1, H170/L82-IgG1 и Fv4-IgG1.
Определение концентрации в плазме антитела против рецептора IL-6 человека с помощью ELISA
Концентрацию антитела против рецептора IL-6 человека в плазме мышей определяли с помощью ELISA. F(abʹ)2-фрагмент антитела против IgG человека (специфичный для γ-цепи) (Sigma) распределяли по иммунопланшету Nunc-ImmunoPlate MaxiSorp (Nalge Nunc International) и допускали нахождение в течение ночи при 4°C для приготовления планшетов с иммобилизованным фрагментом антитела против IgG человека. Готовили образцы для калибровочных кривых, имеющие концентрации в плазме, составляющие 0,8, 0,4, 0,2, 0,1, 0,05, 0,025 и 0,0125 мкг/мл, и образцы плазмы мышей, разведенные в 100 раз или более. 200 мкл 20 нг/мл SR344 добавляли к 100 мкл образцов для калибровочной кривой и образцов плазмы, и затем допускали нахождение образцов в течение 1 часа при комнатной температуре. Впоследствии образцы распределяли по планшетам с иммобилизованным фрагментом антитела против IgG человека, и допускали нахождение планшетов в течение 1 часа при комнатной температуре. Затем добавляли биотинилированное антитело против рецептора IL-6 человека (R&D) для взаимодействия в течение 1 часа при комнатной температуре. Впоследствии добавляли стрептавидин-PolyHRP80 (Stereospecific Detection Technologies) для взаимодействия в течение 1 часа при комнатной температуре, и проводили хромогенную реакцию, используя однокомпонентный субстрат для HRP при использовании в микролунках TMP (BioFX Laboratories) в качестве субстрата. После остановки реакции с помощью 1 н. серной кислоты (Showa Chemical) определяли оптическую плотность при 450 нм с использованием считывающего устройства для микропланшетов. Концентрацию в плазме мышей рассчитывали, исходя из оптической плотности калибровочной кривой, используя аналитическое программное обеспечение SOFTmax PRO (Molecular Devices). Зависимость от времени концентрации в плазме после внутривенного введения, определенной с помощью этого способа, продемонстрирована на фиг. 28.
Определение концентрации SR344 в плазме с помощью электрохемилюминесценции
Концентрацию SR344 в плазме мышей определяли с помощью электрохемилюминесценции. Готовили образцы для калибровочной кривой SR344, приведенного к концентрациям, составляющим 2000, 1000, 500, 250, 125, 62,5 и 31,25 пг/мл, и образцы плазмы мыши, разведенные в 50 раз или больше. Образцы смешивали с раствором моноклонального антитела против рецептора IL-6 человека (R&D), меченного рутением с использованием N-гидроксисукцимидного эфира Sulfo-Tag (Meso Scale Discovery), биотинилированного антитела против рецептора IL-6 человека (R&D), и WT-IgG1, и допускали взаимодействие в течение ночи при 37°C. Конечная концентрация WT-IgG1 составляла 333 мкг/мл, что превышало концентрацию антитела против рецептора IL-6 человека, содержащегося в образцах, с целью связывания почти всех молекул SR344 в образцах с WT-IgG1. Впоследствии образцы распределяли по планшету со стрептавидином MA400 PR (Meso Scale Discovery), и допускали взаимодействие в течение 1 часа при комнатной температуре, и выполняли промывку. Сразу же после распределения буфера Т для считывания (×4) (Meso Scale Discovery) проводили измерение с помощью считывающего устройства Sector PR 400 (Meso Scale Discovery). Концентрацию SR344 рассчитывали, исходя из зависимости калибровочной кривой, используя аналитическое программное обеспечение SOFTmax PRO (Molecular Devices). Зависимость от времени концентрации SR344 в плазме после внутривенного введения, определенной с помощью этого способа, продемонстрирована на фиг. 29.
Эффекты зависящего от pH связывания
Что касается зависимости от времени концентрации антител WT-IgG1 и H54/L28-IgG1, которые не демонстрируют зависящее от pH связывание, и H170/L82-IgG1 и Fv4-IgG1, которые демонстрируют зависящее от pH связывание, зависимости от времени концентрации были примерно идентичными для WT-IgG1, H54/L28-IgG1 и Fv4-IgG1, в то время как H170/L82-IgG1 элиминировался слегка быстрее. Данные зависимости от времени концентрации в плазме анализировали с помощью программного обеспечения для фармакокинетического анализа WinNonlin (Pharsight). Периоды полужизни WT-IgG1, H54/L28-IgG1, Fv4-IgG1 и H170/L28-IgG1 составляли 21,0, 28,8, 26,2 и 7,5 дней, соответственно.
Как описывается в примере 2, когда антигеном является растворимый антиген, введенное антитело связывается с антигеном в плазме и сохраняется в плазме в форме комплекса антиген-антитело. Обычно, в противоположность очень длительному периоду времени сохранения в плазме антитела (скорость элиминации является очень низкой) благодаря действию FcRn, период времени сохранения в плазме антигена является коротким (скорость элиминации является высокой). Таким образом, связавшийся с антителом антиген имеет увеличенный период времени сохранения в плазме, схожий с таковым антитела (скорость элиминации является очень низкой). Схожим образом, когда антиген гуманизированного антитела против рецептора IL-6, SR344 (растворимый рецептор IL-6 человека), вводили сам по себе, SR344 очень быстро элиминировался (период полужизни: 0,2 дня). Однако в случае одновременного введения SR344 с обычным антителом, WT-IgG1 или H54/L28-IgG1, которое не демонстрирует зависящее от pH связывание, скорость элиминации SR344 была значительно снижена, а период времени сохранения в плазме SR344 был увеличен (время полужизни в плазме: 5,3 дней для WT-IgG1, 6,3 дней для H54/L28-IgG1). Это обусловлено тем, что почти все молекулы SR344 были связаны антителами, вводимыми вместе с ними, и поэтому связанные антителами SR344 имели увеличенный период времени сохранения в плазме, схожий с таковым антитела, благодаря действию FcRn, описанному выше.
Однако в случае одновременного введения SR344 с антителом H170/L82-IgG1 или Fv4-IgG1, которое демонстрирует зависящее от pH связывание, элиминация SR344 была значимо быстрой (время полужизни в плазме: 1,3 дня для H170/L82-IgG1, 0,6 дней для Fv4-IgG1), по сравнению со случаем одновременного введения с WT-IgG1 или H54/L28-IgG1. Эта тенденция была особенно отчетливой для Fv4-IgG1. Поскольку аффинность Fv4-IgG1 при pH 7,4 эквивалента аффинности WT-IgG1 и H54/L28-IgG1 или превышает ее, полагали, что почти все молекулы SR344 были связаны с Fv4-IgG1. Даже несмотря на то, что Fv4-IgG1 демонстрирует эквивалентное или слегка более длительное сохранение в плазме и более медленную элиминацию по сравнению с WT-IgG1 и H54/L28-IgG1, элиминация SR344, связанного с Fv4-IgG1, была очень быстрой. Объяснением этого может служить идея технического решения настоящего изобретения, представленная на фиг. 4. В случае обычных антител, которые не демонстрируют зависящее от pH связывание, комплекс антитело-растворимый антиген поступает в эндосомы посредством пиноцитоза в плазме и связывается с FcRn, экспрессируемым в эндосомах, в кислотных условиях внутри эндосом. Поскольку комплекс антитело-растворимый антиген, связанный с FcRn, перемещается как таковой на клеточную поверхность и снова возвращается в плазму, связанный с антителом антиген имеет увеличенный период времени сохранения в плазме, схожий с таковым антитела (элиминация является очень медленной). С другой стороны, в случае антител, которые демонстрируют зависящее от pH связывание, антиген подвергается диссоциации от антитела в кислотных условиях внутри эндосом, и поэтому только антитело связывается с FcRn и снова возвращается в плазму. Поскольку подвергшийся диссоциации от антитела антиген подвергается деградации в лизосомах без возвращения в плазму, элиминация антигена является очень быстрой по сравнению со случаем антител, которые не демонстрируют зависящее от pH связывание. А именно, в случае одновременного введения SR344 с антителом WT-IgG1 или H54/L28-IgG1, которое не демонстрирует зависящее от pH связывание, элиминация SR344 является медленной до степени, схожей с таковой антитела, поскольку SR344 связывается с WT-IgG1 или H54/L28-IgG1 как в плазме, так и в эндосомах. Напротив, в случае одновременного введения SR344 с антителом H170/L82-IgG1 или Fv4-IgG1, которое демонстрирует зависящее от pH связывание, элиминация SR344 является очень быстрой, поскольку SR344 подвергается диссоциации от антитела в интраэндосомальной среде с низким pH. Т.е., поскольку антитела H170/L28-IgG1 и Fv4-IgG1, которые демонстрируют зависящее от pH связывание, подвергаются диссоциации от SR344 в интраэндосомальной среде с низким pH, полагают, что большинство из H170/L82-IgG1 или Fv4-IgG1, снова возвратившихся в плазму с помощью FcRn, не являются связанными с SR344. Таким образом, было установлено, как продемонстрировано на фиг. 4, что демонстрирующее зависящее от pH связывание антитело при подвергании диссоциации от антигена в интраэндосомальной среде с низким pH и возвращении в плазму с помощью FcRn, при этом не будучи связанным с антигеном, может снова связываться с антигеном в плазме. Также было установлено, что при повторении этого процесса антитело, демонстрирующее зависящее от pH связывание, может многократно связываться с множеством антигенов. Это согласуется с представленными в примере 7 данными Biacore, показывающими, что антитела с pH-зависимостью могут многократно связываться с антигенами. Таким образом, путем усиления зависимости от pH связывания антитела с антигеном может быть увеличено число раз повторного связывания с антигенами.
Когда антигеном является растворимый антиген, и антиген связывается с антителом в нейтральных условиях в плазме, но подвергается диссоциации от антитела в эндосомах, и антитело возвращается в плазму с помощью FcRn, антитело может снова связываться с антигеном в нейтральных условиях в плазме. Таким образом, антитело, обладающее способностью к диссоциации от антигена в кислотных условиях внутри эндосом, может многократно связываться с антигенами. По сравнению с тем, когда связанный с антителом антиген не подвергается диссоциации от антитела в эндосомах (т.е., в плазму возвращается связанный с антителом антиген), если связанный с антителом антиген подвергается диссоциации от антитела в эндосомах, скорость элиминации антигена из плазмы увеличивается, поскольку антиген перемещается в лизосомы и подвергается деградации. Следовательно, скорость элиминации антигена из плазмы можно использовать в качестве показателя для определения того, может ли антитело связываться с антигеном многократно. Определение скорости элиминации антигена из плазмы можно выполнить, например, путем введения антигена и антитела in vivo и затем определения концентрации антигена в плазме после введения, как показано в примерах.
Антитело, демонстрирующее зависящее от pH связывание, может повторно связываться с множеством антигенов в противоположность случаю обычного антитела, не демонстрирующего зависящее от pH связывание. Таким образом, количество вводимого антитела может быть значительно снижено, а интервалы между введениями могут быть значительно увеличены.
Повторное связывание с множеством антигенов этого механизма основано на зависящем от pH взаимодействии антигена с антителом. Следовательно, не зависимо от типа антигена, если может быть создано антитело, демонстрирующее зависящее от pH связывание, которое связывается с антигеном при pH 7,4 в плазме, но подвергается диссоциации от антигена при кислотном pH внутри эндосом, такое антитело может повторно связываться с множеством антигенов. В результате, техническое решение настоящего изобретения полезно тем, что оно применимо не только к антителам против рецептора IL-6, IL-6 и рецептора IL-31, но вообще к любому антителу против любого антигена, независимо от типа антигена.
--->
Список последовательностей
<110> CHUGAI SEIYAKU KABUSHIKI KAISHA
<120> Антигенсвязывающая молекула, способная к многократному
связыванию двух или более молекул антигена
<130> C1-A0801Y2P
<140> PCT/JP2009/057309
<141> 2009-04-10
<150> JP 2008-104147
<151> 2008-04-11
<150> JP 2008-247713
<151> 2008-09-26
<150> JP 2009-068744
<151> 2009-03-19
<160> 70
<170> PatentIn version 3.4
<210> 1
<211> 119
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 1
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser Asp Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Gly Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Ala Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 2
<211> 107
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 2
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Tyr Gly Ser Glu Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gly Gln Gly Asn Arg Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu
100 105
<210> 3
<211> 119
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 3
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala His Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Gly Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Ala Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 4
<211> 119
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 4
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His His
20 25 30
His Ala His Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr His Tyr Asn Pro His Leu
50 55 60
Lys Gly Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Ala Tyr Tyr Cys
85 90 95
Ala Arg Val Leu Ala Arg Ile Thr Ala Met Asp His Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 5
<211> 119
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 5
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly His Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro His Leu
50 55 60
Lys Gly Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Ala Tyr Tyr Cys
85 90 95
Ala Arg Phe Leu Ala Arg Ile Thr Ala His Asp His Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 6
<211> 107
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 6
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Gln Ala Ser Gln His Ile Ser Ser His
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Tyr Gly Ser His Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gly Gln Gly Asn Arg Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu
100 105
<210> 7
<211> 107
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 7
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Ser His
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Tyr Gly Ser His Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gly Gln Gly Asn Arg Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu
100 105
<210> 8
<211> 107
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 8
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Ser His
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Tyr Gly Ser His Leu His His Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gly Gln Gly His Arg Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu
100 105
<210> 9
<211> 448
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 9
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Arg Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Ser Ile Thr Ser Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Arg Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Thr Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Met Leu Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Arg Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Ser Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 10
<211> 214
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 10
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 11
<211> 119
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 11
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser Asp Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Gly Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Ala Tyr Tyr Cys
85 90 95
Ala Arg Val Leu Ala Arg Ile Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 12
<211> 107
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 12
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Tyr Gly Ser Glu Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Ser Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu
100 105
<210> 13
<211> 1404
<212> ДНК
<213> Искусственный
<220>
<223> Искусственно синтезированная нуклеотидная последовательность
<400> 13
atgggatgga gctgtatcat cctcttcttg gtagcaacag ctacaggtgt ccactcccag 60
gtccaactgc aggagagcgg tccaggtctt gtgaaaccta gcgagaccct gagcctgacc 120
tgcgccgtgt ctggctactc aattagcgac gatcatgcct ggagctgggt tcgccagcca 180
cctggagaag gtcttgagtg gattggatac attagttata gtggaatcac aaactataat 240
ccatctctca aaggcagagt gacaatatcg agagacacca gcaagaacca gttcagcctg 300
aaactcagca gcgtgacagc cgccgacacc gcggcttatt attgtgcaag atccctagct 360
cggactacgg ctatggacta ctggggtgaa ggcaccctcg tcacagtctc ctcagcctcc 420
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 480
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 540
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 600
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 660
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 720
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 780
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 840
acatgcgtgg tggtggacgt gagccacgaa gaccctgagg tcaagttcaa ctggtacgtg 900
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 960
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 1020
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1080
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga tgagctgacc 1140
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1200
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1260
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1320
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1380
agcctctccc tgtctccggg taaa 1404
<210> 14
<211> 321
<212> ДНК
<213> Искусственный
<220>
<223> Искусственно синтезированная нуклеотидная последовательность
<400> 14
gacatccaga tgacccagag cccaagcagc ctgagcgcca gcgtgggtga cagcgtgacc 60
atcacctgtc aagccagcca ggacatcagc agttacctga attggtacca gcagaagcca 120
ggaaaggctc cagagctgct gatctactac ggctccgaac tgcactctgg tgtgccaagc 180
agattcagcg gtagcggtag cggtaccgac ttcaccttca ccatcagcag cctcgaggca 240
gaggacgccg ctacctacta ctgcgggcag ggtaaccggc ttccatacac gttcggccaa 300
gggaccaagg tggaaatcga a 321
<210> 15
<211> 15
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 15
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 16
<211> 24
<212> ДНК
<213> Искусственный
<220>
<223> Искусственно синтезированная последовательность праймера
<400> 16
ttcccaccag cctgtccgcc tctg 24
<210> 17
<211> 24
<212> ДНК
<213> Искусственный
<220>
<223> Искусственно синтезированная последовательность праймера
<400> 17
cgtgaagcca aaggccccac tccc 24
<210> 18
<211> 32
<212> ДНК
<213> Искусственный
<220>
<223> Искусственно синтезированная последовательность праймера
<400> 18
tagaattcca ccatgctggc cgtcggctgc gc 32
<210> 19
<211> 57
<212> ДНК
<213> Искусственный
<220>
<223> Искусственно синтезированная последовательность праймера
<400> 19
attgcggccg cttatcagtg gtgatgatga tgatgtggta ccgaagaaga atcttgc 57
<210> 20
<211> 1131
<212> ДНК
<213> Искусственный
<220>
<223> Искусственно синтезированная нуклеотидная последовательность
<400> 20
gaattccacc atgctggccg tcggctgcgc gctgctggct gccctgctgg ccgcgccggg 60
ggcggcgctg gccccggggg gctgccctgc acaggaggtg gcgagaggtg tgctgaccag 120
tctgccagga gacagcgtga ctctgacctg cccaggggga gagccggaag acaatgccac 180
tgttcactgg gttctcagga agccagctgt aggctcccac ctcagcagat gggctggcgt 240
gggaaggagg ctgctgctga ggtcggtgca gctccatgac tctggaaact attcatgcta 300
ccgggccggc cgcccggctg gaactgtgca cttgctggtg gatgttcccc ccgaggagcc 360
ccagctctcc tgcttccgga agagccccct cagcaacgtt gtttgtgagt ggggtcctcg 420
gagcacccca tctccgacga ccaaggctgt gctgttggtg aggaagtttc agaacagtcc 480
ggccgaagac ttccaggagc cgtgccagta ttcccaggag tcccagaagt tctcctgcca 540
gttggcagtc ccggagggag acagctcttt ctacatagtg tccatgtgcg tcgccagtag 600
tgtcgggagc aagctcagca aaactcagac ctttcagggt tgtggaatct tgcagcctga 660
tccgcctgcc aacatcacag tcactgccgt ggccagaaac ccccgctggc tcagtgtcac 720
ctggcaagac ccccactcct ggaactcatc tttctacaga ctacggtttg agctcagata 780
tcgagctgaa cggtcaaaga cattcacaac atggatggtc aaggacctcc agcatcactg 840
tgtcatccac gacgcctgga gcggcctgag gcacgtggtg cagcttcgtg cccaggagga 900
gttcgggcaa ggcgagtgga gcgagtggag cccggaggcc atgggcacgc cttggacaga 960
atccaggagt cctccagctg agaacgaggt gtccaccccc acgcaggcac ctactactaa 1020
taaagatgat gataatattc tctccagaga ttctgcaaat gcgacaagcc tcccagtgca 1080
agattcttct tcggtaccac atcatcatca tcaccactga taagcggccg c 1131
<210> 21
<211> 449
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 21
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly His Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro His Leu
50 55 60
Gln Asp Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Ala Tyr Tyr Cys
85 90 95
Ala Arg Phe Leu Ala Arg Ile Thr Ala His Asp His Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 22
<211> 449
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 22
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly His Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro His Leu
50 55 60
Gln Asp Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Leu Ala Arg Ile Thr Ala His Asp His Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 23
<211> 449
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 23
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 24
<211> 449
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 24
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Thr Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 25
<211> 214
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 25
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Gln Ala Ser Arg Asp Ile Ser Ser His
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Tyr Gly Ser His Leu Leu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gly Gln Gly Asn Arg Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 26
<211> 214
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 26
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Gln Ala Ser Arg Asp Ile Ser Ser His
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Tyr Gly Ser His Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gly Gln Gly Asn Arg Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 27
<211> 214
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 27
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Gln Ala Ser Thr Asp Ile Ser Ser His
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Tyr Gly Ser His Leu Leu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gly Gln Gly Asn Arg Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 28
<211> 326
<212> БЕЛОК
<213> Homo sapiens
<400> 28
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 29
<211> 324
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 29
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro
<210> 30
<211> 324
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 30
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Ser Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro
<210> 31
<211> 326
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 31
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Ala His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 32
<211> 324
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 32
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Ser Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Ala His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro
<210> 33
<211> 443
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 33
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu
210 215 220
Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu
290 295 300
Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440
<210> 34
<211> 443
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 34
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Ser Cys Val Glu
210 215 220
Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu
290 295 300
Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440
<210> 35
<211> 443
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 35
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Ser Cys Val Glu
210 215 220
Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu
290 295 300
Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440
<210> 36
<211> 445
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 36
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly His Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro His Leu
50 55 60
Gln Asp Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Leu Ala Arg Ile Thr Ala His Asp His Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu
210 215 220
Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu
290 295 300
Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 37
<211> 443
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 37
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly His Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro His Leu
50 55 60
Gln Asp Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Leu Ala Arg Ile Thr Ala His Asp His Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Ser Cys Val Glu
210 215 220
Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu
290 295 300
Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440
<210> 38
<211> 445
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 38
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Thr Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu
210 215 220
Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu
290 295 300
Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 39
<211> 443
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 39
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Thr Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Ser Cys Val Glu
210 215 220
Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu
290 295 300
Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Ala
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440
<210> 40
<211> 6
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 40
His Asp His Ala Trp Ser
1 5
<210> 41
<211> 16
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 41
His Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro His Leu Gln Asp
1 5 10 15
<210> 42
<211> 16
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 42
Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu Gln Gly
1 5 10 15
<210> 43
<211> 16
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 43
Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Thr Leu Gln Gly
1 5 10 15
<210> 44
<211> 10
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 44
Phe Leu Ala Arg Ile Thr Ala His Asp His
1 5 10
<210> 45
<211> 10
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 45
Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr
1 5 10
<210> 46
<211> 30
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 46
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser
20 25 30
<210> 47
<211> 30
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 47
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser
20 25 30
<210> 48
<211> 14
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 48
Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp Ile Gly
1 5 10
<210> 49
<211> 32
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 49
Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser Leu Lys
1 5 10 15
Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Ala Tyr Tyr Cys Ala Arg
20 25 30
<210> 50
<211> 32
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 50
Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser Leu Lys
1 5 10 15
Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 51
<211> 32
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 51
Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 52
<211> 11
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 52
Trp Gly Glu Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 53
<211> 11
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 53
Gln Ala Ser Arg Asp Ile Ser Ser His Leu Asn
1 5 10
<210> 54
<211> 11
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 54
Gln Ala Ser Thr Asp Ile Ser Ser His Leu Asn
1 5 10
<210> 55
<211> 7
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 55
Tyr Gly Ser His Leu Leu Ser
1 5
<210> 56
<211> 7
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 56
Tyr Gly Ser His Leu Glu Ser
1 5
<210> 57
<211> 9
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 57
Gly Gln Gly Asn Arg Leu Pro Tyr Thr
1 5
<210> 58
<211> 23
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 58
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys
20
<210> 59
<211> 15
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 59
Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile Tyr
1 5 10 15
<210> 60
<211> 32
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 60
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Phe Thr Ile Ser Ser Leu Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys
20 25 30
<210> 61
<211> 10
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 61
Phe Gly Gln Gly Thr Lys Val Glu Ile Glu
1 5 10
<210> 62
<211> 449
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 62
Glu Val Gln Leu Val Glu Ser Gly Gly Lys Leu Leu Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Ala Met Ser Trp Phe Arg Gln Ser Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Glu Ile Ser Ser Gly Gly Ser Tyr Thr Tyr Tyr Pro Asp Thr Val
50 55 60
Thr Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Glu Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Leu Trp Gly Tyr Tyr Ala Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 63
<211> 213
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 63
Gln Ile Val Leu Ile Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Arg Leu Leu Ile Tyr
35 40 45
Asp Thr Ser Asn Leu Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 64
<211> 449
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 64
Glu Val Gln Leu Val Glu Ser Gly Gly Lys Leu Leu Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Ala Met Ser Trp Phe Arg Gln Ser Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Glu Ile Ser Ser Gly Gly Ser Tyr Thr Tyr His Pro His Thr Val
50 55 60
Thr Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Glu Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Leu Trp Gly His Tyr Ala Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 65
<211> 213
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 65
Gln Ile Val Leu Ile Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Arg Leu Leu Ile Tyr
35 40 45
Asp Thr Ser Asn His Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln His Trp Ser Gly His Pro Tyr Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 66
<211> 213
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 66
Gln Ile Val Leu Ile Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Arg Leu Leu Ile Tyr
35 40 45
Asp Thr Ser His His Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln His Trp Ser Gly His Pro Tyr Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 67
<211> 456
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 67
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Asn Ser Gly Gly Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Asp Val Val Pro Ala Ala Met Ser Phe Tyr Tyr Gly Met
100 105 110
Asp Val Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly Lys Gly
450 455
<210> 68
<211> 214
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 68
Leu Pro Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Thr Val
20 25 30
Tyr Trp Tyr Gln Gln Glu Pro Gly Gln Ala Pro Val Leu Val Val Tyr
35 40 45
Asp Asp Thr Asp Arg Pro Ala Gly Ile Pro Glu Arg Phe Ser Gly Ala
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Thr Asp His
85 90 95
Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Asn Pro Thr Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Gly Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 69
<211> 456
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 69
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
His Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Asn Ser Gly Gly Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Asp Val Val Pro Ala Ala Met Ser Phe Tyr Tyr Gly Met
100 105 110
Asp Val Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly Lys Gly
450 455
<210> 70
<211> 449
<212> БЕЛОК
<213> Искусственный
<220>
<223> Искусственно синтезированная пептидная последовательность
<400> 70
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser Asp Asp
20 25 30
Gln Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Gly Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Ala Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, СПОСОБНАЯ МНОГОКРАТНО СВЯЗЫВАТЬСЯ С МНОЖЕСТВОМ АНТИГЕННЫХ МОЛЕКУЛ | 2018 |
|
RU2757786C2 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, СПОСОБНАЯ К МНОГОКРАТНОМУ СВЯЗЫВАНИЮ ДВУХ ИЛИ БОЛЕЕ МОЛЕКУЛ АНТИГЕНА | 2021 |
|
RU2841525C2 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, ИНДУЦИРУЮЩАЯ ИММУННЫЙ ОТВЕТ НА АНТИГЕН-МИШЕНЬ | 2012 |
|
RU2722829C2 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА ДЛЯ УСКОРЕНИЯ ИСЧЕЗНОВЕНИЯ АНТИГЕНА ЧЕРЕЗ FcγRIIB | 2013 |
|
RU2736349C2 |
| СПЕЦИФИЧНАЯ К ТКАНИ-МИШЕНИ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА | 2013 |
|
RU2743463C2 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА ДЛЯ УСКОРЕНИЯ ИСЧЕЗНОВЕНИЯ АНТИГЕНА ЧЕРЕЗ FcγRIIB | 2013 |
|
RU2812226C1 |
| БИБЛИОТЕКА ЗАВИСИМЫХ ОТ КОНЦЕНТРАЦИИ ИОНОВ СВЯЗЫВАЮЩИХ МОЛЕКУЛ | 2012 |
|
RU2732151C2 |
| ИММУНОАКТИВИРУЮЩАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА | 2015 |
|
RU2722788C2 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА ДЛЯ УСКОРЕНИЯ ЭЛИМИНАЦИИ АНТИГЕНОВ | 2012 |
|
RU2772771C1 |
| ТРИФУНКЦИОНАЛЬНАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА | 2015 |
|
RU2753882C2 |
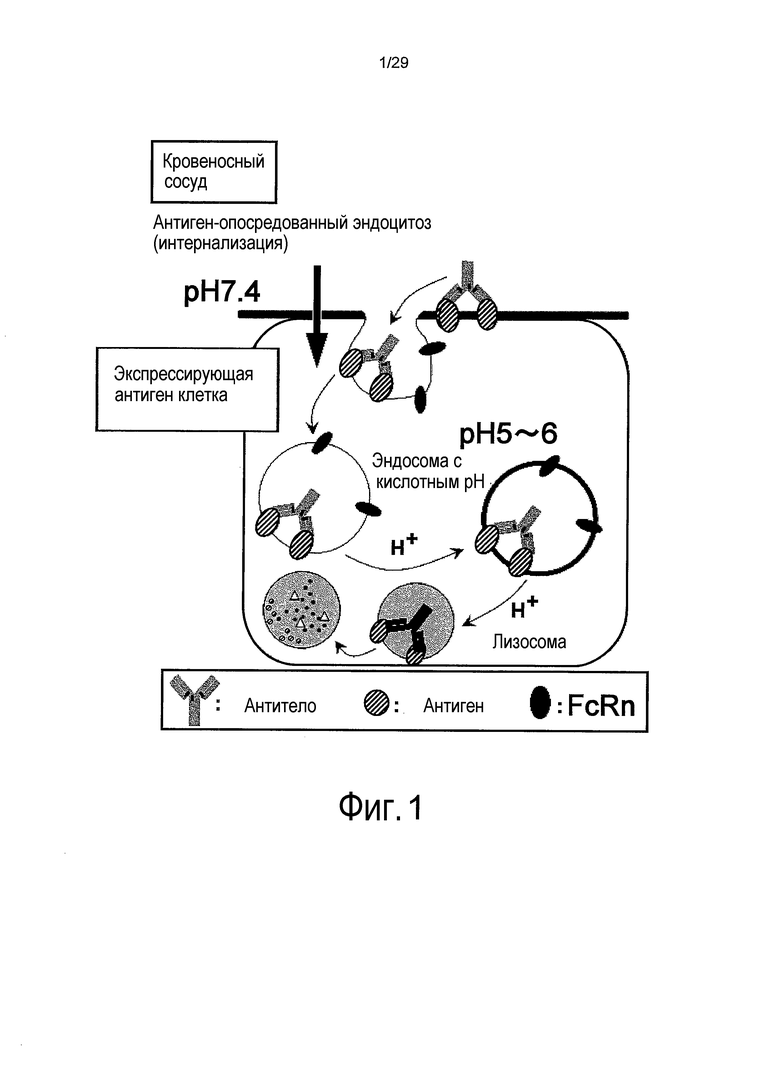
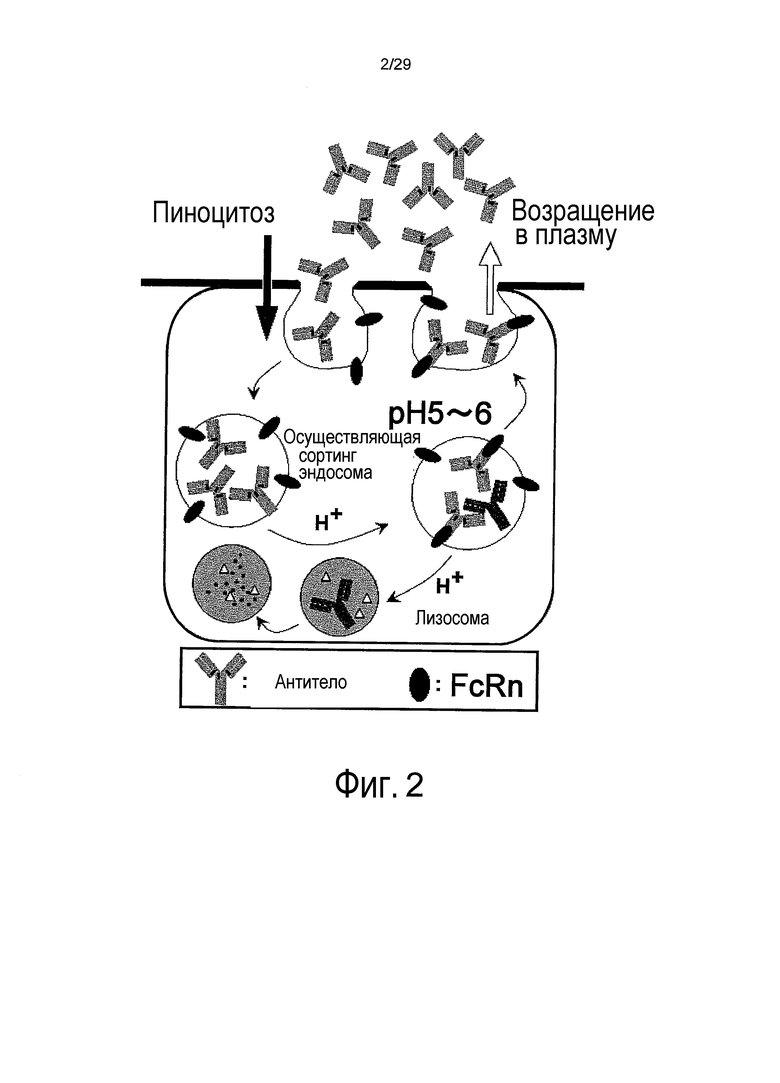
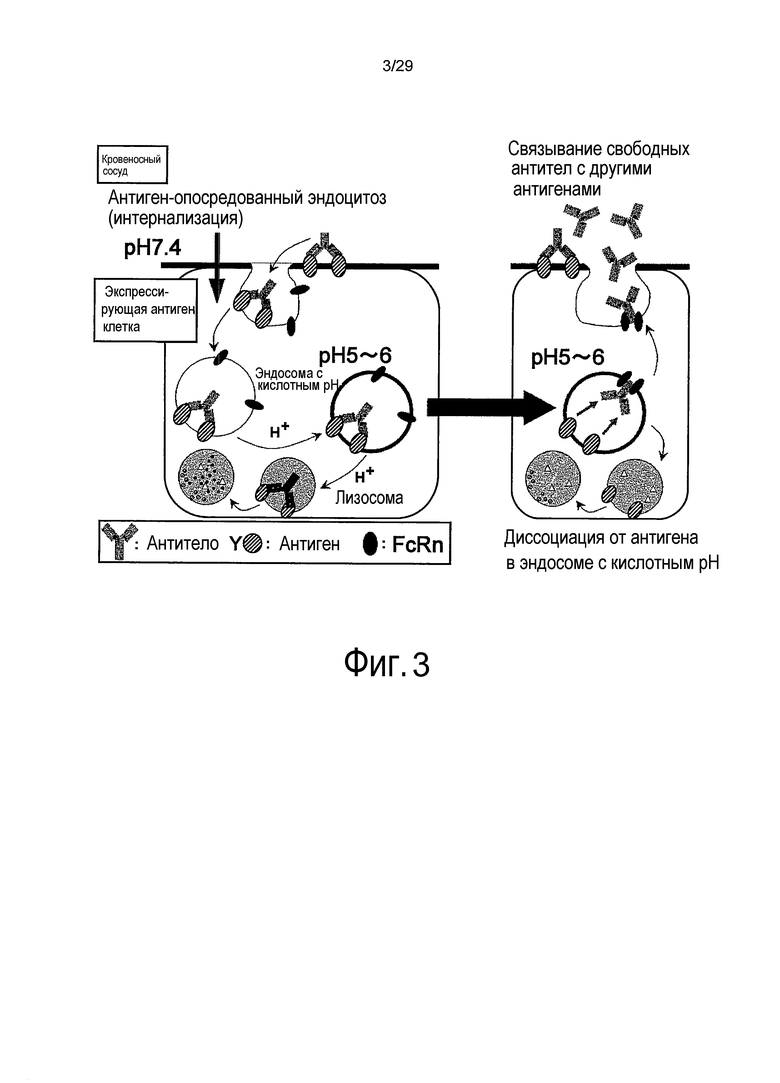
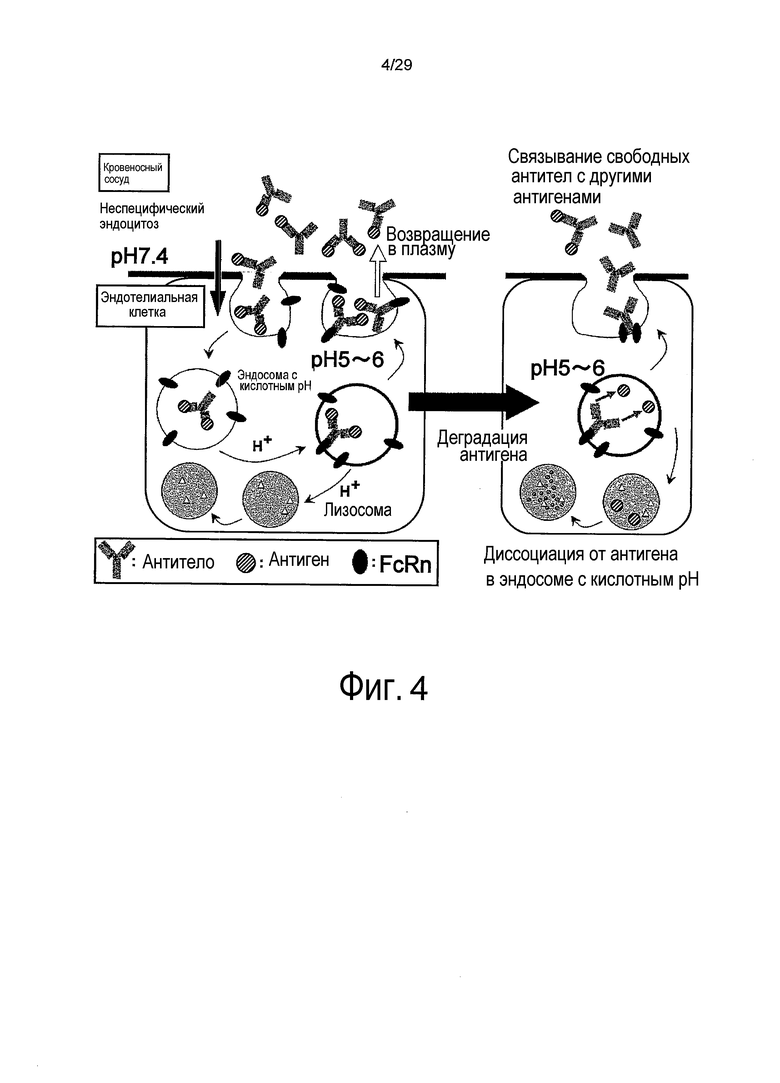
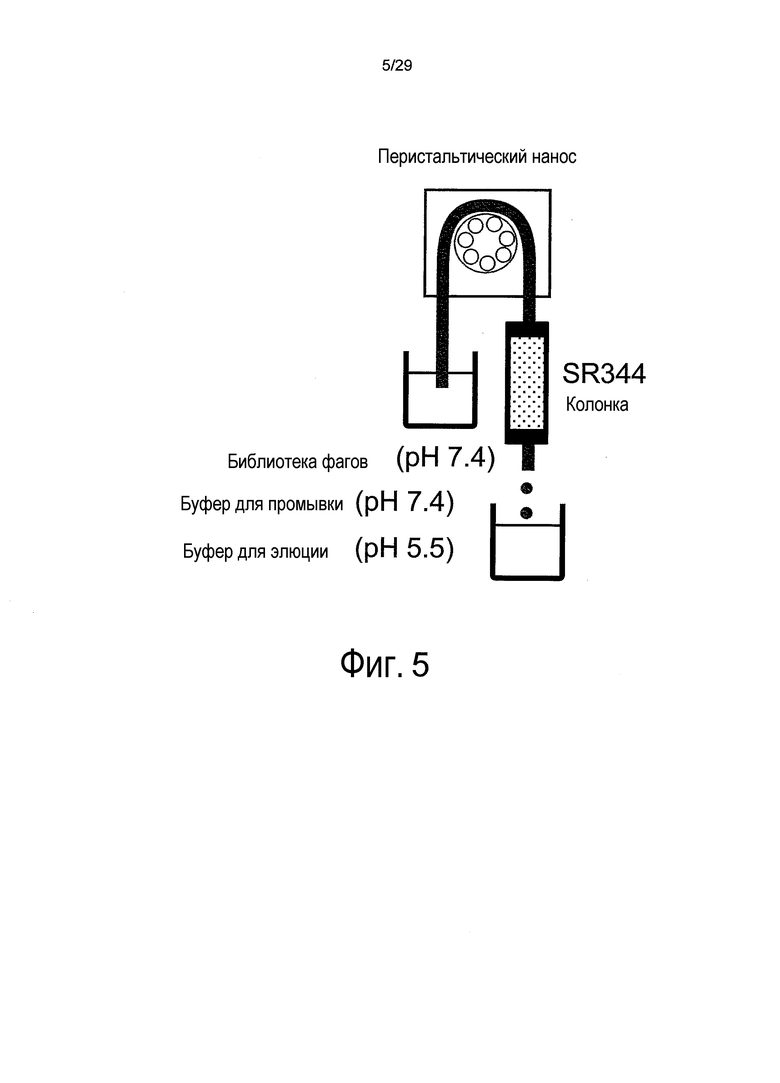
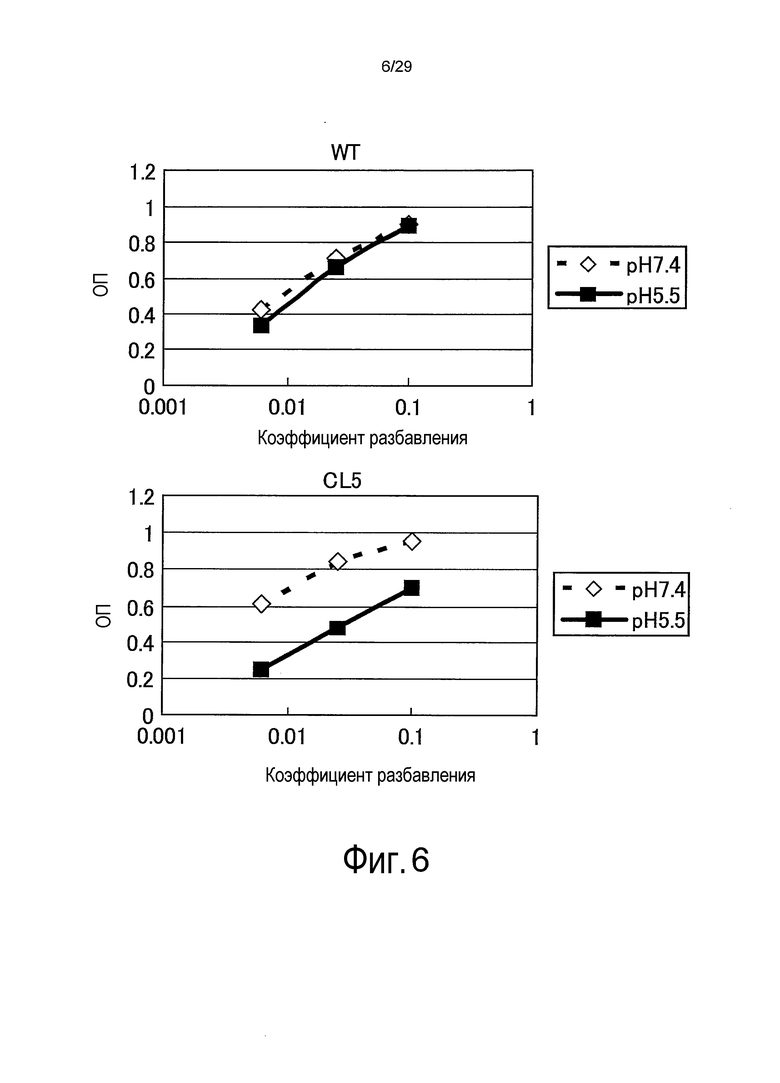
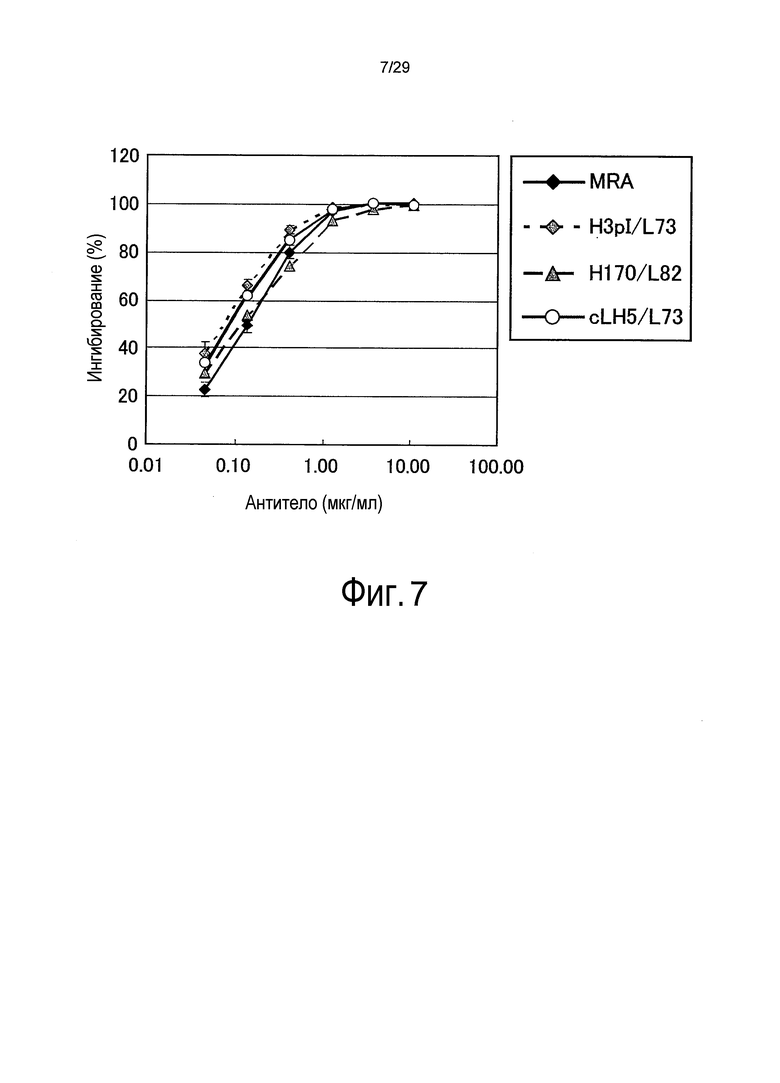
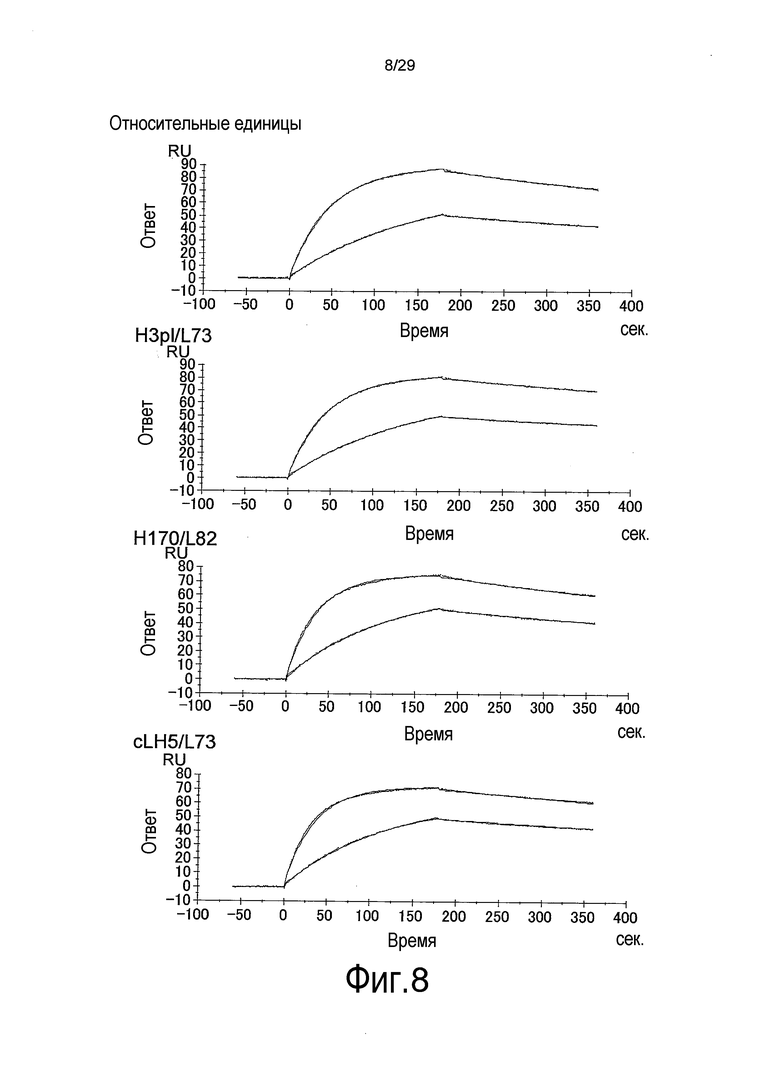
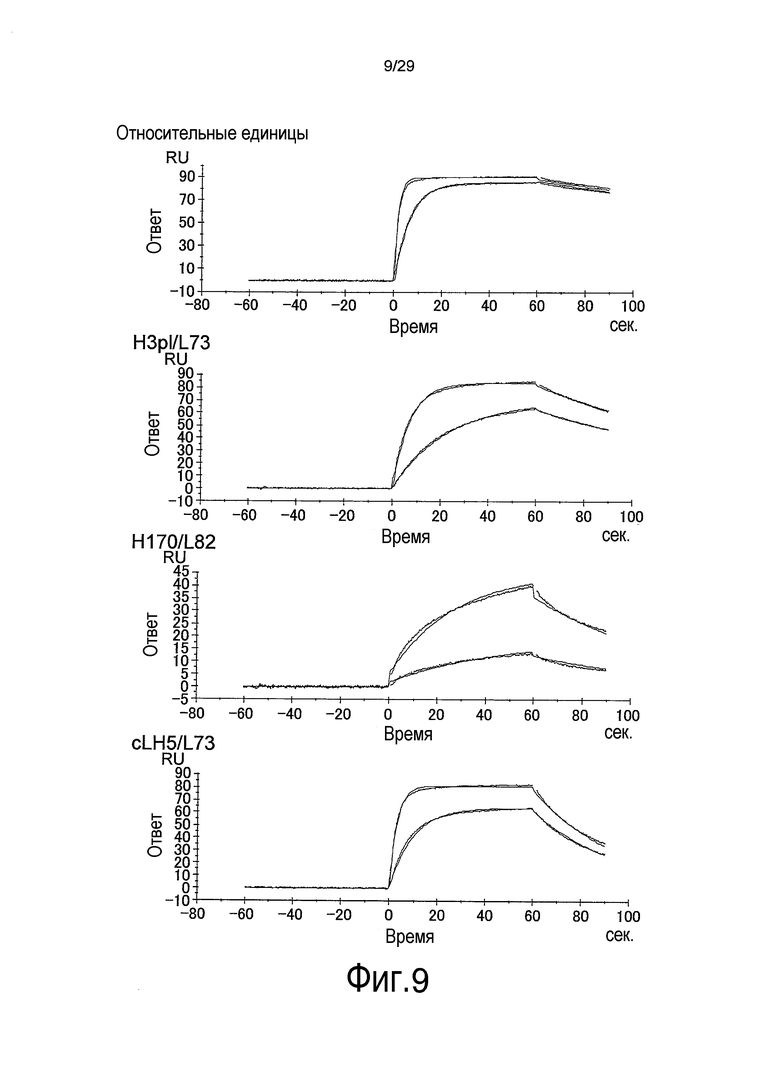
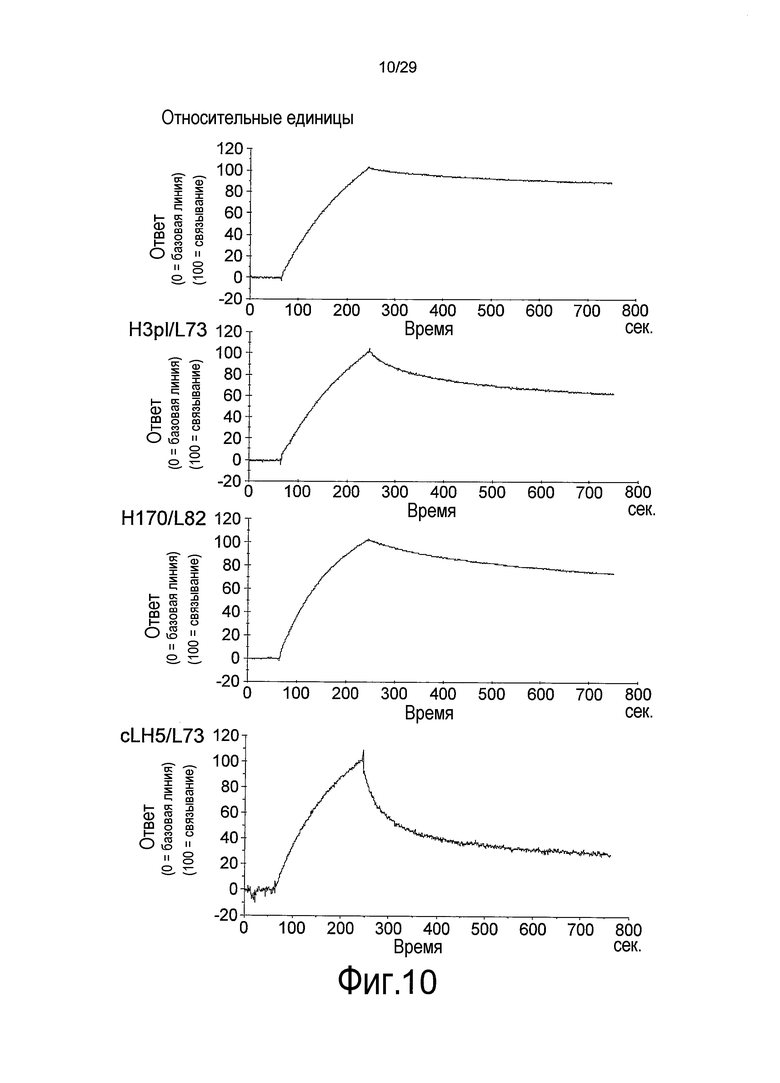
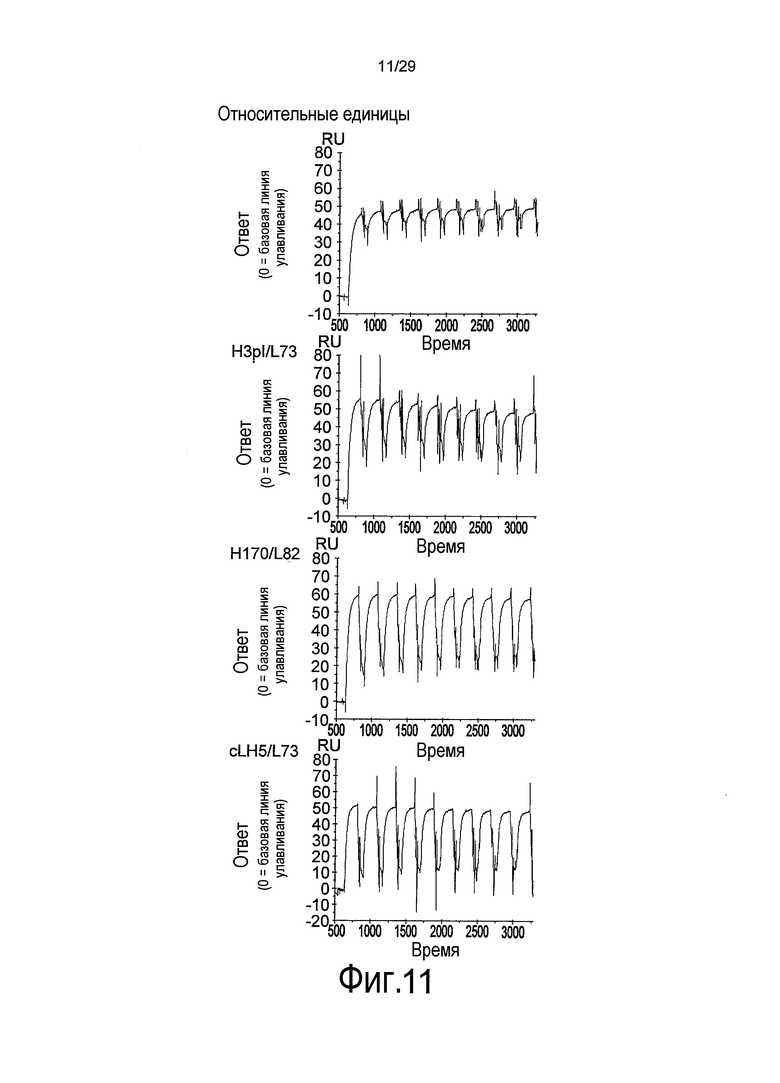
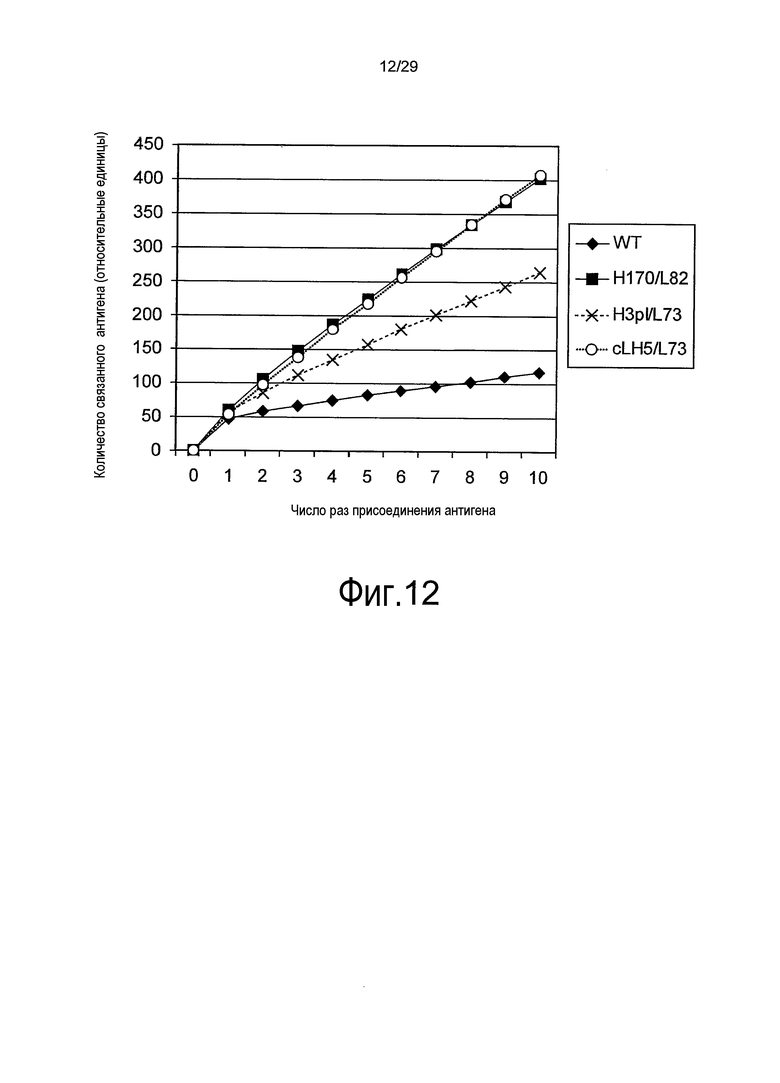
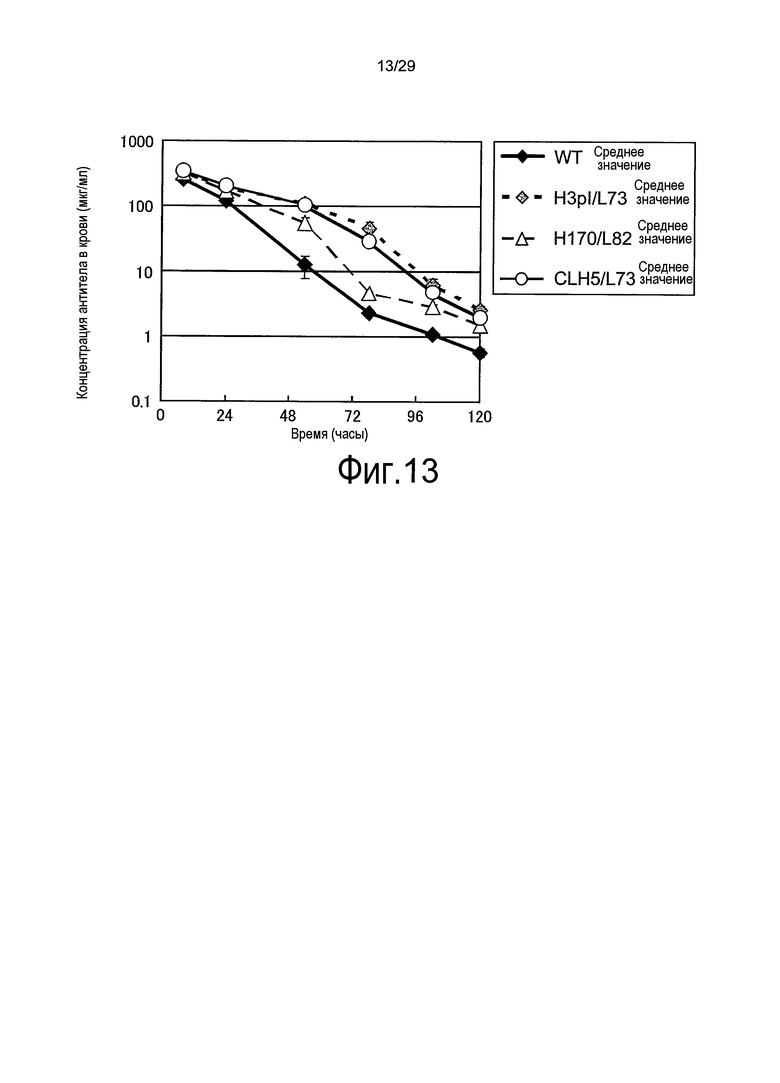
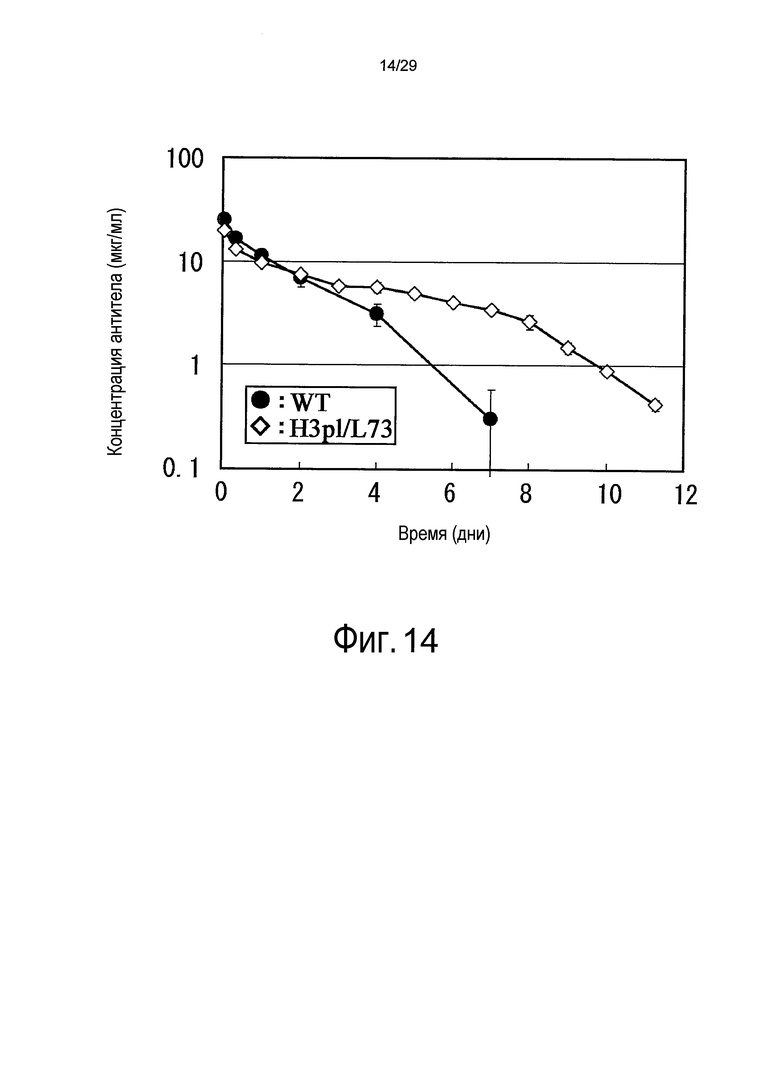
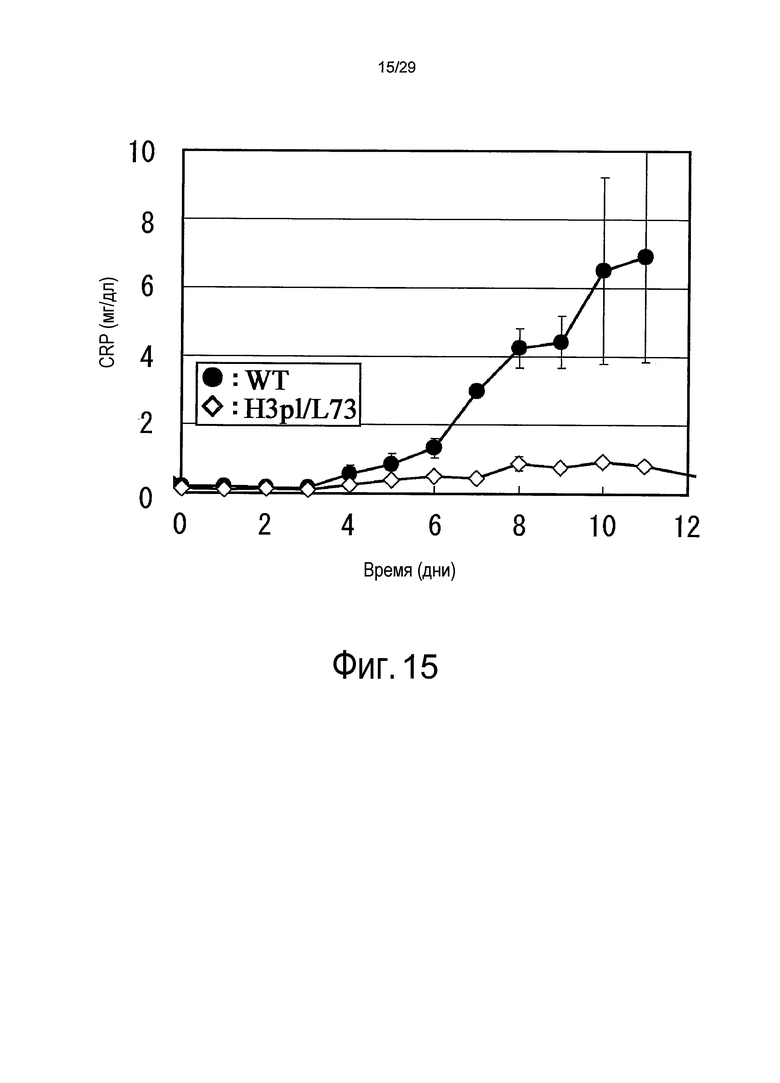
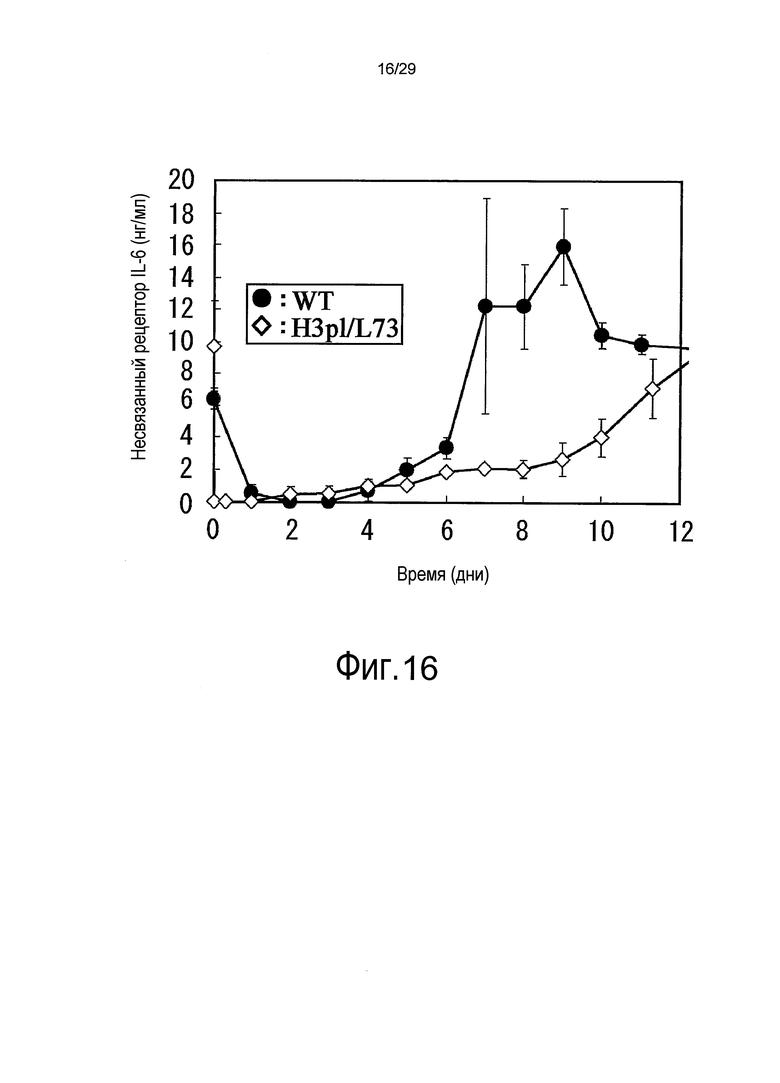
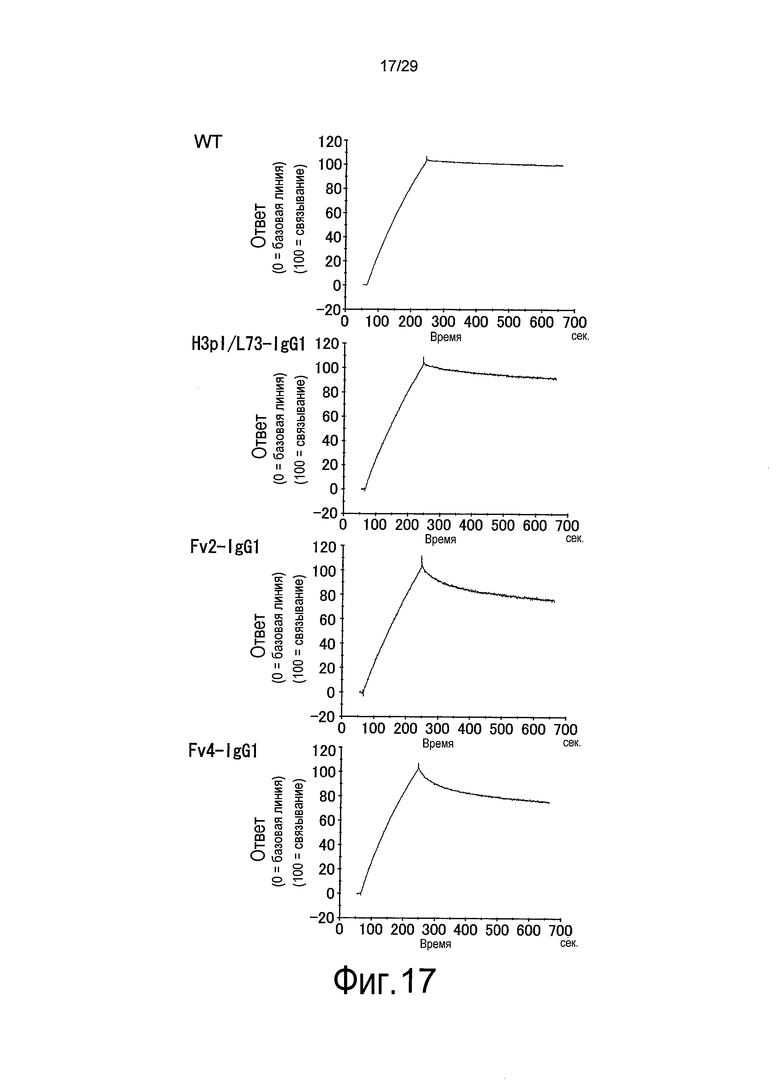
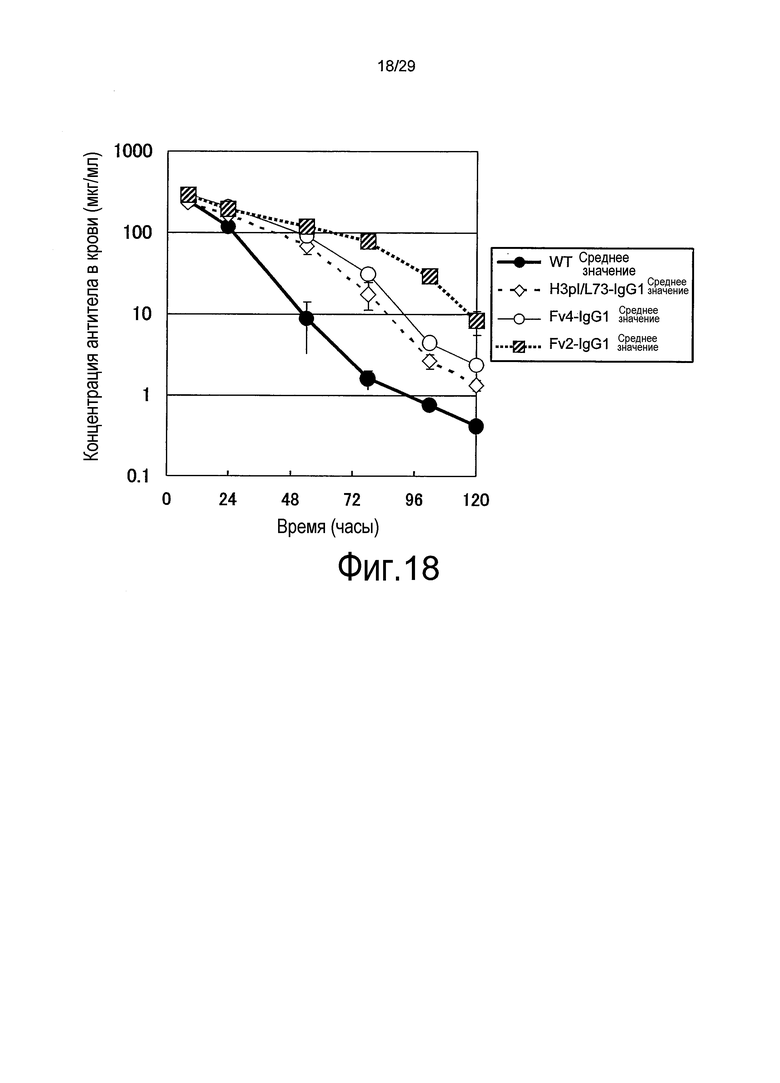
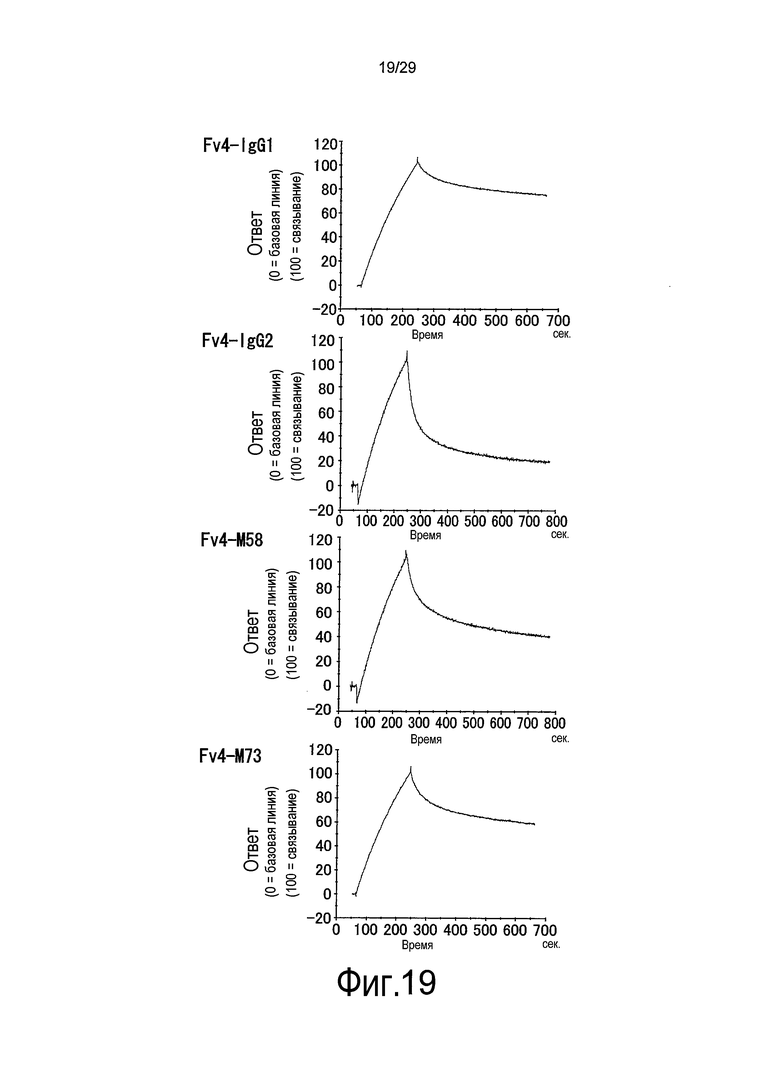
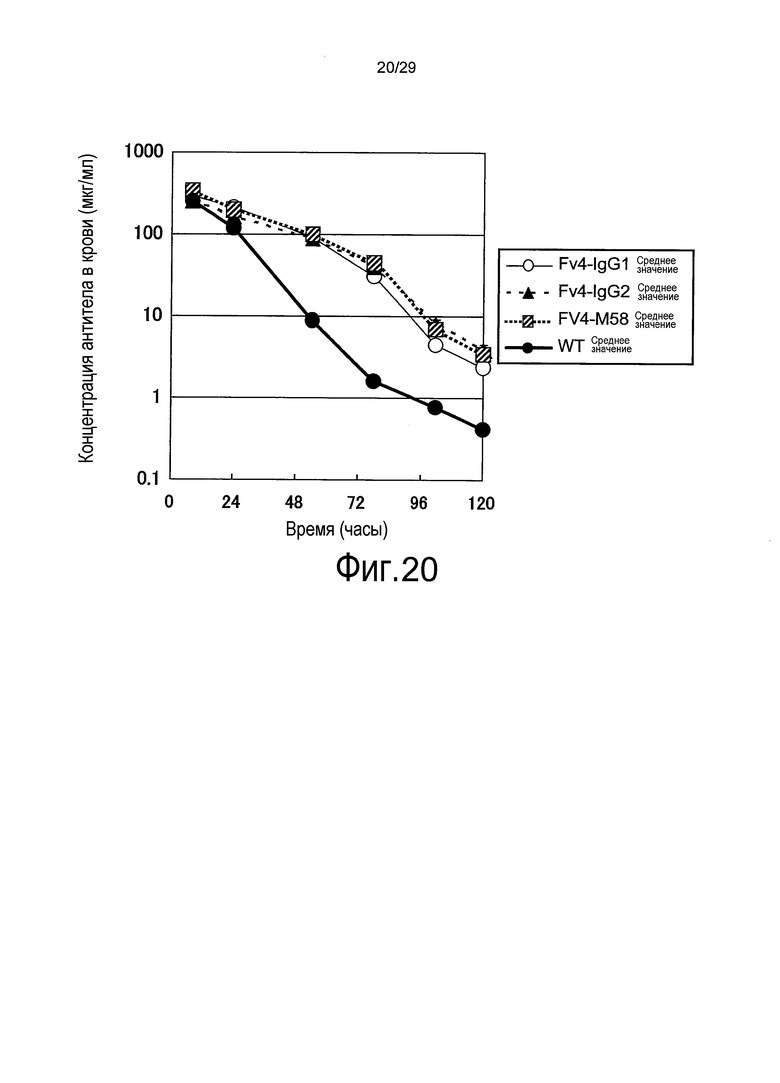
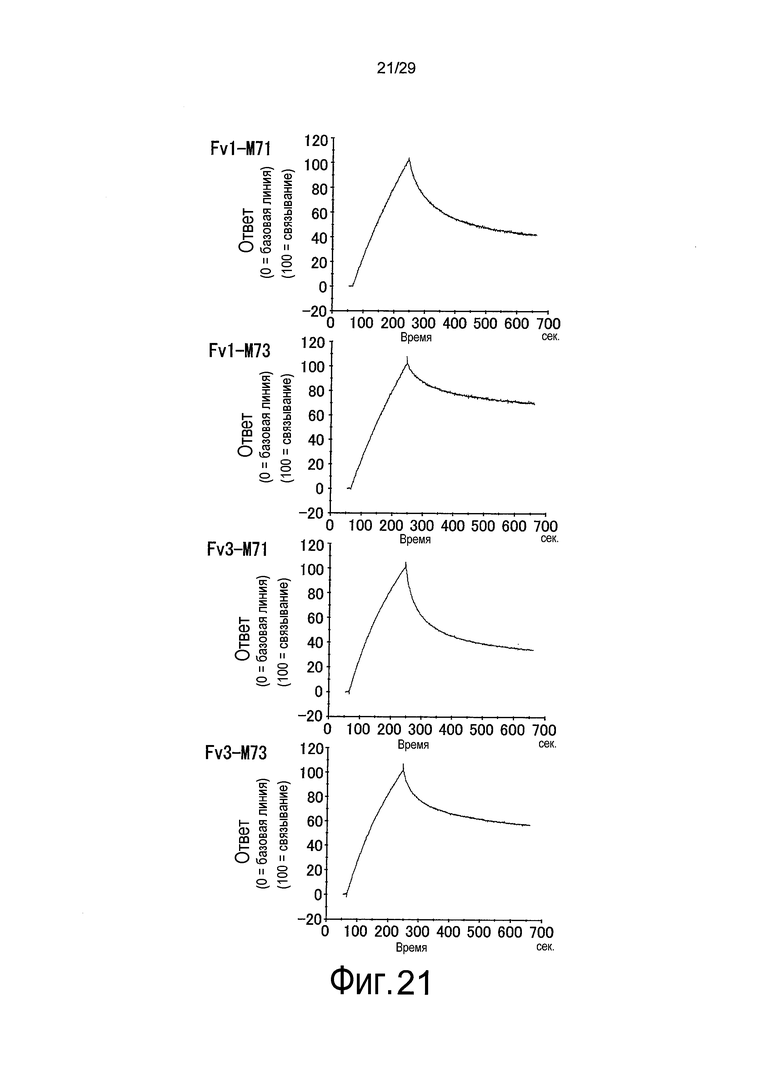
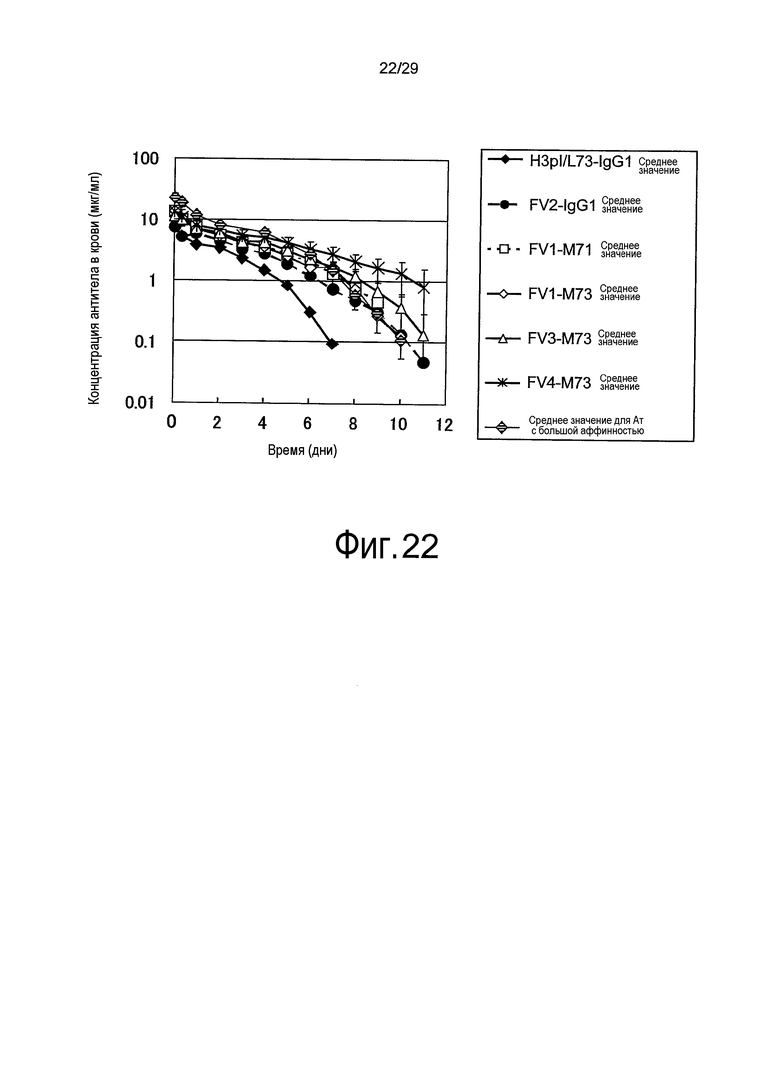
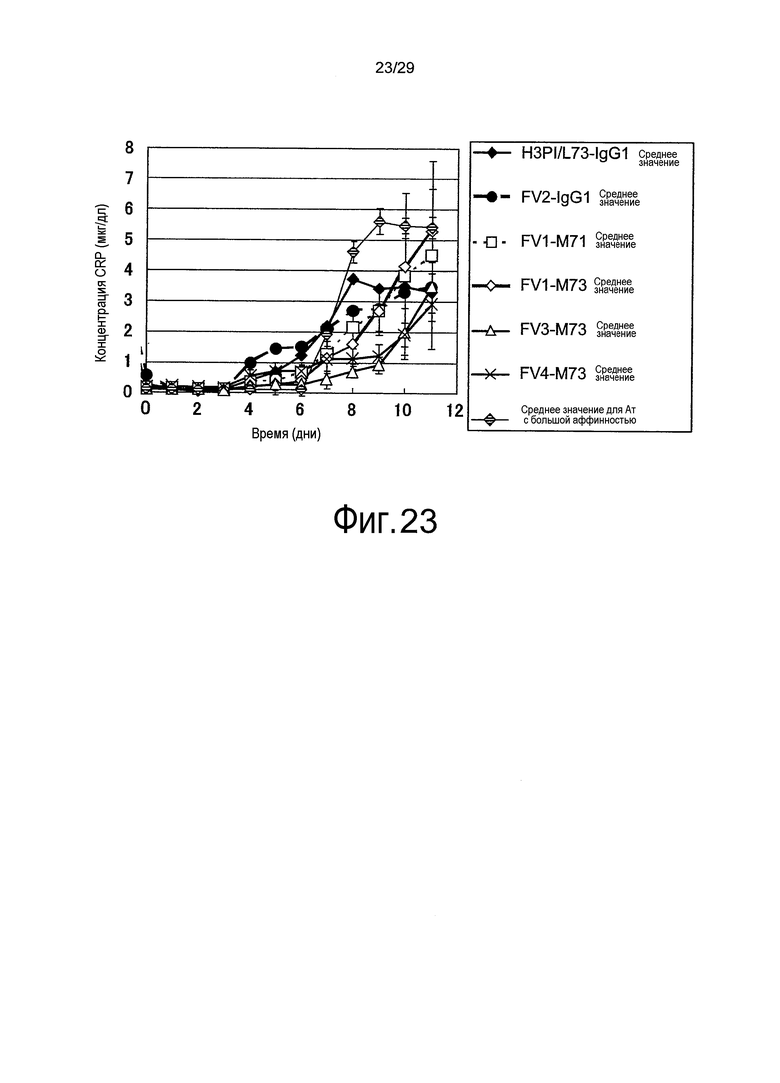
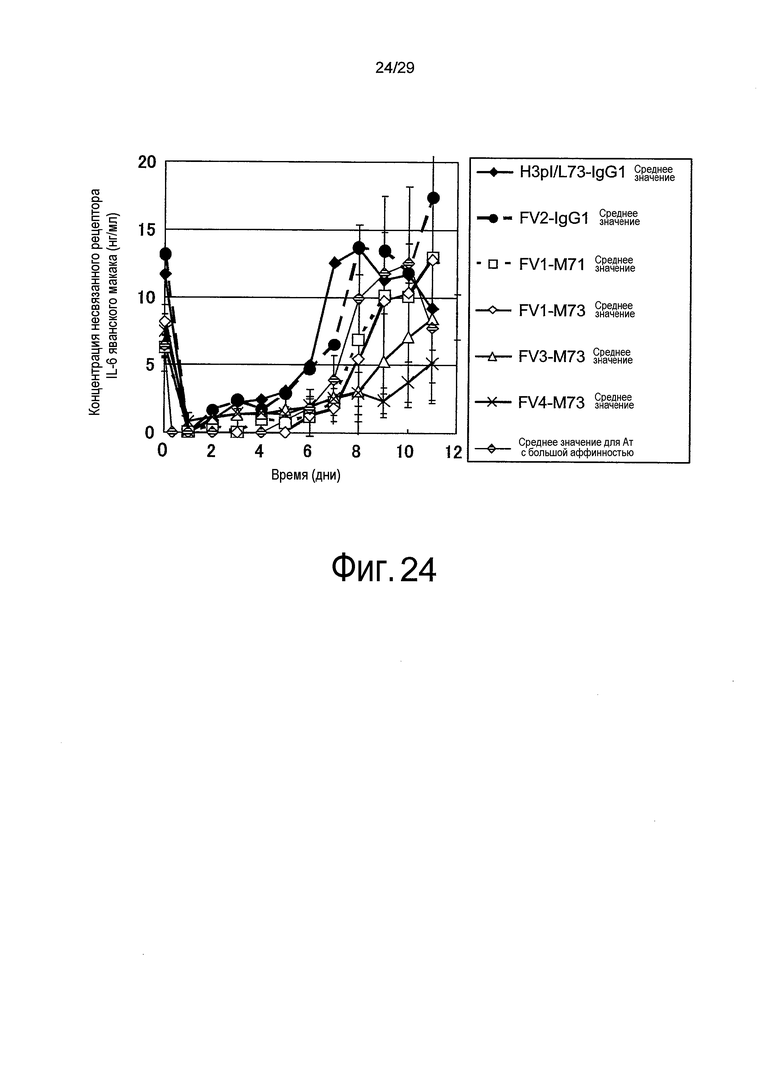
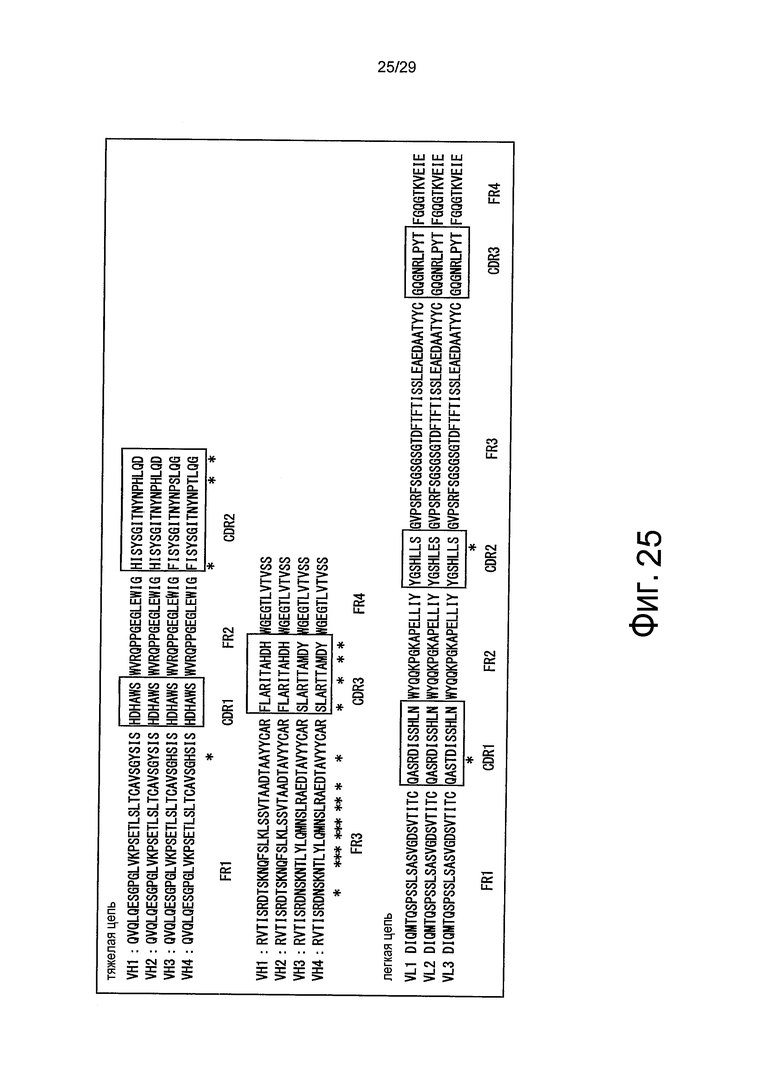
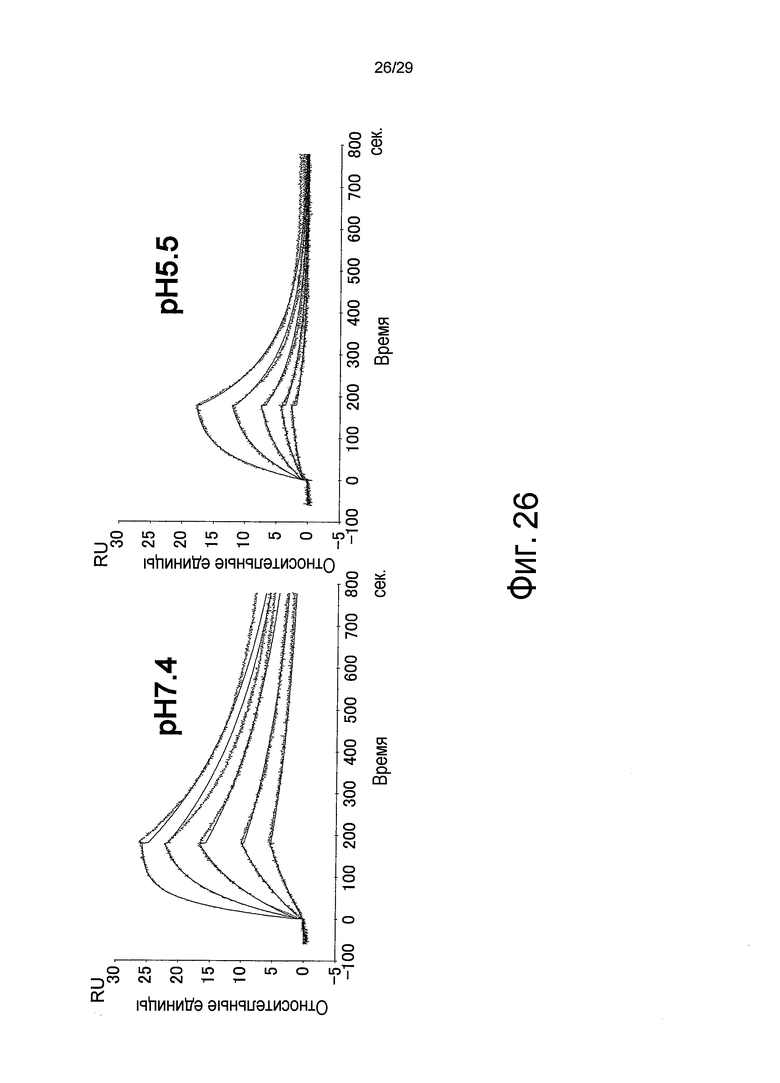
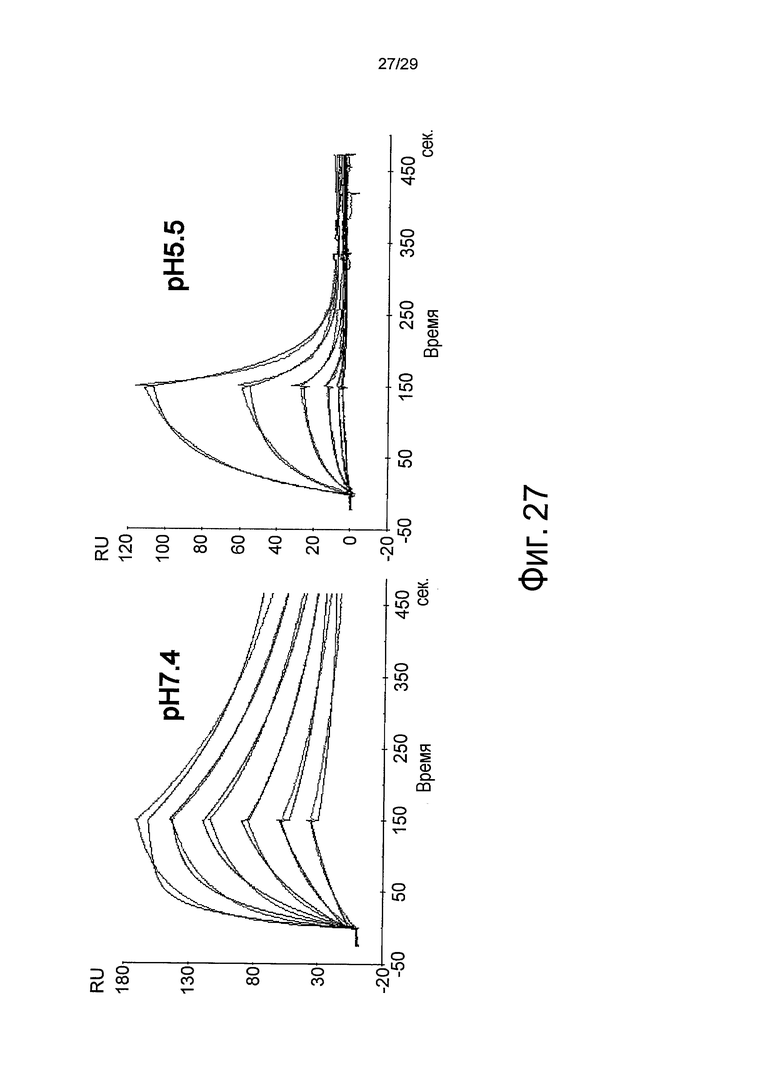
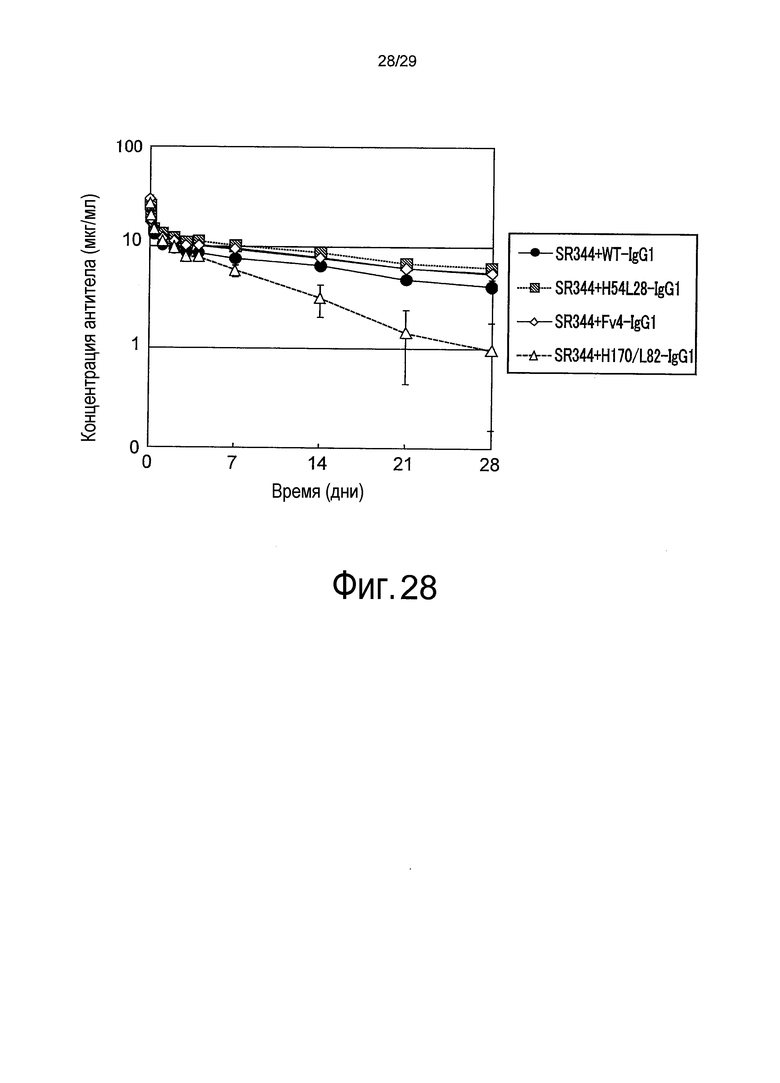
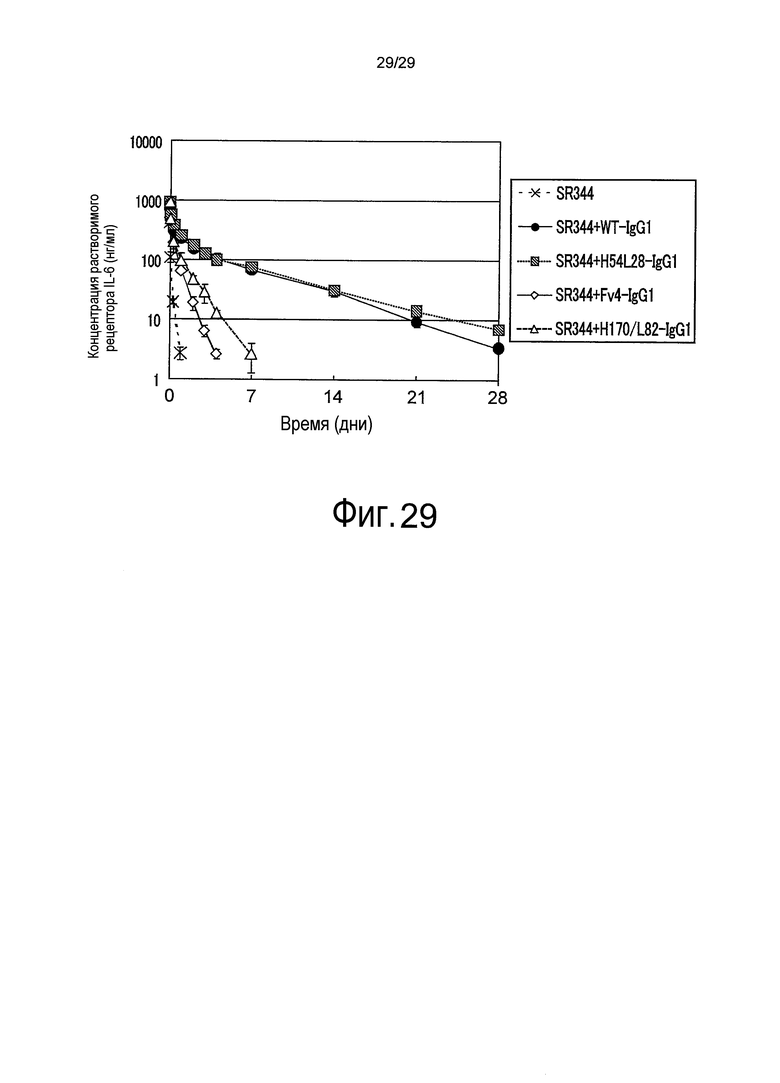
Настоящее изобретение относится к области иммунологии. Предложен способ улучшения фармакокинетики антитела, включающий замену по меньшей мере одной аминокислоты вариабельной области указанного антитела на гистидин. Данное изобретение может найти дальнейшее применение в получении антител с улучшенными фармакокинетическими свойствами. 8 з.п. ф-лы, 29 ил., 17 табл., 18 пр.
1. Способ улучшения фармакокинетики антитела, включающий замену по меньшей мере одной аминокислоты вариабельной области указанного антитела на гистидин, где эта замена гистидином увеличивает значение KD(pH 5,8)/KD(pH 7,4) по сравнению со значением KD(pH 5,8)/KD(pH 7,4) до замены гистидином, где указанное значение KD(pH 5,8)/KD(pH 7,4) определяется как соотношение KD для антигена при pH 5,8 и KD для антигена при pH 7,4, где гистидином замещается по меньшей мере одна аминокислота в положении, выбранном из группы, состоящей из H27, H31, H32, H33, H35, H50, H58, H59, H61, H62, H63, H64, H65, H99, H100b и H102 в вариабельной области тяжелой цепи согласно нумерации Kabat и L24, L27, L28, L32, L53, L54, L56, L90, L92 и L94 в вариабельной области легкой цепи согласно нумерации Kabat, и где значение KD(pH 5,8)/KD(pH 7,4), определяемое как соотношение между KD для антигена при рН 5,8 и KD для антигена при pH 7,4, составляет 2 или больше.
2. Способ по п. 1, где значение KD(pH 5,8)/KD(pH 7,4) составляет 10 или больше.
3. Способ по п. 1 или 2, в котором значение KD(pH 5,8)/KD(pH 7,4) составляет 40 или больше.
4. Способ по любому из пп. 1-3, где указанное антитело обладает антагонистической активностью.
5. Способ по любому из пп. 1-4, где указанное антитело связывается с мембранным антигеном или растворимым антигеном.
6. Способ по любому из пп. 1-5, где гистидин замещает по меньшей мере одну аминокислоту в положении, выбранном из группы, состоящей из H27, H31, H32, H35 и H61 в вариабельной области тяжелой цепи согласно нумерации Kabat.
7. Способ по любому из пп. 1-6, где антитело, в котором по меньшей мере одна аминокислота вариабельной области заменена гистидином, сохраняет 10% или более антигенсвязывающей активности при pH 7.4 по сравнению с антителом до замены гистидином.
8. Способ по любому из пп. 1-7, где число аминокислотных замен на гистидин равно одному или двум.
9. Способ по любому из пп. 1-8, где антитело включает область, которая связывается с FcRn человека.
| ITO W | |||
| et al., "The His-probe method: Effects of histidine residues introduced into the complementarity-determining regions of antibodies on antigen-antibody interactions at different pH values", FEBS letters, 1992, 309(1): 85-88 | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| TAWFIK D | |||
| S | |||
| et al., "pH on-off switching of antibody-hapten binding by site-specific | |||

