Область техники
Настоящее изобретение относится к антигенсвязывающим молекулам, содержащим антигенсвязывающий домен, антигенсвязывающая активность которого варьирует в зависимости от концентрации специфического для ткани-мишени соединения; к способам получения и способам скрининга антигенсвязывающих молекул; и к фармацевтическим композициям, содержащим антигенсвязывающие молекулы.
Уровень техники
Антитела привлекают внимание в качестве фармацевтических средств, поскольку фармацевтические препараты являются высокостабильными в плазме и имеют мало побочных эффектов. В частности, на рынке доступен ряд антительных фармацевтических препаратов IgG-типа, и многие фармацевтические препараты в настоящее время находятся на стадии разработки (непатентные документы 1 и 2).
В качестве лекарственных средств против злокачественной опухоли с использованием антительных фармацевтических средств на настоящий момент одобрены ритуксан против антигена CD20, цетуксимаб против антигена EGFR, герцептин против антигена HER2 и т.п. (непатентный документ 3). Эти молекулы антител связываются с антигенами, экспрессируемыми на злокачественных клетках, и проявляют цитотоксическую активность против злокачественных клеток посредством ADCC и т.п. Известно, что такая цитотоксическая активность посредством ADCC и т.д. зависит от количества антигенов, экспрессируемых на клетках, на которые нацелены терапевтические антитела (непатентный документ 4); таким образом, высокий уровень экспрессии является предпочтительным с точки зрения эффектов терапевтических антител. Однако даже если уровень экспрессии антигена является высоким, когда антигены экспрессируются в нормальных тканях, цитотоксическая активность, опосредуемая ADCC и т.д., будет проявляться против нормальных клеток и, таким образом, побочные эффекты станут существенной проблемой. Таким образом, антигены, на которые нацелены терапевтические антитела, используемые в качестве лекарственных средств от злокачественной опухоли, предпочтительно представляют собой антигены, специфически экспрессирующиеся в злокачественных клетках. Например, молекулы антител против антигена EpCAM, который известен в качестве антигена злокачественной опухоли, считаются перспективными в качестве лекарственных средств от злокачественной опухоли. Однако известно, что антиген EpCAM также экспрессируется в поджелудочной железе, и было описано, что на практике введение антител против EpCAM в клинических испытаниях вызывает панкреатит в качестве побочного эффекта вследствие цитотоксической активности в отношении поджелудочной железы (непатентный документ 5).
После успеха фармацевтических препаратов на основе антител, которые проявляют цитотоксическую активность посредством активности ADCC, было описано второе поколение усовершенствованных молекул антител, которые проявляют выраженную цитотоксическую активность путем усиления активности ADCC посредством удаления фукозы из цепей сахаров N-типа в нативной Fc-области IgG1 человека (непатентный документ 6), усиления активности ADCC путем усиления связывания с FcγRIIIa посредством замены аминокислот в нативной Fc-области IgG1 человека (непатентный документ 7) и т.п. В качестве фармацевтических препаратов на основе антител, которые проявляют цитотоксическую активность против злокачественных клеток через механизм, отличный от упомянутой выше активности ADCC, опосредуемой NK-клетками, также описаны усовершенствованные молекулы антител, которые проявляют более выраженную цитотоксическую активность, такие как конъюгат антитело-лекарственное средство (ADC), в котором антитело конъюгировано с лекарственным средством, обладающим мощной цитотоксической активностью (непатентный документ 8), и низкомолекулярное антитело, которое проявляет токсическую активность против злокачественных клеток путем привлечения T-клеток к злокачественным клеткам.
Такие молекулы антител, обладающие более выраженной цитотоксической активностью, могут проявлять цитотоксическую активность против злокачественных клеток, которые не обладают значительной экспрессией антигена, но, с другой стороны, они проявляют сходную цитотоксическую активность против нормальных тканей с низкой экспрессией антигена. В действительности, по сравнению с цетуксимабом, который представляет собой природный IgG1 человека против антигена EGFR, EGFR-BiTE, который представляет собой биспецифическое антитело против CD3 и EGFR, может проявлять мощную цитотоксическую активность против злокачественных клеток путем привлечения T-клеток к злокачественным клеткам и может проявлять противоопухолевые эффекты. С другой стороны, поскольку EGFR также экспрессируется в нормальных тканях, когда EGFR-BiTE вводили яванским макакам, возникали тяжелые побочные эффекты (непатентный документ 10). Более того, было показано, что биватузумаб мертанзин, ADC, полученный путем связывания мертанзина с антителом против CD44v6, которое на высоком уровне экспрессируется в злокачественных клетках, вызывает тяжелую кожную токсичность и токсичность в печени в клинической практике, поскольку CD44v6 также экспрессируется в нормальных тканях (непатентный документ 11).
Когда антитела, которые могут проявлять мощную цитотоксическую активность против злокачественных клеток, обладающих низкой экспрессией антигена, используют как есть, антиген-мишень должен экспрессироваться в высокой степени специфическим для злокачественной опухоли образом. Однако, поскольку HER2 и EGFR, которые являются антигенами-мишенями герцептина и цетуксимаба, соответственно, также экспрессируются в нормальных тканях, полагают, что количество антигенов злокачественной опухоли, экспрессируемых в высокой степени специфическим для злокачественной опухоли образом ограничено. Таким образом, хотя возможно усилить цитотоксическую активность против злокачественной опухоли, побочные эффекты вследствие цитотоксических действий против нормальных тканей, могут стать проблемой.
Более того, недавно было показано, что ипилимумаб, который усиливает иммунитет опухоли путем ингибирования CTLA4, который участвует в иммуносупрессии в злокачественной опухоли, продлевает общую выживаемость метастатической меланомы (непатентный документ 12). Однако, поскольку ипулимумаб ингибирует CTLA4 системно, хотя иммунитет опухоли повышается, становится проблемой появление подобных аутоиммунному заболеванию тяжелых побочных эффектов вследствие системной активации иммунной системы (непатентный документ 13).
С другой стороны, в качестве антительных фармацевтических препаратов против заболеваний, помимо злокачественной опухоли, известны антительные фармацевтические препараты, которые проявляют терапевтические эффекты путем ингибирования воспалительных цитокинов при воспалительных/аутоиммунных заболеваниях (непатентный документ 14). Например, ремикейд и хумира, которые нацелены на TNF, и актемра, которая нацелена на IL-6R, проявляют высокие терапевтические эффекты против ревматоидного артрита, но, с другой стороны, системная нейтрализация этих цитокинов приводила к тому, что в качестве побочных эффектов наблюдали инфекцию (непатентный документ 15).
В качестве способов, которые можно использовать для антительных фармацевтических препаратов второго поколения, были разработаны различные способы. В то время как были описаны способы улучшения эффекторных функций, антигенсвязывающей способности, фармакокинетики и стабильности, или способы снижения иммуногенных рисков (непатентный документ 16), почти нет сообщений о способах, которые обеспечивают специфическое для ткани-мишени действие антительных фармацевтических препаратов для преодоления таких побочных эффектов. Например, что касается очагов повреждения, таких как злокачественные ткани и воспаленные ткани, были описаны pH-зависимые антитела, которые используют кислые условия в этих тканях-мишенях (патентные документы 1 и 2). Однако снижение pH (т.е. увеличение концентрации ионов водорода) в злокачественных тканях и воспаленных тканях по сравнению с нормальными тканями является небольшим, и, поскольку трудно получить антитела, которые действуют путем обнаружения небольшого увеличения концентрации ионов водорода, которые имеют чрезвычайно малую молекулярную массу, и также поскольку условия кислых значений pH могут быть найдены в нормальных тканях, таких как область резорбции кости остеокластами, и в тканях, отличных от представляющего интерес очага повреждения, использование условий pH в качестве специфического для очага повреждения фактора окружающей среды было расценено как имеющее множество проблем. С другой стороны, были описаны способы получения антител, которые проявляют антигенсвязывающую активность только после их расщепления протеазой, экспрессируемой в очагах повреждения, таких как злокачественные ткани и воспаленные ткани (патентный документ 3). Однако поскольку расщепление антител протеазами является необратимым, когда антитела, которые были расщеплены в очаге повреждения, входят в кровоток и возвращаются в нормальные ткани, они также могут связываться с антигенами в нормальных тканях и это считается проблемой. Более того, также полагают, что специфичность к злокачественной опухоли таких протеаз также является проблемой, которую необходимо решать. Таким образом, способы, которые обеспечивают обратимое действие в областях воспаления или злокачественной опухоли (области повреждения) без системного действия в нормальных тканях и крови для проявления эффективности лекарственного средства, одновременно избегая побочных эффектов, не известны.
[Документы уровня техники]
[Патентные документы]
[Патентный документ 1] WO 2003/105757
[Патентный документ 2] WO 2012/033953
[Патентный документ 3] WO 2010/081173
[Непатентные документы]
[Непатентный документ 1] Monoclonal antibody successes in the clinic. Janice M Reichert, Clark J Rosensweig, Laura B Faden & Matthew C Dewitz, Nat. Biotechnol. (2005) 23, 1073-1078
[Непатентный документ 2] The therapeutic antibodies market to 2008. Pavlou AK, Belsey MJ., Eur. J. Pharm. Biopharm. (2005) 59 (3), 389-396
[Non-patent document 3] Monoclonal antibodies: versatile platforms for cancer immunotherapy. Weiner LM, Surana R, Wang S., Nat. Rev. Immunol. (2010) 10 (5), 317-327
[Непатентный документ 4] Differential responses of human tumor cell lines to anti-p185HER2 monoclonal antibodies. Lewis GD, Figari I, Fendly B, Wong WL, Carter P, Gorman C, Shepard HM, Cancer Immunol. Immunotherapy (1993) 37, 255-263
[Непатентный документ 5] ING-1, a monoclonal antibody targeting Ep-CAM in patients with advanced adenocarcinomas. de Bono JS, Tolcher AW, Forero A, Vanhove GF, Takimoto C, Bauer RJ, Hammond LA, Patnaik A, White ML, Shen S, Khazaeli MB, Rowinsky EK, LoBuglio AF, Clin. Cancer Res. (2004) 10 (22), 7555-7565
[Непатентный документ 6] Non-fucosylated therapeutic antibodies as next-generation therapeutic antibodies. Satoh M, Iida S, Shitara K., Expert Opin. Biol. Ther. (2006) 6 (11), 1161-1173
[Непатентный документ 7] Optimizing engagement of the immune system by anti-tumor antibodies: an engineer's perspective. Desjarlais JR, Lazar GA, Zhukovsky EA, Chu SY., Drug Discov. Today (2007) 12 (21-22), 898-910
[Непатентный документ 8] Antibody-drug conjugates: targeted drug delivery for cancer. Alley SC, Okeley NM, Senter PD., Curr. Opin. Chem. Biol. (2010) 14 (4), 529-537
[Непатентный документ 9] BiTE: Teaching antibodies to engage T-cells for cancer therapy. Baeuerle PA, Kufer P, Bargou R., Curr. Opin. Mol. Ther. (2009) 11 (1), 22-30
[Непатентный документ 10] T cell-engaging BiTE antibodies specific for EGFR potently eliminate KRAS- and BRAF-mutated colorectal cancer cells. Lutterbuese R, Raum T, Kischel R, Hoffmann P, Mangold S, Rattel B, Friedrich M, Thomas O, Lorenczewski G, Rau D, Schaller E, Herrmann I, Wolf A, Urbig T, Baeuerle PA, Kufer P., Proc. Natl. Acad. Sci. U.S.A. (2010) 107 (28), 12605-12610
[Непатентный документ 11] Phase I trial with the CD44v6-targeting immunoconjugate bivatuzumab mertansine in head and neck squamous cell carcinoma. Riechelmann H, Sauter A, Golze W, Hanft G, Schroen C, Hoermann K, Erhardt T, Gronau S., Oral Oncol. (2008) 44 (9), 823-829
[Непатентный документ 12] Ipilimumab in the treatment of melanoma. Trinh VA, Hwu WJ., Expert Opin. Biol. Ther., (2012) Apr 14 (doi:10.1517/14712598.2012.675325)
[Непатентный документ 13] IPILIMUMAB - A NOVEL IMMUNOMODULATING THERAPY CAUSING AUTOIMMUNE HYPOPHYSITIS: A CASE REPORT AND REVIEW. Juszczak A, Gupta A, Karavitaki N, Middleton MR, Grossman A., Eur. J. Endocrinol. (2012) Apr 10 (doi: 10.1530/EJE-12-0167)
[Непатентный документ 14] The Japanese experience with biologic therapies for rheumatoid arthritis. Takeuchi T, Kameda H., Nat. Rev. Rheumatol. (2010) 6 (11), 644-652
[Непатентный документ 15] Current evidence for the management of rheumatoid arthritis with biological disease-modifying antirheumatic drugs: a systematic literature review informing the EULAR recommendations for the management of RA. Nam JL, Winthrop KL, van Vollenhoven RF, Pavelka K, Valesini G, Hensor EM, Worthy G, Landewe R, Smolen JS, Emery P, Buch MH., Ann. Rheum. Dis. (2010) 69 (6), 976-986
[Непатентный документ 16] Antibody engineering for the development of therapeutic antibodies. Kim SJ, Park Y, Hong HJ., Mol. Cells. (2005) 20 (1), 17-29
Сущность изобретения
[Проблемы, решаемые с помощью изобретения]
Настоящее изобретение было осуществлено ввиду описанных выше обстоятельств. Задачей настоящего изобретения является предоставления фармацевтических композиций, которые пригодны для лечения заболеваний, происходящих из тканей-мишеней, и их активных ингредиентов. Другой задачей является предоставление способов скрининга фармацевтических композиций и активных ингредиентов, а также способов их получения.
[Способы решения проблем]
Авторы настоящего изобретения провели тщательные исследования для решения описанных выше задач. В результате они получили антигенсвязывающие молекулы, содержащие антигенсвязывающий домен, антигенсвязывающая активность которого варьирует в зависимости от концентрации специфического для ткани-мишени соединения. Более того, авторы настоящего изобретения обнаружили, что антигенсвязывающие молекулы или фармацевтические композиции, содержащие антигенсвязывающие молекулы, пригодны для лечения заболеваний, которые происходят из ткани-мишени, и что они также пригодны для лечения заболеваний, происходящих из тканей-мишеней, которое включает введение антигенсвязывающих молекул. Также они открыли, что антигенсвязывающие молекулы пригодны для изготовления фармацевтических препаратов для лечения заболеваний, которые происходят из тканей-мишеней. Более того, авторы настоящего изобретения разработали способы скрининга и способы получения антигенсвязывающих молекул, и, тем самым, осуществили настоящее изобретение.
Более конкретно, настоящее изобретение относится к следующему:
[1] Антигенсвязывающая молекула, содержащая антигенсвязывающий домен, антигенсвязывающая активность которого варьирует в зависимости от концентрации специфического для ткани-мишени соединения.
[2] Антигенсвязывающая молекула согласно [1], где тканью-мишенью является злокачественная ткань.
[3] Антигенсвязывающая молекула согласно [2], где соединение, специфическое для злокачественной ткани, является метаболитом, специфическим для злокачественной клетки, метаболитом, специфическим для иммунной клетки, которая инфильтрировала злокачественную ткань, или метаболитом, специфичным к клетке стромы в злокачественной ткани.
[4] Антигенсвязывающая молекула согласно [1], где ткань-мишень представляет собой воспаленную ткань.
[5] Антигенсвязывающая молекула согласно [4], где соединение, специфическое для воспаленной ткани, представляет собой метаболит, специфический для иммунной клетки, которая инфильтрировала воспаленную ткань, или метаболит, специфический для нормальной клетки, которая была повреждена в воспаленной ткани.
[6] Антигенсвязывающая молекула согласно [1], где соединение представляет собой по меньшей мере одно соединение, выбранное из нуклеозида, имеющего структуру пуринового кольца, аминокислоты и ее метаболита, липида и ее метаболита, первичного метаболита гликометаболизма, и никотинамида и его метаболита.
[7] Антигенсвязывающая молекула согласно [6], где соединение представляет собой по меньшей мере одно соединение, выбранное из аденозина, аденозинтрифосфата, инозина, аланина, глутаминовой кислоты, аспарагиновой кислоты, кинуренина, простагландина E2, янтарной кислоты, лимонной кислоты и 1-этилметилникотинамида.
[8] Антигенсвязывающая молекула согласно любому из [1]-[7], где антиген представляет собой молекулу мембранного типа.
[9] Антигенсвязывающая молекула согласно любому из [1]-[8], которая представляет собой антигенсвязывающую молекулу, которая обладает нейтрализующей активностью.
[10] Антигенсвязывающая молекула согласно любому из [1]-[9], которая представляет собой антигенсвязывающую молекулу, которая обладает цитотоксической активностью.
[11] Антигенсвязывающая молекула согласно любому из [1]-[10], которая содержит Fc-область.
[12] Антигенсвязывающая молекула согласно [11], где Fc-область представляет собой Fc-область, содержащуюся в константной области SEQ ID NO:5, 6, 7 или 8.
[13] Антигенсвязывающая молекула согласно [11], где Fc-область содержит измененную FcγR-связывающую Fc-область, которая обладает более высокой активностью связывания Fcγ-рецептора, чем активность связывания Fcγ-рецептора у нативной Fc-области IgG человека.
[14] Антигенсвязывающая молекула согласно [13], где по меньшей мере одна или несколько аминокислот, выбранных из группы, состоящей из аминокислот в положениях 221, 222, 223, 224, 225, 227, 228, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 246, 247, 249, 250, 251, 254, 255, 256, 258, 260, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 278, 279, 280, 281, 282, 283, 284, 285, 286, 288, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 311, 313, 315, 317, 318, 320, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 339, 376, 377, 378, 379, 380, 382, 385, 392, 396, 421, 427, 428, 429, 434, 436 и 440 в соответствии с нумерацией EU в аминокислотной последовательности измененной FcγR-связывающей Fc-области отличаются от аминокислот нативной Fc-области IgG человека.
[15] Антигенсвязывающая молекула согласно [14], которая содержит по меньшей мере одну или несколько аминокислот, выбранных из группы состоящей из:
Lys или Tyr в качестве аминокислоты в положении 221;
Phe, Trp, Glu или Tyr в качестве аминокислоты в положении 222;
Phe, Trp, Glu или Lys в качестве аминокислоты в положении 223;
Phe, Trp, Glu или Tyr в качестве аминокислоты в положении 224;
Glu, Lys или Trp в качестве аминокислоты в положении 225;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 227;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 228;
Ala, Glu, Gly или Tyr в качестве аминокислоты в положении 230;
Glu, Gly, Lys, Pro или Tyr в качестве аминокислоты в положении 231;
Glu, Gly, Lys или Tyr в качестве аминокислоты в положении 232;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 233;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 234;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 235;
Ala, Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 236;
Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 237;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 238;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 239;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 240;
Asp, Glu, Leu, Arg, Trp или Tyr в качестве аминокислоты в положении 241;
Leu, Glu, Leu, Gln, Arg, Trp или Tyr в качестве аминокислоты в положении 243;
His в качестве аминокислоты в положении 244;
Ala в качестве аминокислоты в положении 245;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 246;
Ala, Phe, Gly, His, Ile, Leu, Met, Thr, Val или Tyr в качестве аминокислоты в положении 247;
Glu, His, Gln или Tyr в качестве аминокислоты в положении 249;
Glu или Gln в качестве аминокислоты в положении 250;
Phe в качестве аминокислоты в положении 251;
Phe, Met или Tyr в качестве аминокислоты в положении 254;
Glu, Leu или Tyr в качестве аминокислоты в положении 255;
Ala, Met или Pro в качестве аминокислоты в положении 256;
Asp, Glu, His, Ser или Tyr в качестве аминокислоты в положении 258;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 260;
Ala, Glu, Phe, Ile или Thr в качестве аминокислоты в положении 262;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 263;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 264;
Ala, Leu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 265;
Ala, Ile, Met или Thr в качестве аминокислоты в положении 266;
Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 267;
Asp, Glu, Phe, Gly, Ile, Lys, Leu, Met, Pro, Gln, Arg, Thr, Val или Trp в качестве аминокислоты в положении 268;
Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 269;
Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 270;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 271;
Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 272;
Phe или Ile в качестве аминокислоты в положении 273;
Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 274;
Leu или Trp в качестве аминокислоты в положении 275;
Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 276;
Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp в качестве аминокислоты в положении 278;
Ala в качестве аминокислоты в положении 279;
Ala, Gly, His, Lys, Leu, Pro, Gln, Trp или Tyr в качестве аминокислоты в положении 280;
Asp, Lys, Pro или Tyr в качестве аминокислоты в положении 281;
Glu, Gly, Lys, Pro или Tyr в качестве аминокислоты в положении 282;
Ala, Gly, His, Ile, Lys, Leu, Met, Pro, Arg или Tyr в качестве аминокислоты в положении 283;
Asp, Glu, Leu, Asn, Thr или Tyr в качестве аминокислоты в положении 284;
Asp, Glu, Lys, Gln, Trp или Tyr в качестве аминокислоты в положении 285;
Glu, Gly, Pro или Tyr в качестве аминокислоты в положении 286;
Asn, Asp, Glu или Tyr в качестве аминокислоты в положении 288;
Asp, Gly, His, Leu, Asn, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 290;
Asp, Glu, Gly, His, Ile, Gln или Thr в качестве аминокислоты в положении 291;
Ala, Asp, Glu, Pro, Thr или Tyr в качестве аминокислоты в положении 292;
Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 293;
Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 294;
Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 295;
Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr или Val в качестве аминокислоты в положении 296;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 297;
Ala, Asp, Glu, Phe, His, Ile, Lys, Met, Asn, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 298;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 299;
Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp в качестве аминокислоты в положении 300;
Asp, Glu, His или Tyr в качестве аминокислоты в положении 301;
Ile в качестве аминокислоты в положении 302;
Asp, Gly или Tyr в качестве аминокислоты в положении 303;
Asp, His, Leu, Asn или Thr в качестве аминокислоты в положении 304;
Glu, Ile, Thr или Tyr в качестве аминокислоты в положении 305;
Ala, Asp, Asn, Thr, Val или Tyr в качестве аминокислоты в положении 311;
Phe в качестве аминокислоты в положении 313;
Leu в качестве аминокислоты в положении 315;
Glu или Gln в качестве аминокислоты в положении 317;
His, Leu, Asn, Pro, Gln, Arg, Thr, Val или Tyr в качестве аминокислоты в положении 318;
Asp, Phe, Gly, His, Ile, Leu, Asn, Pro, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 320;
Ala, Asp, Phe, Gly, His, Ile, Pro, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 322;
Ile в качестве аминокислоты в положении 323;
Asp, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 324;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 325;
Ala, Asp, Glu, Gly, Ile, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 326;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 327;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 328;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 329;
Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 330;
Asp, Phe, His, Ile, Leu, Met, Gln, Arg, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 331;
Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 332;
Ala, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Ser, Thr, Val или Tyr в качестве аминокислоты в положении 333;
Ala, Glu, Phe, Ile, Leu, Pro или Thr в качестве аминокислоты в положении 334;
Asp, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 335;
Glu, Lys или Tyr в качестве аминокислоты в положении 336;
Glu, His или Asn в качестве аминокислоты в положении 337;
Asp, Phe, Gly, Ile, Lys, Met, Asn, Gln, Arg, Ser или Thr в качестве аминокислоты в положении 339;
Ala или Val в качестве аминокислоты в положении 376;
Gly или Lys в качестве аминокислоты в положении 377;
Asp в качестве аминокислоты в положении 378;
Asn в качестве аминокислоты в положении 379;
Ala, Asn или Ser в качестве аминокислоты в положении 380;
Ala или Ile в качестве аминокислоты в положении 382;
Glu в качестве аминокислоты в положении 385;
Thr в качестве аминокислоты в положении 392;
Leu в качестве аминокислоты в положении 396;
Lys в качестве аминокислоты в положении 421;
Asn в качестве аминокислоты в положении 427;
Phe или Leu в качестве аминокислоты в положении 428;
Met в качестве аминокислоты в положении 429;
Trp в качестве аминокислоты в положении 434;
Ile в качестве аминокислоты в положении 436; и
Gly, His, Ile, Leu или Tyr в качестве аминокислоты в положении 440
в соответствии с нумерацией EU в аминокислотной последовательности измененной FcγR-связывающей Fc-области.
[16] Антигенсвязывающая молекула согласно [11], где Fc-область модифицирована так, что существует более высокая доля Fc-областей, связанных с цепью сахаров с дефицитом фукозы в составе цепи сахаров, связанной в положении 297, в соответствии с нумерацией EU, Fc-области, или так, что существует более высокая доля Fc-областей с присоединенным биссекторным N-ацетилглюкозамином.
[17] Антигенсвязывающая молекула согласно любому из [11] и [13]-[16], где активность связывания FcRn области Fc в условиях диапазона кислых значений pH усилена по сравнению с активностью связывания FcRn Fc-области SEQ ID NO:5, 6, 7 или 8.
[18] Антигенсвязывающая молекула согласно [17], где Fc-область представляет собой Fc-область с заменой по меньшей мере одной или нескольких аминокислот, выбранных из группы, состоящей из аминокислот в положениях 238, 244, 245, 249, 250, 251, 252, 253, 254, 255, 256, 257, 258, 260, 262, 265, 270, 272, 279, 283, 285, 286, 288, 293, 303, 305, 307, 308, 309, 311, 312, 314, 316, 317, 318, 332, 339, 340, 341, 343, 356, 360, 362, 375, 376, 377, 378, 380, 382, 385, 386, 387, 388, 389, 400, 413, 415, 423, 424, 427, 428, 430, 431, 433, 434, 435, 436, 438, 439, 440, 442, и 447, в соответствии с нумерацией EU, в аминокислотной последовательности Fc-области, содержащейся в константной области SEQ ID NO:5, 6, 7 или 8.
[19] Антигенсвязывающая молекула согласно [18], где Fc-область содержит по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из:
Leu в качестве аминокислоты в положении 238;
Leu в качестве аминокислоты в положении 244;
Arg в качестве аминокислоты в положении 245;
Pro в качестве аминокислоты в положении 249;
Gln или Glu в качестве аминокислоты в положении 250;
Arg, Asp, Glu или Leu в качестве аминокислоты в положении 251;
Phe, Ser, Thr или Tyr в качестве аминокислоты в положении 252;
Ser или Thr в качестве аминокислоты в положении 254;
Arg, Gly, Ile или Leu в качестве аминокислоты в положении 255;
Ala, Arg, Asn, Asp, Gln, Glu, Pro или Thr в качестве аминокислоты в положении 256;
Ala, Ile, Met, Asn, Ser или Val в качестве аминокислоты в положении 257;
Asp в качестве аминокислоты в положении 258;
Ser в качестве аминокислоты в положении 260;
Leu в качестве аминокислоты в положении 262;
Lys в качестве аминокислоты в положении 270;
Leu или Arg в качестве аминокислоты в положении 272;
Ala, Asp, Gly, His, Met, Asn, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 279;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 283;
Asn в качестве аминокислоты в положении 285;
Phe в качестве аминокислоты в положении 286;
Asn или Pro в качестве аминокислоты в положении 288;
Val в качестве аминокислоты в положении 293,
Ala, Glu, Gln или Met в качестве аминокислоты в положении 307;
Ala, Glu, Ile, Lys, Leu, Met, Ser, Val или Trp в качестве аминокислоты в положении 311;
Pro в качестве аминокислоты в положении 309;
Ala, Asp или Pro в качестве аминокислоты в положении 312;
Ala или Leu в качестве аминокислоты в положении 314;
Lys в качестве аминокислоты в положении 316;
Pro в качестве аминокислоты в положении 317;
Asn или Thr в качестве аминокислоты в положении 318;
Phe, His, Lys, Leu, Met, Arg, Ser или Trp в качестве аминокислоты в положении 332;
Asn, Thr или Trp в качестве аминокислоты в положении 339;
Pro в качестве аминокислоты в положении 341;
Glu, His, Lys, Gln, Arg, Thr или Tyr в качестве аминокислоты в положении 343;
Arg в качестве аминокислоты в положении 375;
Gly, Ile, Met, Pro, Thr или Val в качестве аминокислоты в положении 376;
Lys в качестве аминокислоты в положении 377;
Asp, Asn или Val в качестве аминокислоты в положении 378;
Ala, Asn, Ser или Thr в качестве аминокислоты в положении 380;
Phe, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 382;
Ala, Arg, Asp, Gly, His, Lys, Ser или Thr в качестве аминокислоты в положении 385;
Arg, Asp, Ile, Lys, Met, Pro, Ser или Thr в качестве аминокислоты в положении 386;
Ala, Arg, His, Pro, Ser или Thr в качестве аминокислоты в положении 387;
Asn, Pro или Ser в качестве аминокислоты в положении 389;
Asn в качестве аминокислоты в положении 423;
Asn в качестве аминокислоты в положении 427;
Leu, Met, Phe, Ser или Thr в качестве аминокислоты в положении 428;
Ala, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val или Tyr в качестве аминокислоты в положении 430;
His или Asn в качестве аминокислоты в положении 431;
Arg, Gln, His, Ile, Lys, Pro или Ser в качестве аминокислоты в положении 433;
Ala, Gly, His, Phe, Ser, Trp или Tyr в качестве аминокислоты в положении 434;
Arg, Asn, His, Ile, Leu, Lys, Met или Thr в качестве аминокислоты в положении 436;
Lys, Leu, Thr или Trp в качестве аминокислоты в положении 438;
Lys в качестве аминокислоты в положении 440;
Lys в качестве аминокислоты в положении 442; и
Ile, Pro или Thr в качестве аминокислоты в положении 308;
как указано с помощью нумерации EU в аминокислотной последовательности Fc-области, содержащейся в константной области SEQ ID NO:5, 6, 7 или 8.
[20] Антигенсвязывающая молекула согласно любому из [1]-[19], где антигенсвязывающий домен представляет собой мультиспецифический или мультипаратопный антигенсвязывающий домен.
[21] Антигенсвязывающая молекула согласно [20], где антиген, связываемый по меньшей мере одним из антигенсвязывающих доменов, представляет собой молекулу мембранного типа, экспрессируемую на мембране злокачественной клетки, и антиген, связываемый по меньшей мере одним из антигенсвязывающих доменов, представляет собой молекулу мембранного типа, экспрессируемую на мембране эффекторной клетки.
[22] Антигенсвязывающая молекула согласно [21], где эффекторная клетка представляет собой NK-клетку, макрофаг или T-клетку.
[23] Антигенсвязывающая молекула согласно [21] или [22], где молекула мембранного типа, экспрессируемая на мембране эффекторной клетки, представляет собой TCR-образующий полипептид, CD2, CD3, CD28, CD44, CD16, CD32, CD64 или NKG2D.
[24] Антигенсвязывающая молекула согласно [20], где антиген, связываемый по меньшей мере одним из антигенсвязывающих доменов, представляет собой молекулу мембранного типа, экспрессируемую на мембране злокачественной клетки, и антиген, связываемый по меньшей мере одним из антигенсвязывающих доменов, представляет собой цитотоксическое вещество.
[25] Антигенсвязывающая молекула согласно любому из [20]-[24], где антигенсвязывающая молекула представляет собой фрагмент антитела.
[26] Антигенсвязывающая молекула согласно любому из [1]-[24], где антигенсвязывающая молекула представляет собой антитело.
[27] Антигенсвязывающая молекула согласно любому из [1]-[7], где антиген представляет собой растворимую молекулу.
[28] Антигенсвязывающая молекула согласно [27], которая представляет собой антигенсвязывающую молекулу, обладающую нейтрализующей активностью.
[29] Антигенсвязывающая молекула согласно [27] или [28], которая содержит Fc-область.
[30] Антигенсвязывающая молекула согласно [29], где Fc-область представляет собой Fc-область, содержащуюся в константной области SEQ ID NO:5, 6, 7 или 8.
[31] Антигенсвязывающая молекула согласно [29], где активность связывания Fc-области с FcRn в условиях диапазона кислых значений pH усилена по сравнению с активностью связывания с FcRn Fc-области, содержащейся в константной области SEQ ID NO:5, 6, 7 или 8.
[32] Антигенсвязывающая молекула согласно [31], где Fc-область представляет собой Fc-область с заменой по меньшей мере одной или нескольких аминокислот, выбранных из группы, состоящей из аминокислот в положениях 238, 244, 245, 249, 250, 251, 252, 253, 254, 255, 256, 257, 258, 260, 262, 265, 270, 272, 279, 283, 285, 286, 288, 293, 303, 305, 307, 308, 309, 311, 312, 314, 316, 317, 318, 332, 339, 340, 341, 343, 356, 360, 362, 375, 376, 377, 378, 380, 382, 385, 386, 387, 388, 389, 400, 413, 415, 423, 424, 427, 428, 430, 431, 433, 434, 435, 436, 438, 439, 440, 442 и 447, в соответствии с нумерацией EU, в аминокислотной последовательности Fc-области, содержащейся в константной области SEQ ID NO:5, 6, 7 или 8.
[33] Антигенсвязывающая молекула согласно [32], где Fc-область содержит по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из:
Leu в качестве аминокислоты в положении 238;
Leu в качестве аминокислоты в положении 244;
Arg в качестве аминокислоты в положении 245;
Pro в качестве аминокислоты в положении 249;
Gln или Glu в качестве аминокислоты в положении 250;
Arg, Asp, Glu или Leu в качестве аминокислоты в положении 251;
Phe, Ser, Thr или Tyr в качестве аминокислоты в положении 252;
Ser или Thr в качестве аминокислоты в положении 254;
Arg, Gly, Ile или Leu в качестве аминокислоты в положении 255;
Ala, Arg, Asn, Asp, Gln, Glu, Pro или Thr в качестве аминокислоты в положении 256;
Ala, Ile, Met, Asn, Ser или Val в качестве аминокислоты в положении 257;
Asp в качестве аминокислоты в положении 258;
Ser в качестве аминокислоты в положении 260;
Leu в качестве аминокислоты в положении 262;
Lys в качестве аминокислоты в положении 270;
Leu,или Arg в качестве аминокислоты в положении 272;
Ala, Asp, Gly, His, Met, Asn, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 279;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 283;
Asn в качестве аминокислоты в положении 285;
Phe в качестве аминокислоты в положении 286;
Asn или Pro в качестве аминокислоты в положении 288;
Val в качестве аминокислоты в положении 293,
Ala, Glu, Gln или Met в качестве аминокислоты в положении 307;
Ala, Glu, Ile, Lys, Leu, Met, Ser, Val или Trp в качестве аминокислоты в положении 311;
Pro в качестве аминокислоты в положении 309;
Ala, Asp или Pro в качестве аминокислоты в положении 312;
Ala или Leu в качестве аминокислоты в положении 314;
Lys в качестве аминокислоты в положении 316;
Pro в качестве аминокислоты в положении 317;
Asn или Thr в качестве аминокислоты в положении 318;
Phe, His, Lys, Leu, Met, Arg, Ser или Trp в качестве аминокислоты в положении 332;
Asn, Thr или Trp в качестве аминокислоты в положении 339;
Pro в качестве аминокислоты в положении 341;
Glu, His, Lys, Gln, Arg, Thr или Tyr в качестве аминокислоты в положении 343;
Arg в качестве аминокислоты в положении 375;
Gly, Ile, Met, Pro, Thr или Val в качестве аминокислоты в положении 376;
Lys в качестве аминокислоты в положении 377;
Asp, Asn или Val в качестве аминокислоты в положении 378;
Ala, Asn, Ser или Thr в качестве аминокислоты в положении 380;
Phe, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 382;
Ala, Arg, Asp, Gly, His, Lys, Ser или Thr в качестве аминокислоты в положении 385;
Arg, Asp, Ile, Lys, Met, Pro, Ser или Thr в качестве аминокислоты в положении 386;
Ala, Arg, His, Pro, Ser или Thr в качестве аминокислоты в положении 387;
Asn, Pro или Ser в качестве аминокислоты в положении 389;
Asn в качестве аминокислоты в положении 423;
Asn в качестве аминокислоты в положении 427;
Leu, Met, Phe, Ser или Thr в качестве аминокислоты в положении 428;
Ala, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val или Tyr в качестве аминокислоты в положении 430;
His или Asn в качестве аминокислоты в положении 431;
Arg, Gln, His, Ile, Lys, Pro или Ser в качестве аминокислоты в положении 433;
Ala, Gly, His, Phe, Ser, Trp или Tyr в качестве аминокислоты в положении 434;
Arg, Asn, His, Ile, Leu, Lys, Met или Thr в качестве аминокислоты в положении 436;
Lys, Leu, Thr или Trp в качестве аминокислоты в положении 438;
Lys в качестве аминокислоты в положении 440;
Lys в качестве аминокислоты в положении 442; и
Ile, Pro или Thr в качестве аминокислоты в положении 308;
как указано с помощью нумерации EU, в аминокислотной последовательности Fc-области, содержащейся в константной области SEQ ID NO:5, 6, 7 или 8.
[34] Антигенсвязывающая молекула согласно [29], где активность связывания Fc-области с FcRn в условиях диапазона нейтральных значений pH усилена по сравнению с активностью связывания с FcRn Fc-области, содержащейся в константной области SEQ ID NO:5, 6, 7 или 8.
[35] Антигенсвязывающая молекула согласно [34], где Fc-область представляет собой Fc-область с заменой по меньшей мере одной или нескольких аминокислот, выбранных из группы, состоящей из аминокислот в положениях 237, 248, 250, 252, 254, 255, 256, 257, 258, 265, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434 и 436 в соответствии с нумерацией EU, в аминокислотной последовательности Fc-области, содержащейся в константной области SEQ ID NO:5, 6, 7 или 8.
[36] Антигенсвязывающая молекула согласно [35], где Fc-область содержит по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из:
Met в качестве аминокислоты в положении 237;
Ile в качестве аминокислоты в положении 248;
Ala, Phe, Ile, Met, Gln, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 250;
Phe, Trp или Tyr в качестве аминокислоты в положении 252;
Thr в качестве аминокислоты в положении 254;
Glu в качестве аминокислоты в положении 255;
Asp, Asn, Glu или Gln в качестве аминокислоты в положении 256;
Ala, Gly, Ile, Leu, Met, Asn, Ser, Thr или Val в качестве аминокислоты в положении 257;
His в качестве аминокислоты в положении 258;
Ala в качестве аминокислоты в положении 265;
Ala или Glu в качестве аминокислоты в положении 286;
His в качестве аминокислоты в положении 289;
Ala в качестве аминокислоты в положении 297;
Ala в качестве аминокислоты в положении 303;
Ala в качестве аминокислоты в положении 305;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr в качестве аминокислоты в положении 307;
Ala, Phe, Ile, Leu, Met, Pro, Gln или Thr в качестве аминокислоты в положении 308;
Ala, Asp, Glu, Pro или Arg в качестве аминокислоты в положении 309;
Ala, His или Ile в качестве аминокислоты в положении 311;
Ala или His в качестве аминокислоты в положении 312;
Lys или Arg в качестве аминокислоты в положении 314;
Ala, Asp или His в качестве аминокислоты в положении 315;
Ala в качестве аминокислоты в положении 317;
Val в качестве аминокислоты в положении 332;
Leu в качестве аминокислоты в положении 334;
His в качестве аминокислоты в положении 360;
Ala в качестве аминокислоты в положении 376;
Ala в качестве аминокислоты в положении 380;
Ala в качестве аминокислоты в положении 382;
Ala в качестве аминокислоты в положении 384;
Asp или His в качестве аминокислоты в положении 385;
Pro в качестве аминокислоты в положении 386;
Glu в качестве аминокислоты в положении 387;
Ala или Ser в качестве аминокислоты в положении 389;
Ala в качестве аминокислоты в положении 424;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 428;
Lys в качестве аминокислоты в положении 433;
Ala, Phe, His, Ser, Trp или Tyr в качестве аминокислоты в положении 434; и
His, Ile, Leu, Phe, Thr или Val в качестве аминокислоты в положении 436
как указано с помощью нумерации EU, в аминокислотной последовательности Fc-области SEQ ID NO:5, 6, 7 или 8.
[37] Антигенсвязывающая молекула согласно любому из [29] и [31]-[36], где Fc-область обладает более высокой активностью связывания с ингибиторным Fcγ-рецептором, чем с активирующим Fcγ-рецептором.
[38] Антигенсвязывающая молекула согласно [37], где ингибиторный Fcγ-рецептор представляет собой FcγRIIb.
[39] Антигенсвязывающая молекула согласно [37] или [38], где активирующий Fcγ-рецептор представляет собой FcγRIa человека, FcγRIIa (R) человека, FcγRIIa (H) человека, FcγRIIIa (V) человека или FcγRIIIa (F) человека.
[40] Антигенсвязывающая молекула согласно любому из [37]-[39], где аминокислота в положении 238 или 328 (нумерация EU) Fc-области включает аминокислоту, которая отличается от аминокислоты нативной Fc-области IgG человека.
[41] Антигенсвязывающая молекула согласно [40], где аминокислота в положении 238, указанном с помощью нумерации EU, в Fc-области, представляет собой Asp, или аминокислота в положении 328 представляет собой Glu.
[42] Антигенсвязывающая молекула согласно [40] или [41], которая содержит по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из:
Asp в качестве аминокислоты в положении 233;
Trp или Tyr в качестве аминокислоты в положении 234;
Ala, Asp, Glu, Leu, Met, Phe, Trp или Tyr в качестве аминокислоты в положении 237;
Asp в качестве аминокислоты в положении 239;
Ala, Gln или Val в качестве аминокислоты в положении 267;
Asn, Asp или Glu в качестве аминокислоты в положении 268;
Gly в качестве аминокислоты в положении 271;
Ala, Asn, Asp, Gln, Glu, Leu, Met, Ser или Thr в качестве аминокислоты в положении 326;
Arg, Lys или Met в качестве аминокислоты в положении 330;
Ile, Leu или Met в качестве аминокислоты в положении 323; и
Asp в качестве аминокислоты в положении 296 в соответствии с нумерацией EU, в аминокислотной последовательности Fc-области.
[43] Антигенсвязывающая молекула согласно любому из [27]-[42], где антигенсвязывающая молекула представляет собой антитело.
[44] Способ получения антигенсвязывающей молекулы согласно любому из [1]-[43], который включает выбор антигенсвязывающего домена, антигенсвязывающая активность которого варьирует в зависимости от концентрации специфического для ткани-мишени соединения.
[45] Способ скрининга антигенсвязывающей молекулы согласно любому из [1]-[43], который включает выбор антигенсвязывающего домена, антигенсвязывающая активность которого варьирует в зависимости от концентрации специфического для ткани-мишени соединения.
[46] Фармацевтическая композиция, содержащая антигенсвязывающую молекулу согласно любому из [1]-[43].
Краткое описание чертежей
На фиг. 1 показано, что антитело с низкомолекулярным переключающим фактором не связывается с антигенами в нормальных условиях, когда низкомолекулярные соединения не присутствуют, однако связывается с антигенами в ткани-мишени, где низкомолекулярные соединения присутствуют в высокой концентрации.
На фиг. 2 показано, что низкомолекулярные соединения функционируют в качестве переключателя, путем размещения между антителом с низкомолекулярным переключающим фактором и антигеном. Если низкомолекулярное соединение отсутствует, взаимодействие антитело-антиген является недостаточным и антитело не может связываться с антигеном, однако если низкомолекулярное соединение присутствует, антитело может связываться с антигеном вследствие наличия низкомолекулярного соединения, помещенного между антителом и антигеном.
Фиг. 3 представляет собой фигуру, демонстрирующую результат ELISA для связывания антитела с IL-6 человека. На вертикальной оси показаны величины поглощения, которые оценивают активность связывания с IL-6 человека каждого из антител в присутствии или в отсутствие каждого из низкомолекулярных соединений.
На фиг. 4 представлена сенсограмма, демонстрирующая взаимодействие между 4 мкмоль/л IL-6 человека и A11 в присутствии или в отсутствие 100 мкмоль/л кинуренина.
На фиг. 5 представлен график, который оценивает изменение ответа при связывании с A11, иммобилизованном на сенсорном чипе CM5, когда взаимодействию позволяют протекать в течение 60 секунд с 1 мкмоль/л IL-6 в качестве анализируемого соединения. На вертикальной оси показано изменение ответа (RU) до и после взаимодействия с IL-6 и на горизонтальной оси показана концентрация кинуренина (мкмоль/л), содержащегося в растворе в этот момент времени.
На фиг. 6 представлен график, который оценивает ответ на H01, иммобилизованный на сенсорном чипе CM5, когда взаимодействию позволяют протекать в течение 60 секунд с 1 мкмоль/л IL-6 в качестве анализируемого соединения. На вертикальной оси показано изменение ответа (RU) до и после взаимодействия с IL-6 и на горизонтальной оси показана концентрация кинуренина (мкмоль/л), содержащегося в растворе в этот момент времени.
На фиг. 7 представлен график, который оценивает ответ на IL-6, иммобилизованный на сенсорном чипе CM5, когда взаимодействию позволяют протекать в течение 60 секунд с 0,1 мкмоль/л A11 в качестве анализируемого соединения. На вертикальной оси показано изменение ответа (RU) до и после взаимодействия с A11 и на горизонтальной оси показана концентрация кинуренина (мкмоль/л), содержащегося в растворе в этот момент времени.
На фиг. 8 представлен график, полученный путем позволения A11 взаимодействовать с IL-6, иммобилизованным на сенсорном чипе CM5, в присутствии 100 мкмоль/л кинуренина, а затем наблюдения диссоциации A11 от IL6 в присутствии буфера, содержащего 100 мкмоль/л кинуренина или в присутствии буфера, который не содержит кинуренина. На фигуре на вертикальной оси показаны величины, нормализованные путем определения количества A11, связавшегося в присутствии 100 мкмоль/л кинуренина, как 100, и на горизонтальной оси представлен отрезок времени (в секундах) от начала взаимодействия.
На фиг. 9 представлена сенсограмма, полученная путем позволения 800, 400, 200, 100, 50 или 25 нмоль/л кинуренина взаимодействовать с IL-6, иммобилизованном на сенсорном чипе. На вертикальной оси показано изменение количества IL-6, связанного кинуренином (RU) (ответ в начале эксперимента по взаимодействию определяли как 0), и на горизонтальной оси показан отрезок времени от начала эксперимента по взаимодействию.
На фиг. 10 представлена структура 2'-аденозин-PEG-пептида, который представляет собой аналог аденозина, использованный для иммунизации кроликов.
На фиг. 11 представлена структура 5'-аденозин-PEG-пептида, который представляет собой аналог аденозина, использованный для иммунизации кроликов.
На фиг. 12 представлена структура 2'-аденозин-PEG-биотина, полученного путем замены биотина пептидной частью аналога аденозина, используемого для иммунизации кроликов.
На фиг. 13 представлена структура 5'-аденозин-PEG-биотина, полученного путем замены биотина пептидной частью аналога аденозина, используемого для иммунизации кроликов.
На фиг. 14 представлен график, где на вертикальной оси представлена величина (N_связывание_100), полученная путем деления величины связывания при взаимодействии каждого антитела с 2'-аденозин-PEG-биотином на уровень улавливания (RU) каждого антитела, и на горизонтальной оси представлена величина (N_стабильность_100), полученная путем деления величины, полученной через 60 секунд после диссоциации 2'-аденозин-PEG-биотина от каждого антитела, после взаимодействия с 2'-аденозин-PEG-биотином, на уровень улавливания (RU) каждого антитела.
На фиг. 15A представлены сенсограммы, полученные в анализах на основе поверхностного плазмонного резонанса, на которых показано, что клон SMB0002 связывается (взаимодействует с) аденозином. На сенсограммах показаны взаимодействия между SMB0002 и антигеном при 7,81, 31,3, 125 и 500 нМ в возрастающем порядке.
На фиг. 15B представлены сенсограммы, полученные в анализах на основе поверхностного плазмонного резонанса, которые демонстрируют, что клон SMB0002 связывается (взаимодействует) с ATP. На сенсограммах показаны взаимодействия между SMB0002 и антигеном при 78,1, 313, 1250 и 5000 нМ в возрастающем порядке.
На фиг. 15C представлены сенсограммы, полученные в анализах на основе поверхностного плазмонного резонанса, которые демонстрируют, что клон SMB0089 связывается (взаимодействует) с аденозином. На сенсограммах показаны взаимодействия между SMB0089 и антигеном при 7,81, 31,3, 125 и 500 нМ в возрастающем порядке.
На фиг. 15D представлены сенсограммы, полученные в анализах на основе поверхностного плазмонного резонанса, которые демонстрируют, что клон SMB0089 связывается (взаимодействует) с ATP. На сенсограммах показаны взаимодействия между SMB00089 и антигеном при 78,1, 313, 1250 и 5000 нМ в возрастающем порядке.
На фиг. 15E представлены сенсограммы, полученные в анализах на основе поверхностного плазмонного резонанса, которые демонстрируют, что клон SMB0104 связывается (взаимодействует) с аденозином. На сенсограммах показаны взаимодействия между SMB0104 и антигеном при 7,81, 31,3 и 500 нМ в возрастающем порядке.
На фиг. 15F представлены сенсограммы, полученные в анализах на основе поверхностного плазмонного резонанса, которые демонстрируют, что клон SMB0104 связывается (взаимодействует) с ATP. На сенсограммах показаны взаимодействия между SMB0104 и антигеном при 78,1, 313, 1250 и 5000 нМ в возрастающем порядке.
На фиг. 16 представлены сенсограммы, полученные в анализах на основе поверхностного плазмонного резонанса, которые демонстрируют, что клон SMB0171 связывается (взаимодействует) с ATP. На сенсограммах показаны взаимодействия между SMB0171 и антигеном при 5 и 50 мкМ в возрастающем порядке.
На фиг. 17 представлены результаты конкурентного ELISA, которые демонстрируют, что клон SMB0002 связывается с аденозином и ATP.
На фиг. 18 представлен график, который оценивает способность ATP к ингибированию связывания ATNLSA1-4_D12 с меченными биотином антигенами (смесь 5'-аденозин-PEG-биотина и ATP-PEG-биотина).
На фиг. 19 представлена концепция рационально сконструированной библиотеки антител, которая позволяет получить антитела с переключением аденозином/ATP против любого антигена, где библиотека получена из участков вариабельных областей антител, которые контактируют с антигенами, так что аденозин или ATP располагается между антителом и антигеном.
На фиг. 20 представлена концепция библиотеки антител из иммунизированного аденозином кролика, которая обеспечивает антитела с переключением аденозином/ATP против любого антигена, и в которой аденозин или ATP располагается между антителом и антигеном.
Фиг. 21 представляет собой фигуру, демонстрирующую результат ELISA для связывания антитела с IL-6 человека. На вертикальной оси представлена активность связывания каждого антитела с IL-6 человека в зависимости от присутствия или отсутствия аминокислот или метаболитов аминокислот (кинуренин, триптофан, фенилаланин, антраниловая кислота, 3-гидроксикинуренин и кинуреновая кислота), показанная в качестве величин поглощения при длине волны 450 нм.
Фиг. 22 представляет собой фигуру, демонстрирующую результат ELISA для связывания антитела с IL-6 человека. На вертикальной оси представлена активность связывания антитела I6NMSC1-3_#03 с IL-6 человека в зависимости от присутствия или отсутствия каждого низкомолекулярного соединения (ATP, аденозин, инозин, PGE2, янтарная кислота, молочная кислота, кинуренин и коктейль низкомолекулярных соединений), показанная в качестве величин удельной активности, вычисленных из величин поглощения при длине волны 450 нм.
Фиг. 23 представляет собой фигуру, демонстрирующую результат ELISA для связывания антитела с IL-6 человека. На вертикальной оси представлена активность связывания антитела I6NMSC1-3_#17 с IL-6 человека в зависимости от присутствия или отсутствия каждого низкомолекулярного соединения (ATP, аденозин, инозин, PGE2, янтарная кислота, молочная кислота, кинуренин и коктейль низкомолекулярных соединений), показанная в качестве величин удельной активности, вычисленных из величин поглощения при длине волны 450 нм.
Фиг. 24 представляет собой фигуру, демонстрирующую результат ELISA для связывания антитела с HSA. На вертикальной оси представлена активность связывания антитела HSNMSC1-4_#22 с HSA в зависимости от присутствия или отсутствия каждого низкомолекулярного соединения (ATP, аденозин, инозин, PGE2, янтарная кислота, молочная кислота, кинуренин и коктейль низкомолекулярных соединений), показанная в качестве величин поглощения при длине волны 450 нм.
Фиг. 25 представляет собой фигуру, демонстрирующую результат ELISA, проведенного для клона I6DL2C5-4_076, который был получен из библиотеки рационально сконструированных антител против IL-6 человека, в присутствии или в отсутствие ATP и/или аденозина при 1 мМ. На вертикальной оси представлена величина поглощения, которая оценивает активность связывания антитела с IL-6 человека. Результаты, полученные с использованием фага-помощника M13KO7, представлены в качестве отрицательного контроля.
Фиг. 26 представляет собой фигуру, демонстрирующую результат ELISA, проведенного для клона HSDL3C5-4_015, который был получен из рационально сконструированной библиотеки антител против сывороточного альбумина человека в присутствии или в отсутствие ATP и/или аденозина при 1 мМ. На вертикальной оси представлена величина поглощения, которая оценивает активность связывания антитела с сывороточным альбумином человека. Результаты, полученные с использованием фага-помощника M13KO7, представлены в качестве отрицательного контроля.
Фиг. 27 представляет собой фигуру, демонстрирующую результат ELISA, проведенного для клонов 6RAD2C1-4_011 и 6RAD2C1-4_076, которые были получены из рационально сконструированной библиотеки антител против рецептора IL-6 человека, в присутствии или в отсутствие ATP и/или аденозина (обозначено как ADO) при 1 мМ, и в присутствии или в отсутствие коктейля низкомолекулярных соединений (SC). На вертикальной оси представлены величины поглощения, которые оценивают активность связывания антитела с рецептором IL-6 человека. Результаты, полученные с использованием фага-помощника M13KO7, представлены в качестве отрицательного контроля.
Фиг. 28 представляет собой фигуру, демонстрирующую результат ELISA для связывания клона 6RNMSC1-2_F02 с IL-6R человека. На вертикальной оси представлены величины поглощения, которые оценивают активность связывания антитела с IL-6R человека в присутствии или в отсутствие каждого низкомолекулярного соединения.
На фиг. 29 представлена фигура, демонстрирующая результат ELISA для связывания клона 6RNMSC1-3_G02 с IL-6R человека. На вертикальной оси представлены величины поглощения, которые оценивают активность связывания антитела с IL-6R человека в присутствии или в отсутствие каждого низкомолекулярного соединения.
Фиг. 30 представляет собой фигуру, демонстрирующую результат ELISA для связывания антитела с IL-6R человека. На вертикальной оси представлены величины поглощения, которые оценивают активность связывания антитела с IL-6R человека в присутствии или в отсутствие каждой аминокислоты или метаболита аминокислоты.
На фиг. 31 представлены сенсограммы, демонстрирующие взаимодействие между 6RNMSC1-2_F02 и 1 мкмоль/л IL-6R в присутствии 100 мкмоль/л кинуренина, в присутствии 10 ммоль/л ATP, и в отсутствие кинуренина и ATP. Сплошная линия указывает на взаимодействие в присутствии кинуренина, точечная линия указывает на взаимодействие в присутствии ATP, и пунктирной линией указано взаимодействие в их отсутствие.
На фиг. 32 представлен график, полученный путем позволения 6RNMSC1-2_F02 взаимодействовать с IL-6R, иммобилизованном на сенсорном чипе CM5 в присутствии 100 мкмоль/л кинуренина, а затем наблюдения диссоциации 6RNMSC1-2_F02 от IL-6R в присутствии буфера, содержащего 100 мкмоль/л кинуренина, или в присутствии буфера, который не содержит кинуренина. На фигуре на вертикальной оси представлены величины, нормализованные путем определения количества 6RNMSC1-2_F02, связавшегося в присутствии 100 мкмоль/л кинуренина, как 100, и на горизонтальной оси представлен отрезок времени (в секундах) от начала взаимодействия. Сплошной линией указана диссоциация 6RNMSC1-2_F02 от IL-6R в присутствии кинуренина, и пунктирной линией показана диссоциация 6RNMSC1-2_F02 от IL-6R в отсутствие кинуренина.
На фиг. 33 представлен график, полученный путем позволения 5 мкг/л 6RNMSC1-2_F02 взаимодействовать в качестве анализируемого соединения в течение 180 секунд, и оценки ответа на IL-6R, иммобилизованный на сенсорном чипе CM5. На вертикальной оси представлено изменение ответа (RU) до и после взаимодействия 6RNMSC1-2_F02, и на горизонтальной оси представлена концентрация (мкмоль/л) кинуренина, содержащегося в растворе.
На фиг. 34 представлена фигура, демонстрирующая оценку связывания антител с IL-6R человека мембранного типа с помощью FCM. На верхней панели представлены результаты, полученные в присутствии кинуренина, и на нижней панели представлены результаты, полученные в отсутствие кинуренина. На горизонтальной оси представлена интенсивность флуоресценции и на вертикальной оси представлено количество клеток.
На фиг. 35A представлена активность ADCC антител, которые связываются с антигенами в присутствии низкомолекулярных соединений, в отношении клеток, экспрессирующих антигены. На ней показана активность ADCC клона 6RNMSC1-2_F02, который связывается с hIL-6R в присутствии кинуренина, в отношении клеток BaF, экспрессирующих hIL-6R, в присутствии (треугольники) или в отсутствие (круги) кинуренина. Незакрашенные треугольники и круги демонстрируют измеренные величины, и закрашенные треугольники и круги демонстрируют средние значения.
На фиг. 35B представлена активность ADCC антител, которые связываются с антигенами в присутствии низкомолекулярных соединений, в отношении клеток, которые экспрессируют антиген. На ней показана активность ADCC MRA, которое связывается с hIL-6R, независимо от присутствия кинуренина, в отношении клеток BaF, экспрессирующих hIL-6R, в присутствии (треугольники) или в отсутствие (круги) кинуренина. Незакрашенные треугольники и круги демонстрируют измеренные величины, и закрашенные треугольники и круги демонстрируют средние значения.
На фиг. 36 представлена активность ADCC антител, которые связываются с антигенами в присутствии низкомолекулярных соединений, в отношении клеток, экспрессирующих антиген. На ней представлена активность ADCC клона 6RNMSC1-2_F02 в отношении клеток BaF, экспрессирующих hIL-6R, в присутствии (треугольники) или в отсутствие (круги) клона 6RNMSC1-2_F02, который связывается с hIL-6R в присутствии кинуренина. На горизонтальной оси представлена концентрация кинуренина и на вертикальной оси представлена активность ADCC (%). Представлены средние значения и стандартные отклонения для активности ADCC.
Фиг. 37 представляет собой фигуру, демонстрирующую результат ELISA для связывания клона 6RNMSC1-2_F02 в сыворотке мыши с IL-6R человека. На вертикальной оси представлены величины поглощения, которые оценивают активность связывания антитела с IL-6R человека в присутствии или в отсутствие кинуренина.
Фиг. 38 представляет собой фигуру, демонстрирующую результат ELISA, проведенного с клоном I6RLSA1-6_011, который был получен из рационально сконструированной библиотеки антител, против IL-6 человека в присутствии или в отсутствие ATP и аденозина при 10 мМ. На вертикальной оси представлена величина поглощения, которая оценивает активность связывания антитела с IL-6 человека. Результаты, полученные с использованием клона, полученного из рационально сконструированной библиотеки антител, и демонстрирующие активность связывания с IL-6 человека независимо от присутствия низкомолекулярных соединений, представлены в качестве положительного контроля. Результаты, полученные с использованием фага-помощника M13KO7, представлены в качестве отрицательного контроля.
Фиг. 39 представляет собой фигуру, демонстрирующую результат ELISA, проведенного с клоном 6RRLSA1-6_037 и 6RRLSA1-6_045, которые были получены из рационально сконструированной библиотеки антител против рецептора IL-6 человека, в присутствии или в отсутствие ATP и аденозина при 10 мМ. На вертикальной оси представлена величина поглощения, которая оценивает активность связывания антител с рецептором IL-6 человека. Результаты, полученные с использованием фага-помощника M13KO7, представлены в качестве отрицательного контроля.
Фиг. 40 представляет собой фигуру, демонстрирующую результат ELISA, проведенного для 96 клонов, полученных пэннингом рационально сконструированной библиотеки антител четыре раза против IgA-Fc человека с использованием мультивалентного фагового дисплея антител. Величины поглощения, которые оценивают активность связывания антител с IgA-Fc человека в отсутствие ATP и аденозина, представлены на вертикальной оси, и величины поглощения, которые оценивают активность связывания антител с IgA-Fc человека в присутствии ATP и аденозина, представлены на горизонтальной оси.
Фиг. 41 представляет собой фигуру, демонстрирующую результат ELISA, проведенного для 96 клонов, полученных пэннингом рационально сконструированной библиотеки антител четыре раза против IgA-Fc человека с использованием одновалентного фагового дисплея антител. Величины поглощения, которые оценивают активность связывания антител с IgA-Fc человека в отсутствие ATP и аденозина, представлены на вертикальной оси, и величины поглощения, которые оценивают активность связывания антител с IgA-Fc человека в присутствии ATP и аденозина, представлены на горизонтальной оси.
Фиг. 42 представляет собой фигуру, демонстрирующую результат ELISA, проведенного для клона IADL3C5-4_048, полученного из рационально сконструированной библиотеки антител против IgA-Fc человека в присутствии или в отсутствие ATP и аденозина при 1 мМ. На вертикальной оси представлена величина поглощения, которая оценивает активность связывания антитела с IgA-Fc человека. Результаты, полученные с использованием клона, полученного из рационально сконструированной библиотеки антител и демонстрирующего активность связывания с IgA-Fc человека независимо от присутствия низкомолекулярных соединений, представлены в качестве положительного контроля. Результаты, полученные с использованием фага-помощника M13KO7, представлены в качестве отрицательного контроля.
На фиг. 43 представлен график, демонстрирующий уровень связывания (ответ в виде связывания (RU)), когда каждый клон в концентрации 1 мкМ подвергали взаимодействию в течение 120 секунд с IL-6R, иммобилизованном на сенсорном чипе CM5 в присутствии или в отсутствие каждого из низкомолекулярных соединений при 1 мМ.
На фиг. 44A представлена активность ADCC антител, которые связываются с антигенами в присутствии низкомолекулярных соединений, в отношении клеток, экспрессирующих антиген. На этой фигуре представлена активность ADCC клона 6RAD2C1-4_030, который связывается с hIL-6R в присутствии ATP, в отношении клеток CHO, экспрессирующих hIL-6R, в присутствии (треугольники) или в отсутствие (круги) ATP. Незакрашенные треугольники и круги демонстрируют измеренные величины, и закрашенные треугольники и круги демонстрируют средние значения.
На фиг. 44B представлена активность ADCC антител, которые связываются с антигенами в присутствии низкомолекулярных соединений, в отношении клеток, экспрессирующих антиген. На этой фигуре представлена активность ADCC MRA, которое связывается с hIL-6R независимо от присутствия или отсутствия ATP, в отношении клеток CHO, экспрессирующих hIL-6R, в присутствии (треугольники) или в отсутствие (круги) ATP. Незакрашенные треугольники и круги демонстрируют измеренные величины, и закрашенные треугольники и круги демонстрируют средние значения.
На фиг. 44C представлена активность ADCC антител, которые связываются с антигенами в присутствии низкомолекулярных соединений, в отношении клеток, экспрессирующих антиген. На этой фигуре представлена активность ADCC клона 6RAD2C1-4_011, который связывается с hIL-6R, в присутствии ATP, в отношении клеток CHO, экспрессирующих hIL-6R, в присутствии (треугольники) или в отсутствие (круги) ATP. Незакрашенные треугольники и круги демонстрируют измеренные величины, и закрашенные треугольники и круги демонстрируют средние значения.
Фиг. 45 представляет собой фигуру, демонстрирующую результат ELISA, проведенного для клона HSADSA1-6_020, полученного из рационально сконструированной библиотеки антител против 7HSA, в присутствии или в отсутствие ATP и аденозина при 10 мМ. На вертикальной оси представлена величина поглощения, которая оценивает активность связывания антитела с HSA. Результаты, полученные с использованием клона, полученного из рационально сконструированной библиотеки антител и демонстрирующего активность связывания с HSA независимо от присутствия низкомолекулярных соединений, представлены в качестве положительного контроля. Результаты, полученные с использованием фага-помощника M13KO7, представлены в качестве отрицательного контроля.
[Способы осуществления изобретения]
Определения и подробное описание ниже предоставлены для облегчения понимания настоящего изобретения, проиллюстрированного в настоящем описании.
Аминокислоты
В рамках настоящего изобретения аминокислоты описаны с помощью однобуквенного или трехбуквенного кодов или обоих из них, например, Ala/A, Leu/L, Arg/R, Lys/K, Asn/N, Met/M, Asp/D, Phe/F, Cys/C, Pro/P, Gln/Q, Ser/S, Glu/E, Thr/T, Gly/G, Trp/W, His/H, Tyr/Y, Ile/I или Val/V.
Изменение аминокислот
Для изменения аминокислот в аминокислотной последовательности антигенсвязывающей молекулы можно соответствующим образом использовать известные способы, такие как способы сайт-направленного мутагенеза (Kunkel et al. (Proc. Natl. Acad. Sci. USA (1985) 82, 488-492)) и перекрывающаяся ПЦР. Более того, также можно использовать несколько известных способов в качестве способов изменения аминокислот для замены на неприродные аминокислоты (Annu. Rev. Biophys. Biomol. Struct. (2006) 35, 225-249; и Proc. Natl. Acad. Sci. U.S.A. (2003) 100 (11), 6353-6357). Например, является пригодным использование бесклеточной системы трансляции (Clover Direct (Protein Express)), включающей тРНК, в которой неприродная аминокислота связана с комплементарной амбер-супрессорной тРНК одного из стоп-кодонов, кодона UAG (амбер-кодон).
В настоящем описании значение термина "и/или" при описании участка изменения аминокислот, включает каждую комбинацию, где "и" и "или" пригодным образом скомбинированы. В частности, например, "аминокислоты в положениях 33, 55 и/или 96 являются замещенными" включают следующие варианты аминокислотных изменений:
аминокислота(ы) в (a) положении 33, (b) положении 55, (c) положении 96, (d) положениях 33 и 55, (e) положениях 33 и 96, (f) положениях 55 и 96, и (g) положениях 33, 55 и 96.
Более того, в настоящем описании в качестве выражения, демонстрирующего изменение аминокислот, можно соответствующим образом использовать выражение, которое демонстрирует, до и после номера, указывающего на конкретное положение, однобуквенные или трехбуковенные коды для аминокислот до и после изменения, соответственно. Например, изменение N100bL или Asn100bLeu, используемое при замене аминокислоты, содержащейся в вариабельной области антитела, указывает на замену Asn в положении 100b (в соответствии с нумерацией Kabat) на Leu. Иными словами, номер показывает положение аминокислоты в соответствии с нумерацией Kabat, однобуквенный или трехбуквенный код аминокислоты до номера указывает на аминокислоту до замены, и однобуквенный или трехбуквенный код аминокислоты после номера указывает на аминокислоту после замены. Аналогично, изменение P238D или Pro238Asp, используемое при замене аминокислоты Fc-области, содержащейся в константной области антитела, указывает на замену Pro в положении 238 (в соответствии с нумерацией EU) на Asp. Иными словами, номер демонстрирует положение аминокислоты в соответствии с нумерацией EU, однобуквенный или трехбуквенный код аминокислоты до номера указывает на аминокислоту до замены, и однобуквенный или трехбуквенный код аминокислоты после номера указывает на аминокислоту после замены.
Антигены
В рамках настоящего изобретения, "антигены" конкретно не ограничены их структурой, при условии, что они содержат эпитопы, с которыми связываются антигенсвязывающие домены. Иными словами, антигены могут представлять собой неорганические или органические вещества. Другие антигены включают, например, приведенные ниже молекулы: 17-IA, 4-1BB, 4Dc, 6-кето-PGF1a, 8-изо-PGF2a, 8-оксо-dG, рецептор аденозина A1, A33, ACE, ACE-2, активин, активин A, активин AB, активин B, активин C, активин RIA, активин RIA ALK-2, активин RIB ALK-4, активин RIIA, активин RIIB, ADAM, ADAM10, ADAM12, ADAM15, ADAM17/TACE, ADAM8, ADAM9, ADAMTS, ADAMTS4, ADAMTS5, адрессин, aFGF, ALCAM, ALK, ALK-1, ALK-7,альфа-1-антитрипсин, антагонист альфа-V/бета-1, ANG, Ang, APAF-1, APE, APJ, APP, APRIL, AR, ARC, ART, артемин, анти-Id, ASPARTIC, предсердный натрийуретический пептид, интегрин av/b3, Axl, b2M, B7-1, B7-2, B7-H, стимулирующий B-лимфоцииты фактор (BlyS), BACE, BACE-1, Bad, BAFF, BAFF-R, Bag-1, BAK, Bax, BCA-1, BCAM, Bcl, BCMA, BDNF, b-ECGF, bFGF, BID, Bik, BIM, BLC, BL-CAM, BLK, BMP, BMP-2 BMP-2a, остеогенин BMP-3, BMP-4 BMP-2b, BMP-5, BMP-6 Vgr-1, BMP-7 (OP-1), BMP-8 (BMP-8a, OP-2), BMPR, BMPR-IA (ALK-3), BMPR-IB (ALK-6), BRK-2, RPK-1, BMPR-II (BRK-3), BMP, b-NGF, BOK, бомбезин, нейротрофический фактор костей, BPDE, BPDE-ДНК, BTC, фактор комплемента 3 (C3), C3a, C4, C5, C5a, C10, CA125, CAD-8, кальцитонин, cAMP, карциноэмбриональный антиген (CEA), ассоциированный со злокачественной опухолью антиген, катепсин A, катепсин B, катепсин C/DPPI, катепсин D, катепсин E, катепсин H, катепсин L, катепсин O, катепсин S, катепсин V, катепсин X/Z/P, CBL, CCI, CCK2, CCL, CCL1, CCL11, CCL12, CCL13, CCL14, CCL15, CCL16, CCL17, CCL18, CCL19, CCL2, CCL20, CCL21, CCL22, CCL23, CCL24, CCL25, CCL26, CCL27, CCL28, CCL3, CCL4, CCL5, CCL6, CCL7, CCL8, CCL9/10, CCR, CCR1, CCR10, CCR10, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CD1, CD2, CD3, CD3E, CD4, CD5, CD6, CD7, CD8, CD10, CD11a, CD11b, CD11c, CD13, CD14, CD15, CD16, CD18, CD19, CD20, CD21, CD22, CD23, CD25, CD27L, CD28, CD29, CD30, CD30L, CD32, CD33 (p67 белок), CD34, CD38, CD40, CD40L, CD44, CD45, CD46, CD49a, CD52, CD54, CD55, CD56, CD61, CD64, CD66e, CD74, CD80 (B7-1), CD89, CD95, CD123, CD137, CD138, CD140a, CD146, CD147, CD148, CD152, CD164, CEACAM5, CFTR, cGMP, CINC, ботулинический токсин, токсин Clostridium perfringens, CKb8-1, CLC, CMV, CMV UL, CNTF, CNTN-1, COX, C-Ret, CRG-2, CT-1, CTACK, CTGF, CTLA-4, PD1, PDL1, LAG3, TIM3, галектин-9, CX3CL1, CX3CR1, CXCL, CXCL1, CXCL2, CXCL3, CXCL4, CXCL5, CXCL6, CXCL7, CXCL8, CXCL9, CXCL10, CXCL11, CXCL12, CXCL13, CXCL14, CXCL15, CXCL16, CXCR, CXCR1, CXCR2, CXCR3, CXCR4, CXCR5, CXCR6, цитокератиновый ассоциированный с опухолью антиген, DAN, DCC, DcR3, DC-SIGN, регуляторный фактор комплемента (фактор, ускоряющий распад), дез-(1-3)-IGF-I (IGF-1 головного мозга), Dhh, дигоксин, DNAM-1, ДНК-аза, Dpp, DPPIV/CD26, Dtk, ECAD, EDA, EDA-A1, EDA-A2, EDAR, EGF, EGFR (ErbB-1), EMA, EMMPRIN, ENA, рецептор эндотелина, энкефалиназа, eNOS, Eot, эотаксин 1, EpCAM, эфрин B2/EphB4, EPO, ERCC, E-селектин, ET-1, фактор IIa, фактор VII, фактор VIIIc, фактор IX, активирующий фибробласты белок (FAP), Fas, FcR1, FEN-1, ферритин, FGF, FGF-19, FGF-2, FGF3, FGF-8, FGFR, FGFR-3, фибрин, FL, FLIP, Flt-3, Flt-4, фоликулостимулирующий гормон, фракталкин, FZD1, FZD2, FZD3, FZD4, FZD5, FZD6, FZD7, FZD8, FZD9, FZD10, G250, Gas6, GCP-2, GCSF, GD2, GD3, GDF, GDF-1, GDF-3 (Vgr-2), GDF-5 (BMP-14, CDMP-1), GDF-6 (BMP-13, CDMP-2), GDF-7 (BMP-12, CDMP-3), GDF-8 (миостатин), GDF-9, GDF-15 (MIC-1), GDNF, GDNF, GFAP, GFRa-1, GFR-альфа 1, GFR-альфа 2, GFR-альфа 3, GITR, глюкагон, Glut4, гликопротеин IIb/IIIa (GPIIb/IIIa), GM-CSF, gp130, gp72, GRO, рилизинг-фактор гормона роста, гаптен (NP-cap или NIP-cap), HB-EGF, HCC, гликопротеин оболочки HCMV gB, гликопротеин оболочки HCMV gH, HCMV UL, гемопоэтический фактор роста (HGF), Hep B gp120, гепараназа, Her2, Her2/neu (ErbB-2), Her3 (ErbB-3), Her4 (ErbB-4), гликопротеин gB вируса простого герпеса (HSV), гликопротеин gD HSV, HGFA, высокомолекулярный ассоциированный с меланомой антиген (HMW-MAA), gp120 HIV, петля V3 gp 120 IIIB HIV, HLA, HLA-DR, HM1,24, HMFG PEM, HRG, Hrk, сердечный миозин человека, цитомегаловирус человека (HCMV), гормон роста человека (HGH), HVEM, I-309, IAP, ICAM, ICAM-1, ICAM-3, ICE, ICOS, IFNg, Ig, рецептор IgA, IgE, IGF, IGF-связывающий белок, IGF-1R, IGFBP, IGF-I, IGF-II, IL, IL-1, IL-1R, IL-2, IL-2R, IL-4, IL-4R, IL-5, IL-5R, IL-6, IL-6R, IL-8, IL-9, IL-10, IL-12, IL-13, IL-15, IL-18, IL-18R, IL-21, IL-23, IL-27, интерферон (INF)-альфа, INF-бета, INF-гамма, ингибин, iNOS, A-цепь инсулина, B-цепь инсулина, инсулиноподобный фактор роста 1, интегрин альфа 2, интегрин альфа 3, интегрин альфа 4, интегрин альфа 4/бета 1, интегрин альфа 4/бета 7, интегрин альфа 5 (альфа V), интегрин альфа 5/бета 1, интегрин альфа 5/бета 3, интегрин альфа 6, интегрин бета 1, интегрин бета 2, интерферон-гамма, IP-10, I-TAC, JE, калликреин 2, калликреин 5, калликреин 6, калликреин 11, калликреин 12, калликреин 14, калликреин 15, калликреин L1, калликреин L2, калликреин L3, калликреин L4, KC, KDR, фактор роста кератиноцитов (KGF), ламинин 5, LAMP, LAP, LAP (TGF-1), латентный TGF-1, латентный TGF-1 bp1, LBP, LDGF, LECT2, lefty, антиген Lewis-Y, ассоциированный с Lewis-Y антиген, LFA-1, LFA-3, Lfo, LIF, LIGHT, липопротеин, LIX, LKN, Lptn, L-селектин, LT-a, LT-b, LTB4, LTBP-1, белок поверхности легких, лютеинизирующий гормон, рецептор лимфотоксина-бета, Mac-1, MAdCAM, MAG, MAP2, MARC, MCAM, MCAM, MCK-2, MCP, M-CSF, MDC, Mer, металлопротеиназы, рецептор MGDF, MGMT, MHC (HLA-DR), MIF, MIG, MIP, MIP-1-альфа, MK, MMAC1, MMP, MMP-1, MMP-10, MMP-11, MMP-12, MMP-13, MMP-14, MMP-15, MMP-2, MMP-24, MMP-3, MMP-7, MMP-8, MMP-9, MPIF, Mpo, MSK, MSP, муцин (Muc1), MUC18, мюллерово ингибиторное вещество, Mug, MuSK, NAIP, NAP, NCAD, N-C адгерин, NCA 90, NCAM, NCAM, неприлизин, нейротрофин -3, -4 или -6, нейротурин, фактор роста нервов (NGF), NGFR, NGF-бета, nNOS, NO, NOS, Npn, NRG-3, NT, NTN, OB, OGG1, OPG, OPN, OSM, OX40L, OX40R, p150, p95, PADPr, паратиреоидный гормон, PARC, PARP, PBR, PBSF, PCAD, P-кадгерин, PCNA, PDGF, PDGF, PDK-1, PECAM, PEM, PF4, PGE, PGF, PGI2, PGJ2, PIN, PLA2, плацентарная щелочная фосфатаза (PLAP), PlGF, PLP, PP14, проинсулин, прорелаксин, белок C, PS, PSA, PSCA, простатспецифический мембранный антиген (PSMA), PTEN, PTHrp, Ptk, PTN, R51, RANK, RANKL, RANTES, RANTES, A-цепь релаксина, B-цепь релаксина, ренин, F респираторно-синцитиального вируса (RSV), RSV Fgp, Ret, ревматоидный фактор, RLIP76, RPA2, RSK, S100, SCF/KL, SDF-1, SERINE, сывороточный альбумин, sFRP-3, Shh, SIGIRR, SK-1, SLAM, SLPI, SMAC, SMDF, SMOH, SOD, SPARC, Stat, STEAP, STEAP-II, TACE, TACI, TAG-72 (ассоциированный с опухолью гликопротеин-72), TARC, TCA-3, T-клеточный рецептор (например, T-клеточный рецептор альфа/бета), TdT, TECK, TEM1, TEM5, TEM7, TEM8, TERT, PLAP-подобная щелочная фосфатаза семенников, TfR, TGF, TGF-альфа, TGF-бета, панспецифический TGF-бета, TGF-бета RI (ALK-5), TGF-бета RII, TGF-бета RIIb, TGF-бета RIII, TGF-бета 1, TGF-бета 2, TGF-бета 3, TGF-бета 4, TGF-бета 5, тромбин, Ck-1 тимуса, тиреостимулирующий гормон, Tie, TIMP, TIQ, тканевой фактор, TMEFF2, Tmpo, TMPRSS2, TNF, TNF-альфа, TNF-альфа/бета, TNF-бета 2, TNFc, TNF-RI, TNF-RII, TNFRSF10A (TRAIL R1 Apo-2, DR4), TNFRSF10B (TRAIL R2 DR5, KILLER, TRICK-2A, TRICK-B), TNFRSF10C (TRAIL R3 DcR1, LIT, TRID), TNFRSF10D (TRAIL R4 DcR2, TRUNDD), TNFRSF11A (RANK ODF R, TRANCE R), TNFRSF11B (OPG OCIF, TR1), TNFRSF12 (TWEAK R FN14), TNFRSF13B (TACI), TNFRSF13C (BAFF R), TNFRSF14 (HVEM ATAR, HveA, LIGHT R, TR2), TNFRSF16 (NGFR p75NTR), TNFRSF17 (BCMA), TNFRSF18 (GITR AITR), TNFRSF19 (TROY TAJ, TRADE), TNFRSF19L (RELT), TNFRSF1A (TNF RI CD120a, p55-60), TNFRSF1B (TNF RII CD120b, p75-80), TNFRSF26 (TNFRH3), TNFRSF3 (LTbR TNF RIII, TNFC R), TNFRSF4 (OX40 ACT35, TXGP1 R), TNFRSF5 (CD40 p50), TNFRSF6 (Fas Apo-1, APT1, CD95), TNFRSF6B (DcR3 M68, TR6), TNFRSF7 (CD27), TNFRSF8 (CD30), TNFRSF9 (4-1BB CD137, ILA), TNFRSF21 (DR6), TNFRSF22 (DcTRAIL R2 TNFRH2), TNFRST23 (DcTRAIL R1 TNFRH1), TNFRSF25 (DR3 Apo-3, LARD, TR-3, TRAMP, WSL-1), TNFSF10 (лиганд Apo-2 TRAIL, TL2), TNFSF11 (лиганд TRANCE/RANK ODF, лиганд OPG), TNFSF12 (лиганд TWEAK Apo-3, лиганд DR3), TNFSF13 (APRIL TALL2), TNFSF13B (BAFF BLYS, TALL1, THANK, TNFSF20), TNFSF14 (лиганд LIGHT HVEM, LTg), TNFSF15 (TL1A/VEGI), TNFSF18 (лиганд GITR лиганда AITR, TL6), TNFSF1A (TNF-a конектин, DIF, TNFSF2), TNFSF1B (TNF-b LTa, TNFSF1), TNFSF3 (LTb TNFC, p33), TNFSF4 (лиганд OX40 gp34, TXGP1), TNFSF5 (лиганд CD40 CD154, gp39, HIGM1, IMD3, TRAP), TNFSF6 (Fas-лиганд лиганда Apo-1, лиганд APT1), TNFSF7 (CD27-лиганд CD70), TNFSF8 (CD30-лиганд CD153), TNFSF9 (лиганд 4-1BB лиганда CD137), TP-1, t-PA, Tpo, TRAIL, TRAIL R, TRAIL-R1, TRAIL-R2, TRANCE, рецептор трансферрина, TRF, Trk, TROP-2, TLR1 (Toll-подобный рецептор 1), TLR2, TLR3, TLR4, TLR5, TLR6, TLR7, TLR8, TLR9, TLR10, TSG, TSLP, ассоциированный с опухолью антиген CA125, ассоциированный с опухолью антиген, экспрессирующий ассоциированные с Lewis-Y углеводы, TWEAK, TXB2, Ung, uPAR, uPAR-1, урокиназа, VCAM, VCAM-1, VECAD, VE-кадгерин, VE-кадгерин-2, VEFGR-1 (flt-1), VEGF, VEGFR, VEGFR-3 (flt-4), VEGI, VIM, вирусные антигены, VLA, VLA-1, VLA-4, интегрин VNR, фактор фон Виллебранда, WIF-1, WNT1, WNT2, WNT2B/13, WNT3, WNT3A, WNT4, WNT5A, WNT5B, WNT6, WNT7A, WNT7B, WNT8A, WNT8B, WNT9A, WNT9A, WNT9B, WNT10A, WNT10B, WNT11, WNT16, XCL1, XCL2, XCR1, XCR1, XEDAR, XIAP, XPD, HMGB1, IgA, Aβ, CD81, CD97, CD98, DDR1, DKK1, EREG, Hsp90, IL-17/IL-17R, IL-20/IL-20R, окисленный LDL, PCSK9, прекалликреин, RON, TMEM16F, SOD1, хромогранин A, хромогранин B, белок тау, VAP1, высокомолекулярный кининоген, IL-31, IL-31R, Nav1.1, Nav1.2, Nav1.3, Nav1.4, Nav1.5, Nav1.6, Nav1.7, Nav1.8, Nav1.9, EPCR, C1, C1q, C1r, C1s, C2, C2a, C2b, C3, C3a, C3b, C4, C4a, C4b, C5, C5a, C5b, C6, C7, C8, C9, фактор B, фактор D, фактор H, пропердин, склеростин, фибриноген, фибрин, протромбин, тромбин, тканевой фактор, фактор V, фактор Va, фактор VII, фактор VIIa, фактор VIII, фактор VIIIa, фактор IX, фактор IXa, фактор X, фактор Xa, фактор XI, фактор XIa, фактор XII, фактор XIIa, фактор XIII, фактор XIIIa, TFPI, антитромбин III, EPCR, тромбомодулин, TAPI, tPA, плазминоген, плазмин, PAI-1, PAI-2, GPC3, синдекан-1, синдекан-2, синдекан-3, синдекан-4, LPA и S1P; и рецепторы для гормонов и факторов роста. Предпочтительными антигенами являются антигены, которые экспрессируются в злокачественных клетках, иммунных клетках, стромальных клетках и т.п., присутствующих в злокачественных тканях или воспалительных тканях.
Хотя рецепторы приведены в качестве примеров упомянутых выше антигенов, когда эти рецепторы существуют в растворимых формах в биологических жидкостях, их можно использовать в качестве антигенов, которые связываются с антигенсвязывающей молекулой по настоящему изобретению, которая содержит антигенсвязывающий домен, антигенсвязывающая активность которого варьирует в зависимости от концентрации специфичного к ткани-мишени соединения. Примером неограничивающего варианта осуществления такого растворимого рецептора является растворимый IL-6R, который представляет собой белок, состоящий из аминокислот в положениях 1-357 в полипептидной последовательности IL-6R SEQ ID NO:1, как описано в Mullberg et al. (J. Immunol. (1994) 152 (10), 4958-4968).
Молекулы мембранного типа, экспрессируемые на клеточных мембранах, и растворимые молекулы, секретируемые из клеток наружу, входят в примеры упомянутых выше антигенов. Когда антигенсвязывающая молекула по настоящему изобретению, которая содержит антигенсвязывающий домен, антигенсвязывающая активность которого варьирует в зависимости от концентрации специфичного к ткани-мишени соединения, связывается с растворимой молекулой, секретируемой из клеток, предпочтительно, чтобы антигенсвязывающая молекула обладала нейтрализующей активностью, как описано ниже.
Жидкости, в которых существуют растворимые молекулы, не ограничены, и растворимые молекулы могут существовать в биологических жидкостях или, более конкретно, во всех жидкостях, заполняющих пространство между тканями и клетками или сосуды в организмах. В неограничивающем варианте осуществления растворимые молекулы, с которыми связываются антигенсвязывающие молекулы по настоящему изобретению, могут присутствовать во внеклеточной жидкости. У позвоночных внеклеточная жидкость является общим термином для плазмы, интерстициальной жидкости, лимфы, плотной соединительной ткани, цереброспинальной жидкости, спинномозговой жидкости, жидкости пунктата, синовиальной жидкости или сходных компонентов в кости и хряще, альвеолярной жидкости (жидкость бронхоальвеолярного лаважа), перитонеальной жидкости, плевральной жидкости, перикардиального выпота, жидкости кисты, водянистой влаги (внутриглазная жидкость) или сходных трансцеллюлярных жидкостей (различные жидкости в полостях желез и жидкости в полости пищеварительного тракта и другие жидкости полостей тела, продуцированные в результате активного транспорта/секреторной активности клеток).
Когда антигенсвязывающая молекула по настоящему изобретению, содержащая антигенсвязывающий домен, антигенсвязывающая активность которого варьирует в зависимости от концентрации специфичного к ткани-мишени соединения, связывается с молекулой мембранного типа, экспрессируемой на клеточной мембране, подходящие примеры антигенсвязывающей молекулы включают антигенсвязывающие молекулы, которые обладают цитотоксической активностью, связываются с цитотоксическим веществом или обладают способностью связываться с цитотоксическим веществом, как описано ниже. Более того, антигенсвязывающие молекулы, обладающие нейтрализующей активностью вместо свойств наличия цитотоксической активности, связывающиеся с цитотоксическим веществом или обладающие способностью связываться с цитотоксическим веществом; или в дополнение к этим свойствам, также являются пригодными примерами неограничивающего варианта осуществления.
Эпитопы
"Эпитоп" означает антигенную детерминанту в антигене и относится к антигенному центру, с которым связывается антигенсвязывающий домен антигенсвязывающей молекулы, описанной в настоящем описании. Таким образом, например, эпитоп может быть определен в соответствии с его структурой. Альтернативно эпитоп может быть определен в соответствии с антигенсвязывающей активностью антигенсвязывающей молекулы, которая распознает эпитоп. Когда антиген представляет собой пептид или полипептид, эпитоп может быть указан с помощью аминокислотных остатков, образующих эпитоп. Альтернативно, когда эпитоп представляет собой цепь сахара, эпитоп может быть указан с помощью его определенной структуры цепи сахаров.
Линейный эпитоп представляет собой эпитоп, который содержит эпитоп, в котором распознается его первичная аминокислотная последовательность. Такой линейный эпитоп, как правило, содержит по меньшей мере три и наиболее часто по меньшей мере пять, например, от приблизительно 8 до приблизительно 10 или 6-20 аминокислот в конкретной последовательности.
В противоположность линейному эпитопу, "конформационный эпитоп" представляет собой эпитоп, в котором первичная аминокислотная последовательность, содержащая эпитоп, не является единственной детерминантой распознаваемого эпитопа (например, первичная аминокислотная последовательность конформационного эпитопа не обязательно распознается определяющим эпитоп антителом). Конформационные эпитопы могут содержать большее количество аминокислот по сравнению с линейными эпитопами. Антитело, распознающее конформационный эпитоп, распознает трехмерную структуру пептида или белка. Например, когда молекула белка сворачивается и формирует трехмерную структуру, аминокислоты и/или основные цепи полипептида, которые образуют коноформационный эпитоп, располагаются рядом друг с другом, и эпитоп становится распознаваемым для антитела. Способы определения конформаций эпитопов включают, например, рентгеновскую кристаллографию, двумерный ядерный магнитный резонанс, сайт-специфическое спиновое мечение и электронный парамагнитный резонанс, он они не ограничиваются ими. См., например, Epitope Mapping Protocols in Methods in Molecular Biology (1996), Vol. 66, Morris (ed.).
Структуру антигенсвязывающего домена, который связывается с эпитопом, называют паратопом. Эпитоп и паратоп стабильно связываются через воздействие водородных связей, электростатических сил, ванд-дер-ваальсовых сил, гидрофобных связей и т.п. между эпитопом и паратопом. Эту прочность связывания между эпитопом и паратопом называют аффинностью. Общая суммарная прочность связывания, когда связывается множество антигенов и множество антигенсвязывающих молекул, называется авидностью. Когда антитело, содержащее множество антигенсвязывающих доменов (т.е. поливалентное антитело), или сходное с ним связывается с множеством эпитопов, аффинность действует синергично, и, таким образом, авидность становится более высокой, чем аффинность.
Активность связывания
Примеры способа оценки связывания эпитопа исследуемой антигенсвязывающей молекулой, содержащей антигенсвязывающий домен для IL-6R, описаны ниже. В соответствии с примерами ниже, также можно соответствующим образом выполнять способы оценки связывания эпитопа исследуемой антигенсвязывающей молекулой, содержащей антигенсвязывающий домен, с антигеном, отличным от IL-6R.
Например, то, что исследуемая антигенсвязывающая молекула, содержащая антигенсвязывающий домен для IL-6R, распознает линейный эпитоп в молекуле IL-6R, можно подтвердить, например, как описано ниже. Для указанной выше цели синтезируют линейный пептид, содержащий аминокислотную последовательность, образующую внеклеточный домен IL-6R. Пептид можно синтезировать химически или его можно получать способами генетической инженерии с использованием области, кодирующей аминокислотную последовательность, соответствующую внеклеточному домену в кДНК IL-6R. Затем исследуемую антигенсвязывающую молекулу, содержащую антигенсвязывающий домен для IL-6R, оценивают в отношении ее активности связывания с линейным пептидом, содержащим аминокислотную последовательность, образующую внеклеточный домен. Например, иммобилизованный линейный пептид можно использовать в качестве антигена в ELISA для оценки активности связывания антигенсвязывающей молекулы с пептидом. Альтернативно активность связывания с линейным пептидом можно оценивать, исходя из уровня, на котором линейный пептид ингибирует связывание антигенсвязывающей молекулы с клетками, экспрессирующими IL-6R. Эти исследования могут продемонстрировать активность связывания антигенсвязывающей молекулы с линейным пептидом.
Наличие распознавания исследуемой антигенсвязывающей молекулой, содержащей антигенсвязывающий домен для IL-6R, конформационного эпитопа можно оценивать следующим образом. Для указанной выше цели получают экспрессирующие IL-6R клетки. Наличие распознавания исследуемой антигенсвязывающей молекулой, содержащей антигенсвязывающий домен IL-6R, конформационного эпитопа, можно определять, когда она прочно связывается с экспрессирующими IL-6R клетками при контакте, но по существу не связывается с иммобилизованным линейным пептидом, содержащим аминокислотную последовательность, образующую внеклеточный домен IL-6R. В рамках настоящего изобретения "по существу не связывает" означает, что активность связывания составляет 80% или менее, как правило, 50% или менее, предпочтительно 30% или менее, и особенно предпочтительно 15% или менее по сравнению с активностью связывания с клетками, экспрессрующими IL-6R человека.
Способы оценки активности связывания исследуемой антигенсвязывающей молекулы, содержащей антигенсвязывающий домен для IL-6R, с экспрессирующими IL-6R клетками, включают, например, способы, описанные в Antibodies: A Laboratory Manual (Ed Harlow, David Lane, Cold Spring Harbor Laboratory (1988) 359-420). В частности, оценку можно проводить на основе принципа ELISA или активированной флуоресценцией сортировки клеток (FACS) с использованием в качестве антигена экспрессирующих IL-6R клеток.
В формате ELISA активность связывания исследуемой антигенсвязывающей молекулы, содержащей антигенсвязывающий домен для IL-6R, с экспрессирующими IL-6R клетками можно оценивать количественно путем сравнения уровней сигнала, генерируемого ферментативной реакцией. В частности, исследуемый полипептидный комплекс добавляют в планшет для ELISA, на котором иммобилизованы эксперессирующие IL-6R клетки. Затем антигенсвязывающую молекулу, связанную с клетками, выявляют с использованием меченного ферментом антитела, которое распознает исследуемую антигенсвязывающую молекулу. Альтернативно, когда используют FACS, приготавливают серии разведений исследуемой антигенсвязывающей молекулы и можно определять титр связывания антитела с экспрессирующими IL-6R клетками для сравнения активности связывания исследуемой антигенсвязывающей молекулы с экспрессирующими IL-6R клетками.
Связывание исследуемой антигенсвязывающей молекулы с антигеном, экспрессируемым на поверхности клеток, суспендированных в буфере или сходных с ним, можно выявлять с использованием проточного цитометра. Известные проточные цитометры включают, например, следующие устройства:
FACSCanto™ II
FACSAria™
FACSArray™
FACSVantage™ SE
FACSCalibur™ (все являются торговыми названиями BD Biosciences)
EPICS ALTRA HyPerSort
Cytomics FC 500
EPICS XL-MCL ADC EPICS XL ADC
Cell Lab Quanta/Cell Lab Quanta SC (все являются торговыми названиями Beckman Coulter).
Предпочтительные способы оценки активности связывания исследуемой антигенсвязывающей молекулы, содержащей антигенсвязывающий домен для IL-6R, с антигеном, включают, например, следующий способ. Сначала экспрессирующие IL-6R клетки подвергают реакции с исследуемой антигенсвязывающей молекулой, а затем их окрашивают меченным FITC вторичным антителом, которое распознает антигенсвязывающую молекулу. Исследуемую антигенсвязывающую молекулу надлежащим образом разбавляют подходящим буфером с получением молекулы с желаемой концентрацией. Например, молекулу можно использовать в концентрации в диапазоне от 10 мкг/мл до 10 нг/мл. Затем определяют интенсивность флуоресценции и количество клеток с использованием FACSCalibur (BD). Интенсивность флуоресценции, полученную с помощью анализа с использованием программного обеспечения CELL QUEST (BD), т.е. среднее геометрическое значение, отражает количество антитела, связанного с клетками. Следовательно, активность связывания исследуемой антигенсвязывающей молекулы, которую отражает количество связанной исследуемой антигенсвязывающей молекулы, можно определять путем определения геометрического среднего значения.
То, обладает ли исследуемая антигенсвязывающая молекула, содержащая антигенсвязывающий домен для IL-6R, общим эпитопом с другой антигенсвязывающей молекулой, можно оценивать на основе конкуренции между двумя молекулами за один и тот же эпитоп. Конкуренцию между антигенсвязывающими молекулами можно выявлять с помощью анализа перекрестного блокирования или сходных с ним. Например, предпочтительным анализом перекрестного блокирования является конкурентный анализ ELISA.
В частности, в анализе перекрестного блокирования белок IL-6R, иммобилизованный на лунках микропланшета для титрования, предварительно инкубируют в присутствии или в отсутствие являющейся кандидатом конкурентной антигенсвязывающей молекулы, а затем к ним добавляют антигенсвязывающую молекулу. Количество исследуемой антигенсвязывающей молекулы, связавшейся с белком IL-6R в лунках, непрямо коррелирует со связывающей способностью являющейся кандидатом конкурентной антигенсвязывающей молекулы, которая конкурирует за связывание с тем же эпитопом. Следовательно, чем более высокой является аффинность конкурентной антигенсвязывающей молекулы к тому же эпитопу, тем более низкой является активность связывания исследуемой антигенсвязывающей молекулы с покрытыми белком IL-6R лунками.
Количество исследуемой антигенсвязывающей молекулы, связанной с лунками через IL-6R, можно легко определить путем предварительного мечения антигенсвязывающей молекулы. Например, меченную биотином антигенсвязывающую молекулу определяют с использованием конъюгата авидин/пероксидаза и соответствующего субстрата. В частности, анализ перекрестного блокирования, в котором используются ферментные метки, такие как пероксидаза, называют "конкурентным анализом ELISA". Антигенсвязывающую молекулу также можно метить другими агентами для мечения, которые обеспечивают выявление или измерение. В частности, известны радиоактивные метки, флуоресцентные метки и т.п.
Когда являющаяся кандидатом конкурентная антигенсвязывающая молекула может блокировать связывание исследуемой антигенсвязывающей молекулой, содержащей антигенсвязывающий домен для IL-6R, по меньшей мере на 20%, предпочтительно по меньшей мере на 20-50%, и более предпочтительно по меньшей мере на 50% по сравнению с активностью связывания в контрольном эксперименте, исследованном в отсутствие конкурентной антигенсвязывающей молекулы, то определяют, что исследуемая антигенсвязывающая молекула по существу связывается с тем же эпитопом, с которым связывается конкурентная антигенсвязывающая молекула, или конкурирует за связывание с тем же эпитопом.
Когда структура эпитопа, с которым связывается исследуемая антигенсвязывающая молекула, содержащая антигенсвязывающий домен для IL-6R, уже идентифицирована, тогда то, имеют ли исследуемые и контрольные антигенсвязывающие молекулы общий эпитоп, можно оценивать путем сравнения активности связывания двух антигенсвязывающих молекул с пептидом, полученным путем внесения аминокислотных мутаций в пептид, формирующий эпитоп.
Для измерения указанной выше активности связывания, например, активность связывания исследуемых и контрольных антигенсвязывающих молекул в отношении линейного пептида, в который внесена мутация, сравнивают в описанном выше формате ELISA. Помимо способов ELISA, активность связывания с мутантным пептидом, связанным с колонкой, можно определять путем пропускания исследуемых и контрольных антигенсвязывающих молекул через колонку, а затем количественного определения антигенсвязывающей молекулы, элюированной в элюирующем растворе. Способы адсорбции мутантного пептида на колонку, например, в форме слитого пептида с GST, известны.
Альтернативно, когда идентифицированный эпитоп представляет собой конформационный эпитоп, тогда то, обладают ли исследуемые и контрольные антигенсвязывающие молекулы общим эпитопом, можно оценивать следующим способом. Сначала получают экспрессирующие IL-6R клетки и клетки, экспрессирующие IL-6R с мутацией, внесенной в эпитоп. Исследуемые и контрольные антигенсвязывающие молекулы добавляют в суспензию клеток, полученную суспендированием этих клеток в соответствующем буфере, таком как PBS. Затем клеточные суспензии надлежащим образом промывают буфером, и к ним добавляют меченное FITC антитело, которое распознает исследуемые и контрольные антигенсвязывающие молекулы. Интенсивность флуоресценции и количество клеток, окрашенных меченым антителом, определяют с использованием FACSCalibur (BD). Исследуемые и контрольные антигенсвязывающие молекулы надлежащим образом разбавляют с использованием подходящего буфера и используют в желаемых концентрациях. Например, их можно использовать в концентрации в диапазоне от 10 мкг/мл до 10 нг/мл. Интенсивность флуоресценции, определенная с помощью анализа с использованием программного обеспечения CELL QUEST (BD), т.е. геометрическое среднее значение, отражает количество меченого антитела, связанного с клетками. Следовательно, активность связывания исследуемых и контрольных антигенсвязывающих молекул, которую отражает количество связавшегося меченого антитела, можно определять путем измерения геометрического среднего значения.
В описанном выше способе то, что антигенсвязывающая молекула "по существу не связывается с клетками, экспрессирующими мутантный IL-6R", можно оценивать, например, следующим способом. Сначала исследуемые и контрольные антигенсвязывающие молекулы, связанные с клетками, экспрессирующими мутантный IL-6R, окрашивают меченым антителом. Затем определяют интенсивность флуоресценции клеток. Когда используют FACSCalibur для выявления флуоресценции проточной цитометрией, определенную интенсивность флуоресценции можно анализировать с использованием программного обеспечения CELL QUEST. Из геометрических средних значений в присутствии и в отсутствие полипептидного комплекса, можно вычислять сравнительное значение (ΔGeo-Mean) по формуле 1, ниже, для определения соотношения увеличения интенсивности флуоресценции в результате связывания антигенсвязывающей молекулой.
Формула 1:
ΔGeo-Mean=Geo-Mean (в присутствии полипептидного комплекса)/ Geo-Mean (в отсутствие полипептидного комплекса)
Геометрическое среднее сравнительное значение (значение ΔGeo-Mean для мутантной молекулы IL-6R), определенное с помощью описанного выше анализа, которое отражает количество исследуемой антигенсвязывающей молекулы, связанной с клетками, экспрессирующими мутантный IL-6R, сравнивают со сравнительным значением ΔGeo-Mean, которое отражает количество исследуемой антигенсвязывающей молекулы, связавшейся с экспрессирующими IL-6R клетками. В этом случае, особенно предпочтительно, чтобы концентрации исследуемой антигенсвязывающей молекулы, используемой для определения сравнительных значений ΔGeo-Mean для экспрессирующих IL-6R клеток и клеток, экспрессирующих мутантный IL-6R, были скорректированы так, чтобы они были равными или по существу равными. В качестве контрольной антигенсвязывающей молекулы используют антигенсвязывающую молекулу, для которой подтверждено, что она распознает эпитоп в IL-6R.
Если сравнительное значение ΔGeo-Mean исследуемой антигенсвязывающей молекулы для клеток, экспрессирующих мутантный IL-6R, меньше, чем сравнительное значение ΔGeo-Mean исследуемой антигенсвязывающей молекулы для экспрессирующих IL-6R клеток, по меньшей мере на 80%, предпочтительно на 50%, более предпочтительно на 30%, и особенно предпочтительно на 15%, тогда исследуемая антигенсвязывающая молекула "по существу не связывается с клетками, экспрессирующими мутантный IL-6R". Формула для определения значения Geo-Mean (геометрического среднего значения) описана в руководстве пользователя программного обеспечения CELL QUEST (BD biosciences). Если сравнение показывает, что сравнительные величины по существу эквивалентны, то может быть определено, что эпитоп для исследуемой и контрольной антигенсвязывающих молекул является одним и тем же.
Ткань-мишень
Термин "ткань-мишень", как используют в рамках изобретения, относится к ткани, содержащей клетки, содержащие антигены, с которыми связываются антигенсвязывающие молекулы по настоящему изобретению зависимым от соединений образом. Она представляет собой ткань, которая обеспечивает положительные фармакологические эффекты для организма, содержащего ткань, когда антигенсвязывающие молекулы связываются с молекулой мембранного типа, экспрессируемой на клетках, или связывается с растворимой молекулой, присутствующей в ткани. В этом случае выражение "положительные фармакологические эффекты" относится к эффектам, которые облегчают, смягчают, ослабляют или излечивают симптомы, обеспечиваемые патологическими участками, содержащими ткань-мишень, для организма, содержащего ткань. Когда симптомы вызваны злокачественными опухолями, такими как рак, неограничивающим вариантом осуществления механизма, который обеспечивает такой фармакологический эффект, является, например, цитотоксическая активность и ингибирование роста злокачественных клеток, и стимуляция иммунитета в злокачественных тканях. В случае воспалительных заболеваний, примеры такого неограничивающего варианта осуществления механизма включают иммуносупрессию и активность блокирования действий воспалительных цитокинов в воспаленных тканях.
Соединение, специфическое для злокачественной опухоли
Термин "соединение, специфическое для злокачественной ткани (специфическое для злокачественной ткани соединение)", как используют в рамках изобретения, относится к соединению, по-разному присутствующему в злокачественных тканях по сравнению с незлокачественными тканями. В рамках настоящего изобретения термин "злокачественная опухоль", как правило, используют для описания злокачественных новообразований, которые могут быть метастазирующими или неметастазирующими. Неограничивающими примерами карцином, развивающихся из эпителиальных тканей, таких как кожа или пищеварительный тракт, включают опухоль головного мозга, рак кожи, рак головы и шеи, рак пищевода, рак легкого, рак желудка, рак двенадцатиперстной кишки, рак молочной железы, рак предстательной железы, рак шейки матки, рак эндометрия, рак поджелудочной железы, рак печени, рак прямой кишки, рак ободочной и прямой кишки, рак толстого кишечника, рак мочевого пузыря и рак яичника. Неограничивающие примеры сарком, развивающихся из неэпителиальных (интерстициальных) тканей, таких как мышцы, включают остеосаркому, хондросаркому, рабдомиосаркому, лейомиосаркому, липосаркому и ангиосаркому. Неограничивающие примеры гематологической злокачественной опухоли, происходящей из гематологических органов, включают злокачественные лимфомы, включая лимфому Ходжкина и неходжкинскую лимфому; лейкоз, включая острый миелоцитарный лейкоз или хронический миелоцитарный лейкоз, и острый лимфоцитарный лейкоз или хронический лимфоцитарный лейкоз; и множественную миелому. Термин "новообразование", широко используемый в настоящем описании, относится к любой вновь сформировавшейся опухоли пораженной ткани. В рамках настоящего изобретения новообразования вызывают образование опухолей, которые частично характеризуются ангиогенезом. Новообразования могут быть доброкачественными, такими как гемангиома, глиома или тератома, или злокачественными, такими как карцинома, саркома, глиома, астроцитома, нейробластома или ретинобластома.
Термин "злокачественная ткань" относится к ткани, содержащей по меньшей мере одну злокачественную клетку. Таким образом, поскольку злокачественные ткани содержат злокачественные клетки и кровеносные сосуды, они относятся ко всем типам клеток, участвующим в образовании опухолевой массы, содержащей злокачественные клетки и эндотелиальные клетки. В рамках настоящего изобретения, "опухолевая масса" относится к очагам опухолевой ткани. Термин "опухоль", как правило, используют для обозначения доброкачественного новообразования или злокачественного новообразования.
Например, в нескольких вариантах осуществления специфические для злокачественной ткани соединения могут представлять собой соединения, определяемые качественными свойствами злокачественных тканей, такими как присутствие в злокачественных тканях, но отсутствие в незлокачественных тканях, или отсутствие в злокачественных тканях, но присутствие в незлокачественных тканях. В других вариантах осуществления специфические для злокачественной ткани соединения могут представлять собой соединения, определяемые количественными свойствами злокачественных тканей, такими как присутствие в злокачественных тканях в концентрации, отличающейся (например, более высокая концентрация или более низкая концентрация) от концентрации в незлокачественных тканях. Например, специфические для злокачественной ткани соединения присутствуют на разных уровнях в произвольных концентрациях. Как правило, специфические для злокачественной ткани соединения могут присутствовать в концентрации, повышенной по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 100%, по меньшей мере на 110%, по меньшей мере на 120%, по меньшей мере на 130%, по меньшей мере на 140%, по меньшей мере на 150%, по меньшей мере в 2 раза, по меньшей мере в 5 раз, по меньшей мере в 10 раз, по меньшей мере в 50 раз, по меньшей мере в 100 раз, по меньшей мере в 103 раз, по меньшей мере в 104 раз, по меньшей мере в 105 раз, по меньшей мере в 106 раз или более или вплоть до бесконечности (т.е., когда соединение отсутствует в незлокачественных тканях). Альтернативно, они могут присутствовать, как правило, в концентрации, сниженной по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 100% (т.е. отсутствуют). Предпочтительно, специфические для злокачественной ткани дифференциально присутствуют в статистически значимо отличающихся концентрациях (т.е. при определении с использованием либо t-критерия Велша, либо рангового суммарного критерия Вилкоксона, значение p составляет менее 0,05 и/или величина составляет менее 0,10). Примеры неограничивающего варианта осуществления специфического для злокачественной опухоли соединения включают соединения, которые являются специфическими для злокачественной ткани метаболитами, продуцируемыми вследствие метаболической активности, характерной для злокачественных клеток, иммунных клеток или стромальных клеток, содержащихся в злокачественных тканях, таких как ткани, описанные ниже (специфические для злокачественной ткани метаболиты, специфические для злокачественных клеток метаболиты, метаболиты, специфические для иммунных клеток, которые инфильтрировали в злокачественные ткани и специфические для стромальных клеток злокачественной опухоли метаболиты).
Специфические для злокачественной ткани метаболиты
Термин "метаболизм" относится к химическим изменениям, которые происходят в биологических тканях, и включает "анаболизм" и "катаболизм". Анаболизм относится к биосинтезу или накоплению молекул, а катаболизм относится к деградации молекул. "Метаболиты" представляют собой промежуточные соединения или продукты, которые появляются в результате метаболизма. "Первичные метаболиты" относятся к метаболитам, прямо вовлеченным в процесс роста или пролиферации клеток или организмов. "Вторичные метаболиты" относятся к продуктам, которые непрямо вовлечены в такие процессы роста или пролиферации и являются продуктами, такими как пигменты или антибиотики, которые продуцируются в результате метаболизма, который осуществляет биосинтез веществ, которые не вовлечены прямо в биологические явления, общие для клеток и организмов. Метаболиты могут представлять собой метаболиты "биополимеров" или они могут представлять собой метаболиты "низкомолекулярных соединений". "Биополимеры" представляют собой полимеры, содержащие один или несколько типов повторяющихся элементов. Биополимеры, как правило, встречаются в биологических системах, и их примеры включают клетки, образующие организм, и межклеточные матриксы, которые прилегают к ним, молекулы, имеющие молекулярную массу приблизительно 5000 или более, которые формируют структуры, такие как интерстициальные матриксы, в частности, полисахариды (углеводы и т.п.), пептиды (этот термин используют в значении, включающем полипептиды и белки), и полинуклеотиды, и аналогично их аналоги, такие как соединения, состоящие из или включающие аналоги аминокислот или неаминокислотные группы. "Низкомолекулярные соединения" относятся к природным химическим веществам, отличным от "биополимеров", которые существуют in vivo. Подходящие примеры неограничивающего варианта осуществления специфического для злокачественной ткани метаболита, описанного в настоящем описании, включают специфические для злокачественных клеток низкомолекулярные метаболиты (Eva Gottfried, Katrin Peter and Marina P. Kreutz, From Molecular to Modular Tumor Therapy (2010) 3 (2), 111-132). Кроме того, также включены метаболиты, которые на высоком уровне продуцируются иммунными клетками, которые инфильтрируют в злокачественные ткани, и метаболиты, которые на высоком уровне продуцируются стромальными клетками, которые поддерживают выживание и/или рост злокачественных клеток (стромальные клетки злокачественной опухоли или ассоциированные со злокачественной опухолью стромальные фибробласты (CAF)). Инфильтрирующие иммунные клетки представляют собой, например, дендритные клетки, ингибиторные дендритные клетки, ингибиторные T-клетки, истощенные T-клетки и происходящие из миеломы супрессорные клетки (MDSC). Более того, метаболиты по настоящему изобретению включают соединения, высвобожденные из клеток наружу клеток, когда клетки, находящиеся в злокачественных тканях (злокачественные клетки, иммунные клетки или стромальные клетки), погибают вследствие апоптоза, некроза или сходных с ними.
Для идентификации специфических для злокачественных клеток метаболитов можно пригодным образом использовать метаболомные анализы, сфокусированные на определении метаболического профиля, в дополнение к анализам на уровне транскриптома (например, Dhanasekaran et al. (Nature (2001) 412, 822-826), Lapointe et al. (Proc. Natl. Acad. Sci. U.S.A. (2004) 101, 811-816) или Perou et al. (Nature (2000) 406, 747-752)) и анализам на уровне протеома (например, Ahram et al. (Mol. Carcinog. (2002) 33, 9-15), Hood et al. (Mol. Cell. Proteomics (2005) 4, 1741-1753)). Более конкретно, для идентификации метаболитов в исследуемых образцах можно соответствующим образом использовать определение метаболического профиля, в котором используется высокоэффективная жидкостная хроматография (ВЭЖХ), ядерный магнитный резонанс (ЯМР) (Brindle et al. (J. Mol. Recognit. (1997) 10, 182-187), масс-спектрометрия (Gates and Sweeley (Clin. Chem. (1978) 24, 1663-1673) (GC/MS и LC/MS)) и ELISA или сходные с ними по отдельности и/или в комбинации.
Эти исследования установили гетерогенность в сформированных опухолях, являющуюся результатом изменения градиента концентрации факторов роста и метаболитов (глюкоза, кислород или сходные с ними), которая обеспечивает рост злокачественной опухоли в условиях низкого давления кислорода (Dang and Semenza (Trends Biochem. Sci. (1999) 24, 68-72)). В этих исследованиях также используют модели на клеточных линиях для понимания изменения каскада утилизации энергии на различных уровнях злокачественности опухолей (Vizan et al. (Cancer Res. (2005) 65, 5512-5515)). Примеры неограничивающего варианта осуществления технических компонентов платформы метаболомики включают экстракцию образца, разделение, спектроскопический анализ, нормализацию данных, описание специфических для класса метаболитов, картирование каскадов, подтверждение и функциональную охарактеризацию метаболитов-кандидатов, описанные Lawton et al. (Pharmacogenomics (2008) 9, 383). Эти способы позволяют идентификацию специфических для злокачественной опухоли метаболитов в желаемых злокачественных тканях.
Примеры неограничивающих вариантов осуществления специфических для злокачественной ткани соединений или специфических для злокачественной ткани метаболитов, используемых в рамках настоящего изобретения, предпочтительно включают по меньшей мере одно соединение, выбранное из соединений ниже. По меньшей мере одно соединение означает, что в дополнение к случаям, когда антигенсвязывающая активность одного и того же антигенсвязывающего домена, описанного ниже, зависит от одного типа специфических для злокачественной опухоли соединений или метаболитов, включаются случаи, когда она зависит от нескольких типов специфических для злокачественной ткани соединений или метаболитов.
(1) Первичные метаболиты цикла Кребса или гликолитической системы, такие как молочная кислота, янтарная кислота и лимонная кислота
Предпочтительные примеры неограничивающего варианта осуществления специфического для злокачественной ткани соединения, в частности, специфического для злокачественной клетки метаболита, используемого в рамках настоящего изобретения, включают первичные метаболиты, такие как молочная кислота, янтарная кислота и лимонная кислота, которые продуцируются в результате метаболизма глюкозы и присутствуют при более высоких концентрациях в злокачественных тканях по сравнению с окружающими незлокачественными тканями. Общеизвестно, что фенотип гликолитической системы, который характеризуется активацией ферментов гликолитической системы (каскад Эмбдена-Мейергофа), таких как пируваткиназа, гексокиназа и лактатдегидрогеназа (LDH), является характеристикой солидных опухолей, называемой эффектом Варбурга.
Иными словами в опухолях высокая экспрессия изоформы M2 пируваткиназы, которая необходима для анаэробного гликолиза, но не изоформы M1, считается функционирующей преимущественно для роста опухолевых клеток in vivo (Christofk et al. (Nature (2008) 452, 230-233). Пировиноградная кислота, продуцируемая пируваткиназой, ингибируется по принципу отрицательной обратной связи молочной кислотой, продуцируемой лактатдегидрогеназой (LDH) в результате равновесной реакции в анаэробных условиях. Поскольку ингибирование по принципу отрицательной обратной связи вызывает стимуляцию дыхания в митохондриях (цикл Кребса) и ингибирование роста клеток, считают, что активация LDH, гексокиназы и переносчика глюкозы (GLUT) играет важную роль в пролиферации злокачественных клеток (Fantin et al. (Cancer Cell (2006) 9, 425-434)). Глюкоза метаболизируется гликолитической системой и конечный метаболит молочная кислота транспортируется вместе с протонами в окружение опухоли и, в результате, считают, что значение pH тканей, окружающих опухоль, становится кислотным. Известно, что молочная кислота, которая является конечным продуктом гликолитического пути, а также янтарная кислота и лимонная кислота, продуцируемые в результате стимуляции дыхания в митохондриях, накапливаются в злокачественных тканях (Teresa et al. (Mol. Cancer (2009) 8, 41-59)). Примеры неограничивающих вариантов осуществления специфических для злокачественной ткани соединений, в частности, специфических для злокачественных клеток метаболитов, используемых в рамках настоящего изобретения, предпочтительно включают такие первичные метаболиты, как молочная кислота, янтарная кислота и лимонная кислота, продуцируемые в результате метаболизма по гликолитическому пути. Более того, известно, что янтарная кислота, которая присутствует в клетках в высокой концентрации, просачивается наружу клеток при гибели клеток (Nature Immunology, (2008) 9, 1261-1269). Таким образом, полагают, что концентрация янтарной кислоты увеличена в злокачественных клетках, в которых гибель клеток происходит часто.
(2) Аминокислоты, такие как аланин, глутаминовая кислота и аспарагиновая кислота
Помимо упомянутого выше метаболизма глюкозы, известно, что метаболизм аминокислот изменен в опухолевых клетках, для которых требуется непрерывное поступление основных аминокислот и неосновных аминокислот, которые необходимы для биосинтеза биополимеров в анаэробных условиях. Глутамин, который содержит два атома азота в его боковой цепи, действует в качестве переносчика азота и является аминокислотой, которая наиболее широко распространена в организме. Опухолевые клетки, в которых скорость захвата аминглутамина в клетки увеличена, называют функционирующими в качестве аминглутаминовой ловушки. Такое увеличение захвата аминглутамина и активности преобразования в глутаминовую кислоту и молочную кислоту называют "глутаминолизом", и он считается характеристикой трансформированных (опухолевых) клеток (Mazurek and Eigenbrodt (Anticancer Res. (2003) 23, 1149-1154); и Mazurek et al. (J. Cell. Physiol. (1999) 181, 136-146)). В результате, у пациентов со злокачественной опухолью происходит увеличение концентрации глутаминовой кислоты, одновременно со снижением уровня аминглутамина в плазме (Droge et al. (Immunobiology (1987) 174, 473-479)). Более того, наблюдали корреляцию между концентрациями 13C-меченой янтарной кислоты, 13C-меченого аланина, 13C-меченой глутаминовой кислоты и 13C-меченой лимонной кислоты в исследованиях метаболизма радиоактивно 13C-меченой глюкозы в тканях рака легкого. Подходящие примеры неограничивающего варианта осуществления специфических для злокачественной опухоли соединений, используемых в рамках настоящего изобретения, включают аланин, глутаминовую кислоту и аспарагиновую кислоту, которые накапливаются в высоких концентрациях в злокачественных тканях посредством такого глутаминолиза и т.п.
(3) Метаболит аминокислоты, такой как кинуренин
Индоламин-2,3-диоксигеназа (IDO) представляет собой метаболизирующий триптофан фермент, который на высоком уровне экспрессируется во многих злокачественных опухолях, таких как меланома, рак толстого кишечника и рак почки (Uyttenhove et al. (Nat. Med. (2003) 9, 1269-127)); и известно, что он имеет две изоформы (Lob et al. (Cancer Immunol. Immunother. (2009) 58, 153-157)). IDO катализирует преобразование триптофана в кинуренин (показан как соединение 1), и она является первым ферментом в каскаде никотинамиднуклеотидов (NAD) de novo. Более того, в глиоме, которая не экспрессирует IDO, кинуренин образуется из триптофана с помощью триптофан-2,3-диоксигеназы (TDO) в печени (Opitz et al. (Nature (2011) 478, 7368, 197-203)). IDO также экспрессируется в дендритных клетках, инфильтрированных в злокачественные ткани, и дендритные клетки также продуцируют кинуренин (J. Immunol. (2008) 181, 5396-5404). IDO также экспрессируется в супрессорных клетках миелоидного происхождения (MDSC) в злокачественных тканях, и MDSC также продуцируют кинуренин (Yu et al. (J. Immunol. (2013) 190, 3783-3797)).
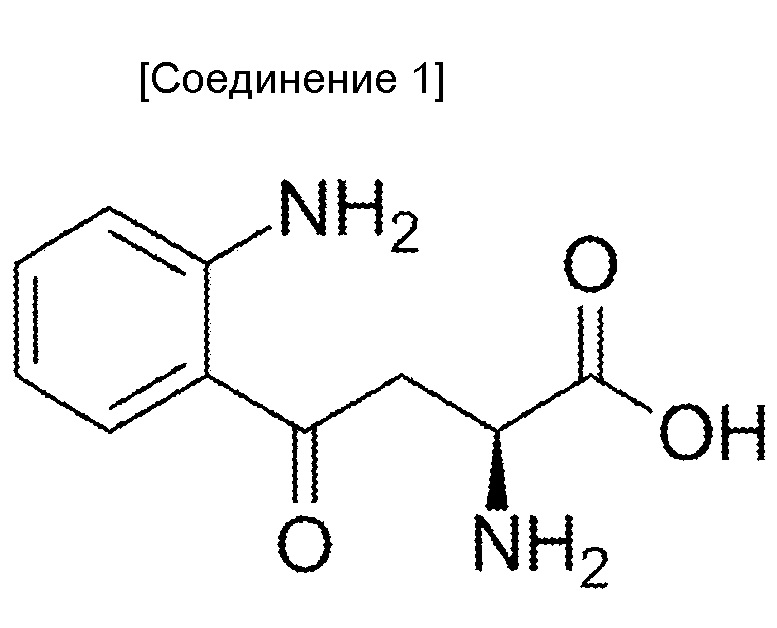
Известно, что кинуренин подавляет тот же тип T-клеточного ответа (Frumento et al. (J. Exp. Med. (2002) 196, 459-468); и было сделано предположение о механизме, в котором опухолевые клетки ускользают от противоопухолевых иммунных ответов посредством такого ингибирования, и пролиферация клеток глиомы усиливается по механизму аутокринной пролиферации, в котором кинуренин действует в качестве эндогенного лиганда для рецептора арильных углеводородов, экспрессируемых на глиомах (Optiz et al. (упоминается выше)). Кинуренин конвертируется в антраниловую кислоту (показана как соединение 2) посредством кинуренидазы и в 3-гидроксикинуренин (показан как соединение 3) посредством кинуренин-3-гидроксигидроксилазы. Как антраниловая кислота, так и 3-гидроксикинуренин, конвертируются в 3-гидроксиантраниловую кислоту, предшественник NAD.
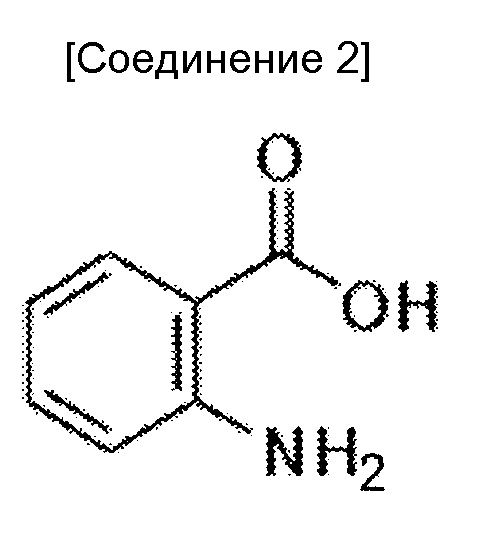

Кинуренин конвертируется в кинуреновую кислоту (показана как соединение 4) посредством кинуренинаминотрансферазы. Примеры неограничивающих вариантов осуществления специфических для злокачественной ткани соединений, в частности, специфических для злокачественной клетки метаболитов, используемых в рамках настоящего изобретения, предпочтительно включают такие метаболиты аминокислот, как кинуренин и его метаболиты, такие как антраниловая кислота, 3-гидроксикинуренин и кинуреновая кислота.
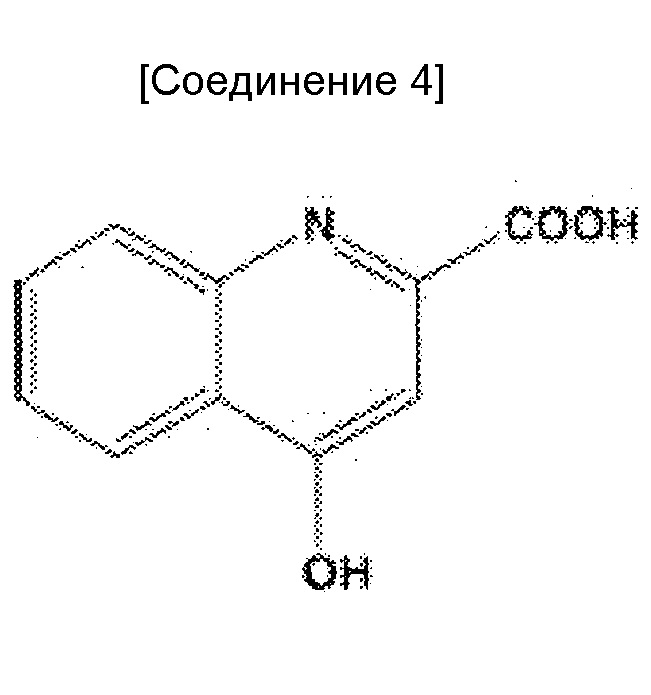
(4) Метаболиты арахидоновой кислоты, такие как простагландин E2
Простагландин E2 (PGE2) (соединение 5) представляет собой метаболит арахидоновой кислоты, называемый простаноидом, который включает тромбоксан и простагландин, синтезируемые посредством циклооксигеназы (COX)-1/2 (Warner and Mitchell (FASEB J. (2004) 18, 790-804)). PGE2 стимулирует пролиферацию клеток рака толстого кишечника и подавляет их апоптоз (Sheng et al. (Cancer Res. (1998) 58, 362-366)). Известно, что экспрессия циклооксигеназы изменяется во многих злокачественных клетках. Более конкретно, в то время как COX-1 экспрессируется конститутивно практически во всех тканях, было обнаружено, что COX-2 в основном индуцируется определенными типами воспалительных цитокинцитокинов и генов злокачественной опухоли в опухолях (Warner and Mitchell (упоминается выше)). Кроме того, было описано, что сверхэкспрессия COX-2 связана с плохим прогнозом рака молочной железы (Denkert et al. (Clin. Breast Cancer (2004) 4, 428-433)), и быстрым прогрессированием заболевания в случае рака яичника (Denker et al. (Mod. Pathol. (2006) 19, 1261-1269)). Ингибиторные T-клетки, которые инфильтрировали в злокачественные ткани, также продуцируют простагландин E2 (Curr. Med. Chem. (2011) 18, 5217-5223). Известно, что низкомолекулярные соединения, такие как метаболиты арахидоновой кислоты простагландин и лейкотриен, действуют в качестве стимулирующего фактора, который регулирует аутокринный и/или паракринный рост злокачественной опухоли (Nat. Rev. Cancer (2012) 12 (11) 782-792). Примеры неограничивающего варианта осуществления специфического для злокачественной ткани соединения, используемого в рамках настоящего изобретения, в частности специфических для злокачественных клеток метаболитов и специфических для иммунных клеток метаболитов, которые инфильтрировали в злокачественные ткани, предпочтительно включают такие метаболиты арахидоновой кислоты, как простагландин E2. Помимо простагландина E2, продукция тромбоксана A2 (TXA2) усилена в злокачественных тканях, таких как ткани рака ободочной и прямой кишки (J. Lab. Clin. Med. (1993) 122, 518-523), и тромбоксан A2 может быть соответственно представлен в качестве неограничивающего варианта осуществления метаболита арахидоновой кислоты по настоящему изобретению.
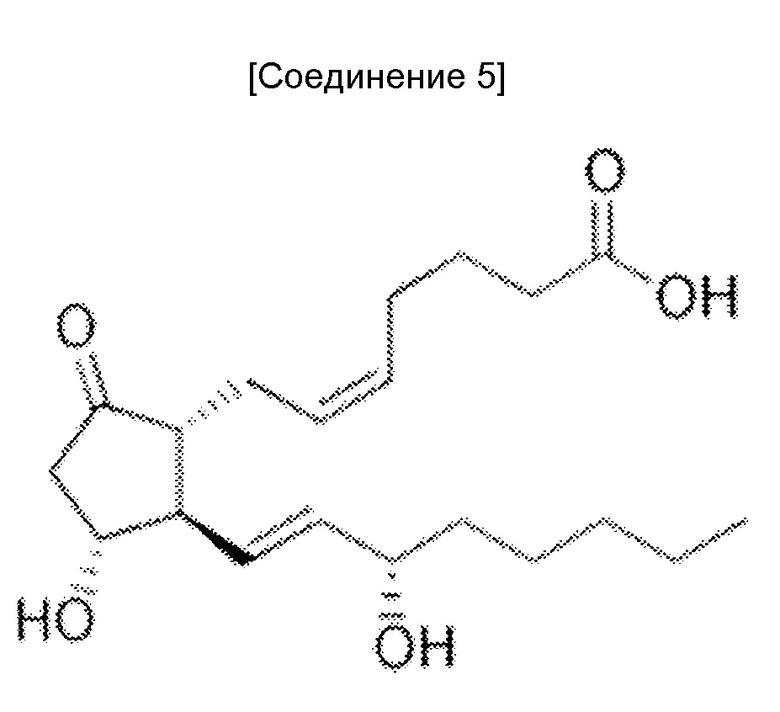
(5) Нуклеозиды, содержащие структуру пуринового кольца, такие как аденозин, аденозинтрифосфат (ATP), аденозиндифосфат (ADP) и аденозинмонофосфат (AMP)
Известно, что, когда злокачественные клетки претерпевают клеточную гибель, большое количество ATP в клетке просачивается наружу клеток. Таким образом, концентрация ATP является значительно более высокой в злокачественных тканях, чем в нормальных тканях (PLoS One. (2008) 3, e2599). Множество типов клеток высвобождают адениннуклеотиды в форме ATP, ADP и AMP. Метаболизм происходит с помощью внеклеточного фермента на клеточной поверхности, такого как внеклеточная 5'-нуклеотидаза (экто-5'-нуклеотидаза) (CD73) (Resta and Thompson (Immunol. Rev. (1998) 161, 95-109) и Sadej et al. (Melanoma Res. (2006) 16, 213-222)). Аденозин представляет собой пуриновый нуклеозид, который конститутивно существует в низкой концентрации во внеклеточной среде, однако сообщалось, что в гипоксических тканях, встречающихся в солидных опухолях, происходит значительное повышение внеклеточной концентрации аденозина (Blay and Hoskin (Cancer Res. (1997) 57, 2602-2605). CD73 экспрессируется на поверхностси иммунных клеток и опухолей (Kobie et al. (J. Immunol. (2006) 177, 6780-6786)), и было обнаружено, что его активность увеличивается при раке молочной железы (Canbolat et al. (Breast Cancer Res. Treat. (1996) 37, 189-193)), раке желудка (Durak et al. (Cancer Lett. (1994) 84, 199-202)), раке поджелудочной железы (Flocke and Mannherz (Biochim. Biophys. Acta (1991) 1076, 273-281) и глиобластоме (Bardot et al. (Br. J. Cancer (1994) 70, 212-218)). Было предположено, что накопление аденозина в злокачественных тканях может быть вызвано увеличением внутриклеточной продукции аденозина посредством дефосфорилфосфорилирования AMP с помощью 5'-нуклеотидазы в цитоплазме (Headrick and Willis (Biochem. J. (1989) 261, 541-550)). Более того, ингибиторные T-клетки и т.п., которые инфильтрировали в злокачественные ткани, также экспрессируют ATP-азу и продуцируют аденозин (Proc. Natl. Acad. Sci. (2006) 103 (35), 13132-13137; Curr. Med. Chem. (2011) 18, 5217-5223). Считается, что продуцированный аденозин обеспечивает в злокачественной ткани иммунодепрессивную среду через рецепторы аденозина, такие как рецептор A2A (Curr. Med. Chem. (2011),18, 5217-23). Примеры неограничивающего варианта осуществления специфического для злокачественной ткани соединения, используемого в рамках настоящего изобретения, предпочтительно включают ATP, ADP, AMP и аденозин, которые накапливаются в высокой концентрации в злокачественных тканях посредством такого метаболизма пуриновых нуклеотидов, таких как ATP. Более того, поскольку аденозин деградирует до инозина с помощью аденозиндезаминазы, инозин накапливается в высокой концентрации.
(6) Мочевая кислота
Мочевая кислота является продуктом метаболического каскада пуриновых нуклеозиднуклеозидов in vivo, и она высвобождается наружу клеток, например, в интерстициальное пространство и кровь. В последние годы было обнаружено, что она высвобождается из погибших клеток, которые присутствуют в очагах повреждения, таких как злокачественные ткани (Nat. Med. (2007) 13, 851-856). Примеры неограничивающего варианта осуществления специфических для злокачественной опухоли соединений, используемых в рамках настоящего изобретения, предпочтительно включают такую мочевую кислоту, которая накапливается в высокой концентрации в злокачественных тканях вследствие метаболизма пуриновых нуклеотидов, таких как ATP.
(7) 1-метилникотинамид
Известно, что фермент никотинамид-N-метилтрансфераза на высоком уровне экспрессируется в нескольких злокачественных тканях человека. Когда этот фермент продуцирует стабильный метаболит 1-этилметилникотинамид из никотинамида, метильная группа S-аденозилтиометионина (SAM), которая служит в качестве донора метильной группы, расходуется; таким образом, было предположено, что высокая экспрессия никотинамид-N-этилметилтрансферазы участвует в образовании опухоли посредством механизма, который нарушает способность к метилированию ДНК, сопровождающую снижение концентрации SAM в злокачественных клетках (Ulanovskaya et al. (Nat. Chem. Biol. (2013) 9 (5) 300-306)). Известно, что стабильный метаболит этого фермента, 1-этилметилникотинамид, секретируется наружу злокачественных клеток (Yamada et al. (J. Nutr. Sci. Vitaminol. (2010) 56, 83-86)), и предпочтительные примеры неограничивающего варианта осуществления специфических для злокачественной ткани соединений, используемых в рамках настоящего изобретения, включают 1-метилникотинамид и т.п., которые накапливаются в высоких концентрациях в злокачественных тканях посредством метаболизма никотинамида.
Специфические для воспаленной ткани соединения
Термин "соединение, специфическое для воспаленной ткани (специфическое для воспаленной ткани соединение)", как используют в рамках изобретения, относится к соединению, которое присутствует на отличающемся уровне в воспаленных тканях по сравнению с невоспаленными тканями. В рамках настоящего изобретения подходящие примеры "воспаленных тканей" включают:
суставы с ревматоидным артритом или остеоартритом;
легкие (альвеолы) с бронхиальной астмой или COPD;
органы пищеварительного тракта при воспалительном заболевании кишечника, болезни Крона или язвенном колите;
фиброзные ткани при фиброзе печени, почки или легкого;
ткани, подвергающиеся реакции отторжения при трансплантации органа;
кровеносные сосуды и сердце (миокард) при артериосклерозе или сердечной недостаточности;
висцеральный жир при метаболическом синдроме;
кожные ткани при атопическом дерматите или другом дерматите; и
спинномозговые нервы при грыже межпозвоночного диска или хронической боли в пояснично-крестцовой области спины.
Метаболиты, специфические для воспаленной ткани
"Специфический для воспаленной ткани метаболит" относится к метаболитам, на высоком уровне продуцируемым иммунными клетками, которые инфильтрировали в воспаленные ткани, и к метаболитам, на высоком уровне продуцируемым, в частности, в нормальных клетках, которые повреждены в воспаленных тканях. Примеры инфильтрирующих иммунных клеток включают эффеторные T-клетки, зрелые дендритные клетки, нейтрофилы, содержащие гранулы клетки (тучные клетки) и базофилы. Более того, метаболиты в рамках настоящего изобретения включают соединения, которые высвободились из клеток наружу клеток, когда клетки, которые присутствуют в воспаленных тканях (иммунные клетки и нормальные клетки) погибают посредством апоптоза, некроза и т.п.
Примеры неограничивающего варианта осуществления специфических для воспаленной ткани соединений или специфических для воспаленной ткани метаболитов, используемых в рамках настоящего изобретения предпочтительно включают по меньшей мере одно соединение, выбранное из приведенных ниже соединений. По меньшей мере одно соединение включает случаи, когда антигенсвязывающая активность одного и того же антигенсвязывающего домена, описанного ниже, зависит от типа специфического для воспаленной ткани соединения или метаболита, а также случаи, когда она зависит от нескольких типов специфических для воспаленной ткани соединений или метаболитов.
(1) Метаболиты арахидоновой кислоты, такие как простагландин E2
Известно, что концентрация PGE2 является высокой при ревматоидном артрите и остеоартрите (Eur. J. Clin. Pharmacol. (1994) 46, 3-7.; Clin. Exp. Rheumatol. (1999) 17, 151-160; Am. J. Vet. Res. (2004) 65, 1269-1275). Примеры неограничивающего варианта осуществления специфических для воспаленной ткани соединений, в частности, специфических для воспаленной ткани метаболитов и метаболитов, специфических для иммунных клеток, которые инфильтрируют в воспаленные ткани, используемых в рамках настоящего изобретения, предпочтительно включают такие метаболиты арахидоновой кислоты, как простагландин E2.
(2) Нуклеозиды, содержащие структуру пуринового кольца, такие как аденозин, аденозинтрифосфат (ATP), аденозиндифосфат (ADP) и аденозинмонофосфат (AMP)
Известно, что концентрация ATP является высокой в альвеолах легких, где происходит воспаление, вызванное бронхиальной астмой (Nat. Med. (2007) 13, 913-919). Также известно, что концентрация ATP является высокой в альвеолах легких, где происходит воспаление, вызванное COPD (Am. J. Respir. Crit. Care Med. (2010) 181, 928-934). Более того, было выявлено, что концентрация аденозина является высокой в суставной жидкости пациентов с ревматоидным артритом (Journal of Pharmaceutical и Biomedical Analysis (2004) 36, 877-882). Более того, известно, что концентрация ATP является высокой в тканях, где происходит реакция отторжения вследствие GVHD (Nat. Med. (2010) 16, 1434-1438). Известно, что концентрация аденозина повышена в фиброзных тканях печени, почки и легкого (FASEB J. (2008) 22, 2263-2272; J. Immunol. (2006) 176, 4449-4458; J. Am. Soc. Nephrol. (2011) 22 (5), 890-901; PLoS ONE J. (2010) 5 (2), e9242). Более того, было выявлено, что концентрация ATP увеличена в фиброзных тканях пациентов с фиброзом легких (Am. J. Respir. Crit. Care Med. (2010) 182, 774-783). Примеры неограничивающего варианта осуществления специфического для воспаленной ткани соединения, используемого в рамках настоящего изобретения, соответственно, включают ATP, ADP, AMP, аденозин и т.п., которые накапливаются в высокой концентрации в воспаленных тканях посредством метаболизма таких пуриновых нуклеотидов, как ATP. Кроме того, инозин накапливается в высокой концентрации вследствие деградации аденозина аденозиндезаминазой с образованием инозина.
(3) Мочевая кислота
Мочевая кислота является продуктом метаболического каскада пуриновых нуклеозиднуклеозидов in vivo, и она высвобождается наружу клеток, например, в интерстициальное пространство и кровь. В последние годы было выявлено, что мочевая кислота, высвобожденная из клеток, претерпевающих некроз, стимулирует воспалительный ответ (J. Clin. Invest. (2010) 120 (6), 1939-1949). Примеры неограничивающего варианта осуществления специфических для воспаленной ткани соединений, используемых в рамках настоящего изобретения, соответственно, включают такую мочевую кислоту, которая накапливается в высокой концентрации в воспаленных тканях вследствие метаболизма пуриновых нуклеотидов, таких как ATP.
Антигенсвязывающий домен
В рамках настоящего изобретения "антигенсвязывающий домен" может представлять собой любую структуру, при условии, что она связывается с представляющим интерес антигеном. Такие домены предпочтительно включают, например:
вариабельные области тяжелых цепей и легких цепей антител;
модуль приблизительно из 35 аминокислот, называемый A-доменом, который содержится в мембранном белке клеток авимере in vivo (международная публикация № WO 2004/044011, международная публикация № WO 2005/040229);
аднектин, содержащий домен 10Fn3, который связывается с белковой частью фибронектина - гликопротеина, экспрессирующегося на клеточной мембране (международная публикация № WO 2002/032925);
аффитело, которое состоит из трехспирального пучка из 58-аминокислот на основе каркаса связывающего IgG домена белка A (международная публикация № WO 1995/001937);
Смоделированные белки анкиринового повтора (DARPin), которые представляют собой область, экспонированную на молекулярной поверхности анкириновых повторов (AR), имеющих структуру, в которой субъединица, состоящая из поворота, содержащего 33 аминокислотных остатка, двух антипараллельных спиралей и петли, многократно сложена в стопку (международная публикация № WO 2002/020565);
антикалины и т.п., которые представляют собой домены, состоящие из четырех петель, которые поддерживают одну сторону структуры бочки, состоящей из восьми расположенных по кругу антипараллельных нитей, которые являются высоко консервативными среди молекул липокалина, таких как ассоциированный с желатином липокалин нейтрофилов (NGAL) (международная публикация № WO 2003/029462); и
вогнутая область, образованная структурой параллельных слоев внутри структуры в форме подковы, образованной уложенными в стопку повторами модуля лейцин-богатого повтора (LRR) вариабельного рецептора лимфоцитов (VLR), который не имеет иммуноглобулиновой структуры и используется в системе приобретенного иммунитета у бесчелюстных позвоночных, таких как минога и миксина (международная публикация № WO 2008/016854). Предпочтительные антигенсвязывающие домены по настоящему изобретению включают, например, антигенсвязывающие домены, имеющие вариабельные области тяжелых цепей и легких цепей антител. Предпочтительные примеры антигенсвязывающих доменов включают "одноцепочечный Fv (scFv)", "одноцепочечное антитело", "Fv", "одноцепочечный Fv 2 (scFv2)", "Fab" и "F(ab')2".
Антигенсвязывающие домены антигенсвязывающих молекул по настоящему изобретению могут связываться с идентичным эпитопом. Такой идентичный эпитоп может присутствовать, например, в белке, содержащем аминокислотную последовательность SEQ ID NO:1. Альтернативно, каждый из антигенсвязывающих доменов антигенсвязывающих молекул по настоящему изобретению может связываться с отличающимся эпитопом. В рамках настоящего изобретения, отличающийся эпитоп может присутствовать, например, в белке, содержащем аминокислотную последовательность SEQ ID NO:1.
Специфичность
"Специфический" означает, что одна из молекул, которая специфически связывается, по существу не связывается с молекулами, отличными от одной или множества молекул-партнеров, с которыми она связывается. Более того, "специфический" также используют, когда антигенсвязывающий домен является специфичным к конкретному эпитопу среди множества эпитопов в антигене. Когда эпитоп, связываемый антигенсвязывающим доменом, содержится в множестве различных антигенов, антигенсвязывающие молекулы, содержащие антигенсвязывающий домен, могут связываться с различными антигенами, которые имеют эпитоп. В рамках настоящего изобретения, выражение "по существу не связываются" определяют в соответствии со способом, описанным в упомянутом выше разделе об активности связывания, и оно относится к активности связывания молекулы, которая специфически связывает молекулу, отличную от молекулы-партнера, где активность связывания составляет не более 80%, обычно не более 50%, предпочтительно не более 30% или особенно предпочтительно не более 15% от активности связывания с ее молекулой-партнером.
Цитотоксическая активность
В неограничивающем варианте осуществления настоящее изобретение относится к антигенсвязывающим молекулам, которые содержат антигенсвязывающий домен, антигенсвязывающая активность которого варьирует в зависимости от концентрации специфического для злокачественной ткани соединения, и которые обладают цитотоксической активностью против клеток, экспрессирующих молекулу мембранного типа на их клеточной мембране; и к фармацевтическим композициям, содержащим эти антигенсвязывающие молекулы в качестве активного ингредиента. В рамках настоящего изобретения цитотоксическая активность включает, например, активность в виде антителозависимой клеточно-опосредуемой цитотоксичности (ADCC), активность в виде комплементзависимой цитотоксичности (CDC) и цитотоксическую активность T-клеток. В рамках настоящего изобретения активность CDC относится к цитотоксической активности системы комплемента. С другой стороны, активность ADCC относится к активности иммунных клеток, обеспечивающей повреждение клеток-мишеней, когда иммунные клетки и т.п. связываются с Fc-областью антигенсвязывающих молекул, содержащих антигенсвязывающий домен, который связывается с молекулой мембранного типа, экспрессируемой на мембране клеток-мишеней, через Fcγ-рецептор, экспрессируемый на иммунных клетках. Наличие у представляющей интерес антигенсвязывающей молекулы активности ADCC или активности CDC можно определять с использованием известных способов (например, Current Protocols in Immunology, Chapter 7. Immunologic studies in humans, Editor, Coligan et al., (1993)).
В частности, сначала получают эффекторные клетки, раствор комплемента и клетки-мишени.
(1) Получение клеток-мишеней
Селезенку извлекают из мыши CBA/N и т.п. и клетки селезенки диспергируют в среде RPMI1640 (Invitrogen). Клетки промывают в той же среде, содержащей 10% эмбриональную телячью сыворотку (FBS, HyClone), а затем получают эффекторые клетки, доводя концентрацию клеток селезенки до 5×106 /мл.
(2) Получение раствора комплемента
Комплемент детенышей кролика (CEDARLANE) разбавляют в 10 раз в культуральной среде (Invitrogen), содержащей 10% FBS, с получением раствора комплемента.
(3) Получение клеток-мишеней
Клетки-мишени можно подвергать радиоактивному мечению путем культивирования клеток, экспрессирующих антиген, с 0,2 мКи 51Cr-хромата натрия (GE Healthcare Bio-Sciences) в среде DMEM, содержащей 10% FBS, в течение одного часа при 37°C. После радиоактивного мечения клетки три раза промывают в среде RPMI1640, содержащей 10% FBS, и клетки-мишени можно получать, доводя концентрацию клеток до 2×105 /мл.
Активность ADCC или активность CDC можно измерять способом, описанным ниже. В случае измерения активности ADCC по 50 мкл клеток-мишеней и антигенсвязывающей молекулы добавляют в 96-луночный планшет с U-образным дном (Becton Dickinson), и им позволяют реагировать в течение 15 минут при комнатной температуре. Затем 100 мкл эффекторных клеток добавляют в планшет и этот планшет помещают в инкубатор с диоксидом углерода на четыре часа. Конечная концентрация антигенсвязывающей молекулы может быть выбрана, например, из 0 мкг/мл или 10 мкг/мл. После инкубации 100 мкл супернатанта собирают из каждой лунки и измеряют радиоактивность с помощью гамма-счетчика (COBRAII AUTO-GAMMA, MODEL D5005, Packard Instrument Company). Цитотоксическую активность (%) можно вычислять с использованием измеренных величин в соответствии с уравнением: (A-C)/(B-C)×100. A представляет собой радиоактивность (cpm) в каждом образце, B представляет собой радиоактивность (cpm) в образце, в который добавлен 1% NP-40 (Nacalai Tesque), и C представляет собой радиоактивность (cpm) в образце, содержащем только клетки-мишени.
Между тем, в случае измерения активности CDC, 50 мкл клеток-мишеней и 50 мкл антигенсвязывающей молекулы добавляют в 96-луночный планшет с плоским дном (Becton Dickinson) и им позволяют реагировать в течение 15 минут на льду. Затем в планшет добавляют 100 мкл раствора комплемента, и этот планшет помещают в инкубатор с диоксидом углерода на четыре часа. Конечная концентрация антигенсвязывающей молекулы может быть выбрана, например, из 0 мкг/мл или 3 мкг/мл. После инкубации из каждой лунки собирают 100 мкл супернатанта, и радиоактивность измеряют с помощью гамма-счетчика. Цитотоксическую активность можно вычислять аналогично определению активности ADCC.
В качестве антигенсвязывающих молекул по настоящему изобретению, обладающих цитотоксической активностью, можно соответственно использовать описанные ниже модифицированные антигенсвязывающие молекулы, с которыми связаны цитотоксические вещества, такие как химиотерапевтические средства, токсические пептиды или радиоактивные химические вещества. Такие модифицированные антигенсвязывающие молекулы (далее обозначаемые как "конъюгат антигенсвязывающая молекула-лекарственное средство") можно получать путем химической модификации полученных антигенсвязывающих молекул. Способы, которые уже разработаны в области конъюгатов антитело-лекарственное средство и т.п., можно соответствующим образом использовать в качестве способа модификации антигенсвязывающих молекул. Более того, модифицированную антигенсвязывающую молекулу со связанным с ней токсическим пептидом можно получать путем экспрессии в соответствующей клетке-хозяине слитого гена, полученного путем связывания гена, кодирующего токсический пептид, в рамке считывания с геном, кодирующим антигенсвязывающую молекулу по настоящему изобретению, а затем выделения молекулы из культурального раствора клеток.
Нейтрализующая активность
В неограничивающем варианте осуществления настоящее изобретение относится к фармацевтической композиции, которая индуцирует иммунный ответ, содержащей в качестве активного ингредиента антигенсвязывающую молекулу, которая содержит антигенсвязывающий домен, антигенсвязывающая активность которого варьирует в зависимости от концентрации специфического для злокачественной ткани соединения, и которая обладает нейтрализующей активностью против молекулы мембранного типа. В другом неограничивающем варианте осуществления настоящее изобретение относится к фармацевтической композиции, которая индуцирует иммунный ответ, содержащей в качестве активного ингредиента антигенсвязывающую молекулу, которая содержит антигенсвязывающий домен, антигенсвязывающая активность которого варьирует в зависимости от концентрации специфического для злокачественной ткани соединения, и которая обладает нейтрализующей активностью против молекулы мембранного типа в дополнение к цитотоксической активности против клеток, экспрессирующих молекулу мембранного типа на их клеточной мембране. Как правило, нейтрализующая активность относится к активности ингибирования биологической активности лиганда, который обладает биологической активностью в отношении клеток, такого как вирусы и токсины. Таким образом, вещество, обладающее нейтрализующей активностью, относится к веществу, которое связывается с лигандом или рецептором, с которым связывается лиганд, и ингибирует связывание между лигандом и рецептором. Рецептор, связывание которого с лигандом блокируется нейтрализующей активностью, не способен осуществлять биологическую активность через рецептор. Когда антигенсвязывающая молекула представляет собой антитело, антитело, обладающее такой нейтрализующей активностью, обычно называют нейтрализующим антителом. Нейтрализующую активность исследуемого вещества можно измерять путем сравнения биологической активности в присутствии лиганда в условиях, когда исследуемое вещество присутствует или отсутствует.
Подходящим примером основного лиганда для рецептора IL-6 является IL-6, который представлен в SEQ ID NO:27. Рецептор IL-6 рецептор, который представляет собой мембранный белок I типа, N-конец которого образует внеклеточный домен, образует гетеротетрамер с рецептором gp130, который был индуцирован посредством IL-6 к димеризации (Heinrich et al. (Biochem. J. (1998) 334, 297-314)). Образование гетеротетрамера активирует Jak, ассоциированную с рецептором gp130. Jak осуществляет аутофосфорилирование и фосфорилирование рецептора. Участки фосфорилирования рецептора и Jak служат в качестве участков связывания для молекул, принадлежащих семейству Stat, имеющих SH2, таких как Stat3, и для MAP-киназы, PI3/Akt и других белков и адаптеров, имеющих SH2. Далее, Stat, который связался с рецептором gp130, фосфорилируется посредством Jak. Фосфорилированный Stat димеризуется и перемещается в ядро и регулирует транскрипцию генов-мишеней. Jak и Stat также могут быть вовлечены в каскад передачи сигнала через рецепторы других классов. Нарушение регуляции каскада передачи сигнала IL-6 наблюдают при воспалении и патологических состояниях аутоиммунных заболеваний, и при злокачественных опухолях, таких как рак предстательной железы и множественная миелома. Stat3, который может действовать в качестве онкогена, конститутивно активирован во многих злокачественных опухолях. При раке предстательной железы и множественной миеломе существует перекрестное действие между каскадом передачи сигнала от рецептора IL-6 и каскадом передачи сигнала от представителей семейства рецептора эпидермального фактора роста (EGFR) (Ishikawa et al. (J. Clin. Exp. Hematopathol. (2006) 46 (2), 55-66)).
Такие каскады внутриклеточной передачи сигнала отличаются для каждого типа клеток; таким образом, соответствующая молекула-мишень может быть выбрана в зависимости от каждой из представляющих интерес клеток-мишеней, и молекула-мишень не ограничивается упомянутыми выше факторами. Активность нейтрализации можно оценивать путем измерения активации сигнала in vivo. Более того, активацию сигналов in vivo также можно выявлять с использованием в качестве индикатора индуцирующего транскрипцию действия гена-мишени, который находится ниже каскада передачи сигнала in vivo. Изменение транскрипционной активности гена-мишени можно обнаруживать по принципу репортерного анализа. В частности, репортерный ген, такой как зеленый флуоресцентный белок (GFP) или люцифераза, помещают ниже фактора транскрипции или промоторной области гена-мишени; и изменение транскрипционной активности можно измерять в значениях активности репортера путем измерения активности репортера. Можно соответственно использовать коммерчески доступные наборы для измерения активации сигнала in vivo (например, Mercury Pathway Profiling Luciferease System (Clontech)).
Более того, в качестве способа измерения активности нейтрализации на лиганде рецептора в семействе рецепторов EGF и т.п., который действует на каскад передачи сигнала, который, как правило, действует в направлении усиления пролиферации клеток, активность нейтрализации у антигенсвязывающей молекулы можно оценивать путем измерения активности пролиферации клеток-мишеней. Например, является пригодным использование следующего способа в качестве способа измерения или оценки ингибиторных эффектов на основе активности нейтрализации антитела против HB-EGF в отношении пролиферации клеток, пролиферация которых стимулирована факторами роста семейства EGF, такими как HB-EGF. В качестве способа оценки и измерения активности ингибирования пролиферации клеток в пробирке используют способ, который измеряет включение живыми клетками [3H]-меченного тимидина, добавленного в культуральную среду, в качестве показателя способности к репликации ДНК. В качестве более удобного способа используют способ исключения красителя, в котором под микроскопом измеряют способности клетки высвобождать краситель, такой как трипановый синий, наружу клетки, или способ MTT. В последнем используется способность живых клеток конвертировать 3-(4,5-диэтилметилтиазол-2-ил)-2,5-дифенилтетразолтетразолий бромид (MTT), который является солью тетразолтетразолия, в синий продукт формазан. Более конкретно, исследуемое антитело добавляют вместе с лигандом в культуральный раствор исследуемой клетки; и после прохождения определенного периода времени в культуру добавляют раствор MTT и его оставляют стоять в течение определенного периода времени, чтобы позволить клетке включить MTT. В результате, MTT, который представляет собой желтое соединение, преобразуется в синее соединение сукцинатдегидрогеназой в митохондриях клетки. После этого синий продукт растворяют для окрашивания, его поглощение измеряют и используют в качестве индикатора количества жизнеспособных клеток. Помимо MTT, также являются коммерчески доступными такие реагенты, как MTS, XTT, WST-1 и WST-8 (Nacalai Tesque и т.п.) и их можно соответственно использовать. Для измерения активности связывающее антитело, которое имеет тот же изотип, что и антитело против HB-EGF, но не обладает активностью ингибирования пролиферации клеток, можно использовать в качестве контрольного антитела таким же образом, как и антитело против HB-EGF, и считается, что антитело против HB-EGF обладает активностью, когда оно демонстрируют более выраженную активность ингибирования пролиферации клеток, чем контрольное антитело.
В качестве клеток для оценки активности можно соответственно использовать, например, клетки, демонстрирующие стимулированную HB-EGF пролиферацию, такие как клеточная линия RMG-1, которая представляет собой клеточную линию рака яичника; и также можно соответственно использовать клетки мыши Ba/F3, трансформированные вектором, в котором ген, кодирующий hEGFR/mg-CSFR, который является слитым белком внеклеточного домена EGFR человека, слитым в рамке считывания с внутриклеточным доменом рецептора G-CSF мыши, связан так, чтобы обеспечить экспрессию. Таким образом, специалисты в данной области могут соответствующим образом выбирать клетки для оценки активности для измерения активности пролиферации клеток, упомянутой выше.
Антитело
В рамках настоящего изобретения "антитело" относится к природному иммуноглобулину или иммуноглобулину, полученному частичным или полным синтезом. Антитела можно выделять из природных источников, таких как природная плазма и сыворотка, или из культуральных супернатантов продуцирующих антитела гибридом. Альтернативно антитела можно частично или полностью синтезировать с использованием способов, таких как генетическая рекомбинация. Предпочтительные антитела включают, например, антитела изотипа иммуноглобулинов или принадлежащего ему подкласса. Известные иммуноглобулины человека включают антитела следующих девяти классов (изотипов): IgG1, IgG2, IgG3, IgG4, IgA1, IgA2, IgD, IgE и IgM. Из этих изотипов антитела по настоящему изобретению включают IgG1, IgG2, IgG3 и IgG4. Ряд аллотипических последовательностей константных областей IgG1 человека, IgG2 человека, IgG3 человека и IgG4 человека вследствие генетического полиморфизма описан в "Sequences of proteins of immunological interest", NIH Publication No.91-3242. Любую из таких последовательностей можно использовать в рамках настоящего изобретения. В частности, для последовательности IgG1 человека, аминокислотная последовательность положений 356-358, как указано с помощью нумерации EU, может быть либо DEL, либо EEM. Несколько аллотипических последовательностей вследствие генетических полиморфизмов описаны в "Sequences of proteins of immunological interest", NIH Publication No. 91-3242 для константной области Igκ (каппа) человека и константной области Igλ (лямбда) человека, и любую из этих последовательностей можно использовать в рамках настоящего изобретения.
Способы получения антитела с желаемой активностью связывания известны специалистам в данной области. Ниже приведен пример, в котором описан способ получения антитела, которое связывается с IL-6R (антитело против IL-6R). Антитела, которые связываются с антигеном, отличным от IL-6R, также можно получать в соответствии с примером, описанным ниже.
Антитела против IL-6R можно получать в качестве поликлональных или моноклональных антител с использованием известных способов. Продуцируемые антитела против IL-6R предпочтительно представляют собой моноклональные антитела, происходящие из млекопитающих. Такие происходящие из млекопитающих моноклональные антитела включают антитела, продуцируемые гибридомами или клетками-хозяевами, трансформированными экспрессирующим вектором, несущим ген антитела, способами генетической инженерии. Моноклональные антитела по настоящему изобретению включают "гуманизированные антитела" или "химерные антитела".
Продуцирующие моноклональные антитела гибридомы можно получать с использованием известных способов, например, как описано ниже. В частности, млекопитающих иммунизируют общепринятыми способами иммунизации с использованием белка IL-6R в качестве сенсибилизирующего антигена. Полученные иммунные клетки подвергают слиянию с известными родительскими клетками общепринятыми способами слияния клеток. Затем гибридомы, продуцирующие антитело против IL-6R, можно отбирать путем скрининга продуцирующих моноклональное антитело клеток с использованием общепринятых способов скрининга.
В частности, моноклональные антитела получают так, как описано ниже. Сначала можно экспрессировать ген IL-6R, нуклеотидная последовательность которого представлена в SEQ ID NO:2, для получения белка IL-6R, представленного в SEQ ID NO:1, который будет использован в качестве сенсибилизирующего антигена для получения антитела. Для этого, последовательность гена, кодирующего IL-6R, встраивают в известный экспрессирующий вектор, и подходящие клетки-хозяева трансформируют этим вектором. Желаемый белок IL-6R человека очищают из клеток-хозяев или их культуральных супернатантов известными способами. Для получения растворимого IL-6R из культуральных супернатантов, например, белок, состоящий из аминокислот в положениях 1-357 в полипептидной последовательности IL-6R SEQ ID NO:1, такой как описана в Mullberg et al. (J. Immunol. (1994) 152 (10), 4958-4968), экспрессируют в качестве растворимого IL-6R вместо белка IL-6R SEQ ID NO:1. Также в качестве сенсибилизирующего антигена можно использовать очищенный природный белок IL-6R.
Очищенный белок IL-6R можно использовать в качестве сенсибилизирующего антигена для иммунизации млекопитающих. Также в качестве сенсибилизирующего антигена можно использовать неполный пептид IL-6R. В этом случае, неполный пептид можно получать путем химического синтеза на основе аминокислотной последовательности IL-6R человека, или путем встраивания неполного гена IL-6R в экспрессирующий вектор для экспрессии. Альтернативно неполный пептид можно получать путем деградации белка IL-6R протеазой. Длина и область неполного пептида IL-6R не ограничены конкретными вариантами осуществления. Предпочтительная область может быть произвольным образом выбрана из аминокислотной последовательности в положениях аминокислот 20-357 аминокислотной последовательности SEQ ID NO:1. Количество аминокислот, образующих пептид, подлежащий применению в качестве сенсибилизирующего антигена, предпочтительно составляет по меньшей мере пять или более, шесть или более, или семь или более. Более конкретно, в качестве сенсибилизирующего антигена можно использовать пептид из 8-50 остатков, более предпочтительно, из 10-30 остатков.
Альтернативно, для сенсибилизирующего антигена можно использовать слитый белок, полученный путем слияния желаемого неполного полипептида или пептида белка IL-6R с отличающимся полипептидом. Например, предпочтительно в качестве сенсибилизирующих антигенов используют Fc-фрагменты антител и пептидные метки. Векторы для экспрессии таких слитых белков можно конструировать путем слияния в рамке считывания генов, кодирующих два или более желаемых полипептидных фрагментов, и встраивания слитого гена в экспрессирующий вектор, как описано выше. Способы получения слитых белков описаны в Molecular Cloning 2nd ed. (Sambrook, J et al., Molecular Cloning 2nd ed., 9.47-9.58 (1989) Cold Spring Harbor Lab. Press). Способы получения IL-6R для применения в качестве сенсибилизирующего антигена и способы иммунизации с использованием IL-6R конкретно описаны в WO 2003/000883, WO 2004/022754, WO 2006/006693 и т.п.
Отсутствуют конкретное ограничение на млекопитающих, подлежащих иммунизации сенсибилизирующим антигеном. Однако предпочтительно выбирать млекопитающих, учитывая их совместимость с родительскими клетками, используемыми для слияния клеток. Как правило, предпочтительно используют грызунов, таких как мыши, крысы и хомячки, кроликов и обезьян.
Указанных выше животных иммунизируют сенсибилизирующим антигеном известными способами. Как правило, выполняемые способы иммунизации включают, например, внутрибрюшинное или подкожное инъекционное введение сенсибилизирующего антигена млекопитающим. В частности, сенсибилизирующий антиген соответствующим образом разбавляют PBS (фосфатно-солевой буфер), физиологическим солевым раствором или сходными с ними. Если желательно, общепринятый адъювант, такой как полный адъювант Фрейнда, смешивают с антигеном и смесь эмульгируют. Затем сенсибилизирующий антиген вводят млекопитающим несколько раз с интервалами 4-21 суток. При иммунизации сенсибилизирующим антигеном можно использовать подходящие носители. В частности, когда в качестве сенсибилизирующего антигена используют низкомолекулярный неполный пептид, иногда желательно связывать сенсибилизирующий антигенный пептид с белком-носителем, таким как альбумин или гемоцианин лимфы улитки, для иммунизации.
Альтернативно гибридомы, продуцирующие желаемое антитело, можно получать с использованием способов иммунизации ДНК, как описано ниже. Иммунизация ДНК представляет собой способ иммунизации, который обеспечивает стимуляцию иммунной системы путем экспрессии сенсибилизирующего антигена у животного, имунизированного в результате введения векторной ДНК, сконструированной для обеспечения экспрессии гена, кодирующего антигенный белок, у животного. По сравнению с общепринятыми способами иммунизации, в которых белковый антиген вводят животным, подлежащим иммунизации, иммунизация ДНК является лучшей в том, что:
- может быть обеспечена стимуляция иммунной системы при сохранении структуры мембранного белка, такого как IL-6R; и
- отсутствует необходимость в очистке антигена для иммунизации.
Для получения моноклонального антитела по настоящему изобретению с использованием иммунизации ДНК, сначала ДНК, экспрессирующую белок IL-6R, вводят животному, подлежащему иммунизации. ДНК, кодирующую IL-6R, можно синтезировать известными способами, такими как ПЦР. Полученную ДНК встраивают в соответствующий экспрессирующий вектор, а затем его вводят животному, подлежащему иммунизации. Предпочтительно используемые экспрессирующие векторы включают, например, коммерчески доступные экспрессирующие векторы, такие как pcDNA3.1. Векторы можно вводить в организм с использованием общепринятых способов. Например, иммунизацию ДНК проводят с использованием генной пушки для введения покрытых экспрессирующим вектором частиц золота в клетки организма животного, подлежащего иммунизации. Антитела, которые распознают IL-6R, также можно получать способами, описанными в WO 2003/104453.
После иммунизации млекопитающего, как описано выше, увеличение титра связывающего IL-6R антитела подтверждают в сыворотке. Затем иммунные клетки собирают от млекопитающего, а затем подвергают слиянию клеток. В частности, предпочтительно в качестве иммунных клеток используют спленоциты.
В качестве клетки, подлежащей слиянию с упомянутыми выше иммунными клетками, используют клетки миеломы млекопитающих. Клетки миеломы предпочтительно содержат подходящий селективный маркер для скрининга. Селективный маркер придает клеткам характеристики для их выживания (или гибели) в конкретных условиях культивирования. В качестве селективных маркеров известны дефицит гипоксантин-гуанин-фосфорибозилтрансферазы (далее сокращенно обозначаемый как дефицит HGPRT) и дефицит тимидинкиназы (далее сокращенно обозначаемый как дефицит TK). Клетки с дефицитом HGPRT или TK обладают чувствительностью к гипоксантину-аминоптерину-тимидину (далее сокращенно обозначаемой чувствительностью HAT). Чувствительные к HAT клетки не могут синтезировать ДНК в селективной среде HAT, и, таким образом, погибают. Однако когда клетки слиты с нормальными клетками, они могут продолжать синтезировать ДНК с использованием пути спасения нормальных клеток, и, таким образом, они могут расти в селективной среде HAT.
Селекцию клеток с дефицитом HGPRT и TK можно проводить в среде, содержащей 6-тиогуанин, 8-азагуанин (далее сокращенно обозначаемый как 8AG) или 5'-бромдезоксиуридин, соответственно. Нормальные клетки погибают, поскольку они включают аналоги пиримидина в их ДНК. Между тем, клетки, которые являются дефицитными по этим ферментам, могут выживать в селективной среде, поскольку они не могут включать эти аналоги пиримидина. Кроме того, селективный маркер, называемый устойчивостью к G418, обеспечиваемый геном устойчивости к неомицину, обеспечивает устойчивость к 2-дезоксистрептаминовым антибиотикам (аналоги гентамицина). Известны различные типы миеломных клеток, которые пригодны для слияния клеток.
Например, предпочтительно можно использовать миеломные клетки, включающие следующие клетки:
P3(P3x63Ag8,653) (J. Immunol. (1979) 123 (4), 1548-1550);
P3x63Ag8U.1 (Current Topics in Microbiology и Immunology (1978)81, 1-7);
NS-1 (C. Eur. J. Immunol. (1976)6 (7), 511-519);
MPC-11 (Cell (1976) 8 (3), 405-415);
SP2/0 (Nature (1978) 276 (5685), 269-270);
FO (J. Immunol. Methods (1980) 35 (1-2), 1-21);
S194/5.XX0.BU.1 (J. Exp. Med. (1978) 148 (1), 313-323);
R210 (Nature (1979) 277 (5692), 131-133), и т.д.
Слияние клеток между иммуноцитами и клетками миеломы по существу проводят с использованием известных способов, например, способа Kohler and Milstein et al. (Methods Enzymol. (1981) 73: 3-46).
Более конкретно, слияние клеток можно проводить, например, в общепринятой культуральной среде в присутствии агента, обеспечивающего слияние клеток. Агенты, обеспечивающие слияние клеток, включают, например, полиэтиленгликоль (PEG) и вирус Сендай (HVJ). Если необходимо, также можно добавлять вспомогательное вещество, такое как диметилсульфоксид, для повышения эффективности слияния.
Соотношение иммунных клеток к клеткам миеломы можно определять самостоятельно, предпочтительно, например, оно составляет одну клетку миеломы на каждые от одного до десяти иммуноцитов. Культуральные среды, используемые для слияния клеток, включают, например, среды, которые пригодны для выращивания клеточных линий, такие как среда RPMI1640 и среда MEM, и другие общепринятые культуральные среды, используемые для этого типа клеточной культуры. Кроме того, предпочтительно в культуральную среду можно добавлять сывороточные добавки, такие как эмбриональная телячья сыворотка (FCS).
Для слияния клеток заданные количества указанных выше иммунных клеток и миеломных клеток смешивают в описанной выше культуральной среде. Затем добавляют раствор PEG (например, средняя молекулярная масса составляет приблизительно от 1000 до 6000), предварительно нагретый до приблизительно 37°C, в концентрации, как правило, от 30% до 60% (масс./об.). Затем эту смесь осторожно перемешивают для получения желаемых слитых клеток (гибридом). Затем к клеткам постепенно добавляют подходящую культуральную среду, упомянутую выше, и смесь многократно центрифугируют для удаления супернатанта. Таким образом, можно удалить агенты слияния клеток и т.п., которые являются неблагоприятными для роста гибридомы.
Селекцию полученных таким образом гибридом можно проводить путем культивирования с использованием общепринятой селективной среды, например, среды HAT (культуральная среда, содержащая гипоксантин, аминоптерин и тимидин). Клетки, отличные от желаемых гибридом (неслитые клетки), можно уничтожать путем продолжения культивирования в описанной выше среде HAT в течение достаточного периода времени. Как правило, период составляет от нескольких суток до нескольких недель. Затем, гибридомы, продуцирующие желаемое антитело, подвергают скринингу и клонируют по отдельности общепринятыми способами лимитирующих разведений.
Селекцию полученных таким образом гибридом можно проводить с использованием селективной среды на основе селективного маркера, которым обладает миелома, используемая для слияния клеток. Например, селекцию клеток с дефицитом HGPRT или TK можно проводить путем культивирования с использованием среды HAT (культуральная среда, содержащая гипоксантин, аминоптерин и тимидин). В частности, когда для слияния клеток используют чувствительные к HAT миеломные клетки, клетки, успешно слитые с нормальными клетками, могут селективно пролиферировать в среде HAT. Клетки, отличные от желаемых гибридом (неслитые клетки), можно уничтожать путем продолжения культивирования в описанной выше среде HAT в течение достаточного периода времени. В частности, селекцию желаемых гибридом можно проводить путем культивирования, как правило, в течение от нескольких суток до нескольких недель. Затем гибридомы, продуцирующие желаемое антитело, подвергают скринингу и по отдельности клонируют общепринятыми способами лимитирующих разведений.
Желаемые антитела можно предпочтительно отбирать и по отдельности клонировать способами скрининга на основе известной реакции антиген/антитело. Например, связывающее IL-6R моноклональное антитело может связываться с IL-6R, экспрессированным на поверхности клетки. Такое моноклональное антитело можно подвергать скринингу с помощью активированной флуоресценцией сортировки клеток (FACS). FACS представляет собой систему, которая оценивает связывание антитела с клеточной поверхностью путем анализа клеток, контактирующих с флуоресцентным антителом с использованием лазерного луча и измерения флуоресценции, испускаемой индивидуальными клетками.
Для скрининга гибридом, которые продуцируют моноклональные антитело по настоящему изобретению, с помощью FACS, сначала получают экспрессирующие IL-6R клетки. Клетки, предпочтительно используемые для скрининга, представляют собой клетки млекопитающих, в которых IL-6R экспрессируется вынужденным образом. В качестве контроля можно проводить селективное выявление активности антитела, которое связывается с IL-6R клеточной поверхности, с использованием в качестве клеток-хозяев нетрансформированных клеток млекопитающих. В частности, гибридомы, продуцирующие моноклональное антитело против IL-6R, можно выделять путем селекции гибридом, которые продуцируют антитело, которое связывает с клетками, в которых экспрессируется IL-6R вынужденным образом, но не с клетками-хозяевами.
Альтернативно активность связывания антитела с иммобилизованными экспрессирующими IL-6R клетками можно оценивать на основе принципа ELISA. Например, экспрессирующие IL-6R клетки иммобилизуют на лунках планшета для ELISA. Культуральные супернатанты гибридом контактируют с иммобилизованными клетками в лунках, и выявляют антитела, которые связываются с иммобилизованными клетками. Когда моноклональные антитела происходят из мыши, антитела, связанные с клетками, можно выявлять с использованием антитела против иммуноглобулина мыши. Селекцию гибридом, продуцирующих желаемое антитело, обладающее антигенсвязывающей способностью, проводят с помощью описанного выше скрининга, и их можно клонировать способом лимитирующих разведений и т.п.
Продуцирующие моноклональное антитело гибридомы, полученные таким образом, можно пассировать в общепринятой культуральной среде и хранить в жидком азоте в течение длительного периода времени.
Описанные выше гибридомы культивируют общепринятым способом, и желаемые моноклональные антитела можно получать из культуральных супернатантов. Альтернативно гибридомы вводят совместимым млекопитающим и выращивают в них, и моноклональные антитела получают из асцитной жидкости. Первый из этих способов пригоден для получения антител с высокой чистотой.
Предпочтительно также можно использовать антитела, кодируемые генами антител, которые клонированы из антителопродуцирующих клеток, таких как описанные выше гибридомы. Клонированный ген антитела встраивают в подходящий вектор, и его вводят в клетку-хозяина для экспрессии антитела, кодируемого геном. Способы выделения генов антител, встраивания генов в векторы и трансформации клеток-хозяев, уже разработаны, например, Vandamme et al. (Eur. J. Biochem. (1990) 192(3), 767-775). Способы продуцирования рекомбинантных антител также известны, как описано ниже.
Например, кДНК, кодирующую вариабельную область (V-область) антитела против IL-6R, получают из клеток гибридомы, экспрессирующих антитело против IL-6R. Для этой цели сначала из гибридом экстрагируют тотальную РНК. Способы, используемые для экстракции мРНК из клеток, включают, например:
- способ ультрацентрифугирования с гуанидином (Biochemistry (1979) 18(24), 5294-5299), и
- способ AGPC (Anal. Biochem. (1987) 162(1), 156-159)
Экстрагированные мРНК можно очищать с использованием набора для очистки мРНК mRNA Purification Kit(GE Healthcare Bioscience) и т.п. Альтернативно также коммерчески доступны наборы для экстракции тотальной мРНК непосредственно из клеток, такие как набор для очистки мРНК QuickPrep mRNA Purification Kit (GE Healthcare Bioscience). мРНК можно получать из гибридом с использованием таких наборов. кДНК, кодирующие V-область антитела, можно синтезировать из полученных мРНК с использованием обратной транскриптазы. кДНК можно синтезировать с использованием набора для синтеза первой цепи кДНК с обратной транскриптазой AMV Reverse Transcriptase First-strand cDNA Synthesis Kit (Seikagaku Co.) и т.п. Более того, для синтеза и амплификации кДНК можно соответствующим образом использовать набор для амплификации кДНК SMART RACE cDNA amplification kit (Clontech) и способ 5'-RACE на основе ПЦР (Proc. Natl. Acad. Sci. U.S.A. (1988) 85(23), 8998-9002; Nucleic Acids Res. (1989) 17(8), 2919-2932). В таком способе синтеза кДНК на оба конца кДНК можно вносить подходящие участки для ферментов рестрикции, описанные ниже.
Представляющий интерес фрагмент кДНК очищают из полученного продукта ПЦР, а затем его лигируют в векторную ДНК. Таким образом, конструируют рекомбинантный вектор и вводят в E. coli и т.п. После селекции колоний из образующих колонии E. coli можно получать желаемый рекомбинантный вектор. Затем исследуют, имеет ли рекомбинантный вектор представляющую интерес нуклеотидную последовательность кДНК, известным способом, таким как способ с терминацией цепи дидезоксинуклеотидами.
Способ 5'-RACE, в котором используются праймеры для амплификации гена вариабельной области, удобно использовать для выделения гена, кодирующего вариабельную область. Сначала конструируют библиотеку кДНК 5'-RACE с помощью синтеза кДНК с использованием РНК, экстрагированных из клеток гибридом в качестве матрицы. Для синтеза библиотеки кДНК 5'-RACE соответствующим образом используют коммерчески доступный набор, такой как набор для амплификации кДНК SMART RACE.
Ген антитела амплифицируют способом ПЦР с использованием полученной библиотеки кДНК 5'-RACE в качестве матрицы. Праймеры для амплификации гена антитела мыши можно конструировать на основе известных последовательностей генов антител. Нуклеотидные последовательности праймеров варьируют, в зависимости от подкласса иммуноглобулинов. Таким образом, предпочтительно, чтобы подкласс был определен заранее с использованием коммерчески доступного набора, такого как набор для изотипирования моноклональных антител мыши Iso Strip mouse monoclonal antibody isotyping kit (Roche Diagnostics).
В частности, например, праймеры, которые обеспечивают амплификацию генов, кодирующих тяжелые цепи γ1, γ2a, γ2b и γ3 и легкие цепи κ и λ, используют для выделения генов, кодирующих IgG мыши. Как правило, для амплификации гена вариабельной области gG используют праймер, который гибридизуется с областью константной области вблизи вариабельной области, в качестве 3'-праймера. Между тем, в качестве 5'-праймера используют праймер, прилагаемый к набору для конструирования библиотеки кДНК 5'-RACE.
Продукты ПЦР, амплифицированные таким образом, используют для переформировывания иммуноглобулинов, состоящих из комбинации тяжелых и легких цепей. Селекцию желаемого антитела можно проводить с использованием активности связывания IL-6R переформированного иммуноглобулина в качестве индикатора. Например, когда задачей является выделение антитела против IL-6R, более предпочтительно, чтобы связывание антитела с IL-6R было специфическим. Скрининг связывающего IL-6R антитела можно проводить, например, с помощью следующих стадий:
(1) контактирование экспрессирующей IL-6R клетки с антителом, содержащим V-область, кодируемую кДНК, выделенной из гибридомы;
(2) выявление связывания антитела с эксперессирующей IL-6R клеткой; и
(3) селекция антитела, которое связывается с экспрессирующей IL-6R клеткой.
Способы выявления связывания антитела с экспрессирующими IL-6R клетками известны. В частности, связывание антитела с экспрессирующими IL-6R клетками можно выявлять с помощью описанных выше способов, таких как FACS. Для оценки активности связывания антитела соответствующим образом используют иммобилизованные образцы экспрессирующих IL-6R клеток.
Предпочтительные способы скрининга антител, в которых в качестве индикатора используется активность связывания, также включают способы пэннинга с использованием фаговых векторов. Способы скрининга с использованием фаговых векторов являются преимущественными, когда гены антител выделяют из библиотек подклассов тяжелых цепей и легких цепей из популяции клеток, экспрессирующей поликлональное антитело. Гены, кодирующие вариабельные области тяжелых цепей и легких цепей, можно связывать соответствующей линкерной последовательностью с получением одноцепочечного Fv (scFv). Фаги, представляющие scFv на их поверхности, можно получать встраиванием гена, кодирующего scFv, в фаговый вектор. Фаги контактируют с представляющим интерес антигеном. Затем ДНК, кодирующую scFv, обладающие представляющей интерес активностью связывания, можно выделять путем сбора фагов, связанных с антигеном. Этот процесс можно повторять при необходимости для увеличения содержания scFv, обладающих представляющий интерес активностью связывания.
После выделения кДНК, кодирующей V-область представляющего интерес антитела против IL-6R, кДНК расщепляют ферментами рестрикции, которые распознают участки рестрикции, встроенные на обоих концах кДНК. Предпочтительные ферменты рестрикции распознают и расщепляют нуклеотидную последовательность, которая встречается в нуклеотидной последовательности гена антитела с низкой частотой. Более того, предпочтительно в представляющий интерес вектор вводят участок рестрикции для фермента, который образует липкий конец, для встраивания одной копии расщепленного фрагмента в правильной ориентации. Кодирующую кДНК V-область антитела против IL-6R расщепляют, как описано выше, и ее встраивают в соответствующий экспрессирующий вектор для конструирования экспрессирующего вектора для антитела. В этом случае, если ген, кодирующий константную область антитела (C-область) и ген, кодирующий описанную выше V-область, слить в рамке считывания, то получат химерное антитело. В рамках настоящего изобретения "химерное антитело" означает, что происхождение константной области отличается от вариабельной области. Таким образом, в дополнение к гетерохимерным антителам мыши/человека, химерные антитела по настоящему изобретению включают аллохимерные антитела человека/человека. Экспрессирующий вектор для химерного антитела можно конструировать путем встраивания гена V-области в экспрессирующий вектор, который уже имеет константную область. В частности, например, последовательность, распознаваемая первым ферментом рестрикции, который расщепляет выше гена V-области, можно соответствующим образом помещать на 5'-конец экспрессирующего вектора, содержащего ДНК, кодирующую желаемую константную область антитела. Экспрессирующий вектор для химерного антитела конструируют путем слияния в рамке считывания двух генов, расщепленных одной и той же комбинацией ферментов рестрикции.
Для получения моноклонального антитела против IL-6R, гены антител встраивают в экспрессирующий вектор так, чтобы гены экспрессировались под контролем регулирующей экспрессию области. Регулирующая экспрессию область для экспрессии антитела включает, например, энхансеры и промоторы. Более того, на N-конец можно присоединять подходящую сигнальную последовательность, так чтобы экспрессируемое антитело секретировалось наружу клеток. В примерах ниже в качестве сигнальной последовательности используют пептид, имеющий аминокислотную последовательность MGWSCIILFLVATATGVHS (SEQ ID NO:3). Между тем, можно присоединять другие подходящие сигнальные последовательности. Экспрессированный полипептид расщепляется на С-конце описанной выше последовательности, и полученный полипептид секретируется из клеток в качестве зрелого полипептида. Затем подходящие клетки-хозяева трансформируют экспрессирующим вектором и получают рекомбинантные клетки, экспрессирущие ДНК, кодирующую антитело IL-6R.
ДНК, кодирующие тяжелую цепь (H-цепь) и легкую цепь (L-цепь) антитела по отдельности встраивают в различные экспрессирующие векторы для экспрессии гена антитела. Молекулу антитела, имеющую H- и L-цепи, можно экспрессировать путем котрансфекции одной и той же клетки-хозяина векторами, в которые соответственно встроены гены H-цепи и L-цепи. Альтернативно клетки-хозяева можно трансформировать одним экспрессирующим вектором, в который встроены ДНК, кодирующие H- и L-цепи (см. WO 1994/011523).
Существуют различные известные комбинации клетка-хозяин/экспрессирующий вектор для получения антител путем введения выделенных генов антител в соответствующих хозяев. Все из этих экспрессирующих систем пригодны для выделения антигенсвязывающих доменов по настоящему изобретению. Подходящие эукариотические клетки, используемые в качестве клеток-хозяев, включают клетки животных, клетки растений и клетки грибов. В частности, клетки животных включают, например, следующие клетки.
(1) клетки млекопитающих: CHO (линия клеток яичника китайского хомяка), COS (линия клеток почки обезьяны), миелома (Sp2/0, NS0 и т.п.), BHK (клеточная линия почки детенышей хомячка), Hela, Vero, HEK293 (линия клеток почки эмбриона человека с расщепленной аденовирусной ДНК (Ad)5), Freestyle293, клетки PER.C6 (линия клеток сетчатки эмбриона человека, трансформированных генами E1A и E1B аденовируса 5 типа (Ad5)), и т.п. (Current Protocols in Protein Science (май 2001 года, раздел 5.9, таблица 5.9.1));
(2) клетки земноводных: ооциты Xenopus и т.п.; и
(3) клетки насекомых: sf9, sf21, Tn5 и т.п.
Кроме того, в качестве клетки растений известна экспрессирующая система для генов антител с использованием клеток, происходящих из рода Nicotiana, таких как Nicotiana tabacum. Для трансформации клеток растений можно соответствующим образом использовать культивированные клетки каллуса.
Более того, в качестве клеток грибов можно использовать следующие клетки:
- дрожжи: род Saccharomyces, как например, Saccharomyces serevisiae, и род Pichia, как например, Pichia pastoris; и
- нитчатые грибы: род Aspergillus, как например, Aspergillus niger.
Более того, также известны экспрессирующие системы для генов антител, в которых используются прокариотические клетки. Например, при использовании бактериальных клеток, в рамках настоящего изобретения можно подходящим образом использовать клетки E. coli, клетки Bacillus subtilis и т.п. Экспрессирующие векторы, несущие представляющие интерес гены антитела, вводят в клетки путем трансфекции. Трансфицированные клетки культивируют in vitro, и из культуры трансформированных клеток можно получать желаемое антитело.
В дополнение к описанным выше клеткам-хозяевам, также для получения рекомбинантного антитела можно использовать трансгенных животных. Следовательно, антитело можно получать из животного, которому введен ген, кодирующий представляющее интерес антитело. Например, ген антитела можно конструировать в качестве слитого гена путем встраивания в рамке считывания в ген, который кодирует белок, специфически продуцируемый в молоке. В качестве белка, секретируемого в молоке, можно использовать, например β-казеин козы и т.п. Фрагменты ДНК, содержащие слитый ген, в который встроен ген антитела, инъецируют в эмбрион козы, а затем этот эмбрион вводят в самку козы. Желаемые антитела можно получать в качестве белка, слитого с белком молока, из молока, продуцируемого трансгенной козой, родившейся от козы, являющейся реципиентом эмбриона (или ее потомства). Кроме того, для увеличения объема молока, содержащего желаемое антитело, продуцируемого трансгенной козой, трансгенной козе при необходимости можно вводить гормоны, Ebert, K. M. et al., Bio/Technology (1994) 12 (7), 699-702).
Когда антигенсвязывающую молекулу, описанную в настоящем описании, вводят человеку, антигенсвязывающий домен, происходящий из генетически рекомбинантного антитела, которое является искусственно измененным для снижения гетерологической антигенности против человека и т.п., можно соответствующим образом использовать в качестве антигенсвязывающего домена антигенсвязывающей молекулы. Такие генетически рекомбинантные антитела включают, например, гуманизированные антитела. Эти измененные антитела получают соответствующим образом известными способами.
Вариабельная область антитела, используемая для получения антигенсвязывающего домена антигенсвязывающей молекулы, описанной в настоящем описании, как правило, образована тремя определяющими комплементарность областями (CDR) которые разделены четырьмя каркасными областями (FR). CDR представляет собой область, которая по существу определяет специфичность связывания антитела. Аминокислотные последовательности CDR являются в высокой степени разнообразными. С другой стороны, образующие FR аминокислотные последовательности часто обладают высокой идентичностью, даже среди антител с отличающейся специфичностью связывания. Таким образом, как правило, специфичность связывания определенного антитела можно вносить в другое антитело путем пересадки CDR.
Гуманизированное антитело также называют переформированным антителом человека. В частности, известны гуманизированные антитела, полученные пересадкой CDR антитела не являющегося человеком животного, такого как антитело мыши, в антитело человека и т.п. Также известны стандартные способы генной инженерии для получения гуманизированных антител. В частности, например, в качестве способа пересадки CDR антитела мыши в FR человека известна перекрывающаяся ПЦР с удлинением. В перекрывающейся ПЦР с удлинением, нуклеотидную последовательность, кодирующую CDR антитела мыши, подлежащую пересадке, добавляют к праймерам для синтеза FR антитела человека. Праймеры получают для каждой из четырех FR. Является общепризнанным, что при пересадке CDR мыши в FR человека, выбор FR человека, который имеет высокую идентичность FR мыши, является преимущественным для сохранения функции CDR. Таким образом, как правило, предпочтительно использовать FR человека, содержащую аминокислотную последовательность, которая обладает высокой идентичностью с аминокислотной последовательностью FR, соседней с CDR мыши, подлежащей пересадке.
Нуклеотидные последовательности, подлежащие лигированию, конструируют так, чтобы они были соединены друг с другом в рамке считывания. FR человека синтезируют по отдельности с использованием соответствующих праймеров. В результате получают продукты, в которых ДНК, кодирующая CDR мыши, связана с индивидуальными ДНК, кодирующими FR. Нуклеотидные последовательности, кодирующие CDR мыши, каждого продукта конструируют так, чтобы они перекрывались друг с другом. Затем проводят синтез комплементарной цепи для гибридизации перекрывающихся CDR-областей продуктов, синтезированных с использованием гена антитела человека в качестве матрицы. С помощью этой реакции FR человека лигируют через последовательности CDR мыши.
Полноразмерный ген V-области, в котором в конечном итоге лигированы три CDR и четыре FR, амплифицируют с использованием праймеров, которые гибридизуются с его 5'- или 3'-концом, в которые добавлены подходящие последовательности, распознаваемые ферментом рестрикции. Экспрессирующий вектор для гуманизированного антитела можно получать путем встраивания ДНК, полученной, как описано выше, и ДНК, которая кодирует C-область антитела человека, в экспрессирующий вектор, так чтобы они лигировались в рамке считывания. После трансфекции рекомбинантным вектором хозяина для получения рекомбинантных клеток, рекомбинантные клетки культивируют и ДНК, кодирующую гуманизированное антитело, экспрессируют с получением гуманизированного антитела в клеточной культуре (см., публикацию патента Европы № EP 239400 и международную публикацию патента № WO 1996/002576).
Путем качественного или количественного определения и оценки антигенсвязывающей активности гуманизированного антитела, полученного, как описано выше, можно пригодным образом отобрать FR антитела человека, которые позволяют CDR образовывать подходящий антигенсвязывающий центр при лигировании через CDR. Аминокислотные остатки в FR можно заменять при необходимости, так чтобы CDR переформированного анитетела человека образовывали соответствующий антигенсвязывающий центр. Например, мутации аминокислотной последовательности можно вносить в FR с использованием способа ПЦР, который используется для пересадки CDR мыши в FR человека. Более конкретно, в праймеры, которые гибридизуются с FR, можно вносить частичные мутации нуклеотидной последовательности. Мутации нуклеотидной последовательности вносятся в FR, синтезированные с использованием таких праймеров. Мутантные последовательности FR, имеющие желаемые характеристики, можно отбирать путем измерения и оценки активности мутанта антитела с замененной аминокислотой в отношении связывания с антигеном с помощью упомянутого выше способа (Cancer Res. (1993) 53: 851-856).
Альтернативно желаемые антитела человека можно получать путем иммунизации трансгенных животных, имеющих полный набор генов антител человека (см. WO 1993/012227; WO 1992/003918; WO 1994/002602; WO 1994/025585; WO 1996/034096; WO 1996/033735) путем иммунизации ДНК.
Более того, также известны способы получения антител человека путем пэннинга с использованием библиотек антител человека. Например, V-область антитела человека экспрессируют в качестве одноцепочечного антитела (scFv) на поверхности фага способом фагового дисплея. Можно отбирать фаги, экспрессирующие scFv, которые связываются с антигеном. Последовательность ДНК, кодирующую V-область антитела человека, которая связывается с антигеном, можно определять путем анализа генов выбранных фагов. Определяют последовательность ДНК scFv, которая связывается с антигеном. Получают Экспрессирующий вектор путем слияния последовательности V-области в рамке считывания с последовательностью C-области желаемого антитела человека, и встраивания ее в соответствующий экспрессирующий вектор. Экспрессирующий вектор вводят в клетки, пригодные для экспрессии, такие как клетки, описанные выше. Антитело человека можно получать путем экспрессии гена, кодирующего антитело человека, в клетках. Эти способы уже известны (см. WO 1992/001047; WO 1992/020791; WO 1993/006213; WO 1993/011236; WO 1993/019172; WO 1995/001438; WO 1995/015388).
В дополнение к способам, описанным выше, для выделения генов антител можно соответствующим образом использовать способы клонирования B-клеток (идентификация каждой кодирующей антитело последовательности, ее клонирование и выделение; использование при конструировании экспрессирующего вектора для получения каждого антитела (IgG1, IgG2, IgG3 или IgG4, в частности); и т.п.), как описано, например Bernasconi et al. (Science (2002) 298: 2199-2202) или в WO 2008/081008.
Система нумерации EU и система нумерации Kabat
В соответствии со способами, используемыми в рамках настоящего изобретения, положения аминокислот, приписываемые CDR и FR антитела, указаны в соответствии с нумерацией по Kabat (Sequences of Proteins of Immunological Interest (National Institute of Health, Bethesda, Md., 1987 и 1991)). В рамках настоящего изобретения, когда антигенсвязывающая молекула представляет собой антитело или антигенсвязывающий фрагмент, аминокислоты вариабельной области указаны в соответствии с нумерацией по системе Kabat, в то время как аминокислоты константной области указаны в соответствии с системой нумерации EU на основе положений аминокислот по Kabat.
Антигенсвязывающие домены, зависимые от специфического для ткани-мишени соединения
Для получения антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен), антигенсвязывающая активность которого варьирует в зависимости от концентрации специфического для ткани-мишени соединения или, более конкретно, антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен), зависимого от специфического для ткани-мишени соединения, можно соответствующим образом использовать способы, описанные в представленном выше разделе об активности связывания. В качестве неограничивающего варианта осуществления ниже представлены некоторые конкретные примеры способов. Например, для подтверждения того, что антигенсвязывающая активность антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен) в присутствии специфического для ткани-мишени соединения становится более высокой, чем антигенсвязывающая активность антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен) в отсутствие соединения, сравнивают антигенсвязывающую активность антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен) в присутствии и в отсутствие специфического для ткани-мишени соединения или в присутствии высоких и низких концентраций соединения. В другом неограничивающем варианте осуществления, например, для подтверждения того, что антигенсвязывающая активность антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен) в присутствии высокой концентрации специфического для ткани-мишени соединения становится более высокой, чем антигенсвязывающая активность антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен) в присутствии низкой концентрации соединения, сравнивают антигенсвязывающую активность антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен) в присутствии высоких и низких концентраций специфического для ткани-мишени соединения.
Более того, в рамках настоящего изобретения выражение "антигенсвязывающая активность в присутствии специфического для ткани-мишени соединения является более высокой, чем антигенсвязывающая активность в отсутствие соединения" альтернативно может быть сформулировано как "антигенсвязывающая активность антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен) в отсутствие специфического для ткани-мишени соединения является более низкой, чем антигенсвязывающая активность в присутствии соединения". Более того, в рамках настоящего изобретения" антигенсвязывающая активность антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен) в отсутствие специфического для ткани-мишени соединения является более низкой, чем антигенсвязывающая активность в присутствии соединения" альтернативно может быть описано как "антигенсвязывающая активность антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен) в отсутствие специфического для ткани-мишени соединения является более слабой, чем антигенсвязывающая активность в присутствии соединения".
Более того, в рамках настоящего изобретения выражение "антигенсвязывающая активность в присутствии высокой концентрации специфического для ткани-мишени соединения является более высокой, чем антигенсвязывающая активность в присутствии низкой концентрации соединения" альтернативно может быть сформулировано как "антигенсвязывающая активность антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен) в присутствии низкой концентрации специфического для ткани-мишени соединения является более низкой, чем антигенсвязывающая активность в присутствии высокой концентрации соединения". В рамках настоящего изобретения "антигенсвязывающая активность антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен) в присутствии низкой концентрации специфического для ткани-мишени соединения является более низкой, чем антигенсвязывающая активность в присутствии высокой концентрации соединения" альтернативно может быть описано как "антигенсвязывающая активность антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен) в присутствии низкой концентрации специфического для ткани-мишени соединения является более слабой, чем антигенсвязывающая активность в присутствии высокой концентрации соединения".
Условия измерения антигенсвязывающей активности, отличные от концентрации специфического для ткани-мишени соединения, конкретно не ограничены, и они могут быть соответствующим образом выбраны специалистами в данной области. Например, ее можно измерять в условиях буфера HEPES и 37°C. Например, для измерения можно использовать Biacore (GE Healthcare) и т.п. Когда антиген представляет собой растворимую молекулу, активность антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен) в отношении связывания с растворимой молекулой можно определять путем нанесения антигена в качестве анализируемого соединения на чип, на который иммобилизован антигенсвязывающий домен (или антигенсвязывающая молекула, содержащая домен). Альтернативно, когда антиген представляет собой молекулу мембранного типа, активность связывания с молекулой мембранного типа можно определять путем нанесения антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен) в качестве анализируемого соединения на чип, на который иммобилизован антиген.
При условии, что антигенсвязывающая активность антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен), содержащегося в антигенсвязывающих молекулах по настоящему изобретению в отсутствие специфического для ткани-мишени соединения является более слабой, чем антигенсвязывающая активность в присутствии специфического для ткани-мишени соединения, соотношение между антигенсвязывающей активностью в отсутствие соединения и антигенсвязывающей активностью в присутствии соединения конкретно не ограничено. Однако величина KD (в отсутствие соединения)/KD (в присутствии соединения), которая представляет собой соотношение константы диссоциации (KD) в отношении антигена в отсутствие специфического для ткани-мишени соединения и KD в присутствии соединения, предпочтительно равно 2 или более, более предпочтительно 10 или более, и еще более предпочтительно 40 или более. Верхний предел величины KD (в отсутствие соединения)/KD (в присутствии соединения) конкретно не ограничен и он может представлять собой любую величину, например, 400, 1000 или 10000 при условии, что она может быть обеспечена с помощью технологий, известных специалистам в данной области. Когда антигенсвязывающую активность не наблюдают в отсутствие специфического для ткани-мишени соединения значение верхнего предела равно бесконечности.
При условии, что антигенсвязывающая активность антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен), содержащегося в антигенсвязывающих молекулах по настоящему изобретению в присутствии низкой концентрации специфического для ткани-мишени соединения является более слабой, чем антигенсвязывающая активность в присутствии высокой концентрации специфического для ткани-мишени соединения, соотношение между антигенсвязывающей активностью в присутствии низкой концентрации соединения и антигенсвязывающей активностью в присутствии высокой концентрации соединения конкретно не ограничено. Однако величина KD (в присутствии низкой концентрации соединения)/KD (в присутствии высокой концентрации соединения), которая представляет собой соотношение константы диссоциации (KD) в отношении антигена в присутствии низкой концентрации специфического для ткани-мишени соединения и KD в присутствии высокой концентрации соединения, предпочтительно равно 2 или более, более предпочтительно 10 или более, и еще более предпочтительно 40 или более. Верхний предел величины KD (в присутствии низкой концентрации соединения)/KD (в присутствии высокой концентрации соединения) конкретно не ограничен, и он может представлять собой любую величину, например, 400, 1000 или 10000, при условии, что она может быть обеспечена технологиями, известными специалистам в данной области. Когда антигенсвязывающую активность не наблюдают в присутствии низкой концентрации специфического для ткани-мишени соединения, величина верхнего предела равна бесконечности.
Для величины антигенсвязывающей активности, если антиген представляет собой растворимую молекулу, можно использовать константу диссоциации (KD); и если антиген представляет собой молекулу мембранного типа, можно использовать кажущуюся константу диссоциации (кажущаяся KD). Константу диссоциации (KD) и кажущуюся константу диссоциации (кажущаяся KD) можно определять способами, известными специалистам в данной области, например, с использованием Biacore (GE Healthcare), графика Скэтчарда, проточного цитометра и т.п.
В качестве другого индикатора, который демонстрирует соотношение между антигенсвязывающей активностью антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен) по настоящему изобретению в отсутствие специфического для ткани-мишени соединения и антигенсвязывающей активностью в присутствии соединения, например, можно, соответственно, использовать константу скорости диссоциации kd. Когда константу скорости диссоциации (kd) используют вместо константы диссоциации (KD) в качестве индикатора, который демонстрирует соотношение активностей связывания, величина kd (в отсутствие соединения)/kd (в присутствии соединения), которая представляет собой соотношение между kd (константа скорости диссоциации) для антигена в отсутствие специфического для ткани-мишени соединения и kd в присутствии соединения, предпочтительно равна 2 или более, более предпочтительно 5 или более, еще более предпочтительно 10 или более, и еще более предпочтительно 30 или более. Верхний предел величины kd (в отсутствие соединения)/kd (в присутствии соединения) конкретно не ограничен и он может представлять собой любую величину, например, 50, 100 или 200, при условии, что она может быть обеспечена с использованием обычных технических знаний специалистов в данной области. Когда антигенсвязывающую активность не наблюдают в отсутствие специфического для ткани соединения, отсутствует диссоциация и величина верхнего предела становится равной бесконечности.
В качестве другого индикатора, который демонстрирует соотношение между антигенсвязывающей активностью антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен) по настоящему изобретению в присутствии низкой концентрации специфического для ткани-мишени соединения и антигенсвязывающей активностью в присутствии высокой концентрации соединения, например, можно соответственно использовать константу скорости диссоциации kd. Когда константу скорости диссоциации (kd) используют вместо константы диссоциации (KD) в качестве индикатора, демонстрирующего соотношение активностей связывания, величина kd (в присутствии низкой концентрации соединения)/kd (в присутствии высокой концентрации соединения), которая представляет собой соотношение между kd (константа скорости диссоциации) в отношении антигена в присутствии низкой концентрации специфического для ткани-мишени соединения и kd в присутствии высокой концентрации соединения, предпочтительно равна 2 или более, более предпочтительно 5 или более, еще более предпочтительно 10 или более, и еще более предпочтительно 30 или более. Верхний предел величины kd (в присутствии низкой концентрации соединения)/kd (в присутствии высокой концентрации соединения) конкретно не ограничен, и он может представлять собой любую величину, например, 50, 100 или 200, при условии, что она может быть обеспечена с использованием обычных технических знаний специалистов в данной области. Когда антигенсвязывающую активность не наблюдают в присутствии низкой концентрации специфического для ткани-мишени соединения, отсутствует диссоциация, и величина верхнего предела становится равной бесконечности.
Для величины антигенсвязывающей активности, если антиген представляет собой растворимую молекулу, можно использовать константу скорости диссоциации (kd); и если антиген представляет собой молекулу мембранного типа можно использовать кажущуюся константу скорости диссоциации (кажущаяся kd). Константу скорости диссоциации (kd) и кажущуюся константу скорости диссоциации (кажущаяся kd) можно определять способами, известными специалистам в данной области, например, с использованием Biacore (GE Healthcare), проточного цитометра или сходных с ними. В рамках настоящего изобретения при измерении антигенсвязывающей активности антигенсвязывающего домена (или антигенсвязывающей молекулы, содержащей домен) при определенной концентрации специфического для ткани-мишени соединения, условия, отличные от концентрации соединения, предпочтительно являются одинаковыми.
Например, в одном варианте осуществления, предусматриваемом в рамках настоящего изобретения, антигенсвязывающий домен (или антигенсвязывающую молекулу, содержащую домен) с более низкой антигенсвязывающей активностью в отсутствие специфического для ткани-мишени соединения, чем в присутствии соединения, можно получать путем скрининга антигенсвязывающих доменов (или антигенсвязывающих молекул), который включает стадии:
(a) определения антигенсвязывающей активности антигенсвязывающих доменов (или антигенсвязывающих молекул) в отсутствие специфического для ткани-мишени соединения;
(b) определения антигенсвязывающей активности антигенсвязывающих доменов (или антигенсвязывающих молекул) в присутствии специфического для ткани-мишени соединения; и
(c) отбора антигенсвязывающего домена (или антигенсвязывающей молекулы) с более низкой антигенсвязывающей активностью в отсутствие специфического для ткани-мишени соединения, чем в присутствии соединения.
Например, в одном варианте осуществления, предусматриваемом в рамках настоящего изобретения, антигенсвязывающий домен (или антигенсвязывающую молекулу, содержащую домен) с более низкой антигенсвязывающей активностью в присутствии низкой концентрации специфического для ткани-мишени соединения, чем в присутствии высокой концентрации соединения, можно получать путем скрининга антигенсвязывающих доменов (или антигенсвязывающих молекул), который включает стадии:
(a) определения антигенсвязывающей активности антигенсвязывающих доменов (или антигенсвязывающих молекул) в присутствии низкой концентрации специфического для ткани-мишени соединения;
(b) определения антигенсвязывающей активности антигенсвязывающих доменов (или антигенсвязывающих молекул) в присутствии высокой концентрации специфического для ткани-мишени соединения; и
(c) отбора антигенсвязывающего домена (или антигенсвязывающей молекулы) с более низкой антигенсвязывающей активностью в присутствии низкой концентрации специфического для ткани-мишени соединения, чем в присутствии высокой концентрации соединения.
Более того, в одном варианте осуществления, предусматриваемом в рамках настоящего изобретения, антигенсвязывающий домен (или антигенсвязывающую молекулу, содержащую домен) с более низкой антигенсвязывающей активностью в отсутствие специфического для ткани-мишени соединения, чем в присутствии соединения, можно получать путем скрининга антигенсвязывающих доменов (или антигенсвязывающих молекул) или их библиотеки, который включает стадии:
(a) контактирования антигенсвязывающих доменов (или антигенсвязывающих молекул) или их библиотеки с антигеном в присутствии специфического для ткани-мишени соединения;
(b) помещения антигенсвязывающих доменов (или антигенсвязывающих молекул), которые связываются с антигеном на указанной стадии (a), в условия отсутствия соединения;
(c) выделения антигенсвязывающего домена (или антигенсвязывающей молекулы), который диссоциировал на стадии (b).
Более того, в одном варианте осуществления, предусматриваемом в рамках настоящего изобретения, антигенсвязывающий домен (или антигенсвязывающую молекулу, содержащую домен) с более низкой антигенсвязывающей активностью в присутствии низкой концентрации специфического для ткани-мишени соединения, чем в присутствии высокой концентрации соединения, можно получать путем скрининга антигенсвязывающих доменов (или антигенсвязывающих молекул) или их библиотеки, который включает стадии:
(a) контактирования антигенсвязывающих доменов (или антигенсвязывающих молекул) или их библиотеки с антигеном в присутствии высокой концентрации специфического для ткани-мишени соединения;
(b) помещение антигенсвязывающих доменов (или антигенсвязывающих молекул), которые связываются с антигеном на стадии (a), в условия присутствия низкой концентрации соединения;
(c) выделения антигенсвязывающего домена (или антигенсвязывающей молекулы), который диссоциировал на указанной стадии (b).
Альтернативно в одном варианте осуществления, предусматриваемом в рамках настоящего изобретения, антигенсвязывающий домен (или антигенсвязывающую молекулу, содержащую домен) с более низкой антигенсвязывающей активностью в отсутствие специфического для ткани-мишени соединения, чем в присутствии соединения, можно получать путем скрининга антигенсвязывающих доменов (или антигенсвязывающих молекул) или их библиотеки, который включает стадии:
(a) контактирования библиотеки антигенсвязывающих доменов (или антигенсвязывающих молекул) с антигеном в отсутствие специфического для ткани-мишени соединения;
(b) отбора антигенсвязывающих доменов (или антигенсвязывающих молекул), которые не связываются с антигеном на указанной стадии (a);
(c) позволения антигенсвязывающим доменам (или антигенсвязывающим молекулам), отобранным на указанной стадии (b) связаться с антигеном в присутствии соединения; и
(d) выделения антигенсвязывающего домена (или антигенсвязывающей молекулы), который связывается с антигеном на указанной стадии (c).
Альтернативно, в одном варианте осуществления, предусматриваемом в рамках настоящего изобретения, антигенсвязывающий домен (или антигенсвязывающую молекулу, содержащую домен) с более низкой антигенсвязывающей активностью в присутствии низкой концентрации специфического для ткани-мишени соединения, чем в присутствии высокой концентрации соединения, можно получать путем скрининга антигенсвязывающих доменов (или антигенсвязывающих молекул) или их библиотеки, который включает стадии:
(a) контактирования библиотеки антигенсвязывающих доменов (или антигенсвязывающих молекул) с антигеном в присутствии низкой концентрации специфического для ткани-мишени соединения;
(b) отбора антигенсвязывающих доменов (или антигенсвязывающих молекул), которые не связываются с антигеном на указанной стадии (a);
(c) позволения антигенсвязывающим доменам (или антигенсвязывающим молекулам), отобранным на указанной стадии (b) связаться с антигеном в присутствии высокой концентрации соединения; и
(d) выделения антигенсвязывающего домена (или антигенсвязывающей молекулы), который связывается с антигеном на указанной стадии (c).
Более того, в одном варианте осуществления, предусматриваемом в рамках настоящего изобретения, антигенсвязывающий домен (или антигенсвязывающую молекулу, содержащую домен) с более низкой антигенсвязывающей активностью в отсутствие специфического для ткани-мишени соединения, чем в присутствии соединения, можно получать способом скрининга, включающим стадии:
(a) контактирования библиотеки антигенсвязывающих доменов (или антигенсвязывающих молекул) с колонкой с иммобилизованным на ней антигеном в присутствии специфического для ткани-мишени соединения;
(b) элюирования антигенсвязывающего домена (или антигенсвязывающей молекулы), который связывается с колонкой на указанной стадии (a), из колонки в отсутствие соединения; и
(c) выделения антигенсвязывающего домена (или антигенсвязывающей молекулы), элюированного на указанной стадии (b).
Более того, в одном варианте осуществления, предусматриваемом в рамках настоящего изобретения, антигенсвязывающий домен (или антигенсвязывающую молекулу, содержащую домен) с более низкой антигенсвязывающей активностью в присутствии низкой концентрации специфического для ткани-мишени соединения, чем в присутствии высокой концентрации соединения, можно получать способом скрининга, включающим стадии:
(a) контактирования библиотеки антигенсвязывающих доменов (или антигенсвязывающих молекул) с колонкой с иммобилизованным на ней антигеном в присутствии высокой концентрации специфического для ткани-мишени соединения;
(b) элюирования антигенсвязывающего домена (или антигенсвязывающей молекулы), который связывается с колонкой на стадии (a), из колонки в присутствии низкой концентрации соединения; и
(c) выделения антигенсвязывающего домена (или антигенсвязывающей молекулы), элюированного на стадии (b).
Более того, в одном варианте осуществления, предусматриваемом в рамках настоящего изобретения, антигенсвязывающий домен (или антигенсвязывающую молекулу, содержащую домен) с более низкой антигенсвязывающей активностью в отсутствие специфического для ткани-мишени соединения, чем в присутствии соединения, можно получать способом скрининга, включающим стадии:
(a) позволения библиотеке антигенсвязывающих доменов (или антигенсвязывающих молекул) пройти через колонку с иммобилизованным на ней антигеном в отсутствие специфического для ткани-мишени соединения;
(b) сбора антигенсвязывающего домена (или антигенсвязывающей молекулы), элюированного без связывания с колонкой на стадии (a);
(c) позволения антигенсвязывающему домену (или антигенсвязывающей молекулы), собранному на стадии (b), связаться с антигеном в присутствии соединения; и
(d) выделения антигенсвязывающего домена (или антигенсвязывающей молекулы), который связывается с антигеном на указанной стадии (c).
Более того, в одном варианте осуществления, предусматриваемом в рамках настоящего изобретения, антигенсвязывающий домен (или антигенсвязывающую молекулу, содержащую домен) с более низкой антигенсвязывающей активностью в присутствии низкой концентрации специфического для ткани-мишени соединения, чем в присутствии высокой концентрации соединения, можно получать способом скрининга, включающим стадии:
(a) позволения библиотеке антигенсвязывающих доменов (или антигенсвязывающих молекул) пройти через колонку с иммобилизованным на ней антигеном в присутствии низкой концентрации специфического для ткани-мишени соединения;
(b) сбора антигенсвязывающего домена (или антигенсвязывающей молекулы), элюированного без связывания с колонкой на указанной стадии (a);
(c) позволения антигенсвязывающему домену (или антигенсвязывающей молекуле), собранному на указанной стадии (b), связаться с антигеном в присутствии высокой концентрации соединения; и
(d) выделения антигенсвязывающего домена (или антигенсвязывающей молекулы), который связывается с антигеном на указанной стадии (c).
Более того, в одном варианте осуществления, предусматриваемом в рамках настоящего изобретения, антигенсвязывающий домен (или антигенсвязывающую молекулу, содержащую домен) с более низкой антигенсвязывающей активностью в отсутствие специфического для ткани-мишени соединения, чем в присутствии соединения, можно получать способом скрининга, включающим стадии:
(a) контактирования антигена с библиотекой антигенсвязывающих доменов (или антигенсвязывающих молекул) в присутствии специфического для ткани-мишени соединения;
(b) получения антигенсвязывающего домена (или антигенсвязывающей молекулы), который связывается с антигеном на указанной стадии (a);
(c) помещения антигенсвязывающего домена (или антигенсвязывающей молекулы), полученного на указанной стадии (b), в условия отсутствия соединения; и
(d) выделения антигенсвязывающего домена (или антигенсвязывающей молекулы), антигенсвязывающая активность которого на указанной стадии (c) является более слабой, чем антигенсвязывающая активность эталона, выбранного на указанной стадии (b).
Более того, в одном варианте осуществления, предусматриваемом в рамках настоящего изобретения, антигенсвязывающий домен (или антигенсвязывающую молекулу, содержащую домен) с более низкой антигенсвязывающей активностью в присутствии низкой концентрации специфического для ткани-мишени соединения, чем в присутствии высокой концентрации соединения, можно получать способом скрининга, включающим стадии:
(a) контактирования антигена с библиотекой антигенсвязывающих доменов (или антигенсвязывающих молекул) в присутствии высокой концентрации специфического для ткани-мишени соединения;
(b) получения антигенсвязывающего домена (или антигенсвязывающей молекулы), который связывается с антигеном на указанной стадии (a);
(c) помещения антигенсвязывающего домена (или антигенсвязывающей молекулы), полученного на указанной стадии (b), в условия присутствия низкой концентрации соединения; и
(d) выделения антигенсвязывающего домена (или антигенсвязывающей молекулы), антигенсвязывающая активность которого на указанной стадии (c) является более слабой, чем антигенсвязывающая активность эталона, выбранного на указанной стадии (b).
Упомянутые выше стадии можно повторять два или более раз. Таким образом, настоящее изобретение относится к антигенсвязывающему домену (или антигенсвязывающей молекуле, содержащей домен) с более низкой антигенсвязывающей активностью в отсутствие специфического для ткани-мишени соединения, чем в присутствии соединения, или к антигенсвязывающему домену (или антигенсвязывающей молекуле, содержащей домен) с более низкой антигенсвязывающей активностью в присутствии низкой концентрации специфического для ткани-мишени соединения, чем в присутствии высокой концентрации соединения, полученному способами скрининга, которые дополнительно включают стадию повторения стадий с (a) по (c) или с (a) по (d) два или более раз в упомянутых выше способах скрининга. Количество повторов стадий с (a) по (c) или с (a) по (d) конкретно не ограничено, и, как правило, оно составляет десять или менее.
В способах скрининга по настоящему изобретению специфическое для ткани-мишени соединение может представлять собой соединение, определяемое количественной величиной специфичности к ткани-мишени, такой как присутствие в ткани мишени в концентрации (например, в высокой концентрации или в низкой концентрации), отличающейся от концентрации в не являющихся мишенью тканях. Например, специфическое для ткани-мишени соединение присутствует на отличающемся уровне при любых концентрациях. Однако, как правило, специфическое для ткани-мишени соединение может присутствовать в концентрации, увеличенной по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 100%, по меньшей мере на 110%, по меньшей мере на 120%, по меньшей мере на 130%, по меньшей мере на 140%, по меньшей мере на 150%, по меньшей мере в 2 раза, по меньшей мере в 5 раз, по меньшей мере в 10 раз, по меньшей мере в 50 раз, по меньшей мере в 100 раз, по меньшей мере в 103 раз, по меньшей мере в 104 раз, по меньшей мере в 105 раз, по меньшей мере в 106 раз или более или вплоть до бесконечности (когда соединение отсутствует в не являющихся мишенью тканях).
Пороговое значение, различающее низкие и высокие концентрации, может быть выбрано соответствующим образом согласно изобретению. Например, в неограничивающем варианте осуществления порогового значения для ATP или аденозина, пороговое значение для условий низкой концентрации может быть выбрано соответственно из величин 10 нМ, 1 нМ, 100 пМ, 10 пМ, 1 пМ и 0 M. В зависимости от заданного порогового значения, условия высокой концентрации могут быть выбраны соответствующим образом из величины, выбранной из величины, по меньшей мере на 110%, по меньшей мере на 120%, по меньшей мере на 130%, по меньшей мере на 140%, по меньшей мере на 150%, по меньшей мере в два раза, по меньшей мере в пять раз, по меньшей мере в 10 раз, по меньшей мере в 50 раз, по меньшей мере в 100 раз, по меньшей мере в 103 раз, по меньшей мере в 104 раз, по меньшей мере в 105 раз, и по меньшей мере в 106 раз превышающей величину каждой пороговой величины. Более того, в неограничивающем варианте осуществления для PGE2 пороговое значение для условий низкой концентрации может быть соответственно выбрано из величин 10 пМ, 1 пМ, 100 фМ, 10 фМ, 1 фМ и 0 M. В зависимости от заданного порогового значения, условия высокой концентрации могут быть выбраны соответственно из величины, выбранной из величины, по меньшей мере на 110%, по меньшей мере на 120%, по меньшей мере на 130%, по меньшей мере на 140%, по меньшей мере на 150%, по меньшей мере в два раза, по меньшей мере в пять раз, по меньшей мере в 10 раз, по меньшей мере в 50 раз, по меньшей мере в 100 раз, по меньшей мере в 103 раз, по меньшей мере 104 раз, по меньшей мере 105 раз и по меньшей мере в 106 раз превышающей величину каждого порогового значения. Более того, в неограничивающем варианте осуществления для кинуренина, пороговое значение для условий низкой концентрации может быть выбрано соответственно из величин 10 мкМ, 1 мкМ, 100 нМ, 10 нМ, и 1 нМ и 0 M. В зависимости от заданного порогового значения, условия высокой концентрации могут быть выбраны соответственно из величины, выбранной из величины, по меньшей мере на 110%, по меньшей мере на 120%, по меньшей мере на 130%, по меньшей мере на 140%, по меньшей мере на 150%, по меньшей мере в два раза, по меньшей мере в пять раз, по меньшей мере в 10 раз, по меньшей мере в 50 раз, по меньшей мере в 100 раз, по меньшей мере в 103 раз, по меньшей мере в 104 раз, по меньшей мере в 105 раз, и по меньшей мере в 106 раз превышающей величину каждой пороговой величины.
Антигенсвязывающую активность антигенсвязывающего домена (или антигенсвязывающей молекулы) можно измерять способом, известным специалистам в данной области, и условия, отличные от концентрации специфического для ткани-мишени соединения, могут быть соответственно выбраны специалистом в данной области. Антигенсвязывающую активность антигенсвязывающего домена (или антигенсвязывающей молекулы) можно оценивать в качестве константы диссоциации (KD), кажущейся константы диссоциации (кажущаяся KD), константы скорости диссоциации (kd), кажущейся константы скорости диссоциации (кажущаяся kd), и т.д. Их можно определять способами, известными специалистам в данной области, например, с использованием Biacore (GE Healthcare), графика Скэтчарда, FACS и т.п.
В рамках настоящего изобретения стадия отбора антитела или антигенсвязывающего домена с более высокой антигенсвязывающей активностью в присутствии специфического для ткани-мишени соединения, чем в отсутствие соединения, имеет то же значение, что и стадия отбора антитела или антигенсвязывающего домена с более низкой антигенсвязывающей активностью в отсутствие специфического для ткани-мишени соединения, чем в присутствии соединения.
В рамках настоящего изобретения стадия отбора антитела или антигенсвязывающего домена с более высокой антигенсвязывающей активностью в присутствии высокой концентрации специфического для ткани-мишени соединения, чем в присутствии низкой концентрации соединения, имеет то же значение, что и стадия отбора антитела или антигенсвязывающего домена с более низкой антигенсвязывающей активностью в отсутствие специфического для ткани-мишени соединения, чем в присутствии соединения.
При условии, что антигенсвязывающая активность в отсутствие специфического для ткани-мишени соединения является более низкой, чем антигенсвязывающая активность в присутствии соединения, различие между антигенсвязывающей активностью в присутствии соединения и антигенсвязывающей активностью в отсутствие соединения конкретно не ограничено, однако предпочтительно антигенсвязывающая активность в присутствии соединения относительно антигенсвязывающей активности в отсутствие соединения является более высокой в два раза или более, более предпочтительно в 10 раз или более, и еще более предпочтительно в 40 раз или более. Верхний предел различия между антигенсвязывающими активностями конкретно не ограничен, и, при условии, что он может быть достигнут способами, известными специалистам в данной области, возможна любая величина, такая как в 400 раз, в 1000 раз или в 10000 раз. В отсутствие специфического для ткани-мишени соединения, когда антигенсвязывающую активность не наблюдают, этот верхний предел становится равным бесконечности.
Антигенсвязывающие домены (или антигенсвязывающие молекулы, содержащие домены) по настоящему изобретению, которые подлежат скринингу упомянутыми выше способами, могут представлять собой любые антигенсвязывающие домены (или антигенсвязывающие молекулы); и, например, можно проводить скрининг упомянутых выше антигенсвязывающих доменов (или антигенсвязывающих молекул). Например, можно проводить скрининг антигенсвязывающих доменов (или антигенсвязывающих молекул), имеющих встречающиеся в природе последовательности, и можно проводить скрининг антигенсвязывающих доменов (или антигенсвязывающих молекул) с аминокислотными последовательностями с заменой.
Библиотека
В соответствии с определенным вариантом осуществления антигенсвязывающий домен (или антигенсвязывающую молекулу, содержащую этот домен) по настоящему изобретению можно получать из библиотеки, в основном содержащей множество антигенсвязывающих молекул, имеющих последовательности, отличающиеся друг от друга, в которых по меньшей мере один аминокислотный остаток, который изменяет активность связывания антигенсвязывающей молекулы с антигеном, зависимым от специфического для ткани-мишени соединения, содержится в антигенсвязывающем домене. Примеры соединения включают (1) первичные метаболиты цикла Кребса или гликолитического каскада, такие как лактоза, янтарная кислота или лимонная кислота, (2) аминокислоты, такие как аланин, глутаминовая кислота или аспарагин, (3) кинуренин и его аминокислотные метаболиты, такие как антраниловая кислота, 3-гидроксикинуренин и кинуреновая кислота, (4) метаболиты арахидоновой кислоты, такие как простагландин E2, и (5) нуклеозиды, содержащие структуру пуринового кольца, такие как аденозин, аденозинтрифосфат (ATP), аденозиндифосфат (ADP) и аденозинмонофосфат (AMP). Ниже приведены примеры такой библиотеки, в основном, содержащей множество антигенсвязывающих молекул, имеющих отличающиеся друг от друга последовательности, в которых в антигенсвязывающем домене содержится по меньшей мере один аминокислотный остаток, который изменяет активность связывания антигенсвязывающей молекулы с антигенами, зависимыми от аденозина и/или ATP, которые являются специфическими для ткани-мишени соединениями.
В рамках настоящего изобретения "библиотека" относится к множеству антигенсвязывающих молекул или множеству слитых полипептидов, содержащих антигенсвязывающие молекулы, или нуклеиновых кислот или полинуклеотидов, кодирующих их последовательности. Последовательности множества антигенсвязывающих молекул или множества слитых полипептидов, содержащих антигенсвязывающие молекулы, в библиотеке не идентичны, а отличаются друг от друга.
В рамках настоящего изобретения выражение "последовательности отличаются друг от друга" в выражении "множество антигенсвязывающих молекул, последовательности которых отличаются друг от друга", означает, что последовательности антигенсвязывающих молекул в библиотеке отличаются друг от друга. В частности, в библиотеке количество последовательностей, отличающихся друг от друга, отражает количество независимых клонов с различными последовательностями, и также может называться "размером библиотеки". Размер библиотеки для обычной библиотеки фагового дисплея находится в диапазоне от 106 до 1012. Размер библиотеки может быть увеличен вплоть до 1014 с использованием известных способов, таких как рибосомный дисплей. Однако истинное количество фаговых частиц, используемых в селекции способом пэннинга фаговой библиотеки, как правило, в 10-10000 раз превышает размер библиотеки. Эту избыточную множественность также называют "количеством эквивалентов библиотеки", и она означает, что существует от 10 до 10000 индивидуальных клонов, которые обладают одной и той же аминокислотной последовательностью. Таким образом, в рамках настоящего изобретения выражение "последовательности отличаются друг от друга" означает, что последовательности независимых антигенсвязывающих молекул в библиотеке, исключая эквиваленты библиотеки, отличаются друг от друга. Более конкретно, указанное выше означает, что существует от 106 до 1014 антигенсвязывающих молекул, последовательности которых отличаются друг от друга, предпочтительно от 107 до 1012 молекул, более предпочтительно от 108 до 1011 молекул, и особенно предпочтительно от 108 до 1010 молекул, последовательности которых отличаются друг от друга.
В рамках настоящего изобретения выражение "множество" в выражении "библиотека в основном состоит из множества антигенсвязывающих молекул" в случае, например, антигенсвязывающих молекул, слитых полипептидов, полинуклеотидных молекул, векторов или вирусов по настоящему изобретению, главным образом, относится к группе из двух или более типов вещества. Например, когда два или более вещества отличаются друг от друга конкретной характеристикой, это означает, что существует два или более типов вещества. Такие примеры могут включать, например, мутантные аминокислоты, наблюдаемые в конкретных аминокислотных положениях аминокислотной последовательности. Например, когда существует две или более антигенсвязывающих молекул по настоящему изобретению, последовательности которых являются по существу одинаковыми или предпочтительно одинаковыми, за исключением нестрогих остатков или за исключением конкретных мутантных аминокислот в гипервариабельных положениях, экспонированных на поверхности, существует множество антигенсвязывающих молекул по настоящему изобретению. В другом примере, когда существует две или более полинуклеотидных молекулы, последовательности которых являются по существу одинаковыми или предпочтительно одинаковыми, за исключением нуклеотидов, кодирующих нестрогие остатки, или нуклеотидов, кодирующих мутантные аминокислоты гипервариабельных положений, экспонированных на поверхности, существует множество полинуклеотидных молекул по настоящему изобретению.
Кроме того, в настоящем описании выражение "в основном состоит из" в выражении "библиотека в основном состоит из множества антигенсвязывающих молекул" отражает количество антигенсвязывающих молекул, антигенсвязывающая активность которых варьирует, в зависимости от концентрации специфического для ткани-мишени соединения, среди независимых клонов с отличающимися последовательностями в библиотеке. Особенно предпочтительно, чтобы в библиотеке существовало по меньшей мере 104 антигенсвязывающих молекул, имеющих такую активность связывания. Более предпочтительно, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, содержащей по меньшей мере 105 антигенсвязывающих молекул, обладающих такой активностью связывания. Еще более предпочтительно, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, содержащей по меньшей мере 106 антигенсвязывающих молекул, обладающих такой активностью связывания. Особенно предпочтительно, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, содержащей по меньшей мере 107 антигенсвязывающих молекул, обладающих такой активностью связывания. Еще более предпочтительно, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, содержащей по меньшей мере 108 антигенсвязывающих молекул, обладающих такой активностью связывания. Альтернативно это также предпочтительно может быть выражено как соотношение количества антигенсвязывающих молекул, в которых антигенсвязывающая активность антигенсвязывающего домена варьирует в зависимости от присутствия или отсутствия аденозина и/или ATP относительно количества независимых клонов, имеющих различные последовательности, в библиотеке. В частности, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в которой антигенсвязывающие молекулы, имеющие такую активность связывания, составляют от 0,1% до 80%, предпочтительно от 0,5% до 60%, более предпочтительно от 1% до 40%, еще более предпочтительно от 2% до 20%, и особенно предпочтительно от 4% до 10% независимых клонов с отличающимися последовательностями в библиотеке. В случае слитых полипептидов, полинуклеотидных молекул или векторов, могут быть возможными сходные выражения, в которых используются количества молекул или отношение к общему количеству молекул. В случае вирусов, также могут быть возможными сходные выражения, в которых используется количество вирионов или отношение к общему количеству вирионов.
Аминокислоты, которые изменяют антигенсвязывающую активность антигенсвязывающего домена в зависимости от присутствия или отсутствия аденозина и/или ATP
Антигенсвязывающие домены или антитела по настоящему изобретению, подвергаемые скринингу описанными выше способами скрининга, можно получать любым способом. Можно использовать заранее существующие антитела, заранее существующие библиотеки (фаговые библиотеки и т.д.), антитела или библиотеки, полученные из гибридом, полученных путем иммунизации животных, или из B-клеток иммунизированных животных, и антитела или библиотеки, полученные из иммунных клеток, таких как B-клетки, животных, иммунизированных конъюгатом, в котором аденозин или ATP пригодным образом связан с адъювантом, таким как высокоиммуногенный пептид эпитопа T-клеток. Неограничивающий пример пептида эпитопа T-клеток пригодным образом включает происходящий из столбнячного токсина хелперный пептид p30 (представленный в SEQ ID NO:4, и также обозначаемый как фрагмент C (FrC)).
Примеры аминокислот, которые изменяют антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от присутствия или отсутствия и аденозина и/или ATP, как описано выше, включают аминокислоты, которые образуют мотив, связывающий аденозин и/или ATP. Положения аминокислот, где в антигенсвязывающем домене содержатся упомянутые выше аминокислоты, не ограничиваются каким-либо конкретным положением. При условии, что антигенсвязывающая активность антигенсвязывающего домена изменяется в зависимости от присутствия или отсутствия аденозина и/или ATP, возможно любое положение в вариабельной области тяжелой цепи или вариабельной области легкой цепи, образующей антигенсвязывающий домен. Более конкретно, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном содержащей антигенсвязывающие молекулы, имеющие отличающиеся друг от друга последовательности, в которых аминокислоты, которые изменяют антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от присутствия или отсутствия аденозина и/или ATP, содержатся в антигенсвязывающем домене тяжелой цепи. В неограничивающем варианте осуществления антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном содержащей антигенсвязывающие молекулы, имеющие отличающиеся друг от друга последовательности, в которых аминокислоты, которые изменяют антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от присутствия или отсутствия аденозина и/или ATP, содержатся в CDR1, CDR2 и/или CDR3 тяжелой цепи. В другом неограничивающем варианте осуществления антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном содержащей антигенсвязывающие молекулы, имеющие отличающиеся друг от друга последовательности, в которых аминокислоты, которые изменяют антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от присутствия или отсутствия аденозина и/или ATP, содержатся в FR1, FR2, FR3 и/или FR4 тяжелой цепи.
Более того, в одном варианте осуществления настоящего изобретения, антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном содержащей антигенсвязывающие молекулы, обладающие отличающимися друг от друга последовательностями, в которых аминокислоты, которые изменяют антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от присутствия или отсутствия аденозина и/или ATP, содержатся в антигенсвязывающем домене тяжелой цепи и/или легкой цепи. В неограничивающем варианте осуществления антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном содержащей антигенсвязывающие молекулы, имеющие отличающиеся друг от друга последовательности, в которых аминокислоты, которые изменяют антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от присутствия или отсутствия аденозина и/или ATP, содержатся в CDR1, CDR2 и/или CDR3 тяжелой цепи и/или легкой цепи. В другом неограничивающем варианте осуществления антигенсвязывающие домены по настоящему изобретению можно получать из библиотеки, в основном содержащей антигенсвязывающие молекулы, имеющие отличающиеся друг от друга последовательности, в которых аминокислоты, которые изменяют антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от присутствия или отсутствия аденозина и/или ATP, содержатся в FR1, FR2, FR3 и/или FR4 тяжелой цепи и/или легкой цепи.
В неограничивающем варианте осуществления примеры таких аминокислот включают любые одну или несколько аминокислот, выбранные из аминокислот в положениях 52, 52a, 53, 96, 100a и 100c, содержащихся в вариабельной области тяжелой цепи. Также в неограничивающем варианте осуществления примеры таких аминокислот включают одну или несколько аминокислот, выбранных из аминокислот, включающих Ser в положении 52, Ser в положении 52a, Arg в положении 53, Gly в положении 96, Leu в положении 100a, и Trp в положении 100c, содержащиеся в вариабельной области тяжелой цепи.
В качестве каркасной последовательности вариабельных областей легкой цепи и/или тяжелой цепи антигенсвязывающей молекулы можно использовать любую каркасную последовательность при условии, что в антигенсвязывающем домене тяжелой цепи и/или легкой цепи содержатся аминокислоты, которые изменяют антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от присутствия или отсутствия аденозина и/или ATP. Происхождение каркасных последовательностей не ограничено, и их можно получать из организма человека или любых не являющихся человеком организмов. Такие организмы предпочтительно включают мышей, крыс, морских свинок, хомячков, песчанок, кошек, кроликов, собак, коз, овец, животных семейства бычьи, лошадей, верблюдов и организмы, выбранные из не являющихся человеком приматов. В особенно предпочтительном варианте осуществления каркасные последовательности вариабельной области легкой цепи и/или тяжелой цепи антигенсвязывающей молекулы предпочтительно имеют каркасные области человека эмбрионального типа. Таким образом, в одном варианте осуществления настоящего изобретения, если каркасные последовательности целиком представляют собой последовательности человека, полагают, что антигенсвязывающая молекула по настоящему изобретению индуцирует малый иммуногенный ответ или не индуцирует его при введении человеку (например, для лечения заболеваний). В указанном выше значении выражение "содержащий последовательность эмбрионального типа" в рамках настоящего изобретения означает, что часть каркасных последовательностей по настоящему изобретению идентична части каркасных последовательностей человека эмбрионального типа. Например, когда последовательность FR2 тяжелой цепи антигенсвязывающей молекулы по настоящему изобретению представляет собой комбинацию последовательностей FR2 тяжелой цепи отличающихся каркасных последовательностей человека эмбрионального типа, такая молекула также является антигенсвязывающей молекулой, "содержащей последовательность эмбриональной линии" в рамках настоящего изобретения. Даже когда каркасные последовательности антигенсвязывающих молекул по настоящему изобретению представляют собой последовательности с заменами, они являются "антигенсвязывающими молекулами, содержащими последовательность эмбрионального типа" по настоящему изобретению. Примеры таких последовательностей с заменами включают, в частности, последовательности, в которых аминокислоты части каркасных последовательностей человека эмбрионального типа заменены аминокислотами, которые изменяют антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от присутствия или отсутствия аденозина и/или ATP.
Предпочтительные примеры каркасных областей включают, например, последовательности полностью человеческих каркасных областей человека, известные в настоящее время, которые включены в web-сайт V-Base (http://vbase.mrc-cpe.cam.ac.uk/) или другие. Эти последовательности каркасных областей можно соответствующим образом использовать в качестве последовательности эмбрионального типа, содержащейся в антигенсвязывающей молекуле по настоящему изобретению. Последовательности эмбрионального типа могут быть подразделены на категории в соответствии с их сходством (Tomlinson et al. (J. Mol. Biol. (1992) 227, 776-798); Williams and Winter (Eur. J. Immunol. (1993) 23, 1456-1461); Cox et al. (Nat. Genetics (1994) 7, 162-168)). Соответствующие последовательности эмбрионального типа могут быть выбраны из Vκ, которые подразделяются на семь подгрупп; Vλ, которые подразделяются на десять подгрупп; и VH, которые подразделяются на семь подгрупп.
Полностью человеческие последовательности VH предпочтительно включают, но не ограничиваются ими, например, последовательности VH:
подгруппы VH1 (например, VH1-2, VH1-3, VH1-8, VH1-18, VH1-24, VH1-45, VH1-46, VH1-58 и VH1-69);
подгруппы VH2 (например, VH2-5, VH2-26 и VH2-70);
подгруппы VH3 (VH3-7, VH3-9, VH3-11, VH3-13, VH3-15, VH3-16, VH3-20, VH3-21, VH3-23, VH3-30, VH3-33, VH3-35, VH3-38, VH3-43, VH3-48, VH3-49, VH3-53, VH3-64, VH3-66, VH3-72, VH3-73 и VH3-74);
подгруппы VH4 (VH4-4, VH4-28, VH4-31, VH4-34, VH4-39, VH4-59 и VH4-61);
подгруппы VH5 (VH5-51);
подгруппы VH6 (VH6-1); и
подгруппы VH7 (VH7-4 и VH7-81).
Они также описаны в известных документах (Matsuda et al. (J. Exp. Med. (1998) 188, 1973-1975)) и т.п., и, таким образом, специалисты в данной области могут соответствующим образом моделировать антигенсвязывающие молекулы по настоящему изобретению, исходя из информации об этих последовательностях. Также предпочтительно использовать другие полностью человеческие каркасные области или каркасные субобласти.
Полностью человеческие последовательности VK предпочтительно включают, но не ограничиваются ими, например:
A20, A30, L1, L4, L5, L8, L9, L11, L12, L14, L15, L18, L19, L22, L23, L24, O2, O4, O8, O12, O14 и O18, выделенные в подгруппу Vk1;
A1, A2, A3, A5, A7, A17, A18, A19, A23, O1 и O11, выделенные в подгруппу Vk2;
A11, A27, L2, L6, L10, L16, L20 и L25, выделенные в подгруппу Vk3;
B3, выделенная в подгруппу Vk4;
B2 (также обозначаемая как Vk5-2), выделенная в подгруппу Vk5; и
A10, A14 и A26, выделенные в подгруппу VK6.
(Kawasaki et al. (Eur. J. Immunol. (2001) 31, 1017-1028); Schable and Zachau (Biol. Chem. Hoppe Seyler (1993) 374, 1001-1022); Brensing-Kuppers et al. (Gene (1997) 191, 173-181)).
Полностью человеческие последовательности VL предпочтительно включают, но не ограничиваются ими, например:
V1-2, V1-3, V1-4, V1-5, V1-7, V1-9, V1-11, V1-13, V1-16, V1-17, V1-18, V1-19, V1-20 и V1-22, выделенные в подгруппу VL1;
V2-1, V2-6, V2-7, V2-8, V2-11, V2-13, V2-14, V2-15, V2-17 и V2-19, выделенные в подгруппу VL1;
V3-2, V3-3 и V3-4, выделенные в подгруппу VL3;
V4-1, V4-2, V4-3, V4-4 и V4-6, выделенные в подгруппу VL4; и
V5-1, V5-2, V5-4 и V5-6, выделенные в подгруппу VL5 (Kawasaki et al. (Genome Res. (1997) 7, 250-261)).
Обычно эти каркасные последовательности отличаются друг от друга одним или несколькими аминокислотными остатками. Эти каркасные последовательности можно использовать в комбинации с "по меньшей мере одним аминокислотным остатком, который изменяет антигенсвязывающую активность антигенсвязывающего домена в зависимости от присутствия или отсутствия аденозина и/или ATP" по настоящему изобретению. Другие примеры полностью человеческих каркасных областей, используемых в комбинации с "по меньшей мере одним аминокислотным остатком, который изменяет антигенсвязывающую активность антигенсвязывающего домена в зависимости от присутствия или отсутствия аденозина и/или ATP" по настоящему изобретению включают, но не ограничиваются ими, например, KOL, NEWM, REI, EU, TUR, TEI, LAY и POM (например, Kabat et al. (1991), выше, Wu et al. (J. Exp. Med. (1970) 132, 211-250)).
Без связи с конкретной теорией, полагают, что одна из причин для того, чтобы ожидать, что использование последовательностей эмбрионального типа предотвратит неблагоприятные иммунные ответы у большинства индивидуумов, состоит в следующем. В результате процесса созревания аффинности в ходе нормальных иммунных ответов, часто в вариабельных областях иммуноглобулинов происходят соматические мутации. Такие мутации чаще всего происходят в области CDR, последовательности которых являются гипервариабельными, но также затрагивают остатки каркасных областей. Такие мутации каркасной области не существуют в генах эмбрионального типа, но также с меньшей вероятностью будут иммуногенными у пациентов. С другой стороны в нормальной человеческой популяции присутствует большинство каркасных последовательностей, экспрессируемых с генов эмбрионального типа. В результате иммунной толерантности, ожидается, что эти каркасные области эмбрионального типа будут иметь низкую иммуногенность у пациентов или не иметь иммуногенности у пациентов. Для максимизации возможности иммунной толерантности можно выбирать гены, кодирующие вариабельную область, из группы часто встречающихся функциональных генов эмбрионального типа.
Для получения антигенсвязывающих молекул по настоящему изобретению, в которых описанные выше последовательности вариабельной области, последовательности вариабельной области тяжелой или легкой цепей, последовательности CDR или каркасные последовательности содержат аминокислоты, которые изменяют антигенсвязывающую активность антигенсвязывающего домена в зависимости от присутствия или отсутствия аденозина и/или ATP, можно соответствующим образом использовать известные способы, такие как сайт-направленный мутагенез (Kunkel et al. (Proc. Natl. Acad. Sci. USA (1985) 82, 488-492)) и перекрывающаяся ПЦР.
Например, библиотека, которая содержит множество антигенсвязывающих молекул по настоящему изобретению, последовательности которых отличаются друг от друга, можно конструировать путем комбинирования вариабельных областей тяжелой цепи, полученных в качестве рандомизированной библиотеки последовательностей вариабельной области с вариабельной областью легкой цепи, выбранной в качестве последовательности CDR и/или каркасной последовательности, исходно содержащей по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающего домена в зависимости от присутствия или отсутствия аденозина и/или ATP.
Альтернативно последовательность вариабельной области тяжелой цепи и/или легкой цепи, выбранную в качестве последовательности CDR и/или каркасной последовательности, исходно содержащей по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающего домена в зависимости от присутствия или отсутствия аденозина и/или ATP, как упоминалось выше, можно конструировать так, чтобы она содержала различные аминокислотные остатки, отличные от указанного выше аминокислотного остатка(ов). В рамках настоящего изобретения такие остатки обозначают как "нестрогие остатки". Количество и положение нестрогих остатков конкретно не ограничено, при условии, что антигенсвязывающая активность антигенсвязывающей молекулы по настоящему изобретению варьирует в зависимости от концентрации специфического для ткани соединения. В частности, последовательности CDR и/или последовательности FR тяжелой цепи и/или легкой цепи могут содержать один или несколько нестрогих остатков. Можно идентифицировать нестрогие остатки и остатки, которые могут быть заменены на другие аминокислоты для получения библиотеки путем внесения мутаций или с помощью анализа кристаллической структуры комплексов, образовавшихся между антителом и аденозином и/или ATP. Например, из анализа кристаллической структуры комплексов, образовавшихся между антителом и аденозином и/или ATP, можно идентифицировать остатки в антителе, которые не вовлечены в связывание с аденозином и/или ATP. Можно выбирать аминокислоты, которые поддерживают связывание с соединениями на соответствующем уровне, даже когда остатки, которые идентифицированы, как не вовлеченные в связывание с аденозином и/или ATP, заменены на другие аминокислоты. Таким образом, можно сконструировать библиотеку, которая имеет выбранные аминокислоты для выбранных остатков. В этом случае можно конструировать библиотеку, в основном содержащую множество антигенсвязывающих молекул, чтобы она представляла собой ансамбль антигенсвязывающих молекул, в котором остатки, идентифицированные как не вовлеченные в связывание с аденозином и/или ATP, замещены аминокислотами, которые отличаются друг от друга. Следовательно, комбинация индивидуальных нестрогих остатков, замещенных аминокислотами, которые отличаются друг от друга, может обеспечить разнообразие последовательности в антигенсвязывающих молекулах, содержащих нестрогие остатки.
Антигенсвязывающие молекулы можно конструировать так, чтобы они включали остатки, где по меньшей мере один из остатков, идентифицированных как вовлеченный в связывание с аденозином и/или ATP, становится любым остатком, выбранным из этого остатка и других остатков, которые отличаются от этого остатка. В неограничивающем варианте осуществления примеры аминокислот, идентифицированных как вовлеченные в связывание с аденозином и/или ATP, могут включать одну или несколько аминокислот, выбранных из аминокислот в положениях 52, 52a, 53, 96, 100a и 100c в вариабельной области тяжелой цепи. В неограничивающем варианте осуществления примеры таких аминокислот включают одну или несколько аминокислот, выбранных из аминокислот, включающих Ser в положении 52, Ser в положении 52a, Arg в положении 53, Gly в положении 96, Leu в положении 100a и Trp в положении 100c, содержащихся в вариабельной области тяжелой цепи. Например, когда Leu в положении 100a, упомянутом выше, идентифицирован как вовлеченный в связывание с аденозином и/или ATP, аминокислотный остаток в положении 100a в антигенсвязывающих молекулах, включенных в библиотеку, может представлять собой любой аминокислотный остаток, выбранный из нестрогих остатков His, Met, Leu, Arg, Trp или Tyr, в дополнение к Leu.
В неограничивающем варианте осуществления примеры нестрогих остатков могут включать аминокислоты в положениях 31, 32, 33, 35, 50, 55, 56, 57, 58, 59, 95, 96, 97, 98, 99, 100, 100a и 100b, содержащихся в вариабельной области тяжелой цепи. В другом неограничивающем варианте осуществления примеры таких аминокислот могут включать аминокислоты в положениях 26, 27, 27a, 27b, 27c, 28, 29, 31, 32, 50, 51, 52, 53, 54, 55, 89, 90, 91, 92, 93, 94, 95a, 96 и 97, содержащихся в вариабельной области легкой цепи.
В неограничивающем варианте осуществления примеры упомянутых выше нестрогих остатков могут включать следующие аминокислоты, содержащиеся в вариабельной области тяжелой цепи:
Asp, Gly, Asn, Ser, Arg или Thr в качестве аминокислоты в положении 31;
Ala, Phe, His, Asn, Ser или Tyr в качестве аминокислоты в положении 32;
Ala, Glu, Asp, Gly, Phe, Ile, His, Lys, Met, Leu, Asn, Gln, Pro, Ser, Arg, Trp, Val, Tyr или Thr в качестве аминокислоты в положении 33;
His, Ser, Thr, Tyr или Asn в качестве аминокислоты в положении 35;
Ala, Glu, Asp, Gly, Phe, Ile, His, Lys, Met, Leu, Asn, Gln, Pro, Arg, Thr, Trp, Val, Tyr или Ser в качестве аминокислоты в положении 50;
Ala, Glu, Asp, Gly, Leu, Thr, Ser, Arg или Asn в качестве аминокислоты в положении 55;
Ala, Glu, Asp, Gly, Phe, Ile, His, Lys, Met, Leu, Gln, Pro, Ser, Thr, Trp, Val или Tyr в качестве аминокислоты в положении 56;
Ala, Lys, Arg, Thr или Ile в качестве аминокислоты в положении 57;
Asp, Gly, Phe, His, Ser, Thr, Tyr или Asn в качестве аминокислоты в положении 58;
Leu или Tyr в качестве аминокислоты в положении 59;
Ala, Ile, Lys, Met, Leu, Arg, Trp, Val, Tyr или Phe в качестве аминокислоты в положении 95;
Ala, Asp, Asn или Ser в качестве аминокислоты в положении 96;
Ala, Asp, Gly, Ile, His, Lys, Met, Leu, Asn, Ser, Val, Tyr или Arg в качестве аминокислоты в положении 97;
Ala, Glu, Asp, Gly, Phe, Ile, His, Met, Leu, Asn, Gln, Pro, Ser, Arg, Thr, Trp, Val, Tyr или Lys в качестве аминокислоты в положении 98;
Ala, Glu, Asp, Phe, His, Lys, Asn, Gln, Ser, Arg, Trp, Val, Tyr или Gly в качестве аминокислоты в положении 99;
Ala, Glu, Gly, Phe, Ile, His, Lys, Met, Leu, Asn, Gln, Pro, Ser, Arg, Thr, Trp, Val, Tyr или Asp в качестве аминокислоты в положении 100;
Ala, Phe, Ile, His, Lys, Met, Arg, Trp, Val или Tyr в качестве аминокислоты в положении 100a; или
Ala, Glu, Asp, Gly, Phe, Ile, His, Lys, Met, Leu, Gln, Pro, Ser, Arg, Thr, Trp, Val, Tyr или Asn в качестве аминокислоты в положении 100b.
В неограничивающем варианте осуществления примеры упомянутых выше нестрогих остатков могут включать следующие аминокислоты, содержащиеся в вариабельной области легкой цепи:
Ala, Ser или Thr в качестве аминокислоты в положении 26;
Thr или Ser в качестве аминокислоты в положении 27;
Gly, Asn, Thr или Ser в качестве аминокислоты в положении 27a;
Asn или Asp в качестве аминокислоты в положении 27b;
Ile или Val в качестве аминокислоты в положении 27c;
Asp или Gly в качестве аминокислоты в положении 28;
Ala, Asp, Phe, Ser, Arg, Thr, Tyr или Gly в качестве аминокислоты в положении 29;
Glu, Asp, Lys или Asn в качестве аминокислоты в положении 31;
Ala, Asp, Ser, Thr или Tyr в качестве аминокислоты в положении 32;
Asp, Gly, Lys, Asn, Gln, Ser, Arg, Tyr или Glu в качестве аминокислоты в положении 50;
Asp, Gly, Lys, Asn, Thr или Val в качестве аминокислоты в положении 51;
Ala, Asp, Asn, Thr или Ser в качестве аминокислоты в положении 52;
Glu, Asp, His, Asn, Gln, Ser, Tyr или Lys в качестве аминокислоты в положении 53;
Lys или Arg в качестве аминокислоты в положении 54;
Leu или Pro в качестве аминокислоты в положении 55;
Ala, Gly, Phe, Leu, Asn, Gln, Thr, Val, Tyr или Ser в качестве аминокислоты в положении 89;
Ala, Leu, Thr, Val или Ser в качестве аминокислоты в положении 90;
Ala, Asp, Phe, His, Lys, Asn, Ser, Arg, Thr, Trp, Val или Tyr в качестве аминокислоты в положении 91;
Glu, Asp, Ser, Arg, Thr, Val, Tyr или Ala в качестве аминокислоты в положении 92;
Ala, Asp, Ile, Asn, Ser, Arg, Thr, Val, Tyr или Gly в качестве аминокислоты в положении 93;
Ala, Asp, Gly, Ile, Asn, Arg, Thr или Ser в качестве аминокислоты в положении 94;
Ala, Glu, Asp, Gly, Phe, Ile, His, Lys, Met, Leu, Gln, Pro, Ser, Arg, Thr, Trp, Val, Tyr или Asn в качестве аминокислоты в положении 95;
Ala, Glu, Asp, Gly, Ile, His, Lys, Leu, Gln, Pro, Ser, Arg, Thr, Tyr или Asn в качестве аминокислоты в положении 95a;
Ala, Asp, Gly, Phe, His, Lys, Leu, Asn, Gln, Pro, Ser, Thr, Trp, Tyr или Val в качестве аминокислоты в положении 96; или
Ala, Gly, Ile, Met, Leu, Ser или Val в качестве аминокислоты в положении 97.
В рамках настоящего изобретения "нестрогий остаток" относится к варьированию аминокислотных остатков, присутствующему в гипервариабельных положениях аминокислот вариабельных областей легкой цепи и тяжелой цепи, в которых существует несколько различных аминокислот, когда сравнивают аминокислотные последовательности известных и/или нативных антител или антигенсвязывающих доменов. Гипервариабельные положения, как правило, расположены в областях CDR. В одном варианте осуществления данные, предоставленные в Kabat, Sequences of Proteins of Immunological Interest (National Institute of Health Bethesda Md., 1987 and 1991) являются пригодными для определения гипервариабельных положений в известных и/или нативных антителах. Более того, в базах данных в Интернете (http://vbase.mrc-cpe.cam.ac.uk/, и http://www.bioinf.org.uk/abs/index.html) предоставлено множество собранных последовательностей легких цепей и тяжелых цепей человека и их положения. Информация о последовательностях и положениях является пригодной для определения гипервариабельных положений в рамках настоящего изобретения. В соответствии с настоящим изобретением, когда определенное аминокислотное положение имеет предпочтительно от приблизительно 2 до приблизительно 20, предпочтительно от приблизительно 3 до приблизительно 19, предпочтительно от приблизительно 4 до приблизительно 18, предпочтительно от 5 до 17, предпочтительно от 6 до 16, предпочтительно от 7 до 15, предпочтительно от 8 до 14, предпочтительно от 9 до 13, и предпочтительно от 10 до 12 возможных вариантов аминокислотных остатков, положение можно считать гипервариабельным. В некоторых вариантах осуществления определенное положение аминокислоты может иметь предпочтительно по меньшей мере приблизительно 2, предпочтительно по меньшей мере приблизительно 4, предпочтительно по меньшей мере приблизительно 6, предпочтительно по меньшей мере приблизительно 8, предпочтительно приблизительно 10, и предпочтительно приблизительно 12 возможных вариантов аминокислотных остатков.
Библиотека по настоящему изобретению, которая содержит множество антигенсвязывающих молекул, имеющих отличающиеся друг от друга последовательности, можно конструировать путем комбинирования вариабельных областей тяжелой цепи, продуцированных в качестве рандомизированной библиотеки последовательностей вариабельных областей с упомянутыми выше вариабельными областями легкой цепи, в которые внесен по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающих доменов в зависимости от присутствия или отсутствия аденозина и/или ATP. Аналогично, библиотека по настоящему изобретению, которая содержит множество антигенсвязывающих молекул, имеющих отличающиеся друг от друга последовательности, также можно получать путем комбинирования вариабельных областей тяжелой цепи, в которые внесен по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающих доменов в зависимости от присутствия или отсутствия аденозина и/или ATP, и которые имеют другие аминокислотные остатки, сконструированные в качестве нестрогих остатков.
Когда вариабельные области тяжелой цепи, продуцированные в качестве рандомизированной библиотеки вариабельных областей, и вариабельные области легкой цепи, в которые внесен по меньшей мере один аминокислотный остаток, который изменяет антигенсвязывающую активность антигенсвязывающей молекулы в зависимости от концентрации специфического для ткани-мишени соединения, комбинируют, как описано выше, последовательности вариабельных областей легкой цепи можно конструировать так, чтобы они содержали нестрогие остатки, аналогично тому, как описано выше. Количество и положение таких нестрогих остатков конкретно не ограничивается конкретными вариантами осуществления при условии, что антигенсвязывающая активность антигенсвязывающий молекулы по настоящему изобретению варьирует в зависимости от присутствия или отсутствия аденозина и/или ATP. В частности, последовательности CDR и/или последовательности FR тяжелой цепи и/или легкой цепи могут содержать один или несколько нестрогих остатков.
Предпочтительные вариабельные области тяжелой цепи, подлежащие комбинированию, включают, например, рандомизированные библиотеки вариабельной области. Для получения рандомизированной библиотеки вариабельной области известные способы комбинируют соответствующим образом. В неограничивающем варианте осуществления настоящего изобретения в качестве рандомизированной библиотеки вариабельной области предпочтительно можно использовать иммунную библиотеку, сконструированную на основе генов антител, происходящих из лимфоцитов животных, иммунизированных конкретным антигеном, пациентов с инфекциями, индивидуумов с повышенным титром антител в крови в результате вакцинации, пациентов со злокачественной опухолью или пациентов с аутоиммунным заболеванием.
В другом неограничивающем варианте осуществления настоящего изобретения, в качестве рандомизированной библиотеки вариабельной области также предпочтительно можно использовать синтетическую библиотеку, полученную путем замены последовательностей CDR V-генов в геномной ДНК или функциональных реконструированных V-генов набором синтетических олигонуклеотидов, содержащих последовательности, кодирующие наборы кодонов соответствующей длины. В этом случае, поскольку в последовательности CDR3 тяжелой цепи наблюдают разнообразие, также можно заменять только последовательность CDR3. Критерием возникновения разнообразия аминокислот в вариабельной области антигенсвязывающей молекулы является то, что разнообразие возникает в аминокислотных остатках в экспонированных на поверхности положениях антигенсвязывающей молекулы. Экспонированное на поверхности положение относится к положению, которое считается способным экспонироваться на поверхности и/или контактировать с антигеном, исходя из структуры, группы структур и/или смоделированной структуры антигенсвязывающей молекулы. Как правило, такие положения представляют собой CDR. Предпочтительно, экспонированные на поверхности положения определяют с использованием координат из трехмерной модели антигенсвязывающей молекулы с использованием компьютерной программы, такой как программа InsightII (Accelrys). Экспонированные на поверхности положения можно определять с использованием алгоритмов, известных в данной области (например, Lee and Richards (J. Mol. Biol. (1971) 55, 379-400); Connolly (J. Appl. Cryst. (1983) 16, 548-558)). Определение экспонированных на поверхности положений можно проводить с использованием программного обеспечения, пригодного для моделирования белка, и информации о трехмерной структуре, полученной для антитела. Программное обеспечение, которое можно использовать для этих целей, предпочтительно включает программное обеспечение SYBYL Biopolymer Module (Tripos Associates). Главным образом или предпочтительно, когда алгоритм требует введения пользователем параметра размера, "размер" образца, который используют в вычислении, устанавливают как радиус, равный приблизительно 1,4 Ангстрем или менее. Более того, способы определения экспонированных на поверхности областей и участков с использованием программного обеспечения для персональных компьютеров описаны Pacios (Comput. Chem. (1994) 18 (4), 377-386; J. Mol. Model. (1995) 1, 46-53).
Более того, в неограничивающем варианте осуществления настоящего изобретения аминокислоты вариабельной области, включающей область CDR и/или каркасную область, можно изменять соответствующим образом для повышения стабильности антитела. В неограничивающем варианте осуществления примеры таких аминокислот могут включать аминокислоты в положениях 1, 5, 10, 30, 48 и 58. Более конкретно, примеры могут включать Gln в положении 1, Gln в положении 5, Asp в положении 10, Asn в положении 30, Leu в положении 48, и Asn в положении 58. Для повышения стабильности антитела эти аминокислоты можно заменять соответствующими аминокислотами, содержащимися в последовательности эмбрионального типа. В неограничивающем варианте осуществления примером такой последовательности эмбрионального типа может быть последовательность VH3-21. В этом случае, Gln в положении 1 может быть заменен на Glu, Gln в положении 5 может быть заменен на Val, Asp в положении 10 может быть заменен на Gly, Asn в положении 30 может быть заменен на Ser, Leu в положении 48 может быть заменен на Val, и Asn в положении 58 может быть заменен на Tyr.
В другом неограничивающем варианте осуществления настоящего изобретения наивную библиотеку, сконструированную из генов антител, происходящих из лимфоцитов здоровых индивидуумов, и состоящую из наивных последовательностей, которые представляют собой последовательности антитела, которые не имеют смещенности в их репертуаре, можно особенно предпочтительно использовать в качестве рандомизированной библиотеки вариабельных областей (Gejima et al. (Human Antibodies (2002) 11, 121-129); Cardoso et al. (Scand. J. Immunol. (2000) 51, 337-344)). В настоящем описании "аминокислотная последовательность, содержащая наивную последовательность" относится к аминокислотной последовательности, полученной из такой наивной библиотеки.
Fc-область
Fc-область содержит аминокислотную последовательность, происходящую из константной области тяжелой цепи антитела. Fc-область представляет собой часть константной области тяжелой цепи антитела, которая включает N-конец шарнирной области, который представляет собой участок расщепления папаином в положении в области аминокислоты 216 (указанном с помощью нумерации EU), и шарнирную область, домен CH2 и домен CH3. Fc-области также могут быть получены из IgG1 человека; однако они не ограничиваются каким-либо конкретным подклассом IgG. Предпочтительные примеры Fc-областей включают Fc-области, обладающие активностью связывания FcRn в диапазоне кислых значений pH, как описано ниже. Предпочтительные примеры Fc-областей включают Fc-области, обладающие активностью связывания Fcγ-рецептора, как описано ниже. В неограничивающем варианте осуществления примеры таких Fc-областей включают Fc-области IgG1 (SEQ ID NO:5), IgG2 (SEQ ID NO:6), IgG3 (SEQ ID NO:7) или IgG4 (SEQ ID NO:8) человека.
Fcγ-рецептор
Fcγ-рецептор (также называемый FcγR) относится к рецептору, способному связываться с Fc-областью моноклональных антител IgG1, IgG2, IgG3 или IgG4, и включает всех представителей, принадлежащих семейству белков, по существу кодируемых генами Fcγ-рецепторов. У человека это семейство включает FcγRI (CD64), включая изоформы FcγRIa, FcγRIb и FcγRIc; FcγRII (CD32), включая изоформы FcγRIIa (включая аллотип H131 и R131, т.е., FcγRIIa(H) и FcγRIIa(R))), FcγRIIb (включая FcγRIIb-1 и FcγRIIb-2), и FcγRIIc; и FcγRIII (CD16) включая изоформу FcγRIIIa (включая аллотип V158 и F158, т.е., FcγRIIIa(V) и FcγRIIIa(F))) и FcγRIIIb (включая аллотип FcγRIIIb-NA1 и FcγRIIIb-NA2); а также неидентифицированные FcγR человека, изоформы FcγR и их аллотипы, однако семейство не ограничивается этими примерами. Не ограничиваясь этим, FcγR включает FcγR человека, мыши, крысы, кролика и обезьяны. FcγR может происходить из любого организма. FcγR мыши включает, но не ограничивается этими примерами, FcγRI (CD64), FcγRII (CD32), FcγRIII (CD16) и FcγRIII-2 (FcγRIV, CD16-2), а также неидентифицированные FcγR мыши, изоформы FcγR и их аллотипы. Предпочтительные примеры таких Fcγ-рецепторов включают, например, FcγRI (CD64), FcγRIIa (CD32), FcγRIIb (CD32), FcγRIIIa (CD16) и/или FcγRIIIb (CD16) человека. Полинуклеотидная последовательность и аминокислотная последовательность FcγRI человека представлены в SEQ ID NO:9 (NM_00056.3) и 10 (NP_000557.1), соответственно; полинуклеотидная последовательность и аминокислотная последовательность FcγRIIa (аллотип H131) представлены в SEQ ID NO:11 (BC020823.1) и 12 (AAH20823.1), соответственно (аллотип R131 представляет собой последовательность, в которой аминокислота в положении 166 SEQ ID NO:12 заменена на Arg); полинуклеотидная последовательность и аминокислотная последовательность FcγIIB представлены в SEQ ID NO:13 (BC146678.1) и 14 (AAI46679.1), соответственно; полинуклеотидная последовательность и аминокислотная последовательность FcγRIIIa представлены в SEQ ID NO:15 (BC033678.1) и 16 (AAH33678.1), соответственно; и полинуклеотидная последовательность и аминокислотная последовательность FcγRIIIb представлены в SEQ ID NO:17 (BC128562.1) и 18 (AAI28563.1), соответственно (номер доступа RefSeq представлен в каждом случае в скобках). То, обладает ли Fcγ-рецептор активностью связывания с Fc-областью моноклонального антитела IgG1, IgG2, IgG3 или IgG4, можно оценивать с помощью скрининга ALPHA (анализ усиленной за счет эффекта близости люминесценции), способа BIACORE на основе поверхностного плазмонного резонанса (SPR) и других способов (Proc. Natl. Acad. Sci. USA (2006) 103(11), 4005-4010), в дополнение к описанным выше форматам FACS и ELISA.
В FcγRI (CD64), включая FcγRIa, FcγRIb, и FcγRIc, и FcγRIII (CD16), включающий изоформы FcγRIIIa (включая аллотипы V158 и F158) и FcγRIIIb (включая аллотипы FcγRIIIb-NA1 и FcγRIIIb-NA2), α-цепь, которая связывается с Fc-областью IgG, ассоциирована с общей γ-цепью, имеющей ITAM, ответственный за передачу внутриклеточного активирующего сигнала. Между тем, цитоплазматический домен FcγRII (CD32), включая изоформы FcγRIIa (включая аллотипы H131 и R131) и FcγRIIc содержит ITAM. Эти рецепторы экспрессируются на многих иммунных клетках, таких как макрофаги, тучные клетки и антигенпредставляющие клетки. Активирующий сигнал, передаваемый при связывании этих рецепторов с Fc-частью IgG, приводит к усилению фагоцитарной активности макрофагов, продукции воспалительных цитокинов, дегрануляции тучных клеток и усилению функции антигенпредставляющих клеток. Fcγ-рецепторы, обладающие способностью передавать активирующий сигнал, как описано выше, также называют активирующими Fcγ-рецепторами.
Между тем, внутрицитоплазматический домен FcγRIIb (включая FcγRIIb-1 и FcγRIIb-2) содержит ITIM, ответственный за передачу ингибиторных сигналов. Связывание между FcγRIIb и B-клеточным рецептором (BCR) на B-клетках подавляет активирующий сигнал от BCR, что приводит к подавлению продукции антитела через BCR. Связывание FcγRIII и FcγRIIb на макрофагах подавляет фагоцитарную активность и продукцию воспалительных цитокинов. Fcγ-рецепторы, обладающие способностью передавать ингибирующий сигнал, как описано выше, также в настоящем описании называют ингибиторным Fcγ-рецептором.
Активность связывания Fc-области с FcγR
Как упоминалось выше, Fc-области, обладающие активностью связывания с Fcγ-рецептором, являются примерами Fc-областей, содержащихся в антигенсвязывающих молекулах по настоящему изобретению. Неограничивающий вариант осуществления такой Fc-области включает Fc-область IgG1 (SEQ ID NO:5), IgG2 (SEQ ID NO:6), IgG3 (SEQ ID NO:7) или IgG4 (SEQ ID NO:8) человека. Наличие у Fcγ-рецептора активности связывания с Fc-областью моноклонального антитела IgG1, IgG2, IgG3 или IgG4 можно оценивать с помощью скрининга ALPHA (анализ усиленной за счет эффекта близости люминесценции), способа BIACORE, который основан на явлении поверхностного плазмонного резонанса (SPR) и т.п. (Proc. Natl. Acad. Sci. USA (2006) 103 (11), 4005-4010), в дополнение к описанным выше форматам FACS и ELISA.
Скрининг ALPHA проводят с помощью технологии ALPHA на основе принципа, описанного ниже, где используется два типа гранул: донорные и акцепторные гранулы. Люминесцентный сигнал выявляется только когда молекулы, связанные с донорными гранулами, биологически взаимодействуют с молекулами, связанными с акцепторными гранулами, и когда эти два типа гранул находятся вблизи друг друга. Возбуждаемый лазерным пучком фотосенсибилизатор в донорных гранулах преобразует кислород вокруг гранулы в возбужденный синглетный кислород. Когда синглетный кислород распространяется среди донорных гранул и достигает акцепторных гранул, расположенных близко, в акцепторных гранулах индуцируется хемилюминесцентная реакция. Эта реакция в конечном итоге приводит к испусканию света. Если молекулы, связанные с донорными гранулами, не взаимодействуют с молекулами, связанными с акцепторными гранулами, синглетный кислород, продуцируемый донорными гранулами, не достигает акцепторных гранул и хемилюминесцентная реакция не происходит.
Например, меченную биотином антигенсвязывающую молекулу, содержащую Fc-область, иммобилизуют на донорных гранулах, и Fcγ-рецептор, меченный глутатион-S-трансферазой (GST), иммобилизуют на акцепторных гранулах. В отсутствие антигенсвязывающей молекулы, содержащей конкурирующий вариант Fc-области, Fcγ-рецептор взаимодействует с антигенсвязывающей молекулой, содержащей нативную Fc-область, что приводит к индукции сигнала в области 520-620 нм. Антигенсвязывающая молекула, имеющая немеченый вариант Fc-области, конкурирует с антигенсвязывающей молекулой, содержащей нативную Fc-область, для взаимодействия с Fcγ-рецептором. Относительную аффинность связывания можно определять путем количественного определения снижения флуоресценции в результате конкуренции. Способы биотинилирования антигенсвязывающих молекул, таких как антитела, с использованием сульфо-NHS-биотина и т.п., известны. Соответствующие способы добавления GST-метки к Fcγ-рецептору включают способы, которые вовлекают слияние полипептидов, кодирующих Fcγ и GST в рамке считывания, экспрессию слитого гена с использованием клеток, в которые введен вектор, с которым ген функционально связан, а затем очистку с использованием колонки с глутатионом. Индуцированный сигнал предпочтительно можно анализировать, например, путем аппроксимации его к односторонней модели конкуренции на основе нелинейного регрессионного анализа с использованием программного обеспечения, такого как GRAPHPAD PRISM (GraphPad, San Diego).
Одно из веществ при наблюдении их взаимодействия иммобилизуют в качестве лиганда на тонкую золотую пленку на сенсорном чипе. Когда свет падает на заднюю поверхность сенсорного чипа, так чтобы общее отражение происходило на поверхности контакта между тонкой золотой пленкой и стеклом, интенсивность отраженного света частично снижается в определенной области (сигнал SPR). Другое из веществ при выявлении взаимодействия инжектируют в качестве анализируемого соединения на поверхность сенсорного чипа. Когда анализируемое соединение связывается с лигандом, масса иммобилизованной молекулы лиганда возрастает. Это изменяет индекс рефракции растворителя на поверхности сенсорного чипа. Изменение индекса рефракции вызывает сдвиг положения сигнала SPR (с другой стороны, диссоциация вызывает сдвиг сигнала обратно в исходное положение). В системе Biacore величину сдвига, упомянутую выше (т.е. изменение массы на поверхности сенсорного чипа) наносят на график по вертикальной оси, и, таким образом, изменение массы с течением времени показывают в качестве данных измерения (сенсограмма). Параметры кинетики (константа скорости ассоциации (ka) и константу скорости диссоциации (kd)), определяют из кривых сенсограммы, и аффинность (KD) определяют из соотношения этих двух констант. В способе BIACORE, предпочтительно используют анализ ингибирования. Примеры такого анализа ингибирования описаны в Proc. Natl. Acad. Sci USA (2006) 103 (11): 4005-4010.
Fc-области с измененным связыванием Fcγ-рецептора (FcγR)
В дополнение к Fc-области IgG1 (SEQ ID NO:5), IgG2 (SEQ ID NO:6), IgG3 (SEQ ID NO:7) или IgG4 (SEQ ID NO:8) человека, в качестве Fc-области в рамках настоящего изобретения можно соответственно использовать Fc-область с измененным связыванием FcγR, которая обладает более высокой активностью связывания Fcγ-рецептора, чем Fc-область нативного IgG человека. В рамках настоящего изобретения, "Fc-область нативного IgG человека" относится к Fc-области, в которой цепь сахара, связанная в положении 297 (нумерация EU) Fc-области IgG1, IgG2, IgG3 или IgG4 человека, представленных в качестве SEQ ID NO:5, 6, 7 или 8, представляет собой содержащую фукозу цепь сахаров. Такие Fc-области с измененным связыванием FcγR можно получать путем изменения аминокислот Fc-области нативного IgG человека. То, является ли активность связывания с FcγR у Fc-области с измененным связыванием с FcγR более высокой, чем у Fc-области нативного IgG человека, можно определять соответствующим образом с использованием способов, описанных в упомянутом выше разделе об активности связывания.
В рамках настоящего изобретения "изменение аминокислот" или "аминокислотное изменение" Fc-области включает изменение аминокислотной последовательности, которое отличается от исходной Fc-области. Исходная Fc-область может представлять собой любую Fc-область при условии, что вариант, модифицированный из исходной Fc-области, может связываться с Fcγ-рецептором человека в диапазоне нейтральных значений pH. Более того, также предпочтительно в качестве Fc-области по настоящему изобретению можно использовать Fc-область, измененную от исходной Fc-области, которая уже была изменена. "Исходная Fc-область" может относиться к самому полипептиду, к композиции, содержащей исходную Fc-область, или к аминокислотной последовательности, кодирующей исходную Fc-область. Исходные Fc-области могут содержать известные Fc-области, полученные посредством рекомбинации, кратко описанные в разделе "Антитела". Источник исходных Fc-областей не ограничен, и они могут быть получены из человека или не являющихся человеком организмов. Такие организмы предпочтительно включают мышей, крыс, морских свинок, хомячков, песчанок, кошек, кроликов, собак, коз, овец, животных семейства бычьих, лошадей, верблюдов и организмы, выбранные из не являющихся человеком приматов. В другом варианте осуществления исходные Fc-области также можно получать из яванских макаков, мартышек, макаков-резус, шимпанзе или людей. Исходные Fc-области можно получать предпочтительно из IgG1 человека; однако они не ограничиваются каким-либо конкретным подклассом IgG. Это означает, что Fc-область, соответствующую IgG1, IgG2, IgG3 (или IgG4 человека, можно использовать соответствующим образом в качестве исходной Fc-области, и в рамках настоящего изобретения это также означает, что Fc-область произвольного класса или подкласса IgG, происходящую из любых организмов, описанных выше, предпочтительно можно использовать в качестве исходной Fc-области. Примеры встречающихся в природе вариантов IgG или измененных форм описаны в опубликованных документах (Curr. Opin. Biotechnol. (2009) 20 (6): 685-91; Curr. Opin. Immunol. (2008) 20 (4), 460-470; Protein Eng. Des. Sel. (2010) 23 (4): 195-202; международные публикации № WO 2009/086320; WO 2008/092117; WO 2007/041635; и WO 2006/105338); однако они не ограничиваются примерами.
Примеры изменений включают изменения с одной или несколькими мутациями, например, мутациями путем замены различными аминокислотными остатками аминокислот исходных Fc-областей, путем встраивания одного или нескольких аминокислотных остатков в исходные Fc-области, или путем делеции одной или нескольких аминокислот из исходной Fc-области. Предпочтительно, аминокислотные последовательности измененных Fc-областей содержат по меньшей мере часть аминокислотной последовательности ненативной Fc-области. Такие варианты обязательно обладают идентичностью или сходством последовательности менее 100% с их исходной Fc-областью. В предпочтительном варианте осуществления варианты обладают идентичностью или сходством аминокислотной последовательности от приблизительно 75% до менее чем 100%, более предпочтительно от приблизительно 80% до менее чем 100%, еще более предпочтительно от приблизительно 85% до менее чем 100%, еще более предпочтительно от приблизительно 90% до менее чем 100%, и наиболее предпочтительно от приблизительно 95% до менее чем 100% с аминокислотной последовательностью их исходной Fc-области. В неограничивающем варианте осуществления настоящего изобретения по меньшей мере одна аминокислота отличается между измененной Fc-областью по настоящему изобретению и ее исходной Fc-областью. Аминокислотное отличие между измененной Fc-областью по настоящему изобретению и ее исходной Fc-областью также может быть предпочтительно установлено на основе аминокислотных отличий в описанных выше конкретных положениях аминокислот в соответствии с нумерацией EU. Примеры способов получения таких вариантов представлены в разделе "Изменение аминокислот".
Включенную в антигенсвязывающие молекулы по настоящему изобретению, Fc-область с измененным связыванием FcγR, которая обладает более высокой активностью связывания с Fcγ-рецептором, чем Fc-область нативного IgG человека (Fc-область с измененным связыванием с FcγR) можно получать любым способом. В частности, Fc-область с измененным связыванием FcγR можно получать путем изменения аминокислот иммуноглобулина человека IgG-типа, использованного в качестве исходной Fc-области. Предпочтительные Fc-области иммуноглобулинов IgG-типа для изменения включают, например Fc-области IgG человека, представленные в SEQ ID NO:5, 6, 7 или 8 (IgG1, IgG2, IgG3 или IgG4, соответственно, и их варианты).
Аминокислоты в любых положениях можно изменять на другие аминокислоты при условии, что активность связывания с Fcγ-рецептором является более высокой, чем у Fc-области нативного IgG человека. Когда антигенсвязывающая молекула содержит Fc-область IgG1 человека в качестве Fc-области человека, она предпочтительно содержит изменение, которое обеспечивает эффект более высокой активности связывания Fcγ-рецептора, чем у Fc-области нативного IgG человека, в которой цепь сахара, связанная в положении 297 (нумерация EU) представляет собой содержащую фукозу цепь сахара. Такие изменения аминокислот описаны, например, в международных публикациях, таких как WO2007/024249, WO2007/021841, WO2006/031370, WO2000/042072, WO2004/029207, WO2004/099249, WO2006/105338, WO2007/041635, WO2008/092117, WO2005/070963, WO2006/020114, WO2006/116260 и WO2006/023403.
Примеры таких аминокислот, которые могут быть изменены, включают по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из положений 221, 222, 223, 224, 225, 227, 228, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 246, 247, 249, 250, 251, 254, 255, 256, 258, 260, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 278, 279, 280, 281, 282, 283, 284, 285, 286, 288, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 311, 313, 315, 317, 318, 320, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 339, 376, 377, 378, 379, 380, 382, 385, 392, 396, 421, 427, 428, 429, 434, 436 и 440 (нумерация EU). Fc-область (Fc-область с измененным связыванием FcγR), обладающая более высокой активностью связывания Fcγ-рецептора, чем Fc-область нативного IgG человека, можно получать путем изменения этих аминокислот.
Примеры особенно предпочтительных изменений для применения в рамках настоящего изобретения включают по меньшей мере одно или несколько изменений аминокислот, выбранных из группы, состоящей из:
Lys или Tyr вместо аминокислоты в положении 221;
Phe, Trp, Glu или Tyr вместо аминокислоты в положении 222;
Phe, Trp, Glu или Lys вместо аминокислоты в положении 223;
Phe, Trp, Glu или Tyr вместо аминокислоты в положении 224;
Glu, Lys или Trp вместо аминокислоты в положении 225;
Glu, Gly, Lys или Tyr вместо аминокислоты в положении 227;
Glu, Gly, Lys или Tyr вместо аминокислоты в положении 228;
Ala, Glu, Gly или Tyr вместо аминокислоты в положении 230;
Glu, Gly, Lys, Pro или Tyr вместо аминокислоты в положении 231;
Glu, Gly, Lys или Tyr вместо аминокислоты в положении 232;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 233;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 234;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 235;
Ala, Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 236;
Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 237;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 238;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp или Tyr вместо аминокислоты в положении 239;
Ala, Ile, Met или Thr вместо аминокислоты в положении 240;
Asp, Glu, Leu, Arg, Trp или Tyr вместо аминокислоты в положении 241;
Leu, Glu, Leu, Gln, Arg, Trp или Tyr вместо аминокислоты в положении 243;
His вместо аминокислоты в положении 244;
Ala вместо аминокислоты в положении 245;
Asp, Glu, His или Tyr вместо аминокислоты в положении 246;
Ala, Phe, Gly, His, Ile, Leu, Met, Thr, Val или Tyr вместо аминокислоты в положении 247;
Glu, His, Gln или Tyr вместо аминокислоты в положении 249;
Glu или Gln вместо аминокислоты в положении 250;
Phe вместо аминокислоты в положении 251;
Phe, Met или Tyr вместо аминокислоты в положении 254;
Glu, Leu или Tyr вместо аминокислоты в положении 255;
Ala, Met или Pro вместо аминокислоты в положении 256;
Asp, Glu, His, Ser или Tyr вместо аминокислоты в положении 258;
Asp, Glu, His или Tyr вместо аминокислоты в положении 260;
Ala, Glu, Phe, Ile или Thr вместо аминокислоты в положении 262;
Ala, Ile, Met или Thr вместо аминокислоты в положении 263;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Trp или Tyr вместо аминокислоты в положении 264;
Ala, Leu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 265;
Ala, Ile, Met или Thr вместо аминокислоты в положении 266;
Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp или Tyr вместо аминокислоты в положении 267;
Asp, Glu, Phe, Gly, Ile, Lys, Leu, Met, Pro, Gln, Arg, Thr, Val или Trp вместо аминокислоты в положении 268;
Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 269;
Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Gln, Arg, Ser, Thr, Trp или Tyr вместо аминокислоты в положении 270;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 271;
Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 272;
Phe или Ile вместо аминокислоты в положении 273;
Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 274;
Leu или Trp вместо аминокислоты в положении 275;
Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 276;
Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp вместо аминокислоты в положении 278;
Ala вместо аминокислоты в положении 279;
Ala, Gly, His, Lys, Leu, Pro, Gln, Trp или Tyr вместо аминокислоты в положении 280;
Asp, Lys, Pro или Tyr вместо аминокислоты в положении 281;
Glu, Gly, Lys, Pro или Tyr вместо аминокислоты в положении 282;
Ala, Gly, His, Ile, Lys, Leu, Met, Pro, Arg или Tyr вместо аминокислоты в положении 283;
Asp, Glu, Leu, Asn, Thr или Tyr вместо аминокислоты в положении 284;
Asp, Glu, Lys, Gln, Trp или Tyr вместо аминокислоты в положении 285;
Glu, Gly, Pro или Tyr вместо аминокислоты в положении 286;
Asn, Asp, Glu или Tyr вместо аминокислоты в положении 288;
Asp, Gly, His, Leu, Asn, Ser, Thr, Trp или Tyr вместо аминокислоты в положении 290;
Asp, Glu, Gly, His, Ile, Gln или Thr вместо аминокислоты в положении 291;
Ala, Asp, Glu, Pro, Thr или Tyr вместо аминокислоты в положении 292;
Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 293;
Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 294;
Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 295;
Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr или Val вместо аминокислоты в положении 296;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 297;
Ala, Asp, Glu, Phe, His, Ile, Lys, Met, Asn, Gln, Arg, Thr, Val, Trp или Tyr вместо аминокислоты в положении 298;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr вместо аминокислоты в положении 299;
Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp вместо аминокислоты в положении 300;
Asp, Glu, His или Tyr вместо аминокислоты в положении 301;
Ile вместо аминокислоты в положении 302;
Asp, Gly или Tyr вместо аминокислоты в положении 303;
Asp, His, Leu, Asn или Thr вместо аминокислоты в положении 304;
Glu, Ile, Thr или Tyr вместо аминокислоты в положении 305;
Ala, Asp, Asn, Thr, Val или Tyr вместо аминокислоты в положении 311;
Phe вместо аминокислоты в положении 313;
Leu вместо аминокислоты в положении 315;
Glu или Gln вместо аминокислоты в положении 317;
His, Leu, Asn, Pro, Gln, Arg, Thr, Val или Tyr вместо аминокислоты в положении 318;
Asp, Phe, Gly, His, Ile, Leu, Asn, Pro, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 320;
Ala, Asp, Phe, Gly, His, Ile, Pro, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 322;
Ile вместо аминокислоты в положении 323;
Asp, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Thr, Val, Trp или Tyr вместо аминокислоты в положении 324;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 325;
Ala, Asp, Glu, Gly, Ile, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 326;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Thr, Val, Trp или Tyr вместо аминокислоты в положении 327;
Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 328;
Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 329;
Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 330;
Asp, Phe, His, Ile, Leu, Met, Gln, Arg, Thr, Val, Trp или Tyr вместо аминокислоты в положении 331;
Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 332;
Ala, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Ser, Thr, Val или Tyr вместо аминокислоты в положении 333;
Ala, Glu, Phe, Ile, Leu, Pro или Thr вместо аминокислоты в положении 334;
Asp, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Val, Trp или Tyr вместо аминокислоты в положении 335;
Glu, Lys или Tyr вместо аминокислоты в положении 336;
Glu, His или Asn вместо аминокислоты в положении 337;
Asp, Phe, Gly, Ile, Lys, Met, Asn, Gln, Arg, Ser или Thr вместо аминокислоты в положении 339;
Ala или Val вместо аминокислоты в положении 376;
Gly или Lys вместо аминокислоты в положении 377;
Asp вместо аминокислоты в положении 378;
Asn вместо аминокислоты в положении 379;
Ala, Asn или Ser вместо аминокислоты в положении 380;
Ala или Ile вместо аминокислоты в положении 382;
Glu вместо аминокислоты в положении 385;
Thr вместо аминокислоты в положении 392;
Leu вместо аминокислоты в положении 396;
Lys вместо аминокислоты в положении 421;
Asn вместо аминокислоты в положении 427;
Phe или Leu вместо аминокислоты в положении 428;
Met вместо аминокислоты в положении 429;
Trp вместо аминокислоты в положении 434;
Ile вместо аминокислоты в положении 436; и
Gly, His, Ile, Leu или Tyr вместо аминокислоты в положении 440;
как указано с помощью нумерации EU в Fc-области. Количество аминокислот, подлежащих изменению, конкретно не ограничено; и аминокислота может быть изменена в одном участке или аминокислоты могут быть изменены в двух или более участках. Примеры комбинаций для изменений аминокислот в двух или более участках включают изменения, описанные в таблице 1 (таблицы с 1-1 по 1-3).
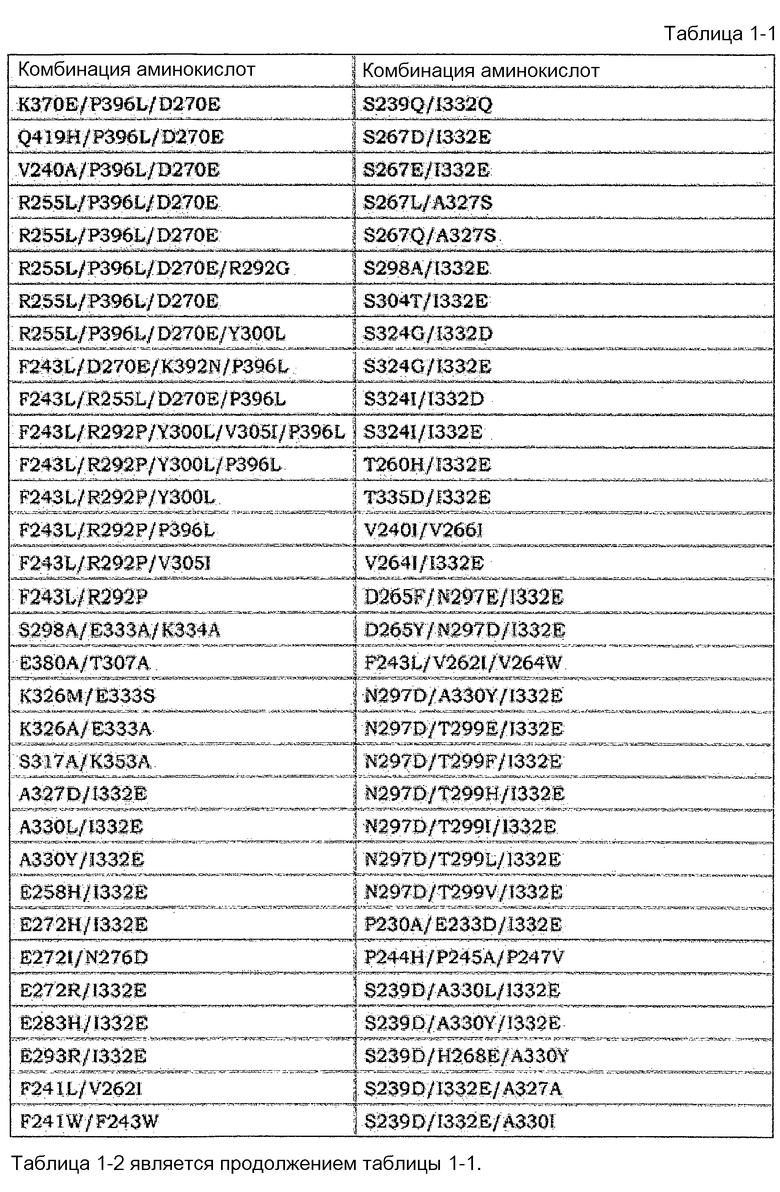
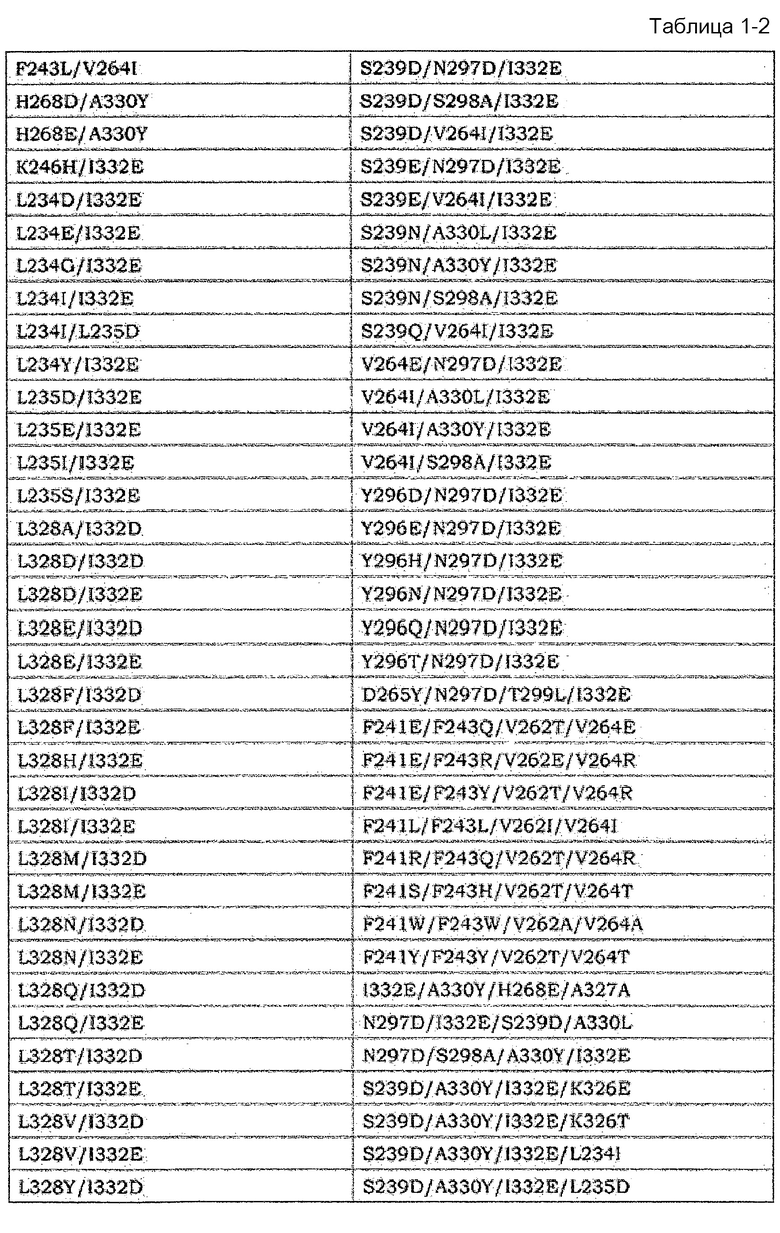

В качестве условий pH для измерения активности связывания связывающего Fcγ-рецептор домена и Fcγ-рецептора, содержащегося в антигенсвязывающей молекуле по настоящему изобретению, можно соответственно использовать условия диапазона кислых значений pH или условия диапазона нейтральных значений pH. Диапазон кислых значений pH или диапазон нейтральных значений pH в качестве условий для измерения активности связывания связывающего Fcγ-рецептор домена и Fcγ-рецептора, содержащегося в антигенсвязывающей молекуле по настоящему изобретению, как правило, указывается как от pH 5,8 до pH 8,0. Предпочтительно, он представляет собой диапазон, указываемый произвольными величинами pH от 6,0 до pH 7,4; и, предпочтительно, его выбирают из pH 6,0, pH 6,1, pH 6,2, pH 6,3, pH 6,4, pH 6,5, pH 6,6, pH 6,7, pH 6,8, pH 6,9, pH 7,0, pH 7,1, pH 7,2, pH 7,3 и pH 7,4; и особенно предпочтительно он представляет собой pH от 6,15 до 7,4, что является близким к pH в злокачественных тканях (Vaupel et al., Cancer Res. (1989) 49, 6449-6665). Что касается температуры, используемой в качестве условий измерения, аффинность связывания между связывающим Fcγ-рецептор доменом и Fcγ-рецептором человека можно оценивать при любой температуре от 10°C до 50°C. Предпочтительно, для определения аффинности связывания между связывающим Fcγ-рецептор доменом человека и Fcγ-рецептором используют температуру от 15°C до 40°C. Более предпочтительно, для определения аффинности связывания между связывающим Fcγ-рецептор доменом и Fcγ-рецептором можно сходным образом использовать любую температуру от 20°C до 35°C, такую как любая из 20°C, 21°C, 22°C, 23°C, 24°C, 25°C, 26°C, 27°C, 28°C, 29°C, 30°C, 31°C, 32°C, 33°C, 34°C или 35°C. В одном варианте осуществления настоящего изобретения неограничивающим примером является температура 25°C.
В рамках настоящего изобретения "Fc-область с измененным связыванием FcγR имеет более высокую активность связывания Fcγ-рецептора, чем нативная Fc-область" означает, что активность связывания Fcγ-рецептора человека у Fc-области с измененным связыванием FcγR с любым из Fcγ-рецепторов человека: FcγRI, FcγRIIa, FcγRIIb, FcγRIIIa и/или FcγRIIIb является более высокой, чем активность связывания нативной Fc-области с этими Fcγ-рецепторами человека. Например, это означает, что, исходя из описанного выше аналитического способа, по сравнению с активностью связывания у антигенсвязывающей молекулы, содержащей нативную Fc-область IgG человека в качестве контроля, активность связывания у антигенсвязывающей молекулы, содержащей Fc-область с измененным связыванием FcγR, составляет 105% или более, предпочтительно 110% или более, 115% или более, 120% или более, 125% или более, особенно предпочтительно 130% или более, 135% или более, 140% или более, 145% или более, 150% или более, 155% или более, 160% или более, 165% или более, 170% или более, 175% или более, 180% или более, 185% или более, 190% или более, 195% или более или превышает в 2 раза или более, в 2,5 раза или более, в 3 раза или более, в 3,5 раза или более, в 4 раза или более, в 4,5 раза или более, в 5 раз или более, в 7,5 раз или более, в 10 раз или более, в 20 раз или более, в 30 раз или более, в 40 раз или более, в 50 раз или более, в 60 раз или более, в 70 раз или более, в 80 раз или более, в 90 раз или более или в 100 раз или более. Исходную Fc-область можно использовать в качестве нативной Fc-области, и также можно использовать нативные Fc-области антител того же подкласса.
В рамках настоящего изобретения Fc-область нативного IgG человека, в которой цепь сахаров, связанная с аминокислотой в положении 297 (нумерация EU) представляет собой содержащую фукозу цепь сахаров, пригодным образом используют в качестве нативной Fc-области IgG человека, подлежащей применению в качестве контроля. То, является ли цепь сахаров, связанная с аминокислотой в положении 297 (нумерация EU) содержащей фукозу цепью сахаров, можно определять с использованием способа, описанного в непатентном документе 6. Например, с помощью такого способа, как указано ниже, возможно определить то, является ли цепь сахаров, связанная с нативной Fc-областью IgG человека, содержащей фукозу цепью сахаров. Посредством реакции N-гликозидазы F (Roche diagnostics) с нативным IgG человека, подлежащим исследованию, цепь сахаров диссоциирует от нативного IgG человека (Weitzhandler et al. (J. Pharma. Sciences (1994) 83, 12, 1670-1675)). Далее высушенный концентрат реакционного раствора, из которого были удалены белки посредством реакции с этанолом (Schenk et al. (J. Clin. Investigation (2001) 108 (11) 1687-1695)), метят флуоресцентной меткой 2-аминопиридином или 2-аминобензамидом (Bigge et al. (Anal. Biochem. (1995) 230 (2) 229-238)). Реагенты удаляют с помощью твердофазной экстракции с использованием кассеты с целлюлозой, и флуоресцентно меченную модифицированную посредством 2-AP или 2-AB цепь сахаров анализируют с помощью нормально-фазовой хроматографии. Путем исследования выявленных пиков на хроматограмме можно определить, является ли цепь сахаров, связанная с нативной Fc-областью IgG человека, содержащей фукозу цепью сахаров.
В качестве антигенсвязывающей молекулы, содержащей нативную Fc-область антитела того же подкласса, которое используют в качестве контроля, можно соответственно использовать антигенсвязывающую молекулу, имеющую Fc-область моноклонального IgG-антитела. Структуры таких Fc-областей представлены в SEQ ID NO:5 (A добавлен на N-конец последовательности с номером доступа RefSeq AAC82527.1), SEQ ID NO:6 (A добавлен на N-конец последовательности с номером доступа RefSeq AAB59393.1), SEQ ID NO:7 (номер доступа RefSeq CAA27268.1) и SEQ ID NO:8 (A добавлен на N-конец последовательности с номером доступа RefSeq AAB59394.1). Более того, когда антигенсвязывающую молекулу, содержащую Fc-область конкретного изотипа антитела, используют в качестве исследуемого вещества, эффект антигенсвязывающей молекулы, содержащей исследуемую Fc-область, в отношении активности связывания Fcγ-рецептора, исследуют с использованием в качестве контроля антигенсвязывающей молекулы, имеющей Fc-область моноклонального IgG-антитела этого конкретного изотипа. Таким путем пригодным образом выбирают антигенсвязывающие молекулы, содержащие Fc-область, для которой продемонстрировано, что ее активность связывания Fcγ-рецептора является высокой.
Fc-области, обладающие активностью селективного связывания с Fcγ-рецептором
Примеры связывающих Fcγ-рецептор доменов, пригодных для применения в рамках настоящего изобретения, включают связывающие Fcγ-рецептор домены, обладающие более высокой активностью связывания с конкретным Fcγ-рецептором, чем с другими Fcγ-рецепторами (связывающие Fcγ-рецептор домены, обладающие активностью селективного связывания с Fcγ-рецептором). Когда антитело используют в качестве антигенсвязывающей молекулы (когда Fc-область используют в качестве связывающего Fcγ-рецептор домена), одна молекула антитела может связывать только одну молекулу Fcγ-рецептора. Таким образом, одна антигенсвязывающая молекула не может связываться с другими активирующими FcγR в связанном с ингибиторным Fcγ-рецептором состоянии и не может связываться с другими активирующими Fcγ-рецепторами или ингибиторными Fcγ-рецепторами в связанном с активирующим Fcγ-рецептором состоянии.
Fc-области с более высокой активностью связывания с активирующим Fcγ-рецептором, чем с ингибирующим Fcγ-рецептором
Как описано выше, предпочтительные активирующие Fcγ-рецепторы включают FcγRI (CD64), в том числе FcγRIa, FcγRIb и FcγRIc; FcγRIIa; и FcγRIII (CD16), в том числе FcγRIIIa (включая аллотипы V158 и F158) и FcγRIIIb (включая аллотипы FcγRIIIb-NA1 и FcγRIIIb-NA2). Между тем, предпочтительные примеры ингибиторных Fcγ-рецепторов включают FcγRIIb (в том числе FcγRIIb-1 и FcγRIIb-2).
В рамках настоящего изобретения примером случая, где активность связывания с определенным Fcγ-рецептором является более высокой, чем активность связывания с другим Fcγ-рецептором, является случай, где активность связывания с активирующим Fcγ-рецептором, является более высокой, чем активность связывания с ингибиторным Fcγ-рецептором. В этом случае, активность связывания Fc-рецептора с любым из Fcγ-рецепторов человека: FcγRIa, FcγRIIa, FcγRIIIa и/или FcγRIIIb считается более высокой, чем активность связывания с FcγRIIb. Например, это означает, что, исходя из описанного выше способа анализа, активность связывания антигенсвязывающей молекулы, содержащей Fc-область, с любым из Fcγ-рецепторов человека: FcγRIa, FcγRIIa, FcγRIIIa и/или FcγRIIIb, составляет 105% или более, предпочтительно 110% или более, 120% или более, 130% или более, 140% или более, особенно предпочтительно 150% или более, 160% или более, 170% или более, 180% или более, 190% или более, 200% или более, 250% или более, 300% или более, 350% или более, 400% или более, 450% или более, 500% или более, 750% или более по сравнению с активностью связывания с FcγRIIb или превышает ее в 10 раз или более, в 20 раз или более, в 30 раз или более, в 40 раз или более, в 50 раз или более, в 60 раз, в 70 раз, в 80 раз, в 90 раз или в 100 раз. Fc-область с более высокой активностью связывания с активирующими Fcγ-рецепторами, чем с ингибиторными Fcγ-рецепторами, может быть подходящим образом включена в антигенсвязывающие молекулы по настоящему изобретению, антигенсвязывающий домен которых связывается с молекулой мембранного типа. Известно, что IgG1-антитела, содержащие такие Fc-области, усиливают активность ADCC, упомянутую ниже. Таким образом, антигенсвязывающие молекулы, содержащие Fc-область, также пригодны в качестве антигенсвязывающих молекул, подлежащих включению в фармацевтические композиции по настоящему изобретению.
В неограничивающем варианте осуществления настоящего изобретения примеры Fc-области, обладающей более высокой активностью связывания с активирующими Fcγ-рецепторами, чем с ингибиторными Fcγ-рецепторами (или обладающей активностью селективного связывания с ингибиторными Fcγ-рецепторами) предпочтительно включают Fc-области, в которых по меньшей мере одна или несколько аминокислот, выбранных из группы, состоящей из аминокислот в положениях 221, 222, 223, 224, 225, 227, 228, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 246, 247, 249, 250, 251, 254, 255, 256, 258, 260, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 278, 279, 280, 281, 282, 283, 284, 285, 286, 288, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 311, 313, 315, 317, 318, 320, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 339, 376, 377, 378, 379, 380, 382, 385, 392, 396, 421, 427, 428, 429, 434, 436, и 440, указанных с помощью нумерации EU, упомянутой выше, изменены на аминокислоты, отличающиеся от аминокислот нативной Fc-области.
В неограничивающем варианте осуществления настоящего изобретения примеры Fc-области, обладающей более высокой активностью связывания с активирующими Fcγ-рецепторами, чем с ингибиторными Fcγ-рецепторами (или обладающей активностью селективного связывания с ингибиторными Fcγ-рецепторами) предпочтительно включают Fc-области, в которых множество аминокислот, указанных в таблицах с 1-1 по 1-3, изменены на аминокислоты, отличающиеся от аминокислот нативной Fc-области.
Fc-области, активность связывания которых с ингибиторным Fcγ-рецептором является более высокой, чем активность связывания с активирующим Fcγ-рецептором
В рамках настоящего изобретения, примером случая, когда активность связывания с определенным Fcγ-рецептором является более высокой, чем активность связывания с другим Fcγ-рецептором, является случай, когда активность связывания с ингибиторным Fcγ-рецептором является более высокой, чем активность связывания с активирующим Fcγ-рецептором. В этом случае активность связывания Fc-области с FcγRIIb считается более высокой, чем активность связывания с любым из Fcγ-рецепторов человека: FcγRIa, FcγRIIa, FcγRIIIa и/или FcγRIIIb. Например, это означает, что, исходя из описанного выше способа анализа, активность связывания антигенсвязывающей молекулы, содержащей Fc-область, с FcγRIIb составляет 105% или более, предпочтительно 110% или более, 120% или более, 130% или более, 140% или более, в частности предпочтительно 150% или более, 160% или более, 170% или более, 180% или более, 190% или более, 200% или более, 250% или более, 300% или более, 350% или более, 400% или более, 450% или более, 500% или более, 750% или более по сравнению с активностью связывания с любым из Fcγ-рецепторов человека: FcγRIa, FcγRIIa, FcγRIIIa и/или FcγRIIIb, или превышает в 10 раз или более, в 20 раз или более, в 30 раз или более, в 40 раз или более, в 50 раз, в 60 раз, в 70 раз, в 80 раз, в 90 раз или в 100 раз или более. Fc-область с более высокой активностью связывания с ингибиторными Fcγ-рецепторами, чем с активирующими Fcγ-рецепторами, может быть подходящим образом включена в антигенсвязывающие молекулы по настоящему изобретению, антигенсвязывающий домен которых связывается с растворимой молекулой.
В неограничивающем варианте осуществления настоящего изобретения примеры Fc-области, обладающей более высокой активностью связывания с ингибирующими Fcγ-рецепторами, чем с активирующими Fcγ-рецепторами (или обладающей активностью селективного связывания с ингибиторными Fcγ-рецепторами) предпочтительно включают Fc-области, в которых из аминокислот указанной выше Fc-области аминокислоты в положениях 238 и 328, указанных с помощью нумерации EU, изменены на аминокислоты, отличающиеся от аминокислот нативной Fc-области.
В неограничивающем варианте осуществления настоящего изобретения примеры Fc-области, обладающей более высокой активностью связывания с ингибиторными Fcγ-рецепторами, чем с активирующими Fcγ-рецепторами (или обладающей активностью селективного связывания с ингибиторными Fcγ-рецепторами) предпочтительно включают Fc-области, измененные в любой одной или нескольких аминокислотах в указанной выше Fc-области, как указано с помощью нумерации EU: аминокислота в положении 238 (указано с помощью нумерации EU) изменена на Asp; и аминокислота в положении 328 (указано с помощью нумерации EU) изменена на Glu. Более того, в качестве Fc-областей, обладающих активностью селективного связывания с ингибиторными Fcγ-рецепторами можно пригодным образом выбирать Fc-области или изменения, описанные в US 2009/0136485.
В другом неограничивающем варианте осуществления настоящего изобретения предпочтительные примеры включают Fc-области, измененные в любой или нескольких аминокислотах в указанной выше Fc-области, как указано с помощью нумерации EU: аминокислота в положении 238 (указано с помощью нумерации EU) изменена на Asp; и аминокислота в положении 328 (указано с помощью нумерации EU) измена на Glu.
В другом неограничивающем варианте осуществления настоящего изобретения предпочтительные примеры включают Fc-области, которые имеют одно или несколько изменений, проиллюстрированных в PCT/JP2012/054624: замена Pro в положении 238 (указано с помощью нумерации EU) на Asp; изменение аминокислоты в положении 237 (указано с помощью нумерации EU) на Trp; изменение аминокислоты в положении 237 (указано с помощью нумерации EU) на Phe; изменение аминокислоты в положении 267 (указано с помощью нумерации EU) на Val; изменение аминокислоты в положении 267 (указано с помощью нумерации EU) на Gln; изменение аминокислоты в положении 268 (указано с помощью нумерации EU) на Asn; изменение аминокислоты в положении 271 (указано с помощью нумерации EU) на Gly; изменение аминокислоты в положении 326 (указано с помощью нумерации EU) на Leu; изменение аминокислоты в положении 326 (указано с помощью нумерации EU) на Gln; изменение аминокислоты в положении 326 (указано с помощью нумерации EU) на Glu; изменение аминокислоты в положении 326 (указано с помощью нумерации EU) на Met; изменение аминокислоты в положении 239 (указано с помощью нумерации EU) на Asp; изменение аминокислоты в положении 267 (указано с помощью нумерации EU) на Ala; изменение аминокислоты в положении 234 (указано с помощью нумерации EU) на Trp; изменение аминокислоты в положении 234 (указано с помощью нумерации EU) на Tyr; изменение аминокислоты в положении 237 (указано с помощью нумерации EU) на Ala; изменение аминокислоты в положении 237 (указано с помощью нумерации EU) на Asp; изменение аминокислоты в положении 237 (указано с помощью нумерации EU) на Glu; изменение аминокислоты в положении 237 (указано с помощью нумерации EU) на Leu; изменение аминокислоты в положении 237 (указано с помощью нумерации EU) на Met; изменение аминокислоты в положении 237 (указано с помощью нумерации EU) на Tyr; изменение аминокислоты в положении 330 (указано с помощью нумерации EU) на Lys; изменение аминокислоты в положении 330 (указано с помощью нумерации EU) на Arg, изменение аминокислоты в положении 233 (указано с помощью нумерации EU) на Asp, изменение аминокислоты в положении 268 (указано с помощью нумерации EU) на Asp, изменение аминокислоты в положении 268 (указано с помощью нумерации EU) на Glu, изменение аминокислоты в положении 326 (указано с помощью нумерации EU) на Asp, изменение аминокислоты в положении 326 (указано с помощью нумерации EU) на Ser, изменение аминокислоты в положении 326 (указано с помощью нумерации EU) на Thr, изменение аминокислоты в положении 323 (указано с помощью нумерации EU) на Ile, изменение аминокислоты в положении 323 (указано с помощью нумерации EU) на Leu, изменение аминокислоты в положении 323 (указано с помощью нумерации EU) на Met, изменение аминокислоты в положении 296 (указано с помощью нумерации EU) на Asp, изменение аминокислоты в положении 326 (указано с помощью нумерации EU) на Ala, изменение аминокислоты в положении 326 (указано с помощью нумерации EU) на Asn, и изменение аминокислоты в положении 330 (указано с помощью нумерации EU) на Met.
Fc-области с модифицированными цепями сахаров
Fc-области, содержащиеся в антигенсвязывающих молекулах, предусматриваемых в рамках настоящего изобретения, могут включать Fc-области, которые модифицированы так, чтобы композиция Fc-областей с которыми связана цепь сахаров, имела более высокий процент Fc-областей, с которыми связана цепь сахаров с дефицитом фукозы, или более высокий процент Fc-областей, к которым добавлен биссекторный N-ацетилглюкозамин. Известно, что удаление остатка фузозы из N-ацетилглюкозамина на восстанавливающем конце комплексных цепей сахаров с N-гликозидной связью, связанных с Fc-областью антитела, повышает аффинность в отношении FcγRIIIa (непатентный документ 6). Известно, что для IgG1-антител, содержащих такие Fc-области, активность ADCC, упомянутая ниже, усиливается; таким образом, антигенсвязывающие молекулы, содержащие такие Fc-области, также являются пригодными в качестве антигенсвязывающих молекул, предназначенных для содержания в фармацевтических композициях по настоящему изобретению. Примеры антител с удаленным остатком фукозы из N-ацетилглюкозамина на восстанавливающем конце комплексных цепей сахаров с N-гликозидной связью, связанных с Fc-областями антител, представляют собой антитела, такие как:
антитела, модифицированные посредством гликозилирования (например, WO 1999/054342); и
антитела, связанные с цепями сахаров с дефицитом фукозы (например, WO 2000/061739, WO 2002/031140 и WO 2006/067913).
Более конкретно, для получения антител, связанных с цепями сахаров с дефицитом фукозы (например, WO 2000/061739, WO 2002/031140 и WO 2006/067913) в качестве другого неограничивающего варианта осуществления антител с удаленным остатком фукозы из N-ацетилглюкозамина на восстанавливающем конце комплексных цепей сахаров с N-гликозидной связью, связанных с Fc-областями антител, получают клетки-хозяева, имеющие низкую способность присоединять фукозу к цепям сахаров, путем изменения активности образования структуры цепи сахаров полипептида, подлежащего гликозилированию. Антитела, которые лишены фукозы на их цепях сахаров, можно собирать из культуры клеток-хозяев путем экспрессии желаемого гена антитела в клетках-хозяевах. Неограничивающие пригодные примеры активности в отношении образования структуры цепи сахаров в полипептиде включают активность переносчика или фермента, выбранного из группы, состоящей из фукозилтрансферазы (EC 2.4.1.152), переносчика фукозы (SLC35C1), GMD (GDP-манноза-4,6-дегидратаза) (EC 4.2.1.47), Fx (GDP-кето-6-дезоксиманноза-3,5-эпимераза, 4-редуктаза) (EC 1.1.1.271) и GFPP (GDP-β-L-фукозопирофосфорилаза (EC 2.7.7.30). При условии, что эти ферменты или переносчики могут проявлять их активность, их структуры не обязательно должны быть точно установленными. В рамках настоящего изобретения, белки, которые проявляют такую активность, обозначают как "функциональные белки". В неограничивающем варианте осуществления способы изменения этой активности включают удаление этой активности. Для получения клеток-хозяев с дефицитом этой активности, можно соответственно использовать известные способы, такие как способ разрушения генов этих функциональных белков, чтобы они стали неспособными функционировать (например, WO2000/061739, WO2002/031140 и WO2006/067913). Клетки-хозяева с дефицитом такой активности можно получать, например, способом, который разрушает гены этих функциональных белков, эндогенных для клеток CHO, клеток BHK, клеток NS0, клеток SP2/0, клеток миеломы YO, клеток миеломы мыши P3X63, клеток PER, клеток PER.C6, клеток HEK293, клеток гибридомы и т.п., так чтобы гены были неспособны функционировать.
Известны антитела, которые имеют цепь сахаров, содержащую биссекторный GlcNAc (WO2002/079255 и т.д.). В неограничивающем варианте осуществления получают клетки-хозяева, экспрессирующие ген, который кодирует функциональный белок, обладающий активностью GnTIII (β-1,4-маннозилгликопротеин-4-β-N-ацетилглюкозаминилтрансфераза) (EC 2.4.1.94) или активностью GalT (β-1,4-галактозилтрансфераза) (EC 2.4.1.143), для получения антител, которые имеют биссекторные содержащие GlcNAc цепи сахаров. В другом подходящем неограничивающем варианте осуществления получают клетки-хозяева, которые экспрессируют, в дополнение к упомянутым выше функциональным белкам, ген, кодирующий функциональный белок, обладающий активностью ManII человека (маннозидаза II) (3.2.1.114), ген, кодирующий функциональный белок, обладающий активностью GnTI (β-1,2-ацетилглюкозаминилтрансфераза I) (EC 2.4.1.94), ген, кодирующий функциональный белок, обладающий активностью GnTII (β-1,2-ацетилглюкозаминилтрансфераза II) (EC 2.4.1.143), ген, кодирующий функциональный белок, обладающий активностью ManI (манноидаза) (EC 3.2.1.113) и α-1,6-фукозилтрансферазы (EC 2.4.1.68) (WO2004/065540).
Антитела с удаленным остатком фукозы из N-ацетилглюкозамина на восстанавливающем конце комплексных цепей сахаров с N-гликозидной связью, связанных с Fc-областями антитела, и антитела, обладающие цепями сахаров, содержащими биссекторный GlcNAc, можно получать, соответственно, путем трансфекции экспрессирующего вектора, содержащего ген антитела, в клетки-хозяева с низкой способностью присоединять фукозу к цепям сахаров, и в клетки-хозяева, обладающие активностью образования биссекторных содержащих структуру GlcNAc цепей сахаров. Способы получения этих антител можно применять в способах получения антигенсвязывающих молекул, содержащих измененные Fc-области, которые модифицированы так, чтобы композиция Fc-областей с присоединенной цепью сахаров по настоящему изобретению имела высокий процент Fc-областей с присоединенной цепью сахаров с дефицитом фукозы или высокий процент Fc-областей с присоединенным биссекторным N-ацетилглюкозамином. Композиция Fc-областей с присоединенными к ней цепями сахаров, содержащимися в антигенсвязывающих молекулах по настоящему изобретению, продуцированных такими способами получения, можно оценивать способом, описанным в разделе "Fc-области с измененным связыванием Fcγ-рецептора (FcγR)" выше.
Мультиспецифические антигенсвязывающие молекулы и мультипаратопные антигенсвязывающие молекулы
Антигенсвязывающая молекула, содержащая по меньшей мере два антигенсвязывающих домена, где по меньшей мере один из антигенсвязывающих доменов связывается с первым эпитопом в молекуле антигена и по меньшей мере один другой из антигенсвязывающих доменов связывается со вторым эпитопом в молекуле антигена, называют "мультиспецифической антигенсвязывающей молекулой" с точки зрения ее реакционной специфичности. Когда два типа антигенсвязывающих доменов, содержащихся в одной антигенсвязывающей молекуле, допускают связывание с двумя различными эпитопами антигенсвязывающей молекулы, эту молекулу называют "биспецифической антигенсвязывающей молекулой". Когда три типа антигенсвязывающих доменов, содержащихся в одной антигенсвязывающей молекуле, допускают связывание с тремя различными эпитопами антигенсвязывающей молекулой, эту антигенсвязывающую молекулу называют "триспецифической антигенсвязывающей молекулой".
Паратоп в антигенсвязывающем домене, который связывается с первым эпитопом в молекуле антигена и паратоп в антигенсвязывающем домене, который связывается со вторым эпитопом, который структурно отличается от первого эпитопа, имеют различные структуры. Таким образом, антигенсвязывающая молекула, содержащая по меньшей мере два антигенсвязывающих домена, где по меньшей мере один из антигенсвязывающих доменов связывается с первым эпитопом в молекуле антигена и по меньшей мере один другой из антигенсвязывающих доменов связывается со вторым эпитопом в молекуле антигена, называют "мультипаратопной антигенсвязывающей молекулой" с точки зрения специфичности ее структуры. Когда два типа антигенсвязывающих доменов, содержащихся в одной антигенсвязывающей молекуле, допускает связывание с двумя различными эпитопами антигенсвязывающей молекулой, эту молекулу называют "бипаратопной антигенсвязывающей молекулой". Когда три типа антигенсвязывающих доменов, содержащихся в одной антигенсвязывающей молекуле, допускают связывание с тремя различными эпитопами антигенсвязывающей молекулой, эту молекулу называют "трипаратопной антигенсвязывающей молекулой".
Поливалентные мультиспецифические или мультипаратопные антигенсвязывающие молекулы, содержащие один или несколько антигенсвязывающих доменов, и способы их получения описаны в непатентных документах, таких как Conrath et al. (J.Biol.Chem. (2001) 276 (10) 7346-7350), Muyldermans (Rev. Mol. Biotech. (2001) 74, 277-302), и Kontermann R.E. (2011) Bispecific Antibodies (Springer-Verlag) и в патентных документах, таких как WO 1996/034103 и WO 1999/023221. Антигенсвязывающие молекулы по настоящему изобретению можно получать с использованием мультиспецифических или мультипаратопных антигенсвязывающих молекул и способов их получения, описанных в этих документах.
Биспецифические антитела и способы их получения
В одном варианте осуществления биспецифические антитела и способы их получения упомянуты ниже в качестве примеров упомянутых выше мультиспецифических или мультипаратопных антигенсвязывающих молекул и способов их получения. Биспецифические антитела представляют собой антитела, содержащие два типа вариабельных областей, которые специфически связываются с различными эпитопами. Биспецифические антитела IgG-типа можно секретировать из гибридной гибридомы (квадромы), полученной путем слияния двух типов гибридом, которые продуцируют IgG-антитела (Milstein et al., Nature (1983) 305, 537-540).
Когда биспецифическое антитело получают с использованием рекомбинантных способов, таких как способы, описанные в упомянутом выше разделе, касающемся антител, можно использовать способ, который вводит гены, кодирующие тяжелые цепи, содержащие два типа представляющих интерес вариабельных областей, в клетки для их коэкспрессии. Однако при рассмотрении только комбинации тяжелых цепей, такой способ будет обеспечивать смесь (i) комбинации пары тяжелых цепей, в которых одна из тяжелых цепей содержит вариабельную область, которая связывается с первым эпитопом, а другая тяжелая цепь содержит вариабельную область, которая связывается со вторым эпитопом, (ii) комбинации пары тяжелых цепей, которые включают только тяжелые цепи, содержащие вариабельную область, которая связывается с первым эпитопом, и (iii) комбинации пары тяжелых цепей, которые включают только тяжелые цепи, содержащие вариабельную область, которая связывается со вторым эпитопом, которые присутствуют в молекулярном соотношении 2:1:1. Трудно очистить антигенсвязывающие молекулы, содержащие желаемую комбинацию тяжелых цепей, из смеси трех типов комбинаций тяжелых цепей.
При получении биспецифических антител с использованием таких рекомбинантных способов, биспецифические антитела, содержащие гетеромерную комбинацию тяжелых цепей, можно предпочтительно секретировать путем добавления соответствующих аминокислотных замен в CH3-домены, составляющие тяжелые цепи. В частности, этот способ проводят путем замены аминокислотой, имеющей более крупную боковую цепь (выступ (что означает "выпуклая часть")) аминокислоты в CH3-домене одной из тяжелых цепей, и замены аминокислотой, имеющей меньшую боковую цепь (углубление (что означает "полость")) аминокислоты в CH3-домене другой тяжелой цепи, так чтобы выступ размещался в полости. Это обеспечивает образование гетеромерной тяжелой цепи и одновременно ингибирует образование гомомерной тяжелой цепи (международная публикация № WO 1996027011; Ridgway et al., Protein Engineering (1996) 9, 617-621; Merchant et al., Nature Biotechnology (1998) 16, 677-681).
Более того, также существуют известные способы получения биспецифического антитела с использованием способов контроля ассоциации полипептидов или ассоциации образованных полипептидом гетеромерных мультимеров для ассоциации между тяжелыми цепями. В частности, способы контроля образования тяжелых цепей можно использовать для получения биспецифического антитела (международная публикация № WO 2006/106905), в котором аминокислотные остатки, образующие поверхность контакта между тяжелыми цепями, изменяют для ингибирования ассоциации между тяжелыми цепями, имеющими ту же последовательность, и для обеспечения образования тяжелых цепей с различными последовательностями. Такие способы можно использовать для получения биспецифических антител.
В неограничивающем варианте осуществления настоящего изобретения два полипептида, образующих Fc-область, происходящие из биспецифического антитела, описанного выше, можно соответствующим образом использовать в качестве Fc-области, содержащейся в антигенсвязывающей молекуле. Более конкретно, предпочтительно использовать два полипептида, образующих Fc-область, которые содержат Cys в качестве аминокислоты в положении 349 и Trp в качестве аминокислоты в положении 366 в соответствии с нумерацией EU в аминокислотной последовательности одного из полипептидов; и Cys в качестве аминокислоты в положении 356, Ser в качестве аминокислоты в положении 366, Ala в качестве аминокислоты в положении 368, и Val в качестве аминокислоты в положении 407, как указано с помощью нумерации EU, в аминокислотной последовательности другого полипептида.
В другом неограничивающем варианте осуществления настоящего изобретения два полипептида, которые составляют Fc-область, и которые содержат Asp в качестве аминокислоты в положении 409 в соответствии с нумерацией EU в аминокислотной последовательности одного из полипептидов, и Lys в качестве аминокислоты в положении 399 в соответствии с нумерацией EU в аминокислотной последовательности другого полипептида, можно соответственно использовать в качестве Fc-области. В упомянутом выше варианте осуществления аминокислота в положении 409 может представлять собой Glu вместо Asp, и аминокислота в положении 399 может представлять собой Arg вместо Lys. Более того, в дополнение к аминокислоте Lys в положении 399, Asp можно соответственно добавлять в качестве аминокислоты в положении 360 или Asp можно соответственно добавлять в качестве аминокислоты в положении 392.
В другом неограничивающем варианте осуществления настоящего изобретения, два полипептида, которые составляют Fc-область, и которые содержат Glu в качестве аминокислоты в положении 370 в соответствии с нумерацией EU в аминокислотной последовательности одного из полипептидов, и Lys в качестве аминокислоты в положении 357 в соответствии с нумерацией EU в аминокислотной последовательности другого полипептида, можно соответственно использовать в качестве Fc-области.
В другом неограничивающем варианте осуществления настоящего изобретения, два полипептида, которые составляют Fc-область, и которые содержат Glu в качестве аминокислоты в положении 439 в соответствии с нумерацией EU в аминокислотной последовательности одного из полипептидов, и Lys в качестве аминокислоты в положении 356 в соответствии с нумерацией EU в аминокислотной последовательности другого полипептида, можно соответственно использовать в качестве Fc-области.
В другом неограничивающем варианте осуществления настоящего изобретения любой из указанных ниже вариантов осуществления для упомянутых выше комбинаций можно соответственно использовать в качестве Fc-области:
(i) два полипептида, которые составляют Fc-область и которые содержат Asp в качестве аминокислоты в положении 409 и Glu в качестве аминокислоты в положении 370 в соответствии с нумерацией EU в аминокислотной последовательности одного из полипептидов, и Lys в качестве аминокислоты в положении 399 и Lys в качестве аминокислоты в положении 357 в соответствии с нумерацией EU в аминокислотной последовательности другого полипептида (в этом варианте осуществления аминокислота в положении 370 в соответствии с нумерацией EU может представлять собой Asp вместо Glu, и аминокислоту Asp в положении 392 можно использовать вместо аминокислоты Glu в положении 370 в соответствии с нумерацией EU);
(ii) два полипептида, которые составляют Fc-область и которые содержат Asp в качестве аминокислоты в положении 409 и Glu в качестве аминокислоты в положении 439 в соответствии с нумерацией EU в аминокислотной последовательности одного из полипептидов; и Lys в качестве аминокислоты в положении 399 и Lys в качестве аминокислоты в положении 356 в соответствии с нумерацией EU в аминокислотной последовательности другого полипептида (в этом варианте осуществления аминокислоту Asp в положении 360, аминокислоту Asp в положении 392 или аминокислоту Asp в положении 439 можно использовать вместо аминокислоты Glu в положении 439 в соответствии с нумерацией EU);
(iii) два полипептида, которые составляют Fc-область и которые содержат Glu в качестве аминокислоты в положении 370, и Glu в качестве аминокислоты в положении 439 в соответствии с нумерацией EU в аминокислотной последовательности одного из полипептидов, и Lys в качестве аминокислоты в положении 357, и Lys в качестве аминокислоты в положении 356 в соответствии с нумерацией EU в аминокислотной последовательности другого полипептида; или
два полипептида, которые составляют Fc-область и которые содержат Asp аминокислота в положении 409, Glu в качестве аминокислоты в положении 370, и Glu в качестве аминокислоты в положении 439 в соответствии с нумерацией EU в аминокислотной последовательности одного из полипептидов; и Lys в качестве аминокислоты в положении 399, Lys в качестве аминокислоты в положении 357, и Lys в качестве аминокислоты в положении 356 в соответствии с нумерацией EU в аминокислотной последовательности другого полипептида (в этом варианте осуществления аминокислота в положении 370 может не быть замещена на Glu, и более того, когда аминокислота в положении 370 не замещена на Glu, аминокислота в положении 439 может представлять собой Asp вместо Glu или аминокислоту Asp в положении 392 можно использовать вместо аминокислоты Glu в положении 439, в соответствии с нумерацией EU).
Кроме того, в другом неограничивающем варианте осуществления настоящего изобретения также может быть пригодным использование двух полипептидов, которые составляют Fc-область и которые содержат Lys в качестве аминокислоты в положении 356 в соответствии с нумерацией EU в аминокислотной последовательности одного из полипептидов, и Arg в качестве аминокислоты в положении 435 и Glu в качестве аминокислоты в положении 439 в соответствии с нумерацией EU в аминокислотной последовательности другого полипептида.
В другом неограничивающем варианте осуществления настоящего изобретения также может быть пригодным использование двух полипептидов, которые составляют Fc-область и которые содержат Lys в качестве аминокислоты в положении 356, и Lys в качестве аминокислоты в положении 357 в соответствии с нумерацией EU в аминокислотной последовательности одного из полипептидов, и Glu в качестве аминокислоты в положении 370, Arg в качестве аминокислоты в положении 435, и Glu в качестве аминокислоты в положении 439 в соответствии с нумерацией EU в аминокислотной последовательности другого полипептида.
Более того, в дополнение к упомянутому выше способу связывания гетерологичных тяжелых цепей, также для получения мультиспецифических или мультипаратопных антигенсвязывающих молекул, предусматриваемых в рамках настоящего изобретения, можно использовать технологию CrossMab, которая известна в качестве технологии связывания гетерологичных легких цепей, в которой легкую цепь, образующую вариабельную область, которая связывается с первым эпитопом, и легкую цепь, образующую вариабельную область, которая связывается со вторым эпитопом, соответственно связывают с тяжелой цепью, образующей вариабельную область, которая связывается с первым эпитопом, и тяжелой цепью, образующей вариабельную область, которая связывается со вторым эпитопом (Scaefer et al. (Proc. Natl. Acad. Sci. U.S.A. (2011) 108, 11187-11192)). Более того, также для получения мультиспецифических или мультипаратопных антигенсвязывающих молекул, предусматриваемых в рамках настоящего изобретения, можно использовать обмен плечами Fab, который известен как технология для связывания гетерологичных тяжелых цепей, в которой тяжелую цепь, образующую вариабельную область, которая связывается с первым эпитопом, и тяжелую цепь, образующую вариабельную область, которая связывается со вторым эпитопом, с использованием этих гетерологичных тяжелых цепей IgG4 обменивают между собой (Labrijn et al. (Proc. Natl. Acad. Sci. U.S.A. (2013) 110, 5145-5150), WO2008119353).
Эффекторные клетки
В рамках настоящего изобретения термин "эффекторные клетки" можно использовать в наиболее широком значении, включающем T-клетки (CD4+ (хелперный лимфоцитарные) T-клетки и/или CD8+ (цитотоксические) T-клетки, многоядерные лейкоциты (нейтрофилы, эозинофилэозинофилы, базофилы, тучные клетки), моноциты, макрофаги, гистиоциты или лейкоциты, такие как натуральные киллерные клетки (NK-клетки), NK-подобные T-клетки, клетки Купфера, клетки Лангерганса или активируемые лимфокинами киллерные клетки (LAK-клетки), B-лимфоциты или антигенпредставляющие клетки, такие как дендритные клетки или макрофаги. Предпочтительные примеры эффекторных клеток включают CD8+ (цитотоксические) T-клетки, NK-клетки или макрофаги. В качестве антигена, с которым связывается по меньшей мере один антигенсвязывающий домен, содержащийся в антигенсвязывающей молекуле по настоящему изобретению, можно использовать молекулы мембранного типа, экспрессируемые на клеточной мембране эффекторных клеток. Неограничивающие примеры предпочтительной молекулы мембранного типа могут представлять собой CD3, CD2, CD28, CD44, CD16, CD32, CD64 или NKG2D, активирующие NK-клетки лиганды или полипептиды, составляющие TCR.
Цитотоксические вещества
Чтобы антигенсвязывающие молекулы по настоящему изобретению связывались со злокачественными клетками и проявляли цитотоксическую активность, цитотоксические вещества можно связывать с антигенсвязывающими молекулами. Цитотоксические вещества могут представлять собой химиотерапевтические средства, проиллюстрированные ниже, или соединения, описанные в Curr Opin Chem Biol (2010) 14, 529-37 и WO 2009/140242; и эти соединения связаны с антигенсвязывающими молекулами подходящими линкерами и т.п. Когда антигенсвязывающие молекулы по настоящему изобретению используют в качестве фармацевтических композиций, эти цитотоксические вещества можно связывать с антигенсвязывающими молекулами перед введением, или их можно вводить до, после или одновременно с введением антигенсвязывающих молекул индивидуумам (исследуемым индивидуумам, пациентам и т.п.).
Описанные ниже модифицированные антигенсвязывающие молекулы, с которыми связаны цитотоксические вещества, такие как химиотерапевтические средства, токсические пептиды или радиоактивные химические вещества, также можно предпочтительно использовать в качестве антигенсвязывающих молекул по настоящему изобретению, обладающих цитотоксической активностью. Такие модифицированные антигенсвязывающие молекулы (далее в настоящем описании обозначаемые как конъюгат антигенсвязывающая молекула-лекарственное средство) можно получать путем химической модификации полученных антигенсвязывающих молекул. В качестве способов модификации антигенсвязывающих молекул можно использовать соответственно способы, которые уже являются установившимися в области конъюгатов антитело-лекарственное средство и т.п. Более того, модифицированную антигенсвязывающую молекулу, с которой связан токсический пептид, можно получать путем экспрессии в соответствующих клетках-хозяевах слитого гена, полученного путем связывания гена, кодирующего токсический пептид, в рамке считывания с геном, кодирующим антигенсвязывающую молекулу по настоящему изобретению, а затем выделения его из клеточной культуры.
Примеры химиотерапевтических средств, связанных с антигенсвязывающим молекулами по настоящему изобретению, могут включать: азарибин, анастрозол, азацитидин, блеомицин, бортезомиб, бриостатин-1, бусульфан, камптотецин, 10-гидроксикамптотецин, кармустин, целебрекс, хлорамбуцил, цисплатин, иринотекан, карбоплатин, кладрибин, циклофосфамид, цитарабин, дакарбазин, доцетаксел, актиномициндактиномицин, дауномицин глюкуронид, даунорубицин, дексаметазон, диэтилстилбестрол, доксорубицин, доксорубицин глюкуронид, эпирубицин, этинилэстрадиол, эстрамустин, этопозид, этопозид глюкуронид, флоксиуридин, флударабинфлударабин, флутамид, фторурацил, флуоксиместерон, гемцитабин, гидроксипрогестерон капроат, гидроксимочевину, идарубицин, ифосфамид, лейковорин, ломустин, майтанзиноид, мехорэтамин, медроксипрогестерона ацетат, магестрола ацетат, мелфалан, меркаптопурин, метотрексат, митоксантрон, митрамицин, митомицин, митотан, фенилбутират, преднизон, прокарбазин, паклитаксел, пентостатин, семустин, стрептозоцин, тамоксифен, таксаны, таксол, тестостерона пропионат, талидомид, тиогуанин, тиотепа, тенипозид, топотекан, урацил мустард, винбластин, винорелбин и винкристин.
В рамках настоящего изобретения предпочтительные химиотерапевтические средства представляют собой низкомолекулярные химиотерапевтические средства. Низкомолекулярные химиотерапевтические средства маловероятно будут препятствовать функции антигенсвязывающих молекул, даже после их связывания с антигенсвязывающими молекулами по настоящему изобретению. В рамках настоящего изобретения низкомолекулярные химиотерапевтичские средства обычно имеют молекулярную массу от 100 до 2000, предпочтительно от 200 до 1000. Все химиотерапевтические средства, проиллюстрированные в настоящем описании, представляют собой низкомолекулярные химиотерапевтические средства. Химиотерапевтические средства по настоящему изобретению включают пролекарства, которые конвертируются в активные химиотерапевтические средства in vivo. Активация пролекарства может представлять собой ферментативное конвертирование или неферментативное конвертирование.
Более того, цитотоксические вещества, которые связаны с антигенсвязывающими молекулами по настоящему изобретению, включают, например, токсические пептиды (токсины), такие как экзотоксин A Pseudomonas, сапорин-s6, дифтерийный токсин, токсин книдарий; радиоактивный йод и фотосенсибилизаторы. Подходящие примеры токсических пептидов включают следующие:
A-цепь дифтерийного токсина (Langone et al. (Methods in Enzymology (1983) 93, 307-308));
Экзотоксин Pseudomonas (Nature Medicine (1996) 2, 350-353);
цепь рицина (A-цепь рицина) (Fulton et al. (J. Biol. Chem. (1986) 261, 5314-5319), Sivam et al. (Cancer Res. (1987) 47, 3169-3173), Cumber et al. (J. Immunol. Metehods (1990) 135, 15-24), Wawrzynczak et al. (Cancer Res. (1990) 50, 7519-7562), и Gheeite et al. (J. Immunol. Methods (1991) 142, 223-230));
Дегликозилированная A-цепь рицина (Thorpe et al. (Cancer Res. (1987) 47, 5924-5931));
A-цепь абрина (Wawrzynczak et al. (Br. J. Cancer (1992) 66, 361-366), Wawrzynczak et al. (Cancer Res. (1990) 50, 7519-7562), Sivam et al. (Cancer Res. (1987) 47, 3169-3173) и Thorpe et al. (Cancer Res. (1987) 47, 5924-5931));
Гелонин (Sivam et al. (Cancer Res. (1987) 47, 3169-3173), Cumber et al. (J. Immunol. Methods (1990) 135, 15-24), Wawrzynczak et al. (Cancer Res., (1990) 50, 7519-7562) и Bolognesi et al. (Clin. exp. Immunol. (1992) 89, 341-346));
PAP-s; противовирусный белок из семян лаконоса (Bolognesi et al. (Clin. exp. Immunol. (1992) 89, 341-346));
бриодин (Bolognesi et al. (Clin. exp. Immunol. (1992) 89, 341-346));
сапорин (Bolognesi et al. (Clin. exp. Immunol. (1992) 89, 341-346));
момордин (Cumber et al. (J. Immunol. Methods (1990) 135, 15-24); Wawrzynczak et al. (Cancer Res. (1990) 50, 7519-7562); и Bolognesi et al. (Clin. exp. Immunol. (1992) 89, 341-346));
моморкохин (Bolognesi et al. (Clin. exp. Immunol. (1992) 89, 341-346));
диантин 32 (Bolognesi et al. (Clin. exp. Immunol. (1992) 89, 341-346));
диантин 30 (Stirpe F., Barbieri L. (FEBS letter (1986) 195, 1-8));
модессин (Stirpe F., Barbieri L. (FEBS letter (1986) 195, 1-8));
вискумин (Stirpe F., Barbieri L. (FEBS letter (1986) 195, 1-8));
волкесин (Stirpe F., Barbieri L. (FEBS letter (1986) 195, 1-8));
додекандрин (Stirpe F., Barbieri L. (FEBS letter (1986) 195, 1-8));
тритин (Stirpe F., Barbieri L. (FEBS letter (1986) 195, 1-8));
луффин (Stirpe F., Barbieri L. (FEBS letter (1986) 195, 1-8)); и
трихокирин (Casellas et al. (Eur. J. Biochem. (1988) 176, 581-588), и Bolognesi et al. (Clin. exp. Immunol., (1992) 89, 341-346)).
Антигенсвязывающая молекула
В рамках настоящего изобретения, термин "антигенсвязывающая молекула, содержащая антигенсвязывающий домен, антигенсвязывающая активность которого в присутствии специфического для ткани-мишени соединения является более высокой, чем в отсутствие специфического для ткани-мишени соединения" используют в наиболее широком значении; и, в частности, он включает различные типы молекул при условии, что ни проявляют антигенсвязывающую активность. Молекулы, в которых антигенсвязывающий домен связан с Fc-областью, включают, например, антитела. Антитела могут включать единичные моноклональные антитела (включая антитела-агонисты и антитела-антагонисты), антитела человека, гуманизированные антитела, химерные антитела и т.п. Альтернативно при использовании в качестве фрагментов антител, они предпочтительно включают антигенсвязывающие домены и антигенсвязывающие фрагменты (например, Fab, F(ab')2, scFv и Fv). Каркасные молекулы, где трехмерные структуры, такие как уже известная стабильная белковая структура α/β-бочонок, используются в качестве каркаса (основы) и только некоторые части структур преобразованы в библиотеки для конструирования антигенсвязывающих доменов, также включены в антигенсвязывающие молекулы по настоящему изобретению.
Антигенсвязывающая молекула по настоящему изобретению может содержать по меньшей мере некоторые части Fc-области, которые опосредуют связывание с Fcγ-рецептором и/или FcRn. В неограничивающем варианте осуществления антигенсвязывающая молекула включает, например, антитела и слитые белки Fc. Слитый белок относится к химерному полипептиду, содержащему полипептид, имеющий первую аминокислотную последовательность, который связан с полипептидом, имеющим вторую аминокислотную последовательность, которые не связаны в природе. Например, слитый белок может содержать аминокислотную последовательность по меньшей мере части Fc-области (например, часть Fc-области, ответственная за связывание с Fcγ-рецептором или часть Fc-области, ответственная за связывание с FcRn). Аминокислотные последовательности могут находиться в отдельных белках, которые переносят вместе в слитый белок, или, как правило, они могут находиться в одном белке; однако они включены в слитый полипептид в новом порядке. Слитые белки можно получать, например, посредством химического синтеза или способами генетической рекомбинации для экспрессии полинуклеотида, кодирующего области пептидов в желаемом порядке.
Соответствующие домены по настоящему изобретению могут быть связаны через линкеры или прямо через полипептидную связь. Линкеры включают произвольные пептидные линкеры, которые можно вносить способами генетической инженерии, синтетические линкеры и линкеры, описанные, например, в Holliger et al., Protein Engineering (1996) 9(3), 299-305. Однако пептидные линкеры являются предпочтительными в рамках настоящего изобретения. Длина пептидных линкеров конкретно не ограничена, и ее могут надлежащим образом выбирать специалисты в данной области в зависимости от назначения. Длина предпочтительно составляет пять аминокислот или более (без конкретных ограничений, верхний предел обычно составляет 30 аминокислоты или менее, предпочтительно 20 аминокислоты или менее), и особенно предпочтительно 15 аминокислот.
Например, такие пептидные линкеры предпочтительно включают:
Ser
Gly·Ser
Gly·Gly·Ser
Ser·Gly·Gly
Gly·Gly·Gly·Ser (SEQ ID NO:19)
Ser·Gly·Gly·Gly (SEQ ID NO:20)
Gly·Gly·Gly·Gly·Ser (SEQ ID NO:21)
Ser·Gly·Gly·Gly·Gly (SEQ ID NO:22)
Gly·Gly·Gly·Gly·Gly·Ser (SEQ ID NO:23)
Ser·Gly·Gly·Gly·Gly·Gly (SEQ ID NO:24)
Gly·Gly·Gly·Gly·Gly·Gly·Ser (SEQ ID NO:25)
Ser·Gly·Gly·Gly·Gly·Gly·Gly (SEQ ID NO:26)
(Gly·Gly·Gly·Gly·Ser (SEQ ID NO:21))n
(Ser·Gly·Gly·Gly·Gly (SEQ ID NO:22))n
где n представляет собой целое число, равное 1 или более. Длину или последовательности пептидных линкеров могут соответствующим образом выбирать специалисты в данной области, в зависимости от назначения.
Обычно для сшивания пептидов используют синтетические линкеры (химические сшивающие агенты), и например:
N-гидроксисукцинимид (NHS),
дисукцинимидилсуберат (DSS),
бис(сульфосукцинимидил)суберат (BS3),
дитиобис(сукцинимидил пропионат) (DSP),
дитиобис(сульфосукцинимидилпропионат) (DTSSP),
этиленгликоль бис(сукцинимидилсукцинат) (EGS),
этиленгликоль бис(сульфосукцинимидилсукцинат) (сульфо-EGS),
дисукцинимидилтартрат (DST), дисульфосукцинимидилтартрат (сульфо-DST),
бис[2-(сукцинимидоксикарбонилокси)этил]сульфон (BSOCOES),
и бис[2-(сульфосукцинимидоксикарбонилокси)этил]сульфон (сульфо-BSOCOES). Эти сшивающие агенты являются коммерчески доступными.
Когда используют множество линкеров для связывания соответствующих доменов, все они могут быть одного типа или они могут быть различных типов. В дополнение к линкерам, проиллюстрированным выше, также можно соответствующим образом использовать линкеры с пептидными метками, такими как His-метка, HA-метка, myc-метка и FLAG-метка. Более того, также можно соответствующим образом использовать образование водородных связей, образование дисульфидных связей, образование ковалентных связей, ионное взаимодействие и свойства связывания друг с другом в результате их комбинации. Например, можно использовать аффинность между CH1 и CL антитела, и также можно использовать Fc-области, происходящие из описанных выше биспецифических антител для гетероассоциации Fc-областей. Более того, также можно соответствующим образом использовать дисульфидные связи, образующиеся между доменами.
Для связывания соответствующих доменов пептидной связью, полинуклеотиды, кодирующие домены, связывают вместе в рамке считывания. Известные способы связывания полинуклеотидов в рамке считывания включают способы, такие как лигирование фрагментов рестрикции, ПЦР со слиянием и перекрывающаяся ПЦР. Такие способы можно соответствующим образом использовать отдельно или в комбинации для получения антигенсвязывающих молекул по настоящему изобретению. В рамках настоящего изобретения термины "связанный" и "слитый", или "связывание" и "слияние" используют взаимозаменяемо. Эти термины означают, что два или более элементов или компонентов, таких как полипептиды, слиты вместе с образованием единой структуры любыми способами, включая описанные выше способы химического связывания и способы генетической рекомбинации. Когда два или более доменов, элементов или компонентов представляют собой полипептиды, связывание в рамке считывания означает связывание двух или более элементов рамок считывания с образованием более длинной непрерывной рамки считывания при сохранении правильных рамок считывания полипептидов. Когда две молекулы Fab используют в качестве антигенсвязывающего домена, в качестве предпочтительной антигенсвязывающей молекулы по настоящему изобретению можно использовать антитело, которое представляет собой антигенсвязывающую молекулу по настоящему изобретению, в которой антигенсвязывающий домен связан в рамке считывания с константной областью, включающей Fc-область, через пептидную связь, но не через линкер.
Низкомолекулярное антитело
Антитела, используемые в рамках настоящего изобретения, не ограничиваются полноразмерными молекулами антител, и они могут представлять собой низкомолекулярные антитела (миниантитела) и их модифицированные продукты. Низкомолекулярное антитело включает фрагмент антитела, который лишен части полноразмерного антитела (например, целое антитело, такое как целый IgG); и оно конкретно не ограничено при условии, что оно обладает антигенсвязывающей активностью. Низкомолекулярное антитело по настоящему изобретению конкретно не ограничено при условии, что оно является частью полноразмерного антитела, но предпочтительно содержит вариабельную область тяжелой цепи (VH) и/или вариабельную область легкой цепи (VL). Аминокислотная последовательность VH или VL может иметь замену(ы), делецию(и), вставку(и) и/или инсерцию(и). Более того, при условии наличия антигенсвязывающей активности, VH и/или VL могут быть частично удалены. Вариабельная область может быть химеризованной или гуманизированной. Конкретные примеры фрагментов антител включают Fab, Fab', F(ab')2 и Fv. Конкретные примеры низкомолекулярных антител включают Fab, Fab', F(ab')2, Fv, scFv (одноцепочечный Fv), диантитело и sc(Fv)2 (одноцепочечный (Fv)2). Мультимеры этих антител (например, димеры, тримеры, тетрамеры и полимеры) также включены в низкомолекулярные антитела по настоящему изобретению.
Фрагменты антител можно получать путем обработки антитела ферментом, таким как папаин и пепсин. Альтернативно гены, кодирующие эти фрагменты антител, можно конструировать, встраивать в экспрессирующие векторы, а затем экспрессировать в подходящих клетках-хозяевах (см., например, Co et al., (J. Immunol. (1994) 152, 2968-2976); Better and Horwitz (Methods in Enzymology (1989) 178, 476-496), Plueckthun and Skerra (Methods in Enzymology (1989) 178, 476-496); Lamoyi (Methods in Enzymology (1989) 121, 652-663); Rousseaux (Methods in Enzymology (1989) 121, 663-669); и Bird, et al., TIBTECH (1991) 9, 132-137).
Диантитело относится к двухвалентному низкомолекулярному антителу, сконструированному посредством слияния генов (Hollinger et al., (Proc. Natl. Acad. Sci. USA 90, 6444-6448 (1993)); EP 404,097; WO 1993/11161 и т.п.). Диантитело представляет собой димер, состоящий из двух полипептидных цепей. Как правило, в каждой полипептидной цепи, составляющей димер, VL и VH связаны с помощью линкера в одной и той же цепи. Линкер в диантителе, как правило, является достаточно коротким, чтобы предотвратить связывание между VL и VH. В частности, аминокислотные остатки, составляющие линкер, представляют собой, например, приблизительно пять остатков. Линкер между VL и VH, которые кодируются одной и той же полипептидной цепью, является слишком коротким, чтобы образовать одноцепочечный фрагмент вариабельной области, и между полипептидными цепями образуется димер. В результате диантитела имеют два антигенсвязывающих центра.
scFv можно получать путем связывания V-области H-цепи и V-области L-цепи антитела. В scFv V-область H-цепи и V-область L-цепи лигированы через линкер, предпочтительно пептидный линкер (Huston, et al., Proc. Natl. Acad. Sci. U.S.A. (1988) 85, 5879-5883). V-область H-цепи и V-область L-цепи scFv могут происходить из любого из антител, описанных в настоящем описании. Пептидный линкер для лигирования V-областей конкретно не ограничен; и, например, в качестве линкера можно использовать любой одноцепочечный пептид, состоящий из 3-25 остатков и т.п., или пептидные линкеры, описанные ниже, и т.п. Для лигирования V-областей можно использовать способы ПЦР, такие как способы, описанные выше. ДНК, кодирующую scFv, можно амплифицировать способом ПЦР с использованием в качестве матрицы либо целой ДНК, либо частичной ДНК, кодирующей желаемую аминокислотную последовательность, которая выбрана из последовательности ДНК, кодирующей H-цепь или V-область H-цепи упомянутого выше антитела, и ДНК, кодирующей L-цепь или V-область L-цепи упомянутого выше антитела; и с использованием пары праймеров, имеющих последовательности, соответствующие последовательностям двух концов. Далее, ДНК, имеющую желаемую последовательность, можно получать путем проведения реакции ПЦР с использованием комбинации ДНК, кодирующей часть, представляющую собой пептидный линкер, и пары праймеров, имеющих последовательности, сконструированные так, чтобы оба конца ДНК лигировались с H-цепью и L-цепью, соответственно. После конструирования кодирующей scFv ДНК можно получать экспрессирующие векторы, имеющие ДНК, и рекомбинантные клетки, трансформированные экспрессирующим вектором, в соответствии с общепринятыми способами. Более того, scFv можно получать путем культивирования полученных рекомбинантных клеток для экспрессии ДНК, кодирующей scFv.
sc(Fv)2 представляет собой низкомолекулярное антитело, полученное путем связывания двух VH и двух VL с линкерами и т.п. с получением единой цепи (Hudson et al. (J. Immunol. Methods 1999; 231: 177-189)). sc(Fv)2 можно получать, например, путем связывания scFv с помощью линкера.
Более того, являются предпочтительными антитела, в которых два VH и два VL расположены в порядке VH, VL, VH и VL, начиная с N-конца одноцепочечного полипептида ([VH]-линкер-[VL]-линкер-[VH]-линкер-[VL]). Порядок двух VH и двух VL конкретно не ограничивается указанным выше расположением, и они могут быть расположены в любом порядке. Примеры включают следующие расположения:
- [VL]-линкер-[VH]-линкер-[VH]-линкер-[VL]
- [VH]-линкер-[VL]-линкер-[VL]-линкер-[VH]
- [VH]-линкер-[VH]-линкер-[VL]-линкер-[VL]
- [VL]-линкер-[VL]-линкер-[VH]-линкер-[VH]
- [VL]-линкер-[VH]-линкер-[VL]-линкер-[VH]
Линкер, сходный с линкером, описанным в разделе "антигенсвязывающие молекулы", выше, можно использовать в качестве линкера для связывания вариабельных областей антитела. Особенно предпочтительный вариант осуществления sc(Fv)2 в рамках настоящего изобретения включает, например, следующие sc(Fv)2:
- [VH]-пептидный линкер (15 аминокислот)-[VL]-пептидный линкер (15 аминокислот)-[VH]-пептидный линкер (15 аминокислот)-[VL]
Как правило, для связывания четырех вариабельных областей антитела требуется три линкера. Используемые линкеры могут быть одного типа или различных типов. Примеры неограничивающего варианта осуществления низкомолекулярного антитела в рамках настоящего изобретения включают диантитело или sc(Fv)2, где паратопы отличаются друг от друга; один из паратопов связывается с эпитопом в молекуле мембранного типа, которая связывается с мембраной злокачественной клетки; и другой паратоп связывается с эпитопом в молекуле мембранного типа, экспрессируемой на мембране эффекторных клеток. В упомянутом выше диантителе или sc(Fv)2 активность связывания одного паратопа, который связывается с эпитопом в молекуле мембранного типа, которая связывается с мембраной злокачественной клетки, может зависеть от специфического для злокачественной ткани соединения, причем активность связывания одного из паратопов с эпитопом в молекуле мембранного типа, которая связывается с мембраной эффекторной клетки, может зависеть от специфического для злокачественной ткани соединения, или активность связывания обоих паратопов может зависеть от специфического для злокачественной ткани соединения.
Неограничивающий вариант осуществления низкомолекулярного антитела в рамках настоящего изобретения включает, например, диантитело или sc(Fv)2, где паратопы отличаются друг от друга; один из паратопов связывается с эпитопом в молекуле мембранного типа, которая связывается с мембраной злокачественной клетки; и другой паратоп связывается с эпитопом в цитотоксическом веществе. В диантителе или sc(Fv)2, упомянутых выше, активность связывания одного из паратопов, который связывается с эпитопом в молекуле мембранного типа, которая связывается с мембраной злокачественной клетки, может зависеть от специфического для злокачественной ткани соединения, активность связывания другого паратопа, который связывается с эпитопом в цитотоксическом веществе, может зависеть от специфического для злокачественной ткани соединения, или активность связывания обоих паратопов может зависеть от специфического для злокачественной ткани соединения.
Такое низкомолекулярное антитело можно получать путем обработки антитела ферментом, таким как папаин или пепсин, для получения фрагментов антител, или путем конструирования ДНК, которые кодируют эти фрагменты антител или низкомолекулярные антитела, встраивания их в экспрессирующие векторы, а затем экспрессии их в соответствующих клетках-хозяевах (см., например, Co, M. S. et al., J. Immunol. (1994) 152, 2968-2976; Better, M. and Horwitz, A. H., Methods Enzymol. (1989) 178, 476-496; Pluckthun, A. and Skerra, A., Methods Enzymol. (1989) 178, 497-515; Lamoyi, E., Methods Enzymol. (1986) 121, 652-663; Rousseaux, J. et al., Methods Enzymol. (1986) 121, 663-669; и Bird, R. E. and Walker, B. W., Trends Biotechnol. (1991) 9, 132-137).
FcRn
В отличие от Fcγ-рецептора, принадлежащего семейству иммуноглобулинов, FcRn, в частности, FcRn человека, структурно сходен с полипептидами главного комплекса гистосовместимости (MHC) класса I, проявляя 22%-29% идентичность последовательности с молекулами MHC класса I (Ghetie el al., Immunol. Today (1997) 18 (12): 592-598). FcRn экспрессируется в качестве гетеродимера, состоящего из растворимой β- или легкой цепи (β2-микроглобулин) в комплексе с трансмембранной α- или тяжелой цепью. Подобно MHC, α-цепь FcRn содержит три внеклеточных домена (α1, α2 и α3) и ее короткий цитоплазматический домен заякоривает белок на клеточной поверхности. α1- и α2-домены взаимодействуют с FcRn-связывающим доменом Fc-области антитела (Raghavan et al., Immunity (1994) 1: 303-315).
FcRn экспрессируется в материнской плаценте и желточном мешке млекопитающих, и он вовлечен в перенос IgG от матери к плоду. Кроме того, в тонком кишечнике новорожденных грызунов, где FcRn экспрессируется, FcRn вовлечен в перенос материнских IgG через эпителий щеточной каемки из проглоченного молозива или молока. FcRn экспрессируется во множестве других тканей и системах эндотелиальных клеток различных видов. FcRn также экспрессируется у взрослого человека в эндотелии, кровеносных сосудах мышц и синусоидных капиллярах печени. Полагают, что FcRn участвует в поддержании концентрации IgG в плазме путем опосредования рециклирования IgG в сыворотку при связывании с IgG. Как правило, связывание FcRn с молекулами IgG строго зависит от pH. Оптимальное связывание наблюдают при кислом диапазоне pH ниже 7,0.
FcRn человека, предшественником которого является полипептид, имеющий сигнальную последовательность SEQ ID NO:28 (полипептид с сигнальной последовательностью представлен в SEQ ID NO:29), образует комплекс с β2-микроглобулином человека in vivo. Растворимый FcRn человека в комплексе с β2-микроглобулином получают с использованием общепринятых способов рекомбинантной экспрессии. Fc-области по настоящему изобретению можно оценивать в отношении их активности связывания с таким растворимым FcRn человека в комплексе с β2-микроглобулином. В рамках настоящего изобретения, если нет иных указаний, FcRn человека относится к форме, способной связываться с Fc-областью по настоящему изобретению. Примеры включают комплекс между FcRn человека и β2-микроглобулином человека.
Активность связывания Fc-области с FcRn, в частности, FcRn человека
Активность связывания Fc-области по настоящему изобретению с FcRn, в частности FcRn человека, можно измерять способами, известными специалистам в данной области, как описано в разделе "Активность связывания" выше. Специалисты в данной области могут соответствующим образом определить условия, отличные от pH. Антигенсвязывающую активность и активность связывания FcRn человека антигенсвязывающей молекулы можно оценивать, основываясь на константе диссоциации (KD), кажущейся константе диссоциации (KD), константе скорости диссоциации (kd), кажущейся константе скорости диссоциации (kd) и т.п. Их можно измерять способами, известными специалистам в данной области. Например, можно использовать Biacore (GE healthcare), график Скэтчарда или проточный цитометр.
Когда измеряют активность связывания FcRn человека Fc-области по настоящему изобретению, условия, отличные от pH, конкретно не ограничены, и их могут соответствующим образом выбирать специалисты в данной области. Измерения можно проводить, например, при 37°C с использованием буфера MES, как описано в международной публикации № WO 2009125825. Альтернативно активность связывания FcRn человека Fc-области по настоящему изобретению можно измерять способами, известными специалистам в данной области, и ее можно измерять с использованием, например, Biacore (GE Healthcare) и т.п. Активность Fc-области в отношении связывания по настоящему изобретению с FcRn человека можно оценивать путем нанесения в качестве анализируемого соединения FcRn человека, Fc-области или антигенсвязывающей молекулы по настоящему изобретению, содержащей Fc-область, на чип, на который иммобилизована Fc-область, антигенсвязывающая молекула по настоящему изобретению, содержащая Fc-область, или FcRn человека.
Диапазон нейтральных значений pH в качестве условий, где Fc-область, содержащаяся в антигенсвязывающей молекуле по настоящему изобретению, имеет активность связывания FcRn, как правило, означает от pH6,7 до pH10,0. Предпочтительно, диапазон нейтральных значений pH представляет собой диапазон, указанный с помощью произвольных величин pH от pH7,0 до pH8,0, и предпочтительно он выбран из pH7,0, 7,1, 7,2, 7,3, 7,4, 7,5, 7,6, 7,7, 7,8, 7,9 и 8,0, и особенно предпочтительно pH7,4, что близко к значению pH плазмы (крови) in vivo. Когда аффинность связывания между FcRn-связывающим доменом человека и FcRn человека при pH7,4 является слишком низкой, чтобы ее оценить, вместо pH7,4 можно использовать pH7,0. В рамках изобретения диапазон кислых значений pH в качестве условий, где Fc-область, содержащаяся в антигенсвязывающей молекуле по настоящему изобретению, имеет активность связывания FcRn, как правило, означает от pH4,0 до pH6,5. Предпочтительно, диапазон кислых значений pH означает от pH5,5 до pH6,5, особенно предпочтительно от pH5,8 до pH6,0, что близко к pH в ранней эндосоме in vivo. Что касается температуры, используемой в качестве условий измерения, аффинность связывания между доменом, связывающим FcRn человека, и FcRn человека можно оценивать при любой температуре от 10°C до 50°C. Предпочтительно, аффинность связывания между доменом, связывающим FcRn человека, и FcRn человека можно определять при от 15°C до 40°C. Более предпочтительно, аффинность связывания между доменом, связывающим FcRn человека, и FcRn человека, можно определять аналогичным образом при произвольной температуре от 20°C до 35°C, такой как любая температура из 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 и 35°C. В одном варианте осуществления настоящего изобретения температура включает, но не ограничивается ими, например, 25°C.
Согласно Journal of Immunology (2009) 182: 7663-7671, активность связывания нативного IgG1 человека с FcRn человека в диапазоне кислых значений pH (pH 6,0) составляет KD 1,7 мкМ, и эта активность практически не поддается выявлению в диапазоне нейтральных значений pH. Таким образом, в предпочтительном варианте осуществления можно проводить скрининг антигенсвязывающих молекул, содержащих Fc-область, у которой активность связывания FcRn человека в диапазоне кислых значений pH составляет 20 мкМ (KD) или сильнее. В более предпочтительном варианте осуществления можно проводить скрининг антигенсвязывающих молекул, содержащих Fc-область, у которой активность связывания FcRn человека в диапазоне кислых значений pH составляет 2,0 мкМ (KD) или сильнее. В еще более предпочтительном варианте осуществления можно проводить скрининг антигенсвязывающих молекул, содержащих Fc-область, у которой активность связывания FcRn человека в диапазоне кислых значений pH составляет 0,5 мкМ (KD) или сильнее. Упомянутые выше величины KD определяют способом, описанным в Journal of Immunology (2009) 182: 7663-7671 (посредством иммобилизации антигенсвязывающей молекулы на чип и нанесения FcRn человека в качестве анализируемого соединения).
Fc-область, обладающая активностью связывания FcRn в условиях диапазона кислых значений pH
Fc-область, обладающая активностью связывания FcRn в условиях диапазона кислых значений pH, также можно предпочтительно использовать в качестве Fc-области, содержащейся в антигенсвязывающей молекуле, предусматриваемой в рамках настоящего изобретения. Как правило, известно, что IgG-антитела обладают длительным временем удержания в плазме вследствие связывания с FcRn. Связывание между IgG и FcRn наблюдается только в кислых условиях (pH 6,0) и связывание практически не наблюдается в нейтральных условиях (pH 7,4). IgG-антитела неспецифически включаются в клетки, однако они возвращаются на клеточную поверхность посредством связывания с FcRn в эндосоме в кислых условиях эндосом, а затем диссоциируют от FcRn в нейтральных условиях в плазме. Когда в Fc-область IgG вносят мутации для устранения связывания FcRn в условиях диапазона кислых значений pH, антитела не рециклируют из эндосомы в плазму. Таким образом, время удержания антигена в плазме значительно снижается. Способ увеличения связывания FcRn в условиях диапазона кислых значений pH описан в качестве способа увеличения времени удержания IgG-антител в плазме. Увеличение связывания FcRn в условиях диапазона кислых значений pH путем внесения аминокислотных замен в Fc-область IgG-антитела может увеличить эффективность рециклирования из эндосомы в плазму и в результате, время удержания в плазме увеличивается.
Настоящее изобретение не ограничивается конкретной теорией, однако, например, когда антигенсвязывающая молекула, предусматриваемая в рамках настоящего изобретения, связывается с антигеном мембранного типа, экспрессируемым на злокачественных клетках, содержащихся в злокачественных тканях, может быть возможным непрерывное подавление пролиферации злокачественных клеток, как описано ниже. Даже после повреждения злокачественных клеток, экспрессирующих молекулу мембранного типа, с которой связалась антигенсвязывающая молекула по настоящему изобретению в присутствии высокой концентрации специфического для злокачественной ткани соединения, вследствие цитотоксической активности, опосредуемой антигенсвязывающей молекулой, антиген все еще может связываться с антигенсвязывающим доменом в антигенсвязывающей молекуле. Из антигенсвязывающих молекул, неспецифически включенных в клетки, антигенсвязывающие молекулы, которые высвобождают антиген в присутствии низкой концентрации специфического для злокачественной ткани соединения, возвращаются на клеточную поверхность посредством связывания с FcRn в эндосоме в кислых условиях внутри эндосомы, а затем диссоциируют от FcRn в нейтральных условиях в плазме. В присутствии высокой концентрации специфического для злокачественной ткани соединения антигенсвязывающие молекулы по настоящему изобретению, рециклировавшие таким образом, могут вновь связываться с их антигенами, которые представляют собой молекулы мембранного типа, экспрессируемые на злокачественных клетках.
Настоящее изобретение не ограничивается конкретной теорией, однако, например, когда растворимый антиген, связываемый антигенсвязывающими молекулами, предусматриваемыми в рамках настоящего изобретения, представляет собой лиганд, который положительно регулирует активацию воспалительных клеток или пролиферацию клеток-мишеней, содержащихся в ткани-мишени, может быть возможным подавление пролиферации клеток-мишеней или активация воспалительных клеток, как описано ниже. Антигенсвязывающие молекулы по настоящему изобретению, связываемые с растворимой молекулой, т.е. ее антигеном, неспецифически включаются в клетки в присутствии высокой концентрации специфических для ткани-мишени соединений. За этим следует высвобождение антигена в присутствии низкой концентрации специфических для ткани-мишени соединений, антигенсвязывающие молекулы возвращаются на клеточную поверхность посредством связывания с FcRn в эндососме в кислых условиях внутри эндосомы, а затем антигенсвязывающие молекулы диссоциируют от FcRn в нейтральных условиях в плазме. Антигенсвязывающие молекулы по настоящему изобретению, рециклировавшие таким образом, могут вновь связываться с растворимыми молекулами, т.е. их антигеном, в присутствии высокой концентрации специфических для ткани-мишени соединений. С другой стороны, антигены, которые диссоциировали от антигенсвязывающих молекул в присутствии низкой концентрации специфических для ткани-мишени соединений, деградируют в лизосоме. Концентрация растворимого антигена снижается по мере того, как он проходит через стадию рециклирования. Таким образом, считается, что пролиферация злокачественных клеток или активация воспалительных клеток могут подавляться.
В рамках настоящего изобретения предпочтительные Fc-области имеют активность связывания FcRn в условиях диапазона кислых значений pH. Когда Fc-область изначально обладает активностью связывания FcRn в условиях диапазона кислых значений pH, этот домен можно использовать как есть. Когда домен обладает слабой активностью связывания FcRn в условиях диапазона кислых значений pH или не обладает ей, Fc-область, обладающую желаемой активностью связывания FcRn, можно получать путем изменения аминокислот антигенсвязывающей молекулы. Fc-области, имеющие желаемую или усиленную активность связывания FcRn в условиях диапазона кислых значений pH, также можно пригодным образом получать путем изменения аминокислот Fc-области. Аминокислотные модификации Fc-области, которые приводят к такой желаемой активности связывания, можно выявлять путем сравнения активности связывания FcRn в условиях диапазона кислых значений pH до и после изменения аминокислот. Специалисты в данной области могут соответствующим образом изменять аминокислоты с использованием известных способов, сходных с упомянутыми выше способами, используемыми для изменения активности связывания Fcγ-рецептора.
Fc-области, содержащиеся в антигенсвязывающих молекулах по настоящему изобретению, которые обладают активностью связывания FcRn в условиях диапазона кислых значений pH, можно получать любым способом. В частности, FcRn-связывающие домены, обладающие активностью связывания FcRn или усиленной активностью связывания FcRn в условиях диапазона кислых значений pH, можно получать путем изменения аминокислот иммуноглобулина человека IgG-типа, используемого в качестве исходной Fc-области. Предпочтительные Fc-области иммуноглобулина IgG-типа для изменения включают, например, Fc-области IgG человека (IgG1, IgG2, IgG3 и IgG4, и их варианты). При условии, что Fc-область обладает активностью связывания FcRn в условиях диапазона кислых значений pH или может увеличивать активность связывания FcRn человека в условиях диапазона кислых значений pH, аминокислоты в любом положении можно изменять на другие аминокислоты. Когда антигенсвязывающая молекула содержит Fc-область IgG1 человека в качестве Fc-области, предпочтительно, чтобы полученная Fc-область содержала изменение, которое приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека. Аминокислоты, которые позволяют такое изменение, включают, например, аминокислоты в положениях 252, 254, 256, 309, 311, 315, 433 и/или 434 в соответствии с нумерацией EU и связанные с ними аминокислоты в положениях 253, 310, 435 и/или 426, как описано в WO 1997/034631. Подходящие примеры включают аминокислоты в положениях 238, 252, 253, 254, 255, 256, 265, 272, 286, 288, 303, 305, 307, 309, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 386, 388, 400, 413, 415, 424, 433, 434, 435, 436, 439 и/или 447, как указано в соответствии с нумерацией EU, как описано в WO 2000/042072. Аналогично, подходящие примеры аминокислот, которые позволяют такое изменение, включают аминокислоты в положениях 251, 252, 254, 255, 256, 308, 309, 311, 312, 385, 386, 387, 389, 428, 433, 434 и/или 436 в соответствии с нумерацией EU, как описано в WO 2002/060919. Более того, аминокислоты, которые позволяют такое изменение, включают, например, аминокислоты в положениях 250, 314 и 428 в соответствии с нумерацией EU, как описано в WO2004/092219. Кроме того, подходящие примеры аминокислот, которые позволяют такое изменение, включают аминокислоты в положениях 238, 244, 245, 249, 252, 256, 257, 258, 260, 262, 270, 272, 279, 283, 285, 286, 288, 293, 307, 311, 312, 316, 317, 318, 332, 339, 341, 343, 375, 376, 377, 378, 380, 382, 423, 427, 430, 431, 434, 436, 438, 440 и/или 442, как описано в WO 2006/020114. Более того, подходящие примеры аминокислот, которые позволяют такое изменение, включают аминокислоты в положениях 251, 252, 307, 308, 378, 428, 430, 434 и/или 436 в соответствии с нумерацией EU, как описано в WO 2010/045193. Изменение этих аминокислот усиливает связывание Fc-области иммуноглобулин IgG-типа с FcRn в условиях диапазона кислых значений pH.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления изменения, которое приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, включает по меньшей мере одно или несколько аминокислотных изменений, выбранных из группы, состоящей из:
Arg или Leu в качестве аминокислоты в положении 251;
Phe, Ser, Thr или Tyr в качестве аминокислоты в положении 252;
Ser или Thr в качестве аминокислоты в положении 254;
Arg, Gly, Ile или Leu в качестве аминокислоты в положении 255;
Ala, Arg, Asn, Asp, Gln, Glu или Thr в качестве аминокислоты в положении 256;
Ile или Thr в качестве аминокислоты в положении 308;
Pro в качестве аминокислоты в положении 309;
Glu, Leu или Ser в качестве аминокислоты в положении 311;
Ala или Asp в качестве аминокислоты в положении 312;
Ala или Leu в качестве аминокислоты в положении 314;
Ala, Arg, Asp, Gly, His, Lys, Ser или Thr в качестве аминокислоты в положении 385;
Arg, Asp, Ile, Lys, Met, Pro, Ser или Thr в качестве аминокислоты в положении 386;
Ala, Arg, His, Pro, Ser или Thr в качестве аминокислоты в положении 387;
Asn, Pro или Ser в качестве аминокислоты в положении 389;
Leu, Met, Phe, Ser или Thr в качестве аминокислоты в положении 428;
Arg, Gln, His, Ile, Lys, Pro или Ser в качестве аминокислоты в положении 433;
His, Phe или Tyr в качестве аминокислоты в положении 434; и
Arg, Asn, His, Lys, Met или Thr в качестве аминокислоты в положении 436, как указано в соответствии с нумерацией EU. Между тем, количество аминокислот, подлежащих изменению, конкретно не ограничено; и аминокислоту можно изменять только в одном участке или можно изменять аминокислоты в двух или более участках.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающим вариантом осуществления изменения, которое приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, могут быть изменения, включающие Ile в качестве аминокислоты в положении 308, Pro в качестве аминокислоты в положении 309 и/или Glu в качестве аминокислоты в положении 311 в соответствии с нумерацией EU. Другой неограничивающий вариант осуществления этого изменения может включать Thr в качестве аминокислоты в положении 308, Pro в качестве аминокислоты в положении 309, Leu в качестве аминокислоты в положении 311, Ala в качестве аминокислоты в положении 312, и/или Ala в качестве аминокислоты в положении 314. Более того, другой неограничивающий вариант осуществления этого изменения может включать Ile или Thr в качестве аминокислоты в положении 308, Pro в качестве аминокислоты в положении 309, Glu, Leu или Ser в качестве аминокислоты в положении 311, Ala в качестве аминокислоты в положении 312, и/или Ala или Leu в качестве аминокислоты в положении 314. Другой неограничивающий вариант осуществления этого изменения может включать Thr в качестве аминокислоты в положении 308, Pro в качестве аминокислоты в положении 309, Ser в качестве аминокислоты в положении 311, Asp в качестве аминокислоты в положении 312, и/или Leu в качестве аминокислоты в положении 314.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления изменения, которое приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, может представлять собой изменения, включающие Leu в качестве аминокислоты в положении 251, Tyr в качестве аминокислоты в положении 252, Ser или Thr в качестве аминокислоты в положении 254, Arg в качестве аминокислоты в положении 255, и/или Glu в качестве аминокислоты в положении 256 в соответствии с нумерацией EU.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления изменения, которое приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, может представлять собой изменения, включающие Leu, Met, Phe, Ser или Thr в качестве аминокислоты в положении 428, Arg, Gln, His, Ile, Lys, Pro или Ser в качестве аминокислоты в положении 433, His, Phe или Tyr в качестве аминокислоты в положении 434, и/или Arg, Asn, His, Lys, Met или Thr в качестве аминокислоты в положении 436 в соответствии с нумерацией EU. Другой неограничивающий вариант осуществления этого изменения может включать His или Met в качестве аминокислоты в положении 428, и/или His или Met в качестве аминокислоты в положении 434.
Когда в качестве Fc-области присутствует Fc-область IgG1
человека, неограничивающий вариант осуществления изменения, которое приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, может представлять собой изменения, включающие Arg в качестве аминокислоты в положении 385, Thr в качестве аминокислоты в положении 386, Arg в качестве аминокислоты в положении 387, и/или Pro в качестве аминокислоты в положении 389 в соответствии с нумерацией EU. Другой неограничивающий вариант осуществления этого изменения может включать Asp в качестве аминокислоты в положении 385, Pro в качестве аминокислоты в положении 386, и/или Ser в качестве аминокислоты в положении 389.
Более того, когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления изменения, которое приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, включает по меньшей мере одно или несколько аминокислотных изменений, выбранных из группы, состоящей из: Gln или Glu в качестве аминокислоты в положении 250; и Leu или Phe в качестве аминокислоты в положении 428 в соответствии с нумерацией EU. Количество аминокислоты, подлежащих изменению, конкретно не ограничено; и аминокислоту можно изменять только в одном участке или аминокислоты можно изменять в двух участках.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления изменения, которое приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, может представлять собой изменения, включающие Gln в качестве аминокислоты в положении 250, и/или Leu или Phe в качестве аминокислоты в положении 428 в соответствии с нумерацией EU. Другой неограничивающий вариант осуществления этого изменения может включать Glu в качестве аминокислоты в положении 250, и/или Leu или Phe в качестве аминокислоты в положении 428.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления изменения, которое приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека включает по меньшей мере два или более изменений аминокислот, выбранных из группы, состоящей из:
Asp или Glu в качестве аминокислоты в положении 251;
Tyr в качестве аминокислоты в положении 252;
Gln в качестве аминокислоты в положении 307;
Pro в качестве аминокислоты в положении 308;
Val в качестве аминокислоты в положении 378;
Ala в качестве аминокислоты в положении 380;
Leu в качестве аминокислоты в положении 428;
Ala или Lys в качестве аминокислоты в положении 430;
Ala, His, Ser или Tyr в качестве аминокислоты в положении 434; и
Ile в качестве аминокислоты в положении 436, как указано в соответствии с нумерацией EU. Количество аминокислоты, подлежащих изменению, конкретно не ограничено; и аминокислоту можно изменять только в двух участках или аминокислоты можно изменять в трех или более участках.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления модификации, которая приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, может представлять собой изменения, включающие Gln в качестве аминокислоты в положении 307, и Ala или Ser в качестве аминокислоты в положении 434 в соответствии с нумерацией EU. Другой неограничивающий вариант осуществления этого изменения может включать Pro в качестве аминокислоты в положении 308, и Ala в качестве аминокислоты в положении 434. Более того, другой неограничивающий вариант осуществления этого изменения может включать Tyr в качестве аминокислоты в положении 252, и Ala в качестве аминокислоты в положении 434. Другой отличающийся неограничивающий вариант осуществления этого изменения может включать Val в качестве аминокислоты в положении 378, и Ala в качестве аминокислоты в положении 434. Другой отличающийся неограничивающий вариант осуществления этого изменения может включать Leu в качестве аминокислоты в положении 428, и Ala в качестве аминокислоты в положении 434. Другой отличающийся неограничивающий вариант осуществления этого изменения может включать Ala в качестве аминокислоты в положении 434, и Ile в качестве аминокислоты в положении 436. Более того, другой неограничивающий вариант осуществления этого изменения может включать Pro в качестве аминокислоты в положении 308, и Tyr в качестве аминокислоты в положении 434. Кроме того, другой неограничивающий вариант осуществления этого изменения может включать Gln в качестве аминокислоты в положении 307, и Ile в качестве аминокислоты в положении 436.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления изменения, которое приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, может представлять собой изменения, включающие любую из Gln в качестве аминокислоты в положении 307, Ala в качестве аминокислоты в положении 380, и Ser в качестве аминокислоты в положении 434 в соответствии с нумерацией EU. Другой неограничивающий вариант осуществления этого изменения может включать Gln в качестве аминокислоты в положении 307, Ala в качестве аминокислоты в положении 380, и Ala в качестве аминокислоты в положении 434. Более того, другой неограничивающий вариант осуществления этого изменения может включать Tyr в качестве аминокислоты в положении 252, Pro в качестве аминокислоты в положении 308, и Tyr в качестве аминокислоты в положении 434. Отличающийся неограничивающий вариант осуществления этого изменения может включать Asp в качестве аминокислоты в положении 251, Gln в качестве аминокислоты в положении 307, и His в качестве аминокислоты в положении 434.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления изменения, которое приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, включает изменение по меньшей мере двух или более аминокислот, выбранных из группы, состоящей из:
Leu в качестве аминокислоты в положении 238;
Leu в качестве аминокислоты в положении 244;
Arg в качестве аминокислоты в положении 245;
Pro в качестве аминокислоты в положении 249;
Tyr в качестве аминокислоты в положении 252;
Pro в качестве аминокислоты в положении 256;
Ala, Ile, Met, Asn, Ser или Val в качестве аминокислоты в положении 257;
Asp в качестве аминокислоты в положении 258;
Ser в качестве аминокислоты в положении 260;
Leu в качестве аминокислоты в положении 262;
Lys в качестве аминокислоты в положении 270;
Leu или Arg в качестве аминокислоты в положении 272;
Ala, Asp, Gly, His, Met, Asn, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 279;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Arg, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 283;
Asn в качестве аминокислоты в положении 285;
Phe в качестве аминокислоты в положении 286;
Asn или Pro в качестве аминокислоты в положении 288;
Val в качестве аминокислоты в положении 293;
Ala, Glu или Met в качестве аминокислоты в положении 307;
Ala, Ile, Lys, Leu, Met, Val или Trp в качестве аминокислоты в положении 311;
Pro в качестве аминокислоты в положении 312;
Lys в качестве аминокислоты в положении 316;
Pro в качестве аминокислоты в положении 317;
Asn или Thr в качестве аминокислоты в положении 318;
Phe, His, Lys, Leu, Met, Arg, Ser или Trp в качестве аминокислоты в положении 332;
Asn, Thr или Trp в качестве аминокислоты в положении 339;
Pro в качестве аминокислоты в положении 341;
Glu, His, Lys, Gln, Arg, Thr или Tyr в качестве аминокислоты в положении 343;
Arg в качестве аминокислоты в положении 375;
Gly, Ile, Met, Pro, Thr или Val в качестве аминокислоты в положении 376;
Lys в качестве аминокислоты в положении 377;
Asp или Asn в качестве аминокислоты в положении 378;
Asn, Ser или Thr в качестве аминокислоты в положении 380;
Phe, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr в качестве аминокислоты в положении 382;
Asn в качестве аминокислоты в положении 423;
Asn в качестве аминокислоты в положении 427;
Ala, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val или Tyr в качестве аминокислоты в положении 430;
His или Asn в качестве аминокислоты в положении 431;
Phe, Gly, His, Trp или Tyr в качестве аминокислоты в положении 434;
Ile, Leu или Thr в качестве аминокислоты в положении 436;
Lys, Leu, Thr или Trp в качестве аминокислоты в положении 438;
Lys в качестве аминокислоты в положении 440; и
Lys в качестве аминокислоты в положении 442 в соответствии с нумерацией EU. Количество аминокислоты, подлежащих изменению, конкретно не ограничено и могут быть изменены аминокислоты только в двух участках и могут быть изменены аминокислоты в трех или более участках.
Когда в качестве Fc-области присутствует Fc-область IgG1 человека, неограничивающий вариант осуществления изменения, которое приводит к эффекту усиления связывания FcRn в условиях диапазона кислых значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека, может представлять собой изменения, включающие Ile в качестве аминокислоты в положении 257, и Ile в качестве аминокислоты в положении 311 в соответствии с нумерацией EU. Другой неограничивающий вариант осуществления этого изменения может включать Ile в качестве аминокислоты в положении 257, и His в качестве аминокислоты в положении 434. Другой неограничивающий вариант осуществления этого изменения может включать Val в качестве аминокислоты в положении 376, и His в качестве аминокислоты в положении 434.
Fc-области, обладающие активностью связывания FcRn в условиях диапазона нейтральных значений pH
Более того, в другом неограничивающем варианте осуществления можно проводить скрининг антигенсвязывающих молекул, содержащих Fc-область с характеристикой наличия активности связывания FcRn человека в диапазоне нейтральных значений pH вместо упомянутой выше характеристики наличия активности связывания FcRn человека в диапазоне кислых значений pH. В предпочтительном варианте осуществления можно проводить скрининг антигенсвязывающих молекул, содержащих Fc-область, у которой активность связывания FcRn человека в диапазоне нейтральных значений pH составляет 40 мкМ (KD) или сильнее. В более предпочтительном варианте осуществления можно проводить скрининг антигенсвязывающих молекул, содержащих Fc-область, у которой активность связывания FcRn человека в диапазоне нейтральных значений pH равна 15 мкМ (KD) или сильнее.
Более того, в другом неограничивающем варианте осуществления можно проводить скрининг антигенсвязывающих молекул, содержащих Fc-область с характеристикой наличия активности связывания FcRn человека в диапазоне нейтральных значений pH вместо упомянутой выше характеристики наличия активности связывания FcRn человека в диапазоне кислых значений pH. В предпочтительном варианте осуществления можно проводить скрининг антигенсвязывающих молекул, содержащих Fc-область, у которой активность связывания FcRn человека в диапазоне нейтральных значений pH составляет 40 мкМ (KD) или сильнее. В более предпочтительном варианте осуществления можно проводить скрининг антигенсвязывающих молекул, содержащих Fc-область, у которой активность связывания FcRn человека в диапазоне нейтральных значений pH равна 15 мкМ (KD) или сильнее.
В рамках настоящего изобретения предпочтительные Fc-области имеют активность связывания FcRn человека в диапазоне кислых значений pH и/или в диапазоне нейтральных значений pH. Когда Fc-область первоначально обладает активностью связывания FcRn человека в диапазоне кислых значений pH и/или в диапазоне нейтральных значений pH, ее можно использовать как есть. Когда Fc-область имеет слабую активность связывания FcRn человека или не имеет активности связывания FcRn человека в диапазоне кислых значений pH и/или в диапазоне нейтральных значений pH, антигенсвязывающие молекулы, содержащие Fc-область, имеющую желаемую активность связывания FcRn человека, можно получать путем изменения аминокислот Fc-области, содержащейся в антигенсвязывающих молекулах. Fc-области, имеющие желаемую активность связывания FcRn человека в диапазоне кислых значений pH и/или в диапазоне нейтральных значений pH, также можно пригодным образом получать путем изменения аминокислот Fc-области человека. Альтернативно антигенсвязывающие молекулы, содержащие Fc-область, имеющую желаемую активность связывания FcRn человека, можно получать путем изменения аминокислот Fc-области, которые первоначально обладают активностью связывания FcRn человека в диапазоне кислых значений pH и/или в диапазоне нейтральных значений pH. Аминокислотные изменения Fc-области человека, которые приводят к такой желаемой активности связывания, могут быть выявлены путем сравнения активности связывания FcRn человека в диапазоне кислых значений pH и/или в диапазоне нейтральных значений pH до и после изменения аминокислот. Специалисты в данной области могут соответствующим образом изменить аминокислоты с использованием известных способов.
В рамках настоящего изобретения "изменение аминокислот" или "аминокислотное изменение" Fc-области включает изменение аминокислотной последовательности, которое отличается от исходной Fc-области. Исходная Fc-область может представлять собой любую Fc-область при условии, что вариант, модифицированный из исходной Fc-области, может связываться с FcRn человека в диапазоне кислых значений pH (т.е. исходная Fc-область не обязательно должна обладать активностью связывания с FcRn человека в диапазоне нейтральных значений pH). Примеры исходных Fc-областей предпочтительно включают Fc-области IgG-антител, т.е. нативные Fc-области. Более того, также в качестве измененной Fc-области по настоящему изобретению предпочтительно можно использовать Fc-область, модифицированную из исходной Fc-области, которая уже модифицирована. "Исходная Fc-область" может относиться к самому полипептиду, композиции, содержащей исходную Fc-область или полинуклеотидной последовательности, кодирующей исходную Fc-область. Исходные Fc-области могут содержать Fc-область известного IgG-антитела, полученную путем рекомбинации, кратко описанной в разделе "Антитела". Источник исходных Fc-областей не ограничен, и их можно получать из человека или любых не являющихся человеком организмов. Такие организмы предпочтительно включают мышей, крыс, морских свинок, хомячков, песчанок, кошек, кроликов, собак, коз, овец, животных семейства бычьи, лошадей, верблюдов и организмов, выбранных из не являющихся человеком приматов. В другом варианте осуществления исходные Fc-области также можно получать из яванских макаков, мартышек, макаков-резус, шимпанзе или людей. Исходные Fc-области можно предпочтительно получать из IgG1 человека; однако они не ограничиваются каким-либо конкретным классом IgG. Это означает, Fc-область, соответствующую IgG1 (SEQ ID NO:5), IgG2 (SEQ ID NO:6), IgG3 (SEQ ID NO:7) или IgG4 (SEQ ID NO:8) человека можно соответствующим образом использовать Fc-область IgG1, IgG2, IgG3 или IgG4 человека, и в рамках настоящего изобретения это также означает, что предпочтительно в качестве исходной Fc-области можно использовать Fc-область произвольного класса или подкласса IgG, происходящую из любых организмов, описанных выше. Примеры встречающихся в природе вариантов или измененных форм IgG описаны в опубликованных документах (Curr. Opin. Biotechnol. (2009) 20 (6): 685-91; Curr. Opin. Immunol. (2008) 20 (4), 460-470; Protein Eng. Des. Sel. (2010) 23 (4): 195-202; международных публикациях № WO 2009/086320; WO 2008/092117; WO 2007/041635 и WO 2006/105338); однако они не ограничиваются этими примерами.
Примеры изменений включают изменения с одной или несколькими мутациями, например, мутациями путем замены другими аминокислотными остатками аминокислот исходных Fc-областей, путем встраивания одного или нескольких аминокислотных остатков в аминокислоты исходных Fc-областей, или путем делеции одной или нескольких аминокислот из аминокислот исходных Fc-областей. Предпочтительно, аминокислотные последовательности измененных Fc-областей включают по меньшей мере часть аминокислотной последовательности ненативной Fc-области. Такие варианты должны обладать идентичностью или сходством последовательности с их исходной Fc-областью, меньшими чем 100%. В предпочтительном варианте осуществления варианты обладают идентичностью или сходством аминокислотной последовательности приблизительно от 75% до менее чем 100%, более предпочтительно приблизительно от 80% до менее чем 100%, еще более предпочтительно приблизительно от 85% до менее чем 100%, еще более предпочтительно, приблизительно от 90% до менее чем 100%, и еще более предпочтительно приблизительно от 95% до менее чем 100% с аминокислотной последовательностью их исходной Fc-области. В неограничивающем варианте осуществления настоящего изобретения по меньшей мере одна аминокислота отличается между измененной Fc-областью и ее исходной Fc-областью. Аминокислотное отличие между модифицированной Fc-областью по настоящему изобретению и ее исходной Fc-областью также может быть предпочтительно указано на основе аминокислотных отличий в описанных выше конкретных положениях аминокислотных остатков, как указано с помощью нумерации EU. Способы получения таких вариантов проиллюстрированы в разделе "Изменения аминокислот".
Fc-области, содержащиеся в антигенсвязывающих молекулах по настоящему изобретению, которые обладают активностью связывания FcRn человека в диапазоне нейтральныйх значений pH, можно получать любым способом. В частности, можно проводить скрининг антигенсвязывающих молекул, содержащих Fc-область, у которой активность связывания FcRn человека в диапазоне нейтральных значений pH составляет 20 мкМ (KD) или сильнее; в более предпочтительном варианте осуществления Fc-область, у которой активность связывания FcRn человека в диапазоне нейтральных значений pH составляет 2,0 мкМ (KD) или сильнее; и в еще более предпочтительном варианте осуществления Fc-область, у которой активность связывания FcRn человека в диапазоне нейтральных значений pH составляет 0,5 мкМ (KD) или сильнее в результате изменения аминокислот иммуноглобулина IgG-типа человека, используемого в качестве исходной Fc-области. Предпочтительные Fc-области иммуноглобулинов IgG-типов для изменения включают, например, Fc-области IgG человека, таких как IgG1, IgG2, IgG3 и IgG4, представленные в SEQ ID NO:5, 6, 7 и 8, соответственно, и их варианты.
Когда антигенсвязывающая молекула содержит Fc-область IgG1 человека в качестве Fc-области, пригодные примеры аминокислот, которые можно изменять для достижения упомянутых выше желаемых эффектов в отношении связывания FcRn в условиях диапазона нейтральных значений pH путем изменения аминокислот иммуноглобулина человека IgG-типа в качестве исходной Fc-области, включают аминокислоты в положениях 238, 252, 253, 254, 255, 256, 265, 272, 286, 288, 303, 305, 307, 309, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 386, 388, 400, 413, 415, 424, 433, 434, 435, 436, 439 и/или 447 в соответствии с нумерацией EU, как описано в WO 2000/042072. Аналогично, подходящие примеры аминокислот, которые позволяют такое изменение, включают аминокислоты в положениях 251, 252, 254, 255, 256, 308, 309, 311, 312, 385, 386, 387, 389, 428, 433, 434 и/или 436 в соответствии с нумерацией EU, как описано в WO 2002/060919. Более того, аминокислоты, которые позволяют такое изменение, включают, например, аминокислоты в положениях 250, 314 и 428 в соответствии с нумерацией EU, как описано в WO2004/092219. Более того, подходящие примеры аминокислот, которые позволяют такое изменение, включают аминокислоты в положениях 251, 252, 307, 308, 378, 428, 430, 434 и/или 436 в соответствии с нумерацией EU, как описано в WO 2010/045193. Изменение этих аминокислот усиливает связывание Fc-области иммуноглобулина человека IgG-типа с FcRn в условиях диапазона нейтральных значений pH.
Fc-области, обладающие активностью связывания FcRn человека в диапазоне нейтральных значений pH, также можно получать путем изменения аминокислот иммуноглобулина человека IgG-типа, используемого в качестве исходной Fc-области. Fc-области иммуноглобулинов IgG-типа, подходящие для изменения, включают, например, Fc-области IgG человека (такого как IgG1, IgG2, IgG3 и IgG4, соответственно, соответствующие SEQ ID NO:5, 6, 7 и 8, их измененные формы). Аминокислоты в любых положениях можно изменять на другие аминокислоты, при условии, что полученные области обладают активностью связывания FcRn человека в диапазоне нейтральных значений pH или увеличенной активностью связывания FcRn человека в нейтральном диапазоне. Когда антигенсвязывающая молекула содержит Fc-область IgG1 человека в качестве Fc-области человека, предпочтительно, чтобы полученная область содержала изменение, которое приводит к эффекту усиления связывания FcRn человека в диапазоне нейтральных значений pH по сравнению с активностью связывания исходной Fc-области IgG1 человека. Аминокислоты, которые допускают такие изменения, включают, например, аминокислоты в следующих положениях: 221-225, 227, 228, 230, 232, 233-241, 243-252, 254-260, 262-272, 274, 276, 278-289, 291-312, 315-320, 324, 325, 327-339, 341, 343, 345, 360, 362, 370, 375-378, 380, 382, 385-387, 389, 396, 414, 416, 423, 424, 426-438, 440 и 442 (указанные согласно нумерации EU). Изменение этих аминокислот усиливает связывание с FcRn человека Fc-области иммуноглобулина IgG-типа в диапазоне нейтральных значений pH.
Среди описанных выше, для применения в рамках настоящего изобретения выбирают подходящие изменения, которые усиливают связывание FcRn человека в диапазоне нейтральных значений pH. Особенно предпочтительные аминокислоты для измененных Fc-областей включают, например, аминокислоты в положениях 237, 248, 250, 252, 254, 255, 256, 257, 258, 265, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434 и 436 (указанные согласно нумерации EU). Активность связывания FcRn человека Fc-области, включенной в антигенсвязывающую молекулу, можно увеличивать в диапазоне нейтральных значений pH путем замены по меньшей мере одной аминокислоты другой аминокислотой.
Особенно предпочтительные изменения в Fc-области включают, например, замены:
Met вместо аминокислоты в положении 237;
Ile вместо аминокислоты в положении 248;
Ala, Phe, Ile, Met, Gln, Ser, Val, Trp или Tyr вместо аминокислоты в положении 250;
Phe, Trp или Tyr вместо аминокислоты в положении 252;
Thr вместо аминокислоты в положении 254;
Glu вместо аминокислоты в положении 255;
Asp, Asn, Glu или Gln вместо аминокислоты в положении 256;
Ala, Gly, Ile, Leu, Met, Asn, Ser, Thr или Val вместо аминокислоты в положении 257;
His вместо аминокислоты в положении 258;
Ala вместо аминокислоты в положении 265;
Ala или Glu вместо аминокислоты в положении 286;
His вместо аминокислоты в положении 289;
Ala вместо аминокислоты в положении 297;
Ala вместо аминокислоты в положении 303;
Ala вместо аминокислоты в положении 305;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr вместо аминокислоты в положении 307;
Ala, Phe, Ile, Leu, Met, Pro, Gln или Thr вместо аминокислоты в положении 308;
Ala, Asp, Glu, Pro или Arg вместо аминокислоты в положении 309;
Ala, His или Ile вместо аминокислоты в положении 311;
Ala или His вместо аминокислоты в положении 312;
Lys или Arg вместо аминокислоты в положении 314;
Ala, Asp или His вместо аминокислоты в положении 315;
Ala вместо аминокислоты в положении 317;
Val вместо аминокислоты в положении 332;
Leu вместо аминокислоты в положении 334;
His вместо аминокислоты в положении 360;
Ala вместо аминокислоты в положении 376;
Ala вместо аминокислоты в положении 380;
Ala вместо аминокислоты в положении 382;
Ala вместо аминокислоты в положении 384;
Asp или His вместо аминокислоты в положении 385;
Pro вместо аминокислоты а в положении 386;
Glu for вместо аминокислоты в положении 387;
Ala или Ser вместо аминокислоты в положении 389;
Ala вместо аминокислоты в положении 424;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 428;
Lys вместо аминокислоты в положении 433;
Ala, Phe, His, Ser, Trp или Tyr вместо аминокислоты в положении 434; и
His, Ile, Leu, Phe, Thr или Val вместо аминокислоты в положении 436 в системе нумерации EU. Между тем количество измененных аминокислот конкретно не ограничено; такие аминокислотные изменения включают единичное изменение аминокислоты и изменение аминокислот в двух или более участках. Эти комбинации изменений аминокислот включают, например, комбинации, описанные в таблицах с 2-1 по 2-33.


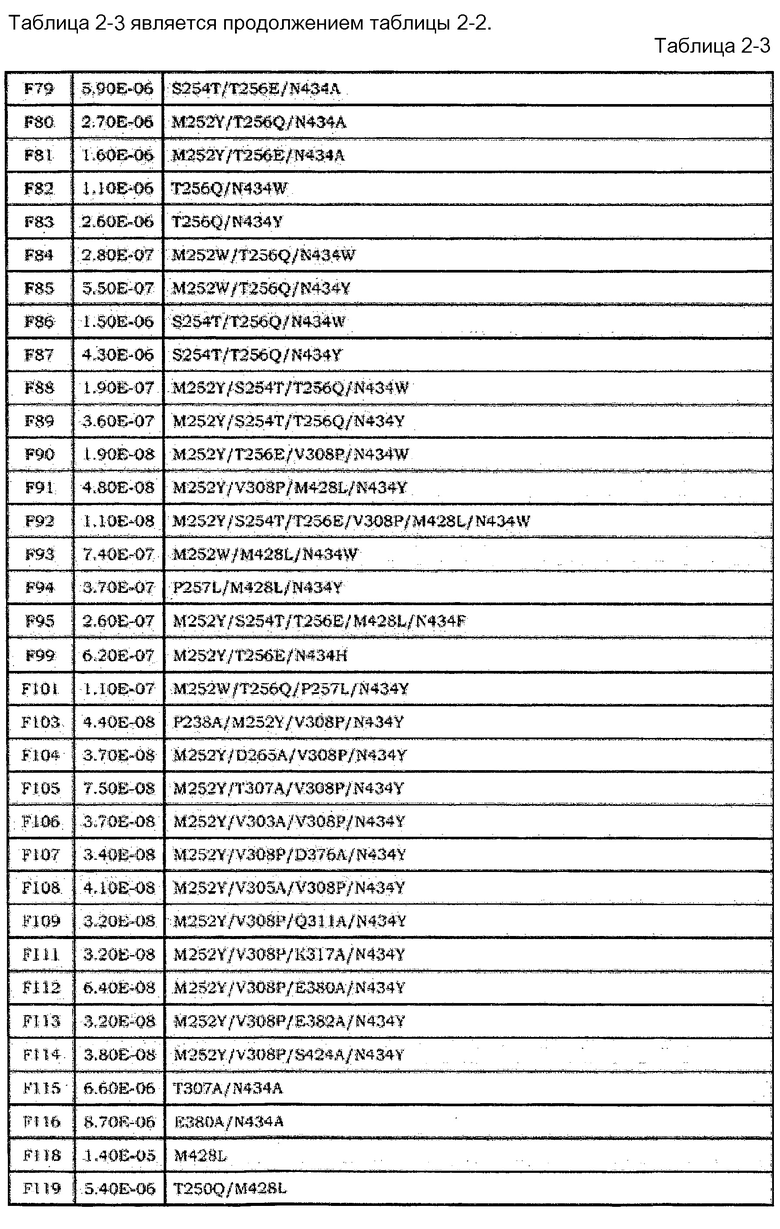

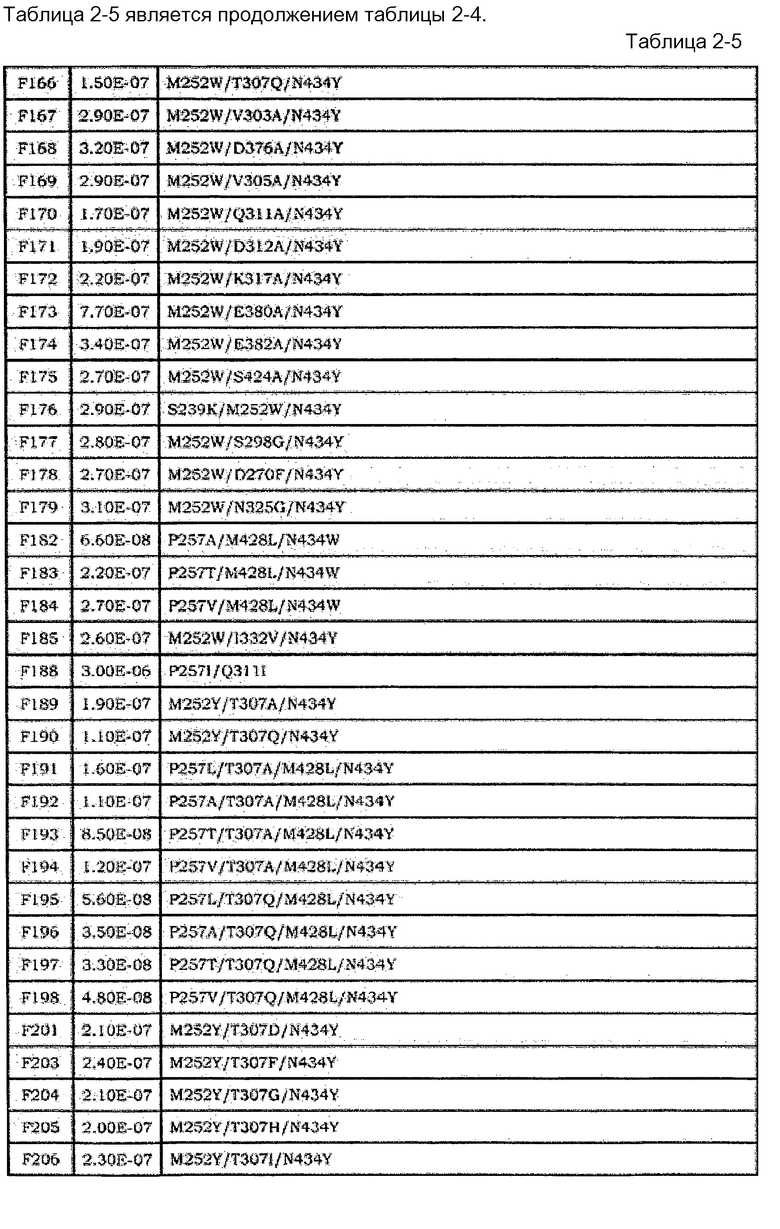










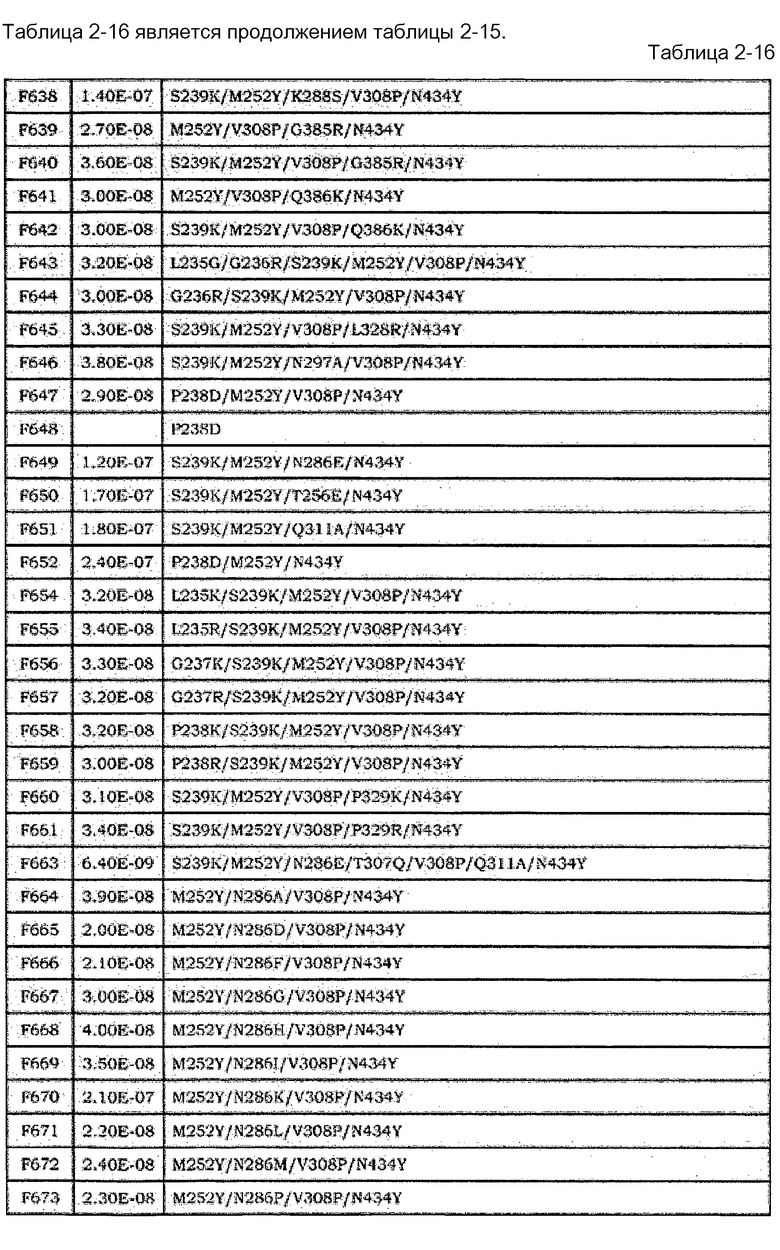


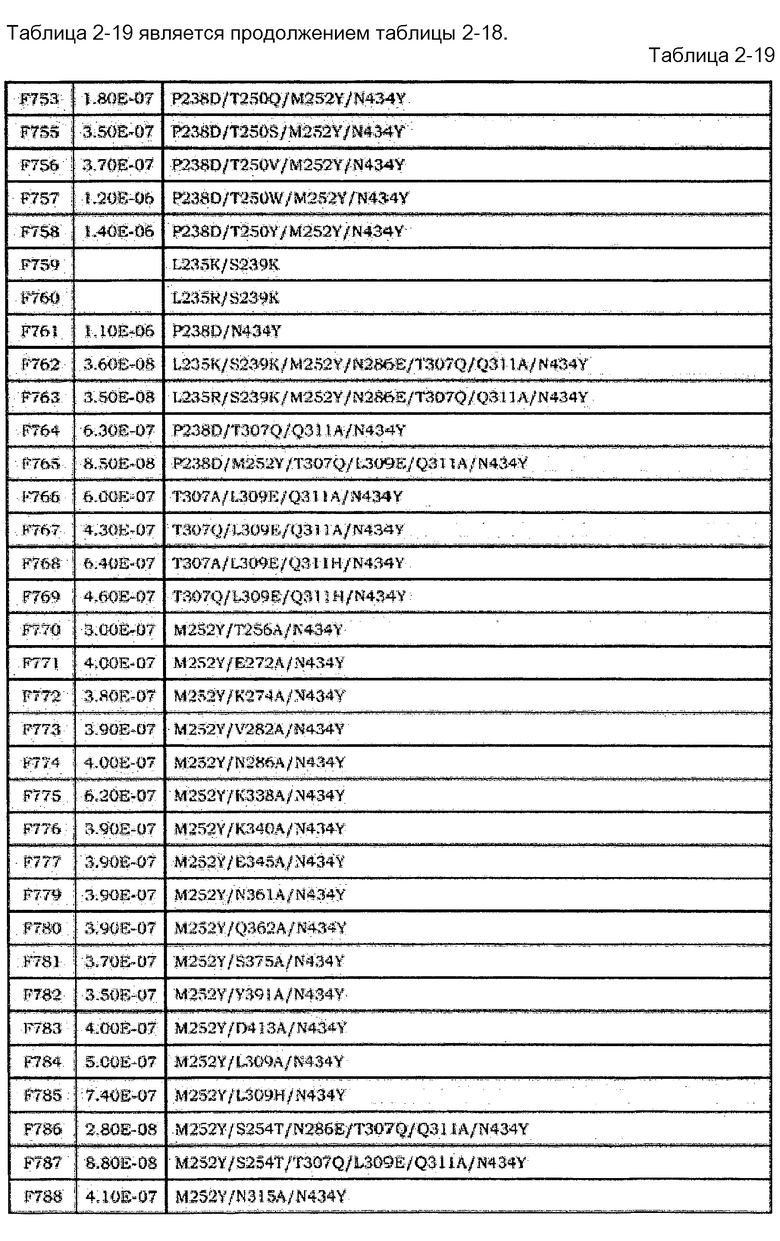
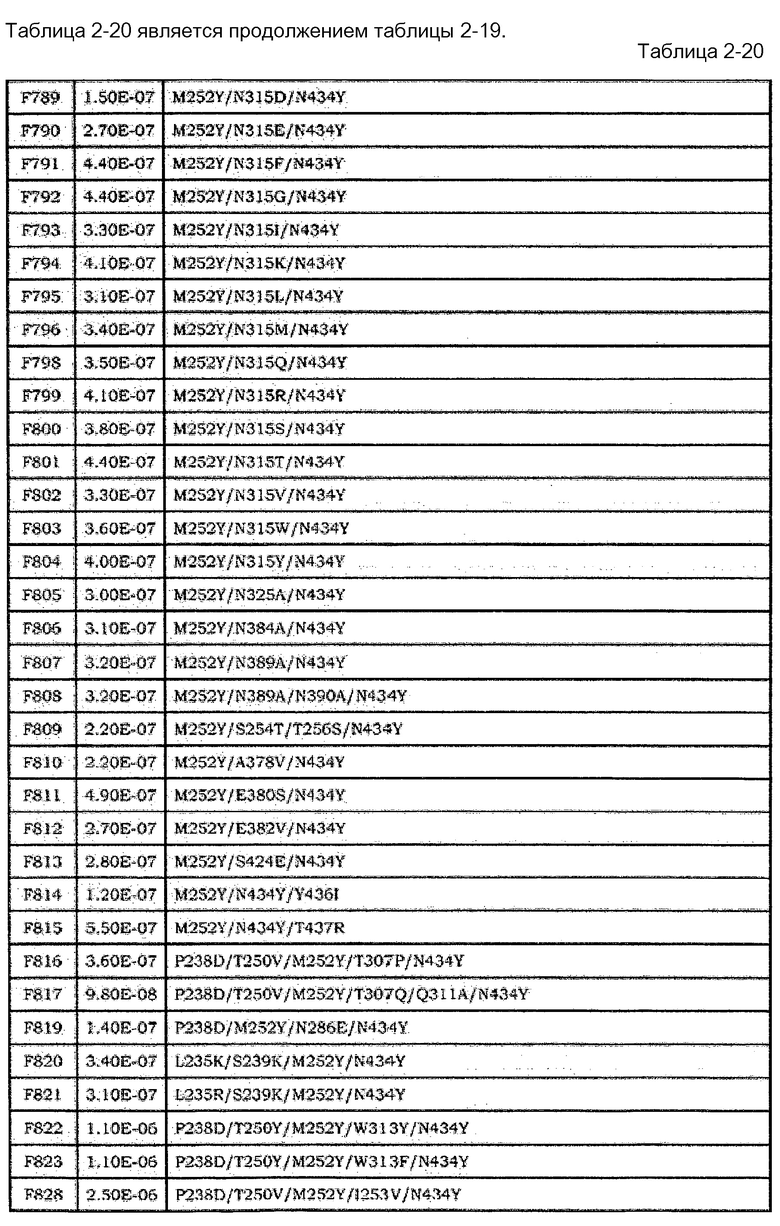
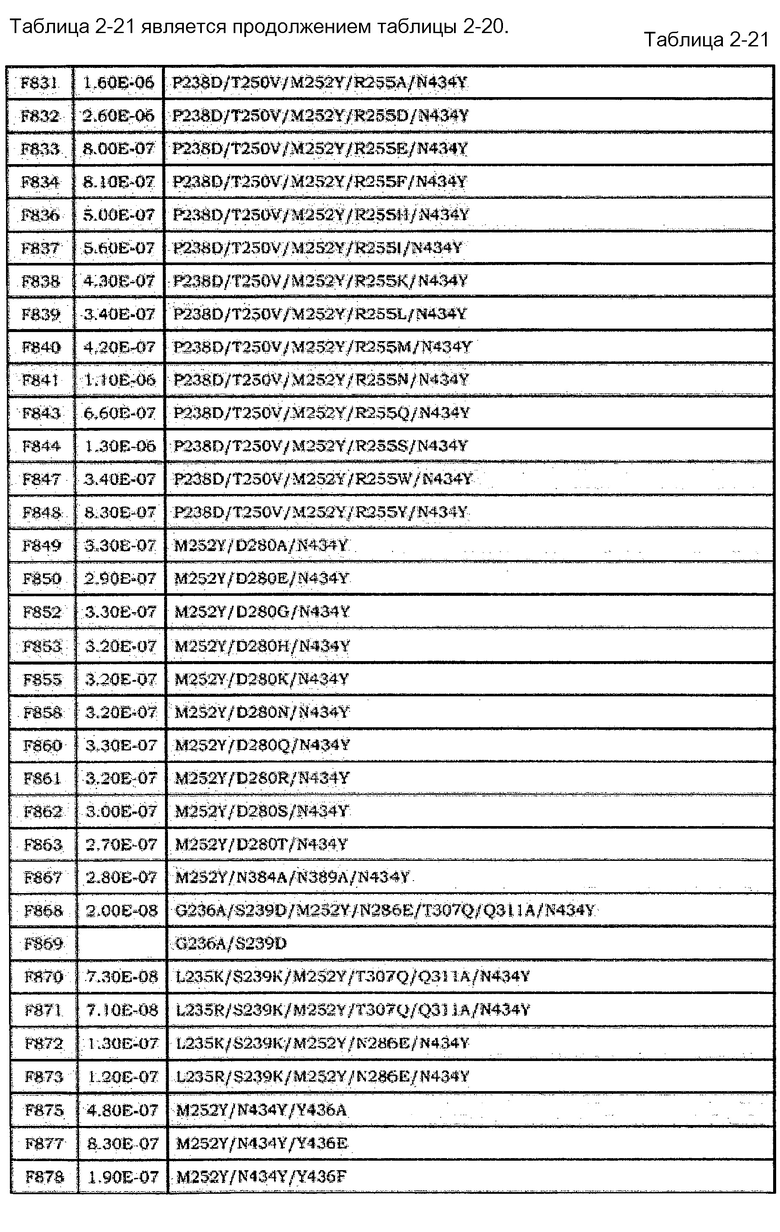


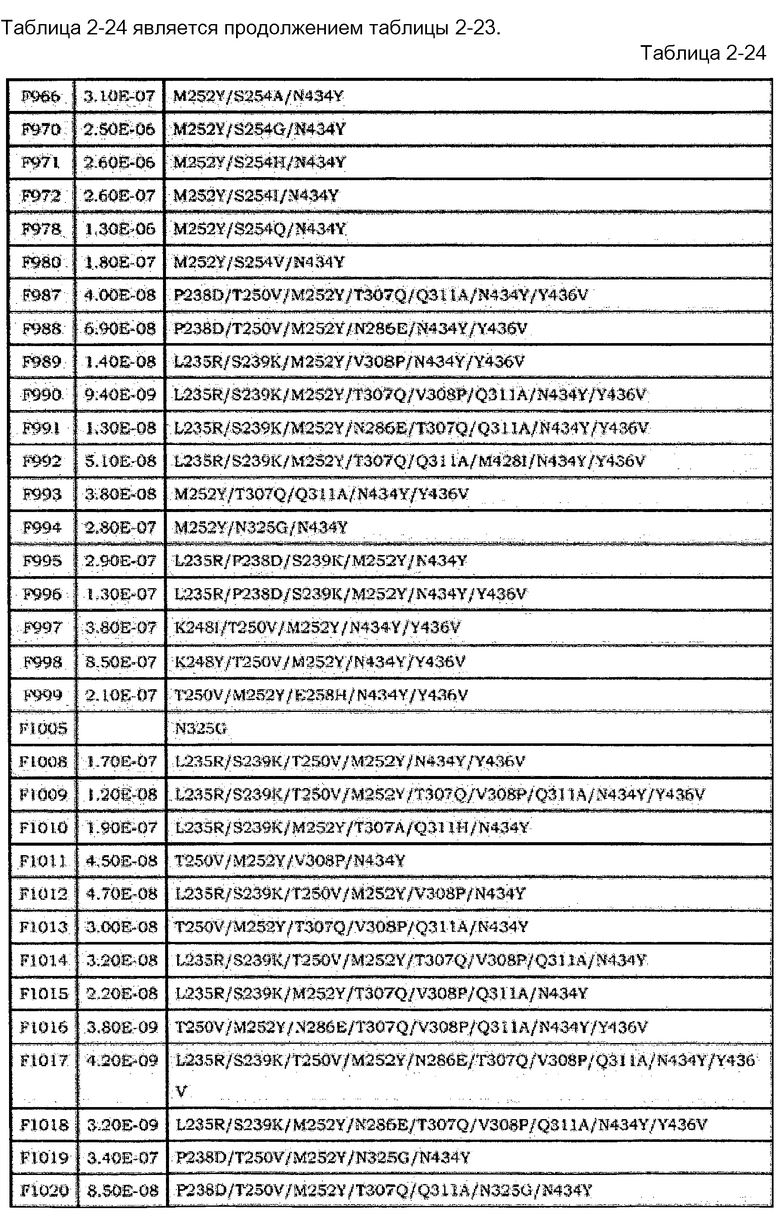
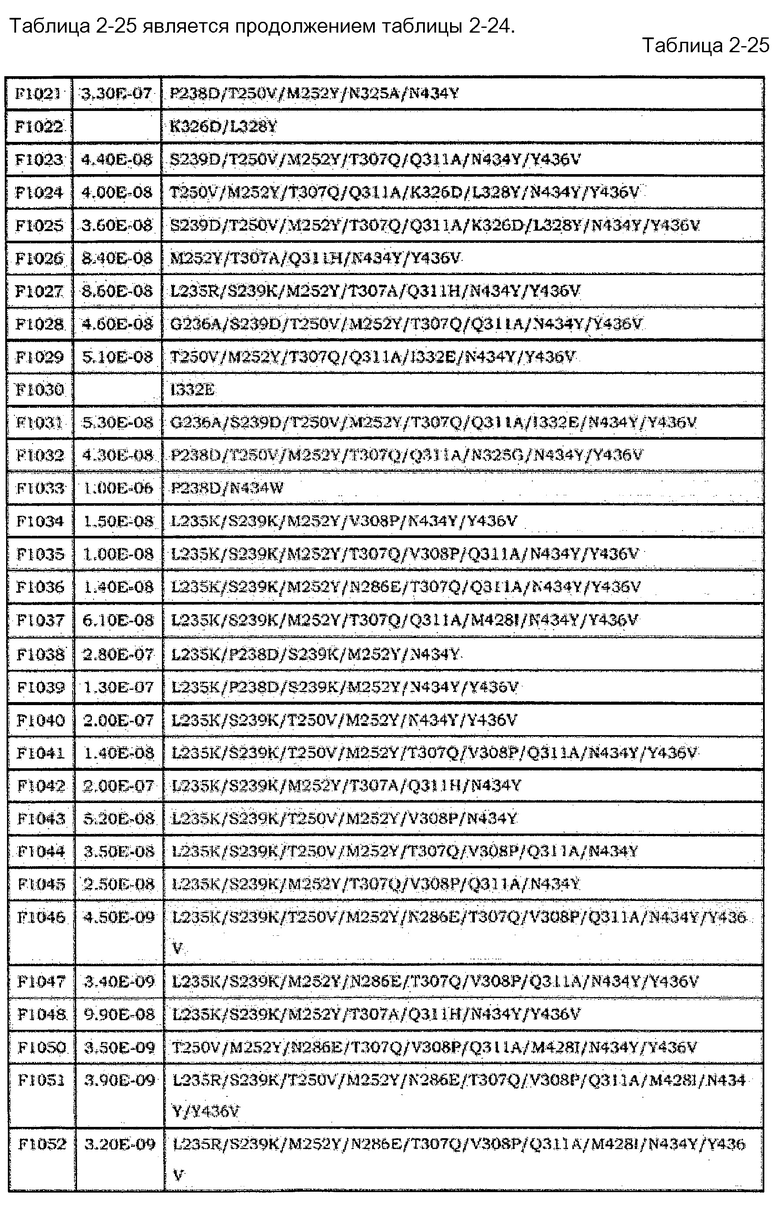
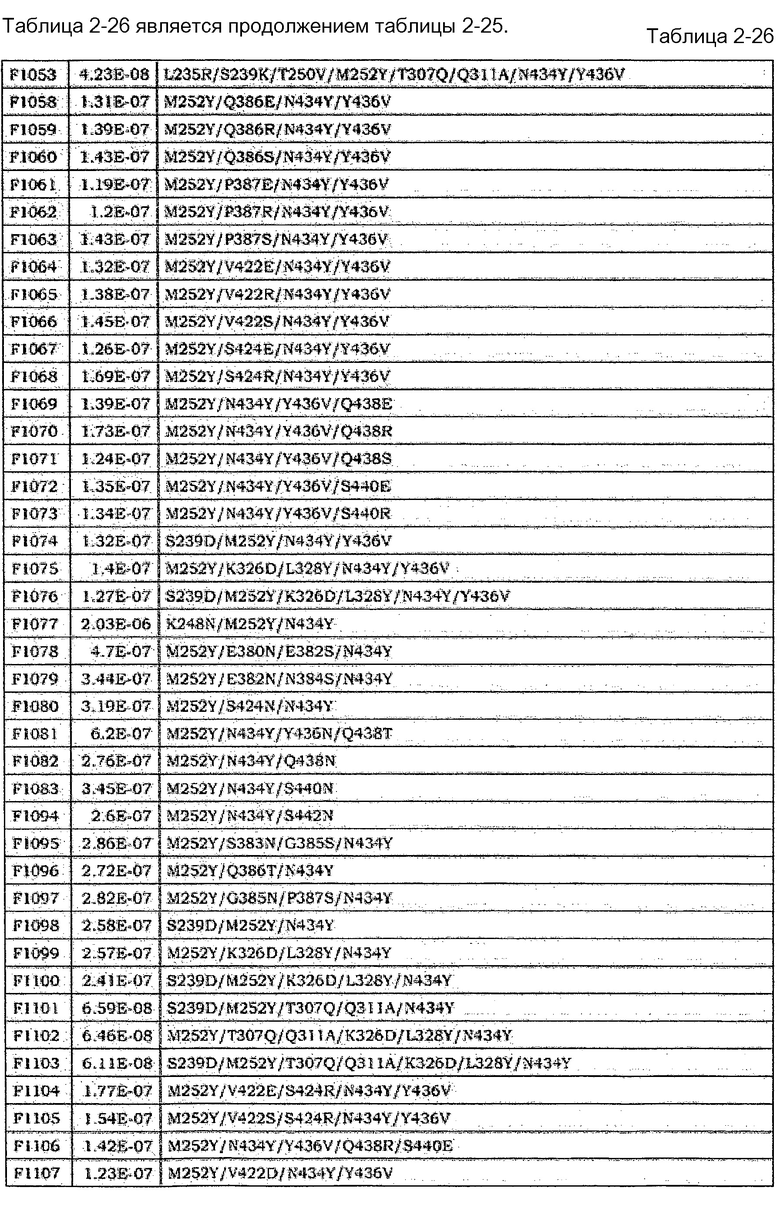




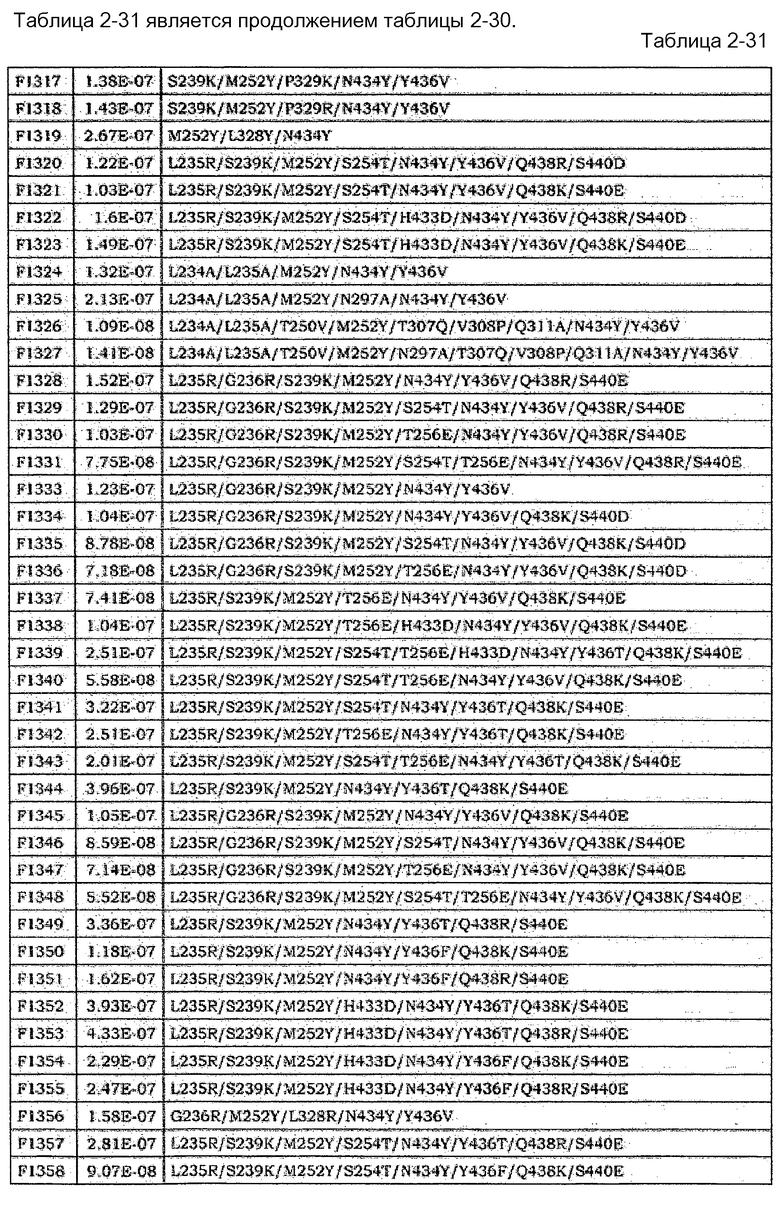
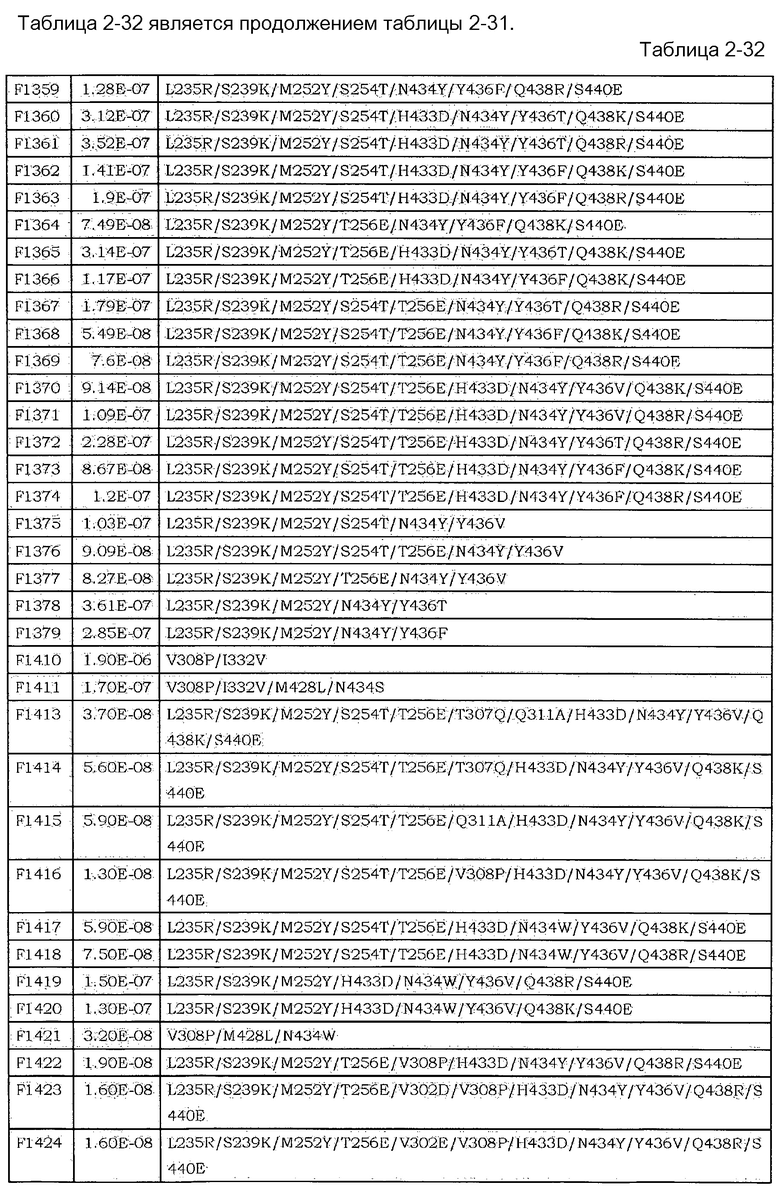

Гетерокомплекс, содержащий четыре молекулы, включая две молекулы FcRn и одну молекулу активирующего Fcγ-рецептора
Кристаллографические исследования FcRn с IgG-антителами продемонстрировали, что комплекс FcRn-IgG состоит из одной молекулы IgG на две молекулы FcRn, и полагают, что эти две молекулы связываются в области поверхности контакта доменов CH2 и CH3, расположенных по обеим сторонам Fc-области IgG (Burmeister et al. (Nature (1994) 372, 336-343)). Между тем, как продемонстрировано в примере 3 PCT/JP2012/058603, было показано, что Fc-область антитела способна образовывать комплекс, содержащий четыре молекулы, включая две молекулы FcRn и одну молекулу активирующего рецептора Fcγ (PCT/JP2012/058603). Это образование гетерокомплекса представляет собой явление, которое было выявлено в результате анализа свойств антигенсвязывающих молекул, содержащих Fc-область, обладающую активностью связывания FcRn в условиях диапазона нейтральных значений pH.
Хотя настоящее изобретение не ограничивается конкретным принципом, можно считать, что антигенсвязывающие молекулы, введенные in vivo, оказывают описанные ниже эффекты на фармакокинетику in vivo (удержание в плазме) антигенсвязывающих молекул и иммунный ответ (иммуногенность) на введенные антигенсвязывающие молекулы в результате образования гетерокомплексов, содержащих четыре молекулы, включающие Fc-область, содержащуюся в антигенсвязывающих молекулах, две молекулы FcRn и одну молекулу активирующего Fcγ-рецептора. В дополнение к различным типам активирующих Fcγ-рецепторов, на иммунных клетках экспрессируется FcRn. Полагают, что образование таких тетрамерных комплексов на иммунных клетках антигенсвязывающими молекулами ускоряет включение антигенсвязывающих молекул в иммунные клетки путем увеличения аффинности в отношении иммунных клеток и путем обеспечения ассоциации внутриклеточных доменов для усиления сигнала интернализации. Указанное выше также применимо для антигенпредставляющих клеток и вероятности того, что антигенсвязывающие молекулы, возможно, включаются в антигенпредставляющие клетки путем образования тетрамерных комплексов на клеточной мембране антигенпредставляющих клеток. Как правило, антигенсвязывающие молекулы, включенные в антигенпредставляющие клетки, деградируют в лизосомах антигенпредставляющих клеток и представляются T-клеткам. В результате время удержания в плазме антигенсвязывающих молекул может снижаться, поскольку включение антигенсвязывающих молекул в антигенпредставляющие клетки ускоряется посредством образования описанных выше тетрамерных комплексов на клеточной мембране антигенпредставляющих клеток. Аналогично, может индуцироваться (усиливаться) иммунный ответ.
По этой причине понятно, что, когда антигенсвязывающую молекулу, обладающую сниженной способностью образовывать такие тетрамерные комплексы, вводят in vivo, время удержания антигенсвязывающих молекул в плазме может увеличиваться, и индукция иммунного ответа in vivo будет подавляться. Предпочтительные варианты осуществления таких антигенсвязывающих молекул, которые ингибируют образование этих комплексов на иммунных клетках, включающих антигенпредставляющие клетки, представляют собой, например, три варианта осуществления, описанных ниже.
Антигенсвязывающие молекулы, которые ингибируют образование гетерокомплексов
(Вариант осуществления 1) Антигенсвязывающая молекула, содержащая Fc-область, обладающую активностью связывания FcRn в условиях диапазона нейтральных значений pH и активность связывания которой с активирующим FcγR является более низкой, чем активность связывания нативной Fc-области с активирующим FcγR
Антигенсвязывающая молекула согласно варианту осуществления 1 образует тримерный комплекс посредством связывания с двумя молекулами FcRn; однако она не образует какой-либо комплекс, содержащий активирующий FcγR. Fc-область, активность связывания которой с активирующим FcγR является более низкой, чем активность связывания нативной Fc-области с активирующим FcγR, можно получать путем изменения аминокислот нативной Fc-области, как описано выше. То, является ли активность связывания измененной Fc-области с активирующим FcγR более низкой, чем активность связывания нативной Fc-области с активирующим FcγR, можно соответствующим образом исследовать с использованием способов, описанных в разделе "активность связывания" выше.
Предпочтительные активирующие Fcγ-рецепторы включают FcγRI (CD64), который включает FcγRIa, FcγRIb, и FcγRIc; FcγRIIa (включая аллотипы R131 и H131); и FcγRIII (CD16), который включает изоформы FcγRIIIa (включая аллотипы V158 и F158) и FcγRIIIb (включая аллотипы FcγRIIIb-NA1 и FcγRIIIb-NA2).
В рамках настоящего изобретения "активность связывания варианта Fc-области с активирующим Fcγ-рецептором является более низкой, чем активность связывания нативной Fc-области с активирующим Fcγ-рецептором" означает, что активность связывания варианта Fc-области с любым из Fcγ-рецепторов человека (FcγRI, FcγRIIa, FcγRIIIa и/или FcγRIIIb) является более низкой, чем активность связывания нативной Fc-области с этими Fcγ-рецепторами человека. Например, это означает, что, исходя из описанного выше способа анализа, активность связывания антигенсвязывающей молекулы, содержащей вариант Fc-области, по сравнению с активностью связывания антигенсвязывающей молекулы, содержащей нативную Fc-область в качестве контроля, составляет 95% или менее, предпочтительно 90% или менее, 85% или менее, 80% или менее, 75% или менее, и особенно предпочтительно 70% или менее, 65% или менее, 60% или менее, 55% или менее, 50% или менее, 45% или менее, 40% или менее, 35% или менее, 30% или менее, 25% или менее, 20% или менее, 15% или менее, 10% или менее, 9% или менее, 8% или менее, 7% или менее, 6% или менее, 5% или менее, 4% или менее, 3% или менее, 2% или менее или 1% или менее. В качестве нативной Fc-области можно использовать исходную Fc-область, и также можно использовать Fc-области антител дикого типа отличающихся изотипов.
Между тем, активность связывания нативной формы с активирующим FcγR предпочтительно представляет собой активность связывания с Fcγ-рецептором для IgG1 человека. Помимо проведения описанных выше изменений, активность связывания с Fcγ-рецептором может быть снижена путем изменения изотипа IgG2 человека, IgG3 человека или IgG4 человека. Альтернативно, помимо проведения упомянутых выше изменений, активность связывания с Fcγ-рецептором также может быть снижена путем экспрессии антигенсвязывающей молекулы, содержащей Fc-область, обладающую активностью связывания с Fcγ-рецептором в хозяевах, которые не присоединяют цепи сахаров, таких как Escherichia coli.
В качестве антигенсвязывающей молекулы, содержащей контрольную Fc-область, можно соответственно использовать антигенсвязывающую молекулу, обладающую Fc-областью моноклонального IgG-антитела. Структуры таких Fc-областей представлены в SEQ ID NO:5 (A добавлен на N-конец последовательности с номером доступа RefSeq AAC82527.1), SEQ ID NO:6 (A добавлен на N-конец последовательности с номером доступа RefSeq AAB59393.1), SEQ ID NO:7 (номер доступа RefSeq CAA27268.1) и SEQ ID NO:8 (A добавлен на N-конец последовательности с номером доступа RefSeq AAB59394.1). Более того, когда антигенсвязывающую молекулу, содержащую Fc-область конкретного изотипа антитела, используют в качестве исследуемого вещества, эффект антигенсвязывающей молекулы, содержащей исследуемую Fc-область, в отношении активности связывания Fcγ-рецептора, исследуют с использованием в качестве контроля антигенсвязывающей молекулы, имеющей Fc-область моноклонального IgG-антитела этого конкретного изотипа. Таким путем пригодным образом выбирают антигенсвязывающие молекулы, содержащие Fc-область, для которой продемонстрировано, что ее активность связывания Fcγ-рецептора является высокой.
В неограничивающем варианте осуществления настоящего изобретения предпочтительные примеры Fc-областей, активность связывания которых с активирующим FcγR является более низкой, чем активность связывания нативной Fc-области с активирующим FcγR, включают Fc-области с изменением одной или нескольких аминокислот в любом из положений 234, 235, 236, 237, 238, 239, 270, 297, 298, 325, 328 и 329, как указано с помощью нумерации EU, в аминокислотах описанной выше Fc-области так, чтобы она отличалась от нативной Fc-области. Изменения в Fc-области не ограничиваются описанным выше примером, и они могут представлять собой, например, модификации, такие как дегликозилгликозилирование (N297A и N297Q), IgG1-L234A/L235A, IgG1-A325A/A330S/P331S, IgG1-C226S/C229S, IgG1-C226S/C229S/E233P/L234V/L235A, IgG1-L234F/L235E/P331S, IgG1-S267E/L328F, IgG2-V234A/G237A, IgG2-H268Q/V309L/A330S/A331S, IgG4-L235A/G237A/E318A и IgG4-L236E, описанные в Cur. Opin. in Biotech. (2009) 20 (6), 685-691; изменения, такие как G236R/L328R, L235G/G236R, N325A/L328R и N325L/L328R, описанные в WO 2008/092117; вставки аминокислот в положениях 233, 234, 235 и 237 в соответствии с нумерацией EU; и изменения в положениях, описанных в WO 2000/042072.
В неограничивающем варианте осуществления настоящего изобретения примеры предпочтительной Fc-области включают Fc-области, имеющие одно или несколько из следующих изменений, как указано с помощью нумерации EU, в упомянутой выше Fc-области:
Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Lys, Met, Phe, Pro, Ser, Thr или Trp в качестве аминокислоты в положении 234;
Ala, Asn, Asp, Gln, Glu, Gly, His, Ile, Lys, Met, Pro, Ser, Thr, Val или Arg в качестве аминокислоты в положении 235;
Arg, Asn, Gln, His, Leu, Lys, Met, Phe, Pro или Tyr в качестве аминокислоты в положении 236;
Ala, Asn, Asp, Gln, Glu, His, Ile, Leu, Lys, Met, Pro, Ser, Thr, Val, Tyr или Arg в качестве аминокислоты в положении 237;
Ala, Asn, Gln, Glu, Gly, His, Ile, Lys, Thr, Trp или Arg в качестве аминокислоты в положении 238;
Gln, His, Lys, Phe, Pro, Trp, Tyr или Arg в качестве аминокислоты в положении 239;
Ala, Arg, Asn, Gln, Gly, His, Ile, Leu, Lys, Met, Phe, Ser, Thr, Trp, Tyr или Val в качестве аминокислоты в положении 265;
Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Lys, Phe, Pro, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 266;
Arg, His, Lys, Phe, Pro, Trp или Tyr в качестве аминокислоты в положении 267;
Ala, Arg, Asn, Gln, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val в качестве аминокислоты в положении 269;
Ala, Arg, Asn, Gln, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val в качестве аминокислоты в положении 270;
Arg, His, Phe, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 271;
Arg, Asn, Asp, Gly, His, Phe, Ser, Trp или Tyr в качестве аминокислоты в положении 295;
Arg, Gly, Lys или Pro в качестве аминокислоты в положении 296;
Ala в качестве аминокислоты в положении 297;
Arg, Gly, Lys, Pro, Trp или Tyr в качестве аминокислоты в положении 298;
Arg, Lys или Pro в качестве аминокислоты в положении 300;
Lys или Pro в качестве аминокислоты в положении 324;
Ala, Arg, Gly, His, Ile, Lys, Phe, Pro, Thr, Trp, Tyr или Val в качестве аминокислоты в положении 325;
Arg, Gln, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val в качестве аминокислоты в положении 327;
Arg, Asn, Gly, His, Lys или Pro в качестве аминокислоты в положении 328;
Asn, Asp, Gln, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Ser, Thr, Trp, Tyr, Val или Arg в качестве аминокислоты в положении 329;
Pro или Ser в качестве аминокислоты в положении 330;
Arg, Gly или Lys в качестве аминокислоты в положении 331; или
Arg, Lys или Pro в качестве аминокислоты в положении 332.
(Вариант осуществления 2) Антигенсвязывающая молекула, содержащая Fc-область, обладающую активностью связывания FcRn в условиях диапазона нейтральных значений pH и активность связывания которой с ингибиторным FcγR превышает активность связывания с активирующим Fcγ-рецептором
Путем связывания с двумя молекулами FcRn и одной молекулой ингибирующего FcγR, антигенсвязывающая молекула согласно варианту осуществления 2 может образовывать комплекс, содержащий эти четыре молекулы. Однако, поскольку одна антигенсвязывающая молекула может связываться только с одной молекулой FcγR, одна антигенсвязывающая молекула в состоянии, связанном с ингибиторным FcγR, не может связываться с другими активирующими FcγR. Более того, было описано, что антигенсвязывающая молекула, которая включается в клетки в состоянии, связанном с ингибиторным FcγR, рециклируют на клеточную мембрану и, таким образом, ускользает от деградации внутри клеток (Immunity (2005) 23, 503-514). Более конкретно, считается, что антигенсвязывающие молекулы, обладающие активностью селективного связывания с ингибиторным FcγR, не могут образовывать гетерокомплексы, содержащие активирующий FcγR и две молекулы FcRn, которые вызывают иммунный ответ.
Предпочтительные активирующие Fcγ-рецепторы включают FcγRI (CD64), который включает FcγRIa, FcγRIb, и FcγRIc; FcγRIIa (включая аллотипы R131 и H131); и FcγRIII (CD16), который включает изоформы FcγRIIIa (включая аллотипы V158 и F158) и FcγRIIIb (включая аллотипы FcγRIIIb-NA1 и FcγRIIIb-NA2). Между тем, примеры предпочтительных ингибиторных Fcγ-рецепторов включают FcγRIIb (включая FcγRIIb-1 и FcγRIIb-2).
В рамках настоящего изобретения "активность связывания с ингибиторным FcγR является более высокой, чем активность связывания с активирующим Fcγ-рецептором", означает, что активность связывания варианта Fc-области с FcγRIIb является более высокой, чем активность связывания с любым из Fcγ-рецепторов человека: FcγRI, FcγRIIa, FcγRIIIa и/или FcγRIIIb. Например, это означает, что, исходя из описанного выше способа анализа, активность связывания с FcγRIIb антигенсвязывающей молекулы, содержащей вариант Fc-области, по сравнению с активностью связывания с любым из Fcγ-рецепторов человека: FcγRI, FcγRIIa, FcγRIIIa и/или FcγRIIIb составляет 105% или более, предпочтительно 110% или более, 120% или более, 130% или более, 140% или более, и в частности предпочтительно 150% или более, 160% или более, 170% или более, 180% или более, 190% или более, 200% или более, 250% или более, 300% или более, 350% или более, 400% или более, 450% или более, 500% или более, 750% или более, или превышает 10 раз или более, в 20 раз или более, в 30 раз или более, в 40 раз или более, в 50 раз или более.
Наиболее предпочтительно, активность связывания с FcγRIIb является более высокой, чем каждая из активностей связывания с FcγRIa, FcγRIIa (включая аллотипы R131 и H131) и FcγRIIIa (включая аллотипы V158 и F158). FcγRIa демонстрирует выраженную высокую аффинность в отношении нативного IgG1; таким образом, полагают, что связывание является насыщенным in vivo вследствие присутствия большого количества эндогенного IgG1. По этой причине может быть возможным ингибирование образования комплекса, даже если активность в отношении FcγRIIb является более высокой, чем активность связывания с FcγRIIa и FcγRIIIa, и является более низкой, чем активность связывания с FcγRIa.
В качестве контрольной антигенсвязывающей молекулы, содержащей Fc-область, предпочтительно можно использовать антигенсвязывающие молекулы, имеющие Fc-область моноклонального IgG-антитела. Структуры таких Fc-областей представлены в SEQ ID NO:5 (A добавлен на N-конец последовательности с номером доступа RefSeq AAC82527.1), SEQ ID NO:6 (A добавлен на N-конец последовательности с номером доступа RefSeq AAB59393.1), SEQ ID NO:7 (номер доступа RefSeq CAA27268.1) и SEQ ID NO:8 (A добавлен на N-конец последовательности с номером доступа RefSeq AAB59394.1). Более того, когда антигенсвязывающую молекулу, содержащую Fc-область конкретного изотипа антитела, используют в качестве исследуемого вещества, эффект антигенсвязывающей молекулы, содержащей исследуемую Fc-область, в отношении активности связывания Fcγ-рецептора, исследуют с использованием в качестве контроля антигенсвязывающей молекулы, имеющей Fc-область моноклонального IgG-антитела этого конкретного изотипа. Таким путем пригодным образом выбирают антигенсвязывающие молекулы, содержащие Fc-область, для которой продемонстрировано, что ее активность связывания с Fcγ-рецептором является высокой.
В неограничивающем варианте осуществления настоящего изобретения предпочтительные примеры Fc-областей, обладающих активностью селективного связывания с ингибиторным FcγR, включают Fc-области, в которых среди аминокислот описанной выше Fc-области аминокислота в положении 238 или 328, как указано с помощью нумерации EU, изменена на аминокислоту, отличающуюся от аминокислоты в нативной Fc-области. Более того, в качестве Fc-области, обладающей активностью селективного связывания с ингибиторным Fcγ-рецептором, можно соответственно выбирать Fc-области или изменения, описанные в US 2009/0136485.
В неограничивающем варианте осуществления настоящего изобретения предпочтительным примером является Fc-область, имеющая одно или несколько из следующих изменений, как указано с помощью нумерации EU, в упомянутой выше Fc-области: аминокислота в положении 238 представляет собой Asp или аминокислота в положении 328 представляет собой Glu.
В другом неограничивающем варианте осуществления настоящего изобретения примеры предпочтительной Fc-области включают Fc-области, имеющие замену Pro в положении 238 в соответствии с нумерацией EU на Asp и имеющие одно или несколько из изменений: изменение аминокислоты в положении 237 в соответствии с нумерацией EU на Trp, аминокислота в положении 237 в соответствии с нумерацией EU представляет собой Phe, аминокислота в положении 267 в соответствии с нумерацией EU представляет собой Val, аминокислота в положении 267 в соответствии с нумерацией EU представляет собой Gln, аминокислота в положении 268 в соответствии с нумерацией EU представляет собой Asn, аминокислота в положении 271 в соответствии с нумерацией EU представляет собой Gly, аминокислота в положении 326 в соответствии с нумерацией EU представляет собой Leu, аминокислота в положении 326 в соответствии с нумерацией EU представляет собой Gln, аминокислота в положении 326 в соответствии с нумерацией EU представляет собой Glu, аминокислота в положении 326 в соответствии с нумерацией EU представляет собой Met, аминокислота в положении 239 в соответствии с нумерацией EU представляет собой Asp, аминокислота в положении 267 в соответствии с нумерацией EU представляет собой Ala, аминокислота в положении 234 в соответствии с нумерацией EU представляет собой Trp, аминокислота в положении 234 в соответствии с нумерацией EU представляет собой Tyr, аминокислота в положении 237 в соответствии с нумерацией EU представляет собой Ala, аминокислота в положении 237 в соответствии с нумерацией EU представляет собой Asp, аминокислота в положении 237 в соответствии с нумерацией EU представляет собой Glu, аминокислота в положении 237 в соответствии с нумерацией EU представляет собой Leu, аминокислота в положении 237 в соответствии с нумерацией EU представляет собой Met, аминокислота в положении 237 в соответствии с нумерацией EU представляет собой Tyr, аминокислота в положении 330 в соответствии с нумерацией EU представляет собой Lys, аминокислота в положении 330 в соответствии с нумерацией EU представляет собой Arg, аминокислота в положении 233 в соответствии с нумерацией EU представляет собой Asp, аминокислота в положении 268 в соответствии с нумерацией EU представляет собой Asp, аминокислота в положении 268 в соответствии с нумерацией EU представляет собой Glu, аминокислота в положении 326 в соответствии с нумерацией EU представляет собой Asp, аминокислота в положении 326 в соответствии с нумерацией EU представляет собой Ser, аминокислота в положении 326 в соответствии с нумерацией EU представляет собой Thr, аминокислота в положении 323 в соответствии с нумерацией EU представляет собой Ile, аминокислота в положении 323 в соответствии с нумерацией EU представляет собой Leu, аминокислота в положении 323 в соответствии с нумерацией EU представляет собой Met, аминокислота в положении 296 в соответствии с нумерацией EU представляет собой Asp, аминокислота в положении 326 в соответствии с нумерацией EU представляет собой Ala, аминокислота в положении 326 в соответствии с нумерацией EU представляет собой Asn, и аминокислота в положении 330 в соответствии с нумерацией EU представляет собой Met.
(Вариант осуществления 3) Антигенсвязывающая молекула, содержащая Fc-область, в которой один из двух полипептидов, составляющих Fc-область, обладает активностью связывания FcRn в условиях диапазона нейтральных значений pH, а другой полипептид не обладает активностью связывания FcRn в условиях диапазона нейтральных значений pH
Посредством связывания с одной молекулой FcRn и одной молекулой FcγR, антигенсвязывающая молекула согласно варианту осуществления 3 может образовывать тримерный комплекс; однако она не образует какой-либо гетерокомплекс, содержащий четыре молекулы, включая две молекулы FcRn и одну молекулу FcγR. В качестве Fc-области, в которой один из двух полипептидов, составляющих Fc-область, обладает активностью связывания FcRn в условиях диапазона нейтральных значений pH, а другой полипептид не обладает какой-либо активностью связывания FcRn в условиях диапазона нейтральных значений pH, которой обладает антигенсвязывающая молекула согласно варианту осуществления 3, можно соответствующим образом использовать Fc-области, происходящие из биспецифических антител. Биспецифические антитела представляют собой два типа антител, обладающие специфичностью в отношении различных антигенов. Биспецифические антитела IgG-типа можно секретировать из гидрибных гибридом (квадром), полученных путем слияния двух типов гибридом, продуцирующих IgG-антитела (Milstein et al. (Nature (1983) 305, 537-540).
Когда антигенсвязывающую молекулу согласно варианту осуществления 3 получают с использованием рекомбинантных способов, таких как способы, описанные в упомянутом выше разделе "Антитела", можно использовать способ, в котором гены, кодирующие полипептиды, которые составляют два типа представляющих интерес Fc-областей, трансфицируют в клетки для их коэкспрессии. Однако продуцированные Fc-области будут представлять собой смесь, в которой следующие компоненты существуют в молекулярном соотношении 2:1:1: Fc-область, в которой один из двух полипептидов, составляющих Fc-область, обладает активностью связывания FcRn в условиях диапазона нейтральных значений pH, а другой полипептид не обладает никакой активностью связывания FcRn в условиях диапазона нейтральных значений pH; Fc-область, в которой оба из двух полипептидов, составляющих Fc-область, обладают активностью связывания FcRn в условиях диапазона нейтральных значений pH; и Fc-область, в которой оба из двух полипептидов, составляющих Fc-область, не обладают активностью связывания FcRn в условиях диапазона нейтральных значений pH. Трудно очищать антигенсвязывающие молекулы, содержащие желаемую комбинацию Fc-областей, из трех типов IgG.
При получении антигенсвязывающих молекул согласно варианту осуществления с использованием таких рекомбинантных способов, антигенсвязывающие молекулы, содержащие гетеромерную комбинацию тяжелых цепей, можно предпочтительно секретировать путем добавления соответствующих аминокислотных замен в CH3-домены, составляющие Fc-области. В частности, этот способ проводят путем замены аминокислотой, имеющей более крупную боковую цепь (выступ (что означает "выпуклая часть")) аминокислоты в CH3-домене одной из тяжелых цепей, и замены аминокислотой, имеющей меньшую боковую цепь (углубление (что означает "полость")) аминокислоты в CH3-домене другой тяжелой цепи, так чтобы выступ размещался в полости. Это обеспечивает образование гетеромерной тяжелой цепи и одновременно ингибирует образование гомомерной тяжелой цепи (WO 1996027011; Ridgway et al., Protein Engineering (1996) 9, 617-621; Merchant et al., Nature Biotechnology (1998) 16, 677-681).
Более того, также существуют известные способы получения биспецифического антитела с использованием способов контроля ассоциации полипептидов или ассоциации образованных полипептидом гетеромерных мультимеров для ассоциации между двумя полипептидами, которые составляют Fc-область. В частности, способы контроля образования тяжелых цепей можно использовать для получения биспецифического антитела (WO 2006/106905), в котором аминокислотные остатки, образующие поверхность контакта между двумя полипептидами, которые составляют Fc-область, изменяют для ингибирования ассоциации между Fc-областями, имеющими ту же последовательность, и для обеспечения образования полипептидных комплексов, образованных двумя Fc-областями с различными последовательностями. В частности, в качестве неограничивающего варианта осуществления для получения антигенсвязывающей молекулы согласно варианту осуществления 3 по настоящему изобретению можно использовать способы, описанные в представленном выше разделе, касающемся биспецифических антител и способов их получения.
Ожидается, что эти антигенсвязывающие молекулы согласно вариантам осуществления 1-3 способны снижать иммуногенность и увеличивать время удержания в плазме по сравнению с антигенсвязывающими молекулами, способными образовывать тетрамерные комплексы.
Способы получения антигенсвязывающих доменов
Настоящее изобретение относится к способам получения антигенсвязывающих доменов, антигенсвязывающая активность которых в присутствии специфического для ткани-мишени соединения является более высокой, чем антигенсвязывающая активность в отсутствие соединения.
Более конкретно, настоящее изобретение относится к способу получения антигенсвязывающего домена, который включает стадии (a)-(e), ниже:
(a) определение антигенсвязывающей активности антигенсвязывающего домена в отсутствие специфического для ткани-мишени соединения;
(b) определение антигенсвязывающей активности антигенсвязывающего домена в присутствии специфического для ткани-мишени соединения;
(c) отбор антигенсвязывающего домена, антигенсвязывающая активность которого в отсутствие специфического для ткани-мишени соединения является более низкой, чем в присутствии соединения;
(d) культивирование клеток, трансфицированных вектором, с которым полинуклеотид, кодирующий антигенсвязывающий домен, отобранный на стадии (c), функционально связан; и
(e) сбор антигенсвязывающего домена из культуральной среды клеток, культивированных на стадии (d).
Настоящее изобретение также относится к способу получения антигенсвязывающего домена, который включает стадии (a)-(e) ниже:
(a) определение антигенсвязывающей активности антигенсвязывающего домена в присутствии низкой концентрации специфического для ткани-мишени соединения;
(b) определение антигенсвязывающей активности антигенсвязывающего домена в присутствии высокой концентрации специфического для ткани-мишени соединения;
(c) отбор антигенсвязывающего домена, антигенсвязывающая активность которого в присутствии низкой концентрации специфического для ткани-мишени соединения является более низкой, чем в присутствии высокой концентрации соединения;
(d) культивирование клеток, трансфицированных вектором, с которым полинуклеотид, кодирующий антигенсвязывающий домен, отобранный на стадии (c), функционально связан; и
(e) сбор антигенсвязывающего домена из культуральной среды клеток, культивированных на стадии (d).
Более того, настоящее изобретение относится к способу получения антигенсвязывающего домена, который включает стадии (a)-(e), ниже:
(a) контактирование антигенсвязывающих доменов или их библиотеки с антигеном в присутствии специфического для ткани-мишени соединения;
(b) помещение антигенсвязывающих доменов, которые связались с антигеном на указанной стадии (a), в условия отсутствия соединения;
(c) выделение антигенсвязывающего домена, который диссоциировал на указанной стадии (b);
(d) культивирование клеток, трансфицированных вектором, с которым полинуклеотид, кодирующий антигенсвязывающий домен, отобранный на стадии (c), функционально связан; и
(e) сбор антигенсвязывающего домена из культуральной среды клеток, культивированных на стадии (d).
Кроме того, настоящее изобретение относится к способу получения антигенсвязывающего домена, который включает стадии (a)-(e), ниже:
(a) контактирование антигенсвязывающих доменов или их библиотеки с антигеном в присутствии высокой концентрации специфического для ткани-мишени соединения;
(b) помещение антигенсвязывающих доменов, которые связываются с антигеном на указанной стадии (a), в условия присутствия низкой концентрации соединения;
(c) выделение антигенсвязывающего домена, который диссоциирует на указанной стадии (b);
(d) культивирование клеток, трансфицированных вектором, с которым полинуклеотид, кодирующий антигенсвязывающий домен, отобранный на стадии (c), функционально связан; и
(e) сбор антигенсвязывающего домена из культуральной среды клеток, культивированных на стадии (d).
Настоящее изобретение относится к способу получения антигенсвязывающего домена, который включает стадии (a)-(f), ниже:
(a) контактирования библиотеки антигенсвязывающих доменов с антигеном в отсутствие специфического для ткани-мишени соединения;
(b) выбор антигенсвязывающих доменов, которые не связываются с антигеном на указанной стадии (a);
(c) позволение антигенсвязывающим доменам, выбранным на указанной стадии (b), связаться с антигеном в присутствии соединения;
(d) выделение антигенсвязывающего домена, который связывается с антигеном на указанной стадии (c);
(e) культивирование клеток, трансфицированных вектором, с которым полинуклеотид, кодирующий антигенсвязывающий домен, отобранный на стадии (d), функционально связан; и
(f) сбор антигенсвязывающего домена из культуральной среды клеток, культивированных на стадии (e).
Настоящее изобретение относится к способу получения антигенсвязывающего домена, который включает стадии с (a) по (f), ниже:
(a) контактирование библиотеки антигенсвязывающих доменов с антигеном в присутствии низкой концентрации специфического для ткани-мишени соединения;
(b) выбор антигенсвязывающих доменов, которые не связываются с антигеном на указанной стадии (a);
(c) позволение антигенсвязывающим доменам, выбранным на указанной стадии (b), связаться с антигеном в присутствии высокой концентрации соединения;
(d) выделение антигенсвязывающего домена, который связывается с антигеном на указанной стадии (c);
(e) культивирование клеток, трансфицированных вектором, с которым полинуклеотид, кодирующий антигенсвязывающий домен, отобранный на стадии (d), функционально связан; и
(f) сбор антигенсвязывающего домена из культуральной среды клеток, культивированных на стадии (e).
Настоящее изобретение относится к способу получения антигенсвязывающего домена, который включает стадии (a)-(e), ниже:
(a) контактирование библиотеки антигенсвязывающих доменов с колонкой с иммобилизованным на ней антигеном в присутствии специфического для ткани-мишени соединения;
(b) элюирование антигенсвязывающих доменов, которые связываются с колонкой на указанной стадии (a), из колонки в отсутствие соединения;
(c) выделение антигенсвязывающего домена, элюированного на указанной стадии (b);
(d) культивирование клеток, трансфицированных вектором, с которым полинуклеотид, кодирующий антигенсвязывающий домен, отобранный на стадии (c), функционально связан; и
(e) сбор антигенсвязывающего домена из культуральной среды клеток, культивированных на стадии (d).
Настоящее изобретение относится к способу получения антигенсвязывающего домена, который включает стадии (a)-(e), ниже:
(a) контактирование библиотеки антигенсвязывающих доменов с колонкой с иммобилизованным на ней антигеном в присутствии высокой концентрации специфического для ткани-мишени соединения;
(b) элюирование антигенсвязывающих доменов, которые связываются с колонкой на указанной стадии (a), из колонки в присутствии низкой концентрации соединения;
(c) выделение антигенсвязывающего домена, элюированного на указанной стадии (b);
(d) культивирование клеток, трансфицированных вектором, с которым полинуклеотид, кодирующий антигенсвязывающий домен, отобранный на стадии (c), функционально связан; и
(e) сбор антигенсвязывающего домена из культуральной среды клеток, культивированных на стадии (d).
Настоящее изобретение относится к способу получения антигенсвязывающего домена, который включает стадии (a)-(f), ниже:
(a) позволение библиотеке антигенсвязывающих доменов пройти через колонку с иммобилизованным на ней антигеном в отсутствие специфического для ткани-мишени соединения;
(b) сбор антигенсвязывающих доменов, которые элюировались без связывания с колонкой на стадии (a);
(c) позволение антигенсвязывающим доменам, собранным на стадии (b), связаться с антигеном в присутствии соединения;
(d) выделение антигенсвязывающего домена, который связывается с антигеном на стадии (c);
(e) культивирование клеток, трансфицированных вектором, с которым полинуклеотид, кодирующий антигенсвязывающий домен, отобранный на стадии (d), функционально связан; и
(f) сбор антигенсвязывающего домена из культуральной среды клеток, культивированных на стадии (e).
Настоящее изобретение относится к способу получения антигенсвязывающего домена, который включает стадии (a)-(f), ниже:
(a) позволение библиотеке антигенсвязывающих доменов пройти через колонку с иммобилизованным на ней антигеном в присутствии низкой концентрации специфического для ткани-мишени соединения;
(b) сбор антигенсвязывающих доменов, которые элюируются без связывания с колонкой на указанной стадии (a);
(c) позволение антигенсвязывающим доменам, собранным на указанной стадии (b), связаться с антигеном в присутствии высокой концентрации соединения;
(d) выделение антигенсвязывающего домена, который связывается с антигеном на указанной стадии (c);
(e) культивирование клеток, трансфицированных вектором, с которым полинуклеотид, кодирующий антигенсвязывающий домен, отобранный на стадии (d), функционально связан; и
(f) сбор антигенсвязывающего домена из культуральной среды клеток, культивированных на стадии (e).
Более того, настоящее изобретение относится к способу получения антигенсвязывающего домена, который включает стадии (a)-(f), ниже:
(a) контактирование антигена с библиотекой антигенсвязывающих доменов в присутствии специфического для ткани-мишени соединения;
(b) получение антигенсвязывающих доменов, которые связываются с антигеном на стадии (a);
(c) помещение антигенсвязывающего домена, полученного на стадии (b), в условия отсутствия соединения;
(d) выделение антигенсвязывающего домена, антигенсвязывающая активность которого на стадии (c) является более слабой, чем у эталона, выбранного на стадии (b);
(e) культивирование клеток, трансфицированных вектором, с которым полинуклеотид, кодирующий антигенсвязывающий домен, отобранный на стадии (d), функционально связан; и
(f) сбор антигенсвязывающего домена из культуральной среды клеток, культивированных на стадии (e).
Настоящее изобретение относится к способу получения антигенсвязывающего домена, который включает стадии (a)-(f), ниже:
(a) контактирование антигена с библиотекой антигенсвязывающих доменов в присутствии высокой концентрации специфического для ткани-мишени соединения;
(b) получение антигенсвязывающих доменов, которые связываются с антигеном на стадии (a);
(c) помещение антигенсвязывающих доменов, полученных на стадии (b), в условия присутствия низкой концентрации соединения;
(d) выделение антигенсвязывающего домена, антигенсвязывающая активность которого на стадии (c) является более слабой, чем у эталона, выбранного на стадии (b);
(e) культивирование клеток, трансфицированных вектором, с которым полинуклеотид, кодирующий антигенсвязывающий домен, отобранный на стадии (d), функционально связан; и
(f) сбор антигенсвязывающего домена из культуральной среды клеток, культивированных на стадии (e).
Термины "клетка", "клеточная линия" и "клеточная культура" используют в настоящем описании синонимично, и такие обозначения могут включать всех потомков клетки или клеточной линии. Таким образом, например, термины "трансформант" и "трансформированные клетки" включают культуры и первичные клетки-мишени, происходящие из них, не зависимо количества пассирований. Более того понятно, что все потомки могут не быть точно идентичными по содержанию ДНК вследствие преднамеренных или случайных мутаций. Также могут быть включены мутантные потомки, которые обладают по существу той же функцией или биологической активностью, например, в отношении которых проводили скрининг первоначально трансформированных клеток. Если подразумевается другое обозначение, это будет очевидно из контекста описания. Клетки, которые являются пригодными для применения, соответственно выбирают из клеток, описанных в разделе "Антитела", выше.
При указании на экспрессию кодирующей последовательности, термин "последовательность контроля" относится к нуклеотидным последовательностям ДНК, которые необходимы для экспрессии функционально связанной кодирующей последовательности в конкретном организме-хозяине. Последовательности контроля, пригодные, например, для прокариот, включают промотор, необязательную последовательность оператора, участок связывания рибосом и, возможно, другие последовательности, которые все еще предстоит изучить. Известно, что эукариотические клетки используют промоторы, сигналы полиаденилирования и энхансеры для экспрессии кодирующей последовательности.
Для нуклеиновой кислоты термин "функционально связанный" означает, что нуклеиновая кислота имеет функциональную взаимосвязь с другой последовательностью нуклеиновой кислоты. Например, ДНК препоследовательности или секреторного лидера функционально связана с ДНК полипептида, если он экспрессируется в качестве белка-предшественника, который участвует в секреции полипептида. Промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию последовательности. Участок связывания рибосом функционально связан с кодирующей последовательностью, если он расположен так, чтобы способствовать трансляции. Обычно выражение "функционально связанный" означает, что связанные последовательности ДНК являются смежными и, в случае секреторной лидерной последовательности, являются смежными и находятся в рамке считывания. Однако энхансеры не должны быть смежными. Связывание осуществляют посредством лигирования в подходящих участках рестрикции. Если такие участки не существуют, используют синтетические олигонуклеотидные адапторы или линкеры в соответствии с общепринятой практикой. Более того, связанные нуклеиновые кислоты можно получать с помощью упомянутой выше технологии перекрывающейся ПЦР.
"Лигирование" относится к процессу формирования фосфодиэфирной связи между двумя фрагментами нуклеиновой кислоты. Для лигирования двух фрагментов концы этих фрагментов должны быть совместимыми друг с другом. В некоторых случаях эти концы обладают совместимостью сразу после расщепления эндонуклеазой. Однако может быть необходимо, чтобы сначала липкие концы, обычно образующиеся при расщеплении эндонуклеазой, были преобразованы в тупые концы, чтобы они стали совместимыми для лигирования. Для преобразования в тупые концы ДНК обрабатывают в подходящем буфере в течение по меньшей мере 15 минут при 15°C приблизительно 10 единицами фрагмента Кленова ДНК-полимеразы I или ДНК-полимеразой T4 в присутствии четырех дезоксирибонуклеотидтрифосфатов в соответствующем буферном растворе. Далее ДНК очищают экстракцией фенолом-хлороформом и преципитацией этанолом, или очисткой на диоксиде кремния. Фрагменты ДНК, подлежащие лигированию, добавляют в эквимолярных количествах к раствору. Этот раствор содержит ATP и буфер лигазы, лигазу, такую как ДНК-лигаза T4 в количестве приблизительно 10 единиц на 0,5 мкг ДНК. Если ДНК подлежит лигированию с вектором, вектор сначала линеаризуют посредством расщепления соответствующей эндонуклеазой(ами) рестрикции. Затем линеаризованный фрагмент обрабатывают бактериальной щелочной фосфатазой или фосфатазой кишечника теленка, тем самым препятствуя самолигированию фрагмента на стадии лигирования.
В способах получения по настоящему изобретению, антигенсвязывающий домен, который обладает более высокой антигенсвязывающей активностью в присутствии специфического для ткани-мишени соединения, чем в его отсутствие, который может быть отобран с помощью способа, описанного в представленном выше разделе "Антигенсвязывающий домен, зависимый от соединения, специфического для ткани-мишени", является выделенным. Например, когда антигенсвязывающий домен, выделенный таким образом, отобран из библиотеки нуклеотид, кодирующий антигенсвязывающий домен выделяют обычно амплификацией генов из вируса, такого как фаг, как описано в разделе "Примеры", ниже. Более того, когда антигенсвязывающий домен или антитело, выделенные таким образом, являются отобранными из культуральной среды клеток, таких как гибридом, ген антитела и т.п. можно выделять обычной амплификацией генов из клеток, как показано в разделе "Антитела", выше.
Способы получения антигенсвязывающих молекул
Настоящее изобретение относится к способам получения антигенсвязывающих молекул, антигенсвязывающая активность которых в присутствии специфического для ткани-мишени соединения является более высокой, чем антигенсвязывающая активность в отсутствие соединения.
Более конкретно, настоящее изобретение относится к способу получения антигенсвязывающих молекул, который включает стадии:
(a) определение антигенсвязывающей активности антигенсвязывающего домена в отсутствие специфического для ткани-мишени соединения;
(b) определение антигенсвязывающей активности антигенсвязывающего домена в присутствии специфического для ткани-мишени соединения;
(c) отбор антигенсвязывающего домена с более низкой антигенсвязывающей активностью в отсутствие специфического для ткани-мишени соединения, чем в присутствии соединения;
(d) связывание полинуклеотида, кодирующего антигенсвязывающий домен, отобранный на стадии (c), с полинуклеотидом, кодирующим полипептид, содержащий Fc-область;
(e) культивирование клеток, в которые введен вектор, с которым полинуклеотид, полученный на стадии (d), функционально связан; и
(f) сбор антигенсвязывающих молекул из культуральной среды клеток, культивированных на стадии (e).
Настоящее изобретение также относится к способу получения антигенсвязывающей молекулы, который включает стадии:
(a) определение антигенсвязывающей активности антигенсвязывающего домена в присутствии низкой концентрации специфического для ткани-мишени соединения;
(b) определение антигенсвязывающей активности антигенсвязывающего домена в присутствии высокой концентрации специфического для ткани-мишени соединения;
(c) отбор антигенсвязывающего домена с более низкой антигенсвязывающей активностью в присутствии низкой концентрации специфического для ткани-мишени соединения, чем в присутствии высокой концентрации соединения;
(d) связывание полинуклеотида, кодирующего антигенсвязывающий домен, отобранный на стадии (c), с полинуклеотидом, кодирующим полипептид, содержащий Fc-область;
(e) культивирование клеток, в которые введен вектор, с которым полинуклеотид, полученный на стадии (d) функционально связан; и
(f) сбор антигенсвязывающих молекул из культуральной среды клеток, культивированных на стадии (e).
Более того, настоящее изобретение относится к способу получения антигенсвязывающей молекулы, который включает стадии:
(a) контактирование антигенсвязывающих доменов или их библиотеки с антигеном в присутствии специфического для ткани-мишени соединения;
(b) помещение антигенсвязывающих доменов, которые связываются с антигеном на указанной стадии (a) в условия отсутствия соединения;
(c) выделение антигенсвязывающего домена, который диссоциирует на указанной стадии (b);
(d) связывание полинуклеотида, кодирующего антигенсвязывающий домен, отобранный на стадии (c), с полинуклеотидом, кодирующим полипептид, содержащий Fc-область;
(e) культивирование клеток, в которые введен вектор, с которым полинуклеотид, полученный на стадии (d), функционально связан; и
(f) сбор антигенсвязывающей молекулы из культуральной среды клеток, культивированных на стадии (e).
Кроме того, настоящее изобретение относится к способу получения антигенсвязывающей молекулы, который включает стадии:
(a) контактирование антигенсвязывающих доменов или их библиотеки с антигеном в присутствии высокой концентрации специфического для ткани-мишени соединения;
(b) помещение антигенсвязывающих доменов, которые связываются с антигеном на указанной стадии (a) в условия присутствия низкой концентрации соединения;
(c) выделение антигенсвязывающего домена, который диссоциирует на указанной стадии (b);
(d) связывание полинуклеотида, кодирующего антигенсвязывающий домен, отобранный на стадии (c) с полинуклеотидом, кодирующим полипептид, содержащий Fc-область;
(e) культивирование клеток, в которые введен вектор, с которым полинуклеотид, полученный на стадии (d), функционально связан; и
(f) сбор антигенсвязывающих молекул из культуральной среды клеток, культивированных на стадии (e).
Настоящее изобретение относится к способу получения антигенсвязывающей молекулы, который включает стадии:
(a) контактирование библиотеки антигенсвязывающих доменов с антигеном в отсутствие специфического для ткани-мишени соединения;
(b) выбор антигенсвязывающих доменов, которые не связываются с антигеном на указанной стадии (a);
(c) позволение антигенсвязывающим доменам, отобранным на указанной стадии (b), связаться с антигеном в присутствии соединения;
(d) выделение антигенсвязывающего домена, который связывается с антигеном на указанной стадии (c);
(e) связывание полинуклеотида, кодирующего антигенсвязывающий домен, отобранный на стадии (d), с полинуклеотидом, кодирующим полипептид, содержащий Fc-область;
(f) культивирование клеток, в которые введен вектор, с которым полинуклеотид, полученный на стадии (e), функционально связан; и
(g) сбор антигенсвязывающих молекул из культуральной среды клеток, культивированных на стадии (f).
Настоящее изобретение относится к способу получения антигенсвязывающей молекулы, который включает стадии:
(a) контактирования библиотеки антигенсвязывающих доменов с антигеном в присутствии низкой концентрации специфического для ткани-мишени соединения;
(b) отбор антигенсвязывающих доменов, которые не связываются с антигеном на указанной стадии (a);
(c) позволение антигенсвязывающим доменам, отобранным на указанной стадии (b), связаться с антигеном в присутствии высокой концентрации соединения;
(d) выделение антигенсвязывающего домена, который связывается с антигеном на указанной стадии (c);
(e) связывание полинуклеотида, кодирующего антигенсвязывающий домен, отобранный на стадии (d), с полинуклеотидом, кодирующим полипептид, содержащий Fc-область;
(f) культивирование клеток, в которые введен вектор, с которым полинуклеотид, полученный на стадии (e), функционально связан; и
(g) сбор антигенсвязывающих молекул из культуральной среды клеток, культивированных на стадии (f).
Настоящее изобретение относится к способу получения антигенсвязывающей молекулы, который включает стадии:
(a) контактирования библиотеки антигенсвязывающих доменов с колонкой с иммобилизованным на ней антигеном в присутствии специфического для ткани-мишени соединения;
(b) элюирование антигенсвязывающих доменов, которые связываются с колонкой на указанной стадии (a), из колонки в отсутствие соединения;
(c) выделение антигенсвязывающего домена, элюированного на указанной стадии (b);
(d) связывание полинуклеотида, кодирующего антигенсвязывающий домен, отобранный на стадии (c), с полинуклеотидом, кодирующим полипептид, содержащий Fc-область;
(e) культивирование клеток, в которые введен вектор, с которым полинуклеотид, полученный на стадии (d), функционально связан; и
(f) сбор антигенсвязывающих молекул из культуральной среды клеток, культивированных на стадии (e).
Настоящее изобретение относится к способу получения антигенсвязывающей молекулы, который включает стадии:
(a) контактирование библиотеки антигенсвязывающих доменов с колонкой с иммобилизованным на ней антигеном в присутствии высокой концентрации специфического для ткани-мишени соединения;
(b) элюирование антигенсвязывающих доменов, которые связываются с колонкой на указанной стадии (a), из колонки в присутствии низкой концентрации соединения;
(c) выделение антигенсвязывающего домена, элюированного на указанной стадии (b);
(d) связывание полинуклеотида, кодирующего антигенсвязывающий домен, отобранный на стадии (c), с полинуклеотидом, кодирующим полипептид, содержащий Fc-область;
(e) культивирование клеток, в которые введен вектор, с которым полинуклеотид, полученный на стадии (d), функционально связан; и
(f) сбор антигенсвязывающих молекул из культуральной среды клеток, культивированных на стадии (e).
Настоящее изобретение относится к способу получения антигенсвязывающей молекулы, который включает стадии:
(a) позволение библиотеке антигенсвязывающих доменов пройти через колонку с иммобилизованным на ней антигеном в отсутствие специфического для ткани-мишени соединения;
(b) сбор антигенсвязывающих доменов, которые элюируются без связывания с колонкой на указанной стадии (a);
(c) позволение антигенсвязывающим доменам, собранным на стадии (b), связаться с антигеном в присутствии соединения;
(d) выделение антигенсвязывающего домена, который связывается с антигеном на стадии (c);
(e) связывание полинуклеотида, кодирующего антигенсвязывающий домен, отобранный на стадии (d), с полинуклеотидом, кодирующим полипептид, содержащий Fc-область;
(f) культивирование клеток, в которые введен вектор, с которым полинуклеотид, полученный на стадии (e), функционально связан; и
(g) сбор антигенсвязывающих молекул из культуральной среды клеток, культивированных на стадии (f).
Настоящее изобретение относится к способу получения антигенсвязывающей молекулы, который включает стадии:
(a) позволение библиотеке антигенсвязывающих доменов пройти через колонку с иммобилизованным на ней антигеном в присутствии низкой концентрации специфического для ткани-мишени соединения;
(b) сбор антигенсвязывающих доменов, которые элюируются без связывания с колонкой на указанной стадии (a);
(c) позволение антигенсвязывающим доменам, собранным на указанной стадии (b), связаться с антигеном в присутствии высокой концентрации соединения;
(d) выделение антигенсвязывающего домена, который связывается с антигеном на указанной стадии (c);
(e) связывание полинуклеотида, кодирующего антигенсвязывающий домен, отобранный на стадии (d), с полинуклеотидом, кодирующим полипептид, содержащий Fc-область;
(f) культивирование клеток, в которые введен вектор, с которым полинуклеотид, полученный на стадии (e), функционально связан; и
(g) сбор антигенсвязывающих молекул из культуральной среды клеток, культивированных на стадии (f).
Более того, настоящее изобретение относится к способу получения антигенсвязывающей молекулы, который включает стадии:
(a) контактирования библиотеки антигенсвязывающих доменов с антигеном в присутствии специфического для ткани-мишени соединения;
(b) получение антигенсвязывающих доменов, которые связываются с антигеном на указанной стадии (a);
(c) помещение антигенсвязывающих доменов, полученных на указанной стадии (b), в условия отсутствия соединения;
(d) выделение антигенсвязывающего домена, антигенсвязывающая активность которого на указанной стадии (c) является более слабой, чем у эталона, выбранного на стадии (b);
(e) связывание полинуклеотида, кодирующего антигенсвязывающий домен, отобранный на стадии (d), с полинуклеотидом, кодирующим полипептид, содержащий Fc-область;
(f) культивирование клеток, в которые введен вектор, с которым полинуклеотид, полученный на стадии (e), функционально связан; и
(g) сбор антигенсвязывающих молекул из культуральной среды клеток, культивированных на стадии (f).
Настоящее изобретение относится к способу получения антигенсвязывающей молекулы, который включает стадии:
(a) контактирования библиотеки антигенсвязывающих доменов с антигеном в присутствии высокой концентрации специфического для ткани-мишени соединения;
(b) получение антигенсвязывающих доменов, которые связываются с антигеном на указанной стадии (a);
(c) помещение антигенсвязывающих доменов, полученных на стадии (b), в условия присутствия низкой концентрации соединения;
(d) выделение антигенсвязывающего домена, антигенсвязывающая активность которого на стадии (c) является более слабой, чем у эталона, отобранного на стадии (b);
(e) связывание полинуклеотида, кодирующего антигенсвязывающий домен, отобранный на стадии (d), с полинуклеотидом, кодирующим полипептид, содержащий Fc-область;
(f) культивирование клеток, в которые введен вектор, с которым полинуклеотид, полученный на стадии (e) функционально связан; и
(g) сбор антигенсвязывающих молекул из культуральной среды клеток, культивированных на стадии (f).
Неограничивающий вариант осуществления Fc-области, полинуклеотидная последовательность которой связана с полинуклеотидом, кодирующим антигенсвязывающий домен, представляет собой, например, Fc-область, содержащуюся в константной области антитела IgG1 (SEQ ID NO:5), IgG2 (SEQ ID NO:6), IgG3 (SEQ ID NO:7) или IgG4 (SEQ ID NO:8) человека. Fc-область представляет собой часть константной области тяжелой цепи антитела, начинающаяся с N-конца шарнирной области, который соответствует участку расщепления папаином в области аминокислота вблизи положения 216 в соответствии с нумерацией EU, и содержит шарнирную область, домен CH2 и домен CH3. Fc-область можно получать из IgG1 человека, однако она не ограничивается конкретным подклассом IgG.
Неограничивающий вариант осуществления Fc-области, полинуклеотидная последовательность которой связана с полинуклеотидом, кодирующим антигенсвязывающий домен, включает, например, Fc-область, у которой активность связывания с Fcγ-рецептором превышает активность связывания с Fcγ-рецептором у Fc-области нативного IgG человека. Примеры таких Fc-областей включают Fc-области, в которых по меньшей мере одна или несколько аминокислот, выбранных из группы, состоящей из аминокислот в положениях 221, 222, 223, 224, 225, 227, 228, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 246, 247, 249, 250, 251, 254, 255, 256, 258, 260, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 278, 279, 280, 281, 282, 283, 284, 285, 286, 288, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 311, 313, 315, 317, 318, 320, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 339, 376, 377, 378, 379, 380, 382, 385, 392, 396, 421, 427, 428, 429, 434, 436 и 440 в соответствии с нумерацией EU, отличаются от соответствующих аминокислотных остатков в соответствии с нумерацией EU в Fc-области, содержащейся в константной области антитела SEQ ID NO:5, 6, 7 или 8.
Более того, неограничивающий вариант осуществления упомянутой выше Fc-области включает, например, Fc-области, содержащие по меньшей мере одно или несколько изменений аминокислот, выбранных из группы, состоящей из:
изменение аминокислоты в положении 221 на Lys или Tyr;
изменение аминокислоты в положении 222 на Phe, Trp, Glu или Tyr;
изменение аминокислоты в положении 223 на Phe, Trp, Glu или Lys;
изменение аминокислоты в положении 224 на Phe, Trp, Glu или Tyr;
изменение аминокислоты в положении 225 на Glu, Lys или Trp;
изменение аминокислоты в положении 227 на Glu, Gly, Lys или Tyr;
изменение аминокислоты в положении 228 на Glu, Gly, Lys или Tyr;
изменение аминокислоты в положении 230 на Ala, Glu, Gly или Tyr;
изменение аминокислоты в положении 231 на Glu, Gly, Lys, Pro или Tyr;
изменение аминокислоты в положении 232 на Glu, Gly, Lys или Tyr;
изменение аминокислоты в положении 233 на Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 234 на Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 235 на Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 236 на Ala, Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 237 на Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 238 на Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 239 на Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 240 на Ala, Ile, Met или Thr;
изменение аминокислоты в положении 241 на Asp, Glu, Leu, Arg, Trp или Tyr;
изменение аминокислоты в положении 243 на Leu, Glu, Leu, Gln, Arg, Trp или Tyr;
изменение аминокислоты в положении 244 на His;
изменение аминокислоты в положении 245 на Ala;
изменение аминокислоты в положении 246 на Asp, Glu, His или Tyr;
изменение аминокислоты в положении 247 на Ala, Phe, Gly, His, Ile, Leu, Met, Thr, Val или Tyr;
изменение аминокислоты в положении 249 на Glu, His, Gln или Tyr;
изменение аминокислоты в положении 250 на Glu или Gln;
изменение аминокислоты в положении 251 на Phe;
изменение аминокислоты в положении 254 на Phe, Met или Tyr;
изменение аминокислоты в положении 255 на Glu, Leu или Tyr;
изменение аминокислоты в положении 256 на Ala, Met или Pro;
изменение аминокислоты в положении 258 на Asp, Glu, His, Ser или Tyr;
изменение аминокислоты в положении 260 на Asp, Glu, His или Tyr;
изменение аминокислоты в положении 262 на Ala, Glu, Phe, Ile или Thr;
изменение аминокислоты в положении 263 на Ala, Ile, Met или Thr;
изменение аминокислоты в положении 264 на Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Trp или Tyr;
изменение аминокислоты в положении 265 на Ala, Leu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 266 на Ala, Ile, Met или Thr;
изменение аминокислоты в положении 267 на Asp, Glu, Phe, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 268 на Asp, Glu, Phe, Gly, Ile, Lys, Leu, Met, Pro, Gln, Arg, Thr, Val или Trp;
изменение аминокислоты в положении 269 на Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 270 на Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Gln, Arg, Ser, Thr, Trp или Tyr;
изменение аминокислоты в положении 271 на Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 272 на Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 273 на Phe или Ile;
изменение аминокислоты в положении 274 на Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 275 на Leu или Trp;
изменение аминокислоты в положении 276 на Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 278 на Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp;
изменение аминокислоты в положении 279 на Ala;
изменение аминокислоты в положении 280 на Ala, Gly, His, Lys, Leu, Pro, Gln, Trp или Tyr;
изменение аминокислоты в положении 281 на Asp, Lys, Pro или Tyr;
изменение аминокислоты в положении 282 на Glu, Gly, Lys, Pro или Tyr;
изменение аминокислоты в положении 283 на Ala, Gly, His, Ile, Lys, Leu, Met, Pro, Arg или Tyr;
изменение аминокислоты в положении 284 на Asp, Glu, Leu, Asn, Thr или Tyr;
изменение аминокислоты в положении 285 на Asp, Glu, Lys, Gln, Trp или Tyr;
изменение аминокислоты в положении 286 на Glu, Gly, Pro или Tyr;
изменение аминокислоты в положении 288 на Asn, Asp, Glu или Tyr;
изменение аминокислоты в положении 290 на Asp, Gly, His, Leu, Asn, Ser, Thr, Trp или Tyr;
изменение аминокислоты в положении 291 на Asp, Glu, Gly, His, Ile, Gln или Thr;
изменение аминокислоты в положении 292 на Ala, Asp, Glu, Pro, Thr или Tyr;
изменение аминокислоты в положении 293 на Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 294 на Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 295 на Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 296 на Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr или Val;
изменение аминокислоты в положении 297 на Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 298 на Ala, Asp, Glu, Phe, His, Ile, Lys, Met, Asn, Gln, Arg, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 299 на Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr;
изменение аминокислоты в положении 300 на Ala, Asp, Glu, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val или Trp;
изменение аминокислоты в положении 301 на Asp, Glu, His или Tyr;
изменение аминокислоты в положении 302 на Ile;
изменение аминокислоты в положении 303 на Asp, Gly или Tyr;
изменение аминокислоты в положении 304 на Asp, His, Leu, Asn или Thr;
изменение аминокислоты в положении 305 на Glu, Ile, Thr или Tyr;
изменение аминокислоты в положении 311 на Ala, Asp, Asn, Thr, Val или Tyr;
изменение аминокислоты в положении 313 на Phe;
изменение аминокислоты в положении 315 на Leu;
изменение аминокислоты в положении 317 на Glu или Gln;
изменение аминокислоты в положении 318 на His, Leu, Asn, Pro, Gln, Arg, Thr, Val или Tyr;
изменение аминокислоты в положении 320 на Asp, Phe, Gly, His, Ile, Leu, Asn, Pro, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 322 на Ala, Asp, Phe, Gly, His, Ile, Pro, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 323 на Ile;
изменение аминокислоты в положении 324 на Asp, Phe, Gly, His, Ile, Leu, Met, Pro, Arg, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 325 на Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 326 на Ala, Asp, Glu, Gly, Ile, Leu, Met, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 327 на Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 328 на Ala, Asp, Glu, Phe, Gly, His, Ile, Lys, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 329 на Asp, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 330 на Cys, Glu, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 331 на Asp, Phe, His, Ile, Leu, Met, Gln, Arg, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 332 на Ala, Asp, Glu, Phe, Gly, His, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 333 на Ala, Asp, Glu, Phe, Gly, His, Ile, Leu, Met, Pro, Ser, Thr, Val или Tyr;
изменение аминокислоты в положении 334 на Ala, Glu, Phe, Ile, Leu, Pro или Thr;
изменение аминокислоты в положении 335 на Asp, Phe, Gly, His, Ile, Leu, Met, Asn, Pro, Arg, Ser, Val, Trp или Tyr;
изменение аминокислоты в положении 336 на Glu, Lys или Tyr;
изменение аминокислоты в положении 337 на Glu, His или Asn;
изменение аминокислоты в положении 339 на Asp, Phe, Gly, Ile, Lys, Met, Asn, Gln, Arg, Ser или Thr;
изменение аминокислоты в положении 376 на Ala или Val;
изменение аминокислоты в положении 377 на Gly или Lys;
изменение аминокислоты в положении 378 на Asp;
изменение аминокислоты в положении 379 на Asn;
изменение аминокислоты в положении 380 на Ala, Asn или Ser;
изменение аминокислоты в положении 382 на Ala или Ile;
изменение аминокислоты в положении 385 на Glu;
изменение аминокислоты в положении 392 на Thr;
изменение аминокислоты в положении 396 на Leu;
изменение аминокислоты в положении 421 на Lys;
изменение аминокислоты в положении 427 на Asn;
изменение аминокислоты в положении 428 на Phe или Leu;
изменение аминокислоты в положении 429 на Met;
изменение аминокислоты в положении 434 на Trp;
изменение аминокислоты в положении 436 на Ile; и
изменение аминокислоты в положении 440 на Gly, His, Ile, Leu или Tyr;
в соответствии с нумерацией EU, в аминокислотных остатках Fc-области, содержащейся в константной области антитела SEQ ID NO:5, 6, 7 или 8. Количество измененных аминокислот конкретно не ограничено; может быть изменена аминокислота только в одном участке или могут быть изменены аминокислоты в двух или более участках. Комбинации изменения аминокислот в двух или более участках включают, например, изменения, описанные в таблице 1 (с таблицы 1-1 по таблицу 1-3).
Неограничивающий вариант осуществления Fc-области полинуклеотидная последовательность которой связана с полинуклеотидом, кодирующим антигенсвязывающий домен, представляет собой, например, Fc-область, обладающую активностью связывания с ингибиторным Fcγ-рецептором, которая превышает активность связывания с активирующим Fcγ-рецептором. В частности, неограничивающий вариант осуществления таких Fc-областей представляет собой Fc-область, активность связывания который с FcγRIIb является более высокой, чем активность связывания с любым из Fcγ-рецепторов человека: FcγRIa, FcγRIIa, FcγRIIIa и/или FcγRIIIb.
Неограничивающий вариант осуществления упомянутой выше Fc-области предпочтительно включает, например, Fc-область, в которой аминокислота в положении 238 или 328 в соответствии с нумерацией EU в Fc-области, содержащейся в константной области антитела SEQ ID NO:5, 6, 7 или 8, изменена на аминокислоту, отличающуюся от аминокислоты в нативной Fc-области. Предпочтительным примером таких Fc-областей является Fc-область, обладающая одним или несколькими из следующих изменений: изменение аминокислоты в положении 238 на Asp, и изменение аминокислоты в положении 328 на Glu, в соответствии с нумерацией EU, в упомянутой выше Fc-области.
В другом неограничивающем варианте осуществления упомянутой выше Fc-области, предпочтительные примеры включают Fc-области, обладающие одним или несколькими из изменений, проиллюстрированных в PCT/JP2012/054624: замена Pro в положении 238 в соответствии с нумерацией EU на Asp, изменение аминокислоты в положении 237 в соответствии с нумерацией EU на Trp, изменение аминокислоты в положении 237 в соответствии с нумерацией EU на Phe, изменение аминокислоты в положении 267 в соответствии с нумерацией EU на Val, изменение аминокислоты в положении 267 в соответствии с нумерацией EU на Gln, изменение аминокислоты в положении 268 в соответствии с нумерацией EU на Asn, изменение аминокислоты в положении 271 в соответствии с нумерацией EU на Gly, изменение аминокислоты в положении 326 в соответствии с нумерацией EU на Leu, изменение аминокислоты в положении 326 в соответствии с нумерацией EU на Gln, изменение аминокислоты в положении 326 в соответствии с нумерацией EU на Glu, изменение аминокислоты в положении 326 в соответствии с нумерацией EU на Met, изменение аминокислоты в положении 239 в соответствии с нумерацией EU на Asp, изменение аминокислоты в положении 267 в соответствии с нумерацией EU на Ala, изменение аминокислоты в положении 234 в соответствии с нумерацией EU на Trp, изменение аминокислоты в положении 234 в соответствии с нумерацией EU на Tyr, изменение аминокислоты в положении 237 в соответствии с нумерацией EU на Ala, изменение аминокислоты в положении 237 в соответствии с нумерацией EU на Asp, изменение аминокислоты в положении 237 в соответствии с нумерацией EU на Glu, изменение аминокислоты в положении 237 в соответствии с нумерацией EU на Leu, изменение аминокислоты в положении 237 в соответствии с нумерацией EU на Met, изменение аминокислоты в положении 237 в соответствии с нумерацией EU на Tyr, изменение аминокислоты в положении 330 в соответствии с нумерацией EU на Lys, изменение аминокислоты в положении 330 в соответствии с нумерацией EU на Arg, изменение аминокислоты в положении 233 в соответствии с нумерацией EU на Asp, изменение аминокислоты в положении 268 в соответствии с нумерацией EU на Asp, изменение аминокислоты в положении 268 в соответствии с нумерацией EU на Glu, изменение аминокислоты в положении 326 в соответствии с нумерацией EU на Asp, изменение аминокислоты в положении 326 в соответствии с нумерацией EU на Ser, изменение аминокислоты в положении 326 в соответствии с нумерацией EU на Thr, изменение аминокислоты в положении 323 в соответствии с нумерацией EU на Ile, изменение аминокислоты в положении 323 в соответствии с нумерацией EU на Leu, изменение аминокислоты в положении 323 в соответствии с нумерацией EU на Met, изменение аминокислоты в положении 296 в соответствии с нумерацией EU на Asp, изменение аминокислоты в положении 326 в соответствии с нумерацией EU на Ala, изменение аминокислоты в положении 326 в соответствии с нумерацией EU на Asn, и изменение аминокислоты в положении 330 в соответствии с нумерацией EU на Met.
Неограничивающий вариант осуществления Fc-области, полинуклеотидная последовательность которой связана с полинуклеотидом, кодирующим антигенсвязывающий домен, включает Fc-области, обладающие активностью связывания с FcRn в диапазоне кислых значений pH. Аминокислоты, которые претерпевают такое изменение, включают, например, аминокислоты в положениях 252, 254, 256, 309, 311, 315, 433 и/или 434 в соответствии с нумерацией EU, и аминокислоты в положениях 253, 310, 435 и/или 426, которые комбинированы с упомянутыми выше аминокислотами, как описано в WO 1997/034631. Предпочтительные примеры включают аминокислоты в положениях 238, 252, 253, 254, 255, 256, 265, 272, 286, 288, 303, 305, 307, 309, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 386, 388, 400, 413, 415, 424, 433, 434, 435, 436, 439 и/или 447 (нумерация EU), как описано в WO 2000/042072. Аналогично, предпочтительные примеры аминокислот, которые претерпевают такое изменение, включают аминокислоты в положениях 251, 252, 254, 255, 256, 308, 309, 311, 312, 385, 386, 387, 389, 428, 433, 434 и/или 436 в соответствии с нумерацией EU, как описано в WO 2002/060919. Более того, аминокислоты, которые претерпевают такое изменение, представляют собой, например, аминокислоты в положениях 250, 314 и 428 в соответствии с нумерацией EU, как описано в WO2004/092219. Кроме того, предпочтительные примеры аминокислот, которые претерпевают такое изменение, включают аминокислоты в положениях 238, 244, 245, 249, 252, 256, 257, 258, 260, 262, 270, 272, 279, 283, 285, 286, 288, 293, 307, 311, 312, 316, 317, 318, 332, 339, 341, 343, 375, 376, 377, 378, 380, 382, 423, 427, 430, 431, 434, 436, 438, 440 и/или 442, как описано в WO 2006/020114. Более того, предпочтительные примеры аминокислот, которые претерпевают такое изменение, включают аминокислоты в положениях 251, 252, 307, 308, 378, 428, 430, 434 и/или 436 в соответствии с нумерацией EU, как описано в WO 2010/045193.
Неограничивающий вариант осуществления упомянутой выше Fc-области включает, например, Fc-области, имеющие по меньшей мере одно или несколько изменений аминокислот, выбранных из группы, состоящей из:
изменение аминокислоты в положении 251 на Arg или Leu;
изменение аминокислоты в положении 252 на Phe, Ser, Thr или Tyr;
изменение аминокислоты в положении 254 на Ser или Thr;
изменение аминокислоты в положении 255 на Arg, Gly, Ile или Leu;
изменение аминокислоты в положении 256 на Ala, Arg, Asn, Asp, Gln, Glu или Thr;
изменение аминокислоты в положении 308 на Ile или Thr;
изменение аминокислоты в положении 309 на Pro;
изменение аминокислоты в положении 311 на Glu, Leu или Ser;
изменение аминокислоты в положении 312 на Ala или Asp;
изменение аминокислоты в положении 314 на Ala или Leu;
изменение аминокислоты в положении 385 на Ala, Arg, Asp, Gly, His, Lys, Ser или Thr;
изменение аминокислоты в положении 386 на Arg, Asp, Ile, Lys, Met, Pro, Ser или Thr;
изменение аминокислоты в положении 387 на Ala, Arg, His, Pro, Ser или Thr;
изменение аминокислоты в положении 389 на Asn, Pro или Ser;
изменение аминокислоты в положении 428 на Leu, Met, Phe, Ser или Thr;
изменение аминокислоты в положении 433 на Arg, Gln, His, Ile, Lys, Pro или Ser;
изменение аминокислоты в положении 434 на His, Phe или Tyr; и
изменение аминокислоты в положении 436 на Arg, Asn, His, Lys, Met или Thr,
в соответствии с нумерацией EU, в аминокислотных остатках Fc-области, содержащейся в константной области антитела SEQ ID NO:5, 6, 7 или 8. Количество аминокислот, подлежащих изменению, конкретно не ограничено; может быть изменена аминокислота только в одном участке или могут быть изменены аминокислоты в двух или более участках.
Другой неограничивающий вариант осуществления Fc-области, активность связывания которой с FcRn в диапазоне кислых значений pH является более сильной, чем активность связывания у исходной Fc-области IgG1 человека, включает Fc-области, в которых аминокислота в положении 308 представляет собой Ile, аминокислота в положении 309 представляет собой Pro, и/или аминокислота в положении 311 представляет собой Glu, в соответствии с нумерацией EU, в аминокислотных остатках Fc-области, содержащейся в константной области антитела SEQ ID NO:5, 6, 7 или 8. Другой неограничивающий вариант осуществления этой Fc-области может включать Fc-области, содержащие Thr вместо аминокислоты в положении 308, Pro вместо аминокислоты в положении 309, Leu вместо аминокислоты в положении 311, Ala вместо аминокислоты в положении 312, и/или Ala вместо аминокислоты в положении 314. Более того, другой неограничивающий вариант осуществления этого изменения может включать Fc-области, содержащие Ile или Thr вместо аминокислоты в положении 308, Pro вместо аминокислоты в положении 309, Glu, Leu или Ser вместо аминокислоты в положении 311, Ala вместо аминокислоты в положении 312, и/или Ala или Leu вместо аминокислоты в положении 314. Другой неограничивающий вариант осуществления этого изменения может включать Fc-области, содержащие Thr вместо аминокислоты в положении 308, Pro вместо аминокислоты в положении 309, Ser вместо аминокислоты в положении 311, Asp вместо аминокислоты в положении 312, и/или Leu вместо аминокислоты в положении 314.
Другой неограничивающий вариант осуществления Fc-области, активность связывания которой с FcRn в диапазоне кислых значений pH является более сильной, чем активность связывания у исходной Fc-области IgG1 человека, включает, например, Fc-области, содержащие Leu вместо аминокислоты в положении 251, Tyr вместо аминокислоты в положении 252, Ser или Thr вместо аминокислоты в положении 254, Arg вместо аминокислоты в положении 255, и/или Glu вместо аминокислоты в положении 256, в соответствии с нумерацией EU, в аминокислотных остатках Fc-области, включенной в константную область антитела SEQ ID NO:5, 6, 7 или 8.
Другой неограничивающий вариант осуществления Fc-области, активность связывания которой с FcRn в диапазоне кислых значений pH является более сильной, чем активность связывания у исходной Fc-области IgG1 человека, включает, например, Fc-области, содержащие Leu, Met, Phe, Ser или Thr вместо аминокислоты в положении 428, Arg, Gln, His, Ile, Lys, Pro или Ser вместо аминокислоты в положении 433, His, Phe или Tyr вместо аминокислоты в положении 434, и/или Arg, Asn, His, Lys, Met или Thr вместо аминокислоты в положении 436, в соответствии с нумерацией EU, в аминокислотных остатках Fc-области, включенной в константную область антитела SEQ ID NO:5, 6, 7 или 8. Более того, другой неограничивающий вариант осуществления этого изменения включает Fc-области, содержащие His или Met вместо аминокислоты в положении 428 и/или His или Met вместо аминокислоты в положении 434.
Другой неограничивающий вариант осуществления Fc-области, активность связывания которой с FcRn в диапазоне кислых значений pH является более сильной, чем активность связывания исходной Fc-области IgG1 человека, может представлять собой, например, изменения, включающие Arg вместо аминокислоты в положении 385, Thr вместо аминокислоты в положении 386, Arg вместо аминокислоты в положении 387, и/или Pro вместо аминокислоты в положении 389, в соответствии с нумерацией EU, в аминокислотных остатках Fc-области, включенной в константную область антитела SEQ ID NO:5, 6, 7 или 8. Другой неограничивающий вариант осуществления этого изменения включает Fc-области, содержащие Asp вместо аминокислоты в положении 385, Pro вместо аминокислоты в положении 386, и/или Ser вместо аминокислоты в положении 389.
Другой неограничивающий вариант осуществления Fc-области, активность связывания которой с FcRn в диапазоне кислых значений pH является более сильной, чем активность связывания у исходной Fc-области IgG1 человека, включает Fc-области, содержащие по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из
Gln или Glu вместо аминокислоты в положении 250; и
Leu или Phe вместо аминокислоты в положении 428, в соответствии с нумерацией EU,
в аминокислотных остатках Fc-области, содержащейся в константной области антитела SEQ ID NO:5, 6, 7 или 8.
Другой неограничивающий вариант осуществления Fc-области, активность связывания которой с FcRn в диапазоне кислых значений pH является более сильной, чем активность связывания у исходной Fc-области IgG1 человека, включает, например, Fc-области, содержащие Gln вместо аминокислоты в положении 250, и/или Leu или Phe вместо аминокислоты в положении 428, в соответствии с нумерацией EU, в аминокислотных остатках Fc-области, содержащейся в константной области антитела SEQ ID NO:5, 6, 7 или 8. Другой неограничивающий вариант осуществления этого изменения может включать Glu вместо аминокислоты в положении 250, и/или Leu или Phe вместо аминокислоты в положении 428.
Другой неограничивающий вариант осуществления Fc-области, активность связывания которой с FcRn в диапазоне кислых значений pH является более сильной, чем активность связывания у исходной Fc-области IgG1 человека, включает Fc-области, содержащие по меньшей мере две или более аминокислот, выбранных из группы, состоящей из:
Asp или Glu вместо аминокислоты в положении 251;
Tyr вместо аминокислоты в положении 252;
Gln вместо аминокислоты в положении 307;
Pro вместо аминокислоты в положении 308;
Val вместо аминокислоты в положении 378;
Ala вместо аминокислоты в положении 380;
Leu вместо аминокислоты в положении 428;
Ala или Lys вместо аминокислоты в положении 430;
Ala, His, Ser или Tyr вместо аминокислоты в положении 434; и
Ile вместо аминокислоты в положении 436;
в соответствии с нумерацией EU, в аминокислотных остатках Fc-области, содержащейся в константной области антитела SEQ ID NO:5, 6, 7 или 8.
Другой неограничивающий вариант осуществления Fc-области, активность связывания которой с FcRn в диапазоне кислых значений pH является более сильной, чем активность связывания у исходной Fc-области IgG1 человека, включает, например, Fc-области, содержащие Gln вместо аминокислоты в положении 307, и Ala или Ser вместо аминокислоты в положении 434, в соответствии с нумерацией EU, в аминокислотных остатках Fc-области, содержащейся в константной области антитела SEQ ID NO:5, 6, 7 или 8. Другой неограничивающий вариант осуществления этой Fc-области включает Fc-области, содержащие Pro вместо аминокислоты в положении 308, и Ala вместо аминокислоты в положении 434. Более того, другой неограничивающий вариант осуществления этой Fc-области включает Fc-области, содержащие Tyr вместо аминокислоты в положении 252, и Ala вместо аминокислоты в положении 434. Другой неограничивающий вариант осуществления этой Fc-области включает Fc-области, содержащие Val вместо аминокислоты в положении 378, и Ala вместо аминокислоты в положении 434. Другой неограничивающий вариант осуществления этой Fc-области включает изменения, включающие Leu вместо аминокислоты в положении 428, и Ala вместо аминокислоты в положении 434. Другой неограничивающий вариант осуществления этой Fc-области включает Fc-области, содержащие Ala вместо аминокислоты в положении 434, и Ile вместо аминокислоты в положении 436. Более того, другой неограничивающий вариант осуществления этого изменения включает Fc-области, содержащие Pro вместо аминокислоты в положении 308, и Tyr вместо аминокислоты в положении 434. Кроме того, другой неограничивающий вариант осуществления этого изменения включает Fc-области, содержащие Gln вместо аминокислоты в положении 307, и Ile вместо аминокислоты в положении 436.
Другой неограничивающий вариант осуществления Fc-области, активность связывания которой с FcRn в диапазоне кислых значений pH является более сильной, чем активность связывания у исходной Fc-области IgG1 человека, включает Fc-области, содержащие любой из Gln вместо аминокислоты в положении 307, Ala вместо аминокислоты в положении 380, и Ser вместо аминокислоты в положении 434, в соответствии с нумерацией EU, в аминокислотных остатках Fc-области, содержащейся в константной области антитела SEQ ID NO:5, 6, 7 или 8. Другой неограничивающий вариант осуществления этой Fc-области включает Fc-области, содержащие Gln вместо аминокислоты в положении 307, Ala вместо аминокислоты в положении 380, и Ala вместо аминокислоты в положении 434. Более того, другой неограничивающий вариант осуществления этой Fc-области включает Fc-области, содержащие Tyr вместо аминокислоты в положении 252, Pro вместо аминокислоты в положении 308, и Tyr вместо аминокислоты в положении 434. Другой неограничивающий вариант осуществления этой Fc-области включает Fc-области, содержащие Asp вместо аминокислоты в положении 251, Gln вместо аминокислоты в положении 307, и His вместо аминокислоты в положении 434.
Другой неограничивающий вариант осуществления Fc-области, активность связывания которой с FcRn в диапазоне кислых значений pH является более сильной, чем активность связывания у исходной Fc-области IgG1 человека, включает по меньшей мере одно или несколько изменений аминокислот, выбранных из группы, состоящей из:
изменение аминокислоты в положении 238 на Leu;
изменение аминокислоты в положении 244 на Leu;
изменение аминокислоты в положении 245 на Arg;
изменение аминокислоты в положении 249 на Pro;
изменение аминокислоты в положении 252 на Tyr;
изменение аминокислоты в положении 256 на Pro;
изменение аминокислоты в положении 257 на Ala, Ile, Met, Asn, Ser или Val;
изменение аминокислоты в положении 258 на Asp;
изменение аминокислоты в положении 260 на Ser;
изменение аминокислоты в положении 262 на Leu;
изменение аминокислоты в положении 270 на Lys;
изменение аминокислоты в положении 272 на Leu или Arg;
изменение аминокислоты в положении 279 на Ala, Asp, Gly, His, Met, Asn, Gln, Arg, Ser, Thr, Trp или Tyr;
изменение аминокислоты в положении 283 на Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Arg, Ser, Thr, Trp или Tyr;
изменение аминокислоты в положении 285 на Asn;
изменение аминокислоты в положении 286 на Phe;
изменение аминокислоты в положении 288 на Asn или Pro;
изменение аминокислоты в положении 293 на Val;
изменение аминокислоты в положении 307 на Ala, Glu или Met;
изменение аминокислоты в положении 311 на Ala, Ile, Lys, Leu, Met, Val или Trp;
изменение аминокислоты в положении 312 на Pro;
изменение аминокислоты в положении 316 на Lys;
изменение аминокислоты в положении 317 на Pro;
изменение аминокислоты в положении 318 на Asn или Thr;
изменение аминокислоты в положении 332 на Phe, His, Lys, Leu, Met, Arg, Ser или Trp;
изменение аминокислоты в положении 339 на Asn, Thr или Trp;
изменение аминокислоты в положении 341 на Pro;
изменение аминокислоты в положении 343 на Glu, His, Lys, Gln, Arg, Thr или Tyr;
изменение аминокислоты в положении 375 на Arg;
изменение аминокислоты в положении 376 на Gly, Ile, Met, Pro, Thr или Val;
изменение аминокислоты в положении 377 на Lys;
изменение аминокислоты в положении 378 на Asp или Asn;
изменение аминокислоты в положении 380 на Asn, Ser или Thr;
изменение аминокислоты в положении 382 на Phe, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val, Trp или Tyr;
изменение аминокислоты в положении 423 на Asn;
изменение аминокислоты в положении 427 на Asn;
изменение аминокислоты в положении 430 на Ala, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Gln, Arg, Ser, Thr, Val или Tyr;
изменение аминокислоты в положении 431 на His или Asn;
изменение аминокислоты в положении 434 на Phe, Gly, His, Trp или Tyr;
изменение аминокислоты в положении 436 на Ile, Leu или Thr;
изменение аминокислоты в положении 438 на Lys, Leu, Thr или Trp;
изменение аминокислоты в положении 440 на Lys; и
изменение аминокислоты в положении 442 на Lys;
в соответствии с нумерацией EU, в аминокислотных остатках Fc-области, содержащейся в константной области антитела SEQ ID NO:5, 6, 7 или 8. Количество аминокислот, подлежащих изменению, конкретно не ограничено, и могут быть изменены аминокислоты только в двух участках и могут быть изменены аминокислоты в трех или более участках.
Другой неограничивающий вариант осуществления Fc-области, активность связывания которой с FcRn в диапазоне кислых значений pH является более сильной, чем активность связывания исходной Fc-области IgG1 человека, включает Fc-области, содержащие Ile вместо аминокислоты в положении 257, и Ile вместо аминокислоты в положении 311, в соответствии с нумерацией EU, в аминокислотных остатках Fc-области, содержащейся в константной области антитела SEQ ID NO:5, 6, 7 или 8. Другой неограничивающий вариант осуществления этой Fc-области включает Fc-области, содержащие Ile вместо аминокислоты в положении 257 и His вместо аминокислоты в положении 434. Другой неограничивающий вариант осуществления этой Fc-области включает Fc-области, содержащие Val вместо аминокислоты в положении 376 и His вместо аминокислоты в положении 434.
Неограничивающий вариант осуществления Fc-области, полинуклеотидная последовательность которой связана с полинуклеотидом, кодирующим антигенсвязывающий домен, включает, например, Fc-области, обладающие активностью связывания с FcRn человека в диапазоне нейтральных значений pH. Примеры Fc-областей, обладающих активностью связывания с FcRn человека в диапазоне нейтральных значений pH, включают Fc-области, в которых по меньшей мере одна или несколько аминокислот в положениях, выбранных из группы, состоящей из положений 221-225, 227, 228, 230, 232, 233-241, 243-252, 254-260, 262-272, 274, 276, 278-289, 291-312, 315-320, 324, 325, 327-339, 341, 343, 345, 360, 362, 370, 375-378, 380, 382, 385-387, 389, 396, 414, 416, 423, 424, 426-438, 440 и 442, в соответствии с нумерацией EU, заменены в аминокислотных остатках Fc-области, включенной в константную область антитела SEQ ID NO:5, 6, 7 или 8.
Другой неограничивающий вариант осуществления упомянутой выше Fc-области, обладающей активностью связывания с FcRn в диапазоне нейтральных значений pH, включает Fc-области, в которых аминокислоты в положениях 237, 248, 250, 252, 254, 255, 256, 257, 258, 265, 286, 289, 297, 298, 303, 305, 307, 308, 309, 311, 312, 314, 315, 317, 332, 334, 360, 376, 380, 382, 384, 385, 386, 387, 389, 424, 428, 433, 434, и 436, в соответствии с нумерацией EU, заменены в аминокислотных остатках Fc-области, содержащейся в константной области антитела SEQ ID NO:5, 6, 7 или 8. Путем замены по меньшей мере одной аминокислоты, выбранной из этих аминокислот, отличающейся аминокислотой, Fc-область, включенную в антигенсвязывающую молекулу, можно связывать с FcRn человека в диапазоне нейтральных значений pH.
Другой неограничивающий вариант осуществления упомянутой выше Fc-области, обладающей активностью связывания с FcRn в диапазоне нейтральных значений pH, включает Fc-области, содержащие по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из:
Met вместо аминокислоты в положении 237;
Ile вместо аминокислоты в положении 248;
Ala, Phe, Ile, Met, Gln, Ser, Val, Trp или Tyr вместо аминокислоты в положении 250;
Phe, Trp или Tyr вместо аминокислоты в положении 252;
Thr вместо аминокислоты в положении 254;
Glu вместо аминокислоты в положении 255;
Asp, Asn, Glu или Gln вместо аминокислоты в положении 256;
Ala, Gly, Ile, Leu, Met, Asn, Ser, Thr или Val вместо аминокислоты в положении 257;
His вместо аминокислоты в положении 258:
Ala вместо аминокислоты в положении 265;
Ala или Glu вместо аминокислоты в положении 286;
His вместо аминокислоты в положении 289;
Ala вместо аминокислоты в положении 297;
Ala вместо аминокислоты в положении 303;
Ala вместо аминокислоты в положении 305;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Met, Asn, Pro, Gln, Arg, Ser, Val, Trp или Tyr вместо аминокислоты в положении 307;
Ala, Phe, Ile, Leu, Met, Pro, Gln или Thr вместо аминокислоты в положении 308;
Ala, Asp, Glu, Pro или Arg вместо аминокислоты в положении 309;
Ala, His или Ile вместо аминокислоты в положении 311;
Ala или His вместо аминокислоты в положении 312;
Lys или Arg вместо аминокислоты в положении 314;
Ala, Asp или His вместо аминокислоты в положении 315;
Ala вместо аминокислоты в положении 317;
Val вместо аминокислоты в положении 332;
Leu вместо аминокислоты в положении 334;
His вместо аминокислоты в положении 360;
Ala вместо аминокислоты в положении 376;
Ala вместо аминокислоты в положении 380;
Ala вместо аминокислоты в положении 382;
Ala вместо аминокислоты в положении 384;
Asp или His вместо аминокислоты в положении 385;
Pro вместо аминокислоты в положении 386;
Glu вместо аминокислоты в положении 387;
Ala или Ser вместо аминокислоты в положении 389;
Ala вместо аминокислоты в положении 424;
Ala, Asp, Phe, Gly, His, Ile, Lys, Leu, Asn, Pro, Gln, Ser, Thr, Val, Trp или Tyr вместо аминокислоты в положении 428;
Lys вместо аминокислоты в положении 433;
Ala, Phe, His, Ser, Trp или Tyr вместо аминокислоты в положении 434; и
His, Ile, Leu, Phe, Thr или Val вместо аминокислоты в положении 436;
в соответствии с нумерацией EU. Количество аминокислот, подлежащих изменению, конкретно не ограничено, и может быть изменена аминокислота только в одном участке или могут быть изменены аминокислоты в двух или более участках. Комбинации этих изменений аминокислот включают, например, изменения, описанные в таблицах с 2-1 по 2-33.
Неограничивающий вариант осуществления Fc-области, полинуклеотидная последовательность которой связана с полинуклеотидом, кодирующим антигенсвязывающий домен, включает, например, Fc-области, активность связывания которых с активирующим FcγR, является более низкой, чем активность связывания нативной Fc-области с активирующим FcγR. Другой неограничивающий вариант осуществления Fc-области предпочтительно включает, например, Fc-области, в которых одна или несколько аминокислот в положениях 234, 235, 236, 237, 238, 239, 270, 297, 298, 325, 328, и 329 в соответствии с нумерацией EU изменены на аминокислоты, которые отличаются от аминокислот нативной Fc-области SEQ ID NO:5, 6, 7 или 8. Изменения в Fc-области не ограничиваются описанным выше примером, и они могут представлять собой, например, такие изменения, как дегликозилгликозилирование (N297A и N297Q), IgG1-L234A/L235A, IgG1-A325A/A330S/P331S, IgG1-C226S/C229S, IgG1-C226S/C229S/E233P/L234V/L235A, IgG1-L234F/L235E/P331S, IgG1-S267E/L328F, IgG2-V234A/G237A, IgG2-H268Q/V309L/A330S/A331S, IgG4-L235A/G237A/E318A и IgG4-L236E, описанные в Cur. Opin. in Biotech. (2009) 20 (6), 685-691; изменения, такие как G236R/L328R, L235G/G236R, N325A/L328R и N325L/L328R, описанные в WO 2008/092117; вставки аминокислот в положениях 233, 234, 235 и 237 в соответствии с нумерацией EU; и изменения в положениях, описанных в WO 2000/042072.
Другой неограничивающий вариант осуществления упомянутой выше Fc-области, активность связывания которой с активирующим FcγR является более низкой, чем активность связывания нативной Fc-области с активирующим FcγR, включает Fc-области, содержащие по меньшей мере одну или несколько аминокислот, выбранных из группы, состоящей из:
Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Lys, Met, Phe, Pro, Ser, Thr или Trp в качестве аминокислоты в положении 234;
Ala, Asn, Asp, Gln, Glu, Gly, His, Ile, Lys, Met, Pro, Ser, Thr, Val или Arg в качестве аминокислоты в положении 235;
Arg, Asn, Gln, His, Leu, Lys, Met, Phe, Pro или Tyr в качестве аминокислоты в положении 236;
Ala, Asn, Asp, Gln, Glu, His, Ile, Leu, Lys, Met, Pro, Ser, Thr, Val, Tyr или Arg в качестве аминокислоты в положении 237;
Ala, Asn, Gln, Glu, Gly, His, Ile, Lys, Thr, Trp или Arg в качестве аминокислоты в положении 238;
Gln, His, Lys, Phe, Pro, Trp, Tyr или Arg в качестве аминокислоты в положении 239;
Ala, Arg, Asn, Gln, Gly, His, Ile, Leu, Lys, Met, Phe, Ser, Thr, Trp, Tyr или Val в качестве аминокислоты в положении 265;
Ala, Arg, Asn, Asp, Gln, Glu, Gly, His, Lys, Phe, Pro, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 266;
Arg, His, Lys, Phe, Pro, Trp или Tyr в качестве аминокислоты в положении 267;
Ala, Arg, Asn, Gln, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val в качестве аминокислоты в положении 269;
Ala, Arg, Asn, Gln, Gly, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val в качестве аминокислоты в положении 270;
Arg, His, Phe, Ser, Thr, Trp или Tyr в качестве аминокислоты в положении 271;
Arg, Asn, Asp, Gly, His, Phe, Ser, Trp или Tyr в качестве аминокислоты в положении 295;
Arg, Gly, Lys или Pro в качестве аминокислоты в положении 296;
Ala в качестве аминокислоты в положении 297;
Arg, Gly, Lys, Pro, Trp или Tyr в качестве аминокислоты в положении 298;
Arg, Lys или Pro в качестве аминокислоты в положении 300;
Lys или Pro в качестве аминокислоты в положении 324;
Ala, Arg, Gly, His, Ile, Lys, Phe, Pro, Thr, Trp, Tyr или Val в качестве аминокислоты в положении 325;
Arg, Gln, His, Ile, Leu, Lys, Met, Phe, Pro, Ser, Thr, Trp, Tyr или Val в качестве аминокислоты в положении 327;
Arg, Asn, Gly, His, Lys или Pro в качестве аминокислоты в положении 328;
Asn, Asp, Gln, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Ser, Thr, Trp, Tyr, Val или Arg в качестве аминокислоты в положении 329;
Pro или Ser в качестве аминокислоты в положении 330;
Arg, Gly или Lys в качестве аминокислоты в положении 331; или
Arg, Lys или Pro в качестве аминокислоты в положении 332;
в соответствии с нумерацией EU. Количество аминокислот, подлежащих изменению, конкретно не ограничено, и можно изменять аминокислоту только в одном участке или можно изменять аминокислоты в двух или более участках.
В неограничивающем варианте осуществления настоящего изобретения два полипептида, образующих Fc, которые происходят из биспецифического антитела, как описано выше можно соответственно использовать в качестве Fc-области, подлежащей включению в антигенсвязывающую молекулу. Более конкретно, является предпочтительным использование двух полипептидов, которые составляют Fc-область и которые содержат Cys в качестве аминокислоты в положении 349 и Trp в качестве аминокислоты в положении 366 в соответствии с нумерацией EU в аминокислотной последовательности одного из полипептидов; и Cys в качестве аминокислоты в положении 356, Ser в качестве аминокислоты в положении 366, Ala в качестве аминокислоты в положении 368, и Val в качестве аминокислоты в положении 407, как указано с помощью нумерации EU, в аминокислотной последовательности другого полипептида.
В другом неограничивающем варианте осуществления настоящего изобретения, два полипептида, которые составляют Fc-область и которые содержат Asp в качестве аминокислоты в положении 409 в соответствии с нумерацией EU в аминокислотной последовательности одного из полипептидов, и Lys в качестве аминокислоты в положении 399 в соответствии с нумерацией EU в аминокислотной последовательности другого полипептида, можно соответственно использовать в качестве Fc-области. В указанном выше варианте осуществления аминокислота в положении 409 может представлять собой Glu вместо Asp, и аминокислота в положении 399 может представлять собой Arg вместо Lys. Более того, в дополнение к аминокислоте Lys в положении 399, Asp можно соответственно добавлять в качестве аминокислоты в положении 360, или Asp можно соответственно добавлять в качестве аминокислоты в положении 392.
В другом неограничивающем варианте осуществления настоящего изобретения, два полипептида, которые составляют Fc-область, и которые содержат Glu в качестве аминокислоты в положении 370 в соответствии с нумерацией EU в аминокислотной последовательности одного из полипептидов, и Lys в качестве аминокислоты в положении 357 в соответствии с нумерацией EU в аминокислотной последовательности другого полипептида, можно соответственно использовать в качестве Fc-области.
В другом неограничивающем варианте осуществления настоящего изобретения, два полипептида, которые составляют Fc-область и которые содержат Glu в качестве аминокислоты в положении 439 в соответствии с нумерацией EU в аминокислотной последовательности одного из полипептидов, и Lys в качестве аминокислоты в положении 356 в соответствии с нумерацией EU в аминокислотной последовательности другого полипептида, можно соответственно использовать в качестве Fc-области.
В другом неограничивающем варианте осуществления настоящего изобретения любую из комбинаций упомянутых выше вариантов осуществления, как показано ниже, можно соответственно использовать в качестве Fc-области:
(i) два полипептида, которые составляют Fc-область и которые содержат Asp в качестве аминокислоты в положении 409 и Glu в качестве в положении 370 в соответствии с нумерацией EU в аминокислотной последовательности одного из полипептидов, и Lys в качестве в положении 399 и Lys в качестве в положении 357 в соответствии с нумерацией EU в аминокислотной последовательности другого полипептида (в этом варианте осуществления аминокислота в положении 370 в соответствии с нумерацией EU может представлять собой Asp вместо Glu, и аминокислоту Asp в положении 392 в соответствии с нумерацией EU можно использовать вместо аминокислоты Glu в положении 370 в соответствии с нумерацией EU);
(ii) два полипептида, которые составляют Fc-область и которые содержат Asp в качестве аминокислоты в положении 409 и Glu в качестве аминокислоты в положении 439 в соответствии с нумерацией EU в аминокислотной последовательности одного из полипептидов; и Lys в качестве аминокислоты в положении 399 и Lys в качестве аминокислоты в положении 356 в соответствии с нумерацией EU в аминокислотной последовательности другого полипептида (в этом варианте осуществления аминокислоту Asp в положении 360 в соответствии с нумерацией EU, аминокислоту Asp в положении 392 в соответствии с нумерацией EU или аминокислоту Asp в положении 439 в соответствии с нумерацией EU можно использовать вместо аминокислоты Glu в положении 439 в соответствии с нумерацией EU);
(iii) два полипептида, которые составляют Fc-область и которые содержат Glu в качестве аминокислоты в положении 370 и Glu в качестве аминокислоты в положении 439 в соответствии с нумерацией EU в аминокислотной последовательности одного из полипептидов, и Lys в качестве аминокислоты в положении 357 и Lys в качестве аминокислоты в положении 356 в соответствии с нумерацией EU в аминокислотной последовательности другого полипептида; или
два полипептида, которые составляют Fc-область и которые содержат аминокислоту Asp в положении 409, Glu в качестве аминокислоты в положении 370, и Glu в качестве аминокислоты в положении 439 в соответствии с нумерацией EU в аминокислотной последовательности одного из полипептидов; и Lys в качестве аминокислоты в положении 399, Lys в качестве аминокислоты в положении 357, и Lys в качестве аминокислоты в положении 356 в соответствии с нумерацией EU в аминокислотной последовательности другого полипептида (в этом варианте осуществления аминокислота в положении 370 в соответствии с нумерацией EU может не быть заменена на Glu, и более того, когда аминокислота в положении 370 не заменена на Glu, аминокислота в положении 439 может представлять собой Asp вместо Glu или аминокислоту Asp в положении 392 можно использовать вместо аминокислоты Glu в положении 439).
Кроме того, в другом неограничивающем варианте осуществления настоящего изобретения также можно соответственно использовать два полипептида, которые составляют Fc-область и которые содержат Lys в качестве аминокислоты в положении 356 в соответствии с нумерацией EU в аминокислотной последовательности одного из полипептидов, и Arg в качестве аминокислоты в положении 435 и Glu в качестве аминокислоты в положении 439 в соответствии с нумерацией EU в аминокислотной последовательности другого полипептида.
В другом неограничивающем варианте осуществления настоящего изобретения, также можно соответственно использовать два полипептида, которые составляют Fc-область и которые содержат Lys в качестве аминокислоты в положении 356 и Lys в качестве аминокислоты в положении 357 в соответствии с нумерацией EU в аминокислотной последовательности одного из полипептидов, и Glu в качестве аминокислоты в положении 370, Arg в качестве аминокислоты в положении 435, и Glu в качестве аминокислоты в положении 439 в соответствии с нумерацией EU в аминокислотной последовательности другого полипептида.
Антигенсвязывающие молекулы по настоящему изобретению выделяют из культуральной среды клеток, трансформированных желаемым экспрессирующим вектором, в котором полинуклеотид, кодирующий антигенсвязывающий домен, и полинуклеотид, кодирующий полипептид, содержащий Fc-область, которые связаны описанным выше образом, функционально связаны.
Когда Fc-область, содержащаяся в антигенсвязывающей молекуле по настоящему изобретению, представляет собой Fc-область, которая модифицирована так, чтобы процент Fc-областей, с которыми связана цепь сахаров с дефицитом фукозы или биссекторный N-ацетилглюкозамин, был более высоким, упомянутые выше трансформированные клетки-хозяева, которые соответственно используют, представляют собой клетки-хозяева, которые обладают низкой способностью присоединять фукозу к цепи сахаров в результате модификации активности для формирования структуры цепи сахаров полипептида, подлежащего модификации цепью сахаров (например, WO 2000/061739, WO 2002/031140, и WO 2006/067913). В неограничивающем варианте осуществления таких клеток-хозяев, можно соответственно использовать клетки-хозяева с дефицитом активности фермента или переносчика, выбранного из группы, состоящей из фукозилтрансферазы (EC 2.4.1.152), переносчика фукозы (SLC35C1), GMD (GDP-манноза-4,6-дегидратаза) (EC 4.2.1.47), Fx (GDP-кето-6-дезоксиманноза-3,5-эпимераза, 4-редуктаза) (EC 1.1.1.271) и GFPP (GDP-β-L-фукозопирофосфорилаза (EC 2.7.7.30) (например, WO 2000/061739, WO 2002/031140 и WO 2006/067913). Клетки-хозяева с дефицитом такой активности можно получать, например, способом, который разрушает гены этих функциональных белков, эндогенных для клеток CHO, клеток BHK, клеток NS0, клеток SP2/0, клеток миеломы YO, клеток миеломы мыши P3X63, клеток PER, клеток PER.C6, клеток HEK293, клеток гибридомы и т.п., так чтобы гены были неспособны функционировать.
Когда Fc-область, содержащаяся в антигенсвязывающей молекуле по настоящему изобретению, представляет собой Fc-область, имеющую цепь сахаров, содержащую биссекторный GlcNAc, описанные выше трансформированные клетки, которые соответственно используют, представляют собой клетки-хозяева, экспрессирующие ген, кодирующий функциональный белок, обладающий активностью GnTIII (β-1,4-маннозилгликопротеин-4-β-N-ацетилглюкозаминилтрансфераза) (EC 2.4.1.94) или активностью GalT (β-1,4-галактозилтрансфераза) (EC 2.4.1.143), для получения антител, которые имеют биссекторные содержащие GlcNAc цепи сахаров (WO2002/079255 и т.п.). В другом подходящем неограничивающем варианте осуществления пригодным образом используют клетки-хозяева, которые экспрессируют, в дополнение к упомянутым выше функциональным белкам, ген, кодирующий функциональный белок, обладающий активностью ManII человека (маннозидаза II) (3.2.1.114), ген, кодирующий функциональный белок, обладающий активностью GnTI (β-1,2-ацетилглюкозаминилтрансфераза I) (EC 2.4.1.94), ген, кодирующий функциональный белок, обладающий активностью GnTII (β-1,2-ацетилглюкозаминилтрансфераза II) (EC 2.4.1.143), ген, кодирующий функциональный белок, обладающий активностью ManI (манноидаза) (EC 3.2.1.113) и α-1,6-фукозилтрансферазы (EC 2.4.1.68) (WO2004/065540).
Антигенсвязывающие молекулы по настоящему изобретению получают с использованием способов, которые аналогичны способам получения антител, такими как выделение из культуральной среды упомянутых выше клеток, которые описаны в разделе "Антитела", выше. Неограничивающий вариант осуществления упомянутых выше полипептидов, содержащих Fc-область, включает, например, константную область антитела SEQ ID NO:5, 6, 7 или 8. Неограничивающий вариант осуществления антигенсвязывающих молекул по настоящему изобретению представляет собой, например, полноразмерную молекулу антитела.
Фармацевтические композиции
Настоящее изобретение относится к фармацевтическим композициям, содержащим антигенсвязывающую молекулу, которая не действует системно в крови или нормальных клетках, а действует на очаги повреждения, такие как злокачественная опухоль и области воспаления, проявляя эффективность лекарственного средства, одновременно избегая побочных эффектов. Антигенсвязывающая молекула, содержащаяся в фармацевтической композиции по настоящему изобретению, связывается с антигеном, экспрессируемым в злокачественных клетках, иммунных клетках, стромальных клетках и т.п. в злокачественных тканях; антигеном, секретируемым в злокачественных тканях; или антигеном, экспрессируемым иммунными клетками и т.п. в воспаленных тканях; и антигеном, секретируемым в воспаленных тканях; и она не может связываться с антигенами, экспрессируемыми в нормальных тканях; таким образом, побочные эффекты вследствие цитотоксической активности, нейтрализующей активности и т.п. против нормальной ткани устраняются; и в то же время обеспечиваются мощные цитотоксические эффекты, эффекты подавления роста и усиливающее иммунитет действие на злокачественные опухоли, или иммунодепрессивные эффекты против воспалительных клеток в воспаленных тканях. Например, биспецифическая или бипаратопная антигенсвязывающая молекула, содержащая антигенсвязывающий домен, который связывается с EGFR, экспрессируемым на злокачественных клетках, и антигенсвязывающий домен, который связывается с CD3, экспрессируемым на T-клетках в зависимости от специфического для злокачественной ткани соединения, не связывается с EGFR, экспрессируемым в нормальных тканях, но связывается с EGFR, экспрессируемым на злокачественных клетках; тем самым, проявляя мощные противоопухолевые эффекты, одновременно избегая побочных эффектов. В частности, в то время как антигенсвязывающая молекула связывается с CD3, экспрессируемым на T-клетках вблизи злокачественных клеток, в зависимости от специфического для злокачественной ткани соединения, молекула не связывается с CD3, экспрессируемым на T-клетках, которые не находятся вблизи злокачественных клеток. Таким образом, молекула активирует T-клетки вблизи злокачественных клеток, проявляя мощные противоопухолевые эффекты, и одновременно избегая побочных эффектов.
Такие антигенсвязывающие молекулы, которые связываются с антигеном в тканях-мишенях, но не в других нормальных тканях и крови, проявляют эффективность лекарственного средства, одновременно избегая побочных эффектов. Антигенсвязывающие молекулы, предусматриваемые в рамках настоящего изобретения, которые связываются с антигеном с использованием низкомолекулярного соединения, присутствующего в высоких концентрациях в тканях-мишенях in vivo в качестве переключающего фактора, а именно, антигенсвязывающие молекулы с низкомолекулярным переключающим фактором, не связываются с антигеном в нормальных условиях, где низкомолекулярное соединение отсутствуют, но могут связываться с антигеном в тканях-мишенях, где низкомолекулярное соединение присутствует в высоких концентрациях.
Неограничивающий вариант осуществления таких антигенсвязывающих молекул с низкомолекулярным переключающим фактором включает специфические для злокачественной ткани или специфические для воспаленной ткани зависимые от соединения антигенсвязывающие молекулы; и специфическое для злокачественной опухоли или специфическое для воспаленной ткани соединение, такое как аденозин, аденозин-5'-трифосфат (ATP), инозин, кинуренин, простагландин E2 (PGE2), янтарная кислота и молочная кислота, которые присутствуют в злокачественных тканях или воспаленных тканях в высоких концентрациях и способны функционировать в качестве переключающего фактора, обеспечивают функцию переключения вследствие размещения между антигенсвязывающей молекулой по настоящему изобретению (паратопом, содержащимся в ней) и антигеном (эпитопом, содержащимся в нем). В отсутствие соединения, взаимодействие между паратопом в антигенсвязывающей молекуле по настоящему изобретению и эпитопом в антигене не является достаточным, чтобы антигенсвязывающая молекула по настоящему изобретению была способна связаться с антигеном. В присутствии соединения соединение располагается между паратопом в антигенсвязывающей молекуле по настоящему изобретению и эпитопом в антигене; и антигенсвязывающая молекула, которая связалась с антигеном в ткани-мишени, такой как злокачественная ткань или воспаленная ткань, где соединение присутствует в высоких концентрациях, может проявлять эффективность лекарственного средства в отношении клеток, экспрессирующих антиген. Более того, поскольку это связывание переключающего соединения является обратимым, связывание антигенсвязывающей молекулы по настоящему изобретению с антигеном с помощью этих переключающих соединений можно контролировать обратимым образом. Таким образом, антигенсвязывающие молекулы по настоящему изобретению, которые могут проявлять эффективность лекарственного средства в очаге повреждения, таком как злокачественная ткань или воспаленная ткань, путем связывания с патогенными клетками, такими как злокачественные клетки или иммунные клетки в злокачественной ткани или воспаленной ткани, или путем связывания с антигеном, секретируемым в злокачественной ткани или воспаленной ткани, являются пригодными в качестве фармацевтических композиций. Фармацевтические композиции по настоящему изобретению могут содержать фармацевтически приемлемый носитель.
В рамках настоящего изобретения фармацевтические композиции, как правило, относятся к фармацевтическим средствам для лечения, или предупреждения, или исследования и диагностики заболевания. Более того, в рамках настоящего изобретения, выражение "фармацевтическая композиция, содержащая антигенсвязывающую молекулу, антигенсвязывающая активность которой варьирует в зависимости от концентрации специфического для ткани-мишени соединения", может быть переформулировано как "способ лечения заболевания, который включает введение индивидууму, подлежащему лечению, антигенсвязывающей молекулы, антигенсвязывающая активность которой варьирует в зависимости от концентрации специфического для ткани-мишени соединения" или может быть переформулировано как "применение антигенсвязывающей молекулы, антигенсвязывающая активность которой варьирует в зависимости от концентрации специфического для ткани-мишени соединения для получения фармацевтического препарата для лечения заболевания". Более того, выражение "фармацевтическая композиция, содержащая антигенсвязывающую молекулу, антигенсвязывающая активность которой варьирует в зависимости от концентрации специфического для ткани-мишени соединения" можно переформулировать как "применение антигенсвязывающей молекулы, антигенсвязывающая активность которой варьирует в зависимости от концентрации специфического для ткани-мишени соединения, для лечения заболевания".
Фармацевтические композиции по настоящему изобретению могут быть составлены способами, известными специалистам в данной области. Например, их можно использовать парентерально в форме инъекций стерильных растворов или суспензий, включающих воду или другую фармацевтически приемлемую жидкость. Например, такие композиции могут быть составлены путем смешения в форме единичной дозы, требуемой повсеместно одобренной практикой производства лекарственных средств, путем комбинирования надлежащим образом с фармакологически приемлемыми носителями или средами, в частности, со стерильной водой, физиологическим раствором, растительным маслом, эмульгатором, суспензией, поверхностно-активным веществом, стабилизатором, вкусовой добавкой, эксципиентом, носителем, консервантом, связующим веществом и т.п. В таких составах количество активного ингредиента корректируют так, чтобы получить соответствующее количество в заданном диапазоне.
Стерильные композиции для инъекций могут быть составлены с использованием носителей, таких как дистиллированная вода для инъекций, в соответствии со стандартной практикой составления.
Водные растворы для инъекций включают, например, физиологический солевой раствор и изотонические растворы, содержащие декстрозу или другие адъюванты (например, D-сорбит, D-маннозу, D-маннит и хлорид натрия). Также их можно использовать в комбинации с соответствующими солюбилизаторами, например, спиртами (этанол и т.п.), полиспиртами (пропиленгликоль, полиэтиленгликоль и т.п.), неионными поверхностно-активными веществами (полисорбат 80(TM), HCO-50 и т.п.).
Масла включают кунжутное масло и соевые масла. В комбинации с солюбилизаторами можно использовать бензилбензоат и/или бензиловый спирт. Также можно комбинировать буферы (например, фосфатный буфер натрий-ацетатный буфер), успокаивающие средства (например, прокаина гидрохлорид), стабилизаторы (например, бензиловый спирт и фенол) и/или антиоксиданты. Полученными инъекционными препаратами заполняют подходящие ампулы.
Фармацевтические композиции по настоящему изобретению предпочтительно вводят парентерально. Например, вводят композиции в дозированной форме для инъекций, трансназального введения, введения через легкие или чрескожного введения. Например, их можно вводить системно или локально путем внутривенной инъекции, внутримышечной инъекции, внутрибрюшинной инъекции, подкожной инъекции и т.п.
Способы введения можно надлежащим образом выбирать, учитывая возраст и симптомы пациента. Доза фармацевтической композиции, содержащей антигенсвязывающую молекулу, может составлять, например, от 0,0001 до 1,000 мг/кг для каждого введения. Альтернативно доза может составлять, например, от 0,001 до 100000 мг на пациента. Однако настоящее изобретение не ограничивается числовыми величинами, описанными выше. Дозы и способы введения варьируют, в зависимости от массы тела пациента, возраста, симптомов и т.п. Специалисты в данной области могут установить подходящие дозы и способы введения, учитывая факторы, описанные выше.
Аминокислоты, содержащиеся в аминокислотных последовательностях по настоящему изобретению, могут быть посттрансляционно модифицированными (например, модификация N-концевого глутамина в пироглутаминовую кислоту путем пироглутамилирования хорошо известна специалистам в данной области). Естественно, такие посттрансляционно модифицированные аминокислоты включены в аминокислотные последовательности по настоящему изобретению.
Все цитированные в описании документы уровня техники включены в настоящее описание в качестве ссылок.
Примеры
[Пример 1] Концепция антител, которые связываются с антигенами через низкомолекулярные соединения, служащие в качестве переключающего фактора, которые присутствуют в тканях-мишенях в высоких концентрациях
Для обеспечения эффективности лекарственного средства, одновременно избегая неблагоприятных эффектов, существует потребность в технологии разработки лекарственных средств, которые действуют в очагах повреждения, таких как злокачественная опухоль или области воспаления, без системного действия на нормальные ткани или кровь. Молекулы антител, которые могут связываться с антигенами, экспрессируемыми на злокачественных клетках, но не способны связываться с антигенами, экспрессируемыми в нормальных тканях, после введения, могут проявлять мощные цитотоксические эффекты против злокачественной опухоли, одновременно избегая неблагоприятных эффектов на нормальные ткани в результате цитотокического действия. Например, антигенсвязывающие молекулы, которые были изменены относительно описанной выше EGFR-BiTE (непатентный документ 9), которая не может связываться с EGFR, экспрессированным в нормальных тканях, но способна связываться с EGFR, экспрессируемым на злокачественных клетках, может проявлять мощный противоопухолевый эффект, одновременно избегая побочных эффектов. Между тем BiTE проявляет противоопухолевый эффект путем привлечения и активации T-клеток через CD3 (непатентный документ 8); и, если бы было возможно сообщить EGFR-BiTE свойство связывания с CD3, экспрессируемым на T-клетках вблизи злокачественных клеток, но не с CD3, экспрессируемым на T-клетках не в близи злокачественных клеток, EGFR-BiTE, измененная так, чтобы иметь это свойство, могла бы активировать T-клетки в злокачественной опухоли и, таким образом, могла бы проявлять мощные противоопухолевые эффекты, одновременно избегая неблагоприятных эффектов.
Однако это не ограничивается только антительными фармацевтическими препаратами против злокачественной опухоли. Когда молекула антитела связывается с цитокинами и ингибирует их в синовиальной жидкости воспаленных суставов при ревматоидном артрите, но не ингибирует системно цитокины, молекула может проявлять мощные терапевтические эффекты против воспалительных/аутоиммунных заболеваний, таких как ревматоидный артрит, одновременно избегая увеличенных рисков инфекции вследствие системной нейтрализации цитокинов.
Как описано выше, антитела, которые связываются с антигенами в злокачественных тканях, но не с антигенами в других тканях, таких как нормальные ткани и кровь, могут проявлять эффективность лекарственного средства, одновременно избегая неблагоприятных эффектов. Однако, идеальные антитела, имеющие такие свойства, не были описаны до настоящего времени. Между тем, как показано на фиг. 1, молекулы антител, которые связываются с антигенами через низкомолекулярные соединения в качестве переключающего фактора, который присутствует в высоких концентрациях в злокачественных тканях in vivo (т.е. антитела с низкомолекулярными переключающими факторами), не связываются с антигенами в условиях отсутствия таких низкомолекулярных соединений; и они могут связываться с антигенами в тканях-мишенях, где низкомолекулярные соединения присутствуют в высоких концентрациях.
При разработке таких антител с низкомолекулярными переключающими факторами, сначала проводили поиск низкомолекулярных соединений, которые присутствуют в высоких концентрациях в злокачественной опухоли, и рассматривали их как пригодные для применения в качестве переключающего фактора. Результат показал, что перспективными в качестве переключающих факторов были аденозин, аденозинтрифосфат (аденозин-5'-трифосфат (ATP)), инозин, кинуренин, простагландин E2 (PGE2), янтарная кислота и молочная кислота. Каждое из этих низкомолекулярных соединений либо продуцируется злокачественными клетками, либо высвобождается из злокачественных клеток после гибели клеток, либо продуцируется иммунными клетками и т.д., инфильтрирующими ткани злокачественной опухоли, и, таким образом, они присутствуют в высоких концентрациях в злокачественных тканях; однако они присутствуют в более низких концентрациях в нормальных тканях и крови по сравнению со злокачественными тканями. Если бы эти низкомолекулярные соединения можно было расположить в комплексе между антителом и антигеном так, как показано на фиг. 2, низкомолекулярные соединения могли бы осуществлять функцию переключающего фактора. В частности, в отсутствие низкомолекулярных соединений взаимодействие антиген-антитело является недостаточным и антитело не может связываться с его антигеном. Между тем, в присутствии низкомолекулярного соединения антитело может связываться с его антигеном через низкомолекулярное соединение, располагающееся между антителом и антигеном. Иными словами, в присутствии низкой концентрации низкомолекулярных соединений взаимодействие антиген-антитело является недостаточным и антитело не может связываться с его антигеном, в то время как в присутствии высокой концентрации низкомолекулярных соединений антитело может связываться с его антигеном через низкомолекулярное соединение, располагающееся между антителом и антигеном. Более того, связывание низкомолекулярных соединений в качестве переключателя является обратимым, и регуляция связывания антигена с низкомолекулярным переключающим фактором также является обратимой.
В этом контексте, изначально, авторы настоящего изобретения предприняли попытку выделить антитела с низкомолекулярными переключающими факторами против IL-6 (Br. J. Haematol. (2011) 152 (5), 579-92), о котором сообщалось, что он вовлечен в рост злокачественных клеток.
[Пример 2] Получение антител, которые связываются с IL-6 человека в присутствии низкомолекулярных соединений, из библиотеки антител человека с использованием технологий фагового дисплея
(2-1) Конструирование библиотеки фагового дисплея наивных антител человека
Библиотеку фагового дисплея антител человека, состоящую из множества фагов, которые презентируют Fab-домены антител человека, последовательности которых отличаются друг от друга, конструировали с использованием в качестве матрицы полиA-РНК, полученную из PBMC человека, коммерчески доступную поли-A РНК человека и т.п. в соответствии со способом, известным специалистам в данной области.
(2-2) Получение антител, которые связываются с IL-6 человека в присутствии низкомолекулярных соединений, из библиотеки способом пэннинга гранул
Библиотеку фагового дисплея наивных антител человека, сконструированную, как описано в (2-1), подвергали скринингу в отношении антител, которые проявляют антигенсвязывающую активность в присутствии низкомолекулярных соединений, в частности, путем сбора фагов, экспонирующих антитела, которые в присутствии низкомолекулярных соединений проявляют антигенсвязывающую активность в отношении антигенов, уловленных гранулами. Фаги собирали из суспензии фагов, элюированной с гранул в отсутствие низкомолекулярных соединений. В этом способе получения использованный антиген представлял собой меченный биотином IL-6 человека.
Фаги, продуцированные в E. coli, содержащих фагмидный вектор, сконструированный для фагового дисплея, очищали общепринятым способом. Затем получали суспензию фаговой библиотеки путем диализа фагов против TBS. Далее, к суспензии фаговой библиотеки добавляли BSA до конечной концентрации 4%. Пэннинг проводили с использованием магнитных гранул с иммобилизованным на них антигеном. Использованные магнитные гранулы представляли собой покрытые нейтравидином гранулы (Sera-Mag SpeedBeads NeutrAvidin-coated) или покрытые стрептавидином гранулы (Dynabeads M-280 Streptavidin).
Для эффективного выделения антител с низкомолекулярным переключающим фактором, которые зависят от низкомолекулярных соединений, которые могут выступать в качестве переключателя в злокачественных тканях, проводили пэннинг для увеличения содержания антител, которые связываются с антигенами в присутствии смешанного раствора низкомолекулярных соединений (аденозин, аденозинтрифосфат (аденозин-5’-трифосфат (ATP)), инозин, кинуренин, простагландин E2 (PGE2), янтарная кислота и молочная кислота (далее обозначается как коктейль низкомолекулярных соединений (SC)), но не в отсутствие SC.
В частности, вместе с 250 пмоль меченного биотином антигена, SC, содержащий натриевую соль аденозинтрифосфата (ATP-Na), аденозин, инозин, янтарную кислоту и молочную кислоту в конечной концентрации 1 мМ, простагландин E2 (PGE2) в конечной концентрации 1 мкМ и кинуренин в конечной концентрации 100 мкМ, который был доведена до pH 7,4 с помощью NaOH, контактировали с полученной суспензией фаговой библиотеки в течение 60 минут при комнатной температуре. Затем к суспензии фаговой библиотеки добавляли блокированные BSA магнитные гранулы, и комплексу антиген-фаг позволяли связываться с магнитными гранулами при комнатной температуре в течение 15 минут. После промывания один раз SC/TBS (TBS, содержащий SC), гранулы объединяли с 0,5 мл раствора трипсина в концентрации 1 мг/мл. Сразу после суспендирования гранул при комнатной температуре в течение 15 минут, суспензию фагов собирали с выделенных гранул с использованием магнитного стенда. Собранную суспензию фагов добавляли к 10 мл клеток E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инфицировали фагом путем инкубации упомянутых выше E. coli при осторожном перемешивании при 37°C в течение одного часа. Инфицированные E. coli высевали в чашку 225 мм × 225 мм. Затем фаги собирали из культуральной среды посеянных E. coli с получением жидкого исходного раствора с фаговой библиотекой.
Первый раунд пэннинга проводили для сбора фагов, которые способны связываться в присутствии низкомолекулярных соединений, в то время как второй и последующий раунды пэннинга проводили для увеличения содержания фагов, которые способны связываться с антигенами в присутствии SC. В частности, полученную суспензию фаговой библиотеки смешивали с 40 пмоль меченного биотином антигена, SC и NaOH, и контактировали с низкомолекулярными соединениями и антигенами в течение 60 минут при комнатной температуре. Добавляли блокированные посредством BSA магнитные гранулы и позволяли им связаться с комплексом антиген-фаг в течение 15 минут при комнатной температуре. Гранулы промывали 1 мл SC/TBST и SC/TBS. К суспензии гранул добавляли 0,5 мл TBS при комнатной температуре, а затем сразу суспензию фагов собирали с выделенных гранул с использованием магнитного стенда. После повторения этой обработки две отдельно элюированных суспензии фагов смешивали. Затем полученные гранулы объединяли с 0,5 мл TBS и перемешивали при комнатной температуре в течение пяти минут. Суспензию фага собирали с выделенных гранул с использованием магнитного стенда. Посредством добавления 5 мкл трипсина в концентрации 100 мг/мл к собранной суспензии фагов, белок pIII (происходящий из фага-помощника белок pIII), который не экспонирует Fab, отщепляли от фагов, и устраняли способность фагов, которые не экспонируют Fab, инфицировать E. coli. Фаги, собранные из трипсинизированной суспензии фагов, добавляли к 10 мл E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубировали при 37°C в течение одного часа при осторожном перемешивании для инфицирования фагом. Инфицированные E. coli высевали в чашку 225 мм × 225 мм. В этот момент времени два типа инфицированных E. coli, полученных посредством второго раунда пэннинга, смешивали в равных количествах. Затем фаги собирали из культуральной среды посеянных E. coli с получением суспензии фаговой библиотеки. Пэннинг проводили три раза для выделения антител, которые обладают антигенсвязывающей активностью в присутствии SC.
(2-3) Получение антител, которые связываются с IL-6 человека в присутствии низкомолекулярных соединений, из библиотеки с использованием способа отрицательной селекции
Сконструированную библиотеку фагового дисплея наивных антител человека подвергали скринингу в отношении антител, которые проявляют антигенсвязывающую активность в присутствии низкомолекулярных соединений. В качестве первой стадии скрининга сконструированную библиотеку фагового дисплея наивных антител человека контактировали с меченным биотином антигеном со стрептавидином в отсутствие низкомолекулярных соединений для устранения фагов, экспонирующих антитела, которые обладают антигенсвязывающей активностью даже в отсутствие низкомолекулярных соединений. Затем, пэннинг проводили в присутствии низкомолекулярных соединений аналогичным образом. Таким образом, проводили скрининг антител, которые обладают антигенсвязывающей активностью в присутствии низкомолекулярных соединений. В качестве антигена использовали меченный биотином IL-6.
Фаги продуцировали в E. coli, сохранивших сконструированную фагмиду фагового дисплея. Продуцированные фаги очищали общепринятым способом, а затем получали суспензию фаговой библиотеки путем диализа фагов против TBS. Затем к суспензии фаговой библиотеки добавляли BSA в конечной концентрации 4%. Использованные магнитные гранулы представляли собой покрытые нейтравидином гранулы (Sera-Mag SpeedBeads NeutrАвидин-coated) или покрытые стрептавидином гранулы (Dynabeads M-280 Streptavidin). Пэннинг проводили с использованием магнитных гранул с иммобилизованным на них антигеном.
Вместе с 250 пмоль меченного биотином антигена, SC, содержащий ATP-Na, аденозин, инозин, янтарную кислоту и молочную кислоту в конечной концентрации 1 мМ, PGE2 в конечной концентрации 1 мкМ и кинуренин в конечной концентрации 100 мкМ, значение pH которого было доведено до 7,4 с помощью NaOH, добавляли и контактировали с полученной суспензией фаговой библиотеки в течение 60 минут при комнатной температуре. Затем к суспензии фаговой библиотеки добавляли блокированные BSA магнитные гранулы, и комплексу антиген-фаг позволяли связываться с магнитными гранулами при комнатной температуре в течение 15 минут. После промывания один раз SC/TBS, гранулы объединяли с 0,5 мл раствора трипсина в концентрации 1 мг/мл. Сразу после суспендирования гранул при комнатной температуре в течение 15 минут суспензию фагов собирали с выделенных гранул с использованием магнитного стенда. Собранную суспензию фагов добавляли к 10 мл клеток E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубировали при 37°C в течение одного часа при осторожном перемешивании для инфицирования фагом. Инфицированные E. coli высевали в чашку 225 мм × 225 мм. Затем фаги собирали из культуральной среды посеянных E. coli с получением жидкого исходного раствора с фаговой библиотекой.
Первый раунд пэннинга проводили для сбора фагов, которые способны связываться в присутствии SC, в то время как второй и последующий раунды пэннинга проводили для увеличения содержания фагов, которые способны связываться с антигенами в присутствии SC. В частности, 250 пмоль биотинилированного антигена добавляли к блокированным посредством BSA гранулам с нейтравидином Sera-Mag для связывания при комнатной температуре в течение 15 минут. Гранулы промывали три раза посредством TBS. К гранулам добавляли суспензию фаговой библиотеки, блокированной посредством BSA, и им позволяли связываться с ней при комнатной температуре в течение одного часа. Фаги, которые не связываются с антигенами или гранулы собирали путем выделения гранул с использованием магнитного стенда. К собранным фагам добавляли 40 пмоль меченного биотином антигена, SC и NaOH. Таким образом, фаговую библиотеку контактировали с низкомолекулярными соединениями в SC при комнатной температуре в течение 60 минут. Затем блокированные посредством BSA магнитные гранулы добавляли к смеси меченого антигена, SC и фаговой библиотеки, и позволяли им связаться с комплексом антиген-фаг в течение 15 минут при комнатной температуре. Гранулы промывали 1 мл SC/TBST и SC/TBS. Затем к смеси добавляли 0,5 мл раствора трипсина в концентрации 1 мг/мл. После перемешивания смешанной суспензии при комнатной температуре в течение 20 минут, фаги собирали с гранул, разделенных с использованием магнитного стенда. Собранные фаги добавляли к 10 мл E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубировали при 37°C в течение одного часа при осторожном перемешивании для инфицирования фагом. Инфицированные E. coli высевали в чашку 225 мм × 225 мм. Пэннинг проводили три раза для выделения антител, которые обладают антигенсвязывающей активностью в присутствии SC.
(2-4) Оценка активности связывания в присутствии низкомолекулярных соединений с помощью ELISA фагов
Культуральные супернатанты, содержащие фаги, собирали в соответствии с общепринятым способом (Methods Mol. Biol. (2002) 178, 133-145) из единичных колоний E. coli, полученных способом, описанным выше. Собранные культуральные супернатанты обрабатывали ультрафильтрацией с использованием NucleoFast 96 (MACHEREY-NAGEL). 100 мкл собранных культуральных супернатантов добавляли в каждую лунку NucleoFas96 и центрифугировали (4500 g в течение 45 минут) для удаления оставшейся жидкой части. В каждую лунку добавляли 100 мкл H2O, и вновь центрифугировали с помощью NucleoFast 96 (4500 g в течение 30 минут) для промывания. Добавляли 100 мкл TBS, а затем NucleoFast 96 позволяли стоять в течение пяти минут при комнатной температуре. Наконец, из супернатанта каждой лунки собирали суспензию фага.
После добавления TBS или SC/TBS очищенные фаги подвергали ELISA по следующей методике. 96-луночный микропланшет для титрования StreptaWell (Roche) покрывали в течение ночи посредством 100 мкл TBS, содержащего меченный биотином антиген. После удаления антигена путем промывания каждой лунки планшета посредством TBST, лунки блокировали 250 мкл смеси 2% обезжиренное молоко-TBS в течение одного часа или более. Смесь 2% обезжиренное молоко-TBS удаляли, а затем в каждую лунку добавляли полученные очищенные фаги. Планшету позволяли стоять при 37°C в течение одного часа для обеспечения связывания экспонирующих антитело фагов с антигеном в присутствии или в отсутствие SC в каждой лунке. После того, как каждую лунку промывали посредством TBST или SC/TBST, в нее добавляли конъюгированное с HRP антитело против M13 (Amersham Pharmacia Biotech), разбавленное TBS или SC/TBS, и планшет инкубировали в течение одного часа. После промывания посредством TBST или SC/TBST, в каждую лунку добавляли простой раствор TMB (ZYMED), и хромогенную реакцию в растворе завершали добавлением серной кислоты. Затем оценивали развитие окраски путем измерения поглощения при 450 нм.
ELISA фагов для 96 выделенных клонов выявил клон "I6NMSC1-3_A11", который обладает активностью связывания с IL-6 человека в качестве антигена в присутствии коктейля низкомолекулярных соединений.
[Пример 3] Оценка антител, которые связываются с антигенами в присутствии низкомолекулярных соединений
(3-1) Экспрессия и очистка антител, которые связываются с IL-6 человека
Амплифицировали гены из клона I6NMSC1-3_A11, который был оценен как обладающий антигенсвязывающей активностью в присутствии SC в ELISA фагов, как описано в примере 2, с использованием специфических праймеров (SEQ ID NO:110 и 112). Нуклеотидные последовательности генов анализировали (последовательности тяжелой цепи и легкой цепи представлены в SEQ ID NO:30 и 31, соответственно). Ген, кодирующий вариабельную область I6NMSC1-3_A11, встраивали в плазмиду для экспрессии в клетках животных IgG1/лямбда человека, в то время как каждый из генов, кодирующих вариабельные области известного антитела против IL-6 человека CLB8-F1 (тяжелая цепь и легкая цепь представляют собой SEQ ID NO:32 и 33, соответственно) и вариабельные области антитела против глипикана 3 человека GC413 (тяжелая цепь и легкая цепь представляют собой SEQ ID NO:34 и 35, соответственно) в качестве отрицательного контроля, встраивали в плазмиду для экспрессии в клетках животных IgG1/каппа человека. Антитела экспрессировали с использованием способа, описанного ниже. FreeStyle 293-F (Invitrogen), которые происходят из клеток почки эмбриона человека, суспендировали при плотности клеток 1,33×106 клеток/мл в среде для экспрессии FreeStyle 293 Expression Medium (Invitrogen) и распределяли аликвотами объемом 3 мл в каждую ячейку планшета с 6 ячейками. Плазмидную ДНК трансфицировали в клетки с помощью липофекции. Из культуральных супернатантов после культивирования в течение четырех суток в инкубаторе с CO2 (37°C, 8% CO2, 90 об/мин), антитела очищали способом, известным специалистам в данной области в данной области, с использованием rProtein A Sepharose™ Fast Flow (Amersham Biosciences). Поглощение растворов очищенных антител измеряли при 280 нм с использованием спектрофотометра. Из величин, полученных с помощью измерения, концентрации очищенных антител вычисляли с использованием коэффициента экстинкции, определенного способом PACE (Protein Science (1995) 4, 2411-2423).
(3-2) Идентификация низкомолекулярных соединений, необходимых для связывания полученных антител с IL-6 человека
Три типа антител: выделенные I6NMSC1-3_A11 (далее сокращенно обозначается как A11) и CLB8-F1 и GC413 в качестве контролей подвергали ELISA в девяти условиях, описанных в таблице 3. Между тем, каждое низкомолекулярное соединение соответственно получали в концентрациях, представленных в таблице 3, с использованием буферов, указанных в таблице 4. В качестве антигена использовали меченный биотином IL-6 человека.
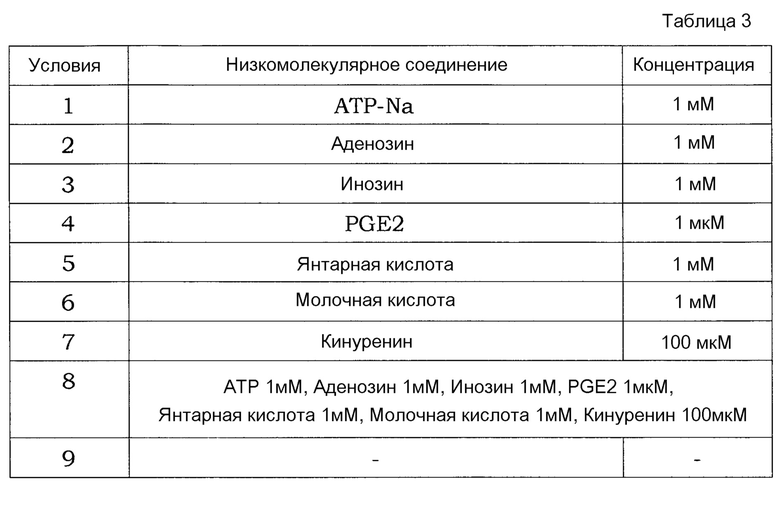

Сначала 96-луночный микропланшет для титрования StreptaWell (Roche) покрывали при комнатной температуре в течение одного часа или более 100 мкл PBS, содержащего меченный биотином антиген. После промывания буфером для промывания для удаления не связавшегося антигена из планшета, каждую лунку блокировали в течение одного часа или более посредством 250 мкл блокирующего буфера. Блокирующий буфер удаляли из каждой лунки. Очищенные IgG приготавливали в концентрации 2,5 мкг/мл в буфере для образца, содержащем низкомолекулярные соединения в конечных концентрациях, представленных в таблице 3, и каждый из них распределяли аликвотами объемом 100 мкл в каждую лунку планшета. Планшету позволяли стоять при комнатной температуре в течение одного часа, чтобы обеспечить связывание каждого IgG с антигеном в каждой лунке. После промывания буфером для промывания, содержащим низкомолекулярные соединения в конечных концентрациях, представленных в таблице 3, в каждую лунку добавляли конъюгированное с HRP антитело против IgG человека (BIOSOURCE), разбавленное буфером для образца, содержащим те же низкомолекулярные соединения. Планшет инкубировали в течение одного часа. После промывания буфером для промывания, содержащим каждое низкомолекулярное соединение, в каждую лунку добавляли простой раствор TMB (ZYMED). Хромогенную реакцию в растворе в каждой лунке завершали добавлением серной кислоты. Затем оценивали развитие окраски путем измерения поглощения при 450 нм.
Результат измерения представлен на фиг.3. Результат показал, что поглощение CLB8-F1 было постоянным, независимо от типа или присутствия низкомолекулярного соединения, в то время как поглощение I6NMSC1-3_A11 было значительно более низким в условиях 9 (без низкомолекулярных соединений) по сравнению с поглощением в условиях 8 (полный раствор коктейля низкомолекулярных соединений). Аналогично ELISA фагов, этот результат показал, что I6NMSC1-3_A11 имело свойство, состоящее в том, что его связывание антигена изменяется в зависимости от присутствия низкомолекулярных соединений. Между тем, I6NMSC1-3_A11 продемонстрировало эквивалентное поглощение в условиях 7 (в присутствии 100 мкМ кинуренина) с поглощением в условиях 8; однако поглощение было значительно более низким в других условиях. Этот результат демонстрирует, что I6NMSC1-3_A11 представляет собой антитело, которое связывается с IL-6 человека в качестве антигена в присутствии кинуренина, но не в отсутствие кинуренина.
[Пример 4] Оценка эффекта кинуренина на связывание IL6 человека с помощью поверхностного плазмонного резонанса
(4-1) Оценка кинуренина в отношении его функции переключения при связывании IL-6 человека
С использованием Biacore T200 (GE Healthcare), A11 анализировали в отношении его взаимодействия с IL-6 человека (Kamakura Techno-Science, Inc.) в реакции антиген-антитело. На сенсорный чип CM5 (GE Healthcare) иммобилизовывали соответствующее количество белка A/G (Invitrogen) способом присоединения аминов. Представляющие интерес антитела улавливали на чипе, чтобы обеспечить реакцию с IL-6 в качестве антигена. Два типа используемых подвижных буферов представляли собой 10 ммоль/л ACES, 150 ммоль/л NaCl, 0,05% (масс./об.) Tween20, 100 мкмоль/л кинуренина, pH 7,4, и 10 ммоль/л ACES, 150 ммоль/л NaCl, 0,05% (масс./об.) Tween20, pH 7,4. Взаимодействие с IL-6 в качестве антигена оценивали при 37°C. Буфер, использованный для разбавления IL-6, представлял тот же подвижный буфер, который описан выше.
Разбавленный раствор IL-6 человека и подвижный буфер в качестве пустого образца инжектировали при скорости потока 5 мкл/мин в течение трех минут для обеспечения взаимодействия IL-6 человека с A11, уловленном на сенсорном чипе. Затем инжектировали подвижный буфер при скорости потока 5 мкл/мин в течение трех минут. После наблюдения диссоциации IL-6 человека от антитела инжектировали 10 ммоль/л глицина-HCl (pH 1,5) при скорости потока 30 мкл/мин в течение 30 секунд для регенерации сенсорного чипа. Константу диссоциации KD (M) A11 вычисляли для IL-6 человека на основе константы скорости ассоциации ka (1/М с) и константы скорости диссоциации kd (1/с), обе из которых являются параметрами кинетики, вычисленными из сенсограммы, полученной при измерении. Каждый параметр вычисляли с использованием оценочного программного обеспечения Biacore T200 Evaluation Software (GE Healthcare).
Сенсограммы для взаимодействия между A11 и 4 мкмоль/л IL-6 человека, полученные путем измерения в присутствии или в отсутствие 100 мкмоль/л кинуренина, представлены на фиг.4. Как показано на фиг.4, A11 связывалось с IL-6 в присутствии 100 мкмоль/л кинуренина; однако в отсутствие кинуренина связывание с IL-6 не выявлялось. Это демонстрирует, что A11 обладает свойством, состоящим в том, что оно связывается с IL-6 через кинуренин в качестве переключающего фактора. Между тем, константа диссоциации KD A11 составила 1,0 E-6 моль/л в присутствии 100 мкмоль/л кинуренина.
(4-2) Оценка эффекта концентрации кинуренина на связывание IL-6 человека
Затем оценивали эффект концентрации кинуренина на реакцию антиген-антитело между A11 и IL-6 человека с использованием Biacore T200 (GE Healthcare). Использованный подвижный буфер представлял собой 10 ммоль/л ACES, 150 ммоль/л NaCl, 0,05% (масс./об.) Tween20, pH 7,4. Реакцию антиген-антитело между A11 и IL-6 человека оценивали при 25°C. A11 иммобилизовывали на сенсорный чип CM5 способом присоединения аминов, и в качестве анализируемого соединения IL-6 разбавляли до 1 мкмоль/л посредством 10 ммоль/л ACES, 150 ммоль/л NaCl, 0,05% (масс./об.) Tween20, pH 7,4, содержащего кинуренин в различных концентрациях, и позволяли им взаимодействовать в течение 60 секунд для выявления изменений в уровне связывания. Результат представлен на фиг. 5. Этот результат продемонстрировал, что чем выше концентрация кинуренина в качестве переключателя, тем больше IL-6 связывается с A11.
Далее тот же эксперимент, который описан выше, проводили для оценки эффекта концентрации кинуренина на реакцию антиген-антитело между IL-6 человека и связывающим IL-6 человека антителом H01 (тяжелая цепь и легкая цепь представляют собой SEQ ID NO:36 и 37, соответственно), иммобилизованном на сенсорный чип CM5, которое было получено из библиотеки и служило в качестве контроля для функции переключения A11 кинуренином. Результат представлен на фиг. 6. Этот результат подтвердил, что для контрольного антитела против IL-6 H01, происходящего из библиотеки, его связывание с IL-6 не изменялось, даже если концентрация кинуренина изменялась.
Затем оценивали эффект концентрации кинуренина в качестве переключающего фактора в отношении двухвалентного связывания A11 с IL-6 с использованием Biacore T200 (GE Healthcare). Использованный подвижный буфер представлял собой 10 ммоль/л ACES, 150 ммоль/л NaCl, 0,05% (масс./об.) Tween20, pH 7,4. Реакцию антиген-антитело между A11 и IL-6 человека оценивали при 25°C. IL-6 иммобилизовывали на сенсорный чип CM5 способом присоединения аминов, и в качестве анализируемого соединения A11 разбавляли до 0,1 мкмоль/л посредством 10 ммоль/л ACES, 150 ммоль/л NaCl, 0,05% (масс./об.) Tween20, pH 7,4, содержащего различные концентрации кинуренина, и позволяли им взаимодействовать в течение 60 секунд для выявления изменений в количестве двухвалентного связывания A11 с IL-6. Результат представлен на фиг.7. В этой аналитической системе ожидалось, что A11 будет связываться двухвалентным образом, поскольку IL-6 иммобилизован на сенсорном чипе. В случае такой аналитической системы, где A11 распознает IL-6 двухвалентным образом, также было выявлено, что количество A11, связавшегося с IL-6, возрастает при увеличении концентрации кинуренина. Этот результат продемонстрировал, что A11 обладает свойством, состоящим в том, что, обладая двухвалентным связыванием, оно также связывается с IL-6 через кинуренин в качестве переключающего фактора.
(4-3) Эффект кинуренина в качестве переключателя на диссоциацию антител от IL-6 человека
С использованием Biacore T200 (GE Healthcare), A11, связавшееся с IL-6 в присутствии кинуренина, исследовали для оценки того, диссоцирует ли оно в отсутствие кинуренин зависимым от концентрации кинуренина образом. Использованный подвижный буфер представлял собой 10 ммоль/л ACES, 150 ммоль/л NaCl, 0,05% (масс./об.) Tween20, pH 7,4, и 10 ммоль/л ACES, 150 ммоль/л NaCl, 0,05% (масс./об.) Tween20, pH 7,4, 100 мкмоль/л кинуренина. Анализ проводили при 25°C. IL-6 иммобилизовывали на сенсорный чип CM5 способом присоединения аминов, и в качестве анализируемого соединения A11 разбавляли до 0,1 мкмоль/л посредством 10 ммоль/л ACES, 150 ммоль/л NaCl, 0,05% (масс./об.) Tween20, pH 7,4, содержащего 100 мкмоль/л кинуренина, и позволяли им взаимодействовать в течение 60 секунд. Затем проводили мониторинг диссоциации IL-6 с помощью каждого типа подвижного буфера. Для сравнения степени диссоциации между соответствующими условиями подвижного буфера, количества IL-6, связавшегося с A11, нормализовывали и сравнивали, принимая величину в присутствии 100 мкмоль/л кинуренина за 100. Сенсограмма, на которой представлено взаимодействие между A11 и IL-6 после нормализации, представлена на фиг. 8. Результат, показанный на фиг. 8 демонстрирует, что A11 обладает свойством, состоящим в том, что оно связывается с IL-6 в присутствии кинуренина, а затем быстро диссоциирует от IL-6 в отсутствие кинуренина. В частности, было продемонстрировано, что опосредуемая кинуренином регуляция связывания антитела с IL-6 человека является полностью обратимой.
Эти результаты показали, что A11 представляет собой антитело, которое связывается с IL-6 в присутствии кинуренина через кинуренин в качестве переключающего фактора, но диссоциирует от IL-6 в отсутствие кинуренина. Также было подтверждено, что является возможной полная регуляция A11 по типу включение/выключение, так чтобы оно не обладало активностью связывания с IL-6 человека в отсутствие кинуренина. Ожидалось, что функция переключения будет достигнута аналогично тому, как показано на фиг. 2.
(4-4) Оценка кинуренина в отношении его связывания с IL-6 человека
Взаимодействие между IL-6 (Kamakura Techno-Science, Inc.) и кинуренином анализировали с использованием Biacore T200 (GE Healthcare). На сенсорный чип CM5 (GE Healthcare) иммобилизовывали приблизительно 5000 RU IL-6 способом присоединения аминов, и 800, 400, 200, 100, 50 или 25 нмоль/л кинуренина позволяли взаимодействовать с IL-6. Использованный подвижный буфер представлял собой 10 ммоль/л ACES, 150 ммоль/л NaCl, 0,05% (масс./об.) Tween20, pH 7,4. Все измерения для взаимодействия, описанного выше, проводили при 25°C. Кинуренин разбавляли с использованием подвижного буфера. Полученная сенсограмма, демонстрирующая взаимодействие между IL-6 и кинуренином, представлена на фиг. 9.
Приблизительно 5000 RU IL-6 иммобилизовывали в описанном выше эксперименте. Молекулярные массы IL-6 и кинуренина составляли приблизительно 20000 г/моль и приблизительно 200 г/моль, соответственно. Таким образом, взаимодействие ожидалось максимум при приблизительно 50 RU кинуренина. Однако в условиях измерения, описанных выше, явное взаимодействие с IL-6 не обнаруживалось, даже когда кинуренину позволяли взаимодействовать при максимальной концентрации 800 нмоль/л.
Исходя из результата примера, описанного выше, было оценено, что KD кинуренина для образования комплекса, состоящего из A11, IL-6 и кинуренин, составляет от нескольких десятков нМ до нескольких нМ. Это также указывает на то, что, если гипотетически кинуренин взаимодействует прямо с IL-6, взаимодействие однозначно наблюдалось бы, когда кинуренину позволяли взаимодействовать при 800 нмоль/л. Результат, описанный выше, указывает на возможность того, что кинуренин не взаимодействует прямо с IL-6, а взаимодействует с A11 или с комплексом A11-IL-6 в концентрации несколько десятков нМ.
[Пример 5] Получение антител против аденозина с помощью клонирования B-клеток кролика
(5-1) Моделирование иммуногена для конструирования связывающей аденозин библиотеки
Иммуногены, использованные у кроликов, представляли собой 2'-аденозин-PEG-хелперный пептид столбнячного токсина p30 (2'-аденозин-PEG-пептид), представленный на фиг. 10 и 5'-аденозин-PEG-хелперный пептид p30 столбнячного токсина (5'-аденозин-PEG-пептид), представленный на фиг. 11. Хелперный пептид p30 столбнячного токсина состоит из аминокислотной последовательности FNNFTVSFWLRVPKVSASHLE (SEQ ID NO:4), и он представляет собой пептид, идентифицированный в качестве эпитопа T-клеточного рецептора, экспрессируемого на хелперных T-клетках (Eur. J. Immunol. (1989) 19, 2237-2242). Известно, что пептид активирует продукцию антител (J. Immunol. (1992) 149, 717-721). Когда он связан с аденозином, пептид служит в качестве адъюванта и, таким образом, ожидается, что он будет усиливать продукцию антител против аденозина. Связывание между аденозином и хелперным пептидом p30 столбнячного токсина моделировали через PEG, так чтобы эпитопы антител против аденозина не включали хелперный пептид p30 столбнячного токсина. Аденозин представляет собой метаболит ATP, и, поскольку фосфатфосфатные группы ATP связаны с 5'-гидроксильной группой аденозина, антитела, которые не распознают 5'-гидроксильную группу аденозина в качестве эпитопа, также могут связываться с ATP, в дополнение к аденозину. Следовательно, было бы легче получить антитела, которые могут связываться как с аденозином, так и с ATP, с использованием в качестве иммуногена конструкции 5'-аденозин-PEG-хелперный пептид p30 столбнячного токсина, в то время как было бы легче получить антитела, которые связывается с аденозином, но не с ATP, с использованием в качестве иммуногена конструкции 2'-аденозин-PEG-хелперный пептид p30 столбнячного токсина. По этой причине было получено два типа иммуногенов, которые содержат хелперный пептид p30 столбнячного токсина, связанный с 2'- или 5'-положением аденозина, аналогично тому, как описано в (5-2).
Кроме того, 2'-аденозин-PEG-биотин (фиг. 12) и 5'-аденозин-PEG-биотин (фиг. 13), в котором биотин конъюгирован вместо хелперного пептида p30 столбнячного токсина получали, как описано ниже. Путем оценки связывания с этими двумя типами конструкции аденозин-PEG-биотин, антитела можно исследовать, чтобы продемонстрировать, что их эпитопы не включают хелперный пептид p30 столбнячного токсина.
(5-2) Синтез иммуногенов для получения связывающей аденозин библиотеки
2'-Аденозин-PEG-пептид (конъюгат аденозин 2'-PEG-пептид или 2'-(PEG-пептид)аденозин) и 2'-аденозин-PEG-биотин (конъюгат аденозин 2'-PEG-биотин или 2'-(PEG-биотин)аденозин) синтезировали, как описано ниже. Синтезированный 2'-аденозин-PEG-пептид и 2'-аденозин-PEG-биотин анализировали или фракционировали в условиях, описанных ниже.
Условия анализа LCMS описаны ниже.
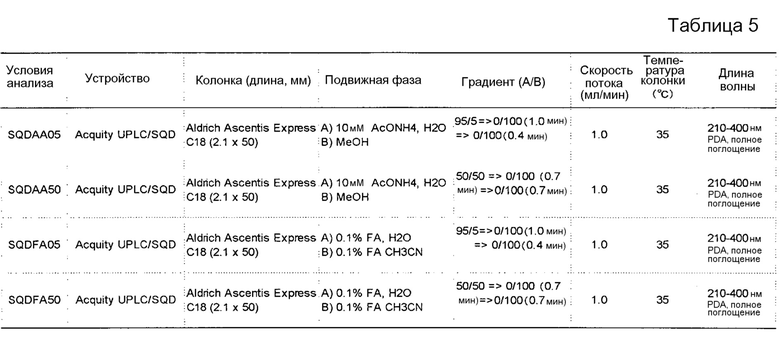
Ниже описаны условия препаративной ВЭЖХ.
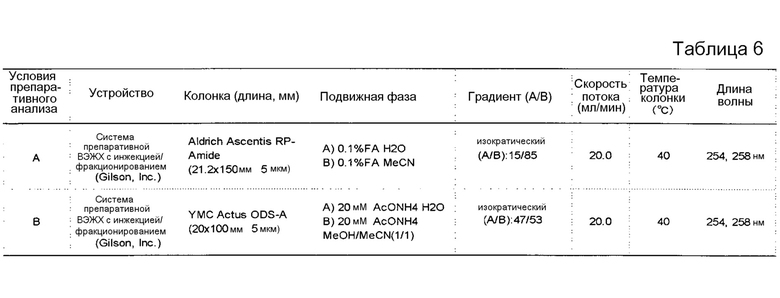
(5-2-1) Синтез соединения 006 (Boc-Phe-Asn-Asn-Phe-Thr (tBu)-Val-Ser (tBu)-Phe-Trp (Boc)-Lue-Arg (Pbf)-Val-Pro-Lys (Boc)-Val-Ser (tBu)-Ala-Ser (tBu)-His (Trt)-Leu-Glu (tBu)-OH)
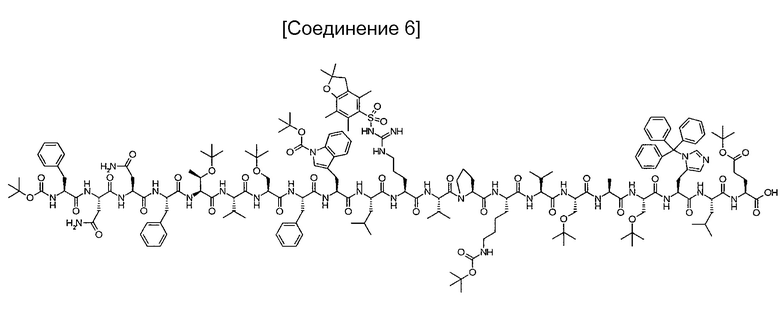
Пептидный синтез проводили способом Fmoc с использованием устройства для синтеза пептидов (Multipep RS; Intavis). Все аминокислоты аминокислоты с Fmoc приобретали от WATANABE CHEMICAL INDUSTRIES, LTD. Подробное описание методики обработки представлено в руководстве, прилагаемом к устройству для синтеза.
Fmoc-Glu(tBu)-OH, связанный на его C-конце с 2-хлортритилсмолой (250 мг/колонка, 30 колонок, 11,7 ммоль), раствор N,N-диметилформамида, содержащжий различные Fmoc-аминокислоты (0,6 моль/л) и 1-гидрокси-7-азабензотриазолтриазол (0,375 моль/л), и раствор диизопропилкарбодиимида в N,N-диметилформамиде (10% об./об.) загружали в устройство для синтеза. Реакцию синтеза проводили с использованием в качестве раствора для удаления Fmoc раствора N,N-диметилформамида (20% об./об.), содержащего пиперидинпиперидин и 5% (масс./об.) мочевину. После промывания смолы N,N-диметилформамидом проводили удаление защитной группы Fmoc, а затем проводили один цикл реакции конденсации с Fmoc-аминокислотой. Этот цикл повторяли для удлинения пептидов на поверхности смолы. После удлинения смолу промывали трифторэтанолом. Пептиды отщепляли от смолы добавлением трифторэтанола/дихлорметанэтана (=1/1). Таким образом, получили соединение 006 (7,2 г) в виде неочищенного продукта.
LCMS(ESI)m/z = 1185(M+3H)3+
Время удержания: 1,24 минуты (условия анализа: SQDAA05)
(5-2-2) Синтез соединения 007
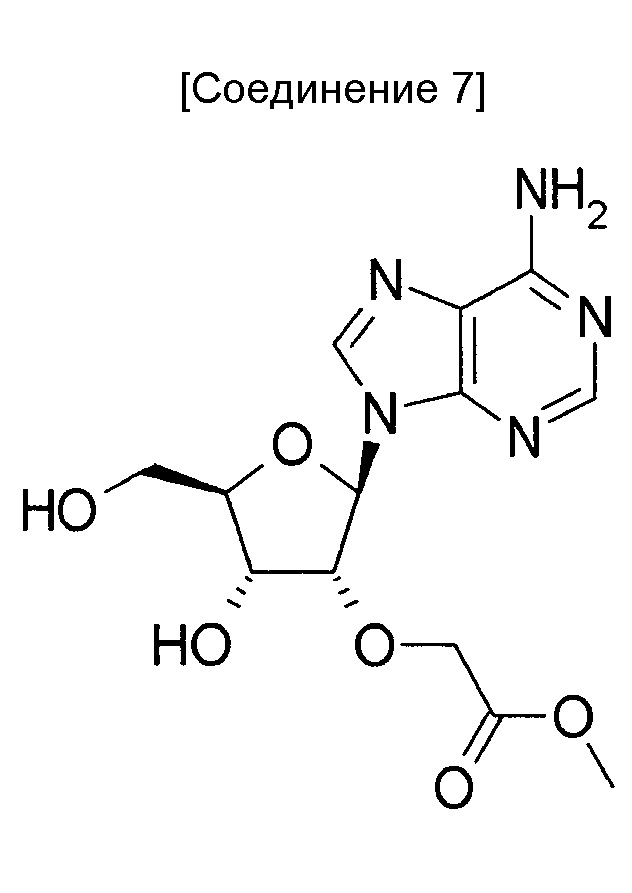
Суспензию аденозина (2,00 г, 7,48 ммоль) в N,N-диметилформамиде (40 мл) охлаждали до 0°C, и к ней добавляли 60% гидрид натрия (0,42 г, 10,48 моль). Реакционную смесь перемешивали в течение одного часа при 0°C. После добавления метилбромацетата (0,76 мл, 8,01 ммоль), полученную реакционную смесь перемешивали в течение пяти часов при комнатной температуре, и к ней добавляли уксусную кислоту (1 мл) и метанол (3 мл). Реакционную смесь концентрировали при пониженном давлении. Полученный осадок очищали нормально-фазовой хроматографией на силикагеле (дихлорметан/метанол). Таким образом, было получено соединение 007 (0,93 г, 37%).
LCMS(ESI) m/z=340(M+H)+
Время удержания: 0,27 минуты (условия анализа: SQDFA05)
(5-2-3) Синтез соединения 008
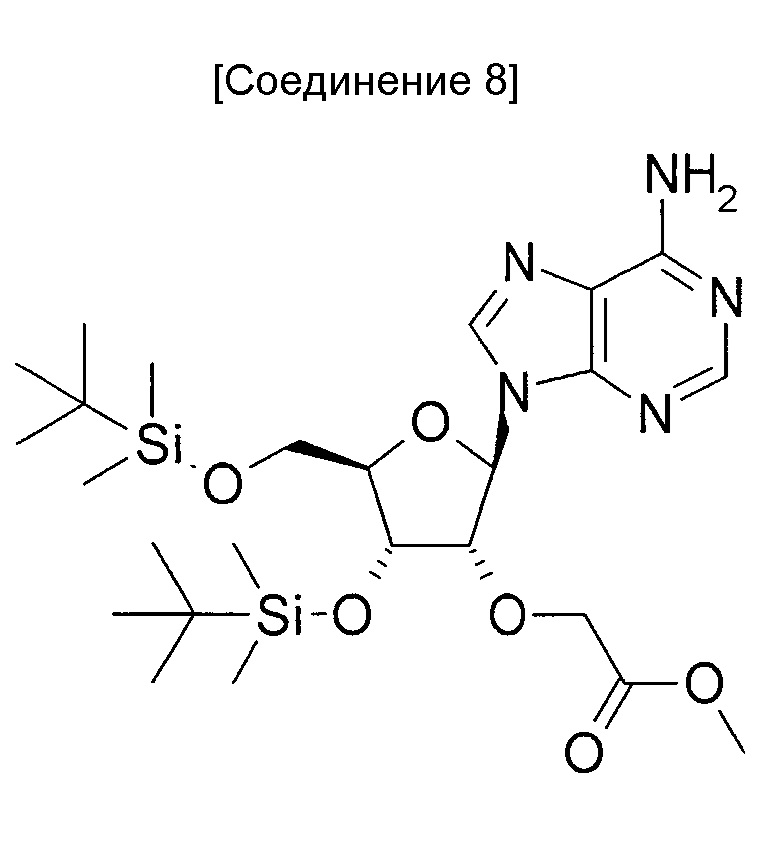
Трет-бутилдиметилсилилхлорид (999 мг, 6,63 моль) и имидазол (722 мг, 10,61 моль) добавляли к раствору соединения 007 (900 мг, 2,65 ммоль) в пиридине. Реакционную смесь перемешивали в течение четырех часов при комнатной температуре и экстрагировали этилацетатом/водой. Экстрагированный органический слой промывали насыщенным раствором хлорида натрия и сушили над безводным сульфатом натрия. После фильтрации органический слой концентрировали при пониженном давлении. Полученный осадок очищали нормально-фазовой колоночной хроматографией на силикагеле (дихлорметан/метанол). Таким образом, было получено соединение 008 (1,17 г, 78%).
LCMS(ESI)m/z = 568(M+H)+
Время удержания: 1,10 минуты (условия анализа: SQDFA05)
(5-2-4) Синтез соединения 009
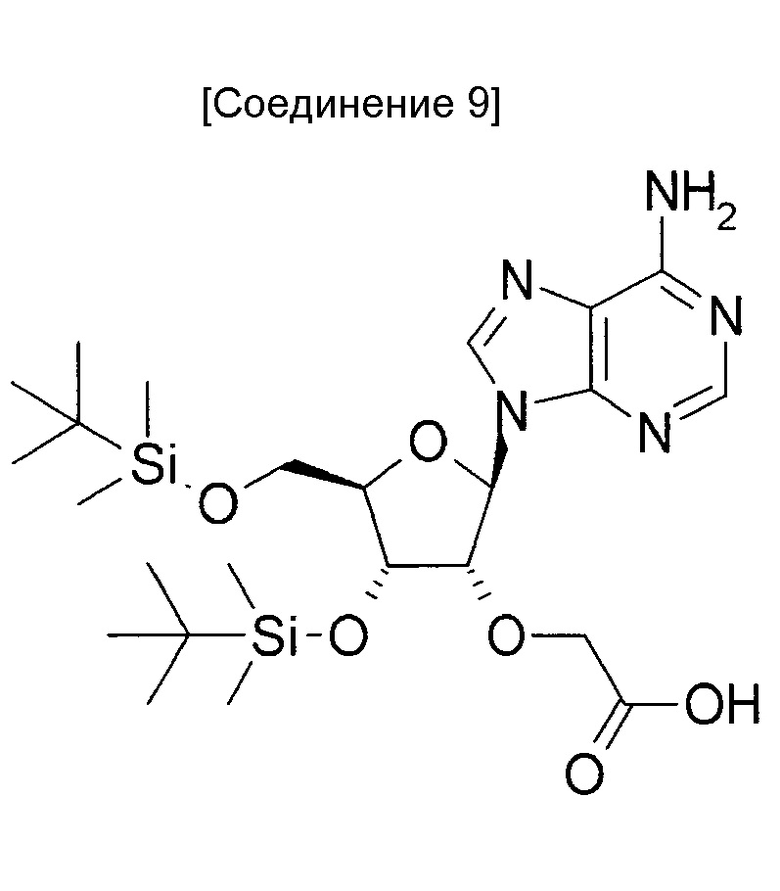
Гидроксид лития (61 мг, 2,5 моль), растворенный в воде (0,17 мл) добавляли к раствору соединения 008 (290 мг, 0,51 ммоль) в метаноле (0,34 мл)/ тетрагидрофуране (0,34 мл). Реакционную смесь перемешивали в течение 30 минут при комнатной температуре. Смесь нейтрализовывали 1 M хлористоводородной кислотой и концентрировали при пониженном давлении. Концентрированный осадок экстрагировали этилацетатом/водой. Полученный органический слой промывали насыщенным раствором хлорида натрия и сушили над безводным сульфатом натрия. После фильтрации органический слой концентрировали при пониженном давлении. Таким образом, было получено соединение 009 (319 мг, 90%).
LCMS(ESI)m/z = 552(M-H)-
Время удержания: 0,97 минуты (условия анализа: SQDFA05)
(5-2-5) Синтез соединений 010 и 011
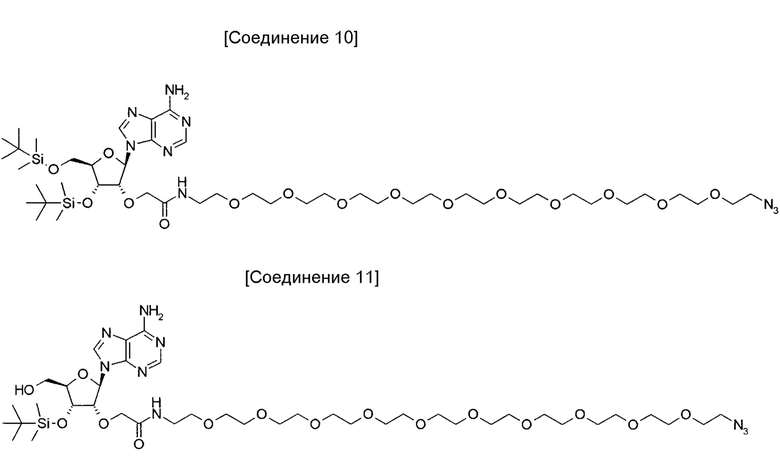
1-гидроксибензотриазолтриазол (75 мг, 0,553 моль) и гидрохлорид 1-этил-3-(3-диметиламинопропил)карбодиимида (106 мг, 0,553 моль) добавляли к раствору соединения 009 (255 мг, 0,460 ммоль) в N,N-диметилформамиде (1,5 мл), и их перемешивали в течение трех минут при комнатной температуре. К реакционной смеси добавляли O-(2-аминоэтил)-O'-2-азидоэтил)нонаэтиленгликоль (291 мг, 0, 53 ммоль) и ее перемешивали в течение трех часов при комнатной температуре. Реакционную смесь концентрировали при пониженном давлении и полученный осадок очищали обращено-фазовой хроматографией на силикагеле (10 мМ водный раствор ацетата аммония/метанол. Получали соединения 010 (177 мг, 4%) и 011 (72 мг, 19%).
Соединение 010
LCMS(ESI)m/z = 1063(M+H)+
Время удержания: 0,98 минуты (условия анализа: SQDFA05)
Соединение 011
LCMS(ESI)m/z = 949(M+H)+
Время удержания: 0,67 минуты (условия анализа, SQDFA05)
(5-2-6) Синтез соединения 012
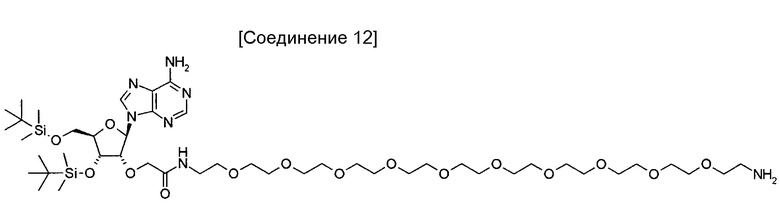
10% палладий на угле (34 мг) добавляли к раствору соединения 010 (170 мг, 0,160 ммоль) в этаноле (1 мл). Реакционную смесь перемешивали в течение двух часов в атмосфере водорода, и вновь добавляли к ней 10% палладий на угле (34 мг). Реакционную смесь перемешивали в течение двух часов в атмосфере водорода для завершения реакции. Фильтрат реакционного раствора концентрировали при пониженном давлении. Получили соединение 012 (34 мг, 95%).
LCMS(ESI)m/z = 1037(M+H)+
Время удержания: 0,70 минуты (условия анализа: SQDFA05)
(5-2-7) Синтез соединений 013 и 014
Соединение 006 (354 мг, 0,110 ммоль), 1-гидроксибензотриазолтриазол (13 мг, 0,100 моль) и гидрохлорид 1-этил-3-(3-диметиламинопропил)карбодиимида (19 мг, 0,100 моль) добавляли к раствору соединения 012 (86 мг, 0,083 ммоль) в N,N-диметилформамиде (1,5 мл), и их перемешивали в течение двух часов при комнатной температуре. Фильтрат реакционной смеси очищали с помощью условий препаративного анализа A, описанных в таблице 6. Получили смесь соединений 013 и 014 (72 мг).
Соединение 013
LCMS(ESI)m/z=1525(M+3H)3+, 1144(M+4H)4+
Время удержания: 1,13 минуты (условия анализа: SQDAA50)
Соединение 014
LCMS(ESI)m/z = 1444(M+3H)3+, 1083(M+4H)4+
Время удержания: 1,02 минуты (условия анализа: SQDAA50)
(5-2-8) Синтез 2'-аденозин-PEG-пептида (конъюгат аденозин 2'-PEG-пептид или 2'-(PEG-пептид)аденозин) (соединение 015)
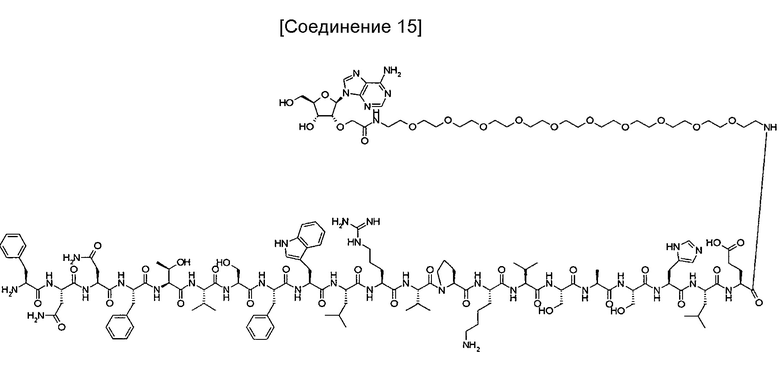
Трифторуксусную кислоту (16 мл), дихлорметан (8 мл), воду (1,3 мл) и тетраизопропилсилан (1,3 мл) добавляли к смеси соединений 013 и 014 (42 мг), и перемешивали в течение шести часов при комнатной температуре. Осадок, полученный концентрированием реакционной смеси при пониженном давлении, очищали с использованием условий препаративного анализа B, описанных в таблице 6. Таким образом, получили соединение 015 (10 мг).
LCMS(ESI)m/z = 1090(M+3H)3+, 818(M+4H)4+
Время удержания: 0,52 минуты (условия анализа: SQDAA50)
(5-2-9) Синтез соединения 016
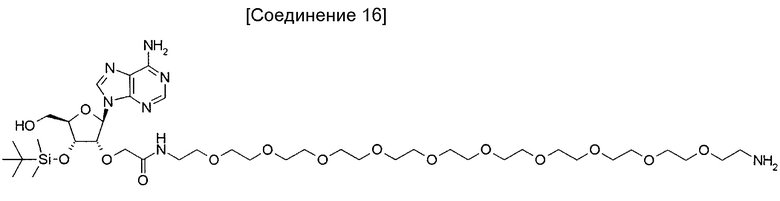
К раствору соединения 011 (70 мг, 0,074 ммоль) в этаноле (1 мл) добавляли 10% палладий на угле (34 мг) и реакционную смесь перемешивали в течение пяти часов в атмосфере водорода. Фильтрат реакционной смеси концентрировали при пониженном давлении. Таким образом, получили соединение 016 (58 мг, 85%).
LCMS(ESI)m/z = 923(M+H)+
Время удержания: 0,50 минуты (условия анализа: SQDFA05)
(5-2-10) Синтез соединения 017
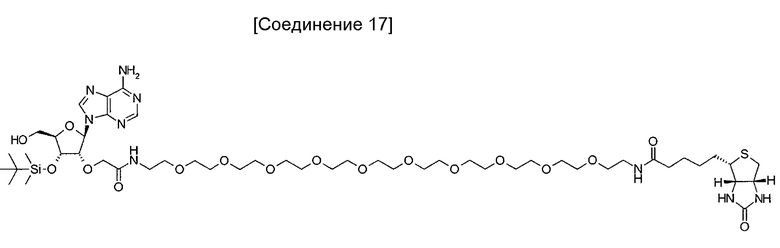
D-биотин-N-сукцинимидил (24 мг, 0,069 ммоль) и триэтиламин (13 мкл, 0,094 моль) добавляли к раствору соединения 016 (58 мг, 0,063 ммоль) в N,N-диметилформамиде (1 мл) и перемешивали в течение двух часов при комнатной температуре. Затем после добавления D-биотин-N-сукцинимидила (5 мг, 0,015 ммоль) реакция завершалась через 1,5 часа при перемешивании при комнатной температуре. Реакционную смесь очищали обращено-фазовой колоночной хроматографией на силикагеле (10 мМ водный раствор ацетата аммония/метанол). Получили соединение 017 (50 мг, 69%).
LCMS(ESI)m/z = 1149(M+H)+
Время удержания: 1,04 минуты (условия анализа: SQDFA05)
(5-2-11) Синтез 2'-аденозин-PEG-биотина (конъюгат аденозин 2'-PEG-биотин или 2’-(PEG-биотин)аденозин) (соединение 018)
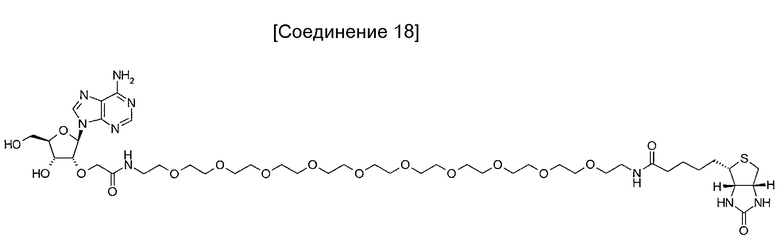
Раствор 1 M фторида тетра-н-бутиламмония в тетрагидрофуране (65 мкл, 0,065 ммоль) добавляли к раствору соединения 017 (62 мг, 0,054 ммоль) в тетрагидрофуране (2 мл) и перемешивали при комнатной температуре в течение одного часа. Затем добавляли 1 M раствор фторида тетра-н-бутиламмония в тетрагидрофуране (20 мкл, 0,020 ммоль) и реакцию завершали перемешиванием при комнатной температуре в течение одного часа. Реакционную смесь концентрировали при пониженном давлении и осадок очищали обращено-фазовой колоночной хроматографией на силикагеле (0,11% водный раствор муравьиной кислоты/0,11% муравьиная кислота в ацетонитриле). Получили соединение 018 (12 мг, 21%).
LCMS(ESI)m/z = 1035(M+H)+
Время удержания: 0,71 минуты (условия анализа: SQDAA05)
Более того, также посредством той же реакции синтезировали 5'-аденозин-PEG-пептид и 5'-аденозин-PEG-биотин.
(5-3) Получение связывающих аденозин антител у животных и скрининга антител
Кроликов иммунизировали 2'-аденозин-PEG-пептидом и/или 5'-аденозин-PEG-пептидом общепринятым способом. Клетки-кандидаты с активностью связывания аденозина отбирали из суспензии клеток, полученной из крови иммунизированных кроликов, с использованием autoMACS Pro Separator и FACSAria (BD), которые используют активность связывания аденозин-PEG-биотина и экспрессию IgG у кролика в качестве показателей. Затем проводили скрининг с антителами, секретированными в культуральные супернатанты отобранных клеток. Во время скрининга проводили ELISA для оценки наличия активности связывания с аденозин-PEG-биотином. Также ELISA проводили для оценки того, подавляет ли аденозин при добавлении в комбинации с аденозин-PEG-биотином на уровне, в 1000 раз или более превышающем уровень аденозин-PEG-биотина, связывание аденозин-PEG-биотина. Вариабельные области H-цепи и L-цепи выделяли способом ПЦР из клеток, отобранных с использованием в качестве индикатора наличия активности связывания аденозин-PEG-биотина, а также подавления связывания с аденозин-PEG-биотином аденозином, добавленным в комбинации с аденозин-PEG-биотином. Полученные вариабельные области экспрессировались в комбинации с константной областью тяжелой цепи и константной областью легкой цепи IgG1 человека.
(5-4) Получение B-клеток для получения связывающей аденозин иммунной библиотеки
Клетки получали из селезенок кроликов, иммунизированных 2'-аденозин-PEG-пептидом столбнячного токсина и 5'-аденозин-PEG-пептидом столбнячного токсина. Из суспензий клеток отбирали клетки-кандидаты с активностью связывания аденозина с использованием autoMACS Pro Separator и FACSAria (BD) с наличием связывания аденозин-PEG-биотина, а также экспрессией IgG или IgM кролика в качестве индикаторов. Отобранные клетки промывали PBS(-) и полученные клеточные осадки использовали для конструирования иммунных библиотек.
[Пример 6] Оценка клонов, полученных клонированием B-клеток кролика
(6-1) Оценка клонов, полученных клонированием B-клеток кролика, в отношении активности связывания с 2'-аденозин-PEG-биотином
Клоны, полученные клонированием B-клеток кролика, оценивали в отношении активности связывания с аденозином способом SPR. Реакцию антиген-антитело между клонами и 2'-аденозин-PEG-биотин кинетически анализировали с использованием Biacore 4000 (GE Healthcare). На сенсорный чип CM5 (GE Healthcare) иммобилизовывали соответствующее количество белка A/G (Invitrogen) способом присоединения аминов. Представляющие интерес антитела улавливали на чипе. Затем после взаимодействия 100 нмоль/л 2'-аденозин-PEG-биотина в качестве анализируемого соединения в течение 60 секунд проводили мониторинг и измерение диссоциации анализируемого соединения в течение 60 секунд. Использованный подвижный буфер представлял собой HBS-P+ (GE Healthcare). Все измерения проводили при 25°C. Анализируемое соединение разбавляли с использованием подвижного буфера.
Соответствующие антитела сравнивали в отношении активности связывания с 2'-аденозин-PEG-биотином с использованием в качестве индикатора величины (N_связывание_100), полученной делением величины связывания при взаимодействии с 2'-аденозин-PEG-биотином на величину улавливания (RU) для каждого антитела, и величины (N_стабильность_100), полученной делением величины диссоциации 2'-аденозин-PEG-биотина от каждого антитела в течение 60 секунд после взаимодействия с 2'-аденозин-PEG-биотином на величину улавливания (RU) для каждого антитела. Что касается антител, для которых величина улавливания составляла 1500 RU или менее, их связывание не достаточно поддавалось обнаружению и, таким образом, они были исключены из исследуемых образцов. Результат представлен на фиг. 14. Результат, представленный на фиг. 14, демонстрирует, что способ клонирования B-клеток обеспечивал связывающие аденозин клоны с различной аффинностью.
(6-2) Оценка связывающих 2'-аденозин-PEG-биотин клонов в отношении их активность связывания с аденозином и ATP, и анализ последовательности клонов
Клоны, для которых было продемонстрировано, что они связываются с 2'-аденозин-PEG-биотином, анализировали в отношении их связывания с аденозином и ATP способом SPR и конкурентного ELISA.
(6-2-1) Оценка с использованием SPR связывающих 2'-аденозин-PEG-биотин клонов в отношении их связывания с аденозином и ATP
С использованием Biacore T200 (GE Healthcare), антитела SMB0002, SMB0089 и SMB0104, полученные способом клонирования B-клеток, анализировали в отношении взаимодействия с аденозином и ATP в реакции антиген-антитело. На сенсорный чип CM5 (GE Healthcare) иммобилизовывали соответствующее количество белка A/G (Invitrogen) способом присоединения аминов. Представляющие интерес антитела улавливали на чипе для обеспечения взаимодействия с аденозином или ATP в качестве антигена. Использованный подвижный буфер представлял собой 10 ммоль/л ACES, 150 ммоль/л NaCl, 0,05% (масс./об.) Tween20, pH 7,4. Все измерения проводили при 25°C. Антигены разбавляли с использованием подвижного буфера.
Что касается SMB0002, SMB0089, и SMB0104, разбавленные растворы антигенов и подвижный буфер в качестве пустого образца инжектировали при скорости потока 20 мкл/мин в течение двух минут для обеспечения взаимодействия каждого антигена с антителами, уловленными на сенсорном чипе. Затем инжектировали подвижный буфер при скорости потока 20 мкл/мин в течение трех минут и наблюдали диссоциацию антигенов от антител. Далее инжектировали 10 ммоль/л глицин-HCl (pH 1,5) при скорости потока 30 мкл/мин в течение 30 секунд для регенерации сенсорного чипа. Константу скорости ассоциации ka (1/М с) и константу скорости диссоциации kd (1/s), обе из которых являются параметрами кинетики, вычисляли из сенсограмм, полученных при измерении. Константу диссоциации KD (M) вычисляли, исходя из этих констант. Каждый параметр вычисляли с использованием оценочного программного обеспечения Biacore T200 Evaluation Software (GE Healthcare).
Результат показал, что множество клонов, включающих SMB0002, SMB0089 и SMB0104, связывались как с аденозином, так и с ATP. Сенсограммы, исследованные для оценки каждого клона в отношении его связывания при концентрациях аденозина 500, 125, 31,3 и 7,81 нМ или при ATP концентрациях 5000, 1250, 313, и 78,1 нМ представлены на фиг.15. Как показано на фиг.15, было продемонстрировано, что SMB0002, SMB0089 и SMB0104 связываются как с аденозином, так и с ATP. KD для SMB0002, SMB0089 и SMB0104 составляли 9,3E-9, 6,9E-9, и 4,1E-8 (моль/л) для аденозина, и 1,0Е-5, 8,8E-7, и 1,4E-7 (моль/л) для ATP, соответственно.
Аналогичным образом с использованием Biacore 4000 (GE Healthcare), антитело SMB0171, полученное клонированием B-клеток, анализировали в отношении его взаимодействия с аденозином и ATP в реакции антиген-антитело. На сенсорный чип CM5 (GE Healthcare) иммобилизовывали соответствующее количество белка A/G (Invitrogen) способом присоединения аминов. Представляющие интерес антитела улавливали на чипе, чтобы обеспечить взаимодействие с аденозином или ATP в качестве антигена. Использованный подвижный буфер представлял собой HBS-P+ (GE Healthcare). Все измерения проводили при 25°C. Антигены разбавляли с использованием подвижного буфера.
Что касается SMB0171, разбавленные растворы антигена и подвижный буфер в качестве пустого образца инжектировали при скорости потока 10 мкл/мин в течение одной минуты, чтобы обеспечить взаимодействие каждого антигена с антителами, уловленными на сенсорном чипе. Затем, инжектировали подвижный буфер при скорости потока 10 мкл/мин в течение трех минут и исследовали диссоциацию антитела от антигенов. Затем, инжектировали 10 ммоль/л глицина-HCl (pH 1,5) при скорости потока 30 мкл/мин в течение 30 секунд для регенерации сенсорного чипа. Константу скорости ассоциации ka (1/М с) и константу скорости диссоциации kd (1/c), обе из которых являются параметрами кинетики, вычисляли из сенсограмм, полученных путем измерения. Исходя из этих констант вычисляли константу диссоциации KD (M). Каждый параметр вычисляли с использованием оценочного программного обеспечения Biacore 4000 Evaluation Software (GE Healthcare).
Результат продемонстрировал, что SMB0171 связывается с ATP. Сенсограммы, полученные при оценке каждого клона в отношении связывания при концентрациях ATP 50 и 5 мкМ, представлены на фиг. 16. Как показано на фиг. 16, было продемонстрировано, что SMB0171 связывается с ATP. KD SMB0171 для ATP составила 5,9E-6 (моль/л).
(6-2-2) Оценка связывающих 2’-аденозин-PEG-биотин клонов в отношении их связывания с аденозином и ATP в конкурентном ELISA
Антитела, для которых было показано связывание с 2’-аденозин-PEG-биотином, разбавляли до 1 мкг/мл посредством PBS и добавляли в каждую лунку 384-луночного MAXISorp (Nunc). Для иммобилизации антител планшету позволяли стоять в течение одного часа или более при комнатной температуре. После удаления антител, разбавленных PBS, из каждой лунки, добавляли TBS, содержащий 1% BSA, и планшету позволяли стоять в течение одного часа или более. Затем TBS (pH 7,4), содержавший 1% BSA, удаляли из планшета. В планшет добавляли 2'-аденозин-PEG-биотин, разбавленный до 50 нМ посредством PBS, смесь 2'-аденозин-PEG-биотина и аденозин, разбавленный до 50 нМ и 500 мкМ соответственно посредством PBS, смесь 2'-аденозин-PEG-биотина и ATP, разбавленных до 50 нМ и 500 мкМ, соответственно, с PBS или PBS отдельно. Планшету позволяли стоять при комнатной температуре в течение одного часа, а затем промывали три раза 80 мкл PBS, содержавшего 0,05% Tween-20. Затем в каждую лунку добавляли стрептавидин-HRP (Thermo fisher scientific), разбавленный в 20000 раз посредством PBS, и планшету позволяли стоять в течение одного часа или более при комнатной температуре. Планшет промывали три раза 80 мкл PBS, содержавшего 0,05% Tween-20, а затем в каждую лунку добавляли хромогенный субстрат (субстрат пероксидазы ABTS). Планшет инкубировали в течение одного часа, а затем развитие окраски в растворе в каждой лунке оценивали путем измерения поглощения при 405 нм с использованием SpectraMax от Molecular Device.
Как показано на фиг. 17, результат показал, что связывание SMB0002 с 2'-аденозин-PEG-биотином ингибировалось добавлением избыточных количеств аденозина и ATP. Таким образом, было показано, что клоны антител связываются не только с 2'-аденозин-PEG-биотином, но также как с аденозином, так и с ATP.
(6-2-3) Анализ последовательности клонов, идентифицированных как аденозин- и АТФ- связывающие, с использованием SPR
Аминокислотные последовательности клонов, для которых было продемонстрировано, что они связываются как с аденозином, так и с ATP, представлены в таблице 7.

[Пример 7] Получение антител, которые связываются с аденозином и/или ATP, из библиотеки антител человека с использованием способов фагового дисплея(7-1)
(7-1) Конструирование библиотеки фагового дисплея наивных антител человека
Библиотеку фагового дисплея антител человека, состоящую из множества фагов, которые презентируют Fab-домены антител человека, последовательности которых отличались друг от друга, конструировали с использованием в качестве матрицы полиA РНК, полученной из PBMC человека, коммерчески доступной полиA РНК человека и т.п. в соответствии со способом, известным специалистам в данной области.
(7-2) Получение антител, которые связываются с аденозином и/или ATP, из библиотеки способом пэннинга гранул
Библиотеку фагового дисплея наивных антител человека, сконструированную, как описано в (7-1), подвергали скринингу в отношении антител, которые проявляют антигенсвязывающую активность, в частности, путем сбора фагов, которые экспонируют антитела с активностью связывания с антигенами, уловленными гранулами. В качестве антигенов использовали биотинилированный ATP, 2’-аденозин-PEG-биотин и 5’-аденозин-PEG-биотин.
Фаги, продуцированные в E. coli, содержащих фагмидный вектор, сконструированный для фагового дисплея очищали общепринятым способом, а затем подвергали диализу против TBS с получением суспензии фаговой библиотеки. Затем в суспензию добавляли BSA в конечной концентрации 4%. Пэннинг проводили с использованием магнитных гранул с иммобилизованным на них антигеном. Использованные магнитные гранулы представляли собой покрытые нейтравидином гранулы (Sera-Mag SpeedBeads NeutrАвидин-coated) и покрытые стрептавидином гранулы (Dynabeads M-280 Streptavidin).
Далее к полученной суспензии фаговой библиотеки добавляли 250 пмоль биотинилированного ATP, 2'-аденозин-PEG-биотина и 5'-аденозин-PEG-биотина. Таким образом, суспензию фаговой библиотеки контактировали с аденозином и ATP в течение 60 минут при комнатной температуре. Затем к суспензии фаговой библиотеки добавляли блокированные BSA магнитные гранулы и комплекс фага с аденозином и/или ATP позволяли связаться с магнитными гранулами при комнатной температуре в течение 15 минут. Гранулы промывали один раз посредством TBS. Затем гранулы объединяли с 0,5 мл раствора трипсина в концентрации 1 мг/мл. Гранулы суспендировали при комнатной температуре в течение 15 минут, и сразу после этого суспензию фага собирали с выделенных гранул с использованием магнитного стенда. Собранную суспензию фагов добавляли к 10 мл клеток E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубировали при 37°C в течение одного часа при осторожном перемешивании для инфицирования фагом. Инфицированные E. coli высевали в чашку 225 мм × 225 мм. Затем фаги собирали из культуральной среды посеянных E. coli с получением жидкого исходного раствора с фаговой библиотекой.
Второй раунд пэннинга проводили, чтобы также увеличить содержание фагов, которые способны связываться с аденозином и/или ATP. Полученную суспензию фаговой библиотеки контактировали с аденозином и ATP в течение 60 минут при комнатной температуре путем добавления 50 пмоль каждого из биотинилированного ATP, 2'-аденозин-PEG-биотина и 5'-аденозин-PEG-биотина. Затем к суспензии фаговой библиотеки добавляли блокированные посредством BSA магнитные гранулы, и комплексу фага с аденозином и/или ATP позволяли связываться с магнитными гранулами при комнатной температуре в течение 15 минут. Гранулы промывали три раза посредством TBST и два раза посредством TBS. Затем гранулы объединяли с 0,5 мл раствора трипсина в концентрации 1 мг/мл. Гранулы суспензировали при комнатной температуре в течение 15 минут, и сразу после этого суспензию фага собирали с выделенных гранул с использованием магнитного стенда. Собранную суспензию фагов добавляли к 10 мл клеток E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубировали при 37°C в течение одного часа при осторожном перемешивании для инфицирования фагом. Инфицированные E. coli высевали в чашку 225 мм × 225 мм. Затем фаги собирали из культуральной среды посеянных E. coli с получением жидкого исходного раствора с фаговой библиотекой.
По той же методике пэннинг проводили три раза для получения антител, которые способны связываться с аденозином и/или ATP. На четвертом раунде пэннинга каждое из промывания TBST и промывания TBS проводили пять раз.
(7-3) Оценка активности связывания аденозина и ATP с помощью ELISA фагов
Из единичных колоний E. coli, полученных пэннингом, как описано в представленном выше примере, культуральные супернатанты, содержащие фаги, собирали в соответствии с общепринятым способом (Method Mol. Biol. (2002) 178, 133-145). Собранные культуральные супернатанты обрабатывали ультрафильтрацией с использованием NucleoFast 96 (MACHERY-NAGEL). 100 мкл культуральных супернатантов добавляли в каждую лунку NucleoFast 96 и центрифугирвоали при 4500 g в течение 45 минут оставшейся жидкой части. После добавления 100 мкл H2O NucleoFast 96 промывали центрифугированием при 4500 g в течение 30 минут. После добавления 100 мкл TBS NucleoFast 96 позволяли стоять в течение пяти минут при комнатной температуре. Затем из супернатантов собирали суспензии фагов.
Очищенные фаги, к которым добавляли TBS, подвергали ELISA по следующей методике. 96-луночный микропланшет для титрования StreptaWell (Roche) покрывали при комнатной температуре в течение одного часа посредством 100 мкл TBS, содержавшего меченные биотином антигены (смесь равных количеств 2'-аденозин-PEG-биотина, 5'-аденозин-PEG-биотина и ATP-PEG-биотина). Антигены удаляли из каждой лунки планшета промыванием TBST (TBS, содержащий 0,1% Tween20), а затем лунки блокировали 250 мкл смеси 2% обезжиренное молоко/TBS в течение одного часа или более. 2% обезжиренное молоко/TBS удаляли, а затем в каждую лунку добавляли полученные очищенные фаги. Планшету позволяли стоять при комнатной температуре в течение одного часа, чтобы позволить экспонированному на фаге антителу связаться с антигенами в каждой лунке. После промывания TBST, в каждую лунку добавляли конъюгированное с HRP антитело против M13 (Amersham Pharmacia Biotech), разбавленное TBS. Планшет инкубировали в течение одного часа. После промывания TBST в каждую лунку добавляли простой раствор TMB (ZYMED). Хромогенную реакцию в растворе каждой лунки останавливали добавлением серной кислоты. Затем оценивали развитие окраски путем измерения поглощения при 450 нм.
Из 192 кулонов, подвергнутых ELISA фагов, получили 106 клонов, которые обладали способностью связываться с любым одним, или двумя, или всеми тремя из 2'-аденозин-PEG-биотина, 5'-аденозин-PEG-биотина и ATP-PEG-биотина.
Далее, для подтверждения того, в отношении какого из антигенов: 2'-аденозин-PEG-биотин, 5'-аденозин-PEG-биотин и ATP-PEG-биотин, эти клоны обладали активностью связывания, очищенные фаги, разбавленные TBS, подвергали ELISA по следующей методике. 96-луночный микропланшет для титрования StreptaWell (Roche) покрывали при комнатной температуре в течение одного часа посредством 100 мкл TBS, содержащего меченный биотином антиген (2'-аденозин-PEG-биотин, 5'-аденозин-PEG-биотин или ATP-PEG-биотин). После удаления антигенов промыванием каждой лунки планшета TBST, лунки блокировали 250 мкл смеси 2% обезжиренное молоко/TBS в течение одного часа или более. 2% обезжиренное молоко/TBS удаляли, а затем в каждую лунку добавляли полученные очищенные фаги. Планшету позволяли стоять при комнатной температуре в течение одного часа, чтобы обеспечить связывание экспонирующих антитело фагов с антигенами в каждой лунке. После промывания TBST в каждую лунку добавляли конъюгированное с HRP антитело против M13 (Amersham Pharmacia Biotech), разбавленное TBS. Планшет инкубировали в течение одного часа. После промывания TBST, в каждую лунку добавляли простой раствор TMB (ZYMED). Хромогенную реакцию в растворе каждой лунки останавливали добавлением серной кислоты. Затем, оценивали развитие окраски путем измерения поглощения при 450 нм. Результат ELISA фагов представлен в таблице 8, ниже.
Среди клонов, подвергнутых ELISA фагов, был продемонстрирован клон, который связывается с двумя или более типами антигенов. Его подвергали амплификации генов со специфическими праймерами с использованием фрагмента антитела в качестве матрицы. Нуклеотидную последовательность гена анализировали. Этот клон обладал способностью связываться как с 5'-аденозин-PEG-биотином, так и с ATP-PEG-биотином, и он был назван ATNLSA1-4_D12. Последовательность вариабельной области тяжелой цепи антитела ATNLSA1-4_D12 представлена в SEQ ID NO:46 и его последовательность вариабельной области легкой цепи представлена в SEQ ID NO:47.
(7-4) Оценка активности связывания аденозина и ATP с помощью конкурентного ELISA фагов
На основе структур 5'-аденозин-PEG-биотина и ATP-PEG-биотина оставалась возможность того, что клон ATNLSA1-4_D12 (вариабельная область тяжелой цепи, SEQ ID NO:46; легкая цепь: SEQ ID NO:47), для которого по результатам ELISA фагов было показано, что оно обладает способностью связываться как с 5'-аденозин-PEG-биотином, так и с ATP-биотином, распознает биотиновую метку или часть PEG. Таким образом, чтобы продемонстрировать, что ATNLSA1-4_D12 не является антителом, которое распознает биотиновую метку или PEG, исследовали, ингибируется ли связывание антигена аденозином или ATP, с помощью ELISA фагов с использованием ATNLSA1-4_D12 и связывающего IL-6R клона PF1 (тяжелая цепь: SEQ ID NO:48; легкая цепь: SEQ ID NO:49), полученного в качестве отрицательного контроля. Каждый из ATNLSA1-4_D12 и PF1 разбавляли TBS и подвергали ELISA по следующей методике.
96-луночный микропланшет для титрования StreptaWell (Roche) покрывали при комнатной температуре в течение одного часа 100 мкл TBS, содержащего меченные биотином антигены (смесь 5'-аденозин-PEG-биотина и ATP-PEG-биотина). После удаления антигенов промыванием каждой лунки планшета TBST, лунки блокировали 250 мкл смеси 2% обезжиренное молоко/TBS в течение одного часа или более. 2% обезжиренное молоко/TBS удаляли, а затем в каждую лунку добавляли полученные очищенные фаги. Планшету позволяли стоять при комнатной температуре в течение одного часа, чтобы обеспечить связывание экспонирующих антитело фагов с антигенами в каждой лунке. Затем в лунки добавляли, TBS, который не содержит антигена или который содержит серийные разведения ATP от равного количества до вплоть до разведения в 10000 относительно количества антигена. Для конкуренции иммобилизованного антигена с ATP планшету позволяли стоять при комнатной температуре в течение одного часа. Затем после промывания TBST в каждую лунку добавляли конъюгированное с HRP антитело против M13 (Amersham Pharmacia Biotech), разбавленное TBS. Планшет инкубировали в течение одного часа. После промывания TBST в каждую лунку добавляли простой раствор TMB (ZYMED). Хромогенную реакцию в растворе каждой лунки останавливали добавлением серной кислоты. Затем, оценивали развитие окраски путем измерения поглощения при 450 нм.
Результат измерения представлен на фиг. 18. Было продемонстрировано, что чем более высокой является концентрация ATP, тем меньшей является степень развития окраски для ATNLSA1-4_D12 в присутствии избыточного количества ATP. Таким образом, было продемонстрировано, что связывание между ATNLSA1-4_D12 и его антигеном ингибируется зависимым от концентрации ATP образом. Между тем, в контрольном эксперименте с PF1 в качестве отрицательного контроля, его связывание антигена не обнаруживалось независимо от концентрации ATP. Указанные выше данные демонстрируют, что ATNLSA1-4_D12 представляет собой антитело, которое обладает способностью связываться с ATP, но не распознает биотиновую метку или PEG.
(7-5) Экспрессия и очистка антител, которые связываются с ATP и аденозином
С использованием специфических праймеров амплифицировали гены из клона ATNLSA1-4_D12, который был оценен как обладающий активностью связывания с ATP и аденозина с помощью ELISA фагов, описанного в примере 7. Нуклеотидные последовательности генов анализировали (последовательности тяжелой цепи и легкой цепи представлены в SEQ ID NO:46 и 47, соответственно). Ген, кодирующий вариабельную область ATNLSA1-4_D12 встраивали в плазмиду для экспрессии в клетках животных IgG1/лямбда человека. Антитело экспрессировали с использованием способа, описанного ниже. Клетки, происходящие из клеток почки эмбриона человека, FreeStyle 293-F (Invitrogen) суспендировали при плотности клеток 1,33×106 клеток/мл в среде для экспрессии FreeStyle 293 Expression Medium (Invitrogen) и распределяли аликвотами объемом 3 мл в каждую ячейку планшета с 6 ячейками. Сконструированную плазмиду вводили в клетки способом липофекции. После культивирования в течение четырех суток в инкубаторе с CO2 (37°C, 8% CO2, 90 об./мин.), антитело очищали из культуральных супернатантов способом, известным специалистам в данной области, с использованием rProtein A SepharoseTM Fast Flow (Amersham Biosciences). Поглощение очищенных растворов антител измеряли при 280 нм с использованием спектрофотометра. Из величин, полученных путем измерения, вычисляли концентрацию очищенного антитела с использованием коэффициента экстинкции, определенного способом PACE (Protein Science (1995) 4, 2411-2423).
(7-6) Оценка связывающего ATP и аденозин антитела в отношении связывания ATP и аденозина с использованием поверхностного плазмонного резонанса
Biacore T200 (GE Healthcare) использовали для анализа взаимодействия D12, в котором константная область IgG связана с вариабельной областью клона ATNLSA1-4_D12 с активностью связывания ATP и аденозина в реакции антиген-антитело. На сенсорный чип CM5 или CM4 (GE Healthcare) иммобилизовывали соответствующее количество белка A (Life technologies) способом присоединения аминов. Представляющее интерес антитело улавливали на чипе для обеспечения взаимодействия с ATP (Wako), аденозином (Wako) или ADP (аденозиндифосфат) (Wako) в качестве антигена. Использованный подвижный буфер представлял собой 50 мМ Tris-HCl (Takara, T903), 500 мМ NaCl, 0,01% (масс./об.) Tween20. Антигену позволяли взаимодействовать в течение 30 секунд при скорости потока 30 мкл/мин и диссоциировали в течение 30 секунд. Взаимодействие с антигеном оценивали при 15°C. Антиген разбавляли с использованием того же подвижного буфера.
Константу диссоциации KD (M) вычисляли, исходя из константы скорости ассоциации ka (1/М с) и константы скорости диссоциации kd (1/с), обе из которых являются параметрами кинетики, вычисленным из сенсограммы, полученной при измерении. Альтернативно константу диссоциации KD (M) вычисляли с использованием стационарного анализа. Каждый параметр вычисляли с использованием оценочного программного обеспечения Biacore T200 Evaluation Software (GE Healthcare).
Для вычисления KD в отношении аденозина ответ в виде связывания оценивали при различных концентрациях аденозина в присутствии или в отсутствие 20 мкмоль/л ADP. Кроме того, ответ в виде связывания отдельно оценивали в присутствии 20 мкмоль/л ADP. Ответ (R) для специфического связывания аденозина получали путем вычитания величины ответа в виде связывания в присутствии ADP отдельно из ответа в виде связывания на различные концентрации аденозина в присутствии of ADP, а затем путем вычитания полученной величины, которая предположительно соответствует компонентам неспецифического связывания, из величины ответа в виде связывания на аденозин в отсутствие ADP. Из кривой, на которой концентрация аденозина нанесена на график по оси X и R, вычисленный по формуле 2, нанесен на график по оси Y, величину KD в отношении аденозина определяли способом наименьших квадратов с использованием функции Solver в Office Excel 2007 (Microsoft).
(Формула 2)
R=Rmax × конц./(KD + конц.)
В формуле 2, конц. соответствует концентрации аденозина (моль/л), в то время как Rmax соответствует величине ответа, ожидаемой для максимального связывания аденозина с антителом. Измеренные величины ответа получали с использованием Scrubber2 (BioLogics. Inc).
KD для D12, определенная с помощью измерения, описанного выше, составила 8,5 мкмоль/л для ATP, 0,25 мкмоль/л для ADP или 1100 мкмоль/л для аденозина. Этот результат демонстрирует, что D12 обладает активностью связывания с ATP, ADP и аденозином; и он также указывает на то, что D12 обладает активностью связывания с AMP (аденозинмонофосфат) и cAMP (циклический аденозинмонофосфат).
[Пример 8] Конструирование библиотеки с использованием антител против ATP/аденозина для получения антител с переключением ATP/аденозином
Известно, что в злокачественных тканях и воспаленных тканях, является высокой не только концентрация аденозина, но также и концентрация ATP. Таким образом, является полезным использование антител, для которых как аденозин, так и ATP (обозначается как ATP/аденозин в этом примере) могут служить в качестве переключающего фактора (в частности, антитела, которые связываются с антигенами, когда аденозин или ATP присутствует в высокой концентрации), а также антител, для которых либо аденозин, либо ATP, отдельно служит в качестве переключающего фактора. ATNLSA1-4_D12, описанное в примере 7-4, представляет собой антитело, которое связывается с ATP/аденозином. Как показано на фиг. 19, полагают, что ATP/аденозин размещается между антителом и его антигеном-мишенью, и, таким образом, антитело содержит вариабельную область антитела, которая контактирует с антигеном-мишенью. Таким образом, авторы настоящего изобретения предположили, что можно сконструировать синтетические библиотеки антител, которые позволят выделить антитела с переключением ATP/аденозином, активность связывания которых с произвольными антигенами изменяется в зависимости от присутствия ATP/аденозина, путем объединения в качестве библиотеки сегментов вариабельных областей антитела, которые способны контактировать с антигеном-мишенью и сохранять связывание ATP/аденозина.
Анализировали кристаллическую структуру комплекса между ATP и антителом с переключением ATP/аденозином ATNLSA1-4_D12, полученным из библиотеки антител человека, как описано в примере 7-4. Результат анализа кристаллической структуры выявил способ распознавания аденозина (или ATP) антителом, а также идентифицировал аминокислотные остатки, которые считаются по существу не вовлеченными в связывание аденозина (или ATP) в вариабельной области антитела. Аминокислотные остатки, которые были идентифицированы как значительно вовлеченные в связывание аденозина (ATP), представляют собой Ser52, Ser52a, Arg53, Gly96, Leu100a и Trp100c (нумерация Kabat) в тяжелой цепи.
При конструировании такой библиотеки положения, которые удовлетворяют по меньшей мере одному из условий, описанных ниже, выбирали в качестве подходящих для конструирования библиотеки.
Условия 1: положение, которое не вовлечено значительно в связывание ATP или, если оно вовлечено в связывание, положение, имеющее аминокислоту, отличную от последовательности дикого типа, которая не ингибирует связывание ATP;
Условие 2: положение, которое является полиморфным в определенной степени с точки зрения частоты встречаемости аминокислот; и
Условие 3: положение, которое не является необходимым для образования канонической структуры.
В областях, содержащихся как в тяжелой цепи, так и в легкой цепи, последовательности ATNLSA1-4_D12 и которые удовлетворяют условиям, описанным выше, аминокислоты в областях CDR1 и CDR2, которые имеют частоту встречаемости 2% или более в последовательности эмбрионального тип, а также аминокислоты в области CDR3, которые имеют частоту встречаемости 1% или более в последовательности эмбрионального типа, подвергали всестороннему замещению. Эти замены объединяли для конструирования множества вариантов ATNLSA1-4_D12.
Участки изменения в тяжелой цепи (в таблице, положения, указанные как "Kabat", в соответствии с нумерацией Kabat), а также аминокислоты до изменения (в таблице, аминокислоты, обозначаемые как "природная последовательность") в этих участках и аминокислоты после изменения (в таблице, аминокислоты, обозначаемые как "измененные аминокислоты"), представлены в таблице 9.
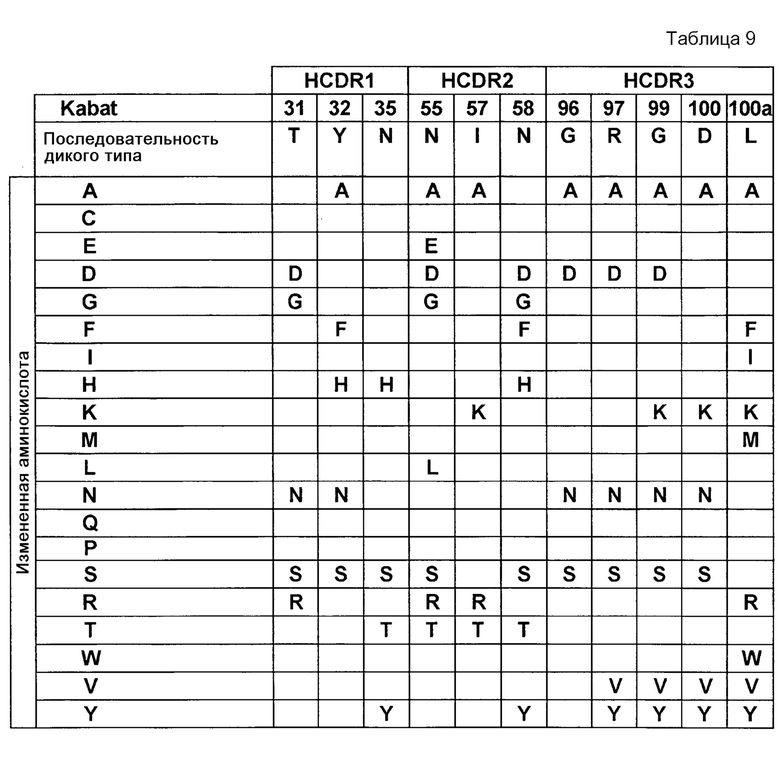
Участки изменения в легкой цепи (в таблице, положения, указанные как "Kabat", в соответствии с нумерацией Kabat), а также аминокислоты до изменения (в таблице, аминокислоты, обозначаемые как "природная последовательность") в этих участках и аминокислоты после изменения (в таблице, аминокислоты, обозначаемые как "измененные аминокислоты"), представлены в таблице 10.
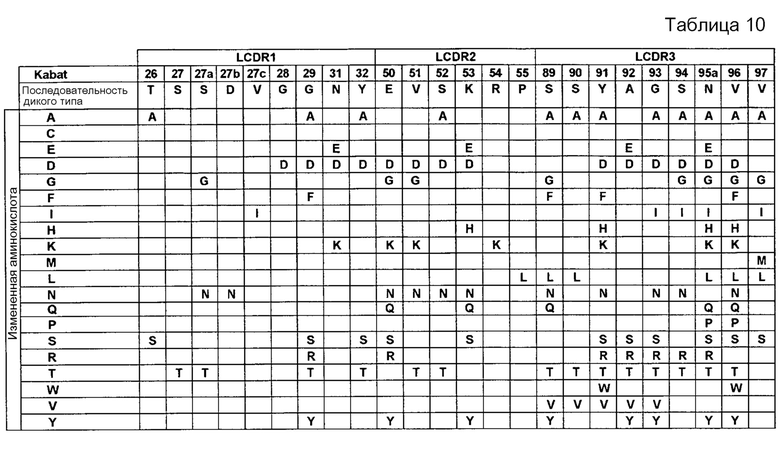
Каждый вариант, экспрессированный и очищенный способом, описанным в примере 7-1, анализировали в отношении его связывания ATP и аденозина тем же способом, который описан в примере 7-6, с использованием Biacore. Исходя из результата анализа аффинность каждого варианта в отношении ATP вычисляли в качестве величины KD. Участки в тяжелой цепи, где изменение не снижает способность связывания ATP до менее чем 1/5 от способьности связывания ATNLSA1-4_D12 (в частности, где величина KD составляет менее 42,5 мкмоль/л), и участки в легкой цепи, где способность связывать ATP является более высокой, чем у ATNLSA1-4_D12 (в частности, где величина KD меньше 8,5 мкмоль/л), оценивали как приемлемые для изменения. Аминокислоты, замещенные в этих участках, считали подходящими для включения в библиотеку (нестрогие остатки для введения в библиотеку).
Исходя из результатов оценки способности каждого варианта связывать ATP, было спрогнозировано, что способность связывать ATP будет снижаться при объединении каждого участка для конструирования библиотеки. Таким образом, в участки вблизи положений, которые, как ожидалось, вовлечены в связывание ATP, вносили замены, и различные варианты, полученные комбинированием этих замен, всесторонне анализировали для исследования того, возможно ли идентифицировать изменения, которые, как ожидается, будут обеспечивать усиление способности связывать ATP. Такие участки изменения (положения, указанные как "Kabat", в соответствии с нумерацией Kabat в таблице), и аминокислоты до изменения (аминокислоты, обозначаемые как "последовательность дикого типа" в таблице) и аминокислоты после изменения (аминокислоты, обозначаемые как "измененные аминокислоты" в таблице) в этих участках представлены в таблице 11.
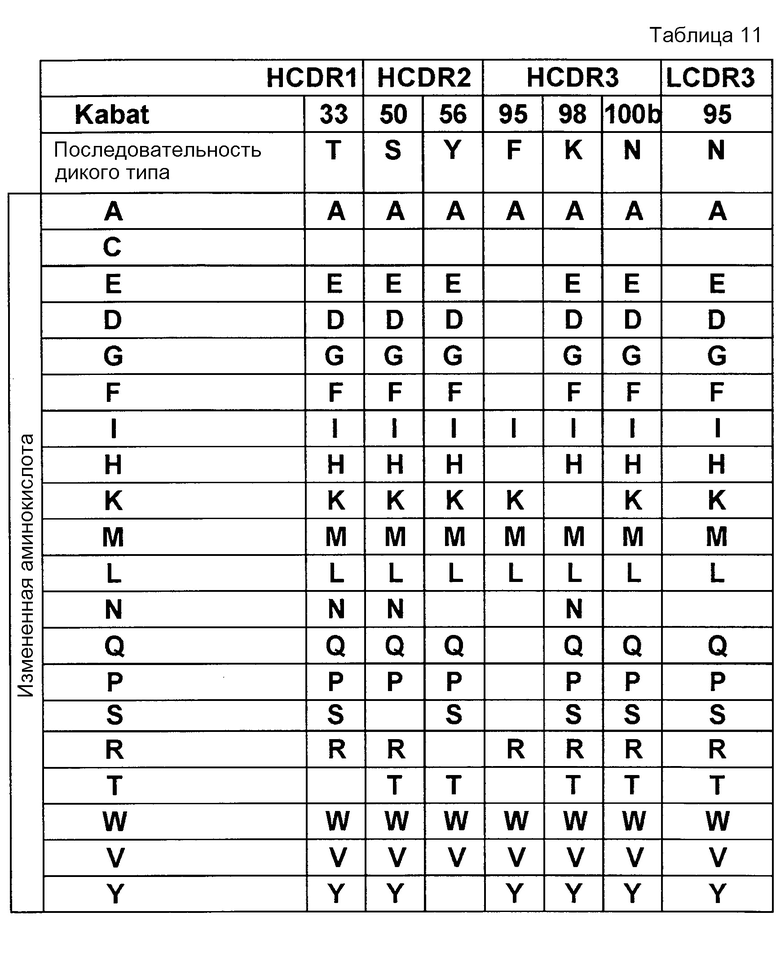
Каждый вариант, экспрессированный и очищенный способом, описанным в примере 7-1, анализировали в отношении его связывания ATP и аденозина тем же способом с использованием Biacore, который описан в примере 7-6. Результат анализа показал, что связывание ATP и аденозина, как ожидалось, усиливалось путем изменений в положениях 56 и 100b и т.п. в соответствии с нумерацией Kabat (например, аминокислотное изменение, такое как Tyr56His и Asn100bLeu). Было определено, что аминокислоты, замещенные в этих участках, могут быть включены в библиотеку (нестрогие остатки для внесения в библиотеку).
В областях CDR ATNLSA1-4_D12, репертуары аминокислот, включающие аминокислоты, отобранные с помощью описанного выше анализа варианта в качестве пригодных для включения в библиотеку (нестрогие аминокислотные остатки для внесения в библиотеку) и аминокислоты до изменения аминокислот (в частности, аминокислоты, включенные в природную последовательность ATNLSA1-4_D12), и участки, содержащие такие репертуары, моделировали для конструирования библиотеки для получения антител с переключением ATP/аденозином. Библиотеку конструировали так, чтобы в репертуаре аминокислот частота встречаемости аминокислот была одинаковой для каждой аминокислоты (например, когда существует десять типов аминокислот в репертуаре аминокислот, каждая аминокислота встречается с частотой 10%).
Участки, содержащие репертуары аминокислот в тяжелой цепи (положения, указанные как "Kabat", в соответствии с нумерацией Kabat в таблице) и репертуары аминокислот в этих участках представлены в таблице 12. Участки, содержащие репертуары аминокислот в легкой цепи (положения, указанные как "Kabat", в соответствии с нумерацией Kabat в таблице) и репертуары аминокислот в этих участках представлены в таблице 13.
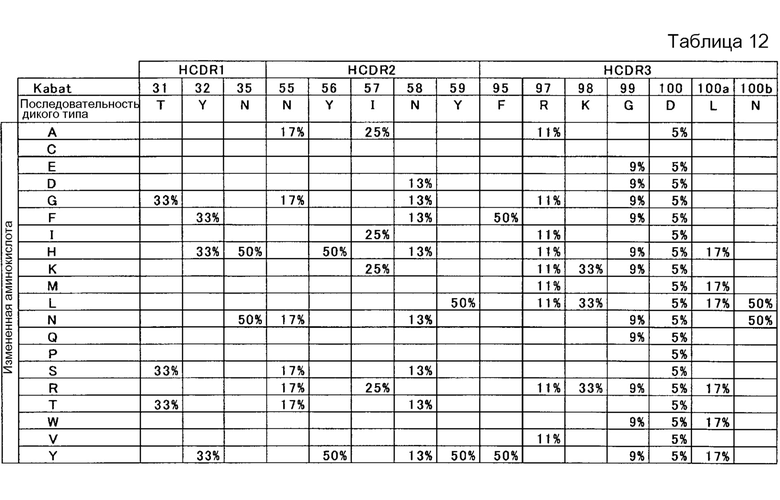
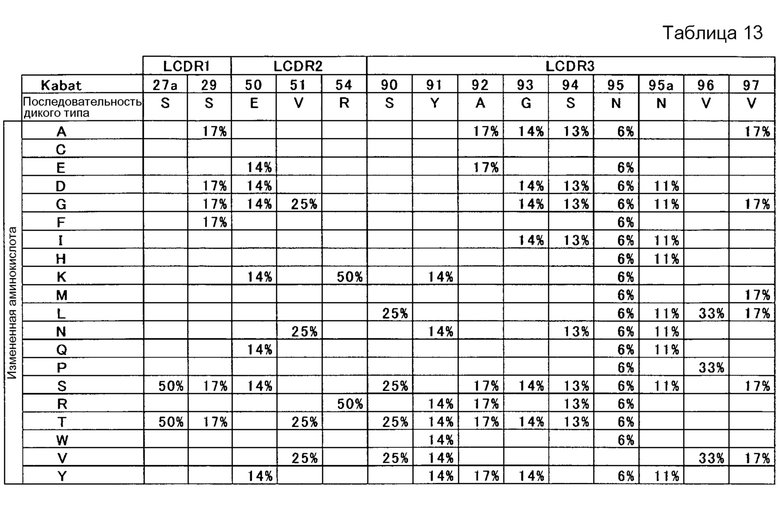
Результат анализа последовательности указывает на то, что каркасная область ATNLSA1-4_D12 происходила из последовательности эмбрионального типа VH3-21. Далее, для повышения стабильности антитела последовательность каркасной области ATNLSA1-4_D12 восстанавливали до последовательности эмбрионального типа VH3-21 путем внесения в каркасную последовательность ATNLSA1-4_D12 изменений Gln01Glu, Gln05Val, Asp10Gly, Asn30Ser, Leu48Val и Asn58Tyr (числа соответствуют номерам по Kabat). Варианты ATNLSA1-4_D12, экспрессированные и очищенные способом, описанным в примере 7-1, измеряли в отношении их Tm способом DSC. Измерение с помощью DSC проводили способом, известным специалистам в данной области. Tm варианта, которая является результатом добавления этих изменений к ATNLSA1-4_D12, значительно улучшалась от 74,37°C до 81,44°C, и наблюдали стабилизацию структуры. Иногда является предпочтительным использование высокостабильных каркасных областей для библиотек антител и, таким образом, каркасную последовательность, в которую были внесены изменения, описанные выше, использовали в качестве каркасной последовательность библиотеки. Каркасная область, использованная для библиотеки, представлена в таблице 14.

Гены синтезировали так, чтобы они содержали соответствующие последовательности в библиотеке, сконструированной, как описано выше (DNA2.0), и библиотеку генов амплифицировали с помощью праймеров, которые способны амплифицировать VH и VL, соответственно, с использованием коллекции (библиотеки) соответствующих генов в качестве матрицы. Последовательности праймеров, использованные для амплификации VL, представлены в SEQ ID NO:102 и 103, в то время как последовательности праймеров, использованных для амплификации VH, представлены в SEQ ID NO:104 и 105. Амплифицированную рационально сконстрированную библиотеку генов вариабельных областей тяжелой цепи и легкой цепи антитела человека встраивали в соответствующий фагмидный вектор, содержащий как происходящую из IgG последовательность CH1 человека, так и происходящую из IgG человека последовательность константной области легкой цепи. Фагмидный вектор вводили E. coli электропорацией для конструирования рационально сконструированной библиотеки, которая презентирует Fab-домены, содержащие вариабельную область-константную область антитела человека, и из которой можно выделять антитела, которые способны связываться с антигенмми через аденозин или ATP в качестве переключающего фактора. Ожидается, что такая рационально сконструированная библиотека с различными H-цепями и L-цепями, которые обладают активностью связывания аденозина или ATP, будет пригодной в качестве библиотеки, содержащей антитела человека, которые с расположенным между антителом и антигеном аденозином (или ATP), как показано на фиг. 19, позволят эффективно получить антитела с переключением аденозином/ATP против любого произвольного антигена. Более того, как описано выше, поскольку ATNLSA1-4_D12 связывается не только с аденозином и ATP, но также и с ADP, было предсказано, что оно обладает активностью связывания с AMP и cAMP, которые структурно сходны с ATP, ADP и аденозином. Это указывает на то, что такие библиотеки являются пригодными для выделения антител с переключением, активность связывания которых с произвольными антигенами-мишенями изменяется в зависимости от присутствия любого одного или нескольких из низкомолекулярных соединений: ATP, ADP, AMP, cAMP и аденозина.
[Пример 9] Конструирование иммунной библиотеки для получения антител с переключением аденозином/ATP, содержащей репертуары антител против ATP/аденозина
Множество библиотек фагового дисплея антител кролика, которые презентируют Fab-домены, содержащие последовательности антител кролика, конструировали с использованием в качестве матрицы мРНК, собранной из популяции B-клеток, отобранной с использованием MACS и FACS, как описано в примере 5-4, которая экспрессируют антитела, связывающие аденозин-PEG-биотин. Способ конструирования осуществляли в соответствии с Rader (Methods Mol. Biol. (2009) 525, 101-28).
Более конкретно, кДНК получали обратной транскрипцией с использованием в качестве матрицы мРНК, собранной из 600000 клеток описанных выше B-клеток, которые были отобраны из девяти иммунизированных кроликов. С использованием кДНК в качестве матрицы последовательность вариабельной области тяжелой цепи и последовательность вариабельной области-константной области легкой цепи амплифицировали способом ПЦР с использованием праймеров, представленных в таблице 15, в подходящих условиях.
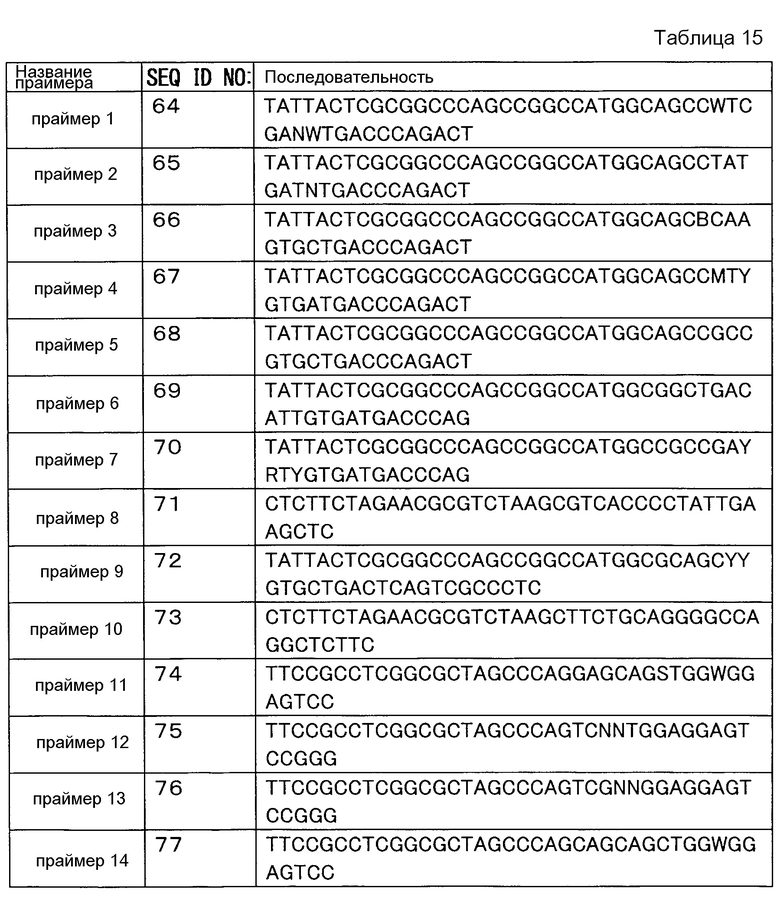
Комбинацию амплифицированной библиотеки генов вариабельных областей тяжелой цепи антител кролика и библиотеки генов вариабельной области-константной области легкой цепи антител кролика встраивали в соответствующий фагмидный вектор, содержащий происходящую из IgG кролика последовательность CH1. Фагмидный вектор вводили в E. coli путем электропорации для конструирования библиотеки фагового дисплея антител кролика (далее, библиотеки из иммунизированных аденозином кроликов), которая презентирует Fab-домены, содержащие вариабельную область-константную область антител кролика, и из которой можно выделять антитела, которые способны связываться с антигенами через аденозин или ATP в качестве переключающего фактора. Ожидается, что такая иммунная библиотека для аденозина, которая состоит из различных H-цепей и L-цепей, которые проявляют свойство связывания аденозина, будет пригодной в качестве иммунной библиотеки, где аденозин (или ATP) располагается между антителом и антигеном, как показано на фиг.20, которая позволит выделить антитела с переключением аденозином/ATP против любого произвольного антигена.
[Пример 10] Получение антител, которые связываются с антигенами в присутствии аденозина и ATP, из библиотеки антител с использованием способов фагового дисплея
(10-1) Получение антител, которые связываются с антигенами в присутствии низкомолекулярных соединений, из библиотеки с использованием смеси аденозина и ATP
Антитела, которые проявляют антигенсвязывающую активность в присутствии аденозина и/или ATP, получали из сконструированной библиотеки фагового дисплея из иммунизированных аденозином кроликов и библиотеки фагового дисплея рационально сконструированных антител. Для получения антител фаги, экспонирующие антитела, которые проявляют способность связываться с антигенами, уловленными гранулами в присутствии аденозина и ATP, объединяли, а затем фаги собирали из элюата с гранул в отсутствие аденозина и ATP.
Фаги продуцировали в E. coli, содержащих фагмидный вектор, сконструированный для фагового дисплея. В культуральную среду E. coli, в которой проводили продуцирование фага добавляли 2,5 M NaCl/10% PEG для осаждения фагов. Фракцию осажденных фагов разбавляли TBS для получения суспензии библиотеки. Затем в суспензию библиотеки фагов добавляли BSA в конечной концентрации 4%. Пэннинг проводили с использованием магнитных гранул с иммобилизованным на них антигеном. Использованные магнитные гранулы представляли собой покрытые нейтравидином гранулы (Sera-Mag SpeedBeads NeutrАвидин-coated) и покрытые стрептавидином гранулы (Dynabeads M-280 Streptavidin).
Кждый из 500 пмоль меченного биотином антигена и ATP-Na и аденозина в конечной концентрации 1 мМ добавляли к суспензии фаговой библиотеки. Суспензию фаговой библиотеки контактировали с антигеном, аденозином и ATP при комнатной температуре в течение 60 минут. К суспензии фаговой библиотеки добавляли блокированные BSA магнитные гранулы и комплексу антиген-фаг позволяли связываться с магнитными гранулами при комнатной температуре в течение 15 минут. Гранулы промывали один раз посредством ATP и TBS с растворенным аденозином. Затем гранулы объединяли с 0,5 мл 1 мг/мл трипсина. Гранулы суспендировали при комнатной температуре в течение 15 минут, а затем сразу суспензию фага собирали с выделенных гранул с использованием магнитного стенда. Собранную суспензию фагов добавляли к 10 мл клеток E. coli cells штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубировали при 37°C при осторожном перемешивании в течение одного часа для инфицирования фагом. Инфицированные E. coli высевали в чашку 225 мм × 225 мм. Затем фаги собирали из культуральной среды посеянных E. coli с получением жидкого исходного раствора с фаговой библиотекой.
Первый раунд пэннинга проводили для сбора фагов, которые способны связывать антиген в присутствии аденозина и ATP, в то время как второй и последующий раунды пэннинга проводили для увеличения содержания фагов, которые способны связывать антиген только в присутствии аденозина и ATP. В частности, каждый из 40 пмоль меченного биотином антигена и аденозина и ATP в конечной концентрации 1 мМ добавляли к полученной суспензии фаговой библиотеки. Таким образом, фаговую библиотеку контактировали с антигеном, аденозином и ATP в течение 60 минут при комнатной температуре. Добавляли блокированные посредством BSA магнитные гранулы и комплексу антиген-фаг позволяли связываться с магнитными гранулами в течение 15 минут при комнатной температуре. Гранулы промывали 1 мл TBST с растворенным аденозином и ATP (далее обозначается как (аденозин+ATP)/TBST), аденозином, и TBS с растворенным аденозином и ATP (далее обозначаемый как (аденозин+ATP)/TBS). Затем, гранулы объединяли с 0,5 мл TBS. Гранулы суспендировали при комнатной температуре, а затем сразу суспензию фага собирали с выделенных гранул с использованием магнитного стенда. После повторения этой обработки две по отдельности элюированных суспензии фагов объединяли вместе. Белок pIII (происходящий из фага-помощника белок pIII), который не экспонирует Fab, отщепляли от фагов добавлением 5 мкл 100 мг/мл трипсина собранной суспензии фагов для устранения способностси фагов, которые не экспонируют Fab, инфицировать E. coli. Фаги, собранные из трипсинизированной суспензии фагов, добавляли к 10 мл клеток E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубировали при 37°C при осторожном перемешивании в течение одного часа для инфицирования фагом. Инфицированные E. coli высевали в чашку 225 мм × 225 мм. Затем фаги собирали из культуральной среды посеянных E. coli с получением суспензии фаговой библиотеки. Пэннинг проводили три раза для выделения антител, которые обладают антигенсвязывающей активностью в присутствии аденозина и ATP.
(10-2) Получение антител, которые связываются с антигенами в присутствии аденозина и ATP, из библиотеки антител с использованием способа отрицательной селекции
Библиотеку фагового дисплея антител, происходящую из кроликов, иммунизированных аденозином, или библиотеку фагового дисплея рационально сконструированных антител подвергали скринингу в отношении антител, которые проявляют антигенсвязывающую активность в присутствии аденозина и/или ATP. В качестве первой стадии скрининга библиотеку фагового дисплея антител контактировали с меченным биотином антигеном-стрептавидином в отсутствие аденозина и ATP для устранения фагов, экспонирующих антитела, которые обладают антигенсвязывающей активностью даже в отсутствие аденозина и ATP. Затем проводили пэннинг аналогичным образом в присутствии аденозина и ATP для скрининга антител, которые проявляют антигенсвязывающую активность в присутствии аденозин и ATP.
Фаги продуцировали в E. coli, содержащих сконструированные фагмиды фагового дисплея. В культуральную среду E. coli, в которой происходило продуцирование фага, добавляли 2,5 M NaCl/10% PEG для осаждения фагов. Осажденную фракцию фагов разбавляли TBS для получения суспензии библиотеки. Затем к суспензии фаговой библиотеки добавляли BSA до конечной концентрации 4%. Пэннинг проводили с использованием магнитных гранул с иммобилизованным на них антигеном. Использованные магнитные гранулы представляли собой покрытые нейтравидином гранулы (Sera-Mag SpeedBeads NeutrАвидин-coated) и покрытые стрептавидином гранулы (Dynabeads M-280 Streptavidin).
Вместе с 250 пмоль меченного биотином антигена, к полученной суспензии фаговой библиотеки добавляли смесь аденозина и ATP в конечной концентрации 1 мМ. Таким образом, суспензию фаговой библиотеки контактировали с антигеном, аденозином и ATP в течение 60 минут при комнатной температуре. Затем к суспензии фаговой библиотеки добавляли блокированные BSA магнитные гранулы, и им позволяли связаться с комплексом антиген-фаг при комнатной температуре в течение 15 минут. Гранулы промывали один раз (аденозин+ATP)/TBS. Затем гранулы объединяли с 0,5 мл раствора трипсина в концентрации 1 мг/мл. Гранулы суспензировали при комнатной температуре в течение 15 минут, и сразу после этого суспензию фага собирали с выделенных гранул с использованием магнитного стенда. Собранную суспензию фагов добавляли к 10 мл клеток E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубировали при 37°C при осторожном перемешивании в течение одного часа для инфицирования фагом. Инфицированные E. coli высевали в чашку 225 мм × 225 мм. Затем фаги собирали из культуральной среды посеянных E. coli с получением жидкого исходного раствора с фаговой библиотекой.
Первый раунд пэннинга проводили для сбора фагов, которые способны связываться в присутствии аденозина и ATP, в то время как второй и последующий раунды пэннинга проводили для увеличения содержания фагов, которые связываться с антигеном только в присутствии аденозина и ATP. В частности, 250 пмоль биотинилированного антигена добавляли к блокированным посредством BSA гранулам с нейтравидином Sera-Mag, и им позволяли связываться при комнатной температуре в течение 15 минут. Гранулы промывали три раза посредством TBS. Суспензию фаговой библиотеки, подвергнутую блокированию посредством BSA, добавляли к промытым гранулам и им позволяли связываться при комнатной температуре в течение одного часа. Фаги, которые не связываются с антигенами или гранулами, собирали путем выделения гранул с использованием магнитного стенда. К собранным фагам добавляли каждый из сорока пмоль меченного биотином антигеном и аденозина и ATP в конечной концентрации 1 мМ. Таким образом, фаговую библиотеку контактировали с антигеном, аденозином и ATP в течение 60 минут при комнатной температуре. К смеси меченого антигена, аденозина, ATP и фаговой библиотеки добавляли блокированные посредством BSA магнитные гранулы, и позволяли им связаться с комплексом антиген-фаг в течение 15 минут при комнатной температуре. Гранулы промывали 1 мл (аденозин+ATP)/TBST и (аденозин+ATP)/TBS. Затем к смеси добавляли 0,5 мл раствора трипсина в концентрации 1 мг/мл. Смешанную суспензию перемешивали при комнатной температуре в течение 20 минут, а затем фаги собирали с гранул, которые были разделены с использованием магнитного стенда. Собранные фаги добавляли к 10 мл клеток E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубировали при 37°C при осторожном перемешивании в течение одного часа для инфицирования фагом. Инфицированные E. coli высевали в чашку 225 мм × 225 мм. Пэннинг проводили три раза для выделения антител, которые обладают антигенсвязывающей активностью в присутствии аденозина и ATP.
(10-3) Получение антител, которые связываются с антигенами в присутствии аденозина и ATP, из библиотеки антител с использованием способа чередующегося пэннинга
Библиотеку фагового дисплея, происходящую из кроликов, иммунизированных аденозином, или библиотеку фагового дисплея рационально сконструированных антител подвергают скринингу в отношении антител, которые проявляют антигенсвязывающую активность в присутствии аденозина и/или ATP. В качестве первой стадии скрининга библиотеку фагового дисплея антител контактируют с биотинилированным аденозином и ATP-нейтравидином в присутствии немеченых антигенов для сбора библиотеки фагового дисплея антител, которые связываются с аденозином и/или ATP в присутствии антигена. Затем библиотеку фагового дисплея контактируют с биотинилированным антигеном-стрептавидином в присутствии аденозина и ATP для сбора антител, которые связываются с антигеном в присутствии аденозина и ATP. Таким образом, проводят скрининг антител, которые обладают антигенсвязывающей активностью в присутствии аденозина и ATP, путем проведения пэннинга, описанного выше, чередующимся образом.
Фаги продуцируют в E. coli, содержащих фагмидный вектор, сконструированный для фагового дисплея. В культуральную среду E. coli, в которой происходит продукция фага добавляют 2,5 M NaCl/10% PEG для осаждения фагов. Фракцию осажденных фагов разбавляют TBS для получения суспензии библиотеки. Затем к суспензии фаговой библиотеки добавляют BSA в конечной концентрации 4%. Пэннинг проводят с использованием магнитных гранул с иммобилизованным на них антигеном. Используемые магнитные гранулы представляют собой покрытые нейтравидином гранулы (Sera-Mag SpeedBeads NeutrAvidin-coated) и покрытые стрептавидином гранулы (Dynabeads M-280 Streptavidin).
Вместе с 1000 пмоль немеченого антигена, 250 пмоль биотинилированного ATP, 2'-аденозин-PEG-биотина и 5'-аденозин-PEG-биотина добавляют к полученной суспензии фаговой библиотеки. Таким образом, суспензию фаговой библиотеки контактируют с антигеном, аденозином и ATP при комнатной температуре в течение 60 минут. Затем к суспензии фаговой библиотеки добавляют блокированные BSA магнитные гранулы, и комплексу фага с антигеном, и аденозином и/или ATP позволяют связываться с магнитными гранулами при комнатной температуре в течение 15 минут. Гранулы промывают один раз TBS, содержащим 1000 пмоль антигена. Затем гранулы объединяют с 0,5 мл раствора трипсина в концентрации 1 мг/мл. Гранулы суспендируют при комнатной температуре в течение 15 минут, а затем сразу суспензию фагов собирают с выделенных гранул с использованием магнитного стенда. Собранную суспензию фагов добавляют к 10 мл клеток E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубируют при 37°C при осторожном перемешивании в течение одного часа для инфицирования фагом. Инфицированные E. coli высевают на планшет 225 мм × 225 мм. Затем фаги собирают из культуральной среды посеянных E. coli с получением суспензии фаговой библиотеки.
Второй раунд пэннинга проводят для увеличения содержания фагов, которые способны связываться с биотинилированным антигеном в присутствии аденозина и ATP. В частности, 40 пмоль биотинилированного антигена и аденозин и ATP в конечной концентрации 1 мМ добавляют к полученному раствору с фаговой библиотекой. Таким образом, суспензию фаговой библиотеки контактируют с антигеном, а также аденозином и ATP, в течение 60 минут при комнатной температуре. Затем к раствору фаговой библиотеки добавляют блокированные посредством BSA магнитные гранулы, и комплексу фага с антигеном, а также аденозином и/или ATP, позволяют связывааться с магнитными гранулами при комнатной температуре в течение 15 минут. Гранулы промывают три раза TBST, содержащим аденозин и ATP, в конечной концентрации 1 мМ, и два раза TBS, содержащим аденозин и ATP в конечной концентрации 1 мМ. Затем гранулы комбинируют с 0,5 мл раствора трипсина в концентрации 1 мг/мл. Гранулы суспендируют при комнатной температуре в течение 15 минут, а затем сразу раствор с фагом собирают с выделенных гранул с использованием магнитного стенда. Раствор с собранным фагом добавляют к 10 мл клеток E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубируют при 37°C при осторожном перемешивании в течение одного часа для инфицирования фагом. Инфицированные E. coli высевают в планшет 225 мм × 225 мм. Затем фаги собирают из культуральной среды посеянных E. coli с получением раствора фаговой библиотеки.
При последующих четных раундах пэннинг проводят аналогично пэннингу второго раунда. Однако на четвертом и последующих раундах пэннинга, количество промываний гранул посредством как (аденозин+ATP)/TBST, так и (аденозин+ATP)/TBS увеличивают до пяти раз.
Третий раунд пэннинга проводят, чтобы также увеличить содержание фагов, которые способны связывать биотинилированный аденозин и ATP в присутствии антигена. В частности, вместе с 250 пмоль биотинилированного ATP, 2'-аденозин-PEG-биотина и 5'-аденозин-PEG-биотина, к полученному раствору с фаговой библиотекой добавляют 1000 пмоль немеченого антигена. Таким образом, раствор с фаговой библиотекой контактируют с антигеном, а также с аденозином и ATP, в течение 60 минут при комнатной температуре. Затем к раствору с фаговой библиотекой добавляют блокированные посредством BSA магнитные гранулы, и комплексу фага с антигеном, а также аденозином и/или ATP, позволяют связаться с магнитными гранулами при комнатной температуре в течение 15 минут. Гранулы промывают три раза TBST, содержащим 1000 пмоль антигена, и два раза TBS, содержащим 1000 пмоль антигена. Затем гранулы объединяют с 0,5 мл раствора 1 мг/мл трипсина. Гранулы суспендируют при комнатной температуре в течение 15 минут, а затем сразу раствор собирают с выделенных гранул с использованием магнитного стенда. Собранный раствор с фагом добавляют к 10 мл клеток E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубируют при 37°C при осторожном перемешивании в течение одного часа для инфицирования фагом. Инфицированные E. coli высевают в чашку 225 мм × 225 мм. Затем фаги собирают из культуральной среды посеянных E. coli для получения раствора с фаговой библиотекой.
При последующих нечетных раундах пэннинг проводят аналогично пэннингу третьего раунда. Однако на четвертом и последующих раундах пэннига, количество промывания гранул как TBST, содержащим антиген, так и TBS, содержащим антиген, увеличивают до пяти раз. Альтернативно, независимо от того, нечетным или четным является раунд, пэннинг на третьем и последующих раунда всегда проводят аналогично пэннингу третьего раунда. Однако на четвертом и последующих раундах пэннинга количество промываний гранул как TBST, содержащим антиген, так и TBS, содержащим антиген, увеличивают до пяти раз.
(10-4) Оценка активности связывания в присутствии или в отсутствие аденозина и/или ATP с помощью ELISA фагов
Содержащие фаг культуральные супернатанты собирали в соответствии с общепринятым способом (Methods Mol. Biol. (2002) 178, 133-145) из единичных колоний E. coli, полученных способом, описанным выше. Собранные культуральные супернатанты обрабатывали ультрафильтрацией с использованием NucleoFast 96 (MACHERY-NAGEL). 100 мкл культуральных супернатантов добавляли в каждую лунку NucleoFast 96 и центрифугировали (4500g в течение 45 минут) для удаления оставшейся жидкой части. После добавления 100 мкл H2O NucleoFast 96 промывали центрифугированием (4500 g в течение 30 минут). Наконец, добавляли 100 мкл TBS и NucleoFast 96 позволяли стоять в течение пяти минут при комнатной температуре. Суспензию фагов собирали из супернатанта в каждой лунке NucleoFast 96.
Очищенные фаги, к которым добавляли TBS или (аденозин+ATP)/TBS, подвергали ELISA по следующей методике. 96-луночный микропланшет для титрования StreptaWell (Roche) покрывали в течение ночи посредством 100 мкл TBS, содержащего меченный биотином антиген. После удаления антигена путем промывания каждой лунки планшета посредством TBST, лунки блокировали 250 мкл смеси 2% обезжиренное молоко/TBS в течение одного часа или более. 2% обезжиренное молоко/TBS удаляли, а затем в каждую лунку добавляли полученные очищенные фаги. Планшету позволяли стоять при 37°C в течение одного часа для обеспечения связывания экспонирующих антитело фагов с антигеном в каждой лунке в присутствии или в отсутствие аденозина и/или ATP. После промывания TBST или (аденозин+ATP)/TBST, в каждую лунку добавляли конъюгированное с HRP антитело против M13 (Amersham Pharmacia Biotech), разбавленное TBS или (аденозин+ATP)/TBS. Планшет инкубировали в течение одного часа. После промываний TBST или (аденозин+ATP)/TBST, в каждую лунку добавляли простой раствор TMB (ZYMED). Хромогенную реакцию в растворе каждой лунки останавливали добавлением серной кислоты. Затем, оценивали развитие окраски путем измерения поглощения при 450 нм. Результат показал, что для трех типов антигенов: IL6 человека, рецептор IL6 человека и HAS (сывороточный альбумин человека), существовало множество типов антител, связывающихся в присутствии низкомолекулярных соединений. Также из библиотеки наивных антител человека были получены антитела, которые связываются в присутствии ATP. Между тем, антитела с переключением против IL6 человека, рецептора IL6 человека и HAS можно было получить с большей эффективностью. Результат ELISA фагов представлен в таблице 16.
(10-5) Оценка в отношении способности связываться антител с переключением, антигенсвязывающая активность которых изменяется в зависимости от присутствия аденозина и ATP, и анализ последовательности
Гены амплифицировали с использованием специфических праймеров (SEQ ID NO:111 и 112) из клонов, которые были оценены как обладающие антигенсвязывающей активностью в присутствии аденозин или ATP, исходя из результатов ELISA фагов, описанных в (10-4). Нуклеотидные последовательности генов анализировали, и результат показал, что получено множество антител, которые связываются с антигенами: IL6 человека, HSA и IL6R человека, и обладают последовательностями, отличающимся друг от друга. Аминокислотные последовательности I6DL2C1-4_076 (антитело к IL6 человека), HSDL3C5-4_015 (антитело к HSA) и 6RAD2C1-4_011 и 6RAD2C1-4_076 (антитела к IL-6R человека) представлены в таблице 17.
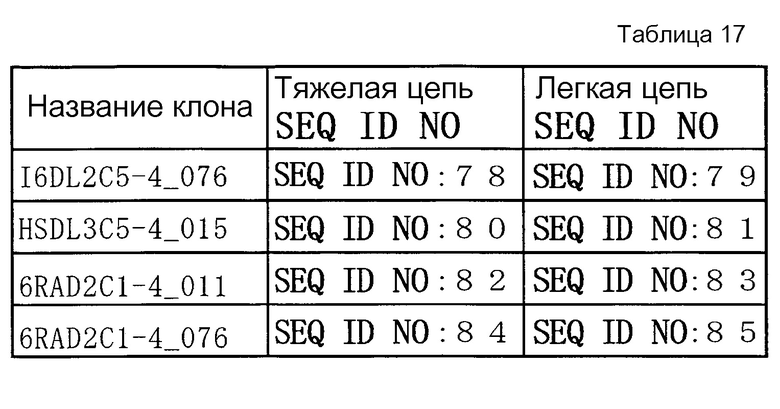
(10-6) Идентификация низкомолекулярных соединений, требуемых для связывания с антигеном полученных антител
Каждое из полученных антител I6DL2C1-4_076, HSDL3C5-4_015, 6RAD2C1-4_011 и 6RAD2C1-4_076 подвергали ELISA. Использованные низкомолекулярные соединения представляли собой 1 мМ ATP, аденозин и их смесь. Использованные антигены представляли собой меченные биотином IL6 человека, IL6R человека и HSA.
Сначала 96-луночный микропланшет для титрования StreptaWell (Roche) покрывали при комнатной температуре в течение одного часа или более 100 мкл TBS, содержащего меченный биотином антиген. После промывания посредством TBST для удаления не связавшегося меченного биотином антигена из планшета каждую лунку блокировали 250 мкл смеси 2% обезжиренное молоко/TBS в течение одного часа или более. 2% обезжиренное молоко/TBS удаляли из каждой лунки, а затем в планшет добавляли 50 мкл экспонирующего антитело фага. Планшету позволяли стоять при комнатной температуре в течение одного часа, чтобы позволить связывание каждого фага с меченным биотином антигеном в каждой лунке в присутствии или в отсутствие ATP и/или аденозина. После промывания TBST с или без ATP и/или аденозина, в каждую лунку добавляли конъюгированное с HRP антитело против M13 (Amersham Pharmacia Biotech), разбавленное TBS или (аденозин и/или ATP)/TBS. Планшет инкубировали в течение одного часа. После промывания TBST с низкомолекулярным соединением или без него в каждую лунку добавляли простой раствор TMB (ZYMED). Хромогенную реакцию в растворе каждой лунки останавливали добавлением серной кислоты. Затем, оценивали развитие окраски путем измерения поглощения при 450 нм (фиг. 25, 26 и 27).
[Пример 11] Активность связывания антител, полученных, как описано в примере 2, с IL-6 человека в присутствии метаболитов аминокислот, отличных от кинуренина
Антитело I6NMSC1-3_A11, полученное, как описано в примере 2-4, которое связывается с IL-6 человека в присутствии низкомолекулярных соединений, представляет собой антитело, которое связывается с IL-6 человека в присутствии кинуренина, как описано в примере 3-2. Кинуренин представляет собой метаболит триптофана, который конвертируется в антраниловую кислоту кинурениназой; в 3-гидроксикинуренин кинуренин-3-гидроксигидроксилазой; и в кинуреновую кислоту кинуренинаминотрансферазой (Stefan Lob et. al., Nat Rev Cancer. (2009) 9 (6), 445-452). Метаболиты аминокислот, такие как метаболиты триптофана, оценивали в отношении того, являются ли они пригодными в качестве неограничивающего варианта осуществления специфических для злокачественной ткани соединений по настоящему изобретению, в частности специфических для злокачественных клеток метаболитов по настоящему изобретению.
I6NMSC1-3_A11, описанное в примере 3-2, которое обладает антигенсвязывающей активностью в присутствии кинуренина, известное антитело против IL-6 CLB8-F1 и GC413 в качестве отрицательного контроля подвергали ELISA при семи условиях, описанных в таблице 18. Между тем, каждую аминокислоту и ее метаболиты получали соответствующим образом в концентрациях, представленных в таблице 18, с использованием буферов, указанных в таблице 4. Использованный антиген представлял собой меченный биотином IL-6 человека.
Сначала 96-луночный микропланшет для титрования StreptaWell (Roche) покрывали при комнатной температуре в течение одного часа или более 100 мкл PBS, содержащего меченный биотином антиген. После промывания буфером для промывания для удаления не связавшегося антигена из планшета, каждую лунку блокировали в течение одного часа или более посредством 250 мкл блокирующего буфера. Блокирующий буфер удаляли из каждой лунки, а затем очищенные IgG приготавливали в концентрации 2,5 мкг/мл в буфере для образца, содержащем низкомолекулярные соединения в конечных концентрациях, представленных в таблице 18, и каждый из них распределяли аликвотами объемом 100 мкл в планшет. Планшету позволяли стоять при комнатной температуре в течение одного часа, чтобы обеспечить связывание каждого IgG с антигеном в каждой лунке. После промывания буфером для промывания, содержащим аминокислоты или метаболиты аминокислот в конечных концентрациях, представленных в таблице 18, в каждую лунку добавляли конъюгированное с HRP антитело против IgG человека (BIOSOURCE), разбавленное буфером для образца, содержащим те же аминокислоты и метаболиты аминокислот. Планшет инкубировали в течение одного часа. После промывания буфером для промывания, содержащим каждую аминокислоту или метаболит аминокислоты, в каждую лунку добавляли простой раствор TMB (ZYMED). Хромогенную реакцию в растворе каждой лунки останавливали добавлением серной кислоты. Затем, оценивали развитие окраски путем измерения поглощения при 450 нм. Композиции использованных буферов представлены в таблице 4.
Результат измерения представлен на фиг.21. поглощение для CLB8-F1 было постоянным независимо от типа и присутствия низкомолекулярных соединений. Между тем, поглощение для I6NMSC1-3_A11 было значительно более низким в условиях 7 (в отсутствие низкомолекулярных соединений) по сравнению с условиями 1 (раствор кинуренина). Более того, поглощение для I6NMSC1-3_A11 было высоким в условиях 2 (раствор триптофана) и условиях 5 (раствор 3-гидроксикинуренина), а также в условиях 1. Это демонстрирует, что I6NMSC1-3_A11 представляет собой антитело, которое связывается с IL-6 человека в качестве антигена не только в присутствии кинуренина, но также в присутствии аминокислоты (триптофан) в качестве предшественника кинуренина или в присутствии метаболита кинуренина.
Эти данные указывают на то, что один и тот же способ можно использовать для получения антител, которые связываются с представляющим интерес антигеном не только в присутствии одного типа метаболита аминокислоты, но также в присутствии множества различных типов аминокислот или метаболитов аминокислот.
[Пример 12] Получение антител, которые связываются с IL-6 человека в присутствии низкомолекулярных соединений, из библиотеки антител человека с использованием технологии фагового дисплея
(12-1) Получение антител, которые связываются с IL-6 человека в присутствии низкомолекулярных соединений, из библиотеки с использованием пэннинга гранул или способа отрицательной селекции
Тем же способом, который описан в разделах 2-2 и 2-3, библиотеку фагового дисплея наивных антител человека, сконструированную, как описано в пример 2-1, подвергали скринингу в отношении антител, которые проявляют антигенсвязывающую активность в присутствии низкомолекулярных соединений.
(12-2) Оценка активности связывания в присутствии низкомолекулярных соединений с помощью ELISA фагов
Культуральные супернатанты, содержащие фаги, получали из единичных колоний E. coli, полученных тем же способом, который описан в примере 2-4. Фаги подвергали ELISA. Путем проведения ELISA фагов с использованием 768 выделенных клонов, вновь получали клоны "I6NMSC1-3_#03" и "I6NMSC1-3_#17", которые проявляли активность связывания с IL-6 человека в качестве антигена в присутствии коктейля низкомолекулярных соединений.
(12-3) Экспрессия и очистка антител, которые связываются с IL-6 человека
Амплифицировали гены из клонов I6NMSC1-3_#03 и I6NMSC1-3_#17, которые были оценены как обладающие антигенсвязывающей активностью в присутствии SC с помощью ELISA фагов, с использованием специфических праймеров (SEQ ID NO:110 и 112); и анализировали их нуклеотидные последовательности. Последовательности тяжелой цепи и легкой цепи I6NMSC1-3_#03 представляют собой последовательности SEQ ID NO:50 и 51, соответственно. Между тем, последовательности тяжелой цепи и легкой цепи I6NMSC1-3_#17 представляют собой последовательности SEQ ID NO:52 и 53, соответственно. Последовательность гена, кодирующую вариабельную область I6NMSC1-3_#17, встраивали в плазмиду для экспрессии в клетках животных IgG1/лямбда человека, в то время как последовательность гена, кодирующую вариабельную область I6NMSC1-3_#03, известное антитело против IL-6 человека CLB8-F1 (последовательности тяжелой цепи и легкой цепи представлены в SEQ ID NO:32 и 33, соответственно) или антитело против глипикана-3 человека GC413 (последовательности тяжелой цепи и легкой цепи представлены в SEQ ID NO:34 и 35, соответственно) в качестве отрицательного контроля встраивали в плазмиду для экспрессии в клетках животных для IgG1/каппа человека. Экспрессированные антитела очищали способом, описанным в примере 3.
(12-4) Идентификация низкомолекулярных соединений, необходимых для связывания антитела I6NMSC1-3_#03 с IL-6 человека
I6NMSC1-3_#03 подвергали ELISA в девяти условиях, описанных в таблице 3. Между тем каждое низкомолекулярное соединение соответственно получали в концентрациях, представленных в таблице 3 с использованием буферов, указанных в таблице 4. Использованный антиген представлял собой меченный биотином IL-6 человека.
Сначала 96-луночный микропланшет для титрования StreptaWell (Roche) покрывали при комнатной температуре в течение одного часа или более 100 мкл PBS, содержавшего меченный биотином антиген. После промывания буфером для промывания для удаления не связавшегося антигена из планшета, каждую лунку блокировали в течение одного часа или более 250 мкл блокирующего буфера. После удаления блокирующего буфера из каждой лунки очищенные IgG приготавливали в концентрации 2,5 мкг/мл в буфере для образца, содержащем низкомолекулярные соединения в конечных концентрациях, представленных в таблице 3, и каждый из них распределяли аликвотами объемом 100 мкл в планшет. Планшету позволяли стоять при комнатной температуре в течение одного часа для обеспечения связывания каждого IgG с антигеном в каждой лунке. После промывания промывочным буфером, содержащим низкомолекулярные соединения в конечных концентрациях, представленных в таблице 3, в каждую лунку добавляли конъюгированное с HRP антитело против IgG человека (BIOSOURCE), разбавленное буфером для образца, содержащим те же низкомолекулярные соединения. Планшет инкубировали в течение одного часа. После промывания буфером для промывания, содержащим каждое низкомолекулярное соединение, в каждую лунку добавляли простой раствор TMB (ZYMED). Хромогенную реакцию в растворе каждой лунки останавливали добавлением серной кислоты. Затем, оценивали развитие окраски путем измерения поглощения при 450 нм. Состав использованного буфера представлен в таблице 4.
Результат измерения представлен на фиг. 22. Результат показал, что поглощение для I6NMSC1-3_#03 было значительно более низким в условиях 9 (без низкомолекулярного соединения) по сравнению с поглощением в условиях 8 (раствор полного коктейля низкомолекулярных соединений). Аналогично результату ELISA фагов, этот результат подтвердил, что I6NMSC1-3_#03 обладает свойством, состоящим в том, что его связывание с антигеном изменяется в зависимости от наличия или отсутствия низкомолекулярных соединений. Более того, поглощение для I6NMSC1-3_#03 было сравнимым между условиями 7 (100 мкМ кинуренина) и условиями 8; однако поглощение было более низким в других условиях. Это продемонстрировало, что подобно I6NMSC1-3_A11, описанному в примере 3, I6NMSC1-3_#03 представляло собой антитело, которое связывается с IL-6 человека в качестве антигена в присутствии кинуренинапа. I6NMSC1-3_#03 обладает аминокислотной последовательностью, отличающейся от I6NMSC1-3_A11, что демонстрирует, что способ описанный выше, можно использовать для выделения различных типов антител, которые связываются с антигенами в присутствии низкомолекулярных соединений.
(12-5) Идентификация низкомолекулярных соединений, необходимых для связывания антитела I6NMSC1-3_#17 с IL-6 человека
Три типа антител: полученное I6NMSC1-3_#17, контрольное CLB8-F1 и отрицательный контроль GC413 подвергали ELISA в девяти условиях, описанных в таблице 3. Каждое низкомолекулярное соединение получали в соответствующей концентрации, представленной в таблице 3, с использованием буферов, представленных в таблице 4. Использованный антиген представлял собой меченный биотином IL-6 человека.
Сначала 96-луночный микропланшет для титрования StreptaWell (Roche) покрывали при комнатной температуре в течение одного часа или более 100 мкл PBS, содержащего меченный биотином антиген. После промывания буфером для промывания для удаления не связавшегося антигена из планшета, каждую лунку блокировали в течение одного часа или более 250 мкл блокирующего буфера. После удаления блокирующего буфера из каждой лунки получали очищенные IgG в концентрации 0,15 мкг/мл в буфере для образца, содержащем низкомолекулярные соединения в конечных концентрациях, представленных в таблице 3, и каждый из них распределяли аликвотами объемом 100 мкл в планшет. Планшету позволяли стоять при комнатной температуре в течение одного часа, чтобы позволить связывание каждого IgG с антигеном в каждой лунке. После промывания буфером для промывания, содержащим низкомолекулярные соединения в конечных концентрациях, представленных в таблице 3, в каждую лунку добавляли конъюгированное с HRP антитело против IgG человека (BIOSOURCE), разбавленное буфером для образца, содержащим те же низкомолекулярные соединения. Планшет инкубировали в течение одного часа. После промывания буфером для промывания, содержащим каждое низкомолекулярное соединение, в каждую лунку добавляли простой раствор TMB (ZYMED). Хромогенную реакцию в растворе каждой лунки останавливали добавлением серной кислоты. Затем, оценивали развитие окраски путем измерения поглощения при 450 нм. Композиция использованного буфера представлена в таблице 4.
Результат измерения представлен на фиг.23. Результат показал, что поглощение для CLB8-F1 было одинаковым независимо от типа и присутствия или отсутствия низкомолекулярного соединения, в то время как поглощение для I6NMSC1-3_#17 было значительно более низким в условиях 9 (без низкомолекулярных соединений) по сравнению с поглощением в условиях 8 (раствор полного коктейля низкомолекулярных соединений). Аналогично результату ELISA фагов, этот результат подтвердил, что I6NMSC1-3_#17 обладает свойством, состоящим в том, что его связывание с антигеном изменяется в зависимости от присутствия или отсутствия низкомолекулярных соединений. Более того, поглощение для I6NMSC1-3_#17 в условиях 1 (1 мМ ATP-Na) и условиях 5 (1 мМ янтарная кислота) было сравнимым с поглощением в условиях 8; однако в других условиях поглощение было более низким. Этот результат указывает на то, что I6NMSC1-3_#17 представляет собой антитело, которое связывается с IL-6 человека в качестве антигена в присутствии либо ATP-Na, либо янтарной кислоты. Известно, что ATP высвобождается из злокачественных клеток. Также известно, что янтарная кислота накапливается внутри и снаружи клеток специфическим для злокачественных клеток образом. Явление, состоящее в том, что даже в аэробных условиях злокачественные клетки осуществляют метаболизм зависимым от гликолиза образом, а не окислительным фосфорилированием, известен как эффект Варбурга. Как гликолиз, так и окислительное фосфорилирование постоянно ограничены в злокачественных опухолях ишемического типа вследствие постоянной недостаточности кровоток, и, таким образом, такие злокачественные опухоли получают энергию в худших условиях. Известно, что злокачественные опухоли ишемического типа осуществляют метаболизм энергии в зависимости от фумаратного дыхания, что приводит к накоплению янтарной кислоты в качестве метаболита фумарата (Cancer Res. (2009) 69 (11), 4918-4925).
Это продемонстрировало, что с использованием такого способа возможно получить антитела, которые связываются с антигенами в присутствии низкомолекулярного соединения, отличного от кинуренина. Также это показало, что возможно получить антитела, которые связываются с антигенами в присутствии низкомолекулярных соединений, таких как ATP-Na и янтарная кислота, которые обладают общим признаком, состоящим в том, что они имеют множество отрицательных зарядов, но структурно отличаются друг от друга.
[Пример 13] Получение антител, которые связываются с сывороточным альбумином человека (далее также обозначаемый как HSA) в присутствии низкомолекулярных соединений способами фагового дисплея из библиотеки антител человека
(13-1) Получение антител, которые связываются с HSA в присутствии низкомолекулярных соединений, из библиотеки пэннингом гранул
Библиотеку фагового дисплея наивных антител человека, сконструированную, как описано в примере 2, подвергали скринингу в отношении антител, которые проявляют активность связывания HSA в присутствии низкомолекулярных соединений, в частности, путем сбора фагов, экспонирующих антитела, которые в присутствии низкомолекулярных соединений проявляют активность связывания с HSA, уловленным на гранулы. Фаги собирали из суспензии фагов, элюированной с гранул в отсутствие низкомолекулярных соединений. В этом способе получения, в качестве антигена использовали меченный биотином HSA.
Фаги, продуцированные в E. coli, содержащих фагмидный вектор, сконструированный для фагового дисплея, очищали общепринятым способом. Затем получали суспензию фаговой библиотеки посредством диализа против TBS. Далее к суспензии фаговой библиотеки добавляли обезжиренное молоко в конечной концентрации 3%. Пэннинг проводили с использованием магнитных гранул с иммобилизованным на них антигеном. Использованные магнитные гранулы представляли собой покрытые нейтравидином гранулы (Sera-Mag SpeedBeads NeutrAvidin-coated) и покрытые стрептавидином гранулы (Dynabeads M-280 Streptavidin).
Для эффективного получения антител с низкомолекулярным переключающим фактором, которые зависят от низкомолекулярных соединений, которые могут служить в качестве переключающего фактора в злокачественных тканях, проводили пэннинг для увеличения содержания антител, которые связываются с антигенами в присутствии смеси таких низкомолекулярных соединений (аденозин, аденозинтрифосфат (аденозин-5’-трифосфат (ATP)), инозин, кинуренин, простагландин E2 (PGE2), янтарная кислота и молочная кислота (далее обозначаемая как коктейль низкомолекулярных соединений (SC)), но не связываются с антигенами в отсутствие SC.
В частности, вместе с 250 пмоль меченного биотином антигена, SC, содержащую натриевую соль аденозинтрифосфата (ATP-Na), аденозин, инозин, янтарную кислоту и молочную кислоту в конечной концентрации 1 мМ, простагландин E2 (PGE2) в конечной концентрации 1 мкМ, и кинуренин в конечной концентрации 100 мкМ, которые были доведены до pH 7,4 посредством NaOH, добавляли и контактировали с полученной суспензией фаговой библиотеки в течение 60 минут при комнатной температуре. Затем к суспензии фаговой библиотеки добавляли блокированные обезжиренным молоком магнитные гранулы, и комплексу антиген-фаг позволяли связываться с магнитными гранулами при комнатной температуре в течение 15 минут. Гранулы промывали один раз посредством SC/TBS (TBS, содержащий SC). Затем гранулы объединяли с 0,5 мл раствора трипсина в концентрации 1 мг/мл. Гранулы суспензировали при комнатной температуре в течение 15 минут, и сразу после этого суспензию фага собирали с выделенных гранул с использованием магнитного стенда. Собранную суспензию фагов добавляли к 10 мл клеток E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубировали при 37°C в течение одного часа при осторожном перемешивании для инфицирования фагом. Инфицированные E. coli высевали в чашку 225 мм × 225 мм. Затем фаги собирали из культуральной среды посеянных E. coli с получением жидкого исходного раствора с фаговой библиотекой.
Первый раунд пэннинга проводили для сбора фагов, которые способны связываться в присутствии низкомолекулярных соединений, в то время как второй и последующий раунды пэннинга проводили для увеличения содержания фагов, которые способны связываться с антигеном в присутствии низкомолекулярных соединений. В частности, 40 пмоль меченного биотином антигена, SC и NaOH добавляли к полученной суспензии фаговой библиотеки. Таким образом, фаговую библиотеку контактировали с антигеном и низкомолекулярными соединениями в течение 60 минут при комнатной температуре. Добавляли блокированные обезжиренным молоком магнитные гранулы и им позволяли связывать комплекс антиген-фаг в течение 15 минут при комнатной температуре. Гранулы промывали 1 мл SC/TBST и SC/TBS. Затем гранулы объединяли с 0,5 мл TBS. Гранулы суспендировали при комнатной температуре, и сразу после этого суспензию фага собирали с выделенных гранул с использованием магнитного стенда. После повторения этой обработки две элюированных по отдельности суспензии фагов объединяли друг с другом. Затем полученные гранулы объединяли с 0,5 мл TBS и перемешивали при комнатной температуре в течение пяти минут. Суспензию фага собирали с выделенных гранул с использованием магнитного стенда. Белок pIII (происходящий из фага-помощника белок pIII), который не экспонирует Fab, отщепляли от фагов добавлением 5 мкл 100 мг/мл трипсина к собранной суспензии фага для устранения способности фагов, которые не экспонируют Fab, инфицировать E. coli. Фаги, собранные из трипсинизированной суспензии фагов, добавляли к 10 мл клеток E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубировали при осторожном перемешивании при 37°C в течение одного часа для инфицирования фагом. Инфицированные E. coli высевали в чашку 225 мм × 225 мм. В этот момент времени два типа инфицированных E. coli, полученных на двух раундах пэннинга, смешивали в равных количествах. Затем фаги собирали из культуральной среды посеянных E. coli с получением суспензии фаговой библиотеки. Пэннинг проводили три раза с получением антител, которые обладают антигенсвязывающей активностью в присутствии низкомолекулярных соединений.
(13-2) Получение антител, которые связываются с HSA в присутствии низкомолекулярных соединений, из библиотеки с использованием способа отрицательной селекции
Сконструированную библиотеку фагового дисплея наивных антител человека подвергали скринингу в отношении антител, которые проявляют активность связывания HSA в присутствии низкомолекулярных соединений. В качестве первой стадии скрининга библиотеку фагового дисплея наивных антител человека контактировали с меченным биотином антигеном со стрептавидином в отсутствие низкомолекулярных соединений для устранения фагов, экспонирующих антитела, которые обладают активностью связывания HSA даже в отсутствие низкомолекулярных соединений. Затем, пэннинг проводили в присутствии низкомолекулярных соединений аналогичным образом. Таким образом, проводили скрининг антител, которые обладают активностью связывания HSA в присутствии низкомолекулярных соединений. Меченный биотином HSA использовали в качестве антигена.
Фаги продуцировали в E. coli, содержащих фагмидный вектор, сконструированный для фагового дисплея. Продуцированные фаги очищали общепринятым способом, а затем получали суспензию фаговой библиотеки путем диализа фагов против TBS. Затем к суспензии фаговой библиотеки добавляли обезжиренное молоко в конечной концентрации 3%. Использованные магнитные гранулы представляли собой покрытые нейтравидином гранулы (Sera-Mag SpeedBeads NeutrАвидин-coated) и покрытые стрептавидином гранулы (Dynabeads M-280 Streptavidin). Пэннинг проводили с использованием меченного биотином HSA, иммобилизованного на магнитных гранулах.
Вместе с 250 пмоль меченного биотином HSA, SC, содержащий ATP-Na, аденозин, инозин, янтарную кислоту и молочную кислоту в конечной концентрации 1 мМ, PGE2 в конечной концентрации 1 мкМ, и кинуренин в конечной концентрации 100 мкМ, который был доведен до pH 7,4 с помощью NaOH, добавляли и контактировали с полученным раствором с фаговой библиотекой в течение 60 минут при комнатной температуре. Затем к раствору с фаговой библиотекой добавляли блокированные обезжиренным молоком магнитные гранулы и позволяли им связываться с комплексом фага с меченным биотином HSA в течение 15 минут при комнатной температуре. Гранулы промывали один раз посредством SC/TBS. Затем гранулы объединяли с 0,5 мл раствора трипсина в концентрации 1 мг/мл. Гранулы суспензировали при комнатной температуре в течение 15 минут, и сразу после этого суспензию фага собирали с выделенных гранул с использованием магнитного стенда. Собранную суспензию фагов добавляли к 10 мл клеток E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубировали при осторожном перемешивании при 37°C в течение одного часа для инфицирования фагом. Инфицированные E. coli высевали в чашку 225 мм × 225 мм. Затем фаги собирали из культуральной среды посеянных E. coli с получением раствора фаговой библиотеки.
Первый раунд пэннинга проводили для сбора фагов, которые способны связываться в присутствии низкомолекулярных соединений, в то время как второй и последующий раунды пэннинга проводили для увеличения содержания фагов, которые способны связываться с меченным биотином HSA в присутствии низкомолекулярных соединений. В частности, 250 пмоль меченного биотином HSA добавляли к блокированным обезжиренным молоком гранулам с нейтравидином Sera-Mag и позволяли им связываться в течение 15 минут при комнатной температуре. Гранулы промывали три раза посредством TBS; и к гранулам добавляли блокированный обезжиренным молоком раствор с фаговой библиотекой, и позволяли им связываться при комнатной температуре в течение одного часа. Гранулы выделяли с использованием магнитного стенда для сбора фагов, которые не связались с меченным биотином HSA или гранулами. К собранным фагам добавляли сорок пмоль меченного биотином HSA, SC и NaOH. Таким образом, фаговую библиотеку контактировали с меченным биотином HSA и низкомолекулярными соединениями в SC в течение 60 минут при комнатной температуре. Затем к смеси меченного биотином HSA, SC и фаговой библиотеки добавляли блокированные обезжиренным молоком гранулы, и комплексу меченного биотином HSA и фага позволяли связываться с магнитными гранулами в течение 15 минут при комнатной температуре. Гранулы промывали 1 мл SC/TBST и SC/TBS. Затем к смеси добавляли 0,5 мл раствора трипсина в концентрации 1 мг/мл. Смешанный раствор перемешивали в течение 20 минут при комнатной температуре, а затем фаги собирали с гранул, разделенных с использованием магнитного стенда. Собранные фаги добавляли к 10 мл клеток E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубировали при осторожном перемешивании при 37°C в течение одного часа для инфицирования фагом. Инфицированные E. coli высевали в чашку 225 мм × 225 мм. Пэннинг проводили три раза с получением антител, которые обладают активностью связывания с меченным биотином HSA в присутствии низкомолекулярных соединений.
(13-3) Оценка активности связывания в присутствии низкомолекулярных соединений с помощью ELISA фагов
Культуральные супернатанты, содержащие фаги собирали в соответствии с общепринятым способом (Methods Mol. Biol. (2002) 178, 133-145) из единичных колоний E. coli, полученных способом, описанным выше. Собранные культуральные супернатанты обрабатывали ультрафильтрацией с использованием NucleoFast 96 (MACHEREY-NAGEL). В каждую лунку добавляли 100 мкл культуральных супернатантов, и NucleoFast 96 центрифугировали (4500 g в течение 45 минут) для удаления оставшейся жидкой части. В каждую лунку добавляли сто мкл H2O и вновь центрифугировали с помощью NucleoFast 96 (4500 g в течение 30 минут) для промывания. Добавляли 100 мкл TBS, а затем NucleoFast 96 позволяли стоять в течение пяти минут при комнатной температуре. Наконец, собирали раствор фага, содержащийся в супернатанте каждой лунки.
Очищенные фаги, к которым были добавлены TBS или SC/TBS, подвергали ELISA по следующей методике. 96-луночный микропланшет для титрования StreptaWell (Roche) покрывали в течение ночи посредством 100 мкл TBS, содержащего меченный биотином HSA. Меченный биотином HSA удаляли промыванием каждой лунуки планшета TBST, а затем лунки блокировали 250 мкл смеси 2% обезжиренное молоко/TBS в течение одного часа или более. 2% обезжиренное молоко/TBS удаляли, а затем в каждую лунку добавляли полученные очищенные фаги. Планшету позволяли стоять при комнатной температуре в течение одного часа, чтобы позволить связывание экспонирующих антитело фагов с меченным биотином HSA в каждой лунке в присутствии или в отсутствие SC. После промывания TBST или SC/TBST, в каждую лунку добавляли конъюгированное с HRP антитело против M13 (Amersham Pharmacia Biotech), разбавленное посредством TBS или SC/TBS. Планшет инкубировали в течение одного часа. После промывания TBST или SC/TBST, в каждую лунку добавляли простой раствор TMB (ZYMED). Хромогенную реакцию в растворе каждой лунки останавливали добавлением серной кислоты. Затем, оценивали развитие окраски путем измерения поглощения при 450 нм.
Клон HSNMSC1-4_#22, который обладает активностью связывания с HSA в качестве антигена в присутствии коктейля низкомолекулярных соединений, получали путем проведения ELISA фагов с использованием 782 выделенных клонов.
(13-4) Экспрессия и очистка связывающих HSA антител
Амплифицировали гены из клона HSNMSC1-4_#22, который был оценен как обладающий активностью связывания с меченным биотином HSA в присутствии SC, с помощь ELISA фагов, описанного в (13-3), с использованием специфических праймеров (SEQ ID NO:110 и 112). Нуклеотидные последовательности генов анализировали (последовательности тяжелой цепи и легкой цепи представлены в SEQ ID NO:54 и 55, соответственно). Гены, кодирующие вариабельные области HSNMSC1-4_#22, встраивали в плазмиду для экспрессии в клетках животных IgG1/лямбда человека. Между тем гены, кодирующие вариабельные области отрицательного контроля в виде антитела против глипикана 3 человека GC413 (тяжелая цепь и легкая цепь представляют собой SEQ ID NO:34 и 35, соответственно) встраивали в плазмиду для экспрессии в клетках животных IgG1/каппа человека. Экспрессированные нтитела очищали способом, описанным в примере 3.
(13-5) Идентификация низкомолекулярных соединений, необходимых для связывания полученных антител с HSA
Два полученных типа антител: HSNMSC1-4_#22 и GC413, подвергали ELISA в девяти условиях, описанных в таблице 3. Между тем, каждое низкомолекулярное соединение соответственно получали в концентрациях, представленных в таблице 3, с использованием буферов, указанных в таблице 19. В качестве антигена использовали меченный биотином HSA.

Сначала 96-луночный микропланшет для титрования StreptaWell (Roche) покрывали при комнатной температуре в течение одного часа или более 100 мкл PBS, содержащего меченный биотином HSA. После промывания буфером для промывания для удаления не связавшегося меченного биотином HSA из планшета, каждую лунку блокировали в течение одного часа или более 250 мкл блокирующего буфера. Блокирующий буфер удаляли из каждой лунки, а затем очищенные IgG приготавливали в концентрации 2,5 мкг/мл в буфере для образца, содержащем низкомолекулярные соединения в конечных концентрациях, представленных в таблице 3, и каждый из них распределяли аликвотами объемом 100 мкл в планшет. Планшету позволяли стоять при комнатной температуре в течение одного часа, чтобы позволить связывание каждого IgG с меченным биотином HSA в каждой лунке. После промывания буфером для промывания, содержащим низкомолекулярные соединения в конечных концентрациях, представленных в таблице 3, в каждую лунку добавляли конъюгированное с HRP антитело против IgG человека (BIOSOURCE), разбавленное буфером для образца, содержащим те же низкомолекулярные соединения. Планшет инкубировали в течение одного часа. После промывания буфером для промывания, содержащим каждое низкомолекулярное соединение, в каждую лунку добавляли простой раствора TMB (ZYMED). Хромогенную реакцию в растворе каждой лунки останавливали добавлением серной кислоты. Затем, оценивали развитие окраски путем измерения поглощения при 450 нм. Композиция использованного буфера представлена в таблице 19.
Результат измерения представлен на фиг. 24. Результат показал, что поглощение для HSNMSC1-4_#22 было значительно более низким в условиях 9 (без низкомолекулярных соединений) по сравнению с поглощением в условиях 8 (раствор полного коктейля низкомолекулярных соединений). Аналогично ELISA фагов, этот результат подтвердил, что HSNMSC1-4_#22 обладает свойством, состоящим в том, что его связывание антигена изменяется в зависимости от наличия или отсутствия низкомолекулярных соединений. Между тем, поглощение для HSNMSC1-4_#22 было одинаковым между условиями 2 (1 мМ аденозин) и условиями 8, однако было значительно более низким в других условиях. Этот результат продемонстрировал, что HSNMSC1-4_#22 представляет собой антитело, которое связывается с HSA в качестве антигена в присутствии аденозина. Таким образом, было продемонстрировано, что такой способ можно использовать для выделения антител, которые связываются с антигенами в присутствии низкомолекулярных соединений, отличных от кинуренина.
[Пример 14] Получение антител, которые связываются с рецептором IL-6 человека (hIL-6R) в присутствии низкомолекулярных соединений, из библиотеки антител человека с использованием способов фагового дисплея
(14-1) Получение антител, которые связываются с hIL-6R в присутствии низкомолекулярных соединений, из библиотеки наивных антител человека, способом пэннинга гранул
Библиотеку фагового дисплея наивных антител человека, сконструированную, как описано в примере 2, подвергали скринингу в отношении антител, которые проявляют активность связывания hIL-6R в присутствии низкомолекулярных соединений, в частности, путем сбора фагов, экспонирующих антитела, которые в присутствии низкомолекулярных соединений проявляют активность связывания с hIL-6R, уловленном на гранулы. Фаги собирали из элюита фага, элюированного с гранул в отсутствие низкомолекулярных соединений. В этом способе получения в качестве антигена использовали меченный биотином hIL-6R.
Фаги, продуцированные в E. coli содержащих фагмидный вектор, сконструированный для фагового дисплея, очищали общепринятым способом. Затем получали раствор с фаговой библиотекой путем диализа фагов против TBS. Далее, к раствору с фаговой библиотекой добавляли BSA в конечной концентрации 4%. Пэннинг проводили с использованием магнитных гранул с иммобилизованным на них антигеном. Использованные магнитные гранулы представляли собой покрытые нейтравидином гранулы (Sera-Mag SpeedBeads NeutrАвидин-coated) и покрытые стрептавидином гранулы (Dynabeads M-280 Streptavidin).
Для эффективного получения антител с низкомолекулярным переключателем, которые зависят от низкомолекулярных соединений, которые могут служить в качестве переключающего фактора в злокачественных тканях, проводили пэннинг, описанный в (2-2), для увеличения содержания антител, которые связываются с антигенами в присутствии SC, но не связываются с антигенами в отсутствие SC.
В частности, вместе с 250 пмоль меченного биотином антигена, SC, полученный, как описано в (2-2), добавляли и контактировали с полученным раствором с фаговой библиотекой при комнатной температуре в течение 60 минут. Затем раствор с фаговой библиотекой добавляли к блокированным BSA магнитным гранулам, и комплексу антиген-фаг позволяли связываться с магнитными гранулами в течение 15 минут при комнатной температуре. Гранулы промывали один раз посредством SC/TBS (TBS, содержащий SC). Затем, гранулы объединяли с 0,5 мл раствора трипсина в концентрации 1 мг/мл. Гранулы суспендировали при комнатной температуре в течение 15 минут, и сразу после этого раствор фага собирали с выделенных гранул с использованием магнитного стенда. Собранный раствор фагов добавляли к 10 мл клеток E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубировали при осторожном перемешивании при 37°C в течение одного часа для инфицирования фагом. Инфицированные E. coli высевали в чашку 225 мм × 225 мм. Затем фаги собирали из культуральной среды посеянных E. coli с получением жидкого исходного раствора с фаговой библиотекой.
Пэннинг проводили, как описано в (2-2), за исключением добавления 10 мкл 100 мг/мл трипсина для отщепления белка pIII (происходящий из хелперного фага белок pIII), который не экспонирует Fab, чтобы устранить способность фагов, которые не экспонируют Fab, инфицировать E. coli.
(14-2) Получение антител, которые связываются с hIL-6R в присутствии низкомолекулярных соединений, из библиотеки наивных антител человека с использованием способа отрицательной селекции
Сконструированную библиотеку фагового дисплея наивных антител человека подвергали скринингу в отношении антител, которые проявляют активность связывания hIL-6R в присутствии низкомолекулярных соединений. В качестве первой стадии скрининга библиотеку фагового дисплея наивных антител человека контактировали с меченным биотином антигеном со стрептавидином в отсутствие низкомолекулярных соединений для устранения фагов, экспонирующих антител, которые обладают активностью связывания hIL-6R даже в отсутствие низкомолекулярных соединений. Затем пэннинг проводили в присутствии низкомолекулярных соединений аналогичным образом для скрининга антител, которые обладают активностью связывания hIL-6R в присутствии низкомолекулярных соединений. Использованный антиген представлял собой меченный биотином hIL-6R. Затем получали раствор с фаговой библиотекой способом, описанным в (2-3), с использованием меченного биотином hIL-6R в качестве антигена.
(14-3) Оценка активности связывания в присутствии низкомолекулярных соединений с помощью ELISA фагов
Культуральные супернатанты, содержащие фаги, собирали в соответствии с общепринятым способом (Methods Mol. Biol. (2002) 178, 133-145) из единичных колоний E. coli, полученных, как описано в (14-2). Фаги, очищенные способом, описанным в (2-4), подвергали ELISA по следующей методике. 96-луночный микропланшет для титрования StreptaWell (Roche) покрывали в течение ночи посредством 100 мкл TBS, содержащего меченный биотином hIL-6R. После удаления меченного биотином hIL-6R посредством промывания каждой лунки планшета посредством TBST, лунки блокировали 250 мкл смеси 2% обезжиренное молоко/TBS в течение одного часа или более. 2% обезжиренное молоко/TBS удаляли, а затем в каждую лунку добавляли полученные очищенные фаги. Планшету позволяли стоять при комнатной температуре в течение одного часа, чтобы позволить связывание экспонирующих антитело фагов с меченным биотином hIL-6R в каждой лунке в присутствии или в отсутствие SC. После промывания TBST или SC/TBST, в каждую лунку добавляли конъюгированное с HRP антитело против M13 (Amersham Pharmacia Biotech), разбавленное TBS или SC/TBS. Планшет инкубировали в течение одного часа. После промывания TBST или SC/TBST, в каждую лунку добавляли простой раствор TMB (ZYMED). Хромогенную реакцию в растворе каждой лунки останавливали добавлением серной кислоты. Затем, оценивали развитие окраски путем измерения поглощения при 450 нм.
Клоны 6RNMSC1-2_F02 и 6RNMSC1-3_G02, которые обладают активностью связывания с hIL-6R в качестве антигена в присутствии коктейля низкомолекулярных соединений, получали путем проведения ELISA фагов с использованием 960 выделенных клонов.
(14-4) Экспрессия и очистка антител, которые связываются с hIL-6R
Гены амплифицировали с использованием специфических праймеров (SEQ ID NO:110 и 112) из клонов 6RNMSC1-2_F02 и 6RNMSC1-3_G02, которые были оценены как обладающие активностью связывания с меченным биотином hIL-6R в присутствии SC с помощью ELISA фагов, описанного в (14-3). Нуклеотидные последовательности генов анализировали (6RNMSC1-2_F02: последовательности тяжелой цепи и легкой цепи представлены в SEQ ID NO:86 и 87, соответственно; 6RNMSC1-3_G02: последовательности тяжелой цепи и легкой цепи представлены в SEQ ID NO:88 и 89, соответственно). Гены, кодирующие вариабельные области 6RNMSC1-2_F02 и 6RNMSC1-3_G02, и вариабельные области отрицательного контроля в виде антитела против глипикана-3 человека GC413 (тяжелая цепь и легкая цепь представляют собой SEQ ID NO:34 и 35, соответственно) встраивали в плазмиду для экспрессии в клетках животных IgG1/каппа человека. Экспрессированные антитела очищали способом, описанным в примере 3.
(14-5) Идентификация низкомолекулярных соединений, необходимых для связывания полученных антител с hIL-6R
Три типа полученных антител: 6RNMSC1-2_F02, и 6RNMSC1-3_G02, и GC413 подвергали ELISA в девяти условиях, описанных в таблице 3. Между тем, каждое низкомолекулярное соединение соответственно получали в концентрациях, представленных в таблице 3 с использованием буферов, указанных в таблице 19. Использованный антиген представлял собой меченный биотином hIL-6R.
Сначала, 96-луночный микропланшет для титрования StreptaWell (Roche) покрывали при комнатной температуре в течение одного часа или более 100 мкл PBS, содержащего меченный биотином hIL-6R. После промывания буфером для промывания для удаления не связавшегося меченного биотином hIL-6R из планшета, каждую лунку блокировали в течение одного часа или более 250 мкл блокирующего буфера. После удаления блокирующего буфера из каждой лунки, очищенные IgG приготавливали в концентрации 2,5 мкг/мл в буфере для образца, содержащем низкомолекулярные соединения в конечных концентрациях, представленных в таблице 3, и каждый из них распределяли в планшет аликвотами объемом 100 мкл. Планшету позволяли стоять при комнатной температуре в течение одного часа, чтобы позволить связывание каждого IgG с меченным биотином hIL-6R в каждой лунке. После промывания буфером для промывания, содержащим низкомолекулярные соединения в конечных концентрациях, представленных в таблице 3, в каждую лунку добавляли конъюгированное с HRP антитело против IgG человека (BIOSOURCE), разбавленное буфером для образца, содержащим низкомолекулярные соединения. Планшет инкубировали в течение одного часа. После промывания буфером для промывания, содержащим каждое низкомолекулярное соединение, в каждую лунку добавляли простой раствор TMB (ZYMED). Хромогенную реакцию в растворе каждой лунки останавливали добавлением серной кислоты. Затем оценивали развитие окраски путем измерения поглощения при 450 нм. Композиция использованного буфера представлена в таблице 19.
Результат измерения представлен на фиг.28 и 29. Когда использовали 6RNMSC1-2_F02 или 6RNMSC1-3_G02, поглощение было значительно более низким в условиях 9 (в отсутствие низкомолекулярных соединений) по сравнению с условиями 8 (раствор полного коктейля низкомолекулярных соединений). Этот результат подтвердил, что 6RNMSC1-2_F02 и 6RNMSC1-3_G02 обладают свойством, что их связывание антигена изменяется в зависимости от присутствия или отсутствия низкомолекулярных соединений. Между тем, когда использовали 6RNMSC1-2_F02, поглощение было одинаковым в условиях 7 (100 мкМ кинуренин) и в условиях 8, однако поглощение было значительно более низким в других условиях, что демонстрирует, что 6RNMSC1-2_F02 представляет собой антитело, которое связывается с hIL-6R в качестве антигена в присутствии кинуренина (фиг. 28). Более того, когда использовали 6RNMSC1-3_G02, поглощение было одинаковым между условиями 1 (1 мМ ATP-Na) и условиями 8, однако поглощение было значительно более низким в других условиях, что демонстрирует, что 6RNMSC1-3_G02 представляет собой антитело, которое связывается с hIL-6R в качестве антигена в присутствии ATP (фиг. 29). Таким образом, было продемонстрировано, что способ, описанный выше, можно использовать для выделения одновременно множества антител, антигенсвязывающая активность которыйх изменяется в присутствии различных низкомолекулярных соединений.
[Пример 15] Охарактеризция антитела 6RNMSC1-2_F02
(15-1) Оценка с помощью ELISA активности связывания hIL6R в присутствии аминокислот и метаболитов аминокислот, отличных от кинуренина
Антитело 6RNMSC1-2_F02, полученное, как описано в примере 14, которое связывается с hIL-6R в присутствии низкомолекулярных соединений, представляет собой антитело, которое связывается с hIL-6R в присутствии кинуренина. Метаболиты аминокислот, такие как метаболиты триптофана, описанные в примере 11, оценивали в отношении того, являются ли они предпочтительными в качестве неограничивающего варианта осуществления соединений, специфических для злокачественной опухоли, в частности, специфических для злокачественной клетки метаболитов, для применения в рамках настоящего изобретения.
6RNMSC1-2_F02, описанное в примере 14, которое обладает антигенсвязывающей активностью в присутствии кинуренина, и отрицательный контроль GC413 подвергали ELISA в семи условиях, описанных в таблице 18. Между тем, каждое низкомолекулярное соединение соответственно получали в концентрациях, представленных в таблице 18, с использованием буферов, указанных в таблице 4. Использованный антиген представлял собой меченный биотином hIL-6R. ELISA проводили с использованием способа, описанного в примере 11.
Результат измерения представлен на фиг.30. Когда использовали 6RNMSC1-2_F02 поглощение было значительно более низким в условиях 7 (в отсутствие низкомолекулярных соединений) по сравнению с условиями 1 (раствор кинуренина). Аналогично, 6RNMSC1-2_F02 продемонстрировало то же высокое поглощение в условиях 5 (раствор 3-гидроксикинуренина), что и в условиях 1, что указывает на то, что оно представляет собой антитело, которое связывается с hIL-6R в качестве антигена не только в присутствии кинуренина, но также в присутствии метаболита кинуренина. Более того, поглощение для 6RNMSC1-2_F02 было значительно более низким в других условиях, что указывает на то, что 6RNMSC1-2_F02 представляет собой антитело, которое не связывается с hIL-6R в качестве антигена в присутствии триптофана, предшественника кинуренина. Уровень экспрессии IDO - фермента, который метаболизирует триптофан с образованием кинуренина, повышен в микроокружении злокачественной опухоли. Таким образом, ожидается, что антитела, которые связываются с антигенами в присутствии кинуренина или его метаболитом, но не в присутствии триптофана, являются важными в качестве антител, которые связываются с антигенами только в микроокружении злокачественной опухоли. Это указывает на то, что тот же способ можно использовать для получения антител, которые связываются с представляющим интерес антигеном не только в присутствии одного метаболита аминокислоты, но также в присутствие множества структурно различающихся метаболитов аминокислот.
(15-2) Оценка кинуренина в отношении его эффекта на связывание рецептора IL6 человека с помощью поверхностного плазмонного резонанса
Biacore T200 (GE Healthcare) использовали для анализа взаимодействия 6RNMSC1-2_F02 с рецептором IL6 человека (IL-6R) в реакции антиген-антитело. На сенсорный чип CM5 (GE Healthcare) иммобилизовывали соответствующее количество белка A (Invitrogen) способом присоединения аминов. Представляющее интерес антитело улавливали на чипе, чтобы позволить взаимодействие с IL-6R в качестве антигена. Использованный подвижный буфер представлял собой 20 ммоль/л ACES, 150 ммоль/л NaCl, 0,05% (масс./об.) Tween20, pH 7,4. Взаимодействие с антигеном IL-6R анализировали при 25°C. Буферы, использованные для разбавления IL-6R, представляли собой подвижный буфер и буфер, полученный добавлением 100 мкмоль/л кинуренина в подвижный буфер, и, кроме того, буфер, полученный в качестве контроля добавлением 10 ммоль/л ATP к подвижному буферу.
Разбавленный раствор IL-6R и подвижный буфер в качестве пустого образца инжектировали при скорости потока 10 мкл/мин в течение одной минуты, чтобы позволить взаимодействие IL-6R с 6RNMSC1-2_F02, уловленном на сенсорном чипе. Затем инжектировали подвижный буфер при скорости потока 10 мкл/мин в течение одной минуты. После наблюдения диссоциации IL-6R от антитела инжектировали 10 ммоль/л глицина-HCl (pH 1,5) при скорости потока 30 мкл/мин в течение 30 секунд для регенерации сенсорного чипа. Константу диссоциации KD (M) 6RNMSC1-2_F02 вычисляли для IL-6R, исходя из константы скорости ассоциации ka (1/М с) и константы скорости диссоциации kd (1/с), обе из которых являются параметрами кинетики, вычисленными из сенсограммы, полученной путем измерения. Каждый параметр вычисляли с использованием оценочного программного обеспечения Biacore T200 Evaluation Software (GE Healthcare).
Сенсограммы, полученные с помощью этого измерения взаимодействия между 6RNMSC1-2_F02 и 1 мкмоль/л IL-6R в присутствии 100 мкмоль/л кинуренина и в присутствии или в отсутствие 10 ммоль/л ATP, представлены в на фиг.31. Как показано на фиг. 31, 6RNMSC1-2_F02 связывается с IL-6R в присутствии 100 мкмоль/л кинуренина; однако его связывание с IL-6R не поддавалось обнаружению в отсутствие кинуренина. Это демонстрирует, что 6RNMSC1-2_F02 обладает свойством, состоящим в том, что оно связывается с IL-6R через кинуренин в качестве переключающего фактора. Между тем константа диссоциации KD 6RNMSC1-2_F02 составляла 1,5 мкмоль/л в присутствии 100 мкмоль/л кинуренина.
(15-3) Эффект кинуренина в качестве переключающего фактора на диссоциацию антител от IL-6R
Biacore T200 (GE Healthcare) использовали для оценки того, диссоциирует ли 6RNMSC1-2_F02, который связывается с IL-6R в присутствии кинуренина, зависимым от концентрации кинуренина образом в присутствии кинуренина. Использованные подвижные буферы представляли собой 20 ммоль/л ACES, 150 ммоль/л NaCl, 0,05% (масс./об.) Tween20, pH 7,4, и 20 ммоль/л ACES, 150 ммоль/л NaCl, 0,05% (масс./об.) Tween20, pH 7,4, 100 мкмоль/л кинуренина, и анализ проводили при 25°C. IL-6R иммобилизовывали на сенсорный чип CM5 способом присоединения аминов; и 6RNMSC1-2_F02 разбавляли до 5 мкг/мл посредством 20 ммоль/л ACES, 150 ммоль/л NaCl, 0,05% (масс./об.) Tween20, pH 7,4, содержащих 100 мкмоль/л кинуренина, и его подвергали взаимодействию в качестве анализируемого соединения в течение 180 секунд. Затем проводили мониторинг диссоциации IL-6R в каждых условиях подвижного буфера. Для сравнения степени диссоциации между соответствующими условиями подвижного буфера, величины нормализовывали, принимая количество 6RNMSC1-2_F02, связавшегося с IL-6R в присутствии 100 мкмоль/л кинуренина, за 100, и сравнивали. Сенсограмма, которая отражает взаимодействие между 6RNMSC1-2_F02 и IL-6R после нормализации, представлена на фиг. 32. Результат, показанный на фиг.32, демонстрирует, что 6RNMSC1-2_F02 обладает свойством, состоящим в том, что он связывается с IL-6R в присутствии кинуренина, а затем быстро диссоциирует от IL-6R в отсутствие кинуренина. В частности, было продемонстрировано, что опосредуемая кинуренином регуляция связывания антитела с IL-6R является обратимой.
(15-4) Оценка эффекта концентрации кинуренина на связывание IL-6R
Далее использовали Biacore T200 (GE Healthcare) для оценки эффекта концентрации кинуренина на реакцию антиген-антитело между 6RNMSC1-2_F02 и IL-6R. Использованный подвижный буфер представлял собой 20 ммоль/л ACES, 150 ммоль/л NaCl, 0,05% (масс./об.) Tween20, pH 7,4. Реакцию антиген-антитело между 6RNMSC1-2_F02 и IL-6R человека анализировали при 25°C. IL-6R иммобилизовывали на сенсорный чип CM5 способом присоединения аминов; и 6RNMSC1-2_F02 разбавляли до 1 мкг/мл посредством 20 ммоль/л ACES, 150 ммоль/л NaCl, 0,05% (масс./об.) Tween20, pH 7,4, содержащего различные концентрации кинуренина, и им позволяли взаимодействовать в качестве анализируемого соединения в течение 180 секунд для выявления изменений в уровне связывания. Результат представлен на фиг.33. Этот результат продемонстрировал, что чем более высокой является концентрация кинуренина, служащего в качестве переключающего фактора, тем выше количество 6RNMSC1-2_F02, связавшегося с IL-6R.
Между тем, поскольку IL-6R иммобилизован на сенсорном чипе в этой системе анализа, полагают, что 6RNMSC1-2_F02 связывается двухвалентным образом. Также было выявлено, что в такой системе анализа, где 6RNMSC1-2_F02 распознает IL-6R двухвалентным образом, количество 6RNMSC1-2_F02, связавшегося с IL-6R, возрастет с увеличением концентрации кинуренина. Этот результат продемонстрировал, что 6RNMSC1-2_F02 обладает свойством, состоящим в том, что он связывается IL-6R через кинуренин в качестве переключающего фактора также посредством двухвалентного связывания.
Эти результаты продемонстрировали, что 6RNMSC1-2_F02 представляет собой антитело, которое связывается с IL-6R в присутствии кинуренина, с кинуренином в качестве переключающего фактора, но диссоциирует от IL-6R в отсутствие кинуренина. Более того, было подтверждено, что возможно иметь полный контроль 6RNMSC1-2_F02 по принципу включения/выключения, так что оно не демонстрирует активности связывания IL-6R в отсутствие кинуренина. Ожидалось, что функция переключения будет достигнута так, как показано на фиг. 2.
(15-5) Эффект кинуренина на активность ADCC 6RNMSC1-2_F02
Гены, кодирующие вариабельные области 6RNMSC1-2_F02, определенные, как описано в примере 14, встраивали в плазмиду для экспрессии в клетках животных IgG1/каппа человека, которое содержит последовательность константной области тяжелой цепи антитела, содержащую последовательность SEQ ID NO:90, и последовательность константной области легкой цепи каппа, содержащую последовательность SEQ ID NO:91. Соответствующие гены, кодирующие вариабельные области известного антитела против IL-6R человека, MRA (последовательности тяжелой цепи и легкой цепи представлены в SEQ ID NO:92 и 93, соответственно), также встраивали в экспрессирующую плазмиду клеток животных для IgG1/каппа человека, которое имеет описанные выше константные области (SEQ ID NO:90 и 91). Антитела экспрессировали с использованием способа, описанного ниже. FreeStyle 293-F (Invitrogen), которые происходили из клеток почки эмбриона человека, суспендировали при плотности клеток 1,33×106 клеток/мл в среде для экспрессии FreeStyle 293 Expression Medium (Invitrogen), и распределяли аликвотами объемом 3 мл в каждую ячейку планшета с 6 ячейками. Плазмидную ДНК трансфицировали в клетки с помощью липофекции. Из культуральных супернатантов после культивирования в течение четырех суток в инкубаторе с CO2 (37°C, 8% CO2, 90 об/мин), антитела очищали способом, известным специалистам в данной области, с использованием rProtein A Sepharose™ Fast Flow (Amersham Biosciences). Поглощение очищенных растворов антител измеряли при 280 нм с использованием спектрофотометра. Из величин, полученных путем измерения, концентрации очищенных антител вычисляли с использованием коэффициента экстинкции, определенного способом PACE (Protein Science (1995) 4, 2411-2423).
Антитело 6RNMSC1-2_F02, которое обладает антигенсвязывающей активностью в присутствии кинуренина, как описано в примере 14, оценивали в отношении его связывания с растворимым hIL-6R. В качестве первой стадии оценки активности ADCC 6RNMSC1-2_F02 в отношении экспрессирующих hIL-6R клеток, 6RNMSC1-2_F02 оценивали в отношении того, обладает ли оно способностью связываться с hIL-6R мембранного типа, экспрессированным в экспрессирующих hIL-6R клетках. В частности, связывание 6RNMSC1-2_F02 с клетками линии BaF/hIL-6R (WO2012/073992) оценивали и анализировали с использованием проточного цитометра. Соответствующее количество клеток BaF/hIL-6R получали и блокировали посредством PBS, содержащего 2% FBS, на льду в течение одного часа или более. Супернатант удуаляли из блокированных клеток центрифугированием, и добавляли 100 мкл 6RNMSC1-2_F02 или контрольного антитела MRA (последовательности тяжелой цепи и легкой цепи представлены в SEQ ID NO:92 и 93, соответственно) в двух условиях: в присутствии или в отсутствие конечной концентрации кинуренина при 100 мкМ. На этой стадии антитела контактировали с hIL-6R клеточной мембраны на льду в течение 30 минут. Комплекс антитело-клетка промывали буфером для промывания, содержащим кинуренин, или буфером для промывания, который не содержит кинуренина. Затем, в присутствии или в отсутствие кинуренина комплекс контактировали со вторичным антителом (Beckman Coulter IM1627), которое распознает константную область антитела. После инкубации в течение 30 минут с антителом на льду клетки вновь промывали буфером для промывания, а затем ресуспендировали в PBS/2% FBS. Связывание 6RNMSC1-2_F02 с полученными клетками оценивали и анализировали с использованием проточного цитометра BD FACS cant II (BD).
Результат анализа показан на фиг. 34. В случае контрольного антитела MRA флуоресценция поддавалась обнаружению независимо от присутствия или отсутствия кинуренина. Напротив, для 6RNMSC1-2_F02 впервые наблюдали сдвиг флуоресценции в присутствии 100 мкМ кинуренина; однако флуоресценция не поддавалась обнаружению в отсутствие кинуренина. Это демонстрирует, что 6RNMSC1-2_F02 представляет собой антитело, которое обладает способностью связываться с hIL-6R, экспрессированным на клеточной мембране в присутствии кинуренина.
Как правило, природные антитела связываются в области их Fab прямо с антигенами на клетках-мишенях, и их Fc связывается с FcγR на эффекторных клетках, что приводит к индукции цитотоксической активности (активность ADCC) эффекторных клеток против клеток-мишеней. В этом контексте оценку того, достигается ли активность ADCC против клеток, экспрессирующих hIL-6R, при связывании 6RNMSC1-2_F02 с hIL-6R в присутствии кинуренина, оценивали способом, описанным ниже.
Использовали вариант 6RNMSC1-2_F02 с увеличенной эффектоной активностью (последовательности тяжелой цепи и легкой цепи представлены в SEQ ID NO:94 и 91, соответственно). При различных концентрациях 6RNMSC1-2_F02, активность ADCC против экспрессирущих hIL-6R клеток анализировали в присутствии или в отсутствие кинуренина в соответствии со способом, описанным в справочном примере 1. Результат анализа представлен на фиг.35.
Результат анализа подтвердил, что в присутствии кинуренина 6RNMSC1-2_F02 индуцировал активность ADCC против клеток, экспрессирующих hIL-6R, зависимым от концентрации антитела образом. Данные демонстрируют, что связывание антител с антигеном через кинуренин индуцирует активность ADCC против экспрессирующих антиген клеток и, с точки зрения функционирования через противоопухолевую активность, такую как активность ADCC, антитела, которые связываются с антигенами в присутствии низкомолекулярных соединений в качестве переключающего фактора, также могут регулироваться присутствием низкомолекулярных соединений, служащих в качестве переключающего фактора.
Более того, существует различие в концентрации кинуренина между нормальными и опухолевыми тканями, и таким образом является предпочтительным, чтобы антитело проявляло активность ADCC против экспрессирующих антиген опухолевых клеток при концентрации кинуренина в опухолевых тканях, в то время как при нормальной концентрации кинуренина в тканях, активность ADCC ослабляется или не проявляется. Затем, 6RNMSC1-2_F02 оценивали в отношении активности ADCC против экспрессирующих hIL-6R клеток при различных концентрациях кинуренина в соответствии со способом, описанным в справочном примере 2. Результат анализа представлен на фиг.36. Результат анализа продемонстрировал, что 6RNMSC1-2_F02 обладало зависимой от концентрации кинуренина активностью ADCC против экспрессирующих hIL-6R. Более того, в противоположность приблизительно 10% активности ADCC при концентрации 4-6 мкМ, которая считается концентрацией кинуренина в нормальных тканях, активность ADCC при концентрации 30-40 мкМ, которая считается концентрацией кинуренина в опухолевых тканях, составляла приблизительно 25%.
Исходя из этих результатов, в случае 6RNMSC1-2_F02 активность ADCC против экспрессирущих hIL-6R клеток была слабой в нормальных тканях, где концентрация кинуренина является низкой; и в опухолевых тканях, где его концентрация является высокой, активность ADCC у 6RNMSC1-2_F02 против экспрессирующих hIL-6R клеток была более высокой. Это указывает на то, что путем введения антитела, которое использует кинуренин в качестве переключающего фактора, цитотоксичность нормальных тканей, экспрессирующих антиген-мишень, может быть снижена без снижения фармацевтического эффекта против опухолевых тканей, экспрессирущих антиген-мишень.
(15-6) Оценка полученного антитела в отношении его активности связывания с hIL-6R в сыворотке мыши с помощью ELISA для IgG
Антитело 6RNMSC1-2_F02, полученное, как описано в примере 14, которое связывается с hIL-6R в присутствии низкомолекулярных соединений, представляет собой антитело, которое связывается с hIL-6R в присутствии кинуренина. До сих пор 6RNMSC1-2_F02 оценивали в отношении его антигенсвязывающей способности в буферах, таких как PBS и TBS. Считается, что в сыворотке мыши существует множество неизвестных низкомолекулярных соединений, включая аминокислоты, и нельзя исключать возможности того, что такие низкомолекулярные соединения будут влиять на связывание 6RNMSC1-2_F02 с антигеном. Таким образом, 6RNMSC1-2_F02 оценивали в отношении его антигенсвязывающей способности в сыворотке мыши.
Как описано в примере 14, антитело 6RNMSC1-2_F02, которое обладает антигенсвязывающей активностью в присутствии кинуренина, и известное антитело против hIL-6R MRA подвергали ELISA в двух условиях, описанных в таблице 20. Использованный антиген представлял собой меченный биотином hIL-6R.
Сначала 96-луночный микропланшет для титрования StreptaWell (Roche) покрывали при комнатной температуре в течение одного часа или более 100 мкл PBS, содержащего меченный биотином антиген. После промывания буфером для промывания для удаления не связавшегося антигена из планшета, каждую лунку блокировали в течение одного часа или более 250 мкл блокирующего буфера. После удаления блокирующего буфера из каждой лунки каждый из очищенных IgG приготавливали в концентрации 2,5 мкг/мл в условиях 2, представленных в таблице 20, и распределяли аликвотами объемом 100 мкл в планшет. Планшету позволяли стоять при комнатной температуре в течение одного часа, чтобы позволить связывание каждого IgG с антигеном в каждой лунке. После промывания буфером для промывания, содержащим 100 мкМ кинуренин, в каждую лунку добавляли конъюгированное с HRP антитело против IgG человека (BIOSOURCE), разбавленное буфером для образца, содержащим кинуренин. Планшет инкубировали в течение одного часа. После промывания буфером, содержащим кинуренин, в каждую лунку добавляли простой раствор TMB (ZYMED). Хромогенную реакцию в растворе каждой лунки останавливали добавлением серной кислоты. Затем оценивали развитие окраски путем измерения поглощения при 450 нм.
Результат измерения представлен на фиг. 37. В случае MRA поглощение было одинаковым независимо от присутствия или отсутствия кинуренина. Напротив, когда использовали 6RNMSC1-2_F02, поглощение было значительно более низким в условиях 1 (сыворотка мыши без кинуренина) по сравнению с условиями 2 (сыворотка мыши в присутствии кинуренина), представленными в таблице 20. Это указывает на то, что 6RNMSC1-2_F02 представляет собой антитело, которое связывается с hIL-6R в качестве антигена в присутствии кинуренина без влияния неизвестных молекул в сыворотке мыши.
[Пример 16] Получение антител, которые связываются с антигенами в отсутствие аденозина или ATP, из библиотеки антител с использованием способов фагового дисплея
(16-1) Получение антител, связывание которых с антигеном ингибируется в присутствии низкомолекулярных соединений, из библиотеки с использованием смеси аденозина и ATP
Антитела, которые связываются с антигенами-мишенями в присутствии низкомолекулярных соединений, служащих в качестве переключающего фактора, получали, как описано в примерах выше. В эксперименте, описанном в этом примере, авторы настоящего изобретения предприняли попытку получить антитела, которые связываются с антигенами-мишенями в отсутствие низкомолекулярных соединений.
Антитела, которые проявляют антигенсвязывающую активность в отсутствие аденозина и/или ATP, но способность к связыванию которых снижется в присутствии аденозина и/или ATP, получали из сконструированной библиотеки фагового дисплея рационально сконструированных антител. В качестве первой стадии выделения антител фаговоую библиотеку антител контактировали с биотинилированным аденозином и ATP-нейтравидином для сбора из библиотеки фагового дисплея антител, которые связываются с аденозином и/или ATP. Затем библиотеку фагового дисплея антител контактировали с биотинилированным антигеном-стрептавидином в отсутствие аденозина и ATP для сбора антител, которые связываются с антигенами в отсутствие аденозина и ATP. Пэннинг проводили чередующимся образом, описанным выше, для скрининга антител, которые обладают активностью связывания с антигеном и аденозином и/или ATP. В присутствии аденозина и ATP, ожидалось, что связывание антител с такими свойствами с антигеном будет ингибироваться связыванием с антителами аденозина и/или ATP.
Фаги продуцировали в E. coli, содержащих фагмидный вектор, сконструированный для фагового дисплея. В культуральную среду E. coli, в которой проводили продукцию фага добавляли 2,5 M NaCl/10% PEG для преципитации фага. Фракцию преципитированных фагов разбавляли TBS для получения раствора с фаговой библиотекой. Затем к раствору с фаговой библиотекой добавляли BSA в конечной концентрации 4%. Пэннинг проводили с использованием магнитных гранул с иммобилизованным на них антигеном. Использованные магнитные гранулы представляли собой покрытые нейтравидином гранулы (Sera-Mag SpeedBeads NeutrАвидин-coated) и покрытые стрептавидином гранулы (Dynabeads M-280 Streptavidin).
К полученному раствору с фаговой библиотекой добавляли 500 пмоль биотинилированного ATP, 2'-аденозин-PEG-биотин и 5'-аденозин-PEG-биотин. Таким образом, раствор с фаговой библиотекой контактировали с аденозином и ATP в течение 60 минут при комнатной температуре. Затем к раствору с фаговой библиотекой добавляли блокированные BSA магнитные гранулы и комплексу фага с аденозином и/или ATP позволяли связываться с магнитными гранулами при комнатной температуре в течение 15 минут. Гранулы промывали один раз TBS, а затем к гранулам добавляли 0,5 мл раствора трипсина в концентрации 1 мг/мл.
Гранулы суспендировали при комнатной температуре в течение 15 минут, и сразу после этого с выделенных гранул собирали раствор с фагами с использованием магнитного стенда. Собранный раствор с фагами добавляли к 10 мл клеток E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубировали при осторожном перемешивании при 37°C в течение одного часа для инфицирования фагом. Инфицированные E. coli высевали в чашку 225 мм × 225 мм. Затем фаги собирали из культуральной среды посеянных E. coli для получения раствора с фаговой библиотекой.
Второй раунд пэннинга проводили для увеличения содержания фагов, способных связываться с биотинилированным антигеном в отсутствие аденозина и ATP. В частности, 250 пмоль биотинилированного антигена добавляли к полученному раствору с фаговой библиотекой. Таким образом, раствор с фаговой библиотекой контактировали с антигеном в течение 60 минут при комнатной температуре. Затем к раствору с фаговой библиотекой добавляли блокированные BSA магнитные гранулы, и комплексу антиген-фаг позволяли связываться с магнитными гранулами при комнатной температуре в течение 15 минут. Гранулы промывали два раза посредством TBST и один раз посредством TBS. Затем к гранулам добавляли 0,5 мл раствора трипсина в концентрации 1 мг/мл. Гранулы суспендировали при комнатной температуре в течение 15 минут, и сразу после этого раствор с фагом собирали с выделенных гранул с использованием магнитного стенда. Собранный раствор с фагом добавляли к 10 мл клеток E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубировали при осторожном перемешивании при 37°C в течение одного часа для инфицирования фагом. Инфицированные E. coli высевали в чашку 225 мм × 225 мм. Затем фаги собирали из культуральной среды посеянных E. coli с получением раствора с фаговой библиотекой.
На последующих нечетных раундах пэннинг проводили аналогичным образом в качестве пэннинга первого раунда. Однако количество промываний гранулы TBST и TBS увеличивали до трех раз и двух раз, соответственно.
При последующих четных раундах пэннинг проводили аналогичным образом в качестве пэннинга второго раунда. Однако на четвертом и последующих раундах пэннинга, количество биотинилированного антигена снижали до 40 пмоль, и количество промываний гранул TBST и TBS увеличивали до трех раз и двух раз, соответственно.
(16-2) Оценка активности связывания в присутствии низкомолекулярных соединений с помощью ELISA фагов
Культуральные супернатанты, содержащие фаги, собирали в соответствии с общепринятым способом (Methods Mol. Biol. (2002) 178, 133-145) из единичных колоний E. coli полученных способом, описанным выше. Собранные культуральные супернатанты ультрацентрифугировали с использованием NucleoFast 96 (MACHEREY-NAGEL). В каждую лунку добавляли 100 мкл культуральных супернатантов, и NucleoFast 96 центрифугировали (4500 g в течение 45 минут) для удаления оставшейся жидкой части. В каждую лунку добавляли 100 мкл H2O и вновь NucleoFast 96 промывали центрифугированием (4500 g в течение 30 минут). Добавляли 100 мкл TBS, а затем NucleoFast 96 позволяли стоять при комнатной температуре в течение пяти минут. Наконец, раствор фага собирали из супернатанта в каждой лунке.
Очищенные фаги, к которым были добавлены TBS или ATP и аденозин/TBS, подвергали ELISA по следующей методике. 96-луночный микропланшет для титрования StreptaWell (Roche) покрывали в течение ночи посредством 100 мкл TBS, содержащего меченный биотином антиген. После удаления антигена из каждой лунки планшета промыванием TBST, лунки блокировали 250 мкл смеси 2% обезжиренное молоко/TBS в течение одного часа или более. 2% обезжиренное молоко/TBS удаляли, а затем в каждую лунку добавляли полученные очищенные фаги. Планшету позволяли стоять при 37°C в течение одного часа, чтобы позволить связаться экспонирующим антитело фага с антигенами в каждой лунке в присутствии или в отсутствие 10 мМ аденозина и ATP. После промывания TBST или 10 мМ (ATP и аденозин)/TBST, в каждую лунку добавляли конъюгированное с HRP антитело против M13 (Amersham Pharmacia Biotech), разбавленное TBS или 10 мМ (ATP и аденозин)/TBS. Планшет инкубировали в течение одного часа. После промывания TBST или 10 мМ (ATP и аденозин)/TBST, в каждую лунку добавляли простой раствор TMB (ZYMED). Хромогенную реакцию в растворе каждой лунки останавливали добавлением серной кислоты. Затем, оценивали развитие окраски путем измерения поглощения при 450 нм.
ELISA фагов проводили с использованием 96 выделенных клонов с получением из библиотеки рационально сконструированных антител клона "I6RLSA1-6_011", который обладал антигенсвязывающей активностью с IL-6 человека в отсутствие ATP и аденозина, клона "HSADSA1-6_020", который обладал антигенсвязывающей активностью с сывороточным альбумином человека (HSA) в отсутствие ATP и аденозина, а также клонов "6RRLSA1-6_037" и "6RRLSA1-6_045", которые обладали антигенсвязывающей активностью в отношении рецептора IL-6 человека в отсутствие ATP и аденозина (фиг. 38, 39 и 45).
(16-3) Анализ последовательности антител, для которых аденозин и ATP служат в качестве переключающего фактора
Гены амплифицировали с использованием специфических праймеров (SEQ ID NO:111 и 112) из клонов, которые были оценены как обладающие антигенсвязывающей активностью в отсутствие аденозина и ATP, исходя из результата ELISA фагов, описанного в (16-2). Нуклеотидные последовательности генов анализировали. Исходя из результата анализа, аминокислотные последовательности представлены в таблице 21 ниже.
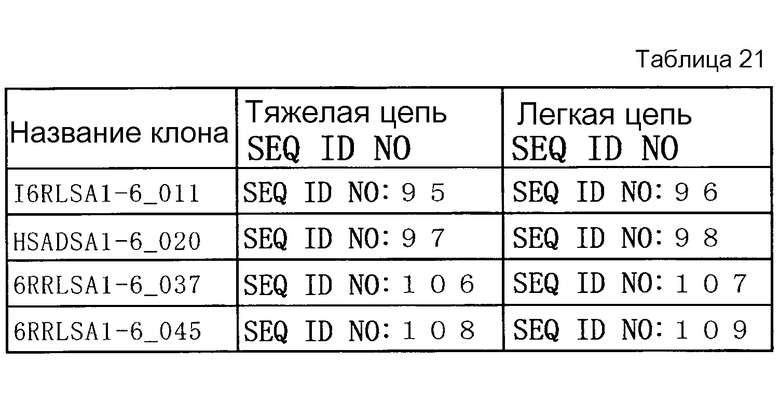
[Пример 17] Получение антител, которые связываются с антигенами в присутствии аденозина и ATP, из библиотеки антител с использованием технологии мультивалентного фагового дисплея
(17-1) Получение антител, которые связываются с антигенами в присутствии низкомолекулярных соединений, из библиотеки с использованием мультивалентного дисплея
Мультивалентный дисплей антител на фаге использовали для получения антител, которые обладают антигенсвязывающей активностью в присутствии аденозина и/или ATP, или из библиотеки фагового дисплея рационально сконструированных антител. При получении антител из библиотеки вероятность получения возрастает, когда соотношение антигенсвязывающей способности между условиями присутствия и отсутствия низкомолекулярных соединений является более высоким. Таким образом, проводили пэннинг на основе усиления кажущейся способности к связыванию для эффективного сбора антител со способностью к связыванию в присутствии низкомолекулярных соединений. Более конкретно, кажущуюся связывающую способность усиливали с помощью эффекта авидности (эффект связывания поливалентного антигена), позволяя фагам экспонировать антитела поливалентным образам. Сначала библиотеку фагового дисплея рационально сконстрированных антител контактировали с биотинилировнными антигенами в присутствии аденозина и ATP для сбора библиотеки фагового диспелея антител, которые связываются с антигенами в присутствии аденозина и ATP. Затем, в соответствии со способом, описанным в Rondot (Nat. Biotechnol. (2001) 19, 75-78), E. coli инфицировали собранной библиотекой фагового дисплея антител, а затем инфицировали фагами-помощниками с дефицитом гена, кодирующего pIII, с получением библиотеки фагового дисплея антител, где антитела присутствуют на всех pIII. Библиотеку мультивалентного фагового дисплея контактировали с биотинилированным антигеном со стрептавидином в присутствии аденозина и ATP. После сбора библиотеки фаги элюировали с гранул в отсутствие аденозина и ATP и собирали в элюате. Этот цикл получения и пэннинга фагов проводили несколько раз для скрининга антител, которые обладают антигенсвязывающей активностью только в присутствии аденозина и/или ATP.
E. coli, содержащие сконструированную фагмиду фагового дисплея, инфицировали фагом-помощником M13KO7 и культивировали в течение ночи при 30°C для получения одновалентной библиотеки фагового дисплея антител. После продуцирования фага в культуральную среду E. coli добавляли 2,5 M NaCl/10% PEG для преципитации фагов. Фракцию преципитированного фага разбавляли TBS для получения раствора с фаговой библиотекой. Затем к раствору с фаговой библиотекой добавляли BSA в конечной концентрации 4%. Пэннинг проводили с использованием магнитных гранул с иммобилизованным на них антигеном. Использованные магнитные гранулы представляли собой покрытые нейтравидином гранулы (Sera-Mag SpeedBeads NeutrАвидин-coated) и покрытые стрептавидином гранулы (Dynabeads M-280 Streptavidin).
Раствор с фаговой библиотекой контактировали с антигеном, аденозином и ATP при комнатной температуре в течение 60 минут путем добавления к полученной фаговой библиотеке 500 пмоль меченного биотином IgA-Fc человека (SEQ ID NO:99) в качестве антигена и ATP-Na и аденозина в конечной концентрации 1 мМ. К раствору с фаговой библиотекой добавляли блокированные BSA магнитные гранулы, и комплексу антиген-фаг позволяли связываться с магнитными гранулами при комнатной температуре в течение 15 минут. Гранулы промывали один раз посредством TBS, в котором были растворены ATP и аденозин. Затем к гранулам добавляли 0,5 мл раствора трипсина в концентрации 1 мг/мл. Гранулы суспендировали при комнатной температуре в течение 15 минут, и сразу после этого раствор с фагами собирали с выделенных гранул с использованием магнитного стенда. Собранный раствор фага добавляли к 10 мл клеток E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубировали при осторожном перемешивании при 37°C в течение одного часа для инфицирования фагом. Инфицированные E. coli высевали в чашку 225 мм × 225 мм. Затем фагу-помощнику M13KO7 или M13KO7ΔpIII (называемый гиперфагом) (PROGEN Biotechnik) позволяли инфицировать культуральную среду посеянных E. coli, и фаги собирали из супернатанта культуры, инкубированной в течение ночи при 30°C, для получения раствора с одновалентной библиотекой фагового дисплея антител или мультивалентной библиотекой фагового дисплея, соответственно.
Первый раунд пэннинга проводили для сбора фагов, которые способны связывать антиген в присутствии аденозина и ATP, в то время как второй и последующий раунды пэннинга проводили для увеличения содержания фагов, которые способны связывать антиген только в присутствии аденозина и ATP. В частности, каждый из 250 пмоль меченного биотином антигена и аденозина и ATP в конечной концентрации 1 мМ добавляли к полученному раствору с фаговой библиотекой. Таким образом, фаговую библиотеку контактировали с антигеном, аденозином и ATP при комнатной температуре в течение 60 минут. Добавляли блокированные посредством BSA магнитные гранулы и позволяли им связываться с комплексом фаг-антиген в течение 15 минут при комнатной температуре. Гранулы промывали 1 мл TBST, в котором были растворены аденозин и ATP (далее обозначаемый как (аденозин+ATP)/TBST), и TBS, в котором были растворены аденозин и ATP (далее обозначаемый как (аденозин+ATP)/TBS). Затем к гранулам добавляли 0,5 мл TBS. Гранулы суспендировали при комнатной температуре, и сразу после этого раствор фага собирали из выделенных гранул с использованием магнитного стенда. После повторения этой обработки, два элюированных по отдельности раствора с фагами объединяли. Белок pIII (происходящий из фага-помощника белок pIII), который не экспонирует Fab, отщепляли от фагов добавлением 5 мкл l 100 мг/мл трипсина к собранному раствору с фагами для устранения способности фагов, которые не экспонируют Fab, инфицировать E. coli. Фаги, собранные из трипсинизированного раствора с фагами добавляли к 10 мл клеток E. coli штамма ER2738 на логарифмической фазе роста (OD600=0,4-0,7). E. coli инкубировали при осторожном перемешивании при 37°C в течение одного часа для инфицирования фагом. Инфицированные E. coli высевали в чашку 225 мм × 225 мм. Затем аналогично тому, как на первом раунде пэннинга, фаги собирали из культуральной среды посеянных E. coli для получения раствора с одновалентной библиотекой фагового дисплея антител и мультивалентной библиотекой фагового дисплея антител. Пэннинг проводили три раза с получением антител, которые обладают антигенсвязывающей активностью в присутствии аденозина и ATP. Между тем, на третьем и последующих раундах пэннинга биотинилированный антиген использовали в концентрации 40 пмоль.
(17-2) Оценка активности связывания в присутствии аденозина и/или ATP с помощью ELISA фагов
Культуральные супернатанты, содержащие фаги, собирали в соответствии с общепринятым способом (Methods Mol. Biol. (2002) 178, 133-145) из единичных колоний E. coli, полученных способом, описанным выше. Собранные культуральные супернатанты ультрацентрифугировали с использованием NucleoFast 96 (MACHERY-NAGEL). 100 мкл культуральных супернатантов добавляли в каждую лунку NucleoFast 96 и центрифугировали (4500 g в течение 45 минут) для удаления оставшейся жидкой части. Добавляли 100 мкл H2O, а затем NucleoFast 96 промывали центрифугированием (4500 g в течение 30 минут). Наконец, добавляли 100 мкл TBS, и NucleoFast 96 позволяли стоять в течение пяти минут при комнатной температуре. Раствор с фагом, содержащийся в супернатанте в каждой лунке, собирали.
Очищенные фаги, к которым добавляли TBS или (аденозин+ATP)/TBS, подвергали ELISA по следующей методике. 96-луночный микропланшет для титрования StreptaWell (Roche) покрывали в течение ночи посредством 100 мкл TBS, содержащего меченный биотином антиген. После удаления антигена из каждой лунки планшета посредством промывания TBST, лунки блокировали 250 мкл смеси 2% обезжиренное молоко/TBS в течение одного часа или более. 2% обезжиренное молоко/TBS удаляли, а затем в каждую лунку добавляли полученные очищенные фаги. Планшету позволяли стоять в течение одного часа для обеспечения связывания экспонирующих антитело фагов с антигеном в каждой лунке в присутствии или в отсутствие аденозина и ATP. После промывания TBST или (аденозин+ATP)/TBST, в каждую лунку добавляли конъюгированное с HRP антитело против M13 (Amersham Pharmacia Biotech), разбавленное TBS или (аденозин+ATP)/TBS. Планшет инкубировали в течение одного часа. После промывания TBST или (аденозин+ATP)/TBST, в каждую лунку добавляли простой раствор TMB (ZYMED). Хромогенную реакцию в растворе каждой лунки останавливали добавлением серной кислоты. Затем, оценивали развитие окраски путем измерения поглощения при 450 нм. Результат показывает, что было получено большее количество антител, которые обладают активностью связывания в присутствии низкомолекулярных соединений из мультивалентной библиотеки фагового дисплея антител (фиг. 40 и 41). Эти данные указывают на то, что антитела, которые обладают активностью связывания в присутствии низкомолекулярных соединений, можно получать более эффективно с использованием способа мультивалентной библиотеки фагового дисплея антител. Результат ELISA фагов представлен в таблице 22 ниже.
(17-3) Оценка связывающей способности антител, которые используют аденозин и ATP в качестве переключающего фактора, и анализ последовательности
Гены амплифицировали с использованием специфических праймеров (SEQ ID NO:111 и 112) из клонов, которые были оценены как обладающие антигенсвязывающей активностью в присутствии аденозина и ATP, исходя из результата ELISA фагов, описанного в (17-2). Нуклеотидные последовательности генов анализировали. В результате, получили клон "2IADL3C5-4_048 (тяжелая цепь, SEQ ID NO:100; легкая цепь, SEQ ID NO:101)", который проявляет антигенсвязывающую активность в присутствии аденозина и ATP (фиг. 42).
[Пример 18] Охарактеризация зависимых от ATP/аденозина, полученных из библиотеки
(18-1) Получение зависимых от ATP/аденозина антител, полученных из библиотеки
Гены амплифицировали с использованием специфических праймеров из клонов 6RAD2C1-4_001, 6RAD2C1-4_005, 6RAD2C1-4_011, 6RAD2C1-4_026, 6RAD2C1-4_030, 6RAD2C1-4_042, 6RAD2C1-4_076, 6RDL3C1-4_085 и 6RDL3C5-4_011, которые были получены, как описано в примере 10, и были оценены как обладающие активностью связывания с меченным биотином hIL-6R в присутствии ATP или аденозина; и их нуклеотидные последовательности анализировали (таблица 23).

Вариабельные области 6RAD2C1-4_001, 6RAD2C1-4_005, 6RAD2C1-4_011, 6RAD2C1-4_026, 6RAD2C1-4_030, 6RAD2C1-4_042, 6RAD2C1-4_076, 6RDL3C1-4_085 и 6RDL3C5-4_011 встраивали в плазмиду для экспрессии в клетках животных IgG1/каппа, которое имеет последовательность константной области тяжелой цепи антитела SEQ ID NO:90 и последовательность константной области легкой цепи каппа SEQ ID NO:91. Антитела экспрессировали с использованием способа, описанного ниже. FreeStyle 293-F (Invitrogen), которые происходили из клеток почки эмбриона человека, суспендировали при плотности клеток 1,33×106 клеток/мл в среде для экспрессии FreeStyle 293 Expression Medium (Invitrogen), и распределяли аликвотами объемом 3 мл в каждую ячейку планшета с 6 ячейками. Плазмидную ДНК трансфицировали в клетки с помощью липофекции. Из культуральных супернатантов после культивирования в течение четырех суток в инкубаторе с CO2 (37°C, 8% CO2, 90 об./мин.), антитела очищали способом, известным специалистам в данной области в данной области с использованием rProtein A Sepharose™ Fast Flow (Amersham Biosciences). Поглощение очищенных растворов антител измеряли при 280 нм с использованием спектрофотометра. Из величин, полученных путем измерения, вычисляли концентрации очищенных антител с использованием коэффициента экстинкции, определенного способом PACE (Protein Science (1995) 4, 2411-2423).
(18-2) Оценка эффекта различных низкомолекулярных соединений на связывание рецептора IL6 человека с помощь поверхностного плазмонного резонанса
Biacore T200 (GE Healthcare) использовали для оценки эффекта различных низкомолекулярных соединений на реакцию антиген-антитело IL-6R с 9 зависимыми от ATP/аденозина антителами (6RAD2C1-4_001, 6RAD2C1-4_005, 6RAD2C1-4_011, 6RAD2C1-4_026, 6RAD2C1-4_030, 6RAD2C1-4_042, 6RAD2C1-4_076, 6RDL3C1-4_085, и 6RDL3C5-4_011), полученными из библиотеки. Использовали подвижный буфер: 20 ммоль/л ACES, 150 ммоль/л NaCl, 0,05% (масс./об.) Tween20, pH 7,4. Анализ проводили при 25°C. IL-6R иммобилизовывали на сенсорный чип CM5 способом присоединения аминов; и антителам позволяли взаимодействовать в качестве анализируемого соединения в течение 120 секунд, и наблюдали изменения величины связывания. Для разбавления антител использовли подвижный буфер или подвижный буфер, содержащий любой из ATP, ADP, AMP, cAMP и аденозина (ADO). Конечную концентрацию каждого низкомолекулярного соединения доводили до 1 мМ и 1 мкМ, соответственно. Между тем в условиях 1 мМ ATP анализ проводили с серией пошаговых концентраций антител. Константу диссоциации KD (моль/л) каждого колона для IL-6R вычисляли из графика равновесной величины против концентрации антитела. Параметры вычисляли с использованием оценочного программного обеспечения Biacore T200 Evaluation Software (GE Healthcare). Константа диссоциации KD каждого клона в присутствии 1 мМ ATP представлена в таблице 24.
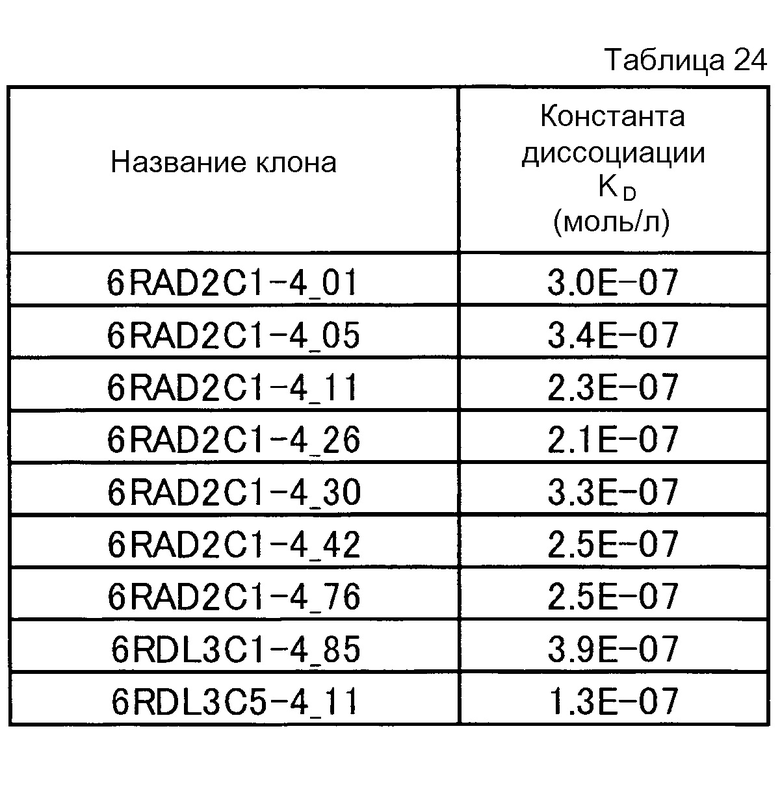
Количество каждого клона, который связывается с IL-6R, при определении с помощью этого анализа в присутствии или в отсутствие 1 мМ низкомолекулярных соединений, представлено на фиг. 43. Как показано на фиг.43, каждый клон связывается с IL-6R в присутствии 1 мМ ATP, однако их связывание не поддавалось обнаружению в отсутствие ATP. Это демонстрирует, что каждый клон обладает свойством, состоящим в том, что он связывается с IL-6R через ATP в качестве переключающего фактора. В низкомолекулярных соединениях, помимо ATP, связывание всех клонов наблюдалось в присутствии ADP, и было показано, что некоторые клоны связываются в присутствии AMP и cAMP. Связывание IL-6R не поддавалось обнаружению в присутствии ADO.
Это демонстрирует, что антитела, которые связываются с антигенами-мишенями в присутствии любого одного или нескольких из ATP, ADP, AMP и cAMP, можно получать с использованием рационально сконструированных библиотек. Как описано в этом примере, пэннинг проводили в присутствии как ATP, так и ADO, которые связываются с антителом ATNLSA1-4_D12, использованным в качестве эталона при конструировании библиотек. Результат показал, что антитела, которые прочно связываются с антигенами-мишенями, были выделены в присутствии ATP, который прочно связывается с ATNLSA1-4_D12, но не в присутствии ADO, связывается с ATNLSA1-4_D12 слабее, чем ATP. Антитела, которые связываются с антигенами только в зависимости от желаемого низкомолекулярного соединения, можно получать путем выделения антигенсвязывающих антител посредством контактирования антигенов с библиотеками в присутствии только низкомолекулярного соединения. Например, антитела, которые связываются в присутствии ADO, можно эффективно получать из библиотек путем проведения пэннинга в присутствии ADO отдельно.
(18-3) Эффект ATP на активность ADCC полученных антител
Полученные антитела 6RAD2C1-4_030 и 6RAD2C1-4_011 анализировали способом, описанным ниже, для исследования того, опосредуется ли активность ADCC против клеток, экспрессирующих hIL-6R, связыванием антител с hIL-6R в присутствии аденозинтрифосфата (ATP). Эту оценку проводили с использованием варианта 6RAD2C1-4_030 с увеличенной эффекторной активностью (вариабельная область тяжелой цепи антитела, SEQ ID NO:119; вариабельная область легкой цепи антитела, SEQ ID NO:120; константная область тяжелой цепи антитела, SEQ ID NO:90, константная область легкой цепи антитела, SEQ ID NO:91) и варианта 6RAD2C1-4_011 с увеличенной эффекторной активностью (вариабельная область тяжелой цепи антитела, SEQ ID NO:82; вариабельная область легкой цепи антитела, SEQ ID NO:83; константная область тяжелой цепи антитела, SEQ ID NO:90; константная область легкой цепи антитела, SEQ ID NO:91), полученного, как описано в примере 18-1, а также известного антител против IL-6R человека MRA (вариабельная область тяжелой цепи антитела, SEQ ID NO:92; вариабельная область легкой цепи антитела, SEQ ID NO:92; константная область тяжелой цепи антитела, SEQ ID NO:90; константная область легкой цепи антитела, SEQ ID NO:91), полученного, как описано в примере 15-5. При различных концентрациях антитела в присутствии или в отсутствие ATP, 6RAD2C1-4_030, 6RAD2C1-4_011 и MRA оценивали в отношении активности ADCC против экспрессирующих hIL-6R клеток в соответствии со способом, описанным в справочном примере 3. Результат анализа представлен на фиг. 44.
Результат анализа подтвердил зависимую от концентрации антитела активность ADCC у 6RAD2C1-4_030 и 6RAD2C1-4_011 в присутствии ATP. Эти данные демонстрируют, что активность ADCC индуцируется против экспрессирующих антиген клеток посредством связывания антиген-антитело, опосредуемого не только кинуренином, но также ATP; и, таким образом, выявлено, что для антител, которые связываются антигенами в присутствии низкомолекулярных соединений в качестве переключающего фактора, их функция, состоящая в противоопухолевой активности, такой как активность ADCC, также может регулироваться присутствием низкомолекулярных соединений, служащих в качестве переключающего фактора.
Эти результаты указывают на то, что выраженная индукция активности ADCC против экспрессирующих hIL-6R клеток ожидается в опухолевых тканях, где концентрация ATP является высокой, и слабая индукция активности ADCC ожидается в нормальных тканях, где концентрация ATP является низкой. Исходя из указанного выше, путем введения антител, для которых ATP служит в качестве переключающего фактора, цитотоксичность против нормальных тканей, экспрессирующих антиген-мишень, может быть снижена без снижения фармцевтического эффекта на опухолевые ткани, экспрессирующие антиген-мишень.
[Справочный пример 1] Активность ADCC у исследуемых антител с использованием мононуклеарных клеток периферической крови человека в качестве эффекторных клеток
Антитела, которые связывают антигены зависимым от кинуренина образом, оценивали в отношении их активности ADCC против экспрессирующих антиген клеток при различных концентрациях антитела в соответствии со способом, описанным ниже. Мононуклеарные клеткимононуклеарные клетки периферической крови человека (далее обозначаемые как PBMC человека) использовали в качестве эффекторных клеток для измерения активности ADCC каждого исследуемого антитела следующим образом.
(1) Получение раствора PBMC человека
Шприцы, предварительно заполненные 200 мкл раствора гепарина в концентрации 1000 единиц/мл (Novo-Heparin, 5000 единиц на инъекцию; Novo Nordisk) использовали для сбора 50 мл периферической крови от здоровых добровольцев (взрослые мужчины), являющихся сотрудниками Chugai Pharmaceutical Co. Ltd. Периферическую кровь разбавляли в два раза посредством PBS(-) и разделяли на четыре равных части, каждую из которых переносили в предварительно центрифугированную пробирку для разделения лейкоцитов Leucosep (Greiner Bio-One), содержавшую 15 мл Ficoll-Paque PLUS. Пробирки для разделения, содержавшие аликвоту периферической крови, центрифугировали при 2150 об/мин в течение десяти минут при комнатной температуре. Затем из пробирок собирали полученные фракции мононуклеарных клеток. Клетки в каждой фракции промывали один раз RPMI-1640 (nacalai tesque), дополненной 10% FBS (далее обозначаемой как 10%FBS/RPMI), а затем суспендировали при плотности клеток 1×107 клеток/мл в 10% FBS/RPMI. Суспензии клеток использовали в качестве раствора PBMC человека в последующих экспериментах.
(2) Получение клеток-мишеней
0,74 МБк Cr-51 добавляли к 3×106 клеток BaF/hIL6R (Mihara et al., (Int. Immunopharmacol. (2005) 5, 1731-40), которые представляют собой клетки Ba/F3, экспрессирущие рецептор IL-6 человека. Затем клетки инкубировали в инкубаторе с 5% CO2 при 37°C в течение 1 часа. После промывания 3 раза 10% FBS/RPMI клетки суспендировали при плотности клеток 2×105 клеток/мл в 10% FBS/RPMI. Суспензию клеток использовали в качестве клеток-мишеней в последующих экспериментах.
(3) Получение раствора кинуренина
L-Кинуренин (sigma) разбавляли до 5 мМ посредством PBS(-), а затем его концентрацию доводили до 400 мкМ с использованием 10% FBS/RPMI. Раствор миспользовали в качестве раствора кинуренина в последующих экспериментах.
(4) Анализ высвобождения хрома (ADCC)
Активность ADCC оценивали, исходя из удельной скорости высвобождения хрома, определенной с помощью анализа высвобождения хрома. Сначала растворы антител, полученные в различных концентрациях (0, 004, 0,4 и 40 мкг/мл) добавляли в количестве 50 мкл в каждую лунку 96-луночного планшета с круглым дном. Затем клетки-мишени, полученные, как описано в (2), высевали в лунки в количестве 50 мкл (1×104 клеток/лунка). Более того, раствор кинуренина, полученный, как описано в (3) добавляли в лунки в количестве 50 мкл, и планшету позволяли стоять при комнатной температуре в течение 15 минут. Затем в каждую лунку добавляли раствор с PBMC человека, полученный, как описано в (1), в количестве 50 мкл (5×105 клеток/лунка). Планшету позволяли стоять в инкубаторе с 5% CO2 при 37°C в течение 4 часов, с последующим центрифугированием. 100 мкл культурального супернатанта из каждой лунки измеряли в отношении радиоактивности с использованием гамма-счетчика. Удельную скорость высвобождения хрома определяли, исходя из представленного ниже уравнения.
Скорость высвобождения хрома (%)= A-C)×100/(B-C)
В этом уравнении "A" представляет собой среднюю радиоактивность (cpm) 100 мкл культурального супернатанта в каждой лунке. "B" представляет собой среднюю радиоактивность (cpm) 100 мкл культурального супернатанта в лунке, содержащей клетки-мишени, 50 мкл 4% водного раствора NP-40 (Nonidet P-40; Nacalai Tesques) и 100 мкл 10% FBS/RPMI. Более того, "C" представляет собой среднюю радиоактивность (cpm) 100 мкл культурального супернатанта в лунке, содержащей клетки-мишени, 150 мкл 10% FBS/RPMI или 100 мкл 10% FBS/RPMI, и 50 мкл раствора кинуренина. Исследование проводили в двух экземплярах. Среднюю удельную скорость высвобождения хрома (%), которая отражает активность ADCC каждого исследуемого антитела, вычисляли на основе анализа, описанного выше.
[Справочный пример 2] Активность ADCC каждого исследуемого антитела с использованием мононуклеарных клеток периферической крови человека в качестве эффекторных клеток
Антитела, которые связываются с антигенами зависимым от кинуренина образом, оценивали в отношении их активности ADCC против экспрессирующих антиген клеток при различных концентрациях кинуренина в соответствии со способом, описанным ниже. С использованием мононуклеарных клеток периферической крови человека в качестве эффекторных клеток, активность ADCC каждого исследуемого антитела анализировали следующим образом. Раствор PBMC человека и клетки-мишени получали тем же способом, который описан в справочном примере 1.
(1) Получение растворов кинуренина
L-кинуренин (sigma) разбавляли до 5 мМ посредством PBS(-), а затем его концентрации доводили до 400, 133, 44, 14,8 и 4,9 мкМ с использованием 10% FBS/RPMI. Растворы использовали в качестве растворов кинуренина в последующих экспериментах.
(2) Анализ высвобождения хрома (ADCC)
Активность ADCC оценивали на основе удельной скорости высвобождения хрома, определенной в анализе высвобождения хрома. Сначала раствор антител, приготовленный в концентрации 200 мкг/мл, добавляли в количестве 50 мкл в каждую лунку 96-луночного планшета с круглым дном. Затем клетки-мишени, полученные, как описано выше, высевали в лунки в количестве 50 мкл (1×104 клеток/лунка). Более того, раствор кинуренина, полученный в каждой концентрации, как описано в (1), добавляли в лунки в количестве 50 мкл, и планшету позволяли стоять при комнатной температуре в течение 15 минут. Затем в каждую лунку добавляли раствор с PBMC человека, полученный, как описано выше, в количестве 50 мкл (5×105 клеток/лунка). Планшету позволяли стоять в инкубаторе с 5% CO2 при 37°C в течение 4 часов, с последующим центрифугированием. 100 мкл культурального супернатанта из каждой лунки планшета измеряли в отношении радиоактивности с использованием гамма счетчика. Удельную скорость высвобождения хрома определяли, исходя из уравнения, описанного в справочном примере 1.
[Справочный пример 3] Активность ADCC для исследования антител с использованием линии NK-клеток человека NK92 в качестве эффекторных клеток
Антитела, которые связываются с антигенами зависимым от ATP образом, оценивали в отношении их активности ADCC против экспрессирующих антиген клеток в различных концентрациях антитела в соответствии со способом, описанным ниже. Активность ADCC каждого исследуемого антитела анализировали с использованием в качестве эффекторных клеток NK92-CD16(V), полученных посредством искусственной экспрессии FcgRIIIa человека в линии NK-клеток человека NK92 следующим образом.
(1) Получение NK92-CD16(V)
NK92-CD16(V) суспендировали при плотности клеток 1×107 клеток/мл в RPMI/10% FBS. Суспензию клеток использовали в качестве раствора NK92-CD16(V) в последующих экспериментах.
(2) Получение клеток-мишеней
0,74 МБк Cr-51 добавляли к 3×106 клеток CHO/hIL6R, которые представляют собой клетки CHO, экспрессирующие рецептор IL-6 человека. Затем клетки инкубировали в инкубаторе с 5% CO2 при 37°C в течение 1 часа. После промывания 3 раза 10% FBS/RPMI, клетки суспендировали при плотности клеток 2×105 клеток/мл в 10% FBS/RPMI. Суспензию клеток использовали в качестве клеток-мишеней в последующих экспериментах.
(3) Получение раствора ATP
ATP (sigma) разбавляли до 100 мМ посредством 10% FBS/RPMI, а затем его концентрацию доводили до 4 мМ. Раствор использовали в качестве раствора ATP в последующих экспериментах.
(4) Анализ высвобождения хрома (ADCC)
Активность ADCC оценивали на основе удельной скорости высвобождения хрома, определенной в анализе высвобождения хрома. Сначала раствор антител, приготовленный в различных концентрациях (0,04, 0,4, 4 или 40 мкг/мл), добавляли в количестве 50 мкл в каждую лунку 96-луночного планшета с круглым дном. Затем клетки-мишени, полученные, как описано в (2), высевали в лунки в количестве 50 мкл (1×104 клеток/лунка). Более того, раствор ATP, полученный в каждой концентрации, как описано в (3), добавляли в лунки в количестве 50 мкл, и планшету позволяли стоять при комнатной температуре в течение 15 минут. Затем в каждую лунку добавляли раствор с NK92-CD16(V) человека, полученный, как описано в (1), в количестве 50 мкл (5×105 клеток/лунка). Планшету позволяли стоять в инкубаторе с 5% CO2 при 37°C в течение 4 часов, с последующим центрифугированием. 100 мкл культурального супернатанта из каждой лунки планшета измеряли в отношении радиоактивности с использованием гамма счетчика. Удельную скорость высвобождения хрома определяли, исходя из уравнения, описанного в справочном примере 1.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CHUGAI SEIYAKU KABUSHIKI KAISHA
<120> СПЕЦИФИЧНАЯ К ТКАНИ-МИШЕНИ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА
<130> C1-A1204Y1P
<140> PCT/JP2013/064975
<141> 2013-05-30
<150> JP 2012-123781
<151> 2012-05-30
<150> JP 2012-177311
<151> 2012-08-09
<160> 126
<170> PatentIn version 3.5
<210> 1
<211> 468
<212> PRT
<213> Homo sapiens
<400> 1
Met Leu Ala Val Gly Cys Ala Leu Leu Ala Ala Leu Leu Ala Ala Pro
1 5 10 15
Gly Ala Ala Leu Ala Pro Arg Arg Cys Pro Ala Gln Glu Val Ala Arg
20 25 30
Gly Val Leu Thr Ser Leu Pro Gly Asp Ser Val Thr Leu Thr Cys Pro
35 40 45
Gly Val Glu Pro Glu Asp Asn Ala Thr Val His Trp Val Leu Arg Lys
50 55 60
Pro Ala Ala Gly Ser His Pro Ser Arg Trp Ala Gly Met Gly Arg Arg
65 70 75 80
Leu Leu Leu Arg Ser Val Gln Leu His Asp Ser Gly Asn Tyr Ser Cys
85 90 95
Tyr Arg Ala Gly Arg Pro Ala Gly Thr Val His Leu Leu Val Asp Val
100 105 110
Pro Pro Glu Glu Pro Gln Leu Ser Cys Phe Arg Lys Ser Pro Leu Ser
115 120 125
Asn Val Val Cys Glu Trp Gly Pro Arg Ser Thr Pro Ser Leu Thr Thr
130 135 140
Lys Ala Val Leu Leu Val Arg Lys Phe Gln Asn Ser Pro Ala Glu Asp
145 150 155 160
Phe Gln Glu Pro Cys Gln Tyr Ser Gln Glu Ser Gln Lys Phe Ser Cys
165 170 175
Gln Leu Ala Val Pro Glu Gly Asp Ser Ser Phe Tyr Ile Val Ser Met
180 185 190
Cys Val Ala Ser Ser Val Gly Ser Lys Phe Ser Lys Thr Gln Thr Phe
195 200 205
Gln Gly Cys Gly Ile Leu Gln Pro Asp Pro Pro Ala Asn Ile Thr Val
210 215 220
Thr Ala Val Ala Arg Asn Pro Arg Trp Leu Ser Val Thr Trp Gln Asp
225 230 235 240
Pro His Ser Trp Asn Ser Ser Phe Tyr Arg Leu Arg Phe Glu Leu Arg
245 250 255
Tyr Arg Ala Glu Arg Ser Lys Thr Phe Thr Thr Trp Met Val Lys Asp
260 265 270
Leu Gln His His Cys Val Ile His Asp Ala Trp Ser Gly Leu Arg His
275 280 285
Val Val Gln Leu Arg Ala Gln Glu Glu Phe Gly Gln Gly Glu Trp Ser
290 295 300
Glu Trp Ser Pro Glu Ala Met Gly Thr Pro Trp Thr Glu Ser Arg Ser
305 310 315 320
Pro Pro Ala Glu Asn Glu Val Ser Thr Pro Met Gln Ala Leu Thr Thr
325 330 335
Asn Lys Asp Asp Asp Asn Ile Leu Phe Arg Asp Ser Ala Asn Ala Thr
340 345 350
Ser Leu Pro Val Gln Asp Ser Ser Ser Val Pro Leu Pro Thr Phe Leu
355 360 365
Val Ala Gly Gly Ser Leu Ala Phe Gly Thr Leu Leu Cys Ile Ala Ile
370 375 380
Val Leu Arg Phe Lys Lys Thr Trp Lys Leu Arg Ala Leu Lys Glu Gly
385 390 395 400
Lys Thr Ser Met His Pro Pro Tyr Ser Leu Gly Gln Leu Val Pro Glu
405 410 415
Arg Pro Arg Pro Thr Pro Val Leu Val Pro Leu Ile Ser Pro Pro Val
420 425 430
Ser Pro Ser Ser Leu Gly Ser Asp Asn Thr Ser Ser His Asn Arg Pro
435 440 445
Asp Ala Arg Asp Pro Arg Ser Pro Tyr Asp Ile Ser Asn Thr Asp Tyr
450 455 460
Phe Phe Pro Arg
465
<210> 2
<211> 1407
<212> ДНК
<213> Homo sapiens
<400> 2
atgctggccg tcggctgcgc gctgctggct gccctgctgg ccgcgccggg agcggcgctg 60
gccccaaggc gctgccctgc gcaggaggtg gcgagaggcg tgctgaccag tctgccagga 120
gacagcgtga ctctgacctg cccgggggta gagccggaag acaatgccac tgttcactgg 180
gtgctcagga agccggctgc aggctcccac cccagcagat gggctggcat gggaaggagg 240
ctgctgctga ggtcggtgca gctccacgac tctggaaact attcatgcta ccgggccggc 300
cgcccagctg ggactgtgca cttgctggtg gatgttcccc ccgaggagcc ccagctctcc 360
tgcttccgga agagccccct cagcaatgtt gtttgtgagt ggggtcctcg gagcacccca 420
tccctgacga caaaggctgt gctcttggtg aggaagtttc agaacagtcc ggccgaagac 480
ttccaggagc cgtgccagta ttcccaggag tcccagaagt tctcctgcca gttagcagtc 540
ccggagggag acagctcttt ctacatagtg tccatgtgcg tcgccagtag tgtcgggagc 600
aagttcagca aaactcaaac ctttcagggt tgtggaatct tgcagcctga tccgcctgcc 660
aacatcacag tcactgccgt ggccagaaac ccccgctggc tcagtgtcac ctggcaagac 720
ccccactcct ggaactcatc tttctacaga ctacggtttg agctcagata tcgggctgaa 780
cggtcaaaga cattcacaac atggatggtc aaggacctcc agcatcactg tgtcatccac 840
gacgcctgga gcggcctgag gcacgtggtg cagcttcgtg cccaggagga gttcgggcaa 900
ggcgagtgga gcgagtggag cccggaggcc atgggcacgc cttggacaga atccaggagt 960
cctccagctg agaacgaggt gtccaccccc atgcaggcac ttactactaa taaagacgat 1020
gataatattc tcttcagaga ttctgcaaat gcgacaagcc tcccagtgca agattcttct 1080
tcagtaccac tgcccacatt cctggttgct ggagggagcc tggccttcgg aacgctcctc 1140
tgcattgcca ttgttctgag gttcaagaag acgtggaagc tgcgggctct gaaggaaggc 1200
aagacaagca tgcatccgcc gtactctttg gggcagctgg tcccggagag gcctcgaccc 1260
accccagtgc ttgttcctct catctcccca ccggtgtccc ccagcagcct ggggtctgac 1320
aatacctcga gccacaaccg accagatgcc agggacccac ggagccctta tgacatcagc 1380
aatacagact acttcttccc cagatag 1407
<210> 3
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 3
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser
<210> 4
<211> 21
<212> PRT
<213> Clostridium sp.
<400> 4
Phe Asn Asn Phe Thr Val Ser Phe Trp Leu Arg Val Pro Lys Val Ser
1 5 10 15
Ala Ser His Leu Glu
20
<210> 5
<211> 330
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 5
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 6
<211> 326
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 6
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 7
<211> 377
<212> PRT
<213> Homo sapiens
<400> 7
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr Cys Pro
100 105 110
Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg
115 120 125
Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys
130 135 140
Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
145 150 155 160
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
165 170 175
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
180 185 190
Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys Trp Tyr
195 200 205
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
210 215 220
Gln Tyr Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu His
225 230 235 240
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
245 250 255
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln
260 265 270
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
275 280 285
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
290 295 300
Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu Asn Asn
305 310 315 320
Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu
325 330 335
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Ile
340 345 350
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Phe Thr Gln
355 360 365
Lys Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 8
<211> 327
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 8
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 9
<211> 1125
<212> ДНК
<213> Homo sapiens
<220>
<221> CDS
<222> (1)..(1125)
<400> 9
atg tgg ttc ttg aca act ctg ctc ctt tgg gtt cca gtt gat ggg caa 48
Met Trp Phe Leu Thr Thr Leu Leu Leu Trp Val Pro Val Asp Gly Gln
1 5 10 15
gtg gac acc aca aag gca gtg atc act ttg cag cct cca tgg gtc agc 96
Val Asp Thr Thr Lys Ala Val Ile Thr Leu Gln Pro Pro Trp Val Ser
20 25 30
gtg ttc caa gag gaa acc gta acc ttg cac tgt gag gtg ctc cat ctg 144
Val Phe Gln Glu Glu Thr Val Thr Leu His Cys Glu Val Leu His Leu
35 40 45
cct ggg agc agc tct aca cag tgg ttt ctc aat ggc aca gcc act cag 192
Pro Gly Ser Ser Ser Thr Gln Trp Phe Leu Asn Gly Thr Ala Thr Gln
50 55 60
acc tcg acc ccc agc tac aga atc acc tct gcc agt gtc aat gac agt 240
Thr Ser Thr Pro Ser Tyr Arg Ile Thr Ser Ala Ser Val Asn Asp Ser
65 70 75 80
ggt gaa tac agg tgc cag aga ggt ctc tca ggg cga agt gac ccc ata 288
Gly Glu Tyr Arg Cys Gln Arg Gly Leu Ser Gly Arg Ser Asp Pro Ile
85 90 95
cag ctg gaa atc cac aga ggc tgg cta cta ctg cag gtc tcc agc aga 336
Gln Leu Glu Ile His Arg Gly Trp Leu Leu Leu Gln Val Ser Ser Arg
100 105 110
gtc ttc acg gaa gga gaa cct ctg gcc ttg agg tgt cat gcg tgg aag 384
Val Phe Thr Glu Gly Glu Pro Leu Ala Leu Arg Cys His Ala Trp Lys
115 120 125
gat aag ctg gtg tac aat gtg ctt tac tat cga aat ggc aaa gcc ttt 432
Asp Lys Leu Val Tyr Asn Val Leu Tyr Tyr Arg Asn Gly Lys Ala Phe
130 135 140
aag ttt ttc cac tgg aat tct aac ctc acc att ctg aaa acc aac ata 480
Lys Phe Phe His Trp Asn Ser Asn Leu Thr Ile Leu Lys Thr Asn Ile
145 150 155 160
agt cac aat ggc acc tac cat tgc tca ggc atg gga aag cat cgc tac 528
Ser His Asn Gly Thr Tyr His Cys Ser Gly Met Gly Lys His Arg Tyr
165 170 175
aca tca gca gga ata tct gtc act gtg aaa gag cta ttt cca gct cca 576
Thr Ser Ala Gly Ile Ser Val Thr Val Lys Glu Leu Phe Pro Ala Pro
180 185 190
gtg ctg aat gca tct gtg aca tcc cca ctc ctg gag ggg aat ctg gtc 624
Val Leu Asn Ala Ser Val Thr Ser Pro Leu Leu Glu Gly Asn Leu Val
195 200 205
acc ctg agc tgt gaa aca aag ttg ctc ttg cag agg cct ggt ttg cag 672
Thr Leu Ser Cys Glu Thr Lys Leu Leu Leu Gln Arg Pro Gly Leu Gln
210 215 220
ctt tac ttc tcc ttc tac atg ggc agc aag acc ctg cga ggc agg aac 720
Leu Tyr Phe Ser Phe Tyr Met Gly Ser Lys Thr Leu Arg Gly Arg Asn
225 230 235 240
aca tcc tct gaa tac caa ata cta act gct aga aga gaa gac tct ggg 768
Thr Ser Ser Glu Tyr Gln Ile Leu Thr Ala Arg Arg Glu Asp Ser Gly
245 250 255
tta tac tgg tgc gag gct gcc aca gag gat gga aat gtc ctt aag cgc 816
Leu Tyr Trp Cys Glu Ala Ala Thr Glu Asp Gly Asn Val Leu Lys Arg
260 265 270
agc cct gag ttg gag ctt caa gtg ctt ggc ctc cag tta cca act cct 864
Ser Pro Glu Leu Glu Leu Gln Val Leu Gly Leu Gln Leu Pro Thr Pro
275 280 285
gtc tgg ttt cat gtc ctt ttc tat ctg gca gtg gga ata atg ttt tta 912
Val Trp Phe His Val Leu Phe Tyr Leu Ala Val Gly Ile Met Phe Leu
290 295 300
gtg aac act gtt ctc tgg gtg aca ata cgt aaa gaa ctg aaa aga aag 960
Val Asn Thr Val Leu Trp Val Thr Ile Arg Lys Glu Leu Lys Arg Lys
305 310 315 320
aaa aag tgg gat tta gaa atc tct ttg gat tct ggt cat gag aag aag 1008
Lys Lys Trp Asp Leu Glu Ile Ser Leu Asp Ser Gly His Glu Lys Lys
325 330 335
gta att tcc agc ctt caa gaa gac aga cat tta gaa gaa gag ctg aaa 1056
Val Ile Ser Ser Leu Gln Glu Asp Arg His Leu Glu Glu Glu Leu Lys
340 345 350
tgt cag gaa caa aaa gaa gaa cag ctg cag gaa ggg gtg cac cgg aag 1104
Cys Gln Glu Gln Lys Glu Glu Gln Leu Gln Glu Gly Val His Arg Lys
355 360 365
gag ccc cag ggg gcc acg tag 1125
Glu Pro Gln Gly Ala Thr
370
<210> 10
<211> 374
<212> PRT
<213> Homo sapiens
<400> 10
Met Trp Phe Leu Thr Thr Leu Leu Leu Trp Val Pro Val Asp Gly Gln
1 5 10 15
Val Asp Thr Thr Lys Ala Val Ile Thr Leu Gln Pro Pro Trp Val Ser
20 25 30
Val Phe Gln Glu Glu Thr Val Thr Leu His Cys Glu Val Leu His Leu
35 40 45
Pro Gly Ser Ser Ser Thr Gln Trp Phe Leu Asn Gly Thr Ala Thr Gln
50 55 60
Thr Ser Thr Pro Ser Tyr Arg Ile Thr Ser Ala Ser Val Asn Asp Ser
65 70 75 80
Gly Glu Tyr Arg Cys Gln Arg Gly Leu Ser Gly Arg Ser Asp Pro Ile
85 90 95
Gln Leu Glu Ile His Arg Gly Trp Leu Leu Leu Gln Val Ser Ser Arg
100 105 110
Val Phe Thr Glu Gly Glu Pro Leu Ala Leu Arg Cys His Ala Trp Lys
115 120 125
Asp Lys Leu Val Tyr Asn Val Leu Tyr Tyr Arg Asn Gly Lys Ala Phe
130 135 140
Lys Phe Phe His Trp Asn Ser Asn Leu Thr Ile Leu Lys Thr Asn Ile
145 150 155 160
Ser His Asn Gly Thr Tyr His Cys Ser Gly Met Gly Lys His Arg Tyr
165 170 175
Thr Ser Ala Gly Ile Ser Val Thr Val Lys Glu Leu Phe Pro Ala Pro
180 185 190
Val Leu Asn Ala Ser Val Thr Ser Pro Leu Leu Glu Gly Asn Leu Val
195 200 205
Thr Leu Ser Cys Glu Thr Lys Leu Leu Leu Gln Arg Pro Gly Leu Gln
210 215 220
Leu Tyr Phe Ser Phe Tyr Met Gly Ser Lys Thr Leu Arg Gly Arg Asn
225 230 235 240
Thr Ser Ser Glu Tyr Gln Ile Leu Thr Ala Arg Arg Glu Asp Ser Gly
245 250 255
Leu Tyr Trp Cys Glu Ala Ala Thr Glu Asp Gly Asn Val Leu Lys Arg
260 265 270
Ser Pro Glu Leu Glu Leu Gln Val Leu Gly Leu Gln Leu Pro Thr Pro
275 280 285
Val Trp Phe His Val Leu Phe Tyr Leu Ala Val Gly Ile Met Phe Leu
290 295 300
Val Asn Thr Val Leu Trp Val Thr Ile Arg Lys Glu Leu Lys Arg Lys
305 310 315 320
Lys Lys Trp Asp Leu Glu Ile Ser Leu Asp Ser Gly His Glu Lys Lys
325 330 335
Val Ile Ser Ser Leu Gln Glu Asp Arg His Leu Glu Glu Glu Leu Lys
340 345 350
Cys Gln Glu Gln Lys Glu Glu Gln Leu Gln Glu Gly Val His Arg Lys
355 360 365
Glu Pro Gln Gly Ala Thr
370
<210> 11
<211> 951
<212> ДНК
<213> Homo sapiens
<220>
<221> CDS
<222> (1)..(951)
<400> 11
atg act atg gag acc caa atg tct cag aat gta tgt ccc aga aac ctg 48
Met Thr Met Glu Thr Gln Met Ser Gln Asn Val Cys Pro Arg Asn Leu
1 5 10 15
tgg ctg ctt caa cca ttg aca gtt ttg ctg ctg ctg gct tct gca gac 96
Trp Leu Leu Gln Pro Leu Thr Val Leu Leu Leu Leu Ala Ser Ala Asp
20 25 30
agt caa gct gct ccc cca aag gct gtg ctg aaa ctt gag ccc ccg tgg 144
Ser Gln Ala Ala Pro Pro Lys Ala Val Leu Lys Leu Glu Pro Pro Trp
35 40 45
atc aac gtg ctc cag gag gac tct gtg act ctg aca tgc cag ggg gct 192
Ile Asn Val Leu Gln Glu Asp Ser Val Thr Leu Thr Cys Gln Gly Ala
50 55 60
cgc agc cct gag agc gac tcc att cag tgg ttc cac aat ggg aat ctc 240
Arg Ser Pro Glu Ser Asp Ser Ile Gln Trp Phe His Asn Gly Asn Leu
65 70 75 80
att ccc acc cac acg cag ccc agc tac agg ttc aag gcc aac aac aat 288
Ile Pro Thr His Thr Gln Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn
85 90 95
gac agc ggg gag tac acg tgc cag act ggc cag acc agc ctc agc gac 336
Asp Ser Gly Glu Tyr Thr Cys Gln Thr Gly Gln Thr Ser Leu Ser Asp
100 105 110
cct gtg cat ctg act gtg ctt tcc gaa tgg ctg gtg ctc cag acc cct 384
Pro Val His Leu Thr Val Leu Ser Glu Trp Leu Val Leu Gln Thr Pro
115 120 125
cac ctg gag ttc cag gag gga gaa acc atc atg ctg agg tgc cac agc 432
His Leu Glu Phe Gln Glu Gly Glu Thr Ile Met Leu Arg Cys His Ser
130 135 140
tgg aag gac aag cct ctg gtc aag gtc aca ttc ttc cag aat gga aaa 480
Trp Lys Asp Lys Pro Leu Val Lys Val Thr Phe Phe Gln Asn Gly Lys
145 150 155 160
tcc cag aaa ttc tcc cat ttg gat ccc acc ttc tcc atc cca caa gca 528
Ser Gln Lys Phe Ser His Leu Asp Pro Thr Phe Ser Ile Pro Gln Ala
165 170 175
aac cac agt cac agt ggt gat tac cac tgc aca gga aac ata ggc tac 576
Asn His Ser His Ser Gly Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr
180 185 190
acg ctg ttc tca tcc aag cct gtg acc atc act gtc caa gtg ccc agc 624
Thr Leu Phe Ser Ser Lys Pro Val Thr Ile Thr Val Gln Val Pro Ser
195 200 205
atg ggc agc tct tca cca atg ggg gtc att gtg gct gtg gtc att gcg 672
Met Gly Ser Ser Ser Pro Met Gly Val Ile Val Ala Val Val Ile Ala
210 215 220
act gct gta gca gcc att gtt gct gct gta gtg gcc ttg atc tac tgc 720
Thr Ala Val Ala Ala Ile Val Ala Ala Val Val Ala Leu Ile Tyr Cys
225 230 235 240
agg aaa aag cgg att tca gcc aat tcc act gat cct gtg aag gct gcc 768
Arg Lys Lys Arg Ile Ser Ala Asn Ser Thr Asp Pro Val Lys Ala Ala
245 250 255
caa ttt gag cca cct gga cgt caa atg att gcc atc aga aag aga caa 816
Gln Phe Glu Pro Pro Gly Arg Gln Met Ile Ala Ile Arg Lys Arg Gln
260 265 270
ctt gaa gaa acc aac aat gac tat gaa aca gct gac ggc ggc tac atg 864
Leu Glu Glu Thr Asn Asn Asp Tyr Glu Thr Ala Asp Gly Gly Tyr Met
275 280 285
act ctg aac ccc agg gca cct act gac gat gat aaa aac atc tac ctg 912
Thr Leu Asn Pro Arg Ala Pro Thr Asp Asp Asp Lys Asn Ile Tyr Leu
290 295 300
act ctt cct ccc aac gac cat gtc aac agt aat aac taa 951
Thr Leu Pro Pro Asn Asp His Val Asn Ser Asn Asn
305 310 315
<210> 12
<211> 316
<212> PRT
<213> Homo sapiens
<400> 12
Met Thr Met Glu Thr Gln Met Ser Gln Asn Val Cys Pro Arg Asn Leu
1 5 10 15
Trp Leu Leu Gln Pro Leu Thr Val Leu Leu Leu Leu Ala Ser Ala Asp
20 25 30
Ser Gln Ala Ala Pro Pro Lys Ala Val Leu Lys Leu Glu Pro Pro Trp
35 40 45
Ile Asn Val Leu Gln Glu Asp Ser Val Thr Leu Thr Cys Gln Gly Ala
50 55 60
Arg Ser Pro Glu Ser Asp Ser Ile Gln Trp Phe His Asn Gly Asn Leu
65 70 75 80
Ile Pro Thr His Thr Gln Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn
85 90 95
Asp Ser Gly Glu Tyr Thr Cys Gln Thr Gly Gln Thr Ser Leu Ser Asp
100 105 110
Pro Val His Leu Thr Val Leu Ser Glu Trp Leu Val Leu Gln Thr Pro
115 120 125
His Leu Glu Phe Gln Glu Gly Glu Thr Ile Met Leu Arg Cys His Ser
130 135 140
Trp Lys Asp Lys Pro Leu Val Lys Val Thr Phe Phe Gln Asn Gly Lys
145 150 155 160
Ser Gln Lys Phe Ser His Leu Asp Pro Thr Phe Ser Ile Pro Gln Ala
165 170 175
Asn His Ser His Ser Gly Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr
180 185 190
Thr Leu Phe Ser Ser Lys Pro Val Thr Ile Thr Val Gln Val Pro Ser
195 200 205
Met Gly Ser Ser Ser Pro Met Gly Val Ile Val Ala Val Val Ile Ala
210 215 220
Thr Ala Val Ala Ala Ile Val Ala Ala Val Val Ala Leu Ile Tyr Cys
225 230 235 240
Arg Lys Lys Arg Ile Ser Ala Asn Ser Thr Asp Pro Val Lys Ala Ala
245 250 255
Gln Phe Glu Pro Pro Gly Arg Gln Met Ile Ala Ile Arg Lys Arg Gln
260 265 270
Leu Glu Glu Thr Asn Asn Asp Tyr Glu Thr Ala Asp Gly Gly Tyr Met
275 280 285
Thr Leu Asn Pro Arg Ala Pro Thr Asp Asp Asp Lys Asn Ile Tyr Leu
290 295 300
Thr Leu Pro Pro Asn Asp His Val Asn Ser Asn Asn
305 310 315
<210> 13
<211> 876
<212> ДНК
<213> Homo sapiens
<220>
<221> CDS
<222> (1)..(876)
<400> 13
atg gga atc ctg tca ttc tta cct gtc ctt gcc act gag agt gac tgg 48
Met Gly Ile Leu Ser Phe Leu Pro Val Leu Ala Thr Glu Ser Asp Trp
1 5 10 15
gct gac tgc aag tcc ccc cag cct tgg ggt cat atg ctt ctg tgg aca 96
Ala Asp Cys Lys Ser Pro Gln Pro Trp Gly His Met Leu Leu Trp Thr
20 25 30
gct gtg cta ttc ctg gct cct gtt gct ggg aca cct gca gct ccc cca 144
Ala Val Leu Phe Leu Ala Pro Val Ala Gly Thr Pro Ala Ala Pro Pro
35 40 45
aag gct gtg ctg aaa ctc gag ccc cag tgg atc aac gtg ctc cag gag 192
Lys Ala Val Leu Lys Leu Glu Pro Gln Trp Ile Asn Val Leu Gln Glu
50 55 60
gac tct gtg act ctg aca tgc cgg ggg act cac agc cct gag agc gac 240
Asp Ser Val Thr Leu Thr Cys Arg Gly Thr His Ser Pro Glu Ser Asp
65 70 75 80
tcc att cag tgg ttc cac aat ggg aat ctc att ccc acc cac acg cag 288
Ser Ile Gln Trp Phe His Asn Gly Asn Leu Ile Pro Thr His Thr Gln
85 90 95
ccc agc tac agg ttc aag gcc aac aac aat gac agc ggg gag tac acg 336
Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn Asp Ser Gly Glu Tyr Thr
100 105 110
tgc cag act ggc cag acc agc ctc agc gac cct gtg cat ctg act gtg 384
Cys Gln Thr Gly Gln Thr Ser Leu Ser Asp Pro Val His Leu Thr Val
115 120 125
ctt tct gag tgg ctg gtg ctc cag acc cct cac ctg gag ttc cag gag 432
Leu Ser Glu Trp Leu Val Leu Gln Thr Pro His Leu Glu Phe Gln Glu
130 135 140
gga gaa acc atc gtg ctg agg tgc cac agc tgg aag gac aag cct ctg 480
Gly Glu Thr Ile Val Leu Arg Cys His Ser Trp Lys Asp Lys Pro Leu
145 150 155 160
gtc aag gtc aca ttc ttc cag aat gga aaa tcc aag aaa ttt tcc cgt 528
Val Lys Val Thr Phe Phe Gln Asn Gly Lys Ser Lys Lys Phe Ser Arg
165 170 175
tcg gat ccc aac ttc tcc atc cca caa gca aac cac agt cac agt ggt 576
Ser Asp Pro Asn Phe Ser Ile Pro Gln Ala Asn His Ser His Ser Gly
180 185 190
gat tac cac tgc aca gga aac ata ggc tac acg ctg tac tca tcc aag 624
Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr Thr Leu Tyr Ser Ser Lys
195 200 205
cct gtg acc atc act gtc caa gct ccc agc tct tca ccg atg ggg atc 672
Pro Val Thr Ile Thr Val Gln Ala Pro Ser Ser Ser Pro Met Gly Ile
210 215 220
att gtg gct gtg gtc act ggg att gct gta gcg gcc att gtt gct gct 720
Ile Val Ala Val Val Thr Gly Ile Ala Val Ala Ala Ile Val Ala Ala
225 230 235 240
gta gtg gcc ttg atc tac tgc agg aaa aag cgg att tca gcc aat ccc 768
Val Val Ala Leu Ile Tyr Cys Arg Lys Lys Arg Ile Ser Ala Asn Pro
245 250 255
act aat cct gat gag gct gac aaa gtt ggg gct gag aac aca atc acc 816
Thr Asn Pro Asp Glu Ala Asp Lys Val Gly Ala Glu Asn Thr Ile Thr
260 265 270
tat tca ctt ctc atg cac ccg gat gct ctg gaa gag cct gat gac cag 864
Tyr Ser Leu Leu Met His Pro Asp Ala Leu Glu Glu Pro Asp Asp Gln
275 280 285
aac cgt att tag 876
Asn Arg Ile
290
<210> 14
<211> 291
<212> PRT
<213> Homo sapiens
<400> 14
Met Gly Ile Leu Ser Phe Leu Pro Val Leu Ala Thr Glu Ser Asp Trp
1 5 10 15
Ala Asp Cys Lys Ser Pro Gln Pro Trp Gly His Met Leu Leu Trp Thr
20 25 30
Ala Val Leu Phe Leu Ala Pro Val Ala Gly Thr Pro Ala Ala Pro Pro
35 40 45
Lys Ala Val Leu Lys Leu Glu Pro Gln Trp Ile Asn Val Leu Gln Glu
50 55 60
Asp Ser Val Thr Leu Thr Cys Arg Gly Thr His Ser Pro Glu Ser Asp
65 70 75 80
Ser Ile Gln Trp Phe His Asn Gly Asn Leu Ile Pro Thr His Thr Gln
85 90 95
Pro Ser Tyr Arg Phe Lys Ala Asn Asn Asn Asp Ser Gly Glu Tyr Thr
100 105 110
Cys Gln Thr Gly Gln Thr Ser Leu Ser Asp Pro Val His Leu Thr Val
115 120 125
Leu Ser Glu Trp Leu Val Leu Gln Thr Pro His Leu Glu Phe Gln Glu
130 135 140
Gly Glu Thr Ile Val Leu Arg Cys His Ser Trp Lys Asp Lys Pro Leu
145 150 155 160
Val Lys Val Thr Phe Phe Gln Asn Gly Lys Ser Lys Lys Phe Ser Arg
165 170 175
Ser Asp Pro Asn Phe Ser Ile Pro Gln Ala Asn His Ser His Ser Gly
180 185 190
Asp Tyr His Cys Thr Gly Asn Ile Gly Tyr Thr Leu Tyr Ser Ser Lys
195 200 205
Pro Val Thr Ile Thr Val Gln Ala Pro Ser Ser Ser Pro Met Gly Ile
210 215 220
Ile Val Ala Val Val Thr Gly Ile Ala Val Ala Ala Ile Val Ala Ala
225 230 235 240
Val Val Ala Leu Ile Tyr Cys Arg Lys Lys Arg Ile Ser Ala Asn Pro
245 250 255
Thr Asn Pro Asp Glu Ala Asp Lys Val Gly Ala Glu Asn Thr Ile Thr
260 265 270
Tyr Ser Leu Leu Met His Pro Asp Ala Leu Glu Glu Pro Asp Asp Gln
275 280 285
Asn Arg Ile
290
<210> 15
<211> 765
<212> ДНК
<213> Homo sapiens
<220>
<221> CDS
<222> (1)..(765)
<400> 15
atg tgg cag ctg ctc ctc cca act gct ctg cta ctt cta gtt tca gct 48
Met Trp Gln Leu Leu Leu Pro Thr Ala Leu Leu Leu Leu Val Ser Ala
1 5 10 15
ggc atg cgg act gaa gat ctc cca aag gct gtg gtg ttc ctg gag cct 96
Gly Met Arg Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
20 25 30
caa tgg tac agg gtg ctc gag aag gac agt gtg act ctg aag tgc cag 144
Gln Trp Tyr Arg Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
35 40 45
gga gcc tac tcc cct gag gac aat tcc aca cag tgg ttt cac aat gag 192
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu
50 55 60
agc ctc atc tca agc cag gcc tcg agc tac ttc att gac gct gcc aca 240
Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr
65 70 75 80
gtt gac gac agt gga gag tac agg tgc cag aca aac ctc tcc acc ctc 288
Val Asp Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu
85 90 95
agt gac ccg gtg cag cta gaa gtc cat atc ggc tgg ctg ttg ctc cag 336
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
100 105 110
gcc cct cgg tgg gtg ttc aag gag gaa gac cct att cac ctg agg tgt 384
Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys
115 120 125
cac agc tgg aag aac act gct ctg cat aag gtc aca tat tta cag aat 432
His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn
130 135 140
ggc aaa ggc agg aag tat ttt cat cat aat tct gac ttc tac att cca 480
Gly Lys Gly Arg Lys Tyr Phe His His Asn Ser Asp Phe Tyr Ile Pro
145 150 155 160
aaa gcc aca ctc aaa gac agc ggc tcc tac ttc tgc agg ggg ctt gtt 528
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Val
165 170 175
ggg agt aaa aat gtg tct tca gag act gtg aac atc acc atc act caa 576
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
180 185 190
ggt ttg tca gtg tca acc atc tca tca ttc ttt cca cct ggg tac caa 624
Gly Leu Ser Val Ser Thr Ile Ser Ser Phe Phe Pro Pro Gly Tyr Gln
195 200 205
gtc tct ttc tgc ttg gtg atg gta ctc ctt ttt gca gtg gac aca gga 672
Val Ser Phe Cys Leu Val Met Val Leu Leu Phe Ala Val Asp Thr Gly
210 215 220
cta tat ttc tct gtg aag aca aac att cga agc tca aca aga gac tgg 720
Leu Tyr Phe Ser Val Lys Thr Asn Ile Arg Ser Ser Thr Arg Asp Trp
225 230 235 240
aag gac cat aaa ttt aaa tgg aga aag gac cct caa gac aaa tga 765
Lys Asp His Lys Phe Lys Trp Arg Lys Asp Pro Gln Asp Lys
245 250
<210> 16
<211> 254
<212> PRT
<213> Homo sapiens
<400> 16
Met Trp Gln Leu Leu Leu Pro Thr Ala Leu Leu Leu Leu Val Ser Ala
1 5 10 15
Gly Met Arg Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
20 25 30
Gln Trp Tyr Arg Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
35 40 45
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu
50 55 60
Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr
65 70 75 80
Val Asp Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu
85 90 95
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
100 105 110
Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys
115 120 125
His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn
130 135 140
Gly Lys Gly Arg Lys Tyr Phe His His Asn Ser Asp Phe Tyr Ile Pro
145 150 155 160
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Val
165 170 175
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
180 185 190
Gly Leu Ser Val Ser Thr Ile Ser Ser Phe Phe Pro Pro Gly Tyr Gln
195 200 205
Val Ser Phe Cys Leu Val Met Val Leu Leu Phe Ala Val Asp Thr Gly
210 215 220
Leu Tyr Phe Ser Val Lys Thr Asn Ile Arg Ser Ser Thr Arg Asp Trp
225 230 235 240
Lys Asp His Lys Phe Lys Trp Arg Lys Asp Pro Gln Asp Lys
245 250
<210> 17
<211> 702
<212> ДНК
<213> Homo sapiens
<220>
<221> CDS
<222> (1)..(702)
<400> 17
atg tgg cag ctg ctc ctc cca act gct ctg cta ctt cta gtt tca gct 48
Met Trp Gln Leu Leu Leu Pro Thr Ala Leu Leu Leu Leu Val Ser Ala
1 5 10 15
ggc atg cgg act gaa gat ctc cca aag gct gtg gtg ttc ctg gag cct 96
Gly Met Arg Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
20 25 30
caa tgg tac agc gtg ctt gag aag gac agt gtg act ctg aag tgc cag 144
Gln Trp Tyr Ser Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
35 40 45
gga gcc tac tcc cct gag gac aat tcc aca cag tgg ttt cac aat gag 192
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu
50 55 60
agc ctc atc tca agc cag gcc tcg agc tac ttc att gac gct gcc aca 240
Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr
65 70 75 80
gtc aac gac agt gga gag tac agg tgc cag aca aac ctc tcc acc ctc 288
Val Asn Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu
85 90 95
agt gac ccg gtg cag cta gaa gtc cat atc ggc tgg ctg ttg ctc cag 336
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
100 105 110
gcc cct cgg tgg gtg ttc aag gag gaa gac cct att cac ctg agg tgt 384
Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys
115 120 125
cac agc tgg aag aac act gct ctg cat aag gtc aca tat tta cag aat 432
His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn
130 135 140
ggc aaa gac agg aag tat ttt cat cat aat tct gac ttc cac att cca 480
Gly Lys Asp Arg Lys Tyr Phe His His Asn Ser Asp Phe His Ile Pro
145 150 155 160
aaa gcc aca ctc aaa gat agc ggc tcc tac ttc tgc agg ggg ctt gtt 528
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Val
165 170 175
ggg agt aaa aat gtg tct tca gag act gtg aac atc acc atc act caa 576
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
180 185 190
ggt ttg gca gtg tca acc atc tca tca ttc tct cca cct ggg tac caa 624
Gly Leu Ala Val Ser Thr Ile Ser Ser Phe Ser Pro Pro Gly Tyr Gln
195 200 205
gtc tct ttc tgc ttg gtg atg gta ctc ctt ttt gca gtg gac aca gga 672
Val Ser Phe Cys Leu Val Met Val Leu Leu Phe Ala Val Asp Thr Gly
210 215 220
cta tat ttc tct gtg aag aca aac att tga 702
Leu Tyr Phe Ser Val Lys Thr Asn Ile
225 230
<210> 18
<211> 233
<212> PRT
<213> Homo sapiens
<400> 18
Met Trp Gln Leu Leu Leu Pro Thr Ala Leu Leu Leu Leu Val Ser Ala
1 5 10 15
Gly Met Arg Thr Glu Asp Leu Pro Lys Ala Val Val Phe Leu Glu Pro
20 25 30
Gln Trp Tyr Ser Val Leu Glu Lys Asp Ser Val Thr Leu Lys Cys Gln
35 40 45
Gly Ala Tyr Ser Pro Glu Asp Asn Ser Thr Gln Trp Phe His Asn Glu
50 55 60
Ser Leu Ile Ser Ser Gln Ala Ser Ser Tyr Phe Ile Asp Ala Ala Thr
65 70 75 80
Val Asn Asp Ser Gly Glu Tyr Arg Cys Gln Thr Asn Leu Ser Thr Leu
85 90 95
Ser Asp Pro Val Gln Leu Glu Val His Ile Gly Trp Leu Leu Leu Gln
100 105 110
Ala Pro Arg Trp Val Phe Lys Glu Glu Asp Pro Ile His Leu Arg Cys
115 120 125
His Ser Trp Lys Asn Thr Ala Leu His Lys Val Thr Tyr Leu Gln Asn
130 135 140
Gly Lys Asp Arg Lys Tyr Phe His His Asn Ser Asp Phe His Ile Pro
145 150 155 160
Lys Ala Thr Leu Lys Asp Ser Gly Ser Tyr Phe Cys Arg Gly Leu Val
165 170 175
Gly Ser Lys Asn Val Ser Ser Glu Thr Val Asn Ile Thr Ile Thr Gln
180 185 190
Gly Leu Ala Val Ser Thr Ile Ser Ser Phe Ser Pro Pro Gly Tyr Gln
195 200 205
Val Ser Phe Cys Leu Val Met Val Leu Leu Phe Ala Val Asp Thr Gly
210 215 220
Leu Tyr Phe Ser Val Lys Thr Asn Ile
225 230
<210> 19
<211> 4
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 19
Gly Gly Gly Ser
1
<210> 20
<211> 4
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 20
Ser Gly Gly Gly
1
<210> 21
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 21
Gly Gly Gly Gly Ser
1 5
<210> 22
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 22
Ser Gly Gly Gly Gly
1 5
<210> 23
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 23
Gly Gly Gly Gly Gly Ser
1 5
<210> 24
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 24
Ser Gly Gly Gly Gly Gly
1 5
<210> 25
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 25
Gly Gly Gly Gly Gly Gly Ser
1 5
<210> 26
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 26
Ser Gly Gly Gly Gly Gly Gly
1 5
<210> 27
<211> 107
<212> PRT
<213> Homo sapiens
<400> 27
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 28
<211> 365
<212> PRT
<213> Homo sapiens
<400> 28
Met Gly Val Pro Arg Pro Gln Pro Trp Ala Leu Gly Leu Leu Leu Phe
1 5 10 15
Leu Leu Pro Gly Ser Leu Gly Ala Glu Ser His Leu Ser Leu Leu Tyr
20 25 30
His Leu Thr Ala Val Ser Ser Pro Ala Pro Gly Thr Pro Ala Phe Trp
35 40 45
Val Ser Gly Trp Leu Gly Pro Gln Gln Tyr Leu Ser Tyr Asn Ser Leu
50 55 60
Arg Gly Glu Ala Glu Pro Cys Gly Ala Trp Val Trp Glu Asn Gln Val
65 70 75 80
Ser Trp Tyr Trp Glu Lys Glu Thr Thr Asp Leu Arg Ile Lys Glu Lys
85 90 95
Leu Phe Leu Glu Ala Phe Lys Ala Leu Gly Gly Lys Gly Pro Tyr Thr
100 105 110
Leu Gln Gly Leu Leu Gly Cys Glu Leu Gly Pro Asp Asn Thr Ser Val
115 120 125
Pro Thr Ala Lys Phe Ala Leu Asn Gly Glu Glu Phe Met Asn Phe Asp
130 135 140
Leu Lys Gln Gly Thr Trp Gly Gly Asp Trp Pro Glu Ala Leu Ala Ile
145 150 155 160
Ser Gln Arg Trp Gln Gln Gln Asp Lys Ala Ala Asn Lys Glu Leu Thr
165 170 175
Phe Leu Leu Phe Ser Cys Pro His Arg Leu Arg Glu His Leu Glu Arg
180 185 190
Gly Arg Gly Asn Leu Glu Trp Lys Glu Pro Pro Ser Met Arg Leu Lys
195 200 205
Ala Arg Pro Ser Ser Pro Gly Phe Ser Val Leu Thr Cys Ser Ala Phe
210 215 220
Ser Phe Tyr Pro Pro Glu Leu Gln Leu Arg Phe Leu Arg Asn Gly Leu
225 230 235 240
Ala Ala Gly Thr Gly Gln Gly Asp Phe Gly Pro Asn Ser Asp Gly Ser
245 250 255
Phe His Ala Ser Ser Ser Leu Thr Val Lys Ser Gly Asp Glu His His
260 265 270
Tyr Cys Cys Ile Val Gln His Ala Gly Leu Ala Gln Pro Leu Arg Val
275 280 285
Glu Leu Glu Ser Pro Ala Lys Ser Ser Val Leu Val Val Gly Ile Val
290 295 300
Ile Gly Val Leu Leu Leu Thr Ala Ala Ala Val Gly Gly Ala Leu Leu
305 310 315 320
Trp Arg Arg Met Arg Ser Gly Leu Pro Ala Pro Trp Ile Ser Leu Arg
325 330 335
Gly Asp Asp Thr Gly Val Leu Leu Pro Thr Pro Gly Glu Ala Gln Asp
340 345 350
Ala Asp Leu Lys Asp Val Asn Val Ile Pro Ala Thr Ala
355 360 365
<210> 29
<211> 119
<212> PRT
<213> Homo sapiens
<400> 29
Met Ser Arg Ser Val Ala Leu Ala Val Leu Ala Leu Leu Ser Leu Ser
1 5 10 15
Gly Leu Glu Ala Ile Gln Arg Thr Pro Lys Ile Gln Val Tyr Ser Arg
20 25 30
His Pro Ala Glu Asn Gly Lys Ser Asn Phe Leu Asn Cys Tyr Val Ser
35 40 45
Gly Phe His Pro Ser Asp Ile Glu Val Asp Leu Leu Lys Asn Gly Glu
50 55 60
Arg Ile Glu Lys Val Glu His Ser Asp Leu Ser Phe Ser Lys Asp Trp
65 70 75 80
Ser Phe Tyr Leu Leu Tyr Tyr Thr Glu Phe Thr Pro Thr Glu Lys Asp
85 90 95
Glu Tyr Ala Cys Arg Val Asn His Val Thr Leu Ser Gln Pro Lys Ile
100 105 110
Val Lys Trp Asp Arg Asp Met
115
<210> 30
<211> 451
<212> PRT
<213> Homo sapiens
<400> 30
Gln Val Gln Leu Gln Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ala Val Thr Gly Asp Asp Trp Tyr Phe Asp Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 31
<211> 214
<212> PRT
<213> Homo sapiens
<400> 31
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Val Tyr
35 40 45
Asp Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Asp His
85 90 95
Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 32
<211> 447
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 32
Glu Val Gln Leu Val Glu Ser Gly Gly Lys Leu Leu Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Ala Met Ser Trp Phe Arg Gln Ser Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Glu Ile Ser Ser Gly Gly Ser Tyr Thr Tyr Tyr Pro Asp Thr Val
50 55 60
Thr Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Glu Met Ser Ser Leu Arg Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Leu Trp Gly Tyr Tyr Ala Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 33
<211> 213
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 33
Gln Ile Val Leu Ile Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Arg Leu Leu Ile Tyr
35 40 45
Asp Thr Ser Asn Leu Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Gly Tyr Pro Tyr Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 34
<211> 443
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 34
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Ile Arg Gln Pro Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
130 135 140
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
180 185 190
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
195 200 205
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440
<210> 35
<211> 219
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 35
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Arg Asn Thr Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala
35 40 45
Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Asn
85 90 95
Thr His Val Pro Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 36
<211> 453
<212> PRT
<213> Homo sapiens
<400> 36
Gln Val Gln Leu Val Leu Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Met Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Gln Ser Gly Asp Thr His Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Val Ser Thr Gly Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Gly Ser Asp Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Leu Ile Thr Ala Ala Gly Pro Pro Pro Phe Glu His
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 37
<211> 213
<212> PRT
<213> Homo sapiens
<400> 37
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Met Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Thr Met Pro Cys Ser Gly Asn Gly Leu Gly Asp Lys Phe Val
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly His Ser Pro Val Ala Val Ile Tyr
35 40 45
Gln Asp Ala Lys Arg Pro Ser Gly Val Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Gly Ser Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala
65 70 75 80
Met Asp Glu Ala Asp Tyr Tyr Cys Gln Ala Trp Asp Ser Gly Thr Ala
85 90 95
Val Phe Gly Thr Gly Thr Arg Leu Ser Ile Leu Gly Gln Pro Lys Ala
100 105 110
Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala
115 120 125
Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala
130 135 140
Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val
145 150 155 160
Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser
165 170 175
Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr
180 185 190
Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala
195 200 205
Pro Thr Glu Cys Ser
210
<210> 38
<211> 446
<212> PRT
<213> Oryctolagus cuniculus
<400> 38
Cys Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr
1 5 10 15
Pro Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Met Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Gly Ala Asp Ser Ser Thr Trp Tyr Pro Ser Trp Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Met
65 70 75 80
Thr Ser Leu Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Gly
85 90 95
Arg Phe Val Gly Tyr Thr Asn Ala Phe Asp Pro Trp Gly Pro Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 39
<211> 220
<212> PRT
<213> Oryctolagus cuniculus
<400> 39
Ala Gln Val Leu Thr Gln Thr Pro Ser Ser Val Ser Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Ser Cys Gln Ser Ser Gln Ser Val Trp Asn Asn
20 25 30
Asn Tyr Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Phe Asp Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Arg Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Gly Val
65 70 75 80
Gln Cys Glu Asp Ala Ala Thr Tyr Tyr Cys His Gly Ser Tyr Ala Asn
85 90 95
Ser Gly Trp Tyr Asp Asn Ala Phe Gly Gly Gly Thr Glu Val Val Val
100 105 110
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
115 120 125
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
130 135 140
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
145 150 155 160
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
180 185 190
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
195 200 205
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<210> 40
<211> 452
<212> PRT
<213> Oryctolagus cuniculus
<400> 40
Cys Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr
1 5 10 15
Pro Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Glu Gly Leu Glu Tyr Ile
35 40 45
Gly Phe Ile Asn Thr Gly Gly Ser Ser Tyr Tyr Ala Pro Trp Ala Ile
50 55 60
Gly Arg Leu Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Ile
65 70 75 80
Thr Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Val
85 90 95
Lys Ser Tyr Val Asn Ser Asn Gly Tyr Phe Ile Phe Ser Arg Leu Asp
100 105 110
Leu Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro
450
<210> 41
<211> 218
<212> PRT
<213> Oryctolagus cuniculus
<400> 41
Ala Gln Val Leu Thr Gln Thr Ala Ser Ser Val Ser Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Ser Cys Gln Ser Ser Lys Ser Val Tyr Asn Asn
20 25 30
Asn Phe Leu Ser Trp Tyr Gln Gln Lys Leu Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Tyr Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Lys Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Asp Leu
65 70 75 80
Glu Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Gly Tyr Ser Gly
85 90 95
Ile Pro Ile Asn Ala Phe Gly Gly Gly Thr Glu Val Val Val Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 42
<211> 446
<212> PRT
<213> Oryctolagus cuniculus
<400> 42
Cys Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr
1 5 10 15
Pro Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Ser Asn
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Gly Gly Ser Gly Asp Thr Gly Tyr Ala Ser Trp Ala Asn
50 55 60
Gly Arg Phe Thr Val Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Met
65 70 75 80
Thr Ser Leu Thr Ala Ala Asp Thr Ala Thr Tyr Phe Cys Val Arg His
85 90 95
Ser Val Gly Ala Ser Trp Trp Val Phe Asn Ile Trp Gly Pro Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
<210> 43
<211> 219
<212> PRT
<213> Oryctolagus cuniculus
<400> 43
Ala Gln Val Leu Thr Gln Thr Pro Ser Ser Val Ser Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Asn Cys Gln Ser Ser Gln Ser Val Tyr Ser Gly
20 25 30
Asn Phe Phe Ala Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Asp Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Lys Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Gly Val
65 70 75 80
Gln Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Gln Gly Thr Tyr Tyr Asn
85 90 95
Ser Gly Trp Ser Asn Val Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 44
<211> 443
<212> PRT
<213> Oryctolagus cuniculus
<400> 44
Cys Gln Ser Leu Glu Glu Ser Gly Gly Arg Leu Val Met Pro Gly Gly
1 5 10 15
Ser Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Asn Tyr
20 25 30
Asn Ile Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile
35 40 45
Gly Phe Ile Asp Ser Gly Gly Ser Ala Tyr Tyr Ala Asn Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Thr Ser Thr Thr Val Asp Leu Lys Met
65 70 75 80
Thr Ser Leu Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Gly
85 90 95
Gly Val Asn Val Asp Tyr Tyr Ile Trp Gly Pro Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
130 135 140
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
180 185 190
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
195 200 205
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440
<210> 45
<211> 217
<212> PRT
<213> Oryctolagus cuniculus
<400> 45
Ala Gln Val Leu Thr Gln Thr Pro Ser Pro Val Ser Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Asn Cys Gln Ala Ser Gln Ser Val Tyr Ser Asn
20 25 30
Asn Tyr Leu Ala Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Arg Ala Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Lys Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Asp Val
65 70 75 80
Val Cys Asp Asp Ala Ala Ser Tyr Tyr Cys Gln Gly Tyr Tyr Tyr Gly
85 90 95
Gly Ile Gly Pro Phe Gly Gly Gly Thr Glu Val Val Val Lys Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 46
<211> 122
<212> PRT
<213> Homo sapiens
<400> 46
Gln Val Gln Leu Gln Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Ile Asn Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Gly Arg Lys Gly Asp Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 47
<211> 110
<212> PRT
<213> Homo sapiens
<400> 47
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Val Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Ser Tyr Ala Gly Ser
85 90 95
Asn Asn Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 48
<211> 119
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 48
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser Asp Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Lys Gly Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Ala Tyr Tyr Cys
85 90 95
Ala Arg Val Leu Ala Arg Ile Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 49
<211> 107
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 49
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Ser Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Glu Leu Leu Ile
35 40 45
Tyr Tyr Gly Ser Glu Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Glu Ala
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gly Gln Gly Asn Arg Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Glu
100 105
<210> 50
<211> 449
<212> PRT
<213> Homo sapiens
<400> 50
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ile Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Gly Pro Leu Val Ser Asp Ala Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 51
<211> 219
<212> PRT
<213> Homo sapiens
<400> 51
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Leu Ser Leu Leu Asn Arg
20 25 30
Asp Gly Lys Thr Phe Leu Tyr Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Glu Val Ser Ser Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Gln Ile
65 70 75 80
Asn Arg Val Glu Ala Asp Asp Ala Gly Val Tyr Tyr Cys Met Gln Gly
85 90 95
Leu His Leu Pro Arg Thr Phe Gly Leu Arg Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 52
<211> 450
<212> PRT
<213> Homo sapiens
<400> 52
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Arg Gly Met Gly Gly Asp Ala Phe Asp Ile Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 53
<211> 216
<212> PRT
<213> Homo sapiens
<400> 53
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Thr Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Ser Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr His Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Asn Gly Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 54
<211> 456
<212> PRT
<213> Homo sapiens
<400> 54
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Tyr Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Gly Met Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Asn Asp Ala Ser Asn Gln Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Arg Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Ser Glu Asp Thr Gly Leu Tyr Tyr Cys
85 90 95
Ala Lys Glu Gly Ser Ser Pro Lys Thr Pro Thr Ser Thr Trp Ser Ser
100 105 110
Leu Glu Ser Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
115 120 125
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
130 135 140
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
145 150 155 160
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
165 170 175
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
180 185 190
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
195 200 205
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215 220
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
355 360 365
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
435 440 445
Ser Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 55
<211> 217
<212> PRT
<213> Homo sapiens
<400> 55
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile His Asp Val Ser Asn Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ser
85 90 95
Thr Thr Pro Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu Gly
100 105 110
Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
115 120 125
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
130 135 140
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
145 150 155 160
Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys
165 170 175
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser
180 185 190
His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu
195 200 205
Lys Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 56
<211> 30
<212> PRT
<213> Homo sapiens
<400> 56
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Phe Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser
20 25 30
<210> 57
<211> 14
<212> PRT
<213> Homo sapiens
<400> 57
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
1 5 10
<210> 58
<211> 32
<212> PRT
<213> Homo sapiens
<400> 58
Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 59
<211> 11
<212> PRT
<213> Homo sapiens
<400> 59
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 60
<211> 22
<212> PRT
<213> Homo sapiens
<400> 60
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys
20
<210> 61
<211> 16
<212> PRT
<213> Homo sapiens
<400> 61
Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu Met Ile Tyr
1 5 10 15
<210> 62
<211> 32
<212> PRT
<213> Homo sapiens
<400> 62
Gly Val Pro Asp Arg Phe Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser
1 5 10 15
Leu Thr Val Ser Gly Leu Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys
20 25 30
<210> 63
<211> 10
<212> PRT
<213> Homo sapiens
<400> 63
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
1 5 10
<210> 64
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<220>
<221> дополнительный признак
<222> (38)..(38)
<223> n представляет собой a, c, g или t
<400> 64
tattactcgc ggcccagccg gccatggcag ccwtcganwt gacccagact 50
<210> 65
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<220>
<221> дополнительный признак
<222> (39)..(39)
<223> n представляет собой a, c, g или t
<400> 65
tattactcgc ggcccagccg gccatggcag cctatgatnt gacccagact 50
<210> 66
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 66
tattactcgc ggcccagccg gccatggcag cbcaagtgct gacccagact 50
<210> 67
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 67
tattactcgc ggcccagccg gccatggcag ccmtygtgat gacccagact 50
<210> 68
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 68
tattactcgc ggcccagccg gccatggcag ccgccgtgct gacccagact 50
<210> 69
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 69
tattactcgc ggcccagccg gccatggcgg ctgacattgt gatgacccag 50
<210> 70
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 70
tattactcgc ggcccagccg gccatggccg ccgayrtygt gatgacccag 50
<210> 71
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 71
ctcttctaga acgcgtctaa gcgtcacccc tattgaagct c 41
<210> 72
<211> 55
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 72
tattactcgc ggcccagccg gccatggcgc agcyygtgct gactcagtcg ccctc 55
<210> 73
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 73
ctcttctaga acgcgtctaa gcttctgcag gggccaggct cttc 44
<210> 74
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 74
ttccgcctcg gcgctagccc aggagcagst ggwggagtcc 40
<210> 75
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<220>
<221> дополнительный признак
<222> (25)..(26)
<223> n представляет собой a, c, g или t
<400> 75
ttccgcctcg gcgctagccc agtcnntgga ggagtccggg 40
<210> 76
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<220>
<221> дополнительный признак
<222> (26)..(27)
<223> n представляет собой a, c, g или t
<400> 76
ttccgcctcg gcgctagccc agtcgnngga ggagtccggg 40
<210> 77
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 77
ttccgcctcg gcgctagccc agcagcagct ggwggagtcc 40
<210> 78
<211> 122
<212> PRT
<213> Homo sapiens
<400> 78
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Gly His
20 25 30
Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Thr His Ile Phe Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Arg Arg Trp Ala Met Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 79
<211> 110
<212> PRT
<213> Homo sapiens
<400> 79
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Asp Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Gly Gly Ser Lys Lys Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Leu Lys Ser Tyr Ala
85 90 95
Glu Gly Pro Met Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 80
<211> 122
<212> PRT
<213> Homo sapiens
<400> 80
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn His Ile Asn Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly His Arg Arg Asp Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 81
<211> 110
<212> PRT
<213> Homo sapiens
<400> 81
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Gly Gly Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Val Thr Tyr Ser Ile
85 90 95
Ala Asp Pro Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 82
<211> 122
<212> PRT
<213> Homo sapiens
<400> 82
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Gly His
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Ser Tyr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Ala Leu Gly Gln Met Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 83
<211> 110
<212> PRT
<213> Homo sapiens
<400> 83
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Asp Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Ser Val Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Ser Arg Ser Ser Ser
85 90 95
Met Asn Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 84
<211> 122
<212> PRT
<213> Homo sapiens
<400> 84
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Gly Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Ser Tyr Ile Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Ala Leu Gly Gln Arg Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 85
<211> 109
<212> PRT
<213> Homo sapiens
<400> 85
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Asp Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Gln Thr Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Ser Arg Ser Ile Lys
85 90 95
His Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 86
<211> 123
<212> PRT
<213> Homo sapiens
<400> 86
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ala Pro Val Val Ala Arg Pro Arg Gly Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 87
<211> 112
<212> PRT
<213> Homo sapiens
<400> 87
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Ser Pro Val Thr Leu Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asp Gly Asn Thr Tyr Leu Ser Trp Leu Gln Gln Arg Pro Gly Gln Pro
35 40 45
Pro Arg Leu Leu Ile His Glu Ile Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ala Gly Thr Asn Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ala
85 90 95
Thr Gln Phe Pro Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 88
<211> 122
<212> PRT
<213> Homo sapiens
<400> 88
Glu Val Gln Leu Val Gln Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Val Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Val Pro Ser Ser Gly Tyr Pro Gly Arg Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 89
<211> 106
<212> PRT
<213> Homo sapiens
<400> 89
Asn Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Ser
20 25 30
Leu Asn Trp Tyr Gln Gln Ile Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Ser Tyr Tyr Cys Gln Gln Ser Tyr Ser Ala Trp Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 90
<211> 328
<212> PRT
<213> Homo sapiens
<400> 90
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 91
<211> 107
<212> PRT
<213> Homo sapiens
<400> 91
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 92
<211> 119
<212> PRT
<213> Homo sapiens
<400> 92
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Arg Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Ser Ile Thr Ser Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Arg Gly Leu Glu Trp
35 40 45
Ile Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Thr Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Met Leu Arg Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Arg Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Ser Leu Val Thr Val Ser Ser
115
<210> 93
<211> 107
<212> PRT
<213> Homo sapiens
<400> 93
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 94
<211> 328
<212> PRT
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 94
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Asp Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Glu Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 95
<211> 122
<212> PRT
<213> Homo sapiens
<400> 95
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Gly Tyr
20 25 30
Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Ser His Ile Ser Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Gly Arg Lys Tyr Arg Met Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 96
<211> 110
<212> PRT
<213> Homo sapiens
<400> 96
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Thr Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Thr Val Thr Gly Ile
85 90 95
Trp Ser Val Gly Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 97
<211> 122
<212> PRT
<213> Homo sapiens
<400> 97
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn His Ala Thr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Gly Lys Lys Gly Arg Tyr Leu Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 98
<211> 110
<212> PRT
<213> Homo sapiens
<400> 98
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Asp Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Tyr Val Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Leu Thr Thr Asp Ser
85 90 95
Leu Asn Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 99
<211> 233
<212> PRT
<213> Homo sapiens
<400> 99
Cys His Pro Arg Leu Ser Leu His Arg Pro Ala Leu Glu Asp Leu Leu
1 5 10 15
Leu Gly Ser Glu Ala Asn Leu Thr Cys Thr Leu Thr Gly Leu Arg Asp
20 25 30
Ala Ser Gly Val Thr Phe Thr Trp Thr Pro Ser Ser Gly Lys Ser Ala
35 40 45
Val Gln Gly Pro Pro Glu Arg Asp Leu Cys Gly Cys Tyr Ser Val Ser
50 55 60
Ser Val Leu Pro Gly Cys Ala Glu Pro Trp Asn His Gly Lys Thr Phe
65 70 75 80
Thr Cys Thr Ala Ala Tyr Pro Glu Ser Lys Thr Pro Leu Thr Ala Thr
85 90 95
Leu Ser Lys Ser Gly Asn Thr Phe Arg Pro Glu Val His Leu Leu Pro
100 105 110
Pro Pro Ser Glu Glu Leu Ala Leu Asn Glu Leu Val Thr Leu Thr Cys
115 120 125
Leu Ala Arg Gly Phe Ser Pro Lys Asp Val Leu Val Arg Trp Leu Gln
130 135 140
Gly Ser Gln Glu Leu Pro Arg Glu Lys Tyr Leu Thr Trp Ala Ser Arg
145 150 155 160
Gln Glu Pro Ser Gln Gly Thr Thr Thr Phe Ala Val Thr Ser Ile Leu
165 170 175
Arg Val Ala Ala Glu Asp Trp Lys Lys Gly Asp Thr Phe Ser Cys Met
180 185 190
Val Gly His Glu Ala Leu Pro Leu Ala Phe Thr Gln Lys Thr Ile Asp
195 200 205
Arg Leu Ala Gly Lys Gly Gly Gly Gly Ser Gly Leu Asn Asp Ile Phe
210 215 220
Glu Ala Gln Lys Ile Glu Trp His Glu
225 230
<210> 100
<211> 122
<212> PRT
<213> Homo sapiens
<400> 100
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Gly His
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn His Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Arg Lys Phe Met Met Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 101
<211> 110
<212> PRT
<213> Homo sapiens
<400> 101
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Gln Asn Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Leu Arg Ser Ser Asn
85 90 95
Leu Ser Pro Gly Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 102
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 102
actcgcggcc cagccggcca tggcg 25
<210> 103
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 103
taggacggtc agcttggtac ctccgcc 27
<210> 104
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 104
gcgcagccgg cgctagcc 18
<210> 105
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 105
tgggcccttg gtcgacgc 18
<210> 106
<211> 122
<212> PRT
<213> Homo sapiens
<400> 106
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Gly His Arg His Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Lys Lys Gly Asn Arg Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 107
<211> 110
<212> PRT
<213> Homo sapiens
<400> 107
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Ser Thr Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Thr Thr Thr Tyr Ala
85 90 95
Lys Asn Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 108
<211> 122
<212> PRT
<213> Homo sapiens
<400> 108
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Gly Tyr
20 25 30
Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn His Arg Ser Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Gly Lys Arg Phe Asp Arg Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 109
<211> 110
<212> PRT
<213> Homo sapiens
<400> 109
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Thr Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Lys Thr Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Ser Arg Arg Tyr Arg
85 90 95
Arg Ser Leu Ser Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 110
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 110
cgcaacgcaa ttaatgtgag 20
<210> 111
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 111
gcgtcacact ttgctatg 18
<210> 112
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Искусственно синтезированная последовательность
<400> 112
tgagttccac gacaccgtca c 21
<210> 113
<211> 122
<212> PRT
<213> Homo sapiens
<400> 113
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Gly His
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Arg Leu His Asp Arg Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 114
<211> 110
<212> PRT
<213> Homo sapiens
<400> 114
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Asp Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Gln Val Ser Lys Lys Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Ser Arg Ser Tyr Gly
85 90 95
Ala Gly Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 115
<211> 122
<212> PRT
<213> Homo sapiens
<400> 115
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr His
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Ser Tyr Ile His Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Gly Lys Leu Gly Glu Arg Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 116
<211> 110
<212> PRT
<213> Homo sapiens
<400> 116
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Asp Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Gln Val Ser Lys Lys Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Val Arg Ser Asp Gly
85 90 95
His Gly Pro Met Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 117
<211> 122
<212> PRT
<213> Homo sapiens
<400> 117
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Gly Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Ala His Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Arg Leu Gly Asp Arg Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 118
<211> 110
<212> PRT
<213> Homo sapiens
<400> 118
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Ala Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Lys Val Ser Lys Lys Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Leu Arg Ala Ala Thr
85 90 95
Thr Gly Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 119
<211> 122
<212> PRT
<213> Homo sapiens
<400> 119
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Gly Tyr Ala Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Gly Ala Leu Asn Thr Leu Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 120
<211> 110
<212> PRT
<213> Homo sapiens
<400> 120
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Asp Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Gln Val Ser Lys Lys Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Thr Arg Thr Ala Asn
85 90 95
Ala Gly Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 121
<211> 122
<212> PRT
<213> Homo sapiens
<400> 121
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Ser Tyr Ala Ser Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Arg Leu Asn Ser His Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 122
<211> 110
<212> PRT
<213> Homo sapiens
<400> 122
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Phe Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Gln Gly Ser Lys Lys Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Thr Arg Ala Thr Thr
85 90 95
Lys Gly Val Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 123
<211> 122
<212> PRT
<213> Homo sapiens
<400> 123
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Gly His
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Asn Tyr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Gly Arg Leu His Asp Arg Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 124
<211> 110
<212> PRT
<213> Homo sapiens
<400> 124
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Asp Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Gln Val Ser Lys Lys Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Ser Arg Ser Tyr Gly
85 90 95
Ala Gly Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 125
<211> 122
<212> PRT
<213> Homo sapiens
<400> 125
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Gly Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Thr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Arg Ser Gly Tyr Ile Phe Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe Gly Lys Leu Asn His Met Asn Trp Val Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 126
<211> 110
<212> PRT
<213> Homo sapiens
<400> 126
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Thr Ser Thr Asp Val Gly Thr Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Gln Val Ser Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Phe Cys Ser Thr Arg Ala Ile Thr
85 90 95
Arg Gly Val Ala Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, ИНДУЦИРУЮЩАЯ ИММУННЫЙ ОТВЕТ НА АНТИГЕН-МИШЕНЬ | 2012 |
|
RU2722829C2 |
| АНТИ-CD137 АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2829536C2 |
| СОДЕРЖАЩИЙ ЛЕКАРСТВЕННОЕ СРЕДСТВО ПЕРЕНОСЧИК В КЛЕТКУ ДЛЯ ФОРМИРОВАНИЯ ИММУННОГО КОМПЛЕКСА | 2012 |
|
RU2739792C1 |
| АНТИТЕЛА, БЛОКИРУЮЩИЕ CD73 | 2020 |
|
RU2819204C2 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА ДЛЯ УСКОРЕНИЯ ИСЧЕЗНОВЕНИЯ АНТИГЕНА ЧЕРЕЗ FcγRIIB | 2013 |
|
RU2812226C1 |
| АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА ДЛЯ УСКОРЕНИЯ ИСЧЕЗНОВЕНИЯ АНТИГЕНА ЧЕРЕЗ FcγRIIB | 2013 |
|
RU2736349C2 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2022 |
|
RU2824285C2 |
| ИММУНОАКТИВИРУЮЩАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА | 2015 |
|
RU2722788C2 |
| АНТИ-PCSK9 АНТИТЕЛО, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2016 |
|
RU2739208C2 |
| СОБАЧЬИ АНТИТЕЛА С МОДИФИЦИРОВАННЫМИ ПОСЛЕДОВАТЕЛЬНОСТЯМИ CH2-CH3 | 2014 |
|
RU2815059C2 |
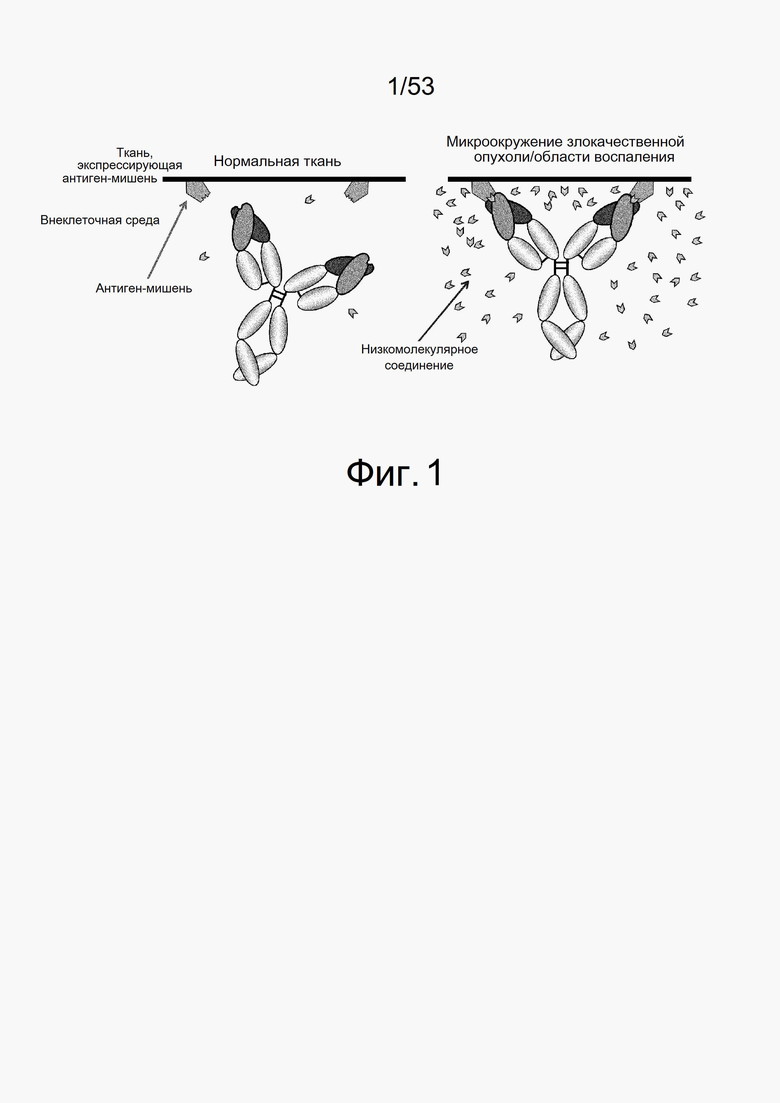
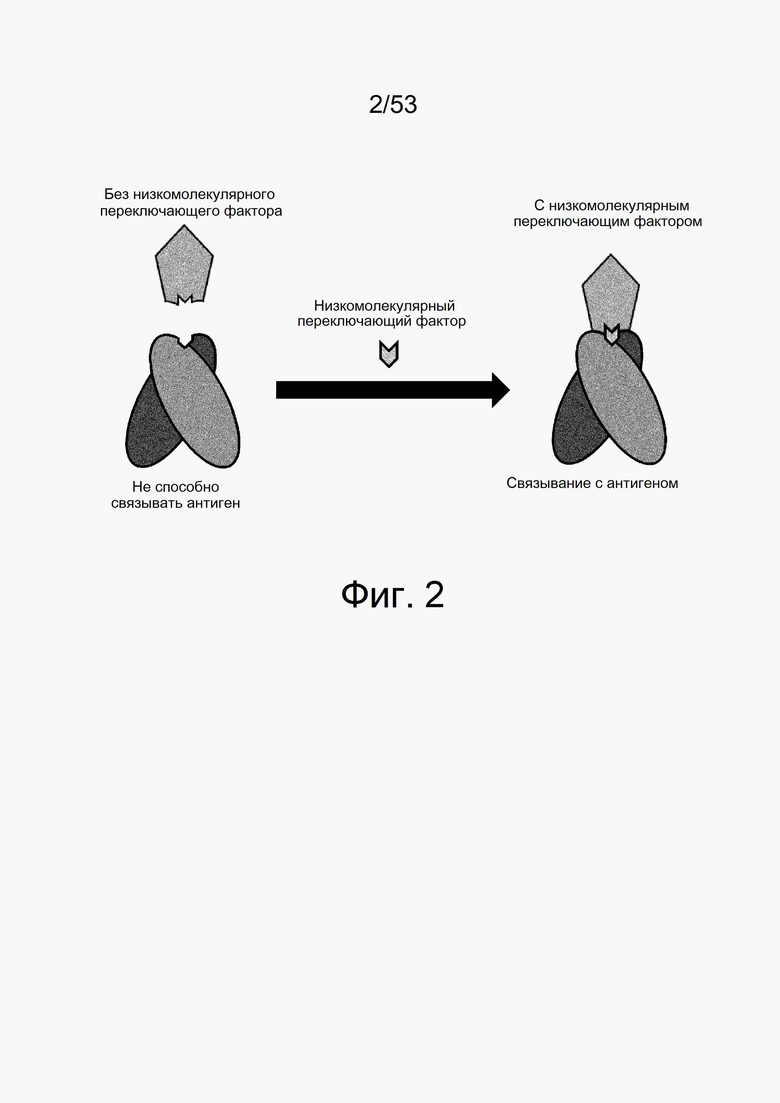
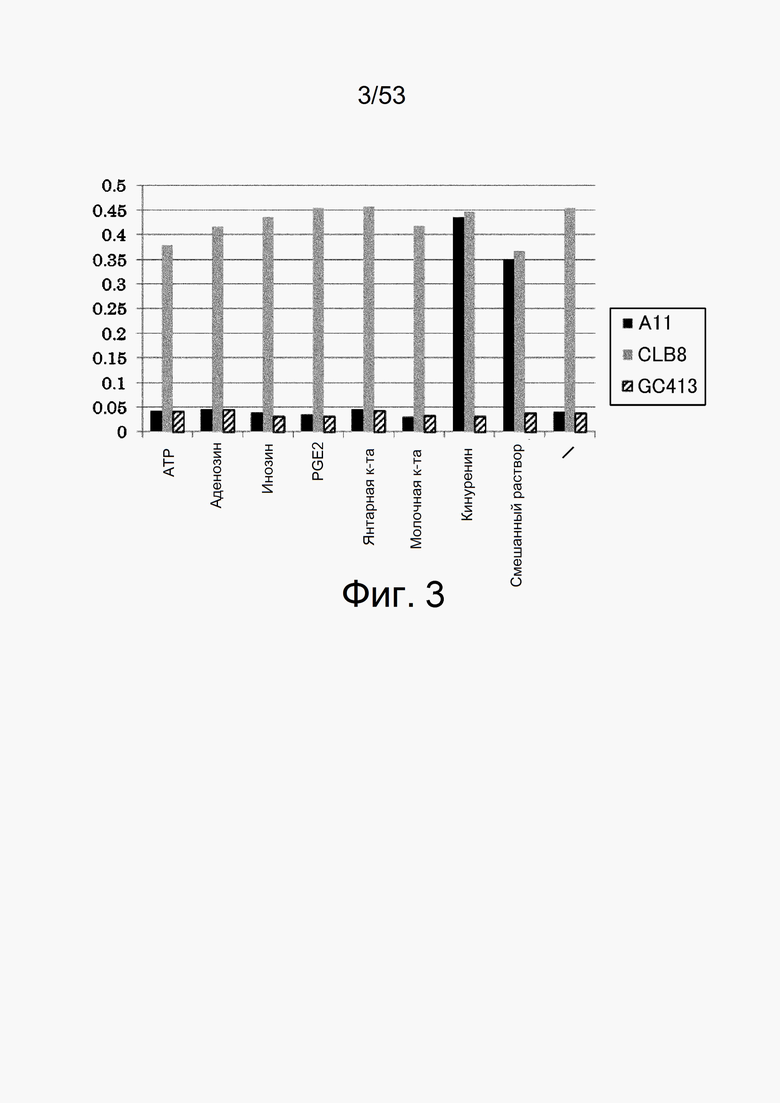
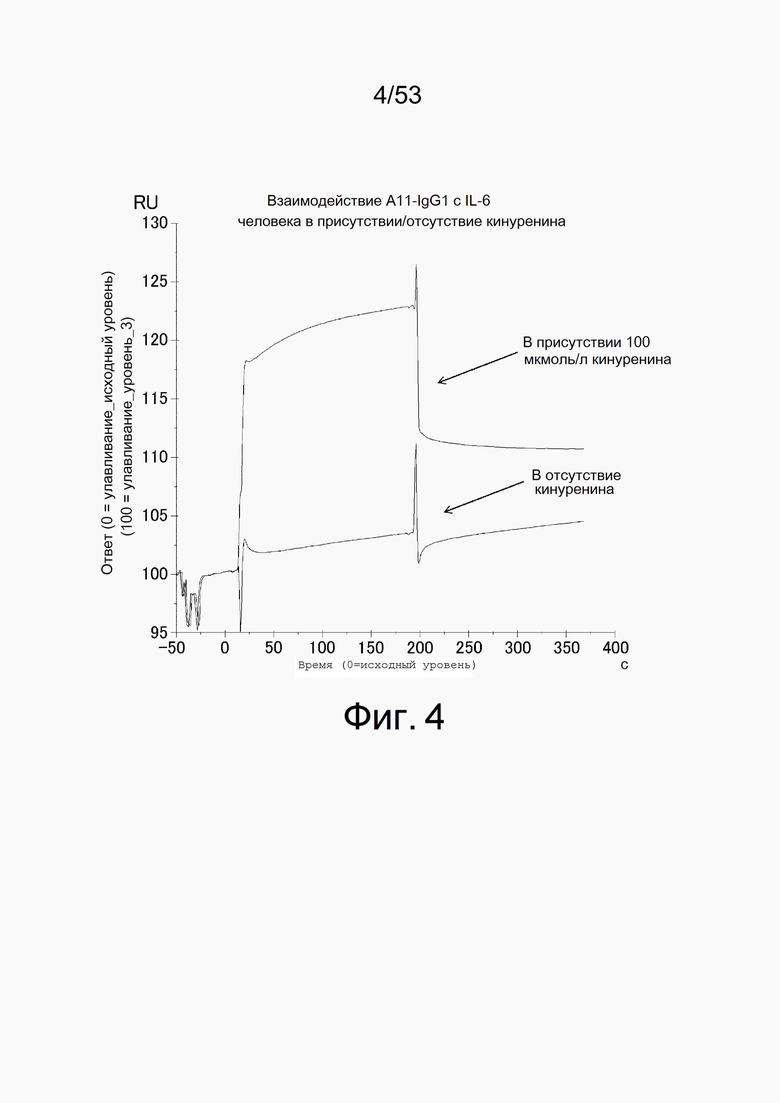
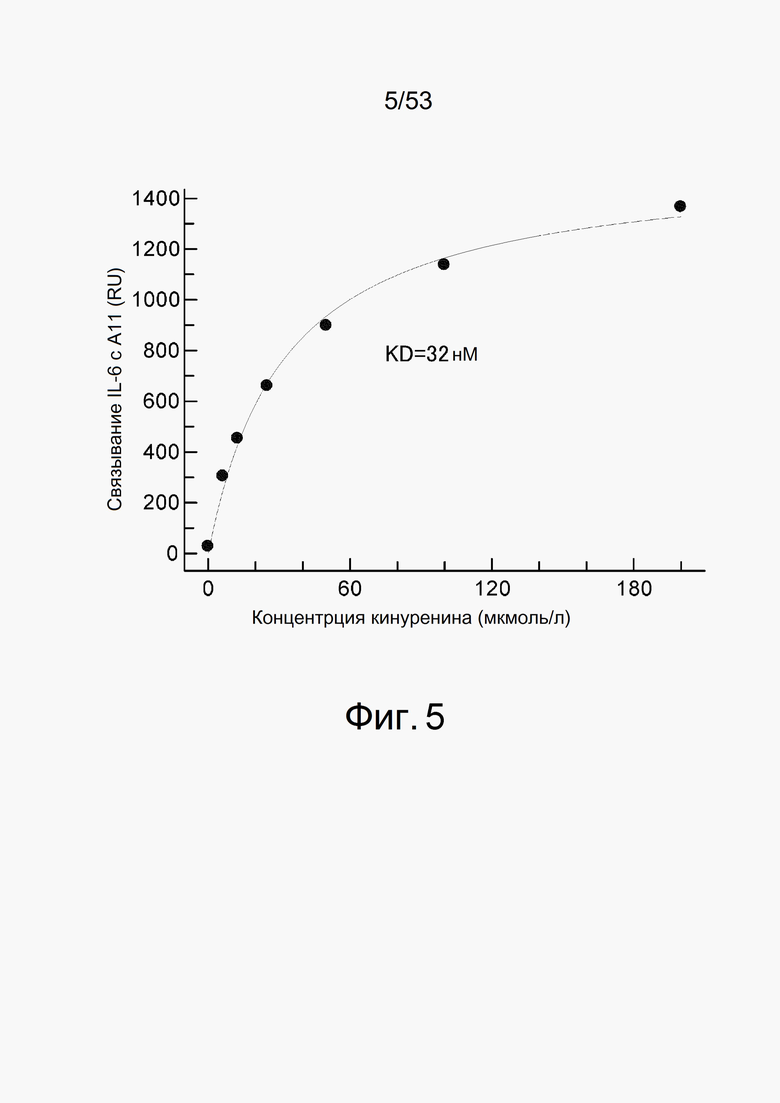
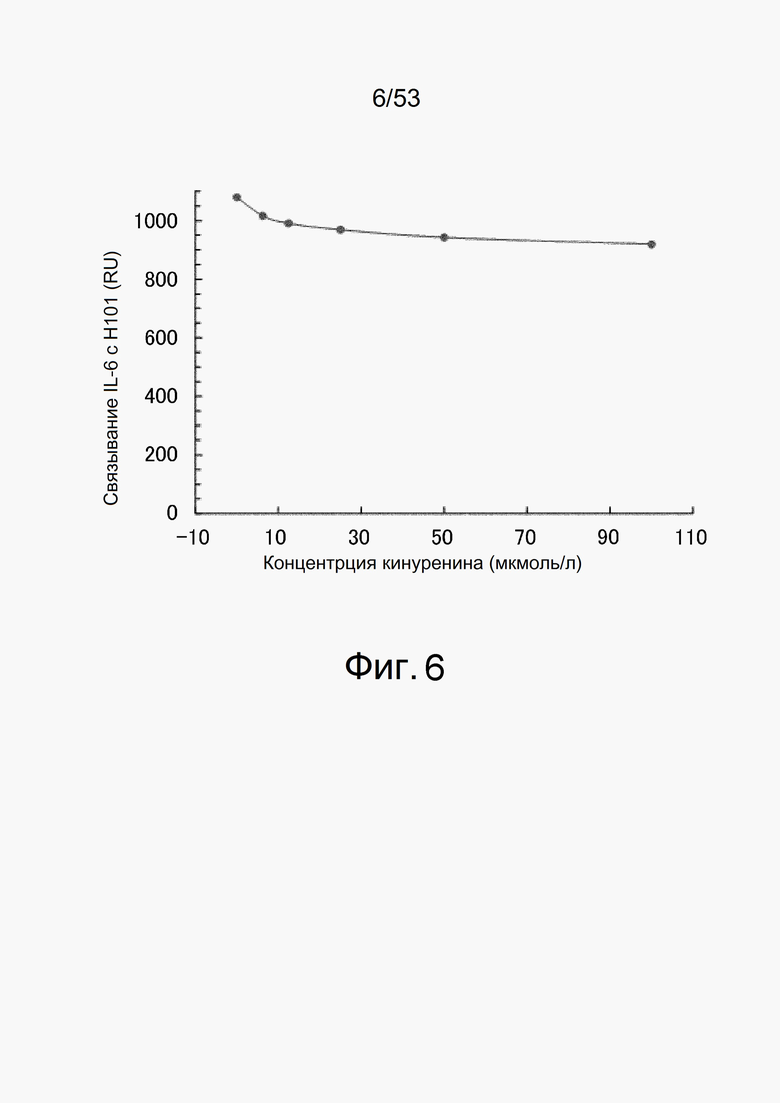
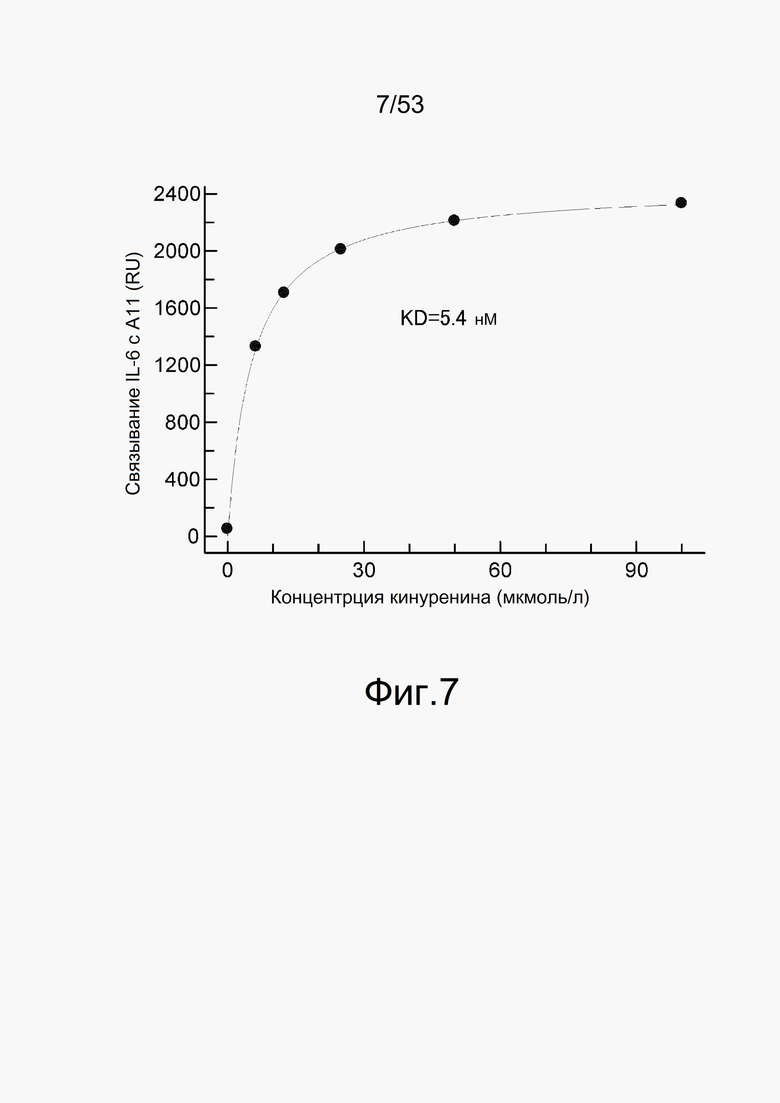
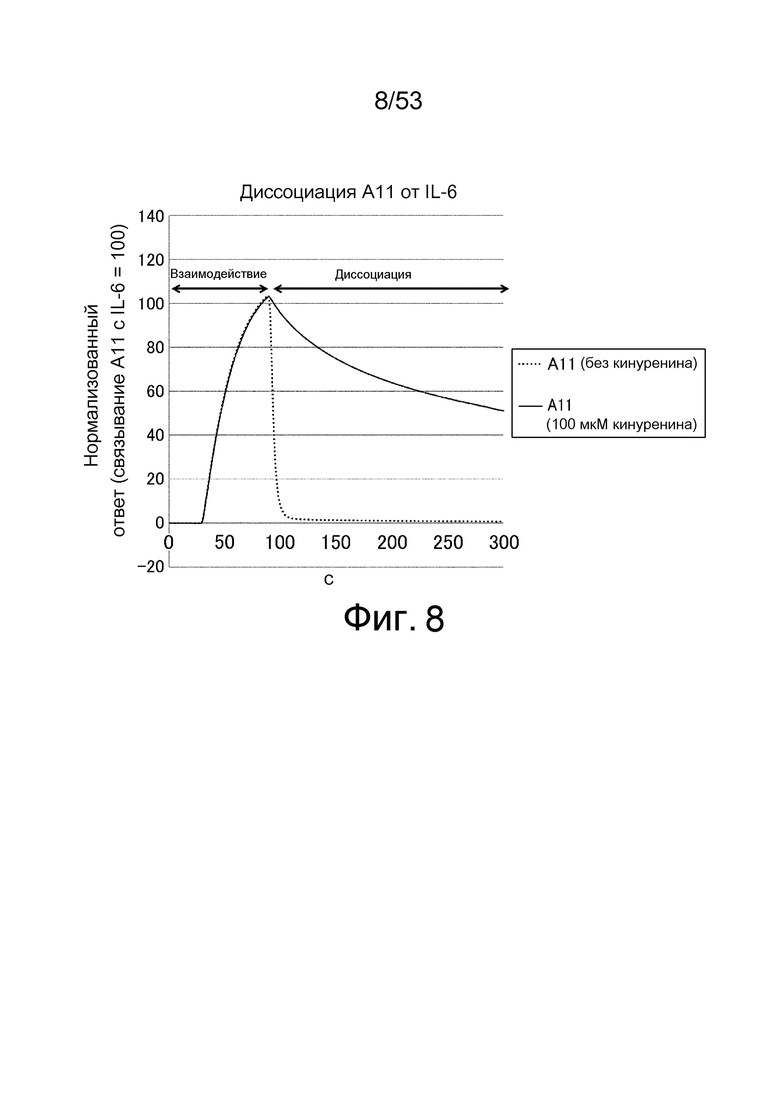
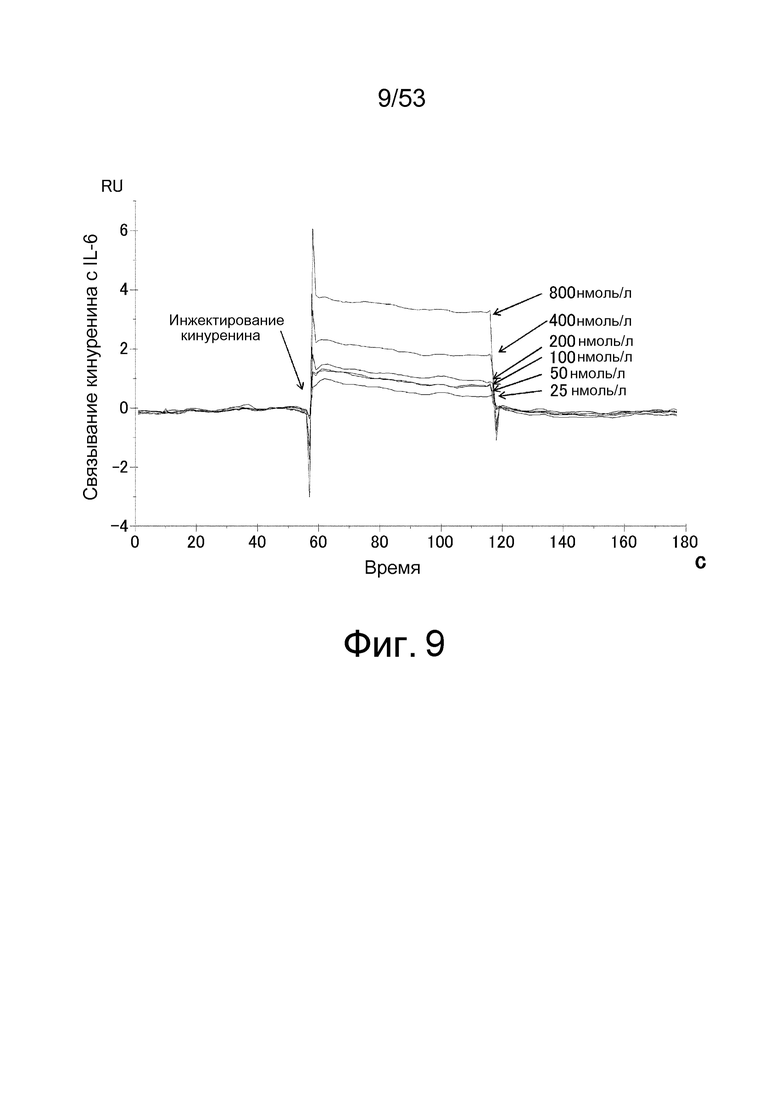
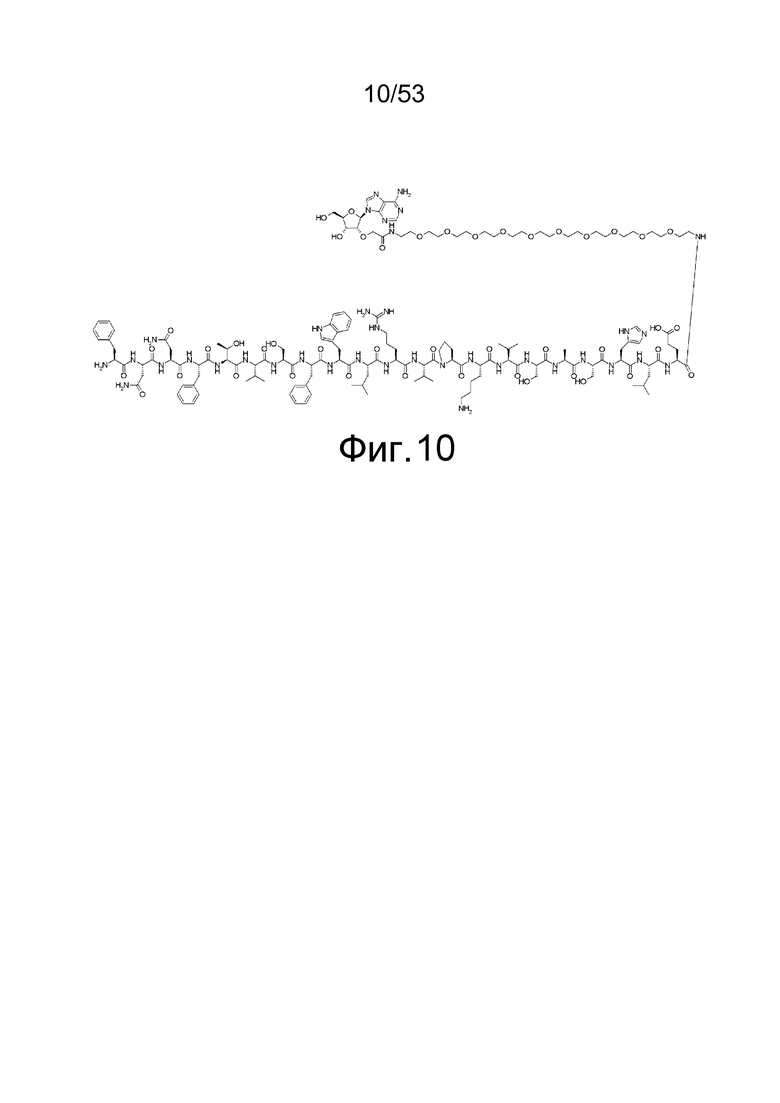
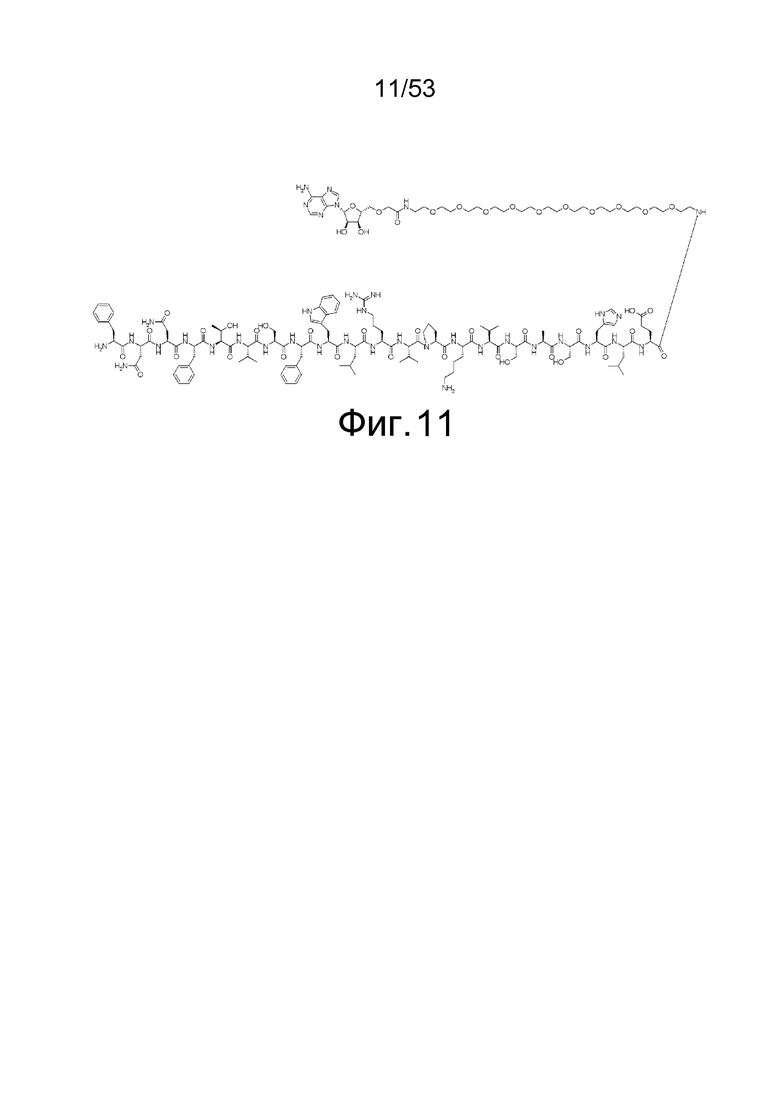
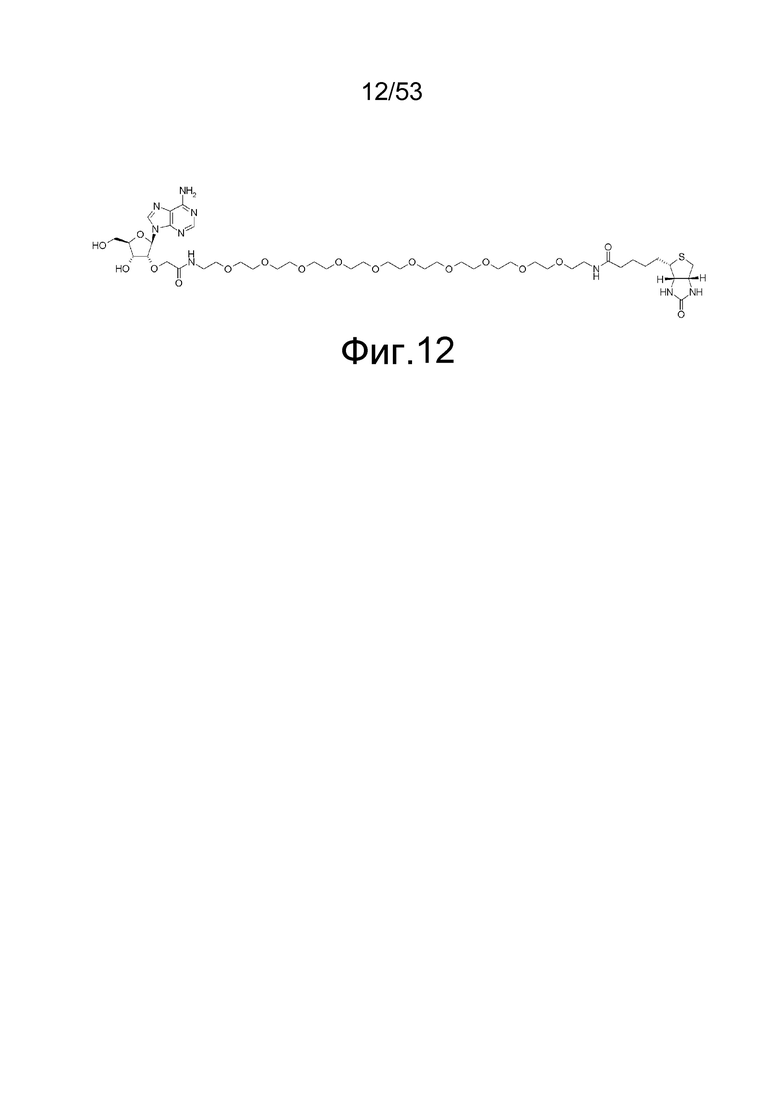
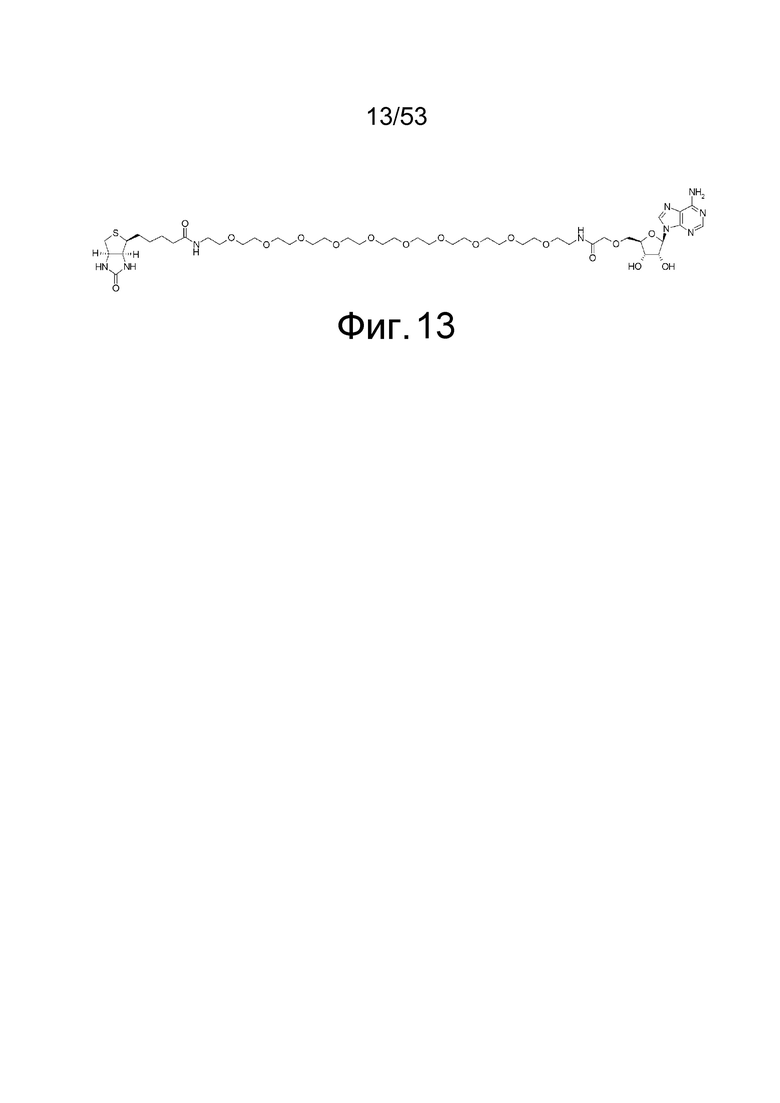
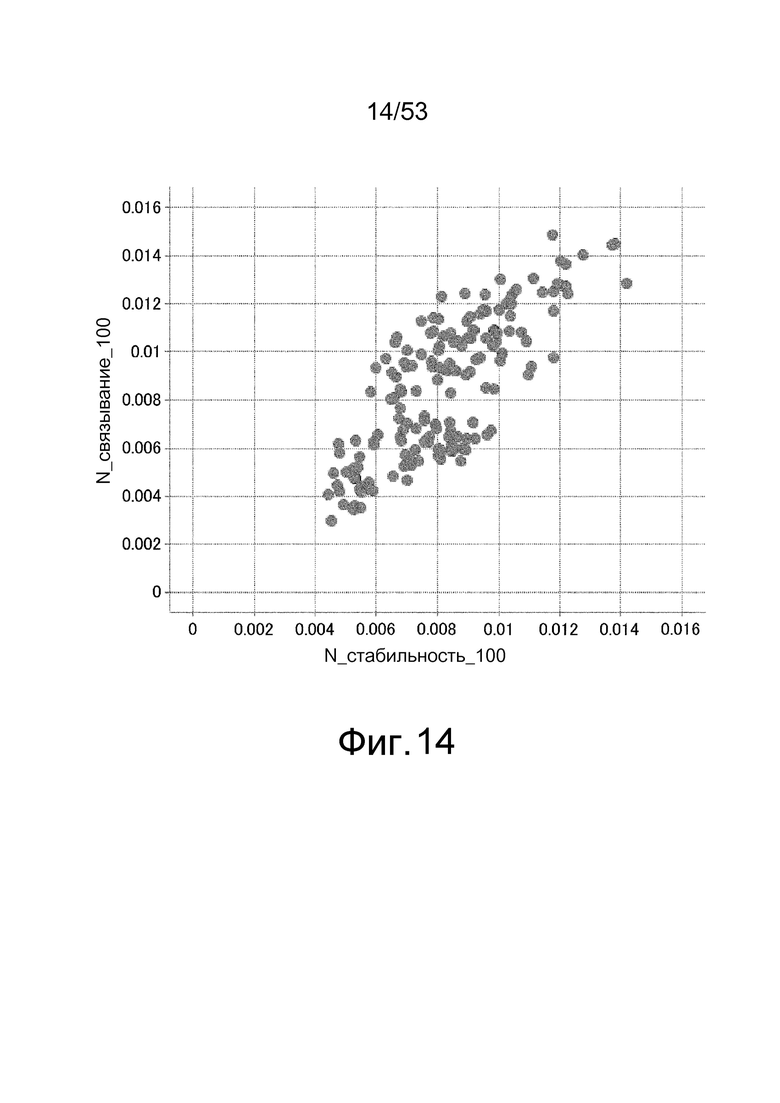
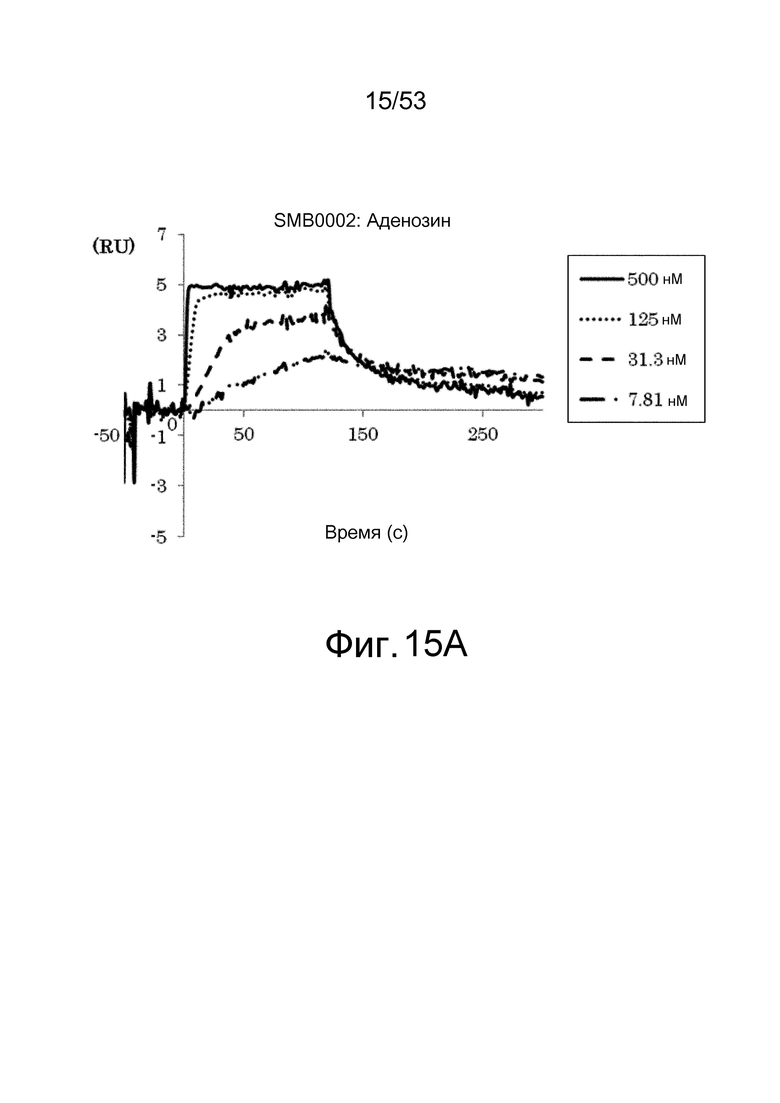

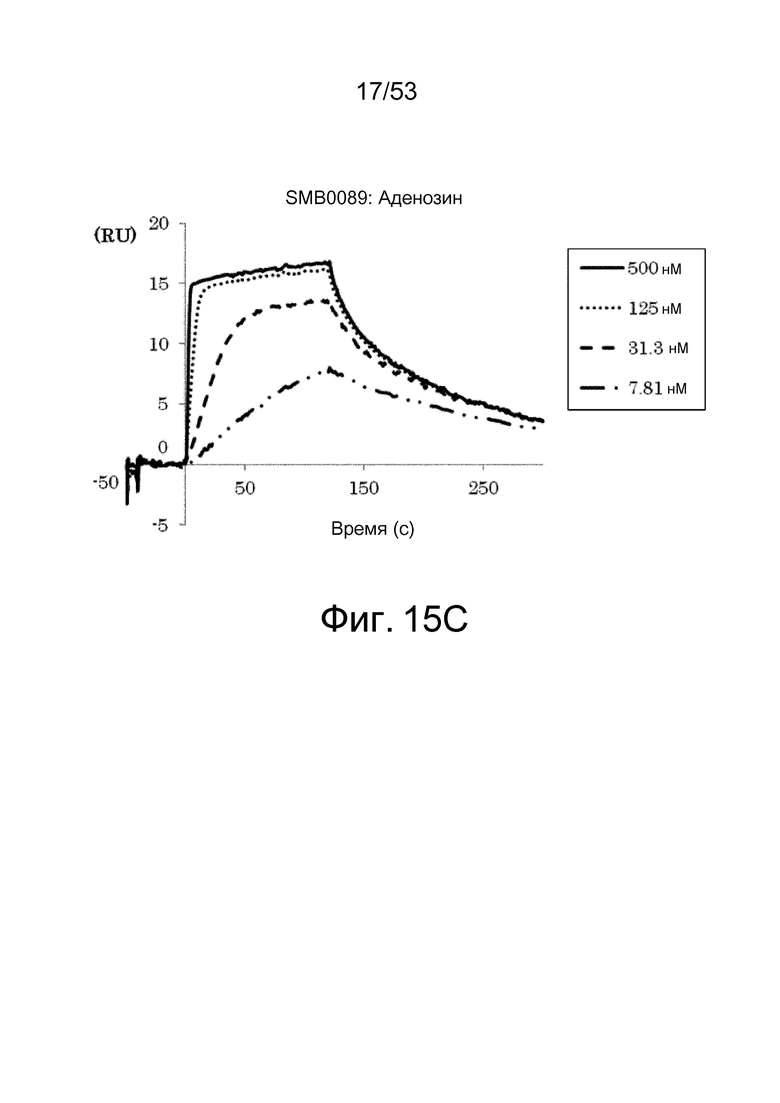
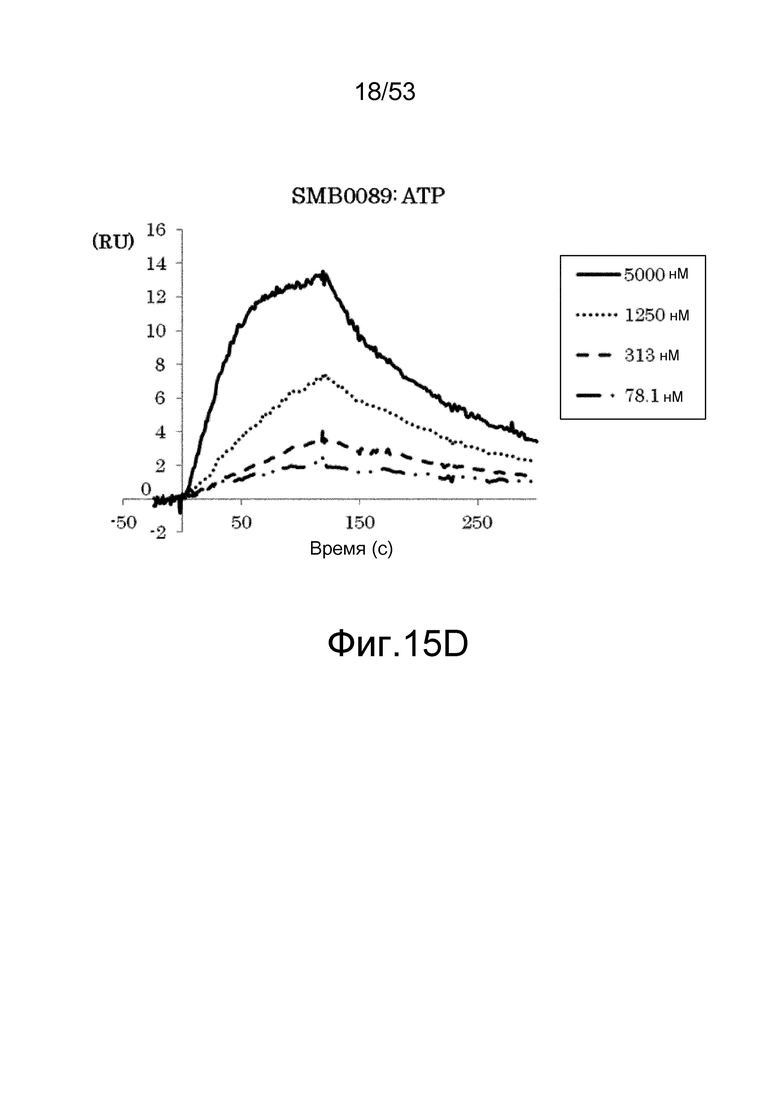

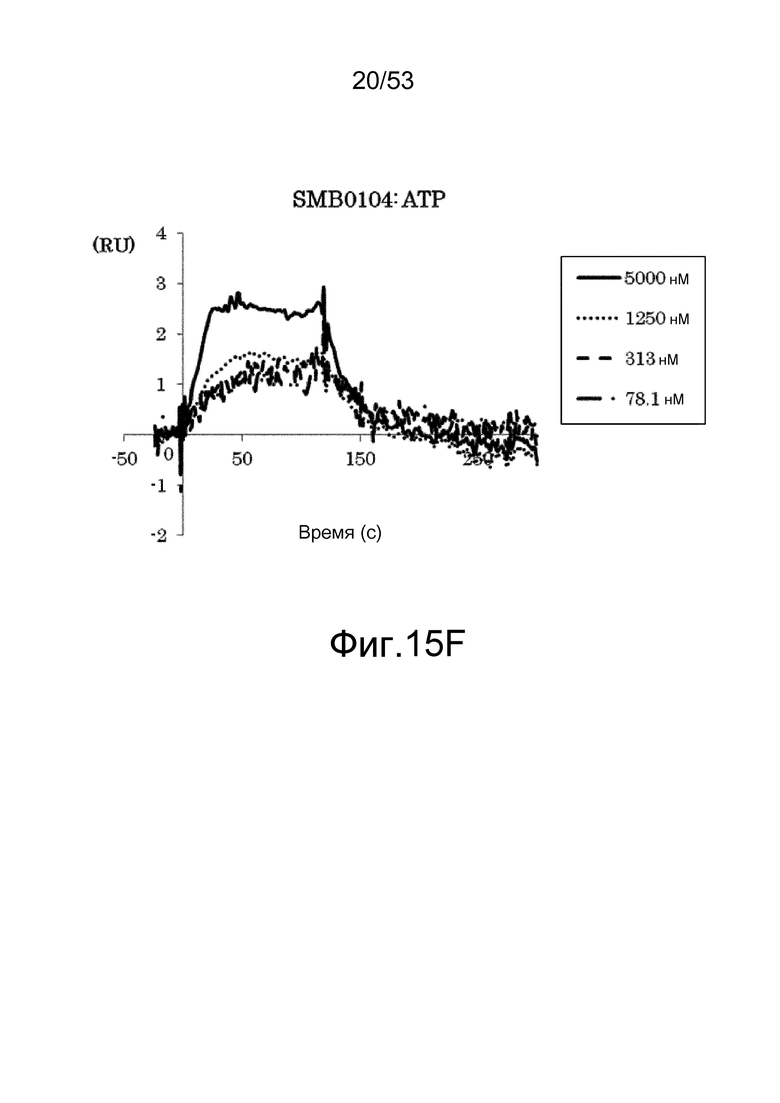
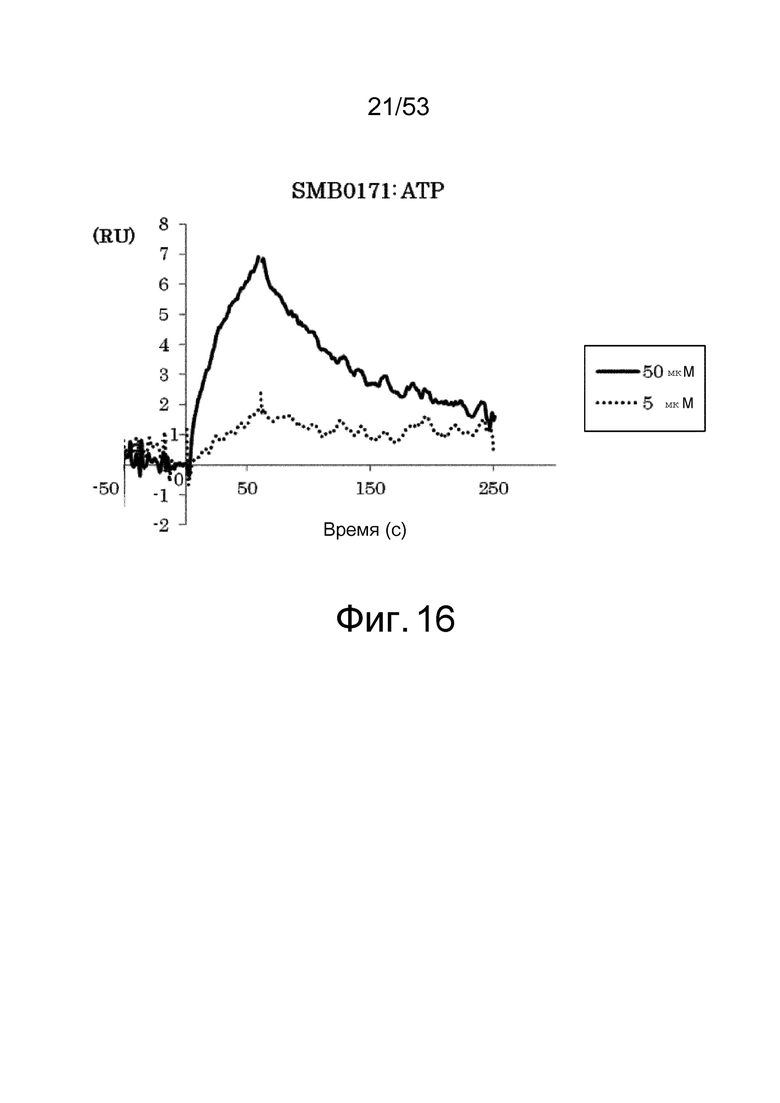

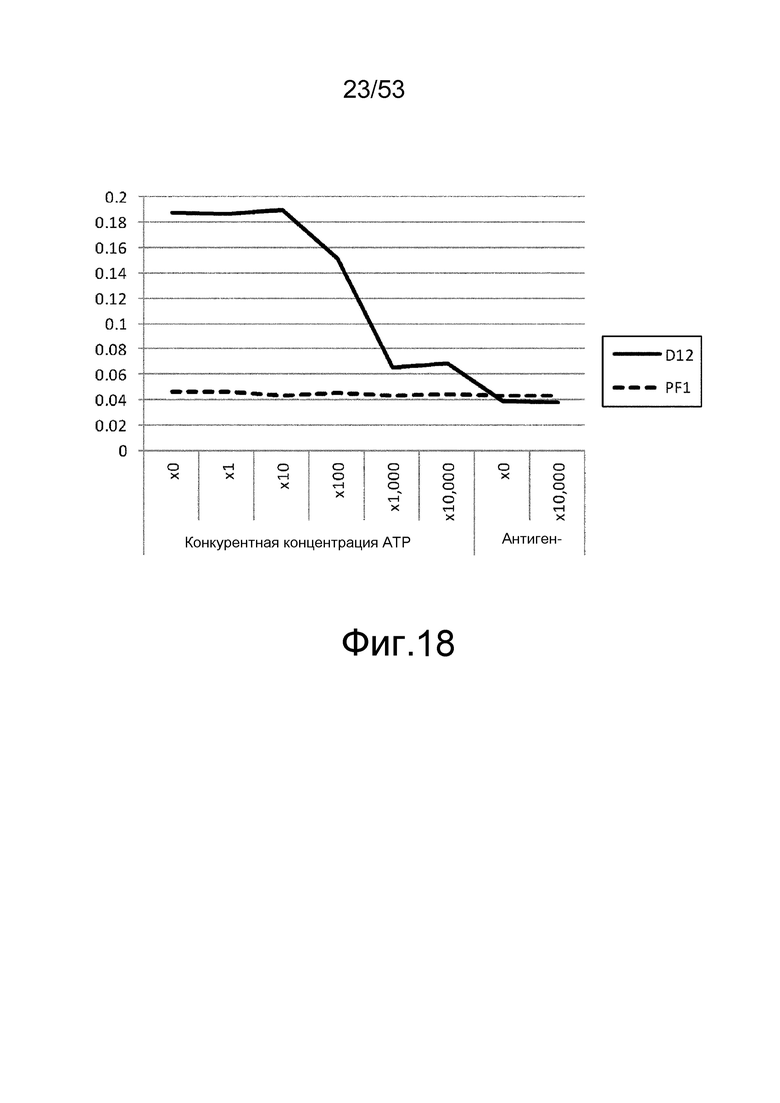
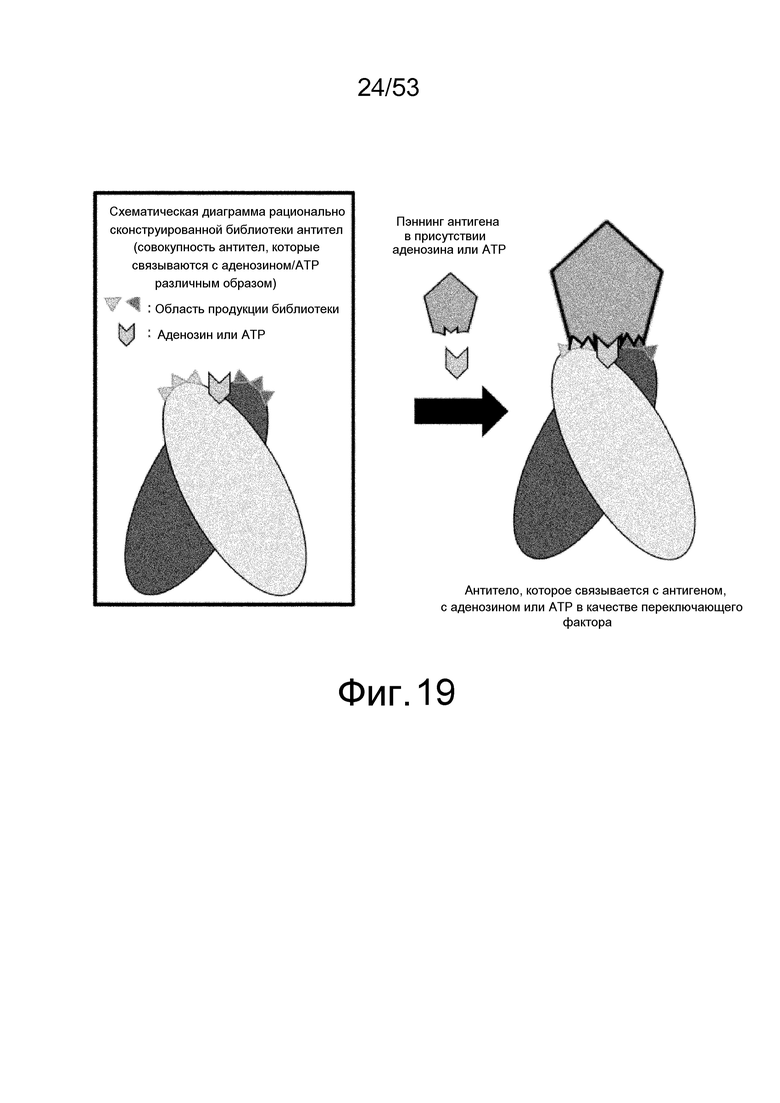

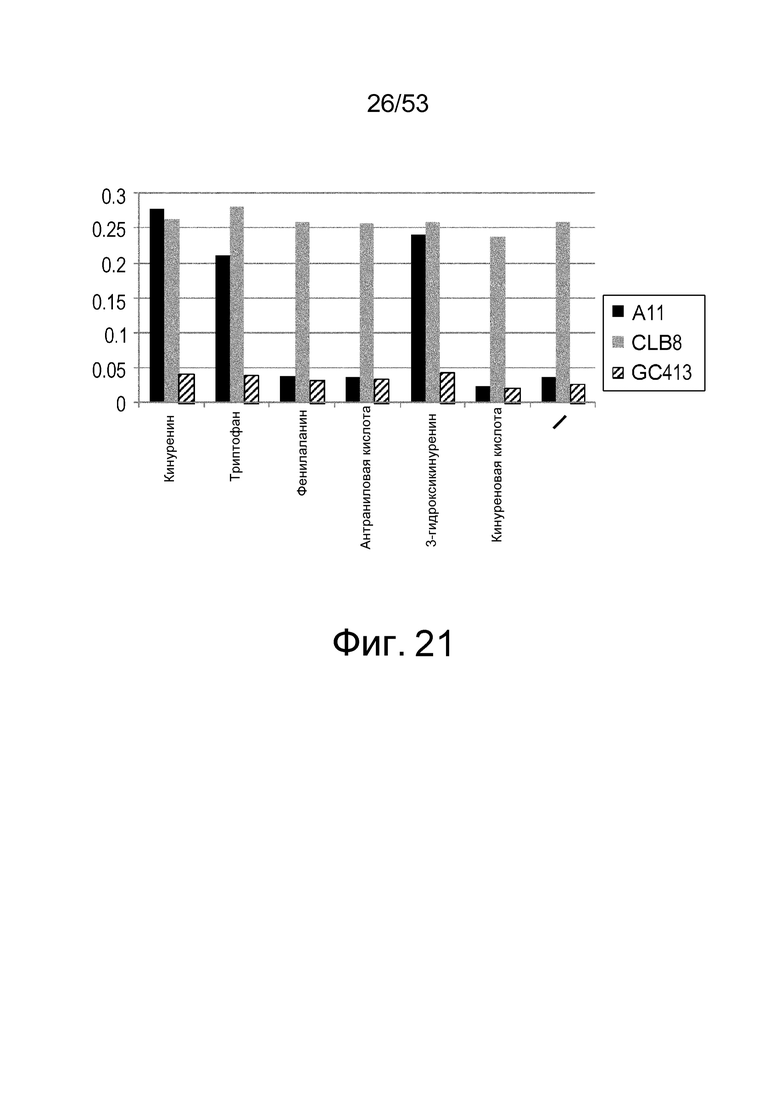
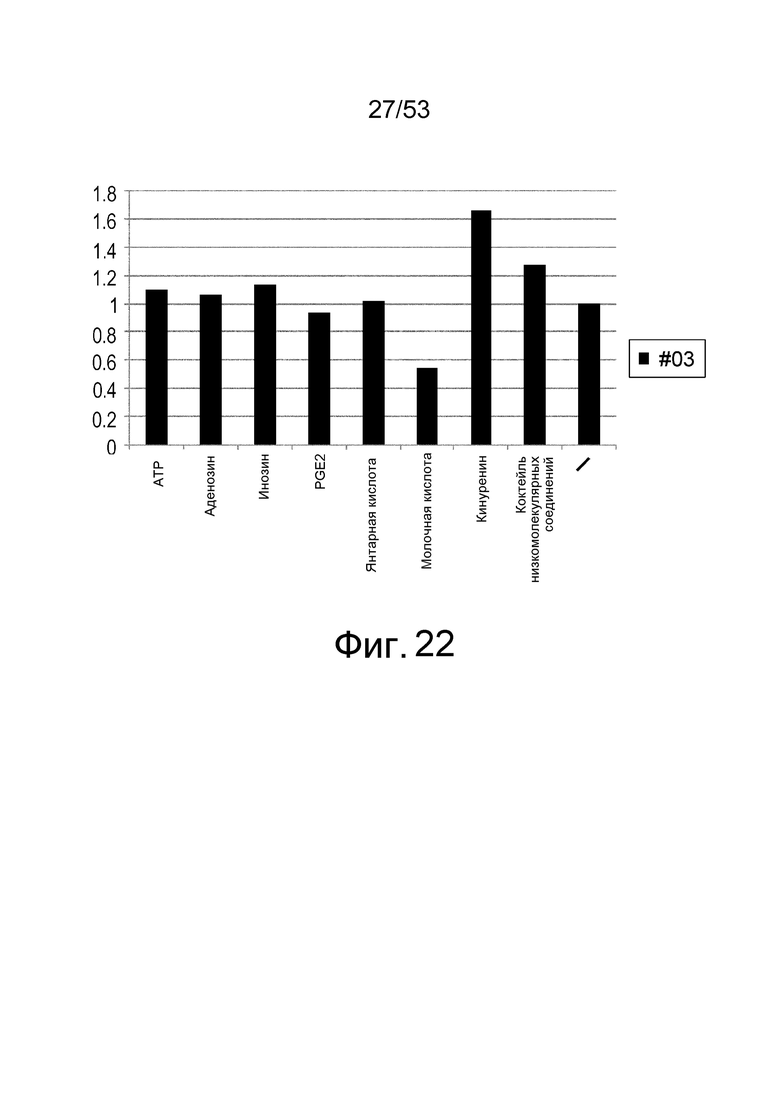
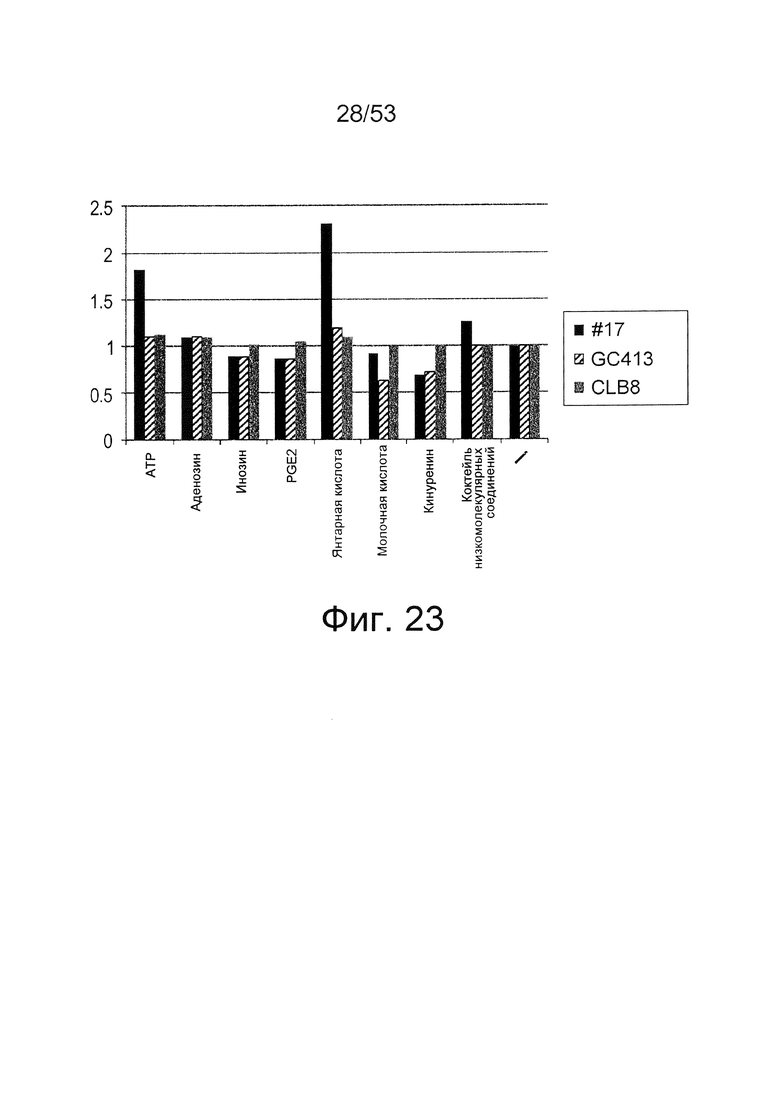
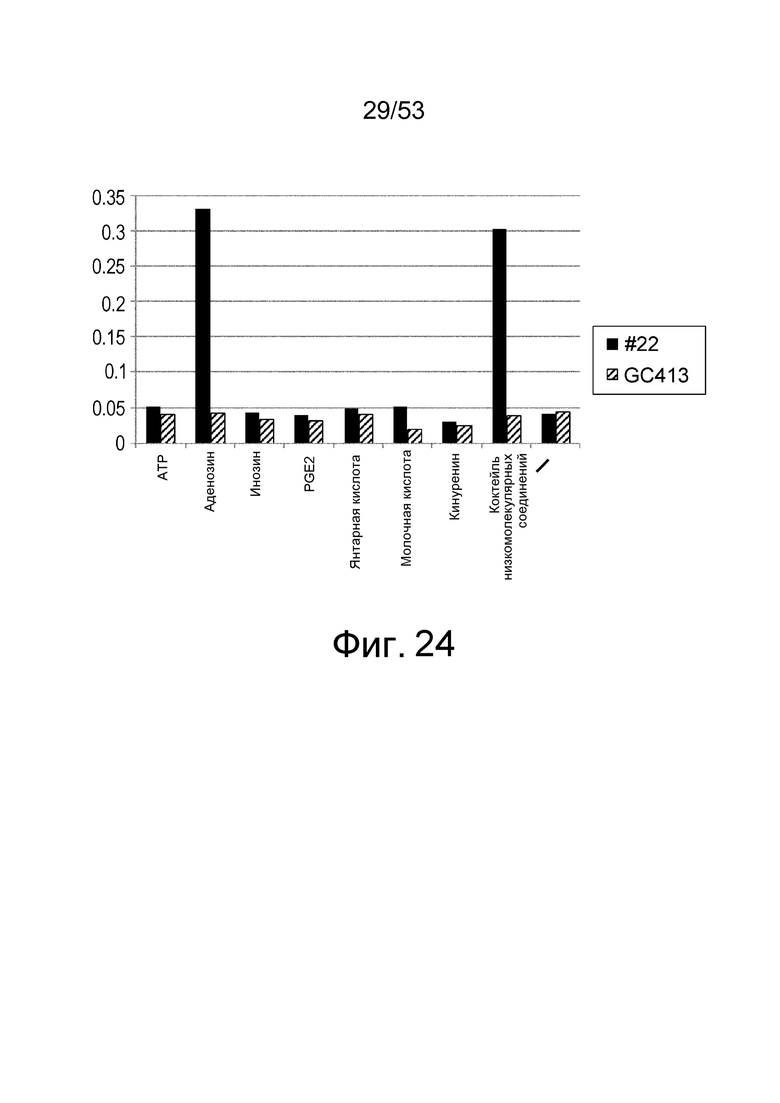
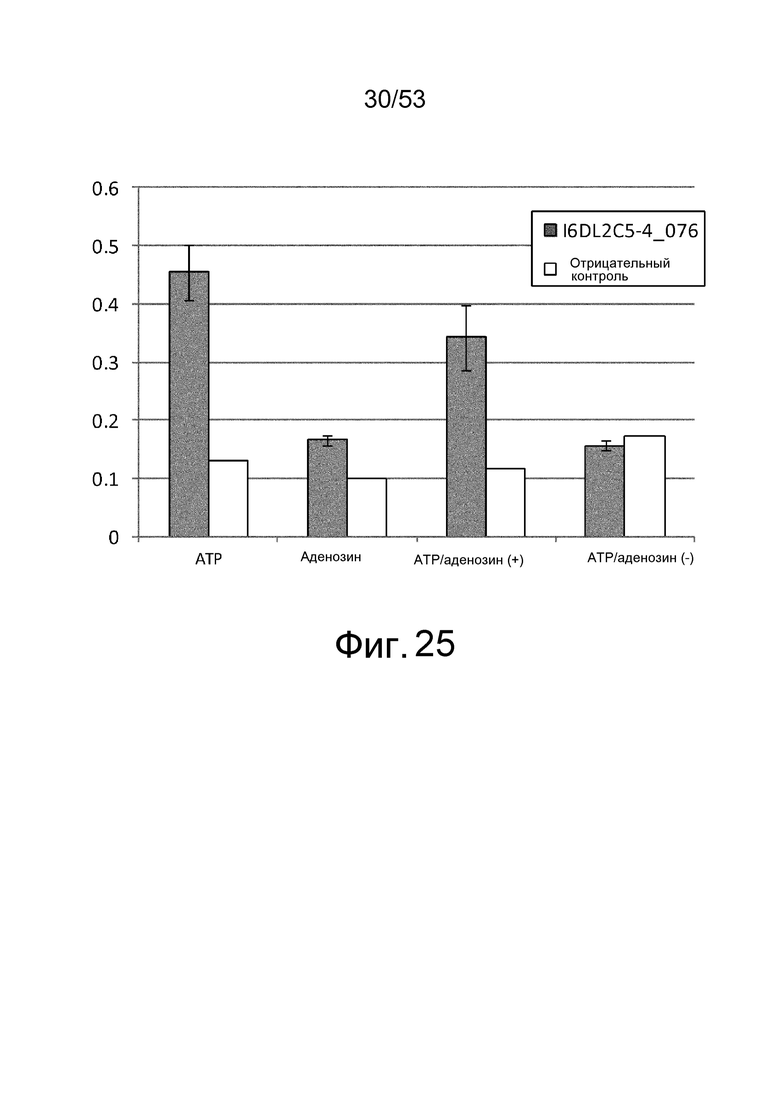
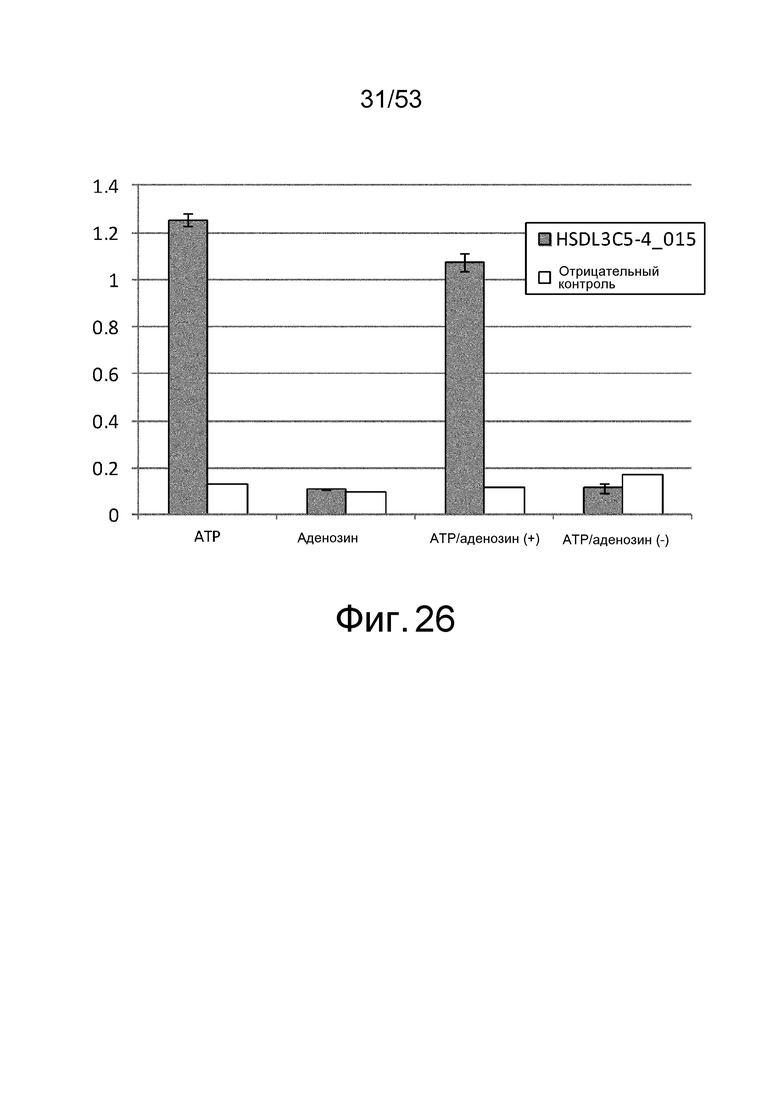
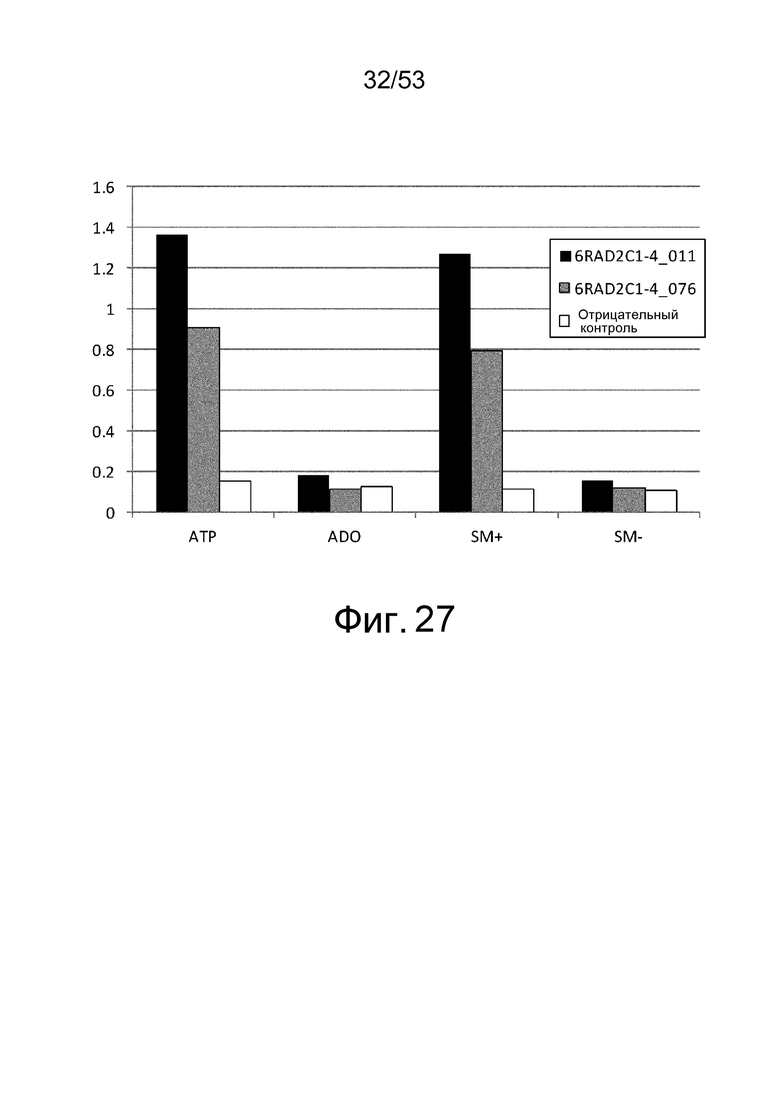
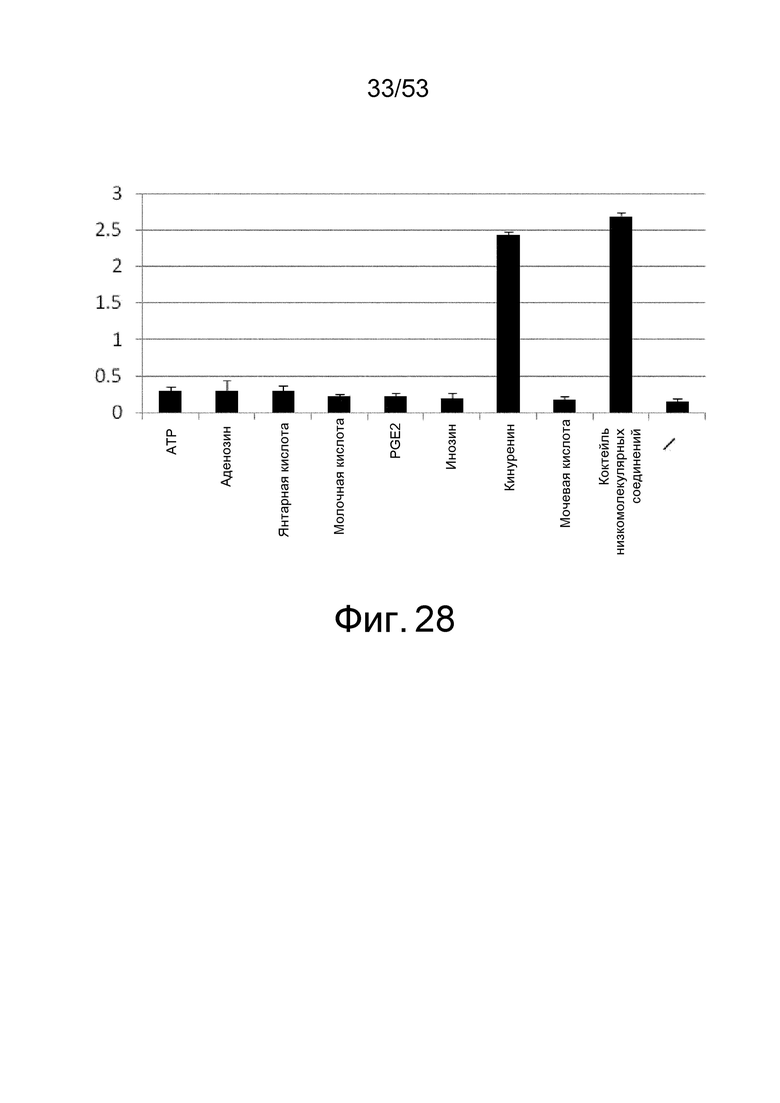

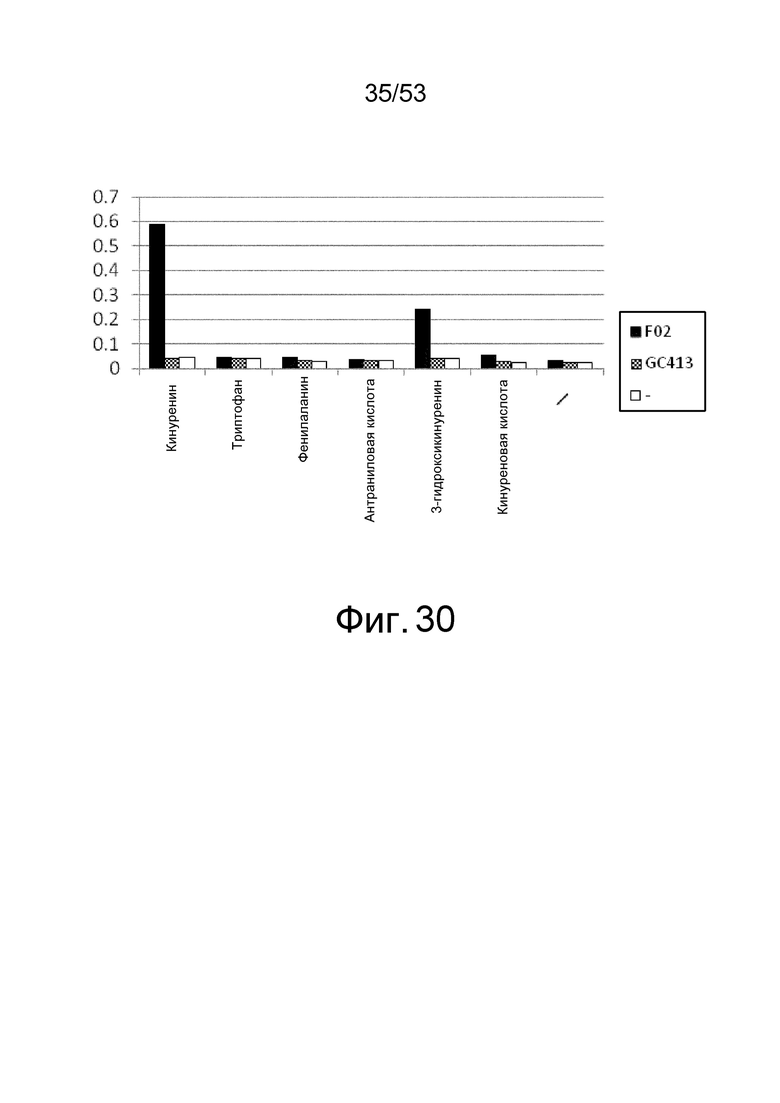
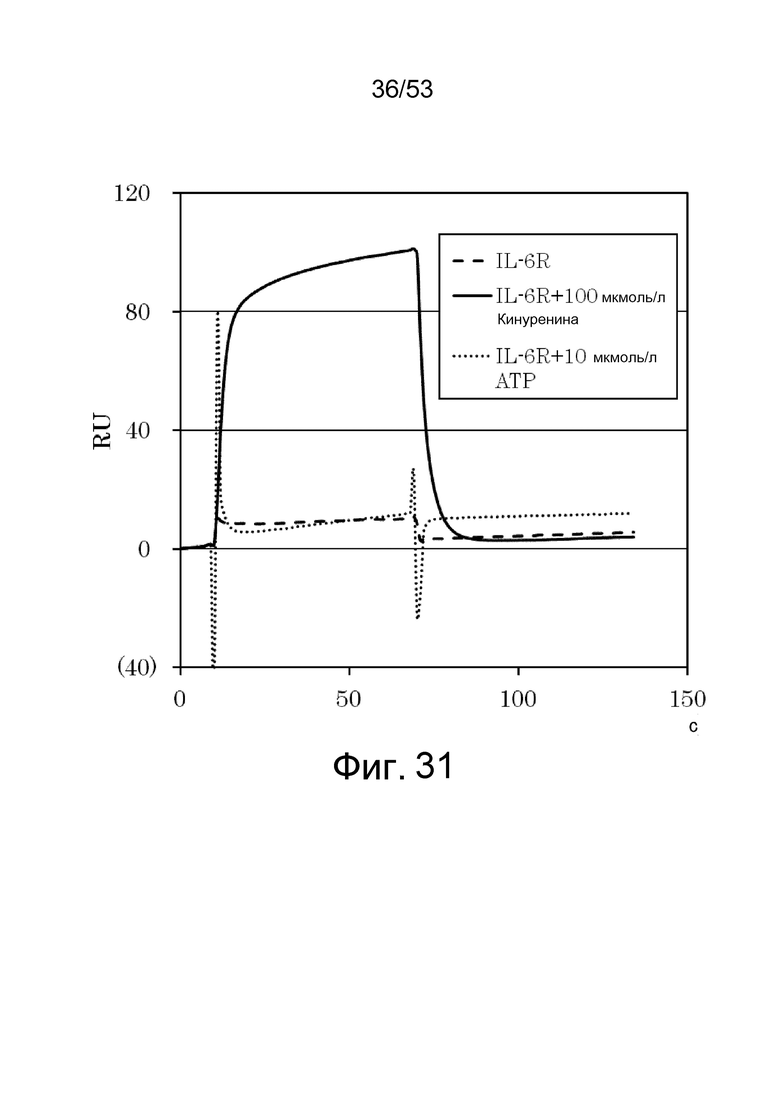
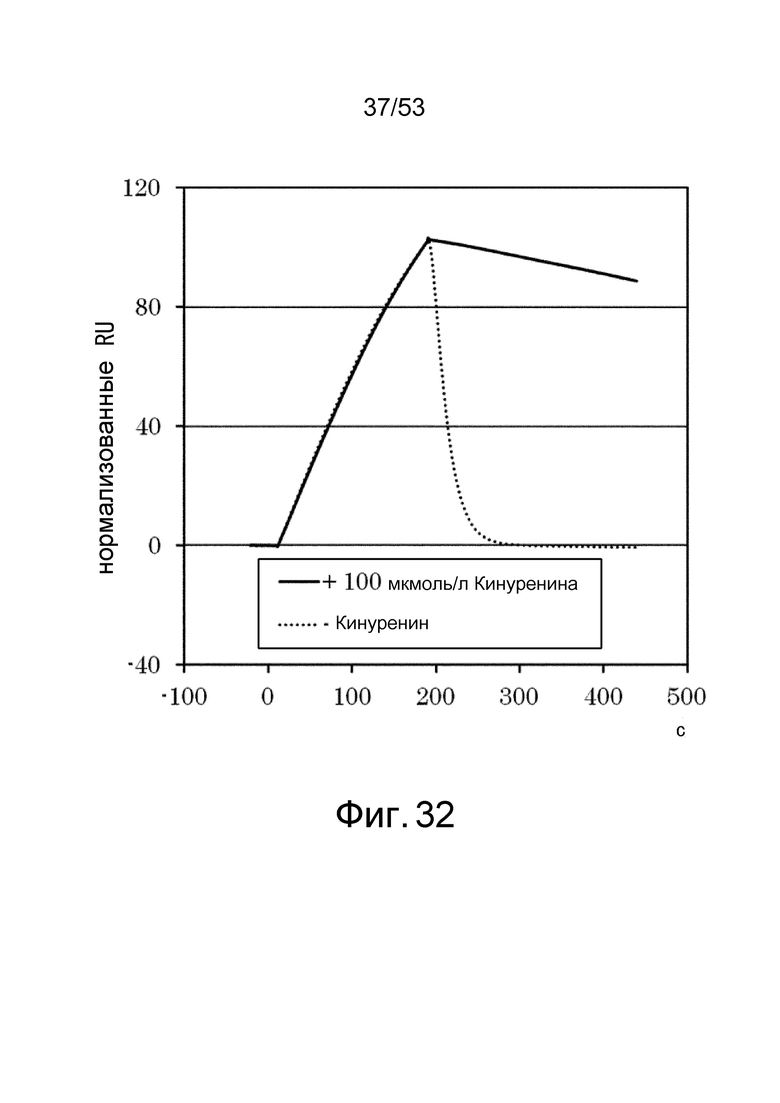
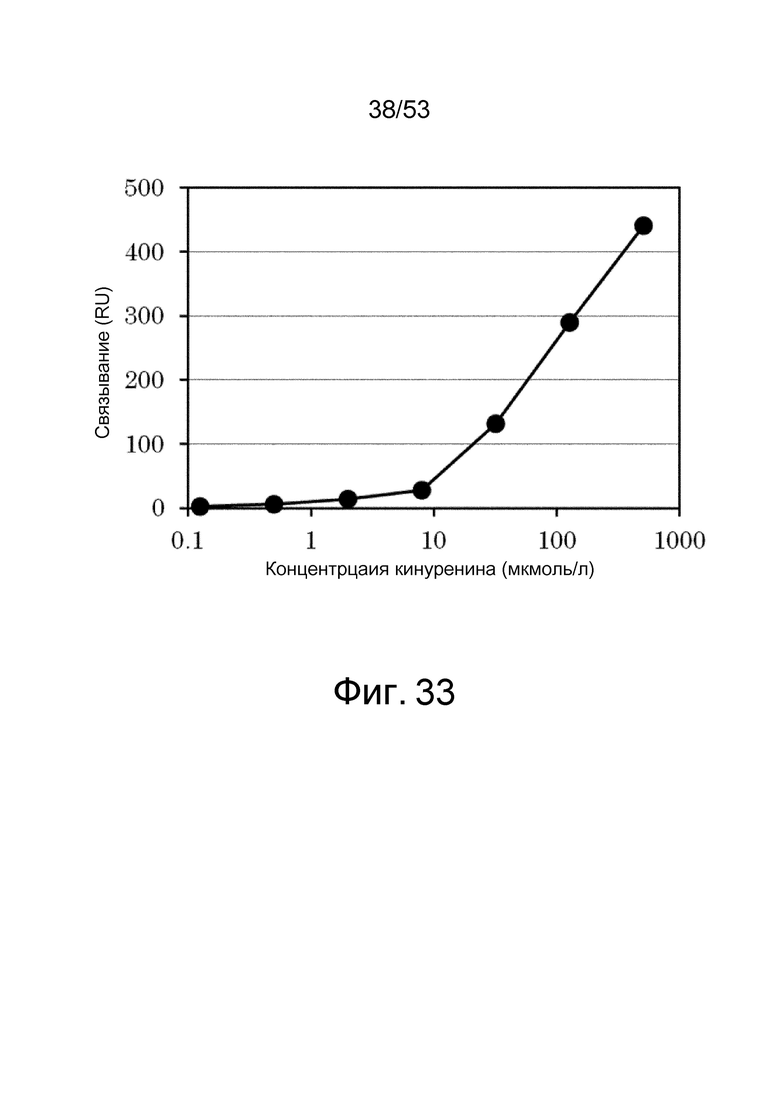
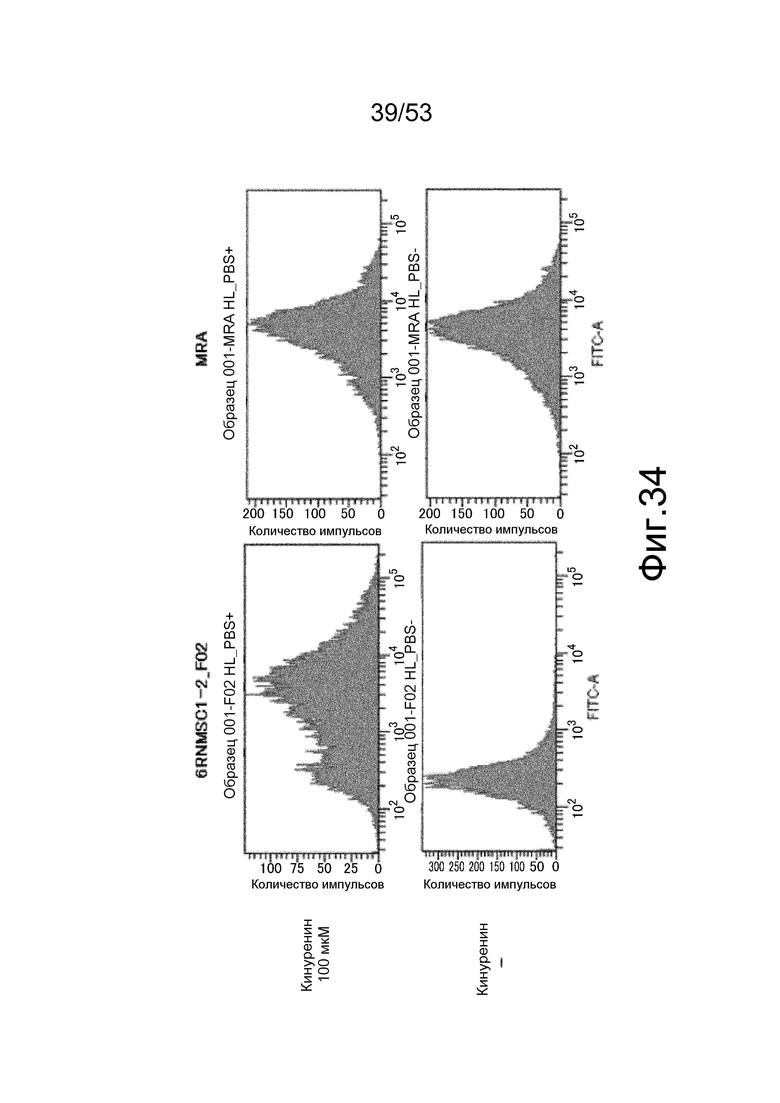
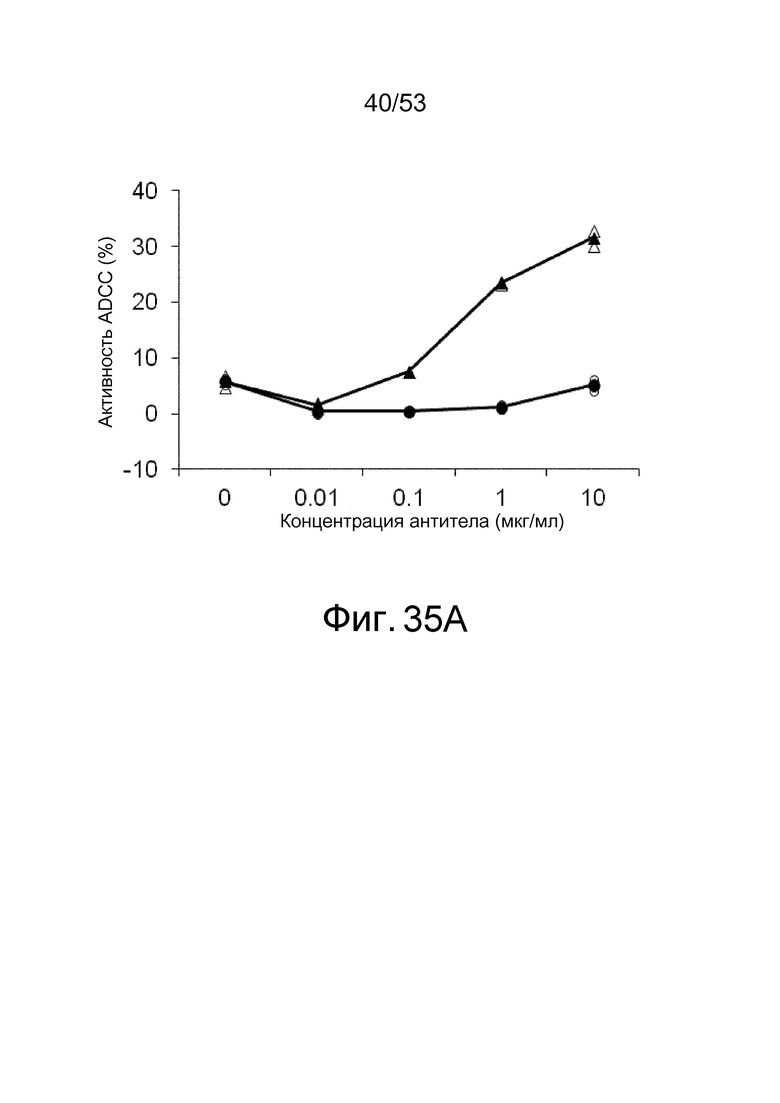
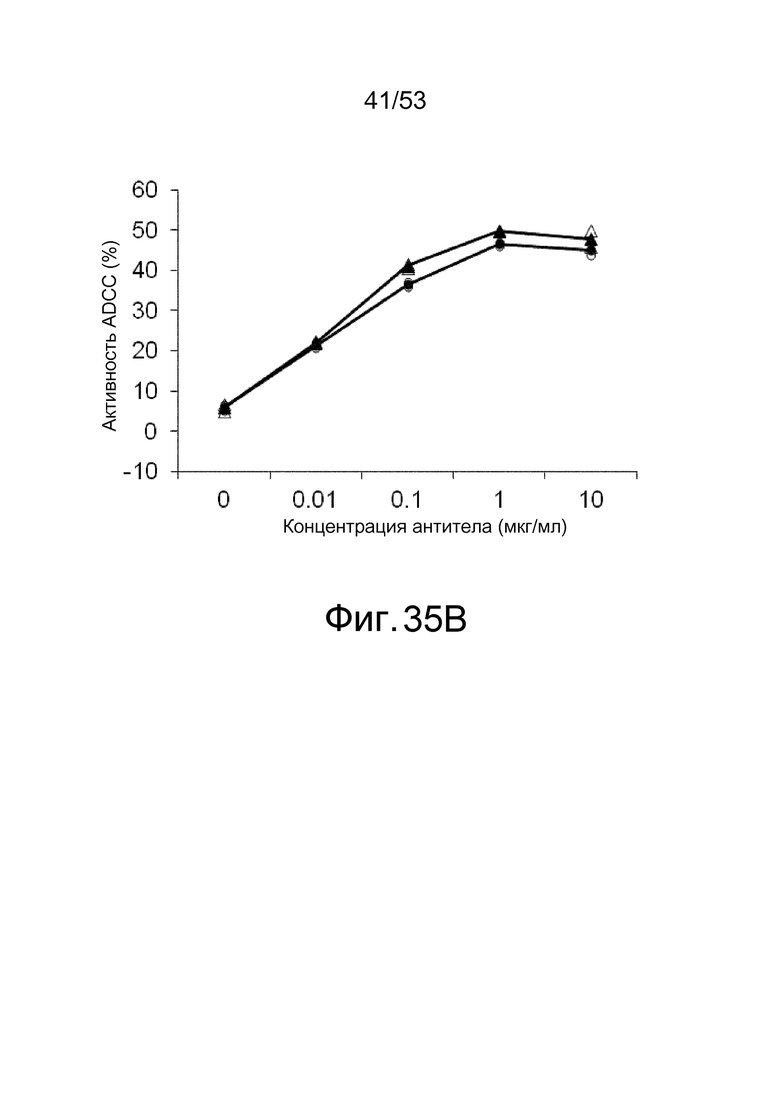
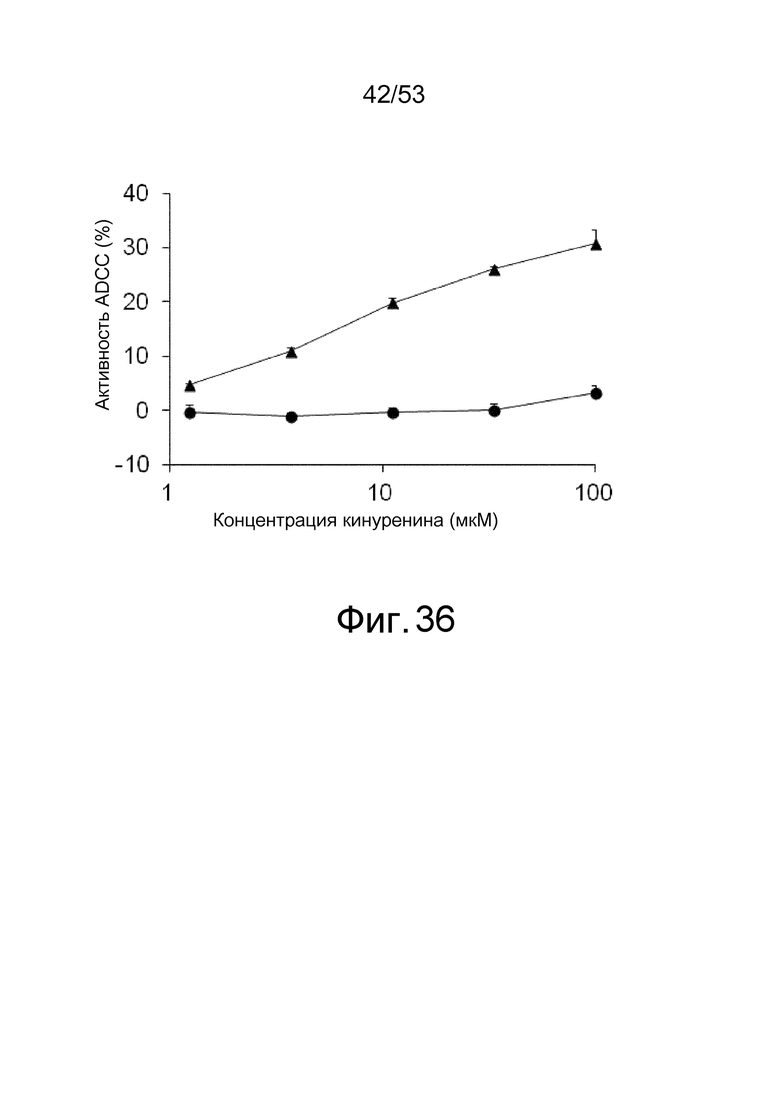
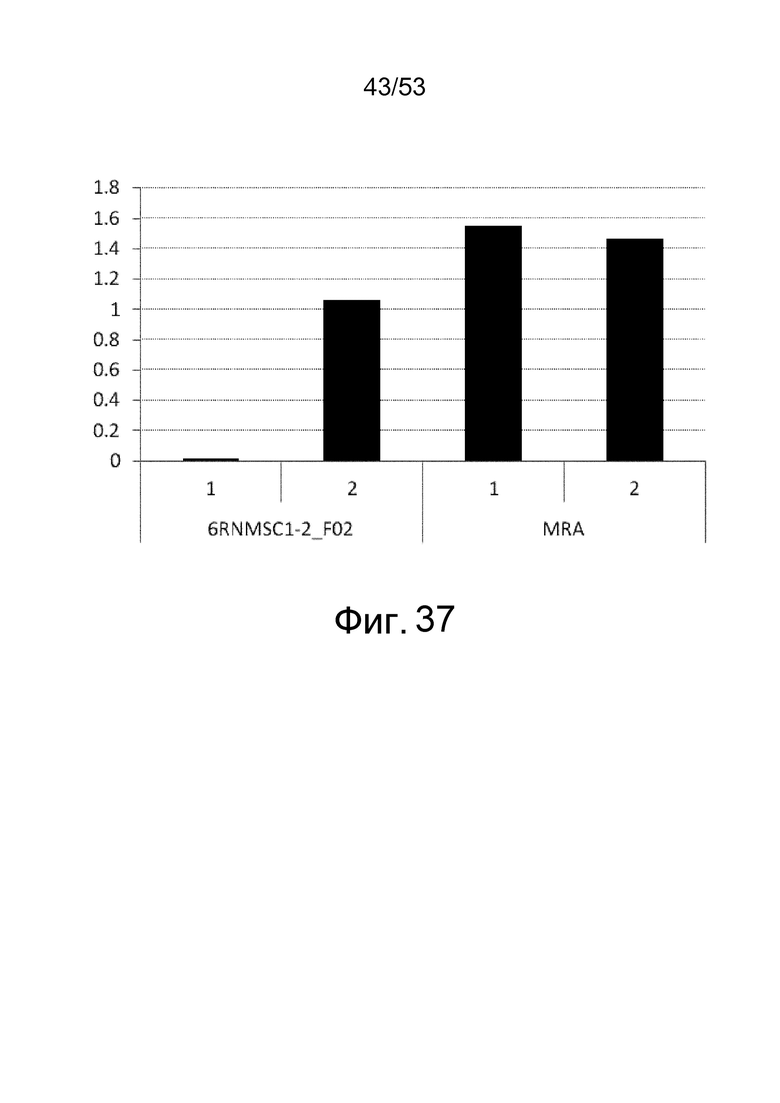
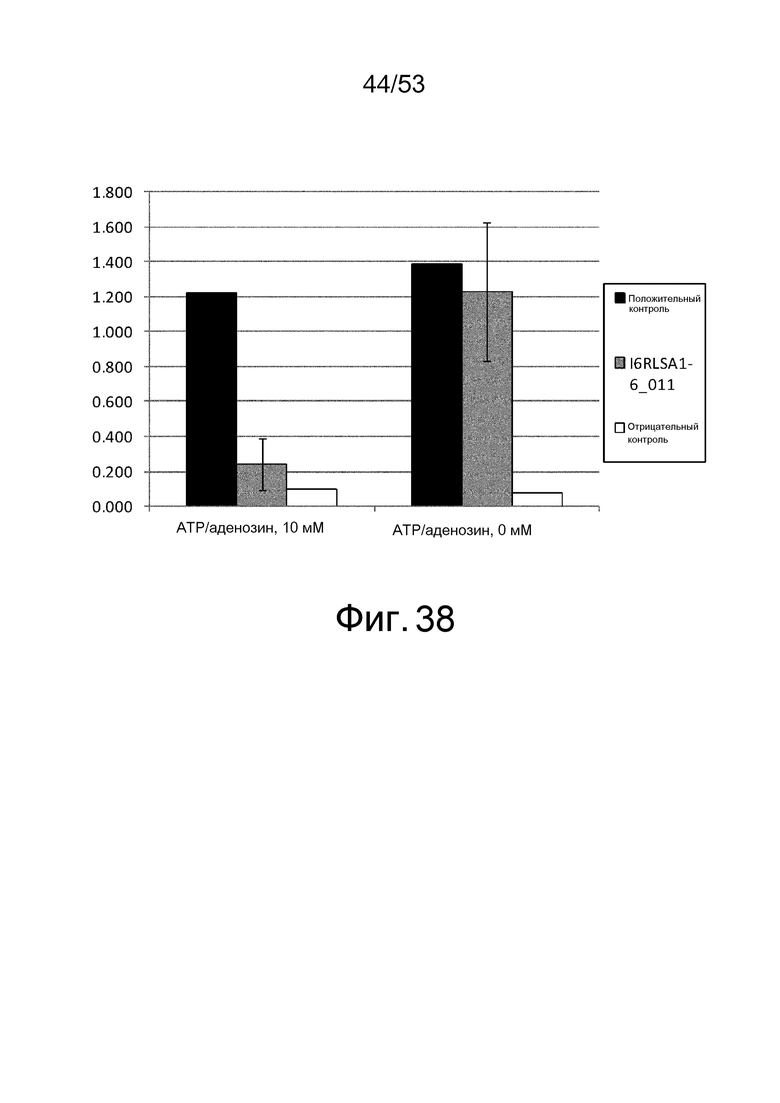
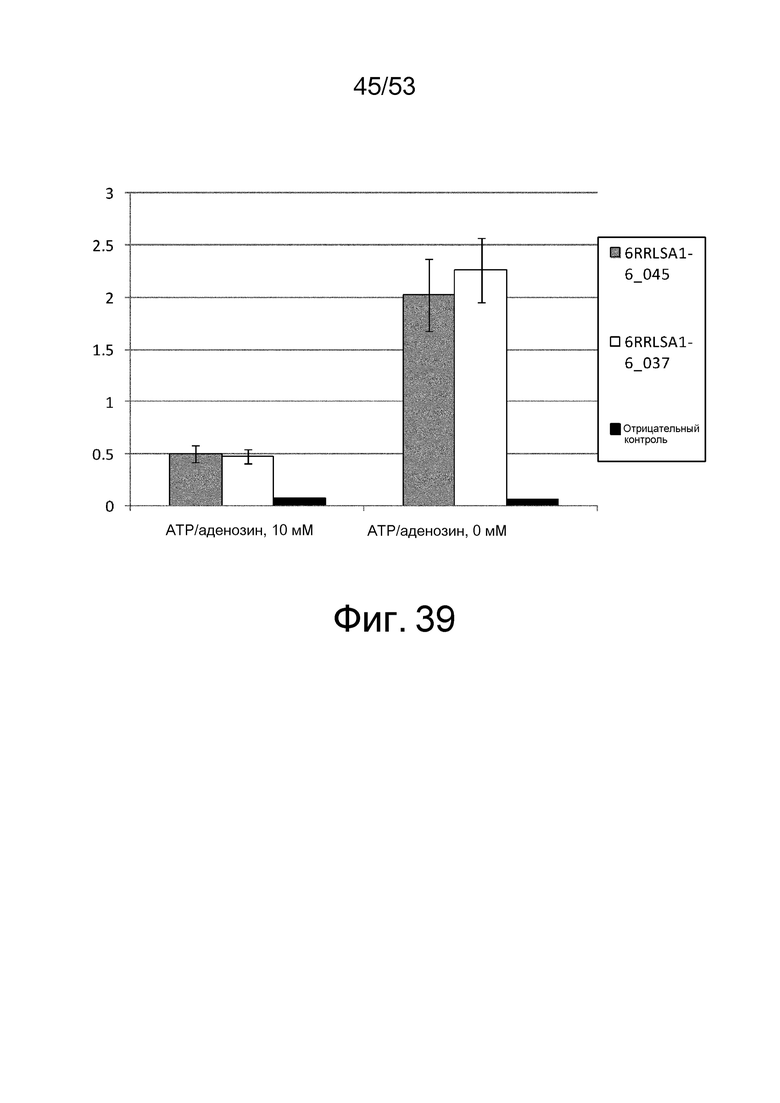
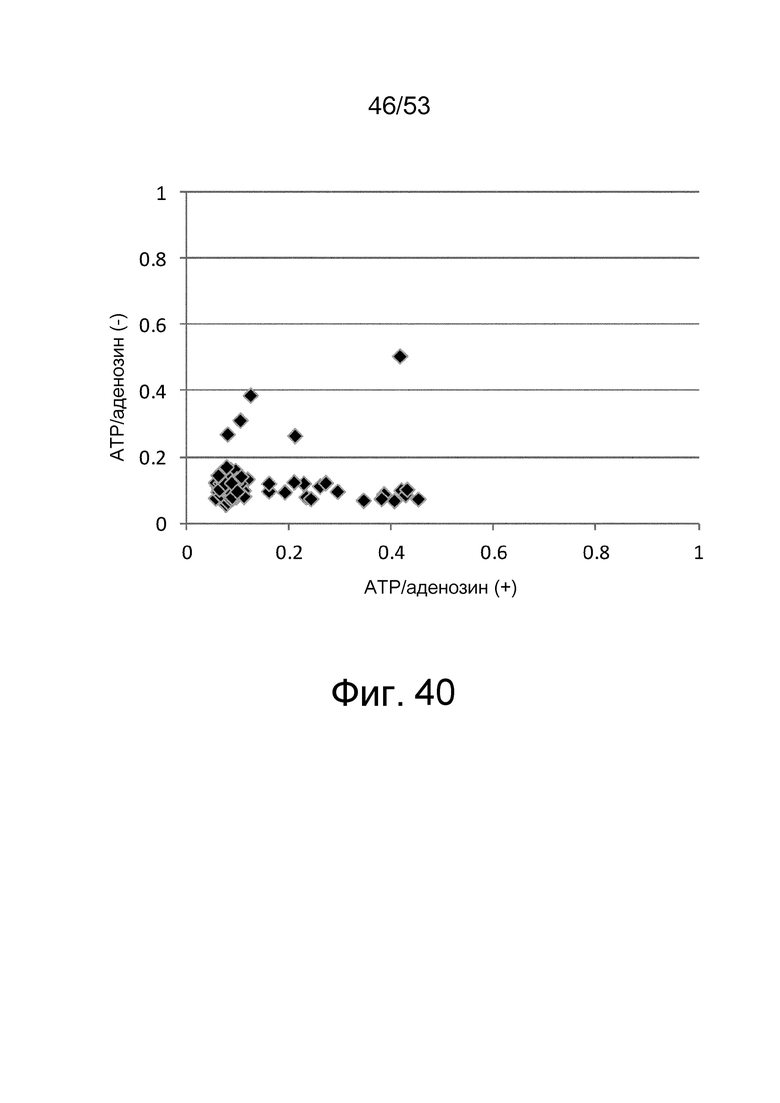

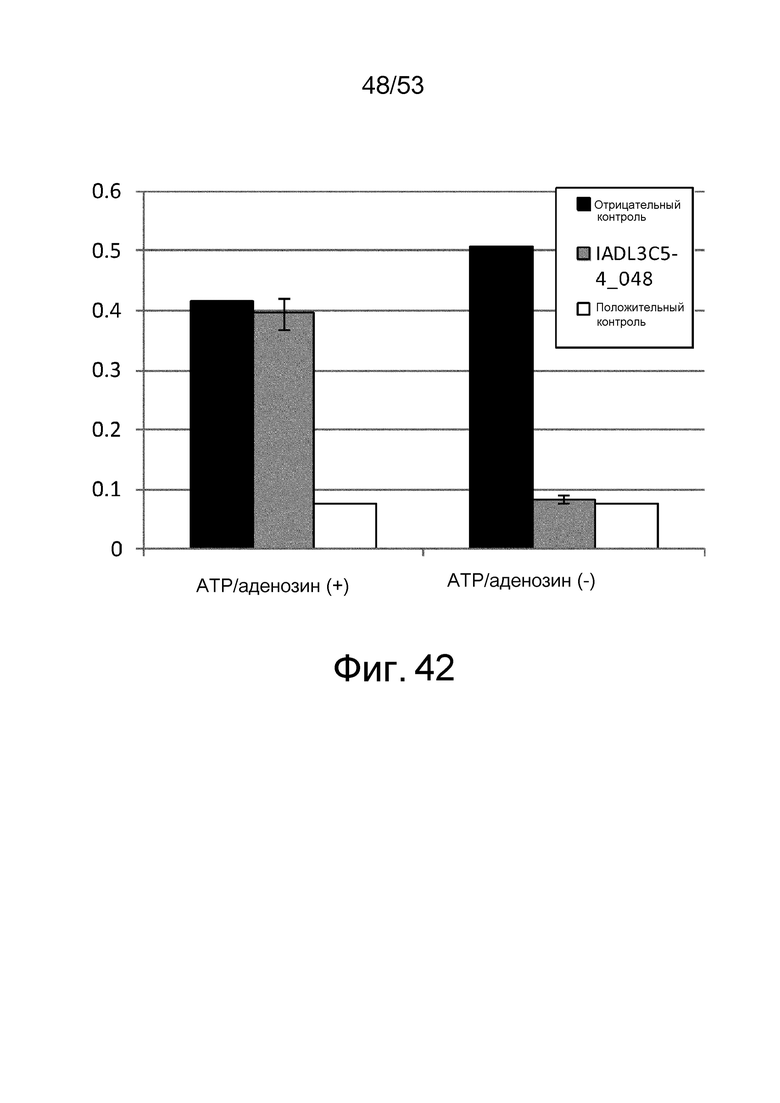
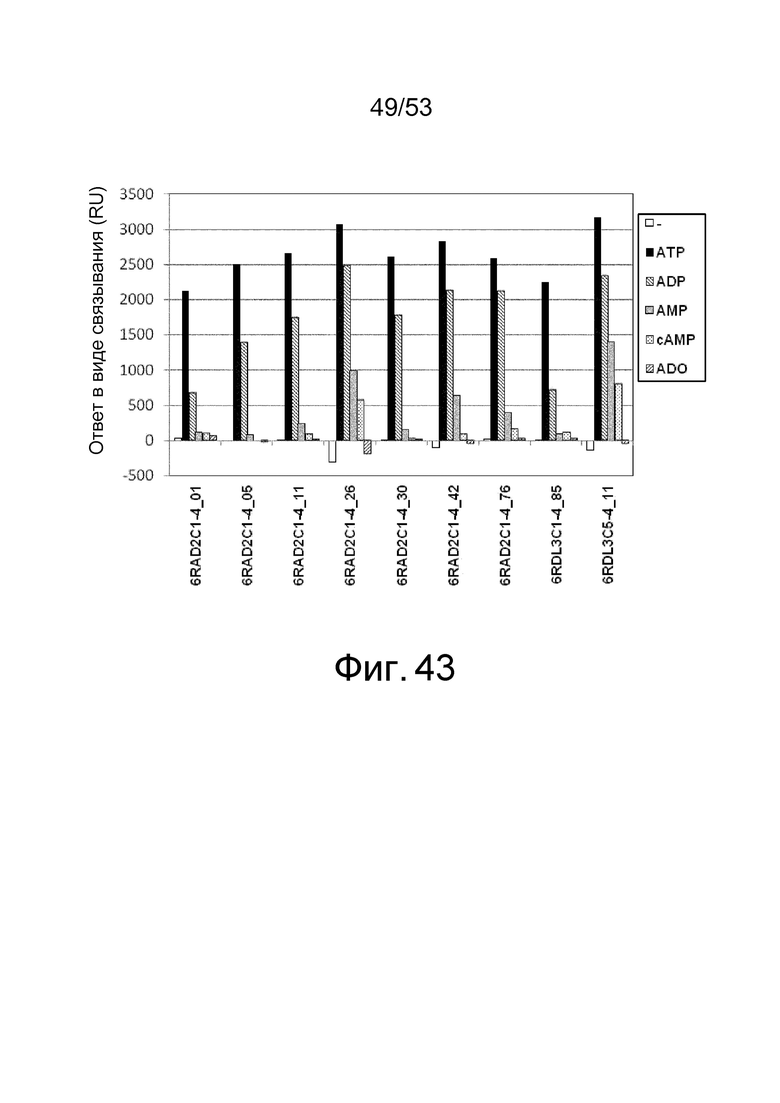
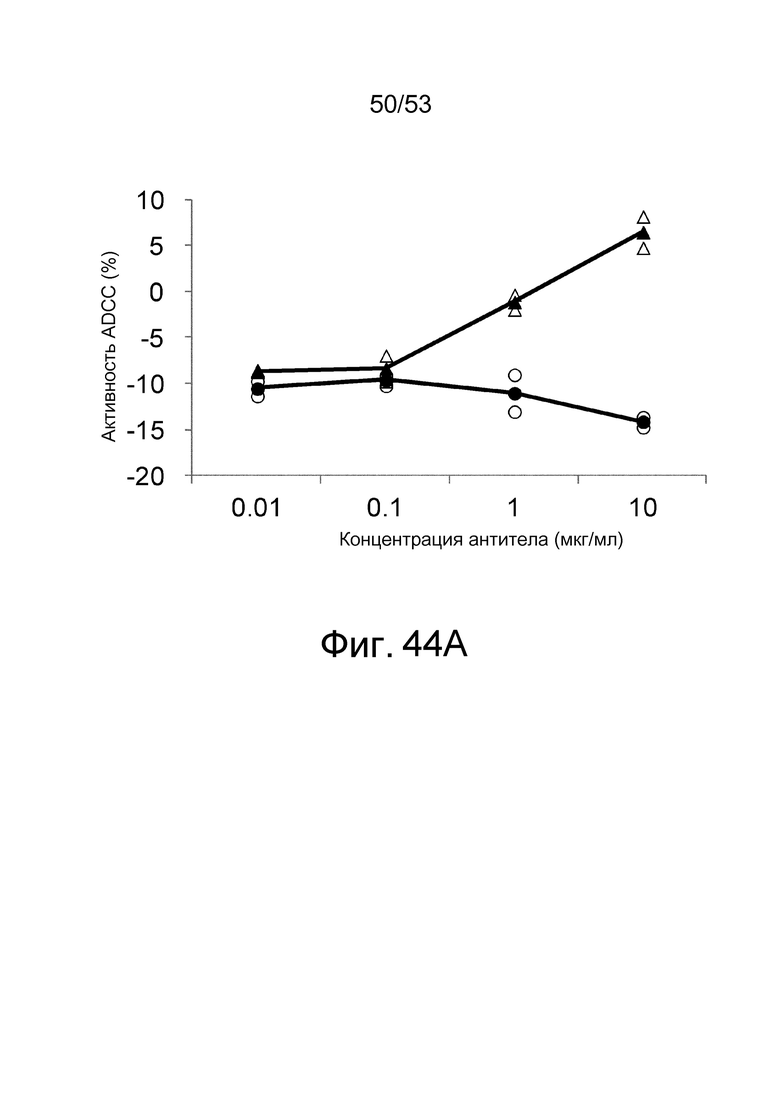

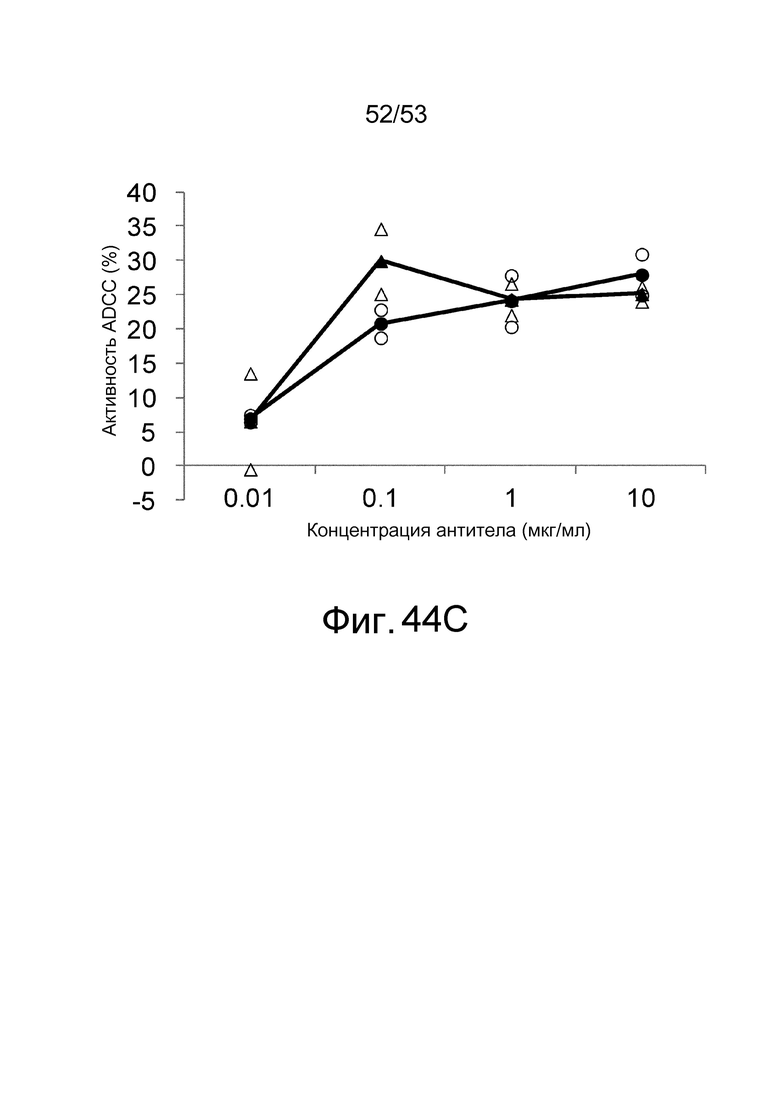
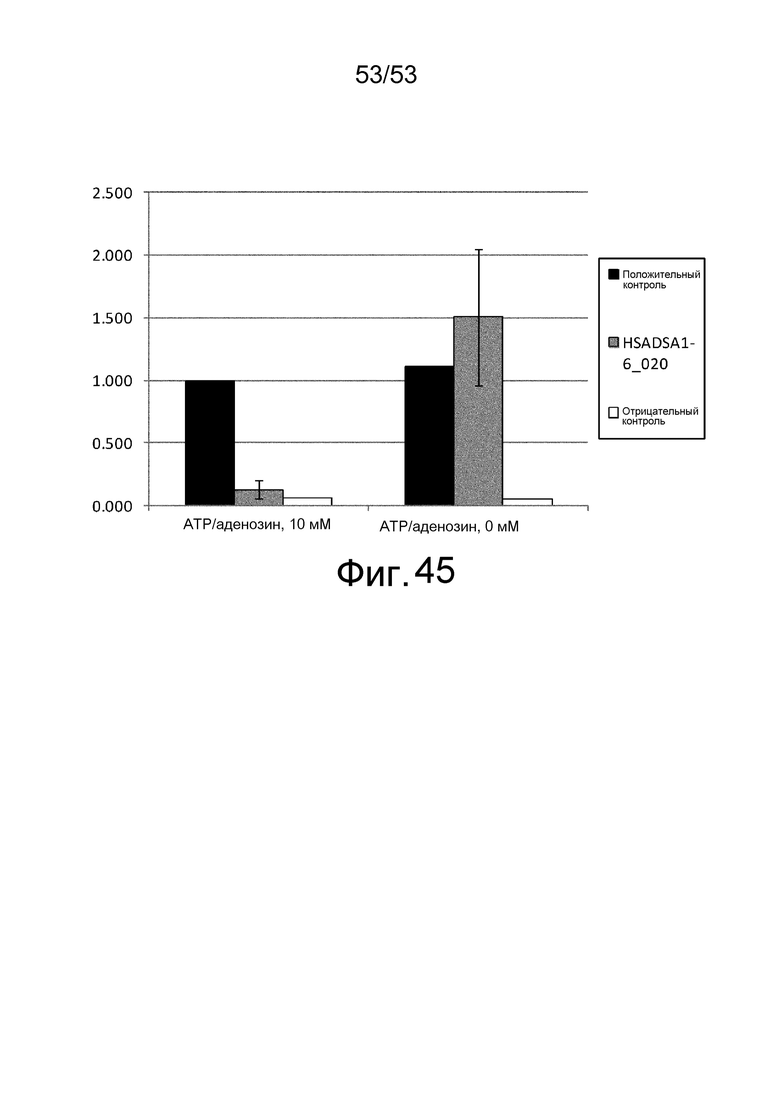
Данная группа изобретений относится к иммунологии. Предложены способы скрининга антигенсвязывающего домена, антигенсвязывающая активность которого варьирует в зависимости от концентрации специфичного к ткани-мишени соединения. Такие способы позволяют получать антигенсвязывающие домены, в частности антигенсвязывающие домены антител, обладающие большей специфичностью для злокачественной ткани, и могут быть пригодны для изготовления фармацевтических препаратов. 3 н. и 13 з.п. ф-лы, 24 табл., 18 пр., 45 ил.
1. Способ скрининга антигенсвязывающего домена, который включает:
(a1) определение антигенсвязывающей активности антигенсвязывающего домена с антигеном в присутствии специфичного для злокачественной ткани соединения при первой концентрации;
(а2) определение антигенсвязывающей активности антигенсвязывающего домена с антигеном в присутствии специфичного для злокачественной ткани соединения при второй концентрации, где вторая концентрация является отличной от первой; и
(а3) отбор антигенсвязывающего домена, антигенсвязывающая активность которого в присутствии соединения при первой концентрации отличается от связывания антигенсвязывающей активности домена с антигеном в присутствии соединения при второй концентрации.
2. Способ по п.1, где вторая концентрация выше, чем первая концентрация, и антигенсвязывающий домен, отобранный на стадии (а3), является антигенсвязывающим доменом, активность связывания с антигеном которого в присутствии соединения при второй концентрации выше, чем активность связывания антигенсвязывающего домена с антигеном в присутствии соединения при первой концентрации.
3. Способ по п.2, где первая концентрация равна нулю.
4. Способ скрининга антигенсвязывающего домена, который включает:
(a1) контактирование антигенсвязывающего домена или библиотеки антигенсвязывающих доменов с антигеном в присутствии соединения, специфичного для злокачественной ткани, при первой концентрации;
(а2) воздействие на антигенсвязывающий домен, который связывается с антигеном на стадии (a1), второй концентрацией соединения, специфичного для злокачественной ткани, где вторая концентрация отличается от первой концентрации;
(а3) отбор антигенсвязывающих доменов, которые отделяются от антигена на стадии (а2).
5. Способ по п.4, где вторая концентрация ниже, чем первая концентрация.
6. Способ по п.5, где вторая концентрация равна нулю.
7. Способ скрининга антигенсвязывающего домена, который включает:
(a1) контактирование антигенсвязывающего домена или библиотеки антигенсвязывающих доменов с антигеном в присутствии соединения, специфичного для злокачественной ткани, при первой концентрации;
(а2) отбор антигенсвязывающих доменов, которые не связываются с антигеном на указанной стадии (a1);
(а3) контактирование антигенсвязывающего домена, отобранного на стадии (а2), с антигеном в присутствии соединения, специфичного для злокачественной ткани, при второй концентрации, где вторая концентрация отличается от первой; и
(а4) отбор антигенсвязывающих доменов, которые связываются с антигеном на стадии (а3).
8. Способ по п.7, где вторая концентрация выше, чем первая концентрация.
9. Способ по п.8, где первая концентрация равна нулю.
10. Способ по любому из пп.1-9, где соединение, специфичное для злокачественной ткани, является метаболитом, специфичным для злокачественной клетки, метаболитом, специфичным для иммунной клетки, которая инфильтрирована в злокачественную ткань, или метаболитом, специфичным для клетки стромы в злокачественной ткани.
11. Способ по любому из пп.1-9, где соединение, специфичное для злокачественной ткани, представляет собой по меньшей мере одно соединение, выбранное из нуклеозида, имеющего структуру пуринового кольца, аминокислоты и ее метаболита, липида и его метаболита, первичного метаболита гликометаболизма, и никотинамида и его метаболита.
12. Способ по п.11, где соединение, специфичное для злокачественной ткани, представляет собой по меньшей мере одно соединение, выбранное из аденозина, аденозинтрифосфата, инозина, аланина, глутаминовой кислоты, аспарагиновой кислоты, кинуренина, простагландина Е2, янтарной кислоты, лимонной кислоты и 1-метилникотинамида.
13. Способ по любому из пп.1-12, где антиген представляет собой молекулу мембранного типа.
14. Способ по любому из пп.1-12, где антиген представляет собой растворимую молекулу.
15. Способ по любому из пп.1-14, где антигенсвязывающий домен представляет собой фрагмент антитела.
16. Способ по любому из пп.1-15, где антигенсвязывающий домен содержит вариабельную область тяжелой цепи антитела и вариабельную область легкой цепи антитела.
| МНОГОПОЛЯРНАЯ ЭЛЕКТРОЛИЗНАЯ ВАННА ДЛЯ ПОЛУЧЕНИЯ ЖИДКИХ МЕТАЛЛОВ ЭЛЕКТРОЛИЗОМ РАСПЛАВОВ И СПОСОБ УСТАНОВКИ ЭЛЕКТРОЛИЗНЫХ ВАНН | 2004 |
|
RU2275443C2 |
| LUTTERBUESE RALF et al., "T Cell-Engaging BiTE Antibodies Specific for EGFR Potently Eliminate KRAS- and BRAF-Mutated Colorectal Cancer Cells", Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(28): 12605-12610; | |||
| LUTTRELL B.M., HENNIKER A.J | |||
| "Reaction coupling of chelation and | |||

