ОБЛАСТЬ ТЕХНИКИ
[0001] Настоящее изобретение относится к грибковым и бактериальным эндофитам растений, повышающим выживаемость семян и/или усиливающим здоровье растений, приводя к общему улучшению сельскохозяйственных характеристик растений при нормальных и стрессовых условиях. Изобретение также относится к таким эндофитам в изолированном состоянии.
УРОВЕНЬ ТЕХНИКИ
[0002] Грибы и бактерии представляют собой повсеместно распространенные микроорганизмы. Термин "эндофит" впервые предложен de Вагу [1866] для обозначения микробов, способных бессимптомно колонизировать растительные ткани [Stone et al., 2000]. О существовании эндофитов известно на протяжении уже более чем столетие [Freeman 1904], и, по-видимому, в каждом отдельно выбранном из 300000 видов растений-хозяев обитает от нескольких до сотен эндофитов [Tan и Zou, 2001]. Эндофиты являются микроорганизмами, связанными с живыми тканями растений-хозяев преимущественно симбиотическими или мутуалистическими отношениями. Многие из них способны повышать устойчивость растений к абиотическим стрессам факторам или могут использоваться растением для защиты от патогенных грибов и бактерий [Singh et al. 2011]. Некоторые из этих микроорганизмов оказались полезными для сельского хозяйства, лесного хозяйства и садоводства, а также, как и растения для производства важных медицинских соединений.
[0003] Эндофиты в значительной степени детерминируют процессы регуляции в растительных клетках и целом геноме растений, в том числе, жизненный цикл растения: (i) процессы, происходящие до и после прорастания семян (миковитализм) [Vujanovic и Vujanovic 2007], (ii) поглощение питательных веществ растением и механизмы стимуляции роста (микогетеротропизм) [Smith и Read 2008] и (iii) невосприимчивость растений к стрессу, обусловленному воздействием окружающей среды и индуцированной системной устойчивости к болезням и вредителям (микосимбиотицизм) [Wallin 1927; Margulis, 1991]. Они могут играть важную роль для выработки биомассы растениями, поглощения СО2 и/или изменения урожайности и, следовательно, имеют важное значение в регуляции экосферы, обеспечивая здоровье растений и продовольственную безопасность. Кроме того, они могут быть важными детекторами (биоиндикаторами) изменений окружающей среды, так, например, изменения структуры и биомассы эндофитных сообществ могут предвещать изменения не только в путях поступления нутриентов (N, Р, К), передачи энергии в пищевых цепях и биогеохимических циклах, но и в изменениях устойчивости к ультрафиолету (УФ-В), повышенной температуре, засухе и засолению, которые влияют на формирование и общую стабильность растительных экосистем. Несмотря на обилие и, вероятно, значимость во всех наземных экосистемах, почти ничего не известно о составе эндофитов семян или спермосферы, взаимосвязей между ними и их общей ответной реакции на изменения окружающей среды.
[0004] Спермосфера представляет собой быстро меняющуюся и микробиологически динамичную зону почвы, окружающую прорастающие семена [Nelson, 2004], тогда как ризосфера является микробиологически активной зоной грунтовой массы, окружающей корни растений [Smith и Read 2008]. Ризосфера поддерживает микогетеротрофию, или симбиорические отношения микориза-растение. Спермосфера, с другой стороны, способствует формированию миковитальности или взаимоотношений между эндофитными грибами и семенами растений, увеличивающих силу, энергию и дружность прорастания семян, значение чего может быть довольно прогнозируемым. Грибковые эндофиты отличаются от микоризы тем, что могут колонизировать не только корни, но и другие органы растений, включая семена [Vujanovic et al. 2000; Hubbard et al. 2011]. Они принадлежат к многоклеточным грибам отделов Ascomycota и Basidiomycota, и форма осуществляемой ими колонизации симбиотических структур отличается от той, которую формируют одноклеточные или ценоцитные представители отдела Glomeromycota, известной как везикулярного-арбускулярный микоризный симбиоз [Abdellatif et al. 2009]. Эндофитные бактерии были также найдены в практически каждом изученном растении, где они занимают экологическую нишу, подобную нише грибов, как, например, в здоровых внутренних тканях. Хотя большинство бактерий-эндофитов, по-видимому, начинают свой жизненный цикл в ризосфере или филосфере; некоторые из них могут передаваться с семенами [Ryan et al. 2008].
[0005] Прорастание семян является чрезвычайно важным этапом жизненного цикла для размножения и выживания растений при оптимальных и стрессовых условиях окружающей среды. Колонизация микробами-эндофитами на стадии семени является особенно критичной, потому что семя исполняет роль органа размножения цветковых растений, важного для их восстановления и распространения [Baskin и Baskin 2004] и по причине играемой микобионтами и симбиотически ассоциированными бактериями (бактобиотами) роли потенциальных активаторов запуска прорастания в природной - невозмущенной, возмущенной или загрязненной - среде обитания [Miihlmann и Peintner 2000; Adriaensen et al. 2006; White и Torres 2010]. Таким образом, разработка способов, посредством которых можно повысить всхожесть и препятствовать ее ограничению воздействием болезней, жарой или засухой, является важной. Использование эндофитных симбионтов является перспективным способом, с помощью которого может быть повышена всхожесть семян. [Vujanovic et al. 2000; Vujanovic и Vujanovic 2006; Vujanovic и Vujanovic 2007]. Было высказано предположение, что стрессоустойчивость растений можно обеспечить при помощи отношений между микобионтом и сменами, известных как миковитальность - явления, обнаруженного у семейства Orchidaceae [Vujanovic 2008] и при помощи бактовитальности, которая является формой бактосимбиоза, путем использования различных штаммов эндофитов, действующих различными способами.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0006] Эндофиты могут приносить пользу растениям-хозяевам, таким как пшеница, ячмень, бобовые, рапс, деревья, кусты или травянистые растения различными способами, в том числе и путем бактовитальности, миковитальности и микогетеротрофии, повышая их устойчивость к экологическим стрессам, как это уже было показано в данном документе. Пренатальный уход в земледелии, как было уже показано на шести эндофитных штаммах, определяет намного больше, чем просто жизнеспособность, здоровье и силу семян или проростков. Он также определяет, чего ожидать до и во время процесса прорастания, образования проростков, и, в последствии, от продуктивности урожая или урожайности.
[0007] Ряд параметров симбиотической эффективности (выход из состояния покоя, прорастание, рост и урожайность) оценивались при взаимодействии соответствующего эндофитного штамма(ов) из Saskatchewan Microbial Collection and Database (SMCD) с культурой(ами) in vitro в фитотроне, теплице и полевых условиях.
[0008] Также были проверены способность бактериальных эндофитов повышать жизнеспособность семян. Как в случае с грибными, так и с бактериальными эндофитами, улучшенная жизнестойкость семян, может повышать устойчивость растений к абиотическим и биотическим стрессовым факторам на стадии прорастания, и до созревания растения благодаря микогетеротрофии.
[0009] Соответственно, в настоящем изобретении предложен изолированный штамм эндофита Streptomyces sp., или его культура, которая хранится в International Depositary Authority of Canada (IDAC, Национальная микробиологическая лаборатория. Агентство общественного здравоохранения Канады. 1015 Арлингтон стрит, Виннипег, Манитоба, Канада R3E 3R2) шифр доступа 081111-06, или который содержит 16S рДНК с последовательностью, представленной в SEQ ID NO: 6; изолированный штамм эндофита Paraconyothirium sp., или его культуру, которая хранится ID АС под шифром доступа 081111-03, или который содержит ITS рДНК, с последовательностью, представленной в SEQ ID NO: 5; изолированный эндофит Pseudeurotium sp. или его культуру, которая хранится под шифром доступа ID АС 081111-02, или которая содержит ITS рДНК с последовательностью, представленной в SEQ ID NO: 4; изолированный эндофит Penicillium sp. или его культуру, которая хранится под шифром доступа ID АС 081111-01, или который содержит ITS рДНК с последовательностью, представленной в SEQ ID NO: 03; изолированную культуру Cladosporium sp., которая хранится под шифром доступа ID АС 200312-06, или которая содержит ITS рДНК с последовательностью, представленной в SEQ ID NO: 1, и/или выделенный эндофит Cladosporium sp. или его культуру, которая хранится под шифром доступа ID АС 200312-05, или которая содержит ITS рДНК с последовательностью, представленной в SEQ ID NO: 2.
[0010] Также в данном документе предлагается композиция, содержащая изолированный эндофит или культуру, представленную в данном документе, или их комбинацию, или их смесь с носителем.
[0011] Дополнительно в данном документе предлагается семя, содержащее эндофиты или культуры, описанные в данном документе. В одном из вариантов реализации изобретения семя покрыто эндофитами. В другом варианте реализации изобретения семя культивируется или высаживается рядом с эндофитами таким образом, чтобы эндофиты могли колонизировать семя.
[0012] В настоящем изобретении также предлагаются способы повышения жизнеспособности семени и улучшения здоровья растения, и повышения урожайности при нормальных и стрессовых условиях. Следовательно, в нем предлагается способ улучшения жизнеспособности семени и здоровья растения и/или повышения урожайности растения, включающий следующие этапы: инокуляцию семени эндофитами или культурой, описанной в данном документе, или их комбинацией, или их смесью с композицией, описанной в данном документе; и выращивание из семени растения первого поколения.
[0013] В одном из вариантов реализации изобретения, способ включает следующие этапы: инокуляция семени изолированным эндофитом Streptomyces sp. или его культурой, которая хранится под шифром ID АС 081111-06, или которая содержит 16S рДНК с последовательностью, представленной в SEQ ID NO: 6. В варианте реализации изобретения, способ повышает всхожесть семени, уменьшает время, необходимое для достижения энергии прорастания, уменьшает гидротермальное время, необходимое для прорастания, повышает силу прорастания семени, увеличивает сырой вес проростков, повышает частоту образования клубней и активность бактерий рода Rhizobium, и/или повышает урожайности проростков. В другом варианте реализации изобретения способ включает уменьшение последствий воздействия стресса, такого как засуха, тепловой стресс и/или биотический стресс, такой как инфицирование патогенным грибом рода Fusarium.
[0014] В другом варианте реализации изобретения, способ включает следующие этапы: инокуляция семени изолированным эндофитом Paraconyothirium sp. или его культурой, которая хранится под шифром ID АС 081111-03, или которая содержит ITS рДНК с последовательностью, представленной в SEQ ID NO: 5. В варианте реализации изобретения, способ увеличивает всхожесть семени, уменьшает время, необходимое для достижения энергии прорастания, уменьшает гидротермальное время, необходимое для прорастания, повышает силу прорастания семени, увеличивает сырой вес проростков, и/или увеличивает урожайность проростков. В другом варианте реализации изобретения, способ включает уменьшение последствий стресса, такого как засуха, тепловой стресс и/или биотический стресс, такой как инфицирование патогенным грибом рода Fusarium.
[0015] В еще одном варианте реализации изобретения, способ включает следующие этапы: инокуляцию семени изолированным эндофитом Pseudeurotium sp. или его культурой, которая хранится в ID АС под шифром 081111-02, или которая содержит ITS рДНК с последовательностью, представленной в SEQ ID NO: 4. В варианте реализации изобретения, способ уменьшает время, необходимое для достижения энергии прорастания, уменьшает гидротермальное время, необходимое для прорастания, увеличивает силу прорастания семени и/или увеличивает сырой вес проростков. В другом варианте реализации изобретения, способ включает уменьшение последствий стресса, такого как засуха и/или тепловой стресс.
[0016] В дополнительном варианте реализации изобретения, способ включает следующие этапы: инокуляцию семени изолированным эндофитом Pseudeurotium sp. или его культурой, которая хранится в ID АС под шифром 081111-01, или которая содержит ITS рДНК с последовательностью, представленной в SEQ ID NO: 3. В варианте реализации изобретения, способ повышает всхожесть семени, уменьшает время, необходимое для достижения энергии прорастания, снижает гидротермальное время, необходимое для прорастания, повышает силу прорастания семени и/или повышает урожайность проростков. В другом варианте реализации изобретения способ включает усиленную стратификацию, прерывание состояния покоя и увеличение стрессоустойчивости путем модуляции экспрессии генов гормоновэнт-кауриновой кислоты (КАО), репрессора роста побегов (RSG), абсцисовой кислоты (АБК (ABA)), гибберелиновой кислоты (TK(GA)), генов 14-3-3 или оксида азота (NO) и/или генов стрессоустойчивости супероксиддисмутазы (СОД (SOD)), марганец-супероксиддисмутазы (МпСОД), генов пролина (Pro) и MYB, снижения последствий воздействия стресса, такого как засуха, тепловой стресс и/или биотического стресса, такого как инфицирование патогенным грибом рода Fusarium.
[0017] В дополнительном варианте реализации изобретения, способ включает следующие этапы: инокуляцию семени изолированным эндофитом Cladosporium sp. или его культурой, которая хранится в ID АС под шифром 200312-06, или которая содержит ITS рДНК с последовательностью, представленной в SEQ ID NO: 1. В варианте реализации изобретения, способ уменьшает время, необходимое для достижения энергии прорастания, уменьшает гидротермальное время, необходимое для прорастания, увеличивает силу прорастания семени и/или увеличивает сырой вес проростков. В варианте реализации изобретения, способ включает уменьшение последствий стресса, такого как засуха и/или тепловой стресс.
[0018] В еще одном варианте реализации изобретения, способ включает следующие этапы: инокуляцию семени изолированным эндофитом Cladosporium sp. или его культурой, которая хранится в ID АС под шифром 200312-05, или которая содержит ITS рДНК с последовательностью, представленной в SEQ ID NO: 2. В варианте реализации изобретения, способ включает уменьшение последствий стресса, такого как засуха, и/или тепловой стресс.
[0019] В варианте реализации изобретения семя покрыто эндофитами, культивируются с эндофитами, или высаживаются рядом с эндофитами. В конкретном варианте реализации семя высаживают рядом с эндофитами, на расстоянии около 4 см от эндофита.
[0020] В качестве растения может быть использовано любое растение. В одном из вариантов реализации изобретения, использованное растение представляет собой злаковые (пшеница или ячмень), бобовые (горох, чечевица или нут), лен, рапс, хвойное дерево (ель или сосна), широколиственное дерево (ива или тополь), кустарник (карагана или терескен) или травянистое растение (овсяница или волоснец).
[0021] В ином аспекте, в данном документе предлагается способ улучшения здоровья растений и/или урожайности растений, включающий в себя обработку растительного посадочного материала или самого растения эндофитом или его культурой, описанной в данном документе, или их комбинацией, смесью или композицией, описанной в данном документе; и культивирование растительного посадочного материала до растение первого поколения, или предоставление возможности растению расти.
[0022] В варианте реализации изобретения, растительный посадочный материал - это любая генеративная/половая (семя, генеративная почка или цветок) или вегетативная/бесполая (стебель, черенок, корень, луковица, корневище, клубень, вегетативная почка или лист) часть растения, из которой можно культивировать новое растение.
[0023] В варианте реализации изобретения изолированным эндофитом, или культурой такого эндофита, являлся изолированный штамм эндофита Streptomyces sp. или его культура, которая хранится в International Depositary Authority of Canada (IDAC, Национальная микробиологическая лаборатория. Агентство общественного здравоохранения Канады. 1015 Арлингтон стрит, Виннипег, Манитоба, Канада, R3E 3R2) код доступа 081111-06, или который содержит 16S рДНК с последовательностью, представленной в SEQ ID NO: 6; изолированный штамм эндофита Paraconyothirium sp. или его культура, которая хранится под шифром доступа ID АС 081111-03, или которая содержит ITS рДНК с последовательностью, представленной в SEQ ID NO: 5; изолированный эндофит Pseudeurotium sp. или его культура, которая хранится под шифром доступа ID АС 081111-02, или которая содержит ITS рДНК с последовательностью, представленной в SEQ ID NO: 4; изолированный эндофит Penicillium sp. или его культура, которая хранится под шифром доступа IDAC 081111-01, или которая содержит ITS рДНК с последовательностью, представленной в SEQ ID NO: 3; изолированный эндофит Cladosporium sp. или его культура, которая хранится под шифром доступа ID АС 200312-06, или которая содержит ITS рДНК с последовательностью, представленной в SEQ ID NO: 1; и/или изолированный эндофит Cladosporium sp. или его культура, которая хранится под шифром доступа ID АС 200312-05, или которая содержит ITS рДНК с последовательностью, представленной в SEQ ID NO: 2.
[0024] В варианте реализации изобретения, способы стимулируют процессы развития ландшафта и ремедиации. Следовательно, в одном из вариантов реализации изобретения предложен способ уменьшения загрязнения почвы, включающий: обработку растительного посадочного материала или растения эндофитами, или культурой, описанной в данном документе, или их комбинацией, или их смесью, или композицией, описанной в данном документе; и культивирование растительного посадочного материала до растения первого поколения или предоставление возможности растению расти. В одном варианте реализации изобретения, загрязнителем почвы являлются углеводороды, нефть или другие химические вещества, соли или металлы, такие как свинец, кадмий или радиоизотопы.
[0025] В другом варианте реализации изобретения, способы уменьшают последствия воздействия стрессовых факторов, таких как засуха, повышенная температура и/или биотических стрессовых факторов.
[0026] В качестве растения может быть использовано любое растение. В одном из вариантов реализации изобретения, растение является злаковым (пшеница или ячмень), бобовым (горох, чечевица или нут), льном, рапсом, хвойным деревом (ель или сосна), широколиственным деревом (ива или тополь), кустарником (карагана илитерескен) или травянистым растением (овсяница или волоснец).
[0027] Другие свойства и преимущества данного изобретения станут очевидными из следующего подробного описания. Однако следует учитывать, что подробное описание и конкретные примеры, раскрывающие отдельные предпочтительные варианты реализации данного изобретения, приведены только в иллюстративных целях и различные изменения в сущности и объеме изобретения будут явными для специалистов в данной области техники из приведенного детального описания и соответствующих графических материалов и условных обозначений графических материалов.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0028] Раскрытие изобретения далее будет описано со ссылкой на графические материалы, где:
[0029] Фигура 1 иллюстрирует фенотипическое проявление эндофитных грибов штаммов SMCD 2204 2004f, 2206, 2208, 2210 и бактериального штамма SMCD 2215; после 10 дней роста на КДА при 21°С.
[0030] Фигура 2 иллюстрирует предполагаемое филогенетическое дерево Cladosporium spp. SMCD 2204 и SMCD 2204F, созданное методом присоединения соседей, исходя из ITS рДНК. Числа на узловых точках отображают значения статистического бутстрэпа, полученные при 1000 повторений; показаны только значения >70%. Планка отображает 0,01 нуклеотидных замен на сайт (положение нуклеотида).
[0031] Фигура 3 иллюстрирует предполагаемое филогенетическое дерево Penicillium sp. SMCD 2206, созданное методом присоединения соседей на основании ITS рДНК. Числа на узловых точках отображают значения статистического бутстрэпа, полученные при 1000 повторений; показаны только значения >70%. Планка отображает 0,01 нуклеотидных замен на сайт (положение нуклеотида).
[0032] Фигура 4 иллюстрирует предполагаемое филогенетическое дерево Pseudeurotium sp. SMCD 2208, созданное методом присоединения соседей на основании ITS рДНК. Числа на узловых точках отображают значения статистического бутстрэпа, полученные при 1000 повторений; показаны только значения >70%. Планка отображает 0,01 нуклеотидных замен на сайт (положение нуклеотида).
[0033] Фигура 5 иллюстрирует предполагаемое филогенетическое дерево Coniothyrium штамма SMCD 2210, созданное методом присоединения соседей на основании ITS рДНК. Числа на узловых точках отображают значения статистического бутстрэпа, полученные при 1000 повторений; показаны только значения >70%. Планка отображает 0,05 нуклеотидных замен на сайт (положение нуклеотида).
[0034] Фигура 6 показывает предполагаемое филогенетическое дерево штамма Streptomyces sp SMCD 2215, созданное методом присоединения соседей на основании 16S рДНК. Числа на узловых точках отображают значения статистического бутстэпа, полученные при 1000 повторений; показаны только значения >60%. Планка отображает 0,05 нуклеотидных замен на сайт (положение нуклеотида).
[0035] Фигура 7 иллюстрирует правую часть разделенной чашки (растение с микробным партнером): проявление фенотипических признаков здоровой пшеницы, корни которой во время роста контактировали с бактериальным матом; и левую часть разделенной чашки (растение без микробного партнера): массовое формирование корневых волосков пшеницей, обусловленное взаимосвязями между грибком и растением, сформированными в левой части разделенной чашки.
[0036] Фигура 8(А) и (С) иллюстрирует прерывистую колонизацию тканей корней пшеницы (эпидермиса и коры) SMCD2206, в сравнении с (В) и (D), которые иллюстрируют патогенную равномерную/непрерывную клеточную колонизацию корней пшеницы грибом Fusarium graminearum, в том числе в зоне сосудистого цилиндра.
[0037] Фигура 9 иллюстрирует индекс Ireg - уровень отклонения (неравномерности) формы клеток эндофитов (SMCD).
[0038] Фигура 10 иллюстрирует индекс Idir - уровень изменений направления при колонизации живой клетки растения-хозяина.
[0039] Фигура 11 иллюстрирует эндофитные гифы в корне проростка пшеницы (А- SMCD 2204; В- SMCD 2206; С- SMCD 2210 и D- SMCD-2215), визуализированые при помощи окрашивания лактофуксином и флуоресцентной микроскопии. Симбиотические структуры/органы: D) бактериальный эндофит SMCD 2215 формирует, в основном, изогнутые межклеточные филаменты, тогда так эндофитный гриб (Фигуры справа) формирует: MCD 2204 - внутриклеточные завитые структуры и арбускулы, SMCD 2206 - внутриклеточные везикулы, и SMCD 2110 - внутриклеточные узелки.
[0040] Фигура 12 иллюстрирует внешний вид прорастающих симбиотических проростков пшеницы после 10 дней на влажной фильтровальной бумаге при 21°С.
[0041] Фигура 13 иллюстрирует длину листьев прорастающих проростков симбиотической пшеницы после 10 дней на влажной фильтровальной бумаге при 21°С.
[0042] Фигура 14 отображает способ инокуляции in vitro (А). Агарная пластинка, площадью 5 мм, вырезанная из края материнской колонии, была помещена покрытой гифами стороной вниз в центр 60 мм чашки Петри, содержащей картофельно-декстрозный агар (КДА). Далее, пять семян со стерилизованной поверхностью помещали вблизи от агарной пластинки, на расстоянии, соответствующем 48 часам роста гиф, и проращивали в темноте. Влияние трех разных способов поверхностной стерилизации семян на их всхожесть (В). Столбцы, отмеченные одной или двумя звездочками (*), имеют значимые или очень значимые отличия от такого же эндофита, выращенного в контрольных условиях (р≤0,05 или р≤0,01, соответственно; ANOVA, с последующим post-hoc LSD анализом). Планки погрешностей отображают стандартную ошибку среднего (SE).
[0043] Фигура 15 иллюстрирует темпы роста свободноживущих эндофитов SMCD 2204, 2206, 2208, 2210 и 2215 in vitro на картофельно-декстрозном агаре (КДА) в условиях теплового стресса (36°С), засухи (8% полиэтиленгликоль (ПЭГ) 8000), в контрольных условиях на протяжении пяти дней и при одновременном действии высокой температуры (36°С) и засухи (8% ПЭГ) в течение шести дней. Столбцы, отмеченные одной или двумя звездочками (*), имеют значимые или очень значимые отличия от такого же эндофита, выращенного в контрольных условиях (р≤0,05 или р≤0,01, соответственно; ANOVA, с последующим post-hoc LSD анализом). Планки погрешностей отображают стандартную ошибку среднего (SE).
[0044] Фигура 16 иллюстрирует процент всхожести и сырой вес проростков при инициальных экспериментах, в которых поверхность семян стерилизовали 5% раствором гипохлорита натрия в течение 3 мин. Процент всхожести семян пшеницы in vitro измерялся после трех дней выращивания на картофельно-декстрозном агаре (КДА) при воздействии теплового стресса (36°С), засухи (8% полиэтиленгликоля (ПЭГ) 8000) и в контрольных условиях (А, В и С). Ось у нормировалась относительно показателя процента всхожести семян, выращенных при таких же условиях, но поверхность которых стерилизовалась 5% гипохлоритом натрия в течение 1 мин. Сырой вес проростков in vitro измерялся на 7-й день выращивания на КДА при воздействии теплового стресса, засухи и в контрольных условиях (D, Е и F). Столбцы, отмеченные одной (*) или двумя звездочками (**) имеют значимые или очень значимые отличия от контроля, не содержащего эндофита (р≤0,05 или р≤0,01, соответственно; ANOVA, с последующим post-hoc LSD анализом). Планки погрешностей отображают стандартную ошибку среднего (SE).
[0045] Фигура 17 иллюстрируют процент всхожести семян пшеницы, культивированных в течение некоторого времени вместе с эндофитами, способствующими наиболее эффективному формированию устойчивости к абиотическим стрессам (SMCD 2206, 2210 и 2215), в сравнении с не колонизированными и не пребывающими в стрессовых условиях семенами (позитивный контроль) и с не колонизированными, но пребывающими в стрессовых условиях семенами (негативный контроль). Энергия прорастания (EG) зависит от срока времени, в днях (ось х), при котором достигается 50% всхожесть (ось у). Символы " ", "х", "ο", "Δ", и "
", "х", "ο", "Δ", и " " отображают позитивный контроль, семена, обработанные SMCD 2206, семена обработанные SMCD 2210, семена обработанные SMCD 2215 и негативный контроль, соответственно. Показатели для теплового стресса и засухи соответствовали 36°С и 8% полиэтиленгликоля (ПЭГ) 8000, соответственно. Планки погрешностей отображают стандартную ошибку среднего (SE). Примечание: семена, используемые при определении EG, были получены на второй стадии экспериментов, и, следовательно, стерилизовались 5% раствором гипохлорита натрия в течение одной, а не трех минут.
" отображают позитивный контроль, семена, обработанные SMCD 2206, семена обработанные SMCD 2210, семена обработанные SMCD 2215 и негативный контроль, соответственно. Показатели для теплового стресса и засухи соответствовали 36°С и 8% полиэтиленгликоля (ПЭГ) 8000, соответственно. Планки погрешностей отображают стандартную ошибку среднего (SE). Примечание: семена, используемые при определении EG, были получены на второй стадии экспериментов, и, следовательно, стерилизовались 5% раствором гипохлорита натрия в течение одной, а не трех минут.
[0046] Фигура 18 иллюстрирует отношение между гидротермальным временем (НТТ), необходимым для достижения 50% всхожести при воздействии жары и засухи по отдельности и 5% всхожести при воздействии комбинации жары и засухи (ось х), и процентом всхожести, достигнутым через семь дней (ось у). Значения всхожести через 7 дней и НТТ получены, исходя из результатов второй стадии экспериментов. Символы "", "υ" and " " изображают семена, которые подвергались тепловому шоку (36°С), засухе (8% полиэтиленгликоль (ПЭГ) 8000) или обоим, тепловому шоку и засухе, соответственно. Значения R-квадрата, ассоциированного с линиями тренда, равняются 0,96, 0,80 и 0,18 для семян, подвергшихся высокой температуре, засухе или высокой температуре и засухе, соответственно. Примечание: Семена, используемые при определении процента всхожести на седьмые сутки и гидротермального времени (НТТ) были получены на второй стадии экспериментов, и, следовательно, обрабатывались 5% раствором гипохлорита натрия в течение одной, а не трех минут.
" изображают семена, которые подвергались тепловому шоку (36°С), засухе (8% полиэтиленгликоль (ПЭГ) 8000) или обоим, тепловому шоку и засухе, соответственно. Значения R-квадрата, ассоциированного с линиями тренда, равняются 0,96, 0,80 и 0,18 для семян, подвергшихся высокой температуре, засухе или высокой температуре и засухе, соответственно. Примечание: Семена, используемые при определении процента всхожести на седьмые сутки и гидротермального времени (НТТ) были получены на второй стадии экспериментов, и, следовательно, обрабатывались 5% раствором гипохлорита натрия в течение одной, а не трех минут.
[0047] Фигура 19 иллюстрирует, что у семян, обработанных или инокулированных штаммами SMCD, наблюдались улучшения всех тестируемых параметров прорастания семян, в том числе эффективности силы прорастания семян (SGV).
[0048] Фигура 20 иллюстрирует соотношения значений эффективности засухоустойчивости (DTE) сортов пшеницы (А) и ячменя (В), не обработанных (Е-) и обработанных (Е+) эндофитами, на основании значений усредненного симбиотического эффекта при использовании всех протестированных выделенных SMCD, при сборе урожая моделировался тепловой стресс в условиях теплицы.
[0049] Фигура 21 иллюстрирует (А) повышение урожайности пшеницы модификаторами-эндофитами (Е+) (SMCD 2206, SMCD 2210 и 2215), по сравнению с контролем (Е-) (урожайность г/3 горшка). (В) повышение урожайности инокулянтами-эндофитами (SMCD 2206, SMCD 2210 и 2215) у двурядного ячменя (Ва) и шестирядного ячменя (Bb) (урожайность: 3 растения/горшок).
[0050] Фигура 22 иллюстрирует (А) шестирядный ячмень сорта AC Metcalfe, слева направо: Засуха (Е-), Засуха и SMCD 2206 (Е+), Контроль (Е-), Контроль и SMCD 2206 (Е+); (В) сорт Wheat-Unity, слева направо: Засуха (Е-), Засуха и SMCD 2215 (Е+), Контроль (Е-), Контроль и SMCD 2215 (Е+); (С) сорт Wheat-Verona, слева направо: Засуха (Е-), Засуха и SMCD 2215 (Е+), Контроль (Е-), Контроль и SMCD 2215 (Е+); (D) твердая пшеница-TEAL, слева направо: Засуха (Е-), Засуха и SMCD 2210 (Е+), Контроль (Е-), Контроль и SMCD 2210 (Е+).
[0051] Фигура 23 иллюстрирует сухой вес стеблей (А) нута, (В) чечевицы и (С) гороха, выращенных в симбиозе с SMCD эндофитами (Е+) при воздействии теплового стресса в условиях фитотрона. Колонки, отмеченные одной (*) или двумя звездочками (**), имеют значимые или очень значимые отличия, в сравнении с подвергшимся стрессу контролем без эндофита (р≤0,05 или р≤0,01, соответственно; ANOVA, с последующим post-hoc LSD анализом).
[0052] Фигура 24 иллюстрирует сухой вес стручков (А) нута, (В) чечевицы и (С) гороха, выращенных в симбиозе с SMCD эндофитами (Е+) при воздействии теплового стресса в условиях фитотрона. Колонки, отмеченные одной (*) или двумя звездочками (**), имеют значимые или очень значимые отличия, в сравнении с подвергшимся стрессу контролем без эндофита (р≤0,05 или р≤0,01, соответственно; ANOVA, с последующим post-hoc LSD анализом).
[0053] Фигура 25 иллюстрирует сухой вес корней (А) нута, (В) чечевицы и (С) гороха, выращенных в симбиозе с SMCD эндофитами (Е+) при воздействии теплового стресса в условиях фитотрона. Колонки, отмеченные одной (*) или двумя звездочками (**), имеют значимые или очень значимые отличия, в сравнении с подвергшимся стрессу контролем без эндофита (р≤0,05 или р≤0,01, соответственно; ANOVA, с последующим post-hoc LSD анализом).
[0054] Фигура 26 иллюстрирует сухой вес стеблей (А) нута, (В) чечевицы и (С) гороха, выращенных в теплице в условиях засухи. Колонки, отмеченные одной (*) или двумя звездочками (**), имеют значимые или очень значимые отличия, в сравнении с подвергшимся стрессу контролем без эндофита (Е-) (р≤0,05 или р≤0,01, соответственно; ANOVA, с последующим post-hoc LSD анализом).
[0055] Фигура 27 иллюстрирует сухой вес стручков (А) нута, (В) чечевицы и (С) гороха, при взаимодействии с эндофитами (Е+) в теплице в условиях засухи. Колонки, отмеченные одной (*) или двумя звездочками (**), имеют значимые или очень значимые отличия в сравнении с подвергшимся стрессу контролем без эндофита (р≤0,05 или р≤0,01, соответственно; ANOVA, с последующим post-hoc LSD анализом).
[0056] Фигура 28 иллюстрирует сухой вес корней (А) нута, (В) чечевицы и (С) гороха, выращенных в теплице в условиях засухи. Колонки, отмеченные одной (*) или двумя звездочками (**), имеют значимые или очень значимые отличия в сравнении с подвергшимся стрессу контролем без эндофита (р≤0,05 или р≤0,01, соответственно; ANOVA, с последующим post-hoc LSD анализом).
[0057] Фигура 29 иллюстрирует: А. цветущие растения нута Vanguard, плодоносящие стручки при воздействии засухи в теплице - левое растение не симбиотическое (Е-), правое растение в симбиотических отношениях с представителями штамма SMCD 2215 (Е+); В и С, растения нута Vanguard, плодоносящие стручки при воздействии засухи в теплице - (В) в не симбиотических и (С) в симбиотических отношениях с представителями штамма SMCD 2215.
[0058] Фигура 30 иллюстрирует образование клубеньков на корнях разных сортов гороха при воздействии теплового стресса в условиях фитотрона: Hendel (сверху) и Golden (снизу), инокулированный (слева) и неинокулированный (справа) с SMCD 2215. Примечание: во всех образцах был обнаружен природный инфекционный агент Rhizobium sp.из семян гороха.
[0059] Фигура 31 в виде функции от времени иллюстрирует значительное увеличение энергии прорастания (≥50%) семян Glamis (чечевицы) эндофитами SMCD2206 и SMCD 2215 в условиях теплового стресса и засухи in vitro.
[0060] Фигура 32 в виде функции от времени иллюстрирует значительное увеличение энергии прорастания (≥50%) семян Handel (горох) эндофитами SMCD2206 и SMCD 2215 в условиях теплового стресса и засухи in vitro.
[0061] Фигура 33 иллюстрирует повышение эндофитными модификаторами (SMCD 2206 и SMCD 2210) урожайности льна в условиях засухи в теплице. Разные буквы над колонками отображают наличие статистически достоверных отличий между образцами (р<0,05, критерий Краскела - Уоллиса).
[0062] Фигура 34 отображает улучшение урожайности рапса эндофитными инокулянтами (SMCD 2206, SMCD 2210 и SMCD 2215) в условиях засухи в теплице. Разные буквы над колонками отображают наличие статистически достоверных отличий между образцами (р<0,05, критерий Краскела - Уоллиса).
[0063] Фигура 35 иллюстрирует выживаемость семян пшеницы, преинокулированных in vitro (чашки Петри в верхнем ряду) и проростков пшеницы, преинокулированных в условиях теплицы (чашки Петри в нижнем ряду) зндофитом SMCD 2206, показывая растения, которые выросли здоровыми, и с патогенными с Fusarium avenaceum и Fusarium graminearum - с отображением симптомов болезни и смерти растений.
[0064] Фигура 36 иллюстрирует выращенные в зернах пшеницы инокулянты Fusarium.
[0065] Фигура 37 иллюстрирует то, что выпревание в послевсходовый период в условиях теплицы было предотвращено эндофитом SMCD 2206.
[0066] Фигура 38 иллюстрирует увеличение биомассы (надземная a-d и корни e-f) пшеницы при наличии эндофита SMCD 2206, в сравнении с необработанным растением, (а) контрольное растение (Е-), (b) инокулированное растение (Е+), (с) контрольное цветущее растение, (d) инокулированное цветущее растение, (е) контрольное растение (Е-, слева) в сравнении с инокулированным SMCD 2206 растением (Е+, справа), и (f) флюоресцентная микроскопия корней пшеницы, колонизированных SMCD 2206 (Е+).
[0067] Фигура 39 иллюстрируетбиомассу наземных частей растения/на одно растение (слева) и биомассу подземных (корни) частей растения /на одно растение (справа) в контрольных (Е-) и растений, инокулированных SMCD (Е+) против F. graminearum и F. avenaceum. Вертикальные планки погрешностей на точках значенийотображают стандартную ошибку среднего.
[0068] Фигура 40 иллюстрирует длину корней контрольных растений (CDC Teal) без эндофитов SMCD в сравнении с растениями, инокулированными штаммами SMCD. Планки на точках значений отображают стандартную ошибку среднего.
[0069] Фигура 41 иллюстрирует сухую массу растения/на одно растение (сорт TEAL) при использовании метода двойной предварительной инокуляции: а) эндофит SMCD + Fusarium avenaceum (F.av), и b) эндофит SMCD + Fusarium graminearum (F.gr). Вертикальные планки погрешностей на точках значений отображают стандартную ошибку среднего.
[0070] Фигура 42 иллюстрирует сравнение размеров колосков пшеницы TEAL, в присутствии патогена (негативный контроль) и при отсутствии патогена (негативный контроль). Фигура слева - слева направо: i) растение+ F.gr, ii) растение + F.av, и (iii) растение; фигура справа- слева направо: i) растение; ii) растение + эндофит; iii) растение + эндофит + F. av; и iv) растение + эндофит + F.gr.
[0071] Фигура 43 иллюстрирует штамм-специфичный паттерн прорастания симбиотических семян, описанный как миковитальностъ: Handel + 6% ПЭГ - Контроль (А), Handel + 6% ПЭГ + SMCD 2204 (В); Handel + 6% ПЭГ + SMCD 2204F (С), Handel + 6% ПЭГ + SMCD 2206 (D), Handel + 6% ПЭГ + SMCD 2210 (Е), Handel + 6% ПЭГ + SMCD 2215 (F) после 7 дней при 21°С в темноте.
[0072] Фигура 44 иллюстрирует экспрессию связанных с (А) СОД и (В) MnCOД генов в Handel, подвергшихся действию ПЭГ, в присутствии эндофитов и без них.
[0073] Фигура 45 иллюстрирует экспрессию генов, связанных с пролином у Handel, подвергшихся действию ПЭГ, с эндофитами и без них.
[0074] Фигура 46 иллюстрирует in vitro прорастание семян пшеницы после трех дней пребывания на картофельно-декстрозном агаре (КДА). Холодная стратификация была усовершенствована путем удерживания семян в холодильной камере при 4°С в течение 48 часов. При непрямой и прямой обработке эндофитами, семена проращивали на приблизительно 4 см расстоянии или при непосредственном контакте с эндофитом, соответственно. А) Процент всхожести в сравнении с энергией прорастания (50% прорастание). В) эффективность прорастания семян пшеницы, подвергшихся холодной и биологической стратификации. Эффективность рассчитывалась путем вычитания из процента всхожести обработанных семян процента всхожести контрольных семян.
[0075] Фигура 47 иллюстрирует специфичные паттерны экспрессии генов гиббереллина (TaGA3ox2 и 14-3-3) и АБК (TaNCED2 и ТаАВА8'ОН1) в колеоризе прорастающих семян пшеницы в течение трех дней холодной и биологической стратификации. Уровень экспрессии генов рассчитывался как 2-Δ CT.
[0076] Фигура 48 иллюстрирует соотношение уровней экспрессии генов (2-Δ CT) гиббереллина (TaGA3ox2 и 14-3-3) и АБК (TaNCED2 и ТаАВА8'ОН1) в колеоризе прорастающих семян пшеницы в течение трех дней холодной и биологической стратификации.
[0077] Фигура 49 иллюстрирует отличительные паттерны экспрессии регуляторных генов гормонов RSG и КАО и генов резистивности MYB 1 and MYB 2 в колеоризе прорастающих семян пшеницы в течение трех дней холодной и биологической стратификации. Уровень экспрессии генов рассчитывался как 2-ΔCT.
[0078] Фигура 50 иллюстрирует выход первичного корня в прорастающем зерне пшеницы (А) инвертированное флюоресцентное (В) и флюоресцентное изображение флюоресценции DAF-2DA при взаимодействии с NO в клетках первичного корня (С) проростка AC Avonlea через 5 мин. после обработки [Nakatsubo et al. 1998] экссудатом грибка SMCD 2206. Флюоресцентная реакция в контрольных клетках зародышевого первичного корня обнаружена не была. Масштаб = 25 мкм; Масштаб = 50 мкм.
[0079] Фигура 51 иллюстрирует значения интенсивности флуоресценции DAF-2T через 5 мин. после обработки первичных корней из проростков пшеницы AC Avonlea эксудатом из грибков SMCD 2206 эксудатом из грибков вместе с поглотителем NO cPTIO, и стерильной водой. Сегменты первичного корня инкубировали в течение 30 мин в детекционном буфере (10 мМ Tris-Hcl, рН 7,4, 10 мМ KCl), содержащем 15 мкМ DAF-2DA (Sigma-Aldrich) или без 1 мМ 2-(4-карбоксифенил)-4,4,5,5-тетраметилимидазолин-1-оксил-3-оксида (cPTIO) в качестве поглотителя NO. Средние значения флуоресценции представлены как отношение интенсивности флуоресценции на 5 мин к интенсивности флуоресценции в момент времени 0. Разные буквы отображают наличие статистически достоверных отличий между образцами (р<0,05, критерий Краскела - Уоллиса).
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Новые штаммы, композиции и семена
[0080] Авторы настоящего изобретения выделили 6 новых штаммов эндофитов, которые улучшают жизнеспособность семян и здоровье растений, и урожайности при нормальных и/или стрессовых условиях. Эти эндофиты хранятся в следующих организациях: International Depository Authority of Canada - IDAC (хранение исходных штаммов - IDAC, Национальная микробиологическая лаборатория, Агентство общественного здравоохранения Канады, 1015 Арлингтон Стрит, Виннипег, Манитоба, Канада, R3E 3R2; квитанции о приеме и подтверждении жизнеспособности в Приложении A) Saskatchewan Microbial Collection and Database - SMCD (копии сохраняемых штаммов) Штаммы:
(a) IDAC 081111-06 = SMCD 2215;
(b) IDAC 081111-03 = SMCD 2210;
(c) IDAC 081111-02 = SMCD 2208;
(d) IDAC 081111-01 = SMCD 2206;
(e) IDAC 200312-06 = SMCD 2204; и
(f) IDAC 200312-05 = SMCD 2204F.
[0081] Соответственно, настоящее изобретение предлагает изолированный штамм эндофита Streptomyces sp., или его культуру, которая хранится под шифром IDAC 081111-06, или которая содержит 16S рДНК с последовательностью, представленной на SEQ ID NO: 6; изолированный штамм эндофита Paraconyothirium sp., или его культуру, которая хранится под шифром IDAC 081111-03, или которая содержит ITS рДНК с последовательностью, представленной на SEQ ID NO: 5; изолированный штамм эндофита Pseudeurotium sp., или его культуру, которая хранится под шифром IDAC 081111-02, или которая содержит ITS рДНК с последовательностью, показанной на SEQ ID NO: 4; изолированный штамм эндофита Penicillium sp., или его культуру, которая хранится под шифром IDAC 081111-01, или которая содержит ITS рДНК с последовательностью, представленной на SEQ ID NO: 3; изолированную культуру Cladosporium sp., которая хранится под шифром IDAC 200312-06, или которая содержит ITS рДНК с последовательностью, представленной на SEQ ID NO: 1; и/или изолированного эндофита Cladosporium sp. или его культуру, которая хранится под шифром IDAC 200312-05 или которая содержит ITS рДНК с последовательностью, представленной на SEQ ID NO: 2; или их комбинации, или смеси.
[0082] Термин "эндофит", используемый в данном документе, относится к грибам или бактериальным организмам, которые могут жить в симбиозе с растением внутри его, и также упоминается в данном документе как "эндосимбионт". Грибной эндофит может пребывать в форме спор, гиф или мицелия. Бактериальный эндофит может пребывать в форме клетки или группы клеток. Термин "эндофит", используемый в данном документе, включает дочерние штаммы, представленные в данном документе.
[0083] Также в данном документе предлагается композиция, содержащая изолированный эндофит или культуру, представленную в данном документе, или их комбинацию, или их смесь с носителем. Типичные носители включают инертный материал (на неуглеродной основе), используемый для поддержания и доставки плотно размещенного активного ингредиента к цели, и, необязательно, вспомогательные соединения, но не ограничиваются этим; способствуют и поддерживают выполнение функций активным ингредиентом, защищая его от УФ-излучения; обеспечивают устойчивость цели к дождю; удерживают влагу или защищают от высыхания; и/или способствуют распространению и рассредоточению биопестицидов при использовании стандартного сельскохозяйственного оборудование, так как это было описано Hynes и Boyetchko (2006, Soil Biology & Biochemistry 38: 845-84).
[0084] В другом варианте реализации изобретения композиция содержит по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5 или 6 штаммов или культур, приведенных в данном документе.
[0085] Также в данном документе предлагается композиция, содержащая изолированный эндофит или его культуру, представленные в документе, или их комбинацию, или их смесь с носителем.
[0086] В одном из вариантов реализации изобретения, семена инокулировали путем инокуляции в почву. В другом варианте реализации изобретения, семя покрыто эндофитом или его культурой. В еще одном варианте реализации изобретения эндофит, или его культура, вводились в семена путем распыления, инъекции, инокуляции, прививания, покрытия ими семян или обработкой ими семян.
Способы
[0087] Ниже предлагается способ повышения жизнеспособности семян, улучшения здоровья растений и/или их урожайности, включающий в себя инокуляцию семян эндофитом или его культурой, описанными в данном документе, или их комбинацией, или их смесью с композицией, описанной в данном документе; и культивирование из семени растения первого поколения.
[0088] Фраза "инокуляция семян", используемая в данном документе, относится к нанесению, заражению, совместной посадке или покрытию семян эндофитами. Методы инокуляции семян хорошо известны в данной области техники, например, как описано Hynes и Boyetchko (2006, Soil Biology & Biochemistry 38: 845-84).
[0089] Термин "усиление жизнестойкости семени", который используется в данном документе, относится к пренатальному уходу за растением, приводящему к улучшению всхожести семени, и его способности к формированию растения в нормальных и/или стрессовых условиях, и включает любой один или более из приведенных ниже эффектов, но не ограничивается ими: выведение из состояния покоя, обеспечение стратификации семени, увеличение всхожести, изменение экспрессии генов, уменьшение времени, необходимого для достижения энергии прорастания, защита от биотических стрессов, защита от абиотических стрессов, уменьшение гидротермального времени, необходимого для прорастания, увеличение силы прорастания семени, увеличение эффективности прорастания семени, увеличение равномерности прорастания семени, увеличение эффективности устойчивости к высоким температурам/засухе, увеличение веса проростков и урожайности проростков. Эффективность устойчивости к засухе/высоким температурам (DTE/THE) является термином противоположным по значению (антонимом) к чувствительности.
[0090] Энергия прорастания определяется как 50% всхожесть, по отношению к общему числу тестируемых семян. Сила прорастания семян отображает разницу между процентом всхожести обработанных семян и процентом всхожести необработанных семян. Понятие гипотермального времени постулирует, что каждое отдельное зерно начинает прорастать тогда, когда сумма температуры и водного потенциала достигла, в течении некоторого периода времени, уровня достаточного для начала прорастания. Эффективность прорастания определяется как процентное соотношение обработанных семян, прорастающих по истечению определенного периода времени после посадки, к числу семян, тестируемых в качестве необработанного контроля. Биологическая стратификация определяется как выведение семян из состояния покоя симбионтом путем стимуляции их прорастания. Однородность прорастания семян является максимальным процентом всхожести семян, достигнутым в течение минимального времени инкубации.
[0091] Термин "улучшение здоровья и/или урожайности растений", использованный в данном документе, относится к общему улучшению сельскохозяйственных характеристик (например, здоровья и продуктивности) приобретенных растением, и включает в себя любой один или более из приведенных ниже эффектов, но не ограничиваясь ими: изменение экспрессии генов RSG, КАО, ABA, GA, 14-3-3 или NO для улучшения гормональной активности растения, изменение экспрессии генов MYB, Pro, СОД или MnCOД для повышения стрессоустойчивости, например, устойчивости к абиотическим и биотическим стрессам, увеличения веса различных органов, таких как корень, стебель, листья и стручки, повышения активности бактерий рода Rhizobium, и частоты образования клубеньков, и улучшения характеристик семян первого или последующих поколений, в том числе, одной или более из приведенных ниже характеристик, но не ограничиваясь ими: веса семян в последующем поколении и энергии прорастания в последующем поколении.
[0092] Гены гормонов КАО, RSG, ABAs, GAs, 14-3-3 и наборы праймеров к ним соответствуют описанным Zhang et al. [2007]. Гены стрессоустойчивости СОД, МпСОД, Pro и MYB и наборы праймеров к ним приведены в Таблице 6 и Таблице 9 (SEQ ID NO: 8-19).
[0093] Термин "снижение" или "повышение", используемый в данном документе, относится к снижению или повышению характеристик семени, обработанного эндофитами или у выросшего из него растения, в сравнении с необработанным семенем или выросшим из него растением. Например, снижения в характеристиках могут иметь значения по меньшей мере 5%, 10%, 15%, 25%, 50%, 75%, 100% или 200% или ниже, чем в необработанном контроле, и повешения могут иметь значения по меньшей мере 5%, 10%, 15%, 25%, 50%, 75%, 100% или 200% или более, чем в необработанном контроле.
[0094] В одном из вариантов реализации изобретения, растение культивируют в условиях абиотического или биотического стресса.
[0095] Термин "абиотический стресс", который используется в данном документе, относится к стрессу под воздействием неживой природы, который, как правило, влияет на жизнеспособность семени и здоровью растения и включает в себя тепловой стресс и засуху, но не ограничивается ими. В одном из вариантов реализации изобретения, абиотический стресс являлся тепловым стрессом. В другом варианте реализации изобретения, абиотический стресс являлся засухой, осмотическим стрессом или засолением. Термин "биотический стресс", который используется в данном документе, относится к стрессу под воздействием живой природы, который, как правило, влияет на жизнеспособность семени и здоровье растения, и включает в себя микробные инфекции растений, но не ограничивается ими. В одном из вариантов реализации изобретения, биотическим стрессом является инфицирование патогенным грибом рода Fusarium.
[0096] В одном из вариантов реализации изобретения, способ включает следующие элементы: инокуляцию семени выделенным эндофитом Streptomyces sp. или культурой его штамма, которая хранится под шифром IDAC 081111-06 или которая содержит 16S рДНК, с последовательностью, представленную в SEQ ID NO: 6. В варианте реализации изобретения, способ увеличивает всхожесть семян, уменьшает время, необходимое для достижения энергии прорастания, снижает гидротермальное время, необходимое для прорастания, повышает силу прорастания семян, увеличивает сырой вес проростков, повышает активность бактерий рода Rhizobium и частоту образования клубеньков и/или повышает урожайность проростков. В одном из вариантов реализации изобретения, способ включает: уменьшение последствий воздействия стрессовых факторов, таких как засуха, высокие температуры и/или биотический стресс.
[0097] В другом варианте реализации изобретения, способ включает следующие этапы: инокуляцию семени изолированным эндофитом Paraconyothirium sp. или его культурой, которая хранится под шифром IDAC 081111-03, или же которая содержит ITS рДНК с последовательностью, представленной в SEQ ID NO: 5. В варианте реализации изобретения, способ увеличивает всхожесть семени, уменьшает время, необходимое для достижения энергии прорастания, уменьшает гидротермальное время, необходимое для прорастания, повышает силу прорастания семени, увеличивает сырой вес проростков и/или увеличивает урожайность проростков. В другом варианте реализации изобретения, способ включает: уменьшение последствий действия стрессовых факторов, таких как засуха, повышенная температура и/или биотический стресс.
[0098] В еще одном варианте реализации изобретения, способ включает следующие этапы: инокуляцию семени изолированным эндофитом Pseudeurotium sp. или его культурой, которая хранится в IDAC под шифром 081111-02, или же которая содержит ITS рДНК с последовательностью, представленной в SEQ ID NO: 4. В варианте реализации изобретения, способ уменьшает время, необходимое для достижения энергии прорастания, снижает гидротермальное время, необходимое для прорастания, увеличивает силу прорастания семени и/или увеличивает сырой вес проростков. В другом варианте реализации изобретения, способ включает уменьшение последствий стресса, такого как засуха и/или тепловой стресс.
[0099] В дополнительном варианте реализации изобретения, способ включает следующие этапы: инокуляцию семени изолированным эндофитом Penicillium sp. или его культурой, которая хранится под шифром IDAC 081111-01, или же которая содержит ITS рДНК с последовательностью, представленной в SEQ ID NO: 3. В варианте реализации изобретения, способ повышает всхожесть семени, уменьшает время, необходимое для достижения энергии прорастания, снижает гидротермальное время, необходимое для прорастания, повышает силу прорастания семени и/или повышает урожайность проростков. В другом варианте реализации изобретения, способ включает: уменьшение последствий действия стрессовых факторов, таких как засуха, повышенная температура и/или биотический стресс. В другом варианте реализации изобретения, способ включает: усиление стратификации, вывод семян из состояния покоя и увеличение стрессоустойчивости путем изменения экспрессии генов гормонов КАО, RSG, ABAs, GAs, 14-3-3 или NO и/или экспрессии генов стрессоустойчивости СОД, МnСОД, Pro или MYB, уменьшения последствий действия стрессовых факторов, таких как засуха, повышенная температура и/или биотический стресс.
[00100] В еще одном варианте реализации изобретения, способ включает следующие этапы: инокуляцию семени изолированным эндофитом Cladosporium sp. или его культурой, которая хранится под шифром IDAC 200312-06, или же которая содержит ITS рДНК с последовательностью, представленной в SEQ ID NO: 1. В варианте реализации изобретения, способ уменьшает время, необходимое для достижения энергии прорастания, уменьшает гидротермальное время, необходимое для прорастания, увеличивает силу прорастания семени и/или увеличивает сырой вес проростков. В варианте реализации изобретения, способ включает уменьшение последствий стресса, такого как засуха и/или тепловой стресс.
[00101] В еще одном варианте реализации изобретения, способ включает следующие этапы: инокуляцию семени изолированным эндофитом Cladosporium sp. или его культурой, которая хранится под шифром IDAC 200312-05, или же которая содержит ITS рДНК с последовательностью, представленной в SEQ ID NO: 2. В варианте реализации изобретения, способ включает уменьшение последствий стресса, такого как засуха, и/или тепловой стресс.
[00102] Термин "растение", используемый в данном документе, относится к представителям царства Plantae и включает в себя растения на всех этапах жизненного цикла, включая семена, но не ограничиваясь ими. В одном из вариантов реализации изобретения, использованное растение являлось злаком (пшеница или ячмень), бобовым (горох, чечевица или нут), льном или рапсом.
[00103] В варианте реализации изобретения семя покрыто эндофитами, культивируются с эндофитами, или высаживаются рядом с эндофитами. В конкретном варианте реализации семя высаживают рядом с эндофитами, на расстоянии около 4 см от эндофита.
[00104] В ином аспекте изобретения, в данном документе предлагается способ улучшения здоровья растений и/или урожайности растений, включающий в себя обработку растительного посадочного материала или самого растения эндофитом или его культурой, описанной в данном документе, или их комбинацией, смесью или композицией, описанной в данном документе; и культивирование растительного посадочного материала до растения первого поколения или предоставление возможности растению вырасти.
[00105] Термин "посадочный материал", используемый в данном документе, относится к любой генеративной/половой или вегетативной/бесполой части растения, из которой можно культивировать новое растение. В варианте реализации изобретения, растительным посадочным материалом являются генеративные - семя, генеративная почка или цветок и вегетативные - стебель, черенок, корень, луковица, корневище, клубень, вегетативная почка или части листьев.
[00106] В варианте реализации в качестве изолированного эндофита или его культуры использовался изолированный штамм эндофита Streptomyces sp.или его культура, которая хранится International Depositary Authority of Canada (IDAC, Национальная микробиологическая лаборатория. Агентство общественного здравоохранения Канады. 1015 Арлингтон Стрит, Виннипег, Манитоба, Канада, R3E 3R2) код доступа 081111-06, или которая содержит 16S рДНК с последовательностью, представленной в SEQ ID NO: 6; изолированный штамм эндофита Paraconyothirium sp., или его культура, которая хранится под шифром доступа IDAC 081111-03, или которая содержит 1ST рДНК с последовательностью, представленной в SEQ ID NO: 5; изолированный штамм эндофита Pseudeurotium sp. или его культура, которая хранится под шифром доступа IDAC 081111-02 или которая содержит 1ST рДНК с последовательностью, представленной в SEQ ID NO: 4; изолированный эндофит Penicillium sp. или его культура, которая хранится под шифром доступа IDAC 081111-01 или которая содержит 1ST рДНК с последовательностью, представленной в SEQ ID NO: 3; изолированная культура Cladosporium sp. которая хранится под шифром доступа IDAC 200312-06 или которая содержит 1ST рДНК с последовательностью, представленной в SEQ ID NO: 1, и/или изолированный эндофит Cladosporium sp. или его культура, которая хранится под шифром доступа IDAC 200312-05 или которая содержит 1ST рДНК с последовательностью, прелставленной в SEQ ID NO: 2.
[00107] В другом варианте реализации изобретения, способы уменьшают последствия воздействия стрессовых факторов, таких как засуха, повышенная температура и/или биотических стрессовых факторов.
[00108] В варианте реализации изобретения, способы стимулируют процессы развития ландшафта и ремедиации.
[00109] Соответственно, в одном из вариантов реализации изобретения описывается способ фиторемедиации или фиторекламации загрязненных участков, включающий в себя обработку растительного посадочного материала, или растения эндофитами, или культурами, описанными в данном документе, или сочетанием их смеси, или композицией, описанной в данном документе, и культивирование растительного посадочного материала до растений первого поколения или предоставление возможности растению вырасти; восстанавливая или ликвидируя таким образом загрязненный участок.
[00110] Термин "фиторемедиация", который используется в данном документе, относиться к применению растений для удаления, уменьшения или нейтрализации веществ, отходов или опасных материалов с участка для того, чтобы предотвратить или минимизировать любое неблагоприятное воздействие на окружающую среду. Термин "фиторекламация", который используется в данном документе, относится к применению растений для восстановления земли с нарушенным покровом до первоначального состояния, или же для продуктивного использования в других целях.
[00111] В одном из вариантов реализации изобретения, участком является почва, такая как почва на полигоне хранения отходов. В одном из вариантов реализации изобретения к числу веществ, отходов или опасных материалов относились углеводороды, нефть или другие химические вещества, соли или металлы, такие как свинец, кадмий или радиоизотопы.
[00112] Фраза "обработка растительного посадочного материала или растений", используемый в данном документе, относится к применению эндофитов, или их культур, отдельно или с любым твердым или жидким средством доставки в растительный посадочный материал, или в растение, или в части указанного растения. В варианте реализации изобретения, обработка включает в себя внекорневое введение эндофита или внесение его в почву, или его комбинация с любым твердым или жидким средством доставки на любых стадиях выращивания растения.
[00113] В качестве растения может быть использовано любое растение. В одном из вариантов реализации изобретения, использованное растение являлось злаком (пшеница или ячмень), бобовым (горох, чечевица или нут), льном, рапсом, хвойным деревом (ель или сосна), широколиственным деревом (ива, тополь), кустарником (карагана или терескен) или травянистым растением (овсяница или волоснец).
[00114] Приведенное выше описание в общих чертах описывает данную заявку. Более полное понимание может быть достигнуто при рассмотрении следующих конкретных примеров. Данные примеры приведены исключительно в целях иллюстрации и не ограничивают объем настоящего изобретения. Изменение формы и замена эквивалентов рассматриваются как условия, которые могут быть предложены или становятся целесообразными. Хотя в данном документе и были использованы конкретные термины, данные термины предназначены для описательных целей, а не для целей ограничения.
[00115] Приведенные ниже примеры не являются ограничивающими и предназначены для иллюстрации изобретения.
ПРИМЕРЫ
ПРИМЕРЫ (1-14)
[00116] Состояние покоя и прорастание находятся в зависимости от нескольких факторов и процессов. Для обеспечения успешного формирования проростков, важно контролировать основные процессы или условия. Роль генетических свойств растений, гормонов и различных тканей семени были изучены довольно основательно. Настоящие примеры являются примерами исследований взаимоотношений эндофит-семя, переходящими к корневой симбиотической стадии при созревании растения.
Пример 1
Таксономия
[00117] International Depository Authority of Canada - IDAC (хранит исходные штаммы) и Saskatchewan Microbial Collection и Database - SMCD (хранит копии штаммов) Штаммы: IDAC 081111-06 = SMCD 2215; IDAC 081111-03 = SMCD 2210; IDAC 081111-02 = SMCD 2208; IDAC 081111-01 = SMCD 2206; IDAC 200312-06 = SMCD 2204; IDAC 200312-05 = SMCD 2204F (Фигуры 1-6 и Таблица 1).
[00118] Штамм SMCD 2215 был изначально выделен из эндофитной бактерии Phyalocephala sensu lato растительный эндофитный SMCD гриб. Классификация, согласно Labeda et al. [2012]. Это филогенетическое исследование изучает почти все описанные виды (615 таксонов) семейства Streptomycetaceae исходя из геннетических последовательностей 16S рДНК и демонстрирует видовое разнообразие этого семейства, в котором было обнаружено наличие 130 клад, подтвержденных статистически.
[00119] Полученные из последовательностей 16S рДНК данные подтверждают, что штамм SMCD 2215 Streptomyces sp. можно отнести к отдельной неизвестной кладе, согласно Labeda et al [2012], но к отдельному orStreptomyces lividans. виду.
Пример 2
Симбиотическая взаимосвязь между микрорганизмом и растением и уровень их совместимости
[00120] Уровень совместимости между микроорганизмами и растениями оценивали при помощи немного модифицированного метода Abdellatif et al. [2009]. На разделенной пополам 10 см чашке с агаром, не содержащей питательных веществ (Фигура 7), производили оценку здоровья растений и формирования им корневых волосков - абсорбентов воды и минеральных веществ, при совместном культивировании, с или без микробных партнеров. На Фигуре 7, левая часть каждой из разделенных чашек отображает культуру с микробным партнером, а правая часть каждой из разделенных чашек отображает культуру без микробного партнера. Эксперимент был воспроизведен дважды в трех повторностях.
[00121] Как показано в левых частях каждой разделенной чашки, здоровые растительные ткани формировались даже тогда, когда корни растений были выращены непосредственно на плотных бактериальных матах. Биомасса корневых волосков увеличилась примерно вдвое, по сравнению с правыми частями каждой из разделенных чашек, где микробный партнер отсутствовал (см. на левые части).
[00122] Эффективность образованной растением симбиотической связи зависит от типа распределения эндофитов в эндодерме корней. Типичная колонизация корней эндофитами прерывиста и неполная, с меньшим <50%, в сравнении с колонизацией патогенными грибами, количеством занятых клеток (Таблица 2) которая характеризуется равномерностью/непрерывностью (частота: 60-80%) колонизации клеток (Фигура 8).
[00123] Производительность эндофита не должна оцениваться исключительно путем измерения производства биомассы потому, что основой значительного повышения урожайности является эффективность колонизации растений эндофитами. Это можно оценить, характеризуя их связь с растительными клетками, тканями или органами (т.е. семенами и первичными корнями) с применением математических Индексов, которые были разработаны [Abdellatif et al. 2009] и применялись в данном исследовании (Фигура 9 и Фигура 10).
[00124] Эти Индексы основываются на следующих наблюдениях: Эндофитные симбиоты имеют паттерны колонизации первичного корня (корней)(регулярность или уровень отклонений форм эндофитных клеток - Ireg и их направления-Idir при колонизации живых клеток), отличные от тех, что присущи мертвым клеткам первичного корня (которые, обычно, остаются колонизированными настоящими сапрофитами).
[00125] Высокие значения индексов Ireg и Idir являются признаком мутуалистических (взаимовыгодных) отношений симбионт-растение. В заключение, результаты показывают, что симбиотические связи микроорганизм-растение характеризуются высоким уровнем совместимости между двумя партнерами, что приводит к сбалансированной (<50% колонизованных клетках коры) и прерывистой колонизации корней микробными эндофитами, измеренной при помощи математических индексов [Abdellatif et al. 2009]. Это мутуалистическое партнерство дополнительно характеризуется прямым воздействием эндофитных микроорганизмов на здоровый рост (бакто- и микозависимость), когда растению затруднительно использовать микробных партнеров исключительно в качестве источника питательных веществ или энергии для роста. Кроме того, наблюдалось увеличение биомассы корневых волосков благодаря воздействию эндофитов, и оно обнаруживалось даже в корнях, которые находились в отдаленных частях разделенных чашек, где не было микробных партнеров, что указывает на вероятный системный эффект усиления роста эндофитами.
Пример 3
Симбиотические органы эндофитов на пшенице
[00126] Каждая таксономическая группа эндофитов устанавливает уникальный тип миковитализма, впоследствии формируя различные симбиотические органы. Характеристика миковитализма проводилась при помощи Abdellatif et al. [2009] методики, состоящей из in vitro ко-культивирования семени и микроорганизма, оценивающей ранние этапы симбиотических отношений микроорганизм-растение. Разнообразие микробных симбиотических органов, сформированных в проростках пшеницы при участии SMCD 2204, 2206, 2210 и 2215, проиллюстрировано на Фигуре 11.
[00127] Подводя итог всему вышесказанному, результаты показывают, что формирование каждым эндофитом особого типа симбиотических органов в корнях пшеницы, вероятно, связано с их различными симбиотическими функциями. Равномерная частота колонизации, очаговые паттерны колонизации, увеличение септирования гиф в живых клетках корня, а также формирование арбускул, узлов, завитков и пузырьков -предполагаемых функциональных симбиотических органов - могут быть признаками местной специализации грибных эндофитов и стимулировать формирование миковитальность и микогетгротрофность у растений. Бактовитальность была представлена, в основном, скрученными межклеточными филаментами, образованными микроорганизмом Streptomyces.
[00128] Симбиоз, сформированный на уровне семени, приводил к увеличению размеров проростков пшеницы через 10 дней после совместной инокуляции (Фигура 12 и Фигура 13).
Пример 4
Эндофиты улучшают всхожесть семени пшеницы, подвергшегося воздействию теплового стресса и засухи
[00129] Прорастание семени является чрезвычайно важной стадией для выживания растения и своевременного формирования проростков, особенно в стрессовых условиях. Было высказано предположение, что эндофиты могут улучшать всхожесть семян, пребывающих под воздействием теплового стресса и засухи. Модель гидротермального времени (НТТ) прорастания - это концептуальная модель, которая является полезной для прогнозирования времени и энергии прорастания (EG) при заданном наборе условий. НТТ и EG применяются для определения повышения жароустойчивости или засухоустойчивости пшеницы, вызванного одним или более совместимыми эндофитами. Проверяемые эндофиты привели к резкому увеличению процента всхожести, улучшению показателей EG и НТТ, и к снижению чувствительности пшеницы к повышенной температуре и засухе, что было определено измерениями сырого веса проростков. При колонизации наиболее успешным эндофитом, значения тестируемых параметров в семенах пшеницы, подвергшихся тепловому стрессу напоминали значения этих параметров у семян, не подвергшихся стрессу.
Материалы и методы
Модель гидротермального времени прорастания и энергии прорастания
[00130] Модель гипотермального (НТТ) времени [Gummerson 1986] постулирует, что каждое отдельное зерно начинает прорастать при соблюдении двух условий. Во-первых, сумма средних суточных температур, превышающая минимальное кардинальное значения (Tmin), накопленных в течение некоторого периода времени, должна преодолеть пороговое значение (θT), измеряемого в градус-днях. Во-вторых, семена должны накопить достаточный водный потенциал (θH) на градус-день. Таким образом, НТТ (θHT) может быть выражена как:

[00131] согласно с Köchy и Tielbörger [2007],

где t отображает время, в днях, и

при постоянных условиях окружающей среды, при условии, что ψсубстрата равна или ниже оптимальной для прорастания семян температуры. В уравнении 3, ψсубстрата и ψmin обозначают водный потенциал субстрата и минимальный водный потенциал, при котором прорастание возможно, в МПа, соответственно. Согласно Bradford [2002], уравнения 2 и 3 можно подставить в уравнение 1 с получением:

[00132] Тем не менее, в настоящем исследовании, температура превышала оптимальную для прорастания пшеницы температуру [рассмотрено McMaster (2009)], в связи с чем необходимо учитывать максимальную температуру (Tmax), выше которой прорастание невозможно. Таким образом, уравнение 2 было превращено в:

в котором Tmin≤Тсубстрата≤Tmax. Если уравнение 5 заменить уравнением 2, в уравнении 4, получим следующие результаты:
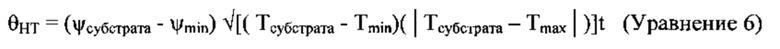
в котором Tmin≤Тсубстрата≤Tmax.
[00133] Энергия прорастания (EG) может быть определена несколькими способами, в том числе по соотношению процента семян, прорастающих после определенного периода времени после посадки, к общему количеству тестируемых семян [Ruan et al. 2002; Dongdong et al. 2009], или достижению 50% всхожести [Allen 1958]. Для того, чтобы совместить EG с моделью прорастания НТТ используется последнее определение, это означает, что EG равна t в уравнении 2.
Оценка параметров
[00134] Оценка Tmin и Tmax пшеницы была основана как на информации, полученной из литературы, так и на собственных наблюдениях изобретателей. McMaster [2009] обобщил данные, взятые из Friend et al. [1962], Сао и Moss [1989], и Jame et al. [1998] указывающие на существование криволинейной зависимости между скоростью развития пшеницы и температурой. Поскольку прорастание и развитие пшеницы не происходит при температуре ниже 0°С или выше 40°С, Tmin и Tmax были присвоены значения 0°С и 40°С, соответственно.
[00135] Параметр ψmin оценивался in vitro на прорастающих семенах пшеницы, выращенных картофельно-декстрозном агаре (КДА, Difco), содержащем полиэтиленгликоль (ПЭГ) 8000 в диапазоне концентраций (Amresco Inc.). Активность воды (aw) КДА без добавок и КДА, содержащего 8%, 12% и 16% ПЭГ была измерена с помощью AquaLab 4ТЕ, Series 4 Quick Start, Decagon Devices. Активность воды была конвертирована в водный потенциал (ψ), используя соотношение, адаптированное Bloom и Richard [2002]:
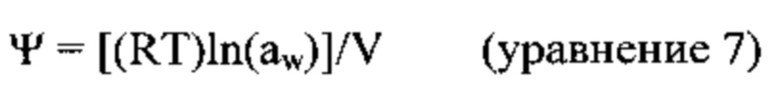
в котором R это универсальная газовая константа (8,314 Дж моль-1 К-1), Т - это температура в °К, и V - это парциальный молярный объем воды (18 мл/моль). Для преобразования единиц измерения, 1 Дж/мл = 1 МПа = 10 бар. Водный потенциал равен нулю для свободной поверхности воды или насыщенной среды; все другие значения являются отрицательными.
[00136] Активность воды в КДА и КДА, содержащем 8%, 12% и 16% ПЭГ была 0,9974, 0,9890, 0,9863 и 0,9825, соответственно. Эти значения эквивалентны -0,35, -1,51, -1,88 и -2,41 МПа, соответственно, что согласуется с литературными данными [Leone et al. 1994].
Растительные и грибные материалы
[00137] В качестве растительных материалов использовали сорт твердой пшеницы АС Avonlea, который имеет низкую устойчивость к экологическому стрессу [SaskSeed guide 2008]. Семена, которые использовались на первом этапе экспериментов, были произведены Paterson Grain в 2008 году, в полевых условиях, и не были сертифицированы на отсутствие в них микробов. Семена, которые использовались на второй стадии экспериментов, были произведены Agriculture and Agri-Food Canada (AAFC) Seed Increase Unit Research Farm в 2006 году в тепличных условиях, и были сертифицированны на отсутствие в них микробов. Поверхность семян пшеницы стерилизовалась 95% этанолом в течение 10 с, семена промывали в стерильной дистиллированной воде в течение 10 с, погружали на 3 мин (первый этап экспериментов с несертифицированными на отсутствие микробов семенами), или на 1 мин (второй этап экспериментов со сертифицированными на отсутствие микробов семенами) в 5% гипохлорит натрия (Javex), промывали три раза в стерильной дистиллированной воде и КДА для их прорастания [Abdellatif et al. 2009]. Третий способ стерилизации семян, включающий в себя выдерживание в хлоре (газообразном) в течение 3 часов (полученного путем смешивания в стакане 25 мл 6% гипохлорита натрия с 1,0 мл концентрированной соляной кислоты) в закрытой пластмассовой коробке, помещенной в вытяжной шкаф [Rivero et al. 2011] также проверялся. Процент всхожести семян, подвергнутых разным протоколам стерилизации и размещенных на КДА на три дня, проиллюстрирован на Фигуре 14 В. Только 3-минутное погружение в гипохлорит натрия приводило к значительному снижению всхожести (р≤0,01). Поверхностная стерилизация семян была осуществлена для устранения микробов, которые могут конкурировать с исследуемыми эндофитами. Кроме того, микроорганизмы, присутствующие на поверхности семян, могли распространиться за пределы чашки и появляющихся проростков, ингибируя рост растений. Все семена, используемые в исследовании, рассматривались как не содержащие микроорганизмов, на основании отсутствия непреднамеренного микробного роста на чашке.
[00138] Четыре митоспоровых эндофитных гриба отдела Ascomycota (систематизированных согласно Kiffer and Morelet [2000]): SMCD 2204, SMCD 2206, SMCD 2208 и SMCD 2210, а также выделенные нитевидные грамположительные бактериальные актиномицеты (Actinomycetes) SMCD 2215; совместимые с Triticum turgidum L. [Abdellatif et al. 2009] были использованы в данном исследовании. Эндофиты выращивались на КДА в течение по меньшей мере, трех дней при комнатной температуре в темноте, до использования в эксперименте.
Эндофиты как свободноживущие организмы
[00139] Агарные пластинки (5 мм), вырезанные из краев материнской колонии, были размещены в центре 90 мм чашки Петри, содержащей либо КДА без добавок, либо с добавлением 8% ПЭГ (модель засухи). Чашки Петри герметизировали парафильмом (Pechiney Plastic Packaging) для поддержания стерильности и помещали в настольный инкубатор (Precision Thermo Scientific, модель 3522) при 23°C, или в условиях теплового стресса, при 36°С, в темноте. Диаметр колоний измерялся через 24, 48, 72, 96 часов и на пятый и шестой день. Изменения в диаметре были использованы для расчета скорости роста колоний. Ростовые параметры измерялись как минимум с тремя повторностями для каждого изолята.
Свойства эндофитов увеличивать жаро- и засухоустойчивость пшеницы
[00140] Каждый изолят наносился на семена пшеницы по отдельности, перед прорастанием согласно способу, описанному в Abdellatif et al. [2010] и проиллюстрированному на Фигуре 14А. Вкратце, пять поверхностно-стерилизованных семян были размещены на расстоянии от 5 мм2агарной пластины, эквивалентному 48 часам роста гиф, помещенной покрытой гифами стороной вниз в центр 60 мм чашки Петри. Для обеспечения потребностей медленно растущих штаммов, агарную пластинку с колониями эндофитов помешали в чашку Петри в период от одного до четырех дней, предшествующих их введению в семена. Проростки проращивали в течение одной недели при воздействии абиотических стрессов, и в контрольных условиях.
[00141] Засуха была индуцирована при помощи КДА, содержащего 8% ПЭГ. Тепловой стресс был индуцирован в настольном инкубатора в условиях темноты; каждые 2 часа температуру поднимали на 2°С, таким образом она была постепенно увеличена с 28°С до 36°С.На начальных этапах экспериментов, производилось измерение процента всхожести на третьи сутки и сырого веса спустя одну неделю. Каждый эксперимент включал шесть чашек Петри и был трижды независимо повторен. В последующих экспериментах процент всхожести оценивался каждые 24 часа в течение семи дней. Каждый эксперимент включал 10 чашек Петри и повторяли либо два раза (сочетание засухи и теплового стресса) или три раза (тепловой стресс, засуха и контрольные условия).
[00142] Устойчивая внутренняя колонизация корней пшеницы предполагаемыми эндофитами была подтверждена при помощи повторной изоляции эндофитных организма из корней, поверхность которых, для предотвращения роста внешних микроорганизмов, стерилизовалась с использованием модифицированной процедуры из Larran et al. [2002]. Фрагмент корня (~0,5 см) подвергали поверхностной стерилизации в 95% этаноле в течение 10 с, корень промывали в стерильной дистиллированной воде в течение 10 с, погружали на 20 с в 5% раствор гипохлорита натрия (Javex), промывали три раза в стерильной дистиллированной воде и помещали на КПА в чашке Петри диаметром 60 мм. Чашку Петри герметизировали парафильмом и инкубировали в темноте при комнатной температуре в период от четырех до семи дней до исследования.
Анализ статистических данных
[00143] Темпы роста колонии свободноживущих эндофитных организмов, выращенных в условиях теплового стресса или засухи, сравнивались со значением данного параметра тех же организмов, выращенных в контрольных условиях с использованием дисперсионного анализа (ANOVA) с последующим применением post-hoc теста наименьшей существенной разницы (LSD) Фишера Данные, касающееся значений процента всхожести, до статистического анализа подвергались арксинус-преобразованию [McDonald 2009]. Статистические различия между значениями процента всхожести после трех и после семь дней, и значениями сырого веса в течении семи дней были оценены с применением однофакторного анализа ANOVA для сопоставления всех воздействий. Впоследствии, для оценки значимости различий между безэндофитным контролем и семенами, обработанными каждым из эндофитов применялся post-hoc LSD анализ. Уровень статистической значимости различий между EG и НТТ, необходимых для достижения 50% всхожести колонизированных эндофитами и контрольных семян, были оценены путем вычисления EG при каждом из трех независимых повторов эксперимента. Полученные данные были подвергнуты ANOVA и post-hoc LSD анализу. Значения Р менее 0,05 и 0,01 оценивались как значимые и очень значимые, соответственно. Статистический анализ проводился программой SPSS Inc. 2011.
Результаты
[00144] В каждой секции, результаты были систематизированы согласно типу стрессовых факторов: тепловой стресс, засуха, тепловой стресс в комбинации с засухой или отсутствие стресса. В каждом типе стресса результаты, касающиеся растительных материалов, были представлены согласно проростками и/или измеренными характеристикам проростков: процент прорастания на третий и седьмой день, сырой вес на седьмой день, EG и НТТ.
Свободноживущие эндофиты
[00145] Фенотипы SMCD 2206, 2210 и 2215 не были изменены жарой (36°С), в то время как SMCD 2204 и 2208 не росли при 36°С. Темпы роста колоний SMCD 2206 и 2210 снизились при 36°С, по сравнению с нестрессовыми условиями (р≤0,01), в то время как темпы роста SMCD 2215 при 36°С повысились (с≤0,05) (Фигура 15). При 36°С SMCD 2215 рос наиболее быстро, а за ним, в порядке убывания, следуют 2206 и 2210 (Фигура 15).
[00146] Морфология SMCD 2204, 2206, 2208 и 2215 не была заметно изменена засухой (8% ПЭГ). Однако, когда SMCD 2210 подвергся воздействию засухи, этот организм потерял свой "шерстистый" внешний вид, и, вместо этого приобрел "блестящий" или "слизистый" внешний вид. Темпы роста колоний SMCD 2204 2206 и 2208 были снижены засухой (р≤0,01, р≤0,01, р≤0,05 соответственно), в то время как темпы роста колонии всех остальных эндофитов осталась неизменными (Фигура 15). При воздействии засухи самым высоким был темп роста SMCD 2204, а за ним, в порядке убывания, следуют: 2206, 2210, 2208 и 2215 (Фигура 15).
[00147] При одновременном воздействии 36°С жары и засухи (8% ПЭГ), SMCD 2204 и 2208 перестали расти, в то время как SMCD 2206, 2210 и 2215 росли значительно медленнее, чем в контрольных условиях (р≤0,01) (Фигура 15). В контрольных условиях, SMCD 2204 рос наиболее быстро, за ним, в порядке убывания, следовали: SMCD 2206, 2210, 2208 и 2215 (Фигура 15).
Реакция колонизированной эндофитами пшеницы на высокие температуры
[00148] При 36°С, колонизация микроорганизмами SMCD 2206 и 2215 привела к увеличению всхожести по истечению трех дней (р≤0,05 и р≤0,01, соответственно; Фигура 16А), в то время как SMCD 2204, 2208 и 2210 не вызвали изменений значения этого параметра (p>0,1; Фигура 16А). По истечению шести дней, проросло 63% и 56% семян, культивированных вместе с SMCD 2204 и 2208, соответственно. Эти значения статистически не отличались (р>0,1) от 59% всхожести, достигнутой не колонизированным контролем. Напротив, эндосимбионты SMCD 2206, 2210 и 2215 стимулировали прорастание, по результатам семи дней (р≤0,01; Фигура 17).
[00149] При воздействии 36°С, сырой вес проростков пшеницы, культивированных-вместе с 2204, 2206, 2208 и 2210 не изменился, в то время как SMCD 2215 значительно повысил значение этого параметра (р≤0,01, соответственно; Фигура 16D).
[00150] EG семян, культивированных при 36°С вместе с грибным эндофитом SMCD 2210 (р≤0,05; Таблица 3, Фигура 17) было выше, чем у семян, не содержащих эндофитов. Тем не менее, SMCD 2204, 2206, 2208 и 2215 не изменили значение EG (р>0,1; Таблица 3), в сравнении с контролем. SMCD 2210 увеличил EG в наибольшей степени, за ним следовали SMCD 2206 и 2215 (Таблица 3). SMCD 2210 снизил время, необходимое для прорастания 50% семян всего лишь до 2 дней.
[00151] При воздействии теплового стресса, НТТ, необходимая для прорастания, была уменьшена у семян пшеницы, колонизированных SMCD 2210 (р≤0,05; Таблица 3), но не была изменена каким-либо другим из проверяемых эндофитов (р>0,1; Таблица 3). Семенам, не содержащим эндофитов, для достижения 50% всхожести, потребовалось на 50 МПа°С дней больше, чем семенам, которые были колонизированы SMCD 2210 (наиболее результативный из тестируемых зндофитов) (Таблица 3). Была обнаружена четкая прямая негативная корреляция между НТТ, необходимой для достижения 50% прорастания, и процентом всхожести по истечении семи дней, проведенных при воздействии теплового стресса (Фигура 18).
Ответ колонизированной эндофитами пшеницы на засуху
[00152] При воздействии засухи в течение трех дней, процент всхожести культивированных вместе с SMCD 2208 семян пшеницы снизился, в сравнении с не содержащими эндофитов семенами (р≤0,01; Фигура 16 В), тогда как SMCD 2204, 2206, 2210 и 2215 не изменили значение этого параметра (р>0,1; Фигура 16 В). Обработка семян эндофитами SMCD 2206, 2210 и 2215 привела к повышению всхожести через семь дней (р≤0,01, р≤0,05 и р≤0,01, соответственно; Фигура 17). В отличие от этого, 65 и 67% семян культивированных вместе с SMCD 2204 и 2208 проросли по истечению семи дней. Ни один из этих значений статистически значимо не отличалось от значения 59%, достигнутого не колонизированными семенами, проросшими в сходных условиях (р>0,1). В условиях засухи, по истечению семи дней SMCD 2208 и 2210 снизили сырой вес (р≤0,05 и р≤0,01, соответственно; Фигура 16Е). Ни один из остальных микобионтов не изменил значение этого параметра (р>0,1; Фигура 16Е).
[00153] EG снизилась в семенах пшеницы, культивированных со всеми тестируемыми эндофитамы, в сравнении с семенами, не содержащими эндофиты (0,05<р≤0,1 для SMCD 2204 и 2208, и р≤0,05 для 2206, 2210 и 2215; Таблица 3). SMCD 2206 улучшил значение EG в наибольшей степени, уменьшая время, необходимое для достижения 50% прорастания, которое было достигнуто через 2,6 дня (Таблица 3; Фигура 17).
[00154] В условиях засухи, НТТ, необходимая для прорастания уменьшилась у семян пшеницы, обработанных всеми тестируемыми эндофитами (Таблица 3). В то время, когда не колонизированным семенам для достижения 50% прорастания требовалось 80 МПа °С дней, семенам, колонизированные эндофитом SMCD 2206 (наиболее продуктивный из проверяемых эндофитов) требовалось всего лишь 34 МПа°С дней, снижая потребность на 46 МПа °С дней (Таблица 3). Была обнаружена заметная отрицательная линейная корреляция между НТТ, необходимым для достижения 50% прорастания и процентом всхожеси на седьмые сутки под воздействием засухи (Фигура 18). Тем не менее, значение связанной с этой линейной зависимостью R было меньшим, чем в случае с найденной при тепловом стрессе корреляцией. Диапазоны НТТ, необходимых для достижения 50% всхожести, разнятся при воздействии теплового стресса и засухи, являются уникальными для семян, подвергшихся засухе и тепловому стрессу, и имеют значения от 34 до 44 МПа °С дней и от 80 до 94 МПа °С дней, соответственно (Фигура 18; Таблица 3). Диапазоны значений процента всхожести по истечению семи дней схожи для семян, подвергнутых засухе и семян, которые подвергались воздействию тепла, хотя значения процента всхожести подвергнутых тепловому стрессу семян охватывают несколько больший диапазон (Фигура 18).
Реакция колонизированной эндофитами пшеницы на действие комбинации жары и засухи
[00155] Очень немногие семена пшеницы проросли при совместном воздействии засухи (8% ПЭГ) и теплового стресса (36°С) (Фигура 17). Колонизация эндофитами SMCD 2210 и 2215 по истечении нескольких дней увеличила процент всхожести (р≤0,01; Фигура 17). С другой стороны, SMCD 2204, 2206 и 2208 не смогли улучшить значение этого параметра (р>0,1). Семена, культивированные совместно с SMCD 2215 (наиболее благотворный по результатам тестирования улучшения этого параметра микроорганизм) достигли 24% значение всхожести, что в четыре раза выше уровня, достигнутого их аналогами - свободноживущими эндофитами (Фигура 17).
[00156] Так как ни неколонизированные семена, ни семена, колонизированные любым из эндофитов, не достигли всхожести в 50% в течение семи дней, EG не может быть определена, и НТТ была рассчитана для 5%, а не 50% всхожести. Время, необходимое для достижения 5% всхожести, было в диапазоне от 24 часов до четырех дней. Ни один из тестируемых эндофитов не уменьшил время, необходимое для достижения 5% всхожести, или значения НТТ (р>0,1). В целом, НТТ, необходимое для достижения 5% всхожести, варьировалась в диапазоне от 11 до 43 МПа °С дней (НТТсредее=23,9) (Фигура 18; Таблица 3).
[00157] Диапазон значений НТТ для семян, подвергнутых тепловому стрессу и засухе был уникальным, если сравнивать его с значениями НТТ, полученными при воздействии повышенной температуры или засухи по отдельности. Была обнаружена линейная отрицательная взаимосвязь между необходимым НТТ и процентом всхожести при комбинированном воздействии теплового стресса и жары. Тем не менее, значение связанного с этой линейной зависимостью параметра R2 было меньшим, чем в случае с корреляцией, обнаруженной при воздействии теплового стресса и засухи по отдельности (Фигура 18).
Реакция колонизированной эндофитами пшеницы на контрольные условия
[00158] По истечению трех дней, SMCD 2215 значительно увеличивал всхожесть семян при нестрессовых условиях, по сравнению с неколонизированными семенами (р≤0,01) (Фигура 16С). SMCD 2206, 2208 и 2210 позитивно влияли на процент всхожести, в то время ка SMCD 2204 не изменил его значение. В нестрессовых условиях, SMCD 2204, 2210 и 2215 увеличили сырой вес за семь дней (р≤0,05 и р≤0,01, соответственно). Тем не менее, SMCD 2206 и 2208 не имел никакого влияния на изменение сырого веса, при сравнении с неколонизированными саженцами (Фигура 16F).
[00159] В контрольных условиях, значения параметров EG и НТТ были слегка улучшены эндофитами SMCD 2206 и 2215 (Таблица 3). Совместное культивирование семян пшеницы с разными изолятами связывают с относительно небольшим изменением в значениях параметров EG и НТТ.
Пример 5
Эндофиты повышали урожайность пшеницы и ячменя в условиях сильной засухи
[00160] Краткое описание: В связи с изменением климата и ростом численности населения, усовершенствование методов, повышающих устойчивость сельскохозяйственных культур к стрессовым условиям, является чрезвычайно важным. Инокуляция тремя симбиотическими эндофитами, которые являются аборигенными для канадских прерий, приводит к повышению устойчивости пшеницы и ячменя к засухам и тепловому стрессу, а также к увеличению веса семян и урожая зерновых. Использование в этой сфере подобных грибных и бактериальных эндофитов создает потенциал для увеличения энергии прорастания семян (SGV = разница между общим процентном Е- прорастающих семян и Е + прорастающих семян) (Фигура 19, Фигура 20 А и В), а также для повышения урожайности в потенциально стрессовых условиях (Таблица 4; Фигура 21А, Ва, и Bb). Данные подтверждают, что штаммы SMCD повышают жизнеспособность семян и силу растений (Фигура 22A-D). В целом, результаты показывают, что пренатальный уход за семенами с применением эндофитных микроорганизмов, в частности SMCD штаммов, обеспечивает превосходную урожайность пшеницы и ячменя, благодаря улучшениям физиологических параметров.
Материалы и методы
[00161] Семена пшеницы и ячменя были произведены на экспериментальных участках Университета Саскачевана (University of Saskatchewan) и в Crop Science Field Laboratory (Saskatoon). Поверхность визуально здоровых семян стерилизовали в 95% этаноле в течение 10 с, семена промывали в стерильной дистиллированной воде в течение 10 с, погружали в 5% раствор гипохлорита натрия (Javex) на 1 мин, а затем промывали три раза в стерильной дистиллированной воде.
[00162] Эндофитные изоляты, используемые в данном исследовании, были первоначально выделены из корней твердых сортов пшеницы Triticum turgidum L., выращенной на полевых участках в провинции Саскачеван, Канада [Vujanovic 2007b]: SMCD 2204, 2206, 2208, 2210, 2215. Все полученные эндофитные изоляты культивировали на картофельно-декстозном агаре (КДА, Difco), в отсутствие растения-хозяина. До использования в эксперементе, изоляты выращивались на КДА в темноте при комнатной температуре (23°С) в течение трех дней.
[00163] Экспериментальная инокуляция производилась в вегетационных сосудах. Каждый из имеющийся эндофитных изолятов наносился на семена злаков (пшеницы и ячменя) до их прорастания, согласно способу, описанному в Abdellatif et al. [2010]. Вкратце, пять семян, поверхность которых была стерилизована, были размещены на расстоянии, эквивалентному 48 часам роста гиф, от агарной пластинки площадью 5 мм2, расположенной покрытой гифами стороной вниз, в центре 2 л пластикового культурального сосуда, наполненного 300 г (сухой вес) автоклавированной влажной грунтовой смеси Sunshine mix 4. Затем семена и агарная пластинка покрывалась 3,5-4,0 см слоем грунтовой смеси Sunshine mix 4. Семена, обработанные эндофитом одного типа, рассаживали по двенадцати сосудам, в каждый сосуд помещали по 5 семян. Горшки, содержащие растения, помещали в теплицу, для создания условий засухи и осуществления контроля обработки. Горшки были размещены блоками, в произвольном порядке.
[00164] Индукцию засухи производили в период с мая по сентябрь, когда максимальная суточная температура в теплице колебалась в диапазоне от 18 до 26°С. В солнечные дни, освещение осуществлялось естественным солнечным светом, а в пасмурные дни или зимние дни, имеющие укороченный фотопериод, в качестве дополнительного источника освещения применялись 1000 Вт натриевые лампы, подвешенные к потолку на высоте около 2 м над растениями. В первом эксперименте, подвергшиеся засухе и контрольные (хорошо поливаемые) растения выращивали при 25% массовой влажности почвы и 100% влагоемкости, соответственно. В ходе эксперимента контрольные растения поливались до доведения значения влагоемкости почвы до 100%, три раза в неделю, в то время как растения, которые подвергались засухе, поливались раз в неделю, до доведения значения влагоемкости почвы до 100%. Такой режим засухи был установлен для имитирования естественного цикла протекания засух, которые могут возникать в североамериканских прериях во время вегетации [Chipanshi et al. 2006].
[00165] Производили сбор зрелых колосков, сухие зерна взвешивали в лаборатории при помощи весов Mettler Toledo PG802-S.
Результаты и их обсуждение
Увеличение силы прорастания семян пшеницы (SGV)
[00166] В контрольных условиях in vitro, семена пшеницы, обработанные SMCD (2204, 2206, 2208, 2210, 2215) прорастали быстрее, более равномерно и с гораздо более высоким значением SGE (эффективности прорастания семян). SGV семян, инокулированных SMCD (Е+), был выше на 15-40%, по сравнению с необработанными (Е-) семенами (Фигура 19), демонстрируя тем самым эффективность регулирования SMCD периода покоя семян и увеличения энергии прорастания семян. Также было показано позитивное воздействие штаммов SMCD на урожайность пшеницы и ячменя при сильной засухе.
[00167] Как правило, ячмень имел более высокую чувствительность (низкие показатели DTE (Drought Tolerance Efficacy)) и более низкую урожайность, чем у пшеницы (Таблица 4), возможно, из-за того, что условия острой засухи в теплице являются более подходящими для пшеницы. В частности, двурядный ячмень CDC Kendall, не содержащий эндофита (Е-), имел более высокую, чем у других представителей ячменя, чувствительность к засухе. Тем не менее, обработка эндофитами (Е+) оказывает сильный положительный эффект на урожайность всех генотипов (Таблица 4). Приобретенная устойчивость колебалась в диапазоне от низкой устойчивости у CDC Kendall до высокой устойчивости у New Dale, тогда как приобретенная устойчивость пшеницы была неизменно высокой.
[00168] На стадии зрелости пшеницы и ячменя эндофиты SMCD вызывали у них сильное возрастание значений таких показателей засухоустойчивости, как эффективность DTE и урожайность. Применение SMCD на Avonlea, наиболее чувствительном к засухе сорту пшеницы из обнаруженных (DTE=16,1), привело к сильному возрастанию урожайности (77%) в условиях засухи, в сравнении с контролем, или стандартным поливом. Пшеница сорта Carberry извлекла наибольшую выгоду из эндофитов в контрольных (нормальных) условиях, в то время как CDC Utmost VB и BW 423 проявляли себя одинаково хорошо и в условиях засухи, и в контроле.
[00169] В заключение, сочетание засухоустойчивых генотипов с совместимыми к ним эндофитными микробными симбионтами растений SMCD 2206, 2210, SMCD и SMCD 2215 приводит к увеличению засухоустойчивости, важного для обеспечения продовольственной безопасности аспекта. Не желая ограничиваться теорией, это говорит о том, что взаимосвязь с симбионтами была наиболее выгодной для наиболее восприимчивых к засухе (низкие значения DTE) сортов пшеницы (Фигура 19А) и ячменя (Фигура 19В) при воздействии на них засухи.
[00170] Единственным обнаруженным исключением являлся шестирядный ячмень сорта Legacy, продемонстрировавший чрезвычайно низкий DTE=1,1. Несмотря на то, что в контрольных условиях в присутствие эндофитов он продемонстрировал позитивное повышение урожайности на 26,9%, при засухе симбиоз привел к увеличению его урожайности всего лишь на 5%. Поэтому, этот сорт был исключен из модели для ячменя, представленной на Фигуре 19В.
Влияние отдельных штаммов SMCD на продуктивность пшеницы и ячменя
[00171] Отдельные штаммы SMCD имели положительное влияние на среднюю урожайность каждого сорта, хотя фактические значения этой величины зависели от конкретной комбинации штамм-сорт. Фигура 21 иллюстрирует результаты, полученные в условиях засухи в теплице (Фигура 21: А - Пшеница; Ва - Ячмень (двурядный), и Bb - ячмень (шестирядный)).
[00172] Ранний контакт семян с совместимыми с ними SMCD изолятами является предпосылкой для защиты от засухи сельскохозяйственных культур, результатом чего является увеличение урожайности, или продуктивности, зерновых. SMCD 2206, как правило, вызывал наиболее сильные улучшения у большинства сортов. Тем не менее, наиболее сильные индивидуальные улучшения были обусловлены специфичными взаимоотношениями штамм-сорт, например Пшеница-РТ580 и Ячмень-CDC Copeland предпочитали SMCD 2210; тогда как Пшеница-ВШ423 и РТ580, также как и CDC Kendall демонстрировали наибольшую продуктивность и устойчивость к засухе при из инокуляции SMCD 2215.
[00173] Результаты подчеркивают важность миковитализма для семян пшеницы и ячменя, пребывающих в стрессовых условиях, помогая селекционерам в создании высокопродуктивных сортов, способных выдерживать условия засухи значительно лучше, чем любой из сортов без помощи эндофитов (Фигура 22: A-D). Исходя из продемонстрированной эффективности SMCD штаммов в полевых условиях, селекционеры, для защиты урожая, смогут создать симбиотические растительные продукты, и агро-бизнес будет получать выгоду от получения гарантированного уровня позитивной прибыли от урожая, независимо от изменений условий окружающей среды.
Пример 6
Эксперементальное воспроизведение теплового стресса на бобовых в фитотроне
[00174] Этот эксперимент был проведен в условиях фитотрона. Все сорта семян, инокулированных и не инокулированных эндофитами (SMCD 2204F, SMCD 2206, SMCD 2210 и SMCD 2215), высаживали в вегетационные сосуды, содержащие грунтовую смесь. Детальная информация о подходах, применяемых при инокуляции растений эндофитами были описаны ранее в Примере 5. Сосуды, содержащие растения, которые будут подвергнуты тепловому стрессу, были помещены в ростовую камеру фитотрона Conviron PGR15 (Controlled Environments Ltd.), используя метод случайных блоков. Для теплового стресса была выбрана температура около 33°С.Растения подвергались действию этой температуры в течение 8 часов, после чего растения подвергались воздействию температуры в 21°С в течение от 16 часов до 10 дней. После воздействия теплового шока, температуру изменяли до значения в 16°С на 8 часов и 21°С на 16 часов.
Результаты
[00175] Вкратце, результаты показывают, что эффективность придания жароустойчивости каждым из тестируемых эндофитов зависит от конкретного растения, или хозяина эндофита (А-нут, В-чечевица и С-горох), и что увеличение общей биомассы зависит об отдельных конкретных органов растения, таких как: стручок (Фигура 23), стебель (Фигура 24) и корень (Фигура 25), на которые тепловой стресс действует по-разному.
[00176] SMCD 2215, в большинстве случаев, увеличивал биомассу стебля и стручка у гороха, и биомассу корня у нута. SMCD 2206 вызывал увеличение биомассы стебля и стручка у чечевицы, и биомассы корней у нута, гороха и чечевицы. SMCD 2210 увеличивал в основном биомассу стебля и стручков у нута, и биомассу корней у гороха. SMCD 2204F увеличивал биомассу стручков у большинства тестируемых культур (у нута, гороха и чечевицы). Наиболее продуктивная комбинация эндофит-сорт (Е+) увеличивала биомассу стручков, стеблей и корней на уровне 300%, в сравнении с подвергшимся тепловому стрессу контролю, не содержащему эндофитов (Е-).
[00177] Стебель: Следующие эндофиты продемонстрировали наиболее продуктивную ответную реакцию на тепловой стресс: Нут: Amit: SMCD 2210. Vanguard: SMCD 2204F; Горох: Golden: SMCD 2215. Handel: SMCD 2215; и Чечевица: Glamis: SMCD 2206. Sedley: SMCD 2206.
[00178] Стручки: Следующие эндофиты продемонстрировали наиболее продуктивную ответную реакцию на тепловой стресс: Нут: Amit: SMCD 2210. Vanguard: SMCD 2204F; Горох: Golden: SMCD 2204F. Handel: SMCD 2215; Чечевица: Glamis: SMCD 2206. Sedley: SMCD 2204F.
[00179] Корень: Следующие эндофиты продемонстрировали наиболее продуктивную ответную реакцию на тепловой стресс: Нут: Amit: SMCD 2215. Vanguard: SMCD 2206; SMCD 2215; Горох: Golden: SMCD 2210; SMCD 2215. Handel: SMCD 2206; Чечевица: Glamis: SMCD 2206; Sedley: SMCD 2204F.
Пример 7
Эксперимент по воспроизведению в теплице засухи у бобовых
[00180] В данном исследовании использовались семена шести сортов [Amit, Vanguard (нут), Golden, Handel (горох) и Glamis, Sedley (чечевица)] и эндофиты SMCD 2204, SMCD 2204F, SMCD 2206, SMCD 2210, и SMCD 2215. Данные эксперименты проводились в теплице. После инокуляции эндофитами и посева семян, для имитации сильной засухи, вегетационные сосуды оставили без воды на 14 дней, как было предложено Charlton et.al. [2008] и согласно методике и условиям, изложенным Gan et al. [2004].
Результаты
[00181] В целом, результаты показывают, что каждый штамм SMCD положительно влияет на некоторые сельскохозяйственные параметры: формирование стручков или урожайность (Фигура 27), биомассу стеблей (Фигура 26) и корней (Фигура 28) у нута (А), гороха (В), чечевицы (С) в условиях засухи. В целом, сорта сельскохозяйственных культур, после их колонизации симбиотическими эндофитами (Е+), становились более устойчивыми к засухе, чем к тепловому стрессу. Уровень эффективности тестируемых эндофитов в обеспечении засухоустойчивости варьировал в разных органах растений: выход стручков был значительно увеличен эндофитом SMCD 2204 у Glamis, SMCD 2204F у Vanguard, SMCD 2206 у Sedley, SMCD 2210 у Golden и SMCD 2215 у Handel.
[00182] Стебель: Следующие эндофиты продемонстрировали наилучший ответ на засуху: Нут: Amit: SMCD 2204F, Vanguard: SMCD 2206; Горох: Golden: SMCD 2204, Handel: SMCD 2204; SMCD 2210; SMCD 2215; Чечевица: Glamis: SMCD 2204F; SMCD 2206. Sedley: SMCD 2204F; SMCD 2206.
[00183] Стручки: Следующие эндофиты продемонстрировали наилучший ответ на засуху: Нут: Amit: SMCD 2204; SMCD 2210. Vanguard: SMCD 2204; SMCD 2206; SMCD 2215; Горох: Golden: SMCD 2210; SMCD2215. Handel: SMCD 2204F; SMCD 2206; SMCD 2215; Чечевица: Glamis: SMCD 2204F; SMCD 2206. Sedley: SMCD 2210; SMCD2215.
[00184] Корень: Следующие эндофиты продемонстрировали наилучший ответ на засуху: Нут: Amit: SMCD 2204; SMCD 2215. Vanguard: SMCD 2204F; SMCD 2206; Горох: Golden: SMCD 2204F; SMCD2215. Handel: SMCD 2204F; Чечевица: Glamis: SMCD 2204F; SMCD 2206; SMCD 2210. Sedley: SMCD 2206; SMCD 2210.
Пример 8
Streptomyces sp.SMCD 2215 увеличивал активность бактерий рода Rhizobium и частоту образования клубеньков у гороха, пребывающего под воздействием теплового стресса.
[00185] Как недавно было отмечено, другие BVijxuStreptomyces, S. lydicusWYECIO [Tokala et al. 2002], Streptomyces sp. nov. SMCD2215 колонизируют корни молодых проростков гороха из семян, полученных из растений, выращенных в контрольных условиях. Это приводит к специфичным изменениям, таким как усиление цветения и урожайности стручков (Фигура 29) и увеличению формирования клубеньков на корнях бактерией рода Rhizobium sp.(Фигура 30), природным эндофитным колонизатором семян гороха, открытым в этом исследовании (Таблица 5). Вегетативные гифы Streptomyces sp. nov. SMCD2215 колонизировали клетки развивающихся клубеньков, что было обнаружено при культивировании на чашках (КДА), флуоресцентной микроскопии (Carl Zeiss Axioskop 2) и ПЦР амплификации (BioRad) [Schrey and Tarkka 2008]
Пример 9
Эндофиты увеличивают устойчивость бобовых к абиотическим стрессам путем увеличения живучести семян
[00186] Бобовые культуры относят группе, состоящей из более чем шестидесяти разных зернобобовых культур со всего мира. Семена бобовых играют важную роль в питании людей. Основными ограничивающими факторами при выращивании бобовых являются биотические и абиотические стрессовые факторы, такие как засуха, жара, холод и засоленность. Последние исследования показывают, что взаимодействие между эндофитным микроорганизмом и растением является эффективным предопределяющим фактором адаптации растений.
[00187] В этом исследовании выдвигается гипотеза, согласно которой эндофиты увеличивают скорость и равномерность прорастания семян in vitro в оптимальных и стрессовых условиях. Целью было, во-первых, произвести оценку присущей эндофитам способности запускать прорастание и, во-вторых, оценить эффективность формирования вызванной эндофитами жаро- и засухоустойчивости у представителей бобовых.
Материалы и методы
[00188] Два сорта представителей бобовых, Glamis (чечевица) и Handel (горох), культивировали совместно с SMCD 2206 и SMCD 2215, штаммами симбиотических эндофитных грибов и бактерий, соответственно. Способность эндофитов индуцировать стрессоустойчивость у представителей сортов Golden (Фигура 31) и Handel (Фигура 32) проверялась путем in vitro проращивания семян при моделировании условий засухи (6% PEG) и высокой температуры (33°С).
[00189] Поверхность семян стерилизовали 95% этанолом в течение 20 секунд, дважды промывали стерильной дистиллированной водой в течение 10 секунд, после чего помещали на 2 минуты в 3% раствор гипохлорита натрия (Javex). В заключение, семена промывали стерильной дистиллированной водой 4 раза. Инокуляцию семян проводили на среде КДА, содержащей или не содержащей эндофиты, в темноте и при комнатной температуре [Abdellatif et al. 2009]. Микроорганизмы выращивались на КДА в течение по меньшей мере трех дней в темноте и при комнатной температуре, до использования в эксперименте. Оценка способности эндофитов к индуцированию формирования стрессоустойчивости производилась при помощи измерения энергии прорастания, которая служит для фиксации временных рамок прорастания и которая определяется как количество дней, необходимых для достижения прорастания 50% семян.
Результаты
[00190] Настоящее исследование характеризует индивидуальные свойства грибных и бактериальных эндофитов к индукции формирования у бобовых засухо- и жароустойчивости, специфичные к комбинаци бактериальный или грибной штамм-сорт растения-абиотический стресс. Для лучшего понимания механизмов формирования индицированной эндофитами стрессоустойчивости бобовых, в данном исследовании были использованы методы протеомного и молекулярного анализа.
[00191] Штаммы SMCD значительно повысили частоту прорастания бобовых in vitro в стандартных условиях (Фигура 33). В стрессовых условиях, оба эндофита (SMCD 2206 и SMCD 2215) повысили частоту прорастания, в сравнении с не колонизированными семенами. Частота прорастания варьировала от 70-100% в обработанных эндофитами семенах до 60-80% в контроле, что означает, что тестируемые эндофиты имеют потенциал для увеличения энергии прорастания семян (SGV) на >15%. Наибольшая частота прорастания (100%) была выявлена у Glamis (чечевицы), которая вызывалась обоими эндофитами, SMCD 2206 и SMCD 2215, при сильной засухе или тепловом стрессе. При совместной инокуляции штаммами SMCD, энергия прорастания (прорастание >50% семян) у Glamis достигалась за 2 дня в условиях засухи и за 3 в условиях теплого стресса. Подобные результаты были достигнуты и горохом Handel, если не учитывать того, что этот сорт сам по себе является более устойчивым к тепловому шоку, чем Glamis (чечевица).
Пример 10
Эндофиты повышали урожайности льна и рапса в условиях острой засухи при экспериментах в теплице.
[00192] Целью данного исследования было применение трех случайно выбранных изолятов (SMCD 2206, SMCD 2210 and SMCD 2215 для проверки эффективности их влияния на формирование урожая льна и рапса в условиях засухи.
Материалы и методы
[00193] План эксперимента, процедуры, которые проводились с семенами льна (Bethun и Sorel) и рапса (1768S), способы введения эндофитов (SMCD 2206, SMCD 2210 и SMCD 2215), условия засухи и способы оценки урожайности соответствуют показанным в Примере 5, с незначительными изменениями. Вкратце, контрольные растения поливались до доведения значения влагоемкости почвы до 100%, три раза в неделю, тогда как подвергаемые засухе растения поливались раз в неделю до доведения значения влагоемкости почвы до 100%. Такой режим засухи был установлен для имитирования естественного цикла протекания засух, которые могут возникать во время периода вегетации в канадских прериях, во время которых осадки могут не выпадать в течение семи дней подряд, или даже более того.
Результаты и их обсуждение
[00194] Сильная засуха привела к снижению урожая не симбиотического льна, тогда как инокуляция эндофитами SMCD 2206 и SMCD 2210 привела к сильному повышению урожайности льна в таких же условиях. В частности, в условиях засухи, SMCD 2206 поддерживал почти 100% урожайность у Bethun, в то время как SMCD 2210 обеспечивал 50% урожайность, в сравнении с не подвергшимся стрессу контролем в условиях теплицы (Фигура 34). В случае с рапсом, повышение урожайности было зафиксировано в комбинации с SMCD 2210 (>100%), за ним следует SMCD 2206 (~50%) и SMCD 2215 (~30%), в сравнении с не подвергшимся стрессу контролем (Фигура 35).
[00195] Способность к защите от Fusarium avenaceum и F. graminearum также была проверена в условиях теплицы. Автоклавированные семена инфицировали инокулятом Fusaria инкубировали в темноте в течение 7 дней при 25°С (Фигура 36), после чего инокулировали эндофитами, выращеными на чашках Петри, как описано в Abdellatif et al. [2009].
[00196] Грунтовая смесь была мнокулирована двадцатью семенами, содержащими Fusarium. Состав грунтовой смеси: 55-65% канадский сфагнум торф мох, перлит, и известняк, смешанные с песком. Стандартные условия теплицы включали 8 часовое освещение чередующееся с 16 часовым фотопериодом (1000 люкс) приотносительной влажности 70% и постоянной температуре 25°С±2°С.
[00197] Обработка растений производилась следующим образом:
O1: Необработанные растения (контроль)
O2: Растение + эндофит
O3: Растение + патоген, Fusarium avenaceum
O4: Растение + патоген, Fusarium graminearum
O5: Растение + эндофитный гриб + Fusarium avenaceum
O6: Растение + эндофит + Fusarium graminearum
[00198] Каждую обработку повторяли в вегетационных сосудах, и проростки поливали три раза в неделю при контролируемых условиях. Колонизацию корней эндофитами проверяли с помощью флуоресцентного микроскопа, для разграничения симбиотических и патогенных отношений эндофит-пшеница. [Abdellatif et al. 2009].
[00199] Фигуры 37-40 иллюстрируют позитивное влияние эндофитов на противодействие проростков последствиям стресса (Фигура 37), формирование биомассы листьев и корней (Фигура 38 и Фигура 39), фазу цветения и формирования колосьев (Фигура 38, Фигура 39 и Фигура 40). Все тестированные эндофиты индуцировали рост нормально сформированных листьев, по сравнению с контролем, так же как и рост нормально сформированных цветков в присутствии эндофитов.
[00200] Для подтверждения способности эндофитов стимулировать рост зрелых растений в присутствии патогенного гриба рода Fusarium, продуктивность стадии цветения, результатом которой было формирование колосьев, оценивалась на более поздних стадиях роста.
[00201] Гистограммы на Фигуре 41 иллюстрируют продуктивность эндофитов в увеличении биомассы, или сухого веса, колосьев пшеницы после двойной инокуляции (зндофитом SMCD и патогенным грибом рода Fusarium).
[00202] Урожайность пшеницы в присутствии эндофитов и патогенного гриба рода Fusarium была значительно повышена применением эндофитов всех штаммов, в сравнении с пшеницей, инфицированной F. graminearum и F. avenaceum, но не обработанной эндофитами (Е-) (Фигура 41). Растения, обработанные только патогеном, имели значительно меньший размер колосьев, по сравнению с контрольными растениями и растениями, обработанными эндофитами (Е+) (Фигура 42).
Пример 11
Эндофито-опосредованная регуляция экспрессии генов устойчивости к абиотическому стрессу у бобовых
Реферат
[00203] Геномные и белковые механизмы позитивного влияния эндофитов на устойчивость растений-хозяев к абиотическим стрессам изучены недостаточно. Одна из современных теорий утверждает, что симбиотические растения защищаются от оксидативного стресса, вызванного такими стрессовыми факторами, как повышенная температура, засуха и засоленность, продукцией антиоксидантных молекул. Целью данного исследования является прояснение механизмов защитного симбиоза гороха, нута и чечевицы путем оценки экспрессии генов Pro, СОД, и МnСОД, запускаемой взаимодействием между хозяином и эндофитом. Результаты данных исследований показали наличие опосредованной эндофитами регуляции экспресии генов в растениях, инокулированных эндофитами. Данные гены играют важную роль и обеспечивают защиту хозяина путем усиления устойчивости к испытываемым абиотическим стрессорам.
Материалы и методы
[00204] С целью проведения анализа были собраны листья с нормальных и подвергнутых стрессу семян шести сортов (Amit, Vanguard [нут] (Фигура 43), Golden, Handel [горох] и Glamis, Sedley [чечевица]), обработанных и необработанных эндофитами.
[00205] Для амплификации генов, таких как гены пролина (Pro), СОД и МnСОД, применялся ПЦР в реальном времени, с использованием праймеров с последовательностями, указанными в SEQ ID NOs: 8-15 (Таблица 6), при этом стрессовые белки, как правило, играют особую роль в защите цитоплазмы от обезвоживания и в защите растений путем снижения токсичности, вызванной высокой концентрацией ионов. ПЦР проводили при следующих условиях: 3 минуты при 95°С (активация фермента), 40 циклов по 30 секунд при 95°С (денатурация) и 30 секунд при 60°С (отжиг/удлинение). В конечном счете, анализ кривой плавления производился в диапазоне температур от 65° до 95°С с шагом в 0,5°С, каждый шаг длился 5 секунд, чтобы убедится в наличии единственного продукта и отсутствии димеров праймеров. Количественный анализ относительно контрольного гена производился путем вычитания СТ контрольного гена от СТ таргетного гена (ΔCT). Для определения относительных уровней экспрессии, полученную разность числа циклов делили на значение калибратора - нормализованное значение таргетного гена, полученное значение (ΔΔCT) является экспонентой с основанием 2 (из-за удвоения функции ПЦР).
Результаты
[00206] Было проанализировано экспрессию разных генов во время воздействия засухи. Таблица 6 отображает гены, которые подверглись анализу. Некоторые из полученных результатов относятся к представителям сорта Handel, подвергшимся воздействию 6% ПЭГ.
СОД и МnСОД
[00207] В целом, СОД играет основную роль в осуществлении антиоксидантных защитных механизмов. В настоящем исследовании был обнаружен очень высокий уровень экспрессии СОД в нормальных (Е-, контроль) листьях, подвергшееся воздействию 6% ПЭГ (почти 200-кратное повышение экспрессии). Эндофиты играли очень важную роль в уменьшении этого стресса. В особенности, SMCD 2215, за ним следовали SMCD 2210, SMCD 2204 и SMCD 2206. Эти симбионты резко снижали стресс путем снижения экспрессии всего лишь в 9 и 24 раза (Фигура 44А).
[00208] MnCOД - это одна из форм СОД. В контрольных листьях наблюдалось 16-кратное увеличение экспрессии генов, тогда как SMCD 2215 подавлял стресс и снижал изменение уровня экспрессии с 16-кратного до 2-кратного, за ним следовали SMCD 2206, SMCD 2210 и SMCD 2204 (Фигура 44В).
Пролин
[00209] Пролин является важным для первичного метаболизма. Биосинтез пролина у растений контролируется активностью двух генов P5CS. Экспрессия этого гена оценивалась у содержащего эндофиты гороха сорта Handel, пребывающего в условиях засухи. Как и предполагалось, ген P5CS активировался, и его экспрессия увеличилась в 5 раз в листьях, полученных из растений, которые подвергались воздействию ПЭГ. В то же время в листьях, собранных с растений, семена которых были ассоциированы с SMCD 2206, экспрессия увеличивалась в 2,8 раза, за ним следовал SMCD 2215 с 3,4-кратным повышением экспрессии генов, кодирующих пролин (Фигура 45). Данные результаты подтвердили, что эндофиты играют значительную роль в обеспечении стрессоустойчивости путем изменения экспрессии генов пролина, в сравнении с подвергшимися стрессу неинокулированными растениями.
Пример 12
Паттерны экспрессии генов в колеоризе пшеницы при холодной и биологической стратификации
Реферат
[00210] Пшеница представляет собой одну из широко используемых основных сельскохозяйственных культур в мире. Однако, за последние два десятилетия всемирная продукция пшеницы уменьшилась на 5,5%, и предсказано последующее уменьшение, в связи с глобальным потеплением. Таким образом, нахождение условий и методов, которые могли бы позволить увеличить всхожесть семян, является очень важным. Холодная стратификация представляет собой широко известный способ выведения семян из состояния покоя и стимуляции их прорастания. Биологическая стратификация грибными эндофитами также может стимулировать прорастание семян многих зерновых культур. Колеориза представляет собой одну из наиболее активных тканей семени, а также - часть семени, которая первой выходит наружу при прорастании. Для оценки эффективности метода стратификации, был определен процент всхожести семян пшеницы, подвергшихся холодной и биологической стратификации, а также был определен уровень экспрессии генов гибберелина и абсцисовой кислоты в колеоризе. Как холодная, как и биологическая стратификация значительно (Р<0,05) повышают уровень и эффективность прорастания. Расстояние между грибным эндофитом и семенами является определяющим для биологической стратификации фактором, так как семена, пребывающие в непосредственном контакте с грибным эндофитом, имеют наивысший процент прорастания (до 86%). Повышенная экспрессия гена GA3ox2 в колеоризе пшеницы была обнаружена на протяжении периода прорастания путем выявления устойчивой продукции биоактивной молекулы GA3. Экспрессия гена 14-3-3 была наиболее низкой после прямой обработки эндофитом. Экспрессия гена биосинтеза абсцизовой кислоты (АБК), TaNCED2, была значительно выше в семенах, которые подвергались холодной стратификации, что отображает роль абсцизовой кислоты как гормона стрессовой адаптации. В колеоризе также было обнаружено повышение экспрессии гена ТаАВА8'OH1. В целом, это исследование дает молекулярно-биологические доказательства важности колеоризы для прорастающих семян пшеницы. Сравнивая холодный и биологический способы стратификации, можно сделать вывод, что всхожесть семян может быть заметно повышена путем применения грибных эндофитов, и что расстояние между зероном и эндофитом является ключевым для миковитальности фактором.
Материалы и методы
Семена пшеницы
[00211] В данном исследовании применялись семена твердой пшеницы сорта АС Avonlea, имеющие низкую устойчивость к стрессорным условиям окружающей среды. Эти семена были произведены Agriculture and Agri-Food Canada Seed Increase Unit Research Farm в 2006 году в тепличных условиях, и были рекомендованы как не содержащие микроорганизмов. Семена помещали в стерильные пакеты с застежкой и хранили при 4°С до последующего применения.
Сравнение протоколов стерилизации семян
[00212] Для стерилизации поверхности семян пшеницы были предложены различные способы. В данном документе четыре широко признанных способа стерилизации семян сравнивали между собой для определения наиболее подходящего протокола, с помощью которого можно было бы эффективно стерилизовать поверхность семян, не ухудшая качества и жизнеспособности семян пшеницы этого сорта. В первом способе, поверхность семян стерилизовали 95% этанолом в течение 10 с, с последующей трехразовой, по 1 мин, промывкой стерильной дистиллированной водой [Zhang et al., 2007. ВМС Genetics 8]. Второй протокол являлся стерилизацией отбеливателем, при которой поверхность семян стерилизуют в 5% раствором гипохлорита натрия в течение 3 мин с последующим тщательным трехразовым, по 1 мин, ополаскиванием семян стерильной дистиллированной водой. В соответствии с третьим протоколом, поверхность семян стерилизовали 95% этанолом в течение 10 с, семена промывали стерильной дистиллированной водой, а затем погружали на 3 мин в 5% раствор гипохлорита натрия, промывали три раза в стерильной дистиллированной воде и помещали на картофельно-декстрозный агар (КДА) для обеспечения их прорастания [Abdellatif et al. 2009]. Четвертый способ являлся стерилизацией семян парами хлора (газообразного) [Desfeux et al., 2000]. Под вытяжным шкафом, небольшой стакан, содержащий 20 мл отбеливателя, помещали в 5-литровую коробку с газоотводными коннекторами. Семена пшеницы помещали в 96-луночный планшет и удерживали его в коробке с газоотводными коннекторами. Затем, для создания газообразного хлора, в стакан с отбеливателем добавляли 3 мл концентрированной соляной кислоты. Для удержания семян в контакте с газообразным хлором, крышку не открывали в течение 4 часов. После стерилизации, 96-луночный планшет помещали в ламинарный бокс на 1 час, для рассеивания остатков газообразного хлора. Затем стерилизованные семена трижды промывали стерильной дистиллированной водой и высеивали на чашки с КДА. Сравнение этих способов стерилизации показало, что стерилизация газообразным хлором является самым эффективным способом, обеспечивающим 80% всхожесть семян без контаминации, при том, что контрольные семена имели более высокий уровень контаминации (Таблица 7). И хотя способы стерилизации этанолом и отбеливателем успешно снижали контаминацию, при этом они сильно повлияли на всхожесть семян. Таким образом, протокол стерилизации газообразным хлором является весьма эффективным способом стерилизации семян пшеницы, и именно он был выбран для стерилизации семян, необходимой для осуществления экспериментов, проведенных в данном исследовании.
Холодная и биологическая стратификация
[00213] Для проведения холодной стратификации, поверхностно стерилизованные семена удерживали на влажной фильтровальной бумаге в холодильной комнате при 4°С в течение 48 часов [Mukhopadhyay et al., 2004; Wu et al., 2008]. Через 2 дня, семена, прошедшие холодную стратификацию, перемещали в условия комнатной температуры и быстро промывали стерильной дистиллированной водой, после чего помещали на чашки с картофельно-декстрозным агаром (КДА). Для проведения биологической стратификации, стерилизованные семена инкубировали в присутствии SMCD 2206. Грибной эндофит выращивали на КДА в темноте при комнатной температуре в течение по меньшей мере трех дней до применения. Для оценки эффективности стратификации, семена пшеницы проращивали в непосредственном контакте с грибным эндофитом, а также на некотором расстоянии от него. (5 мм2) агарную пластинку с эндофитом, отделенную от краев материнской колонии, помещали в центр 90 см чашки Петри с КДА. Затем 10 поверхностно стерилизованных семян помещали на периферии чашки Петри, на расстоянии около 4 см, окружая таким образом агарную пластинку с грибами. Все чашки Петри запечатывались 5 слоями пленки Parafilm® (Pechiny Plastic Packaging) во избежание биологической контаминации и распространения летучих газообразных соединений. Влияние прямого контакта грибного эндофита выяснялось путем размещения 3 мм2 агарной пластинки между двумя соседними поверхностно стерилизованными семенами и 5 мм2 пластинки в центр КДА. Все процедуры были выполнены в трех повторностях на чашках с КДА, в каждую чашку Петри помещали десять поверхностно стерилизованных семян. Чашки Петри инкубировали в настольном инкубаторе при комнатной температуре (~20°С) в темноте. Время инкубации записывали и добавляли в базу данных, изоляцию колеоризы проводили через 24, 48 и 72 часа.
Процент всхожести
[00214] Появление ранних первичных корней тщательно отслеживалось. Процент всхожести рассчитывался путем оценки количества проросших семян на 10 семян пшеницы на каждой чашке с КДА. Всхожесть в 50% оценивалась как энергия прорастания. Эффективность прорастания при различных способах обработки оценивалась при помощи следующего уравнения:
Эффективность = % прорастания при обработке - % прорастания в контроле
[Уравн. 1]
[00215] Уровни всхожести наблюдались для всех обработанных образцов и на всех повторностях. Для оценки общей жизнеспособности образцов на день 2 и день 3 уровни всхожести контролировались начиная с дня 1. Чашки с КДА сохраняли запечатанными в течение всего периода сбора данных.
Выделение колеоризы
[00216] После оценки уровней прорастания, чашки с КДА немедленно переносились в стерильный ламинарный бокс для изоляции колеоризы. Семена пшеницы аккуратно расчленяли под микроскопом, слои колеоризы отделяли при помощи стерильной иглы и скальпеля. Выделенную колеоризу хранили в очищенных от РНКаз стерильных микроцентрифужных пробирках. Семена, полученные в результате всех биологических повторностей были объединены, и для получения оптимального для экстракции РНК количества растительного материала было выделено около 20-30 колеориз.
Экстракция РНК и синтез ДНК
[00217] Во избежание деградации растительного материала, выделение РНК проводили каждый день, сразу после выделения колеоризы. Для выделения РНК использовали около 20 мг образцов колеоризы. Выделение общей РНК производили при помощи Aurum™ Total RNA Mini Kits, согласно инструкциям производителя (Bio-Rad Laboratories). Концентрацию РНК измеряли спектрофотометрическим методом на Nanodrop (Thermo Scientific). Синтез кДНК производился сразу после выделения РНК при помощи iScript cDNA Synthesis Kit, согласно инструкции от производителя (Bio-Rad Laboratories). Для синтеза кДНК брали 600 нг аликвоты РНК. Обратная транскрипция производилась в течение 30 минут при 42°С, с конечной денатурацией в течение 5 минут при 85°С.
Количественный ПЦР в реальном времени
[00218] Экспрессия функциональных генов гибберелина и абсцизовой кислоты измерялась при помощи относительного количественного ПЦР в реальном времени (QRT-PCR). Различные катаболические и биосинтетические гены были отобраны для оценки их соответствующей роли в холодной и биологической стратификации. Фрагмент гена актина пшеницы длинной 131 п.н использовался в качестве внутреннего контроля [Nakamura et al., 2010]. QRT-PCR производился при помощи термоциклера MJ-Mini Gradient Thermal Cycler (Bio-Rad Laboratories), согласно инструкции от производителя. Условия проведения ПЦР: 1 цикл при 95°С в течение 1 минуты и 40 циклов по 20 секунд при 94°С, 60°С в течение 30 секунд, и 72°С в течение 1 минуты. Для ПЦР в реальном времени применялись полученные образцы кДНК, все реакции проводились в трех повторах, с двумя отрицательными контроллями. Каждые 25 мкл реакционной смеси содержали 18 мкл iQ™ SYBR® Green supermix (Bio-Rad Laboratories), 10 пМоль соответствующих прямых и обратных праймеров, 2,5 мкл бычьего сывороточного альбумина и 25 нг образцов кДНК. Количественный анализ выполнялся согласно Zhang et al. [2007]. Уровни экспрессии были рассчитаны с использованием значения порогового цикла (СТ), определенного согласно установленной вручную базовой линии. Разницу между значениями Ct гена-мишени и актина (Cttarget-Ctactin) оценивали как Ct, после чего уровень экспрессии рассчитывался как 2-Ct. Средние значения 2-Ct были использованы для оценки разницы уровней экспрессии в контроле и образцах, подвергшихся стратификации. Для проверки наличия амплификонов и специфичности их последовательностей, анализ кривых плавления и электрофорез в агарозном геле производился после прохождения каждой QRT-PCR.
Секвенирование
[00219] Ампликоны актина и различных генов GA и ABA очищали при помощи BioBasic PCR Purification Kit (Bio Basic Inc.). Для каждой обработки, очищенные ампликоны были отправлены для секвенирования в Plant Biotechnology Institute (NRC-PBI). Последовательности генов были определены путем проведения анализа Basic Local Alignment Search Tool (BLAST) (http://blast.ncbi.nlm.nih.gov).
Анализ статистических данных
[00220] Однофакторный дисперсионный анализ данных, касающихся процента всхожести и экспрессии генов, производился при помощи программного обеспечения IBM SPSS Statistics, версия 19. Различия между контрольной и подвергаемой стратификации группами оценивались при помощи post-hoc теста Дункана.
Результаты и их обсуждение
Процент всхожести и эффективность прорастания
[00221] Как холодная, так и биологическая стратификация значительно повысили уровень всхожести при всех трех типах обработки, демонстрируя более высокий процент всхожести, чем у контроля (Фигура 46А; Таблица 8). После прямой обработки эндофитами каждый день наблюдалось увеличение процента всхожести, а также его увеличение на 60% в период с дня 1 по день 3. На протяжении всего периода прорастания (3 дня) у них была отмечена значительно (Р<0,05) более высокая всхожесть, чем при трех других типах обработки. Только биологическая стратификация привела к более чем 50% всхожести после второго дня. Интерестно отметить, что непрямая обработка эндофитами не привела к прорастанию после первого дня, но вызвала примечательную 50% всхожесть после второго дня. Воздействие холодной стратификации не привело к появлению существенных отличий от контроля после первого дня, но затем постепенно увеличивало их, продемонстрировав появление значительных различий после второго и третьего дней. Паттерн увеличения прорастания также отображается значениями R2. В то время, как в контроле значение R2 соответствовало 0,40, при холодной стратификация и прямой обработкой эндофитами оно равнялось 0,60 и 0,75, соответственно. С другой стороны, по причине его 50% возрастания с первого по второй день, непрямая обработка эндофитами продемонстрировала наивысшее значение R2, равное 0,93, что в около 2,5 раза выше, чем в контроле. Энергия прорастания является критическим параметром, определяющим способность семян к прерывания состояния покоя и старту прорастания. Энергия прорастания рассчитывается как процент всхожести семян через определенный период времени, или как количество дней, необходимых для достижения 50% всхожести. Прямая стратификация эндофитами показала наивысшую эффективность, за ней следовали непрямая и холодная стратификация (Фигура 46В). Так как при непрямой обработке эндофитами не наблюдалось прорастание семян после первого дня, эффективность прорастания была отрицательной. В целом, стратификация показала чрезвычайно положительный результат, приведя к достижению 50% всхожести через 48 часов.
[00222] Стратификация играет важную экологическую роль при выходе из состояния первичного покоя и в повышении всхожести семян [Bewley and Black 1982; Probert et al., 1989]. Ослабление состояния покоя семян и улучшение всхожести путем холодной стратификации были достигнуты у многих видов, включая злаковые [Schutz и Rave 1999], шелковицу [Koyuncu 2005], сосну [Carpita et al., 1983], табак [Wu et al., 2008], рис [Mukhopadhyay et al. 2004], и яблоню [Bogatek и Lewak 1988]. При помощи холодной стратификации также была увеличена всхожесть 33 видов однолетних сорняков и было предположено, что стратификация способна свести к нулю отличия между всхожестью семян у разных популяций [Milberg и Andersson 1998]. Тем не менее, имеется мало информации о влиянии холодной стратификации на прорастание семян пшеницы. Данное исследование показало, что для реализации эффекта холодной стратификации необходим период инициации и, поэтому, всхожесть семян существенно не отличалась от контроля в первый день. Тем не менее, она продемонстрировала значительное влияние на прорастание, начиная со второго дня, и увеличила процент всхожести на целых 20%, в сравнении с контролем. При подборе длительности холодной стратификации в данном исследовании руководствовались данными предыдущих отчетов, которые показали, что период в 48 часов эффективен для холодной стратификации табака [Wu et al., 2008] и риса [Mukhopadhyay et al. 2004]. Более ранние исследования показали, что эффект от холодной стратификации пропорционально зависит об длительности ее воздействия [Baskin et al. 1992; Cavieres и Arroyo, 2000]. Полученные нами данные подтверждают это, и позволяют предвидеть, что для достижения максимальной всхожести пшеницы может потребоваться немного больший период стратификации (~4 дней).
[00223] В нескольких отчетах говорится о повышении всхожести семян в результате применения грибковых эндофитов [Vujanovic 2007b; Hubbard et al. 2012; Vujanovic and Vujanovic 2007]. Настоящее исследование поддерживает концепцию "миковитализма", сутью которой является увеличение жизненной силы путем грибной колонизации. Широко известным является свойство грибных эндофитов производить летучие соединения, которые влияют на продолжительность растительных фенофаз [Mitchell et al., 2009; Strobel et al., 2004]. Таким образом, эндофиты могут быть способными воздействовать на прорастание семян, даже если они не находятся в прямом контакте с семенами, и эта черта является особенно ценной в полевых условиях. В данном исследовании также проверяется то, как физическое расстояние при биологической стратификации может влиять на всхожесть. Полученные результаты показывают, что семена, находящееся в непосредственном контакте с грибным эндофитом, получали несомненно больше выгоды, чем их аналоги. Прямой контакт с эндофитом был причиной наиболее высокого процента всхожести и эффективности прорастания в каждый день периода прорастания. Подобно прямому контакту с эндофитом, семена, которые находились в 4 см от эндофита, также имели значительно более высокий уровень всхожести, чем контроль. Тем не менее, расстояние действительно повлияло на процент всхожести и эффективность прорастания, и у не пребывающих в прямом контакте семян значение процента всхожести было на 14-27% меньше, чем у пребывающих в прямом контакте. Кроме того, прорастание не наблюдалось в первый день, за чем следовало резкое возрастание его значения (50%) на второй день. Всхожесть семян является чрезвычайно сложным процессом и лежащие в основе этого явления механизмы относительно слабо изучены [Nonogaki et al., 2010]. Так, не ясно, каким образом грибные эндофиты облегчают выход семян из состояния покоя и запускают прорастание семян. Учитывая способность грибов производить широкий спектр веществ, усиливающих рост растений, представляется возможным, что эти вещества действуют более эффективно при непосредственной близости. Следовательно, семена, прошедшие прямую обработку эндофитами, имеют значительно более высокую всхожесть, чем при других способах обработки. Напротив, семена, прошедшее непрямую обработку эндофитами, демонстрировали высокую эффективность прорастания через 48 часов, этот период должен был позволить им накопить достаточное количество веществ, стимулирующих рост. Существует разница (6,6%) между значениями процента всхожести у контрольных и прошедших непрямую обработку эндофитами семян, однако она не существенна.
Уровни экспрессии генов гибберелинов и абсцизовой кислоты
[00224] Гены ОА3-оксидазы 2 и 14-3-3 были выбраны как гены биосинтеза ГК и негативный регулятор сигнального пути биосинтеза ГК, соответственно [Ji et al., 2011; Zhang et al., 2007]. Ген NCED является широко известным из-за выполнения им важной роли в сигнальных путях биосинтеза АБК, тогда как ген АБК 8'-гидроксилазы принимает участие в катаболических сигнальных путях [Ji et al., 2011]. Анализ количественного ПЦР в реальном времени показал, что значения дифференциальной экспрессии (Фигура 47) и соотношение уровней экспрессии (Фигура 48) различных функциональных генов значительно различались (Р<0,05) при разных способах обработки. Достаточные для детекции уровни экспрессии всех генов, за исключением гена 14-3-3 в третий день, выявлялись каждый день. Уровни экспрессии всех генов в первый день снизились, в сравнении с контролем. Экспрессия гена биосинтеза ГК, TaGA3ox2, при холодной стратификации была значительно выше, чем при биологической стратификации. С другой стороны, не было обнаружено значимых различий между уровнями экспрессии гена 14-3-3 при обработке холодом и эндофитами, хотя уровень его экспрессии при холодной стратификации был немного выше, чем при эндофитной. Уровни транскрипции TaNCED2, гена биосинтеза АБК, не отличались у контрольной и подвергнувшейся холодной стратификации групп, но значительно повышались при обработке эндофитами. При всех трех способах стратификации обнаруживалось снижение экспрессии гена АБК 8'-гидроксидазы, ТаАВА8'ОН1, при этом самый низкий уровень экспрессии наблюдался при холодной стратификации. Паттерны экспрессии при прямой и непрямой обработке эндофитами не отличались. На второй день, экспрессия TaGA3ox2 была значительно снижена по сравнению с контролем, при всех типах стратификации. При холодной стратификации и непрямой обрабоке эндофитами экспрессия не отличалась, самый низкий уровень экспрессии был зафиксирован в колеоризах, которые подвергались прямой обработке эндофитами. Значимых различий между уровнями транскрипции 14-3-3 при всех четырех типах обработки обнаружено не было, хотя при холодной стратификации экспрессия была немного выше. Экспрессия генов TaNCED2 при обработке эндофитами была значительно ниже, чем при холодной стратификации или в контроле. Аналогичным образом, при стратификации было обнаружено значительное снижение экспрессии гена ТаАВА8'ОН1, по сравнению с контролем. Наиболее низкий уровень экспрессии был зафиксирован при непрямой обработке эндофитами. Уровни экспрессии гена TaGA3ox2 при разных типах обработки также значимо отличались на третий день. При холодной стратификации его экспрессия была приблизительно в десять раз выше, чем в контроле, тогда как при обработке эндофитами обеими способами она значимо не отличалась. Напротив, экспрессия генов TaNCED2 и ТаАВА8'ОН1 снизилась при стратификации всех типов, с наиболее низкими уровнями экспрессии при прямой и непрямой обработке эндофитами, соответственно. Экспрессия гена 14-3-3 на третий день не наблюдалась.
[00225] Соотношения уровней экспрессии генов биосинтеза ГК и АБК, TaGA3ox2:TaNCED2, при разных типах обработки значимо не отличались в первый день, но позже они неуклонно возрастали (Фигура 48). На второй день после непрямой обработки эндофитами было выявлено наивысшее его значение, которое было выше, чем при других типах обработки в около 5-10 раз; тем не менее, значения этого параметра на третий день были равны при всех типах обработки. Напротив, соотношение экспрессии генов биосинтеза ГК и катаболических генов (TaGA3ox2:14-3-3) в первый день было наивысшим при прямой обработке эндофитами, за ней следовала непрямая обработка, холодная стратификация и контроль, что очень похоже на тенденцию с процентом их всхожести. На первый день при всех типах обработки обнаруживались сходные паттерны соотношения экспрессии генов биосинтеза ГК и генов катаболизма АБК, TaGA3ox2:TaABA1, но, тем не менее, на второй день, при непрямой обработке эндофитами, оно было значительно выше, чем при других типах обработки. На первый день, при холодной и непрямой стратификации эндофитами обнаруживались равные уровни их экспрессии, а у контроля она была незначительной. Значения соотношения между генами биосинтеза АБК и генами ее катаболизма (TaNCED2:TaABAl) при разных типах обработки не отличалось в течение всего периода прорастания, хотя холодная стратификация привела к повышению уровня стратификации в первый день.
[00226] Гены, кодирующие ферменты биосинтеза и катаболизма ГК и АБК, имели различные паттерны, в зависимости от накопления транскриптов [Hedden and Phillips, 2000]. Паттерны экспрессии генов GA3ox1 были изучены во множестве видов растений, включая арабидопсис [Phillips et al., 1995], рис [Oikawa et al., 2004]и пшеницу [Zhang et al., 2007]. В то время как другие гены биосинтеза ГК, такие как GA-20ox, связывают с ростом вегетативных тканей и цветов, GA3ox (GA3ox2 или GA4H) экспрессируются исключительно в семенах при их прорастании и, предположительно, играют в этом ключевую роль [Phillips et al., 1995; Yamaguchi et al., 1998; Hedden and Phillips, 2000]. Подобно предыдущим отчетам, это исследование также продемонстрировало высокий уровень экспрессии гена GA3ox2 в колеоризе пшеницы во время ее прорастания. Потенциально, не ограничиваясь рамками теории, это говорит о стабильной выработке биологически активной формы ГК - молекулы GA3, в колеоризе пшеницы при прорастании. С другой стороны, в колеоризе также была выявлена низкая экспрессия гена 14-3-3, негативного регулятора биосинтеза ГК. С постепенным вырастанием проростков и увеличением содержания эндогенной ГК, уровень транскрипции 14-3-3 тоже уменьшался, до окончательного его снижения на второй день. Интересно, что в контроле наблюдался наивысший уровень 14-3-3, за ним следовали холодная стратификация, прямая и непрямая обработка эндофитами, что отразилось на их прорастании. Эти результаты соответствуют предыдущим отчетам Zhang et al. [2007], которые показали, что гены биосинтеза ГК и катаболические гены тесно ваимосвязаны с содержанием ГК и ростом побегов.
[00227] Система экспрессии генов пути АБК была исследована в широком спектре представителей злаков и бобовых, таких как рис [Oliver et al., 2007], пшеница [Ji et al., 2011; Nakamura et al., 2010], боб [Qin and Zeevart, 1999]. Текущие результаты указывают на то, что экспрессия гена TaNCED2 не сильно различалась между группами, подвергавшимися разным способам обработки, за исключением групп контроля и холодной стратификации в первый день. Абсцизовая кислота играет ключевую роль в сигнальных путях адаптации растений к стрессу [Nakamura et al., 2010]. Поскольку семена, подвергшиеся холодной стратификации, пребывали при 4°С в течение 48 часов до их инкубации при комнатной температуре, содержание абсцизовой кислоты, возможно, было выше. С другой стороны, повышенная экспрессия TaNCED2 в контроле может привести к усилению синтеза АБК и, таким образом, к замедлению уровня всхожести. Последние работы подтверждают, что катаболизм АБК происходит, преимущественно, в колеоризе [Millar et al., 2006; Okamoto et al., 2006]. Более того, Barrero et al. [2009] сообщили об активации и наивысшей экспрессии АВА8'ОН-1 в колеоризе ячменя. Подобно этим данным, в данном исследовании высокий уровень экспрессии гена ТаАВА8'ОН1 был отмечен в колеоризе пшеницы. Значение соотношения уровней генов биосинтеза ГК и АБК тесно связано с процентом всхожести. Несмотря на то, что значение TaGA3ox2:TaNCED2 значительно не отличалось после первого дня, при непрямой обработке эндофитами оно было наивысшим на второй день, по причине его значительного увеличения. С другой стороны, все три типа стратификации привели к значительному увеличению TaGA3ox2:TaNCED2 на третий день, что отобразилось на их всхожести.
[00228] Механизмы, лежащие в основе биологической стратификации, все еще относительно неизвестны, но они могли бы помочь понять, как происходят взаимодействия между растением и грибом на ранних стадиях прорастания. Роль грибных эндофитов, как биостимуляторов, является общепризнанной [Arnold et al., 2001; Hubbard et al. 2011; Saikkonen et al., 1998; Khan et al. 2012]. В этом исследовании мы продемонстрировали, что грибные эндофиты могут значительно стимулировать прорастание семян, и сила такой миковитальности пропорциональна расстоянию между семенем и грибным эндофитом. Более того, эффект от опосредованной грибными эндофитами биологической стратификации является значительно более выраженным, чем от предварительной обработки холодом. Предыдущие исследования показали, что степень инициации прорастания пропорциональна длительности холодной стратификации [Cavieres и Arroyo, 2000b], учитывая это, в последущих исследованиях период холодной стратификации может быть продлен (>48 часов), для увеличения всхожести семян пшеницы. Несмотря на то, что холодная стратификация увеличивает уровень транскрипции генов биосинтеза АБК, грибные эндофиты не стимулируют непосредственно экспрессию генов фитогормонов в колеоризе. Однако эти исследования уделили особое внимание определению экспрессии четырех генов в колеоризе.
[00229] Ни одно из исследований не сравнивало степени всхожести после холодной и биологической стратификации и не производило оценку экспрессии генов биосинтеза и катаболизма ГК и АБК в колеоризе пшеницы. Ранее колеориза была представлена как высокоактивный компонент прорастающих семян [Barrero et al., 2009]. В соответствии с этим примером, в колеоризе прорастающих семян пшеницы была также продемонстрирована высокая экспрессия различных функциональных генов. Прорастание семян в значительной степени может быть усилено путем применения грибных эндофитов: 1) путем непрямой миковитальности, или при отсутствии контакта между семенами и эндофитами на тестируемом расстоянии (например, в этом примере использовалось расстояние равное 4 см и 2 см) путем прямой миковитальности, или вслучае достижения эндофитом зерна.
Пример 13
Стратификация эндофитами воздействует на гормональные регуляторы (RSG and КАО) и гены резистентности MYB
[00230] Стратификация представляет собой воздействие на семена холодными и влажными условиями, с целью прерывания состояния покоя и усиления всхожести. Так как в настоящее время стратификация ограничена применением в этой роли абиотических факторов, это исследование направлено на достижение более широкого признания роли биотических факторов, с использованием миковитальности, или модели симбиоза семя-гриб. Это подтверждает существование как холодной, так и биологической стратификации. Всхожесть семян пшеницы, подверженных холодной стратификации при 4°С, сравнивалась со всхожестью семян, которые были инокулированы при комнатной температуре. Семена были инокулированы эндофитами штамма SMCD2206. Производилась оценка изменений в паттернах экспрессии генов-регуляторов стимуляции роста семян (RSG и КАО), фитогормональных гибберелинов (ГКы); и приобретенных генов устойчивости (MYB) в условиях абиотического и биотического воздействия на ранних этапах прерывания периода покоя и прорастания. Измерения в клетках колеоризы производили при помощи qRT-PCR (как это было описано в Примере 12). Полученные результаты показали, что экспрессия генов RSG и КАО (Фигура 49), кодирующих ферменты биосинтеза ГК, и генов резистентности MYB (Фигура 49) увеличивается в инокулированных семенах. Таким образом, миковитальность вызывает эффект перепрограммирования процессов, происходящих в семенах пшеницы до и после ее прорастания, приводя к учащению прерывания периода покоя и увеличению всхожести, эффективно способствуя пренатальному уходу за семенами злаковых культур.
Материалы и методы
Образцы РНК
[00231] Это исследование является продолжением Примера 12. Те же материалы (пшеница SMCD 2206) и способы in vitro, а также выделенные пробы РНК применялись для оценки экспрессии генов-регуляторов фитогормонов RSG и КАО и генов резистентности MYB при помощи qRT-PCR.
[00232] До экстракции РНК, пробирки, содержащие ткани колеоризы сохраняли в жидком азоте, в который они помещались сразу после выделения тканей колеоризы, для сохранения клеток и предотвращения денатурации РНК. Для экстракции общей РНК из растительных клеток применялся Aurum™ Total RNA Mini Kit (Bio-Rad Laboratories), что подразумевает, что для каждой пробы было необходимо не менее 20 мг растительных тканей. Все этапы экстракции проводились очень быстро, и в течение всего процесса пробы находились на льду, так какрнк очень быстро денатурирует при комнатной температуре. Свежевыделенная общая PHKS непосредственно загружалась в пробирки с предварительно смешанными реагентами для синтеза РНК, полученными из iScript cDNA Synthesis Kit (Bio-Rad Laboratories). Обратная транскрипция производилась в термоцикл ере в течение 30 минут при 42°С, с завершающей денатурацией при 85°С в течение 5 минут.Концентрация ДНК измерялась при помощи Nanodrop (Thermo Scientific), и доводилась до 100 нг/мкл.
Количественный RT-PCR и анализ статистических данных
[00233] Количественный ПЦР в реальном времени (QRT-PCR) производился при помощи MiniOpticon™ Real-Time PCR Detection System (Bio-Rad Laboratories) с применением iQ™ SYBR® Green supermix kit (Bio-Rad Laboratories). Для нормализации данных QRT-PCR, ген актина (фрагмент длиной 131 п. н.) был выбран в качестве контрольного гена и выполнял функцию внутреннего контроля, для предотвращения получения ошибочной оценки экспрессии генов при низкой концентрации кДНК [Zhang et al. 2007; Nicot 2005]. Праймеры генов КАО и RSG, взятые из Zhang et al. [2007], тестировались в данном эксперименте, в то время как оригинальные праймеры создавались для MYB1 и MYB2, исходя из общедоступных последовательностей Triticum aestivum (http://compbio.dfci.harvard.edu/cgi-bin/tgi/geneprod_search.pl) в лаборатории компьютерной биологии и функциональной геномики (Гарвардский университет). По-новому спроектированные праймеры MBY (Таблица 9):
[00234] мРНК транскрипционного фактора МуЬ2 (158 п.н), содержащая последовательности, представленные в SEQ ID NO: 16 и SEQ ID NO: 17 и мРНК транскрипционного фактора Myb1 (152 п.н), содержащая последовательности, представленные в SEQ ID NO: 18 и SEQ ID NO: 19 (Таблица 9).
[00235] 100 нг/мкл пробы кДНК повторно разбавляли до 10 нг/мкл, для каждой 25 мкл реакциионной пробы использовали по 2 мкл раствора кДНК. Кроме того, для получения 25 мкл реакционной смеси использовали 12,5 мкл раствора iQ™ SYBR® Green supermix, 8,5 мл стерильной milli-Q воды, и по 1 мкл каждого прямого и обратного праймера (10 пмоль). Протокол работы термоциклера: 10 минут при 95°С, 40 циклов по 20 секунд при 94°С,30 секунд при 60°С и 72°С в течении 1 минуты. QRT-PCR выполнялся в трех повторениях для всех образцов кДНК, полученных при всех типах обработки, с двумя негативными контролями в каждой реакции. Уровни экспрессии генов отсылались к количественным кривым, построенными при помощи программного обеспечения CFX Manager™ Software (Bio-Rad Laboratories). Количественное значение цикла (Cq) из записанных измерений уровня флуоресценции устанавливалось вручную, исходя из базовой линии. В данном исследовании применялся статистический способ относительного количественного определения [Gizinger 2002]. Соотношения значения данного целевого гена к значению гена эндогенного контроля использовали для сравнения проб, подвергшихся разным типам обработки. Количественный анализ (ΔCT) производился путем вычитания из значения Cq целевого гена значения Cq контрольного гена. Далее от ΔCT вычиталось значение калибратора с получением соответствующих значений ΔΔСТ, которые были преобразованы в log2 (функция двоичного логарифма ПЦР) для получения относительных уровней экспрессии генов [Jurado et al., 2010]. Продукты амплификации генов RSG, КАО и MYB очищали при помощи BioBasic PCR Purification Kit (Bio Basic Inc.) и отсылали для секвенирования в Plant Biotechnology Institute (NRC-PBI). Последовательности генов были определены путем проведения анализа Basic Local Alignment Search Tool (BLAST) (http://blast.ncbi.nlm.nih.gov). Последовательности высоко идентичных или похожих генов соответствующих различным гомологичным организмам собирались и выравнивались при помощи программы MEGA5 (Molecular Evolutionary Genetics Analysis). Филогенетическое дерево было создано при помощи статистического способа присоединения соседей на основе выровненных генов.
Пример 14
Было показано регулирующее действие оксида азота (NO) на муковитализм на ранних стадиях прорастания семян.
[00236] Оксид азота (NO) является очень реактивной сигнальной молекулой, общей для грибных, животных и растительных систем. NO также известен тем, что он является сигнальной молекулой, вовлеченной в гормональную сигнализацию эукариотических клеток [Guo et al. 2003] и осуществление ответа растений на биотический и абиотический стресс [Hayat et al. 2010]. Есть доказательства того, что аккумуляция NO, повышение активации СОД и пролина способствуют сохранению О" и аккумуляции Н2О2 в листьях пшеницы при солевом стрессе, хотя информация о грибных эндофитах и их действии на прорастание (миковитализм) почти отсутствует. В данном исследовании, возникновение NO на ранних этапах прорастания семян пшеницы AC Avonlea исследовали в течение трех дней в присутствии эндофита SMCD 2206 на КДА, сосредотачивая внимание на ответе первичного корня на молекулы, диффундирующие от грибов. NO в первичном корне (ранний корневой орган) культур проростков визуализировали при помощи флуоресцентной микроскопии с применением специфического зонда 4,5-diaminofluoresce в диацетате; оценка производилась после пятиминутного воздействия грибным экссудатом, достаточного для индукции значительного накопления NO [Calcagno et al. 2012]. Так, экссудат SMCD 2206 вызвал выработку значительного количества NO в тканях корня пшеницы; не ограничиваясь теорией, возможно, что такая выработка регулируется путем молекулярного диалога при симбиозе у пшеницы.
Материалы и методы
[00237] Накопление NO в тканях первичного корня анализировали в прорастающих семенах пшеницы AC Avonlea (in vitro, подход представлен в Примере 12) с помощью проникающей в клетку NO-специфичной пробы DAF-2DA, согласно Calcagno et al. [2012] которая превращается в флуоресцентное производное триазола DAF-2T при реакции с NO. Образование DAF-2T визуализировали при помощи флуоресцентной микроскопии (Carl Zeiss Axioscop 2). Проростки AC Avonlea оценивали через 5 мин после обработки экссудатом грибов SMCD 2206 при помощи следующей процедуры, предложенной Nakatsubo et al. [1998]
[00238] Специфичность ответа на эндофит SMCD 2206 подтвердилась отсутствием ответа в неинокулированных клетках первичного корня. Анализ был повторен тремя независимыми биологическими повторностями.
Результаты и их обсуждение
[00239] Обработка семян грибным экссудатом может имитировать - в какой-то степени - приближение гиф эндофитов во время пресимбиотической фазы взаимодействия, как это было предложено для микоризы AM при сокультивировании с корнями арабидопсиса [Calcagno et al. 2010]. Грибной экссудат, следовательно, можно с уверенностью применять для проверки того, вызывают ли диффундирующие от грибов сигнальные молекулы накопление NO в тканях хозяина-пшеницы (Фигура 51) во время прохождения повышающих миковитальность событий на ранних этапах прорастания.
[00240] Доказательство на клеточном уровне, следовательно, предполагает, что накопление NO является новым компонентом сигнального пути, который приводит к формированию связанного с миковитализмом симбиоза семян пшеницы (Фигура 51). Это открытие имеет как теоретическую, так и практическую ценность для усовершенствования техник пренатального ухода за растениями с использованием эндофитных симбионтов.
Пример 15
Изучение еффектов эндофитов в фиторемедиации и фиторекламации
[00241] Фиторемедиация является перспективной экотехнологией. Было показано, что она является экономически выгодной для восстановления почв, загрязненных углеводородами/нефтью, солями и тяжелыми металлами. В данном исследовании будет изучено влияние хвойных (Picea или Pinus) и лиственных (Salix или Populus) деревьев, кустарников (Caragana или Krascheninnikovia) и травянистых растений (Festuca или Elymuss), инфицированных (Е+) и не инфицированных (Е-) эндофитными организмами (через инфицирование посадочного материала, семени или корня и путем колонизации) (SMCD 2204 2206, 2208, 2210 и 2215) на разложение, преобразование или деградацию нефтяных углеводородов на загрязненных нефтью почвах. Растения будут выращиваться в вегетационных сосудах, содержащих загрязненную и не загрязненную нефтью почву. Растения будут инокулированны и будут инкубироваться в течение 6 месяцев с использованием парникового способа, предложенного Soleimani et al. (2010). Незасеянные горшки будут использованы в качестве контроля. В конце эксперимента, колонизация корней растений (Abdellatif et al. 2009), гидрофобность почвы (Chau 2012), общее содержание гидрокарбонатов нефти (TPHs) и полициклических ароматических гидрокарбонатов (PAHs) будет проанализировано (Germida et al. 2010). Различия в биомассе корней и побегов у Е+ и Е- растений, а также эффективность фотосинтеза в листьях будут сопоставлены (Hubbard et al. 2012) с выведением РАН и ТРН в ризосфере растений. Незасеянные горшки будут использоваться в качестве контроля при расчете эффективности разложения нефтяных углеводородов симбиотическими (Е+) растениями (Soleimani et al. 2010). Инфицированные растения будут разлагать, преобразовывать или вызывать деградацию углеводородов и солей, поглощать, накапливать, убирать или устранить тяжелые металлы и радиоактивные изотопы в загрязненном участке почвы или окружающей среды.
[00242] Хотя настоящее изобретение было описано со ссылкой на то, что в настоящее время рассматривается в качестве примеров, следует понимать, что настоящее изобретение не ограничено раскрытыми примерами. Наоборот, изобретение охватывает различные модификации и эквивалентные схемы, попадающие в рамки сущности и объема изобретения, как определено в прилагаемой формуле изобретения.
[00243] Все публикации, патенты и заявки на патенты, указанные в данном документе, включены в качестве ссылки во всей полноте в той же мере, как если бы каждая отдельная публикация, патент или патентная заявка были конкретно и индивидуально указаны как включенные посредством ссылки во всей своей полноте.




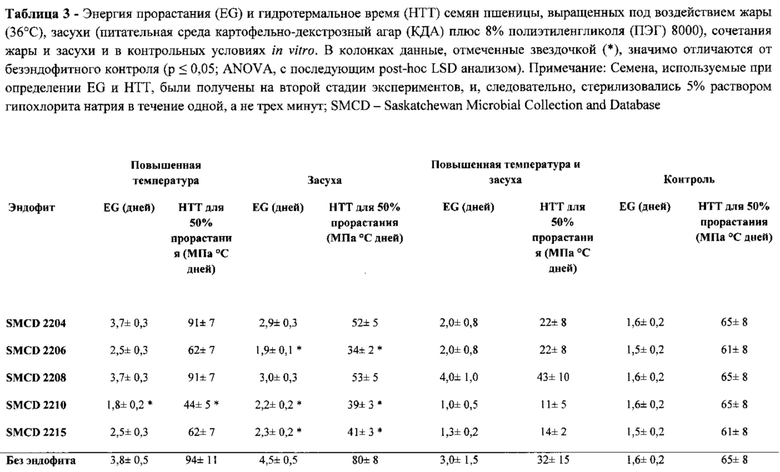



Природный нодулятор Rhizobium во взаимодействии со Streptomyces SMCD2215
16S F (Golden) Rhizobium sp.






Ссылки
Abdellatif et al. 2009. Mycological Research, 113: 782-791.
Abdellatif et al. 2010. Can JPlant Pathol, 32: 468-480.
Adriaensen et al. 2006. Mycorrhiza, 16: 553-558.
Agius et al. 2006. PNAS, 103: 11796-11801.
Ali et al. 1994. Annals of Applied Biology, 125: 367-375.
Allen 1958. Forest Chron, 34: 266-298.
Armas et al. 2004. Ecology, 85: 2682-2686.
Arnold et al. 2001. Mycological Research, 105: 1502-1507.
Bacon and White 2000. In: Bacon CW and White JFJ (Eds), Microbial endophytes. Marcel Dekker Inc; New York, NY, USA. 237-263.
Bae et al. 2009. J Exp Bot 60: 3279-3295.
Baird et al. 2010. Mycorrhiza. 20: 541-549.
Barrero et al. 2009. Plant Physiology, 150: 1006-1021.
Baskin et al. 1992. International Journal of Plant Sciences, 153: 239-243.
Baskin and Baskin 2004. Sci. Res., 14: 1-16.
Bewley and Black 1982. Physiology and Biochemistry of Seeds. 2. Viability, Dormancy, and Environmental Control. Springer-Verlag, Berlin.
Bloom and Richard 2002. ASAE Paper No 027010. ASAE, St. Joseph, MI.
Bogatek and Lewak 1988. Physiologia Plantarum, 73: 406-411.
Boyko and Kovalchuk 2008. Environmental and Molecular Mutagenesis, 49: 61-72.
Bradford 2002. In: J. Kigel J, Galili G (eds), Seed Develop and Germin. Marcel Dekker Inc, New York, pp. 351-396.
Calcagno et al. 2012. Mycorrhiza, 22: 259-69.
Cao and Moss. 1989. Crop Sci, 29: 1018-1021.
Carpita et al. 1983. Physiologia Plantarum 59: 601-606.
Cavieres and Arroyo, 2000. Plant Ecology 149: 1-8.
Cavieres and Arroyo, 2000b. Gayana Botanica 64: 40-45.
Charlton et.al. 2008. Metabolomics, 4: 312-327.
Chauetal. 2012. Fungal Biology, 116: 1212-1218.
Chipanshi et al. 2006. Clim Res, 30: 175-187.
Choi and Sano, 2007. Molecular Genetics and Genomics, 277: 589-600.
Davitt et al. 2010. New Phytol, 188: 824-834.
de Bary 1866. Vol. II. Hofmeister's Handbook of Physiological Botany. Leipzig, Germany.
Desfeux et al., 2000. Plant Physiology, 123: 895-904.
Dong-dong et al. 2009. J Zhejiang Univ-Sci B, 9: 964-968.
Farquhar and Richards 1984. Australian Journal Plant Physiology 11: 539-552.
Farquhar et al. 1989 In: Jones HG, Flowers TJ and Jones MB (Eds) Plants under stress. Cambridge University Press, Cambridge, pp 47-69.
Farquhar et al. 1989b. Annual Review of Plant Physiology and Plant Molecular Biology. 40: 503-537.
Finnegan et al. 1998. Plant Molecular Biology 95: 5824-5829.
Freeman 1904. Philosophical Transactions of the Royal Society London (Biology) 196: 1-27.
Friend et al. 1962. Can J Bot, 40: 1299-1311.
Gan et al. 2004. Can. J Plant Sci, 84: 697-704.
Germida et al. 2010. Field-scale assessment of phytoremediation at a former oil tank battery in Bruderheim, Alberta. World Congress of Soil Science, Soil Solutions for a Changing World, 1-6 August 2010. Brisbane, Australia. Available on-line at: http://www.iuss.org/19th%20WCSS/Symposium/pdf/0694.pdf
Gizinger 2002. Experimental Hematology, 30: 503-512.
Gomall et al. 2010. Phil. Trans. R. Soc. B, 365, 2973-2989.
Grant et al. 2009. Tree Physiology, 29: 1-17.
Gummerson 1986. J Exp Bot, 37: 729-741.
Gundel et al. 2010. Evol Appl, 3: 538-546.
Guo et al. 2003. Science, 302: 100-103.
Hayat et al. 2010. Nitric Oxide in Plant Physiology, Issue 58, Willey-VCH Verlag, Germany.
Hedden and Phillips, 2000. Trends in Plant Science, 5: 523-530.
Hubbard et al. 2011. In: Wheat: Genetics, Crops and Food Production. Nova Science Publishers Hauppauge, NY, USA. pp. 333-345.
Hubbard et al. 2012. Botany, 90(2): 137-149.
Jame et al. 1998. Agric Forest Meteorol 92: 241-249.
Ji et al., 2011. Plant Physiology, 156: 647-662.
Johannes et al. 2009. Plos Genetics 5: el000530.
Johannes et al. 2011. Genetics, 188: 215-227.
Johnson et al. 1990. Crop Science, 30: 338-343.
Jost et al. 2001. Nucleic Acids Research, 29: 4452-4461.
Jumpponen and Trappe 1998. New Phytologist, 140: 295-310.
Jurado et al., 2010. Food Microbiology, 27: 50-57.
Kane 2011. Environmental and Experimental Botany, 71: 337-344.
Kang et al. 2008. International Journal of Sustainable Development and World Ecology, 15: 440-447.
Karavata & Manetas 1999. Photosynthetica, 36: 41-49.
Khan et al. 2010. Pakistan Journal of Botany, 42: 259-267.
Khan et al. 2012. BMC Microbiol, 12; 12:3.
Kiffer and Morelet 2000. Science Publisher Inc, Enfield, NH, Plymouth.
Köchy and Tielborger 2007. Basic Appl Ecol 8: 171-182.
Koyuncu 2005. Acta Biologica Cracoviensia Series Botanica, 47: 23-26.
Labeda et al 2012. Antonie van Leeuwenhoek, 101: 73-104.
Lang-Mladek et al. 2010. Molecular Plant, 3: 594-602.
Larran et al. 2002. World Journal of Microbial Biotechnology, 18: 683-686.
Leone et al. 1994. Physiol Plantarum, 92: 21-30.
Li et al. 2008. Ecological Research, 23: 927-930.
Li et al. 2011. Agronomy Journal, 103: 1619-1628.
Lu et al. 2007. Plant Biology, 49: 1599-1607.
Lucht et al. 2002. Nature Genetics, 30: 311-314.
Madsood et al. 2005. Engineering Applications of Artificial Intelligence, 18: 115-125.
Margulis, 1991. In Symbiosis as a Source of Evolutionary Innovation, L. Margulis and R. Fester, ed. The MIT Press: Cambridge, pp. 1-14.
Marquez et al. 2007. Science, 315: 513-515.
McCormick MC, Siegel (eds.) 1999. Prenatal Care: Effectiveness and implementation. Cambridge University Press UK.
McDonald 2009. Handbook of biological statistics. 2nd ed. Sparky House Publishing, Baltimore, Maryland.
McMaster 2009. In: Carver BF (ed), Wheat, science and trade, Wiley-Blackwell, Iowa, USA, pp. 31-55.
Milberg and Andersson 1998. Plant Ecology, 134: 225-234. Millar et al., 2006. Plant Journal, 45: 942-954.
Miransari et al. 2011. Applied Microbiology and Biotechnology, 92: 875-885. Mitchell et al., 2009. Microbiology-SGM, 156: 270-277.
Mühlmann and Peintner 2000. Mycorrhiza, 18: 171-180.
Mukhopadhyay et al., 2004. PNAS, 101: 6309-6314.
Nakatsubo et al. 1998. FEBS Lett, 427: 263-266.
Nakamura et al., 2010. Euphytica, 171: 111-120.
Nelson, 2004. Annu Rev Phytopathol, 42: 271-309.
Nicot 2005. Journal of Experimental Botany, 56: 2907-2914.
Nonogaki et al., 2010. Plant Science, 179: 574-581.
Oikawa et al., 2004. Plant Molecular Biology, 55: 687-700.
Okamoto et al., 2006. Plant Physiology, 141: 97-107.
Oliver et al., 2007. Plant and Cell Physiology, 48: 1319-1330.
Penterman et al. 2007. PNAS, 104: 6752-6757.
Phillips et al., 1995. Plant Physiology, 108: 1049-1057.
Probert et al., 1989. Journal of Experimental Botany, 40: 293-301.
Qin and Zeevart, 1999. PNAS, 96: 15354-15361.
Reynolds et al. 2007 Journal of Experimental Botany, 58: 177-186.
Richards et al. 2002. Crop Science, 42: 111-121.
Ries et al. 2000. Nature, 406: 98-101.
Rivero et al. 2011. International Conference on Arabidopsis Research. June 22-25, Madison USA.
Ruan et al. 2002. Seed Sci Technol, 30: 61-67.
Ryan et al. 2008. FEMS Microbiol Lett, 278: 1-9.
Saikkonen et al., 1998. Annual Review of Ecology and Systematics, 29: 319-343.
Saze 2008. Seminars in Cell and Developmental Biology, 19: 527-536.
Schrey and Tarkka 2008. Antonie van Leeuwenhoek, 94: 11-19.
Schutz and Rave 1999. Ecology, 144: 215-230.
Semenov and Shewry 2011. Scientific Reports, 1: 66-71.
Sinclair et al. 1984. Bioscience, 34: 36-40.
Singh et al. 2011. Plant Signal Behav, 6: 175-191.
Smith and Read 2008. Mycorrhizal symbiosis, Third Edition. Elsevier Ltd. Mycorrhizas in acholorophyllous plants (mycoheterotrophs). Chapter 13: 458-507.
Solaiman et al. 2010. Australian Journal of Soil Research, 48: 546-554.
Soleimani et al. 2010. Chemosphere, 81: 1084-1090.
Stone et al., 2000. In: Bacon, C.W. and White, J.F. eds., Microbial Endophytes, Marcel Dekker: New York Chap. 1: 3-29.
Strobel et al., 2004. Journal of Natural Products, 67: 257-268.
Sun et al. 2010. Journal of Plant Physiolog, 167: 1009-1017.
Tan and Zou, 2001. Nat Prod Rep, 18: 448-45.
Tokala et al. 2002. Appl Environ Microbiol, 68:2161-2171.
Vaughn et al. 2007. PloS Biology, 5: 1617-1629.
Verhoeven et al. 2010. New Phytologis, 185: 1108-1118.
Vujanovic et al. 2000. Annals of Botany, 86: 79-86.
Vujanovic and Brisson 2002. Mycological Progress. 1: 147-154.
Vujanovic and Vujanovic 2006. Floriculture, Ornamental and Plant Biotech, 63: 563-569.
Vujanovic and Vujanovic 2007. Symbiosis, 44: 93-99.
Vujanovic 2007b. Can J Plant Pathol, 29: 451-451.
Vujanovic 2008. 19th International Conference on Arabidopsis. Research Proceedings- ICAR13, July 23-27, Montreal, QC, Canada.
Waller et al. 2005. PNAS, 102: 13386-13391.
Wallin 1927. Symbionticism and the Origin of Species. London: Bailliere, Tindall and Cox.
Wang et al. 2011. Journal of Experimental Botany, 62: 1951-1960.
Whalley et al. 2006. Plant and Soil, 280: 279-290.
White and Torres 2010. Physiol. Plant, 138: 440-446.
Wu et al. 2008. Plant Physiology, 148: 1953-1963.
Wu and Sardo 2010. Lichtfouse E. (Ed.), Sociology, Organic Farming, Climate Change and Soil Science. Sustainable Agriculture Reviews. 3: DOI 10.1007/978-90-481-3333-8_3.
Yamaguchi et al. 1998. Plant Cell, 10: 2115-2126
Yang et al., 2002. Planta, 215: 645-652.
Zadoks et al. 1974. Weed Research, 14:415-421.
Zhang et al., 2007. BMC Genet, 2007, 8: 40.
Zhang et al. 2010. Journal of Cereal Science, 52: 263-269.
Zhang et al. 2011. African Journal of Microbiology Research, 5: 5540-5547.
Zhao et al. 2007. Journal of Plant Nutrition, 30: 947-963.
Zhong et al. 2009. African Journal of Biotechnology, 8: 6201-6207.
Zhu et al. 2007. Current Biology, 17: 54-59.
Foresight. The future of food and farming: challenges and choices for global sustainability. Final Project Report. London: The Government Office for Science, UK, 2011.
IPCC Climate Change 2007. In: Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt KB, Tignor M and Miller HL (Eds). Cambridge University Press, Cambridge, UK.
Saskatchewan Ministry of Agriculture 2008. Varieties of Grain Crops. SaskSeed guide. Regina, SK, Canada.
--->
Перечень последовательностей
<110> University of Saskatchewan
<120> ENDOPHYTIC MICROBIAL SYMBIONTS IN PLANT PRENATAL CARE
<130> 13764-P41875PC00
<160> 19
<170> PatentIn version 3.5
<210> 1
<211> 581
<212> DNA
<213> Cladosporium sp.
<400> 1
cctatagctg actgcggagg gacattacaa gtgaccccgg tctaaccacc gggatgttca 60
taaccctttg ttgtccgact ctgttgcctc cggggcgacc ctgccttcgg gcgggggctc 120
cgggtggaca cttcaaactc ttgcgtaact ttgcagtctg agtaaactta attaataaat 180
taaaactttt aacaacggat ctcttggttc tggcatcgat gaagaacgca gcgaaatgcg 240
ataagtaatg tgaattgcag aattcagtga atcatcgaat ctttgaacgc acattgcgcc 300
ccctggtatt ccggggggca tgcctgttcg agcgtcattt caccactcaa gcctcgcttg 360
gtattgggca acgcggtccg ccgcgtgcct caaatcgacc ggctgggtct tctgtcccct 420
aagcgttgtg gaaactattc gctaaagggt gttcgggagg ctacgccgta aaacaacccc 480
atttctaagg ttgacctcgg atcaggtagg gatacccgct gaacttaagc atatcaataa 540
gcggaggaaa agaaaccaac agggattgcc ccagtaacga a 581
<210> 2
<211> 571
<212> DNA
<213> Cladosporium sp.
<400> 2
tcgatctagc tcatagtgac tgcggaggga cattacaagt gaccccggtc taaccaccgg 60
gatgttcata accctttgtt gtccgactct gttgcctccg gggcgaccct gccttcgggc 120
gggggctccg ggtggacact tcaaactctt gcgtaacttt gcagtctgag taaacttaat 180
taataaatta aaacttttaa caacggatct cttggttctg gcatcgatga agaacgcagc 240
gaaatgcgat aagtaatgtg aattgcagaa ttcagtgaat catcgaatct ttgaacgcac 300
attgcgcccc ctggtattcc ggggggcatg cctgttcgag cgtcatttca ccactcaagc 360
ctcgcttggt attgggaaac gcggaccgac gcgtgcctca aatcgaccgg cagggtcttc 420
tgtcccctaa acgttgtgaa aattattcga taaaggatgt tccgtgctac attgtgaata 480
gaaccgcatt tataacattg attataaact aattacgact acatggtaag atagatatat 540
caaggaactt cctctaaatg accaagaaac c 571
<210> 3
<211> 592
<212> DNA
<213> Penicillium sp.
<400> 3
tcgacggcgt atcctagtga ctgcggagga tcattaccga gtgagggccc tctgggtcca 60
acctcccacc cgtgtttaat ttaccttgtt gcttcggcgg gcccgcctta actggccgcc 120
ggggggctta cgcccccggg cccgcgcccg ccgaagacac cctcgaactc tgtctgaaga 180
ttgtagtctg agtgaaaata taaattattt aaaactttca acaacggatc tcttggttcc 240
ggcatcgatg aagaacgcag cgaaatgcga tacgtaatgt gaattgcaaa ttcagtgaat 300
catcgagtct ttgaacgcac attgcgcccc ctggtattcc ggggggcatg cctgtccgag 360
cgtcattgct gccctcaagc acggcttgtg tgttgggccc cgtcctccga tcccggggga 420
cgggcccgaa aggcagcggc ggcaccgcgt ccggtcctcg agcgtatggg gctttgtcac 480
ccgctctgta ggcccggccg gcgcttgccg atcaacccaa atttttatcc aggttgacct 540
cggatcaggt agggataccc gctgaactta agcatatcaa taagcggagg aa 592
<210> 4
<211> 605
<212> DNA
<213> Pseudeurotium sp.
<400> 4
taactgattt ggcggactgg cggaaggaca ttaaagagac gttgcccttc ggggtatacc 60
tcccaccctt tgtttacctt ttcctttgtt gctttggcgg gcccgtcctc ggaccaccgg 120
tttcggctgg tcagtgcccg ccagaggacc taaaactctg tttgttcata ttgtctgagt 180
actatataat agttaaaact ttcaacaacg gatctcttgg ttctggcatc gatgaagaac 240
gcagcgaaat gcgataagta atgtgaattg cagaattcag tgaatcatcg aatctttgaa 300
cgcacattgc gccccctggt attccggggg gcatgcctgt tcgagcgtca ttacaaccct 360
caagctctgc ttggtattgg gctctgccgg tcccggcagg ccttaaaatc attggcggtg 420
ccattcggct tcaagcgtag taattcttct cgctttggag acccgggtgc gtgcttgcca 480
tcaaccccca attttttcag gttgacctcg gatcaggtag ggatacccgc tgaacttaag 540
catatcaata agcggaggaa aagaaaccaa cagggattgt cccaataacg aatttataaa 600
taata 605
<210> 5
<211> 579
<212> DNA
<213> Paraconyothirium sp.
<400> 5
tcgagagttc ggactaagtg cctgatccga ggtcaagacg gtaatgttgc ttcgtggacg 60
cgggccacgc ccccccgcag acgcaattgt gctgcgcgag aggaggcaag gaccgctgcc 120
aatgaatttg gggcgagtcc gcgcgcgaag gcgggacaga cgcccaacac caagcagagc 180
ttgagggtgt agatgacgct cgaacaggca tgccccatgg aataccaagg ggcgcaatgt 240
gcgttcaaag attcgatgat tcactgaatt ctgcaattca cactacttat cgcatttcgc 300
tgcgttcttc atcgatgcca gagccaagag atccattgtt gaaagttgta acgattgttt 360
gtatcagaac aggtaatgct agatgcaaaa aaggttttgt taagttccag cggcaggttg 420
ccccgccgaa ggagaacgaa aggtgctcgt aaaaaaagga tgcaggaatg cggcgcgtga 480
gggtgttacc cctaccaccc gggagagaac ccccgagggc cgcgaccgca cctggttgag 540
atggataatg atccttccgc aggttcacct acggaaacc 579
<210> 6
<211> 1160
<212> DNA
<213> Streptomyces sp.
<400> 6
ccgggggcac tccactgcgt atgtgtgacg agtagaccgc tgcgcttagc tgaggtctga 60
tgaaatgtag aacacttaac aaaaatatgc ccggatggat atacttttca acgacagggc 120
tgcgattgga tgatctcctt tgaaacacag aactagtcac ggcgacgaat actcaacttc 180
gacccccccc ctttctggag gcgcgtctta gtcccctcct tgatggagct gccccgtgct 240
cggcggccgg agtcggcggt gttttccgct gtacctgaga cgctggacca actccttcgg 300
gaggcagcag tggggaatat tgcacaatgg gcgcaagcct gatgcagcga cgccgcgtga 360
gggatgacgg ccttcgggtt gtaaacctct ttcagcaggg aagaagcgca agtgacggta 420
cctgcagaag aagcgccggc taactacgtg ccagcagccg cggtaatacg tagggcgcaa 480
gcgttgtccg gaattattgg gcgtaaagag ctcgtaggcg gcttgtcacg tcgattgtga 540
aagcccgagg cttaacctcg ggtctgcagt cgatacgggc aggctagagt gtggtagggg 600
agatcggaat tcctggtgta gcggtgaaat gcgcagatat caggaggaac accggtggcg 660
aaggcggatc tctgggccat tactgacgct gaggagcgaa agcgtgggga gcgaacagga 720
ttagataccc tggtagtcca cgccgtaaac ggtgggaact aggtgttggc gacattccac 780
gtcgtcggtg ccgcagctaa cgcattaagt tccccgcctg gggagtacgg ccgcaaggct 840
aaaactcaaa ggaattgacg ggggcccgca caagcggcgg agcatgtggc ttaattcgac 900
gcaacgcgaa gaaccttacc aaggcttgac atacaccgga aacatccaga gatgggtgcc 960
cccttgtggt cggcgtacag gtcgtgcatg gctgtcgtca gctcgtgtcg tgagatgttg 1020
ggtaagtccc gcaacgagcg caaccttgtt ctggtgctgc cagcatgccc ttcgggtgat 1080
gggacttcac cacggagacc gcggctccac tccgacgagg tgggggacga cgtcagtcat 1140
catgccctaa tgtctggctg 1160
<210> 7
<211> 1176
<212> DNA
<213> Rhizobium sp.
<400> 7
ggaagggggg cggcttacca tgcaagtcga gcgccccgca aggggagcgg cagacgggtg 60
agtaacgcgt gggaatctac ccttgactac ggaataacgc agggaaactt gtgctaatac 120
cgtatgtgtc cttcgggaga aagatttatc ggtcaaggat gagcccgcgt tggattagct 180
agttggtggg gtaaaggcct accaaggcga cgatccatag ctggtctgag aggatgatca 240
gccacattgg gactgagaca cggcccaaac tcctacggga ggcagcagtg gggaatattg 300
gacaatgggc gcaagcctga tccagccatg ccgcgtgagt gatgaaggcc ctagggttgt 360
aaagctcttt caccggagaa gataatgacg gtatccggag aagaagcccc ggctaacttc 420
gtgccagcag ccgcggtaat acgaaggggg ctagcgttgt tcggaattac tggccgtaaa 480
gcgcacgtag gcggatcgat cagtcagggg tgaaatccca gggctcaacc ctggaactgt 540
ctttgatact gtcgatctgg agaacttcct gctcgagtga tttacccaca tggcgagcac 600
cggcaccccg tttcgacatg caaaaaatga tgcccaggct tatgtttgac ctggctgcta 660
cggctctctt cggcgtggac cccggcctcc tatccccgga gatgccaccc atggacgccg 720
cagtctccat ggatatatca tggaggtggg ttttctccga ctcatgatgc cggcttcttg 780
ctggaagttg atgaagcaac taaacatcag ccctgagaga aagcttcgca tgccgcgcag 840
ggtgctccga gtgttcgtct ggagatgatg aaatagacga agatcatctc atgtcatgtt 900
ggtaacgacg agaacaagat ggtgtggatt ttgtgtcttc catcctccat gaccctgacg 960
atgctgatga tgacgtggtt catgctatga tgactcgata ctggtcgctg caagcggata 1020
cagttgggac ctaccgctaa catggttctt tctacaacct ccccccaaac cgcataggat 1080
cgtggtcaat cattcggcac gaacctcttc ccccattgcc tccaactagt ttatcgctct 1140
agagttgggg agccctgtgt gacctttcgt acgcga 1176
<210> 8
<211> 24
<212> DNA
<213> Pea-Pisum sativum
<400> 8
ccacattacc tgtatcggat gaca 24
<210> 9
<211> 22
<212> DNA
<213> Pea-Pisum sativum
<400> 9
gagcccagaa caggagctaa ca 22
<210> 10
<211> 25
<212> DNA
<213> Pea-Pisum sativum
<400> 10
gcagaaaaac cctatcctcc gtgct 25
<210> 11
<211> 20
<212> DNA
<213> Pea-Pisum sativum
<400> 11
gctccaaagc tccgtagtcg 20
<210> 12
<211> 20
<212> DNA
<213> Pea-Pisum sativum
<400> 12
ctgtactcgc tgttggggtg 20
<210> 13
<211> 20
<212> DNA
<213> Pea-Pisum sativum
<400> 13
gcatggatat ggaagccgtg 20
<210> 14
<211> 20
<212> DNA
<213> Pea-Pisum sativum
<400> 14
aatggccgaa agcattgcca 20
<210> 15
<211> 24
<212> DNA
<213> Pea-Pisum sativum
<400> 15
aaggacggtg atgccgatgg actc 24
<210> 16
<211> 21
<212> DNA
<213> Wheat-Triticum aestivum
<400> 16
acatcaagcg cggcaacttc a 21
<210> 17
<211> 23
<212> DNA
<213> Wheat-Triticum aestivum
<400> 17
gagccgcttc ttgaggtggg tgt 23
<210> 18
<211> 20
<212> DNA
<213> Wheat-Triticum aestivum
<400> 18
ccagggagga cggacaacga 20
<210> 19
<211> 20
<212> DNA
<213> Wheat-Triticum aestivum
<400> 19
ctctgcgccg tctcgaagga 20
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| УЛУЧШЕННЫЕ ЭНДОФИТОМ СЕЯНЦЫ С УВЕЛИЧЕННОЙ УСТОЙЧИВОСТЬЮ К ВРЕДИТЕЛЯМ | 2007 |
|
RU2453601C2 |
| ЭНДОФИТНЫЕ ГРИБЫ EPICHLOE/NEOTYPHODIUM SPP. ДЛЯ ИСКУССТВЕННОГО ЗАРАЖЕНИЯ SECALE SPP. И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2683379C2 |
| ВЫДЕЛЕННЫЙ ШТАММ CLONOSTACHYS ROSEA ДЛЯ ПРИМЕНЕНИЯ В КАЧЕСТВЕ АГЕНТА БИОЛОГИЧЕСКОЙ ЗАЩИТЫ | 2014 |
|
RU2689608C2 |
| ШТАММ MYCOSPHAERELLA SP., КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ БОРЬБЫ С ФУЗАРИОЗОМ | 2012 |
|
RU2705286C2 |
| ГЕНЫ-МИШЕНИ ДЛЯ НАПРАВЛЕННОГО ВОЗДЕЙСТВИЯ НА АЗОТФИКСАЦИЮ ДЛЯ УЛУЧШЕНИЯ КАЧЕСТВ РАСТЕНИЙ | 2018 |
|
RU2805085C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ БОРЬБЫ С ФУЗАРИОЗОМ | 2019 |
|
RU2736382C1 |
| ШТАММ MICROBACTERIUM, КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ БОРЬБЫ С ФУЗАРИОЗОМ | 2012 |
|
RU2634386C2 |
| Способ стимулирования роста шестого зародышевого корня у пшеницы | 2021 |
|
RU2769730C1 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ БОРЬБЫ С ФУЗАРИОЗОМ | 2020 |
|
RU2777606C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ УЛУЧШЕНИЯ ПРИЗНАКОВ РАСТЕНИЙ | 2016 |
|
RU2775712C2 |






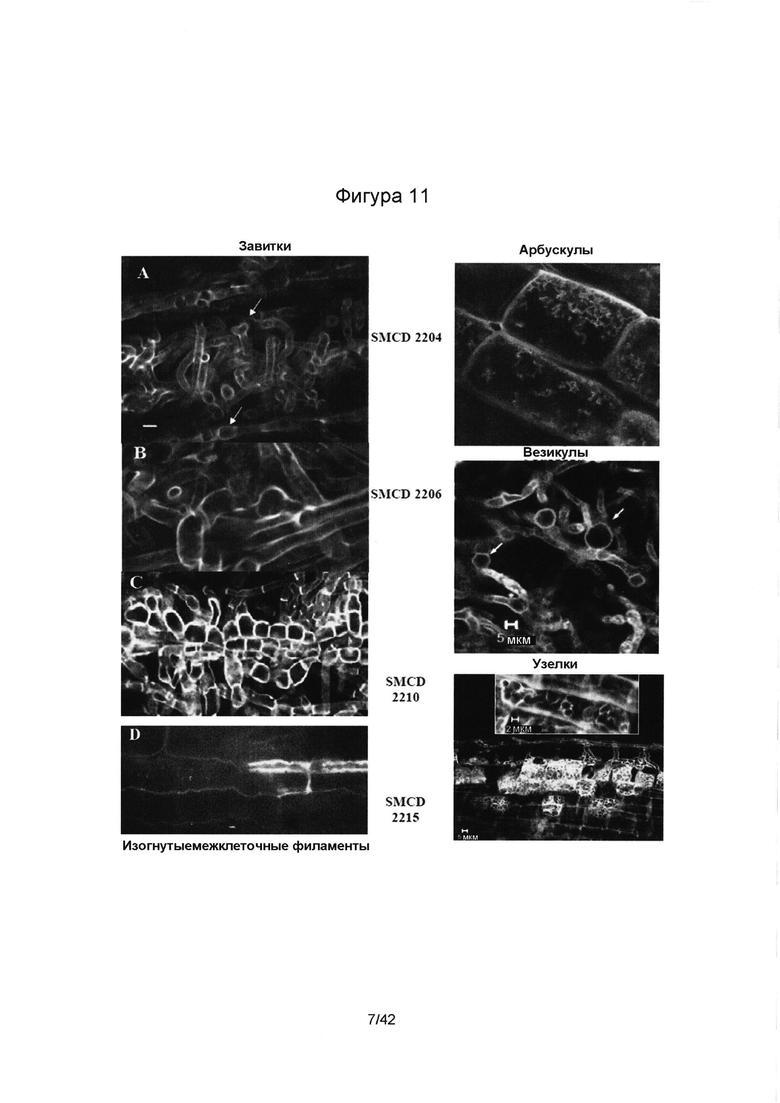


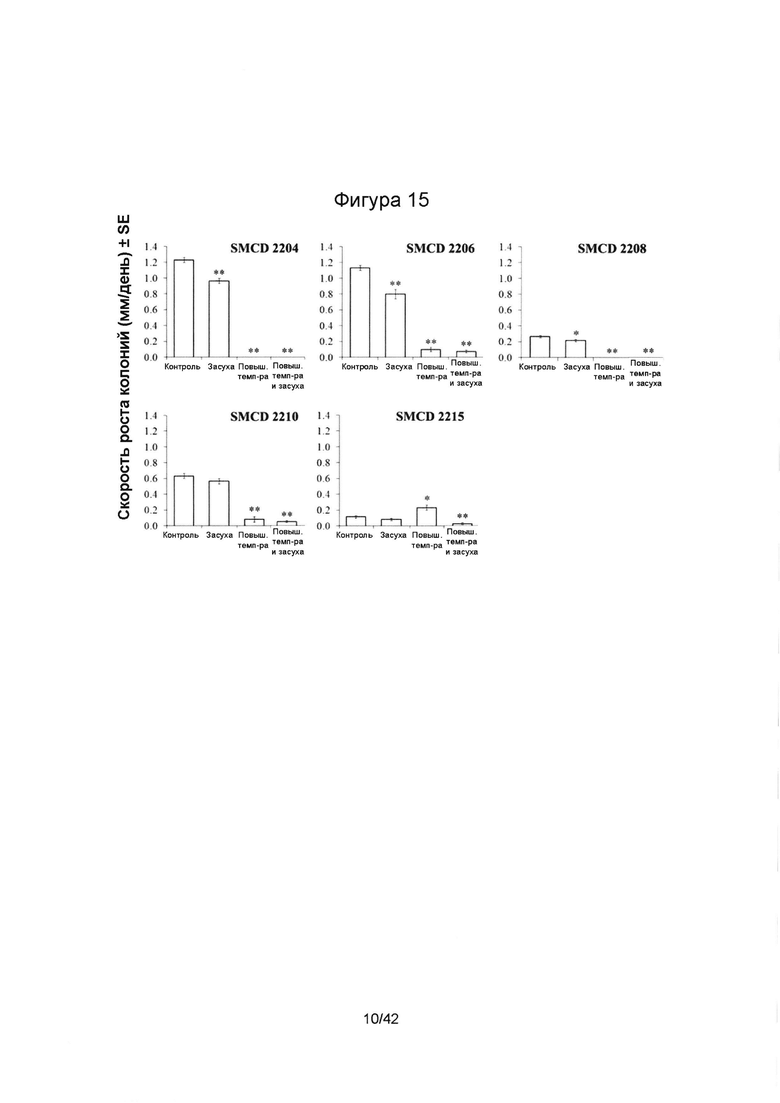


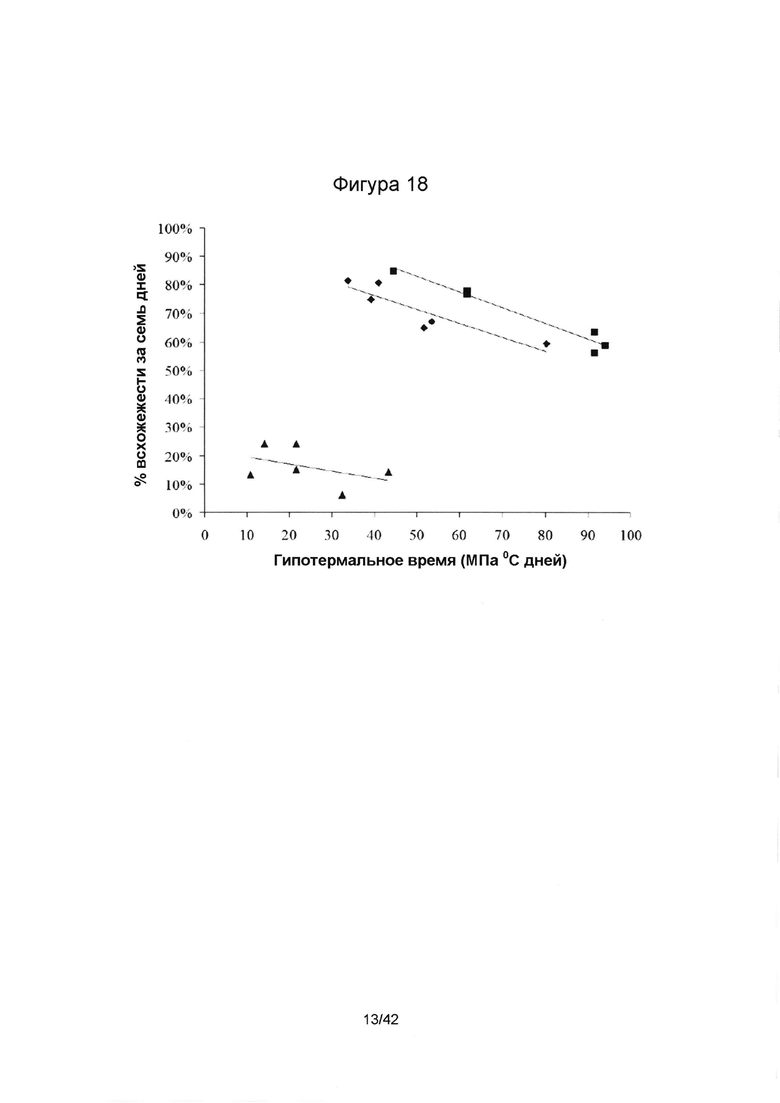




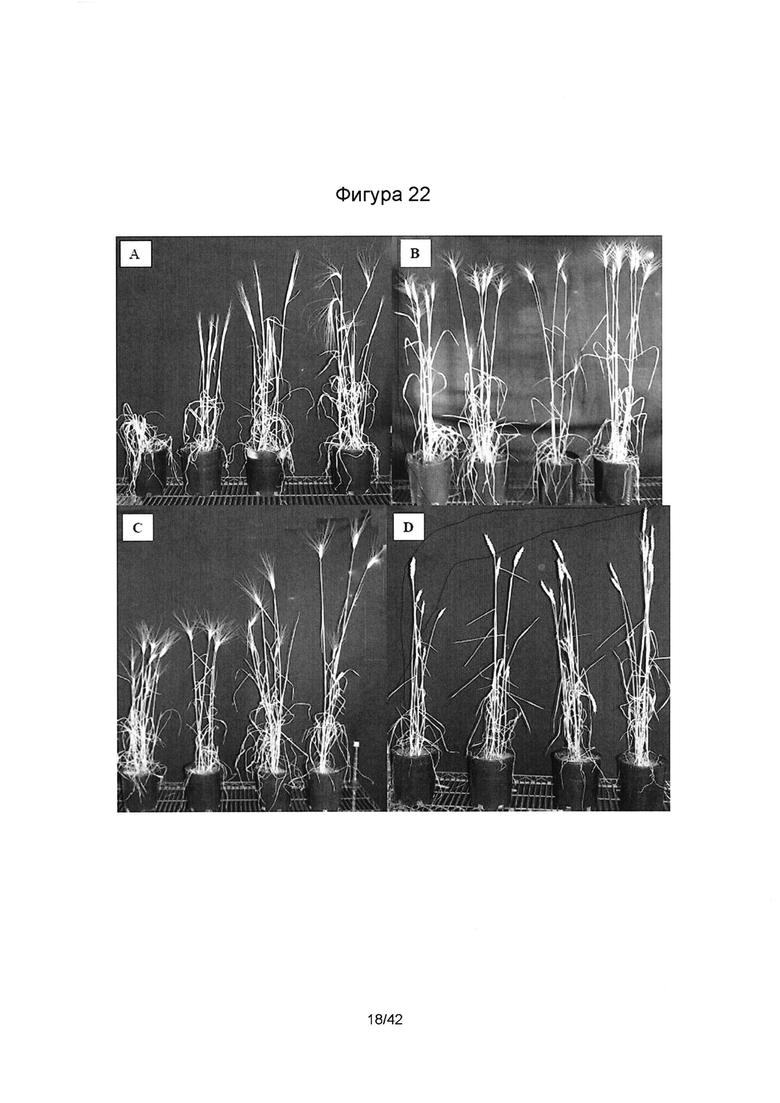
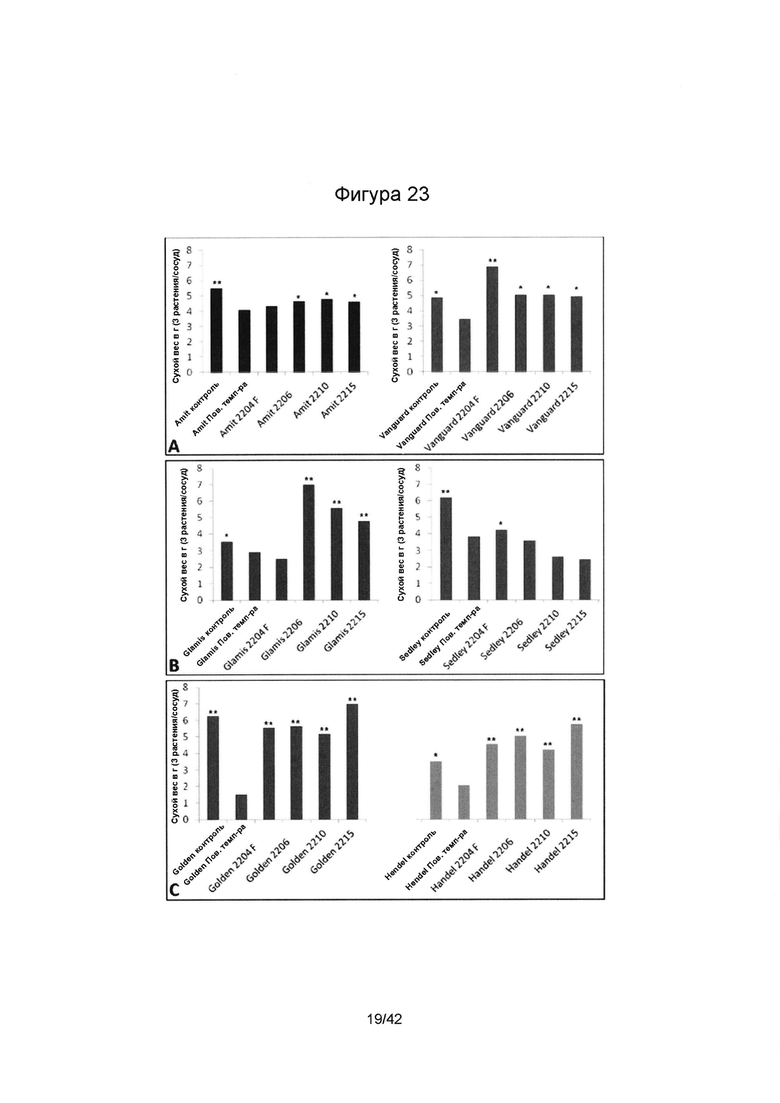

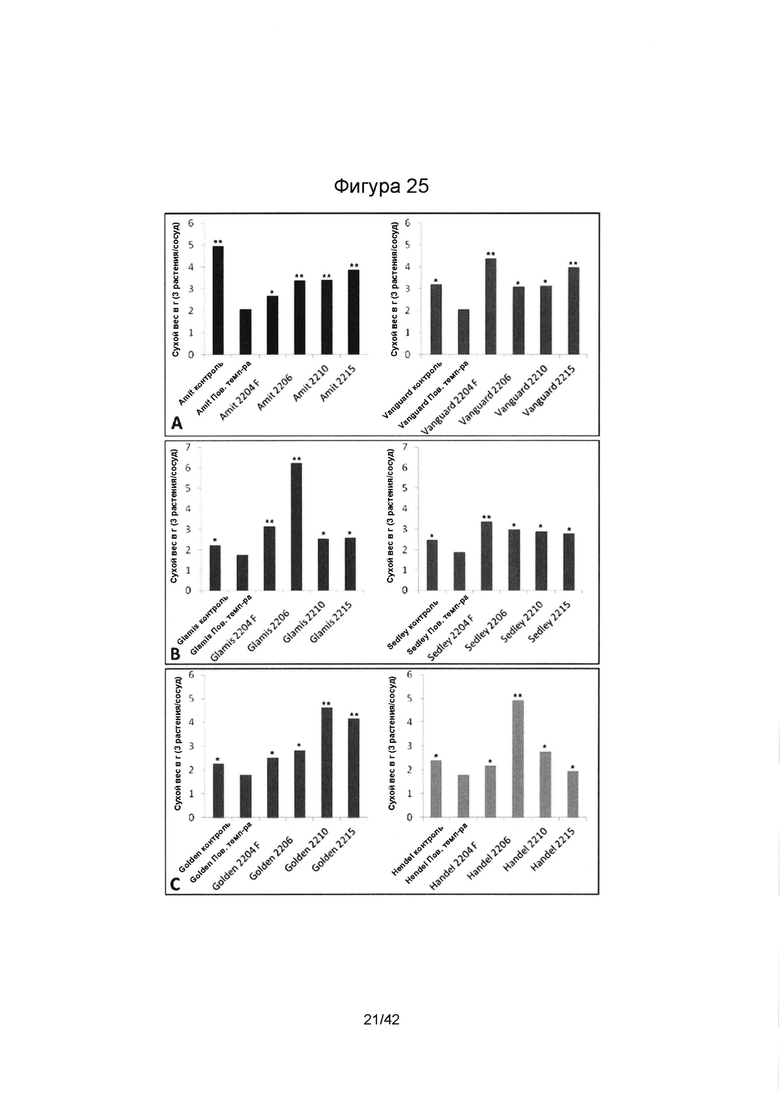


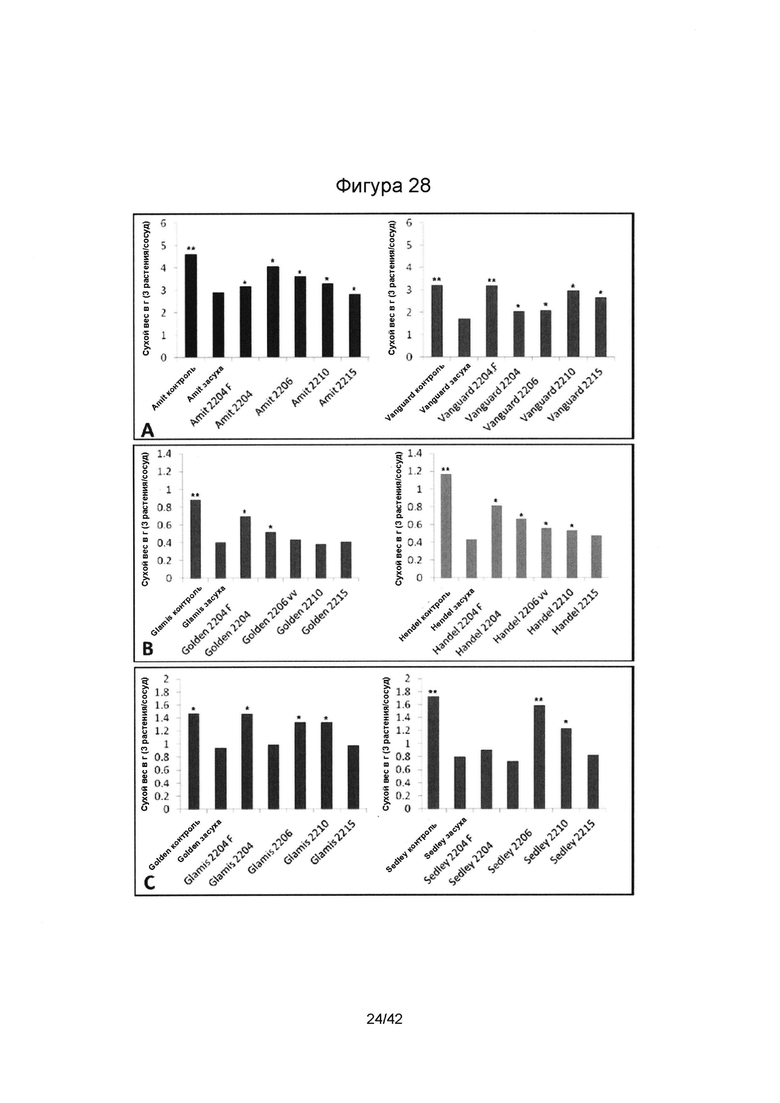



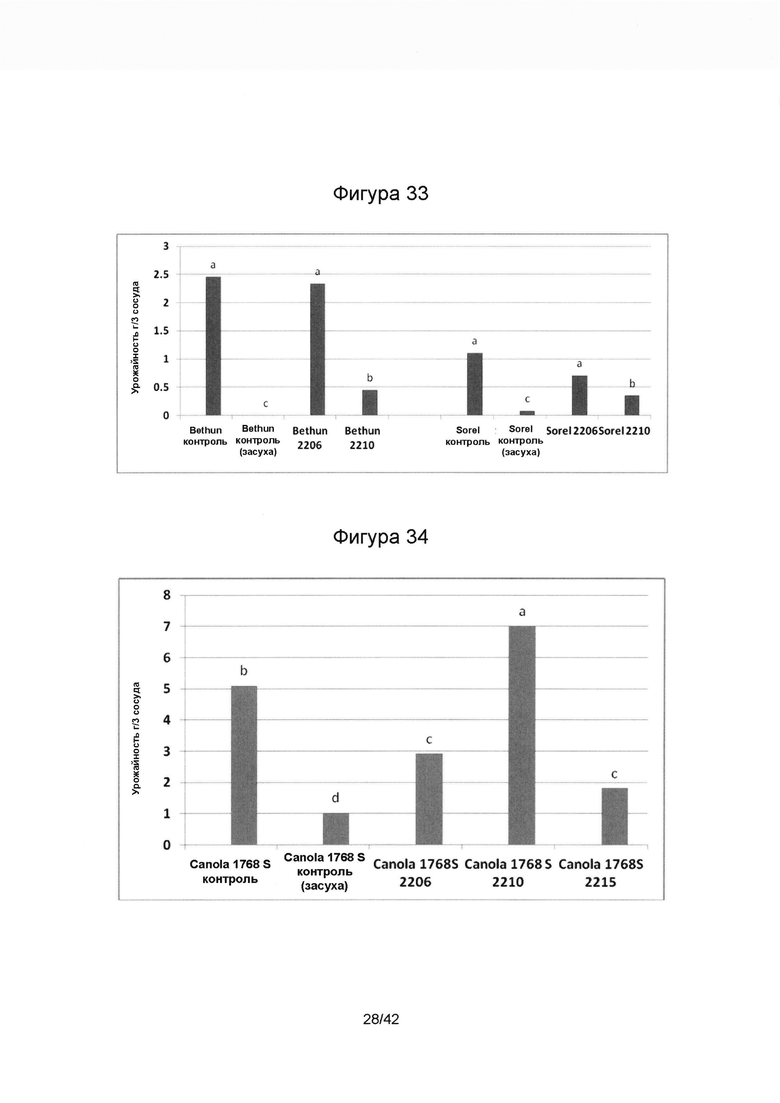
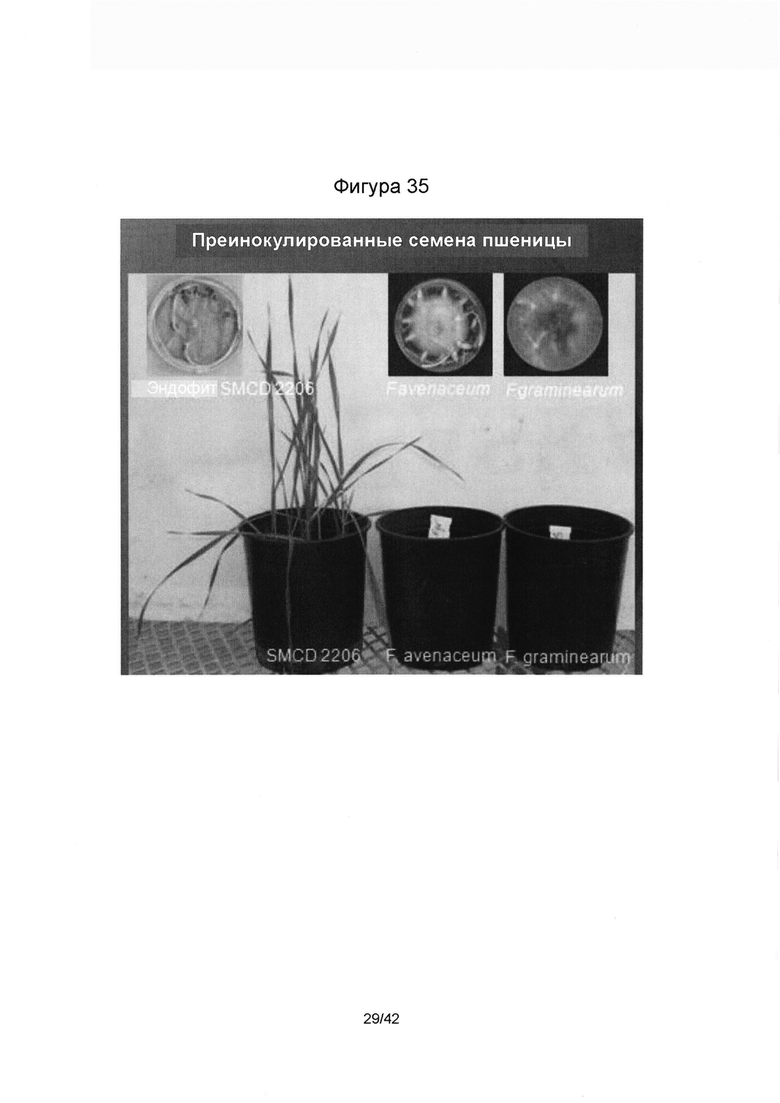



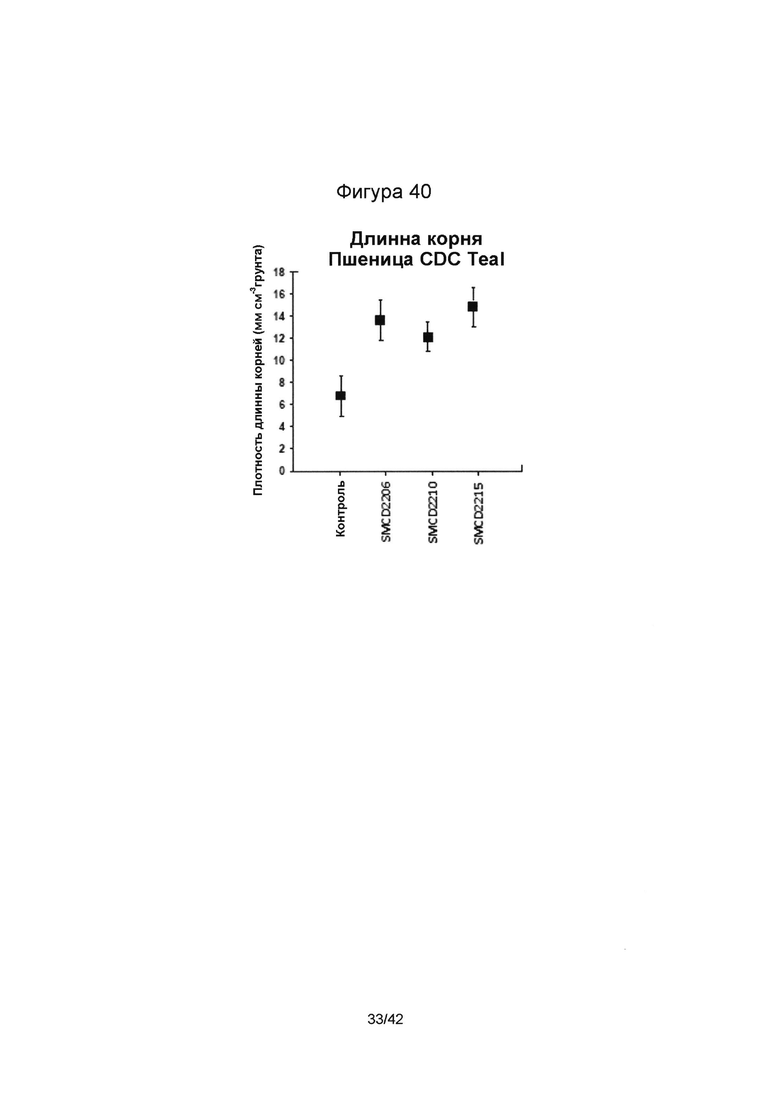

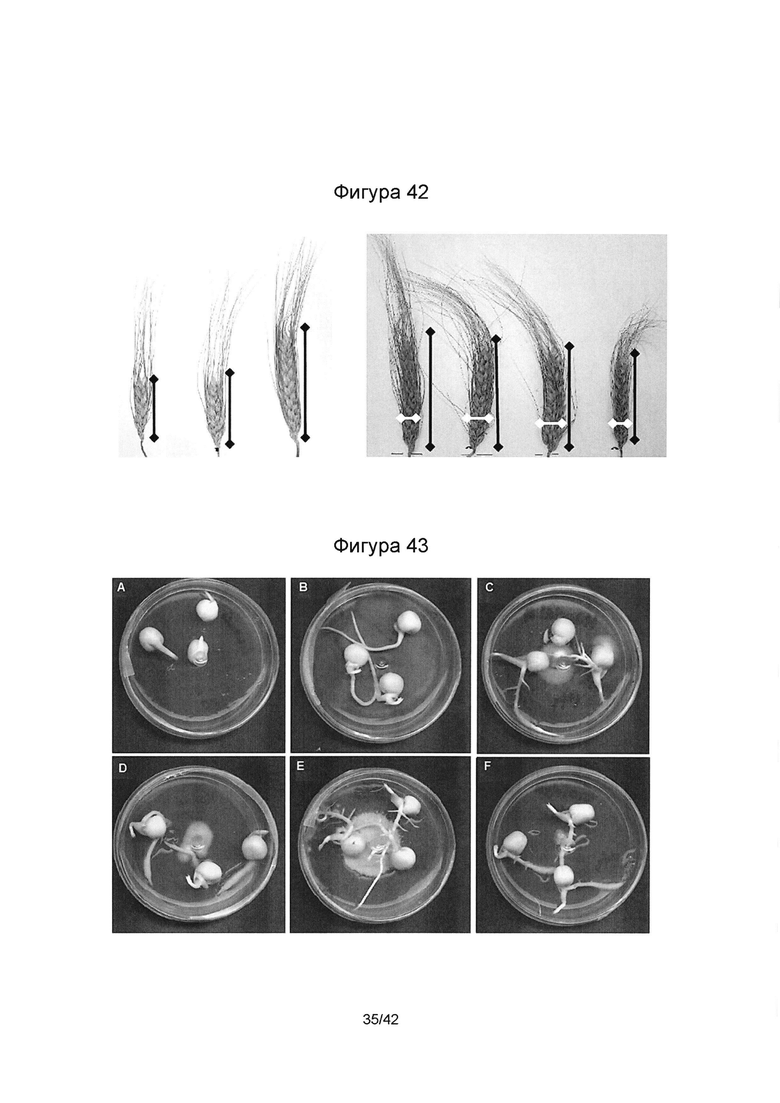







Изобретение относится к области биохимии, в частности к применению штамма Streptomyces sp. или культуры Streptomyces sp. для повышения биомассы корневых волосков у зерновой культуры, для увеличения длины листа у зерновой культуры, для повышения урожайности зерновой культуры, а также для повышения устойчивости зерновой культуры к патогену Fusarium. Изобретение позволяет эффективно улучшать сельскохозяйственные характеристики зерновой культуры. 4 н.п. ф-лы, 51 ил., 9 табл., 15 пр.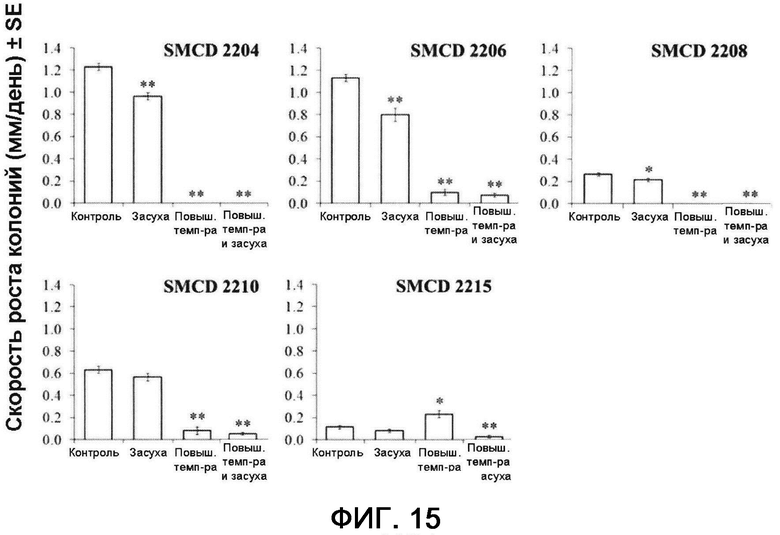
1. Применение штамма Streptomyces sp., депонированного в IDАС под номером IDАС 081111-06, или культуры Streptomyces sp., где вышеуказанный штамм и культура содержат 16S рДНК с последовательностью, представленной в SEQ ID NO:6, для повышения биомассы корневых волосков у зерновой культуры, где зерновая культура представляет собой пшеницу, где применение включает обработку семени злака указанным штаммом или указанной культурой и культивирование из семени злака растения первого поколения.
2. Применение штамма Streptomyces sp., депонированного в IDАС под номером IDАС 081111-06, или культуры Streptomyces sp., где вышеуказанный штамм и культура содержат 16S рДНК с последовательностью, представленной в SEQ ID NO:6, для увеличения длины листа у зерновой культуры, где зерновая культура представляет собой пшеницу, где применение включает обработку семени злака указанным штаммом или указанной культурой и культивирование из семени злака растения первого поколения.
3. Применение штамма Streptomyces sp., депонированного в IDАС под номером IDАС 081111-06, или культуры Streptomyces sp., где вышеуказанный штамм и культура содержат 16S рДНК с последовательностью, представленной в SEQ ID NO:6, для повышения урожайности зерновой культуры, где зерновая культура представляет собой пшеницу, где применение включает обработку семени злака указанным штаммом или указанной культурой и культивирование из семени злака растения первого поколения.
4. Применение штамма Streptomyces sp., депонированного в IDАС под номером IDАС 081111-06, или культуры Streptomyces sp., где вышеуказанный штамм и культура содержат 16S рДНК с последовательностью, представленной в SEQ ID NO:6, для повышения устойчивости зерновой культуры, представляющей собой пшеницу, к патогену Fusarium, где применение включает обработку семени злака указанным штаммом или указанной культурой и культивирование из семени растения первого поколения.
| MICHELLE HUBBARD et al., Fungal endophytes improve wheat seed germination under heat and drought stress, Botany, 9 February 2012, Vol | |||
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |
| Способ приготовления строительного изолирующего материала | 1923 |
|
SU137A1 |
| WO 2013016361 A2, 31.01.2013 | |||
| US 6312940 B1, 06.11.2001 | |||
| US 7601346 B1, 13.10.2009 | |||
| УЛУЧШЕННЫЕ ЭНДОФИТОМ СЕЯНЦЫ С УВЕЛИЧЕННОЙ УСТОЙЧИВОСТЬЮ К ВРЕДИТЕЛЯМ | 2007 |
|
RU2453601C2 |

