Область техники
Изобретение относится к физиологии растений, сельскому хозяйству и биотехнологии и может быть использовано при выращивании сельскохозяйственных злаковых культур с большим числом зародышевых корней, прорастающих на ранних стадиях развития, а также в сельском хозяйстве и агротехнической практике при обработке семян до или после посева для стимулирования образования шестого зародышевого корня с целью повышения способности поглощать воду и питательные элементы из среды, в биотехнологии, когда наличие дополнительного проросшего зародышевого корня является необходимостью или преимуществом.
Уровень техники
Корневая система выполняет функции закрепления растения в почве, запасания питательных веществ, поглощения воды и минеральных элементов. Влага и питательные элементы распределены в почве неравномерно, и развитая корневая система дает растению дополнительные преимущества в освоении больших объемов почвы, обеспечении ресурсами, необходимыми для роста и развития растений. Хорошо развитая, мощная корневая система является важной характеристикой сельскохозяйственных культур, в значительной степени определяющей продуктивность растений. В связи с современными изменениями климата, увеличением площадей засушливых почв, обеднением почв при одновременной интенсификации сельскохозяйственного производства назрела необходимость более эффективного использования ресурсов почв (Ilina et al. 2017). Адаптироваться к изменчивым условиям окружающей среды растениям помогают сформированные в процессе эволюции защитные механизмы, отвечающие за формирование архитектуры корневой системы (Bao et al. 2014). Разработка способов стимуляции этих адаптивных механизмов - наиболее перспективный путь к повышению урожайности сельскохозяйственных растений (Grossman and Rice 2012; Kano et al. 2011; Lynch 1995).
Пшеница является самой важной продовольственной культурой в России. По оценкам экспертов, в ближайшее время потребности человечества в пшенице увеличатся на 50-60%, при этом цена на злаковые увеличится на 29% (https://www.ipcc.ch/). Максимальная продуктивность пшеницы реализуется лишь в оптимальных условиях, а действительная продуктивность растений в значительной степени определяется способностью адаптироваться к широкому диапазону условий окружающей среды. Раннее развитие мощной корневой системы позволяет успешно обеспечивать потребности растения в воде и минеральном питании даже в условиях их недостатка. Однако механизмы, регулирующие формирование корневой системы у пшеницы, остаются малоисследованными (Kano et al. 2011; Meister et al. 2014; Salvi et al. 2016). Формирование архитектуры корней лучше исследовано у двудольных растений, для которых характерна стержневая корневая система, представленная главным корнем, сохраняющимся в течение всей жизни растения, и системой боковых корней разного порядка. Для однодольных характерна мочковатая корневая система, и в этом случае особое значение имеют зародышевые (или первичные) корни, которые развиваются из корневых примордиев зародыша на ранних стадиях развития растений. Число и скорость появления зародышевых корней имеют особое значение для адаптации растений к условиям дефицита воды, так как прорастающие семена и молодые проростки особенно чувствительны к недостатку влаги (Cane et al. 2014; Golan et al. 2018; Manschadi et al. 2008). Существует некоторая генетически-детерминированная вариабильность в числе прорастающих зародышевых корней, в частности у пшеницы это число может варьировать от трех до шести, в зависимости от сорта (Araki and Iijima 2001; Golan et al. 2018; Robertson et al. 1979). Первым из прорастающего зерна появляется первичный корень, за ним следует одна или две пары зародышевых корней, у некоторых сортов может появляться шестой зародышевый корень (Atkinson et al. 2015; Iannucci et al. 2017; Ma et al. 2017; Maccaferri et al. 2016; Ren et al. 2012; Sanguineti et al. 2007). Число развившихся зародышевых корней у пшеницы определяется как генетическими факторами, так и условиями окружающей среды. В работе (Shorinola et al. 2019) показано, что у растений твердой пшеницы первый, второй и третий зародышевые корни вносят больший вклад в поглощение воды в условиях достаточного количества влаги в почве, однако, роль четвертого, пятого и шестого корней заметно возрастает в условиях дефицита воды и питательных элементов, так как их наличие значительно увеличивает поглощающую поверхность корней.
У диких видов пшеницы обычно развиваются три зародышевых корня, хотя пять корневых примордиев отчетливо наблюдаются в зародыше (Golan et al. 2018). Два из пяти примордиев могут не развиться в корни или прорасти только после эпизода засухи, в случае повреждения или гибели первых трех зародышевых корней. Шестой зародышевый корень крайне редко появляется при нормальных условиях, но вырастает у 55% сеянцев в ходе восстановления после стресса дегидратации (Golan et al. 2018; Shorinola et al. 2019). У культурных видов пшеницы, включая сорта твердой гексаплоидной пшеницы (Triticum durum) и мягкой тетраплоидной пшеницы (Triticum aestivum), часто пять корней формируются на самых ранних стадиях развития растений, хотя этот параметр варьирует, особенно у гексаплоидных сортов. Шесть зародышевых корней чаще прорастают у сортов Т. durum ( 2014; Golan et al. 2018).
2014; Golan et al. 2018).
Зародышевые корни могут расти на глубину до 180 см (Kirkegaard and Lilley 2007), в то время как узловые корни располагаются в поверхностном слое почвы. Зародышевые корни функционируют на протяжении всей жизни растения, извлекая воду и минералы из более глубоких слоев почвы (Morozov 1950; Troughton and Whittington 1970). Большее число зародышевых корней обеспечивает большую площадь поверхности и большую биомассу корней, и более длинную корневую систему (Golan et al. 2018). Кроме того, узловые корни, лежащие в самом верхнем слое почвы, более подвержены повреждению в условиях засухи, из чего следует, что большее число зародышевых корней связано с повышенной устойчивостью растений к засухе (Reynolds and Tuberosa 2008; Richards 2008). Недавнее исследование растений твердой пшеницы с разной архитектурой корневой системы показало, что в условиях дефицита влаги генотипы с глубокой корневой системой производят на 35% больше урожая зерна, и масса тысячи зерен этих растений на 9% выше по сравнению с генотипами с неглубокой корневой системой (El Hassouni et al. 2018). В другом исследовании было показано, что количество корней проростков пшеницы, культивируемых в лабораторных условиях, коррелирует с урожайностью растений в полевых условиях (Liu et al. 2013). Следовательно, большое количество развитых зародышевых корней является потенциально ценным сельскохозяйственным признаком, и разработка способов регуляции развития зародышевых корней является важной задачей прикладной биологии растений.
Несмотря на то, что многие исследования были нацелены на идентификацию Локусов Количественных Признаков (ЛКП, Quantitative Trait Loci, QTLs), определяющих архитектуру корней у пшеницы (Atkinson et al. 2015; Hamada et al. 2012; Iannucci et al. 2017; Ma et al. 2017; Maccaferri et al. 2016; Manschadi et al. 2008; Ren et al. 2012; Sanguineti et al. 2007; Shorinola et al. 2019), генетические факторы, определяющие количество образованных зародышевых корней, до сих пор не идентифицированы. Из-за того, что формирование корневой системы регулируется большим числом генов, то есть имеет полигенный характер, получение растений с заданными характеристиками корневой системы представляет собой сложную задачу (Flowers and Yeo 1995; Roy et al. 2014). Более того, количество развитых зародышевых корней может значительно различаться у отдельных растений одного сорта, и на этот параметр могут влиять как эндогенные факторы, так и условия окружающей среды. Данные о физиологических и экологических факторах, влияющих на развитие зародышевых корней, также весьма ограничены. Было показано, что старение семян может влиять на способность образовывать зародышевые корни, в то время как свежие семена дают больше зародышевых корней (Meyer 1976). Вес семян положительно коррелирует со средним числом корней, но в пределах каждого уровня плоидности были обнаружены образцы, имеющие одинаковый вес семян и значительно отличающиеся средние значения числа корней (Robertson et al. 1979). Число корней увеличивается с увеличением размера зародыша (Manner 1965; Meyer 1976; Williams 1960), и определяется факторами, присутствующими в прорастающем эмбрионе, а не в эндосперме (Golan et al. 2018). Было показано, что преждевременный сбор семян, концентрация сахарозы в среде проращивания и температура влияют на появление зародышевых корней (Huang et al. 1991; Meyer and Alston 1978; Meyer 1976). Также было показано, что рост шестого корня инициируется в стрессовых условиях после гибели ранее сформированных зародышевых корней с помощью еще не известных регуляторных механизмов (Golan et al. 2018; Shorinola et al. 2019). В связи с тем, что регуляция формирования шестого корня имеет полигенную природу, селекцию по данному признаку значительно затруднена (Golan et al. 2018).
Развитие корневой системы, сложный, до настоящего времени не полностью понятный процесс, в регуляцию которого вовлечены множество регуляторных систем, включая системы гормонов. Известно, что растительные гормоны ауксины являются основными регулятором развития боковых корней (Reed et al. 1998). Обработка растений ауксинами может приводит к увеличению числа боковых и вспомогательных корней (патент RU 2271098 C2), хотя и не во всех случаях (Ilina et al. 2018). Роль гормонов жасмонатов, их взаимодействие с ауксинами и цитокининами в регуляции развития корневой системы до сих пор до конца не понятна (da Costa et al. 2013; Gutierrez et al. 2012).
Жасмонаты - группа соединений родственных жасмоновой кислоте (1R,2R)-3-оксо-2-(2Z)-пент-2-енилциклопентанэтановая кислота [(1R,2R)-3-Oxo-2-(2Z)-2-pentenyl-cyclopentaneacetic acid] (Фиг. 1). Жасмонаты являются важными сигнальными компонентами, координирующими реакцию растений в стрессовых условиях (Wasternack and Strnad 2018). Биосинтез жасмонатов из α-линоленовой кислоты инициируется в хлоропластах, где опосредованное липоксигеназой окисление приводит к образованию 13-гидроперокси-9,11,15-октадекатриеновой кислоты (13-ГПОТ, 13-НРОТ) (Feussner and Wasternack 2002). 13-ГПОТ далее превращается в (9S, 13S)-12-оксофитодиеновую кислоту (ОФДК, OPDA) в результате последовательного воздействия алленоксидсинтазы и алленоксидциклазы (Schaller and Stintzi 2009). Затем ОФДК транспортируется в пероксисомы, где восстанавливается с помощью оксофитодиеноатредуктазы (OPDA REDUCTASE, OPR) (Stintzi and Browse 2000), затем происходят три цикла (3-окисления боковой цепи молекулы с образованием жасмоновой кислоты (ЖК, JA) (Miersch and Wasternack 2000). ЖК может быть дополнительно модифицирована в цитоплазме путем образования конъюгата с аминокислотой изолейцином (Иле, Ile), что дает начало биоактивному сигнальному лиганду ЖК-Иле (JA-Ile), который отвечает за регуляцию большинства ЖК-зависимых процессов (Fonseca et al. 2009; Thines et al. 2007). ЖК-Иле воспринимается рецептором CORONATINE INSENSITIVE 1 (COI1), который является компонентом убиквитин Е3 лигазного комплекса Skp1-Cullin-F-box (SCF-COI1). После связывания с ЖК-Иле комплекс SCF-COI1 направляет репрессорные белки JAZ (Jasmonate ZIM domain) на деградацию на 26S протеасомах, тем самым инициируя экспрессию жасмонат-зависимых генов (Devoto et al. 2002; Yan et al. 2009). Метилжасмонат (Фиг. 2) является одним из наиболее часто используемых производных жасмоновой кислоты, это соединение отличается большей растворимостью по сравнению с другими жасмонатами, легко проникает сквозь мембраны и в растительных тканях деметилируется, превращаясь в жасмоновую кислоту (Fattorini et al. 2009).
У однодольных, особенно у пшеницы, жасмонатная система остается недостаточно охарактеризованной (Lyons et al. 2013).
Благодаря высокой биологической активности, жасмонаты нашли практическое применение в сельском хозяйстве, биотехнологии, производстве косметических средств и медицине (подробно рассмотрено в обзорной статье (Ghasemi Pirbalouti et al. 2014)). В первую очередь применение этих соединений связано с их способностью регулировать защитные ответы растений в условиях биотических стрессов, и, в связи с этим, несколькими патентами закреплено применение жасмонатов и их производных для защиты растений от насекомых и фитопатогенов. Примерами могут служить патенты US Pat. No. 5118711, US Pat. No. 8992653, US Pat. No. 8115053 B2, US 8507756 B2. Роль жасмонатов в регуляции роста и развития растений менее понятна (Savchenko et al. 2019; Savchenko et al. 2014; Wasternack and Stmad 2018). Вызываемые жасмонатами эффекты могут по-разному проявляться у разных видов растений, например, ингибирование роста одних растений/тканей и стимулирование ростовых процессов у других растений, других тканей или органов. Патент US 20150230462 A1 описывает применение жасмонатов для стимулирования прорастания семян и повышения их устойчивости к воздействию стрессовых факторов. В патенте U.S. Pat. No. 6114284 заявлено, что эфиры (эстеры) жасмоновой кислоты в сочетании с гиббереллином ускоряют рост и развитие растений, а патент U.S. Pat. No. 5436226 раскрывает применение жасмонатов для ингибирования прорастания и потемнения клубней.
Данное изобретение основывается на открытом нами феномене влияния жасмонатов, эндогенно продуцируемых или экзогенно добавленных к прорастающим семенам, на формирование архитектуры корневой системы пшеницы (Pigolev et al. 2021).
Новизна изобретения заключается в том, что никто ранее не использовал жасмонаты и их производные в качестве индуктора роста шестого зародышевого корня у пшеницы.
Задача изобретения заключается в разработке способа изменения архитектуры корневой системы путем стимулирования роста шестого зародышевого корня у пшеницы. Такое изменение корневой системы растения неизбежно скажется на способности поглощать воду и питательные элементы с самых ранних стадий развития и в течение всей жизни, в связи с тем, что зародышевые корни первыми появляются при прорастании семян и функционируют в течение всей жизни, а у взрослых растений - при различных условиях окружающей среды, так как именно эти корни проникают на большую глубину по сравнению с другими корнями. Технический результат заключается в увеличении числа растений с проросшим шестым зародышевым корнем, как следствие, в увеличении числа растений с архитектурой корневой системы, характеризующейся большим числом развитых зародышевых корней, путем обработки семян фитогормонами жасмонатного ряда или путем изменения эндогенного содержания жасмонатов в результате генетической модификации растения. Способ характеризуется надежностью и воспроизводимостью эффектов, основан на экзогенной обработке растений известными доступными натуральными нетоксичными соединениями, обладающими выраженной биологической активностью, или альтернативно, на изменении эндогенного содержания гормонов жасмонатов посредством изменения экспрессии гена пути биосинтеза жасмонатов. Результат может быть использован в сельском хозяйстве, в биотехнологии, в полевых и лабораторных условиях, когда дополнительное число зародышевых корней является желательным признаком или предоставляет какие-либо преимущества. Обработка семян может проводиться путем опрыскивания или вымачивания семян перед посадкой, посредством полива почвы раствором, содержащим производные жасмоновой кислоты, или добавлением этих соединений в синтетическую среду, среду проращивания. Генетическая модификация растений, ведущая к изменению содержания жасмонатов в прорастающих зернах посредством сверхэкспрессии гена пути биосинтеза жасмонатов, может осуществляться в результате сверхэкспрессии чужеродного гена или собственного гена растения, кодирующего один из ферментов пути биосинтеза жасмонатов, или изменения активности промоторов, регулирующих экспрессию генов ферментов биосинтеза жасмонатов.
Существует множество запатентованных способов обработки семян, направленных на стимулирование роста прорастающих семян растений или защиты от неблагоприятных факторов внешней среды. Примерами могут служить патенты Патент РФ №2249584 и Патент РФ №2130264, которые предполагают использование сложных компонентов, таких как, например, водный экстракт из листьев каланхоэ, ингибиторы развития почвенных вредителей и возбудителей грибковых и бактериальных заболеваний, гуматы, комплексы штаммов бактерий. Использование таких сложных компонентов связано с тем, что биохимический состав смесей для обработки может значительно варьировать, что, в свою очередь, может негативно сказаться на воспроизводимости ожидаемых эффектов. Описаны и более простые составы, и индивидуальные соединения, обладающие рострегулирующей активностью, такие как производные мочевины, пирролидоны и амиды карбоновых кислот (Murthy et al. 1998), но все из описанных способов характеризуются наличием существенных недостатков и лишены специфичности по отношению к индуцируемым эффектам. Например, тидиазурон (N-фенил-N'-1,2,3-тиадиазол-5-илмочевина) демонстрирует уникальное сочетание свойств, характерных для ауксина и цитокинина, однако, в некоторых случаях он может индуцировать ингибирование ферментов растений и, тем самым, вызывать обратный эффект, а также при повышенном содержании тяжелых металлов в почве он способствует накоплению их в тканях растений. Было описано использование веществ, таких как 1-(4-нитрофенил)-4-N-этиламинопирролидон-2, 1-(4-нитрофенил)-4-N-бутиламинопирролидон-2, N-фениламид, 3-N-фениламино-4-амино-(4-нитрофенил)-бутановой кислоты в качестве регуляторов роста для улучшения посевных качеств семян озимой пшеницы, активации прорастания и увеличения продуктивности растений (Музыченко et al. 2005). Описывается применение полигексаметиленгуанидин-гидрохлорида в качестве стимулятора роста и развития зерновых культур для повышения всхожести семян кукурузы и пшеницы, ускорения их роста и развития, снижения поражаемости фитопатогенами и увеличения урожайность зерновых культур. Однако, данные соединения по строению далеки от природных фитогормонов и к тому же зачастую обладают невысокой растворимостью в воде, что ограничивает их применение.
В отличие от всех перечисленных способов, предлагаемый способ отличается высокой специфичностью по отношению к индукции шестого зародышевого корня, наличие которого повышает способность растений добывать воду и минеральное питание в широком диапазоне условий. Применяемое вещество является естественным фитогормоном, который метаболизируется в растительных тканях, вещество растворимо в воде, его производство в промышленных масштабах возможно.
Применение генно-модифицированных организмов в настоящее время в России ограничено (регулируется законами Федеральный закон от 05.07.1996 N 86-ФЗ, (ред. от 03.07.2016) N 96-ФЗ от 12 июля 2000 г.). Однако известно множество патентов по трансгенным растениям и в России, и за рубежом, включая предыдущий патент автора данной заявки (US 20120011599 А1/ЕР 2403329 А4) и патенты по созданию трансгенной пшеницы (примеры - RU 2646108 C1, RU 2377303 C2). Наличие большого числа зарубежных патентов в области трансгенных растений доказывает тот факт, что в зарубежных странах не только уже созданы солидные базы запатентованных данных в этой области, но и наблюдается активное пополнение этих баз. Более того, организмы, полученные с помощью современного метода генетической модификации CRISPR-CAS9 и вариаций этого метода, использование которых позволяет не вносить чужеродные последовательности в модифицируемый организм, а лишь изменить внутренние последовательности генома, рассматриваются как нетрансгенные. Данное изобретение также предлагает новый способ стимулирования роста шестого зародышевого корня у пшеницы путем изменения эндогенного уровня жасмонатов в растениях в результате активации экспрессии гена пути биосинтеза этого гормона. Сущность изобретения
Одним из аспектов изобретения является индукция роста шестого зародышевого корня у пшеницы путем экзогенной обработки семян жасмонатами, полученными из растительных объектов, синтетическими жасмонатами или их аналогами, сохраняющими способность индуцировать жасмонат-зависимые ответы в растениях.
Другим аспектом изобретения является экзогенная обработка жасмонатами или их производными, путем опрыскивания или вымачивания семян перед посадкой, посредством полива почвы с использованием растворов, содержащих жасмонаты или их производные, или добавлением этих соединений в синтетическую среду, среду проращивания.
Другим аспектом изобретения является экзогенная обработка жасмонатами семян пшеницы любых сортов и генотипов, включая тетраплоидные и гексаплоидные сорта и виды.
Другим аспектом изобретения является способ индукции роста шестого зародышевого корня у пшеницы путем изменения эндогенного уровня жасмонатов путем изменения экспрессии генов пути биосинтеза жасмонатов, включая гены, кодирующие локализованные в хлоропластах ферменты пути биосинтеза жасмонатов, липоксигеназу, алленоксидсинтазу, алленоксидциклазу, локализованные в пероксисомах оксофитодиеноатредуктазу, а также локализованные в цитоплазме ферменты JAR (JASMONATE RESISTANT), отвечающие за конъюгацию жасмоновой кислоты и аминокислоты изолейцина, что приводит к формированию наиболее активной формы жасмоната, или путем изменения промоторов вышеуказанных генов.
Другим аспектом изобретения является способ индукции роста шестого зародышевого корня у пшеницы посредством изменения эндогенного уровня жасмонатов путем экспрессии генов пути биосинтеза жасмонатов из любых растительных объектов, включая пшеницу, или путем экспрессии модифицированных последовательностей генов, кодирующих функциональный белок, способный осуществлять превращения, характерные для реакций пути биосинтеза жасмонатов.
Другим аспектом изобретения является способ индукции роста шестого зародышевого корня у пшеницы путем создания трансгенных растений с измененным профилем экспрессии генов пути биосинтеза жасмонатов посредством получения трансгенных растений, включая методы биобаллистики и агротрансформации.
Другим аспектом изобретения является способ индукции роста шестого зародышевого корня у пшеницы путем создания трансгенных растений, в которых экспрессия перенесенного гена контролируется конститутивными или индуцируемыми промоторами.
Другим аспектом изобретения является способ индукции роста шестого зародышевого корня у пшеницы путем создания геномно-редактированных растений с измененным эндогенным уровнем жасмонатов в результате редактирования нуклеотидной последовательности генов, кодирующих функциональные белки, способные осуществлять превращения, характерные для реакций пути биосинтеза жасмонатов или редактирования последовательности нативных промотора(-ов) генов пути биосинтеза жасмонатов растения, обеспечивающее изменение уровня их эндогенной экспрессии.
Другим аспектом изобретения является способ индукции роста шестого зародышевого корня у пшеницы, сочетающий изменение эндогенно уровня жасмонатов и экзогенную обработку жасмонатами.
Краткое описание чертежей
Формула жасмоновой кислоты иллюстрируется Фиг. 1.
Формула метилжасмоната иллюстрируется Фиг. 2.
Прорастающие семена Сар-60 без шестого зародышевого корня и с развитым шестым зародышевым корнем иллюстрируются Фиг. 3. Прорастающие семена без шестого зародышевого корня обозначены буквами А и Б, с развитым шестым зародышевым корнем - буквами В, Г и Д.
Описание изобретения
Формирование органов на ранних стадиях развития растений, особенно корневой системы, определяет их выживание и дальнейший рост, развитие, продуктивность. Детальный анализ развития полученных нами растений с измененным содержанием жасмонатов (Pigolev et al. 2018) показал, что семена некоторых трансгенных линий характеризуются более высокой частотой прорастания шестого зародышевого корня (Pigolev et al. 2021). Применение экзогенной обработки растений жасмонатами вызывало схожий эффект (Pigolev et al. 2021). Таким образом сущность предлагаемого способа влияния на корнеобразование пшеницы основана на способности растительных гормонов жасмонатов регулировать формирование шестого зародышевого корня. Примеры осуществления способа.
Пример 1. Влияние метилжасмоната на образование шестого зародышевого корня у растений гексаплоидного сорта Саратовская-60 (Сар-60).
Сравнивали процент растений с проросшим шестым зародышевым корнем среди растений мягкой гексаплоидной пшеницы сорта Сар-60, обработанных и необработанных метилжасмонатом. Перед началом опыта все семена пшеницы были поверхностно стерилизованы обработкой 2% раствором гипохлорита натрия в течение 20 минут, с перемешиванием и последующей отмывкой водой (4 раза по 5 минут). После этого семена были помещены в квадратные чашки Петри размером 10 на 10 см (Sarstedt), с двумя слоями фильтровальной бумаги на дне. В каждую чашку было добавлено 15-17 мл раствора метил жасмоната или автоклавированной дистилированной H2O. В среднем в каждой чашке было от 30 до 40 семян. При прорастании семян в эксперименте поддерживали следующие условия: 1) Температура в комнате была 21-23 градуса; 2) Для освещения использовали лампу мощностью 10 Wt (LED, Uniel (Russia), спектр подходящий для фотосинтеза), которая находилась на расстоянии 45 см от чашек, интенсивность света на уровне чашек составляла 2-5 мкм, со световым режимом - день-ночь 12:12 часов. Метилжасмонат для приготовления растворов растворяли в воде, подогретой до 30 градусов с интенсивным перемешиванием. По мере роста проростков пшеницы в чашки с водой и низкой концентрацией метилжасмоната, начиная где-то с 4 или 5 дня эксперимента, при необходимости добавляли воду. Подсчет корней производили через 5 дней после начала эксперимента, если семена прорастали на воде или в присутствии жасмоната в концентрации 20-50 мкм. При более высоких концентрациях метилжасмоната (100-500 мкм), когда корни были очень короткими, им давали подрасти еще один день. Подсчет растений с шестым корнем производили при помощи USB микроскопа Andonstar ADSM301. Все эксперименты был выполнены минимум в 3-х биологических повторностях. Для оценки статистической значимости и достоверности различий использовался критерий Стьюдента для парных сравнений и Анова и пост-хок критерий Тьюки для множественных сравнений. В ряде случаев после подсчета корней, эксперимент не заканчивали, и проростки растений переносили в пластиковые коробки с водой на дне слоем 1-1,5 см, достаточным для того, чтобы покрыть корни, и контролировали появление других корней вплоть до 14-го дня с начала эксперимента. В экспериментах, где изменялось время обработки метилжасмонатом, после обозначенного времени обработки, семена доставали из чашек, тщательно промывали и перекладывали в чашки с водой. Шестой зародышевый корень идентифицировали по характерному положению (между 4 и 5 зародышевыми корнями, над первым зародышевым корнем) и по характерному времени появления на четвертый-пятый день у необработанных и на пятый-шестой день у обработанных жасмонатами растений (Фиг. 3).
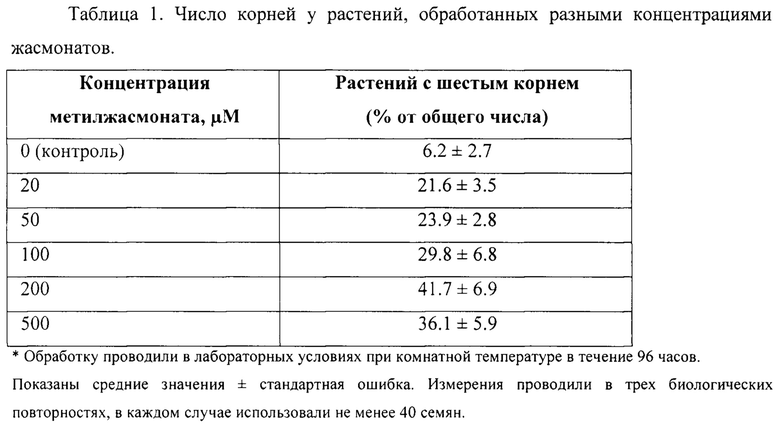
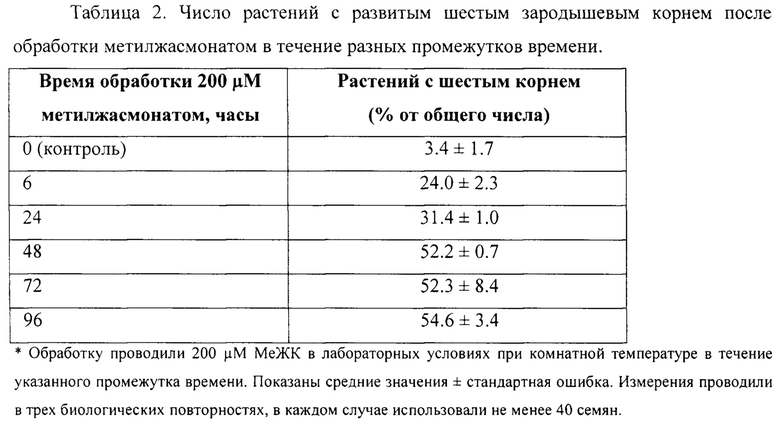
Разные концентрации метилжасмоната наносили на влажную салфетку с прорастающими семенами Сар-60 или обработку 200 μМ метилжасмонатом осуществляли в течение разных промежутков времени. Результаты анализа приведены в Таблицах 1 и 2.
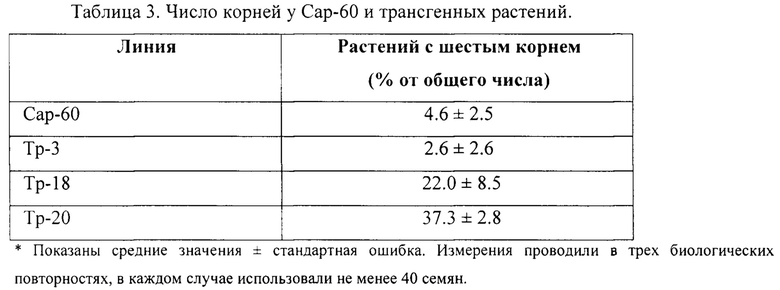
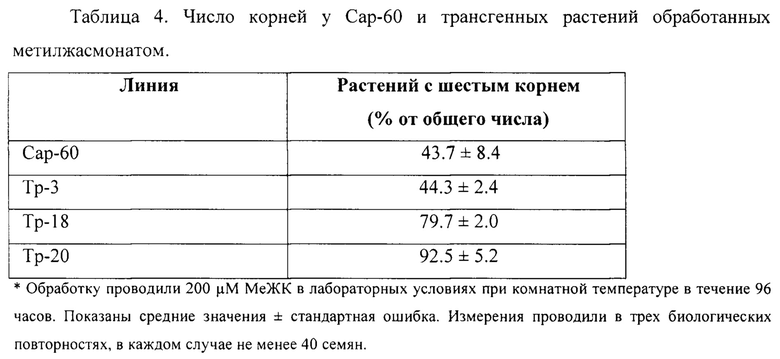
Пример 2. Стимулирование появления шестого зародышевого корня вследствие изменения эндогенного уровня жасмонатов. Анализ проводили на контрольных и трансгенных растениях Сар-60 с измененным уровнем жасмонатов в результате конститутивной экспрессии гена одного из ферментов в пути биосинтеза жасмонатов, 12-оксофитодиеноатредуктазы AtOPR3 из Arabidopsis thaliana. Создание и характеристика растений описаны нами ранее (Pigolev et al. 2018). Процент прорастающих семян с развитым шестым зародышевым корнем был выше среди растений трансгенных линий Тр-18 и Тр-20.
Пример 3. Стимулирование появления шестого зародышевого корня вследствие изменения эндогенного уровня жасмонатов и экзогенной обработки метилжасмонатом. Растения Сар-60 и трансгенные растения обрабатывали метилжасмонатом. Число растений с развитым шестым зародышевым корнем было очень высоко у растений Тр-18 и Тр-20 после обработки метилжасмонатом и составляло 79.7% в случае Тр-18 и 92.5% в случае Тр-20.
Промышленная воспроизводимость Формирование зародышевых корней является важной морфологической, физиологической и агрономической характеристикой пшеницы. Прорастание шестого зародышевого корня крайне редко наблюдается у растений пшеницы. Применение описанного способа, основанного на обработке семян фитогормонами жасмонатного ряда или путем изменения эндогенного содержания жасмонатов в результате генетической модификации растений позволяет значительно увеличить вероятность прорастания шестого зародышевого корня и, таким образом, увеличить число растений с проросшим шестым зародышевым корнем. Способ характеризуется надежностью и воспроизводимостью эффектов, что подтверждено в результате проведения большого числа экспериментов на протяжении нескольких лет исследований. Так как фенотип числа корней, сформированных в лабораторных условиях, воспроизводится при выращивании растений в почве в полевых условиях (Richard et al. 2015; Watt et al. 2013), данный способ может быть использован в сельском хозяйстве и биотехнологическом производстве, в полевых и лабораторных условиях, когда дополнительное число зародышевых корней является желательным признаком или предоставляет какие-либо преимущества. Экзогенная обратка растений подразумевает применение доступных натуральных нетоксичных соединений, обладающих выраженной биологической активностью. Применяемые вещества являются естественными фитогормонами. Производное жасмоновой кислоты метилжасмонат легко метаболизируется в растительных тканях, вещество растворимо в воде, его производство в промышленных масштабах возможно. Обработка семян может проводиться разными способами, в том числе путем опрыскивания или вымачивания семян перед посадкой, посредством полива почвы раствором, содержащим производные жасмоновой кислоты, или добавлением этих соединений в синтетическую среду, среду проращивания. Генетическая модификация растений, ведущая к изменению содержания жасмонатов в прорастающих зернах посредством изменения экспрессии гена пути биосинтеза жасмонатов, может осуществляться в результате сверхэкспрессии чужеродного гена или собственного гена растения, кодирующего один из ферментов пути биосинтеза жасмонатов или изменения активности промотора генов жасмонатного пути.
Источники информации
Araki, Н. and Iijima, М. (2001) Deep rooting in winter wheat: rooting nodes of deep roots in two cultivars with deep and shallow root systems. Plant Prod Sci 4: 215-219.
Atkinson, J.A., Wingen, L.U., Griffiths, M., Pound, M.P., Gaju, O., Foulkes, M.J., et al. (2015) Phenotyping pipeline reveals major seedling root growth QTL in hexaploid wheat. J Exp Bot 66: 2283-2292.
Bao, Y., Aggarwal, P., Robbins, N.E., 2nd, Sturrock, C.J., Thompson, M.C., Tan, H.Q., et al. (2014) Plant roots use a patterning mechanism to position lateral root branches toward available water. Proc Natl Acad Sci USA 111: 9319-9324.
Cane, M.A., Maccaferri, M., Nazemi, G., Salvi, S., Francia, R., Colalongo, C., et al. (2014) Association mapping for root architectural traits in durum wheat seedlings as related to agronomic performance. Mol Breed 34: 1629-1645.
da Costa, C.T., de Almeida, M.R., Ruedell, С.М., Schwambach, J., Maraschin, F.S. and Fett-Neto, A.G. (2013) When stress and development go hand in hand: main hormonal controls of adventitious rooting in cuttings. Frontiers in plant science 4: 133.
Fattorini, L., Falasca, G., Kevers, C, Rocca, L.M., Zadra, C. and Altamura, M.M. (2009) Adventitious rooting is enhanced by methyl jasmonate in tobacco thin cell layers. Planta 231: 155-168.
Flowers, T.J. and Yeo, A.R. (1995) Breeding for salinity resistance in crop plants - where next.. Functional Plant Biology 22: 875-884.
Ghasemi Pirbalouti, A., Sajjadi, S.E. and Parang, K. (2014) A review (research and patents) on jasmonic acid and its derivatives. Arch Pharm (Weinheim) 347: 229-239.
Golan, G., Hendel, E., Mendez Espitia, G.E., Schwartz, N. and Peleg, Z. (2018) Activation of seminal root primordia during wheat domestication reveals underlying mechanisms of plant resilience. Plant Cell Environ 41: 755-766.
Grossman, J.D. and Rice, K.J. (2012) Evolution of root plasticity responses to variation in soil nutrient distribution and concentration. Evol Appl 5: 850-857.
Gutierrez, L., Mongelard, G., Flokova, K., Pacurar, D.I., Novak, O., Staswick, P., et al. (2012) Auxin controls Arabidopsis adventitious root initiation by regulating jasmonic acid homeostasis. Plant Cell 24: 2515-2527.
Iannucci, A., Marone, D., Russo, M.A., De Vita, P., Miullo, V., Ferragonio, P., et al. (2017) Mapping QTL for Root and Shoot Morphological Traits in a Durum Wheat x T. dicoccum Segregating Population at Seedling Stage. Int J Genomics 2017: 6876393.
Ilina, E.L., Kiryushkin, A.S., Semenova, V.A., Demchenko, N.P., Pawlowski, K. and Demchenko, K.N. (2018) Lateral root initiation and formation within the parental root meristem of Cucurbita pepo: is auxin a key player? Ann Bot 122: 873-888.
Ilina, E.L., Kiryushkin, A.S., Tsyganov, V.E., Pawlowski, K. and Demchenko, K.N. (2017) Molecular, genetic and hormonal outlook in root branching. Agric. Biol. 52,: 856-868.
Kano, М., Inukai, Y., Kitano, H., Kitano, H. and Yamauch, A. (2011) Root plasticity as the key root trait for adaptation to various intensities of drought stress in rice. Plant and Soil 342: 117-128.
Lynch, J. (1995) Root Architecture and Plant Productivity. Plant Physiol 109: 7-13.
Ma, J., Luo, W., Zhang, H., Zhou, X.H., Qin, N.N., Wei, Y.M., et al. (2017) Identification of quantitative trait loci for seedling root traits from Tibetan semi-wild wheat (Triticum aestivum subsp.tibetanum). Genome 60: 1068-1075.
Maccaferri, M., El-Feki, W., Nazemi, G., Salvi, S., Сапе, M.A., Colalongo, M.C., et al. (2016) Prioritizing quantitative trait loci for root system architecture in tetraploid wheat. J Exp Bot 67: 1161-1178.
Manschadi, A.M., Hammer, G.L., Christopher, J.T. and de Voil, P. (2008) Genotypic variation in seedling root architectural traits and implications for drought adaptation in wheat (Triticum aestivum L.). Plant and Soil 2008: 115-129.
Meister, R., Rajani, M.S., Ruzicka, D. and Schachtman, D.P. (2014) Challenges of modifying root traits in crops for agriculture. Trends Plant Sci 19: 779-788.
Murthy, B.N.S., Murch, S.J. and Saxena, P.K. (1998) Thidiazuron: A potent regulator ofin vitro plant morphogenesis. In Vitro Cellular & Developmental Biology - Plant 34: 267.
Pigolev, A., Miroshnichenko, D., Dolgov, S. and Savchenko, T. (2021) Regulation of Sixth Seminal Root Formation by Jasmonate in Triticum aestivum L. Plants 10: 219.
Pigolev, A.V., Miroshnichenko, D.N., Pushin, A.S., Terentyev, V.V., Boutanayev, A.M., Dolgov, S.V., et al. (2018) Overexpression of Arabidopsis OPR3 in Hexaploid Wheat (Triticum aestivum L.) Alters Plant Development and Freezing Tolerance. Int J Mol Sci 19.
Reed, R.C., Brady, S.R. and Muday, G.K. (1998) Inhibition of auxin movement from the shoot into the root inhibits lateral root development in Arabidopsis. Plant Physiol 118: 1369-1378. Ren, Y., He, X., Liu, D., Li, J., Zhao, X., Li, В., et al. (2012) Major quantitative trait loci for seminal root morphology of wheat seedlings. Molecular Breeding 30: 139-148.
Richard, C.A., Hickey, L.T., Fletcher, S., Jennings, R., Chenu, K. and Christopher, J.T. (2015) High-throughput phenotyping of seminal root traits in wheat. Plant Methods 11: 13.
Robertson, B.M., Waines, J.G. and Gill, B.S. (1979) Genetic Variability for Seedling Root Numbers in Wild and Domesticated Wheats. Crop Science 19 843-847.
Roy, S.J., Negrao, S. and Tester, M. (2014) Salt resistant crop plants. Curr Opin Biotechnol 26: 115-124.
Salvi, S., Giuliani, S., Ricciolini, C., Carraro, N., Maccaferri, M., Presterl, Т., et al. (2016) Two major quantitative trait loci controlling the number of seminal roots in maize co-map with the root developmental genes rtcs and rum1. J Exp Bot 67: 1149-1159.
Sanguineti, M.C., Li, S., Maccaferri, M., Corneti, S., Rotondo, F., Chiari, Т., et al. (2007) Genetic dissection of seminal root architecture in elite durum wheat germplasm. Annals of Applied Biology 151: 291-305.
Savchenko, T.V., Rolletschek, H. and Dehesh, K. (2019) Jasmonates-Mediated Rewiring of Central Metabolism Regulates Adaptive Responses. Plant Cell Physiol 60: 2613-2620. Savchenko, T.V., Zastrijnaja, O.M. and Klimov, V.V. (2014) Oxylipins and plant abiotic stress resistance. Biochemistry (Mosc) 79: 362-375.
Shorinola, O., Kaye, R., Golan, G., Peleg, Z., Kepinski, S. and Uauy, C. (2019) Genetic Screening for Mutants with Altered Seminal Root Numbers in Hexaploid Wheat Using a High-Throughput Root Phenotyping Platform. G3 (Bethesda) 9: 2799-2809.
Wasternack, C. and Strnad, M. (2018) Jasmonates: News on Occurrence, Biosynthesis, Metabolism and Action of an Ancient Group of Signaling Compounds. Int J Mol Sci 19.
Watt, M., Moosavi, S., Cunningham, S.C., Kirkegaard, J.A., Rebetzke, G.J. and Richards, R.A. (2013) A rapid, controlled-environment seedling root screen for wheat correlates well with rooting depths at vegetative, but not reproductive, stages at two field sites. Ann Bot 112: 447-455.
Музыченко, Г.Ф., Ненько, Н.И., Бурлака, С.Д., Сибирякова, М.А. and Копань, А.С.(2005) Эффективность новых производных 4-N-Х-аминопирролидонов-2, обладающих рострегулирующей и антистрессовой активностью. Агрохимия 5: 71-75.
| название | год | авторы | номер документа |
|---|---|---|---|
| ЭНДОФИТНЫЕ МИКРОБНЫЕ СИМБИОНТЫ В ПРЕНАТАЛЬНОМ УХОДЕ ЗА РАСТЕНИЯМИ | 2013 |
|
RU2723946C2 |
| СПОСОБ ВЫЯВЛЕНИЯ СОБЫТИЙ РЕДАКТИРОВАНИЯ ГЕНА GBSSI У ЗЕРНОВЫХ КУЛЬТУР С ПОМОЩЬЮ НАБОРА ОЛИГОНУКЛЕОТИДНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ | 2023 |
|
RU2817377C1 |
| СПОСОБ ВЫЯВЛЕНИЯ СОБЫТИЙ РЕДАКТИРОВАНИЯ ГЕНА GBSSI У ЗЕРНОВЫХ КУЛЬТУР С ПОМОЩЬЮ НАБОРА ОЛИГОНУКЛЕОТИДНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ | 2023 |
|
RU2834229C1 |
| СПОСОБ ВЫЯВЛЕНИЯ СОБЫТИЙ РЕДАКТИРОВАНИЯ ГЕНА ISA1 У ЗЕРНОВЫХ КУЛЬТУР С ПОМОЩЬЮ НАБОРА ОЛИГОНУКЛЕОТИДНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ | 2023 |
|
RU2833966C1 |
| СПОСОБ ВЫЯВЛЕНИЯ СОБЫТИЙ РЕДАКТИРОВАНИЯ ГЕНА ISA1 У ЗЕРНОВЫХ КУЛЬТУР С ПОМОЩЬЮ НАБОРА ОЛИГОНУКЛЕОТИДНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ | 2023 |
|
RU2833963C1 |
| АКТИВАТОР СИГНАЛЬНОГО ПУТИ ЖАСМОНОВОЙ КИСЛОТЫ | 2016 |
|
RU2728472C2 |
| СПОСОБ ПОДДЕРЖАНИЯ РОСТА РАСТЕНИЙ В УСЛОВИЯХ НЕДОСТАТКА ВЛАГИ | 2022 |
|
RU2795856C1 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ ПШЕНИЦЫ С БИАЛЛЕЛЬНЫМИ МУТАЦИЯМИ В ПРОТОМОТОРНОЙ ОБЛАСТИ ГЕНА VRN-A1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2020 |
|
RU2772575C2 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ ПШЕНИЦЫ С НУКЛЕОТИДНОЙ ВСТАВКОЙ В ПРОМОТОРНОЙ ОБЛАСТИ ГЕНА VRN-A1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2020 |
|
RU2772578C2 |
| ПРИМЕНЕНИЕ ЛИПОХИТООЛИГОСАХАРИДОВ И/ИЛИ ХИТООЛИГОСАХАРИДОВ В КОМБИНАЦИИ С МИКРООРГАНИЗМАМИ, ПРИДАЮЩИМИ РАСТВОРИМОСТЬ ФОСФАТАМ, ДЛЯ УСИЛЕНИЯ РОСТА РАСТЕНИЙ | 2012 |
|
RU2658362C1 |

Способ относится к области биотехнологии, в соответствии с которым стимулируется рост шестого зародышевого корня при прорастании путем изменения активности жасмонатной гормональной системы. Для этого семена обрабатывают жасмонатами или их синтетическими аналогами, сохраняющими способность индуцировать жасмонат-зависимые ответы в растениях, или изменяют эндогенное содержание жасмонатов путем генетической модификации растений. Экзогенная обработка жасмонатами или их производными осуществляется путем опрыскивания или вымачивания семян перед посадкой, или посредством полива почвы с использованием растворов, содержащих жасмонаты или их производные, или добавлением этих соединений в синтетическую среду, среду проращивания. Для изменения эндогенного уровня жасмонатов в растения привносятся функциональные гены пути биосинтеза жасмонатов из любых растительных объектов или модифицированные последовательности генов, кодирующих функциональный белок, способный осуществлять превращения, характерные для реакций пути биосинтеза жасмонатов. Экспрессия перенесенного гена контролируется конститутивным или индуцируемым промоторами. Генетическая модификация растений осуществляется с использованием биобаллистикого метода доставки векторов или систем редактирования генома. Геномное редактирование используется для редактирования нуклеотидных последовательностей генов ферментов пути биосинтеза жасмонатов или последовательностей нативных промоторов генов пути биосинтеза жасмонатов растения, обеспечивающее изменение уровня эндогенной экспрессии генов ферментов пути биосинтеза жасмонатов. 5 з.п. ф-лы, 3 ил., 4 табл., 3 пр.
1. Способ индукции шестого зародышевого корня у пшеницы путем изменения активности жасмонатной системы растения, отличающийся тем, что стимулируется появление шестого зародышевого корня на ранних стадиях развития растения.
2. Способ по п. 1, где указанное растение является белозерной и краснозерной разновидностью пшеницы мягкой (Т. aestivum L.), пшеницы плотноколосой (Т. compaction Host.), пшеницы спельта (Т. spelta L.), пшеницы шарозерной (T. shaerococcum Perciv.), пшеницы твердой (Т. durum Desf), пшеницы полба (Т. dicoccum (Schrank) Schubl.), пшеницы польской (Т. polonicum L.) и пшеницы тучной (T: turgidum L.).
3. Способ по п. 1, отличающийся тем, что индукция роста шестого зародышевого корня у пшеницы осуществляется путем экзогенной обработки семян жасмонатами, полученными из растительных объектов, синтетическими жасмонатами или их аналогами, сохраняющими способность индуцировать жасмонат-зависимые ответы в растениях.
4. Способ по п. 1, отличающийся тем, что экзогенная обработка жасмонатами или их производными осуществляется путем опрыскивания или вымачивания семян перед посадкой, или посредством полива почвы с использованием растворов, содержащих жасмонаты или их производные, или добавлением этих соединений в синтетическую среду, среду проращивания.
5. Способ по п. 1, отличающийся тем, что активность гормональной жасмонатной системы растения изменяется путем генетических модификаций, приводящих к изменению эндогенного уровня жасмонатов в растениях.
6. Способ по п. 5, отличающийся тем, что растение с измененным эндогенным уровнем жасмонатов представляет собой трансгенное растение, содержащее привнесенные функциональные гены ферментов пути биосинтеза жасмонатов, включая гены, кодирующие локализованные в хлоропластах ферменты пути биосинтеза жасмонатов, липоксигеназу, алленоксидсинтазу, алленоксидциклазу, локализованные в пероксисомах оксофитодиеноатредуктазу, а также локализованные в цитоплазме ферменты JAR (JASMONATE RESISTANT), отвечающие за конъюгацию жасмоновой кислоты и аминокислоты изолейцина.
| СИДОРОВ А.В., и др., Результаты селекции яровой пшеницы на увеличение числа и степени развития зародышевых корней, Вестник КрасГАУ, 2015, N3, с.77-82 | |||
| ШАКИРОВА Ф.М., и др., Влияние предобработки метилжасмонатом на устойчивость проростков пшеницы к солевому стрессу, Агрохимия, 2010, N7, с.26-32 | |||
| WASTERNACK, C., et al, Jasmonates: News on |

