Область техники, к которой относится настоящее изобретение
Настоящее изобретение, в целом, относится к эндофиту эпихлое, который формирует стабильную симбиотическую ассоциацию с рожью (Secale cereale).
Предшествующий уровень техники настоящего изобретения
Растение Secale cereale, известное под названием рожь, выращивают во всем мире, обычно для производства зерна. Зерно главным образом используют для изготовления муки и хлеба и для прямого потребления, в частности, в тех странах, для которых исторически характерно потребление ржаного хлеба. Вегетативные части S. cereale могут быть использованы в качестве соломы или переработаны на силос для использования в качестве корма для животных включительно для загонного выпаса.
Как отмечалось выше, рожь в основном выращивают на зерно. Таким образом, на протяжении культивирования необходимо обеспечение эффективной защиты от вредителей для производства достаточного количества зерна приемлемого качества. Рожь обычно считается озимой сельскохозяйственной культурой, посев которой производится осенью. Чередование выращиваемых культур с другими зерновыми культурами или полевыми культурами можно использовать для уменьшения размножения вредителей и/или возбудителей грибковых заболеваний, присутствующих в почве, или которые размножаются в послеуборочных растительных остатках.
Основные вредители ржи включают без ограничений тли; трипсы; проволочники и личинки хрущей; личинки комаров-долгоножек (Tipula spp.); муху озимую (Delia coarctata); минирующие мушки (Agromyza spp.); шведскую овсяную муху (Oscinella frit); жужелицу (Zabrus tenebrioides); маковую седельную галлицу (Haplodiplosis marginata); пьявицу красногрудую (Oulema melanopus, О. gallaeciand); нематод; и слизней.
Известные методы борьбы с вредителями ржи включают некоторые или все следующие методы: использование устойчивых к вредителям культиваров, оптимизацию времени посева и посев здоровых семян, эффективное чередование выращиваемых культур, уничтожение и/или захоронение, или удаление послеуборочных растительных остатков (пожнивные остатки). Дополнительные методы борьбы с вредителями, которые могут понадобиться, включают использование различных пестицидов на растениях и/или семенах. В некоторых случаях может понадобиться одновременное применение двух или более действующих веществ для борьбы с вредителями.
Однако использование многих пестицидов может быть проблематичным из-за известных проблем, связанных с химическими препаратами, используемыми для таких целей. Многие пестициды являются токсичными и могут быть опасными для здоровья человека и животных, потребляющих протравленные сельскохозяйственные культуры (Casida и Quistad, 1998). В частности, накопление пестицидов в организме людей и животных может приводить к серьезным проблемам со здоровьем, в частности, для субъектов на протяжении раннего этапа развития. Например, воздействие пестицидов может быть связано с респираторными нарушениями, злокачественными опухолями, возникающими в процессе развития, и, как было показано, имеет долгосрочные последствия для развития умственных способностей Zejda et al. (1993).
Может оказаться затруднительным контролировать использование пестицидов в различных условиях окружающей среды, что может приводить к нежелательному распространению токсичных веществ, например, вследствие сноса пестицидов при опрыскивании или выщелачивании почвы. В дополнение к этому, по ряду причин вредители могут приобрести устойчивость к пестицидам включительно из-за ненадлежащего использования и обращения, что может отрицательно сказаться на урожайности сельскохозяйственных (зерновых) культур. В соответствии с этим, существует необходимость в средствах борьбы с вредителями, исключающих применение пестицидов.
Целью настоящего изобретения является получение по меньшей мере одного штамма эндофитного гриба эпихлое, который при комбинации по меньшей мере с одним видом Secale spp. обеспечивает по меньшей мере некоторый уровень защиты растения Secale spp. от вредителей, и/или предоставление пользователям подходящего выбора.
Ссылки на описания патентов, других сторонних документов или источников информации предоставляются здесь в качестве контекста для обсуждения характеристик настоящего изобретения. Если не указано иное, такие ссылки на сторонние документы или источники информации в любой юрисдикции не следует толковать как относящиеся к уровню техники данного изобретения или общеизвестной информации в данной области техники.
Краткое раскрытие настоящего изобретения
Согласно первому аспекту настоящее изобретение относится к выделенному штамму эндофитного гриба эпихлое, выбранному из группы, состоящей из AR3039 (NRRL# 50716), AR3046 (NRRL# 50576), AR3049 (NRRL#50577), AR3050 (NRRL# 50578), AR3064 (NRRL# 50718), AR3067 (NRRL# 50719), AR3068 (NRRL# 50720), AR3073 (NRRL# 50721), AR3074 (NRRL# 50722), AR3076 (NRRL# 50723) и AR3078 (NRRL# 50724), и их комбинаций. Предпочтительно настоящее изобретение относится к выделенным штаммам AR3039, AR3046, AR3050, AR3067, AR3068, AR3074 и AR3078, предпочтительно AR3046, AR3050 или AR3068.
Согласно одному варианту осуществления выделенный штамм является биологически чистым.
Согласно второму аспекту настоящее изобретение относится к зараженному эндофитным грибом растению Secale spp., причем Secale spp. не является естественным хозяином эндофита и, причем растение и эндофит формируют стабильную симбиотическую ассоциацию, которая обеспечивает прохождение растением нормального жизненного цикла.
Согласно одному варианту осуществления эндофит передается от первого поколения Secale spp. второму поколению Secale spp. путем вертикальной передачи. Предпочтительно вертикальная передача от первого поколения растения-хозяина второму поколению растения-хозяина осуществляется через семена.
Согласно одному варианту осуществления эндофит представляет собой эндофит, выделенный из Elymus spp. Предпочтительно эндофит выделен из Elymus mutabilis. Предпочтительно эндофит представляет собой эндофит эпихлое. Предпочтительно эндофит эпихлое представляет собой эндофитный штамм эпихлое, выбранный из группы, состоящей из AR3039 (NRRL# 50716), AR3046 (NRRL# 50576), AR3049 (NRRL#50577), AR3050 (NRRL# 50578), AR3064 (NRRL# 50718), AR3067 (NRRL# 50719), AR3068 (NRRL# 50720), AR3073 (NRRL# 50721), AR3074 (NRRL# 50722), AR3076 (NRRL# 50723) и AR3078 (NRRL# 50724), или их комбинаций. Предпочтительно настоящее изобретение относится к эндофитным штаммам AR3039, AR3046, AR3050, AR3067, AR3068, AR3074 и AR3078, предпочтительно AR3046, AR3050 или AR3068.
Согласно одному варианту осуществления у зараженного эндофитным грибом растения Secale spp. не проявляются внешние признаки заражения.
Согласно одному варианту осуществления растение Secale spp., зараженное эндофитным грибом, характеризуется морфологически нормальным фенотипом.
Согласно одному варианту осуществления растение Secale spp., зараженное эндофитным грибом, продуцирует лолиновые алкалоиды и/или перамин.
Согласно одному варианту осуществления растение Secale spp., зараженное эндофитным грибом, обладает повышенной устойчивостью к одному или нескольким вредителям или повышенной устойчивостью по меньшей мере к одной болезни растений, или устойчивостью и к вредителям, и болезни по сравнению с растением Secale spp., которое не заражено эндофитным грибом.
Согласно одному варианту осуществления растение Secale spp., зараженное эндофитным грибом, обладает повышенной устойчивостью к одному или нескольким вредителям, причем один или несколько вредителей выбраны из группы, состоящей из: (1) видов тли (Aphididae), представленных Rhopalosiphum padi, Schizaphis graminum, Rhopalosiphum maidis, Metopoliphium dirhodum, Sitobion spp., Sitobion avenae, Sitobion fragariae, Diuraphis noxis; (2) насекомых-вредителей культурных и диких злаков (Agromyzidae; Anthomyiidae, Chloropidae, Cephidae и Cecidomyiidae), представленных Oscinella frit, Oscinella pusilla, Mayetiola destructor, Cerodontha spp., Cerodontha australis, Cerodontha angustipennis, Formia fumigata, Meromyze americana, Haplodiplosis marginata, Chlorops pumilionis, Tipula spp. Chromatomyia fuscula, Cephus pygmaeus, Chromatomyia fuscula, Contarinia tritici; (3) видов трипсов (Thripidae), представленных Limothrips cerealium, Limothrips denticornis, Aptinothrips rufus, Stenothrips graminum; (4) видов кузнечиков и сверчков (Acrididae и Gryllidae), представленных Locusta migratoria, Phaulacridium marginale, Phaulacridium. vittatum, Melanoplus spp., Teleogryllus commodus; (5) видов клопов (Lygaeidae), представленных Nyssius huttoni, Blissus leucopertus leucopertus; (6) видов долгоносиков (Curculionidae), представленных Sphenophorus spp.; (7) видов гусениц, поражающих злаковые растения, и гусениц озимой совки (Noctuidae), представленных Pseudaletia unipuncta, Spodoptera spp., Mythimna separata; Persectania aversa, Agrostis ipsilon; (8) видов листоедов (Chysomelidae), представленных Oulema melanopus; (9) видов личинок хрущей (Scarabaeidae), представленных Popillia japonica, Costelytra zealandica, Phyllopertha spp., Rhizotrogus majalis, Anisoplia segetum; (10) видов мучнистых червецов (Pseudococcidae и Coccidae), представленных Phenacoccus hordei, Balanococcus poae, Ripersella rumicis, Porphyrophora tritici; (11) видов проволочников (Elateridae), представленных Conoderus spp., Limonius spp.; видов жуков (Carabidae), представленных Zabrus tenebrioides; (13) видов клещей (Eriophyidae и Penthaleidae), представленных Penthaleus spp., Halotydeus destructor, Aceria spp.; (14) видов вредителей запасов продовольствия (Curculionidae, Silvanidae, Pyralidae, Gelechiidae, Tenebrionidae, Bostrichidae), представленных Sitophilus oryzae, Sitophilus granarius, Sitotroga cerealella, Rhyzopertha dominica, Cryptolestes spp., Oryzaephilus surinamensis, Cadra cautella, Plodia interpunctella, Tribolium confusum, Tribolium castaneum, Lasioderma erricorne; (15) видов цикад-пенниц (Cercopoidea), представленных Philaenus spumarius (16) видов нематод, представленных корневой нематодой (Pratylenchus spp., в частности, P. thornei, P. crenatus, P. neglectus и Р. penetrans), овсяной цистообразующей нематодой (Heterodera spp. и Punctodera spp., в частности, H. avenae, H. latipons, H. hordecalis, H. fllipjevi, H. moni, H. bifenestra, H. pakistanensis и P. punctata), клубеньковой нематодой (Meloidogyne spp., в частности, M. chitwoodi, M. naasi, M. artiellia, M. microtyla, M. ottersoni, M. graminicola, M. graminis, M. kikuyensis и M. spartinae), стеблевой нематодой (Ditylenchus spp., в частности, D. dipsicai и D. radicicola); пшеничной нематодой (Anguina tritici); (16) видов слизней (Dereceras reticulatum и Arion spp., в частности, A. hortensis agg. и A. subfuscus). Согласно одному варианту осуществления вредителями являются нематоды, предпочтительно корневые нематоды (Pratylenchus spp.), или минирующие мухи, Cerodontha australis (Diptera: Agromyzidae), также известный как минер влагалища листа пшеницы.
Согласно одному варианту осуществления растение Secale spp., зараженное эндофитным грибом, обладают повышенной устойчивостью к болезням растений, причем болезнь растений вызвана патогеном растений, выбранным из группы, состоящей из вируса желтой карликовости ячменя (Leteovirus), почвообитающего вируса мозаики пшеницы (Furovirus) и вируса полосатой мозаики пшеницы (Tritimovirus), Xanthomonas campestris, Pseudomonas syringae, Colletotrichum graminicola, Glomerella graminicola [телеоморфа], Alternaria spp., Cladosporium herbarum, Mycosphaerella tassiana [телеоморфа], Epicoccum spp., Sporobolomyces spp., Stemphylium spp., Bipolaris sorokiniana, Cochliobolus sativus [телеоморфа], Fusarium spp., Tilletia caries, Tilletia tritici, Tilletia laevis, Tilletia foetida, Hymenula cerealis, Cephalosporium gramineum, Helminthosporium sativum, Cochliobolus sativus [телеоморфа], Coprinus sychromorbidus, Dilophospora alopecuri, Tilletia controversa, Claviceps purpurea, Sphacelia segetum [анаморфа], Fusarium culmorum, Pseudoseptoria donacis, Selenophoma donacis, Neovossia indica, Tilletia indica, Puccinia recondita, Aecidium clematidis [анаморфа], Cercosporidium graminis, Scolicotrichum graminis, Phaeosphaeria herpotrichoides, Leptosphaeria herpotrichoides, Ustilago tritici, Microdochium nivale, Fusarium nivale, Monographella nivalis [телеоморфа], Erysiphe graminis, Pythium aphanidermatum, Pythium arrhenomanes, Pythium debaryanum, Pythium graminicola, Pythium ultimum, Gibberella zeae, Fusarium graminearum [анаморфа], Septoria secalis, Septoria tritici, Mycosphaerella graminicola [телеоморфа], Rhizoctonia cerealis, Rhizoctonia solani, Rhizoctonia zeae, Blumeria spp., Ceratobasidium cereale [телеоморфа], Myriosclerotinia borealis, Sclerotinia borealis, Typhula idahoensis, Typhula incarnate, Typhula ishikariensis, Typhula ishikariensis var. canadensis, Stagonospora nodorum, Septoria nodorum, Phaeosphaeria nodorum [телеоморфа], Leptosphaeria nodorum, Urocystis occulta, Puccinia graminis, Aspergillus spp., Nigrospora spp., Penicillium spp., Rhizopus spp., Pseudocercosporella herpotrichoides, Tapesia acuformis [телеоморфа], Uredo glumarum [анаморфа], Pyrenophora tritici-repentis, Drechslera tritici-repentis [анаморфа], Helminthosporium tritici-repentis, Puccinia triticina, Pythium spp., Rhynchosporium secalis, Puccinia striiformis, Gaeumannomyces graminis и Fusarium pseudograminearum.
Предпочтительно патоген растений представляет собой Puccinia recondita, Puccinia triticina, Puccinia graminis, Fusarium spp., Pythium spp., Rhynchosporium secalis, Puccinia striiformis, Gaeumannomyces graminis, Rhizoctonia solani или Fusarium pseudograminearum.
Согласно одному варианту осуществления растение Secale spp., зараженное эндофитным грибом, выбрано из группы, состоящей из Secale cereale, Secale montanum, Secale strictum, Secale sylvestre и Secale vavilovii. Предпочтительно Secale spp. представляет собой Secale cereale.
Согласно третьему аспекту настоящее изобретение относится к способу получения стабильной комбинации растения-хозяина/эндофитного гриба эпихлое, предусматривающий искусственное заражение растения Secale spp. по меньшей мере одним эндофитным грибом, который формирует стабильную комбинацию с инокулированным растением, причем у растения-хозяина не проявляются внешние признаки заражения эндофитом.
Согласно одному варианту осуществления стабильная комбинация растения и гриба является достаточно стабильной для обеспечения вертикальной передачи эндофита. Согласно одному варианту осуществления вертикальная передача осуществляется через побеги, в частности, через цветочные побеги в семена или через черенки. Предпочтительно вертикальная передача от первого поколения растения-хозяина второму поколению растения-хозяина осуществляется через семена.
Согласно одному варианту осуществления вертикальная передача эндофита приводит к вертикальной передаче эндофита от первого поколения второму поколению растений-хозяев. Предпочтительно вертикальная передача от первого поколения растения-хозяина второму поколению растения-хозяина осуществляется через семена.
Согласно одному варианту осуществления указанный способ дополнительно предусматривает стадию отбора растения-хозяина Secale из популяции зараженных растений-хозяев, у которого не проявляются внешние признаки заражения эндофитным грибом.
Согласно четвертому аспекту настоящее изобретение относится к способу обеспечения по меньшей мере некоторого уровня защиты растения-хозяина Secale spp. от вредителей, предусматривающему искусственное заражение растения Secale spp. по меньшей мере одним эндофитным грибом эпихлое, причем комбинация эндофитного гриба и растения Secale обеспечивает продукцию по меньшей мере одного алкалоида в количестве, достаточном для обеспечения по меньшей мере некоторого уровня защиты растения-хозяина от вредителей.
Согласно пятому аспекту настоящее изобретение относится к способу обеспечения защиты от вредителей растения-хозяина Secale spp., предусматривающему искусственное заражение растения Secale spp. по меньшей мере одним эндофитным грибом эпихлое, причем комбинация эндофитного гриба и растения Secale обеспечивает продукцию по меньшей мере одного алкалоида, который обеспечивает указанную защиту растения-хозяина от вредителей.
Согласно одному варианту осуществления четвертого и пятого аспектов по меньшей мере один алкалоид представляет собой алкалоид, выбранный из группы, состоящей из перамина, N-ацетилнорлолина, лолина, N-формиллолина, N-ацетиллолина и N-метиллолина.
Согласно одному варианту осуществления четвертого и пятого аспектов по меньшей мере один алкалоид представляет собой лолин или перамин, или оба алкалоида.
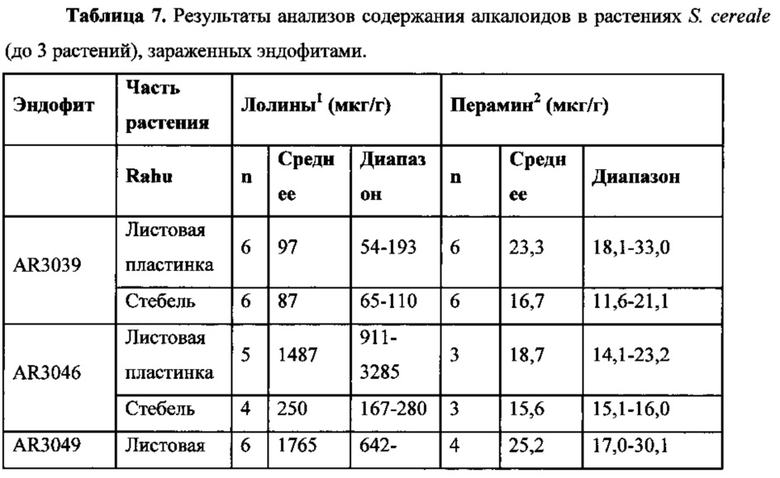
Согласно одному варианту осуществления четвертого и пятого аспектов количество продуцируемого лолина составляет по меньшей мере 25 мкг/г.
Согласно одному варианту осуществления четвертого и пятого аспектов количество продуцируемого лолина приведено в Таблице 7 для лолина.
Согласно одному варианту осуществления четвертого и пятого аспектов количество продуцируемого лолина находится в диапазоне по меньшей мере от 25 мкг/г до приблизительно 3660 мкг/г.
Согласно одному варианту осуществления четвертого и пятого аспектов количество продуцируемого перамина составляет по меньшей мере 1 мкг/г.
Согласно одному варианту осуществления четвертого и пятого аспектов количество продуцируемого перамина приведено в Таблице 7 для перамина.
Согласно одному варианту осуществления четвертого и пятого аспектов количество продуцируемого перамина находится в диапазоне по меньшей мере от 1 мкг/г до приблизительно 45 мкг/г.
Согласно одному варианту осуществления указанный способ дополнительно предусматривает стадию отбора комбинации эндофитного гриба и растения Secale, которая обеспечивает продукцию по меньшей мере одного алкалоида в количестве, достаточном для обеспечения по меньшей мере некоторого уровня защиты растения-хозяина от вредителей.
Согласно шестому аспекту настоящее изобретение относится к семени Secale spp., зараженному эндофитным грибом эпихлое. Предпочтительно Семя растения Secale spp. представляет собой семя растения Secale spp., выбранного из группы, состоящей из Secale cereale, Secale montanum, Secale strictum, Secale sylvestre и Secale vavilovii. Более предпочтительно семя представляет собой семя растения Secale cereale.
Согласно одному варианту осуществления любого из третьего, четвертого, пятого и шестого аспектов настоящего изобретения, которые изложены выше, Secale spp.выбран из группы, состоящей из Secale cereale, Secale montanum, Secale strictum, Secale sylvestre и Secale vavilovii. Предпочтительно Secale spp. представляет собой Secale cereale.
Согласно одному варианту осуществления любого из третьего, четвертого, пятого и шестого аспектов настоящего изобретения, которые изложены выше, по меньшей мере один эндофитный гриб представляет собой эндофит, выделенный из Elymus spp.
Предпочтительно эндофитный гриб выделен из Elymus mutabilis. Предпочтительно эндофитный гриб представляет собой эндофит эпихлое. Предпочтительно эндофит эпихлое выбран из группы, состоящей из AR3039 (NRRL# 50716), AR3046 (NRRL# 50576), AR3049 (NRRL#50577), AR3050 (NRRL# 50578), AR3064 (NRRL# 50718), AR3067 (NRRL# 50719), AR3068 (NRRL# 50720), AR3073 (NRRL# 50721), AR3074 (NRRL# 50722), AR3076 (NRRL# 50723) и AR3078 (NRPvL# 50724), и их комбинаций. Предпочтительно настоящее изобретение относится к выделенным штаммам AR3039, AR3046, AR3050, AR3067, AR3068, AR3074 и AR3078, предпочтительно AR3046, AR3050 или AR3068.
Другие аспекты настоящего изобретения будут понятны из следующего ниже описания, которое приводится только в качестве примера и со ссылкой на прилагаемые графические материалы.
Краткое описание Фигур
Настоящее изобретение описано далее только в качестве примера и со ссылкой на прилагаемые графические материалы, в которых:
На фигуре 1 показана дендрограмма филогенетических взаимоотношений на основе размеров аллелей SSR, которые перечислены в таблице 2.
На фигуре 2 показаны четыре культивара Rahu растения Secale céréale; один незараженный (Е-); и три зараженных AR3046. Слева направо: морфологически нормальное, карликовое и низкорослое растение. У Е- и AR3046-зapaжeнныx нормальных и карликовых растений появляются цветки, а у карликового нет.
На фигуре 3 показаны результаты по определению количества слюны или числа личинок слюнявых пенниц/растение, наблюдаемых на культиваре Rahu растений Secale cereale, зараженных эндофитом AR3046 (Rahu AR3046), и на незараженных растениях (Rahu Nil). Также показано количество слюны или число личинок слюнявых пенниц/растение, наблюдаемых на растениях овсяницы луговой (Festuca pratensis syn. Lolium pratense syn. Schedonorus pratensis), зараженных характерным для нее эндофитом, Neotyphodium uncinatum (MF E+), и на незараженных растениях (MF Е-).
На фигуре 4 показано, что определенные эндофитные штаммы, которые статистически значимо (Р≤0,05) ингибировали рост мицелия Fusarium graminearum в двойной культуре (данные из неопубликованного исследования Stuart Card, AgResearch Ltd).
На фигуре 5 показаны штаммы эндофитных грибов зерновых растений, которые статистически значимо (Р≤0,05) ингибировали рост мицелия Rhizoctonia solani в двойной культуре (данные из неопубликованного исследования Stuart Card, AgResearch Ltd).
Подробное раскрытие настоящего изобретения
Определения
Следующие определения представлены для лучшего понимания настоящего изобретения и в качестве руководства для специалистов в данной области техники при практическом воплощении настоящего изобретения.
Если не указано иное, следует понимать, что все технические и научные термины, использованные в настоящем документе, имеют те же значения, которые понятны специалисту в соответствующей области техники, к которой относится настоящее изобретение. Примеры определений общепринятых терминов в ботанике, микробиологии, молекулярной биологии и биохимии можно найти в источниках: Biology of Plants, Raven et al. (eds.), W.H. Freeman and Company, (2005); Plant Physiology, Taiz et al. (eds.), Sinauer Associates, Incorporated, (2010); Botany: An Introduction to Plant Biology, J.D. Mauseth, Jones & Bartlett Learning, (2003); Methods for General and Molecular Microbiology, 3rd Edition, C.A. Reddy, et al. (eds.), ASM Press, (2008); Encyclopedia of Microbiology, 2nd ed., Joshua Lederburg, (ed.), Academic Press, (2000); Microbiology By Cliffs Notes, I. Edward Alcamo, Wiley, (1996); Dictionary of Microbiology and Molecular Biology, Singleton et al. (2d ed.) (1994); Biology of Microorganisms 11th ed., Brock et al., Pearson Prentice Hall, (2006); Biodiversity of Fungi: Inventory and Monitoring Methods, Mueller et al., Academic Press, (2004); Genes IX, Benjamin Lewin, Jones & Bartlett Publishing, (2007); The Encyclopedia of Molecular Biology, Kendrew et al. (eds.), Blackwell Science Ltd., (1994); Molecular Biology and Biotechnology: a Comprehensive Desk Reference, Robert A. Meyers (ed.), VCH Publishers, Inc., (1995); Symbioses of grasses with seedborne fungal endophytes. Schardl CL et al. (2004) Annual Review of Plant Biology 55: 315-340; и Chemotype diversity of эпихлое, fungal symbionts of grasses, Schardl CL, Young CA, Faulkner JR, Florea S, Pan J (2012) Fungal Ecology 331-344 (Schardl et al., 2012).
Также предполагается, что настоящее изобретение может быть осуществлено на практике с использованием стандартных протоколов и методик, применяемых в ботанике, микробиологии, молекулярной биологии и биохимии, которые известны в данной области техники и которые описаны, например, в источниках: Methods of Studying Root Systems, vol. 33,Wolfgang  , Springer-Verlag, (1979); Root methods: A Handbook, Albert L. Smit Springer, (2000); Biodiversity of Fungi: Inventory and Monitoring Methods, Mueller et al., Academic Press, (2004); Environmental Microbiology: Methods and Protocols, J.F.T. Spencer et al., Humana Press, (2004); Environmental Microbiology, P.D. Sharma, Alpha Science International, (2005); Environmental Microbiology, J.R. Leadbetter, Gulf Professional Publishing, (2005), Molecular Cloning: A Laboratory Manual, Maniatis et al., Cold Spring Harbor Laboratory Press, (1982); Molecular Cloning: A Laboratory Manual (2 ed.), Sambrook et al., Cold Spring Harbor Laboratory Press, (1989); Guide to Molecular Cloning Techniques Vol.152, S.L. Berger and A.R. Kimmerl (Eds.), Academic Press Inc., (1987); Biotechnology of Endophytic Fungi of Grasses. 1994 Bacon and White (Eds.) и других широкодоступных справочных материалах, относящихся к области техники, к которой относится настоящее изобретение, причем все из них включены в настоящий документ во всей их полноте посредством ссылки.
, Springer-Verlag, (1979); Root methods: A Handbook, Albert L. Smit Springer, (2000); Biodiversity of Fungi: Inventory and Monitoring Methods, Mueller et al., Academic Press, (2004); Environmental Microbiology: Methods and Protocols, J.F.T. Spencer et al., Humana Press, (2004); Environmental Microbiology, P.D. Sharma, Alpha Science International, (2005); Environmental Microbiology, J.R. Leadbetter, Gulf Professional Publishing, (2005), Molecular Cloning: A Laboratory Manual, Maniatis et al., Cold Spring Harbor Laboratory Press, (1982); Molecular Cloning: A Laboratory Manual (2 ed.), Sambrook et al., Cold Spring Harbor Laboratory Press, (1989); Guide to Molecular Cloning Techniques Vol.152, S.L. Berger and A.R. Kimmerl (Eds.), Academic Press Inc., (1987); Biotechnology of Endophytic Fungi of Grasses. 1994 Bacon and White (Eds.) и других широкодоступных справочных материалах, относящихся к области техники, к которой относится настоящее изобретение, причем все из них включены в настоящий документ во всей их полноте посредством ссылки.
Используемый в настоящем документе термин «растение» охватывает целые растения и все части растения из всех стадий жизненного цикла растения, включая без ограничений вегетативные и репродуктивные клетки и ткани, черенки, семена, зародыши, побеги, стебли, листья, влагалища листа и листовые пластинки, соцветия, корни, пыльники, лигулы, палисадную ткань, мезофилл, эпидермис, ушки листа, верхнюю цветковую чешую, нижнюю цветковую чешую и боковые побеги.
Используемый в настоящем документе термин «эпихлое» относится к совокупной группе эндофитных грибов, состоящей из двух родов эндофитных грибов: представителей анаморф формального рода Neotyphodium и представителей телеоморф рода  .
.
Используемый в настоящем документе термин «эндофит эпихлое» относится к эндофиту группы «эпихлое», которая известна в данной области, или, как было показано в настоящем документе, формирует симбиотическую ассоциацию с растением-хозяином.
Используемый в настоящем документе термин «обеспечение по меньшей мере некоторого уровня защиты от вредителей» охватывает поддающееся измерению уменьшение степени, выраженности и/или продолжительности воздействия вредителя на растение Secale spp. Предпочтительно поддающееся измерению уменьшение - это статистически значимое уменьшение с Р-значением 0,05 или меньше.
Используемый в настоящем документе термин «уровень, достаточный для обеспечения защиты от вредителей» в отношении уровней алкалоидов означает любой уровень алкалоида, продуцируемого при симбиозе растения и эндофита, который является достаточным для обеспечения поддающегося измерению уменьшения степени, выраженности или продолжительности зараженности вредителями, заражения или негативного воздействия на растение-хозяина Secale spp., которое заражено эндофитным грибом согласно настоящему изобретению. Предпочтительно алкалоид представляет собой перамин или лолин, или производное лолина. Предпочтительно поддающееся измерению уменьшение - это статистически значимое уменьшение с Р-значением 0,05 или меньше.
Используемый в настоящем документе термин «статистически значимый» относится к вероятности того, что результат или взаимоотношение обусловлены чем-то, кроме случайности. С помощью проверки статистических гипотез, которые известны и используются в данной области техники, можно оценить статистическую значимость результата. При проверке статистических гипотез используют «Р-значение», которое, как известно в данной области техники, является вероятностью того, что результат измерений обусловлен только случайностью. В целом, в данной области техники принято, что уровни значимости 5% (0,05) или ниже считаются статистически значимыми.
Используемый в настоящем документе термин «повышенная защита от вредителей» относится к обеспечиваемому уровню защиты растения Secale spp. от вредителей в симбиотической ассоциации с эндофитным грибом эпихлое, при котором уменьшается степень, выраженность и/или продолжительность зараженности, заражения или негативного воздействия вредителей на растение, обусловленного наличием и/или активностью определенного вредителя по сравнению со степенью, выраженностью и/или продолжительностью заряженности, заражения и/или негативного воздействия на растение Secale spp. такого же вредителя в отсутствие эндофитного гриба (контрольное растение) и/или растения Secale spp., имеющего другой эндофитный гриб.
Используемые в настоящем документе термины «искусственное заражение» и «искусственная инокуляция» охватывают любую инокуляцию растения, в частности, растения Secale spp., в частности, Secale cereale, эндофитным грибом с образованием симбиотической ассоциации растения и гриба, которая не встречается в природе.
Используемый в настоящем документе термин «in planta» в контексте эндофитных грибов относится к эндофиту, который живет в симбиозе с растением-хозяином.
Используемый в настоящем документе термин «стабильный симбиоз растения и гриба» относится к симбиотической ассоциации, которая длится на протяжении жизненного цикла растения, причем у растения не проявляются внешние признаки заражения эндофитом. При «стабильной симбиотической комбинации» растение-хозяин заражено эндофитом в первом поколении и производит семена, которые при прорастании дают второе поколение растений-хозяев, которые также заражены эндофитом.
Используемый в настоящем документе термин «нормальный жизненный цикл» относится к нормальному репродуктивному циклу Secale, который включает рост первого поколения растения с производством семян, которые при прорастании дают второе поколение растение.
Используемый в настоящем документе термин «не проявляются внешние признаки заражения эндофитом» в отношении растения-хозяина, содержащего эндофитный гриб, означает, что растение-хозяин, по существу, характеризуется морфологически нормальным фенотипом, который известен в данной области техники для растения-хозяина. При этом используемый в настоящем документе термин «по существу, морфологически нормальный фенотип» растения-хозяина относится к данным по характерной морфологии растения-хозяина, которые известны и, в целом, приняты в данной области техники в отношении определенного растения-хозяина для определенного комплекс условий роста.
Используемый в настоящем документе термин «нормальный фенотип» растения-хозяина относится к характерной морфологии, показателям роста и другим фенотипическим признакам растения-хозяина, которые проявляются на протяжении жизненного цикла растения-хозяина, включая данные по репродуктивному циклу растения-хозяина и семенам растения-хозяина, которые известны и, в целом, приняты в данной области техники для определенного растения-хозяина в отсутствие эндофита.
Используемый в настоящем документе термин «атипичный фенотип» касательно растения-хозяина относится к данным по морфологии, показателям роста или другим фенотипическим признакам растения-хозяина на любой стадии жизненного цикла растения-хозяина, включая репродуктивный цикл растения-хозяина и семена растения-хозяина, которые отличаются от известных и, в целом, принятых в данной области техники и которые считаются характерными, или попадают в пределы наблюдаемых диапазонов, в целом, характерных для этого растения-хозяина. Используемый в настоящем документе термин «атипичный фенотип» касательно растения-хозяина может включать низкорослые растения или карликовые растения, или растения с визуально заметными внешними признаками заражения эндофитом или растения, которые не могут полноценно пройти нормальный репродуктивный цикл через стадию формирования семян, но не ограничивается ими.
Используемый в настоящем описании термин «предусматривающий» означает «состоящий по меньшей мере частично из». Согласно настоящему описанию при толковании утверждений, в которых использован этот термин, необходимо учесть обязательное наличие всех признаков, предваряемых этим термином в каждом утверждении, но также могут быть учтены другие признаки. Родственные термины, такие как «включающий» и «состоящий из», должны толковаться подобным образом.
Подразумевается, что ссылка на диапазон чисел, раскрытых в настоящем документе (например, от 1 до 10), также охватывает ссылку на все рациональные числа в пределах этого диапазона (например, 1, 1,1, 2, 3, 3,9, 4, 5, 6, 6,5, 7, 8, 9 и 10), а также любой диапазон рациональных чисел в пределах этого диапазона (например, от 2 до 8, от 1,5 до 5,5 и от 3,1 до 4,7) и, таким образом, в настоящем документе в явной форме раскрыты все поддиапазоны всех диапазонов, в явной форме раскрытых в настоящем документе. Это всего лишь конкретные примеры, и все возможные комбинации числовых значений между наиболее низким значением и наиболее высоким из приведенных значений, должны подобным образом рассматриваться, как установленные в явной форме в настоящей заявке.
Подробное раскрытие
Многие злаковые холодного сезона вегетации (Роасеае, подсемейство Pooideae) характеризуются наличием передающихся с семенами эндофитных грибов эпихлое, которые известны своими биозащитными свойствами и особенно продукцией алкалоидов, действие которых направлено против вредителей, как, например, лолины (Zhang et al., 2010) и перамин (Koulman et al., 2007). Грибы эпихлое (вид Neotyphodium), размножающиеся бесполым путем, главным образом или исключительно передаются вертикальным путем, тогда как в половых структурах (строма) родственных видов могут развиваться споры (аскоспоры), передаваемые горизонтальным путем (Zhang et al, 2010).
Большинство видов Neotyphodium считаются близкородственными видам рода . Многие виды Neotyphodium могли эволюционировать от по механизму, включающему межвидовую гибридизацию (Tsai et al., 1994). Согласно данным молекулярной филогенетики некоторые авторы считают, что размножающиеся бесполым путем виды Neotyphodium эволюционировали либо от отдельных видов , либо от гибридов, имеющих по меньшей мере два общих предка, принадлежащих виду (Tsai et al., 1994; Moon et al., 2004). В современной таксономии считается, что представители анаморфной формы рода Neotyphodium очень близкородственными представителями, принадлежащими роду телеоморфных грибов (Glenn et al., 1996). Согласно предыдущим кодексам ботанической номенклатуры к роду анаморфоных грибов относятся грибы, формирующие споры для бесполого размножения или размножающиеся вегетативным путем, а к роду телеоморфных грибов относятся грибы, размножающиеся половым путем. В настоящее время согласно кодексу ботанической номенклатуры одному присваивается одно название (Miller et al. 2011). В целом, два рода Neotyphodium и известны в данной области техники как эндофиты «эпихлое».
Симбиотические ассоциации между грибами эпихлое и травянистыми растениями-хозяевами широко распространены, и данные молекулярной филогенетики позволяют предположить, что видовая специфичность, наблюдаемая при этих симбиотических ассоциациях, обусловлена совместной эволюцией этих групп растений и эндофитных грибов (Schardl et al., 2008).
Никакие из современных культурных зерновых растений не заражены эндофитами эпихлое по природе, хотя такая возможность существует для некоторых родственных видов дикого типа (Marshall et al., 1999). Не имея намерения ограничиваться какой-либо теорией, авторы настоящего изобретения полагают, что на протяжении эволюция современных зерновых растений применение сельскохозяйственных методы, таких как хранение семян, могло привести к потере исторических ассоциаций, если они существовали (Welty et al., 1987).
Установление стабильного симбиоза растения и гриба между эндофитным грибом эпихлое и растением-хозяином, которое не является природным хозяином для гриба, является и проблематичным, и непредсказуемым (Simpson и Mace, 2012).
Считается, что при формировании такого симбиоза существует необходимость в успешной интеграции многих переменных параметров между партнерами, которые могут включать несовместимость на экологическом, биохимическом и/или молекулярном уровнях (Christensen et al., 2000). В настоящем описании подробно раскрыт большой объем исследовательской работы, включая обширную экспериментальную работу методом проб и ошибок, что было необходимо для разработки успешных протоколов и методик, с помощью которых были установлены стабильные симбиотические ассоциации между определенными штаммами эндофитного гриба эпихлое и растениями-хозяевами Secale spp., которые не являются природными хозяевами для таких грибов.
Неожиданно авторами настоящего изобретения было установлено, что для установления стабильного симбиоза растения и гриба между некоторыми эндофитными грибами эпихлое и растениями-хозяевами Secale spp. можно использовать искусственную инокуляцию. С использованием способов согласно настоящему изобретению, описанных в настоящем документе, авторы настоящего изобретения смогли получить зараженные растения-хозяева Secale spp., которые формируют стабильную симбиотическую ассоциацию с заражающим грибом, которая обеспечивает прохождение растением нормального жизненного цикла и/или приводит к тому, что у зараженного растения не проявляются внешние признаки заражения эндофитом. В дополнение к этому, авторы настоящего изобретения обнаружили, что установление таких симбиотических ассоциаций может оказывать по меньшей мере некоторое полезное действие на растение-хозяин с точки зрения продукции растением по меньшей мере одного лолинового алкалоида, производного лолинового алкалоида или перамина, или их комбинации, которая может обеспечивать по меньшей мере некоторый уровень защиты растения-хозяина от вредителей.
Авторы настоящего изобретения неожиданно установили, что определенные изоляты эндофитного гриба, взятые из диких родственных видов зерновых растений, подходят для установления стабильного симбиоза растения и гриба с Secale spp. растениями-хозяевами, в частности, Secale cereale. В частности, авторы настоящего изобретения установили симбиотические ассоциации, в результате которых получают комбинацию эндофитного гриба и растения-хозяина Secale spp., которая может обеспечивать по меньшей мере некоторый уровень повышенной защиты от вредителей по сравнению с контрольными растениями Secale spp.; т.е. такими же растениями Secale spp., которые не заражены таким же штаммом симбиотического гриба эпихлое. Не имея намерения ограничиваться какой-либо теорией, авторы настоящего изобретения полагают, что достижение повышенной защиты от вредителей должно ожидаться при комбинациях эндофитного гриба и растения-хозяина, что обеспечивает продукцию определенных алкалоидов, в частности, лолина, производных лолина и/или перамина. В соответствии с этим, авторы настоящего изобретения полагают, что продукция лолинового(ых) и/или пераминового(ых) алкалоида(ов) эндофитным грибом эпихлое или при комбинации эндофитного гриба/растения-хозяина обеспечивает по меньшей мере некоторый уровень повышенной защиты растения-хозяина от вредителей. В частности, согласно настоящему изобретению, исходя из использования эндофитных штаммов и способов, раскрытых в настоящем документе, авторы настоящего изобретения установили, растения-хозяева Secale spp., в частности, растения-хозяева Secale cereale, зараженные определенными штаммами эндофитного гриба эпихлое, обладают повышенной защитой против нематод по сравнению с контрольными растениями Secale.
В общих чертах симбиотические ассоциации, формируемые между растениями-хозяевами и их эндофитными грибами эпихлое, основаны на сложных и внутренних биологических взаимодействиях, которые приводят к высокой степени видовой специфичности, характерной и для эндофита, и хозяина (Simpson и Mace, 2012).
В результате продолжительной научно-исследовательской программы заявители настоящего изобретения идентифицировали в первый раз гриб эпихлое, способный формировать стабильный симбиоз растения и гриба с растениями-хозяевами Secale spp. Заявители настоящего изобретения дополнительно идентифицировали эндофит, который при симбиозе с растениями-хозяевами Secale spp., в частности, растениями-хозяевами Secale cereale, способный придавать зараженному растению-хозяину способность продуцировать один или несколько алкалоидов, которые, как известно, обеспечивают по меньшей мере некоторый уровень повышенной защиты растения от вредителей, по сравнению с незараженным контрольным растением. В частности, один или несколько алкалоидов могут представлять собой лолин, производные лолина или пераминовые алкалоиды.
В соответствии с этим, согласно одному аспекту настоящее изобретение относится к выделенному штамму эндофитного гриба эпихлое, выбранному из группы, состоящей из AR3039 (NRRL# 50716), AR3046 (NRRL# 50576), AR3049 (NRRL#50577), AR3050 (NRRL# 50578), AR3064 (NRRL# 50718), AR3067 (NRRL# 50719), AR3068 (NRRL# 50720), AR3073 (NRRL# 50721), AR3074 (NRRL# 50722), AR3076 (NRRL# 50723) и AR3078 (NRRL# 50724), и их комбинаций. Согласно одному варианту осуществления выделенный штамм является биологически чистым. Предпочтительно настоящее изобретение относится к выделенным штаммам AR3039, AR3046, AR3050, AR3067, AR3068, AR3074 и AR3078, предпочтительно AR3046.
Перечисленные выше штаммы эндофитных грибов были депонированы согласно Будапештскому договору для целей патентной процедуры 13 октября 2011 года в Департаменте сельского хозяйства, Службе сельскохозяйственных исследований среднего запада США, Национальном центре для исследований по применению сельскохозяйственной продукции с адресом 1815 North University Street, Пеория, Иллинойс, 61604-3902, США, что касается штаммов:
AR3046 (NRRL# 50576), AR3049 (NRRL#50577) и AR3050 (NRRL# 50578) и 6 марта 2012, что касается штаммов AR3039 (NRRL# 50716), AR3064 (NRRL# 50718), AR3067 (NRRL# 50719), AR3068 (NRRL# 50720), AR3073 (NRRL# 50721), AR3074 (NRRL# 50722), AR3076 (NRRL# 50723) и AR3078 (NRRL# 50724).
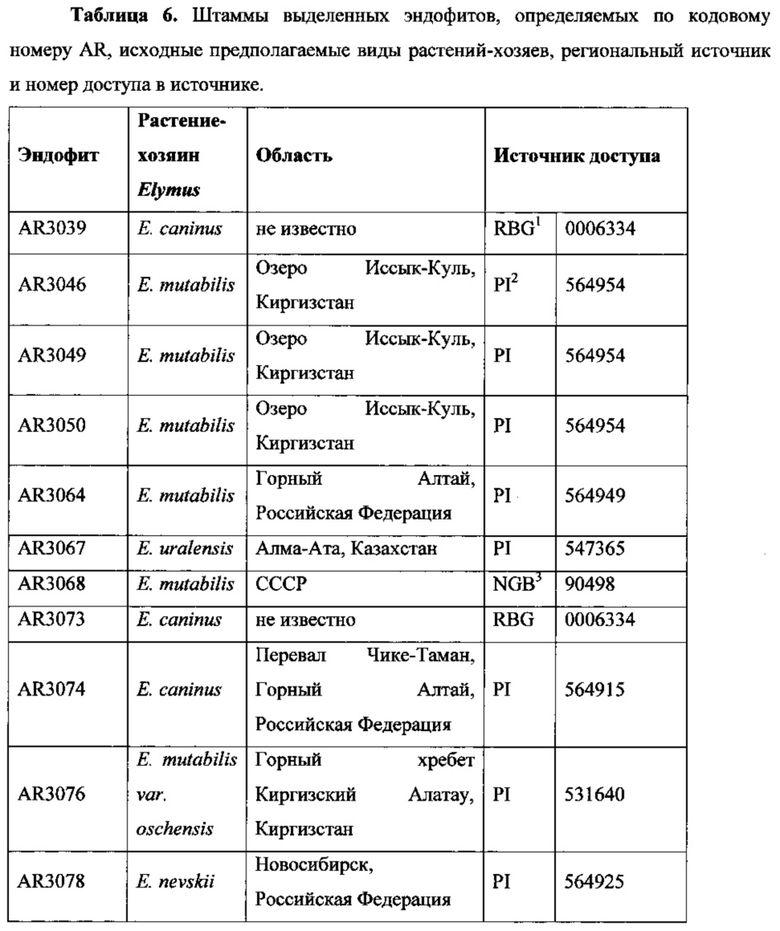
Эндофиты были вьщелены из Elymus spp., включая Ε. mutabilis, полученных из областей Киргизстана, Казахстана, СССР и Российской Федерации, как показано в Таблице 6.
Перечисленные выше эндофиты были выделены из зараженных эндофитами растений Elymus spp., включая Ε. mutabilis, после поверхностной стерилизации ткани растений, как было описано ранее (Christensen et al., 2002).
После выделения выделенный и/или биологически чистый эндофитный гриб можно культивировать с использованием стандартных методов, которые известны в данной области техники и которые раскрыты в настоящем документе, включительно в примерах.
Согласно одному варианту осуществления эндофитный гриб культивируют на антибиотикосодержащем картофельно-декстрозном агаре (ABPDA) при температуре от 20°С до 25°С, предпочтительно от 21°С до 23°С. Оптимальной температурой для роста эндофитного гриба является 22°С. Существует возможность роста эндофитного гриба при температурах выше или ниже этого диапазона, хотя рост может подавляться или может полностью прекратиться. Согласно одному варианту осуществления эндофитный гриб культивируют в темноте.
Настоящее изобретение также относится к способу защиты растения Secale spp. от вредителей, предусматривающий искусственное заражение растения Secale spp. по меньшей мере одним эндофитным грибом эпихлое, причем эндофитный гриб in planta обеспечивает продукцию достаточного количества по меньшей мере одного алкалоида для обеспечения по меньшей мере некоторого уровня защиты растения-хозяина от вредителей. Согласно одному варианту осуществления по меньшей мере один алкалоид представляет собой алкалоид, выбранный из группы, состоящей из перамина, N-ацетилнорлолина, лолина, N-формиллолина, N-ацетиллолина и N-метиллолина.
Инокуляцию можно осуществить с использованием проростков Secale spp., культивированных приблизительно в течение двух недель. Предпочтительно проростки культивируют от 4 до 9 суток.
Вне этого диапазона эффективность формирования этими проростками ассоциаций с грибами все еще высока, но в некоторых случаях они могут быть недозрелыми или перезрелыми для формирования симбиоза с эндофитным грибом. Необходимо, чтобы семена не содержали нецелевых грибов и бактерий во избежание контаминации проростков микроорганизмами.
Согласно одному варианту осуществления искусственную инокуляцию можно осуществить с использованием основной инокуляции проростков Secale spp. Для эффективного установления ассоциации между грибом-симбионтом и растением-хозяином Secale spp. эндофит необходимо инокулировать в меристему растения-хозяина путем разрезания ткани растения и введения мицелия культивированного гриба.
Специалистам в данной области техники известно, что естественная устойчивость и защита травянистых злаковых растений-хозяев от вредителей обеспечивается эндофитами эпихлое, живущими в симбиозе с растениями, и что эта комбинация может обеспечивать некоторую защиту от вредителей. В частности, известно, что лолиновые алкалоиды и алкалоид перамин обеспечивают некоторый уровень такой защиты в отсутствие заметной или известной токсичности для млекопитающих или людей, употребляющих злаки, или в отсутствие токсичности продуктов, образующихся непосредственно после употребления злаков.
Долины представляют собой группу родственных биоактивных природных веществ, которые обладают различными химическими и биологическими свойствами. Полины представляют собой алкалоиды, т.е. органические соединения содержащие атомы азотистых оснований и с химической точки зрения относятся к насыщенным 1-аминопирролизидинам с внутренним эфирным мостиком, соединяющим два углеводородных кольца (от С-2 до С-7). Внутренний эфирный мостик, который не характерен для органических соединений, считается отличительной особенностью этой группы соединений. Конкретные примеры лолинов включают норлолин, причем к его производным по первой аминогруппе можно отнести лолин (с метальной группой), N-метиллолин (с двумя метальными группами, NML), N-ацетилнорлолин (с ацетильной группой, NANL), N-ацеталлолин (с метальной группой и ацетильной группой, NAL) и N-формиллолин (с формильной группой, NFL) (Schardl et al., 2007; Schardl et al., 2012).
В целом, известно, что долины относятся к обладающим пестацидным и сдерживающим эффектом соединениям, продуцируемым злаковыми травами, зараженными эндофитными грибами-симбионтами эпихлое ( / Neotyphodium spp.). Было показано, что лолины повышают устойчивость злаковых растений-хозяев от поедания вредителями (Bush et al., 1997). Биологическое действие определенных лолинов, направленное против определенных вредителей, может в некоторой степени варьировать. Также предполагалось, наличие лолинов может обеспечивать некоторый уровень защиты растений-хозяев от стрессорных факторов окружающей среды, включая засуху и конкуренцию за пространство (Malinowski и Belesky, 2000).
Лолиновые алкалоиды могут продуцироваться при симбиотической комбинации либо эндофитным грибом, либо растением-хозяином. Важное значение имеет обеспечиваемая такой комбинацией продукция лолиновых алкалоидов, причем эта продукция обусловлена комбинацией в результате наличия эндофитного гриба на растении или внутри тканей растения, в частности, в результате наличия грибных гиф между растительными клетками. Исторически было обнаружено, что воспроизведение условий, характерных для симбиоза для обеспечения продукции лолиновых алкалоидов in vitro является чрезвычайно трудной задачей (Porter 1994). Таким образом, до относительно недавнего времени не было известно, продуцируются ли лолиновые алкалоиды в результате образования этих симбиотических ассоциаций эндофитным грибом самим по себе, или они синтезируются в растении в ответ на заражение. Только относительно недавно в работе Blankenship et al. (2001) было продемонстрировано, что эндофит Neotyphodium uncinatum может продуцировать лолины в ростовой среде с определенным химическим составом. Эта работа дает основания предполагать, что эндофит также является продуцентом лолинов при симбиозе с его природными травянистыми растениями-хозяевами (Blankenship et al., 2001). С помощью прямого химического анализа встречающегося в природе эпихлое также был продемонстрирован этот эффект (Schardl et al., 2007).
Перамин (пирролопиразиновый алкалоид) представляет собой биоактивный алкалоид, продуцируемый в результате формирования некоторых комбинаций эндофитов и растений (Schardl et al., 2012). Было показано, что продукция перамина зависит от функционирования по меньшей мере одного гена, принадлежащего эндофиту (Tanaka et al., 2005). Было показано, что перамин сдерживает поедание растений некоторыми насекомыми, которые вызывают повреждение растений, и может обеспечивать защиту от поражения зараженных эндофитами растений некоторыми насекомыми (Rowan и Latch, 1994).
Настоящее изобретение дополнительно относится к способу получения стабильной комбинации растения-хозяина и эндофитного гриба эпихлое, предусматривающему искусственное заражение растения Secale spp. по меньшей мере одним эндофитным грибом, который формирует стабильную комбинацию с инокулированным растением, причем у растения-хозяина не проявляются внешние признаки заражения эндофитом. Это означает, что после установления комбинации зараженное растение-хозяин Secale spp. имеет морфологически нормальный фенотип, характерный для растений Secale spp., который известен в данной области и который предположительно будет получен с учетом условий роста, в которых была обнаружена такая комбинация.
В определенных условиях эндофитные грибы, которые являются облигатными симбионтами одного вида или штамма растения-хозяина, могут быть введены в растение-хозяин другого вида или штамма, с формированием комбинации, которая обычно не встречается в природе. Однако такая комбинации может быть нестабильной и может приводить к образованию растений-хозяев, имеющих атипичный фенотип, т.е., атипичные морфологические и/или физиологические признаки по сравнению с растениями-хозяевами того же штамма или вида, которые либо не подвергались заражению, либо которые содержат природный симбионт. Атипичные фенотипические признаки могут включать карликовые растения (Simpson и Mace, 2012), растения с видимым ростом эпифитов (Christensen et al., 2012), колонизацию сосудистых пучков (Christensen et al., 2001) и локализованную гибель клеток (Christensen, 1995).
Заявитель патента впервые получил стабильную симбиотическую комбинацию между растениями-хозяевами Secale spp. и эндофитными грибами эпихлое, что приводит к образованию стабильной комбинации растения и гриба, которая, как показано, не оказывает атипичное действие в результате заражения эндофитом. Стабильная симбиотическая комбинация, к которой относится настоящее изобретение, может характеризоваться морфологически нормальным фенотипом и полноценным, и нормальным репродуктивным циклом.
Согласно одному варианту осуществления формирование стабильной симбиотической комбинации является достаточным условием обеспечения вертикальной передачи эндофита. Согласно одному варианту осуществления вертикальная передача осуществляется через цветочные побеги и впоследствии через образующиеся семена. Согласно одному варианту осуществления вертикальная передача эндофита осуществляется от первого поколения второму поколению растений-хозяев. Предпочтительно вертикальная передача от первого поколения растения-хозяина второму поколению растения-хозяина осуществляется через семена.
Согласно одному варианту осуществления растение-хозяин представляет собой растение Secale spp., выбранное из группы, состоящей из Secale cereale, Secale montanum, Secale strictum, Secale sylvestre и Secale vavilovii. Предпочтительно растение-хозяин представляет собой Secale cereale.
Настоящее изобретение дополнительно относится к эндофитам, способным при заражении обеспечивать возможность продукции растением-хозяином Secale spp. в составе комбинации эндофита и растения продуцировать один или несколько алкалоидов из лолина, производных лолина и/или пераминовых алкалоидов.
Настоящее изобретение дополнительно относится к семени растения Secale spp., которое заражено по меньшей мере одним эндофитным грибом эпихлое. Предпочтительно эндофит эпихлое выделен из Elymus spp. Предпочтительно эндофитный гриб выделен из Elymus mutabilis. Предпочтительно эндофит эпихлое выбран из группы, состоящей из AR3039 (NRRL# 50716), AR3046 (NRRL# 50576), AR3049 (NRRL#50577), AR3050 (NRRL# 50578), AR3064 (NRRL# 50718), AR3067 (NRRL# 50719), AR3068 (NRRL# 50720), AR3073 (NRRL# 50721), AR3074 (NRRL# 50722), AR3076 (NRRL# 50723) и AR3078 (NRRL# 50724), и их комбинаций. Предпочтительно настоящее изобретение относится к выделенным штаммам AR3039, AR3046, AR3050, AR3067, AR3068, AR3074 и AR3078, предпочтительно AR3046.
Предпочтительно Secale spp. семя представляет собой семя растения Secale cereale, Secale montanum, Secale strictum, Secale sylvestre или Secale vavilovii. Более предпочтительно семя представляет собой семя растения Secale cereale.
Различные аспекты настоящего изобретения далее будут проиллюстрированы в неограничивающих примерах посредством ссылки.
ПРИМЕРЫ
ПРИМЕР 1
Определение штаммов эндофитных грибов
Свыше 580 образцов семян растений Elymus spp. были получены из различных источников и, если позволяло количество семян, вплоть до приблизительно 50 отдельных семян или проростков исследовали на предмет заражения эндофитом. Жизнеспособность эндофитов, содержащихся во влагалище листа проростков, выращенных до стадии двух или более боковых побегов, определяли по методу Simpson et al. (2012). Было показано, что приблизительно 6% образцов были способны давать по меньшей мере один проросток, содержащий жизнеспособный эндофит, что было дополнительно исследовано в рамках следующих примеров.
ПРИМЕР 2
Определение генетических вариантов штаммов эндофитных грибов
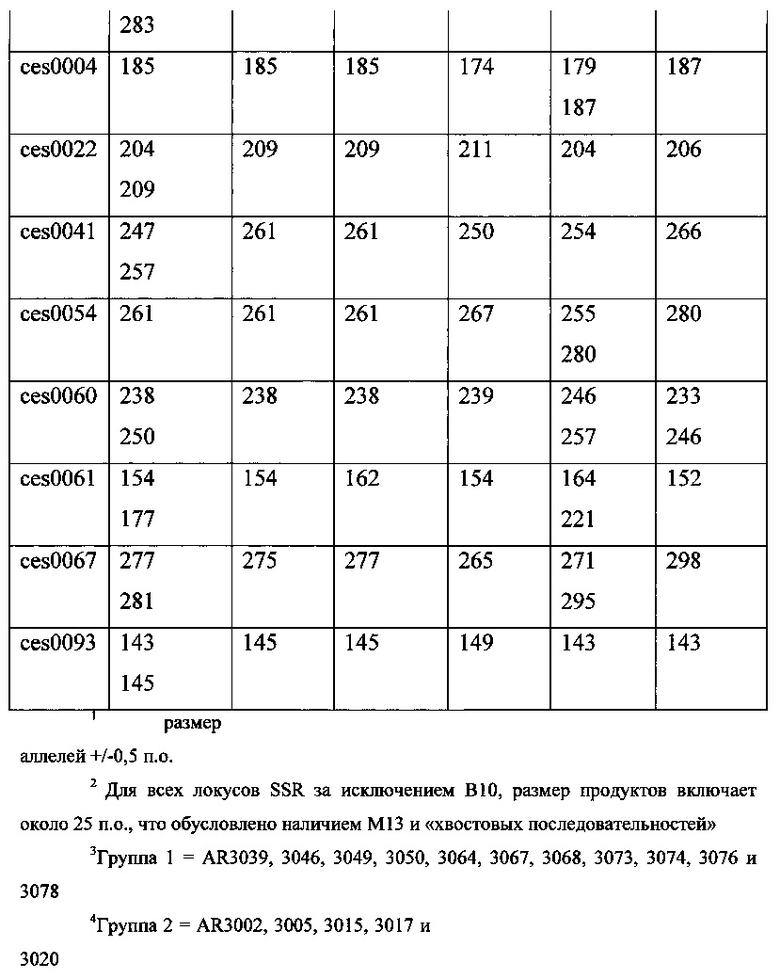
Для того чтобы сосредоточиться на выборе эндофитных штаммов из тех Elymus spp., содержащих жизнеспособный эндофит, который представлял дополнительный интерес, были частично охарактеризованы эндофиты вплоть до 6 растений каждого образца и дифференцированы в отношении генетической изменчивости с помощью «фингерпринтинга» ДНК, исходя из данных генотипирования, полученных от вплоть до 8 выбранных маркерных локусов простых повторяющихся последовательностей (SSR) с использованием последовательностей праймеров из Таблицы 1. Эти последовательности праймеров, как было ранее показано, в целом, принимают участие в амплификации полиморфных последовательностей ДНК эндофита эпихлое с момента, когда эндофиты находятся in planta.
Образцы основного побега с массой сырой ткани приблизительно 100 мг использовали для выделения общей геномной ДНК (растение + эндофит) после процедуры выделения ДНК растений с помощью набора FastDNA согласно рекомендациям производителя (Bio 101, Виста, Калифорния).
Амплификацию SSR проводили с использованием пар олигонуклеотидных праймеров с использованием одного из двух протоколов полимеразной цепной реакции (ПЦР) (Таблица 1). Согласно обоим протоколам ПЦР проводили с использованием термоциклера iCycler (BioRad, Геркулес, Калифорния, США).
Использовали протокол 1, который был описан в следующем источнике (Moon et al., 1999), за исключением того, что была использована температура отжига 60°С. Согласно этому протоколу прямые праймеры были мечены на 5'-конце флуорофором 6-FAM™ (Applied Biosystems, Foster City, Калифорния).
Согласно протоколу 2 прямые праймеры синтезировали с использованием хвостовой последовательности М13 длиной 21 нуклеотид на 5'-конце (5'-TGTAAAACGACGGCCAGT-3') (SEQ ID NO: 1), для облегчения общего мечения ПЦР-продуктов с использованием 6-РАМ™-меченого М13 праймера (Schuelke, 2000). Обратные праймеры синтезировали с использованием последовательности 5'-GTTTCTT-3' (SEQ ID NO: 2) на 5'-конце для обеспечения для обеспечения нематричного аденилирования на 3'-конце ПЦР-продукта (Brownstein et al., 1996). Был использован объем реакционной смеси для ПЦР 10 мкл, содержащей приблизительно 10 нг общей геномной ДНК, 2,5 мМ хлорида магния, 1х Буфер для ПЦР, 0,05 мМ каждого дНТФ, 0,0375 мкл прямого праймера, 0,15 мкл обратного праймера, 0,15 мкл флуоресцентно меченого праймера М13 и 0,75 Е ДНК-полимеразы Platinum Taq (Invitrogen, Карлсбад, Калифорния). ПЦР проводили с использованием следующего протокола: (1) 94°С в течение 4:00 минут, (2) 30 циклов: 94°С в течение 30 секунд, 55°С в течение 30 секунд и 72°С в течение 30 секунд, (3) 8 циклов: 94°С в течение 30 секунд, 53°С в течение 30 секунд и 72°С в течение 30 секунд, (4) 72°С в течение 30 минут (согласно Schuelke 2000).
ПЦР-продукты анализировали с помощью капиллярного электрофореза на анализаторе генов ABI 3100 с использованием 22 см набора капилляров из полимера РОР-7™ (Applied Biosystems). GS500 LIZ (Applied Biosystems) использовали в качестве внутреннего стандарта. Электроферограммы анализировали с использованием ABI Prism GeneScan (v 3.7, Applied Biosystems), и данные генотипировнаия оценивали с использованием ABI Prism Genotyper (v 3.7, Applied Biosystems).
Растения, исследованные, как описано выше, затем дополнительно были охарактеризованы путем осуществления химических анализов. Шесть зараженных проростков дополнительно исследовали на наличие алкалоидов, связанных с наличием эндофитов, таких как индольные дитерпены, алкалоиды спорыньи, перамин и лолины. Представители каждого генотипа грибов были отобраны для более подробного генотипирования с использованием праймеров, подобранных для полиморфных локусов SSR.
Результаты в таблице 2 и на фигуре 2 показывают, что эндофиты согласно настоящему изобретению относятся к одной кладе, обозначенной Группа 1 в таблице 2, и могут быть охарактеризованы как имеющие по меньшей мере большинство аллелей SSR одинакового размера для локусов SSR, приведенных в таблице 1 и таблице 2. Другие эндофиты из таблицы 2, таблицы 3 и на фигуре 2 приведены с целью иллюстрации частоты встречаемости и взаимоотношений разных типов эндофитов, которые не соответствовали требованиям настоящего изобретения.



ПРИМЕР 3
Выделение штаммов эндофитных грибов
Грибы были выделены из большого числа зараженных эндофитами растений после поверхностной стерилизации ткани растений, в целом, как известно в данной области техники, в частности, как описано Christensen et al. (2002). Боковые побеги отделяли от растений путем отрезания от основания и нарезания до приблизительно 5 см до поверхностной стерилизации. Нарезанные на части боковые побеги подвергали поверхностной стерилизации путем быстрой промывки 96% этанолом и пропитки в течение 1 минуты в 10% раствора отбеливающего средства с последующим промыванием два раза стерильной водой. Боковые побеги нарезали на части в поперечном направлении; оделяли кольца листовых влагалищ и помещали на картофельно-декстрозный агар, содержащий 5 мкг/мл тетрациклина (ABPDA). Чашки Петри инкубировали в темноте при 22-25°С в течение 3-5 недель. Культуры можно субкультивировать на той же самой среде.
Культуры исследовали на предмет скорости роста колоний, морфологии колоний, способности формировать конидии, диапазона размеров конидий, последовательности генов β-тубулина (tub2) (Moon et al. 2004) и других отличительных признаков, все из которых были учтены для селекции эндофитов с целью дополнительного изучения.
Выбранные культуры, полученные и иногда субкультивированные аналогично этому примеру, использовали для исследования инокуляции и возможного перенесения заражения проростками растений Secale cereale, как описано ниже.
ПРИМЕР 4
Характеристики эндофитов
Характеристики при выращивании in vitro на PDA были сопоставимыми с характеристиками гриба Neotyphodium (Christensen et al., 1993; Glenn et al., 1996), который характеризуется медленным или умеренно медленным ростом, что изменяется через 4 недели выращивания на PDA. Колонии, выделенные из агара, были белого цвета, опушенными, слегка или сильно извитыми, с войлочной текстурой, с большим количеством гиф воздушного мицелия. Края колоний имели коричневый или кремовый цвет. Клетки-конидиеносцы были одиночными, располагались перпендикулярно гифам, были более широкими у основания и сужающимися к верхушке. Фиалиды были прозрачными, гладкими, ладьевидными или в форме полумесяца, длиной 2,05-14,96 мкм × шириной 1,37-8,19 мкл. Ни один из изолятов не был стерильным. Отдельные характеристики для каждого штамма приведены в Таблице 3.


ПРИМЕР 5
Применение последовательности гена β-тубулина для определения ваимоотношений между эндофитами и известными видами
К эндофитным грибам злаковых трав можно отнести виды или виды Neotyphodium. Некоторые, но не все виды Neotyphodium несомненно являются гибридами двух или нескольких видов , которые описаны главным образом путем сравнения выбранных последовательностей генов (Moon et al., 2004).
Наличие многих аллелей для некоторых локусов, использованных для анализа SSR эндофитов (как в приведенном выше ПРИМЕРЕ 2), могут свидетельствовать о том, что клады, к которым они относятся, по-видимому, имеют гибридное происхождение.
Последовательности генов tub2 трех примеров эндофитов, AR3039, AR3046 и AR3064, согласно настоящей заявке были исследованы с использованием незначительно измененного метода Moon et al. (2004) для определения вероятности происхождения эндофитов от одного или нескольких установленных видов . Подобным образом также были исследованы четыре других эндофита из явно различающихся клад для установления их взаимоотношений с известными видами .
Праймеры и условия, использованные для ПЦР-амплификаций, были модифицированы таким образом, чтобы повысить возможность определения многих аллелей tub2 во всех образцах, для которых гибридное происхождение считается вероятным или возможным. Были выбраны следующие последовательности праймеров: прямой праймер TCG GCC ТСА CGA CGC АСА AC (RJ251-F) (SEQ ID NO: 29) и обратный праймер ССС ATA CAT ТАС АСС ТТТ CTG GCG (RJ252-R) (SEQ ID NO: 30) таким образом, чтобы получить ПЦР-продукты генов эндофитов из Elymus spp., соответствующие, по существу, и включающие соответствующие описанные в литературе последовательности и используемые для определения взаимоотношений между эндофитами и Neotyphodium и их гибридами (Moon et al., 2004). ПЦР проводили с использованием первоначального этапа 95° (3 минуты); 35 циклов 95° (30 секунд), 62° (30 секунд), 72° (45 секунд); конечный этап 72° (5 минут).
ПЦР-продукты секвенировали по методу Сенгера (1997) на анализаторе ДНК ABI PRISM 3700 (Applied Biosystems). Были принято, что последовательности, для которых были получены многие перекрывающиеся пики, имеют гибридное происхождение, тогда как последовательности, для которых не были получены такие пики, имеют не гибридное происхождение.
Клонирование ПЦР-ампликонов, когда было установлено несколько последовательностей, осуществляли с использованием набора TOPO® ТА Cloning® (с использованием вектора pCR®2.1-TOPO®) (Invitrogen). Секвенировали вплоть до 10 клонов, которые описаны выше, и выравнивали с использованием модуля Align X из Vector NTI Advance 11 (Invitrogen), с последовательностями отдельных аллелей tub2.
Отдельные аллели tub1 были отнесены к наиболее близкородственным негибридным видам , как указано далее: последовательности выравнивали с использованием Dialign-TX (Subramanian et al., 2008) и составляли древовидные схемы максимальной вероятности, исходя из данных этого выравнивания, с использованием пакета программ Phylip (Felsenstein, 2005). Для составления окончательной дендрограммы выполняли программу DNAML с использованием 1000 начальных загрузок и трех этапов рандомизации порядка ввода последовательностей на набор, и сокращали до получения одного консенсусного древа с использованием подхода по принципу большинства. Дендрограмму получали с использованием программы Dendroscope (Huson et al., 2007) для установления наиболее близкородственных связей между известными видами с выбранными эндофитами.
Были установлены две последовательности эндофитов AR3039, AR3046 и AR3064 согласно настоящей заявке tub2, свидетельствующие о том, что эти эндофиты являются гибридами Е. bromicola и Е. amarillans. До настоящего времени не было сообщений об этом гибриде, и его можно считать новым видом, который обозначен Е. bromicola x amarillans.
Согласно анализу SSR эндофитные штаммы AR3049, AR3050, AR3067, AR3068, AR3073, AR3074 и AR3078 тесно сочетались с AR3039, AR3046 и AR3064 (Фигуре 1), и весьма вероятно их также можно отнести к Е. bromicola x amarillans. Трое из других эндофитов по данным сравнительных исследований сочетались только с Е. bromicola и один другой эндофит сочетался с Е. yangzii, причем это было характерно для других групп эндофитов с общими наборами генотипов SSR, наблюдаемых для источников Elymus spp.
В целом, данные по SSR и β-тубулину позволяют предположить, что эндофитные штаммы AR3039, AR3046, AR3049, AR3050, AR3064, AR3067, AR3068, AR3073, AR3074 и AR3078 формируют характерную группу из вновь описанных эндофитов, которая может считаться функциональной кладой видов грибов, которые могли быть перенесены от их природного растения-хозяина к промежуточному растению-хозяину Secale в результате искусственно созданной стабильной симбиотической комбинации растения Secale и эндофита, при этом зараженное растение-хозяин Secale характеризуется нормальным ростом и нормальным жизненным циклом.
ПРИМЕР 6
Инокуляция эндофитных грибов эпихлое в ткани злаковых растений Secale
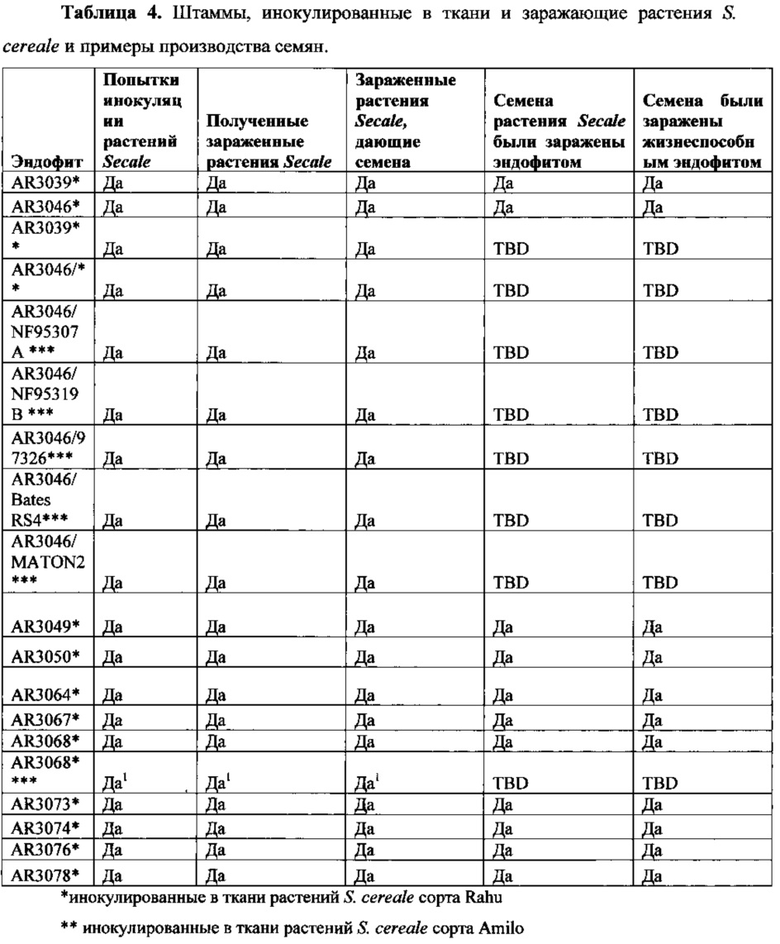
Семена Secale cereale, культиваров Rahu, Amilo и «KWS», и коллекции культиваров S. cereale, полученных от Др. Марка Ньюэлла, Нобелевский фонд Сэмюеля Робертса (NF95307A, NF95319B, 97326, Bates RS4 или MATON2), подвергали поверхностной стерилизации и инокулировали, как описано Latch и Christensen (1985). Семена подвергали поверхностной стерилизации путем погружения в 50%-ный раствор серной кислоты на 15 минут с последующим промыванием пять раз водопроводной водой и погружением в 10%-ный раствора бытового отбеливающего средства (Janola) на 15 минут с последующим промыванием два раза стерильной водой. Семена высушивали в шкафу с ламинарным потоком воздуха на стерильной Ватманской фильтровальной бумаге до размещения на чашки Петри с 4%-ным водным раствором агара. Семена на чашках оставляли прорастать в темноте при 22-25°С в течение 4-9 суток, и полученные в результате этиолированные проростки инокулировали до возвращения в темноту в инкубатор на 7 суток. После этой инкубации чашки помещали под флуоресцентный источник света видимой части спектра по меньшей мере на 7 суток до извлечения проростков и высаживания их в коммерчески доступную почвенную смесь и выращивания их в теплице. Растения выращивали в течение около 6 недель до идентификации зараженных вариантов. Зараженные растения идентифицировали по методу Simpson et al. (2012). Растения дополнительно выращивали в теплице для изучения фенотипа зараженного растения в сравнении с характерным незараженным растением и, в частности, для определения возможности формирования соцветий и семенных коробочек.
Представленные в кратком виде результаты успешной инокуляции обозначены словом «да» включены в Таблицу 4 для этих эндофитных штаммов, если по меньшей мере некоторые из инокулированных растений характеризовались, по существу, нормальным фенотипом и были способны пройти нормальный жизненный цикл (Фигура 2). Собирали семена растений, которые указаны в Таблице 4.

ПРИМЕР 7
Продукция алкалоидов в зараженных эндофитным грибом природных материнских растениях
Было проанализировано содержание лолиновых алкалоидов с помощью принятых методов с незначительной модификацией (Kennedy и Bush, 1983; Yates et al., 1989) и перамина (Rasmussen et al., 2012) в листовых пластинках и псевдостеблях растений Elymus mutabilis, Elymus caninus, Elymus uralensis, Elymus nevskii и Elymus mutabilis var. oschensis, зараженных определенными эндофитными штаммами. Результаты показаны в Таблице 5. Общее количество лолиновых алкалоидов было большим в листьях исследованных растений, чем в псевдостеблях. Не имея намерения ограничиваться какой-либо теорией, авторы настоящего изобретения полагают, что общее количество алкалоидов, присутствующих в исследованных растениях, соответствует диапазону, достаточному для обеспечения по меньшей мере некоторого уровня защиты растения-хозяина от вредителей (Wilkinson et al., 2000). Эти результаты и данные по β-тубулину, представленные в других местах в настоящем документе, полученные in planta, показывают, что ряд эндофитных штаммов в составе клады, определяемой по SSR, может придавать симбиотической комбинации способность продуцировать измеримое количество лолиновых алкалоидов или перамина, или обоих алкалоидов.


ПРИМЕР 8
Источник и географическое происхождение эндофитов, выбранных из Elymus spp.
В Таблице 6 перечислен ряд выделенных эндофитных штаммов, раскрытых в настоящем документе, номера доступа в источнике, из которого они были получены, предполагаемые виды исходного растения-хозяина и региональный источник доступа. В основном, растения-хозяева Elymus spp. были получены из Центральной Азии.

ПРИМЕР 9
Продукция алкалоидов в растениях Secale cereale
Проростки Secale cereale культивара Rahu инокулировали эндофитами, которые перечислены в таблице 7, выращивали в теплице, и проверяли наличие эндофитов и фенотипически нормальных растений. Содержание лолиновых алкалоидов (Kennedy и Bush, 1983; Yates et al., 1989) или перамина (Garthwaite et al., 1994) анализировали отдельно в листовых пластинках и стеблях. Результаты, показанные в Таблице 7, относятся к диапазонам концентраций и для лолиновых алкалоидов, и перамина. Не имея намерения ограничиваться какой-либо теорией, авторы настоящего изобретения полагают, что общее количество лолиновых алкалоидов и перамина, присутствующих в исследованных растениях, соответствует диапазону, достаточному для обеспечения по меньшей мере некоторого уровня защиты растения-хозяина от вредителей (Rowan 1993; Wilkinson et al., 2000; Bacetty et al., 2009a 2009b).

ПРИМЕР 10
Комбинации Elymus mutabilis и эндофитов, обладающие биологической активностью против вредителей зерновых культур
В отсутствие контрольных растений Е. mutabilis, не содержащих эндофитов, была использована овсяница луговая в присутствии и в отсутствии ее природного эндофита (Neotyphodium uncinatum), поскольку известно, что он продуцирует лолиновые алкалоиды и обладает активностью против тлей.
Влияние на тлей
Тля Rhopalosiphum padi является основным вредителем зерновых растений, поскольку она является переносчиком вируса желтой карликовости ячменя.
В биотесте пищевого предпочтения, проведенном с использованием боковых побегов в чашках Петри, количество тлей Rhopalosiphum padi на Elymus mutabilis, зараженном AR3050, было аналогичным количеству тлей на овсянице луговой, зараженной ее природным эндофитом Neotyphodium uncinatum, и статистически значимо меньшим, чем количество тлей на контрольном растении - овсянице луговой, не содержащей эндофита (Таблица 8).

Результаты этого испытания дают основания предполагать, что лолиновые алкалоиды, продуцируемые в результате симбиотической ассоциации, формируемой между Е. mutabilis и AR3050, могут сдерживать размножение питающихся на растениях тлей, как видно в случае с луговыми травами (Wilkinson et al., 2000).
Влияние на клещей Aceria
Aceria spp., в особенности A. tosichella, представляют собой клещей, которые являются переносчиками вируса полосатой мозаики пшеницы в Австралии. Вид клещей, использованный в этих испытаниях, был определен как Aceria spp., предположительно A. tosichella.
Из-за отсутствия контрольных растений Е. mutabilis без эндофита и неизвестного влияния генотипа растений на встречаемость клещей Aceria влияние лолиновых алкалоидов на клещей оценивали на шести зараженных эндофитами и шести не содержащих эндофитов растениях овсяницы луговой. Количество клещей Aceria подсчитывали на трех боковых побегах каждого растения. Статистически значимо большее количество клещей встречалось на не содержащих эндофитов растениях, чем на зараженных эндофитами растениях (Число/боковой побег: 74 для Е+ cf 454 для Е-. Р<0,001). Результаты приведены ниже в Таблице 9.
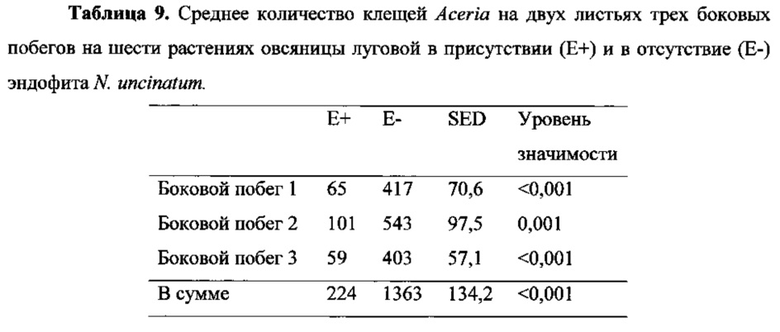
Как показано в описанном выше испытании, результаты в этом испытании с клещами предполагают, что лолиновые алкалоиды отвечают за отпугивание питающихся на растениях клещей, аналогично влиянию на тлей, что было показано для луговых трав (Wilkinson et al., 2000).
Влияние на светло-коричневую яблонную моль
Известно, что это насекомое обладает чувствительностью к биологически активным веществам, продуцируемым эндофитами плевела. В настоящей работе его использовали, чтобы показать присутствие биологически активных веществ в растении Е. mutabilis, зараженном AR3046.
Было установлено выраженное влияние AR3046, когда растительный материал Е. mutabilis, взятый из растений, зараженных AR3046, был включен в искусственный корм и скормлен личинкам светло-коричневой яблонной моли. Среднее количество личинок в процентах, которые устойчиво употребляли корм, и личинок, которые начали употреблять корм в пределах первых 24 часов после получения новорожденными личинками исследуемого корма, составляло: 88% для среднего для трех других видов Elymus, незараженных эндофитами, по сравнению с 4% для личинок, корм которых содержал зараженное AR3046 растение Е. mutabilis. К 10-м суткам испытания 60% личинок, питающихся не содержащим эндофитов растительным материалом, прошли первую личиночную стадию по сравнению только с 13% личинок, питающихся AR3046, прошедшими первую личиночную стадию. Результаты этого испытания приведены ниже в Таблице 10.

Результаты в этом испытании с молью явно предполагают, что AR3046 продуцирует лолиновые алкалоиды, которые отвечают за сдерживание личинок питающейся растительным соком светло-коричневой яблонной моли.
ПРИМЕР 11
Защита зараженных эндофитом растений Secale от вредителей
Был проведен эксперимент для исследования способности эндофита обеспечивать защиту Secale от корневых нематод (Pratylenchus spp.) с использованием боковых побегов культивара Rahu растения Secale cereale в присутствии или в отсутствие эндофита AR3046. Одиннадцать отдельных боковых побегов растений Е+ и E-Rahu, выбранных из 4 независимых материнских растений на воздействие (2-3 клона на материнское растение), пересаживали в глубокие контейнеры для стимуляции роста корней, размером 5×5×12 см, содержащие 100 г полевой почвы, характеризующейся природным заражением 30 Pratylenchus spp. на 100 г. Дополнительные 30 личинок разведенного в лабораторных условиях Pratylenchus penetrans добавляли в каждый контейнер для стимуляции роста корней, таким образом, чтобы обеспечить доступ 60 нематодам к каждому растению. Растения инкубировали в освещенных шкафах для роста при 20°С в течение 30 суток. Затем растения вынимали из почвы, корни промывали, затем очищали с использованием 1,5%-ного раствора хлорида натрия в течение 3 минут. Присутствующие в пределах корневой системы нематоды были окрашены с использованием раствора анилинового синего в глицерине. Корни затем исследовали под микроскопом, и подсчитывали число нематод в пределах корневых систем. Заражение эндофитом не влияло на массу корней.
Наличие эндофита AR3046 приводило к статистически значимому уменьшению (Р<0,05) числа нематод на корневую систему (Таблица 11).
Как показано в описанных выше исследованиях сдерживающего эффекта, оказываемого на насекомых, результаты этого исследования с нематодами дают веские основания предполагать, что лолиновые алкалоиды, продуцируемые AR3046, отвечают за обеспечение по меньшей мере некоторого уровня защиты зараженных растений Secale от вредителей путем сдерживания колонизации нематодами корней зараженных эндофитами растений Secale. Это согласуется с данными по лолинам, которые, как показано, оказывают сдерживающий и пестицидный эффект на нематод (Bacetty et al., 2009а 2009b).

ПРИМЕР 12
Влияние заражения сорта «Rahu» растения Secale cereale эндофитом AR3046 на Cerodontha australis
Злаковые травы и зерновые растения в Новой Зеландии и Австралии часто поражены личинками мухи-минера листьев, Cerodontha australis (Diptera: Agromyzidae), также известной под названием минер влагалища листа пшеницы. Личинки этой мухи питаются внутренними тканями целого ряда злаковых трав и зерновых растений, вызывая повреждение, которые могут приводить к разрушению боковых побегов. Мухи-минеры оставляют видимую бороздку в листьях.
В этом эксперименте растения S. cereale сорта Rahu, зараженные продуцирующим лолины эндофитом AR3046, или незараженные (Nil) растения, заражали С australis. Растения овсяницы луговой, зараженные ее природным эндофитом Neotyphodium uncinatum, были включены в эксперимент, поскольку этот эндофит также продуцирует лолиновые алкалоиды.
Способ
Сравнивали шесть растений Secale каждого сорта «Rahu», зараженных AR3046, и растений овсяницы луговой, зараженных N. uncinatum, с их не содержащими эндофитов вариантами (Nil) в отношении оказываемого ими влияния на заражение С australis. Растения случайным образом разделяли на параллельные группы, состоящие из одного растения на каждое воздействие, и четыре куколки мухи-минера листьев, выделенных из растений плевела многолетнего, растущих теплице помещали у основания каждого растения (между боковыми побегами). В дополнение к этому, одну взрослую женскую особь и одну взрослую мужскую особь мухи, которые вылупились из собранных куколок, и помещали на растения параллельных групп 1-5, и одну мужскую особь на растения параллельной группы 6. Растения каждой параллельной группы из четырех растений накрывали мелкосетчатой тканью, которую помещали поверх решетчатого короба. Короба снимали с растений через 2 недели после их установки. Эксперимент проводили в теплице с максимальной суточной температурой 22°С, и растения в случае необходимости вручную поливали растения.
Фиксировали число боковых побегов и число и длину мух-минеров в личиночной стадии в каждом листе на 4 и на 6 неделю после помещения куколок и взрослых особей на растения, а также фиксировали наличие повреждения, наносимого мухами-минерами в личиночной стадии или куколками в каждом стебле через 6 недель.
Полученные данные подвергали логарифмическому преобразованию (In) и преобразованию путем извлечения квадратного корня из пропорциональных данных до анализа с применением программы Genstat v16 с помощью дисперсионного анализа, блокированного по параллельным группам.
Результаты
Только одно из шести растений Rahu, зараженных AR3046, было поражено мухами в сравнении со всеми шестью растениями Rahu Nil. Количество боковых побегов/растение было статистически значимо меньшим (Р<0,001), и относительное число боковых побегов на зараженных AR3046 растениях Rahu (Р<0,002), пораженных С.australis, было меньшим, чем на растениях Rahu Nil и через 4, и через 6 недель испытаний (Таблица 12). В дополнение к этому, использование AR3046 приводило к уменьшению числа мин/растение (Р<0,001) (мины представляют собой вертикальные бороздки, оставляемые личинками по мере поедания внутренних тканей растений) и числа оснований боковых побегов (т.е. нижняя часть каждого бокового побега), которые были повреждены или в которых содержались личинки или куколки.
В целом растения овсяницы луговой были сильнее поражены мухами, причем все растения были атакованы. Заражение растений овсяницы луговой N. uncinatum приводило к уменьшению относительного числа боковых побегов, пораженных С. australis на 4 (Р<0,007) и на 6 неделю (Р<0,002) (Таблица 12). В дополнение к этому, было повреждено статистически значимо меньшее количество (Р<0,05) оснований боковых побегов содержащих эндофитов растений, или в них содержалось статистически значимо меньшее количество личинок, чем в растениях овсяницы луговой.
Таблица 12. В таблице ниже приведено среднее число жизнеспособных боковых побегов, число поврежденных боковых побегов, относительное число поврежденных боковых побегов и число мин/растение при воздействии Cerodontha australis на 4 и 6 неделю после помещения куколок и взрослых особей на растения Secale cereale сорта «Rahu» и растения овсяницы луговой в присутствии (MF N. unc.) и в отсутствие (MF Nil) эндофита. Среднее число боковых побегов с повреждением и или куколками в основании боковых побегов приведено для 6 недели испытаний. Данные по средним числам подвергали логарифмическому преобразованию (ln) и преобразованию путем извлечения квадратного корня из пропорциональных данных.


Вывод
Заражение растений Secale сорта Rahu продуцирующим лолин эндофитом, AR3046, приводило к статистически значимому уменьшению степени поражения мухой-минером листьев Cerodontha australis. Заражение растений овсяницы луговой другими видами продуцирующих лолин эндофитов также приводило к уменьшению относительного числа боковых побегов, поврежденных личинками мухи, и числа боковых побегов с повреждением основания.
Результаты, полученные в этом испытании, дают веские основания предполагать это лолиновые алкалоиды, продуцируемые AR3046, отвечают за обеспечение по меньшей мере некоторого уровня защиты зараженных растений Secale spp. от вредителей путем уменьшения степени поражения С. australis зараженных эндофитами растений Secale spp. Это согласуется с данными по лолинам, которые, как показано, оказывают сдерживающий и пестицидный эффект на других насекомых и вредителей растений (Schardl et. al., 2007; Bacetty et al., 2009a 2009b).
ПРИМЕР 13
Влияние заражения «Rahu» Secale эндофитом AR3046 на слюнявые пенницы
Был проведен эксперимент, в котором сравнивали влияние заражения растения Secale cereale сорта Rahu грибом AR3046 и заражения овсяницы луговой ее природным продуцирующем лолины эндофитом N. uncinatum наряду с соответствующими незараженными контрольными растениями, на питающееся ксилемой насекомое-вредитель Philaenus spumarius (слюнявая пенница), который предположительно является чувствительным к лолиновым алкалоидам. Растения клонировали таким образом, что для каждого эксперимента с разными насекомыми использовали шесть генетически идентичных растений зараженного Rahu и зараженной и незараженной овсяницы луговой, но только три из шести растений Rahu, не содержащих эндофитов, растения были одинаковыми.
Были проведены два эксперимента, первый с использованием зрелых личинок слюнявой пенницы, которые достаточно быстро превратились во взрослых особей, и второй с использованием менее зрелых личинок. В обоих случаях три личинки слюнявых пенниц были помещены на каждое растение. В первом эксперименте растения были покрыты коробами из ацетататного волокна, которые не совсем устойчивы к воздействию насекомых, тогда как во втором эксперименте использовали нейлоновые покрытия, устойчивые к воздействию насекомых. Число имеющихся личинок слюнявой пенницы и число «сгустков» слюны небольшого, среднего или большого объема, которую личинки слюнявых пенниц выделяют по мере поедания корма, определяли регулярно на протяжении каждого эксперимента. Заражение растения Rahu Secale эндофитом AR3046, приводило к уменьшению и числа «сгустков» слюны, и объема слюны, продуцируемой личинками слюнявой пенницы, в сравнении с растениями, не содержащими эндофитов (На фигуре 3).
В первом эксперименте на растениях, зараженных AR3046, число личинок слюнявых пенниц было статистически значимо меньшим в сравнении с растениями, не содержащими эндофитов, на протяжении всего эксперимента, тогда как во втором эксперименте уменьшение числа личинок было статистически значимым только в начале эксперимента. В отличие от этих результатов заражение растений овсяницы луговой эндофитами не оказывало никакого влияния на число или пищевое поведение личинок слюнявой пеннице.
Результаты этого исследования дают веские основания предполагать, что лолиновые алкалоиды, продуцируемые AR3046, отвечают за обеспечение по меньшей мере некоторого уровня защиты зараженного растения Secale от вредителей путем уменьшения степени поражения личинками слюнявых пенниц зараженных эндофитами растений Secale. Это согласуется с данными по долинам, которые, как показано, оказывают сдерживающий и пестицидный эффект на других насекомых и вредителей растений (Schardl et. al., 2007; Bacetty et al., 2009a 2009b).
ПРИМЕР 14
Исследования, связанные с болезнями растений
Ниже приведены результаты, полученные in vitro, показывающие влияние выбранных эндофитов на патогенные грибы злаковых культур, включая подавление развития целого ряда патогенных и сапротрофных грибов. Например, ряд эндофитных штаммов статистически значимо (Р≤0,05) подавлял рост мицелия Fusarium graminearum и Rhizoctonia solani (Фиг. 4 и Фиг. 5). Эти два патогена являются возбудителями заболевания фузариоза колоса и черной парши, соответственно, оба представляют собой опасные заболевания зерновых культур. Эти эндофиты обладают потенциалом для обеспечения защиты против многих заболеваний злаковых культур. Не имея намерения ограничиваться какой-либо теорией, авторы настоящего изобретения полагают, что хотя никакого(их) механизма(ов) действия не было установлено до настоящего времени для объяснения этого подавления, но вероятным механизмом может быть антибиоз, опосредованный продукцией неизвестных вторичных метаболитов.
Результаты исследований продемонстрировали, что ряд эндофитных штаммов статистически значимо (Р≤0,05) подавлял рост мицелия Fusarium graminearum (Фиг. 4) и Rhizoctonia solani (Фиг. 5).
Хотя настоящее изобретение было описано в качестве примера и со ссылкой на конкретные варианты осуществления, следует понимать, что в пределах объема настоящего изобретения могут быть внесены модификации и/или улучшения.
В дополнение к этому, если признаки или аспекты изобретения описаны в терминах групп Маркуша, то для специалистов в данной области техники представляется очевидным, что в изобретении описан таким образом любой индивидуальный представитель или подгруппа представителей формулы Маркуша.
Специалисту в данной области техники понятно, что настоящее изобретение, которое изложено и описано в настоящем документе, не ограничивается исключительно описанными аспектами, вариантами осуществления и примерами, но также охвачено в пределах сущности и объема настоящего изобретения, тех изменений и модификаций настоящего изобретения, которые представляются очевидными специалисту в данной области техники (включая среднего специалиста техники в данной области техники), в свете описаний, представленных в настоящем документе, и известного уровня техники.
ПРИМЕНЕНИЕ В ПРОМЫШЛЕННЫХ ЦЕЛЯХ
Все эндофитные штаммы эпихлое, симбиозы растения и гриба, семена, образующиеся в результате таких симбиозов, и способы получения таких симбиозов согласно настоящему изобретению, которые раскрыты в настоящем документе, могут применяться в промышленных целях для производства растений, которые используются для потребления людьми или животными.
СПИСОК ЛИТЕРАТУРЫ
Bacetty АА, Snook ME, Glenn AE, Noe JP, Hill N, Culbreath A, Timper P, Bacon CW (2009a) Toxicity of endophyte-infected tall fescue alkaloids and grass metabolites on Pratylenchus scribneri. Phytopathology 99: 1336-1345
Bacetty AA, Snook ME, Glenn AE, Noe JP, Nagabhyru P, Bacon CW (2009b) Chemotaxis disruption in Pratylenchus Scribneri by tall fescue root extracts and alkaloids. Journal of Chemical Ecology 35: 844-850
Blankenship ID, Spiering M J, Wilkinson HH, Fannin FF, Bush LP, Schardl CL (2001). Production of loline alkaloids by the grass endophyte, Neotyphodium uncinatum, in defined media. Phytochemistry 58: 395-401
Brownstein M J, Carpten JD, Smith JR (1996) Modulation of non-templated nucleotide addition by Taq DNA polymerase: Primer modifications that facilitate genotyping. BioTechniques 20: 1004-1010
Bush LP, Wilkinson HH, Schardl CL (1997) Bioprotective Alkaloids of Grass-Fungal Endophyte Symbioses. Plant Physiology 114: 1-7
Casida JE, Quistad GB (1998) Golden Age of Insecticide Research: Past, Present, or Future? Annual Review of Entomology 43: 1-16.
Christensen MJ (1995) Variation in the ability of Acremonium endophytes of Lolium perenne, Festuca arundinacea and F. pratensis to form compatible associations in the 3 grasses. Mycological Research 99: 466-470
Christensen MJ, Bennett RJ, Schmid J (2002) Growth of  / Neotyphodium and p-endophytes in leaves of Lolium and Festuca grasses. Mycological Research 106: 93-106
/ Neotyphodium and p-endophytes in leaves of Lolium and Festuca grasses. Mycological Research 106: 93-106
Christensen MJ, Bennett RJ, Schmid J (2001) Vascular bundle colonisation by Neotyphodium endophytes in natural and novel associations with grasses. Mycological Research 105: 1239-1245
Christensen MJ, Leuchtmann A, Rowan DD, Tapper В A (1993) Taxonomy of Acremonium Endophytes of Tall Fescue (Festuca-Arundinacea), Meadow Fescue (F-Pratensis) and Perennial Rye-Grass (Lolium-Perenne). Mycological Research 97: 1083-1092
Christensen MJ, Simpson WR, Al Samarrai Τ (2000) Infection of tall fescue and perennial ryegrass plants by combinations of different Neotyphodium endophytes. Mycological Research 104: 974-978
Christensen MJ, Saulsbury K, Simpson WR (2012) Conspicuous epiphytic growth of an interspecific hybrid Neotyphodium sp.endophyte on distorted host inflorescences. Fungal Biology 116: 42-48
Felsenstein, J. (2005) PHYLIP (Phylogeny Inference Package) version 3.6. Distributed by the author. Department of Genome Sciences, University of Washington, Seattle
Garthwaite I, Sprosen J, Briggs L, Collin R, Towers N (1994) Food quality on the farm: Immunological detection of mycotoxins in New Zealand pastoral agriculture. Food & Agricultural Immunology 6: 123-129
Glenn AE, Bacon CW, Price R, Hanlin RT (1996) Molecular phylogeny of Acremonium and its taxonomic implications. Mycologia 88: 369-383
Huson D, Richter D, Rausch C, Dezulian T, Franz M, Rupp R (2007) Dendroscope: An interactive viewer for large phylogenetic trees. BMC bioinformatics 8: 460
Kennedy CW, Bush LP (1983) Effect of environment and management factors on the accumulation of N-acetyl and iV-formyl loline alkaloids in tall fescue. Crop Science 23: 547-552
Koulman A, Lane GA, Christensen MJ, Fraser К, Tapper ΒΑ (2007) Peramine and other fungal alkaloids are exuded in the guttation fluid of endophyte-infected grasses. Phytochemistry 68: 355-360
Latch GCM, Christensen MJ (1985) Artificial Infection of Grasses with Endophytes. Annals of Applied Biology 107: 17-24
Malinowski DP, Belesky DP (2000) Adaptations of endophyte-infected cool-season grasses to environmental stresses: Mechanisms of drought and mineral stress tolerance. Crop Science: 40: 923-940
Marshall D, Tunali B, Nelson LR (1999) Occurrence of fungal endophytes in species of wild triticum. Crop Science 39: 1507-1512
Miller JS, Funk VA, Wagner WL, Barrie F, Hoch PC, Herendeen Ρ (2011) Outcomes of the 2011 botanical nomenclature section at the XVIII International Botanical Congress. PhytoKeys 5: 1-3
Moon CD, Tapper ВA, Scott В (1999) Identification of endophytes in planta by a microsatellite-based PCR fingerprinting assay with automated analysis. Applied and Environmental Microbiology 65: 1268-1279
Moon CD, Craven KD, Leuchtmann A, Clements SL, Schardl CL (2004). Prevalence of interspecific hybrids amongst asexual fungal endophytes of grasses. Molecular Ecology 13 (6): 1455-1467
Porter JK (1994). Chemical constituents of grass endophytes. In: Bacon, C.W., White Jr., J.F (Eds), Biotechnology of Endophytic Fungi of Grasses. CRC, Boca Raton, FL, pp. 103-123
Rasmussen S, Lane G A, Mace W, Parsons A J, Fraser К, Xue H (2012) The use of genomics and metabolomics methods to quantify fungal endosymbionts and alkaloids in grasses. Methods in Molecular Biology 860: 213-226
Rowan DD (1993) Lolitrems, peramine and paxilline: mycotoxins of the ryegrass/endophyte interaction. Agriculture, Ecosystems and Environment 44:, 103-122
Rowan DD, Latch GCM (1994) Utilization of endophyte-infected perennial ryegrasses for increased insect resistance. In: Bacon CW, White Jr. JF (eds), Biotechnology of Endophyte Fungi of Grasses. CRC Press, Boca Raton, Fonda, pp. 169-183
Sanger F, Nicklen S, Coulson AR (1977), DNA sequencing with chain-terminating inhibitors, Proceedings of the National Academy of Sciences USA 74 (12): 5463-5467
Schardl CL, Craven KD, Speakman S, Stromberg A, Lindstrom A, Yoshida R (2008). A novel test for host-symbiont codivergence indicates ancient origin of fungal endophytes in grasses. Syst Biol. 57: 483-498
Schardl CL, Grossman RB, Nagabhyru P, Faulkner JR, Mallik UP (2007) Loline alkaloids: Currencies of mutualism. Phytochemistry 68: 980-996
Schardl CL, Young CA, Faulkner JR, Florea S, Pan J (2012) Chemotypic diversity of эпихлое fungal symbionts of grasses. Fungal Ecology 5: 331-344
Schuelke M (2000) An economic method for the fluorescent labelling of PCR fragments. Nature Biotechnology 18: 233-234
Simpson WR, Mace WJ (2012) Novel associations between эпихлое endophytes and grasses: Possibilities and outcomes. In 'Эпихлое, endophytes of cool season grasses: Implications, utilization and biology.' (Eds СA Young, GE Aiken, RL McCulley, JK Strickland, CL Schardl.) pp.35-39. (The Samuel Roberts Noble Foundation: Ardmore, Oklahoma)
Simpson WR, Schmid J, Singh J, Faville M J, Johnson RD (2012) A morphological change in the fungal symbiont Neotyphodium lolii induces dwarfing in its host plant Lolium perenne. Fungal Biology 116: 234-240
Subramanian AR, Kaufmann M, Morgenstern В (2008) DIALIGN-TX: greedy and progressive approaches for segment-based multiple sequence alignment. Algorithms for Molecular Biology 3: article 6
Tanaka A, Tapper BA, Popay A, Parker EJ, Scott В (2005) A symbiosis expressed non-ribosomal peptide synthetase from a mutualistic fungal endophyte of perennial ryegrass confers protection to the symbiotum from insect herbivory. Molecular Microbiology 57: 1036-1050
Tsai HF, Liu JS, Staben С, Christensen MJ, Latch GC, Siegel MR, Schardl CL
(1994). Evolutionary diversification of fungal endophytes of tall fescue grass by hybridization with species. Proc. Natl. Acad. Sci. USA 91 (7): 2542-2546
Welty RE, Azevedo MD, Cooper TM (1987) Influence of moisture content, temperature, and length of storage on seed germination and survival of endophytic fungi in seeds of tall fescue and perennial ryegrass. Phytopathology 77: 893-900
Wilkinson HH, Siegel MR, Blankenship ID, Mallory AC, Bush LP, Schardl CL (2000). Contribution of fungal loline alkaloids to protection from aphids in a grass-endophyte mutualism. Molecular Plant Microbe Interactions 13: 1027-1033
Yates SG, Fenster JC, Bartelt RJ (1989) Assay of tall fescue seed extracts, fractions and alkaloids using the large milkweed bug. Journal of Agriculture and Food Chemistry 37: 354-357
Zejda JE, McDuffie HH, Dosman JA (1993) Epidemiology of health and safety risks in agriculture and related industries - Practical applications for rural physicians. Western Journal of Medicine 158: 56-63
Zhang, DX, Nagabhyru P, Blankenship JD, Schardl CL (2010) Are loline alkaloid levels regulated in grass endophytes by gene expression or substrate availability? Plant Signaling and Behavior 5(11): 1419-22
ОПИСАНИЕ ДЕПОНИРОВАННЫХ МИКРООРГАНИЗМОВ СОГЛАСНО БУДАПЕШТСКОМУ ДОГОВОРУ
Были депонированы следующие биологические образцы согласно условиям Будапештского договора о международном признании депонирования микроорганизмов для целей патентной процедуры

Ниже прилагаются удостоверения для депонируемых экземпляров и заявления, касающиеся жизнеспособности, для перечисленных выше депонированных микроорганизмов.






















| название | год | авторы | номер документа |
|---|---|---|---|
| УЛУЧШЕННЫЕ ЭНДОФИТОМ СЕЯНЦЫ С УВЕЛИЧЕННОЙ УСТОЙЧИВОСТЬЮ К ВРЕДИТЕЛЯМ | 2007 |
|
RU2453601C2 |
| МИКРООРГАНИЗМЫ - СТИМУЛЯТОРЫ РОСТА РАСТЕНИЙ И ИХ ИСПОЛЬЗОВАНИЕ | 2012 |
|
RU2664863C2 |
| ШТАММ MYCOSPHAERELLA SP., КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ БОРЬБЫ С ФУЗАРИОЗОМ | 2012 |
|
RU2705286C2 |
| ШТАММ MICROBACTERIUM, КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ БОРЬБЫ С ФУЗАРИОЗОМ | 2012 |
|
RU2634386C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ БОРЬБЫ С ФУЗАРИОЗОМ | 2019 |
|
RU2736382C1 |
| ЭНДОФИТНЫЕ МИКРОБНЫЕ СИМБИОНТЫ В ПРЕНАТАЛЬНОМ УХОДЕ ЗА РАСТЕНИЯМИ | 2013 |
|
RU2723946C2 |
| ВЫДЕЛЕННЫЙ ШТАММ CLONOSTACHYS ROSEA ДЛЯ ПРИМЕНЕНИЯ В КАЧЕСТВЕ АГЕНТА БИОЛОГИЧЕСКОЙ ЗАЩИТЫ | 2014 |
|
RU2689608C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ БОРЬБЫ С ФУЗАРИОЗОМ | 2020 |
|
RU2777606C2 |
| Штамм бактерий Bacillus subtilis с комплексной биологической активностью | 2019 |
|
RU2733132C1 |
| БИОАКТИВНЫЕ КОМПОЗИЦИИ И МЕТАБОЛИТЫ CHROMOBACTERIUM | 2011 |
|
RU2580105C2 |




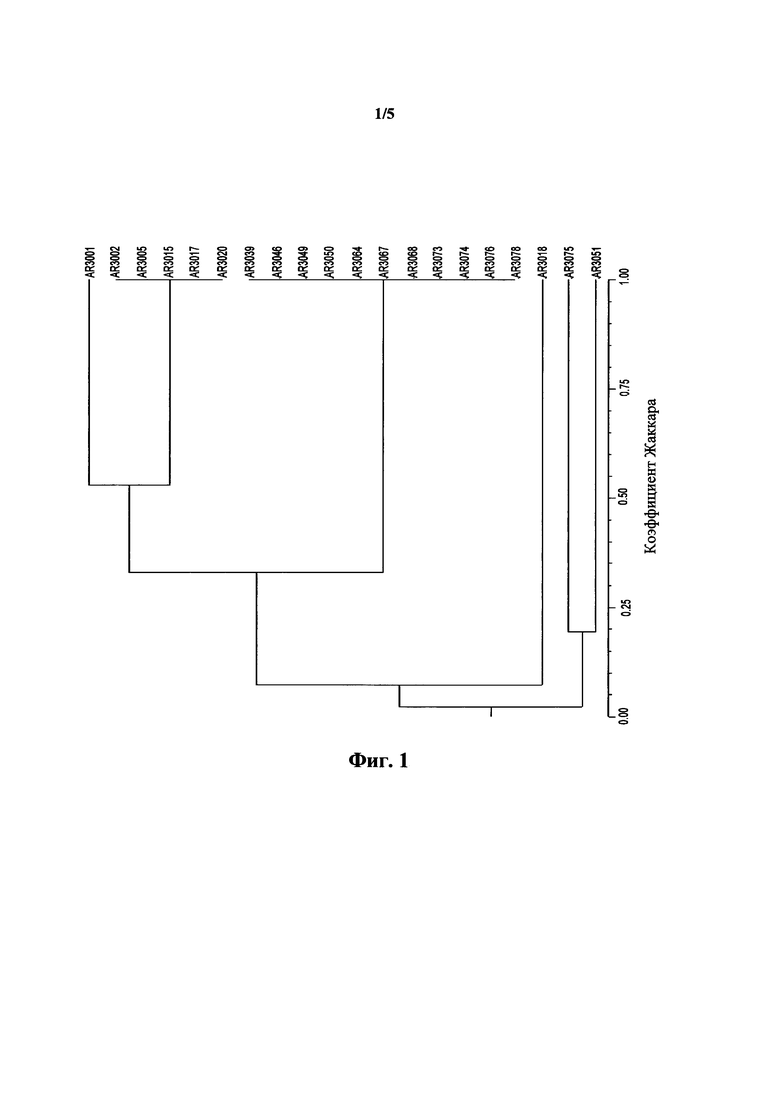



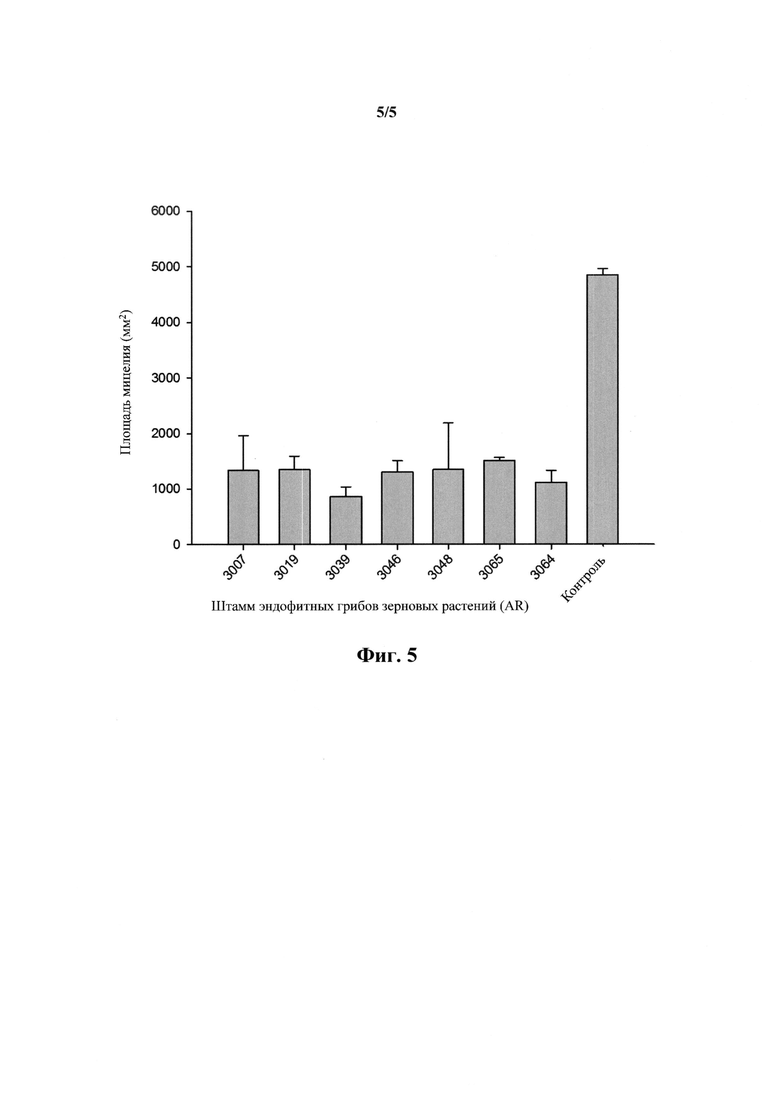
Группа изобретений относится к биотехнологии и сельскому хозяйству. Предложены штаммы эндофитного гриба эпихлое Epichloe/Neotyphodium spp., который представляет собой NRRL# 50716, NRRL# 50576, NRRL#50577, NRRL# 50578, NRRL# 50718, NRRL# 50719, NRRL# 50720, NRRL# 50721, NRRL# 50722, NRRL# 50723 или NRRL# 50724, или их комбинации, для искусственного заражения растения Secale spp. или семени Secale spp. Также предложены зараженное указанным эндофитным грибом эпихлое растение Secale spp., причем растение Secale spp. не является естественным хозяином эндофита, способ получения стабильной комбинации растения-хозяина и эндофитного гриба эпихлое, способ защиты растения-хозяина Secale spp. от вредителей и семя растения Secale spp., зараженное эндофитным грибом эпихлое. Изобретения позволяют получать стабильную комбинацию растения Secale spp. и гриба эпихлое, которое обладает повышенной устойчивостью к одному или нескольким вредителям или повышенной устойчивостью к болезни растений, или устойчивостью и к вредителям, и болезни по сравнению с Secale spp., которое не заражено эндофитным грибом, причем растение и эндофит формируют стабильную симбиотическую ассоциацию, которая обеспечивает прохождение растением нормального жизненного цикла. 16 н. и 42 з.п. ф-лы, 5 ил., 12 табл., 14 пр.
1. Выделенный штамм эндофитного гриба эпихлое Epichloe / Neotyphodium spp., который представляет собой NRRL# 50716 для искусственного заражения растения Secale spp. или семени Secale spp.
2. Выделенный штамм эндофитного гриба эпихлое Epichloe / Neotyphodium spp., который представляет собой NRRL# 50576 для искусственного заражения растения Secale spp. или семени Secale spp.
3. Выделенный штамм эндофитного гриба эпихлое Epichloe / Neotyphodium spp., который представляет собой NRRL# 50577 для искусственного заражения растения Secale spp. или семени Secale spp.
4. Выделенный штамм эндофитного гриба эпихлое Epichloe / Neotyphodium spp., который представляет собой NRRL# 50578 для искусственного заражения растения Secale spp. или семени Secale spp.
5. Выделенный штамм эндофитного гриба эпихлое Epichloe / Neotyphodium spp., который представляет собой NRRL# 50718 для искусственного заражения растения Secale spp. или семени Secale spp.
6. Выделенный штамм эндофитного гриба эпихлое Epichloe / Neotyphodium spp., который представляет собой NRRL# 50719 для искусственного заражения растения Secale spp. или семени Secale spp.
7. Выделенный штамм эндофитного гриба эпихлое Epichloe / Neotyphodium spp., который представляет собой NRRL# 50720 для искусственного заражения растения Secale spp. или семени Secale spp.
8. Выделенный штамм эндофитного гриба эпихлое Epichloe / Neotyphodium spp., который представляет собой NRRL# 50721 для искусственного заражения растения Secale spp. или семени Secale spp.
9. Выделенный штамм эндофитного гриба эпихлое Epichloe / Neotyphodium spp., который представляет собой NRRL# 50722 для искусственного заражения растения Secale spp. или семени Secale spp.
10. Выделенный штамм эндофитного гриба эпихлое Epichloe / Neotyphodium spp., который представляет собой NRRL# 50723 для искусственного заражения растения Secale spp. или семени Secale spp.
11. Выделенный штамм эндофитного гриба эпихлое Epichloe / Neotyphodium spp., который представляет собой NRRL# 50724 для искусственного заражения растения Secale spp. или семени Secale spp.
12. Комбинация выделенных штаммов эндофитного гриба эпихлое Epichloe / Neotyphodium spp. для искусственного заражения растения Secale spp. или семени Secale spp., которые выбраны из группы, состоящей из штаммов по пп. 1-11.
13. Выделенный штамм по любому из пп. 1-11, который является биологически чистым.
14. Искусственно зараженное эндофитным грибом эпихлое выбранным из группы, состоящей из NRRL 50716, NRRL# 50576, NRRL#50577, NRRL# 50578, NRRL# 50718, NRRL# 50719, NRRL# 50720, NRRL# 50721, NRRL# 50722, NRRL 50723 и NRRL# 50724, или их комбинаций, растение Secale spp., которое обладает повышенной устойчивостью к одному или нескольким вредителям или повышенной устойчивостью к болезни растений, или устойчивостью и к вредителям, и болезни по сравнению с Secale spp., которое не заражено эндофитным грибом, причем указанное растение Secale spp. не является естественным хозяином эндофита и, причем растение и эндофит формируют стабильную симбиотическую ассоциацию, которая обеспечивает прохождение растением нормального жизненного цикла.
15. Растение Secale spp. по п. 14, причем эндофит передается от первого поколения Secale spp. второму поколению Secale spp. путем вертикальной передачи.
16. Растение Secale spp. по п. 15, причем вертикальная передача от первого поколения растения-хозяина второму поколению растения-хозяина осуществляется через семена.
17. Растение Secale spp. по любому из пп. 14-16, причем эндофит представляет собой эндофит, выделенный из Elymus spp.
18. Растение Secale spp. по п. 17, причем эндофит выделен из Elymus mutabilis.
19. Растение Secale spp. по любому из пп. 14-16, причем эндофит представляет собой эндофитный штамм эпихлое, выбранный из группы, состоящей из NRRL#50716, NRRL# 50576, NRRL# 50578, NRRL# 50719, NRRL# 50720, NRRL# 50722 и NRRL# 50724.
20. Растение Secale spp. по любому из пп. 14-16, причем эндофит представляет собой NRRL#50576, NRRL# 50578 или NRRL# 50720.
21. Растение Secale spp. по любому из пп. 14-16, причем у зараженного эндофитным грибом растения Secale spp. не проявляются внешние признаки заражения.
22. Растение Secale spp. по любому из пп. 14-16, причем зараженное эндофитным грибом растение Secale spp. характеризуется морфологически нормальным фенотипом.
23. Растение Secale spp. по любому из пп. 14-16, причем зараженное эндофитным грибом растение Secale spp. продуцирует лолиновые алкалоиды и/или перамин.
24. Растение Secale spp. по любому из пп. 14-16, причем зараженное эндофитным грибом растение Secale spp. обладает повышенной устойчивостью к одному или нескольким вредителям, причем вредители выбраны из группы, состоящей из: (1) видов тли (Aphididae) Rhopalosiphum padi, Schizaphis graminum, Rhopalosiphum maidis, Metopoliphium dirhodum, Sitobion spp., Sitobion avenae, Sitobion fragariae, Diuraphis noxis; (2) видов насекомых-вредителей культурных и диких злаков (Agromyzidae; Anthomyiidae, Chloropidae, Cephidae и Cecidomyiidae) Oscinella frit, Oscinella pusilla, Mayetiola destructor, Cerodontha spp., Cerodontha australis, Cerodontha angustipennis, Formia fumigata, Meromyze americana, Haplodiplosis marginata, Chlorops pumilionis, Tipula spp. Chromatomyia fuscula, Cephus pygmaeus, Chromatomyia fuscula, Contarinia tritici; (3) видов трипсов (Thripidae) Limothrips cerealium, Limothrips denticornis, Aptinothrips rufus, Stenothrips graminum; (4) видов саранчовых и сверчков (Acrididae и Gryllidae) Locusta migratoria, Phaulacridium marginale, Phaulacridium vittatum, Melanoplus spp., Teleogryllus commodus; (5) видов клопов (Lygaeidae) Nyssius huttoni, Blissus leucopertus leucopertus; (6) видов долгоносиков (Curculionidae) Sphenophorus spp.; (7) видов гусениц, поражающих злаковые растения, и гусениц озимой совки (Noctuidae) Pseudaletia unipuncta, Spodoptera spp., Mythimna separata; Persectania aversa, Agrostis ipsilon; (8) видов листоедов (Chysomelidae) Oulema melanopus; (9) видов личинок хрущей (Scarabaeidae) Popillia japonica, Costelytra zealandica, Phyllopertha spp., Rhizotrogus majalis, Anisoplia segetum; (10) видов мучнистых червецов (Pseudococcidae и Coccidae) Phenacoccus hordei, Balanococcus poae, Riper sella rumicis, Porphyrophora tritici; (11) видов проволочников (Elateridae) Conoderus spp., Limonius spp.; (12) видов жуков (Carabidae) Zabrus tenebrioides; (13) видов клещей (Eriophyidae и Penthaleidae) Penthaleus spp., Halotydeus destructor, Aceria spp.; (14) видов вредителей запасов продовольствия (Curculionidae, Silvanidae, Pyralidae, Gelechiidae, Tenebrionidae, Bostrichidae) Sitophilus oryzae, Sitophilus granarius, Sitotroga cerealella, Rhyzopertha dominica, Cryptolestes spp., Oryzaephilus surinamensis, Cadra cautella, Plodia interpunctella, Tribolium confusum, Tribolium castaneum, Lasioderma erricorne; (15) видов цикад-пенниц (Cercopoidea) Philaenus spumarius, (16) видов нематод: корневых нематод Pratylenchus spp., P. thornei, P. crenatus, P. neglectus, P. penetrans, овсяных цистообразующих нематод Heterodera spp., Punctodera spp., H. avenae, H. latipons, H. hordecalis, H.filipjevi, H. mani, H. bifenestra, H. pakistanensis, P. punctata, клубеньковых нематод Meloidogyne spp., M. chitwoodi, M. naasi, M. artiellia, M. microtyla, M. ottersoni, M. graminicola, M. graminis, M. kikuyensis, M. spartinae, стеблевых нематод Ditylenchus spp., D. dipsicai, D. radicicola; пшеничных нематод Anguina tritici; (17) видов слизней Deroceras reticulatum, Arion spp., A. hortensis agg., A. subfuscus и минирующих мух Cerodontha australis (Diptera: Agromyzidae), также известных как минер влагалища листа пшеницы.
25. Растение Secale spp. по любому из пп. 14-16, причем зараженное эндофитным грибом растение Secale spp. обладает повышенной устойчивостью по меньшей мере к одной болезни растений, причем болезнь растений вызвана патогеном растений, выбранным из группы, состоящей из вируса желтой карликовости ячменя (Leteovirus), почвообитающего вируса мозаики пшеницы (Furovirus) и вируса полосатой мозаики пшеницы (Tritimovirus), Xanthomonas campestris, Pseudomonas syringae, Colletotrichum graminicola, Glomerella graminicola [телеоморфа], Alternaria spp., Cladosporium herbarum, Mycosphaerella tassiana [телеоморфа], Epicoccum spp., Sporobolomyces spp., Stemphylium spp., Bipolaris sorokiniana, Cochliobolus sativus [телеоморфа], Fusarium spp., Tilletia caries, Tilletia tritici, Tilletia laevis, Tilletia foetida, Hymenula cerealis, Cephalosporium gramineum, Helminthosporium sativum, Cochliobolus sativus [телеоморфа], Coprinus sychromorbidus, Dilophospora alopecuri, Tilletia controversa, Claviceps purpurea, Sphacelia segetum [анаморфа], Fusarium culmorum, Pseudoseptoria donacis, Selenophoma donacis, Neovossia indica, Tilletia indica, Puccinia recondita, Aecidium clematidis [анаморфа], Cercosporidium graminis, Scolicotrichum graminis, Phaeosphaeria herpotrichoides, Leptosphaeria herpotrichoides, Ustilago tritici, Microdochium nivale, Fusarium nivale, Monographella nivalis [телеоморфа], Erysiphe graminis, Pythium aphanidermatum, Pythium arrhenomanes, Pythium debaryanum, Pythium graminicola, Pythium ultimum, Gibberella zeae, Fusarium graminearum [анаморфа], Septoria secalis, Septoria tritici, Mycosphaerella graminicola [телеоморфа], Rhizoctonia cerealis, Rhizoctonia solani, Rhizoctonia zeae, Blumeria spp., Ceratobasidium cereale [телеоморфа], Myriosclerotinia borealis, Sclerotinia borealis, Typhula idahoensis, Typhula incarnate, Typhula ishikariensis, Typhula ishikariensis var. canadensis, Stagonospora nodorum, Septoria nodorum, Phaeosphaeria nodorum [телеоморфа], Leptosphaeria nodorum, Urocystis occulta, Puccinia graminis, Aspergillus spp., Nigrospora spp., Penicillium spp., Rhizopus spp., Pseudocercosporella herpotrichoides, Tapesia acuformis [телеоморфа], Uredo glumarum [анаморфа], Pyrenophora tritici-repentis, Drechslera tritici-repentis [анаморфа], Helminthosporium tritici-repentis, Puccinia triticina, Pythium spp., Rhynchosporium secalis, Puccinia striiformis, Gaeumannomyces graminis и Fusarium pseudograminearum.
26. Растение Secale spp. по п. 25, причем патогеном растений является Puccinia recondita, Puccinia triticina, Puccinia graminis, Fusarium spp., Pythium spp., Rhynchosporium secalis, Puccinia striiformis, Gaeumannomyces graminis, Rhizoctonia solani или Fusarium pseudograminearum.
27. Растение Secale spp. по любому из пп. 14-16, причем растение Secale spp. выбрано из группы, состоящей из Secale cereale, Secale montanum, Secale strictum, Secale sylvestre и Secale vavilovii.
28. Растение Secale spp. по любому из пп. 14-16, причем растение Secale spp. представляет собой Secale cereale.
29. Способ получения стабильной комбинации растения-хозяина/эндофитного гриба эпихлое, предусматривающий искусственное заражение растения Secale spp. по меньшей мере одним эндофитным грибом эпихлое, который формирует стабильную симбиотическую комбинацию с инокулированным растением, причем у растения-хозяина не проявляются внешние признаки заражения эндофитом, где эндофит выбран из группы, состоящей из NRRL# 50716, NRRL# 50576, NRRL#50577, NRRL# 50578, NRRL# 50718, NRRL# 50719, NRRL# 50720, NRRL# 50721, NRRL# 50722, NRRL# 50723 и NRRL# 50724, или их комбинаций.
30. Способ по п. 29, причем стабильная комбинация растения/гриба является достаточно стабильной для обеспечения вертикальной передачи эндофита.
31. Способ по п. 30, причем вертикальная передача осуществляется через побеги или черенки.
32. Способ по п. 30 или 31, причем вертикальная передача осуществляется через цветочные побеги в семена.
33. Способ по любому из пп. 29-31, причем вертикальная передача эндофита приводит к вертикальной передаче эндофита от первого поколения второму поколению растений-хозяев.
34. Способ по любому из пп. 29-31, причем вертикальная передача от первого поколения растения-хозяина второму поколению растения-хозяина осуществляется через семена.
35. Способ по любому из пп. 29-31, причем указанный способ дополнительно предусматривает стадию отбора растения-хозяина Secale spp. из популяции зараженных растений-хозяев, у которого не проявляются внешние признаки заражения эндофитным грибом.
36. Способ по любому из пп. 29-31, причем растение Secale spp. выбрано из группы, состоящей из Secale cereale, Secale montanum, Secale strictum, Secale sylvestre и Secale vavilovii.
37. Способ по любому из пп. 29-31, причем растение Secale spp. представляет собой Secale cereale.
38. Способ по любому из пп. 29-31, причем по меньшей мере один эндофитный гриб представляет собой эндофит, выделенный из Elymus spp.
39. Способ по любому из пп. 29-31, причем эндофитный гриб выделен из Elymus mutabilis.
40. Способ по любому из пп. 29-39, причем эндофит представляет собой NRRL#50716, NRRL# 50576, NRRL# 50578, NRRL# 50719, NRRL# 50720, NRRL# 50722 или NRRL# 50724.
41. Способ по любому из пп. 29-39, причем эндофит представляет собой NRRL#50576, NRRL# 50578 или NRRL# 50720.
42. Способ обеспечения защиты растения-хозяина Secale spp. от вредителей, предусматривающий искусственное заражение растения Secale spp. по меньшей мере одним эндофитным грибом эпихлое выбранным из группы, состоящей из NRRL# 50716, NRRL# 50576, NRRL#50577, NRRL 50578, NRRL# 50718, NRRL# 50719, NRRL# 50720, NRRL# 50721, NRRL 50722, NRRL# 50723 и NRRL# 50724, или их комбинаций, причем комбинация эндофитного гриба и растения Secale обеспечивает продукцию по меньшей мере одного алкалоида в количестве, достаточном для обеспечения защиты растения-хозяина от вредителей.
43. Способ по п. 42, причем по меньшей мере один алкалоид представляет собой алкалоид, выбранный из группы, состоящей из перамина, N-ацетилнорлолина, лолина, N-формиллолина, N-ацетиллолина и N-метиллолина.
44. Способ по п. 42 или 43, причем по меньшей мере один алкалоид представляет собой лолин или перамин, или оба алкалоида.
45. Способ по п. 42 или 43, причем указанный способ дополнительно предусматривает стадию отбора комбинации эндофитного гриба и растения Secale, которая обеспечивает продукцию по меньшей мере одного алкалоида в количестве, достаточном для обеспечения защиты растения-хозяина от вредителей.
46. Способ по п. 42 или 43, причем Secale spp. выбрано из группы, состоящей из Secale cereale, Secale montanum, Secale strictum, Secale sylvestre и Secale vavilovii.
47. Способ по п. 42 или 45, причем Secale spp. представляет собой Secale cereale.
48. Способ по п. 42 или 45, причем по меньшей мере один эндофитный гриб представляет собой эндофит, выделенный из Elymus spp.
49. Способ по п. 42 или 45, причем эндофитный гриб выделен из Elymus mutabilis.
50. Способ по любому из пп. 42-49, причем эндофит представляет собой NRRL#50716, NRRL# 50576, NRRL# 50578, NRRL# 50719, NRRL# 50720, NRRL# 50722 или NRRL# 50724.
51. Способ по любому из пп. 42-49, причем эндофит представляет собой NRRL# 50576, NRRL# 50578 или NRRL# 50720.
52. Семя растения Secale spp., искусственно зараженное эндофитным грибом эпихлое, которое обладает повышенной устойчивостью к одному или нескольким вредителям или повышенной устойчивостью к болезни растений, или устойчивостью и к вредителям, и болезни по сравнению с Secale spp., которое не заражено эндофитным грибом, где эндофит эпихлое выбран из группы, состоящей из NRRL# 50716, NRRL# 50576, NRRL#50577, NRRL# 50578, NRRL# 50718, NRRL# 50719, NRRL# 50720, NRRL# 50721, NRRL# 50722, NRRL# 50723 и NRRL# 50724, или их комбинаций.
53. Семя растения Secale spp. по п. 52, причем указанное семя представляет собой семя Secale spp., выбранного из группы, состоящей из Secale cereale, Secale montanum, Secale strictum, Secale sylvestre и Secale vavilovii.
54. Семя растения Secale spp. по п. 52 или 53, причем указанное семя представляет собой семя растения Secale cereale.
55. Семя растения Secale spp. по п. 52 или 53, причем по меньшей мере один эндофитный гриб представляет собой эндофит, выделенный из Elymus spp.
56. Семя растения Secale spp.по п. 52 или 53, причем эндофитный гриб выделен из Elymus mutabilis.
57. Семя растения Secale spp. по любому из пп. 52-56, причем эндофит представляет собой NRRL#50716, NRRL# 50576, NRRL# 50578, NRRL# 50719, NRRL# 50720, NRRL# 50722 или NRRL# 50724.
58. Семя растения Secale spp. по любому из пп. 52-57, причем эндофит представляет собой NRRL#50576, NRRL# 50578 или NRRL# 50720.
| WO 2008100892 A2, 21.08.2008 | |||
| US 2012144533 A1, 07.06.2012 | |||
| WO 2004082384 A2, 30.09.2004 | |||
| СУДОВОЙ ЗАБОРТНЫЙ ТРАП | 0 |
|
SU213616A1 |
| JOHNSON L | |||
| "The Exploitation of Epichloe Endophytes for Agricultural Advantage", COST-Action "Endophytes in Biotechnology and Agriculture": Working Group 1-4 Meeting: Endophytes: From discovery to application, Trento/S | |||
| Michele, Italy November 12-14, 2012, p.1-41, Найдено в Интернет [27.01.2018] URL: http://endophytes.eu/trento2012/Trento_2012_JohnsonLinda | |||
| БЛАГОВЕЩЕНСКАЯ Е.Ю | |||
| "ЭНДОФИТНЫЕ ГРИБЫ ЗЛАКОВ", АВТОРЕФЕРАТ диссертации на соискание ученой степени кандидата биологических наук, Москва 2006, 26 с. | |||

