ОБЛАСТЬ ТЕХНИКИ
Настоящая группа изобретений относится к биологическим методам борьбы с фитопатогенными болезнями. В частности, изобретение относится к штаммам, биопрепаратам, способам их получения и способам, которые могут использоваться в борьбе с фузариозом сельскохозяйственных культур.
УРОВЕНЬ ТЕХНИКИ
Постоянный рост численности населения вызывает ряд различных социальных и экономических проблем. В частности, одной из подобных проблем считается обеспечение населения качественной сельскохозяйственной продукцией. При этом одной из основных сельскохозяйственных культур, возделываемых на территории Российской федерации и на территории Южного федерального округа (ЮФО), в частности, является озимая пшеница.
Среди актуальных проблем в сельскохозяйственной отрасли известное место занимают грибковые болезни, главной в ЮФО из которых является фузариоз колоса озимой пшеницы. Заболевание наблюдается в большинстве регионов, где выращивается пшеница и обусловливает потери 25-30% урожая [Гагкаева Т.Ю., Ганнибал Ф.Б., Гаврилова О.П. Зараженность зерна пшеницы грибами Fusarium и Alternaria на юге России в 2010 году // Защита и карантин растений. 2012. №1.; Гагкаева Т.Ю., Гаврилова О. П. Зараженность зерна грибами Fusarium в Краснодарском и Ставропольском краях // Защита и карантин растений. 2014. №3].
Возбудителями выступают грибы рода Fusarium, которые находятся в различных частях растения, как в колосковом стержне и чешуях, так и в семенах, особенно в зародышевой части семян.
На настоящий момент времени заражение грибами рода Fusarium сельскохозяйственных культур приобрело характер пандемии и при благоприятных условиях развивается в большей части случаев и представляет серьезную опасность для здоровья населения.
Особую опасность грибы данного рода представляют из-за продукции микотоксинов трихотецинового ряда, которые накапливаются в семенах растений и представляют угрозу, как для людей при приеме в пищу, так и в сельском хозяйстве при кормлении скота [Гагкаева Т.Ю., Гаврилова О.П., Левитин М.М. Биоразнообразие и ареалы основных токсинопродуцирующих грибов рода Fusarium // Биосфера. 2014. №1.].
По химическому составу данная группа микотоксинов подразделяется на 2 группы: А (Т-2 и НТ-2 токсины, диацетоксисцирпенол и их производные); В (дезоксиниваленол (ДОН), ниваленол (НИВ) и их производные.
Существуют различные способы защиты от фитопатогенных грибов, при этом наиболее часто применяемым из них является использование фунгицидов. Однако, эффективность применения подобных веществ постоянно снижается. Так, например, в ряде работ было показано, что эффективность применения химических средств по борьбе с фитопатогенными грибами значительно уменьшается по причине постепенного развития резистентности к действию различных фунгицидных веществ. [Becker R., Hettwer U., Karlovsky P., Deising H.B. et al. Adaptation of Fusarium graminearum to tebuconazole yielded descendants diverging for levels of fitness, fungicide resistance, virulence, and mycotoxin production // Phytopath., 2010, №100, p. 444-453; Чекмарев В.В., Кобыльская Г.В., Бучнева Г.Н., Корабельская О.И. Резистентность грибов рода Fusarium к протравителям семян // Защита и карантин растений. 2011. №3]. По данным других авторов следует, что максимальный эффект, которого можно достигнуть при применении фунгицидов составляет не более 7 5%, однако этого удается достичь, если применять комплексы из нескольких фунгицидных препаратов [Санин С.С., Мотовилин А.А., Корнева Л.Г., Жохова Т.П., Полякова Т.М., Акимова Е.А. Химическая защита пшеницы от болезней при интенсивном зернопроизводстве // Защита и карантин растений. 2011. №8; Гасич Е.Л., Хлопунова Л.Б., Гагкаева Т.Ю., Дмитриев А.П. Действие фунгицидов на развитие гибеллиноза пшеницы при искусственном заражении в лабораторных условиях // Защита и карантин растений. 2015. №1].
В настоящее же время, вектор проводимых исследований сместился с поиска химических веществ на поиск и применение бактерий-антагонистов и их метаболитов, проявляющих антагонистические свойства к различным грибковым возбудителям болезней сельскохозяйственных культур [Сидоренко Олег Дмитриевич Перспективы использования биологических препаратов на основе микроорганизмов // Известия ТСХА. 2012. №6].
Одной из перспективных групп микроорганизмов, отличающихся как способностью эффективно противостоять фитопатогенам, так и проявлять фитостимулирующие свойства, является группа почвенных бактерий-антагонистов класса Bacilli.
В последнее время, бактерии данного семейства рассматриваются в качестве перспективных агентов биологического контроля болезней растений, а также в качестве организмов, проявляющих фитостимулирующую активность в силу их широкой распространенности, природного антагонизма ко многим фитопатогенным грибам и способности продуцировать вещества гормональной природы.
Зарубежный опыт указывает на высокую эффективность применения биопрепаратов на основе бацилл для биологического контроля патогенных грибов рода Fusarium. В частности, показана их эффективность на кукурузе, огурцах и картофеле [Cavaglieri, L., Orlando, J. R. M. I., Rodriguez, M. I., Chulze, S., & Etcheverry, M. (2005). Biocontrol of Bacillus subtilis against Fusarium verticillioides in vitro and at the maize root level. Research in Microbiology, 156(5), 748-754.; Sadfi, N., Cherif, M., Fliss, I., Boudabbous, A., & Antoun, H. (2001). Evaluation of bacterial isolates from salty soils and Bacillus thuringiensis strains for the biocontrol of Fusarium dry rot of potato tubers. Journal of Plant Pathology, 101-117.; Chen, F., Wang, M., Zheng, Y., Luo, J., Yang, X., & Wang, X. (2010). Quantitative changes of plant defense enzymes and phytohormone in biocontrol of cucumber Fusarium wilt by Bacillus subtilis B579. World Journal of Microbiology and Biotechnology, 26(4), 675-684].
Механизм действия заключается в колонизации бактериями корней растений в период прорастания семян и подавление патогенных грибов за счет выделения хитинолитических ферментов, сидерофоров и специфических антимикробных пептидов.
Однако почвенные бактерии обладают высокой чувствительностью к локальным почвенно-климатическим условиям - типу почв, температурному режиму, влажности, составу местных микробных сообществ. Этим обусловлена неэффективность или слабая эффективность применения штаммов, выделенных в определенных областях вне этих регионов. Если вносимые штаммы являются чужеродными, они быстро подавляются уже имеющимся в почве микробным сообществом и теряют способность влиять на патогенные грибы. Помимо этого, способ, основанный на применении штаммов и видов бактерий-антагонистов, нехарактерных для данных био- и агроценозов, является экологически небезопасным по причине того, что может быть нарушен естественный баланс между различными группами почвенных микроорганизмов, что в свою очередь может привести к снижению плодородия почвы и урожайности сельскохозяйственных культур.
Таким образом, целесообразно выделять штаммы из почв тех регионов, где планируется их применение, (см. например Pii Y. et al. The interaction between iron nutrition, plant species and soil type shapes the rhizosphere microbiome // Plant Physiology and Biochemistry. - 2016. - T. 99. - C. 39-48. Ma J. et al. Bacterial diversity and composition in major fresh produce growing soils affected by physiochemical properties and geographic locations // Science of the Total Environment. -2016. - T. 563. - C. 199-209.  S.,
S.,  R., Baldrian P. Drivers of microbial community structure in forest soils // Applied microbiology and biotechnology. - 2018. - T. 102. - №. 10. - C. 4331-4338. и др.).
R., Baldrian P. Drivers of microbial community structure in forest soils // Applied microbiology and biotechnology. - 2018. - T. 102. - №. 10. - C. 4331-4338. и др.).
Из уровня техники известно изобретение, которое относится к биотехнологии, микробиологии и сельскому хозяйству и может быть использовано для получения бактериального препарата против болезней растений, вызываемых фитопатогенными грибами рода Fusarium, Microdochium, Pyrenophora и Puccinia. Штамм Bacillus subtilis BZR 517 обладает фунгицидной активностью в отношении фитопатогенных бактерий и грибов. Штамм Bacillus subtilis депонирован в Коллекции ГНУ ВНИИСХМ Россельхозакадемии под регистрационным номером RCAM01728 (Патент РФ №2552146 С1, опубл. 10.06.2015).
Также из уровня техники известно изобретение, которое относится к сельскому хозяйству. Способ получения супрессивного компоста по отношению к возбудителю фузариоза растений Fusarium oxysporum и заключается в приготовлении компоста, полученного из сельскохозяйственных отходов, выделении из компоста штаммов микроорганизмов, приготовлении на основе выделенных штаммов биопрепарата, внесении биопрепарата в компост, оценке супрессивности компоста, причем проводят оценку супрессивных свойств штаммов по 4 механизмам супрессивности, оценку взаимной антагонистической активности штаммов, осуществляют двукратное внесение биопрепарата в компост, проводят оценку удобрительных свойств супрессивного компоста. Изобретение позволяет получить компосты, обладающие одновременно удобрительными свойствами и стабильными супрессивными свойствами по отношению к возбудителю фузариоза растений Fusarium oxysporum (Патент РФ №2629776 С1, опубл. 04.09.2017).
Кроме того, из уровня техники известно изобретение, которое относится к композиции и способам, предоставленных для комбинации нового штамма Bacillus amyloliquefaciens RTI301 и нового штамма Bacillus subtilis. RTI477, комбинации, обладающей активностью, стимулирующей рост, и активностью против патогенов растений. Композиции, содержащие штаммы RTI301 и RTI477, полезны для улучшения роста растений и/или обеспечения защиты от патогенной инфекции при нанесении на корни растений, семена, каллусную ткань, трансплантаты и черенки. Синергетические результаты наблюдаются для комбинации штаммов, и комбинация штаммов полезна для увеличения урожайности сельскохозяйственных культур, включая сою и кукурузу. Композиции, содержащие комбинацию штаммов, могут применяться отдельно или в комбинации с другими микробными, биологическими или химическими инсектицидами, фунгицидами, нематоцидами, бактериоцидами, гербицидами, экстрактами растений, регуляторами роста растений и удобрениями. (Патент US 9622484 В2, опубл. 18.04.2017)
Решения, описанные в источниках RU 2552146 C1, RU 2629776 C1, US 9622484 В2 являются ближайшими решениями к исследуемому объекту, однако разрабатываемый объект отличается по таким признакам, как штаммы бактерий-антагонистов, способ культивирования, форма хранения препарата, культура по отношению к которой возможно применение, кроме того заявленная группа изобретений имеет ряд существенных преимуществ.
Патент RU 2478290 С2 является наиболее близким к исследуемому объекту. В нем предложен способ получения биопрепарата, который содержит биомассу Bacillus amyloliquefaciens Всероссийская коллекция промышленных микроорганизмов (далее ВКПМ) В-11008 и гуматы при следующем соотношении компонентов, в об. %: биомасса Bacillus amyloliquefaciens ВКПМ В-11008 - вегетативных клеток и спор 1,24÷1,30×1010 КОЕ/мл культуральной жидкости и содержанием спор 94% от общего количества КОЕ - 99,0, гуматы - 1,0. В качестве питательной среды для получения заявляемого биопрепарата используют дешевую среду на основе отходов зернового производства следующего состава, г/л: отруби пшеничные - 40,0; кукурузный экстракт - 0,5; MgSO4 - 0,9; KH2PO4 - 0,5; CaCO3 - 1,0; рН 7,0±0,2. Указанный состав среды обеспечивает высокий уровень накопления биомассы В. amyloliquefaciens ВКПМ В-11008 (5,6÷5,9 г/л), высокий титр жизнеспособных клеток и спор (1,24÷1,28×1010 КОЕ/мл) и интенсивность спорообразования (содержание спор - 94,0% от общего количества КОЕ). Штамм данного вида был проверен на следующих представителях рода: Fusarium: F. graminearum, F. solani, F. oxysporum, F. sambucinum. Данный штамм показал высокий уровень антагонистической активности. Неясной остается проверка действия данного штамма на фитопатогенные грибы. Так предлагается следующий метод: посевы с штаммом бактерий инкубируют при 28°С в течение 3 сут. К выросшей культуре В. amyloliquefaciens ВКПМ В-11008 подсевают штрихом суточные суспензии тест-культур фитопатогенных бактерий (титр суспензий 108 КОЕ/мл). Затем чашки инкубируют при температуре 28°С в течение 24 ч и снимают результаты. В реальных условиях данные группы находятся в соперничестве за колонизацию ризоплана, и более правильным является следующий способ оценки антагонистической активности: одновременный посев культуры гриба и бактерии на плотной среде. Также не проверена возможность применения данного препарата на озимой пшенице. Также фитопатогенные грибы в данном способе не выделяются в ходе работы, а берутся уже известные культуры фитопатогенных грибов из различных коллекций, что может сильно сказаться на эффективности действия данного препарата в различных регионах и странах.
РАСКРЫТИЕ ГРУППЫ ИЗОБРЕТЕНИЙ
Задача, решаемая заявленной группой изобретений, состоит в получении новых штаммов споробразующих бактерий-антагонистов с антифузариозной активностью, в разработке биопрепарата на основе данных штаммов и способа его изготовления, а также способа биологической защиты сельскохозяйственных культур от фузариоза.
Технический результат заявленной группы изобретений заключается в расширении арсенала штаммов споробразующих бактерий-антагонистов с антифузариозной активностью, а также композиций биопрепаратов, направленных на более эффективную борьбу с фузариозом. Кроме того, технический результат заявленного способа изготовления биопрепарата заключается в легкости его воспроизводимости, технологичности и стабильности, возможности использования простой по составу, недорогой среды, получая высокий выход готового продукта, стабильность полученного биопрепарата при хранении.
Указанные технические результаты достигаются путем выделения новых 10 штаммов споробразующих бактерий-антагонистов с антифузариозной активностью. Новые штаммы депонированы 7 ноября 2017 г. (национальное патентное депонирование) и 18 ноября 2019 г. (международное патентное депонирование) в Биоресурсном Центре Всероссийской Коллекции Промышленных Микроорганизмов (БРЦ ВКПМ) НИЦ «Курчатовский институт» - ГосНИИгенетика (адрес: 117545, г. Москва, 1-ый Дорожный проезд, 1) - далее ВКПМ.
ВКПМ осуществляет депонирование для целей патентной процедуры в российской коллекции, уполномоченной на депонирование для целей патентной процедуры и удовлетворяет соответствующим для таких коллекций требованиям, а также гарантирует поддержание жизнеспособности депонируемых объектов в течение по меньшей мере срока действия патента, в том числе является Международным органом по депонированию согласно Будапештскому договору о международном признании депонирования микроорганизмов.
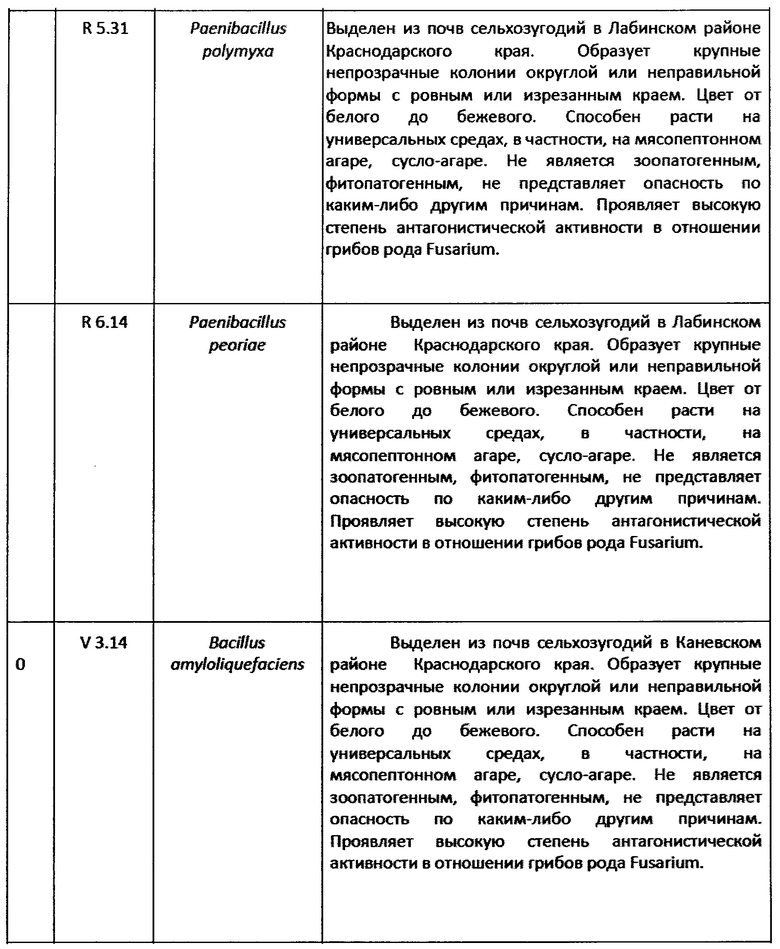
1. Штамм Paenibacillus jamilae K 1.14 (ВКПМ В-13016).
2. Штамм Paenibacillus peoriae О 1.27 (ВКПМ В-13017).
3. Штамм Paenibacillus peoriae О 2.11 (ВКПМ В-13018).
4. Штамм Paenibacillus peoriae R 3.13 (ВКПМ В-13019).
5. Штамм Paenibacillus peoriae R 4.5 (ВКПМ В-13020).
6. Штамм Bacillus amyloliquefaciens R 4.6 ВКПМ B-13021).
7. Штамм Paenibacillus jamilae R 4.24 (ВКПМ B-13022).
8. Штамм Paenibacillus polymyxa R 5.31 (ВКПМ B-13023).
9. Штамм Paenibacillus peoriae R 6.14 (ВКПМ B-13048).
10. Штамм Bacillus amyloliquefaciens V 3.14 (ВКПМ B-13024).
Кроме того, технические результаты достигаются за счет композиции биопрепарата и разработанного способа получения биопрепарата, содержащего 10 вышеуказанных штаммов споробразующих бактерий-антагонистов с антифузариозной активностью.
Биопрепарат для защиты сельскохозяйственных культур от фузариоза, включающий, комбинацию 10 вышеуказанных штаммов, содержащий не менее 108 КОЕ/г жизнеспособных бактерий-антагонистов, при этом остаточная влажность сухого биопрепарата не более 10%.
Способ изготовления биопрепарата на основе комбинации штаммов спорообразующих бактерий-антагонистов с антифузариозной активностью путем их твердофазного культивирования на питательном субстрате, включающий подготовку субстрата путем гидратирования в воде, термообработку субстрата, инокуляцию термообработанного гидратированного субстрата, инкубацию, гомогенизацию субстрата, размягченного при термообработке и в процессе ферментации и последующую сушку.
Способ защиты сельскохозяйственных культур от фузариоза, включает выделение грибов рода Fusarium из образцов, отобранных на полях, подверженных опасности фузариоза; отбор штаммов спорообразующих бактерий-антагонистов, наиболее активно подавляющих штаммы грибов рода Fusarium, распространенных на целевой территории; приготовление вышеуказанного биопрепарата вышеуказанным способом, обработку биопрепаратом посевного материала.
Разработчиками изготовлены партии биопрепарата массой 10 и 50 кг и проведено его тестирование в производственных условиях на пшенице сорта «Гром» на площади 30 га с использованием установки ПС10, а также на кукурузе на площади 50 га. Осенью 2018 года проведен полномасштабный опыт на озимой пшенице сортов «Гром» и «Таня» на площади 200 га. По первым двум опытам проведена серия полевых и лабораторных анализов за весь период вегетации, а исследования посевов озимой пшеницы осенью 2018 года проведены до стадии осеннего кущения. По итогам проведенных лабораторных исследований зарегистрировано снижение численности фитопатогенных грибов p. Fusarium:
1) в ризосфере озимой пшеницы в фазу кущения (снижение в 1,98 раза);
2) в зерне озимой пшеницы (снижение более чем в 5 раз);
3) в ризосфере кукурузы в фазу всходов (в 2,64 раза);
4) в ризосфере кукурузы в фазу молочной спелости (в 1,82 раза);
5) в стеблях озимой пшеницы в фазу всходов (в 2,28 раз на орошаемых полях и в 1,41 раза на полях без ирригации);
6) в ризосфере озимой пшеницы в фазу всходов (в 5 раз на орошаемых полях и в 1,26 раза на полях без ирригации);
7) в стеблях озимой пшеницы в фазу кущения (в 1,83 раз на орошаемых полях и в 2,25 раза на полях без ирригации).
На ранних этапах развития растений обработка семян озимой пшеницы биопрепаратом приводит к протекторному эффекту как в растениях, так и в прилегающей к корням почве, что указывает на колонизацию поверхности корня бактериями, которые препятствуют внедрению патогена. К фазе кущения процент растений, в тканях которых присутствовали грибы p. Fusarium снижается по сравнению с фазой всходов. Таким образом, препарат сохраняет свое защитное действие к фазе кущения, что говорит в пользу активной колонизации растений пшеницы бактериями биопрепарата. Это обеспечивает долговременную активную защиту растений, и при наступлении неблагоприятных внешних условий (подмокание, задержка снега в понижениях рельефа) может препятствовать развитию корневых гнилей.
Самый мощный эффект зарегистрирован для наиболее ценного параметра - в зерне озимой пшеницы, полученной от растений, выращенных из обработанных биопрепаратом семян наблюдалось более чем пятикратное снижение зараженности фузариозом. Значительное снижение численности фитопатогенных грибов р. Fusarium отмечено также в ризосфере озимой пшеницы в фазу всходов. Наиболее активное проникновение грибов в растения происходит в течение самых ранних этапов развития пшеницы - сразу же после прорастания. Применение биопрепарата позволяет обеспечить контроль численности патогена именно в этот критический период, и результаты опыта показывают, что в вариантах, где в фазу всходов численность грибов в ризосфере была снижена, впоследствии оказывается значительно ниже и процент растений, пораженных грибами.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фигуре 1 показан первичный отбор бактерий, при этом высветленные участки показывают отсутствие роста гриба.
На фигуре 2 показано определение антагонизма между исследуемыми бактериями (по краям) и грибом p. Fusarium (в центре), где видны широкие зоны подавления роста.
На фигуре 3 показано определение антагонизма между исследуемыми бактериями (по краям стекла) и грибом p. Fusarium (в центре), где видно, что гриб растет на тонком слое агара, нанесенном на предметное стекло, но не может приблизиться к колониям бактерий.
На фигуре 4 показано изучение способности к биопленкообразованию у штаммов бацилл с высокой фунгицидной активностью.
На фигуре 5 показан рост пшеницы в контроле (А) и после обработки штаммами Ra 3.12 (Б) и Ra 3.13 (В).
На фигуре 6 показано прорастание семян пшеницы в контроле (сверху) и на газоне Ra 2.6 (снизу) на среде КСА.
На фигуре 7 показано изменение длины побега у растений, вырастающих из зараженных семян при их обработке химическим протравителем и биопрепаратом.
На фигуре 8 показан рост фитопатогенной микрофлоры в контроле (А) и его подавление препаратами (Б, В, Г, Д).
На фигуре 9 показаны колонии штаммов из смеси препаратов на корнях проростков пшеницы.
На фигуре 10 показаны результаты ПЦР-анализа зараженных проб. G - результат амплификации с праймерами G1 (Таблица 7), О - результат амплификации с праймерами 02 (Таблица 8).
На фигуре 11 показана численность грибов p. Fusarium в ризосфере озимой пшеницы в фазу кущения.
На фигуре 12 показана численность грибов p. Fusarium в ризосфере озимой пшеницы в фазу созревания.
На фигуре 13 показана численность грибов p. Fusarium в зерне озимой пшеницы.
На фигуре 14 показано действие биопрепарата Ф-1 на зараженность зерна озимой пшеницы грибами p. Fusarium. Лишь в одной из опытных проб наблюдается заражение. (видны характерные колонии Fusarium graminearum розового цвета).
На фигуре 15 показана численность грибов p. Fusarium в ризосфере кукурузы в фазу всходов.
На фигуре 16 показана численность грибов p. Fusarium в ризосфере кукурузы в фазу молочной спелости.
На фигуре 17 показана проверка эффективности ПЦР при заданных параметрах амплификации.
На фигуре 18 показаны результаты ПЦР-анализа избранных проб с родоспецифичными праймерами F2.
Пробы 1-8 - Анализ 8 колоний с посевов пшеницы
Пробы 9-15 - Анализ 7 колоний с посевов кукурузы
16 - отрицательный контроль, 17 - положительный контроль (чистая культура гриба из коллекции), 18 - маркер длин ДНК 50 bp+ (Евроген).
На фигуре 19 представлены результаты амплификации проб с видоспецифичными праймерами на F. culmorum (А) и F.graminearum(Б).
На фигуре 20 редставлены результаты амплификации проб с видоспецифичными праймерами на F. oxysporum (А) и F. sporotrichoides(Б).
На фигуре 21 показаны результаты амплификации проб с видоспецифичными праймерами на F. роае(А) и F. avenaceum(Б).
На фигуре 22 показан процент пораженности растений фитопатогенными грибами p. Fusarium на орошаемых полях (всходы).
На фигуре 23 показан процент пораженности растений фитопатогенными грибами p. Fusarium на орошаемых полях (всходы).
На фигуре 24 показана численность грибов p. Fusarium в ризосферной почве на орошаемых полях, КОЕ/г почвы.
На фигуре 25 показана численность грибов p. Fusarium в ризосферной почве на полях без ирригации, КОЕ/г почвы.
На фигуре 26 показан процент пораженности растений фитопатогенными грибами p. Fusarium на орошаемых полях (осеннее кущение).
На фигуре 27 показан процент пораженности растений фитопатогенными грибами p. Fusarium на полях без ирригации (осеннее кущение).
На фигуре 28 показана численность грибов p. Fusarium в ризосферной почве в фазу осеннего кущения, КОЕ/г почвы.
ОСУЩЕСТВЛЕНИЕ ГРУППЫ ИЗОБРЕТЕНИЙ
Материалы для исследования были получены в ходе двух полевых экспедиций. В ходе экспедиций были отобраны пробы почвы и растений из хозяйства "Воля" Каневского р-на Кранодарского края, а также хозяйств Лабинского р-на "Рассвет"(в т.ч. секции Упорная, Каладжинская, Переправная), "Отрадненское", "Мостовское", "Курганинское" и "Родина". С указанных полей было отобрано 30 образцов почвы, а также образцы растений (озимая пшеница) и пожнивных остатков, являющихся резервуаром инфекции (кукуруза, сахарная свекла).
В ходе разработки способа по получению биопрепарата из этих материалов были выделены 5 штаммов грибов p. Fusarium, а также 1040 штаммов бактерий, проявляющих антагонизм к данным штаммам. Эти культуры послужили материалом для дальнейших исследований антагонистической активности и отбора наиболее активных штаммов.
Выделение культур фитопатогенных грибов p. Fusarium производилось методом посева образцов пораженных растений на плотные питательные среды сусло-агар и агар Сабуро с добавлением стрептомицина.
Выделение культур бактерий-антагонистов производилось методом посева предварительно пастеризованных образцов пораженных растений и почвы на сусло-агар с предварительным посевом на него штаммов грибов p. Fusarium. Отбирались лишь те колонии, которые проявляли видимый антагонизм по отношению к культуре фитопатогена.
Оценка антагонистической активности выделенных культур производилась методом совместного культивирования в условиях роста на чашках Петри и в микрокультуре на стеклах.
Выделенные фитопатогенные грибы были подвергнуты идентификации в ВКПМ.
Десять отобранных наиболее активных штаммов бактерий-антагонистов были подвергнуты идентификации на основании анализа ДНК во ВКПМ, и депонированы в этой коллекции.
Тестирование влияния штаммов на растениях производилось путем инокуляции семян культурой испытуемого штамма с последующим определением всхожести, энергии прорастания, а также показателей роста и развития растений в присутствии и при отсутствии фитопатогенных грибов. Вегетационные опыты по изучению стимулирующего эффекта бактерий-антагонистов производились в условиях климат-камеры и теплицы, а опыты с фитопатогенами - в контролируемых условиях фитотрона.
Получение штаммов микроорганизмов.
Выделение и отбор фитопатогенных грибов и бактерий-антагонистов, проявляющих к ним антагонистическое действие.
Грибы рода Fusarium были выделены со следующих полей: Хозяйство "Воля" - гриб Fus 1, Каладжинская (поле №67, 39Га), гриб с индексом Рас 6.1; Упорная (поле №154, 43 Га), гриб с индексом Рас 6.2.1; Упорная (поле №154, 43 Га), гриб с индексом Рас 6.2.2; 154, 43 Га), гриб с индексом Рас 6.2.2; Отрадненское (поле №68, 68 Га) гриб с индексом Отр 1, Переправная (поле №6, 117 Га) гриб Fus 5.
Первичный скрининг, направленный на выделение штаммов, проявляющих антагонизм, использовалась культура гриба Fusarium graminearum (Fus1).
Выделение штаммов-антагонистов осуществлялось по следующей схеме: 10 г почвы растирали в фарфоровой ступке резиновым пестиком, затем добавляли 100 мл стерильной воды.
Для выделения из почвенных образцов штаммов-антагонистов, относящихся к классу Bacilli, производилась пастеризация данной почвенной суспензии 20 минут при температуре 90°С. При этом сохранялись только споры бактерий-антагонистов данного класса.
На плотную питательную среду Сусло агар газоном засевалась культура гриба, относительно которой производился отбор штаммов бактерий-антагонистов. Поверх культуры гриба наносилась полученная почвенная суспензия (в объеме 50 мкл) и растиралась с помощью шпателя Дригальского до полного высыхания поверхности среды. Инкубировали 3 суток при температуре 25°С.
Отбор производился по принципу есть антагонизм/нет антагонизма. Отбирались все бактерии, демонстрировавшие зону подавления роста гриба вокруг своих колоний (Фиг. 1). Всем штаммам присваивались названия на основании почвенного образца, из которого данный штамм был выделен.
Дальнейший этап работы заключался в определении степени выраженности выделенных на первом этапе 1040 штаммов по отношению к другим 4 видам рода Fusarium.
В центр чашки с плотной питательной средой Сусло агар наносили крючком биомассу исследуемого гриба, по краям чашки -петлей исследуемые бактерии. Инкубировали 7 суток при температуре 25°С. После инкубации измеряли размер зоны подавления роста между грибом и бациллами (Фиг. 2).
В результате данного этапа работы, было выявлено, что из 1040 первично выделенных штаммов наибольшую степень антагонизма проявляли 212 штаммов. В ходе оценки степени подавления роста всех 5 штаммов грибов из 212 штаммов было получено 37 штаммов с наибольшей антагонистической активностью. Степень антагонистической активности данных штаммов.
Также были проведены исследования с помощью метода микрокультуры на стеклах (Фиг. 3), подтвердившие высокую антагонистическую активность выделенных культур.
Отбор штаммов, наиболее перспективных для биотехнологического использования.
После выделения 37 наиболее активных культур они были исследованы на предмет наличия свойств, позволяющих этим штаммам быть использованными для разработки способа получения комплексного противофузариозного биопрепарата.
К числу таких свойств относились:
А) Способность штаммов к формированию устойчивых биопленок, что важно для эффективности колонизации корней проростков пшеницы, а также имеет значение для технологического процесса твердофазной ферментации при наработке препаративной формы.
Б) Способность культур к росту при повышенной температуре, что позволяет подавлять развитие посторонних бактерий-антагонистов при наработке препаративной формы.
Биопленкообразование является одним из наиболее важных генетически детерминированных процессов в жизни бактерий-антагонистов. В протекании многих болезней человека участвуют бактерии, пребывающие в состояния биопленок. Благодаря «чувству кворума» в этой структуре, они решают жизненно важные задачи для своего выживания в тяжелых условиях внешних среды. Осуществляется горизонтальный перенос генов, повышается резистентность к антибиотикам, происходит кооперирование с другими видами. Все это помогает им в постоянной жесткой борьбе, как в организме человека, так и в почве, на корнях растений. Поиск активных штаммов-биопленкообразователей может являться перспективным для использования в биотехнологии, так как образование биопленки тесно связано с колонизацией корневой системы растения. Выявление генетических закономерностей данного процесса у микробиоты, а также изучение морфологии, разнообразия и специфичности свойств биопленок у отдельных видов и штаммов может играть ключевую роль как в разработке лекарственных средств нацеленных против них, так и в создании биопрепаратов для растений, использующих эти образования.
Бактерий рода Bacillus, выделенных из ризосферы озимой пшеницы, выращивали на агаризованной среде LB. Были получены чистые культуры, которые были исследованы на способность к биопленкообразованию. Была использована стандартная методика культивирования и окрашивания генциановым фиолетовым клеток (Stepanovic et al., 2000).
Контроль представляет собой не инокулированную среду.
Культивирование и окрашивание проводили по следующей методике:
1. В 3 лунки на 1 культуру вносили по 200 мкл бактериальной суспензии. Негативный контроль - питательный бульон без культуры.
2. Инкубировали планшет 24 часа, при 30 С°.
3. Удаляли бульон, промыть 3 раза 250 мкл стерильным физиологическим раствором.
4. Закрепляли биопленку 200 мкл абсолютным этанолом.
5. Через 15 минут удаляли этанол, и давали высохнуть.
6. Окрашивали 5 минут, 200 мкл 1% кристаллическим фиолетовым.
7. Промывали под проточной водой.
8. Просушивали, добавляли 200 мкл этанола, инкубировали планшет 5 минут.
9. Измеряли оптическую плотность при длине волны 57 0 нм на FLUOstarOmega.
Статистическую обработку проводили по критерию Стьюдента. Все представленные результаты статистически достоверны.
В результате проведенного скрининга удалось установить наиболее перспективные штаммы для дальнейшего исследования генетических основ биопленкообразования (Фиг. 4.).
Для оценки способности к росту при повышенных температурах тестируемые штаммы высевались на плотную среду LB и культивировлись при температурах от 25 до 42°С.Для этого исследуемые штаммы высевались на твердую питательную среду Сусло-агар и помещались на 3 суток в термостаты с температурой 25, 36 и 42°С. Раз в сутки снимались данные о степени роста штаммов (Фиг. 5). Данные представлены в таблице 1.
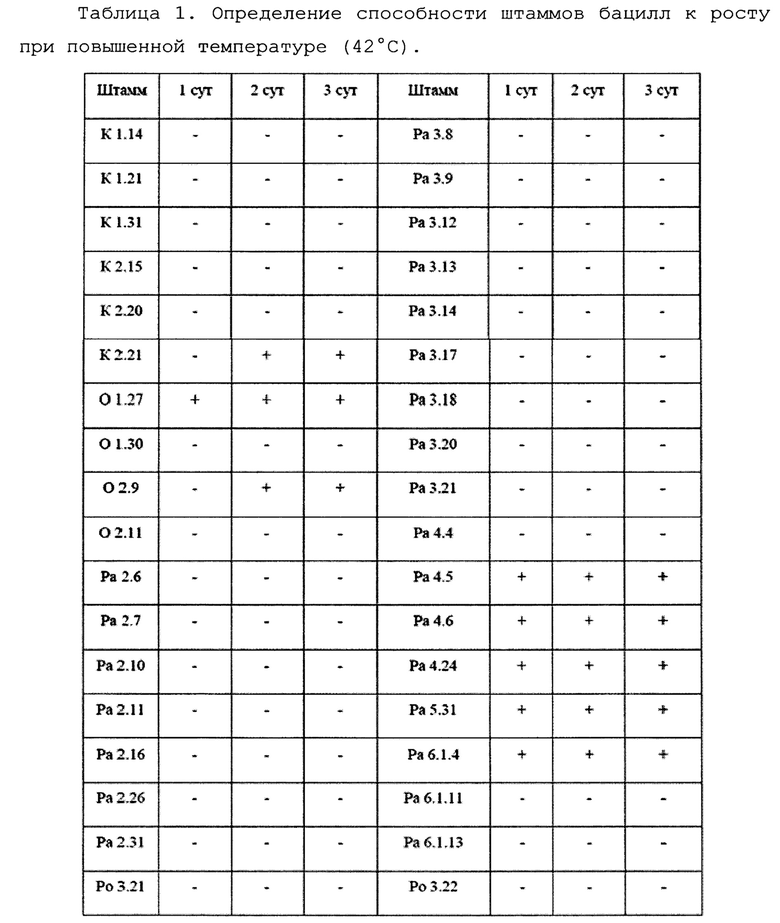
При окончательном отборе 10 наиболее эффективных штаммов учитывалась степень антагонистической активности по отношению к разным штаммам фитопатогенных грибов p. Fusarium, а также уровень биопленкообразования и устойчивость к высоким температурам.
Идентификация отобранных штаммов бактерий-антагонистов и фитопатогенных грибов использовавшихся для их тестирования.
Отобранные по комплексу признаков 10 лучших штаммов бактерий-антагонистов, а также культуры фитопатогенных грибов p. Fusarium были переданы во ВКПМ для проведения их идентификации до уровня вида.
При этом были получены следующие сведения:
По бактериям:
K 1.14, R 4.24 - Paenibacillus jamilae
О 1.27, О 2.11, R 3.13, R 4.5, R 6.14 - Paenibacillus peoriae 18
R 5.31 - Paenibacillus polymyxa
R 4.6, V 3.14 - Bacillus amyloliquefaciens
Таким образом, была проведена молекулярно-генетическая идентификация 10 штаммов спорообразующих бактерий-антагонистов и 6 штаммов фитопатогенных грибов, из которых 5 относятся к роду Fusarium.
Бактериальные культуры были депонированы во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) по процедуре национального патентного депонирования.
Проверка фунгицидной активности 10 отобранных лучших культур в модельных опытах на растениях.
Фунгицидная активность как отдельных препаратов, так и смеси 10 препаратов была проверена в системе пшеница/гриб/бактерия в условиях теплицы.
В присутствии растений взаимбдействие фитопатогенов и их антагонистов может отличаться от их прямого взаимодействия в условиях микробиологического эксперимента.
Для успеха защитного действия на растения бактерии-антагонисты должны обладать рядом свойств, наиболее важные среди которых:
- отсутствие фитопатогенного действия на растения самих бактерий-антагонистов;
- способность к эффективному подавлению фитопатогенной флоры на поверхности прорастающих семян
- способность к колонизации корневой системы растений-продуцентов
- способность к эндофитному росту (не обязательное, но желательное условие).
Тестирование влияния штаммов бактерий-антагонистов на всхожесть, энергию прорастания, а также показатели роста и развития растений в присутствии и при отсутствии фитопатогенных грибов.
На первом этапе исследований была поставлена задача проверить отсутствие фитопатогенного действия на растения самих бактерий-антагонистов (при отсутствии фитопатогенных грибов).
Фитопатогенными являются организмы, подавляющие нормальное развитие растения и приводящие к его гибели. Фитопатогенность может быть не абсолютной и не приводить к гибели растения, но значительно ослаблять и приводить к потере или значительному снижению продуктивности.
Для оценки влияния бактерий-антагонистов на начальные стадии роста и развития растений поверхностно стерилизованные семена обрабатывали бактериальной суспензией из расчета 3×107 КОЕ на 10 г семян и помещались на чашки Петри и в пластиковые контейнеры. В качестве подложки для прорастающих семян в чашках Петри использовали двойной слой фильтровальной бумаги, увлажненной водопроводной водой, в пластиковых контейнерах -песок, увлажненный специальным раствором для выращивания пшеницы.
В отношении тестированных штаммов (Таблица 2) не было отмечено признаков фитопатогенности на ранних стадиях развития. Ни один из протестированных штаммов не показал резкого подавления развития, хотя для некоторых отмечалась небольшая задержка роста на чашках Петри, при этом рост на песке не показывал видимых отличий от контроля. Ни в одном случае не было отмечено очагов некроза тканей.
Некоторые штаммы демонстрировали эффект стимуляции роста. При этом для одних отмечалось увеличение вегетативной массы побегов, для других - корневой системы (Фиг. 6).
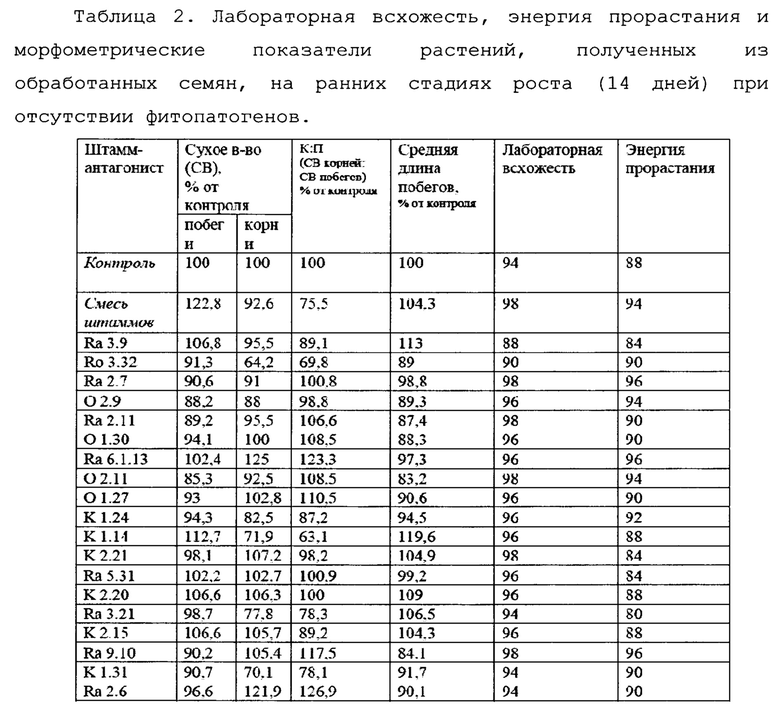
Для уверенности в отсутствии негативного влияния бактерий-антагонистов на всхожесть семян применяли метод высева семян на газон тестируемых микроорганизмов. В этом случае концентрация бактерий-антагонистов многократно превышала обычно применяемую и позволяла оценить возможный негативный эффект в случае передозировки препарата.
Проращивание на бактериальном газоне для тестируемых штаммов показало, что прорастание в большинстве случае задерживается, однако всхожесть не уменьшается, а проростки не несут признаков нарушения развития.
На следующем этапе работ был поставлен опыт в условиях теплицы. Растения были выращены из семян, обработанных тестируемыми штаммами бактерий и их смесью.
Было установлено, что в условиях теплицы растения, обработанные смесью штаммов, демонстрируют нормальный рост и развитие, что говорит об отсутствии угнетающего действия бацилл на растения пшеницы.
На завершающем этапе исследований было проведено исследование всхожести, энергии прорастания и показателей роста и развития растений в присутствии фитопатогенов. Для этого использовались семена пшеницы сорта "Гром", обработанные смесью 10 вышеуказанных штаммов (биопрепарат с рабочим названием Ф-1) в сочетании со стандартным протравителем "Шансил-Трио", а также семена, обработанные только препаратом 23 Шансил-Трио, и необработанные семена в качестве контроля. Как показал микробиологический анализ, данные семена в значительной мере заражены грибами p. Fusarium (Таблица 3).
Для этого был поставлен опыт на растениях пшеницы в условиях песчаной культуры (N=210 растений на один вариант опыта). Данные приведены в таблице 3.
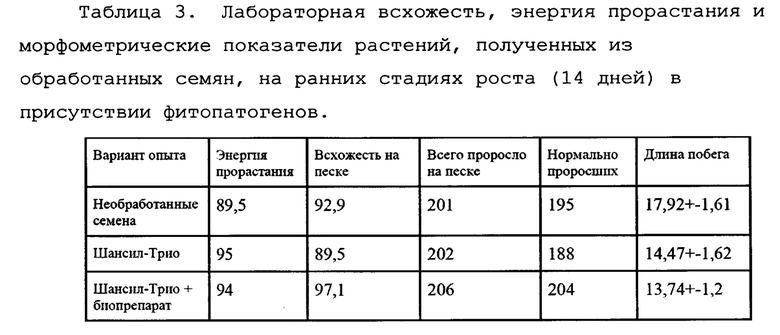
Как видно из таблицы 3, оба варианта обработки семян препаратами приводили к повышению энергии прорастания, а также уменьшением длины растений по сравнению с контролем. Это связано с угнетением фитопатогенных грибов p. Fusarium, которые, как известно, продуцируют гибберелины, что приводит к быстрому вытягиванию растений.
Однако, обработка биопрепаратом приводила к более значительному эффекту, достоверно отличающемуся от контроля (Фиг. 7.).
Кроме того, значительно больше становился процент нормально проросших и развившихся растений, как по сравнению с необработанным контролем, так и с химически обработанными семенами (4й столбик).
Оценка фунгицидной активности препаратов при обработке семян.
Десять тонн семян пшеницы сорта "Гром", обработанных ранее препаратом Шансил Трио были обработаны 100 л водной суспензии биопрепарата Ф-1 с помощью протравливателя ПС-10. Контрольным образцом служили семена необработанные, исследовали также семена обработанные только препаратом Шансил Трио по стандартной технологии также с помощью протравливателя ПС-10.
Примерно 1 грамм семян (21 семя) помещали в пробирку и заливали 10 мл стерильной воды. Интенсивно встряхивали в течение 10 мин. Водную фазу поверхностно на чашки с твердой питательной средой КСА, без добавления антибиотика ципрофлоксацина и с добавлением ципрофлоксацина в концентрации 10 мкг/мл с целью подавления роста бактерий-антагонистов.
Способность к эффективному подавлению фитопатогенной флоры на поверхности прорастающих семян может значительно снизить угнетающий эффект фитопатогенов и дать преимущество обработанным семенам уже на начальных стадиях роста.
Семена обрабатывали либо препаратом "Шансил трио", либо препаратом "Шансил трио" и препаратом на основе полученных бактерий-антагонистов, либо оставляли без обработки.
1 грамм семян (21 семя) из каждой группы помещали в пробирку и заливали 10 мл стерильной воды. Интенсивно встряхивали в течение 10 мин. Водную фазу поверхностно высевали на чашки с твердой питательной средой КСА, без добавления ципрофлоксацина и с добавлением ципрофлоксацина в концентрации 10 мкг/мл с целью подавления роста бактерий-антагонистов.
Чашки инкубировались 2 сут.при температуре 25°С, затем был произведен подсчет бактериальных колоний. После чашки инкубировались еще 5 сут.при той же температуре для выявления роста грибов.
Колонии грибов были промикроскопированы с целью определения рода гриба.
Полученные данные представлены в таблице 4.
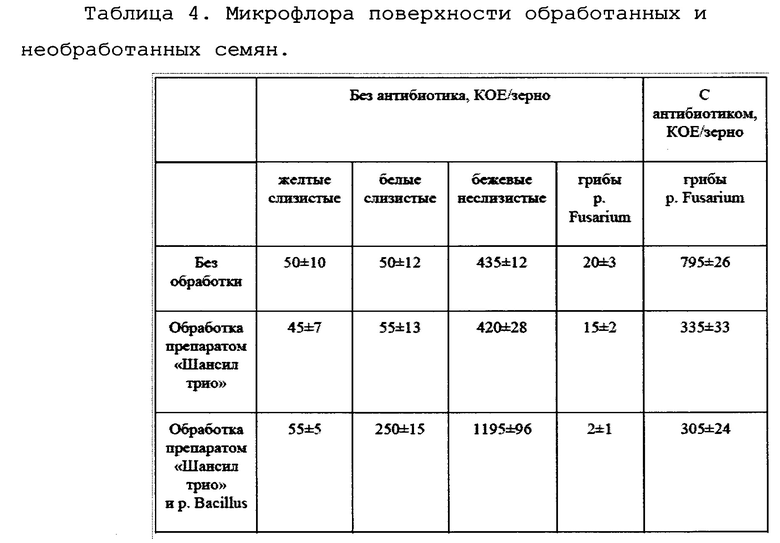
Всего на чашках выделялось три морфологически отличных друг от друга типа колоний. К первому типу относились желтоватые слизистые колонии, бактерии относятся у неспорообразующим микроорганизмам. Белые слизистые и бежевые неслизистые колонии принадлежали спорообразующим бактериям р. Bacillus разных видов. Обсемененность бактериями семян без обработки и обработанных препаратом «Шансил трио» достоверно не отличалась друг от друга, однако, на семенах, обработанных препаратом «Шансил трио» и препаратом бацилл, было зафиксировано значительно большее количество бактерий р. Bacillus, что связано с обработкой препаратом бацилл.
На чашках с совместным ростом бактерий-антагонистов и грибов отмечается единичный рост грибов p. Fusarium (по данным микроскопирования). На чашках с антибиотиком, подавляющим рост бактерий-антагонистов, отмечается значительное количество бактериальных колоний, причем на обработанных семенах количество КОЕ гриба значительно меньше, больше, чем в два раза.
Следует отметить, что смыв с семян, обработанных только препаратом «Шансил Трио», содержал значительное количество спор не фитопатогенных грибов p. Mucor
Таким образом, исследования микрофлоры семян показали следующее:
1. На поверхности необработанных семян содержится достаточное число (около 500 КОЕ/семя) «диких» спорообразующих бактерий-антагонистов, в основном бацилл.
2. Эти бациллы способны подавлять рост грибов p. Fusarium, так как в случае их уничтожения с помощью антибиотика грибов вырастает примерно в 40 раз больше. То есть когда производится смыв с поверхности семян грибов и живых бактерий-антагонистов водой, а затем посев на чашку, бактерии убивают большинство грибов.
3. Стандартная протравка Шансил Трио убивает примерно половину грибов (см. столбец 5).
4. Добавление выращенных нами бацилл (это в сущности те же, что и на зерне, но мы выбрали и вырастили штаммы с максимальной антигрибковой активностью) значительно, примерно в 10 раз, снижает число грибов (см. столбец 4).
5. Обработка нашим препаратом в дозе 7 кг на 10 тонн пшеницы увеличивает число находящихся на поверхности семян бацилл примерно в три раза. Очевидно, что 2/3 бацилл на поверхности обработанных семян принадлежат штаммам заявленного препарата.
Следует отметить, что обработка семян биопрепаратом снижала численность грибов на поверхности семян в 2,6 раза (см. столбец 5). При совместном же росте на питательной среде число колоний грибов снижается значительно сильнее (см. столбец 4). Это связано с тем, что бактерии биопрепарата активны в вегетативном состоянии и начинают активно работать только при прорастании семян, а значит, прорастании грибов и бацилл и их взаимодействии.
Это можно наблюдать в следующем опыте, когда семена обрабатывали препаратами бактерий-антагонистов и раскладывали на твердой питательной среде КСА, давая им прорасти.
Эффективность подавления грибов оценивали по количеству очагов естественной фитопатогенной флоры.
Во всех случаях число очагов плесневения резко снижалось по сравнению с контролем. В условиях роста на бактериальном газоне наблюдалось резкое подавление роста имеющейся фитопатогенной микрофлоры (Фиг. 8).
Таким образом, доказано, что заявленный биопрепарат работает. Он не является чужеродным для агроэкосистемы пшеничного поля и усиливает естественную фунгицидную активность бактериальной микрофлоры пшеницы.
Способность к колонизации корневой системы растений-продуцентов желательна для обеспечения постоянного присутствия защитных микроорганизмов в зоне роста корневой системы.
Не все бактерии способны выживать на корневой системе растения или вблизи нее. Причиной гибели микроорганизмов может стать реакция иммунной системы растений. Для ряда штаммов-антагонистов показана хорошая приживаемость на корнях (Фиг. 9).
При колонизации корней бактерии вытесняют грибы из ризосферы и с ризопланы, что затрудняет их проникновение внутрь растения. Активное противодействие бактерий-антагонистов колонизации растений фитопатогенными грибами рассматривается как дополнительное преимущество, поскольку позволяет надеяться на подавление роста грибов p. Fusarium во внутренних полостях растения. Известно, что основным местом развития Fusarium graminearum - возбудителя твердой головни пшеницы - являются полые стебли злаков. Внутри стебля может накапливаться большая масса гриба, которая после уборки урожая остается на пожнивных остатках и является основной причиной распространения болезни в следующем году. Уменьшение количества грибов p. Fusarium внутри стебля наряду с работой по обезвреживанию пожнивных остатков поможет снизить общую зараженность территорий.
Семена пшеницы обрабатывались одновременно суспензией грибов p. Fusarium и бактериями-антагонистами. В качестве контроля использовали растения, обработанные только грибами р. Fusarium. Растения выращивали в условиях теплицы в сосудах с естественной почвой в течение 4 недель. После этого растения извлекались из сосудов, отмывались от остатков почвы. Стебли, листья и корни поверхностно стерилизовались и помещались на среду, способствующую росту грибов.
Из эксплантов всех органов растений - корней, стеблей и листьев, обработанных только грибами p. Fusarium, наблюдалось интенсивное прорастание гриба. В случае растений, обработанных одновременно грибами p. Fusarium и штаммом-антагонистом, роста грибов p. Fusarium не отмечалось.
Таким образом, на данном этапе было установлено, что как отдельные штаммы бактерий-антагонистов, составляющие основу разрабатываемого препарата, так и смесь 10 отобранных наиболее перспективных штаммов, не демонстрируют фитопатогенного действия на растения, проявляют антагонистическую активность по отношению к фитопатогенным Fusarium sp., а также препятствуют колонизации растений патогенами.
Разработка способа изготовления биопрепарата с антифузариозной активностью на основе твердофазной ферментации спорообразующих бацилл на растительном субстрате.
Ниже представлено описание низкозатратного способа изготовления небольших партий биопрепарата спорообразующих бактерий-антагонистов с антифузариозной активностью на основе твердофазной ферментации спорообразующих бацилл на растительном субстрате с применением стандартного лабораторного оборудования.
Твердофазная ферментация дает возможность добиваться высоких титров целевых штаммов без применения сложного специализированного оборудования. Предыдущие исследования показали, что оптимальными субстратами для спорообразующих бацилл являются гидратированные соевые бобы, прошедшие термообработку автоклавированием при 121°С. Максимальный титр целевых штаммов бактерий-антагонистов достигался при выращивании при температурах 37°С - 42°С и рН 6,8. Наилучшим способом сушки биопрепаратов, полученных твердофазным способом, является сушка на воздухе. Помимо сои в качестве субстрата может использоваться горох и подсолнечный шрот.
Полученные результаты послужили основой для разработки низкозатратной технологии для производства небольших количеств препарата на основе спорообразующих бацилл при помощи неспециализированного бытового и стандартного лабораторного оборудования персоналом, не имеющим специального образования.
Главным преимуществом данного способа является ее легкая воспроизводимость, технологичность и стабильность, использование простой по составу, недорогой среды, высокий выход готового продукта, стабильность полученного препарата при хранении.
Экспериментальная оценка эффективности выращивания споробразующих бактерий-антагонистов с антифузариозной активностью с применением лабораторного и бытового оборудования.
В качестве субстрата для твердофазной ферментации были выбраны соевые бобы, эффективность которых для выращивания бацилл была показана ранее (Chistyakov et al., 2015). По содержанию белка, жира, фосфатидов и некоторых других питательных веществ соевые бобы значительно превосходят многие масличные и злаковые культуры. В зависимости от места и условий произрастания сои содержание питательных веществ 30
может меняться в значительных пределах (белок от 29 до 50,3%; жир - от 13,5 до 25,4%, а сумма белка и жира - от 52 до 65%).
Суть разработанного способа твердофазного культивирования состоит в эксплуатации способности технологически значимых штаммов спорообразующих бактерий-антагонистов образовывать на поверхности питательных субстратов биопленки. Формируя биопленки, бактерии образуют квазимногоклеточное сообщество, обладающее повышенной, по сравнению с планктонной формой, устойчивостью к деструктивным факторам среды. Кроме того, бактерии целевых штаммов, образовавшие равномерно покрывающую субстрат биопленку, подавляют развитие гнилостной микрофлоры. Эти особенности создают для твердофазной ферментации предпосылки технологической пластичности, недостижимые при жидкофазной ферментации.
Однако, разрабатывая реальную технологию твердофазного культивирования, необходимо учитывать, что в производственных условиях риск контаминации патогенной микрофлорой значительно выше, чем в лабораторных условиях. Эффективным средством предотвращения контаминации является культивирование бактерий-антагонистов при повышенной температуре.
В ходе исследований была проведена проверка возможности воспроизведения оптимальных параметров роста использованных в изобретении бактерий-антагонистов при температуре 42°С. На основе полученных данных была проведена корректировка способа культивирования целевых штаммов бактерий-антагонистов, а также набора оборудования.
Для разработки технологии использовали 10 выделенных наиболее эффективных штаммов спорообразующих бактерий-антагонистов (Таблица 5).
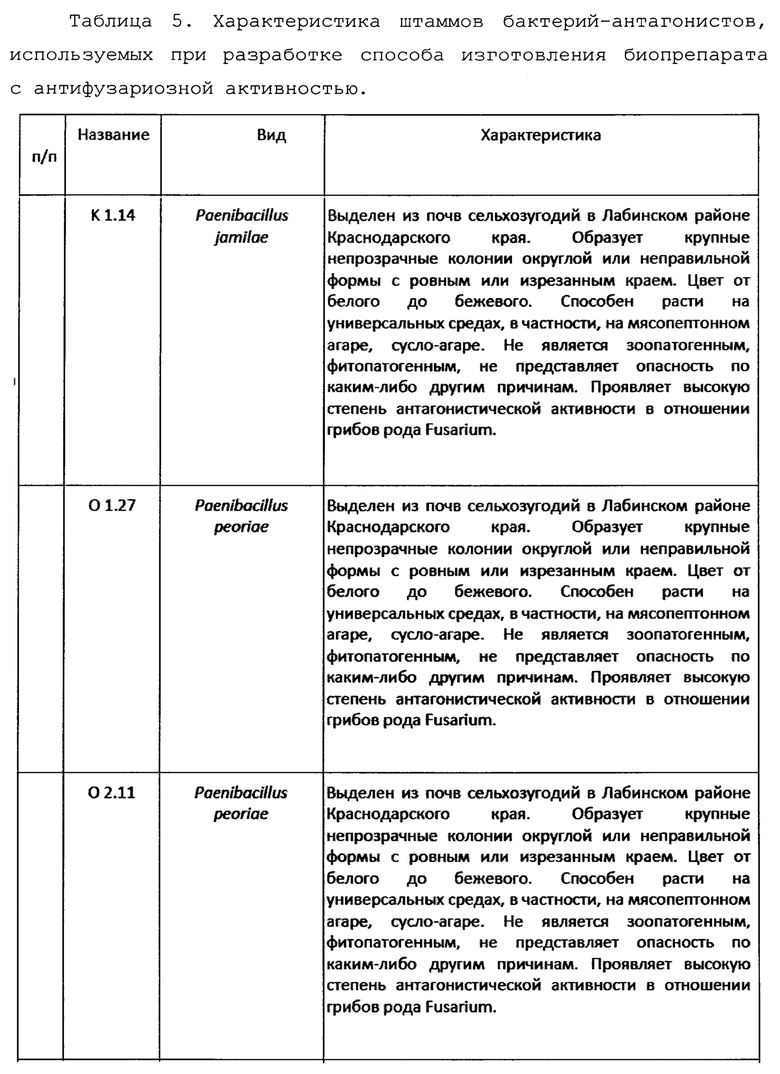
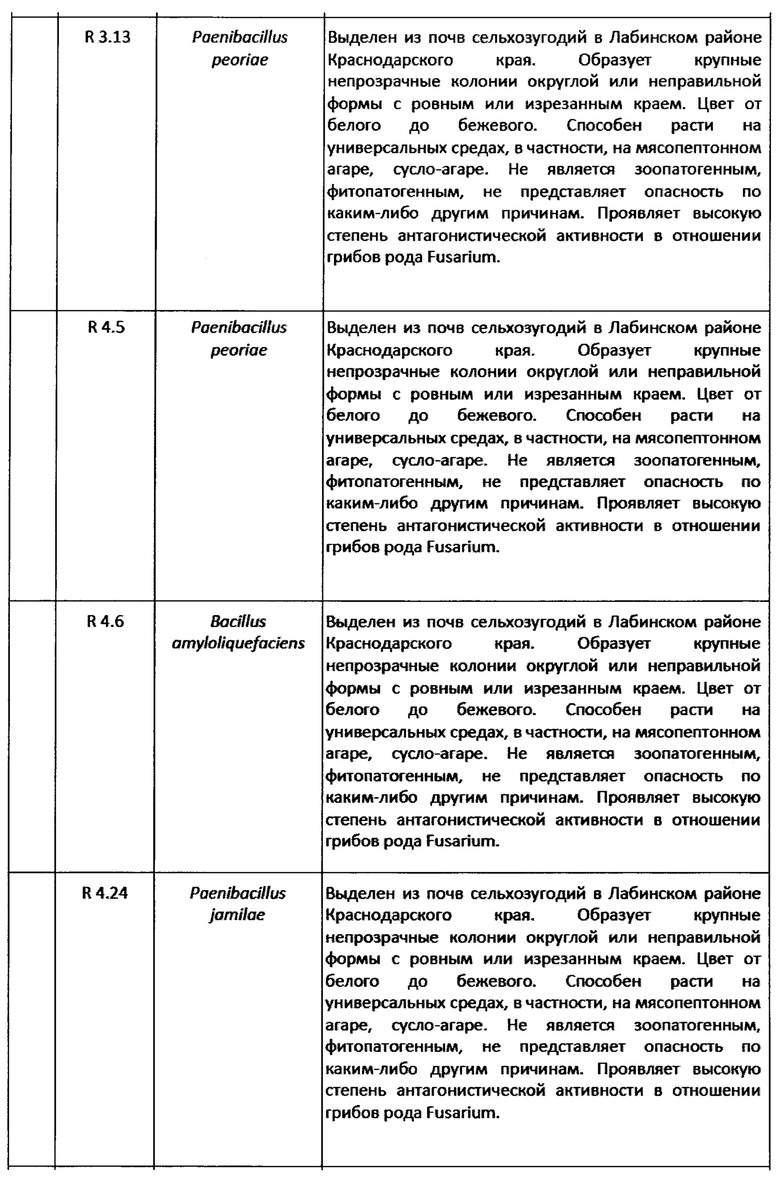
Согласно заключению ВКПМ, работа с данными штаммами не требует специальных мер предосторожности. Подготовка сырья.
Субстратом для твердофазного культивирования служила соя (сорт «Лань»), гидратированная в воде комнатной температуры в течение 12 часов. Объем воды, необходимый для отмывки соевых бобов, определяли эмпирически, визуально контролируя отсутствие на их поверхности пыли и грязи. Оптимальным объемом воды для замачивания 1 кг субстрата является 1 л.
Термообработка субстрата.
Термообработку субстрата для выращивания биопрепарата проводили в автоклаве («Sanyo» 3781(b), Япония) 20 минут при 120°С. Итоговая влажность субстрата составляла при обработке в автоклаве 60%.
При термообработке соевых бобов ключевым фактором является соотношение температуры и времени обработки. Температура и продолжительность термообработки субстрата обеспечивающая полную стерилизацию при термообработке, была подобрана на основании предварительных опытов с количествами субстрата по 1 кг и составила 20 минут при 120°С.Эффективность термообработки оценивали по отсутствию мезофильных аэробных и факультативно анаэробных микроорганизмов в центре пробы. Из центра пробы брали по три образца массой 10 г (ГОСТ 26669-85). Определение КМАФАнМ проводили согласно ГОСТ 10444.15-94.
Исходная величина рН соевого субстрата составила 7,0-7,2.
Инокуляция.
Для инокуляции 1 кг термообработанных гидратированных соевых бобов использовали одну чашку Петри с агаром LB или Сабуро с выросшим на ее поверхности газоном целевого штамма. При этом на 1 кг субстрата приходится более 1,0 - 3,0×108 КОЕ адресного штамма. При подборе времени инокуляции главный параметр - равномерность перемешивания инокулята и субстрата.
Инкубация.
Использование твердофазной ферментации позволяет достичь высокой технологической гибкости. При сохранении указанных выше параметров инокуляции, ферментация может проводиться на любых поддонах из пищевых материалов, в том числе на пластиковых, эмалированных подносах и лотках и на одноразовых поддонах, изготовленных из полиэтиленовой пленки.
В проведенной серии экспериментов твердофазное выращивание осуществляли в бытовых пластиковых подносах и поддонах из гофрокартона, выстеленных полиэтиленовой пленкой, в слое субстрата толщиной 2-3 см продолжительность выращивания 4 8 ч.
Выращивание проводили в термостате (ТС-80, Россия), который позволяет проводить инкубацию при температуре 42±0,2°С.
Рост культур оценивали по содержанию жизнеспособных клеток (КОЕ), определяемого путем высева серийных разведений культуры на плотную среду (МП7А). Опыты проводили в трех повторностях. Гомогенизация.
Гомогенизация соевых бобов, размягченных при термообработке, и становящихся еще мягче в процессе ферментации, не представляет технологической проблемы. Бытовая электромясорубка Kenwood MG 700 мощностью 2000 Вт позволяет гомогенизировать до 3 кг субстрата в течение минуты. Таким образом, скорость гомогенизации ограничивается только скоростью работы персонала. Максимально простой и низкозатратный вариант гомогенизации может быть осуществлен при помощи ручной мясорубки, например, "Мясорубки ручной МА-С», "РПП" (г. Ровно, Украина).
Сушка.
Сушка биопрепарата осуществлялась при температуре 45°С.
После гомогенизации измельченный препарат распределяли тонким слоем (1-3 см) на пластиковых подносах и высушивали до влажности 10%.
Определение жизнеспособности бактерий-антагонистов.
Выживаемость культур оценивали по содержанию жизнеспособных клеток (КОЕ). Для подсчета количества КОЕ/г (колониеобразующих единиц/г) использовали стандартную методику параллельных разведений в физиологическом растворе и глубинного посева на плотные агаризованные среды.
Из каждого разведения делали по три таких посева. Чашки с посевами инкубировали в термостате при 37°С в течение 48 часов. Для каждой серии посевов подсчитывали среднее число колоний, выросших на трех чашках.
Одним из свойств бацилл служит способность к росту в виде биопленки. Согласно современным представлениям, биопленки состоят из живых и мертвых клеток, клеточных фрагментов и внеклеточных полимеров, закрепленных на поверхности (Bridier et al., 2017). Особенностью биопленок является образование бактериями слизи. Образующие ее внеклеточные полимерные вещества обеспечивают защиту от агрессивных агентов, дегидратации, ультрафиолетового излучения (Ngo Thi Na, Naumann, 2009; Ипполитов и др., 2002). Все эти особенности обеспечивают высокую экологическую и технологическую пластичность культур бацилл, полученных методом твердофазной ферментации.
Результаты исследований подтверждают, что использование соевых питательных субстратов в комплексе с экономичным способом твердофазного культивирования может стать основой создания низкозатратных технологий производства биопрепаратов.
Результаты по интенсивности роста исследованных микроорганизмов по окончании твердофазной ферментации на соевом субстрате в течение 24 часов представлены в таблице б. Оптимальные температуры роста различных штаммов были подобраны ранее в опытах на чашках Петри.
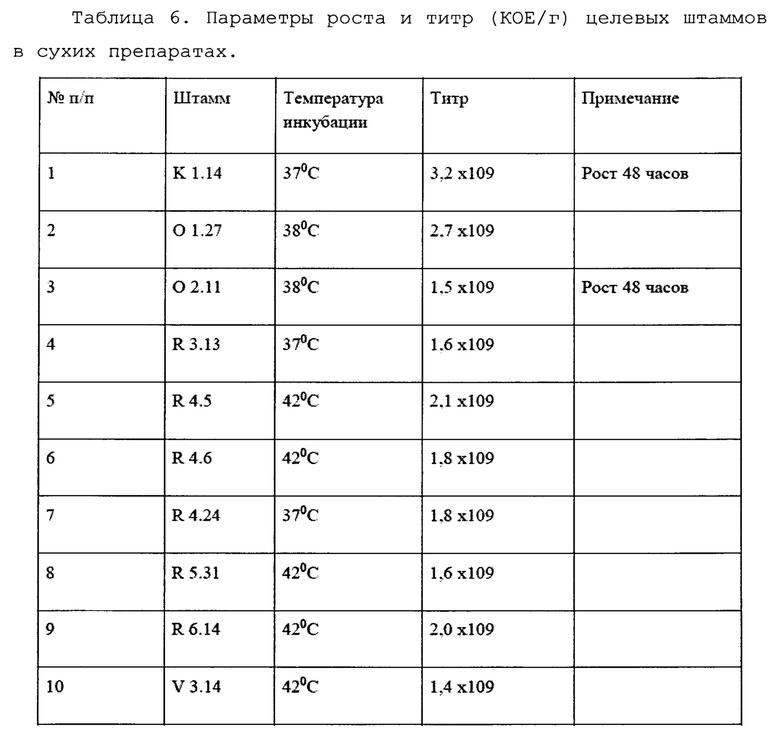
Пример осуществления способа, обеспечивающего получение с помощью стандартного лабораторного оборудования в течение семи дней 0,1 кг сухого биопрепарата (остаточная влажность не более 10%) с титром бактерий-антагонистов не менее 108 КОЕ/г на основе 10 заявленных штаммов спорообразующих бактерий-антагонистов.
Взвешивается необходимый объем соевых бобов.
Бобы промываются проточной водопроводной водой.
Гидратация - бобы сои замачиваются на 12 часов в воде в эмалированной емкости при температуре (22±1)°С.
Бобы сои стерилизуются в автоклаве при 120°С в течение 20 минут.
Для получения инокулята используется маточная культура. Сухая маточная культура должна быть получена из специализированной микробиологической лаборатории. Культура оживляется по стандартной методике переноса в бульон Сабуро, МПБ либо LB..
Проавтоклавированные бобы переносятся в емкость для инокулирования и охлаждаются до 60°С. Для инокуляции 1 кг термообработанных гидратированных соевых бобов использовали одну чашку Петри с агаром LB или Сабуро с выросшим на ее поверхности газоном целевого штамма. При этом на 1 кг субстрата приходится более 1,0 - 3,0×108 КОЕ адресного штамма.
Инокулированные соевые бобы раскладываются на поддонах (толщина слоя 10 см), закрываются полиэтиленовой пленкой. Поддоны с инокулированными бобами сои помещаются в инкубатор. Процесс инкубации должен длиться в течение 24-48 часов при температуре 37-42°С.
После каждой инкубации термостат-инкубатор подвергают влажной уборке с применением моющих средств и, затем, обрабатывают 3% перекисью водорода и высушивают. Пластиковые поддоны после инкубации обрабатывают таким же образом.
Ферментированный субстрат измельчается при помощи электрической либо ручной мясорубки.
Измельченный препарат распределяется тонким слоем на пластиковых подносах и высушивается при температуре 450°С до влажности 8-10%.
Высушенный препарат измельчают при помощи бытовой мельницы. Комплексный препарат получают смешиванием равных навесок отдельных препаратов.
Условия хранения биопрепаратов существенно влияют на жизнеспособность бактериальных клеток, входящих в состав препарата, и, соответственно, на срок его годности. Содержание влаги в препарате должно быть минимальным, что имеет первостепенное значение для сохранения жизнеспособности бактерий-антагонистов.
Готовый биопрепарат должен храниться в холодильнике при температуре (4±2)°С и влажности не более 10%.
Отработка молекулярно-генетических методов анализа результатов с целью последующего скрининга эффективности биопрепарата в полевых опытах.
Для количественного анализа степени зараженности образцов патогенными грибками был проведен анализ полимеразной цепной реакции (ПЦР-анализ) в режиме "реального времени". На первом этапе работ был произведен подбор родоспецифичных и видоспецифичных праймеров. Затем был проведен ПЦР-анализ с последующим электрофорезом и визуализацией в агарозном геле, для подтверждения наличия в пробах целевых фрагментов, а также предварительной идентификации видов Fusarium. Затем был проведен ПЦР-анализ в режиме "реального времени".
Подбор праймеров.
Высокочувствительная детекция патогена до заражения растений или до развития симптомов болезни важна для предотвращения их инфицирования или сдерживания развития и распространения заболевания а, следовательно, и для уменьшения экономических потерь. Диагностические процедуры должны обладать достаточно высокой чувствительностью. До открытия методов амплификации нуклеиновых кислот, в частности ПЦР, для обнаружения конкретного организма использовались методы, основанные на применении специфичных гибридизационных зондов. Однако, из-за низкой чувствительности такие методы зачастую приводили к получению «ложноотрицательных» результатов (Ward et al., 2004). Благодаря высокой чувствительности метод ПЦР дает возможность детекции даже следовых количеств ДНК патогена (Ward et al., 2004). Однако, высокая чувствительность метода может приводить к получению «ложноположительного» результата. Поэтому, для обеспечения достоверности получаемых результатов необходимо строгое соблюдение правил постановки реакции, использование контролей, ограждение образцов и реактивов от возможной ДНК-контаминации через аэрозоли, а также наличие отдельных зон для процедур пробоподготовки и анализа продуктов ПЦР (Kwok, Higushi, 1989).
На практике, для принятия решений о проведении мер по защите урожая, методы более чувствительные, чем традиционная ПЦР, вряд ли потребуются, поскольку пороговый уровень инфекции может быть легко детектирован с помощью ПЦР.
Для молекулярной диагностики фитопатогенов в качестве мишеней часто используются консервативные гены, нуклеотидные последовательности которых у близкородственных микроорганизмов зачастую отличаются лишь несколькими нуклеотидными заменами (single-nucleotide polymorphism, SNP).
Нуклеотидные последовательности для исследуемых штаммов, полученные при секвенировании вариабельных участков генов, кодирующих 16S рРНК, представлены в перечне последовательностей SEQ ID NO 1 - SEQ ID NO 10.
В настоящее время чаще всего используютсятабли:
1) Регионы ITS рибосомальной РНК (рРНК). В геноме грибов каждая копия рРНК представлена тремя генами, разделенными внутренними транскрибируемыми регионами (internal transcribed spacers, ITS),
По ней существуют обширные БД, высокая копийность генов рРНК в любом геноме обеспечивает высокую чувствительность ПЦР. ITS содержат консервативные и вариабельные области, что позволяет проводить классификацию грибов на разных таксономических уровнях.
Однако, нуклеотидные последовательности рРНК не всегда обладают достаточным уровнем вариабельности для различения отдельных видов (Tooleyetal., 1996).
2) гены «домашнего хозяйства»: гены бета-тубулина, актина, фактора элонгации трансляции 1-альфа (Абрамова, 2013); гены фосфатпермеазы (РНО), ген большой субъединицы АТФ-цитратлиазы (acl1).
Ген фактора элонгации трансляции 1-альфа (tef1α) широко используется в филогенетических исследованиях фузариевых грибов.
Преимущества заключаются в том, что его нуклеотидные последовательности высокоинформативны на уровне вида, его неортологические копии не были обнаружены для видов рода Fusarium, на основе последовательностей данного гена была создана база данных FusariumID, ген tef1α представлен в геноме единственной копией (Kristensen et al., 2005, 2007).
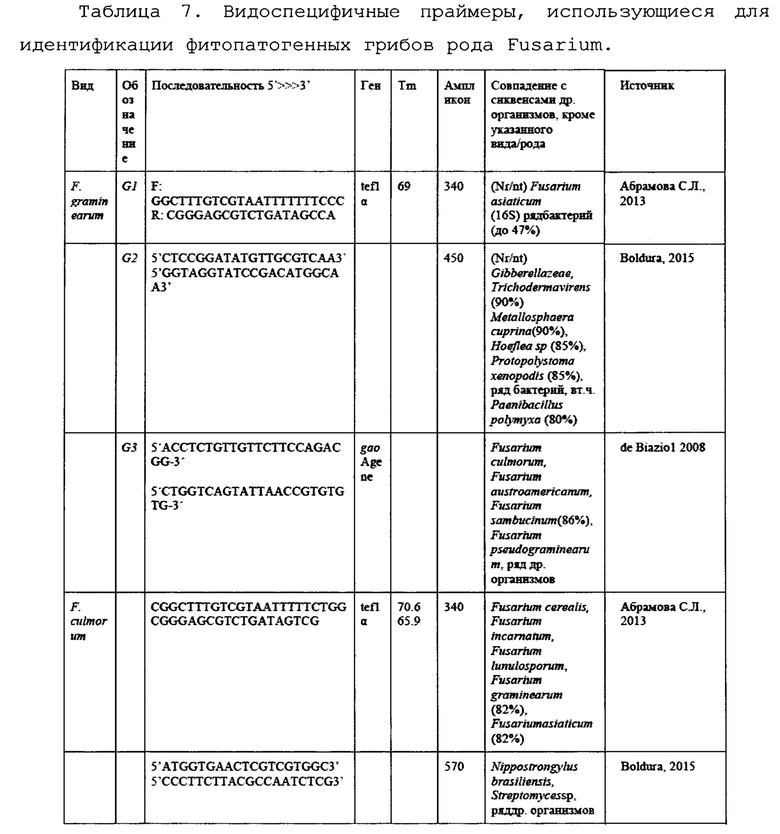
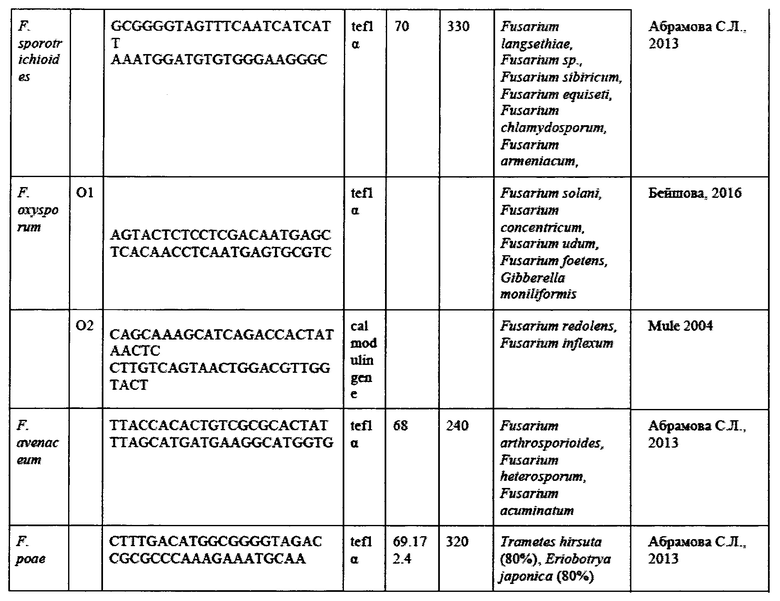
На основании анализа литературы были подобраны следующие праймеры для анализа (Таблица 7).
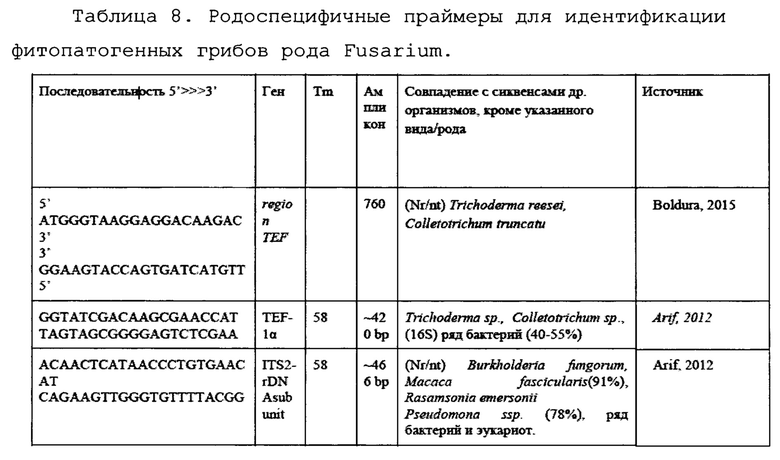
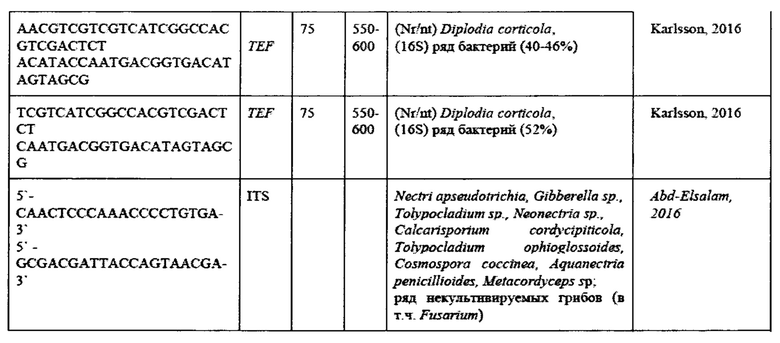
Проверка эффективности ПЦР для идентификации патогенов.
Методом ПЦР было установлено, что в зараженных образцах присутствуют два вида Fusarium: F. graminearum (образцы 2 и 3, фиг. 20) и F. oxysporum (образцы 4-9, Фиг. 20). Поэтому в количественном анализе в дальнейшем использовали видоспецифичные праймеры для этих двух видов, а также родоспецифичные для определения присутствия патогенов других видов.
Таким образом, была проведена работа по подбору праймеров для оценки присутствия фитопатогенов в тканях растений молекулярно-генетическими методами.
Ниже представлены опытные испытания заявленного биопрепарата.
Опыт №1. Влияние препарата на посевы озимой пшеницы, весна-лето 2018.
Учет зараженности ризосферной почвы грибами p. Fusarium Для изучения влияния обработки семян озимой пшеницы биопрепаратом Ф-1 осенью 2017. была проведена обработка 10 т семян с помощью ПС-10. Отбор проб был произведен в фазу кущения в апреле 2018 г. Опытное и контрольное поля были разбиты на 30 секторов под контролем GPS, и в каждом секторе были отобраны пробы растений. Отбор производили по методу конверта из 5 точек сектора для получения репрезентативной картины. В условиях лаборатории производился посев корней пшеницы с ризосферной почвой на среду Чапека, с последующим учетом и идентификацией грибов p. Fusarium. Идентификацию патогенов вели по морфологии колоний, с подтверждением при помощи световой микроскопии. Также отбирали биомассу патогенов для подтверждения принадлежности к p. Fusarium с помощью молекулярно-генетических методов. Данные по численности фитопатогенных грибов представлены в таблице 9.
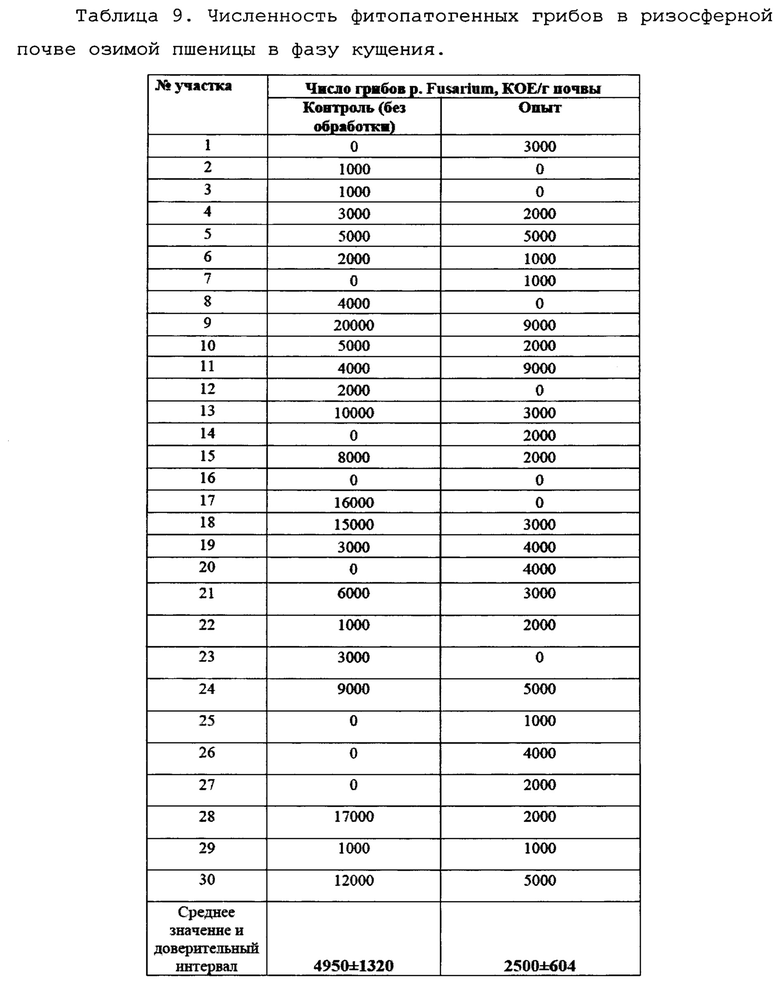
Как можно видеть, численность фитопатогенных грибов на опытном поле снижалась практически вдвое.
Видно также, что на обработанном участке отсутствуют пробы с содержанием грибов рода Fusarium более 9000 КОЕ/г.Это говорит о том, что испытываемый препарат хотя и не уничтожает фитопатоген полностью, но позволяет достаточно эффективно держать его под контролем.
Для статистической обработки данных был использован критерий Пирсона (Х-квадрат). В результате получено значение р=0,03501, что говорит о статистической значимости различий. Графическое представление данных приведено на фиг. 11.
Численность фитопатогенных грибов в ризосферной почве озимой пшеницы в фазу созревания представлена в таблице 10.
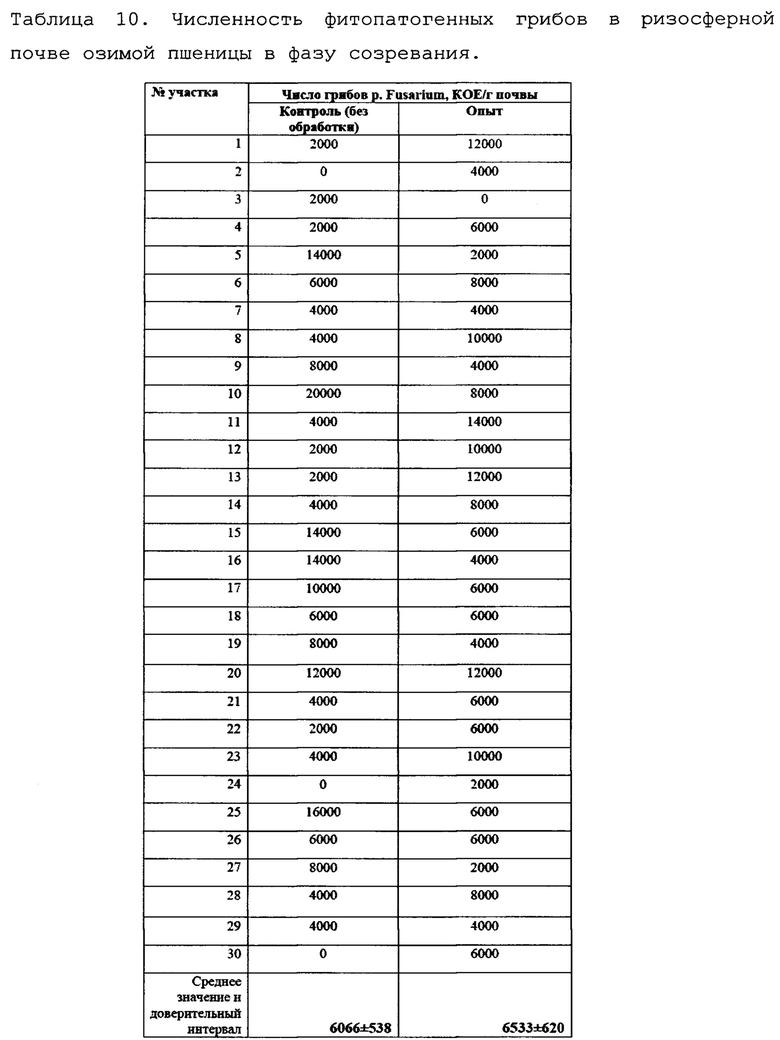
Как можно видеть, численность фитопатогенных грибов в ризосфере озимой пшеницы на опытном и контрольном поле к фазе созревания оказывалась примерно на одном уровне, что может быть связано с прекращением вегетации растений и низким количеством осадков. Данные представлены на фиг. 12.
Оценка влияния обработки биопрепаратом на естественное микробное сообщество почвы.
В весенний период было проведено исследование влияния обработки препаратом на общую численность плесневых грибов в исследуемых образцах. Данные по этому опыту приведены в таблице 11.
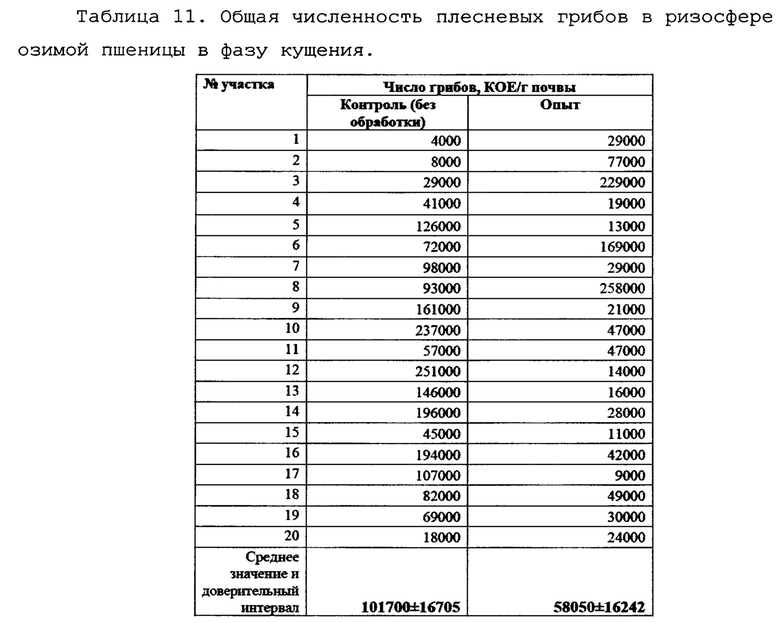
Из данных таблицы 11 видно, что обработка препаратом снижает численность плесневых грибов, то есть фунгицидное действие биопрепарата Ф-1 не ограничивается только грибами p. Fusarium и потенциально может служить инструментом для контроля и других грибковых инфекций.
Работы по исследованию влияния обработки семян озимой пшеницы биопрепаратом Ф-1 на зараженность растений грибами р. Fusarium.
В ходе изучения влияния биопрепарата Ф-1 на озимую пшеницу и ее пораженность фузариозом был произведен сбор колосьев пшеницы для определения морфометрических показателей растений (приводится в описании ниже). Было определено содержание грибов p. Fusarium в зерне озимой пшеницы. Для этого был произведен микробиологический анализ смывов с зерна пшеницы, отобранной на опытном и контрольном поле. Для смыва отбиралось 10 г зерна, которые обрабатывали 10 мл стерильной воды для смыва спор грибов, затем данные смывы высевали на питательную среду. Данные по численности грибов на зерне пшеницы приведены в таблице 12.
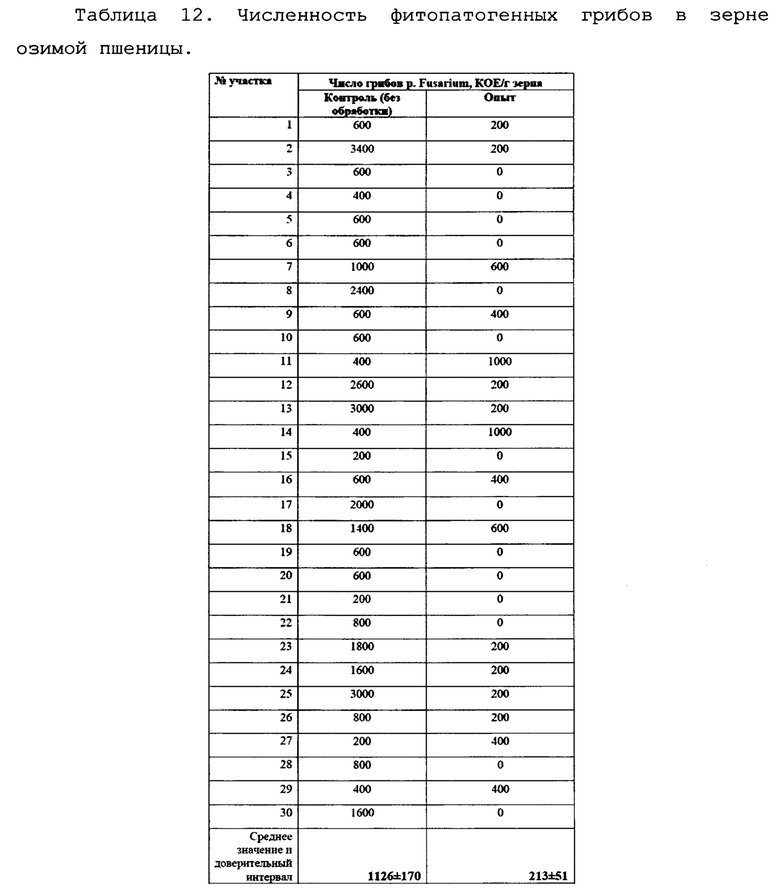
Как можно видеть, зерно в опытном варианте оказалось в гораздо меньшей степени заражено грибами p. Fusarium, по сравнению с контролем (Фиг. 13).
Различия с контролем составили 5,28 раз, кроме того, во многих пробах зерна с опытного поля фитопатогенные грибы отсутствовали полностью, чего не наблюдалось в зерне с контрольного поля. Также снизилась и общая численность грибов (Фиг. 14).
Работы по исследованию влияния биопрепарата Ф-1 на морфометрические показатели растений пшеницы.
Для оценки влияния биопрепарата Ф-1 на рост и развитие растений пшеницы в полевых условиях был проведен анализ основных морфобиометрических показателей растений пшеницы. Полученные результаты сведены в таблицу 13.
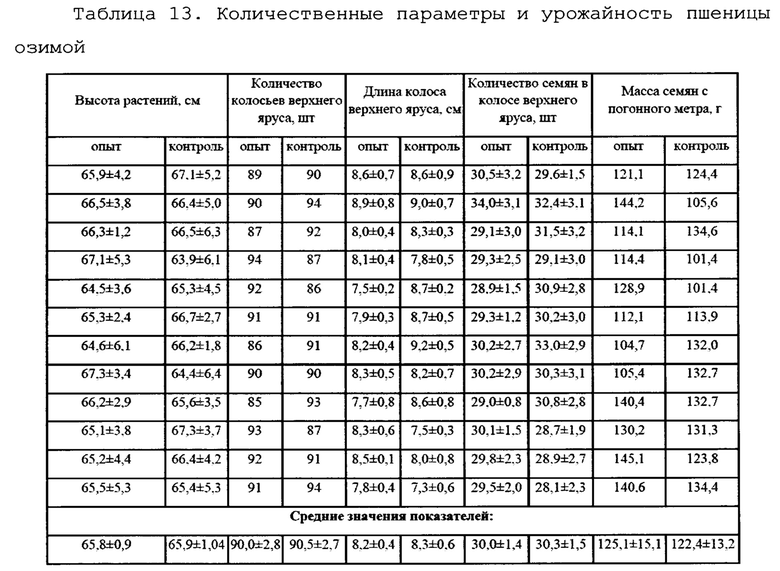
По итогам анализа и статистической обработки полученных данных установлено, что обработка биопрепаратом не повлияла достоверно ни на один из показателей развития растений, а также не повлияла на урожайность (оценивалась масса семян с погонного метра). Эти данные совпадают с данными, полученными в ходе предварительных опытов в 2017 году в условиях теплицы и фитотрона.
Опыт №2. Влияние препарата на посевы кукурузы, весна-лето 2018.
Работы по исследованию влияния обработки семян кукурузы биопрепаратом Ф-1 на зараженность почв и растений грибами р. Fusarium.
Весной 2018 года был заложен опыт по влиянию обработки семян кукурузы на пораженность растений грибами p. Fusarium. Семена кукурузы вручную обрабатывали биопрепаратом Ф-1, после чего был произведен посев. Отборы проб были проведены в фазу всходов и в фазу молочной спелости перед уборкой кукурузы на силос.
Проводили отбор ризосферной почвы с корней растений. Данные по микробиологическому анализу почвы представлены ниже.
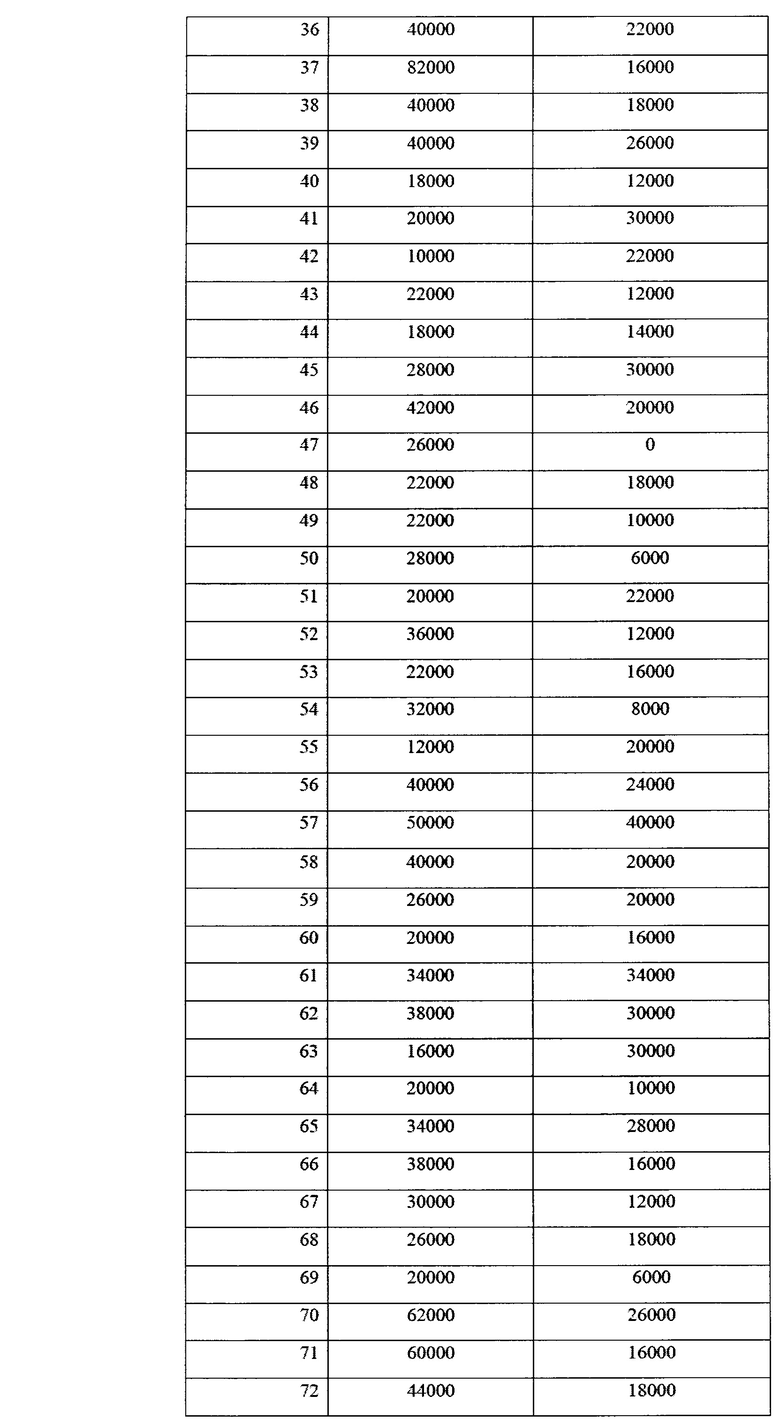
Учет зараженности ризосферной почвы грибами p. Fusarium представлен в таблице 14.
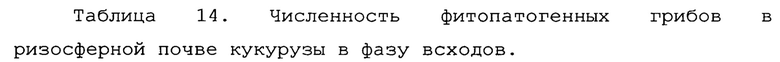
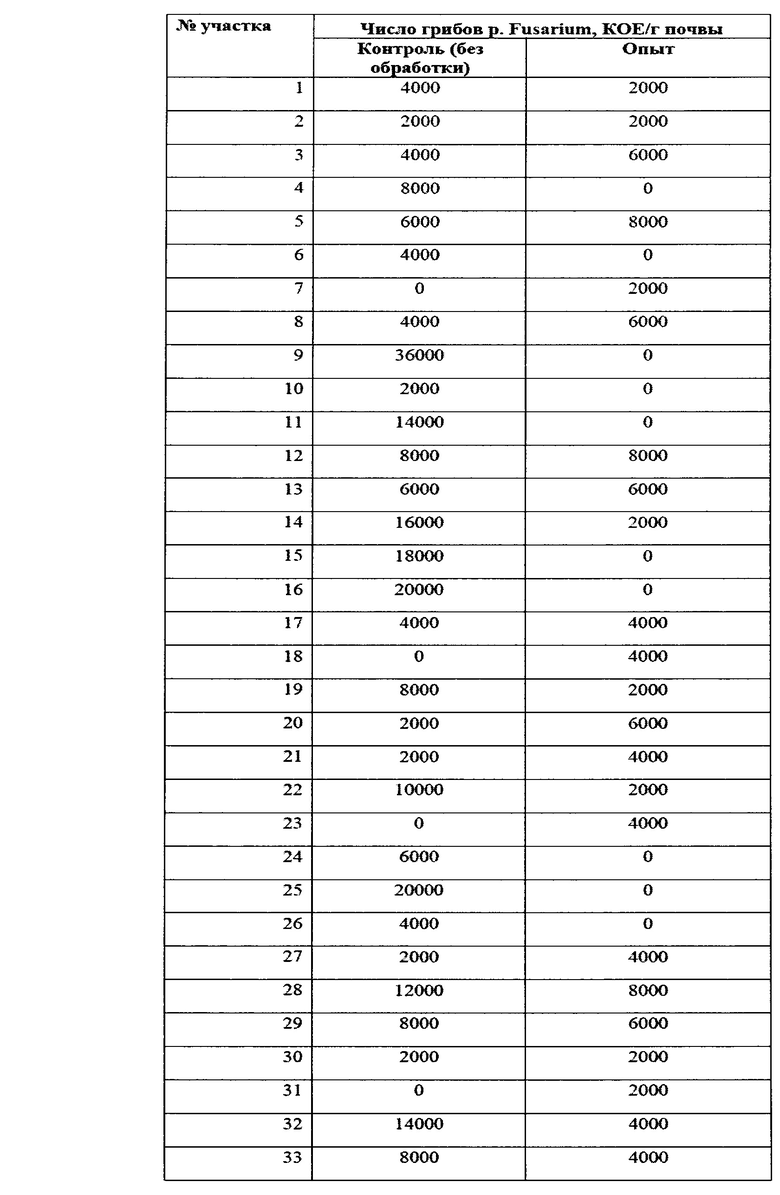
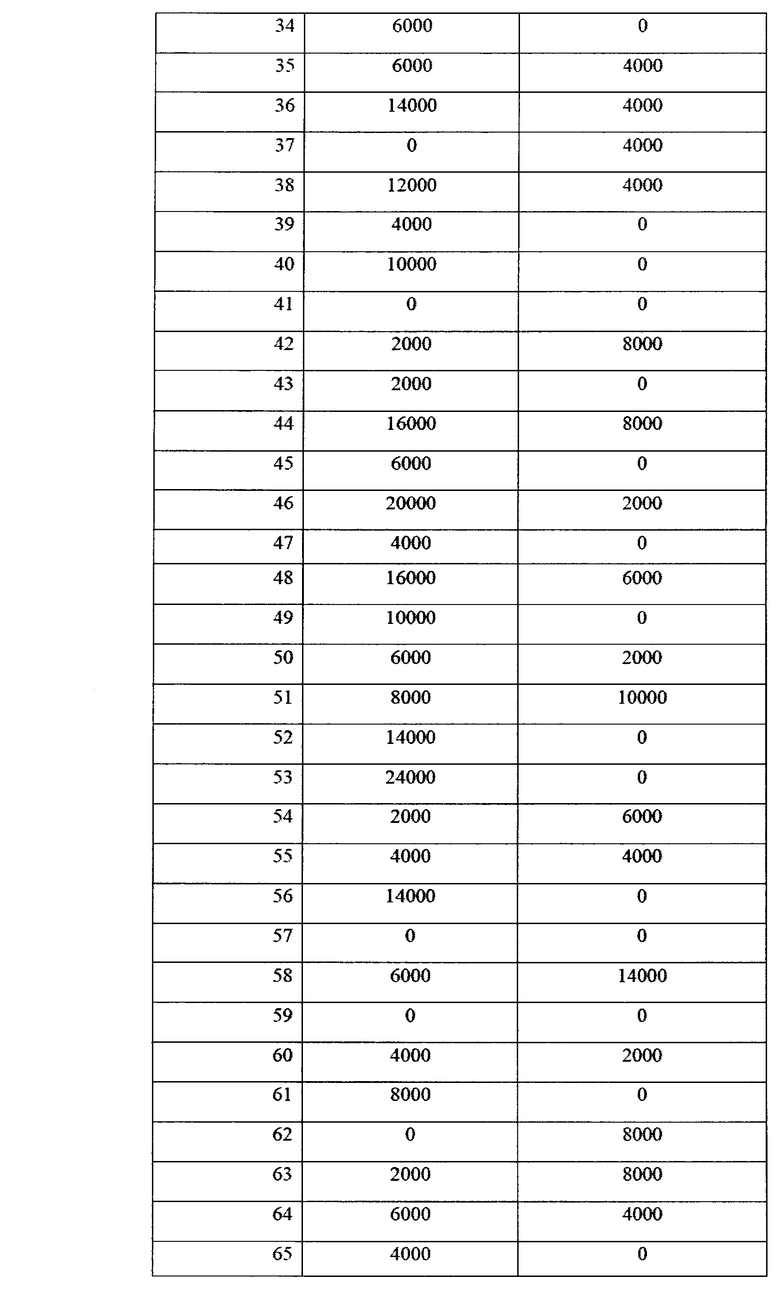
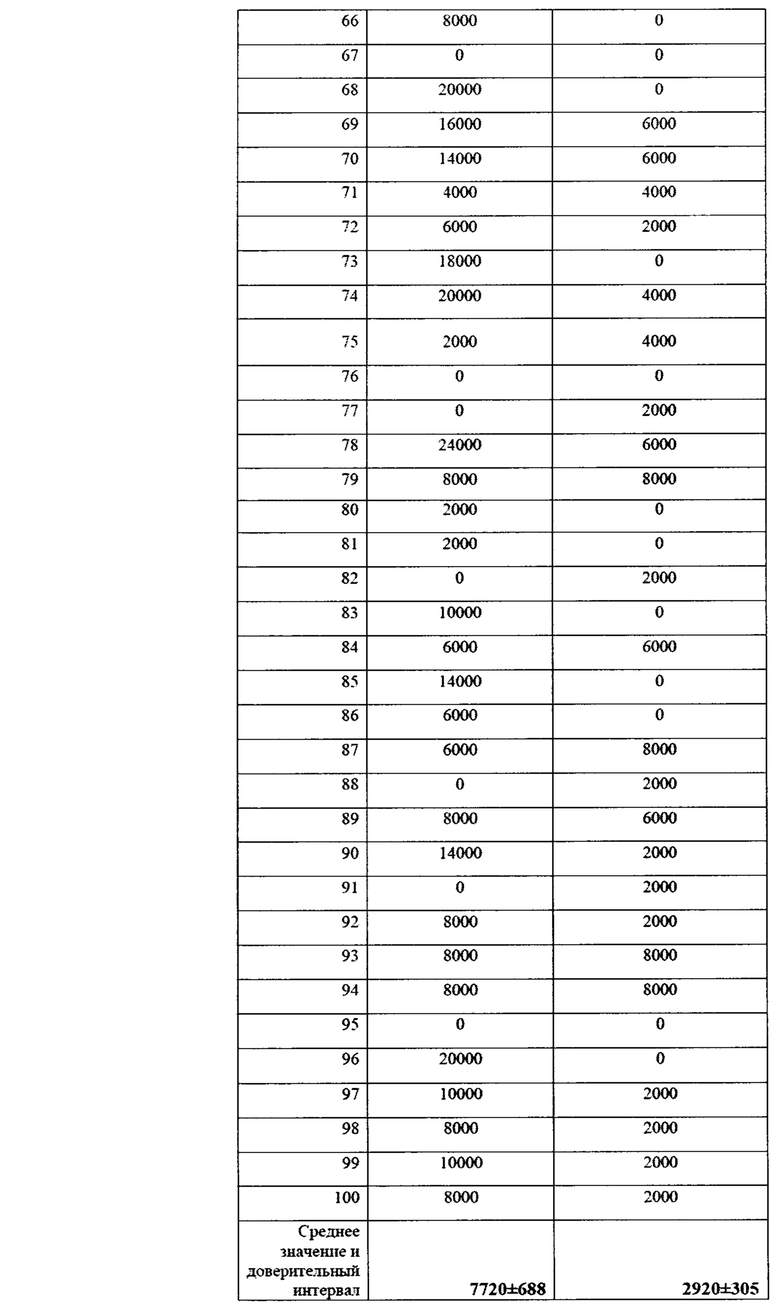
Как видно из данных таблицы 14, в фазу всходов численность грибов p. Fusarium в опыте была ниже, чем в контроле в 2,64 раза. Патогенные грибы не обнаруживались в 37% опытных площадок на опытном поле, и в 16% площадок на контрольном варианте. Также, в опытном варианте наблюдалось 2 площадки с численностью фитопатогенных грибов выше 10000 КОЕ/г почвы, тогда как в контрольном варианте - 31 площадка. В комплексе это говорит об эффективном контроле над численностью фитопатогенных грибов в прикорневой зоне при применении биопрепарата Ф-1.
Данные по численности представлены также в виде диаграммы на фиг. 15.
Таким образом, можно видеть статистически достоверные различия в численности фитопатогенных грибов между контролем и опытом.
Следующий отбор проб проводили в фазу молочной спелости растений, непосредственно перед уборкой кукурузы на силос.
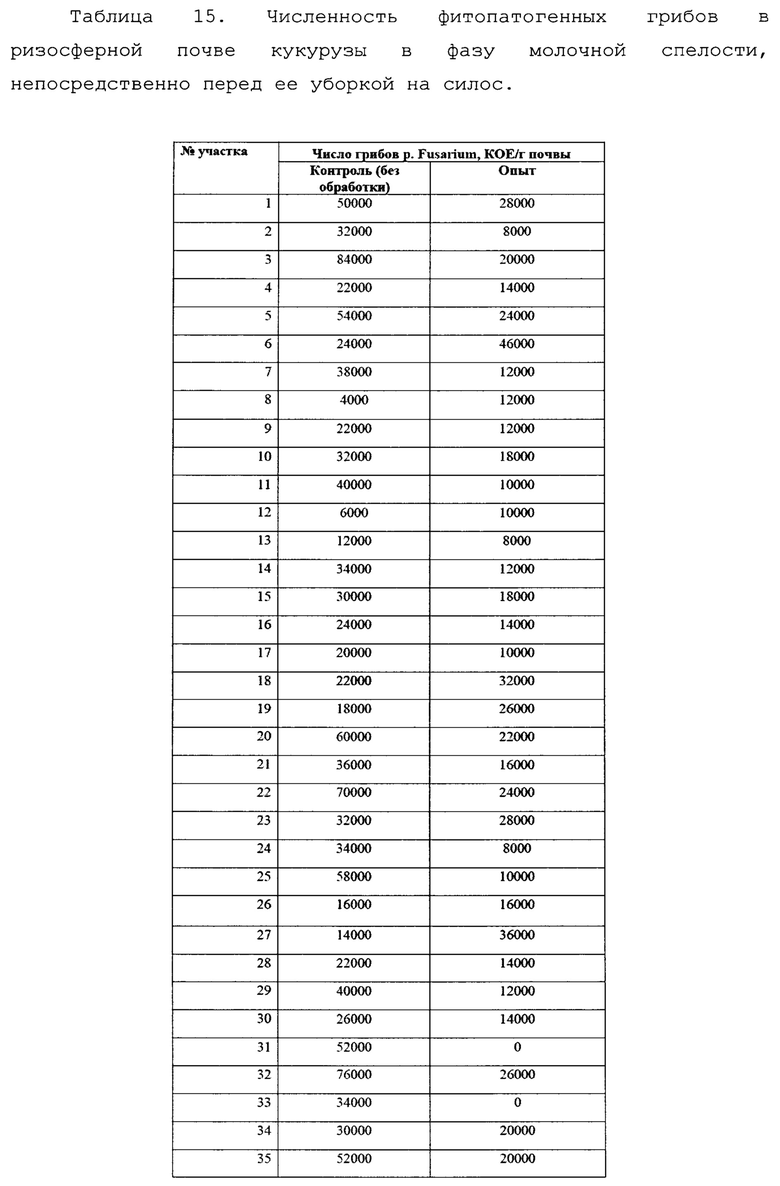
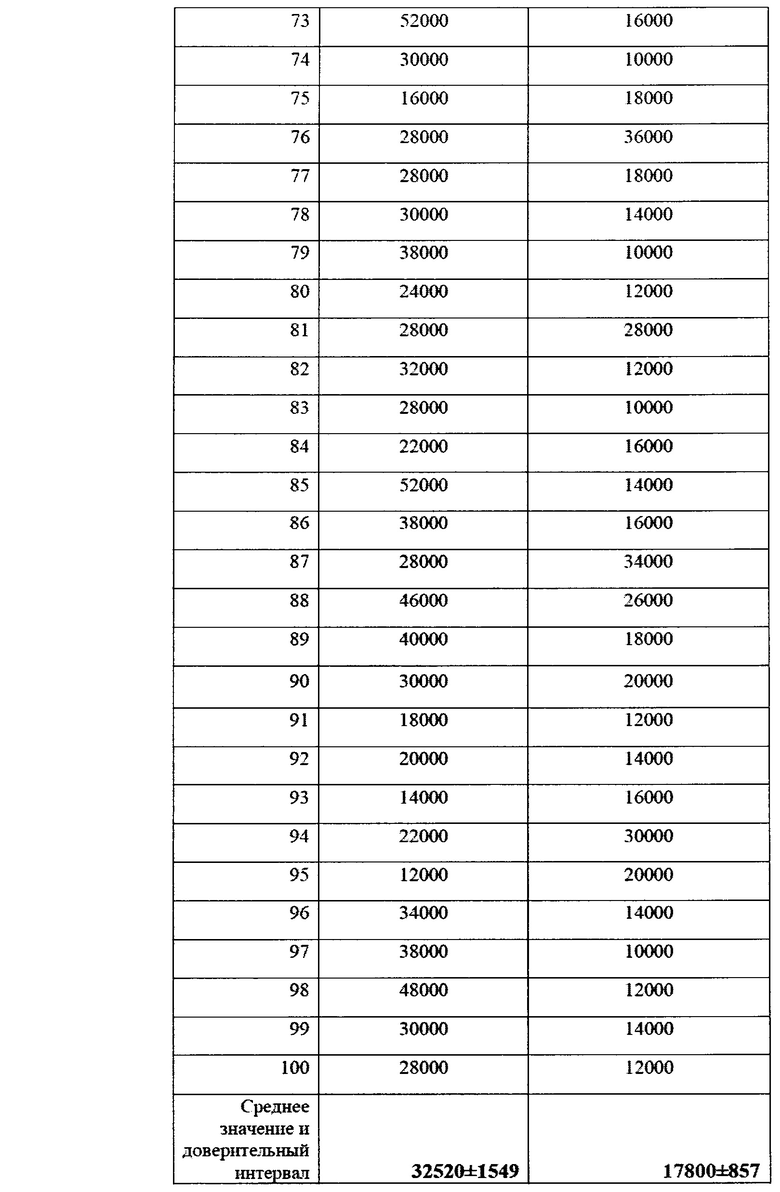
В данную фазу развития растений наблюдалось существенное увеличение численности фитопатогенов в прикорневой зоне, по сравнению с фазой всходов. Тем не менее, в контрольном варианте численность фитопатогенных грибов p. Fusarium была примерно в два раза выше, чем в опыте.
Эти различия также оказались статистически достоверными (Фиг. 16).
Как и в случае с пшеницей, после периода засухи эффект от применения биопрепарата сглаживался, однако не столь значительно, что связано с более мощной корневой системой кукурузы и сохранением влаги в более глубоких слоях почвы. Это позволило различиям сохраниться и в летний период.
Исследование влияния биопрепарата Ф-1 на морфометрические показатели растений кукурузы.
Для оценки влияния биопрепарата Ф-1 на рост и развитие растений кукурузы в полевых условиях был проведен анализ основных морфобиометрических показателей растений кукурузы. Среди таких параметров учитывали длину и массу початков на момент отбора, а также на каждом участке измеряли высоту 5-ти растений. Полученные результаты сведены в таблицах 16 и 17.
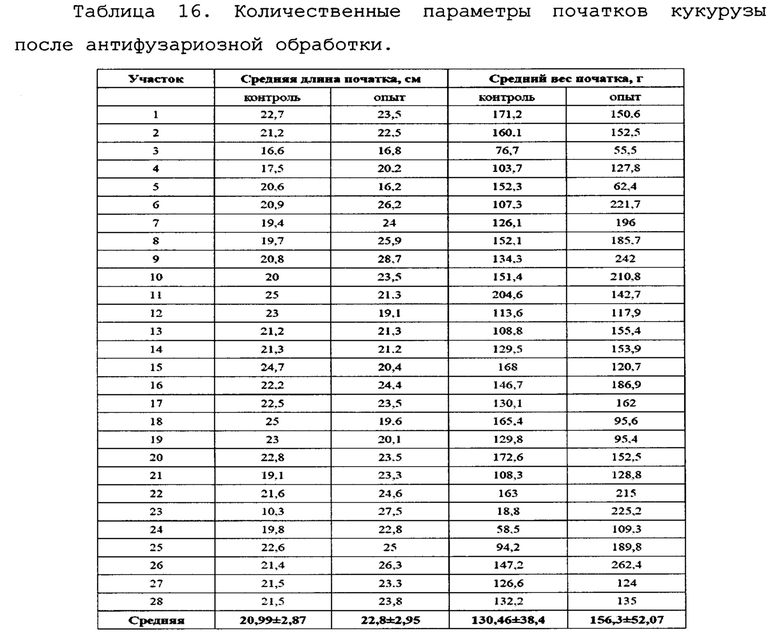
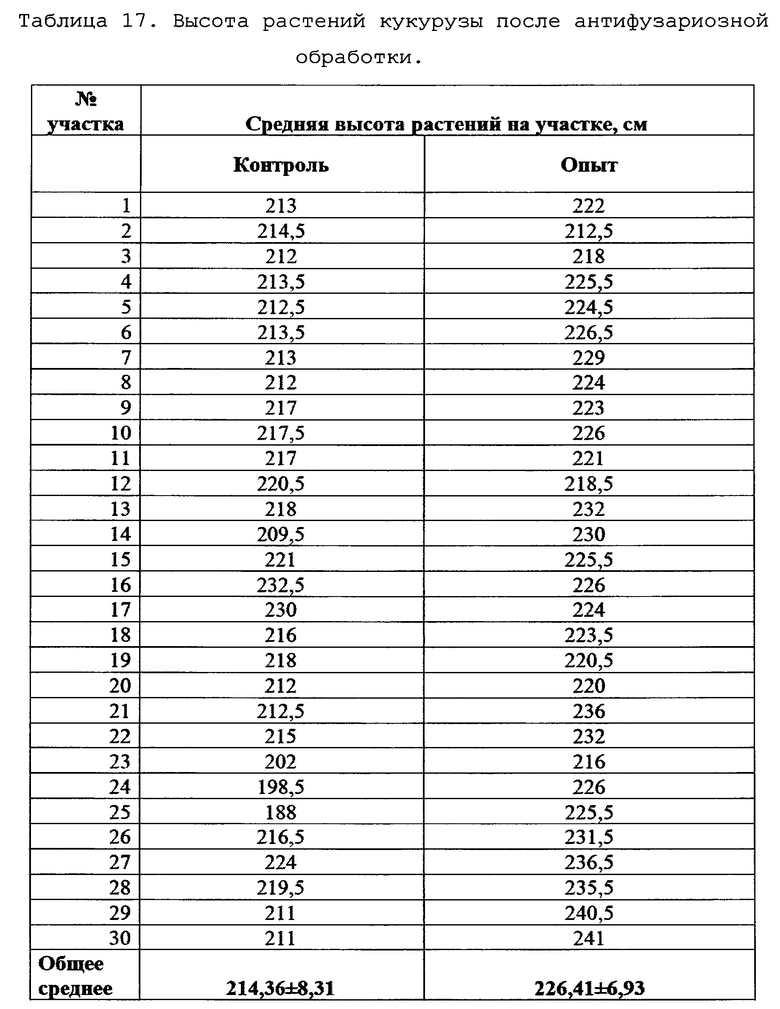
Как можно видеть из данных, приведенных в таблицах 16 и 17, различия основных морфобиометрических параметров растений кукурузы не достигают уровня статистической значимости, то есть, биопрепарат Ф-1 не оказывает негативного влияния на рост и развитие растений.
Исследования по подтверждению принадлежности учитываемых фитопатогенных грибов к p. Fusarium с помощью молекулярно-генетических методов.
Для точной идентификации культур, рост которых наблюдался в чашках Петри, использовали также метод полимеразной цепной реакции (ПЦР).
Выделение ДНК проводили с помощью набора для выделения М-SorbTube с магнитными частицами (Синтол, Россия). Культуру грибов выращивали на плотном питательном агаре в течение 15 часов. Биомассу смывали с чашек стерильным физиологическим раствором, центрифугировали при 3000 об./мин в течение 15 минут. Осадок замораживали в жидком азоте и механически растирали в ступке для разрушения клеточных стенок грибов. Процедуру повторяли трижды. Затем добавляли раствор лизирующего буфера и пробирки встряхивали и инкубировали при 75°С в течение 10 минут. После лизиса пробирки центрифугировали при 3000 об./мин в течение 1 мин и жидкую фракцию, содержащую ДНК, переносили в пробирку, содержащую частицы магнитного сорбента. Затем добавляли 500 мкл осаждающего раствора, содержимое пробирок тщательно перемешивали и оставляли на 5 минут при комнатной температуре. Пробирки центрифугировали при 16000 об./мин в течение 5 минут и помещали в магнитный штатив. Жидкая фракция удалялась. Осадок магнитных частиц с сорбированной ДНК промывали 3 раза осаждающим раствором путем ресуспендирования и центрифугирования. Затем пробирки оставляли открытыми для высушивания осадка. После этого этапа ДНК элюировали с магнитных частиц с помощью элюирующего буфера и хранили при -80°С до анализа.
Общее количество ДНК в пробе (тотальная ДНК) оценивали при помощи спектрофотометра NanoDrop 2000 с «Thermo Scientific» (USA).
Амплификация ДНК.
Для каждого образца готовили смесь для ПЦР: H2O (DD): 18,3 мкл; 25 мМ раствор нуклеотидов 10xdNTP - 2 мкл; 10х DreamTaq ПЦР-буфер (20 мМ MgC12) (ThermoFisher) - 2,5 мкл; DreamTaq полимераза - 0,2 мкл (ThermoFisher), матрица ДНК 1 мкл (5 нг мкл-1) и 0,5 мкл прямого (F) праймера и обратного (R) праймера (0,5 мкМ).
Протокол амплификации был следующим:
1. Первая денатурация при 95°С - 5 мин;
2. Денатурация при 95°С - 30 сек;
3. Отжиг праймера при 58-70°С - 30 сек (оптимальная температура подбиралась для каждой пары праймеров по отдельности);
4. Элонгация при 72°С - 1,5 мин.;
5. Цикл 2-4 повторялся 35 раз;
6. Финальная элонгация при 72°С - 3 мин.
Эффективность ПЦР проверяли при помощи контрольного набора реактивов PCR Control Set (Евроген, Россия). В пробы для отрицательного контроля вносили все реактивы и праймеры, кроме матрицы ДНК.
Для установления родовой принадлежности грибов использовались родоспецифичные праймеры (Таблица 18) на ген фактора элонгации трансляции 1-альфа (tef1α) и ген региона ITS (внутреннего транскрибируемого регионама) рибосомальной РНК (рРНК). Праймеры были подобраны на предыдущем этапе исследования.
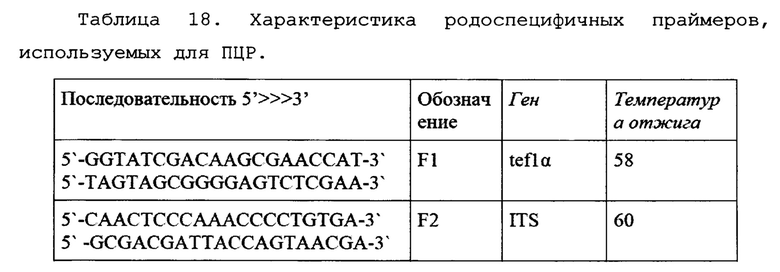
Для установления видовой специфичности были использованы видоспецифичные праймеры (Таблица 19) на основные виды представителей рода Fusarium, поражающих исследуемые поля.
Праймеры были подобраны и проверены на предыдущем этапе исследования.
На фиг. 17 видно, что реакция для штамма Fus1 идет с праймерами G1 для F. graminearum при температуре 62°С, реакция для штамма Рас 6.2.1 - с праймерами O2 для F. oxysporum, что соответствует данным о их генетической идентификации, проведенной на первом этапе работ.
На фиг. 18 можно видеть, что в пробах, содержащих грибы рода Fusarium, образуется специфический продукт амплификации размером 340 пар нуклеотидов. Из 15 колоний, по морфологии похожих на колонии грибов рода Fusarium, лишь 9 оказались таковыми по данным анализа ДНК. Только такие колонии учитывались в статистических расчетах. Это делает тесты более громоздкими, но значительно повышает их точность.
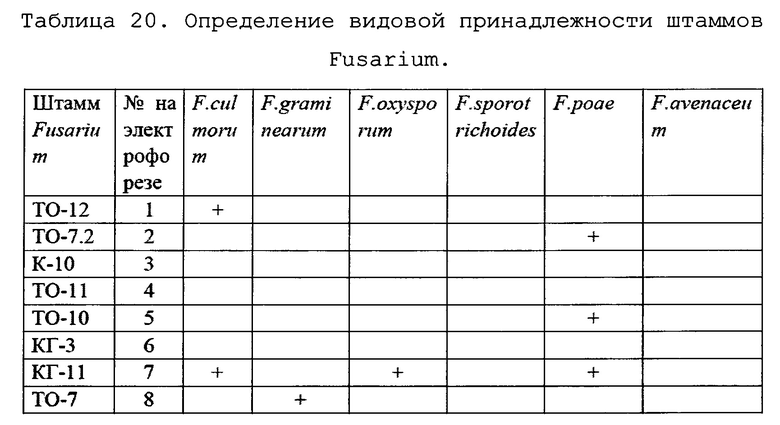
Кроме того, была проведена идентификация видов патогенных грибов, выделенных из посевов, также методом ПЦР.
В результате амплификации с видоспецифичными праймерами были получены следующие результаты (Таблица 20):
На фиг. 19-21 представлены результаты гель-электрофореза в агарозном геле, проведенного для визуализации результатов ПЦР-амплификации.
На указанных фигурах можно видеть выраженные полосы флуоресценции, соответствующие амплифицированным фрагментам длиной 250-400 п. н. Слабо выраженные полосы меньшего размера относятся к так называемым неспецифическим продуктам ПЦР и не учитываются.
Таким образом, можно заключить, что:
• Штамм ТО-12 относится к виду F. culmorum;
• Штамм ТО-7 относится к виду F. graminearum;
• Штаммы ТО-7.2 и ТО-10 относятся к виду F. poae;
• Штамм КГ11 демонстирует реакцию на видоспецифичные праймеры к F. culmorum, F. oxysporum и F. poae.
Для его дальнейшей идентификации необходимы дополнительные исследования. Однако можно предположить, что поскольку в нашей тест-системе праймеры на F. oxysporum, в отличие от двух других, также вызвавших реакцию амплификации, комплементарны гену кальмодулина, вероятно, данный штамм несет мутацию в гене tef1α или получил значительный его участок от близкородственного вида путем горизонтального переноса, но относится, тем не менее, к виду F.oxysporum. Представителей видов F. avenaceum и F. sporotrichoides среди выделенных штаммов не найдено.
Опыт №3. Влияние препарата на посевы озимой пшеницы, осень 2018.
Работы по исследованию влияния обработки семян озимой пшеницы биопрепаратом Ф-1 на пораженность растений грибами р. Fusarium осенью 2018 года (всходы).
Для изучения влияния обработки семян озимой пшеницы биопрепаратом Ф-1 осенью 2018 года была проведена обработка 61 т семян с помощью ПС-10.
Данные семена использовали для посева на орошаемых полях (поле 40, бригада 3, 92 га, предшественник - картофель), а также на полях без ирригации (поле 94, 90 га, бригада 6), предшественник - кукуруза. В качестве контроля были выбраны поля, где для посева применяли семена пшеницы, обработанные только химическими фунгицидами - орошаемое поле 39, 87 га, бригада 3, предшественник - сахарная свекла и поле без ирригации 91, 91 га, бригада 6, предшественник - кукуруза.
Отбор проб был произведен в фазу всходов в ноябре 2018 г. и в фазу кущения в декабре 2018. Отбор образцов производили по методу конверта, в 5 независимых повторностях. Каждое поле делили на 12 секторов и в каждом секторе отбирали по 5 образцов, всего по 60 образцов с поля.
В условиях лаборатории производился посев фрагментов растений пшеницы, (участки стебля без корневой системы) на среду Чапека, с последующим учетом и идентификацией грибов р. Fusarium. На каждую чашку помещалось 10 фрагментов стебля длиной 1 см. Идентификацию патогенов вели по морфологии колоний, с подтверждением при помощи световой микроскопии. Также отбирали биомассу патогенов для подтверждения принадлежности к p. Fusarium с помощью молекулярно-генетических методов. Данные по пораженности растений пшеницы фитопатогенными грибов представлены в виде % фрагментов растений, давших рост колоний грибов p. Fusarium и приведены в таблицах 21 и 22 а также на фигурах 22 и 23.
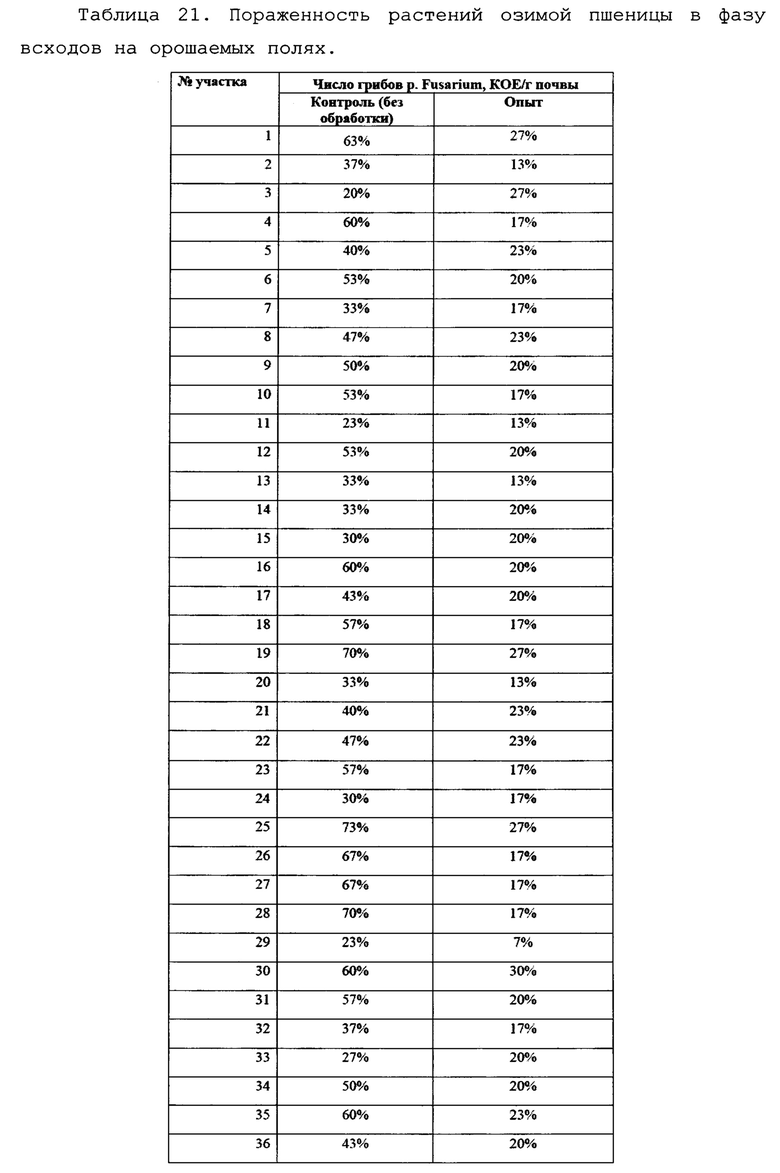
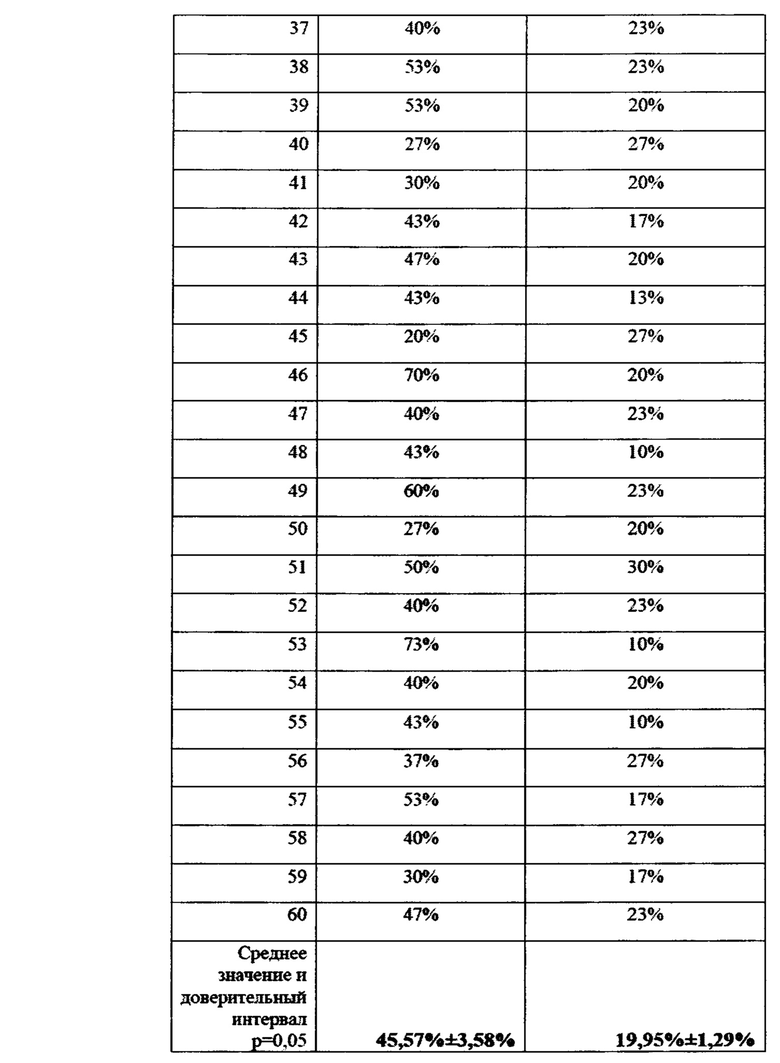
По данным анализа наблюдается достоверное снижение пораженности растений пшеницы фитопатогенными грибами р. Fusarium в 2,28 раз. По видимому, обработка семян пшеницы биопрепаратом затрудняет и замедляет проникновение гриба в ткани растения за счет формирования биопленки бактерий-антагонистов на поверхности прорастающего корня.
Данные по средней пораженности растений представлены графически на фиг. 22
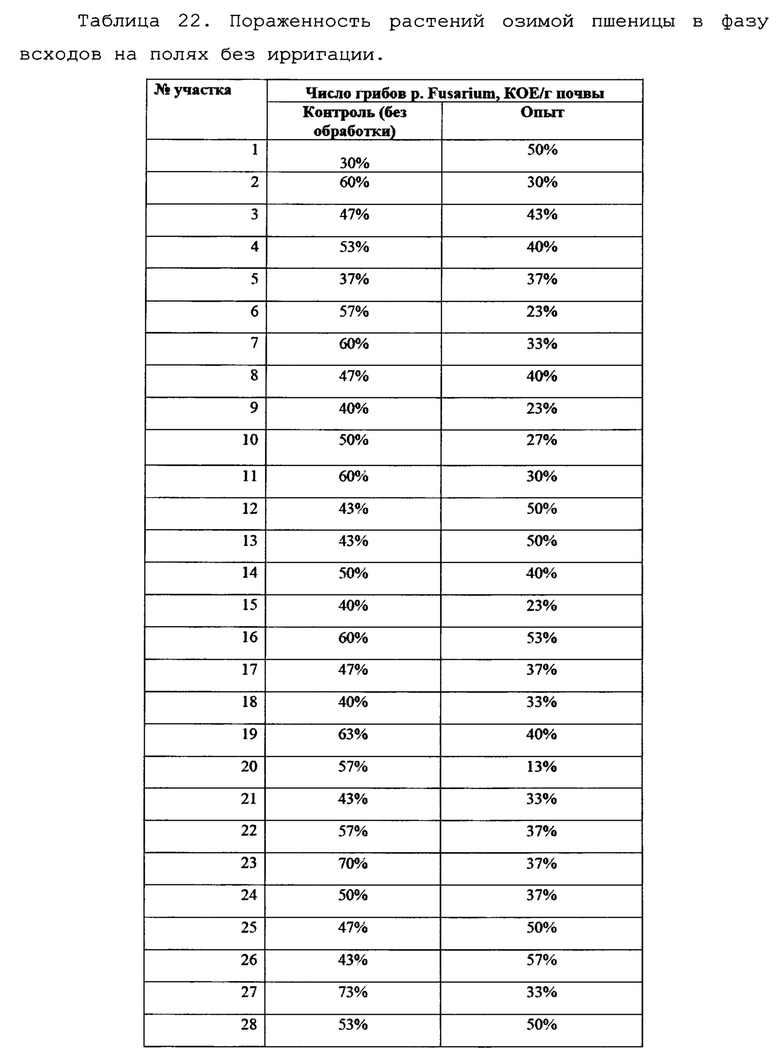
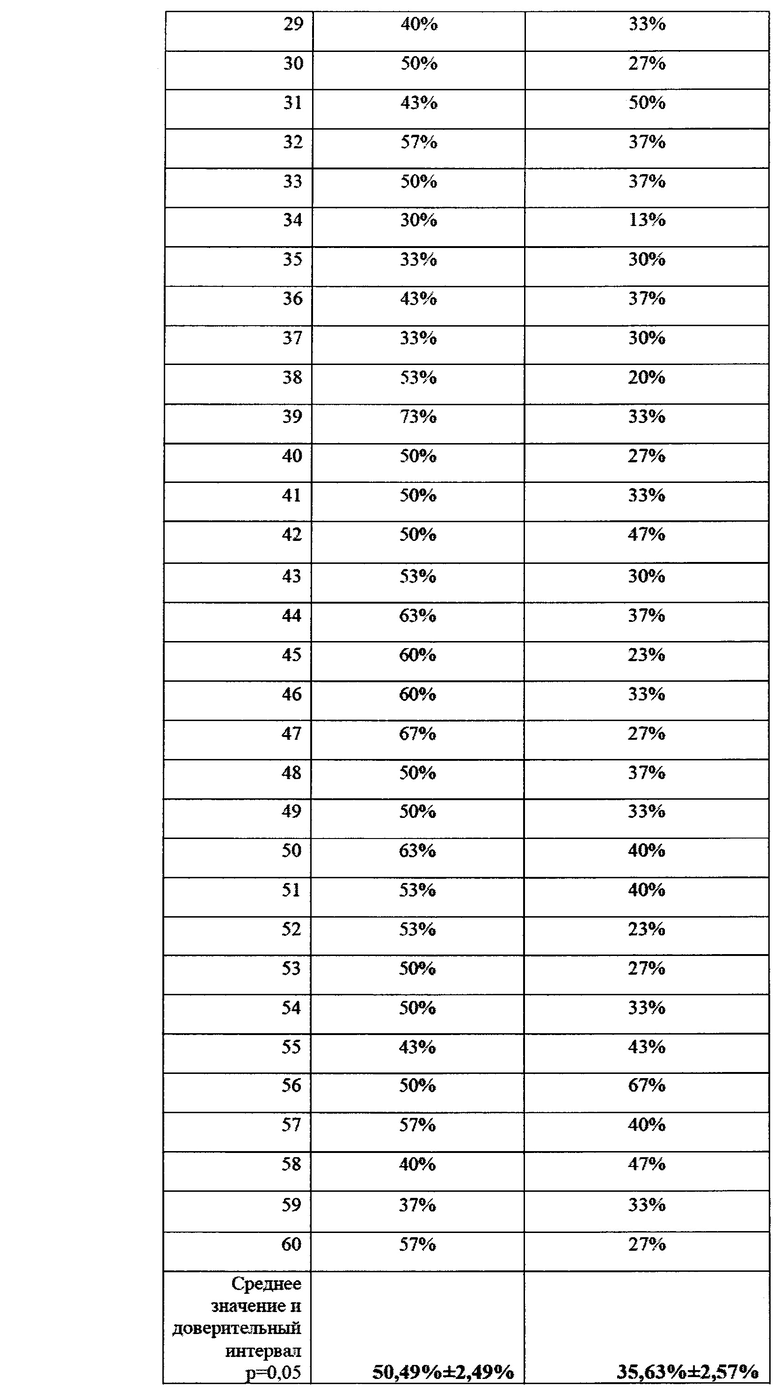
Как видно из таблицы 22, на полях без ирригации снижение степени пораженности растений грибами p. Fusarium при применении биопрепарата Ф-1 было ниже (в 1,41 раз), однако различия также были статистически достоверны. По-видимому, такая разница может объясняться различающимися культурами-предшественниками на полях 4 0 и 39, тогда как на полях без ирригации №94 и №91 предшественник был одинаковый (кукуруза).
Культура-предшественник в значительной степени влияет на исходный инфекционный фон, и это может оказывать влияние на наблюдаемую эффективность препарата. Данные по средней пораженности растений представлены графически на фиг. 23.
Исследование влияния обработки семян озимой пшеницы биопрепаратом Ф-1 на зараженность ризосферной почвы грибами р. Fusarium осенью 2018 года (всходы).
Отбор производили по методу конверта из 5 точек каждого сектора поля для получения репрезентативной картины. Получали смешанную пробу для оценки численности грибов p. Fusarium в прилегающей к корням почве. Всего с каждого поля отбирали по 12 смешанных почвенных проб. В условиях лаборатории производился посев корней пшеницы с ризосферной почвой на среду Чапека, с последующим учетом и идентификацией грибов р. Fusarium. Данные приведены в таблицах 23 и 24, а также на фигурах 24 и 25.

Данные, приведенные в таблице 23, показывают наличие очень существенной разницы в численности грибов p. Fusarium в почве. Несмотря на большой размах варьирования численности грибов в почве опытного поля, заметно значительно более низкое их количество по сравнению с контролем. Это косвенно показывает, что бактерии-антагонисты колонизируют корни растений и контролируют численность грибов p. Fusarium в непосредстенно примыкающем к корням слое почвы. Данные представлены в виде диаграммы на фиг. 24.
Сходная картина наблюдалась на полях без ирригации, хотя и в меньшем масштабе (Таблица 24).
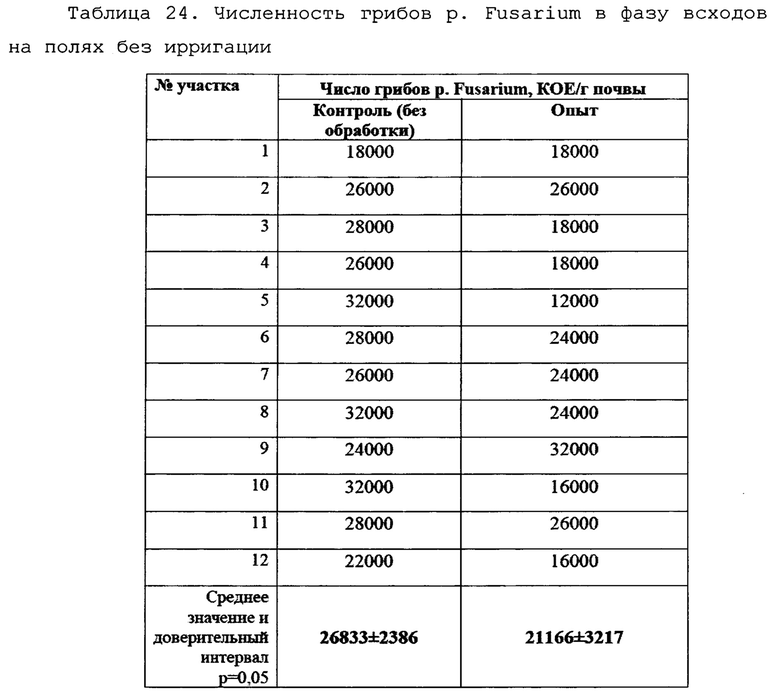
На полях без ирригации численность грибов p. Fusarium в ризосферной почве в опыте была на 22% ниже, чем контроле. Данные различия были статистически достоверны (р=0,02). Средние значения численности фитопатогенных грибов графически представлены на фиг. 25.
В целом, данные полученные в ходе анализа ризосферной почвы хорошо соотносятся с данными, полученными для фрагментов стеблей растений пшеницы. Это указывает на то, что бактерии, входящие в состав биопрепарата активно колонизируют поверхность корня и препятствуют проникновению инфекции в растения.
Исследование влияния биопрепарата Ф-1 на морфометрические показатели растений пшеницы (всходы).
Для оценки влияния биопрепарата Ф-1 на рост и развитие растений пшеницы в полевых условиях осенью 2018 года был проведен анализ основных морфобиометрических показателей растений пшеницы. Полученные результаты сведены в таблицу 25.
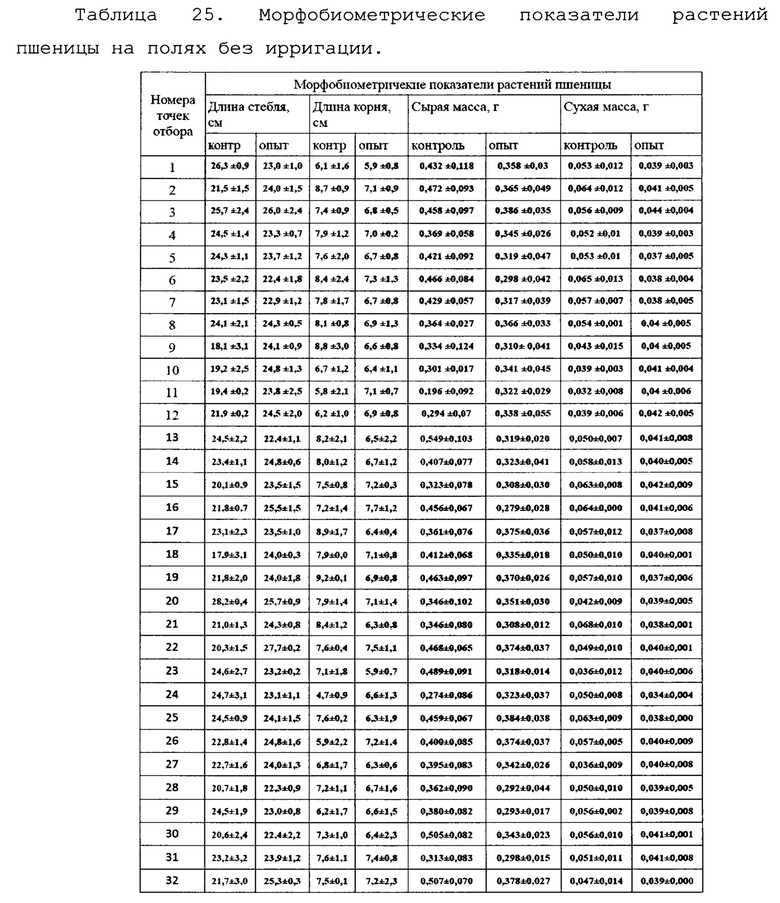
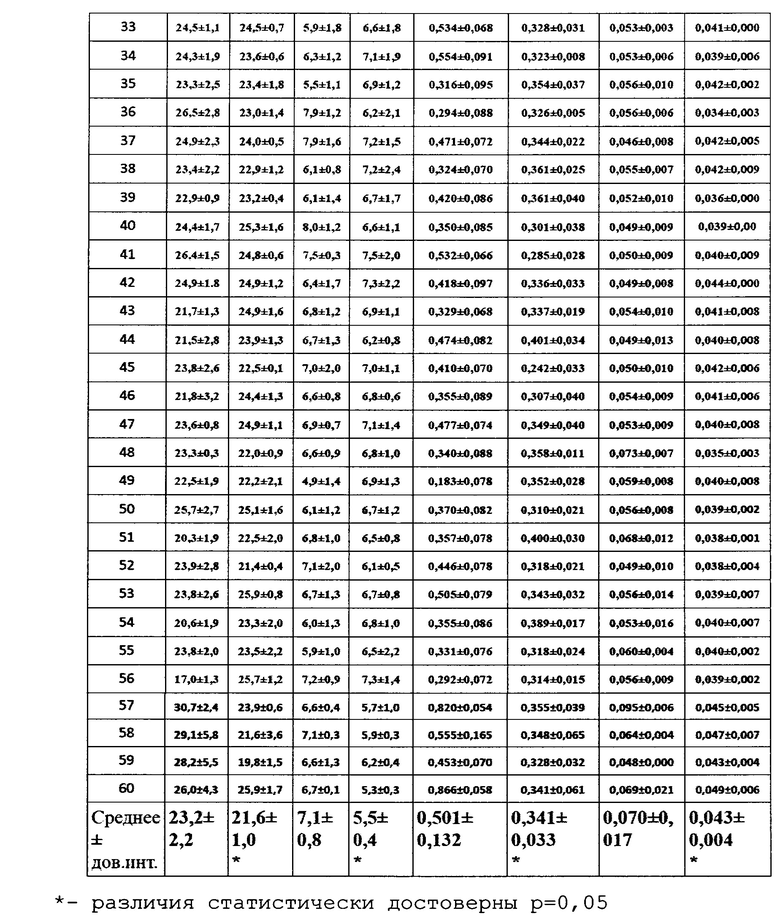
На полях с ирригацией отличия между опытом и контролем также были достоверны, и более ярко выражены по сравнению с полями без ирригации. На это мог повлиять более высокий инфекционный фон, связанный с высокой влажностью в течение предыдущего сельскохозяйственного сезона.
Исследование влияния обработки семян озимой пшеницы биопрепаратом Ф-1 на пораженность растений грибами p. Fusarium осенью 2018 года представлены в таблице 26.
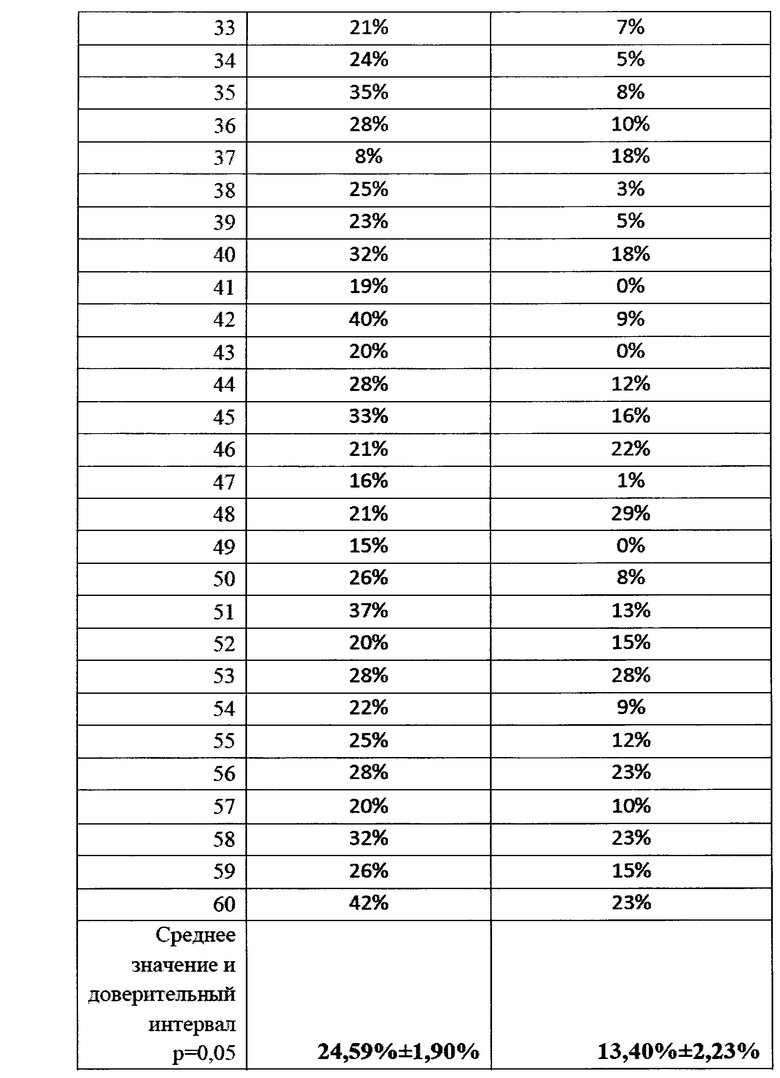
По данным анализа встречаемости грибов p. Fusarium в тканях растений отмечено достоверное снижение пораженности растений пшеницы фитопатогенными грибами p. Fusarium в 1,83 раза. Следует отметить, что по сравнению с фазой всходов (45,57% ± 3,58% в контроле, и 19,95% ± 1,29% в опыте) встречаемость фитопатогенов в фазу кущения стала ниже. По-видимому, в ходе развития растений пшеницы, часть проникающих грибов элиминируется за счет активного иммунитета растений. При этом обработка биопрепаратом может способствовать этому процессу. Важно, что процент зараженных растений оказывается ниже еще в процессе прорастания, и эта тенденция сохраняется по мере развития растений и перехода их в фазу кущения. Графически встречаемость грибов p. Fusarium в тканях растений отражена на диаграмме (Фиг. 26).
На полях без ирригации наблюдается сходная картина (Таблица 27).
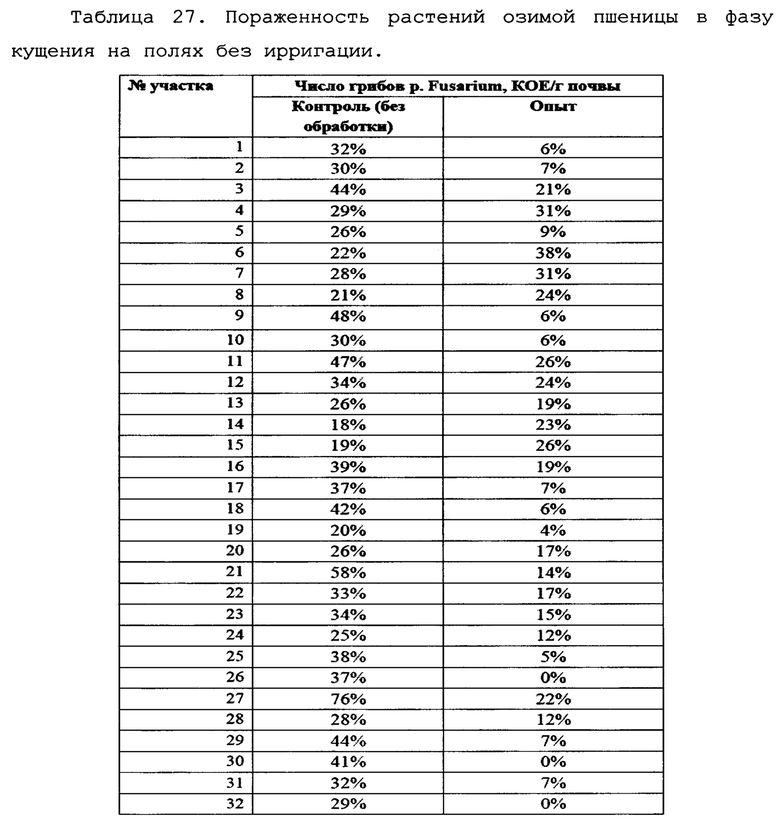
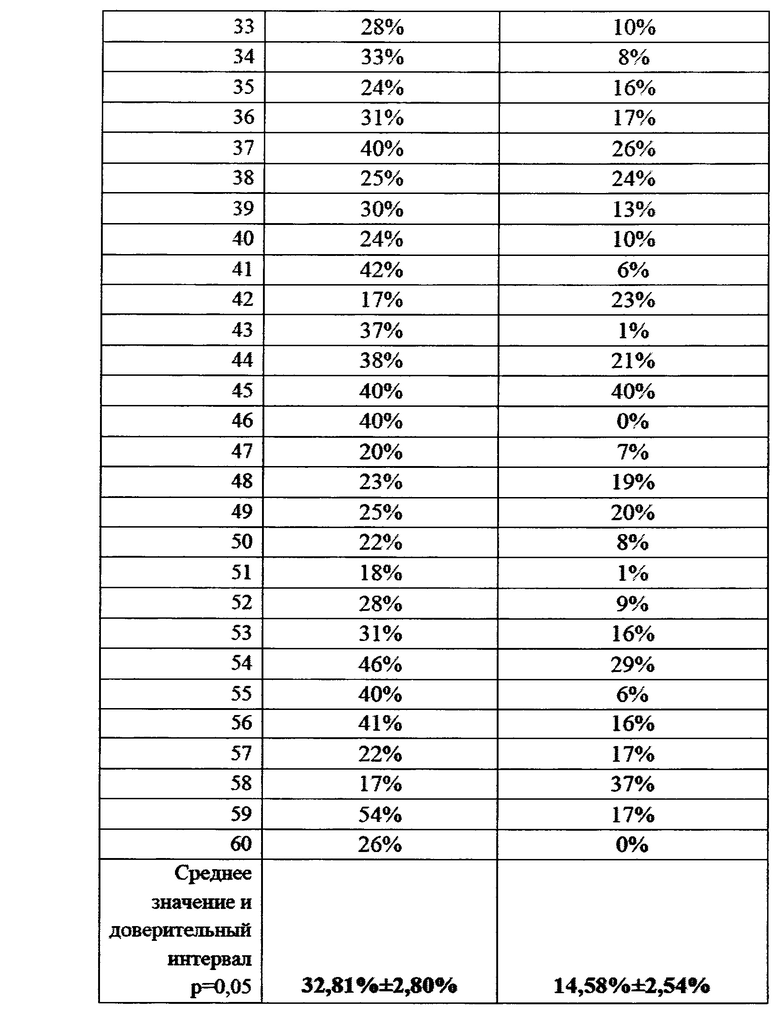
На полях без ирригации встречаемость грибов p. Fusarium в тканях растений в фазу кущения в опыте была ниже чем в контроле в 2,25 раза. Так же как и на орошаемых полях, к фазе кущения процент растений, в тканях которых присутствовали грибы p. Fusarium снижается. В контроле снижение идет с 50,49% до 32,81%, а в опыте с 35,63% до 14,58%.
Таким образом, биопрепарат сохраняет свое защитное действие к фазе кущения, что говорит в пользу активной колонизации растений пшеницы бактериями биопрепарата (Фиг. 27).
Исследование влияния обработки семян озимой пшеницы биопрепаратом Ф-1 на зараженность ризосферной почвы грибами р. Fusarium осенью 2018 года (осеннее кущение) представлено в таблице 28.
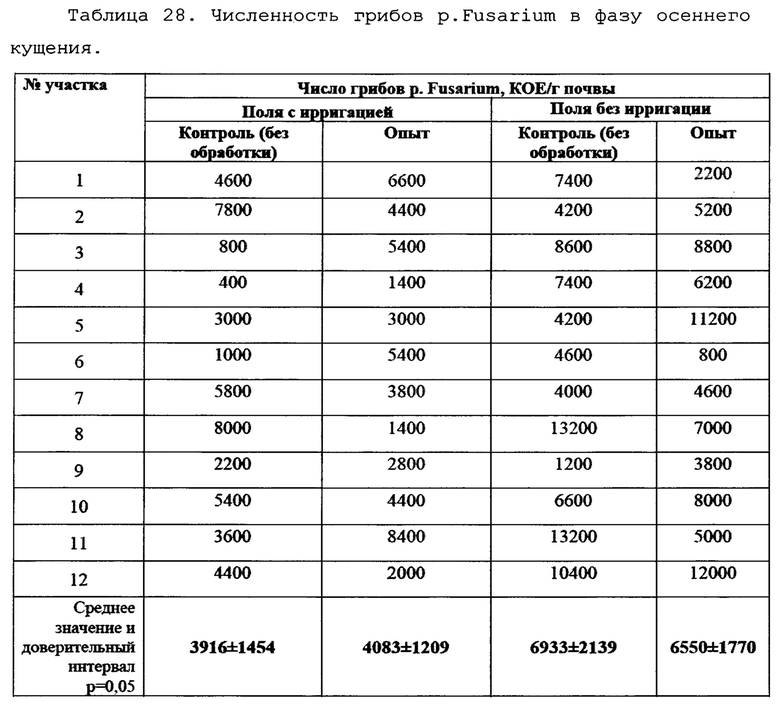
Исследование ризосферной почвы в фазу кущения показало (Фиг. 28), что на данном этапе развития растений численность грибов p. Fusarium не отличалась в почвах контрольных и опытных полей. Кроме того, по сравнению с фазой всходов, наблюдалось существенное падение их численности в ризосфере пшеницы. Одним из объяснений может быть установление низких температур и снегопады в ноябре (отбор проводился в декабре, после схода снега).
По-видимому, наиболее активное проникновение грибов в растения происходит в течение самых ранних этапов развития пшеницы - сразу же после прорастания, а на более поздних этапах численность грибов в ризосфере снижается. При этом важно обеспечить контроль численности патогена именно в этот критический период, и результаты опыта показывают, что в вариантах, где в фазу всходов численность грибов в ризосфере была снижена, впоследствии оказывается ниже процент растений, в которые грибы успешно внедрились.
Исследование влияния биопрепарата Ф-1 на морфометрические показатели растений пшеницы (осеннее кущение).
В фазу осеннего кущения определялись следующие морфобиометрические показатели растений: длина стебля, длина корня, сырая и сухая масса растения, а также количество стеблей на растение. Данные приведены в таблицах 2 9 и 30.
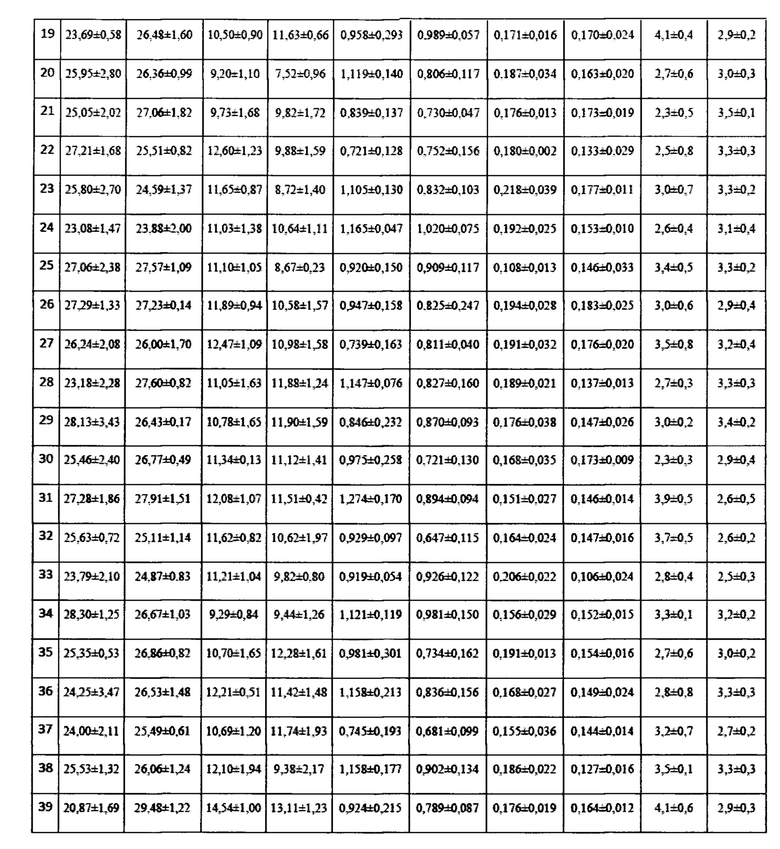
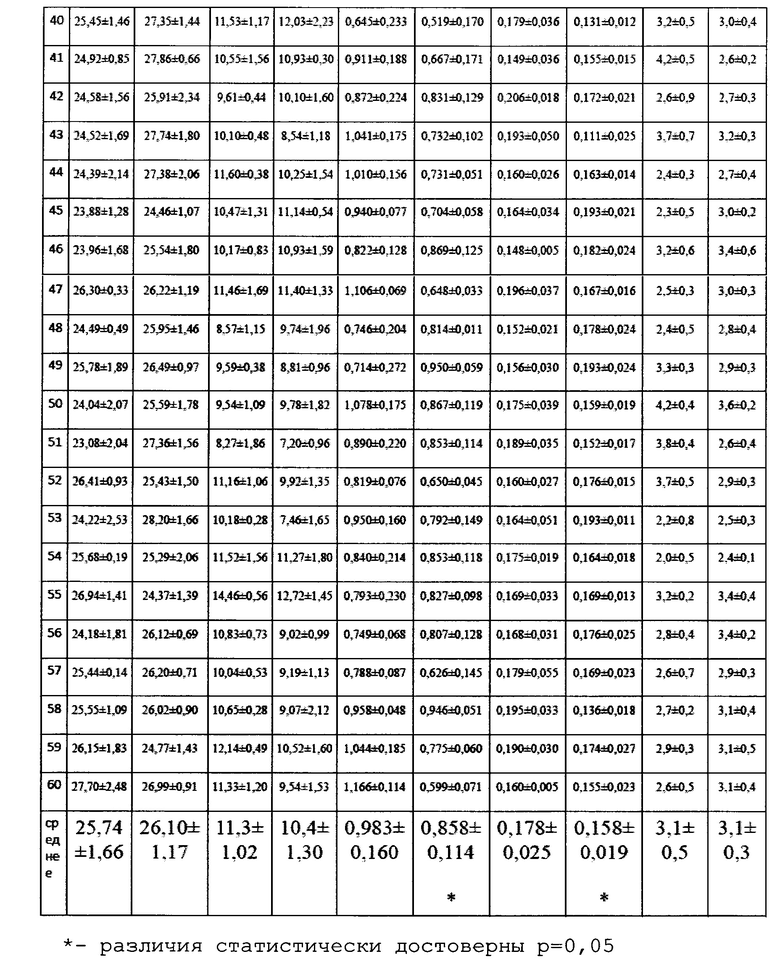
К фазе кущения растения на опытных и контрольных полях без ирригации выравнивались по длине побегов и корней, различия по массе сохранялись, но также становились менее выраженными. Это связано, что в фазу всходов даже незначительное количество фитогормонов, продуцируемых грибами, оказывает на растение существенное влияние, а по мере роста и развития растений вклад экзогенных фитогормонов ослабевает.
При этом по числу стеблей на растение контрольное и опытное поля оказались совершенно одинаковыми, на данный признак обработка не повлияла.
На полях с ирригацией разница между контрольным и опытным полем также сглаживается, но все еще более заметна, по сравнению с не орошаемыми полями (Таблица 30).
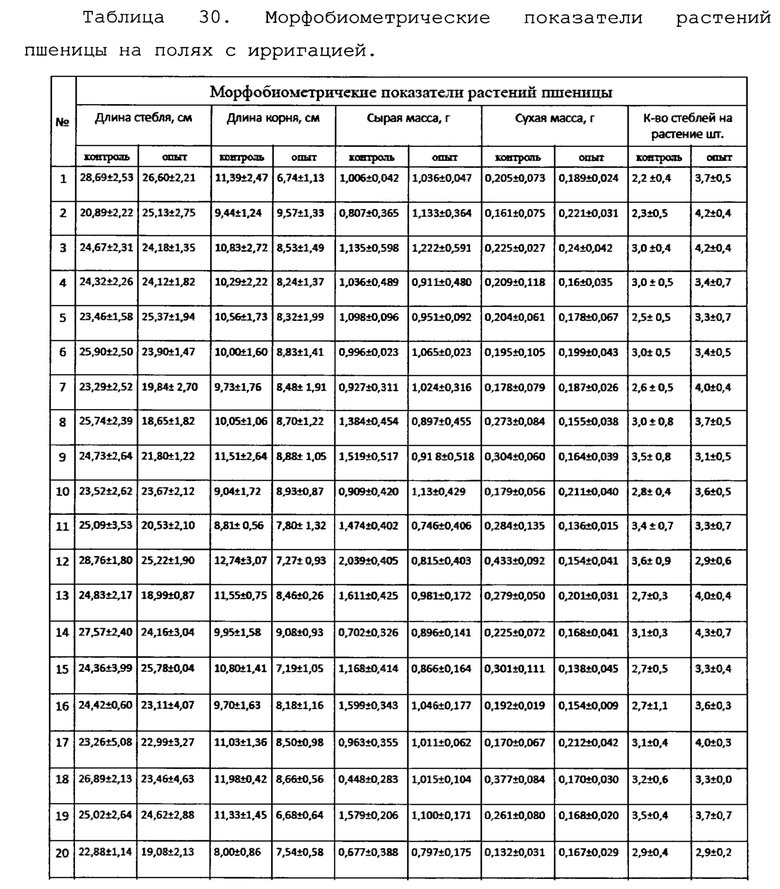
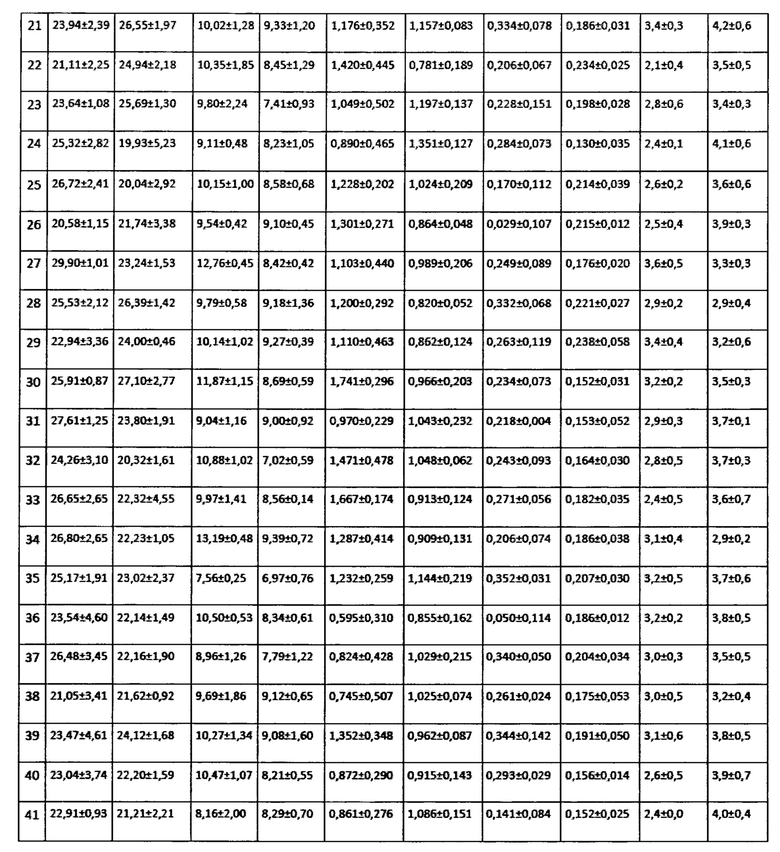
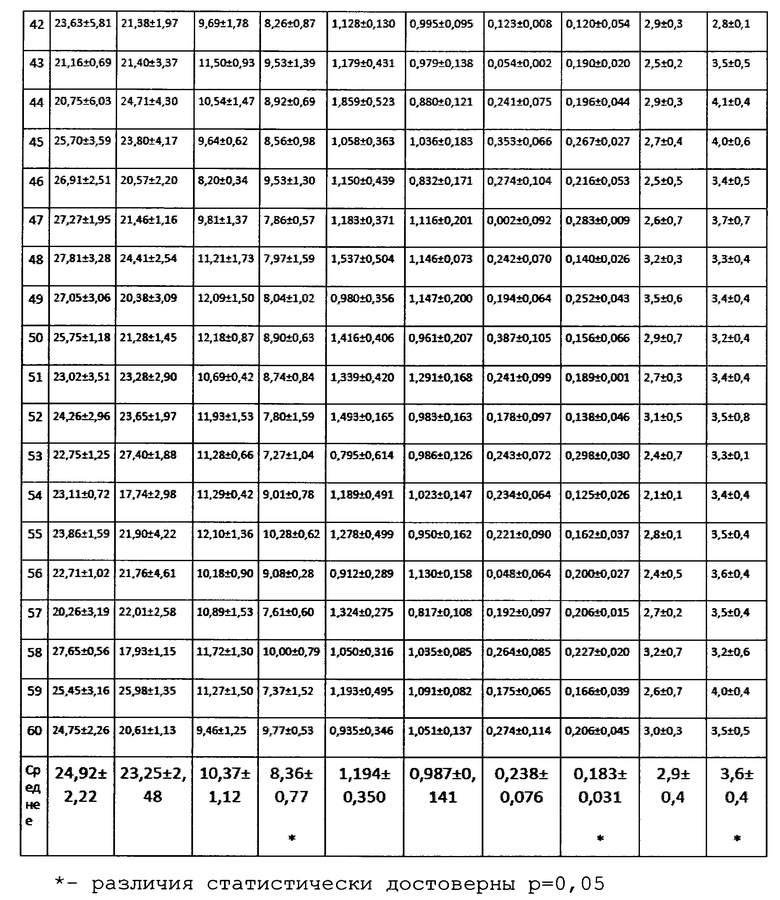
Растения на контрольном и опытном полях различались по длине корня и сухой массе (в контроле несколько больше, чем в опыте). Однако при этом наблюдалось достоверное различие в числе побегов на растение - в опыте 3,6 побега на растение, в контроле 2,9. Данный показатель потенциально может оказывать положительное влияние на урожайность, однако требуется учет продуктивности боковых побегов пшеницы.
Таким образом, в результате исследовательских работ было выделено 104 0 культур спорообразующих бактерий p. Bacillus из почв полей, принадлежащих Заказчику и корневых систем растений озимой пшеницы. Также было выделено 5 штаммов грибов р. Fusarium, доминирующих на полях Заказчика, из которых 3 штамма принадлежали к виду Fusarium graminearum и 2 к виду Fusarium oxysporum, и кроме того был выделен фитопатогенный гриб Plectosphaerella cucumerina.
При этом была произведена оценка антагонистической активности выделенных культур и была проверена методом совместного культивирования в условиях роста на чашках Петри и в микрокультуре на стеклах. В результате было отобрано сначала 212 штаммов, проявляющих антагонизм, затем из них отобрано 37 самых активных культур. Из этих культур было выбрано 10 штаммов, в наибольшей степени отвечающих цели исследований, то есть разработке биопрепарата и способа получения биопрепарата.
При этом были выделены штаммы бактерий-антагонистов и фитопатогенных грибов, которые были идентифицированы специалистами ВКПМ. Штаммы бактерий-антагонистов были переданы в ВКПМ для депонирования по процедуре национального патентного депонирования.
При этом было проведено тестирование влияния штаммов на растения, которое производилось путем обработки семян культурой испытуемого штамма с последующим определением всхожести, энергии прорастания, а также показателей роста и развития растений в присутствии и при отсутствии фитопатогенных грибов. Вегетационные опыты по изучению стимулирующего эффекта бактерий-антагонистов были проведены в климат-камере и теплице, в песчаной и почвенной культуре. В результате было показано отсутствие негативных эффектов для растений, нейтральное и позитивное действие на показатели их роста и развития.
При этом был разработан способ получения биопрепарата, содержащего 10 штаммов споробразующих бактерий-антагонистов с антифузариозной активностью.
Пробная партия препарата была испытана в реальных условиях на 10 т пшеницы сорта "Гром" с использованием установки ПС10.
Обработка зерна показала эффективность разработанного биопрепарата по сравнению с химическим фунгицидом "Шансил Трио" в условиях лабораторного опыта.
Разработанный биопрепарат Ф-1 проявил способность снижать численность грибов рода Fusarium в следующих вариантах опыта:
1) в ризосфере озимой пшеницы в фазу кущения (снижение в 1,98 раза)
2) в зерне озимой пшеницы (снижение более чем в 5 раз)
3) в ризосфере кукурузы в фазу всходов (в 2,64 раза)
4) в ризосфере кукурузы в фазу молочной спелости (в 1,82 раза)
5) в стеблях озимой пшеницы в фазу всходов (в 2,28 раз на орошаемых полях ив 1,41 раза на полях без ирригации)
6) в ризосфере озимой пшеницы в фазу всходов (в 5 раз на орошаемых полях и в 1,26 раза на полях без ирригации)
7) в стеблях озимой пшеницы в фазу кущения (в 1,83 раз на орошаемых полях и в 2,25 раза на полях без ирригации)
При этом самый мощный эффект зарегистрирован для наиболее ценного параметра - в зерне озимой пшеницы, обработанной биопрепаратом Ф-1 наблюдалось более чем пятикратное снижение зараженности фузариозом.
На ранних этапах развития растений обработка семян озимой пшеницы биопрепаратом Ф-1 приводит к протекторному эффекту как в растениях, так и в прилегающей к корням ризосферной почве, что указывает на колонизацию ризопланы бактериями, которые препятствуют внедрению патогена. Следствием колонизации корневой системы бактериями является то, что в фазу кущения процент растений, в тканях которых присутствовали грибы р. Fusarium в опыте значительно ниже, чем в контроле: в 1,83 раза на орошаемых полях и в 2,25 раз на полях без ирригации.
--->
Перечень последовательностей
<110> Общество с ограниченной ответственностью «Агрофирма «Урожайная»
(Obshchestvo s ogranichennoi otvetstvennostiu «Agrofirma «Urozhainaia»)
<120> Штаммы, биопрепарат, способ получения биопрепарата и способ биологической защиты сельскохозяйственных культур от фузариоза
<140> RU 2019139147
<141> 2019-12-02
<160> 10
<210> 1
<211> 844
<212> ДНК
<213> Paenibacillus peoriae R 3.13
<400> 1
cccaggcgaa tcttaatgta gttaacttcg gcaccaaggg tatcgaaacc cctaacacct 60
agcattcatc gtttacggcg tggactacca gggtatctaa tcctgtttgc tccccacgct 120
ttcgcgcctc agcgtcagtt acagcccaga gagtcgcctt cgccactggt gttcctccac 180
atatctacgc atttcaccgc tacacgtgga attccactct cctcttctgc actcaagctc 240
cccagtttcc agtgcgaccc gaagttgagc ctcgggatta aacaccagac ttaaagagcc 300
gcctgcgcgc gctttacgcc caataattcc ggacaacgct tgccccctac gtattaccgc 360
ggctgctggc acgtagttag ccggggcttt cttctcaggt accgtcactc ttgtagcagt 420
tactctacaa gacgttcttc cctggcaaca gagctttacg atccgaaaac cttcatcact 480
cacgcggcgt tgctccgtca gctttcgccc attgcggaag attccctact gctgccyccc 540
gtagagtctg ggccgtgtct cagtcccagt gtggccgatc accctctcag gtcggctacg 600
catcgtcgcc ttggtaggcc tttaccccac caactagcta atgcgccgcm gccatccaca 660
agtgacagat tgctccgtct ttcctccttc tcccatgcag gaaaaggatg tatcgggtat 720
tagctaccgt ttccggtagt tatccctgtc ttgtgggcag gttgcctacg tgttactcac 780
ccgtccgccg ctagttrttt agaagcaagc twtctaatya ccccgckcga cttgcatgya 840
ttag 844
<210> 2
<211> 701
<212> ДНК
<213> Paenibacillus peoriae R 4.5
<400> 2
agttaacttc ggcaccaagg gtatcgaaac ccctaacacc tagcattcat cgtttacggc 60
gtggactacc agggtatcta atcctgtttg ctccccacgc tttcgcgcct cagcgtcagt 120
tacagcccag agagtcgcct tcgccactgg tgttcctcca catctctacg catttcaccg 180
ctacacgtgg aattccactc tcctcttctg cactcaagct ccccagtttc cagtgcgacc 240
cgaagttgag cctcgggatt aaacaccaga cttaaagagc cgcctgcgcg cgctttacgc 300
ccaataattc cggacaacgc ttgcccccta cgtattaccg cggctgctgg cacgtagtta 360
gccggggctt tcttctcagg taccgtcacy yywrkagcag ttactctmym arrcgttctt 420
ccctggcaac agagctttac gatccgaaaa ccttcatcac tcacgcggcg ttgctccgtc 480
agctttcgcc cattgcggaa gattccctac tgctgcctcc cgtagagtct gggccgtgtc 540
tcagtcccag tgtggccgat caccctctca ggtcggctac gcatcgtcgc cttggtagsc 600
ctttacccca ccaactagct aatgcgccgc mgccatccca caagtgacag attrctcccs 660
ctttcctcct tctcccatgc aggaaaarga tgtatcgggt a 701
<210> 3
<211> 867
<212> ДНК
<213> Bacillus amyloliquefaciens R 4.6
<400> 3
agmcggatgc ttaatgcgtt agctgcagca ctaaggggcg gaaaccccct aacacttagc 60
actcatcgtt tacggcgtgg actaccaggg tatctaatcc tgttcgctcc ccacgctttc 120
gctcctcagc gtcagttaca gaccagagag tcgccttcgc cactggtgtt cctccacatc 180
tctacgcatt tcaccgctac acgtggaatt ccactctcct cttctgcact caagttcccc 240
agtttccaat gaccctcccc ggttgagccg ggggctttca catcagactt aagaaaccgc 300
ctgcgagccc tttacgccca ataattccgg acaacgcttg ccacctacgt attaccgcgg 360
ctgctggcac gtagttagcc gtggctttct ggttaggtac cgtcaaggtg ccgccctatt 420
tgaacggcac ttgttcttcc ctaacaacag agctttacga tccgaaaacc ttcatcactc 480
acgcggcgtt gctccgtcag actttcgtcc attgcggaag attccctact gctgcctccc 540
gtaggagtct gggccgtgtc tcagtcccag tgtggccgat caccctctca ggtcggctac 600
gcatcgtcgc cttggtgagc cgttacctca ccaactagct atgcgccgcg ggtccwtctg 660
taagtggtac cgaagccacc ttttatgtct gaccatgcgg ttcaacaacc atccggtatt 720
agccccggtt tcccggagtt atcycrkctt acaggcaggt tacccacgtg ttactcaccc 780
gtcgcgctta catcaggagy agcttccatc tgwccgctcg acttgcatgt attaggcacg 840
ccgcagcgtt ysgtccttga ccgggat 867
<210> 4
<211> 770
<212> ДНК
<213> Paenibacillus jamilae R 4.24
<400> 4
tccccaggcg gaatcttaat gtgttaactt cggcaccaag ggtatcgaaa cccctaacac 60
ctagcattca tcgtttacgg cgtggactac cagggtatct aatcctgttt gctccccacg 120
ctttcgcgcc tcagcgtcag ttacagccca gagagtcgcc ttcgccactg gtgttcctcc 180
acatctctac gcatttcacc gctacacgtg gaattccact ctcctcttct gcactcaagc 240
tccccagttt ccagtgcgac ccgaagttga gcctcgggat taaacaccag acttaaagag 300
ccgcctgcgc gcgctttacg cccaataatt ccggacaacg cttgccccct acgtattacc 360
gcggctgctg gcacgtagtt agccggggct ttcttctcag gtaccgtcac yyywrkagca 420
gttactctmy marrcgttct tccctggcaa cagagcttta cgatccgaaa accttcatca 480
ctcacgcggc gttgctccgt cagctttcgc ccattgcgga agattcccta ctgctgcctc 540
ccgtagagtc tgggccgtgt ctcagtccca gtgtggccga tcaccctctc aggtcggcta 600
cgcatcgtcg ccttggtags ctttacccca ccaactagct aatgcgccgc mgccatccac 660
aagtgacaga ttrctccgcc tttcctcctt ctcccatgca ggaaargatg tatcgggtat 720
tagctaccgt ttccggtagt tatccctgtc ttgtrggcag rttgcctacg 770
<210> 5
<211> 467
<212> ДНК
<213> Paenibacillus polymyxa R 5.31
<400> 5
tccccaggcg atsttaatgt gttaacttcg gcaccaaggg tatcgaaacc cctaacacct 60
agcattcatc gtttacggcg tggactacca gggtatctaa tcctgtttgc tccccacgct 120
ttcgcgcctc agcgtcagtt acagcccaga gagtcgcctt cgccactggt gttcctccac 180
atctctacgc atttcaccgc tacacgtgga attccactct cctcttctgc actcaagctc 240
cccagtttcc agtgcgaccc gaagttgagc ctcgggatta aacaccagac ttaaagagcc 300
gcctgcgcgc gctttacgcc caataattcc ggacaacgct tgccccctac gtattaccgc 360
ggctgctggc acgtagttag ccggggcttt cttctcaggt accgtcacyy ctagagcagt 420
tactctctma gacgttcttc cctggsaaaa magctttacg atcggaa 467
<210> 6
<211> 857
<212> ДНК
<213> Paenibacillus peoriae R6.14
<400> 6
atgtgttaac ttcggcacca agggtatcga aacccctaac acctagcatt catcgtttac 60
ggcgtggact accagggtat ctaatcctgt ttgctcccca cgctttcgcg cctcagcgtc 120
agttacagcc cagagagtcg ccttcgccac tggtgttcct ccacatctct acgcatttca 180
ccgctacacg tggaattcca ctctcctctt ctgcactcaa gctccccagt ttccagtgcg 240
acccgaagtt gagcctcggg attaaacacc agacttaaag agccgcctgc gcgcgcttta 300
cgcccaataa ttccggacaa cgcttgcccc ctacgtatta ccgcggctgc tggcacgtag 360
ttagccgggg ctttcttctc aggtaccgtc actcttgtag cagttactct acaagacgtt 420
cttccctggc aacagagctt tacgatccga aaaccttcat cactcacgcg gcgttgctcc 480
gtcaggcttt cgcccattgc ggaagattcc ctactgctgc ctcccgtagg agtctgggcc 540
gtgtctcagt cccagtgtgg ccgatcaccc tctcaggtcg gctacgcatc gtcgccttgg 600
taggccttta ccccaccaac tagctaatgc gccgcaggcc catccacaag tgacagattg 660
ctccgtcttt cctccytctc ccatgcagga aaggatgtat cgggtattag ctaccgtttc 720
cggtagttat ccctgtcttg tgggcaggtt gcctacgtgt tactcacccg tccgccgcta 780
ggttrtttag aagcaagctt ctamayaacc ccgctcgact tgcatgtatt aggcacgccg 840
ccgcgttcgt cctgagc 857
<210> 7
<211> 843
<212> ДНК
<213> Bacillus amyloliquefaciens V 3.14
<400> 7
gcgttagctg cagcactaag gggcggaaacc ccctaacact tagcactcat cgtttacggc 60
gtggactacc agggtatcta atcctgttcgc tccccacgct ttcgctcctc agcgtcagtt 120
acagaccaga gagtcgcctt cgccactggtg ttcctccaca tctctacgca tttcaccgct 180
acacgtggaa ttccactctc ctcttctgcac tcaagttccc cagtttccaa tgaccctccc 240
cggttgagcc gggggctttc acatcagactt aagaaaccgc ctgcgagccc tttacgccca 300
ataattccgg acaacgcttg ccacctacgta ttaccgcggc tgctggcacg tagttagccg 360
tggctttctg gttaggtacc gtcaaggtgcc gccctatttg aacggcactt gttcttccct 420
aacaacagag ctttacgatc cgaaaaccttc atcactcacg cggcgttgct ccgtcagact 480
ttcgtccatt gcggaagatt ccctactgctg cctcccgtag gagtctgggc cgtgtctcag 540
tcccagtgtg gccgatcacc ctctcaggtcg gctacgcatc gtcgccttgg tgagccgtta 600
cctcaccaac tagctaatgc gccgcgggtcc atctgtaagt ggtagccgaa gccacctttt 660
atgtctgaac catgcggttc agacaaccatc cggtattagc cccggtttcc cggagttatc 720
ccagtcttac aggcaggtta cccacgtgtta ctcacccgtc cgccgctcac atcaggagca 780
agctcccatc tgtccgctcg acttgcatggt attaggcacg ccgccagcgt tcgttctgag 840
cag 843
<210> 8
<211> 682
<212> ДНК
<213> Paenibacillus peoriae O 1.27
<400> 8
ttaatgyagt taacttcggc accaagggta tcgaaacccc taacacctag cattcatcgt 60
ttacggcgtg gactaccagg gtatctaatc ctgtttgctc cccacgcttt cgcgcctcag 120
cgtcagttac agcccagaga gtcgccttcg ccactggtgt tcctccacat ctctacgcat 180
ttcaccgcta cacgtggaat tccactctcc tcttctgcac tcaagctccc cagtttccag 240
tgcgacccga agttgagcct cgggattaaa caccagactt aaagagccgc ctgcgcgcgc 300
tttacgccca ataattccgg acaacgcttg ccccctacgt attaccgcgg ctgctggcac 360
gtagttagcc ggggctttct tctcaggtac cgtcactctt rtagcagtta ctctacaaga 420
cgttcttccc tggcaacaga gctttacgat ccgaaaacct tcatcactca cgcggcgttg 480
ctccgtcagc tttcgcccat tgcggaagat tccctactgc tgcctcccgt agagtctggg 540
ccgtgtctca gtcccagtgt ggccgatcac cctctcaggt cggctacgca tcgtcgcctt 600
ggtagscctt taccccacca actagctaat gcgccgcmgs catccmcaag tgacagattg 660
ctccgtcttt cctccttctc cc 682
<210> 9
<211> 851
<212> ДНК
<213> Paenibacillus jamilae K 1.14
<400> 9
atgtgttaac ttcggcacca agggtatcga aacccctaac acctagcatt catcgtttac 60
ggcgtggact accagggtat ctaatcctgt ttgctcccca cgctttcgcg cctcagcgtc 120
agttacagcc cagagagtcg ccttcgccac tggtgttcct ccacatctct acgcatttca 180
ccgctacacg tggaattcca ctctcctctt ctgcactcaa gctcyccagt ttccagtgcg 240
acccgaagtt gagcctcggg attaaacacc agacttaaag agccgcctgc gcgcgcttta 300
cgcccaataa ttccggacaa cgcttgcccc ctacgtatta ccgcggctgc tggcacgtag 360
ttagccgggg ctttcttctc aggtaccgtc acyyywrkag cagttactct mymarrcgtt 420
cttccctggc aacagagctt tacgatccga aaaccttcat cactcacgcg gcgttgctcc 480
gtcaggcttt cgcccattgc ggaagattcc ctactgctgc ctcccgtagg agtctgggcc 540
gtgtctcagt cccagtgtgg ccgatcaccc tctcaggtcg gctacgcatc gtcgccttgg 600
taggccttta ccccaccaac tagctatgcg ccgcaggccc atccacaagt gacagattgc 660
tccgcctttc ctccttctcc catgcaggaa aggatgtatc gggtattagc taccgtttcc 720
ggtagttatc cctgtcttgt gggcaggttg cctacgtgtt actcacccgt ccgccgctag 780
gttarttaga agcaagcttc taatyaaccy ccgctcgact tgcatgtata acacgccgcc 840
gcgtcgtctg a 851
<210> 10
<211> 852
<212> ДНК
<213> Paenibacillus jamilae O 2.11
<400> 10
atgtgttaac ttcggcacca agggtatcga aacccctaac acctagcatt catcgtttac 60
ggcgtggact accagggtat ctaatcctgt ttgctcccca cgctttcgcg cctcagcgtc 120
agttacagcc cagagagtcg ccttcgccac tggtgttcct ccacatctct acgcatttca 180
ccgctacacg tggaattcca ctctcctctt ctgcactcaa gctccccagt ttccagtgcg 240
acccgaagtt gagcctcggg attaaacacc agacttaaag agccgcctgc gcgcgcttta 300
cgcccaataa ttccggacaa cgcttgcccc ctacgtatta ccgcggctgc tggcacgtag 360
ttagccgggg ctttcttctc aggtaccgtc actcttgtag cagttactct acaagacgtt 420
cttccctggc aacagagctt tacgatccga aaaccttcat cactcacgcg gcgttgctcc 480
gtcaggcttt cgcccattgc ggaagattcc ctactgctgc ctcccgtagg agtctgggcc 540
gtgtctcagt cccagtgtgg ccgatcaccc tctcaggtcg gctacgcatc gtcgccttgg 600
taggccttta ccccaccaac tagctatgcg ccgcaggccc atccacaagt gacagattrc 660
tccgtctttc ctccytctcc catgcaggaa aggatrtatc gggtattagc taccgtttcc 720
ggtagttatc cctgtcttgt ggcaggttgc ctacgtgtta ctcacccgtc cgccgctggr 780
ttrtktagaa gcaagcttct amayaayccc gctcgacttg catgtattaa cacgccgtag 840
cgttcgtcct ga 852
<---


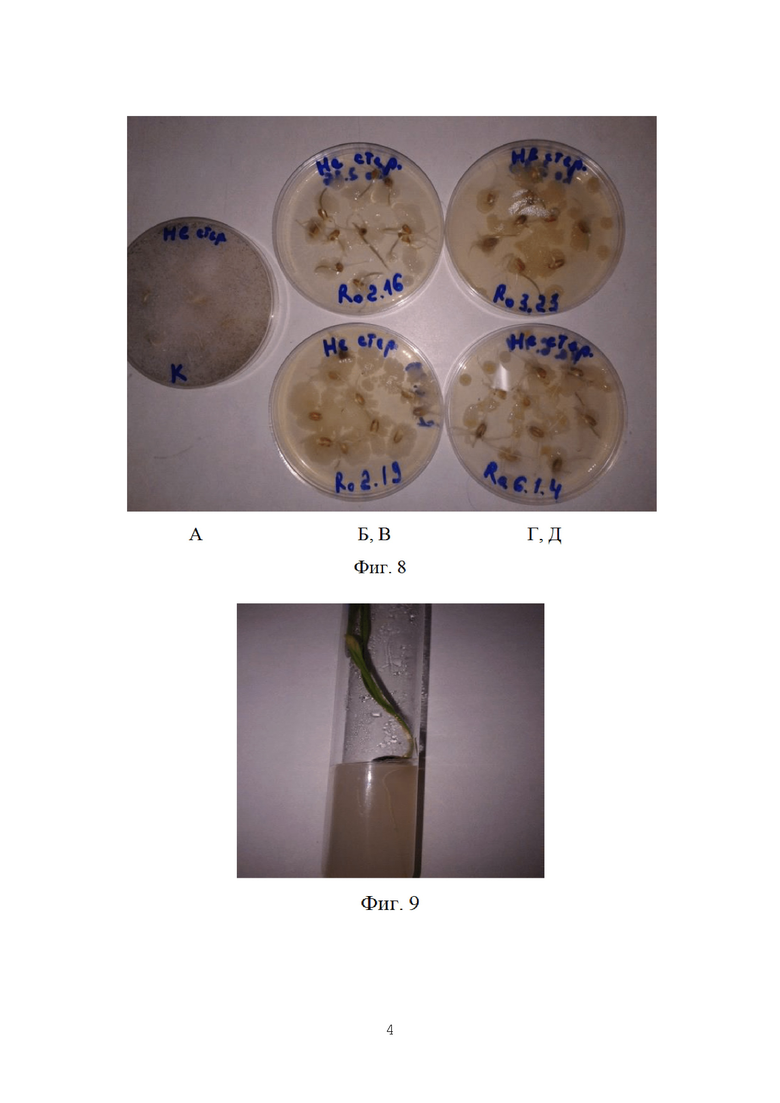
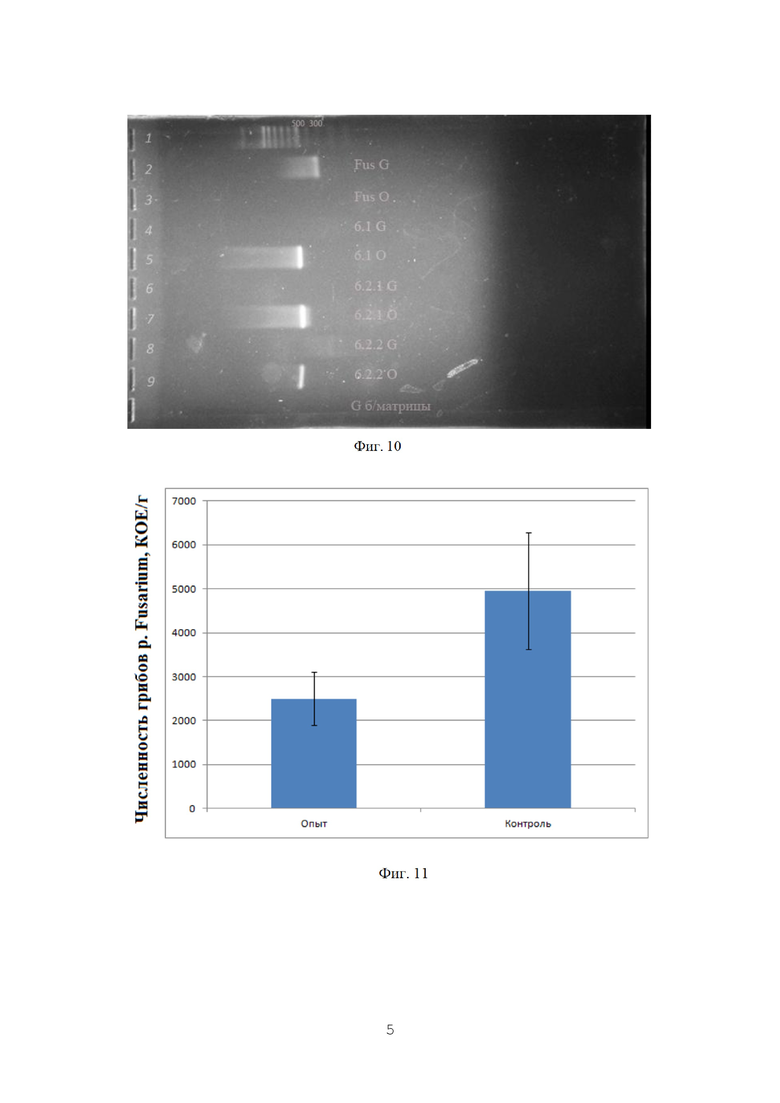
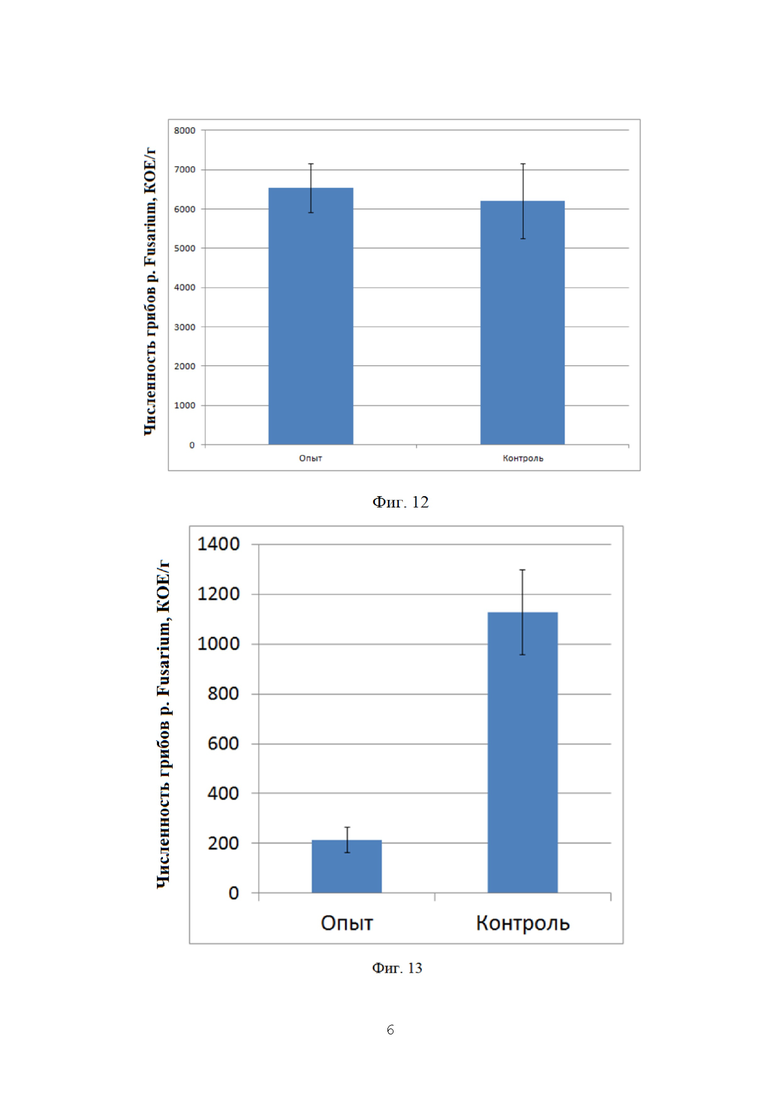
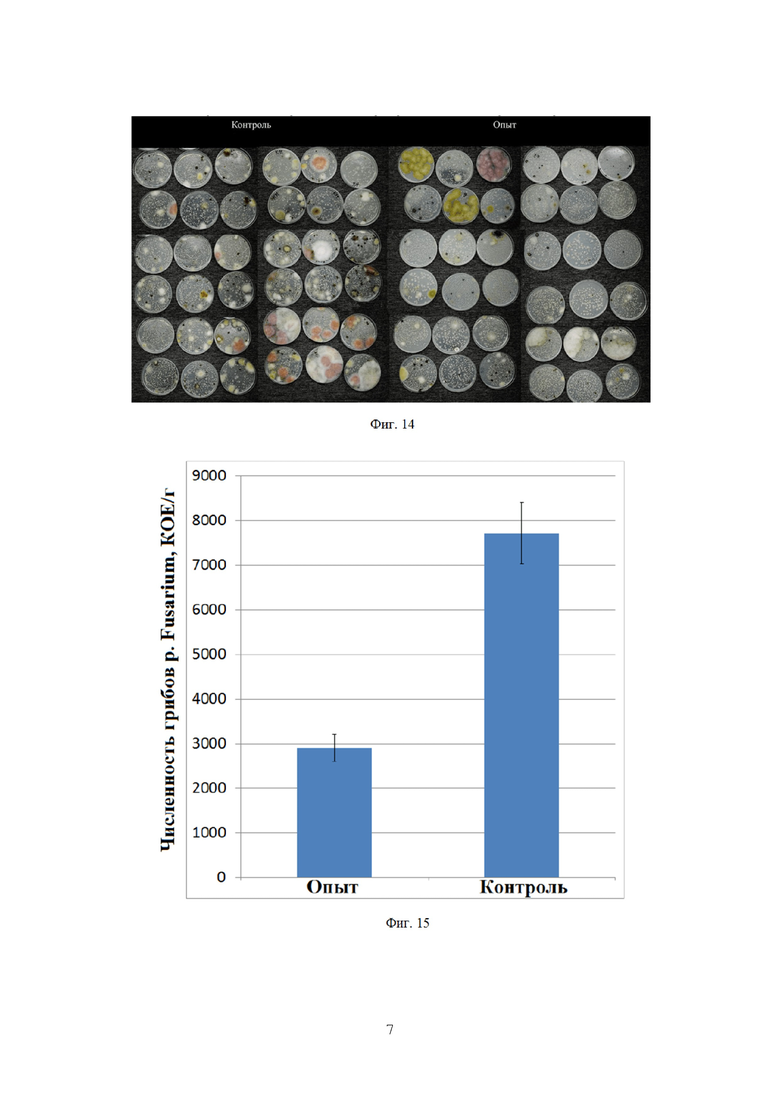
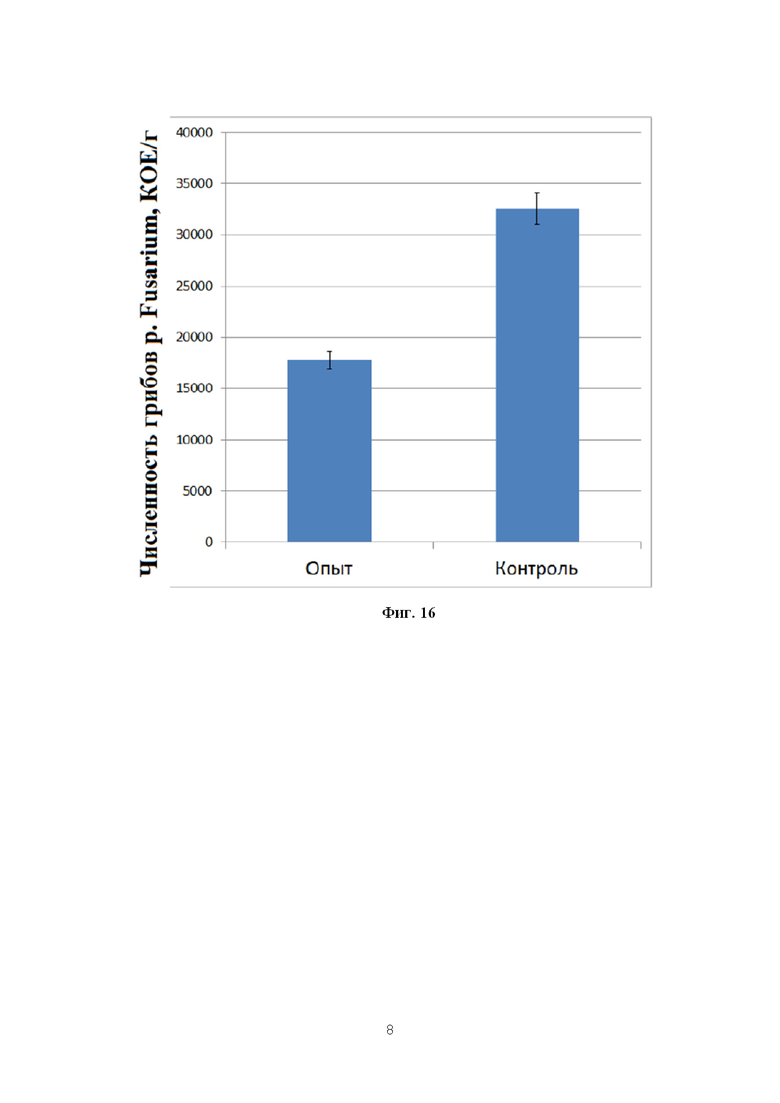

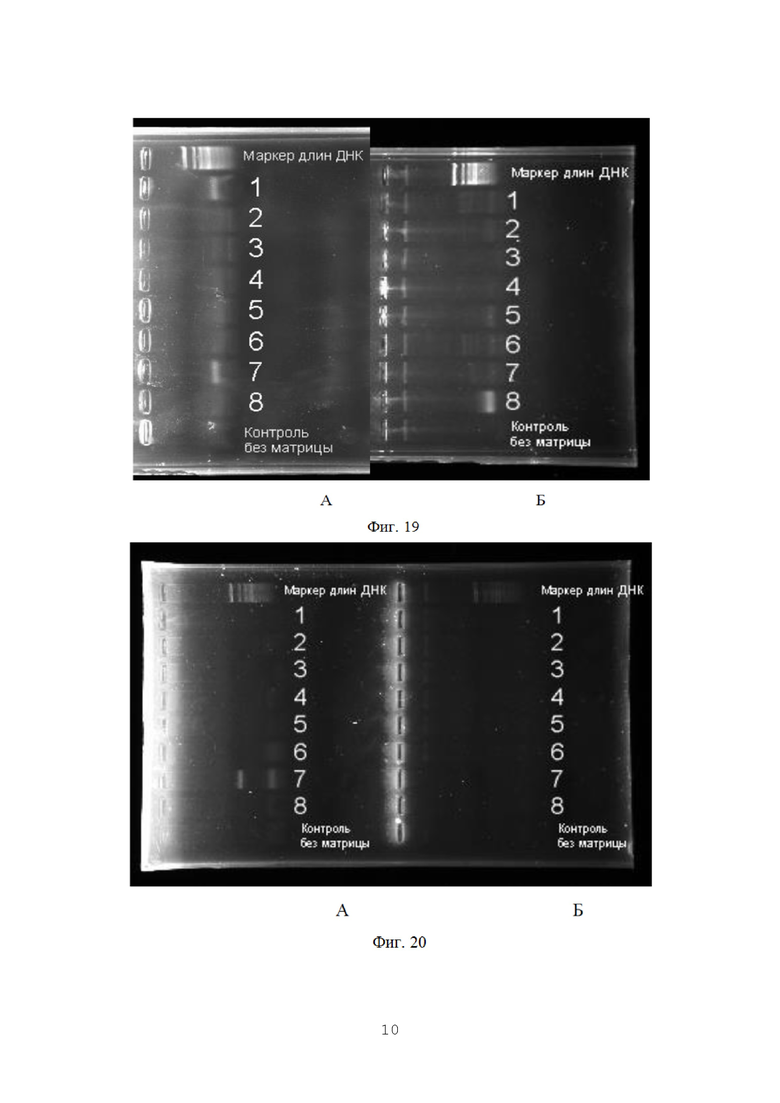
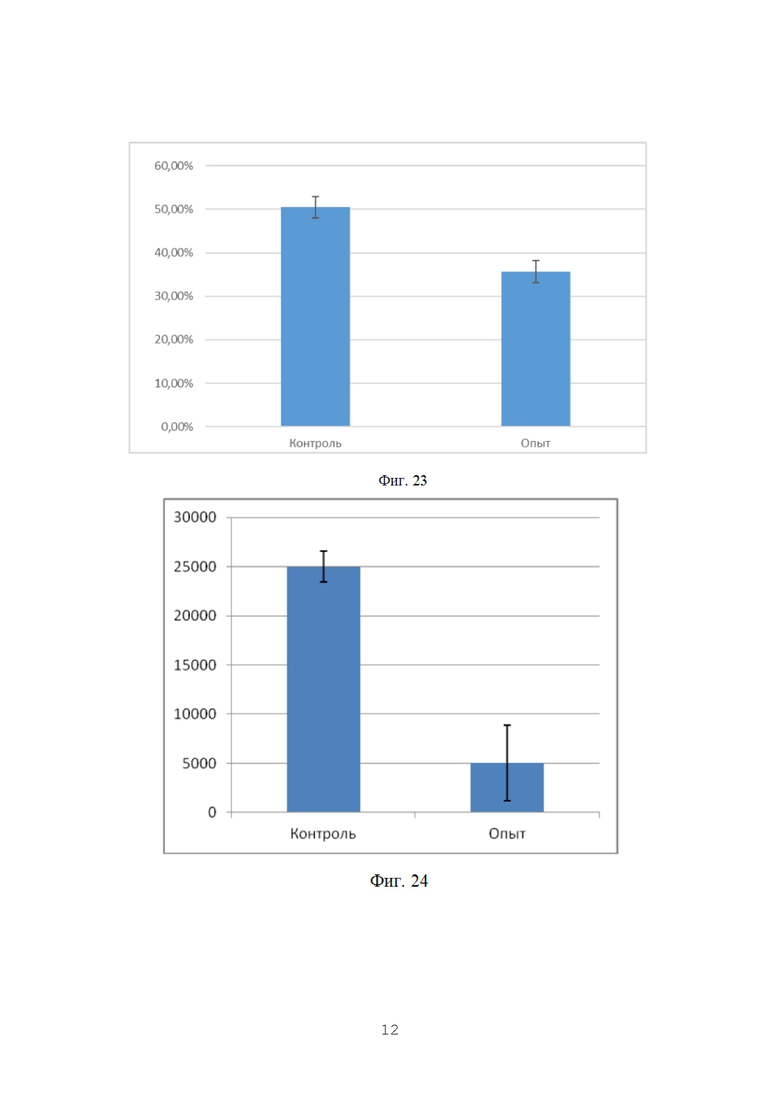
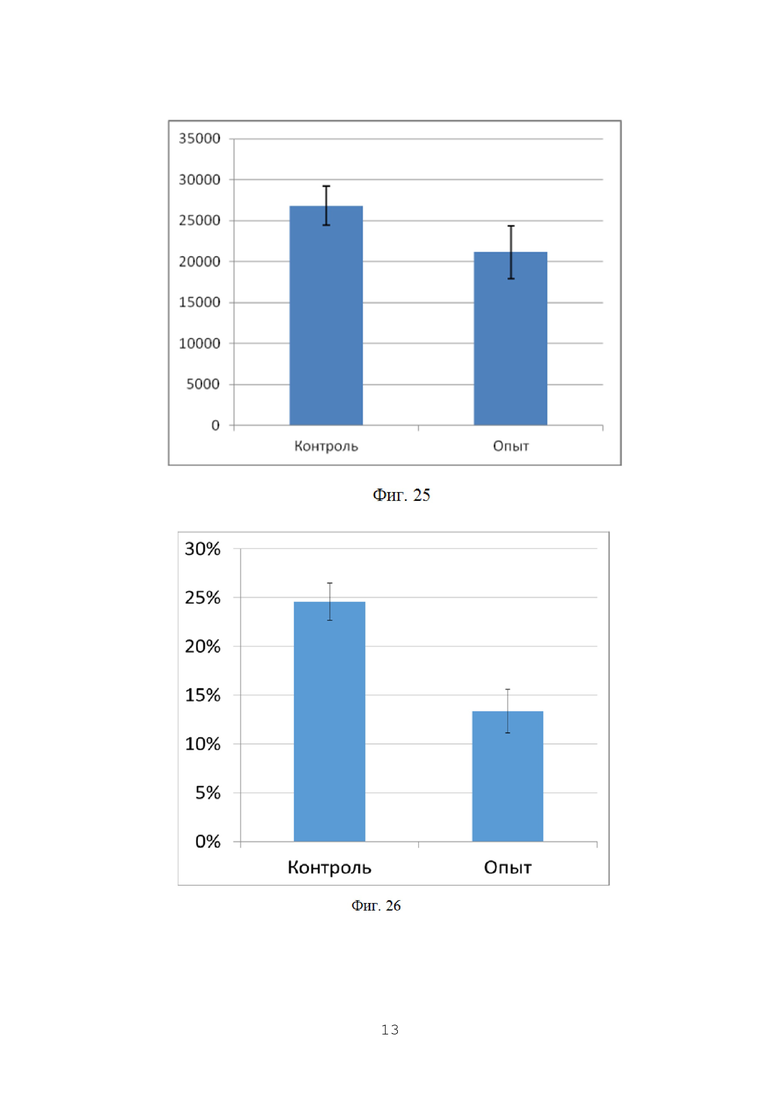
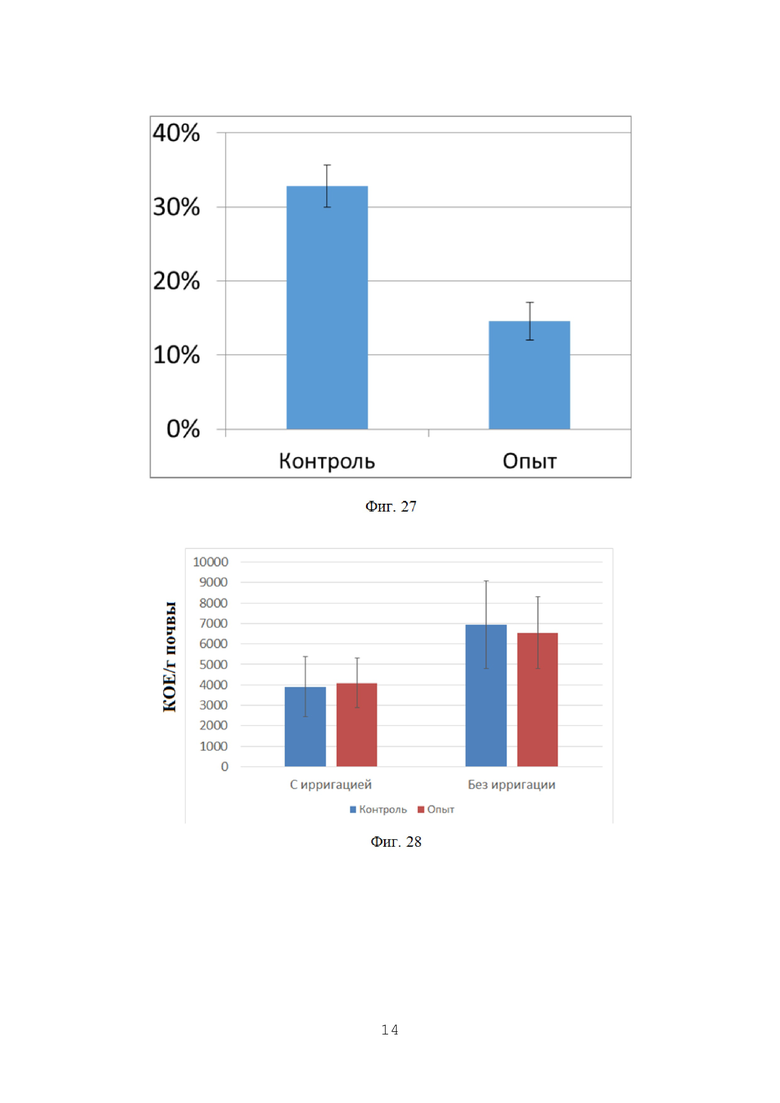
Группа изобретений относится к области биотехнологии. Предложены штаммы споробразующих бактерий-антагонистов с антифузариозной активностью (варианты), биопрепарат для защиты сельскохозяйственных культур от фузариоза, способ изготовления биопрепарата и способ защиты сельскохозяйственных культур от фузариоза. Биопрепарат включает комбинацию предложенных штаммов, при этом содержит не менее 108 КОЕ/г жизнеспособных бактерий-антагонистов и имеет остаточную влажность не более 10%. Способ изготовления биопрепарата включает твердофазное культивирование предложенных штаммов на питательном субстрате. Способ защиты сельскохозяйственных культур включает выделение грибов рода Fusarium из отобранных на полях образцов, отбор наиболее активно подавляющих штаммы грибов рода Fusarium штаммов спорообразующих бактерий-антагонистов, приготовление биопрепарата и обработку биопрепаратом посевного материала. Изобретения обеспечивают расширение арсенала штаммов споробразующих бактерий-антагонистов с антифузариозной активностью и биопрепаратов, направленных на эффективную борьбу с фузариозом, а также обеспечивают при изготовлении биопрепарата возможность использования простой по составу среды с высоким выходом готового продукта и стабильностью полученного биопрепарата при хранении. 13 н.п. ф-лы, 28 ил., 30 табл.
1. Штамм Paenibacillus jamilae K 1.14 споробразующих бактерий-антагонистов с антифузариозной активностью, депонированный во Всероссийской коллекции промышленных микроорганизмов под номером В-13016.
2. Штамм Paenibacillus peoriae О 1.27 споробразующих бактерий-антагонистов с антифузариозной активностью, депонированный во Всероссийской коллекции промышленных микроорганизмов под номером В-13017.
3. Штамм Paenibacillus peoriae О 2.11 споробразующих бактерий-антагонистов с антифузариозной активностью, депонированный во Всероссийской коллекции промышленных микроорганизмов под номером В-13018.
4. Штамм Paenibacillus peoriae R 3.13 споробразующих бактерий-антагонистов с антифузариозной активностью, депонированный во Всероссийской коллекции промышленных микроорганизмов под номером В-13019.
5. Штамм Paenibacillus peoriae R 4.5 споробразующих бактерий-антагонистов с антифузариозной активностью, депонированный во Всероссийской коллекции промышленных микроорганизмов под номером В-13020.
6. Штамм Bacillus amyloliquefaciens R 4.6 споробразующих бактерий-антагонистов с антифузариозной активностью, депонированный во Всероссийской коллекции промышленных микроорганизмов под номером В-13021.
7. Штамм Paenibacillus jamilae R 4.24 споробразующих бактерий-антагонистов с антифузариозной активностью, депонированный во Всероссийской коллекции промышленных микроорганизмов под номером В-13022.
8. Штамм Paenibacillus polymyxa R 5.31 споробразующих бактерий-антагонистов с антифузариозной активностью, депонированный во Всероссийской коллекции промышленных микроорганизмов под номером В-13023.
9. Штамм Paenibacillus peoriae R 6.14 споробразующих бактерий-антагонистов с антифузариозной активностью, депонированный во Всероссийской коллекции промышленных микроорганизмов под номером В-13048.
10. Штамм Bacillus amyloliquefaciens V 3.14 споробразующих бактерий-антагонистов с антифузариозной активностью, депонированный во Всероссийской коллекции промышленных микроорганизмов под номером В-13024.
11. Биопрепарат для защиты сельскохозяйственных культур от фузариоза, включающий, комбинацию штаммов по пп. 1-10, содержащий не менее 108 КОЕ/г жизнеспособных бактерий-антагонистов, при этом остаточная влажность сухого биопрепарата не более 10%.
12. Способ изготовления биопрепарата на основе комбинации штаммов спорообразующих бактерий-антагонистов с антифузариозной активностью по пп. 1-10 путем их твердофазного культивирования на питательном субстрате, включающий подготовку субстрата путем гидратирования в воде, термообработку субстрата, инокуляцию термообработанного гидратированного субстрата, инкубацию, гомогенизацию субстрата, размягченного при термообработке и в процессе ферментации и последующую сушку.
13. Способ защиты сельскохозяйственных культур от фузариоза, включающий выделение грибов рода Fusarium из образцов, отобранных на полях, подверженных опасности фузариоза; отбор штаммов спорообразующих бактерий-антагонистов, наиболее активно подавляющих штаммы грибов рода Fusarium, распространенных на целевой территории; приготовление биопрепарата по п. 11, способом по п. 12, обработку биопрепаратом посевного материала.
| Способ получения цианистых соединений | 1924 |
|
SU2018A1 |
| CN 109266589 A, 25.01.2019 | |||
| KR 20140115022 A, 30.09.2014 | |||
| US 6602500 B1, 05.08.2003 | |||
| ШТАММ БАКТЕРИЙ Bacillus amyloliquefaciens subsp. plantarum BS89 В КАЧЕСТВЕ СРЕДСТВА ПОВЫШЕНИЯ ПРОДУКТИВНОСТИ РАСТЕНИЙ И ИХ ЗАЩИТЫ ОТ БОЛЕЗНЕЙ | 2015 |
|
RU2599416C1 |

