Изобретение относится к молекулярной генетике, а именно к способам определения полиморфизма генов, в частности гена дистрофина (DMD), обуславливающего наличие стресс-синдрома у свиней, и может быть использовано в племенном свиноводстве.
Важнейшим условием устойчивого развития свиноводства является селекционно-племенной потенциал отрасли. Научное обеспечение этой деятельности считается одним из наиболее перспективных направлений, на которых должны быть сосредоточены усилия [Отраслевая программа «Развитие свиноводства в Российской Федерации на 2013-2015 годы». М., 2013. С. 28.]. Большой вклад в развитие современного свиноводства в РФ внес импорт зарубежного племенного материала. Такой завоз был сопряжен с неконтролируемым вводом и распространением в популяциях племенного скота в России наследственных аномалий, способных нанести существенный вред развитию племенного животноводства и поставить под угрозу биологическую безопасность страны [Молекулярные методы диагностики заболеваний и наследственных дефектов сельскохозяйственных животных / Е.А. Гладырь, Н.А. Зиновьева, Л.К. Эрнст и др. // Зоотехния, 2009. №8. С. 26-27].
Элиминация рецессивных наследственных аномалий в популяциях сельскохозяйственных животных требует разработки и внедрения систем ДНК-идентификации животных - носителей мутантных аллелей. Использование тест-систем ДНК-диагностики позволит очистить популяции животных от скрытых носителей и тем самым за счет современных методов управления генетическими ресурсами повысить эффективность свиноводства. Чувствительность свиней к стрессам представляет определенную проблему в свиноводстве [Генетические аномалии в свиноводстве / О.В. Костюнина, В.Р. Харзинова, Е.А. Гладырь, Н.А. Зиновьева, Е.И. Сизарева // Таврийский научный вестник. - 2011. - №76(2). - С. 190-193]. Стрессы обусловлены рядом факторов, такими как генетическая предрасположенность животных, поведение обслуживающего персонала, скученность при транспортировке и содержании, сезон года и другими [The fatigued pig syndrome / M. Ritter, M. Ellis, M. Benjamin, E. Berg, P. DuBois, J. Marchant-Forde, A. Green, P. Matzat, P. Mormede, Т. Moyer, K. Pfalzgraf, M. Siemens, J. Sterle, T. Whiting, B. Wolter, and A. Johnson // Journal of Animal Science. 2005. 83(Suppl. 1):258. (Abstr.)] К примеру, в 2006 году потери свиноводческой промышленности США в результате транспортного стресса составили примерно 46 млн. долл. [Transport losses in market weight pigs: I. A review of definitions, incidence and economic impact / M.J. Ritter, M. Ellis, N.L. Berry, S.E. Curtis, L, Anil, M. Benjamin, D. Butler, C. Dewey, B. Driessen, P. DuBois, J. Hill, J. Marchant-Forde, P. Matzat, J. McGlone, P. Mormede, T. Moyer, K. Pfalzgraf, J. Salak-Johnson, J. Sterle, C. Stull, T. Whiting, B. Wolter, S. R. Niekamp, A.K.. Johnson // Professional Animal Scientist. 2009. 25:404-414.
У чувствительных к стрессу свиней помимо нарушений в конституции могут проявляться симптомы, приводящие к гибели животных. Известно, что одним из генетических факторов является мутация HAL-1843 в гене рианодинового рецептора 1, RYR1, которая приводит к возникновению злокачественной гипертермии у свиней и пороку PSE (бледное, дряблое, экссудативное мясо). Эта мутация, приводящая к аминокислотной замене (аргинин - цистеин) в 615-й позиции RYR1, была обнаружена в 1991 году и считается ответственной за большинство потерь, обусловленных стрессом [The fatigued pig syndrome / M. Ritter, M. Ellis, M. Benjamin, E. Berg, P. DuBois, J. Marchant-Forde, A. Green, P. Matzat, P. Mormede, T. Moyer. K. Pfalzgraf, M. Siemens, J. Sterle, T. Whiting, B. Wolter, and A. Johnson // Journal of Animal Science. - 2005. - 83(Suppl. 1):258. (Abstr.); Identification of a mutation in porcine ryanodine receptor associated with malignant hyperthermia / J. Fujii, K. Otsu, F. Zorzato. S. de Leon, V.K. Khanna, J.E. Weiler, P.J.  D.H. MacLennan // Science. - 1991. - №253. - P. 448-451; The effects of halothane sensitivity on carcass composition and meat quality in HAL-1843 normal pigs / C. P. Allison, R. C. Johnson, M. E. Doumit // Journal of Animal Science. - 2005. - №83. - P. 871-678.]. Однако не все фенотипические проявления стресса обусловлены данной мутацией.
D.H. MacLennan // Science. - 1991. - №253. - P. 448-451; The effects of halothane sensitivity on carcass composition and meat quality in HAL-1843 normal pigs / C. P. Allison, R. C. Johnson, M. E. Doumit // Journal of Animal Science. - 2005. - №83. - P. 871-678.]. Однако не все фенотипические проявления стресса обусловлены данной мутацией.
В 2012 году коллектив американских ученых во главе с Nonneman D.J. обнаружил симптомы, характерные для классического стресса, у животных, свободных от мутации HAL-1843 в гене RYR1 [A defect in dystrophin causes a novel porcine stresssyndrome / Dan J., Nonneman, Tami Brown-Brandl, Shuna A. Jones, Ralph T.Wiedmann and Gary A. Rohrer // BMCGenomics, 2012. №13. P. 233].
Проведенные полногеномные исследования позволили обнаружить статистически значимую ассоциацию региона на половой хромосоме X (SSCX), локализованного в области 25.1-27.7 Мб гена дистрофина (DMD), со стресс-синдромом. Причиной мутацией является нуклеотидная замена (85890_783; ss410758971, экзон 41), которая приводит к замене аргинина на триптофан в позиции 1958 (R1958W) аминокислотной последовательности DMD. Ген дистрофина кодирует 15-й повтор биспиральной области центрального стержня спектрина и таким образом обуславливает функции белка дистрофина, являющегося важной составляющей структуры мышечной ткани. Дистрофии обеспечивает структурную устойчивость ассоциированного с ним гликопротеинового комплекса (ДАГ-комплекса), расположенного на клеточной мембране [Dystrophin. More thanjust the sum of its parts / E. Le Rumeur,S.J. Winder, J.F. Hubert // Biochim Biophys Acta. - 2010. - №1804. - P. 1713-1722.].
По некоторым данным, мутация R1958W приводит к уменьшению экспрессии дистрофина в диафрагме, поясничной и длиннейшей мышце спины [The physiological response of protease inhibition in dystrophic muscle/K. Hollinger, J.T. Selsby // Acta Physiol, 2013. №208. P. 234-244.].
Из-за локализации гена на Х-хромосоме стресс-синдром, как правило, встречается в основном у самцов, унаследовавших Х-хромосому с дефектным аллелем от матери. Наибольшая восприимчивость к стрессу у свиней отмечается в возрасте двух-трех месяцев [A defect in dystrophin causes a novel porcine stresssyndrome / D.J., Nonneman, T. Brovvn-Brandl, S.A. Jones, R.T.Wiedmann, G.A. Rohrer // BMC Genomics, 2012. №13. P. 233].
В качестве прототипа заявленного способа, наиболее близким по технической сущности и достигаемому результату, можно считать прямое секвенирование продуктов ПЦР [A defect in dystrophin causes a novel porcine stresssyndrome / D.J., Nonneman, T. Brown-Brandl, S.A. Jones, R.T.Wiedmann, G.A. Rohrer // BMC Genomics, 2012. №13. P. 233], использование которого требует наличие дорогостоящего оборудования (ДНК-секвенаторов). Отличие нашего способа заключается в выявлении полиморфизма DMD с использованием метода ПЦР с последующим рестрикционным гидролизом образующегося фрагмента (анализа ПДРФ - полиморфизма длин амплифицированных фрагментов).
Следует отметить, что простота нашего способа определения полиморфизма DMD на основе ПЦР-ПДРФ, отсутствие необходимости использования дорогостоящего оборудования позволит использовать его в селекционной практике с целью идентификации животных, являющихся скрытыми носителями.
Анализ последовательности DMD в области мутации показал, что последняя приводит к исключению сайта рестрикции эндонуклеазы Aci I (последовательность узнавания 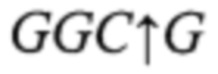 ), поэтому в качестве базового метода для моделирования тест-системы определения полиморфизма DMD был выбран метод полимеразной цепной реакции с последующим анализом полиморфизма длин рестрицированных фрагментов (ПЦР-ПДРФ), что является отличием заявленного изобретения от известных способов диагностики аллелей DMD. Такой подход позволяет проводить идентификацию результатов с помощью метода электрофореза в агарозном геле без использования дорогостоящего оборудования, что обеспечит относительно невысокую стоимость разрабатываемого способа.
), поэтому в качестве базового метода для моделирования тест-системы определения полиморфизма DMD был выбран метод полимеразной цепной реакции с последующим анализом полиморфизма длин рестрицированных фрагментов (ПЦР-ПДРФ), что является отличием заявленного изобретения от известных способов диагностики аллелей DMD. Такой подход позволяет проводить идентификацию результатов с помощью метода электрофореза в агарозном геле без использования дорогостоящего оборудования, что обеспечит относительно невысокую стоимость разрабатываемого способа.
Суть разработанной тест-системы заключается в амплификации фрагмента гена DMD, содержащего мутацию, длиной 234 п.о. с последующим рестрикционным гидролизом образующегося фрагмента эндонуклеазой AciI. Это приводит к образованию фрагмента длиной 202 п.о., специфичного для нормального аллеля (нуклеотид С), в то время как мутантному аллелю (нуклеотид Т) соответствует нерестрицированный фрагмент длиной 234 п.о. Животные - носители стресс-синдрома имеют генотипы ТТ (свинки) и Т0 (хрячки), свинки - носители гетерозиготного генотипа ТС, а животные - не носители - генотип СС (свинки) и С0 (хрячки), что позволяет дифференцировать мутантные и немутантные аллели DMD методом электрофореза в агарозном геле.
Способ отличается тем, что с применением нескольких технически простых и не требующих дорогостоящих реактивов, оборудования, затрат сил и времени методов, возможно выявление мутантного аллеля Т гена DMD, что позволит применить данный метод в селекции животных.
При создании настоящего изобретения задача состояла в разработке способа обнаружения аллеля Т гена DMD, обуславливающего наличие стресс-синдрома, с целью идентификации скрытых носителей и разработки программ их использования в селекции без снижения репродуктивных способностей свиней.
Задача нашего изобретения - создание дешевого способа идентификации полиморфизма в гене DMD, обуславливающего наличие стресс-синдрома у свиней, и может быть использовано в племенном свиноводстве.
Технический результат изобретения достигается тем, что предложен способ диагностики полиморфизма в гене DMD, обуславливающего наличие стресс-синдрома у свиней, для использования в селекции свиней, включающий метод полимеразной цепной реакции с последующим гидролизом продуктов реакции с использованием эндонуклеазы рестрикиции AciI. Это приводит к образованию фрагментов длиной 202 п.о. и 32 п.о., специфичных для нормального аллеля (нуклеотид С), в то время как мутантному аллелю (нуклеотид Т) соответствует нерестрицированный фрагмент длиной 234 п.о., а идентификация генотипов животных осуществляется по результатам электрофореза продуктов ПЦР-ПДРФ в агарозном геле. Это обеспечит снижение трудоемкости, увеличение производительности, возможности использования реагентов отечественного производства, относительно невысокую стоимость разрабатываемого способа.
Способ отличается тем, что с применением нескольких технически простых и не требующих дорогостоящих реактивов, затрат сил и времени методов, возможно выявление мутантного аллеля Т гена DMD, что позволит применить данный метод в селекции животных.
Сущность изобретения - определение полиморфизма гена DMD, обуславливающего наличие стресс-синдрома у свиней, методом ПЦР-ПДРФ.
Разрабатываемый способ базируется на определении нуклеотидной трансверсии С→Т в позиции 1958 гена DMD (R1958W). С этой целью был выбран участок гена DMD с мутацией, обуславливающей наличие стресс-синдрома. В электронной базе была найдена нуклеотидная последовательность интересующего нас участка ДНК (номер GenBank NW_003612750).
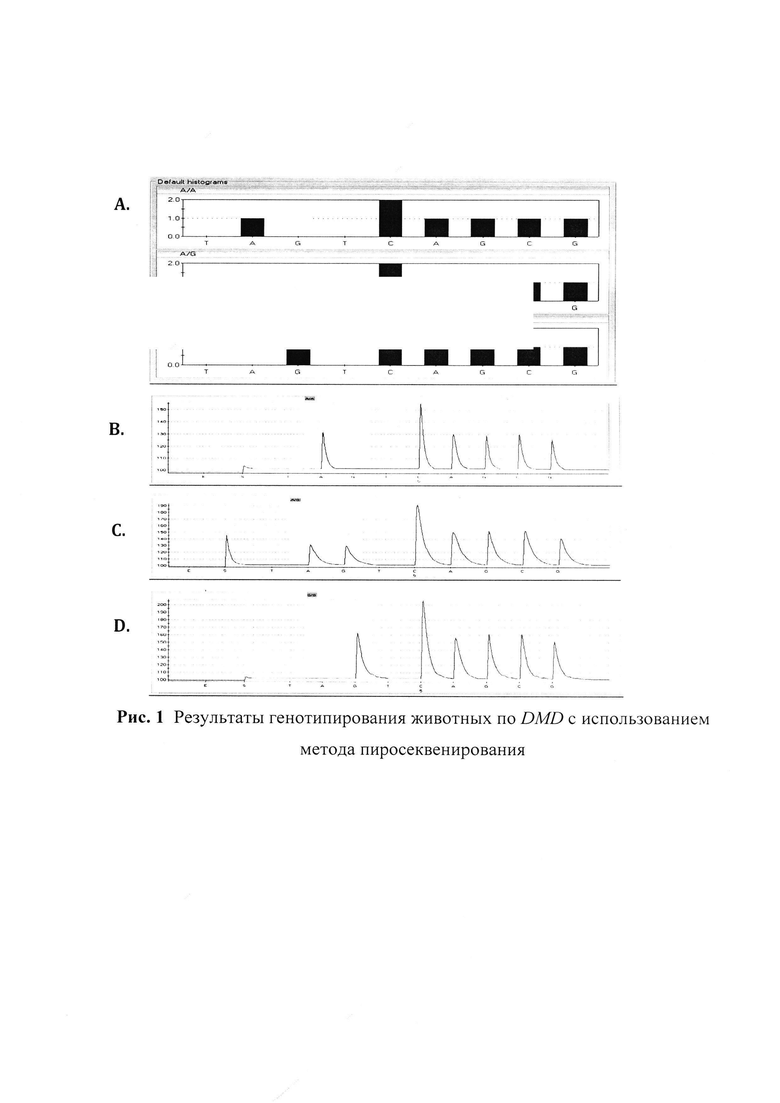
Для создания серии референтных образцов с известными генотипами по DMD, (n=48) из проб ткани (ушной выщип, n=24 (хряки) и n=24 (свиноматки) выделяли ДНК: по 24 образца каждого из видов биоматериала методом экстракции перхлоратом с использованием магнитных частиц (ООО «Изоген», Россия) и по 24 образца - с использованием колонок Nexttec (Nexttec Biotechnologie GmbH, Германия) в соответствии с рекомендациями производителей. Референтные генотипы генерировали посредством прямого определения последовательности в области мутации методом пиросеквенирования. С этой целью проводили амплификацию фрагмента длиной 234 п.о., содержащего мутацию, с использованием праймеров DMD_1Pyro,и DMD_2Pyro_Bio (мечен биотином) с последующим отжигом зонда DMD_Zond и пиросеквенированием. Для пиросеквенирования использовали базовую последовательность TA/GTCAGCG, где A/G - мутируемый нуклеотид, Т, Т - контрольные нуклеотиды. Были теоретически смоделированы гистограммы возможных генотипов DMD и получены результаты генотипирования животных (рис. 1). Где А - теоретически смоделированные гистограммы в зависимости от генотипа по DMD, ось X - базовая последовательность нуклеотидов, ось Y - условные единицы (соответствуют теоретически рассчитанной силе люминесценции); В, С, D - результирующие пикограммы последовательности DMD в области исследуемой мутации, ось X - базовая последовательность нуклеотидов, ось Y - условные единицы (регистрируемая прибором сила люминесценции);
В результате проведенного генотипирования была создана серия референтных образцов (n=48), в том числе 5 образцов с генотипом ТТ (стресс-чувствительный) и 43 образца с генотипом СС (стресс-устойчивый).
Определение полиморфизма DMD предложенным способом выполняли следующим образом:
1. Исходя из локализации мутации были подобраны два праймера, фланкирующие определяемую мутацию, при этом праймер DMD_1 является прямым, а праймер DMD_2 - обратным:
DMD_ 1 5' gccaactcagatccagctcagc3'
DMD_2 5'agcccaatgtgaggagaagct3'
Продукт амплификации праймеров DMD_1 и DMD_2 имеет длину 234 п.о. Схема представлена в приложении 1 на рисунке 2А. Где DMD_1, DMD_2 - праймеры, фланкирующие фрагмент; c/t - локализация исследуемой мутации; AciI - эндонуклеаза; Б: электрофореграмма продуктов PCR в 3% агарозном геле: дорожки 1, 2, 3, 4, 6, 9, 10, 11, 12, 14 - генотип Т0 либо ТТ (носитель); дорожка 5, 7, 13 - генотип СТ (гетерозигота, носитель); дорожка 15 - генотип СС либо С0 (не носитель); дорожка 8 - маркер молекулярной массы Fast Ruler Low Ranger; длины рестрикционных фрагментов в п.о. указаны слева от фотографии.
2. Реакции проводили в 15 мкл реакционной смеси следующего состава: 1×ПЦР буфер (16.6 мМ (NH4)2SO4, 67.7 мМ Трис-HCl, рН=8.8, 0.1% (v/v) Tween 20, 1.5 мМ MgCl2), 0,2 мМ дНТФ, 10 пмоль каждого из праймеров, 2 мМ MgCl2, 1 Ед Taq-полимеразы и 1 мкл ДНК при следующем температурно-временном режиме: 1 цикл - 95°С 5 мин, 44 цикла последовательно - 94°С - 45 с., 62°С - 45 с., 72°С - 45 с., заключительная элонгация при 72°С - 8 мин;
3. По завершении ПЦР к реакционной смеси добавляли 0,2 мкл эндонуклеазы AciI (Neb, Великобритания) и 1,5 мкл буфера IX NEB-буфер CutSmart и инкубировали при 37°С в течение 10-12 ч.
4. Определение аллелей DMD осуществляли методом гель-электрофореза, нанося по 5 мкл продуктов гидролиза в 3% агарозный гель, электрофоретически разделяли в 1х ТАЕ буфере 20 мин при 100 V и детектировали под ультрафиолетовым светом (УФ), при этом генотипу СС или С0 соответствует фрагмент длиной 202 п.о., генотипу СТ - два фрагмента длиной 234 и 202 п.о. (прил. рис. 2Б). Генотипу Т0 или ТТ (в зависимости от пола животного) соответствует наличие одного фрагмента длиной 234 п.о.
5. Результативность разработанной тест-системы оценивали посредством сравнения результатов генотипирования референтных образцов.
Пример. Контрольное использование предложенного способа определения полиморфизма DMD было апробировано на 100 свиньях породы дюрок, 100 свиньях породы крупная белая и 500 свиньях породы ландрас. Исследование выявило отсутствие аллеля Т у свиней пород крупная белая и дюрок, у свиней породы ландрас частота генотипов СТ и Т0 составила 11,33 и 0,39%%, соответственно.
Таким образом, предложенный способ может быть использован в генетике сельскохозяйственных животных для выявления полиморфизма в гене DMD обуславливающего наличие стресс-синдрома, с целью проведения тестирования племенного поголовья на наличие аллеля Т в гене DMD у племенных свиней различного происхождения с целью профилактики распространения мутации в стадах.
DMD_1 5’gccaactcagatccagctcagc3’
DMD_2 5’agcccaatgtgaggagaagct3’

Изобретение относится к области молекулярной генетики и предназначено для определения полиморфизма С→Т в позиции 1958 (R1958W) гена DMD, обуславливающего наличие стресс-синдрома у свиней. Проводят амплификацию фрагмента гена, содержащего мутацию, с использованием двух праймеров с последующим гидролизом продуктов ПЦР эндонуклеазой Aci I. Идентификация генотипов животных осуществляется по результатам электрофореза продуктов ПЦР-ПДРФ в агарозном геле. Фрагмент длиной 202 п.о. специфичен для нормального аллеля (нуклеотид С). Фрагмент длиной 234 п.о. соответствует мутантному аллелю (нуклеотид Т). Изобретение обеспечивает создание дешевого способа идентификации полиморфизма в гене DMD, обуславливающего наличие стресс-синдрома у свиней. 2 ил., 1 пр.
Способ определения полиморфизма С→Т в позиции 1958 (R1958W) гена DMD, обуславливающего наличие стресс-синдрома у свиней, включающий амплификацию фрагмента гена, содержащего мутацию, с использованием двух праймеров с последующим гидролизом продуктов ПЦР эндонуклеазой Aci I, это приводит к образованию фрагмента длиной 202 п.о., специфичного для нормального аллеля (нуклеотид С), в то время как мутантному аллелю (нуклеотид Т) соответствует нерестрицированный фрагмент длиной 234 п.о., а идентификация генотипов животных осуществляется по результатам электрофореза продуктов ПЦР-ПДРФ в агарозном геле.
| NONNEMAN D.J | |||
| et al | |||
| A defect in dystrophin causes a novel porcine stress syndrome | |||
| BMC Genomics | |||
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ СТРЕССОВОЙ ЧУВСТВИТЕЛЬНОСТИ У СВИНОМАТОК | 2010 |
|
RU2456957C2 |
| Способ отбора свиней по устойчивости к стресс-синдрому | 1980 |
|
SU923485A1 |

