Изобретение относится к области биотехнологии и касается разработки способа получения секретируемой в дрожжах, полностью функциональной фосфолипазы А2.
Фосфолипаза А2 (Pla2) катализирует гидролиз сложноэфирной связи во втором положении глицерофосфолипидов (фосфолипидов) - соединений, являющихся основными компонентами мембран всех живых клеток и по этой причине содержащихся во многих продуктах животного и растительного происхождения. Многообразие молекулярных форм фосфолипидов определяется типом фосфатной группы (холин, этаноламин, инозит или серии) и строением остатков жирных кислот.
Pla2 является одним из востребованных ферментов на рынке ферментных препаратов (ФП). В пищевой промышленности ее применяют для улучшения качества майонеза, дегуммирования растительных масел, изготовления хлебобулочных изделий, молочных продуктов, а также при производстве сыра [Бакланов 2008;  & Dennis, 1982]. Другими областями применения Pla2 являются текстильная промышленность, кормопроизводство, а также синтез искусственных фосфолипидов, востребованных, в частности, в фармакологии и парфюмерии [De Maria et al., 2007; Hoogevest & Wendel, 2014; Liu et al., 2015]. Интенсивное использование Pla2 ожидают в топливно-энергетической промышленности для получения биодизеля [Cesarini et al., 2015].
& Dennis, 1982]. Другими областями применения Pla2 являются текстильная промышленность, кормопроизводство, а также синтез искусственных фосфолипидов, востребованных, в частности, в фармакологии и парфюмерии [De Maria et al., 2007; Hoogevest & Wendel, 2014; Liu et al., 2015]. Интенсивное использование Pla2 ожидают в топливно-энергетической промышленности для получения биодизеля [Cesarini et al., 2015].
Из числа ФП, содержащих Pla2 наиболее известны следующие:
• ФП Maxapal® А2 (DSM), получаемый микробиологическим синтезом с использованием рекомбинантного штамма грибов Aspergillus niger,
• ФП «DENAZYME PLA2» или «PLA2 Nagase» (Nagase Corporation), основу которого составляет Pla2 Streptomyces violaceoruber, получаемый микробиологическим синтезом с использованием продуцентов рода Streptomyces;
• ФП «Lecitase L10» (Novozymes A/S), получаемый очисткой из поджелудочной железы свиней.
ФП Pla2 получают с использованием природных или рекомбинантных штаммов-продуцентов. Однако, независимо от происхождения, коммерческие ФП Pla2 характеризуются высокой себестоимостью, которая обусловлена низкой продукцией, проистекающей из особенностей биосинтеза этого фермента, в частности, из его способности связывать и гидролизовать фосфолипиды клеточных мембран, вызывая их разрушение, приводящее к гибели продуцирующих клеток.
Преимущества природных продуцентов Pla2 обусловлены их относительно хорошей адаптацией к биосинтезу этого токсичного фермента [Sugiyama et. al. 2002; WO 2004/097012].
Один из механизмов адаптации иллюстрируют грибные продуценты Pla2. Большинство из них синтезируют секретируемые Pla2 в виде незрелых про-ферментов с пониженной активностью. Их активация осуществляется внутриклеточно на поздних стадиях секреции под действием специализированных протеиназ. В результате из клеток в культуральную среду выходят зрелые, полностью активные Pla2 [WO 2004/097012]. Такая стратегия биосинтеза токсичных белков, способных в случае преждевременной активации нанести повреждения клеткам-продуцентам, широко распространена в природе, а также является традиционной для использования в биотехнологии.
Известен также родственный способ продукции токсичных белков. От предыдущего он отличается тем, что продуцирующие клетки не обеспечивают внутриклеточного созревания и секретируют во внеклеточную среду про-ферменты, которые остаются в неактивном или слабоактивном состоянии в течение неопределенно долгого времени до момента активации. В этом случае про-ферменты называют зимогенами1, (1 Зимоген (zymogen) [греч. zyme - закваска и genes - порождающий, рождающийся] - функционально неактивный или слабоактивный профермент, который активируется при посттрансляционном удалении из молекулы фрагмента, способствующего поддержанию латентного состояния (напр., зимоген пепсиногена превращается в пищеварительный фермент пепсин при расщеплении на соответствующие пептиды)) и их активация осуществляется за пределами клеток под действием других ферментов, либо автокаталитически при изменении внешних условий (например, рН). В последнем случае говорят про самоактивируемый зимоген. В частности, в форме неактивных зимогенов продуцируются факторы свертывания крови животных [Stubbs & Bode 1994]. Получение фосфолипаз А2 в форме природных или искусственных зимогенов до сих пор не было известно.
Еще одним способом природной адаптации организмов к биосинтезу Pla2 считается изменение фосфолипидного состава клеточных мембран и исключение из них компонентов, к которым продуцируемый фермент проявляет наибольшее сродство. В результате адаптации снижается токсическое действие Pla2 на клетку-хозяина и, как следствие, появляется возможность увеличения продукции фермента [Sugiyama et al., 2002].
Даже при наличии адаптации использование природных штаммов для производства Pla2 сопряжено с экономическими ограничениями. Как правило, для достижения высокого выхода фермента штамм-продуцент необходимо культивировать с использованием специального оборудования и комплексных сред, содержащих, в том числе, жировой компонент [Valero, 2012]. При этом организм-хозяин, как правило, синтезирует множество изоформ не только целевой Pla2, но и других липаз, которые изменяют специфичность и селективность ФП и способны ухудшать его свойства, а глубокая очистка целевого фермента представляется экономически неоправданной.
В этой связи оптимальным способом увеличения производства высококачественных ФП является разработка рекомбинантных технологий, обеспечивающих экспрессию целевой Pla2 в клетках специально подобранного реципиентного микроорганизма.
Будем различать рекомбинантные производные природных ферментов, аминокислотная последовательность которых идентична природным прототипам и не содержит целенаправленно-введенных искусственных модификаций, и рекомбинантные производные природных ферментов, аминокислотная последовательность которых была искусственно модифицирована в результате генно-инженерных манипуляций и не идентична природным прототипам. Будем называть первые РПМ(-), а вторые РПМ(+) природных ферментов.
Также будем различать специфическую активность Pla2, выявляемую в специальных условиях, например, при постоянном рН, в присутствии высокой концентрации ионов кальция Са2+ на лецитиновом субстрате, солюбилизированного с использованием растворителя ПАВ «Тритон Х-100», а также специфическую активность, выявляемую на яичном желтке, не содержащем никаких посторонних добавок. Первую будем называть «специальной», а вторую - «собственной» активностью Pla2. Некоторые Pla2 обладают высокой «специальной» и низкой «собственной» активностью (Чеперегин с соавт. 2019). В то же время действие Pla2 на клетки в процессе биосинтеза и перспективы промышленного применения определяются уровнем «собственной», а не «специальной» активности, и полностью функциональная РПМ(+) Pla2 должна обладать показателями «специальной» и «собственной» активности, сходными с показателями природного фермента.
В настоящее время дрожжи удерживают статус эффективных и продвинутых эукариотических систем экспрессии генов [Gasser et al. 2013; Singh et al. 2016; Zahrl et al. 2017; Raveendran et al. 2018]. Они обладают низким уровнем продукции собственных секретируемых белков, но при этом способны обеспечить эффективную секрецию в культуральную среду целевых рекомбинантных продуктов.
Известно, что, не обладая собственными фосфолипазами А2, дрожжи способны продуцировать РПМ(-) природных ферментов. В частности, известно о продукции в дрожжах РПМ(-) Pla2 индийской кобры Naja naja naja [Lefkowitz et al. 1999] и РПМ(-) Pla2 штамма бактерий 2917 S. violaceoruber [Liu et al. 2015]. Однако у этих РПМ(-) Pla2 зафиксирована низкая «специальная» активность, 50-80 ед/мг [Lefkowitz et al. 1999] и 170 ед/мг [Liu et al., 2015], что существенно ниже показателей высокоактивных Pla2, синтезируемых природными продуцентами, штаммом S.violaceoruber А-2688 - 1000 ед/мг [Sugiyama et. al. 2002; http://wayback.archive-it.org/7993/20171031055659/ https://www.fda.gov/downloads/Food/IngredientsPackadngLabeling/GRAS/NoticeInventon,/UCM263914.pdf] или штаммами грибов - 648 ед/мг и выше [WO 2004/097012], составляющих основу коммерческих препаратов. Кроме того, показано, что РПМ(-) Pla2 штамма бактерий 2917 S. violaceoruber обладает близким к нулю показателем «собственной» активности [Чеперегин с соавт. 2019]. В этой связи, даже несмотря на высокую продукцию, составлявшую 30 и 200 мг/л, соответственно, указанные РПМ(-) Pla2 должны быть отнесены к низкоактивным ферментам, а их потенциальное промышленное применение -к компромиссным технологиям.
Известно об эффективной экспрессии в дрожжах высокоактивных РПМ(+) Pla2 [Чеперегин с соавт. 2019]. Однако вследствие внесенных изменений эти РПМ(+) приобретают высокую «собственную» активность только в присутствии дополнительных количеств ионов кальция Са2+ [Чеперегин с соавт. 2019]. В отсутствие дополнительных количеств кальция эти РПМ(+) Pla2 являются низкоактивными ферментами, для применения которых требуются компромиссные технологии.
Примеров разработки на основе дрожжей продуцентов РПМ(-) или РПМ(+) Pla2, обладающих высокой «собственной» активностью, в открытых источниках информации не обнаружено. В то же время именно они востребованы для промышленного производства.
Природная Pla2 бактерий S. violaceoruber является уникальной ферментом, не имеющим «загрязняющей» липазной активности, в связи с чем ее использование снижает потери при дегуммировании масел [Liu et al., 2015]. Из источников информации известны две природные Pla2 S. violaceoruber: высокоактивная Pla2 штамма А-2688 [Sugiyama et. al., 2002], называемая далее Pla22 [Чеперегин с соавт. 2019], и низкоактивная Pla2 штамма 2917 [Liu et al., 2015]. Информация об экспрессии в дрожжах раскрыта для низкоактивной Pla2 [Liu et al., 2015], а также для РПМ(+) Pla22 [Чеперегин с соавт. 2019], на базе которой разработан негликозилированный вариант Pla22ng, несущий мутации Ser31Ala, Ser82Ala и Asn108Gln в сайтах N-гликозилирования [Чеперегин с соавт. 2019].
Последовательности вариантов Pla22 и Pla22ng фосфолипазы А2 используют в качестве основы для разработки заявляемого изобретения. Препарат природной высокоактивной фосфолипазы А2 штамма S. violaceoruber А-2688 [Sugiyama et. al., 2002], производят в форме коммерческого препарата «Pla2 Nagase».
Задача заявляемой группы изобретений состоит в разработке способа получения в дрожжах Saccharomyces cerevisiae секретируемой полностью функциональной фосфолипазы А2.
Задача решена путем:
- разработки способа получения секретируемой полностью функциональной фосфолипазы А2, осуществляемого путем получения и введения в клетки дрожжей Saccharomyces cerevisiae генетической конструкции, кодирующей белок-предшественник, включающий последовательность нативной, содержащей сайты N-гликозилирования, или мутантной, не содержащей сайтов N-негликозилирования, фосфолипазы А2 штамма S. violaceoruber А-2688, содержащей инсерцию интеина между остатком глицина в позиции 75 и остатком серина в позиции 76, а также имеющей на N-конце дополнительный дипептид, состоящий из аминокислотных остатков серина и глицина, с последующим биосинтезом этого белка-предшественника и его автокаталитическим процессингом с образованием целевого продукта;
- получения генетической конструкции, кодирующей белок-предшественник, содержащий инсерцию интеина PRP8 штамма Penicillium chrysogenum ВКПМ F-3 в составе нативной фосфолипазы А2 штамма Streptomyces violaceoruber А-2688, для осуществления заявляемого способа;
- получения генетической конструкции, кодирующей белок-предшественник, содержащий инсерцию интеина PRP8 штамма Penicillium chrysogenum ВКПМ F-3, несущего мутацию Cys11Tyr, в составе нативной фосфолипазы А2 штамма Streptomyces violaceoruber А-2688, для осуществления заявляемого способа;
- получения генетической конструкции, кодирующей белок-предшественник, содержащий инсерцию интеина PRP8 штамма Neosartorya aurata NRRL 4378 в составе фосфолипазы А2 штамма Streptomyces violaceoruber А-2688, несущей мутации Ser31Ala, Ser82Ala и Asn108Gln, для осуществления заявляемого способа;
- получения генетической конструкции, кодирующей белок-предшественник, содержащий инсерцию интеина PRP8 штамма Penicillium chrysogenum ВКПМ F-3, несущего мутацию Cys11Tyr, в составе фосфолипазы А2 штамма Streptomyces violaceoruber А-2688, несущей мутации Ser31Ala, Ser82Ala и Asn108Gln, для осуществления заявляемого способа.
Способ получения секретируемой полностью функциональной фосфолипазы А2
Решение задачи включает следующие этапы:
- используя методы молекулярной биологии и генетической инженерии, клонируют структурный ген интеина PRP8 штамма P. chrysogenum (Int4), интеина PRP8 штамма P. chrysogenum, несущего мутацию Cys11Tyr (Int4Y), и интеина PRP8 штамма N. aurata NRRL 4378 (Int5);
- с использованием структурного гена, кодирующего РПМ(-) Pla22 [Чеперегин с соавт. 2019], получают генетические конструкции SEQ ID N1 и SEQ ID N2, кодирующие белок - предшественник для получения фосфолипазы А2 путем автокаталитического процессинга, представляющий собой гликозилированную фосфолипазу А2 штамма S. violaceoruber А-2688, содержащую инсерцию интеина Int4 или интеина Int4Y, соответственно, между остатком глицина в позиции 75 и остатком серина в позиции 76 и имеющую на N-конце дополнительный дипептид, состоящий из аминокислотных остатков серина и глицина;
- с использованием структурного гена, кодирующего РПМ(+) Pla22ng [Чеперегин с соавт. 2019], получают генетические конструкции SEQ ID N3 и SEQ ID N4, кодирующие белок - предшественник для получения фосфолипазы А2 путем автокаталитического процессинга, представляющий собой негликозилированную фосфолипазу А2 штамма А-2688 S. violaceoruber, несущую мутации S31A, S82A и N108Q и содержащую инсерцию интеина Int5 или интеина Int4Y, соответственно, между остатком глицина в позиции 75 и остатком серина в позиции 76 и имеющую на N-конце дополнительный дипептид, состоящий из аминокислотных остатков серина и глицина;
- полученные генетические конструкции SEQ ID N1, SEQ ID N2, SEQ ID N3 и SEQ ID N4 вводят в клетки дрожжей S. cerevisiae. Для этого используют реципиентные штаммы, векторы, промоторы, лидерные области, которые идентичны или аналогичны описанным ранее [Чеперегин с соавт. 2019, RU 2460795 и RU 2522479] и не являются существенными признаками изобретения. В результате получают штаммы-продуценты заявляемых белков-предшественников;
- осуществляют культивирование каждого из полученных штаммов дрожжей, содержащих генетические конструкции SEQ ID N1, SEQ ID N2, SEQ ID N3 и SEQ ID N4, для чего используют условия, обеспечивающие экспрессию этих конструкций, идентичные или аналогичные описанным ранее [Чеперегин с соавт. 2019, RU 2460795 и RU 2522479], не являющиеся существенными признаками изобретения. В результате культивирования каждого штамма получают культуральную жидкость, содержащую секретированный заявляемый белок-предшественник, который в результате автокаталитического процессинга образует целевой продукт; анализ активности которого доказывает, что он представляет собой полностью функциональную фосфолипазу А2.
Изобретение проиллюстрировано следующими фигурами:
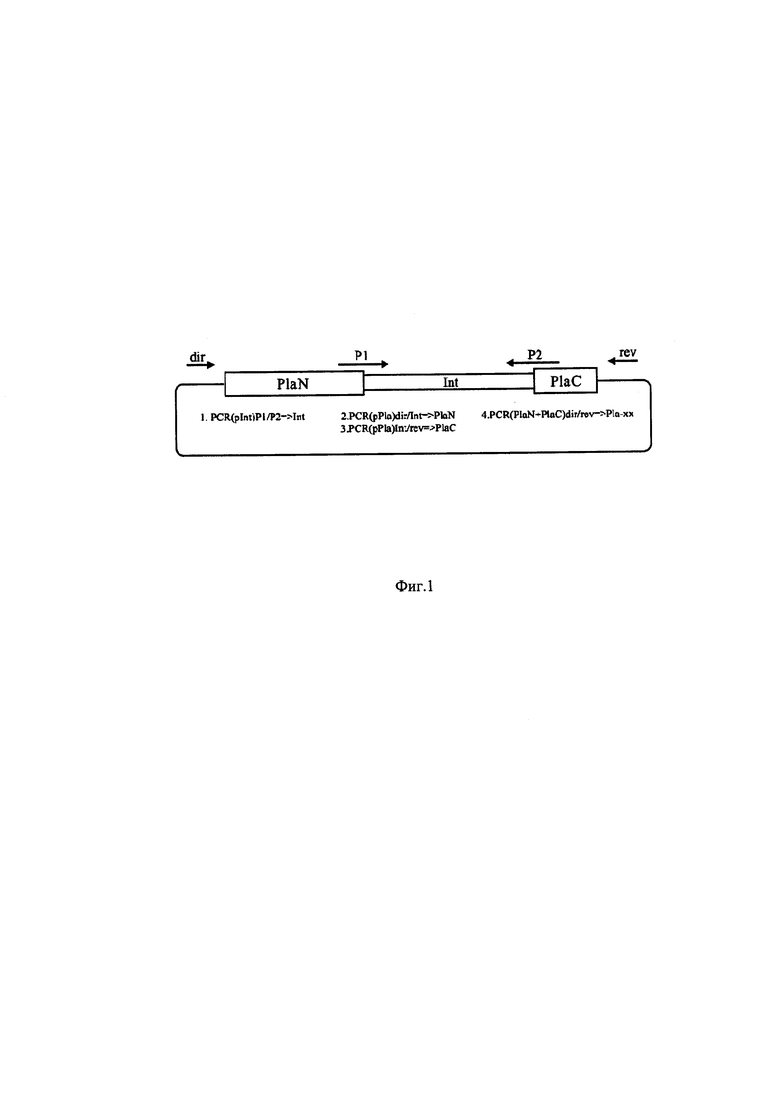
Фиг. 1 - Схема конструирования фрагментов ДНК, кодирующих заявляемые белки-предшественники. Условные обозначения: pINT - плазмида pInt4, pInt4Y или pInt5; INT -последовательность ДНК, кодирующая интеин Int4, Int4Y или Int5; PlaN и PlaC - последовательности ДНК, кодирующие N-концевую (с 1 по 75 остаток) и С-концевую (с 76 по 122 остаток) части фосфолипазы А2 в составе заявляемых белков-предшественников; pPla - плазмиды pUC18x-Pla22 или pUC18x-Pla22ng; Pla-хх - последовательность ДНК, кодирующая заявляемый белок-предшественник
Фиг. 2. ПААГ-электрофореграмма белков культуральной жидкости дрожжей штаммов D-Pla22ng(Int5) и D-Pla22ng(Int4Y) до (-) и после (+) процессинга, проведенного в течение 69 часов при комнатной температуре. Показано положение белковых полос соответствующих белкам-предшественникам (Pla22ng(Int)), интеинам Int5 и Int4Y, а также зрелой фосфолипазе А2 (Pla22ng).
Пример 1. Клонирование структурного гена интеина Int4
Фрагмент ДНК, кодирующий мини-интеин PRP8 P. chrysogenum (GeneBank АМ042015.1), называемый Int4, получают с использованием ПЦР. ПЦР осуществляют с использованием ДНК-полимеразы Phusion (ThermoFisher). Матрицей служит тотальная геномная ДНК штамма P. chrysogenum ВКПМ F-3, очищенная по стандартной методике [Kaiser et. al., 1994]. Фрагмент ДНК, кодирующий Int4 амплифицируют с использованием праймеров:
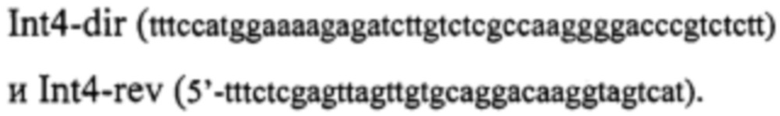
После открывания концов с использованием рестриктаз NcoI и XhoI амплифицированный фрагменты ДНК клонируют в лабораторном векторе pUC19mx, производном стандартного вектора pUC19, содержащем модифицированный полилинкер в окружении сайтов узнавания рестриктаз NcoI и XhoI.
В результате клонирования получают плазмиду pInt4, содержащую последовательность структурного гена интеина Int4.
Пример 2. Клонирование структурного гена интеина Int4Y
Фрагмент ДНК, кодирующий мутантный вариант мини-интеина PRP8 P. chrysogenum (GeneBank АМ042015.1), называемый Int4Y, клонируют, как описано в примере 1, но для амплификации используют праймеры:
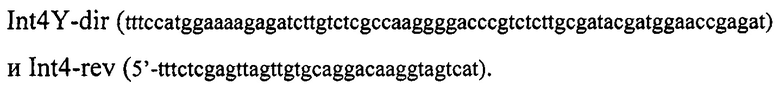
В результате клонирования получают плазмиду pInt4Y, содержащую последовательность структурного гена мутантного интеина Int4Y, содержащего замену Cys11Tyr.
Пример 3. Синтез структурного гена интеина Int5
Фрагмент ДНК SEQ ID N1, кодирующий интеин Nau PRP8 штамма N. aurata NRRL 4378, называемый Int5, получают в виде синтетического BamHI/XhoI фрагмента ДНК. Этот фрагмент клонируют в лабораторной плазмиде pUC18x. В результате получают плазмиду pInt5, содержащую последовательность структурного гена интеина Int5.
Пример 4. Конструирование гена Pla22
Фрагмент ДНК, заключающий структурный ген фосфолипазы А2, называемый Pla22, кодирующий аминокислотную последовательность зрелой фосфолипазы А2 штамма S. violaceoruber А-2688 (GenBank АЕМ88445), получают с использованием ПЦР. Матрицей для ПЦР служит последовательность синтетического гена Pla2 [Чеперегин с соавт. 2019]. Фрагмент ДНК, содержащий этот ген, клонирован по сайтам BamHI/XhoI в векторе pUC18x, являющемся производным стандартного вектора pUC18 и содержащем в составе модифицировонного полилинкера сайт узнавания рестриктазы XhoI вместо сайта EcoRI.
Искомый фрагмент ДНК получают в 2 этапа. На первом этапе с помощью ПЦР получают 2 перекрывающихся фрагмента ДНК:
- фрагмент 1 получают, используя праймер  и
и 
- фрагмент 2 получают, используя праймер  и
и 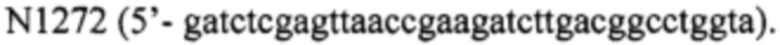
Амплифицированные фрагменты ДНК очищают из геля и на следующем этапе проводят ПЦР-опосредованное лигирование очищенных фрагментов ДНК. С этой целью проводят ПЦР с использованием смеси фрагментов 1 и 2 в качестве матрицы. Праймерами для амплификации служат N168 и N1272. Амплифицированный фрагмент ДНК элюируют из агарозного геля и после открывания концов с использованием рестриктаз BamHI и XhoI клонируют в плазмиде pUC18x, расщепленной по тем же сайтам. В результате клонирования получают плазмиду pUC18x-Pla22, в составе которой нуклеотидную последовательность клонированного гена подтверждают секвенированием.
Полученная плазмида pUC18x-Pla22 содержит BamHI/XhoI фрагмент ДНК, кодирующий вариант Pla22 фосфолипазы А2, аминокислотная последовательность которой заключает остаток треонина в позиции 46 (замена Ser46Thr) и остатки фенилаланина и глицина на С-конце в позициях 121 и 122, соответственно (замена Leu121(PheGly)). Плазмиду pUCT8x-Pla22 используют для получения гена мутантного варианта фосфолипазы и для инсерции последовательностей ДНК, кодирующих интеины.
Пример 5. Конструирование гена Pla22ng
Фрагмент ДНК, заключающий структурный ген Pla22ng, кодирующий мутантный вариант фосфолипазы, из аминокислотной последовательности которой исключают три сайта N-гликозилирования, получают с помощью ПЦР. В качестве матрицы для ПЦР используют ДНК плазмиды pUC18x-Pla22. ПЦР осуществляют в 2 этапа.
На первом этапе последовательность мутантного гена получают в виде 4 фрагментов ДНК, полученных с помощью ПЦР и имеющих попарно перекрывающиеся концы:
- фрагмент 1 получают, используя праймер  и
и 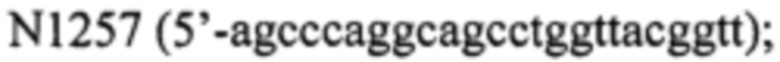
- фрагмент 2 получают, используя праймер  и
и 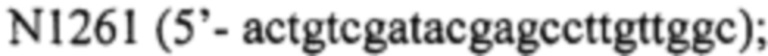
- фрагмент 3 получают, используя праймер 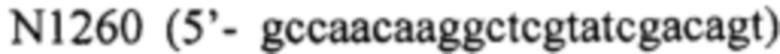 и
и 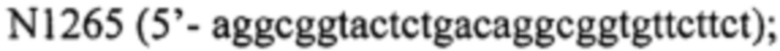
- фрагмент 4 получают, используя праймер 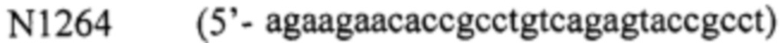 и
и 
Амплифицированные фрагменты ДНК очищают из геля, используя с этой целью набор Qiagen (Qiagen, cat. №28706).
На следующем этапе проводят ПЦР-опосредованное лигирование 4 очищенных фрагментов ДНК. С этой целью проводят ПЦР с использованием смеси фрагментов 1, 2, 3 и 4 в качестве матрицы. Праймерами для амплификации служат N1346 и N169. Амплифицированный фрагмент ДНК элюируют из агарозного геля и после открывания концов с использованием рестриктаз BamHI и XhoI клонируют в плазмиде pUC18x (Пример 4), расщепленной по тем же сайтам. В результате клонирования получают плазмиду pUC18x-Pla22ng, в составе которой нуклеотидную последовательность клонированного гена подтверждают секвенированием.
Полученная плазмида pUC18x-Pla22ng содержит BamHI/XhoI фрагмент ДНК, кодирующий мутантную фосфолипазу, содержащую 3 замены: Ser31Ala, Ser82Ala и Asn108Gln, инактивирующие все 3 потенциальных сайта N-гликозилирования. Мутантный вариант фосфолипазы называют Pla22ng.
Плазмиду pUC18x-Pla22ng используют для инсерции последовательностей ДНК, кодирующих интеины.
Пример 6. Получение фрагментов ДНК, кодирующих заявляемые интеин-содеожащие белки-предшественники
Заявляемые белки-предшественники нативной, содержащей сайты N-гликозилирования, фосфолипазы А2, содержащие инсерции интеинов Int4 или Int4Y, называют Pla22(Int4) и Pla22(Int4Y), соответственно. Для получения последовательностей ДНК, кодирующих эти белки, используют плазмиду pUC18x-Pla22.
Заявляемые белки-предшественники мутантной, не содержащей сайтов N-негликозилирования, фосфолипазы А2, содержащие инсерции интеинов Int5 или Int4Y, называют Pla22ng(Int5) и Pla22ng(Int4Y), соответственно. Для получения последовательностей ДНК, кодирующих эти белки, используют плазмиду pUC18x-Pla22ng.
Для конструирования фрагментов ДНК, кодирующих заявляемые белки-предшественники, содержащие интеиновые инсерции, используют технологию ПЦР-опосредованного лигирования перекрывающихся последовательностей ДНК по схеме Фиг. 1. Для этого используют универсальные праймеры для амплификации dir 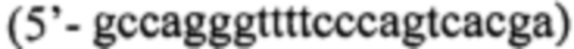 и rev
и rev 
Кроме них, для получения последовательностей ДНК, кодирующих белки-предшественники Pla22(Int4), Pla22(Int4Y) и Pla22ng(Int4Y), используют праймеры Р1 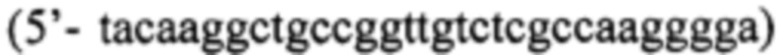 и Р2
и Р2 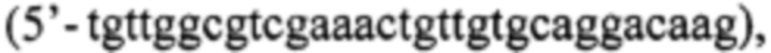 а для получения последовательности ДНК, кодирующей белок-предшественник Pla22ng(Int5) используют праймеры Р1
а для получения последовательности ДНК, кодирующей белок-предшественник Pla22ng(Int5) используют праймеры Р1 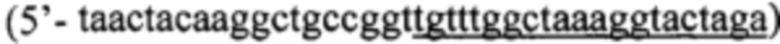 и Р2
и Р2  Для ПЦР используют ДНК-полимеразу Phusion (ThermoFisher Scientific, F-#F530S).
Для ПЦР используют ДНК-полимеразу Phusion (ThermoFisher Scientific, F-#F530S).
В результате ПЦР-лигирования получают BamHI/XhoI фрагменты ДНК, заключающие последовательности ДНК SEQ ID N1, SEQ ID N2, SEQ ID N3 и SEQ ID N4, кодирующие заявляемые белки-предшественники Pla22(Int4), Pla22(Int4Y), Pla22ng(Int5) и Pla22ng(Int4Y), соответственно.
Эти фрагменты ДНК используют для конструирования экспрессионных плазмид, штаммов-продуцентов и получения соответствующих вариантов фосфолипазы А2.
Пример 7. Конструирование экспрессионных плазмид для биосинтеза заявляемых белков-предшественников в дрожжах S. cerevisiae
Экспрессионные плазмиды конструируют путем лигирования трех фрагментов ДНК.
Фрагментом 1 служит XhoI/HindIII фрагмент ДНК лабораторного бирепликонного вектора pPDX3 [RU 2460795]. Данный фрагмент содержит, начиная от сайта XhoI: область терминации транскрипции гена CYC1 дрожжей S. cerevisiae; фрагмент ДНК, обеспечивающий репликацию и селекцию плазмид pPDX3 в клетках Е. coli; фрагмент ДНК эндогенной 2-мкм плазмиды дрожжей, обеспечивающий способность плазмид pPDX3 поддерживаться в клетках дрожжей S. cerevisiae в эписомном многокопийном состоянии; структурные гены URA3 и PGK1 дрожжей S. cerevisiae, обеспечивающие селективное поддержание плазмид pPDX3 в штаммах, имеющих соответствующие хромосомные мутации [RU 2460795, RU 2515913 и др.].
Фрагментом 2 является «регуляторный» HindIII/BglII фрагмент ДНК, кодирующий промотор GAL1, используемый для экспрессии, и лидерную область matHH, направляющую секрецию вариантов фосфолипазы А2 в дрожжах S. cerevisiae.
Для конструирования «регуляторного» фрагмента ДНК используют плазмиду pUC18x-GAL1matHH-GH, содержащую ген зрелого соматропина, слитый с фрагментом ДНК, кодирующим лидерный полипептид, включающий последовательности сигнального пептида mat и удвоенной про-области белка HSP150 дрожжей S. cerevisiae [RU 2460795].
Конструирование проводят путем замещения NcoI/XhoI фрагмента ДНК плазмиды pUC18x-GAL1matHH-GH, кодирующего соматропин, на синтетический двуцепочечный фрагмент ДНК, заключающий сайт узнавания рестриктазы BglII и полученный в результате отжига двух олигонуклеотидов:

В результате клонирования получают плазмиду pUC18x-GAL1matHH-(BglII), в составе которой в последовательности ДНК, кодирующей лидерный полипептид matHH, сконструирован сайт рестриктазы BglII, формируемый кодоном аргинина, входящим в сайт узнавания протеиназы Кех2, и кодоном серина, следующим непосредственно за кодоном аргинина.
Плазмида pUC18x-GAL1matHH-(BglII) служит источником «регуляторного» HindIII/BglII фрагмента ДНК (Фрагмента 2).
Фрагментом 3 служит BamHI/XhoI фрагмент ДНК заключающий последовательность ДНК SEQ ID N1 или SEQ ID N2 или SEQ ID N3 или SEQ ID N4, кодирующий заявляемый белок-предшественник Pla22(Int4), Pla22(Int4Y), Pla22ng(Int5) или Pla22ng(Int4Y), соответственно.
В частности,
- плазмиду pPDX3-Pla22(Int4) получают путем лигирования фрагментов 1 и 2, а также фрагмента 3, заключающего последовательность ДНК SEQ ID N1;
- плазмиду pPDX3-Pla22(Int4Y) получают путем лигирования фрагментов 1 и 2, а также фрагмента 3, заключающего последовательность ДНК SEQ ID N2;
- плазмиду pPDX3-Pla22ng(Int5) получают путем лигирования фрагментов 1 и 2, а также фрагмента 3, заключающего последовательность ДНК SEQ ID N3;
- плазмиду pPDX3-Pla22ng(Int4Y) получают путем лигирования фрагментов 1 и 2, а также фрагмента 3, заключающего последовательность ДНК SEQ ID N4.
Полученные экспрессионные плазмиды серии pPDX3 содержат в своем составе последовательности ДНК, кодирующие заявляемые белки-предшественники, слитые с последовательностями ДНК, кодирующими промотор GAL1 дрожжей S.cerevisiae и лидерный полипептид matHH, заключающий в своем составе удвоенную (НН) про-области белка HSP150 дрожжей S. cerevisiae. Эти плазмиды используют для трансформации клеток реципиентных штаммов дрожжей S. cerevisiae и наработки соответствующих образцов фосфолипазы А2.
Пример 8. Конструирование штаммов-продуцентов заявляемых белков-предшественников с использованием лабораторного реципиентного штамма beta-0 S. cerervisiae
Для конструирования штаммов-продуцентов клетки лабораторного реципиентного штамма S. cerervisiae beta-0 [Чеперегин с соавт. 2019] трансформируют с использованием полученных экспрессионных плазмид серии pPDX3. Трансформацию осуществляют с использованием литий ацетата как описано [Gietz & Schiestl 2007]. В результате трансформации получают штаммы-продуценты серии В. В частности,
- в результате трансформации клеток лабораторного реципиентного штамма beta-0 S. cerervisiae плазмидой pPDX3-Pla22(Int4) получают штамм B-Pla22(Int4), который используют для биосинтеза белка-предшественника Pla22(Int4).
- в результате трансформации клеток лабораторного реципиентного штамма beta-0 S. cerervisiae плазмидой pPDX3-Pla22(Int4Y) получают штамм B-Pla22(Int4Y), который используют для биосинтеза белка-предшественника Pla22(Int4Y).
- в результате трансформации клеток лабораторного реципиентного штамма beta-0 S. cerervisiae плазмидой pPDX3-Pla22ng(Int5) получают штамм B-Pla22ng(Int5), который используют для биосинтеза белка-предшественника Pla22ng(Int5).
- в результате трансформации клеток лабораторного реципиентного штамма beta-0 S. cerervisiae плазмидой pPDX3-Pla22ng(Int4Y) получают штамм B-Pla22ng(Int4Y), который используют для биосинтеза белка-предшественника Pla22ng(Int4Y).
Пример 9. Конструирование штаммов-продуцентов заявляемых белков-предшественников с использованием лабораторного реципиентного штамма S. cerervisiae D721W
Для конструирования штаммов-продуцентов клетки лабораторного реципиентного штамма S. cerervisiae D721W [RU 2460795] трансформируют с использованием полученных экспрессионных плазмид серии pPDX3. Трансформацию осуществляют по методу Ito [Ito et al. 1983]. В результате трансформации получают штаммы-продуценты серии D.
В частности,
- в результате трансформации клеток лабораторного реципиентного штамма D721W S. cerervisiae плазмидой pPDX3-Pla22(Int4) получают штамм D-Pla22(Int4), который используют для биосинтеза белка-предшественника Pla22(Int4).
- в результате трансформации клеток лабораторного реципиентного штамма D721W S. cerervisiae плазмидой pPDX3-Pla22(Int4Y) получают штамм D-Pla22(Int4Y), который используют для биосинтеза белка-предшественника Pla22(Int4Y).
- в результате трансформации клеток лабораторного реципиентного штамма D721W S. cerervisiae плазмидой pPDX3-Pla22ng(Int5) получают штамм D-Pla22ng(Int5), который используют для биосинтеза белка-предшественника Pla22ng(Int5).
- в результате трансформации клеток лабораторного реципиентного штамма D721W S. cerervisiae плазмидой pPDX3-Pla22ng(Int4Y) получают штамм D-Pla22ng(Int4Y), который используют для биосинтеза белка-предшественника Pla22ng(Int4Y).
Пример 10. Получение заявляемых белков-предшественников и продуктов их процессинга Для получения заявляемых белков-предшественников осуществляют культивирование сконструированных штаммов-продуцентов. С этой целью:
- штаммы-продуценты серии В культивируют на жидкой среде YPDG (дрожжевой экстракт - 1%; бактопептон - 2%; глюкоза - 2%; галактоза - 2%; вода - остальное) в течение 96 ч до стационарной фазы роста с использованием ротационного шейкера со скоростью 250 об/мин при температуре 22°С.
- штаммы-продуценты серии D культивируют на жидкой среде 223 (дрожжевой экстракт - 2%; бактопептон - 2%; глюкоза - 3%; вода - остальное) в течение 96 ч до стационарной фазы роста с использованием ротационного шейкера со скоростью 250 об/мин при температуре 22°С.
По завершении культивирования штаммов-продуцентов обеих серий получают образцы культуральной жидкости, которые освобождают от клеток дрожжей путем центрифугирования в течение 10 мин с ускорением 16000 g и последующего переноса образцов над осадочной жидкости в подходящие чистые емкости.
В результате получают свободные от клеток дрожжей обработанные образцы культуральной жидкости, каждый из которых содержит один из заявляемых белков-предшественников, Pla22(Int4) или Pla22(Int4Y) или Pla22ng(Int5) или Pla22ng(Int4Y), который не обязательно очищают и концентрируют, однако обязательно подвергают автокаталитическому процессингу.
На фиг. 2 приведены результаты автокаталитического процессинга белков-предшественников Pla22ng(Int5) и Pla22ng(Int4Y), продуцированных штаммами D-Pla22ng(Int5) и D-Pla22ng(Int4Y), соответственно, который осуществляют при комнатной температуре в течение 69 часов.
В результате автокаталитического процессинга получают образцы целевого продукта.
Пример 11. Оценка «специальной» и «собственной» активности образцов целевого продукта
Определение «специальной» активности образцов целевого продукта проводят с использованием рН-титратора по методике «Инструментального определения активности в стандартных условиях с использованием рН-титратора» [Чеперегин с соавт. 2019]. Для калибровки используют образец коммерческого препарата «Pla2 Nagase». В результате измерения получают данные о величине «специальной» активности каждого образца целевого продукта, выраженной в единицах активности коммерческого препарата «Pla2 Nagase». Другими словами, получают данные об относительной активности каждого образца целевого продукта и контрольного препарата «Pla2 Nagase».
На основании полученных данных для каждого образца целевого продукта готовят контрольный образец «Pla2 Nagase», расчетное значение «специальной» активности которого равно активности соответствующего образца целевого продукта. В результате формируют экспериментальные пары, состоящие из образца целевого продукта и разведенного контрольного образца «Pla2 Nagase.
В каждой экспериментальной паре проводят повторное измерение «специальной» активности. Результаты измерения активности образцов целевого продукта выражают в виде доли от активности соответствующего контрольного образца «Pla2 Nagase» и заносят в табл. 1.
Затем в каждой экспериментальной паре проводят измерение «собственной» активности разведенного контрольного образца «Pla2 Nagase» и образца целевого продукта. Измерение проводят по методике «Инструментального определения активности на яичном желтке» [Чеперегин с соавт. 2019]. Как и в случае измерения «специальной» активности, результаты измерения «собственной» активности образца целевого продукта выражают в виде доли от активности соответствующего контрольного образца «Pla2 Nagase» и заносят в табл. 1.
Результаты, приведенные в табл. 1 подтверждают, что заявляемый способ обеспечивает получение образцов целевого продукта, обладающих показателями «специальной» и «собственной» активности, сходными с показателями коммерческого препарата Pla2-Nagase. Наличие у образцов целевого продукта показателей «специальной» и «собственной» активности, сходных с показателями коммерческого препарата Pla2-Nagase, доказывает, что целевой продукт является полностью функциональной фосфолипазой А2.

Преимущества заявляемого способа:
- микробиологический синтез целевого продукта осуществляют с использованием дрожжей Saccharomyces cerevisiae, традиционно признаваемых безопасными для получения ферментов для пищевой промышленности, к которым относится фосфолипаза А2;
- способ позволяет получать нативную фосфолипазу А2, то есть содержащую сайты N-гликозилирования, или мутантную, то есть не содержащую сайтов N-гликозилирования;
- при осуществлении способа могут быть применены разные интеины, в том числе мутантные;
- способ не требует больших трудозатрат, поскольку целевой продукт является секретируемым, а процесс образования целевого продукта из его предшественника - протекает самопроизвольно;
- способ не нуждается в использовании протеаз, поскольку процессинг белка-предшественника осуществляется на основе автокаталитического механизма;
- способ обеспечивает получение целевого продукта, ключевые показатели которого, характеризующие не только «специальную», но и «собственную» активность, находятся на уровне показателей коммерческого препарата «Pla2-Nagase» и, таким образом, соответствуют показателям природной высокоактивной фосфолипазы А2.
Литература
Бакланов К.В. Совершенствование технологии высококалорийных майонезов - М: 2008, с. 23
Чеперегин С.Э., Санникова Е.П., Малышева А.В., Клебанов Ф.А., Козлов Д.Г. (2019) Высокоактивные модифицированные варианты рекомбинантной фосфолипазы А2 Streptomyces violaceoruber для эффективного биосинтеза в дрожжах. Биотехнология, 2019, Т. 35, №3, С. 30-41. doi: 10.21519/0234-2758-2019-35-3-30-41
Cesarini S. Moving towards a Competitive Fully Enzymatic Biodiesel Process / S. Cesarini, F.I.J. Pastor, P.M. Nielsen, P. Diaz. // Sustainability. - 2015. - V.7 (6). - P. 7884-7903. doi: 10.3390/su7067884
De Maria L., Vind J.,  , Svendsen A., Patkar S. (2007). Phospholipases and their industrial applications. Appl Microbiol Biotechnol, 74: 290-300. DOI 10.1007/s00253-006-0775-x
, Svendsen A., Patkar S. (2007). Phospholipases and their industrial applications. Appl Microbiol Biotechnol, 74: 290-300. DOI 10.1007/s00253-006-0775-x
Gasser В., Prielhofer R., Marx H., et al. Pichia pastoris: protein production host and model organism for biomedical research. Future Microbiol., 2013, 8(2), 191-208. doi: 10.2217/fmb. 12.133.v
Gietz R.D., Schiestl R.H. Quick and easy yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat Protoc., 2007, 2 (1), 35-37. doi: 10.1038/nprot.2007.14
Hoogevest P., Wendel A. (2014). The use of natural and synthetic phospholipids as pharmaceutical excipients. Eur J Lipid Sci Technol., 116(9): 1088-1107. doi: 10.1002/ejlt.201400219.
Ito H., Fukuda Y., Murata K., Kimura A. Transformation of intact yeast cells treated with alkali cations. J Bacteriol., 1983,153(1): 163-168.
Kaiser et. al. (1994) Methods in Yeast Genetics. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY
Lefkowitz L.J., Deems R.A., Dennis E.A. (1999). Expression of Group IA Phospholipase A2 in Pichia pastoris: Identification of a Phosphatidylcholine Activator Site Using Site-Directed Mutagenesis. Biochemistry, 38, 14174-14184.
Liu A., Yu X.-W., Sha C, Xu Y. (2015). Streptomyces violaceoruber phospholipase A2: expression in Pichia pastoris, properties, and application in oil degumming. Appl Biochem Biotechnol, 17'5(6): 3195-3206. DOI 10.1007/s 12010-015-1492-7
 A. and Dennis, E.A. (1982) Acyl and phosphoryl migration in lysophospholipids: importance in phospholipids synthesis and phospholipase activity, Biochemistry, 21: 1743-1750.
A. and Dennis, E.A. (1982) Acyl and phosphoryl migration in lysophospholipids: importance in phospholipids synthesis and phospholipase activity, Biochemistry, 21: 1743-1750.
Raveendran S., Parameswaran В., Ummalyma S.B., Abraham A., Mathew A.K., Madhavan A., Rebello S., Pandey A. Applications of Microbial Enzymes in Food Industry. Food Technol Biotechnol, 2018, 56(1), 16-30. doi: 10.17113/ftb.56.01.18.5491.
Stubbs M.T., Bode W. Coagulation factors and their inhibitors. Curr Opin Struct Biol., 1994,4(6):823-832. doi:10.1016/0959-440x(94)90263-1
Singh R., Kumar M., Mittal A., Mehta P.K. Microbial enzymes: industrial progress in 21st century. 3 Biotech., 2016, 6(2), 174. doi: 10.1007/s13205-016-0485-8.
Sugiyama M., Ohtan K., Izuhara M., Koike Т., Suzuki K., Imamura S., Misaki H. (2002). A Novel Prokaryotic Phospholipase A2. Characterization, gene cloning, and solution structure. The Journal of Biological Chemistry, 277(22): 20051-20058.
Valero F. (2012). Heterologous expression systems for lipases: A review. In Lipases and Phospholipases: Methods and Protocols, Methods in Molecular Biology; Sandoval, G., Ed.; Springer Science Business Media: New York, NY, USA, 2012; Volume 861, pp. 161-178.
Zahrl R.J.,  , Mattanovich D., Gasser B. Systems biotechnology for protein production in Pichia pastoris. FEMS Yeast Res., 2017, 17(7), fox068. doi: 10.1093/femsyr/fox068
, Mattanovich D., Gasser B. Systems biotechnology for protein production in Pichia pastoris. FEMS Yeast Res., 2017, 17(7), fox068. doi: 10.1093/femsyr/fox068
--->
Перечень последовательностей
<110> Федеральное государственное бюджетное учреждение «Государственный научно-исследовательский институт генетики и селекции промышленных микроорганизмов» Национального исследовательского центра «Курчатовский институт» (НИЦ «Курчатовский институт» - ГосНИИгенетика)
<120> Способ получения секретируемой полностью функциональной фосфолипазы А2 в дрожжах Saccharomyces cerevisiae, белок-предшественник для осуществления этого способа (варианты)
<160> 4
<210> 1
<211> 852
<212> DNA
<213> Artificial sequence for the synthesis of recombinant phospholipase A2 of Streptomyces violaceoruber strain А-2688
<220>
<221> CDS
<222> (1)…(846)
<223> Sequence encodes precursor of the recombinant phospholipase A2
<221> misc_feature
<222> (1)…(6)
<223> Sequence encodes additional dipeptide conjugated to the N-end of the recombinant phospholipase A2
<221> misc_feature
<222> (1)…(241)
<223> Sequence encodes N-terminal part of the recombinant phospholipase A2
<221> misc_feature
<222> (242)…(702)
<223> Sequence encodes native intein PRP8 of Penicillium chrysogenum
<221> misc_feature
<222> (703)…(846)
<223> Sequence encodes C-terminal part of the recombinant phospholipase A2
<221> misc_feature
<222> (847)…(852)
<223> Sequence encodes recognition site for endonuclease XhoI
<400> 1
TCC GGA GCT CCT GCC GAC AAG CCT CAG GTC CTG GCC AGT TTC ACC CAG 48
Ser Gly Ala Pro Ala Asp Lys Pro Gln Val Leu Ala Ser Phe Thr Gln
1 5 10 15
ACC AGT GCC AGT TCT CAG AAC GCC TGG CTG GCT GCC AAC CGT AAC CAG 96
Thr Ser Ala Ser Ser Gln Asn Ala Trp Leu Ala Ala Asn Arg Asn Gln
20 25 30
AGT GCC TGG GCT GCC TAC GAG TTC GAC TGG AGT ACC GAC TTG TGT ACA 144
Ser Ala Trp Ala Ala Tyr Glu Phe Asp Trp Ser Thr Asp Leu Cys Thr
35 40 45
CAG GCC CCT GAC AAC CCT TTC GGT TTC CCT TTC AAC ACC GCC TGT GCC 192
Gln Ala Pro Asp Asn Pro Phe Gly Phe Pro Phe Asn Thr Ala Cys Ala
50 55 60
CGT CAT GAC TTC GGT TAC CGT AAC TAC AAG GCT GCC GGT TGT CTC GCC 240
Arg His Asp Phe Gly Tyr Arg Asn Tyr Lys Ala Ala Gly Cys Leu Ala
65 70 75 80
AAG GGG ACC CGT CTC TTG CGA TGC GAT GGA ACC GAG ATC AAT GTG GAA 288
Lys Gly Thr Arg Leu Leu Arg Cys Asp Gly Thr Glu Ile Asn Val Glu
85 90 95
GAC GTG CGC GAA GGT GAC CTA CTT CTG GGT CCC GAT GGA GAG CCT CGC 336
Asp Val Arg Glu Gly Asp Leu Leu Leu Gly Pro Asp Gly Glu Pro Arg
100 105 110
CGT GCA TTC AAC ATA GTG AAT GGC ATC GAC CGC CTG TAC CGC ATC AAG 384
Arg Ala Phe Asn Ile Val Asn Gly Ile Asp Arg Leu Tyr Arg Ile Lys
115 120 125
ATC GGC GGT GAG AAA GAG GAC CTT GTG GTG ACG CCG AAC CAT ATT CTG 432
Ile Gly Gly Glu Lys Glu Asp Leu Val Val Thr Pro Asn His Ile Leu
130 135 140
GTG CTT TAT AGA GAG GAT GGT TCC AAG AAT GTG GAG AAG CAA ACG GTG 480
Val Leu Tyr Arg Glu Asp Gly Ser Lys Asn Val Glu Lys Gln Thr Val
145 150 155 160
GAG ATC ACT GCT GCC GAG TTT GCC GCG CTT TCT ACC GAG GAA AGA AGC 528
Glu Ile Thr Ala Ala Glu Phe Ala Ala Leu Ser Thr Glu Glu Arg Ser
165 170 175
CTC TAT AGT GCC TTT ACA TCT CCT AGG GCT GAG AAG GGC GCC GAT GAT 576
Leu Tyr Ser Ala Phe Thr Ser Pro Arg Ala Glu Lys Gly Ala Asp Asp
180 185 190
TCG GCT CAA ACG CAC AGT TTC AAG ATT GAG CAA GTT AGC CTC GAA TCC 624
Ser Ala Gln Thr His Ser Phe Lys Ile Glu Gln Val Ser Leu Glu Ser
195 200 205
GAG AAG ACA GAG TGG GCT GGT TTC CGA GTC GAC AAA GAT CAG CTT TAC 672
Glu Lys Thr Glu Trp Ala Gly Phe Arg Val Asp Lys Asp Gln Leu Tyr
210 215 220
CTG CGT CAT GAC TAC CTT GTC CTG CAC AAC AGT TTC GAC GCC AAC AAG 720
Leu Arg His Asp Tyr Leu Val Leu His Asn Ser Phe Asp Ala Asn Lys
225 230 235 240
AGT CGT ATC GAC AGT GCC TTC TAC GAG GAC ATG AAG CGT GTC TGT ACT 768
Ser Arg Ile Asp Ser Ala Phe Tyr Glu Asp Met Lys Arg Val Cys Thr
245 250 255
GGT TAC ACC GGT GAG AAG AAC ACC GCC TGT AAC AGT ACC GCC TGG ACC 816
Gly Tyr Thr Gly Glu Lys Asn Thr Ala Cys Asn Ser Thr Ala Trp Thr
260 265 270
TAC TAC CAG GCC GTC AAG ATC TTC GGT TAA CTCGAG 852
Tyr Tyr Gln Ala Val Lys Ile Phe Gly ***
275 280
<210> 2
<211> 852
<212> DNA
<213> Artificial sequence for the synthesis of recombinant phospholipase A2 of Streptomyces violaceoruber strain А-2688
<220>
<221> CDS
<222> (1)…(846)
<223> Sequence encodes precursor of the recombinant phospholipase A2
<221> misc_feature
<222> (1)…(6)
<223> Sequence encodes additional dipeptide conjugated to the N-end of the recombinant phospholipase A2
<221> misc_feature
<222> (1)…(241)
<223> Sequence encodes N-terminal part of the recombinant phospholipase A2
<221> misc_feature
<222> (242)…(702)
<223> Sequence encodes mutant intein PRP8 of Penicillium chrysogenum, enclosing mutation Cys11Tyr
<221> misc_feature
<222> (703)…(846)
<223> Sequence encodes C-terminal part of the recombinant phospholipase A2
<221> misc_feature
<222> (847)…(852)
<223> Sequence encodes recognition site for endonuclease XhoI
<400> 2
TCC GGA GCT CCT GCC GAC AAG CCT CAG GTC CTG GCC AGT TTC ACC CAG 48
Ser Gly Ala Pro Ala Asp Lys Pro Gln Val Leu Ala Ser Phe Thr Gln
1 5 10 15
ACC AGT GCC AGT TCT CAG AAC GCC TGG CTG GCT GCC AAC CGT AAC CAG 96
Thr Ser Ala Ser Ser Gln Asn Ala Trp Leu Ala Ala Asn Arg Asn Gln
20 25 30
AGT GCC TGG GCT GCC TAC GAG TTC GAC TGG AGT ACC GAC TTG TGT ACA 144
Ser Ala Trp Ala Ala Tyr Glu Phe Asp Trp Ser Thr Asp Leu Cys Thr
35 40 45
CAG GCC CCT GAC AAC CCT TTC GGT TTC CCT TTC AAC ACC GCC TGT GCC 192
Gln Ala Pro Asp Asn Pro Phe Gly Phe Pro Phe Asn Thr Ala Cys Ala
50 55 60
CGT CAT GAC TTC GGT TAC CGT AAC TAC AAG GCT GCC GGT TGT CTC GCC 240
Arg His Asp Phe Gly Tyr Arg Asn Tyr Lys Ala Ala Gly Cys Leu Ala
65 70 75 80
AAG GGG ACC CGT CTC TTG CGA TAC GAT GGA ACC GAG ATC AAT GTG GAA 288
Lys Gly Thr Arg Leu Leu Arg Tyr Asp Gly Thr Glu Ile Asn Val Glu
85 90 95
GAC GTG CGC GAA GGT GAC CTA CTT CTG GGT CCC GAT GGA GAG CCT CGC 336
Asp Val Arg Glu Gly Asp Leu Leu Leu Gly Pro Asp Gly Glu Pro Arg
100 105 110
CGT GCA TTC AAC ATA GTG AAT GGC ATC GAC CGC CTG TAC CGC ATC AAG 384
Arg Ala Phe Asn Ile Val Asn Gly Ile Asp Arg Leu Tyr Arg Ile Lys
115 120 125
ATC GGC GGT GAG AAA GAG GAC CTT GTG GTG ACG CCG AAC CAT ATT CTG 432
Ile Gly Gly Glu Lys Glu Asp Leu Val Val Thr Pro Asn His Ile Leu
130 135 140
GTG CTT TAT AGA GAG GAT GGT TCC AAG AAT GTG GAG AAG CAA ACG GTG 480
Val Leu Tyr Arg Glu Asp Gly Ser Lys Asn Val Glu Lys Gln Thr Val
145 150 155 160
GAG ATC ACT GCT GCC GAG TTT GCC GCG CTT TCT ACC GAG GAA AGA AGC 528
Glu Ile Thr Ala Ala Glu Phe Ala Ala Leu Ser Thr Glu Glu Arg Ser
165 170 175
CTC TAT AGT GCC TTT ACA TCT CCT AGG GCT GAG AAG GGC GCC GAT GAT 576
Leu Tyr Ser Ala Phe Thr Ser Pro Arg Ala Glu Lys Gly Ala Asp Asp
180 185 190
TCG GCT CAA ACG CAC AGT TTC AAG ATT GAG CAA GTT AGC CTC GAA TCC 624
Ser Ala Gln Thr His Ser Phe Lys Ile Glu Gln Val Ser Leu Glu Ser
195 200 205
GAG AAG ACA GAG TGG GCT GGT TTC CGA GTC GAC AAA GAT CAG CTT TAC 672
Glu Lys Thr Glu Trp Ala Gly Phe Arg Val Asp Lys Asp Gln Leu Tyr
210 215 220
CTG CGT CAT GAC TAC CTT GTC CTG CAC AAC AGT TTC GAC GCC AAC AAG 720
Leu Arg His Asp Tyr Leu Val Leu His Asn Ser Phe Asp Ala Asn Lys
225 230 235 240
AGT CGT ATC GAC AGT GCC TTC TAC GAG GAC ATG AAG CGT GTC TGT ACT 768
Ser Arg Ile Asp Ser Ala Phe Tyr Glu Asp Met Lys Arg Val Cys Thr
245 250 255
GGT TAC ACC GGT GAG AAG AAC ACC GCC TGT AAC AGT ACC GCC TGG ACC 816
Gly Tyr Thr Gly Glu Lys Asn Thr Ala Cys Asn Ser Thr Ala Trp Thr
260 265 270
TAC TAC CAG GCC GTC AAG ATC TTC GGT TAA CTCGAG 852
Tyr Tyr Gln Ala Val Lys Ile Phe Gly ***
275 280
<210> 3
<211> 852
<212> DNA
<213> Artificial sequence for the synthesis of recombinant phospholipase A2 of Streptomyces violaceoruber strain А-2688, enclosing mutations Ser31Ala, Ser82Ala и Asn108Gln
<220>
<221> CDS
<222> (1)…(873)
<223> Sequence encodes precursor of the recombinant phospholipase A2
<221> misc_feature
<222> (1)…(6)
<223> Sequence encodes additional dipeptide conjugated to the N-end of the recombinant phospholipase A2
<221> misc_feature
<222> (1)…(241)
<223> Sequence encodes N-terminal part of the recombinant phospholipase A2
<221> misc_feature
<222> (242)…(723)
<223> Sequence encodes mutant intein PRP8 of the strain Neosartorya aurata NRRL 4378
<221> misc_feature
<222> (724)…(867)
<223> Sequence encodes C-terminal part of the recombinant phospholipase A2
<221> misc_feature
<222> (847)…(852)
<223> Sequence encodes recognition site for endonuclease XhoI
<400> 3
TCC GGA GCT CCT GCC GAC AAG CCT CAG GTC CTG GCC AGT TTC ACC CAG 48
Ser Gly Ala Pro Ala Asp Lys Pro Gln Val Leu Ala Ser Phe Thr Gln
1 5 10 15
ACC AGT GCC AGT TCT CAG AAC GCC TGG CTG GCT GCC AAC CGT AAC CAG 96
Thr Ser Ala Ser Ser Gln Asn Ala Trp Leu Ala Ala Asn Arg Asn Gln
20 25 30
GCT GCC TGG GCT GCC TAC GAG TTC GAC TGG AGT ACC GAC TTG TGT ACA 144
Ala Ala Trp Ala Ala Tyr Glu Phe Asp Trp Ser Thr Asp Leu Cys Thr
35 40 45
CAG GCC CCT GAC AAC CCT TTC GGT TTC CCT TTC AAC ACC GCC TGT GCC 192
Gln Ala Pro Asp Asn Pro Phe Gly Phe Pro Phe Asn Thr Ala Cys Ala
50 55 60
CGT CAT GAC TTC GGT TAC CGT AAC TAC AAG GCT GCC GGT TGT CTC GCC 240
Arg His Asp Phe Gly Tyr Arg Asn Tyr Lys Ala Ala Gly Cys Leu Ala
65 70 75 80
AAA GGT ACT AGA TTA TTG AGA TAT GAT GGT TCT GAA ATT GAA GTT CAA 288
Lys Gly Thr Arg Leu Leu Arg Tyr Asp Gly Ser Glu Ile Glu Val Gln
85 90 95
GAT GTC AAA GAA GGT GAT TTG TTA TTG GGT CCA GAT GGA GGT CCA AGA 336
Asp Val Lys Glu Gly Asp Leu Leu Leu Gly Pro Asp Gly Gly Pro Arg
100 105 110
AGA GCT TTT AAC ATA GTT TCA GGT GAA GAT AGG TTG TAT AGA GTT AAG 384
Arg Ala Phe Asn Ile Val Ser Gly Glu Asp Arg Leu Tyr Arg Val Lys
115 120 125
ATT GAC GGT TCT GTT GAG GAT TTA GTT GTT ACT CCT AAT CAT ATC TTG 432
Ile Asp Gly Ser Val Glu Asp Leu Val Val Thr Pro Asn His Ile Leu
130 135 140
GTC TTT CAT AGA GAA CAG AAA GCT AGG GAT AAA GAA GAT GAT CAA TTG 480
Val Phe His Arg Glu Gln Lys Ala Arg Asp Lys Glu Asp Asp Gln Leu
145 150 155 160
CCA GAA TCT TAT GAC ACA GTT GAA ATG ACA GCA GCT GAG TTT GCT GCT 528
Pro Glu Ser Tyr Asp Thr Val Glu Met Thr Ala Ala Glu Phe Ala Ala
165 170 175
TTA TCT GCT GAA GAT AGA TCA AGA TAT AGA GCA TTC AGG TCT CCA TCT 576
Leu Ser Ala Glu Asp Arg Ser Arg Tyr Arg Ala Phe Arg Ser Pro Ser
180 185 190
TTC GAT TTG TCT GAA AAA GCT GTA CCT ACT AAT CAC AGA TTT GCT ATT 624
Phe Asp Leu Ser Glu Lys Ala Val Pro Thr Asn His Arg Phe Ala Ile
195 200 205
AAG GAT ATT AGA TTA GAA TTG GAG ACT ACT GAA TGG GCA GGA TTT AGA 672
Lys Asp Ile Arg Leu Glu Leu Glu Thr Thr Glu Trp Ala Gly Phe Arg
210 215 220
GTC GAT AAA GAT CAA TTG TAT TTG AGG CAT GAT TAC TTA GTT TTG CAT 720
Val Asp Lys Asp Gln Leu Tyr Leu Arg His Asp Tyr Leu Val Leu His
225 230 235 240
AAT AGT TTC GAC GCC AAC AAG GCT CGT ATC GAC AGT GCC TTC TAC GAG 768
Asn Ser Phe Asp Ala Asn Lys Ala Arg Ile Asp Ser Ala Phe Tyr Glu
245 250 255
GAC ATG AAG CGT GTC TGT ACT GGT TAC ACC GGT GAG AAG AAC ACC GCC 816
Asp Met Lys Arg Val Cys Thr Gly Tyr Thr Gly Glu Lys Asn Thr Ala
260 265 270
TGT CAG AGT ACC GCC TGG ACC TAC TAC CAG GCC GTC AAG ATC TTC GGT 864
Cys Gln Ser Thr Ala Trp Thr Tyr Tyr Gln Ala Val Lys Ile Phe Gly
275 280 285
TAA CTCGAG 873
***
<210> 4
<211> 852
<212> DNA
<213> Artificial sequence for the synthesis of recombinant phospholipase A2 of Streptomyces violaceoruber strain А-2688, enclosing mutations Ser31Ala, Ser82Ala и Asn108Gln
<220>
<221> CDS
<222> (1)…(846)
<223> Sequence encodes precursor of the recombinant phospholipase A2
<221> misc_feature
<222> (1)…(6)
<223> Sequence encodes additional dipeptide conjugated to the N-end of the recombinant phospholipase A2
<221> misc_feature
<222> (1)…(241)
<223> Sequence encodes N-terminal part of the recombinant phospholipase A2
<221> misc_feature
<222> (242)…(702)
<223> Sequence encodes mutant intein PRP8 of Penicillium chrysogenum, enclosing mutation Cys11Tyr
<221> misc_feature
<222> (703)…(846)
<223> Sequence encodes C-terminal part of the recombinant phospholipase A2
<221> misc_feature
<222> (847)…(852)
<223> Sequence encodes recognition site for endonuclease XhoI
<400> 4
TCC GGA GCT CCT GCC GAC AAG CCT CAG GTC CTG GCC AGT TTC ACC CAG 48
Ser Gly Ala Pro Ala Asp Lys Pro Gln Val Leu Ala Ser Phe Thr Gln
1 5 10 15
ACC AGT GCC AGT TCT CAG AAC GCC TGG CTG GCT GCC AAC CGT AAC CAG 96
Thr Ser Ala Ser Ser Gln Asn Ala Trp Leu Ala Ala Asn Arg Asn Gln
20 25 30
GCT GCC TGG GCT GCC TAC GAG TTC GAC TGG AGT ACC GAC TTG TGT ACA 144
Ala Ala Trp Ala Ala Tyr Glu Phe Asp Trp Ser Thr Asp Leu Cys Thr
35 40 45
CAG GCC CCT GAC AAC CCT TTC GGT TTC CCT TTC AAC ACC GCC TGT GCC 192
Gln Ala Pro Asp Asn Pro Phe Gly Phe Pro Phe Asn Thr Ala Cys Ala
50 55 60
CGT CAT GAC TTC GGT TAC CGT AAC TAC AAG GCT GCC GGT TGT CTC GCC 240
Arg His Asp Phe Gly Tyr Arg Asn Tyr Lys Ala Ala Gly Cys Leu Ala
65 70 75 80
AAG GGG ACC CGT CTC TTG CGA TAC GAT GGA ACC GAG ATC AAT GTG GAA 288
Lys Gly Thr Arg Leu Leu Arg Tyr Asp Gly Thr Glu Ile Asn Val Glu
85 90 95
GAC GTG CGC GAA GGT GAC CTA CTT CTG GGT CCC GAT GGA GAG CCT CGC 336
Asp Val Arg Glu Gly Asp Leu Leu Leu Gly Pro Asp Gly Glu Pro Arg
100 105 110
CGT GCA TTC AAC ATA GTG AAT GGC ATC GAC CGC CTG TAC CGC ATC AAG 384
Arg Ala Phe Asn Ile Val Asn Gly Ile Asp Arg Leu Tyr Arg Ile Lys
115 120 125
ATC GGC GGT GAG AAA GAG GAC CTT GTG GTG ACG CCG AAC CAT ATT CTG 432
Ile Gly Gly Glu Lys Glu Asp Leu Val Val Thr Pro Asn His Ile Leu
130 135 140
GTG CTT TAT AGA GAG GAT GGT TCC AAG AAT GTG GAG AAG CAA ACG GTG 480
Val Leu Tyr Arg Glu Asp Gly Ser Lys Asn Val Glu Lys Gln Thr Val
145 150 155 160
GAG ATC ACT GCT GCC GAG TTT GCC GCG CTT TCT ACC GAG GAA AGA AGC 528
Glu Ile Thr Ala Ala Glu Phe Ala Ala Leu Ser Thr Glu Glu Arg Ser
165 170 175
CTC TAT AGT GCC TTT ACA TCT CCT AGG GCT GAG AAG GGC GCC GAT GAT 576
Leu Tyr Ser Ala Phe Thr Ser Pro Arg Ala Glu Lys Gly Ala Asp Asp
180 185 190
TCG GCT CAA ACG CAC AGT TTC AAG ATT GAG CAA GTT AGC CTC GAA TCC 624
Ser Ala Gln Thr His Ser Phe Lys Ile Glu Gln Val Ser Leu Glu Ser
195 200 205
GAG AAG ACA GAG TGG GCT GGT TTC CGA GTC GAC AAA GAT CAG CTT TAC 672
Glu Lys Thr Glu Trp Ala Gly Phe Arg Val Asp Lys Asp Gln Leu Tyr
210 215 220
CTG CGT CAT GAC TAC CTT GTC CTG CAC AAC AGT TTC GAC GCC AAC AAG 720
Leu Arg His Asp Tyr Leu Val Leu His Asn Ser Phe Asp Ala Asn Lys
225 230 235 240
GCT CGT ATC GAC AGT GCC TTC TAC GAG GAC ATG AAG CGT GTC TGT ACT 768
Ala Arg Ile Asp Ser Ala Phe Tyr Glu Asp Met Lys Arg Val Cys Thr
245 250 255
GGT TAC ACC GGT GAG AAG AAC ACC GCC TGT CAG AGT ACC GCC TGG ACC 816
Gly Tyr Thr Gly Glu Lys Asn Thr Ala Cys Gln Ser Thr Ala Trp Thr
260 265 270
TAC TAC CAG GCC GTC AAG ATC TTC GGT TAA CTCGAG 852
Tyr Tyr Gln Ala Val Lys Ile Phe Gly ***
275 280
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Генетическая конструкция, кодирующая предшественник белка YB-1 человека, штамм Escherichia coli - продуцент предшественника белка YB-1 человека, способ микробиологического синтеза этого предшественника | 2019 |
|
RU2728237C1 |
| СПОСОБ УПРАВЛЕНИЯ РИТМОМ СЕРДЦА И СОКРАЩЕНИЕМ ОТДЕЛЬНЫХ КАРДИОМИОЦИТОВ ПРИ ПОМОЩИ ТЕРМОГЕНЕТИКИ | 2022 |
|
RU2802995C1 |
| МАТЕРИАЛЫ И МЕТОДЫ, ИСПОЛЬЗУЕМЫЕ ДЛЯ ЛЕЧЕНИЯ РЕСПИРАТОРНЫХ ЗАБОЛЕВАНИЙ У СОБАК | 2020 |
|
RU2811752C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ БИОЛОГИЧЕСКОГО ПОЛУЧЕНИЯ ЛАКТАТА ИЗ С1-СОЕДИНЕНИЙ С ПРИМЕНЕНИЕМ ТРАНСФОРМАНТОВ ЛАКТАТ ДЕГИДРОГЕНАЗЫ | 2014 |
|
RU2710714C2 |
| Вакцина против герпеса | 2019 |
|
RU2731073C1 |
| ИММУНОИНДУЦИРУЮЩЕЕ СРЕДСТВО | 2016 |
|
RU2744843C2 |
| Антитела против белка р17 ВИЧ-1 субтипа А | 2019 |
|
RU2727673C1 |
| ВИРУС ГРИППА, СПОСОБНЫЙ ИНФИЦИРОВАТЬ СОБАЧЬИХ, И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2802222C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРЕДОТВРАЩЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2016 |
|
RU2714205C2 |
| СПОСОБ ПРОДУЦИРОВАНИЯ ПОЧЕЧНЫХ КЛЕТОК-ПРЕДШЕСТВЕННИКОВ | 2016 |
|
RU2730861C2 |

Разработан способ получения в дрожжах Saccharomyces cerevisiae секретируемой полностью функциональной фосфолипазы А2, включающей последовательность фосфолипазы А2 штамма Streptomyces violaceoruber А-2688, в том числе несущей мутации S31A, S82A и N108Q, и содержащей на N-конце дополнительный дипептид, состоящий из аминокислотных остатков серина и глицина. Заявляемый способ включает биосинтез предшественника целевого белка, представляющего собой вариант фосфолипазы А2, содержащей инсерцию интеина между остатком глицина в позиции 75 и остатком серина в позиции 76. Также изобретение включает в себя четыре белка-предшественника, которые кодируются последовательностями SEQ ID N: 1-4 соответственно. Удаление инсерции интеина и превращение белка-предшественника в активный целевой белок происходит путем автокаталитического процессинга. 5 н.п. ф-лы, 11 пр., 1 табл., 2 ил.
1. Способ получения секретируемой полностью функциональной фосфолипазы А2, осуществляемый путем получения и введения в клетки дрожжей Saccharomyces cerevisiae генетической конструкции, кодирующей белок-предшественник, включающий последовательность нативной, содержащей сайты N-гликозилирования, или мутантной, не содержащей сайтов N-гликозилирования, фосфолипазы А2 штамма Streptomyces violaceoruber А-2688, содержащей инсерцию интеина между остатком глицина в позиции 75 и остатком серина в позиции 76, а также имеющей на N-конце дополнительный дипептид, состоящий из аминокислотных остатков серина и глицина, с последующим биосинтезом этого белка-предшественника и его автокаталитическим процессингом с образованием целевого продукта.
2. Белок-предшественник для осуществления способа по п. 1, кодируемый последовательностью SEQ ID N: 1 и содержащий инсерцию интеина PRP8 штамма Penicillium chrysogenum ВКПМ F-3 в составе нативной фосфолипазы А2 штамма Streptomyces violaceoruber А-2688.
3. Белок-предшественник для осуществления способа по п. 1, кодируемый последовательностью SEQ ID N: 2 и содержащий инсерцию интеина PRP8 штамма Penicillium chrysogenum ВКПМ F-3, несущего мутацию Cys11Tyr, в составе нативной фосфолипазы А2 штамма Streptomyces violaceoruber А-2688.
4. Белок-предшественник для осуществления способа по п. 1, кодируемый последовательностью SEQ ID N: 3 и содержащий инсерцию интеина PRP8 штамма Neosartorya aurata NRRL 4378 в составе мутантной фосфолипазы А2 штамма Streptomyces violaceoruber А-2688, несущей мутации Ser31Ala, Ser82Ala и Asn108Gln.
5. Белок-предшественник для осуществления способа по п. 1, кодируемый последовательностью SEQ ID N: 4 и содержащий инсерцию интеина PRP8 штамма Penicillium chrysogenum ВКПМ F-3, несущего мутацию Cys11Tyr, в составе фосфолипазы А2 Streptomyces violaceoruber А-2688, несущей мутации Ser31Ala, Ser82Ala и Asn108Gln.
| ЧЕПЕРЕГИН С.Э | |||
| и др, Высокоактивные модифицированные варианты рекомбинантной фосфолипазы А2 Streptomyces violaceoruber для эффективного биосинтеза в дрожжах, БИОТЕХНОЛОГИЯ, 2019, т.35, н.3, стр.30-41 | |||
| ВЕЛЬКОВ В.В., С-реактивный белок и липопротеин-ассоциированная фосфолипаза А2: новые факты и новые возможности для диагностики и стратификации |

