Изобретение относится к области биотехнологии и иммунологии, в частности к антителам к белкам ВИЧ.
Матриксный белок р17 вируса иммунодефицита человека первого типа (ВИЧ-1) играет значимую роль в патогенезе ВИЧ-инфекции и последующем развитии синдрома приобретенного иммунодефицита (СПИДа). Данный белок является одним из продуктов протеолиза белка-предшественника Gag и выполняет структурообразующую функцию в вирионе. Известно, что белок р17 входит в состав вирусной оболочки, покрытой снаружи двуслойной липидной мембраной [Ganser-Pornillos В.K., Yeager М., Pornillos О. Assembly and architecture of HIV // Adv Exp Med Biol. 2012; 726:441-465]. Однако, по-видимому, часть молекул белка р17 может быть экспонирована на поверхности вир иона, поскольку известны антитела к белку р17, обладающие вируснейтрализующей активностью [Kageyama S., Katsumoto Т., Taniguchi K., et al., Neutralization of human immunodeficiency virus type 1 (HIV-1) with antibody from carriers' plasma against HIV-1 protein pi7 // Acta Virol. 1996; 40(4): 195-200; Papsidero L.D., Sheu M., Ruscetti F.W. Human immunodeficiency virus type 1-neutralizing monoclonal antibodies which react with p17 core protein: characterization and epitope mapping // J of Virology 1989; 63(1): 267-272; US 5185147].
Помимо содержащегося в вирионе матриксного белка р17 также существует экстраклеточный белок р17, который выделяется ВИЧ-инфицированными клетками в течение всего жизненного цикла вируса. Связываясь с клеточными рецепторами, в частности, с CXCR1, CXCR2, экстраклеточный белок р17 взаимодействует с клетками иммунной системы: В-клетками и активированными Т-клетками, моноцитами, NK-клетками, плазмацитоидными дендритными клетками. На примере моноцитов и активированных лимфоцитов периферической крови было показано, что белок p17 вызывает секрецию клетками провоспалительных цитокинов интерферона-гамма и фактора некроза опухоли-альфа, а также усиливает секрецию МСР-1 [Fiorentini S., Marini Е., Caracciolo S., Caruso A. Functions of the HIV-1 matrix protein p17 // New Microbiol. 2006, 29(1): 1-10; Caccun F., Marsico S., Fiorentini S., Caruso A., Giagulli C. HIV-1 Matrix Protein p17 and its Receptors // Current Drag Targets, 2016, 17(1): 23-32].
Модулируя пролиферацию клеток и цитокиновое микроокружение, р17 способствует репликации вируса [De Francesco М.А., Caruso A., Fallacara F., Canaris A.D., Dima F., Poiesi C., Licenziati S., Corulli M., Martinelli F., Fiorentini S., Turano A. HIV p17 enhances lymphocyte proliferation and HIV-1 replication after binding to a human serum factor // AIDS. 1998, 2(3):245-252]. Таким образом, экстраклеточный p17 может действовать как специфический вирусный цитокин, создающий подходящую среду, способствующую репликации вируса. Белок р17 обнаруживается в лимфатических узлах больных даже при длительном проведении высокоэффективной антиретровирусной терапии (ВААРТ), сопровождающейся снижением титров циркулирующего в крови вируса; возможно, именно он участвует в восстановлении репликации вируса, наблюдаемой при прекращении или в перерывах между курсами ВААРТ [Popovic М., Tenner-Racz K., Pelser С., et al. Persistence of HIV-1 structural proteins and glycoproteins in lymph nodes of patients under highly active antiretroviral therapy // Proc Natl Acad Sci USA 2005; 102(41): 14807-14812]. Также известны мутантные формы белка р17, обладающие клоногенной активностью по отношению к В-клеткам и участвующие в образовании лимфом, которые являются частыми осложнениями при ВИЧ-инфекции [Dolcetti R., Giagulli С., Не W., et al. Role of HIV-1 matrix protein p17 variants in lymphoma pathogenesis // Proc Natl Acad Sci USA. 2015, 112(46): 14331-1436]. Клинические исследования показали, что наличие высокого уровня антител к белку р17 коррелирует с медленным развитием СПИДа, в то время как уровень антител в крови снижается при переходе от бессимптомного течения инфекции к СПИДу. Поэтому представляется перспективным рассмотрение белка р17 в качестве мишени для противовирусной терапии.
Ранее белок р17 ВИЧ или его фрагменты использовали для создания иммуногенов, которые могут стать основой для терапевтических вакцин [US 185147, US 6111068, US 5480967, ЕА 006308, ЕА 201000829, RU 2337922, RU 2306950, RU 2010133961, RU 2412946], однако примеры практического применения таких иммуногенов или вакцин, с терапевтической или профилактической целью, отсутствуют.
Были предложены антитела к белку р17: мышиные моноклональные антитела MBS-3 [RU 2337922] и МВА-1 [RU 2012146085], блокирующие влияние белка р17 на продукцию провоспалительных цитокинов мононуклеарными клетками периферической крови человека; мышиное моноклональное антитело, ингибирующее образование белка р17 из его предшественника белка Gag [WO 9119000]; и одноцепочечное антитело, которое, экспрессируясь внутри клеток, эффективно ингибирует репликацию ВИЧ-1 при последующем заражении [Tewari D., Goldstein S.L., Notkins A.L., Zhou P. cDNA Encoding a Single-Chain Antibody to HIV p17 with Cytoplasmic or Nuclear Retention Signals Inhibits HIV-1 Replication // J Immunol, 1998, 161(5):2642-2647]. Также известны моноклональное мышиное антитело 32/1.24.89 (Абкам, Великобритания), которое в концентрации менее 1 мкг/мл является эффективным ингибитором репликации вируса ВИЧ-1, и моноклональное антитело 32/5.8.42 (Абкам, Великобритания), которое также обладает вируснейтрализующей активностью, но при более высокой концентрации [Papsidero L.D., Sheu М., Ruscetti F.W. Human immunodeficiency virus type 1-neutralizing monoclonal antibodies which react with p17 core protein: characterization and epitope mapping // Journal of Virology; 1989; 63(1):267-272].
Однако вышеперечисленные антитела были разработаны против белков р17 и Gag ВИЧ-1 субтипа В, преобладающего в США и странах Западной Европы, в то время как в России и странах Ближнего Зарубежья преобладает субтип А, при этом последовательности аминокислот в белках р17 субтипов А и В различаются [Masharsky А.Е., Klimov N.A., Kozlov А.Р. Molecular cloning and analysis of full-length genome of HIV type 1 strains prevalent in countries of the former Soviet Union // AIDS Res Hum Retroviruses. 2003, 19(10):933-939]. В частности, сравнение последовательностей аминокислот белков р17 ВИЧ-1 субтипов А и В из базы данных UniProt (Expasy/UniProt KB/Р05888/Р24736-1) с помощью программы BLAST [www.ncbi.nlm.nih.gov/pubmedVBLAST] показывает, что последовательности данных белков различаются на 25%. Для использования в нашей стране в качестве терапевтических антител против ВИЧ-1 необходимо, чтобы такие антитела эффективно связывали белок р17 субтипа А.
Технической задачей, на решение которой направлено настоящее изобретение, является расширение спектра антител, связывающих белок р17 вируса иммунодефицита человека первого типа, в частности субтипа А.
Технический результат достигался созданием моноклонального антитела мыши к белку р17 российского изолята субтипа А ВИЧ-1, специфически связывающего белок р17, а также созданием химерного антитела, которое специфически и с высокой аффинностью связывается с р17.
Мышиное моноклональное антитело изотипа IgG2κ содержит гипервариабельные участки вариабельной области тяжелой цепи по SEQ ID NO: 3-5 и гипервариабельные участки вариабельной области легкой цепи по SEQ ID NO: 6-8, изоэлектрическая точка антитела в диапазоне pi 6,4-6,8.
Химерное моноклональное антитело, содержит тяжелую цепь по SEQ ID NO: 11 и легкую цепь по SEQ ID NO: 12, имеет молекулярную массу 160 кДа, специфически связывается с белком р17, при этом в исследованном диапазоне концентрации количество связавшегося с антигеном химерного моноклонального антитела зависит от концентрации антитела. Сравнительный анализ последовательностей CDR тяжелой и легкой цепей показал отсутствие гомологий между известными антителами к р17 и заявленными антителами.
Технология получения мышиного антитела включала в себя иммунизацию животных антигеном, представляющим собой рекомбинантный белок р17 российского изолята субтипа А ВИЧ-1. В качестве антигенов для отбора позитивных продуцентов моноклональных антител при постановке иммуноферментного анализа (ИФА) использовали тот же белок, что и для иммунизации. В дальнейшем в последовательностях легкой и тяжелой цепей полученного мышиного моноклонального антитела была произведена замена константных областей на константные области иммуноглобулина G человека с получением химерного (мышь-человек) антитела. Химерное антитело к р17 ВИЧ-1 было наработано в трансфицированных культивируемых клетках китайского хомячка линии СНО-K1, очищено и проанализировано.
Изобретение иллюстрируется следующими графическими материалами:
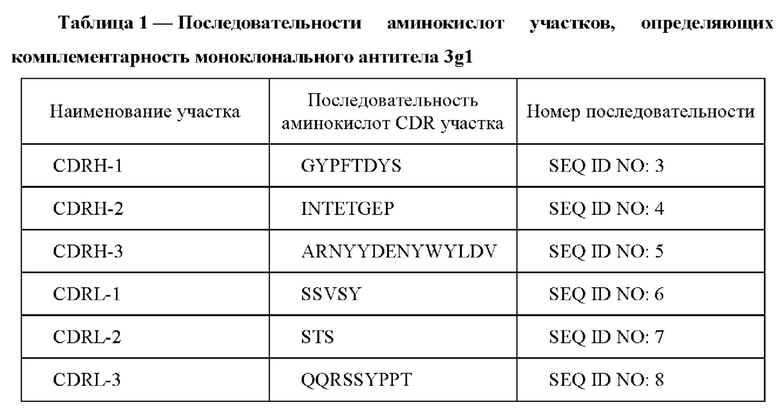
На Фиг. 1 представлена электрофореграмма очищенного химерного антитела 3gl-him в 4-20% полиакриламидном геле: 1 - образец очищенного антитела 3gl-him (восстанавливающие условия); 2 - маркеры молекулярного веса; 3 - образец очищенного антитела 3gl-him (невосстанавливающие условия);
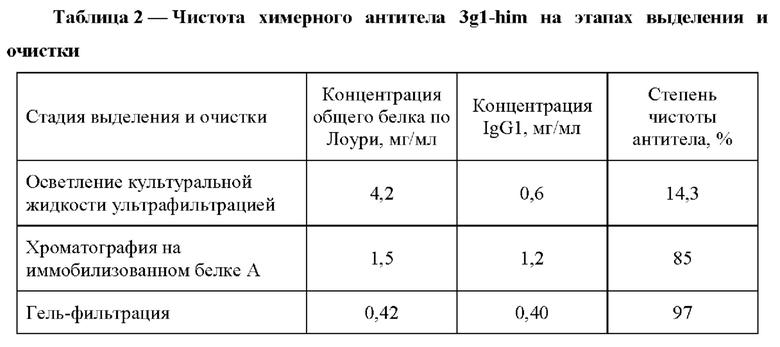
На Фиг. 2 представлен Вестерн-блот рекомбинантного белка р17 российского изолята субтипа А ВИЧ-1 после электрофореза и последующей инкубации с химерным антителом 3gl-him: 4 - в восстанавливающих условиях, 5 - маркеры молекулярного веса; 6 - в не восстанавливающих условиях.
Сущность и промышленная применимость изобретения поясняется следующими примерами:
Пример 1. Получение белка р17 российского изолята субтипа А ВИЧ-1.
Рекомбинантный белок р17 российского изолята субтипа А ВИЧ-1 получали, как описано ранее [Аль-Шехадат Р.И., Духовлинов И.В., Кобатов А.И., Климов Н.А., Козлов А.П. Получение рекомбинантного белка вируса р17 иммунодефицита человека субтипа А // Биотехнология, 2010, 6:19-26]. Последовательность данного белка содержит на N-конце 6-гистидиновый таг, используемый для очистки белка с помощью металлохелатной хроматографии. Чистота очищенного белка р17 составила 97%.
Пример 2. Иммунизация животных и получение гибридомы, продуцирующей антитело к белку р17.
Мышей линии Balb/c иммунизировали очищенным рекомбинантным белком р17 разработанным нами ранее методом с применением интерлейкина-1β в качестве адъюванта [RU 2412946]. Иммунизированных животных подвергали эвтаназии и из их селезенок и периферических лимфоузлов выделяли лимфоциты. Слияние полученных лимфоцитов с клетками миеломы мыши Sp2/0 проводили с помощью полиэтиленгликоля по стандартной методике. После селекции на селективной питательной среде, содержащей гипоксантин, аминоптерин и тимидин (HAT), из 5300 ячеек с растущими клетками отбирали образцы культуральной жидкости для скрининга антител к р17 человека с помощью твердофазного иммуноферментного анализа (ИФА). Для отбора стабильных клонов позитивные продуценты моноклональных антител подвергали дальнейшему клонированию. В результате нескольких циклов клонирования и скрининга был отобран клон гибридных клеток s3gl, стабильно секретирующих антитело 3gl к белку р17 ВИЧ-1.
Пример 3. Очистка моноклонального антитела и его свойства.
Антитело 3gl из асцитной жидкости мышей линии Balb/c с внутрибрюшинно привитой гибридомой s3gl выделяли с помощью гель-проникающей и ионообменной хроматографии для его дальнейшей характеризации.
По результатам электрофореза в 4-20% полиакриламидном геле с ДДС натрия молекулярный вес мышиного моноклонального антитела 3gl в нативных (невосстанавливающих) условиях соответствует ожидаемому для нативного IgG мыши (160 кДа), в восстанавливающих условиях антитело диссоциирует на тяжелую (52 кДа) и легкую (28 кДа) цепи. С помощью имму но ферментного набора ISO-2 kit 072K4849 (Sigma, США) был установлен изотип тяжелой цепи антитела 3gl - G2, и легкой цепи - κ. С помощью ИФА была проверена специфичность антитела. Так было установлено, что антитело 3gl не связывается с белками сыворотки крови и селезенки мышей, а также с белками сыворотки крови человека и растворимыми белками гомогената культивируемых клеток яичников китайского хомячка (СНО). Изоэлектрическая точка антитела 3gl находится в диапазоне рН 6,4-6,8.
Пример 4. Синтез и секвенирование кДНК, кодирующих вариабельные области легкой и тяжелой цепей моноклонального антитела 3gl.
Из клеток гибрид омы 3gl выделяли РНК, на матрице которой путем обратной транскрипции и амплификации с помощью известных наборов синтетических праймеров (Immunogenetics Information System http://www.imgt.org) были амплифицированы фрагменты генов, кодирующие вариабельные области тяжелой (VH) и легкой (VL) цепей антитела. Полученные фрагменты генов были клонированы в вектор pALTA (Евроген, Россия) и секвенированы с внешних праймеров (М13). Последовательности фрагментов генов, кодирующих вариабельные области VH и VL моноклонального антитела 3gl, представлены на SEQ ID NO: 1 и SEQ ID NO: 2 соответственно. Анализ полученных последовательностей производили с помощью программы IMGT V-Quest (IMGT, the international ImMunoGeneTics information system http://www.imgt.org), что позволило выделить участки гипервариабельных доменов CDR, определяющие комплементарность антитела антигену (Таблица 1).
Последовательности аминокислот SEQ ID NO: 3-8 с помощью программы BLAST (National Center for Biotechnology Information http://www.ncbi.nlm.nih.gov) сравнивали с последовательностями вариабельных областей тяжелых и легких цепей известных моноклональных антител к белку p17 ВИЧ-1. Участков гомологии не обнаружено.
Пример 5. Конструирование химерного антитела 3gl-him.
С помощью полимеразной цепной реакции последовательности ДНК, кодирующие вариабельные области тяжелой и легкой цепей моноклонального антитела 3gl, были состыкованы с последовательностями ДНК, кодирующими сигнальный пептид легкой цепи иммуноглобулина мыши, а также константную область тяжелой цепи IgG1 человека и константную область легкой каппа цепи иммуноглобулина G человека соответственно. Сконструированное химерное антитело получило название 3gl-him.
Полученная последовательность нуклеотидов, кодирующая тяжелую цепь химерного (мышь-человек) антитела 3gl-him, представлена в SEQ ID NO: 9, последовательность аминокислот тяжелой цепи химерного антитела - в SEQIDNO: 11.
Полученная последовательность нуклеотидов, кодирующая легкую цепь химерного антитела 3gl-him, представлена в SEQ ID NO: 10, последовательность аминокислот легкой цепи химерного антитела - в SEQ ID NO: 12.
Полученные в примере 4 последовательности ДНК, кодирующие тяжелую и легкую цепи химерного антитела, удлиняли путем постановки ПЦР с праймерами, содержавшими сайты рестрикции XbaI и NotI, обрабатывали данными рестриктазами и лигировали с рестрицированными XbaI и NotI плазмидными векторами серии pTVK4. В результате были получены плазмиды pTVK4g/Anti-P17-him-H и pTVK4d/ Anti-P17-him-L.
Плазмида pTVK4g/Anti-P17-him-H несет последовательность ДНК, кодирующую тяжелую цепь химерного антитела против белка р17, промотор цитомегаловируса, сайт полиаденилирования, ген устойчивости к генетицину и элемент UCOE из генома мыши, длиной 2990 нуклеотидов, обеспечивающий преимущественную интеграцию вектора в транскрипционно активные участки хроматина [Jia J. Hou С, Hughes B.S., Smede M., Leung K.M., Levine K., Rigby S., Gray P.P., Munro T.P. High-throughput ClonePix FL analysis of mAb-expressing clones using the UCOE expression system // New Biotechnology, 2014, 31(3):214-220; Nair A.R., Xie Jinger X., Hermiston T.W. Effect of different UCOE-promoter combinations in creation of engineered cell lines for the production of Factor VIII // BMC Research Notes, 2011, 4:178]. Плазмида pTVK4d/Anti-P17-him-L несет последовательность ДНК, кодирующую легкую цепь химерного антитела против белка р17, промотор цитомегаловируса, сайт полиаденилирования, ген дигидрофолатредуктазы, обеспечивающий устойчивость к метотрексату, и элемент UCOE из 5'-нетранслируемой области гена RPS3 мыши, длиной 2990 нуклеотидов.
Пример 6. Получение штамма клеток СНО - продуцента химерного антитела против белка р17 ВИЧ.
Для постоянной трансфекции использовали клетки линии яичников китайского хомячка, дефицитные по гену дигидрофолатредуктазы (CHOdnfr-), ранее адаптированные к суспензионному росту в бессывороточных условиях [Biaggio R.T., Abreu-Neto M.S., Covas D.T., Swiech K. Serum-free suspension culturing of human cells: adaptation, growth, and cryo-preservation // Bioprocess BiosystEng., 2015, 38(8): 1495-1507].
Клетки культивировали в среде CDM4CHO (HyClone, США) с добавлением 4 мМ аланилглутамина, 0,1 мМ гипоксантина, 0,016 мМ тимидина и 0,1% Pluronic F-68 (Sigma, США) в микробиореакторах типа TubeSpin 50 (ТРР, Швейцария) на орбитальном шейкере Sanyo MIR-S100C (200 об/мин), который был помещен в CO2-инкубатор, обеспечивающий содержание в воздухе 5% CO2 и температуру 37°С.
Трансфекцию проводили с помощью липосомального реагента «Freestyle» (Life Technologies, США) по инструкции производителя, используя смесь из 3,6 мкг плазмиды pTVK4g/Anti-P17-him-H и 2,4 мкг плазмиды pTVK4d/ Anti-P17-him-L на 5⋅106 клеток. Через 48 часов после трансфекции клетки переводили на среду без гипоксантина/тимидина с добавлением 5 мкг/мл метотрексата. В последующие дни среду меняли с интервалом 3 суток. Выросшие клетки затем пересевали на среду, содержавшую 0,2 мг/мл генетицина. Через 15 суток клетки переносили в колбы Эрленмейера в культуральную среду для последующего наращивания, после чего клетки общего пула, показавшие устойчивый рост на селективной среде, были расклонированы в 40 96-луночных планшетов фирмы Nunc с клеточной концентрацией 100 клеток на планшет. Через месяц культивирования с периодической сменой среды и адаптацией к росту в среде с генетицином в микробиореакторах типа Tube Spin 50 (ТРР, Швейцария) на орбитальном шейкере Sanyo MIR-S100C (200 об/мин), который был помещен в CO2-инкубатор (5% CO2, 37°С), было обнаружено 30 клонов, из них 12 клонов были положительными по продукции IgG1 человека, определяемых методом твердофазного ИФА. Положительные клоны пересевали и вновь исследовали продукцию ими IgG1 человека на 5-е сутки культивирования. По результатам скрининга было отобрано 4 клона с продукцией IgG1 более 200 мкг/мл в культуральной жидкости. Дальнейшее изучение стабильности и продуктивности клонов позволило выбрать лучший из них с продукцией IgG1 человека до 500 мкг/мл. Этот клон был помещен на хранение в криобанк и использован для культивирования в биореакторе в 1 л среды CDM4CHO (HyClone, США), в которую добавляли 4 мМ аланил-глутамина (Applichem, Германия), 0,1% Pluronic F-68 (Sigma, США) и 5% EX-CELL® CD Hydrolysate Fusion (HyClone, США). Исходная концентрация клеток составляла 0,6⋅106 клеток/мл. В первые трое суток культивирования поддерживали температуру 37°С, затем ее понижали до 32°С. Дальнейшее культивирование проводили при 32°С. Начиная с четвертых суток, в биореактор ежедневно добавляли питательные добавки Cell Boost 7а и Cell Boost 7b (HyClone, США) по 2% и 0,2% от первоначального объема культуры соответственно. Культивирование продолжали до 14 суток, достигнув максимальной плотности клеток более 9⋅106 клеток/мл и концентрации химерного антитела 700 мкг/мл.
Пример 7. Очистка химерного антитела 3gl-him и определение его свойств.
Очистку химерного антитела 3gl-him из культуральной жидкости проводили в три этапа, указанные в Таблице 2, последовательно используя ячейку для ультрафильтрации с с диаметром пор 0,22 мкм (GE-Healthcare Life Sciences, США), сорбент HITrap мембраной Protein A HP (GE-Healthcare Life Sciences, США) и колонку для гель-фильтрации BioSil SEC 125-5, 300×7,8 мм (Bio-Rad, США). Концентрацию антитела определяли методом ИФА по связыванию с сорбированным на планшете белком р17.
Анализ очищенного антитела 3gl-him методом электрофореза в 4-20% полиакриламидном геле с ДДС натрия показал, что оно характеризуется кажущейся молекулярной массой приблизительно 160 кДа в невосстанавливающих условиях, а в восстанавливающих условиях антитело диссоциирует на тяжелую и легкую цепи с кажущимися молекулярными массами 55 и 25 кДа - соответственно, что соответствует показателям для тяжелой и легкой цепей иммуноглобулина G1 (Фиг. 1).
Проверку специфичности антитела проверяли методом Вестерн-блота. Сначала проводили электрофорез рекомбинантного белка р17 российского изолята субтипа А ВИЧ-1 в восстанавливающих и в невосстанавливающих условиях и электроперенос белка из геля на нитроцеллюлозную мембрану. Нитроцеллюлозную пластину с перенесенным антигеном помещали на ночь в фосфатно-солевой буфер рН 7,2, содержащий 1% БСА, затем инкубировали с полученным антителом к р17 3gl-him в концентрации 20 мкг/мл в течение 60 мин. Детекцию проводили, инкубируя пластину с раствором поликлональных антител козы против Fc фрагмента IgG человека (Sigma), коньюгированных с пероксидазой хрена (разведение 1:4000) в течение 1 часа. Окраску проводили субстратным раствором. Результаты Вестерн-блота представлены на Фиг. 2. Полученное химерное антитело 3gl-him специфично связывает белок р17 российского изолята субтипа А ВИЧ-1.
--->
Перечень последовательностей
<110> Федеральное государственное унитарное предприятие «Государственный научно-исследовательский институт особо чистых биопрепаратов» Федерального медико-биологического агентства
Federal State Unitary Enterprise «State Research Institute of Highly Pure Biopreparations» of the Federal Medical and Biological Agency
<120> Антитела против белка р17 ВИЧ-1 субтипа А
<130>
<160> 12
<210> 1
<211> 364
<212> ДНК
<213> Mus musculus
<220>
<222> (1)…(364)
<223> ДНК, кодирующая вариабельную область тяжелой цепи мышиного моноклонального антитела 3g1
<400> 1
CAG ATC CAC TTA TTG CAG TCT GGA CCT GAG CTG AAG AAG CCT GGA GAG 48
Gln Ile His Leu Leu Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
ACA GTC AAG ATC TCC TGC AAG GCT TCT GGT TAT CCC TTC ACA GAC TAT 96
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Pro Phe Thr Asp Tyr
20 25 30
TCA ATG CAC TGG GTG AAG CAG GCT CCA GGA AAG GGT TTA AAG TGG ATG 144
Ser Met His Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met
35 40 45
GGC TGG ATA AAC ACT GAG ACT GGT GAG CCA GCA TAT GCA GAT GAC TTC 192
Gly Trp Ile Asn Thr Glu Thr Gly Glu Pro Ala Tyr Ala Asp Asp Phe
50 55 60
AAG GGA CGG TTT GCC TTC TCT TTG GAA ACC TCT GCC AGC ACT GCC TAT 240
Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
TTG CAG ATC AAC ATC CTC AAA AAT GAG GAC ACG GCT ACA TAT TTC TGT 288
Leu Gln Ile Asn Ile Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys
85 90 95
GCC CGG AAT TAC TAC GAT GAA AAC TAC TGG TAC CTC GAT GTC TGG GGC 336
Ala Arg Asn Tyr Tyr Asp Glu Asn Tyr Trp Tyr Leu Asp Val Trp Gly
100 105 110
GCA GGG ACC ACG GTC ACC GTC TCC TCС G 364
Ala Gly Thr Thr Val Thr Val Ser Ser
115 120 121
<210> 2
<211> 319
<212> ДНК
<213> Mus musculus
<220>
<222> (1)…(319)
<223> ДНК, кодирующая вариабельную область легкой цепи мышиного моноклонального антитела 3g1
<400> 2
CAA ATT GTT CTC ACC CAG TCT CCA GCA ATC ATG TCT ACA TCT CCA GGG 48
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Thr Ser Pro Gly
1 5 10 15
GAG AAG GTC ACC ATA ACC TGC AGT GCC AGC TCA AGT GTA AGT TAC ATG 96
Glu Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
CAC TGG TTC CAG CAG AAG CCA GGC ACT TCT CCC AAA CTC TGG ATT TAT 144
His Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Trp Ile Tyr
35 40 45
AGC ACA TCC AAC CTG GCT TCT GGA GTC CCT GCT CGC TTC AGT GGC AGT 192
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
GGA TCT GGG ACC TCT TAC TCT CTC ACA ATC AGC CGA ATG GAG GCT GAA 240
GGC TGG ATA AAC ACT GAG ACT GGT GAG CCA GCA TAT GCA GAT GAC TTC 192
Gly Trp Ile Asn Thr Glu Thr Gly Glu Pro Ala Tyr Ala Asp Asp Phe
50 55 60
AAG GGA CGG TTT GCC TTC TCT TTG GAA ACC TCT GCC AGC ACT GCC TAT 240
Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
TTG CAG ATC AAC ATC CTC AAA AAT GAG GAC ACG GCT ACA TAT TTC TGT 288
Leu Gln Ile Asn Ile Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys
85 90 95
GCC CGG AAT TAC TAC GAT GAA AAC TAC TGG TAC CTC GAT GTC TGG GGC 336
Ala Arg Asn Tyr Tyr Asp Glu Asn Tyr Trp Tyr Leu Asp Val Trp Gly
100 105 110
GCA GGG ACC ACG GTC ACC GTC TCC TCС G 364
Ala Gly Thr Thr Val Thr Val Ser Ser
115 120 121
<210> 2
<211> 319
<212> ДНК
<213> Mus musculus
<220>
<222> (1)…(319)
<223> ДНК, кодирующая вариабельную область легкой цепи мышиного моноклонального антитела 3g1
<400> 2
CAA ATT GTT CTC ACC CAG TCT CCA GCA ATC ATG TCT ACA TCT CCA GGG 48
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Thr Ser Pro Gly
1 5 10 15
GAG AAG GTC ACC ATA ACC TGC AGT GCC AGC TCA AGT GTA AGT TAC ATG 96
Glu Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
CAC TGG TTC CAG CAG AAG CCA GGC ACT TCT CCC AAA CTC TGG ATT TAT 144
His Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Trp Ile Tyr
35 40 45
AGC ACA TCC AAC CTG GCT TCT GGA GTC CCT GCT CGC TTC AGT GGC AGT 192
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
GGA TCT GGG ACC TCT TAC TCT CTC ACA ATC AGC CGA ATG GAG GCT GAA 240
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Met Glu Ala Glu
65 70 75 80
GAT GCT GCC ACT TAT TAC TGC CAG CAA AGG AGT AGT TAC CCA CCC ACG 288
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Ser Tyr Pro Pro Thr
85 90 95
TTC GGT GCT GGG ACC AAG CTG GAG CTG AAA C 319
Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105 106
<210> 3
<211> 8
<212> белок
<213> Mus musculus
<220>
<222> (1)…(8)
<223> Последовательность аминокислот участка CDRH-1 мышиного моноклонального антитела 3g1
<400> 3
Gly Tyr Pro Phe Thr Asp Tyr Ser
1 5 8
<210> 4
<211> 8
<212> белок
<213> Mus musculus
<220>
<222> (1)…(8)
<223> Последовательность аминокислот участка CDRH-2 мышиного моноклонального антитела 3g1
<400> 4
Ile Asn Thr Glu Thr Gly Glu Pro
1 5 8
<210> 5
<211> 14
<212> белок
<213> Mus musculus
<220>
<222> (1)…(14)
<223> Последовательность аминокислот участка CDRH-3 мышиного моноклонального антитела 3g1
<400> 5
Ala Arg Asn Tyr Tyr Asp Glu Asn Tyr Trp Tyr Leu Asp Val
1 5 10 14
<210> 6
<211> 5
<212> белок
<213> Mus musculus
<220>
<222> (1)…(5)
<223> Последовательность аминокислот участка CDRL-1 мышиного моноклонального антитела 3g1
<400> 6
Ser Ser Val Ser Tyr
1 5
<210> 7
<211> 3
<212> белок
<213> Mus musculus
<220>
<222> (1)…(3)
<223> Последовательность аминокислот участка CDRL-2 мышиного моноклонального антитела 3g1
<400> 7
Ser Thr Ser
1 3
<210> 8
<211> 9
<212> белок
<213> Mus musculus
<220>
<222> (1)…(9)
<223> Последовательность аминокислот участка CDRL-3 мышиного моноклонального антитела 3g1
<400> 8
Gln Gln Arg Ser Ser Tyr Pro Pro Thr
1 5 9
<210> 9
<211> 1422
<212> ДНК
<213> искусственная последовательность
<220>
<222> (1)…(1422)
<223> Последовательность нуклеотидов, кодирующая тяжелую цепь химерного антитела 3g1-him
<400> 9
ATG GAT TTC CAG GTG CAG ATC TTC AGC TTT CTC CTG ATT AGC GCT AGC 48
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
GTG ATC ATT AGC AGG GGC CAG ATC CAC TTA TTG CAG TCT GGA CCT GAG 96
Val Ile Ile Ser Arg Gly Gln Ile His Leu Leu Gln Ser Gly Pro Glu
20 25 30
CTG AAG AAG CCT GGA GAG ACA GTC AAG ATC TCC TGC AAG GCT TCT GGT 144
Leu Lys Lys Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly
35 40 45
TAT CCC TTC ACA GAC TAT TCA ATG CAC TGG GTG AAG CAG GCT CCA GGA 192
Tyr Pro Phe Thr Asp Tyr Ser Met His Trp Val Lys Gln Ala Pro Gly
50 55 60
AAG GGT TTA AAG TGG ATG GGC TGG ATA AAC ACT GAG ACT GGT GAG CCA 240
Lys Gly Leu Lys Trp Met Gly Trp Ile Asn Thr Glu Thr Gly Glu Pro
65 70 75 80
GCA TAT GCA GAT GAC TTC AAG GGA CGG TTT GCC TTC TCT TTG GAA ACC 288
Ala Tyr Ala Asp Asp Phe Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr
85 90 95
TCT GCC AGC ACT GCC TAT TTG CAG ATC AAC ATC CTC AAA AAT GAG GAC 336
Ser Ala Ser Thr Ala Tyr Leu Gln Ile Asn Ile Leu Lys Asn Glu Asp
100 105 110
ACG GCT ACA TAT TTC TGT GCC CGG AAT TAC TAC GAT GAA AAC TAC TGG 384
Thr Ala Thr Tyr Phe Cys Ala Arg Asn Tyr Tyr Asp Glu Asn Tyr Trp
115 120 125
TAC CTC GAT GTC TGG GGC GCA GGG ACC ACG GTC ACC GTC TCC TCA GCC 432
Tyr Leu Asp Val Trp Gly Ala Gly Thr Thr Val Thr Val Ser Ser Ala
130 135 140
TCC ACC AAG GGC CCA TCG GTC TTC CCC CTG GCA CCC TCC TCC AAG AGC 480
Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser
145 150 155 160
ACC TCT GGG GGC ACA GCG GCC CTG GGC TGC CTG GTC AAG GAC TAC TTC 528
Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe
165 170 175
CCC GAA CCG GTG ACG GTG TCG TGG AAC TCA GGC GCC CTG ACC AGC GGC 576
Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly
180 185 190
TG CAC ACC TTC CCG GCT GTC CTA CAG TCC TCA GGA CTC TAC TCC CTC 624
Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu
195 200 205
AGC AGC GTG GTG ACC GTG CCC TCC AGC AGC TTG GGC ACC CAG ACC TAC 672
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr
210 215 220
ATC TGC AAC GTG AAT CAC AAG CCC AGC AAC ACC AAG GTG GAC AAG AAG 720
Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys
225 230 235 240
GTT GAG CCC AAA TCT TGT GAC AAA ACT CAC ACA TGC CCA CCG TGC CCA 768
Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
245 250 255
GCA CCT GAA CTC CTG GGG GGA CCG TCA GTC TTC CTC TTC CCC CCA AAA 816
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
260 265 270
CCC AAG GAC ACC CTC ATG ATC TCC CGG ACC CCT GAG GTC ACA TGC GTG 864
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
275 280 285
GTG GTG GAC GTG AGC CAC GAA GAC CCT GAG GTC AAG TTC AAC TGG TAC 912
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
290 295 300
GTG GAC GGC GTG GAG GTG CAT AAT GCC AAG ACA AAG CCG CGG GAG GAG 960
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
305 310 315 320
CAG TAC AAC AGC ACG TAC CGT GTG GTC AGC GTC CTC ACC GTC CTG CAC 1008
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
325 330 335
CAG GAC TGG CTG AAT GGC AAG GAG TAC AAG TGC AAG GTC TCC AAC AAA 1056
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
340 345 350
GCC CTC CCA GCC CCC ATC GAG AAA ACC ATC TCC AAA GCC AAA GGG CAG 1104
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
355 360 365
CCC CGA GAA CCA CAG GTG TAC ACC CTG CCC CCA TCC CGG GAG GAG ATG 1152
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
370 375 380
ACC AAG AAC CAG GTC AGC CTG ACC TGC CTG GTC AAA GGC TTC TAT CCC 1200
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
385 390 395 400
AGC GAC ATC GCC GTG GAG TGG GAG AGC AAT GGG CAG CCG GAG AAC AAC 1248
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
405 410 415 TAC AAG ACC ACG CCT CCC GTG CTG GAC TCC GAC GGC TCC TTC TTC CTC 1296
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
420 425 430
TAT AGC AAG CTC ACC GTG GAC AAG AGC AGG TGG CAG CAG GGG AAC GTC 1344
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
435 440 445
TTC TCA TGC TCC GTG ATG CAT GAG GCT CTG CAC AAC CAC TAC ACG CAG 1392
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
450 455 460
AAG AGC CTC TCC CTG TCC CCG GGT AAA TGA 1422
Lys Ser Leu Ser Leu Ser Pro Gly Lys ***
465 470 474
<210> 10
<211> 708
<212> ДНК
<213> искусственная последовательность
<220>
<222> (1)…(708)
<223> Последовательность нуклеотидов, кодирующая легкую цепь химерного антитела 3g1-him
<400> 10
ATG GAT TTC CAG GTG CAG ATC TTC AGC TTT CTC CTG ATT AGC GCT AGC 48
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
GTG ATC ATT AGC AGG GGC CAA ATT GTT CTC ACC CAG TCT CCA GCA ATC 96
Val Ile Ile Ser Arg Gly Gln Ile Val Leu Thr Gln Ser Pro Ala Ile
20 25 30
ATG TCT ACA TCT CCA GGG GAG AAG GTC ACC ATA ACC TGC AGT GCC AGC 144
Met Ser Thr Ser Pro Gly Glu Lys Val Thr Ile Thr Cys Ser Ala Ser
35 40 45
TCA AGT GTA AGT TAC ATG CAC TGG TTC CAG CAG AAG CCA GGC ACT TCT 192
Ser Ser Val Ser Tyr Met His Trp Phe Gln Gln Lys Pro Gly Thr Ser
50 55 60
CCC AAA CTC TGG ATT TAT AGC ACA TCC AAC CTG GCT TCT GGA GTC CCT 240
Pro Lys Leu Trp Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro
65
GCT CGC TTC AGT GGC AGT GGA TCT GGG ACC TCT TAC TCT CTC ACA ATC 288
Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile
85 90 95
AGC CGA ATG GAG GCT GAA GAT GCT GCC ACT TAT TAC TGC CAG CAA AGG 336
Ser Arg Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Arg
100 105 110
AGT AGT TAC CCA CCC ACG TTC GGT GCT GGG ACC AAG CTG GAG CTG AAA 384
Ser Ser Tyr Pro Pro Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
115 120 125
CGG ACC GTG GCC GCA CCT AGC GTG TTC ATC TTC CCA CCT TCC GAC GAG 432
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
130 135 140
CAG CTG AAG TCC GGC ACC GCT TCT GTC GTG TGC CTG CTG AAC AAC TTC 480
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
145 150 155 160
TAC CCC CGC GAG GCC AAG GTG CAG TGG AAG GTG GAC AAC GCC CTG CAG 528
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
165 170 175
TCC GGC AAC TCC CAG GAA TCC GTG ACC GAG CAG GAC TCC AAG GAC AGC 576
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
180 185 190
ACC TAC TCC CTG TCC TCC ACC CTG ACC CTG AGC AAG GCC GAC TAC GAG 624
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
195 200 205
AAG CAC AAG GTG TAC GCC TGC GAA GTG ACC CAC CAG GGC CTG TCT AGC 672
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
210 215 220
CCC GTG ACC AAG TCT TTC AAC CGG GGC GAG TGC TGA 708
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys ***
225 230 235 236
<210> 11
<211> 473
<212> белок
<213> искусственная последовательность
<220>
<222> (1)…(473)
<223> Последовательность аминокислот тяжелой цепи химерного антитела 3g1-him
<400> 11
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Ile Ser Arg Gly Gln Ile His Leu Leu Gln Ser Gly Pro Glu
20 25 30
Leu Lys Lys Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly
35 40 45
Tyr Pro Phe Thr Asp Tyr Ser Met His Trp Val Lys Gln Ala Pro Gly
50 55 60
Lys Gly Leu Lys Trp Met Gly Trp Ile Asn Thr Glu Thr Gly Glu Pro
65 70 75 80
Ala Tyr Ala Asp Asp Phe Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr
85 90 95
Ser Ala Ser Thr Ala Tyr Leu Gln Ile Asn Ile Leu Lys Asn Glu Asp
100 105 110
Thr Ala Thr Tyr Phe Cys Ala Arg Asn Tyr Tyr Asp Glu Asn Tyr Trp
115 120 125
Tyr Leu Asp Val Trp Gly Ala Gly Thr Thr Val Thr Val Ser Ser Ala
130 135 140
Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser
145 150 155 160
Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe
165 170 175
Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly
180 185 190
Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu
195 200 205
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr
210 215 220
Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys
225 230 235 240
Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
245 250 255
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
260 265 270
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
275 280 285
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
290 295 300
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
305 310 315 320
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
325 330 335
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
340 345 350
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
355 360 365
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
370 375 380
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
385 390 395 400
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
405 410 415
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
420 425 430
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
435 440 445
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
450 455 460
Lys Ser Leu Ser Leu Ser Pro Gly Lys
465 470 473
<210> 12
<211> 235
<212> белок
<213> искусственная последовательность
<220>
<222> (1)…(235)
<223> Последовательность аминокислот легкой цепи химерного антитела 3g1-him
<400> 12
Met Asp Phe Gln Val Gln Ile Phe Ser Phe Leu Leu Ile Ser Ala Ser
1 5 10 15
Val Ile Ile Ser Arg Gly Gln Ile Val Leu Thr Gln Ser Pro Ala Ile
20 25 30
Met Ser Thr Ser Pro Gly Glu Lys Val Thr Ile Thr Cys Ser Ala Ser
35 40 45
Ser Ser Val Ser Tyr Met His Trp Phe Gln Gln Lys Pro Gly Thr Ser
50 55 60
Pro Lys Leu Trp Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro
65 70 75 80
Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile
85 90 95
Ser Arg Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Arg
100 105 110
Ser Ser Tyr Pro Pro Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
115 120 125
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
130 135 140
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
145 150 155 160
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
165 170 175
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
180 185 190
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
195 200 205
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
210 215 220
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230 235
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ШТАММ КЛЕТОК CHO-SE-9/4 - ПРОДУЦЕНТ ХИМЕРНОГО АНТИТЕЛА ПРОТИВ ЭРИТРОПОЭТИНА ЧЕЛОВЕКА И ХИМЕРНОЕ АНТИТЕЛО, ПРОДУЦИРУЕМОЕ ДАННЫМ ШТАММОМ | 2019 |
|
RU2717038C1 |
| СПОСОБ УПРАВЛЕНИЯ РИТМОМ СЕРДЦА И СОКРАЩЕНИЕМ ОТДЕЛЬНЫХ КАРДИОМИОЦИТОВ ПРИ ПОМОЩИ ТЕРМОГЕНЕТИКИ | 2022 |
|
RU2802995C1 |
| Вакцина против герпеса | 2019 |
|
RU2731073C1 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРЕДОТВРАЩЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2016 |
|
RU2714205C2 |
| БИОСЕНСОРНАЯ СИСТЕМА ДЛЯ БЫСТРОГО ОБНАРУЖЕНИЯ ОПРЕДЕЛЯЕМЫХ КОМПОНЕНТОВ | 2016 |
|
RU2717658C2 |
| СПОСОБ ПРОДУЦИРОВАНИЯ ПОЧЕЧНЫХ КЛЕТОК-ПРЕДШЕСТВЕННИКОВ | 2016 |
|
RU2730861C2 |
| МАТЕРИАЛЫ И МЕТОДЫ, ИСПОЛЬЗУЕМЫЕ ДЛЯ ЛЕЧЕНИЯ РЕСПИРАТОРНЫХ ЗАБОЛЕВАНИЙ У СОБАК | 2020 |
|
RU2811752C2 |
| АНТИТЕЛА, ИНДУЦИРУЮЩИЕ ИММУННУЮ ТОЛЕРАНТНОСТЬ, ИНДУЦИРОВАННЫЕ ЛИМФОЦИТЫ И ТЕРАПЕВТИЧЕСКИЙ АГЕНТ/СПОСОБ КЛЕТОЧНОЙ ТЕРАПИИ С ИСПОЛЬЗОВАНИЕМ ИНДУЦИРОВАННЫХ ЛИМФОЦИТОВ | 2019 |
|
RU2816592C2 |
| АНТИТЕЛО К CD73 ЧЕЛОВЕКА | 2017 |
|
RU2754058C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2019 |
|
RU2781542C2 |
Изобретение относится к области биотехнологии и иммунологии, в частности к антителам к белкам ВИЧ, и представляет собой химерное (мышь-человек) моноклональное антитело, эффективно связывающее матриксный белок р17 вируса иммунодефицита человека первого типа субтипа А. Изобретение может быть эффективно использовано для профилактики и терапии ВИЧ-инфекции. 1 з.п. ф-лы, 2 ил., 2 табл., 7 пр.
1. Моноклональное антитело к белку р17 вируса иммунодефицита человека 1 типа субтипа А, содержащее гипервариабельные участки тяжелой цепи с последовательностями SEQ ID NO: 3-5 и гипервариабельные участки легкой цепи с последовательностями SEQ ID NO: 6-8.
2. Антитело по п. 1, имеющее последовательность тяжелой цепи по SEQ ID NO: 11 и последовательность легкой цепи по SEQ ID NO: 12.
| ИЗОЛИРОВАННЫЕ ПОЛИПЕПТИДЫ НА ОСНОВЕ НЕЙТРАЛИЗУЮЩЕГО ЭПИТОПА БЕЛКА p17 ВИРУСА ВИЧ, ИСПОЛЬЗУЕМЫЕ В КАЧЕСТВЕ ВАКЦИН, А ТАКЖЕ НЕЙТРАЛИЗУЮЩИЕ АНТИ-p17-АНТИТЕЛА, СПЕЦИФИЧЕСКИ РАСПОЗНАЮЩИЕ УКАЗАННЫЙ НЕЙТРАЛИЗУЮЩИЙ ЭПИТОП | 2002 |
|
RU2337922C9 |
| СПОСОБ ПОЛУЧЕНИЯ АНТИТЕЛ К БЕЛКУ р17 ВИЧ | 2009 |
|
RU2412946C1 |
| RU 2017104770 A, 14.08.2018 | |||
| ВАКЦИНА ПРОТИВ ВИЧ/СПИД | 2010 |
|
RU2475264C2 |
| Аппарат для производства углекислого аммония | 1925 |
|
SU6308A1 |

