Группа изобретений относится к области биотехнологии и иммунологии, в частности к вакцинам на основе плазмидной ДНК против вирусов герпеса.
Герпесвирусные инфекции широко распространены по всему миру и имеют постоянную тенденцию к неуклонному росту. Около 90% населения планеты являются носителями одного или обоих вирусов герпеса человека 1 и 2 типов (ВГЧ-1 и ВГЧ-2). Распространенность вируса герпеса человека 3 типа (вирус ветряной оспы, ВГЧ-3) зависит от популяции и варьирует от 61% до 100%, около 90% для 4 типа (вирус Эпштейна-Барр, ВГЧ-4), от 64 до 83% для 6 типа (ВГЧ-6), около 85% для 7 типа (ВГЧ-7). Вирус герпеса человека 5 типа (цитомегаловирус, ВГЧ-5) получил наибольшее распространение среди детей дошкольного возраста. В России ежегодно регистрируют более 120 млн. случаев герпетической инфекции.
Вирусы герпеса человека обладают способностью вовлекать в инфекционный процесс различные органы и системы, что и обусловливает многообразие вызываемых ими заболеваний: от простых кожно-слизистых до угрожающих жизни генерализованных инфекций. При этом вирусы герпеса являются серьезной проблемой для иммунокомпрометированных людей: больных ВИЧ или туберкулезом, пожилых людей.
Особенностью вируса герпеса является механизм его взаимодействия с организмом человека. В отличие от большинства вирусов герпес, попадая в клетки, персистирует в виде экстрахромосомных кольцевых молекул ДНК (эписом), поэтому герпетические инфекции рассматриваются как пожизненные. Возможными стратегиями борьбы с последствиями заражения вирусом герпеса являются борьба с рецидивами с помощью противовирусных препаратов, а также вакцинация как единственный способ обеспечить надежный иммунный ответ организма. Вакцинация способна предотвращать проникновение вируса в клетки организма, так как подготовленный иммунитет уничтожает вирус до передачи его генетического материала в клетки.
Использующиеся в данный момент методы вакцинации подразумевают введение в организм живых или ослабленных форм вируса герпеса, что приводит к реакции иммунитета и снижает риск развития тяжелой формы заболевания. Однако введение такого типа препарата может спровоцировать затяжной рецидив герпетической инфекции, вызвать энцефалит, пневмонию или гепатит. Среди разработок, касающихся вакцин на основе живых вирусов, как правило, 2 типа, следует отметить изобретения по заявке РТ2516629 и патентам RU 2575067 и RU 2236256. ВРТ2516629 описан дефектный ВГЧ-2 с высоким уровнем экспрессии гликопротеина D ВГЧ-2. Дополнительно в дефектном вирусе могут экспрессироваться иммуномодулирующие гены, такие как ИЛ15 и/или основные антигены ВГЧ-1 или ВГЧ-2, например gB или gC. В RU 2575067 предлагается дефектный по репликации доминантно-негативный рекомбинантный ВГЧ-2, вакцина на его основе и способ иммунизации, а в RU 2236256 - вакцина на основе мутантного вируса герпеса.
Вакцинные композиции на основе ВГЧ-1 или ВГЧ-2, инактивированных формалином или гамма-излучением, и иммунокомпетентного вещества (ИФН и комбинация двух независимых аминокислот) предложены ФГУП СПбНИИВС ФМБА России (RU 2552341) и ООО «РусГен» (RU 2291690). Композиция может дополнительно содержать фармацевтически приемлемый носитель и аминокислоты.
Вместе с тем указанные разработки не перспективны для внедрения в медицинскую практику в связи с небольшими преимуществами перед уже используемыми вакцинами. В настоящее время одобренными к применению в России вакцинами против ВГЧ-1 и ВГЧ-2 являются препараты Витагерпавак и Герповакс, основанные на аттенуированных штаммах вирусов. Также существуют основанные на том же принципе вакцины против ВГЧ-3 под торговыми названиями Varivax (Varicella Virus Vaccine Live) и Zostavax (Merck&Co, Inc). В то же время инактивированных вакцин против вирусов герпеса человека 4-8 типов не существует.
При использовании препарата Varivax отмечена высокая частота развития лихорадки, для которой характерно повышение температуры от 37,7 до 39°С, а также возникновение сходной с симптомом вируса герпеса 3 типа сыпи. К наиболее частым побочным эффектам препарата Zostavax относятся диарея, повышение чувствительности к респираторным инфекциям, ринит, жар, головная боль и астения.
В силу данных ограничений и возможности развития побочных эффектов современные исследования сосредоточены на разработке вакцин на основе рекомбинантных белков. В частности, предлагается использовать для создания иммунного ответа полипептиды на фармацевтически приемлемом носителе (RU 2585961) и антигены (US 2017275339, US 2017216428).
Однако лечение герпесвирусных инфекций с использованием указанных инактивированных вакцин сопряжено с риском возникновения рецидива инфекции, возникновения энцефалита, пневмонии, гепатита и других осложнений. Так, вакцины Витагерпавак и Герповакс нельзя использовать на активной стадии герпеса, вакцинацию можно начинать не ранее чем через 2 недели после исчезновения внешних проявлений. Также эти вакцины нельзя использовать при любых острых заболеваниях, инфекционных и неинфекционных, тяжелых декомпенсированных соматических заболеваниях (инфаркт, гепатит, цирроз печени, нефрит), злокачественных опухолях любой локализации, беременности и аллергии на антибиотики аминогликозидного ряда.
На сегодняшний день не существует одобренных к применению на людях рекомбинантных вакцин, основанных на отдельных антигенных детерминантах вирусов, против различных вирусов герпеса человека. Все существующие кандидаты на роль таких вакцин находятся на разных этапах доклинических и клинических испытаний. В частности, известны следующие кандидатные вакцины против различных типов вирусов герпеса человека, в данный момент проходящие клинические испытания:
- GEN-003, субъединичная вакцина против ВГЧ-2 на основе вирусных белков ICP4 и gD2;
- CMV-MVA Triplex, ДНК-вакцина, кодирующая три антигена ВГЧ-5 (Nakamura R., NCT02506933);
- Shingrix, субъединичная вакцина против ВГЧ-3 компании GlaxoSmithKline. В данный момент находится на фазе III испытаний.
Однако сейчас нет ни одной вакцины, направленной на лечение сразу нескольких типов вирусов герпеса человека. Таким образом, становится актуальной задача разработки кандидатной вакцины нового поколения против вирусов герпеса человека. Решением этой задачи может быть разработка поливалентной ДНК-вакцины на основе эпитопов высокоиммуногенных вирусных белков, специфичных для вирусов герпеса человека всех типов, которая будет обеспечивать защиту от всех патогенных для человека типов вируса герпеса без риска возникновения осложнений.
Биоинформатический подход при дизайне ДНК-вакцин позволяет предсказать, какие МНС-лиганды образуются в результате презентации иммунными клетками какого-либо белка [Backert L., Kohlbacher О. Immunoinformatics and Epitope Prediction in the Age of Genomic Medicine // Genome Medicine, 2015, 7: 119]. Используя данный подход, была смоделирована белковая последовательность, которая представляет собой полипептид, специально собранный из иммуногенных эпитопов вирусов герпеса человека для эффективной презентации антигенов иммунопротеасомой на основе базы данных МНС-лигандов SYFPEITHI [Kloetzel P.M. Antigen Processing by the Proteasome // Nature Reviews. Molecular Cell Biology, 2001, 2(3): 179-87]. Предполагается, что презентированные эпитопы могут распознаваться Т-лимфоцитами, то есть экспрессирующийся в клетках человека белок по настоящему изобретению вызовет активацию специализированных Т-клеточных клонов, а именно цитотоксических Т-лимфоцитов, в результате чего образуется эффективный иммунный ответ [Patronov A., Doytchinova I.T-Cell Epitope Vaccine Design by Immunoinformatics // Open Biology, 2013, 3(1): 120139].
Задачей, на решение которой направлено настоящее изобретение, являлось создание вакцины на основе плазмидной ДНК, кодирующей полипептид, представляющий антигены всех восьми основных типов ВГЧ.
Задача решалась созданием плазмиды, несущей искусственную последовательность ДНК, кодирующую рекомбинантный слитый белок, схема которого приведена на Фиг. 1, содержащий антигенные детерминанты следующих вирусных белков: gpD и gpC ВГЧ-1 и ВГЧ-2; gpE ВГЧ-3; gp350 и LMP2 ВГЧ-4; димepagH/gL и пентамераgH/gL/UL128/UL130/UL131A ВГЧ-5; IEP1, рр85, gH и МСР ВГЧ-6; gp65 ВГЧ-7; LANA1, K12, gpB и K8.1 ВГЧ-8. При введении в организм заявленной плазмидной ДНК происходит экспрессия рекомбинантного слитого белка, активирующего гуморальное и клеточное звенья иммунитета.
Заявляемая последовательность была синтезирована методом ПНР с использованием перекрывающихся последовательностей олигонуклеотидов, предварительно синтезированных химическим путем с помощью синтезатора олигонуклеотидов. Синтетическая ДНК, кодирующая рекомбинантный слитый белок по настоящему изобретению, имеет последовательность SEQ ID NO: 2 с оптимизированным для синтеза в человеческих клетках кодонным составом.
Полученная ДНК была вставлена по сайтам рестрикции XhoI и HindIII в экспрессионный вектор pcDNA3.1. В результате был получен эукариотический экспрессионный вектор, схема которого приведена на Фиг. 2, содержащий последовательность SEQ ID NO: 2 и селективный маркер устойчивости к ампициллину.
При трансформации клеток E.coli BL21 [DE3] описанным вектором был получен штамм-продуцент плазмидной ДНК. Из биомассы штамма была выделена плазмидная ДНК, кодирующая рекомбинантный слитый белок по настоящему изобретению. Она представляет собой двуцепочечную кольцевую молекулу ДНК длиной 9509 п.н.
Изобретение иллюстрируется следующими графическими материалами:
На Фиг. 1 представлена аминокислотная последовательность рекомбинантного слитого белка, содержащего основные иммуногенные эпитопы ВГЧ 1-8 типов. Составные элементы белка (С-конец каждого элемента обозначен цифрой): 1 - эпитоп гликопротеина gD ВГЧ-1/2; 2 - эпитоп гликопротеина gC ВГЧ-1/2; 3 - эпитоп белка LANA1 ВГЧ-8; 4 - эпитоп белка рр85 ВГЧ-6; 5 - эпитоп белка МСР ВГЧ-6; 6 - эпитоп гликопротеина gp350 ВГЧ-4; 7 - эпитоп пентамерного гликопротеина gH/gL/UL128/UL130/UL131A ВГЧ-5; 8 - эпитоп белка МСР ВГЧ-6; 9 - эпитоп белка K8.1 ВГЧ-8; 10 - эпитоп белка МСР ВГЧ-6; 11 - эпитоп белка K8.1 ВГЧ-8; 12 - эпитоп гликопротеина gpE ВГЧ-3; 13 - эпитоп белка МСР ВГЧ-6; 14 - эпитоп гликопротеина gpB ВГЧ-8; 15 - эпитоп гликопротеина gpEBГЧ-3; 16 - эпитоп белка LANA1 ВГЧ-8; 17 - эпитоп гликопротеина gH/gLBГЧ-5;18 - эпитоп гликопротеина gH/gLBГЧ-5; 19 - эпитоп гликопротеина gpHBГЧ-6; 20 - эпитоп гликопротеина gpBBГЧ-8; 21 - эпитоп гликопротеина gp65BГЧ-7; 22 -эпитоп гликопротеина gpHBГЧ-6; 23 - эпитоп белка Iep1 ВГЧ-6; 24 - эпитоп белка K12 ВГЧ-8; 25 - эпитоп белка LANA1 ВГЧ-8; 26 - эпитоп белка LМР2 ВГЧ-4; 27 - эпитоп гликопротеина Gp350BГЧ-4.
На Фиг. 2 представлена карта плазмидной ДНК, кодирующей рекомбинантный слитый белок по настоящему изобретению.
На Фиг. 3 представлена электрофореграмма фрагментов ДНК. 1 - положительный контроль, ПНР на плазмидной ДНК; 2 - результат ПЦР с РНК, выделенной из трансфицированных клеток; 3 - отрицательный контроль; М - маркер длины ДНК GeneRuler 100 bp DNA Ladder
Сущность и промышленная применимость изобретения поясняется следующими примерами.
Пример 1. Дизайн нуклеотидной последовательности гена, кодирующего рекомбинантный слитый белок.
Последовательность рекомбинантного слитого белка по настоящему изобретению была собрана из иммуногенных эпитопов вирусов герпеса человека 3-8 типов специально для эффективной презентации антигенов иммунопротеасомой на основе базы данных МНС-лигандов SYFPEITHI в программе EpiToolKit 2.0. К этой последовательности с N-конца были присоединены эпитопы гликопротеинов gD и gC ВГЧ-1/2 через последовательности 2А-пептидов. Схема сборки рекомбинантного слитого белка приведена на Фиг. 1, его аминокислотная последовательность представлена в SEQ ID NO: 1
После моделирования последовательность аминокислотных остатков рекомбинантного слитого белка переведена в нуклеотидную последовательность в программе UniproUGENEc учетом кодонной оптимизации для экспрессии в клетках человека. Данная кодирующая последовательность, представленная в SEQ ID NO: 2, была синтезирована методом ПЦР с использованием перекрывающихся последовательностей олигонуклеотидов, предварительно синтезированных химическим путем с помощью синтезатора нуклеиновых кислот.
Пример 2. Получение плазмидной ДНК, кодирующей рекомбинантный слитый белок.
Искусственный ген, полученный в примере 1, вводили в плазмидный экспрессионный вектор pDNA3.1 для последующей экспрессии в эукариотических клетках. Для этого провели реакцию рестрикции по сайтам XhoI и HindIII с последующим лигированием в вектор при +20°С в течение 2 часов. Карта полученного экспрессионного вектора представлена на Фиг. 2.
Пример 3. Конструирование прокариотического штамма для амплификации плазмидной ДНК.
Плазмидной ДНК, полученной в примере 2, трансформировали клетки Escherichia coli штамма DH10B методом электропорации с использованием устройства Bio-Rad Gene Pulser. После трансформации клетки инкубировали в SOC-среде (2% бакто-триптон, 0,5% дрожжевой экстракт, 10 мМ NaCl, 2,5 мМ KСl, 10 мМ MgCl2, 10 мМ MgSO4, 20 мМ глюкоза) в течение 40 мин при +37°С.
Бактериальные колонии, содержащие вектор, отбирали по признаку устойчивости к антибиотику ампициллину на селективной питательной среде (LB-агар, 50 мг/мл антибиотика). Выросшие на такой среде колонии проверяли на наличие гена интереса.
Из выросших клонов выделяли плазмидную ДНК щелочным методом с использованием ацетата аммония высокой концентрации. Раствор, содержащий целевую плазмидную ДНК, отделяли от осадка центрифугированием, после чего ДНК-конструкцию осаждали из супернатанта добавлением изопропилового спирта.
Полученный осадок растворяли в физиологическом растворе и обрабатывали рибонуклеазой А для гидролиза молекул РНК. Целевую ДНК-конструкцию затем переосаждали из раствора с помощью смеси 96% этанола и 10 М ацетата аммония в соотношении 3:1. Осадок перерастворяли в физиологическом растворе. Полученный таким образом раствор плазмидной ДНК затем очищали с помощью хроматографических методов.
Пример 4. Определение специфической активности плазмидной ДНК. Активным компонентом разрабатываемой ДНК-вакцины против вируса герпеса человека является плазмида, несущая ген, кодирующий рекомбинантный слитый белок. После введения в организм плазмидная ДНК неспецифически проникает в клетки, находящиеся в месте инъекции (миоциты, дендритные клетки), а затем транспортируется в ядро, где происходит экспрессия целевого гена. Соответствующий ему белок презентируется на молекулах главного комплекса гистосовместимости (HLA) как I, так и II класса антигенпрезентирующих клеток (АПК) наивным Т-лимфоцитам. Данное взаимодействие обеспечивает запуск как клеточного, так и гуморального иммунного ответа.
В качестве показателя специфической активности ДНК-вакцины против вируса герпеса человека использовалась способность плазмидной ДНК, являющейся действующим веществом ДНК-вакцины, проникать в клетки млекопитающих и запускать экспрессию целевого гена. Экспрессия целевого гена оценивалась на уровне синтеза молекулы РНК.
Тест на определение специфической активности включает следующие этапы:
1) трансфекция клеток млекопитающих плазмидной ДНК, полученной в Примере 3;
2) выделение общей РНК из трансфицированных клеток;
3) синтез молекулы кДНК с помощью реакции обратной транскрипции;
4) проведение полимеразной цепной реакции, амплификация фрагмента кДНК с использованием праймеров, специфичных к участку последовательности гена, кодирующего заявленный рекомбинантный слитый белок;
5) анализ результатов амплификации с помощью горизонтального электрофореза в агарозном геле.
По результатам проведенного анализа в геле должна быть видна полоса, соответствующая участку гена размером 513 п. н.
Исследование проводили на линии клеток 293 (клетки почки эмбриона человека, трансфицированные ДНК аденовируса 5 типа).
Предварительно размороженные и стабильные в культуре клетки линии 293 трансфицировали плазмидной ДНК, полученной в примере 3, в концентрации 12,5 мкг на 2⋅106 клеток. Трансфекцию проводили на монослое клеток кальций-фосфатным методом. Через 48 ч после трансфекции, клетки снимали с пластика, выделяли РНК с помощью реагента TRI Reagent® (Sigma Aldrich) в соответствии с протоколом производителя. Выделенную РНК обрабатывали ДНКазой (Sigma) для удаления примесей остаточной плазмидной ДНК и исключения ложноположительного результата исследования, проводили контроль качества РНК с помощью горизонтального элекрофореза в агарозном геле.
Синтезировали молекулы кДНК в реакции обратной транскрипции с использованием полученной РНК в качестве матрицы. Синтез молекулы кДНК с помощью реакции обратной транскрипции проводили с помощью набора Thermo Scientific RevertAid Н Minus First Strand cDNA Synthesis Kit (ThermoFisher Scientific) в соответствии с протоколом, рекомендуемым производителем. В качестве праймеров были использованы случайные гексамерные праймеры, входящие в состав набора. Полученные в ходе реакции обратной транскрипции кДНК амплифицировали с помощью полимеразной цепной реакции с праймерами, специфическими к участку гена, кодирующего рекомбинантный слитый белок по настоящему изобретению, с последующим горизонтальным электрофорезом в агарозном геле.
Полученный результат анализировали с помощью горизонтального электрофореза в агарозном геле. Результаты исследования представлены на Фиг. 3. После проведения трансфекции, выделения РНК, реакции обратной транскрипции и ПЦР с использованием праймеров, специфичных к участку гена, кодирующего рекомбинантный слитый белок по настоящему изобретению, на электрофореграмме детектируется полоса с ожидаемым размером 513 п.н. Наличие данной полосы свидетельствует о проникновении плазмидной ДНК в клетки и запуске процесса транскрипции с образованием молекулы РНК целевого гена, служащей матрицей для последующего синтеза белковой молекулы.
Пример 5. Исследование механизма действия in vivo.
Исследования специфической активности проводили на самках мышей линии Balb/c массой 18-22 г, возраст 3 месяца (ФГУП ПЛЖ Рапполово). Группу отрицательного контроля не подвергали иммунизации. Исследуемую группу иммунизировали внутримышечно плазмидной ДНК в дозе 6250 мкг/кг (125 мкг или 500 мкл на животное). Иммунизации проводили двукратно с интервалом 14 дней. Через две недели после последней иммунизации животные были умерщвлены методом декапитации с тотальным забором крови и выделением спленоцитов животных исследованных групп.
Пример 5.1. Исследование гуморального иммунного ответа проводили с помощью иммуноферментного анализа. Выявляли антитела, специфичные к пептидам рекомбинантного слитого белка по настоящему изобретению, в сыворотке крови иммунизированных мышей. Присутствие антител свидетельствует о развитии специфического иммунного ответа и подтверждает синтез белка с мРНК.
Исследовали синтез антител, специфичных к пептидам рекомбинантного слитого белка по настоящему изобретению, с помощью твердофазного иммуноферментного анализа. В качестве антигена, сорбированного на пластик использовали пул из 3 пептидов из рекомбинантного слитого белка по настоящему изобретению.
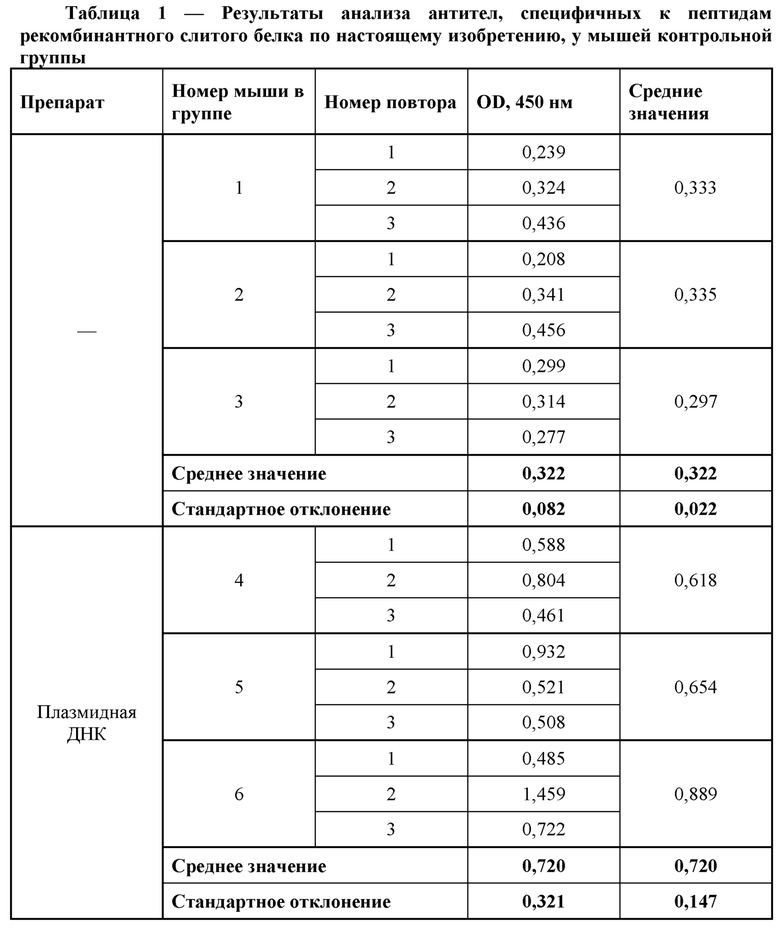
Результаты анализа антител, специфичных к пептидам рекомбинантного слитого белка по настоящему изобретению, обнаруженных в сыворотке крови мышей, иммунизированных плазмидной ДНК, и мышей контрольной группы представлены в Таблице 1.
Уровень антител к специфическим пептидам герпесвирусов у иммунизированных животных невысок, однако почти в два раза выше, чем в контрольной группе. Таким образом, подтвержден предполагаемый механизм действия плазмидной ДНК против вируса герпеса человека, заключающийся в проникновении плазмидных ДНК в клетки, экспрессии закодированных в них генов и запуске специфического иммунного ответа у исследованных животных.
Пример 5.2. Индукцию специфического клеточного иммунного ответа оценивали с помощью окрашивания поверхностных маркеров CD3, CD4, CD8 и внутриклеточного окрашивания цитокинов ИФН-гамма и ФНО-альфа после in vitro стимуляции спленоцитов пептидами из рекомбинантного слитого белка по настоящему изобретению.
Методика внутриклеточного окрашивания цитокинов основана на способности CD4 и CD8 Т-клеток синтезировать цитокины в ответ на стимуляцию «знакомым» антигеном. Данный метод включает в себя стимуляцию выделенных спленоцитов специфическими антигенами, выявление с помощью антител поверхностных клеточных маркеров, фиксацию клеток и пермеабилизацию клеточных мембран с последующим внутриклеточным окрашиванием цитокинов антителами.
При исследовании специфической активности из селезенок мышей, иммунизированных плазмидной ДНК, и мышей контрольной группы выделяли спленоциты методом лизиса эритроцитов 0,86% хлоридом аммония.
Специфическая стимуляция спленоцитов проводилась пулом, содержащим 3 пептида из рекомбинантного слитого белка по настоящему изобретению. Стимуляция проводилась в присутствии брефелдина A (Sigma) и монензина (Sigma). Брефельдин и монензин необходимы для остановки работы аппарата Гольджи, вследствие чего синтезируемые цитокины не выделяются из клетки.
Для окрашивания цитокинов и клеточных маркеров использовали следующие антитела (eBioscience):
- anti -mouse CD3e-biotin,
- anti-mouse CD4 (L3T4)-phycoerythrin-Cy5 (PE-Cy5),
- anti-mouse CD8a- phycoerythrin-Cy5 (PE-Cy5),
- anti-mouse IFNγ - fluorescein isothiocyanate (FITC),
- anti-mouse TNFα - Phycoerythrin (PE).
В случае биотинилированных антител в качестве хромогенного субстрата использовали стрептавидин, конъюгированный с Phycoerythrin-PE-TexasRed. Анализ внутриклеточного окрашивания цитокинов проводили на проточном цитофлуориметре EPICS XL (Beckman Coulter).
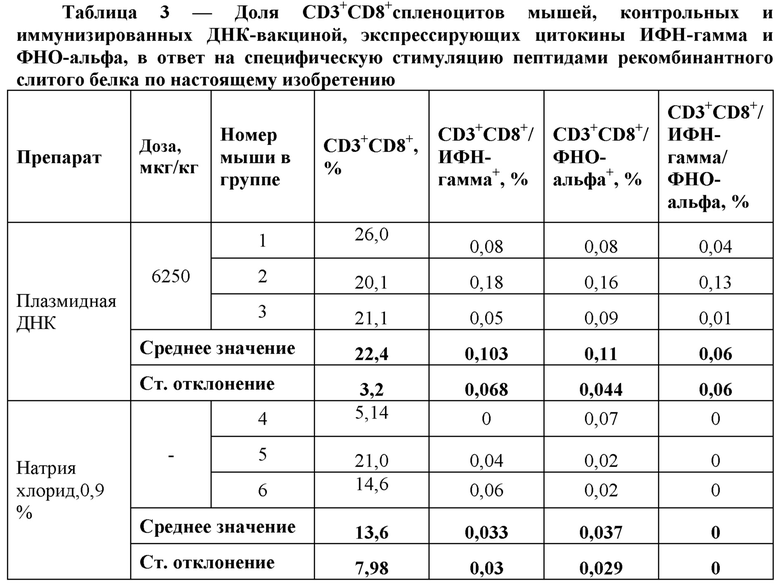
Результаты окрашивания поверхностных маркеров CD3, CD4, CD8 и внутриклеточного окрашивания цитокинов ИФН-гамма и ФНО-альфа после in vitro стимуляции спленоцитов пептидами рекомбинантного слитого белка по настоящему изобретению представлены в Таблицах 2 и 3.
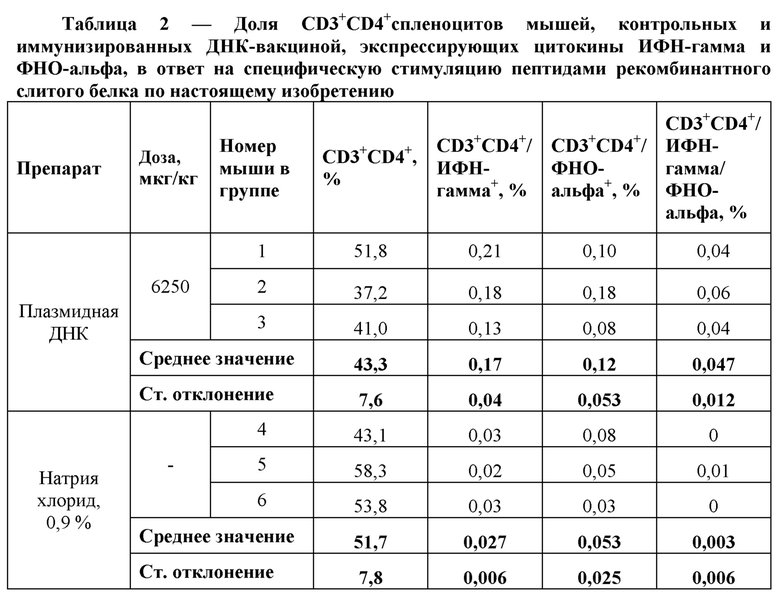
Анализ полученных данных свидетельствует о повышении синтеза цитокинов ИФН-гамма и ФНО-альфа CD3/CD4 и CD3/CD8 спленоцитами, выделенными из селезенок мышей, иммунизированных ДНК-вакциной, по сравнению с контрольной группой животных.
Таким образом, показано, что плазмидная ДНК по настоящему изобретению проникает в клетки линии 293, с нее транскрибируются молекулы РНК целевого гена, служащие матрицей для последующего синтеза белковой молекулы. У иммунизированных животных детектируется повышение оптической плотности относительно показателей контрольной группы в ИФА, что свидетельствует о развитии гуморального иммунного ответа. Механизм действия ДНК-вакцины против вируса герпеса человека заключается в проникновении плазмидных ДНК в клетки, экспрессии закодированных в них генов и запуске специфического иммунного и гуморального клеточного ответа.
--->
Перечень последовательностей
<110> Федеральное государственное унитарное предприятие
«Государственный научно-исследовательский институт особо чистых
биопрепаратов» Федерального медико-биологического агентства
Federal State Unitary Enterprise «State Research Institute of Highly Pure
Biopreparations» of the Federal Medical and Biological Agency
<120> Вакцина против герпеса
<130>
<160> 2
<210> 1
<211> 1383
<212> белок
<213> Искусственная последовательность
<220>
<222> (1)…(1383)
<223> Последовательность аминокислот рекомбинантного слитого
белка, содержащего иммуногенные эпитопы ВГЧ 8 основных типов.
<400> 1
Met Gly Val Lys Val Leu Phe Ala Leu Ile Cys Ile Ala Val Ala Glu
1 5 10 15
Ala Gly Cys Cys Ala Cys Cys Ala Thr Gly Gly Gly Cys Gly Thr Gly
20 25 30
Ala Ala Gly Gly Thr Gly Cys Thr Gly Thr Thr Cys Gly Cys Cys Cys
35 40 45
Thr Gly Ala Thr Cys Thr Gly Cys Ala Thr Thr Gly Cys Cys Gly Thr
50 55 60
Gly Gly Cys Cys Gly Ala Ala Gly Cys Cys Lys Tyr Ala Leu Ala Asp
65 70 75 80
Pro Ser Leu Lys Met Ala Asp Pro Asn Arg Phe Arg Gly Lys Asn Leu
85 90 95
Pro Val Leu Asp Arg Leu Thr Asp Pro Pro Gly Val Lys Arg Val Tyr
100 105 110
His Ile Gln Pro Ser Leu Glu Asp Pro Phe Gln Pro Pro Ser Ile Pro
115 120 125
Ile Thr Val Tyr Tyr Ala Val Leu Glu Arg Ala Cys Arg Ser Val Leu
130 135 140
Leu His Ala Pro Ser Glu Ala Pro Gln Ile Val Arg Gly Ala Ser Asp
145 150 155 160
Glu Ala Arg Lys His Thr Tyr Asn Leu Thr Ile Ala Trp Tyr Arg Met
165 170 175
Gly Asp Asn Cys Ala Ile Pro Ile Thr Val Met Glu Tyr Thr Glu Cys
180 185 190
Pro Tyr Asn Lys Ser Leu Gly Val Cys Pro Ile Arg Thr Gln Pro Arg
195 200 205
Trp Ser Tyr Tyr Asp Ser Phe Ser Ala Val Ser Glu Asp Asn Leu Gly
210 215 220
Phe Leu Met His Ala Pro Ala Phe Glu Thr Ala Gly Thr Tyr Leu Arg
225 230 235 240
Leu Val Lys Ile Asn Asp Trp Thr Glu Ile Thr Gln Phe Ile Leu Glu
245 250 255
His Arg Ala Arg Ala Ser Cys Lys Tyr Ala Leu Pro Leu Arg Ile Pro
260 265 270
Pro Ala Ala Cys Leu Thr Ser Lys Ala Tyr Gln Gln Gly Val Thr Val
275 280 285
Asp Ser Ile Gly Met Leu Pro Arg Phe Ile Pro Glu Asn Gln Arg Thr
290 295 300
Val Ala Leu Tyr Ser Leu Lys Ile Ala Gly Trp His Gly Pro Lys Pro
305 310 315 320
Pro Tyr Thr Ser Thr Leu Leu Pro Pro Glu Leu Ser Asp Thr Thr Asn
325 330 335
Ala Thr Gln Pro Glu Leu Val Pro Glu Asp Pro Glu Asp Ser Ala Leu
340 345 350
Leu Glu Asp Pro Ala Gly Thr Val Ser Ser Gln Ile Pro Pro Asn Trp
355 360 365
His Ile Pro Ser Ile Gln Asp Val Ala Pro His His Ala Pro Ala Ala
370 375 380
Pro Ser Asn Pro Gly Ser Gly Ala Thr Asn Phe Ser Leu Leu Lys Gln
385 390 395 400
Ala Gly Asp Val Glu Glu Asn Pro Gly Pro Ser Pro Gly Arg Thr Ile
405 410 415
Thr Val Gly Pro Arg Gly Asn Ala Ser Asn Ala Ala Pro Ser Val Pro
420 425 430
Arg Asn Arg Ser Ala Pro Arg Thr Thr Pro Thr Pro Pro Gln Pro Arg
435 440 445
Lys Ala Thr Lys Ser Lys Ala Ser Thr Ala Lys Pro Ala Pro Pro Pro
450 455 460
Lys Thr Gly Pro Pro Lys Thr Ser Ser Glu Pro Val Arg Cys Asn Arg
465 470 475 480
His Asp Pro Leu Ala Arg Tyr Gly Ser Arg Val Gln Ile Arg Cys Arg
485 490 495
Phe Pro Asn Ser Thr Arg Thr Glu Ser Arg Leu Gln Ile Trp Arg Tyr
500 505 510
Ala Thr Ala Thr Asp Ala Glu Ile Gly Thr Ala Pro Ser Leu Glu Glu
515 520 525
Val Met Val Asn Val Ser Ala Pro Pro Gly Gly Gln Leu Val Tyr Asp
530 535 540
Ser Ala Pro Asn Arg Thr Asp Pro His Val Ile Trp Ala Glu Gly Ala
545 550 555 560
Gly Pro Gly Ala Ser Pro Arg Leu Tyr Ser Val Val Gly Pro Leu Gly
565 570 575
Arg Gln Arg Leu Ile Ile Glu Glu Leu Thr Leu Glu Thr Gln Gly Met
580 585 590
Tyr Tyr Trp Val Trp Gly Arg Thr Asp Arg Pro Ser Ala Tyr Gly Thr
595 600 605
Trp Val Arg Val Arg Val Phe Arg Pro Pro Ser Leu Thr Ile His Pro
610 615 620
His Ala Val Leu Glu Gly Gln Pro Phe Lys Ala Thr Cys Thr Ala Ala
625 630 635 640
Thr Tyr Tyr Pro Gly Asn Arg Ala Glu Phe Val Trp Phe Glu Asp Gly
645 650 655
Arg Arg Val Phe Asp Pro Ala Gln Ile His Thr Gln Thr Gln Glu Asn
660 665 670
Pro Asp Gly Phe Ser Thr Val Ser Thr Val Thr Ser Ala Ala Val Gly
675 680 685
Gly Gln Gly Pro Pro Arg Thr Phe Thr Cys Gln Leu Thr Trp His Arg
690 695 700
Asp Ser Val Ser Phe Ser Arg Arg Asn Ala Ser Gly Thr Ala Ser Val
705 710 715 720
Leu Pro Arg Pro Thr Ile Thr Met Glu Phe Thr Gly Asp His Ala Val
725 730 735
Cys Thr Ala Gly Cys Val Pro Glu Gly Val Thr Phe Ala Trp Phe Leu
740 745 750
Gly Asp Asp Ser Ser Pro Ala Glu Lys Val Ala Val Ala Ser Gln Thr
755 760 765
Ser Cys Gly Arg Pro Gly Thr Ala Thr Ile Arg Ser Thr Leu Pro Val
770 775 780
Ser Tyr Glu Gln Thr Glu Tyr Ile Cys Arg Leu Ala Gly Tyr Pro Asp
785 790 795 800
Gly Ile Pro Val Leu Glu His His Gly Ser His Gln Pro Pro Pro Arg
805 810 815
Asp Pro Thr Glu Arg Gln Val Ile Arg Ala Val Glu Gly Gly Ser Gly
820 825 830
Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro
835 840 845
Gly Pro Cys Cys Leu Cys Leu Thr Val Phe Leu Asp Gly Gly Asp Gly
850 855 860
Asn Lys Thr Leu Arg Asp Asn Ser Tyr Met Pro Leu Ile Ala Leu Ser
865 870 875 880
Leu His Glu Asn Gly Ala Asp Asn Lys Val Trp Ile Arg His His Val
885 890 895
Gly Ile Glu Lys Pro Asn Pro Ser Glu Gly Glu Ala Ala Asp Asn Lys
900 905 910
Val Trp Gly Ser Ala Lys Pro Gly Asn Gly Ser Tyr Phe Ala Ser Val
915 920 925
Lys Thr Glu Met Leu Gly Asn Glu Ile Asp Ala Asp Asn Val His Leu
930 935 940
Val Tyr Ile Leu Ser Lys Gln Asn Gln Gln His Leu Ile Pro Gln Trp
945 950 955 960
Ala Leu Arg Gln Ile Ala Asp Phe Ala Asp Asn Lys Val Trp Asp Pro
965 970 975
Ser Arg Tyr Asn Ile Ser Phe Glu Ala Leu Leu Gly Ile Tyr Ser Ala
980 985 990
Asp Asn Val Thr Trp Arg Leu Ala Ala Gly Ser Pro Ser Ser Ala Asp
995 1000 1005
Asn Val His Leu Phe Lys Ser Leu Ile Tyr Ile Asn Glu Asn Thr Lys
1010 1015 1020
Ile Leu Glu Val Glu Ala Asp Asn Val Thr Trp Ala Leu Ile Ser Ala
1025 1030 1035 1040
Phe Ser Gly Ser Ala Asp Asn Val Thr Trp Ser Ala Gln Glu Asp Leu
1045 1050 1055
Gly Asp Asp Thr Gly Ile His Val Ile Ala Asp His Asn Lys Val Thr
1060 1065 1070
Val Trp Ser Leu Leu Ser Ile Met Thr Leu Ala Ala Met His Ser Lys
1075 1080 1085
Leu Ser Pro Ala Asp Asn Val Thr Trp Met Leu Met Ile Ile Ile Val
1090 1095 1100
Ile Ala Ile Ala Asp Asn Val Thr Trp Phe His Met Trp Asn Tyr His
1105 1110 1115 1120
Ser His Val Phe Ser Val Gly Asp Ala Asp Asn Val Thr Trp Pro Val
1125 1130 1135
Val Ser Thr His Glu Gln Ile Ala Asp Asn Val Thr Trp Phe Ala Leu
1140 1145 1150
Asp Pro Tyr Asn Glu Ala Asp Asn Lys Val Trp Ala Lys Arg Ala Leu
1155 1160 1165
Trp Thr Pro Asp Gln Ile Thr Asp Ile Thr Ser Leu Arg Asp Asn Lys
1170 1175 1180
Thr Leu Val Asp Glu Glu Tyr Arg Phe Ile Ser Asp Ala Thr Phe Val
1185 1190 1195 1200
Asp Glu Thr Ala Asp Asn Lys Val Trp Ser Ser Met Lys Val Asn Val
1205 1210 1215
Asn Gly Val Ala Asp Asn Lys Val Trp Thr Thr Tyr Ala Trp Arg Trp
1220 1225 1230
Ser Ala Asp Asn Lys Val Trp Gly Ile Tyr Tyr Ile Arg Val Val Glu
1235 1240 1245
Val Arg Gln Met Gln Tyr Asp Asn Ala Asp Asn Lys Cys Leu Ala Met
1250 1255 1260
Leu Val Leu Leu Ala Glu Ile Ala Asp Glu Gly Asn Lys Val Trp Cys
1265 1270 1275 1280
Ile Gln Ser Ile Gly Ala Ser Val Ala Asp Asn Lys Val Trp Trp Arg
1285 1290 1295
Leu Gly Ala Ile Pro Pro Leu Val Ala Asp Asn Lys Cys Leu Pro Glu
1300 1305 1310
Ser Ser Gln Arg Pro Pro Leu Ala Asp Asn Val His Leu Thr Tyr Gly
1315 1320 1325
Pro Val Phe Met Cys Leu Gly Gly Leu Leu Thr Met Val Ala Gly Ala
1330 1335 1340
Asp Asn Val His Leu Asp Asp Arg Thr Thr Leu Gln Leu Ala Gln Asn
1345 1350 1355 1360
Pro Val Tyr Ile Pro Glu Thr Tyr Pro Tyr Ile Lys Trp Asp Asn Ser
1365 1370 1375
Ser Trp Asp Phe Ile Thr Val
1380 1383
<210> 1
<211> 4155
<212> ДНК
<213> Искусственная последовательность
<220>
<222> (1)…(4155)
<223> Последовательность нуклеотидов ДНК, кодирующей
рекомбинантный слитый белок, содержащий иммуногенные эпитопы ВГЧ 8
основных типов.
<400> 1
ATG GGA GTG AAA GTA CTC TTT GCC TTG ATT TGC ATT GCT GTT GCT GAA 48
Met Gly Val Lys Val Leu Phe Ala Leu Ile Cys Ile Ala Val Ala Glu
1 5 10 15
GCG GGC TGC TGC GCC TGT TGT GCA ACA GGC GGG GGC TGC GGG ACA GGG 96
Ala Gly Cys Cys Ala Cys Cys Ala Thr Gly Gly Gly Cys Gly Thr Gly
20 25 30
GCC GCA GGA GGG ACC GGC TGT ACT GGC ACC ACC TGT GGG TGT TGC TGC 144
Ala Ala Gly Gly Thr Gly Cys Thr Gly Thr Thr Cys Gly Cys Cys Cys
35 40 45
ACG GGT GCT ACT TGC ACT GGT TGT GCT ACA ACT GGG TGC TGT GGC ACA 192
Thr Gly Ala Thr Cys Thr Gly Cys Ala Thr Thr Gly Cys Cys Gly Thr
50 55 60
GGT GGA TGC TGC GGC GCA GCA GGC TGC TGT AAA TAT GCT CTG GCG GAC 240
Gly Gly Cys Cys Gly Ala Ala Gly Cys Cys Lys Tyr Ala Leu Ala Asp
65 70 75 80
CCA TCT CTG AAG ATG GCC GAC CCC AAC CGC TTC CGC GGC AAG AAC CTC 288
Pro Ser Leu Lys Met Ala Asp Pro Asn Arg Phe Arg Gly Lys Asn Leu
85 90 95
CCT GTC TTG GAC AGG CTG ACA GAT CCA CCA GGG GTG AAG AGG GTG TAC 336
Pro Val Leu Asp Arg Leu Thr Asp Pro Pro Gly Val Lys Arg Val Tyr
100 105 110
CAC ATT CAG CCT TCC TTG GAG GAC CCC TTC CAG CCG CCA TCT ATT CCC 384
His Ile Gln Pro Ser Leu Glu Asp Pro Phe Gln Pro Pro Ser Ile Pro
115 120 125
ATC ACT GTC TAC TAT GCG GTG CTT GAG CGG GCC TGC AGG AGC GTG CTG 432
Ile Thr Val Tyr Tyr Ala Val Leu Glu Arg Ala Cys Arg Ser Val Leu
130 135 140
CTC CAT GCT CCA AGT GAA GCA CCT CAG ATC GTG AGG GGG GCT TCA GAT 480
Leu His Ala Pro Ser Glu Ala Pro Gln Ile Val Arg Gly Ala Ser Asp
145 150 155 160
GAG GCC CGG AAG CAC ACC TAC AAC CTG ACC ATT GCC TGG TAC AGG ATG 528
Glu Ala Arg Lys His Thr Tyr Asn Leu Thr Ile Ala Trp Tyr Arg Met
165 170 175
GGC GAC AAC TGT GCC ATT CCT ATT ACA GTG ATG GAA TAT ACT GAA TGT 576
Gly Asp Asn Cys Ala Ile Pro Ile Thr Val Met Glu Tyr Thr Glu Cys
180 185 190
CCC TAT AAT AAG AGC CTG GGG GTC TGC CCC ATT CGG ACG CAG CCT CGC 624
Pro Tyr Asn Lys Ser Leu Gly Val Cys Pro Ile Arg Thr Gln Pro Arg
195 200 205
TGG TCC TAC TAC GAC AGC TTC TCC GCT GTC TCA GAA GAC AAC TTG GGC 672
Trp Ser Tyr Tyr Asp Ser Phe Ser Ala Val Ser Glu Asp Asn Leu Gly
210 215 220
TTT CTG ATG CAT GCC CCA GCT TTT GAA ACA GCA GGT ACA TAC CTG CGG 720
Phe Leu Met His Ala Pro Ala Phe Glu Thr Ala Gly Thr Tyr Leu Arg
225 230 235 240
CTG GTA AAG ATC AAT GAC TGG ACA GAA ATC ACC CAG TTC ATT CTG GAA 768
Leu Val Lys Ile Asn Asp Trp Thr Glu Ile Thr Gln Phe Ile Leu Glu
245 250 255
CAC CGC GCC AGG GCC AGC TGC AAG TAC GCC CTC CCA CTG CGC ATC CCG 816
His Arg Ala Arg Ala Ser Cys Lys Tyr Ala Leu Pro Leu Arg Ile Pro
260 265 270
CCG GCC GCC TGC TTG ACA AGC AAA GCC TAC CAG CAA GGA GTC ACA GTG 864
Pro Ala Ala Cys Leu Thr Ser Lys Ala Tyr Gln Gln Gly Val Thr Val
275 280 285
GAT TCC ATT GGG ATG CTT CCC CGG TTC ATC CCA GAG AAC CAG AGA ACT 912
Asp Ser Ile Gly Met Leu Pro Arg Phe Ile Pro Glu Asn Gln Arg Thr
290 295 300
GTT GCC CTG TAC AGC TTG AAA ATT GCT GGC TGG CAT GGC CCC AAA CCT 960
Val Ala Leu Tyr Ser Leu Lys Ile Ala Gly Trp His Gly Pro Lys Pro
305 310 315 320
CCC TAC ACC TCC ACC CTC CTT CCT CCA GAA CTG AGT GAC ACA ACC AAC 1008
Pro Tyr Thr Ser Thr Leu Leu Pro Pro Glu Leu Ser Asp Thr Thr Asn
325 330 335
GCC ACC CAG CCC GAG CTG GTC CCT GAG GAC CCA GAA GAT TCT GCG CTG 1056
Ala Thr Gln Pro Glu Leu Val Pro Glu Asp Pro Glu Asp Ser Ala Leu
340 345 350
TTG GAA GAT CCT GCC GGA ACA GTT TCC AGC CAA ATC CCC CCT AAC TGG 1104
Leu Glu Asp Pro Ala Gly Thr Val Ser Ser Gln Ile Pro Pro Asn Trp
355 360 365
CAC ATC CCT AGC ATC CAG GAT GTG GCC CCT CAT CAC GCG CCC GCT GCA 1152
His Ile Pro Ser Ile Gln Asp Val Ala Pro His His Ala Pro Ala Ala
370 375 380
CCA AGC AAT CCT GGC AGT GGT GCC ACA AAT TTT TCT TTG CTG AAA CAG 1200
Pro Ser Asn Pro Gly Ser Gly Ala Thr Asn Phe Ser Leu Leu Lys Gln
385 390 395 400
GCC GGG GAC GTG GAG GAG AAC CCC GGG CCA TCC CCA GGC CGC ACA ATC 1248
Ala Gly Asp Val Glu Glu Asn Pro Gly Pro Ser Pro Gly Arg Thr Ile
405 410 415
ACG GTG GGT CCT AGA GGC AAT GCA AGC AAC GCA GCT CCC AGT GTG CCA 1296
Thr Val Gly Pro Arg Gly Asn Ala Ser Asn Ala Ala Pro Ser Val Pro
420 425 430
AGA AAC AGA AGC GCC CCG AGG ACC ACT CCC ACT CCA CCC CAG CCA AGG 1344
Arg Asn Arg Ser Ala Pro Arg Thr Thr Pro Thr Pro Pro Gln Pro Arg
435 440 445
AAG GCC ACC AAG TCC AAG GCT AGC ACA GCC AAG CCC GCC CCC CCA CCC 1392
Lys Ala Thr Lys Ser Lys Ala Ser Thr Ala Lys Pro Ala Pro Pro Pro
450 455 460
AAG ACC GGC CCT CCA AAA ACA TCT TCT GAG CCC GTC AGG TGC AAC AGG 1440
Lys Thr Gly Pro Pro Lys Thr Ser Ser Glu Pro Val Arg Cys Asn Arg
465 470 475 480
CAT GAC CCT CTG GCC CGC TAC GGC TCT CGG GTG CAG ATC CGC TGC CGC 1488
His Asp Pro Leu Ala Arg Tyr Gly Ser Arg Val Gln Ile Arg Cys Arg
485 490 495
TTT CCA AAC AGC ACC AGG ACG GAG TCC AGG CTC CAG ATA TGG AGA TAT 1536
Phe Pro Asn Ser Thr Arg Thr Glu Ser Arg Leu Gln Ile Trp Arg Tyr
500 505 510
GCC ACT GCC ACG GAT GCC GAG ATC GGC ACT GCA CCC TCT CTT GAA GAG 1584
Ala Thr Ala Thr Asp Ala Glu Ile Gly Thr Ala Pro Ser Leu Glu Glu
515 520 525
GTC ATG GTC AAC GTG TCT GCT CCT CCT GGA GGA CAG CTT GTC TAT GAC 1632
Val Met Val Asn Val Ser Ala Pro Pro Gly Gly Gln Leu Val Tyr Asp
530 535 540
TCC GCC CCC AAC CGG ACT GAT CCC CAC GTG ATC TGG GCC GAA GGA GCA 1680
Ser Ala Pro Asn Arg Thr Asp Pro His Val Ile Trp Ala Glu Gly Ala
545 550 555 560
GGG CCC GGC GCC TCC CCC AGA CTC TAC TCA GTG GTT GGT CCC CTG GGC 1728
Gly Pro Gly Ala Ser Pro Arg Leu Tyr Ser Val Val Gly Pro Leu Gly
565 570 575
CGG CAG CGC CTG ATC ATC GAA GAA CTC ACG CTG GAA ACT CAA GGC ATG 1776
Arg Gln Arg Leu Ile Ile Glu Glu Leu Thr Leu Glu Thr Gln Gly Met
580 585 590
TAC TAC TGG GTG TGG GGA AGA ACA GAC AGA CCT TCT GCC TAT GGA ACT 1824
Tyr Tyr Trp Val Trp Gly Arg Thr Asp Arg Pro Ser Ala Tyr Gly Thr
595 600 605
TGG GTC CGA GTG CGG GTC TTC CGG CCC CCC AGC CTT ACC ATC CAC CCC 1872
Trp Val Arg Val Arg Val Phe Arg Pro Pro Ser Leu Thr Ile His Pro
610 615 620
CAT GCA GTC CTG GAG GGG CAG CCC TTT AAA GCA ACC TGC ACC GCG GCC 1920
His Ala Val Leu Glu Gly Gln Pro Phe Lys Ala Thr Cys Thr Ala Ala
625 630 635 640
ACC TAT TAT CCT GGG AAC CGA GCA GAA TTT GTG TGG TTC GAG GAT GGT 1968
Thr Tyr Tyr Pro Gly Asn Arg Ala Glu Phe Val Trp Phe Glu Asp Gly
645 650 655
AGG AGA GTG TTT GAT CCA GCC CAG ATT CAT ACT CAG ACA CAG GAG AAC 2016
Arg Arg Val Phe Asp Pro Ala Gln Ile His Thr Gln Thr Gln Glu Asn
660 665 670
CCT GAC GGC TTC AGC ACT GTG TCC ACA GTC ACT TCA GCT GCT GTG GGC 2064
Pro Asp Gly Phe Ser Thr Val Ser Thr Val Thr Ser Ala Ala Val Gly
675 680 685
GGC CAG GGC CCG CCC CGC ACC TTC ACC TGC CAG CTG ACG TGG CAC AGA 2112
Gly Gln Gly Pro Pro Arg Thr Phe Thr Cys Gln Leu Thr Trp His Arg
690 695 700
GAT TCA GTC TCC TTC AGT AGA AGA AAT GCC AGT GGC ACC GCA TCT GTT 2160
Asp Ser Val Ser Phe Ser Arg Arg Asn Ala Ser Gly Thr Ala Ser Val
705 710 715 720
CTG CCC AGG CCC ACA ATA ACA ATG GAG TTC ACT GGA GAC CAC GCC GTG 2208
Leu Pro Arg Pro Thr Ile Thr Met Glu Phe Thr Gly Asp His Ala Val
725 730 735
TGC ACA GCT GGA TGT GTG CCC GAG GGC GTG ACC TTT GCC TGG TTT TTG 2256
Cys Thr Ala Gly Cys Val Pro Glu Gly Val Thr Phe Ala Trp Phe Leu
740 745 750
GGT GAC GAC AGC AGC CCT GCT GAG AAG GTG GCA GTG GCC AGC CAG ACC 2304
Gly Asp Asp Ser Ser Pro Ala Glu Lys Val Ala Val Ala Ser Gln Thr
755 760 765
AGC TGT GGC AGG CCT GGT ACT GCT ACC ATC AGA TCC ACT CTT CCA GTC 2352
Ser Cys Gly Arg Pro Gly Thr Ala Thr Ile Arg Ser Thr Leu Pro Val
770 775 780
AGC TAT GAG CAG ACT GAG TAC ATC TGC CGG CTT GCT GGC TAC CCA GAT 2400
Ser Tyr Glu Gln Thr Glu Tyr Ile Cys Arg Leu Ala Gly Tyr Pro Asp
785 790 795 800
GGG ATC CCT GTG CTG GAG CAC CAC GGC AGC CAC CAG CCA CCT CCT CGG 2448
Gly Ile Pro Val Leu Glu His His Gly Ser His Gln Pro Pro Pro Arg
805 810 815
GAC CCC ACC GAG AGG CAG GTG ATT AGA GCT GTA GAA GGC GGT TCT GGG 2496
Asp Pro Thr Glu Arg Gln Val Ile Arg Ala Val Glu Gly Gly Ser Gly
820 825 830
GAA GGG CGG GGC TCC CTG CTG ACT TGT GGA GAT GTT GAG GAG AAT CCC 2544
Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu Glu Asn Pro
835 840 845
GGA CCC TGC TGC CTG TGC CTC ACC GTG TTC CTG GAT GGA GGC GAT GGC 2592
Gly Pro Cys Cys Leu Cys Leu Thr Val Phe Leu Asp Gly Gly Asp Gly
850 855 860
AAC AAG ACT CTG AGA GAC AAC AGT TAC ATG CCC CTC ATT GCA CTC AGC 2640
Asn Lys Thr Leu Arg Asp Asn Ser Tyr Met Pro Leu Ile Ala Leu Ser
865 870 875 880
CTG CAC GAG AAT GGC GCG GAT AAC AAA GTT TGG ATC CGG CAC CAT GTG 2688
Leu His Glu Asn Gly Ala Asp Asn Lys Val Trp Ile Arg His His Val
885 890 895
GGA ATT GAA AAA CCA AAT CCA TCA GAG GGA GAA GCC GCT GAC AAC AAG 2736
Gly Ile Glu Lys Pro Asn Pro Ser Glu Gly Glu Ala Ala Asp Asn Lys
900 905 910
GTC TGG GGG TCA GCA AAG CCA GGA AAC GGG AGC TAC TTC GCC TCT GTC 2784
Val Trp Gly Ser Ala Lys Pro Gly Asn Gly Ser Tyr Phe Ala Ser Val
915 920 925
AAG ACA GAG ATG CTG GGA AAT GAG ATT GAT GCT GAC AAT GTT CAC CTG 2832
Lys Thr Glu Met Leu Gly Asn Glu Ile Asp Ala Asp Asn Val His Leu
930 935 940
GTT TAT ATC CTC TCA AAG CAG AAT CAG CAG CAC CTG ATT CCA CAG TGG 2880
Val Tyr Ile Leu Ser Lys Gln Asn Gln Gln His Leu Ile Pro Gln Trp
945 950 955 960
GCA CTT AGA CAA ATT GCC GAC TTT GCT GAC AAT AAG GTG TGG GAT CCC 2928
Ala Leu Arg Gln Ile Ala Asp Phe Ala Asp Asn Lys Val Trp Asp Pro
965 970 975
AGC CGC TAT AAC ATC TCC TTT GAG GCC CTT CTT GGG ATT TAC AGT GCT 2976
Ser Arg Tyr Asn Ile Ser Phe Glu Ala Leu Leu Gly Ile Tyr Ser Ala
980 985 990
GAT AAT GTT ACA TGG AGG TTG GCT GCC GGC AGC CCC TCC AGT GCA GAC 3024
Asp Asn Val Thr Trp Arg Leu Ala Ala Gly Ser Pro Ser Ser Ala Asp
995 1000 1005
AAT GTG CAC CTC TTC AAG TCT CTC ATC TAC ATC AAC GAA AAC ACC AAA 3072
Asn Val His Leu Phe Lys Ser Leu Ile Tyr Ile Asn Glu Asn Thr Lys
1010 1015 1020
ATC CTG GAG GTG GAG GCT GAT AAT GTC ACC TGG GCT TTG ATC AGT GCC 3120
Ile Leu Glu Val Glu Ala Asp Asn Val Thr Trp Ala Leu Ile Ser Ala
1025 1030 1035 1040
TTC TCT GGA AGC GCA GAT AAT GTG ACC TGG TCT GCA CAG GAA GAC CTG 3168
Phe Ser Gly Ser Ala Asp Asn Val Thr Trp Ser Ala Gln Glu Asp Leu
1045 1050 1055
GGT GAT GAC ACC GGC ATT CAC GTC ATT GCA GAT CAC AAC AAG GTC ACG 3216
Gly Asp Asp Thr Gly Ile His Val Ile Ala Asp His Asn Lys Val Thr
1060 1065 1070
GTC TGG AGC CTC CTC TCC ATC ATG ACC TTG GCA GCC ATG CAC AGC AAG 3264
Val Trp Ser Leu Leu Ser Ile Met Thr Leu Ala Ala Met His Ser Lys
1075 1080 1085
CTG TCC CCT GCA GAT AAC GTG ACA TGG ATG CTC ATG ATC ATT ATT GTC 3312
Leu Ser Pro Ala Asp Asn Val Thr Trp Met Leu Met Ile Ile Ile Val
1090 1095 1100
ATC GCC ATA GCT GAC AAT GTA ACC TGG TTC CAC ATG TGG AAC TAC CAT 3360
Ile Ala Ile Ala Asp Asn Val Thr Trp Phe His Met Trp Asn Tyr His
1105 1110 1115 1120
TCT CAT GTC TTT TCC GTG GGG GAT GCA GAC AAT GTG ACT TGG CCC GTG 3408
Ser His Val Phe Ser Val Gly Asp Ala Asp Asn Val Thr Trp Pro Val
1125 1130 1135
GTC TCT ACT CAT GAA CAG ATC GCA GAT AAT GTC ACC TGG TTT GCA CTG 3456
Val Ser Thr His Glu Gln Ile Ala Asp Asn Val Thr Trp Phe Ala Leu
1140 1145 1150
GAC CCT TAC AAT GAA GCT GAT AAT AAA GTG TGG GCC AAA AGA GCC CTG 3504
Asp Pro Tyr Asn Glu Ala Asp Asn Lys Val Trp Ala Lys Arg Ala Leu
1155 1160 1165
TGG ACC CCA GAC CAG ATT ACT GAC ATC ACA TCA CTG AGG GAC AAT AAA 3552
Trp Thr Pro Asp Gln Ile Thr Asp Ile Thr Ser Leu Arg Asp Asn Lys
1170 1175 1180
ACC CTG GTG GAC GAG GAA TAC AGA TTT ATT TCT GAC GCC ACA TTT GTT 3600
Thr Leu Val Asp Glu Glu Tyr Arg Phe Ile Ser Asp Ala Thr Phe Val
1185 1190 1195 1200
GAT GAG ACC GCC GAC AAT AAA GTC TGG AGT TCC ATG AAG GTG AAC GTG 3648
Asp Glu Thr Ala Asp Asn Lys Val Trp Ser Ser Met Lys Val Asn Val
1205 1210 1215
AAT GGA GTT GCA GAC AAC AAG GTT TGG ACT ACT TAT GCA TGG CGG TGG 3696
Asn Gly Val Ala Asp Asn Lys Val Trp Thr Thr Tyr Ala Trp Arg Trp
1220 1225 1230
TCA GCC GAC AAC AAA GTG TGG GGC ATC TAT TAC ATT CGC GTG GTG GAA 3744
Ser Ala Asp Asn Lys Val Trp Gly Ile Tyr Tyr Ile Arg Val Val Glu
1235 1240 1245
GTG CGC CAG ATG CAG TAT GAT AAT GCT GAT AAC AAA TGT CTG GCT ATG 3792
Val Arg Gln Met Gln Tyr Asp Asn Ala Asp Asn Lys Cys Leu Ala Met
1250 1255 1260
CTG GTG CTC CTG GCA GAG ATA GCA GAT GAA GGT AAT AAG GTA TGG TGC 3840
Leu Val Leu Leu Ala Glu Ile Ala Asp Glu Gly Asn Lys Val Trp Cys
1265 1270 1275 1280
ATC CAG AGC ATT GGA GCT TCT GTG GCG GAC AAC AAA GTC TGG TGG CGC 3888
Ile Gln Ser Ile Gly Ala Ser Val Ala Asp Asn Lys Val Trp Trp Arg
1285 1290 1295
CTT GGA GCC ATC CCC CCG TTG GTG GCT GAC AAC AAG TGC CTG CCT GAG 3936
Leu Gly Ala Ile Pro Pro Leu Val Ala Asp Asn Lys Cys Leu Pro Glu
1300 1305 1310
AGC TCC CAG AGG CCA CCG CTG GCC GAT AAC GTT CAT CTC ACA TAT GGG 3984
Ser Ser Gln Arg Pro Pro Leu Ala Asp Asn Val His Leu Thr Tyr Gly
1315 1320 1325
CCT GTT TTC ATG TGC CTT GGT GGC CTG CTG ACC ATG GTG GCT GGG GCA 4032
Pro Val Phe Met Cys Leu Gly Gly Leu Leu Thr Met Val Ala Gly Ala
1330 1335 1340
GAC AAC GTC CAC TTG GAT GAT AGA ACC ACA CTG CAG CTC GCT CAG AAC 4080
Asp Asn Val His Leu Asp Asp Arg Thr Thr Leu Gln Leu Ala Gln Asn
1345 1350 1355 1360
CCA GTG TAT ATT CCT GAA ACC TAC CCT TAT ATC AAG TGG GAC AAC TCC 4128
Pro Val Tyr Ile Pro Glu Thr Tyr Pro Tyr Ile Lys Trp Asp Asn Ser
1365 1370 1375
TCC TGG GAC TTC ATC ACC GTC TGA TAA 4155
Ser Trp Asp Phe Ile Thr Val * *
1380 1383
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ПОМОЩЬЮ КОРИНЕБАКТЕРИЙ С ПРИМЕНЕНИЕМ СИСТЕМЫ РАСЩЕПЛЕНИЯ ГЛИЦИНА | 2015 |
|
RU2713298C2 |
| Химерные белки на основе утрофина и дистрофина человека и их применение для лечения миодистрофии Дюшенна | 2021 |
|
RU2767335C1 |
| МАТЕРИАЛЫ И МЕТОДЫ, ИСПОЛЬЗУЕМЫЕ ДЛЯ ЛЕЧЕНИЯ РЕСПИРАТОРНЫХ ЗАБОЛЕВАНИЙ У СОБАК | 2020 |
|
RU2811752C2 |
| УНИВЕРСАЛЬНЫЙ АНТИТЕЛООПОСРЕДОВАННЫЙ БИОСЕНСОР | 2016 |
|
RU2746486C2 |
| ИММУНОИНДУЦИРУЮЩЕЕ СРЕДСТВО | 2016 |
|
RU2744843C2 |
| КОНСТРУКЦИИ НУКЛЕИНОВОЙ КИСЛОТЫ И ВЕКТОРЫ ДЛЯ ГЕНОТЕРАПИИ ДЛЯ ПРИМЕНЕНИЯ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ВИЛЬСОНА И ДРУГИХ СОСТОЯНИЙ | 2015 |
|
RU2745567C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРЕДОТВРАЩЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2016 |
|
RU2714205C2 |
| ВИРУС ГРИППА, СПОСОБНЫЙ ИНФИЦИРОВАТЬ СОБАЧЬИХ, И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2802222C2 |
| СПОСОБ УПРАВЛЕНИЯ РИТМОМ СЕРДЦА И СОКРАЩЕНИЕМ ОТДЕЛЬНЫХ КАРДИОМИОЦИТОВ ПРИ ПОМОЩИ ТЕРМОГЕНЕТИКИ | 2022 |
|
RU2802995C1 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ БИОЛОГИЧЕСКОГО ПОЛУЧЕНИЯ ЛАКТАТА ИЗ С1-СОЕДИНЕНИЙ С ПРИМЕНЕНИЕМ ТРАНСФОРМАНТОВ ЛАКТАТ ДЕГИДРОГЕНАЗЫ | 2014 |
|
RU2710714C2 |

Группа изобретений относится к области биотехнологии и иммунологии. Рекомбинантный слитый белок из иммуногенных эпитопов вирусов герпеса человека 1-8 типов для иммунизации против герпеса характеризуется аминокислотной последовательностью SEQ ID NO: 1. ДНК, кодирующая белок, с нуклеотидной последовательностью SEQ ID NO: 2. Плазмидная ДНК для иммунизации против герпеса представляет собой плазмидный экспрессионный вектор pcDNA3.1, содержащий указанную ДНК. Группа изобретений позволяет выработать гуморальный и клеточный иммунный ответ к вирусам герпеса. 3 н.п. ф-лы, 3 табл., 7 пр., 3 ил.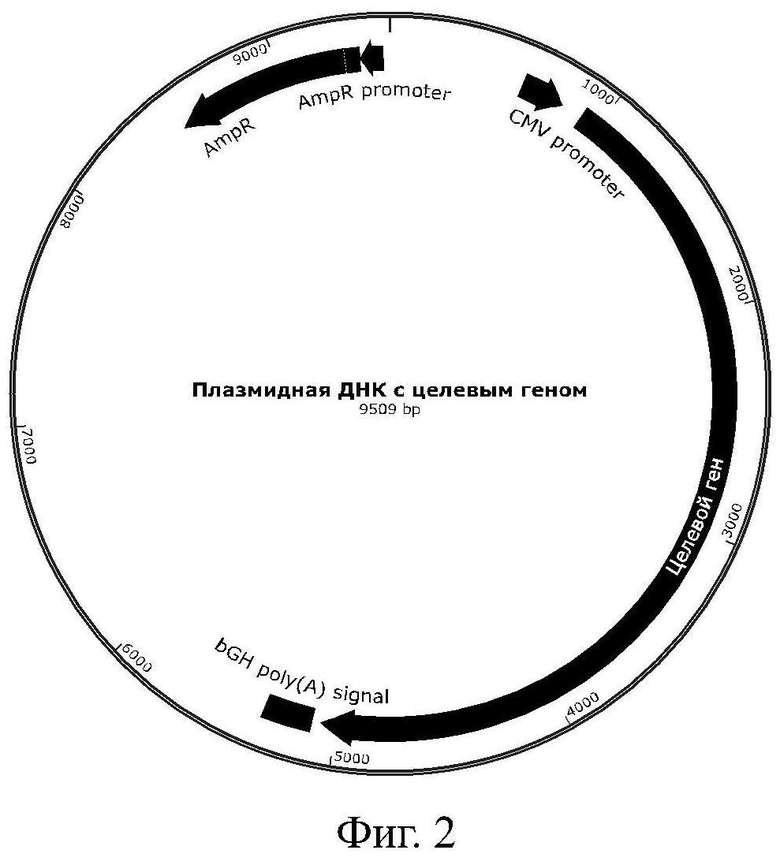
1. Рекомбинантный слитый белок из иммуногенных эпитопов вирусов герпеса человека 1-8 типов для иммунизации против герпеса, характеризующийся аминокислотной последовательностью по SEQ ID NO: 1.
2. ДНК, кодирующая белок по п. 1, нуклеотидная последовательность которой представлена в SEQ ID NO: 2.
3. Плазмидная ДНК для иммунизации против герпеса, представляющая собой плазмидный экспрессионный вектор pcDNA3.1, содержащий ДНК по п. 2.
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ПОЛУЧЕНИЯ АНТИГЕРПЕТИЧЕСКОЙ ПОЛИВАКЦИНЫ И ЛЕКАРСТВЕННОЙ ФОРМЫ НА ЕЕ ОСНОВЕ | 2014 |
|
RU2552341C1 |
| ФАРМАЦЕВТИЧЕСКАЯ ПРОТИВОГЕРПЕТИЧЕСКАЯ КОМПОЗИЦИЯ | 2004 |
|
RU2291690C2 |
| ЧЕПУРНОВА Н.С | |||
| и др., Перспективы применения противогерпетических вакцин, Современные проблемы науки и образования, издательский дом Академия естествознания, Пенза, N 5, 2015, с.327. | |||

