РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Эта заявка испрашивает приоритет предварительной заявки США № 62/806076, поданной 15 февраля 2019 г., содержание которой полностью включено в данный документ посредством ссылки.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Данная заявка включает перечень последовательностей, полностью включенный в данный документ посредством ссылки. Указанная ASCII-копия, созданная 13 февраля 2020 г., имеет название 131698-05420_SL.txt и размер 388,896 байт.
ОБЛАСТЬ ТЕХНИКИ
[0003] Данное изобретение относится к области генной терапии, в том числе к доставке экзогенных последовательностей ДНК в целевую клетку, ткань, орган или организм.
УРОВЕНЬ ТЕХНИКИ
[0004] Целью генной терапии является улучшение клинических исходов у пациентов, страдающих либо генетическими мутациями, либо приобретенными заболеваниями, вызываемыми аберрациями в профиле генной экспрессии. Генная терапия включает лечение или предотвращение медицинских состояний, обусловленных дефектными генами или аномальной регуляцией, или экспрессией, например, недостаточной экспрессией или избыточной экспрессией, которые могут приводить к расстройству, заболеванию, злокачественному новообразованию и т.п. Например, лечение, предотвращение или облегчение заболевания или расстройства, вызываемого дефектным геном, может быть осуществлено путем доставки пациенту корректирующего генетического материала, приводящей к терапевтической экспрессии указанного генетического материала у указанного пациента. Генная терапия основана на обеспечении транскрипционной кассеты с активным генным продуктом (иногда называемым трансгеном), например, что может приводить к положительному эффекту приобретения функции, отрицательному эффекту потери функции или другому исходу, такому как, например, онколитический эффект. Лечение моногенных расстройств у человека может осуществляться путем доставки и экспрессии нормального гена в целевых клетках. Доставка и экспрессия корректирующего гена в целевые клетки пациента могут быть осуществлены с применением многочисленных способов, в том числе с применением сконструированных вирусов и вирусных векторов для доставки генов. Среди множества доступных векторов вирусного происхождения (например, из рекомбинантного ретровируса, рекомбинантного лентивируса, рекомбинантного аденовируса и т.п.) рекомбинантный аденоассоциированный вирус (rAAV) набирает популярность в качестве универсального вектора для генной терапии.
[0005] Аденоассоциированные вирусы (AAV) принадлежат к семейству Parvoviridae, более конкретно, они составляют род Dependoparvovirus. Геном AAV состоит из линейной одноцепочечной молекулы ДНК, которая содержит приблизительно 4,7 тысячи пар основ (т.п.о) и состоит из двух основных открытых рамок считывания (ORF), кодирующих неструктурный белок Rep (репликация) и структурный белок Cap (капсид). В гене cap была идентифицирована вторая ORF, которая кодирует белок (AAP) активирующий сборку. ДНК, фланкирующие кодирующие участки AAV, представляют собой две последовательности действующих в цис-положении инвертированных концевых повторов (ITR) длиной приблизительно 145 нуклеотидов, с прерывистыми палиндромными последовательностями, которые могут укладываться в энергетически стабильные шпилечные структуры, функционирующие в качестве праймеров при репликации ДНК. Наряду с ролью в репликации ДНК указанные последовательности ITR, как было показано, вовлечены в интеграцию вирусной ДНК в клеточный геном, «спасение» из генома хозяина или плазмиды, и заключение в капсид вирусной нуклеиновой кислоты в зрелых вирионах (Muzyczka, (1992) Curr. Top. Micro. Immunol. 158:97-129).
[0006] Векторы, происходящие из ААV (т.е. рекомбинантные ААV (rААV) или ААV-векторы), являются перспективным способом доставки генетического материала, поскольку (i) они способны инфицировать (трансдуцировать) широкий спектр типов неделящихся и делящихся клеток, в том числе миоциты и нейроны; (ii) они лишены вирусных структурных генов, что обеспечивает уменьшение ответов клетки-хозяина на вирусную инфекцию, например, интерферон-опосредованных ответов; (iii) вирусы дикого типа считаются непатогенными для человека; (iv) в отличие от ААV дикого типа, который способен к интеграции в геном клетки-хозяина, у дефектных по репликации ААV-векторов отсутствует ген rep, и они обычно персистируют в виде эписом, что ограничивает риск инсерционного мутагенеза или генотоксичности; и (v) по сравнению с другими векторными системами, ААV-векторы в целом считаются относительно слабыми иммуногенами и, соответственно, не вызывают значимого иммунного ответа (см. ii), обеспечивая таким образом персистенцию векторной ДНК и, потенциально, долгосрочную экспрессию терапевтических трансгенов. AAV-векторы могут также быть получены и введены в составы с высокими титрами, и доставлены путем внутриартериальных, внутривенных, или внутрибрюшинных инъекций, что обеспечивает распределение вектора и перенос генов в значимые мышечные области посредством однократной инъекции у грызунов (Goyenvalle et al., 2004; Fougerousse et al., 2007; Koppanati et al., 2010; Wang et al., 2009) и собак. В клиническом исследовании лечения спинальной мышечной дистрофии типа 1 AAV-векторы доставляли системно с целью воздействовать на головной мозг, что приводило к очевидным клиническим улучшениям.
[0007] Однако применение частиц AAV в качестве вектора для доставки генов имеет ряд серьезных недостатков. Один существенный недостаток, ассоциированный с rAAV, заключается в ограниченной емкости вирусной упаковки, составляющей приблизительно 4,5 т.п.о. гетерологичной ДНК (Dong et al., 1996; Athanasopoulos et al., 2004; Lai et al., 2010). В результате применение AAV-векторов было ограничено емкостью для кодирования белка менее чем 150 кДа. Второй недостаток заключается в том, что ввиду распространенности инфекции AAV дикого типа в популяции, кандидаты для генной терапии rAAV должны проходить скрининг на присутствие нейтрализующих антител, элиминирующих вектор из организма пациента. Третий недостаток связан с иммуногенностью капсида, препятствуещего повторному введению пациентам, которые не были отстранены от начального лечения. Иммунная система пациента может отвечать на вектор, который фактически выступает в роли «бустерной» прививки, стимулируя иммунную систему, генерирующую высокие титры антител против AAV, которые исключают возможность лечения в будущем. В некоторых недавних сообщениях выражены опасения, касающиеся иммуногенности в ситуациях, подразумевающих применение высоких доз. Другой заметный недостаток заключается в относительно медленном начале опосредованной AAV генной экспрессии, принимая во внимание то, что одноцепочечная ДНК AAV должна быть преобразована в двухцепочечную ДНК до экспрессии гетерологичного гена. Хотя были предприняты попытки обойти указанную проблему путем конструирования двухцепочечных ДНК-векторов, указанная стратегия дополнительно ограничивает размер трансгенной экспрессионной кассеты, которая может быть интегрирована в AAV-вектор (McCarty, 2008; Varenika et al., 2009; Foust et al., 2009).
[0008] Кроме того, стандартные вирионы AAV с капсидами получают путем введения плазмиды или плазмид, содержащих геном AAV, гены rep и гены cap (Grimm et al., 1998). При введении указанных хелперных плазмид в транс-положение происходит «спасение» генома AAV (т.е. высвобождение с последующей амплификацией) из генома хозяина и его дальнейшее заключение в капсид (вирусные капсиды) с получением биологически активных AAV-векторов. Однако, как было обнаружено, такие заключенные в капсиды вирусные AAV-векторы неэффективно трансдуцируют определенные типы клеток и тканей. Указанные капсиды также индуцируют иммунный ответ.
[0009] Соответственно, применение векторов на основе аденоассоциированного вируса (AAV-векторов) для генной терапии ограничено однократным введением пациентам (из-за иммунного ответа у пациентов), ограниченным диапазоном трансгенного генетического материала, подходящего для доставки в AAV-векторах ввиду минимальной емкости вирусной упаковки (приблизительно 4,5 т.п.о.) ассоциированного капсида AAV, а также медленной AAV-опосредованной генной экспрессией. Применение клинической генной терапии с использованием rAAV дополнительно затруднено вариабельностью среди пациентов, которую нельзя предсказать на основании дозовой зависимости в сингенных моделях на мышах или других видах моделей.
[0010] Рекомбинантные бескапсидные AAV-векторы могут быть получены в виде выделенной линейной молекулы нуклеиновой кислоты, содержащей экспрессируемые участки трансгена и участки промотора, фланкированные двумя последовательностями инвертированных концевых повторов (ITR) AAV дикого типа, включая сайты связывания Rep и сайты концевого разрешения (TRS). Указанные рекомбинантные AAV-векторы не содержат последовательностей, кодирующих капсидный белок AAV, и могут быть одноцепочечными, двуцепочечными или дуплексными, где один или оба конца ковалентно связаны посредством двух палиндромных последовательностей ITR дикого типа (например, WO2012/123430, патент США 9598703). Они позволяют избежать многих проблем AAV-опосредованной генной терапии благодаря значительно большей трансгенной емкости, быстрому началу трансгенной экспрессии, и действительному распознаванию иммунной системой пациента молекул ДНК как вируса, который необходимо удалить. Однако постоянная экспрессия трансгена может быть желательной не во всех случаях, а канонические ITR AAV дикого типа могут не поддаваться оптимизации для использования в качестве зкДНК. Поэтому, все еще имеется значительная неудовлетворенная потребность в контролируемых рекомбинантных ДНК-векторах, а также в улучшенных характеристиках получения и/или экспрессии.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0011] Описанное в данном документе изобретение относится к улучшенному получении невирусного бескапсидного ДНК-вектора с ковалентно-замкнутыми концами (называемом в данном документе «вектором с ДНК с замкнутыми концами», или «зкДНК-вектором»). ЗкДНК-векторы, которые получены способами описаными в данном документе, представляют собой бескапсидные линейные дуплексные молекулы ДНК, образованные непрерывной цепью комплементарной ДНК с ковалентно замкнутыми концами (линейная, непрерывная и не заключенная в капсид структура), которые содержат последовательность инвертированного 5'-концевого повтора (ITR) и последовательность инвертированного 3' ITR, которые отличаются друг от друга или являются асимметричными.
[0012] Описанная технология относится к получению вектора зкДНК или вектора AAV в клетке (например, клетке насекомого, клетке млекопитающего) или в бесклеточной системе с единственным видом белка Rep. В частности, данное раскрытие частично основано на неожиданном открытии того, что достаточно одного только Rep78 или Rep68 для получения вектора зкДНК или вектора AAV в клетке. Это усовершенствованный и более эффективный способ получения вектора зкДНК, чем описанный в предшествующем уровне техники, в котором векторы AAV или зкДНК продуцируются в клетках (например, клетках насекомых), требующих два белка Rep; например, по меньшей мере, один малый белок Rep (например, Rep52 или Rep40) и по меньшей мере один большой белок Rep (например, Rep78 или Rep68). Таким образом, в предшествующем уровне техники описано, что получение векторов зкДНК или векторов AAV осуществляется с использованием двух белков Rep, либо кодируемых на отдельных конструктах нуклеиновых кислот, каждый из которых функционально связан с промотором, либо двух белков Rep, кодируемых на одном конструкте нуклеиновой кислоты с двумя сайты инициации, оперативно связанные с одним промотором.
[0013] Соответственно, один аспект описанной здесь технологии относится к конструкту нуклеиновой кислоты для получения ДНК-векторов, например, зкДНК-векторов и других рекомбинантных парвовирусных (например, аденоассоциированных вирусов) векторов в клетках (например, клетках насекомых, клетках млекопитающих) и безклеточных систем, в которых, например, клетки насекомых или бесклеточная система содержат первую нуклеотидную последовательность, кодирующую единственный парвовирусный белок Rep, в которой нуклеотидная последовательность не имеет открытой рамки считывания (ORF) и не имеет функционального кодона инициации ниже первого инициирующего кодона и/или отсутствуют альтернативные сайты сплайсинга, предотвращающие пропуск экзона, тем самым обеспечивая трансляцию только одного парвовирусного белка Rep (например, белка Rep78 или Rep 68) без трансляции дополнительных белков Rep на более позднем инициирующем кодоне (например, любого одного или нескольких из Rep52 или Rep 40) в клетках насекомых или бесклеточной системе. То есть нуклеиновая кислота, кодирующая Rep78, также не продуцирует белок Rep52, и аналогично, нуклеиновая кислота, кодирующая Rep68, не продуцирует белок Rep40. Кроме того, никакой другой Rep белок не присутствует и не экспрессируется в системе.
[0014] В некоторых вариантах реализации способы и композиции, описанные в данном документе для использования, единственный белок Rep может быть использован для получения любого вектора зкДНК, включая, помимо прочего, вектор зкДНК, содержащий асимметричные ITRы, как описано в международной патентной заявке PCT/US18/49996 поданной 7 сентября 2018 г. (см., например, Примеры 1-4); вектор зкДНК для редактирования генов, как описано в Международной патентной заявке PCT/US18/64242, поданной 6 декабря 2018 г. (см., например, Примеры 1-7), или вектор зкДНК для получения антител или слитых белков, как описано в Международной патентной заявке PCT/US19/18016, поданной 14 февраля 2019 г. (например, см. Примеры 1-4), все из которых полностью включены в данный документ посредством ссылки. В некоторых вариантах реализации также предполагается, что способы и композиции, описанные в данном документе, с использованием единственного белка Rep, могут быть использованы в синтетическом получении вектора зкДНК, например, в бесклеточной или не принадлежащей насекомым системе продуцирования зкДНК, как раскрыто в Международной заявке PCT/US19/14122, поданной 18 января 2019 г., полностью включенной в данный документ посредством ссылки, в которой единственный белок Rep может быть использован для лигирования содержащихся в нем олигонуклеотидов ITR с помощью белка.
[0015] Описанная здесь технология относится к усовершенствованному способу получения вектора зкДНК, содержащего по меньшей мере одну модифицированную последовательность инвертированного концевого повтора (ITR) AAV и экспрессируемый трансген. Согласно описанию в данном документе, зкДНК-векторы могут быть получены способами, описанными в данном документе, в эукариотических клетках и, соответственно, не содержать прокариотических модификаций ДНК и загрязнения бактериальным эндотоксином из клеток насекомых.
[0016] Аспекты изобретения относятся к способам и композициям для получения векторов зкДНК и векторов AAV с использованием единственного белка Rep, как описано в данном документе. Другие варианты реализации относятся к зкДНК-вектору, полученному с применением способа и композиций согласно данному изобретению.
[0017] В одном аспекте невирусные бескапсидные векторы ДНК с ковалентно-замкнутыми концами, полученные с применением способов описанных в данном документе, предпочтительно представляют собой линейные дуплексные молекулы и могут быть получены из векторного полинуклеотида, который кодирует гетерологичную нуклеиновую кислоту, функционально расположенную между двумя последовательностями разных инвертированных концевых повторов (ITR) (например, ITR AAV), причем по меньшей мере один из ITR содержит сайт концевого разрешения и сайт связывания белка репликации (RPS) (иногда называемый сайтом связывания репликативного белка), например, Rep-связывающий сайт, а один из ITR содержит делецию, вставку или замену относительно другого ITR. То есть один из ITR асимметричен по отношению к другому ITR. В одном варианте реализации по меньшей мере один из ITR представляет собой ITR AAV, например, ITR AAV дикого типа или модифицированный ITR AAV. В одном варианте реализации по меньшей мере один из ITR является модифицированным ITR относительно другого ITR, то есть зкДНК содержит ITR, которые являются асимметричными относительно друг друга. В одном варианте реализации по меньшей мере один из ITR является нефункциональным ITR.
[0018] Согласно некоторым вариантам реализации указанный зкДНК-вектор, полученный способами и композициями, описанными в данном документе, содержит: (1) экспрессионную кассету, содержащую цис-регуляторный элемент, промотор и по меньшей мере один трансген; или (2) промотор, функционально связанный по меньшей мере с одним трансгеном, и (3) две самокомплементарные симметричные последовательности, например, симметричные модифицированные ITR, фланкирующие указанную экспрессионную кассету, причем указанный зкДНК-вектор не ассоциирован с капсидным белком. В некоторых вариантах реализации зкДНК-вектор содержит две самокомплементарные последовательности, присутствующие в геноме AAV, при этом по меньшей мере один ITR содержит функциональный Rep-связывающий элемент (RBE) (также иногда называемый в данном документе «RBS») и сайт концевого разрешения (trs) AAV или функциональный вариант RBE, и один или нескольких цис-регуляторных элементов, функционально связанных с трансгеном. В некоторых вариантах реализации зкДНК-вектор содержит дополнительные компоненты для регуляции экспрессии трансгена, например, регуляторные переключатели, которые описаны в данном документе в разделе, озаглавленном «Регуляторные переключатели», для контроля и регулирования экспрессии трансгена, и может включать регуляторный переключатель, например, «аварийный выключатель», обеспечивающий контролируемую клеточную гибель клетки, содержащей зкДНК-вектор.
[0019] В некоторых вариантах реализации две самокомплементарные последовательности могут быть последовательностями ITR из любого известного парвовируса, например, депендовируса, такого как AAV (например, AAV1-AAV12). Может быть использован любой серотип AAV, включая, без ограничений, модифицированную последовательность ITR AAV2, которая сохраняет Rep-связывающий сайт (RBS), такой как 5'-GCGCGCTCGCTCGCTC-3' (SEQ ID NO: 531), и сайт концевого разрешения (trs) в дополнение к переменной палиндромной последовательности, позволяющей сформировать вторичную структуру шпильки. В некоторых вариантах реализации ITR представляет собой синтетическую последовательность ITR, которая сохраняет функциональный Rep-связывающий сайт (RBS), такой как 5'-GCGCGCTCGCTCGCTC-3' (SEQ ID NO: 531), и сайт концевого разрешения (TRS), в дополнение к вариабельной палиндромной последовательности, позволяющей формировать вторичную структуру шпильки. Согласно некоторым примерам, модифицированная последовательность ITR сохраняет последовательность RBS, trs, и структуру и положение связывающего элемента Rep, образующего концевую петлевую часть одной из шпилечных вторичных структур ITR, как в соответствующей последовательности AAV2 ITR дикого типа.
[0020] Типичными последовательностями ITR для использования в векторах зкДНК, полученных описанными здесь способами и композициями, могут быть любая одна или несколько из Таблиц 2-10A и 10B, или SEQ ID NO: 2, 52, 101-499 и 545-547, или частичные последовательности ITR, показанные на Фиг. 26А-26В. В некоторых вариантах реализации векторы зкДНК, полученные способами и композициями, описанными в данном документе, не имеют ITR, содержащего любую последовательность, выбранную из SEQ ID NO: 500-529.
[0021] Согласно некоторым вариантам реализации зкДНК-вектор, полученный способами и композициями, описанными в данном документе, может содержать ITR с модификацией в ITR, соответствующей любой из модификаций в последовательностях ITR или частичных последовательностях ITR, представленных в любой одной или более из таблиц 2, 3, 4, 5, 6, 7, 8, 9, 10A и 10B данного документа.
[0022] В качестве типичного примера в данном изобретении представлен ДНК-вектор с замкнутыми концами, полученный способами и композициями, описанными в данном документе, содержит промотор, функционально связанный с трансгеном, при этом зкДНК лишена капсидных белков и: (а) продуцируется из зкДНК-плазмиды (например, см. Примеры 1-2 и/или Фиг. 1A-B), которая кодирует мутированную правую сторону ITR AAV2, имеющую то же число внутримолекулярных дуплексных пар оснований, что и SEQ ID NO:2, или мутированную левую сторону ITR AAV2, имеющую такое же число внутримолекулярных дуплексных пар оснований, что и SEQ ID NO:51 в конфигурации вторичной шпильки (предпочтительно, за исключением делеции любой AAA- или TTT-концевой петли в этой конфигурации по сравнению с такими референсными последовательностями), и (b) идентифицируется как зкДНК с использованием анализа для идентификации зкДНК методом электрофореза в агарозном геле, в нативном геле и денатурирующих условиях, указанных в Примере 1. Примеры таких модифицированных последовательностей ITR представлены в Таблицах 2, 3, 4, 5, 6, 7, 8, 9, 10A и 10B данного документа.
[0023] Технология, описанная в данном документе, также относится к получению зкДНК-вектору, который может быть использован для доставки и кодирования одного или более трансгенов в целевой клетке, например, если указанный зкДНК-вектор содержит мультицистронную последовательность, или если указанный трансген и его природный геномный контекст (например, трансген, интроны и эндогенные нетранслируемые участки) вместе включены в указанный зкДНК-вектор. Указанные трансгены могут представлять собой кодирующие белок транскрипты, некодирующие транскрипты, или и то, и другое. Указанный зкДНК-вектор, полученный способами и композициями, описанными в данном документе, может содержать несколько кодирующих последовательностей и неканонический сайт инициации трансляции или более одного промотора для экспрессии кодирующих белок транскриптов, некодирующих транскриптов или обоих. Указанный трансген может содержать последовательность, кодирующую более одного белка, или может представлять собой последовательность некодирующего транскрипта. Указанная экспрессионная кассета может содержать, например, более 4000 нуклеотидов, 5000 нуклеотидов, 10000 нуклеотидов или 20000 нуклеотидов, или 30000 нуклеотидов, или 40000 нуклеотидов, или 50000 нуклеотидов, или любой диапазон значений в пределах между примерно 4000-10000 нуклеотидов или 10000-50000 нуклеотидов, или более 50000 нуклеотидов. Указанные зкДНК-векторы, полученные способами и композициями, описанными в данном документе, не имеют ограничений по размеру, характерных для заключенных в капсиды AAV-векторов, что позволяет доставлять экспрессионную кассету значительного размера для обеспечения эффективной экспрессии трансгенов. В некоторых вариантах реализации вектор зкДНК, полученный способами и композициями, описанными в данном документе, лишен прокариот-специфичного метилирования.
[0024] Кассета экспрессии вектора зкДНК, полученная способами и композициями, описанными в данном документе, также может содержать внутренний сайт входа в рибосому (IRES) и/или элемент 2А. Цис-регуляторные элементы включают, не ограничиваясь перечисленными, промотор, рибопереключатель, инсулятор, miR-регулируемый элемент, посттранскрипционный регуляторный элемент, тканеспецифический и клеточноспецифический промотор и энхансер. Согласно некоторым вариантам реализации указанный ITR может действовать в качестве промотора для трансгена. Согласно некоторым вариантам реализации указанный зкДНК-вектор содержит дополнительные компоненты для регуляции экспрессии трансгена. Например, дополнительный регуляторный компонент может представлять собой регуляторный переключатель согласно описанию в данном документе, в том числе, но не ограничиваясь указанным, «аварийный выключатель», который способен убивать инфицированную зкДНК клетку, при необходимости, и другие индуцируемые и/или репрессируемые элементы.
[0025] Описанная здесь технология дополнительно обеспечивает новые способы эффективного получения вектора зкДНК или другого вектора AAV, который может избирательно экспрессировать один или несколько трансгенов. ЗкДНК-вектор, полученный способами и композициями, описанными в данном документе, может поглощаться клетками-хозяевами, а также транспортироваться в ядро в отсутствие капсида AAV. Кроме того, зкДНК-векторы, полученные способами и композициями, описанными в данном документе, не имеют капсида и, соответственно, избегают иммунного ответа, который может возникать в ответ на капсид-содержащие векторы.
[0026] В одном варианте реализации свободный от капсида невирусный ДНК-вектор (зкДНК-вектор) получают из плазмиды (называемой здесь «зкДНК-плазмида»), содержащей матрицу конструкта для экспрессии полинуклеотидов, которая содержит в следующем порядке: первый 5'-инвертированный концевой повтор (например, AAV ITR); кассету экспрессии; и 3' ITR (например, AAV ITR), где по меньшей мере один из 5' и 3' ITR является модифицированным ITR, или когда оба 5' и 3' ITR являются модифицированными, причем они имеют модификации отличные друг от друга и не являются одной и той же последовательностью. В таком варианте реализации вектор зкДНК получают способом, который проиллюстрирован в примерах и показан на Фиг. 4A-4D, где для получения требуется только единственный белок Rep.
[0027] ЗкДНК-вектор, согласно описанию в данном документе, может быть получен несколькими способами, которые будут понятны обычному специалисту в данной области техники после прочтения данного документа. Например, полинуклеотидная матрица экспрессионного конструкта, используемая для генерирования зкДНК-векторов по данному изобретению, может представлять собой зкДНК-плазмиду (например, см. Таблицу 12 или Фиг. 10В), зкДНК-бакмиду и/или зкДНК-бакуловирус. В одном варианте реализации зкДНК-плазмида содержит сайт рестрикционного клонирования для возможной вставки (например, SEQ ID NO: 7), функционально расположенный между ITR, где может быть вставлена экспрессионная кассета, содержащая, например, промотор, функционально связанный с трансгеном, например, репортерным геном и/или терапевтическим геном. В некоторых вариантах реализации зкДНК-векторы получают из полинуклеотидной матрицы (например, зкДНК-плазмиды, зкДНК-бакмиды, зкДНК-бакуловируса), содержащей ITR, модифицированную по сравнению с соответствующей фланкирующей ITR AAV3 или последовательностью ITR AAV2 дикого типа, причем модификация представляет собой одно или несколько из делеции, вставки и/или замены.
[0028] Согласно некоторым аспектам раскрытие предлагает способ получения вектора зкДНК в клетке насекомого (например, клетках Sf9, Sf21, клетках Trichoplusia ni и клетках High Five) или клетках млекопитающих (например, HEK293, Huh-7, HeLa, HepG2, HeplA, 911, CHO, COS, MeWo, NIH3T3, A549, HT1080, моноциты и зрелые и незрелые дендритные клетки); способ, включающий культивирование клетки насекомого или клетки млекопитающего, содержащей первую нуклеотидную последовательность, кодирующую единственный парвовирусный белок Rep, где в первой нуклеотидной последовательности отсутствует функциональный кодон инициации ниже первого кодона инициации и отсутствуют альтернативные сайты сплайсинга, предотвращающие пропуск экзона, что позволяет трансляцию только одного белка Rep (например, Rep78) без трансляции дополнительных белков Rep на более поздний кодон инициации (например,, любой один или несколько Rep52 или Rep40) или сплайсированный вариант полноразмерного (например, Rep68) в ячейку.
[0029] В соответствии с некоторыми другими аспектами в описании предложен способ получения вектора зкДНК в клетке насекомого (например, клетках Sf9, Sf21, Trichoplusia ni и клетках High Five) или клетках млекопитающих (например, HEK293, Huh-7, HeLa, HepG2, HeplA, 911, CHO, COS, MeWo, NIH3T3, A549, HT1080, моноциты, а также зрелые и незрелые дендритные клетки); способ, включающий культивирование клетки насекомого или клетки млекопитающего, содержащей первую нуклеотидную последовательность, кодирующую однозначный парвовирусный белок Rep, при этом первая нуклеотидная последовательность лишена функционального инициирующего кодона, расположенного ниже первого инициирующего кодона, и содержит делецию на карбокси-конце срощенной последовательности (например, любая часть или полная длина c-концевого интрона/пропущенного экзона), тем самым обеспечивая трансляцию только сигнального белка Rep (например, Rep68) без трансляции дополнительных белков Rep в более позднем инициирующем кодоне (например, любой один или несколько из Rep52 или Rep40) или полноразмерный белок Rep72 в клетке.
[0030] В соответствии с некоторыми другими аспектами в описании предложен способ получения вектора зкДНК в клетке насекомого (например, клетки Sf9, Sf21, Trichoplusia ni, клетки High Five) или клетке млекопитающего (например, HEK293, Huh-7, HeLa, HepG2, HeplA, 911, CHO, COS, MeWo, NIH3T3, A549, HT1080, моноциты и зрелые и незрелые дендритные клетки); способ, включающий культивирование клетки насекомого или клетки млекопитающего, содержащей первую нуклеотидную последовательность, кодирующую один или два белка Rep (например, белок Rep 78 и/или Rep68), при этом в первой нуклеотидной последовательности отсутствует функциональный кодон инициации после первого кодона инициации и интактные альтернативные сайты сплайсинга, тем самым обеспечивая трансляцию только белка Rep78 и/или Rep68 без трансляции дополнительных белков Rep на более позднем инициирующем кодоне (например, любого одного или нескольких из Rep52 или Rep40).
[0031] Клетка, описанная в способах выше, может дополнительно содержать вторую нуклеотидную последовательность, содержащую по меньшей мере одну последовательность инвертированного концевого повтора (ITR) AAV, фланкирующую гетерологичную последовательность при условиях, что когда первая последовательность экспрессируется с получением Rep78 и/или Rep68, зкДНК продуцируется посредством белка Rep78 и/или Rep68 без присутствия Rep52 или Rep40. Затем вектор зкДНК может быть извлечен из клетки. Согласно некоторым вариантам реализации нуклеотидная последовательность, содержащая по меньшей мере один AAV, является частью экспрессионного конструкта. Согласно некоторым вариантам реализации гетерологичная последовательность включает терапевтическую нуклеиновую кислоту. Согласно некоторым вариантам реализации терапевтическая нуклеиновая кислота является частью экспрессионного конструкта. Согласно некоторым вариантам реализации клетка дополнительно содержит нуклеиновую кислоту, которая служит маркером. Согласно некоторым вариантам реализации нуклеиновая кислота, которая служит маркером, является частью экспрессионного конструкта.
[0032] В пермиссивной клетке-хозяине, в присутствии, например, единственного белка Rep, полинуклеотидная матрица, содержащая по меньшей мере один модифицированный ITR, реплицируется, продуцируя зкДНК-векторы. Получение зкДНК-вектора проходит в два этапа: (i) вырезание («высвобождение») матрицы из остова матрицы (например, зкДНК-плазмиды, зкДНК-бакмиды, зкДНК-бакуловирусного генома и т.п.) посредством белков Rep; и (ii) опосредованной Rep репликации вырезанного зкДНК-вектора. Единственный Rep белок, необходимый для стадий (i) и (ii) эксцизии и репликации, может быть любым Rep-белком, описанным в данном документе. Белки Rep и сайты связывания Rep у различных серотипов AAV хорошо известны специалистам в данной области техники.
[0033] Рядовой специалист в данной области техники понимает, что следует выбирать белок Rep из серотипа, который связывается с последовательностью нуклеиновой кислоты и реплицирует ее, на основе, по меньшей мере, одного функционального ITR. Например, если компетентный по репликации ITR относится к серотипу ААV2, соответствующий Rep будет принадлежать серотипу ААV, который работает с указанным серотипом, таким как ITR ААV2 с Rep ААV2 или ААV4, но не с Rep AAV5, который неспособен на это. После репликации (например, после стадии (ii)) зкДНК-вектор с ковалентно замкнутыми концами продолжает накапливаться в пермиссивных клетках, и зкДНК-вектор предпочтительно является достаточно стабильным во времени в присутствии белка Rep в стандартных условиях репликации, например, для накопления в количестве, составляющем по меньшей мере 1 пг/клетку, предпочтительно, по меньшей мере 2 пг/клетку, предпочтительно, по меньшей мере 3 пг/клетку, более предпочтительно, по меньшей мере 4 пг/клетку, еще более предпочтительно, по меньшей мере 5 пг/клетку.
[0034] Соответственно, один аспект изобретения относится к способу, включающему следующие этапы: a) инкубация популяции клеток-хозяев (например, клеток насекомых), несущих указанный матричный полинуклеотидный экспрессионный конструкт (например, зкДНК-плазмиду, зкДНК-бакмиду и/или зкДНК-бакуловирус), которая не содержит последовательностей, кодирующих вирусный капсид, в присутствии одного белка Rep в эффективных условиях и на протяжении времени, достаточного для индукции продуцирования зкДНК-вектора в клетках-хозяевах, причем указанные клетки-хозяева не содержат последовательностей, кодирующих вирусный капсид; и b) сбор и выделение указанного зкДНК-вектора из клеток-хозяев. Присутствие единственного белка Rep индуцирует репликацию векторного полинуклеотида с модифицированным ITR для продуцирования зкДНК-вектора в клетке-хозяине. Однако вирусные частицы (например, вирионы AAV) при этом не экспрессируются. Таким образом, отсутствуют ограничения по размеру, накладываемые вирионами. Предполагается, что если последовательность нуклеиновой кислоты, кодирующая белок Rep, кодирует большой белок Rep, например, белок Rep78 или Rep 68, то кодон инициации для меньших белков Rep модифицируется таким образом, что только большой белок Rep экспрессируется в клетка.
[0035] Согласно некоторым аспектам в раскрытии представлена клетка насекомого (например, клетки Sf9, Sf21, Trichoplusia ni и клетки High Five) или клетка млекопитающего (например, HEK293, Huh-7, HeLa, HepG2, HeplA, 911, CHO, COS, MeWo, NIH3T3, A549, HT1080, моноциты и зрелые и незрелые дендритные клетки); клетка насекомого или клеточная линия млекопитающего, содержащая первую нуклеотидную последовательность, кодирующую единственный парвовирусный белок Rep, где в первой нуклеотидной последовательности отсутствует функциональный кодон инициации ниже первого кодона инициации и отсутствуют альтернативные сайты сплайсинга, предотвращающие пропуск экзона, тем самым обеспечивая трансляцию только единственного белка Rep (например, Rep78) без трансляции дополнительных белков Rep на более поздний кодон инициации (например, любой один или несколько из Rep52 или Rep40) или сплайсированный вариант полноразмерного (например, Rep68) в камере.
[0036] Согласно некоторым другим аспектам в описании представлена клетка насекомого (например, клетки Sf9, Sf21, Trichoplusia ni и клетки High Five) или клетка млекопитающего (например, HEK293, Huh-7, HeLa, HepG2, HeplA, 911, CHO, COS, MeWo, NIH3T3, A549, HT1080, моноциты и зрелые и незрелые дендритные клетки); клетка насекомого или клетка млекопитающего, содержащая первую нуклеотидную последовательность, кодирующую единственный парвовирусный белок Rep, где первая нуклеотидная последовательность не имеет функционального инициирующего кодона ниже первого инициирующего кодона и содержит делецию карбоксиконцевой сращенной последовательности (например, любой части или полноразмерный c-концевой интрон/пропущенный экзон), тем самым делая возможной трансляцию только сигнального белка Rep (например, Rep68) без трансляции дополнительных белков Rep на более поздний кодон инициации (например, любой из или более Rep52 или Rep40) или полноразмерный белок Rep72 в клетке.
[0037] Согласно некоторым другим аспектам в описании представлена клетка насекомого (например, Sf9, Sf21, клетки Trichoplusia ni, клетки High Five) или клетка млекопитающего (например, HEK293, Huh-7, HeLa, HepG2, HeplA, 911, CHO, COS, MeWo, NIH3T3, A549, HT1080, моноциты и зрелые и незрелые дендритные клетки); клетка насекомого или клеточная линия млекопитающего, содержащая первую нуклеотидную последовательность, кодирующую один или два белка Rep (например, белок Rep78 и/или Rep68), где в первой нуклеотидной последовательности отсутствует функциональный кодон инициации ниже первого кодона инициации и интактные альтернативные сайты сплайсинга, тем самым обеспечивая трансляцию только белка Rep78 и/или Rep68, без трансляции дополнительных белков Rep на более поздний кодон инициации (например, любой один или несколько из Rep52 или Rep40)
[0038] Описанная выше клетка может дополнительно содержать вторую нуклеотидную последовательность, содержащую по меньшей мере одну последовательность инвертированного концевого повтора (ITR) AAV, фланкирующую гетерологичную последовательность при условиях, что, когда первая последовательность экспрессируется для продуцирования Rep78 и/или Rep68, зкДНК продуцируется посредством белка Rep78 и/или Rep68 без присутствия Rep52 или Rep40. Затем вектор зкДНК может быть извлечен из клетки. Согласно некоторым вариантам реализации нуклеотидная последовательность, содержащая по меньшей мере один AAV, является частью экспрессионного конструкта. Согласно некоторым вариантам реализации гетерологичная последовательность включает терапевтическую нуклеиновую кислоту. Согласно некоторым вариантам реализации терапевтическая нуклеиновая кислота является частью экспрессионного конструкта. Согласно некоторым вариантам реализации клетка дополнительно содержит нуклеиновую кислоту, которая служит маркером. Согласно некоторым вариантам реализации нуклеиновая кислота, которая служит маркером, является частью экспрессионного конструкта.
[0039] Согласно некоторым аспектам в описании представлена бесклеточная система, содержащая первую нуклеотидную последовательность, кодирующую один парвовирусный белок Rep, где в нуклеотидной последовательности отсутствует функциональный кодон инициации ниже первого кодона инициации и/или отсутствуют альтернативные сайты сплайсинга, предотвращающие пропуск экзона, тем самым обеспечивая трансляцию только единственного парвовирусного белка Rep (например, белка Rep78 или Rep 68) без трансляции дополнительных белков Rep на более поздний кодон инициации (например, любой один или несколько Rep52 или Rep 40) в бесклеточной системе. Согласно некоторым вариантам реализации нуклеиновая кислота, кодирующая Rep78, также не продуцирует белок Rep52 или Rep40. Согласно некоторым вариантам реализации нуклеиновая кислота, кодирующая Rep68, не продуцирует белок Rep52 или Rep40. Согласно некоторым вариантам реализации клетка насекомого, клетка млекопитающего или бесклеточная система не экспрессируют какой-либо другой белок Rep.
[0040] ЗкДНК-вектор, полученный способами описанными в данном документе, используя единственный Rep белок, выделяется из клеток-хозяев, а его присутствие может быть подтверждено путем расщепления ДНК, выделенной из клетки-хозяина, рестрикционным ферментом, имеющим единственный сайт распознавания на зкДНК-векторе, и анализа расщепленного материала ДНК на денатурирующем и неденатурирующем гелях для подтверждения присутствия характерных полос линейной и непрерывной ДНК по сравнению с линейной и прерывистой ДНК.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0041] Фиг. 1А иллюстрирует типичную структуру вектора зкДНК, полученного с использованием одного белка Rep в соответствии со способами и композициями, описанными в данном документе. Согласно указанному варианту реализации указанный пример зкДНК-вектора содержит экспрессионную кассету, содержащую промотор CAG, WPRE и BGHpA. Открытая рамка считывания (ORF), кодирующая трансген люциферазы, вставляется в сайт клонирования (R3/R4) между промотором CAG и WPRE. Экспрессионная кассета фланкирована двумя инвертированными концевыми повторами (ITR) ITR ААV2 дикого типа перед (на 5'-конце) и модифицированным ITR после (на 3'-конце) экспрессионной кассеты, таким образом, два ITR, фланкирующие экспрессионную кассету, являются асимметричными относительно друг друга. Рядовой специалист в данной области техники поймет, что можно использовать любой ITR. В иллюстративных целях, ITR в конструктах зкДНК на этой Фигуре и в Примерах этого документа представляют собой модифицированный ITR и ITR WT. Однако, данный документ охватывает зкДНК-векторы, содержащие последовательность гетерологичной нуклеиновой кислоты, расположенные между двумя последовательностями инвертированных концевых повторов (ITR), причем последовательности ITR могут быть асимметричной парой ITR или симметричной или по существу симметричной парой ITR, как эти термины определены в данном документе. ЗкДНК вектор, раскрытый в данном документе, может содержать последовательности ITR выбранные из любых из: (i) по меньшей мере, одного ITR дикого типа и по меньшей мере одного модифицированного инвертированного концевого повтора ААV (mod-ITR) (например, асимметричных модифицированных ITR); (ii) двух модифицированных ITR, причем пара mod-ITR имеет различную трехмерную пространственную организацию относительно друг друга (например, асимметричных модифицированных ITR), или (iii) пары симметричных или по существу симметричных ITR WT-WT, причем каждый из ITR-WT имеет одинаковую трехмерную пространственную организацию, или (iv) пары симметричных или по существу симметричных модифицированных ITR, причем каждый из mod-ITR имеет одинаковую трехмерную пространственную организацию. В некоторых вариантах реализации способы данного раскрытия включают использование единственного белка rep для получения вектора зкДНК, который составлен в композицию, которая включает систему доставки, такую как, но не ограничиваясь этим, липосомная система доставки наночастиц.
[0042] Фиг. 1B иллюстрирует типичную структуру вектора зкДНК, полученного с использованием единственного белка Rep в соответствии со способами и композициями, описанными в данном документе, где вектор зкДНК включает кассету экспрессии, содержащую промотор CAG, WPRE и BGHpA. Открытая рамка считывания (ORF), кодирующая трансген люциферазы, вставляется в сайт клонирования между промотором CAG и WPRE. Экспрессионная кассета фланкирована двумя инвертированными концевыми повторами (ITR) модифицированным ITR перед (на 5'-конце) и ITR дикого типа после (на 3'-конце) экспрессионной кассеты. Как показано на Фиг. 1A, квалифицированный специалист может легко выбрать последовательности ITR, которые будут асимметричной парой ITR или симметричной или по существу симметричной парой ITR, как эти термины определены в данном документе.
[0043] Фиг. 1C иллюстрирует типичную структуру вектора зкДНК, полученного с использованием единственного белка Rep в соответствии со способами и композициями, описанными в данном документе, где вектор зкДНК включает кассету экспрессии, содержащую энхансер/промотор, открытую рамку считывания (ORF) для вставки трансгена, посттранскрипционный элемент (WPRE) и сигнал полиА. Открытая рамка считывания (ORF) позволяет вставить трансген в сайт клонирования между промотором CAG и WPRE. Экспрессионная кассета фланкирована двумя инвертированными концевыми повторами (ITR), которые являются асимметричными относительно друг друга - модифицированным ITR слева от (на 5'-конце) и модифицированным ITR справа от (на 3'-конце) экспрессионной кассеты, причем 5'-ITR и 3'-ITR оба являются модифицированными ITR, но имеют разные модификации (т.е. они имеют неодинаковые модификации). Как показано на Фиг. 1A, квалифицированный специалист может легко выбрать последовательности ITR, которые будут асимметричной парой ITR или симметричной или по существу симметричной парой ITR, как эти термины определены в данном документе.
[0044] На Фиг. 2А представлена Т-образная структура стебель-петля левого ITR дикого типа AAV2 (SEQ ID NO: 538) с обозначенными A-A'-плечом, B-B'-плечом, C-C'-плечом, двумя сайтами связывания Rep (RBE и RBE'), а также с указанным сайтом концевого разрешения (trs). RBE содержит ряд из 4 дуплексных тетрамеров, предположительно, взаимодействующих с Rep 78 либо с Rep 68. Кроме того, RBE' также предположительно взаимодействует с комплексом Rep, собранным на ITR дикого типа или мутированном ITR в указанного конструкта. Участки D и D' содержат сайты связывания транскрипционных факторов и другие консервативные структуры. Фиг. 2B демонстрирует предполагаемую катализируемую Rep никующую и лигирующую активности левого ITR дикого типа (SEQ ID NO: 539), включая Т-образную структуру стебель-петля левого ITR дикого типа AAV2 с обозначенными A-A'-плечом, B-B'-плечом, C-C'-плечом, двумя сайтами связывания Rep (RBE и RBE'), а также с указанным сайтом концевого разрешения (TRS), и участок D и D', содержащий несколько сайтов связывания транскрипционных факторов и другую консервативную структуру.
[0045] На Фиг. 3А представлена первичная структура (полинуклеотидная последовательность) (слева) и вторичная структура (справа) RBE-содержащих частей A-A'-плеча, и C-C'- и B-B'-плеча левого ITR AAV2 дикого типа (SEQ ID NO: 540). Фиг. 3B демонстрирует типичный пример последовательности мутированного ITR (также называемого модифицированным ITR) для левого ITR. Представлена первичная структура (слева) и предсказанная вторичная структура (справа) RBE-участка A-A'-плеча, C-плечо и B-B'-плечо типичного мутированного левого ITR (ITR-1, левый) (SEQ ID NO: 113). Фиг. 3C демонстрирует первичную структуру (слева) и вторичную структуру (справа) RBE-содержащей части петли A-A' и B-B' и C-C' плеч правого ITR AAV2 дикого типа (SEQ ID NO: 541). Фиг. 3D демонстрирует типичный пример правого модифицированного ITR. Представлены первичная структура (слева) и прогнозируемая вторичная структура (справа) RBE-содержащей части A-A' плеча, B-B' и C плеч типичного примера мутантного правого ITR (ITR-1, правый) (SEQ ID NO: 114). Может быть использована любая комбинация левого и правого ITR (например, AAV2 ITR или другого вирусного серотипа или синтетические ITR) при условии, что левый ITR асимметричен или отличается от правого ITR. Каждая из полинуклеотидных последовательностей на Фиг. 3А-3D относится к последовательностям, используемым в плазмиде или бакмиде/бакуловирусном геноме, применяемых для продуцирования зкДНК, как описано в данном документе. Также каждая из Фиг. 3A-3D включает соответствующие предполагаемые вторичные структуры зкДНК, полученные на основании конфигураций зкДНК-вектора в плазмиде или бакмиде/бакуловирусном геноме и предсказанных значений свободной энергии Гиббса.
[0046] На Фиг. 4A приведено схематическое изображение предшествующего процесса получения инфицированных бакуловирусом клеток насекомых (BIIC), которые подходят для получения зкДНК способом, схематически представленным на Фиг. 4B. В этих вариантах реализации две бакмиды генерируются путем транспонирования плазмиды зкДНК или Rep-плазмиды (кодирующей единственный белок Rep) в вектор экспрессии бакуловируса для создания бакмиды вектора зкДНК (т.е. Бакмиды-1) и одной бакмиды Rep (Rep-Бакмиды), которые используются для трансфекции клеток насекомых с получением клеток насекомых с введенным бакуловирусом, BIIC-1 и BICC-2 (единственный Rep), соответственно. Фиг. 4B представляет собой схему типичного способа получения зкДНК с использованием клеток насекомых (например, BICC-2), содержащих Rep-Бакмиду, содержащую последовательность нуклеиновой кислоты для единственного белка Rep, а Фиг. 4С иллюстрирует биохимический метод и процесс подтверждения продукции вектора зкДНК с использованием описанной здесь методологии единственного Rep-белка. На Фиг. 4D и Фиг. 4Е представлены схематические изображения, описывающие процесс идентификации присутствия зкДНК в ДНК, собранной из клеточных осадков, полученных в процессе продуцирования зкДНК в соответствии с Фиг. 4В. Фиг. 4E демонстрирует ДНК, имеющую прерывистую структуру. ЗкДНК может быть разрезана рестрикционной эндонуклеазой, имеющей один сайт распознавания на зкДНК-векторе, с образованием двух фрагментов ДНК разного размера (1 т.п.о. и 2 т.п.о.) как в нейтральных, так и в денатурирующих условиях. На Фиг. 4E также продемонстрирована зкДНК, имеющая линейную и непрерывную структуру. Указанный зкДНК-вектор может быть разрезан рестрикционной эндонуклеазой с образованием двух фрагментов ДНК, которые мигрируют в виде фрагментов размером 1 т.п.о. и 2 т.п.о. в нейтральных условиях, однако в денатурирующих условиях указанные цепи остаются соединенными и образуют одиночные цепи, которые мигрируют в виде фрагментов размером 2 т.п.о. и 4 т.п.о. На Фиг. 4D схематически представлены ожидаемые полосы для типичной зкДНК, нерасщепленной или расщепленной рестрикционной эндонуклеазой, а затем подвергнутой электрофорезу либо на нативном, либо на денатурирующем геле. На крайней схеме слева показан нативный гель и несколько полос, указывающих на то, что в дуплексной и неразрезанной форме зкДНК существует по меньшей мере в мономерном и димерном состояниях, которые проявляются в виде быстрее мигрирующего мономера меньшего размера и медленнее мигрирующего димера с размером, в 2 раза большим, чем у мономера. Второе слева схематическое изображение демонстрирует, что при разрезании зкДНК рестрикционной эндонуклеазой, исходные полосы исчезают, а появляются полосы мигрирующие быстрее (например, меньшего размера), которые соответствуют ожидаемым размерам фрагментов, остающихся после расщепления. В денатурирующих условиях исходная дуплексная ДНК является одноцепочечной и мигрирует в виде объекта вдвое большего размера по сравнению с наблюдаемым на нативном геле, поскольку комплементарные цепи ковалентно связаны. Соответственно, на второй справа схеме расщепленная зкДНК демонстрирует распределение полос, аналогичное наблюдаемому на нативном геле, однако указанные полосы мигрируют как фрагменты, размер которых в два раза больше их эквивалентов на нативном геле. Крайняя справа схема демонстрирует, что неразрезанная зкДНК в денатурирующих условиях мигрирует в виде одноцепочечного раскрытого кольца, и, соответственно, наблюдаемые полосы имеют в два раза больший размер, чем наблюдаемые в нативных условиях, когда кольцо не раскрыто. На указанной фигуре «т.п.о.» используется для указания относительного размера нуклеотидных молекул, основанного, в зависимости от контекста, либо на длине нуклеотидной цепи (например, для одноцепочечных молекул, наблюдаемых в денатурирующих условиях), либо на числе пар оснований (например, для двуцепочечных молекул, наблюдаемых в нативных условиях).
[0047] Фиг. 5 демонстрирует иллюстративное изображение прогона в денатурирующем геле примеров зкДНК-векторов с (+) или без (-) расщепления эндонуклеазами (EcoRI для зкДНК-конструктов 1 и 2; BamH1 для зкДНК-конструктов 3 и 4; SpeI для зкДНК-конструктов 5 и 6; и XhoI для зкДНК-конструктов 7 и 8). Размеры для полос, помеченных звездочкой, определены и приведены в нижней части чертежа.
[0048] На Фиг. 6A показаны результаты анализа экспрессии белка in vitro, измеряющего активность люциферазы (ось y, RQ (Luc)) в клетках HEK293 через 48 часов после трансфекции 400 нг (черный), 200 нг (серый) или 100 нг (белый) конструктом, идентифицированным на оси абсцисс (конструкт-1, конструкт-3, конструкт-5, конструкт-7 (Таблица 12). На Фиг. 6B показана активность люциферазы (ось y, RQ (Luc)), измеренная в клетках HEK293 через 48 часов после трансфекции 400 нг (черный), 200 нг (серый) или 100 нг (белый) конструктами, идентифицированными на оси x (конструкт-2, конструкт-4, конструкт-6, конструкт-8) (Таблица 12). Также представлены активности люциферазы, измеренные в клетках HEK293, обработанных Fugene без каких-либо плазмид («Fugene»), или в необработанных клетках HEK293 («Необработанные»).
[0049] Фиг. 7A показывает жизнеспособность клеток HEK293 (ось y) через 48 часов после трансфекции 400 нг (черный), 200 нг (серый) или 100 нг (белый) конструктами, идентифицированными на оси x (конструкт-1, конструкт-3, конструкт-5, конструкт-7). Фиг. 7B показывает жизнеспособность клеток HEK293 (ось y) через 48 часов после трансфекции 400 нг (черный), 200 нг (серый) или 100 нг (белый) конструктами, идентифицированной на оси x (конструкт-2, конструкт-4, конструкт-6, конструкт-8).
[0050] Фиг. 8A представляет собой типичную Rep-бакмиду в плазмиде pFBDLSR, содержащей последовательности нуклеиновых кислот для модифицированного белка Rep78, где модифицированный белок Rep 78 представляет собой модификацию аминокислотного остатка 225 (Met) SEQ ID NO: 530, причем аминокислотный остаток 225 заменен на глицин (Gly) (например, M225G или Met225Gly) или треонин (Thr) (например, M225T или Met225Thr). Эта иллюстративная Rep-бакмида содержит: фрагмент промотора IE1 (SEQ ID NO: 66); нуклеотидную последовательность Rep78, кодирующую модифицированный белок Rep78, в котором отсутствует функциональный инициирующий кодон ниже первого инициирующего кодона, тем самым обеспечивая трансляцию одного белка Rep78. Как будет понятно специалисту в данной области, можно модифицировать эту модифицированную бакмиду Rep78 или модифицированную плазмиду Rep78 нуклеиновой кислотой, кодирующей любой единственный белок Rep (например, Rep68, Rep52, Rep40), которая была модифицирована так, чтобы иметь единственный инициирующий кодон и поэтому кодирует единственный белок Rep. На Фиг. 8B приведено схематическое изображение примера зкДНК-плазмиды-1, с wt-L ITR (левым ITR дикого типа), промотором CAG, трансгеном люциферазы, WPRE и последовательностью полиаденилирования; и mod-R (модифицированным правым) ITR.
[0051] На Фиг. 9A показана предсказанная структура с наименьшей энергией, содержащей RBE части плеча A-A' и C-C' плеча типичного модифицированного левого ITR («ITR-2 (Левый)» SEQ ID NO: 101), а на Фиг. 9B показана предсказанная структура с наименьшей энергией содержащей RBE части плеча A-A' и C-C' плеча типичного модифицированного правого ITR («ITR-2 (Правый)» SEQ ID NO: 102). Предполагается, что они образуют структуру с одним плечом (C-C') и одной непарной петлей. Прогнозируемые значения их свободных энергий Гиббса для развертывания составляют -72,6 ккал/моль.
[0052] На Фиг. 10A показана предсказанная структура с наименьшей энергией содержащей RBE части плеча A-A' и B-B' плеча типичного модифицированного левого ITR («ITR-3 (Левый)» SEQ ID NO: 103), а на Фиг. 10B показана предсказанная структура с наименьшей энергией содержащей RBE части плеча A-A' и B-B' плеча типичного модифицированного правого ITR («ITR-3 (Правый)» SEQ ID NO: 104). Предполагается, что они образуют структуру с одним плечом (B-B') и одной непарной петлей. Прогнозируемые значения их свободных энергий Гиббса для развертывания составляют -74,8 ккал/моль.
[0053] На Фиг. 11A показана предсказанная структура с наименьшей энергией содержащей RBE части плеча A-A' и C-C' плеча типичного модифицированного левого ITR («ITR-4 (Левый)» SEQ ID NO: 105), а на Фиг. 11B показана предсказанная структура с наименьшей энергией, содержащей RBE части плеча A-A' и C-C' плеча типичного модифицированного правого ITR («ITR-4 (Правый)» SEQ ID NO: 106). Предполагается, что они образуют структуру с одним плечом (C-C') и одной непарной петлей. Прогнозируемые значения их свободных энергий Гиббса для развертывания составляют -76,9 ккал/моль.
[0054] На Фиг. 12A показана предсказанная структура с наименьшей энергией, содержащей RBE части плеча A-A' и частей C-C' и B-B' типичного модифицированного левого ITR, которая показывает комплементарные пары оснований частей C-B 'и C'-B («ITR-10 (Левый)» SEQ ID NO: 107), а на Фиг. 12B показана предсказанная структура с наименьшей энергией содержащей RBE части плеча A-A' и частей C-C' и B-B' типичного модифицированного левого ITR, которая показывает комплементарные пары оснований частей C-B 'и C'-B («ITR-10 (Правый)» SEQ ID NO: 108). Предполагается, что они образуют структуру с одним плечом (часть C'-B и C-B'или часть B'-C и B-C') и одной непарной петлей. Прогнозируемые значения их свободных энергий Гиббса для развертывания составляют -83,7 ккал/моль.
[0055] На Фиг. 13A показана предсказанная структура с наименьшей энергией, содержащей RBE части плеча A-A' и частей C-C' и B-B' типичного модифицированного левого ITR («ITR-17 (Левый)» SEQ ID NO: 109); на Фиг. 13B показана предсказанная структура с наименьшей энергией содержащей RBE части плеча A-A' и частей C-C' и B-B' типичного модифицированного левого ITR («ITR-17 (Правый)» SEQ ID NO: 110). Предполагается, что как ITR-17 (левый), так и ITR-17 (правый) образуют структуру с одним плечом (B-B') и одной непарной петлей. Прогнозируемые значения их свободных энергий Гиббса для развертывания составляют -73,3 ккал/моль.
[0056] На Фиг. 14A показана предсказанная структура с наименьшей энергией, содержащей RBE части плеча A-A' типичного модифицированного левого ITR («ITR-6 (Левый)» SEQ ID NO: 111), а на Фиг. 14B показана предсказанная структура с наименьшей энергией содержащей RBE части плеча A-A' типичного модифицированного правого ITR («ITR-6 (Правый)» SEQ ID NO: 112). Предполагается, что как ITR-6 (левый), так и ITR-6 (правый) образуют структуру с одним плечом. Прогнозируемые значения их свободных энергий Гиббса для развертывания составляют -54,4 ккал/моль.
[0057] На Фиг. 15A показана предсказанная структура с наименьшей энергией, содержащей RBE части плеча A-A', плеча C и плеча B-B' типичного модифицированного левого ITR («ITR-1 (Левый)» SEQ ID NO: 113), а на Фиг. 15B показана предсказанная структура с наименьшей энергией содержащей RBE части плеча A-A', плечо C и плечо B-B' типичного модифицированного правого ITR («ITR-1 (Правый)» SEQ ID NO: 114). Предполагается, что как ITR-1 (левый), так и ITR-1 (правый) образуют структуру с двумя плечами, одно из которых является усеченным. Прогнозируемые значения их свободных энергий Гиббса для развертывания составляют -74,7 ккал/моль.
[0058] На Фиг. 16A показана предсказанная структура с наименьшей энергией, содержащей RBE части плеча A-A', плеча C и плеча B-B' типичного модифицированного левого ITR («ITR-5 (Левый)» SEQ ID NO: 545), а на Фиг. 16B показана предсказанная структура с наименьшей энергией содержащей RBE части плеча A-A', плечо B-B' и плечо C типичного модифицированного правого ITR («ITR-5 (Правый)» SEQ ID NO: 116). Предполагается, что как ITR-5 (левый), так и ITR-5 (правый) образуют структуру с двумя плечами, одно из которых (например, плечо C') является усеченным. Прогнозируемые значения их свободных энергий Гиббса для развертывания составляют -73,4 ккал/моль.
[0059] На Фиг. 17A показана предсказанная структура с наименьшей энергией, содержащей RBE части плеча A-A', плеча C-C' и плеча B-B' типичного модифицированного левого ITR («ITR-7 (Левый)» SEQ ID NO: 117), а на Фиг. 17B показана предсказанная структура с наименьшей энергией содержащей RBE части плеча A-A', плечо C-C' и плечо B-B' типичного модифицированного правого ITR («ITR-7 (Правый)» SEQ ID NO: 118). Предполагается, что как ITR-17 (левый), так и ITR-17 (правый) образуют структуру с двумя плечами, одно из которых (например, плечо B-B') является усеченным. Прогнозируемые значения их свободных энергий Гиббса для развертывания составляют -89,6 ккал/моль.
[0060] На Фиг. 18A показана предсказанная структура с наименьшей энергией содержащей RBE части плеча A-A', плеча C-C' и плеча B-B' типичного модифицированного левого ITR («ITR-8 (Левый)» SEQ ID NO: 119), а на Фиг. 18B показана предсказанная структура с наименьшей энергией содержащей RBE части плеча A-A', плечо C-C' и плечо B-B' типичного модифицированного правого ITR («ITR-8 (Правый)» SEQ ID NO: 120). Предполагается, что как ITR-8 (левый), так и ITR-8 (правый) образуют структуру с двумя плечами, одно из которых является усеченным. Прогнозируемые значения их свободных энергий Гиббса для развертывания составляют -86,9 ккал/моль.
[0061] На Фиг. 19A показана предсказанная структура с наименьшей энергией, содержащей RBE части плеча A-A', плеча C-C' и плеча B-B' типичного модифицированного левого ITR («ITR-9 (Левый)» SEQ ID NO: 121), а на Фиг. 19B показана предсказанная структура с наименьшей энергией содержащей RBE части плеча A-A', плеча C-C' и плеча B-B' типичного модифицированного правого ITR («ITR-9 (Правый)» SEQ ID NO: 122). Предполагается, что как ITR-9 (левый), так и ITR-9 (правый) образуют структуру с двумя плечами, одно из которых является усеченным. Прогнозируемые значения их свободных энергий Гиббса для развертывания составляют -85,0 ккал/моль.
[0062] На Фиг. 20A показана предсказанная структура с наименьшей энергией, содержащей RBE части плеча A-A', плеча C-C' и плеча B-B' типичного модифицированного левого ITR («ITR-11 (Левый)» SEQ ID NO: 123), а на Фиг. 20B показана предсказанная структура с наименьшей энергией содержащей RBE части плеча A-A', плеча C-C' и плеча B-B' типичного модифицированного правого ITR («ITR-11 (Правый)» SEQ ID NO: 124). Предполагается, что как ITR-11 (левый), так и ITR-11 (правый) образуют структуру с двумя плечами, одно из которых является усеченным. Прогнозируемые значения их свободных энергий Гиббса для развертывания составляют -89,5 ккал/моль.
[0063] На Фиг. 21A показана предсказанная структура с наименьшей энергией, содержащей RBE части плеча A-A', плеча C-C' и плеча B-B' типичного модифицированного левого ITR («ITR-12 (Левый)» SEQ ID NO: 125), а на Фиг. 21B показана предсказанная структура с наименьшей энергией содержащей RBE части плеча A-A', плеча C-C' и плеча B-B' типичного модифицированного правого ITR («ITR-12 (Правый)» SEQ ID NO: 126). Предполагается, что как ITR-12 (левый), так и ITR-12 (правый) образуют структуру с двумя плечами, одно из которых является усеченным. Прогнозируемые значения их свободных энергий Гиббса для развертывания составляют -86,2 ккал/моль.
[0064] На Фиг. 22A показана предсказанная структура с наименьшей энергией, содержащей RBE части плеча A-A', плеча C-C' и плеча B-B' типичного модифицированного левого ITR («ITR-13 (Левый)» SEQ ID NO: 127), а на Фиг. 22B показана предсказанная структура с наименьшей энергией содержащей RBE части плеча A-A', плеча C-C' и плеча B-B' типичного модифицированного правого ITR («ITR-13 (Правый)» SEQ ID NO: 128). Предполагается, что как ITR-13 (левый), так и ITR-13 (правый) образуют структуру с двумя плечами, одно из которых (например, плечо B-B') является усеченным. Прогнозируемые значения их свободных энергий Гиббса для развертывания составляют -82,9 ккал/моль.
[0065] На Фиг. 23A показана предсказанная структура с наименьшей энергией, содержащей RBE части плеча A-A', плеча C-C' и плеча B-B' типичного модифицированного левого ITR («ITR-14 (Левый)» SEQ ID NO: 129), а на Фиг. 23B показана предсказанная структура с наименьшей энергией содержащей RBE части плеча A-A', плеча C-C' и плеча B-B' типичного модифицированного правого ITR («ITR-14 (Правый)» SEQ ID NO: 130). Предполагается, что как ITR-14 (левый), так и ITR-14 (правый) образуют структуру с двумя плечами, одно из которых (например, плечо B-B') является усеченным. Прогнозируемые значения их свободных энергий Гиббса для развертывания составляют -80,5 ккал/моль.
[0066] На Фиг. 24A показана предсказанная структура с наименьшей энергией, содержащей RBE части плеча A-A', плеча C-C' и плеча B-B' типичного модифицированного левого ITR («ITR-15 (Левый)» SEQ ID NO: 131), а на Фиг. 24B показана предсказанная структура с наименьшей энергией содержащей RBE части плеча A-A', плеча C-C' и плеча B-B' типичного модифицированного правого ITR («ITR-15 (Правый)» SEQ ID NO: 132). Предполагается, что как ITR-15 (левый), так и ITR-15 (правый) образуют структуру с двумя плечами, одно из которых (например, плечо B-B') является усеченным. Прогнозируемые значения их свободных энергий Гиббса для развертывания составляют -77,2 ккал/моль.
[0067] На Фиг. 25A показана предсказанная структура с наименьшей энергией, содержащей RBE части плеча A-A', плеча C-C' и плеча B-B' типичного модифицированного левого ITR («ITR-16 (Левый)» SEQ ID NO: 133), а на Фиг. 25B показана предсказанная структура с наименьшей энергией содержащей RBE части плеча A-A', плеча C-C' и плеча B-B' типичного модифицированного правого ITR («ITR-16 (Правый)» SEQ ID NO: 134). Предполагается, что как ITR-16 (левый), так и ITR-16 (правый) образуют структуру с двумя плечами, одно из которых (например, плечо B-B') является усеченным. Прогнозируемые значения их свободных энергий Гиббса для развертывания составляют -73,9 ккал/моль.
[0068] На Фиг. 26А показаны предсказанные структуры RBE-содержащей части плеча A-A' и модифицированного плеча B-B' и/или модифицированного плеча C-C' типичных примеров модифицированных правых ITR, приведенных в Таблице 10A. Фиг. 26B демонстрирует прогнозируемые структуры RBE-содержащей части плеча A-A' и модифицированного плеча C-C' и/или модифицированного плеча B-B' примеров модифицированных правых ITR, приведенных в Таблице 10B. Показанные структуры являются предсказанными структурами с наименьшей свободной энергией. Обозначение условными цветами: красный = вероятность >99%; оранжевый = вероятность 99% - 95%; бежевый = вероятность 95-90%; темно-зеленый 90% - 80%; ярко-зеленый = 80% - 70%; голубой = 70% - 60%; темно-синий 60% - 50%; и розовый = < 50%.
[0069] Фиг. 27 демонстрирует люциферазную активность клеток насекомых Sf9 GlycoBac, трансфицированных выбранными асимметричными мутантными вариантами ITR из Таблиц 10A и 10B. Вектор зкДНК имел ген люциферазы, фланкированный ITR дикого типа, и модифицированный асимметричный ITR, выбранный из Таблиц 10A или 10B. «ITR-50 R no rep» является известным спасаемым мутантом без коинфекции Rep- содержащим бакуловирусом. «Ложные» условия представлены только реагентами для трансфекции, без донорной ДНК.
[0070] Фиг. 28 показывает нативный агарозный гель (1% агарозы, 1x буфер TAE) репрезентативных неочищенных экстрактов зкДНК из культур клеток насекомых Sf9, трансфицированных зкДНК-плазмидами, содержащими левый wt-ITR с другим ITR, выбранным из различных мутантных правых ITR, раскрытых в таблице 10A. На каждую дорожку вносили 2 мкг общего экстракта. Слева направо: полоса 1) маркеры 1 т.п.о. плюс, дорожка 2) правый ITR-18, дорожка 3) правый ITR-49 дорожка 4) правый ITR-19, дорожка 5) правый ITR-20, дорожка 6) правый ITR-21, дорожка 7) правый ITR-22, дорожка 8) правый ITR-23, дорожка 9) правый ITR-24, дорожка 10) Правый ITR-25, дорожка 11) правый ITR-26, дорожка 12) правый ITR-27, дорожка 13), правый ITR-28, дорожка 14) правый ITR-50, дорожка 15) 1 т.п.о. плюс маркеры.
[0071] Фиг. 29 показывает денатурирующий гель (0,8% щелочной агарозы) репрезентативных конструктов из библиотеки мутантов ITR. Вектор зкДНК получают из плазмидных конструктов, содержащих левую ITR дикого типа с другой ITR, выбранной из различных мутантных правых ITR, описанных в таблице 10А. Слева направо, дорожка 1) маркеры 1 т.п.о. плюс, дорожка 2) правый нерасщепленный ITR-18, дорожка 3) правый расщепленный ITR-18, дорожка 4) правый неразрезанный ITR-19, дорожка 5) правый расщепленный ITR- 19, дорожка 6) правый неразрезанный ITR-21, полоса 7) правый расщепленный ITR-21, дорожка 8) правый расщепленный ITR-25, дорожка 9) правый расщепленный ITR-25. Экстракты обрабатывали эндонуклеазой рестрикции EcoRI. Ожидается, что каждая мутантная зкДНК будет иметь единственный сайт распознавания EcoRI, дающий два характерных фрагмента, ~ 2000 п.о. и ~ 3000 п.о., которые в денатурирующих условиях будут иметь длину ~ 4000 и ~ 6000 п.о., соответственно. Необработанные экстракты зкДНК имеют размер ~ 5000 п.о., и ожидается, что они будут мигрировать в денатурирующих условиях на ~ 11000 п.о.
[0072] Фиг. 30 показывает активность люциферазы in vitro в клетках HEK293 мутантов ITR правого ITR-18, правого ITR-19, правого ITR-21 и правого ITR-25, и ITR-49, где левый ITR в векторе зкДНК представляет собой WT ITR. «Ложные» условия представлены только реагентами для трансфекции, без донорной ДНК, а необработанные являются негативным контролем.
[0073] Фиг. 31 представляет собой таблицу, показывающую различные свойства и активности (например, связывание ДНК, никирование ДНК, геликазную активность, АТФазную активность и активность Zn-пальцев) различных видов Rep-белков (например, Rep78 дикого типа, Rep68 дикого типа, Rep52 дикого типа и Rep40 дикого типа) и модифицированные виды Rep68, например, в которых аминокислота белка Rep78 модифицирована до любого из Y156, K340H, Met→ Gly (M225G). При модификации Met→ Gly (M225G) Rep78 сохраняет все свойства и активности белка Rep78 дикого типа.
[0074] Фиг. 32A и 32B представляют собой неденатурирующие гели, показывающие присутствие высокостабильных ДНК-векторов и характерные полосы, подтверждающие присутствие вектора высокостабильной ДНК с замкнутыми концами (зкДНК), полученного с использованием единственного белка Rep, с использованием описанных здесь способов. На Фиг. 32A более высокие количества вектора зкДНК продуцируются с использованием нуклеиновой кислоты модифицированного Rep78 с модификацией Rep78 в Met→ Gly (M225G) (дорожка 1) или Rep в Met→Thr (M225T) (дорожка 2) по сравнению с получением при использовании нуклеиновой кислоты, кодирующей Rep78 дикого типа (дорожка 5), в котором нуклеиновая кислота экспрессирует как белок Rep78/68, так и белок Rep52/40. Вектор зкДНК не был получен со связывающими/никирующими мутантами Rep78, содержащими модификации Gly (Y156F) (дорожка 3) или Thr (Y156F) (дорожка 4). На Фиг. 32B, мутанты Rep68 Met→ Gly (M225G) и Rep68 Met→Thr (M225T) также продуцировали вектор зкДНК на уровнях, равных или превышающих количество вектора зкДНК, полученного с использованием нуклеиновой кислоты модифицированного Rep78 с модификацией Rep78 в Met→ Gly (M225G) или Rep Met→Thr (M225T). DLSR: плазмидный конструкт, экспрессирующий длинный (Rep78) и короткий (Rep52) белок Rep в тандеме; pIE78: полноразмерная последовательность Rep78 дикого типа; Rep78 M→G: полноразмерный Rep78, содержащий одиночную мутацию M225G; Rep78M→Т: полноразмерный Rep78, содержащий одиночную мутацию M225T; Rep78Y156F: полноразмерный Rep78, имеющий единственную мутацию в никазном домене.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
I. Определения
[0075] Если в данном документе не указано иное, научные и технические термины, используемые применительно к данной заявке, будут иметь значения, общепринятые для специалистов в области техники, к которой относится данное изобретение. Следует понимать, что данное изобретение не ограничено конкретной методологией, протоколами и реагентами, и т.п., описанными в данном документе, и, соответственно, предусматривает различные варианты. Используемая в данном документе терминология служит только для описания конкретных вариантов реализации, и не предназначена для ограничения объема данного изобретения, который определяется исключительно формулой изобретения. Определения общепринятых терминов иммунологии и молекулярной биологии можно найти в следующих источниках: The Merck Manual of Diagnosis and Therapy, 19-е изд., опубликовано Merck Sharp & Dohme Corp., 2011 (ISBN 978-0-911910-19-3); Robert S. Porter et al. (ред.), Fields Virology, 6-е изд., опубликовано Lippincott Williams & Wilkins, Philadelphia, PA, USA (2013), Knipe, D.M. and Howley, P.M. (ред.), The Encyclopedia of Molecular Cell Biology and Molecular Medicine, опубликовано Blackwell Science Ltd., 1999-2012 (ISBN 9783527600908); и Robert A. Meyers (ред.), Molecular Biology and Biotechnology: a Comprehensive Desk Reference, опубликовано VCH Publishers, Inc., 1995 (ISBN 1-56081-569-8); Immunology, Werner Luttmann, опубликовано Elsevier, 2006; Janeway's Immunobiology, Kenneth Murphy, Allan Mowat, Casey Weaver (ред.), Taylor & Francis Limited, 2014 (ISBN 0815345305, 9780815345305); Lewin's Genes XI, опубликовано Jones & Bartlett Publishers, 2014 (ISBN-1449659055); Michael Richard Green and Joseph Sambrook, Molecular Cloning: A Laboratory Manual, 4е изд., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., USA (2012) (ISBN 1936113414); Davis et al., Basic Methods in Molecular Biology, Elsevier Science Publishing, Inc., New York, USA (2012) (ISBN 044460149X); Laboratory Methods in Enzymology: DNA, Jon Lorsch (ред.) Elsevier, 2013 (ISBN 0124199542); Current Protocols in Molecular Biology (CPMB), Frederick M. Ausubel (ред.), John Wiley and Sons, 2014 (ISBN 047150338X, 9780471503385), Current Protocols in Protein Science (CPPS), John E. Coligan (ред.), John Wiley and Sons, Inc., 2005; и Current Protocols in Immunology (CPI) (John E. Coligan, ADA M Kruisbeek, David H Margulies, Ethan M Shevach, Warren Strobe, (ред.) John Wiley and Sons, Inc., 2003 (ISBN 0471142735, 9780471142737), содержание которых в полном объеме включено в данный документ посредством ссылок.
[0076] В данном документе термины «гетерологичная нуклеотидная последовательность» и «трансген» используются взаимозаменяемо и относятся к представляющей интерес нуклеиновой кислоте (отличной от нуклеиновой кислоты, кодирующей полипептид капсида), которая встроена в зкДНК-вектор и может быть доставлена и экспрессирована зкДНК-вектором согласно описанию в данном документе. Представляющие интерес трансгены включают, не ограничиваясь перечисленным, нуклеиновые кислоты, кодирующие полипептиды, предпочтительно, терапевтические (например, для применения в медицинских, диагностических или ветеринарных целях) или иммуногенные полипептиды (например, для вакцин). Согласно некоторым вариантам реализации представляющие интерес нуклеиновые кислоты включают нуклеиновые кислоты, которые транскрибируются в терапевтическую РНК. Трансгены, включенные для использования в зкДНК-векторах по изобретению, включают, без ограничений, экспрессирующие или кодирующие один или несколько полипептидов, пептидов, рибозимов, аптамеров, пептидных нуклеиновых кислот, миРНК, РНКи, микроРНК, днкРНК (длинных некодирующих РНК), антисмысловых олиго- или полинуклеотидов, антител, антигенсвязывающих фрагментов, или любую их комбинацию.
[0077] В данном документе термины «экспрессионная кассета» и «транскрипционная кассета» используются взаимозаменяемо и относятся к линейному отрезку нуклеиновых кислот, который содержит трансген, функционально связанный с одним или несколькими промоторами или другими регуляторными последовательностями, достаточными для направления транскрипции указанного трансгена, но не содержит кодирующие капсид последовательности, другие векторные последовательности или участки инвертированных концевых повторов. Экспрессионная кассета может дополнительно содержать одну или более действующих в цис-положении последовательностей (например, промоторов, энхансеров или репрессоров), один или более интронов, и один или более посттранскрипционных регуляторных элементов.
[0078] В данном документе термин «концевой повтор» или «TR» включает любой вирусный концевой повтор или любую синтетическую последовательность, которая содержит по меньшей мере одну минимальную требуемую точку начала репликации и участок, содержащий палиндромную шпилечную структуру. Связывающая Rep последовательность («RBS») (также называемая RBE (связывающий Rep элемент)) и сайт концевого разрешения («TRS») вместе образуют «минимальную требуемую точку начала репликации» и, соответственно, TR содержит по меньшей мере одну RBS и по меньшей мере один TRS. Каждый из TR, обратно-комплементарных друг относительно друга в пределах определенного отрезка полинуклеотидной последовательности, как правило, называют «инвертированным концевым повтором» или «ITR». В контексте вируса, ITR опосредуют репликацию, упаковку вируса, интеграцию и высвобождение провируса. Как было неожиданно обнаружено авторами данного изобретения, концевые повторы, не являющиеся обратно комплементарными по всей их длине, могут, тем не менее, выполнять традиционные функции ITR и, таким образом, термин ITR в данном документе относится к КП в геноме зкДНК или зкДНК-векторе, который способен опосредовать репликацию зкДНК-вектора. Специалисту в данной области техники будет понятно, что в зкДНК-векторах сложной конфигурации может присутствовать более двух пар ITR или асимметричных ITR. Указанный ITR может представлять собой AAV ITR или ITR не из AAV, или может происходить из AAV ITR или из ITR не из AAV. Например, указанный ITR может происходить из семейства Parvoviridae, куда входят парвовирусы и депендовирусы (например, парвовирус собачьих, парвовирус крупного рогатого скота, парвовирус мышей, парвовирус свиньи, парвовирус человека B-19), или шпилька SV40, которая служит в качестве точки начала репликации SV40, может быть использована как ITR, который дополнительно может быть модифицирован путем усечения, замены, делеции, вставки и/или добавления. Семейство вирусов Parvoviridae состоит из двух подсемейств: Parvovirinae, инфицирующих позвоночных, и Densovirinae, инфицирующих беспозвоночных. Депендопарвовирусы включают вирусное семейство аденоассоциированных вирусов (ААV), которые способны к репликации у позвоночных животных-хозяев, включая, без ограничений, человека, виды приматов, бычьих, собачьих, лошадей и овец. Для удобства в данном документе ITR, расположенный в 5' направлении (выше) относительно экспрессионной кассеты в зкДНК-векторе, называется «5' ITR» или «левым ITR», а ITR, расположенный в 3' направлении (ниже) экспрессионной кассеты в зкДНК-векторе, называется «3' ITR» или «правым ITR».
[0079] «ITR дикого типа» или «WT-ITR» относится к последовательности встречающейся в природе ITR-последовательности ААV или другого депендовируса, которая сохраняет, например, активность связывания Rep и никующую способность Rep. Нуклеотидная последовательность WT-ITR из любого серотипа AAV может незначительно отличаться от канонической встречающейся в природе последовательности из-за вырожденности генетического кода или дрейфа, и, соответственно, последовательности WT-ITR, предусмотренные для применения в данном документе, включают последовательности WT-ITR, образованные в результате встречающихся в природе изменений, происходящих в ходе процесса продуцирования (например, ошибки репликации).
[0080] В данном документе термин «по существу симметричные WT-ITR» или «пара по существу симметричных WT-ITR» относится к паре WT-ITR в составе одного зкДНК-генома или зкДНК-вектора, где оба ITR представляют собой ITR дикого типа, которые имеют обратно-комплементарную последовательность по всей длине. Например, ITR может считаться последовательностью дикого типа, даже если он содержит один или более нуклеотидов, отличающихся от нуклеотидов в канонической встречающейся в природе последовательности, при условии, что указанные изменения не влияют на свойства и общую трехмерную структуру последовательности. Согласно некоторым аспектам, указанные отличающиеся нуклеотиды представляют собой консервативные изменения последовательностей. Неограничивающий пример представлен последовательностью, по меньшей мере на 95%, 96%, 97%, 98% или 99% идентичной канонической последовательности (по оценке, например, с помощью BLAST при параметрах, устанавливаемых по умолчанию), а также имеет симметричную трехмерную пространственную организацию относительно другого WT-ITR, так что их трехмерная структура имеет одинаковую форму в геометрическом пространстве. Указанный по существу симметричный WT-ITR содержит такие же петли A, C-C' и B-B' в трехмерном пространстве. Может быть функционально подтверждено, что по существу симметричный WT-ITR является WT, путем определения наличия в нем функционального сайта связывания Rep (RBE или RBE') и сайта концевого разрешения (trs), который спаривается с соответствующим белком Rep. Можно дополнительно проверить другие функции, включая экспрессию трансгена в пермиссивных условиях.
[0081] В данном документе выражения «модифицированный ITR» или «mod-ITR» или «мутантный ITR» используются в данном документе взаимозаменяемо и относятся к ITR, который содержит мутацию по меньшей мере в одном или более нуклеотидах по сравнению с WT-ITR из того же серотипа. Указанная мутация может приводить к изменению в одной или более из участков A, C, C', B, B' в ITR, и может приводить к изменению трехмерной пространственной организации (т.е. трехмерной структуры в геометрическом пространстве) по сравнению с трехмерной пространственной организацией WT-ITR из того же серотипа.
[0082] Используемый в данном документе термин «асимметричные ITR», также называемый «асимметричными парами ITR», относится к паре ITR в одном геноме зкДНК или зкДНК-векторе, которые не являются обратными комплементами по их полной длине. В качестве одного неограничивающего примера, в асимметричной паре ITR не имеют симметричной трехмерной пространственной организации со своим ITR-партнером, и их трехмерные структуры имеют разные формы в геометрическом пространстве. Другими словами, асимметричная пара ITR имеет разные общие геометрические структуры, т.е. они имеют разную организацию своих A, C-C'- и B-B'-петель в трехмерном пространстве (например, один ITR может иметь короткое плечо C-C' и/или короткое плечо B-B' по сравнению с родственным ITR). Различие в последовательностях между двумя ITR может быть связано с вставкой, делецией, усечением или точечной мутацией одного или нескольких нуклеотидов. В одном варианте реализации, один ITR асимметричной пары ITR может быть последовательностью ITR ААV дикого типа, а другой ITR - модифицированным ITR, как определено в данном документе (например, последовательностью ITR не-дикого типа или синтетической). В другом варианте реализации ни один из ITR асимметричной пары ITR не является последовательностью ААV дикого типа, и указанные два ITR представляют собой модифицированные ITR, которые имеют разные формы в геометрическом пространстве (т.е. разную общую геометрическую структуру). В некоторых вариантах реализации один mod-ITR асимметричной пары ITR может иметь короткое C-C'-плечо, а другой ITR может иметь другую модификацию (например, одно плечо или короткое B-B'-плечо и т.д.), так что они имеют различную трехмерную пространственную организацию по сравнению с парным ему асимметричным mod-ITR.
[0083] В данном документе термин «симметричный ITR» относится к паре ITR в составе одного зкДНК-генома или зкДНК-вектора, мутированных или модифицированных относительно последовательностей депендовирусного ITR дикого типа, и обратно-комплементарных по всей длине. Ни один из ITR не является последовательностью ITR ААV2 дикого типа (т.е. они представляют собой модифицированные ITR, также называемые мутантными ITR), и могут отличаться от последовательности ITR дикого типа вследствие вставки, делеции, замены, усечения или точечной мутации нуклеотидов. Для удобства в данном документе ITR, расположенный в 5' направлении (выше) относительно экспрессионной кассеты в зкДНК-векторе, называется «5' ITR» или «левым ITR», а ITR, расположенный в 3' направлении (ниже) экспрессионной кассеты в зкДНК-векторе, называется «3' ITR» или «правым ITR».
[0084] Используемый в данном документе термин «по существу симметричные модифицированные ITR» или «по существу симметричная пара mod-ITR» относится к паре модифицированных ITR в одном геноме зкДНК или зкДНК-векторе, которые оба имеют обратно комплементарные последовательности по всей своей длине. Например, модифицированный ITR может считаться по существу симметричным, даже имея некоторые нуклеотидные последовательности, отклоняющиеся от обратно комплементарной последовательности, если указанные изменения не влияют на свойства и общую форму. Один неограничивающий пример представлен последовательностью, которая по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98% или 99% идентична канонической последовательности (по оценке в BLAST при использовании параметров, устанавливаемых по умолчанию), а также имеет симметричную трехмерную пространственную организацию в отношении родственного модифицированного ITR, так что их трехмерные структуры имеют одинаковую форму в геометрическом пространстве. Иными словами, пара по существу симметричных модифицированных ITR содержит одинаково организованные в трехмерном пространстве петли A, C-C' и B-B'. Согласно некоторым вариантам реализации ITR из пары mod-ITR могут содержать разные обратно-комплементарные нуклеотидные последовательности, но все же иметь одинаковую симметричную трехмерную пространственную организацию - то есть оба ITR содержат мутации, которые приводят к одной и той же общей трехмерной форме. Например, один ITR (например, 5' ITR) в паре mod-ITR может быть взят из одного серотипа, а другой ITR (например, 3' ITR) может быть взят из другого серотипа, однако оба ITR могут содержать одинаковую соответствующую мутацию (например, в том случае, если 5'ITR содержит делецию в участке C, родственный модифицированный 3'ITR из другого серотипа содержит делецию в аналогичном положении в участке C'), таким образом, что ITR в паре модифицированных ITR имеют одинаковую симметричную трехмерную пространственную организацию. В таких вариантах реализации каждый ITR в паре модифицированных ITR может принадлежать к разным серотипам (например, AAV1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 и 12), таким как комбинация ААV2 и ААV6, причем модификация одного ITR отображается в соответствующем положении родственного ITR другого серотипа. Согласно одному варианту реализации пара по существу симметричных модифицированных ITR относится к паре модифицированных ITR (mod-ITR) при условии, что различие нуклеотидных последовательностей указанных ITR не влияет на свойства или общую форму, и они имеют по существу одинаковую форму в трехмерном пространстве. Неограничивающий пример представлен mod-ITR, последовательность которого по меньшей мере на 95%, 96%, 97%, 98% или 99% идентична каноническому mod-ITR по оценке с применением стандартных способов, хорошо известных в данной области техники, таких как BLAST (Basic Local Alignment Search Tool) или BLASTN, при параметрах, устанавливаемых по умолчанию, а также имеет симметричную трехмерную пространственную организацию, так что их трехмерная структура имеет одинаковую форму в геометрическом пространстве. ITR в паре по существу симметричных mod-ITR содержит одинаковые петли A, C-C'и B-B' в трехмерном пространстве, например, если модифицированный ITR в паре по существу симметричных mod-ITR содержит делецию в плече C-C', то родственный mod-ITR содержит соответствующую делецию в плече C-C', а также имеет аналогичную трехмерную структуру остальных петель A и B-B', принимающую такую же форму в геометрическом пространстве, что и у родственного mod-ITR.
[0085] Термин «фланкирование» касается относительного положения одной последовательности нуклеиновой кислоты по отношению к другой последовательности нуклеиновой кислоты. В общем, в последовательности ABC A и C фланкируют B. То же самое верно для расположения AxBxC. Таким образом, фланкирующая последовательность предшествует или следует за фланкируемой последовательностью, но не обязательно должна быть смежной с, или непосредственно прилегающей к, фланкируемой последовательности. В одном варианте реализации термин фланкирование относится к концевым повторам на каждом конце линейного дуплексного зкДНК-вектора.
[0086] Используемый в данном документе термин «геном зкДНК» относится к экспрессионной кассете, которая дополнительно включает по меньшей мере один участок инвертированного концевого повтора. ЗкДНК-геном может дополнительно содержать одну или более спейсерных участков. В некоторых вариантах реализации геном зкДНК включен в виде межмолекулярного дуплексного полинуклеотида ДНК в плазмиду или вирусный геном.
[0087] Используемый в данном документе термин «спейсерный участок зкДНК» относится к промежуточной последовательности, которая разделяет функциональные элементы в указанном зкДНК-векторе или геноме зкДНК. Согласно некоторым вариантам реализации спейсерные участки зкДНК удерживают два функциональных элемента на требуемом расстоянии для оптимальной функциональности. Согласно некоторым вариантам реализации спейсерные участки зкДНК обеспечивают или повышают генетическую стабильность зкДНК-генома в составе, например, плазмиды или бакуловируса. Согласно некоторым вариантам реализации спейсерные участки зкДНК облегчают подготовленные генетические манипуляции с зкДНК-геномом, обеспечивая удобное расположение для сайтов клонирования и т.п. Например, согласно определенным аспектам олигонуклеотидный «полилинкер», содержащий несколько сайтов рестрикции эндонуклеазами или последовательность не из открытой рамки считывания, сконструированный так, чтобы он не содержал известных сайтов связывания белка (например, транскрипционного фактора), может быть размещен в зкДНК-геноме для разделения цис-действующих факторов, например, путем вставки 6-мера, 12-мера, 18-мера, 24-мера, 48-мера, 86-мера, 176-мера и т.п. между сайтом концевого разрешения и расположенным в направлении 5' регуляторным элементом транскрипции. Аналогичным образом, спейсер может быть встроен между сигнальной последовательностью полиаденилирования и 3'-сайтом концевого разрешения.
[0088] В данном документе термины «сайт связывания Rep», «связывающий Rep элемент», «RBE» и «RBS» используются взаимозаменяемо и относятся к сайту связывания для белка Rep (например, AAV Rep 78 или AAV Rep 68), который после связывания белком Rep позволяет белку Rep осуществлять сайт-специфическую эндонуклеазную активность в отношении последовательности, включающей RBS. Последовательность RBS вместе с ее обратным комплементом образуют один RBS. Последовательности RBS известны в данной области техники и включают, например, 5'-GCGCGCTCGCTCGCTC-3′ (SEQ ID NO: 531), представляющую собой последовательность RBS, идентифицированную в AAV2. Любая известная последовательность RBS может применяться во вариантах реализации данного изобретения, в том числе другие известные последовательности RBS AAV и другие известные встречающиеся в естественной среде или синтетические последовательности RBS. Без связи с какой-либо теорией, считается, что нуклеазный домен белка Rep связывается с дуплексной последовательностью нуклеотидов GCTC и, соответственно, на дуплексном олигонуклеотиде 5'-(GCGC)(GCTC)(GCTC)(GCTC)-3' (SEQ ID NO: 531) происходит прямое связывание и стабильная сборка двух известных белков Rep AAV. Кроме того, растворимые агрегированные конформеры (т.е., неопределенное число взаимосвязанных белков Rep) диссоциируют и связываются с олигонуклеотидами, которые содержат сайты связывания Rep. Каждый белок Rep взаимодействует и с азотистыми основаниями, и с фосфодиэфирным остовом на каждой цепи. Взаимодействия с азотистыми основаниями обеспечивают специфичность в отношении последовательности, тогда как взаимодействия с фосфодиэфирным остовом неспецифичны или менее специфичны в отношении последовательностей и стабилизируют комплекс белка с ДНК.
[0089] Используемые в данном документе термины «сайт концевого разрешения» и «TRS» используются взаимозаменяемо и относятся к участку, в котором Rep образует тирозин-фосфодиэфирную связь с 5'-тимидином, с образованием 3'-ОН, который служит субстратом для удлинения ДНК с помощью клеточной ДНК-полимеразы, например, ДНК-полимеразой дельта или ДНК-полимеразой эпсилон. Как вариант, комплекс Rep-тимидин может принимать участие в скоординированной реакции лигирования. Согласно некоторым вариантам реализации TRS охватывает, как минимум, неспаренное тимидиновое основание. Согласно некоторым вариантам реализации никирующую эффективность TRS по меньшей мере отчасти можно контролировать за счет расстояния между ним и RBS в пределах одной молекулы. Если акцепторный субстрат представляет собой комплементарный ITR, итоговый продукт представляет собой внутримолекулярный дуплекс. TRS последовательности известны в данной области техники, и включают, например, 5'-GGTTGA-3' (SEQ ID NO: 45), представляющую собой гексануклеотидную последовательность, идентифицированную в AAV2. Согласно вариантам реализации данного изобретения может применяться любая известная последовательность TRS, в том числе другие известные последовательности TRS AAV и другие известные природные или синтетические последовательности TRS, такие как AGTT (SEQ ID NO: 46), GGTTGG (SEQ ID NO: 47), AGTTGG (SEQ ID NO: 48), AGTTGA (SEQ ID NO: 49); и другие мотивы, такие как RRTTRR (SEQ ID NO: 50).
[0090] Используемый в данном документе термин «зкДНК-плазмида» относится к плазмиде, которая содержит зкДНК-геном в виде межмолекулярного дуплекса.
[0091] Используемый в данном документе термин «зкДНК-бакмида» относится к геному инфекционного бакуловируса, включающему геном зкДНК в виде межмолекулярного дуплекса, способному размножаться в E. coli в виде плазмиды и, соответственно, выполнять роль челночного вектора для бакуловируса.
[0092] В данном документе термин «зкДНК-бакуловирус» относится к бакуловирусу, который содержит геном зкДНК-геном в виде межмолекулярного дуплекса в составе генома бакуловируса.
[0093] В данном документе термины «инфицированная зкДНК-бакуловирусом клетка насекомого» и «зкДНК-BIIC» используются взаимозаменяемо и относятся к клетке-хозяину беспозвоночного животного (включая, без ограничений, клетку насекомого (например, клетку Sf9)), инфицированную зкДНК-бакуловирусом.
[0094] В данном документе термин «зкДНК» относится к бескапсидной замкнутой линейной двухцепочечной (дц) дуплексной ДНК для невирусного переноса генов, синтетической или другой. Подробное описание зкДНК приведено в международной заявке PCT/US2017/020828, поданной 3 марта 2017 г., содержание которой полностью явным образом включено в данный документ посредством ссылки. Некоторые способы получения зкДНК-вектора, содержащего различные последовательности и конфигурации инвертированных концевых повторов (ITR), с использованием клеточных методов описаны в примере 1 в международной заявке PCT/US18/49996, поданной 7 сентября 2018 г., и международной заявке PCT/US2018/064242, поданной 6 декабря 2018 г., каждая из которых полностью включена в данный документ посредством ссылки. Некоторые способы получения синтетических зкДНК-векторов содержащие различные последовательности и конфигурации ITR, описаны, например, в международной заявке PCT/US2019/14122, поданной 18 января 2019 г., содержание которой полностью включено в данный документ посредством ссылки.
[0095] Используемый в данном документе термин «ДНК-вектор с замкнутыми концами» относится к бескапсидному ДНК-вектору с по меньшей мере одним ковалентно замкнутым концом, причем по меньшей мере часть вектора имеет структуру внутримолекулярного дуплекса.
[0096] Используемые в данном документе термины «зкДНК-вектор» и «зкДНК» используются взаимозаменяемо и относятся к ДНК-вектору с замкнутыми концами, содержащему по меньшей мере один концевой палиндром. Согласно некоторым вариантам реализации указанная зкДНК содержит два ковалентно замкнутых конца.
[0097] В данном документе термин «нкДНК» или «никированная зкДНК» относится к ДНК с замкнутыми концами, содержащей разрыв или пропуск размером 1-100 пар оснований в участке «стебля» или спейсерном участке 5', расположенном выше открытой рамки считывания (например, промотор и трансген для экспрессии).
[0098] В данном документе термины «пропуск» и «разрыв» используются взаимозаменяемо и относятся к прерванной части синтетического ДНК-вектора согласно данному изобретению, создавая отрезок части одноцепочечной ДНК в двухцепочечной зкДНК. Длина пропуска может составлять от одной 1 пары оснований до 100 пар оснований в одной цепи дуплексной ДНК. Типичные пропуски, разработанные и созданные с применением способов, описанных в данном документе, и синтетические векторы, полученные указанными способами, могут иметь длину, например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59 или 60 пар оснований. Примеры пропусков в данном описании могут иметь длину от 1 пар оснований до 10 пар оснований, от 1 до 20 пар оснований, от 1 до 30 пар оснований.
[0099] Как определено в данном документе, «репортеры» относятся к белкам, которые могут использоваться для обеспечения детектируемых показаний. Репортеры обычно продуцируют измеримый сигнал, например, флуоресцентный, цветовой или люминесцентный. Кодирующие последовательности репортерных белков кодируют белки, присутствие которых в клетке или в организме легко наблюдать. Например, флуоресцентные белки заставляют клетку флуоресцировать при возбуждении светом с конкретной длиной волны, люциферазы заставляют клетку катализировать реакцию, которая генерирует свет, а ферменты, такие как β-галактозидаза, преобразуют субстрат в окрашенный продукт. Типичные примеры репортерных полипептидов, пригодных для экспериментальных или диагностических целей, включают, без ограничений, β-лактамазу, β-галактозидазу (LacZ), щелочную фосфатазу (ЩФ), тимидинкиназу (TK), зеленый флуоресцентный белок (ЗФБ) и другие флуоресцентные белки, хлорамфениколацетилтрансферазу (ХАТ), люциферазу и другие, хорошо известные в данной области техники.
[00100] Используемый в данном документе термин «эффекторный белок» относится к полипептиду, который обеспечивает детектируемые показания, либо, например, как репортерный полипептид, либо, более предпочтительно, как полипептид, убивающий клетку, например, токсин, или агент, придающий клетке чувствительность к убиванию определенным агентом или при отсутствии определенного агента. Эффекторные белки включают любой белок или пептид, который прямо нацелен на ДНК и/или РНК клетки-хозяина или повреждает их. Например, эффекторные белки могут включать, не ограничиваясь перечисленными, рестрикционную эндонуклеазу, которая нацелена на последовательность ДНК клетки-хозяина (геномный или внехромосомный элемент), протеазу, расщепляющую полипептид-мишень, необходимый для выживания клетки, ингибитор ДНК-гиразы и токсин рибонуклеазного типа. В некоторых вариантах реализации экспрессия эффекторного белка, контролируемая синтетическим биологическим контуром, как описано в данном документе, может принимать участие в качестве фактора в другом синтетическом биологическом контуре, с расширением таким образом диапазона и сложности отклика системы биологического контура.
[00101] Регуляторы транскрипции относятся к активаторам и репрессорам транскрипции, которые либо активируют, либо подавляют транскрипцию гена, представляющего интерес. Промоторы представляют собой участки нуклеиновой кислоты, которые инициируют транскрипцию конкретного гена. Активаторы транскрипции обычно связываются поблизости от транскрипционных промоторов и рекрутируют РНК-полимеразу, чтобы прямо инициировать транскрипцию. Репрессоры связываются с промоторами транскрипции и стерически препятствуют инициации транскрипции РНК-полимеразой. Другие регуляторы транскрипции могут служить либо активатором, либо репрессором в зависимости от места их связывания и условий в клетке и окружающей среде. Неограничивающие примеры классов регуляторов транскрипции включают, без ограничений, гомеодоменные белки, белки с мотивом «цинковый палец», белки с мотивом «крылатая спираль» (forkhead-белки) и белки с мотивом «лейциновая застежка».
[00102] При использовании в данном документе, «белок-репрессор» или «белок-индуктор» представляет собой белок, который связывается с элементом регуляторной последовательности и подавляет или активирует, соответственно, транскрипцию последовательностей, функционально связанных с указанным элементом регуляторной последовательности. Предпочтительные репрессорные и индукторные белки согласно описанию в данном документе чувствительны к присутствию или отсутствию по меньшей мере одного входного агента или фактора внешней среды. Предпочтительные белки, как описано в данном документе, являются модульными по форме и содержат, например, отделяемые ДНК-связывающие и связывающие вводимые агенты, или откликающиеся на них элементы или домены.
[00103] Используемый в данном документе термин «носитель» включает любые и все возможные растворители, дисперсионные среды, основы, покрытия, разбавители, антибактериальные и противогрибковые агенты, изотонические и замедляющие абсорбцию агенты, буферы, растворы-носители, суспензии, коллоиды и т.п. Применение таких сред и агентов для фармацевтически активных веществ хорошо известно в данной области техники. Также вспомогательные активные ингредиенты могут быть включены в указанные композиции. Выражение «фармацевтически приемлемый» относится к молекулярным объектам и композициям, которые не приводят к токсической, аллергической или аналогичной нежелательной реакции при введении хозяину.
[00104] Используемый в данном документе термин «домен, реагирующий на вводимый агент», представляет собой домен транскрипционного фактора, который связывает или иначе отвечает на условие или вводимый агент, обеспечивая реакцию присоединенного к нему ДНК-связывающего слитого домена на наличие указанного условия или входного воздействия. Согласно одному варианту реализации присутствие указанного условия или входного агента приводит к конформационному изменению в отвечающем на входной агент домене, или в белке, с которым он слит, которое модифицирует модулирующую транскрипцию активность транскрипционного фактора.
[00105] Термин «in vivo» относится к анализам или способам, которые реализуют на организме или в организме, таком как многоклеточное животное. Согласно некоторым описанным здесь аспектам, может быть указано, что способ или применение реализуют «in vivo», если используется одноклеточный организм, такой как бактерия. Термин «ex vivo» относится к способам и применению, которые реализуют с использованием живой клетки с интактной мембраной, находящейся вне организма многоклеточного животного или растения, например, эксплантаты, культивируемые клетки, в том числе первичные клетки и линии клеток, трансформированные линии клеток, и экстрагированные ткани или клетки, включая, в том числе, клетки крови. Термин «in vitro» относится к анализам и способам, которые не требуют присутствия клетки с интактной мембраной, например, клеточных экстрактов, и могут относиться к введению программируемой синтетической биологической цепи в бесклеточной системе, такой как среда, не содержащая клеток или клеточных систем, таких как клеточные экстракты.
[00106] Термин «промотор» в данном документе относится к любой последовательности нуклеиновой кислоты, которая регулирует экспрессию другой последовательности нуклеиновой кислоты за счет управления транскрипцией указанной последовательности нуклеиновой кислоты, которая может представлять собой гетерологичный целевой ген, кодирующий белок или РНК. Промоторы может быть конститутивными, индуцируемыми, репрессируемыми, тканеспецифическими или представлять собой любую комбинацию перечисленного. Промотор представляет собой контрольный участок последовательности нуклеиновой кислоты, в которой осуществляется контроль инициации и скорости транскрипции остальной части последовательности нуклеиновой кислоты. Промотор может также содержать генетические элементы, с которыми могут связываться регуляторные белки и молекулы, такие как РНК-полимераза и другие транскрипционные факторы. Согласно некоторым вариантам реализации описанных в данном документе аспектов промотор может управлять экспрессией транскрипционного фактора, который регулирует экспрессию самого промотора, или экспрессию другого промотора, используемого в другом модульном компоненте синтетических биологических цепей, описанных в данном документе. В последовательности промотора расположен сайт инициации транскрипции, а также связывающие белки домены, отвечающие за связывание РНК-полимеразы. Эукариотические промоторы часто, однако не всегда содержат «TATA»-боксы и «CAT»-боксы. Различные промоторы, в том числе индуцируемые промоторы, могут применяться для управления экспрессией трансгенов в зкДНК-векторах согласно описанию в данном документе.
[00107] Термин «энхансер» в данном документе относится к действующей в цис-положении (цис-действующей) регуляторной последовательности (например, размером 50-1500 пар оснований), которая связывает один или более белков (например, белки-активаторы или транскрипционный фактор) для усиления транскрипционной активации последовательности нуклеиновой кислоты. Энхансеры могут быть расположены на расстоянии до 1 000 000 пар оснований в 5'-направлении от сайта начала гена или в 3'-направлении от сайта начала гена, который они регулируют. Энхансер может быть расположен в пределах интронного участка или в экзонном участке неродственного гена.
[00108] Можно сказать, что промотор управляет экспрессией или управляет транскрипцией последовательности нуклеиновой кислоты, которую он регулирует. Выражения «функционально связанный», «функционально расположенный», «с функциональной связью», «под контролем» и «под транскрипционным контролем» указывают, что промотор находится в корректном функциональном положении и/или ориентации относительно последовательности нуклеиновой кислоты, которую он регулирует, для контроля инициации транскрипции и/или экспрессии этой последовательности. «Инвертированный промотор» в данном документе относится к промотору, последовательность нуклеиновой кислоты в котором располагается в обратной ориентации, так что цепь, которая была смысловой, становится некодирующей цепью, и наоборот. Последовательности инвертированных промоторов могут применяться, согласно различным вариантам реализации, для регуляции состояния переключателя. Кроме того, согласно различным вариантам реализации промотор может применяться в сочетании с энхансером.
[00109] Промотор может быть представлен промотором, в естественных условиях, ассоциированным с геном или последовательностью, а также может быть получен путем выделения некодирующих 5' последовательностей, расположенных в 5' направлении от кодирующего сегмента и/или экзона определенного гена или последовательности. Такой промотор может называться «эндогенным». Аналогичным образом, согласно некоторым вариантам реализации энхансер может быть представлен энхансером, в естественных условиях ассоциированным с последовательностью нуклеиновой кислоты, расположенной либо в 5'-3', либо в 3'-5' направлении от указанной последовательности.
[00110] Согласно некоторым вариантам реализации кодирующий сегмент нуклеиновой кислоты расположен под контролем «рекомбинантного промотора» или «гетерологичного промотора»; оба указанных термина относятся к промотору, в норме не ассоциированному с кодируемой последовательностью нуклеиновой кислоты, с которой промотор функционально связан в естественной среде. Рекомбинантный или гетерологичный энхансер относится к энхансеру, в норме не ассоциированному с определенной последовательностью нуклеиновой кислоты в естественной среде. Такие промоторы или энхансеры могут включать промоторы или энхансеры других генов; промоторы или энхансеры, выделенные из любой другой прокариотической, вирусной или эукариотической клетки; и синтетические промоторы или энхансеры, которые не являются «встречающимися в природе», т.е., содержат другие элементы других транскрипционных регуляторных участков, и/или мутации, которые изменяют экспрессию, полученные с помощью способов генетического конструирования, которые известны в данной области техники. Наряду с получением последовательностей нуклеиновых кислот промоторов и энхансеров синтетическим путем, последовательности промоторов могут быть получены с применением технологии рекомбинантного клонирования и/или амплификации нуклеиновой кислоты, в том числе ПЦР, применительно к синтетическим биологическим цепям и модулям согласно описанию в данном документе (см., например, патент США №4683202, патент США №5928906, каждый из которых включен в данный документ посредством ссылки). Кроме того, предусматривается также возможность применения контрольных последовательностей, которые направляют транскрипцию и/или экспрессию последовательностей в неядерных органеллах, таких как митохондрии, хлоропласты и т.п.
[00111] Как описано в данном документе, «индуцибельный промотор» представляет собой промотор, который характеризуется инициацией или усилением транскрипционной активности в присутствии, под влиянием, или при контакте с, индуктором или индуцирующим агентом. «Индуктор» или «индуцирующий агент» согласно определению в данном документе может быть эндогенным или в норме экзогенным соединением или белком, который вводят таким образом, чтобы он был активен в отношении индукции транскрипционной активности индуцируемого промотора. Согласно некоторым вариантам реализации указанный индуктор или индуцирующий агент, т.е., химическое вещество, соединение или белок, сам может быть результатом транскрипции или экспрессии последовательности нуклеиновой кислоты (т.е., индуктор может представлять собой индукторный белок, экспрессируемый другим компонентом или модулем), который сам может быть под контролем или представлять собой индуцируемый промотор. Согласно некоторым вариантам реализации индуцируемый промотор индуцируется в отсутствие определенных агентов, таких как репрессор. Примеры индуцируемых промоторов включают, не ограничиваясь перечисленными, тетрациклин, металлотионин, экдизон, вирусы млекопитающих (например, поздний аденовирусный промотор; длинный концевой повтор вируса опухоли молочной железы мышей (MMTV-LTR)) и другие отвечающие на стероиды промоторы, промоторы, отвечающие на рапамицин и т.п.
[00112] Термин «субъект» в данном документе относится к человеку или животному, для которого предложено лечение, в том числе профилактическое лечение, зкДНК-вектором в соответствии с данным изобретением. Обычно указанное животное представляет собой позвоночное, такое как, не ограничиваясь перечисленными, примат, грызун, домашнее животное или дичь. Приматы включают, не ограничиваясь перечисленными, шимпанзе, яванского макака, паукообразных обезьян и макак, например, макака-резус. Грызуны включают мышей, крыс, сурков, хорьков, кроликов и хомяков. Домашние и дикие животные включают, не ограничиваясь перечисленными, коров, лошадей, свиней, оленей, бизонов, буйволов, кошачьих, например, домашних кошек, собачьих, например, собак, лисиц, волков, виды птиц, например, курицу, эму, страуса, и рыб, например, форель, сома и лосося. Согласно определенным вариантам реализации аспектов, описанных в данном документе, указанный субъект представляет собой млекопитающее, например, примата или человека. Субъект может быть мужского или женского пола. Кроме того, субъект может представлять собой младенца или ребенка. Согласно некоторым вариантам реализации указанный субъект может представлять собой новорожденного или нерожденного субъекта, например, субъект находится in utero. Предпочтительно, указанный субъект представляет собой млекопитающее. Указанное млекопитающее может представлять собой человека, не человекообразного примата, мышь, крысу, собаку, кошку, лошадь или корову, однако указанные примеры не являются ограничивающими. Млекопитающие, отличные от человека, могут быть целесообразным образом использованы в качестве субъектов в животных моделях заболеваний и расстройств. Кроме того, описанные в данном документе способы и композиции могут применяться для одомашненных животных и/или домашних питомцев. Субъект-человек может быть любого возраста, пола, принадлежать к любой расе или этнической группе, например, представлять собой европеоида (белая раса), азиата, африканца, чернокожего, афроамериканца, афро-европейца, латиноамериканца, иметь ближневосточное происхождение и т.п. Согласно некоторым вариантам реализации субъект может представлять собой пациента или другого субъекта в клинических условиях. Согласно некоторым вариантам реализации указанный субъект уже проходит лечение.
[00113] В данном контексте термин «антитело» используется в самом широком смысле и охватывает различные структуры антител, в том числе, но не ограничиваясь перечисленными, моноклональные антитела, поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела) и фрагменты антител, при условии, что они демонстрируют требуемую антигенсвязывающую активность. «Фрагмент антитела» относится к молекуле, отличной от интактного антитела, которая содержит часть интактного антитела, которая связывает тот же антиген, с которым связывается интактное антитело. Согласно одному варианту реализации антитело или его фрагмент содержит цепь иммуноглобулина или фрагмент антитела и по меньшей мере одну последовательность вариабельного домена иммуноглобулина. Примеры антител или их фрагментов включают, не ограничиваясь перечисленными, Fv, scFv, Fab-фрагмент, Fab', F(ab')2, Fab'-SH, однодоменное антитело (dAb), тяжелую цепь, легкую цепь, тяжелую и легкую цепь, полное антитело (например, включающее все из Fc, Fab, тяжелых цепей, легких цепей, вариабельных участков и т.п.), биспецифическое антитело, диатело, линейное антитело, одноцепочечное антитело, интратело, моноклональное антитело, химерное антитело, мультиспецифическое антитело или мультимерное антитело. Антитело или его фрагмент может относиться к любому классу, в том числе, но не ограничиваясь перечисленными, IgA, IgD, IgE, IgG и IgM, и любому их подклассу, в том числе, но не ограничиваясь перечисленными, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Кроме того, антитело может быть получено от любого млекопитающего, например, приматов, людей, крыс, мышей, лошадей, коз и т.д. Согласно одному варианту реализации указанное антитело представляет собой антитело человека или гуманизированное антитело. Согласно некоторым вариантам реализации указанное антитело представляет собой модифицированное антитело. Согласно некоторым вариантам реализации компоненты антитела могут экспрессироваться отдельно, таким образом, чтобы после экспрессии белковых компонентов происходила самосборка антитела. Согласно некоторым вариантам реализации указанное антитело «гуманизировано» для снижения иммуногенных реакций у человека. Согласно некоторым вариантам реализации указанное антитело обладает требуемой функцией, например, функцией взаимодействия с нужным белком и его ингибирование для лечения заболевания или симптома заболевания. Согласно одному варианту реализации указанное антитело или его антигенсвязывающий фрагмент содержит каркасный участок или участок Fc.
[00114] В данном документе термин «антигенсвязывающий домен» молекулы антитела относится к части молекулы антитела, например, молекулы иммуноглобулина (Ig), которая принимает участие в связывании антигена. Согласно вариантам реализации указанный сайт связывания антигена образован остатками аминокислот вариабельных (V) участков тяжелой (H) и легкой (L) цепей. Три высоконеоднородных участка в вариабельных участках тяжелой и легкой цепей, называемые гипервариабельными участками, располагаются между более консервативными фланкирующими отрезками, называемыми «каркасными участками» (FR). FR представляют собой аминокислотные последовательности, которые в естественных условиях расположены между гипервариабельными участками в иммуноглобулинах и примыкают к ним. Согласно вариантам реализации в молекуле антитела три гипервариабельных участка легкой цепи и три гипервариабельных участка тяжелой цепи расположены друг относительно друга в трехмерном пространстве таким образом, что они образуют антигенсвязывающую поверхность, комплементарную трехмерной поверхности связываемого антигена. Указанные три гипервариабельные участки каждой из тяжелой и легкой цепей называются «определяющими комплементарность участками» или «CDR». Каркасный участок и CDR были определены и описаны, например, в источниках: Kabat, E. A., et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242, и Chothia, C. et al. (1987) J. Mol. Biol. 196:901-917. Каждая вариабельная цепь (например, вариабельная тяжелая цепь и вариабельная легкая цепь), как правило, состоит из трех CDR и четырех FR, расположенных в последовательности аминокислот, в направлении от аминоконца к карбоксильному концу, в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4.
[00115] В данном документе термин «полноразмерное антитело» относится к молекуле иммуноглобулина (Ig) (например, IgG-антителу), например, встречающейся в природе и образующейся в результате нормальных процессов рекомбинации фрагментов иммуноглобулиновых генов.
[00116] В данном документе термин «функциональный фрагмент антитела» относится к фрагменту, который связывается с тем же антигеном, что и распознаваемый интактным (например, полноразмерным) антителом. Термины «фрагмент антитела» или «функциональный фрагмент» также включают выделенные фрагменты, состоящие из вариабельных участков, такие как «Fv»-фрагменты, состоящие из вариабельных участков тяжелых и легких цепей, или рекомбинантных одноцепочечных полипептидных молекул, в которых легкие и тяжелые вариабельные участки связаны пептидным линкером («scFv-белки»). Согласно некоторым вариантам реализации фрагмент антитела не содержит частей антител без антигенсвязывающей активности, таких как Fc-фрагменты или отдельные остатки аминокислот.
[00117] В данном документе «последовательность вариабельного домена иммуноглобулина» относится к последовательности аминокислот, которая может образовывать структуру вариабельного домена иммуноглобулина. Например, указанная последовательность может включать полностью или частично последовательность аминокислот встречающегося в природе вариабельного домена. Например, указанная последовательность может включать или не включать одну, две или более N- или C-концевых аминокислот, или может включать другие изменения, которые совместимы с образованием структуры белка.
[00118] Термины «полинуклеотид» и «нуклеиновая кислота», используемые в данном документе взаимозаменяемо, относятся к полимерной форме нуклеотидов любой длины, будь то рибонуклеотиды либо дезоксирибонуклеотиды. Таким образом, этот термин включает одно-, двух- или многоцепочечные ДНК или РНК, геномную ДНК, кДНК, гибриды ДНК-РНК или полимер, включающий пуриновые и пиримидиновые основания или другие природные, химически или биохимически модифицированные, неприродные или дериватизированные нуклеотидные основания. «Олигонуклеотид» обычно относится к полинуклеотидам, содержащим от примерно 5 до примерно 100 нуклеотидов одно- или двухцепочечной ДНК. Однако для целей данного изобретения не существует верхнего предела длины олигонуклеотида. Олигонуклеотиды также известны как «олигомеры» или «олиго» (oligos) и могут быть выделены из генов или химически синтезированы способами, известными в данной области. Термины «полинуклеотид» и «нуклеиновая кислота» следует понимать как включающие, применительно к описываемым вариантам реализации, одноцепочечные (такие как смысловые или антисмысловые) и двухцепочечные полинуклеотиды. ДНК может находиться в форме, например, антисмысловых молекул, плазмидной ДНК, дуплексов ДНК-ДНК, предварительно конденсированной ДНК, продуктов ПЦР, векторов (P1, PAC, BAC, YAC, искусственные хромосомы), экспрессионных кассет, химерных последовательностей, хромосомной ДНК, или производных и комбинаций указанных групп. ДНК может находиться в форме миникольца, плазмиды, бакмиды, минигена, служебной ДНК (линейного ковалентно замкнутого ДНК-вектора), линейной дуплексной ДНК с замкнутыми концами (CELiD или зкДНК), ДНК Doggybone (dbDNA ™), гантелеобразную ДНК, минималистичного вектора для иммунологически определенной генной экспрессии (MIDGE), вирусного вектора или невирусных векторов. РНК может находиться в форме короткой интерферирующей РНК (киРНК), дцРНК - субстрата Dicer, малой шпилечной РНК (мшРНК), aсимметричной интерферирующей РНК (аиРНК), микроРНК (миРНК), мРНК, рРНК, тРНК, вирусной РНК (вРНК) и их комбинаций. Нуклеиновые кислоты включают нуклеиновые кислоты, содержащие известные аналоги нуклеотидов или модифицированные остатки остова или связи, которые являются синтетическими, встречающимися в природе и не встречающимися в природе, которые имеют связывающие свойства, аналогичные свойствам референсной нуклеиновой кислоты. Примеры таких аналогов и/или модифицированных остатков включают, без ограничения, фосфотиоаты, фосфородиамидат-морфолиновый олигомер (морфолино), фосфорамидаты, метилфосфонаты, хиральные метилфосфонаты, 2'-O-метил рибонуклеотиды, закрытую нуклеиновую кислоту (LNA™) и пептидные нуклеиновые кислоты (ПНК). За исключением конкретно указанных ограничений термин охватывает известные аналоги природных нуклеотидов, которые имеют связывающие свойства, аналогичные свойствам референсной нуклеиновой кислоты. Если не указано иное, подразумевается также, что конкретная последовательность нуклеиновой кислоты также охватывает консервативно модифицированные варианты (например, вырожденные замены кодонов), аллели, ортологи, однонуклеотидные полиморфизмы (SNP) и комплементарные последовательности, а также последовательность, указанная явным образом.
[00119] «Нуклеотиды» содержат сахар дезоксирибозу (ДНК) или рибозу (РНК), основание и фосфатную группу. Нуклеотиды соединены между собой указанными фосфатными группами.
[00120] «Основания» включают пурины и пиримидины, которые дополнительно включают природные соединения аденин, тимин, гуанин, цитозин, урацил, инозин и природные аналоги, а также синтетические производные пуринов и пиримидинов, которые включают, но не ограничиваются ими, модификации, которые вводят новые реакционноспособные группы, такие как, но не ограничиваясь ими, амины, спирты, тиолы, карбоксилаты и алкилгалогениды.
[00121] Под «гибридизуемым» или «комплементарным» или «по существу комплементарным» подразумевается, что нуклеиновая кислота (например, РНК) включает последовательность нуклеотидов, которая позволяет ей нековалентно связываться, т.е. формировать пары оснований по Уотсону-Крику и/или пары оснований G/U, подвергаться «отжигу» или «гибридизироваться» с другой нуклеиновой кислотой специфичным для последовательности, антипараллельным образом (т.е. нуклеиновая кислота специфически связывается с комплементарной нуклеиновой кислотой) в соответствующих in vitro и/или in vivo условиях температуры и ионной силы раствора. Как известно в данной области техники, стандартное спаривание оснований по Уотсону-Крику включает спаривание аденина (A) с тимидином (T), спаривание аденина (A) с урацилом (U) и спаривание гуанина (G) с цитозином (C). Кроме того, в данной области техники также известно, что при гибридизации двух молекул РНК (например, дцРНК) гуаниновое (G) основание спаривается с урацилом (U). Например, спаривание оснований G/U частично отвечает за вырожденность (т.е. избыточность) генетического кода в контексте спаривания оснований анти-кодонов тРНК с кодонами в мРНК. В контексте данного изобретения, гуанин (G) связывающего белок сегмента (дуплекса дцРНК) нацеливающей на ДНК молекулы РНК по данному изобретению считается комплементарным урацилу (U), и наоборот. Таким образом, когда пара оснований G/U может быть получена в данном нуклеотидном положении со связывающим белок сегментом (дуплекс дцРНК) нацеливающей на ДНК молекулы РНК по данному изобретению, это положение не рассматривается как некомплементарное, но вместо этого считается комплементарным.
[00122] Используемый в данном документе термин «конструкт нуклеиновой кислоты» относится к молекуле нуклеиновой кислоты, одноцепочечной или двухцепочечной, которая выделена из встречающегося в природе гена, или модифицирована так, чтобы она содержала сегменты нуклеиновых кислот способом, не существующим в природе, или является синтетической. Термин «конструкт нуклеиновой кислоты» является синонимом термина «экспрессионная кассета», когда конструкт нуклеиновой кислоты содержит контрольные последовательности, необходимые для экспрессии кодирующей последовательности по данному изобретению. «Экспрессионная кассета» включает кодирующую последовательность ДНК, функционально связанную с промотором.
[00123] В данном документе выражения «терапевтическое средство с нуклеиновой кислотой», «терапевтическая нуклеиновая кислота» и «ТНК» используются взаимозаменяемо и относятся к любому варианту терапевтического средства с использованием нуклеиновых кислот в качестве активного компонента терапевтического агента для лечения заболевания или расстройства. В данном документе указанные выражения относятся к терапевтическим средствам на основе РНК и терапевтическим средствам на основе ДНК. Неограничивающие примеры терапевтических средств на основе РНК включают мРНК, антисмысловую РНК и олигонуклеотиды, рибозимы, аптамеры, интерферирующие РНК (РНКи), дцРНК - субстрата Dicer, малую шпилечную РНК (мшРНК), асимметричные интерферирующие РНК (аиРНК), микроРНК (миРНК). Неограничивающие примеры терапевтических средств на основе ДНК включают миникольцо ДНК, миниген, вирусную ДНК (например, геном лентивируса или AAV) или невирусные синтетические ДНК-векторы, линейную дуплексную ДНК с замкнутыми концами (зкДНК/CELiD), плазмиды, бакмиды, ДНК-векторы Doggybone (dbDNA™), минималистичный вектор для иммунологически определенной генной экспрессии (MIDGE)-вектор, невирусный служебный ДНК-вектор (линейный ковалентно замкнутый ДНК-вектор) или гантелеобразный минимальный ДНК-вектор («гантелеобразная ДНК»).
[00124] Термины «пептид», «полипептид» и «белок» используются в данном документе взаимозаменяемо и относятся к полимерной форме аминокислот любой длины, которая может включать кодируемые и некодируемые аминокислоты, химически или биохимически модифицированные или дериватизированные аминокислоты, и полипептиды, имеющие модифицированные пептидные остовы.
[00125] В данном документе термины «синтетический AAV-вектор» и «получение AAV-вектора синтетическим путем» относятся к AAV-вектору и способам его получения синтетическим путем в полностью бесклеточной среде.
[00126] В данном документе термин «содержащий» («включающий») или «содержит» («включает») используется в отношении композиций, способов и их соответствующего компонента (компонентов), существенных для указанного способа или композиции, но допускающих включение не указанных элементов, существенных или нет.
[00127] В данном документе термин «состоящий по существу из» относится к элементам, необходимым для определенного варианта реализации. Указанный термин допускает присутствие элементов, не влияющих существенным образом на основную и новую или функциональную характеристику (характеристики) указанного варианта реализации.
[00128] Термин «состоящий из» относится к композициям, способам и их соответствующим компонентам согласно описанию в данном документе, которые не включают каких-либо элементов, не указанных в приведенном описании варианта реализации.
[00129] В данном описании и прилагаемой формуле изобретения термины в единственном числе включают их эквиваленты во множественном числе, если иное явным образом не следует из контекста. Соответственно, например, «способ» включает один или более способов, и/или этапов типа, описанного в данном документе, и/или таких, которые будут очевидными для специалистов в данной области техники после прочтения данного описания, и т.п. Аналогичным образом термин «или» включает «и», если из контекста явным образом не следует иное. Хотя при практической реализации или тестировании данного изобретения могут применяться способы и материалы, аналогичные или эквивалентные описанным в данном документе, ниже описаны подходящие способы и материалы. Выражение «например» в данном документе указывает на неограничивающий пример. Соответственно, выражение «например» синонимично выражению «к примеру».
[00130] За исключением рабочих примеров или, если указано иное, все числа, выражающие количества ингредиентов или условий реакции, используемые в данном документе, следует понимать как модифицированные термином «приблизительно» во всех случаях. Термин «приблизительно» при использовании применительно к процентам может означать ±1%. Данное изобретение дополнительно подробно разъяснено на приведенных ниже примерах, однако они не предназначены для ограничения объема данного изобретения.
[00131] Следует понимать, что данное изобретение не ограничено конкретной методологией, протоколами и реагентами, и т.п., описанными в данном документе, и, соответственно, предусматривает различные варианты. Используемая в данном документе терминология служит только для описания конкретных вариантов реализации, и не предназначена для ограничения объема данного изобретения, который определяется исключительно формулой изобретения.
II. Белки инициатора репликации (Rep)
[00132] Как описано в данном документе, описанная здесь технология относится к композиции и улучшенным способам получения ДНК-векторов, например, вектора зкДНК, как описано в данном документе, или вектора AAV с единственным видом белка Rep. В соответствии с некоторыми аспектами в описании предложен способ получения ДНК-вектора, например, вектора зкДНК, как описано в данном документе, или вектора AAV с использованием одного белка Rep, где белок Rep не является Rep52 или Rep40. Согласно некоторым вариантам реализации, единственным белком Rep является Rep78. Согласно некоторым вариантам реализации, единственным белком Rep является Rep68. Это усовершенствованный и более эффективный метод получения вектора зкДНК, который дает более высокий выход вектора зкДНК, чем способы, описанные в предшествующем уровне техники, в которых используются два белка Rep, включающие Rep78 или 68 и Rep52 или 40 (например, Rep78 и Rep 52, см. Фиг. 32). Действительно, до данного изобретения считалось, что для продуцирования частиц AAV должны присутствовать два Rep-белка, один длинный (например, Rep78 или Rep 68) и один короткий (например, Rep52 или Rep40). В частности, считалось, что Rep78 и Rep52 должны присутствовать либо в виде отдельных единиц, либо с использованием одной кодирующей последовательности для белков Rep78 и Rep52 для получения частиц AAV.
[00133] Соответственно, один аспект технологии, описанной в данном документе, относится к способу получения вектора ДНК, например вектора зкДНК, как описано в данном документе, или вектора AAV с использованием единственного белка Rep, в отличие от использования двух белков Rep. Согласно некоторым вариантам реализации, единственным белком Rep является Rep78. Согласно некоторым вариантам реализации, единственным белком Rep является Rep68. Согласно некоторым вариантам реализации белок Rep может быть Rep78 и Rep68, но не Rep52 или Rep40.
[00134] Другой аспект технологии, описанной в данном документе, относится к композиции, содержащей конструкт нуклеиновой кислоты, который содержит первую нуклеотидную последовательность, кодирующую один парвовирусный белок Rep, где нуклеотидная последовательность не имеет открытой рамки считывания (ORF) и не содержит функционального инициирующего кодона, расположенного ниже первого инициирующего кодона и/или в нем отсутствуют альтернативные сайты сплайсинга, предотвращающие пропуск экзонов, тем самым обеспечивая трансляцию одного парвовирусного белка Rep (например, белка Rep78 или Rep68) без трансляции дополнительных белков Rep (например, любого одного или нескольких из Rep52 или Rep40) в клетках насекомых или бесклеточной системе. То есть нуклеиновая кислота, кодирующая Rep78, также не продуцирует белок Rep52, и аналогично, нуклеиновая кислота, кодирующая Rep68, не продуцирует белок Rep40. Кроме того, никакой другой белок Rep не присутствует и не экспрессируется в системе конструкта нуклеиновой кислоты для получения ДНК-векторов, например, зкДНК-векторов и других рекомбинантных парвовирусных (например, аденоассоциированных вирусов) векторов в клетках (например, клетках насекомых, клетках млекопитающих) и бесклеточных системах, например, клетках насекомых или бесклеточных системах.
Rep-белки в общем
[00135] Rep-гены воспроизводят вирусный геном. В нуклеиновой кислоте дикого типа, кодирующей Rep78 или Rep68, событие сплайсинга в открытой рамке считывания Rep либо Rep78, либо Rep68 приводит к появлению двух белков Rep при трансляции: Rep52 и Rep40, соответственно. То есть белок Rep78 и белок Rep68 кодируются одной нуклеиновой кислотой, которая подвергается дифференциальному сплайсингу с продуцированием Rep78 и Rep68. Точно так же белок Rep52 и белок Rep40 кодируются одной нуклеиновой кислотой, которая подвергается дифференциальному сплайсингу с продуцированием белков Rep52 и Rep40. Rep78 представляет собой полноразмерный белок, продуцируемый из исходного первого сайта инициации трансляции, тогда как Rep52 является продуктом трансляции из нижележащего внутреннего «второго (AUG)» сайта инициации трансляции. Следовательно, когда экспрессируется полноразмерный геном AAV дикого типа, обычно присутствуют все четыре вида Rep-белков (например, Rep78, Rep68, Rep52 и Rep40) в основном из-за двух разных сайтов инициации трансляции, а также из-за того, что сайты альтернативного сплайсинга около карбоксильного конца. Каждый белок Rep имеет различные функциональные возможности, например, никирование ДНК, связывание ДНК, геликазную активность, лигазную и АТФазную активность. Функциональность данного белка Rep дополнительно описана на Фиг. 31. Ранее сообщалось, что и Rep78, и Rep52 белки необходимы для получения вектора AAV или вектора зкДНК в различных системах, например, клетках насекомых и клеточных системах млекопитающих. Однако, как обсуждается в данном документе, авторы изобретения демонстрируют, что только один белок Rep или, альтернативно, по меньшей мере, комбинация длинных белков Rep (Rep78 и Rep68), но не коротких белков Rep (Rep52 и Rep40), может использоваться для получения вектора AAV или вектора зкдНК. Единственный вид Rep-белка, используемый в композициях и способе, описанных в данном документе, включает все три функции: никирование ДНК, связывание ДНК и функциональное лигирование ДНК. В некоторых вариантах реализации единственный белок Rep дополнительно содержит функциональные группы геликазы и АТФазы.
[00136] В некоторых вариантах реализации единственный вид белка Rep, используемый в композициях и способах, описанных в данном документе, представляет собой белок Rep AAV2, если ITR относится к серотипу 2 (например, AAV2). В альтернативных вариантах реализации единственный белок Rep может происходить из любого из 42 серотипов AAV или, более предпочтительно, из белка Rep AAV1, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11 или белка AAV12 Rep. В некоторых вариантах реализации единственный белок Rep, охватываемый для использования в раскрытых здесь способах и композициях, соответствует белку Rep парвовируса животных, если ITR относится к серотипу 2 (например, AAV2). Белок Rep работает как часть системы с ITR, для связывания с ITR и инициации репликации с конечным разрешением и катализированием образования векторной молекулы зкДНК с замкнутыми концами.
[00137] В некоторых вариантах реализации единственный белок Rep, используемый в композициях и способах, описанных в данном документе, представляет собой Rep78. В альтернативных вариантах реализации единственный белок Rep, используемый в композициях и способах, описанных в данном документе является Rep68. В альтернативных вариантах реализации единственный вид Rep-белка представляет собой Rep52 или Rep40, который был модифицирован таким образом, чтобы содержать функциональность Rep78 или Rep68, например, чтобы иметь ДНК-связующую, ДНК-никирующую, геликазную и АТФазную активность. Альтернативно, в некоторых вариантах реализации белок Rep, используемый в композиции и способе, описанных в данном документе, может представлять собой комбинацию длинных белков Rep (например, Rep78 и Rep68) без Rep52 или Rep40, коротких белков Rep.
[00138] Другой аспект описанной здесь технологии относится к конструкту нуклеиновой кислоты, кодирующий единственный белок Rep, в котором нуклеиновая кислота не индуцирует и не позволяет экспрессию второго белка Rep. Соответственно, в одном аспекте конструкт нуклеиновой кислоты, кодирующий единственный белок Rep, модифицирован так, что в ней отсутствует функциональный кодон инициации для другого белка Rep.
[00139] В одном варианте реализации присутствие единственного вида Rep (например, при отсутствии других видов) определяется специфическими мутациями, которые предотвращают трансляцию p19 Rep, и отсутствием других видов Rep на вестерн-блоттинге с использованием анти-Rep антител известных в данной области техники.
[00140] Конструкты нуклеиновых кислот, кодирующие модифицированные белки Rep
[00141] В одном варианте реализации единственный вид белка Rep кодируется нуклеотидной последовательностью, кодирующей модифицированный белок Rep, например, он может кодировать модифицированный белок Rep78, при этом нуклеотидная последовательность не имеет функционального кодона инициации для кодирования Rep52 белка, а также отсутствуют сайты сплайсинга для пропуска экзонов для получения Rep68 или Rep40. Например, модифицированная нуклеотидная последовательность Rep78 включает модификацию или мутацию в кодоне инициации для Rep52, так что кодон инициации (например, AUG) для Rep52 изменен и больше не кодирует метионин, а кодирует другую аминокислоту. В некоторых вариантах реализации кодон инициации (Met) для Rep52 в последовательности нуклеиновой кислоты Rep78 мутирован для кодирования глицина (например, AUG мутирован в один из: GGU, GGC, GGA, GGG, который кодирует Gly) или аминокислоту треонин (например, AUG мутирован в один из ACT, ACC, ACA и ACG, который кодирует Thr).
Модифицированные Rep белки
[00142] В некоторых вариантах реализации модифицированная нуклеотидная последовательность Rep78 может кодировать модифицированный белок Rep78, который содержит модификацию аминокислотного остатка 225 (Met) SEQ ID NO: 530, в котором аминокислотный остаток 225 заменен на глицин (Gly) (например, M225G или Met225Gly) или треонин (Thr) (например, M225T или Met225Thr). В одном варианте реализации мутированный белок Rep78 содержит последовательность SEQ ID NO: 530 или включает последовательность, содержащую по меньшей мере 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичности последовательности с SEQ ID NO: 530, в которой аминокислота в положении 225 не является Met, и где модифицированный белок Rep имеет, по меньшей мере, функциональность связывания ДНК и никирования ДНК, а кодирующий его ген не способствует получению второго белка Rep. Специалист в данной области сможет создать точечную мутацию, используя, например, сайт-направленный мутагенез. Чтобы оценить, была ли мутация в нуклеотидной последовательности сгенерирована правильно, можно выполнить выравнивание последовательности с модифицированным белком Rep (т.е. белком Rep, содержащим точечную мутацию) по сравнению с белком Rep дикого типа.
[00143] В одном варианте реализации нуклеотидная последовательность, кодирующая единственный белок Rep, используемый в композициях и способах, раскрытых в данном документе, включает последовательность контроля экспрессии, например, промотор, цис-регуляторные элементы или регуляторный переключатель, как описано в данном документе, расположенную перед кодоном инициации нуклеотидной последовательности, кодирующей парвовирусный белок Rep78, в которой последовательность нуклеиновой кислоты не имеет функционального инициирующего кодона для Rep52. В одном варианте реализации нуклеотидная последовательность, кодирующая единственный белок Rep, используемый в композициях и способах, раскрытых в данном документе, включает последовательность контроля экспрессии перед кодоном инициации нуклеотидной последовательности, кодирующей парвовирусный белок Rep78, в которой последовательность нуклеиновой кислоты не имеет функциональных сайтов сплайсингу для кодирования Rep68.
[00144] То есть в некоторых вариантах реализации нуклеиновая кислота, кодирующая Rep78, имеет только один инициирующий кодон, что позволяет осуществлять трансляцию только белка Rep78 или белка Rep68. В таких вариантах реализации нуклеиновая кислота Rep78 имеет функциональный первый инициирующий кодон, обеспечивающий трансляцию белка Rep78, но модифицированный (или нефункциональный) инициирующий кодон ниже исходного инициирующего кодона, что приводит к тому, что Rep52 не экспрессируется.
[00145] Во всех случаях не используются никакие другие векторы, которые кодируют другой Rep. Также не используется белок Rep, который уже присутствует в клетках насекомых или клетках млекопитающих, используемых в способах создания ДНК-векторов, например, векторах зкДНК или векторах AAV, в соответствии со способами, описанными в данном документе.
[00146] В одном варианте реализации единственный белок Rep, используемый в композициях и способах, раскрытых в данном документе, относится к семейству парвовирусов. В другом варианте реализации единственный белок Rep, используемый в композициях и способах, раскрытых в данном документе, предпочтительно происходит из подсемейства Dependovirus вируса Rep. В другом варианте реализации единственный Rep-белок, используемый в композициях и способах, раскрытых в данном документе, более предпочтительно является AAV Rep.
[00147] В одном варианте реализации нуклеотидная последовательность по изобретению включает последовательность контроля экспрессии, кодирующую белок Rep 68 AAV, где последовательность нуклеиновой кислоты не имеет функционального инициирующего кодона для Rep40, но имеет делецию в последовательности интрона на его карбокси-конце, результатом чего является Rep68. В другом варианте реализации последовательность нуклеиновой кислоты имеет делецию в интронной последовательности полноразмерного Rep78 и не имеет других функциональных сайтов сплайсинга, что приводит к получению транскрипта, который способен быть транслирован только в Rep68. То есть в некоторых вариантах реализации нуклеиновая кислота, кодирующая Rep68, имеет только один инициирующий кодон, что позволяет осуществлять трансляцию только белка Rep68 с удаленной с-концевой последовательностью интрона. В таких вариантах реализации нуклеиновая кислота Rep68 имеет функциональный первый инициирующий кодон, обеспечивающий трансляцию белка Rep68, но инициирующий кодон, расположенный ниже исходного инициирующего кодона, модифицирован или нефункционален мутацией (например, M225G или M225T), которая приводит к тому, что Rep40 не экспресируется. Альтернативно, нуклеиновая кислота, кодирующая Rep68, модифицирована так, что второй инициирующий кодон модифицирован или нефункционален мутацией (например, M225G или M225T), но расположенные ниже c-концевые сайты сплайсинга являются функциональными и позволяют экспрессию белка Rep78 и белка Rep68.
[00148] Последовательность, практически идентичная нуклеотидной последовательности SEQ. ID NO: 530 представляет собой последовательность, которая имеет по меньшей мере 60%, 70%, 80% или 90% идентичности с SEQ ID NO: 530.
III. Подробное описание способа получения зкДНК-вектора с использованием одного белка Rep
A. Получение в общем
[00149] Как описано в данном документе, вектор зкДНК может быть получен способом с использованием только одного белка Rep, в отличие от более чем одного, например, двух белков Rep. Соответственно, один аспект данного изобретения относится к способу, включающему следующие этапы: a) инкубация популяции клеток-хозяев (например, клеток насекомых), несущих указанный матричный полинуклеотидный экспрессионный конструкт (например, зкДНК-плазмиду, зкДНК-бакмиду и/или зкДНК-бакуловирус), который не содержит последовательностей, кодирующих вирусный капсид, в присутствии белка Rep в эффективных условиях и на протяжении времени, достаточного для индукции продуцирования зкДНК-вектора в клетках-хозяевах, причем указанные клетки-хозяева не содержат последовательностей, кодирующих вирусный капсид; и b) сбор и выделение указанного зкДНК-вектора из клеток-хозяев. Присутствие единственного белка Rep индуцирует репликацию векторного полинуклеотида с модифицированным ITR для продуцирования зкДНК-вектора в клетке-хозяине. Однако вирусные частицы (например, вирионы AAV) при этом не экспрессируются. Таким образом, отсутствуют ограничения по размеру, такие как естественным образом присутствующие в AAV-векторах или других вирусных векторах.
[00150] Присутствие зкДНК-вектора, выделенного из клеток-хозяев, может быть подтверждено путем расщепления ДНК, выделенной из клетки-хозяина, рестрикционным ферментом, имеющим один сайт распознавания на зкДНК-векторе, и анализа расщепленного ДНК-материала на неденатурирующем геле для подтверждения присутствия характерных полос линейной и непрерывной ДНК по сравнению с линейной и прерывистой ДНК.
[00151] В еще одном аспекте изобретение предусматривает применение линий клеток-хозяев, имеющих стабильно интегрированную в собственный геном полинуклеотидную матрицу экспрессионного ДНК-вектора (зкДНК-матрицу), при продуцировании невирусного ДНК-вектора, например, как описано в: Lee, L. et al. (2013) Plos One 8(8): e69879. Предпочтительно Rep добавляют в клетки-хозяева с величиной МЗ (множественность заражения), равной приблизительно 3. Если линия клеток-хозяев представляет собой линию клеток млекопитающего, например, клетки HEK293, то указанные линии клеток могут содержать стабильно интегрированную полинуклеотидную векторную матрицу, а второй вектор, например, герпесвирусный, может применяться для введения в клетки белка Rep, обеспечивающего вырезание и амплификацию зкДНК в присутствии Rep и хелперного вируса.
[00152] Согласно одному варианту реализации клетки-хозяева, используемые для получения зкДНК-векторов, описанных в данном документе, представляют собой клетки насекомых, и для доставки как полинуклеотида, который кодирует белок Rep, так и матричный полинуклеотидный экспрессионный конструкт невирусного ДНК-вектора для зкДНК используют бакуловирус, например, согласно описанию на Фиг. 4A-4C и в Примере 1. Согласно некоторым вариантам реализации клетку-хозяина модифицируют таким образом, чтобы она экспрессировала единственный белок Rep.
[00153] Затем зкДНК-вектор собирают и выделяют из клеток-хозяев. Время сбора и извлечения зкДНК-векторов, описанных в данном документе, из клеток может быть выбрано и оптимизировано так, чтобы обеспечить получение указанных зкДНК-векторов с высоким выходом. Например, время сбора может быть выбрано с учетом жизнеспособности клеток, морфологии клеток, роста клеток и т.д. Согласно одному варианту реализации клетки культивируют в достаточных условиях и собирают через достаточное время после инфекции бакуловирусом, чтобы получить зкДНК-векторы, но до начала гибели большинства клеток вследствие токсичности бакуловируса. Указанные ДНК-векторы могут быть выделены с применением наборов для очистки плазмид, таких как наборы без эндотоксина для выделения плазмид (Endo-Free Plasmid Kits) от Qiagen. Другие способы, разработанные для выделения плазмид, также могут быть адаптированы для ДНК-векторов. Как правило, могут быть использованы любые способы очистки нуклеиновых кислот.
[00154] Указанные ДНК-векторы могут быть очищены любыми способами очистки ДНК, известными специалистам в данной области техники. Согласно одному варианту реализации очищенные зкДНК-векторы получают в виде молекул ДНК. Согласно другому варианту реализации очищенные зкДНК-векторы получают в виде экзосом или микрочастиц.
[00155] Присутствие зкДНК-вектора может быть подтверждено путем расщепления векторной ДНК, выделенной из клеток, рестрикционным ферментом, имеющим один сайт распознавания на ДНК-векторе, и анализа как расщепленного, так и нерасщепленного ДНК-материала с применением гель-электрофореза для подтверждения присутствия характерных полос линейной и непрерывной ДНК, отличных от полос линейной и прерывистой ДНК. Фиг. 4C и Фиг. 4E иллюстрируют один вариант реализации идентификации присутствия зкДНК-векторов с замкнутыми концами, продуцируемых описанными в данном документе способами. Фиг. 5 изображает гель, подтверждающий получение зкДНК из множества плазмидных конструктов с использованием способа, описанного в примерах.
B. зкДНК-плазмида
[00156] зкДНК-плазмида представляет собой плазмиду, используемую для последующего получения зкДНК-вектора. В некоторых вариантах реализации зкДНК-плазмида может быть сконструирована с применением известных методик для обеспечения по меньшей мере нижеперечисленного, в виде функционально связанных компонентов, в направлении транскрипции: (1) 5'-ITR; (2) экспрессионной кассеты, содержащей цис-регуляторный элемент, например, промотор, индуцибельный промотор, регуляторный переключатель, энхансеры и т.п.; и (3) 3'-ITR, причем последовательность 3'-ITR является ассимметричной к последовательности 5'-ITR. Согласно некоторым вариантам реализации указанная экспрессионная кассета, фланкированная ITR, содержит сайт клонирования для введения экзогенной последовательности. Указанная экспрессионная кассета заменяет кодирующие участки rep и cap геномов AAV.
[00157] В одном аспекте зкДНК-вектор получают из плазмиды, называемой в данном документе «зкДНК-плазмида», кодирующей, в указанном порядке: первый инвертированный концевой повтор (ITR) аденоассоциированного вируса (AAV), экспрессионную кассету, содержащую трансген, и мутированный или модифицированный ITR AAV, причем указанная зкДНК-плазмида не содержит кодирующих последовательностей капсидного белка AAV. В альтернативных вариантах реализации такая зкДНК-плазмида кодирует, в указанном порядке: первый (или 5') модифицированный или мутированный ITR ААV, экспрессионную кассету, содержащую трансген, и второй (или 3') ITR ААV дикого типа, причем указанная зкДНК-плазмида лишена кодирующих последовательностей капсидного белка ААV, а 5'- и 3'-ITR являются ассимметричными друг относительно друга. В альтернативных вариантах реализации зкДНК-плазмида кодирует, в указанном порядке: первый (или 5') модифицированный или мутированный ITR ААV, экспрессионную кассету, содержащую трансген, и второй (или 3') мутированный или модифицированный ITR ААV, причем указанная зкДНК-плазмида лишена кодирующих последовательностей капсидного белка ААV, а модифицированные 5'- и 3'-ITR отличаются и не имеют одинаковых модификаций.
[00158] Согласно дополнительному варианту реализации система зкДНК-плазмиды лишена кодирующих последовательностей вирусного капсидного белка (т.е. не содержит генов капсида AAV, а также генов капсидов других вирусов). Кроме того, согласно конкретному варианту реализации зкДНК-плазмида также лишена кодирующих последовательностей белка Rep AAV. Соответственно, в предпочтительном варианте реализации зкДНК-плазмида лишена функциональных генов cap ААV и rep ААV, GG-3′ для AAV2), а также вариабельной палиндромной последовательности, обеспечивающей образование шпильки.
[00159] ЗкДНК-плазмида по данному изобретению может быть получена с использованием природных нуклеотидных последовательностей геномов любых серотипов AAV, хорошо известных в данной области техники. В одном варианте реализации, остов плазмиды зкДНК получают из генома ААV1, ААV2, ААV3, ААV4, AAV5, AAV5, AAV7, AAV8, AAV9, AAV10, AAV 11, AAV12, AAVrh8, AAVrh10, AAV-DJ и AAV-DJ8. Например, NCBI: NC 002077; NC 001401; NC001729; NC001829; NC006152; NC 006260; NC 006261; из указателя вирусов Kotin and Smith («The Springer Index of Viruses»), доступного по ссылке, поддерживаемой Springer (веб-адрес: oesys.springer.de/viruses/database/mkchapter.asp?virID=42.04.) (примечание: ссылки на указатель URL или базу данных подразумевают содержание доступного по ссылке ресурса URL или базы данных на фактическую дату подачи данной заявки). В конкретном варианте реализации остов зкДНК-плазмиды получают из генома AAV2. В другом конкретном варианте реализации, остов зкДНК-плазмиды представляет собой синтетический остов, сконструированный методами генной инженерии с включением на 5'- и 3'-концах ITR, происходящих из одного из указанных геномов AAV.
[00160] ЗкДНК-плазмида может необязательно включать селектируемый или селективный маркер для использования при создании линии клеток, продуцирующей зкДНК-вектор. Согласно одному варианту реализации указанный селективный маркер может быть инсертирован ниже (то есть в направлении 3') относительно последовательности 3'ITR. Согласно другому варианту реализации указанный селективный маркер может быть инсертирован выше (т.е., в направлении 5') относительно последовательности 5' ITR. Подходящие селективные маркеры включают, например, маркеры, придающие устойчивость к лекарственному средству. Селекционными маркерами могут быть, например, гены устойчивости к бластицидину S, канамицину, генетицину и т.п. Согласно предпочтительному варианту реализации селективный маркер для отбора по чувствительности к лекарственному средству представляет собой ген устойчивости к бластицидину S.
[00161] Типичную зкДНК (например, rAAV0) продуцируют из плазмиды рAAV. Способ получения вектора rAAV может включать: (а) доставку в клетку-хозяина плазмиды rAAV, как описано выше, при этом как клетка-хозяин, так и плазмида не содержат генов, кодирующих капсидный белок, (b) культивирование указанной клетки-хозяина в условиях, обеспечивающих продуцирование генома зкДНК; и (c) сбор клеток и выделение генома AAV, продуцированного указанными клетками.
C. Примеры способа получения зкДНК-векторов из зкДНК-плазмид
[00162] В данном документе также предложены способы получения бескапсидных зкДНК-векторов, в частности, способ с достаточно высоким выходом для обеспечения достаточного количества вектора для экспериментов in vivo.
[00163] Согласно некоторым вариантам реализации способ получения зкДНК-вектора включает следующие этапы: (1) введение конструкта нуклеиновых кислот, содержащего экспрессионную кассету и две ассимметричных последовательности ITR, в клетку-хозяина (например, клетки Sf9), (2) необязательно, получение клональной линии клеток, например, с применением селективного маркера, присутствующего в плазмиде, (3) введение гена кодирующего Rep (путем трансфекции или инфекции бакуловирусом, несущим указанный ген) в указанную клетку насекомого; и (4) сбор клеток и очистка зкДНК-вектора. Конструкт нуклеиновых кислот, содержащий экспрессионную кассету и две последовательности ITR, описанных выше, для получения бескапсидного AAV-вектора, может находиться в форме cfAAV-плазмиды, или бакмиды, или бакуловируса, полученных из cfAAV-плазмиды согласно приведенному ниже описанию. Указанный конструкт нуклеиновых кислот может быть введен в клетку-хозяина путем трансфекции, вирусной трансдукции, стабильной интеграции или других способов, известных в данной области техники.
D. Клеточные линии:
[00164] Линии клеток-хозяев, используемые для продуцирования зкДНК-вектора, могут включать линии клеток насекомых, полученные из Spodoptera frugiperda, такие как клетки Sf9, Sf21 или клетки Trichoplusia ni, или линии клеток других беспозвоночных животных, позвоночных животных или других эукариот, в том числе клетки млекопитающих. Также могут быть использованы другие линии клеток, известные специалисту в данной области техники, такие как HEK293, Huh-7, HeLa, HepG2, HeplA, 911, CHO, COS, MeWo, NIH3T3, A549, HT1080, моноциты, а также зрелые и незрелые дендритные клетки. Линии клеток-хозяев могут быть трансфицированы для стабильной экспрессии зкДНК-плазмиды для продуцирования зкДНК-вектора с высоким выходом.
[00165] ЗкДНК-плазмиды могут быть введены в клетки Sf9 путем транзиентной трансфекции с использованием реагентов (например, липосом, фосфата кальция) или физических методов (например, электропорации), известных в данной области. Как вариант, могут быть созданы стабильные линии клеток Sf9, которые стабильно интегрировали зкДНК-плазмиду в геном. Такие стабильные линии клеток могут быть созданы путем включения селективного маркера в зкДНК-плазмиду согласно описанию выше. Если зкДНК-плазмида, используемая для трансфекции линии клеток, включает селективный маркер, такой как антибиотик, клетки, которые были трансфицированы зкДНК-плазмидой и интегрировали зкДНК-плазмиду в геном, могут быть отобраны путем добавления антибиотика в среду для роста клеток. Затем устойчивые клоны клеток могут быть выделены с помощью методов разведения отдельных клеток или переноса колоний и размножены.
E. Выделение и очистка зкДНК-векторов
[00166] Примеры способа получения и выделения зкДНК-векторов представлены на Фиг. 4A-4E и в конкретных примерах ниже. ЗкДНК-векторы согласно описанию в данном документе могут быть получены из клеток-продуцентов, экспрессирующих белок (белки) AAV Rep, дополнительно трансформированных зкДНК-плазмидой, зкДНК-бакмидой или зкДНК-бакуловирусом. Плазмиды, подходящие для получения зкДНК-векторов, включают плазмиды, показанные на Фиг. 8A (подходят для получения Rep BIIC), Фиг. 8B (плазмида, используемая для получения зкДНК-вектора).
[00167] В одном аспекте полинуклеотид кодирует белок Rep AAV (Rep78 или 68), доставляемый в клетку-продуцент в плазмиде (Rep-плазмиде), бакмиде (Rep-бакмиде) или бакуловирусе (Rep-бакуловирусе). Rep-плазмида, Rep-бакмида и Rep-бакуловирус могут быть получены описанными выше способами.
[00168] Способы получения зкДНК-вектора, который представляет собой пример зкДНК-вектора, описаны в данном документе. Экспрессионные конструкты, используемые для получения зкДНК-векторов согласно данному изобретению, могут представлять собой плазмиду (например, зкДНК-плазмиды), бакмиду (например, зкДНК-бакмиду) и/или бакуловирус (например, зкДНК-бакуловирус). В неограничивающем примере зкДНК-вектор может быть получен из клеток, коинфицированных зкДНК-бакуловирусом и Rep-бакуловирусом. Белки Rep продуцированные с Rep-бакуловируса, могут реплицировать зкДНК-бакуловирус с получением зкДНК-векторов. Альтернативно, векторы зкДНК могут быть получены из клеток, стабильно трансфицированных конструктов, содержащей последовательность, кодирующую один белок Rep AAV (например, Rep78, Rep68 или Rep52), доставленный в Rep-плазмидах, Rep-бакмидах или Rep-бакуловирусе. ЗкДНК-бакуловирусом можно транзиентно трансфицировать клетки, он может быть реплицирован белком Rep и может продуцировать зкДНК-векторы.
[00169] Бакмидой (например, зкДНК-бакмидой) могут быть трансфицированы пермиссивные клетки насекомых, такие как Sf9, Sf21, клетки Tni (Trichoplusia ni), клетки High Five, и указанная бакмида продуцирует зкДНК-бакуловирус, который представляет собой рекомбинантный бакуловирус, включающий последовательности, содержащие ассимметричные ITR и экспрессионную кассету. Затем клетки насекомых могут быть инфицированы зкДНК-бакуловирусом для получения нового поколения рекомбинантного бакуловируса. Необязательно, указанный этап может быть повторен один или более раз для получения большего количества рекомбинантного бакуловируса.
[00170] Время сбора и извлечения зкДНК-векторов, описанных в данном документе, из клеток может быть выбрано и оптимизировано так, чтобы обеспечить получение указанных зкДНК-векторов с высоким выходом. Например, время сбора может быть выбрано с учетом жизнеспособности клеток, морфологии клеток, роста клеток и т.п. Как правило, клетки могут быть собраны по прошествии достаточного времени после инфицирования бакуловирусом для продуцирования зкДНК-векторов (например, зкДНК-векторов), но до того, как большинство клеток начнут умирать вследствие токсичности вируса. ЗкДНК-векторы можно выделить из клеток Sf9 с использованием наборов для очистки плазмид, таких как наборы ENDO-FREE PLASMID® от Qiagen. Другие методы, разработанные для выделения плазмид, также могут быть адаптированы для зкДНК-векторов. Как правило, можно использовать любые известные в данной области способы очистки нуклеиновых кислот, а также коммерчески доступные наборы для экстракции ДНК.
[00171] Как вариант, очистка может быть проведена путем щелочного лизиса клеточного осадка, центрифугирования полученного лизата и выполнения хроматографического разделения. В качестве одного неограничивающего примера, указанный процесс может быть осуществлен путем загрузки супернатанта на ионообменную колонку (например, SARTOBIND Q®), которая удерживает нуклеиновые кислоты, с последующей элюцией (например, 1,2 M раствором NaCl), и проведения дополнительной хроматографической очистки на гель-фильтрационной колонке (например, 6 Fast Flow GE). Затем выделяют бескапсидный вектор AAV, например, путем осаждения.
[00172] Согласно некоторым вариантам реализации зкДНК-векторы могут также быть очищены в форме экзосом, или микрочастиц. В данной области техники известно, что многие типы клеток высвобождают не только растворимые белки, но и сложные белковые/нуклеиновокислотные грузы путем шеддинга мембранных микровезикул (Cocucci et al, 2009; EP 10306226.1). Такие везикулы включают микровезикулы (также называемые микрочастицами) и экзосомы (также называемые нановезикулами); и первые, и вторые содержат белки и РНК в качестве груза. Микровезикулы образуются в результате прямого отпочковывания от плазматической мембраны, а экзосомы высвобождаются во внеклеточную среду при слиянии мультивезикулярных эндосом с плазматической мембраной. Соответственно, содержащие зкДНК-вектор микровезикулы и/или экзосомы могут быть выделены из клеток, трансдуцированных с зкДНК-плазмидой, или бакмидой, или бакуловирусом, полученными с применением зкДНК-плазмиды.
[00173] Микровезикулы можно выделить, выполнив фильтрацию или ультрацентрифугирование культуральной среды при 20000 x g, а экзосомы - при 100000 x g. Оптимальная продолжительность ультрацентрифугирования может быть определена экспериментально и зависит от конкретного типа клеток, из которых выделяют везикулы. Предпочтительно, культуральную среду сначала очищают низкоскоростным центрифугированием (например, при 2000 x g в течение 5-20 минут) и подвергают центрифугированию с использованием, например, спин-колонки AMICON® (Millipore, Уотфорд, Великобритания). Микровезикулы и экзосомы могут быть дополнительно очищены с помощью FACS или MACS с применением специфических антител, распознающих специфические поверхностные антигены, присутствующие на микровезикулах и экзосомах. Другие методы очистки микровезикул и экзосом включают, не ограничиваясь перечисленными, иммунопреципитацию, аффинную хроматографию, фильтрацию и магнитные гранулы, покрытые специфическими антителами или аптамерами. После очистки везикулы промывают, например, физиологическим раствором с фосфатным буфером. Одним из преимуществ использования микровезикул или экзосом для доставки зкДНК-содержащих везикул является то, что эти везикулы могут быть нацелены на клетки различных типов путем включения в их мембраны белков, распознаваемых специфическими рецепторами на соответствующих типах клеток. (см. также EP 10306226).
[00174] Другой аспект данного изобретения относится к способам очистки зкДНК-векторов из линий клеток-хозяев, которые стабильно интегрировали конструкт зкДНК в геном. Согласно одному варианту реализации очищенные зкДНК-векторы получают в виде молекул ДНК. Согласно другому варианту реализации очищенные зкДНК-векторы получают в виде экзосом или микрочастиц.
[00175] На Фиг. 5 показан гель, подтверждающий получение зкДНК с нескольких конструктов зкДНК-плазмид с применением способа, описанного в разделе Примеры. ЗкДНК подтверждается характерным рисунком полос в геле, как описано со ссылкой на Фиг. 4D в Примерах. Другие характеристики процесса продуцирования зкДНК и промежуточных продуктов суммированы на Фиг. 6A и 6B и Фиг. 7A и 7B, как описано в Примерах.
IV. зкДНК вектор
[00176] Как описано в данном документе, способы и композиции, в которых используется единственный белок Rep, применимы для получения бескапсидной молекулы зкДНК с векторами с ковалентно замкнутыми концами (зкДНК). В некоторых вариантах реализации эти векторы зкДНК могут быть продуцированы в пермиссивных клетках-хозяевах, которые содержат единственный белок Rep, и получены из конструктов экспрессии (например, плазмиды зкДНК, бакмиды зкДНК, бакуловируса зкДНК или интегрированной клеточной линии), содержащий гетерологичный ген (трансген), расположенный между двумя последовательностями инвертированных концевых повторов (ITR), где последовательности ITR могут быть асимметричной парой ITR или симметричной или по существу симметричной парой ITR, как эти термины определены в данном документе. ЗкДНК-вектор содержащий NLS как описано в данном документе, может содержать последовательности ITR, которые выбраны из любых из: (i) по меньшей мере одного ITR дикого типа (WT) и по меньшей мере одного модифицированного инвертированного концевого повтора (mod-ITR) AAV (например, асимметричных модифицированных ITR); (ii) двух модифицированных ITR, причем пара mod-ITR имеет различную трехмерную пространственную организацию друг относительно друга (например, асимметричных модифицированных ITR), или (iii) пары симметричных или по существу симметричных ITR WT-WT, в которой каждый ITR-WT имеет одинаковую трехмерную пространственную организацию, или (iv) пары симметричных или по существу симметричных модифицированных ITR, в которой каждый mod-ITR имеет одинаковую трехмерную пространственную организацию, при этом способы по данному изобретению могут дополнительно включать систему доставки, такую как, без ограничений, система доставки на основе липосомных наночастиц.
[00177] ЗкДНК-вектор предпочтительно является дуплексным, например, самокомплементарным, по меньшей мере в части молекулы, такой как экспрессионная кассета (например, зкДНК не является двухцепочечной кольцевой молекулой). ЗкДНК-вектор имеет ковалентно замкнутые концы и, таким образом, устойчив к расщеплению экзонуклеазой (например, экзонуклеазой I или экзонуклеазой III), например, в течение часа при 37°С.
[00178] ЗкДНК-вектор, полученный в соответствии со способами и композициями с использованием единственного белка Rep, описанными в данном документе, не имеет ограничений, связанных с упаковкой, накладываемых ограниченным пространством внутри вирусного капсида. ЗкДНК-векторы представляют собой жизнеспособную эукариотически-продуцируемую альтернативу прокариотически-продуцируемым векторам плазмидной ДНК, в отличие от инкапсулированных геномов AAV. Это позволяет вставлять контрольные элементы, например, регуляторные переключатели, как описано в данном документе, большие трансгены, множественные трансгены и т.д.
[00179] В одном варианте реализации зкДНК-вектор, полученный в соответствии со способами и композициями с использованием единственного белка Rep, описанными в данном документе, содержит, в направлении от 5' к 3': первый инвертированный концевой повтор (ITR) аденоассоциированного вируса (AAV), нуклеотидную последовательность, представляющую интерес (например, экспрессионную кассету, как описано в данном документе), и второй ITR AAV, причем первый ITR и второй ITR являются асимметричными относительно друг друга, т.е. они отличаются друг от друга. В качестве типичного варианта реализации первая ITR может быть ITR дикого типа, а вторая ITR может быть мутированной или модифицированной ITR. В некоторых вариантах реализации первая ITR может быть мутированной или модифицированной ITR, а вторая ITR - ITR дикого типа. В другом варианте реализации первый ITR и второй ITR оба являются модифицированными, но представляют собой разные последовательности, или имеют разные модификации, или не являются идентично модифицированными ITR. Другими словами, ITR асимметричны в том смысле, что любые изменения в одном ITR не отражаются в другом ITR; или, альтернативно, если ITR отличаются друг от друга. Типичные примеры асимметричных ITR в зкДНК-векторе и для использования при получении зкДНК-плазмиды описаны ниже в разделе, озаглавленном «ITR».
[00180] Последовательности ITR дикого типа или мутированные или иным образом модифицированные, представленные в данном документе, представляют собой последовательности ДНК, включенные в экспрессионный конструкт (например, зкДНК-плазмиду, зкДНК-бакмиду, зкДНК-бакуловирус) для продуцирования зкДНК-вектора. Соответственно, последовательности ITR, фактически содержащиеся в указанном зкДНК-векторе, продуцируемом с зкДНК-плазмиды или другого экспрессионного конструкта, могут быть или не быть идентичными последовательностям ITR по данному изобретению, что обусловлено естественными изменениями, происходящими в процессе получения (например, ошибкой репликации).
[00181] Согласно некоторым вариантам реализации зкДНК-вектор, полученный в соответствии со способами и композициями с использованием единственного белка Rep, описанными в данном документе, содержит экспрессионную кассету с трансгеном, который может представлять собой, например, регуляторную последовательность, последовательность, кодирующую нуклеиновую кислоту (например, такую как miR или антисмысловая последовательность), или последовательность, кодирующую полипептид (например, такую как трансген). Согласно одному варианту реализации указанный трансген может быть функционально связан с одной или более регуляторными последовательностями, которые обеспечивают или контролируют экспрессию трансгена. Согласно одному варианту реализации указанный полинуклеотид содержит первую последовательность ITR и вторую последовательность ITR, причем представляющая интерес нуклеотидная последовательность фланкирована первой и второй последовательностями ITR, и указанные первая и вторая последовательности ITR асимметричны друг другу.
[00182] В одном варианте реализации в каждом из указанных аспектов экспрессионная кассета расположена между двумя ITR, которые содержат, в следующем порядке, вместе с чем-либо одним или более из перечисленного: промотор, функционально связанный с трансгеном, посттранскрипционный регуляторный элемент; и сигнал полиаденилирования и терминации. В одном варианте реализации промотор является регулируемым - индуцибельным или репрессируемым. Промотор может представлять собой любую последовательность, которая облегчает транскрипцию трансгена. В одном варианте реализации указанный промотор представляет собой промотор CAG (например, SEQ ID NO: 03) или его вариант. Указанный посттранскрипционный регуляторный элемент представляет собой последовательность, которая модулирует экспрессию трансгена, в неограничивающем примере - любую последовательность, образующую третичную структуру, которая усиливает экспрессию трансгена.
[00183] В одном варианте реализации указанный посттранскрипционный регуляторный элемент содержит WPRE (например, SEQ ID NO: 08). В одном варианте реализации указанный сигнал полиаденилирования и терминации содержит поли(A) BGH (например, SEQ ID NO: 09). Дополнительно может быть использован любой цис-регуляторный элемент, известный в данной области техники, или их комбинация, например, энхансерная 5'-последовательность позднего сигнала полиаденилирования SV40 (USE) или другие элементы посттранскрипционного процессинга, в том числе, но не ограничиваясь перечисленными, ген тимидинкиназы вируса простого герпеса или вируса гепатита B (HBV). Согласно одному варианту реализации длина экспрессионной кассеты в 5'-3' направлении больше известной максимальной длины последовательности, заключаемой в капсид вириона AAV. Согласно одному варианту реализации указанная длина составляет более 4,6 т.п.о., или более 5 т.п.о., или более 6 т.п.о., или более 7 т.п.о. Различные примеры экспрессионных кассет представлены в данном документе.
[00184] Указанная экспрессионная кассета в зкДНК векторе, полученном в соответствии со способами и композициями с использованием единственного белка Rep, описанным в данном документе, может содержать более 4000 нуклеотидов, 5000 нуклеотидов, 10000 нуклеотидов или 20000 нуклеотидов, или 30000 нуклеотидов, или 40000 нуклеотидов, или 50000 нуклеотидов, или любой диапазон значений в пределах между примерно 4000-10000 нуклеотидов или 10000-50000 нуклеотидов, или более 50000 нуклеотидов. В некоторых вариантах реализации экспрессионная кассета может содержать трансген или нуклеиновую кислоту имеющий длину в диапазоне от 500 до 50000 нуклеотидов. В некоторых вариантах реализации экспрессионная кассета может содержать трансген или нуклеиновую кислоту имеющий длину в диапазоне от 500 до 75000 нуклеотидов. В некоторых вариантах реализации экспрессионная кассета может содержать трансген или нуклеиновую кислоту имеющий длину в диапазоне от 500 до 10000 нуклеотидов. В некоторых вариантах реализации экспрессионная кассета может содержать трансген или нуклеиновую кислоту имеющий длину в диапазоне от 1000 до 10000 нуклеотидов. В некоторых вариантах реализации экспрессионная кассета может содержать трансген или нуклеиновую кислоту имеющий длину в диапазоне от 500 до 5000 нуклеотидов. Указанные зкДНК-векторы не имеют ограничений по размеру, характерных для заключенных в капсиды AAV-векторов, что позволяет доставлять экспрессионную кассету значительного размера для обеспечения эффективной экспрессии трансгенов. Согласно некоторым вариантам реализации указанный зкДНК-вектор не содержит специфического для прокариот метилирования.
[00185] В некоторых вариантах реализации кассета экспрессии в векторе зкДНК, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, также может содержать внутренний сайт входа в рибосому (IRES) и/или элемент 2A. Цис-регуляторные элементы включают, не ограничиваясь перечисленными, промотор, рибопереключатель, инсулятор, miR-регулируемый элемент, посттранскрипционный регуляторный элемент, тканеспецифический и клеточноспецифический промотор и энхансер. Согласно некоторым вариантам реализации указанный ITR может действовать в качестве промотора для трансгена. В некоторых вариантах реализации указанный зкДНК-вектор содержит дополнительные компоненты для регуляции экспрессии указанного трансгена, например, один или несколько регуляторных переключателей, которые описаны в данном документе в разделе «Регуляторные переключатели», для контроля и регуляции экспрессии указанного трансгена, и может содержать, при необходимости, регуляторный переключатель, который представляет собой «аварийный выключатель», позволяющий обеспечивать контролируемую смерть клетки, содержащей зкДНК-вектор.
[00186] На Фиг. 1А-1С приведены схематические изображения неограничивающих типичных примеров зкДНК-векторов, или соответствующая последовательность зкДНК-плазмид. ЗкДНК-векторы являются бескапсидными и могут быть получены из плазмиды, кодирующей, в указанном порядке: первый ITR, экспрессируемую трансгенную кассету и второй ITR, при этом по меньшей мере одна из последовательностей первого и/или второго ITR мутирована относительно соответствующей последовательности ITR AAV2 дикого типа. Указанная экспрессируемая трансгенная кассета предпочтительно включает в указанном порядке, что-либо одно или более из следующего: энхансер/промотор, ORF-репортер (трансген), посттранскрипционный регуляторный элемент (например, WPRE) и сигнал полиаденилирования и терминации (например, поли(А) BGH).
[00187] Кассета экспрессии в векторе зкДНК, полученном в соответствии со способами и композициями с использованием одного белка Rep, как раскрыто в данном документе, может содержать любой интересующий трансген. Трансгены, представляющие интерес, включают, не ограничиваясь перечисленными, нуклеиновые кислоты, кодирующие полипептиды, или некодирующие нуклеиновые кислоты (например, РНКи, miR и т.п.) предпочтительно терапевтические (например, для применения в медицинских, диагностических или ветеринарных целях) или иммуногенные полипептиды (например, для вакцин) полипептиды. Согласно некоторым вариантам реализации трансгены в экспрессионной кассете кодируют один или более полипептидов, пептидов, рибозимов, пептидных нуклеиновых кислот, киРНК, РНКи, антисмысловых олигонуклеотидов, антисмысловых полинуклеотидов, антител, антигенсвязывающих фрагментов или любой комбинации перечисленного. Согласно некоторым вариантам реализации указанный трансген представляет собой терапевтический ген или маркерный белок. Согласно некоторым вариантам реализации указанный трансген представляет собой агонист или антагонист. Согласно некоторым вариантам реализации указанный антагонист представляет собой миметик антитела или фрагмент антитела, или его антигенсвязывающий фрагмент, например, нейтрализующее антитело или фрагмент антитела и т.п. Согласно некоторым вариантам реализации указанный трансген кодирует антитело, в том числе полноразмерное антитело или фрагмент антитела согласно определению в данном документе. Согласно некоторым вариантам реализации указанное антитело представляет собой антигенсвязывающий домен или последовательность вариабельного домена иммуноглобулина согласно определению в данном документе.
[00188] В частности, указанный трансген может кодировать один или более терапевтических агентов, в том числе, но не ограничиваясь перечисленными, например, белок (белки), полипептид(ы), пептид(ы), фермент(ы), антитела, антигенсвязывающие фрагменты, а также их варианты и/или активные фрагменты для применения при лечении, профилактике и/или облегчении одного или более симптомов заболеваний, дисфункции, повреждения и/или расстройства. Примеры трансгенов описаны в данном документе в разделе, названном «Способ лечения».
[00189] Существует множество структурных признаков зкДНК-векторов, полученных в соответствии со способами и композициями с использованием единого белка Rep, как раскрыто в данном документе, которые отличают их от экспрессионных векторов на основе плазмид. ЗкДНК-векторы могут обладать одним или более из следующих признаков: отсутствие оригинальной (т.е. не инсертированной) бактериальной ДНК, отсутствие прокариотической точки начала репликации, автономность, т.е., они не требуют каких-либо последовательностей, кроме двух ITR, включая связывающие Rep сайты и сайты концевого разрешения (RBS и TRS), и экзогенной последовательности между ITR, присутствие последовательностей ITR, которые образуют шпильки, эукариотического происхождения (т.е., они продуцируются в эукариотических клетках), и отсутствие метилирования ДНК бактериального типа или, по сути, любого другого метилирования, воспринимаемого как аномальное хозяином-млекопитающим. В целом, предпочтительно, чтобы предложенные векторы не содержали какой-либо прокариотической ДНК, однако предусматривается возможность вставки какой-либо прокариотической ДНК в виде экзогенной последовательности, в качестве неограничивающего примера, в промоторном или энхансерном участке. Другой важный признак, отличающий зкДНК-векторы от плазмидных экспрессионных векторов, заключается в том, что зкДНК-векторы представляют собой одноцепочечную линейную ДНК с замкнутыми концами, тогда как плазмиды всегда представлены двуцепочечной ДНК.
[00190] ЗкДНК-векторы, полученных в соответствии со способами и композициями с использованием единого белка Rep, как раскрыто в данном документе, предпочтительно имеют линейную и непрерывную структуру, а не прерывистую структуру, при определении с помощью анализа расщепления рестриктазой (Фиг. 4D). Считается, что линейная и непрерывная структура более устойчива к атаке клеточными эндонуклеазами, а также с меньшей вероятностью подвергается рекомбинации и вызывает мутагенез. Соответственно, зкДНК-вектор, имеющий линейную и непрерывную структуру, представляет собой предпочтительный вариант реализации. Непрерывный линейный одноцепочечный зкДНК-вектор с внутримолекулярным дуплексом может иметь ковалентно связанные концы без последовательностей, кодирующих белки капсида AAV. Указанные зкДНК-плазмиды структурно отличаются от плазмид (включая зкДНК-плазмиды, описанные в данном документе), которые представляют собой кольцевые дуплексные молекулы нуклеиновых кислот бактериального происхождения. Комплементарные цепи плазмид могут быть разделены после денатурации с получением двух молекул нуклеиновой кислоты, тогда как, напротив, зкДНК-векторы, содержащие комплементарные цепи, представлены единственной молекулой ДНК и, соответственно, даже после денатурации остаются в виде единственной молекулы. В некоторых вариантах реализации зкДНК-векторы, как описано в данном документе, могут быть получены без метилирования оснований ДНК по прокариотическому типу, в отличие от плазмид. Соответственно, зкДНК-векторы и зкДНК-плазмиды отличаются как структурой (в частности, линейной или кольцевой), так и способами, применяемыми для получения и очистки указанных отличающихся объектов (см. ниже), а также характером метилирования ДНК, которое происходит по прокариотическому типу в зкДНК-плазмидах и по эукариотическому типу в зкДНК-векторе.
[00191] Несколько преимуществ использования зкДНК-вектора, как описано в данном документе, по сравнению с экспрессионными векторами на основе плазмид, включают, без ограничений, следующие: 1) плазмиды содержат последовательности бактериальной ДНК и подвергаются специфическому прокариотическому метилированию, например, метилированию с образованием 6-метиладенозина и 5-метилцитозина, тогда как бескапсидные последовательности AAV-вектора имеют эукариотическое происхождение и не подвергаются специфическому прокариотическому метилированию; в результате бескапсидные AAV-векторы будут с меньшей вероятностью индуцировать воспалительные и иммунные ответы по сравнению с плазмидами; 2) если плазмиды требуют наличия гена устойчивости в процессе получения, то зкДНК-векторы - нет; 3) если кольцевая плазмида не доставляется в ядро при введении в клетку и требует избыточной нагрузки, чтобы избежать разложения клеточными нуклеазами, то зкДНК-векторы содержат вирусные цис-элементы, т.е. ITR, придающие устойчивость к нуклеазам, и могут быть сконструированы для нацеливания и доставки в ядро. Выдвинуто предположение, что минимальными определяющими элементами, обязательными для функции ITR, являются Rep-связывающий сайт (RBS; 5'-GCGCGCTCGCTCGCTC-3' (SEQ ID NO: 531) для AAV2) и сайт концевого разрешения (TRS; 5'-AGTTGG-3' (SEQ ID NO: 48) для AAV2), плюс вариабельная палиндромная последовательность, обеспечивающая образование шпильки; и 4) зкДНК-векторы не характеризуются часто обнаруживаемой в происходящих из прокариот плазмидах чрезмерной представленностью CpG-динуклеотидов, которые, по имеющимся данным, связываются с представителем Toll-подобного семейства рецепторов, вызывая опосредованный T-клетками иммунный ответ. Напротив, трансдукция бескапсидными AAV-векторами согласно описанию в данном документе может быть эффективно нацелена на типы клеток и тканей, которые трудно трансдуцировать стандартными вирионами AAV с применением различных реагентов для доставки.
V. ITR
[00192] Вектор зкДНК, полученный в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, содержит гетерологичный ген, расположенный между двумя последовательностями инвертированных концевых повторов (ITR), которые отличаются друг от друга (т.е. являются асимметричными ITR). В некоторых вариантах реализации по меньшей мере один из ITR модифицирован путем делеции, вставки и/или замены по сравнению с последовательностью ITR дикого типа (например, ITR AAV); и по меньшей мере один из ITR содержит функциональный сайт связывания Rep (RBS; например, 5'-GCGCGCTCGCTCGCTC-3' для AAV2, SEQ ID NO: 531) и функциональный сайт терминального разрешения (TRS; например, 5'-AGTT-3', SEQ ID NO: 46.) В одном варианте реализации, по меньшей мере один из указанных ITR является нефункциональным ITR. В одном варианте реализации не каждый из разных или модифицированных ITR является ITR дикого типа из разных серотипов.
[00193] Хотя ITR, приведенные для иллюстрации в данном документе в описании и Примерах, представляют собой ITR AAV2, специалисту в данной области техники известно, что, как было указано выше, можно использовать ITR из любого известного парвовируса, например, депендовируса, такого как AAV (например, из генома AAV1, AAV2, AAV3, AAV4, AAV5, AAV 5, AAV7, AAV8, AAV9, AAV10, AAV 11, AAV12, AAVrh8, AAVrh10, AAV-DJ и AAV-DJ8. Например, NCBI: NC 002077; NC 001401; NC001729; NC001829; NC006152; NC 006260; NC 006261), химерные ITR или ITR из любых синтетических AAV. Согласно некоторым вариантам реализации указанный AAV способен инфицировать теплокровных животных, например, представлен аденоассоциированными вирусами птиц (AAAV), крупного рогатого скота (BAAV), собачьих, лошадей и овец. Согласно некоторым вариантам реализации ITR взят из парвовируса B19 (номер доступа GenBank: NC 000883), мелкого вируса мышей (MVM) (номер доступа GenBank NC 001510); парвовируса гусей (номер доступа в GenBank NC 001701); парвовируса змей 1 (номер доступа GenBank NC 006148).
[00194] Согласно некоторым вариантам реализации указанная последовательность ITR в зкДНК векторе, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, может быть взята из вирусов семейства Parvoviridae, которое включает два подсемейства: Parvovirinae, которые инфицируют позвоночных, и Densovirinae, которые инфицируют насекомых. Подсемейство Parvovirinae (называемое парвовирусами) включает род Dependovirus, представители которого, в большинстве случаев, нуждаются в коинфицировании хелперным вирусом, таким как аденовирус или вирус герпеса, для продуктивной инфекции. Род Dependovirus включает аденоассоциированный вирус (AAV), который обычно инфицирует людей (например, серотипы 2, 3A, 3B, 5 и 6) или приматов (например, серотипы 1 и 4), и родственные вирусы, которые инфицируют других теплокровных животных (например, аденоассоциированные вирусы крупного рогатого скота, собачьих, лошадей, овец). Общее описание парвовирусов и других представителей семейства Parvoviridae приведено в источнике: Kenneth I. Berns, "Parvoviridae: The Viruses and Their Replication," глава 69 издания «FIELDS VIROLOGY» (3 издание, 1996).
[00195] Специалисту в данной области техники известно, что последовательности ITR имеют общую структуру, содержащую двухцепочечную структуру Холлидея, которая, как правило, представляет собой T-образную или Y-образную шпилечную структуру (см., например, Фиг. 2A и Фиг. 3A), при этом каждый ITR образован двумя палиндромными плечами или петлями (B-B' и C-C'), погруженными в палиндромное плечо большего размера (A-A'), и одноцепочечную последовательность D (при этом порядок указанных палиндромных последовательностей определяет ориентацию ITR), и, соответственно, можно легко определить соответствующие модифицированные последовательности ITR из любого серотипа AAV для применения в зкДНК-векторе или зкДНК-плазмиде на основе типичных последовательностей AAV2 ITR приведенных в данном документе. См., например, описание структурного анализа и сравнения последовательностей ITR из разных серотипов AAV (AAV1-AAV6) в источниках Grimm et al., J. Virology, 2006; 80(1); 426-439; Yan et al., J. Virology, 2005; 364-379; Duan et al., Virology 1999; 261; 8-14.
[00196] Конкретные изменения и мутации в ITR подробно описаны в данном документе, но в контексте «измененных» или «мутированных» или «модифицированных» ITR, эти определения указывают на то, что нуклеотиды были вставлены, удалены и/или замещены относительно принадлежащей к дикому типу, референсной или исходной последовательности ITR, и могут быть изменены относительно другого фланкирующего ITR в векторе зкДНК, имеющем два фланкирующих ITR. Измененный или мутированный ITR может представлять собой сконструированный ITR. В данном документе «сконструированный» относится к аспекту, который подвергается манипуляциям человека. Например, полипептид считается «рекомбинантным», когда по меньшей мере один аспект полипептида, например, его последовательность, был подвергнут манипуляциям со стороны человека для создания отличий от аспекта, в котором он существует в природе.
[00197] В некоторых вариантах реализации ITR в векторе зкДНК, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, может быть синтетическим. Согласно одному варианту реализации синтетический ITR основан на последовательностях ITR более чем из одного серотипа AAV. Согласно другому варианту реализации синтетический ITR не включает последовательностей на основе AAV. Согласно еще одному варианту реализации, синтетический ITR сохраняет описанную выше структуру ITR, хотя и содержит только часть или не содержит последовательности, происходящей из AAV. В некоторых аспектах синтетический ITR может взаимодействовать преимущественно с Rep дикого типа или с Rep определенного серотипа, или, в некоторых случаях, не будет распознаваться Rep дикого типа и будет распознаваться только мутированным Rep.
[00198] Последовательности ITR устроены однотипно и содержат двуцепочечную структуру Холлидея, которая, как правило, представляет собой T-образную или Y-образную шпилечную структуру (см., например, Фиг. 2A и Фиг. 3A), при этом каждый ITR образован двумя палиндромными плечами или петлями (B-B' и C-C'), встроенными в палиндромное плечо большего размера (A-A'), и одноцепочечной последовательностью D (где порядок указанных палиндромных последовательностей определяет направление ориентации ITR). Рядовой специалист в данной области техники может легко определить последовательности ITR или модифицированного ITR из любого серотипа AAV для применения в зкДНК-векторе или зкДНК-плазмиде, на основании примеров последовательностей ITR AAV2, приведенных в данном документе. См., например, сравнение последовательностей ITR из различных серотипов AAV (AAV1-AAV6 и AAV птиц (AAAV) и AAV крупного рогатого скота (BAAV)), описанное в Grimm et al., J. Virology, 2006; 80(1); 426-439; где приведены % идентичности левого ITR AAV2 и левых ITR из других серотипов: AAV-1 (84%), AAV-3 (86%), AAV-4 (79%), AAV-5 (58%) AAV-6 (левый ITR) (100%) и AAV-6 (правый ITR) (82%).
[00199] Соответственно, в то время как ITR AAV2 используются в качестве иллюстративных ITR в векторе зкДНК, полученном в соответствии со способами и композициями с использованием одного белка Rep, как раскрыто в данном документе, вектор зкДНК может быть получен с ITR любого известного серотипа AAV или на его основе, включая, например, AAV серотип 1 (AAV1), AAV серотип 2 (AAV2), AAV серотип 4 (AAV4), AAV серотип 5 (AAV5), AAV серотип 6 (AAV6), AAV серотип 7 (AAV7), AAV серотип 8 (AAV8), AAV серотип 9 (AAV9), AAV серотип 10 (AAV10), AAV серотип 11 (AAV11) или AAV серотип 12 (AAV12). Специалист в данной области техники сможет определить соответствующую последовательность в других серотипах известными способами. Например, путем определения того, находится ли изменение в участке A, A', B, B', C, C' или D, и определения соответствующего участка в другом серотипе. Можно использовать BLAST® (Basic Local Alignment Search Tool) или другие программы выравнивания для определения гомологии с настройками по умолчанию для определения соответствующей последовательности. Согласно данному изобретению также предложены популяции и совокупности зкДНК-векторов, содержащих ITR из комбинации разных серотипов AAV - то есть один ITR может быть взят из одного серотипа AAV, а другой ITR может быть взят из другого серотипа. Не связываясь какой-либо теорией, укажем, что в одном варианте реализации один ITR может быть взят из последовательности ITR AAV2 или быть на ней основан, а другой ITR зкДНК-вектора может быть взят из или быть основан на любой одной или нескольких последовательностях ITR AAV серотипа 1 (AAV1), AAV серотипа 4 (AAV4), AAV серотипа 5 (AAV5), AAV серотипа 6 (AAV6), AAV серотипа 7 (AAV7), AAV серотипа 8 (AAV8), AAV серотипа 9 (AAV9), AAV серотипа 10 (AAV10), AAV серотипа 11 (AAV11) или AAV серотипа 12 (AAV12).
[00200] Любой парвовирусный ITR может использоваться как ITR или в качестве базового ITR для модификации. Предпочтительно, указанный парвовирус представляет собой депендовирус. Более предпочтительно указанный парвовирус представляет собой AAV. Выбор серотипа может быть основан на тканевом тропизме серотипа. AAV2 обладает тропизмом к широкому ряду тканей, AAV1 преимущественно нацелен на нервную ткань и скелетные мышцы, а AAV5 преимущественно нацелен на нервную ткань, пигментированный эпителий сетчатки и фоторецепторы. AAV6 преимущественно нацелен на скелетные мышцы и легкие. AAV8 преимущественно нацелен на печень, скелетные мышцы, сердце и ткани поджелудочной железы. AAV9 преимущественно направлен на печень, скелетные мышцы и ткани легких. В одном варианте реализации модифицированный ITR основан на ITR AAV2. Например, он выбран из группы, состоящей из: SEQ ID NO: 2 и SEQ ID NO: 52. В одном варианте каждого из этих аспектов векторный полинуклеотид содержит пару ITR, выбранных из группы, состоящей из: SEQ ID NO: 1 и SEQ ID NO: 52; и SEQ ID NO: 2 и SEQ ID NO: 51. В одном варианте каждого из этих аспектов векторный полинуклеотид или невирусные, бескапсидные ДНК-векторы с ковалентно замкнутыми концами содержат пару различных ITR, выбранных из группы, состоящей из: SEQ ID NO: 101 и SEQ ID NO. : 102; SEQ ID NO: 103 и SEQ ID NO: 104, SEQ ID NO: 105 и SEQ ID NO: 106; SEQ ID NO: 107 и SEQ ID NO: 108; SEQ ID NO: 109 и SEQ ID NO: 110; SEQ ID NO: 111 и SEQ ID NO: 112; SEQ ID NO: 113 и SEQ ID NO: 114; и SEQ ID NO: 115 и SEQ ID NO: 116. В некоторых вариантах реализации модифицированный ITR выбран из любой из ITR или частичных последовательностей ITR SEQ ID NO: 2, 52, 63, 64, 101-499 или 545-547.
[00201] Согласно некоторым вариантам реализации зкДНК-вектор, полученный в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, может содержать ITR с модификацией в ITR, соответствующей любой из модификаций в последовательностях ITR или частичных последовательностях ITR, представленных в любой одной или более из таблиц 2, 3, 4, 5, 6, 7, 8, 9, 10A и 10B данного документа, или последовательностей представленных на Фиг 26A или 26B.
[00202] В некоторых вариантах реализации вектор зкДНК, полученный в соответствии со способами и композициями с использованием единственного Rep-белка, как раскрыто в данном документе, может образовывать внутримолекулярную дуплексную вторичную структуру. В неограничивающем примере вторичная структура первого ITR и асимметричного второго ITR представлена в контексте ITR дикого типа (см., например, фиг. 2A, 3A, 3C) и/или модифицированных структур ITR (см., например, Фиг. 2B и Фиг. 3B, 3D). Вторичные структуры выводят или предсказывают на основании последовательностей ITR плазмиды, используемой для получения зкДНК-вектора. Примерные вторичные структуры модифицированных ITR, в которых удалена часть структуры стебель-петля, показаны на Фиг. 9A-25B и Фиг. 26A-26B, а также показаны в Таблицах 10A и 10B. Типичные вторичные структуры модифицированных ITR, содержащих единственный стебель и две петли, приведены на Фиг. 9A-13B. Примерная вторичная структура модифицированного ITR с одним стеблем и одной петлей показана на Фиг. 14. Согласно некоторым вариантам реализации указанная вторичная структура может быть выведена, как показано в данном документе, с использованием термодинамических методов, основанных на правилах ближайшего соседа, которые предсказывают стабильность структуры, количественно определяемую изменением свободной энергии укладки. Например, указанная структура может быть предсказана путем нахождения структуры с наименьшей свободной энергией. Согласно некоторым вариантам реализации для предсказания структуры ITR может быть использован алгоритм, описанный в источнике: Reuter, J. S., & Mathews, D. H. (2010) RNAstructure: software for RNA secondary structure prediction and analysis. BMC Bioinformatics. 11, 129, и реализованный в программном обеспечении RNAstructure (доступно в сети Интернет по адресу: «rna.urmc.rochester.edu/RNAstructureWeb/index.html”). Указанный алгоритм может также включать как параметры изменения свободной энергии при 37°C, так и параметры изменения энтальпии, полученные из экспериментальной литературы, что позволяет прогнозировать стабильность конформации при произвольной температуре. С применением программного обеспечения для анализа структуры РНК могут быть предсказаны некоторые из модифицированных структур ITR в виде модифицированных T-образных структур типа «петля-на-стебле» с расчетной свободной энергией Гиббса(ΔG) для разворачивания в физиологических условиях, представленными на Фиг. 3A-3D. С применением программного обеспечения для анализа структуры РНК предсказаны три типа модифицированных ITR, имеющие более высокую свободную энергию Гиббса для разворачивания, чем ITR дикого типа AAV2 (-92, 9 ккал/моль); указанные ITR представлены следующими: (a) модифицированные ITR со структурой с единственным плечом/единственной неспаренной петлей согласно данному изобретению, согласно прогнозу имеющие свободную энергию Гиббса для разворачивания в диапазоне от -85 до -70 ккал/моль. (b) предсказано, что модифицированные ITR, имеющие структуру с единственной шпилькой, описанные в данном документе, будут иметь свободную энергию Гиббса для разворачивания в диапазоне от -70 до -40 ккал/моль. (c) предсказано, что модифицированные ITR, имеющие структуру с двумя плечами, описанные в данном документе, будут иметь свободную энергию Гиббса для разворачивания в диапазоне от -90 до -70 ккал/моль. Без связи с какой-либо теорией, структуры с более высокой свободной энергией Гиббса будут легче разворачиваться для репликации белками репликации Rep 68 или Rep 78. Таким образом, модифицированные ITR, имеющие более высокую свободную энергию Гиббса для разворачивания - например, структура с одним плечом/одной непарной петлей, структура с одной шпилькой, усеченная структура - демонстрируют тенденцию к более эффективной репликации, чем ITR дикого типа.
[00203] В одном варианте реализации левый ITR вектора зкДНК, полученный в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, модифицирован или мутирован по отношению к структуре ITR AAV дикого типа (wt), а правый ITR является диким типа AAV ITR. Согласно одному варианту реализации правый ITR зкДНК-вектора модифицирован относительно структуры AAV ITR дикого типа, а левый ITR симметричен (обратно-комплементарен) ему и содержит такие же мутации. Согласно такому варианту реализации модификация ITR (например, левого или правого ITR) может быть получена путем делеции, вставки или замены одного или более нуклеотидов ITR дикого типа, происходящего из генома AAV.
[00204] Используемые согласно данному изобретению ITR могут быть разрешаемыми и неразрешаемыми, и выбранные для применения в зкДНК-векторах ITR предпочтительно представляют собой последовательности AAV, причем предпочтительными являются последовательности из серотипов 1, 2, 3, 4, 5, 6, 7, 8 и 9. Разрешаемые AAV ITR не обязательно представлены последовательностью ITR дикого типа (например, эндогенная последовательность AAV ITR или последовательность AAV ITR дикого типа может быть изменена путем вставки, делеции, усечения и/или введения миссенс-мутаций), при условии, что указанный концевой повтор опосредует требуемые функции, например, репликацию, упаковку, интеграцию вируса и/или «спасение» провируса, и т.п. Как правило, но не обязательно, ITR взяты из одного серотипа AAV, например, обе последовательности ITR зкДНК-вектора взяты из AAV2. ITR могут быть синтетическими последовательностями, которые функционируют как инвертированные концевые повторы AAV, такими как «последовательность с двойной D», как описано в патенте США No. № 5478745, Samulski et al. Хотя это не обязательно, ITR могут быть от одного и того же парвовируса, например, обе последовательности ITR происходят от AAV2.
[00205] Согласно одному варианту реализации зкДНК, полученный в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, может включать структуру ITR, которая мутирована относительно одного из ITR дикого типа как раскрыто в данном документе, однако при этом мутированный или модифицированный ITR сохраняет функциональный сайт связывания Rep (RBE или RBE') и сайт концевого разрешения (trs). Согласно одному варианту реализации указанный мутированный зкДНК ITR включает функциональный сайт белка репликации (RPS-1), и при продуцировании используют репликативно-компетентный белок, который связывает сайт RPS-1.
[00206] В одном варианте реализации по меньшей мере одна из ITR в векторе зкДНК, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, представляет собой дефектную ITR в отношении связывания Rep и/или репликации. Согласно одному варианту реализации указанный дефект представлен по меньшей мере 30%, согласно другим вариантам реализации - по меньшей мере 35%..., 50%..., 65%..., 75%..., 85%..., 90%..., 95%..., 98%..., недостаточностью функции относительно ITR дикого типа или ее полным отсутствием, или недостаточностью функции, соответствующей какому-либо промежуточному значению. Клетки-хозяева не экспрессируют вирусные капсидные белки и полинуклеотидная векторная матрица не содержит каких-либо последовательностей, кодирующих вирусный капсид. Согласно одному варианту реализации указанные полинуклеотидные векторные матрицы и клетки-хозяева, которые не содержат генов капсида AAV и итогового белка, также не кодируют или экспрессируют генов капсидов других вирусов. Кроме того, согласно конкретному варианту реализации указанная молекула нуклеиновой кислоты также не содержит последовательностей, кодирующих белок Rep AAV.
[00207] Согласно некоторым вариантам реализации структурный элемент ITR может быть представлен любым структурным элементом, который вовлечен в функциональное взаимодействие ITR с единственным большим белком Rep (например, Rep 78 или Rep 68). Согласно некоторым вариантам реализации указанный структурный элемент обеспечивает селективность взаимодействия ITR с единственным большим белком Rep, т.е., определяет по меньшей мере частично, какой белок Rep функционально взаимодействует с указанным ITR. Согласно другим вариантам реализации указанный структурный элемент физически взаимодействует с единственным большим белком Rep при связывании указанного белка Rep с ITR. Каждый структурный элемент может представлять собой, например, вторичную структуру ITR, нуклеотидную последовательность ITR, промежуточную последовательность между двумя или более элементами; или комбинацию любых вышеперечисленных элементов. Согласно одному варианту реализации указанные структурные элементы выбраны из группы, состоящей из плеча A и плеча A', плеча B и плеча B', плеча C и плеча C', плеча D, связывающего Rep сайта (RBE) и RBE' (т.е. последовательности, комплементарной RBE), и сайта концевого разрешения (trs).
[00208] Более конкретно, способность структурного элемента ITR в векторе зкДНК, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, функционально взаимодействовать с конкретным единственным белком Rep, например, большим белком Rep или малым белком Rep, можно изменить, изменив структурный элемент. Например, нуклеотидная последовательность структурного элемента может быть модифицирована по сравнению с последовательностью ITR дикого типа. Согласно одному варианту реализации структурный элемент (например, плечо A, плечо A', плечо B, плечо B', плечо C, плечо C', плечо D, RBE, RBE' и trs) ITR может быть удален и заменен структурным элементом дикого типа из другого парвовируса. Например, заменяющая структура может быть взята из AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAV12, AAV13, парвовируса змей (например, парвовируса королевского питона), парвовируса крупного рогатого скота, парвовируса коз, парвовируса птиц, парвовируса собачьих, парвовируса лошадиных, парвовируса креветок, свиного парвовируса или AAV насекомых. Например, ITR может представлять собой ITR AAV2, и плечо A или A', или RBE могут быть заменены структурным элементом из AAV5. В другом примере, ITR может представлять собой ITR AAV5, а плечи C или C', RBE и trs могут быть заменены структурным элементом из AAV2. В другом примере ITR AAV может представлять собой ITR AAV5, в котором плечи B и B' заменены плечами B и B' ITR AAV2.
[00209] Исключительно для примера в Таблице 1 приведены иллюстративные модификации по меньшей мере одного нуклеотида (например, делеция, вставка и/или замена) в участках модифицированных ITR, где X указывает на модификацию по меньшей мере одной нуклеиновой кислоты (например, делецию, вставку и/или замену) в указанном участке относительно соответствующего ITR дикого типа. Согласно некоторым вариантам реализации при любой модификации по меньшей мере одного нуклеотида (например, делеции, вставки и/или замены) в любом из участков C и/или C', и/или B, и/или B' сохраняется три последовательных нуклеотида T (т.е. ТТТ) по меньшей мере в одной концевой петле. Например, если указанная модификация приводит к чему-либо из: ITR с единственным плечом (например, с единственным плечом C-C' или единственным плечом B-B'), или с модифицированным плечом C-B' или плечом C'-B, или ITR с двумя плечами, содержащему по меньшей мере одно усеченное плечо (например, усеченное плечо C-C' и/или усеченное плечо B-B'), то по меньшей мере в указанном единственном плече, или по меньшей мере в одном из плеч ITR с двумя плечами (у которого одно плечо может быть усеченным) сохраняются три последовательных нуклеотида T (т.е. TTT) по меньшей мере в одной концевой петле. Согласно некоторым вариантам реализации усеченное плечо C-C' и/или усеченное плечо B-B' содержит три последовательных нуклеотида T (т.е. TTT) в концевой петле.
Таблица 1: Примеры комбинаций модификаций по меньшей мере одного нуклеотида (например, делеции, вставки и/или замены) в разных участках или плечах B-B' и C-C' ITR (X указывает на модификацию нуклеотида, например, добавление, делецию или замену по меньшей мере одного нуклеотида в указанном участке).
[00210] Согласно некоторым вариантам реализации модифицированный ITR для применения в зкДНК векторе, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, может содержать любую из комбинаций модификаций, представленных в Таблице 1, и также модификацию по меньшей мере одного нуклеотида в любом одном или более участках, выбранных из: участка между A' и C, между C и C', между C' и B, между B и B' и между B' и A. Согласно некоторым вариантам реализации при любой модификации по меньшей мере одного нуклеотида (например, делеции, вставки и/или замене) в участке C, или C', или B, или B', все же сохраняется концевая петля структуры «петля-на-стебле». Согласно некоторым вариантам реализации при любой модификации по меньшей мере одного нуклеотида (например, делеции, вставки и/или замене) между C и C' и/или B и B' сохраняются три последовательных нуклеотида T (т.е., TTT) по меньшей мере в одной концевой петле. В альтернативных вариантах реализации любая модификация по меньшей мере одного нуклеотида (например, делеции, вставки и/или замене) между C и C' и/или B и B' сохраняет три последовательных нуклеотида «A» (т.е. AAA) по меньшей мере в одной концевой петле. В некоторых вариантах реализации модифицированный ITR для применения в зкДНК векторе, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, может содержать любую одну из комбинаций модификаций, представленных в Таблице 1, а также модификацию по меньшей мере одного нуклеотида (например, делецию, вставку и/или замену) в любом из одного или нескольких участков, выбранных из: A', A и/или D. Например, в некоторых вариантах реализации модифицированный ITR для использования по данному изобретению может содержать любую из комбинаций модификаций, представленных в Таблице 1, а также модификацию по меньшей мере одного нуклеотида (например, делецию, вставку и/или замену) в участке А. Согласно некоторым вариантам реализации модифицированный ITR для применения в зкДНК векторе, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, может содержать любую из комбинаций модификаций, представленных в Таблице 1, а также модификацию по меньшей мере одного нуклеотида (например, делецию, инсерцию и/или замену) в участке A'. Согласно некоторым вариантам реализации модифицированный ITR для применения в данном документе может содержать любую из комбинаций модификаций, представленных в Таблице 1, а также модификацию по меньшей мере одного нуклеотида (например, делецию, вставку и/или замену) в участке A и/или A'. Согласно некоторым вариантам реализации модифицированный ITR для применения в данном документе может содержать любую из комбинаций модификаций, представленных в Таблице 1, а также модификацию по меньшей мере одного нуклеотида (например, делецию, вставку и/или замену) в участке D.
[00211] Согласно одному варианту реализации нуклеотидная последовательность структурного элемента может быть модифицирована (например, путем модификации 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, или 20, или более нуклеотидов, или любого числа нуклеотидов в промежуточном диапазоне) с получением модифицированного структурного элемента. В одном варианте реализации конкретные модификации ITR в векторе зкДНК, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, проиллюстрированы здесь примерами (например, SEQ ID NO: 2, 52, 63, 64, 101-499, или 545-547). Согласно некоторым вариантам реализации ITR в зкДНК векторе, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, может быть модифицирован (например, путем модификации 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, или 20, или более нуклеотидов, или любого числа нуклеотидов в промежуточном диапазоне). В других вариантах реализации ITR в зкДНК векторе, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, может иметь по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или более, идентичности последовательности с одной из модифицированных ITR SEQ ID NOS: 469-499 или 545-547, или RBE-содержащего отдела плеча A-A' и плеч C-C' и B-B' SEQ ID NO: 101-134 или 545-547.
[00212] Согласно некоторым вариантам реализации модифицированный ITR в зкДНК векторе, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, может, например, включать удаление или делецию конкретного плеча полностью, например, полное или частичное удаление или делецию плеча A-A', или полное или частичное удаление или делеция плеча B-B', или полное или частичное удаление или делеция плеча C-C', или, как вариант, удаление 1, 2, 3, 4, 5, 6, 7, 8, 9 или более пар оснований, образующих стебель петли, при условии сохранения итоговой петли, которой завершается стебель (например, единственного плеча) (например, см. ITR-6). Согласно некоторым вариантам реализации модифицированный ITR может включать удаление 1, 2, 3, 4, 5, 6, 7, 8, 9 или более пар оснований из плеча B-B'. Согласно некоторым вариантам реализации модифицированный ITR может включать удаление 1, 2, 3, 4, 5, 6, 7, 8, 9 или более пар оснований из плеча C-C'. Согласно некоторым вариантам реализации модифицированный ITR может включать удаление 1, 2, 3, 4, 5, 6, 7, 8, 9 или более пар оснований из плеча C-C' и удаление 1, 2, 3, 4, 5, 6, 7, 8, 9 или более пар оснований из плеча B-B'. Предусмотрена любая комбинация удаления пар оснований, например, 6 пар оснований могут быть удалены в плече C-C 'и 2 пары оснований в плече B-B'. В качестве иллюстративного примера, Фиг. 13A-13B демонстрируют пример модифицированного ITR с делецией по меньшей мере 7 пар оснований из каждой из части C и части C', заменой нуклеотида в петле между участком C и C', и делецией по меньшей мере одной пары оснований из каждого из участка B и участка B', так что модифицированный ITR содержит два плеча, по меньшей мере одно из которых (например, C-C') является усеченным. Обратим внимание, что в указанном примере, поскольку модифицированный ITR содержит по меньшей мере одну делецию пар оснований в каждом из участков B и B', плечо B-B' также усечено по сравнению с WT ITR.
[00213] Согласно некоторым вариантам реализации 1, 2, 3, 4, 5, 6, 7, 8, 9 или более комплементарных пар оснований удаляют как из C-части, так и из C'-части плеча C-C', таким образом, что плечо C-C' становится усеченным. Иначе говоря, при удалении основания из C-части плеча C-C', удаляют комплементарную пару оснований из C'-части, с усечением таким образом плеча C-C'. Согласно таким вариантам реализации удаляют 2, 4, 6, 8 или более пар оснований из плеча C-C', таким образом, что плечо C-C' становится усеченным. Согласно альтернативным вариантам реализации удаляют 1, 2, 3, 4, 5, 6, 7, 8, 9 или более пар оснований из C-части плеча C-C' таким образом, чтобы сохранилась только C'-часть плеча. Согласно альтернативным вариантам реализации удаляют 1, 2, 3, 4, 5, 6, 7, 8, 9 или более пар оснований из C'-части плеча C-C' таким образом, чтобы сохранилась только C-часть плеча.
[00214] Согласно некоторым вариантам реализации удаляют 1, 2, 3, 4, 5, 6, 7, 8, 9 или более комплементарных пар оснований как из B-части, так и из B'-части плеча B-B' таким образом, чтобы плечо B-B' становится усеченным. Иначе говоря, при удалении основания из B-части плеча B-B' удаляют комплементарную пару оснований из B'-части, с усечением таким образом плеча B-B'. Согласно таким вариантам реализации удаляют 2, 4, 6, 8 или более пар оснований из плеча B-B' таким образом, что плечо B-B' становится усеченным. Согласно альтернативным вариантам реализации удаляют 1, 2, 3, 4, 5, 6, 7, 8, 9 или более пар оснований из B-части плеча B-B' таким образом, чтобы сохранилась только B'-часть плеча. Согласно альтернативным вариантам реализации удаляют 1, 2, 3, 4, 5, 6, 7, 8, 9 или более пар оснований из B'-части плеча B-B' таким образом, чтобы сохранилась только B-часть плеча.
[00215] Согласно некоторым вариантам реализации модифицированный ITR в зкДНК векторе, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, может содержать от 1 до 50 (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49 или 50) делеций нуклеотидов относительно полноразмерной последовательности ITR дикого типа. Согласно некоторым вариантам реализации модифицированный ITR может содержать от 1 до 30 делеций нуклеотидов относительно полноразмерной последовательности WT ITR. Согласно некоторым вариантам реализации модифицированный ITR содержит от 2 до 20 делеций нуклеотидов относительно полноразмерной последовательности ITR дикого типа.
[00216] Согласно некоторым вариантам реализации модифицированный ITR образует две противопоставленных продольно-симметричных «петли-на-стебле», например, петля C-C' имеет длину, отличную от длины петли B-B'. Согласно некоторым вариантам реализации одна из противопоставленных продольно-симметричных петель-на-стебле модифицированного ITR содержит часть стебля C-C' и/или B-B' с длиной в диапазоне от 8 до 10 пар оснований, и петлевую часть (например, между C-C' или между B-B'), которая содержит от 2 до 5 неспаренных дезоксирибонуклеотидов. Согласно некоторым вариантам реализации одна из продольно-симметричных петель-на-стебле модифицированного ITR содержит часть стебля C-C' и/или B-B' длиной менее 8 или менее 7, 6, 5, 4, 3, 2, 1 пар оснований и петлевую часть (например, между C-C' или между B-B'), которая содержит 0-5 нуклеотидов. Согласно некоторым вариантам реализации модифицированный ITR с продольно-асимметричной петлей-на-стебле содержит часть стебля C-C' и/или B-B' длиной менее чем 3 пары оснований.
[00217] В некоторых вариантах реализации модифицированный ITR в зкДНК векторе, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, не содержит каких-либо нуклеотидных делеций в RBE-содержащей части участков A или A', чтобы не мешать репликации ДНК (например, связыванию с RBE белка Rep или никированию в сайте концевого разрешения). Согласно некоторым вариантам реализации модифицированный ITRв зкДНК векторе, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, содержит одну или более делеций в участке B, B', C и/или C согласно описанию в данном документе. Несколько неограничивающих примеров модифицированных ITR показаны на Фиг. 9A-26B.
[00218] В некоторых вариантах реализации модифицированный ITR в зкДНК векторе, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, может включать делецию плеча B-B', так что плечо C-C' остается, например, см. типичные ITR -2 (левый) и ITR-2 (правый), показанные на Фиг. 9A-9B, и ITR-4 (левый) и ITR-4 (правый) (Фиг. 11A-11B). Согласно некоторым вариантам реализации модифицированный ITR может включать делецию плеча C-C', таким образом, сохраняется плечо B-B', например, см. типичные ITR-3 (левый) and ITR-3 (правый) приведенные на Фиг. 10A-10B. Согласно некоторым вариантам реализации модифицированный ITR может включать делецию плеча B-B' и плеча C-C', таким образом, чтобы сохранялась единственная петля-на-стебле, например, см. типичные ITR-6 (левый) и ITR-6 (правый), показанные на Фиг. 14A-14B, и ITR-21 (левый) и ITR-37 (правый). В некоторых вариантах реализации модифицированный ITR в зкДНК векторе, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, может включать делецию участка C', так что остается усеченная C-петля и плечо B-B', например см. типичные ITR-1 (левый) и ITR-1 (правый), показанные на Фиг. 15A-15B. Аналогичным образом, в некоторых вариантах реализации модифицированный ITR может содержать делецию участка C, так что остается усеченная C'-петля и плечо B-B', например, см. типичные ITR-5 (левый) и ITR-5 (правый), показанные на Фиг. 16A-16B.
[00219] Согласно некоторым вариантам реализации модифицированный ITR в зкДНК векторе, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, может включать делецию пар оснований в любой одной или более из: C-части, C'-части, B-части или B'-части, таким образом, чтобы между частями C-B' и частями C'-B происходило спаривание комплементарных оснований с получением единственного плеча, например, см. ITR-10 (правый) и ITR-10 (левый) (Фиг. 12А-12В).
[00220] Согласно некоторым вариантам реализации наряду с модификацией в одном или более нуклеотидов в участке C, C', B и/или B', модифицированный ITR для применения в зкДНК векторе, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, может содержать модификацию (например, делецию, замену или добавление) по меньшей мере 1, 2, 3, 4, 5, 6 нуклеотидов в любом одном или более участков, выбранных из: участок между A' и C, между C и C', между C' и B, между B и B' и между B' и A. Например, нуклеотид A между B' и C в модифицированном правом ITR может быть заменен на G, C или A или удален, или может быть добавлен один или более нуклеотидов; нуклеотид T между C' и B в модифицированном левом ITR может быть заменен на G, C или A, или удален, или может быть добавлен один или более нуклеотидов.
[00221] В некоторых вариантах реализации вектор зкДНК, полученный в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, не имеет модифицированного ITR, состоящего из нуклеотидной последовательности, выбранной из любой из: SEQ ID NO: 550-557. В некоторых вариантах реализации вектор зкДНК, полученный в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, не имеет модифицированного ITR, содержащего нуклеотидную последовательность, выбранную из любой из: SEQ ID NO: 550-557.
[00222] В некоторых вариантах реализации зкДНК-вектор содержит регуляторный переключатель, как описано в данном документе, и выбранный модифицированный ITR имеющий нуклеотидную последовательность, выбранную из любых из группы, состоящей из: SEQ ID NO: 550-557.
[00223] В другом варианте реализации структура структурного элемента ITR в векторе зкДНК, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, может быть изменена. Например, в указанном структурном элементе изменена высота стебля и/или число нуклеотидов в петле. Например, высота стебля может составлять примерно 2, 3, 4, 5, 6, 7, 8 или 9 нуклеотидов или более или соответствовать любому диапазону значений. В одном варианте реализации стебель может иметь высоту от примерно 5 нуклеотидов до примерно 9 нуклеотидов и функционально взаимодействовать с Rep. В другом варианте реализации стебель может иметь высоту около 7 нуклеотидов и функционально взаимодействовать с Rep. В другом примере петля может иметь 3, 4, 5, 6, 7, 8, 9 или 10 нуклеотидов или более, или любой диапазон указанных значений.
[00224] В другом варианте реализации, число сайтов связывания GAGY или связанных с GAGY сайтов связывания в пределах RBE или расширенного RBE может быть увеличено или уменьшено. Согласно одному примеру RBE или расширенный RBE может содержать 1, 2, 3, 4, 5 или 6 или более сайтов связывания GAGY, или любое их количество в диапазоне указанных значений. Каждый сайт связывания GAGY может независимым образом представлять собой точную последовательность GAGY или последовательность, аналогичную GAGY, при условии, что эта последовательность является достаточной для связывания белка Rep.
[00225] Согласно другому варианту реализации промежуточная последовательность между двумя элементами (такими как, без ограничений, RBE и шпилька) может быть изменена (например, увеличена или уменьшена), для изменения функционального взаимодействия с единственным большим белком Rep. Например, указанный промежуток может составлять приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или 21 нуклеотидов или более, или любой диапазон указанных значений.
[00226] ЗкДНК-вектор, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, может включать структуру ITR, которая модифицирована относительно структуры ITR AAV2 дикого типа, раскрытой в данном документе, но все еще сохраняет функциональную часть RBE, trs и RBE'. На Фиг. 2А и Фиг. 2B продемонстрирован один из возможных механизмов работы сайта trs в части структуры ITR дикого типа зкДНК-вектора. В некоторых вариантах реализации зкДНК-вектор содержит одну или несколько функциональных полинуклеотидных последовательностей ITR, которые содержат Rep-связывающий сайт (RBS; 5'-GCGCGCTCGCTCGCTC-3' (SEQ ID NO: 531) для AAV2) и сайт концевого разрешения (TRS; 5'-AGTT (SEQ ID NO: 46)). В некоторых вариантах реализации по меньшей мере один ITR (дикого типа или модифицированный ITR) является функциональным. В альтернативных вариантах реализации, в тех случаях, когда зкДНК-вектор содержит два модифицированных ITR, которые отличаются друг от друга или асимметричны друг относительно друга, по меньшей мере один модифицированный ITR является функциональным и по меньшей мере один модифицированный ITR является нефункциональным.
[00227] В некоторых вариантах реализации вектор зкДНК, полученный в соответствии со способами и композициями с использованием единственного Rep-белка, как раскрыто в данном документе, не имеет модифицированного ITR, выбранного из какой-либо последовательности, состоящей из или состоящей по существу из: SEQ ID NO: 500-529, как предусмотрено в данном документе. Согласно некоторым вариантам реализации зкДНК-вектор не содержит ITR, который выбран из любой последовательности, выбранной из SEQ ID NO: 500-529.
[00228] В некоторых вариантах реализации модифицированный ITR (например, левый или правый) вектора зкДНК, полученного в соответствии со способами и композициями с использованием единственного Rep-белка, как раскрыто в данном документе, имеет модификации внутри петлевого плеча, усеченного плеча или спейсера. Типичные последовательности ITR, имеющие модификации в петлевом плече, усеченном плече или спейсере, перечислены в Таблице 2.
[00229] В некоторых вариантах реализации модифицированный ITR (например, левый или правый) вектора зкДНК, полученного в соответствии со способами и композициями с использованием единственного Rep-белка, как раскрыто в данном документе, имеет модификации в пределах петлевого плеча и усеченного плеча. Типичные последовательности ITR, имеющие модификации в петлевом плече и усеченном плече, перечислены в Таблице 3.
[00230] В некоторых вариантах реализации модифицированный ITR (например, левый или правый) вектора зкДНК, полученного в соответствии со способами и композициями с использованием единственного Rep-белка, как раскрыто в данном документе, имеет модификации внутри петлевого плеча и спейсера. Типичные последовательности ITR, имеющие модификации в петлевом плече и спейсере, перечислены в Таблице 4.
[00231] В некоторых вариантах реализации модифицированный ITR (например, левый или правый) вектора зкДНК, полученного в соответствии со способами и композициями с использованием единственного Rep-белка, как раскрыто в данном документе, имеет модификации внутри усеченного плеча и спейсера. Типичные последовательности ITR, имеющие модификации в усеченном плече и спейсере, перечислены в Таблице 5.
[00232] В некоторых вариантах реализации модифицированный ITR (например, левый или правый) вектора зкДНК, полученного в соответствии со способами и композициями с использованием единственного Rep-белка, как раскрыто в данном документе, имеет модификации в петлевом плече, усеченном плече и спейсере. Типичные последовательности ITR, имеющие модификации в петлевом плече, усеченном плече и спейсере, перечислены в Таблице 6.
[00233] В некоторых вариантах реализации модифицированный ITR (например, левый или правый) вектора зкДНК, полученного в соответствии со способами и композициями с использованием единственного Rep-белка, как раскрыто в данном документе, модифицирован таким образом, чтобы он имел самую низкую энергию разворачивания («низкоэнергетическая структура»). Низкая энергия будет иметь меньшую свободную энергию Гиббса по сравнению с ITR дикого типа. Типичные последовательности ITR, которые модифицированы до низкой (то есть с пониженной) энергией для разворачивания, представлены здесь в Таблице 7-9.
[00234] В некоторых вариантах реализации модифицированный ITR (например, левый или правый) вектора зкДНК, полученного в соответствии со способами и композициями с использованием единственного Rep-белка, как раскрыто в данном документе, выбран из любого белка или их комбинации, показанных в Таблицах 2-9, 10A или 10B.
[00235] Таблица 2: Последовательности ITR с изменениями в петлевом плече, усеченном плече или спейсере. Они включают последовательность RBS GCGCGCTCGCTCGCTC (SEQ ID NO: 531) на 5'-конце и комплементарную последовательность RBE' GAGCGAGCGAGCGCGC (SEQ ID NO: 536) на самом 3'-конце.
Таблица 3: Модифицированные последовательности ITR с модификациями в петлевом плече и усеченном плече
Таблица 4: Последовательности ITR с изменениями в петлевом плече и спейсере
Таблица 5: Последовательности ITR с изменениями в усеченном плече и спейсере
Таблица 6: Последовательности ITR с изменениями в петлевом плече, усеченном плече и спейсере
[00236] Как раскрыто в данном документе модифицированный ITR в векторе зкДНК, полученный в соответствии со способами и композициями с использованием единственного белка Rep, раскрытого в данном документе, может быть создан, чтобы включать делецию, вставку или замену одного или нескольких нуклеотидов из ITR дикого типа, полученного из AAV генома. Модифицированный ITR может быть получен путем генетической модификации во время намножения в плазмиде в Escherichia coli или в виде генома бакуловируса в клетках Spodoptera frugiperda или другими биологическими методами, например in vitro с использованием полимеразной цепной реакции или химического синтеза.
[00237] В некоторых вариантах реализации модифицированный ITR в векторе зкДНК, полученный в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, может включать делецию, вставку или замену одного или нескольких нуклеотидов из ITR дикого типа AAV2 (слева) (SEQ ID NO: 51) или ITR дикого типа AAV2 (справа) (SEQ ID NO: 1). В частности, один или несколько нуклеотидов удалены, вставлены или заменены из B-C' или C-C' в Т-образной структуре стебель-петля. Кроме того, модифицированный ITR не включает в себя модификацию Rep-связывающих элементов (RBE) и сайта конечного разрешения (trs) ITR дикого типа AAV2, хотя RBE' (TTT) может присутствовать или отсутствовать в зависимости от того, подверглась ли матрица одному циклу репликации, тем самым преобразовав триплет AAA в комплементарный RBE' - TTT.
[00238] Приведены примеры трех типов модифицированных ITR: (1) модифицированный ITR, имеющий структуру с наименьшей энергией, включающую одно плечо и одну непарную петлю («структура с одним плечом/одной непарной петлей»); (2) модифицированный ITR, имеющий структуру с наименьшей энергией с одной шпилькой («структура с одной шпилькой»); и (3) модифицированный ITR, имеющий структуру с наименьшей энергией с двумя плечами, одно из которых усечено («усеченная структура»).
[00239] Модифицированный ITR с одним плечем/структурой с одинарной непарной петлей.
[00240] ITR дикого типа можно модифицировать для образования вторичной структуры, содержащей одно плечо и одну непарную петлю (т.е. «структура с одним плечом/одной непарной петлей»). Свободная энергия Гиббса (ΔG) для разворачивания структуры может находиться в диапазоне от -85 ккал/моль до -70 ккал/моль. Приведены типичные структуры модифицированных ITR.
[00241] Предполагается, что модифицированные ITR формируют структуру с одним плечом/одной непарной петлей, которая может включать делецию, вставку или замену одного или более нуклеотидов относительно ITR дикого типа в последовательностях, образующих плечо B и B' и/или плечо C и C'. Модифицированный ITR может быть получен путем генетической модификации или биологического и/или химического синтеза.
[00242] Например, ITR-2, левый и правый, представленные на Фиг. 9A-9B (SEQ ID NO: 101 и 102) созданы таким образом, чтобы включать делецию двух нуклеотидов из плеча C-C' и делецию 16 нуклеотидов из плеча B-B' в ITR дикого типа AAV2. Три нуклеотида, оставшиеся в плече B-B' модифицированного ITR, не образуют комплементарного спаривания. Таким образом, левый и правый ITR-2 имеют структуру с наименьшей энергией с одним плечом C-C' и одним непарной петлей. Свободная энергия Гиббса для разворачивания структуры составляет около -72,6 ккал/моль.
[00243] Левый и правый ITR-3, представленные на Фиг. 10A и 10B (SEQ ID NO: 103 и 104) созданы таким образом, чтобы включать 19 нуклеотидных делеций в плече C-C' из ITR дикого типа AAV2. Три нуклеотида, оставшиеся в плече B-B' модифицированного ITR, не образуют комплементарного спаривания. Таким образом, левый и правый ITR-3 имеют структуру с наименьшей энергией с одним плечом B-B' и одной непарной петлей. Свободная энергия Гиббса для разворачивания структуры составляет около -74,8 ккал/моль.
[00244] Левый и правый ITR-4, представленные на Фиг. 11A и 11B (SEQ ID NO: 105 и 106) созданы таким образом, чтобы включать 19 нуклеотидных делеций в плече B-B' из ITR дикого типа AAV2. Три нуклеотида, оставшиеся в плече B-B' модифицированного ITR, не образуют комплементарного спаривания. Таким образом, левый и правый ITR-4 имеют структуру с наименьшей энергией с одним плечом C-C' и одной непарной петлей. Свободная энергия Гиббса для разворачивания структуры составляет около -76,9 ккал/моль.
[00245] Правый и левый ITR-10, представленные на Фиг. 12A и 12B (SEQ ID NO: 107 и 108) созданы таким образом, чтобы включать 8 нуклеотидных делеций в плече B-B' из ITR дикого типа AAV2. Нуклеотиды, оставшиеся в плечах B-B' и C-C', образуют новые дополнительные связи между мотивами B и C' (левый ITR-10) или между мотивами C и B' (правый ITR-10). Таким образом, левый и правый ITR-10 имеют структуру с наименьшей энергией с одним плечом B-C' или C-B' и одной непарной петлей. Свободная энергия Гиббса для разворачивания структуры составляет около -83,7 ккал/моль.
[00246] Левый и правый ITR-17, представленные на Фиг. 13A и 13B (SEQ ID NO: 109 и 110) созданы таким образом, чтобы включать 14 нуклеотидных делеций в C-C' плече из ITR дикого типа AAV2. Восемь нуклеотидов, оставшихся в плече C-C', не образуют дополнительных связей. В результате, левый и правый ITR-17 имеют структуру с наименьшей энергией с одним плечом B-B' и одной непарной петлей. Свободная энергия Гиббса для разворачивания структуры составляет около -73,3 ккал/моль.
[00247] Левые и правые последовательности ITR дикого типа (вверху) и различные левые и правые модифицированные ITR (внизу), которые, как предполагается, формируют структуру с одним плечом/одной непарной петлей, выровнены и представлены ниже в Таблице 7.
[00248] Таблица 7: Выравнивание wt-ITR и модифицированных ITR (ITR-2, ITR-3, ITR-4, ITR-10 и ITR-17) с одноцепочечной/одинарной непарной петлей.
(SEQ ID NO)
ITR-2
(SEQ:101)
GCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGCAAAGCCCGGGCGTCGGGCGACCTTTGG
:::::::::::::::::::::::::::::::: :: ::::::::: :::
GCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGAAA--CCCGGGCGT---GCG--------
10 20 30 40
70 80 90
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
:::::::::::::::::::::::
--------CCTCAGTGAGCGAGCGAGCGCGC
50 60 70
ITR-2
(SEQ:102)
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACG--CCCGGGCGGC
:::::::::::::::::::::::: : :::::: ::::::::::
GCGCGCTCGCTCGCTCACTGAGGC------------GCACGCCCGGGTTTCCCGGGCGGC
10 20 30 40
60 70 80
CTCAGTGAGCGAGCGAGCGCGC
::::::::::::::::::::::
CTCAGTGAGCGAGCGAGCGCGC
50 60 70
ITR-3
(SEQ:103)
GCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGCAAAGCCCGGGCGTCGGGCGACCTTTGG
:::::::::::::::::::::::::: :::::::::::::::
GCGCGCTCGCTCGCTCACTGAGGCCG-------------------TCGGGCGACCTTTGG
10 20 30 40
70 80 90
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
:::::::::::::::::::::::::::::::
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
50 60 70
ITR-3
(SEQ:104)
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCCGGGCGGCCT
:::::::::::::::::::::::::::::::::::::::::::::::: ::::
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACG--------GCCT
10 20 30 40 50
70 80
CAGTGAGCGAGCGAGCGCGC
::::::::::::::::::::
CAGTGAGCGAGCGAGCGCGC
60 70
ITR-4
(SEQ:105)
GCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGCAAAGCCCGGGCGTCGGGCGACCTTTGG
:::::::::::::::::::::::::::::::::::::::::::::::::
GCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGCAAAGCCCGGGCGTCGG-----------
10 20 30 40
70 80 90
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
:::::::::::::::::::::::
--------CCTCAGTGAGCGAGCGAGCGCGC
50 60 70
ITR-4
(SEQ:106)
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCCGGGCGGCCT
:::::::::::::::::::::::::: : : :: : :::::::::::::
GCGCGCTCGCTCGCTCACTGAGGCCG--------ACGCCCGGGCTTTGCCCGGGCGGCCT
10 20 30 40 50
70 80
CAGTGAGCGAGCGAGCGCGC
::::::::::::::::::::
CAGTGAGCGAGCGAGCGCGC
60 70
ITR-10
(SEQ:107)
GCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGCAAAGCCCGGGCGTCGGGCGACCTTTGG
::::::::::::::::::::::::::::::::::::::::::::::::::: :::
GCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGCAAAGCCCGGGCGTCGGGC----TTT--
10 20 30 40 50
70 80 90
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
:::::::::::::::::::::::::::::
--GCCCGGCCTCAGTGAGCGAGCGAGCGCGC
60 70 80
ITR-10
(SEQ:108)
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAG---GTCGCCCGAC----GCCCGG
::::::::::::::::::::::::::::: :::: : :::::: ::::::
GCGCGCTCGCTCGCTCACTGAGGCCGGGC----AAAGCCCGACGCCCGGGCTTTGCCCGG
10 20 30 40 50
60 70 80
GCGGCCTCAGTGAGCGAGCGAGCGCGC
:::::::::::::::::::::::::::
GCGGCCTCAGTGAGCGAGCGAGCGCGC
60 70 80
ITR-17
(SEQ:109)
gcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcgacctttgg
:::::::::::::::::::::::::: ::: :::::::::::::::::
GCGCGCTCGCTCGCTCACTGAGGCCG-------AAA-------CGTCGGGCGACCTTTGG
10 20 30 40
70 80 90
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
:::::::::::::::::::::::::::::::
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
50 60 70
ITR-17
(SEQ:110)
gcgcgctcgctcgctcactgaggccgggcgaccaaaggtcgcccgacgcccgggcggcct
:::::::::::::::::::::::::::::::::::::::::::::::: ::::::
gcgcgctcgctcgctcactgaggccgggcgaccaaaggtcgcccgacgttt---cggcct
10 20 30 40 50
70 80
CAGTGAGCGAGCGAGCGCGC
::::::::::::::::::::
CAGTGAGCGAGCGAGCGCGC
60 70
[00249] Модифицированный ITR со структурой с единственной шпилькой
[00250] ITR дикого типа можно модифицировать, чтобы получить структуру с наименьшей энергией, включающую структуру с единственной шпилькой. Свободная энергия Гиббса (ΔG) для разворачивания структуры может находиться в диапазоне от -70 ккал/моль до -40 ккал/моль. Типичные структуры модифицированных ITR представлены на Фиг. 14A и 14B.
[00251] Предполагается, что модифицированные ITR формируют структуру с единственной шпилькой, которая может включать делецию, вставку или замену одного или более нуклеотидов относительно ITR дикого типа в последовательностях, образующих плечо B и B' и/или плечо C и C'. Модифицированный ITR может быть получен путем генетической модификации или биологического и/или химического синтеза.
[00252] Например, левый и правый ITR-6, представленные на Фиг. 14A-14B (SEQ ID NO: 111 и 112), включают делеции 40 нуклеотидов в плечах B-B' и C-C' по сравнению с ITR AAV2 дикого типа. Предполагается, что нуклеотиды, оставшиеся в модифицированном ITR, образуют структуру с единственной шпилькой. Свободная энергия Гиббса для разворачивания структуры составляет около -54,4 ккал/моль.
[00253] Последовательности ITR дикого типа и ITR-6 (как левого, так и правого) выровнены и представлены ниже в Таблице 8.
[00254] Таблица 8: Выравнивание wt-ITR и модифицированного ITR-6 со структурой с единственной шпилькой
(SEQ ID NO)
ITR-6
(SEQ:111)
GCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGCAAAGCCCGGGCGTCGGGCGACCTTTGG
:::::::::::::::::::::::: ::::::
GCGCGCTCGCTCGCTCACTGAGGC---------AAAGCC---------------------
10 20 30
70 80 90
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
:::::::::::::::::::::
----------TCAGTGAGCGAGCGAGCGCGC
40 50
ITR-6 (SEQ:544)
, GCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGCGTCGGGCGACCTTTGGTCGCCCGGCCT
:::::::::::::::::::::::: :::: :::
, GCGCGCTCGCTCGCTCACTGAGGCC-------------------TTTG---------CCT
10 20
20 10
, CAGTGAGCGAGCGAGCGCGC (SEQ ID NO: 543)
::::::::::::::::::::
, CAGTGAGCGAGCGAGCGCGC (SEQ ID NO: 544)
40 50
[00255] Модифицированный ITR с усеченной структурой
[00256] ITR дикого типа можно модифицировать, чтобы иметь структуру с наименьшей энергией, состоящую из двух плеч, одно из которых усечено. Их свободная энергия Гиббса (ΔG) для разворачивания находится в диапазоне от -90 до -70 ккал/моль. Таким образом, их свободные энергии для разворачивания по Гиббсу ниже, чем ITR дикого типа AAV2.
[00257] Модифицированные ITR могут включать делецию, вставку или замену одного или более нуклеотидов относительно ITR дикого типа в последовательностях, образующих плечо B и B' и/или плечо C и C'. В некоторых вариантах реализации модифицированный ITR может, например, включать удаление всей конкретной петли, например, петли A-A', петли B-B' или петли C-C', или, альтернативно, удаление 1, 2, 3, 4, 5, 6, 7, 8, 9 или более пар оснований, образующих стебель петли, при условии, что последняя петля на конце стебля все еще присутствует. Модифицированный ITR может быть получен путем генетической модификации или биологического и/или химического синтеза.
[00258] Типичные структуры модифицированных ITR с усеченной структурой представлены на Фиг. 15А-15В.
[00259] Предполагается, что последовательности различных модифицированных ITR образуют усеченную структуру, выровнены с последовательностью ITR дикого типа и представлены ниже в Таблице 9.
[00260] Таблица 9: Выравнивание wt-ITR и модифицированных ITR (ITR-5, ITR-7, ITR-8, ITR-9, ITR-11, ITR-12, ITR-13, ITR-14, ITR-1 и ITR-16) с усеченной структурой.
(SEQ ID NO)
ITR-5
(SEQ:545)
gcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcgacctttgg
:::::::::::::::::::::::: ::::::::::::::::::::::::
gcgcgctcgctcgctcactgaggc------------gcccgggcgtcgggcgacctttgg
10 20 30 40
70 80 90
tcgcccggcctcagtgagcgagcgagcgcgc
:::::::::::::::::::::::::::::::
tcgcccggcctcagtgagcgagcgagcgcgc (SEQ ID NO: 545)
50 60 70
ITR-5
(SEQ:116)
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCCGGGCGGCCT
:::::::::::::::::::::::::::::::::::::::::::::::::::::::: :::
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCCGGGCG-CCT
10 20 30 40 50
70 80
CAGTGAGCGAGCGAGCGCGC
::::::::::::::::::::
CAGTGAGCGAGCGAGCGCGC
60 70
ITR-7
(SEQ:117)
gcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcgacctttgg
:::::::::::::::::::::::::::::::::::::::::::::::::::::: :: :
gcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcgac--tttg
10 20 30 40 50
70 80 90
tcgcccggcctcagtgagcgagcgagcgcgc
:::::::::::::::::::::::::::::::
tcgcccggcctcagtgagcgagcgagcgcgc
60 70 80
ITR-7
(SEQ:118)
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCCGGGC-----
:::::::::::::::::::::::::::::::: :: ::::::::::::::::::
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACAAA--GTCGCCCGACGCCCGGGCTTTGC
10 20 30 40 50
60 70 80
------GGCCTCAGTGAGCGAGCGAGCGCGC
:::::::::::::::::::::::::
CCGGGCGGCCTCAGTGAGCGAGCGAGCGCGC
60 70 80
ITR-8
(SEQ:119)
gcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcgacctttgg
::::::::::::::::::::::::::::::::::::::::::::::::::::: :::
gcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcga--ttt--
10 20 30 40 50
70 80 90
tcgcccggcctcagtgagcgagcgagcgcgc
:::::::::::::::::::::::::::::::
tcgcccggcctcagtgagcgagcgagcgcgc
60 70 80
ITR-8
(SEQ:120)
gcgcgctcgctcgctcactgaggccgggcgaccaaaggtcgcccgacgcccgggc-----
::::::::::::::::::::::::::::::: ::: :::::::::::::::::
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGA--AAA--TCGCCCGACGCCCGGGCTTTGC
10 20 30 40 50
60 70 80
------GGCCTCAGTGAGCGAGCGAGCGCGC
:::::::::::::::::::::::::
CCGGGCGGCCTCAGTGAGCGAGCGAGCGCGC
60 70 80
ITR-9
(SEQ:121)
GCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGCAAAGCCCGGGCGTCGGGCGACCTTTGG
:::::::::::::::::::::::::::::::::::::::::::::::::::: ::
GCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGCAAAGCCCGGGCGTCGGGCG----TT--
10 20 30 40 50
70 80 90
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
:::::::::::::::::::::::::::::::
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
60 70 80
ITR-9
(SEQ:122)
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCCGGGC-----
::::::::::::::::::::::::::::::: :: ::::::::::::::::
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGA--AA----CGCCCGACGCCCGGGCTTTGC
10 20 30 40 50
60 70 80
------GGCCTCAGTGAGCGAGCGAGCGCGC
CCGGGCGGCCTCAGTGAGCGAGCGAGCGCGC
60 70 80
ITR-11
(SEQ:123)
GCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGCAAAGCCCGGGCGTCGGGCGACCTTTGG
:::::::::::::::::::::::::::::::: :: :::::::::::::::::::::::
GCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGAAA--CCCGGGCGTCGGGCGACCTTTGG
10 20 30 40 50
70 80 90
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
:::::::::::::::::::::::::::::::
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
60 70 80
ITR-11
(SEQ:124)
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCCGGG------
::::::::::::::::::::::::::::::::::::::::::::::::::::::
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCCGGGTTTCCC
70 80 90 100 110 120
60 70 80
---CGGCCTCAGTGAGCGAGCGAGCGCGC
::::::::::::::::::::::::::
GGGCGGCCTCAGTGAGCGAGCGAGCGCGC
130 140 150
ITR-12
(SEQ:125)
GCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGCAAAGCCCGGGCGTCGGGCGACCTTTGG
::::::::::::::::::::::::::::::: ::: ::::::::::::::::::::::
GCGCGCTCGCTCGCTCACTGAGGCCGCCCGG--AAA-CCGGGCGTCGGGCGACCTTTGG
10 20 30 40 50
70 80 90
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
:::::::::::::::::::::::::::::::
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
60 70 80
ITR-12
(SEQ:126)
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCCGG-------
:::::::::::::::::::::::::::::::::::::::::::::::::::::
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCCGGTTTCCGG
10 20 30 40 50 60
60 70 80
GCGGCCTCAGTGAGCGAGCGAGCGCGC
:::::::::::::::::::::::::::
GCGGCCTCAGTGAGCGAGCGAGCGCGC
70 80
ITR-13
(SEQ:127)
GCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGCAAAGCCCGGGCGTCGGGCGACCTTTGG
:::::::::::::::::::::::::::::: ::: :::::::::::::::::::::
GCGCGCTCGCTCGCTCACTGAGGCCGCCCG---AAA---CGGGCGTCGGGCGACCTTTGG
10 20 30 40 50
70 80 90
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
:::::::::::::::::::::::::::::::
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
60 70 80
ITR-13
(SEQ:128)
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCCG-----GGC
:::::::::::::::::::::::::::::::::::::::::::::::::::: :::
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCCGTTTCGGGC
10 20 30 40 50 60
60 70 80
GGCCTCAGTGAGCGAGCGAGCGCGC
:::::::::::::::::::::::::
GGCCTCAGTGAGCGAGCGAGCGCGC
70 80
ITR-14
(SEQ:129)
GCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGCAAAGCCCGGGCGTCGGGCGACCTTTGG
::::::::::::::::::::::::::::: :::: :::::::::::::::::::
GCGCGCTCGCTCGCTCACTGAGGCCGCCC----AAAG----GGCGTCGGGCGACCTTTGG
10 20 30 40 50
70 80 90
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
:::::::::::::::::::::::::::::::
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
60 70 80
ITR-14
(SEQ:130)
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCC---GGGCGG
::::::::::::::::::::::::::::::::::::::::::::::::::: ::::::
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCCTTTGGGCGG
10 20 30 40 50 60
60 70 80
CCTCAGTGAGCGAGCGAGCGCGC
:::::::::::::::::::::::
CCTCAGTGAGCGAGCGAGCGCGC
70 80
ITR-15
(SEQ:131)
GCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGCAAAGCCCGGGCGTCGGGCGACCTTTGG
:::::::::::::::::::::::::::: :::: ::::::::::::::::::
GCGCGCTCGCTCGCTCACTGAGGCCGCC-----AAAG-----GCGTCGGGCGACCTTTGG
10 20 30 40 50
70 80 90
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
:::::::::::::::::::::::::::::::
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
60 70 80
ITR-15
(SEQ:132)
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCCG-GGCGGCC
:::::::::::::::::::::::::::::::::::::::::::::::::: :::::::
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCTTTGGCGGCC
10 20 30 40 50 60
60 70 80
TCAGTGAGCGAGCGAGCGCGC
:::::::::::::::::::::
TCAGTGAGCGAGCGAGCGCGC
70 80
ITR-16
(SEQ:133)
GCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGCAAAGCCCGGGCGTCGGGCGACCTTTGG
::::::::::::::::::::::::::: ::::: ::::::::::::::::
GCGCGCTCGCTCGCTCACTGAGGCCGC------AAAGC------GTCGGGCGACCTTTGG
10 20 30 40
70 80 90
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
:::::::::::::::::::::::::::::::
TCGCCCGGCCTCAGTGAGCGAGCGAGCGCGC
50 60 70
ITR-16
(SEQ:134)
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCCGGGCGGCCT
::::::::::::::::::::::::::::::::::::::::::::::::: : ::::::
GCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCTTTG-CGGCCT
10 20 30 40 50
70 80
CAGTGAGCGAGCGAGCGCGC
::::::::::::::::::::
CAGTGAGCGAGCGAGCGCGC
60 70
[00261] Дополнительные типичные модифицированные ITR в каждом из вышеуказанных классов для использования в векторе зкДНК, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, представлены в таблицах 10A и 10B. Предсказанная вторичная структура правых модифицированных ITR в таблице 10A показана на Фиг. 26A, и предсказанная вторичная структура левых модифицированных ITR в Таблице 10B показана на Фиг. 26B.
[00262] Таблица 10A и Таблица 10B показывают типичные правые и левые модифицированные ITR в векторе зкДНК, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе.
[00263] Таблица 10А: Типичные правые модифицированные ITR. Эти типичные модифицированные правые ITR могут содержать RBE GCGCGCTCGCTCGCTC-3′ (SEQ ID NO: 531), спейсер ACTGAGGC (SEQ ID NO: 532), комплемент спейсера GCCTCAGT (SEQ ID NO: 535) и RBE' (т.е. комплемент RBE) GAGCGAGCGAGCGCGC (SEQ ID NO: 536).
[00264] ТАБЛИЦА 10В: Типичные модифицированные левые ITR в векторе зкДНК, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе. Эти примеры модифицированных левых ITR могут содержать RBE GCGCGCTCGCTCGCTC-3′ (SEQ ID NO: 531), спейсер ACTGAGGC (SEQ ID NO: 532), комплемент спейсера GCCTCAGT (SEQ ID NO: 535) и комплемент RBE (RBE') GAGCGAGCGAGCGCGC (SEQ ID NO: 536).
[00265] Согласно вариантам реализации данного изобретения зкДНК-вектор, полученный в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, не содержит модифицированных ITR, имеющих нуклеотидную последовательность, выбранную из любой последовательности из группы SEQ ID No: 550, 551, 552, 553, 553, 554, 555, 556 и 557.
[00267] В той степени, в которой вектор зкДНК, полученный в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, имеет модифицированный ITR, который имеет одну из модификаций в участке B, B', C или C', как описано в SEQ ID NO: 550-557, как определено в одном или нескольких пунктах формулы данной заявки, или в любом изобретении, которое будет определено в измененной формуле изобретения, которая в будущем может быть подана в этой заявке или в любом патенте, полученном на ее основе, и в той степени, в которой законы любой соответствующей страны или стран, к которым применяются те или иные требования, настоящим оставляется право отказать в указанном раскрытии из пунктов формулы данной заявки или любого патента, полученного из нее, в той степени, в которой это необходимо для предотвращения признания недействительной данной заявки или любого другого патента, полученного на основании этого.
[00267] Например, без ограничений, оставляется право отказать в рассмотрении любого из следующих предметов из любого пункта формулы данной заявки, в данное время или с поправками, внесенными в будущем, или любого патента, полученного на его основе:
[00268] A. модифицированный ITR, выбранный из любой группы, состоящей из: SEQ ID NO: 2, 52, 63, 64, 113, 114, 550, 551; 552, 553, 553, 554, 555, 556, 557, используемые в векторе зкДНК, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как описано в данном документе, без регуляторного переключателя
[00269] B. указанные выше модифицированные ITR в A., в векторе зкДНК, полученном в соответствии со способами и композициями с использованием единственного белка Rep, как описано в данном документе, без регуляторной последовательности, в котором гетерологичная нуклеиновая кислота кодирует ABCA4, USA2A var1, VEGFR CEP290, фактор VIII BDD (FVIII), фактор VIII, vWF_His, vWF, лецитин-холестерин-ацетилтрансферазу, PAH, G6PC или CFTR
VI. Регуляторные элементы.
[00270] Композиция, используемая в способах получения зкДНК-вектора, например, вектора зкДНК, как описано в данном документе, или вектора AAV, содержит последовательность нуклеиновой кислоты, кодирующую единственный модифицированный белок Rep, может дополнительно содержать регуляторный элемент, например, цис-регуляторный элемент, как описано в этом документе, перед или функционально связанной с нуклеиновой кислотой, кодирующей единственный модифицированный белок Rep. Например, нуклеотидная последовательность, кодирующая модифицированный белок Rep, например, кодирующая модифицированный белок Rep 78, но не содержащая функциональный кодон инициации для кодирования белка Rep 52 или сайты сплайсинга для пропуска экзона для получения Rep 68 или Rep40, является функционально связанной с регуляторным элементом, например, цис-регуляторным элементом.
[00271] В одном варианте реализации нуклеотидная последовательность, кодирующая единственный белок Rep, используемый в композициях и способах, раскрытых в данном документе, включает последовательность контроля экспрессии, например, промотор, цис-регуляторные элементы или регуляторный переключатель, как описано в данном документе, расположенные выше кодона инициации нуклеотидной последовательности, кодирующей парвовирусный белок Rep78, в которой последовательность нуклеиновой кислоты не имеет функционального инициирующего кодона для Rep52 и/или сайтов сплайсинга для пропуска экзона для получения Rep 68 или Rep40. В одном варианте реализации нуклеотидная последовательность, кодирующая единственный белок Rep, используемый в композициях и способах, раскрытых в данном документе, включает последовательность контроля экспрессии перед кодоном инициации нуклеотидной последовательности, кодирующей парвовирусный белок Rep78, в которой последовательность нуклеиновой кислоты не имеет функциональных сайтов сплайсингу для кодирования Rep68.
[00272] Аналогично, вектор зкДНК, полученный в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, может быть получен из экспрессионных конструктов, которые дополнительно содержат конкретную комбинацию цис-регуляторных элементов. Цис-регуляторные элементы включают, не ограничиваясь перечисленными, промотор, рибопереключатель, инсулятор, miR-регулируемый элемент, посттранскрипционный регуляторный элемент, тканеспецифический и клеточноспецифический промотор и энхансер. Согласно некоторым вариантам реализации указанный ITR может действовать в качестве промотора для трансгена. Согласно некоторым вариантам реализации указанный зкДНК-вектор содержит дополнительные компоненты для регуляции экспрессии трансгена, например, регуляторные переключатели согласно описанию в данном документе, для регулирования экспрессии трансгена, или «аварийный выключатель», который способен убить клетку, содержащую указанный зкДНК-вектор.
[00273] ЗкДНК-вектор, полученный в соответствии со способами и композициями с использованием единственного белка Rep, как раскрыто в данном документе, может быть получен из экспрессирующих конструктов, которые дополнительно содержат специфическую комбинацию цис-регуляторных элементов, таких как посттранскрипционный регуляторный элемент WHP (WPRE) (например, SEQ ID NO: 8) и поли(А) BGH (SEQ ID NO: 9). Подходящие экспрессионные кассеты для применения в экспрессионных конструктах не имеют налагаемых вирусным капсидом ограничений упаковки. Экспрессионные кассеты по данному изобретению включают промотор, который может влиять на общие уровни экспрессии, а также на клеточную специфичность. Для экспрессии трансгена они могут включать высокоактивный немедленный ранний промотор вирусного происхождения. Экспрессионные кассеты могут содержать тканеспецифические эукариотические промоторы для ограничения трансгенной экспрессии конкретными типами клеток и снижения токсических эффектов и иммунных ответов, обусловленных нерегулируемой эктопической экспрессией.
[00274] В предпочтительных вариантах реализации промоторы или регуляторные элементы для использования при экспрессии модифицированного единственного Rep-белка или в экспрессионной кассете вектора зкДНК, полученного описанными здесь способами, могут содержать синтетический регуляторный элемент, такой как промотор CAG (SEQ ID NO: 3). Промотор CAG содержит (i) ранний энхансерный элемент цитомегаловируса (CMV), (ii) промотор, первый экзон и первый интрон гена бета-актина курицы, и (iii) акцептор сплайсинга гена бета-глобина кролика. Альтернативно, промоторы или регуляторные элементы для использования при экспрессии модифицированного единственного Rep-белка или в экспрессионной кассете вектора зкДНК, полученного описанными здесь способами, могут содержать промотор альфа-1-антрипсина (AAT) (SEQ ID NO: 4 или SEQ ID NO: 74), печень-специфический (LP1) промотор (SEQ ID NO: 5 или SEQ ID NO: 16) или промотор фактора удлинения-1-альфа (EF1a) человека (например, SEQ ID NO: 6 или SEQ ID NO: 15). В некоторых вариантах реализации промоторы или регуляторные элементы для использования при экспрессии модифицированного единственного Rep-белка или в экспрессионной кассете вектора зкДНК, полученного описанными здесь способами, выбраны из одного или нескольких конститутивных промоторов, например ретровируса. промотор LTR вируса саркомы Рауса (RSV) (необязательно с энхансером RSV) или немедленный ранний промотор цитомегаловируса (CMV) (необязательно с энхансером CMV, например, SEQ ID NO: 22). Альтернативно, индуцибельный промотор, нативный промотор для трансгена, тканеспецифический промотор или различные промоторы, известные в данной области, могут быть функционально связаны с нуклеиновой кислотой, кодирующей модифицированный единственный белок Rep, или в кассете экспрессии вектора зкДНК, полученного способами, раскрытыми в данном документе.
[00275] Подходящие промоторы, в том числе описанные выше, могут происходить из вирусов и, соответственно, могут называться вирусными промоторами, или они могут происходить из любого организма, в том числе прокариотических или эукариотических организмов. Для управления экспрессией с помощью любой РНК-полимеразы (например, pol I, pol II, pol III) могут применяться подходящие промоторы. Типичные промоторы, которые могут быть функционально связаны с нуклеиновой кислотой, кодирующей модифицированный единственный белок Rep, или c кассетой экспрессии вектора зкДНК, полученной описанными здесь способами, включают, без ограничений, ранний промотор SV40, промотор из длинного концевого повтора вируса опухоли молочной железы мышей (LTR); аденовирусный большой поздний промотор (Ad MLP); промотор вируса простого герпеса (HSV), промотор цитомегаловируса (CMV), такой как участок немедленного раннего промотора CMV (CMVIE), промотор вируса саркомы Рауса (RSV), промотор U6 малой ядерной РНК человека (U6, например, SEQ ID NO: 18) (Miyagishi et al., Nature Biotechnology 20, 497-500 (2002)), усиленный промотор U6 (например, Xia et al., Nucleic Acids Res. 2003 Sep. 1; 31(17)), промотор H1 человека (H1) (например, SEQ ID NO: 19), промотор CAG, промотор альфа-1-антитрипсина человека (HAAT) (например, SEQ ID NO: 21), и т.п. Согласно вариантам реализации указанные промоторы изменяют на содержащем интрон 3′-конце так, чтобы они включали один или более сайтов расщепления нуклеазами. Согласно некоторым вариантам реализации ДНК, содержащая сайт(ы) расщепления нуклеазой, является чужеродной для ДНК промотора.
[00276] Промотор может содержать одну или более специфических регуляторных последовательностей транскрипции для дополнительного усиления экспрессии, и/или для пространственного и/или временнóго изменения экспрессии. Промотор может также включать дистальные энхансерные или репрессорные элементы, которые могут быть локализованы на расстоянии нескольких тысяч пар оснований от сайта старта транскрипции. Промотор может происходить из источников, включающих вирусы, бактерии, грибы, растения, насекомые и животные. Промотор может регулировать экспрессию генного компонента конститутивно или дифференциально в отношении клетки, ткани или органа, где происходит экспрессия, или в отношении стадии развития, на которой происходит экспрессия, или в ответ на внешние стимулы, такие как физиологические стрессы, патогены, ионы металлов или индуцирующие агенты. Репрезентативные примеры промоторов, которые могут быть функционально связаны с нуклеиновой кислотой, кодирующей модифицированный единственный белок Rep, или c кассетой экспрессии вектора зкДНК, полученной описанными здесь способами, включают, без ограничений, промотор бактериофага Т7, промотор бактериофага T3, промотор SP6, оператор-промотор lac, промотор tac, поздний промотор SV40, ранний промотор SV40, промотор RSV-LTR, промотор CMV IE, ранний промотор SV40 или поздний промотор SV40 и промотор CMV IE, а также промоторы перечисленые ниже. Такие промоторы и/или энхансеры можно применять для экспрессии любого представляющего интерес гена, например, молекул для редактирования генов, донорной последовательности, терапевтических белков и т.п. Например, указанный вектор может содержать промотор, который функционально связан с последовательностью нуклеиновой кислоты, кодирующей терапевтический белок. Указанный промотор, функционально связанный с кодирующей последовательностью терапевтического белка, может представлять собой промотор вируса обезьян 40 (SV40), промотор вируса опухоли молочной железы мышей (MMTV), промотор вируса иммунодефицита человека (ВИЧ), например, промотор длинного концевого повтора (LTR) вируса иммунодефицита крупного рогатого скота (BIV), промотор вируса Молони, промотор вируса лейкоза птиц (ALV), промотор цитомегаловируса (CMV), такой как ранний промотор CMV, промотор вируса Эпштейна-Барр (EBV) или промотор вируса саркомы Рауса (RSV). Согласно некоторым вариантам реализации промотор также может представлять собой промотор гена человека, такого как убиквитин С человека (hUbC), актин человека, миозин человека, гемоглобин человека, мышечный креатин человека или металлотионеин человека. Промотор также может представлять собой тканеспецифичный промотор, такой как специфический для печени промотор, такой как промотор альфа-1-антитрипсина человека (HAAT), природный или синтетический. В одном варианте реализации доставка в печень может быть достигнута с использованием эндогенного ApoE-специфического нацеливания композиции, содержащей зкДНК-вектор, на гепатоциты посредством рецептора липопротеина низкой плотности (LDL), присутствующего на поверхности гепатоцитов.
[00277] Согласно одному варианту реализации используемый промотор представляет собой нативный промотор гена, кодирующий терапевтический белок. Промоторы и другие регуляторные последовательности соответствующих генов, кодирующих терапевтические белки, известны и были охарактеризованы. Используемый участок промотора может дополнительно включать одну или несколько дополнительных регуляторных последовательностей (например, нативных), например, энхансеров (например, SEQ ID NO: 22 и SEQ ID NO: 23).
[00278] Неограничивающие примеры подходящих промоторов для использования при экспрессии модифицированного единственного Rep-белка или вектора зкДНК, полученного описанными здесь способами, включают промотор CAG, например (SEQ ID NO: 3), промотор HAAT (SEQ ID NO: 21), промотор EF1-α человека (SEQ ID NO: 6) или фрагмент промотора EF1a (SEQ ID NO: 15), промотор IE2 (например, SEQ ID NO: 20) и промотор EF1-α крысы (SEQ ID NO: 24).
[00279] Последовательности полиаденилирования: В некоторых вариантах реализации последовательность, кодирующая последовательность полиаденилирования, может быть функционально связана с нуклеиновой кислотой, кодирующей модифицированный единственный белок Rep, или вектор зкДНК, полученный способами, описанными в данном документе, для стабилизации экспрессируемой мРНК, и/или для помощи в ядерном экспорте и трансляции. В одном варианте реализации конструкт, содержащаяий нуклеиновую кислоту, кодирующую единственный модифицированный белок Rep, или вектор зкДНК, полученный описанными здесь способами, не включает последовательность полиаденилирования. Согласно другим вариантам реализации конструкт, содержащий нуклеиновую кислоту, кодирующую единственный модифицированный белок Rep, или вектор зкДНК, полученный описанными здесь способами включает по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 10, по меньшей мере 15, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50 или более адениновых динуклеотидов. В некоторых вариантах реализации указанная последовательность полиаденилирования содержит примерно 43 нуклеотида, примерно 40-50 нуклеотидов, примерно 40-55 нуклеотидов, примерно 45-50 нуклеотидов, примерно 35-50 нуклеотидов, или в любом диапазоне между указанными значениями.
[00280] Конструкт, содержащий нуклеиновую кислоту, кодирующую единственный модифицированный белок Rep, или вектор зкДНК, полученный описанными здесь способами, может включать последовательность полиаденилирования, известную в данной области техники, или ее вариант, например, встречающуюся в природе последовательность, выделенную из бычьей BGHpA (например, SEQ ID NO: 74), или из вирусной SV40pA (например, SEQ ID NO: 10), или синтетическую последовательность (например, SEQ ID NO: 27). Некоторые экспрессионные кассеты также включают последовательность 5'-энхансера позднего поли(А)-сигнала (USE) SV40. Согласно некоторым вариантам реализации указанные USE могут применяться в комбинации с SV40pA или гетерологичным поли(A)-сигналом.
[00281] Указанные экспрессионные кассеты могут также включать посттранскрипционный элемент для повышения экспрессии трансгена. В некоторых вариантах реализации для повышения экспрессии трансгена применяют посттранскрипционный регуляторный элемент (WPRE) вируса гепатита сурков (WHP) (например, SEQ ID NO: 8). Могут применяться другие элементы посттранскрипционного процессинга, такие как посттранскрипционный элемент гена тимидинкиназы вируса простого герпеса или вируса гепатита B (HBV). К трансгенам могут быть присоединены секреторные последовательности, например, последовательности VH-02 и VK-A26, например, SEQ ID NO: 25 и SEQ ID NO: 26.
VI. Регуляторные переключатели
[00282] Молекулярный регуляторный переключатель - это переключатель, который генерирует измеряемое изменение состояния в ответ на сигнал. Такие регуляторные переключатели можно успешно комбинировать с конструктом, содержащим нуклеиновую кислоту, кодирующую модифицированный единственный белок Rep, или вектор зкДНК, полученный описанными здесь способами, для контроля выхода вектора зкДНК. В некоторых вариантах реализации конструкт, содержащая нуклеиновую кислоту, кодирующую один модифицированный белок Rep, или вектор зкДНК, полученный описанными здесь способами, содержит регуляторный переключатель, который служит для точной настройки экспрессии одного белка Rep или трансгена в зкДНК векторе. Например, он может служить для обеспечения функции биологического сдерживания зкДНК-вектора. Согласно некоторым вариантам реализации указанный переключатель представляет собой «двухпозиционный» переключатель, который сконструирован таким образом, чтобы запускать или останавливать (т.е. прекращать) экспрессию представляющего интерес гена в зкДНК контролируемым и регулируемым образом. Согласно некоторым вариантам реализации указанный переключатель может включать «аварийный выключатель», который может давать клетке, содержащей указанный зкДНК-вектор, команду для запуска механизма запрограммированной клеточной смерти после активации переключателя.
[00283] A. Бинарные регуляторные переключатели
[00284] Согласно некоторым вариантам реализации указанный зкДНК-вектор содержит регулятор, который может служить для контролируемой модуляции экспрессии трансгена. Согласно такому варианту реализации указанная экспрессионная кассета, локализованная между ITR зкДНК-вектора, может дополнительно содержать регуляторный участок, например, промотор, цис-элемент, репрессор, энхансер и т.п., функционально связанную с представляющим интерес геном, причем указанный регуляторный участок регулирует один или более кофакторов или экзогенных агентов. Соответственно, согласно одному варианту реализации транскрипция и экспрессия представляющего интерес гена с зкДНК-вектора будет происходить только тогда, когда в клетке присутствуют указанные один или более кофакторов или экзогенных агентов. Согласно другому варианту реализации один или более кофакторов или экзогенных агентов могут применяться для дерепрессии транскрипции и экспрессии представляющего интерес гена.
[00285] Любые регуляторные участки нуклеиновых кислот, известные специалисту в данной области техники, могут применяться в зкДНК-векторе, разработанном так, чтобы содержать регуляторный переключатель. В неограничивающем примере регуляторные участки могут модулировать низкомолекулярные переключатели или индуцируемые или репрессируемые промоторы. Неограничивающими примерами индуцибельных промоторов являются индуцируемые гормонами или индуцируемые металлами промоторы. Другие примеры индуцируемых промоторных/энхансерных элементов включают, не ограничиваясь перечисленными, индуцируемый RU486 промотор, индуцируемый экдизоном промотор, индуцируемый рапамицином промотор и промотор металлотионеина. Предусмотрено применение классических переключателей на основе тетрациклина или других антибиотиков, в том числе описанных в источнике: (Fussenegger et al., Nature Biotechnol. 18: 1203-1208 (2000)).
[00286] B. Низкомолекулярные регуляторные переключатели
[00287] Различные известные из уровня техники регуляторные переключатели на основе малых молекул известны в данной области техники и могут быть скомбинированы с зкДНК-векторами согласно описанию в данном документе с образованием контролируемого регуляторным переключателем зкДНК-вектора. Согласно некоторым вариантам реализации указанный регуляторный переключатель может быть выбранным из чего-либо одного или из комбинации следующего: ортогональная пара лиганд/ядерный рецептор, например, вариант ретиноидного рецептора/LG335 и GRQCIMFI, наряду с искусственным промотором, контролирующим экспрессию функционально связанного трансгена, например, согласно описанию в источнике: Taylor, et al. BMC Biotechnology 10 (2010): 15; сконструированные стероидные рецепторы, например, модифицированный рецептор прогестерона с усечением на C-конце, который не способен связывать прогестерон, но связывает RU486 (мифепристон) (Патент США № 5364791); рецептор экдизона из Drosophila и их экдистероидные лиганды (Saez, et al., PNAS, 97(26)(2000), 14512-14517; или переключатель, управляемый антибиотиком триметопримом (TMP), согласно описанию в источнике: Sando R 3rd; Nat Methods. 2013, 10(11):1085-8.
[00288] Другие регуляторные переключатели на основе малых молекул, известные специалисту в данной области техники, также предусмотрены для применения для контроля трансгенной экспрессии зкДНК и включают, не ограничиваясь перечисленными, описанные в источнике: Buskirk et al., Cell; Chem and Biol., 2005; 12(2); 151-161; чувствительный к абсцизовой кислоте положительный переключатель; такой как описанный в источнике: Liang, F.-S., et al., (2011) Science Signaling, 4(164); чувствительные к экзогенному L-аргинину положительные переключатели, такие как описанные в источнике: Hartenbach, et al. Nucleic Acids Research, 35(20), 2007, синтетические чувствительные к желчным кислотам положительные переключатели, такие как описанные в источнике: Rössger et al., Metab Eng. 2014, 21: 81-90; чувствительные к биотину положительные переключатели, такие как описанные в источнике: Weber et al., Metab. Eng. 2009 Mar; 11(2): 117-124; регуляторные переключатели с двойным входным сигналом, чувствительные к пищевым добавкам бензоата/ванилина, такие как описанные в источнике: Xie et al., Nucleic Acids Research, 2014; 42(14);e116; чувствительные к 4-гидрокситамоксифену переключатели, такие как описанные в источнике: Giuseppe et al., Molecular Therapy, 6(5), 653 - 663; и чувствительные к флавоноиду (флоретину) регуляторные переключатели, такие как описанные в источнике: Gitzinger et al., Proc. Natl. Acad. Sci. U S A. 2009 Jun 30; 106(26): 10638-10643.
[00289] Согласно некоторым вариантам реализации регуляторный переключатель для контроля трансгена или экспрессируемый зкДНК-вектором представляет собой переключатель для активации пролекарств, такой как описанные в патентах США 8771679 и 6339070.
[00290] Примеры регуляторных переключателей для применения в зкДНК-векторах включают, не ограничиваясь перечисленными, представленные в Таблице 11.
[00291] C. Регуляторные переключатели «с кодом доступа»
[00292] Согласно некоторым вариантам реализации указанный регуляторный переключатель может представлять собой «переключатель с кодом доступа» или «цепь с кодом доступа». Переключатели с кодом доступа позволяют точно настроить контроль экспрессии трансгена с зкДНК-вектора при возникновении конкретных условий - то есть для реализации трансгенной экспрессии и/или репрессии должна выполняться комбинация условий. Например, для осуществления экспрессии трансгена должны возникнуть по меньшей мере условия A и B. Регуляторный переключатель с кодом доступа может требовать выполнения любого количества условий, например, для осуществления трансгенной экспрессии должны возникнуть по меньшей мере 2 условия, или по меньшей мере 3, или по меньшей мере 4, или по меньшей мере 5, или по меньшей мере 6 или по меньшей мере 7 или более условий. Согласно некоторым вариантам реализации должны возникнуть по меньшей мере 2 условия (например, условия A, B); согласно некоторым вариантам реализации должны возникнуть по меньшей мере 3 условия (например, A, B и C, или A, B и D). Согласно неограничивающему примеру для осуществления генной экспрессии с зкДНК, содержащей регуляторный переключатель с кодом доступа «ABC», должны выполняться условия A, B и C. Условия A, B и C могут быть следующими; условие A представлено наличием состояния или заболевания, условие B представляет собой гормональный ответ, а условие C представляет собой ответ на трансгенную экспрессию. В неограничивающем типичном примере, когда трансген представлен инсулином, условие A выполняется в том случае, если субъект страдает диабетом, условие B - высокий уровень сахара в крови, а условие C - уровень эндогенного инсулина при его недостаточной экспрессии. Как только уровень сахара снижается или достигается требуемый уровень инсулина, трансген (например, инсулин) «отключается» до тех пор, пока снова не возникнут указанные 3 условия, «включающие» его повторно. В другом типичном примере, в том случае, если трансген представлен EPO, условием A является наличие хронической болезни почек (ХПН), условие B выполняется при гипоксических состояниях в почках субъекта, условие C выполняется при нарушении рекрутинга эритропоэтин-продуцирующих клеток (EPC) в почках; или, как вариант, нарушении активации HIF-2. Как только повышается уровень кислорода или достигается требуемый уровень EPO, трансген (например, EPO) «отключается» до тех пор, пока снова не возникнут указанные 3 условия, «включающие» его повторно.
[00293] Регуляторные переключатели с кодом доступа подходят для точной настройки экспрессии трансгена с зкДНК-вектора. Например, регуляторный переключатель с кодом доступа может быть модульным, то есть он содержит несколько переключателей, например, тканеспецифический индуцируемый промотор, который включается только в присутствии определенного уровня метаболита. Согласно такому варианту реализации для осуществления трансгенной экспрессии с зкДНК-вектора индуцируемый агент должен присутствовать (условие A) в клетках требуемого типа (условие B), а уровень метаболита должен быть равен, быть выше или быть ниже определенного порогового значения (условие C). Согласно альтернативным вариантам реализации указанный регуляторный переключатель с кодом доступа может быть разработан таким образом, чтобы трансгенная экспрессия «включалась» при выполнении условий A и B, но «выключалась» при выполнении условия C. Такой вариант реализации полезен, когда выполнение условия C является прямым результатом экспрессии трансгена - то есть условие C используется в качестве положительной обратной связи для отключения трансгенной экспрессии с зкДНК-вектора, после того как указанный трансген оказал достаточный требуемый терапевтический эффект.
[00294] Согласно некоторым вариантам реализации регуляторный переключатель с кодом доступа, предусмотренный для применения в зкДНК-векторе, раскрыт в WO2017/059245, содержание которой полностью включено в данный документ посредством ссылки, в которой описан переключатель, называемый «переключателем с кодом доступа» («Passcode switch») или «цепью с кодом доступа» («Passcode circuit») или «аварийным выключателем с кодом доступа» («Passcode kill switch»), который представляет собой синтетическую биологическую цепь, задействующую гибридные транскрипционные факторы (ТФ) для конструирования комплексных требований к условиям среды для выживания клеток. Регуляторные переключатели с кодом доступа описанные в WO2017/059245, в частности, подходят для применения в зкДНК-векторах, поскольку являются модульными и настраиваемыми, как в отношении условий окружающей среды, контролирующих активацию цепи, так и в отношении выходных модулей, контролирующих судьбу клеток. Кроме того, в частности, «цепь с кодом доступа» подходит для применения в зкДНК-векторах, так как без надлежащих молекул «кода доступа» он позволяет трансгенной экспрессии происходить только при выполнении необходимых заранее заданных условий. В том случае, если с клеткой произойдет что-либо непредусмотренное или по какой-то причине больше не будет нужна трансгенная экспрессия, может быть запущен соответствующий «аварийный выключатель» (т.е. переключатель-блокатор).
[00295] Согласно некоторым вариантам реализации регуляторный переключатель с кодом доступа или «цепь с кодом доступа», предусмотренный для применения в зкДНК-векторе, содержит гибридные транскрипционные факторы (ТФ) для расширения диапазона и сложности сигналов среды, используемых для задания условий биологического сдерживания. В отличие от переключателя-блокатора, который запускает клеточную смерть при выполнении заранее заданного условия, «цепь с кодом доступа» обеспечивает выживание клеток или трансгенную экспрессию в присутствии конкретного «кода доступа» и может быть легко перепрограммирована, чтобы обеспечивать трансгенную экспрессию и/или выживание клеток только при выполнении заранее заданного условия среды или присутствии кода доступа.
[00296] Согласно одному аспекту система «с кодом доступа», которая ограничивает рост клеток присутствием заранее заданного набора из по меньшей мере двух выбранных агентов, включает одну или более конструктов нуклеиновых кислот, кодирующих модули экспрессии, включающие: i) модуль экспрессии токсина, который кодирует токсин, токсичный для клетки-хозяина, отличающийся тем, что последовательность, кодирующая токсин, функционально связана с промотором P1, репрессия которого происходит при связывании первого гибридного репрессорного белка hRP1; ii) модуль экспрессии первого гибридного репрессорного белка, который кодирует первый гибридный репрессорный белок hRP1, отличающийся тем, что экспрессию hRP1 контролирует логический И-элемент, образуемый двумя гибридными транскрипционными факторами hTF1 и hTF2, связывание или активность которых зависят от агентов A1 и A2, соответственно, таким образом, что для экспрессии hRP1 необходимы оба агента A1 и A2, при этом в отсутствие A1 или A2 экспрессия hRP1 недостаточна для репрессии модуля промотора токсина P1 и продуцирования токсина, так что клетка-хозяин погибает. В указанной системе все гибридные факторы, hTF1, hTF2 и hRP1, содержат чувствительный к среде модуль из одного транскрипционного фактора и модуль распознавания ДНК из другого транскрипционного фактора, что делает связывание соответствующего регуляторного переключателя с кодом доступа чувствительным к присутствию агента среды, A1 или A2, который отличается от агента, с которым соответствующие субъединицы обычно связываются в природе.
[00297] Соответственно, зкДНК-вектор может содержать «регуляторную цепь с кодом доступа», которая требует присутствия и/или отсутствия конкретных молекул для активации выходного модуля. Согласно некоторым вариантам реализации в тех случаях, когда гены, кодирующие клеточные токсины, помещают в выходной модуль, указанная регуляторная цепь с кодом доступа может быть использована не только для регуляции трансгенной экспрессии, но и для создания механизма «аварийного выключателя», в котором цепь убивает клетку в том случае, если указанная клетка ведет себя нежелательным образом (например, покидает конкретную среду, определяемую сенсорными доменами, или дифференцирует в клетку другого типа). В одном неограничивающем примере модульная структура гибридных транскрипционных факторов, архитектура цепи и выходной модуль позволяют переконфигурировать цепь для восприятия других сигналов среды, реакции на сигналы среды иным образом и контроля других функций в клетке, наряду с индуцируемой клеточной смертью, как известно в данной области техники.
[00298] Для регуляторного переключателя с кодом доступа согласно описанию в данном документе могут применяться любые комбинации регуляторных переключателей согласно описанию в данном документе, например, низкомолекулярные переключатели, переключатели на основе нуклеиновых кислот, гибридные переключатели на основе малых молекул и нуклеиновых кислот, переключатели для посттранскрипционной регуляции трансгенов, посттрансляционные регуляторы, контролируемые излучением переключатели, опосредованные гипоксией переключатели и другие регуляторные переключатели, известные специалистам в данной области техники. Регуляторные переключатели, предусмотренные для применения, также описаны в обзорной статье Kis et al., J R Soc Interface. 12: 20141000 (2015), и обобщены в Таблице 1 указанной статьи. В некоторых вариантах реализации регуляторный переключатель для применения в системе с паролем может быть выбран из любых переключателей, приведенных в Таблице 11, или их комбинации.
[00299] D. Регуляторные переключатели на основе нуклеиновых кислот для контроля трансгенной экспрессии.
[00300] Согласно некоторым вариантам реализации регуляторный переключатель для контроля трансгена, экспрессируемого зкДНК, основан на механизме контроля на основе нуклеиновых кислот. Примеры механизмов контроля на основе нуклеиновых кислот известны в данной области техники и предусмотрены для применения. Например, такие механизмы включают рибопереключатели, такие как описанные, например, в US2009/0305253, US2008/0269258, US2017/0204477, WO2018026762A1, патенте США US 9222093 и заявке EP288071, все из которых полностью включены в данный документ посредством ссылки, а также раскрыты в обзорной работе Villa JK et al., Microbiol Spectr. 2018 May; 6 (3), содержание которой полностью включено в данный документ посредством ссылки. Также включены транскрипционные биосенсоры, отвечающие на метаболиты, такие как описанные в источнике: WO2018/075486 и WO2017/147585, содержание которой полностью включено в данный документ посредством ссылки. Другие известные в данной области техники механизмы, предусмотренные для применения, включают сайленсинг трансгена с помощью молекулы миРНК или РНКи (например, miR, мшРНК). Например, зкДНК-вектор может содержать регуляторный переключатель, который кодирует молекулу РНКи, комплементарную трансгену, экспрессируемому зкДНК-вектором. При экспрессии такой РНКи даже в том случае, когда зкДНК-вектор экспрессирует трансген, будет происходить его сайленсинг под действием комплементарной молекулы РНКи, а если РНКи не экспрессируется при экспрессии трансгена зкДНК-вектором, сайленсинг трансгена под действием РНКи не происходит. Такой пример молекулы РНКи, контролирующей генную экспрессию, или используемой в качестве регуляторного переключателя, описан в US2017/0183664. Согласно некоторым вариантам реализации указанный регуляторный переключатель содержит репрессор, который блокирует экспрессию трансгена с зкДНК-вектора. Согласно некоторым вариантам реализации указанный положительный/отрицательный переключатель представляет собой переключатель на основе малой активирующей транскрипцию РНК («Small transcription activating RNA», STAR), например, такую как описанная в источниках: Chappell J. et al., Nat Chem Biol. 2015 Mar;11(3):214-20; и Chappell et al., Microbiol Spectr. 2018 May;6(3). Согласно некоторым вариантам реализации указанный регуляторный переключатель представляет собой переключатель типа «toehold», такой как описанный в источниках: US2009/0191546, US2016/0076083, WO2017/087530, US2017/0204477, WO2017/075486 и Green et al, Cell, 2014; 159(4); 925-939 все из которых полностью включены в данный документ посредством ссылки.
[00301] Согласно некоторым вариантам реализации указанный регуляторный переключатель представляет собой тканеспецифический самоинактивирующийся регуляторный переключатель, например, согласно описанию в US2002/0022018, отличающийся тем, что он преднамеренно отключает трансгенную экспрессию в сайте, где в противном случае трансгенная экспрессия могла бы быть неблагоприятной. Согласно некоторым вариантам реализации регуляторный переключатель представляет собой обратимую с помощью рекомбиназы систему генной экспрессии, например, согласно описанию в US2014/0127162 и патенте США 8324436.
[00302] Согласно некоторым вариантам реализации регуляторный переключатель для контроля представляющего интерес трансгена или гена, экспрессируемого зкДНК-вектором, представляет собой гибрид механизма контроля на основе нуклеиновых кислот и низкомолекулярной регуляторной системы. Такие системы хорошо известны специалистам в данной области техники и предусмотрены для применения в данном документе. Примеры таких регуляторных переключателей включают, не ограничиваясь перечисленными, систему LTRi или систему «Lac-Tet-RNAi», например, согласно описанию в источниках: US2010/0175141, Deans T. et al., Cell., 2007, 130(2); 363-372, WO2008/051854 и патент США 9388425.
[00303] Согласно некоторым вариантам реализации регуляторный переключатель для контроля представляющего интерес трансгена или гена, экспрессируемого зкДНК-вектором, включает циклическую перестановку согласно описанию в патенте США 8338138. Согласно такому варианту реализации указанный молекулярный переключатель является мультистабильным, т.е. способен переключаться между по меньшей мере двумя состояниями, или, как вариант, бистабильным, т.е. находится либо в "выключенном", либо во "включенном" состоянии, например, способен или неспособен испускать свет, способен или неспособен связываться, способен или неспособен катализировать, способен или неспособен переносить электроны, и так далее. Согласно другому аспекту, в молекулярном переключателе используют слитую молекулу, за счет чего указанный переключатель способен переключаться между более чем двумя состояниями. Например, в ответ на конкретное пороговое состояние, демонстрируемое вставочной последовательностью или акцепторной последовательностью, соответствующая другая последовательность слитой молекулы может демонстрировать диапазон состояний (например, диапазон связывающей активности, диапазон ферментного катализа и т.п.). Соответственно, вместо переключения между «включенным» и «выключенным» состоянием слитая молекула может демонстрировать дифференцированный ответ на стимул.
[00304] Согласно некоторым вариантам реализации регуляторный переключатель на основе нуклеиновой кислоты может быть выбран из любого переключателя или комбинации переключателей из Таблицы 11.
[00305] E. Посттранскрипционный и посттрансляционные регуляторные переключатели.
[00306] Согласно некоторым вариантам реализации регуляторный переключатель для контроля представляющего интерес трансгена или гена, экспрессируемого зкДНК-вектором, представляет собой систему посттранскрипционной модификации. Например, такой регуляторный переключатель может представлять собой аптазимный рибопереключатель, чувствительный к тетрациклину или теофиллину, согласно описанию в US2018/0119156, GB201107768, WO2001/064956A3, патенте EP 2707487 и Beilstein et al., ACS Synth. Biol., 2015, 4 (5), pp 526-534; Zhong et al., Elife. 2016 Nov 2;5. pii: e18858. Согласно некоторым вариантам реализации предполагается, что специалист в данной области техники может закодировать как трансген, так и ингибиторную киРНК, которая содержит чувствительный к лиганду аптамер (отрицательный переключатель), в результате чего будет сформирован положительный переключатель, чувствительный к лиганду.
[00307] Согласно некоторым вариантам реализации регуляторный переключатель для контроля представляющего интерес трансгена или гена, экспрессируемого зкДНК-вектором, представляет собой систему посттрансляционной модификации. Согласно альтернативным вариантам реализации представляющий интерес ген или белок экспрессируется в виде пропротеина или пре-пропротеина, или содержит сигнальный элемент ответа (SRE) или дестабилизирующий домен (DD), присоединенный к экспрессируемому белку, что предотвращает корректную укладку и/или активность белка до тех пор, пока не произойдет посттрансляционная модификация. В случае дестабилизирующего домена (DD) или SRE указанный дестабилизирующий домен посттрансляционно расщепляется в присутствии экзогенного агента или малой молекулы. Специалист в данной области техники сможет использовать такие способы контроля согласно описанию в патенте США 8173792 и заявке PCT WO2017180587. Другие переключатели посттранскрипционного контроля, предусмотренные для применения в зкДНК-векторе для контроля активности функционального трансгена, раскрыты в источниках: Rakhit et al., Chem Biol. 2014; 21 (9): 1238-52 и Navarro et al., ACS Chem Biol. 2016; 19; 11 (8): 2101-2104A.
[00308] Согласно некоторым вариантам реализации регуляторный переключатель для контроля представляющего интерес трансгена или гена, экспрессируемого зкДНК-вектором, представляет собой систему посттрансляционной модификации, которая встраивает чувствительные к лиганду интеины в кодирующую последовательность трансгена таким образом, чтобы указанный трансген или экспрессируемый белок оставался ингибированным до сплайсинга. Например, это было продемонстрировано с применением 4-гидрокситамоксифена и тиреоидного гормона (см., например, патенты США 7541450, 9200045; 7192739, Buskirk, et al, Proc Natl Acad Sci U S A. 2004 Jul 20; 101(29): 10505-10510; ACS Synth Biol. 2016 Dec 16; 5(12): 1475-1484; и 2005 Feb; 14(2): 523-532. Согласно некоторым вариантам реализации посттранскрипционный регуляторный переключатель может быть выбран из любого переключателя или их комбинации из Таблицы 11.
[00309] F. Другие примеры регуляторных переключателей
[00310] Любой известный регуляторный переключатель может быть использован в зкДНК-векторе для контроля генной экспрессии трансгена, экспрессируемого зкДНК-вектором, включая переключатели, запускаемые изменениями окружающей среды. Дополнительные примеры включают, не ограничиваясь перечисленными: метод BOC Сузуки и соавторов (Suzuki et al., Scientific Reports 8; 10051 (2018)); расширение генетического кода и нефизиологическая аминокислота; управляемые излучением или ультразвуком положительные/отрицательные переключатели (см., например, Scott S et al., Gene Ther. 2000 Jul;7(13):1121-5; патенты США 5612318; 5571797; 5770581; 5817636; и WO1999/025385A1). Согласно некоторым вариантам реализации указанный регуляторный переключатель контролирует имплантируемая система, например, согласно патенту США 7840263; US2007/0190028A1, где генную экспрессию контролирует одна или более форм энергии, в том числе электромагнитная энергия, которая активирует промоторы, функционально связанные с трансгеном в зкДНК-векторе.
[00311] Согласно некоторым вариантам реализации регуляторный переключатель, предусмотренный для применения в зкДНК-векторе, представляет собой опосредуемый гипоксией или активируемый стрессом переключатель, например, такой как описанные в источниках: WO1999060142A2, патенты США 5834306; 6218179; 6709858; US2015/0322410; Greco et al., (2004) Targeted Cancer Therapies 9, S368, а также элементы FROG, TOAD и NRSE и индуцируемые условиями элементы сайленсинга, в том числе элементы ответа на гипоксию (HRE), элементы ответа на воспаление (IRE) и активируемые сдвиговым напряжением элементы (SSAE), например, согласно описанию в патенте США 9394526. Такой вариант реализации подходит для "включения" экспрессии трансгена с зкДНК-вектора после ишемии или в ишемических тканях и/или в опухолях.
[00312] Согласно некоторым вариантам реализации регуляторный переключатель, предусмотренный для применения в зкДНК-векторе, представляет собой оптогенетический (например, контролируемый светом) регуляторный переключатель, например, такой как один из переключателей, которые рассмотрены в источнике: Poleskaya et al., BMC Neurosci. 2018; 19 (Suppl 1): 12, и также предусмотрены для применения согласно данному изобретению. Согласно таким вариантам реализации зкДНК-вектор может содержать генетические элементы, которые являются светочувствительными и могут регулировать трансгенную экспрессию в ответ на свет в видимом диапазоне длин волн (например, синий свет, ближняя ИК-область спектра). зкДНК-векторы, содержащие оптогенетические регуляторные переключатели, подходят для экспрессии трансгена в местоположениях в организме, доступных для таких источников света, например, в коже, глазах, мышцах и т.п., и могут также быть использованы при экспрессии зкДНК-векторов во внутренних органах и тканях, куда световой сигнал может быть доставлен подходящим способом (например, с помощью имплантируемого устройства согласно описанию в данном документе). Такие оптогенетические регуляторные переключатели включают применение светочувствительных элементов или индуцируемых светом транскрипционных эффекторов (LITE) (например, согласно описанию в 2014/0287938), системы Light-On (например, согласно описанию в источнике: Wang et al., Nat Methods. 2012 Feb 12;9(3):266-9); которая, как сообщалось, позволяет in vivo контролировать экспрессию трансгена инсулина, системы Cry2/CIB1 (например, согласно описанию в источнике: Kennedy et al., Nature Methods; 7, 973-975 (2010); и системы FKF1/GIGANTEA (например, согласно описанию в источнике: Yazawa et al., Nat Biotechnol. 2009 Oct;27(10):941-5).
[00313] G. «Аварийные выключатели»
[00314] Другие варианты реализации данного изобретения относятся к зкДНК-вектору, содержащему «аварийный выключатель». «Аварийный выключатель» согласно описанию в данном документе позволяет убить клетку, содержащую указанный зкДНК-вектор, или обеспечить ее программируемую клеточную смерть в качестве способа окончательного удаления введенного зкДНК-вектора из системы субъекта. Специалисту в данной области техники будет понятно, что применение «аварийных выключателей» в зкДНК-векторах согласно данному изобретению, как правило, сопряжено с нацеливанием зкДНК-вектора на ограниченное число клеток, потеря которого является приемлемой для субъекта, или на тип клеток, апоптоз которых желателен (например, на раковые клетки). Во всех аспектах «аварийный выключатель» согласно описанию в данном документе разработан таким образом, чтобы обеспечить быстрый и надежный киллинг клетки, содержащей зкДНК-вектор, в отсутствие входящего сигнала выживания или другого заданного условия. Другими словами, «аварийный выключатель», кодируемый зкДНК-вектором согласно данному документу, может ограничивать выживание клетки, содержащей зкДНК-вектор, средой, задаваемой специфическими входными сигналами. Такие «аварийные выключатели» служат для обеспечения функции биологического сдерживания, если требуется либо удалить зкДНК-вектор из субъекта, либо обеспечить отсутствие экспрессии с него кодируемого трансгена. Соответственно, «аварийные выключатели» представляют собой синтетические биологические цепи в зкДНК-векторе, сопрягающие сигналы внешней среды с обусловленным выживанием клетки, содержащей указанный зкДНК-вектор. Согласно некоторым вариантам реализации разные зкДНК-векторы может быть разработаны так, что они имеют разные «аварийные выключатели». Это обеспечивает возможность контролировать, киллинг каких клеток, экспрессирующих трансген, происходит при использовании коктейлей зкДНК-векторов.
[00315] Согласно некоторым вариантам реализации зкДНК-вектор может содержать «аварийный выключатель», который представляет собой модульную цепь биологического сдерживания. Согласно некоторым вариантам реализации «аварийный выключатель», предусмотренный для применения в зкДНК-векторе, описан в WO2017/059245, где описан переключатель, называемый «аварийным блокиратором», который содержит по меньшей мере две репрессируемые последовательности, расположенные взаимно ингибирующим образом, так что сигнал окружающей среды репрессирует активность второй молекулы в конструкте (например, связывающий малые молекулы транскрипционный фактор используют, чтобы обеспечить состояние «выживания» за счет репрессии продуцирования токсина). В клетках, содержащих зкДНК-вектор, содержащий «аварийный блокиратор», после потери сигнала окружающей среды цепь окончательно переключается в состояние «блокировки», при котором происходит дерепрессия токсина, приводящая к продуцированию токсина, что убивает клетку. Согласно другому варианту реализации предложена синтетическая биологическая цепь, называемая «цепью с кодом доступа» или «аварийным выключателем с кодом доступа», где использованы гибридные транскрипционные факторы (ТФ) для конструирования комплексных требований к условиям среды для выживания клеток. «Аварийные блокираторы» и «аварийные выключатели с кодом доступа», описанные в WO2017/059245, в частности, подходят для применения в зкДНК-векторах, поскольку они являются модульными и настраиваемыми, как в отношении условий окружающей среды, контролирующих активацию цепи, так и в отношении выходных модулей, контролирующих судьбу клеток. При правильном подборе токсинов, которые включают, не ограничиваясь указанным, эндонуклеазу, например, EcoRI, «цепи с кодами доступа», присутствующие в зкДНК-векторе, которые можно применять не только для уничтожения клетки-хозяина, содержащей вектор зкДНК, но также для разрушения ее генома и сопутствующих плазмид.
[00316] Другие «аварийные выключатели», известные специалисту в данной области техники, предусмотрены для применения в зкДНК-векторе согласно описанию в данном документе, например, согласно описанию в US2010/0175141; US2013/0009799; US2011/0172826; US2013/0109568, а также «аварийные выключатели», описанные в источниках: Jusiak et al, Reviews in Cell Biology and Molecular Medicine; 2014; 1-56; Kobayashi et al., PNAS, 2004; 101; 8419-9; Marchisio et al., Int. Journal of Biochem and Cell Biol., 2011; 43; 310-319; и Reinshagen et al., Science Translational Medicine, 2018, 11.
[00317] Соответственно, согласно некоторым вариантам реализации указанный зкДНК-вектор может содержать конструкт нуклеиновой кислоты «аварийного выключателя», который содержит нуклеиновую кислоту, кодирующую эффекторный токсин или репортерный белок, причем экспрессию эффекторного токсина (например, белка смерти) или репортерного белка контролирует заранее заданное условие. Например, заранее заданным условием может быть присутствие агента окружающей среды, такого как, например, экзогенный агент, без которого клетка по умолчанию будет экспрессировать эффекторный токсин (например, белок смерти) и погибнет. Согласно альтернативным вариантам реализации заранее заданным условием является присутствие двух или более агентов окружающей среды, например, клетка выживет только при поступлении двух или более необходимых экзогенных агентов, и без какого-либо из них клетка, содержащая зкДНК-вектор, погибнет.
[00318] Согласно некоторым вариантам реализации зкДНК-вектор модифицируют так, чтобы он содержал «аварийный выключатель» для уничтожения клеток, содержащих указанный зкДНК-вектор, для эффективного прекращения in vivo экспрессии трансгена, экспрессируемого указанным зкДНК-вектором (например, терапевтического гена, белка или пептида, и т.е.). В частности, также зкДНК-вектор генетически сконструирован для экспрессии белка-переключателя, который не функционирует в клетках млекопитающих в нормальных физиологических условиях. Клетки, экспрессирующие белок-переключатель, будут уничтожены только после введения лекарственного средства или воздействия условия внешней среды, специфически нацеленных на указанный белок-переключатель, и, соответственно, тогда будет прекращена экспрессия терапевтического белка или пептида. Например, сообщалось, что клетки, экспрессирующие HSV-тимидинкиназу, могут погибать при введении таких лекарственных средств, как ганцикловир и цитозиндезаминаза. См., например, Dey and Evans, Suicide Gene Therapy by Herpes Simplex Virus-1 Thymidine Kinase (HSV-TK), в источнике: Targets in Gene Therapy, под редакцией You (2011); и Beltinger et al., Proc. Natl. Acad. Sci. USA 96(15):8699-8704 (1999). Согласно некоторым вариантам реализации зкДНК-вектор может содержать «аварийный выключатель» киРНК, называемый DISE (смерть, вызываемая элиминацией гена выживания, «Death Induced by Survival gene Elimination») (Murmann et al., Oncotarget. 2017; 8:84643-84658. Induction of DISE in ovarian cancer cells in vivo).
[00319] Согласно некоторым аспектам «аварийный блокиратор» представляет собой биологическую цепь или систему, придающую клеточному ответу чувствительность к заранее определенному условию, такому как отсутствие агента в среде, где растут клетки, например, экзогенного агента. Такая цепь или система может содержать нуклеиновую кислоту, содержащую экспрессионные модули, которые образуют аварийную регуляторную цепь, чувствительную к заранее заданному условию, при этом указанный конструкт содержит:
i) модуль экспрессии первого репрессорного белка, отличающегося тем, что указанный первый репрессорный белок связывает нуклеиновую кислоту связывающего элемента первого репрессорного белка, и репрессирует транскрипцию с кодирующей последовательности, содержащей указанный связывающий элемент первого репрессорного белка, при этом репрессорная активность первого репрессорного белка чувствительна к ингибированию первым экзогенным агентом, присутствие или отсутствие которого определяет заранее заданное условие;
ii) модуль экспрессии второго репрессорного белка, отличающегося тем, что второй репрессорный белок связывает нуклеиновую кислоту связывающего элемента второго репрессорного белка и репрессирует транскрипцию с кодирующей последовательности, содержащей указанный связывающий элемент второго репрессорного белка, при этом второй репрессорный белок отличается от первого репрессорного белка; и
iii) модуль экспрессии эффектора, содержащий последовательность нуклеиновой кислоты, кодирующую эффекторный белок, функционально связанный с генетическим элементом, содержащим связывающий элемент второго репрессорного белка, так что экспрессия второго репрессорного белка вызывает репрессию экспрессии эффектора с модуля экспрессии эффектора, при этом второй модуль экспрессии содержит элемент, связывающий нуклеиновую кислоту первого репрессорного белка, который обеспечивает репрессию транскрипции второго репрессорного белка, если указанный элемент связан первым репрессорным белком; соответствующие модули образуют регуляторную цепь таким образом, что в отсутствие первого экзогенного агента, первый репрессорный белок продуцируется с модуля экспрессии первого репрессорного белка и репрессирует транскрипцию с модуля экспрессии второго репрессорного белка, так что подавление экспрессии эффектора вторым репрессорным белком устраняется, что обеспечивает экспрессию эффекторного белка, но в присутствии первого экзогенного агента активность первого репрессорного белка ингибирована, что разрешает экспрессию второго репрессорного белка, который поддерживает экспрессию эффекторного белка в «выключенном» состоянии, таким образом, первый экзогенный агент в цепи необходим для поддержания экспрессии эффекторного белка в «выключенном» состоянии, и удаление или отсутствие первого экзогенного агента приводит к экспрессии эффекторного белка по умолчанию.
[00320] Согласно некоторым вариантам реализации указанный эффектор представляет собой токсин или белок, который индуцирует программу клеточной смерти. Может быть использован любой белок, токсичный для клетки-хозяина. Согласно некоторым вариантам реализации указанный токсин убивает только те клетки, в которых происходит его экспрессия. Согласно другим вариантам реализации указанный токсин убивает и другие клетки того же организма-хозяина. Для использования в «аварийном блокираторе» подходят любые из множества продуктов, которые приводят к клеточной смерти. В частности, целесообразно использование агентов, которые ингибируют репликацию ДНК, трансляцию белка или другие процессы или, например, разрушают нуклеиновую кислоту клетки-хозяина. Для идентификации эффективного механизма уничтожения клеток-хозяев при активации цепи, было протестировано несколько генов токсинов, которые непосредственно повреждают ДНК или РНК клетки-хозяина. Тестировали эндонуклеазу ecoRI21, ингибитор ДНК-гиразы ccdB22 и токсин рибонуклеазного типа mazF23, поскольку они подробно описаны, являются нативными для E. coli и обеспечивают диапазон механизмов уничтожения. Чтобы повысить надежность цепи и обеспечить независимый способ обуславливаемой цепью смерти клеток, указанная система может быть дополнительно адаптирована для экспрессии, например, нацеленной протеазы или нуклеазы, которая дополнительно взаимодействует с репрессором, который поддерживает ген смерти в "выключенном" состоянии. При утрате или отмене сигнала выживания репрессия гена смерти устраняется еще эффективнее за счет, например, активного разложения репрессорного белка или его транскрипта. В неограничивающем примере использовали протеазу mf -Lon не только для разложения LacI, но и для нацеливания на важные белки для их разложения. Метка разложения mf-Lon pdt#1 может быть присоединена к 3'-концу пяти важных генов, белковые продукты которых, в частности, чувствительны к разложению mf-Lon20; жизнеспособность клеток измеряли после удаления ATc. Среди протестированных важных целевых генов ген биосинтеза пептидогликана murC обеспечивал наиболее выраженный и быстрый фенотип клеточной смерти (доля выживших клеток < 1 x 10-4 не более чем через 6 часов).
[00321] В данном документе термин «заранее заданный входной сигнал» относится к агенту или состоянию, которое известным образом влияет на активность полипептида транскрипционного фактора. Обычно такие агенты способны связывать полипептид транскрипционного фактора и/или изменять его конформацию, тем самым модифицируя активность указанного полипептида транскрипционного фактора. Примеры заранее заданных входных сигналов включают, не ограничиваясь перечисленными, входные агенты среды, не требуемые для выживания определенного организма-хозяина (т.е. в отсутствие синтетической биологической цепи согласно описанию в данном документе). Условия, которые могут обеспечивать заранее заданный входной сигнал, включают, например, температуру, например, если активность одного или более факторов чувствительна к температуре, присутствие или отсутствие света, в том числе света с длинами волн определенного спектра, и концентрацию газа, соли, металла или минерала. Входные агенты среды включают, например, малую молекулу, биологические агенты, такие как феромоны, гормоны, факторы роста, метаболиты, питательные вещества и т.п., и их аналоги; концентрации химических веществ, побочные продукты среды, ионы металлов и другие такие молекулы или агенты; уровни света; температуру; механическое напряжение или давление; или электрические сигналы, такие как токи и напряжения.
[00322] Согласно некоторым вариантам реализации используют репортеры для количественной оценки силы или активности сигнала, принимаемого модулями или программируемыми синтетическими биологическими цепями согласно данному изобретению. Согласно некоторым вариантам реализации репортеры могут быть слиты внутри рамки с другими кодирующими последовательностями для идентификации локализации белка в клетке или организме. Люциферазы могут применяться в качестве эффекторных белков в различных вариантах реализации данного изобретения, например, для изменения низких уровней генной экспрессии, поскольку клетки, как правило, практически не имеют или совершенно не имеют фоновой люминесценции в отсутствие люциферазы. Согласно другим вариантам реализации ферменты, которые продуцируют окрашенные субстраты, можно количественно оценивать с применением спектрофотометров или других позволяющих измерять поглощение инструментов, в том числе планшет-ридеров. Как и люциферазы, такие ферменты, как β-галактозидаза, могут применяться для измерения низких уровней генной экспрессии, поскольку они имеют тенденцию усиливать слабые сигналы. Согласно некоторым вариантам реализации эффекторный белок может представлять собой фермент, который может разлагать или иным образом уничтожать определенный токсин. Согласно некоторым вариантам реализации эффекторный белок может представлять собой фермент-одоратор, который превращает субстрат в продукт, имеющий запах. Согласно некоторым вариантам реализации эффекторный белок может представлять собой фермент, который фосфорилирует или дефосфорилирует либо малые молекулы, либо другие белки, или фермент, который метилирует или деметилирует другие белки или ДНК.
[00323] Согласно некоторым вариантам реализации эффекторный белок может представлять собой рецептор, лиганд, или литический белок. Рецепторы, как правило, имеют три домена: внеклеточный домен для связывания лигандов, таких как белки, пептиды или малые молекулы, трансмембранный домен и внутриклеточный или цитоплазматический домен, который часто может участвовать в каком-либо явлении при передаче сигнала, таком как фосфорилирование. Согласно некоторым вариантам реализации в качестве эффекторных белков используют последовательности генов транспортеров, каналов или насосов. Неограничивающие примеры и последовательности эффекторных белков для применения с «аварийными выключателями» согласно описанию в данном документе можно найти в Реестре стандартных биологических компонентов во всемирной сети Интернет по адресу: parts.igem.org.
[00324] В данном документе «белок-модулятор» представляет собой белок, который модулирует экспрессию с целевой последовательности нуклеиновой кислоты. Белки-модуляторы включают, например, транскрипционные факторы, в том числе транскрипционные активаторы и репрессоры, а также белки которые связываются или модифицируют транскрипционный фактор и влияют на его активность. Согласно некоторым вариантам реализации белок-модулятор включает, например, протеазу, которая разлагает белковый фактор, вовлеченный в регуляцию экспрессии с последовательности целевой нуклеиновой кислоты. Предпочтительные белки-модуляторы включают модульные белки, в которых, например, связывающие ДНК и связывающие входной агент, или ответные элементы или домены разделены и могут быть перенесены, так что, например, слияние связывающего ДНК домена первого белка-модулятора с отвечающим на входной агент доменом второго белка-модулятора приводит к образованию нового белка, который связывает последовательность ДНК, распознаваемую первым белком, но чувствителен к входному агенту, на который в норме отвечает второй белок. Соответственно, в данном документе термин «полипептид-модулятор», более конкретно - «репрессорный полипептид» включает, наряду с указанными полипептидами, например, «(репрессорный) полипептид LacI», варианты или производные таких полипептидов, которые отвечают на другой входной агент или вариант входного агента. Соответственно, в случае полипептида LacI предусмотрены мутанты или варианты LacI, которые связываются с агентами, не являющимися лактозой или IPTG. В данной области техники известен широкий спектр таких агентов.
[00325] Таблица 11. Примеры регуляторных переключателей. bВозможность включения эффектором; отличная от удаления эффектора, который обеспечивает выключенное состояние. cВозможность выключения эффектором; отличная от удаления эффектора, который обеспечивает включенное состояние. dЛиганд или другой физический стимул (например, температура, электромагнитное излучение, электричество), который стабилизирует переключатель либо во включенном, либо в выключенном состоянии. eОтносится к референсному числу, указанному в источнике: Kis et al., J R Soc Interface. 12:20141000 (2015), содержание которого вместе с цитируемыми источниками включено в данный документ посредством ссылки.
VII. Фармацевтические композиции
[00326] Согласно другому аспекту предложены фармацевтические композиции. Указанная фармацевтическая композиция содержит зкДНК-вектор согласно описанию в данном документе и фармацевтически приемлемый носитель или разбавитель.
[00327] ДНК-векторы согласно описанию в данном документе могут быть включены в фармацевтические композиции, подходящие для введения субъекту для доставки in vivo в клетки, ткани или органы субъекта. Как правило, указанная фармацевтическая композиция содержит зкДНК-вектор согласно описанию в данном документе и фармацевтически приемлемый носитель. Например, зкДНК-векторы описанные в данном документе могут быть включены в фармацевтическую композицию, подходящую для требуемого пути терапевтического введения (например, парентерального введения). Также предусмотрена пассивная трансдукция ткани путем внутривенной или внутриартериальной инфузии под высоким давлением, а также внутриклеточной инъекции, такой как внутриядерная микроинъекция или внутрицитоплазматическая инъекция. Фармацевтические композиции для терапевтических целей могут быть введены в состав раствора, микроэмульсии, дисперсии, липосом или другой упорядоченной структуры, подходящей для высокой концентрации зкДНК-вектора. Стерильные растворы для инъекций могут быть приготовлены путем введения соединения зкДНК-вектора в необходимом количестве в соответствующий буфер с одним ингредиентом или комбинацией ингредиентов, перечисленных выше, при необходимости, с последующей стерилизующей фильтрацией.
[00328] Фармацевтически активные композиции, содержащие зкДНК-вектор, могут быть введены в состав для доставки трансгена в нуклеиновой кислоте в клетки реципиента, что приводит к терапевтической экспрессии в них указанного трансгена. Указанная композиция также может включать фармацевтически приемлемый носитель.
[00329] ЗкДНК-вектор согласно описанию в данном документе может быть введен в фармацевтическую композицию, подходящую для местного, системного, внутриамниотического, интратекального, интракраниального, внутриартериального, внутривенного, внутрилимфатического, внутрибрюшинного, подкожного, трахеального, внутритканевого (например, внутримышечного, внутрисердечного, внутрипеченочного, внутри почечного, интрацеребрального), интратекального, интравезикального, конъюнктивальное (например, внеорбитального, интраорбитального, ретроорбитального, интраретинального, субретинального, хориоидального, субхориоидального, интрастромального, интракамерального и интравитреального), интракохлеарного и мукозального (например, перорального, ректального, назального) введение. Также предусмотрена пассивная трансдукция ткани путем внутривенной или внутриартериальной инфузии под высоким давлением, а также внутриклеточной инъекции, такой как внутриядерная микроинъекция или внутрицитоплазматическая инъекция.
[00330] Фармацевтические композиции для терапевтических целей, как правило, должны быть стерильными и стабильными в условиях изготовления и хранения. Указанная композиция может быть введена в состав раствора, микроэмульсии, дисперсии, липосом или другой упорядоченной структуры, подходящей для высокой концентрации зкДНК-вектора. Стерильные растворы для инъекций могут быть приготовлены путем введения соединения зкДНК-вектора в необходимом количестве в соответствующий буфер с одним ингредиентом или комбинацией ингредиентов, перечисленных выше, при необходимости, с последующей стерилизующей фильтрацией.
[00331] В данной области техники известны различные методы и способы доставки нуклеиновых кислот в клетки. Например, нуклеиновые кислоты, такие как зкДНК, могут быть введены в состав липидных наночастиц (ЛНЧ), липидоидов, липосом, липидных наночастиц, липоплексов или наночастиц типа ядро/оболочка. Как правило, ЛНЧ состоят из молекул нуклеиновой кислоты (например, зкДНК), одного или более ионизируемых или катионных липидов (или их солей), одного или более неионогенных или нейтральных липидов (например, фосфолипида), молекулы, которая предотвращает агрегацию (например, ПЭГ или ПЭГ-липидного конъюгата), и необязательно стерола (например, холестерина).
[00332] Другой способ доставки нуклеиновых кислот, таких как зкДНК, в клетку, заключается в конъюгировании нуклеиновой кислоты с лигандом, который интернализуется указанной клеткой. Например, указанный лиганд может связывать рецептор на поверхности клетки и интернализоваться посредством эндоцитоза. Указанный лиганд может быть ковалентно связан с нуклеотидом в нуклеиновой кислоте. Примеры конъюгатов для доставки нуклеиновых кислот в клетку описаны, например, в WO2015/006740, WO2014/025805, WO2012/037254, WO2009/082606, WO2009/073809, WO2009/018332, WO2006/112872, WO2004/090108, WO2004/091515 и WO2017/177326.
[00333] Нуклеиновые кислоты, такие как зкДНК, могут также быть доставлены в клетку путем трансфекции. Подходящие способы трансфекции включают, не ограничиваясь перечисленными, опосредованную липидами трансфекцию, опосредованную катионным полимером трансфекцию или осаждение фосфатом кальция. Реагенты для трансфекции хорошо известны в данной области техники и включают, не ограничиваясь перечисленными, TurboFect Transfection Reagent (Thermo Fisher Scientific), Pro-Ject Reagent (Thermo Fisher Scientific), TRANSPASS™ P Protein Transfection Reagent (New England Biolabs), CHARIOT™ Protein Delivery Reagent (Active Motif), PROTEOJUICE™ Protein Transfection Reagent (EMD Millipore), реагент 293fectin, липофектамин - LIPOFECTAMINE ™ 2000, LIPOFECTAMINE ™ 3000 (Thermo Fisher Scientific), LIPOFECTAMINE ™ (Thermo Fisher Scientific), липофектин - LIPOFECTIN™ (Thermo Fisher Scientific), DMRIE-C, CELLFECTIN™ (Thermo Fisher Scientific), OLIGOFECTAMINE™ (Thermo Fisher Scientific), LIPOFECTACE™, FUGENE™ (Roche, Базель, Швейцария), FUGENE™ HD (Roche), TRANSFECTAM™(трансфектам, Promega, Мэдисон, Висконсин), TFX-10™ (Promega), TFX-20™ (Promega), TFX-50™ (Promega), TRANSFECTIN™ (BioRad, Геркулес, Калифорния), SILENTFECT™ (Bio-Rad), Effectene™ (Qiagen, Валенсия, Калифорния), DC-chol (Avanti Polar Lipids), GENEPORTER™ (Gene Therapy Systems, Сан-Диего, Калифорния), DHARMAFECT 1™ (Dharmacon, Лафайет, Колорадо), DHARMAFECT 2™ (Dharmacon), DHARMAFECT 3™ (Dharmacon), DHARMAFECT 4™ (Dharmacon), ESCORT™ III (Sigma, Сент-Луис, Миссури) и ESCORT™ IV (Sigma Chemical Co.). Нуклеиновые кислоты, такие как зкДНК, также могут быть доставлены в клетку с помощью способов микрофлюидики, известных специалистам в данной области техники.
[00334] Способы невирусной доставки нуклеиновых кислот in vivo или ex vivo включают электропорацию, липофекцию (см. патент США №5049386; 4946787; и коммерчески доступные реагенты, такие как трансфектам (Transfectam™) и липофектин (Lipofectin™)), микроинъекцию, биолистику, виросомы, липосомы (см., например, Crystal, Science 270:404-410 (1995); Blaese et al., Cancer Gene Ther. 2:291-297 (1995); Behr et al., Bioconjugate Chem. 5:382-389 (1994); Remy et al., Bioconjugate Chem. 5:647-654 (1994); Gao et al., Gene Therapy 2:710-722 (1995); Ahmad et al., Cancer Res. 52:4817-4820 (1992); патенты США № 4186183, №4217344, №4235871, №4261975, №4485054, №4501728, №4774085, №4837028 и №4946787), иммунолипосомы, конъюгаты нуклеиновых кислот с поликатионами или липидами, «голую» ДНК и усиленное агентами поглощение ДНК. Сонопорация с использованием, например, системы Sonitron 2000 (Rich-Mar) также может быть использована для доставки нуклеиновых кислот.
[00335] ЗкДНК-векторы согласно описанию в данном документе могут также быть введены прямо в организм для трансдукции клеток in vivo. Введение осуществляют любым из путей, обычно используемых для приведения молекулы в максимальный контакт с клетками крови или тканей, в том числе, но не ограничиваясь перечисленным, инъекцию, инфузию, местное нанесение и электропорацию. Подходящие способы введения таких нуклеиновых кислот доступны и хорошо известны специалистам в данной области техники, и хотя для введения конкретной композиции можно использовать более одного пути, конкретный путь часто может обеспечить более быструю и более эффективную реакцию по сравнению с другим путем введения.
[00336] Способы введения зкДНК-вектора из нуклеиновых кислот согласно описанию в данном документе могут включать доставку в гематопоэтические стволовые клетки, например, способами, описанными, например, в патенте США № 5928638.
[00337] ЗкДНК-векторы в соответствии с данным изобретением могут быть добавлены в липосомы для доставки в клетку или целевой орган субъекта. Липосомы представляют собой везикулы, имеющие по меньшей мере один липидный бислой. Липосомы часто используются в качестве носителей для доставки лекарственных/терапевтических средств в контексте фармацевтических разработок. Они работают за счет слияния с клеточной мембраной и перегруппировки ее липидной структуры для доставки лекарственного средства или активного фармацевтического ингредиента (АФИ). Липосомальные композиции для такой доставки состоят из фосфолипидов, в частности, соединений, содержащих фосфатидилхолиновую группу, хотя указанные композиции могут также включать другие липиды.
[00338] Согласно некоторым аспектам данного изобретения предложен липосомальный состав, который включает одно или более соединений с функциональной группой полиэтиленгликоля (ПЭГ) (так называемые «пегилированные соединения»), которые могут снижать иммуногенность/антигенность, обеспечивать гидрофильность и гидрофобность указанному соединению (соединениям) и снижение частоты его дозирования. В других случаях, липосомальная композиция просто содержит полимер полиэтиленгликоля (ПЭГ) в качестве дополнительного компонента. В таких аспектах молекулярная масса ПЭГ или функциональной группы ПЭГ может составлять от 62 Да до примерно 5000 Да.
[00339] В некоторых аспектах данное изобретение относится к композиции липосом, которая доставляет АФИ с профилем пролонгированного или контролируемого высвобождения на протяжении периода времени от нескольких часов до недель. Согласно некоторым родственным аспектам указанный липосомальный состав может включать водные камеры, которые связаны липидными бислоями. Согласно другим родственным аспектам указанный липосомальный состав инкапсулирует АФИ с компонентами, которые претерпевают физический переход при повышенной температуре, который высвобождает АФИ в течение периода от нескольких часов до недель.
[00340] Согласно некоторым аспектам указанный липосомальный состав содержит сфингомиелин и один или более липидов согласно описанию в данном документе. В некоторых аспектах липосомная композиция содержит оптисомы.
[00341] В некоторых аспектах данное изобретение предусматривает липосомальную композицию, которая включает один или несколько липидов, выбранных из: натриевой соли N-(карбонилметоксиполиэтиленгликоль 2000)-1,2-дистеароил-sn-глицеро-3-фосфоэтаноламина, (дистеароил-sn-глицерофосфоэтаноламина), MPEG (метоксиполиэтиленгликоль)-конъюгированного липида, HSPC (гидрогенизированного соевого фосфатидилхолина); ПЭГ (полиэтиленгликоля); DSPE (дистеароил-sn-глицерофосфоэтаноламина); DSPC (дистеароилфосфатидилхолина); DOPC (диолеоилфосфатидилхолина); DPPG (дипальмитоилфосфатидилглицерина); EPC (яичного фосфатидилхолина); DOPS (диолеоилфосфатидилсерина); POPC (пальмитоилолеоилфосфатидилхолина); SM (сфингомиелина); MPEG (метоксиполиэтиленгликоля); DMPC (димиристоилфосфатидилхолина); DMPG (димиристоилфосфатидилглицерина); DSPG (дистеароилфосфатидилглицерина); DEPC (диэрукоилфосфатидилхолина); DOPE (диолеоил-sn-глицерофосфоэтаноламина), холестерилсульфата (CS), дипальмитоилфосфатидилглицерина (DPPG), DOPC (диолеоил-sn-глицерофосфатидилхолина) или любой их комбинации.
[00342] В некоторых аспектах данное изобретение предусматривает липосомальную композицию, содержащую фосфолипид, холестерин и пегилированный липид в молярном соотношении 56:38:5. Согласно некоторым аспектам общее содержание липидов в липосомальном составе составляет 2-16 мг/мл. Согласно некоторым аспектам данного изобретения предложен липосомальный состав, содержащий липид, содержащий фосфатидилхолиновую функциональную группу, липид, содержащий этаноламиновую функциональную группу, и пегилированный липид. Согласно некоторым аспектам данного изобретения предложен липосомальный состав, содержащий липид, содержащий фосфатидилхолиновую функциональную группу, липид, содержащий этаноламиновую функциональную группу, и пегилированный липид, в молярном соотношении 3:0,015:2, соответственно. Согласно некоторым аспектам данного изобретения предложен липосомальный состав, содержащий липид, содержащий фосфатидилхолиновую функциональную группу, холестерин и пегилированный липид. Согласно некоторым аспектам данного изобретения предложен липосомальный состав, содержащий липид, содержащий фосфатидилхолиновую функциональную группу и холестерин. Согласно некоторым аспектам указанный пегилированный липид представлен PEG-2000-DSPE. Согласно некоторым аспектам данного изобретения предложен липосомальный состав, содержащий DPPG, соевый ФХ, конъюгат липида с MPEG-DSPE и холестерин.
[00343] Согласно некоторым аспектам данного изобретения предложен липосомальный состав, содержащий один или более липидов, содержащих фосфатидилхолиновую функциональную группу, и один или более липидов, содержащих этаноламиновую функциональную группу. Согласно некоторым аспектам данного изобретения предложен липосомальный состав, содержащий что-либо одно или более из: липидов, содержащих фосфатидилхолиновую функциональную группу, липидов, содержащих этаноламиновую функциональную группу, и стеролов, например, холестерина. Согласно некоторым аспектам указанный липосомальный состав содержит DOPC/ DEPC; и DOPE.
[00344] Согласно некоторым аспектам данного изобретения предложен липосомальный состав, дополнительно содержащий одно или более фармацевтических вспомогательных веществ, например, сахарозу и/или глицин.
[00345] Согласно некоторым аспектам данного изобретения предложен липосомальный состав, имеющий униламеллярную или мультиламеллярную структуру. В некоторых аспектах данное изобретение относится к липосомной композиции, которая включает мультивезикулярные частицы и/или частицы на основе пены. В некоторых аспектах данное изобретение предусматривает композицию липосом, размер которых больше, чем у обычных наночастиц, и составляет примерно от 150 до 250 нм. В некоторых аспектах липосомальная композиция представляет собой лиофилизированный порошок.
[00346] Согласно некоторым аспектам данного изобретения предложен липосомальный состав, который получают и нагружают зкДНК-векторами, раскрытыми или описанными в данном документе, путем добавления слабого основания к смеси, содержащей выделенную зкДНК, вне липосомы. Указанное добавление повышает значение pH вне липосом до приблизительно 7,3 и стимулирует вход АФИ в липосому. Согласно некоторым аспектам данного изобретения предложен липосомальный состав, имеющий значения pH, кислотные внутри липосом. В таких случаях значение pH внутри липосом может составлять 4-6,9 и более предпочтительно - pH 6,5. Согласно другим аспектам данного изобретения предложен липосомальный состав, полученный с применением технологии внутрилипосомальной стабилизации лекарственных средств. В таких случаях используют полимерные или неполимерные высокозаряженные анионы и внутрилипосомальные захватывающие агенты, например, полифосфат или октасульфат сахарозы.
[00347] В других аспектах данное изобретение предлагает липосомную композицию, содержащую фосфолипиды, лецитин, фосфатидилхолин и фосфатидилэтаноламин.
[00348] Реагенты для доставки, такие как липосомы, нанокапсулы, микрочастицы, микросферы, липидные частицы, везикулы и т.п., можно использовать для введения композиций по данному изобретению в подходящие клетки-хозяева. В частности, композиции нуклеиновых кислот могут быть составлены для доставки в инкапсулированном виде в липидной частице, липосоме, везикуле, наносфере, наночастице, частице золота и т.п. Такие составы могут быть предпочтительными для введения фармацевтически приемлемых композиций нуклеиновых кислот, раскрытых в данном документе.
[00349] Различные способы доставки, известные в данной области, или их модификации, могут быть использованы для доставки зкДНК-векторов in vitro или in vivo. Например, согласно некоторым вариантам реализации зкДНК-векторы доставляют путем транзиентного проникновения через клеточную мембрану с использованием механической, электрической, ультразвуковой, гидродинамической или лазерной энергии для облегчения входа ДНК в целевые клетки. Например, зкДНК-вектор может быть доставлен путем транзиентного разрыва клеточной мембраны путем продавливания клетки через канал ограниченного размера или другими способами, известными в данной области техники. В некоторых случаях прямо вводят зкДНК-вектор по отдельности в виде депротеинизированной ДНК в клетки кожи, вилочковой железы, сердечной мышцы, скелетных мышц или печени.
[00350] В некоторых случаях зкДНК-вектор доставляют с помощью генной пушки. Сферические частицы золота или вольфрама (диаметром 1-3 мкм), покрытые бескапсидными AAV-векторами, могут быть разогнаны до высокой скорости с помощью сжатого газа для проникновения в клетки целевой ткани.
[00351] Согласно некоторым вариантам реализации для доставки зкДНК-векторов используют электропорацию. Электропорация вызывает временную дестабилизацию клеточной мембраны клеток целевой ткани в результате введения в ткань пары электродов, так что молекулы ДНК из среды вокруг дестабилизированной мембраны могут проникать в цитоплазму и нуклеоплазму указанной клетки. Электропорацию использовали in vivo для многих типов тканей, таких как кожа, легкие и мышцы.
[00352] В некоторых случаях зкДНК-вектор доставляют путем гидродинамической инъекции, которая представляет собой простой и высокоэффективный способ прямой внутриклеточной доставки любых водорастворимых соединений и частиц во внутренние органы и скелетные мышцы во всей конечности.
[00353] В некоторых случаях зкДНК-векторы доставляют с помощью ультразвука, создавая наноскопические поры в мембране для облегчения внутриклеточной доставки частиц ДНК в клетки внутренних органов или опухолей, поэтому размер и концентрация плазмидной ДНК имеют большое значение для эффективности системы. В некоторых случаях зкДНК-векторы доставляют путем магнитофекции с использованием магнитных полей для концентрации частиц, содержащих нуклеиновую кислоту, в целевых клетках.
[00354] В некоторых случаях могут быть использованы химические системы доставки, например, с использованием наномерных комплексов, которые включают уплотнение отрицательно заряженной нуклеиновой кислоты поликатионными наномерными частицами, в составе катионных липосом/мицелл или катионных полимеров. Катионные липиды, используемые в указанном способе доставки, включают, без ограничений, моновалентные катионные липиды, поливалентные катионные липиды, гуанидин-содержащие соединения, соединения-производные холестерина, катионные полимеры (например, полиэтиленимин, поли-L-лизин, протамин, другие катионные полимеры), и гибриды липид-полимер.
[00355] А. Экзосомы:
[00356] В некоторых вариантах реализации зкДНК-вектор, как раскрыто в данном документе, доставляют путем упаковки в экзосому. Экзосомы представляют собой небольшие мембранные везикулы эндоцитарного происхождения, которые высвобождаются во внеклеточную среду после слияния мультивезикулярных телец с плазматической мембраной. Их поверхность состоит из липидного бислоя клеточной мембраны донорной клетки, они содержат цитозоль из клетки, которая продуцировала экзосому, и экспонируют на поверхности мембранные белки родительской клетки. Экзосомы продуцируются различными типами клеток, в том числе эпителиальными клетками, В- и Т-лимфоцитами, тучными клетками (MC), а также дендритными клетками (DC). Согласно некоторым вариантам реализации предусмотрены для применения экзосомы с диаметром от 10 нм до 1 мкм, от 20 нм до 500 нм, от 30 нм до 250 нм, от 50 до 100 нм. Экзосомы могут быть выделены для доставки в целевые клетки либо с применением донорных клеток, либо путем введения в них конкретных нуклеиновых кислот. Различные подходы, известные в данной области техники, могут применяться для получения экзосом, содержащих бескапсидные AAV-векторы согласно данному изобретению.
[00357] B. Микрочастицы/наночастицы
[00358] Согласно некоторым вариантам реализации зкДНК-вектор согласно описанию в данном документе доставляют на липидной наночастице. Обычно липидные наночастицы содержат ионизируемый аминолипид (например, гептатриаконта-6, 9, 28, 31-тетраен-19-ил 4- (диметиламино)бутаноат, DLin-MC3-DMA, фосфатидилхолин(1,2-дистеароил-sn-глицеро-3-фосфохолин, DSPC), холестерин и липид оболочки (полиэтиленгликоль- димиристоилглицерин, ПЭГ-ДМГ), например, как описано в источнике: Tam et al. (2013). Advances in Lipid Nanoparticles for siRNA delivery. Pharmaceuticals 5(3): 498-507.
[00359] В некоторых вариантах реализации липидная наночастица имеет средний диаметр от примерно 10 до примерно 1000 нм. В некоторых вариантах реализации липидная наночастица имеет диаметр менее 300 нм. В некоторых вариантах реализации липидная наночастица имеет диаметр от примерно 10 до примерно 300 нм. В некоторых вариантах реализации липидная наночастица имеет диаметр менее 200 нм. В некоторых вариантах реализации липидная наночастица имеет диаметр от примерно 25 до примерно 200 нм. В некоторых вариантах реализации препарат липидных наночастиц (например, композиция, содержащая множество липидных наночастиц) имеет распределение по размерам, при котором средний размер (например, диаметр) составляет от примерно 70 нм до примерно 200 нм, и более типично средний размер составляет примерно 100 нм или меньше.
[00360] Различные липидные наночастицы, известные в данной области, могут быть использованы для доставки зкДНК-вектора, раскрытого в данном документе. Например, различные способы доставки с применением липидных наночастиц описаны в патентах США №9404127, №9006417 и №9518272.
[00361] Согласно некоторым вариантам реализации a зкДНК-вектор согласно описанию в данном документе доставляют на золотой наночастице. Как правило, нуклеиновая кислота может быть ковалентно связана с золотой наночастицей или нековалентно связана с золотой наночастицей (например, связана заряд-зарядным взаимодействием), например, согласно описанию в источнике: Ding et al. (2014). Gold Nanoparticles for Nucleic Acid Delivery. Mol. Ther. 22(6); 1075-1083. Согласно некоторым вариантам реализации конъюгаты золотых наночастиц и нуклеиновой кислоты получают с использованием способов, описанных, например, в патенте США № 6812334.
[00362] C. Липосомы
[00363] Получение и применение липосом в целом известно специалистам в данной области техники. Были разработаны липосомы с улучшенной стабильностью в сыворотке и временем полужизни в циркуляторной системе (патент США № 5741516). Кроме того, были описаны различные способы получения липосомальных и подобных липосомальным составов в качестве потенциальных носителей лекарственных средств (патенты США №5567434; №5552157; №5565213; №5738868 и №5795587).
[00364] Липосомы успешно применялись с некоторыми типами клеток, которые обычно устойчивы к трансфекции с использованием других процедур. Кроме того, липосомы свободны от ограничений по длине ДНК, типичных для систем доставки на основе вирусов. Липосомы эффективно применялись для введения генов, лекарственных средств, радиотерапевтических агентов, вирусов, транскрипционных факторов и аллостерических эффекторов в различные культивируемые линии клеток и в организм животных. Кроме того, было завершено несколько успешных клинических испытаний для изучения эффективности опосредованной липосомами доставки лекарственных средств.
[00365] Липосомы образуются из фосфолипидов, которые диспергированы в водной среде, и спонтанно образуют мультиламеллярные концентрические бислойные везикулы (также называемые многослойными везикулами (MLV). MLV обычно имеют диаметр от 25 нм до 4 мкм. Обработка MLV ультразвуком приводит к образованию малых однослойных везикул (SUV) с диаметром в диапазоне от 200 до 500 ангстрем, содержащих водный раствор в ядре.
[00366] Согласно некоторым вариантам реализации липосома содержит катионные липиды. Термин «катионный липид» включает липиды и синтетические липиды, имеющие как полярные, так и неполярные домены, которые могут быть положительно заряженными при физиологических значениях pH или около них, и которые связываются с полианионами, такими как нуклеиновые кислоты, и способствуют доставке нуклеиновых кислот в клетки. Согласно некоторым вариантам реализации катионные липиды включают насыщенные и ненасыщенные алкильные и алициклические простые эфиры и сложные эфиры аминов, амидов или их производных. В некоторых вариантах реализации катионные липиды включают неразветвленные, разветвленные алкильные, алкенильные группы, или любую комбинацию вышеуказанного. В некоторых вариантах реализации катионные липиды содержат от 1 до примерно 25 атомов углерода (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 атомов углерода). В некоторых вариантах реализации катионные липиды содержат более 25 атомов углерода. Согласно некоторым вариантам реализации прямоцепочечные или разветвленные алкильные или алкеновые группы содержат шесть или более атомов углерода. Катионный липид может также содержать, согласно некоторым вариантам реализации, одну или более алициклических групп. Неограничивающие примеры алициклических групп включают холестерин и другие стероидные группы. Согласно некоторым вариантам реализации политических липидов с использованием одного или более противоионов. Примеры противоионов (анионов) включают, не ограничиваясь перечисленными, Cl-, Br-, I-, F-, ацетат, трифторацетат, сульфат, нитрит и нитрат.
[00367] Неограничивающие примеры катионных липидов включают звездчатые дендримеры полиэтиленимина и полиамидоамина (PAMAM), липофектин (комбинация DOTMA и DOPE), липофектазу, липофектамин (LIPOFECTAMINE™, например, LIPOFECTAMINE™ 2000), DOPE, цитофектин (Gilead Sciences, Фостер-Сити, Калифорния) и эуфектины (JBL, Сан-Луис-Обиспо, Калифорния). Примеры катионных липосом могут быть получены из N-[1-(2, 3-диолеолокси)-пропил]-N, N,N-триметилхлорида аммония (DOTMA), N-[1-(2, 3-диолеолокси)-пропил]-N,N,N-триметиламмония метилсульфата (DOTAP), 3β-[N-(N′, N′-диметиламиноэтан)карбамоил]холестерина (DC-Chol), 2, 3-диолеилокси-N-[2(сперминкарбоксамидо)этил]-N,N-диметил-1- пропанаминия трифторацетата (DOSPA), 1,2-димиристилоксипропил-3-диметил-гидроксиэтиламмония бромида; и диметилдиоктадециламмония бромида (DDAB). Нуклеиновые кислоты (например, CELiD) могут также быть введены в комплексы, например, с поли(L-лизином) или авидином, и липиды, например, стерил-поли(L-лизин), могут быть включены или не включены в указанную смесь.
[00368] Согласно некоторым вариантам реализации зкДНК-вектор согласно описанию в данном документе доставляют с применением катионного липида, описанного в патенте США №8158601, или полиаминного соединения или липида согласно описанию в патенте США №8034376.
[00369] C. Конъюгаты
[00370] Согласно некоторым вариантам реализации зкДНК-вектор согласно описанию в данном документе конъюгирован (например, ковалентно связан с агентом, который повышает поглощение клетками). «Агент, который повышает поглощение клетками» представляет собой молекулу, которая облегчает транспорт нуклеиновой кислоты через липидную мембрану. Например, нуклеиновая кислота может быть конъюгирована с липофильным соединением (например, холестерином, токоферолом и т.п.), проникающим в клетки пептидом (СРР) (например, пенетратином, ТАТ, Syn1B и т.п.) и полиаминами (например, спермином). Дополнительные примеры агентов, которые повышают поглощение клетками, раскрыты, например, в источнике: Winkler (2013). Oligonucleotide conjugates for therapeutic applications. Ther. Deliv. 4(7); 791-809.
[00371] Согласно некоторым вариантам реализации зкДНК-вектор согласно описанию в данном документе конъюгирован с полимером (например, полимерной молекулой) или молекулой фолата (например, молекулой фолиевой кислоты). В целом, доставка нуклеиновых кислот, конъюгированных с полимерами, известна в данной области техники, например, описана в WO2000/34343 и WO2008/022309. Согласно некоторым вариантам реализации зкДНК-вектор согласно описанию в данном документе конъюгирован с поли(амидным) полимером, например, согласно описанию в патенте США № 8987377. Согласно некоторым вариантам реализации нуклеиновая кислота, описанная в данном документе, конъюгирована с молекулой фолиевой кислоты согласно описанию в патенте США № 8507455.
[00372] Согласно некоторым вариантам реализации зкДНК-вектор согласно описанию в данном документе конъюгирован с углеводом, например, согласно описанию в патенте США № 8450467.
[00373] Е. Нанокапсула
[00374] Как вариант, могут применяться нанокапсульные составы зкДНК-вектора согласно описанию в данном документе. Как правило, нанокапсулы могут захватывать вещества стабильным и воспроизводимым образом. Чтобы избежать побочных эффектов, обусловленных внутриклеточной перегрузки полимерами, такие ультратонкие частицы (размером около 0,1 мкм) необходимо конструировать с использованием полимеров, способных разлагаться in vivo. Предусмотрены для применения биоразлагаемые наночастицы полиалкилцианоакрилата, которые отвечают указанным требованиям.
VIII. Способы доставки зкДНК-векторов
[00375] Согласно некоторым вариантам реализации зкДНК-вектор может быть доставлен в целевую клетку in vitro или in vivo различными подходящими способами. Могут применяться или вводиться зкДНК-векторы по отдельности. ЗкДНК-векторы могут быть доставлены в клетку без помощи реагента для трансфекции или другого физического способа. Как вариант, зкДНК-векторы могут быть доставлены с применением любого известного в данной области техники реагента для трансфекции или другого известного в данной области техники физического способа, который облегчает вход ДНК в клетку, например, липосом, спиртов, богатых полилизином соединений, богатых аргинином соединений, фосфата кальция, микровезикул, микроинъекции, электропорации и т.п.
[00376] Напротив, трансдукция бескапсидными AAV-векторами согласно описанию в данном документе может быть эффективно нацелена на типы клеток и тканей, которые трудно трансдуцировать стандартными вирионами AAV с применением различных реагентов для доставки.
IX. Дополнительные применения зкДНК-векторов
[00377] Композиции и зкДНК-векторы согласно данному описанию можно использовать для доставки трансгена с различными целями. Согласно некоторым вариантам реализации указанный трансген кодирует белок или функциональную РНК, которая предназначена для использования в исследовательских целях, например, для создания соматической модели на трансгенных животных, несущих трансген, например, для изучения функции продукта трансгена. В другом примере указанный трансген кодирует белок или функциональную РНК, которая предназначена для использования с целью создания модели заболевания на животных. Согласно некоторым вариантам реализации указанный трансген кодирует один или более пептидов, полипептидов или белков, которые подходят для лечения, профилактики или облегчения болезненных состояний или расстройств у субъекта - млекопитающего. Указанный трансген может быть перенесен в организм субъекта (например, экспрессироваться у пациента) в количестве, достаточном для лечения заболевания, ассоциированного с пониженной экспрессией, отсутствием экспрессии или дисфункцией указанного гена. Согласно некоторым вариантам реализации указанный трансген может быть перенесен в организм субъекта (например, экспрессироваться у субъекта) в количестве, достаточном для лечения заболевания, ассоциированного с повышенной экспрессией, активностью продукта гена или неадекватной стимулирующей регуляцией гена, который супрессирует или экспрессию которого снижает иным образом указанный трансген.
X. Способы применения
[00378] ЗкДНК-вектор, согласно данному изобретению, также можно использовать в способе доставки представляющей интерес нуклеотидной последовательности в целевую клетку. Указанный способ, в частности, может представлять собой способ доставки представляющего интерес терапевтического гена в клетку нуждающегося в этом субъекта. Данное изобретение позволяет осуществлять экспрессию in vivo полипептида, белка или олигонуклеотида, кодируемого терапевтической экзогенной последовательностью ДНК в клетках субъекта таким образом, чтобы происходила экспрессия указанного полипептида, белка или олигонуклеотида на терапевтических уровнях. Указанные результаты наблюдаются как в in vivo, так и в in vitro режимах доставки зкДНК-вектора.
[00379] Способ доставки представляющей интерес нуклеиновой кислоты в клетку субъекта может включать введение указанному субъекту зкДНК-вектора согласно данному изобретению, содержащего указанную представляющую интерес нуклеиновую кислоту. Кроме того, согласно данному изобретению предложен способ доставки представляющей интерес нуклеиновой кислоты в клетку нуждающегося в этом субъекта, включающий многократное введение зкДНК-вектора согласно данному изобретению, содержащего указанную нуклеиновую представляющую интерес кислоту. Поскольку вектор зкДНК, согласно данному изобретению, не индуцирует иммунного ответа, такая стратегия многократного введения не будет нарушать ответ иммунной системы хозяина против зкДНК-вектора согласно данному изобретению, в отличие от ситуации, которая наблюдается с заключенными в капсиды векторами.
[00380] Нуклеиновую кислоту (кислоты) зкДНК-вектора вводят в достаточных количествах для трансфекции клеток нужной ткани и для обеспечения достаточных уровней переноса и экспрессии генов без нежелательных побочных эффектов. Обычные и фармацевтически приемлемые пути введения включают, без ограничений, внутривенный (например, в липосомном составе), прямую доставку в выбранный орган (например, внутрипортальную доставку в печень), внутримышечный и другие парентеральные пути введения. Пути введения могут быть скомбинированы, если требуется.
[00381] ЗкДНК-вектор не ограничен одним видом зкДНК-вектора. Следовательно, согласно другому аспекту несколько зкДНК-векторов, содержащих разные экзогенные последовательности ДНК, могут быть одновременно или последовательно доставлены в целевые клетку, ткань, орган или организм субъекта. Соответственно, указанная стратегия может обеспечивать экспрессию нескольких генов. Доставка также может осуществляться неоднократно и, что важно для генной терапии в клинических условиях, с последующим увеличением или снижением доз, учитывая отсутствие иммунного ответа хозяина против капсида благодаря отсутствию вирусного капсида. Ожидается, что никакой реакции против капсида возникать не будет из-за отсутствия капсида.
[00382] Согласно данному изобретению также предложен способ лечения заболевания у субъекта, включающий введение в нуждающуюся в этом целевую клетку (в частности, в мышечную клетку или ткань) указанного субъекта терапевтически эффективного количества зкДНК-вектора, необязательно с фармацевтически приемлемым носителем. Хотя указанный зкДНК-вектор может быть введен в присутствии носителя, такой носитель не является необходимым. Предложенный зкДНК-вектор содержит представляющую интерес нуклеотидную последовательность, подходящую для лечения заболевания. В частности, указанный зкДНК-вектор может содержать требуемую экзогенную последовательность ДНК, функционально связанную с контрольными элементами, способными направлять транскрипцию требуемого полипептида, белка или олигонуклеотида, кодируемого указанной экзогенной последовательность ДНК при введении субъекту. Указанный зкДНК-вектор может быть введен любым подходящим путем, как описано выше и в других разделах данного документа.
XI. Способы лечения
[00383] Описанная в данном документе технология также демонстрирует способы получения, а также способы применения раскрытых зкДНК-векторов разнообразными путями, включая, например, варианты применения, методы, диагностические процедуры и/или схемы генной терапии ex situ, in vitro и in vivo.
[00384] Согласно данному изобретению предложен способ лечения заболевания или расстройства у субъекта, включающий введение в нуждающуюся в этом целевую клетку (например, мышечную клетку или ткань, или другой тип пораженных клеток) субъекта терапевтически эффективного количества зкДНК-вектора, необязательно с фармацевтически приемлемым носителем. Хотя указанный зкДНК-вектор может быть введен в присутствии носителя, такой носитель не является необходимым. Предложенный зкДНК-вектор содержит представляющую интерес нуклеотидную последовательность, подходящую для лечения заболевания. В частности, указанный зкДНК-вектор может содержать требуемую экзогенную последовательность ДНК, функционально связанную с контрольными элементами, способными направлять транскрипцию требуемого полипептида, белка или олигонуклеотида, кодируемого указанной экзогенной последовательность ДНК при введении субъекту. Указанный зкДНК-вектор может быть введен любым подходящим путем, как описано выше и в других разделах данного документа.
[00385] Любой трансген может быть доставлен с помощью зкДНК-векторов согласно описанию в данном документе. Трансгены, представляющие интерес, включают нуклеиновые кислоты, кодирующие полипептиды, или некодирующие нуклеиновые кислоты (например, РНКи, miR и т.п.), предпочтительно терапевтические (например, для медицинского, диагностического или ветеринарного применения) или иммуногенные (например, для вакцин) полипептиды.
[00386] Согласно некоторым вариантам реализации трансгены, которые требуется экспрессировать с помощью зкДНК-векторов, описанных в данном документе, экспрессируют или кодируют один или более полипептидов, пептидов, рибозимов, пептидных нуклеиновых кислот, киРНК, РНКи, антисмысловых олигонуклеотидов, антисмысловых полинуклеотидов, антител, антигенсвязывающих фрагментов, или любую комбинацию перечисленного.
[00387] В частности, указанный трансген может кодировать один или более терапевтических агентов, включая, но не ограничиваясь перечисленными, например, белок (белки), полипептид(ы), пептид(ы), фермент(ы), антитела, антигенсвязывающие фрагменты, а также варианты и/или активные фрагменты перечисленного, агонисты, антагонисты, миметики для применения при лечении, профилактике и/или облегчении одного или более симптомов заболевания, дисфункции, повреждения и/или расстройства. Согласно одному аспекту заболевание, дисфункция, травма, повреждение и/или расстройство представляет собой заболевание, дисфункцию, травму, повреждение и/или расстройство у человека.
[00388] Как отмечено в данном документе, указанный трансген может кодировать терапевтический белок или пептид, или последовательность терапевтической нуклеиновой кислоты или терапевтического агента, в том числе, но не ограничиваясь перечисленными, один или более агонистов, антагонистов, антиапоптотических факторов, ингибиторов, рецепторов, цитокинов, цитотоксинов, эритропоэтических агентов, гликопротеинов, факторов роста, рецепторов факторов роста, гормонов, рецепторов гормонов, интерферонов, интерлейкинов, рецепторов интерлейкина, факторов роста нервов, нейроактивных пептидов, рецепторов нейроактивных пептидов, протеаз, ингибиторов протеаз, протеиндекарбоксилаз, протеинкиназ, ингибиторов протеинкиназ, ферментов, связывающих рецепторы белков, транспортных белков или одного или более их ингибиторов, рецепторов серотонина или одного или более ингибиторов его поглощения, серпинов, рецепторов серпинов, супрессоров опухоли, диагностических молекул, химиотерапевтических агентов, цитотоксинов, или любой комбинации перечисленного.
[00389] Согласно некоторым вариантам реализации трансген в экспрессионной кассете, экспрессионного конструкта или зкДНК-векторе, описанном в данном документе, может быть кодон-оптимизирован для клетки-хозяина. В данном документе термин «кодон-оптимизированный» или «оптимизация кодонов» относится к процессу модификации последовательности нуклеиновой кислоты для усиления экспрессии в клетках представляющих интерес позвоночных, например, мыши или человека (например, гуманизированные последовательности), путем замены по меньшей мере одного, более чем одного или значимого числа кодонов нативной последовательности (например, прокариотической последовательности) на кодоны, которые чаще или наиболее часто используются в генах указанного позвоночного. У различных видов наблюдается конкретный систематический перевес определенных кодонов для конкретных аминокислот. Как правило, оптимизация кодонов не изменяет аминокислотную последовательность оригинального транслируемого белка. Оптимизированные кодоны могут быть определены с применением, например, платформы для оптимизации кодонов и индивидуализированного синтеза генов Gene Forge® от Aptagen (Aptagen, Inc.) или другой общедоступной базы данных.
[00390] В данном документе раскрыты композиции и составы зкДНК-векторов, которые включают один или более зкДНК-векторов согласно данному изобретению вместе с одним или несколькими фармацевтически приемлемыми буферами, разбавителями или вспомогательными веществами. Такие композиции могут быть включены в один или более диагностических или терапевтических наборов для диагностики, предотвращения, лечения или облегчения одного или более симптомов заболевания, повреждения, расстройства, травмы или дисфункции. Согласно одному аспекту указанное заболевание, повреждение, расстройство, травма или дисфункция представляют собой заболевание, повреждение, расстройство, травму или дисфункцию у человека.
[00391] Другой аспект технологии, описанной в данном документе, предусматривает способ обеспечения нуждающегося в этом субъекта диагностически или терапевтически эффективным количеством зкДНК-вектора, причем указанный способ включает введение в клетку, ткань или орган нуждающегося в этом субъекта некоторого количества зкДНК-вектора согласно описанию в данном документе в течение периода времени, обеспечивающего возможность экспрессии трансгена с зкДНК-вектора, с обеспечением таким образом субъекта диагностически или терапевтически эффективным количеством белка, пептида, нуклеиновой кислоты, экспрессируемых указанным зкДНК-вектором. Согласно дополнительному аспекту указанный субъект представляет собой человека.
[00392] Согласно другому аспекту технологии, описанной в данном документе, предложен способ диагностики, предотвращения, лечения или облегчения по меньшей мере одного или более симптомов заболевания, расстройства, нарушения функции, повреждения, аномального состояния или травмы у субъекта. В общем и широком смысле указанный способ включает, по меньшей мере, этап введения нуждающемуся в этом субъекту одного или более раскрытых зкДНК-векторов, в количестве и в течение времени, достаточных для диагностики, предотвращения, лечения или облегчения одного или более симптомов заболевания, расстройства, дисфункции, повреждения, аномального состояния или травмы у субъекта. Согласно дополнительному аспекту указанный субъект представляет собой человека.
[00393] Другой аспект представлен применением указанного зкДНК-вектора в качестве инструмента для лечения или снижения одного или более симптомов заболевания или болезненных состояний. Существует ряд наследственных заболеваний, для которых известны дефектные гены, и которые, как правило, делят на два класса: состояния недостаточности, как правило, недостаточности ферментов, которые обычно наследуются по рецессивному типу, и состояния дисбаланса, в которые могут быть вовлечены регуляторные или структурные белки, и которые, как правило, но не всегда, наследуются по доминантному типу. В случае заболеваний с состояниями недостаточности зкДНК-векторы могут применяться для доставки трансгенов с целью введения нормального гена в пораженные ткани для заместительной терапии, а также, согласно некоторым вариантам реализации, для создания моделей заболевания на животных с применением антисмысловых мутаций. В случае болезненных состояний разбалансированности, зкДНК-векторы могут применяться для создания болезненного состояния в модельной системе, которая затем может использоваться для противодействия указанному болезненному состоянию. Таким образом, зкДНК-векторы и способы, раскрытые в данном документе, позволяют лечить генетические заболевания. Согласно данному документу лечение болезненного состояния осуществляют путем частичного или полного устранения недостаточности или дисбаланса, которые вызывают заболевание или делают его более тяжелым.
[00394] В еще одном дополнительном аспекте зкДНК-вектор согласно описанию в данном документе может применяться для доставки гетерологичной нуклеотидной последовательности в ситуациях, когда требуется регуляция уровня трансгенной экспрессии (например, экспрессии трансгенов, кодирующих гормоны или факторы роста, согласно описанию в данном документе).
[00395] Соответственно, согласно некоторым вариантам реализации зкДНК-вектор, описанный в данном документе, может применяться для коррекции аномального уровня и/или функции генного продукта (например, отсутствия или дефекта белка), который приводит к заболеванию или расстройству. Указанный зкДНК-вектор может продуцировать функциональный белок и/или модифицировать уровни указанного белка для облегчения или снижения симптомов, возникающих в результате конкретного заболевания или расстройства, вызываемого отсутствием или дефектом белка или обеспечивать преимущество при указанном заболевании или расстройстве. Например, лечение недостаточности OTC может осуществляться путем продуцирования функционального фермента OTC; лечение гемофилии A и B может осуществляться путем модификации уровней фактора VIII, фактора IX и фактора X; лечение ФКУ может осуществляться путем модификации уровней фермента фенилаланингидроксилазы; лечение болезни Фабри или болезни Гоше может осуществляться путем продуцирования функциональной альфа-галактозидазы или бета-глюкоцереброзидазы, соответственно; лечение MLD или MPSII может осуществляться путем продуцирования функциональной арилсульфатазы A или идуронат-2-сульфатазы, соответственно; лечение кистозного фиброза может осуществляться путем продуцирования функционального регулятора трансмембранной проводимости при кистозном фиброзе; лечение болезни накопления гликогена может осуществляться путем восстановления функции функционального фермента глюкозо-6-фосфатазы; и лечение PFIC может осуществляться путем получения функциональных генов ATP8B1, ABCB11, ABCB4 или TJP2.
[00396] Согласно альтернативным вариантам реализации зкДНК-векторы согласно описанию в данном документе могут применяться для обеспечения клетки антисмысловой нуклеиновой кислотой in vitro или in vivo. Например, если трансген представляет собой молекулу РНКи, экспрессия антисмысловой нуклеиновой кислоты или РНКи в целевой клетке снижает уровень экспрессии конкретного белка указанной клеткой. Соответственно, трансгены, представляющие собой молекулы РНКи или антисмысловые нуклеиновые кислоты, могут быть введены для снижения уровня экспрессии конкретного белка у нуждающегося в этом субъекта. Антисмысловые нуклеиновые кислоты также можно вводить в клетки in vitro для регуляции физиологии клеток, например, для оптимизации систем культивирования клеток или тканей.
[00379] В некоторых вариантах реализации типичные примеры трансгенов, кодируемых зкДНК-вектором, включают, без ограничений: X, лизосомальные ферменты (например, гексозаминидазу A, ассоциированную с болезнью Тея-Сакса, или идуронатсульфатазу, ассоциированную с синдромом Гунтера/MPS II), эритропоэтин, ангиостатин, эндостатин, супероксиддисмутазу, глобин, лептин, каталазу, тирозингидроксилазу, а также цитокины (например, интерферон, β-интерферон, интерферон-γ, интерлейкин-2, интерлейкин-4, интерлейкин 12, гранулоцитарно-макрофагальный колониестимулирующий фактор, лимфотоксин и т.п.), пептидные факторы роста и гормоны (например, соматотропин, инсулин, инсулиноподобные факторы роста 1 и 2, тромбоцитарный фактор роста (ТРФ), эпидермальный фактор роста (ЭФР), фактор роста фибробластов (ФРФ), фактор роста нервов (ФРН), нейротрофический фактор-3 и 4, нейротрофический фактор головного мозга (BDNF), глиальный фактор роста (GDNF), трансформирующие факторы роста α и β; и т.п.), рецепторы (например, рецептор фактора некроза опухоли). В некоторых примерах вариантов реализации указанный трансген кодирует моноклональное антитело, специфическое в отношении одной или более требуемых мишеней. В некоторых примерах вариантов реализации указанный зкДНК-вектор кодирует более одного трансгена. В некоторых примерах вариантов реализации указанный трансген кодирует слитый белок, содержащий два разных представляющих интерес полипептида. Согласно некоторым вариантам реализации указанный трансген кодирует антитело, в том числе полноразмерное антитело или фрагмент антитела согласно определению в данном документе. Согласно некоторым вариантам реализации указанное антитело представляет собой последовательность антигенсвязывающего домена или вариабельного домена иммуноглобулина согласно определению в данном документе. Другие иллюстративные примеры трансгенных последовательностей кодируют продукты суицидных генов (тимидинкиназу, цитозиндезаминазу, дифтерийный токсин, цитохром P450, дезоксицитидинкиназу и фактор некроза опухоли), белки, придающие устойчивость к лекарственным средствам, применяемым в противораковой терапии, и продукты генов-супрессоров опухолей.
[00398] Согласно репрезентативному варианту реализации экспрессируемый указанным зкДНК-вектором трансген может применяться для лечения мышечной дистрофии у нуждающегося в этом субъекта, при этом указанный способ включает: введение эффективного для лечения, облегчения или предотвращения количества зкДНК-вектора, описанного в данном документе, причем указанный зкДНК-вектор содержит гетерологичную нуклеиновую кислоту, кодирующую дистрофин, минидистрофин, микродистрофин, пропептид миостатина, фоллистатин, растворимый рецептор активина II типа, ИФР-1, противовоспалительные полипептиды, такие как доминантный мутант I-каппа-B, саркоспан, утрофин, микродистрофин, ламинин-α2, α-саркогликан, β-саркогликан, γ-саркогликан, Δ-саркогликан, ИФР-1, антитело или фрагмент антитела против миостатина или пропептида миостатина, и/или РНКи, направленная против миостатина. Согласно конкретным вариантам реализации указанный зкДНК-вектор может быть введен в скелетную, диафрагмальную и/или сердечную мышцу согласно описанию в тексте данного документа.
[00399] В некоторых вариантах реализации указанный зкДНК-вектор может применяться для доставки трансгена в скелетную, сердечную или диафрагмальную мышцу, для продуцирования полипептида (например, фермента) или функциональной РНК (например, РНК-интерференции, микроРНК, антисмысловой РНК), которая в норме циркулирует в крови, или для системной доставки в другие ткани, для лечения, облегчения и/или предотвращения расстройства (например, метаболического расстройства, такого как диабет (например, инсулина), гемофилии (например, VIII), мукополисахаридозного расстройства (например, синдрома Слая, синдрома Гурлер, синдрома Шейе, синдрома Гурлер-Шейе, синдрома Гунтера, синдрома Санфилиппо A, B, C, D, синдрома Моркио, синдрома Марото-Лами и т.п.) или расстройства лизосомального накопления (такого как болезнь Гоше [глюкоцереброзидаза], болезнь Помпе [лизосомальная кислая альфа-глюкозидаза] или болезнь Фабри [альфа-галактозидаза A]) или расстройства накопления гликогена (такие как болезнь Помпе [лизосомальная кислая альфа-глюкозидаза]). Другие подходящие белки для лечения, облегчения и/или предотвращения метаболических расстройств описаны выше.
[00400] Согласно другим вариантам реализации зкДНК-вектор согласно описанию в данном документе может применяться для доставки трансгена в способе лечения, облегчения и/или предотвращения метаболического расстройства у нуждающегося в этом субъекта. Иллюстративные примеры метаболических расстройств и трансгенов, кодирующих полипептиды, описаны в данном документе. Необязательно, указанный полипептид секретируется (например, полипептид, который представляет собой секретируемый полипептид в естественном состоянии, или полипептид, сконструированный таким образом, чтобы обеспечить его секрецию, например, за счет достижения функциональной связи с секреторной сигнальной последовательностью, как известно специалистам в данной области техники).
[00401] Согласно другим вариантам реализации зкДНК-вектор согласно описанию в данном документе можно применять для лечения судорог, например, для снижения возникновения, частоты или тяжести судорог. Эффективность терапевтического лечения судорог может быть оценена на основании поведенческих признаков (например, дрожи, глазных или ротовых тиков) и/или электрографическими способами (большинство судорог характеризуются отличительными электрографическими аномалиями). Таким образом, зкДНК-вектор, описанный в данном документе, также можно применять для лечения эпилепсии, для которой характерны множественные припадки во временной перспективе. Согласно одному репрезентативному варианту реализации соматостатин (или его активный фрагмент) вводят в головной мозг с применением зкДНК-вектора согласно описанию в данном документе для лечения опухоли гипофиза. В соответствии с указанным вариантом реализации зкДНК-вектор согласно описанию в данном документе, кодирующий соматостатин (или его активный фрагмент), вводят в гипофиз путем микроинфузии. Аналогичным образом, такое лечение можно применять для лечения акромегалии (аномальной секреции гормона роста гипофизом). Последовательности нуклеиновой кислоты (например, № доступа GenBank J00306) и последовательность аминокислот (например, № доступа GenBank P01166; содержит процессированные активные пептиды соматостатин-28 и соматостатин-14) соматостатинов известны специалистам в данной области техники. Согласно конкретным вариантам реализации указанный зкДНК-вектор может кодировать трансген, который содержит секреторный сигнал, согласно описанию в патенте США № 7071172.
[00402] Другой аспект данного изобретения относится к применению зкДНК-вектора согласно описанию в данном документе для получения антисмысловой РНК, РНКи или другой функциональной РНК (например, рибозима) для системной доставки субъекту in vivo. Соответственно, согласно некоторым вариантам реализации указанный зкДНК-вектор может содержать трансген, который кодирует антисмысловую нуклеиновую кислоту, рибозим (например, согласно описанию в патенте США №5877022), РНК, которые влияют на опосредованный сплайсосомой транс-сплайсинг (см. Puttaraju et al., (1999) Nature Biotech. 17:246; патент США №6013487; патент США №6083702), интерферирующие РНК (РНКи), опосредующие сайленсинг генов (см. Sharp et al., (2000) Science 287:2431), или другие нетранслируемые РНК, такие как «гидовые» РНК (Gorman et al., (1998) Proc. Nat. Acad. Sci. USA 95:4929; патент США № 5869248, выданный Yuan с соавторами); и т.п.
[00403] Согласно некоторым вариантам реализации указанный зкДНК-вектор может дополнительно также содержать трансген, который кодирует репортерный полипептид (например, фермент, такой как зеленый флуоресцентный белок или щелочная фосфатаза). Согласно некоторым вариантам реализации трансген, который кодирует репортерный белок, подходящий для экспериментальных или диагностических целей, выбирают из любого из следующих трансгенов: β-лактамазы, β-галактозидазы (LacZ), щелочной фосфатазы, тимидинкиназы, зеленого флуоресцентного белка (GFP), хлорамфениколацетилтрансферазы (CAT), люциферазы и других генов, хорошо известных в данной области техники. Согласно некоторым аспектам зкДНК-векторы, содержащие трансген, кодирующие репортерный полипептид, могут быть использованы в диагностических целях или в качестве маркеров активности зкДНК-вектора у субъекта, которому их вводят.
[00404] Согласно некоторым вариантам реализации указанный зкДНК-вектор может содержать трансген или гетерологичную нуклеотидную последовательность, который(ая) обладает гомологией в отношении локуса на хромосоме хозяина и рекомбинирует с ним. Указанный подход может быть использован для коррекции генетического дефекта в клетке-хозяине.
XII. Введение
[00405] Согласно конкретным вариантам реализации может быть проведено более одного введения (например, два, три, четыре или более введений) для достижения желаемого уровня экспрессии гена на протяжении периода времени с интервалами различной продолжительности, например, ежедневно, еженедельно, ежемесячно, ежегодно и т.п.
[00406] Примеры способов введения зкДНК-вектора, описанного в данном документе, включают пероральное, ректальное, трансмукозальное, интраназальное, ингаляционное (например, в аэрозоле), буккальное (например, сублингвальное), вагинальное, интратекальное, внутриглазное, чрескожное, интраэндотелиальное введение, введение in utero (или in ovo), парентеральное (например, внутривенное, подкожное, внутрикожное, внутричерепное, внутримышечное [в том числе введение в скелетную, диафрагмальную и/или сердечную мышцу], внутриплевральное, интрацеребральное и внутрисуставное), местное (например, введение на поверхность кожи и слизистых оболочек, в том числе поверхности дыхательных путей, и чрескожное введение), внутрилимфатическое введение и т.п., а также прямую инъекцию в ткань или орган (например, в печень, глаз, скелетную мышцу, сердечную мышцу, диафрагмальную мышцу или головной мозг).
[00407] Введение указанного зкДНК-вектора субъекту может осуществляться на любом участке, включая, без ограничений, участок, выбранный из группы, состоящей из головного мозга, скелетной мышцы, гладкой мышцы, сердца, диафрагмы, эпителия дыхательных путей, печени, почки, селезенки, поджелудочной железы, кожи и глаза. Также может осуществляться введение зкДНК-вектора в опухоль (например, внутрь опухоли или лимфатического узла или рядом с ней). Наиболее подходящий путь в том или ином конкретном случае зависит от природы и тяжести состояния, лечение, облегчение и/или предотвращение которого проводят, и от природы конкретного используемого зкДНК-вектора. Кроме того, зкДНК позволяет вводить более одного трансгена в единственном векторе или в нескольких зкДНК-векторах (например, в коктейле зкДНК).
[00408] А. Диапазоны доз
[00409] Необязательно могут быть использованы анализы in vivo и/или in vitro для определения оптимальных диапазонов дозировок для применения. Точная доза для использования в композиции будет также зависеть от пути введения и серьезности состояния и должна определяться на основании решения специалиста в данной области техники и с учетом обстоятельств каждого субъекта. Эффективные дозы могут быть экстраполированы из кривых доза-ответ, полученных из тест-систем in vitro или на животных моделях.
[00410] ЗкДНК-вектор вводят в достаточных количествах для трансфекции клеток требуемой ткани и обеспечения достаточных уровней переноса генов и экспрессии без чрезмерных нежелательных явлений. Стандартные и фармацевтически приемлемые пути введения включают, не ограничиваясь перечисленными, описанные ниже в разделе «Введение», такие как прямая доставка в выбранный орган (например, интрапортальная доставка в печень), пероральная доставка, ингаляция (в том числе интраназальная и внутритрахеальная доставка), интраокулярная, внутривенная, внутримышечная, подкожная, внутрикожная, внутриопухолевая доставка; и другие парентеральные пути введения. Пути введения могут быть скомбинированы, если требуется.
[00411] Доза количества зкДНК-вектора, необходимая для достижения определенного «терапевтического эффекта», будет варьировать в зависимости от нескольких факторов, в том числе, но не ограничиваясь перечисленными: от пути введения нуклеиновой кислоты, уровня экспрессии гена или РНК, необходимого для достижения терапевтического эффекта, конкретного заболевания или расстройства, лечение которого проводят, и стабильности гена (генов), РНК-продукта (продуктов) или итогового экспрессируемого белка (белков). Специалист в данной области техники легко сможет определить диапазон доз зкДНК-вектора для лечения пациента, имеющего конкретное заболевание или расстройство, на основе вышеупомянутых факторов, а также других факторов, хорошо известных в данной области техники.
[00412] Режим дозирования может быть скорректирован для обеспечения оптимального терапевтического ответа. Например, олигонуклеотид может вводиться неоднократно, например, ежедневно может вводиться несколько доз, или доза может быть пропорционально снижена с учетом диктуемой терапевтической ситуацией необходимости. Специалист в данной области техники сможет легко определить подходящие дозы и схемы введения олигонуклеотидов согласно данному изобретению, как для введения в клетки, так и для введения субъектам.
[00413] «Терапевтически эффективная доза» находится в относительно широком диапазоне значений, который может быть определен в ходе клинических испытаний и зависит от конкретного применения (нервные клетки потребуют очень незначительных количеств, тогда как для системных инъекций потребуются значительные количества). Например, для прямой инъекции in vivo в скелетную или сердечную мышцу человека, величина терапевтически эффективной дозы будет составлять от примерно 1 мкг до 100 г зкДНК-вектора. Если для доставки зкДНК-вектора используются экзосомы или микрочастицы, то терапевтически эффективная доза может быть определена экспериментально, но ожидается, что она должна доставить от 1 мкг до примерно 100 г вектора.
[00414] Приготовление фармацевтически приемлемых вспомогательных веществ и растворов носителей хорошо известно специалистам в данной области, как и разработка подходящих режимов дозирования и лечения для применения конкретных композиций, описанных в данном документе, в различных схемах лечения.
[00415] Для трансфекции in vitro эффективное количество зкДНК-вектора, которое нужно доставить в клетки (1×106 клеток), составляет порядка 0,1-100 мкг зкДНК-вектора, предпочтительно от 1 до 20 мкг, и более предпочтительно от 1 до 15 мкг, или от 8 до 10 мкг. Для зкДНК-векторов большего размера требуются более высокие дозы. Если используются экзосомы или микрочастицы, эффективная доза in vitro может быть определена экспериментально, однако она должна обеспечивать доставку, в общем, такого же количества зкДНК-вектора.
[00416] Лечение может включать введение разовой дозы или нескольких доз. Согласно некоторым вариантам реализации субъекту может быть введено более одной дозы; фактически, дозы могут вводиться многократно по мере необходимости, поскольку зкДНК-вектор не вызывает у хозяина иммунного ответа против капсида ввиду отсутствия вирусного капсида. Следовательно, специалист в данной области может легко определить подходящее количество доз. Количество вводимых доз может составлять, например, примерно 1-100, предпочтительно 2-20 доз.
[00417] Без связи с какой-либо конкретной теорией отметим, что отсутствие типичного противовирусного иммунного ответа при введении зкДНК-вектора согласно описанию в данном документе (т.е. отсутствие капсидных компонентов) позволяет вводить зкДНК-вектор хозяину неоднократно. Согласно некоторым вариантам реализации число введений гетерологичной нуклеиновой кислоты субъекту составляет от 2 до 10 раз (например, 2, 3, 4, 5, 6, 7, 8, 9 или 10 раз). Согласно некоторым вариантам реализации зкДНК-вектор доставляют субъекту более 10 раз.
[00418] Согласно некоторым вариантам реализации дозу зкДНК-вектора вводят субъекту не более одного раза в календарный день (например, за 24-часовой период). Согласно некоторым вариантам реализации дозу зкДНК-вектора вводят субъекту не более одного раза в 2, 3, 4, 5, 6, или 7 календарных дней. Согласно некоторым вариантам реализации дозу зкДНК-вектора вводят субъекту не более одного раза в календарную неделю (например, 7 календарных дней). Согласно некоторым вариантам реализации дозу зкДНК-вектора вводят субъекту не чаще одного раза в две недели (например, один раз за период, равный двум календарным неделям). Согласно некоторым вариантам реализации дозу зкДНК-вектора вводят субъекту не более одного раза в календарный месяц (например, один раз в 30 календарных дней). Согласно некоторым вариантам реализации дозу зкДНК-вектора вводят субъекту не более одного раза в шесть календарных месяцев. Согласно некоторым вариантам реализации дозу зкДНК-вектора вводят субъекту не более одного раза в календарный год (например, за 365 дней или 366 дней в высокосном году).
[00419] В. Единичные лекарственные формы
[00420] В некоторых вариантах реализации фармацевтические композиции могут быть удобно представлены в виде единичной лекарственной формы. Единичная лекарственная форма обычно будет адаптирована к одному или нескольким конкретным путям введения фармацевтической композиции. Согласно некоторым вариантам реализации указанная единичная лекарственная форма адаптирована для введения путем ингаляции. Согласно некоторым вариантам реализации указанная единичная лекарственная форма адаптирована для введения с помощью испарителя. Согласно некоторым вариантам реализации указанная единичная лекарственная форма адаптирована для введения с помощью небулайзера. Согласно некоторым вариантам реализации единичная лекарственная форма адаптирована для введения с помощью генератора аэрозоля. Согласно некоторым вариантам реализации единичная лекарственная форма адаптирована для перорального введения, для буккального введения или для сублингвального введения. Согласно некоторым вариантам реализации единичная лекарственная форма адаптирована для внутривенного, внутримышечного или подкожного введения. Согласно некоторым вариантам реализации единичная лекарственная форма адаптирована для интратекального или интрацеребровентрикулярного введения. Согласно некоторым вариантам реализации фармацевтическая композиция представлена составом для местного введения. Количество активного ингредиента, которое может быть скомбинировано с материалом носителя для получения единичной лекарственной формы, обычно представлено таким количеством указанного соединения, которое обеспечивает терапевтический эффект.
XIII. Различные применения
[00421] Композиции и зкДНК-векторы, представленные в данном документе, можно использовать для доставки трансгена с различными целями, как описано выше. Согласно некоторым вариантам реализации указанный трансген кодирует белок или функциональную РНК, которая предназначена для использования в исследовательских целях, например, для создания соматической модели на трансгенных животных, несущих трансген, например, для изучения функции продукта трансгена. В другом примере указанный трансген кодирует белок или функциональную РНК, которая предназначена для использования с целью создания модели заболевания на животных.
[00422] Согласно некоторым вариантам реализации указанный трансген кодирует один или более пептидов, полипептидов или белков, которые подходят для лечения, облегчения или предотвращения болезненных состояний у субъекта-млекопитающего. Указанный трансген может быть перенесен в организм пациента (например, экспрессироваться у пациента) в количестве, достаточном для лечения заболевания, ассоциированного с пониженной экспрессией, отсутствием экспрессии или дисфункцией указанного гена.
[00423] Согласно некоторым вариантам реализации предусмотрено применение указанных зкДНК-векторов в способах диагностики и скрининга, где трансген транзиентно или стабильно экспрессируют в системе культуры клеток или, как вариант, в модели на трансгенных животных.
[00424] Согласно другому аспекту описанная в данном документе технология обеспечивает способ трансдукции популяции клеток млекопитающих. В общем и широком смысле указанный способ включает, по меньшей мере, этап введения в одну или более клеток популяции композиции, которая содержит эффективное количество одной или более зкДНК согласно описанию в данном документе.
[00425] Кроме того, согласно данному изобретению предложены композиции, а также терапевтические и/или диагностические наборы, которые включают один или более раскрытых зкДНК-векторов или одну или более раскрытых композиций зкДНК, в составах, полученных с использованием одного или более дополнительных ингредиентов, или подготовленных вместе с одной или более инструкциями по их применению.
ПРИМЕРЫ
[00426] Приведенные ниже примеры предназначены для иллюстрации, а не для ограничения.
ПРИМЕР 1: Конструирование зкДНК-векторов
[00427] Описано получение зкДНК-векторов с использованием матрицы полинуклеотидного конструкта. Например, матрица полинуклеотидного конструкта, используемая для получения зкДНК-векторов согласно данному изобретению, может представлять собой зкДНК-плазмиду, зкДНК-бакмиду и/или зкДНК-бакуловирус. Без ограничения теорией, в пермиссивной клетке-хозяине в присутствии, например, Rep, матрица полинуклеотидного конструкта, содержащая два ITR и экспрессионный конструкт, где по меньшей мере один из ITR модифицирован, реплицируется с получением зкДНК-векторов. Получение зкДНК-вектора проходит в два этапа: во-первых, вырезание («спасение») матрицы из остова матрицы (например, зкДНК-плазмиды, зкДНК-бакмиды, генома зкДНК-бакуловируса и т.п.) посредством белков Rep, и, во-вторых, опосредованная Rep репликация вырезанного зкДНК-вектора.
[00428] Примером способа получения зкДНК-векторов является зкДНК-плазмида согласно описанию в данном документе. На Фиг. 1А и 1В матрица полинуклеотидного конструкта каждой из зкДНК-плазмид включает как левый мутированный ITR, так и правый мутированный ITR, с расположенными между указанными ITR следующими последовательностями: (i) энхансера/промотора; (ii) сайта клонирования трансгена; (iii) посттранскрипционного элемента ответа (например, посттранскрипционный регуляторный элемент вируса гепатита сурков (WPRE)); и (iv) сигнала полиаденилирования (например, из гена бычьего гормона роста (BGHpA)). Также между всеми компонентами были введены уникальные сайты распознавания рестрикционными эндонуклеазами (R1-R6) (показаны на Фиг. 1A и Фиг. 1B) для облегчения введения новых генетических компонентов в конкретные сайты конструкта. Сайты ферментов R3 (PmeI) GTTTAAAC (SEQ ID NO: 7) и R4 (PacI) TTAATTAA (SEQ ID NO: 542) встраивают в сайт клонирования для введения открытой рамки считывания трансгена. Указанные последовательности клонировали в плазмиду pFastBac HT B, полученную от ThermoFisher Scientific.
[00429] Вкратце, ряд зкДНК-векторов получали из конструктов зкДНК-плазмид, показанных в Таблице 12, используя способ, показанный на Фиг. 4A-4C. В Таблице 12 указано количество соответствующих полинуклеотидных последовательностей для каждого компонента, включая последовательности, активные в качестве сайта белка репликации (RPS) (например, сайта связывания Rep) на любом конце промотора, функционально связанного с трансгеном. Номера в Таблице 12 относятся к SEQ ID NO в данном документе, соответствующим последовательностям каждого компонента.
[00430] Таблица 12: Типичные зкДНК-конструкты
[00431] Согласно некоторым вариантам реализации конструкт для получения зкДНК-векторов содержит промотор, который представляет собой регуляторный переключатель согласно описанию в данном документе, например, индуцируемый промотор. Другие конструкты использовали для создания векторов зкДНК, например, конструкт-10, конструкт-11, конструкт-12 и конструкт-13 (см., например, Таблицу 14A), которые содержат промотор MND или HLCR, функционально связанный с трансгеном люциферазы.
[00432] Продуцирование зкДНК-бакмид:
[00433] Как изображено на Фиг. 4А, компетентные клетки DH10Bac (компетентные клетки MAX EFFICIENCY® DH10Bac™, Thermo Fisher) трансформировали либо тестируемой, либо контрольной плазмидой согласно протоколу в соответствии с инструкциями производителя. Индуцировали рекомбинацию между плазмидой и бакуловирусным челночным вектором в клетках DH10Bac для получения рекомбинантных зкДНК-бакмид. Рекомбинантные бакмиды отбирали путем скрининга с положительным отбором на основе сине-белого скрининга в E.coli (маркер Φ80dlacZΔM15 обеспечивает α-комплементацию гена β-галактозидазы из бакмидного вектора) на чашке с бактериальным агаром, содержащим X-gal и IPTG с антибиотиками, для отбора трансформантов и поддержания бакмиды и транспозазных плазмид. Белые колонии, образующиеся в результате транспозиции, которая разрушает индикаторный ген β-галактозида, отбирали и культивировали в 10 мл среды.
[00434] Рекомбинантные зкДНК-бакмиды выделяли из E. coli и трансфицировали ими клетки насекомых Sf9 или Sf21 с применением FugeneHD для получения инфекционного бакуловируса. Адгезивные клетки насекомых Sf9 или Sf21 культивировали в 50 мл среды в колбах T25 при 25 °С. Через 4 дня культуральную среду (содержащую вирус P0) отделяли от клеток, фильтровали через фильтр с размером пор 0,45 мкм, отделяя инфекционные бакуловирусные частицы от клеток или клеточного дебриса.
[00435] Необязательно, первое поколение бакуловируса (P0) амплифицировали путем инфицирования наивных клеток насекомых Sf9 или Sf21 в 50-500 мл среды. Клетки культивировали в суспензионных культурах в инкубаторе с орбитальным шейкером при 130 об/мин при 25°С, отслеживая диаметр и жизнеспособность клеток до тех пор, пока клетки не достигнут диаметра 18-19 нм (от диаметра наивных клеток, равного 14-15 нм) и плотности около 4,0E+6 клеток/мл. Через 3-8 суток после инфицирования бакуловирусные частицы P1 в среде собирали после центрифугирования для удаления клеток и дебриса, с последующей фильтрацией через фильтр с размером пор 0,45 мкм.
[00436] ЗкДНК-бакуловирус, содержащий тестируемые конструкты, собирали и определяли инфекционную активность или титр бакуловируса. В частности, 4 × 20 мл культуры клеток Sf9 с плотностью 2,5E+6 клеток/мл обрабатывали бакуловирусом P1 в следующих разведениях: 1/1000, 1/10000, 1/50000, 1/100000, и инкубировали при 25-27°C. Инфективность определяли ежедневно на протяжении 4-5 дней по показателю увеличения диаметра клеток и остановке клеточного цикла, а также изменению жизнеспособности клеток.
[00437] Ссылаясь на Фиг. 4A, «Rep-плазмида», которая содержит единственный белок Rep (см., например, Фиг. 8A), была продуцирована в экспрессионном векторе pFASTBAC™-Dual (ThermoFisher).
[00438] Указанной Rep-плазмидой трансформировали компетентные клетки DH10Bac (MAX EFFICIENCY® DH10Bac™ Competent Cells (Thermo Fisher)), следуя предоставленному производителем протоколу. Индуцировали рекомбинацию между Rep-плазмидой и бакуловирусным челночным вектором в клетках DH10Bac для получения рекомбинантных бакмид («Rep-бакмид»). Рекомбинантные бакмиды отбирали путем положительного отбора, который включал сине-белый скрининг в E. coli (маркер Φ80dlacZΔM15 обеспечивает α-комплементацию гена β-галактозидазы из бакмидного вектора) на чашке с бактериальным агаром, содержащим X-gal и IPTG. Изолированные белые колонии собирали и инокулировали в 10 мл селективной среды (канамицин, гентамицин, тетрациклин в бульоне LB). Рекомбинантные бакмиды (Rep-бакмиды) выделяли из E.coli, и указанными Rep-бакмидами трансфицировали клетки насекомых Sf9 или Sf21 для получения инфекционного бакуловируса.
[00439] Клетки насекомых Sf9 или Sf21 культивировали в 50 мл среды в течение 4 дней и выделяли из культуры инфекционный рекомбинантный бакуловирус («Rep-бакуловирус»). Необязательно, Rep-бакуловирус первого поколения (P0) амплифицировали путем инфицирования необработанных клеток насекомых Sf9 или Sf21 и культивирования в 50-500 мл среды. Через 3-8 дней после инфицирования бакуловирусные частицы P1 в среде собирали, отделяя клетки путем центрифугирования или фильтрации, или используя другой способ фракционирования. Собирали Rep-бакуловирус и определяли инфекционную активность бакуловируса. В частности, 4 x 20 мл культур клеток Sf9 при плотности 2,5x106 клеток/мл обрабатывали бакуловирусом P1 в следующих разведениях: 1/1000, 1/10000, 1/50000, 1/100000, и инкубировали. Инфективность определяли ежедневно на протяжении 4-5 дней по показателю увеличения диаметра клеток и остановке клеточного цикла, а также изменению жизнеспособности клеток.
[00440] Получение и характеризация зкДНК-вектора
[00441] Как показано на Фиг. 4B, затем добавляли культуральную среду для клеток насекомых Sf9, содержащую либо (1) образец, содержащий зкДНК-бакмиду или зкДНК-бакуловирус, либо (2) Rep-бакуловирус, описанные выше, в свежую культуру клеток Sf9 (2,5E+6 клеток/мл, 20 мл) в соотношении 1:1000 и 1:10000, соответственно. Затем клетки культивировали при 130 об/мин при 25 °С. Через 4-5 дней после коинфицирования детектировали диаметр и жизнеспособность клеток. Когда диаметр клеток достигал 18-20 нм при жизнеспособности, составляющей приблизительно 70-80%, клеточные культуры центрифугировали, среду удаляли и собирали клеточные осадки. Клеточные осадки сначала ресуспендировали в достаточном объеме водной среды - либо воды, либо буфера. ЗкДНК-вектор выделяли из клеток и очищали с использованием протокола очистки Qiagen MIDI PLUS™ (Qiagen, обработка 0,2 мг массы клеточного осадка на колонку).
[00442] Значения выхода для зкДНК-векторов, продуцированных и очищенных из клеток насекомых Sf9, первоначально определяли на основании поглощения в УФ-диапазоне при 260 нм. Выходы различных векторов зкДНК, определенные на основе УФ-поглощения, представлены ниже в Таблице 13.
[00443] Таблица 13: Выход векторов зкДНК из типичных конструктов.
(Диаметр в микрометрах)
Жизнеспособность: 53,3%
Диаметр: 18,4
[00444] Оценка зкДНК-векторов может быть проведена путем идентификации с применением электрофореза на агарозном геле в нативных или денатурирующих условиях, как проиллюстрировано на Фиг. 4D, где наблюдается (a) присутствие характеристических полос, мигрирующих с вдвое большим размером на денатурирующих гелях по сравнению с нативными гелями после расщепления рестрикционной эндонуклеазой и гель-электрофоретического анализа, и (b) наличие полос мономера и димера (2×) на денатурирующих гелях для нерасщепленного материала, что характерно для присутствия зкДНК-вектора.
[00445] Структуры выделенных зкДНК-векторов дополнительно анализировали путем расщепления ДНК, полученной из коинфицированных клеток Sf9 (согласно описанию в данном документе) рестрикционными эндонуклеазами, выбранными на основании a) присутствия только одного сайта разрезания в зкДНК-векторах, и b) получения итоговых фрагментов достаточно большого размера, чтобы они были четко видны при фракционировании на 0,8% денатурирующем агарозном геле (>800 п.о.). Как продемонстрировано на Фиг. 4E, линейные ДНК-векторы с прерывистой структурой и зкДНК-вектор с линейной и непрерывной структурой можно различить по размеру продуктов реакции - например, ожидается, что ДНК-вектор с прерывистой структурой будет давать фрагменты размером 1 т.п.о. и 2 т.п.о., а неинкапсидированный вектор с непрерывной структурой - фрагменты размером 2 т.п.о. и 4 т.п.о.
[00446] Таким образом, для того, чтобы качественным образом продемонстрировать, что выделенные зкДНК-векторы имеют ковалентно замкнутые концы, как требует их определение, образцы расщепляли рестрикционной эндонуклеазой, идентифицированной в контексте последовательности конкретного ДНК-вектора, как эндонуклеаза с одним сайтом рестрикции, предпочтительно, обеспечивающая образование двух продуктов расщепления неравного размера (например, 1000 п.о. и 2000 п.о.). После расщепления и электрофореза на денатурирующем геле (который разделяет две комплементарные цепи ДНК), линейная ДНК, не являющаяся ковалентно замкнутой, будет разделяться на фрагменты размером 1000 п.о. и 2000 п.о., в то время как ковалентно замкнутая ДНК (т.е. зкДНК-вектор) будет разделяться на фрагменты в 2 раза большего размера (2000 п.о. и 4000 п.о.), так как две цепи ДНК связаны, а после разворачивания будут иметь вдвое большую длину (хотя и будут одноцепочечными). Кроме того, при расщеплении мономерных, димерных и n-мерных форм ДНК-векторов все они будут разделяться на фрагменты одинаковых размеров из-за связывания концов мультимерных ДНК-векторов (см. Фиг. 4D).
[00447] Фиг. 5 представляет собой иллюстративную картину денатурирующего геля с векторами зкДНК следующим образом: конструкт-1, конструкт-2, конструкт-3, конструкт-4, конструкт-5, конструкт-6, конструкт-7 и конструкт-8 (все описанные в Таблице 12 выше), с (+) или без (-) расщепления эндонуклеазой. Каждый вектор зкДНК от конструкта-1 до конструкта-8 давал две полосы (*) после эндонуклеазной реакции. Размеры их двух полос, определенные на основе размера маркера, представлены в нижней части рисунка. Размеры полос подтверждают, что каждый из векторов зкДНК, полученных из плазмид, содержащих от конструкта-1 до конструкта-8, имеет непрерывную структуру.
[00448] В данном документе фраза «анализ для идентификации ДНК-векторов с помощью электрофореза на агарозном геле в условиях нативного и денатурирующего геля» относится к анализу для оценки наличия замкнутых концов зкДНК путем проведения расщепления рестрикционной эндонуклеазой с последующей электрофоретической оценкой продуктов расщепления. Один пример такого анализа приведен ниже, хотя специалисту в данной области техники будет понятно, что возможно осуществление многих известных в данной области техники вариантов указанного примера. Рестрикционную эндонуклеазу выбирают таким образом, чтобы это был фермент, выполняющий единственный разрез представляющего интерес зкДНК-вектора с образованием продуктов, длина которых составляет приблизительно 1/3× и 2/3× длины ДНК-вектора. Это обеспечивает разделение полос как на нативном, так и на денатурирующем гелях. Перед денатурацией важно удалить буфер из образца. Набор для очистки продуктов ПЦР от Qiagen или обессоливающие «центрифужные колонки», например, колонки GE HEALTHCARE ILUSTRA™ MICROSPIN™ G-25, являются примерами известных в данной области техники средств для расщепления эндонуклеазами. Указанный анализ включает, например, i) расщепление ДНК подходящей рестрикционной эндонуклеазой (эндонуклеазами), 2) внесение, например, в набор для очистки продуктов ПЦР от Qiagen, элюирование дистиллированной водой, iii) добавление 10x денатурирующего раствора (10× = 0,5 М NaOH, 10 мМ EDTA), добавление 10x не забуференного раствора красителя и проведение анализа вместе с маркерами ДНК, полученными путем добавления 10x денатурирующего раствора к 4х, на 0,8-1,0% геле, предварительно инкубированном с 1 мМ EDTA и 200 мМ NaOH для обеспечения однородности концентрации NaOH в геле и гелевой камере, и проведение геля в присутствии 1× денатурирующего раствора (50 мМ NaOH, 1 мМ EDTA). Специалисту в данной области техники будет понятно, какое напряжение следует использовать для проведения электрофореза в зависимости от размеров и желаемого времени получения результатов. После электрофореза гели подсушивают и нейтрализуют в 1× TBE или TAE и переносят в дистиллированную воду или 1× TBE/TAE с 1× SYBR Gold. Затем полосы могут быть визуализированы, например, с применением красителя SYBR® Gold Nucleic Acid Gel Stain от Thermo Fisher (концентрат 10 000X в ДМСО) и эпифлуоресцентного света (синего) или УФ (312 нм).
[00449] Чистота полученного зкДНК-вектора может быть оценена с применением любого известного в данной области техники способа. Согласно одному неограничивающему примеру способа вклад зкДНК-плазмиды в общее поглощение УФ образцом может быть рассчитан путем сравнения интенсивности флуоресцентности зкДНК-вектора и стандарта. Например, если на основании поглощения УФ определено, что на гель загружено 4 мкг зкДНК-вектора, а интенсивность флуоресцентности зкДНК-вектора эквивалентна полосе 2 т.п.о. при известной массе 1 мкг, значит, масса зкДНК-вектора равна 1 мкг, и зкДНК-вектор составляет 25% от общего количества поглощающего УФ материала. Затем строят график зависимости интенсивности полосы на геле от вычисленного введенного количества, которому соответствует полоса - например, если общее количество зкДНК-вектора соответствует 8 т.п.о., а вырезанная сравнительная полоса соответствует 2 т.п.о., то интенсивность указанной полосы на графике будет соответствовать 25% от общего введенного количества, что в данном случае составит 0,25 мкг при введении 1,0 мкг. Используя титрование плазмиды зкДНК-вектора для построения стандартной кривой, рассчитывают количество для полосы зкДНК-вектора по уравнению линии регрессии, которое затем можно использовать для определения процента от общего введенного количества, представленного зкДНК-вектором, или процента чистоты.
ПРИМЕР 2: получение вирусной ДНК в зкДНК клетках
[00450] Векторы зкДНК были также получены из конструктов 11, 12, 13 и 14, показанных в Таблице 14A. ЗкДНК-плазмиды, содержащие конструкты 11-14, были получены методами молекулярного клонирования, хорошо известными в данной области. Плазмиды в Таблице 14A были сконструированы с WPRE, содержащим SEQ ID NO: 8, за которым следует BGHpA, содержащий SEQ ID NO: 9, в 3'-нетранслируемом участке между трансгеном и правым ITR.
Таблица 14A
[00451] Векторный остов для конструктов 11-14 выглядит следующим образом: (i) asymITR-MND -люцифераза-wPRE-BGH-polyA-ITR в pFB-HTb (конструкт-11), (ii) ITR-MND-люцифераза -wPRE-BGH-polyA-asymITR в pFB-HTb (конструкт-12), (iii) asymITR-HLCR-AAT-luc-wPRE(O)-BGH-polyA-ITR в pFB-HTb (конструкт-13); и ITR -HLCR-AAT-luc-wPRE(O)-BGH-polyA-asymITR в pFB-HTb (конструкт-14), каждый конструкт имеет по меньшей мере один асимметричный ITR относительно друг друга. Эти конструкты также содержат одну или несколько из следующих последовательностей: wPRE0 (SEQ ID NO: 72) и последовательность BGH-PolyA (SEQ ID NO: 73), или последовательности, которые по меньшей мере 85%, или по меньшей мере 90%, или по меньшей мере 95%. идентичны указанным последовательностям.
[00452] Затем проводили получение вектора зкДНК в соответствии с процедурой, показанной на Фиг. 4A-4C, например, (а) получение рекомбинантной зкДНК-бакмидной ДНК и трансфекция клеток насекомых рекомбинантной зкДНК-бакмидной ДНК; (b) получение исходного материала P1 (низкого титра), исходного раствора P2 (высокого титра) и определение титра вируса с помощью количественной ПЦР для получения 5 мл, >1E+7 бляшкообразующих или инфекционных единиц «БОЕ» на мл бакуловирусного объема, согласно сертификату анализа бакуловирусного объема. Выделение вектора зкДНК осуществляли путем совместного инфицирования 50 мл клеток насекомых бакуловирусным объемом, для последующих пар инфицирования: Rep-bacmid, как описано в данном документе, и по меньшей мере, одним из следующих конструктов: конструкт-11, конструкт-12, конструкт-13 и конструкт-14. Выделение вектора зкДНК проводили с использованием набора QIAGEN Plasmid Midi Kit для получения очищенного ДНК-материала для дальнейшего анализа. Таблица 14B и Таблица 14C показывают выход (определенный по показателям OD) вектора зкДНК, полученного из конструктов 11-14.
[00453] Таблица 14B: Выход (определенный по показателям OD) типичных векторов зкДНК, полученных из конструктов 11-14.
[00454] В Таблице 14C показано количество материала ДНК, полученного (определено по показателям OD) с использованием конструктов 12 и 14 из Таблицы 14C. Выход материала общей ДНК был приемлемым по сравнению с типичными выходами около 3 мг/л материала ДНК из способа в вышеизложенного Примера 1 (Таблица 13).
ПРИМЕР 3: экспрессирование зкДНК-вектором трансгена люциферазы in vitro
[00455] Конструкты получали путем введения открытой рамки считывания, кодирующей репортерный ген люциферазы, в сайт клонирования плазмидных конструктов зкДНК: конструкт-1, конструкт-3, конструкт-5 и конструкт-7. ЗкДНК-плазмиды (см. выше в Таблице 12), включая кодирующую последовательность люциферазы, называются плазмидным конструктом 1-Luc, c плазмидным конструктом-3-Luc, плазмидном конструктом -5-Luc и плазмидным конструктом 7-Luc соответственно.
[00456] Клетки HEK293 культивировали и трансфицировали 100 нг, 200 нг или 400 нг плазмидных конструктов 1, 3, 5 и 7, с применением FUGENE® (Promega Corp.) в качестве агента для трансфекции. Экспрессию люциферазы из каждой из плазмид определяли на основе активности люциферазы в каждой культуре клеток, и результаты представлены на Фиг. 6А. Люциферазная активность не была обнаружена в необработанных контрольных клетках («Необработанные») или в клетках, обработанных одним фугеном («Fugene»), что подтверждает, что люциферазная активность является результатом экспрессии генов из плазмид. Как показано на Фиг. 6A и Фиг. 6B, устойчивая экспрессия люциферазы была обнаружена в конструктах 1 и 7. Экспрессия из конструкта-7 экспрессировала люциферазу с обнаружением дозозависимого увеличения активности люциферазы.
[00457] Рост и жизнеспособность клеток, трансфицированных каждой из плазмид, также были определены и представлены на Фиг. 7A и Фиг. 7В. Рост клеток и жизнеспособность трансфицированных клеток существенно не различались между разными группами клеток, обработанными разными конструктами.
[00458] Соответственно, активность люциферазы, измеренная в каждой группе и нормализованная на основе роста и жизнеспособности клеток, не отличалась от активности люциферазы без нормализации. ЗкДНК-плазмида с конструктом 1-Luc показала наиболее устойчивую экспрессию люциферазы с нормализацией или без нее.
[00459] Таким образом, данные, представленные на Фиг. 6A, 6B, 7A и 7B демонстрируют, что конструкт-1, содержащий от 5' до 3'- WT-ITR (SEQ ID NO: 51), промотор CAG (SEQ ID NO: 3), сайт клонирования R3/R4 (SEQ ID NO: 7), WPRE (SEQ ID NO: 8), BGHpA (SEQ ID NO: 9) и модифицированный ITR (SEQ ID NO: 2) является эффективным при получении вектора зкДНК, который может экспрессировать белок трансгена с зкДНК вектора.
ПРИМЕР 4: Экспрессия трансгенного белка люциферазы in vivo с зкДНК-векторов.
[00460] In vivo экспрессию белка трансгена с зкДНК-векторов, продуцированных с конструктов 1-8, описанных выше, оценивали на мышах. ЗкДНК-вектор, полученный из конструкта-1 зкДНК-плазмиды (как описано в Таблице 12), был протестирован и продемонстрировал продолжительную и устойчивую трансгенную экспрессию люциферазы в модели на мышах после гидродинамической инъекции конструкта зкДНК без липосом и повторного введения дозы (на день 28) и продолжительность (до дня 42) зкДНК экзогенной люциферазы светлячков. В различных экспериментах экспрессия люциферазы в выбранных векторах зкДНК оценивается in vivo, при этом векторы зкДНК содержат трансген люциферазы и по меньшей мере один модифицированный ITR, выбранный из любого, показанного в Таблицах 10A-10B, или ITR, содержащий по меньшей мере одну последовательность, показанную на Фиг. 26A-26B.
[00461] Экспрессия люциферазы in vivo: самцам мышей CD-1 IGS в возрасте 5-7 недель (Charles River Laboratories) вводят 0,35 мг/кг зкДНК-вектора, экспрессирующего люциферазу, в объеме 1,2 мл путем внутривенного (в/в) гидродинамического введения в хвостовую вену на день 0.. Экспрессию люциферазы оценивают с помощью визуализации IVIS в дни 3, 4, 7, 14, 21, 28, 31, 35 и 42. Вкратце, мышам внутрибрюшинной вводят 150 мг/кг субстрата люциферина, а затем оценивают люминесценцию всего тела с помощью визуализации IVIS®.
[00462] Визуализацию IVIS проводят на день 3, день 4, день 7, день 14, день 21, день 28, день 31, день 35 и день 42, и визуализируют отобранные органы ex vivo после умерщвления на день 42.
[00463] На протяжении исследования животных ежедневно взвешивают и проводят мониторинг общего состояния здоровья и самочувствия. При умерщвлении кровь каждого животного собирают путем терминальной пункции сердца, разделяют на две части и обрабатывают для получения 1) плазмы и 2) сыворотки, причем плазму мгновенно замораживают, а сыворотку используют для панели анализов на ферменты печени, после чего мгновенно замораживают. Кроме того, отбирают печень, селезенки, почки и паховые лимфатические узлы (LN) и визуализируют ex vivo с применением IVIS.
[00464] Экспрессию люциферазы в печени оценивают с применением набора для люциферазного ИФА-анализа MAXDISCOVERY® Luciferase ELISA assay (BIOO Scientific/PerkinElmer), кПЦР на люциферазу образцов печени, гистопатологии образцов печени и/или панели анализов на ферменты печени в сыворотке (VetScanVS2; Abaxis Preventive Care Profile Plus).
ПРИМЕР 5: «Прогонный» скрининг мутантов ITR
[00465] Проводили дополнительные анализы взаимосвязи структуры ITR с образованием зкДНК. Конструировали серию мутантов для изучения влияния специфических структурных изменений на образование зкДНК и способность экспрессировать трансген кодируемый указанной зкДНК. Конструирование мутантов, анализ образования зкДНК и оценка трансгенной экспрессии зкДНК в культуре клеток человека более подробно описаны ниже.
A. Конструирование мутантных ITR
[00466] Библиотеку из 31 плазмид с уникальными кассетами симметричных мутантных AAV ITR типа II разрабатывали in silico, а затем проводили оценку на клетках насекомых Sf9 и клетках эмбриональной почки человека (HEK293). Каждая кассета ITR содержала репортерный ген либо люциферазы (LUC), либо зеленого флуоресцентного белка (GFP) под управлением последовательности промотора p10 для экспрессии в клетках насекомых, и последовательности промотора CAG для экспрессии в клетках млекопитающих. Мутации в последовательности ITR были созданы либо в правом, либо в левом участке ITR. Библиотека содержала 15 правосторонних (RS) и 16 левосторонних (LS) мутантов, раскрытых в Таблицах 10A и 10B и на Фиг. 26A и 26B в данном документе.
[00467] Суспензионные культуры Sf9 культивировали в среде Sf900 III (Gibco) в вентилируемых флаконах объемом 200 мл для культур тканей. Культуры пересевали каждые 48 часов и перед каждым пересевом измеряли количество клеток и показатели роста с помощью счетчика ViCell Counter (Beckman Coulter). Культуры культивировали на шейкере (орбита 1 дюйм (25 мм), 130 об/мин) при 27 °C. Адгезивные культуры клеток HEK293 поддерживали в среде GlutiMax DMEM (модифицированной по Дульбекко среде Игла, Gibco) с 1% фетальной бычьей сыворотки и 0,1% пенициллина-стрептомицина (PenStrep) в культуральных флаконах объемом 250 мл при 37° C с 5% CO2. Культуры обрабатывали трипсином и пересевали каждые 96 часов. Для каждого пересева использовали разведение 1:10 из флакона с 90-100% конфлюэнтностью.
[00468] зкДНК-векторы получали и конструировали согласно описанию в Примере 1 выше. Вкратце, как показано на Фиг. 4B, клетки Sf9, трансдуцированные плазмидными конструктами, оставляли для адгезивного роста в течение 24 часов в стационарных условиях при 27°C. Через 24 часа трансфицированные клетки Sf9 инфицировали Rep вектором через инфицированные бакуловирусом клетки насекомых (BIIC). BIIC предварительно анализировали для определения характеристик инфективности и использовали в итоговом разведении 1:2000. BIIC в разведении 1:100 в среде для клеток насекомых Sf900 добавляли в каждую лунку с предварительно инфицированными клетками. BIIC без Rep-вектора добавляли в подгруппу лунок в качестве отрицательного контроля. Содержимое в планшетах перемешивали путем аккуратного встряхивания на качалочной мешалке для планшетов в течение 2 минут. Клетки культивировали в течение дополнительных 48 часов при 27°C в стационарных условиях. Все экспериментальные и контрольные конструкты анализировали в трех повторах.
[00469] Через 48 часов 96-луночный планшет извлекали из инкубатора, оставляли на непродолжительное время для уравновешивания до комнатной температуры и анализировали экспрессию люциферазы (OneGlo Luciferase Assay (Promega Corporation)). Суммарную люминесценцию измеряли с применением микропланшетного ридера SpectraMax M Series. Данные для повторов усредняли. Результаты представлены на Фиг. 27. Как и ожидалось, три отрицательных контрольных образца (только среда, ложная трансфекция без донорской ДНК, образец, который был обработан в отсутствие клеток с содержащим Rep бакуловирусом) демонстрировали отсутствие значимой экспрессии люциферазы. Устойчивая экспрессия люциферазы наблюдалась в каждом из мутантных образцов, что указывает на то, что для каждого образца трансфекция кодируемым зкДНК трансгеном была успешной и он экспрессировался независимо от мутации.
B. Анализ образования зкДНК
[00470] Чтобы убедиться, что зкДНК, полученная в предыдущем исследовании, имела ожидаемую структуру с замкнутыми концами, проводили эксперименты для получения достаточного количества каждой зкДНК, которая впоследствии могла бы быть протестирована на надлежащую структуру. Вкратце, суспензионные культуры Sf9 трансфицировали ДНК, принадлежащей одной ITR-мутантной плазмиде из библиотеки. Культуры высевали с плотностью 1,25 × 106 клеток/мл в культуральные флаконы Эрленмейера с ограниченным газообменом. Комплексы ДНК:липид для трансфекции получали с использованием реагента для трансфекции FuGene в соответствии с инструкциями производителя. Готовили смеси комплексов и инкубировали таким же образом, как описано выше для люцеферазного анализа на планшетах, с увеличением объемов пропорционально количеству трансфицируемых клеток. Как и в случае анализа с репортерным геном, использовали соотношение 4,5:1 (объем реагента/масса ДНК). Параллельно с экспериментальными культурами готовили ложнотрансфицированные (только реагенты для трансфекции) и необработанные контрольные культуры. После добавления реагентов для трансфекции культуры оставляли для восстановления в течение 10-15 минут при комнатной температуре с легким вращением, после чего переносили в инкубатор с шейкером и температурой 27°C. После 24 часов инкубации при встряхивании проводили подсчет клеток и измеряли показатели роста для всех флаконов (экспериментальных и контрольных) с помощью счетчика ViCell (Beckman Coulter). Все флаконы (за исключением контроля роста) инфицировали BIIC, содержащими Rep-вектор, в конечном разведении 1:5000. Также получали положительный контроль с использованием установленной процедуры двойного инфицирования BIIC для получения зкДНК. Для культуры с двойной инфекцией высевали количество клеток, равное среднему количеству жизнеспособных клеток во всех экспериментальных культурах. Контрольная культура с двойной инфекцией была инфицирована Rep и BIIC с репортерным геном в конечном разведении 1:5000 для каждого конструкта, соответственно. После инфицирования культуры снова помещали в инкубатор в ранее описанные условия со встряхиванием. Количество клеток, показатели роста и жизнеспособности измеряли ежедневно для всех флаконов в течение 3 дней после инфицирования. Измерения в момент времени Т=0 проводили после того, как вновь инфицированным культурам давали возможность восстановиться в течение приблизительно 2 часов в условиях инкубации со встряхиванием. Через 3 дня клетки собирали посредством центрифугирования в течение 15 минут. Супернатант утилизировали, записывали массу осадка и замораживали осадок при -80°C до экстракции ДНК.
[00471] Предполагаемую неочищенную зкДНК экстрагировали из всех флаконов (экспериментальных и контрольных) с использованием набора для очистки Qiagen Plasmid Plus Midi Purification Kit (Qiagen) в соответствии с протоколом производителя для «высокого выхода». Количественное определение элюатов проводили с использованием данных измерения оптической плотности, полученных с помощью NanoDrop OneC (ThermoFisher). Полученные экстракты зкДНК хранили при 4°C.
[00472] Вышеописанные экстракты зкДНК проганяли на нативном агарозном геле (1% агароза, 1x буфер TAE), приготовленному с использованием разведения SYBR Safe Gel Stain (ThermoFisher Scientific) 1:10000, вместе с ДНК-маркерами на 1 т.п.о. TrackIt Plus DNA. Затем гель визуализировали с помощью Gbox Mini Imager в УФ/синем свете. Согласно приведенному ранее описанию, ожидается, что в образцах зкДНК, которые прогнали на нативных гелях, будет две первичных полосы: полоса, соответствующая примерно 5500 п.о., представляющая мономерное вещество, и полоса, соответствующая примерно 11000 п.о., соответствующая димерному веществу. Все мутантные образцы были протестированы и показали ожидаемые полосы мономера и димера на нативных агарозных гелях. Результаты для репрезентативного образца указанных конструктов представлены на Фиг. 28. Предполагаемую неочищенную зкДНК с мутантным ITR и контрольные экстракты, полученные в малом масштабе, дополнительно анализировали с применением парного рестрикционного расщепления и денатурирующего агарозного геля для подтверждения двуцепочечной структуры ДНК, указывающей на зкДНК. Ожидается, что у каждого мутантного зкДНК имеется единственный сайт рестрикции EcoR1; поэтому, если она правильно сформирована, то образует два характерных фрагмента при расщеплении EcoR1. Высокоточную рестрикционную эндонуклеазу EcoRI (New England Biolabs) использовали для расщепления экстракта предполагаемой зкДНК в соответствии с инструкциями производителя. Экстракты ложнотрансфицированных и контрольных культур не анализировали, поскольку спектрофотометрическое количественное определение с применением NanoDrop (ThermoFisher), а также анализ на нативном агарозном геле показал, что в элюатах отсутствуют детектируемый зкДНК/плазмидо-подобный продукт. Расщепленный материал очищали с применением набора для ПЦР Qiagen PCR Clean-up Kit (Qiagen соответствии с инструкциями производителя, за исключением того, что очищенный расщепленный материал элюировали в воде без нуклеаз вместо буфера для элюирования от Qiagen. Щелочной агарозный гель (0,8% щелочной агарозы) уравновешивали в буфере для уравновешивания (1 мМ EDTA, 200 мМ NaOH) в течение ночи при 4°C. 10x денатурирующий раствор (50 мМ NaOH, 1 мМ EDTA) добавляли к образцам очищенных расщепленных зкДНК и соответствующих нерасщепленных зкДНК (в общем 1 мкг), и образцы нагревали при 65°C в течение 10 минут. 10x краситель для нанесения (бромфеноловый синий, 50% глицерина) добавляли к каждому денатурированному образцу и перемешивали. Маркеры Plus DNA TrackIt 1 т.п.о. (ThermoFisher Scientific) также наносили на гель в качестве референсной ДНК. Прогон на геле проводили в течение ~18 часов при 4°C и постоянном напряжении (25 В) с последующим промыванием деионизированной H2O и нейтрализацией в буфере 1xTAE (Tris-ацетат, EDTA), pH 7,6, в течение 20 минут при аккуратном помешивании. Затем гель переносили в раствор 1x TAE/1x SYBR Gold примерно на 1 час при аккуратном помешивании. Затем гель визуализировали с помощью Gbox Mini Imager (Syngene) в УФ/синем свете. Ожидалось, что неразрезанные денатурированные образцы будут мигрировать на уровне примерно 11000 пар оснований, а образцы, обработанные EcoRI - давать две полосы, одну на уровне примерно 4000 пар оснований и другую на уровне примерно 6000 пар оснований.
[00473] Все мутантные образцы в этом эксперименте показали аналогичные результаты. Две значимых полосы были видны на каждой дорожке с обработанными EcoR1 образцами, мигрировавшими на денатурирующем геле с ожидаемыми размерами, что резко отличается от нерасщепленных образцов, которыйе мигрировали с ожидаемым размером примерно 11000 пар оснований. На Фиг. 27 показаны результаты для репрезентативного образца мутантов, где видны две полосы со значениями выше фоновых для каждого расщепленного мутантного образца, в отличие от единственной полосы, наблюдаемой в случае нерасщепленных мутантных образцов. Соответственно, в мутантных образцах, по-видимому, корректно формировалась зкДНК.
A. Функциональная экспрессия в культуре клеток человека
[00474] Для оценки функциональности мутантной ITR зкДНК, полученной в процессе маломасштабного производства, клетки HEK293 трансфицировали некоторыми репрезентативными образцами мутантной зкДНК. Активно делящиеся клетки HEK293 были посеяны в 96-луночные микротитрационные планшеты с плотностью 3x106 клеток на лунку (80% конфлюэнтность) и инкубируют в течение 24 часов в ранее описанных условиях для адгезивных культур HEK293. Через 24 часа проводили трансфекцию 200 нг неочищенной полученной в малом масштабе зкДНК с применением липофектамина (Invitrogen, ThermoFisher Scientific). Комплексы трансфекции получали в соответствии с инструкциями производителя, и 10 мкл общего объема комплекса трансфекции использовали для трансфекции ранее высеянных клеток HEK293. Все экспериментальные и контрольные конструкты анализировали в трех повторах. Трансфицированные клетки инкубировали в описанных ранее условиях в течение 72 часов. Через 72 часа 96-луночный планшет извлекали из инкубатора и ненадолго оставляют для уравновешивания до комнатной температуры. Был проведен OneGlo люциферазный анализ. После 10 минут на орбитальном шейкере измеряли суммарную люминесценцию с применением микропланшетного ридера SpectraMax M Series. Данные для повторов усредняли. Результаты представлены на Фиг. 30. Каждый из протестированных мутантных образцов, который экспрессировал люциферазу в культуре клеток человека, показывает, что зкДНК была корректно образована и экспрессирована для каждого образца в контексте клеток человека.
[00475] ПРИМЕР 6: Конструкты только с Rep78 или Rep68 способны продуцировать зкДНК
[00476] Ген репликации (Rep) AAV кодирует четыре неструктурных или репликационных (Rep) белка из одной и той же открытой рамки считывания. Rep78, Rep68, Rep52 и Rep40 названы по их предполагаемой молекулярной массе, рассчитанной по их подвижности в SDS-PAGE (Mendelson et al., 1986. J Virol. 60: 823-832). Rep78/68 транслируются с мРНК, происходящей от промотора транскрипции с элемента 5 на схеме (P5). Rep78 и Rep68 служат в качестве белков инициатора репликации вируса, которые распознают родственные сайты связывания в вирусной точки начала репликации и "разрывают" точку начала на сайте концевого разрешения. Процес никирования обеспечивает свободную 3'-гидроксильную группу, которая запускает синтез вирусной ДНК. Было показано, что в дополнение к ДНК-связывающей и сайт-специфической эндонуклеазной активности Rep78 и Rep68 обладают геликазной и АТФазной активностями. Белки Rep52/40 транслируются с мРНК, происходящей от промотора транскрипции элемента 19 на схеме (P19). Белки Rep52 и Rep40 опосредуют сборку вируса. Белки Rep68 и Rep40 отличаются от своих более длинных аналогов тем, что они транслируются со сплайсированных мРНК с промоторов P5 и P19 соответственно. Сплайсинг удаляет 92 аминокислотных остатка с карбоксильных концов белков Rep78 и Rep52 и заменяет их 9 аминокислотами, расположенными на С-концах Rep68 и Rep40.
[00477] Были проведены эксперименты, чтобы определить, достаточно ли присутствия Rep78 или Rep68 для образования зкДНК. Добавляли точечную мутацию для устранения трансляции Rep 52 промотором p19 (M->G и M->T), чтобы исследовать влияние делеции Rep52/40 на образование зкДНК, как описано в Примере 1 выше. Таким образом, конструкты, модифицированные точечной мутацией Rep52 (например, аминокислоты 225 M->G и M->T), будут демонстрировать только продукт зкДНК из Rep78. Были созданы два дополнительных конструкта, чтобы определить, обладает ли Rep68 какой-либо активностью при образовании зкДНК. Мутанты Rep68 Met→ Gly (M225G) и Rep68 Met→Thr (M225T) были сконструированы для удаления внутреннего сайта трансляции и с-концевой интронной последовательности (92 аминокислотных остатка и замена на 9 аминокислот, как описано выше). Был получен дополнительный мутант с точечной мутацией в домене активности никазы (Y156F). На Фиг. 32A и B представляют собой неденатурирующие гели, показывающие присутствие высокостабильных ДНК-векторов и характерные полосы, подтверждающие присутствие вектора высокостабильной ДНК с замкнутыми концами (зкДНК), полученного с использованием единственного белка Rep, с использованием описанных здесь способов. Как показано на Фиг. 32A, более высокие количества вектора зкДНК продуцируются с использованием нуклеиновой кислоты модифицированного Rep78 с модификацией Rep78 из Met.→ Gly (M225G) (дорожка 1) или Rep Met→ Thr (M225T) (дорожка 2) по сравнению с продуцированием при использовании нуклеиновой кислоты, кодирующей Rep78 дикого типа (дорожка 5), при котором нуклеиновая кислота экспрессирует как белок Rep78, так и белок Rep52. Вектор зкДНК не был получен со связывающими мутантами Rep78, содержащими модификации Gly (Y156F) (дорожка 3) или Thr (Y156F) (дорожка 4). Фиг. 32B дополнительно показывает, что мутантные Rep68 Met→ Gly (M225G) и Rep68 Met→ Thr (M225T) также продуцировали вектор зкДНК до уровней, равных или превышающих количество вектора зкДНК, полученного с использованием нуклеиновой кислоты модифицированного Rep78 с модификацией Rep78 в Met→ Gly (M225G) или Rep Met→ Thr (M225T) и делецией с-концевого интрона.
[00478] Соответственно, эти эксперименты продемонстрировали, что одного Rep78 или одного Rep68 достаточно для образования зкДНК без присутствия Rep52 или Rep40.
ИСТОЧНИКИ
[00479] Все источники, перечисленные и раскрытые в данном описании и в примерах, в том числе патенты, патентные заявки, международные патентные заявки и публикации включены в данный документ полностью посредством ссылок.
[00480] REP последовательности
SEQ ID NO. 558 представляет собой аминокислотную последовательность Rep 40 из AAV1.
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ala
Pro Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
Asn Gly Tyr Glu Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
Gln Lys Arg Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Glu Phe Phe Arg Trp Ala Gln Asp His Val Thr Glu Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Asn Lys Arg Pro Ala
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
Asp Leu Ala Arg Gly Gln Pro Leu (SEQ ID NO: 558)
SEQ ID NO. 559 представляет собой аминокислотную последовательность Rep 40 из AAV2.
Met Glu Leu Val Gly Trp Leu Val Asp Lys Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Gln Gln
Pro Val Glu Asp Ile Ser Ser Asn Arg Ile Tyr Lys Ile Leu Glu Leu
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
Thr Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Thr Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Asp Phe Phe Arg Trp Ala Lys Asp His Val Val Glu Val
Glu His Glu Phe Tyr Val Lys Lys Gly Gly Ala Lys Lys Arg Pro Ala
Pro Ser Asp Ala Asp Ile Ser Glu Pro Lys Arg Val Arg Glu Ser Val
Ala Gln Pro Ser Thr Ser Asp Ala Glu Ala Ser Ile Asn Tyr Ala Asp
Arg Leu Ala Arg Gly His Ser Leu (SEQ ID NO: 559)
SEQ ID NO. 560 представляет собой аминокислотную последовательность Rep 40 из AAV3A.
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Ser Asn
Pro Pro Glu Asp Ile Thr Lys Asn Arg Ile Tyr Gln Ile Leu Glu Leu
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Glu Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Glu Phe
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Asp Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
Ser Asn Asp Ala Asp Val Ser Glu Pro Lys Arg Glu Cys Thr Ser Leu
Ala Gln Pro Thr Thr Ser Asp Ala Glu Ala Pro Ala Asp Tyr Ala Asp
Leu Ala Arg Gly Gln Pro Phe (SEQ ID NO: 560)
SEQ ID NO. 561 представляет собой аминокислотную последовательность Rep 40 из AAV3B.
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Ser Asn
Pro Pro Glu Asp Ile Thr Lys Asn Arg Ile Tyr Gln Ile Leu Glu Leu
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Glu Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Asp Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
Ser Asn Asp Ala Asp Val Ser Glu Pro Lys Arg Gln Cys Thr Ser Leu
Ala Gln Pro Thr Thr Ser Asp Ala Glu Ala Pro Ala Asp Tyr Ala Asp
Leu Ala Arg Gly Gln Pro Phe (SEQ ID NO: 561)
SEQ ID NO. 562 представляет собой аминокислотную последовательность Rep 40 из AAV4.
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Gln Asn
Pro Pro Glu Asp Ile Ser Ser Asn Arg Ile Tyr Arg Ile Leu Glu Met
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Lys Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
Thr His Glu Phe Tyr Val Arg Lys Gly Gly Ala Arg Lys Arg Pro Ala
Pro Asn Asp Ala Asp Ile Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Gln Pro Ser Thr Ser Asp Ala Glu Ala Pro Val Asp Tyr Ala Asp
Leu Ala Arg Gly Gln Pro Leu (SEQ ID NO: 562)
SEQ ID NO. 563 представляет собой аминокислотную последовательность Rep 40 из AAV5.
Met Ala Leu Val Asn Trp Leu Val Glu His Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asn Gln Glu Ser Tyr Leu Ser Phe Asn Ser Thr
Gly Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Thr Lys
Ile Met Ser Leu Thr Lys Ser Ala Val Asp Tyr Leu Val Gly Ser Ser
Val Pro Glu Asp Ile Ser Lys Asn Arg Ile Trp Gln Ile Phe Glu Met
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Ile Leu Tyr Gly Trp Cys
Gln Arg Ser Phe Asn Lys Arg Asn Thr Val Trp Leu Tyr Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Thr Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Leu Ile Trp Trp Glu Glu Gly Lys Met Thr Asn
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Val Gln Ile Asp Ser Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Val Val Val Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Glu Asp Arg Met Phe Lys Phe
Glu Leu Thr Lys Arg Leu Pro Pro Asp Phe Gly Lys Ile Thr Lys Gln
Glu Val Lys Asp Phe Phe Ala Trp Ala Lys Val Asn Gln Val Pro Val
Thr His Glu Phe Lys Val Pro Arg Glu Leu Ala Gly Thr Lys Gly Ala
Glu Lys Ser Leu Lys Arg Pro Leu Gly Asp Val Thr Asn Thr Ser Tyr
Lys Ser Leu Glu Lys Arg Ala Arg Leu Ser Phe Val Pro Glu Thr Pro
Arg Ser Ser Asp Val Thr Val Asp Pro Ala Pro Leu Arg Pro Leu Asn
Trp Asn Ser Leu Val Gly Pro Ser Trp (SEQ ID NO: 563)
SEQ ID NO. 564 представляет собой аминокислотную последовательность Rep 40 из AAV6.
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ala
Pro Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
Gln Lys Arg Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Glu Phe Phe Arg Trp Ala Gln Asp His Val Thr Glu Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Asn Lys Arg Pro Ala
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
Asp Leu Ala Arg Gly Gln Pro Leu (SEQ ID NO: 564)
SEQ ID NO. 565 представляет собой аминокислотную последовательность Rep 40 из AAV7.
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ser
Leu Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Ser Lys Arg Pro Ala
Pro Asp Asp Ala Asp Ile Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
Asp Leu Ala Arg Gly Gln Pro Leu (SEQ ID NO: 565)
SEQ ID NO. 566 представляет собой аминокислотную последовательность Rep 40 из AAV8.
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ser
Leu Pro Ala Asp Ile Thr Gln Asn Arg Ile Tyr Arg Ile Leu Ala Leu
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Ser Lys Arg Pro Ala
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
Asp Leu Ala Arg Gly Gln Pro Leu (SEQ ID NO: 566)
SEQ ID NO. 567 представляет собой консенсусную аминокислотную последовательность SEQ ID NO: 558-566.
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Xaa Ser
Pro Pro Glu Asp Ile Ser Thr Asn Arg Ile Tyr Arg Ile Leu Ala Leu
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Xaa Pro Ser Thr Ser Asp Ala Glu Ala Pro Val Asp Phe Ala Asp
Leu Ala Arg Gly Gln Pro Leu (SEQ ID NO: 567)
SEQ ID NO. 568 представляет собой аминокислотную последовательность Rep 52 из AAV1.
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ala
Pro Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
Asn Gly Tyr Glu Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
Gln Lys Arg Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Glu Phe Phe Arg Trp Ala Gln Asp His Val Thr Glu Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Asn Lys Arg Pro Ala
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
Asp Arg Tyr Gln Asn Lys Cys Ser Arg His Ala Gly Met Leu Gln Met
Leu Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Asn Phe Asn Ile
Cys Phe Thr His Gly Thr Arg Asp Cys Ser Glu Cys Phe Pro Gly Val
Ser Glu Ser Gln Pro Val Val Arg Lys Arg Thr Tyr Arg Lys Leu Cys
Ala Ile His His Leu Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser Ala
Cys Asp Leu Val Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln (SEQ ID NO: 568)
SEQ ID NO. 569 представляет собой аминокислотную последовательность Rep 52 из AAV2.
Met Glu Leu Val Gly Trp Leu Val Asp Lys Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Gln Gln
Pro Val Glu Asp Ile Ser Ser Asn Arg Ile Tyr Lys Ile Leu Glu Leu
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
Thr Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Thr Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Asp Phe Phe Arg Trp Ala Lys Asp His Val Val Glu Val
Glu His Glu Phe Tyr Val Lys Lys Gly Gly Ala Lys Lys Arg Pro Ala
Pro Ser Asp Ala Asp Ile Ser Glu Pro Lys Arg Val Arg Glu Ser Val
Ala Gln Pro Ser Thr Ser Asp Ala Glu Ala Ser Ile Asn Tyr Ala Asp
Arg Tyr Gln Asn Lys Cys Ser Arg His Val Gly Met Asn Leu Met Leu
Phe Pro Cys Arg Gln Cys Glu Arg Met Asn Gln Asn Ser Asn Ile Cys
Phe Thr His Gly Gln Lys Asp Cys Leu Glu Cys Phe Pro Val Ser Glu
Ser Gln Pro Val Ser Val Val Lys Lys Ala Tyr Gln Lys Leu Cys Tyr
Ile His His Ile Met Gly Lys Val Pro Asp Ala Cys Thr Ala Cys Asp
Leu Val Asn Val Asp Leu Asp Asp Cys Ile Phe Glu Gln (SEQ ID NO: 569)
SEQ ID NO. 570 представляет собой аминокислотную последовательность Rep 52 из AAV3A.
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Ser Asn
Pro Pro Glu Asp Ile Thr Lys Asn Arg Ile Tyr Gln Ile Leu Glu Leu
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Glu Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Glu Phe
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Asp Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
Ser Asn Asp Ala Asp Val Ser Glu Pro Lys Arg Glu Cys Thr Ser Leu
Ala Gln Pro Thr Thr Ser Asp Ala Glu Ala Pro Ala Asp Tyr Ala Asp
Arg Tyr Gln Asn Lys Cys Ser Arg His Val Gly Met Asn Leu Met Leu
Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Ile Ser Asn Val Cys
Phe Thr His Gly Gln Arg Asp Cys Gly Glu Cys Phe Pro Gly Met Ser
Glu Ser Gln Pro Val Ser Val Val Lys Lys Lys Thr Tyr Gln Lys Leu
Cys Pro Ile His His Ile Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser
Ala Cys Asp Leu Ala Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln (SEQ ID NO: 570)
SEQ ID NO. 571 представляет собой аминокислотную последовательность Rep 52 из AAV3B.
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Ser Asn
Pro Pro Glu Asp Ile Thr Lys Asn Arg Ile Tyr Gln Ile Leu Glu Leu
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Glu Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Asp Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
Ser Asn Asp Ala Asp Val Ser Glu Pro Lys Arg Gln Cys Thr Ser Leu
Ala Gln Pro Thr Thr Ser Asp Ala Glu Ala Pro Ala Asp Tyr Ala Asp
Arg Tyr Gln Asn Lys Cys Ser Arg His Val Gly Met Asn Leu Met Leu
Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Ile Ser Asn Val Cys
Phe Thr His Gly Gln Arg Asp Cys Gly Glu Cys Phe Pro Gly Met Ser
Glu Ser Gln Pro Val Ser Val Val Lys Lys Lys Thr Tyr Gln Lys Leu
Cys Pro Ile His His Ile Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser
Ala Cys Asp Leu Ala Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln (SEQ ID NO: 571)
SEQ ID NO. 572 представляет собой аминокислотную последовательность Rep 52 из AAV4.
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Gln Asn
Pro Pro Glu Asp Ile Ser Ser Asn Arg Ile Tyr Arg Ile Leu Glu Met
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Lys Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
Thr His Glu Phe Tyr Val Arg Lys Gly Gly Ala Arg Lys Arg Pro Ala
Pro Asn Asp Ala Asp Ile Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Gln Pro Ser Thr Ser Asp Ala Glu Ala Pro Val Asp Tyr Ala Asp
Arg Tyr Gln Asn Lys Cys Ser Arg His Val Gly Met Asn Leu Met Leu
Phe Pro Cys Arg Gln Cys Glu Arg Met Asn Gln Asn Val Asp Ile Cys
Phe Thr His Gly Val Met Asp Cys Ala Glu Cys Phe Pro Val Ser Glu
Ser Gln Pro Val Ser Val Val Arg Lys Arg Thr Tyr Gln Lys Leu Cys
Pro Ile His His Ile Met Gly Arg Ala Pro Glu Val Ala Cys Ser Ala
Cys Glu Leu Ala Asn Val Asp Leu Asp Asp Cys Asp Met Glu Gln (SEQ ID NO: 572)
SEQ ID NO. 573 представляет собой аминокислотную последовательность Rep 52 из AAV5.
Met Ala Leu Val Asn Trp Leu Val Glu His Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asn Gln Glu Ser Tyr Leu Ser Phe Asn Ser Thr
Gly Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Thr Lys
Ile Met Ser Leu Thr Lys Ser Ala Val Asp Tyr Leu Val Gly Ser Ser
Val Pro Glu Asp Ile Ser Lys Asn Arg Ile Trp Gln Ile Phe Glu Met
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Ile Leu Tyr Gly Trp Cys
Gln Arg Ser Phe Asn Lys Arg Asn Thr Val Trp Leu Tyr Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Thr Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Leu Ile Trp Trp Glu Glu Gly Lys Met Thr Asn
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Val Gln Ile Asp Ser Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Val Val Val Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Glu Asp Arg Met Phe Lys Phe
Glu Leu Thr Lys Arg Leu Pro Pro Asp Phe Gly Lys Ile Thr Lys Gln
Glu Val Lys Asp Phe Phe Ala Trp Ala Lys Val Asn Gln Val Pro Val
Thr His Glu Phe Lys Val Pro Arg Glu Leu Ala Gly Thr Lys Gly Ala
Glu Lys Ser Leu Lys Arg Pro Leu Gly Asp Val Thr Asn Thr Ser Tyr
Lys Ser Leu Glu Lys Arg Ala Arg Leu Ser Phe Val Pro Glu Thr Pro
Arg Ser Ser Asp Val Thr Val Asp Pro Ala Pro Leu Arg Pro Leu Asn
Trp Asn Ser Arg Tyr Asp Cys Lys Cys Asp Tyr His Ala Gln Phe Asp
Asn Ile Ser Asn Lys Cys Asp Glu Cys Glu Tyr Leu Asn Arg Gly Lys
Asn Gly Cys Ile Cys His Asn Val Thr His Cys Gln Ile Cys His Gly
Ile Pro Pro Trp Glu Lys Glu Asn Leu Ser Asp Phe Gly Asp Phe Asp
Asp Ala Asn Lys Glu Gln (SEQ ID NO: 573)
SEQ ID NO. 574 представляет собой аминокислотную последовательность Rep 52 из AAV6.
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ala
Pro Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
Gln Lys Arg Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Glu Phe Phe Arg Trp Ala Gln Asp His Val Thr Glu Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Asn Lys Arg Pro Ala
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
Asp Arg Tyr Gln Asn Lys Cys Ser Arg His Ala Gly Met Leu Gln Met
Leu Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Asn Phe Asn Ile
Cys Phe Thr His Gly Thr Arg Asp Cys Ser Glu Cys Phe Pro Gly Val
Ser Glu Ser Gln Pro Val Val Arg Lys Arg Thr Tyr Arg Lys Leu Cys
Ala Ile His His Leu Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser Ala
Cys Asp Leu Val Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln (SEQ ID NO: 574)
SEQ ID NO. 575 представляет собой аминокислотную последовательность Rep 52 из AAV7.
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ser
Leu Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Ser Lys Arg Pro Ala
Pro Asp Asp Ala Asp Ile Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
Asp Arg Tyr Gln Asn Lys Cys Ser Arg His Ala Gly Met Ile Gln Met
Leu Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Asn Phe Asn Ile
Cys Phe Thr His Gly Val Arg Asp Cys Leu Glu Cys Phe Pro Gly Val
Ser Glu Ser Gln Pro Val Val Arg Lys Lys Thr Tyr Arg Lys Leu Cys
Ala Ile His His Leu Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser Ala
Cys Asp Leu Val Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln (SEQ ID NO: 575)
SEQ ID NO. 576 представляет собой аминокислотную последовательность Rep 52 из AAV8.
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ser
Leu Pro Ala Asp Ile Thr Gln Asn Arg Ile Tyr Arg Ile Leu Ala Leu
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Ser Lys Arg Pro Ala
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
Asp Arg Tyr Gln Asn Lys Cys Ser Arg His Ala Gly Met Leu Gln Met
Leu Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Asn Phe Asn Ile
Cys Phe Thr His Gly Val Arg Asp Cys Ser Glu Cys Phe Pro Gly Val
Ser Glu Ser Gln Pro Val Val Arg Lys Arg Thr Tyr Arg Lys Leu Cys
Ala Ile His His Leu Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser Ala
Cys Asp Leu Val Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln (SEQ ID NO: 576)
SEQ ID NO. 577 представляет собой консенсусную аминокислотную последовательность SEQ ID NOs 568-576.
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Xaa Ser
Pro Pro Glu Asp Ile Ser Thr Asn Arg Ile Tyr Arg Ile Leu Ala Leu
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Asp Pro Ser Thr Ser Asp Ala Glu Ala Pro Val Asp Phe Ala Asp
Arg Tyr Gln Asn Lys Cys Ser Arg His Ala Gly Met Xaa Gln Met Leu
Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Asn Xaa Asn Ile Cys
Phe Thr His Gly Xaa Arg Asp Cys Xaa Glu Cys Phe Pro Gly Val Ser
Glu Ser Gln Xaa Val Val Arg Lys Arg Thr Tyr Xaa Lys Leu Cys Xaa
Ile His His Leu Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser Ala Cys
Asp Leu Val Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln (SEQ ID NO: 577)
SEQ ID NO. 578 представляет собой аминокислотную последовательность Rep 68 из AAV1.
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Ser Trp Val Ala Glu
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Leu Asn Leu Ile
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
Val Gln Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Leu His Ile Leu Val Glu
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
Arg Asp Lys Leu Val Gln Thr Ile Tyr Arg Gly Ile Glu Pro Thr Leu
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Glu Tyr Ile
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Leu Asn
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ala
Pro Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
Asn Gly Tyr Glu Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
Gln Lys Arg Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Glu Phe Phe Arg Trp Ala Gln Asp His Val Thr Glu Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Asn Lys Arg Pro Ala
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
Asp Leu Ala Arg Gly Gln Pro Leu (SEQ ID NO: 578)
SEQ ID NO. 579 представляет собой аминокислотную последовательность Rep 68 из AAV2.
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asn Trp Val Ala Glu
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Leu Asn Leu Ile
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
Thr Glu Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Met His Val Leu Val Glu
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
Arg Glu Lys Leu Ile Gln Arg Ile Tyr Arg Gly Ile Glu Pro Thr Leu
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Gln Tyr Leu
Ser Ala Cys Leu Asn Leu Thr Glu Arg Lys Arg Leu Val Ala Gln His
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Gln Asn
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
Met Glu Leu Val Gly Trp Leu Val Asp Lys Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Gln Gln
Pro Val Glu Asp Ile Ser Ser Asn Arg Ile Tyr Lys Ile Leu Glu Leu
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
Thr Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Thr Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Asp Phe Phe Arg Trp Ala Lys Asp His Val Val Glu Val
Glu His Glu Phe Tyr Val Lys Lys Gly Gly Ala Lys Lys Arg Pro Ala
Pro Ser Asp Ala Asp Ile Ser Glu Pro Lys Arg Val Arg Glu Ser Val
Ala Gln Pro Ser Thr Ser Asp Ala Glu Ala Ser Ile Asn Tyr Ala Asp
Arg Leu Ala Arg Gly His Ser Leu (SEQ ID NO: 579)
SEQ ID NO. 580 представляет собой аминокислотную последовательность Rep 68 из AAV3A.
Met Pro Gly Phe Tyr Glu Ile Val Leu Lys Val Pro Ser Asp Leu Asp
Glu Arg Leu Pro Gly Ile Ser Asn Ser Phe Val Asn Trp Val Ala Glu
Lys Glu Trp Asp Val Pro Pro Asp Ser Asp Met Asp Pro Asn Leu Ile
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Glu Phe Leu
Val Glu Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
Gln Phe Glu Lys Gly Glu Thr Tyr Phe His Leu His Val Leu Ile Glu
Thr Ile Gly Val Lys Ser Met Val Val Gly Arg Tyr Val Ser Gln Ile
Lys Glu Lys Leu Val Thr Arg Ile Tyr Arg Gly Val Glu Pro Gln Leu
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
Asn Lys Val Val Asp Asp Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Asp Gln Tyr Leu
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Gln Asn
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Ser Asn
Pro Pro Glu Asp Ile Thr Lys Asn Arg Ile Tyr Gln Ile Leu Glu Leu
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Glu Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Glu Phe
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Asp Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
Ser Asn Asp Ala Asp Val Ser Glu Pro Lys Arg Glu Cys Thr Ser Leu
Ala Gln Pro Thr Thr Ser Asp Ala Glu Ala Pro Ala Asp Tyr Ala Asp
Leu Ala Arg Gly Gln Pro Phe (SEQ ID NO: 580)
SEQ ID NO. 581 представляет собой аминокислотную последовательность Rep 68 из AAV3B.
Met Pro Gly Phe Tyr Glu Ile Val Leu Lys Val Pro Ser Asp Leu Asp
Glu His Leu Pro Gly Ile Ser Asn Ser Phe Val Asn Trp Val Ala Glu
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Pro Asn Leu Ile
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Glu Phe Leu
Val Glu Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
Gln Phe Glu Lys Gly Glu Thr Tyr Phe His Leu His Val Leu Ile Glu
Thr Ile Gly Val Lys Ser Met Val Val Gly Arg Tyr Val Ser Gln Ile
Lys Glu Lys Leu Val Thr Arg Ile Tyr Arg Gly Val Glu Pro Gln Leu
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
Asn Lys Val Val Asp Asp Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Asp Gln Tyr Leu
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Gln Asn
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Ser Asn
Pro Pro Glu Asp Ile Thr Lys Asn Arg Ile Tyr Gln Ile Leu Glu Leu
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Glu Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Asp Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
Ser Asn Asp Ala Asp Val Ser Glu Pro Lys Arg Gln Cys Thr Ser Leu
Ala Gln Pro Thr Thr Ser Asp Ala Glu Ala Pro Ala Asp Tyr Ala Asp
Leu Ala Arg Gly Gln Pro Phe (SEQ ID NO: 581)
SEQ ID NO. 582 представляет собой аминокислотную последовательность Rep 68 из AAV4.
Met Pro Gly Phe Tyr Glu Ile Val Leu Lys Val Pro Ser Asp Leu Asp
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Ser Trp Val Ala Glu
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Leu Asn Leu Ile
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Glu Phe Leu
Val Glu Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
Gln Phe Glu Lys Gly Asp Ser Tyr Phe His Leu His Ile Leu Val Glu
Thr Val Gly Val Lys Ser Met Val Val Gly Arg Tyr Val Ser Gln Ile
Lys Glu Lys Leu Val Thr Arg Ile Tyr Arg Gly Val Glu Pro Gln Leu
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
Asn Lys Val Val Asp Asp Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Asp Gln Tyr Ile
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Gln Asn
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Gln Asn
Pro Pro Glu Asp Ile Ser Ser Asn Arg Ile Tyr Arg Ile Leu Glu Met
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Lys Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
Thr His Glu Phe Tyr Val Arg Lys Gly Gly Ala Arg Lys Arg Pro Ala
Pro Asn Asp Ala Asp Ile Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Gln Pro Ser Thr Ser Asp Ala Glu Ala Pro Val Asp Tyr Ala Asp
Leu Ala Arg Gly Gln Pro Leu (SEQ ID NO: 582)
SEQ ID NO. 583 представляет собой аминокислотную последовательность Rep 68 из AAV5.
Met Ala Thr Phe Tyr Glu Val Ile Val Arg Val Pro Phe Asp Val Glu
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asp Trp Val Thr Gly
Gln Ile Trp Glu Leu Pro Pro Glu Ser Asp Leu Asn Leu Thr Leu Val
Glu Gln Pro Gln Leu Thr Val Ala Asp Arg Ile Arg Arg Val Phe Leu
Tyr Glu Trp Asn Lys Phe Ser Lys Gln Glu Ser Lys Phe Phe Val Gln
Phe Glu Lys Gly Ser Glu Tyr Phe His Leu His Thr Leu Val Glu Thr
Ser Gly Ile Ser Ser Met Val Leu Gly Arg Tyr Val Ser Gln Ile Arg
Ala Gln Leu Val Lys Val Val Phe Gln Gly Ile Glu Pro Gln Ile Asn
Asp Trp Val Ala Ile Thr Lys Val Lys Lys Gly Gly Ala Asn Lys Val
Val Asp Ser Gly Tyr Ile Pro Ala Tyr Leu Leu Pro Lys Val Gln Pro
Glu Leu Gln Trp Ala Trp Thr Asn Leu Asp Glu Tyr Lys Leu Ala Ala
Leu Asn Leu Glu Glu Arg Lys Arg Leu Val Ala Gln Phe Leu Ala Glu
Ser Ser Gln Arg Ser Gln Glu Ala Ala Ser Gln Arg Glu Phe Ser Ala
Asp Pro Val Ile Lys Ser Lys Thr Ser Gln Lys Tyr Met Ala Leu Val
Asn Trp Leu Val Glu His Gly Ile Thr Ser Glu Lys Gln Trp Ile Gln
Glu Asn Gln Glu Ser Tyr Leu Ser Phe Asn Ser Thr Gly Asn Ser Arg
Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Thr Lys Ile Met Ser Leu
Thr Lys Ser Ala Val Asp Tyr Leu Val Gly Ser Ser Val Pro Glu Asp
Ile Ser Lys Asn Arg Ile Trp Gln Ile Phe Glu Met Asn Gly Tyr Asp
Pro Ala Tyr Ala Gly Ser Ile Leu Tyr Gly Trp Cys Gln Arg Ser Phe
Asn Lys Arg Asn Thr Val Trp Leu Tyr Gly Pro Ala Thr Thr Gly Lys
Thr Asn Ile Ala Glu Ala Ile Ala His Thr Val Pro Phe Tyr Gly Cys
Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp Cys Val Asp Lys
Met Leu Ile Trp Trp Glu Glu Gly Lys Met Thr Asn Lys Val Val Glu
Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg Val Asp Gln Lys
Cys Lys Ser Ser Val Gln Ile Asp Ser Thr Pro Val Ile Val Thr Ser
Asn Thr Asn Met Cys Val Val Val Asp Gly Asn Ser Thr Thr Phe Glu
His Gln Gln Pro Leu Glu Asp Arg Met Phe Lys Phe Glu Leu Thr Lys
Arg Leu Pro Pro Asp Phe Gly Lys Ile Thr Lys Gln Glu Val Lys Asp
Phe Phe Ala Trp Ala Lys Val Asn Gln Val Pro Val Thr His Glu Phe
Lys Val Pro Arg Glu Leu Ala Gly Thr Lys Gly Ala Glu Lys Ser Leu
Lys Arg Pro Leu Gly Asp Val Thr Asn Thr Ser Tyr Lys Ser Leu Glu
Lys Arg Ala Arg Leu Ser Phe Val Pro Glu Thr Pro Arg Ser Ser Asp
Val Thr Val Asp Pro Ala Pro Leu Arg Pro Leu Asn Trp Asn Ser Leu
Val Gly Pro Ser Trp (SEQ ID NO: 583)
SEQ ID NO. 584 представляет собой аминокислотную последовательность Rep 68 из AAV6.
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asn Trp Val Ala Glu
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Leu Asn Leu Ile
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
Val Gln Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Leu His Ile Leu Val Glu
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
Arg Asp Lys Leu Val Gln Thr Ile Tyr Arg Gly Ile Glu Pro Thr Leu
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Glu Tyr Ile
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala His Asp
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Leu Asn
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ala
Pro Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
Gln Lys Arg Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Glu Phe Phe Arg Trp Ala Gln Asp His Val Thr Glu Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Asn Lys Arg Pro Ala
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
Asp Leu Ala Arg Gly Gln Pro Leu (SEQ ID NO: 584)
SEQ ID NO. 585 представляет собой аминокислотную последовательность Rep 68 из AAV7.
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asn Trp Val Ala Glu
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Leu Asn Leu Ile
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
Val Gln Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Leu His Val Leu Val Glu
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
Arg Glu Lys Leu Val Gln Thr Ile Tyr Arg Gly Val Glu Pro Thr Leu
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Glu Tyr Ile
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Leu Asn
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ser
Leu Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Ser Lys Arg Pro Ala
Pro Asp Asp Ala Asp Ile Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
Asp Leu Ala Arg Gly Gln Pro Leu (SEQ ID NO: 585)
SEQ ID NO. 586 представляет собой аминокислотную последовательность Rep 68 из AAV8.
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asn Trp Val Ala Glu
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Arg Asn Leu Ile
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
Val Gln Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Leu His Val Leu Val Glu
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
Arg Glu Lys Leu Gly Pro Asp His Leu Pro Ala Gly Ser Ser Pro Thr
Leu Pro Asn Trp Phe Ala Val Thr Lys Asp Ala Val Met Ala Pro Ala
Gly Gly Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu
Pro Lys Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Glu
Tyr Ile Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala
Gln His Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn
Leu Asn Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala
Arg Tyr Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser
Glu Lys Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn
Ala Ala Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala
Gly Lys Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly
Pro Ser Leu Pro Ala Asp Ile Thr Gln Asn Arg Ile Tyr Arg Ile Leu
Ala Leu Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly
Trp Ala Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly
Pro Ala Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala
Val Pro Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe
Asn Asp Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met
Thr Ala Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys
Val Arg Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr
Pro Val Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly
Asn Ser Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe
Lys Phe Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr
Lys Gln Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr
Glu Val Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Ser Lys Arg
Pro Ala Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro
Ser Val Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp
Phe Ala Asp Leu Ala Arg Gly Gln Pro Leu (SEQ ID NO: 586)
SEQ ID NO. 587 представляет собой аминокислотную последовательность Rep 78 из AAV1.
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Ser Trp Val Ala Glu
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Leu Asn Leu Ile
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
Val Gln Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Leu His Ile Leu Val Glu
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
Arg Asp Lys Leu Val Gln Thr Ile Tyr Arg Gly Ile Glu Pro Thr Leu
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Glu Tyr Ile
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Leu Asn
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ala
Pro Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
Asn Gly Tyr Glu Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
Gln Lys Arg Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Glu Phe Phe Arg Trp Ala Gln Asp His Val Thr Glu Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Asn Lys Arg Pro Ala
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
Asp Arg Tyr Gln Asn Lys Cys Ser Arg His Ala Gly Met Leu Gln Met
Leu Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Asn Phe Asn Ile
Cys Phe Thr His Gly Thr Arg Asp Cys Ser Glu Cys Phe Pro Gly Val
Ser Glu Ser Gln Pro Val Val Arg Lys Arg Thr Tyr Arg Lys Leu Cys
Ala Ile His His Leu Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser Ala
Cys Asp Leu Val Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln (SEQ ID NO: 587)
SEQ ID NO. 588 представляет собой аминокислотную последовательность Rep 78 из AAV2.
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asn Trp Val Ala Glu
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Leu Asn Leu Ile
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
Thr Glu Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Met His Val Leu Val Glu
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
Arg Glu Lys Leu Ile Gln Arg Ile Tyr Arg Gly Ile Glu Pro Thr Leu
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Gln Tyr Leu
Ser Ala Cys Leu Asn Leu Thr Glu Arg Lys Arg Leu Val Ala Gln His
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Gln Asn
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
Met Glu Leu Val Gly Trp Leu Val Asp Lys Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Gln Gln
Pro Val Glu Asp Ile Ser Ser Asn Arg Ile Tyr Lys Ile Leu Glu Leu
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
Thr Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Thr Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Asp Phe Phe Arg Trp Ala Lys Asp His Val Val Glu Val
Glu His Glu Phe Tyr Val Lys Lys Gly Gly Ala Lys Lys Arg Pro Ala
Pro Ser Asp Ala Asp Ile Ser Glu Pro Lys Arg Val Arg Glu Ser Val
Ala Gln Pro Ser Thr Ser Asp Ala Glu Ala Ser Ile Asn Tyr Ala Asp
Arg Tyr Gln Asn Lys Cys Ser Arg His Val Gly Met Asn Leu Met Leu
Phe Pro Cys Arg Gln Cys Glu Arg Met Asn Gln Asn Ser Asn Ile Cys
Phe Thr His Gly Gln Lys Asp Cys Leu Glu Cys Phe Pro Val Ser Glu
Ser Gln Pro Val Ser Val Val Lys Lys Ala Tyr Gln Lys Leu Cys Tyr
Ile His His Ile Met Gly Lys Val Pro Asp Ala Cys Thr Ala Cys Asp
Leu Val Asn Val Asp Leu Asp Asp Cys Ile Phe Glu Gln (SEQ ID NO: 588)
SEQ ID NO. 589 представляет собой аминокислотную последовательность Rep 78 из AAV3A.
Met Pro Gly Phe Tyr Glu Ile Val Leu Lys Val Pro Ser Asp Leu Asp
Glu Arg Leu Pro Gly Ile Ser Asn Ser Phe Val Asn Trp Val Ala Glu
Lys Glu Trp Asp Val Pro Pro Asp Ser Asp Met Asp Pro Asn Leu Ile
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Glu Phe Leu
Val Glu Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
Gln Phe Glu Lys Gly Glu Thr Tyr Phe His Leu His Val Leu Ile Glu
Thr Ile Gly Val Lys Ser Met Val Val Gly Arg Tyr Val Ser Gln Ile
Lys Glu Lys Leu Val Thr Arg Ile Tyr Arg Gly Val Glu Pro Gln Leu
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
Asn Lys Val Val Asp Asp Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Asp Gln Tyr Leu
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Gln Asn
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Ser Asn
Pro Pro Glu Asp Ile Thr Lys Asn Arg Ile Tyr Gln Ile Leu Glu Leu
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Glu Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Glu Phe
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Asp Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
Ser Asn Asp Ala Asp Val Ser Glu Pro Lys Arg Glu Cys Thr Ser Leu
Ala Gln Pro Thr Thr Ser Asp Ala Glu Ala Pro Ala Asp Tyr Ala Asp
Arg Tyr Gln Asn Lys Cys Ser Arg His Val Gly Met Asn Leu Met Leu
Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Ile Ser Asn Val Cys
Phe Thr His Gly Gln Arg Asp Cys Gly Glu Cys Phe Pro Gly Met Ser
Glu Ser Gln Pro Val Ser Val Val Lys Lys Lys Thr Tyr Gln Lys Leu
Cys Pro Ile His His Ile Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser
Ala Cys Asp Leu Ala Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln (SEQ ID NO: 589)
SEQ ID NO. 590 представляет собой аминокислотную последовательность Rep 78 из AAV3B.
Met Pro Gly Phe Tyr Glu Ile Val Leu Lys Val Pro Ser Asp Leu Asp
Glu His Leu Pro Gly Ile Ser Asn Ser Phe Val Asn Trp Val Ala Glu
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Pro Asn Leu Ile
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Glu Phe Leu
Val Glu Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
Gln Phe Glu Lys Gly Glu Thr Tyr Phe His Leu His Val Leu Ile Glu
Thr Ile Gly Val Lys Ser Met Val Val Gly Arg Tyr Val Ser Gln Ile
Lys Glu Lys Leu Val Thr Arg Ile Tyr Arg Gly Val Glu Pro Gln Leu
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
Asn Lys Val Val Asp Asp Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Asp Gln Tyr Leu
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Gln Asn
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Ser Asn
Pro Pro Glu Asp Ile Thr Lys Asn Arg Ile Tyr Gln Ile Leu Glu Leu
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Glu Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Asp Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
Ser Asn Asp Ala Asp Val Ser Glu Pro Lys Arg Gln Cys Thr Ser Leu
Ala Gln Pro Thr Thr Ser Asp Ala Glu Ala Pro Ala Asp Tyr Ala Asp
Arg Tyr Gln Asn Lys Cys Ser Arg His Val Gly Met Asn Leu Met Leu
Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Ile Ser Asn Val Cys
Phe Thr His Gly Gln Arg Asp Cys Gly Glu Cys Phe Pro Gly Met Ser
Glu Ser Gln Pro Val Ser Val Val Lys Lys Lys Thr Tyr Gln Lys Leu
Cys Pro Ile His His Ile Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser
Ala Cys Asp Leu Ala Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln (SEQ ID NO: 590)
SEQ ID NO. 591 представляет собой аминокислотную последовательность Rep 78 из AAV4.
Met Pro Gly Phe Tyr Glu Ile Val Leu Lys Val Pro Ser Asp Leu Asp
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Ser Trp Val Ala Glu
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Leu Asn Leu Ile
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Glu Phe Leu
Val Glu Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
Gln Phe Glu Lys Gly Asp Ser Tyr Phe His Leu His Ile Leu Val Glu
Thr Val Gly Val Lys Ser Met Val Val Gly Arg Tyr Val Ser Gln Ile
Lys Glu Lys Leu Val Thr Arg Ile Tyr Arg Gly Val Glu Pro Gln Leu
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
Asn Lys Val Val Asp Asp Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Asp Gln Tyr Ile
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Gln Asn
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Gln Asn
Pro Pro Glu Asp Ile Ser Ser Asn Arg Ile Tyr Arg Ile Leu Glu Met
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Lys Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
Thr His Glu Phe Tyr Val Arg Lys Gly Gly Ala Arg Lys Arg Pro Ala
Pro Asn Asp Ala Asp Ile Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Gln Pro Ser Thr Ser Asp Ala Glu Ala Pro Val Asp Tyr Ala Asp
Arg Tyr Gln Asn Lys Cys Ser Arg His Val Gly Met Asn Leu Met Leu
Phe Pro Cys Arg Gln Cys Glu Arg Met Asn Gln Asn Val Asp Ile Cys
Phe Thr His Gly Val Met Asp Cys Ala Glu Cys Phe Pro Val Ser Glu
Ser Gln Pro Val Ser Val Val Arg Lys Arg Thr Tyr Gln Lys Leu Cys
Pro Ile His His Ile Met Gly Arg Ala Pro Glu Val Ala Cys Ser Ala
Cys Glu Leu Ala Asn Val Asp Leu Asp Asp Cys Asp Met Glu Gln (SEQ ID NO: 591)
SEQ ID NO. 592 представляет собой аминокислотную последовательность Rep 78 из AAV5.
Met Ala Thr Phe Tyr Glu Val Ile Val Arg Val Pro Phe Asp Val Glu
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asp Trp Val Thr Gly
Gln Ile Trp Glu Leu Pro Pro Glu Ser Asp Leu Asn Leu Thr Leu Val
Glu Gln Pro Gln Leu Thr Val Ala Asp Arg Ile Arg Arg Val Phe Leu
Tyr Glu Trp Asn Lys Phe Ser Lys Gln Glu Ser Lys Phe Phe Val Gln
Phe Glu Lys Gly Ser Glu Tyr Phe His Leu His Thr Leu Val Glu Thr
Ser Gly Ile Ser Ser Met Val Leu Gly Arg Tyr Val Ser Gln Ile Arg
Ala Gln Leu Val Lys Val Val Phe Gln Gly Ile Glu Pro Gln Ile Asn
Asp Trp Val Ala Ile Thr Lys Val Lys Lys Gly Gly Ala Asn Lys Val
Val Asp Ser Gly Tyr Ile Pro Ala Tyr Leu Leu Pro Lys Val Gln Pro
Glu Leu Gln Trp Ala Trp Thr Asn Leu Asp Glu Tyr Lys Leu Ala Ala
Leu Asn Leu Glu Glu Arg Lys Arg Leu Val Ala Gln Phe Leu Ala Glu
Ser Ser Gln Arg Ser Gln Glu Ala Ala Ser Gln Arg Glu Phe Ser Ala
Asp Pro Val Ile Lys Ser Lys Thr Ser Gln Lys Tyr Met Ala Leu Val
Asn Trp Leu Val Glu His Gly Ile Thr Ser Glu Lys Gln Trp Ile Gln
Glu Asn Gln Glu Ser Tyr Leu Ser Phe Asn Ser Thr Gly Asn Ser Arg
Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Thr Lys Ile Met Ser Leu
Thr Lys Ser Ala Val Asp Tyr Leu Val Gly Ser Ser Val Pro Glu Asp
Ile Ser Lys Asn Arg Ile Trp Gln Ile Phe Glu Met Asn Gly Tyr Asp
Pro Ala Tyr Ala Gly Ser Ile Leu Tyr Gly Trp Cys Gln Arg Ser Phe
Asn Lys Arg Asn Thr Val Trp Leu Tyr Gly Pro Ala Thr Thr Gly Lys
Thr Asn Ile Ala Glu Ala Ile Ala His Thr Val Pro Phe Tyr Gly Cys
Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp Cys Val Asp Lys
Met Leu Ile Trp Trp Glu Glu Gly Lys Met Thr Asn Lys Val Val Glu
Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg Val Asp Gln Lys
Cys Lys Ser Ser Val Gln Ile Asp Ser Thr Pro Val Ile Val Thr Ser
Asn Thr Asn Met Cys Val Val Val Asp Gly Asn Ser Thr Thr Phe Glu
His Gln Gln Pro Leu Glu Asp Arg Met Phe Lys Phe Glu Leu Thr Lys
Arg Leu Pro Pro Asp Phe Gly Lys Ile Thr Lys Gln Glu Val Lys Asp
Phe Phe Ala Trp Ala Lys Val Asn Gln Val Pro Val Thr His Glu Phe
Lys Val Pro Arg Glu Leu Ala Gly Thr Lys Gly Ala Glu Lys Ser Leu
Lys Arg Pro Leu Gly Asp Val Thr Asn Thr Ser Tyr Lys Ser Leu Glu
Lys Arg Ala Arg Leu Ser Phe Val Pro Glu Thr Pro Arg Ser Ser Asp
Val Thr Val Asp Pro Ala Pro Leu Arg Pro Leu Asn Trp Asn Ser Arg
Tyr Asp Cys Lys Cys Asp Tyr His Ala Gln Phe Asp Asn Ile Ser Asn
Lys Cys Asp Glu Cys Glu Tyr Leu Asn Arg Gly Lys Asn Gly Cys Ile
Cys His Asn Val Thr His Cys Gln Ile Cys His Gly Ile Pro Pro Trp
Glu Lys Glu Asn Leu Ser Asp Phe Gly Asp Phe Asp Asp Ala Asn Lys
Glu Gln (SEQ ID NO: 592)
SEQ ID NO. 593 представляет собой аминокислотную последовательность Rep 78 из AAV6.
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asn Trp Val Ala Glu
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Leu Asn Leu Ile
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
Val Gln Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Leu His Ile Leu Val Glu
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
Arg Asp Lys Leu Val Gln Thr Ile Tyr Arg Gly Ile Glu Pro Thr Leu
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Glu Tyr Ile
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala His Asp
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Leu Asn
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ala
Pro Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
Gln Lys Arg Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Glu Phe Phe Arg Trp Ala Gln Asp His Val Thr Glu Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Asn Lys Arg Pro Ala
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
Asp Arg Tyr Gln Asn Lys Cys Ser Arg His Ala Gly Met Leu Gln Met
Leu Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Asn Phe Asn Ile
Cys Phe Thr His Gly Thr Arg Asp Cys Ser Glu Cys Phe Pro Gly Val
Ser Glu Ser Gln Pro Val Val Arg Lys Arg Thr Tyr Arg Lys Leu Cys
Ala Ile His His Leu Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser Ala
Cys Asp Leu Val Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln (SEQ ID NO: 593)
SEQ ID NO. 594 представляет собой аминокислотную последовательность Rep 78 из AAV7.
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asn Trp Val Ala Glu
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Leu Asn Leu Ile
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
Val Gln Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Leu His Val Leu Val Glu
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
Arg Glu Lys Leu Val Gln Thr Ile Tyr Arg Gly Val Glu Pro Thr Leu
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Glu Tyr Ile
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Leu Asn
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ser
Leu Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Ser Lys Arg Pro Ala
Pro Asp Asp Ala Asp Ile Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
Asp Arg Tyr Gln Asn Lys Cys Ser Arg His Ala Gly Met Ile Gln Met
Leu Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Asn Phe Asn Ile
Cys Phe Thr His Gly Val Arg Asp Cys Leu Glu Cys Phe Pro Gly Val
Ser Glu Ser Gln Pro Val Val Arg Lys Lys Thr Tyr Arg Lys Leu Cys
Ala Ile His His Leu Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser Ala
Cys Asp Leu Val Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln (SEQ ID NO: 594)
SEQ ID NO. 595 представляет собой аминокислотную последовательность Rep 78 из AAV8.
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asn Trp Val Ala Glu
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Arg Asn Leu Ile
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
Val Gln Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Leu His Val Leu Val Glu
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
Arg Glu Lys Leu Gly Pro Asp His Leu Pro Ala Gly Ser Ser Pro Thr
Leu Pro Asn Trp Phe Ala Val Thr Lys Asp Ala Val Met Ala Pro Ala
Gly Gly Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu
Pro Lys Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Glu
Tyr Ile Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala
Gln His Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn
Leu Asn Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala
Arg Tyr Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser
Glu Lys Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn
Ala Ala Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala
Gly Lys Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly
Pro Ser Leu Pro Ala Asp Ile Thr Gln Asn Arg Ile Tyr Arg Ile Leu
Ala Leu Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly
Trp Ala Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly
Pro Ala Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala
Val Pro Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe
Asn Asp Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met
Thr Ala Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys
Val Arg Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr
Pro Val Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly
Asn Ser Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe
Lys Phe Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr
Lys Gln Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr
Glu Val Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Ser Lys Arg
Pro Ala Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro
Ser Val Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp
Phe Ala Asp Arg Tyr Gln Asn Lys Cys Ser Arg His Ala Gly Met Leu
Gln Met Leu Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Asn Phe
Asn Ile Cys Phe Thr His Gly Val Arg Asp Cys Ser Glu Cys Phe Pro
Gly Val Ser Glu Ser Gln Pro Val Val Arg Lys Arg Thr Tyr Arg Lys
Leu Cys Ala Ile His His Leu Leu Gly Arg Ala Pro Glu Ile Ala Cys
Ser Ala Cys Asp Leu Val Asn Val Asp Leu Asp Asp Cys Val Ser Glu
Gln (SEQ ID NO: 595)
SEQ ID NO. 596 представляет собой консенсусную аминокислотную последовательность Rep78 из SEQ ID NO: 587-595.
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asn Trp Val Ala Glu
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Arg Asn Leu Ile
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
Val Gln Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Leu His Val Leu Val Glu
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
Arg Glu Lys Leu Val Xaa Xaa Ile Tyr Arg Gly Ile Glu Pro Thr Leu
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Glu Tyr Ile
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Leu Asn
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Xaa Ser
Pro Pro Glu Asp Ile Ser Thr Asn Arg Ile Tyr Arg Ile Leu Ala Leu
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
Ala Asp Pro Ser Thr Ser Asp Ala Glu Ala Pro Val Asp Phe Ala Asp
Arg Tyr Gln Asn Lys Cys Ser Arg His Ala Gly Met Xaa Gln Met Leu
Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Asn Xaa Asn Ile Cys
Phe Thr His Gly Xaa Arg Asp Cys Xaa Glu Cys Phe Pro Gly Val Ser
Glu Ser Gln Xaa Val Val Arg Lys Arg Thr Tyr Xaa Lys Leu Cys Xaa
Ile His His Leu Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser Ala Cys
Asp Leu Val Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ДЖЕНЕРЕЙШЕН БИО КО.
<120> МОДУЛЯЦИЯ АКТИВНОСТИ REP БЕЛКА ПРИ ПОЛУЧЕНИИ ДНК С ЗАМКНУТЫМИ КОНЦАМИ (ЗКДНК)
<130> 131698-05420
<140>
<141>
<150> 62/806,076
<151> 2019-02-15
<160> 606
<170> PatentIn версия 3.5
<210> 1
<211> 141
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 1
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg gctttgcccg ggcggcctca gtgagcgagc 120
gagcgcgcag ctgcctgcag g 141
<210> 2
<211> 130
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 2
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg gcggcctcag tgagcgagcg agcgcgcagc 120
tgcctgcagg 130
<210> 3
<211> 1923
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 3
tcaatattgg ccattagcca tattattcat tggttatata gcataaatca atattggcta 60
ttggccattg catacgttgt atctatatca taatatgtac atttatattg gctcatgtcc 120
aatatgaccg ccatgttggc attgattatt gactagttat taatagtaat caattacggg 180
gtcattagtt catagcccat atatggagtt ccgcgttaca taacttacgg taaatggccc 240
gcctggctga ccgcccaacg acccccgccc attgacgtca ataatgacgt atgttcccat 300
agtaacgcca atagggactt tccattgacg tcaatgggtg gagtatttac ggtaaactgc 360
ccacttggca gtacatcaag tgtatcatat gccaagtccg ccccctattg acgtcaatga 420
cggtaaatgg cccgcctggc attatgccca gtacatgacc ttacgggact ttcctacttg 480
gcagtacatc tacgtattag tcatcgctat taccatggtc gaggtgagcc ccacgttctg 540
cttcactctc cccatctccc ccccctcccc acccccaatt ttgtatttat ttatttttta 600
attattttgt gcagcgatgg gggcgggggg gggggggggg cgcgcgccag gcggggcggg 660
gcggggcgag gggcggggcg gggcgaggcg gagaggtgcg gcggcagcca atcagagcgg 720
cgcgctccga aagtttcctt ttatggcgag gcggcggcgg cggcggccct ataaaaagcg 780
aagcgcgcgg cgggcgggag tcgctgcgac gctgccttcg ccccgtgccc cgctccgccg 840
ccgcctcgcg ccgcccgccc cggctctgac tgaccgcgtt actcccacag gtgagcgggc 900
gggacggccc ttctcctccg ggctgtaatt agcgcttggt ttaatgacgg cttgtttctt 960
ttctgtggct gcgtgaaagc cttgaggggc tccgggaggg ccctttgtgc gggggggagc 1020
ggctcggggg gtgcgtgcgt gtgtgtgtgc gtggggagcg ccgcgtgcgg cccgcgctgc 1080
ccggcggctg tgagcgctgc gggcgcggcg cggggctttg tgcgctccgc agtgtgcgcg 1140
aggggagcgc ggccgggggc ggtgccccgc ggtgcggggg gggctgcgag gggaacaaag 1200
gctgcgtgcg gggtgtgtgc gtgggggggt gagcaggggg tgtgggcgcg gcggtcgggc 1260
tgtaaccccc ccctgcaccc ccctccccga gttgctgagc acggcccggc ttcgggtgcg 1320
gggctccgta cggggcgtgg cgcggggctc gccgtgccgg gcggggggtg gcggcaggtg 1380
ggggtgccgg gcggggcggg gccgcctcgg gccggggagg gctcggggga ggggcgcggc 1440
ggcccccgga gcgccggcgg ctgtcgaggc gcggcgagcc gcagccattg ccttttatgg 1500
taatcgtgcg agagggcgca gggacttcct ttgtcccaaa tctgtgcgga gccgaaatct 1560
gggaggcgcc gccgcacccc ctctagcggg cgcggggcga agcggtgcgg cgccggcagg 1620
aaggaaatgg gcggggaggg ccttcgtgcg tcgccgcgcc gccgtcccct tctccctctc 1680
cagcctcggg gctgtccgcg gggggacggc tgccttcggg ggggacgggg cagggcgggg 1740
ttcggcttct ggcgtgtgac cggcggctct agagcctctg ctaaccatgt tttagccttc 1800
ttctttttcc tacagctcct gggcaacgtg ctggttattg tgctgtctca tcatttgtcg 1860
acagaattcc tcgaagatcc gaaggggttc aagcttggca ttccggtact gttggtaaag 1920
cca 1923
<210> 4
<211> 1272
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 4
aggctcagag gcacacagga gtttctgggc tcaccctgcc cccttccaac ccctcagttc 60
ccatcctcca gcagctgttt gtgtgctgcc tctgaagtcc acactgaaca aacttcagcc 120
tactcatgtc cctaaaatgg gcaaacattg caagcagcaa acagcaaaca cacagccctc 180
cctgcctgct gaccttggag ctggggcaga ggtcagagac ctctctgggc ccatgccacc 240
tccaacatcc actcgacccc ttggaatttc ggtggagagg agcagaggtt gtcctggcgt 300
ggtttaggta gtgtgagagg gtccgggttc aaaaccactt gctgggtggg gagtcgtcag 360
taagtggcta tgccccgacc ccgaagcctg tttccccatc tgtacaatgg aaatgataaa 420
gacgcccatc tgatagggtt tttgtggcaa ataaacattt ggtttttttg ttttgttttg 480
ttttgttttt tgagatggag gtttgctctg tcgcccaggc tggagtgcag tgacacaatc 540
tcatctcacc acaaccttcc cctgcctcag cctcccaagt agctgggatt acaagcatgt 600
gccaccacac ctggctaatt ttctattttt agtagagacg ggtttctcca tgttggtcag 660
cctcagcctc ccaagtaact gggattacag gcctgtgcca ccacacccgg ctaatttttt 720
ctatttttga cagggacggg gtttcaccat gttggtcagg ctggtctaga ggtaccggat 780
cttgctacca gtggaacagc cactaaggat tctgcagtga gagcagaggg ccagctaagt 840
ggtactctcc cagagactgt ctgactcacg ccaccccctc caccttggac acaggacgct 900
gtggtttctg agccaggtac aatgactcct ttcggtaagt gcagtggaag ctgtacactg 960
cccaggcaaa gcgtccgggc agcgtaggcg ggcgactcag atcccagcca gtggacttag 1020
cccctgtttg ctcctccgat aactggggtg accttggtta atattcacca gcagcctccc 1080
ccgttgcccc tctggatcca ctgcttaaat acggacgagg acagggccct gtctcctcag 1140
cttcaggcac caccactgac ctgggacagt gaatccggac tctaaggtaa atataaaatt 1200
tttaagtgta taatgtgtta aactactgat tctaattgtt tctctctttt agattccaac 1260
ctttggaact ga 1272
<210> 5
<211> 547
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 5
ccctaaaatg ggcaaacatt gcaagcagca aacagcaaac acacagccct ccctgcctgc 60
tgaccttgga gctggggcag aggtcagaga cctctctggg cccatgccac ctccaacatc 120
cactcgaccc cttggaattt ttcggtggag aggagcagag gttgtcctgg cgtggtttag 180
gtagtgtgag aggggaatga ctcctttcgg taagtgcagt ggaagctgta cactgcccag 240
gcaaagcgtc cgggcagcgt aggcgggcga ctcagatccc agccagtgga cttagcccct 300
gtttgctcct ccgataactg gggtgacctt ggttaatatt caccagcagc ctcccccgtt 360
gcccctctgg atccactgct taaatacgga cgaggacagg gccctgtctc ctcagcttca 420
ggcaccacca ctgacctggg acagtgaatc cggactctaa ggtaaatata aaatttttaa 480
gtgtataatg tgttaaacta ctgattctaa ttgtttctct cttttagatt ccaacctttg 540
gaactga 547
<210> 6
<211> 1179
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 6
ggctccggtg cccgtcagtg ggcagagcgc acatcgccca cagtccccga gaagttgggg 60
ggaggggtcg gcaattgaac cggtgcctag agaaggtggc gcggggtaaa ctgggaaagt 120
gatgtcgtgt actggctccg cctttttccc gagggtgggg gagaaccgta tataagtgca 180
gtagtcgccg tgaacgttct ttttcgcaac gggtttgccg ccagaacaca ggtaagtgcc 240
gtgtgtggtt cccgcgggcc tggcctcttt acgggttatg gcccttgcgt gccttgaatt 300
acttccacct ggctgcagta cgtgattctt gatcccgagc ttcgggttgg aagtgggtgg 360
gagagttcga ggccttgcgc ttaaggagcc ccttcgcctc gtgcttgagt tgaggcctgg 420
cctgggcgct ggggccgccg cgtgcgaatc tggtggcacc ttcgcgcctg tctcgctgct 480
ttcgataagt ctctagccat ttaaaatttt tgatgacctg ctgcgacgct ttttttctgg 540
caagatagtc ttgtaaatgc gggccaagat ctgcacactg gtatttcggt ttttggggcc 600
gcgggcggcg acggggcccg tgcgtcccag cgcacatgtt cggcgaggcg gggcctgcga 660
gcgcggccac cgagaatcgg acgggggtag tctcaagctg gccggcctgc tctggtgcct 720
ggtctcgcgc cgccgtgtat cgccccgccc tgggcggcaa ggctggcccg gtcggcacca 780
gttgcgtgag cggaaagatg gccgcttccc ggccctgctg cagggagctc aaaatggagg 840
acgcggcgct cgggagagcg ggcgggtgag tcacccacac aaaggaaaag ggcctttccg 900
tcctcagccg tcgcttcatg tgactccacg gagtaccggg cgccgtccag gcacctcgat 960
tagttctcga gcttttggag tacgtcgtct ttaggttggg gggaggggtt ttatgcgatg 1020
gagtttcccc acactgagtg ggtggagact gaagttaggc cagcttggca cttgatgtaa 1080
ttctccttgg aatttgccct ttttgagttt ggatcttggt tcattctcaa gcctcagaca 1140
gtggttcaaa gtttttttct tccatttcag gtgtcgtga 1179
<210> 7
<211> 8
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 7
gtttaaac 8
<210> 8
<211> 581
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 8
gagcatctta ccgccattta ttcccatatt tgttctgttt ttcttgattt gggtatacat 60
ttaaatgtta ataaaacaaa atggtggggc aatcatttac atttttaggg atatgtaatt 120
actagttcag gtgtattgcc acaagacaaa catgttaaga aactttcccg ttatttacgc 180
tctgttcctg ttaatcaacc tctggattac aaaatttgtg aaagattgac tgatattctt 240
aactatgttg ctccttttac gctgtgtgga tatgctgctt tatagcctct gtatctagct 300
attgcttccc gtacggcttt cgttttctcc tccttgtata aatcctggtt gctgtctctt 360
ttagaggagt tgtggcccgt tgtccgtcaa cgtggcgtgg tgtgctctgt gtttgctgac 420
gcaaccccca ctggctgggg cattgccacc acctgtcaac tcctttctgg gactttcgct 480
ttccccctcc cgatcgccac ggcagaactc atcgccgcct gccttgcccg ctgctggaca 540
ggggctaggt tgctgggcac tgataattcc gtggtgttgt c 581
<210> 9
<211> 225
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 9
tgtgccttct agttgccagc catctgttgt ttgcccctcc cccgtgcctt ccttgaccct 60
ggaaggtgcc actcccactg tcctttccta ataaaatgag gaaattgcat cgcattgtct 120
gagtaggtgt cattctattc tggggggtgg ggtggggcag gacagcaagg gggaggattg 180
ggaagacaat agcaggcatg ctggggatgc ggtgggctct atggc 225
<210> 10
<211> 213
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 10
taagatacat tgatgagttt ggacaaacca caactagaat gcagtgaaaa aaatgcttta 60
tttgtgaaat ttgtgatgct attgctttat ttgtaaccat tataagctgc aataaacaag 120
ttaacaacaa caattgcatt cattttatgt ttcaggttca gggggaggtg tgggaggttt 180
tttaaagcaa gtaaaacctc tacaaatgtg gta 213
<210> 11
<211> 1386
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 11
atgcagcgcg tgaacatgat catggccgag agccccggcc tgatcaccat ctgcctgctg 60
ggctacctgc tgagcgccga gtgcaccgtg ttcctggacc acgagaacgc caacaagatc 120
ctgaaccgcc ccaagcgcta caacagcggc aagctggagg agttcgtgca gggcaacctg 180
gagcgcgagt gcatggagga gaagtgcagc ttcgaggagg cccgcgaggt gttcgagaac 240
accgagcgca ccaccgagtt ctggaagcag tacgtggacg gcgaccagtg cgagagcaac 300
ccctgcctga acggcggcag ctgcaaggac gacatcaaca gctacgagtg ctggtgcccc 360
ttcggcttcg agggcaagaa ctgcgagctg gacgtgacct gcaacatcaa gaacggccgc 420
tgcgagcagt tctgcaagaa cagcgccgac aacaaggtgg tgtgcagctg caccgagggc 480
taccgcctgg ccgagaacca gaagagctgc gagcccgccg tgcccttccc ctgcggcagg 540
gtgagcgtga gccagaccag caagctgacc cgcgccgagg ccgtgttccc cgacgtggac 600
tacgtgaaca gcaccgaggc cgagaccatc ctggacaaca tcacccagag cacccagagc 660
ttcaacgact tcacccgcgt ggtgggcggc gaggacgcca agcccggcca gttcccctgg 720
caggtggtgc tgaacggcaa ggtggacgcc ttctgcggcg gcagcatcgt gaacgagaag 780
tggatcgtga ccgccgccca ctgcgtggag accggcgtga agatcaccgt ggtggccggc 840
gagcacaaca tcgaggagac cgagcacacc gagcagaagc gcaacgtgat ccgcatcatc 900
ccccaccaca actacaacgc cgccatcaac aagtacaacc acgacatcgc cctgctggag 960
ctggacgagc ccctggtgct gaacagctac gtgaccccca tctgcatcgc cgacaaggag 1020
tacaccaaca tcttcctgaa gttcggcagc ggctacgtga gcggctgggg ccgcgtgttc 1080
cacaagggcc gcagcgccct ggtgctgcag tacctgcgcg tgcccctggt ggaccgcgcc 1140
acctgcctgc gcagcaccaa gttcaccatc tacaacaaca tgttctgcgc cggcttccac 1200
gagggcggca gggacagctg ccagggcgac agcggcggcc cccacgtgac cgaggtggag 1260
ggcaccagct tcctgaccgg catcatcagc tggggcgagg agtgcgccat gaagggcaag 1320
tacggcatct acaccaaggt gagccgctac gtgaactgga tcaaggagaa gaccaagctg 1380
acctaa 1386
<210> 12
<211> 1386
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 12
atgcagcgcg tgaacatgat catggccgag agccccggcc tgatcaccat ctgcctgctg 60
ggctacctgc tgagcgccga gtgcaccgtg ttcctggacc acgagaacgc caacaagatc 120
ctgaaccgcc ccaagcgcta caacagcggc aagctggagg agttcgtgca gggcaacctg 180
gagcgcgagt gcatggagga gaagtgcagc ttcgaggagg cccgcgaggt gttcgagaac 240
accgagcgca ccaccgagtt ctggaagcag tacgtggacg gcgaccagtg cgagagcaac 300
ccctgcctga acggcggcag ctgcaaggac gacatcaaca gctacgagtg ctggtgcccc 360
ttcggcttcg agggcaagaa ctgcgagctg gacgtgacct gcaacatcaa gaacggccgc 420
tgcgagcagt tctgcaagaa cagcgccgac aacaaggtgg tgtgcagctg caccgagggc 480
taccgcctgg ccgagaacca gaagagctgc gagcccgccg tgcccttccc ctgcggcagg 540
gtgagcgtga gccagaccag caagctgacc cgcgccgagg ccgtgttccc cgacgtggac 600
tacgtgaaca gcaccgaggc cgagaccatc ctggacaaca tcacccagag cacccagagc 660
ttcaacgact tcacccgcgt ggtgggcggc gaggacgcca agcccggcca gttcccctgg 720
caggtggtgc tgaacggcaa ggtggacgcc ttctgcggcg gcagcatcgt gaacgagaag 780
tggatcgtga ccgccgccca ctgcgtggag accggcgtga agatcaccgt ggtggccggc 840
gagcacaaca tcgaggagac cgagcacacc gagcagaagc gcaacgtgat ccgcatcatc 900
ccccaccaca actacaacgc cgccatcaac aagtacaacc acgacatcgc cctgctggag 960
ctggacgagc ccctggtgct gaacagctac gtgaccccca tctgcatcgc cgacaaggag 1020
tacaccaaca tcttcctgaa gttcggcagc ggctacgtga gcggctgggg ccgcgtgttc 1080
cacaagggcc gcagcgccct ggtgctgcag tacctgcgcg tgcccctggt ggaccgcgcc 1140
acctgcctgc tgagcaccaa gttcaccatc tacaacaaca tgttctgcgc cggcttccac 1200
gagggcggca gggacagctg ccagggcgac agcggcggcc cccacgtgac cgaggtggag 1260
ggcaccagct tcctgaccgg catcatcagc tggggcgagg agtgcgccat gaagggcaag 1320
tacggcatct acaccaaggt gagccgctac gtgaactgga tcaaggagaa gaccaagctg 1380
acctaa 1386
<210> 13
<211> 1876
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 13
cgcagccacc atggcggggt tttacgagat tgtgattaag gtccccagcg accttgacgg 60
gcatctgccc ggcatttctg acagctttgt gaactgggtg gccgagaagg aatgggagtt 120
gccgccagat tctgacatgg atctgaatct gattgagcag gcacccctga ccgtggccga 180
gaagctgcag cgcgactttc tgacggaatg gcgccgtgtg agtaaggccc cggaggccct 240
tttctttgtg caatttgaga agggagagag ctacttccac atgcacgtgc tcgtggaaac 300
caccggggtg aaatccatgg ttttgggacg tttcctgagt cagattcgcg aaaaactgat 360
tcagagaatt taccgcggga tcgagccgac tttgccaaac tggttcgcgg tcacaaagac 420
cagaaatggc gccggaggcg ggaacaaggt ggtggatgag tgctacatcc ccaattactt 480
gctccccaaa acccagcctg agctccagtg ggcgtggact aatatggaac agtatttaag 540
cgcctgtttg aatctcacgg agcgtaaacg gttggtggcg cagcatctga cgcacgtgtc 600
gcagacgcag gagcagaaca aagagaatca gaatcccaat tctgatgcgc cggtgatcag 660
atcaaaaact tcagccaggt acatggagct ggtcgggtgg ctcgtggaca aggggattac 720
ctcggagaag cagtggatcc aggaggacca ggcctcatac atctccttca atgcggcctc 780
caactcgcgg tcccaaatca aggctgcctt ggacaatgcg ggaaagatta tgagcctgac 840
taaaaccgcc cccgactacc tggtgggcca gcagcccgtg gaggacattt ccagcaatcg 900
gatttataaa attttggaac taaacgggta cgatccccaa tatgcggctt ccgtctttct 960
gggatgggcc acgaaaaagt tcggcaagag gaacaccatc tggctgtttg ggcctgcaac 1020
taccgggaag accaacatcg cggaggccat agcccacact gtgcccttct acgggtgcgt 1080
aaactggacc aatgagaact ttcccttcaa cgactgtgtc gacaagatgg tgatctggtg 1140
ggaggagggg aagatgaccg ccaaggtcgt ggagtcggcc aaagccattc tcggaggaag 1200
caaggtgcgc gtggaccaga aatgcaagtc ctcggcccag atagacccga ctcccgtgat 1260
cgtcacctcc aacaccaaca tgtgcgccgt gattgacggg aactcaacga ccttcgaaca 1320
ccagcagccg ttgcaagacc ggatgttcaa atttgaactc acccgccgtc tggatcatga 1380
ctttgggaag gtcaccaagc aggaagtcaa agactttttc cggtgggcaa aggatcacgt 1440
ggttgaggtg gagcatgaat tctacgtcaa aaagggtgga gccaagaaaa gacccgcccc 1500
cagtgacgca gatataagtg agcccaaacg ggtgcgcgag tcagttgcgc agccatcgac 1560
gtcagacgcg gaagcttcga tcaactacgc agacaggtac caaaacaaat gttctcgtca 1620
cgtgggcatg aatctgatgc tgtttccctg cagacaatgc gagagaatga atcagaattc 1680
aaatatctgc ttcactcacg gacagaaaga ctgtttagag tgctttcccg tgtcagaatc 1740
tcaacccgtt tctgtcgtca aaaaggcgta tcagaaactg tgctacattc atcatatcat 1800
gggaaaggtg ccagacgctt gcactgcctg cgatctggtc aatgtggatt tggatgactg 1860
catctttgaa caataa 1876
<210> 14
<211> 1191
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 14
ttgttcaaag atgcagtcat ccaaatccac attgaccaga tcgcaggcag tgcaagcgtc 60
tggcaccttt cccatgatat gatgaatgta gcacagtttc tgatacgcct ttttgacgac 120
agaaacgggt tgagattctg acacgggaaa gcactctaaa cagtctttct gtccgtgagt 180
gaagcagata tttgaattct gattcattct ctcgcattgt ctgcagggaa acagcatcag 240
attcatgccc acgtgacgag aacatttgtt ttggtagcgg tctgcgtagt tgatcgaagc 300
ttccgcgtct gacgtcgatg gctgcgcaac tgactcgcgc acccgtttgg gctcacttat 360
atctgcgtca ctgggggcgg gtcttttctt ggctccaccc tttttgacgt agaattcatg 420
ctccacctca accacgtgat cctttgccca ccggaaaaag tctttgactt cctgcttggt 480
gaccttccca aagtcatgat ccagacggcg ggtgagttca aatttgaaca tccggtcttg 540
caacggctgc tggtgttcga aggtcgttga gttcccgtca atcacggcgc acatgttggt 600
gttggaggtg acgatcacgg gagtcgggtc tatctgggcc gaggacttgc atttctggtc 660
cacgcgcacc ttgcttcctc cgagaatggc tttggccgac tccacgacct tggcggtcat 720
cttcccctcc tcccaccaga tcaccatctt gtcgacacag tcgttgaagg gaaagttctc 780
attggtccag tttacgcacc cgtagaaggg cacagtgtgg gctatggcct ccgcgatgtt 840
ggtcttcccg gtagttgcag gcccaaacag ccagatggtg ttcctcttgc cgaacttttt 900
cgtggcccat cccagaaaga cggaagccgc atattgggga tcgtacccgt ttagttccaa 960
aattttataa atccgattgc tggaaatgtc ctccacgggc tgctggccca ccaggtagtc 1020
gggggcggtt ttagtcaggc tcataatctt tcccgcattg tccaaggcag ccttgatttg 1080
ggaccgcgag ttggaggccg cattgaagga gatgtatgag gcctggtcct cctggatcca 1140
ctgcttctcc gaggtaatcc ccttgtccac gagccacccg accagctcca t 1191
<210> 15
<211> 141
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 15
aataaacgat aacgccgttg gtggcgtgag gcatgtaaaa ggttacatca ttatcttgtt 60
cgccatccgg ttggtataaa tagacgttca tgttggtttt tgtttcagtt gcaagttggc 120
tgcggcgcgc gcagcacctt t 141
<210> 16
<211> 556
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 16
ccctaaaatg ggcaaacatt gcaagcagca aacagcaaac acacagccct ccctgcctgc 60
tgaccttgga gctggggcag aggtcagaga cctctctggg cccatgccac ctccaacatc 120
cactcgaccc cttggaattt cggtggagag gagcagaggt tgtcctggcg tggtttaggt 180
agtgtgagag gggaatgact cctttcggta agtgcagtgg aagctgtaca ctgcccaggc 240
aaagcgtccg ggcagcgtag gcgggcgact cagatcccag ccagtggact tagcccctgt 300
ttgctcctcc gataactggg gtgaccttgg ttaatattca ccagcagcct cccccgttgc 360
ccctctggat ccactgctta aatacggacg aggacactcg agggccctgt ctcctcagct 420
tcaggcacca ccactgacct gggacagtga atccggacat cgattctaag gtaaatataa 480
aatttttaag tgtataattt gttaaactac tgattctaat tgtttctctc ttttagattc 540
caacctttgg aactga 556
<210> 17
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 17
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 18
<400> 18
000
<210> 19
<400> 19
000
<210> 20
<400> 20
000
<210> 21
<400> 21
000
<210> 22
<400> 22
000
<210> 23
<400> 23
000
<210> 24
<400> 24
000
<210> 25
<400> 25
000
<210> 26
<400> 26
000
<210> 27
<400> 27
000
<210> 28
<400> 28
000
<210> 29
<400> 29
000
<210> 30
<400> 30
000
<210> 31
<400> 31
000
<210> 32
<400> 32
000
<210> 33
<400> 33
000
<210> 34
<400> 34
000
<210> 35
<400> 35
000
<210> 36
<400> 36
000
<210> 37
<400> 37
000
<210> 38
<400> 38
000
<210> 39
<400> 39
000
<210> 40
<400> 40
000
<210> 41
<400> 41
000
<210> 42
<400> 42
000
<210> 43
<400> 43
000
<210> 44
<400> 44
000
<210> 45
<211> 6
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 45
ggttga 6
<210> 46
<211> 4
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 46
agtt 4
<210> 47
<211> 6
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 47
ggttgg 6
<210> 48
<211> 6
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 48
agttgg 6
<210> 49
<211> 6
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 49
agttga 6
<210> 50
<211> 6
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 50
rrttrr 6
<210> 51
<211> 141
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 51
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc t 141
<210> 52
<211> 130
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 52
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcgtcg ggcgaccttt 60
ggtcgcccgg cctcagtgag cgagcgagcg cgcagagagg gagtggccaa ctccatcact 120
aggggttcct 130
<210> 53
<211> 3123
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 53
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggaaacc cgggcgtgcc 60
cgggcgcctc agtgagcgag cgagcgcgca gagagggagt ggccaactcc atcactaggg 120
gttcctgaac agagaaacag gagaatatgg gccaaacagg atatctgtgg taagcagttc 180
ctgccccggc tcagggccaa gaacagttgg aacagcagaa tatgggccaa acaggatatc 240
tgtggtaagc agttcctgcc ccggctcagg gccaagaaca gatggtcccc agatgcggtc 300
ccgccctcag cagtttctag agaaccatca gatgtttcca gggtgcccca aggacctgaa 360
atgaccctgt gccttatttg aactaaccaa tcagttcgct tctcgcttct gttcgcgcgc 420
ttctgctccc cgagctctat ataagcagag ctcgtttagt gaaccgtcag atcgcctgga 480
gacgccatcc acgctgtttt gacttccata gaaggccgcc accatggaag acgccaaaaa 540
cataaagaaa ggcccggcgc cattctatcc gctggaagat ggaaccgctg gagagcaact 600
gcataaggct atgaagagat acgccctggt tcctggaaca attgctttta cagatgcaca 660
tatcgaggtg gacatcactt acgctgagta cttcgaaatg tccgttcggt tggcagaagc 720
tatgaaacga tatgggctga atacaaatca cagaatcgtc gtatgcagtg aaaactctct 780
tcaattcttt atgccggtgt tgggcgcgtt atttatcgga gttgcagttg cgcccgcgaa 840
cgacatttat aatgaacgtg aattgctcaa cagtatgggc atttcgcagc ctaccgtggt 900
gttcgtttcc aaaaaggggt tgcaaaaaat tttgaacgtg caaaaaaagc tcccaatcat 960
ccaaaaaatt attatcatgg attctaaaac ggattaccag ggatttcagt cgatgtacac 1020
gttcgtcaca tctcatctac ctcccggttt taatgaatac gattttgtgc cagagtcctt 1080
cgatagggac aagacaattg cactgatcat gaactcctct ggatctactg gtctgcctaa 1140
aggtgtcgct ctgcctcata gaactgcctg cgtgagattc tcgcatgcca gagatcctat 1200
ttttggcaat caaatcattc cggatactgc gattttaagt gttgttccat tccatcacgg 1260
ttttggaatg tttactacac tcggatattt gatatgtgga tttcgagtcg tcttaatgta 1320
tagatttgaa gaagagctgt ttctgaggag ccttcaggat tacaagattc aaagtgcgct 1380
gctggtgcca accctattct ccttcttcgc caaaagcact ctgattgaca aatacgattt 1440
atctaattta cacgaaattg cttctggtgg cgctcccctc tctaaggaag tcggggaagc 1500
ggttgccaag aggttccatc tgccaggtat caggcaagga tatgggctca ctgagactac 1560
atcagctatt ctgattacac ccgaggggga tgataaaccg ggcgcggtcg gtaaagttgt 1620
tccatttttt gaagcgaagg ttgtggatct ggataccggg aaaacgctgg gcgttaatca 1680
aagaggcgaa ctgtgtgtga gaggtcctat gattatgtcc ggttatgtaa acaatccgga 1740
agcgaccaac gccttgattg acaaggatgg atggctacat tctggagaca tagcttactg 1800
ggacgaagac gaacacttct tcatcgttga ccgcctgaag tctctgatta agtacaaagg 1860
ctatcaggtg gctcccgctg aattggaatc catcttgctc caacacccca acatcttcga 1920
cgcaggtgtc gcaggtcttc ccgacgatga cgccggtgaa cttcccgccg ccgttgttgt 1980
tttggagcac ggaaagacga tgacggaaaa agagatcgtg gattacgtcg ccagtcaagt 2040
aacaaccgcg aaaaagttgc gcggaggagt tgtgtttgtg gacgaagtac cgaaaggtct 2100
taccggaaaa ctcgacgcaa gaaaaatcag agagatcctc ataaaggcca agaagggcgg 2160
aaagatcgcc gtgtaagagc atcttaccgc catttattcc catatttgtt ctgtttttct 2220
tgatttgggt atacatttaa atgttaataa aacaaaatgg tggggcaatc atttacattt 2280
ttagggatat gtaattacta gttcaggtgt attgccacaa gacaaacatg ttaagaaact 2340
ttcccgttat ttacgctctg ttcctgttaa tcaacctctg gattacaaaa tttgtgaaag 2400
attgactgat attcttaact atgttgctcc ttttacgctg tgtggatatg ctgctttata 2460
gcctctgtat ctagctattg cttcccgtac ggctttcgtt ttctcctcct tgtataaatc 2520
ctggttgctg tctcttttag aggagttgtg gcccgttgtc cgtcaacgtg gcgtggtgtg 2580
ctctgtgttt gctgacgcaa cccccactgg ctggggcatt gccaccacct gtcaactcct 2640
ttctgggact ttcgctttcc ccctcccgat cgccacggca gaactcatcg ccgcctgcct 2700
tgcccgctgc tggacagggg ctaggttgct gggcactgat aattccgtgg tgttgtctgt 2760
gccttctagt tgccagccat ctgttgtttg cccctccccc gtgccttcct tgaccctgga 2820
aggtgccact cccactgtcc tttcctaata aaatgaggaa attgcatcgc attgtctgag 2880
taggtgtcat tctattctgg ggggtggggt ggggcaggac agcaaggggg aggattggga 2940
agacaatagc aggcatgctg gggatgcggt gggctctatg gcaggaaccc ctagtgatgg 3000
agttggccac tccctctctg cgcgctcgct cgctcactga ggccgggcga ccaaaggtcg 3060
cccgacgccc gggctttgcc cgggcggcct cagtgagcga gcgagcgcgc agctgcctgc 3120
agg 3123
<210> 54
<211> 3117
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 54
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc tgaacagaga aacaggagaa tatgggccaa acaggatatc 180
tgtggtaagc agttcctgcc ccggctcagg gccaagaaca gttggaacag cagaatatgg 240
gccaaacagg atatctgtgg taagcagttc ctgccccggc tcagggccaa gaacagatgg 300
tccccagatg cggtcccgcc ctcagcagtt tctagagaac catcagatgt ttccagggtg 360
ccccaaggac ctgaaatgac cctgtgcctt atttgaacta accaatcagt tcgcttctcg 420
cttctgttcg cgcgcttctg ctccccgagc tctatataag cagagctcgt ttagtgaacc 480
gtcagatcgc ctggagacgc catccacgct gttttgactt ccatagaagg ccgccaccat 540
ggaagacgcc aaaaacataa agaaaggccc ggcgccattc tatccgctgg aagatggaac 600
cgctggagag caactgcata aggctatgaa gagatacgcc ctggttcctg gaacaattgc 660
ttttacagat gcacatatcg aggtggacat cacttacgct gagtacttcg aaatgtccgt 720
tcggttggca gaagctatga aacgatatgg gctgaataca aatcacagaa tcgtcgtatg 780
cagtgaaaac tctcttcaat tctttatgcc ggtgttgggc gcgttattta tcggagttgc 840
agttgcgccc gcgaacgaca tttataatga acgtgaattg ctcaacagta tgggcatttc 900
gcagcctacc gtggtgttcg tttccaaaaa ggggttgcaa aaaattttga acgtgcaaaa 960
aaagctccca atcatccaaa aaattattat catggattct aaaacggatt accagggatt 1020
tcagtcgatg tacacgttcg tcacatctca tctacctccc ggttttaatg aatacgattt 1080
tgtgccagag tccttcgata gggacaagac aattgcactg atcatgaact cctctggatc 1140
tactggtctg cctaaaggtg tcgctctgcc tcatagaact gcctgcgtga gattctcgca 1200
tgccagagat cctatttttg gcaatcaaat cattccggat actgcgattt taagtgttgt 1260
tccattccat cacggttttg gaatgtttac tacactcgga tatttgatat gtggatttcg 1320
agtcgtctta atgtatagat ttgaagaaga gctgtttctg aggagccttc aggattacaa 1380
gattcaaagt gcgctgctgg tgccaaccct attctccttc ttcgccaaaa gcactctgat 1440
tgacaaatac gatttatcta atttacacga aattgcttct ggtggcgctc ccctctctaa 1500
ggaagtcggg gaagcggttg ccaagaggtt ccatctgcca ggtatcaggc aaggatatgg 1560
gctcactgag actacatcag ctattctgat tacacccgag ggggatgata aaccgggcgc 1620
ggtcggtaaa gttgttccat tttttgaagc gaaggttgtg gatctggata ccgggaaaac 1680
gctgggcgtt aatcaaagag gcgaactgtg tgtgagaggt cctatgatta tgtccggtta 1740
tgtaaacaat ccggaagcga ccaacgcctt gattgacaag gatggatggc tacattctgg 1800
agacatagct tactgggacg aagacgaaca cttcttcatc gttgaccgcc tgaagtctct 1860
gattaagtac aaaggctatc aggtggctcc cgctgaattg gaatccatct tgctccaaca 1920
ccccaacatc ttcgacgcag gtgtcgcagg tcttcccgac gatgacgccg gtgaacttcc 1980
cgccgccgtt gttgttttgg agcacggaaa gacgatgacg gaaaaagaga tcgtggatta 2040
cgtcgccagt caagtaacaa ccgcgaaaaa gttgcgcgga ggagttgtgt ttgtggacga 2100
agtaccgaaa ggtcttaccg gaaaactcga cgcaagaaaa atcagagaga tcctcataaa 2160
ggccaagaag ggcggaaaga tcgccgtgta agagcatctt accgccattt attcccatat 2220
ttgttctgtt tttcttgatt tgggtataca tttaaatgtt aataaaacaa aatggtgggg 2280
caatcattta catttttagg gatatgtaat tactagttca ggtgtattgc cacaagacaa 2340
acatgttaag aaactttccc gttatttacg ctctgttcct gttaatcaac ctctggatta 2400
caaaatttgt gaaagattga ctgatattct taactatgtt gctcctttta cgctgtgtgg 2460
atatgctgct ttatagcctc tgtatctagc tattgcttcc cgtacggctt tcgttttctc 2520
ctccttgtat aaatcctggt tgctgtctct tttagaggag ttgtggcccg ttgtccgtca 2580
acgtggcgtg gtgtgctctg tgtttgctga cgcaaccccc actggctggg gcattgccac 2640
cacctgtcaa ctcctttctg ggactttcgc tttccccctc ccgatcgcca cggcagaact 2700
catcgccgcc tgccttgccc gctgctggac aggggctagg ttgctgggca ctgataattc 2760
cgtggtgttg tctgtgcctt ctagttgcca gccatctgtt gtttgcccct cccccgtgcc 2820
ttccttgacc ctggaaggtg ccactcccac tgtcctttcc taataaaatg aggaaattgc 2880
atcgcattgt ctgagtaggt gtcattctat tctggggggt ggggtggggc aggacagcaa 2940
gggggaggat tgggaagaca atagcaggca tgctggggat gcggtgggct ctatggcagg 3000
aacccctagt gatggagttg gccactccct ctctgcgcgc tcgctcgctc actgaggccg 3060
cccgggaaac ccgggcgtgc gcctcagtga gcgagcgagc gcgcagctgc ctgcagg 3117
<210> 55
<211> 2841
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 55
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggaaacc cgggcgtgcc 60
cgggcgcctc agtgagcgag cgagcgcgca gagagggagt ggccaactcc atcactaggg 120
gttcctgaac agagaaacag gagaatatgg gccaaacagg atatctgtgg taagcagttc 180
ctgccccggc tcagggccaa gaacagttgg aacagcagaa tatgggccaa acaggatatc 240
tgtggtaagc agttcctgcc ccggctcagg gccaagaaca gatggtcccc agatgcggtc 300
ccgccctcag cagtttctag agaaccatca gatgtttcca gggtgcccca aggacctgaa 360
atgaccctgt gccttatttg aactaaccaa tcagttcgct tctcgcttct gttcgcgcgc 420
ttctgctccc cgagctctat ataagcagag ctcgtttagt gaaccgtcag atcgcctgga 480
gacgccatcc acgctgtttt gacttccata gaaggccgcc accatgatca tggccgagag 540
ccctggcctg atcaccatct gcctgctggg ctacctgctg agcgccgagt gcaccgtgtt 600
cctggaccac gagaacgcca acaagatcct gaaccggccc aagagataca acagcggcaa 660
gctggaggag ttcgtgcagg gcaacctgga gagggagtgc atggaggaga agtgcagctt 720
cgaggaggcc agggaagtgt tcgagaacac cgagcggacc accgagttct ggaagcagta 780
cgtggacggc gaccagtgcg agagcaaccc ttgcctgaac ggcggcagct gcaaggacga 840
catcaacagc tacgagtgct ggtgcccttt cggcttcgag ggcaagaact gcgagctgga 900
cgtgacctgc aacatcaaga acggccgctg cgagcagttc tgcaagaaca gcgccgacaa 960
caaagtggtg tgtagctgca ccgagggcta cagactggcc gagaaccaga agagctgcga 1020
gcccgccgtg cccttcccct gcggcagagt gagcgtgtcc cagaccagca agctgaccag 1080
agccgagacc gtgttccccg acgtggacta cgtgaatagc accgaggccg agaccatcct 1140
ggacaacatc acccagagca cccagtcctt caacgacttc accagagttg tgggcggcga 1200
ggacgccaag cccggccagt tcccctggca ggtggtgctg aacggcaaag tggatgcctt 1260
ctgcggcggc agcatcgtga acgagaagtg gatcgtgaca gccgcccact gcgtggagac 1320
cggcgtgaag atcaccgtgg tggccggcga acacaatatc gaggagaccg agcacaccga 1380
gcagaagcgg aacgtcatcc ggattatccc ccaccacaac tacaacgccg ccatcaacaa 1440
gtacaaccac gacatcgccc tgctggagct ggacgagcct ctggtgctga atagctacgt 1500
gacccccatc tgcatcgccg acaaggagta caccaacatc ttcctgaagt tcggcagcgg 1560
ctacgtgtcc ggctggggca gagtgttcca caagggcaga agcgccctgg tgctgcagta 1620
cctgagagtg cccctggtgg acagagccac ctgcctgttg agcaccaagt tcaccatcta 1680
caacaacatg ttctgcgccg gcttccacga gggcggcaga gacagctgcc agggcgacag 1740
cggcggaccc cacgtgaccg aagtggaggg caccagcttc ctgaccggca tcatcagctg 1800
gggcgaggag tgcgccatga agggcaagta cggcatctac accaaagtga gccggtacgt 1860
gaactggatc aaggagaaaa ccaagctgac ctgagagcat cttaccgcca tttattccca 1920
tatttgttct gtttttcttg atttgggtat acatttaaat gttaataaaa caaaatggtg 1980
gggcaatcat ttacattttt agggatatgt aattactagt tcaggtgtat tgccacaaga 2040
caaacatgtt aagaaacttt cccgttattt acgctctgtt cctgttaatc aacctctgga 2100
ttacaaaatt tgtgaaagat tgactgatat tcttaactat gttgctcctt ttacgctgtg 2160
tggatatgct gctttatagc ctctgtatct agctattgct tcccgtacgg ctttcgtttt 2220
ctcctccttg tataaatcct ggttgctgtc tcttttagag gagttgtggc ccgttgtccg 2280
tcaacgtggc gtggtgtgct ctgtgtttgc tgacgcaacc cccactggct ggggcattgc 2340
caccacctgt caactccttt ctgggacttt cgctttcccc ctcccgatcg ccacggcaga 2400
actcatcgcc gcctgccttg cccgctgctg gacaggggct aggttgctgg gcactgataa 2460
ttccgtggtg ttgtctgtgc cttctagttg ccagccatct gttgtttgcc cctcccccgt 2520
gccttccttg accctggaag gtgccactcc cactgtcctt tcctaataaa atgaggaaat 2580
tgcatcgcat tgtctgagta ggtgtcattc tattctgggg ggtggggtgg ggcaggacag 2640
caagggggag gattgggaag acaatagcag gcatgctggg gatgcggtgg gctctatggc 2700
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 2760
ccgggcgacc aaaggtcgcc cgacgcccgg gctttgcccg ggcggcctca gtgagcgagc 2820
gagcgcgcag ctgcctgcag g 2841
<210> 56
<211> 2835
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 56
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc tgaacagaga aacaggagaa tatgggccaa acaggatatc 180
tgtggtaagc agttcctgcc ccggctcagg gccaagaaca gttggaacag cagaatatgg 240
gccaaacagg atatctgtgg taagcagttc ctgccccggc tcagggccaa gaacagatgg 300
tccccagatg cggtcccgcc ctcagcagtt tctagagaac catcagatgt ttccagggtg 360
ccccaaggac ctgaaatgac cctgtgcctt atttgaacta accaatcagt tcgcttctcg 420
cttctgttcg cgcgcttctg ctccccgagc tctatataag cagagctcgt ttagtgaacc 480
gtcagatcgc ctggagacgc catccacgct gttttgactt ccatagaagg ccgccaccat 540
gatcatggcc gagagccctg gcctgatcac catctgcctg ctgggctacc tgctgagcgc 600
cgagtgcacc gtgttcctgg accacgagaa cgccaacaag atcctgaacc ggcccaagag 660
atacaacagc ggcaagctgg aggagttcgt gcagggcaac ctggagaggg agtgcatgga 720
ggagaagtgc agcttcgagg aggccaggga agtgttcgag aacaccgagc ggaccaccga 780
gttctggaag cagtacgtgg acggcgacca gtgcgagagc aacccttgcc tgaacggcgg 840
cagctgcaag gacgacatca acagctacga gtgctggtgc cctttcggct tcgagggcaa 900
gaactgcgag ctggacgtga cctgcaacat caagaacggc cgctgcgagc agttctgcaa 960
gaacagcgcc gacaacaaag tggtgtgtag ctgcaccgag ggctacagac tggccgagaa 1020
ccagaagagc tgcgagcccg ccgtgccctt cccctgcggc agagtgagcg tgtcccagac 1080
cagcaagctg accagagccg agaccgtgtt ccccgacgtg gactacgtga atagcaccga 1140
ggccgagacc atcctggaca acatcaccca gagcacccag tccttcaacg acttcaccag 1200
agttgtgggc ggcgaggacg ccaagcccgg ccagttcccc tggcaggtgg tgctgaacgg 1260
caaagtggat gccttctgcg gcggcagcat cgtgaacgag aagtggatcg tgacagccgc 1320
ccactgcgtg gagaccggcg tgaagatcac cgtggtggcc ggcgaacaca atatcgagga 1380
gaccgagcac accgagcaga agcggaacgt catccggatt atcccccacc acaactacaa 1440
cgccgccatc aacaagtaca accacgacat cgccctgctg gagctggacg agcctctggt 1500
gctgaatagc tacgtgaccc ccatctgcat cgccgacaag gagtacacca acatcttcct 1560
gaagttcggc agcggctacg tgtccggctg gggcagagtg ttccacaagg gcagaagcgc 1620
cctggtgctg cagtacctga gagtgcccct ggtggacaga gccacctgcc tgttgagcac 1680
caagttcacc atctacaaca acatgttctg cgccggcttc cacgagggcg gcagagacag 1740
ctgccagggc gacagcggcg gaccccacgt gaccgaagtg gagggcacca gcttcctgac 1800
cggcatcatc agctggggcg aggagtgcgc catgaagggc aagtacggca tctacaccaa 1860
agtgagccgg tacgtgaact ggatcaagga gaaaaccaag ctgacctgag agcatcttac 1920
cgccatttat tcccatattt gttctgtttt tcttgatttg ggtatacatt taaatgttaa 1980
taaaacaaaa tggtggggca atcatttaca tttttaggga tatgtaatta ctagttcagg 2040
tgtattgcca caagacaaac atgttaagaa actttcccgt tatttacgct ctgttcctgt 2100
taatcaacct ctggattaca aaatttgtga aagattgact gatattctta actatgttgc 2160
tccttttacg ctgtgtggat atgctgcttt atagcctctg tatctagcta ttgcttcccg 2220
tacggctttc gttttctcct ccttgtataa atcctggttg ctgtctcttt tagaggagtt 2280
gtggcccgtt gtccgtcaac gtggcgtggt gtgctctgtg tttgctgacg caacccccac 2340
tggctggggc attgccacca cctgtcaact cctttctggg actttcgctt tccccctccc 2400
gatcgccacg gcagaactca tcgccgcctg ccttgcccgc tgctggacag gggctaggtt 2460
gctgggcact gataattccg tggtgttgtc tgtgccttct agttgccagc catctgttgt 2520
ttgcccctcc cccgtgcctt ccttgaccct ggaaggtgcc actcccactg tcctttccta 2580
ataaaatgag gaaattgcat cgcattgtct gagtaggtgt cattctattc tggggggtgg 2640
ggtggggcag gacagcaagg gggaggattg ggaagacaat agcaggcatg ctggggatgc 2700
ggtgggctct atggcaggaa cccctagtga tggagttggc cactccctct ctgcgcgctc 2760
gctcgctcac tgaggccgcc cgggaaaccc gggcgtgcgc ctcagtgagc gagcgagcgc 2820
gcagctgcct gcagg 2835
<210> 57
<211> 3912
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 57
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggaaacc cgggcgtgcc 60
cgggcgcctc agtgagcgag cgagcgcgca gagagggagt ggccaactcc atcactaggg 120
gttcctggct cagaggctca gaggcacaca ggagtttctg ggctcaccct gcccccttcc 180
aacccctcag ttcccatcct ccagcagctg tttgtgtgct gcctctgaag tccacactga 240
acaaacttca gcctactcat gtccctaaaa tgggcaaaca ttgcaagcag caaacagcaa 300
acacacagcc ctccctgcct gctgaccttg gagctggggc agaggtcaga gacctctctg 360
ggcccatgcc acctccaaca tccactcgac cccttggaat ttcggtggag aggagcagag 420
gttgtcctgg cgtggtttag gtagtgtgag agggtccggg ttcaaaacca cttgctgggt 480
ggggagtcgt cagtaagtgg ctatgccccg accccgaagc ctgtttcccc atctgtacaa 540
tggaaatgat aaagacgccc atctgatagg gtttttgtgg caaataaaca tttggttttt 600
ttgttttgtt ttgttttgtt ttttgagatg gaggtttgct ctgtcgccca ggctggagtg 660
cagtgacaca atctcatctc accacaacct tcccctgcct cagcctccca agtagctggg 720
attacaagca tgtgccacca cacctggcta attttctatt tttagtagag acgggtttct 780
ccatgttggt cagcctcagc ctcccaagta actgggatta caggcctgtg ccaccacacc 840
cggctaattt tttctatttt tgacagggac ggggtttcac catgttggtc aggctggtct 900
agaggtaccg gatcttgcta ccagtggaac agccactaag gattctgcag tgagagcaga 960
gggccagcta agtggtactc tcccagagac tgtctgactc acgccacccc ctccaccttg 1020
gacacaggac gctgtggttt ctgagccagg tacaatgact cctttcggta agtgcagtgg 1080
aagctgtaca ctgcccaggc aaagcgtccg ggcagcgtag gcgggcgact cagatcccag 1140
ccagtggact tagcccctgt ttgctcctcc gataactggg gtgaccttgg ttaatattca 1200
ccagcagcct cccccgttgc ccctctggat ccactgctta aatacggacg aggacagggc 1260
cctgtctcct cagcttcagg caccaccact gacctgggac agtgccgcca ccatggaaga 1320
cgccaaaaac ataaagaaag gcccggcgcc attctatccg ctggaagatg gaaccgctgg 1380
agagcaactg cataaggcta tgaagagata cgccctggtt cctggaacaa ttgcttttac 1440
agatgcacat atcgaggtgg acatcactta cgctgagtac ttcgaaatgt ccgttcggtt 1500
ggcagaagct atgaaacgat atgggctgaa tacaaatcac agaatcgtcg tatgcagtga 1560
aaactctctt caattcttta tgccggtgtt gggcgcgtta tttatcggag ttgcagttgc 1620
gcccgcgaac gacatttata atgaacgtga attgctcaac agtatgggca tttcgcagcc 1680
taccgtggtg ttcgtttcca aaaaggggtt gcaaaaaatt ttgaacgtgc aaaaaaagct 1740
cccaatcatc caaaaaatta ttatcatgga ttctaaaacg gattaccagg gatttcagtc 1800
gatgtacacg ttcgtcacat ctcatctacc tcccggtttt aatgaatacg attttgtgcc 1860
agagtccttc gatagggaca agacaattgc actgatcatg aactcctctg gatctactgg 1920
tctgcctaaa ggtgtcgctc tgcctcatag aactgcctgc gtgagattct cgcatgccag 1980
agatcctatt tttggcaatc aaatcattcc ggatactgcg attttaagtg ttgttccatt 2040
ccatcacggt tttggaatgt ttactacact cggatatttg atatgtggat ttcgagtcgt 2100
cttaatgtat agatttgaag aagagctgtt tctgaggagc cttcaggatt acaagattca 2160
aagtgcgctg ctggtgccaa ccctattctc cttcttcgcc aaaagcactc tgattgacaa 2220
atacgattta tctaatttac acgaaattgc ttctggtggc gctcccctct ctaaggaagt 2280
cggggaagcg gttgccaaga ggttccatct gccaggtatc aggcaaggat atgggctcac 2340
tgagactaca tcagctattc tgattacacc cgagggggat gataaaccgg gcgcggtcgg 2400
taaagttgtt ccattttttg aagcgaaggt tgtggatctg gataccggga aaacgctggg 2460
cgttaatcaa agaggcgaac tgtgtgtgag aggtcctatg attatgtccg gttatgtaaa 2520
caatccggaa gcgaccaacg ccttgattga caaggatgga tggctacatt ctggagacat 2580
agcttactgg gacgaagacg aacacttctt catcgttgac cgcctgaagt ctctgattaa 2640
gtacaaaggc tatcaggtgg ctcccgctga attggaatcc atcttgctcc aacaccccaa 2700
catcttcgac gcaggtgtcg caggtcttcc cgacgatgac gccggtgaac ttcccgccgc 2760
cgttgttgtt ttggagcacg gaaagacgat gacggaaaaa gagatcgtgg attacgtcgc 2820
cagtcaagta acaaccgcga aaaagttgcg cggaggagtt gtgtttgtgg acgaagtacc 2880
gaaaggtctt accggaaaac tcgacgcaag aaaaatcaga gagatcctca taaaggccaa 2940
gaagggcgga aagatcgccg tgtaagagca tcttaccgcc atttattccc atatttgttc 3000
tgtttttctt gatttgggta tacatttaaa tgttaataaa acaaaatggt ggggcaatca 3060
tttacatttt tagggatatg taattactag ttcaggtgta ttgccacaag acaaacatgt 3120
taagaaactt tcccgttatt tacgctctgt tcctgttaat caacctctgg attacaaaat 3180
ttgtgaaaga ttgactgata ttcttaacta tgttgctcct tttacgctgt gtggatatgc 3240
tgctttatag cctctgtatc tagctattgc ttcccgtacg gctttcgttt tctcctcctt 3300
gtataaatcc tggttgctgt ctcttttaga ggagttgtgg cccgttgtcc gtcaacgtgg 3360
cgtggtgtgc tctgtgtttg ctgacgcaac ccccactggc tggggcattg ccaccacctg 3420
tcaactcctt tctgggactt tcgctttccc cctcccgatc gccacggcag aactcatcgc 3480
cgcctgcctt gcccgctgct ggacaggggc taggttgctg ggcactgata attccgtggt 3540
gttgtctgtg ccttctagtt gccagccatc tgttgtttgc ccctcccccg tgccttcctt 3600
gaccctggaa ggtgccactc ccactgtcct ttcctaataa aatgaggaaa ttgcatcgca 3660
ttgtctgagt aggtgtcatt ctattctggg gggtggggtg gggcaggaca gcaaggggga 3720
ggattgggaa gacaatagca ggcatgctgg ggatgcggtg ggctctatgg caggaacccc 3780
tagtgatgga gttggccact ccctctctgc gcgctcgctc gctcactgag gccgggcgac 3840
caaaggtcgc ccgacgcccg ggctttgccc gggcggcctc agtgagcgag cgagcgcgca 3900
gctgcctgca gg 3912
<210> 58
<211> 3906
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 58
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc tggctcagag gctcagaggc acacaggagt ttctgggctc 180
accctgcccc cttccaaccc ctcagttccc atcctccagc agctgtttgt gtgctgcctc 240
tgaagtccac actgaacaaa cttcagccta ctcatgtccc taaaatgggc aaacattgca 300
agcagcaaac agcaaacaca cagccctccc tgcctgctga ccttggagct ggggcagagg 360
tcagagacct ctctgggccc atgccacctc caacatccac tcgacccctt ggaatttcgg 420
tggagaggag cagaggttgt cctggcgtgg tttaggtagt gtgagagggt ccgggttcaa 480
aaccacttgc tgggtgggga gtcgtcagta agtggctatg ccccgacccc gaagcctgtt 540
tccccatctg tacaatggaa atgataaaga cgcccatctg atagggtttt tgtggcaaat 600
aaacatttgg tttttttgtt ttgttttgtt ttgttttttg agatggaggt ttgctctgtc 660
gcccaggctg gagtgcagtg acacaatctc atctcaccac aaccttcccc tgcctcagcc 720
tcccaagtag ctgggattac aagcatgtgc caccacacct ggctaatttt ctatttttag 780
tagagacggg tttctccatg ttggtcagcc tcagcctccc aagtaactgg gattacaggc 840
ctgtgccacc acacccggct aattttttct atttttgaca gggacggggt ttcaccatgt 900
tggtcaggct ggtctagagg taccggatct tgctaccagt ggaacagcca ctaaggattc 960
tgcagtgaga gcagagggcc agctaagtgg tactctccca gagactgtct gactcacgcc 1020
accccctcca ccttggacac aggacgctgt ggtttctgag ccaggtacaa tgactccttt 1080
cggtaagtgc agtggaagct gtacactgcc caggcaaagc gtccgggcag cgtaggcggg 1140
cgactcagat cccagccagt ggacttagcc cctgtttgct cctccgataa ctggggtgac 1200
cttggttaat attcaccagc agcctccccc gttgcccctc tggatccact gcttaaatac 1260
ggacgaggac agggccctgt ctcctcagct tcaggcacca ccactgacct gggacagtgc 1320
cgccaccatg gaagacgcca aaaacataaa gaaaggcccg gcgccattct atccgctgga 1380
agatggaacc gctggagagc aactgcataa ggctatgaag agatacgccc tggttcctgg 1440
aacaattgct tttacagatg cacatatcga ggtggacatc acttacgctg agtacttcga 1500
aatgtccgtt cggttggcag aagctatgaa acgatatggg ctgaatacaa atcacagaat 1560
cgtcgtatgc agtgaaaact ctcttcaatt ctttatgccg gtgttgggcg cgttatttat 1620
cggagttgca gttgcgcccg cgaacgacat ttataatgaa cgtgaattgc tcaacagtat 1680
gggcatttcg cagcctaccg tggtgttcgt ttccaaaaag gggttgcaaa aaattttgaa 1740
cgtgcaaaaa aagctcccaa tcatccaaaa aattattatc atggattcta aaacggatta 1800
ccagggattt cagtcgatgt acacgttcgt cacatctcat ctacctcccg gttttaatga 1860
atacgatttt gtgccagagt ccttcgatag ggacaagaca attgcactga tcatgaactc 1920
ctctggatct actggtctgc ctaaaggtgt cgctctgcct catagaactg cctgcgtgag 1980
attctcgcat gccagagatc ctatttttgg caatcaaatc attccggata ctgcgatttt 2040
aagtgttgtt ccattccatc acggttttgg aatgtttact acactcggat atttgatatg 2100
tggatttcga gtcgtcttaa tgtatagatt tgaagaagag ctgtttctga ggagccttca 2160
ggattacaag attcaaagtg cgctgctggt gccaacccta ttctccttct tcgccaaaag 2220
cactctgatt gacaaatacg atttatctaa tttacacgaa attgcttctg gtggcgctcc 2280
cctctctaag gaagtcgggg aagcggttgc caagaggttc catctgccag gtatcaggca 2340
aggatatggg ctcactgaga ctacatcagc tattctgatt acacccgagg gggatgataa 2400
accgggcgcg gtcggtaaag ttgttccatt ttttgaagcg aaggttgtgg atctggatac 2460
cgggaaaacg ctgggcgtta atcaaagagg cgaactgtgt gtgagaggtc ctatgattat 2520
gtccggttat gtaaacaatc cggaagcgac caacgccttg attgacaagg atggatggct 2580
acattctgga gacatagctt actgggacga agacgaacac ttcttcatcg ttgaccgcct 2640
gaagtctctg attaagtaca aaggctatca ggtggctccc gctgaattgg aatccatctt 2700
gctccaacac cccaacatct tcgacgcagg tgtcgcaggt cttcccgacg atgacgccgg 2760
tgaacttccc gccgccgttg ttgttttgga gcacggaaag acgatgacgg aaaaagagat 2820
cgtggattac gtcgccagtc aagtaacaac cgcgaaaaag ttgcgcggag gagttgtgtt 2880
tgtggacgaa gtaccgaaag gtcttaccgg aaaactcgac gcaagaaaaa tcagagagat 2940
cctcataaag gccaagaagg gcggaaagat cgccgtgtaa gagcatctta ccgccattta 3000
ttcccatatt tgttctgttt ttcttgattt gggtatacat ttaaatgtta ataaaacaaa 3060
atggtggggc aatcatttac atttttaggg atatgtaatt actagttcag gtgtattgcc 3120
acaagacaaa catgttaaga aactttcccg ttatttacgc tctgttcctg ttaatcaacc 3180
tctggattac aaaatttgtg aaagattgac tgatattctt aactatgttg ctccttttac 3240
gctgtgtgga tatgctgctt tatagcctct gtatctagct attgcttccc gtacggcttt 3300
cgttttctcc tccttgtata aatcctggtt gctgtctctt ttagaggagt tgtggcccgt 3360
tgtccgtcaa cgtggcgtgg tgtgctctgt gtttgctgac gcaaccccca ctggctgggg 3420
cattgccacc acctgtcaac tcctttctgg gactttcgct ttccccctcc cgatcgccac 3480
ggcagaactc atcgccgcct gccttgcccg ctgctggaca ggggctaggt tgctgggcac 3540
tgataattcc gtggtgttgt ctgtgccttc tagttgccag ccatctgttg tttgcccctc 3600
ccccgtgcct tccttgaccc tggaaggtgc cactcccact gtcctttcct aataaaatga 3660
ggaaattgca tcgcattgtc tgagtaggtg tcattctatt ctggggggtg gggtggggca 3720
ggacagcaag ggggaggatt gggaagacaa tagcaggcat gctggggatg cggtgggctc 3780
tatggcagga acccctagtg atggagttgg ccactccctc tctgcgcgct cgctcgctca 3840
ctgaggccgc ccgggaaacc cgggcgtgcg cctcagtgag cgagcgagcg cgcagctgcc 3900
tgcagg 3906
<210> 59
<211> 3630
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 59
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggaaacc cgggcgtgcc 60
cgggcgcctc agtgagcgag cgagcgcgca gagagggagt ggccaactcc atcactaggg 120
gttcctggct cagaggctca gaggcacaca ggagtttctg ggctcaccct gcccccttcc 180
aacccctcag ttcccatcct ccagcagctg tttgtgtgct gcctctgaag tccacactga 240
acaaacttca gcctactcat gtccctaaaa tgggcaaaca ttgcaagcag caaacagcaa 300
acacacagcc ctccctgcct gctgaccttg gagctggggc agaggtcaga gacctctctg 360
ggcccatgcc acctccaaca tccactcgac cccttggaat ttcggtggag aggagcagag 420
gttgtcctgg cgtggtttag gtagtgtgag agggtccggg ttcaaaacca cttgctgggt 480
ggggagtcgt cagtaagtgg ctatgccccg accccgaagc ctgtttcccc atctgtacaa 540
tggaaatgat aaagacgccc atctgatagg gtttttgtgg caaataaaca tttggttttt 600
ttgttttgtt ttgttttgtt ttttgagatg gaggtttgct ctgtcgccca ggctggagtg 660
cagtgacaca atctcatctc accacaacct tcccctgcct cagcctccca agtagctggg 720
attacaagca tgtgccacca cacctggcta attttctatt tttagtagag acgggtttct 780
ccatgttggt cagcctcagc ctcccaagta actgggatta caggcctgtg ccaccacacc 840
cggctaattt tttctatttt tgacagggac ggggtttcac catgttggtc aggctggtct 900
agaggtaccg gatcttgcta ccagtggaac agccactaag gattctgcag tgagagcaga 960
gggccagcta agtggtactc tcccagagac tgtctgactc acgccacccc ctccaccttg 1020
gacacaggac gctgtggttt ctgagccagg tacaatgact cctttcggta agtgcagtgg 1080
aagctgtaca ctgcccaggc aaagcgtccg ggcagcgtag gcgggcgact cagatcccag 1140
ccagtggact tagcccctgt ttgctcctcc gataactggg gtgaccttgg ttaatattca 1200
ccagcagcct cccccgttgc ccctctggat ccactgctta aatacggacg aggacagggc 1260
cctgtctcct cagcttcagg caccaccact gacctgggac agtgccgcca ccatgatcat 1320
ggccgagagc cctggcctga tcaccatctg cctgctgggc tacctgctga gcgccgagtg 1380
caccgtgttc ctggaccacg agaacgccaa caagatcctg aaccggccca agagatacaa 1440
cagcggcaag ctggaggagt tcgtgcaggg caacctggag agggagtgca tggaggagaa 1500
gtgcagcttc gaggaggcca gggaagtgtt cgagaacacc gagcggacca ccgagttctg 1560
gaagcagtac gtggacggcg accagtgcga gagcaaccct tgcctgaacg gcggcagctg 1620
caaggacgac atcaacagct acgagtgctg gtgccctttc ggcttcgagg gcaagaactg 1680
cgagctggac gtgacctgca acatcaagaa cggccgctgc gagcagttct gcaagaacag 1740
cgccgacaac aaagtggtgt gtagctgcac cgagggctac agactggccg agaaccagaa 1800
gagctgcgag cccgccgtgc ccttcccctg cggcagagtg agcgtgtccc agaccagcaa 1860
gctgaccaga gccgagaccg tgttccccga cgtggactac gtgaatagca ccgaggccga 1920
gaccatcctg gacaacatca cccagagcac ccagtccttc aacgacttca ccagagttgt 1980
gggcggcgag gacgccaagc ccggccagtt cccctggcag gtggtgctga acggcaaagt 2040
ggatgccttc tgcggcggca gcatcgtgaa cgagaagtgg atcgtgacag ccgcccactg 2100
cgtggagacc ggcgtgaaga tcaccgtggt ggccggcgaa cacaatatcg aggagaccga 2160
gcacaccgag cagaagcgga acgtcatccg gattatcccc caccacaact acaacgccgc 2220
catcaacaag tacaaccacg acatcgccct gctggagctg gacgagcctc tggtgctgaa 2280
tagctacgtg acccccatct gcatcgccga caaggagtac accaacatct tcctgaagtt 2340
cggcagcggc tacgtgtccg gctggggcag agtgttccac aagggcagaa gcgccctggt 2400
gctgcagtac ctgagagtgc ccctggtgga cagagccacc tgcctgttga gcaccaagtt 2460
caccatctac aacaacatgt tctgcgccgg cttccacgag ggcggcagag acagctgcca 2520
gggcgacagc ggcggacccc acgtgaccga agtggagggc accagcttcc tgaccggcat 2580
catcagctgg ggcgaggagt gcgccatgaa gggcaagtac ggcatctaca ccaaagtgag 2640
ccggtacgtg aactggatca aggagaaaac caagctgacc tgagagcatc ttaccgccat 2700
ttattcccat atttgttctg tttttcttga tttgggtata catttaaatg ttaataaaac 2760
aaaatggtgg ggcaatcatt tacattttta gggatatgta attactagtt caggtgtatt 2820
gccacaagac aaacatgtta agaaactttc ccgttattta cgctctgttc ctgttaatca 2880
acctctggat tacaaaattt gtgaaagatt gactgatatt cttaactatg ttgctccttt 2940
tacgctgtgt ggatatgctg ctttatagcc tctgtatcta gctattgctt cccgtacggc 3000
tttcgttttc tcctccttgt ataaatcctg gttgctgtct cttttagagg agttgtggcc 3060
cgttgtccgt caacgtggcg tggtgtgctc tgtgtttgct gacgcaaccc ccactggctg 3120
gggcattgcc accacctgtc aactcctttc tgggactttc gctttccccc tcccgatcgc 3180
cacggcagaa ctcatcgccg cctgccttgc ccgctgctgg acaggggcta ggttgctggg 3240
cactgataat tccgtggtgt tgtctgtgcc ttctagttgc cagccatctg ttgtttgccc 3300
ctcccccgtg ccttccttga ccctggaagg tgccactccc actgtccttt cctaataaaa 3360
tgaggaaatt gcatcgcatt gtctgagtag gtgtcattct attctggggg gtggggtggg 3420
gcaggacagc aagggggagg attgggaaga caatagcagg catgctgggg atgcggtggg 3480
ctctatggca ggaaccccta gtgatggagt tggccactcc ctctctgcgc gctcgctcgc 3540
tcactgaggc cgggcgacca aaggtcgccc gacgcccggg ctttgcccgg gcggcctcag 3600
tgagcgagcg agcgcgcagc tgcctgcagg 3630
<210> 60
<211> 3624
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 60
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc tggctcagag gctcagaggc acacaggagt ttctgggctc 180
accctgcccc cttccaaccc ctcagttccc atcctccagc agctgtttgt gtgctgcctc 240
tgaagtccac actgaacaaa cttcagccta ctcatgtccc taaaatgggc aaacattgca 300
agcagcaaac agcaaacaca cagccctccc tgcctgctga ccttggagct ggggcagagg 360
tcagagacct ctctgggccc atgccacctc caacatccac tcgacccctt ggaatttcgg 420
tggagaggag cagaggttgt cctggcgtgg tttaggtagt gtgagagggt ccgggttcaa 480
aaccacttgc tgggtgggga gtcgtcagta agtggctatg ccccgacccc gaagcctgtt 540
tccccatctg tacaatggaa atgataaaga cgcccatctg atagggtttt tgtggcaaat 600
aaacatttgg tttttttgtt ttgttttgtt ttgttttttg agatggaggt ttgctctgtc 660
gcccaggctg gagtgcagtg acacaatctc atctcaccac aaccttcccc tgcctcagcc 720
tcccaagtag ctgggattac aagcatgtgc caccacacct ggctaatttt ctatttttag 780
tagagacggg tttctccatg ttggtcagcc tcagcctccc aagtaactgg gattacaggc 840
ctgtgccacc acacccggct aattttttct atttttgaca gggacggggt ttcaccatgt 900
tggtcaggct ggtctagagg taccggatct tgctaccagt ggaacagcca ctaaggattc 960
tgcagtgaga gcagagggcc agctaagtgg tactctccca gagactgtct gactcacgcc 1020
accccctcca ccttggacac aggacgctgt ggtttctgag ccaggtacaa tgactccttt 1080
cggtaagtgc agtggaagct gtacactgcc caggcaaagc gtccgggcag cgtaggcggg 1140
cgactcagat cccagccagt ggacttagcc cctgtttgct cctccgataa ctggggtgac 1200
cttggttaat attcaccagc agcctccccc gttgcccctc tggatccact gcttaaatac 1260
ggacgaggac agggccctgt ctcctcagct tcaggcacca ccactgacct gggacagtgc 1320
cgccaccatg atcatggccg agagccctgg cctgatcacc atctgcctgc tgggctacct 1380
gctgagcgcc gagtgcaccg tgttcctgga ccacgagaac gccaacaaga tcctgaaccg 1440
gcccaagaga tacaacagcg gcaagctgga ggagttcgtg cagggcaacc tggagaggga 1500
gtgcatggag gagaagtgca gcttcgagga ggccagggaa gtgttcgaga acaccgagcg 1560
gaccaccgag ttctggaagc agtacgtgga cggcgaccag tgcgagagca acccttgcct 1620
gaacggcggc agctgcaagg acgacatcaa cagctacgag tgctggtgcc ctttcggctt 1680
cgagggcaag aactgcgagc tggacgtgac ctgcaacatc aagaacggcc gctgcgagca 1740
gttctgcaag aacagcgccg acaacaaagt ggtgtgtagc tgcaccgagg gctacagact 1800
ggccgagaac cagaagagct gcgagcccgc cgtgcccttc ccctgcggca gagtgagcgt 1860
gtcccagacc agcaagctga ccagagccga gaccgtgttc cccgacgtgg actacgtgaa 1920
tagcaccgag gccgagacca tcctggacaa catcacccag agcacccagt ccttcaacga 1980
cttcaccaga gttgtgggcg gcgaggacgc caagcccggc cagttcccct ggcaggtggt 2040
gctgaacggc aaagtggatg ccttctgcgg cggcagcatc gtgaacgaga agtggatcgt 2100
gacagccgcc cactgcgtgg agaccggcgt gaagatcacc gtggtggccg gcgaacacaa 2160
tatcgaggag accgagcaca ccgagcagaa gcggaacgtc atccggatta tcccccacca 2220
caactacaac gccgccatca acaagtacaa ccacgacatc gccctgctgg agctggacga 2280
gcctctggtg ctgaatagct acgtgacccc catctgcatc gccgacaagg agtacaccaa 2340
catcttcctg aagttcggca gcggctacgt gtccggctgg ggcagagtgt tccacaaggg 2400
cagaagcgcc ctggtgctgc agtacctgag agtgcccctg gtggacagag ccacctgcct 2460
gttgagcacc aagttcacca tctacaacaa catgttctgc gccggcttcc acgagggcgg 2520
cagagacagc tgccagggcg acagcggcgg accccacgtg accgaagtgg agggcaccag 2580
cttcctgacc ggcatcatca gctggggcga ggagtgcgcc atgaagggca agtacggcat 2640
ctacaccaaa gtgagccggt acgtgaactg gatcaaggag aaaaccaagc tgacctgaga 2700
gcatcttacc gccatttatt cccatatttg ttctgttttt cttgatttgg gtatacattt 2760
aaatgttaat aaaacaaaat ggtggggcaa tcatttacat ttttagggat atgtaattac 2820
tagttcaggt gtattgccac aagacaaaca tgttaagaaa ctttcccgtt atttacgctc 2880
tgttcctgtt aatcaacctc tggattacaa aatttgtgaa agattgactg atattcttaa 2940
ctatgttgct ccttttacgc tgtgtggata tgctgcttta tagcctctgt atctagctat 3000
tgcttcccgt acggctttcg ttttctcctc cttgtataaa tcctggttgc tgtctctttt 3060
agaggagttg tggcccgttg tccgtcaacg tggcgtggtg tgctctgtgt ttgctgacgc 3120
aacccccact ggctggggca ttgccaccac ctgtcaactc ctttctggga ctttcgcttt 3180
ccccctcccg atcgccacgg cagaactcat cgccgcctgc cttgcccgct gctggacagg 3240
ggctaggttg ctgggcactg ataattccgt ggtgttgtct gtgccttcta gttgccagcc 3300
atctgttgtt tgcccctccc ccgtgccttc cttgaccctg gaaggtgcca ctcccactgt 3360
cctttcctaa taaaatgagg aaattgcatc gcattgtctg agtaggtgtc attctattct 3420
ggggggtggg gtggggcagg acagcaaggg ggaggattgg gaagacaata gcaggcatgc 3480
tggggatgcg gtgggctcta tggcaggaac ccctagtgat ggagttggcc actccctctc 3540
tgcgcgctcg ctcgctcact gaggccgccc gggaaacccg ggcgtgcgcc tcagtgagcg 3600
agcgagcgcg cagctgcctg cagg 3624
<210> 61
<211> 4257
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 61
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc ttgtagttaa tgattaaccc gccatgctac ttatctacgt 180
agccatgctc tagagcggcc gcacgcgtag atcttcaata ttggccatta gccatattat 240
tcattggtta tatagcataa atcaatattg gctattggcc attgcatacg ttgtatctat 300
atcataatat gtacatttat attggctcat gtccaatatg accgccatgt tggcattgat 360
tattgactag ttattaatag taatcaatta cggggtcatt agttcatagc ccatatatgg 420
agttccgcgt tacataactt acggtaaatg gcccgcctgg ctgaccgccc aacgaccccc 480
gcccattgac gtcaataatg acgtatgttc ccatagtaac gccaataggg actttccatt 540
gacgtcaatg ggtggagtat ttacggtaaa ctgcccactt ggcagtacat caagtgtatc 600
atatgccaag tccgccccct attgacgtca atgacggtaa atggcccgcc tggcattatg 660
cccagtacat gaccttacgg gactttccta cttggcagta catctacgta ttagtcatcg 720
ctattaccat ggtcgaggtg agccccacgt tctgcttcac tctccccatc tcccccccct 780
ccccaccccc aattttgtat ttatttattt tttaattatt ttgtgcagcg atgggggcgg 840
gggggggggg ggggcgcgcg ccaggcgggg cggggcgggg cgaggggcgg ggcggggcga 900
ggcggagagg tgcggcggca gccaatcaga gcggcgcgct ccgaaagttt ccttttatgg 960
cgaggcggcg gcggcggcgg ccctataaaa agcgaagcgc gcggcgggcg ggagtcgctg 1020
cgacgctgcc ttcgccccgt gccccgctcc gccgccgcct cgcgccgccc gccccggctc 1080
tgactgaccg cgttactccc acaggtgagc gggcgggacg gcccttctcc tccgggctgt 1140
aattagcgct tggtttaatg acggcttgtt tcttttctgt ggctgcgtga aagccttgag 1200
gggctccggg agggcccttt gtgcgggggg gagcggctcg gggggtgcgt gcgtgtgtgt 1260
gtgcgtgggg agcgccgcgt gcggcccgcg ctgcccggcg gctgtgagcg ctgcgggcgc 1320
ggcgcggggc tttgtgcgct ccgcagtgtg cgcgagggga gcgcggccgg gggcggtgcc 1380
ccgcggtgcg gggggggctg cgaggggaac aaaggctgcg tgcggggtgt gtgcgtgggg 1440
gggtgagcag ggggtgtggg cgcggcggtc gggctgtaac ccccccctgc acccccctcc 1500
ccgagttgct gagcacggcc cggcttcggg tgcggggctc cgtacggggc gtggcgcggg 1560
gctcgccgtg ccgggcgggg ggtggcggca ggtgggggtg ccgggcgggg cggggccgcc 1620
tcgggccggg gagggctcgg gggaggggcg cggcggcccc cggagcgccg gcggctgtcg 1680
aggcgcggcg agccgcagcc attgcctttt atggtaatcg tgcgagaggg cgcagggact 1740
tcctttgtcc caaatctgtg cggagccgaa atctgggagg cgccgccgca ccccctctag 1800
cgggcgcggg gcgaagcggt gcggcgccgg caggaaggaa atgggcgggg agggccttcg 1860
tgcgtcgccg cgccgccgtc cccttctccc tctccagcct cggggctgtc cgcgggggga 1920
cggctgcctt cgggggggac ggggcagggc ggggttcggc ttctggcgtg tgaccggcgg 1980
ctctagagcc tctgctaacc atgttttagc cttcttcttt ttcctacagc tcctgggcaa 2040
cgtgctggtt attgtgctgt ctcatcattt gtcgacagaa ttcctcgaag atccgaaggg 2100
gttcaagctt ggcattccgg tactgttggt aaagccacca tggaagacgc caaaaacata 2160
aagaaaggcc cggcgccatt ctatccgctg gaagatggaa ccgctggaga gcaactgcat 2220
aaggctatga agagatacgc cctggttcct ggaacaattg cttttacaga tgcacatatc 2280
gaggtggaca tcacttacgc tgagtacttc gaaatgtccg ttcggttggc agaagctatg 2340
aaacgatatg ggctgaatac aaatcacaga atcgtcgtat gcagtgaaaa ctctcttcaa 2400
ttctttatgc cggtgttggg cgcgttattt atcggagttg cagttgcgcc cgcgaacgac 2460
atttataatg aacgtgaatt gctcaacagt atgggcattt cgcagcctac cgtggtgttc 2520
gtttccaaaa aggggttgca aaaaattttg aacgtgcaaa aaaagctccc aatcatccaa 2580
aaaattatta tcatggattc taaaacggat taccagggat ttcagtcgat gtacacgttc 2640
gtcacatctc atctacctcc cggttttaat gaatacgatt ttgtgccaga gtccttcgat 2700
agggacaaga caattgcact gatcatgaac tcctctggat ctactggtct gcctaaaggt 2760
gtcgctctgc ctcatagaac tgcctgcgtg agattctcgc atgccagaga tcctattttt 2820
ggcaatcaaa tcattccgga tactgcgatt ttaagtgttg ttccattcca tcacggtttt 2880
ggaatgttta ctacactcgg atatttgata tgtggatttc gagtcgtctt aatgtataga 2940
tttgaagaag agctgtttct gaggagcctt caggattaca agattcaaag tgcgctgctg 3000
gtgccaaccc tattctcctt cttcgccaaa agcactctga ttgacaaata cgatttatct 3060
aatttacacg aaattgcttc tggtggcgct cccctctcta aggaagtcgg ggaagcggtt 3120
gccaagaggt tccatctgcc aggtatcagg caaggatatg ggctcactga gactacatca 3180
gctattctga ttacacccga gggggatgat aaaccgggcg cggtcggtaa agttgttcca 3240
ttttttgaag cgaaggttgt ggatctggat accgggaaaa cgctgggcgt taatcaaaga 3300
ggcgaactgt gtgtgagagg tcctatgatt atgtccggtt atgtaaacaa tccggaagcg 3360
accaacgcct tgattgacaa ggatggatgg ctacattctg gagacatagc ttactgggac 3420
gaagacgaac acttcttcat cgttgaccgc ctgaagtctc tgattaagta caaaggctat 3480
caggtggctc ccgctgaatt ggaatccatc ttgctccaac accccaacat cttcgacgca 3540
ggtgtcgcag gtcttcccga cgatgacgcc ggtgaacttc ccgccgccgt tgttgttttg 3600
gagcacggaa agacgatgac ggaaaaagag atcgtggatt acgtcgccag tcaagtaaca 3660
accgcgaaaa agttgcgcgg aggagttgtg tttgtggacg aagtaccgaa aggtcttacc 3720
ggaaaactcg acgcaagaaa aatcagagag atcctcataa aggccaagaa gggcggaaag 3780
atcgccgtgt aattctagag tcggggcggc cggccgcttc gagcagacat gataagatac 3840
attgatgagt ttggacaaac cacaactaga atgcagtgaa aaaaatgctt tatttgtgaa 3900
atttgtgatg ctattgcttt atttgtaacc attataagct gcaataaaca agttaacaac 3960
aacaattgca ttcattttat gtttcaggtt cagggggagg tgtgggaggt tttttaaagc 4020
aagtaaaacc tctacaaatg tggtaaaatc gataagtgcg gaccgagcgg ccgctctaga 4080
gcatggctac gtagataagt agcatggcgg gttaatcatt aactacacct gcaggcagct 4140
gcgcgctcgc tcgctcactg aggccgcccg ggcgtcgggc gacctttggt cgcccggcct 4200
cagtgagcga gcgagcgcgc agagagggag tggccaactc catcactagg ggttcct 4257
<210> 62
<211> 2510
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 62
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc ttgtagttaa tgattaaccc gccatgctac ttatctacgt 180
agccatgctc tagagcggcc gcgctagccc ctaaaatggg caaacattgc aagcagcaaa 240
cagcaaacac acagccctcc ctgcctgctg accttggagc tggggcagag gtcagagacc 300
tctctgggcc catgccacct ccaacatcca ctcgacccct tggaatttcg gtggagagga 360
gcagaggttg tcctggcgtg gtttaggtag tgtgagaggg gaatgactcc tttcggtaag 420
tgcagtggaa gctgtacact gcccaggcaa agcgtccggg cagcgtaggc gggcgactca 480
gatcccagcc agtggactta gcccctgttt gctcctccga taactggggt gaccttggtt 540
aatattcacc agcagcctcc cccgttgccc ctctggatcc actgcttaaa tacggacgag 600
gacactcgag ggccctgtct cctcagcttc aggcaccacc actgacctgg gacagtgaat 660
ccggacatcg attctaaggt aaatataaaa tttttaagtg tataatttgt taaactactg 720
attctaattg tttctctctt ttagattcca acctttggaa ctgaattcta gaccaccatg 780
cagagggtga acatgatcat ggctgagagc cctggcctga tcaccatctg cctgctgggc 840
tacctgctgt ctgctgagtg cactgtgttc ctggaccatg agaatgccaa caagatcctg 900
aacaggccca agagatacaa ctctggcaag ctggaggagt ttgtgcaggg caacctggag 960
agggagtgca tggaggagaa gtgcagcttt gaggaggcca gggaggtgtt tgagaacact 1020
gagaggacca ctgagttctg gaagcagtat gtggatgggg accagtgtga gagcaacccc 1080
tgcctgaatg ggggcagctg caaggatgac atcaacagct atgagtgctg gtgccccttt 1140
ggctttgagg gcaagaactg tgagctggat gtgacctgca acatcaagaa tggcagatgt 1200
gagcagttct gcaagaactc tgctgacaac aaggtggtgt gcagctgcac tgagggctac 1260
aggctggctg agaaccagaa gagctgtgag cctgctgtgc cattcccatg tggcagagtg 1320
tctgtgagcc agaccagcaa gctgaccagg gctgaggctg tgttccctga tgtggactat 1380
gtgaacagca ctgaggctga aaccatcctg gacaacatca cccagagcac ccagagcttc 1440
aatgacttca ccagggtggt ggggggggag gatgccaagc ctggccagtt cccctggcaa 1500
gtggtgctga atggcaaggt ggatgccttc tgtgggggca gcattgtgaa tgagaagtgg 1560
attgtgactg ctgcccactg tgtggagact ggggtgaaga tcactgtggt ggctggggag 1620
cacaacattg aggagactga gcacactgag cagaagagga atgtgatcag gatcatcccc 1680
caccacaact acaatgctgc catcaacaag tacaaccatg acattgccct gctggagctg 1740
gatgagcccc tggtgctgaa cagctatgtg acccccatct gcattgctga caaggagtac 1800
accaacatct tcctgaagtt tggctctggc tatgtgtctg gctggggcag ggtgttccac 1860
aagggcaggt ctgccctggt gctgcagtac ctgagggtgc ccctggtgga cagggccacc 1920
tgcctgagga gcaccaagtt caccatctac aacaacatgt tctgtgctgg cttccatgag 1980
gggggcaggg acagctgcca gggggactct gggggccccc atgtgactga ggtggagggc 2040
accagcttcc tgactggcat catcagctgg ggggaggagt gtgccatgaa gggcaagtat 2100
ggcatctaca ccaaagtctc cagatatgtg aactggatca aggagaagac caagctgacc 2160
taatgactcc atggttcgaa tgctttattt gtgaaatttg tgatgctatt gctttatttg 2220
taaccattat aagctgcaat aaacaagtta acaacaacaa ttgcattcat tttatgtttc 2280
aggttcaggg ggaggtgtgg gaggtttttt aaaactagtg cggccgctct agagcatggc 2340
tacgtagata agtagcatgg cgggttaatc attaactaca aggaacccct agtgatggag 2400
ttggccactc cctctctgcg cgctcgctcg ctcactgagg ccgggcgacc aaaggtcgcc 2460
cgacgcccgg gcggcctcag tgagcgagcg agcgcgcagc tgcctgcagg 2510
<210> 63
<211> 126
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 63
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggaaacc cgggcgtgcc 60
cgggcgcctc agtgagcgag cgagcgcgca gagagggagt ggccaactcc atcactaggg 120
gttcct 126
<210> 64
<211> 120
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 64
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgcccggga aacccgggcg tgcgcctcag tgagcgagcg agcgcgcagc tgcctgcagg 120
<210> 65
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 65
gcgcgctcgc tcgctcactg aggccgcccg ggcgtcgggc gacctttggt cgcccggcct 60
cagtgagcga gcgagcgcgc 80
<210> 66
<211> 141
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 66
aataaacgat aacgccgttg gtggcgtgag gcatgtaaaa ggttacatca ttatcttgtt 60
cgccatccgg ttggtataaa tagacgttca tgttggtttt tgtttcagtt gcaagttggc 120
tgcggcgcgc gcagcacctt t 141
<210> 67
<211> 1876
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 67
cgcagccacc atggcggggt tttacgagat tgtgattaag gtccccagcg accttgacga 60
gcatctgccc ggcatttctg acagctttgt gaactgggtg gccgagaagg aatgggagtt 120
gccgccagat tctgacatgg atctgaatct gattgagcag gcacccctga ccgtggccga 180
gaagctgcag cgcgactttc tgacggaatg gcgccgtgtg agtaaggccc cggaggccct 240
tttctttgtg caatttgaga agggagagag ctacttccac atgcacgtgc tcgtggaaac 300
caccggggtg aaatccatgg ttttgggacg tttcctgagt cagattcgcg aaaaactgat 360
tcagagaatt taccgcggga tcgagccgac tttgccaaac tggttcgcgg tcacaaagac 420
cagaaatggc gccggaggcg ggaacaaggt ggtggatgag tgctacatcc ccaattactt 480
gctccccaaa acccagcctg agctccagtg ggcgtggact aatatggaac agtatttaag 540
cgcctgtttg aatctcacgg agcgtaaacg gttggtggcg cagcatctga cgcacgtgtc 600
gcagacgcag gagcagaaca aagagaatca gaatcccaat tctgatgcgc cggtgatcag 660
atcaaaaact tcagccaggt acatggagct ggtcgggtgg ctcgtggaca aggggattac 720
ctcggagaag cagtggatcc aggaggacca ggcctcatac atctccttca atgcggcctc 780
caactcgcgg tcccaaatca aggctgcctt ggacaatgcg ggaaagatta tgagcctgac 840
taaaaccgcc cccgactacc tggtgggcca gcagcccgtg gaggacattt ccagcaatcg 900
gatttataaa attttggaac taaacgggta cgatccccaa tatgcggctt ccgtctttct 960
gggatgggcc acgaaaaagt tcggcaagag gaacaccatc tggctgtttg ggcctgcaac 1020
taccgggaag accaacatcg cggaggccat agcccacact gtgcccttct acgggtgcgt 1080
aaactggacc aatgagaact ttcccttcaa cgactgtgtc gacaagatgg tgatctggtg 1140
ggaggagggg aagatgaccg ccaaggtcgt ggagtcggcc aaagccattc tcggaggaag 1200
caaggtgcgc gtggaccaga aatgcaagtc ctcggcccag atagacccga ctcccgtgat 1260
cgtcacctcc aacaccaaca tgtgcgccgt gattgacggg aactcaacga ccttcgaaca 1320
ccagcagccg ttgcaagacc ggatgttcaa atttgaactc acccgccgtc tggatcatga 1380
ctttgggaag gtcaccaagc aggaagtcaa agactttttc cggtgggcaa aggatcacgt 1440
ggttgaggtg gagcatgaat tctacgtcaa aaagggtgga gccaagaaaa gacccgcccc 1500
cagtgacgca gatataagtg agcccaaacg ggtgcgcgag tcagttgcgc agccatcgac 1560
gtcagacgcg gaagcttcga tcaactacgc agacaggtac caaaacaaat gttctcgtca 1620
cgtgggcatg aatctgatgc tgtttccctg cagacaatgc gagagaatga atcagaattc 1680
aaatatctgc ttcactcacg gacagaaaga ctgtttagag tgctttcccg tgtcagaatc 1740
tcaacccgtt tctgtcgtca aaaaggcgta tcagaaactg tgctacattc atcatatcat 1800
gggaaaggtg ccagacgctt gcactgcctg cgatctggtc aatgtggatt tggatgactg 1860
catctttgaa caataa 1876
<210> 68
<211> 129
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 68
atcatggaga taattaaaat gataaccatc tcgcaaataa ataagtattt tactgttttc 60
gtaacagttt tgtaataaaa aaacctataa atattccgga ttattcatac cgtcccacca 120
tcgggcgcg 129
<210> 69
<211> 1203
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 69
gccgccacca tggagttggt gggctggctc gtggacaaag gcattacttc ggaaaagcag 60
tggattcagg aggatcaggc atcttacatc tcattcaacg ctgccagtaa ctcgaggtcc 120
cagatcaagg cagcgctgga caacgcggga aagattatga gtctgaccaa aactgctcca 180
gactacctcg ttggtcagca accggtggaa gatatctcca gcaacaggat ctacaagatt 240
ctggagctca acggctacga ccctcaatac gctgcctcag tgttcttggg ttgggccacc 300
aagaaattcg gcaagagaaa cactatctgg ctgttcggcc ccgctaccac tggaaagaca 360
aacatcgcag aagcgattgc tcacacggtg ccattctacg gctgcgtcaa ctggacaaac 420
gagaacttcc cgttcaacga ctgtgtcgat aagatggtta tctggtggga ggaaggaaag 480
atgacggcca aagtggtcga aagcgccaag gcaattctgg gtggctctaa agtgcgcgtc 540
gaccagaagt gcaaatcttc agctcaaatc gatcctaccc ccgttattgt gacatcaaac 600
acgaacatgt gtgccgtgat cgacggaaac agtacaacgt tcgaacacca gcaacctctc 660
caggatcgta tgttcaagtt cgagctcacc cgccgtttgg accatgattt cggcaaggtc 720
actaaacaag aggttaagga cttcttccgc tgggctaaag atcacgttgt ggaggttgaa 780
catgagttct acgtcaagaa aggaggtgct aagaaacgtc cagccccgtc ggacgcagat 840
atctccgaac ctaagagggt gagagagtcg gtcgcacagc caagcacttc tgacgcagaa 900
gcttccatta actacgcaga taggtaccaa aacaagtgca gcagacacgt gggtatgaac 960
ttgatgctgt tcccatgccg ccagtgtgag cgtatgaacc aaaactctaa catctgtttc 1020
acacatggcc agaaggactg cctcgaatgt ttccctgtgt cagagagtca gcccgtctca 1080
gtcgttaaga aagcttacca aaagttgtgc tacatccacc atattatggg taaagtccct 1140
gatgcctgta ccgcttgtga tctggtcaac gtggatttgg acgactgtat tttcgagcaa 1200
taa 1203
<210> 70
<211> 388
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 70
gaacagagaa acaggagaat atgggccaaa caggatatct gtggtaagca gttcctgccc 60
cggctcaggg ccaagaacag ttggaacagc agaatatggg ccaaacagga tatctgtggt 120
aagcagttcc tgccccggct cagggccaag aacagatggt ccccagatgc ggtcccgccc 180
tcagcagttt ctagagaacc atcagatgtt tccagggtgc cccaaggacc tgaaatgacc 240
ctgtgcctta tttgaactaa ccaatcagtt cgcttctcgc ttctgttcgc gcgcttctgc 300
tccccgagct ctatataagc agagctcgtt tagtgaaccg tcagatcgcc tggagacgcc 360
atccacgctg ttttgacttc catagaag 388
<210> 71
<211> 1662
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 71
gccgccacca tggaagacgc caaaaacata aagaaaggcc cggcgccatt ctatccgctg 60
gaagatggaa ccgctggaga gcaactgcat aaggctatga agagatacgc cctggttcct 120
ggaacaattg cttttacaga tgcacatatc gaggtggaca tcacttacgc tgagtacttc 180
gaaatgtccg ttcggttggc agaagctatg aaacgatatg ggctgaatac aaatcacaga 240
atcgtcgtat gcagtgaaaa ctctcttcaa ttctttatgc cggtgttggg cgcgttattt 300
atcggagttg cagttgcgcc cgcgaacgac atttataatg aacgtgaatt gctcaacagt 360
atgggcattt cgcagcctac cgtggtgttc gtttccaaaa aggggttgca aaaaattttg 420
aacgtgcaaa aaaagctccc aatcatccaa aaaattatta tcatggattc taaaacggat 480
taccagggat ttcagtcgat gtacacgttc gtcacatctc atctacctcc cggttttaat 540
gaatacgatt ttgtgccaga gtccttcgat agggacaaga caattgcact gatcatgaac 600
tcctctggat ctactggtct gcctaaaggt gtcgctctgc ctcatagaac tgcctgcgtg 660
agattctcgc atgccagaga tcctattttt ggcaatcaaa tcattccgga tactgcgatt 720
ttaagtgttg ttccattcca tcacggtttt ggaatgttta ctacactcgg atatttgata 780
tgtggatttc gagtcgtctt aatgtataga tttgaagaag agctgtttct gaggagcctt 840
caggattaca agattcaaag tgcgctgctg gtgccaaccc tattctcctt cttcgccaaa 900
agcactctga ttgacaaata cgatttatct aatttacacg aaattgcttc tggtggcgct 960
cccctctcta aggaagtcgg ggaagcggtt gccaagaggt tccatctgcc aggtatcagg 1020
caaggatatg ggctcactga gactacatca gctattctga ttacacccga gggggatgat 1080
aaaccgggcg cggtcggtaa agttgttcca ttttttgaag cgaaggttgt ggatctggat 1140
accgggaaaa cgctgggcgt taatcaaaga ggcgaactgt gtgtgagagg tcctatgatt 1200
atgtccggtt atgtaaacaa tccggaagcg accaacgcct tgattgacaa ggatggatgg 1260
ctacattctg gagacatagc ttactgggac gaagacgaac acttcttcat cgttgaccgc 1320
ctgaagtctc tgattaagta caaaggctat caggtggctc ccgctgaatt ggaatccatc 1380
ttgctccaac accccaacat cttcgacgca ggtgtcgcag gtcttcccga cgatgacgcc 1440
ggtgaacttc ccgccgccgt tgttgttttg gagcacggaa agacgatgac ggaaaaagag 1500
atcgtggatt acgtcgccag tcaagtaaca accgcgaaaa agttgcgcgg aggagttgtg 1560
tttgtggacg aagtaccgaa aggtcttacc ggaaaactcg acgcaagaaa aatcagagag 1620
atcctcataa aggccaagaa gggcggaaag atcgccgtgt aa 1662
<210> 72
<211> 581
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 72
gagcatctta ccgccattta ttcccatatt tgttctgttt ttcttgattt gggtatacat 60
ttaaatgtta ataaaacaaa atggtggggc aatcatttac atttttaggg atatgtaatt 120
actagttcag gtgtattgcc acaagacaaa catgttaaga aactttcccg ttatttacgc 180
tctgttcctg ttaatcaacc tctggattac aaaatttgtg aaagattgac tgatattctt 240
aactatgttg ctccttttac gctgtgtgga tatgctgctt tatagcctct gtatctagct 300
attgcttccc gtacggcttt cgttttctcc tccttgtata aatcctggtt gctgtctctt 360
ttagaggagt tgtggcccgt tgtccgtcaa cgtggcgtgg tgtgctctgt gtttgctgac 420
gcaaccccca ctggctgggg cattgccacc acctgtcaac tcctttctgg gactttcgct 480
ttccccctcc cgatcgccac ggcagaactc atcgccgcct gccttgcccg ctgctggaca 540
ggggctaggt tgctgggcac tgataattcc gtggtgttgt c 581
<210> 73
<211> 225
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 73
tgtgccttct agttgccagc catctgttgt ttgcccctcc cccgtgcctt ccttgaccct 60
ggaaggtgcc actcccactg tcctttccta ataaaatgag gaaattgcat cgcattgtct 120
gagtaggtgt cattctattc tggggggtgg ggtggggcag gacagcaagg gggaggattg 180
ggaagacaat agcaggcatg ctggggatgc ggtgggctct atggc 225
<210> 74
<211> 1177
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 74
ggctcagagg ctcagaggca cacaggagtt tctgggctca ccctgccccc ttccaacccc 60
tcagttccca tcctccagca gctgtttgtg tgctgcctct gaagtccaca ctgaacaaac 120
ttcagcctac tcatgtccct aaaatgggca aacattgcaa gcagcaaaca gcaaacacac 180
agccctccct gcctgctgac cttggagctg gggcagaggt cagagacctc tctgggccca 240
tgccacctcc aacatccact cgaccccttg gaatttcggt ggagaggagc agaggttgtc 300
ctggcgtggt ttaggtagtg tgagagggtc cgggttcaaa accacttgct gggtggggag 360
tcgtcagtaa gtggctatgc cccgaccccg aagcctgttt ccccatctgt acaatggaaa 420
tgataaagac gcccatctga tagggttttt gtggcaaata aacatttggt ttttttgttt 480
tgttttgttt tgttttttga gatggaggtt tgctctgtcg cccaggctgg agtgcagtga 540
cacaatctca tctcaccaca accttcccct gcctcagcct cccaagtagc tgggattaca 600
agcatgtgcc accacacctg gctaattttc tatttttagt agagacgggt ttctccatgt 660
tggtcagcct cagcctccca agtaactggg attacaggcc tgtgccacca cacccggcta 720
attttttcta tttttgacag ggacggggtt tcaccatgtt ggtcaggctg gtctagaggt 780
accggatctt gctaccagtg gaacagccac taaggattct gcagtgagag cagagggcca 840
gctaagtggt actctcccag agactgtctg actcacgcca ccccctccac cttggacaca 900
ggacgctgtg gtttctgagc caggtacaat gactcctttc ggtaagtgca gtggaagctg 960
tacactgccc aggcaaagcg tccgggcagc gtaggcgggc gactcagatc ccagccagtg 1020
gacttagccc ctgtttgctc ctccgataac tggggtgacc ttggttaata ttcaccagca 1080
gcctcccccg ttgcccctct ggatccactg cttaaatacg gacgaggaca gggccctgtc 1140
tcctcagctt caggcaccac cactgacctg ggacagt 1177
<210> 75
<400> 75
000
<210> 76
<400> 76
000
<210> 77
<400> 77
000
<210> 78
<400> 78
000
<210> 79
<400> 79
000
<210> 80
<400> 80
000
<210> 81
<400> 81
000
<210> 82
<400> 82
000
<210> 83
<400> 83
000
<210> 84
<400> 84
000
<210> 85
<400> 85
000
<210> 86
<400> 86
000
<210> 87
<400> 87
000
<210> 88
<400> 88
000
<210> 89
<400> 89
000
<210> 90
<400> 90
000
<210> 91
<400> 91
000
<210> 92
<400> 92
000
<210> 93
<400> 93
000
<210> 94
<400> 94
000
<210> 95
<400> 95
000
<210> 96
<400> 96
000
<210> 97
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 97
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 98
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 98
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacggc ctcagtgagc 60
aagcgagcgc gc 72
<210> 99
<211> 122
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 99
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacggcctc agtgagcgag cgagcgcgca gctgcctgca 120
gg 122
<210> 100
<211> 130
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 100
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg gcggcctcag tgagcgagcg agcgcgcagc 120
tgcctgcagg 130
<210> 101
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 101
gcgcgctcgc tcgctcactg aggccgcccg ggaaacccgg gcgtgcgcct cagtgagcga 60
gcgagcgcgc 70
<210> 102
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 102
gcgcgctcgc tcgctcactg aggcgcacgc ccgggtttcc cgggcggcct cagtgagcga 60
gcgagcgcgc 70
<210> 103
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 103
gcgcgctcgc tcgctcactg aggccgtcgg gcgacctttg gtcgcccggc ctcagtgagc 60
gagcgagcgc gc 72
<210> 104
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 104
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacggc ctcagtgagc 60
gagcgagcgc gc 72
<210> 105
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 105
gcgcgctcgc tcgctcactg aggccgcccg ggcaaagccc gggcgtcggc ctcagtgagc 60
gagcgagcgc gc 72
<210> 106
<211> 72
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 106
gcgcgctcgc tcgctcactg aggccgacgc ccgggctttg cccgggcggc ctcagtgagc 60
gagcgagcgc gc 72
<210> 107
<211> 83
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 107
gcgcgctcgc tcgctcactg aggccgcccg ggcaaagccc gggcgtcggg ctttgcccgg 60
cctcagtgag cgagcgagcg cgc 83
<210> 108
<211> 83
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 108
gcgcgctcgc tcgctcactg aggccgggca aagcccgacg cccgggcttt gcccgggcgg 60
cctcagtgag cgagcgagcg cgc 83
<210> 109
<211> 77
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 109
gcgcgctcgc tcgctcactg aggccgaaac gtcgggcgac ctttggtcgc ccggcctcag 60
tgagcgagcg agcgcgc 77
<210> 110
<211> 77
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 110
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgtt tcggcctcag 60
tgagcgagcg agcgcgc 77
<210> 111
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 111
gcgcgctcgc tcgctcactg aggcaaagcc tcagtgagcg agcgagcgcg c 51
<210> 112
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 112
gcgcgctcgc tcgctcactg aggctttgcc tcagtgagcg agcgagcgcg c 51
<210> 113
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 113
gcgcgctcgc tcgctcactg aggccgcccg ggcgtcgggc gacctttggt cgcccggcct 60
cagtgagcga gcgagcgcgc 80
<210> 114
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 114
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 115
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 115
gcgcgctcgc tcgctcactg aggcgcccgg gcgtcgggcg acctttggtc gcccggcctc 60
agtgagcgag cgagcgcgc 79
<210> 116
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 116
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cgggcgcctc 60
agtgagcgag cgagcgcgc 79
<210> 117
<211> 89
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 117
gcgcgctcgc tcgctcactg aggccgcccg ggcaaagccc gggcgtcggg cgactttgtc 60
gcccggcctc agtgagcgag cgagcgcgc 89
<210> 118
<211> 89
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 118
gcgcgctcgc tcgctcactg aggccgggcg acaaagtcgc ccgacgcccg ggctttgccc 60
gggcggcctc agtgagcgag cgagcgcgc 89
<210> 119
<211> 87
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 119
gcgcgctcgc tcgctcactg aggccgcccg ggcaaagccc gggcgtcggg cgattttcgc 60
ccggcctcag tgagcgagcg agcgcgc 87
<210> 120
<211> 87
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 120
gcgcgctcgc tcgctcactg aggccgggcg aaaatcgccc gacgcccggg ctttgcccgg 60
gcggcctcag tgagcgagcg agcgcgc 87
<210> 121
<211> 85
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 121
gcgcgctcgc tcgctcactg aggccgcccg ggcaaagccc gggcgtcggg cgtttcgccc 60
ggcctcagtg agcgagcgag cgcgc 85
<210> 122
<211> 85
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 122
gcgcgctcgc tcgctcactg aggccgggcg aaacgcccga cgcccgggct ttgcccgggc 60
ggcctcagtg agcgagcgag cgcgc 85
<210> 123
<211> 89
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 123
gcgcgctcgc tcgctcactg aggccgcccg ggaaacccgg gcgtcgggcg acctttggtc 60
gcccggcctc agtgagcgag cgagcgcgc 89
<210> 124
<211> 89
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 124
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cgggtttccc 60
gggcggcctc agtgagcgag cgagcgcgc 89
<210> 125
<211> 87
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 125
gcgcgctcgc tcgctcactg aggccgcccg gaaaccgggc gtcgggcgac ctttggtcgc 60
ccggcctcag tgagcgagcg agcgcgc 87
<210> 126
<211> 87
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 126
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cggtttccgg 60
gcggcctcag tgagcgagcg agcgcgc 87
<210> 127
<211> 85
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 127
gcgcgctcgc tcgctcactg aggccgcccg aaacgggcgt cgggcgacct ttggtcgccc 60
ggcctcagtg agcgagcgag cgcgc 85
<210> 128
<211> 85
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 128
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cgtttcgggc 60
ggcctcagtg agcgagcgag cgcgc 85
<210> 129
<211> 83
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 129
gcgcgctcgc tcgctcactg aggccgccca aagggcgtcg ggcgaccttt ggtcgcccgg 60
cctcagtgag cgagcgagcg cgc 83
<210> 130
<211> 83
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 130
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc ctttgggcgg 60
cctcagtgag cgagcgagcg cgc 83
<210> 131
<211> 81
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 131
gcgcgctcgc tcgctcactg aggccgccaa aggcgtcggg cgacctttgg tcgcccggcc 60
tcagtgagcg agcgagcgcg c 81
<210> 132
<211> 81
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 132
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc tttggcggcc 60
tcagtgagcg agcgagcgcg c 81
<210> 133
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 133
gcgcgctcgc tcgctcactg aggccgcaaa gcgtcgggcg acctttggtc gcccggcctc 60
agtgagcgag cgagcgcgc 79
<210> 134
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 134
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgct ttgcggcctc 60
agtgagcgag cgagcgcgc 79
<210> 135
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 135
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgaagcc cgggctgcct 60
cagtgagcga gcgagcgcgc 80
<210> 136
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 136
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacacc cgggtggcct 60
cagtgagcga gcgagcgcgc 80
<210> 137
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 137
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgac cggtcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 138
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 138
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgca cgtgcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 139
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 139
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc atggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 140
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 140
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgaagac cggtctgcct 60
cagtgagcga gcgagcgcgc 80
<210> 141
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 141
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacaca cgtgtggcct 60
cagtgagcga gcgagcgcgc 80
<210> 142
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 142
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgac atgtcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 143
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 143
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgaagca cgtgctgcct 60
cagtgagcga gcgagcgcgc 80
<210> 144
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 144
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgaaacc cgggttgcct 60
cagtgagcga gcgagcgcgc 80
<210> 145
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 145
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgaagcc atggctgcct 60
cagtgagcga gcgagcgcgc 80
<210> 146
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 146
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacaac cggttggcct 60
cagtgagcga gcgagcgcgc 80
<210> 147
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 147
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacacc atggtggcct 60
cagtgagcga gcgagcgcgc 80
<210> 148
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 148
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgaa cgttcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 149
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 149
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgca attgcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 150
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 150
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgaaaac cggtttgcct 60
cagtgagcga gcgagcgcgc 80
<210> 151
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 151
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgaagaa cgttctgcct 60
cagtgagcga gcgagcgcgc 80
<210> 152
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 152
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgaagca attgctgcct 60
cagtgagcga gcgagcgcgc 80
<210> 153
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 153
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacaaa cgtttggcct 60
cagtgagcga gcgagcgcgc 80
<210> 154
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 154
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgaa atttcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 155
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 155
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgaagca attgctgcct 60
cagtgagcga gcgagcgcgc 80
<210> 156
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 156
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgaaacc atggttgcct 60
cagtgagcga gcgagcgcgc 80
<210> 157
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 157
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacaac atgttggcct 60
cagtgagcga gcgagcgcgc 80
<210> 158
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 158
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgaagac atgtctgcct 60
cagtgagcga gcgagcgcgc 80
<210> 159
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 159
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacaca attgtggcct 60
cagtgagcga gcgagcgcgc 80
<210> 160
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 160
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgaaaaa cgttttgcct 60
cagtgagcga gcgagcgcgc 80
<210> 161
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 161
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgaaaac atgtttgcct 60
cagtgagcga gcgagcgcgc 80
<210> 162
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 162
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgaaaca attgttgcct 60
cagtgagcga gcgagcgcgc 80
<210> 163
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 163
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgaagaa atttctgcct 60
cagtgagcga gcgagcgcgc 80
<210> 164
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 164
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacaaa attttggcct 60
cagtgagcga gcgagcgcgc 80
<210> 165
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 165
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgaaaaa atttttgcct 60
cagtgagcga gcgagcgcgc 80
<210> 166
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 166
gcgcgctcgc tcgctcgctg aggccgggcg accaaaggtc gcccgacgcc cgggcggcct 60
cagcgagcga gcgagcgcgc 80
<210> 167
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 167
gcgcgctcgc tcgctcaatg aggccgggcg accaaaggtc gcccgacgcc cgggcggcct 60
cattgagcga gcgagcgcgc 80
<210> 168
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 168
gcgcgctcgc tcgctcaccg aggccgggcg accaaaggtc gcccgacgcc cgggcggcct 60
cggtgagcga gcgagcgcgc 80
<210> 169
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 169
gcgcgctcgc tcgctcacta aggccgggcg accaaaggtc gcccgacgcc cgggcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 170
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 170
gcgcgctcgc tcgctcactg gggccgggcg accaaaggtc gcccgacgcc cgggcggccc 60
cagtgagcga gcgagcgcgc 80
<210> 171
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 171
gcgcgctcgc tcgctcactg aagccgggcg accaaaggtc gcccgacgcc cgggcggctt 60
cagtgagcga gcgagcgcgc 80
<210> 172
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 172
gcgcgctcgc tcgctcactg agaccgggcg accaaaggtc gcccgacgcc cgggcggtct 60
cagtgagcga gcgagcgcgc 80
<210> 173
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 173
gcgcgctcgc tcgctcgatg aggccgggcg accaaaggtc gcccgacgcc cgggcggcct 60
catcgagcga gcgagcgcgc 80
<210> 174
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 174
gcgcgctcgc tcgctcgcgg aggccgggcg accaaaggtc gcccgacgcc cgggcggcct 60
ccgcgagcga gcgagcgcgc 80
<210> 175
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 175
gcgcgctcgc tcgctcgcta aggccgggcg accaaaggtc gcccgacgcc cgggcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 176
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 176
gcgcgctcgc tcgctcgctg gggccgggcg accaaaggtc gcccgacgcc cgggcggccc 60
cagcgagcga gcgagcgcgc 80
<210> 177
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 177
gcgcgctcgc tcgctcgctg aagccgggcg accaaaggtc gcccgacgcc cgggcggctt 60
cagcgagcga gcgagcgcgc 80
<210> 178
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 178
gcgcgctcgc tcgctcgctg agaccgggcg accaaaggtc gcccgacgcc cgggcggtct 60
cagcgagcga gcgagcgcgc 80
<210> 179
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 179
gcgcgctcgc tcgctcgagg aggccgggcg accaaaggtc gcccgacgcc cgggcggcct 60
cctcgagcga gcgagcgcgc 80
<210> 180
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 180
gcgcgctcgc tcgctcgata aggccgggcg accaaaggtc gcccgacgcc cgggcggcct 60
tatcgagcga gcgagcgcgc 80
<210> 181
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 181
gcgcgctcgc tcgctcgatg gggccgggcg accaaaggtc gcccgacgcc cgggcggcct 60
catcgagcga gcgagcgcgc 80
<210> 182
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 182
gcgcgctcgc tcgctcgatg aagccgggcg accaaaggtc gcccgacgcc cgggcggctt 60
catcgagcga gcgagcgcgc 80
<210> 183
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 183
gcgcgctcgc tcgctcgatg agaccgggcg accaaaggtc gcccgacgcc cgggcggtct 60
catcgagcga gcgagcgcgc 80
<210> 184
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 184
gcgcgctcgc tcgctcgaga aggccgggcg accaaaggtc gcccgacgcc cgggcggcct 60
tctcgagcga gcgagcgcgc 80
<210> 185
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 185
gcgcgctcgc tcgctcgagg gggccgggcg accaaaggtc gcccgacgcc cgggcggccc 60
cctcgagcga gcgagcgcgc 80
<210> 186
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 186
gcgcgctcgc tcgctcgagg aagccgggcg accaaaggtc gcccgacgcc cgggcggctt 60
cctcgagcga gcgagcgcgc 80
<210> 187
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 187
gcgcgctcgc tcgctcgagg gggccgggcg accaaaggtc gcccgacgcc cgggcggccc 60
cctcgagcga gcgagcgcgc 80
<210> 188
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 188
gcgcgctcgc tcgctcgagg aagccgggcg accaaaggtc gcccgacgcc cgggcggctt 60
cctcgagcga gcgagcgcgc 80
<210> 189
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 189
gcgcgctcgc tcgctcgagg agaccgggcg accaaaggtc gcccgacgcc cgggcggtct 60
cctcgagcga gcgagcgcgc 80
<210> 190
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 190
gcgcgctcgc tcgctcgaga gggccgggcg accaaaggtc gcccgacgcc cgggcggccc 60
tctcgagcga gcgagcgcgc 80
<210> 191
<400> 191
000
<210> 192
<400> 192
000
<210> 193
<400> 193
000
<210> 194
<400> 194
000
<210> 195
<400> 195
000
<210> 196
<400> 196
000
<210> 197
<400> 197
000
<210> 198
<400> 198
000
<210> 199
<400> 199
000
<210> 200
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 200
gcgcgctcgc tcgctcgaga aagccgggcg accaaaggtc gcccgacgcc cgggcggctt 60
tctcgagcga gcgagcgcgc 80
<210> 201
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 201
gcgcgctcgc tcgctcgaga agaccgggcg accaaaggtc gcccgacgcc cgggcggcct 60
tctcgagcga gcgagcgcgc 80
<210> 202
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 202
gcgcgctcgc tcgctcgaga gagccgggcg accaaaggtc gcccgacgcc cgggcggctc 60
tctcgagcga gcgagcgcgc 80
<210> 203
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 203
gcgcgctcgc tcgctcgaga ggaccgggcg accaaaggtc gcccgacgcc cgggcggtcc 60
tctcgagcga gcgagcgcgc 80
<210> 204
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 204
gcgcgctcgc tcgctcgaga gaaccgggcg accaaaggtc gcccgacgcc cgggcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 205
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 205
gcgcgctcgc tcgctcaaga gaaccgggcg accaaaggtc gcccgacgcc cgggcggttc 60
tcttgagcga gcgagcgcgc 80
<210> 206
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 206
gcgcgctcgc tcgctcacga gaaccgggcg accaaaggtc gcccgacgcc cgggcggttc 60
tcgtgagcga gcgagcgcgc 80
<210> 207
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 207
gcgcgctcgc tcgctcacta gaaccgggcg accaaaggtc gcccgacgcc cgggcggttc 60
tagtgagcga gcgagcgcgc 80
<210> 208
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 208
gcgcgctcgc tcgctcactg gaaccgggcg accaaaggtc gcccgacgcc cgggcggttc 60
cagtgagcga gcgagcgcgc 80
<210> 209
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 209
gcgcgctcgc tcgctcactg aaaccgggcg accaaaggtc gcccgacgcc cgggcggttt 60
cagtgagcga gcgagcgcgc 80
<210> 210
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 210
gcgcgctcgc tcgctcgcgg gggccgggcg accaaaggtc gcccgacgcc cgggcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 211
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 211
gcgcgctcgc tcgctcaata aaaccgggcg accaaaggtc gcccgacgcc cgggcggttt 60
tattgagcga gcgagcgcgc 80
<210> 212
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 212
gcgcgctcgc tcgctcactg aggccaggcg accaaaggtc gcctgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 213
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 213
gcgcgctcgc tcgctcactg aggccgagcg accaaaggtc gctcgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 214
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 214
gcgcgctcgc tcgctcactg aggccggacg accaaaggtc gtccgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 215
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 215
gcgcgctcgc tcgctcactg aggccgggag accaaaggtc tcccgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 216
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 216
gcgcgctcgc tcgctcactg aggccgggca accaaaggtt gcccgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 217
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 217
gcgcgctcgc tcgctcactg aggccgggcg gccaaaggcc gcccgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 218
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 218
gcgcgctcgc tcgctcactg aggccgggcg aacaaagttc gcccgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 219
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 219
gcgcgctcgc tcgctcactg aggccgggcg acaaaatgtc gcccgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 220
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 220
gcgcgctcgc tcgctcactg aggccaagcg accaaaggtc gcttgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 221
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 221
gcgcgctcgc tcgctcactg aggccaaacg accaaaggtc gtttgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 222
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 222
gcgcgctcgc tcgctcactg aggccaaaag accaaaggtc ttttgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 223
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 223
gcgcgctcgc tcgctcactg aggccaaaaa accaaaggtt ttttgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 224
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 224
gcgcgctcgc tcgctcactg aggccaaaaa gccaaaggct ttttgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 225
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 225
gcgcgctcgc tcgctcactg aggccaaaaa gacaaagtct ttttgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 226
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 226
gcgcgctcgc tcgctcactg aggccaaaaa gaaaaattct ttttgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 227
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 227
gcgcgctcgc tcgctcactg aggccaaaaa aaaaaatttt ttttgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 228
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 228
gcgcgctcgc tcgctcactg aggccgaaaa aaaaaatttt tttcgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 229
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 229
gcgcgctcgc tcgctcactg aggccggaaa gaaaaattct ttccgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 230
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 230
gcgcgctcgc tcgctcactg aggccgggaa gaaaaattct tcccgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 231
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 231
gcgcgctcgc tcgctcactg aggccgggca gaaaaattct gcccgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 232
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 232
gcgcgctcgc tcgctcactg aggccgggcg gaaaaattcc gcccgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 233
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 233
gcgcgctcgc tcgctcactg aggccgggcg aaaaaatttc gcccgacgcc cgggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 234
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 234
gcgcgctcgc tcgctcactg aggccaggcg accaaaggtc gcctgacacc cgggtggcct 60
cagtgagcga gcgagcgcgc 80
<210> 235
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 235
gcgcgctcgc tcgctcactg aggccaggcg accaaaggtc gcctgacgcc atggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 236
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 236
gcgcgctcgc tcgctcactg aggccaggcg accaaaggtc gcctgacgac atgtcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 237
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 237
gcgcgctcgc tcgctcactg aggccaggcg accaaaggtc gcctgacgaa cgttcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 238
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 238
gcgcgctcgc tcgctcactg aggccaggcg accaaaggtc gcctgaagca attgctgcct 60
cagtgagcga gcgagcgcgc 80
<210> 239
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 239
gcgcgctcgc tcgctcactg aggccgggcg gccaaaggcc gcccgacacc cgggtggcct 60
cagtgagcga gcgagcgcgc 80
<210> 240
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 240
gcgcgctcgc tcgctcactg aggccgggcg gccaaaggcc gcccgacgcc atggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 241
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 241
gcgcgctcgc tcgctcactg aggccgggcg gccaaaggcc gcccgacgac atgtcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 242
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 242
gcgcgctcgc tcgctcactg aggccgggcg gccaaaggcc gcccgacgaa cgttcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 243
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 243
gcgcgctcgc tcgctcactg aggccgggcg gccaaaggcc gcccgaagca attgctgcct 60
cagtgagcga gcgagcgcgc 80
<210> 244
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 244
gcgcgctcgc tcgctcactg aggccgggcg acaaaatgtc gcccgacacc cgggtggcct 60
cagtgagcga gcgagcgcgc 80
<210> 245
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 245
gcgcgctcgc tcgctcactg aggccgggcg acaaaatgtc gcccgacgcc atggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 246
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 246
gcgcgctcgc tcgctcactg aggccgggcg acaaaatgtc gcccgacgac atgtcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 247
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 247
gcgcgctcgc tcgctcactg aggccgggcg acaaaatgtc gcccgacgaa cgttcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 248
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 248
gcgcgctcgc tcgctcactg aggccgggcg acaaaatgtc gcccgaagca attgctgcct 60
cagtgagcga gcgagcgcgc 80
<210> 249
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 249
gcgcgctcgc tcgctcactg aggccaaaag accaaaggtc ttttgacacc cgggtggcct 60
cagtgagcga gcgagcgcgc 80
<210> 250
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 250
gcgcgctcgc tcgctcactg aggccaaaag accaaaggtc ttttgacgcc atggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 251
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 251
gcgcgctcgc tcgctcactg aggccaaaag accaaaggtc ttttgacgac atgtcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 252
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 252
gcgcgctcgc tcgctcactg aggccaaaag accaaaggtc ttttgacgaa cgttcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 253
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 253
gcgcgctcgc tcgctcactg aggccaaaag accaaaggtc ttttgaagca attgctgcct 60
cagtgagcga gcgagcgcgc 80
<210> 254
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 254
gcgcgctcgc tcgctcactg aggccaaaaa aaaaaatttt ttttgacacc cgggtggcct 60
cagtgagcga gcgagcgcgc 80
<210> 255
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 255
gcgcgctcgc tcgctcactg aggccaaaaa aaaaaatttt ttttgacgcc atggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 256
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 256
gcgcgctcgc tcgctcactg aggccaaaaa aaaaaatttt ttttgacgac atgtcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 257
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 257
gcgcgctcgc tcgctcactg aggccaaaaa aaaaaatttt ttttgacgaa cgttcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 258
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 258
gcgcgctcgc tcgctcactg aggccaaaaa aaaaaatttt ttttgaagca attgctgcct 60
cagtgagcga gcgagcgcgc 80
<210> 259
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 259
gcgcgctcgc tcgctcactg aggccgggca gaaaaattct gcccgacacc cgggtggcct 60
cagtgagcga gcgagcgcgc 80
<210> 260
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 260
gcgcgctcgc tcgctcactg aggccgggca gaaaaattct gcccgacgcc atggcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 261
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 261
gcgcgctcgc tcgctcactg aggccgggca gaaaaattct gcccgacgac atgtcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 262
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 262
gcgcgctcgc tcgctcactg aggccgggca gaaaaattct gcccgacgaa cgttcggcct 60
cagtgagcga gcgagcgcgc 80
<210> 263
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 263
gcgcgctcgc tcgctcactg aggccgggca gaaaaattct gcccgaagca attgctgcct 60
cagtgagcga gcgagcgcgc 80
<210> 264
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 264
gcgcgctcgc tcgctcacta aggccaggcg accaaaggtc gcctgacgcc cgggcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 265
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 265
gcgcgctcgc tcgctcacta aggccgggcg gccaaaggcc gcccgacgcc cgggcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 266
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 266
gcgcgctcgc tcgctcacta aggccgggcg acaaaatgtc gcccgacgcc cgggcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 267
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 267
gcgcgctcgc tcgctcacta aggccaaaag accaaaggtc ttttgacgcc cgggcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 268
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 268
gcgcgctcgc tcgctcacta aggccaaaaa aaaaaatttt ttttgacgcc cgggcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 269
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 269
gcgcgctcgc tcgctcacta aggccgggca gaaaaattct gcccgacgcc cgggcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 270
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 270
gcgcgctcgc tcgctcgatg gggccaggcg accaaaggtc gcctgacgcc cgggcggcct 60
catcgagcga gcgagcgcgc 80
<210> 271
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 271
gcgcgctcgc tcgctcgatg gggccgggcg gccaaaggcc gcccgacgcc cgggcggcct 60
catcgagcga gcgagcgcgc 80
<210> 272
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 272
gcgcgctcgc tcgctcgatg gggccgggcg acaaaatgtc gcccgacgcc cgggcggcct 60
catcgagcga gcgagcgcgc 80
<210> 273
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 273
gcgcgctcgc tcgctcgatg gggccaaaag accaaaggtc ttttgacgcc cgggcggcct 60
catcgagcga gcgagcgcgc 80
<210> 274
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 274
gcgcgctcgc tcgctcgatg gggccaaaaa aaaaaatttt ttttgacgcc cgggcggcct 60
catcgagcga gcgagcgcgc 80
<210> 275
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 275
gcgcgctcgc tcgctcgatg gggccgggca gaaaaattct gcccgacgcc cgggcggcct 60
catcgagcga gcgagcgcgc 80
<210> 276
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 276
gcgcgctcgc tcgctcgaga gaaccaggcg accaaaggtc gcctgacgcc cgggcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 277
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 277
gcgcgctcgc tcgctcgaga gaaccgggcg gccaaaggcc gcccgacgcc cgggcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 278
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 278
gcgcgctcgc tcgctcgaga gaaccgggcg acaaaatgtc gcccgacgcc cgggcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 279
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 279
gcgcgctcgc tcgctcgaga gaaccaaaag accaaaggtc ttttgacgcc cgggcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 280
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 280
gcgcgctcgc tcgctcgaga gaaccaaaaa aaaaaatttt ttttgacgcc cgggcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 281
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 281
gcgcgctcgc tcgctcgaga gaaccgggca gaaaaattct gcccgacgcc cgggcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 282
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 282
gcgcgctcgc tcgctcgcgg gggccaggcg accaaaggtc gcctgacgcc cgggcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 283
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 283
gcgcgctcgc tcgctcgcgg gggccgggcg gccaaaggcc gcccgacgcc cgggcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 284
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 284
gcgcgctcgc tcgctcgcgg gggccgggcg acaaaatgtc gcccgacgcc cgggcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 285
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 285
gcgcgctcgc tcgctcgcgg gggccaaaag accaaaggtc ttttgacgcc cgggcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 286
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 286
gcgcgctcgc tcgctcgcgg gggccaaaaa aaaaaatttt ttttgacgcc cgggcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 287
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 287
gcgcgctcgc tcgctcgcgg gggccgggca gaaaaattct gcccgacgcc cgggcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 288
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 288
gcgcgctcgc tcgctcaata aaaccaggcg accaaaggtc gcctgacgcc cgggcggttt 60
tattgagcga gcgagcgcgc 80
<210> 289
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 289
gcgcgctcgc tcgctcaata aaaccgggcg gccaaaggcc gcccgacgcc cgggcggttt 60
tattgagcga gcgagcgcgc 80
<210> 290
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 290
gcgcgctcgc tcgctcaata aaaccgggcg acaaaatgtc gcccgacgcc cgggcggttt 60
tattgagcga gcgagcgcgc 80
<210> 291
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 291
gcgcgctcgc tcgctcaata aaaccaaaag accaaaggtc ttttgacgcc cgggcggttt 60
tattgagcga gcgagcgcgc 80
<210> 292
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 292
gcgcgctcgc tcgctcaata aaaccaaaaa aaaaaatttt ttttgacgcc cgggcggttt 60
tattgagcga gcgagcgcgc 80
<210> 293
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 293
gcgcgctcgc tcgctcaata aaaccgggca gaaaaattct gcccgacgcc cgggcggttt 60
tattgagcga gcgagcgcgc 80
<210> 294
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 294
gcgcgctcgc tcgctcacta aggccgggcg accaaaggtc gcccgacacc cgggtggcct 60
tagtgagcga gcgagcgcgc 80
<210> 295
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 295
gcgcgctcgc tcgctcacta aggccgggcg accaaaggtc gcccgacgcc atggcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 296
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 296
gcgcgctcgc tcgctcacta aggccgggcg accaaaggtc gcccgacgac atgtcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 297
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 297
gcgcgctcgc tcgctcacta aggccgggcg accaaaggtc gcccgacgaa cgttcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 298
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 298
gcgcgctcgc tcgctcacta aggccgggcg accaaaggtc gcccgaagca attgctgcct 60
tagtgagcga gcgagcgcgc 80
<210> 299
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 299
gcgcgctcgc tcgctcgatg gggccgggcg accaaaggtc gcccgacacc cgggtggcct 60
catcgagcga gcgagcgcgc 80
<210> 300
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 300
gcgcgctcgc tcgctcgatg gggccgggcg accaaaggtc gcccgacgcc atggcggcct 60
catcgagcga gcgagcgcgc 80
<210> 301
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 301
gcgcgctcgc tcgctcgatg gggccgggcg accaaaggtc gcccgacgac atgtcggcct 60
catcgagcga gcgagcgcgc 80
<210> 302
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 302
gcgcgctcgc tcgctcgatg gggccgggcg accaaaggtc gcccgacgaa cgttcggcct 60
catcgagcga gcgagcgcgc 80
<210> 303
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 303
gcgcgctcgc tcgctcgatg gggccgggcg accaaaggtc gcccgaagca attgctgcct 60
catcgagcga gcgagcgcgc 80
<210> 304
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 304
gcgcgctcgc tcgctcgaga gaaccgggcg accaaaggtc gcccgacacc cgggtggttc 60
tctcgagcga gcgagcgcgc 80
<210> 305
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 305
gcgcgctcgc tcgctcgaga gaaccgggcg accaaaggtc gcccgacgcc atggcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 306
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 306
gcgcgctcgc tcgctcgaga gaaccgggcg accaaaggtc gcccgacgac atgtcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 307
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 307
gcgcgctcgc tcgctcgaga gaaccgggcg accaaaggtc gcccgacgaa cgttcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 308
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 308
gcgcgctcgc tcgctcgaga gaaccgggcg accaaaggtc gcccgaagca attgctgttc 60
tctcgagcga gcgagcgcgc 80
<210> 309
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 309
gcgcgctcgc tcgctcgcgg gggccgggcg accaaaggtc gcccgacacc cgggtggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 310
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 310
gcgcgctcgc tcgctcgcgg gggccgggcg accaaaggtc gcccgacgcc atggcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 311
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 311
gcgcgctcgc tcgctcgcgg gggccgggcg accaaaggtc gcccgacgac atgtcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 312
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 312
gcgcgctcgc tcgctcgcgg gggccgggcg accaaaggtc gcccgacgaa cgttcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 313
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 313
gcgcgctcgc tcgctcgcgg gggccgggcg accaaaggtc gcccgaagca attgctgccc 60
ccgcgagcga gcgagcgcgc 80
<210> 314
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 314
gcgcgctcgc tcgctcaata aaaccgggcg accaaaggtc gcccgacacc cgggtggttt 60
tattgagcga gcgagcgcgc 80
<210> 315
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 315
gcgcgctcgc tcgctcaata aaaccgggcg accaaaggtc gcccgacgcc atggcggttt 60
tattgagcga gcgagcgcgc 80
<210> 316
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 316
gcgcgctcgc tcgctcaata aaaccgggcg accaaaggtc gcccgacgac atgtcggttt 60
tattgagcga gcgagcgcgc 80
<210> 317
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 317
gcgcgctcgc tcgctcaata aaaccgggcg accaaaggtc gcccgacgaa cgttcggttt 60
tattgagcga gcgagcgcgc 80
<210> 318
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 318
gcgcgctcgc tcgctcaata aaaccgggcg accaaaggtc gcccgaagca attgctgttt 60
tattgagcga gcgagcgcgc 80
<210> 319
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 319
gcgcgctcgc tcgctcacta aggccaggcg accaaaggtc gcctgacacc cgggtggcct 60
tagtgagcga gcgagcgcgc 80
<210> 320
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 320
gcgcgctcgc tcgctcacta aggccaggcg accaaaggtc gcctgacgcc atggcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 321
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 321
gcgcgctcgc tcgctcacta aggccaggcg accaaaggtc gcctgacgac atgtcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 322
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 322
gcgcgctcgc tcgctcacta aggccaggcg accaaaggtc gcctgacgaa cgttcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 323
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 323
gcgcgctcgc tcgctcacta aggccaggcg accaaaggtc gcctgaagca attgctgcct 60
tagtgagcga gcgagcgcgc 80
<210> 324
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 324
gcgcgctcgc tcgctcacta aggccgggcg gccaaaggcc gcccgacacc cgggtggcct 60
tagtgagcga gcgagcgcgc 80
<210> 325
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 325
gcgcgctcgc tcgctcacta aggccgggcg gccaaaggcc gcccgacgcc atggcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 326
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 326
gcgcgctcgc tcgctcacta aggccgggcg gccaaaggcc gcccgacgac atgtcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 327
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 327
gcgcgctcgc tcgctcacta aggccgggcg gccaaaggcc gcccgacgaa cgttcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 328
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 328
gcgcgctcgc tcgctcacta aggccgggcg gccaaaggcc gcccgaagca attgctgcct 60
tagtgagcga gcgagcgcgc 80
<210> 329
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 329
gcgcgctcgc tcgctcacta aggccgggcg acaaaatgtc gcccgacacc cgggtggcct 60
tagtgagcga gcgagcgcgc 80
<210> 330
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 330
gcgcgctcgc tcgctcacta aggccgggcg acaaaatgtc gcccgacgcc atggcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 331
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 331
gcgcgctcgc tcgctcacta aggccgggcg acaaaatgtc gcccgacgac atgtcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 332
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 332
gcgcgctcgc tcgctcacta aggccgggcg acaaaatgtc gcccgacgaa cgttcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 333
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 333
gcgcgctcgc tcgctcacta aggccgggcg acaaaatgtc gcccgaagca attgctgcct 60
tagtgagcga gcgagcgcgc 80
<210> 334
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 334
gcgcgctcgc tcgctcacta aggccaaaag accaaaggtc ttttgacacc cgggtggcct 60
tagtgagcga gcgagcgcgc 80
<210> 335
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 335
gcgcgctcgc tcgctcacta aggccaaaag accaaaggtc ttttgacgcc atggcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 336
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 336
gcgcgctcgc tcgctcacta aggccaaaag accaaaggtc ttttgacgac atgtcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 337
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 337
gcgcgctcgc tcgctcacta aggccaaaag accaaaggtc ttttgacgaa cgttcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 338
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 338
gcgcgctcgc tcgctcacta aggccaaaag accaaaggtc ttttgaagca attgctgcct 60
tagtgagcga gcgagcgcgc 80
<210> 339
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 339
gcgcgctcgc tcgctcacta aggccaaaaa aaaaaatttt ttttgacacc cgggtggcct 60
tagtgagcga gcgagcgcgc 80
<210> 340
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 340
gcgcgctcgc tcgctcacta aggccaaaaa aaaaaatttt ttttgacgcc atggcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 341
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 341
gcgcgctcgc tcgctcacta aggccaaaaa aaaaaatttt ttttgacgac atgtcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 342
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 342
gcgcgctcgc tcgctcacta aggccaaaaa aaaaaatttt ttttgacgaa cgttcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 343
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 343
gcgcgctcgc tcgctcacta aggccaaaaa aaaaaatttt ttttgaagca attgctgcct 60
tagtgagcga gcgagcgcgc 80
<210> 344
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 344
gcgcgctcgc tcgctcacta aggccgggca gaaaaattct gcccgacacc cgggtggcct 60
tagtgagcga gcgagcgcgc 80
<210> 345
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 345
gcgcgctcgc tcgctcacta aggccgggca gaaaaattct gcccgacgcc atggcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 346
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 346
gcgcgctcgc tcgctcacta aggccgggca gaaaaattct gcccgacgac atgtcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 347
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 347
gcgcgctcgc tcgctcacta aggccgggca gaaaaattct gcccgacgaa cgttcggcct 60
tagtgagcga gcgagcgcgc 80
<210> 348
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 348
gcgcgctcgc tcgctcacta aggccgggca gaaaaattct gcccgaagca attgctgcct 60
tagtgagcga gcgagcgcgc 80
<210> 349
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 349
gcgcgctcgc tcgctcgatg gggccaggcg accaaaggtc gcctgacacc cgggtggcct 60
catcgagcga gcgagcgcgc 80
<210> 350
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 350
gcgcgctcgc tcgctcgatg gggccaggcg accaaaggtc gcctgacgcc atggcggcct 60
catcgagcga gcgagcgcgc 80
<210> 351
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 351
gcgcgctcgc tcgctcgatg gggccaggcg accaaaggtc gcctgacgac atgtcggcct 60
catcgagcga gcgagcgcgc 80
<210> 352
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 352
gcgcgctcgc tcgctcgatg gggccaggcg accaaaggtc gcctgacgaa cgttcggcct 60
catcgagcga gcgagcgcgc 80
<210> 353
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 353
gcgcgctcgc tcgctcgatg gggccaggcg accaaaggtc gcctgaagca attgctgcct 60
catcgagcga gcgagcgcgc 80
<210> 354
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 354
gcgcgctcgc tcgctcgatg gggccgggcg gccaaaggcc gcccgacacc cgggtggcct 60
catcgagcga gcgagcgcgc 80
<210> 355
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 355
gcgcgctcgc tcgctcgatg gggccgggcg gccaaaggcc gcccgacgcc atggcggcct 60
catcgagcga gcgagcgcgc 80
<210> 356
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 356
gcgcgctcgc tcgctcgatg gggccgggcg gccaaaggcc gcccgacgac atgtcggcct 60
catcgagcga gcgagcgcgc 80
<210> 357
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 357
gcgcgctcgc tcgctcgatg gggccgggcg gccaaaggcc gcccgacgaa cgttcggcct 60
catcgagcga gcgagcgcgc 80
<210> 358
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 358
gcgcgctcgc tcgctcgatg gggccgggcg gccaaaggcc gcccgaagca attgctgcct 60
catcgagcga gcgagcgcgc 80
<210> 359
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 359
gcgcgctcgc tcgctcgatg gggccgggcg acaaaatgtc gcccgacacc cgggtggcct 60
catcgagcga gcgagcgcgc 80
<210> 360
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 360
gcgcgctcgc tcgctcgatg gggccgggcg acaaaatgtc gcccgacgcc atggcggcct 60
catcgagcga gcgagcgcgc 80
<210> 361
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 361
gcgcgctcgc tcgctcgatg gggccgggcg acaaaatgtc gcccgacgac atgtcggcct 60
catcgagcga gcgagcgcgc 80
<210> 362
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 362
gcgcgctcgc tcgctcgatg gggccgggcg acaaaatgtc gcccgacgaa cgttcggcct 60
catcgagcga gcgagcgcgc 80
<210> 363
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 363
gcgcgctcgc tcgctcgatg gggccgggcg acaaaatgtc gcccgaagca attgctgcct 60
catcgagcga gcgagcgcgc 80
<210> 364
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 364
gcgcgctcgc tcgctcgatg gggccaaaag accaaaggtc ttttgacacc cgggtggcct 60
catcgagcga gcgagcgcgc 80
<210> 365
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 365
gcgcgctcgc tcgctcgatg gggccaaaag accaaaggtc ttttgacgcc atggcggcct 60
catcgagcga gcgagcgcgc 80
<210> 366
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 366
gcgcgctcgc tcgctcgatg gggccaaaag accaaaggtc ttttgacgac atgtcggcct 60
catcgagcga gcgagcgcgc 80
<210> 367
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 367
gcgcgctcgc tcgctcgatg gggccaaaag accaaaggtc ttttgacgaa cgttcggcct 60
catcgagcga gcgagcgcgc 80
<210> 368
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 368
gcgcgctcgc tcgctcgatg gggccaaaag accaaaggtc ttttgaagca attgctgcct 60
catcgagcga gcgagcgcgc 80
<210> 369
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 369
gcgcgctcgc tcgctcgatg gggccaaaaa aaaaaatttt ttttgacacc cgggtggcct 60
catcgagcga gcgagcgcgc 80
<210> 370
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 370
gcgcgctcgc tcgctcgatg gggccaaaaa aaaaaatttt ttttgacgcc atggcggcct 60
catcgagcga gcgagcgcgc 80
<210> 371
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 371
gcgcgctcgc tcgctcgatg gggccaaaaa aaaaaatttt ttttgacgac atgtcggcct 60
catcgagcga gcgagcgcgc 80
<210> 372
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 372
gcgcgctcgc tcgctcgatg gggccaaaaa aaaaaatttt ttttgacgaa cgttcggcct 60
catcgagcga gcgagcgcgc 80
<210> 373
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 373
gcgcgctcgc tcgctcgatg gggccaaaaa aaaaaatttt ttttgaagca attgctgcct 60
catcgagcga gcgagcgcgc 80
<210> 374
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 374
gcgcgctcgc tcgctcgatg gggccgggca gaaaaattct gcccgacacc cgggtggcct 60
catcgagcga gcgagcgcgc 80
<210> 375
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 375
gcgcgctcgc tcgctcgatg gggccgggca gaaaaattct gcccgacgcc atggcggcct 60
catcgagcga gcgagcgcgc 80
<210> 376
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 376
gcgcgctcgc tcgctcgatg gggccgggca gaaaaattct gcccgacgac atgtcggcct 60
catcgagcga gcgagcgcgc 80
<210> 377
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 377
gcgcgctcgc tcgctcgatg gggccgggca gaaaaattct gcccgacgaa cgttcggcct 60
catcgagcga gcgagcgcgc 80
<210> 378
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 378
gcgcgctcgc tcgctcgatg gggccgggca gaaaaattct gcccgaagca attgctgcct 60
catcgagcga gcgagcgcgc 80
<210> 379
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 379
gcgcgctcgc tcgctcgaga gaaccaggcg accaaaggtc gcctgacacc cgggtggttc 60
tctcgagcga gcgagcgcgc 80
<210> 380
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 380
gcgcgctcgc tcgctcgaga gaaccaggcg accaaaggtc gcctgacgcc atggcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 381
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 381
gcgcgctcgc tcgctcgaga gaaccaggcg accaaaggtc gcctgacgac atgtcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 382
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 382
gcgcgctcgc tcgctcgaga gaaccaggcg accaaaggtc gcctgacgaa cgttcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 383
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 383
gcgcgctcgc tcgctcgaga gaaccaggcg accaaaggtc gcctgaagca attgctgttc 60
tctcgagcga gcgagcgcgc 80
<210> 384
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 384
gcgcgctcgc tcgctcgaga gaaccgggcg gccaaaggcc gcccgacacc cgggtggttc 60
tctcgagcga gcgagcgcgc 80
<210> 385
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 385
gcgcgctcgc tcgctcgaga gaaccgggcg gccaaaggcc gcccgacgcc atggcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 386
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 386
gcgcgctcgc tcgctcgaga gaaccgggcg gccaaaggcc gcccgacgac atgtcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 387
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 387
gcgcgctcgc tcgctcgaga gaaccgggcg gccaaaggcc gcccgacgaa cgttcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 388
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 388
gcgcgctcgc tcgctcgaga gaaccgggcg gccaaaggcc gcccgaagca attgctgttc 60
tctcgagcga gcgagcgcgc 80
<210> 389
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 389
gcgcgctcgc tcgctcgaga gaaccgggcg acaaaatgtc gcccgacacc cgggtggttc 60
tctcgagcga gcgagcgcgc 80
<210> 390
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 390
gcgcgctcgc tcgctcgaga gaaccgggcg acaaaatgtc gcccgacgcc atggcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 391
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 391
gcgcgctcgc tcgctcgaga gaaccgggcg acaaaatgtc gcccgacgac atgtcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 392
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 392
gcgcgctcgc tcgctcgaga gaaccgggcg acaaaatgtc gcccgacgaa cgttcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 393
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 393
gcgcgctcgc tcgctcgaga gaaccgggcg acaaaatgtc gcccgaagca attgctgttc 60
tctcgagcga gcgagcgcgc 80
<210> 394
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 394
gcgcgctcgc tcgctcgaga gaaccaaaag accaaaggtc ttttgacacc cgggtggttc 60
tctcgagcga gcgagcgcgc 80
<210> 395
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 395
gcgcgctcgc tcgctcgaga gaaccaaaag accaaaggtc ttttgacgcc atggcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 396
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 396
gcgcgctcgc tcgctcgaga gaaccaaaag accaaaggtc ttttgacgac atgtcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 397
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 397
gcgcgctcgc tcgctcgaga gaaccaaaag accaaaggtc ttttgacgaa cgttcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 398
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 398
gcgcgctcgc tcgctcgaga gaaccaaaag accaaaggtc ttttgaagca attgctgttc 60
tctcgagcga gcgagcgcgc 80
<210> 399
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 399
gcgcgctcgc tcgctcgaga gaaccaaaaa aaaaaatttt ttttgacacc cgggtggttc 60
tctcgagcga gcgagcgcgc 80
<210> 400
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 400
gcgcgctcgc tcgctcgaga gaaccaaaaa aaaaaatttt ttttgacgcc atggcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 401
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 401
gcgcgctcgc tcgctcgaga gaaccaaaaa aaaaaatttt ttttgacgac atgtcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 402
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 402
gcgcgctcgc tcgctcgaga gaaccaaaaa aaaaaatttt ttttgacgaa cgttcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 403
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 403
gcgcgctcgc tcgctcgaga gaaccaaaaa aaaaaatttt ttttgaagca attgctgttc 60
tctcgagcga gcgagcgcgc 80
<210> 404
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 404
gcgcgctcgc tcgctcgaga gaaccgggca gaaaaattct gcccgacacc cgggtggttc 60
tctcgagcga gcgagcgcgc 80
<210> 405
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 405
gcgcgctcgc tcgctcgaga gaaccgggca gaaaaattct gcccgacgcc atggcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 406
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 406
gcgcgctcgc tcgctcgaga gaaccgggca gaaaaattct gcccgacgac atgtcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 407
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 407
gcgcgctcgc tcgctcgaga gaaccgggca gaaaaattct gcccgacgaa cgttcggttc 60
tctcgagcga gcgagcgcgc 80
<210> 408
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 408
gcgcgctcgc tcgctcgaga gaaccgggca gaaaaattct gcccgaagca attgctgttc 60
tctcgagcga gcgagcgcgc 80
<210> 409
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 409
gcgcgctcgc tcgctcgcgg gggccaggcg accaaaggtc gcctgacacc cgggtggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 410
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 410
gcgcgctcgc tcgctcgcgg gggccaggcg accaaaggtc gcctgacgcc atggcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 411
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 411
gcgcgctcgc tcgctcgcgg gggccaggcg accaaaggtc gcctgacgac atgtcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 412
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 412
gcgcgctcgc tcgctcgcgg gggccaggcg accaaaggtc gcctgacgaa cgttcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 413
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 413
gcgcgctcgc tcgctcgcgg gggccaggcg accaaaggtc gcctgaagca attgctgccc 60
ccgcgagcga gcgagcgcgc 80
<210> 414
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 414
gcgcgctcgc tcgctcgcgg gggccgggcg gccaaaggcc gcccgacacc cgggtggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 415
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 415
gcgcgctcgc tcgctcgcgg gggccgggcg gccaaaggcc gcccgacgcc atggcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 416
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 416
gcgcgctcgc tcgctcgcgg gggccgggcg gccaaaggcc gcccgacgac atgtcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 417
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 417
gcgcgctcgc tcgctcgcgg gggccgggcg gccaaaggcc gcccgacgaa cgttcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 418
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 418
gcgcgctcgc tcgctcgcgg gggccgggcg gccaaaggcc gcccgaagca attgctgccc 60
ccgcgagcga gcgagcgcgc 80
<210> 419
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 419
gcgcgctcgc tcgctcgcgg gggccgggcg acaaaatgtc gcccgacacc cgggtggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 420
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 420
gcgcgctcgc tcgctcgcgg gggccgggcg acaaaatgtc gcccgacgcc atggcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 421
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 421
gcgcgctcgc tcgctcgcgg gggccgggcg acaaaatgtc gcccgacgac atgtcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 422
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 422
gcgcgctcgc tcgctcgcgg gggccgggcg acaaaatgtc gcccgacgaa cgttcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 423
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 423
gcgcgctcgc tcgctcgcgg gggccgggcg acaaaatgtc gcccgaagca attgctgccc 60
ccgcgagcga gcgagcgcgc 80
<210> 424
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 424
gcgcgctcgc tcgctcgcgg gggccaaaag accaaaggtc ttttgacacc cgggtggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 425
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 425
gcgcgctcgc tcgctcgcgg gggccaaaag accaaaggtc ttttgacgcc atggcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 426
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 426
gcgcgctcgc tcgctcgcgg gggccaaaag accaaaggtc ttttgacgac atgtcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 427
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 427
gcgcgctcgc tcgctcgcgg gggccaaaag accaaaggtc ttttgacgaa cgttcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 428
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 428
gcgcgctcgc tcgctcgcgg gggccaaaag accaaaggtc ttttgaagca attgctgccc 60
ccgcgagcga gcgagcgcgc 80
<210> 429
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 429
gcgcgctcgc tcgctcgcgg gggccaaaaa aaaaaatttt ttttgacacc cgggtggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 430
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 430
gcgcgctcgc tcgctcgcgg gggccaaaaa aaaaaatttt ttttgacgcc atggcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 431
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 431
gcgcgctcgc tcgctcgcgg gggccaaaaa aaaaaatttt ttttgacgac atgtcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 432
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 432
gcgcgctcgc tcgctcgcgg gggccaaaaa aaaaaatttt ttttgacgaa cgttcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 433
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 433
gcgcgctcgc tcgctcgcgg gggccaaaaa aaaaaatttt ttttgaagca attgctgccc 60
ccgcgagcga gcgagcgcgc 80
<210> 434
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 434
gcgcgctcgc tcgctcgcgg gggccgggca gaaaaattct gcccgacacc cgggtggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 435
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 435
gcgcgctcgc tcgctcgcgg gggccgggca gaaaaattct gcccgacgcc atggcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 436
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 436
gcgcgctcgc tcgctcgcgg gggccgggca gaaaaattct gcccgacgac atgtcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 437
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 437
gcgcgctcgc tcgctcgcgg gggccgggca gaaaaattct gcccgacgaa cgttcggccc 60
ccgcgagcga gcgagcgcgc 80
<210> 438
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 438
gcgcgctcgc tcgctcgcgg gggccgggca gaaaaattct gcccgaagca attgctgccc 60
ccgcgagcga gcgagcgcgc 80
<210> 439
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 439
gcgcgctcgc tcgctcaata aaaccaggcg accaaaggtc gcctgacacc cgggtggttt 60
tattgagcga gcgagcgcgc 80
<210> 440
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 440
gcgcgctcgc tcgctcaata aaaccaggcg accaaaggtc gcctgacgcc atggcggttt 60
tattgagcga gcgagcgcgc 80
<210> 441
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 441
gcgcgctcgc tcgctcaata aaaccaggcg accaaaggtc gcctgacgac atgtcggttt 60
tattgagcga gcgagcgcgc 80
<210> 442
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 442
gcgcgctcgc tcgctcaata aaaccaggcg accaaaggtc gcctgacgaa cgttcggttt 60
tattgagcga gcgagcgcgc 80
<210> 443
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 443
gcgcgctcgc tcgctcaata aaaccaggcg accaaaggtc gcctgaagca attgctgttt 60
tattgagcga gcgagcgcgc 80
<210> 444
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 444
gcgcgctcgc tcgctcaata aaaccgggcg gccaaaggcc gcccgacacc cgggtggttt 60
tattgagcga gcgagcgcgc 80
<210> 445
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 445
gcgcgctcgc tcgctcaata aaaccgggcg gccaaaggcc gcccgacgcc atggcggttt 60
tattgagcga gcgagcgcgc 80
<210> 446
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 446
gcgcgctcgc tcgctcaata aaaccgggcg gccaaaggcc gcccgacgac atgtcggttt 60
tattgagcga gcgagcgcgc 80
<210> 447
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 447
gcgcgctcgc tcgctcaata aaaccgggcg gccaaaggcc gcccgacgaa cgttcggttt 60
tattgagcga gcgagcgcgc 80
<210> 448
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 448
gcgcgctcgc tcgctcaata aaaccgggcg gccaaaggcc gcccgaagca attgctgttt 60
tattgagcga gcgagcgcgc 80
<210> 449
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 449
gcgcgctcgc tcgctcaata aaaccgggcg acaaaatgtc gcccgacacc cgggtggttt 60
tattgagcga gcgagcgcgc 80
<210> 450
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 450
gcgcgctcgc tcgctcaata aaaccgggcg acaaaatgtc gcccgacgcc atggcggttt 60
tattgagcga gcgagcgcgc 80
<210> 451
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 451
gcgcgctcgc tcgctcaata aaaccgggcg acaaaatgtc gcccgacgac atgtcggttt 60
tattgagcga gcgagcgcgc 80
<210> 452
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 452
gcgcgctcgc tcgctcaata aaaccgggcg acaaaatgtc gcccgacgaa cgttcggttt 60
tattgagcga gcgagcgcgc 80
<210> 453
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 453
gcgcgctcgc tcgctcaata aaaccgggcg acaaaatgtc gcccgaagca attgctgttt 60
tattgagcga gcgagcgcgc 80
<210> 454
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 454
gcgcgctcgc tcgctcaata aaaccaaaag accaaaggtc ttttgacacc cgggtggttt 60
tattgagcga gcgagcgcgc 80
<210> 455
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 455
gcgcgctcgc tcgctcaata aaaccaaaag accaaaggtc ttttgacgcc atggcggttt 60
tattgagcga gcgagcgcgc 80
<210> 456
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 456
gcgcgctcgc tcgctcaata aaaccaaaag accaaaggtc ttttgacgac atgtcggttt 60
tattgagcga gcgagcgcgc 80
<210> 457
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 457
gcgcgctcgc tcgctcaata aaaccaaaag accaaaggtc ttttgacgaa cgttcggttt 60
tattgagcga gcgagcgcgc 80
<210> 458
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 458
gcgcgctcgc tcgctcaata aaaccaaaag accaaaggtc ttttgaagca attgctgttt 60
tattgagcga gcgagcgcgc 80
<210> 459
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 459
gcgcgctcgc tcgctcaata aaaccaaaaa aaaaaatttt ttttgacacc cgggtggttt 60
tattgagcga gcgagcgcgc 80
<210> 460
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 460
gcgcgctcgc tcgctcaata aaaccaaaaa aaaaaatttt ttttgacgcc atggcggttt 60
tattgagcga gcgagcgcgc 80
<210> 461
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 461
gcgcgctcgc tcgctcaata aaaccaaaaa aaaaaatttt ttttgacgac atgtcggttt 60
tattgagcga gcgagcgcgc 80
<210> 462
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 462
gcgcgctcgc tcgctcaata aaaccaaaaa aaaaaatttt ttttgacgaa cgttcggttt 60
tattgagcga gcgagcgcgc 80
<210> 463
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 463
gcgcgctcgc tcgctcaata aaaccaaaaa aaaaaatttt ttttgaagca attgctgttt 60
tattgagcga gcgagcgcgc 80
<210> 464
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 464
gcgcgctcgc tcgctcaata aaaccgggca gaaaaattct gcccgacacc cgggtggttt 60
tattgagcga gcgagcgcgc 80
<210> 465
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 465
gcgcgctcgc tcgctcaata aaaccgggca gaaaaattct gcccgacgcc atggcggttt 60
tattgagcga gcgagcgcgc 80
<210> 466
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 466
gcgcgctcgc tcgctcaata aaaccgggca gaaaaattct gcccgacgac atgtcggttt 60
tattgagcga gcgagcgcgc 80
<210> 467
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 467
gcgcgctcgc tcgctcaata aaaccgggca gaaaaattct gcccgacgaa cgttcggttt 60
tattgagcga gcgagcgcgc 80
<210> 468
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 468
gcgcgctcgc tcgctcaata aaaccgggca gaaaaattct gcccgaagca attgctgttt 60
tattgagcga gcgagcgcgc 80
<210> 469
<211> 120
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 469
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
cgcacgcccg ggtttcccgg gcggcctcag tgagcgagcg agcgcgcagc tgcctgcagg 120
<210> 470
<211> 122
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 470
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgacgcccg ggctttgccc gggcggcctc agtgagcgag cgagcgcgca gctgcctgca 120
gg 122
<210> 471
<211> 129
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 471
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg gcgcctcagt gagcgagcga gcgcgcagct 120
gcctgcagg 129
<210> 472
<211> 101
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 472
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ctttgcctca gtgagcgagc gagcgcgcag ctgcctgcag g 101
<210> 473
<211> 139
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 473
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgaca aagtcgcccg acgcccgggc tttgcccggg cggcctcagt gagcgagcga 120
gcgcgcagct gcctgcagg 139
<210> 474
<211> 137
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 474
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgaaa atcgcccgac gcccgggctt tgcccgggcg gcctcagtga gcgagcgagc 120
gcgcagctgc ctgcagg 137
<210> 475
<211> 135
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 475
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgaaa cgcccgacgc ccgggctttg cccgggcggc ctcagtgagc gagcgagcgc 120
gcagctgcct gcagg 135
<210> 476
<211> 133
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 476
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcaaag cccgacgccc gggctttgcc cgggcggcct cagtgagcga gcgagcgcgc 120
agctgcctgc agg 133
<210> 477
<211> 139
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 477
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg gtttcccggg cggcctcagt gagcgagcga 120
gcgcgcagct gcctgcagg 139
<210> 478
<211> 137
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 478
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgg tttccgggcg gcctcagtga gcgagcgagc 120
gcgcagctgc ctgcagg 137
<210> 479
<211> 135
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 479
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgcccgt ttcgggcggc ctcagtgagc gagcgagcgc 120
gcagctgcct gcagg 135
<210> 480
<211> 133
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 480
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgccctt tgggcggcct cagtgagcga gcgagcgcgc 120
agctgcctgc agg 133
<210> 481
<211> 131
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 481
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgccttt ggcggcctca gtgagcgagc gagcgcgcag 120
ctgcctgcag g 131
<210> 482
<211> 129
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 482
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgctttg cggcctcagt gagcgagcga gcgcgcagct 120
gcctgcagg 129
<210> 483
<211> 127
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 483
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgggcgacc aaaggtcgcc cgacgtttcg gcctcagtga gcgagcgagc gcgcagctgc 120
ctgcagg 127
<210> 484
<211> 120
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 484
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggaaacc cgggcgtgcg 60
cctcagtgag cgagcgagcg cgcagagagg gagtggccaa ctccatcact aggggttcct 120
<210> 485
<211> 122
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 485
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgt cgggcgacct ttggtcgccc 60
ggcctcagtg agcgagcgag cgcgcagaga gggagtggcc aactccatca ctaggggttc 120
ct 122
<210> 486
<211> 122
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 486
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
ggcctcagtg agcgagcgag cgcgcagaga gggagtggcc aactccatca ctaggggttc 120
ct 122
<210> 487
<211> 129
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 487
cctgcaggca gctgcgcgct cgctcgctca ctgaggcgcc cgggcgtcgg gcgacctttg 60
gtcgcccggc ctcagtgagc gagcgagcgc gcagagaggg agtggccaac tccatcacta 120
ggggttcct 129
<210> 488
<211> 101
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 488
cctgcaggca gctgcgcgct cgctcgctca ctgaggcaaa gcctcagtga gcgagcgagc 60
gcgcagagag ggagtggcca actccatcac taggggttcc t 101
<210> 489
<211> 139
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 489
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacttt gtcgcccggc ctcagtgagc gagcgagcgc gcagagaggg agtggccaac 120
tccatcacta ggggttcct 139
<210> 490
<211> 137
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 490
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgatttt cgcccggcct cagtgagcga gcgagcgcgc agagagggag tggccaactc 120
catcactagg ggttcct 137
<210> 491
<211> 135
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 491
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgtttcg cccggcctca gtgagcgagc gagcgcgcag agagggagtg gccaactcca 120
tcactagggg ttcct 135
<210> 492
<211> 133
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 492
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggctttgcc cggcctcagt gagcgagcga gcgcgcagag agggagtggc caactccatc 120
actaggggtt cct 133
<210> 493
<211> 139
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 493
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggaaacc cgggcgtcgg 60
gcgacctttg gtcgcccggc ctcagtgagc gagcgagcgc gcagagaggg agtggccaac 120
tccatcacta ggggttcct 139
<210> 494
<211> 137
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 494
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccggaaaccg ggcgtcgggc 60
gacctttggt cgcccggcct cagtgagcga gcgagcgcgc agagagggag tggccaactc 120
catcactagg ggttcct 137
<210> 495
<211> 135
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 495
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgaaacggg cgtcgggcga 60
cctttggtcg cccggcctca gtgagcgagc gagcgcgcag agagggagtg gccaactcca 120
tcactagggg ttcct 135
<210> 496
<211> 133
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 496
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccaaagggcg tcgggcgacc 60
tttggtcgcc cggcctcagt gagcgagcga gcgcgcagag agggagtggc caactccatc 120
actaggggtt cct 133
<210> 497
<211> 131
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 497
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc caaaggcgtc gggcgacctt 60
tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca actccatcac 120
taggggttcc t 131
<210> 498
<211> 129
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 498
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc aaagcgtcgg gcgacctttg 60
gtcgcccggc ctcagtgagc gagcgagcgc gcagagaggg agtggccaac tccatcacta 120
ggggttcct 129
<210> 499
<211> 127
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 499
cctgcaggca gctgcgcgct cgctcgctca ctgaggccga aacgtcgggc gacctttggt 60
cgcccggcct cagtgagcga gcgagcgcgc agagagggag tggccaactc catcactagg 120
ggttcct 127
<210> 500
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 500
gcccgctggt ttccagcggg ctgcgggccc gaaacgggcc cgc 43
<210> 501
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 501
cgggcccgtg cgggcccaaa gggcccgc 28
<210> 502
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 502
gcccgggcac gcccgggttt cccgggcg 28
<210> 503
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 503
cgtgcgggcc caaagggccc gc 22
<210> 504
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 504
cgggcgacca aaggtcgccc g 21
<210> 505
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 505
cgcccgggct ttgcccgggc 20
<210> 506
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 506
cgggcgacca aaggtcgccc gacgcccggg ctttgcccgg gc 42
<210> 507
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 507
cgggcgacca aaggtcgccc g 21
<210> 508
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 508
cgcccgggct ttgcccgggc 20
<210> 509
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 509
cgggcgacca aaggtcgccc gacgcccggg cggc 34
<210> 510
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 510
cgggcgacca aaggtcgccc g 21
<210> 511
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 511
cgcccgggct ttgcccgggc 20
<210> 512
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 512
cggggcccga cgcccgggct ttgcccgggc 30
<210> 513
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 513
cgggcgacca aaggtcgccc g 21
<210> 514
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 514
cgcccgggct ttgcccgggc 20
<210> 515
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 515
cgggcccgac gcccgggctt tgcccgggc 29
<210> 516
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 516
cgggcgacca aaggtcgccc g 21
<210> 517
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 517
cgcccgggct ttgcccgggc 20
<210> 518
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 518
gcccgggcaa agcccgggcg 20
<210> 519
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 519
cgggcgacct ttggtcgccc g 21
<210> 520
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 520
gcccgggcaa agcccgggcg tcgggcgacc tttggtcgcc cg 42
<210> 521
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 521
gcccgggcaa agcccgggcg 20
<210> 522
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 522
cgggcgacct ttggtcgccc g 21
<210> 523
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 523
gcccgggcaa agcccgggcg 20
<210> 524
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 524
cgggcgacct ttggtcgccc g 21
<210> 525
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 525
gccgcccggg cgacgggcga cctttggtcg cccg 34
<210> 526
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 526
gcccgggcaa agcccgggcg 20
<210> 527
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 527
cgggcgacct ttggtcgccc g 21
<210> 528
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 528
gcccgggcgt cgggcgacct ttggtcgccc g 31
<210> 529
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 529
cgggcgacct ttggtcgccc g 21
<210> 530
<400> 530
000
<210> 531
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 531
gcgcgctcgc tcgctc 16
<210> 532
<211> 8
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 532
actgaggc 8
<210> 533
<400> 533
000
<210> 534
<400> 534
000
<210> 535
<211> 8
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 535
gcctcagt 8
<210> 536
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 536
gagcgagcga gcgcgc 16
<210> 537
<400> 537
000
<210> 538
<211> 165
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 538
aggaacccct agtgatggag ttggccactc cctctctgcg cgctcgctcg ctcactgagg 60
ccgcccgggc aaagcccggg cgtcgggcga cctttggtcg cccggcctca gtgagcgagc 120
gagcgcgcag agagggagtg gccaactcca tcactagggg ttcct 165
<210> 539
<211> 140
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 539
cccctagtga tggagttggc cactccctct ctgcgcgctc gctcgctcac tgaggccgcc 60
cgggcaaagc ccgggcgtcg ggcgaccttt ggtcgcccgg cctcagtgag cgagcgagcg 120
cgcagagaga tcactagggg 140
<210> 540
<211> 91
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 540
gcgcgctcgc tcgctcactg aggccgcccg ggcaaagccc gggcgtcggg cgacctttgg 60
tcgcccggcc tcagtgagcg agcgagcgcg c 91
<210> 541
<211> 91
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 541
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgcc cgggctttgc 60
ccgggcggcc tcagtgagcg agcgagcgcg c 91
<210> 542
<211> 8
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 542
ttaattaa 8
<210> 543
<211> 80
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 543
cgcgcgagcg agcgagtgac tccggcccgc tggtttccag cgggctgcgg gcccgccgga 60
gtcactcgct cgctcgcgcg 80
<210> 544
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 544
gcgcgctcgc tcgctcactg aggcctttgc ctcagtgagc gagcgagcgc gc 52
<210> 545
<211> 79
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 545
gcgcgctcgc tcgctcactg aggcgcccgg gcgtcgggcg acctttggtc gcccggcctc 60
agtgagcgag cgagcgcgc 79
<210> 546
<211> 77
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 546
gcgcgctcgc tcgctcactg aggccgggcg accaaaggtc gcccgacgtt tcggcctcag 60
tgagcgagcg agcgcgc 77
<210> 547
<211> 77
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 547
gcgcgctcgc tcgctcactg aggccgaaac gtcgggcgac ctttggtcgc ccggcctcag 60
tgagcgagcg agcgcgc 77
<210> 548
<400> 548
000
<210> 549
<400> 549
000
<210> 550
<400> 550
000
<210> 551
<400> 551
000
<210> 552
<400> 552
000
<210> 553
<400> 553
000
<210> 554
<400> 554
000
<210> 555
<400> 555
000
<210> 556
<400> 556
000
<210> 557
<400> 557
000
<210> 558
<211> 312
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 558
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
1 5 10 15
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
20 25 30
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
35 40 45
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ala
50 55 60
Pro Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
65 70 75 80
Asn Gly Tyr Glu Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
85 90 95
Gln Lys Arg Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
100 105 110
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
115 120 125
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
130 135 140
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
145 150 155 160
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
165 170 175
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
180 185 190
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
195 200 205
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
210 215 220
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
225 230 235 240
Glu Val Lys Glu Phe Phe Arg Trp Ala Gln Asp His Val Thr Glu Val
245 250 255
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Asn Lys Arg Pro Ala
260 265 270
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
275 280 285
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
290 295 300
Asp Leu Ala Arg Gly Gln Pro Leu
305 310
<210> 559
<211> 312
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 559
Met Glu Leu Val Gly Trp Leu Val Asp Lys Gly Ile Thr Ser Glu Lys
1 5 10 15
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
20 25 30
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
35 40 45
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Gln Gln
50 55 60
Pro Val Glu Asp Ile Ser Ser Asn Arg Ile Tyr Lys Ile Leu Glu Leu
65 70 75 80
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
85 90 95
Thr Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
100 105 110
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Thr Val Pro
115 120 125
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
130 135 140
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
145 150 155 160
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
165 170 175
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
180 185 190
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
195 200 205
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
210 215 220
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
225 230 235 240
Glu Val Lys Asp Phe Phe Arg Trp Ala Lys Asp His Val Val Glu Val
245 250 255
Glu His Glu Phe Tyr Val Lys Lys Gly Gly Ala Lys Lys Arg Pro Ala
260 265 270
Pro Ser Asp Ala Asp Ile Ser Glu Pro Lys Arg Val Arg Glu Ser Val
275 280 285
Ala Gln Pro Ser Thr Ser Asp Ala Glu Ala Ser Ile Asn Tyr Ala Asp
290 295 300
Arg Leu Ala Arg Gly His Ser Leu
305 310
<210> 560
<211> 311
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 560
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
1 5 10 15
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
20 25 30
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
35 40 45
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Ser Asn
50 55 60
Pro Pro Glu Asp Ile Thr Lys Asn Arg Ile Tyr Gln Ile Leu Glu Leu
65 70 75 80
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
85 90 95
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
100 105 110
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
115 120 125
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
130 135 140
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
145 150 155 160
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
165 170 175
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Glu Pro Thr Pro Val
180 185 190
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
195 200 205
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Glu Phe
210 215 220
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
225 230 235 240
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Asp Val
245 250 255
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
260 265 270
Ser Asn Asp Ala Asp Val Ser Glu Pro Lys Arg Glu Cys Thr Ser Leu
275 280 285
Ala Gln Pro Thr Thr Ser Asp Ala Glu Ala Pro Ala Asp Tyr Ala Asp
290 295 300
Leu Ala Arg Gly Gln Pro Phe
305 310
<210> 561
<211> 311
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 561
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
1 5 10 15
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
20 25 30
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
35 40 45
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Ser Asn
50 55 60
Pro Pro Glu Asp Ile Thr Lys Asn Arg Ile Tyr Gln Ile Leu Glu Leu
65 70 75 80
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
85 90 95
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
100 105 110
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
115 120 125
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
130 135 140
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
145 150 155 160
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
165 170 175
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Glu Pro Thr Pro Val
180 185 190
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
195 200 205
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
210 215 220
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
225 230 235 240
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Asp Val
245 250 255
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
260 265 270
Ser Asn Asp Ala Asp Val Ser Glu Pro Lys Arg Gln Cys Thr Ser Leu
275 280 285
Ala Gln Pro Thr Thr Ser Asp Ala Glu Ala Pro Ala Asp Tyr Ala Asp
290 295 300
Leu Ala Arg Gly Gln Pro Phe
305 310
<210> 562
<211> 311
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 562
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
1 5 10 15
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
20 25 30
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
35 40 45
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Gln Asn
50 55 60
Pro Pro Glu Asp Ile Ser Ser Asn Arg Ile Tyr Arg Ile Leu Glu Met
65 70 75 80
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
85 90 95
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
100 105 110
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
115 120 125
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
130 135 140
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
145 150 155 160
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
165 170 175
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
180 185 190
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
195 200 205
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
210 215 220
Glu Leu Thr Lys Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
225 230 235 240
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
245 250 255
Thr His Glu Phe Tyr Val Arg Lys Gly Gly Ala Arg Lys Arg Pro Ala
260 265 270
Pro Asn Asp Ala Asp Ile Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
275 280 285
Ala Gln Pro Ser Thr Ser Asp Ala Glu Ala Pro Val Asp Tyr Ala Asp
290 295 300
Leu Ala Arg Gly Gln Pro Leu
305 310
<210> 563
<211> 329
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 563
Met Ala Leu Val Asn Trp Leu Val Glu His Gly Ile Thr Ser Glu Lys
1 5 10 15
Gln Trp Ile Gln Glu Asn Gln Glu Ser Tyr Leu Ser Phe Asn Ser Thr
20 25 30
Gly Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Thr Lys
35 40 45
Ile Met Ser Leu Thr Lys Ser Ala Val Asp Tyr Leu Val Gly Ser Ser
50 55 60
Val Pro Glu Asp Ile Ser Lys Asn Arg Ile Trp Gln Ile Phe Glu Met
65 70 75 80
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Ile Leu Tyr Gly Trp Cys
85 90 95
Gln Arg Ser Phe Asn Lys Arg Asn Thr Val Trp Leu Tyr Gly Pro Ala
100 105 110
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Thr Val Pro
115 120 125
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
130 135 140
Cys Val Asp Lys Met Leu Ile Trp Trp Glu Glu Gly Lys Met Thr Asn
145 150 155 160
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
165 170 175
Val Asp Gln Lys Cys Lys Ser Ser Val Gln Ile Asp Ser Thr Pro Val
180 185 190
Ile Val Thr Ser Asn Thr Asn Met Cys Val Val Val Asp Gly Asn Ser
195 200 205
Thr Thr Phe Glu His Gln Gln Pro Leu Glu Asp Arg Met Phe Lys Phe
210 215 220
Glu Leu Thr Lys Arg Leu Pro Pro Asp Phe Gly Lys Ile Thr Lys Gln
225 230 235 240
Glu Val Lys Asp Phe Phe Ala Trp Ala Lys Val Asn Gln Val Pro Val
245 250 255
Thr His Glu Phe Lys Val Pro Arg Glu Leu Ala Gly Thr Lys Gly Ala
260 265 270
Glu Lys Ser Leu Lys Arg Pro Leu Gly Asp Val Thr Asn Thr Ser Tyr
275 280 285
Lys Ser Leu Glu Lys Arg Ala Arg Leu Ser Phe Val Pro Glu Thr Pro
290 295 300
Arg Ser Ser Asp Val Thr Val Asp Pro Ala Pro Leu Arg Pro Leu Asn
305 310 315 320
Trp Asn Ser Leu Val Gly Pro Ser Trp
325
<210> 564
<211> 312
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 564
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
1 5 10 15
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
20 25 30
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
35 40 45
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ala
50 55 60
Pro Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
65 70 75 80
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
85 90 95
Gln Lys Arg Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
100 105 110
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
115 120 125
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
130 135 140
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
145 150 155 160
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
165 170 175
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
180 185 190
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
195 200 205
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
210 215 220
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
225 230 235 240
Glu Val Lys Glu Phe Phe Arg Trp Ala Gln Asp His Val Thr Glu Val
245 250 255
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Asn Lys Arg Pro Ala
260 265 270
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
275 280 285
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
290 295 300
Asp Leu Ala Arg Gly Gln Pro Leu
305 310
<210> 565
<211> 312
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 565
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
1 5 10 15
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
20 25 30
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
35 40 45
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ser
50 55 60
Leu Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
65 70 75 80
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
85 90 95
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
100 105 110
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
115 120 125
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
130 135 140
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
145 150 155 160
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
165 170 175
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
180 185 190
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
195 200 205
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
210 215 220
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
225 230 235 240
Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
245 250 255
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Ser Lys Arg Pro Ala
260 265 270
Pro Asp Asp Ala Asp Ile Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
275 280 285
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
290 295 300
Asp Leu Ala Arg Gly Gln Pro Leu
305 310
<210> 566
<211> 312
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 566
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
1 5 10 15
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
20 25 30
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
35 40 45
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ser
50 55 60
Leu Pro Ala Asp Ile Thr Gln Asn Arg Ile Tyr Arg Ile Leu Ala Leu
65 70 75 80
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
85 90 95
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
100 105 110
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
115 120 125
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
130 135 140
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
145 150 155 160
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
165 170 175
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
180 185 190
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
195 200 205
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
210 215 220
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
225 230 235 240
Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
245 250 255
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Ser Lys Arg Pro Ala
260 265 270
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
275 280 285
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
290 295 300
Asp Leu Ala Arg Gly Gln Pro Leu
305 310
<210> 567
<211> 311
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<220>
<221> Модифицированная аминокислота
<222> (63)..(63)
<223> Любая аминокислота
<220>
<221> Модифицированная аминокислота
<222> (290)..(290)
<223> Любая аминокислота
<400> 567
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
1 5 10 15
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
20 25 30
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
35 40 45
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Xaa Ser
50 55 60
Pro Pro Glu Asp Ile Ser Thr Asn Arg Ile Tyr Arg Ile Leu Ala Leu
65 70 75 80
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
85 90 95
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
100 105 110
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
115 120 125
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
130 135 140
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
145 150 155 160
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
165 170 175
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
180 185 190
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
195 200 205
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
210 215 220
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
225 230 235 240
Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
245 250 255
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
260 265 270
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
275 280 285
Ala Xaa Pro Ser Thr Ser Asp Ala Glu Ala Pro Val Asp Phe Ala Asp
290 295 300
Leu Ala Arg Gly Gln Pro Leu
305 310
<210> 568
<211> 399
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 568
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
1 5 10 15
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
20 25 30
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
35 40 45
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ala
50 55 60
Pro Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
65 70 75 80
Asn Gly Tyr Glu Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
85 90 95
Gln Lys Arg Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
100 105 110
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
115 120 125
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
130 135 140
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
145 150 155 160
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
165 170 175
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
180 185 190
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
195 200 205
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
210 215 220
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
225 230 235 240
Glu Val Lys Glu Phe Phe Arg Trp Ala Gln Asp His Val Thr Glu Val
245 250 255
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Asn Lys Arg Pro Ala
260 265 270
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
275 280 285
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
290 295 300
Asp Arg Tyr Gln Asn Lys Cys Ser Arg His Ala Gly Met Leu Gln Met
305 310 315 320
Leu Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Asn Phe Asn Ile
325 330 335
Cys Phe Thr His Gly Thr Arg Asp Cys Ser Glu Cys Phe Pro Gly Val
340 345 350
Ser Glu Ser Gln Pro Val Val Arg Lys Arg Thr Tyr Arg Lys Leu Cys
355 360 365
Ala Ile His His Leu Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser Ala
370 375 380
Cys Asp Leu Val Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln
385 390 395
<210> 569
<211> 397
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 569
Met Glu Leu Val Gly Trp Leu Val Asp Lys Gly Ile Thr Ser Glu Lys
1 5 10 15
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
20 25 30
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
35 40 45
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Gln Gln
50 55 60
Pro Val Glu Asp Ile Ser Ser Asn Arg Ile Tyr Lys Ile Leu Glu Leu
65 70 75 80
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
85 90 95
Thr Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
100 105 110
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Thr Val Pro
115 120 125
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
130 135 140
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
145 150 155 160
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
165 170 175
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
180 185 190
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
195 200 205
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
210 215 220
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
225 230 235 240
Glu Val Lys Asp Phe Phe Arg Trp Ala Lys Asp His Val Val Glu Val
245 250 255
Glu His Glu Phe Tyr Val Lys Lys Gly Gly Ala Lys Lys Arg Pro Ala
260 265 270
Pro Ser Asp Ala Asp Ile Ser Glu Pro Lys Arg Val Arg Glu Ser Val
275 280 285
Ala Gln Pro Ser Thr Ser Asp Ala Glu Ala Ser Ile Asn Tyr Ala Asp
290 295 300
Arg Tyr Gln Asn Lys Cys Ser Arg His Val Gly Met Asn Leu Met Leu
305 310 315 320
Phe Pro Cys Arg Gln Cys Glu Arg Met Asn Gln Asn Ser Asn Ile Cys
325 330 335
Phe Thr His Gly Gln Lys Asp Cys Leu Glu Cys Phe Pro Val Ser Glu
340 345 350
Ser Gln Pro Val Ser Val Val Lys Lys Ala Tyr Gln Lys Leu Cys Tyr
355 360 365
Ile His His Ile Met Gly Lys Val Pro Asp Ala Cys Thr Ala Cys Asp
370 375 380
Leu Val Asn Val Asp Leu Asp Asp Cys Ile Phe Glu Gln
385 390 395
<210> 570
<211> 400
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 570
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
1 5 10 15
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
20 25 30
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
35 40 45
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Ser Asn
50 55 60
Pro Pro Glu Asp Ile Thr Lys Asn Arg Ile Tyr Gln Ile Leu Glu Leu
65 70 75 80
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
85 90 95
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
100 105 110
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
115 120 125
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
130 135 140
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
145 150 155 160
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
165 170 175
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Glu Pro Thr Pro Val
180 185 190
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
195 200 205
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Glu Phe
210 215 220
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
225 230 235 240
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Asp Val
245 250 255
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
260 265 270
Ser Asn Asp Ala Asp Val Ser Glu Pro Lys Arg Glu Cys Thr Ser Leu
275 280 285
Ala Gln Pro Thr Thr Ser Asp Ala Glu Ala Pro Ala Asp Tyr Ala Asp
290 295 300
Arg Tyr Gln Asn Lys Cys Ser Arg His Val Gly Met Asn Leu Met Leu
305 310 315 320
Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Ile Ser Asn Val Cys
325 330 335
Phe Thr His Gly Gln Arg Asp Cys Gly Glu Cys Phe Pro Gly Met Ser
340 345 350
Glu Ser Gln Pro Val Ser Val Val Lys Lys Lys Thr Tyr Gln Lys Leu
355 360 365
Cys Pro Ile His His Ile Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser
370 375 380
Ala Cys Asp Leu Ala Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln
385 390 395 400
<210> 571
<211> 400
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 571
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
1 5 10 15
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
20 25 30
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
35 40 45
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Ser Asn
50 55 60
Pro Pro Glu Asp Ile Thr Lys Asn Arg Ile Tyr Gln Ile Leu Glu Leu
65 70 75 80
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
85 90 95
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
100 105 110
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
115 120 125
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
130 135 140
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
145 150 155 160
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
165 170 175
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Glu Pro Thr Pro Val
180 185 190
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
195 200 205
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
210 215 220
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
225 230 235 240
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Asp Val
245 250 255
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
260 265 270
Ser Asn Asp Ala Asp Val Ser Glu Pro Lys Arg Gln Cys Thr Ser Leu
275 280 285
Ala Gln Pro Thr Thr Ser Asp Ala Glu Ala Pro Ala Asp Tyr Ala Asp
290 295 300
Arg Tyr Gln Asn Lys Cys Ser Arg His Val Gly Met Asn Leu Met Leu
305 310 315 320
Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Ile Ser Asn Val Cys
325 330 335
Phe Thr His Gly Gln Arg Asp Cys Gly Glu Cys Phe Pro Gly Met Ser
340 345 350
Glu Ser Gln Pro Val Ser Val Val Lys Lys Lys Thr Tyr Gln Lys Leu
355 360 365
Cys Pro Ile His His Ile Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser
370 375 380
Ala Cys Asp Leu Ala Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln
385 390 395 400
<210> 572
<211> 399
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 572
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
1 5 10 15
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
20 25 30
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
35 40 45
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Gln Asn
50 55 60
Pro Pro Glu Asp Ile Ser Ser Asn Arg Ile Tyr Arg Ile Leu Glu Met
65 70 75 80
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
85 90 95
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
100 105 110
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
115 120 125
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
130 135 140
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
145 150 155 160
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
165 170 175
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
180 185 190
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
195 200 205
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
210 215 220
Glu Leu Thr Lys Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
225 230 235 240
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
245 250 255
Thr His Glu Phe Tyr Val Arg Lys Gly Gly Ala Arg Lys Arg Pro Ala
260 265 270
Pro Asn Asp Ala Asp Ile Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
275 280 285
Ala Gln Pro Ser Thr Ser Asp Ala Glu Ala Pro Val Asp Tyr Ala Asp
290 295 300
Arg Tyr Gln Asn Lys Cys Ser Arg His Val Gly Met Asn Leu Met Leu
305 310 315 320
Phe Pro Cys Arg Gln Cys Glu Arg Met Asn Gln Asn Val Asp Ile Cys
325 330 335
Phe Thr His Gly Val Met Asp Cys Ala Glu Cys Phe Pro Val Ser Glu
340 345 350
Ser Gln Pro Val Ser Val Val Arg Lys Arg Thr Tyr Gln Lys Leu Cys
355 360 365
Pro Ile His His Ile Met Gly Arg Ala Pro Glu Val Ala Cys Ser Ala
370 375 380
Cys Glu Leu Ala Asn Val Asp Leu Asp Asp Cys Asp Met Glu Gln
385 390 395
<210> 573
<211> 390
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 573
Met Ala Leu Val Asn Trp Leu Val Glu His Gly Ile Thr Ser Glu Lys
1 5 10 15
Gln Trp Ile Gln Glu Asn Gln Glu Ser Tyr Leu Ser Phe Asn Ser Thr
20 25 30
Gly Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Thr Lys
35 40 45
Ile Met Ser Leu Thr Lys Ser Ala Val Asp Tyr Leu Val Gly Ser Ser
50 55 60
Val Pro Glu Asp Ile Ser Lys Asn Arg Ile Trp Gln Ile Phe Glu Met
65 70 75 80
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Ile Leu Tyr Gly Trp Cys
85 90 95
Gln Arg Ser Phe Asn Lys Arg Asn Thr Val Trp Leu Tyr Gly Pro Ala
100 105 110
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Thr Val Pro
115 120 125
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
130 135 140
Cys Val Asp Lys Met Leu Ile Trp Trp Glu Glu Gly Lys Met Thr Asn
145 150 155 160
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
165 170 175
Val Asp Gln Lys Cys Lys Ser Ser Val Gln Ile Asp Ser Thr Pro Val
180 185 190
Ile Val Thr Ser Asn Thr Asn Met Cys Val Val Val Asp Gly Asn Ser
195 200 205
Thr Thr Phe Glu His Gln Gln Pro Leu Glu Asp Arg Met Phe Lys Phe
210 215 220
Glu Leu Thr Lys Arg Leu Pro Pro Asp Phe Gly Lys Ile Thr Lys Gln
225 230 235 240
Glu Val Lys Asp Phe Phe Ala Trp Ala Lys Val Asn Gln Val Pro Val
245 250 255
Thr His Glu Phe Lys Val Pro Arg Glu Leu Ala Gly Thr Lys Gly Ala
260 265 270
Glu Lys Ser Leu Lys Arg Pro Leu Gly Asp Val Thr Asn Thr Ser Tyr
275 280 285
Lys Ser Leu Glu Lys Arg Ala Arg Leu Ser Phe Val Pro Glu Thr Pro
290 295 300
Arg Ser Ser Asp Val Thr Val Asp Pro Ala Pro Leu Arg Pro Leu Asn
305 310 315 320
Trp Asn Ser Arg Tyr Asp Cys Lys Cys Asp Tyr His Ala Gln Phe Asp
325 330 335
Asn Ile Ser Asn Lys Cys Asp Glu Cys Glu Tyr Leu Asn Arg Gly Lys
340 345 350
Asn Gly Cys Ile Cys His Asn Val Thr His Cys Gln Ile Cys His Gly
355 360 365
Ile Pro Pro Trp Glu Lys Glu Asn Leu Ser Asp Phe Gly Asp Phe Asp
370 375 380
Asp Ala Asn Lys Glu Gln
385 390
<210> 574
<211> 399
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 574
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
1 5 10 15
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
20 25 30
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
35 40 45
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ala
50 55 60
Pro Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
65 70 75 80
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
85 90 95
Gln Lys Arg Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
100 105 110
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
115 120 125
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
130 135 140
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
145 150 155 160
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
165 170 175
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
180 185 190
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
195 200 205
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
210 215 220
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
225 230 235 240
Glu Val Lys Glu Phe Phe Arg Trp Ala Gln Asp His Val Thr Glu Val
245 250 255
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Asn Lys Arg Pro Ala
260 265 270
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
275 280 285
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
290 295 300
Asp Arg Tyr Gln Asn Lys Cys Ser Arg His Ala Gly Met Leu Gln Met
305 310 315 320
Leu Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Asn Phe Asn Ile
325 330 335
Cys Phe Thr His Gly Thr Arg Asp Cys Ser Glu Cys Phe Pro Gly Val
340 345 350
Ser Glu Ser Gln Pro Val Val Arg Lys Arg Thr Tyr Arg Lys Leu Cys
355 360 365
Ala Ile His His Leu Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser Ala
370 375 380
Cys Asp Leu Val Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln
385 390 395
<210> 575
<211> 399
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 575
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
1 5 10 15
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
20 25 30
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
35 40 45
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ser
50 55 60
Leu Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
65 70 75 80
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
85 90 95
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
100 105 110
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
115 120 125
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
130 135 140
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
145 150 155 160
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
165 170 175
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
180 185 190
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
195 200 205
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
210 215 220
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
225 230 235 240
Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
245 250 255
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Ser Lys Arg Pro Ala
260 265 270
Pro Asp Asp Ala Asp Ile Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
275 280 285
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
290 295 300
Asp Arg Tyr Gln Asn Lys Cys Ser Arg His Ala Gly Met Ile Gln Met
305 310 315 320
Leu Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Asn Phe Asn Ile
325 330 335
Cys Phe Thr His Gly Val Arg Asp Cys Leu Glu Cys Phe Pro Gly Val
340 345 350
Ser Glu Ser Gln Pro Val Val Arg Lys Lys Thr Tyr Arg Lys Leu Cys
355 360 365
Ala Ile His His Leu Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser Ala
370 375 380
Cys Asp Leu Val Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln
385 390 395
<210> 576
<211> 399
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 576
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
1 5 10 15
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
20 25 30
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
35 40 45
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ser
50 55 60
Leu Pro Ala Asp Ile Thr Gln Asn Arg Ile Tyr Arg Ile Leu Ala Leu
65 70 75 80
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
85 90 95
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
100 105 110
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
115 120 125
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
130 135 140
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
145 150 155 160
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
165 170 175
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
180 185 190
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
195 200 205
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
210 215 220
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
225 230 235 240
Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
245 250 255
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Ser Lys Arg Pro Ala
260 265 270
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
275 280 285
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
290 295 300
Asp Arg Tyr Gln Asn Lys Cys Ser Arg His Ala Gly Met Leu Gln Met
305 310 315 320
Leu Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Asn Phe Asn Ile
325 330 335
Cys Phe Thr His Gly Val Arg Asp Cys Ser Glu Cys Phe Pro Gly Val
340 345 350
Ser Glu Ser Gln Pro Val Val Arg Lys Arg Thr Tyr Arg Lys Leu Cys
355 360 365
Ala Ile His His Leu Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser Ala
370 375 380
Cys Asp Leu Val Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln
385 390 395
<210> 577
<211> 398
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<220>
<221> Модифицированная аминокислота
<222> (63)..(63)
<223> Любая аминокислота
<220>
<221> Модифицированная аминокислота
<222> (317)..(317)
<223> Любая аминокислота
<220>
<221> Модифицированная аминокислота
<222> (333)..(333)
<223> Любая аминокислота
<220>
<221> Модифицированная аминокислота
<222> (341)..(341)
<223> Любая аминокислота
<220>
<221> Модифицированная аминокислота
<222> (345)..(345)
<223> Любая аминокислота
<220>
<221> Модифицированная аминокислота
<222> (356)..(356)
<223> Любая аминокислота
<220>
<221> Модифицированная аминокислота
<222> (364)..(364)
<223> Любая аминокислота
<220>
<221> Модифицированная аминокислота
<222> (368)..(368)
<223> Любая аминокислота
<400> 577
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
1 5 10 15
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
20 25 30
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
35 40 45
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Xaa Ser
50 55 60
Pro Pro Glu Asp Ile Ser Thr Asn Arg Ile Tyr Arg Ile Leu Ala Leu
65 70 75 80
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
85 90 95
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
100 105 110
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
115 120 125
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
130 135 140
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
145 150 155 160
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
165 170 175
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
180 185 190
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
195 200 205
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
210 215 220
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
225 230 235 240
Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
245 250 255
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
260 265 270
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
275 280 285
Ala Asp Pro Ser Thr Ser Asp Ala Glu Ala Pro Val Asp Phe Ala Asp
290 295 300
Arg Tyr Gln Asn Lys Cys Ser Arg His Ala Gly Met Xaa Gln Met Leu
305 310 315 320
Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Asn Xaa Asn Ile Cys
325 330 335
Phe Thr His Gly Xaa Arg Asp Cys Xaa Glu Cys Phe Pro Gly Val Ser
340 345 350
Glu Ser Gln Xaa Val Val Arg Lys Arg Thr Tyr Xaa Lys Leu Cys Xaa
355 360 365
Ile His His Leu Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser Ala Cys
370 375 380
Asp Leu Val Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln
385 390 395
<210> 578
<211> 536
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 578
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
1 5 10 15
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Ser Trp Val Ala Glu
20 25 30
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Leu Asn Leu Ile
35 40 45
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
50 55 60
Val Gln Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
65 70 75 80
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Leu His Ile Leu Val Glu
85 90 95
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
100 105 110
Arg Asp Lys Leu Val Gln Thr Ile Tyr Arg Gly Ile Glu Pro Thr Leu
115 120 125
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
130 135 140
Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
145 150 155 160
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Glu Tyr Ile
165 170 175
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
180 185 190
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Leu Asn
195 200 205
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
210 215 220
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
225 230 235 240
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
245 250 255
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
260 265 270
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ala
275 280 285
Pro Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
290 295 300
Asn Gly Tyr Glu Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
305 310 315 320
Gln Lys Arg Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
325 330 335
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
340 345 350
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
355 360 365
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
370 375 380
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
385 390 395 400
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
405 410 415
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
420 425 430
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
435 440 445
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
450 455 460
Glu Val Lys Glu Phe Phe Arg Trp Ala Gln Asp His Val Thr Glu Val
465 470 475 480
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Asn Lys Arg Pro Ala
485 490 495
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
500 505 510
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
515 520 525
Asp Leu Ala Arg Gly Gln Pro Leu
530 535
<210> 579
<211> 536
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 579
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
1 5 10 15
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asn Trp Val Ala Glu
20 25 30
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Leu Asn Leu Ile
35 40 45
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
50 55 60
Thr Glu Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
65 70 75 80
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Met His Val Leu Val Glu
85 90 95
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
100 105 110
Arg Glu Lys Leu Ile Gln Arg Ile Tyr Arg Gly Ile Glu Pro Thr Leu
115 120 125
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
130 135 140
Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
145 150 155 160
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Gln Tyr Leu
165 170 175
Ser Ala Cys Leu Asn Leu Thr Glu Arg Lys Arg Leu Val Ala Gln His
180 185 190
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Gln Asn
195 200 205
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
210 215 220
Met Glu Leu Val Gly Trp Leu Val Asp Lys Gly Ile Thr Ser Glu Lys
225 230 235 240
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
245 250 255
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
260 265 270
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Gln Gln
275 280 285
Pro Val Glu Asp Ile Ser Ser Asn Arg Ile Tyr Lys Ile Leu Glu Leu
290 295 300
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
305 310 315 320
Thr Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
325 330 335
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Thr Val Pro
340 345 350
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
355 360 365
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
370 375 380
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
385 390 395 400
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
405 410 415
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
420 425 430
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
435 440 445
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
450 455 460
Glu Val Lys Asp Phe Phe Arg Trp Ala Lys Asp His Val Val Glu Val
465 470 475 480
Glu His Glu Phe Tyr Val Lys Lys Gly Gly Ala Lys Lys Arg Pro Ala
485 490 495
Pro Ser Asp Ala Asp Ile Ser Glu Pro Lys Arg Val Arg Glu Ser Val
500 505 510
Ala Gln Pro Ser Thr Ser Asp Ala Glu Ala Ser Ile Asn Tyr Ala Asp
515 520 525
Arg Leu Ala Arg Gly His Ser Leu
530 535
<210> 580
<211> 535
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 580
Met Pro Gly Phe Tyr Glu Ile Val Leu Lys Val Pro Ser Asp Leu Asp
1 5 10 15
Glu Arg Leu Pro Gly Ile Ser Asn Ser Phe Val Asn Trp Val Ala Glu
20 25 30
Lys Glu Trp Asp Val Pro Pro Asp Ser Asp Met Asp Pro Asn Leu Ile
35 40 45
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Glu Phe Leu
50 55 60
Val Glu Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
65 70 75 80
Gln Phe Glu Lys Gly Glu Thr Tyr Phe His Leu His Val Leu Ile Glu
85 90 95
Thr Ile Gly Val Lys Ser Met Val Val Gly Arg Tyr Val Ser Gln Ile
100 105 110
Lys Glu Lys Leu Val Thr Arg Ile Tyr Arg Gly Val Glu Pro Gln Leu
115 120 125
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
130 135 140
Asn Lys Val Val Asp Asp Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
145 150 155 160
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Asp Gln Tyr Leu
165 170 175
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
180 185 190
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Gln Asn
195 200 205
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
210 215 220
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
225 230 235 240
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
245 250 255
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
260 265 270
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Ser Asn
275 280 285
Pro Pro Glu Asp Ile Thr Lys Asn Arg Ile Tyr Gln Ile Leu Glu Leu
290 295 300
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
305 310 315 320
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
325 330 335
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
340 345 350
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
355 360 365
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
370 375 380
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
385 390 395 400
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Glu Pro Thr Pro Val
405 410 415
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
420 425 430
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Glu Phe
435 440 445
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
450 455 460
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Asp Val
465 470 475 480
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
485 490 495
Ser Asn Asp Ala Asp Val Ser Glu Pro Lys Arg Glu Cys Thr Ser Leu
500 505 510
Ala Gln Pro Thr Thr Ser Asp Ala Glu Ala Pro Ala Asp Tyr Ala Asp
515 520 525
Leu Ala Arg Gly Gln Pro Phe
530 535
<210> 581
<211> 535
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 581
Met Pro Gly Phe Tyr Glu Ile Val Leu Lys Val Pro Ser Asp Leu Asp
1 5 10 15
Glu His Leu Pro Gly Ile Ser Asn Ser Phe Val Asn Trp Val Ala Glu
20 25 30
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Pro Asn Leu Ile
35 40 45
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Glu Phe Leu
50 55 60
Val Glu Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
65 70 75 80
Gln Phe Glu Lys Gly Glu Thr Tyr Phe His Leu His Val Leu Ile Glu
85 90 95
Thr Ile Gly Val Lys Ser Met Val Val Gly Arg Tyr Val Ser Gln Ile
100 105 110
Lys Glu Lys Leu Val Thr Arg Ile Tyr Arg Gly Val Glu Pro Gln Leu
115 120 125
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
130 135 140
Asn Lys Val Val Asp Asp Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
145 150 155 160
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Asp Gln Tyr Leu
165 170 175
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
180 185 190
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Gln Asn
195 200 205
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
210 215 220
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
225 230 235 240
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
245 250 255
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
260 265 270
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Ser Asn
275 280 285
Pro Pro Glu Asp Ile Thr Lys Asn Arg Ile Tyr Gln Ile Leu Glu Leu
290 295 300
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
305 310 315 320
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
325 330 335
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
340 345 350
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
355 360 365
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
370 375 380
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
385 390 395 400
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Glu Pro Thr Pro Val
405 410 415
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
420 425 430
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
435 440 445
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
450 455 460
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Asp Val
465 470 475 480
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
485 490 495
Ser Asn Asp Ala Asp Val Ser Glu Pro Lys Arg Gln Cys Thr Ser Leu
500 505 510
Ala Gln Pro Thr Thr Ser Asp Ala Glu Ala Pro Ala Asp Tyr Ala Asp
515 520 525
Leu Ala Arg Gly Gln Pro Phe
530 535
<210> 582
<211> 535
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 582
Met Pro Gly Phe Tyr Glu Ile Val Leu Lys Val Pro Ser Asp Leu Asp
1 5 10 15
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Ser Trp Val Ala Glu
20 25 30
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Leu Asn Leu Ile
35 40 45
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Glu Phe Leu
50 55 60
Val Glu Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
65 70 75 80
Gln Phe Glu Lys Gly Asp Ser Tyr Phe His Leu His Ile Leu Val Glu
85 90 95
Thr Val Gly Val Lys Ser Met Val Val Gly Arg Tyr Val Ser Gln Ile
100 105 110
Lys Glu Lys Leu Val Thr Arg Ile Tyr Arg Gly Val Glu Pro Gln Leu
115 120 125
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
130 135 140
Asn Lys Val Val Asp Asp Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
145 150 155 160
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Asp Gln Tyr Ile
165 170 175
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
180 185 190
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Gln Asn
195 200 205
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
210 215 220
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
225 230 235 240
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
245 250 255
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
260 265 270
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Gln Asn
275 280 285
Pro Pro Glu Asp Ile Ser Ser Asn Arg Ile Tyr Arg Ile Leu Glu Met
290 295 300
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
305 310 315 320
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
325 330 335
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
340 345 350
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
355 360 365
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
370 375 380
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
385 390 395 400
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
405 410 415
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
420 425 430
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
435 440 445
Glu Leu Thr Lys Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
450 455 460
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
465 470 475 480
Thr His Glu Phe Tyr Val Arg Lys Gly Gly Ala Arg Lys Arg Pro Ala
485 490 495
Pro Asn Asp Ala Asp Ile Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
500 505 510
Ala Gln Pro Ser Thr Ser Asp Ala Glu Ala Pro Val Asp Tyr Ala Asp
515 520 525
Leu Ala Arg Gly Gln Pro Leu
530 535
<210> 583
<211> 549
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 583
Met Ala Thr Phe Tyr Glu Val Ile Val Arg Val Pro Phe Asp Val Glu
1 5 10 15
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asp Trp Val Thr Gly
20 25 30
Gln Ile Trp Glu Leu Pro Pro Glu Ser Asp Leu Asn Leu Thr Leu Val
35 40 45
Glu Gln Pro Gln Leu Thr Val Ala Asp Arg Ile Arg Arg Val Phe Leu
50 55 60
Tyr Glu Trp Asn Lys Phe Ser Lys Gln Glu Ser Lys Phe Phe Val Gln
65 70 75 80
Phe Glu Lys Gly Ser Glu Tyr Phe His Leu His Thr Leu Val Glu Thr
85 90 95
Ser Gly Ile Ser Ser Met Val Leu Gly Arg Tyr Val Ser Gln Ile Arg
100 105 110
Ala Gln Leu Val Lys Val Val Phe Gln Gly Ile Glu Pro Gln Ile Asn
115 120 125
Asp Trp Val Ala Ile Thr Lys Val Lys Lys Gly Gly Ala Asn Lys Val
130 135 140
Val Asp Ser Gly Tyr Ile Pro Ala Tyr Leu Leu Pro Lys Val Gln Pro
145 150 155 160
Glu Leu Gln Trp Ala Trp Thr Asn Leu Asp Glu Tyr Lys Leu Ala Ala
165 170 175
Leu Asn Leu Glu Glu Arg Lys Arg Leu Val Ala Gln Phe Leu Ala Glu
180 185 190
Ser Ser Gln Arg Ser Gln Glu Ala Ala Ser Gln Arg Glu Phe Ser Ala
195 200 205
Asp Pro Val Ile Lys Ser Lys Thr Ser Gln Lys Tyr Met Ala Leu Val
210 215 220
Asn Trp Leu Val Glu His Gly Ile Thr Ser Glu Lys Gln Trp Ile Gln
225 230 235 240
Glu Asn Gln Glu Ser Tyr Leu Ser Phe Asn Ser Thr Gly Asn Ser Arg
245 250 255
Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Thr Lys Ile Met Ser Leu
260 265 270
Thr Lys Ser Ala Val Asp Tyr Leu Val Gly Ser Ser Val Pro Glu Asp
275 280 285
Ile Ser Lys Asn Arg Ile Trp Gln Ile Phe Glu Met Asn Gly Tyr Asp
290 295 300
Pro Ala Tyr Ala Gly Ser Ile Leu Tyr Gly Trp Cys Gln Arg Ser Phe
305 310 315 320
Asn Lys Arg Asn Thr Val Trp Leu Tyr Gly Pro Ala Thr Thr Gly Lys
325 330 335
Thr Asn Ile Ala Glu Ala Ile Ala His Thr Val Pro Phe Tyr Gly Cys
340 345 350
Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp Cys Val Asp Lys
355 360 365
Met Leu Ile Trp Trp Glu Glu Gly Lys Met Thr Asn Lys Val Val Glu
370 375 380
Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg Val Asp Gln Lys
385 390 395 400
Cys Lys Ser Ser Val Gln Ile Asp Ser Thr Pro Val Ile Val Thr Ser
405 410 415
Asn Thr Asn Met Cys Val Val Val Asp Gly Asn Ser Thr Thr Phe Glu
420 425 430
His Gln Gln Pro Leu Glu Asp Arg Met Phe Lys Phe Glu Leu Thr Lys
435 440 445
Arg Leu Pro Pro Asp Phe Gly Lys Ile Thr Lys Gln Glu Val Lys Asp
450 455 460
Phe Phe Ala Trp Ala Lys Val Asn Gln Val Pro Val Thr His Glu Phe
465 470 475 480
Lys Val Pro Arg Glu Leu Ala Gly Thr Lys Gly Ala Glu Lys Ser Leu
485 490 495
Lys Arg Pro Leu Gly Asp Val Thr Asn Thr Ser Tyr Lys Ser Leu Glu
500 505 510
Lys Arg Ala Arg Leu Ser Phe Val Pro Glu Thr Pro Arg Ser Ser Asp
515 520 525
Val Thr Val Asp Pro Ala Pro Leu Arg Pro Leu Asn Trp Asn Ser Leu
530 535 540
Val Gly Pro Ser Trp
545
<210> 584
<211> 536
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 584
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
1 5 10 15
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asn Trp Val Ala Glu
20 25 30
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Leu Asn Leu Ile
35 40 45
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
50 55 60
Val Gln Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
65 70 75 80
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Leu His Ile Leu Val Glu
85 90 95
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
100 105 110
Arg Asp Lys Leu Val Gln Thr Ile Tyr Arg Gly Ile Glu Pro Thr Leu
115 120 125
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
130 135 140
Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
145 150 155 160
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Glu Tyr Ile
165 170 175
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala His Asp
180 185 190
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Leu Asn
195 200 205
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
210 215 220
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
225 230 235 240
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
245 250 255
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
260 265 270
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ala
275 280 285
Pro Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
290 295 300
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
305 310 315 320
Gln Lys Arg Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
325 330 335
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
340 345 350
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
355 360 365
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
370 375 380
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
385 390 395 400
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
405 410 415
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
420 425 430
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
435 440 445
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
450 455 460
Glu Val Lys Glu Phe Phe Arg Trp Ala Gln Asp His Val Thr Glu Val
465 470 475 480
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Asn Lys Arg Pro Ala
485 490 495
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
500 505 510
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
515 520 525
Asp Leu Ala Arg Gly Gln Pro Leu
530 535
<210> 585
<211> 536
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 585
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
1 5 10 15
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asn Trp Val Ala Glu
20 25 30
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Leu Asn Leu Ile
35 40 45
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
50 55 60
Val Gln Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
65 70 75 80
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Leu His Val Leu Val Glu
85 90 95
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
100 105 110
Arg Glu Lys Leu Val Gln Thr Ile Tyr Arg Gly Val Glu Pro Thr Leu
115 120 125
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
130 135 140
Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
145 150 155 160
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Glu Tyr Ile
165 170 175
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
180 185 190
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Leu Asn
195 200 205
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
210 215 220
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
225 230 235 240
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
245 250 255
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
260 265 270
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ser
275 280 285
Leu Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
290 295 300
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
305 310 315 320
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
325 330 335
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
340 345 350
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
355 360 365
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
370 375 380
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
385 390 395 400
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
405 410 415
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
420 425 430
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
435 440 445
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
450 455 460
Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
465 470 475 480
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Ser Lys Arg Pro Ala
485 490 495
Pro Asp Asp Ala Asp Ile Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
500 505 510
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
515 520 525
Asp Leu Ala Arg Gly Gln Pro Leu
530 535
<210> 586
<211> 538
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 586
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
1 5 10 15
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asn Trp Val Ala Glu
20 25 30
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Arg Asn Leu Ile
35 40 45
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
50 55 60
Val Gln Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
65 70 75 80
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Leu His Val Leu Val Glu
85 90 95
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
100 105 110
Arg Glu Lys Leu Gly Pro Asp His Leu Pro Ala Gly Ser Ser Pro Thr
115 120 125
Leu Pro Asn Trp Phe Ala Val Thr Lys Asp Ala Val Met Ala Pro Ala
130 135 140
Gly Gly Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu
145 150 155 160
Pro Lys Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Glu
165 170 175
Tyr Ile Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala
180 185 190
Gln His Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn
195 200 205
Leu Asn Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala
210 215 220
Arg Tyr Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser
225 230 235 240
Glu Lys Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn
245 250 255
Ala Ala Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala
260 265 270
Gly Lys Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly
275 280 285
Pro Ser Leu Pro Ala Asp Ile Thr Gln Asn Arg Ile Tyr Arg Ile Leu
290 295 300
Ala Leu Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly
305 310 315 320
Trp Ala Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly
325 330 335
Pro Ala Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala
340 345 350
Val Pro Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe
355 360 365
Asn Asp Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met
370 375 380
Thr Ala Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys
385 390 395 400
Val Arg Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr
405 410 415
Pro Val Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly
420 425 430
Asn Ser Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe
435 440 445
Lys Phe Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr
450 455 460
Lys Gln Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr
465 470 475 480
Glu Val Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Ser Lys Arg
485 490 495
Pro Ala Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro
500 505 510
Ser Val Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp
515 520 525
Phe Ala Asp Leu Ala Arg Gly Gln Pro Leu
530 535
<210> 587
<211> 623
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 587
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
1 5 10 15
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Ser Trp Val Ala Glu
20 25 30
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Leu Asn Leu Ile
35 40 45
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
50 55 60
Val Gln Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
65 70 75 80
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Leu His Ile Leu Val Glu
85 90 95
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
100 105 110
Arg Asp Lys Leu Val Gln Thr Ile Tyr Arg Gly Ile Glu Pro Thr Leu
115 120 125
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
130 135 140
Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
145 150 155 160
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Glu Tyr Ile
165 170 175
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
180 185 190
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Leu Asn
195 200 205
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
210 215 220
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
225 230 235 240
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
245 250 255
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
260 265 270
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ala
275 280 285
Pro Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
290 295 300
Asn Gly Tyr Glu Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
305 310 315 320
Gln Lys Arg Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
325 330 335
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
340 345 350
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
355 360 365
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
370 375 380
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
385 390 395 400
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
405 410 415
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
420 425 430
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
435 440 445
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
450 455 460
Glu Val Lys Glu Phe Phe Arg Trp Ala Gln Asp His Val Thr Glu Val
465 470 475 480
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Asn Lys Arg Pro Ala
485 490 495
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
500 505 510
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
515 520 525
Asp Arg Tyr Gln Asn Lys Cys Ser Arg His Ala Gly Met Leu Gln Met
530 535 540
Leu Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Asn Phe Asn Ile
545 550 555 560
Cys Phe Thr His Gly Thr Arg Asp Cys Ser Glu Cys Phe Pro Gly Val
565 570 575
Ser Glu Ser Gln Pro Val Val Arg Lys Arg Thr Tyr Arg Lys Leu Cys
580 585 590
Ala Ile His His Leu Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser Ala
595 600 605
Cys Asp Leu Val Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln
610 615 620
<210> 588
<211> 621
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 588
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
1 5 10 15
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asn Trp Val Ala Glu
20 25 30
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Leu Asn Leu Ile
35 40 45
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
50 55 60
Thr Glu Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
65 70 75 80
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Met His Val Leu Val Glu
85 90 95
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
100 105 110
Arg Glu Lys Leu Ile Gln Arg Ile Tyr Arg Gly Ile Glu Pro Thr Leu
115 120 125
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
130 135 140
Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
145 150 155 160
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Gln Tyr Leu
165 170 175
Ser Ala Cys Leu Asn Leu Thr Glu Arg Lys Arg Leu Val Ala Gln His
180 185 190
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Gln Asn
195 200 205
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
210 215 220
Met Glu Leu Val Gly Trp Leu Val Asp Lys Gly Ile Thr Ser Glu Lys
225 230 235 240
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
245 250 255
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
260 265 270
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Gln Gln
275 280 285
Pro Val Glu Asp Ile Ser Ser Asn Arg Ile Tyr Lys Ile Leu Glu Leu
290 295 300
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
305 310 315 320
Thr Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
325 330 335
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Thr Val Pro
340 345 350
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
355 360 365
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
370 375 380
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
385 390 395 400
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
405 410 415
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
420 425 430
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
435 440 445
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
450 455 460
Glu Val Lys Asp Phe Phe Arg Trp Ala Lys Asp His Val Val Glu Val
465 470 475 480
Glu His Glu Phe Tyr Val Lys Lys Gly Gly Ala Lys Lys Arg Pro Ala
485 490 495
Pro Ser Asp Ala Asp Ile Ser Glu Pro Lys Arg Val Arg Glu Ser Val
500 505 510
Ala Gln Pro Ser Thr Ser Asp Ala Glu Ala Ser Ile Asn Tyr Ala Asp
515 520 525
Arg Tyr Gln Asn Lys Cys Ser Arg His Val Gly Met Asn Leu Met Leu
530 535 540
Phe Pro Cys Arg Gln Cys Glu Arg Met Asn Gln Asn Ser Asn Ile Cys
545 550 555 560
Phe Thr His Gly Gln Lys Asp Cys Leu Glu Cys Phe Pro Val Ser Glu
565 570 575
Ser Gln Pro Val Ser Val Val Lys Lys Ala Tyr Gln Lys Leu Cys Tyr
580 585 590
Ile His His Ile Met Gly Lys Val Pro Asp Ala Cys Thr Ala Cys Asp
595 600 605
Leu Val Asn Val Asp Leu Asp Asp Cys Ile Phe Glu Gln
610 615 620
<210> 589
<211> 624
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 589
Met Pro Gly Phe Tyr Glu Ile Val Leu Lys Val Pro Ser Asp Leu Asp
1 5 10 15
Glu Arg Leu Pro Gly Ile Ser Asn Ser Phe Val Asn Trp Val Ala Glu
20 25 30
Lys Glu Trp Asp Val Pro Pro Asp Ser Asp Met Asp Pro Asn Leu Ile
35 40 45
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Glu Phe Leu
50 55 60
Val Glu Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
65 70 75 80
Gln Phe Glu Lys Gly Glu Thr Tyr Phe His Leu His Val Leu Ile Glu
85 90 95
Thr Ile Gly Val Lys Ser Met Val Val Gly Arg Tyr Val Ser Gln Ile
100 105 110
Lys Glu Lys Leu Val Thr Arg Ile Tyr Arg Gly Val Glu Pro Gln Leu
115 120 125
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
130 135 140
Asn Lys Val Val Asp Asp Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
145 150 155 160
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Asp Gln Tyr Leu
165 170 175
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
180 185 190
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Gln Asn
195 200 205
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
210 215 220
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
225 230 235 240
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
245 250 255
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
260 265 270
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Ser Asn
275 280 285
Pro Pro Glu Asp Ile Thr Lys Asn Arg Ile Tyr Gln Ile Leu Glu Leu
290 295 300
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
305 310 315 320
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
325 330 335
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
340 345 350
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
355 360 365
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
370 375 380
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
385 390 395 400
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Glu Pro Thr Pro Val
405 410 415
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
420 425 430
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Glu Phe
435 440 445
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
450 455 460
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Asp Val
465 470 475 480
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
485 490 495
Ser Asn Asp Ala Asp Val Ser Glu Pro Lys Arg Glu Cys Thr Ser Leu
500 505 510
Ala Gln Pro Thr Thr Ser Asp Ala Glu Ala Pro Ala Asp Tyr Ala Asp
515 520 525
Arg Tyr Gln Asn Lys Cys Ser Arg His Val Gly Met Asn Leu Met Leu
530 535 540
Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Ile Ser Asn Val Cys
545 550 555 560
Phe Thr His Gly Gln Arg Asp Cys Gly Glu Cys Phe Pro Gly Met Ser
565 570 575
Glu Ser Gln Pro Val Ser Val Val Lys Lys Lys Thr Tyr Gln Lys Leu
580 585 590
Cys Pro Ile His His Ile Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser
595 600 605
Ala Cys Asp Leu Ala Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln
610 615 620
<210> 590
<211> 624
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 590
Met Pro Gly Phe Tyr Glu Ile Val Leu Lys Val Pro Ser Asp Leu Asp
1 5 10 15
Glu His Leu Pro Gly Ile Ser Asn Ser Phe Val Asn Trp Val Ala Glu
20 25 30
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Pro Asn Leu Ile
35 40 45
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Glu Phe Leu
50 55 60
Val Glu Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
65 70 75 80
Gln Phe Glu Lys Gly Glu Thr Tyr Phe His Leu His Val Leu Ile Glu
85 90 95
Thr Ile Gly Val Lys Ser Met Val Val Gly Arg Tyr Val Ser Gln Ile
100 105 110
Lys Glu Lys Leu Val Thr Arg Ile Tyr Arg Gly Val Glu Pro Gln Leu
115 120 125
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
130 135 140
Asn Lys Val Val Asp Asp Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
145 150 155 160
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Asp Gln Tyr Leu
165 170 175
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
180 185 190
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Gln Asn
195 200 205
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
210 215 220
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
225 230 235 240
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
245 250 255
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
260 265 270
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Ser Asn
275 280 285
Pro Pro Glu Asp Ile Thr Lys Asn Arg Ile Tyr Gln Ile Leu Glu Leu
290 295 300
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
305 310 315 320
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
325 330 335
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
340 345 350
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
355 360 365
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
370 375 380
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
385 390 395 400
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Glu Pro Thr Pro Val
405 410 415
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
420 425 430
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
435 440 445
Glu Leu Thr Arg Arg Leu Asp His Asp Phe Gly Lys Val Thr Lys Gln
450 455 460
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Asp Val
465 470 475 480
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
485 490 495
Ser Asn Asp Ala Asp Val Ser Glu Pro Lys Arg Gln Cys Thr Ser Leu
500 505 510
Ala Gln Pro Thr Thr Ser Asp Ala Glu Ala Pro Ala Asp Tyr Ala Asp
515 520 525
Arg Tyr Gln Asn Lys Cys Ser Arg His Val Gly Met Asn Leu Met Leu
530 535 540
Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Ile Ser Asn Val Cys
545 550 555 560
Phe Thr His Gly Gln Arg Asp Cys Gly Glu Cys Phe Pro Gly Met Ser
565 570 575
Glu Ser Gln Pro Val Ser Val Val Lys Lys Lys Thr Tyr Gln Lys Leu
580 585 590
Cys Pro Ile His His Ile Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser
595 600 605
Ala Cys Asp Leu Ala Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln
610 615 620
<210> 591
<211> 623
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 591
Met Pro Gly Phe Tyr Glu Ile Val Leu Lys Val Pro Ser Asp Leu Asp
1 5 10 15
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Ser Trp Val Ala Glu
20 25 30
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Leu Asn Leu Ile
35 40 45
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Glu Phe Leu
50 55 60
Val Glu Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
65 70 75 80
Gln Phe Glu Lys Gly Asp Ser Tyr Phe His Leu His Ile Leu Val Glu
85 90 95
Thr Val Gly Val Lys Ser Met Val Val Gly Arg Tyr Val Ser Gln Ile
100 105 110
Lys Glu Lys Leu Val Thr Arg Ile Tyr Arg Gly Val Glu Pro Gln Leu
115 120 125
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
130 135 140
Asn Lys Val Val Asp Asp Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
145 150 155 160
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Asp Gln Tyr Ile
165 170 175
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
180 185 190
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Gln Asn
195 200 205
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
210 215 220
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
225 230 235 240
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
245 250 255
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Ser Lys
260 265 270
Ile Met Ser Leu Thr Lys Thr Ala Pro Asp Tyr Leu Val Gly Gln Asn
275 280 285
Pro Pro Glu Asp Ile Ser Ser Asn Arg Ile Tyr Arg Ile Leu Glu Met
290 295 300
Asn Gly Tyr Asp Pro Gln Tyr Ala Ala Ser Val Phe Leu Gly Trp Ala
305 310 315 320
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
325 330 335
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
340 345 350
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
355 360 365
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
370 375 380
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
385 390 395 400
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
405 410 415
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
420 425 430
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
435 440 445
Glu Leu Thr Lys Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
450 455 460
Glu Val Lys Asp Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
465 470 475 480
Thr His Glu Phe Tyr Val Arg Lys Gly Gly Ala Arg Lys Arg Pro Ala
485 490 495
Pro Asn Asp Ala Asp Ile Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
500 505 510
Ala Gln Pro Ser Thr Ser Asp Ala Glu Ala Pro Val Asp Tyr Ala Asp
515 520 525
Arg Tyr Gln Asn Lys Cys Ser Arg His Val Gly Met Asn Leu Met Leu
530 535 540
Phe Pro Cys Arg Gln Cys Glu Arg Met Asn Gln Asn Val Asp Ile Cys
545 550 555 560
Phe Thr His Gly Val Met Asp Cys Ala Glu Cys Phe Pro Val Ser Glu
565 570 575
Ser Gln Pro Val Ser Val Val Arg Lys Arg Thr Tyr Gln Lys Leu Cys
580 585 590
Pro Ile His His Ile Met Gly Arg Ala Pro Glu Val Ala Cys Ser Ala
595 600 605
Cys Glu Leu Ala Asn Val Asp Leu Asp Asp Cys Asp Met Glu Gln
610 615 620
<210> 592
<211> 610
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 592
Met Ala Thr Phe Tyr Glu Val Ile Val Arg Val Pro Phe Asp Val Glu
1 5 10 15
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asp Trp Val Thr Gly
20 25 30
Gln Ile Trp Glu Leu Pro Pro Glu Ser Asp Leu Asn Leu Thr Leu Val
35 40 45
Glu Gln Pro Gln Leu Thr Val Ala Asp Arg Ile Arg Arg Val Phe Leu
50 55 60
Tyr Glu Trp Asn Lys Phe Ser Lys Gln Glu Ser Lys Phe Phe Val Gln
65 70 75 80
Phe Glu Lys Gly Ser Glu Tyr Phe His Leu His Thr Leu Val Glu Thr
85 90 95
Ser Gly Ile Ser Ser Met Val Leu Gly Arg Tyr Val Ser Gln Ile Arg
100 105 110
Ala Gln Leu Val Lys Val Val Phe Gln Gly Ile Glu Pro Gln Ile Asn
115 120 125
Asp Trp Val Ala Ile Thr Lys Val Lys Lys Gly Gly Ala Asn Lys Val
130 135 140
Val Asp Ser Gly Tyr Ile Pro Ala Tyr Leu Leu Pro Lys Val Gln Pro
145 150 155 160
Glu Leu Gln Trp Ala Trp Thr Asn Leu Asp Glu Tyr Lys Leu Ala Ala
165 170 175
Leu Asn Leu Glu Glu Arg Lys Arg Leu Val Ala Gln Phe Leu Ala Glu
180 185 190
Ser Ser Gln Arg Ser Gln Glu Ala Ala Ser Gln Arg Glu Phe Ser Ala
195 200 205
Asp Pro Val Ile Lys Ser Lys Thr Ser Gln Lys Tyr Met Ala Leu Val
210 215 220
Asn Trp Leu Val Glu His Gly Ile Thr Ser Glu Lys Gln Trp Ile Gln
225 230 235 240
Glu Asn Gln Glu Ser Tyr Leu Ser Phe Asn Ser Thr Gly Asn Ser Arg
245 250 255
Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Thr Lys Ile Met Ser Leu
260 265 270
Thr Lys Ser Ala Val Asp Tyr Leu Val Gly Ser Ser Val Pro Glu Asp
275 280 285
Ile Ser Lys Asn Arg Ile Trp Gln Ile Phe Glu Met Asn Gly Tyr Asp
290 295 300
Pro Ala Tyr Ala Gly Ser Ile Leu Tyr Gly Trp Cys Gln Arg Ser Phe
305 310 315 320
Asn Lys Arg Asn Thr Val Trp Leu Tyr Gly Pro Ala Thr Thr Gly Lys
325 330 335
Thr Asn Ile Ala Glu Ala Ile Ala His Thr Val Pro Phe Tyr Gly Cys
340 345 350
Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp Cys Val Asp Lys
355 360 365
Met Leu Ile Trp Trp Glu Glu Gly Lys Met Thr Asn Lys Val Val Glu
370 375 380
Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg Val Asp Gln Lys
385 390 395 400
Cys Lys Ser Ser Val Gln Ile Asp Ser Thr Pro Val Ile Val Thr Ser
405 410 415
Asn Thr Asn Met Cys Val Val Val Asp Gly Asn Ser Thr Thr Phe Glu
420 425 430
His Gln Gln Pro Leu Glu Asp Arg Met Phe Lys Phe Glu Leu Thr Lys
435 440 445
Arg Leu Pro Pro Asp Phe Gly Lys Ile Thr Lys Gln Glu Val Lys Asp
450 455 460
Phe Phe Ala Trp Ala Lys Val Asn Gln Val Pro Val Thr His Glu Phe
465 470 475 480
Lys Val Pro Arg Glu Leu Ala Gly Thr Lys Gly Ala Glu Lys Ser Leu
485 490 495
Lys Arg Pro Leu Gly Asp Val Thr Asn Thr Ser Tyr Lys Ser Leu Glu
500 505 510
Lys Arg Ala Arg Leu Ser Phe Val Pro Glu Thr Pro Arg Ser Ser Asp
515 520 525
Val Thr Val Asp Pro Ala Pro Leu Arg Pro Leu Asn Trp Asn Ser Arg
530 535 540
Tyr Asp Cys Lys Cys Asp Tyr His Ala Gln Phe Asp Asn Ile Ser Asn
545 550 555 560
Lys Cys Asp Glu Cys Glu Tyr Leu Asn Arg Gly Lys Asn Gly Cys Ile
565 570 575
Cys His Asn Val Thr His Cys Gln Ile Cys His Gly Ile Pro Pro Trp
580 585 590
Glu Lys Glu Asn Leu Ser Asp Phe Gly Asp Phe Asp Asp Ala Asn Lys
595 600 605
Glu Gln
610
<210> 593
<211> 623
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 593
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
1 5 10 15
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asn Trp Val Ala Glu
20 25 30
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Leu Asn Leu Ile
35 40 45
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
50 55 60
Val Gln Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
65 70 75 80
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Leu His Ile Leu Val Glu
85 90 95
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
100 105 110
Arg Asp Lys Leu Val Gln Thr Ile Tyr Arg Gly Ile Glu Pro Thr Leu
115 120 125
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
130 135 140
Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
145 150 155 160
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Glu Tyr Ile
165 170 175
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala His Asp
180 185 190
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Leu Asn
195 200 205
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
210 215 220
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
225 230 235 240
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
245 250 255
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
260 265 270
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ala
275 280 285
Pro Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
290 295 300
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
305 310 315 320
Gln Lys Arg Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
325 330 335
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
340 345 350
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
355 360 365
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
370 375 380
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
385 390 395 400
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
405 410 415
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
420 425 430
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
435 440 445
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
450 455 460
Glu Val Lys Glu Phe Phe Arg Trp Ala Gln Asp His Val Thr Glu Val
465 470 475 480
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Asn Lys Arg Pro Ala
485 490 495
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
500 505 510
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
515 520 525
Asp Arg Tyr Gln Asn Lys Cys Ser Arg His Ala Gly Met Leu Gln Met
530 535 540
Leu Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Asn Phe Asn Ile
545 550 555 560
Cys Phe Thr His Gly Thr Arg Asp Cys Ser Glu Cys Phe Pro Gly Val
565 570 575
Ser Glu Ser Gln Pro Val Val Arg Lys Arg Thr Tyr Arg Lys Leu Cys
580 585 590
Ala Ile His His Leu Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser Ala
595 600 605
Cys Asp Leu Val Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln
610 615 620
<210> 594
<211> 623
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 594
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
1 5 10 15
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asn Trp Val Ala Glu
20 25 30
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Leu Asn Leu Ile
35 40 45
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
50 55 60
Val Gln Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
65 70 75 80
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Leu His Val Leu Val Glu
85 90 95
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
100 105 110
Arg Glu Lys Leu Val Gln Thr Ile Tyr Arg Gly Val Glu Pro Thr Leu
115 120 125
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
130 135 140
Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
145 150 155 160
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Glu Tyr Ile
165 170 175
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
180 185 190
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Leu Asn
195 200 205
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
210 215 220
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
225 230 235 240
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
245 250 255
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
260 265 270
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Pro Ser
275 280 285
Leu Pro Ala Asp Ile Lys Thr Asn Arg Ile Tyr Arg Ile Leu Glu Leu
290 295 300
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
305 310 315 320
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
325 330 335
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
340 345 350
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
355 360 365
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
370 375 380
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
385 390 395 400
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
405 410 415
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
420 425 430
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
435 440 445
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
450 455 460
Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
465 470 475 480
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Ser Lys Arg Pro Ala
485 490 495
Pro Asp Asp Ala Asp Ile Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
500 505 510
Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp Phe Ala
515 520 525
Asp Arg Tyr Gln Asn Lys Cys Ser Arg His Ala Gly Met Ile Gln Met
530 535 540
Leu Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Asn Phe Asn Ile
545 550 555 560
Cys Phe Thr His Gly Val Arg Asp Cys Leu Glu Cys Phe Pro Gly Val
565 570 575
Ser Glu Ser Gln Pro Val Val Arg Lys Lys Thr Tyr Arg Lys Leu Cys
580 585 590
Ala Ile His His Leu Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser Ala
595 600 605
Cys Asp Leu Val Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln
610 615 620
<210> 595
<211> 625
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 595
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
1 5 10 15
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asn Trp Val Ala Glu
20 25 30
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Arg Asn Leu Ile
35 40 45
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
50 55 60
Val Gln Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
65 70 75 80
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Leu His Val Leu Val Glu
85 90 95
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
100 105 110
Arg Glu Lys Leu Gly Pro Asp His Leu Pro Ala Gly Ser Ser Pro Thr
115 120 125
Leu Pro Asn Trp Phe Ala Val Thr Lys Asp Ala Val Met Ala Pro Ala
130 135 140
Gly Gly Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu
145 150 155 160
Pro Lys Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Glu
165 170 175
Tyr Ile Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala
180 185 190
Gln His Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn
195 200 205
Leu Asn Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala
210 215 220
Arg Tyr Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser
225 230 235 240
Glu Lys Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn
245 250 255
Ala Ala Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala
260 265 270
Gly Lys Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly
275 280 285
Pro Ser Leu Pro Ala Asp Ile Thr Gln Asn Arg Ile Tyr Arg Ile Leu
290 295 300
Ala Leu Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly
305 310 315 320
Trp Ala Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly
325 330 335
Pro Ala Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala
340 345 350
Val Pro Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe
355 360 365
Asn Asp Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met
370 375 380
Thr Ala Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys
385 390 395 400
Val Arg Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr
405 410 415
Pro Val Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly
420 425 430
Asn Ser Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe
435 440 445
Lys Phe Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr
450 455 460
Lys Gln Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr
465 470 475 480
Glu Val Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Ser Lys Arg
485 490 495
Pro Ala Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro
500 505 510
Ser Val Ala Asp Pro Ser Thr Ser Asp Ala Glu Gly Ala Pro Val Asp
515 520 525
Phe Ala Asp Arg Tyr Gln Asn Lys Cys Ser Arg His Ala Gly Met Leu
530 535 540
Gln Met Leu Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Asn Phe
545 550 555 560
Asn Ile Cys Phe Thr His Gly Val Arg Asp Cys Ser Glu Cys Phe Pro
565 570 575
Gly Val Ser Glu Ser Gln Pro Val Val Arg Lys Arg Thr Tyr Arg Lys
580 585 590
Leu Cys Ala Ile His His Leu Leu Gly Arg Ala Pro Glu Ile Ala Cys
595 600 605
Ser Ala Cys Asp Leu Val Asn Val Asp Leu Asp Asp Cys Val Ser Glu
610 615 620
Gln
625
<210> 596
<211> 622
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<220>
<221> Модифицированная аминокислота
<222> (118)..(119)
<223> Любая аминокислота
<220>
<221> Модифицированная аминокислота
<222> (287)..(287)
<223> Любая аминокислота
<220>
<221> Модифицированная аминокислота
<222> (541)..(541)
<223> Любая аминокислота
<220>
<221> Модифицированная аминокислота
<222> (557)..(557)
<223> Любая аминокислота
<220>
<221> Модифицированная аминокислота
<222> (565)..(565)
<223> Любая аминокислота
<220>
<221> Модифицированная аминокислота
<222> (569)..(569)
<223> Любая аминокислота
<220>
<221> Модифицированная аминокислота
<222> (580)..(580)
<223> Любая аминокислота
<220>
<221> Модифицированная аминокислота
<222> (588)..(588)
<223> Любая аминокислота
<220>
<221> Модифицированная аминокислота
<222> (592)..(592)
<223> Любая аминокислота
<400> 596
Met Pro Gly Phe Tyr Glu Ile Val Ile Lys Val Pro Ser Asp Leu Asp
1 5 10 15
Glu His Leu Pro Gly Ile Ser Asp Ser Phe Val Asn Trp Val Ala Glu
20 25 30
Lys Glu Trp Glu Leu Pro Pro Asp Ser Asp Met Asp Arg Asn Leu Ile
35 40 45
Glu Gln Ala Pro Leu Thr Val Ala Glu Lys Leu Gln Arg Asp Phe Leu
50 55 60
Val Gln Trp Arg Arg Val Ser Lys Ala Pro Glu Ala Leu Phe Phe Val
65 70 75 80
Gln Phe Glu Lys Gly Glu Ser Tyr Phe His Leu His Val Leu Val Glu
85 90 95
Thr Thr Gly Val Lys Ser Met Val Leu Gly Arg Phe Leu Ser Gln Ile
100 105 110
Arg Glu Lys Leu Val Xaa Xaa Ile Tyr Arg Gly Ile Glu Pro Thr Leu
115 120 125
Pro Asn Trp Phe Ala Val Thr Lys Thr Arg Asn Gly Ala Gly Gly Gly
130 135 140
Asn Lys Val Val Asp Glu Cys Tyr Ile Pro Asn Tyr Leu Leu Pro Lys
145 150 155 160
Thr Gln Pro Glu Leu Gln Trp Ala Trp Thr Asn Met Glu Glu Tyr Ile
165 170 175
Ser Ala Cys Leu Asn Leu Ala Glu Arg Lys Arg Leu Val Ala Gln His
180 185 190
Leu Thr His Val Ser Gln Thr Gln Glu Gln Asn Lys Glu Asn Leu Asn
195 200 205
Pro Asn Ser Asp Ala Pro Val Ile Arg Ser Lys Thr Ser Ala Arg Tyr
210 215 220
Met Glu Leu Val Gly Trp Leu Val Asp Arg Gly Ile Thr Ser Glu Lys
225 230 235 240
Gln Trp Ile Gln Glu Asp Gln Ala Ser Tyr Ile Ser Phe Asn Ala Ala
245 250 255
Ser Asn Ser Arg Ser Gln Ile Lys Ala Ala Leu Asp Asn Ala Gly Lys
260 265 270
Ile Met Ala Leu Thr Lys Ser Ala Pro Asp Tyr Leu Val Gly Xaa Ser
275 280 285
Pro Pro Glu Asp Ile Ser Thr Asn Arg Ile Tyr Arg Ile Leu Ala Leu
290 295 300
Asn Gly Tyr Asp Pro Ala Tyr Ala Gly Ser Val Phe Leu Gly Trp Ala
305 310 315 320
Gln Lys Lys Phe Gly Lys Arg Asn Thr Ile Trp Leu Phe Gly Pro Ala
325 330 335
Thr Thr Gly Lys Thr Asn Ile Ala Glu Ala Ile Ala His Ala Val Pro
340 345 350
Phe Tyr Gly Cys Val Asn Trp Thr Asn Glu Asn Phe Pro Phe Asn Asp
355 360 365
Cys Val Asp Lys Met Val Ile Trp Trp Glu Glu Gly Lys Met Thr Ala
370 375 380
Lys Val Val Glu Ser Ala Lys Ala Ile Leu Gly Gly Ser Lys Val Arg
385 390 395 400
Val Asp Gln Lys Cys Lys Ser Ser Ala Gln Ile Asp Pro Thr Pro Val
405 410 415
Ile Val Thr Ser Asn Thr Asn Met Cys Ala Val Ile Asp Gly Asn Ser
420 425 430
Thr Thr Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Phe Lys Phe
435 440 445
Glu Leu Thr Arg Arg Leu Glu His Asp Phe Gly Lys Val Thr Lys Gln
450 455 460
Glu Val Lys Glu Phe Phe Arg Trp Ala Ser Asp His Val Thr Glu Val
465 470 475 480
Ala His Glu Phe Tyr Val Arg Lys Gly Gly Ala Lys Lys Arg Pro Ala
485 490 495
Pro Asp Asp Ala Asp Lys Ser Glu Pro Lys Arg Ala Cys Pro Ser Val
500 505 510
Ala Asp Pro Ser Thr Ser Asp Ala Glu Ala Pro Val Asp Phe Ala Asp
515 520 525
Arg Tyr Gln Asn Lys Cys Ser Arg His Ala Gly Met Xaa Gln Met Leu
530 535 540
Phe Pro Cys Lys Thr Cys Glu Arg Met Asn Gln Asn Xaa Asn Ile Cys
545 550 555 560
Phe Thr His Gly Xaa Arg Asp Cys Xaa Glu Cys Phe Pro Gly Val Ser
565 570 575
Glu Ser Gln Xaa Val Val Arg Lys Arg Thr Tyr Xaa Lys Leu Cys Xaa
580 585 590
Ile His His Leu Leu Gly Arg Ala Pro Glu Ile Ala Cys Ser Ala Cys
595 600 605
Asp Leu Val Asn Val Asp Leu Asp Asp Cys Val Ser Glu Gln
610 615 620
<210> 597
<211> 1530
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 597
atgtccatgg gggcaccgcg gtccctcctc ctggccctgg ctgctggcct ggccgttgcc 60
cgtccgccca acatcgtgct gatctttgcc gacgacctcg gctatgggga cctgggctgc 120
tatgggcacc ccagctctac cactcccaac ctggaccagc tggcggcggg agggctgcgg 180
ttcacagact tctacgtgcc tgtgtctctg tgcacaccct ctagggccgc cctcctgacc 240
ggccggctcc cggttcggat gggcatgtac cctggcgtcc tggtgcccag ctcccggggg 300
ggcctgcccc tggaggaggt gaccgtggcc gaagtcctgg ctgcccgagg ctacctcaca 360
ggaatggccg gcaagtggca ccttggggtg gggcctgagg gggccttcct gcccccccat 420
cagggcttcc atcgatttct aggcatcccg tactcccacg accagggccc ctgccagaac 480
ctgacctgct tcccgccggc cactccttgc gacggtggct gtgaccaggg cctggtcccc 540
atcccactgt tggccaacct gtccgtggag gcgcagcccc cctggctgcc cggactagag 600
gcccgctaca tggctttcgc ccatgacctc atggccgacg cccagcgcca ggatcgcccc 660
ttcttcctgt actatgcctc tcaccacacc cactaccctc agttcagtgg gcagagcttt 720
gcagagcgtt caggccgcgg gccatttggg gactccctga tggagctgga tgcagctgtg 780
gggaccctga tgacagccat aggggacctg gggctgcttg aagagacgct ggtcatcttc 840
actgcagaca atggacctga gaccatgcgt atgtcccgag gcggctgctc cggtctcttg 900
cggtgtggaa agggaacgac ctacgagggc ggtgtccgag agcctgcctt ggccttctgg 960
ccaggtcata tcgctcccgg cgtgacccac gagctggcca gctccctgga cctgctgcct 1020
accctggcag ccctggctgg ggccccactg cccaatgtca ccttggatgg ctttgacctc 1080
agccccctgc tgctgggcac aggcaagagc cctcggcagt ctctcttctt ctacccgtcc 1140
tacccagacg aggtccgtgg ggtttttgct gtgcggactg gaaagtacaa ggctcacttc 1200
ttcacccagg gctctgccca cagtgatacc actgcagacc ctgcctgcca cgcctccagc 1260
tctctgactg ctcatgagcc cccgctgctc tatgacctgt ccaaggaccc tggtgagaac 1320
tacaacctgc tggggggtgt ggccggggcc accccagagg tgctgcaagc cctgaaacag 1380
cttcagctgc tcaaggccca gttagacgca gctgtgacct tcggccccag ccaggtggcc 1440
cggggcgagg accccgccct gcagatctgc tgtcatcctg gctgcacccc ccgcccagct 1500
tgctgccatt gcccagatcc ccatgcctga 1530
<210> 598
<211> 1653
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полинуклеотид
<400> 598
atgccgccac cccggaccgg ccgaggcctt ctctggctgg gtctggttct gagctccgtc 60
tgcgtcgccc tcggatccga aacgcaggcc aactcgacca cagatgctct gaacgttctt 120
ctcatcatcg tggatgacct gcgcccctcc ctgggctgtt atggggataa gctggtgagg 180
tccccaaata ttgaccaact ggcatcccac agcctcctct tccagaatgc ctttgcgcag 240
caagcagtgt gcgccccgag ccgcgtttct ttcctcactg gcaggagacc tgacaccacc 300
cgcctgtacg acttcaactc ctactggagg gtgcacgctg gaaacttctc caccatcccc 360
cagtacttca aggagaatgg ctatgtgacc atgtcggtgg gaaaagtctt tcaccctggg 420
atatcttcta accataccga tgattctccg tatagctggt cttttccacc ttatcatcct 480
tcctctgaga agtatgaaaa cactaagaca tgtcgagggc cagatggaga actccatgcc 540
aacctgcttt gccctgtgga tgtgctggat gttcccgagg gcaccttgcc tgacaaacag 600
agcactgagc aagccataca gttgttggaa aagatgaaaa cgtcagccag tcctttcttc 660
ctggccgttg ggtatcataa gccacacatc cccttcagat accccaagga atttcagaag 720
ttgtatccct tggagaacat caccctggcc cccgatcccg aggtccctga tggcctaccc 780
cctgtggcct acaacccctg gatggacatc aggcaacggg aagacgtcca agccttaaac 840
atcagtgtgc cgtatggtcc aattcctgtg gactttcagc ggaaaatccg ccagagctac 900
tttgcctctg tgtcatattt ggatacacag gtcggccgcc tcttgagtgc tttggacgat 960
cttcagctgg ccaacagcac catcattgca tttacctcgg atcatgggtg ggctctaggt 1020
gaacatggag aatgggccaa atacagcaat tttgatgttg ctacccatgt tcccctgata 1080
ttctatgttc ctggaaggac ggcttcactt ccggaggcag gcgagaagct tttcccttac 1140
ctcgaccctt ttgattccgc ctcacagttg atggagccag gcaggcaatc catggacctt 1200
gtggaacttg tgtctctttt tcccacgctg gctggacttg caggactgca ggttccacct 1260
cgctgccccg ttccttcatt tcacgttgag ctgtgcagag aaggcaagaa ccttctgaag 1320
cattttcgat tccgtgactt ggaagaggat ccgtacctcc ctggtaatcc ccgtgaactg 1380
attgcctata gccagtatcc ccggccttca gacatccctc agtggaattc tgacaagccg 1440
agtttaaaag atataaagat catgggctat tccatacgca ccatagacta taggtatact 1500
gtgtgggttg gcttcaatcc tgatgaattt ctagctaact tttctgacat ccatgcaggg 1560
gaactgtatt ttgtggattc tgacccattg caggatcaca atatgtataa tgattcccaa 1620
ggtggagatc ttttccagtt gttgatgcct tga 1653
<210> 599
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 599
gcgcgctcgc tcgctc 16
<210> 600
<211> 8
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 600
actgaggc 8
<210> 601
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 601
cgggcgacca aaggtcgccc ga 22
<210> 602
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 602
cgcccgggcg 10
<210> 603
<211> 8
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 603
gcctcagt 8
<210> 604
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 604
gagcgagcga gcgcgc 16
<210> 605
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 605
gcccgggcgt cgggcgacct ttggtcgccc g 31
<210> 606
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
олигонуклеотид
<400> 606
gcccgggcaa agcccgggcg 20
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КОНСТРУКЦИИ НУКЛЕИНОВОЙ КИСЛОТЫ И ВЕКТОРЫ ДЛЯ ГЕНОТЕРАПИИ ДЛЯ ПРИМЕНЕНИЯ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ВИЛЬСОНА И ДРУГИХ СОСТОЯНИЙ | 2015 |
|
RU2745567C2 |
| Выделенная нуклеиновая кислота, которая кодирует слитый белок на основе FVIII-BDD и гетерологичного сигнального пептида, и ее применение | 2022 |
|
RU2818229C2 |
| МОЛЕКУЛЫ НУКЛЕИНОВОЙ КИСЛОТЫ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2819144C2 |
| Вакцина на основе AAV5 для индукции специфического иммунитета к вирусу SARS-CoV-2 и/или профилактики коронавирусной инфекции, вызванной SARS-CoV-2 | 2020 |
|
RU2760301C1 |
| МОЛЕКУЛЫ НУКЛЕИНОВОЙ КИСЛОТЫ И ПУТИ ИХ ПРИМЕНЕНИЯ ДЛЯ НЕВИРУСНОЙ ГЕННОЙ ТЕРАПИИ | 2019 |
|
RU2834625C2 |
| Вакцина на основе AAV5 для индукции специфического иммунитета к вирусу SARS-CoV-2 и/или профилактики коронавирусной инфекции, вызванной SARS-CoV-2 | 2020 |
|
RU2783313C1 |
| Кодон-оптимизированная нуклеиновая кислота, которая кодирует белок фактора свёртывания крови IX, и ее применение | 2021 |
|
RU2831751C2 |
| Вакцина на основе AAV5 для индукции специфического иммунитета к вирусу SARS-CoV-2 и/или профилактики коронавирусной инфекции, вызванной SARS-CoV-2 | 2021 |
|
RU2761879C1 |
| ГЕНОТЕРАПЕВТИЧЕСКИЕ КОНСТРУКЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ВИЛЬСОНА | 2020 |
|
RU2807158C2 |
| СОЛЮБИЛИЗИРОВАННЫЕ АПИРАЗЫ, СПОСОБЫ И ПРИМЕНЕНИЕ | 2019 |
|
RU2791992C2 |
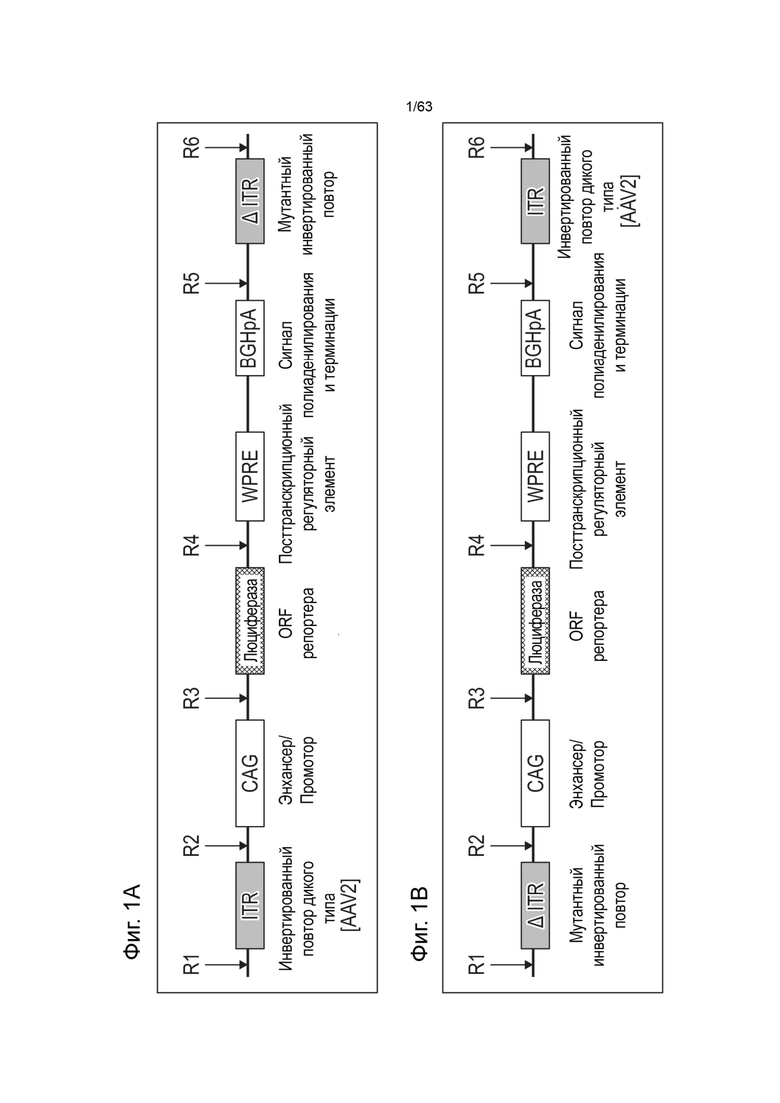
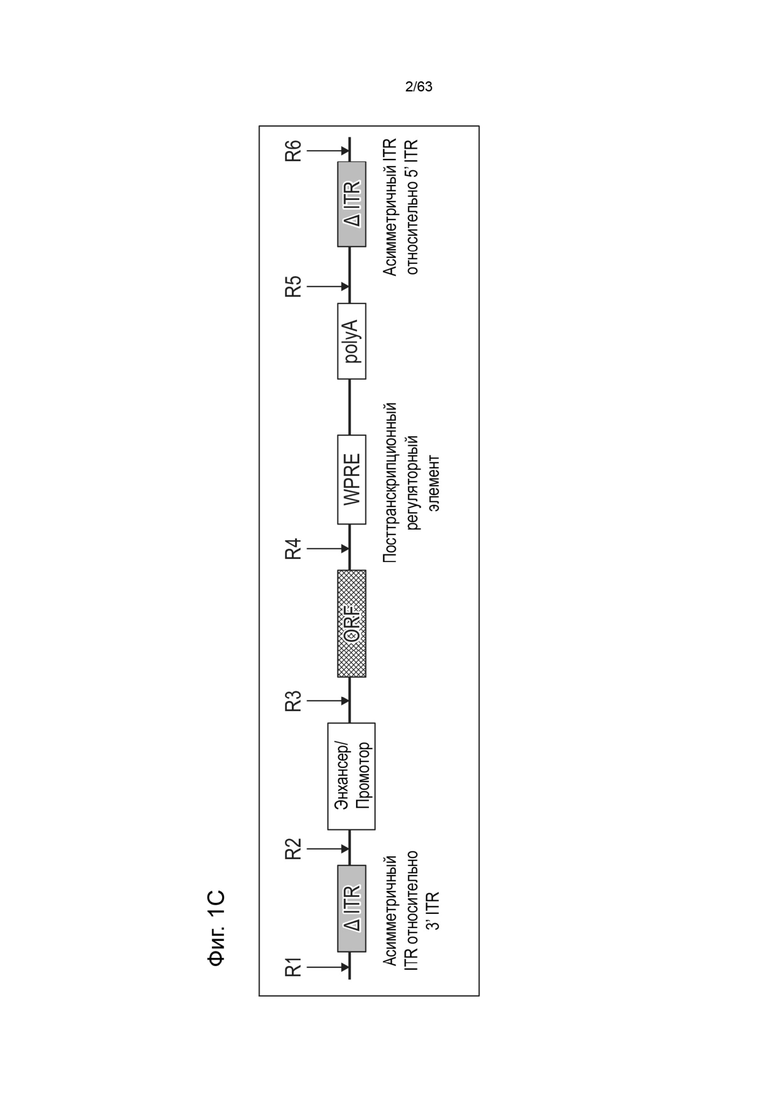
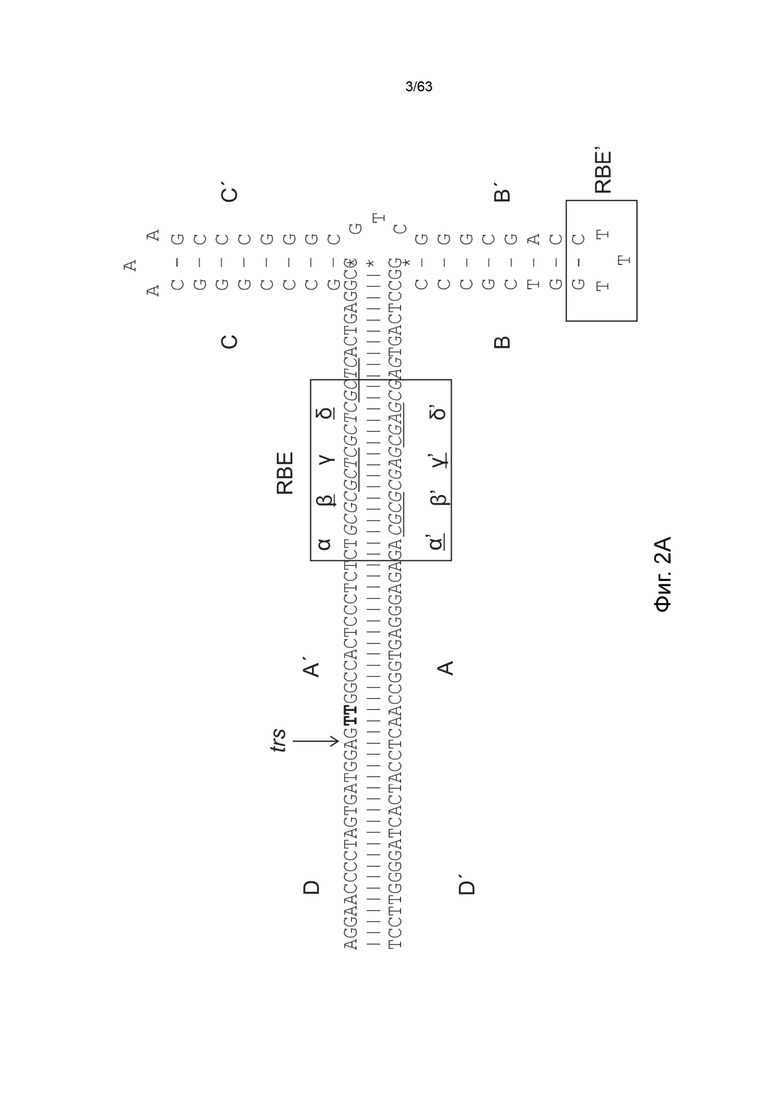
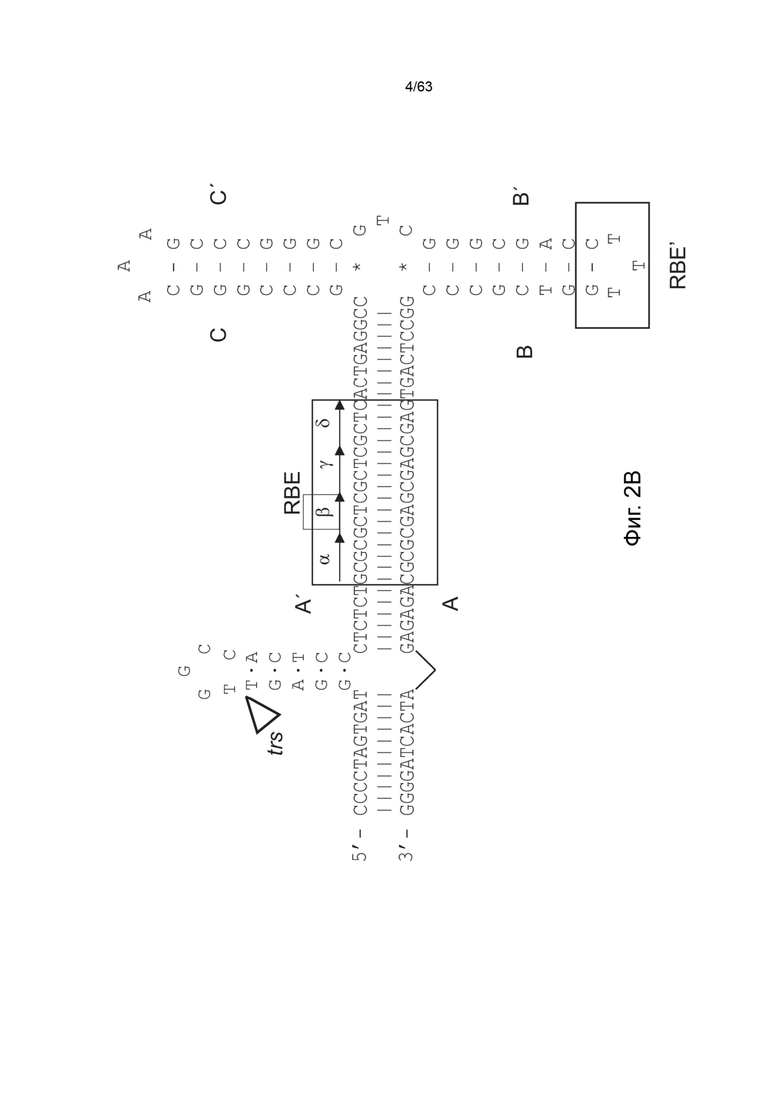
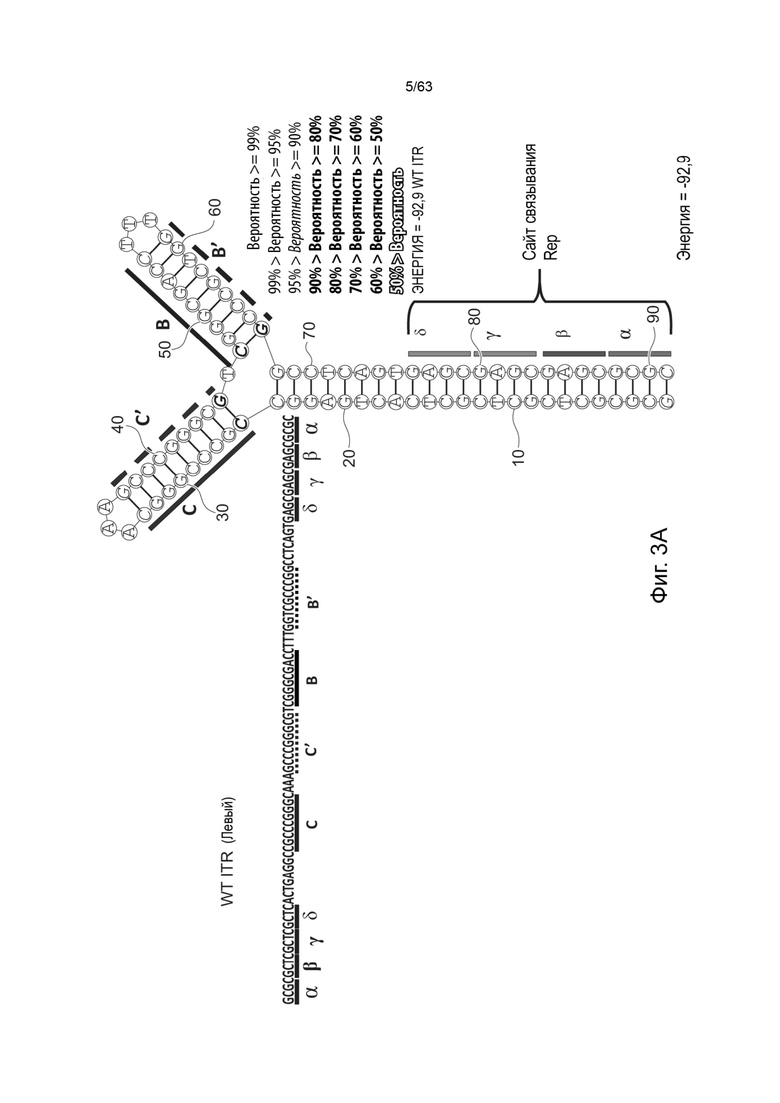
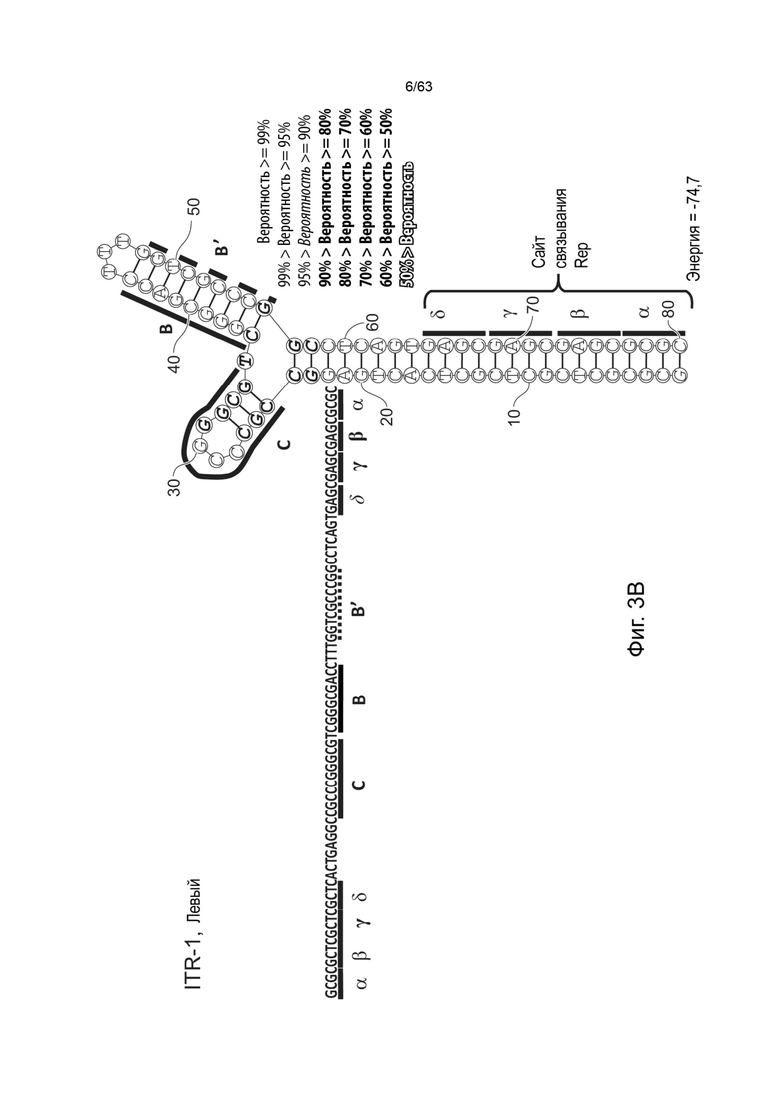
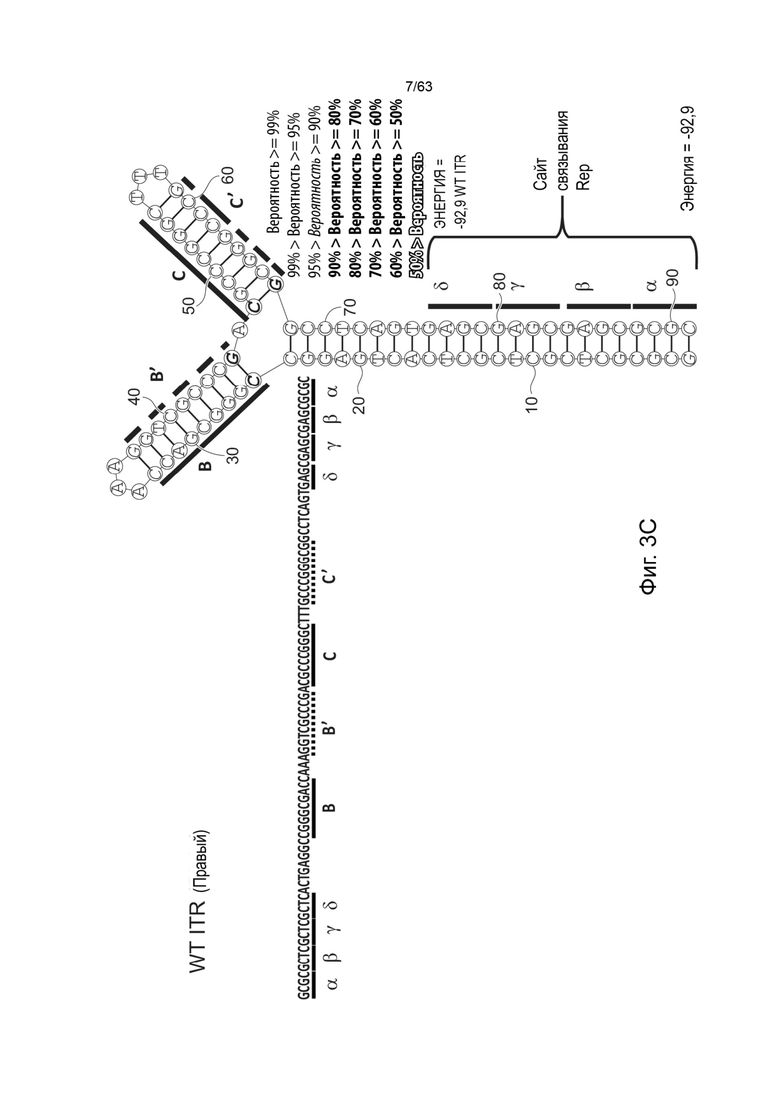
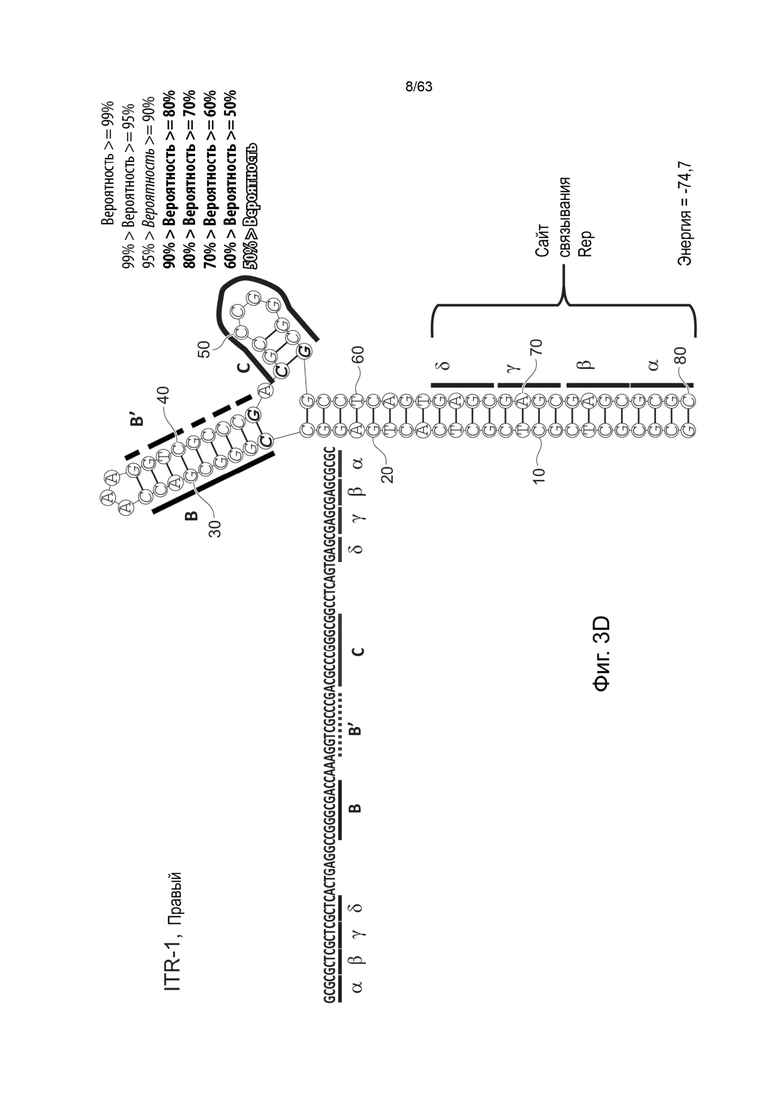
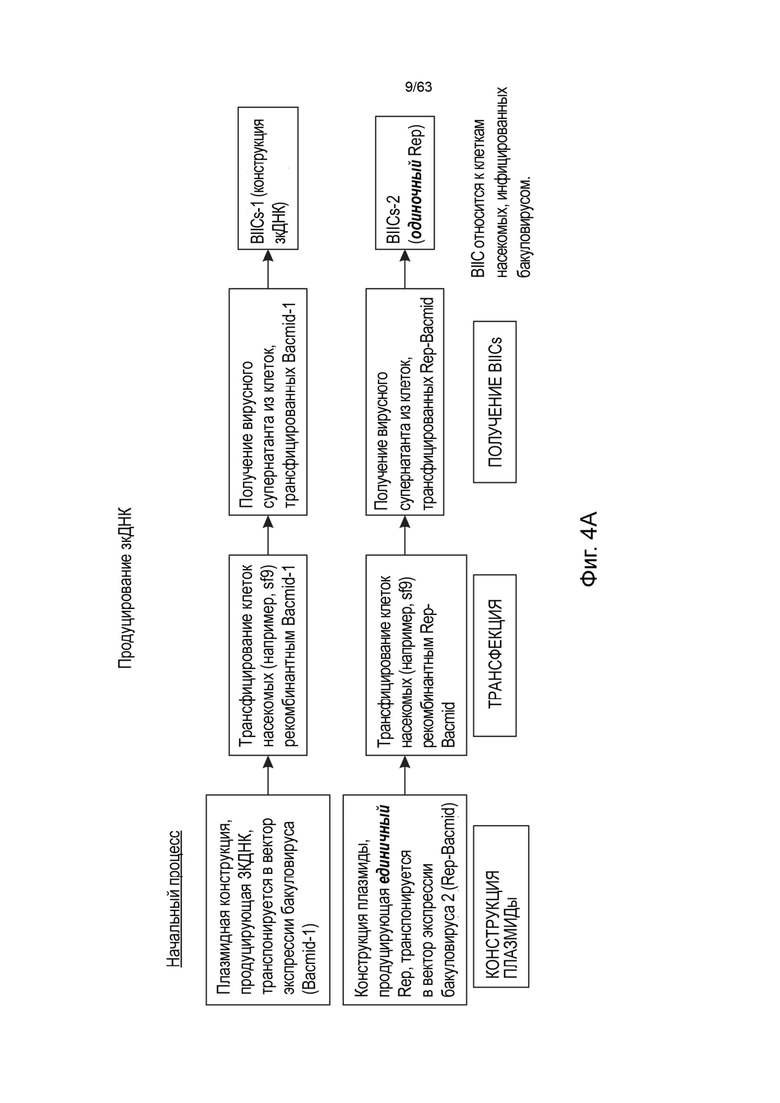
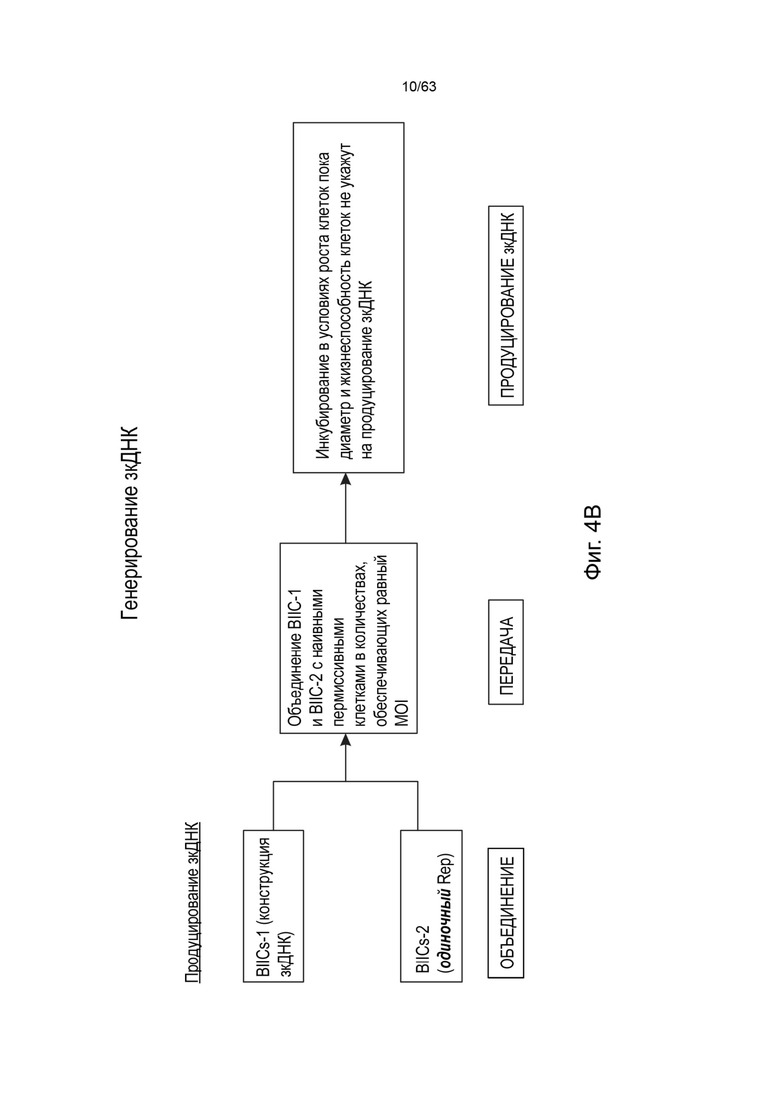
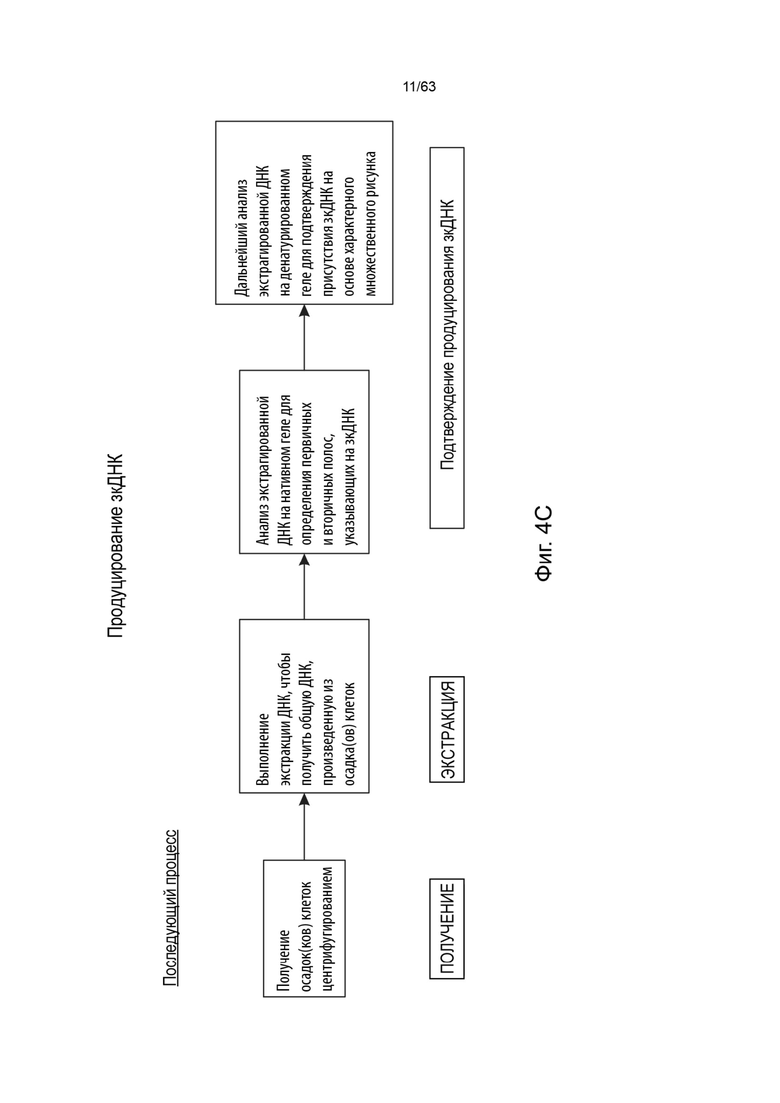
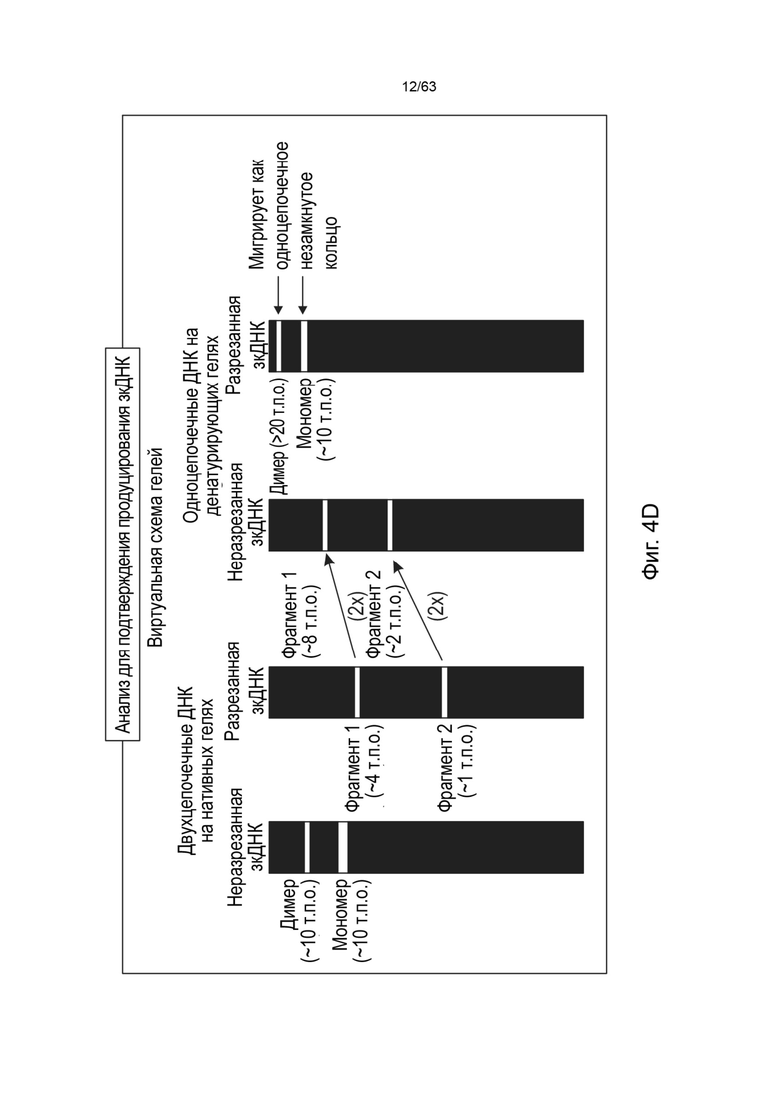
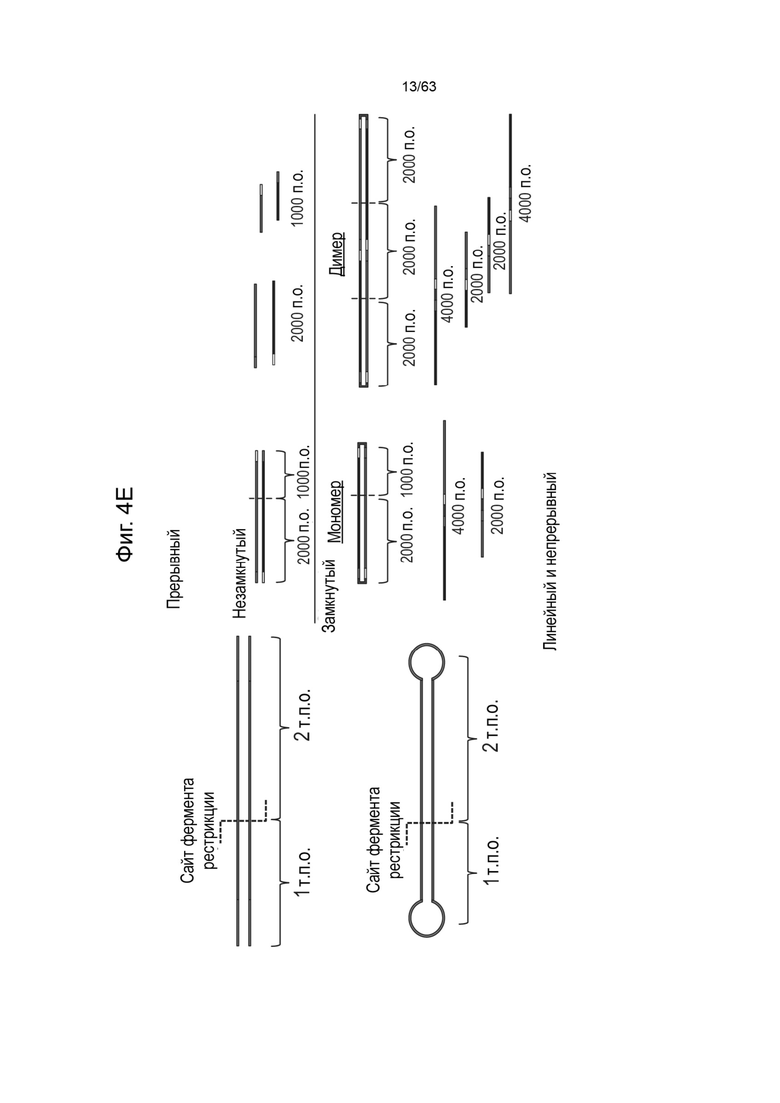
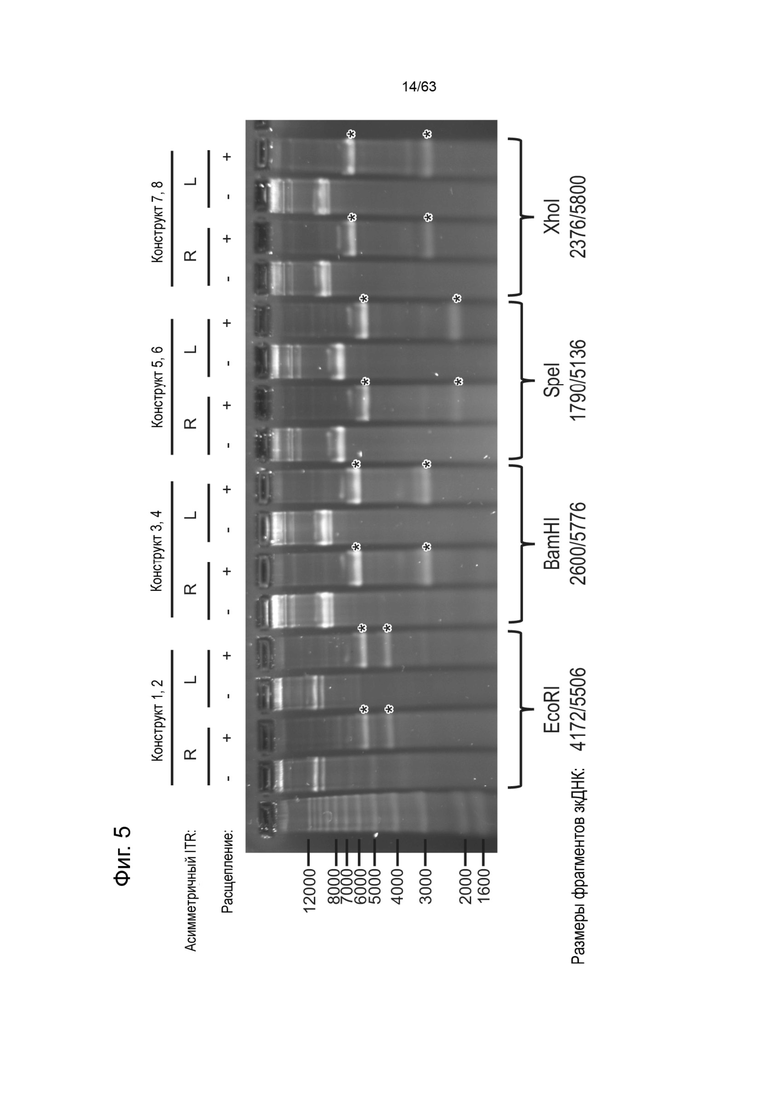
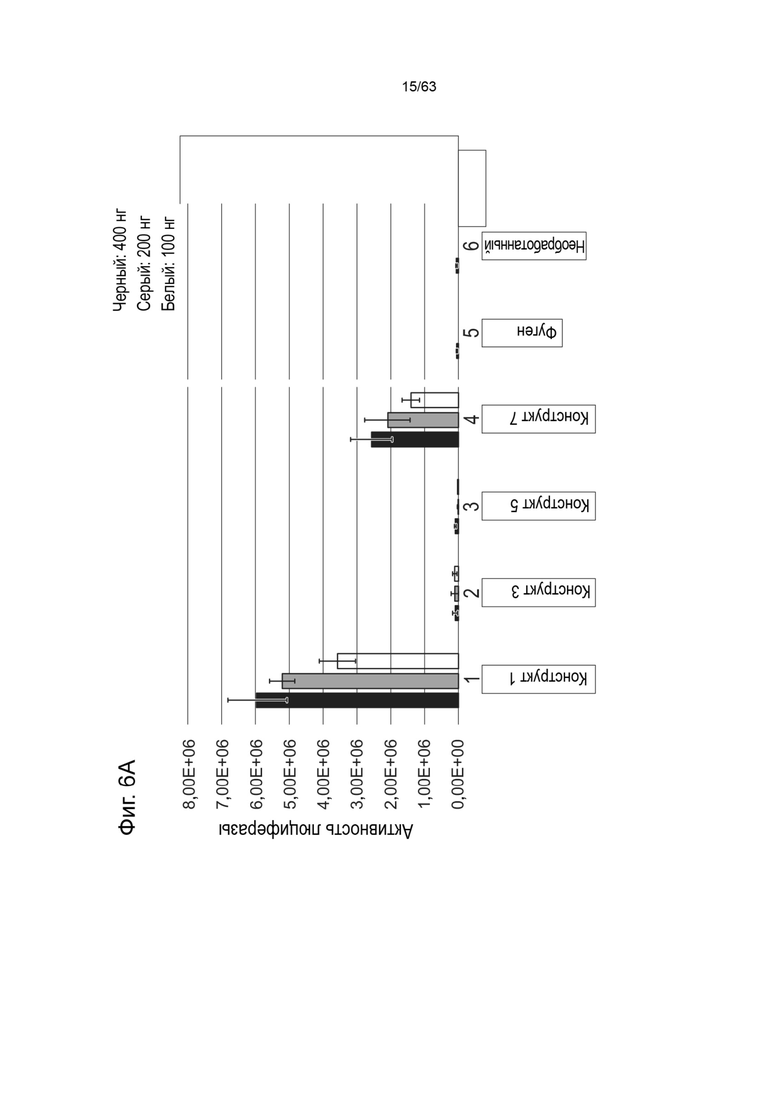
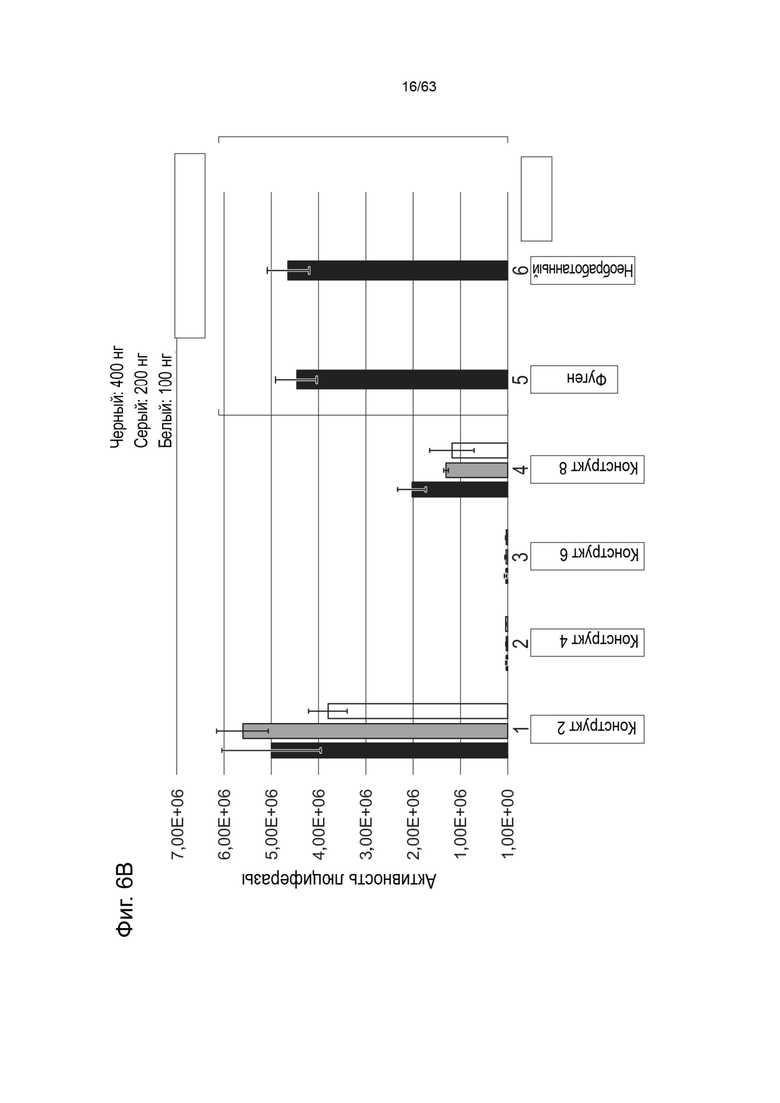
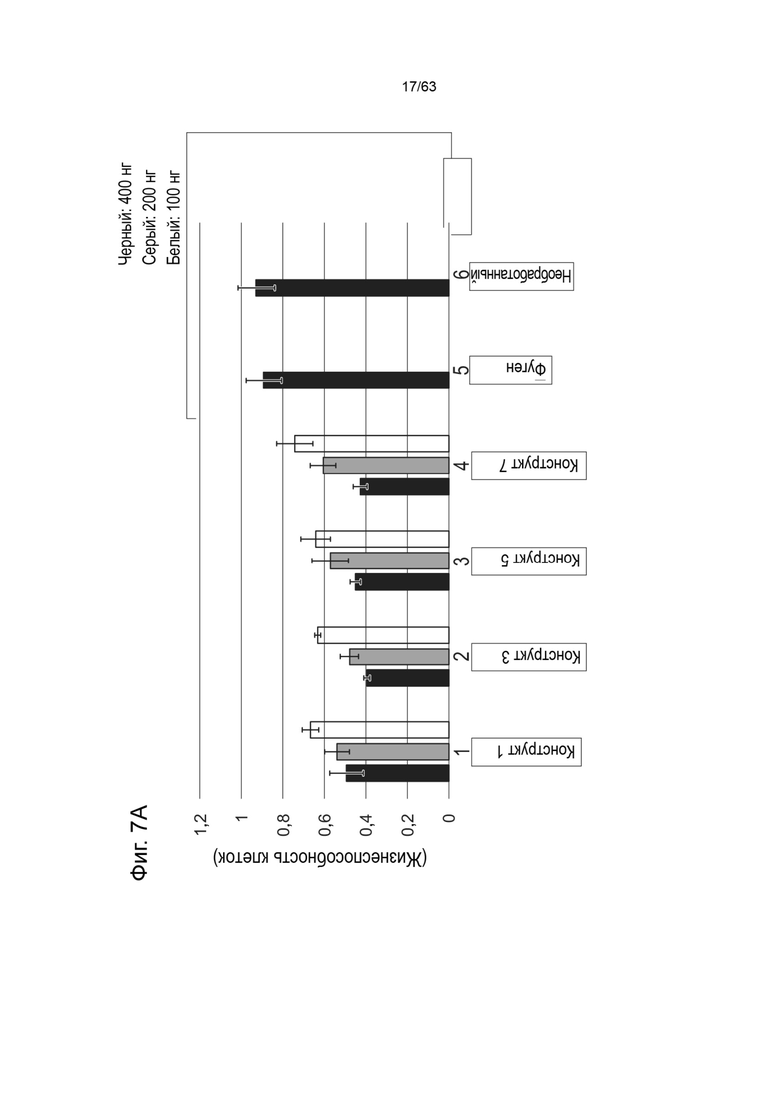
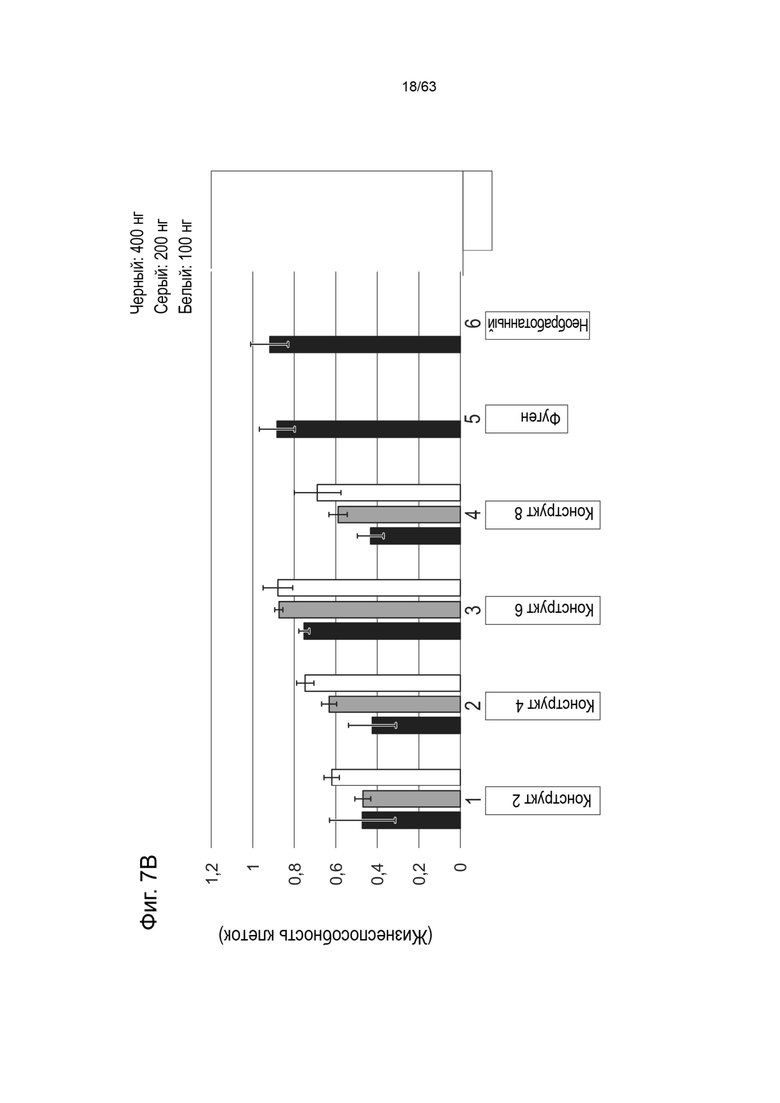
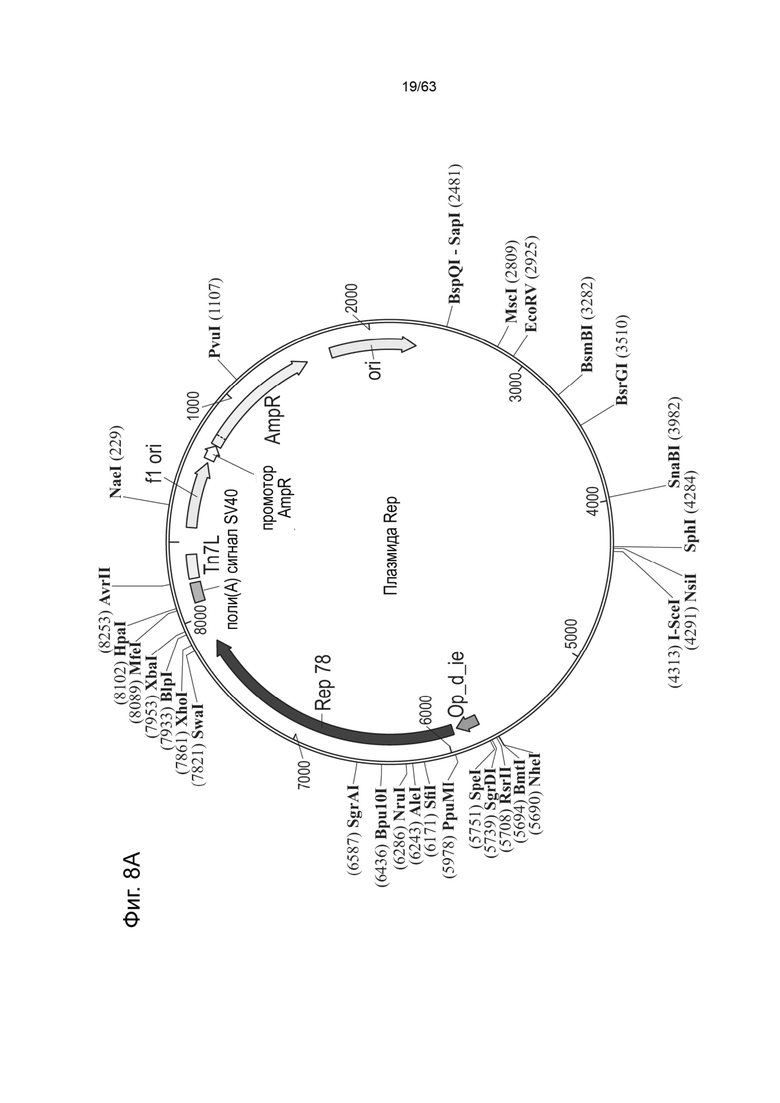
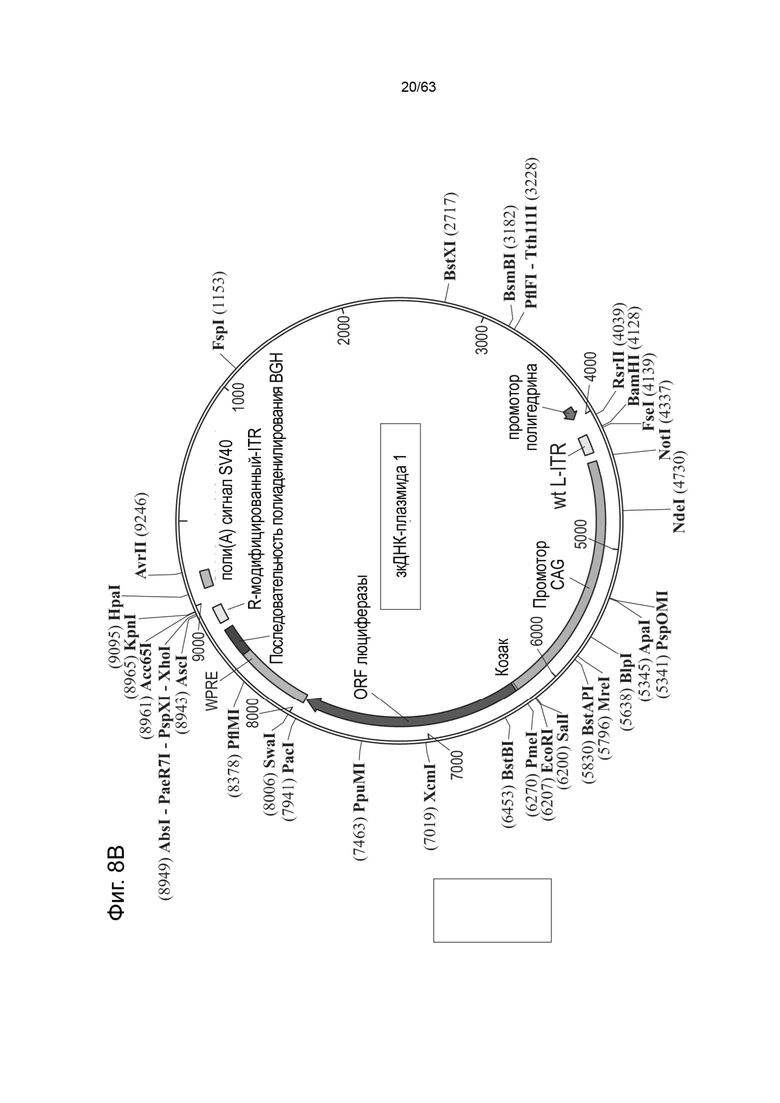
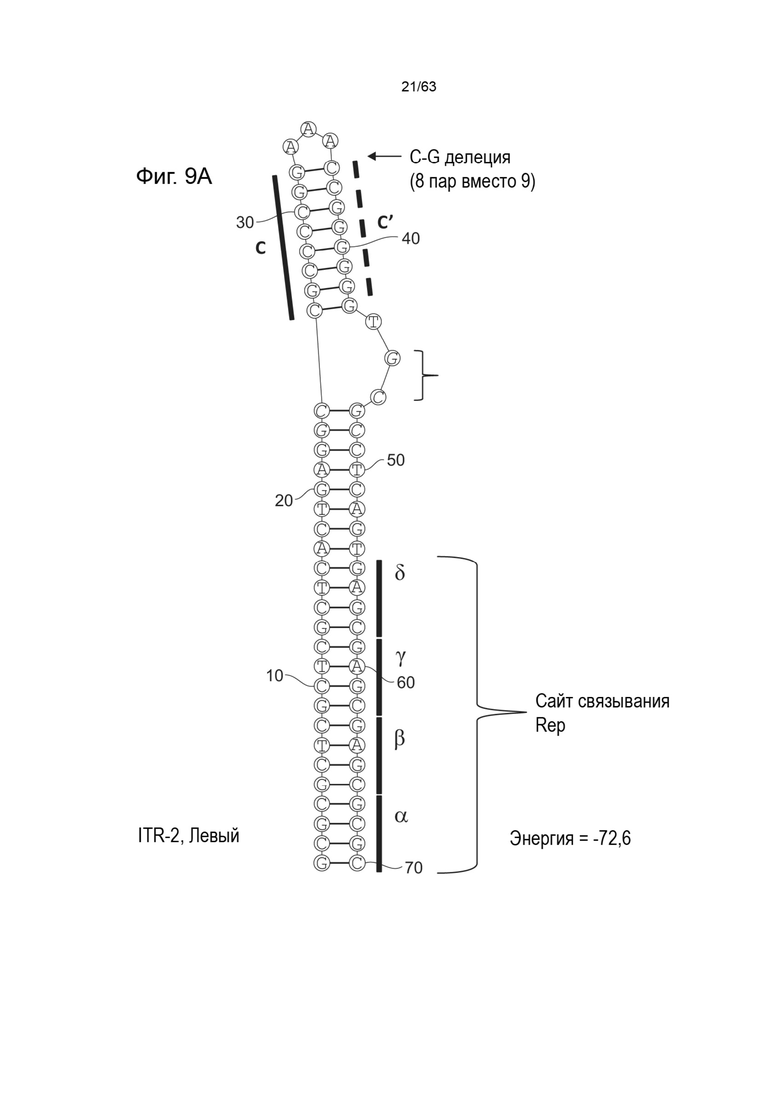
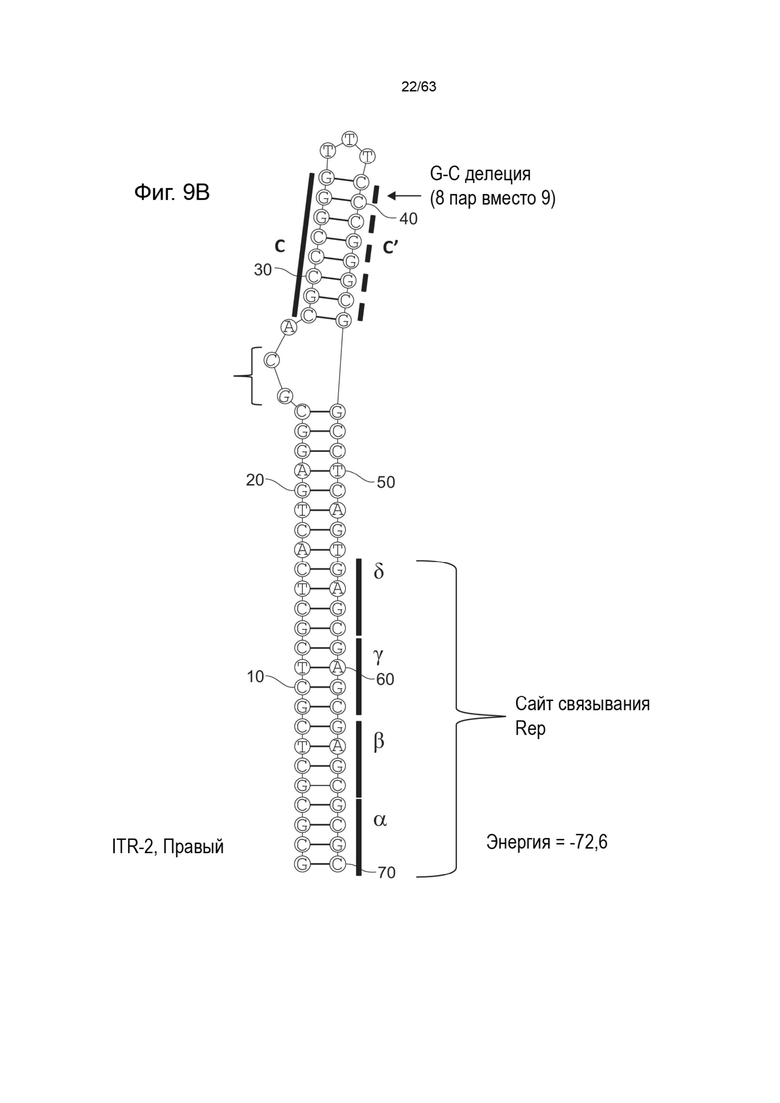
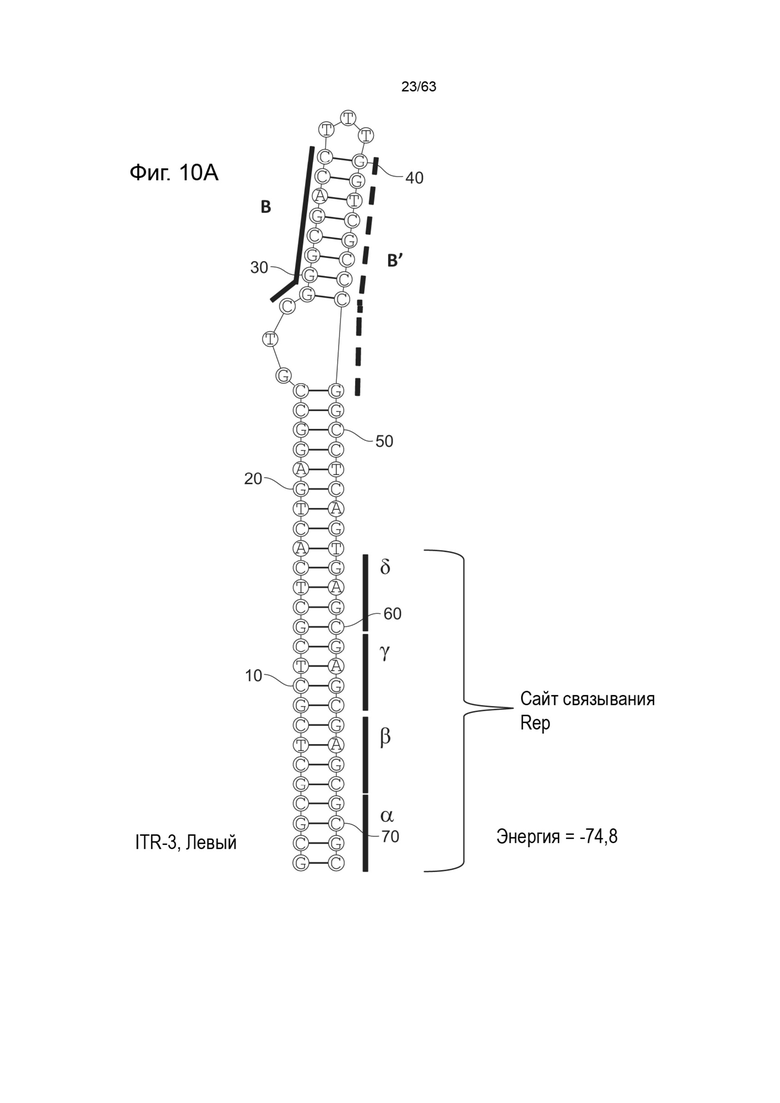
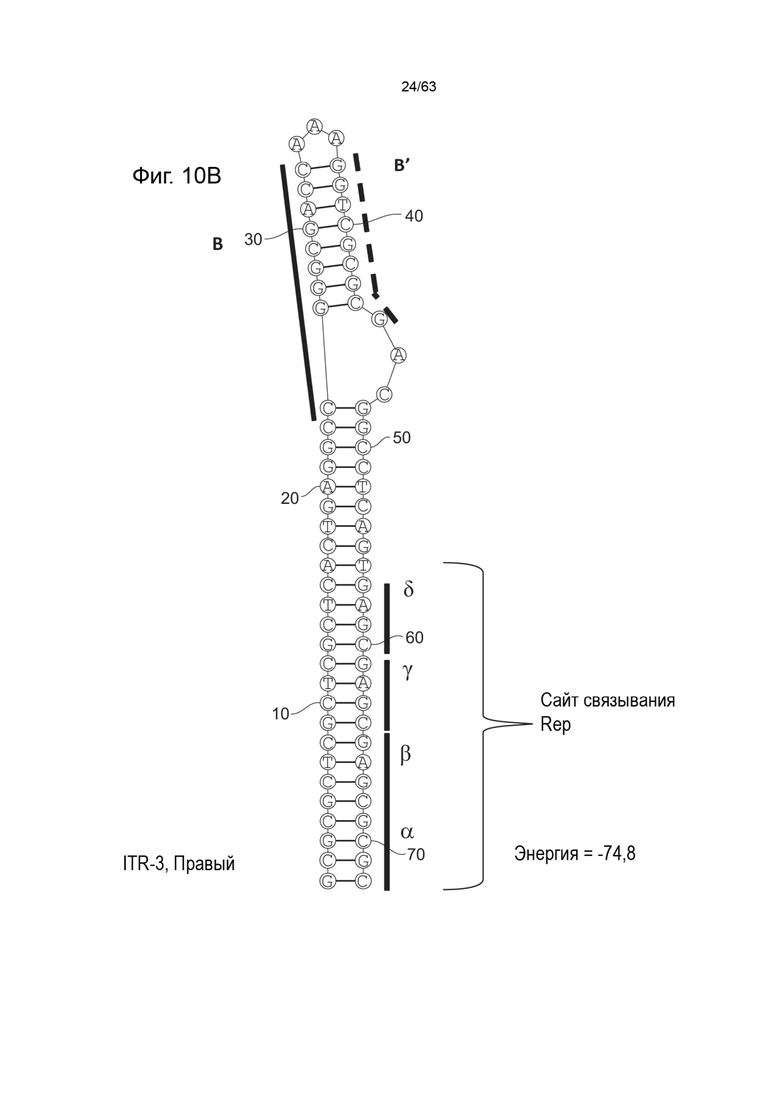
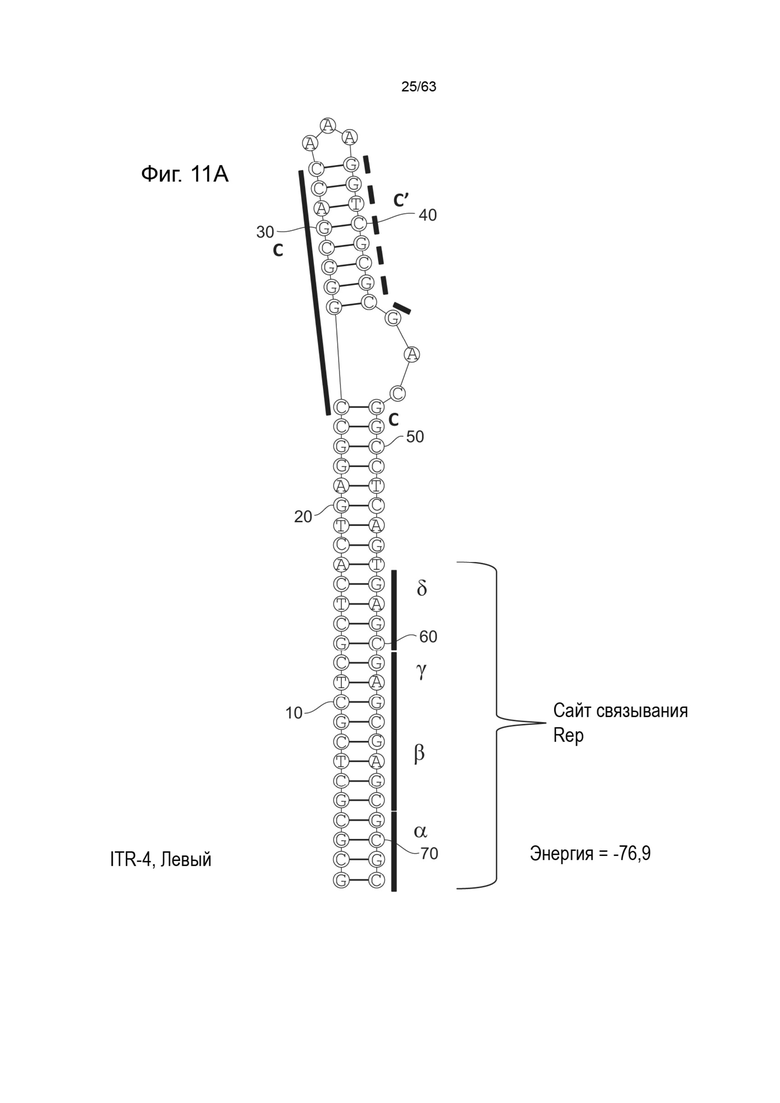
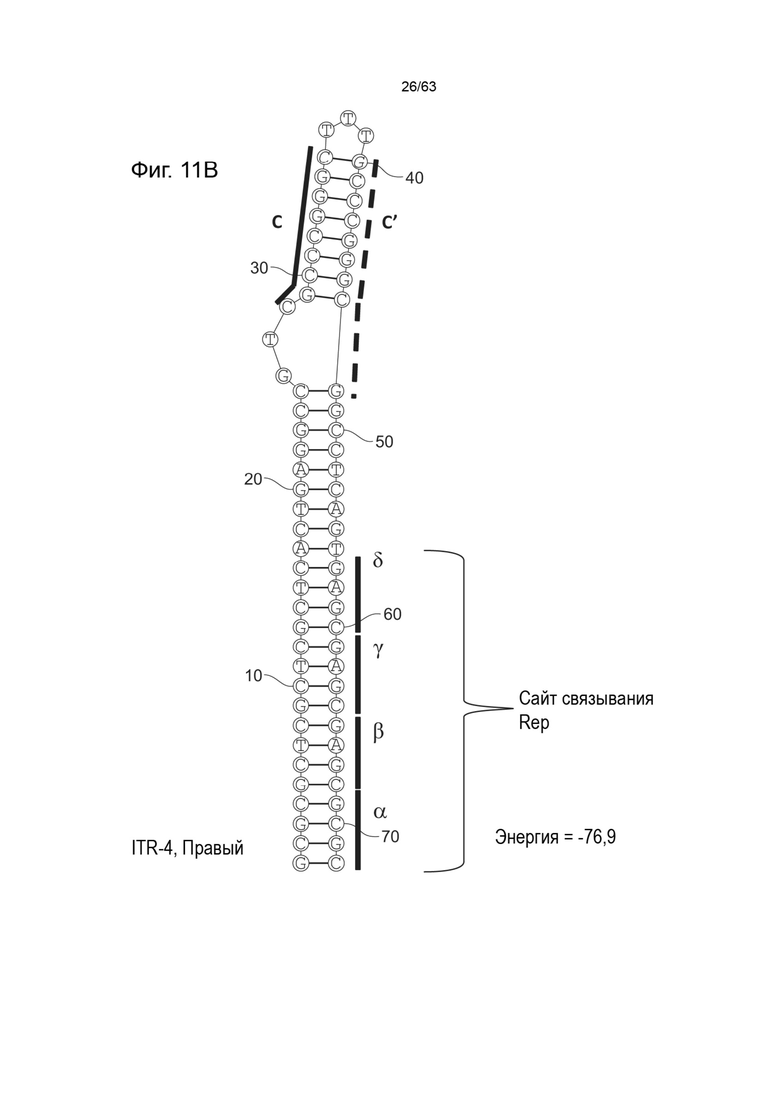
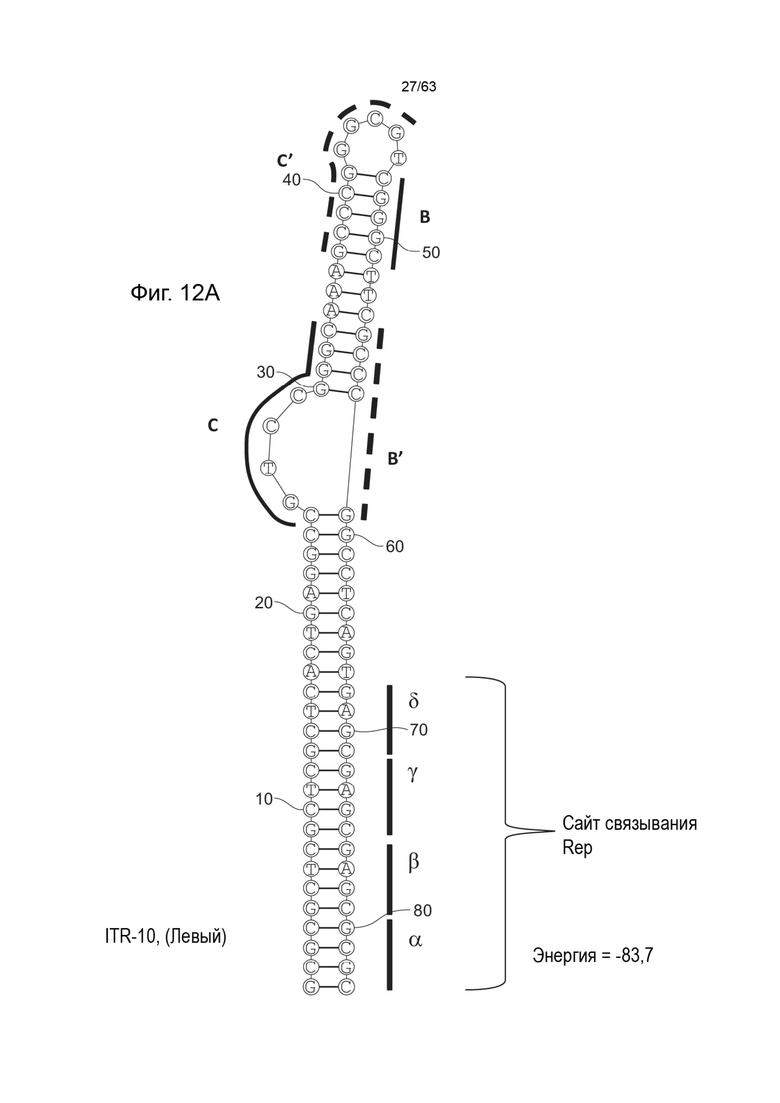
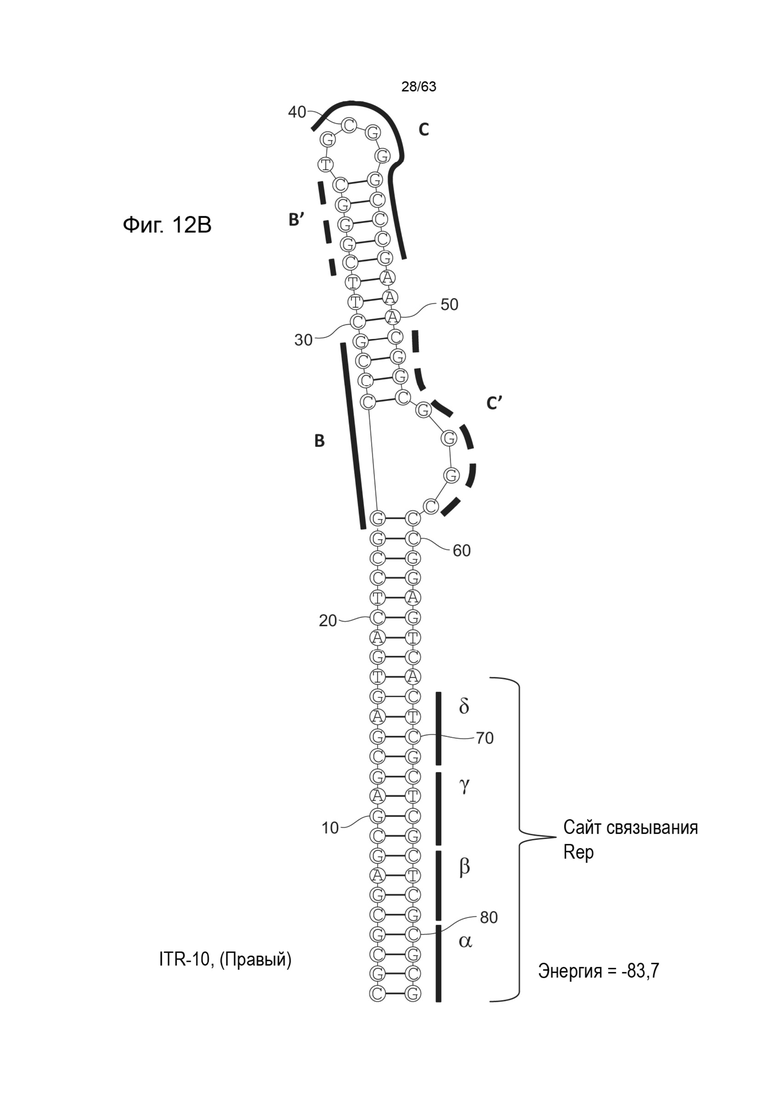
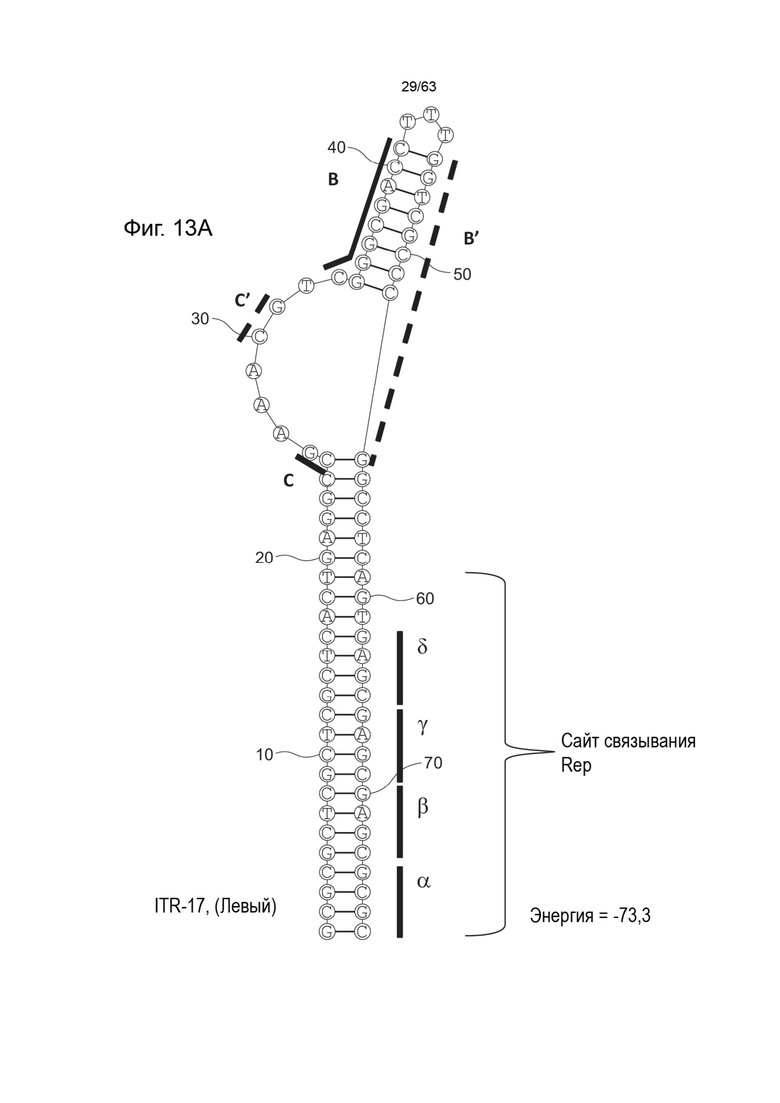
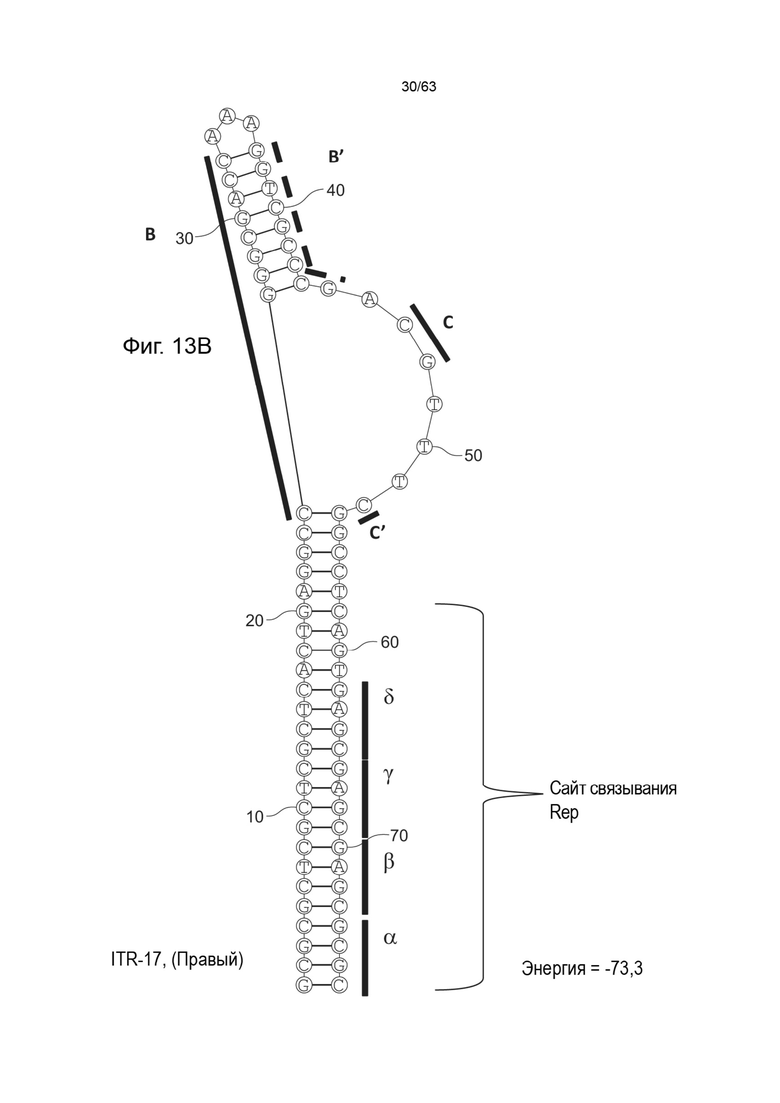
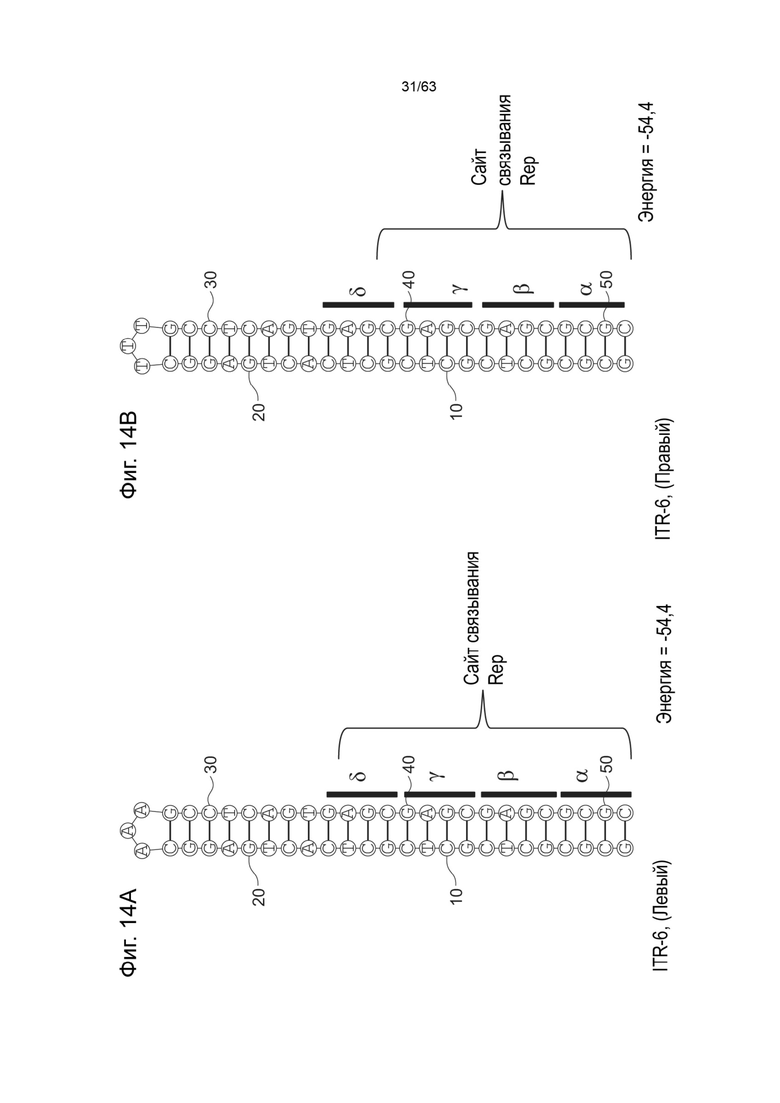
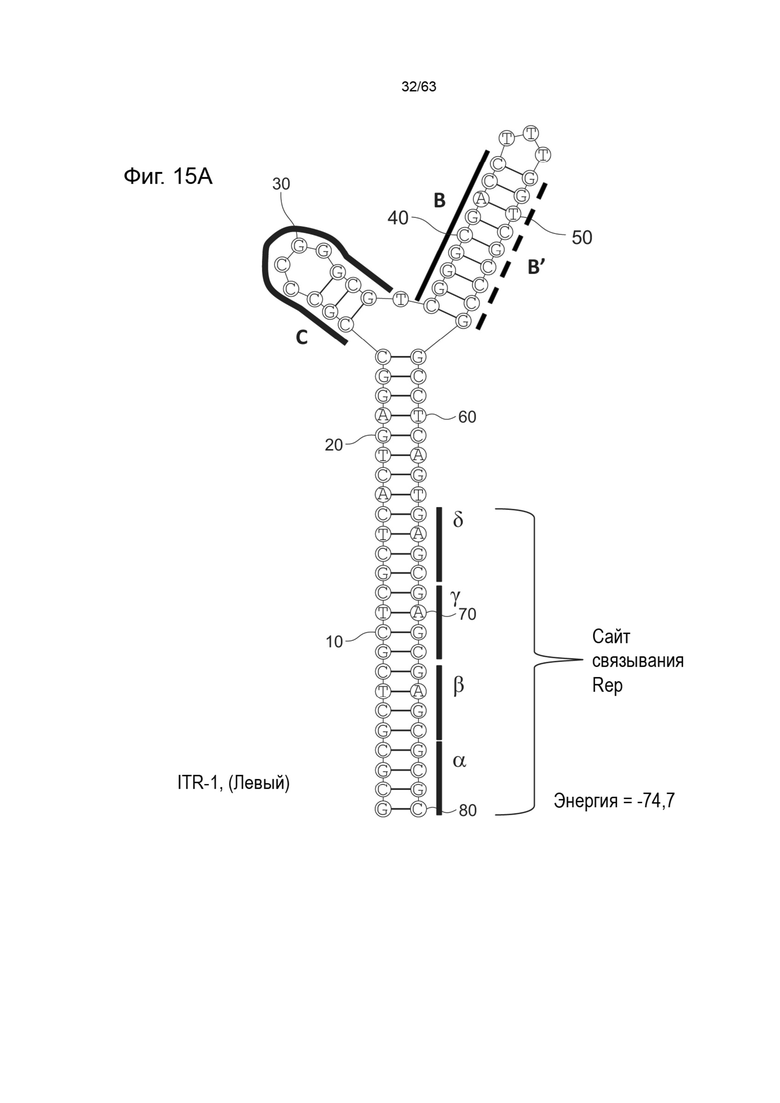
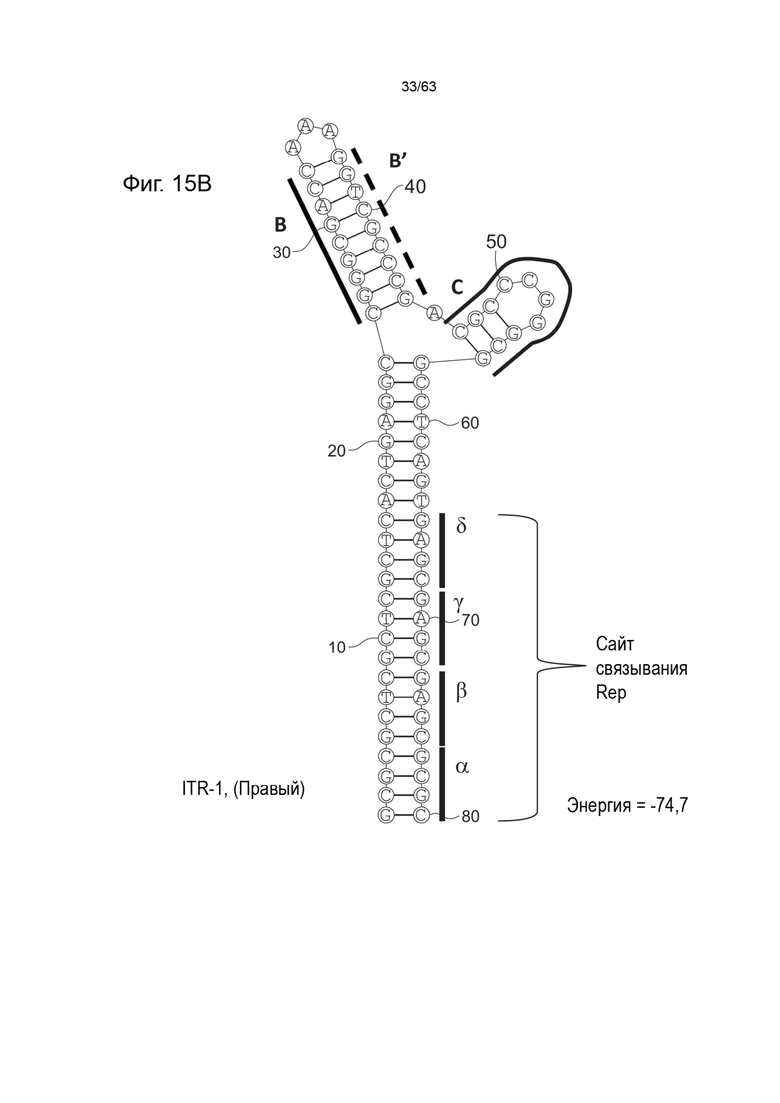
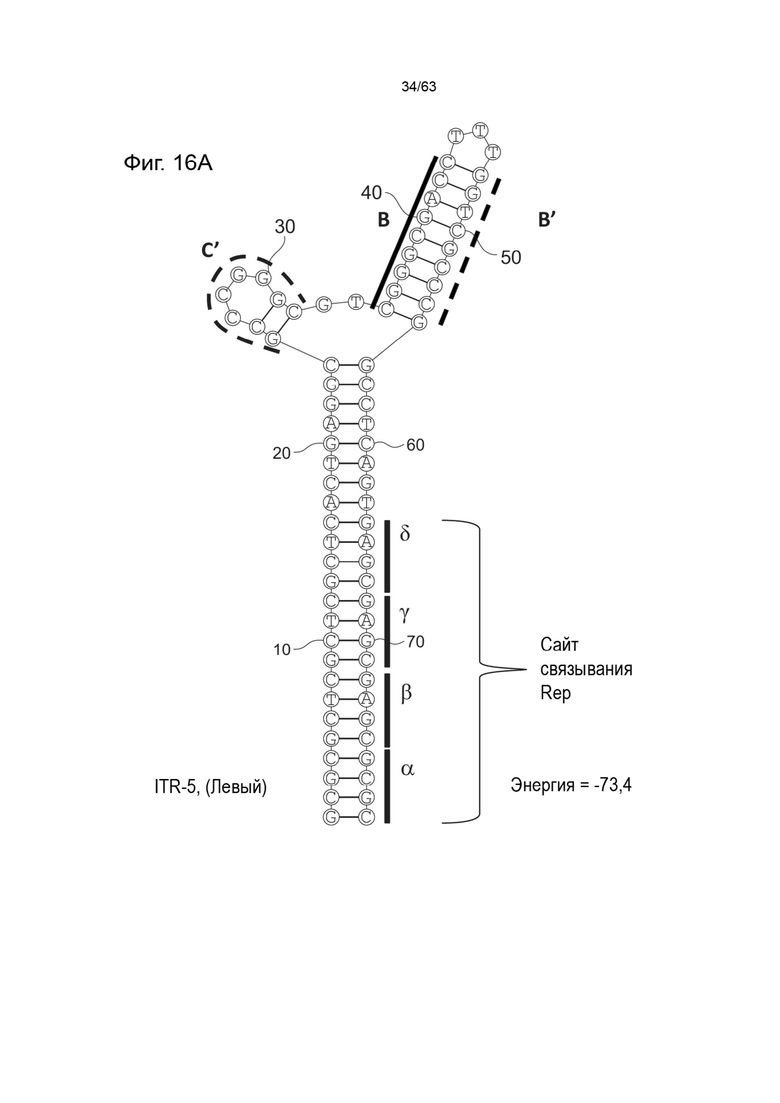
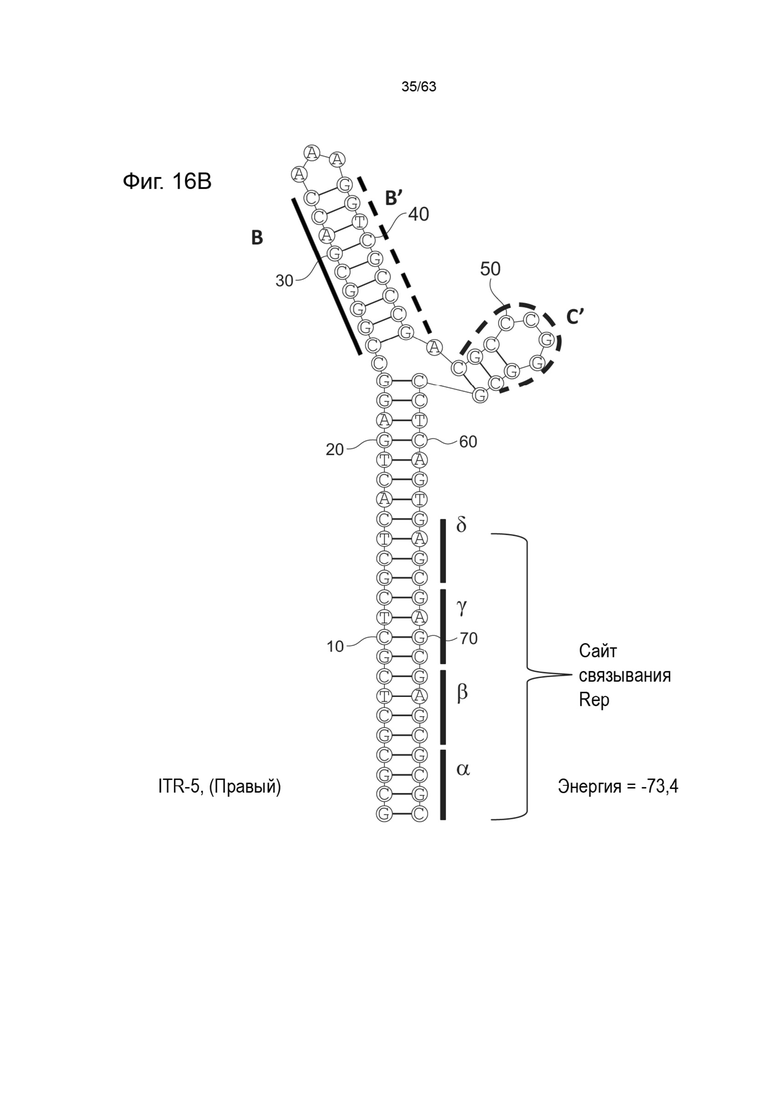
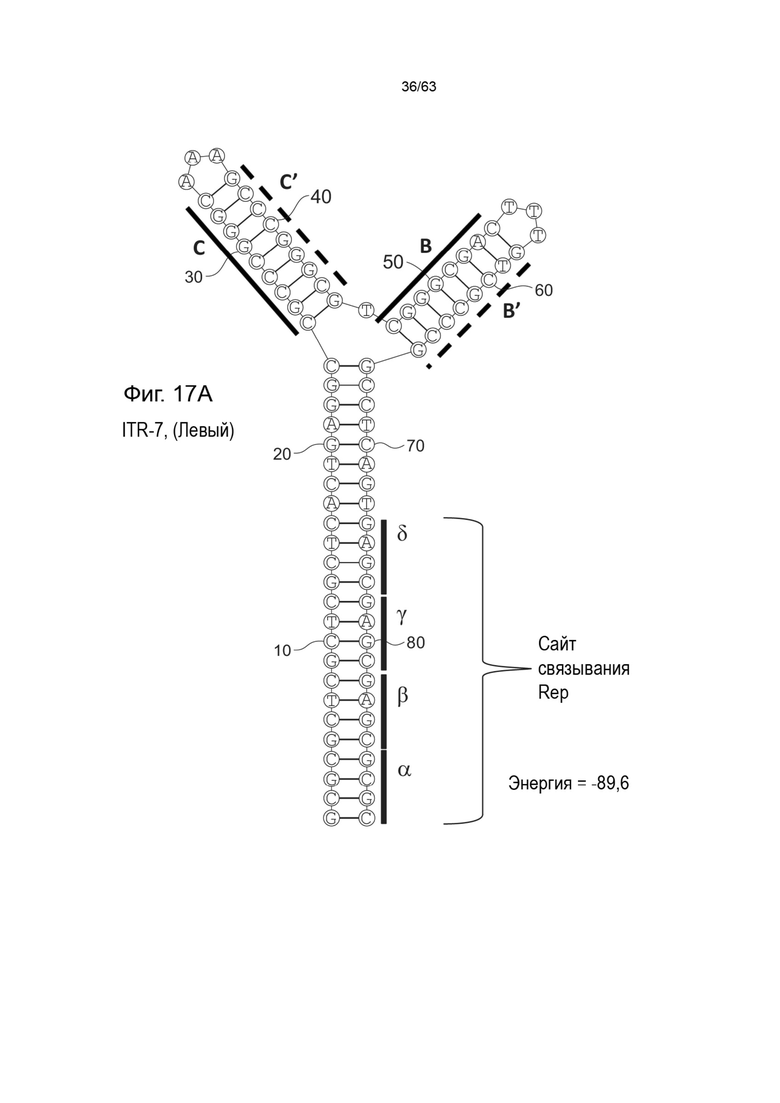
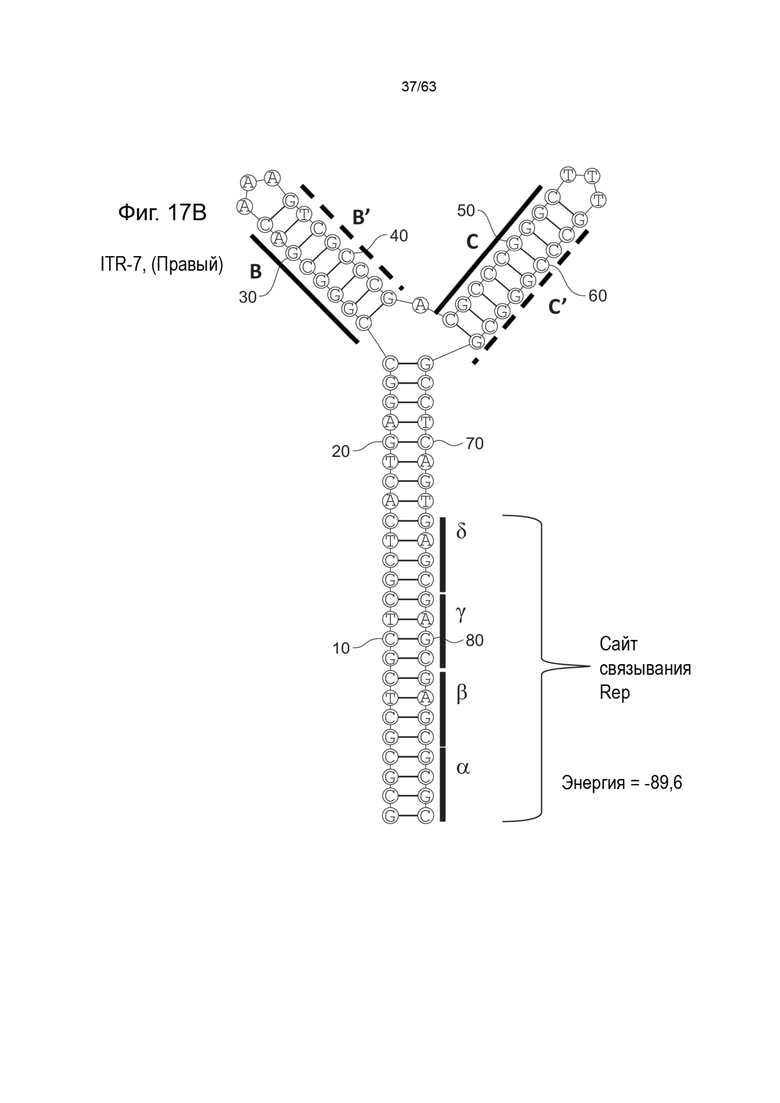
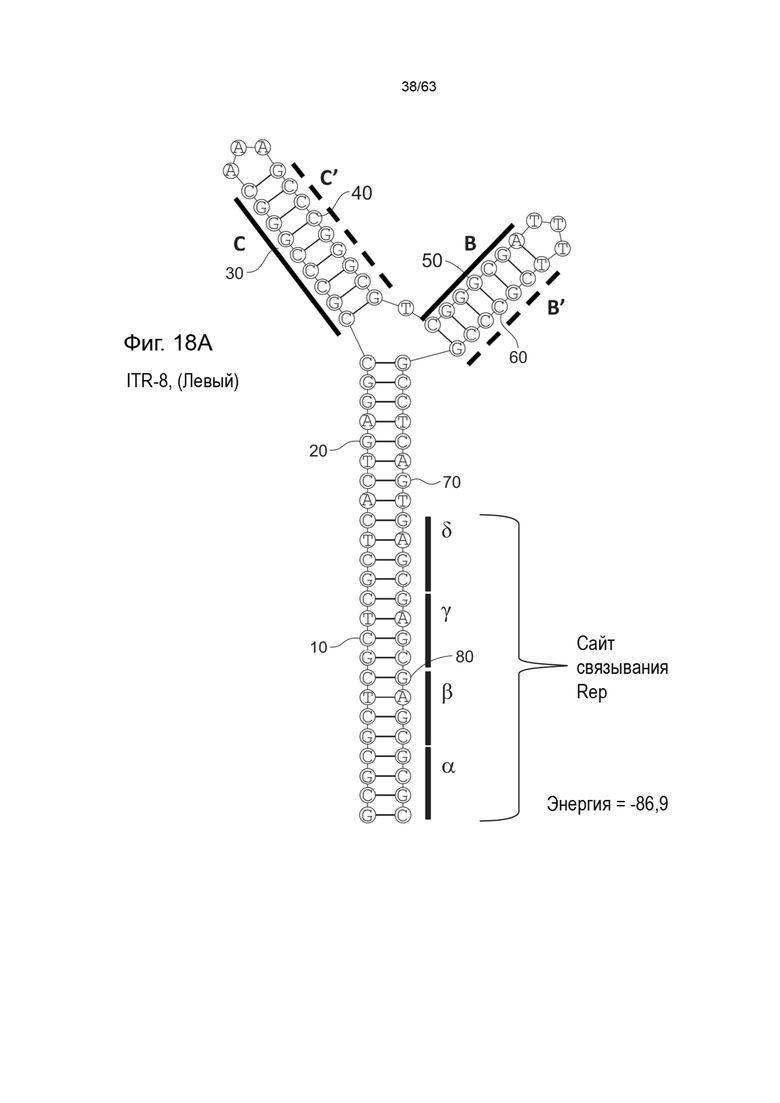
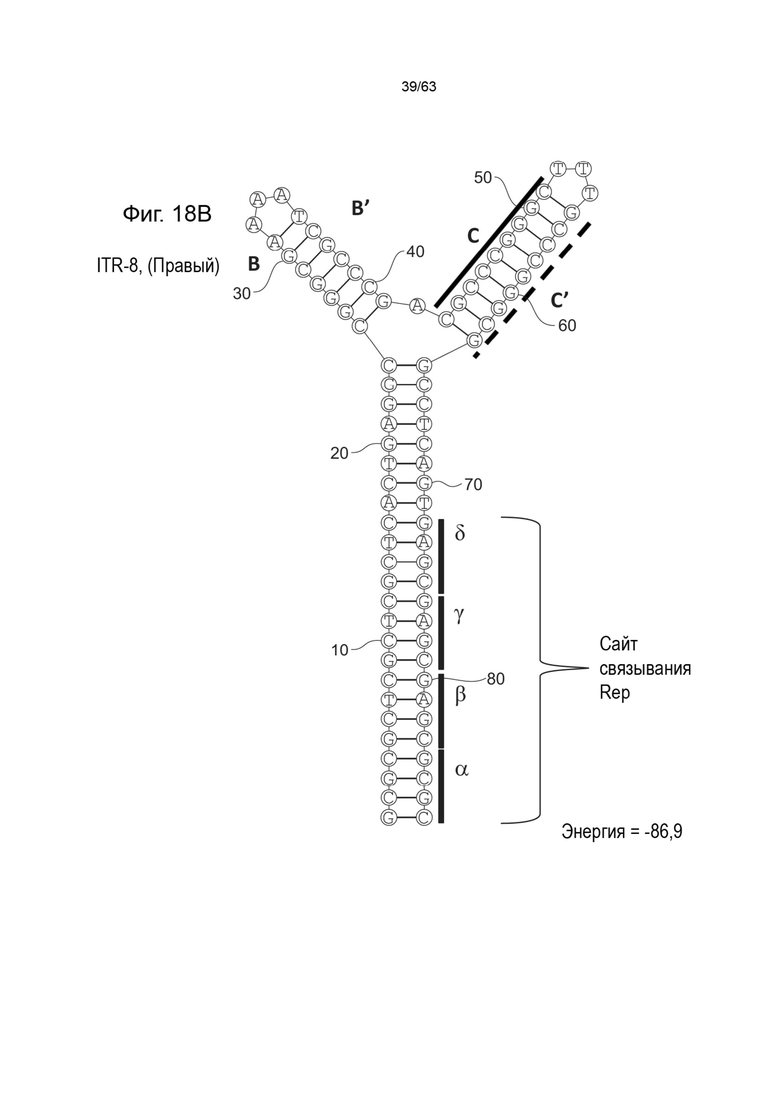
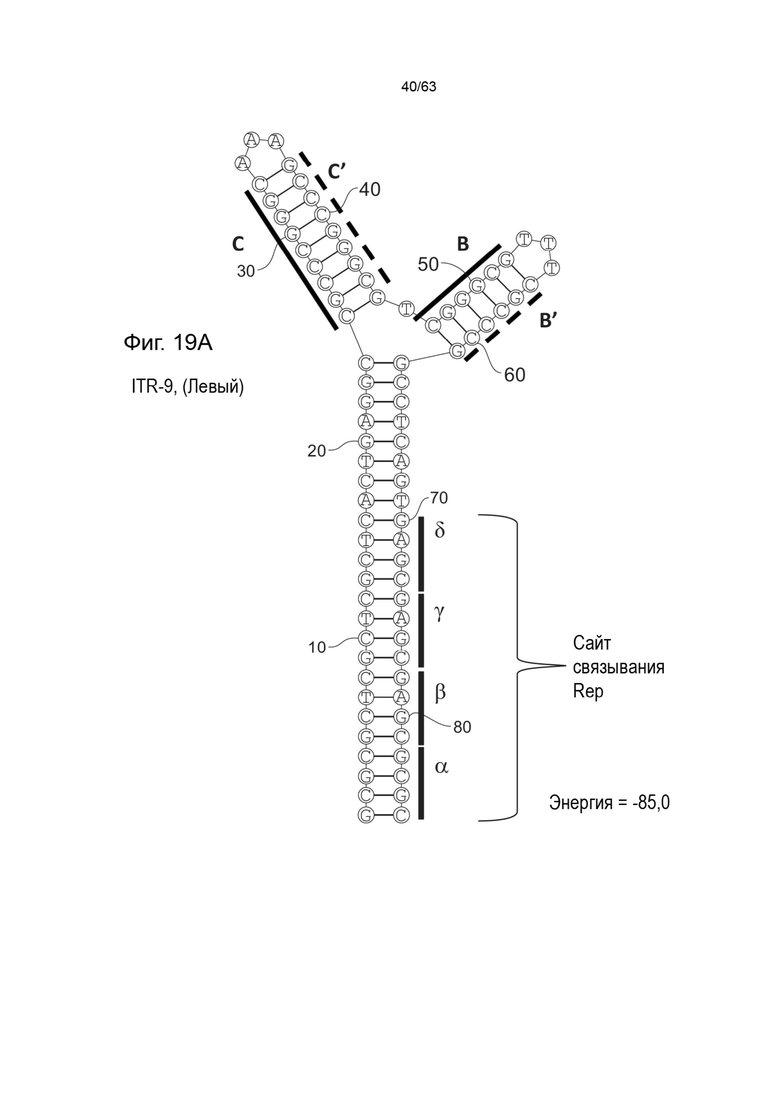
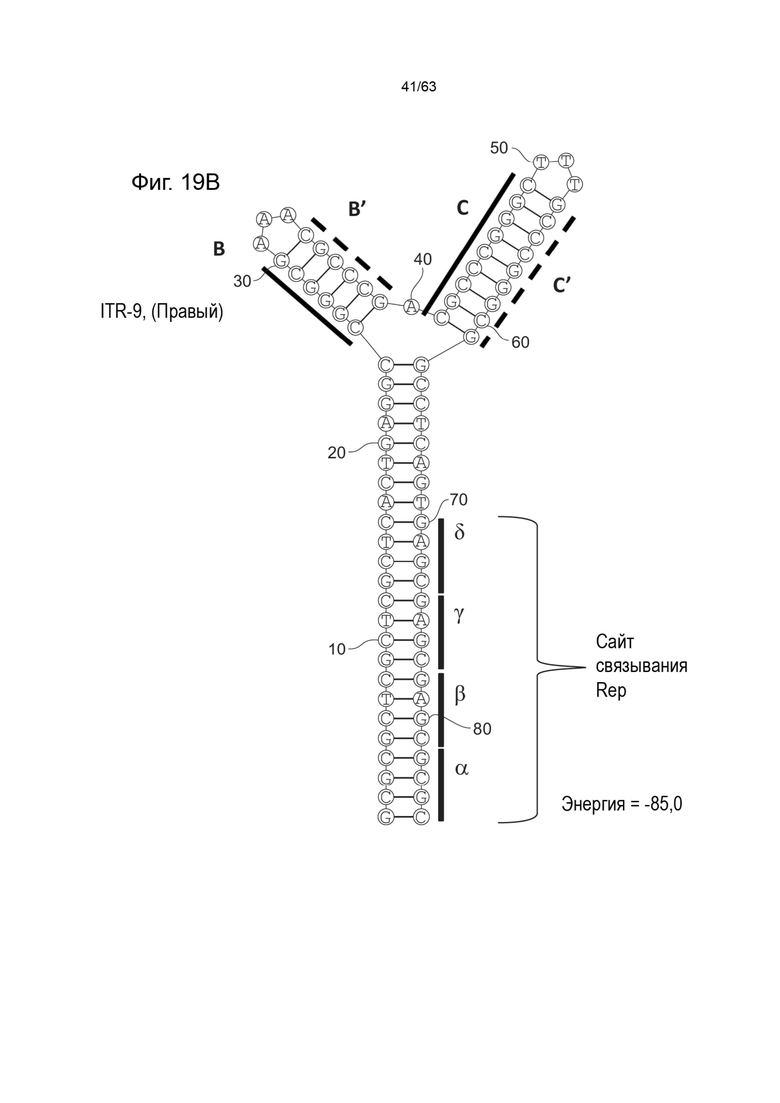
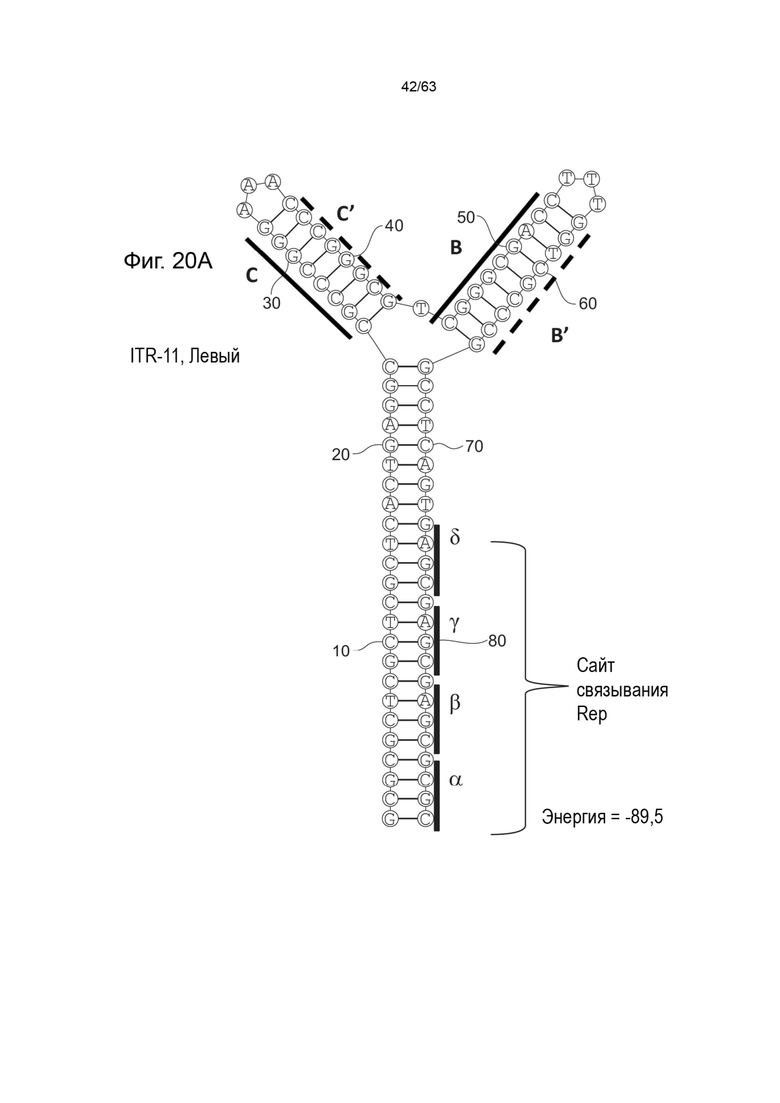
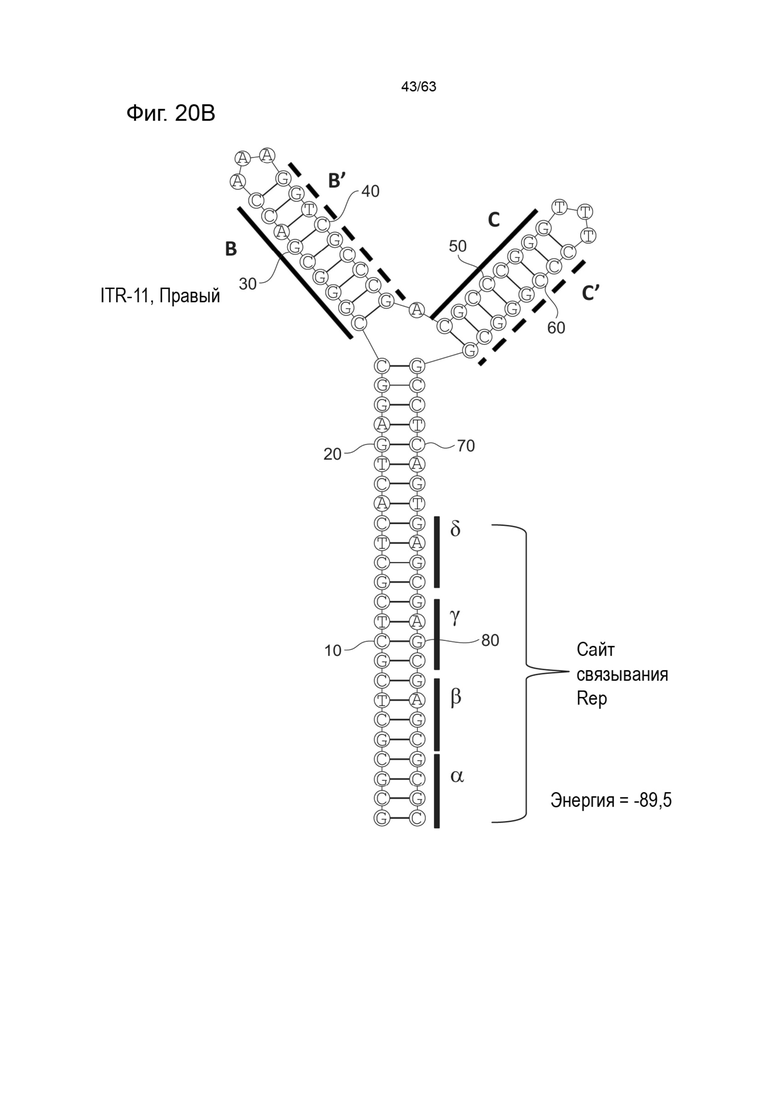
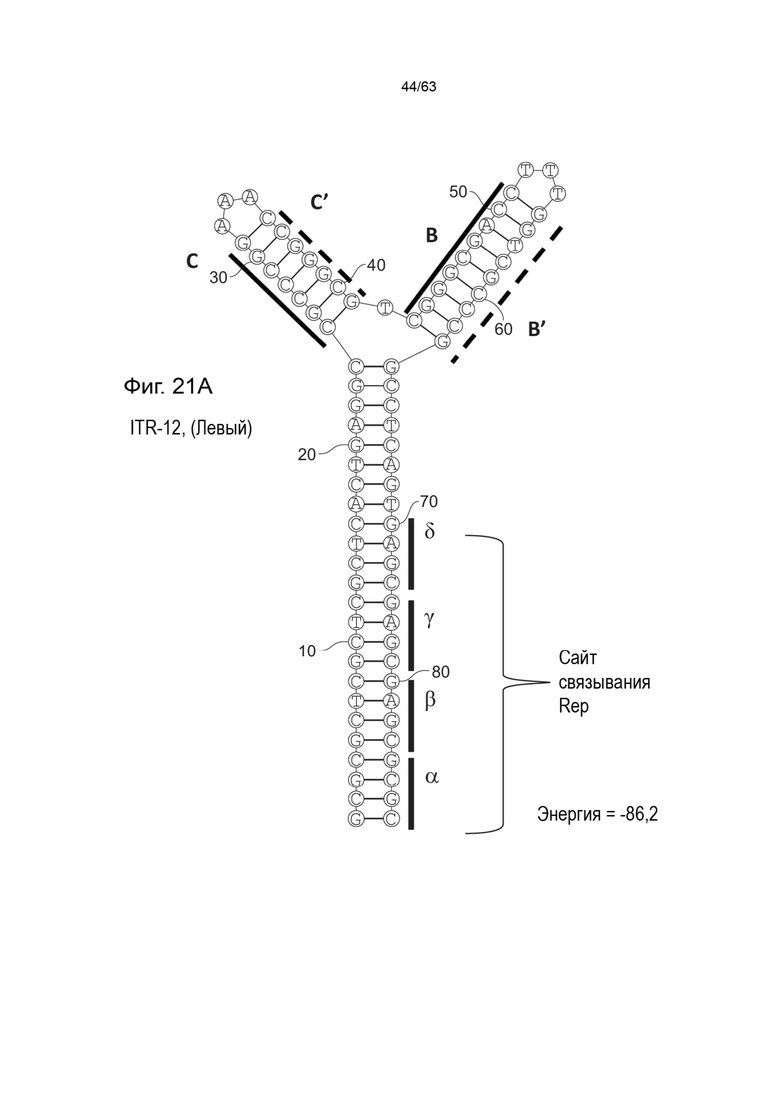
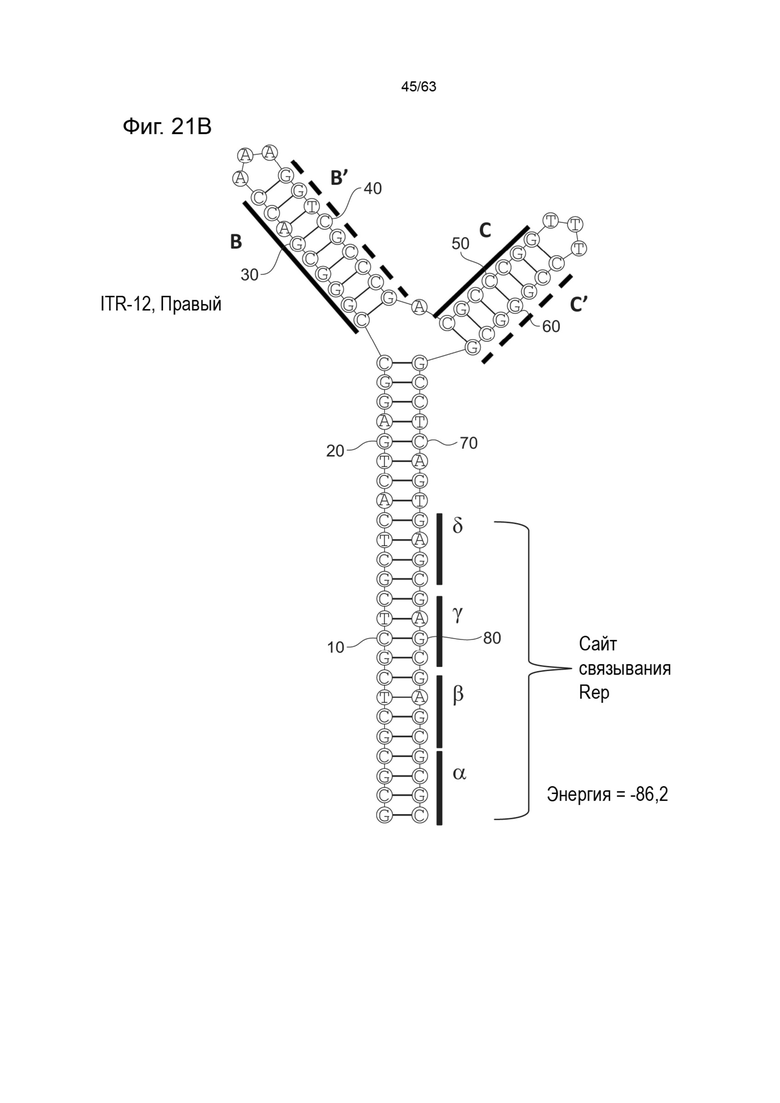
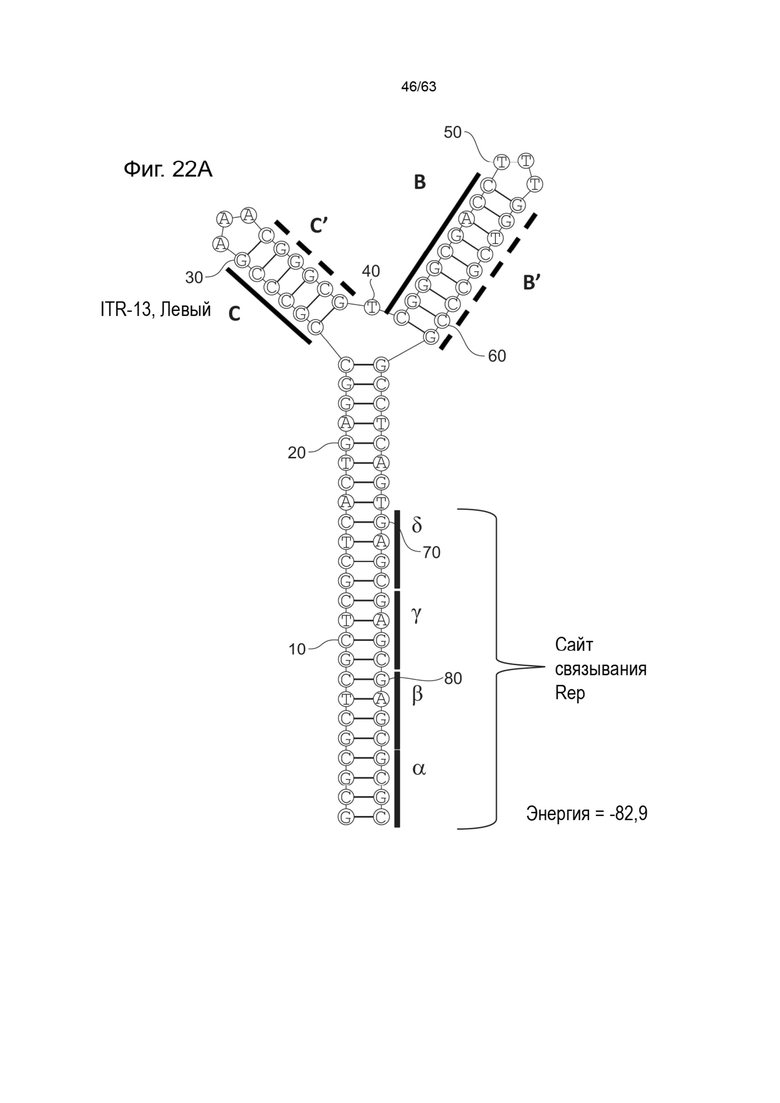
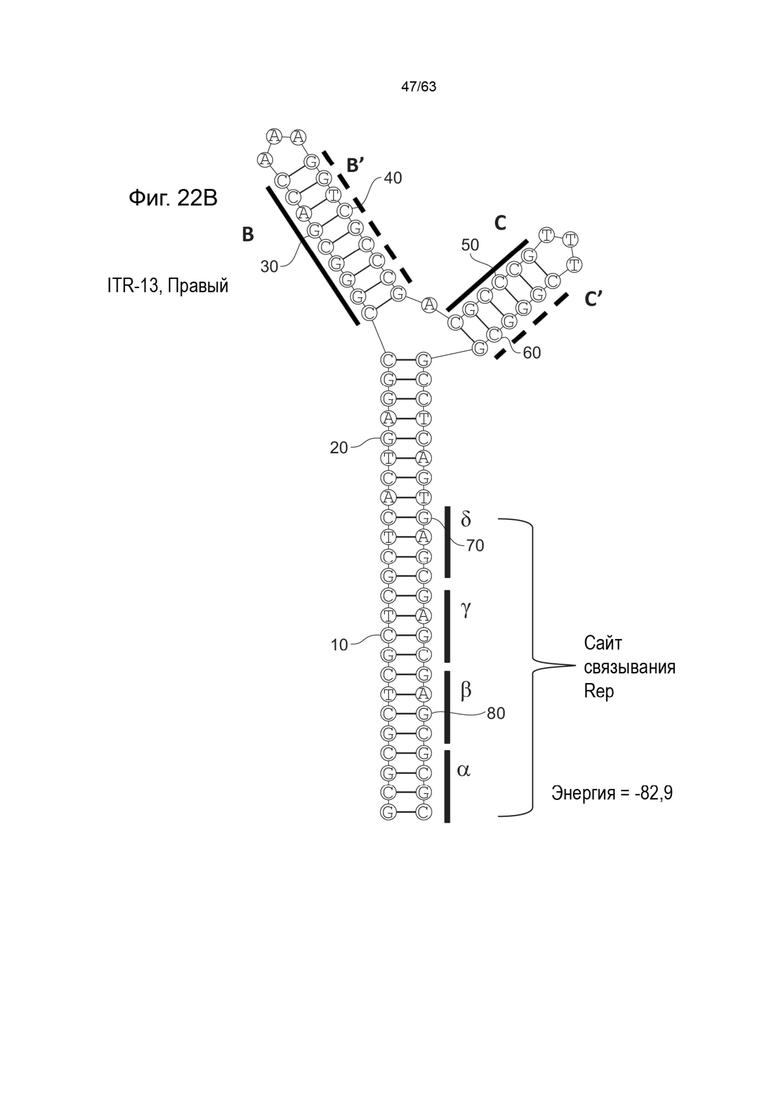
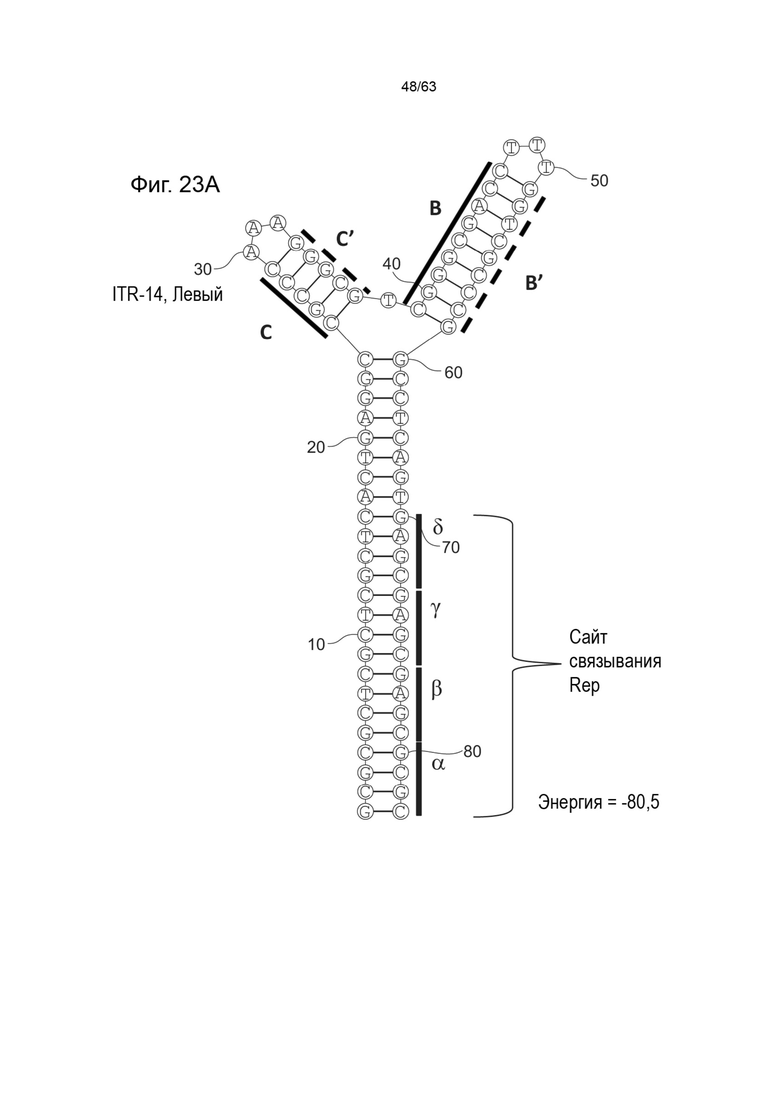
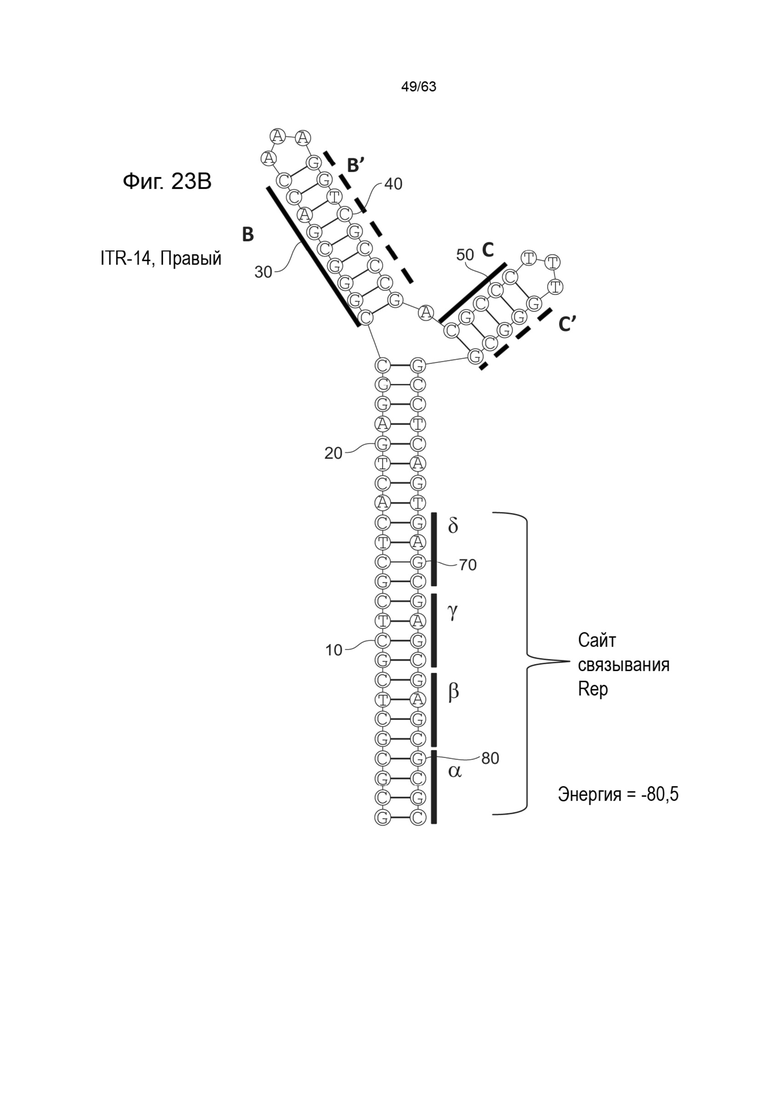
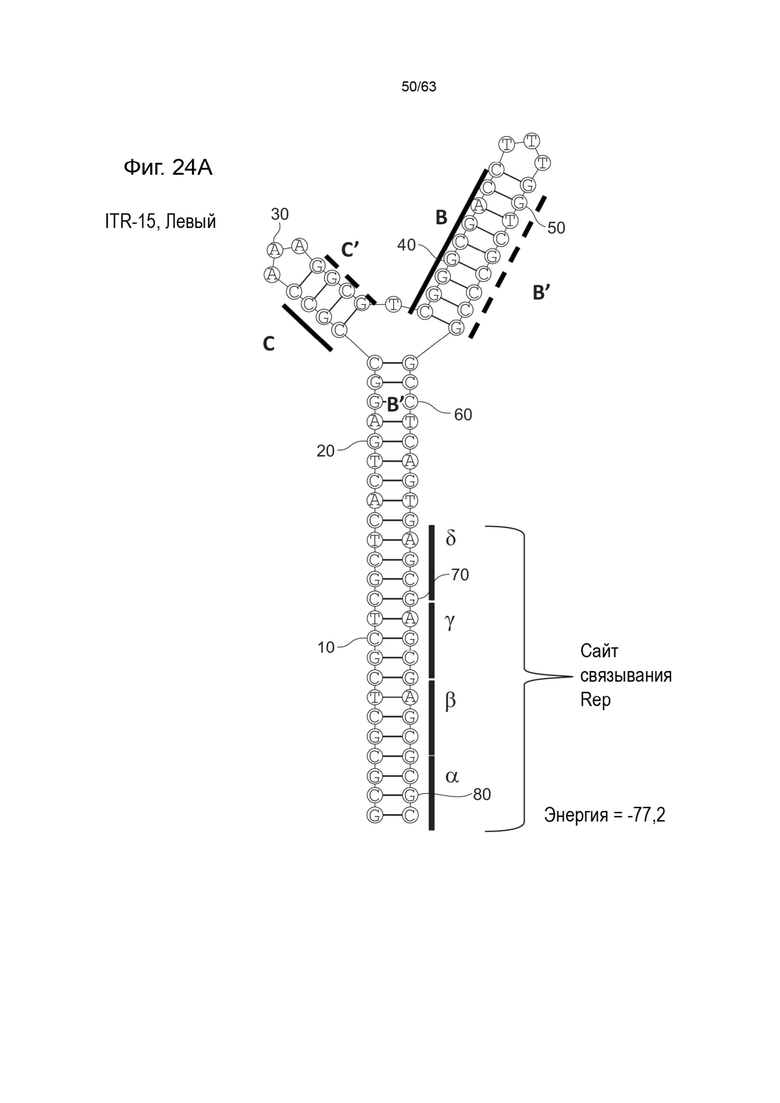
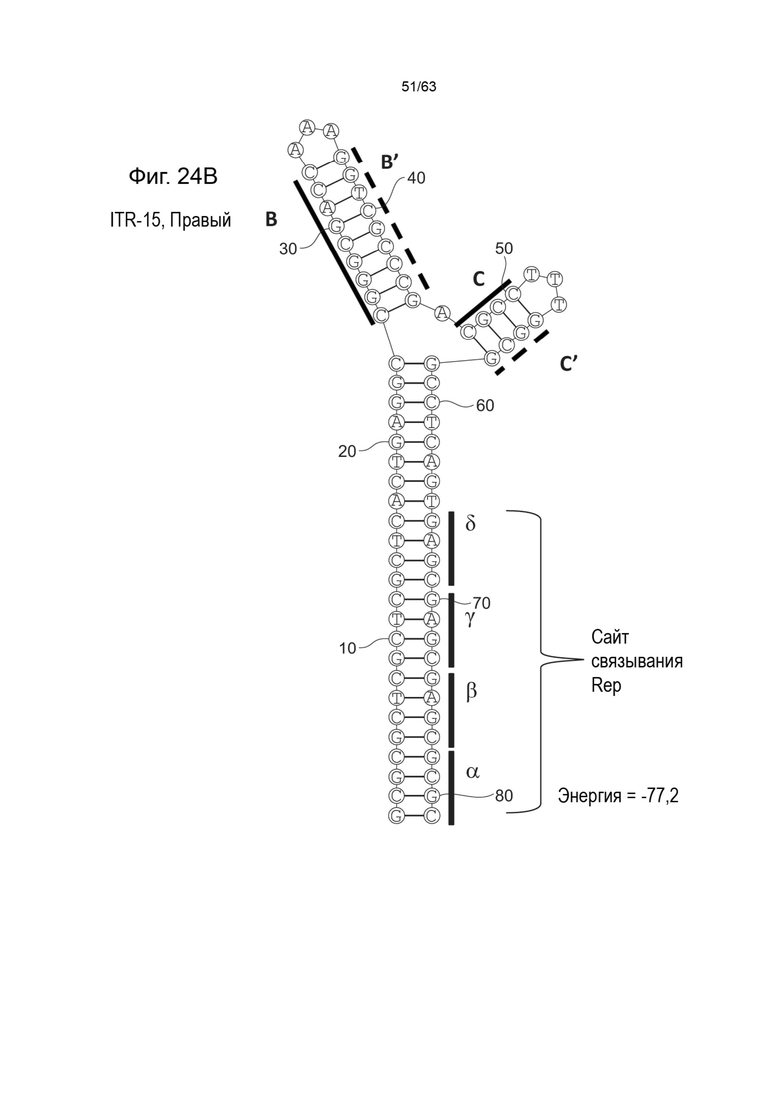
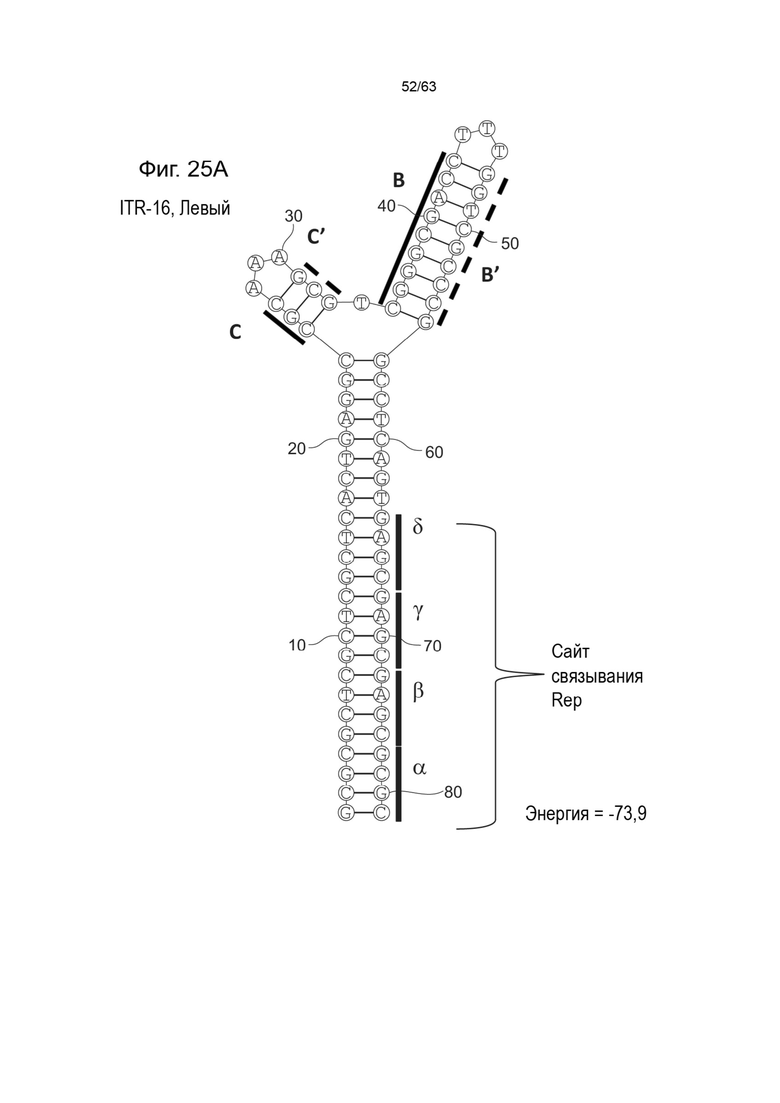
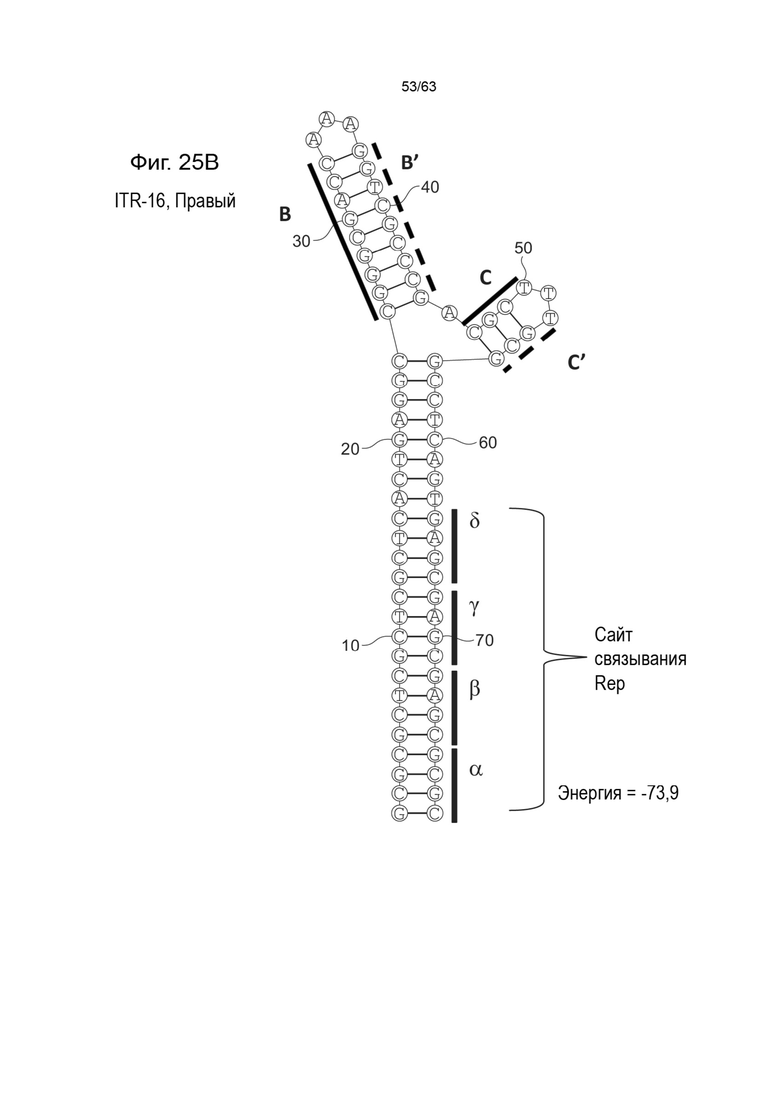
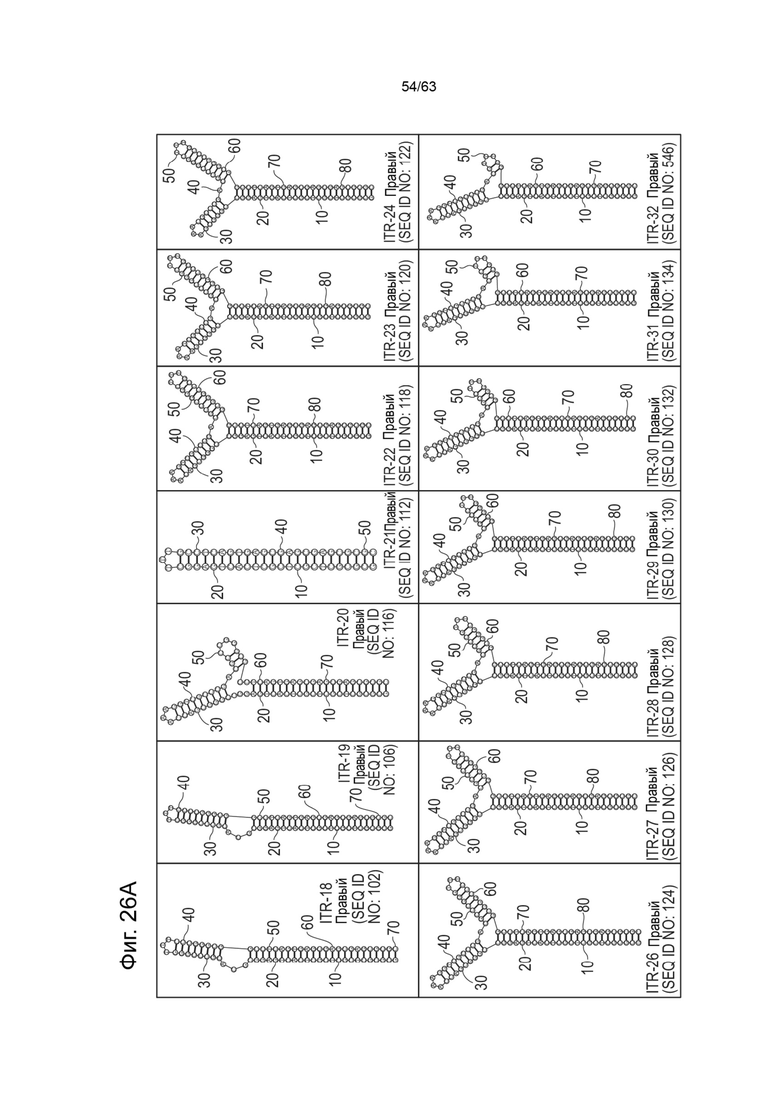
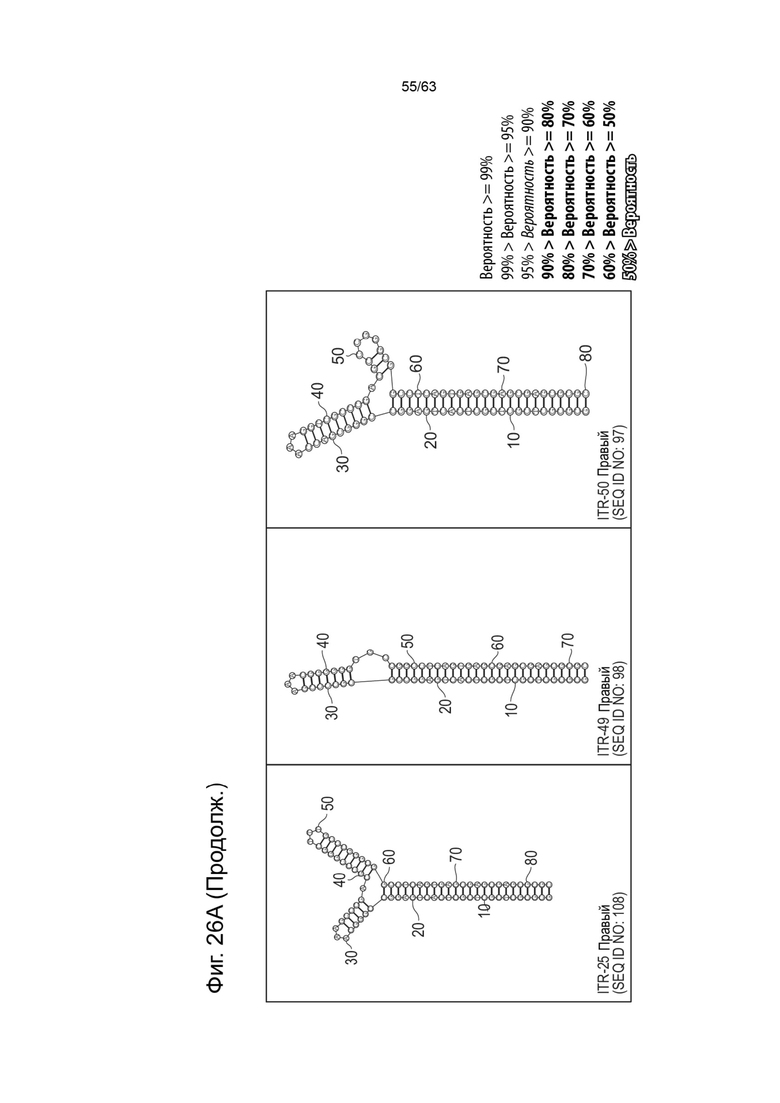
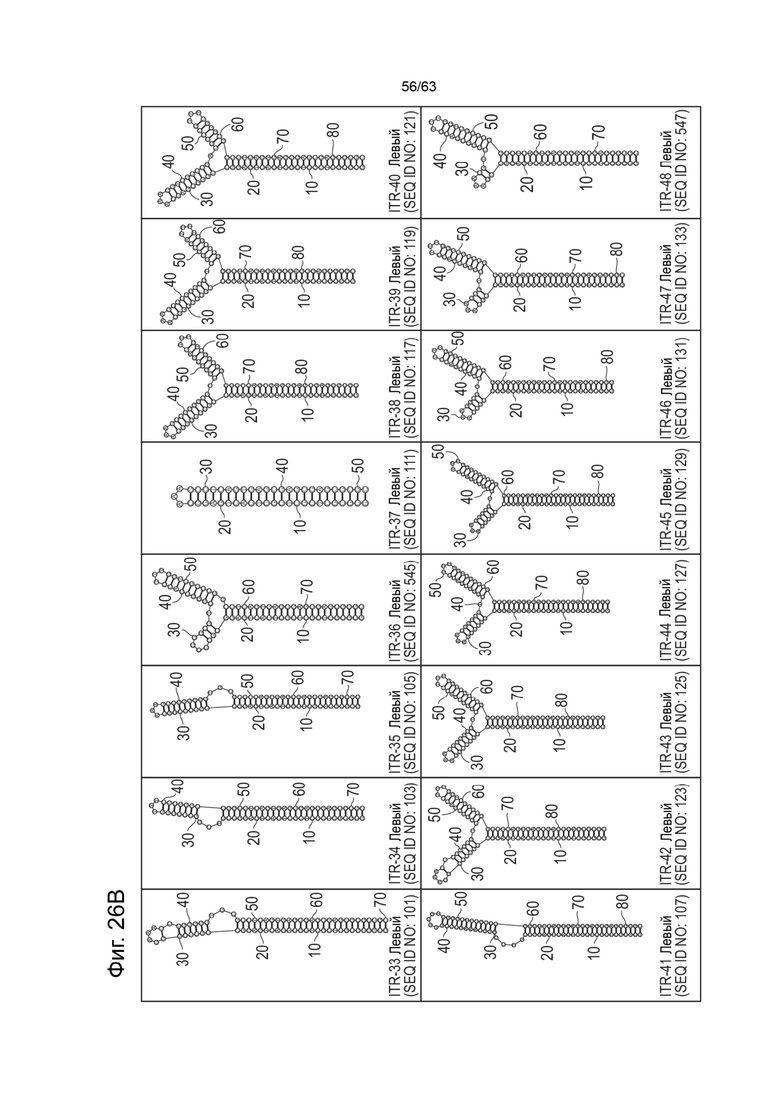
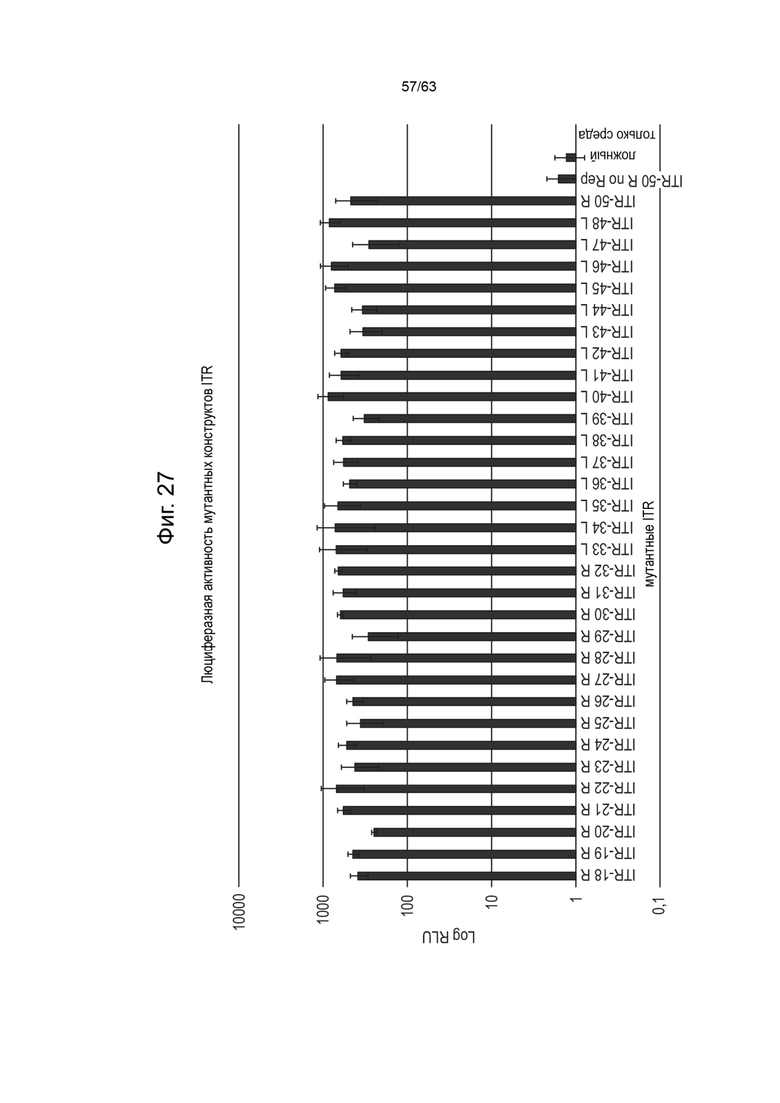

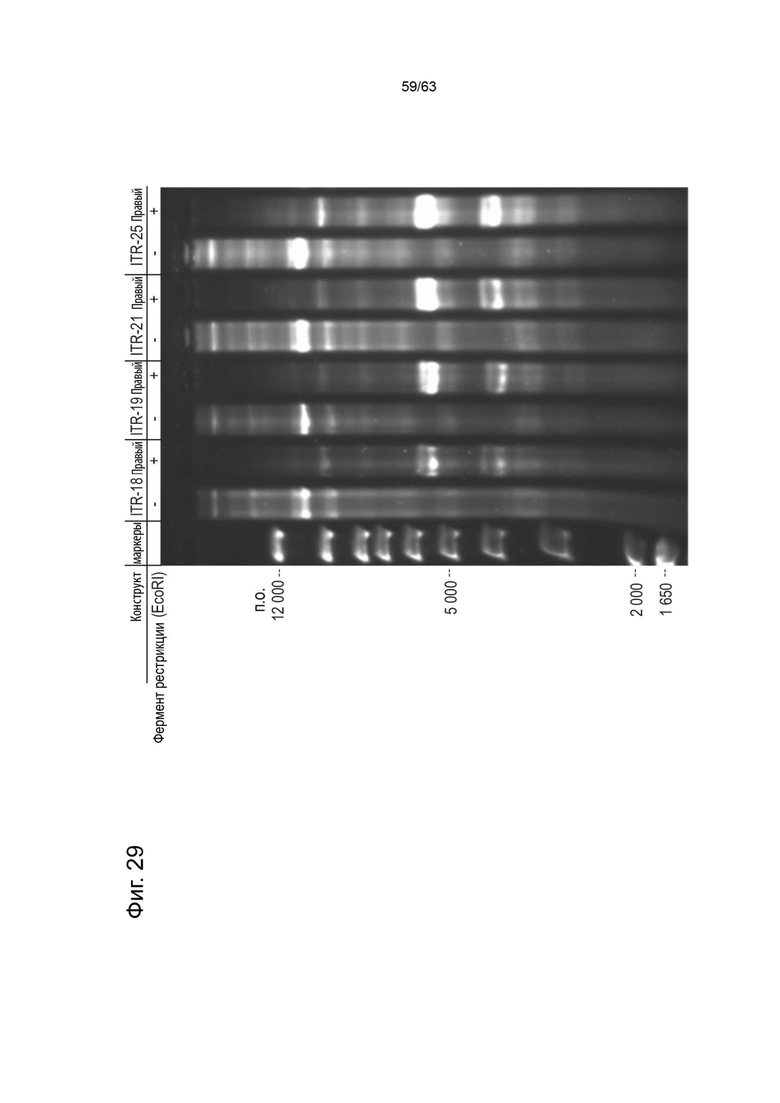
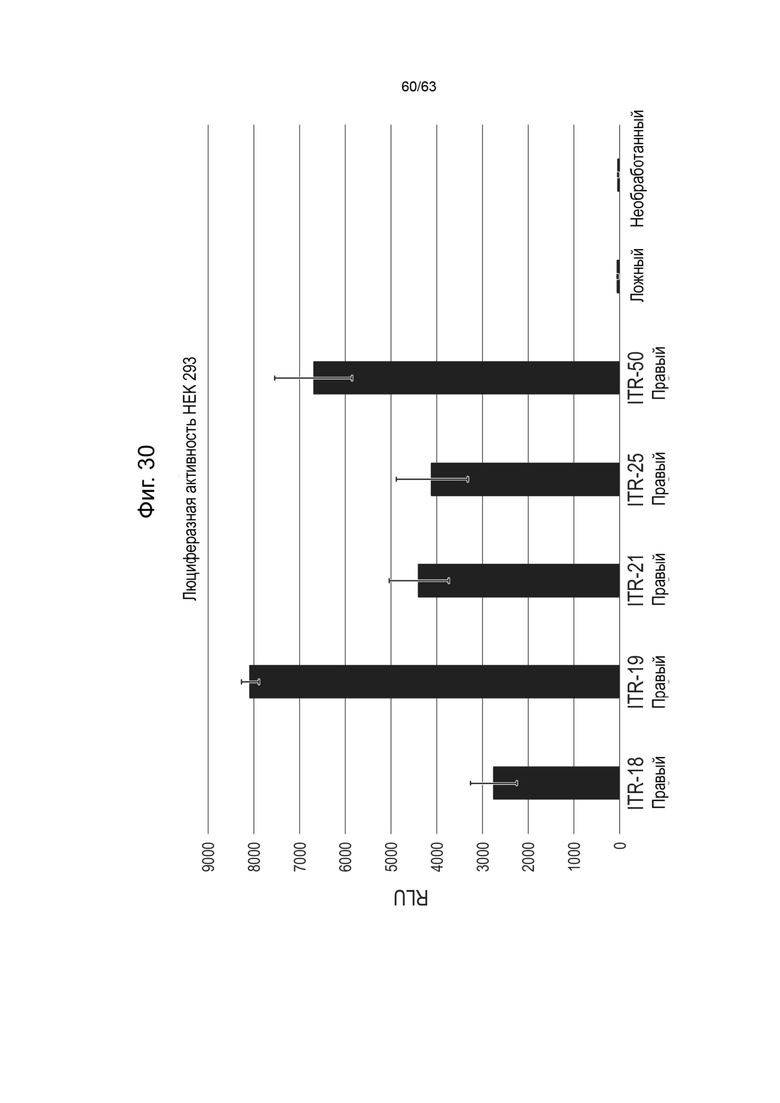
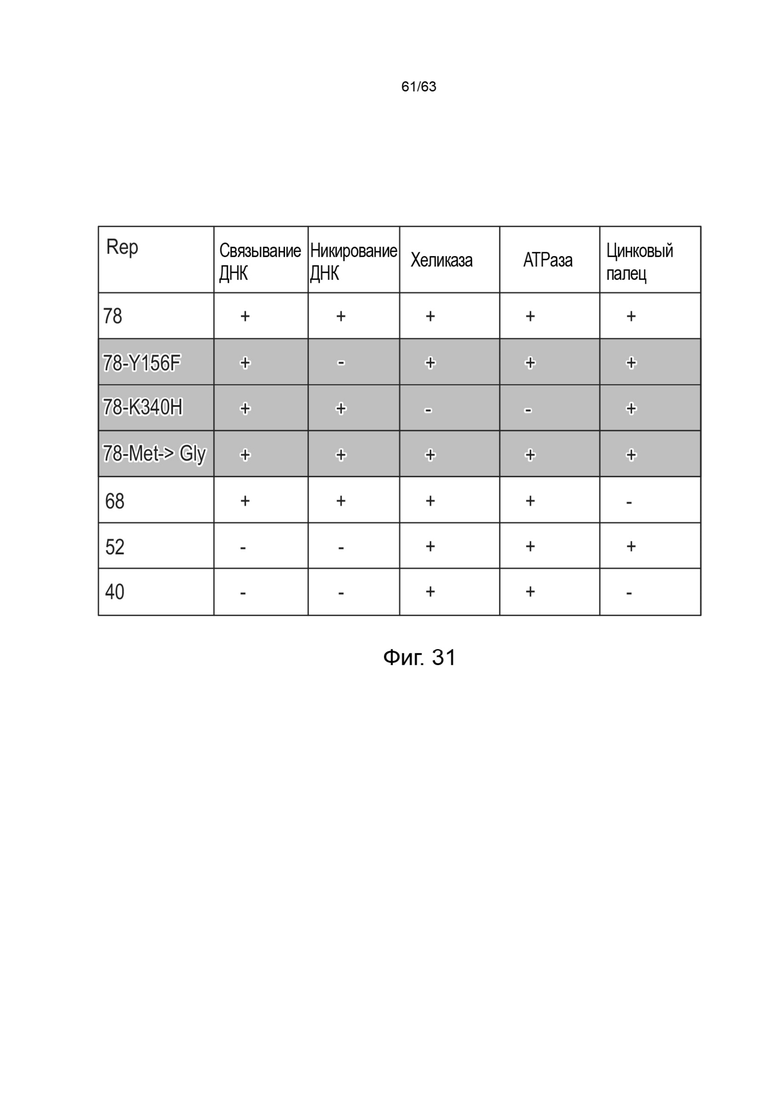
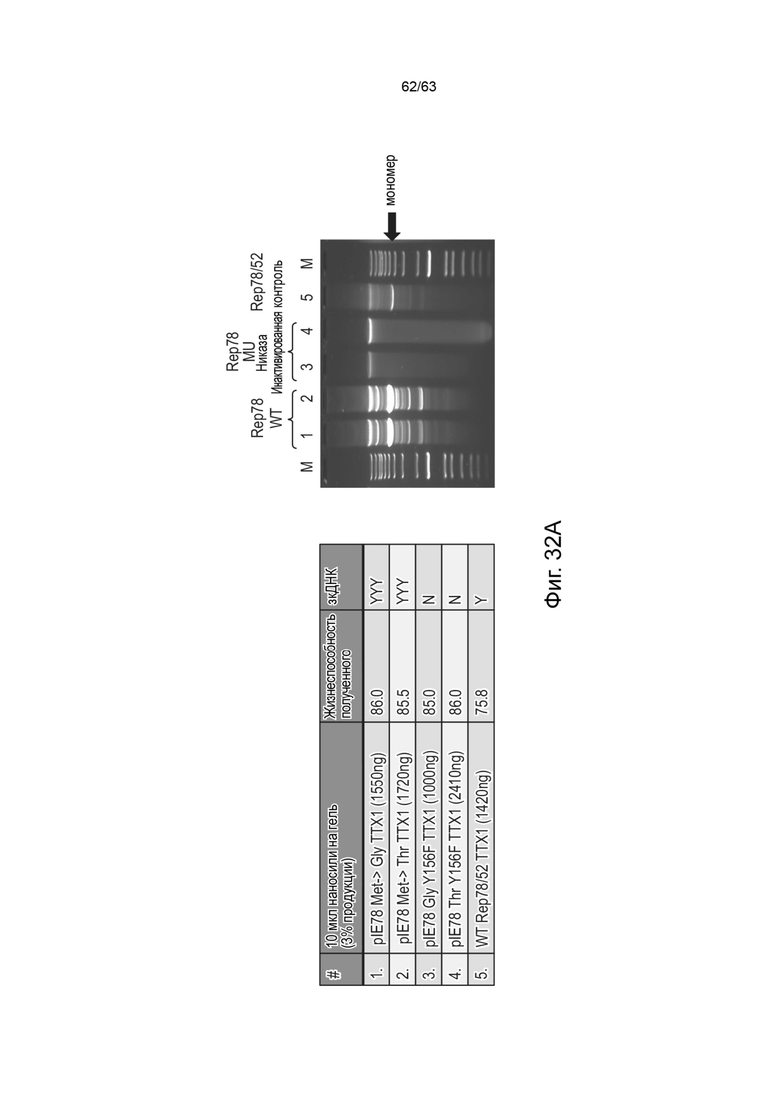
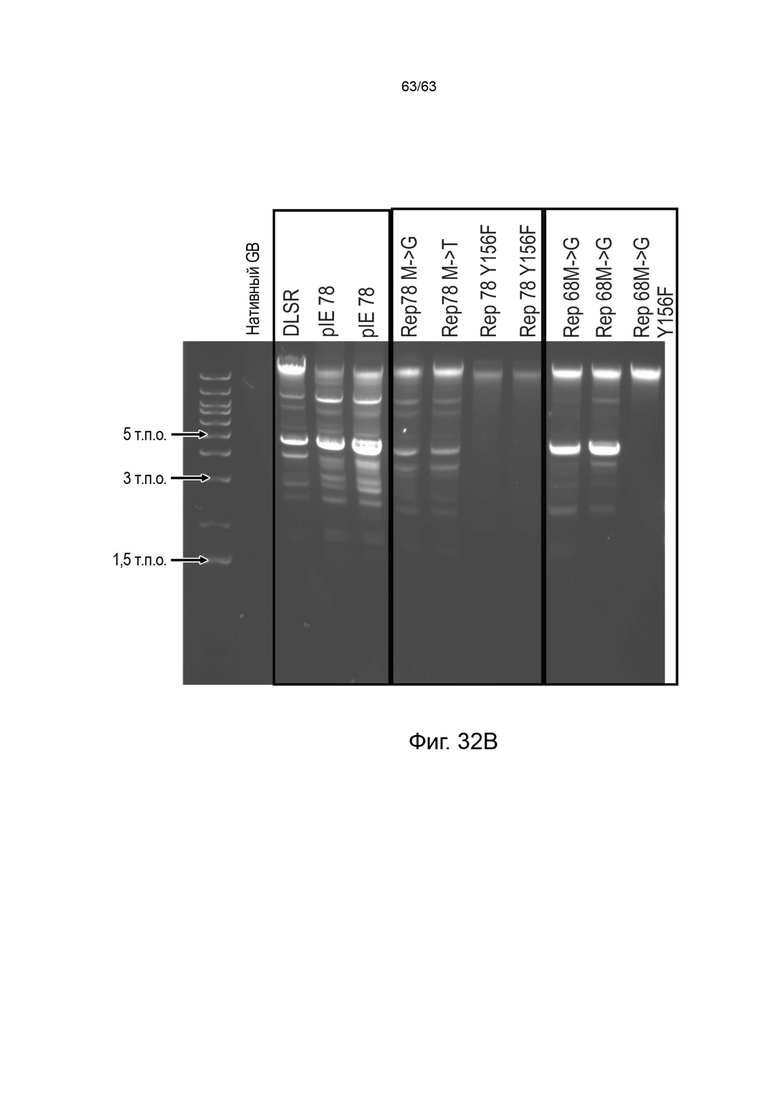
Изобретение относится к области биотехнологии. Описана группа изобретений, включающая ДНК-вектор для доставки полинуклеотидной последовательности (варианты), полинуклеотид для создания ДНК-вектора, способ получения ДНК-вектора, фармацевтическую композицию для генной терапии, набор для получения ДНК-вектора, клетку для получения зкДНК-вектора. В одном варианте реализации ДНК-вектор получен из векторного полинуклеотида, в котором векторный полинуклеотид кодирует последовательность нуклеиновой кислоты, расположенную между первым инвертированным концевым повтором (ITR) и вторым ITR, причем по меньшей мере один из первого ITR и второго ITR содержит последовательность нуклеиновой кислоты, соответствующую сайту связывания белка репликации (Rep) для индуцирования репликации ДНК-вектора в клетке в присутствии единственного вида Rep-белка, обладающего по крайней мере ДНК-связывающей и ДНК-никирующей функциональностью. Изобретение расширяет арсенал средств для доставки полинуклеотидной последовательности. 8 н. и 81 з.п. ф-лы, 64 ил., 16 табл., 6 пр.
1. ДНК-вектор для доставки полинуклеотидной последовательности, полученный из векторного полинуклеотида, в котором векторный полинуклеотид кодирует последовательность нуклеиновой кислоты, расположенную между первым инвертированным концевым повтором (ITR) и вторым ITR, причем по меньшей мере один из первого ITR и второго ITR содержит последовательность нуклеиновой кислоты, соответствующую сайту связывания белка репликации (Rep) для индуцирования репликации ДНК-вектора в клетке в присутствии единственного вида Rep-белка, обладающего, по крайней мере, ДНК-связывающей и ДНК-никирующей функциональностью, причем ДНК-вектор получен способом, включающим стадии:
a) инкубация популяции клеток, несущих векторный полинуклеотид, который лишен кодирующих последовательностей вирусного капсида, в присутствии указанного единственного вида Rep-белка, в условиях, эффективных и в течение времени, достаточного для индуцирования получения ДНК-вектора внутри клеток, причем клетки не содержат последовательностей, кодирующих вирусный капсид, причем не присутствуют другие виды Rep-белков, и причем клетки не контактируют с последовательностью нуклеиновой кислоты, кодирующей второй белок Rep; и
b) сбор и выделение полученного ДНК-вектора из клеток.
2. ДНК-вектор по п. 1, отличающийся тем, что единственный вид белка Rep дополнительно имеет геликазную, лигазную и АТФазную функциональность.
3. ДНК-вектор по п. 1, отличающийся тем, что единственный вид белка Rep представляет собой белок Rep AAV.
4. ДНК-вектор по п. 2, отличающийся тем, что единственный вид белка Rep выбран из группы, состоящей из AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11 и белка Rep AAV12.
5. ДНК-вектор по п. 2, отличающийся тем, что единственный вид белка Rep представляет собой белок Rep68 AAV2.
6. ДНК-вектор по п. 2, отличающийся тем, что единственный вид белка Rep представляет собой белок Rep78 AAV2.
7. ДНК-вектор по п. 6, отличающийся тем, что белок Rep78 кодируется мутантной последовательностью нуклеиновой кислоты Rep78.
8. ДНК-вектор по п. 7, отличающийся тем, что мутантная последовательность нуклеиновой кислоты Rep78 не имеет функционального кодона инициации трансляции для Rep52.
9. ДНК-вектор по п. 7, отличающийся тем, что мутантная последовательность нуклеиновой кислоты Rep78 кодирует мутантный белок Rep78, который содержит мутацию в положении 225 аминокислоты SEQ ID NO: 588.
10. ДНК-вектор по п. 7, отличающийся тем, что аминокислота в положении 225 SEQ ID NO: 588 мутирована в глицин (Gly) или треонин (Thr).
11. ДНК-вектор по п. 6, отличающийся тем, что мутантная последовательность нуклеиновой кислоты Rep78 кодирует мутантный белок Rep78, который содержит последовательность, имеющую по меньшей мере 95% идентичность последовательности с SEQ ID NO: 588, и имеет по меньшей мере функциональность связывания ДНК и никирования ДНК.
12. ДНК-вектор по п. 1, отличающийся тем, что первый ITR и второй ITR каждый представляет собой ITR парвовируса.
13. ДНК-вектор по п. 12, отличающийся тем, что парвовирус представляет собой депендовирус.
14. ДНК-вектор по п. 13, отличающийся тем, что парвовирус представляет собой AAV.
15. ДНК-вектор по п. 14, отличающийся тем, что AAV происходит из серотипа AAV, выбранного из группы, состоящей из AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11 и AAV12.
16. ДНК-вектор по п. 1, отличающийся тем, что ДНК-вектор представляет собой невирусный бескапсидный двухцепочечный ДНК-вектор с ковалентно замкнутыми концами (вектор зкДНК).
17. ДНК-вектор по п. 16, отличающийся тем, что присутствие зкДНК-вектора, выделенного из клеток, может быть подтверждено путем расщепления ДНК, выделенной из клетки, рестрикционным ферментом, имеющим единственный сайт распознавания на ДНК-векторе, и анализа расщепленного ДНК-материала на неденатурирующем геле для подтверждения присутствия характерных полос линейной и непрерывной ДНК по сравнению с линейной и прерывистой ДНК.
18. ДНК-вектор для доставки полинуклеотидной последовательности, полученный из векторного полинуклеотида, причем векторный полинуклеотид кодирует последовательность нуклеиновой кислоты, расположенную между двумя разными инвертированными концевыми повторами (ITR), причем по крайней мере один из указанных двух разных ITR представляет собой функциональный ITR, содержащий функциональный концевой сайт разрешения и сайт связывания белка репликации (Rep) для индуцирования репликации векторного полинуклеотида в клетке и в присутствии единственного вида Rep-белка, обладающего, по крайней мере, ДНК-связывающей и ДНК-никирующей функциональностью, причем ДНК-вектор получен способом, включающим стадии:
a) инкубация популяции клеток, несущих векторный полинуклеотид, который лишен кодирующих последовательностей вирусного капсида, в присутствии единственного вида Rep-белка, в условиях, эффективных и в течение времени, достаточного для индуцирования получения ДНК-вектора внутри клеток, причем клетки не содержат нуклеиновую кислоту, кодирующую Rep52 или Rep40 в клетках, при этом другие виды Rep не присутствуют в клетке, и причем клетки не контактируют с последовательностью нуклеиновой кислоты, кодирующей второй белок Rep; и
b) сбор и выделение вектора ДНК из клеток.
19. Полинуклеотид для создания ДНК-вектора, включающий последовательность нуклеиновой кислоты, кодирующую единственный вид аминокислотной последовательности Rep белка, которая имеет по меньшей мере ДНК-связующую и ДНК-никирующую функциональность, связанную по меньшей мере с одной последовательностью контроля экспрессии.
20. Полинуклеотид по п. 19, отличающийся тем, что единственный вид белка Rep имеет геликазную, лигазную и АТФазную функциональность.
21. Полинуклеотид по п. 19, отличающийся тем, что единственный вид белка Rep представляет собой белок Rep AAV.
22. Полинуклеотид по п. 21, отличающийся тем, что белок Rep AAV выбран из группы, состоящей из белка AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11 и белка AAV12 Rep.
23. Полинуклеотид по п. 21, отличающийся тем, что белок Rep AAV представляет собой белок Rep AAV2.
24. Полинуклеотид по п. 21, отличающийся тем, что белок Rep AAV представляет собой белок Rep78 AAV2.
25. Полинуклеотид по п. 24, отличающийся тем, что белок Rep78 AAV2 кодируется мутантной последовательностью нуклеиновой кислоты Rep78.
26. Полинуклеотид по п. 25, отличающийся тем, что мутантная последовательность нуклеиновой кислоты Rep78 не имеет функционального кодона инициации трансляции для Rep52.
27. Полинуклеотид по п. 25, отличающийся тем, что белок Rep78 содержит мутацию в положении 225 аминокислоты SEQ ID NO: 588.
28. Полинуклеотид по п. 27, отличающийся тем, что аминокислота 225 из SEQ ID NO: 588 мутирована в глицин (Gly) или треонин (Thr).
29. Полинуклеотид по п. 25, отличающийся тем, что мутантная последовательность нуклеиновой кислоты Rep78 кодирует мутантный белок Rep78, который содержит последовательность, имеющую по меньшей мере 95% идентичность последовательности с SEQ ID NO: 588 и имеющую по меньшей мере ДНК-связующую и ДНК-никирующую функциональность.
30. Полинуклеотид по п. 19, отличающийся тем, что по меньшей мере одна последовательность контроля экспрессии кодирует промотор, выбранный из группы, состоящей из: промотор IE, промотор ΔIE и промотор CMV.
31. Полинуклеотид по п. 19, отличающийся тем, что ДНК-вектор представляет собой невирусный бескапсидный двухцепочечный ДНК-вектор с ковалентно замкнутыми концами (вектор зкДНК).
32. Полинуклеотид по п. 31, отличающийся тем, что присутствие зкДНК-вектора, выделенного из клеток, может быть подтверждено путем расщепления ДНК, выделенной из клетки, рестрикционным ферментом, имеющим единственный сайт распознавания на ДНК-векторе, и анализа расщепленного ДНК-материала на неденатурирующем геле для подтверждения присутствия характерных полос линейной и непрерывной ДНК по сравнению с линейной и прерывистой ДНК.
33. Способ получения ДНК-вектора, включающий контактирование клетки с:
(1) последовательностью нуклеиновой кислоты, кодирующей единственный вид белка репликации (Rep), имеющей по меньшей мере ДНК-связующую и ДНК-никирующую функциональность, причем последовательность нуклеиновой кислоты связана по меньшей мере с одной последовательностью контроля экспрессии, причем клетка не экспрессирует какие-либо другие виды белка Rep и не контактирует с другими видами белка Rep;
(2) двухцепочечным ДНК-конструктом, содержащим:
экспрессионную кассету;
первый ITR на 5'-конце перед кассетой экспрессии; и
второй ITR на 3'-конце после кассеты экспрессии; и
(3) сбор ДНК-вектора.
34. Способ по п. 33, отличающийся тем, что единственный вид белка Rep представляет собой белок Rep AAV.
35. Способ по п. 34, отличающийся тем, что белок Rep AAV выбран из группы, состоящей из AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11 и белка AAV12 Rep.
36. Способ по п. 34, отличающийся тем, что белок Rep AAV представляет собой белок Rep68 AAV2.
37. Способ по п. 34, отличающийся тем, что белок Rep AAV представляет собой белок Rep78 AAV2.
38. Способ по п. 37, отличающийся тем, что белок Rep78 AAV2 кодируется мутантной последовательностью нуклеиновой кислоты Rep78.
39. Способ по п. 38, отличающийся тем, что мутантная последовательность нуклеиновой кислоты Rep78 не имеет функционального кодона инициации трансляции для Rep52.
40. Способ по п. 38, отличающийся тем, что мутантная последовательность нуклеиновой кислоты Rep78 кодирует мутантный белок Rep78, который содержит мутацию в положении 225 аминокислоты SEQ ID NO: 588.
41. Способ по п. 40, отличающийся тем, что аминокислота 225 из SEQ ID NO: 588 мутирована в глицин (Gly) или треонин (Thr).
42. Способ по п. 38, отличающийся тем, что мутантная последовательность нуклеиновой кислоты Rep78 кодирует мутантный белок Rep78, который содержит последовательность, имеющую по меньшей мере 95% идентичность последовательности с SEQ ID NO: 588.
43. Способ по п. 33, отличающийся тем, что по меньшей мере одна последовательность контроля экспрессии кодирует промотор, выбранный из группы, состоящей из: промотор ID, промотор ΔIE и промотор CMV.
44. Способ по п. 33, отличающийся тем, что указанный двухцепочечный конструкт ДНК выбран из группы, состоящей из бакмиды, плазмиды, мини-кольца и линейной двухцепочечной молекулы ДНК.
45. Способ по п. 33, отличающийся тем, что первый ITR представляет собой ITR дикого типа.
46. Способ по п. 33, отличающийся тем, что первый ITR и второй ITR являются симметричными или по существу симметричными или асимметричными относительно друг друга.
47. Способ по п. 45, отличающийся тем, что ITR дикого типа содержит SEQ ID NO: 51.
48. Способ по п. 33, отличающийся тем, что второй ITR представляет собой модифицированный ITR.
49. Способ по п. 48, отличающийся тем, что модифицированный ITR содержит SEQ ID NO: 2.
50. Способ по п. 33, отличающийся тем, что первый ITR представляет собой модифицированный ITR.
51. Способ по п. 50, отличающийся тем, что модифицированный ITR содержит SEQ ID NO: 52.
52. Способ по п. 50, отличающийся тем, что второй ITR представляет собой ITR дикого типа.
53. Способ по п. 52, отличающийся тем, что ITR дикого типа содержит SEQ ID NO: 1.
54. Способ по п. 33, отличающийся тем, что первый ITR и второй ITR являются репликационно-компетентными.
55. Способ по п. 33, отличающийся тем, что по меньшей мере один из первого ITR и второго ITR содержит последовательность нуклеиновой кислоты, соответствующую сайту связывания белка Rep для индуцирования репликации ДНК-вектора в клетке.
56. Способ по п. 33, отличающийся тем, что один или оба из первого ITR и второго ITR представляет собой ITR AAV.
57. Способ по п. 33, отличающийся тем, что экспрессионная кассета содержит цис-регуляторный элемент.
58. Способ по п. 57, отличающийся тем, что цис-регуляторный элемент выбирают из группы, состоящей из посттранскрипционного регуляторного элемента и поли(А)-сигнала BGH.
59. Способ по п. 58, отличающийся тем, что указанный посттранскрипционный регуляторный элемент содержит посттранскрипционный регуляторный элемент вируса гепатита сурков (WPRE).
60. Способ по п. 33, отличающийся тем, что экспрессионная кассета дополнительно содержит промотор, выбранный из группы, состоящей из промотора CAG, промотора ААТ, промотора LP1 и промотора EF1a.
61. Способ по п. 33, отличающийся тем, что указанная экспрессионная кассета содержит последовательность нуклеиновой кислоты, выбранную из группы, состоящей из SEQ ID NO: 3, SEQ ID NO: 7, SEQ ID NO: 8 и SEQ ID NO: 9.
62. Способ по п. 33, отличающийся тем, что указанная экспрессионная кассета дополнительно содержит экзогенную последовательность.
63. Способ по п. 62, отличающийся тем, что экзогенная последовательность содержит по меньшей мере 2000 нуклеотидов.
64. Способ по п. 62, отличающийся тем, что указанная экзогенная последовательность кодирует белок.
65. Способ по п. 62, отличающийся тем, что экзогенная последовательность кодирует белок, выбранный из группы, состоящей из: репортерный белок, терапевтический белок, антиген, ген-редактирующий белок и цитотоксический белок.
66. Способ по п. 33, отличающийся тем, что ДНК-вектор имеет линейную и непрерывную структуру.
67. ДНК-вектор для доставки полинуклеотидной последовательности, получаемый способом по п. 33.
68. Фармацевтическая композиция для генной терапии, содержащая ДНК-вектор по п. 67 и эксципиент.
69. Набор для получения ДНК-вектора, содержащий:
экспрессионный конструкт, содержащий по меньшей мере один сайт рестрикции для вставки по меньшей мере одной последовательности нуклеиновой кислоты или регуляторного переключателя или обоих, по меньшей мере один сайт рестрикции, расположенный между асимметричными инвертированными концевыми повторами (асимметричные ITR), в которых по меньшей мере один из асимметричных ITR содержит функциональный концевой сайт разрешения и сайт связывания белка репликации (Rep); а также
вектор, содержащий последовательность нуклеиновой кислоты, кодирующую единственный вид Rep-белка, при этом вектор подходит для экспрессии одного вида Rep-белка в клетке насекомого.
70. Набор по п. 69, дополнительно содержащий популяцию клеток насекомых, которая не содержит последовательностей, кодирующих вирусный капсид, которые в присутствии единственного вида белка Rep могут индуцировать продуцирование ДНК-вектора.
71. Клетка для получения зкДНК-вектора, содержащая последовательность нуклеиновой кислоты, кодирующую единственный вид белка репликации (Rep) AAV, имеющую по меньшей мере ДНК-связующую и ДНК-никирующую функциональность, связанную по меньшей мере с одной последовательностью контроля экспрессии, причем клетка не экспрессирует какие-либо виды белка Rep парвовируса и не контактирует с другими видами белка Rep.
72. Клетка по п. 71, отличающаяся тем, что клетка дополнительно содержит конструкт двухцепочечной ДНК, содержащий экспрессионную кассету, первый ITR на 5'-конце перед экспрессионной кассетой и второй ITR на 3'-конце после экспрессионной кассеты.
73. Клетка по п. 71, отличающаяся тем, что клетка представляет собой клетку насекомого.
74. Клетка по п. 73, отличающаяся тем, что клетка насекомого выбрана из группы, состоящей из клетки Sf9, клетки Sf21, клетки Trichoplusia ni и клетки High Five.
75. Клетка-хозяин по п. 74, отличающаяся тем, что указанная клетка насекомого представляет собой клетку Sf9.
76. Клетка по п. 74, отличающаяся тем, что клетка насекомого представляет собой клетку High Five.
77. Клетка по п. 71, отличающаяся тем, что клетка представляет собой клетку млекопитающего.
78. Клетка по п. 77, отличающаяся тем, что клетка млекопитающего выбрана из группы, состоящей из HEK293, Huh-7, HeLa, HepG2, HeplA, 911, CHO, COS, MeWo, NIH3T3, A549, HT1080, моноцитов и зрелых и незрелых дендритных клеток.
79. Клетка по п. 77, отличающаяся тем, что клетка млекопитающего представляет собой HEK293.
80. Клетка по п. 71, отличающаяся тем, что последовательность нуклеиновой кислоты, кодирующая единственный вид белка Rep AAV, кодирует Rep78 или Rep68.
81. Клетка по п. 80, отличающаяся тем, что последовательность нуклеиновой кислоты не имеет функционального инициирующего кодона для Rep52 или Rep40.
82. Клетка по п. 81, отличающаяся тем, что последовательность нуклеиновой кислоты кодирует белок Rep78.
83. Клетка по п. 82, отличающаяся тем, что последовательность нуклеиновой кислоты кодирует белок Rep68.
84. Клетка по п. 82, отличающаяся тем, что последовательность нуклеиновой кислоты кодирует мутантный белок Rep78, который содержит мутацию в положении 225 аминокислоты SEQ ID NO: 588.
85. Клетка по п. 84, отличающаяся тем, что аминокислота 225 (метионин) из SEQ ID NO: 588 мутирована в глицин (Gly) или треонин (Thr).
86. Клетка по п. 84, отличающаяся тем, что последовательность нуклеиновой кислоты дополнительно включает одну или более модификаций в альтернативных сайтах сплайсинга на карбоксиконце, предотвращая событие сплайсинга, ведущее к получению Rep68, тем самым обеспечивая продукцию только Rep78.
87. Клетка по п. 81, отличающаяся тем, что последовательность нуклеиновой кислоты является полноразмерной и содержит интактные альтернативные сайты сплайсинга на карбоксиконце, что приводит к продукции Rep78 и Rep68.
88. Клетка по п. 81, отличающаяся тем, что последовательность нуклеиновой кислоты содержит делецию карбоксиконцевой интронной последовательности, приводящую к получению только Rep68.
89. Клетка по п. 81, отличающаяся тем, что последовательность нуклеиновой кислоты кодирует белок Rep78, который содержит последовательность, имеющую по меньшей мере 95% идентичность последовательности с SEQ ID NO: 588 и имеющую по меньшей мере ДНК-связующую и ДНК-никирующую функциональность.
| US 20190032083 A1 31.01.2019 | |||
| Lamartina S | |||
| et al | |||
| Способ получения смеси хлоргидратов опийных алкалоидов (пантопона) из опийных вытяжек с любым содержанием морфия | 1921 |
|
SU68A1 |
| Способ и аппарат для получения гидразобензола или его гомологов | 1922 |
|
SU1998A1 |
| Контактный детектор | 1927 |
|
SU7653A1 |
| WO 2019032898 A1 14.02.2019 | |||
| WO 2012123430 A1 20.09.2012 | |||
| WO 2017152149 A1 08.09.2017 | |||
| AAV ВЕКТОРЫ С УСОВЕРШЕНСТВОВАННЫМИ Rep-КОДИРУЮЩИМИ ПОСЛЕДОВАТЕЛЬНОСТЯМИ, ИСПОЛЬЗУЕМЫМИ В СИСТЕМАХ ПРОДУКЦИИ НА ОСНОВЕ КЛЕТОК НАСЕКОМЫХ | 2007 |
|
RU2457252C2 |
| WO 2008016391 A2 | |||
