Область техники
[0001] Настоящее изобретение относится к антисмысловому олигомеру для пропуска экзона, содержащему нуклеотидную последовательность, комплементарную двум или более разным последовательностям в целевом экзоне. В частности, настоящее изобретение относится к антисмысловому олигомеру, который вызывает пропуск экзона 44 в человеческом гене дистрофина, и к фармацевтической композиции, содержащей этот олигомер.
[0002] Мышечная дистрофия Дюшенна (МДД) представляет собой наиболее распространенную форму наследственной прогрессирующей мышечной дистрофии, поражающей одного из приблизительно 3500 новорожденных мальчиков. Хотя двигательные функции редко отличаются от здоровых людей в младенчестве и детстве, мышечная слабость наблюдается у детей в возрасте приблизительно от 4 до 5 лет. Впоследствии мышечная слабость прогрессирует до потери способности передвигаться приблизительно к 12 годам и смерти в результате сердечной или дыхательной недостаточности в возрасте 20-30 лет. МДД является тяжелым нарушением. В настоящее время не существует доступной эффективной терапии МДД, и существует острая необходимость в разработке нового терапевтического агента.
[0003] Как известно, МДД вызвана мутацией в гене дистрофина. Ген дистрофина располагается на X-хромосоме и является огромным по размеру геном, состоящим из 2,2 миллиона пар нуклеотидов ДНК. ДНК транскрибируется с образованием предшественников мРНК, интроны удаляются посредством сплайсинга с образованием мРНК длиной 13993 основания, в которой 79 экзонов соединены вместе. Эта мРНК транслируется с образованием белка дистрофина длиной 3685 аминокислот. Белок дистрофин связан с поддержанием стабильности мембраны в мышечных клетках и необходим для уменьшения хрупкости мышечных клеток. Ген дистрофина у пациентов с МДД содержит мутацию и, следовательно, белок дистрофин, функционирующий в мышечных клетках, редко экспрессируется. Поэтому структура мышечных клеток не может поддерживаться в организме пациентов с МДД, что приводит к входу большого количества ионов кальция в мышечные клетки. Вследствие этого развивается похожий на воспалительный ответ, способствующий развитию фиброза, что затрудняет регенерацию мышечных клеток.
[0004] Мышечная дистрофия Беккера (МДБ) также вызывается мутацией в гене дистрофина. Симптомы включают мышечную слабость, но прогрессия мышечной слабости, как правило, умеренная или слабая по сравнению с МДД. Во многих случаях начало заболевания приходится на взрослый возраст. Различия в клинических симптомах между МДД и МДБ, как полагают, основаны на том, нарушает или нет мутация открытую рамку считывания для аминокислот при трансляции мРНК дистрофина с образованием белка дистрофина (Непатентный документ 1). В частности, при МДД наличие мутации вызывает сдвиг рамки считывания для аминокислот таким образом, что прекращается экспрессия функционального белка дистрофина, в то время как при МДБ образуется функционирующий, хотя и неполноценный, белок дистрофин, поскольку рамка считывания для аминокислот сохраняется, в то время как часть экзонов отсутствует в результате мутации.
[0005] Пропуск экзонов, как ожидают, будет служить способом лечения МДД. Такой способ включает модификацию сплайсинга с целью восстановления рамки считывания для аминокислот в мРНК дистрофина и индукции экспрессии белка дистрофина, обладающего частично восстановленной функцией (Непатентный документ 2). Часть аминокислотной последовательности, являющейся мишенью для пропуска экзона, будет утрачена. По этой причине белок дистрофин, экспрессирующийся при такой обработке, становится короче нормального белка, но поскольку рамка считывания для аминокислот сохраняется, функция стабилизации мышечных клеток частично восстанавливается. Следовательно, ожидается что пропуск экзона приведет к тому, что МДД будет иметь симптомы, похожие на симптомы МДБ, которая является более умеренным заболеванием. Подход, основанный на пропуске экзонов, прошел испытания на животных с применением мышей или собак и в настоящее время изучается в рамках клинических исследований на людях с МДД.
[0006] Пропуск экзонов можно индуцировать посредством связывания антисмысловых нуклеиновых кислот либо, мишенью которых являются либо 5'-, либо 3'-сайты сплайсинга, либо как 5'-, так и 3'-сайты сплайсинга, либо сайты внутри экзона. Экзон включится в мРНК только при узнавании обоих сайтов сплайсинга комплексом сплайсосомы. Таким образом, пропуск экзона можно индуцировать, действуя на сайты сплайсинга с помощью антисмысловых нуклеиновых кислот. Более того, считается, что связывание белка SR, обгащенного серином и аргинином, с экзонным энхансером сплайсинга (ESE, exonic splicing enhancer) необходимо для узнавания экзона механизмом сплайсинга. Соответственно, пропуск экзона можно индуцировать, действуя на ESE.
[0007] Поскольку мутации в гене дистрофина могут различаться у разных пациентов с МДД, антисмысловые нуклеиновые кислоты следует конструировать на основании сайта или типа соответствующей генетической мутации. Существует несколько сообщений об антисмысловых нуклеиновых кислотах, которые индуцируют пропуск экзона, с одной последовательностью в качестве мишени к единственному экзону в гене дистрофина (Патентные документы 1-6 и Непатентные документы 1 и 2). Также сообщалось о том, что при смешивании и функционировании двух типов антисмысловых нуклеиновых кислот, действующих на один и тот же экзон в гене дистрофина (при двойном действии), можно усилить пропускающую экзон активность по сравнению с применением каждой антисмысловой нуклеиновой кислоты по отдельности (Патентный документ 7).
[0008] Однако ни одно из предыдущих сообщений не показывало, что объединенная одноцепочечная антисмысловая нуклеиновая кислота (объединенный тип), действующая на два и более сайтов в одном и том же экзоне, обладает пропускающей экзон активностью (Патентный документ 1).
Документы, относящиеся к предшествующему уровню техники
Патентные документы
[0009] Патентный документ 1: Международная публикация WO 2004/048570
Патентный документ 2: Международная публикация WO 2009/139630
Патентный документ 3: Международная публикация WO 2010/048586
Патентный документ 4: US 2010/0168212
Патентный документ 5: Международная публикация WO 2011/057350
Патентный документ 6: Международная публикация WO 2006/000057
Патентный документ 7: Международная публикация WO 2007/135105
Непатентные документы
[0010] Непатентный документ 1: Annemieke Aartsma-Rus et al., (2002) Neuromuscular Disorders 12: S71-S77
Непатентный документ 2: Wilton S. D., e t al., Molecular Therapy 2007: 15: p. 1288-96
РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
Задачи изобретения
[0011] В свете описанных обстоятельств основной целью настоящего изобретения является получение нового антисмыслового олигомера объединенного типа, который индуцирует пропуск экзона, действуя на две разные нуклеотидные последовательности в одном и том же экзоне в гене дистрофина, и терапевтического агента для лечения мышечной дистрофии, содержащего этот олигомер.
Средства решения задачи
[0012] Авторы настоящего изобретения провели обширные исследования содержания техник, описанных в указанных выше документах, и структуры гена дистрофина и пр. и обнаружили в результате, что антисмысловой олигомер, полученный посредством объединения олигомеров, действующих на два разных сайта в экзоне 44 в гене дистрофина человека, может индуцировать пропуск этого экзона. На основании этого открытия авторы настоящего изобретения создали настоящее изобретение.
[0013] Таким образом, настоящее изобретение заключается в следующем.
[1] Антисмысловой олигомер длиной от 15 до 30 оснований, где
(a) первая часть олигомера, содержащая нуклеотидную последовательность, комплементарную первой нуклеотидной последовательности из 7-15 последовательно расположенных оснований в целевом экзоне; и
(b) вторая часть олигомера, содержащая нуклеотидную последовательность, комплементарную второй нуклеотидной последовательности из 7-15 последовательно расположенных оснований в целевом экзоне,
соединены, где
первая нуклеотидная последовательность и вторая нуклеотидная последовательность не следуют друг за другом и не перекрываются друг с другом, и
антисмысловой олигомер индуцирует пропуск целевого экзона, или его фармацевтически приемлемая соль или гидрат.
[2] Антисмысловой олигомер по п.1, где олигомер из первой и/или из второй части содержит нуклеотидную последовательность комплементарную части нуклеотидной последовательности интрона рядом с целевым экзоном, или его фармацевтически приемлемая соль или гидрат.
[3] Антисмысловой олигомер по п.1 или 2, где целевой экзон является экзоном человеческого гена дистрофина, или его фармацевтически приемлемая соль или гидрат.
[4] Антисмысловой олигомер по п.1 или 2, где первая нуклеотидная последовательность является нуклеотидной последовательностью из 7-15 последовательно расположенных оснований, выбираемых из нуклеотидной последовательности, представленной SEQ ID NO: 1, или его фармацевтически приемлемая соль или гидрат.
[5] Антисмысловой олигомер по любому из п.п.1-3, где вторая нуклеотидная последовательность является нуклеотидной последовательностью из 7-15 последовательно расположенных оснований, выбираемых из нуклеотидной последовательности, представленной SEQ ID NO: 2, или его фармацевтически приемлемая соль или гидрат.
[6] Антисмысловой олигомер по п.1 или 2, где две части олигомера, выбранные из группы, состоящей из следующих пунктов с (c) по (e), соединены:
(c) часть олигомера, состоящая из нуклеотидной последовательности, комплементарной нуклеотидной последовательности из 7-15 последовательно расположенных оснований, выбираемых из нуклеотидной последовательности, представленной SEQ ID NO: 3;
(d) часть олигомера, состоящая из нуклеотидной последовательности, комплементарной нуклеотидной последовательности из 7-15 последовательно расположенных оснований, выбираемых из нуклеотидной последовательности, представленной SEQ ID NO: 4; и
(e) часть олигомера, состоящая из нуклеотидной последовательности, комплементарной нуклеотидной последовательности из 7-15 последовательно расположенных оснований, выбираемых из нуклеотидной последовательности, представленной SEQ ID NO: 5, или его фармацевтически приемлемая соль или гидрат.
[7] Антисмысловой олигомер по п.1 или 2, который состоит из нуклеотидной последовательности, выбираемой из группы, состоящей из SEQ ID No с 6 по 9, или его фармацевтически приемлемая соль или гидрат.
[8] Антисмысловой олигомер по любому из п.п.1-7, который является олигонуклеотидом, или его фармацевтически приемлемая соль или гидрат.
[9] Антисмысловой олигомер по п.8, где остаток сахара и/или область соединения через фосфатную группу по меньшей мере одного нуклеотида, входящего в состав олигонуклеотида, модифицированы, или его фармацевтически приемлемая соль или гидрат.
[10] Антисмысловой олигомер по п.8 или 9, где остаток сахара по меньшей мере одного нуклеотида, входящего в состав олигонуклеотида, является рибозой, в которой 2'-OH-группа заменена любой группой, выбранной из группы, состоящей из OR, R, R'OR, SH, SR, NH2, NHR, NR2, N3, CN, F, Cl, Br и I (где R является алкилом или арилом и R' является алкиленом), или его фармацевтически приемлемая соль или гидрат.
[11] Антисмысловой олигомер по любому из п.п.8-10, где область соединения через фосфатную группу по меньшей мере одного нуклеотида, входящего в состав олигонуклеотида, является любой областью, выбираемой из группы, состоящей из тиофосфатной связи, дитиофосфатной связи, алкилфосфонатной связи, фосфорамидатной связи и боранофосфатной связи, или его фармацевтически приемлемая соль или гидрат.
[12] Антисмысловой олигомер по любому из п.п.1-7, который является морфолино-олигомером, или его фармацевтически приемлемая соль или гидрат.
[13] Антисмысловой олигомер по п.12, который является морфолино-олигомером, или его фармацевтически приемлемая соль или гидрат.
[14] Антисмысловой олигомер по п.12 или 13, в котором 5'-конец имеет любую из приведенных ниже химических формул (1)-(3), или его фармацевтически приемлемая соль или гидрат.
[Формула 1]
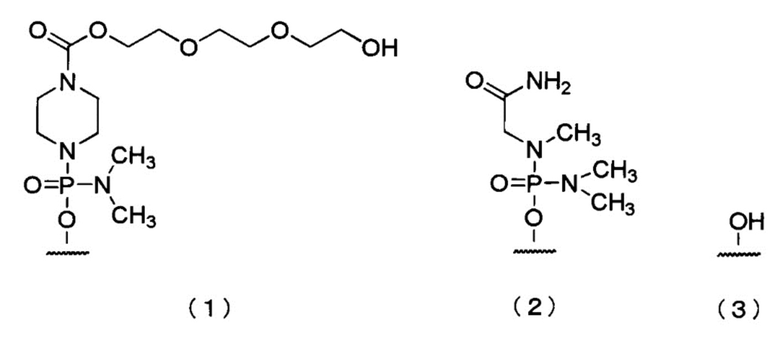
[15] Фармацевтическая композиция для лечения мышечной дистрофии, содержащая в качестве активного ингредиента антисмысловой олигомер по любому из п.п.1-14, или его фармацевтически приемлемую соль или гидрат.
[16] Фармацевтическая композиция по п.15, содержащая фармацевтически приемлемый носитель.
[17] Способ лечения мышечной дистрофии, который включает введение пациенту с мышечной дистрофией антисмыслового олигомера или его фармацевтически приемлемой соли или гидрата по любому из п.п.1-12, или фармацевтической композиции по любому из п.п.1-16.
[18] Способ лечения по п.17, где у пациента с мышечной дистрофией есть мутация (мутации), на которую действует пропуск экзона 44 в гене дистрофина.
[19] Способ лечения по п.17 или 18, где пациентом является человек.
[20] Применение антисмыслового олигомера или его фармацевтически приемлемой соли или гидрата по любому из п.п.1-14 для получения фармацевтической композиции для лечения мышечной дистрофии.
[21] Антисмысловой олигомер или его фармацевтически приемлемая соль или гидрат по любому из п.п.1-14 для применения в лечении мышечной дистрофии.
[22] Антисмысловой олигомер или его фармацевтически приемлемая соль или гидрат по п.21, где у пациента с мышечной дистрофией в указанном лечении есть мутация (мутации), на которую действует пропуск экзона 44 в гене дистрофина.
[23] Антисмысловой олигомер по п.21 или 22 или его фармацевтически приемлемая соль или гидрат, где пациентом является человек.
[24] Способ получения антисмыслового олигомера по п.1, который включает соединение
(a) первой части олигомера, содержащей нуклеотидную последовательность, комплементарную первой нуклеотидной последовательности из 7-15 последовательно расположенных оснований в целевом экзоне; и
(b) второй части олигомера, содержащей нуклеотидную последовательность, комплементарную второй нуклеотидной последовательности из 7-15 последовательно расположенных оснований в целевом экзоне,
с образованием антисмыслового олигомера длиной от 15 до 30 оснований, где
первая нуклеотидная последовательность и вторая нуклеотидная последовательность не следуют друг за другом и не перекрываются друг с другом.
[25] Способ по п.24, также включающий:
измерение эффективности пропуска при использовании полученного антисмыслового олигомера; и
отбор антисмыслового олигомера, который обладает эффективностью пропуска экзона, превышающей референтное значение.
[26] Способ скрининга антисмыслового олигомера, включающий:
(a) отбор
(i) первой части олигомера, содержащей нуклеотидную последовательность, комплементарную первой нуклеотидной последовательности из 7-15 последовательно расположенных оснований в целевом экзоне; и
(ii) второй части олигомера, содержащей нуклеотидную последовательность, комплементарную второй нуклеотидной последовательности из 7-15 последовательно расположенных оснований в целевом экзоне, где
первая нуклеотидная последовательность и вторая нуклеотидная последовательность не следуют друг за другом и не перекрываются друг с другом;
(b) соединение первого и второго олигомеров с образованием антисмыслового олигомера длиной от 15 до 30 оснований;
(c) измерение эффективности пропуска экзона при использовании антисмыслового олигомера, полученного на стадии (b); и
(d) отбор антисмыслового олигомера, который обладает эффективностью пропуска экзона, превышающей референтное значение.
ЭФФЕКТЫ ИЗОБРЕТЕНИЯ
[0014] Антисмысловой олигомер настоящего изобретения может индуцировать пропуск экзона 44 в гене дистрофина человека с высокой эффективностью. Также симптомы мышечной дистрофии Дюшенна можно эффективно облегчить посредством введения фармацевтической композиции настоящего изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0015] Фиг. 1 показывает эффективность пропуска экзона 44 в гене дистрофина человека в линии клеток рабдомиосаркомы человека (в клетках RD).
Фиг. 2 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 3 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 4 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 5 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 6 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 7 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 8 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 9 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 10 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 11 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 12 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 13 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 14 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 15 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 16 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 17 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 18 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 19 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 20 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 21 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 22 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 23 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 24 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 25 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 26 показывает эффективность пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) при соответствующих концентрациях олигомеров.
Фиг. 27 показывает сравнение эффективности пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) объединенной формы и смеси двух олигомерных частей, действующих на разные сайты.
Фиг. 28 показывает сравнение эффективности пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) объединенной формы и смеси двух олигомерных частей, действующих на разные сайты.
Фиг. 29 показывает сравнение эффективности пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) объединенной формы и смеси двух олигомерных частей, действующих на разные сайты.
Фиг. 30 показывает сравнение эффективности пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) каждой части олигомера в отдельности, объединенной формы и смеси двух олигомерных частей, действующих на разные сайты.
Фиг. 31 показывает сравнение эффективности пропуска экзона 44 в гене дистрофина человека в клетках рабдомиосаркомы человека (в клетках RD) каждой части олигомера в отдельности, объединенной формы и смеси двух олигомерных частей, действующих на разные сайты.
Фиг. 32 показывает эффективность пропуска экзона 44 в гене дистрофина человека в фибробластах человека с МДД с делецией экзона 45.
[0016] Вариант осуществления настоящего изобретения
[0017] Здесь и далее приведено подробное описание настоящего изобретения. Варианты осуществления настоящего изобретения, описанные ниже, приведены в качестве примера, лишь для описания настоящего изобретения, но не для ограничения настоящего изобретения следующими вариантами осуществления настоящего изобретения. Настоящее изобретение можно осуществить различными способами, не отступая от сути настоящего изобретения.
Все публикации, опубликованные патентные заявки, патенты и другие патентные документы, процитированные в настоящем описании изобретения, включены в настоящую заявку в полном объеме посредством ссылки. Описание изобретения в настоящей заявке включает посредством ссылки содержание описания изобретения и чертежей из заявки на патент Японии (№ 2014-124157), поданной 17 июня 2014 г., согласно которой испрашивает приоритет настоящая заявка.
[0018] 1. Антисмысловой олигомер
Настоящее изобретение представляет антисмысловой олигомер длиной от 15 до 30 оснований, где
(a) первая часть олигомера, содержащая нуклеотидную последовательность, комплементарную первой нуклеотидной последовательности из 7-15 последовательно расположенных оснований в целевом экзоне; и
(b) вторая часть олигомера, содержащая нуклеотидную последовательность, комплементарную второй нуклеотидной последовательности из 7-15 последовательно расположенных оснований в целевом экзоне,
соединены, где
первая нуклеотидная последовательность и вторая нуклеотидная последовательность не следуют друг за другом и не перекрываются друг с другом, и антисмысловой олигомер индуцирует пропуск целевого экзона, или его фармацевтически приемлемую соль или гидрат.
Здесь и далее «антисмысловой олигомер или его фармацевтически приемлемая соль или гидрат» можно обобщенно называть «антисмысловым олигомером».
[0019] Описанный выше антисмысловой олигомер можно получить с помощью способа получения, включающего соединение
(a) первой части олигомера, содержащей нуклеотидную последовательность, комплементарную первой нуклеотидной последовательности из 7-15 последовательно расположенных оснований в целевом экзоне; и
(b) второй части олигомера, содержащей нуклеотидную последовательность, комплементарную второй нуклеотидной последовательности из 7-15 последовательно расположенных оснований в целевом экзоне,
с образованием антисмыслового олигомера длиной от 15 до 30 оснований, где
первая нуклеотидная последовательность и вторая нуклеотидная последовательность не следуют друг за другом и не перекрываются друг с другом.
Способ получения может также включать
стадию измерения эффективности пропуска при использовании полученного антисмыслового олигомера; и
вторую стадию отбора антисмыслового олигомера, который обладает эффективностью пропуска экзона, превышающей референтное значение.
[0020] На второй стадии описанного выше способа получения эффективность пропуска можно определить следующим образом. мРНК гена, содержащего целевой экзон, выделяют из исследуемых клеток; в мРНК измеряют уровень «А» полинуклеотидной полосы в случае, когда экзон пропущен, и уровень «В» полинуклеотидной полосы в случае, когда экзон не пропущен. Используя эти величины измерений «А» и «В», рассчитывают эффективность согласно следующему уравнению:
Эффективность пропуска (%)=A/(A+B)×100
Альтернативно, расчет эффективности пропуска можно посмотреть в Международной публикации заявки WO2012/029986.
[0021] На второй стадии референтное значение эффективности пропуска составляет 10% или более, 20% или более, 30% или более, 40% или более, 50% или более, 60% или более, 70% или более, 80% или более и 90% или более.
Как упомянуто выше, при соединении нескольких олигомерных частей антисмысловой олигомер, обладающий повышенной пропускающей экзон активностью, можно получить даже если каждая олигомерная часть обладает низкой пропускающей экзон активностью (или не обладает пропускающей экзон активностью).
[0022] Настоящее изобретение также представляет способ скрининга антисмыслового олигомера, включающий:
(a) отбор
(i) первой части олигомера, содержащей нуклеотидную последовательность, комплементарную первой нуклеотидной последовательности из 7-15 последовательно расположенных оснований в целевом экзоне; и
(ii) второй части олигомера, содержащей нуклеотидную последовательность, комплементарную второй нуклеотидной последовательности из 7-15 последовательно расположенных оснований в целевом экзоне, где
первая нуклеотидная последовательность и вторая нуклеотидная последовательность не следуют друг за другом и не перекрываются друг с другом;
(b) соединение первого и второго олигомеров с образованием антисмыслового олигомера длиной от 15 до 30 оснований;
(c) измерение эффективности пропуска экзона при использовании антисмыслового олигомера, полученного на стадии (b); и
(d) отбор антисмыслового олигомера, который обладает эффективностью пропуска экзона, превышающей референтное значение.
[0023] В описанном выше антисмысловом олигомере первую и вторую части олигомера можно соединить таким образом, что либо первая, либо вторая часть олигомера располагается с 5'- или с 3'-стороны другой части. В одном варианте осуществления настоящего изобретения первая часть олигомера расположена с 5'-стороны, а вторая часть олигомера расположена с 3'-стороны при соединении.
Также антисмысловой олигомер может содержать третью часть олигомера, содержащую нуклеотидную последовательность, комплементарную третьей нуклеотидной последовательности из 7-15 расположенных последовательно оснований в целевом экзоне.
[0024] Как применяют в настоящей заявке, термин «соединять» обозначает соединение двух частей олигомера друг с другом напрямую или соединение двух частей олигомера друг с другом через линкер. Если две части олигомера соединены друг с другом напрямую, 3'-конец части олигомера, расположенной с 5'-стороны, и 5'-конец другой части олигомера, расположенной с 3'-стороны, образуют фосфатную связь или группу, показанную ниже. Примеры линкеров включают нуклеиновую кислоту (цепь) длиной от 1 до 5 оснований, а также известные линкеры, обычно применяемые для соединения нуклеиновых кислот или морфолино-производных нуклеиновых кислот, такие как 3-аминопропил, сукцинил, 2,2'-диэтанолсульфонил и длинноцепочечный алкиламино (LCAA, long chain alkylamino).
[Формула 2]
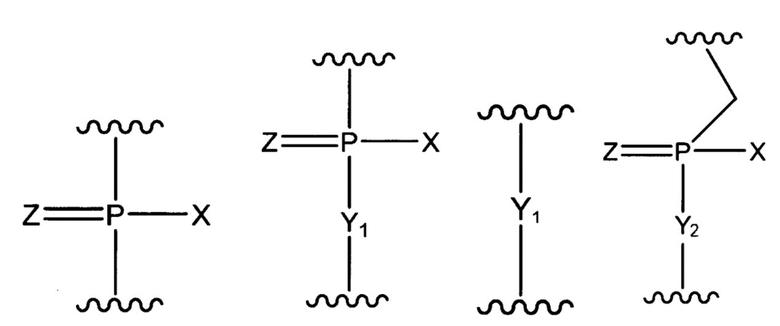
где X представляет -OH, -CH2R1, -O-CH2R1, -S-CH2R1, -NR2R3 или F;
R1 представляет H или алкил;
R2 и R3, которые могут быть одинаковыми или разными, каждый из которых представляет H, алкил, циклоалкил или арил;
Y1 представляет O, S, CH2 или NR1;
Y2 представляет O, S или NR1;
Z представляет O или S.
[0025] Первая и/или вторая часть олигомера могут содержать нуклеотидную последовательность, комплементарную части нуклеотидной последовательности интрона рядом с целевым экзоном. В одном варианте осуществления настоящего изобретения, в котором, например, первая и вторая части олигомера соединены друг с другом таким образом, что первая часть олигомера располагается с 5'-стороны, а вторая часть олигомера располагается с 3'-стороны, 5'-сторона первой части олигомера может содержать нуклеотидную последовательность, комплементарную нуклеотидной последовательности, расположенной рядом с 3'-концом интрона рядом с 5'-стороной целевого экзона, и/или 3'-сторона второй части олигомера может содержать нуклеотидную последовательность, комплементарную нуклеотидной последовательности, расположенной рядом с 5'-концом интрона рядом с 3'-стороной целевого экзона.
Первая и/или вторая часть олигомера может содержать нуклеотидную последовательность, комплементарную части нуклеотидной последовательности экзонного энхансера сплайсинга (ESE, exonic splicing enhancer) целевого экзона.
[0026] Целевой экзон специально не ограничивается. В одном варианте осуществления настоящего изобретения целевой экзон является экзоном в гене человека, и, более того, является экзоном в гене дистрофина человека.
В частности, целевым экзоном является экзон 44 в гене дистрофина человека.
Таким образом, в одном варианте осуществления настоящее изобретение представляет антисмысловой олигомер, который вызывает пропуск экзона 44 в гене дистрофина человека (здесь и далее называемый олигомером настоящего изобретения). Здесь и далее структура антисмыслового олигомера настоящего изобретения будет описана подробнее.
[0027] [Экзон 44 в гене дистрофина человека]
В настоящем изобретении термин «ген» предназначен для обозначения геномного гена, а также включает кДНК, предшественник мРНК и мРНК. Предпочтительно, чтобы ген был предшественником мРНК, т.е. пре-мРНК.
В геноме человека ген дистрофина человека располагается в локусе Xp21.2. Ген дистрофина человека имеет размер, равный 3,0 миллиона пар оснований и является самым большим геном среди известных генов человека. Однако кодирующие области гена дистрофина человека, рассредоточенные в виде 79 экзонов на всем протяжении гена дистрофина человека, составляют лишь 14 т.п.о. (Roberts, RG. et al., Genomics, 16: 536-538 (1993)). Пре-мРНК, которая является транскриптом гена дистрофина человека, подвергается сплайсингу с образованием зрелой мРНК размером 14 т.о. Нуклеотидная последовательность гена дистрофина человека дикого типа известна (№ доступа в GenBank - NM_004006).
Нуклеотидная последовательность экзона 44 в гене дистрофина человека дикого типа представлена SEQ ID NO: 10.
[0028] В одном варианте осуществления настоящего изобретения олигомер настоящего изобретения создан для индукции пропуска экзона 44 в гене дистрофина человека, с модифицированием тем самым белка, кодируемого геном дистрофина типа МДД, в белок дистрофина типа МДБ. Соответственно, экзон 44 в гене дистрофина, который является мишенью пропуска экзона с помощью антисмыслового олигомера, включает как дикий тип, так и мутантные типы.
В частности, мутанты гена дистрофина человека по экзону 44 включают полинуклеотиды, определенные в п. (I) или (II) ниже.
(I) полинуклеотид, который гибридизуется в строгих условиях с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной нуклеотидной последовательности SEQ ID NO: 10; и
(II) полинуклеотид, состоящий из нуклеотидной последовательности, по меньшей мере на 90% идентичной нуклеотидной последовательности SEQ ID NOs: 10.
Как применяют в настоящей заявке, термин «полинуклеотид» предназначен для обозначения ДНК или РНК.
Как применяют в настоящей заявке, термин «полинуклеотид, который гибридизуется в строгих условиях», относится, например, к полинуклеотиду, получаемому при гибридизации колоний, гибридизации бляшек, гибридизации по Саузерну и т.п., с применением в качестве зонда всего полинуклеотида, состоящего из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, например, SEQ ID NO: 10, или его части. Способ гибридизации, который можно использовать, включает способы, описанные, например, в "Sambrook & Russell, Molecular Cloning: A Laboratory Manual Vol. 3, Cold Spring Harbor, Laboratory Press 2001," "Ausubel, Current Protocols in Molecular Biology, John Wiley & Sons 1987-1997" и пр.
Как применяют в настоящей заявке, термин «комплементарная нуклеотидная последовательность» не ограничен только нуклеотидными последовательностями, которые образуют пары по Уотсону-Крику с целевыми нуклеотидными последовательностями, но предназначен также для включения нуклеотидных последовательностей, которые образуют неоднозначные пары оснований. Как применяют в настоящей заявке, термин «пара по Уотсону-Крику» обозначает пару нуклеиновых оснований, в которой водородные связи формируются между аденином и тимином, аденином и урацилом или между гуанином и цитозином, а термин «неоднозначная пара оснований» обозначает пару нуклеиновых оснований, в которой водородные связи формируются между гуанином и урацилом, инозином и урацилом, инозином и аденином или между инозином и цитозином. Как применяют в настоящей заявке, термин «комплементарная нуклеотидная последовательность» относится не только к нуклеотидной последовательности, на 100% комплементарной целевой нуклеотидной последовательности, но также обозначает комплементарную нуклеотидную последовательность, которая может содержать, например, от 1 до 3, 1 или 2 или один нуклеотид, некомплементарный целевой нуклеотидной последовательности.
Как применяют в настоящей заявке, «строгие условия» могут быть любыми из условий «малой строгости», условий «средней строгости» и условий «высокой строгости». Условиями «малой строгости» являются, например, 5×SSC, 5×раствор Денхардта, 0,5% додецилсульфат натрия, 50% формамид при 32°С. Условиями «средней строгости» являются, например, 5×SSC, 5×раствор Денхардта, 0,5% додецилсульфат натрия, 50% формамид при 42°С или 5×SSC, 1% додецилсульфат натрия, 50 мМ Трис-HCl (pH 7,5), 50% формамид при 42°С. Условиями «высокой строгости» являются, например, 5×SSC, 5×раствор Денхардта, 0,5% додецилсульфат натрия, 50% формамид при 50°С или 0,2×SSC, 0,1% додецилсульфат натрия при 65°С. В этих условиях ожидается, что полинуклеотиды с большей гомологией будут с успехом получены при более высоких температурах, хотя в строгость гибридизации вносят вклад разнообразные факторы, включающие температуру, концентрацию зонда, длину зонда, ионную силу, время, концентрацию соли и другие, и специалисты в данной области техники могут соответствующим образом подобрать эти факторы для достижения схожей строгости.
При применении для гибридизации имеющихся в продаже наборов можно использовать, например, Alkphos Direct Labeling and Detection System (GE Healthcare). В этом случае в соответствии с прилагаемым протоколом после культивирования с меченым зондом в течение ночи мембрану отмывают основным буфером для отмывки, содержащим 0,1% (вес/объем) додецилсульфата натрия при 55°С, выявляя тем самым гибридизованные полинуклеотиды. Альтернативно, в случае создания зонда на основе всей нуклеотидной последовательности, комплементарной нуклеотидной последовательности SEQ ID NO: 10, или ее части гибридизацию можно выявить с помощью набора DIG Nucleic Acid Detection Kit (Roche Diagnostics), когда зонд метят дигоксигенином (DIG, digoxigenin), используя имеющийся в продаже реагент (например, PCR Labeling Mix (Roche Diagnostics) и т.п.).
Помимо полинуклеотидов, описанных выше, другие полинуклеотиды, которые могут гибридизоваться, включают полинуклеотиды, идентичные на 90% или более, 91% или более, 92% или более, 93% или более, 94% или более, 95% или более, 96% или более, 97% или более, 98% или более, 99% или более, 99,1% или более, 99,2% или более, 99,3% или более, 99,4% или более, 99,5% или более, 99,6% или более, 99,7% или более, 99,8% или более, или 99,9% или более нуклеотидной последовательности SEQ ID NO: 10, в соответствии с расчетом с использованием программы для поиска гомологии, такой как BLAST, с применением параметров, заданных по умолчанию.
Идентичность нуклеотидных последовательностей можно определить, используя алгоритм BLAST (Basic Local Alignment Search Tool средство поиска основного локального совмещения) Karlin и Altschul (Proc. Natl. Acad. Sci. USA 872264-2268, 1990; Proc. Natl. Acad. Sci. USA 90: 5873, 1993). Были разработаны программы, названные BLASTN и BLASTX, на основе алгоритма BLAST (Altschul SF et al., J. Mol. Biol. 215: 403, 1990). В случае установления нуклеотидной последовательности, используя BLASTN, параметрами являются, например, оценка=100 и длина слова=12. В случае использования программ BLAST и Gapped BLAST (BLAST с использованием пропусков) применяют параметры, заданные по умолчанию для каждой программы.
[0029] Олигомер настоящего изобретения как раз является антисмысловым олигомером длиной от 15 до 30 оснований, в котором соединены две части олигомера, выбранные из группы, состоящей из следующих п. (a) и (b):
(a) часть олигомера, состоящая из нуклеотидной последовательности, комплементарной нуклеотидной последовательности из 7-15 расположенных последовательно оснований, выбираемых из нуклеотидной последовательности, представленной SEQ ID NO: 1; и
(b) часть олигомера, состоящая из нуклеотидной последовательности, комплементарной нуклеотидной последовательности из 7-15 расположенных последовательно оснований, выбираемых из нуклеотидной последовательности, представленной SEQ ID NO: 2.
Например, первая нуклеотидная последовательность может быть нуклеотидной последовательностью из 7-15 расположенных последовательно оснований, выбираемых из нуклеотидной последовательности, представленной SEQ ID NO: 1, и/или вторая нуклеотидная последовательность может быть нуклеотидной последовательностью из 7-15 расположенных последовательно оснований, выбираемых из нуклеотидной последовательности, представленной SEQ ID NO: 2.
Предпочтительно, чтобы олигомер настоящего изобретения был антисмысловым олигомером длиной от 15 до 30 оснований, в котором соединены две части олигомера, выбираемые из группы, состоящей из следующих п. с (c) по (e):
(c) часть олигомера, состоящая из последовательности, комплементарной нуклеотидной последовательности из 7-15 расположенных последовательно оснований, выбираемых из нуклеотидной последовательности, представленной SEQ ID NO: 3;
(d) часть олигомера, состоящая из последовательности, комплементарной нуклеотидной последовательности из 7-15 расположенных последовательно оснований, выбираемых из нуклеотидной последовательности, представленной SEQ ID NO: 4; и
(e) часть олигомера, состоящая из последовательности, комплементарной нуклеотидной последовательности из 7-15 расположенных последовательно оснований, выбираемых из нуклеотидной последовательности, представленной SEQ ID NO: 5.
[0030] В настоящей заявке нуклеотидные последовательности, представленные SEQ ID NO: 1 и 2, являются последовательностями, состоящими из оснований в положениях с -1 до 44 и с 58 до 115, соответственно, с 5'-конца нуклеотидной последовательности экзона 44 (SEQ ID NO: 10) в человеческом гене дистрофина дикого типа.
Нуклеотидная последовательность, представленная SEQ ID NO: 3, является последовательностью, состоящей из оснований в положениях с 18 до 34 с 5'-конца нуклеотидной последовательности экзона 44 (SEQ ID NO: 10) в человеческом гене дистрофина дикого типа. Аналогично, нуклеотидные последовательности, представленные SEQ ID NO: 4 и 5, являются последовательностями, состоящими из оснований в положениях с 61 по 77 и с 88 по 104, соответственно.
[0031] Размер каждой части олигомера с (a) по (e) (здесь и далее также называемые просто частями) составляет от 7 до 15 оснований в длину, предпочтительно, от 8 до 15 оснований в длину, от 9 до 15 оснований в длину, от 10 до 15 оснований в длину, от 10 до 14 оснований в длину, от 10 до 13 оснований в длину или от 11 до 13 оснований в длину. Части с (a) по (e) могут быть одинакового или разного размера.
[0032] Для отбора двух частей олигомера из группы, состоящей из (a) и (b), две части олигомера могут быть комбинацией одинаковых частей олигомера или могут быть комбинацией разных частей олигомера. В частности, две части олигомера могут быть комбинацией (a) и (a) или комбинацией (b) и (b) или могут быть комбинацией (a) и (b).
Для отбора двух частей олигомера из группы, состоящей из п. с (c) по (e), две части олигомера могут быть комбинацией одинаковых частей олигомера или могут быть комбинацией разных частей олигомера. Предпочтительно, чтобы две части соответственно выбирали из разных типов. Например, если (c) выбирают в качестве одной части, предпочтительно, чтобы другая часть была (d) или (e). Аналогично, если выбрана (d) в качестве одной части, предпочтительно, чтобы другая часть была (c) или (e). Также если выбрана (e) в качестве одной части, предпочтительно, чтобы другая часть была (c) или (d).
[0033] Если выбраны части (a) и (b), любая из двух выбранных частей может располагаться с 5'-стороны. Если выбраны части (a) и (b), предпочтительно, чтобы часть (a) присоединялась с 3'-стороны.
Если выбраны две части из п. с (c) по (e), любая из двух выбранных частей может располагаться с 5'-стороны. Если выбраны части (c) и (d), предпочтительно, чтобы часть (c) присоединялась с 3'-стороны. Если выбраны части (d) и (e), предпочтительно, чтобы часть (d) присоединялась с 3'-стороны. Если выбраны части (c) и (e), предпочтительно, чтобы часть (c) присоединялась с 3'-стороны.
[0034] Как применяют в настоящей заявке, термин «соединять» относится к прямому связыванию двух частей, выбранных из (a) и (b), или двух частей, выбранных из п. с (c) по (e). В частности, термин «когда две части соединены» обозначает, что 3'-конец части, расположенной с 5'-стороны, и 5'-конец части, расположенной с 3'-стороны, образуют фосфатную связь или группу, показанную ниже.
[Формула 3]
где X представляет -OH, -CH2R1, -O-CH2R1, -S-CH2R1, -NR2R3 или F;
R1 представляет H или алкил;
R2 и R3, которые могут быть одинаковыми или разными, каждый из которых представляет H, алкил, циклоалкил или арил;
Y1 представляет O, S, CH2 или NR1;
Y2 представляет O, S или NR1;
Z представляет O или S.
[0035] Термин «вызывает пропуск экзона 44 в гене дистрофина человека», предназначен для обозначения того, что в результате связывания олигомера настоящего изобретения с сайтом, соответствующим экзону 44 транскрипта (например, пре-мРНК) гена дистрофина человека, например, связывания в результате сплайсинга нуклеотидной последовательности, соответствующей 5'-концу экзона 46 с 3'-концом экзона 43, у пациентов с МДД с делецией экзона 45, когда транскрипт подвергается сплайсингу, образуется зрелая мРНК, в которой нет сдвига рамки считывания.
[0036] В настоящей заявке описанный выше термин «связывание», как предполагается, означает, что при смешивании олигомера настоящего изобретения с транскриптом гена дистрофина человека, оба гибридизуются в физиологических условиях с образованием двухцепочечной нуклеиновой кислоты. Термин «в физиологических условиях» относится к условиям, заданным для воспроизводства in vivo окружения по показателю рН, состава солей и температуры. Условиями являются, например, 25-40°С, предпочтительно 37°С, рН 5-8, предпочтительно рН 7,4, и 150 мМ концентрация хлорида натрия.
[0037] Наличие или отсутствие пропуска экзона 44 в гене дистрофина человека, можно подтвердить посредством введения олигомера настоящего изобретения в клетку, экспрессирующую дистрофин (например, в клетки рабдомиосаркомы человека), амплификации района, окружающего экзон 44 мРНК гена дистрофина человека, на основе общей РНК из клетки, экспрессирующей дистрофин, с помощью ОТ-ПЦР и выполнения «вложенной» ПЦР или анализа последовательности на амплифицированном с помощью ПЦР продукте.
Эффективность пропуска можно определить следующим образом. мРНК гена дистрофина человека выделяют из исследуемых клеток; в мРНК измеряют уровень «А» полинуклеотидной полосы в случае, когда экзон 44 пропущен, и уровень «В» полинуклеотидной полосы в случае, когда экзон 44 не пропущен. Используя эти величины измерений «А» и «В», рассчитывают эффективность согласно следующему уравнению:
Эффективность пропуска (%)=A/(A+B)×100
Альтернативно, расчет эффективности пропуска можно посмотреть в Международной публикации заявки WO2012/029986.
[0038] Предпочтительно, чтобы антисмысловой олигомер настоящего изобретения вызывал пропуск целевого экзона (напр., экзона 44) с эффективностью, равной 10% или более, 20% или более, 30% или более, 40% или более, 50% или более, 60% или более, 70% или более, 80% или более и 90% или более.
[0039] Антисмысловой олигомер настоящего изобретения включает, например, олигонуклеотид, морфолино-олигомер или олигонуклеотид пептидной нуклеиновой кислоты (ПНК), имеющий длину от 15 до 30 оснований. Предпочтительно, чтобы длина составляла от 16 до 30, от 17 до 30, от 18 до 30, от 19 до 30, от 20 до 30, от 20 до 29, от 20 до 28, от 20 до 27, от 20 до 26, от 21 до 26 или от 22 до 26 оснований, и предпочтительными являются морфолино-олигомеры.
[0040] Описанный выше олигонуклеотид (здесь и далее называемый олигонуклеотидом настоящего изобретения) представляет собой олигомер настоящего изобретения, состоящий из нуклеотидов в качестве составных единиц. Такие нуклеотиды могут быть любыми из рибонуклеотидов, дезоксирибонуклеотидов и модифицированных нуклеотидов.
[0041] Модифицированный нуклеотид относится к нуклеотиду, имеющему полностью и частично модифицированные нуклеиновые основания, остатки сахаров и/или области соединения через фосфатную группу, который является рибонуклеотидом или дезоксирибонуклеотидом.
[0042] Нуклеиновые основания включают, например, аденин, гуанин, гипоксантин, цитозин, тимин, урацил и их модифицированные основания. Примеры таких модифицированных оснований включают, но не ограничиваются псевдоурацилом, 3-метилурацилом, дигидроурацилом, 5-алкилцитозинами (например, 5-метилцитозином), 5-алкилурацилами (например, 5-этилурацилом), 5-галогенурацилами (5-бромурацилом), 6-азапиримидином, 6-алкилпиримидинами (6-метилурацилом), 2-тиоурацилом, 4-тиоурацилом, 4-ацетилцитозином, 5-(карбоксигидроксиметил)урацилом, 5'-карбоксиметиламинометил-2-тиоурацилом, 5-карбоксиметиламинометилурацилом, 1-метиладенином, 1-метилгипоксантином, 2,2-диметилгуанином, 3-метилцитозином, 2-метиладенином, 2-метилгуанином, N6-метиладенином, 7-метилгуанином, 5-метоксиаминометил-2-тиоурацилом, 5-метиламинометилурацилом, 5-метилкарбонилметилурацилом, 5-метилоксиурацилом, 5-метил-2-тиоурацилом, 2-метилтио-N6-изопентениладенином, урацил-5-оксиуксусной кислотой, 2-тиоцитозином, пурином, 2,6-диаминопурином, 2-аминопурином, изогуанином, индолом, имидазолом, ксантином и т.д.
[0043] Модификация остатка сахара может включать, например, модификации в 2'-положении рибозы и модификации других положений сахара. Модификация в 2'-положении рибозы включает замену 2'-ОН рибозы на OR, R, R'OR, SH, SR, NH2, NHR, NR2, N3, CN, F, Cl, Br или I, где R представляет собой алкил или арил, а R' представляет собой алкилен.
Модификация в других положениях сахара включает, например, замену О в 4'-положении рибозы или дезоксирибозы на S, образование мостика между 2'- и 4'-положениями сахара, например, LNA (locked nucleic acid, закрытой нуклеиновой кислоты) или ENA (2'-O,4'-C-ethylene-bridged nucleic acids, нуклеиновых кислот с 2'-О,4'-С-этиленовым мостиком), но не ограничивается ими.
[0044] Модификация области соединения через фосфатную группу включает, например, модификацию с заменой фосфодиэфирной связи на тиофосфатную связь, дитиофосфатную связь, алкилфосфонатную связь, фосфорамидатную связь или боранофосфатную связь (Enya et al., Bioorganic & Medicinal Chemistry, 2008, 18, 9154-9160) (см., например, отечественные повторные публикации в Японии РСТ-заявок с № 2006/129594 и 2006/038608).
[0045] Предпочтительно, чтобы алкилом был неразветвленный или разветвленный алкил, содержащий от 1 до 6 атомов углерода. Конкретные примеры включают метил, этил, н-пропил, изопропил, н-бутил, изобутил, втор-бутил, трет-бутил, н-пентил, изопентил, неопентил, трет-пентил, н-гексил и изогексил. Алкил может необязательно быть замещенным. Примерами таких заместителей являются галоген, алкокси, циано и нитро. Алкил может быть замещен 1-3 такими заместителями.
Предпочтительно, чтобы циклоалкилом был циклоалкил, содержащий 5-12 атомов углерода. Конкретные примеры включают циклопентил, циклогексил, циклогептил, циклооктил, циклодецил и циклододецил.
Галоген включает фтор, хлор, бром и йод.
Алкокси представляет собой неразветвленный или разветвленный алкокси, содержащий 1-6 атомов углерода, такой как метокси, этокси, н-пропокси, изопропокси, н-бутокси, изобутокси, втор-бутокси, трет-бутокси, н-пентилокси, изопентилокси, н-гексилокси, изогексилокси и т.д. Среди прочего, алкокси, содержащий 1-3 атомов углерода, является предпочтительным.
Предпочтительно, чтобы арилом был арил, содержащий 6-10 атомов углерода. Конкретные примеры включают фенил, α-нафтил и β-нафтил. Среди прочих, предпочтительным является фенил. Арил может быть, при желании, замещенным. Примерами таких заместителей являются алкил, галоген, алкокси, циано и нитро. Арил может быть замещен одним-тремя такими заместителями.
В настоящем изобретении предпочтительно, чтобы алкиленом был неразветвленный или разветвленный алкилен, содержащий от 1 до 6 атомов углерода. Конкретные примеры включают метилен, этилен, триметилен, тетраметилен, пентаметилен, гексаметилен, 2-(этил)триметилен и 1-(метил)тетраметилен.
Ацил включает неразветвленный или разветвленный алканоил или ароил. Примеры алканоила включают формил, ацетил, 2-метилацетил, 2,2-диметилацетил, пропионил, бутирил, изобутирил, пентаноил, 2,2-диметилпропионил, гексаноил и т.д. Примеры ароила включают бензоил, толуоил и нафтоил. Ароил может быть при желании замещенным в замещаемых положениях и может быть замещен алкилом (алкилами).
[0046] Предпочтительно, чтобы олигонуклеотидом настоящего изобретения был олигомер настоящего изобретения, содержащий составную единицу, представляемую общей формулой ниже, где группа -ОН в положении 2' рибозы замещена метокси, а областью соединения через фосфатную группу является тиофосфатная связь:
[Формула 4]
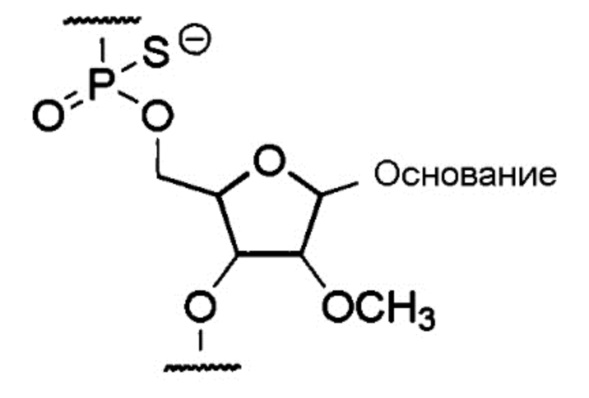
где «основание» представляет собой нуклеиновое основание.
[0047] Олигонуклеотид настоящего изобретения можно легко синтезировать, используя различные автоматизированные синтезаторы (например, AKTA oligopilot plus 10/100 (GE Healthcare)). В альтернативном случае синтез можно также поручить сторонней организации (например, Promega Inc., Takara Co. или Japan Bio Service Co.) и т.д.
[0048] Описанным выше морфолино-олигомером является олигомер настоящего изобретения, содержащий составную единицу, представленную общей формулой ниже:
[Формула 5]
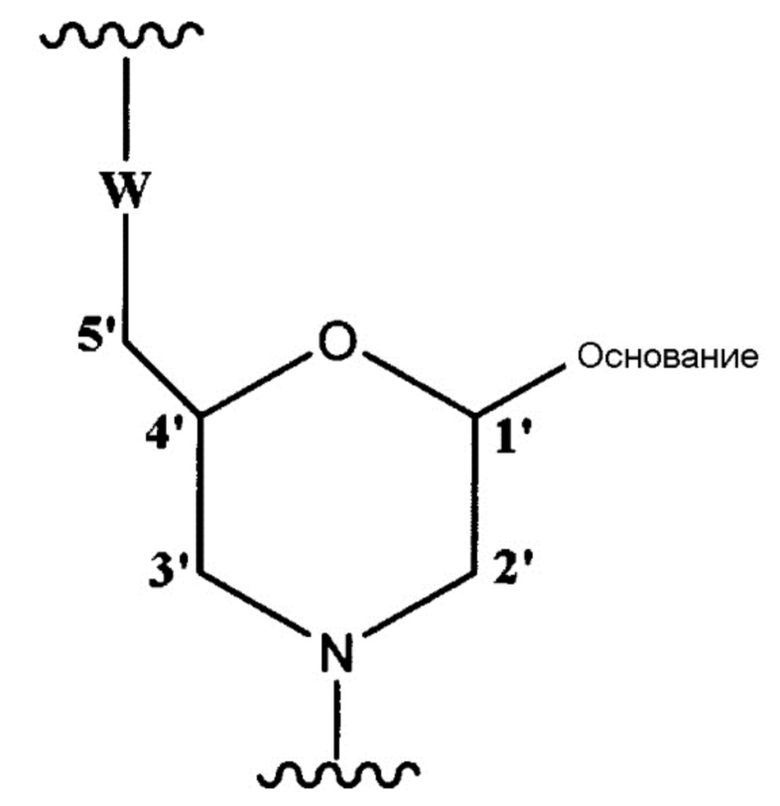
где «основание» имеет то же определенное выше значение, и
W представляет собой группу, представленную любой из следующих групп:
[Формула 6]
где X представляет собой -CH2R1, -O-CH2R1, -S-CH2R1, -NR2R3 или F;
R1 представляет собой Н или алкил;
R2 и R3, которые могут быть одинаковыми или различными, представляют собой каждый Н, алкил, циклоалкил или арил;
Y1 представляет собой O, S, CH2 или NR1;
Y2 представляет собой O, S или NR1;
Z представляет собой O или S.
[0049] Предпочтительно, чтобы морфолино-олигомером был олигомер, содержащий составную единицу, представленную общей формулой ниже, (морфолино-фосфородиамидатный олигомер (здесь далее называемый РМО, phosphorodiamidate morpholino oligomer)).
[Формула 7]
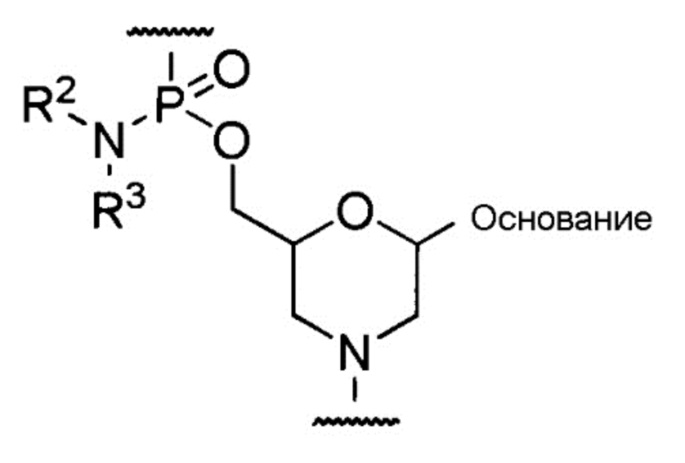
где «основание», R2 и R3 имеют те же определенные выше значения.
[0050] Морфолино-олигомер можно получить в соответствии, например, с WO 1991/009033 или WO 2009/064471. В частности, РМО можно получить с помощью процедуры, описанной в WO 2009/064471 или WO2013/100190.
[0051] [Способ получения PMO]
Вариантом РМО является, например, соединение, представленное общей формулой (I) ниже, (здесь и далее РМО (I)):
[Формула 8]
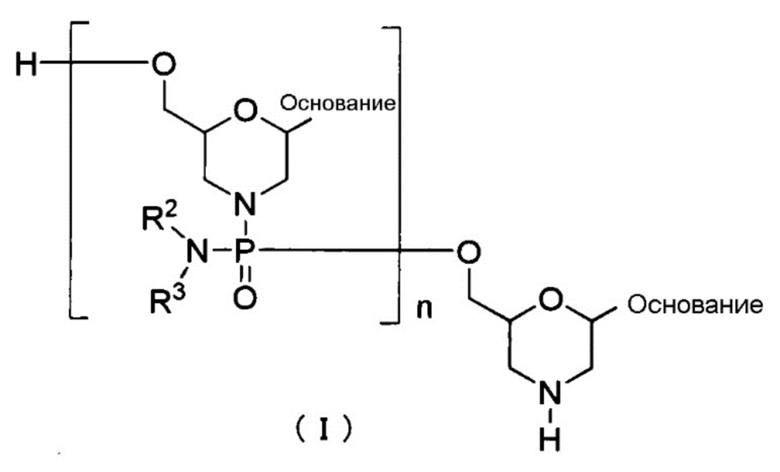
где «основание», R2 и R3 имеют те же определенные выше значения; и,
n является заданным целым числом от 1 до 99, предпочтительно, заданным целым числом от 18 до 28.
[0052] РМО (I) можно получить в соответствии с известным способом, например можно получить посредством выполнения процедур следующих стадий.
Соединения и реагенты, используемые в стадиях ниже, специально не ограничены при условии, что их обычно используют для получения РМО.
[0053] Также все следующие стадии можно выполнить с использованием жидкофазного способа или твердофазного способа (используя руководства или имеющиеся в продаже автоматизированные устройства для твердофазного синтеза). При получении РМО твердофазным способом желательно использовать автоматизированные синтезаторы ввиду простых технических требований и точности синтеза.
[0054] (1) Стадия A:
Соединение, представленное общей формулой (II) ниже (здесь и далее называемое Соединением (II)) реагирует с кислотой с образованием соединения, представленного общей формулой (III) ниже (здесь и далее называемого Соединением (III)):
[Формула 9]
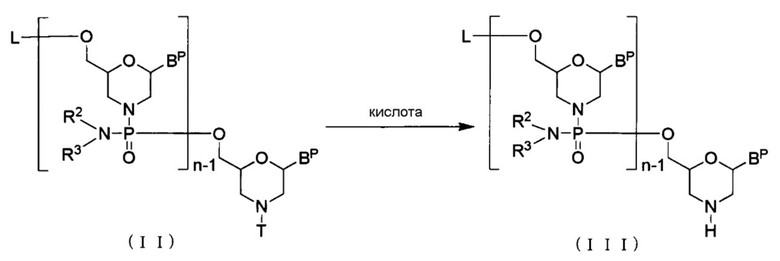
где n, R2 и R3 имеют те же определенные выше значения;
каждое BP независимо представляет собой нуклеиновое основание, которое при желании может быть защищено;
Т представляет собой тритил, монометокситритил или диметокситритил; и
L представляет собой водород, ацил или группу, представленную общей формулой (IV) ниже, (здесь и далее называемую группой (IV).
[Формула 10]
В случае BP «нуклеиновое основание» включает то же нуклеиновое основание, что в случае «основания» при условии, что аминогруппу или гидроксильную группу в нуклеиновом основании, представленном BP, можно защитить.
Такая защитная группа для аминогруппы специально не ограничивается при условии, что она используется в качестве защитной группы для нуклеиновых кислот. Конкретные примеры включают бензоил, 4-метоксибензоил, ацетил, пропионил, бутирил, изобутирил, фенилацетил, феноксиацетил, 4-трет-бутилфеноксиацетил, 4-изопропилфеноксиацетил и (диметиламино)метилен. Конкретные примеры защитной группы для гидроксильной группы включают 2-цианоэтил, 4-нитрофенетил, фенилсульфонилэтил, метилсульфонилэтил и триметилсилилэтил, и фенил, который может быть замещен 1-5 электроноакцепторными группами в оптимальных замещаемых положениях, дифенилкарбамоил, диметилкарбамоил, диэтилкарбамоил, метилфенилкарбамоил, 1-пиролидинилкарбамоил, морфолинокарбамоил, 4-(трет-бутилкарбокси)бензил, 4-[(диметиламино)карбокси]бензил и 4-(фенилкарбокси)бензил (см., например, WO 2009/064471).
«Твердый носитель» специально не ограничивается при условии, что это носитель, применяемый для твердофазной реакции синтеза нуклеиновых кислот. Желательно, чтобы твердый носитель обладал следующими свойствами: например, (i) был труднорастворимым в реагентах, которые можно использовать для синтеза морфолино-производных нуклеиновых кислот, (например, в дихлорметане, ацетонитриле, тетразоле, N-метилимидазоле, пиридине, уксусном ангидриде, лутидине, трифторуксусной кислоте); (ii) был химически устойчивым к реагентам, используемым для синтеза морфолино-производных нуклеиновых кислот; (iii) его можно было химически модифицировать; (iv) его можно было нагрузить желаемыми морфолино-производными нуклеиновых кислот; (v) он обладал прочностью, достаточной, чтобы выдерживать высокое давление на протяжении всех обработок; и (vi) он характеризовался постоянным диапазоном диаметра частиц и их распределением. В частности, он включает поддающийся разбуханию полистирол (например, аминометилполистирольную смолу, поперечно сшитую 1% дивинилбензолом (200-400 меш) (2,4-3,0 ммоль/г) (производства Tokyo Chemical Industry), аминометилированную полистирольную смолу HCl [1% дивинилбензола, 100-200 меш] (производства Peptide Institute, Inc.)), не поддающийся разбуханию полистирол (например, основной носитель (производства GE Healthcare), присоединенный к цепи ПЭГ полистирол (например, NH2-ПЭГ-смолу (производства Watanabe Chemical Co), смолу TentaGel), стекло с контролируемым размером пор (стекло с контролируемым размером пор; CPG) (производства, например, CPG), оксалильное стекло с контролируемым размером пор (см., например, Alul et al., Nucleic Acid Research, Vol. 19, 1527 (1991)), носитель TentaGel-аминополиэтиленгликоль-дериватизированный носитель (например, Wright et al., см., Tetrahedron Letters, Vol. 34, 3373 (1993) и сополимер Poros-полистирол/дивинилбензол.
«Линкер», который можно применять, является известным линкером, обычно используемым для соединения нуклеиновых кислот или морфолино-производных нуклеиновых кислот. Примеры включают 3-аминопропил, сукцинил, 2,2'-диэтанолсульфонил и длинноцепочечный алкиламино (LCAA, long-chain alkyl amino).
[0055] Эту стадию можно выполнить, проводя реакцию между Соединением (II) и кислотой.
[0056] «Кислота», которую можно использовать на этой стадии, включает, например, трифторуксусную кислоту, дихлоруксусную кислоту и трихлоруксусную кислоту. Соответственно, используемая кислота находится в диапазоне, например, от 0,1 моль-эквивалента до 1000 моль-эквивалентов, если исходить из 1 моль Соединения (II), предпочтительно, в диапазоне от 1 моль-эквивалента до 100 моль-эквивалентов, если исходить из 1 моль Соединения (II).
В сочетании с описанной выше кислотой можно использовать органический амин. Органический амин специально не ограничивается и включает, например, триэтиламин. Соответственно, количество используемого органического амина находится в диапазоне, например, от 0,01 моль-эквивалентов до 10 моль-эквивалентов и, предпочтительно, в диапазоне от 0,1 моль-эквивалентов до 2 моль-эквивалентов, если исходить из 1 моль кислоты.
В случае использования на этой стадии соли или смеси кислоты и органического амина соль или смесь включает, например, соль или смесь трифторуксусной кислоты и триэтиламина, а конкретнее, смесь 1 эквивалента триэтиламина и 2 эквивалентов трифторуксусной кислоты.
Кислота, которую можно применять на этой стадии, можно также применять в форме, разведенной в соответствующем растворителе, в концентрации, составляющей от 0,1% до 30%. Растворитель специально не ограничивается, когда речь идет об инертном по отношению к реакции растворителе, и включает, например, дихлорметан, ацетонитрил, спирт (этанол, изопропанол, трифторэтанол и т.д.), воду или их смесь.
[0057] Предпочтительно, чтобы температура описанной выше реакции находилась в диапазоне, например, от 10°С до 50°С, предпочтительнее, в диапазоне от 20°С до 40°С и, наиболее предпочтительно, в диапазоне от 25°С до 35°С.
Время реакции может варьировать в зависимости от вида используемой кислоты и температуры реакции и находится, соответственно, в диапазоне, как правило, от 0,1 минуты до 24 часов, предпочтительно, в диапазоне от 1 минуты до 5 часов.
[0058] После завершения этой стадии можно добавить основание, при необходимости, для нейтрализации кислоты, оставшейся в системе. «Основание» специально не ограничивается и включает, например, диизопропиламин. Основание можно также применять в форме, разведенной в соответствующем растворителе, в концентрации, составляющей от 0,1% (объем/объем) до 30% (объем/объем).
Растворитель, применяемый на этой стадии, специально не ограничивается, когда речь идет об инертном по отношению к реакции растворителе, и включает дихлорметан, ацетонитрил, спирт (этанол, изопропанол, трифторэтанол и т.д.), воду или их смесь. Предпочтительно, чтобы температура описанной выше реакции находилась в диапазоне, например, от 10°С до 50°С, предпочтительнее, в диапазоне от 20°С до 40°С и, наиболее предпочтительно, в диапазоне от 25°С до 35°С.
Время реакции может варьировать в зависимости от вида используемой кислоты и температуры реакции и находится, соответственно, в диапазоне, как правило, от 0,1 минуты до 24 часов, предпочтительно, в диапазоне от 1 минуты до 5 часов.
[0059] В случае Соединения (II) соединение общей формулы (IIa) ниже (в дальнейшем Соединение (IIa)), где n равно 1, а L представляет собой группу (IV), можно получить с помощью следующей процедуры.
[Формула 11]
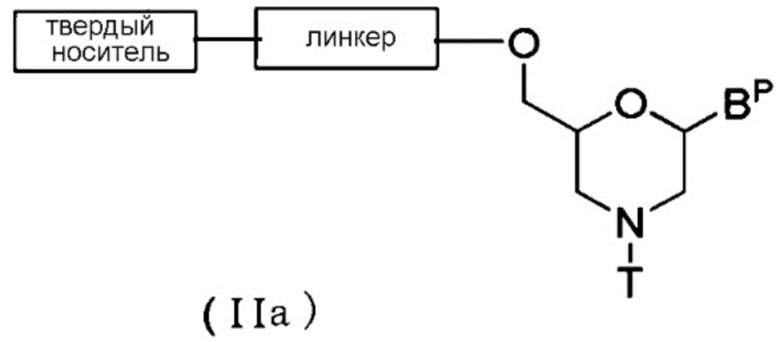
где BP, T, линкер и твердый носитель имеют те же определенные выше значения.
[0060] Стадия 1:
Соединение, представленное общей формулой (V) ниже, реагирует с ацилирующим агентом с образованием соединения, представленного общей формулой (VI) ниже (здесь и далее называемого Соединением (VI)).
[Формула 12]
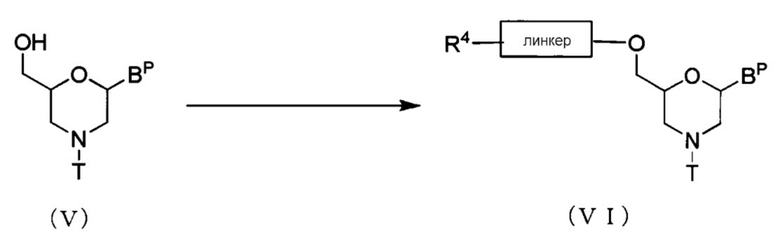
где BP, T и линкер имеют те же определенные выше значения; и
R4 представляет собой гидроксильную галогеновую, карбоксильную или аминогруппу.
[0061] Эту стадию можно выполнить с помощью известных процедур для введения линкеров, используя Соединение (V) в качестве исходного материала.
В частности, соединение, представленное общей формулой (VIa) ниже, можно получить посредством выполнения способа, известного как этерификация, используя Соединение (V) и янтарный ангидрид.
[Формула 13]
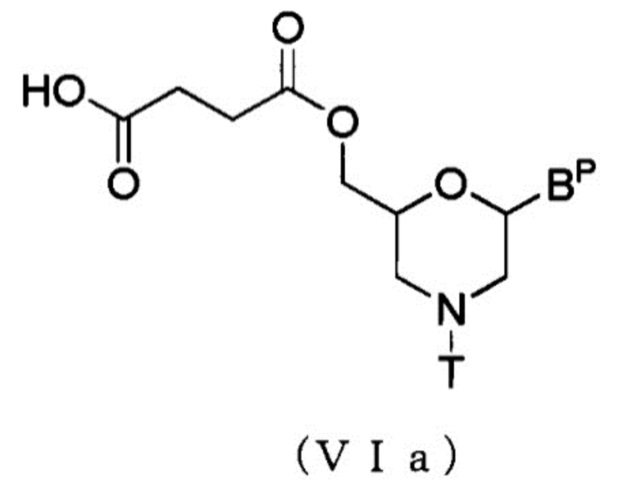
где BP и T имеют те же определенные выше значения.
[0062] Стадия 2:
Соединение (VI) реагирует с твердым носителем с помощью конденсирующего агента с образованием Соединения (IIa).
[Формула 14]
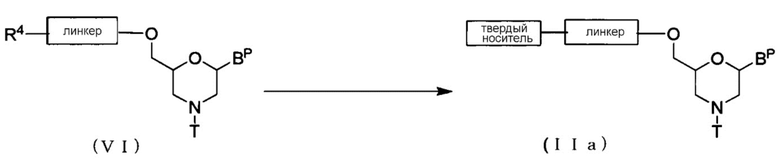
где BP, R4, T, линкер и твердый носитель имеют те же определенные выше значения.
Эту стадию можно выполнить, используя Соединение (VI) и твердый носитель, в соответствии с процессом, известным как реакция конденсации.
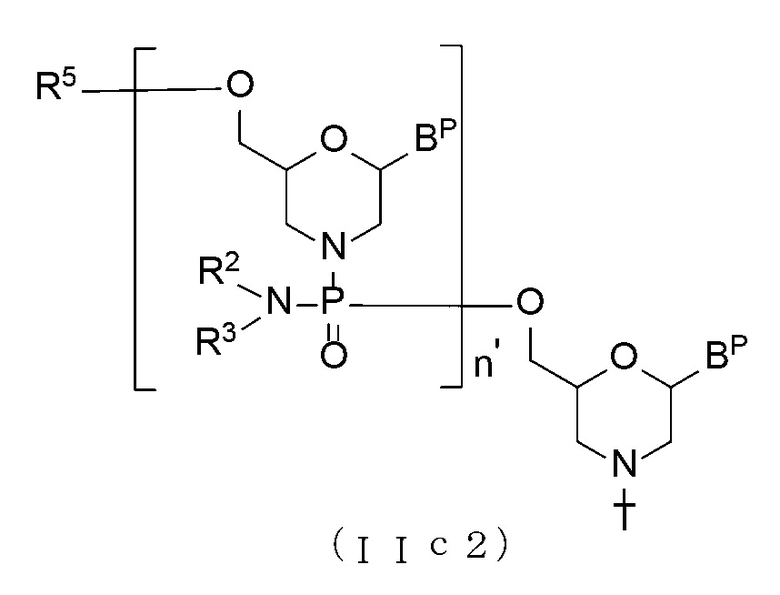
[0063] В случае Соединения (II) соединение, представленное общей формулой (IIa2) ниже, где n составляет от 2 до 99, а L является группой, представленной общей формулой (IV), можно получить, используя Соединение (IIa) в качестве исходного материала и повторяя стадию А и стадию В способа получения РМО, описанного здесь, нужное число раз.
[Формула 15]
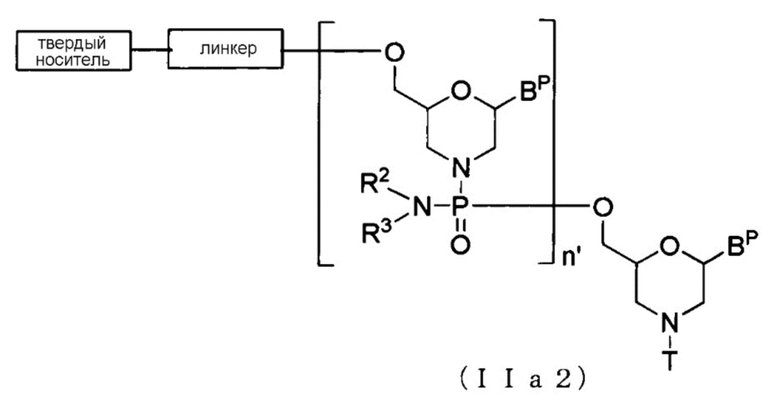
где BP, R2, R3, T, линкер и твердый носитель имеют те же определенные выше значения; и
n' составляет от 1 до 98.
[0064] В соединении (II), соединение с общей формулой (IIb), показанное ниже, где n равно 1, а L является водородом, можно получить при помощи процедуры, описанной, например, в WO 1991/009033.
[Формула 16]
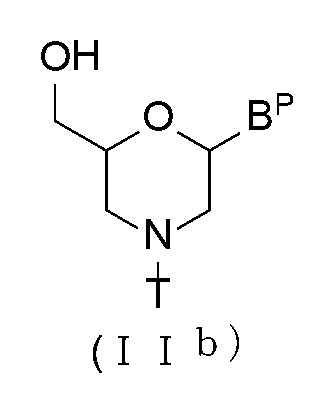
где BP и T имеют те же определенные выше значения.
[0065] В соединении (II), соединение с общей формулой (IIb2), показанное ниже, где n составляет от 2 до 99, а L является водородом, можно получить, используя Соединение (IIb) в качестве исходного материала и повторяя стадию А и стадию В способа получения РМО, описанного в настоящем описании изобретения, нужное число раз.
[Формула 17]
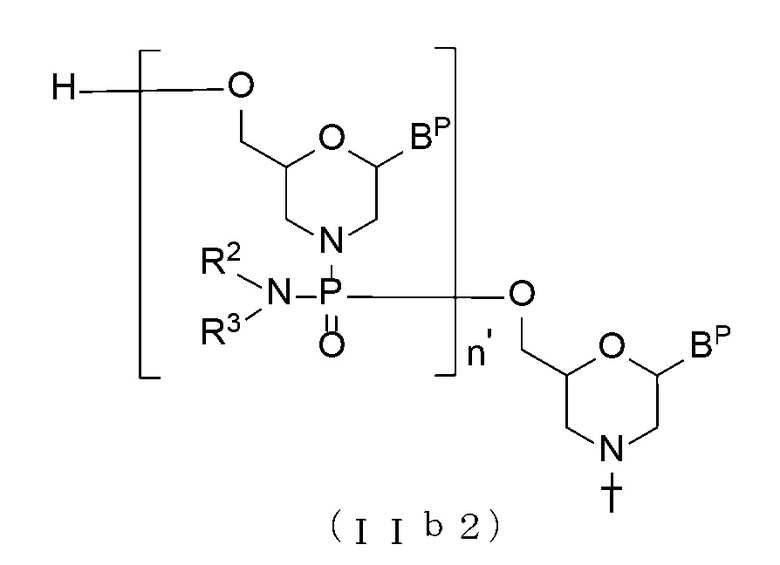
где BP, n', R2, R3 и T имеют те же определенные выше значения.
[0066] В соединении (II), соединение с общей формулой (IIс), показанное ниже, где n равно 1, а L является ацилом, можно получить, проводя процедуру, известную как реакция ацилирования, используя Соединение (IIb).
[Формула 18]
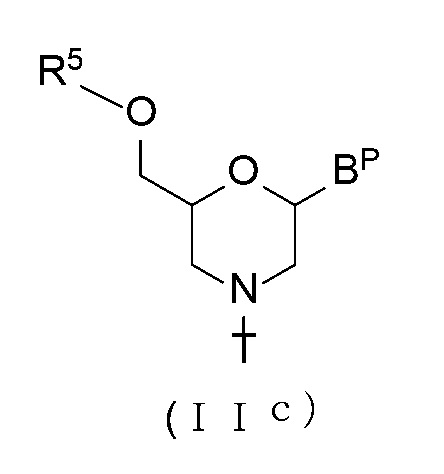
где BP и T имеют те же определенные выше значения; и
R5 представляет собой ацил.
[0067] В соединении (II), соединение с общей формулой (IIс2), показанное ниже, где n составляет от 2 до 99, а L является ацилом, можно получить, используя Соединение (IIс) в качестве исходного материала и повторяя стадию А и стадию В способа получения РМО, описанного здесь, нужное число раз.
[Формула 19]
где BP, n', R2, R3, R5 и T имеют те же определенные выше значения.
[0068] (2) Стадия B
Соединение (III) реагирует с морфолино-мономерным соединением в присутствии основания с образованием соединения, представленного общей формулой (VII) ниже (здесь и далее называемого Соединением (VII)):
[Формула 20]
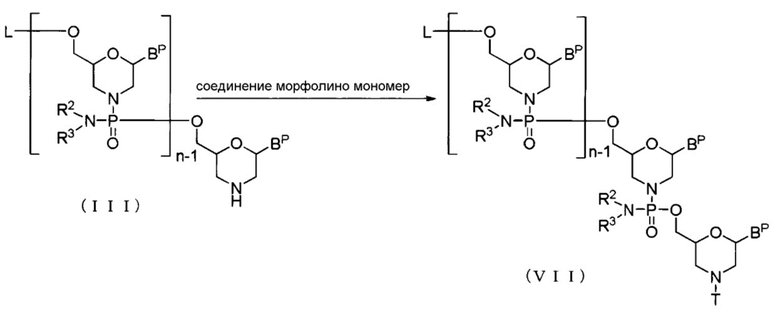
где BP, L, n, R2, R3 и T имеют те же определенные выше значения.
[0069] Эту стадию можно выполнить, проведя реакцию Соединения(III) с морфолино-мономерным соединением в присутствии основания.
[0070] Морфолино-мономерное соединение включает, например, соединения, представленные общей формулой (VIII) ниже:
[Формула 21]
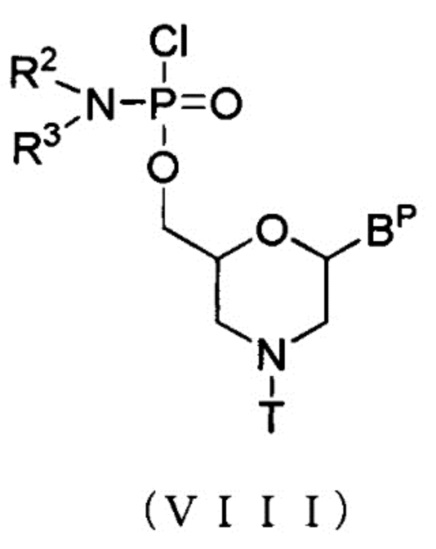
где BP, R2, R3 и T имеют те же определенные выше значения.
«Основание», которое можно применять на этой стадии, включает, например, диизопропиламин, триэтиламин и N-этилморфолин. Применяемое количество основания находится, соответственно, в диапазоне от 1 моль-эквивалента до 1000 моль-эквивалентов, если исходить из 1 моль Соединения (III), предпочтительно, от 10 моль-эквивалентов до 100 моль-эквивалентов, если исходить из 1 моль соединения (III).
Морфолино-мономерное соединение и основание, которые можно применять на этой стадии, можно также использовать в форме, разведенной в соответствующем растворителе, в концентрации, составляющей от 0,1% до 30%. Растворитель специально не ограничивается, когда речь идет об инертном по отношению к реакции растворителе, и включает, например, N,N-диметилимидазолидон, N-метилпиперидон, диметилформамид, дихлорметан, ацетонитрил, тетрагидрофуран или их смесь.
[0071] Предпочтительно, чтобы температура реакции находилась в диапазоне, например, от 0°С до 100°С, предпочтительнее, в диапазоне от 10°С до 50°С.
Время реакции может варьировать в зависимости от вида используемого основания и температуры реакции и находится, соответственно, в диапазоне, как правило, от 1 минуты до 48 часов, предпочтительно, в диапазоне от 30 минут до 24 часов.
[0072] Более того, после завершения этой стадии можно добавить ацилирующий агент в случае необходимости. «Ацилирующий агент» включает, например, уксусный ангидрид, ацетилхлорид и феноксиуксусный ангидрид. Ацилирующий агент можно также использовать в виде разведения соответствующим растворителем в концентрации, составляющей от 0,1% до 30%. Растворитель специально не ограничивается, когда речь идет об инертном по отношению к реакции растворителе, и включает, например, дихлорметан, ацетонитрил, спирт(ы) (этанол, изопропанол, трифторэтанол и т.д.), воду или их смесь.
При необходимости, основание, такое как пиридин, лутидин, коллидин, триэтиламин, диизопропилэтиламин, N-этилморфолин и т.д., можно также применять в комбинации с ацилирующим агентом. Количество ацилирующего агента находится, соответственно, в диапазоне от 0,1 моль-эквивалентов до 10000 моль-эквивалентов и, предпочтительно, в диапазоне от 1 моль-эквивалента до 1000 моль-эквивалентов. Количество основания находится, соответственно, в диапазоне, например, от 0,1 моль-эквивалентов до 100 моль-эквивалентов и, предпочтительно, в диапазоне от 1 моль-эквивалента до 10 моль-эквивалентов, если исходить из 1 моль ацилирующего агента.
Предпочтительно, чтобы температура этой реакции находилась в диапазоне от 10°С до 50°С, предпочтительнее, в диапазоне от 10°С до 50°С, еще более предпочтительно, в диапазоне от 20°С до 40°С и, наиболее предпочтительно, в диапазоне от 25°С до 35°С. Время реакции может варьировать в зависимости от вида используемого ацилирующего агента и температуры реакции и находится, соответственно, в диапазоне, как правило, от 0,1 минуты до 24 часов, предпочтительно, в диапазоне от 1 минуты до 5 часов.
[0073] (3) Стадия C:
В Соединении (VII), полученном на Стадии B, защитную группу удаляют, применяя агент для снятия защиты, с образованием соединения, представленного общей формулой (IX).
[Формула 22]
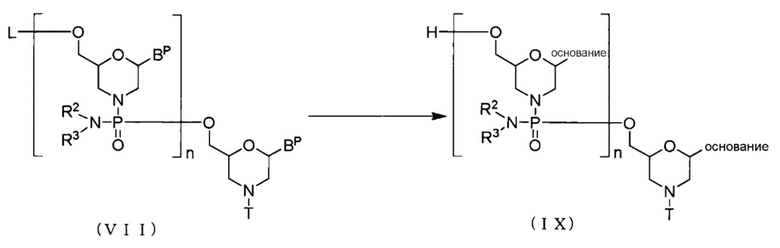
где основание, BP, L, n, R2, R3 и T имеют те же определенные выше значения.
[0074] Эту стадию можно выполнить, проведя реакцию Соединения(VII) с агентом для снятия защиты.
[0075] «Агент для снятия защиты» включает, например, концентрированный водный раствор аммиака и метиламин. «Агент для снятия защиты», применяемый на этой стадии, можно также применять в форме, разведенной, например, водой, метанолом, этанолом, изопропиловым спиртом, ацетонитрилом, тетрагидрофураном, диметилформамидом, N,N-диметилимидазолидоном, N-метилпиперидоном или смесью этих растворителей. Среди прочих предпочтительным является этанол. Применяемое количество агента для снятия защиты находится, соответственно, в диапазоне, например, от 1 моль-эквивалента до 100000 моль-эквивалентов и, предпочтительно, в диапазоне от 10 моль-эквивалентов до 1000 моль-эквивалентов, если исходить из 1 моль соединения (VII).
[0076] Предпочтительно, чтобы температура реакции находилась в диапазоне от 15°С до 75°С, предпочтительнее, в диапазоне от 40°С до 70°С, еще более предпочтительно, в диапазоне от 50°С до 60°С. Время реакции снятия защиты может варьировать в зависимости от вида используемого Соединения (VII), температуры реакции и т.п., и находится, соответственно, в диапазоне, как правило, от 10 минут до 30 часов, предпочтительно, в диапазоне от 30 минут до 24 часов и, еще более предпочтительно, в диапазоне от 5 часов до 20 часов.
[0077] (4) Стадия D:
PMO (I) получают, проводя реакцию Соединения (IX), образованного на стадии С, с кислотой:
[Формула 23]
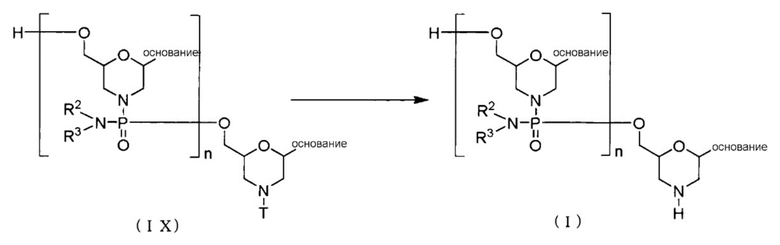
где основание, n, R2, R3 и T имеют те же определенные выше значения.
[0078] Эту стадию можно выполнить, добавив кислоту к Соединению(IX).
[0079] «Кислота», которую можно применять на этой стадии, включает, например, трихлоруксусную кислоту, дихлоруксусную кислоту, уксусную кислоту, фосфорную кислоту, соляную кислоту и т.д. Применяемая кислота используется надлежащим образом, чтобы обеспечивать рН раствора в диапазоне от 0,1 до 4,0 и, предпочтительнее, в диапазоне от 1,0 до 3,0. Растворитель специально не ограничивается, когда речь идет об инертном по отношению к реакции растворителе, и включает, например, ацетонитрил, воду или смесь этих растворителей.
[0080] Соответствующая температура реакции находится в диапазоне от 10°С до 50°С, предпочтительно, в диапазоне от 20°С до 40°С, еще более предпочтительно, в диапазоне от 25°С до 35°С. Время реакции снятия защиты может варьировать в зависимости от вида используемого Соединения (IX), температуры реакции и т.п, и находится, соответственно, в диапазоне от 0,1 минуты до 5 часов, предпочтительно, в диапазоне от 1 минуты до 1 часа и, еще более предпочтительно, в диапазоне от 1 минуты до 30 минут.
[0081] РМО (I) можно получить, проводя с реакционной смесью, полученной на этой стадии, разделение и очистку обычными средствами, таким как экстракция, концентрирование, нейтрализация, фильтрация, разделение центрифугированием, перекристаллизация, хроматография на колонке С8-С18 с обращенной фазой, катионообменная хроматография на колонке, анионообменная хроматография на колонке, гель-фильтрация на колонке, жидкостная хроматография высокого разрешения, диализ, ультрафильтрация и т.д., по отдельности или в сочетании. Таким образом, можно выделить и очистить желаемый РМО (I) (см., например, WO 1991/09033).
При очистке РМО (I) с использованием хроматографии с обращенной фазой в качестве растворителя для элюирования можно применять, например, смесь растворов 20 мМ триэтиламина/ацетатного буфера и ацетонитрила.
При очистке РМО (I) с использованием ионообменной хроматографии в качестве растворителя для элюирования можно применять, например, смесь растворов 1 М солевого раствора и 10 мМ водного раствора гидроксида натрия.
[0082] Описанная выше пептидная нуклеиновая кислота является олигомером настоящего изобретения, имеющим в качестве составной единицы группу, представленную следующей общей формулой:
[Формула 24]
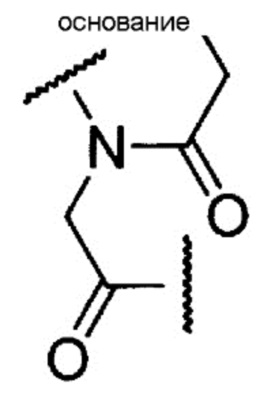
где «основание» имеет то же определенное выше значение.
Пептидо-нуклеиновые кислоты можно получить, в соответствии со следующей литературой.
1) P. E. Nielsen, M. Egholm, R. H. Berg, O. Buchardt Science, 254, 1497 (1991)
2) M. Egholm, O. Buchardt, P. E. Nielsen, R. H. Berg, Jacs., 114, 1895 (1992)
3) K. L. Dueholm, M. Egholm, C. Behrens, L. Christensen, H. F. Hansen, T. Vulpius, K. H. Petersen, R. H. Berg, P. E. Nielsen, O. Buchardt J. Org. Chem., 59, 5767 (1994)
4) L. Christensen, R. Fitzpatrick, B. Gildea, K. H. Petersen, H. F. Hansen, T. Koch, M. Egholm, O. Buchardt, P. E. Nielsen, J.
Coull, R. H. Berg, J. Pept. Sci., 1, 175 (1995)
5) T. Koch, H. F. Hansen, P. Andersen, T. Larsen, H. G. Batz, K. Otteson, H. Orum, J. Pept. Res., 49, 80 (1997)
[0083] В олигомере настоящего изобретения 5'-конец может быть любой из химических структур (1)-(3) ниже и, предпочтительно, является (3)-ОН.
[Формула 25]
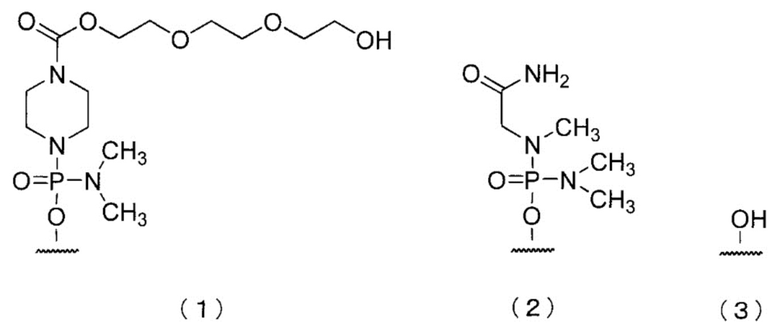
Здесь и далее группы, представленные (1), (2) и (3) выше, называют «группой (1)», «группой (2)» и «группой (3)» соответственно.
[0084] 2. Фармацевтическая композиция
Олигомер настоящего изобретения вызывает пропуск экзона 44 в гене дистрофина. Поэтому ожидается, что состояние мышечной дистрофии можно облегчить посредством введения фармацевтической композиции, содержащей олигомер настоящего изобретения, пациентам с МДД с мутацией, являющейся мишенью пропуска экзона 44, при которой восстанавливается рамка считывания при пропуске экзона 44.
Также процесс получения олигомера настоящего изобретения, длина которого является небольшой, является простым, а стоимость получения олигомера настоящего изобретения можно снизить.
В другом варианте осуществления настоящее изобретение представляет фармацевтическую композицию для лечения мышечной дистрофии, включающую в качестве активного ингредиента олигомер настоящего изобретения, его фармацевтически приемлемую соль или гидрат (здесь и далее называемую композицией настоящего изобретения).
[0085] Примерами фармацевтически приемлемой соли олигомера настоящего изобретения, содержащегося в композиции настоящего изобретения, являются соли щелочных металлов, такие как соли натрия, калия и лития; соли щелочноземельных металлов, такие как соли кальция и магния; соли металлов, такие как соли алюминия, железа, цинка, меди, никеля, кобальта и т.д., аммонийные соли, соли органических аминов, такие как соли трет-октиламина, дибензиламина, морфолина, глюкозамина, алкилового эфира фенилглицина, этилендиамина, N-метилглюкамина, гуанидина, диэтиламина, триэтиламина, дициклогексиламина, N,N'-дибензилэтилендиамина, хлорпрокаина, прокаина, диэтаноламина, N-бензилфенэтиламина, пиперазина, тетраметиламмония, трис(гидроксиметил)аминометана, соли гидрогалогенидов, такие как соли гидрофторатов, гидрохлоридов, гидробромидов и гидроиодидов, соли неорганических кислот, такие как нитраты, перхлораты, сульфаты, фосфаты и т.д., (низший алкан)сульфонаты, такие как метансульфонаты, трифторметансульфонаты и этансульфонаты, арилсульфонаты, такие как бензолсульфонаты и пара-толуолсульфонаты, соли органических кислот, такие как ацетаты, малаты, фумараты, сукцинаты, цитраты, тартраты, оксалаты, малеаты и т.д., и соли аминокислот, такие как соли глицина, лизина, аргинина, орнитина, глютаминовой кислоты и аспарагиновой кислоты. Эти соли можно получить известными способами. В альтернативном случае олигомер настоящего изобретения, содержащийся в композиции настоящего изобретения, может быть в форме его гидрата.
[0086] Способ введения композиции настоящего изобретения специально не ограничивается при условии, что он является фармацевтически приемлемым способом введения, и может быть выбран в зависимости от способа лечения. Ввиду легкости доставки в мышечные ткани предпочтительными являются внутривенное введение, внутриартериальное введение, внутримышечное введение, подкожное введение, пероральное введение, введение в ткани, чрескожное введение и т.д. Также лекарственные формы, подходящие для композиции настоящего изобретения, специально не ограничиваются и включают, например, различные инъекции, пероральные агенты, капли, ингаляции, мази, лосьоны и т.д.
[0087] При введении олигомера настоящего изобретения пациентам с мышечной дистрофией предпочтительно, чтобы композиция настоящего изобретения содержала носитель, способствующий доставке олигомера в мышечные ткани. Такой носитель специально не ограничивается, когда речь идет о фармацевтически приемлемом носителе, и примеры включают катионные носители, такие как катионные липосомы, катионные полимеры и т.д., или носители с использованием оболочки вируса. Катионными липосомами являются, например, липосомы, состоящие из 2-О-(2-диэтиламиноэтил)карабамоил-1,3-О-диолеоилглицерина и фосфолипидов в качестве основных составляющих (в дальнейшем называемые «липосомами А»), Oligofectamine (зарегистрированный товарный знак) (производства Invitrogen Corp.), Lipofectin (зарегистрированный товарный знак) (производства Invitrogen Corp.), Lipofectamine (зарегистрированный товарный знак) (производства Invitrogen Corp.), Lipofectamine 2000 (зарегистрированный товарный знак) (производства Invitrogen Corp.), DMRIE-C (зарегистрированный товарный знак) (производства Invitrogen Corp.), GeneSilencer (зарегистрированный товарный знак) (производства Gene Therapy Systems), TransMessenger (зарегистрированный товарный знак) (производства QIAGEN, Inc.), TransIT TKO (зарегистрированный товарный знак) (производства Mirus) и Nucleofector II (Lonza). Среди прочих предпочтительными являются липосомы А. Примерами катионных полимеров являются JetSI (зарегистрированный товарный знак) (производства Qbiogene, Inc.) и Jet-PEI (зарегистрированный товарный знак) (полиэтиленимин, производства Qbiogene, Inc.). Примером носителей с использованием оболочки вируса является GenomeOne (зарегистрированный товарный знак) (липосома HVJ-E, производства Ishihara Sangyo). В альтернативном случае могут также использоваться медицинские изделия, описанные в патенте Японии с № 2924179, и катионные носители, описанные в отечественных повторных публикациях в Японии РСТ-заявок с № 2006/129594 и 2008/096690.
[0088] Концентрация олигомера настоящего изобретения, содержащегося в композиции настоящего изобретения, может варьировать в зависимости от вида носителя и т.д. и находится, соответственно, в диапазоне от 0,1 нМ до 100 мкМ, предпочтительно, в диапазоне от 1 нМ до 10 мкМ, и, предпочтительнее, в диапазоне от 10 нМ до 1 мкМ. Весовое соотношение олигомера настоящего изобретения, содержащегося в композиции настоящего изобретения, и носителя (носитель/антисмысловой олигомер настоящего изобретения) может варьировать в зависимости от свойств олигомера, типа носителя и т.д. и находится, соответственно, в диапазоне от 0,1 до 100, предпочтительно, в диапазоне от 1 до 50, и, предпочтительнее, в диапазоне от 10 до 20.
[0089] Помимо олигомера настоящего изобретения и носителя, описанного выше, при желании в состав композиции настоящего изобретения могут входить фармацевтически приемлемые добавки. Примерами таких добавок являются вспомогательные средства для эмульгирования (например, жирные кислоты, содержащие 6-22 атомов углерода, и их фармацевтически приемлемые соли, альбумин и декстран), стабилизаторы (например, холестерин и фосфатидная кислота), агенты для придания изотоничности (например, хлорид натрия, глюкоза, мальтоза, лактоза, сахароза, трегалоза) и агенты для контроля рН (например, соляная кислота, серная кислота, фосфорная кислота, уксусная кислота, гидроксид натрия, гидроксид калия и триэтаноламин). Можно применять одну или несколько таких добавок. Содержание добавки в композиции настоящего изобретения составляет 90% по весу или менее, предпочтительно, 70% по весу или менее и, более предпочтительно, 50% по весу или менее.).
[0090] Композицию настоящего изобретения можно получить посредством добавления олигомера настоящего изобретения к дисперсии носителя и надлежащего перемешивания смеси. Добавки можно добавлять на соответствующей стадии либо до, либо после добавления олигомера настоящего изобретения. Содержащий воду растворитель, который можно применять при добавлении олигомера настоящего изобретения, специально не ограничивается, когда речь идет о фармацевтически приемлемом растворителе, и примерами являются вода для инъекции или дистиллированная вода для инъекции, раствор электролитов, такой как физиологический солевой раствор и т.д., и раствор сахара, такой как раствор глюкозы, раствор мальтозы и т.д. В этом случае специалист в данной области техники может соответствующим образом подобрать условия рН и температуры.
[0091] Композицию настоящего изобретения можно приготовить, например, в виде жидкой формы и ее лиофилизированного препарата. Лиофилизированный препарат можно приготовить с помощью лиофилизации композиции настоящего изобретения в жидкой форме обычным способом. Лиофилизацию можно провести, например, посредством надлежащей стерилизации композиции настоящего изобретения, находящейся в жидкой форме, распределения аликвот по контейнерам-флаконам, выполнения предварительного замораживания в течение 2 часов в условиях приблизительно от -40°С до -20°С, первичного высушивания при 0-10°С при пониженном давлении, а затем вторичного высушивания при приблизительно 15-25°С при пониженном давлении. Как правило, лиофилизированный препарат композиции настоящего изобретения можно получить с помощью замещения содержимого флакона азотом и закупоривания.
[0092] Как правило, лиофилизированный препарат композиции настоящего изобретения можно применять после восстановления посредством добавления произвольного подходящего раствора (жидкости для восстановления) и перерастворения препарата. Такая жидкость для перерастворения включает воду для инъекции, физиологический солевой раствор и другие жидкости для инфузии. Объем жидкости для восстановления может варьировать в зависимости от намеченного применения и т.д. и специально не ограничивается, и подходящий объем превышает в 0,5-2 раза объем до лиофилизации или составляет не более 500 мл.
[0093] Желательно контролировать вводимую дозу композиции настоящего изобретения, принимая во внимание следующие факторы: тип и лекарственную форму содержащегося олигомера настоящего изобретения, состояние пациентов, в том числе возраст, вес тела и т.д., путь введения, характеристики и степень заболевания. Суточная доза, рассчитанная как количество антисмыслового олигомера настоящего изобретения, как правило, находится в интервале от 0,1 мг до 10 г/человека и, предпочтительно, от 1 мг до 1 г/человека. Этот интервал числовых значений может иногда меняться в зависимости от типа целевого заболевания, пути введения и молекулы-мишени. Следовательно, в каком-то случае может быть достаточной доза ниже этого интервала, и наоборот, иногда может потребоваться доза выше этого интервала. Композицию можно вводить от одного до нескольких раз в день или с интервалами от одного дня до нескольких дней.
[0094] В другом варианте применения композиции настоящего изобретения представлена фармацевтическая композиция, содержащая вектор, способный экспрессировать олигонуклеотид настоящего изобретения, и описанный выше носитель. Таким экспрессионным вектором может быть вектор, способный экспрессировать множество олигонуклеотидов настоящего изобретения. Композицию можно составлять в форме с фармацевтически приемлемыми добавками, как в случае композиции настоящего изобретения, содержащей олигомер настоящего изобретения. Концентрация экспрессионного вектора, содержащегося в композиции, может варьировать в зависимости от типа носителя и т.д. и находится, соответственно, в диапазоне от 0,1 нМ до 100 мкМ, предпочтительно, в диапазоне от 1 нМ до 10 мкМ, и, предпочтительнее, в диапазоне от 10 нМ до 1 мкМ. Весовое соотношение экспрессионного вектора, содержащегося в композиции, и носителя (носитель/экспрессионный вектор) может варьировать в зависимости от свойств экспрессионного вектора, типа носителя и т.д. и находится, соответственно, в диапазоне от 0,1 до 100, предпочтительно, в диапазоне от 1 до 50, и, предпочтительнее, в диапазоне от 10 до 20. Содержание носителя, входящего в состав композиции, является таким же, как и в случае композиции настоящего изобретения, содержащей олигомер настоящего изобретения, а способ получения фармацевтической композиции, включающей вектор, является таким же, как и в случае композиции настоящего изобретения.
[0095] Здесь и далее будет приведено более подробное описание настоящего изобретения со ссылкой на ПРИМЕРЫ и ТЕСТОВЫЕ ПРИМЕРЫ ниже, но они не предназначены для ограничения настоящего изобретения.
[ПРИМЕРЫ]
[0096] [СПРАВОЧНЫЙ ПРИМЕР 1]
4-{[(2S,6R)-6-(4-бензамидо-2-оксопиримидин-1-ил)-4-тритилморфолин-2-ил]метокси}-4-оксомасляная кислота, нагруженная на аминополистирольную смолу
Стадия 1: Получение 4-{[(2S,6R)-6-(4-бензамидо-2-оксопиримидин-1-ил)-4-тритилморфолин-2-ил]метокси}-4-оксомасляной кислоты
В атмосфере аргона 3,44 г N-{1-[(2R,6S)-6-(гидроксиметил)-4-тритилморфолин-2-ил]-2-оксо-1,2-дигидропиримидин-4-ил}бензамида и 1,1 г 4-диметиламинопиридина (4-DMAP, 4-dimethylaminopyridine) суспендировали в 50 мл дихлорметана, и к суспензии добавляли 0,90 г янтарного ангидрида с последующим перемешиванием при комнатной температуре в течение 3 часов. К реакционной смеси добавляли 10 мл метанола и концентрировали смесь при пониженном давлении. Остаток экстрагировали, используя этилацетат и 0,5 М водный раствор дигидрофосфата калия. Образованный в результате органический слой промывали последовательно 0,5 М водным раствором дигидрофосфата калия, водой и соляным раствором в указанном порядке. Образованный в результате органический слой сушили над сульфатом натрия и концентрировали при пониженном давлении с образованием 4,0 г продукта.
[0097] Стадия 2: Получение 4-{[(2S,6R)-6-(4-бензамидо-2-оксопиримидин-1-ил)-4-тритилморфолин-2-ил]метокси}-4-оксомасляной кислоты, нагруженной на аминополистирольную смолу
После растворения 4,0 г 4-{[(2S,6R)-6-(4-бензамидо-2-оксопиримидин-1(2Н)-ил)-4-тритилморфолин-2-ил]метокси}-4-оксомасляной кислоты в 200 мл пиридина (обезвоженного) к раствору добавляли 0,73 г 4-DMAP и 11,5 г 1-этил-3-(3-диметиламинопропил)карбодиимида гидрохлорида. Затем к смеси добавляли 25,0 г аминополистирольной смолы Primer support 200 amino (производства GE Healthcare Japan Co., Ltd., 17-5214-97), и 8,5 мл триэтиламина с последующим встряхиванием при комнатной температуре в течение 4 дней. После завершения реакции смолу отделяли с помощью фильтрации. Полученную в результате смолу последовательно промывали пиридином, метанолом и дихлорметаном в указанном порядке и сушили при пониженном давлении. К образованной в результате смоле добавляли 200 мл тетрагидрофурана (обезвоженного), 15 мл уксусного ангидрида и 15 мл 2,6-лутидина и смесь встряхивали при комнатной температуре в течение 2 часов. Смолу отделяли с помощью фильтрации, последовательно промывали пиридином, метанолом и дихлорметаном в указанном порядке и сушили при пониженном давлении с образованием 26,7 г продукта.
Величину привеса продукта определяли на основании молярного количества тритила на г смолы посредством измерения поглощения ультрафиолетовых лучей при 409 нм, используя известный способ. Величина привеса смолы составляла 192,2 мкмоль/г.
[0098] Условия измерения УФ
Прибор: U-2910 (Hitachi, Ltd.)
Растворитель: метансульфоновая кислота
Длина волны: 265 нм
Значение ε: 45000
[0099] [СПРАВОЧНЫЙ ПРИМЕР 2]
4-{[(2S, 6R)-6-(5-метил-2,4-диоксопиримидин-1-ил)-4-тритилморфолин-2-ил]метокси}-4-оксомасляная кислота, нагруженная на аминополистирольную смолу
Указанное в заголовке соединение получали схожим с описанным в СПРАВОЧНОМ ПРИМЕРЕ 1 способом, за исключением того, что на этой стадии использовали 1-[(2R,6S)-6-(гидроксиметил)-4-тритилморфолин-2-ил]-5-метилпиримидин-2,4(1H,3H)-дион вместо N-{1-[(2R,6S)-6-(гидроксиметил)-4-тритилморфолин-2-ил]-2-оксо-1,2-дигидропиримидин-4-ил}бензамида, применяемого на Стадии 1 в СПРАВОЧНОМ ПРИМЕРЕ 1.
Количество наносимого продукта определяли по молярному количеству тритила на грамм смолы, измеряя поглощение УФ при 409 нм, используя известный способ. Количество наносимой смолы составляло 164,0 мкмоль/г.
[0100] [СПРАВОЧНЫЙ ПРИМЕР 3]
4-{[(2S,6R)-6-(6-бензамидопурин-9-ил)-4-тритилморфолин-2-ил]метокси}-4-оксомасляная кислота, нагруженная на аминополистирольную смолу
Указанное в заголовке соединение получали схожим с описанным в СПРАВОЧНОМ ПРИМЕРЕ 1 способом, за исключением того, что на этой стадии использовали N-{9-[(2R,6S)-6-(гидроксиметил)-4-тритилморфолин-2-ил]-пурин-6-ил}бензамидо вместо N-{1-[(2R,6S)-6-(гидроксиметил)-4-тритилморфолин-2-ил]-2-оксо-1,2-дигидропиримидин-4-ил}бензамида, применяемого на Стадии 1 в СПРАВОЧНОМ ПРИМЕРЕ 1.
Количество наносимого продукта определяли по молярному количеству тритила на грамм смолы, измеряя поглощение УФ при 409 нм, используя известный способ. Количество наносимой смолы составляло 185,7 мкмоль/г.
[0101] [СПРАВОЧНЫЙ ПРИМЕР 4]
4-{{(2S,6R)-6-(6-2-цианоэтокси}-2-[(2-феноксиацетил)-амино]-пурин-9-ил}-4-тритилморфолин-2-ил]метокси}-4-оксомасляная кислота, нагруженная на аминополистирольную смолу
Указанное в заголовке соединение получали схожим с описанным в СПРАВОЧНОМ ПРИМЕРЕ 1 способом, за исключением того, что на этой стадии использовали N-{6-(2-цианоэтокси)-9-[(2R,6S)-6-(гидроксиметил)-4-тритилморфолин-2-ил]-пурин-2-ил}-2-феноксиацетамидо вместо N-{1-[(2R,6S)-6-(гидроксиметил)-4-тритилморфолин-2-ил]-2-оксо-1,2-дигидропиримидин-4-ил}бензамида, применяемого на Стадии 1 в СПРАВОЧНОМ ПРИМЕРЕ 1.
Количество наносимого продукта определяли по молярному количеству тритила на грамм смолы, измеряя поглощение УФ при 409 нм, используя известный способ. Количество наносимой смолы составляло 164,8 мкмоль/г.
[0102] В соответствии с описаниями в ПРИМЕРАХ 1 ниже, были синтезированы РМО, представленные РМО с № 1-118 в ТАБЛИЦЕ 1. Синтезированный РМО растворяли в воде для инъекции (производства Otsuka Pharmaceutical Factory, Inc.).
[0103] [Таблица 1-1]
[ТАБЛИЦА 1]
3'-конец: ацетилирование
[0104]
[ПРИМЕР 1]
В качестве основания на 5'-конце на колонку с фильтром наносили 0,2 г 4-{[(2S,6R)-6-(4-бензамид-2-оксопиримидин-1(2H)-ил)-4-тритилморфолин-2-ил]метокси}-4-оксомасляной кислоты, нагруженной на аминополистирольную смолу (Справочный пример 1), 4-{[(2S,6R)-6-(5-метил-2,4-диоксопиримидин-1-ил)-4-тритилморфолин-2-ил]-метокси}-4-оксомасляной кислоты, нагруженной на аминополистирольную смолу (Справочный пример 2), 4-{[(2S, 6R)-6-(6-бензамидопурин-9-ил)-4-тритилморфолин-2-ил]-метокси}-4-оксомасляной кислоты, нагруженной на аминополистирольную смолу (Справочный пример 3), или 4-{{(2S, 6R)-6-{6-(2-цианоэтокси)-2-[(2-феноксиацетил)амино]-пурин-9-ил}-4-тритилморфолин-2-ил]-метокси}-4-оксомасляной кислоты, нагруженной на аминополистирольную смолу (Справочный пример 4). Затем начинали синтетический цикл, показанный ниже, используя прибор для синтеза нуклеиновых кислот (AKTA Oligopilot 10 plus). Заданное морфолино-мономерное соединение добавляли на каждом цикле соединения с образованием последовательности оснований, описанной в Таблице 1 (см. Таблицу 2 ниже).
[0105] [ТАБЛИЦА 2]
Примечание: Стадии 1, 2, 7 и 8 проводили снова после финального цикла только в случае ацетилирования 3'-конца.
[0106] Применяемым раствором для снятия защиты был раствор дихлорметана, содержащий 3% (вес/объем) трифторуксусной кислоты. Применяемым раствором для нейтрализации и промывки был раствор, полученный в результате растворения N,N-диизопропилэтиламина до конечной концентрации 10% (объем/объем) и тетрагидрофурана до 5% (объем/объем) в дихлорметане, содержащем 35% (объем/объем) ацетонитрила. Применяемым раствором А для соединения был раствор, полученный в результате растворения морфолино-мономерного соединения в тетрагидрофуране до концентрации 0,10 М. Применяемым раствором В для соединения был раствор, полученный в результате растворения N,N-диизопропилэтиламина до концентрации 20% (объем/объем) и тетрагидрофурана до концентрации 10% (объем/объем) в ацетонитриле. Применяемым раствором для кэпирования был раствор, полученный в результате растворения 20% (объем/объем) уксусного ангидрида и 30% (объем/объем) 2,6-лутидина в ацетонитриле.
[0107] Аминополистирольную смолу, нагруженную синтезированным выше РМО, извлекали из реакционного сосуда и сушили при комнатной температуре в течение по крайней мере 2 часов при пониженном давлении. Высушенный РМО, нагруженный на аминополистирольную смолу, загружали в реакционный сосуд и к нему добавляли 5 мл смеси 28% водного раствора аммиака-этанола (1/4). Смесь перемешивали при 55°С в течение 15 часов. Аминополистирольную смолу отделяли с помощью фильтрации и промывали 1 мл смеси воды-этанола (1/4). Образованный в результате фильтрат концентрировали при пониженном давлении. Образованный в результате остаток растворяли в 10 мл смеси растворителей, состоящей из 20 мМ буфера уксусная кислота - триэтиламин (буфера ТЕАА) и ацетонитрила, (4/1) и фильтровали через мембранный фильтр. Полученный фильтрат очищали с помощью ВЭЖХ с обращенной фазой. Применяли условия, показанные в Таблице 3 ниже.
[0108] [ТАБЛИЦА 3]
[0109] Исследовали каждую фракцию, целевой продукт восстанавливали и концентрировали при пониженном давлении. К концентрированному остатку добавляли 0,5 мл 2М водного раствора фосфорной кислоты и перемешивали смесь в течение 15 минут. Затем добавляли 2 мл 2М водного раствора гидроксида натрия для подщелачивания смеси, а затем фильтровали через мембранный фильтр (0,45 мкм).
Образованный в результате водный раствор, содержащий целевой продукт, очищали с помощью колонки с анионообменной смолой. Применяли условия, показанные в Таблице 4 ниже.
[0110] [ТАБЛИЦА 4]
[0111] Каждую фракцию исследовали (с помощью ВЭЖХ) и целевой продукт получали в виде водного раствора. К образованному в результате водному раствору добавляли 0,1 М фосфатного буфера (рН 6,0) для нейтрализации. Затем полученную смесь обессоливали с помощью обращено-фазовой ВЭЖХ в условиях, описанных в Таблице 5 ниже.
[0112] [ТАБЛИЦА 5]
[0113] Целевой продукт восстанавливали и проводили концентрирование при пониженном давлении. Образованный в результате остаток растворяли в воде. Полученный водный раствор лиофилизировали с образованием целевого соединения в виде белого, похожего на хлопок твердого вещества.
Рассчитанные значения и экспериментальные значения, полученные с помощью ESI-TOF-MS (время-пролетной масс-спектрометрии с ионизацией электрораспылением) представлены в Таблице 6 ниже.
[Талица 6-1]
[ТАБЛИЦА 6]
[0114] [ТЕСТОВЫЙ ПРИМЕР 1]
Исследование in vitro
С помощью набора для нуклеофекции клеточных линий Amaxa Cell Line Nucleofector Kit L на Nucleofector II (Lonza) от 0,1 до 30 мкМ антисмысловых олигомеров из Таблицы 1 трансфицировали 3,5×105 клеток RD (клеточная линия рабдомиосаркомы человека). Применяли программу Program T-030.
После трансфекции клетки культивировали в течение трех суток в 2 мл минимальной питательной среды Игла (EMEM, Eagle's minimal essential medium) (производства Sigma, здесь и далее та же самая), содержащей 10% фетальной бычьей сыворотки (FCS, fetal calf serum) (производства Invitrogen) при 37°C и 5% CO2.
Клетки однократно промывали фосфатно-солевым буфером (производства Nissui, здесь и далее тот же самый) и добавляли к клеткам 350 мкл буфера RLT (производства Qiagen), содержащего 1% 2-меркаптоэтанола (производства Nacalai Tesque). После инкубации клеток при комнатной температуре в течение нескольких минут для проведения лизиса клеток лизат собирали в гомогенизатор QIAshredder (производства Qiagen). Затем лизат центрифугировали при 15000 об/мин в течение 2 минут для получения гомогената. Общую РНК экстрагировали в соответствии с протоколом, приложенным к набору RNeasy Mini Kit (производства Qiagen). Концентрацию экстрагированной общей РНК определяли с помощью NanoDrop ND-1000 (производства LMS).
[0115] Проводили одностадийную ОТ-ПЦР с 400 нг экстрагированной общей РНК с помощью набора для ОТ-ПЦР Qiagen One Step RT-PCR Kit (производства Qiagen). Реакционный раствор готовили в соответствии с протоколом, приложенным к набору. PTC-100 (производства MJ Research) или TaKaRa PCR Thermal Cycler Dice Touch (производства Takara Bio) применяли в качестве термоциклера. Применяли следующую программу ОТ-ПЦР.
50°C, 30 мин: реакция обратной транскрипции
95°C, 15 мин: активация полимеразы, инактивация обратной транскриптазы, тепловая денатурация кДНК
[94°C, 30 секунд; 60°C, 30 секунд; 72°C, 1 минута]×35 циклов: ПЦР-амплификация
72°C, 10 мин: конечная элонгация
[0116] Нуклеотидные последовательности прямого праймера и обратного праймера, применяемых для ОТ-ПЦР, приведены ниже.
Прямой праймер 5'- GCTCAGGTCGGATTGACATT -3' (SEQ ID NO: 125)
Обратный праймер 5'- GGGCAACTCTTCCACCAGTA -3' (SEQ ID NO: 126)
[0117] Продукт реакции, 1 мкл описанной выше ПЦР, анализировали с помощью Bioanalyzer (производства Agilent Technologies, Inc.).
Измеряли уровень «А» полинуклеотидной полосы при пропуске экзона 44, и уровень «В» полинуклеотидной полосы без пропуска экзона 44. На основании этих измерений значений «A» и «B» определяли эффективность пропуска по следующему уравнению:
Эффективность пропуска (%)=A/(A+B)×100
[0118] Результаты эксперимента
Результаты показаны на Фиг.1-26. Этот эксперимент показал, что олигомер настоящего изобретения, полученный посредством соединения коротких частей олигомера, выбранных из оснований в положениях с -1 по 44 (SEQ ID NO: 1) и в положениях с 58 по 115 (SEQ ID NO: 2), соответственно, с 5'-конца нуклеотидной последовательности экзона 44 (SEQ ID NO: 10) в человеческом гене дистрофина дикого типа настоящего изобретения, может эффективно индуцировать пропуск экзона 44.
[0119] [ТЕСТОВЫЙ ПРИМЕР 2]
Исследование in vitro
Эксперимент проводили, как показано в ТЕСТОВОМ ПРИМЕРЕ 1, за исключением того, что 3,5×105 клеток RD (клеточная линия рабдомиосаркомы человека) трансфицировали олигомерами настоящего изобретения PMO № 34, 100, 45, 73, 49 и 47, каждый из которых применяли в индивидуальном виде или в форме, где две части олигомера, входящие в состав каждого олигомера, находились в индивидуальном виде или в виде смеси в концентрации 1, 3 или 10 мкМ с применением набора для нуклеофекции клеточных линий Amaxa Cell Line Nucleofector Kit L на Nucleofector II (Lonza). Применяли программу Program T-030. Комбинации последовательностей для трансфекции указаны ниже.
[ТАБЛИЦА 7]
Комбинация трансфицированных последовательностей
[0120] Результаты эксперимента
Результаты показаны на Фиг.27-31. Этот эксперимент показал, что ни один из PMO № с 110 по 115, PMO № 117 и PMO № 118, действующих на сайт в экзоне 44, не смог самостоятельно вызвать пропуск экзона 44. Этот эксперимент также показал, что по сравнению со смесями двух антисмысловых нуклеиновых кислот, действующих на разные сайты в экзоне 44 (смесью PMO № 114 и PMO № 115; смесью PMO №109 и PMO № 114; смесью PMO № 110 и PMO № 111; смесью PMO № 112 и PMO № 113; смесью PMO № 117 и PMO № 118 и смесью PMO №119 и PMO № 120) олигомеры настоящего изобретения PMO № 34, PMO № 100, PMO № 45, PMO № 73, PMO № 49 и PMO № 47, каждая соответствующая часть олигомера соединена с другой частью, вызывают пропуск экзона 44 с высокой эффективностью.
[0121] [ТЕСТОВЫЙ ПРИМЕР 3]
Исследование in vitro, проведенное на фибробластах человека
Пропускающую экзон 44 активность определяли с использованием клеток GM05112 (человеческих фибробластов, полученных от пациента и МДД с делецией экзона 45, Coriell Institute for Medical Research). В качестве ростовой среды использовали среду Игла, модифицированную по способу Дульбекко, и питательную смесь F-12 (DMEM/F-12, Dulbecco's Modified Eagle Medium:F-12) (Life Technologies), содержащую 10% FCS (HyClone Laboratories, Inc.) и 1% пенициллина/стрептомицина (P/S) (Sigma-Aldrich, Inc.), и культивировали клетки при 37°C и 5% CO2.
[0122] Клетки культивировали во флаконах T225 и к 30 мл ростовой среды добавляли 2,5 мл ретровируса (коэкспрессия ZsGreen1), экспрессирующего человеческий myoD (SEQ ID NO: 127), иполибрен до конечной концентрации 8 мкг/мл (Sigma-Aldrich, Inc.). После инкубации при 32°C в течение 2 дней среду заменяли свежей ростовой средой и продолжали инкубацию при 37°C в течение 3 дней. ZsGreen1-положительные трансформированные MyoD фибробласты собирали при помощи клеточного сортера BD FACSAria Cell Sorter (BD Bioscience). Отобранные клетки суспендировали в среде для дифференцировки (DMEM/F-12, содержащей 2% лошадиной сыворотки (LifeTechnologies), 1% P/S и жидкой добавки инсулина-трансферрина-селенита натрия (ITS Liquid Media Supplement) (Sigma-Aldrich, Inc.)), и рассаживали в количестве 9,4×104 клеток в лунке в покрытый коллагеном 24-луночный планшет. Среду меняли каждые 2-3 дня и продолжали инкубацию до дифференцировки в миотрубочки.
[0123] На 7 день после рассаживания в 24-луночный планшет среду заменяли средой для дифференцировки и добавляли олигомеры PMO № 34, 45, 49 и 73 в конечной концентрации 10 мкМ. После инкубации клеток в течение 2 дней среду заменяли средой для дифференцировки без PMO и инкубировали клетки еще пять дней. Затем клетки собирали для экстрагирования общей РНК с помощью набора RNeasy Mini Kit (QIAGEN). ОТ_ПЦР проводили с 50 нг экстрагированной общей РНК, применяя набор для ОТ-ПЦР QIAGEN OneStep RT-PCR Kit. Реакционный раствор готовили в соответствии с протоколом, приложенным к набору. iCycler (производства Bio-Rad Laboratories) применяли в качестве термоциклера. Применяли следующую программу ОТ-ПЦР.
50°C, 30 мин: реакция обратной транскрипции
95°C, 15 мин: активация полимеразы, инактивация обратной транскриптазы, тепловая денатурация кДНК
[94°C, 1 минута; 60°C, 1 минута; 72°C, 1 минута]×35 циклов: ПЦР-амплификация
72°C, 7 мин: конечная элонгация.
[0124] Нуклеотидные последовательности прямого праймера и обратного праймера, применяемых для ОТ-ПЦР, приведены ниже.
Прямой праймер 5'- GCTCAGGTCGGATTGACATT -3' (SEQ ID NO: 125)
Обратный праймер 5'- GGGCAACTCTTCCACCAGTA -3' (SEQ ID NO: 126)
[0125] 1 мкл ПЦР-продукта анализировали с помощью наборов Experion DNA 1K Analysis Kits (Bio-Rad Laboratories) с помощью установки для электрофореза Experion Electrophoresis Station (Bio-Rad Laboratories). Исследование DNA 1K assay выбирали в программе Experion Software версии 3.2 (Bio-Rad Laboratories) и проводили измерение. Определяли уровень (А) полосы около 317 п.о. и уровень (В) полосы около 465 п.о. (единицы: нмоль/л). Эффективность пропуска (%) определяли по следующему уравнению, применяя Excel 2007 SP3 (Microsoft):
Эффективность пропуска (%)=A/(A+B)×100
[0126] Результаты эксперимента
Результаты показаны на Фиг. 32. Этот эксперимент показал, что антисмысловые олигомеры настоящего изобретения PMO № 34, 45, 49 и 73 смогли вызвать пропуск экзона 44 с высокой эффективностью в клетках, полученных от пациента с МДД с делецией экзона 45.
ПРОМЫШЛЕННАЯ ПРИМЕНИМОСТЬ
[0127] Результаты экспериментов, приведенные в ТЕСТОВЫХ ПРИМЕРАХ, показывают, что олигомеры настоящего изобретения, в которых соединены короткие олигомеры, вызывали пропуск экзона 44 в клетках RD. Таким образом, олигомеры настоящего изобретения являются особенно полезными для лечения МДД.
[0128] Список последовательностей в произвольной форме
[0129]
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
--->
<110> NIPPON SHINYAKU CO., LTD.
NATIONAL CENTER OF NEUROLOGY AND PSYCHIATRY
<120> Антисмысловая нуклеиновая кислота
<130> G1076WO
<150> JP 2014-124157
<151> 2014-06-17
<160> 128
<170> PatentIn версия 3.5
<210> 1
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 1
ggcgatttga cagatctgtt gagaaatggc ggcgttttca ttatg 45
<210> 2
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 2
taatcagtgg ctaacagaag ctgaacagtt tctcagaaag acacaaattc ctgagaat 58
<210> 3
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 3
ttgagaaatg gcggcgt 17
<210> 4
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 4
tcagtggcta acagaag 17
<210> 5
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 5
tctcagaaag acacaaa 17
<210> 6
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 6
gtgtctttct gagccgccat ttctca 26
<210> 7
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 7
tctgttagcc acgccgccat tt 22
<210> 8
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 8
ctgttagcca ctcgccgcca tttc 24
<210> 9
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 9
ttgtgtcttt cttctgttag ccac 24
<210> 10
<211> 148
<212> ДНК
<213> Homo sapiens
<400> 10
gcgatttgac agatctgttg agaaatggcg gcgttttcat tatgatataa agatatttaa 60
tcagtggcta acagaagctg aacagtttct cagaaagaca caaattcctg agaattggga 120
acatgctaaa tacaaatggt atcttaag 148
<210> 11
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 11
ttgtgtcttt ctgtctcaac agatct 26
<210> 12
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 12
tgttagccac tgatctcaac agatct 26
<210> 13
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 13
gttcagcttc tgttctcaac agatct 26
<210> 14
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 14
ttgtgtcttt ctgcgccgcc atttct 26
<210> 15
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 15
ctgagaaact gttcgccgcc atttct 26
<210> 16
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 16
tctcaggaat ttgcgccgcc atttct 26
<210> 17
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 17
tgttagccac tgacgccgcc atttct 26
<210> 18
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 18
gttcagcttc tgtcgccgcc atttct 26
<210> 19
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 19
attctcagga atttgaacgc cgccatttct 30
<210> 20
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 20
atttgtgtct ttctgtttct caacagatct 30
<210> 21
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 21
attctcagga atttgtttct caaca 25
<210> 22
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 22
atttgtgtct ttctgaacgc cgccatttct 30
<210> 23
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 23
tctcaggaat ttgtctcaac agatct 26
<210> 24
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 24
ttgtgtcttt ctgaacgccg ccattt 26
<210> 25
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 25
ttgtgtcttt ctgccgccat ttctca 26
<210> 26
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 26
gtgtctttct gagcgccgcc atttct 26
<210> 27
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 27
ctgagaaact gtttctcaac agatct 26
<210> 28
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 28
atttgtgtct ttcccgccat ttctca 26
<210> 29
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 29
gtgtctttct gagaacgccg ccattt 26
<210> 30
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 30
atttgtgtct ttcaacgccg ccattt 26
<210> 31
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 31
ttgtgtcttt ctcgccgcca tttc 24
<210> 32
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 32
cgccgccatt tctttgtgtc tttctg 26
<210> 33
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 33
tgtgtctttc tgccgccatt tc 22
<210> 34
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 34
tgtgtctttc ccgccatttc 20
<210> 35
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 35
ttagccactg attccgccat ttctca 26
<210> 36
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 36
atttgtgtct ttccgccgcc atttct 26
<210> 37
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 37
tctgttagcc actcgccgcc atttct 26
<210> 38
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 38
tgttagccac tgaccgccat ttctca 26
<210> 39
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 39
tctgttagcc actccgccat ttctca 26
<210> 40
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 40
ttagccactg attcgccgcc atttct 26
<210> 41
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 41
tgttagccac tgaaacgccg ccattt 26
<210> 42
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 42
tctgttagcc actaacgccg ccattt 26
<210> 43
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 43
ttagccactg attaacgccg ccattt 26
<210> 44
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 44
ttgtgtcttt ctgtgttagc cactga 26
<210> 45
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 45
tgttcagctt ctgtgttagc cactga 26
<210> 46
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 46
ttagccactg cgccgccatt 20
<210> 47
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 47
ctgttagcca cgccgccatt 20
<210> 48
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 48
tgttcagctt tagccactga 20
<210> 49
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 49
ctgttagcca ccgccatttc 20
<210> 50
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 50
ttgtgtcttt ctggccattt ctcaac 26
<210> 51
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 51
ctgttagcca ctgcataatg aaaac 25
<210> 52
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 52
ttctgttagc caaaaacgcc gcca 24
<210> 53
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 53
ttagccactg acgccgccat tt 22
<210> 54
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 54
ctgttagcca cgccgccatt tc 22
<210> 55
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 55
actgttcagc gccactgatt 20
<210> 56
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 56
aactgttcag ctttagccac tgatta 26
<210> 57
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 57
ctgagaaact gtttgttagc cactga 26
<210> 58
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 58
aacgccgcca tttgtgtctt tc 22
<210> 59
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 59
tgtgtctttc tgttagccat ttctcaac 28
<210> 60
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 60
cgccgccatt tcttgtgtct ttct 24
<210> 61
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 61
ccgccatttc tcattgtgtc tttctg 26
<210> 62
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 62
ccgccatttc tcatgttagc cactga 26
<210> 63
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 63
ctttctgaga aactgttagc cactga 26
<210> 64
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 64
gccactgacc tctcagcttc tgtta 25
<210> 65
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 65
aacgccgcca tttatttgtg tctttc 26
<210> 66
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 66
aaacgccgcc atttgtgtct ttctga 26
<210> 67
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 67
ctttctgtta gccactgaac 20
<210> 68
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 68
cgccgccatt tcttttctga gaaact 26
<210> 69
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 69
aacgccgcca tttctgagaa actgtt 26
<210> 70
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 70
cccagcttct gttagccact gacc 24
<210> 71
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 71
cttgcttctg ttagccactg accc 24
<210> 72
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 72
cgccgccatt tctgagaaac t 21
<210> 73
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 73
ccgccatttc tcagtgtctt tctgag 26
<210> 74
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 74
tgttagccac tgattgtgtc tttctg 26
<210> 75
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 75
atttgtgtct ttctgttagc cactga 26
<210> 76
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 76
gtctttctga gcgccatttc tc 22
<210> 77
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 77
ttgtgtcttt cctgttagcc ac 22
<210> 78
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 78
tgtctttctg agccgccatt tctc 24
<210> 79
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 79
aaactgttca gcttctgtca aatcgc 26
<210> 80
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 80
acgccgccat ttctcaggaa tttgtg 26
<210> 81
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 81
ctgttcagct ccactgatta 20
<210> 82
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 82
actgttcagc ccactgatta 20
<210> 83
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 83
tgtgtctttc tcgccgccat tt 22
<210> 84
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 84
tgtctttctg acgccatttc tc 22
<210> 85
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 85
cgccgccatt tctttgtgtc tttc 24
<210> 86
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 86
cgccgccatt tttgtgtctt tc 22
<210> 87
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 87
ctgttcagct tgttagccac 20
<210> 88
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 88
ttctgagaaa ctgttagcca c 21
<210> 89
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 89
ctgttcagcc actgatta 18
<210> 90
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 90
ctgagaaact tgttagccac 20
<210> 91
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 91
tgtctttctg accgccattt ct 22
<210> 92
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 92
ctttctgaga acgccatttc tc 22
<210> 93
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 93
ctttctgaga aacgccgcc 19
<210> 94
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 94
tgtctttctg ttagccattt ctc 23
<210> 95
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 95
ctttctgtta gccatttctc 20
<210> 96
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 96
ctttctgaga gccatttctc 20
<210> 97
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 97
ctgttagcca ctccgccatt tc 22
<210> 98
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 98
ctgtgtcttt ctgttagcca c 21
<210> 99
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 99
ctgttcagcg ccactgatta c 21
<210> 100
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 100
ctttctgaga aacgccgcca tttc 24
<210> 101
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 101
tgtctttctg agccgccatt tctca 25
<210> 102
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 102
ctttctgaga accgccattt c 21
<210> 103
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 103
tgttagccac tgtgtcaaat cgcc 24
<210> 104
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 104
tgtctttctg agtgtcaaat cgcc 24
<210> 105
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 105
ccgccatttc tctgtcaaat cgcc 24
<210> 106
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 106
gtgtctttct gagccgccat ttctc 25
<210> 107
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 107
gtgtctttct gaccgccatt tctca 25
<210> 108
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 108
gtgtctttct gaccgccatt tctc 24
<210> 109
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 109
ttcagcttct gtcaaatcgc c 21
<210> 110
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 110
tgttagccac tgtcaaatcg cc 22
<210> 111
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 111
tgtctttctg agtcaaatcg cc 22
<210> 112
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 112
cgccatttct cgtcaaatcg cc 22
<210> 113
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 113
gtgtctttct gccgccattt ctc 23
<210> 114
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 114
ccgccatttc tc 12
<210> 115
<211> 11
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 115
tctgttagcc a 11
<210> 116
<211> 11
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 116
cgccgccatt t 11
<210> 117
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 117
ttgtgtcttt ct 12
<210> 118
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 118
tctgttagcc ac 12
<210> 119
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 119
gtgtctttct gag 13
<210> 120
<211> 13
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 120
ccgccatttc tca 13
<210> 121
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 121
ctgttagcca ct 12
<210> 122
<211> 12
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 122
cgccgccatt tc 12
<210> 123
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 123
gccactgatt 10
<210> 124
<211> 10
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 124
actgttcagc 10
<210> 125
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 125
gctcaggtcg gattgacatt 20
<210> 126
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая нуклеиновая кислота
<400> 126
gggcaactct tccaccagta 20
<210> 127
<211> 963
<212> ДНК
<213> Homo sapiens
<220>
<221> CDS
<222> (1)..(963)
<400> 127
atg gag cta ctg tcg cca ccg ctc cgc gac gta gac ctg acg gcc ccc 48
Met Glu Leu Leu Ser Pro Pro Leu Arg Asp Val Asp Leu Thr Ala Pro
1 5 10 15
gac ggc tct ctc tgc tcc ttt gcc aca acg gac gac ttc tat gac gac 96
Asp Gly Ser Leu Cys Ser Phe Ala Thr Thr Asp Asp Phe Tyr Asp Asp
20 25 30
ccg tgt ttc gac tcc ccg gac ctg cgc ttc ttc gaa gac ctg gac ccg 144
Pro Cys Phe Asp Ser Pro Asp Leu Arg Phe Phe Glu Asp Leu Asp Pro
35 40 45
cgc ctg atg cac gtg ggc gcg ctc ctg aaa ccc gaa gag cac tcg cac 192
Arg Leu Met His Val Gly Ala Leu Leu Lys Pro Glu Glu His Ser His
50 55 60
ttc ccc gcg gcg gtg cac ccg gcc ccg ggc gca cgt gag gac gag cat 240
Phe Pro Ala Ala Val His Pro Ala Pro Gly Ala Arg Glu Asp Glu His
65 70 75 80
gtg cgc gcg ccc agc ggg cac cac cag gcg ggc cgc tgc cta ctg tgg 288
Val Arg Ala Pro Ser Gly His His Gln Ala Gly Arg Cys Leu Leu Trp
85 90 95
gcc tgc aag gcg tgc aag cgc aag acc acc aac gcc gac cgc cgc aag 336
Ala Cys Lys Ala Cys Lys Arg Lys Thr Thr Asn Ala Asp Arg Arg Lys
100 105 110
gcc gcc acc atg cgc gag cgg cgc cgc ctg agc aaa gta aat gag gcc 384
Ala Ala Thr Met Arg Glu Arg Arg Arg Leu Ser Lys Val Asn Glu Ala
115 120 125
ttt gag aca ctc aag cgc tgc acg tcg agc aat cca aac cag cgg ttg 432
Phe Glu Thr Leu Lys Arg Cys Thr Ser Ser Asn Pro Asn Gln Arg Leu
130 135 140
ccc aag gtg gag atc ctg cgc aac gcc atc cgc tat atc gag ggc ctg 480
Pro Lys Val Glu Ile Leu Arg Asn Ala Ile Arg Tyr Ile Glu Gly Leu
145 150 155 160
cag gct ctg ctg cgc gac cag gac gcc gcg ccc cct ggc gcc gca gcc 528
Gln Ala Leu Leu Arg Asp Gln Asp Ala Ala Pro Pro Gly Ala Ala Ala
165 170 175
gcc ttc tat gcg ccg ggc ccg ctg ccc ccg ggc cgc ggc ggc gag cac 576
Ala Phe Tyr Ala Pro Gly Pro Leu Pro Pro Gly Arg Gly Gly Glu His
180 185 190
tac agc ggc gac tcc gac gcg tcc agc ccg cgc tcc aac tgc tcc gac 624
Tyr Ser Gly Asp Ser Asp Ala Ser Ser Pro Arg Ser Asn Cys Ser Asp
195 200 205
ggc atg atg gac tac agc ggc ccc ccg agc ggc gcc cgg cgg cgg aac 672
Gly Met Met Asp Tyr Ser Gly Pro Pro Ser Gly Ala Arg Arg Arg Asn
210 215 220
tgc tac gaa ggc gcc tac tac aac gag gcg ccc agc gaa ccc agg ccc 720
Cys Tyr Glu Gly Ala Tyr Tyr Asn Glu Ala Pro Ser Glu Pro Arg Pro
225 230 235 240
ggg aag agt gcg gcg gtg tcg agc cta gac tgc ctg tcc agc atc gtg 768
Gly Lys Ser Ala Ala Val Ser Ser Leu Asp Cys Leu Ser Ser Ile Val
245 250 255
gag cgc atc tcc acc gag agc cct gcg gcg ccc gcc ctc ctg ctg gcg 816
Glu Arg Ile Ser Thr Glu Ser Pro Ala Ala Pro Ala Leu Leu Leu Ala
260 265 270
gac gtg cct tct gag tcg cct ccg cgc agg caa gag gct gcc gcc ccc 864
Asp Val Pro Ser Glu Ser Pro Pro Arg Arg Gln Glu Ala Ala Ala Pro
275 280 285
agc gag gga gag agc agc ggc gac ccc acc cag tca ccg gac gcc gcc 912
Ser Glu Gly Glu Ser Ser Gly Asp Pro Thr Gln Ser Pro Asp Ala Ala
290 295 300
ccg cag tgc cct gcg ggt gcg aac ccc aac ccg ata tac cag gtg ctc 960
Pro Gln Cys Pro Ala Gly Ala Asn Pro Asn Pro Ile Tyr Gln Val Leu
305 310 315 320
tga 963
<210> 128
<211> 320
<212> PRT
<213> Homo sapiens
<400> 128
Met Glu Leu Leu Ser Pro Pro Leu Arg Asp Val Asp Leu Thr Ala Pro
1 5 10 15
Asp Gly Ser Leu Cys Ser Phe Ala Thr Thr Asp Asp Phe Tyr Asp Asp
20 25 30
Pro Cys Phe Asp Ser Pro Asp Leu Arg Phe Phe Glu Asp Leu Asp Pro
35 40 45
Arg Leu Met His Val Gly Ala Leu Leu Lys Pro Glu Glu His Ser His
50 55 60
Phe Pro Ala Ala Val His Pro Ala Pro Gly Ala Arg Glu Asp Glu His
65 70 75 80
Val Arg Ala Pro Ser Gly His His Gln Ala Gly Arg Cys Leu Leu Trp
85 90 95
Ala Cys Lys Ala Cys Lys Arg Lys Thr Thr Asn Ala Asp Arg Arg Lys
100 105 110
Ala Ala Thr Met Arg Glu Arg Arg Arg Leu Ser Lys Val Asn Glu Ala
115 120 125
Phe Glu Thr Leu Lys Arg Cys Thr Ser Ser Asn Pro Asn Gln Arg Leu
130 135 140
Pro Lys Val Glu Ile Leu Arg Asn Ala Ile Arg Tyr Ile Glu Gly Leu
145 150 155 160
Gln Ala Leu Leu Arg Asp Gln Asp Ala Ala Pro Pro Gly Ala Ala Ala
165 170 175
Ala Phe Tyr Ala Pro Gly Pro Leu Pro Pro Gly Arg Gly Gly Glu His
180 185 190
Tyr Ser Gly Asp Ser Asp Ala Ser Ser Pro Arg Ser Asn Cys Ser Asp
195 200 205
Gly Met Met Asp Tyr Ser Gly Pro Pro Ser Gly Ala Arg Arg Arg Asn
210 215 220
Cys Tyr Glu Gly Ala Tyr Tyr Asn Glu Ala Pro Ser Glu Pro Arg Pro
225 230 235 240
Gly Lys Ser Ala Ala Val Ser Ser Leu Asp Cys Leu Ser Ser Ile Val
245 250 255
Glu Arg Ile Ser Thr Glu Ser Pro Ala Ala Pro Ala Leu Leu Leu Ala
260 265 270
Asp Val Pro Ser Glu Ser Pro Pro Arg Arg Gln Glu Ala Ala Ala Pro
275 280 285
Ser Glu Gly Glu Ser Ser Gly Asp Pro Thr Gln Ser Pro Asp Ala Ala
290 295 300
Pro Gln Cys Pro Ala Gly Ala Asn Pro Asn Pro Ile Tyr Gln Val Leu
305 310 315 320
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИСМЫСЛОВЫЕ НУКЛЕИНОВЫЕ КИСЛОТЫ | 2012 |
|
RU2651468C1 |
| ИНГИБИТОР СИГНАЛА МИОСТАТИНА | 2019 |
|
RU2820270C2 |
| АНТИСМЫСЛОВАЯ НУКЛЕИНОВАЯ КИСЛОТА | 2016 |
|
RU2724554C2 |
| АНТИСМЫСЛОВЫЕ НУКЛЕИНОВЫЕ КИСЛОТЫ | 2015 |
|
RU2730681C2 |
| СОЕДИНЕНИЕ ДЛЯ МОДУЛЯЦИИ ЭКСПРЕССИИ ГЕНА C9ORF72 И ЕГО ПРИМЕНЕНИЕ | 2013 |
|
RU2730677C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ СНИЖЕНИЯ ЭКСПРЕССИИ TAU | 2016 |
|
RU2747734C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ СНИЖЕНИЯ ЭКСПРЕССИИ TAU | 2016 |
|
RU2777570C2 |
| ВАРИАНТЫ HSD17B13 И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2760851C2 |
| Вирус-кодируемые регулируемые агенты, связывающие T-клетки (CATVERT) или NK-клетки (CATVERN), нацеленные на рак | 2020 |
|
RU2827779C2 |
| ОЛИГОНУКЛЕОТИД ДЛЯ ЛЕЧЕНИЯ ПАЦИЕНТОВ С МЫШЕЧНОЙ ДИСТРОФИЕЙ | 2013 |
|
RU2789279C2 |
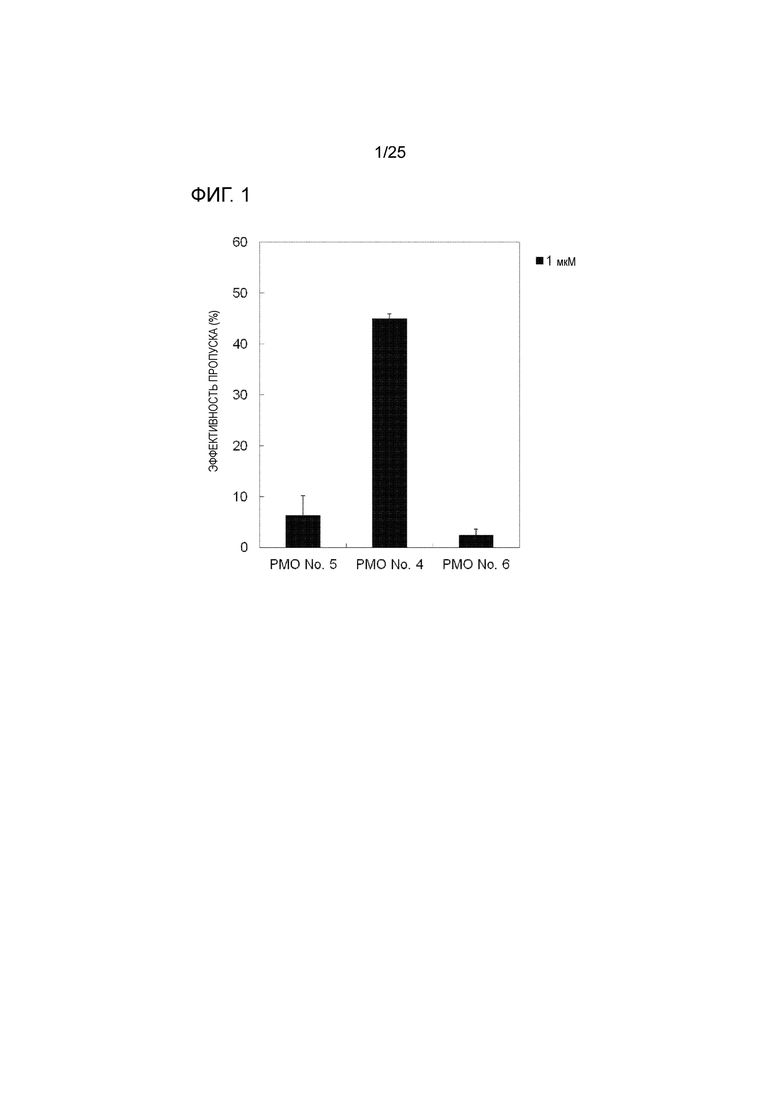
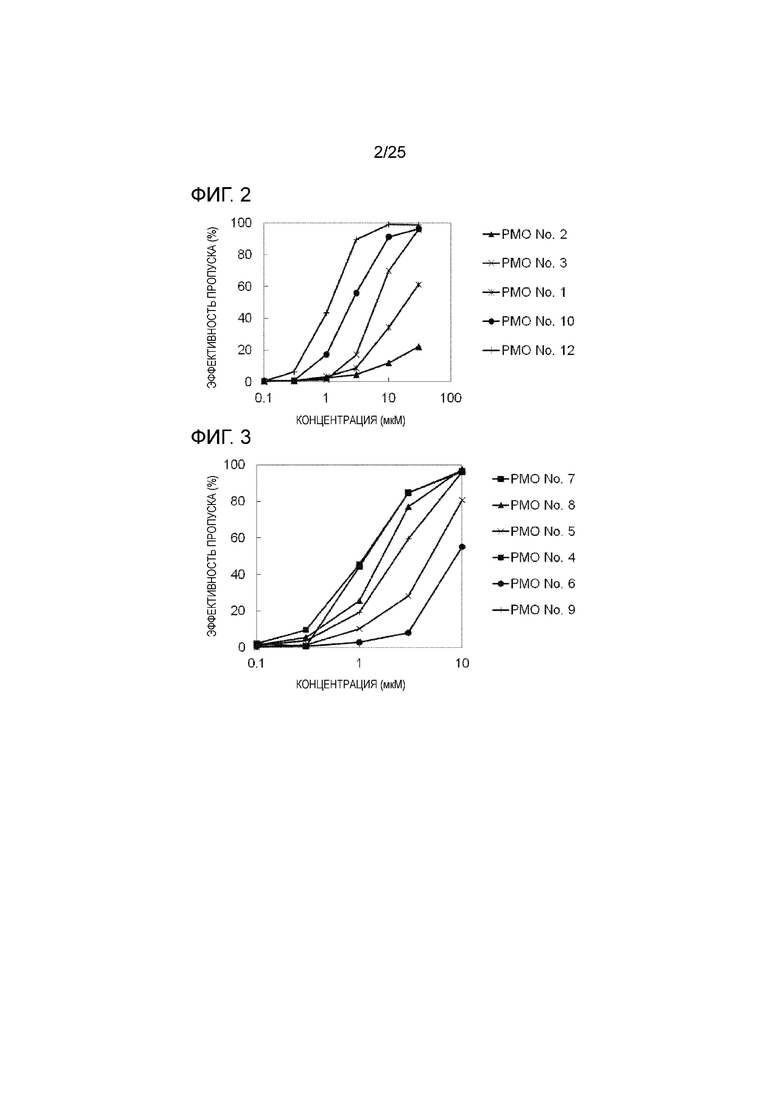
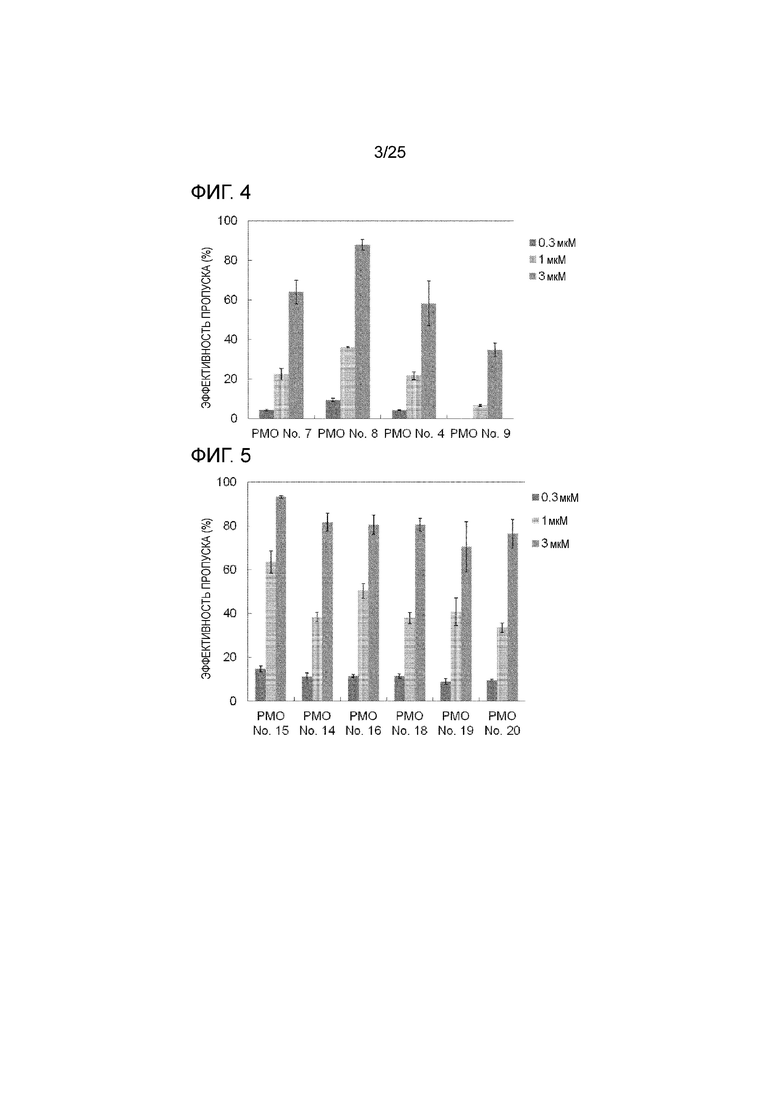
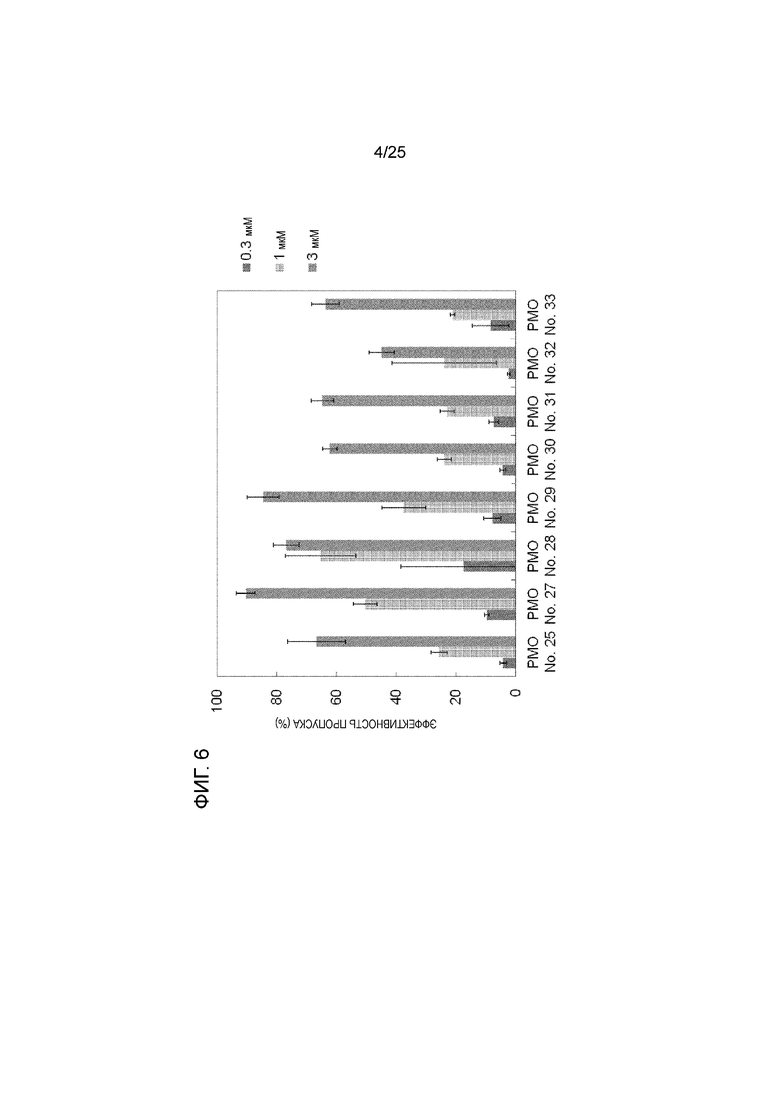
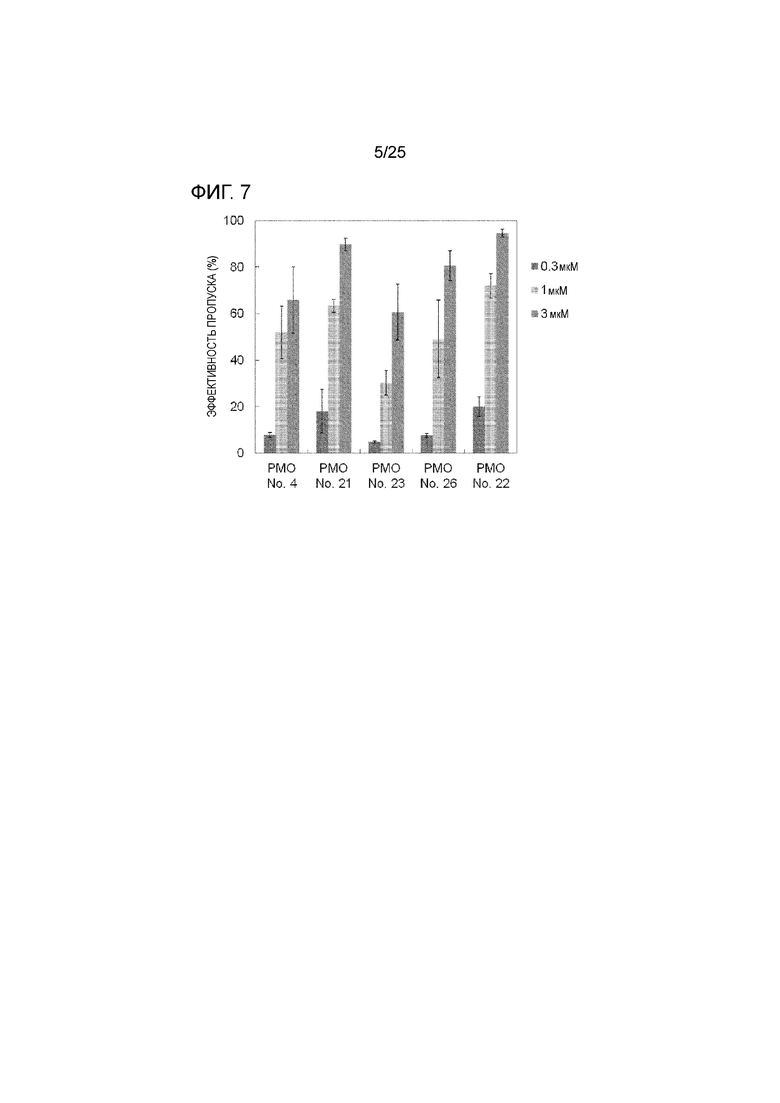
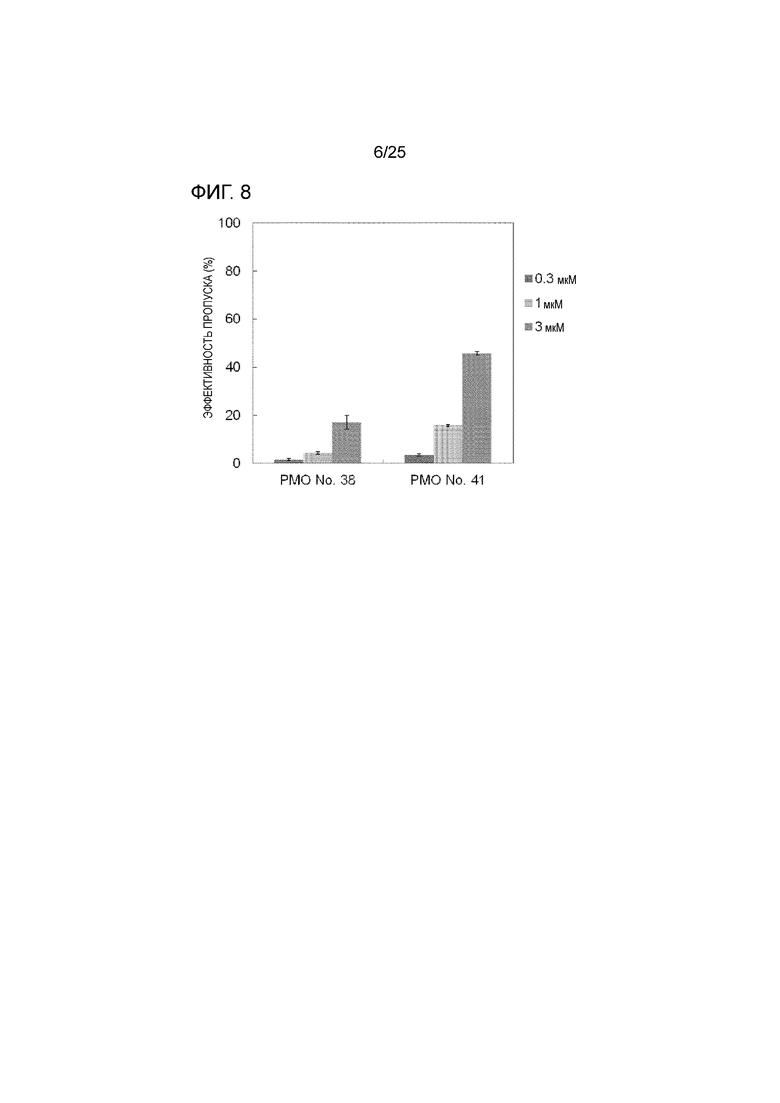
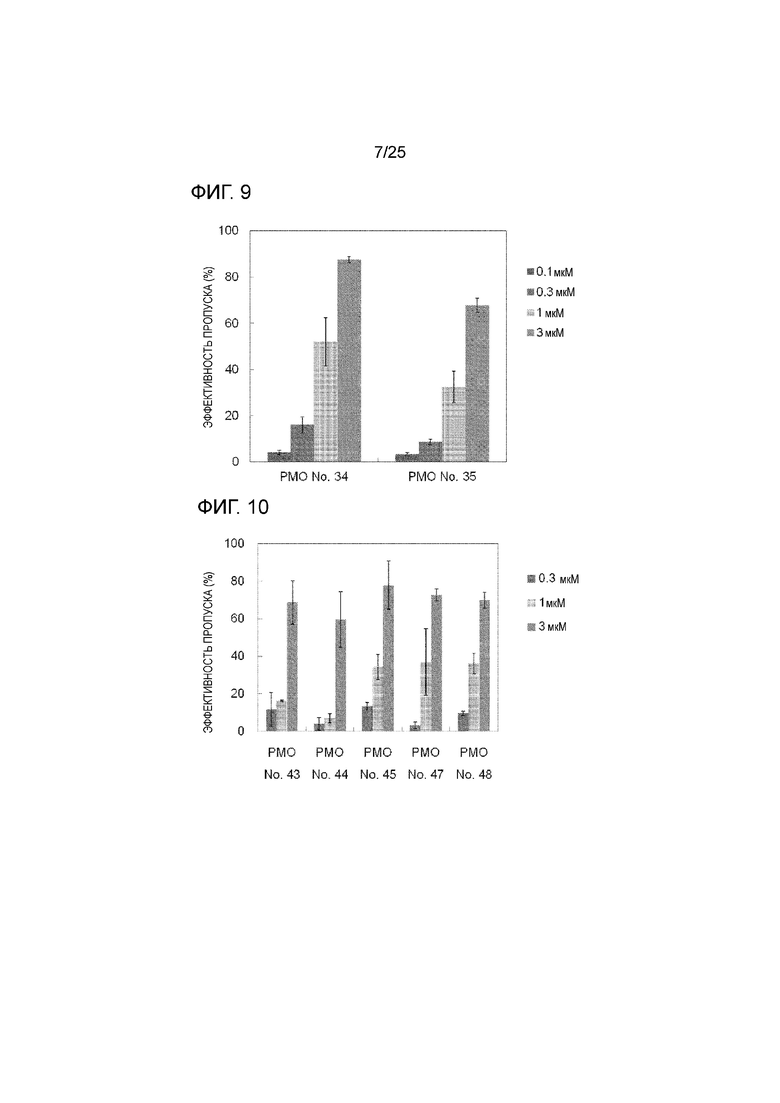
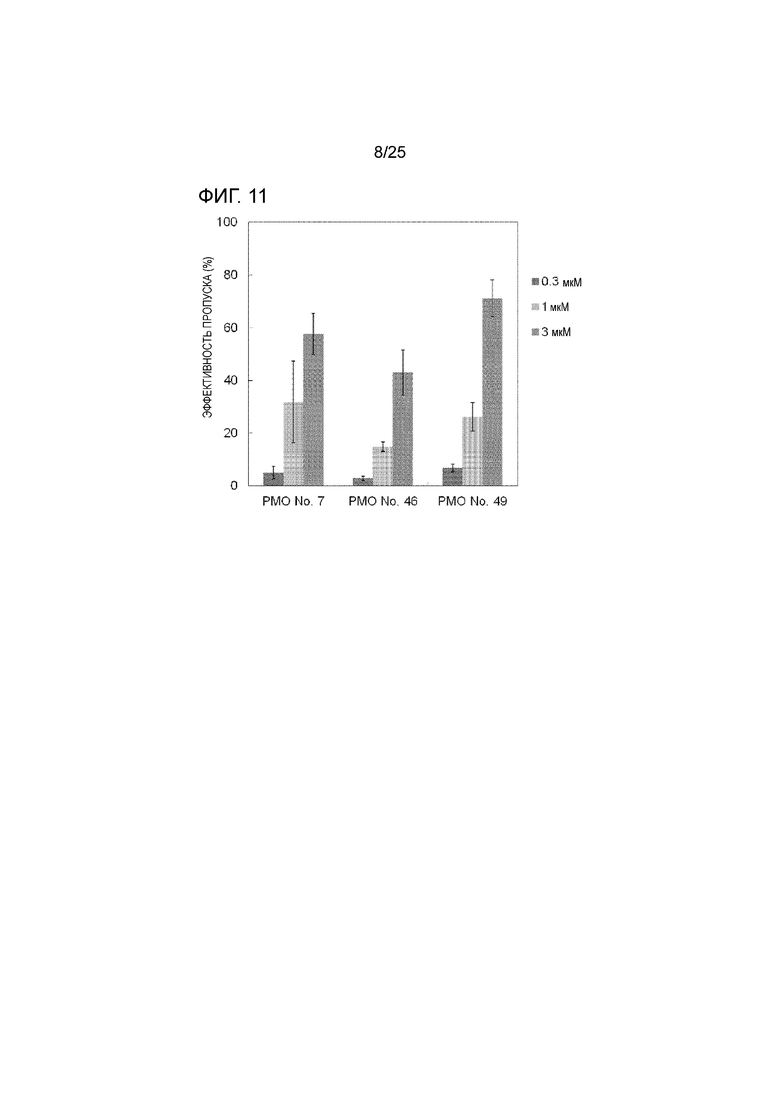
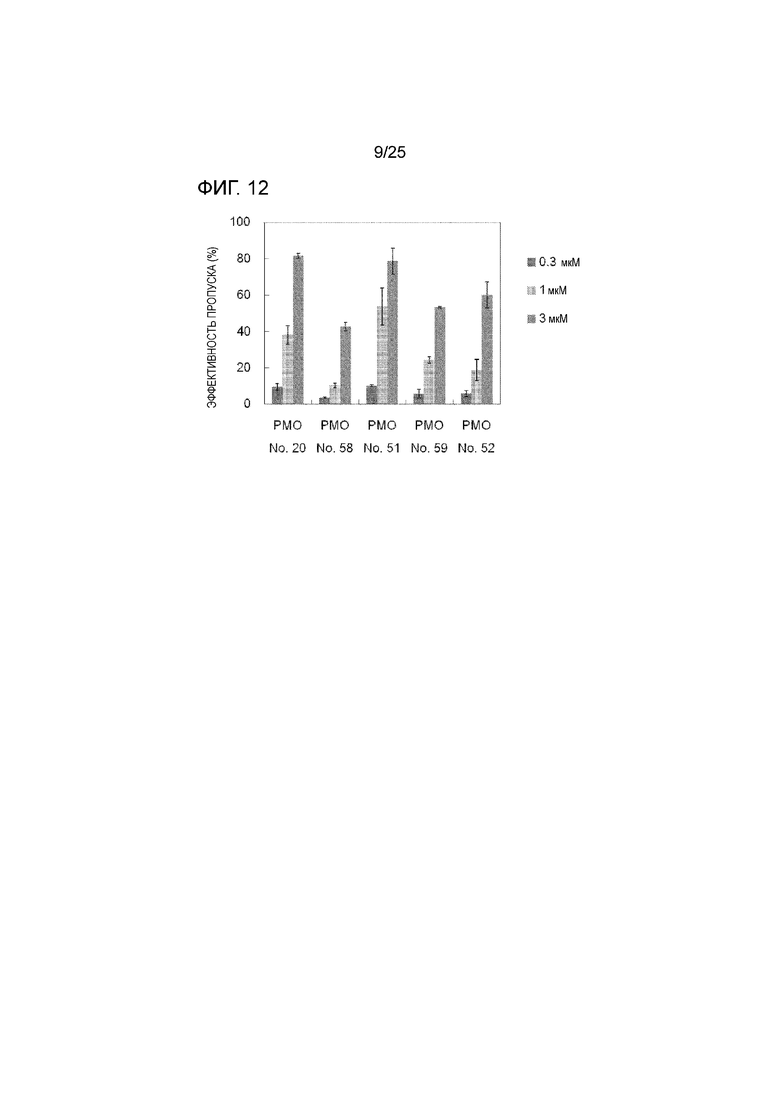
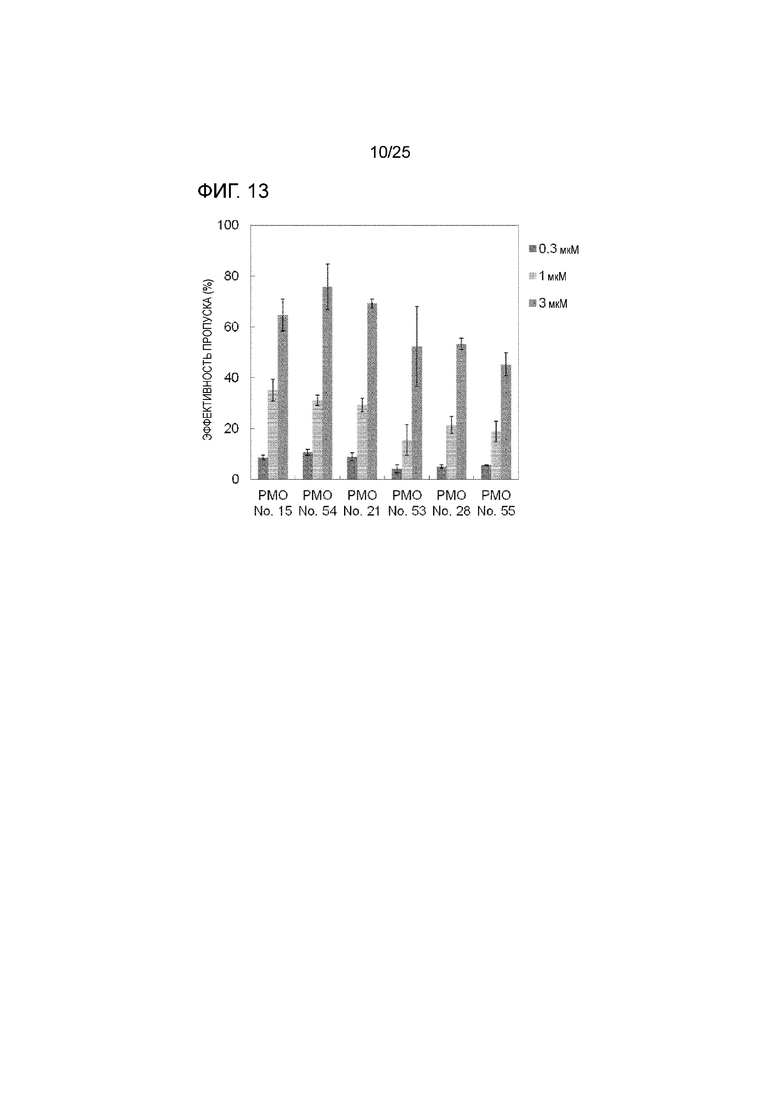
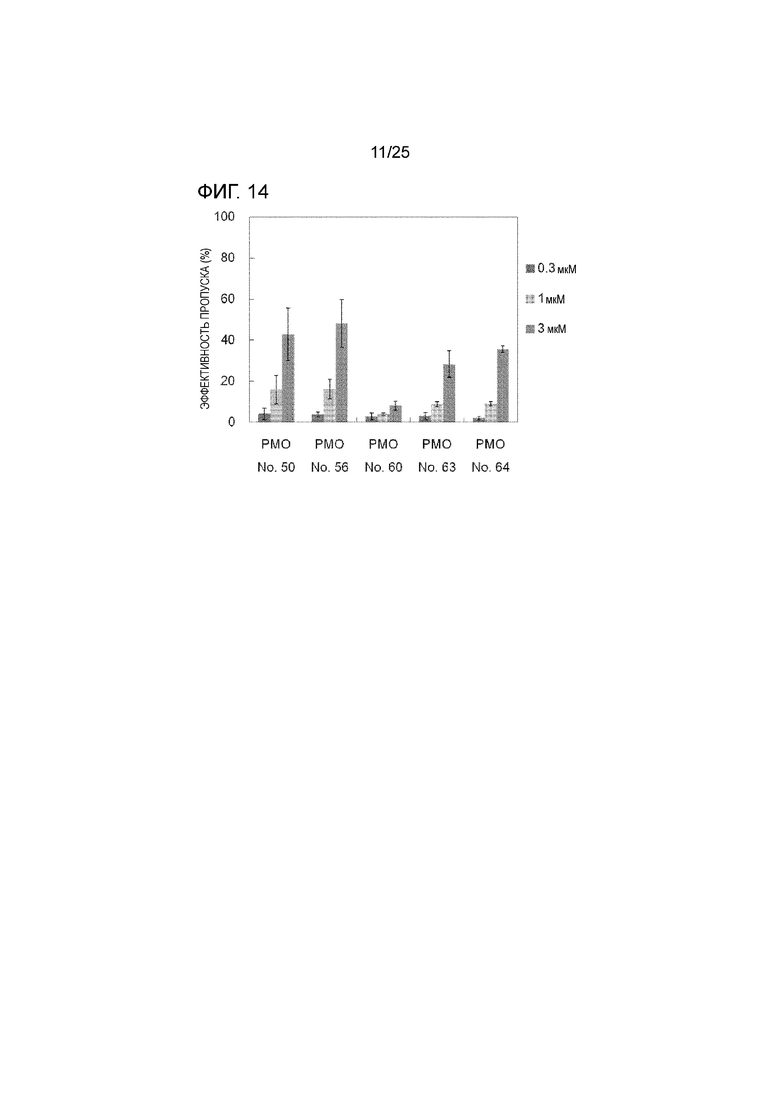
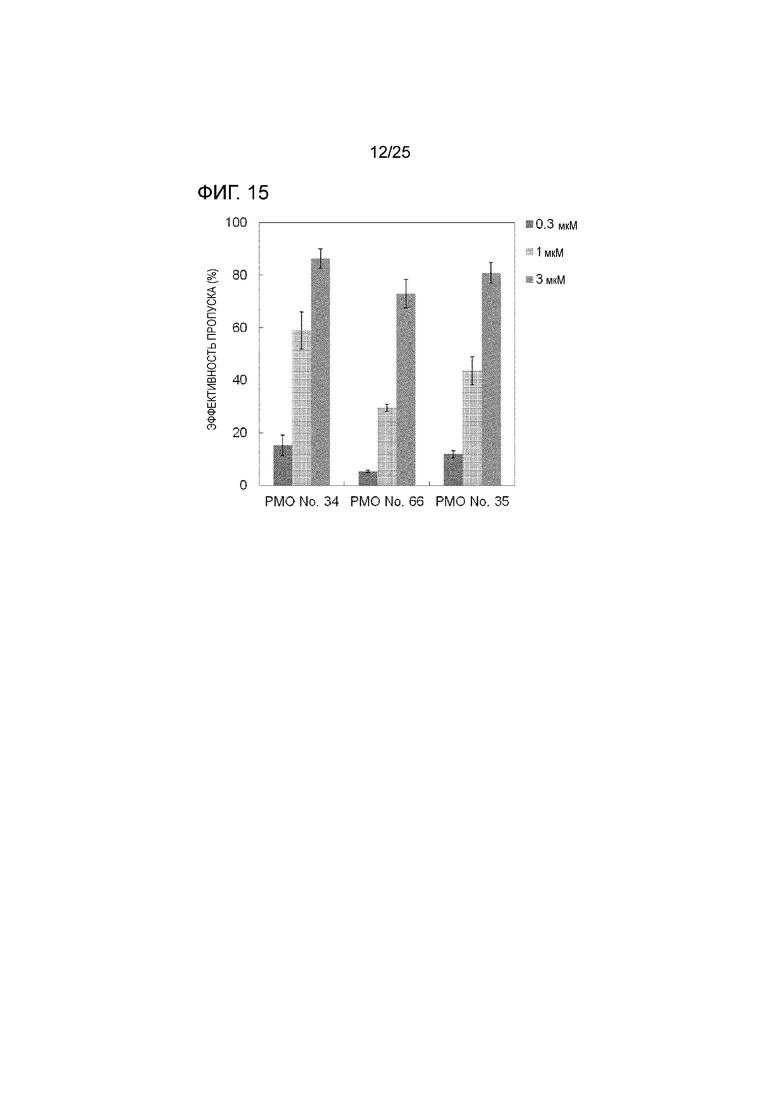
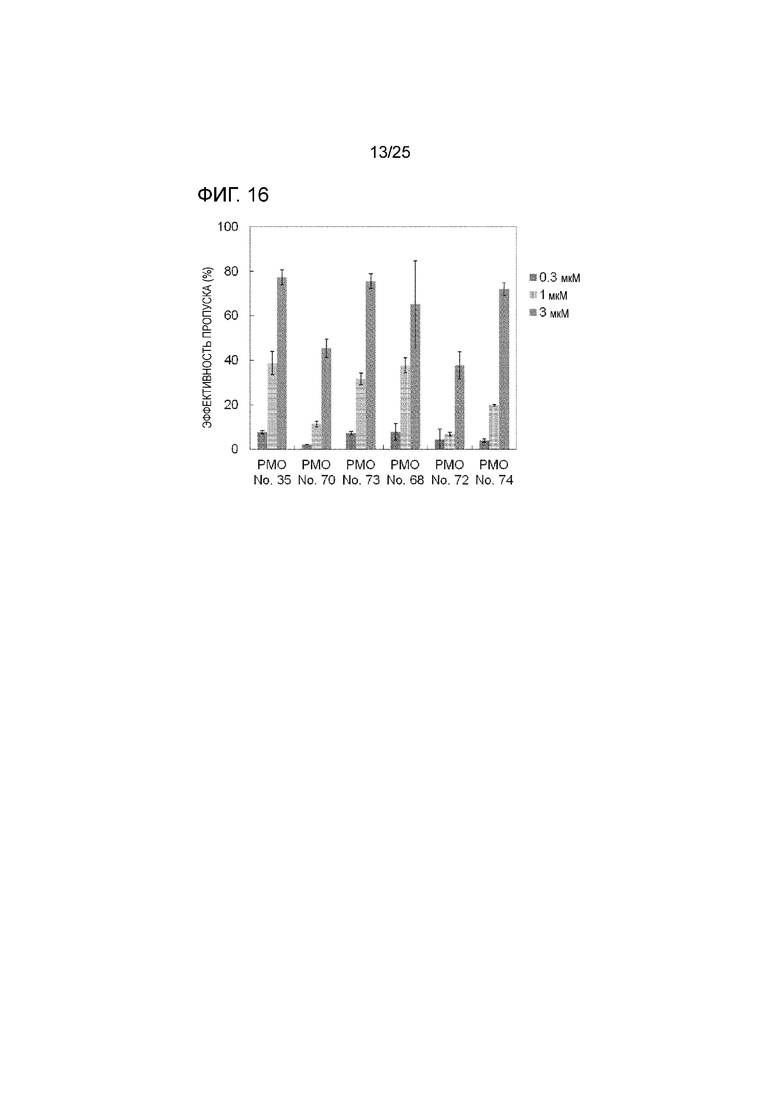
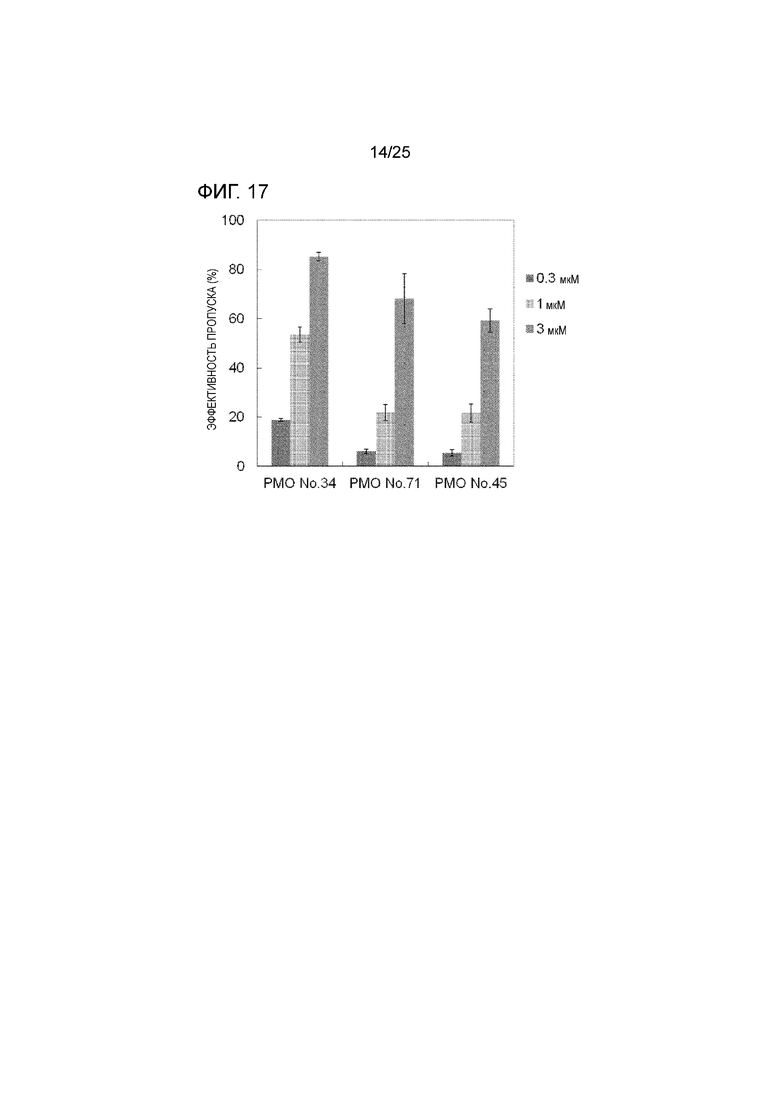
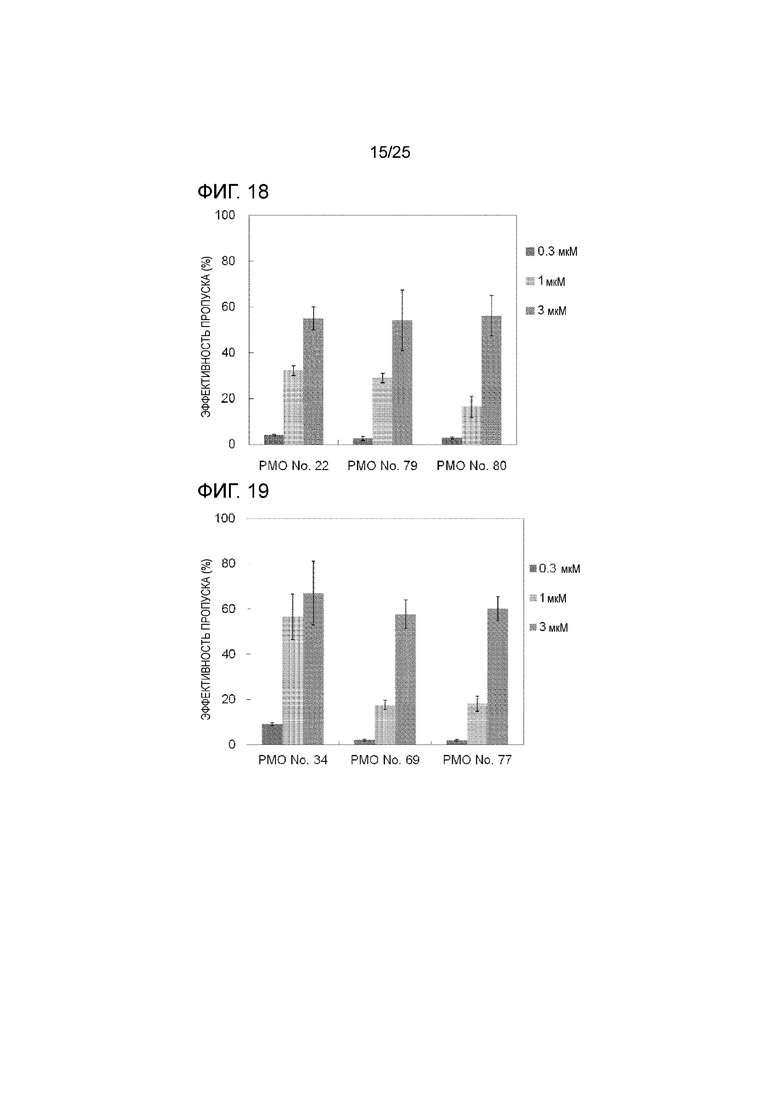
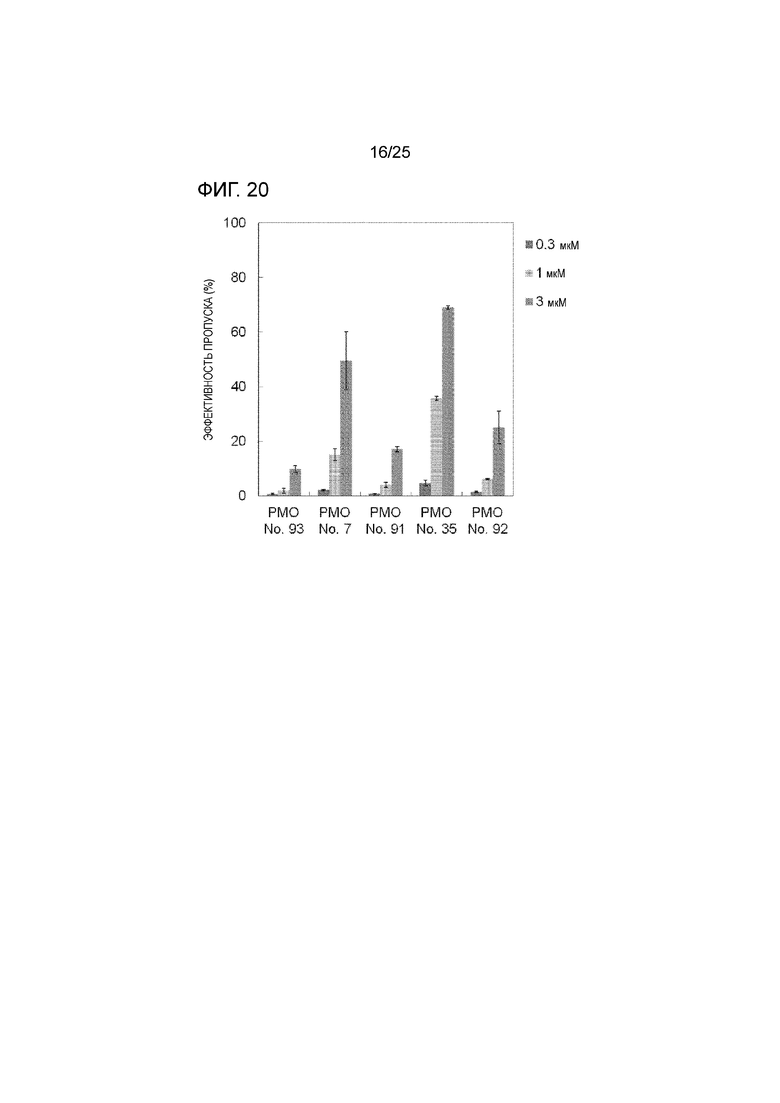
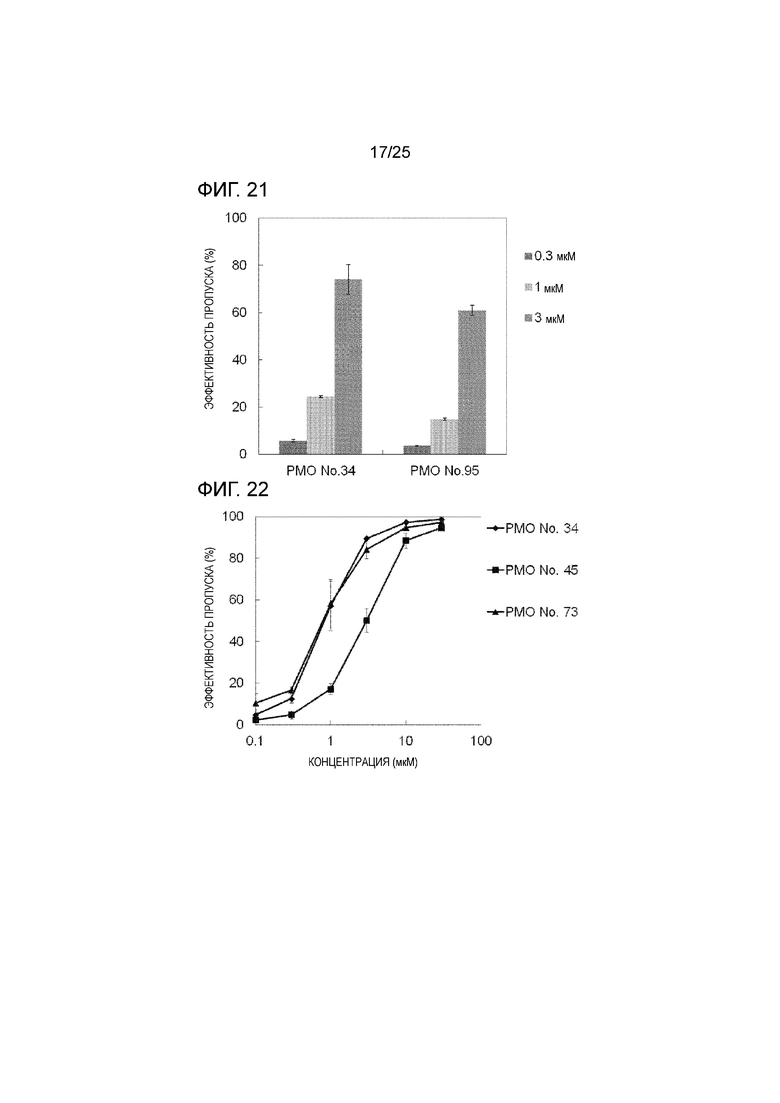
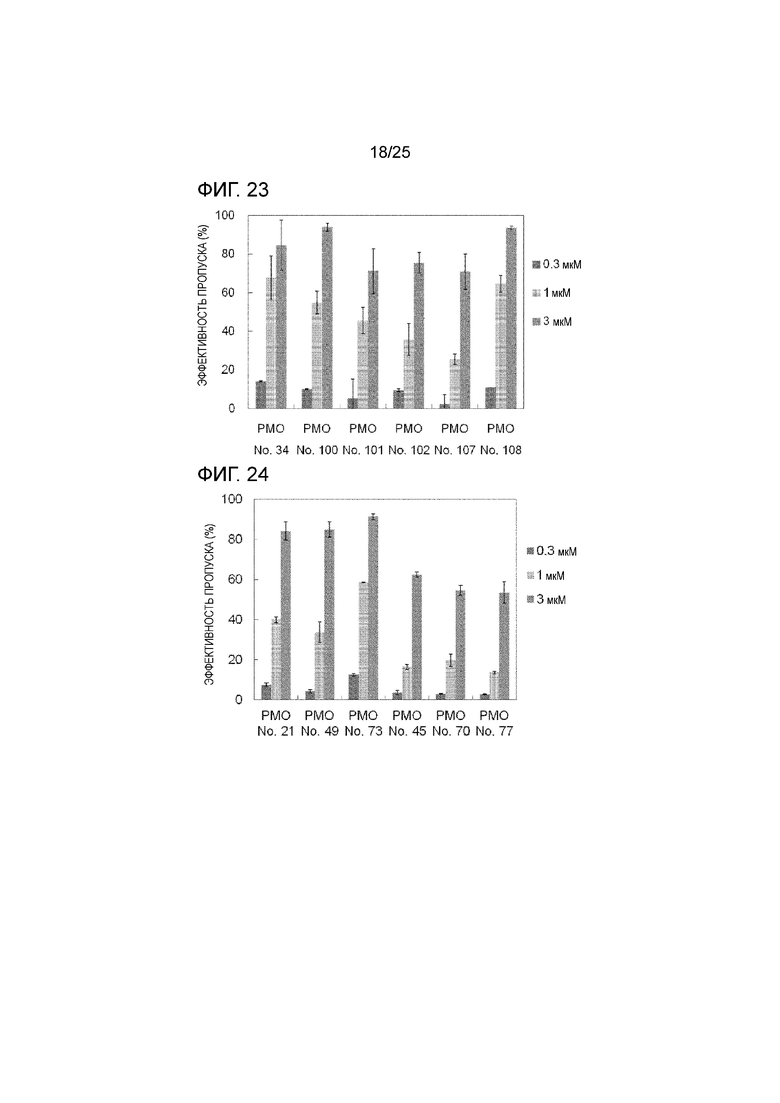
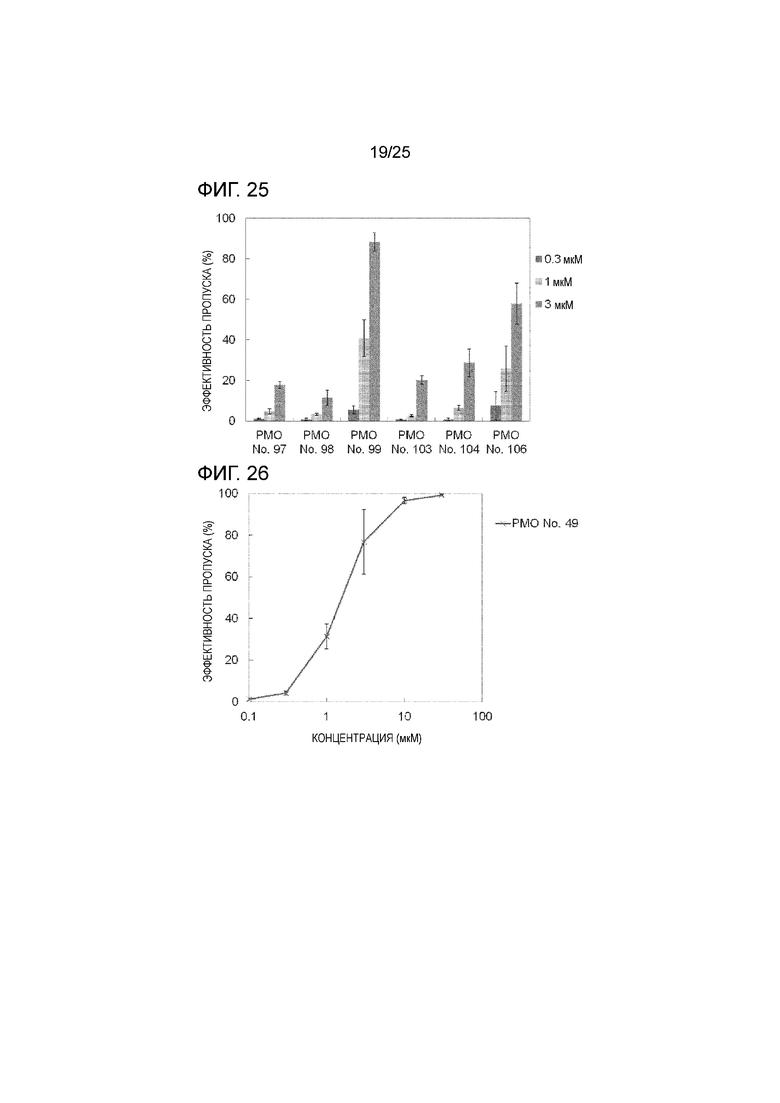
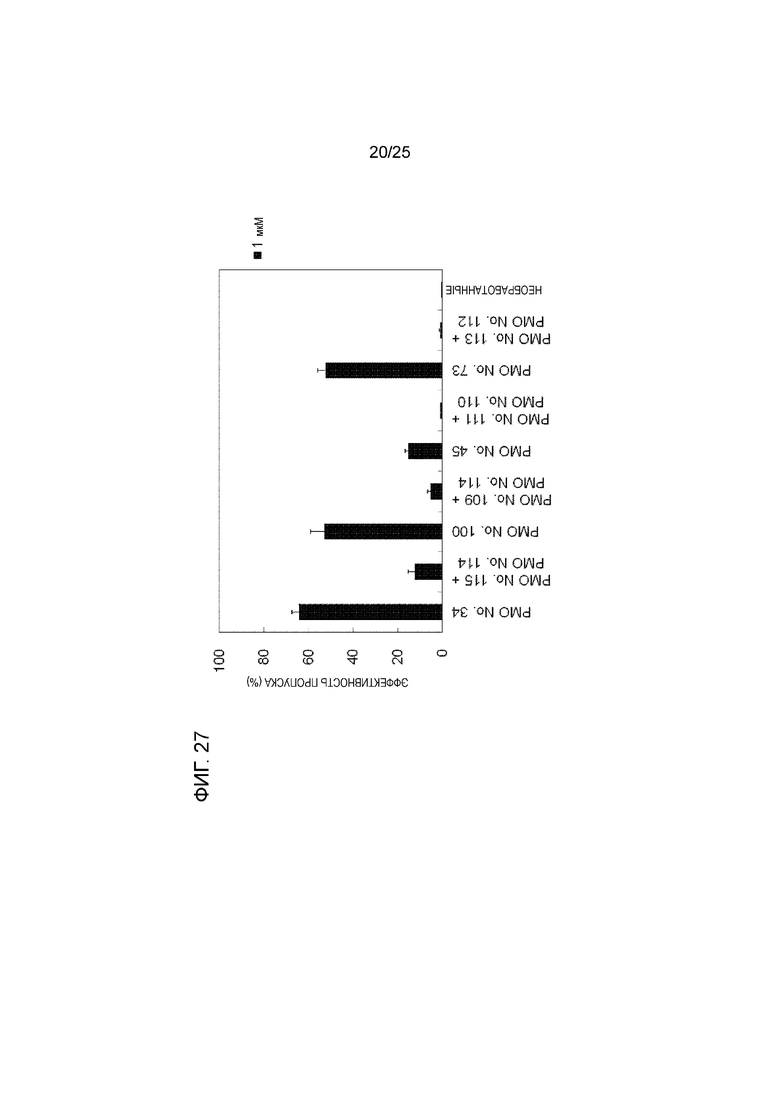
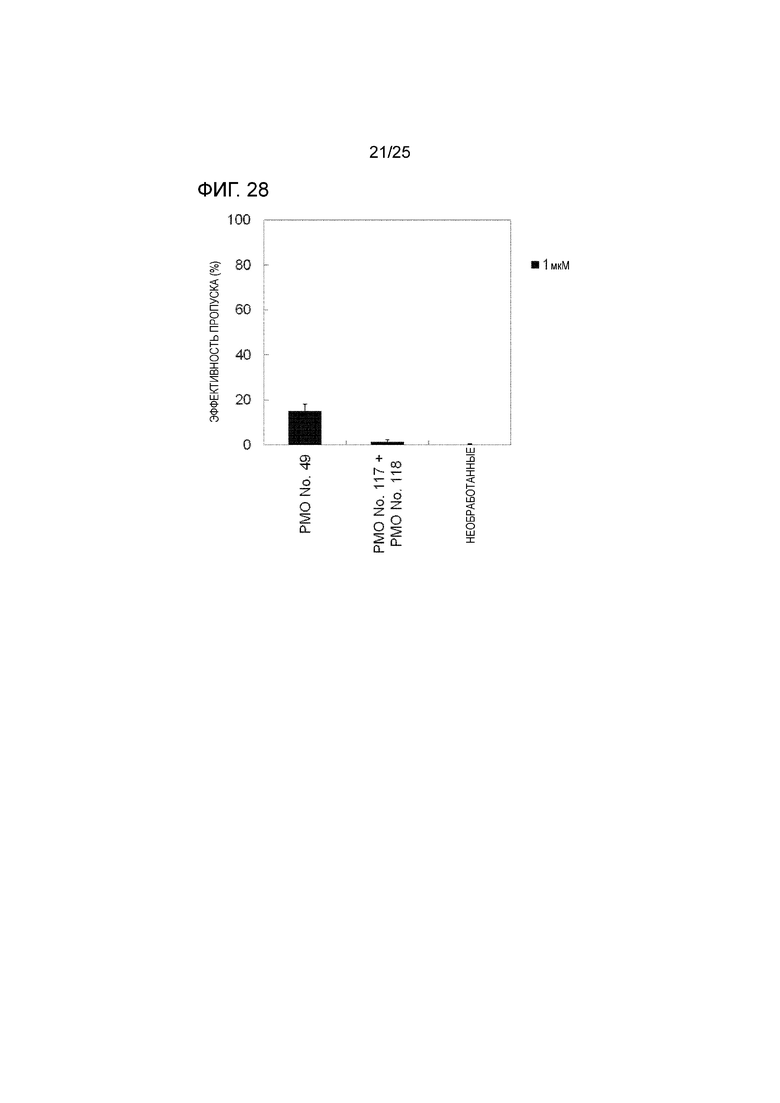
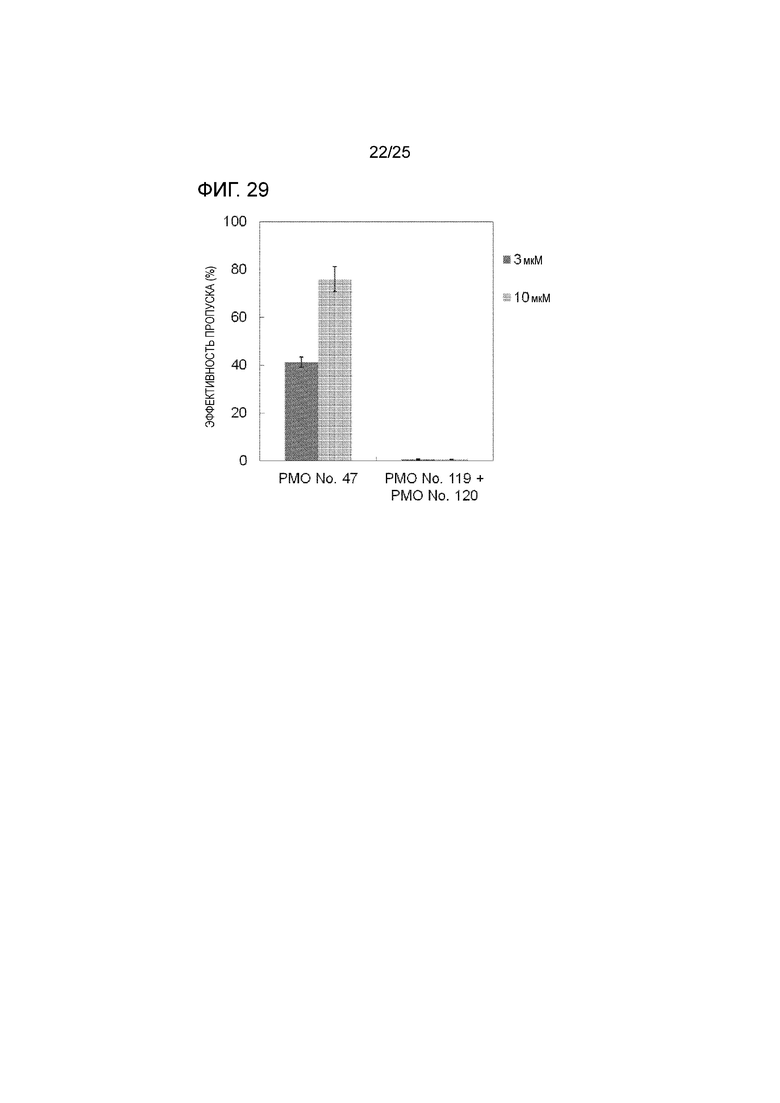
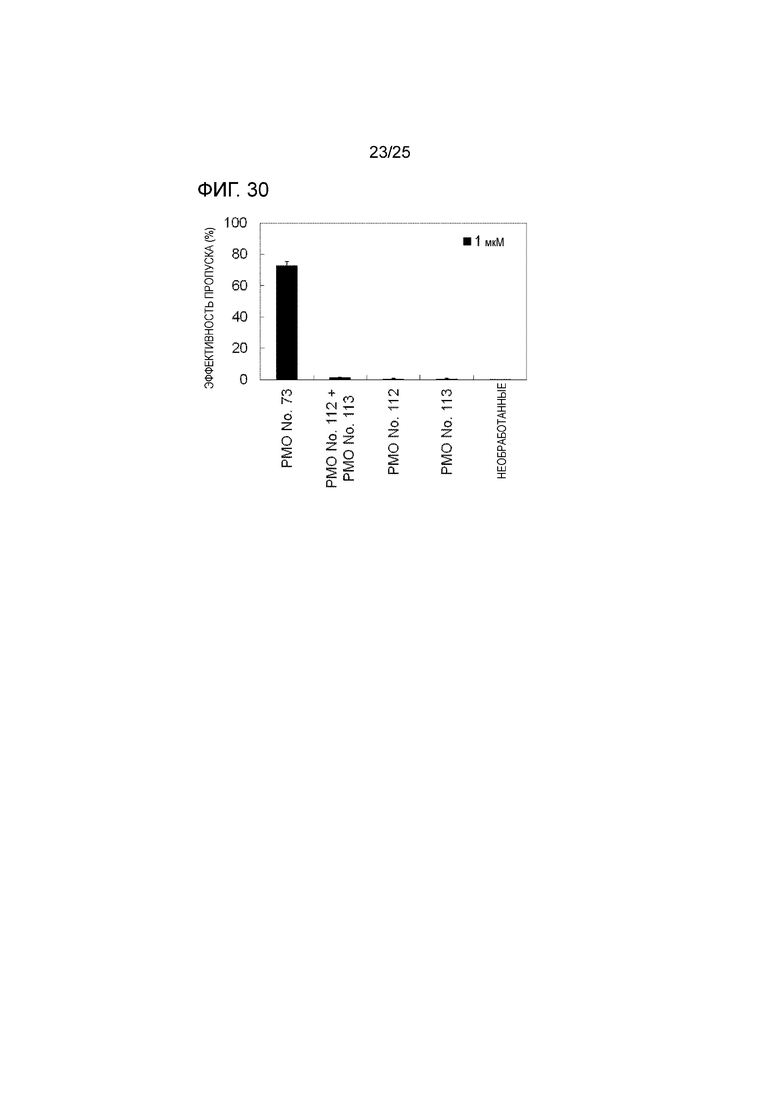

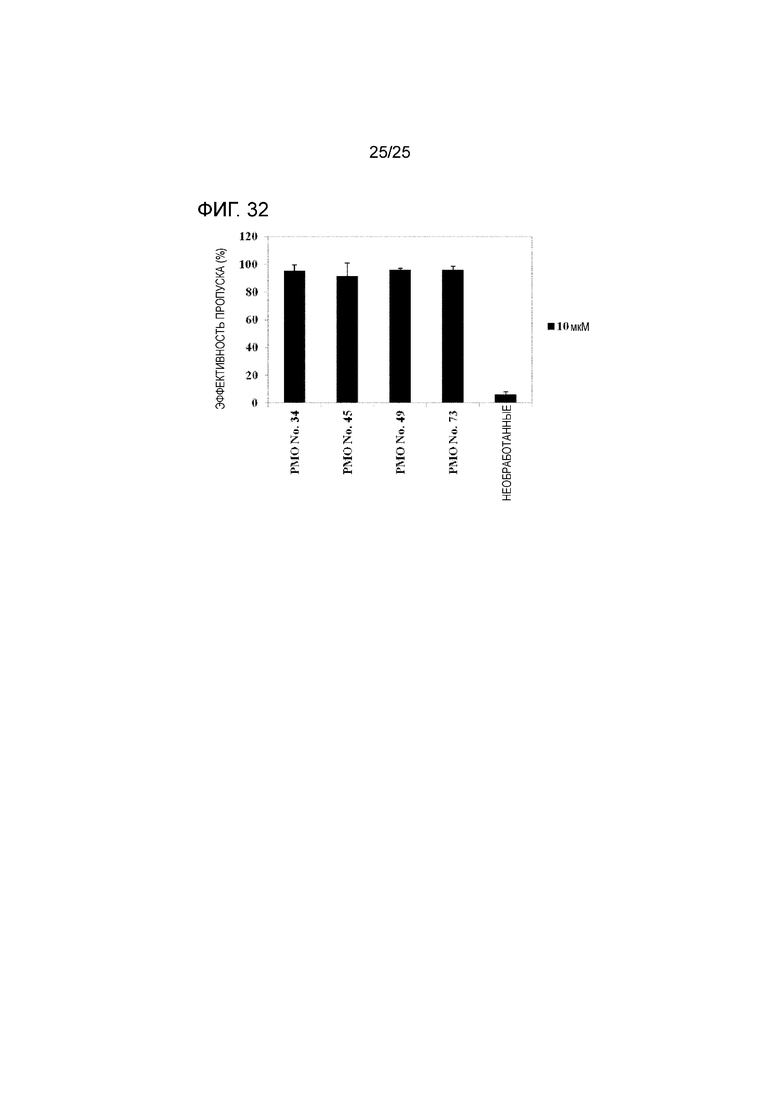
Изобретение относится к области биотехнологии. Предложены антисмысловой олигомер или его фармацевтически приемлемая соль или гидрат, где антисмысловой олигомер индуцирует пропуск экзона 44 гена дистрофина человека. Также предложена фармацевтическая композиция, содержащая указанный олигомер или его фармацевтически приемлемую соль или гидрат. Кроме того, изобретение относится к способу лечения мышечной дистрофии, который включает введение пациенту с мышечной дистрофией указанного олигомера или указанной фармацевтической композиции. Изобретение может быть использовано для получения лекарственного средства, обеспечивающего высокоэффективный пропуск экзона. 5 н. и 12 з.п. ф-лы, 32 ил., 7 табл., 4 пр.
1. Антисмысловой олигомер, состоящий из нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID NO: 11-20, 22, 24-26, 28-33, 35-44, 47, 50, 52-54, 56-63, 65-67, 70, 71, 73, 75-80, 83, 85, 86, 97, 98, 101, 103, 105, 107-110, 112, и 113, или его фармацевтически приемлемая соль или гидрат, где антисмысловой олигомер индуцирует пропуск экзона 44 гена дистрофина человека.
2. Антисмысловой олигомер по п. 1, который является олигонуклеотидом, или его фармацевтически приемлемая соль или гидрат.
3. Антисмысловой олигомер по п. 2, в котором остаток сахара и/или область соединения через фосфатную группу по меньшей мере одного нуклеотида, входящего в состав олигонуклеотида, модифицированы, или его фармацевтически приемлемая соль или гидрат.
4. Антисмысловой олигомер по п. 2 или 3, в котором остаток сахара по меньшей мере одного нуклеотида, входящего в состав олигонуклеотида, является рибозой, в которой 2'-OH-группа заменена любой группой, выбранной из группы, состоящей из OR, R, R'OR, SH, SR, NH2, NHR, NR2, N3, CN, F, Cl, Br и I (где R является алкилом или арилом и R' является алкиленом), или его фармацевтически приемлемая соль или гидрат.
5. Антисмысловой олигомер по любому из пп. 2-4, в котором область соединения через фосфатную группу по меньшей мере одного нуклеотида, входящего в состав олигонуклеотида, является любой областью, выбранной из группы, состоящей из тиофосфатной связи, дитиофосфатной связи, алкилфосфонатной связи, фосфорамидатной связи и боранофосфатной связи, или его фармацевтически приемлемая соль или гидрат.
6. Антисмысловой олигомер по п. 1, который является морфолино-олигомером, или его фармацевтически приемлемая соль или гидрат.
7. Антисмысловой олигомер по п. 6, который является фосфорамидат морфолино-олигомером, или его фармацевтически приемлемая соль или гидрат.
8. Антисмысловой олигомер по п. 6 или 7, в котором 5'-конец имеет любую из приведенных ниже химических формул (1)-(3), или его фармацевтически приемлемая соль или гидрат.
[Формула 26]
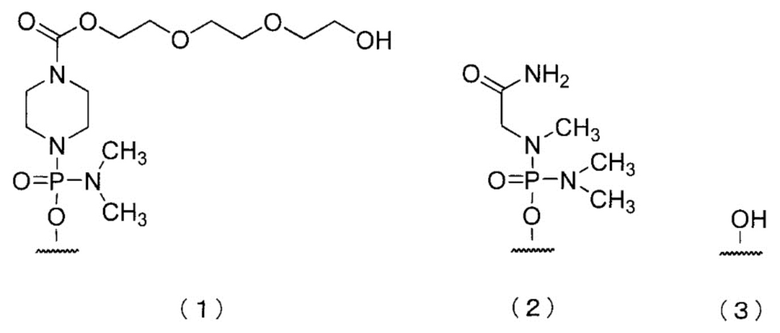
9. Фармацевтическая композиция для лечения мышечной дистрофии, которая содержит в качестве активного ингредиента антисмысловой олигомер по любому из пп. 1-8, или его фармацевтически приемлемую соль или гидрат.
10. Фармацевтическая композиция по п. 9, которая содержит фармацевтически приемлемый носитель.
11. Способ лечения мышечной дистрофии, который включает введение пациенту с мышечной дистрофией антисмыслового олигомера или его фармацевтически приемлемой соли или гидрата по любому пп. 1-8, или фармацевтической композиции по п. 9 или 10.
12. Способ лечения по п. 11, где у пациента с мышечной дистрофией есть мутация (мутации), на которую действует пропуск экзона 44 в гене дистрофина.
13. Способ лечения по п. 11 или 12, где пациентом является человек.
14. Применение антисмыслового олигомера или его фармацевтически приемлемой соли или гидрата по любому из пп. 1-8 для получения фармацевтической композиции для лечения мышечной дистрофии.
15. Антисмысловой олигомер или его фармацевтически приемлемая соль или гидрат по любому из пп. 1-8 для применения в лечении мышечной дистрофии.
16. Антисмысловой олигомер или его фармацевтически приемлемая соль или гидрат по п. 15, где у пациента с мышечной дистрофией в указанном лечении есть мутация (мутации), на которую действует пропуск экзона 44 в гене дистрофина.
17. Антисмысловой олигомер или его фармацевтически приемлемая соль или гидрат по п. 15 или 16, где пациентом является человек.
| WO 2006000057 A1, 05.01.2006 | |||
| WO 2010048586 A1, 29.04.2010 | |||
| AARTSMA-RUS A | |||
| et al., Guidelines for Antisense Oligonucleotide Design and Insight Into Splice-modulating Mechanisms, Molecular Therapy, 2009, vol.17, no.3, pp.548-553 | |||
| ЗЕНКОВА М.А., КАРПОВА Г.Г., Несовершенные комплексы нуклеиновых кислот и их биохимическое проявление, Успехи химии, |

