ОБЛАСТЬ ИЗОБРЕТЕНИЯ
[0001] Областью настоящего изобретения являются фармацевтические композиции для лечения заболеваний и нарушений.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
[0002] Аберрантная активация иммунной системы является отличительной особенностью многих заболеваний и состояний человека. Аутоиммунные заболевания возникают, когда иммунная система организма ошибочно воспринимает аутологичный антиген в качестве чужеродного и атакует собственные ткани организма. Воспалительные заболевания и аллергии могут возникать, когда иммунная система организма ошибочно стимулируется обычными пищевыми антигенами или антигенами окружающей среды. Полипептиды и белки, применяемые для лечения ряда заболеваний человека, зачастую разрушаются, нейтрализуются или иным способом становятся малоэффективными под действием иммунных клеток, которые реагируют на них, как если бы они были чужеродными антигенами.
[0003] Современный способ лечения заболеваний, обусловленных неправильной активацией иммунной системы, включает иммуносупрессию с помощью химических средств, таких как кортикостероиды, или ингибиторов медиаторов воспаления, таких как антигистамины, антитела или цитокины. Эти универсальные способы лечения ассоциированы со значительным повышением частоты заболеваний, например, восприимчивости к заражению, поскольку они подавляют иммунную систему в целом.
[0004] Для некоторых тяжелых случаев аллергии выполняются клинические исследования по индукции "толерантности" к аллергенам путем воздействия медленно возрастающих доз провоцирующего белка с течением времени. В настоящее время эти способы лечения не достигают долгосрочной эффективности и ассоциированы с риском развития тяжелой анафилаксии.
[0005] Существует потребность в новых лекарственных препаратах для лечения данных заболеваний.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В определенных аспектах настоящее изобретение относится к выделенным безъядерным гемопоэтическим клеткам, экспрессирующим антиген. Безъядерные гемопоэтические клетки будут называться в данном документе "EHC-клетки" (или в форме единственного числа "EHC"). В некоторых вариантах осуществления в безъядерных гемопоэтических клетках или клетках EHC отсутствует ядерный материал. Например, EHC-клетки могут представлять собой клетки эритроидного ряда или клетки тромбоцитарного ряда. В некоторых вариантах осуществления EHC-клетки, у которых отсутствует ядерный материал, представляют собой красные кровяные клетки, эритроциты, ретикулоциты или тромбоциты. В некоторых вариантах осуществления безъядерные гемопоэтические клетки или EHC-клетки представляют собой ядерные клетки-предшественники эритроидного ряда или клетки-предшественники тромбоцитарного ряда, которые, например, индуцированы к утрате своего ядерного материала или оказались функционально безъядерными и неспособными к воспроизведению. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC представляет собой циркулирующую в крови клетку, такую как красная кровяная клетка. В некоторых вариантах осуществления экспрессирующую экзогенный антиген EHC культивируют из гемопоэтического предшественника с применением определенных факторов. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC представляет собой клетку тромбоцитарного ряда, такую как тромбоцит. В некоторых вариантах осуществления клетку тромбоцитарного ряда культивируют из гемопоэтического предшественника с применением определенных факторов. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC представляет собой первичную клетку, выделенную из пациента, или для аутологичного, или для аллогенного применения, которую приводят в контакт с антигеном.
[0006] Определенные аспекты настоящего изобретения относятся к экспрессирующим экзогенный антиген EHC-клеткам, которые способны индуцировать иммунологическую толерантность при введении субъекту, например, в форме фармацевтической композиции, содержащей экзогенные антиген-экспрессирующие EHC-клетки. Экзогенный антиген, экспрессируемый EHC-клетками, может быть связан с определенным заболеванием, нарушением или состоянием. Экспрессирующие экзогенный антиген EHC-клетки могут содержать антиген в различной форме, как, например, в виде помещения на поверхности клетки, внутриклеточной экспрессии, внутриклеточной загрузки и конъюгации с поверхностью клетки антигена, представляющего интерес. Экспрессирующие экзогенный антиген EHC-клетки могут лечить заболевания, связанные с аберрантной активацией иммунной системы, более эффективно и/или с меньшим количеством побочных эффектов, чем существующие способы лечения. Например, экспрессирующие экзогенный антиген EHC-клетки могут селективно модулировать иммунную систему, при этом оставляя физиологию иммунной системы в широком смысле практически не нарушенной. В некоторых вариантах осуществления экспрессирующие экзогенный антиген EHC-клетки могут индуцировать разрушение, инактивацию и/или анергию антиген-специфичных T- и B-лимфоцитов. В качестве альтернативы или дополнительно экспрессирующие экзогенный антиген EHC-клетки могут индуцировать пролиферацию антиген-специфичных регуляторных T-лимфоцитов.
[0007] Определенные аспекты настоящего изобретения относятся к экспрессирующим экзогенный антиген EHC-клеткам, которые содержат экзогенный антиген, распознаваемый иммунными клетками при аутоиммунных заболеваниях, таких как, например, рассеянный склероз, сахарный диабет 1 типа, ревматоидный артрит и мембранозный нефрит.
[0008] Определенные аспекты настоящего изобретения относятся к экспрессирующим экзогенный антиген EHC-клеткам, которые содержат экзогенный антиген, распознаваемый иммунными клетками при воспалительных заболеваниях, таких как, например, болезнь Крона, язвенный колит, целиакия или другие идиопатические воспалительные заболевания кишечника.
[0009] Определенные аспекты настоящего изобретения относятся к экспрессирующим экзогенный антиген EHC-клеткам, которые содержат экзогенный антиген, распознаваемый иммунными клетками при заболеваниях, опосредованных несоответствием человеческих лейкоцитарных антигенов (HLA), таких как, например, реакция "трансплантат против хозяина" или отторжение трансплантированных органов.
[0010] Определенные аспекты настоящего изобретения относятся к экспрессирующим экзогенный антиген EHC-клеткам, которые содержат экзогенный антиген, распознаваемый иммунными клетками при аллергических заболеваниях, таких как, например, астма, аллергия на арахис, аллергия на морепродукты, аллергия на пыльцу, аллергия на белок молока, аллергия на укусы насекомых и аллергия на латекс.
[0011] Определенные аспекты настоящего изобретения относятся к экспрессирующим экзогенный антиген EHC-клеткам, которые содержат экзогенный антиген, представляющий собой терапевтический белок, эффективность или активность которого снижена или ослаблена под действием иммунных клеток, такой как, например, фактор свертывания VIII при гемофилии A, фактор свертывания IX при гемофилии B, антитела к фактору некроза опухоли альфа (TNFα) при ревматоидном артрите и других воспалительных заболеваниях, глюкоцереброзидаза при болезни Гоше или аспарагиназа при остром лимфобластном лейкозе (ALL).
[0012] Определенные аспекты настоящего изобретения относятся к экспрессирующим экзогенный антиген EHC-клеткам, которые содержат экзогенный антиген, содержащий полноразмерные полипептиды, усечения и химерные слияния полипептидов, которые a) опосредуют регуляцию комплемента, b) опосредуют связывание и секвестрацию иммунных комплексов, c) являются аутоиммунными антителами или d) являются патогенными частицами. В некоторых вариантах осуществления экзогенный антиген содержит полноразмерные полипептиды, усечения и химерные слияния полипептидов, которые обладают ферментативной активностью в отношении превращения одного низкомолекулярного субстрата в другой низкомолекулярный продукт или одного полипептидного субстрата во второй полипептидный продукт, в том числе, например, расщепления полипептидного субстрата.
[0013] В определенных аспектах настоящего изобретения также представлены способы лечения заболеваний с применением экспрессирующих экзогенный антиген EHC-клеток и их фармацевтических композиций, представленных в данном документе.
[0014] В некоторых аспектах данного документа раскрыт способ индукции иммунологической толерантности. Способ включает введение субъекту-человеку, страдающему от аутоиммунного заболевания, нарушения или состояния или имеющему риск их развития, фармацевтической композиции, содержащей безъядерную гемопоэтическую клетку, экспрессирующую экзогенный антиген, где фармацевтическую композицию вводят в количестве, эффективном для индукции у субъекта иммунологической толерантности в отношении антигена, опосредующего аутоиммунное заболевание, нарушение или состояние.
[0015] В некоторых вариантах осуществления аутоиммунное заболевание выбрано из группы, состоящей из рассеянного склероза, сахарного диабета 1 типа и перечисленных в таблице F.
[0016] В некоторых вариантах осуществления способ дополнительно включает введение фармацевтической композиции по меньшей мере дважды за период лечения, так что аутоиммунное заболевание, нарушение или состояние подвергаются лечению, или их симптом ослабляется.
[0017] В определенных вариантах осуществления способ дополнительно включает введение фармацевтической композиции по меньшей мере дважды за период лечения, так что аутоиммунное заболевание, нарушение или состояние предотвращаются.
[0018] В других вариантах осуществления способ дополнительно включает введение фармацевтической композиции достаточное число раз за период лечения, так что процентная доля антиген-специфичных иммунных клеток значительно снижается во время периода лечения.
[0019] В некоторых вариантах осуществления иммунная клетка представляет собой T-клетку. В некоторых вариантах осуществления иммунная клетка представляет собой B-клетку.
[0020] В некоторых вариантах осуществления снижение концентрации антиген-специфичных иммунных клеток измеряют с помощью проточной цитометрии биологического образца, взятого у субъекта.
[0021] В некоторых вариантах осуществления биологический образец представляет собой биоптат лимфатического узла, образец ткани селезенки или периферической крови.
[0022] В некоторых вариантах осуществления концентрация антиген-специфичных иммунных клеток снижается по меньшей мере на приблизительно 1%, 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99,5%, 99,9%, 99,99% или более 99,99% во время части или на всей протяженности периода лечения.
[0023] В других вариантах осуществления концентрация антиген-специфичных иммунных клеток снижается по меньшей мере на приблизительно 1%, 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99,5%, 99,9%, 99,99% или более 99,99% в пределах приблизительно 1, 5, 10, 15, 20, 30, 40 или 50 минут, или приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22 или 23 часов, или 1, 2, 3, 4, 5 или 6 дней, или приблизительно 1, 2, 3, 4, 5 или 6 недель после введения.
[0024] В некоторых вариантах осуществления фармацевтическую композицию вводят достаточное число раз за период лечения, так что концентрация антиген-специфичных иммунных клеток значительно снижается в течение по меньшей мере приблизительно одной недели, двух недель, трех недель, четырех недель, одного месяца, двух месяцев, трех месяцев, четырех месяцев, пяти месяцев, шести месяцев или более шести месяцев.
[0025] В определенных вариантах осуществления фармацевтическую композицию вводят достаточное число раз за период лечения, так что концентрация антиген-специфичных иммунных клеток значительно снижается в течение периода времени по меньшей мере такого же, как период лечения.
[0026] В некоторых вариантах осуществления фармацевтическую композицию вводят достаточное число раз за период лечения, так что концентрация антиген-специфичных антител в кровотоке значительно снижается во время периода лечения.
[0027] В некоторых вариантах осуществления концентрацию антиген-специфичных антител в кровотоке измеряют с помощью ELISA.
[0028] В некоторых вариантах осуществления концентрация антиген-специфичного циркулирующего в крови антитела снижается по меньшей мере на приблизительно 1%, 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99,5%, 99,9%, 99,99% или более 99,99% во время части или на всей протяженности периода лечения.
[0029] В некоторых вариантах осуществления концентрация антиген-специфичного антитела снижается по меньшей мере на приблизительно 1%, 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99,5%, 99,9%, 99,99% или более 99,99% в пределах приблизительно 1, 5, 10, 15, 20, 30, 40 или 50 минут, или приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22 или 23 часов, или 1, 2, 3, 4, 5 или 6 дней, или приблизительно 1, 2, 3, 4, 5 или 6 недель после введения.
[0030] В определенных вариантах осуществления фармацевтическую композицию вводят достаточное число раз за период лечения, так что концентрация антиген-специфичного циркулирующего в крови антитела значительно снижается в течение по меньшей мере приблизительно одной недели, двух недель, трех недель, четырех недель, одного месяца, двух месяцев, трех месяцев, четырех месяцев, пяти месяцев, шести месяцев или более шести месяцев.
[0031] В определенных вариантах осуществления фармацевтическую композицию вводят достаточное число раз за период лечения, так что концентрация антиген-специфичного циркулирующего в крови антитела значительно снижается в течение периода времени по меньшей мере такого же, как период лечения.
[0032] В некоторых вариантах осуществления фармацевтическую композицию вводят достаточное число раз за период лечения, так что процентная доля антиген-специфичных регуляторных T-клеток значительно повышается во время периода лечения.
[0033] В некоторых вариантах осуществления снижение концентрации антиген-специфичных иммунных клеток измеряют с помощью проточной цитометрии биологического образца, взятого у субъекта.
[0034] В некоторых вариантах осуществления биологический образец представляет собой биоптат лимфатического узла, образец ткани селезенки или периферической крови.
[0035] В определенных вариантах осуществления концентрация антиген-специфичных регуляторных T-клеток повышается по меньшей мере на приблизительно 1%, 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99,5%, 99,9%, 99,99%, или более 99,99% во время части или на всей протяженности периода лечения.
[0036] В определенных вариантах осуществления концентрация антиген-специфичных регуляторных T-клеток повышается по меньшей мере на приблизительно 1%, 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99,5%, 99,9%, 99,99% или более 99,99% в пределах приблизительно 1, 5, 10, 15, 20, 30, 40 или 50 минут, или приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22 или 23 часов, или 1, 2, 3, 4, 5 или 6 дней, или приблизительно 1, 2, 3, 4, 5 или 6 недель после введения.
[0037] В некоторых вариантах осуществления фармацевтическую композицию вводят достаточное число раз за период лечения, так что концентрация антиген-специфичных регуляторных T-клеток значительно повышается в течение по меньшей мере приблизительно одной недели, двух недель, трех недель, четырех недель, одного месяца, двух месяцев, трех месяцев, четырех месяцев, пяти месяцев, шести месяцев или более шести месяцев.
[0038] В некоторых вариантах осуществления фармацевтическую композицию вводят достаточное число раз за период лечения, так что концентрация антиген-специфичных регуляторных T-клеток значительно повышается в течение периода времени по меньшей мере такого же, как период лечения.
[0039] В некоторых вариантах осуществления фармацевтическую композицию вводят достаточное число раз за период лечения, так что один или несколько симптомов аутоиммунного заболевания, нарушения или состояния предотвращаются, снижаются или развиваются с отсрочкой.
[0040] В некоторых вариантах осуществления период лечения не превышает год, шесть месяцев, три месяца, два месяца, один месяц, две недели, одну неделю, три дня, два дня, один день.
[0041] В определенных вариантах осуществления временной интервал между введениями в пределах периода лечения не превышает период, за который число безъядерных гемопоэтических клеток, экспрессирующих экзогенный антиген, снижается до менее чем приблизительно 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% или 95% от числа безъядерных гемопоэтических клеток, экспрессирующих экзогенный антиген, которые присутствовали во введенной фармацевтической композиции.
[0042] В некоторых вариантах осуществления частота введения является достаточной для эффективного снижения концентраций антиген-специфичных иммунных клеток ниже уровня, который ассоциирован с симптомом аутоиммунного заболевания, нарушения или состояния.
[0043] В определенных вариантах осуществления частота введения является достаточной для эффективного снижения концентрации антиген-специфичного циркулирующего в крови антитела ниже уровня, который ассоциирован с симптомом аутоиммунного заболевания, нарушения или состояния.
[0044] В определенных вариантах осуществления частота введения является достаточной для эффективного повышения концентрации антиген-специфичных регуляторных T-клеток выше порогового уровня, который ассоциирован с симптомом аутоиммунного заболевания, нарушения или состояния.
[0045] В некоторых вариантах осуществления безъядерная гемопоэтическая клетка представляет собой клетку эритроидного ряда, клетку тромбоцитарного ряда или их предшественника. В некоторых вариантах осуществления клетка эритроидного ряда представляет собой эритроцит или ретикулоцит. В некоторых вариантах осуществления клетка тромбоцитарного ряда представляет собой тромбоцит.
[0046] В некоторых вариантах осуществления безъядерная гемопоэтическая клетка выделена из донора. В некоторых вариантах осуществления безъядерная гемопоэтическая клетка имеет аутологичное происхождение из субъекта. В некоторых вариантах осуществления безъядерная гемопоэтическая клетка имеет аллогенное происхождение. В некоторых вариантах осуществления безъядерная гемопоэтическая клетка имеет ксеногенное происхождение.
[0047] В некоторых вариантах осуществления безъядерная гемопоэтическая клетка происходит из ядерной клетки-предшественника при помощи способа на основе культуры клеток, который индуцирует удаление ее ядра. В некоторых вариантах осуществления безъядерная гемопоэтическая клетка получена из ядерной клетки-предшественника, над которой проводили химические или физические манипуляции для удаления ее ядра.
[0048] В некоторых вариантах осуществления безъядерная гемопоэтическая клетка получена при помощи облучения или химического разрушения ядра ядерной клетки-предшественника. В некоторых вариантах осуществления химическое разрушение проведено с помощью цитохалазина B. В некоторых вариантах осуществления облучение проведено при помощи по меньшей мере 5 Гр, 7 Гр, 10 Гр, 15 Гр, 25 Гр, 30 Гр, 40 Гр или по меньшей мере 50 Гр.
[0049] В некоторых вариантах осуществления экзогенный антиген представляет собой полипептид, кодируемый экзогенной нуклеиновой кислотой.
[0050] В некоторых вариантах осуществления экзогенный антиген ассоциирован с клеточной мембраной безъядерной гемопоэтической клетки.
[0051] В некоторых вариантах осуществления экзогенный антиген представляет собой слияние или химерный полипептид.
[0052] В некоторых вариантах осуществления слияние или химера содержит по меньшей мере один из S-домена, A-домена или U-домена, где S-домен представляет собой поверхностный домен на безъядерной гемопоэтической клетке, где A-домен представляет собой якорь в клеточной мембране или на ней, где U-домен обращен к внутриклеточной, не подвергающейся воздействию стороне безъядерной гемопоэтической клетки, и где S-домен, A-домен и/или U-домен представляют собой полипептиды различного происхождения.
[0053] В некоторых вариантах осуществления S-домен и/или A-домен содержит по меньшей мере 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или по меньшей мере 500 аминокислот. В некоторых вариантах осуществления S-домен и/или A-домен содержит по меньшей мере 500, 750, или по меньшей мере 1000 аминокислот.
[0054] В некоторых вариантах осуществления экзогенный антиген выбран из группы, состоящей из основного белка миелина, протеолипидного белка, гликопротеина миелина олигодендроцитов, антигена бета-клеток поджелудочной железы, инсулина и перечисленных в таблице F, таблице 6 и таблице 8.
[0055] В некоторых вариантах осуществления безъядерная гемопоэтическая клетка содержит по меньшей мере 10 копий, 100 копий, 1000 копий, 10000 копий, 25000 копий, 50000 копий, 100000 копий, 500000 копий, 1000000 копий или 2000000 копий экзогенного антигена.
[0056] В некоторых вариантах осуществления фармацевтическая композиция дополнительно содержит фармацевтически активное средство.
[0057] В некоторых вариантах осуществления способ дополнительно включает стадию введения фармацевтически активного средства, где фармацевтически активное средство вводят до, после или одновременно с фармацевтической композицией.
[0058] В некоторых вариантах осуществления фармацевтическую композицию вводят внутривенно.
[0059] В некоторых вариантах осуществления фармацевтически активное средство выбрано из биологического средства, низкомолекулярного средства или средства на основе нуклеиновой кислоты.
[0060] В некоторых вариантах осуществления фармацевтическая композиция дополнительно содержит фармацевтически приемлемый носитель.
[0061] В определенных вариантах осуществления способ дополнительно включает стадию выбора лечения для субъекта, страдающего от аутоиммунного заболевания, нарушения или состояния, выбранных из группы, состоящей из тромбоцитопенического акроангиотромбоза, CAPS, APS, тяжелой миастении, синдрома Гудпасчера, мембранозного нефрита, сахарного диабета 1 типа, ревматоидного артрита, рассеянного склероза, болезни Крона или перечисленных в таблице F и таблице G, или имеющего риск их развития.
[0062] В некоторых аспектах данного документа раскрыта фармацевтическая композиция, содержащая безъядерную гемопоэтическую клетку, экспрессирующую экзогенный антиген, где введение эффективного количества фармацевтической композиции способно индуцировать иммунологическую толерантность у субъекта-человека, страдающего от аутоиммунного заболевания, нарушения или состояния или имеющего риск их развития, при введении с помощью способа по любому из предыдущих пунктов.
[0063] В некоторых вариантах осуществления фармацевтическая композиция дополнительно содержит фармацевтически приемлемый носитель.
[0064] В некоторых вариантах осуществления фармацевтическая композиция содержит популяцию гемопоэтических клеток, экспрессирующих экзогенный антиген. В некоторых вариантах осуществления фармацевтическая композиция содержит по меньшей мере 1×103 гемопоэтических клеток, экспрессирующих экзогенный антиген.
[0065] В определенных вариантах осуществления фармацевтической композиции гемопоэтические клетки, экспрессирующие экзогенный антиген, представлены в объеме, составляющем приблизительно 10 нл, 100 нл, 1 мкл, 10 мкл, 100 мкл, 1 мл, 10 мл, 20 мл или 50 мл. В других вариантах осуществления фармацевтической композиции гемопоэтические клетки, экспрессирующие экзогенный антиген, представлены в объеме, составляющем приблизительно 1 мл, 10 мл, 20 мл, 50 мл, 100 мл, 250 мл или 500 мл.
[0066] В некоторых вариантах осуществления фармацевтической композиции композиция составлена для длительного хранения. В некоторых вариантах осуществления фармацевтической композиции композиция заморожена. В некоторых вариантах осуществления фармацевтическая композиция содержит фармацевтически активное средство.
[0067] В определенных вариантах осуществления фармацевтической композиции фармацевтически активное средство выбрано из биологического средства, низкомолекулярного средства или средства на основе нуклеиновой кислоты.
[0068] В некоторых аспектах данного документа раскрыта лекарственная форма, содержащая композиции, описанные в данном документе, составленная в виде жидкой суспензии для внутривенной инъекции.
[0069] В некоторых аспектах данного документа раскрыто медицинское устройство, содержащее контейнер, вмещающий фармацевтические композиции, описанные в данном документе, и аппликатор для внутривенной инъекции фармацевтической композиции субъекту.
[0070] В некоторых аспектах данного документа раскрыт медицинский набор, содержащий фармацевтические композиции, описанные в данном документе, и медицинское устройство для внутривенной инъекции фармацевтической композиции субъекту.
[0071] В некоторых аспектах данного документа раскрыты гемопоэтические клетки, экспрессирующие экзогенный антиген, из фармацевтической композиции, вводимой с помощью любого из способов, описанных в данном документе.
[0072] В некоторых аспектах данного документа раскрыта популяция гемопоэтических клеток, экспрессирующих экзогенный антиген, раскрываемых в данном документе.
[0073] В некоторых вариантах осуществления популяция гемопоэтических клеток, экспрессирующих экзогенный антиген, составлена в виде жидкости.
[0074] В некоторых вариантах осуществления популяция гемопоэтических клеток, экспрессирующих экзогенный антиген, заморожена.
[0075] В некоторых аспектах данного документа раскрыт выделенный антиген, экспрессированный популяцией гемопоэтических клеток, описанной в данном документе.
[0076] В некоторых аспектах данного документа раскрыта экзогенная нуклеиновая кислота, кодирующая экзогенный антиген, описанный в данном документе.
[0077] В некоторых аспектах данного документа раскрыта безъядерная гемопоэтическая клетка, содержащая экзогенный антиген, который содержит по меньшей мере один из S-домена, A-домена или U-домена, где S-домен представляет собой внеклеточный поверхностный домен, A-домен представляет собой якорь, и U-домен расположен внутри клетки, и где безъядерная гемопоэтическая клетка способна индуцировать иммунологическую толерантность при введении субъекту.
[0078] В некоторых вариантах осуществления безъядерной гемопоэтической клетки, раскрытой в данном документе, экзогенный антиген представляет собой слияние или химерный полипептид.
[0079] В некоторых вариантах осуществления безъядерной гемопоэтической клетки, раскрытый в данном документе, S-домен, A-домен и/или U-домен представляют собой полипептиды различного происхождения.
[0080] В определенных вариантах осуществления безъядерной гемопоэтической клетки, раскрытой в данном документе, S-домен и/или A-домен содержат по меньшей мере 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250 или по меньшей мере 500 аминокислот.
[0081] В определенных вариантах осуществления безъядерной гемопоэтической клетки, раскрытой в данном документе, S-домен и/или A-домен содержат по меньшей мере 500, 750 или по меньшей мере 1000 аминокислот.
[0082] В некоторых вариантах осуществления безъядерной гемопоэтической клетки, раскрытой в данном документе, экзогенный антиген выбран из группы, состоящей из основного белка миелина, протеолипидного белка, гликопротеина миелина олигодендроцитов, антигена бета-клеток поджелудочной железы, инсулина и перечисленных в таблице F, таблице 6 и таблице 8.
[0083] В некоторых вариантах осуществления безъядерная гемопоэтическая клетка содержит по меньшей мере 10 копий, 100 копий, 1000 копий, 10000 копий, 25000 копий, 50000 копий, 100000 копий, 500000 копий, 1000000 копий или 2000000 копий экзогенного антигена.
[0084] В определенных вариантах осуществления безъядерная гемопоэтическая клетка представляет собой клетку эритроидного ряда, клетку тромбоцитарного ряда или их предшественника. В некоторых вариантах осуществления клетка эритроидного ряда представляет собой эритроцит или ретикулоцит. В некоторых вариантах осуществления клетка тромбоцитарного ряда представляет собой тромбоцит.
[0085] В некоторых вариантах осуществления безъядерная гемопоэтическая клетка выделена из донора. В некоторых вариантах осуществления безъядерная гемопоэтическая клетка имеет аутологичное происхождение из субъекта. В некоторых вариантах осуществления безъядерная гемопоэтическая клетка имеет аллогенное происхождение. В некоторых вариантах осуществления безъядерная гемопоэтическая клетка имеет ксеногенное происхождение.
[0086] В определенных вариантах осуществления безъядерная гемопоэтическая клетка происходит из ядерной клетки-предшественника при помощи способа на основе культуры клеток, который индуцирует удаление ее ядра.
[0087] В определенных вариантах осуществления безъядерная гемопоэтическая клетка получена из ядерной клетки-предшественника, над которой проводили химические или физические манипуляции для удаления ее ядра.
[0088] В определенных вариантах осуществления безъядерная гемопоэтическая клетка получена при помощи облучения или химического разрушения ядра ядерной клетки-предшественника. В некоторых вариантах осуществления химическое разрушение проведено с помощью цитохалазина B. В некоторых вариантах осуществления облучение проведено при помощи по меньшей мере 5 Гр, 7 Гр, 10 Гр, 15 Гр, 25 Гр, 30 Гр, 40 Гр или по меньшей мере 50 Гр.
[0089] В некоторых вариантах осуществления безъядерной гемопоэтической клетки, раскрытой в данном документе, экзогенный антиген представляет собой полипептид, кодируемый экзогенной нуклеиновой кислотой.
[0090] В некоторых вариантах осуществления безъядерной гемопоэтической клетки, раскрытой в данном документе, клетка происходит из источника, относящегося к человеку.
[0091] В некоторых вариантах осуществления безъядерной гемопоэтической клетки, раскрытой в данном документе, клетка происходит из источника, не относящегося к человеку, выбранного из группы, состоящей из свиньи, шимпанзе, макака, примата, не являющегося человеком, и млекопитающего, не являющегося приматом.
[0092] В некоторых вариантах осуществления безъядерной гемопоэтической клетки, раскрытой в данном документе, полипептидный антиген расположен внутри клетки. В некоторых вариантах осуществления безъядерной гемопоэтической клетки, раскрытой в данном документе, полипептидный антиген расположен вне клетки на поверхности клетки. В некоторых вариантах осуществления безъядерной гемопоэтической клетки, раскрытой в данном документе, полипептидный антиген слит с эндогенным клеточным белком. В некоторых вариантах осуществления безъядерной гемопоэтической клетки, раскрытой в данном документе, полипептидный антиген слит с внутриклеточным участком эндогенного трансмембранного белка. В некоторых вариантах осуществления безъядерной гемопоэтической клетки, раскрытой в данном документе, полипептидный антиген слит с внеклеточным участком эндогенного трансмембранного белка. В некоторых вариантах осуществления безъядерной гемопоэтической клетки, раскрытой в данном документе, полипептидный антиген слит с белком, заякоренным с помощью гликозилфосфатидилинозитола (GPI).
[0093] В некоторых аспектах данного документа раскрыта партия культуры ткани, содержащая безъядерные гемопоэтические клетки, описанные в данном документе.
[0094] В некоторых аспектах данного документа раскрыта популяция безъядерных гемопоэтических клеток, описанных в данном документе.
[0095] В некоторых аспектах данного документа раскрыта фармацевтическая композиция, содержащая популяции клеток, описанные в данном документе.
[0096] В некоторых аспектах данного документа раскрыт способ индукции иммунологической толерантности, включающий введение субъекту, нуждающемуся в этом, фармацевтических композиций, описанных в данном документе, в количестве и/или с частотой, достаточными для индукции иммунологической толерантности у субъекта.
[0097] В некоторых аспектах данного документа раскрыт способ лечения заболевания, обусловленного активацией иммунной системы, включающий введение субъекту, нуждающемуся в этом, фармацевтических композиций, описанный в данном документе, в количестве и/или с частотой, достаточными для лечения заболевания, обусловленного активацией иммунной системы.
[0098] В некоторых вариантах осуществления заболевание выбрано из группы, состоящей из заболевания, опосредованного аутореактивными антителами, аутоиммунного заболевания, воспалительного заболевания, аллергического заболевания, заболевания, опосредованного несоответствием HLA, и заболевания, излечимого с помощью иммуногенного терапевтического белка.
[0099] В некоторых аспектах данного документа раскрыт способ снижения или ослабления активации иммунной системы в ответ на схему лечения терапевтическим белком, включающий введение субъекту, нуждающемуся в этом, фармацевтических композиций, описанных в данном документе, в количестве и/или с частотой, достаточными для значительного снижения или ослабления активации иммунной системы.
[00100] В некоторых вариантах осуществления терапевтический белок выбран из группы, состоящей из перечисленных в таблице I, таблице J и таблице 7.
[00101] В некоторых аспектах данного документа раскрыт вектор экспрессии, содержащий последовательность нуклеиновой кислоты, кодирующую эндогенный белок клетки эритроидного ряда, слитый с одним или несколькими экзогенными полипептидными антигенами, выбранными из группы, состоящей из перечисленных в таблице F, таблице G, таблице H, таблице I, таблице J, таблице 6, таблице 7 и таблице 8.
[00102] В некоторых аспектах данного документа раскрыта матричная РНК, содержащая последовательность нуклеиновой кислоты, кодирующую эндогенный белок клетки эритроидного ряда, слитый с одним или несколькими экзогенными полипептидными антигенами, выбранными из группы, состоящей из перечисленных в таблице F, таблице G, таблице H, таблице I, таблице J, таблице 6, таблице 7 и таблице 8.
[00103] В некоторых аспектах данного документа раскрыт способ индукции иммунологической толерантности, включающий введение субъекту-человеку, страдающему от опосредованного аллергеном заболевания, нарушения или состояния или имеющему риск их развития, фармацевтической композиции, содержащей безъядерную гемопоэтическую клетку, экспрессирующую экзогенный антиген, где фармацевтическую композицию вводят в количестве, эффективном для индукции у субъекта иммунологической толерантности в отношении аллергена, опосредующего заболевание, нарушение или состояние.
[00104] В определенных вариантах осуществления экзогенный антиген выбран из группы, состоящей из Ara h2, 2S альбумина, гиалуронидазы и перечисленных в таблице H.
[00105] В определенных вариантах осуществления опосредованное аллергеном заболевание, нарушение или состояние выбрано из группы, состоящей из аллергии на арахис, аллергии на лесной орех, аллергии на яд насекомых и перечисленных в таблице H.
[00106] В некоторых аспектах данного документа раскрыт способ индукции иммунологической толерантности, включающий введение субъекту-человеку, страдающему от заболевания, нарушения или состояния, опосредованного несоответствием человеческих лейкоцитарных антигенов (HLA), или имеющему риск их развития, фармацевтической композиции, содержащей безъядерную гемопоэтическую клетку, экспрессирующую экзогенный антиген, где фармацевтическую композицию вводят в количестве, эффективном для индукции у субъекта иммунологической толерантности в отношении HLA, опосредующих заболевание, нарушение или состояние.
[00107] В некоторых аспектах данного документа раскрыт способ индукции иммунологической толерантности, включающий введение субъекту-человеку, страдающему от заболевания, нарушения или состояния, которые можно лечить с помощью иммуногенной терапевтической молекулы, или имеющему риск их развития, фармацевтической композиции, содержащей безъядерную гемопоэтическую клетку, экспрессирующую экзогенный антиген, где фармацевтическую композицию вводят в количестве, эффективном для индукции у субъекта иммунологической толерантности в отношении иммуногенных терапевтических молекул, применяемых для лечения заболевания, нарушения или состояния.
[00108] В некоторых вариантах осуществления терапевтическая молекула выбрана из группы, состоящей из Рекомбинанта (фактор VIII), Бенефикса (фактор IX), Хумиры (антитело к TNFα) и перечисленных в таблице I, таблице J и таблице 7.
КРАТКОЕ ОПИСАНИЕ ФИГУР
[00109] На фиг. 1 представлен ряд графиков с результатами проточной цитометрии красных кровяных клеток, приведенных в контакт с флуоресцентно меченным IgG, инкапсулированным внутри липосом. Показаны клетки, которые инкубировали в отсутствие липосом (слева), при низкой дозе липосом (по центру) и высокой дозе липосом (справа). На нижних гистограммах показана процентная доля клеток, которые являются флуоресцирующими.
[00110] На фиг. 2 представлен график уровней экспрессии на клеточной поверхности, оцениваемых с помощью количественной проточной цитометрии. На графике показаны различные рецепторы клеточной поверхности - гликофорин A (треугольники, соединенные сплошной линией), cKIT (квадратики, соединенные пунктирной линий), трансферриновый рецептор (ромбы, соединенные точечным пунктиром) - и экзогенный поверхностный трансген (незаштрихованные кружки) во время хода дифференцировки клетки эритроидного ряда.
[00111] На фиг. 3 A-C, F, I-M, O-Z и AA-AU представлен ряд графиков с результатами проточной цитометрии и вестерн-блот-анализа, которые демонстрируют экспрессию большого массива иллюстративных антигенов на поверхности, в цитоплазме, в виде слияний и в виде интактных белков в трех типах клеток, безъядерных клетках эритроидного ряда, ядерных клетках-предшественниках эритроидного ряда и эритролейкозных клетках.
[00112] На фиг. 3 A-C, F, I-M и O-S показана экзогенная экспрессия поверхностных и цитоплазматических белков в безъядерных культивируемых клетках эритроидного ряда.
[00113] Фиг. 3A - экспрессия гликофорина A с HA-эпитопной меткой на цитоплазматическом C-конце, оцениваемая по экспрессии совместно транслируемого GFP.
[00114] Фиг. 3B - экспрессия гликофорина A с HA-эпитопной меткой на N-конце между лидерной последовательностью и основной частью гена, оцениваемая по окрашиванию антителом к HA.
[00115] Фиг. 3C - экспрессия фрагмента, происходящего из рецептора комплемента 1, размером ~70 кДа с HA эпитопной меткой на N-конце, оцениваемая по окрашиванию антителом к HA.
[00116] Фиг. 3F - экспрессия scFv-антитела в виде N-концевого слияния с гликофорином A, оцениваемая по окрашиванию антителом к HA.
[00117] Фиг. 3I - экспрессия scFv-антитела, слитого с C-концом происходящего из Kell фрагмента из 71 аминокислоты, оцениваемая по окрашивания антителом к HA.
[00118] Фиг. 3J - экспрессия scFv-антитела, слитого с C-концом происходящего из Kell фрагмента из 79 аминокислот, оцениваемая по окрашиванию антителом к HA.
[00119] Фиг. 3K - экспрессия CD55 с HA-эпитопной меткой на внеклеточном N-конце после лидерной последовательности, оцениваемая по окрашиванию антителом к HA.
[00120] Фиг. 3L - экспрессия CD59 с HA-эпитопной меткой на внеклеточном N-конце после лидерных последовательностей, оцениваемая по окрашиванию антителом к HA.
[00121] Фиг. 3M - экспрессия scFv-антитела, слитого с N-концом происходящего из CD55 фрагмента из 37 аминокислот, оцениваемая с помощью вестерн-блот-анализа с антителом к HA.
[00122] Фиг. 3O - цитоплазматическая экспрессия аденозиндезаминазы, слитой с HA-меткой, оцениваемая с помощью вестерн-блот-анализа с антителом к HA. Ожидаемый размер приблизительно 40 кДа.
[00123] Фиг. 3P - цитоплазматическая экспрессия фенилаланингидроксилазы, слитой с HA-меткой, оцениваемая с помощью вестерн-блот-анализа с антителом к HA. Ожидаемый размер приблизительно 33 кДа.
[00124] Фиг. 3Q - цитоплазматическая экспрессия фенилаланингидроксилазы, слитой с аденозиндезаминазой и HA-меткой, оцениваемая с помощью вестерн-блот-анализа с антителом к HA.
[00125] Фиг. 3R - цитоплазматическая экспрессия аденозиндезаминазы, слитой с внутриклеточным C-концом гликофорина A, оцениваемая с помощью вестерн-блот-анализа с антителом к HA. Ожидаемый размер приблизительно 55 кДа.
[00126] Фиг. 3S - цитоплазматическая экспрессия фенилаланингидроксилазы, слитой с внутриклеточным C-концом гликофорина A, оцениваемая с помощью вестерн-блот-анализа с антителом к HA. Ожидаемый размер приблизительно 50 кДа.
[00127] На фиг. 3 T-AO показана экзогенная экспрессия поверхностных и цитоплазматических белков в ядерных культивируемых клетках-предшественниках эритроидного ряда.
[00128] Фиг. 3T - экспрессия гликофорина A с HA-эпитопной меткой на цитоплазматическом C-конце, оцениваемая по экспрессии совместно транслируемого GFP.
[00129] Фиг. 3U - экспрессия гликофорина A с HA-эпитопной меткой на N-конце между лидерной последовательностью и основной частью гена, оцениваемая по окрашиванию антителом к HA.
[00130] Фиг. 3V - сверхэкспрессия рецептора комплемента 1, оцениваемая по окрашиванию антителом к CR1.
[00131] Фиг. 3W - экспрессия фрагмента, происходящего из рецептора комплемента 1, размером ~70 кДа с HA-эпитопной меткой на N-конце, оцениваемая по окрашиванию антителом к HA.
[00132] Фиг. 3X - экспрессия фрагмента, происходящего из рецептора комплемента 1, размером ~210 кДа с HA-эпитопной меткой на N-конце, оцениваемая по окрашиванию антителом к HA.
[00133] Фиг. 3Y - экспрессия фрагмента, происходящего из рецептора комплемента 1, размером ~230 кДа, слитого с N-концом гликофорина A с HA-эпитопной меткой на N-конце, оцениваемая по окрашиванию антителом к HA.
[00134] Фиг. 3Z - экспрессия scFv-антитела в виде N-концевого слияния с гликофорином A, оцениваемая по окрашиванию антителом к HA.
[00135] Фиг. 3AA - экспрессия scFv-антитела, слитого с внеклеточным C-концом Kell, оцениваемая по окрашиванию антителом к HA. Ожидаемый размер приблизительно 108 кДа.
[00136] Фиг. 3AB - экспрессия HA-метки, слитой с внеклеточным C-концом Kell, оцениваемая по окрашиванию антителом к HA.
[00137] Фиг. 3AC - экспрессия происходящего из Kell фрагмента из 71 аминокислоты с HA-меткой на C-конце (внеклеточном), оцениваемая по окрашиванию антителом к HA.
[00138] Фиг. 3AD - экспрессия происходящего из Kell фрагмента из 79 аминокислот с HA-меткой на C-конце, оцениваемая по окрашиванию антителом к HA.
[00139] Фиг. 3AE - экспрессия scFv-антитела, слитого с C-концом происходящего из Kell фрагмента из 71 аминокислоты, оцениваемая по окрашиванию антителом к HA.
[00140] Фиг. 3AF - экспрессия scFv-антитела, слитого с C-концом происходящего из Kell фрагмента из 79 аминокислот, оцениваемая по окрашиванию антителом к HA.
[00141] Фиг. 3AG - экспрессия CD55 с HA-эпитопной меткой на внеклеточном N-конце после лидерной последовательности, оцениваемая по окрашиванию антителом к HA.
[00142] Фиг. 3AH - экспрессия CD59 с HA-эпитопной меткой на внеклеточном N-конце после лидерных последовательностей, оцениваемая по окрашиванию антителом к HA.
[00143] Фиг. 3AI - экспрессия scFv-антитела, слитого с N-концом происходящего из CD55 фрагмента из 37 аминокислот, оцениваемая по окрашиванию антителом к HA.
[00144] Фиг. 3AJ - экспрессия scFv-антитела, слитого с N-концом CD59, оцениваемая по окрашиванию антителом к HA.
[00145] Фиг. 3AK - цитоплазматическая экспрессия аденозиндезаминазы, слитой с HA-меткой, оцениваемая с помощью вестерн-блот-анализа с антителом к HA. Ожидаемый размер приблизительно 40 кДа.
[00146] Фиг. 3AL - цитоплазматическая экспрессия фенилаланингидроксилазы, слитой с HA-меткой, оцениваемая с помощью вестерн-блот-анализа с антителом к HA. Ожидаемый размер приблизительно 33 кДа.
[00147] Фиг. 3AM - цитоплазматическая экспрессия фенилаланингидроксилазы, слитой с аденозиндезаминазой и HA-меткой, оцениваемая с помощью проточной цитометрии по флуоресценции от совместно транслируемого GFP.
[00148] Фиг. 3AN - цитоплазматическая экспрессия аденозиндезаминазы, слитой с внутриклеточным C-концом гликофорина A, оцениваемая с помощью вестерн-блот-анализа с антителом к HA. Ожидаемый размер приблизительно 55 кДа.
[00149] Фиг. 3AO - цитоплазматическая экспрессия фенилаланингидроксилазы, слитой с внутриклеточным C-концом гликофорина A, оцениваемая с помощью вестерн-блот-анализа с антителом к HA. Ожидаемый размер приблизительно 50 кДа.
[00150] На фиг. 3 AP-AU показана экзогенная экспрессия поверхностных и цитоплазматических белков на эритролейкозных клетках K562.
[00151] Фиг. 3AP - сверхэкспрессия рецептора комплемента 1, оцениваемая по окрашиванию антителом к CR1.
[00152] Фиг. 3AQ - экспрессия scFv-антитела в виде N-концевого слияния с гликофорином A, оцениваемая по окрашиванию антителом к HA.
[00153] Фиг. 3AR - экспрессия scFv-антитела, слитого с N-концом происходящего из CD55 фрагмента из 37 аминокислот, оцениваемая по окрашиванию антителом к HA.
[00154] Фиг. 3AS - экспрессия scFv-антитела, слитого с N-концом CD59, оцениваемая по окрашиванию антителом к HA.
[00155] Фиг. 3AT - цитоплазматическая экспрессия аденозиндезаминазы, слитой с HA-меткой, оцениваемая с помощью вестерн-блот-анализа с антителом к HA. Ожидаемый размер приблизительно 40 кДа.
[00156] Фиг. 3AU - цитоплазматическая экспрессия фенилаланингидроксилазы, слитой с HA-меткой, оцениваемая с помощью вестерн-блот-анализа с антителом к HA. Ожидаемый размер приблизительно 33 кДа.
[00157] На фиг. 4 представлен ряд гистограмм с результатами проточной цитометрии, в которых измерена флуоресценция в первичных тромбоцитах, которые были трансфицированы с помощью мРНК, кодирующей флуоресцентный белок (GFP). (A) Нетрансфицированные тромбоциты. (B) Тромбоциты, трансфицированные с помощью 3 мкг мРНК GFP. (4C) Тромбоциты, трансфицированные с помощью 6,8 мкг мРНК GFP.
[00158] На фиг. 5 показана экспрессия белка и ферментативная активность трансгенных клеток эритроидного ряда в культуре. (A) представляет собой вестерн-блот-анализ экзогенно экспрессируемой аденозиндезаминазы, обнаруживаемой с помощью антитела к HA в ходе дифференцировки от ядерных клеток-предшественников ("Diff I D5") вплоть до безъядерных клеток эритроидного ряда ("Diff III D8"). (B) представляет собой столбчатую диаграмму, отражающую инозин, полученный из аденозина с помощью интактных клеток 293Т, экспрессирующих аденозиндезаминазу. (C) представляет собой вестерн-блот-анализ экзогенно экспрессируемой фенилаланингидроксилазы, обнаруживаемой с помощью антитела к HA в разные моменты времени в ходе дифференцировки от ядерных клеток-предшественников ("Diff I D5") вплоть до безъядерных клеток эритроидного ряда ("Diff III D8"). (D) представляет собой столбчатую диаграмму, отражающий тирозин, полученный из фенилаланина с применением лизатов из культивируемых безъядерных клеток эритроидного ряда, экспрессирующих фенилаланингидроксилазу.
[00159] На фиг. 6 показан захват иммунных комплексов и перенос их к макрофагам за счет культивируемых клеток эритроидного ряда, которые сверхэкспрессируют рецептор комплемента 1 (CR1). (A) представляет собой график результатов проточной цитометрии, на котором показан захват флуоресцентных иммунных комплексов (белая гистограмма) и иммунных комплексов без комплемента (зашрихованная гистограмма) за счет культивируемых клеток эритроидного ряда, которые сверхэкспрессируют CR1. (B) представляет собой столбчатую диаграмму на основе данных проточной цитометрии, оценивающую поглощение флуоресцентных иммунных комплексов (заштрихованные столбцы), иммунных комплексов без комплемента (серые столбцы) или при отсутствии иммунных комплексов (черные столбцы) за счет макрофагов (левый набор) или макрофагов, инкубируемых с культивируемыми клетками эритроидного ряда, которые сверхэкспрессируют CR1 (правый набор).
[00160] На фиг. 7 показана активность scFv-антитела, которое связывает поверхностный антиген гепатита B (scFv), на поверхности культивируемой клетки эритроидного ряда. (A) представляет собой гистограмму результатов проточной цитометрии, на которой показано связывание 450 нM антигена (белая гистограмма) или отсутствие связывания антигена (серая гистограмма). (B) представляет собой титриметрический анализ сигнала связывания, оцениваемый с помощью проточной цитометрии в отношении диапазона концентраций антигена. (C-D) представляют собой графики результатов проточной цитометрии кровяных клеток от мышей, которым был инъецирован флуоресцентный антиген и культивируемые клетки эритроидного ряда, которые (C) не экспрессируют или (D) экспрессируют scFv. По оси Y отложена флуоресценция антигена. По оси X отложена флуоресценция культивируемых клеток.
[00161] На фиг. 8 показан специфичный клиренс циркулирующих в крови антител, опосредованный экспрессирующими экзогенный антиген EHC-клетками in vivo. (A) представляет собой набор графиков результатов проточной цитометрии, которые демонстрируют отсутствие связывания (вверху) и связывание (внизу) циркулирующих в крови Dylight650-меченных мышиных антител к HA с помощью CFSE-меченных культивируемых человеческих клеток эритроидного ряда, которые либо не экспрессируют (вверху), либо экспрессируют (внизу) HA-эпитопную метку на своей поверхности, при выделении из крови мыши-реципиента. По оси Х отложена флуоресценция CFSE. По оси Y отложена флуоресценция Dylight650 на антителе к HA. (B) представляет собой данные ELISA с субстратом HA-эпитопной меткой для сравнения уровней антитела к HA в зависимости от времени в плазме крови, собранной от мышей, которым инъецировали антитело к HA (незаштрихованные кружки, сплошная линия), антитело к HA с последующей инъекцией культивируемых человеческих клеток эритроидного ряда, которые не экспрессировали HA-эпитопную метку (пунктирная линия), или антитело к HA с последующей инъекцией культивируемых человеческих клеток эритроидного ряда, которые экспрессировали HA-эпитопную метку (точечная пунктирная линия). (C) представляет собой набор графиков результатов проточной цитометрии, которые демонстрируют отсутствие связывания (вверху) и связывание (внизу) Dylight650-меченного мышиного антитела к биотину с CFSE-меченными первичными человеческими эритроцитами, которые либо не были конъюгированы (вверху), либо были конъюгированы (внизу) с биотином на своей поверхности. По оси Х отложена флуоресценция CFSE. По оси Y отложена флуоресценция Dylight650 на антителе к биотину. (D) представляет собой данные ELISA с субстратом биотином для сравнения уровней антитела к биотину в зависимости от времени в плазме крови, собранной от мышей, которым инъецировали антитело к биотину (незаштрихованные кружки, сплошная линия), антитело к биотину с последующей инъекцией культивируемых человеческих клеток эритроидного ряда, которые не были конъюгированы с биотином (пунктирная линия), или антитело к биотину с последующей инъекцией культивируемых человеческих клеток эритроидного ряда, которые были конъюгированы с биотином (точечная пунктирная линия).
[00162] На фиг. 9 показана скорость клиренса культивируемых человеческих клеток эритроидного ряда у мыши. (A) представляет собой репрезентативную точечную диаграмму результатов проточной цитометрии свежевзятой крови, окрашенной в отношении человеческого гликофорина A (ось Y) и CFSE (ось Х), в которой культивируемые человеческие клетки являются двойными положительными. (B) представляет собой график скорости клиренса в зависимости от времени, представленный как процентная доля двойных положительных клеток, сохраняющихся после того, как мышам NSG инъецировали человеческие красные кровяные клетки (кружки, соединенные сплошной линией), культивируемые безъядерные клетки эритроидного ряда (ромбы, соединенные пунктиром), культивируемые безъядерные клетки эритроидного ряда, которые экспрессируют внутриклеточный экзогенный белок (квадратики, соединенные точечным пунктиром), и культивируемые безъядерные клетки эритроидного ряда, которые экспрессируют поверхностный экзогенный белок (незакрашенные треугольники).
[00163] На фиг. 10 представлена оценка побочных эффектов после инъекции культивируемых человеческих клеток эритроидного ряда в мышь. (A-B) демонстрируют уровни (A) фибринопептида A и (B) фибринопептида B, оцениваемые с помощью ELISA в плазме крови, собранной у мышей через 20 минут (черный), 6 часов (серый) и 48 часов (белый) после инъекции (1) человеческих красных кровяных клеток, (2) культивируемых человеческих клеток эритроидного ряда, (3) культивируемых человеческих клеток эритроидного ряда, экспрессирующих экзогенный цитоплазматический белок, (4) культивируемых человеческих клеток эритроидного ряда, экспрессирующих экзогенный поверхностный трансген, или (5) рекомбинантного белка. (C-D) демонстрируют изображения под микроскопом окрашенных гистологических срезов селезенки мышей, которым инъецировали (C) культивируемые человеческие клетки эритроидного ряда и (D) рекомбинантный белок.
[00164] На фиг. 11 показано отслеживание экспрессии экзогенного белка на культивируемых клетках эритроидного ряда в кровотоке. (A) представляет собой данные проточной цитометрии крови, собранной у мыши, которой инъецировали культивируемые человеческие клетки эритроидного ряда, экспрессирующие экзогенный поверхностный белок, где показан процент культивируемых человеческих клеток эритроидного ряда, которые были HA-положительными, в зависимости от времени. (B) представляет собой данные вестерн-блот-анализа крови, собранной у двух мышей, где одной мыши инъецировали культивируемые человеческие клетки эритроидного ряда, экспрессирующие экзогенный цитоплазматический белок, и где другой мыши инъецировали очищенный рекомбинантно полученный экзогенный белок в отсутствие каких-либо клеток, где показан уровень HA-содержащего белка в крови в зависимости от времени.
[00165] На фиг. 12 представлена оценка размножения и дифференцировки культивируемых человеческих клеток эритроидного ряда. (A) представляет собой график скоростей размножения у отдельных культур in vitro дифференцированных клеток эритроидного ряда, которые содержат трансгены (пунктирная линия и точечная пунктирная линия), и клеток, которые не содержат трансген (сплошная линия). (B) представляет собой график результатов проточной цитометрии маркеров клеточной поверхности GPA и CKIT для отдельных культур культивируемых человеческих клеток эритроидного ряда, которые не содержат (слева) или содержат (справа) трансген. (C) представляет собой график результатов проточной цитометрии культивируемых человеческих клеток эритроидного ряда, которые не содержат (слева) или содержат (справа) трансген, где клетки окрашивали с помощью красителя ДНК DRAQ5 (ось Y) и антитела к гликофорину A (ось Х), при этом на котором идентифицированы отдельные популяции (1) безъядерных клеток, (2) ядерных клеток и (3) ядер.
[00166] Фигура 13A представляет собой схему трех способов, с помощью которых антиген может быть локализован в экспрессирующей экзогенный антиген EHC. B представляет собой схему трех способов, с помощью которых антиген, локализованный в экспрессирующей экзогенный антиген EHC или на ней, может действовать на мишень в кровотоке. C представляет собой схему автокаталитического слияния эндогенного полипептидного якоря с антигеном с использованием механизма SpyTag-SpyCatcher.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[00167] В определенных аспектах настоящего изобретения представлены выделенные клетки, которые получены с помощью способов генной инженерии или модифицированы, чтобы они содержали экзогенные антигены, представляющие интерес. В определенных аспектах выделенные EHC-клетки по настоящему изобретению содержат один или несколько антигенов, которые содержат полипептиды или состоят из них. В некоторых вариантах осуществления антиген представляет собой полноразмерный белок. В некоторых вариантах осуществления антиген составлен из одного или нескольких полипептидов, содержащихся в пределах полноразмерного белка, имеющих любую длину, превышающую приблизительно 7 аминокислот. Полипептиды, содержащие антиген, могут содержать один или несколько иммунных эпитопов, которые могут быть конформационными эпитопами или могут быть линейными эпитопами. Антиген может быть составлен из одного или нескольких полипептидов из одного или нескольких различных белков. В определенных аспектах EHC-клетки по настоящему изобретению содержат один или несколько антигенов, которые содержат углеводы или состоят из них. В определенных аспектах EHC-клетки по настоящему изобретению содержат один или несколько антигенов, которые содержат липиды или состоят из них. В определенных аспектах EHC-клетки по настоящему изобретению содержат один или несколько антигенов, которые содержат один или несколько полипептидов, липидов и/или углеводов и любую их комбинацию или состоят из них. Клетки могут представлять собой циркулирующие в крови клетки, такие как EHC-клетки. EHC-клетки можно культивировать из гемопоэтических предшественников с применением определенных факторов, таких как, например, фактор стволовых клеток, цитокины, такие как IL-3 и IL-6, инсулин, трансферрин, эритропоэтин, гидрокортизон и эстрогены.
[00168] Аспекты настоящего изобретения относятся к способам культивирования EHC-клеток, чтобы они содержали экзогенные антигены, представляющие интерес. Экзогенные антигены, представляющие интерес, можно вводить с помощью целого ряда способов, таких как, например, внутриклеточная экспрессия, экспрессия на поверхности клетки, слияние с эндогенным белком, конъюгация с помощью химических или ферментативных средств с белком клеточной поверхности или физическая загрузка во внутриклеточное пространство. Антигенсодержащие клетки по настоящему изобретению можно применять в качестве терапевтических средств.
[00169] Аспекты настоящего изобретения относятся к применению этих антигенсодержащих клеток в лечении заболеваний, обусловленных активацией иммунной системы, посредством индукции периферической толерантности. В некоторых аспектах индукция периферической толерантности означает делецию или инактивацию антиген-специфичных иммунных клеток, таких как, например, CD8 T-лимфоциты (CD8 T-клетки), CD4 T-лимфоциты (CD4 T-клетки), CD4 регуляторные T-лимфоциты (Treg) или B-лимфоциты (B-клетки). Заболевания, обусловленные активацией иммунной системы, включают аутоиммунные заболевания, такие как, например, рассеянный склероз, сахарный диабет 1 типа, ревматоидный артрит и мембранозный нефрит. Заболевания, обусловленные активацией иммунной системы, также включают воспалительные заболевания, такие как, например, болезнь Крона, язвенный колит, целиакия или другие идиопатические воспалительные заболевания кишечника. Заболевания, обусловленные активацией иммунной системы, также включают аллергические заболевания, такие как, например, астма, аллергия на арахис, аллергия на морепродукты, аллергия на пыльцу, аллергия на белок молока, аллергия на укусы насекомых и аллергия на латекс. Заболевания, обусловленные активацией иммунной системы, также включают активацию иммунной системы в ответ на терапевтический белок, вводимый для лечения основного заболевания, что уменьшает эффективность терапевтического белка, такого как, например, фактор свертывания VIII при гемофилии A, фактор свертывания IX при гемофилии B, антитела к фактору некроза опухоли альфа (TNFa) при ревматоидном артрите и других воспалительных заболеваниях, глюкоцереброзидаза при болезни Гоше или аспарагиназа при остром лимфобластном лейкозе (ALL).
Биология иммунологической толерантности
[00170] У организма возникли сложные механизмы для предотвращения аберрантной активации иммунной системы и аутоиммунных заболеваний, совместно называемые иммунологическая толерантность. Центральная толерантность относится к антиген-специфичной делеции аутореактивных T-клеток и B-клеток во время развития в первичных лимфоидных органах, например, в тимусе и костном мозге. Периферическая толерантность относится к делеции или инактивации зрелых T- и B-лимфоцитов вне первичных лимфоидных органов. Периферическая толерантность включает супрессию аутореактивных лимфоцитов с помощью регуляторных T-клеток (Treg) или индукцию анергии или отсутствия реактивности у антиген-специфичных эффекторных лимфоцитов путем продолжительного воздействия низкими дозами антигена в отсутствие костимулирующих сигналов "опасности". Как активацию Treg, так и анергию лимфоцитов можно индуцировать путем секреции ингибирующих факторов, таких как, например, TGF-бета, IL-10 и IL-4.
[00171] Для активации иммунной системы в ответ на антиген часто требуется вторичный сигнал "опасности", такой как лиганд толл-подобного рецептора, зачастую происходящий из микробных или вирусный патогенов (Matzinger, Annu Rev Immuno 1994). Такие сигналы опасности включают двухнитевую РНК, однонитевую ДНК, липополисахарид, бактериальные липопротеины, флагеллин, зимозан и другие. Антигенпрезентирующие клетки, которые получают как антиген, так и сигнал опасности, помещают на своей поверхности костимулирующие молекулы, подобные CD80 и CD86, в дополнение к антигенным пептидам. T-клетки, которые распознают как антигенный пептид, так и костимулирующие молекулы, становятся активированными. Клетки, которые получают только сигнал антигенного пептида, становятся анергическими.
[00172] Терапевтические стратегии, в которых для индукции иммунологической толерантности используется презентация антигена в отсутствие сигналов опасности, были разработаны для экспериментального лечения многих типов пищевой аллергии. Исследования имеют вид длительного воздействия возрастающих доз аллергена с целью индукции толерантности. В тринадцати исследованиях, начиная с 2007 года, в данном формате протестировали целый ряд широко распространенных пищевых аллергенов, таких как арахис, молоко и яйцо. 50-100% пациентов становились сенсибилизированными, то есть, способными переживать пищевую провокацию без развития анафилаксии. Однако долгосрочная толерантность была менее успешной, при этом только 25-50% пациентов были способны переносить антиген через один месяц после прекращения терапии. См., например, Burks et al., New England Journal of Medicine 2012.
[00173] Полагают, что аллергия опосредована IgE при активации тучных клеток и базофилов. Пероральное введение аллергена в низкой дозе, как, например, непрерывное введение с пищей, индуцирует Treg посредством презентации антигена CD11c+ дендритными клетками и секрецией TGF-бета, IL-10 и IL-4. Пероральное введение при высоких дозах индуцирует антиген-специфичную делецию и анергию T-клеток посредством плазмацитоидных дендритных клеток. В исследованиях на человеке пероральное введение аллергена приводит к снижению IgE, тучных клеток и базофилов, повышению IgG4, TGF-бета, IL-10 и временному повышению числа Treg в начале терапии. См., например, Herzog, Adv Drug Deliv Rev 2013.
[00174] Без желания ограничиваться каким-либо конкретным механизмом, полагают, что периферическая иммунологическая толерантность может индуцироваться аутореактивными антигенами из подвергающихся апоптозу клеток (Griffith and Ferguson, Immunity 2011; Green et al., Nat Rev Immunol 2009). Хотя точные механизмы с молекулярной точки зрения не полностью понятны, собственные белки, такие как HSP90 и другие молекулярные структуры, ассоциированные с повреждениями, облегчают поглощение дендритными клетками. Рецепторы дендритных клеток, например CD205, распознают такие сигналы, кросс-презентируют антиген и индуцируют толерогенные цитокины, а также супрессируют экспрессию костимулирующего белка (Bonifaz, J Exp Med 2002).
[00175] Исследуются терапевтические стратегии, которые эксплуатируют толерогенный потенциал подвергающихся апоптозу клеток для индукции периферической иммунологической толерантности. Как правило, данные стратегии предусматривают химическое присоединение антигенов, представляющих интерес, к поверхности клеток. В исследованиях на мышах, крысах и морских свинках целый ряд белковых антигенов химически прикрепляли к поверхности спленоцитов и лейкоцитов. См., например, Miller et al., J Exp Med 1979; Braley-Mullen et al., Cell Immunol 1980; Luo et al., PNAS 2008; Smarr et al., J Immunol 2011.
[00176] В недавнем клиническом исследовании фазы I на человеке смесь пептидных антигенов, ассоциированных с рассеянным склерозом, химически присоединяли к аутологичным мононуклеарам периферической крови и подвергали реинфузии в пациентов (Lutterotti and Martin, Sci Trans Med 2013). Эти клетки хорошо переносились, и имелись доказательства снижения антиген-специфичных T-клеточных ответов.
[00177] EHC-клетки представляют собой известный источник погибающих клеток. Каждый день огромное число эритроцитов подвергается клиренсу после апоптозоподобной запрограммированной клеточной гибели, называемой эриптоз (у человека более 1% в день, приблизительно 1×1011 клеток). Хотя точные триггерные факторы клиренса эритроцитов остаются неясными, подвергающиеся эриптозу эритроциты характеризуются асимметрией фосфатидилсерина, гетерогенностью мембраны и связыванием аннексина-V, что напоминает апоптотические ядерные клетки.
[00178] EHC-клетки также длительно сохраняются в организме. Эритроциты находятся в крови взрослого человека в течение до 120 дней. В связи с этим, EHC-клетки, которые содержат антиген, представляющий интерес, могут обеспечивать возможность долгосрочного воздействия антигена на хозяина. Как описано выше, хотя точные молекулярные механизмы не полностью понятны, полагают, что долгосрочное воздействие антигена может индуцировать периферическую толерантность посредством презентации антигена в отсутствие костимулирующих сигналов, что приводит к экспансии регуляторных T-клеток, делеции и анергии эффекторных T- и B-клеток и секреции противовоспалительных и протолерогенных цитокинов.
[00179] Индукция периферической толерантности путем использования эритроцитов также была исследована экспериментальным путем. В предварительных изысканиях было показано, что модельный антиген овальбумин индуцирует антиген-специфичную делецию CD8 T-клеток и антиген-специфичную индукцию Treg в случае нековалентного прикрепления к эритроцитам (Kontos et al., PNAS 2013) или осмотической загрузки в эритроциты (Cremel and Godfrin, Int J Pharm 2013).
[00180] Культивируемые EHC-клетки по настоящему изобретению, содержащие экзогенный антиген, представляющий интерес, могут обладать явными преимуществами по сравнению с антигеном, который нековалентно прикреплен к эритроцитам, например, посредством полипептид-связывающего домена. Одним преимуществом может быть то, что биораспределение EHC, содержащей экзогенный антиген, представляющий интерес, является более определенным, чем у полипептидной композиции антигена с нацеливающим доменом. EHC, содержащая экзогенный антиген, представляющий интерес, будет удерживаться в сосудистой системе и в местах, в которых, как правило, пребывают эритроциты, например, селезенке. EHC-клетки, содержащие экзогенный антиген, представляющий интерес, не будут отфильтровываться в почках или выходить в периферическую ткань, что является проблемами, которые могут возникать при введении композиций на основе полипептидного антигена. Доза экзогенного антигена на EHC может быть значительно выше, если клетки, содержащие экзогенный антиген, представляющий интерес, подвергают культивированию, чем если полипептидный антиген напрямую инъецируют в кровяное русло и он распределяется среди приблизительно 10 триллионов эритроцитов в кровяном русле. В некоторых случаях может быть предпочтительным иметь экзогенный антиген, представляющий интерес, удерживаемый во внутриклеточном компартменте EHC. Например, если антиген является иммуногенным, внутриклеточная локализация может быть преимущественной, поскольку она может маскировать иммуногенный антиген от иммунной системы и, таким образом, предотвращать или снижать активацию иммунной системы. Эта конфигурация невозможна в случае композиции на основе полипептидного антигена.
[00181] Культивируемые EHC-клетки, которые экспрессируют экзогенный антиген, представляющий интерес, могут иметь явные преимущества по сравнению с антигеном, который осмотически загрузили в EHC-клетки. Культивируемые EHC-клетки, содержащие экзогенный антиген, представляющий интерес, будут иметь клеточные мембраны и цитоскелет, которые в значительной степени не подверглись изменениям, в отличие от продукта процедуры осмотической загрузки, при которой крупные поры нарушают целостность клеточной мембраны и цитоскелета. Морфология и биофизические характеристики EHC-клеток являются главными факторами, определяющими биораспределение, циркуляцию в кровотоке и взаимодействие данных клеток с сосудистой системой и иммунными клетками (например, Pries et al., Cardiovasc. Res. 1996), и, следовательно, поддержание целостности клетки может иметь решающее значение для сохранения эффективности. Экзогенный антиген, который физически прикреплен к культивируемой EHC, например, посредством прямого слияния с эндогенным цитоплазматическим белком или слияния с эндогенным трансмембранным белком, не будет просачиваться из клетки и будет воздействовать на иммунную систему до тех пор, пока EHC не подвергнутся уничтожению. Проблема просачивания может возникнуть, если клетку приводят в контакт с антигеном с применением процедуры осмотической загрузки, при которой клеточная мембрана может повреждаться.
[00182] Культивируемые EHC-клетки, которые экспрессируют антиген, представляющий интерес, можно вводить напрямую субъекту, нуждающемуся в таком антигене. Отделение и очистка антигена во время производства продукта не требуется. В этом состоит отличие от продукта осмотической загрузки, при которой антиген следует синтезировать и очищать отдельно, а затем объединять с клеткой, и это может обеспечивать значительный выигрыш в затратах и времени при производстве продукта. Число культивируемых EHC-клеток, которые экспрессируют антиген, представляющий интерес, можно пропорционально увеличивать с помощью размножения в культуре. Можно вырабатывать крупные, промышленные партии клеток с получением в значительной степени однородной фармацевтической композиции из EHC-клеток для данного антигена, которую можно применять для универсального лечения многих субъектов. В отличие от этого, осмотическая загрузка обычно ограничивается масштабом "от одного донора-одному субъекту".
Получение клеток
[00183] Экспрессирующие экзогенный антиген безъядерные гемопоэтические клетки по настоящему изобретению можно получать с помощью любого способа, описанного в данном документе. В некоторых вариантах осуществления стадии включают приведение в контакт выделенных, необязательно подвергнутых культивированию клеток, происходящих от гемопоэтических стволовых клеток, с антигеном. Гемопоэтические стволовые клетки служат источником всех типов кровяных клеток, обнаруженных в крови млекопитающих, в том числе миелоидной (моноциты и макрофаги, нейтрофилы, базофилы, эозинофилы, эритроциты, мегакариоциты/тромбоциты, дендритные клетки) и лимфоидной линий (T-клетки, B-клетки, NK-клетки). Гемопоэтические стволовые клетки можно выделять из костного мозга костей взрослого организма, в том числе, например, кости бедра, тазовой кости, ребра или грудины. Клетки можно получать напрямую из тазовой кости, например, путем вымывания клеток из костного мозга с применением аспирации с помощью иглы и шприца. В качестве альтернативы, гемопоэтические стволовые клетки можно выделять из нормальной периферической крови после предварительно обработки с помощью цитокинов, таких как, например, гранулоцитарный колониестимулирующий фактор (G-CSF). G-CSF активирует высвобождение клеток из компартмента костный мозг в периферический кровоток. Другие источники гемопоэтических стволовых клеток включают пуповинную кровь и плаценту.
[00184] Выделенные гемопоэтические стволовые клетки можно культивировать, размножать и дифференцировать ex vivo с получением разнообразного исходного материала для получения экспрессирующих экзогенный антиген EHC-клеток. Например, гемопоэтические стволовые клетки, выделенные из костного мозга, простимулированной цитокинами периферической крови или пуповинной крови можно размножать и дифференцировать ex vivo в зрелые эритроциты (Giarratana et al., Nature Biotech. 23:69-74 (2005); заявка на патент США 2007/0218552). В связи с этим, CD34+ клетки выделяют из костного мозга или периферической или пуповинной крови с применением, например, отбора с магнитными микрогранулами и колонок Mini-MACS (Miltenyi Biotech). В одном примере клетки последовательно культивируют в модифицированной бессывороточной среде, дополненной 1% бычьего сывороточного альбумина (BSA), 120 мг/мл насыщенного железом человеческого трансферрина, 900 нг/мл сульфата двухвалентного железа, 90 нг/мл нитрата трехвалентного железа и 10 мг/мл инсулина, и поддерживали при 37°C с 5% диоксида углерода в атмосфере воздуха. Размножение и дифференцировка клеточной культуры могут происходить в несколько стадий. Например, на стадии первоначального выращивания клетки можно размножать в среде, описанной в данном документе, в присутствии множества факторов роста, в том числе, например, гидрокортизона, фактора стволовых клеток, IL-3 и эритропоэтина. На второй стадии, клетки необязательно можно подвергать совместному культивированию, например, на слое прикрепленных стромальных клеток в присутствии эритропоэтина. На третьей стадии клетки можно культивировать на слое прикрепленных стромальных клеток в культуральной среде в отсутствие экзогенных факторов. Слой прикрепленных стромальных клеток может состоять, например, из мышиных стромальных клеток MS-5. В качестве альтернативы, слой прикрепленных стромальных клеток может состоять из мезенхимальных стромальных клеток, происходящих из костного мозга взрослого организма. Прикрепленные стромальные клетки можно поддерживать на RPMI, дополненной, например, 10% фетальной телячьей сыворотки. В некоторых вариантах осуществления клетки-предшественники эритроидного ряда и клеточные популяции, происходящие из них, не подвергают совместному культивированию с клетками, не относящимися к EHC, например, со слоем прикрепленных стромальных клеток, т.е. их культивируют в отсутствие клеток, не относящихся к EHC. В некоторых вариантах осуществления EHC-клетки, содержащие антиген, культивируют в отсутствие клеток, не относящихся к EHC, и подвергают дифференцировке, так что более 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или более 98% EHC-клеток являются безъядерными, и популяцию безъядерных клеток получают без стадии обогащения, такой как гравитационное разделение, магнитная или флуоресцентная сортировка, облучение, применение яда для ядерных клеток и т.п., для отбора безъядерных клеток.
[00185] В некоторых случаях может быть желательным размножить и подвергнуть частичной дифференцировке CD34+ гемопоэтические стволовые клетки in vitro и обеспечить возможность протекания окончательной дифференцировки в зрелые эритроциты in vivo (см., например, Neildez-Nguyen et al., Nature Biotech. 20:467-472 (2002)). Выделенные CD34+ гемопоэтические стволовые клетки можно размножать in vitro в отсутствие слоя прикрепленных стромальных клеток в среде, содержащей различные факторы, в том числе, например, лиганд Flt3, фактор стволовых клеток, тромбопоэтин, эритропоэтин и инсулиновый фактор роста. Полученные клетки-предшественники эритроидного ряда могут характеризоваться экспрессией CD36 и GPA на поверхности клетки, и их можно переливать в субъекта, где обеспечивается возможность протекания окончательной дифференцировки в зрелые эритроциты.
[00186] В некоторых вариантах осуществления популяция EHC содержит множество безъядерных EHC-клеток, которые содержат полипептид антигена, который сохраняется во время удаления ядра. Полученная выделенная безъядерная EHC, содержащая полипептид антигена, проявляет практически такую же хрупкость осмотической мембраны, что и соответствующая выделенная немодифицированная не подвергнутая культивированию EHC.
[00187] В некоторых вариантах осуществления популяция EHC содержит множество клеток-предшественников эритроцитов на практически той же стадии дифференцировки и/или стадии клеточного цикла, при этом клетки-предшественники содержат рекомбинантную нуклеиновую кислоту, кодирующую антиген. Большинство клеток-предшественников эритроцитов, которые содержат рекомбинантную нуклеиновую кислоту, кодирующую антиген, способны к дифференцировке в зрелые эритроциты, которые сохраняют антиген без сохранения рекомбинантной нуклеиновой кислоты.
[00188] В некоторых вариантах осуществления первичные клетки можно собирать посредством венепункции, пункции капилляров или пункции артерии. Из собранной цельной крови затем можно выделять эритроциты, тромбоциты или другие клетки с применением одной или комбинации методик, в том числе плазмафереза, градиента плотности, Hetastarch, PrepaCyte-CB и центрифугирования.
Аллогенные и аутологичные источники
[00189] В некоторых вариантах осуществления получение экспрессирующей экзогенный антиген EHC включает приведение в контакт выделенных, необязательно подвергнутых культивированию клеток, которые являются аутологичными и/или аллогенными в отношении субъекта, с антигеном. Например, эритроциты, аллогенные в отношении субъекта, включают один или несколько эритроцитов, специфичных по группе крови, или один или несколько эритроцитов универсального донора. В некоторых вариантах осуществления экспрессирующие экзогенный антиген EHC-клетки можно получать посредством слияния эритроцитов, например, слияния эритроцитов, аутологичных для субъекта, с одним или несколькими аллогенными эритроцитами, липосомами и/или искусственными везикулами.
[00190] В определенных вариантах осуществления аутологичное переливание экспрессирующих экзогенный антиген EHC-клеток включает выделение эритроцитов, ретикулоцитов или гемопоэтических стволовых клеток из субъекта, получение соответствующей экспрессирующей экзогенный антиген EHC путем приведения клетки в контакт с антигеном с помощью способов, описанных в данном документе, и введение (например, путем переливания) экспрессирующей экзогенный антиген EHC в того же субъекта.
[00191] В определенных вариантах осуществления аллогенное переливание экспрессирующих экзогенный антиген EHC-клеток включает выделение эритроцитов, ретикулоцитов или гемопоэтических стволовых клеток из донора, получение соответствующей экспрессирующей экзогенный антиген EHC путем приведения клетки в контакт с антигеном с помощью способов, описанных в данном документе, и введение (например, путем переливания) экспрессирующей экзогенный антиген EHC в субъекта, который не является донором. Если для переливания применяют аллогенные клетки, следует обращать внимание на применение совместимой группы крови по системе ABO для предотвращения острой внутрисосудистой гемолитической реакции при переливании крови, которая характеризуется активацией комплемента и лизисом несовместимых эритроцитов. Типы крови по системе ABO определяются на основании присутствия или отсутствия антигенов типа крови A и B, моносахаридных углеводных структур, которые находятся на концах олигосахаридных цепей, ассоциированных с гликопротеинами и гликолипидами на поверхности эритроцитов (обзор в Liu et al., Nat. Biotech. 25:454-464 (2007)). Эритроциты группы O не содержат ни одну из этих антигенных моносахаридных структур. У субъектов с эритроцитами группы A имеются природные антитела к эритроцитам группы B, тогда как у субъектов с эритроцитами группы B имеются антитела к эритроцитам группы A. У субъектов с группой крови AB нет ни одного антитела, а у индивидуумов с группой крови O имеются оба. Субъектам с одним из антитела к A и/или антитела к B нельзя переливать кровь, содержащую соответствующий антиген. Поскольку эритроциты группы O не содержат ни антиген A, ни антиген B, их можно безопасно переливать реципиентам с любой группой крови по системе ABO, например, реципиентам с группой A, B, AB или O. Эритроциты группы O считаются универсальными и их можно применять для переливания субъектам всех типов крови. В отличие от этого, эритроциты группы A можно вливать реципиентам с группами A и AB, эритроциты группы B можно вливать реципиентам с группами B и AB и эритроциты группы AB можно вливать только реципиентам с группой AB. В вариантах осуществления, в которых экспрессирующие экзогенный антиген EHC-клетки получают путем приведения эритроцитов или их предшественников в контакт с антигеном, эритроциты или их предшественники получены из источника, который совпадает по совместимости с реципиентом.
[00192] В некоторых случаях может быть полезным превратить экспрессирующую экзогенный антиген EHC, включающую эритроцит, не относящийся к группе O, в клетку универсального типа крови. Ферментативное удаление иммунодоминантных моносахаридов на поверхности эритроцитов группы A и группы B можно применять для получения популяции экспрессирующих экзогенный антиген EHC-клеток группы, аналогичной группе O (см., например, Liu et al., Nat. Biotech. 25:454-464 (2007)). Экспрессирующие экзогенный антиген EHC-клетки группы B можно превращать с применением α-галактозидазы, происходящей из сырых кофейных зерен. В качестве альтернативы или дополнительно можно применять ферменты α-N-ацетилгалактозаминидазу и α-галактозидазу, происходящие из бактерии E. meningosepticum, для соответственного удаления иммунодоминантных антигенов A и B (Liu et al., Nat. Biotech. 25:454-464 (2007)), если они присутствуют на экспрессирующих экзогенный антиген EHC-клетках. В одном примере эритроцитарную массу, выделенную как описано в данном документе, инкубируют в 200 мМ глицина (pH 6,8) и 3 мМ NaCl в присутствии либо α-N-ацетилгалактозаминидазы, либо α-галактозидазы (приблизительно 300 мкг/мл эритроцитарной массы) в течение 60 минут при 26°C. После обработки красные кровяные клетки промывают с помощью 3-4 промывок в солевом растворе с центрифугированием и проводят типирование по системе ABO в соответствии со стандартными методиками заготовки крови.
[00193] В конкретных вариантах осуществления экспрессирующие экзогенный антиген EHC-клетки, описанные в данном документе, можно получать следующим способом. Во-первых, выделяют клетки-предшественники эритроидного ряда. В качестве альтернативы. эти клетки могут быть аутологичными для пациента или происходить из практически универсальной донорской крови. Например, клетки могут иметь тип O по системе ABO, быть отрицательными по резус-фактору Rh r/r, Duffy -/- и большому антигену K1 из системы Kell. В ходе дифференцировки из клетки-предшественника эритроидного ряда в EHC вводят рекомбинантную нуклеиновую кислоту, кодирующую антиген. Рекомбинантная нуклеиновая кислота, кодирующая антиген, может находиться под контролем промотора, специфичного для клеток эритроидного ряда, такого как промотор GATA-1 (см., например, Repik et al., Clin Exp Immunol 2005, 140:230). Рекомбинантную нуклеиновую кислоту, кодирующую антиген, можно вводить любым способом, известным из уровня техники, например, в виде плазмидной ДНК, вирусной или мРНК. Введение нуклеиновой кислоты можно осуществлять с помощью целого ряда стандартных способов, например, посредством трансфекции, трансдукции или электропорации.
Изменение тромбоцитов и их созревание
[00194] В конкретных вариантах осуществления экспрессирующие экзогенный антиген EHC-клетки, описанные в данном документе, можно получать путем приведения тромбоцитов в контакт с антигеном. Каждый день во взрослом организме вырабатывается 2×1011 красных кровяных клеток, и приблизительно половина этого количества белых клеток и тромбоцитов. У человека практически все продуцирование кровяных клеток происходит в красном костном мозге, который представляет собой иерархическую систему развития, составленную их гемопоэтических стволовых клеток, прогениторных клеток промежуточного уровня и созревающих клеток, коммитированных по каждой линии дифференцировки.
[00195] Хотя морфология всех главных типов кровяных клеток подобна на протяжении начальных стадий их развития, мегакариоциты, клетки, коммитированные на продукцию тромбоцитов, характеризуются явным структурным и функциональным отличием после уровня дифференцировки бластных клеток, они вырастают до размера, который в 10 раз превышает диаметр большинства других клеток костного мозга и кровяных клеток, и содержат до 128 нормальных хромосомных наборов, эти клетки дают начало тромбоцитам крови. После серии нормальных клеточных делений развивающийся предшественник мегакариоцита вступает в уникальный клеточный цикл, отличающийся короткой (приблизительно 1 ч) G1-фазой, типичной (7 ч) S-фазой, очень короткой (~45 минут) G2-фазой, за которой следует фаза эндомитоза (несостоявшаяся M-фаза). После того как у клетки развивается высоко полиплоидное ядро, в ней также развиваются разграничивающие мембраны, необходимые для фрагментации цитоплазмы. Данное событие сопровождается экспрессией гликопротеина GPIIbIIIa (фибриногеновый рецептор тромбоцитов; Papayannopoulou et al., Exp. Hematol., 24: 660-9, 1996) и GPIb (рецептор фактора фон Виллебранда; Kaushansky et al., Nature, 369: 568-571, 1994), гранул, которые содержат АДФ, серотонин, -тромбоглобулин и другие вещества, крайне важные для функционирования зрелых тромбоцитов. Наконец, высоко полиплоидные мегакариоциты подвергаются разделению цитоплазмы, что обеспечивает высвобождение тысяч тромбоцитов (Choi et al., Blood, 85: 402-413, 1995; Cramer et al., Blood, 89: 2336-2346, 1997).
[00196] Подобное всем предшественникам кровяных клеток мегакариоциты происходят из плюрипотентных стволовых клеток костного мозга, которые сохраняют способность к длительному самообновлению или к дифференцировке во все элементы крови. Продукция тромбоцитов частично регулируется сигнальными механизмами, индуцируемыми взаимодействием тромбопоэтина (TPO) и его клеточного рецептора TPOR/MPUc-MPL.
[00197] Тромбопоэтин (TPO) представляет собой гемопоэтический фактор роста, вовлеченный в стимуляцию мегакариоцитопоэза и продукции тромбоцитов. TPO экспрессируется в печени и почке, и, в ответ на запрос в отношении тромбоцитов, его экспрессия также может подвергаться повышающей регуляции в микроокружении костного мозга (Kato et al., Stem Cells, 16: 322-328, 1998; McCarty et al., Blood, 86:3668-3675, 1995). Поскольку экспрессия TPO является по большей части конститутивной, полагают, что уровни TPO регулируются путем изоляции за счет тромбоцитов (Fielder et al., Blood 87: 2154, 1996).
[00198] Ген, кодирующий TPO, подвергли клонированию и определению характеристик (Kuter et al., Proc. Natl. Acad. Sci. USA, 91:11104-11108, 1994; Bartley et al., Cell, 77:1117-1124, 1994; Kaushansky et al., Nature, 369:568-571, 1994; Wendling et al., Nature, 369:571-574, 1994, и de Sauvage et al., Nature, 369:533-538, 1994). кДНК человеческого TPO (hTPO) кодирует полипептид длиной в 353 аминокислоты. Полноразмерный hTPO, секретируемый клетками млекопитающих, после отщепления сигнального пептида состоит из 332 аминокислот. Хотя предсказанная молекулярная масса данного белка составляет 38 кДа, молекулярные массы, которые приведены на основании измерений материала сыворотки или культуральной жидкости рекомбинантных клеток, варьируют от 18 до 85 кДа (гликозилирование и посттрансляционный протеолитический процессинг).
[00199] Рецептор клеточной поверхности для TPO (TPOR/MPL/c-MPL) представляет собой продукт протоонкогена c-mpl, гомолога v-mpl, белка оболочки вируса миелопролиферативного лейкоза (MPLV), который, как показано, индуцирует нарушение, связанное со всеми типами миелоидных клеток (Wendling, Virol., 149:242-246, 1986). Человеческий ген c-mpl кодирует белок из 635 aa с предсказанной молекулярной массой 71 кДа (Vigon et al., Proc. Natl. Acad. Sci. USA, 89:5640-44, 1992; Mignotte et al., Genomics, 20: 5-12, 1994).
[00200] У мышей, переведенных в состояние нуль-мутации относительно экспрессии либо TPO, либо его рецептора (TPOR/MPL/c-MPL), проявлялся фенотип тяжелой тромбоцитопении (Gurney et al., Science, 265: 1445, 1994; Kaushansky et al., J. Clin. Invest., 96: 1683, 1995; de Sauvage et al., J. Exp. Med., 183: 651, 1996).
[00201] Было показано, что многие цитокины (например, фактор стволовых клеток [SCF], IL-1, IL-3, IL-6, IL-11, фактор ингибирования лейкоза [LIF], G-CSF, GM-CSF, M-CSF, эритропоэтин (EPO), kit-лиганд и -интерферон) обладают тромбоцитопоэтической активностью.
Активация тромбоцитов
[00202] Полученные тромбоциты представляют собой небольшие дисковидные клеточные фрагменты, которые подвергаются быстрой трансформации, когда они попадают в участки повреждения сосудов. Они становятся более сферическими и образуют выступающие псевдоподии, их фибриногеновые рецепторы активируются, что приводит к агрегации, и они высвобождают содержимое своих гранул и, в конечном итоге, они образуют пробку, которая отвечает за первичный гемостаз (Siess, W., Physiol. Rev. 69: 58-178, 1989). Активация тромбоцитов также наблюдается при патогенезе нестабильной стенокардии, инфаркта миокарда и инсульта (Packham, M. A., Can J. Physiol Pharmacol. 72: 278-284).
[00203] В активации тромбоцитов задействованы некоторые физиологически активные вещества, такие как коллаген, который расположен на поверхности субэндотелия, тромбин, вырабатываемый каскадной системой свертывания крови, и тромбоксан A2 (TXA2) и АДФ, которые высвобождаются из активированных тромбоцитов. Коллаген связывается с некоторыми мембранными белками тромбоцитов, в том числе интегрином α2 β1, что приводит к активации тромбоцитов в виде высвобождения TXA2 и АДФ (Shattil, S. J., et al., Curr. Opin. Cell Biol. 6: 695-704, 1994). В то же время тромбин, TXA2 и АДФ напрямую активируют рецепторы, сопряженные с G-белком, и индуцируют агрегацию тромбоцитов и высвобождение гранул (Hourani, S. M, and Cusack, N. J., Pharmacol. Rev. 43: 243-298, 1991). Считается, что главные события, вовлеченные в активацию тромбоцитов, являются результатом активации β-изоформ фосфолипазы C (PLC), приводящей к образованию инозитол-1,4,5-трифосфата и диацилглицерина. Тромбоциты в основном содержат две изоформы, PLC-β2 и PLC-β3.
[00204] Рецепторы тромбоцитов, которые опосредуют адгезию и агрегацию тромбоцитов, расположены на двух главных поверхностных гликопротеиновых комплексах тромбоцита. Эти комплексы представляют собой комплекс гликопротеинов Ib-IX, который облегчает адгезию тромбоцитов путем связывания фактора фон Виллебранда (vWF), и комплекс гликопротеинов IIb-IIIa, который связывает тромбоциты в агрегаты путем связывания с фибриногеном. У пациентов с синдромом Бернара-Сулье, врожденным нарушением свертывания крови, наблюдается недостаточная адгезия тромбоцитов вследствие дефицита комплекса гликопротеинов Ib-IX, который связывает vWF, умеренная тромбоцитопения и крупные лимфоцитоподобные тромбоциты.
[00205] Гликопротеин V (GPV) представляет собой главный (≈12000 молекул/тромбоцит) сильно гликозилированный мембранный белок тромбоцитов (мол. масса 82000). Воздействие тромбина на тромбоциты приводит к выделению растворимого фрагмента размером 69 кДа, называемого GPVfl. GPV может нековалентно взаимодействовать с комплексом GPIb-IX, комплексом, образуемым за счет нековалентной ассоциации GPIb (состоит из GPIbα, белка размером 145 кДа, присоединенного дисульфидной связью к GPIbβ, белку размером 24 кДа) с GPIX (белок размером 22 кДа). Было определено, что участки связывания для фактора фон Виллебранда и для тромбина в комплексе GPIb-IX расположены на GPIbα. Поскольку теперь известно, что тромбин активирует тромбоциты с помощью расщепления тромбинового рецептора (Vu et. al., Cell 64:1057-1068 (1990)), рецептора, сопряженного с G-белком, неизвестно, расщепляет ли тромбин GPV случайно как следствие связывания тромбина с GPIbα или играет ли данное расщепление физиологическую роль. GPIBα, GPIBβ и GPIX содержат один или несколько гомологичных богатых лейцином доменов из 24 аминокислот. Эти домены также обнаружены в большом семействе богатых лейцином гликопротеинов (LRG).
[00206] GPV является маркером мегакариоцитной клеточной линии. Моноклональное антитело, специфичное в отношении GPV (SW16), не связывается с красными клетками, лейкоцитами, эндотелиальными клетками или клеточными линиями, такими как HEL или MEG-01, которые, как известно, экспрессируют маркеры тромбоцитов-мегакариоцитов.
[00207] Зрелый GPV состоит из 543 аминокислот, которые составляют один трансмембранный домен, короткий цитоплазматический домен (16 остатков) и крупный внеклеточный домен с 8 потенциальными сайтами N-гликозилирования. Анализ внеклеточного домена выявил присутствие 15 тандемных богатых Leu повторов из 24 аминокислот с гомологией с GPIbα и идентифицировал участок расщепления для тромбина возле C-конца с гомологией с Aα-цепью фибриногена.
Условия культивирования
[00208] Источники получения экзогенных антиген-экспрессирующих EHC-клеток, описанных в данном документе, включают циркулирующие в крови клетки, такие как EHC-клетки. Соответствующий источник клеток можно выделять из субъекта, как описано в данном документе, из происходящих из пациента гемопоэтических или прогениторных клеток эритроидного ряда, происходящих из иммортализованных линий EHC, или происходящих из индуцированных плюрипотентных стволовых клеток, необязательно подвергать культивированию и дифференцировке. Способы получения эритроцитов с применением методики клеточной культуры хорошо известны из уровня техники, например, Giarratana et al., Blood 2011, 118:5071, Huang et al., Mol Ther 2013, предварительная электронная публикация 3 сентября, или Kurita et al., PLOS One 2013, 8:e59890. Протоколы варьируют в зависимости от факторов роста, исходных клеточных линий, периода культивирования и морфологических признаков, которыми характеризуются полученные клетки. Также были созданы системы культивирования для получения крови, которая может служить заменой переливаниям донорской крови (Fibach et al. 1989 Blood 73:100). Недавно CD34+ клетки были дифференцированы до стадии ретикулоцитов, с последующим успешным переливанием в субъекта-человека (Giarratana et al., Blood 2011, 118:5071).
[00209] В данном документе представлены способы культивирования EHC-клеток и экспрессирующих экзогенный антиген EHC-клеток, происходящих из EHC-клеток. EHC-клетки можно культивировать из гемопоэтических прогениторных клеток, в том числе, например, CD34+ гемопоэтических прогениторных клеток (Giarratana et al., Blood 2011, 118:5071), индуцированных плюрипотентных стволовых клеток (Kurita et al., PLOS One 2013, 8:e59890) и эмбриональных стволовых клеток (Hirose et al. 2013 Stem Cell Reports 1:499). Смеси из факторов роста и дифференцировки, которые пригодны для размножения и дифференцировки прогениторных клеток, известны из уровня техники. Примеры соответствующих факторов размножения и дифференцировки включают без ограничений фактор стволовых клеток (SCF), интерлейкин (IL), такой как IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-11, IL-12, CSF, G-CSF, тромбопоэтин (TPO), GM-CSF, эритропоэтин (EPO), Flt3, Flt2, PIXY 321 и фактор ингибирования лейкоза (LIF).
[00210] EHC-клетки можно культивировать из гемопоэтических прогениторных клеток, таких как CD34+ клетки, посредством приведения прогениторных клеток в контакт с определенными факторами в процессе культивирования, включающем множество стадий. Например, EHC-клетки можно культивировать из гемопоэтических прогениторных клеток в способе из трех стадий.
[00211] Первая стадия может включать приведение клеток в культуре в контакт с фактором стволовых клеток (SCF) из расчета 1-1000 нг/мл, эритропоэтином (EPO) из расчета 1-100 Ед/мл и интерлейкином-3 (IL-3) из расчета 0,1-100 нг/мл. Первая стадия необязательно включает приведение клеток в культуре в контакт с лигандом, который связывает и активирует ядерный рецептор гормонов, такой как например, глюкокортикоидный рецептор, эстрогеновый рецептор, прогестероновый рецептор, андрогеновый рецептор или прегнан-x-рецептор. Лиганды для этих рецепторов включают, например, кортикостероид, такой как, например, дексаметазон из расчета 10 нМ - 100 мкМ или гидрокортизон из расчета 10 нМ - 100 мкМ; эстроген, такой как, например, бета-эстрадиол из расчета 10 нМ - 100 мкМ; прогестоген, такой как, например, прогестерон из расчета 10 нМ - 100 мкМ, гидроксипрогестерон из расчета 10 нМ - 100 мкМ, 5a-дигидропрогестерон из расчета 10 нМ - 100 мкМ, 11-дезоксикортикостерон из расчета 10 нМ - 100 мкМ, или синтетический прогестин, такой как, например, хлормадинона ацетат из расчета 10 нМ - 100 мкМ; андроген, такой как, например, тестостерон из расчета 10 нМ - 100 мкМ, дигидротестостерон из расчета 10 нМ - 100 мкМ или андростендион из расчета 10 нМ - 100 мкМ; или лиганд прегнан-x-рецептора, такой как, например, рифампицин из расчета 10 нМ - 100 мкМ, гиперфорин из расчета 10 нМ - 100 мкМ, зверобой (гиперицин) из расчета 10 нМ - 100 мкМ, или витамин E-подобные молекулы, такие как, например, токоферол из расчета 10 нМ - 100 мкМ. Первая стадия также может необязательно включать приведение клеток в культуре в контакт с инсулиноподобной молекулой, такой как, например, инсулин из расчета 1-50 мкг/мл, инсулиноподобный фактор роста 1 (IGF-1) из расчета 1-50 мкг/мл, инсулиноподобный фактор роста 2 (IGF-2) из расчета 1-50 мкг/мл или механозависимый фактор роста из расчета 1-50 мкг/мл. Кроме того, первая стадия может необязательно включать приведение клеток в культуре в контакт с трансферрином из расчета 0,1-5 мг/мл.
[00212] Первая стадия может необязательно включать приведение клеток в культуре в контакт с одним или несколькими интерлейкинами (IL) или факторами роста, такими как, например, IL-1, IL-2, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-11, IL-12, гранулоцитарный колониестимулирующий фактор (G-CSF), макрофагальный колониестимулирующий фактор (M-CSF), гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF), тромбопоэтин, фактор роста фибробластов (FGF), фактор роста тромбоцитов (PDGF), трансформирующий фактор роста бета (TGF-B), фактор некроза опухоли-альфа (TNF-A), фактор роста и развития мегакариоцитов (MGDF), фактор ингибирования лейкоза (LIF) и Flt3-лиганд. Каждый интерлейкин или фактор роста, как правило, может обеспечиваться из расчета концентрации 0,1-100 нг/мл. Первая стадия также может необязательно включать приведение клеток в культуре в контакт с белками или небелковыми молекулами сыворотки, как, например, фетальной бычьей сывороткой (1-20%), человеческой плазмой (1-20%), плазманатом (1-20%), человеческой сывороткой (1-20%), альбумином (0,1-100 мг/мл) или гепарином (0,1-10 Ед/мл).
[00213] Вторая стадия может включать приведение клеток в культуре в контакт с фактором стволовых клеток (SCF) из расчета 1-1000 нг/мл и эритропоэтином (EPO) из расчета 1-100 Ед/мл. Вторая стадия также может необязательно включать приведение клеток в культуре в контакт с инсулиноподобной молекулой, такой как, например, инсулин из расчета 1-50 мкг/мл, инсулиноподобный фактор роста 1 (IGF-1) из расчета 1-50 мкг/мл, инсулиноподобный фактор роста 2 (IGF-2) из расчета 1-50 мкг/мл или механозависимый фактор роста из расчета 1-50 мкг/мл. Кроме того, вторая стадия может необязательно включать приведение клеток в культуре в контакт с трансферрином из расчета 0,1-5 мг/мл. Вторая стадия также может необязательно включать приведение клеток в культуре в контакт с белками или небелковыми молекулами сыворотки крови, как, например, фетальной бычьей сывороткой (1-20%), человеческой плазмой (1-20%), плазманатом (1-20%), человеческой сывороткой (1-20%), альбумином (0,1-100 мг/мл) или гепарином (0,1-10 Ед/мл).
[00214] Третья стадия может включать приведение клеток в культуре в контакт с эритропоэтином (EPO) из расчета 1-100 Ед/мл. Третья стадия может необязательно включать приведение клеток в культуре в контакт с фактором стволовых клеток (SCF) из расчета 1-1000 нг/мл. Кроме того, третья стадия может необязательно включать приведение клеток в культуре в контакт с инсулиноподобной молекулой, такой как, например, инсулин из расчета 1-50 мкг/мл, инсулиноподобный фактор роста 1 (IGF-1) из расчета 1-50 мкг/мл, инсулиноподобный фактор роста 2 (IGF-2) из расчета 1-50 мкг/мл или механозависимый фактор роста из расчета 1-50 мкг/мл. Третья стадия также может необязательно включать приведение клеток в культуре в контакт с трансферрином из расчета 0,1-5 мг/мл. Третья стадия также может необязательно включать приведение клеток в культуре в контакт с белками или небелковыми молекулами сыворотки, как, например, фетальной бычьей сывороткой (1-20%), человеческой плазмой (1-20%), плазманатом (1-20%), человеческой сывороткой (1-20%), альбумином (0,1-100 мг/мл) или гепарином (0,1-10 Ед/мл).
[00215] Процесс культивирования может необязательно включать приведение клеток в контакт с помощью способа, известного из уровня техники, с молекулой, например, молекулой ДНК, молекулой РНК, мРНК, миРНК, микроРНК, lncRNA, shRNA, гормоном или малой молекулой, которая активирует или приводит к нокдауну одного или нескольких генов. Целевые гены могут включать, например, гены, которые кодируют фактор транскрипции, фактор роста или рецептор фактора роста, в том числе без ограничений, например, GATA1, GATA2, CMyc, hTERT, p53, EPO, SCF, инсулин, EPO-R, SCF-R, трансферрин-R, инсулин-R.
[00216] В одном варианте осуществления CD34+ клетки помещают в культуру, содержащую различные количества IMDM, FBS, глутамина, BSA, холотрансферрина, инсулина, дексаметазона, β-эстрадиола, IL-3, SCF и эритропоэтина, на трех отдельных стадиях дифференцировки в течение в целом 22 дней.
[00217] В одном варианте осуществления CD34+ клетки помещают в культуру, содержащую различные количества IMDM, FBS, глутамина, BSA, холотрансферрина, инсулина, дексаметазона, β-эстрадиола, IL-3, SCF и тромбопоэтина, на трех отдельных стадиях дифференцировки в течение в целом 14 дней.
[00218] В одном варианте осуществления CD34+ клетки помещают в культуру, содержащую различные количества IMDM, FBS, глутамина, BSA, холотрансферрина, инсулина, дексаметазона, β-эстрадиола, IL-3, SCF и GCSF, на трех отдельных стадиях дифференцировки в течение в целом 15 дней.
[00219] В определенных вариантах осуществления клетки, которые содержат экзогенный антиген, представляющий интерес, могут быть составлены или происходить из множества циркулирующих в крови клеток, в том числе без ограничений перечисленных в таблице A. В предпочтительном варианте осуществления циркулирующие в крови клетки по настоящему изобретению представляют собой EHC-клетки, такие как, например, ядерные красные кровяные клетки, предшественники красных кровяных клеток или безъядерные красные кровяные клетки. Например, EHC-клетки представляют собой стволовую клетку пуповинной крови, CD34+ клетку, гемопоэтическую стволовую клетку (HSC), клетку колониеобразующей единицы селезенки (CFU-S), общую миелоидную клетку-предшественник (CMP), бластную колониеобразующую клетку, взрывообразующую единицу эритроидного ряда (BFU-E), эритро-мегакариоцитарную клетку-предшественника (MEP), колониеобразующую единицу эритроидного ряда (CFU-E), ретикулоцит, эритроцит, индуцированную плюрипотентную стволовую клетку (iPSC), мезенхимальную стволовую клетку (MSC), полихромный нормобласт, ортохромный нормобласт, или перечисленные в таблице A1, или их комбинацию. В некоторых вариантах осуществления EHC-клетки представляют собой бессмертные или иммортализованные клетки, например, иммортализованные эритробластные клетки, полученные с помощью ретровирусной трансдукции CD34+ гемопоэтических прогениторных клеток для экспрессии Oct4, Sox2, Klf4, cMyc и супрессии TP53 (например, Huang et al., Mol Ther 2013, предварительная электронная публикация 3 сентября).
[00220] В данном документе представлены композиции из эритроцитов, в которых множество эритроцитов экспрессирует экзогенный антиген, представляющий интерес, или его фрагмент. Клетки можно культивировать из происходящих из пациента гемопоэтических или прогениторных клеток эритроидного ряда, происходящих из иммортализованных линий EHC или происходящих из индуцированных плюрипотентных стволовых клеток. Способы получения эритроцитов в клеточной культуре известны из уровня техники, например, Giarratana et al., Blood 2011, 118:5071, Huang et al., Mol Ther 2013, или Kurita et al., PLOS One 2013, 8:e59890. Экзогенные антигены можно вводить путем трансфекции одной или множественных копий генов, трансдукции с помощью вируса или электропорации в присутствии ДНК или РНК. Способы экспрессии экзогенных белков в клетках млекопитающих хорошо известны из уровня техники. Например, экспрессия экзогенного фактора IX в гемопоэтических клетках индуцируется вирусной трансдукцией CD34+ прогениторных клеток, см. Chang et al., Nat Biotechnol 2006, 24:1017.
[00221] Композиции из эритроцитов, описанные в данном документе, можно получать следующим образом. Во-первых, выделяют клетки-предшественники эритроидного ряда. В качестве альтернативы. эти клетки могут быть аутологичными для пациента или происходить из практически универсальной донорской крови. Например, клетки могут иметь тип O по системе ABO, быть отрицательными по резус-фактору Rh r/r, Duffy -/- и большому антигену K1 из системы Kell. В ходе дифференцировки из клетки-предшественника эритроидного ряда в EHC вводят нуклеиновые кислоты, кодирующие экзогенный антиген. Нуклеиновая кислота, кодирующая экзогенный антиген, может находиться под контролем промотора, специфичного для клеток эритроидного ряда, такого как промотор GATA-1 (см., например, Repik et al., Clin Exp Immunol 2005, 140:230). Нуклеиновую кислоту, кодирующую экзогенный антиген, можно вводить любым способом, известным из уровня техники, например, в виде плазмидной ДНК, вирусной или мРНК. Введение нуклеиновой кислоты можно осуществлять с помощью целого ряда стандартных способов, например, посредством трансфекции, трансдукции или электропорации.
[00222] Модификация прогениторных клеток. Нуклеиновые кислоты, такие как ДНК-векторы экспрессии или мРНК для продукции антигена, представляющего интерес, можно вводить в прогениторные клетки, которые можно выделять из исходного источника или получать из размноженных, как описано выше, посредством традиционной технологии генной инженерии, представленной в данном документе. В некоторых случаях векторы экспрессии можно разрабатывать так, что они могут встраиваться в геном клетки с помощью гомологичной или негомологичной рекомбинации посредством способов, известных из уровня техники.
[00223] В некоторых случаях нуклеиновую кислоту, кодирующую полипептид, который может селективно нацеливаться и разрезать геном, например, CRISPR/Cas9, эффекторную нуклеазу, подобную активатору транскрипции (TALEN), или цинк-пальцевую нуклеазу, применяют для управления вставкой загруженной в вектор экспрессии нуклеиновой кислоты в конкретное местоположение в геноме, например, в локус CR1 (1q32.2), локус гемоглобина (11p15.4) или другого белка, ассоциированного с клеткой эритроидного ряда, в том числе без ограничений перечисленных в таблице C.
[00224] В некоторых случаях нуклеиновая кислота представляет собой молекулу РНК, или молекулу ДНК, которая кодирует молекулу РНК, которая осуществляет сайленсинг или подавляет экспрессию целевого гена. Например, молекула может представлять собой малую интерферирующую РНК (siRNA), молекулу антисмысловой РНК или молекулу короткой шпилечной РНК (shRNA).
[00225] Способы переноса векторов экспрессии в прогениторные клетки включают без ограничений опосредованный вирусом перенос генов, опосредованный липосомами перенос, трансформацию, генные пушки, трансфекцию и трансдукцию, например, опосредованный вирусом перенос генов, такой как применение векторов, основанных на ДНК вирусов, таких как аденовирус, аденоассоциированный вирус и вирус герпеса, а также векторов на основе ретровирусов. Примеры методов переноса генов включают, например, депротеинизированную ДНК, осаждение с помощью CaPO4, DEAE-дестран, электропорацию, слияние протопластов, липофекцию и микроинъекцию в клетку.
[00226] Любые подвергнутые генетической модификации прогениторные клетки, описанные в данном документе, можно культивировать в соответствующих условиях, обеспечивающих возможность дифференцировки в зрелые безъядерные красные кровяные клетки, например, с помощью способа культивирования in vitro, описанного в данном документе. Полученные безъядерные красные кровяные клетки проявляют и экспрессируют белки, ассоциированные со зрелыми эритроцитами, например, гемоглобин, гликофорин A, которые можно подтверждать и количественно оценивать с помощью стандартных способов (например, вестерн-блоттинга или FACS-анализа).
Стратегии экспрессии экзогенного антигена
[00227] В данном документе представлены антигены, которые выставляются экспрессирующими экзогенный антиген EHC-клетками. В некоторых вариантах осуществления антиген способен к взаимодействию с мишенью, например, ассоциирует с мишенью или связывается с ней. Антиген может содержать или может состоять фактически из полипептида. В некоторых вариантах осуществления антиген содержит полипептид, углевод, нуклеиновую кислоту, липид, малую молекулу или их комбинацию. В некоторых вариантах осуществления антигены не взаимодействуют с мишенью, но служат в качестве загрузок, подлежащих доставке с помощью экспрессирующей экзогенный антиген EHC в клетку, ткань или другой участок в организме субъекта.
Полипептиды, химеры и слияния антигена
[00228] В некоторых вариантах осуществления антигены содержат полипептиды. Полученные полипептиды могут варьировать в размере от 6 аминокислот до 3000 аминокислот и могут выходить за рамки 6, 10, 15, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 150, 200, 300, 400 или могут выходить за рамки 500 аминокислот. Полученные полипептиды могут варьировать в размере от приблизительно 20 аминокислот до приблизительно 500 аминокислот, от приблизительно 30 аминокислот до приблизительно 500 аминокислот или от приблизительно 40 аминокислот до приблизительно 500 аминокислот.
[00229] В некоторых вариантах осуществления полипептид антигена содержит химерный белок или белок слияния, который может содержать два или более отличающихся белковых домена. Эти химерные антигены являются гетерологичными или экзогенными в том смысле, что различные домены происходят из различных источников, и в связи с этим, не обнаруживаются совместно в природе и могут кодироваться, например, рекомбинантными нуклеиновыми кислотами. Полипептиды антигена можно получать с помощью целого ряда способов, многие из которых хорошо известны из уровня техники и также описаны в данном документе. Например, полипептиды антигена можно получать посредством экстракции (например, из выделенных клеток), посредством экспрессии рекомбинантной нуклеиновой кислоты, кодирующей полипептид антигена, или посредством химического синтеза. Полипептиды антигена можно получать, например, с помощью генной инженерии и векторов экспрессии, кодирующих полипептид, вводимых в клетки-хозяева (например, с помощью трансформации или трансфекции) для экспрессии закодированного полипептида антигена.
[00230] Имеется множество консервативных изменений, которые обычно можно осуществлять в аминокислотной последовательности без изменения активности. Эти изменения называют консервативными заменами или мутациями; то есть аминокислота, принадлежащая к группе аминокислот с определенным размером, зарядом или другой характеристикой, может замещаться другой аминокислотой. Замены для аминокислотной последовательности могут быть выбраны из других представителей класса, к которому принадлежит данная аминокислота. Например, неполярные (гидрофобные) аминокислоты включают аланин, лейцин, изолейцин, валин, пролин, фенилаланин, триптофан, метионин и тирозин. Полярные нейтральные аминокислоты включают глицин, серин, треонин, цистеин, тирозин, аспарагин и глутамин. Положительно заряженные (основные) аминокислоты включают аргинин, лизин и гистидин. Отрицательно заряженные (кислотные) аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту. Ожидается, что такие изменения не будут оказывать значительного воздействия на кажущуюся молекулярную массу, определяемую с помощью электрофореза в полиакриламидном геле или изоэлектрической точки. Консервативные замены также включают замещения оптических изомеров в последовательности на другие оптические изомеры, в частности, D-аминокислоты вместо L-аминокислот для одного или нескольких остатков последовательности. Более того, все аминокислоты в последовательности можно подвергать замене D-изомера L-изомером. Иллюстративные консервативные замены включают без ограничений Lys вместо Arg и наоборот для сохранения положительного заряда; Glu вместо Asp и наоборот для сохранения отрицательного заряда; Ser вместо Thr, так что сохраняется свободная ~ OH-группа; и Gln вместо Asn для сохранения свободной NH2-группы. Более того, точковые мутации, делеции и вставки полипептидных последовательностей или соответствующих последовательностей нуклеиновой кислоты в некоторых случаях можно осуществлять без потери функции фрагмента полипептида или нуклеиновой кислоты. Замены могут включать, например, 1, 2, 3 или более остатков. Любое раскрытие конкретной аминокислотной последовательности или рекомбинантной нуклеиновой кислоты, кодирующей полипептид, или раскрытие ее названия включает любые консервативные замены, точковые мутации, делеции и вставки этих полипептидных последовательностей или соответствующих последовательностей нуклеиновой кислоты и любой последовательности, депонированной в виде белка или гена в базе данных, которые можно осуществлять без потери функции фрагмента полипептида или нуклеиновой кислоты.
[00231] В некоторых вариантах осуществления полипептид антигена ассоциирован с мембраной экспрессирующей экзогенный антиген EHC. В других вариантах осуществления полипептид антигена не ассоциирован с мембраной экспрессирующей экзогенный антиген EHC.
[00232] В одном варианте осуществления соотношение массы липидов к антигену в экспрессирующей экзогенный антиген EHC составляет менее 1:1000, приблизительно 1:1000, приблизительно 1:500, приблизительно 1:250, приблизительно 1:100, приблизительно 1:50, приблизительно 1:25, приблизительно 1:10, приблизительно 1:9, приблизительно 1:8, приблизительно 1:7, приблизительно 1:6, приблизительно 1:5, приблизительно 1:4, приблизительно 1:3, приблизительно 1:2, приблизительно 1:1, приблизительно 2:1, приблизительно 3:1, приблизительно 4:1, приблизительно 5:1, приблизительно 6:1, приблизительно 7:1, приблизительно 8:1, приблизительно 9:1, приблизительно 10:1, приблизительно 25:1, приблизительно 50:1, приблизительно 100:1, приблизительно 250:1, приблизительно 500:1, приблизительно 1000:1, приблизительно 10000:1, приблизительно 100000:1, приблизительно 1000000:1, приблизительно 10000000:1, приблизительно 100000000:1, приблизительно 1000000000:1 или более приблизительно 1000000000:1.
[00233] В одном варианте осуществления соотношение массы полипептидов, не являющихся экзогенным антигеном, к антигену в экспрессирующей экзогенный антиген EHC составляет менее 1:1000, приблизительно 1:1000, приблизительно 1:500, приблизительно 1:250, приблизительно 1:100, приблизительно 1:50, приблизительно 1:25, приблизительно 1:10, приблизительно 1:9, приблизительно 1:8, приблизительно 1:7, приблизительно 1:6, приблизительно 1:5, приблизительно 1:4, приблизительно 1:3, приблизительно 1:2, приблизительно 1:1, приблизительно 2:1, приблизительно 3:1, приблизительно 4:1, приблизительно 5:1, приблизительно 6:1, приблизительно 7:1, приблизительно 8:1, приблизительно 9:1, приблизительно 10:1, приблизительно 25:1, приблизительно 50:1, приблизительно 100:1, приблизительно 250:1, приблизительно 500:1, приблизительно 1000:1, приблизительно 10000:1, приблизительно 100000:1, приблизительно 1000000:1, приблизительно 10000000:1, приблизительно 100000000:1, приблизительно 1000000000:1 или более приблизительно 1000000000:1.
[00234] В определенных вариантах осуществления полипептидный антиген расположен на поверхности и подвержен воздействию окружающей среды вокруг экспрессирующей экзогенный антиген EHC. В некоторых вариантах осуществления полипептидный антиген расположен внутри и обращен к не повергающейся воздействию стороне экспрессирующей экзогенный антиген EHC.
[00235] В определенных вариантах осуществления полипептидный антиген содержит по меньшей мере один из следующих доменов: S-домен (поверхностный), A-домен (якорь) и/или U-домен (не повергающийся воздействию), где S-домен представляет собой поверхностный домен, подвергающийся воздействию окружающей среды вокруг экспрессирующей экзогенный антиген EHC, где A-домен представляет собой якорь, и где U-домен расположен внутри и/или обращен к не подвергающейся воздействию стороне экспрессирующей экзогенный антиген EHC.
[00236] Необязательно полипептид антигена содержит i) один или несколько дополнительных S-доменов, называемых S'-домены, или ii) один или несколько дополнительных U-доменов, называемых U'-домены.
[00237] В некоторых вариантах осуществления S-домен и A-домен образуют часть одной и той же полипептидной цепи.
[00238] В некоторых вариантах осуществления A-домен и U-домен образуют часть одной и той же полипептидной цепи.
[00239] В некоторых вариантах осуществления любой один или несколько из S-, A-, U-домена добавляют к экспрессирующей экзогенный антиген EHC извне.
[00240] В некоторых вариантах осуществления любой один или несколько из S-, A-, U-домена продуцируется внутри экспрессирующей экзогенный антиген EHC.
[00241] В некоторых вариантах осуществления любой один или несколько из S-, A-, U-домена представляет собой полипептид.
[00242] В некоторых вариантах осуществления любой один или несколько из S-, A-, U-домена не представляет собой полипептид.
[00243] Схемы иллюстративных конформаций антигенов внутри экспрессирующих экзогенный антиген EHC-клеток или на них показаны на фигурах 13A, 13B и 13C.
A-домен
[00244] В определенных вариантах осуществления A-домен представляет собой мембранный полипептид. A-домен, например, может представлять собой интегральный мембранный полипептид или ассоциированный с мембраной полипептид.
[00245] A-домен может быть выбран из одного из следующих классов, в том числе без ограничений, например, альфа-спиральные битопические, альфа-спиральные политопические, трансмембранные со структурой бета-бочонок, все альфа монотопические/периферические, все бета-монотопические/периферические, альфа/бета-монотопические/периферические, альфа+бета-монотопические/периферические, пептиды с альфа-спиралью, пептиды с бета-шпилькой, пептиды с бета-спиралью, трансмембранный белок 1 типа (N-конец внеклеточный), трансмембранный белок 2 типа (N-конец внутриклеточный), трансмембранный белок 3 типа, трансмембранный белок 4A типа, трансмембранный белок 4B типа, белок, заякоренный с помощью липида, белок, заякоренный с помощью гликозилфосфатидилинозитола (GPI), белок, заякоренный с помощью пренильной цепи, или пептиды с нерегулярной структурой.
[00246] В определенных вариантах осуществления A-домен является эндогенным, например, эндогенным в отношении EHC, тромбоцита или гемопоэтической клетки. В некоторых вариантах осуществления A-домен является эндогенным в отношении клетки млекопитающего.
[00247] В определенных вариантах осуществления A-домен является экзогенным, например, экзогенным в отношении EHC, тромбоцита или гемопоэтической клетки. В некоторых вариантах осуществления A-домен является экзогенным в отношении клетки млекопитающего.
[00248] A-домен можно выбрать из следующих молекул или их фрагментов, в том числе без ограничения CD1, CD2, CD3, CD4, CD5, CD6, CD7, CD8, CD9, CD10, CD11a, CD11b, CD11c, CD12w, CD13, CD14, CD15, CD16, CDw17, CD18, CD19, CD20, CD21, CD22, CD23, CD24, CD25, CD26, CD27, CD28, CD29, CD30, CD31, CD32, CD33, CD34, CD35, CD36, CD37, CD38, CD39, CD40, CD41, CD42, CD43, CD44, CD45, CD46, CD47, CD48, CD49a, CD49b, CD49c, CD49d, CD49e, CD49f, CD53, CD54, CD55, CD56, CD57, CD58, CD59, CD61, CD62E, CD62L, CD62P, CD63, CD68, CD69, CD71, CD72, CD73, CD74, CD80, CD81, CD82, CD83, CD86, CD87, CD88, CD89, CD90, CD91, CD95, CD96, CD100, CD103, CD105, CD106, CD107, CD107a, CD107b, CD109, CD117, CD120, CD122, CD123, CD127, CD132, CD133, CD134, CD135, CD138, CD141, CD142, CD143, CD144, CD147, CD151, CD152, CD154, CD155, CD156, CD158, CD163, CD165, CD166, CD168, CD184, CDw186, CD195, CD197, CDw199, CD209, CD202a, CD220, CD221, CD235a, CD271, CD279, CD303, CD304, CD309, CD326, Ras-родственного белка 1A, предшественника семапорина 7A, кальций- и интегрин-связывающего белка 1, 55 кДа мембранного белка эритроцитов, флотиллина-1, флотиллина-2, белка, ассоциированного с мембраной клетки эритроидного ряда, фактора инициации трансляции эукариот 2C 2, цитохром-b5-редуктазы, гомолога белка 42 регуляции клеточного деления, белка KIAA1363, Band 3, аннексина VII, аквапорина, экто-АДФ-рибозилтрансферазы 4, антигена системы Kell, LFA-3, представителя 1 семейства 2 переносчиков растворенных веществ, белка LGALS3, переносчика мочевины, полипептида антигена CE по Rh-системе групп крови, Rh-ассоциированного гликопротеина, дематина, антигенов групп крови по системе ABO, аквапорина 3, антигенов группы крови Оберже, Band 3, базиджина, C41, CD44, цис-AB, антигена системы Colton, компонента комплемента 4, CR1, DAF, антигена Diego, антигена системы Duffy, антигена Hh/Бомбей, антигена ii, антигена Индийской системы групп крови, антигена Kell, антигена Kidd, антигена Lewis, антигена Лютеран, антигена системы MNS, антигена группы Cost, антигена группы Er, дематина, стоматина, тропомиозина, переносчика глюкозы, аддуцина, рабфилина, C1-тетрагидрофолатсинтазы, антигена группы Vel, антигена Lan, антигена At, антигена Jr, антигена AnWj, антигена Sd, антигенов Batty, Bilkes, Box, Christiansen, HJK, HOFM, JFV, JONEs, Jensen, Katagiri, Livesay, Milne, Oldeide, Peters, Rasmussen, Reid, REIT, SARA, антигена D по резус-системе групп крови, альдолазы, тропомодулина, аргиназы, креатинкиназы, белка B-Cam, Rap1A, антигена Bennett-Goodspeed, антигенов P-системы, антигенов Rh-системы групп крови, антигенов Xg-системы, белка XK, антигенов системы Yt/Cartwright, CD58, Rh, антигена системы Scianna, Radin, DARC (Duffy), CR1 Knops-McCoy, антигена DAF системы Cromer, Gerbich (GYPC), CD47, гликофорина A, полосы 3 (AE3), GYPB Ss, C4A, C4B Chido/Rodgers, компонента C4 системы комплемента, HLA Bg HLA класса I, RHAG Rh-ассоциированного гликопротеин транспорта аммония, белка водного канала Colton (Co), ацетилхолинэстеразы ACHE Cartwright (Yt), глутатион-трансферазы, гликофорина C, аквапорина, эритробласт-ассоциированного мембранного белка, CD44, синаптобревина 2, рибонуклеазы, цитохрома b двенадцатиперстной кишки, ABO-гликозилтрансфераз, CD59, антигена CD44 Индийской системы (In), рецептора адгезии AnWj, MER2, АДФ-рибозилтрансферазы DOK Dombrock, гипотетического рецептора адгезии SEMA7A JMH, белка UMOD Sda Tamm-Horsfall (уромодулин), белка анионного канала (полоса 3, AE1) по системе Diego (Di), Wright (Wr), переносчика мочевины Kidd (Jk), альфа(1,3) фукозилтрансферазы FUT3 по системе Lewis (Le), нейротелина OK Oka, гипотетической молекулы адгезии, рецептора адгезии LW, секреторной (Se) альфа(1,2)-фукозилтрансферазы FUT2, Hh альфа(1,2)-фукозилтрансферазы FUT1, рецептора адгезии LU Лютеран (Lu), P1-гликозилтрансферазы, гипотетического переносчика нейромедиаторов XK Kx, XG Xg, ранее называемого PBDX, MIC2, гемоглобина, анкирина, спектрина, металлопротеиназы KEL Kell (формы K, k, Kp, Js), антигена Torkildsen, кофермента Q10, Rab 35, Ral A-связывающего белка, связывающего белка блестящей оболочки, белка Lyn B, белка KIaa1741, DC38, кальций-транспортирующей АТФазы, GPIX, GPIba, GPIbb, GPV, GPIb-IX-V, GPVI, GPIa/IIa, GPIIb/IIIa, GPV/IIa.
S-домен
[00249] В некоторых вариантах осуществления S-домен представляет собой белок или полипептид. В других вариантах осуществления S-домен представляет собой нуклеиновую кислоту. В некоторых вариантах осуществления S-домен представляет собой химическое вещество. В определенных вариантах осуществления S-домен представляет собой малую молекулу.
[00250] В некоторых вариантах осуществления S-домен представляет собой полипептид, выбранный из или происходящий из одного или нескольких следующих классов, в том числе без ограничений гибкого линкера, эпитопной метки, фермента, протеазы, нуклеазы, антигена, антителоподобной молекулы, лиганда антитела, фактора роста, цитокина, хемокина, рецептора фактора роста, цитокинового рецептора, хемокинового рецептора, последовательности, распознаваемой ферментом, последовательности, распознаваемой транспептидазой, последовательности, распознаваемой протеазой, расщепляемого домена, интеина, ДНК-связывающего белка и РНК-связывающего белка, регуляторной молекулы комплемента, молекулы системы комплемента, молекулы системы свертывания крови, хелатора, регуляторного домена комплемента, SCR-домена, CCP-домена, иммуноглобулинового или иммуноглобулиноподобного домена, повтора armadillo, лейциновой застежки-молнии, домена смерти, кадгеринового повтора, EF-руки, фосфотирозин-связывающего домена, плекстрин-гомологичного домена, домена SCR гомологии 2, цинковопальцевого домена, циклического пептида, проникающего пептида.
[00251] В некоторых вариантах осуществления S-домен представляет собой молекулу, не относящуюся к полипептидам, например, нуклеиновую кислоту, углевод или малую молекулу. В некоторых вариантах осуществления S-домен представляет собой нуклеиновую кислоту, выбранную из одного или нескольких следующих классов, в том числе без ограничений ДНК-аптамера, РНК-аптамера, siRNA, shRNA, однонитевого РНК-зонда, однонитевого ДНК-зонда, мРНК, химически модифицированного нуклеотида. В некоторых вариантах осуществления S-домен представляет собой малую молекулу, выбранную из одного или нескольких следующих классов, в том числе без ограничений хелатора, DOTA, радионуклида, изотопа, визуализирующего средства, флуоресцентной молекулы, хемилюминесцентной молекулы, газа.
U-домен
[00252] В некоторых вариантах осуществления U-домен представляет собой белок или полипептид. В других вариантах осуществления U-домен представляет собой нуклеиновую кислоту. В некоторых вариантах осуществления U-домен представляет собой химическое вещество. В определенных вариантах осуществления U-домен представляет собой малую молекулу.
[00253] В некоторых вариантах осуществления U-домен представляет собой полипептид, выбранный из или происходящий из одного или нескольких следующих классов, в том числе без ограничений гибкого линкера, эпитопной метки, фермента, протеазы, нуклеазы, антигена, антителоподобной молекулы, лиганда антитела, фактора роста, цитокина, хемокина, рецептора фактора роста, цитокинового рецептора, хемокинового рецептора, последовательности, распознаваемой ферментом, последовательности, распознаваемой транспептидазой, последовательности, распознаваемой протеазой, расщепляемого домена, интеина, ДНК-связывающего белка и РНК-связывающего белка, регуляторной молекулы комплемента, молекулы системы комплемента, молекулы системы свертывания крови, хелатора, регуляторного домена комплемента, SCR-домена, CCP-домена, иммуноглобулинового или иммуноглобулиноподобного домена, повтора armadillo, лейциновой застежки-молнии, домена смерти, кадгеринового повтора, EF-руки, фосфотирозин-связывающего домена, плекстрин-гомологичного домена, домена SCR гомологии 2, цинковопальцевого домена, циклического пептида, проникающего в клетку пептида, киназного домена, фосфатазного домена, белка цитоскелета, белка, который взаимодействует с белком цитоскелета, рецептора, сопряженного с G-белком, тирозинкиназы, ITIM-домена, ITAM-домена.
[00254] В некоторых вариантах осуществления U-домен представляет собой молекулу, не относящуюся к полипептидам, например, нуклеиновую кислоту, углевод или малую молекулу. В некоторых вариантах осуществления U-домен представляет собой нуклеиновую кислоту, выбранную из одного или нескольких следующих классов, в том числе без ограничений ДНК-аптамера, РНК-аптамера, siRNA, shRNA, однонитевого РНК-зонда, однонитевого ДНК-зонда, мРНК, химически модифицированного нуклеотида. В некоторых вариантах осуществления U-домен представляет собой малую молекулу, выбранную из одного или нескольких следующих классов, в том числе без ограничений хелатора, DOTA, радионуклида, изотопа, визуализирующего средства, флуоресцентной молекулы, хемилюминесцентной молекулы, газа.
Примеры полипептидов антигена
[00255] Примеры полипептидов антигена включают следующие: полипептидный антиген содержит гликофорин A с HA-эпитопной меткой на N-конце; полипептидный антиген содержит лидерную последовательность гликофорина A, HA-эпитопную метку и основную последовательность гликофорина A; полипептидный антиген содержит рецептор комплемента 1 (CR1); полипептидный антиген содержит лидерную последовательность CR1, HA-эпитопную метку, основную последовательность CR1; полипептидный антиген содержит лидерную последовательность CR1, HA-эпитопную метку, шесть SCR-доменов LHR-A и LHR-B из CR1, два мембранно-проксимальных SCR-домена CR1, трансмембранную область CR1 и внутриклеточную область CR1; полипептидный антиген содержит лидерную последовательность CR1, HA-эпитопную метку, девять SCR-доменов LHR-A, и LHR-B, и LHR-C из CR1, два мембранно-проксимальных SCR-домена CR1, трансмембранную область CR1 и внутриклеточную область CR1; полипептидный антиген содержит лидерную последовательность CR1, LHR-A из CR1, LHR-B из CR1, LHR-C из CR1, два мембранно-проксимальных SCR-домена CR1, трансмембранную область CR1 и внутриклеточную область CR1; полипептидный антиген содержит лидерную последовательность CR1, LHR-A из CR1, LHR-B из CR1, LHR-C из CR1, два мембранно-проксимальных SCR-домена CR1, трансмембранную область и внутриклеточную область гликофорина A; полипептидный антиген содержит лидерную последовательность гликофорина A, scFv антитела против поверхностного антигена гепатита B (scFv), гибкий линкер (Gly3Ser)2, HA-эпитопную метку и основную часть гликофорина A; полипептидный антиген содержит Kell, гибкий линкер (Gly3Ser)2, HA-эпитопную метку и scFv; полипептидный антиген содержит Kell и HA-эпитопную метку; полипептидный антиген содержит N-терминальный фрагмент Kell из 71 аминокислоты и HA-эпитопную метку; полипептидный антиген содержит N-терминальный фрагмент Kell из 71 аминокислоты, гибкий линкер (Gly3Ser)2 и HA-эпитопную метку; полипептидный антиген содержит N-терминальный фрагмент Kell из 79 аминокислот и HA эпитопную метку; полипептидный антиген содержит N-терминальный фрагмент Kell из 79 аминокислот, гибкий линкер (Gly3Ser)2 и HA-эпитопную метку; полипептидный антиген содержит N-терминальный фрагмент Kell из 71 аминокислоты, гибкий линкер (Gly3Ser)2, scFv и HA-эпитопную метку; полипептидный антиген содержит N-терминальный фрагмент Kell из 79 аминокислот, гибкий линкер (Gly3Ser)2, scFv и HA-эпитопную метку; полипептидный антиген содержит лидерную последовательность CD55, scFv, HA-эпитопную метку и 37 терминальных аминокислот CD55; полипептидный антиген содержит лидерную последовательность CD55, HA-эпитопную метку и основную часть CD55. В одном варианте осуществления полипептидный антиген содержит лидерную последовательность CD59, scFv, HA-эпитопную метку и основную часть CD59; полипептидный антиген содержит лидерную последовательность CD59, и HA-эпитопную метку, и основную часть CD59; полипептидный антиген содержит аденозиндезаминазу и HA-эпитопную метку; полипептидный антиген содержит фенилаланингидроксилазу и HA-эпитопную метку; полипептидный антиген содержит аденозиндезаминазу, гибкий линкер (Gly3Ser)2, фенилаланингидроксилазу и HA-эпитопную метку; полипептидный антиген содержит гликофорин A, аденозиндезаминазу на цитоплазматическом C-конце и HA-эпитопную метку; полипептидный антиген содержит гликофорин A, фенилаланингидроксилазу на цитоплазматическом C-конце и HA-эпитопную метку.
[00256] В определенных вариантах осуществления антиген способен взаимодействовать с макрофагом или взаимодействует с ним. Полипептид антигена может содержать один или несколько из следующих: рецептора комплемента (Rieu et al., J. Cell Biol. 127:2081-2091 (1994)), фагоцитарного рецептора (Brasseur et al., Photochem. Photobiol. 69:345-352 (1999)), трансферринового рецептора (Dreier et al., Bioconjug. Chem. 9:482-489 (1998); Hamblin et al., J. Photochem. Photobiol. 26:4556 (1994)); Fc-рецептора (Rojanasakul et al., Pharm. Res. 11:1731-1733 (1994)) и маннозного рецептора (Frankel et al., Carbohydr. Res. 300:251-258 (1997); Chakrabarty et al., J. Protozool. 37:358-364 (1990)).
[00257] Другие антигены, способные взаимодействовать или взаимодействующие с макрофагами, включают: липопротеины низкой плотности (Mankertz et al., Biochem. Biophys. Res. Commun. 240:112-115 (1997); von Baeyer et al., Int. J. Clin. Pharmacol. Ther. Toxicol. 31:382-386 (1993)), липопротеины очень низкой плотности (Tabas et al., J. Cell Biol. 115:1547-1560 (1991)), остатки маннозы и другие углеводные компоненты (Pittet et al., Nucl. Med. Biol. 22:355-365 (1995)), поликатионные молекулы, такие как поли-L-лизин (Hamblin et al., J. Photochem. Photobiol. 26:45-56 (1994)), липосомы (Bakker-Woudenberg et al., J. Drug Target. 2:363-371 (1994); Betageri et al., J. Pharm. Pharmacol. 45:48-53 (1993)) и 2-макроглобулин (Chu et al., J. Immunol. 152:1538-1545 (1994)).
[00258] В данном документе представлены композиции, содержащие EHC-клетки, содержащие антиген, обладающий функциональными активностями, которые или i) не присутствуют в нативных EHC-клетках той же линии дифференцировки, или ii) присутствуют в нативных EHC-клетках той же линии дифференцировки со сниженными уровнями или со сниженными уровнями активности по сравнению с EHC-клетками, содержащими антиген. Такие функциональные активности включают ингибирование комплемента, клиренс иммунных комплексов, искусственную презентацию антигенов, модуляцию системы свертывания крови, перенос кислорода, доставку лекарственных средств, поглощение цитотоксинов, уклонение от фагоцитоза и продление времени циркуляции в кровотоке.
[00259] В некоторых вариантах осуществления EHC-клетки имеют более высокие уровни полипептида рецептора комплемента, такого как CR1, чем нативные EHC-клетки той же линии дифференцировки за счет содержания антигена CR-1. В альтернативном варианте осуществления EHC-клетки, содержащие антиген, обладают более высокими уровнями полипептида-агониста рецептора комплемента или ассоциированного с комплементом полипептида, чем нативные EHC-клетки той же линии дифференцировки, в том числе без ограничений полипептидов, перечисленных в таблице 6 и таблице 8. Полипептид антигена-рецептора комплемента содержит полипептид человеческого рецептора комплемента-1 (CR1), его вариант или функциональный фрагмент. Полипептид антигена CR1 может происходить из одной или нескольких нативных аллелей CR1, например, аллели A (также называемой аллель F или аллель CR1*1), аллели B (также называемой аллель S или аллель CR1*2), аллели C (также называемой аллель F' или аллель CR1*3) или аллели D (также называемой аллель CR1*4). Последовательности и номера доступа в базе данных для этих нативных форм представлены в таблице 3. В некоторых вариантах осуществления полипептид антигена CR1 содержит домен полипептида CR1. Например, полипептид CR1 может содержать один или несколько доменов с короткими консенсусными повторами (SCR), также называемых модули белков контроля комплемента (CCP) или Суси-домены, например, номер доступа в Genbank AAV65577.1. В одном варианте осуществления полипептид антигена CR1 содержит один или несколько коротких консенсусных повторов (SCR), например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44 или более 44 SCR. В другом варианте осуществления полипептид антигена CR1 содержит один или несколько структурных единиц CR1, длинных гомологичных повторов (LHR), например, LHR-A, LHR-B, LHR-C или LHR-D, например, 1, 2, 3, 4, 5, 6 или более 6 LHR-доменов. В другом варианте осуществления полипептид антигена CR1 может содержать один или несколько внеклеточных доменов CR1, слитых с другим белком клеточной мембраны, например, гликофорином A, гликофорином B, гликофорином C, гликофорином D, антигеном Kell, Band 3, аквапорином 1, glut 1, белком антигена kidd, антигеном резус-системы, в том числе без ограничений компонентами клеточной поверхности, перечисленными в таблице 1 и таблице 6.
[00260] В некоторых вариантах осуществления EHC содержит рекомбинантную нуклеиновую кислоту, кодирующую полипептид антигена-рецептора комплемента, или в качестве альтернативы или в комбинации, полипептид антигена-агониста рецептора комплемента или полипептид ассоциированного с комплементом антигена, в том числе без ограничений полипептиды и агонисты полипептидов, перечисленные в таблице 8. В некоторых вариантах осуществления EHC-клетки дополнительно содержат экзогенный полипептид фактора ускорения распада (CD59, GenBank: CAG46523.1) или экзогенный полипептид мембранного кофакторного белка (CD46, GenBank: BAA12224.1) или их вариант или функциональный фрагмент, или их комбинацию.
[00261] Функции CR1 включают связывание с C3b-содержащими иммунными комплексами и перемещение этих иммунных комплексов из кровотока к макрофагам ретикулоэндотелиальной системы в печени и селезенке. После встречи с клетками ретикулоэндотелиальной системы иммунный комплекс фагоцитируется фагоцитарной клеткой, а красная кровяная клетка сохраняется, чтобы продолжить свое существование в кровотоке. Удаление иммунного комплекса иногда приводит к протеолитическому отщеплению CR1 с поверхности красной кровяной клетки. Для измерения активности связывания можно проводить in vitro анализ связывания EHC-клеток и иммунных комплексов. Для измерения сохранения EHC можно проводить in vitro анализ фагоцитоза с помощью фагоцитарных клеток и EHC-клеток, загруженных иммунными комплексами. Для измерения клиренса in vivo циркулирующих в крови иммунных комплексов в печень можно проводить анализ клиренса и биораспределения с применением иммунных комплексов с радиоактивной меткой.
[00262] Представлены композиции, содержащие EHC-клетки, содержащие антиген, содержащий нативный полипептид на уровне большем, чем уровень у гемопоэтической клетки той же линии дифференцировки, не содержащей полипептид антигена. Например, популяции EHC-клеток содержат антигены, такие как рецептор комплемента 1, с уровнями по меньшей мере приблизительно в 1,1, например, 1,2, 1,3, 1,4, 1,5, 1,6, 1,7, 1,8, 1,9, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100, 150, 200, 250, 300, 350, 400, 450, 500, 600, 700, 800, 900, 1000, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000 или более чем в 10000 раз выше, чем у соответствующих гемопоэтических клеток той же линии дифференцировки, у которых отсутствует полипептид антигена CR1. Уровни CR1 на ретикулоцитах и эритроцитах, как правило, составляют 50-2000 молекул на клетку (Lach-Trifilieff, J Immunol 1999, 162:7549). Представлены композиции, которые содержат популяции EHC-клеток с уровнями CR1, составляющими по меньшей мере приблизительно 2500, 5000, 6000, 7000, 8000, 9000, 10000, 15000, 20000, 25000, 30000, 40000, 50000, 100000, 200000, 300000, 400000, 500000, 600000, 700000, 800000, 900000, 1000000 или более чем 1000000 молекул на клетку. Уровни CR1 EHC-клеток дикого типа и экспрессирующих экзогенный антиген EHC-клеток можно измерять и количественно оценивать, например, с помощью проточной цитометрии с антителами, специфичными в отношении CR1.
[00263] В некоторых вариантах осуществления в данном документе представлены EHC-клетки, содержащие антиген, популяции EHC-клеток, содержащих антиген, и композиции из EHC-клеток, содержащих антиген. В некоторых вариантах осуществления антиген взаимодействует с циркулирующим в крови патогеном, таким как вирус или бактерия. В некоторых вариантах осуществления EHC экспрессирует рекомбинантный ген, кодирующий антитело, scFv или нанотело, специфичное в отношении циркулирующего в крови патогена. Анитело, scFv или нанотело могут экспрессироваться как белок слияния. В других вариантах осуществления антиген в виде антитела, scFv или нанотела или другой антиген с аффинностью к циркулирующим в крови патогенам, загружают в EHC или на нее. Антиген в виде антитела, scFv или нанотела или другой антиген с аффинностью к циркулирующим в крови патогенам может быть локализован внутри клетки или вне клетки. В некоторых вариантах осуществления антиген является специфичным в отношении вирусного или бактериального антигена, такого как поверхностный антиген, антиген оболочки или капсида.
[00264] В определенных вариантах осуществления в данном документе представлены EHC-клетки, содержащие антиген, популяции EHC-клеток, содержащих антиген, и композиции из EHC-клеток, содержащих антиген. В некоторых вариантах осуществления антиген взаимодействует с токсином, предпочтительно чужеродным токсином, таким как происходящий из патогена или же из окружающей среды. В некоторых вариантах осуществления EHC экспрессирует рекомбинантный ген, кодирующий антиген, содержащий аминокислотную последовательность, происходящую от липополисахарид-связывающего белка (LBP), бактерицидного/повышающего проницаемость белка (BPI), компонента P амилоида или катионного белка. Токсин-связывающие антигены могут экспрессироваться как белок слияния. В других вариантах осуществления токсин-связывающие антигены могут быть загружены в EHC или на нее. Токсин-связывающие антигены могут быть локализованы внутри клетки или вне клетки. В некоторых вариантах осуществления токсин-связывающий антиген является специфичным в отношении бактериального токсина, такого как ботулинический токсин или сибиреязвенный токсин.
[00265] Кроме того, экспрессирующие экзогенный антиген EHC-клетки могут экспрессировать антиген, способный усиливать их способность к изоляции мишени. Потенциальные антигены-усилители секвестрации включают полипептидные переносчики, в том числе без ограничений таковые в таблице 1.
[00266] В одном варианте осуществления антиген содержит полипептид, который содержит аминокислотную последовательность, происходящую из антигена Duffy-рецептора для хемокинов (DARC). В одном варианте осуществления EHC экспрессирует рекомбинантный ген, кодирующий аминокислотную последовательность, полученную из антигена Duffy-рецептора для хемокинов (DARC). Антиген DARC может экспрессироваться как полноразмерный белок или его фрагмент. DARC может экспрессироваться как белок слияния. В других вариантах осуществления белок DARC загружен в EHC или на нее. В некоторых вариантах осуществления загруженный DARC дополнительно функционализирован или иным образом модифицирован. Молекула антигена DARC может быть локализована внутри клетки или вне клетки.
[00267] DARC идентифицировали как сильный мультилигандный хемокиновый рецептор. DARC принадлежит к семейству родопсиноподобных семиспиральных трансмембранных белков. Кроме эритроцитов DARC экспрессируется в эндотелиальных клетках посткапиллярных венул, которые представляют собой главный участок переселения лейкоцитов в большинстве тканей. DARC обеспечивает высокоспецифичный участок связывания для CC- и CXC-хемокинов. Полагают, что DARC обладает более высокой аффинностью в отношении ELR-мотива CXC-хемокинов. CXC-хемокины представляют собой хемоаттрактанты нейтрофилов и потенциально могут быть проангиогенными.
[00268] При взаимодействии DARC и CXCL8 была показана константа диссоциации (Kd), составляющая 5 нмоль/л, а участки связывания рецепторов оценивались в 1000-9000 на эритроцит (Hadley, Blood, 1997). В отличие от остальных хемокиновых рецепторов с семью трансмембранными доменами, у DARC отсутствует высококонсервативный мотив сопряжения с G-белком, расположенный во второй цитоплазматической петле (Meny, Immunohematology, 2010). DARC не сопряжен с G-белком и не обладает известным альтернативным механизмом передачи сигнала. Биологическая роль DARC не полностью ясна. Полагают, что DARC является a) мультиспецифичным; b) неспособным инициировать внутриклеточные сигналы, и c) считается, что хемокины, связанные с поверхностью эритроцита, являются не доступными для их обычной мишени, воспалительных клеток (Neote, J Biol Chem, 1993). Эритроциты могут играть роль в регуляции воспалительных процессов благодаря присутствию DARC.
[00269] Сигнальные молекулы воспаления, такие как цитокины, могут стимулировать местное и системное повреждение ткани, если присутствуют в высоких концентрациях. "Цитокиновые взрывы" вовлечены в патогенез бактериального сепсиса, ревматоидного артрита и некоторых других воспалительных заболеваний. Модифицированные EHC-клетки, которые экзогенно экспрессируют естественные цитокиновые рецепторы или синтетические антителоподобные миметики рецепторов, могут изолировать воспалительные цитокины. Иллюстративный хемокиновый рецептор представляет собой DARC. В данном документе представлены EHC-клетки, содержащие антиген, который представляет собой цитокиновый рецептор или хемокиновый рецептор, в том числе без ограничений DARC. Например, EHC-клетки, экспрессирующие антиген DARC (что, тем самым, повышает его количество, присутствующее на нативных эритроцитах), можно применять для модуляции уровней хемокинов в кровотоке и/или в пределах периферических тканей организма. EHC-клетки, содержащие антиген DARC, либо могут маркироваться для разрушения, либо могут медленно высвобождать медиаторы воспаления обратно в кровоток, но при низкой концентрации и диффузном распределении. EHC, содержащая антиген, который содержит хемокиновый или цитокиновый рецептор, может функционировать в качестве резервуара для пептидов сигнальной трансдукции.
[00270] В одном варианте осуществления антиген содержит полипептид, который содержит аминокислотную последовательность, происходящую из антитела. В одном варианте осуществления EHC экспрессирует рекомбинантный ген, кодирующий аминокислотную последовательность, происходящую из антитела. Антиген в виде антитела может экспрессироваться как полноразмерный белок или его фрагмент. Антитело может экспрессироваться как белок слияния. В других вариантах осуществления белок антитела загружен в EHC или на нее. В некоторых вариантах осуществления загруженное антитело дополнительно функционализировано или иным образом модифицировано. Антиген в виде антитела может быть локализован внутри клетки или вне клетки. В одном варианте осуществления антиген содержит аминокислотную последовательность антитела, которая является специфичной в отношении необходимой мишени. В некоторых вариантах осуществления антитело представляет собой scFv. В других вариантах осуществления антитело представляет собой нанотело.
[00271] В определенных вариантах осуществления EHC-клетки содержат антиген, который содержит антитело или его фрагмент, которое является специфичным в отношении мишени и расположено на клеточной поверхности. Например, вариабельный фрагмент (Fv) антитела, специфичного в отношении связывания ботулинического токсина, экспрессируется на поверхности EHC. Антитела, связывающие ботулинический токсин, известны из уровня техники (Amersdorfer, Inf and Immunity, 1997), как и экспрессия Fv-части антитела (Hoedemaeker, Journ of Bio Chemistry, 1997). После связывания токсин удерживается EHC за счет Fv-области, изолируется и переносится посредством кровеносной системы в печень для выведения из организма.
[00272] В одном варианте осуществления антиген содержит полипептид, который содержит аминокислотную последовательность, происходящую из scFv-антитела. В одном варианте осуществления EHC экспрессирует рекомбинантный ген, кодирующий аминокислотную последовательность, происходящую из scFv-антитела. Антиген в виде scFv-антитела может экспрессироваться как полноразмерный белок или его фрагмент. scFv-антитело может экспрессироваться как белок слияния. В других вариантах осуществления scFv-белок загружен в EHC или на нее. Соответствующие полипептиды scFv-антигена, которые могут экспрессироваться EHC-клетками, включают без ограничений перечисленные в таблице 6.
[00273] scFv-антитела были получены, преимущественно, из гибридомы, клеток селезенки иммунизированных мышей и B-лимфоцитов человека. Вариабельная область антитела образуется с помощью нековалентного гетеродимера из вариабельных V(H)- и V(L)-доменов, которые затем можно применять в конструировании рекомбинантного scFv-антитела.
[00274] Получение scFv известно из уровня техники, и сначала требуется выделение мРНК из гибридомы (или также из селезенки, лимфоцитов и костного мозга), с последующей обратной транскрипцией в кДНК, которая служит в качестве матрицы для амплификации гена антитела (ПЦР). С помощью данного способа можно создавать большие библиотеки с различными наборами scFv, происходящих из антител (набор, сопоставимый с таковым исходных антител, из которого моделировали scFv).
[00275] scFv-антиген можно сделать специфичным в отношении любой целевой молекулы, в том числе без ограничений молекул в таблице 4.
[00276] В одном примере scFv-антиген, специфичный в отношении сибиреязвенного токсина, может экспрессироваться на EHC. После введения субъекту, нуждающемуся в этом, эффективная доза популяции EHC, содержащих молекулу антигена, специфичного в отношении сибиреязвенного токсина, может применяться для захвата и изоляции сибиреязвенного токсина. EHC мигрирует в печень, где происходит выведение.
[00277] В определенных вариантах осуществления эритроциты содержат антиген, содержащий происходящее от верблюжьего антитела нанотело, экспрессированное на поверхности клетки. Нанотела обычно имеют размер 12-15 кДа. Они заметно меньше, чем антитела и scFv. Таким образом, нанотела можно легче подвергать трансфекции, и антиген в виде нанотела будет легче экспрессироваться, транслироваться и/или транспортироваться к клеточной поверхности в EHC. В определенных вариантах осуществления антигены в виде нанотел используются для сведения к минимуму иммуногенных эффектов, вызванных специфичным антигеном. Вследствие своего малого размера нанотела будут обладать сниженным иммуногенным потенциалом. В определенных вариантах осуществления используют антигенные нанотела, поскольку они ограничивают изменения в механическом и морфологическом поведении плазматической мембраны EHC. Это может обеспечивать возможность того, что EHC проявляет поведение нормальной циркуляторной красной кровяной клетки. В определенных вариантах осуществления используют антигенные нанотела, поскольку они обладают усиленной способностью распознавать скрытые или необычные эпитопы по сравнению со стандартными антителами. Например, он могут связываться с небольшими ферментативными полостями мишени и модулировать поведение молекулы-мишени.
[00278] В определенных вариантах осуществления EHC-клетки содержат антигенные нанотела со специфичностью в отношении целевых эпитопов молекул в человеческой системе комплемента. Такие EHC-клетки можно вводить субъекту, нуждающемуся в этом, для селективного истощения одного или нескольких сверхактивных факторов системы комплемента. Например, на C5 могут целенаправленно воздействовать EHC-клетки, содержащие антигенные нанотела со специфичностью в отношении целевых эпитопов C5, и они подвергаются клиренсу из системы с помощью EHC-клеток после введения клеток в субъекта. Данный подход пригоден для обеспечения терапевтического эффекта, например, нарушения, обусловленного системой комплемента, такого как пароксизмальная ночная гемоглобинурия. В определенных вариантах осуществления EHC-клетки содержат антигенные нанотела со специфичностью в отношении целевых эпитопов молекул, в том числе без ограничений перечисленных в таблице 4.
[00279] В некоторых вариантах осуществления антиген содержит полипептид, который содержит аминокислотную последовательность, происходящую из одного из следующего: протеаз, нуклеаз, амилазы, лиазы (сахаразы) или гидролазы (ДНКазы, липазы). В одном варианте осуществления EHC экспрессирует рекомбинантный ген, кодирующий аминокислотную последовательность, происходящую из одного из следующих: протеаз, нуклеаз, амилазы, лиазы (сахаразы) или гидролазы (ДНКазы, липазы). Антигенные протеазы, нуклеазы, амилазы, лиазы и гидролазы могут экспрессироваться как полноразмерный белок или его фрагмент. Антигенные протеазы, нуклеазы, амилазы, лиазы и гидролазы могут экспрессироваться как белок слияния. В других вариантах осуществления антигенные протеазы, нуклеазы, амилазы, лиазы или гидролазы загружены в EHC или на нее. В некоторых вариантах осуществления загруженные антигенные протеазы, нуклеазы, амилазы, лиазы или гидролазы дополнительно функционализированы или модифицированы иным образом. Молекула антигенной протеазы, нуклеазы, амилазы, лиазы или гидролазы может быть локализована внутри клетки или вне клетки.
[00280] В определенных вариантах осуществления EHC-клетки содержат антиген, содержащий протеазу, нуклеазу, амилазу, лиазу или гидролазу. EHC, содержащая антиген в виде протеазы, нуклеазы, амилазы, лиазы или гидролазы, способна приводить к разрушению мишени на EHC, независимо от клиренса в кровотоке, например, с помощью макрофагов в печени. В определенных вариантах осуществления EHC-клетки, содержащие антиген, содержащий протеазу, нуклеазу, амилазу, лиазу или гидролазу, можно вводить субъекту, нуждающемуся в этом, для лечения рака посредством селективного разрушения метаболитов, которые необходимы для роста раковой клетки. Например, аспарагиназу применяют для снижения местных уровней аспарагина для лечения острого лимфобластного лейкоза и острого миелоидного лейкоза. Соответствующие антигены, например, могут включать один или оба из двух главных классов ферментов, способных разрушать целевые молекулы, лиазы и гидролазы. В определенных вариантах осуществления представлены EHC-клетки, содержащие антиген, содержащий молекулу, в том числе без ограничений перечисленные в таблице 6.
[00281] В определенных вариантах осуществления эритроциты содержат антиген, содержащий лиазу. В одном варианте осуществления лиаза представляет собой валин-декарбоксилазу. Антиген валин-декарбоксилаза может экспрессироваться во внутриклеточном пространстве EHC. EHC-клетки, содержащие антиген валин-декарбоксилазу, можно вводить субъекту, нуждающемуся в этом, для модуляции уровней валина в крови. EHC-клетки, содержащие антиген валин-декарбоксилазу, пригодны для лечения валинемии, наследственного нарушения, при котором повышены уровни аминокислоты валина в крови. У пораженных индивидуумов, как правило, развивается рвота, отсутствие прибавки массы тела, умственная отсталость и вялость. Валинемию вызывает дефицит фермента валинтрансаминазы, и она характеризуется аутосомным рецессивным характером наследования.
[00282] В определенных вариантах осуществления эритроциты содержат антиген, содержащий гидролазу. В одном варианте осуществления гидролаза представляет собой дезоксирибонуклеазу I (ДНКазу I). Антиген ДНКаза I может экспрессироваться на поверхности EHC. EHC-клетки, содержащие антиген ДНКазу I, можно вводить субъекту, нуждающемуся в этом, для преимущественного расщепления циркулирующей в крови ДНК по фосфодиэфирным связям, прилегающим к пиримидиновому нуклеотиду, что дает полинуклеотиды с фосфатом на 5'-конце, со свободной гидроксильной группой в положении 3'. Обычно продуцируются тетрануклеотиды. EHC-клетки, содержащие антиген ДНКазы I, пригодны для лечения состояний, усугубленных высокими уровнями иммуногенной ДНК в кровотоке, таких как системная красная волчанка (SLE).
[00283] В определенных вариантах осуществления антиген способен реагировать на внешний стимул, например, после связывания с лигандом или контакта со стимулом, где реакция охватывает, например, передвижение, рефолдинг, изменение конформации, образование димера, образование гомодимера, образование гетеродимера, образование мультимера, передачу сигнала, испускание энергии в поддающейся обнаружению форме (например, флуоресценция), функциональное взаимодействие с другим антигеном или функциональное взаимодействие с полипептидом неэкзогенного антигена.
Мишени
[00284] В данном документе представлены экспрессирующие экзогенный антиген EHC-клетки, содержащие полипептид экзогенного антигена, способный к взаимодействию с мишенью. В данном документе дополнительно представлены экспрессирующие экзогенный антиген EHC-клетки, содержащие неполипептидный экзогенный антиген, способный к взаимодействию с мишенью. Экспрессирующие экзогенный антиген EHC-клетки можно вводить субъекту, нуждающемуся в этом, для модуляции количества или концентрации мишени, находящейся в кровеносной системе субъекта. Для взаимодействия с конкретной мишенью может быть выбран соответствующий экзогенный антиген. Соответствующие мишени включают объекты, которые ассоциированы с конкретным заболеванием, нарушением или состоянием. Однако мишени также можно выбирать независимо от конкретного заболевания, нарушения или состояния.
[00285] В некоторых вариантах осуществления мишень представляет собой антитело или антителоподобную молекулу, например аутоиммунное или аутореактивное антитело, или чужеродное антитело, или терапевтическое антитело, в том числе без ограничений, например, антитело против бета-2 гликопротеина 1, антитело против антигена I/i, антитело против NC1-домена коллагена a3(IV), антитело против гликопротеина тромбоцитов, антитело против рецептора фосфолипазы A2, антитело против гликофорина A, B или C эритроцитов или антитело против Rh-антигена эритроцитов.
[00286] В некоторых вариантах осуществления мишень представляет собой молекулу системы комплемента, например, C1, C1r, C1s, C1q, C2, C2a, C2b, C3, C3a, C3b, C4, C4b, C4a, C3bBb, C3bBb3b, C4b2b, C4b2b3b, C5, C5a, C5b, C6, C7, C8, C9, поли-C9, мембраноатакующий комплекс, фактор B, фактор D, пропердин, C3, C3a, C3b, iC3b, C3c, C3dg, C3dk, C3e, Bb, фактор I, C1q, C1r, C1s, C4, C4a, C4b, C2, C4bp, маннозосвязывающий лектин (MBL), MBL-ассоциированную сериновую протеазу 1 (MASP1), MBL-ассоциированную сериновую протеазу 2 (MASP2), C5, C5a, C6, C7, C8, C9, CR1, CR2, CR3, CR4, C3aR, C3eR, фактор ускорения распада (DAF), мембранный кофакторный белок (MCP), CD59, рецептор бета-цепи C3, ингибитор C1, C4-связывающий белок, фактор I, фактор H.
[00287] В некоторых вариантах осуществления мишень представляет собой иммунный комплекс, например иммунный комплекс с IgG, иммунный комплекс с IgA, иммунный комплекс с IgM.
[00288] В некоторых вариантах осуществления мишень представляет собой амилоидную бляшку, например, бляшку, состоящую из бета-амилоида, IAPP (амилина), альфа-синуклеина, PrPSc, гентингтина, кальцитонина, предсердного натрийуретического фактора, аполипопротеина AI, сывороточного амилоида A, медина, пролактина, транстиретина, лизоцима, бета-2-микроглобулина, гельзолина, кератоэпителина, цистатина, легкой цепи AL иммуноглобулина, S-IBM.
[00289] В некоторых вариантах осуществления мишень представляет собой бактерию, например, Enterococcus, Streptococcus или Mycobacteria, Rickettsia, Mycoplasma, Neisseria meningitides, Neisseria gonorrheoeae, Legionella, Vibrio cholerae, Streptococci, Staphylococcus aureus, Staphylococcus epidermidis, Pseudomonas aeruginosa, Corynobacteria diphtheriae, Clostridium spp., энтеротоксигенные Eschericia coli и Bacillus anthracis. Другие патогены, для которых сообщалась бактериемия на некотором уровне, включают следующих: Rickettsia, Bartonella henselae, Bartonella quintana, Coxiella burnetii, хламидия, Mycobacterium leprae, Salmonella; шигелла; Yersinia enterocolitica; Yersinia pseudotuberculosis; Legionella pneumophila; Mycobacterium tuberculosis; Listeria monocytogenes; Mycoplasma spp.; Pseudomonas fluorescens; Vibrio cholerae; Haemophilus influenzae; Bacillus anthracis; Treponema pallidum; Leptospira; Borrelia; Corynebacterium diphtheriae; Francisella; Brucella melitensis; Campylobacter jejuni; Enterobacter; Proteus mirabilis; Proteus и Klebsiella pneumoniae.
[00290] В некоторых вариантах осуществления мишень представляет собой вирус, в том числе без ограничений вирусы, инфицирование которыми приводит к введению генетического материала в клетки-хозяева после связывания с рецепторами клеточной поверхности, вирусы, инфицирование которыми опосредуется рецепторами клеточной поверхности. Неограничивающие примеры этих вирусов могут быть выбраны из Paramyxoviridae (например, Pneumovirus, Morbillivirus, Metapneumovirus, Respirovirus или Rubulavirus), Adenoviridae (например, Adenovirus), Arenaviridae (например, Arenavirus, такой как вирус лимфоцитарного хориоменингита), Arteriviridae (например, вирус репродуктивно-респираторного синдрома свиней или вирус артериита лошадей), Bunyaviridae (например, Phlebovirus или Hantavirus), Caliciviridae (например, вирус Норуолк), Coronaviridae (например, Coronavirus или Torovirus), Filoviridae (например, Эбола-подобные вирусы), Flaviviridae (например, Hepacivirus или Flavivirus), Herpesviridae (например, Simplexvirus, Varicellovirus, Cytomegalovirus, Roseolovirus или Lymphocryptovirus), Orthomyxoviridae (например, вирус гриппа или Thogotovirus), Parvoviridae (например, Parvovirus), Picomaviridae (например, Enterovirus или Hepatovirus), Poxviridae (например, Orthopoxvirus, Avipoxvirus или Leporipoxvirus), Retroviridae (например, Lentivirus или Spumavirus), Reoviridae (например, Rotavirus), Rhabdoviridae (например, Lyssavirus, Novirhabdovirus или Vesiculovirus) и Togaviridae (например, Alphavirus или Rubivirus). Конкретные примеры этих вирусов включают респираторный коронавирус человека, вирусы гриппа A-C, вирусы гепатита A-G и вирусы простого герпеса 1-9.
[00291] В некоторых вариантах осуществления мишенью является паразит, в том числе без ограничений, например, кишечные или переносимые с кровью паразиты, простейшие, трипаносомы; паразитирующие в крови простейшие и паразиты, способные вызывать малярию; кишечные и системные цестоды, в том числе цестоды-тенииды; кишечные кокцидии; кишечные жгутиковые простейшие; филярийные нематоды; желудочно-кишечные и системные нематоды и анкилостомы.
[00292] В некоторых вариантах осуществления мишень представляет собой гриб, в том числе без ограничений, например, Candida albicans, Candida glabrata, Aspergillus, T. glabrata, Candida tropicalis, C. krusei и C. parapsilosis.
[00293] В некоторых вариантах осуществления мишенью является бактериальный токсин, в том числе без ограничений, например, AB-токсин, альфа-токсин, сибиреязвенный токсин, бактериоцин, ботулинический токсин, холестерин-зависимый цитолизин, C3-токсин Clostridium botulinum, токсин A Clostridium difficile, токсин B Clostridium difficile, энтеротоксин Clostridium, альфа-токсин Clostridium perfringens, бета-токсин Clostridium perfringens, фактор жгутикообразования, Cry1Ac, криптофицин, дельта-эндотоксин, дифтерийный токсин, энтеротоксин типа B, эритрогенный токсин, эксфолиатин, гемолизин E, термолабильный энтеротоксин, термостабильный энтеротоксин, гемолизин, лейкоцидин, липополисахарид, листериолизин O, микроцин, лейкоцидин Пантон-Валентина, остров патогенности, фенолрастворимый модулин, пневмолизин, порообразующий токсин, экзотоксин Pseudomonas, RTX-токсин, сакацин, альфа-токсин Staphylococcus aureus, бета-токсин Staphylococcus aureus, дельта-токсин Staphylococcus aureus, стрептолизин, симплокамид A, табтоксин, тетанолизин, тетаноспазмин, тиолактивируемый цитолизин, толаазин, токсин синдрома токсического шока, токсофлавин, димиколат трегалозы, вероцитотоксин и вибриоцин.
[00294] В некоторых вариантах осуществления мишень представляет собой белок приона, в том числе без ограничений, например, PRP, PRPc, PRPsc, PRPres.
[00295] В некоторых вариантах осуществления мишень представляет собой цитокин, или хемокин, или фактор роста, в том числе без ограничений, например, белок, стимулирующий ацилирование, адипокин, альбинтерферон, CCL1, CCL11, CCL12, CCL13, CCL14, CCL15, CCL16, CCL17, CCL18, CCL19, CCL2, CCL20, CCL21, CCL22, CCL23, CCL24, CCL25, CCL26, CCL27, CCL28, CCL3, CCL5, CCL6, CCL7, CCL8, CCL9, колониестимулирующий фактор, CX3CL1, CX3CR1, CXCL1, CXCL10, CXCL11, CXCL13, CXCL14, CXCL15, CXCL16, CXCL17, CXCL2, CXCL3, CXCL5, CXCL6, CXCL7, CXCL9, эритропоэтин, Gc-MAF, гранулоцитарный колониестимулирующий фактор, гранулоцитарно-макрофагальный колониестимулирующий фактор, фактор роста гепатоцитов, семейство IL 10, семейство IL 17, IL1A, IL1B, интерферон, интерферон бета 1a, интерферон бета 1b, интерферон гамма, интерферон I типа, интерферон II типа, интерферон III типа, интерлейкин, семейство 1 интерлейкинов, антагонист рецептора интерлейкина 1, интерлейкин 10, интерлейкин 12, бета-субъединицу интерлейкина 12, интерлейкин 13, интерлейкин 16, интерлейкин 2, интерлейкин 23, альфа-субъединицу интерлейкина 23, интерлейкин 34, интерлейкин 35, интерлейкин 6, интерлейкин 7, интерлейкин 8, интерлейкин-36, фактор ингибирования лейкоза, фактор стимуляции лейкоцитов, лимфокин, лимфотоксин, лимфотоксин альфа, лимфотоксин бета, макрофагальный колониестимулирующий фактор, воспалительный белок макрофагов, фактор активации макрофагов, монокин, миокин, мионектин, никотинамид-фосфорибозилтрансферазу, онкостатин M, опрелвекин, фактор 4 тромбоцитов, провоспалительный цитокин, промегапоэтин, RANKL, фактор стромальных клеток 1, Talimogene laherparepvec, фактор некроза опухоли альфа, фактор некроза опухоли, XCL1, XCL2, XCR1, ангиопоэтин, основной фактор роста фибробластов, бетацеллюлин, костный морфогенетический белок, нейротрофический фактор головного мозга, CCN-белок внутриклеточной передачи сигнала, CTGF, дарбепоэтин альфа, эндоглин, эпидермальный фактор роста, эпоэтин альфа, эпоэтин бета, эритропоэтин, FGF15, FGF15/19, фактор роста фибробластов, фактор роста фибробластов 23, филграстим, фактор созревания глии, гранулоцитарный колониестимулирующий фактор, гранулоцитарно-макрофагальный колониестимулирующий фактор, ростовой фактор дифференцировки-9, эберпрот-P, гемопоэтические факторы роста, гепарин-связывающий EGF-подобный фактор роста, фактор роста гепатоцитов, инсулиноподобный фактор роста, инсулиноподобный фактор роста 1, инсулиноподобный фактор роста 2, фактор роста кератиноцитов, миостатин, фактор роста нервной ткани, нейротрофин-3, нейротрофин-4, онкомодулин, стимулирующий рост костей, палифермин, PDGFB, плацентарный фактор роста, альфа-гранула тромбоцитов, фактор роста тромбоцитов, рецептор фактора роста тромбоцитов, пролиферативный индекс, тромбопоэтин, трансформирующий фактор роста, фактор роста эндотелия сосудов.
[00296] В некоторых вариантах осуществления мишенью является малая молекула, например, химическое вещество, аминокислота, атом, элемент, органическая кислота, <2000 Да, <1000 Да, <500 Да, в том числе без ограничений, например, железо, медь, кальций, калий, этанол, метанол, глицин, аланин, валин, лейцин, изолейцин, серин, цистеин, селеноцистеин, треонин, метионин, пролин, фенилаланин, тирозин, триптофан, гистидин, лизин, аргинин, аспартат, глутамат, аспарагин, глутамин.
[00297] В некоторых вариантах осуществления мишень представляет собой липидный комплекс, протеолипидный комплекс или холестерин, в том числе без ограничений, например, LDL, VLDL, HDL, HDL2B, триглицериды, LP(a), холестерин.
[00298] В некоторых вариантах осуществления мишенью является клетка млекопитающего, в том числе без ограничений, например, человеческая клетка, циркулирующая в крови клетка, иммунная клетка, нейтрофил, эозинофил, базофил, лимфоцит, моноцит, B-клетка, T-клетка, CD4+ T-клетка, CD8+ T-клетка, гамма-дельта T-клетка, регуляторная T-клетка, клетка-натуральный киллер, T-клетка-натуральный киллер, макрофаг, купферова клетка, дендритная клетка, раковая клетка, раковая стволовая клетка, циркулирующая в крови опухолевая клетка, раковая клетка из одного из следующих типов рака, в том числе без ограничений
[00299] острого лимфобластного лейкоза (ALL), острого миелоидного лейкоза (AML), рака анального канала, рака желчевыводящих путей, рака мочевого пузыря, рака костей, рака кишечника, опухолей головного мозга, рака молочной железы, рака с неизвестной первичной локализацией, метастазирования рака в кость, метастазирования рака в головной мозг, метастазирования рака в печень, метастазирования рака в легкое, карциноида, рака шейки матки, хориокарциномы, хронического лимфоцитарного лейкоза (CLL), хронического миелоидного лейкоза (CML), рака толстой кишки, колоректального рака, рака эндометрия, рака глаза, рака желчного пузыря, рака желудочно-кишечного тракта, гестационных трофобластных опухолей (GTT), гистиоцитарного ретикулоэндотелиоза, рака головы и шеи, лимфомы Ходжкина, рака почки, рака гортани, лейкоза, рака печени, рака легкого, лимфомы, меланомного рака кожи, мезотелиомы, рака мужчин, пузырного заноса, рака полости рта и ротоглотки, миеломы, типов рака носовой полости и синусов, рака носоглотки, неходжкинской лимфомы (NHL), рака пищевода, рака яичника, рака поджелудочной железы, рака полового члена, рака предстательной железы, редких типов рака, рака прямой кишки, рака слюнной железы, вторичных раковых опухолей, рака кожи (немеланомного), саркомы мягких тканей, рака желудка, рака яичка, рака щитовидной железы, рака без выявленного первичного очага, рака матки, рака влагалища и рака вульвы.
Экспрессия, конъюгация, загрузка антигена
[00300] В определенных вариантах осуществления полипептидный антиген экспрессируется внутри экспрессирующей экзогенный антиген EHC. Полипептидный антиген может выставляться на поверхности экспрессирующей экзогенный антиген EHC или может пребывать внутри экспрессирующей экзогенный антиген EHC.
[00301] В определенных вариантах осуществления полипептидный антиген конъюгирован с экспрессирующей экзогенный антиген EHC. Обычно полипептидный антиген конъюгирован с поверхностью экспрессирующей экзогенный антиген EHC. Конъюгацию можно осуществлять химическим или ферментативным путем посредством способов, известных из уровня техники и описанных в данном документе. Неполипептидные антигены также можно конъюгировать с экспрессирующей экзогенный антиген EHC. В некоторых вариантах осуществления антиген не конъюгирован с экспрессирующей экзогенный антиген EHC.
[00302] В определенных вариантах осуществления полипептидный антиген загружен в экспрессирующую экзогенный антиген EHC. Неполипептидные антигены также можно загружать внутрь экспрессирующей экзогенный антиген EHC. В некоторых вариантах осуществления антиген не загружен в экспрессирующую экзогенный антиген EHC или на нее.
[00303] В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC содержит антиген, который необязательно экспрессируется за счет рекомбинантной нуклеиновой кислоты, конъюгированной с EHC, загруженной в EHC или на нее, и любой их комбинации. Необязательно экспрессирующая экзогенный антиген EHC содержит терапевтическое средство или другую загрузку.
[00304] В некоторых вариантах осуществления экспрессирующую экзогенный антиген EHC получают с помощью приведения соответствующей выделенной клетки, например, EHC, ретикулоцита, предшественника EHC, тромбоцита или предшественника тромбоцита, в контакт с рекомбинантной нуклеиновой кислотой, кодирующей полипептид антигена. В некоторых вариантах осуществления полипептид антигена закодирован в ДНК, которую приводят в контакт с ядерной клеткой-предшественником эритроидного ряда или ядерной клеткой-предшественником тромбоцита. В некоторых вариантах осуществления полипептид антигена закодирован в РНК, которую приводят в контакт с тромбоцитом, ядерной EHC, ядерной клеткой-предшественником тромбоцита или ретикулоцитом. В некоторых вариантах осуществления антиген представляет собой полипептид, который приводят в контакт с первичным тромбоцитом, ядерной EHC, ядерной клеткой-предшественником тромбоцита, ретикулоцитом или эритроцитом.
[00305] Полипептид антигена может представлять собой экспрессируемый за счет трансгена, введенного в EHC с помощью электропорации, химической трансфекции или трансфекции с помощью полимерных носителей, вирусной трансдукции, механического разрушения мембраны или другого способа; полипептид антигена, который экспрессируется за счет мРНК, которую вводят в клетку с помощью электропорации, химической трансфекции или трансфекции с помощью полимерных носителей, вирусной трансдукции, механического разрушения мембраны или другого способа; полипептид антигена, полученный за счет сверхэкспрессии нативного локуса вследствие введения внешнего фактора, например, активатора транскрипции, репрессора транскрипции или усилителя секреторного пути; и/или полипептид антигена, который синтезирован, экстрагирован или продуцируется производящей клеткой или другой внешней системой и внедрен в EHC.
[00306] В некоторых вариантах осуществления антиген представляет собой полноразмерный белок. В некоторых вариантах осуществления антиген составлен из одного или нескольких полипептидов, содержащихся в пределах полноразмерного белка, имеющих любую длину, превышающую приблизительно 7 аминокислот. Например, полипептиды могут иметь 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или больше 20 аминокислот, например, 30, 40, 50, 60, 70, 80, 90, 100 или больше 100 аминокислот. Полипептиды, содержащие антиген, могут содержать один или несколько иммунных эпитопов, которые могут быть конформационными эпитопами или могут быть линейными эпитопами. Антиген может быть составлен из одного или нескольких полипептидов из одного или нескольких различных белков.
[00307] Антиген, представляющий интерес, может экспрессироваться в циркулирующей в крови клетке в виде слияния с эндогенным клеточным белком, в том числе без ограничений перечисленными в таблице B и таблице C. Слияние с эндогенным белком может быть необходимо, поскольку полагают, что во время естественного процесса дифференцировки и удаления ядра, EHC сбрасывает и избавляется от многих эндогенных белков, требуемых для эритропоэза, но не для функционирования зрелого эритроцита, таких как, например, c-Kit (SCF-рецептор) и трансферрин. См., например, Keerthivasan et al., Stem Cells International 2011; Migliaccio, Haematologica 2010. Белки, которые остаются, включают определенные мембранные белки, такие как, например, гликофорин A, Band 3 и аквапорин; определенные цитоплазматические белки, такие как, например, гемоглобин альфа, гемоглобин бета и аденозиндезаминаза; и белки цитоскелета.
[00308] Антиген, представляющий интерес, может экспрессироваться во внутриклеточном пространстве EHC с помощью целого ряда способов, включающих непосредственную экспрессию трансгена, слияние с эндогенным внутриклеточным белком, таким как, например, гемоглобин, слияние с внутриклеточным доменом эндогенного белка клеточной поверхности, таким как, например, Band 3, гликофорин A, Kell, или слияние со структурным компонентом цитоскелета клетки эритроидного ряда.
[00309] Антиген, представляющий интерес, может экспрессироваться на внеклеточной поверхности EHC с помощью целого ряда способов, включающих непосредственную экспрессию трансгена, если он содержит трансмембранный домен или другой домен прикрепления к мембране, слияния с эндогенным мембранным белком клетки эритроидного ряда или с трансмембранным доменом указанного белка, такого как, например, Band 3, гликофорин A или Kell; или слияния с GPI-линкерным акцепторным пептидов эндогенного GPI-связанного белка поверхности клетки эритроидного ряда, такого как, например, ацетилхолинэстераза, CD55, CD58 или CD59 (см., например, Kooyman et al., Science 1995).
[00310] Антиген, представляющий интерес, может быть конъюгирован с поверхностью культивируемой EHC с помощью различных химических и ферментативных методов, в том числе без ограничений перечисленных в таблице D, таблице D1 и таблице E. Эти способы включают химическую конъюгацию с помощью бифункциональных сшивающих средств, таких как, например, сложный эфир NHS-малеимидное гетеробифункциональное сшивающее средство для соединения группы первичного амина с группой восстановленного тиола. Эти способы также включают ферментативные стратегии, такие как, например, реакцию транспептидазы, опосредованную ферментом сортазой, для соединения одного полипептида, содержащего акцепторную последовательность LPXTG или LPXTA, с полипептидом, содержащим N-терминальную донорную последовательность GGG, см., например, Swee et al., PNAS 2013. Способы также включают комбинированные способы, такие как, например, сортаза-опосредованная конъюгация основ для клик-химии (азида и алкина) c антигеном и клеткой, соответственно, с последующей реакцией циклоприсоединения для химического связывания антигена с клеткой, см., например, Neves et al., Bioconjugate Chemistry, 2013.
[00311] При необходимости домен каталитического связь-образующего полипептида может экспрессироваться на EHC или в ней, или внутри клетки, или вне клетки. Существует множество каталитических связь-образующих полипептидов, в том числе транспептидазы, сортазы и изопептидазы, в том числе происходящие из Spy0128, белка, выделенного из Streptococcus pyogenes.
[00312] Было продемонстрировано, что расщепление аутокаталитической изопептиднаясвязь-образующей субъединицы (CnaB2-домена) из Spy0128 приводит к получению двух отдельных полипептидов, которые сохраняют каталитическую активность со специфичностью в отношении друг друга. Полипептиды в этой системе называются SpyTag и SpyCatcher. После смешивания SpyTag и SpyCatcher подвергаются образованию изопептидной связи между Asp117 на SpyTag и Lys31 на SpyCatcher (Zakeri and Howarth, JACS 2010, 132:4526). Данная реакция совместима с клеточным окружением и высоко специфична в отношении конъюгации белков/пептидов (Zakeri, B.; Fierer, J. O.; Celik, E.; Chittock, E. C.; Schwarz-Linek, U.; Moy, V. T.; Howarth, M. Proc. Natl. Acad. Sci. U.S.A. 2012, 109, E690-E697). Было показано, что SpyTag и SpyCatcher управляют посттрансляционной топологической модификацией эластиноподобного белка. Например, помещение SpyTag на N-конце, а SpyCatcher на C-конце управляет образованием кольцевых эластиноподобных белков (Zhang et al., Journal of the American Chemical Society, 2013).
[00313] Компоненты SpyTag и SpyCatcher можно менять местами, так что система, в которой молекула A слита со SpyTag и молекула B слита со SpyCatcher, функционально эквивалента системе, в которой молекула A слита со SpyCatcher и молекула B слита со SpyTag. Для целей данного документа, когда применяются SpyTag и SpyCatcher, следует понимать, что комплементарная молекула может быть подставлена на ее место.
[00314] Каталитический связь-образующий полипептид, такой как система SpyTag/SpyCatcher, можно применять для прикрепления экзогенного антигена, представляющего интерес, к поверхности EHC. Последовательность полипептида SpyTag может экспрессироваться на внеклеточной поверхности EHC. Полипептид SpyTag, например, может быть слит с N-концом трансмембранного белка 1 типа или 3 типа, например, гликофорином A, слит с C-концом трансмембранного белка 2 типа, например, Kell, вставлен в рамке на внеклеточном конце или внеклеточной петле трансмембранного белка, несколько раз проходящего через мембрану, например, Band 3, слит с GPI-акцепторным полипептидом, например, CD55 или CD59, слит с заякоренным с помощью липидной цепи полипептидом или слит с периферическим мембранным белком. Последовательность нуклеиновой кислоты, кодирующая слияние со SpyTag, может экспрессироваться внутри EHC. Экзогенный антиген, представляющий интерес, может быть слит со SpyCatcher. Последовательность нуклеиновой кислоты, кодирующая слияние со SpyCatcher, может экспрессироваться и секретироваться из той же EHC, которая экспрессирует слияние со SpyTag. В качестве альтернативы, последовательность нуклеиновой кислоты, кодирующая слияние со SpyCatcher, может быть получена экзогенно, например, в продукционной системе на основе клетки бактерии, гриба, насекомого, млекопитающего или в продукционной системе, не содержащей клеток. После реакции полипептидов SpyTag и SpyCatcher будет образовываться ковалентная связь, которая прикрепит экзогенный антиген, представляющий интерес, к поверхности EHC.
[00315] В одном варианте осуществления полипептид SpyTag может экспрессироваться в виде слияния с N-концом гликофорина A под контролем промотора Gata1 в EHC. Экзогенный антиген, представляющий интерес, например экзогенные антигены, перечисленные в таблице F, таблице G, таблице H, таблице I и таблице J, слитые с последовательностью полипептида SpyCatcher, могут экспрессироваться под контролем промотора Gata1 в той же EHC. После экспрессии обоих полипептидов слияния между полипептидами SpyTag и SpyCatcher будет образовываться изопептидная связь, что приведет к образованию ковалентной связи между поверхностью EHC и экзогенным антигеном, представляющим интерес.
[00316] В другом варианте осуществления полипептид SpyTag может экспрессироваться в виде слияния с N-концом гликофорина A под контролем промотора Gata1 в EHC. Экзогенный антиген, представляющий интерес, например экзогенные антигены, перечисленные в таблице F, таблице G, таблице H, таблице I и таблице J, слитые с последовательностью полипептида SpyCatcher, могут экспрессироваться в соответствующей системе экспрессии на основе клетки млекопитающего, например, в клетках HEK293. После экспрессии полипептида слияния со SpyTag на EHC полипептид слияния со SpyCatcher можно приводить в контакт с клеткой. При соответствующих реакционных условиях между полипептидами SpyTag и SpyCatcher будет образовываться изопептидная связь, что приведет к образованию ковалентной связи между поверхностью EHC и экзогенным антигеном, представляющим интерес.
[00317] Каталитический связь-образующий полипептид, такой как система SpyTag/SpyCatcher, можно применять для заякоривания экзогенного антигена, представляющего интерес, во внутриклеточном пространстве EHC. Последовательность полипептида SpyTag может экспрессироваться во внутриклеточном пространстве EHC с помощью целого ряда способов, включающих непосредственную экспрессию трансгена, слияние с эндогенным внутриклеточным белком, таким как, например, гемоглобин, слияние с внутриклеточным доменом эндогенного белка клеточной поверхности, таким как, например, Band 3, гликофорин A, Kell, или слияние со структурным компонентом цитоскелета клетки эритроидного ряда. Последовательность Spy ограничивается прикреплением к концу полипептида, и она может быть интегрирована внутрь последовательности эндогенного полипептида так, чтобы не нарушались трансляция и локализация полипептида. Экзогенный антиген, представляющий интерес, может быть слит со SpyCatcher. Последовательность нуклеиновой кислоты, кодирующая слияние со SpyCatcher, может экспрессироваться внутри той же EHC, которая экспрессирует слияние со SpyTag. После реакции полипептидов SpyTag и SpyCatcher будет образовываться ковалентная связь, которая служит для заякоривания антигена, представляющего интерес, во внутриклеточном пространстве EHC.
[00318] В одном варианте осуществления EHC может экспрессировать SpyTag, слитый с гемоглобином бета, внутри клетки. EHC можно подвергать генетической модификации с помощью генной последовательности, которая включает гемоглобиновый промотор, ген бета-глобина и последовательность SpyTag, так что после трансляции внутриклеточный бета-глобин является слитым со SpyTag на его C-конце. Кроме того, EHC экспрессирует ведомый промотором ген Gata1, который кодирует SpyCatcher, управляющий экспрессией фенилаланингидроксилазы (PAH), так что после трансляции внутриклеточная PAH слита со SpyCatcher на его N-конце. После экспрессии обоих белков слияния связанный со SpyTag бета-глобин соединяется посредством изопептидной связи со связанной со SpyCatcher PAH во внутриклеточном пространстве, что обеспечивает возможность заякоривания PAH с бета-глобином и сохранения во время созревания.
[00319] В другом варианте осуществления полипептид SpyTag может экспрессироваться как слияние с экзогенным антигном, представляющим интерес, в EHC. Полипептид SpyCatcher может экспрессироваться в виде слияния с C-концом (внутриклеточным) гликофорина A в той же EHC. После экспрессии обоих полипептидов слияния между полипептидами SpyTag и SpyCatcher будет образовываться изопептидная связь, что приведет к образованию ковалентной связи между заякоренным в мембране эндогенным полипептидом клетки эритроидного ряда и экзогенным антигеном, представляющим интерес.
[00320] В другом примере экзогенный антиген, представляющий интерес, можно загружать в культивируемую EHC физическим образом (в противоположность экспрессии) с помощью целого ряда способов, в том числе с помощью осмотической загрузки или циклов с гипотоническими-гипертоническими условиями, при которых экзогенный антиген диффундирует через поры, введенные в мембрану EHC (см., например, Cremel and Godfrin, Int J Pharm 2013), и слияния с проникающим в клетку пептидом, таким как происходящий из бактериального токсина, см., например, Kwon et al., J Contr Rel 2009.
[00321] Экзогенный антиген, представляющий интерес, может представлять собой экспрессируемый за счет трансгена, введенного в EHC с помощью электропорации, химической трансфекции или трансфекции с помощью полимерных носителей, вирусной трансдукции, механического разрушения мембраны или другого способа; экзогенный антиген, который экспрессируется за счет мРНК, которую вводят в клетку с помощью электропорации, химической трансфекции или трансфекции с помощью полимерных носителей, вирусной трансдукции, механического разрушения мембраны или другого способа; полипептид экзогенного антигена, полученный за счет сверхэкспрессии нативного локуса вследствие введения внешнего фактора, например, активатора транскрипции, репрессора транскрипции или усилителя секреторного пути; экзогенный антиген, который синтезирован, экстрагирован или продуцируется производящей клеткой или другой внешней системой и внедрен в EHC.
[00322] EHC-клетки по настоящему изобретению необязательно можно загружать материалами (загрузка), такими как пептиды, белки, ДНК, РНК, siRNA и другие макромолекулы, путем применения в отношении клетки контролируемого повреждения в течение заранее определенного количества времени, чтобы вызвать в клеточной мембране нарушения, так что материалы могут быть доставлены внутрь клетки (например, в цитоплазму).
[00323] В предпочтительных вариантах осуществления EHC представляет собой ретикулоцит. Например, в ретикулоциты можно загрузить мРНК, кодирующую экзогенный антиген с помощью контролируемого повреждения клетки. При необходимости мРНК может быть депротеинизированной или модифицированной. Модификация мРНК, которая усиливает стабильность мРНК и/или снижает иммуногенность, включает, например, ARCA: аналог кэпа, предотвращающий его обращение (m27.3'-OGP3G), GP3G (неметилированный аналог кэпа), m7GP3G (монометилированный аналог кэпа), m32.2.7GP3G (триметилированный аналог кэпа), m5CTP (5'-метил-цитидин трифосфат), m6ATP (N6-метил-аденозин-5'-трифосфат), s2UTP (2-тио-уридин-трифосфат) и Ψ (псевдоуридин-трифосфат).
[00324] В другом предпочтительном варианте осуществления EHC представляет собой эритроцит. Например, в эритроциты можно загружать экзогенный антиген с помощью контролируемого повреждения клетки. Повреждение клетки может быть вызвано, например, давлением, обусловленным механической напряжением или усилиями сдвига, что вызывает у клетки деформацию, сокращение, быстрое растяжение, быстрое сжатие или пульсацию с высокой скоростью сдвига. Контролируемое повреждение клетки приводит к попаданию материала (загрузки) в цитоплазму клетки из среды, окружающей клетку.
[00325] Применение контролируемого повреждения клетки, основанного на контролируемой деформации клетки (например, механической деформации клетки, по мере ее прохождения через сужение), приводит к поглощению материала (загрузка) путем диффузии, а не эндоцитоза. После поглощения клеткой при контролируемом повреждении материал (загрузка) присутствует в цитоплазме, а не в эндосомах, что, тем самым, делает материал легкодоступным для клетки. Контролируемое повреждение клетки, например, с помощью контролируемой деформации, сохраняет жизнеспособность клетки (например, более 50%, 70%, или более 90%). В определенных вариантах осуществления контролируемое повреждение клетки, например, с помощью контролируемой деформации, сохраняет состояние клеточной дифференцировки и активности. При необходимости применяют комбинированную обработку, например, контролируемое повреждение с помощью деформации, с последующей или предшествующей, например, электропорацией или другим способом повышения проницаемости клеточной мембраны. Необязательно можно применять поверхностно-активные вещества.
[00326] Способы механической деформации особенно пригодны для клеток, которые неудовлетворительно переносят другие способы повышения проницаемости мембраны, например, демонстрируют сниженную жизнеспособность или другое состояние дифференцировки после осуществления таких способов. Способы механической деформации также пригодны для материала (загрузки), который неудовлетворительно переносит другие способы повышения проницаемости мембраны. В качестве альтернативы или дополнительно при применении альтернативных способов загрузка может вводиться в клетку в недостаточной степени, например, вследствие, например, заряда, гидрофобности или размера загрузки.
[00327] Один иллюстративный способ контролируемого повреждения с помощью деформации и устройств, пригодных для таких способов, описан, например, в PCT-публикации № WO2013059343 INTRACELLULAR DELIVERY, включенной в данный документ посредством ссылки.
[00328] В конкретном варианте осуществления представлена популяция ретикулоцитов, которая была подвергнута контролируемому повреждению клетки с помощью контролируемой деформации. Например, клетки можно сжимать и деформировать с помощью прохождения через микроканал с диаметром, меньшим, чем отдельный ретикулоцит, тем самым вызывая нарушения в клеточной мембране, так что в мембране появляются поры. Клетки продвигаются, например проталкиваются, через каналы или трубки с помощью применения давления. Сжатие и деформация происходят в среде доставки, содержащей загрузку, например, экзогенный полипептид или олигонуклеотид (например, ДНК, РНК, такую как мРНК). Например, среда доставки может содержать экзогенный антиген, перечисленный в таблице F, таблице G, таблице H, таблице I и таблице J, или его кодирующую мРНК. После деформации ретикулоцит захватывает и сохраняет экзогенный материал. После контролируемого повреждения клетки путем сужения, растяжения и/или пульсации с высокой скоростью сдвига клетки необязательно инкубируют в среде доставки, которая содержит материал (загрузку). Клетки можно поддерживать в среде доставки в течение нескольких минут для восстановления, например, для закрытия повреждения, вызванного прохождением через сужение. Это может происходить при комнатной температуре.
[00329] Контролируемое повреждение клетки, применяемое в данном документе, включает: i) трансфекцию, опосредованную вирусом (например, вирусом простого герпеса, аденовирусом, аденоассоциированным вирусом, вирусом осповакцины или вирусом Синдбис), ii) трансфекцию, опосредованную химическим веществом, например, катионным полимером, фосфатом кальция, катионным липидом, полимерами и наночастицами, такими как циклодекстрин, липосомами, катионными липосомами, DEAE-декстраном, полиэтиленимином, дендримером, полибреном, фосфатом кальция, липофектином, DOTAP, липофектамином, CTAB/DOPE, DOTMA; и iii) трансфекцию, опосредованную физическими методами, в том числе прямой инъекцией, доставкой с биолистическими частицами, электропорацией, облучением лазером, сонопорацией, магнитными наночастицами и контролируемой деформацией (например, обжим клетки), примером чего является микроигла, наноигла, фемтошприц, наконечник для атомно-силовой микроскопии (AFM), генной пушкой (например, наночастицы золота), Amaxa Nucleofector, фототрансфекцией (мультифотонный лазер), импалефекцией и магнитофекцией, и другими соответствующими способами, известными из уровня техники. Любой пригодный способ можно применять для получения EHC-клеток, описанных в данном документе, которые содержат один или несколько необходимых ДНК, РНК (например, мРНК) или полипептидов, содержащих антиген.
[00330] Экзогенный антиген, представляющий интерес, можно обнаруживать на EHC по настоящему изобретению. Присутствие экзогенного антигена можно подтверждать и количественно оценивать с применением стандартных способов молекулярной биологии, например, вестерн-блоттинга или FACS-анализа. Экзогенные антигены, присутствующие во внутриклеточной среде, можно количественно оценить после лизиса клеток или с применением флуоресцентного обнаружения.
Производство
[00331] В некоторых вариантах осуществления EHC получают с применением в качестве источника гемопоэтической клетки-предшественника, например, CD34+ клетки, эритроцита, тромбоцита, мегакариоцита или нейтрофила. В некоторых вариантах осуществления гемопоэтическая клетка-предшественник выделена из донора-человека с помощью способа, соответствующего принципам GMP. В некоторых вариантах осуществления исходные клетки берут начало от аутологичного донора. В некоторых вариантах осуществления исходные клетки берут начало от аллогенного донора. У донора можно провести типирование в отношении полиморфизма антигенов клеток крови и/или донора генотипируют в отношении антигенов клеток крови. Донор может быть донором с универсальным типом крови. В некоторых вариантах осуществления донор имеет фенотип Бомбей, т.е. у него не экспрессируется антиген H. В некоторых вариантах осуществления донор имеет тип крови O по системе ABO и является Rh-отрицательным.
[00332] В некоторых вариантах осуществления EHC получают с применением CD34+ гемопоэтических прогениторных клеток, активированных периферических CD34+ клеток или CD34+ клеток, происходящих из костного мозга, в качестве источника исходного материала. В некоторых вариантах осуществления исходные клетки происходят из пуповинной крови, представляют собой индуцированные плюрипотентные стволовые клетки, или представляют собой эмбриональные стволовые клетки.
[00333] Экспрессирующая экзогенный антиген EHC может быть культивируемой. Число культивируемых EHC-клетки можно пропорционально увеличивать от настольного масштаба до масштаба биореактора. Например, EHC-клетки культивируют то тех пор, пока они не достигнут максимальной плотности популяции, например, 1×105, 1×106, 1×107 или более 1×107 EHC-клеток на мл. Необязательно после достижения максимальной плотности популяции EHC-клетки можно переносить в больший объем свежей среды. Экспрессирующие экзогенный антиген EHC-клетки можно культивировать в биореакторе, таком как, например, биореактор типа Wave, биореактор с перемешиванием. Различные конфигурации биореакторов известны из уровня техники, и при необходимости можно выбрать соответствующую конфигурацию. Конфигурации, пригодные для культивирования и/или размножения популяций экспрессирующих экзогенный антиген EHC-клеток, может легко определить специалист в данной области без излишнего проведения экспериментов. Биореактор может быть обогащен кислородом. Биореактор может необязательно содержать один или несколько лопастных мешалок, поток рециркуляции, входящий поток для среды и элементы управления для регуляции притока среды и питательных веществ или для регуляции оттока среды, питательных веществ и отходов.
[00334] В некоторых вариантах осуществления биореактор может содержать популяцию человеческих EHC-клеток, содержащих экзогенный антиген, которые избавляются от своей внутриклеточной ДНК в ходе процесса культивирования. Например, биореактор может содержать популяцию человеческих EHC-клеток, безъядерных EHC-клеток и пиреноцитов после культивирования. В конкретном варианте осуществления человеческие EHC-клетки и безъядерные EHC-клетки содержат экзогенный антиген, и экзогенный антиген сохраняется в безъядерной EHC, тогда как рекомбинантная нуклеиновая кислота, кодирующая экзогенный антиген, не сохраняется в безъядерной клетке. В определенных вариантах осуществления безъядерная EHC, содержащая экзогенный антиген, проявляет практически такую же хрупкость осмотической мембраны, что и соответствующая выделенная немодифицированная не подвергнутая культивированию EHC.
[00335] В одном варианте осуществления популяция экспрессирующих экзогенный антиген EHC-клеток, полученных из EHC-клеток или предшественников EHC в биореакторе, подвергается общему возрастанию количества в более чем 20000 раз за 14 дней или больше. В некоторых вариантах осуществления экзогенный антиген вводят в культивируемый или свежевыделенный предшественник EHC, и после введения рекомбинантной нуклеиновой кислоты, кодирующей экзогенный антиген, популяция экспрессирующих экзогенный антиген EHC-клеток, полученных из предшественников EHC в биореакторе, увеличивается в количестве в биореакторе по сравнению с количеством клеток-предшественников более чем в 20000 раз.
[00336] В некоторых вариантах осуществления биореактор представляет собой биореактор Wave или перемешиватель, работающий от лопастной мешалки. Биореактор может аэрироваться с помощью рассекателя. В одном варианте осуществления биореактор предназначен для однократного использования. В одном варианте осуществления биореактор имеет CIP (система очистки на месте). Конечное количество экспрессирующих экзогенный антиген EHC-клеток, которое можно получить в условиях биореактора, описанного в данном документе, может составлять более 109, 1010, 1011, 1012, 1013 или более 1013 EHC-клеток. Плотность экспрессирующих экзогенный антиген EHC-клеток можно отслеживать во время культивирования путем измерения плотности плеток путем подсчета с помощью гемоцитометра или путем измерения оптической плотности при 600 нм. Необязательно процесс культивирования отслеживают в отношении уровней pH, насыщения кислородом, скорости перемешивания и/или степени рециркуляции.
Способы и свойства
[00337] Идентичность экспрессирующих экзогенный антиген EHC-клеток можно оценивать с помощью in vitro анализов. Например, идентичность экспрессирующих экзогенный антиген EHC-клеток оценивают путем подсчета количества EHC-клеток в популяции, например, с помощью микроскопии, с помощью проточной цитометрии или с помощью гемоцитометрии. В качестве альтернативы или дополнительно идентичность экспрессирующих экзогенный антиген EHC-клеток оценивают с помощью анализа содержания белка в EHC-клетках, например, с помощью проточной цитометрии, вестерн-блот-анализа, иммунопреципитации, флуоресцентной спектроскопии, хемолюминесценции, масс-спектрометрии или спектроскопии поглощения. В одном варианте осуществления оценивают содержание белка, который не является белком клеточной поверхности, например, представляет собой интегральный мембранный белок, гемоглобин, зрелый гемоглобин, фетальный гемоглобин, эмбриональный гемоглобин, белок цитоскелета. В одном варианте осуществления оценивают содержание белка, который представляет собой поверхностный белок, например, маркер дифференцировки, рецептор, корецептор, переносчик, гликопротеин. В одном варианте осуществления белок клеточной поверхности выбран из перечня, включающего без ограничений гликофорин A, CKIT, трансферриновый рецептор, Band3, Kell, CD45, CD46, CD47, CD55, CD59, CR1. В некоторых вариантах осуществления идентичность экспрессирующих экзогенный антиген EHC-клеток оценивают с помощью анализа содержания экзогенного антигена в EHC-клетках, например, с помощью проточной цитометрии, вестерн-блот-анализа, иммунопреципитации, флуоресцентной спектроскопии, хемолюминесценции, масс-спектрометрии или спектроскопии поглощения. Например, идентичность экспрессирующих экзогенный антиген EHC-клеток можно оценить путем оценки содержания мРНК в EHC-клетках, например, с помощью RT-PCR, проточной цитометрии или нозерн-блот-анализа. Идентичность экспрессирующих экзогенный антиген EHC-клеток можно оценить посредством оценки содержания ядерного материала, например, с помощью проточной цитометрии, микроскопии или Саузерн-блот-анализа, с применением, например, ядерного красителя или зонда на основе нуклеиновой кислоты. В качестве альтернативы или дополнительно идентичность экспрессирующих экзогенный антиген EHC-клеток оценивают путем оценки содержания липидов в EHC-клетках, например, с помощью проточной цитометрии, жидкостной хроматографии или с помощью масс-спектрометрии.
[00338] В некоторых вариантах осуществления идентичность экспрессирующих экзогенный антиген EHC-клеток оценивают с помощью определения метаболической активности EHC-клеток, например, посредством масс-спектрометрии, хемолюминесценции, флуоресцентной спектроскопии, спектроскопии поглощения. Метаболическую активность можно оценить с помощью скорости потребления АТФ и/или метаболическую активность оценивают путем измерения уровня 2,3-дифосфоглицерата (2,3-DPG) в экспрессирующей экзогенный антиген EHC. Метаболическую активность можно оценить как скорость метаболизма одного из следующего, в том числе без ограничений ацетилсалициловой кислоты, N-ацетилцистеина, 4-аминофенола, азатиоприна, бунолола, каптоприла, хлорпромазина, дапсона, даунорубицина, дегидроэпиандростерона, диданозина, допамина, эпинефорина, эсмолола, эстрадиола, эстрона, этопозида, галоперидола, героина, инсулина, изопротеренола, изосорбида динитрата, LY 217896, 6-меркаптопурина, мизонидазола, нитроглицерина, норэпинефрина, пара-аминобензойной кислоты. В некоторых вариантах осуществления идентичность экспрессирующих экзогенный антиген EHC-клеток оценивают с помощью расщепления субстрата EHC-клетками, например, посредством масс-спектрометрии, хемолюминесценции, флуоресцентной спектроскопии, спектроскопии поглощения. Субстрат может представлять собой одно из следующего, в том числе без ограничений, ацетазоламид, арбутин, буметамид, креатинин, дарстин, десэтилдорзоламид, дигоксигенина дигитоксид, дигоксин-16′-глюкуронид, эпинефрин, гентамицин, гиппуровую кислоту, метформин, норэпинефрин, п-аминогиппуровую кислоту, папаверин, пенициллин G, феноловый красный, серотонин, сульфосалициловую кислоту, такролимус, тетрациклин, тукарезол и ванкомицин.
[00339] В одном варианте осуществления популяция экспрессирующих экзогенный антиген EHC-клеток дифференцирована из клетки-предшественника. В этом варианте осуществления состояние дифференцировки популяции экспрессирующих экзогенный антиген EHC-клеток оценивают с помощью анализа in vitro. Анализы in vitro включают описанные в данном документе для оценки идентичности EHC-клеток, в том числе без ограничений оценку скорости размножения, количества, содержания или уровня экспрессии белка, содержания или уровня экспрессия мРНК, содержания липидов, расщепления субстрата, каталитической активности или метаболической активности.
[00340] В некоторых вариантах осуществления экспрессирующие экзогенный антиген EHC-клетки культивируют и состояние дифференцировки EHC-клеток оценивают в нескольких моментах времени в ходе процесса культивирования.
[00341] Экспрессирующие экзогенный антиген EHC-клетки можно получать с применением ретикулоцитов в качестве источника исходного материала. Чистоту выделенных ретикулоцитов можно оценить с применением микроскопии, при этом ретикулоциты характеризуются ретикулярной (ситоподобной) сетью из рибосомальной РНК, которая становится видна под микроскопом при определенных видах окрашивания, таких как новым метиленовым синим или бриллиантовым крезиловым синим. Экспрессия трансферринового рецептора (CD71) на поверхности клетки также выше у ретикулоцитов и снижается по мере их созревания до эритроцитов, что обеспечивает возможность обогащения и анализа популяций ретикулоцитов с применением антител к CD71 (см., например, листок-вкладыш для CD71-микрогранул от Miltenyi, No. 130-046-201). В качестве альтернативы, для оценки свойств выделенных ретикулоцитов по сравнению со зрелыми эритроцитами можно применять анализ содержания креатина и гемоглобина A1C и активности ферментов пируваткиназы, аспартатаминотрансферазы и порфобилиногендезаминазы (см., например, Brun et al., Blood 76:2397-2403 (1990)). Например, активность порфобилиногендезаминазы примерно в 9 раз выше, тогда как содержание гемоглобина A1C примерно в 10 раз меньше у ретикулоцитов по сравнению со зрелыми эритроцитами.
[00342] В некоторых вариантах осуществления клетки, пригодные для получения экспрессирующих экзогенный антиген EHC-клеток, дифференцируют ex vivo и/или in vivo из одной или нескольких стволовых клеток. В одном варианте осуществления одна или несколько стволовых клеток представляют собой одну или несколько гемопоэтических стволовых клеток. Можно проводить различные анализы для подтверждения дифференцировки культивируемых гемопоэтических стволовых клеток в ретикулоциты и эритроциты ex vivo, в том числе, например, микроскопический анализ, гематологический анализ, проточную цитометрию, измерения деформируемости, анализ активности ферментов, а также анализ гемоглобина и определение функциональных свойств (Giarratana et al., Nature Biotech. 23:69-74 (2005)). Фенотип культивируемых гемопоэтических стволовых клеток можно оценивать с применением микроскопического исследования клеток, окрашенных, например, с помощью крезилового бриллиантового синего. В ретикулоцитах, например, при таких условиях окрашивания видна ретикулярная сеть из рибосомальной РНК, тогда как эритроциты не окрашиваются. У безъядерных клеток также можно контролировать стандартные гематологические показатели, в том числе средний объем клетки (MCV; фемтолитры (фл)), среднюю концентрацию гемоглобина в клетке (MCHC; %) и среднее содержание гемоглобина в клетке (MCH; пг/клетка) с применением, например, устройства XE2100 (Sysmex, Roche Diagnostics).
[00343] В некоторых вариантах осуществления экспрессирующие экзогенный антиген EHC-клетки оценивают в отношении их основных физических свойств, например, размера, массы, объема, диаметра, плавучести, плотности, и свойств мембраны, например, вязкости, изменения деформируемости и текучести.
[00344] В одном варианте осуществления диаметр экспрессирующих экзогенный антиген EHC-клеток измеряют с помощью микроскопии или с помощью автоматизированных устройств, например, устройства для гематологического анализа. В одном варианте осуществления диаметр экспрессирующих экзогенный антиген EHC-клеток составляет приблизительно 1-20 микрон. В одном варианте осуществления диаметр экспрессирующих экзогенный антиген EHC-клеток по меньшей мере в одном измерении составляет приблизительно 1-20 микрон. В одном варианте осуществления диаметр экспрессирующих экзогенный антиген EHC-клеток составляет менее приблизительно 1 микрона. В одном варианте осуществления диаметр EHC-клеток в одном измерении превышает приблизительно 20 микрон. В одном варианте осуществления диаметр экспрессирующих экзогенный антиген EHC-клеток составляет от приблизительно 1 микрона до приблизительно 20 микрон, от приблизительно 2 микрон до приблизительно 20 микрон, от приблизительно 3 микрон до приблизительно 20 микрон, от приблизительно 4 микрон до приблизительно 20 микрон, от приблизительно 5 микрон до приблизительно 20 микрон, от приблизительно 6 микрон до приблизительно 20 микрон, от приблизительно 5 микрон до приблизительно 15 микрон или от приблизительно 10 микрон до приблизительно 30 микрон.
[00345] В одном варианте осуществления средний объем клетки у экспрессирующих экзогенный антиген EHC-клеток измеряют с применением устройства для гематологического анализа. В одном варианте осуществления значение среднего объема клетки у EHC-клеток составляет более 10 фл, 20 фл, 30 фл, 40 фл, 50 фл, 60 фл, 70 фл, 80 фл, 90 фл, 100 фл, 110 фл, 120 фл, 130 фл, 140 фл, 150 фл или более 150 фл. В одном варианте осуществления средний объем клетки у EHC-клеток составляет менее 30 фл, 40 фл, 50 фл, 60 фл, 70 фл, 80 фл, 90 фл, 100 фл, 110 фл, 120 фл, 130 фл, 140 фл, 150 фл, 160 фл, 170 фл, 180 фл, 190 фл, 200 фл или менее 200 фл. В одном варианте осуществления средний объем клетки у EHC-клеток составляет 80-100 фемтолитров (фл).
[00346] В одном варианте осуществления среднюю плавучую массу экспрессирующих экзогенный антиген EHC-клеток (пг/клетка) измеряют с применением суспензионного микроканального вибратора или двойного суспензионного микроканального вибратора (см., например, Byun et al. PNAS 2013 110(19):7580 и Bryan et al. Lab Chip 2014 14(3):569).
[00347] В одном варианте осуществления плотность в сухом состоянии экспрессирующих экзогенный антиген EHC-клеток измеряют с помощью плавучей массы в анализе обмена H2O-D2O (см., например, Feijo Delgado et al., PLOS One 2013 8(7):e67590).
[00348] В некоторых вариантах осуществления экспрессирующие экзогенный антиген EHC-клетки обладают средним изменением деформируемости мембраны со стандартным отклонением, превышающим 10, 20, 30, 40, 50, 60, 70, 80, 90, 100 или более 100 мрад, измеряемым с помощью интерференционной пространственной световой микроскопии (SLIM) (см., например, Bhaduri et al., Sci Reports 2014, 4:6211).
[00349] В одном варианте осуществления среднюю вязкость мембраны у популяции экспрессирующих экзогенный антиген EHC-клеток измеряют путем определения средней флуоресценции после инкубации с зависящими от вязкости флуорофорами с высоким квантовым выходом (см., например, Haidekker et al. Chem & Biol 2001 8(2):123).
[00350] В одном варианте осуществления текучесть мембраны у экспрессирующих экзогенный антиген EHC-клеток измеряют с помощью флуоресцентной поляризации, например, с помощью считывающего устройства для микропланшетов BMG Labtech POLARstar Omega.
[00351] Например, для измерения деформируемости ретикулоциты можно отделять от ядерных клеток в день 15 культивирования, например, путем пропускания через фильтр для удаления лейкоцитов (например, Leucolab LCG2, Macopharma), и впоследствии анализировать с применением эктацитометрии. Безъядерные клетки суспендируют в 4% растворе поливинилпирролидона и затем подвергают воздействию возрастающего омотического градиента от 60 до 450 мосмоль/л. Изменения в характере лазерной дифракции (индекса деформируемости) клеток регистрируют в зависимости от осмолярности для оценки динамической деформируемости клеточной мембраны. Максимальный индекс деформируемости, полученный при физиологически релевантной осмолярности, связан со средней площадью поверхности эритроцитов.
[00352] В некоторых вариантах осуществления экспрессирующие экзогенный антиген EHC-клетки анализируют на содержание гемоглобина. Анализы на гемоглобин можно применять для оценки фенотипа дифференцированных клеток (Giarratana et al., Nature Biotech. 23:69-74 (2005)). Например, для оценки процентной доли различных фракций гемоглобина можно применять высокоэффективную жидкостную хроматографию (HPLC) с применением анализатора Bio-Rad Variant II Hb (Bio-Rad Laboratories). Равновесие кислорода можно измерять с применением непрерывного способа с помощью двуволнового спектрофотометра (например, Hemox analyzer, TCS). Связывающую способность гемоглобина можно оценить с применением импульсного фотолиза. В данном способе повторное связывание CO с внутриклеточными тетрамерами гемоглобина анализируют при 436 нм после фотолиза с помощью 10-наносекундного импульса при 532 нм.
[00353] При необходимости экспрессирующие экзогенный антиген EHC-клетки, описанные в данном документе, можно очищать после получения. В уровне техники известно множество соответствующих способов очистки. Например, экспрессирующие экзогенный антиген EHC-клетки очищают с помощью центрифугирования, магнитофореза, облучения, акустофореза и химического или физического удаления ядра. В одном варианте осуществления экспрессирующие экзогенный антиген EHC-клетки очищают посредством созревания ex vivo с помощью, например, совместного культивирования со стромальным клетками. В одном варианте осуществления экспрессирующие экзогенный антиген EHC-клетки очищают посредством химической или ферментативной обработки EHC-клеток, например, путем обработки дегликозилирующим ферментом.
[00354] В одном варианте осуществления экспрессирующие экзогенный антиген EHC-клетки очищают посредством блокирования какого-либо остаточного репликативного потенциала экспрессирующих экзогенный антиген EHC-клеток. В одном варианте осуществления экспрессирующие экзогенный антиген EHC-клетки подвергают облучению, например, рентгеновскими лучами, гамма-лучами, бета-частицами, альфа-частицами, нейтронами, протонами, ядрами элементов, УФ-лучами, чтобы повредить остаточные способные к репликации нуклеиновые кислоты.
[00355] Ионизирующая радиация представляет собой энергию, переносимую посредством рентгеновских лучей, гамма-лучей, бета-частиц (быстрые электроны), альфа-частиц (ядро атома гелия), нейтронов, протонов и других тяжелых ионов, таких как ядра аргона, азота, углерода и других элементов. Рентгеновские лучи и гамма лучи представляют собой электромагнитные волны, подобные свету, но их энергия намного превышает таковую света (их длины волн намного короче). Ультрафиолетовый (УФ) свет представляет собой излучение с промежуточной энергией, которое может повреждать клетки, но УФ-свет отличается от упомянутых выше форм электромагнитного излучения тем, что он вызывает не ионизацию (потерю электрона) атомов или молекул, а возбуждение (изменение энергетического уровня электрона). Другие формы излучения – частицы - имеют или отрицательный заряд (электроны), или положительный заряд (протоны, альфа-лучи и другие тяжелые ионы), или электрически нейтральны (нейтроны).
[00356] Ионизация, индуцированная облучением, может действовать напрямую на молекулы клеточного компонента или опосредовано на молекулы воды с образованием радикалов, происходящих из воды. Радикалы реагируют с соседними молекулами за очень короткий промежуток времени, что приводит к расщеплению химических связей или окислению (присоединению атомов кислорода) подвергающихся воздействию молекул. Главным эффектом в клетке является разрывы ДНК. Поскольку ДНК состоит из пары комплементарных двойных нитей, могут происходить разрывы в любой одиночной нити или обеих нитях. Однако считается, что последнее намного важнее с биологической точки зрения. Большинство однонитевых разрывов в норме могут подвергаться репарации благодаря двухнитевой природе молекулы ДНК (две нити комплементарны друг другу, так что интактная нить может служить матрицей для репарации другой поврежденной нити). В случае двухнитевых разрывов, однако, репарация является более сложной, и может происходить ошибочное присоединение разорванных концов. Эта так называемая неправильная репарация приводит к индукции мутаций, хромосомным аберрациям или гибели клетки.
[00357] Делеция сегментов ДНК представляет собой преобладающую форму радиационного повреждения у клеток, которые пережили облучение. Она может быть обусловлена (1) неправильной репарацией двух отдельных двухнитевых разрывов в молекуле ДНК с соединением двух наружных концов и утратой фрагмента между разрывами или (2) процессом очистки (ферментативным отщеплением нуклеотидов - составных элементов ДНК) разорванных концов перед повторным соединением для репарации одного двухнитевого разрыва.
[00358] Виды излучения отличаются не только своими составляющими (электроны, протоны, нейтроны и т.д.), но также своей энергией. Виды излучения, которые обусловливают интенсивную ионизацию по ходу своего пробега (такие как нейтроны), называют излучением с высокой линейной передачи энергии (с высокой LET), физическим параметром, описывающим среднее количество энергии, высвобождаемой на единицу длины пробега. (См. сопровождающую фигуру). Виды излучения с низкой LET приводят лишь к редкой ионизации по ходу своего пробега и, следовательно, почти однородной в пределах клетки. Доза облучения представляет собой количество энергии на единицу биологического материала (например, количество ионизаций на клетку). Таким образом, виды излучения с высокой LET более разрушительны для биологического материала, чем виды излучения с низкой LET - такие как рентгеновские и гамма-лучи - поскольку при одной и той же дозе виды излучения с низкой LET индуцируют в клетке такое же количество радикалов более рассеянно в пределах клетки, тогда как виды излучения с высокой LET - такие как нейтроны и альфа-частицы - передают большую часть своей энергии небольшой области клетки. Локальные повреждения ДНК, обусловленные интенсивной ионизацией за счет видов облучения с высокой LET, сложнее подвергать репарации, чем диффузное повреждение ДНК, обусловленное редкими ионизациями за счет видов излучения с низкой LET.
[00359] В одном варианте осуществления популяция экспрессирующих экзогенный антиген EHC-клеток подвергнута облучению гамма-лучами с применением дозы облучения, составляющей более 1, 5, 10, 15, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100 или более 100 кГр.
[00360] В одном варианте осуществления популяция экспрессирующих экзогенный антиген EHC-клеток подвергнута облучению рентгеновскими лучами с применением дозы облучения, составляющей более 0,1, 0,5, 1, 5, 10, 15, 20, 25, 30, 35, 40, 50, 60, 70, 80, 90, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000 или более 10000 мЗв.
[00361] Чистоту популяции экспрессирующих экзогенный антиген EHC-клеток можно оценить путем измерения однородности популяции. В одном варианте осуществления среднюю ширину распределения по объему у экспрессирующих экзогенный антиген EHC-клеток измеряют с помощью устройства для гематологического анализа. В одном варианте осуществления популяция экспрессирующих экзогенный антиген EHC-клеток характеризуется отношением ретикулоцитов к неретикулоцитам, составляющим более 10, 100, 1000, 104, 105, 106 или более 106. Однородность популяции экспрессирующих экзогенный антиген EHC-клеток можно оценить путем измерения содержания стромальных клеток в популяции. В одном варианте осуществления популяция экспрессирующих экзогенный антиген EHC-клеток характеризуется менее 1 ppb стромальных клеток. В качестве альтернативы или дополнительно однородность популяции экспрессирующих экзогенный антиген EHC-клеток оценивают путем измерения вирусного титра и/или бактериального колониеобразующего потенциала популяции.
[00362] В одном варианте осуществления однородность популяции экспрессирующих экзогенный антиген EHC-клеток оценивают с помощью анализа in vitro. Анализы in vitro включают описанные в данном документе для оценки идентичности EHC-клеток, в том числе без ограничений оценку скорости размножения, количества, содержания или уровня экспрессии белка, содержания или уровня экспрессия мРНК, содержания липидов, расщепления субстрата, каталитической активности или метаболической активности.
[00363] Зрелые эритроциты для применения в получении экспрессирующих экзогенный антиген EHC-клеток можно выделять с применением различных способов, таких как, например, применение устройства для промывки клеток, сепаратора клеток с непрерывным потоком, разделение по градиенту плотности, сортировка флуоресцентно-активированных клеток (FACS), иммуномагнитное истощение от Miltenyi (MACS), или комбинации этих способов (см., например, van der Berg et al., Clin. Chem. 33:1081-1082 (1987); Bar-Zvi et al., J. Biol. Chem. 262:17719-17723 (1987); Goodman et al., Exp. Biol. Med. 232:1470-1476 (2007)).
[00364] Эритроциты можно выделить из цельной крови с помощью простого центрифугирования (см., например, van der Berg et al., Clin. Chem. 33:1081-1082 (1987)). Например, цельную кровь, антикоагулированную с помощью EDTA, можно центрифугировать при 800×g в течение 10 мин при 4°C. Богатую тромбоцитами плазму и лейкоцитарную пленку удаляют и красные кровяные клетки трижды промывают изотоническим физиологическим раствором (NaCl, 9 г/л).
[00365] В качестве альтернативы, эритроциты можно выделять с применением центрифугирования в градиенте плотности с различными средами для разделения, такими как, например, Ficoll, Hypaque, Histopaque, Percoll, Sigmacell или их комбинации. Например, некоторый объем Histopaque-1077 наслаивают на поверхность равного объема Histopaque-1119. Цельную кровь, антикоагулированную с помощью EDTA, которую развели 1:1 в равном объеме изотонического физиологического раствора (NaCl, 9 г/л), наслаивают на поверхность Histopaque и образец центрифугируют при 700×g в течение 30 минут при комнатной температуре. При этих условиях гранулоциты мигрируют к границе раздела 1077/1119, лимфоциты, другие мононуклеарные клетки и тромбоциты остаются на границе раздела плазма/1077, а красные кровяные клетки осаждаются. Красные кровяные клетки трижды промывают изотоническим физиологическим раствором.
[00366] В качестве альтернативы, эритроциты можно выделять посредством центрифугирования с применением ступенчатого градиента Percoll (см., например, Bar-Zvi et al., J. Biol. Chem. 262:17719-17723 (1987)). Например, свежую кровь смешивают с антикоагулирующим раствором, содержащим 75 мМ цитрата натрия и 38 мМ лимонной кислоты, и клетки быстро промывают забуференным Hepes физиологическим раствором. Лейкоциты и тромбоциты удаляют путем поглощения с помощью смеси α-целлюлозы и Sigmacell (1:1). Эритроциты далее отделяют от ретикулоцитов и остаточных белых кровяных клеток путем центрифугирования в 45/75% ступенчатом градиенте Percoll в течение 10 минут при 2500 об/мин на роторе Sorvall SS34. Эритроциты уходят в осадок, тогда как полоса ретикулоцитов располагается на границе раздела 45/75%, а полоса оставшихся белых кровяных клеток на границе раздела 0/45%. Percoll удаляют из эритроцитов с помощью нескольких промывок в забуференном Hepes физиологическом растворе. Другие материалы, которые можно применять для получения градиента плотности для выделения эритроцитов, включают OptiPrep™, 60% раствор йодиксанола в воде (от Axis-Shield, Данди, Шотландия).
[00367] Эритроциты можно отделить от ретикулоцитов, например, с применением проточной цитометрии (см., например, Goodman el al., Exp. Biol. Med. 232:1470-1476 (2007)). В этом случае цельную кровь центрифугируют (550×g, 20 минут, 25°C) для отделения клеток от плазмы. Осадок клеток ресуспендируют в растворе фосфатно-солевого буфера и далее фракционируют на Ficoll-Paque (плотность 1,077), например, с помощью центрифугирования (400×g, 30 минут, 25°C) для отделения эритроцитов от белых кровяных клеток. Полученный осадок клеток ресуспендируют в RPMI, дополненной 10% фетальной бычьей сыворотки, и подвергают сортировке на устройстве для FACS, таком как, например, Becton Dickinson FACSCalibur (BD Biosciences, Франклин Лейкс, Нью-Джерси, США), на основе размера и зернистости.
[00368] Эритроциты можно отделять с помощью иммуномагнитного истощения (см., например, Goodman, el al., (2007) Exp. Biol. Med. 232:1470-1476). В этом случае магнитные гранулы для элиминации клеток, не относящихся к эритроцитам, применяют с антителами, специфичными в отношении типа клетки. Например, эритроциты отделяют от большинства других компонентов крови с применением градиента плотности, описываемого в данном документе, с последующим иммуномагнитным истощением любых остаточных ретикулоцитов. Клетки предварительно обрабатывают человеческой иммунной сывороткой в течение 20 минут при 25°C и затем обрабатывают антителами против антигенов, специфичных для ретикулоцитов, таких как, например, CD71 и CD36. Антитела могут быть напрямую прикреплены к магнитным гранулам или конъюгированы с PE, например, с которым будут реагировать магнитные гранулы с антителом к PE. Комплекс антитело-магнитная гранула способен селективно удалять остаточные ретикулоциты, например, из популяции эритроцитов.
[00369] Эритроциты также можно выделять с применением афереза. Процесс афереза предусматривает забор цельной крови у пациента или донора, разделение компонентов крови с применением центрифугирования или сортировки клеток, изъятие одной или нескольких отделенных частей и переливание оставшихся компонентов обратно пациенту или донору. Для этого в настоящее время применяют целый ряд устройств, таких как, например, устройства Amicus и Alyx от Baxter (Дирфилд, Иллинойс, США), устройство Trima Accel от Gambro BCT (Лейквуд, Колорадо, США) и устройство MCS+9000 от Haemonetics (Брейнтри, Массачуссетс, США). Дополнительные способы очистки могут быть необходимы для достижения соответствующей степени чистоты клеток.
[00370] В некоторых вариантах осуществления экспрессирующие экзогенный антиген EHC-клетки дифференцируют ex vivo и/или in vivo из одного или нескольких ретикулоцитов. Для получения экспрессирующих экзогенный антиген EHC-клеток можно использовать ретикулоциты. Ретикулоциты представляют собой незрелые красные кровяные клетки, и они составляют приблизительно 1% красных кровяных клеток в организме человека. Ретикулоциты развиваются и созревают в костном мозге. После выхода в кровоток ретикулоциты быстро подвергаются терминальной дифференцировке в зрелые эритроциты. Подобно зрелым эритроцитам ретикулоциты не имеют клеточного ядра. В отличие от зрелых эритроцитов ретикулоциты сохраняют способность к осуществлению синтеза белка. В некоторых вариантах осуществления рекомбинантную нуклеиновую кислоту (такую как мРНК), кодирующую экзогенный антиген, вводят в ретикулоциты для получения экспрессирующих экзогенный антиген EHC-клеток.
[00371] Из периферической крови можно выделять ретикулоциты различного возраста на основании отличий в плотности клеток по мере созревания ретикулоцитов. Ретикулоциты можно выделять из периферической крови с применением дифференциального центрифугирования благодаря различным градиентам плотности. Например, для выделения ретикулоцитов можно применять градиенты Percoll (см., например, Noble el al., Blood 74:475-481 (1989)). Стерильные изотонические растворы Percoll с плотностью 1,096 и 1,058 г/мл получают разведением Percoll (Sigma-Aldrich, Сент-Луис, Миссури, США) до конечной концентрации с помощью 10 мМ триэтаноламина, 117 мМ NaCl, 5 мМ глюкозы и 1,5 мг/мл бычьего сывороточного альбумина (BSA). Эти растворы характеризуются осмолярностью от 295 до 310 мосмоль/л. Пять миллилитров, например, первого раствора Percoll (плотность 1,096) вносят в стерильную коническую центрифужную пробирку на 15 мл. Два миллилитра, например, второго раствора Percoll (плотность 1,058) наслаивают поверх первого раствора Percoll с более высокой плотностью. Два-четыре миллилитра цельной крови наслаивают на поверхность содержимого пробирки. Пробирку центрифугируют при 250×g в течение 30 минут в охлаждаемой центрифуге с колебательным держателем пробирок. Ретикулоциты и некоторые белые клетки мигрируют к границе раздела между двумя слоями Percoll. Клетки, расположенные на границе раздела, переносят в новую пробирку и дважды промывают фосфатно-солевым буфером (PBS) с 5 мМ глюкозы, 0,03 мМ азида натрия и 1 мг/мл BSA. Остаточные белые кровяные клетки удаляют с помощью хроматографии в PBS через эксклюзионную колонку.
[00372] В качестве альтернативы, ретикулоциты можно выделять посредством позитивного отбора с применением подхода иммуномагнитного разделения (см., например, Brun et al., Blood 76:2397-2403 (1990)). Этот подход использует то, что большое количество трансферриновых рецепторов, экспрессировано на поверхности ретикулоцитов по сравнению с эритроцитами перед созреванием. Магнитные гранулы, покрытые антителом к трансферриновому рецептору, можно применять для селективного выделения ретикулоцитов из смешанной популяции клеток крови. Антитела к трансферриновому рецептору различных видов млекопитающих, в том числе человека, доступны из коммерческих источников (например, Affinity BioReagents, Голден, Колорадо, США; Sigma-Aldrich, Сент-Луис, Миссури, США). Антитело к трансферриновому рецептору может быть напрямую связано с магнитными гранулами. В качестве альтернативы, антитело к трансферриновому рецептору может быть опосредованно связано с магнитными гранулами через вторичное антитело. Например, мышиное моноклональное антитело 10D2 (Affinity BioReagents, Голден, Колорадо, США) против человеческого трансферрина можно смешать с иммуномагнитными гранулами, покрытыми антителом козы к иммуноглобулину G мыши (Dynal/Invitrogen, Карлсбад, Калифорния, США). Затем иммуномагнитные гранулы инкубируют с истощенной по лейкоцитам фракцией красных кровяных клеток. Гранулы и красные кровяные клетки инкубируют при 22°C при слабом перемешивании в течение 60-90 минут с последующим выделением гранул с прикрепленными ретикулоцитами с применением магнитного поля. Выделенные ретикулоциты можно освободить от магнитных гранул с применением, например, раствора DETACHaBEAD® (от Invitrogen, Карлсбад, Калифорния, США). В качестве альтернативы, ретикулоциты можно выделять из выращиваемых и созревающих in vitro CD34+ гемопоэтических стволовых клеток с применением способов, описанных в данном документе.
[00373] Терминально дифференцированные безъядерные эритроциты можно отделить от других клеток на основе их содержания ДНК. В неограничивающем примере клетки вначале метят с помощью витального красителя ДНК, такого как Hoechst 33342 (Invitrogen Corp.). Hoechst 33342 представляет собой проникающий в клетки ядерный контрастный краситель, который испускает голубую флуоресценцию при связывании с двухнитевой ДНК. Недифференцированные клетки-предшественники, макрофаги или другие ядерные клетки в культуре окрашиваются с помощью Hoechst 33342, при этом безъядерные эритроциты являются Hoechst-отрицательными. Hoechst-положительные клетки можно отделить от безъядерных эритроцитов с применением устройств для сортировки флуоресцентно-активированных клеток или других методик сортировки клеток. Краситель Hoechst можно удалять из выделенных эритроцитов с помощью диализа или других соответствующих способов.
[00374] Популяцию экспрессирующих экзогенный антиген EHC-клеток можно очищать путем снижения содержания ядерного материала в популяции EHC-клеток. Например, повышают скорость удаление ядра в популяции EHC-клеток и/или повышают количество безъядерных экспрессирующих экзогенный антиген EHC-клеток или проводят их обогащение.
[00375] Популяции экспрессирующих экзогенный антиген EHC-клеток можно инкубировать с малой молекулой, например ингибитором актина, например цитохалазином A, B, C, D, E, F, H, J, и затем центрифугировать для удаления ядерного материала. В качестве альтернативы или дополнительно над популяцией экспрессирующих экзогенный антиген EHC-клеток можно проводить механические манипуляции путем пропускания через постепенно уменьшающиеся фильтры с ограничением по размеру для удаления ядерного материала. Популяцию экспрессирующих экзогенный антиген EHC-клеток также можно инкубировать на покрытой фибронектином пластиковой поверхности для повышения скорости удаления ядерного материала. В одном варианте осуществления популяцию экспрессирующих экзогенный антиген EHC-клеток инкубируют в совместной культуре со стромальными клетками, например, макрофагами, для повышения скорости удаления ядерного материала.
[00376] В некоторых вариантах осуществления популяция экспрессирующих экзогенный антиген EHC-клеток более чем на 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 99%, 99,5%, 99,9% или более чем на 99,9% состоит из безъядерных клеток.
[00377] В некоторых вариантах осуществления экспрессирующие экзогенный антиген EHC-клетки не подвергают совместному культивированию с поддерживающими клетками, например, со слоем прикрепленных стромальных клеток. В некоторых вариантах осуществления популяцию экспрессирующих экзогенный антиген EHC-клеток получают путем приведения EHC-клеток в контакт с экзогенным антигеном и дифференцировки EHC-клеток с получением популяции безъядерных клеток, содержащих экзогенный антиген. Популяцию экспрессирующих экзогенный антиген EHC-клеток получают без стадии обогащения, такой как гравитационное разделение, магнитная или флуоресцентная сортировка, облучение, применение яда для ядерных клеток и т.п., для отбора безъядерных клеток.
[00378] В некоторых вариантах осуществления популяция экспрессирующих экзогенный антиген EHC-клеток составлена более чем на 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 99%, 99,5%, 99,9% или более 99,9% экспрессирующими экзогенный антиген EHC-клетками, у которых отсутствует ядерный материал, как определено с помощью анализа для обнаружения ядерного материала, такого как анализы, описанные в данном документе.
[00379] В некоторых вариантах осуществления оценивают присутствие, биологическую активность и/или функционирование экзогенного антигена, такого как полипептид экзогенного антигена, выставляемый экспрессирующими экзогенный антиген EHC-клетками. Множество соответствующих анализов доступны и известны из уровня техники.
[00380] В одном варианте осуществления экзогенный антиген представляет собой полипептид на поверхности экспрессирующей экзогенный антиген EHC. Присутствие экзогенного антигена можно оценить с помощью анализов, в том числе без ограничений проточной цитометрии, вестерн-блоттинга, RT-PCR, нозерн-блоттинга, розеткообразования по Кумбсу, масс-спектрометрии. В одном варианте осуществления экзогенный антиген представляет собой полипептид внутри экспрессирующей экзогенный антиген EHC. Присутствие экзогенного антигена можно оценить с помощью анализов, в том числе без ограничений вестерн-блоттинга, RT-PCR, нозерн-блоттинга, ПЦР, Саузерн-блоттинга, масс-спектрометрии.
[00381] В одном варианте осуществления экзогенный антиген представляет собой нуклеиновую кислоту на поверхности экспрессирующей экзогенный антиген EHC. Присутствие экзогенного антигена можно оценить с помощью анализов, в том числе без ограничений проточной цитометрии, проточной цитометрии с гомологичным флуоресцентным зондом, Саузерн-блоттинга, нозерн-блоттинга, ПЦР. В одном варианте осуществления экзогенный антиген представляет собой нуклеиновую кислоту внутри экспрессирующей экзогенный антиген EHC. Присутствие экзогенного антигена можно оценить с помощью анализов, в том числе без ограничений Саузерн-блоттинга, нозерн-блоттинга, ПЦР.
[00382] В одном варианте осуществления экзогенный антиген представляет собой малую молекулу на поверхности экспрессирующей экзогенный антиген EHC. Присутствие экзогенного антигена можно оценить с помощью анализов, в том числе без ограничений проточной цитометрии, масс-спектрометрии. В одном варианте осуществления экзогенный антиген представляет собой малую молекулу внутри экспрессирующей экзогенный антиген EHC. Присутствие экзогенного антигена можно оценить с помощью анализов, в том числе без ограничений масс-спектрометрии, флуоресцентной спектроскопии.
[00383] В одном варианте осуществления экзогенный антиген представляет собой липид в мембране экспрессирующей экзогенный антиген EHC. Присутствие экзогенного антигена можно оценить с помощью анализов, в том числе без ограничений масс-спектрометрии, проточной цитометрии, определения растворимости мембраны, флуоресцентной поляризации, интерференционной пространственной световой микроскопии.
[00384] В одном варианте осуществления экзогенный антиген является флуоресцентным, или слит с флуоресцентной молекулой, или совместно экспрессируется с рекомбинантной нуклеиновой кислоты (например, в векторе) с флуоресцентным репортерным белком, таким как GFP. Присутствие экзогенного антигена в экспрессирующей экзогенный антиген EHC или на ней можно оценить с помощью анализов, в том числе без ограничений проточной цитометрии, флуоресцентной спектроскопии, спектроскопии поглощения.
[00385] В одном варианте осуществления экзогенный антиген представляет собой газообразную молекулу. Присутствие экзогенного антигена в экспрессирующей экзогенный антиген EHC или на ней можно оценить с помощью анализов, в том числе без ограничений анализов хемолюминесценции, масс-спектроскопии.
[00386] Присутствие экзогенного антигена в экспрессирующей экзогенный антиген EHC или на ней можно оценить с помощью проточной цитометрии в режиме количественного определения с применением калибровочных гранул, таких как коммерчески доступные калибровочные гранулы для цитометрии, чтобы количественно оценить число экзогенных антигенов на отдельной EHC. В качестве альтернативы или дополнительно присутствие экзогенного антигена в экспрессирующей экзогенный антиген EHC или на ней можно оценить с помощью вестерн-блот-анализа в режиме количественного определения с применением стандарта с известной концентрацией, который можно обнаруживать с применением тех же реагентов для обнаружения, что и для экзогенного антигена, и таким образом можно количественно оценить число экзогенных антигенов на отдельной EHC.
[00387] В некоторых вариантах осуществления присутствие двух или больше различных экзогенных антигенов можно оценить с помощью одного и того же или различных способов, или одновременно, или последовательно, или в режиме параллельного выполнения. Например, в одном варианте осуществления экзогенный антиген, находящийся на поверхности, можно оценить с помощью проточной цитометрии с применением антитела, специфичного в отношении экзогенного антигена, а другой экзогенный антиген, на находящийся на поверхности, который является флуоресцентным, можно определить по флуоресцентному сигналу с применением другого канала при проточной цитометрии. В другом примере экзогенный антиген, находящийся на поверхности, можно оценить с помощью проточной цитометрии, а другой экзогенный антиген, не находящийся на поверхности, можно оценить с помощью вестерн-блот-анализа.
[00388] В конкретном варианте осуществления экзогенный антиген сохраняется на экспрессирующей экзогенный антиген EHC после терминальной дифференцировки источника клеток. Например, экспрессирующую экзогенный антиген EHC получают из культивируемой EHC, а экспрессию или присутствие экзогенного антигена оценивают после терминальной дифференцировки клетки с помощью соответствующего способа, например, с помощью проточной цитометрии, вестерн-блот-анализа, иммунопреципитации, флуоресцентной спектроскопии, хемолюминесценции, Саузерн-блот-анализа, нозерн-блот-анализа или спектроскопии поглощения.
[00389] В конкретном варианте осуществления экзогенный антиген сохраняется на экспрессирующей экзогенный антиген EHC после циркуляции в кровотоке in vivo после введения экспрессирующей экзогенный антиген EHC субъекту. Экспрессирующую экзогенный антиген EHC можно инъецировать лабораторному животному или животной модели, такой как мышь, внутривенно, например, через хвостовую вену, или инъецировать человеку внутривенно. Затем кровь собирают и присутствие экзогенного антигена на экспрессирующей экзогенный антиген EHC оценивают с помощью соответствующего анализа, например, с помощью проточной цитометрии, вестерн-блот-анализа, иммунопреципитации, флуоресцентной спектроскопии, хемолюминесценции, Саузерн-блот-анализа, нозерн-блот-анализа или спектроскопии поглощения.
[00390] В некоторых вариантах осуществления биологическую активность экзогенного антигена в экспрессирующей экзогенный антиген EHC или на ней, общую биологическую активность EHC и общую активность популяции EHC-клеток можно оценить с помощью анализов in vitro.
[00391] В некоторых вариантах осуществления активность экспрессирующей экзогенный антиген EHC быстро улучшают с применением модельной линии клеток. Например, библиотека подходящих экзогенных антигенов экспрессируется на модельной линии клеток, например, HEK293T или K562, и активность оценивают посредством соответствующего анализа; затем самый лучший кандидат экзогенного антигена, например, экспрессируемый на самом высоком уровне или демонстрирующий самую высокую активность в соответствующем анализе, экспрессируют, например, в культивируемых EHC-клетках с получением экспрессирующих экзогенный антиген EHC-клеток.
[00392] В одном варианте осуществления активность экспрессирующей экзогенный антиген EHC быстро улучшают с применением культивирования и мышиной модели. Например, библиотеку подходящих экзогенных антигенов экспрессируют в культивируемых мышиных EHC-клетках; активность оценивают на соответствующей мышиной модели заболевания или соответствующей мышиной модельной системе для оценки активности; самый лучший кандидат экзогенного антигена, например, экспрессируемый на самом высоком уровне или демонстрирующий самую высокую активность в соответствующем анализе, затем экспрессируют, например, в культивируемых EHC-клетках с получением экспрессирующих экзогенный антиген EHC-клеток.
[00393] В некоторых случаях экзогенный антиген представляет собой фермент, и активность экзогенного антигена можно оценить с помощью ферментного анализа, при котором обнаруживают исчезновение молекулы специфичного субстрата или обнаруживают появление молекулы специфичного продукта. Такие анализы включают без ограничений колориметрические анализы, масс-спектрометрию, HPLC, флуоресцентные анализы.
[00394] Например, a) экзогенный антиген представляет собой аденозиндезаминазу (ADA), и ферментный анализ обнаруживает превращение аденозина в инозин; b) экзогенный антиген представляет собой фенилаланингидроксилазу (PAH), и анализ обнаруживает превращение фенилаланина в тирозин; c) экзогенный антиген представляет собой фенилаланин-аммоний-лиазу (PAL), и анализ обнаруживает превращение фенилаланина в транс-коричную кислоту; d) экзогенный антиген представляет собой тимидинфосфорилазу (TP), и анализ обнаруживает превращение тимидина в тимин и 2-дезоксирибозу; e) экзогенный антиген представляет собой пурин-нуклеозидфосфорилазу (PNP), и анализ обнаруживает превращение инозина в гипоксантин, аденозина в аденин и гуанозина в гуанин; f) экзогенный антиген представляет собой гомогентизат-1,2-диоксигеназу (HDG), и анализ обнаруживает превращение гомогентизата в малеилацетоацетат; g) экзогенный антиген представляет собой цистатионин-бета-синтазу, и анализ обнаруживает превращение серина и гомоцистеина в цистатионин; h) экзогенный антиген представляет собой оксалатоксидазу, и анализ обнаруживает окисление оксалата.
[00395] В некоторых вариантах осуществления активность экспрессирующей экзогенный антиген EHC оценивают на животной модели заболевания, например мышиной модели, и иммунодефицитной мыши, или NSG-мыши, при этом заболевание представляет собой, например, метаболическое заболевание или дефицит фермента, или модели, которая может демонстрировать эффект экспрессирующей экзогенный антиген EHC, например, мышь, в которую инъецируют субстрат и измеряют продукт превращения, опосредованного экзогенным антигеном.
[00396] В одном варианте осуществления экзогенный антиген представляет собой полипептид рецептора комплемента 1 (CR1), его производное или функциональный фрагмент. Активность экзогенного антигена CR1 можно оценить несколькими способами, включающими, например, специфический захват иммунных комплексов экзогенным антигеном CR1, эффективный перенос иммунных комплексов к макрофагам или клиренс иммунных комплексов из мыши in vivo.
[00397] Функциональность EHC-клеток, сверхэкспрессирующих экзогенный антиген CR1, можно оценить с помощью одного или нескольких процессов: захвата иммунных комплексов на поверхности EHC, содержащей экзогенный антиген CR1, высвобождение иммунных комплексов для макрофагов, при сохранении EHC, содержащей экзогенный антиген CR1, и надлежащая циркуляция в кровотоке EHC-клеток, содержащих экзогенный антиген CR1. Эти три параметра можно оценивать in vitro. Анализы захвата иммунных комплексов описаны в уровне техники, например, Oudin et al., J Immunol 2000, и Schifferli et al., J Immunol 1991. Например, меченые иммунные комплексы инкубируют с EHC-клетками, экспрессирующими нативный CR1, или полипептид экзогенного антигена CR1, или его фрагмент, и количество иммунных комплексов, захватываемых EHC-клетками, оценивают с помощью проточной цитометрии. Анализы переноса в макрофаги описаны в уровне техники, например, Kuhn et al., J Immunol 1998. Например, меченые иммунные комплексы, загруженные на эритроциты, экспрессирующие нативный CR1, или полипептид экзогенного антигена CR1, или его фрагмент, инкубируют с макрофагами. Перенос иммунных комплексов с поверхности эритроцита к макрофагу и поглощение или сохранение эритроцитов макрофагами можно измерить с помощью проточной цитометрии. Надлежащую циркуляцию в кровотоке можно предсказать путем анализа морфологии и деформируемости EHC. Морфологию EHC-клеток, экспрессирующих нативный CR1, или полипептид экзогенного антигена CR1, или его фрагмент, можно оценить визуально с применением стандартных методик микроскопии, описываемых, например, в Giarratana et al., Blood 2011, и Repik et al., Clin Exp Immunol 2005. Деформируемость EHC-клеток, экспрессирующих нативный CR1, или полипептид экзогенного антигена CR1, или его фрагмент, можно оценить с помощью эктацитометрии, также известной как лазерный оптический ротационный анализ клеток (LORCA), описываемой, например, в Giarratana et al., Blood 2011.
[00398] Например, экспрессирующие экзогенный антиген CR1 EHC (EHC содержит полипептид экзогенного антигена CR1) инкубируют с иммунными комплексами, такими как полученные in vitro иммунные комплексы или происходящие из пациента иммунные комплексы. Захват иммунных комплексов экзогенным антигеном CR1 оценивают, например, с помощью проточной цитометрии с применением флуоресцентного маркера в иммунном комплексе или с помощью проточной цитометрии с применением вторичного средства для обнаружения против элемента иммунного комплекса.
[00399] В одном варианте осуществления экспрессирующую экзогенный антиген CR1 EHC сначала инкубируют с иммунными комплексами, а затем инкубируют с макрофагами, такими как первичные макрофаги, первичные моноциты, культивируемые макрофаги, культивируемые моноциты, клетки U937, PMA-активированные клетки U937, клетки AA9, клетки RAW 264.7, клетки J774, клетки THP1, клетки KG-1, клетки NR8383, клетки MV-4-11, клетки 3D4/31, клетки MD, клетки Fcwf-4, клетки DH82. Макрофаги анализируют с помощью, например, проточной цитометрии или радиографии в отношении присутствия иммунных комплексов, перенесенных с помощью экспрессирующей экзогенный антиген CR1 EHC. Перенос захваченных иммунных комплексов от культивируемых EHC-клеток к макрофагам представляет собой стандартных анализ из уровня техники, см., например: Repik et al. 2005 Clin Exp Immunol. 140:230; Li et al. 2010 Infection Immunity 78(7):3129.
[00400] В одном варианте осуществления активность экспрессирующей экзогенный антиген CR1 EHC оценивают на животной модели. Например, можно применять соответствующую мышиную модель, такую как иммунодефицитная мышь или NSG-мышь. Мышиная модель заболевания может включать, например, иммунокомплексное заболевание, такое как волчанка. Мышиные модели включают NZBWF1/J, MRL/MpJ, MRL/MpJ-Fas(lpr), Smn.C3-Fasl/J, NZM2410/Aeg, 129S4-Cd48, Cg-Sle1, NZM-Sle1 Sle2 Sle3/LmoJ и BXSB.129P2. В качестве альтернативы или дополнительно клиническое проявление заболевания можно обеспечивать путем введения в мышь, например, путем инъекции иммунных комплексов. Экспрессирующие экзогенный антиген CR1 EHC-клетки можно инъецировать в любую соответствующую мышь (или другую животную модель) для тестирования одного или нескольких биологических эффектов EHC, например, клиренса инъецированных иммунных комплексов за счет экспрессирующей экзогенный антиген CR1 EHC.
[00401] В одном варианте осуществления экзогенный антиген представляет собой регуляторную молекулу комплемента или характеризуется активностью регуляции комплемента. Такую активность экзогенного антигена можно оценить с помощью анализов как in vitro, так и in vivo. К примеру, активность экзогенного антигена можно оценить с помощью измерения снижения в анализе in vitro активации комплемента, например, в анализе CH50, в котором измеряется комплемент-опосредованный лизис сенсибилизированных эритроцитов овцы, или в анализе AH50, в котором измеряется альтернативный путь комплемент-опосредованного лизиса несенсибилизированных эритроцитов кролика. В качестве альтернативы, активность экзогенного антигена можно оценить с помощью обнаружения расщепления или отсутствия расщепления, которое может приводить или может не приводить к появлению нового эпитопа, рекомбинантного компонента комплемента, который инкубировали с экзогенным антигеном, в том числе без ограничений, например, расщепления рекомбинантного C2 на C2a и C2b, расщепления фактора B на фактор Ba и фактор Bb, расщепления фактора C3b на iC3bH и iC3bL, расщепления C3bBb на C3b и Bb, расщепления C4bBb на C4b и Bb или расщепления фактора C4b на iC4bH и iC4bL. Расщепление или отсутствие расщепления соответствующего рекомбинантного компонента комплемента можно оценить с помощью способов белкового анализа, известных из уровня техники, в том числе без ограничений, например, хроматографии, гель-электрофореза, ELISA и вестерн-блоттинга. Соответствующие анализы для определения активности экзогенного антигена in vivo включают инъекцию экспрессирующей экзогенный антиген EHC в животное, например мышь, и проверку отложения факторов комплемента, например мембраноатакующего комплекса, с помощью гистологического окрашивания.
[00402] В одном варианте осуществления экзогенный антиген способен связывать или захватывать мишень, и активность экзогенного антигена можно оценить с помощью обнаружения захваченной мишени на экзогенном антигене in vitro или in vivo.
[00403] В одном варианте осуществления экспрессирующую экзогенный антиген EHC инкубируют с мишенью in vitro, и захват мишени экзогенным антигеном обнаруживают с применением анализа in vitro, в том числе без ограничений, например, проточной цитометрии, иммуногистохимии, магнитного разделения, радиографии, анализов колониеобразования, микроскопии.
[00404] В одном варианте осуществления экспрессирующую экзогенный антиген EHC инкубируют с мишенью in vitro, и захват мишени экзогенным антигеном обнаруживают с применением анализа совместного культивирования in vitro, в том числе без ограничений, например, анализа поглощения макрофагами опсонизированной экспрессирующей экзогенный антиген EHC, анализа активации T-клеток, анализа стимуляции B-клеток, анализа дегрануляции тучных клеток, анализа инфекционного потенциала.
[00405] В варианте осуществления экспрессирующую экзогенный антиген EHC инкубируют с мишенью in vitro, и высвобождение или скорость диссоциации захваченной мишени измеряют с применением анализа in vitro, в том числе без ограничений, например, проточной цитометрии, иммуногистохимии, магнитного разделения, радиографии, анализов колониеобразования, микроскопии.
[00406] Захват мишени экспрессирующей экзогенный антиген EHC можно оценить в анализе in vivo, например, на животном, в том числе мышиной модели заболевания, при котором мишень естественным образом присутствует в мыши. Соответствующие заболевания включают бактериальные инфекции, вирусные инфекции, грибковые инфекции, иммунокомплексные заболевания, заболевания, обусловленные аутореактивными антителами, гиперлипидемию, гипергликемию. В других мышиных моделях мишень вводят мыши извне, например, с помощью инъекции или путем введения с пищей. В этих анализах захват мишени экспрессирующей экзогенный антиген EHC оценивают либо с помощью исследования животного, например, плазмы, ткани, в отношении снижения или удержания мишени, либо с помощью выделения или сбора экспрессирующей экзогенный антиген EHC из животного, например, из крови, из плазмы, из ткани, и анализируют присутствие мишени на экзогенном антигене с применением анализа in vitro, в том числе без ограничений, например, проточной цитометрии, иммуногистохимии, магнитного разделения, радиографии, анализов колониеобразования, микроскопии.
[00407] В некоторых вариантах осуществления экзогенный антиген способен связывать или захватывать мишень и значительно повышать клиренс мишени in vivo или значительно снижать концентрацию мишени в кровотоке. Активность экзогенного антигена на экспрессирующей экзогенный антиген EHC можно оценить с помощью обнаружения усиленного клиренса мишени in vitro или in vivo.
[00408] В одном варианте осуществления экспрессирующую экзогенный антиген EHC инкубируют с мишенью in vitro, и захват мишени экзогенным антигеном обнаруживают с применением анализа in vitro, в том числе без ограничений, например, проточной цитометрии, иммуногистохимии, магнитного разделения, радиографии, анализов колониеобразования, микроскопии. Затем экспрессирующую экзогенный антиген EHC инкубируют в анализе совместного культивирования с клеткой, которая, как известно, усиливает клиренс, например, макрофагом или моноцитом, и клиренс мишени и экспрессирующей экзогенный антиген EHC оценивают с помощью, например, проточной цитометрии, иммуногистохимии, магнитного разделения, радиографии, анализов колониеобразования, микроскопии.
[00409] В одном варианте осуществления экспрессирующую экзогенный антиген EHC инкубируют с мишенью in vitro, и захват мишени экзогенным антигеном обнаруживают с применением анализа in vitro, в том числе без ограничений, например, проточной цитометрии, иммуногистохимии, магнитного разделения, радиографии, анализов колониеобразования, микроскопии. Затем экспрессирующую экзогенный антиген EHC инкубируют в физической системе, которая имитирует механизм клиренса EHC in vivo, например, в искусственной селезенке, микроканале, наполненной колонке, смоле, тканевом эксплантате, центрифуге, и клиренс мишени и экспрессирующей экзогенный антиген EHC оценивают с помощью, например, проточной цитометрии, иммуногистохимии, магнитного разделения, радиографии, анализов колониеобразования, микроскопии.
[00410] В одном варианте осуществления клиренс мишени за счет экспрессирующей экзогенный антиген EHC оценивают в анализе in vivo, например на животном, в том числе, например, мышиной модели заболевания, в которой мишень естественным образом присутствует в мыши, например, бактериальной инфекции, вирусной инфекции, грибковой инфекции, иммунокомплексного заболевания, заболевания, обусловленного аутореактивными антителами, гиперлипидемии, гипергликемии или, например, мышиной модели, где мишень вводят в мышь извне, например, с помощью инъекции или путем введения с пищей. В этих анализах клиренс мишени за счет экспрессирующей экзогенный антиген EHC оценивают либо с помощью исследования животного, например, плазмы, ткани, в отношении снижения количества мишени, либо с помощью выделения или сбора экспрессирующей экзогенный антиген EHC из животного, например, из крови, из плазмы, из ткани, и анализируют присутствие мишени на экзогенном антигене с применением анализа in vitro, в том числе без ограничений, например, проточной цитометрии, иммуногистохимии, магнитного разделения, радиографии, анализов колониеобразования, микроскопии.
[00411] В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC способна доставить соответствующий экзогенный антиген в конкретный внутриклеточный компартмент, например, в лизосому.
[00412] Например, экзогенный антиген может быть доставлен в лизосомальный компартмент клетки-мишени, например, макрофага. Успешную доставку экзогенного антигена в лизосомальный компартмент клетки-мишени оценивают с помощью микроскопии и обнаружения точечных пятен, соответствующих флуоресцентному экзогенному антигену или флуоресцентному средству для обнаружения экзогенного антигена. В качестве альтернативы или дополнительно успешную доставку экзогенного антигена в лизосомальный компартмент оценивают с помощью микроскопии и совместной локализации флуоресцентного средства для обнаружения экзогенного антигена с флуоресцентным средством для обнаружения известного лизомосного маркера, например, лизотрекером, LAMP-1.
[00413] В некоторых вариантах осуществления экзогенный антиген представляет собой фермент, который может разрушать токсические компоненты, накопившиеся в лизосоме клетки, проявляющей генотип или фенотип лизосомной болезни накопления или полученной от пациента с таким заболеванием. Например, экзогенный антиген способен разрушать токсический материал, накопившийся в клетке, и восстанавливать фенотип клетки, например, предотвращать гибель клетки.
[00414] Популяцию экспрессирующих экзогенный антиген EHC-клеток можно оценить в отношении неспособности EHC-клеток к репликации, способности EHC-клеток к безопасной циркуляции в сосудистой системе и отсутствия иммуногенности у EHC-клеток.
[00415] В некоторых вариантах осуществления безопасность популяции экспрессирующих экзогенный антиген EHC-клеток оценивают путем измерения потенциала воспроизведения популяции EHC-клеток с применением соответствующего анализа in vitro или in vivo. Например, тесты на фактическую неспособность экспрессирующих экзогенный антиген EHC-клеток к саморепликации включают: a) фактическую неспособность образовывать опухоль при инъекции мыши, характеризующейся ослабленным иммунитетом; b) фактическую неспособность образовывать колонию при культивировании на мягком агаре; c) фактическую неспособность включать тимидин в анализе включения тимидина; d) фактическое отсутствие положительного сигнала после трансфекции с помощью ДНК, кодирующей флуоресцентный репортер, например, менее 10%, 1%, 0,1%, 0,01%, 0,001%, 1 ppm, 100 ppb, 10 ppb, 1 ppb, 100 ppt, 10 ppt, 1 ppt или менее 1 ppt положительного сигнала; e) фактическое отсутствие положительного сигнала после окрашивания ядерным красителем, например, менее 10%, 1%, 0,1%, 0,01% и 0,001%, 1 ppm, 100 ppb, 10 ppb, 1 ppb, 100 ppt, 10 ppt, 1 ppt или менее 1 ppt положительного сигнала; f) фактическое отсутствие положительного сигнала после окрашивания на клеточные маркеры гемобластоза, например, CKIT, CD34, EpCam, например, менее 10%, 1%, 0,1%, 0,01%, 0,001%, 1 ppm, 100 ppb, 10 ppb, 1 ppb, 100 ppt, 10 ppt, 1 ppt, или менее 1 ppt положительного сигнала. В определенных вариантах осуществления экспрессирующие экзогенный антиген EHC-клетки получают таким образом, что они не содержат значительного количества реплицирующейся нуклеиновой кислоты.
[00416] В некоторых вариантах осуществления безопасность популяции экспрессирующих экзогенный антиген EHC-клеток оценивают путем измерения способности введенных EHC-клеток к циркуляции в кровотоке in vivo (в кровеносной системе субъекта) без значительной окклюзии сосуда или индукции системы свертывания крови. Необязательно, можно оценить фармакокинетику экспрессирующих экзогенный антиген EHC-клеток в кровотоке.
[00417] В одном варианте осуществления фармакокинетику экспрессирующих экзогенный антиген EHC-клеток в кровотоке оценивают путем внутривенной инъекции EHC в животное, такое как мышь. Мышь может представлять собой иммунодефицитную мышь NSG (nod-SCID-gamma). Мышь подвергают деплеции макрофагов перед инъекцией EHC, например, с помощью внутрибрюшинной инъекции человеческих красных кровяных клеток или с помощью внутривенной инъекции липисом с клодронатом. Экспрессирующие экзогенный антиген EHC-клетки можно метить флуоресцентным красителем, например, CFSE. После инъекции EHC-клеток проводят забор крови, и количество оставшихся экспрессирующих экзогенный антиген EHC-клеток оценивают с помощью, например, проточной цитометрии, вестерн-блот-анализа, и скорость клиренса экспрессирующих экзогенный антиген EHC-клеток выводят на основе эти данных. Человеческие красные кровяные клетки можно инъецировать в ту же животную модель, что и экспрессирующие экзогенный антиген EHC-клетки, и сравнить скорости клиренса EHC-клеток и человеческих красных кровяных клеток.
[00418] В одном варианте осуществления риск активации системы свертывания крови с помощью экспрессирующей экзогенный антиген EHC оценивают с помощью анализа in vitro. В одном варианте осуществления экспрессирующую экзогенный антиген EHC инкубируют с человеческой кровью и активацию системы свертывания крови оценивают путем измерения времени, требуемого для свертывания в присутствии каолина, отрицательно заряженных фосфолипидов и кальция (тест на активированное частичное тромбопластиновое время (aPTT)), см., например, Exner and Rickard, Biomedicine 1977 27(2):62, или путем измерения времени, требуемого для свертывания в присутствии тромбопластина и кальция (тест на протромбиновое время (PT)), см., например, Jaques and Dunlop 1945, Am J Physiol 145:67. Нормальный диапазон для aPTT-теста составляет приблизительно 25-38 секунд. Нормальный диапазон для PT-теста составляет приблизительно 10-12 секунд.
[00419] В одном варианте осуществления любые побочные эффекты, индуцируемые экспрессирующими экзогенный антиген EHC-клетками, оценивают путем инъекции EHC-клеток в животное внутривенно и оценки активации системы свертывания крови. Уровень индукции системы свертывания крови оценивают путем забора крови и оценки уровней компонентов системы свертывания крови в плазме с помощью, например, вестерн-блот-анализа или ELISA. Компоненты системы свертывания крови в основном представляют собой продукты распада фибриногена, например, фибринопептид A и фибринопептид B. Например, уровень индукции системы свертывания крови оценивают с помощью забора крови и оценки уровней активности свертывания в плазме с помощью анализа активации тромбоцитов, например, путем инкубации плазмы с тромбоцитами и оценки активации тромбоцитов с помощью проточной цитометрии, например, путем окрашивания маркеров активации, например, путем окрашивания PAC-1, CD62p или CD40L.
[00420] В одном варианте осуществления любые побочные эффекты, индуцируемые экспрессирующими экзогенный антиген EHC-клетками, оценивают путем внутривенной инъекции EHC-клеток животному и оценки активации путей развития воспаления. Уровень воспаления можно оценить с помощью забора крови и оценки уровней воспалительных цитокинов в плазме с помощью, например, вестерн-блот-анализа или ELISA. В одном варианте осуществления воспалительные цитокины представляют собой интерферон гамма, фактор некроза опухоли альфа или фрагмент IL-12 p70.
[00421] В одном варианте осуществления любые побочные эффекты, индуцируемые экспрессирующими экзогенный антиген EHC-клетками, оценивают путем внутривенной инъекции EHC-клеток животному и оценки состояния тканей, например, печени, селезенки, сердца, легких, мозга, кожи, почек. Состояние тканей можно оценить с помощью общей аутопсии, иссечения ткани, гистологического окрашивания и визуализации с помощью микроскопии.
[00422] В одном варианте осуществления способность экспрессирующей экзогенный антиген EHC к циркуляции в кровотоке in vivo без значительной окклюзии сосуда или активации системы свертывания крови оценивают путем измерения деформируемости EHC-клеток. Деформируемость экспрессирующей экзогенный антиген EHC оценивают с применением анализа in vitro. Например, анализ представляет собой анализ осмотической хрупкости. Механическую хрупкость экспрессирующей экзогенный антиген EHC можно оценить с помощью измерения структурной целостности в ответ на напряжение сдвига в системе сдвигового деформирования типа Куэтта. В одном варианте осуществления деформируемость экспрессирующей экзогенный антиген EHC оценивают с применением эктацитометра. В одном варианте осуществления деформируемость экспрессирующей экзогенный антиген EHC оценивают путем измерения индекса удлинения при заданном давлении с помощью лазерной дифракции с применением устройства лазерно-симулированный оптический ротационный анализатор клеток (LORCA). В одном варианте осуществления деформируемость экспрессирующей экзогенный антиген EHC оценивают путем измерения времени прохождения через серию микронных сужений с заданными размерами при фиксированном давлении в микрофлюидном устройстве. В одном варианте осуществления деформируемость экспрессирующей экзогенный антиген EHC оценивают путем измерения уровня выживаемости при прохождении через серию микронных сужений с заданными размерами при фиксированном давлении в микрофлюидном устройстве. Микрофлюидное устройство можно выбрать из одного из следующих, в том числе без ограничений, полидиметилсилоксанового (PDMS) чипа с микронными сужениями (например, Hoelzle et al. J Vis Exp 2014 91:e51474); чипа с воронковидными сужениями (например, Guo et al. Lab Chip 2012 12:1143); PDMS-чипа со столбиками (например, Zhang et al. PNAS 2012 109(46):18707) или in vitro искусственной селезенки в виде колонки, загруженной микрогранулами (Guillaume DePlaine et al., Blood 2011, 117(8)).
[00423] В одном варианте осуществления способность экспрессирующей экзогенный антиген EHC к циркуляции в кровотоке in vivo без значительной окклюзии сосуда или активации системы свертывания крови оценивают путем измерения окклюзии сосудов EHC-клетками. Окклюзию сосуда экспрессирующей экзогенный антиген EHC можно оценить с применением анализа in vitro. Например, окклюзия сосуда экспрессирующей экзогенный антиген EHC оценивают с применением анализа ex vivo. Экспрессирующую экзогенный антиген EHC инкубируют при соотношении 1:1 с эталонными человеческими красными кровяными клетками и индукцию многоклеточных розеткообразных структур оценивают с помощью светового микроскопа в сравнении с эталонным анализом с несовпадающей по Rh кровью. Окклюзию сосуда экспрессирующей экзогенный антиген EHC оценивают путем измерения адгезии EHC-клеток к человеческим клетками эндотелия сосудов в условиях потока, см., например, Kaul DK, Finnegan E, and Barabino GA (2009) Microcirculation 16(1):97-111. В качестве альтернативы или дополнительно окклюзию сосуда оценивают путем измерения повышения единиц общего периферического сопротивления сосудов (PRU) в анализе перфузии эндотелия сосудов крысы ex vivo, см., например, Kaul, Fabry and Nagel, PNAS 1989, 86:3356. Кроме того, окклюзию сосуда оценивают с помощью витальной микроскопии, см., например, Zennadi et al., 2007 Blood 110(7):2708. Окклюзию сосуда также можно оценить путем измерения скорости течения и адгезии EHC-клеток с применением in vitro проточных камер с градиентом высоты, см., например, Zennadi et al., 2004, Blood 104(12):3774.
[00424] В некоторых вариантах осуществления безопасность популяции экспрессирующих экзогенный антиген EHC-клеток оценивают путем измерения иммуногенности популяции EHC-клеток с применением соответствующего анализа in vitro или in vivo.
[00425] Например, популяция экспрессирующих экзогенный антиген EHC-клеток a) не индуцирует агглютинацию в тесте Кумбса с применением сыворотки от предполагаемого субъекта-реципиента или b) не индуцирует агглютинацию в тесте Кумбса с применением объединенной человеческой сыворотки.
[00426] В одном варианте осуществления популяция экспрессирующих экзогенный антиген EHC-клеток происходит из прогениторной клетки, которая была генотипирована в отношении главнейших антигенов групп крови и совпадает по генотипу антигенов групп крови с реципиентом.
[00427] В одном варианте осуществления популяция экспрессирующих экзогенный антиген EHC-клеток содержит экзогенный антиген или другой экзогенный белок, который характеризуется менее 10%, 1%, 0,1%, 0,01%, 0,001% или менее 0,001% прогнозируемой реактивностью T-клеток, определенной с помощью алгоритма прогнозирования эпитопа для T-клеток in silico.
[00428] В одном варианте осуществления популяция экспрессирующих экзогенный антиген EHC-клеток содержит экзогенный антиген или другой экзогенный белок, который характеризуется менее 10%, 1%, 0,1%, 0,01%, 0,001% или менее 0,001% реактивностью в in vitro анализе активации T-клеток, например, анализе EpiScreen от Antitope Inc.
[00429] Например, экспрессирующие экзогенный антиген EHC-клетки, происходящие из эритроцитов, можно центрифугировать и ресуспендировать в подходящем растворе (например, стандартном растворе AS-3) для инфузии субъектам, нуждающимся в этом. В некоторых вариантах осуществления экспрессирующие экзогенный антиген EHC-клетки, подлежащие инфузии, имеют тот же тип по системе ABO, что и у реципиента, для сведения к минимуму риска иммунных реакций, ассоциированных с инфузией. Экспрессирующие экзогенный антиген EHC-клетки также можно подвергать предварительной обработке для удаления антигенов, специфичных для типа крови, или иным образом снижать антигенность. Способы, пригодные для этой цели, включают без ограничений описанные в публикациях заявок на патент США №№ 20010006772 и 20030207247.
Терапевтические композиции
[00430] Представлены композиции, содержащие EHC-клетки с эффективными уровнями экзогенного антигена, представляющего интерес. Такие композиции содержат множество EHC-клеток, например, 1×103 клеток, или 1×104, 1×105, 1×106, 1×107, 1×108, 1×109, 1×1010, 1×1011, 1×1012 или более 1×1012 клеток. EHC-клетки по настоящему изобретению, например, можно вводить в виде эритроцитарной массы в физиологическом растворе при концентрации, составляющей 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или более 90% соотношения массы и объема (% масса/объем). Время введения пациенту может варьировать от 10 минут до четырех часов или более.
[00431] Культивируемые EHC-клетки по настоящему изобретению можно хранить в подходящем буфере, например, в одобренном FDA антикоагулянтном консервирующем растворе, таком как антикоагулянт цитрат-декстроза A (ACD-A), цитрат-фосфат-декстроза (CPD), цитратфосфат-декстроза-декстроза (CP2D) или цитрат-фосфат-декстроза-аденин (CPDA-1). Композиции можно хранить в течение периода длительностью до 21 дня.
[00432] В качестве альтернативы, культивируемые EHC-клетки по настоящему изобретению можно хранить в растворе одобренной добавки, например, AS-1 (Adsol), AS-3 (Nutricel), AS-5 (Optisol) или AS-7 (SOLX).
[00433] В качестве альтернативы, культивируемые EHC-клетки по настоящему изобретению можно хранить в криозащитном растворе глицерина. Композиции можно заморозить и хранить в течение периода длительностью до 10 лет. Замороженные клетки можно оттаивать и освобождать от глицерина посредством последовательных стадий отмывки, например, с помощью 0,9% хлорида натрия, перед применением.
[00434] Учитывая продолжительность жизни человеческих и мышиных эритроцитов (120 и 50 дней, соответственно (см., например, Khandelwal et al., Transfusion 2007), может быть выгодно стимулировать искусственное старение эритроцитов для повышения фагоцитоза с помощью антигенпрезентирующих клеток. Наиболее важным и физиологическим механизмом удаления эритроцитов из кровотока является опосредованный иммунной системой (Singer et al., PNAS 1986) после появления новых антигенных участков на поверхности RBC-клеток, как, например, экстернализация фосфатидилсерина (Schroit et al., J Biol Chem 1985) или кластеризация белка Band 3 (Kay, PNAS 1975; Turrini et al., J Biol Chem 1991), индуцируемые искусственно с помощью кальциевого ионофора или обработки химическим веществом BS3, соответственно.
[00435] Культивируемые EHC-клетки по настоящему изобретению можно обработать средством для индукции фагоцитоза, таким как, например, кальциевый ионофор или BS3. Множество культивируемых EHC-клеток по настоящему изобретению могут содержать EHC-клетки, которые были обработаны средством индукции фагоцитоза, таким как, например, кальциевый ионофор или BS3, в течение различных промежутков времени, так что после введения субъекту множество культивируемых EHC-клеток по настоящему изобретению подвергаются фагоцитозу с различной скоростью, например, непрерывно в течение одного дня или нескольких дней, а не как болюс.
[00436] В данном документе представлены композиции и фармацевтические композиции, содержащие множество культивируемых EHC-клеток, которые содержат экзогенный антиген, представляющий интерес. Композиции и фармацевтические композиции могут содержать раствор подходящего буфера для хранения, такой как, например, антикоагулянт цитрат-декстроза A. Композиции и фармацевтические композиции, содержащие множество культивируемых EHC-клеток, которые содержат экзогенный антиген, представляющий интерес, могут дополнительно содержать одобренную добавку, такую как, например, Adsol. Композиции и фармацевтические композиции, содержащие множество культивируемых EHC-клеток, которые содержат экзогенный антиген, представляющий интерес, могут дополнительно содержать криозащитный раствор глицерина для хранения в замороженном состоянии.
[00437] В данном документе представлены EHC-клетки, содержащие экзогенный антиген, представляющий интерес, выбранный из одного или нескольких антигенов в таблице F, таблице G, таблице H, таблице I и таблице J, таких как, например, основной белок миелина, протеолипидный белок, гликопротеин миелина олигодендроцитов, антиген бета-клеток поджелудочной железы, инсулин, флагеллин, глютен, 2S альбумин, гиалуронидаза, фактор VIII, фактор IX и антитело к TNFa, аденозиндезаминаза, L-аспарагиназа, расбуриказа, антитимоцитарный глобулин, L-аргиназа, L-метионаза, препроинсулин, проинсулин, инсулин, GAD65, GAD67, IA-2, ΙΑ-2β, тиреоглобулин, тиреоидная пероксидаза, рецептор тиреотропного гормона, гликопротеин миелина олигодендроцитов, протеолипидный белок, коллаген II, коллаген IV, ацетилхолиновый рецептор, матриксный металлопротеин 1 и 3, молекулярный шаперон-белок теплового шока 47, фибриллин-1, PDGF-рецептор α, PDGF-рецептор β, ядерный белок SS-A, конарахин (Ara h 1), аллерген II (Ara h 2), агглютинин арахиса (Ara h 6), α-лактальбумин (ALA), лактотрансферрин, глютеин, низкомолекулярный глютеин, α-глиадин, γ-глиадин, гордеин, секалин, авенин и их комбинации. Множество EHC-клеток, содержащих экзогенный антиген, представляющий интерес, может быть получено в виде композиции или фармацевтической композиции.
[00438] В данном документе представлены векторы экспрессии, кодирующие один или несколько антигенов из таблицы F, таблицы G, таблицы H, таблицы I и таблицы J, таких как, например, основной белок миелина, протеолипидный белок, гликопротеин миелина олигодендроцитов, антиген бета-клеток поджелудочной железы, инсулин, флагеллин, глютен, Ara h2, 2S альбумин, гиалуронидаза, фактор VIII, фактор IX и антитело к TNFa, необязательно слитых с одним из эндогенных белков клеток эритроидного ряда из таблицы C.
[00439] В данном документе представлены фармацевтические композиции, содержащие экспрессирующие экзогенный антиген EHC-клетки, которые пригодны для введения субъекту. Фармацевтические композиции обычно содержат популяцию экспрессирующих экзогенный антиген EHC-клеток и фармацевтически приемлемый носитель в форме, пригодной для введения субъекту. Фармацевтически приемлемые носители частично определяются конкретной композицией, подлежащей введению, а также конкретным способом, применяемым для введения композиции. Соответственно, существует широкий спектр пригодных составов фармацевтических композиций, содержащих популяцию экспрессирующих экзогенный антиген EHC-клеток. (См., например, Remington's Pharmaceutical Sciences, Mack Publishing Co., Easton, Pa. 18th ed. (1990)). Фармацевтические композиции обычно составляют стерильными, практически изотоническими и в полном соответствии с правилами Надлежащей производственной практики (GMP) Управления США по санитарному надзору за качеством пищевых продуктов и медикаментов.
[00440] Фармацевтически приемлемые наполнители включают наполнители, которые, в целом, безопасны, нетоксичны и желательны, в том числе наполнители, которые подходят для применения в ветеринарии, а также для применения в фармацевтических препаратах для человека. Такие наполнители могут быть твердыми, жидкими, мягкими или, в случае аэрозольной композиции, газообразными.
[00441] Примеры носителей или разбавителей включают без ограничений воду, солевой раствор, растворы Рингера, раствор декстрозы и 5% человеческий сывороточный альбумин. Применение таких сред и соединений для фармацевтически активных веществ хорошо известно из уровня техники. Кроме тех случаев, когда какая-либо традиционная среда или соединение не совместимо с экспрессирующими экзогенный антиген EHC-клетками, описанными в данном документе, подразумевается их применение в композиции. В композиции также могут быть включены вспомогательные терапевтические средства. Как правило, фармацевтическую композицию составляют так, чтобы она была совместима с предполагаемым путем введения. Экспрессирующие экзогенный антиген EHC-клетки можно вводить посредством парентерального, местного, внутривенного, перорального, подкожного, внутриартериального, интрадермального, трансдермального, ректального, внутричерепного, внутрибрюшинного, интраназального; внутримышечного пути или в виде средств для ингаляции. Экспрессирующие экзогенный антиген EHC-клетки необязательно можно вводить в комбинации с другими терапевтическими средствами, которые по меньшей мере частично эффективны в лечении заболевания, нарушения или состояния, для лечения которого предназначены экспрессирующие экзогенный антиген EHC-клетки.
[00442] Растворы или суспензии, применяемые для парентерального, интрадермального или подкожного применения, могут включать следующие компоненты: стерильный разбавитель, такой как вода для инъекций, физиологический раствор, нелетучие масла, полиэтиленгликоли, глицерин, пропиленгликоль или другие синтетические растворители; антибактериальные соединения, такие как бензиловый спирт или метилпарабены; антиоксиданты, такие как аскорбиновая кислота или бисульфит натрия; хелатообразующие соединения, такие как этилендиаминтетрауксусная кислота (EDTA); буферы, такие как ацетаты, цитраты или фосфаты, и соединения для регуляции тоничности, такие как хлорид натрия или декстроза. pH можно регулировать с помощью кислот или оснований, таких как соляная кислота или гидроксид натрия. Препарат для парентерального введения может быть заключен в ампулы, одноразовые шприцы или флаконы для многократных доз, сделанные из стекла или пластика.
[00443] Фармацевтические композиции, пригодные для инъекционного применения, включают стерильные водные растворы (если они растворимы в воде) или дисперсии и стерильные порошки для экстемпорального приготовления стерильных инъекционных растворов или дисперсии. В случае внутривенного введения пригодные носители включают физиологический раствор, бактериостатическую воду, Cremophor EL™ (BASF, Парсиппани, Нью-Джерси) или фосфатно-солевой буфер (PBS). В любом случае композиция должна быть стерильной и должна быть жидкой до такой степени, чтобы существовала легкая возможность введения через шприц. Она должна быть стабильной в условиях производства и хранения, и ее следует предохранить от загрязняющего действия микроорганизмов, таких как бактерии и грибы. Носитель может представлять собой растворитель или дисперсионную среду, содержащую, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и т.п.), и их пригодные смеси. Надлежащую текучесть можно поддерживать, например, с помощью применения покрытия, такого как лецитин, с помощью поддержания требуемого размера частиц в случае дисперсии и с помощью применения поверхностно-активных веществ. Предотвращение действия микроорганизмов может достигаться с помощью различных антибактериальных и противогрибковых соединений, например, парабенов, хлорбутанола, фенола, аскорбиновой кислоты, тимеросала и т.п. Во многих случаях было бы предпочтительно включить в композицию соединения для поддержания изотоничности, например, сахара, полиспиртов, таких как маннит, сорбит, хлорида натрия. Пролонгированного всасывания инъекционных композиций можно добиться путем включения в композицию соединения, которое приводит к отсрочке всасывания, например, моностеарата алюминия и желатина.
[00444] Стерильные инъекционные растворы можно получить путем введения экспрессирующих экзогенный антиген EHC-клеток в эффективное количество подходящего растворителя, при необходимости, с одним или комбинацией ингредиентов, перечисленных в данном документе. В целом, дисперсии получают путем введения экспрессирующих экзогенный антиген EHC-клеток в стерильную среду, которая содержит основную дисперсионную среду и любые требуемые прочие ингредиенты. В случае стерильных порошков для приготовления стерильных инъекционных растворов способы получения представляют собой вакуумную сушку и лиофилизацию, которые дают порошок активного ингредиента, а также любого дополнительного требуемого ингредиента из их раствора, предварительно стерилизованного с помощью фильтрации. Экспрессирующие экзогенный антиген EHC-клетки можно вводить в форме инъекции-депо или препарата-импланта, которые можно составлять таким образом, который обеспечивает непрерывное или пульсирующее высвобождение экспрессирующих экзогенный антиген EHC-клеток, их экзогенного антигена(ов) и/или их необязательной загрузки(загрузок).
[00445] Композиции для перорального введения обычно включают инертный разбавитель или съедобный носитель. Они могут быть заключены в желатиновые капсулы или спрессованы в таблетки. С целью введения перорального терапевтического средства экспрессирующие экзогенный антиген EHC-клетки можно включать с наполнителями и применять в форме таблеток, пастилок или капсул. Композиции для перорального введения также можно получать с применением жидких носителей для применения в качестве жидкости для полоскания рта, где соединение в жидком носителе применяют перорально, и полоскают рот и выплевывают или проглатывают. Фармацевтически совместимые связывающие соединения и/или вспомогательные материалы можно включать как часть композиции. Таблетки, гранулы, капсулы, пастилки и т.п. могут содержать любой из следующих ингредиентов или соединения аналогичной природы: связующее, такое как микрокристаллическая целлюлоза, трагакантовая камедь или желатин; наполнитель, такой как крахмал или лактоза, соединение, способствующее распаданию, такое как альгиновая кислота, Primogel или кукурузный крахмал; смазывающее вещество, такой как стеарат магния или Sterotes; скользящее вещество, такое как коллоидный диоксид кремния; подсластитель, такой как сахароза или сахарин, или ароматизатор, такой как мята перечная, метилсалицилат или апельсиновый ароматизатор.
[00446] Для введения посредством ингаляции экспрессирующие экзогенный антиген EHC-клетки доставляют в форме распыляемого аэрозоля из находящегося под давлением контейнера или дозатора, который содержит соответствующий пропеллент, например, газ, такой как диоксид углерода, или ингалятора.
[00447] Системное введение композиций, содержащих экспрессирующие экзогенный антиген EHC-клетки, также можно осуществлять с помощью средств для трансмукозального или трансдермального введения. Для трансмукозального или трансдермального введения в составе используют проникающие вещества, подходящие для барьера, который следует преодолеть. Такие проникающие вещества в целом известны из уровня техники и включают, например, в случае трансмукозального введения, детергенты, соли желчных кислот и производные фузидовой кислоты. Трансмукозальное введение можно осуществлять посредством применения распыляемых составов или суппозиториев. Для трансдермального введения модифицированные красные кровяные клетки составляют в мази, бальзамы, гели или кремы, общеизвестные из уровня техники.
[00448] Экспрессирующие экзогенный антиген EHC-клетки также можно получать как фармацевтические композиции в форме суппозиториев (например, с традиционными основами для суппозиториев, такими как масло какао и другие глицериды) или удерживающих клизм для ректальной доставки.
[00449] В некоторых вариантах осуществления экспрессирующие экзогенный антиген EHC-клетки получают с носителями, которые будут снижать скорость, с которой экспрессирующие экзогенный антиген EHC-клетки элиминируются из организма субъекта. Например, пригодные составы с контролируемым высвобождением включают импланты и микроинкапсулированные системы доставки. Можно применять биоразлагаемые, биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, сложные полиортоэфиры и полимолочная кислота. Способы получения таких составов будут очевидны специалистам в данной области. Материалы также можно приобретать у Alza Corporation и Nova Pharmaceuticals, Inc.
[00450] В одном варианте осуществления фармацевтическую композицию, содержащую экспрессирующие экзогенный антиген EHC-клетки, вводят внутривенно субъекту, который будет получать пользу от фармацевтической композиции. В других вариантах осуществления композицию вводят в лимфатическую систему, например, посредством внутрилимфатической инъекции или посредством внутриузловой инъекции (см., например, Senti et al., 2008 PNAS 105(46):17908) или посредством внутримышечной инъекции, посредством подкожного введения, посредством прямой инъекции в тимус или в печень.
[00451] В одном варианте осуществления фармацевтическую композицию, содержащую экспрессирующие экзогенный антиген EHC-клетки, вводят в виде жидкой суспензии. В одном варианте осуществления фармацевтическую композицию вводят в виде коагулированного состава, который после введения способен формировать депо и в предпочтительном варианте осуществления медленно высвобождать экспрессирующие экзогенный антиген EHC-клетки в кровоток или в предпочтительном варианте осуществления оставаться в форме депо.
[00452] В одном варианте осуществления фармацевтическую композицию, содержащую экспрессирующие экзогенный антиген EHC-клетки, хранят с применением способов и буферных композиций, которые способны поддерживать жизнеспособность экспрессирующих экзогенный антиген EHC-клеток. Например, деоксигенирование перед хранением для поддержания анаэробного состояния, манипуляцию с pH, обогащение предшественниками метаболитов, манипуляцию с осмотическим балансом, повышение объема суспендирующей среды и/или снижение окислительного стресса путем добавления защитных молекул можно применять для поддержания жизнеспособности экспрессирующих экзогенный антиген EHC-клеток. В некоторых исследованиях, использующих комбинацию этих стратегий, сообщалось о поддержании жизнеспособности эритроцитов, что обеспечивает возможность продления хранения после 6 недель (см., например, Yoshida and Shevkoplyas, Blood Transfus 2010 8:220).
[00453] Фармацевтически приемлемые носители или наполнители можно применять для доставки экспрессирующих экзогенный антиген EHC-клеток, описанных в данном документе. Наполнитель относится к инертному веществу, применяемому в качестве разбавителя или среды. Фармацевтически приемлемые носители применяют, как правило, с соединением так, чтобы сделать соединения применимым для терапии или в качестве продукта. В целом, в случае любого вещества фармацевтически приемлемый носитель представляет собой материал, который объединяют с веществом для доставки субъекту субъект. Традиционные фармацевтические носители, водные, порошковые или масляные основы, загустители и т.п. могут быть необходимыми или желательными. В некоторых случаях носитель необходим для доставки, например, для солюбилизации нерастворимого соединения для доставки в жидком состоянии; как буфер для контроля pH вещества для сохранения его активности или разбавитель для предотвращения потери вещества в сосуде для хранения. В других случаях, однако, носитель служит для удобства, например, жидкость для более удобного введения. Фармацевтически приемлемые соли соединений, описанных в данном документе, можно синтезировать в соответствии со способами, известными специалистам в данной области.
[00454] Как правило, фармацевтически приемлемые композиции являются высокоочищенными, чтобы не содержать загрязнителей, являются биосовместимыми и нетоксичными и подходят для введения субъекту. Если вода служит составной частью носителя, вода является высокоочищенной и обработанной, чтобы не содержать загрязнителей, например, эндотоксинов.
[00455] Фармацевтически приемлемый носитель может представлять собой лактозу, декстрозу, сахарозу, сорбит, маннит, крахмал, аравийскую камедь, фосфат кальция, альгинаты, желатин, силикат кальция, микрокристаллическую целлюлозу, поливинилпирролидон, целлюлозу, воду, сироп, метилцеллюлозу, метилгидроксибензоат, пропилгидроксибонзоат, тальк, стеарат магния и/или минеральное масло, но не ограничивается ими. Фармацевтическая композиция может дополнительно включать смазывающее вещество, смачивающее средство, подсластитель, усилитель вкуса, эмульгирующее средство, суспендирующее средство и/или консервант.
[00456] Представлены фармацевтические композиции, содержащие экспрессирующие экзогенный антиген EHC-клетки с эффективными уровнями экзогенных антигенов. Такие композиции содержат множество экспрессирующих экзогенный антиген EHC-клеток, например, 1×103 EHC-клеток или 1×104, 1×105, 1×106, 1×107, 1×108, 1×109, 1×1010, 1×1011, 1×1012 или более 1×1012 EHC-клеток. В конкретных примерах экспрессирующие экзогенный антиген EHC-клетки, полученные из EHC-клеток, можно вводить в виде эритроцитарной массы в физиологическом растворе при концентрации, составляющей 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или более 90% соотношения массы и объема (% масса/объем). Время введения пациенту может варьировать от 10 минут до четырех часов или более.
[00457] В конкретных примерах, экспрессирующие экзогенный антиген EHC-клетки, полученные из EHC-клеток, можно хранить в подходящем буфере, например, в одобренном FDA антикоагулянтном консервирующем растворе, таком как антикоагулянт цитрат-декстроза A (ACD-A), цитрат-фосфат-декстроза (CPD), цитратфосфат-декстроза-декстроза (CP2D) или цитрат-фосфат-декстроза-аденин (CPDA-1). Композиции можно хранить в течение периода длительностью до 21 дня.
[00458] В качестве альтернативы, экспрессирующие экзогенный антиген EHC-клетки, полученные из EHC-клеток, можно хранить в растворе одобренной добавки, например, AS-1 (Adsol), AS-3 (Nutricel), AS-5 (Optisol) или AS-7 (SOLX).
[00459] В качестве альтернативы, экспрессирующие экзогенный антиген EHC-клетки, полученные из EHC-клеток, можно хранить в криозащитном растворе глицерина. Композиции можно заморозить и хранить в течение периода длительностью до 10 лет. Замороженные клетки можно оттаивать и освобождать от глицерина посредством последовательных стадий отмывки, например, с помощью 0,9% хлорида натрия, перед применением.
[00460] В данном документе представлены композиции и фармацевтические композиции, содержащие множество культивируемых EHC-клеток, которые содержат экзогенный антиген. Композиции и фармацевтические композиции могут содержать раствор подходящего буфера для хранения, такой как, например, антикоагулянт цитрат-декстроза A. Композиции и фармацевтические композиции, содержащие множество культивируемых EHC-клеток, которые содержат экзогенный антиген, могут дополнительно содержать одобренную добавку, такую как, например, Adsol. Композиции и фармацевтические композиции, содержащие множество культивируемых EHC-клеток, которые содержат экзогенный антиген, могут дополнительно содержать криозащитный раствор глицерина для хранения в замороженном состоянии.
[00461] В одном варианте осуществления экспрессирующая экзогенный антиген EHC способна формировать множественный сложный комплекс, например, димер, тример, мультимер, с другой экспрессирующей экзогенный антиген EHC.
[00462] В одном варианте осуществления экспрессирующая экзогенный антиген EHC способна формировать множественный сложный комплекс, например, димер, тример, мультимер, с компонентом кровеносной системы, например, эритроцитом, ретикулоцитом, тромбоцитом, макрофагом, лимфоцитом, T-клеткой, B-клеткой, тучной клеткой.
[00463] Дозировку и частоту введения экспрессирующих экзогенный антиген EHC-клеток и содержащих их фармацевтических композиций может определять лечащий врач, исходя из различных факторов, таких как тяжесть заболевания, возраст, пол и пищевой рацион пациента, тяжесть любого воспаления, время введения и других клинических факторов. В одном примере внутривенное введение начинают при дозе, которая является минимально эффективной, и дозу повышают в течение предварительно установленного периода времени вплоть до наблюдения положительного эффекта. Впоследствии выполняют пошаговое повышение дозировки, ограничиваясь уровнями, которые приводят к соответствующему повышению эффекта, при этом учитывая любые побочные эффекты, которые могут возникнуть.
[00464] Неограничивающие примеры соответствующих дозировок могут варьировать в диапазоне, например, от 1×103 до 1×1014, от 1×103 до 1×107, от 1×105 до 1×106, от 1×107 до 1×1011, от 1×108 до 1×109, от 1×1010 до 1×1014, от 1×1011 до 1×1013 или от 5×1011 до 5×1012 экспрессирующих экзогенный антиген EHC-клеток. Конкретные примеры включают приблизительно 1×103, 2×103, 3×103, 4×103, 5×103, 6×103, 7×103, 8×103, 9×103, 1×104, 2×104, 3×104, 4×104, 5×104, 6×104, 7×104, 8×104, 9×104, 1×105, 2×105, 3×105, 4×105, 5×105, 6×105, 7×105, 8×105, 9×105, 1×106, 2×106, 3×106, 4×106, 5×106, 6×106, 7×106, 8×106, 9×106, 1×107, 2×107, 3×107, 4×107, 5×107, 6×107, 7×107, 8×107, 9×107, 1×108, 2×108, 3×108, 4×108, 5×108, 6×108, 7×108, 8×108, 9×108, 1×109, 2×109, 3×109, 4×109, 5×109, 6×109, 7×109, 8×109, 9×109, 1×1010, 2×1010, 3×1010, 4×1010, 5×1010, 6×1010, 7×1010, 8×1010, 9×1010, 1×1011, 2×1011, 3×1011, 4×1011, 5×1011, 6×1011, 7×1011, 8×1011, 9×1011, 1×1012 или более экспрессирующих экзогенный антиген EHC-клеток. Каждую дозу экспрессирующих экзогенный антиген EHC-клеток можно вводить с интервалами, такими как один раз в день, один раз в неделю, два раза в неделю, один раз в месяц или два раза в месяц. Каждая EHC может экспрессировать некий диапазон молекул антигена, например, от приблизительно 100 до 107 или от приблизительно 103 до 106. Конкретные примеры включают приблизительно 1000, 3000, 5000, 1×104, 3×104, 5×104, 1×105, 3×105, 5×105, 1×106, 3×106, 5×106, 1×107 или более молекул экзогенного антигена на EHC.
[00465] "Пропорциональная дозировка EHC" представляет собой число экспрессирующих экзогенный антиген EHC-клеток, вводимых в виде дозы относительно встречающегося в природе количества циркулирующих в крови объектов. Циркулирующие в крови объекты могут представлять собой клетки, например, эритроциты, ретикулоциты или лимфоциты, или мишени, например, антигены, антитела, вирусы, токсины, цитокины и т.д. Единицы определяют как экспрессирующая экзогенный антиген EHC на циркулирующий в крови объект, т. е. SCMRC/CE. Такая единица дозировки может включать 10-7, 10-6, 10-5, 10-4, 10-3, 10-2, 10-1, 1, 10, 102, 103, 104, 105, 106, 107, 108, 109.
[00466] Фармацевтические композиции, описанные в данном документе, содержат экспрессирующую экзогенный антиген EHC и необязательно фармацевтически активное или терапевтическое средство. Терапевтическое средство может представлять собой биологическое средство, низкомолекулярное средство или средство на основе нуклеиновой кислоты.
[00467] Представлены лекарственные формы, которые содержат фармацевтическую композицию, содержащую экспрессирующую экзогенный антиген EHC, описанную в данном документе. В некоторых вариантах осуществления лекарственная форма составлена в виде жидкой суспензии для внутривенной инъекции.
[00468] Представлены медицинские устройства, которые содержат контейнер, вмещающий фармацевтическую композицию, содержащую экспрессирующую экзогенный антиген EHC, описанную в данном документе, и аппликатор для внутривенной инъекции фармацевтической композиции субъекту.
[00469] Представлены медицинские наборы, которые содержат фармацевтическую композицию, содержащую экспрессирующую экзогенный антиген EHC, описанную в данном документе, и медицинское устройство для внутривенной инъекции фармацевтической композиции субъекту.
[00470] Фармацевтически приемлемая суспензия экспрессирующих экзогенный антиген EHC-клеток предпочтительно упакована в объеме, составляющем от приблизительно 10 до приблизительно 250 мл. Упаковка может представлять собой шприц или пакет для внутривенного вливания, пригодный для переливаний. Введение суспензий проводят, например, посредством внутривенной или внутриартериальной инъекции, необязательно с применением капельного вливания из пакета для внутривенного вливания и т.п. Введение, как правило, проводят внутривенно в руку или через центральный катетер. В случае введений, превышающих 50 мл, предпочтительным является применение капельного введения.
Лечение заболеваний
[00471] Представлены способы индукции иммунологической толерантности. Способы включают введение субъекту, нуждающемуся в индукции иммунологической толерантности, фармацевтической композиции из клеток-эритроцитов, которые содержат экзогенный антиген, представляющий интерес, представленных в данном документе, в количестве и/или при частоте введения, достаточных для индукции иммунологической толерантности у субъекта.
[00472] Фармацевтические композиции по настоящему изобретению обеспечивают клетки-эритроциты, которые содержат экзогенный антиген, представляющий интерес, применимые для стимуляции или усиления иммунологической толерантности. Иммунологическая толерантность может применяться для лечения, предотвращения или снижения тяжести заболевания, нарушения или состояния, ассоциированных с активацией иммунной системы.
[00473] Заболевания, обусловленные активацией иммунной системы, включают аутоиммунные заболевания, такие как, например, рассеянный склероз, сахарный диабет 1 типа, ревматоидный артрит и мембранозный нефрит, и перечисленные в таблице F. Заболевания, обусловленные активацией иммунной системы, также включают воспалительные заболевания, такие как, например, болезнь Крона, язвенный колит, целиакия или другие идиопатические воспалительные заболевания кишечника, и перечисленные в таблице G. Заболевания, обусловленные активацией иммунной системы, также включают аллергические заболевания, такие как, например, астма, аллергия на арахис, аллергия на морепродукты, аллергия на пыльцу, аллергия на белок молока, аллергия на укусы насекомых и аллергия на латекс, и перечисленные в таблице H. Заболевания, обусловленные активацией иммунной системы, также включают активацию иммунной системы в ответ на терапевтический белок, вводимый для лечения основного заболевания, что уменьшает эффективность терапевтического белка, такого как, например, фактор свертывания VIII при гемофилии A, фактор свертывания IX при гемофилии B, антитела к фактору некроза опухоли альфа (TNFa) при ревматоидном артрите и других воспалительных заболеваниях, глюкоцереброзидаза при болезни Гоше или аспарагиназа при остром лимфобластном лейкозе (ALL), и перечисленные в таблице I, таблице J, таблице 5 и таблице 7.
[00474] Дополнительно представлены способы лечения заболевания, обусловленного активацией иммунной системы. Способы включают введение субъекту, нуждающемуся в индукции лечения, фармацевтической композиции из клеток-эритроцитов, которые содержат экзогенный антиген, представляющий интерес, представленных в данном документе, в количестве, достаточном для лечения заболевания, обусловленного активацией иммунной системы. Например, субъект, который имеет или с подозрением на наличие заболевания, обусловленного активацией иммунной системы, такого как аутоиммунное заболевание, воспалительное заболевание или аллергическое заболевание, будет получать пользу от представленных способов лечения.
[00475] В некоторых вариантах осуществления пациент страдает от аутоиммунного заболевания или состояния, или от опосредованного аутореактивными антителами заболевания или состояния, при котором иммунная система пациента активируется против эндогенной молекулы, например, белкового антигена, так что иммунная система атакует эндогенную молекулу, индуцирует воспаление, разрушает ткань и иным образом приводит к симптомам аутоиммунных или вызванных аутореактивными антителами заболевания или состояний. Иммунный ответ может запускаться антителами, которые связываются с эндогенной молекулой, или может запускаться гиперактивными T-клетками, которые атакуют клетки, экспрессирующие эндогенную молекулу, или он может запускаться другими иммунными клетками, такими как регуляторные T-клетки, NK-клетки, NKT-клетки или B-клетки. В этих вариантах осуществления антигенный белок или его фрагмент может экспрессироваться на безъядерной гемопоэтической клетке по настоящему изобретению. Популяции этих клеток, при введении один раз или более пациенту, страдающему от заболевания или состояния, будет достаточно для индукции толерантности к антигенному белку, так что он более не индуцирует активацию иммунной системы, и, таким образом, она будет лечить или ослаблять симптомы лежащего в основе заболевания или состояния.
[00476] Например, пациент, страдающий от приобретенного тромбоцитопенического акроангиотромбоза (TTP), имеет опосредованное аберрантными аутореактивными антителами заболевание, при котором вырабатываются антитела против эндогенного белка ADAMTS13, делающие его неэффективным в осуществлении его активности расщепления фактора фон Виллебранда, что приводит к образованию микротромбов в сосудистой системе и являющейся следствием тромбоцитопении. В этом варианте осуществления антиген ADAMTS13 экспрессируется на безъядерной гемопоэтической клетке и вводится пациенту, страдающему от TTP, в количестве, эффективном для индукции толерантности к ADAMTS13, таким образом снижается количество ингибирующих аутореактивных антител к ADAMTS13 в кровотоке и восстанавливается способность организма расщеплять фактор фон Виллебранда, тем самым снижаются симптомы заболевания. В предпочтительном варианте осуществления только антигенный фрагмент ADAMTS13 экспрессируется на безъядерной гемопоэтической клетке. В предпочтительном варианте осуществления полноразмерный ADAMTS13 экспрессируется на безъядерной гемопоэтической клетке. В предпочтительном варианте осуществления полноразмерный ADAMTS13 экспрессируется на безъядерной гемопоэтической клетке и является ферментативно активным, так что введенный клеточный продукт способен одновременно индуцировать толерантность и также терапевтически расщеплять фактор фон Виллебранда.
[00477] В другом примере пациент, страдающий от атипичного гемолитического анемического синдрома (aHUS), имеет аберрантный ответ с аутореактивными антителами против эндогенного белкового фактора комплемента H (CFH), который не дает CFH осуществлять его функцию регуляции комплемента. В результате этого происходит сверхактивация комплемента в сосудистой системе, что приводит к внутрисосудистому гемолизу. В этом варианте осуществления антиген CFH экспрессируется на безъядерной гемопоэтической клетке и вводится пациенту, страдающему от aHUS, в количестве, эффективном для индукции толерантности к CFH, таким образом снижается количество ингибирующих аутореактивных антител к CFH в кровотоке и восстанавливается способность организма ингибировать комплемент, тем самым снижаются симптомы заболевания. В предпочтительном варианте осуществления только антигенный фрагмент CFH экспрессируется на безъядерной гемопоэтической клетке. В предпочтительном варианте осуществления полноразмерный CFH экспрессируется на безъядерной гемопоэтической клетке. В предпочтительном варианте осуществления полноразмерный CFH экспрессируется на безъядерной гемопоэтической клетке и является терапевтически активным, так что введенный клеточный продукт способен одновременно индуцировать толерантность и также терапевтически стимулировать регуляцию комплемента.
[00478] В другом примере пациент, страдающий от рассеянного склероза (MS), имеет аутоиммунный ответ на полипептидный миелин, который покрывает оболочкой нейроны. В результате этого T-клетки атакуют миелин и возникающее воспаление вызывает демиелинизацию нервных волокон и нарушает возможности передачи вдоль нервов электрических сигналов, что приводит к симптомам рассеянного склероза. В этом варианте осуществления антиген миелин экспрессируется на безъядерной гемопоэтической клетке и вводится пациенту, страдающему от MS, в количестве, эффективном для индукции толерантности к антигену миелину, таким образом снижается иммунный ответ против миелина и восстанавливается способность организма к передаче электрических импульсов вниз по покрытым миелином нервным волокнам, тем самым снижаются симптомы заболевания. В предпочтительном варианте осуществления только один или несколько антигенных фрагментов миелина экспрессируются на безъядерной гемопоэтической клетке. В предпочтительном варианте осуществления полноразмерный белок миелин экспрессируется на безъядерной гемопоэтической клетке.
[00479] В другом примере пациент, страдающий от сахарного диабета 1 типа (T1D), имеет аутоиммунный ответ на островковые бета-клетки поджелудочной железы. В результате этого T-клетки уничтожают островковые бета-клетки, снижая или устраняя способность поджелудочной железы вырабатывать и секретировать инсулин, что приводит к симптомам и патологии T1D. В этом варианте осуществления антиген бета-клеток экспрессируется на безъядерной гемопоэтической клетке и вводится пациенту, страдающему от T1D, в количестве, эффективном для индукции толерантности к антигену бета-клеток, таким образом снижается иммунный ответ против бета-клеток и восстанавливается способность поджелудочной железы вырабатывать и секретировать инсулин, тем самым снижаются симптомы заболевания. В предпочтительном варианте осуществления только один или несколько антигенных фрагментов антигена бета-клеток экспрессируются на безъядерной гемопоэтической клетке. В предпочтительном варианте осуществления полноразмерный белок бета-клеток экспрессируется на безъядерной гемопоэтической клетке.
[00480] Дополнительно представлены способы снижения или ослабления активации иммунной системы в ответ на схему лечения терапевтическим белком. Способы включают введение субъекту, нуждающемуся в снижении или ослаблении активация иммунной системы в ответ на схему лечения терапевтическим белком, фармацевтической композиции из клеток-эритроцитов, которые содержат экзогенный антиген, представляющий интерес, представленных в данном документе, в количестве, достаточном для снижения или ослабления активации иммунной системы в ответ на схему лечения терапевтическим белком.
[00481] В некоторых вариантах осуществления пациент страдает от заболевания или состояния, в случае которых терапевтический белок может вводиться для лечения или ослабления симптома заболевания или состояния, но терапевтический белок является иммуногенным, так что у пациента развивается иммунный ответ против терапевтического белка, так что он более не является эффективным в лечении или ослаблении исходного заболевания. Например, иммуногенный терапевтический белок может происходить из источника, не относящегся к человеку, например, крупного рогатого скота, свиньи или от примата, не являющегося человеком, или из источника, не относящегося к млекопитающему, например, бактерий, грибов или растений, или иммуногенный терапевтический белок может происходить из источника, относящегося к человеку, при этом повторяющееся воздействие и дозирование могут быть достаточными для индукции иммуногености. Иммунный ответ может запускаться антителами, которые связываются с иммуногенным терапевтическим белком и ингибируют его функцию (нейтрализующие антитела) или которые связываются с иммуногенным терапевтическим белком и приводят к его клиренсу под действием других иммунных клеток (опсонизирующие антитела). В этих вариантах осуществления иммуногенный терапевтический белок или его антигенный фрагмент может экспрессироваться на безъядерной гемопоэтической клетке по настоящему изобретению. Популяции этих клеток, при введении один раз или более пациенту, страдающему от заболевания, будет достаточно для индукции толерантности к иммуногенному терапевтическому белку, так что он больше не находится под нейтрализующим или опсонизирующим действием иммунной системы. В одном предпочтительном варианте осуществления иммуногенный терапевтический белок, экспрессированный на поверхности безъядерной гемопоэтической клетки по настоящему изобретению, является терапевтически активным на клетке в кровотоке, так что композиция из клетки и белка способна одновременно индуцировать толерантность и лечить или ослаблять симптомы лежащего в основе заболевания или состояния при введении пациенту. В другом предпочтительном варианте осуществления антигенный фрагмент иммуногенного терапевтического белка, экспрессируемого на поверхности безъядерной гемопоэтической клетки по настоящему изобретению, не является терапевтически активным на клетке в кровотоке, так что композиция из клетки и белка способна индуцировать толерантность при введении пациенту, но отдельный состав с иммуногенным терапевтическим белком вводят для лечения или ослабления симптомов лежащего в основе заболевания или состояния.
[00482] Например, для пациента, страдающего от гемофилии A, требуются вливания фактора свертывания VIII (FVIII) для восстановления надлежащей коагуляции. Однако у многих пациентов развиваются нейтрализующие антитела к FVIII, несмотря на то, что он имеет человеческое происхождение, которые делают его терапевтически неэффективным и приводят к риску опасного для жизни кровотечения. В одном варианте осуществления безъядерную гемопоэтическую клетку, экспрессирующую экзогенный FVIII, вводят пациенту, страдающему от гемофилии A, так что (1) уровни циркулирующего в крови активного FVIII восстанавливаются до уровня, необходимого для ослабления симптомов и предотвращения тяжелого неконтролируемого кровотечения, и так что (2) индуцируется толерантность к FVIII.
[00483] В другом примере пациенту, страдающему от ревматоидного артрита, требуются инъекции антитела к TNFa для снижения воспаления, ассоциированного с этим заболеванием. Однако у многих пациентов развиваются нейтрализующие антитела против антитела к TNFa, которые делают терапевтическое антитело неэффективным. В этом случае пациент, как правило, страдает от ухудшения симптомов и либо вынужден повышать дозу антитела к TNFa, либо переключаться на другое антитело к TNFa с другой кодирующей последовательностью аминокислот. В этом варианте осуществления безъядерную гемопоэтическую клетку, экспрессирующую антигенный фрагмент антитела к TNFa, вводят пациенту с ревматоидным артритом, у которого развились нейтрализующие антитела против антитела к TNFa. Композицию вводят с дозой, достаточной для индукции толерантности к антителу к TNFa, что обеспечивает возможность эффективного введения антитела к TNFa со снижением уровней циркулирующего в крови TNFa и, таким образом, со снижением симптомов ревматоидного артрита у пациента.
[00484] В другом примере пациента, страдающего от фенилкетонурии (PKU), лечат с помощью пегилированной фенилаланин-аммоний-лиазы (PAL), фермента, не являющегося человеческим. У пациента развиваются опсонизирующие и нейтрализующие антитела против PAL, что также вызывает аллергическую реакцию после введения терапевтического белка. Иммунный ответ не только делает PAL неэффективной, он также угрожает здоровью пациента. В одном варианте осуществления безъядерные гемопоэтические клетки, экспрессирующие экзогенную PAL, вводят пациенту, страдающему от PKU, в количестве, достаточном для индукции толерантности к PAL. В предпочтительном варианте осуществления PAL, экспрессируемый в клетке, является активным на клетке, и композиция способна снижать уровни циркулирующего в крови фенилаланина и лечить или ослаблять симптомы фенилкетонурии в дополнение к предотвращению опасной иммунной реакции против PAL. В другом предпочтительном варианте осуществления антигенный фрагмент PAL экспрессируется на безъядерной гемопоэтической клетке, и этот фрагмент, экспрессируемый в клетке, не является терапевтически активным, поэтому отельный состав с PAL вводят пациенту для лечения или ослабления симптомов фенилкетонурии. Индуцирующую толерантность композицию клеток можно вводить до введения терапевтического состава с PAL или индуцирующую толерантность композицию клеток можно вводить одновременно с введением терапевтического состава с PAL.
[00485] В некоторых вариантах осуществления пациент страдает от аллергического заболевания, например, аллергии на перхоть животных, черный орех, бразильский орех, орех кешью, съедобный каштан, пылевых клещей, яйцо, грецкий орех, рыбу, фундук, яд насекомых, латекс, молоко, плесень, арахис, пыльцу, траву, морепродукты, сою, лесные орехи или пшеницу. У пациента, страдающего от аллергии, может развиться иммунный ответ после контакта с антигенным фрагментом аллергена, например через пищу, контакт с кожей, инъекцию или воздействие окружающей среды. В иммунный ответ могут вовлекаться антитело IgE, сенсибилизированные тучные клетки, дегрануляция, высвобождение гистамина и анафилаксия, а также канонические иммунные клетки, такие как T-клетки, B-клетки, дендритные клетки, регуляторные T-клетки, NK-клетки, нейтрофилы и NKT-клетки. Аллергическая реакция может вызывать дискомфорт, или она может быть достаточно серьезной, чтобы вызывать смерть, и, таким образом, требуется постоянная бдительность со стороны больного, а также его или ее семьи и сиделок. В этих вариантах осуществления антигенный белок или его фрагмент может экспрессироваться на безъядерной гемопоэтической клетке по настоящему изобретению. Популяции этих клеток, при введении один раз или более пациенту, страдающему от аллергического заболевания или состояния, будет достаточно для индукции толерантности к антигенному белку, так что он более не индуцирует активацию иммунной системы после воздействия, и, таким образом, она будет лечить или ослаблять симптомы лежащего в основе аллергического заболевания или состояния.
[00486] В одном примере пациент, страдающий от аллергии на арахис, имеет иммунный ответ после воздействия антигена арахиса AraH1. В этом варианте осуществления AraH1 экспрессируется на безъядерной гемопоэтической клетке и вводится пациенту, страдающему от аллергии на арахис, в количестве, эффективном для индукции толерантности к антигену AraH1, таким образом снижается аллергический иммунный ответ и восстанавливается способность индивидуума безопасно потреблять арахис, тем самым снижаются симптомы заболевания. В предпочтительном варианте осуществления только один или несколько антигенных фрагментов AraH1 экспрессируются на безъядерной гемопоэтической клетке. В предпочтительном варианте осуществления полноразмерный белок AraH1 экспрессируется на безъядерной гемопоэтической клетке.
[00487] Определенные аспекты настоящего изобретения относятся к EHC-клеткам, которые содержат антиген, распознаваемый иммунными клетками при заболеваниях, опосредованных несоответствием человеческих лейкоцитарных антигенов (HLA), таких как, например, реакция "трансплантат против хозяина" или отторжение трансплантированных органов.
[00488] В некоторых вариантах осуществления пациент страдает от заболевания или состояния, связанных с несоответствием человеческих лейкоцитарных антигенов (HLA), при котором иммунные клетки активируются против HLA-антигенов на ткани и атакуют ткань. Это обычно происходит после аллогенной трансплантации органа или ткани от донора, который не характеризуется точным соответствием, и приводит к медицинскому состоянию отторжения трансплантата, при котором иммунная система пациента атакует чужеродную ткань или орган и вызывает гибель трансплантированного органа или ткани. Другое обычное состояние, обусловленное несоответствием HLA, является реакция "трансплантат против хозяина" (GVHD), при которой пациенту произвели пересадку аллогенного костного мозга от донора, который характеризуется не точным соответствием, и при которой трансплантированные иммунные клетки (трансплантат) становятся активированными и атакуют органы реципиента (хозяин), которые распознаются как чужеродные, что вызывает повреждение тканей и органов хозяина и приводит к серьезным последствиям, в том числе к смерти. Активация иммунной системы, обусловленная несоответствием HLA, как правило, опосредуется T-клетками, но в нее также могут вовлекаться регуляторные T-клетки, NK-клетки, NKT-клетки, B-клетки, антитела, дендритные клетки, моноциты, макрофаги и нейтрофилы. В этих вариантах осуществления антигенная молекула HLA или ее фрагмент может экспрессироваться на безъядерной гемопоэтической клетке по настоящему изобретению. Популяции этих клеток, при введении один раз или более пациенту, страдающему от заболевания или состояния, обусловленного несоответствием HLA, будет достаточно для индукции толерантности к антигенной молекуле HLA, так что она более не индуцирует активацию иммунной системы, и, таким образом, она будет лечить или ослаблять симптомы лежащего в основе заболевания или состояния, обусловленного несоответствием HLA, например, выживания трансплантированного органа или выживания пациента. В предпочтительном варианте осуществления молекула HLA, экспрессируемая на поверхности клетки, также содержит пептид, загруженный в молекулу HLA.
[00489] Клетки-эритроциты, которые содержат экзогенный антиген, который применяется для способов, описанных в данном документе, могут иметь аутологичное происхождение, т.е. происходить из одного и того же субъекта, или могут иметь аллогенное происхождение, т.е. происходить из другого донора клеток.
[00490] Фармацевтические композиции можно вводить субъекту, например, посредством внутривенного переливания или внутримышечной инъекции.
Другие варианты осуществления
[00491] Следует понимать, что настоящее изобретение не ограничивается конкретными способами, реагентами, соединениями, композициями или биологическими системами, которые, конечно же, могут варьировать. Также следует понимать, что терминология, применяемая в данном документе, служит только для цели описания конкретных аспектов и не подразумевается как ограничивающая.
[00492] Все признаки, раскрытые в настоящем описании, можно объединять в любой комбинации. Каждый признак, раскрытый в настоящем описании, можно заменить альтернативным признаком, служащим для такой же, эквивалентной или подобной цели. Таким образом, если явно не заявляется иное, каждый раскрытый признак представляет собой только пример общей серии эквивалентов или подобных признаков.
[00493] В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC, содержащая экзогенный антиген CR1, не получена в мыши и/или не получена из мышиных клеток эритроидного ряда. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC, содержащая экзогенный антиген CR1, не получена в лабораторном животном и/или не получена из клеток эритроидного ряда, происходящих из лабораторного животного. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC получена из мегакариоцитов или тромбоцитов. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC получена из клетки эритроидного ряда, такой как, например, эритроцит или ретикулоцит. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не получена из нейтрофила, эозинофила или базофила. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не получена из моноцита или макрофага. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не получена из CD34+Thy-1+ гемопоэтической стволовой клетки или популяций клеток, обогащенных по CD34+Lin- или CD34+Thy-1+Lin- клеткам. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит экзогенный антиген, содержащий внеклеточный домен корецептора HIV. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит экзогенный антиген, способный связываться с вирусом. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит экзогенный антиген, содержащий CD4. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит экзогенный антиген, содержащий корецептор HIV. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит экзогенный антиген, содержащий CXCR4, CCR5, CCR1, CCR2, CCR3, CCR4, CCR8, CXCR1, CXCR2, CXCR3, CXCR6, GPR15, APJ, CMKLR1, или CX3CR1, или любую их комбинацию. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит экзогенную нуклеиновую кислоту, кодирующую антиген аденозиндезаминазу. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит экзогенный антиген, содержащий аденозиндезаминазу (ADA). В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит экзогенную нуклеиновую кислоту, кодирующую онкоген. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит экзогенный антиген, содержащий онкоген. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит экзогенную нуклеиновую кислоту, кодирующую cdx1, cdx2 или cdx4. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит экзогенный антиген, содержащий cdx1, cdx2, или cdx4, или любую их комбинацию. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит экзогенный антиген, содержащий химерный полипептид, содержащий домен связывания лиганда. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит экзогенный антиген, содержащий S-домен, который способен связывать лиганд. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит экзогенный антиген, содержащий CD3ζ, CD3η, рецептор IL-2, рецептор IL-3, рецептор IL-4, рецептор IL-7, рецептор IL-11, рецептор IL-13, рецептор GM-CSF, рецептор LIF, рецептор CNTF, рецептор онкостатина M, рецептор TGF-β, рецептор EGF, ATR2/neu, HER2/neu, HER3/c-erbB-3, Xmrk, инсулиновый рецептор, рецептор IGF-1, IRR, рецептор PDGF, рецептор CSF-1, c-kit, STK-1/flk-2, рецептор FGF, flg, bek, рецептор NGF, Ror1 и Ror2 или любую их комбинацию. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит экзогенный антиген, содержащий гены E6 или E7 папилломавируса человека. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит экзогенный антиген содержащий опухолевый антиген. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит экзогенный антиген, содержащий глюкоцереброзидазу. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит экзогенный антиген, содержащий аспарагиназу. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит экзогенный антиген, содержащий аргининдеиминазу. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит молекулу слияния, способную стимулировать слияние экспрессирующей экзогенный антиген EHC с клеткой-мишенью, которая i) отличается от и/или ii) действует независимо от экзогенного антигена, где экзогенный антиген способен взаимодействовать с мишенью. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит экзогенный антиген, содержащий синцитин-1. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит светочувствительное синтетическое соединение, такое как, например, соединение, которое может активироваться фотонами, или подвергаемые гашению соединения. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит активируемое молекулярное средство для обнаружения, способное производить обнаруживаемый ответ. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит диагностическое соединение. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит вирус или бактерию. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не получена из аутологичной CD34+ клетки или не содержит ее. В некоторых вариантах осуществления способы лечения и предотвращения с применением экспрессирующих экзогенный антиген EHC-клеток, полученных из клеток эритроидного ряда, описанных в данном документе, не включают стадию обнаружения клетки эритроидного ряда in vivo, например, благодаря средству для обнаружения, которое ассоциировано с клеткой эритроидного ряда. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не получена из плюрипотентной гемопоэтической стволовой клетки донора-человека. В некоторых вариантах осуществления популяция экспрессирующих экзогенный антиген EHC-клеток не подвергнута размножению в биореакторе. В некоторых вариантах осуществления популяция экспрессирующих экзогенный антиген EHC-клеток после размножения и/или дифференцировки не содержит один вид дифференцированных человеческих кровяных клеток. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не представляет собой дифференцированную зрелую человеческую кровяную клетку. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не получена из кровяной клетки, происходящей от универсального донора, например, с типом крови O, отрицательного по Rh-фактору. В некоторых вариантах осуществления экспрессирующую экзогенный антиген-полипептид ADA EHC не применяют для лечения тяжелой комбинированной иммунной недостаточности (ADA-SCID). В некоторых вариантах осуществления способы размножения и дифференцировки экспрессирующих экзогенный антиген EHC-клеток не включают культивирование экспрессирующих экзогенный антиген EHC-клеток в среде, содержащей лиганд миелопролиферативного рецептора (mpl). В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит загрузку, содержащую синтетический аналог трифосфорилированного нуклеозида. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит загрузку, содержащую 2',3'-дидезоксицитидин-5'-трифосфат (ddCTP) и/или 3'-азидо-3'-дезокситимидин-5'-трифосфат (AZT-TP). В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит загрузку, содержащую бисфосфонат. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC получена путем приведения клетки эритроидного ряда в контакт с экзогенным антигеном и необязательно загрузки без лизирования и повторного закрытия пор клеток для включения экзогенного антигена и/или загрузки. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC получена путем приведения клетки эритроидного ряда в контакт с экзогенным антигеном и необязательно загрузки, где приведение в контакт не включает гипотонический диализ. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC получена путем приведения клетки эритроидного ряда в контакт с экзогенным антигеном и необязательно загрузки, где приведение в контакт не включает загрузку экзогенного антигена и/или загрузки в клетку эритроидного ряда или на нее. В некоторых вариантах осуществления экзогенный антиген получен в объекте, который не представляет собой клетку эритроидного ряда, приводимую в контакт, и/или экзогенный антиген выделен из образца, который не содержит клетку эритроидного ряда, приводимую в контакт. Например, в случае полипептидного экзогенного антигена соответствующие объекты включают линию клеток, систему экспрессии in vitro, бактериальную систему экспрессии и т. д.
[00494] В некоторых вариантах осуществления полипептид экзогенного антигена, экспрессируемый EHC, присутствует на поверхности EHC, но не является нековалентно связанным с поверхностью EHC. В некоторых вариантах осуществления нековалентное прикрепление антигена к поверхности EHC не опосредуется антителом, фрагментом антитела, антителоподобным полипептидом или остовом связывания полипептида, не являющимся антителом. В некоторых вариантах осуществления нековалентное прикрепление экзогенного антигена к поверхности EHC не направлено против антигена клетки эритроидного ряда, такого как Band 3 (CD233), аквапорин -1, Glut-1, антиген Kidd, RhAg/Rh50 (CD241), Rh (CD240), Rh30 CE (CD240CE), Rh30D (CD240D), Kx, гликофорин B (CD235b), гликофорин C (CD235c), гликофорин D (CD235d), Kell (CD238), Duffy/DARCi (CD234), CRl (CD35 ), DAF (CD55), глобозид, CD44, ICAM-4 (CD242), Lu/B-CAM (CD239), XGl/XG2 (CD99), EMMPRIN/нейро телин (CD147), JMH, гликозилтрансфераза, Cartwright, Dombrock, C4A/CAB, Scianna, MER2, s томатин, BA-l (CD24), GPIV (CD36), CD108, CD139 и антиген H (CD173) или другой связывающий эритроциты компонент.
[00495] В некоторых вариантах осуществления полипептид экзогенного антигена не получен независимо от EHC и затем конъюгирован с EHC. В некоторых вариантах осуществления полипептид экзогенного антигена не подвергнут ферментативной конъюгации, например, с помощью аутокаталитической реакции образования изопептидной связи, такой как осуществляемая, например, под действием транспептидазы, сортазы и/или изопептидазы. В одном варианте осуществления полипептид экзогенного антигена не подвергнут ферментативной конъюгации с применением сортазы.
[00496] В некоторых вариантах осуществления полипептид экзогенного антигена не подвергнут химической конъюгации, например, с помощью сшивающего средства, такого как карбодиимид (в том числе сортаза-1-этил-3-(3-диметиламинопропил)карбодиимид (EDC)).
[00497] В одном варианте осуществления экзогенный антиген не получен независимо от EHC и затем инкапсулирован с помощью EHC. В одном варианте осуществления инкапсулирование экзогенного антигена не опосредовано гипотоническим диализом EHC в присутствии экзогенного антигена.
[00498] Многие модификации и другие варианты осуществления настоящего изобретения, изложенные в данном документе, легко придут на ум специалисту в области, к которой принадлежат настоящие изобретения, с получением пользы от идей, представленных в вышеизложенном описании и ассоциированных графических материалах. Следовательно, следует понимать, что настоящие изобретения не следует ограничивать конкретными раскрытыми вариантами осуществления, и подразумевается, что модификации и другие варианты осуществления включены в объем прилагаемой формулы изобретения. Хотя в данном документе используются специальные термины, они применяются только в общем и описательном смысле, а не с целью ограничения.
[00499] Применяемые в настоящем описании и прилагаемой формуле изобретения формы единственного числа включают ссылки на множественное число, если содержание явно не предписывает иное.
[00500] Применение альтернативы (например, "или") следует понимать как означающее одну, обе или любую комбинацию этих альтернатив.
[00501] Термин "приблизительно", применяемый в данном документе при ссылке на измеряемую величину, такую как количество, промежуток времени и т.п., подразумевает, что он охватывает отклонения, составляющие ±20% или ±10%, более предпочтительно ±5%, даже более предпочтительно ±1% и еще более предпочтительно ±0,1% от указанного значения, если такие отклонения подходят для осуществления раскрытых способов.
[00502] Применяемый в данном документе любой диапазон концентраций, диапазон процентных долей, диапазон соотношений или диапазон целочисленных значений следует понимать как включающий значение любого целого числа в пределах изложенного диапазона и, при необходимости, его доли (такие как одна десятая и одна сотая целого числа), если не указано иное.
[00503] "Содержать", "содержащий" и "содержит", а также "состоящий из", применяемые в данном документе, являются синонимами с "включать", "включающий", "включает" или "состоять", "состоящий", "состоит" и представляют собой включающие или неограничивающие термины, которые определяют присутствие того, что следует, например компонента, и не исключают или препятствуют присутствию дополнительных неперечисленных компонентов, признаков, элементов, представителей, стадий, известных из уровня техники или раскрытых в нем.
[00504] Подразумевается, что применяемые в данном документе термины "такой как", "например" и т. п. ссылаются на иллюстративные варианты осуществления и не ограничивают объем настоящего раскрытия.
[00505] Если не определено иное, все технические и научные термины, применяемые в данном документе, имеют такое же значение, общепонятное специалисту в области, к которой относится настоящее изобретение. Хотя любые способы и материалы, подобные или эквивалентные описанным в данном документе, можно применять при практическом осуществлении для тестирования настоящего изобретения, предпочтительные материалы и способы описаны в данном документе.
[00506] Все публикации и патентные заявки, процитированные в настоящем описании, включены в данный документ посредством ссылки в своей полноте для всех целей, как если бы каждая индивидуальная публикация или патентная заявка были бы специально и индивидуально указаны, как включенные посредством ссылки для всех целей. Публикации, обсуждаемые в данном документе, представлены исключительно для их раскрытия перед датой подачи настоящей заявки. Ничто в данном документе не следует истолковывать как признание того, что авторы данного изобретения, описанного в данном документе, не имеют право на датирование такого раскрытия задним числом из-за предшествующего изобретения или по какой-либо другой причине.
Определения
[00507] "Введение", "осуществление введения" и его варианты означают введение композиции, такой как экспрессирующая экзогенный антиген EHC, или средства в субъекта и включают одновременное и последовательное введение композиции или средства. Введение композиции или средства в субъекта осуществляется с помощью любого соответствующего пути, в том числе перорально, пульмонально, интраназально, парентерально (внутривенно, внутримышечно, внутрибрюшинно или подкожно), ректально, внутрь лимфатической системы или местно. Введение включает самостоятельное введение и введение при помощи другого человека. Соответствующий путь введения обеспечивает возможность композиции или средству осуществить их предполагаемую функцию. Например, если соответствующий путь является внутривенным, композицию вводят путем введения композиции или средства в вену субъекта. Введение можно осуществлять с помощью любого пригодного пути введения.
[00508] "Якорь", или "якорный домен", или "A-домен" применяется для ссылки на часть полипептида экзогенного антигена, в том числе слияния или химерного полипептида экзогенного антигена, которая находится в контакте с клеточной мембраной EHC. Полипептид экзогенного антигена может взаимодействовать с липидным слоем клеточной мембраны через вставку фосфолипидного хвоста, ковалентное связывание с элементом липидного слоя, ионную связь, водородную связь или через домен один или несколько раз проходящего через мембрану трансмембранного полипептида, которые пересекают один или несколько липидных слоев клеточной мембраны.
[00509] Применяемый в данном документе термин "антитело" охватывает иммуноглобулин, вне зависимости от того, является ли он природным или частично или полностью получен синтетическим путем, и его фрагменты. Термин также относится к любому белку со связывающим доменом, который является гомологичным связывающему домену иммуноглобулина. Эти белки могут происходить из природных источников или могут быть получены частично или полностью синтетическим путем. Кроме того, "антитело" включает полипептид, содержащий каркасную область из гена иммуноглобулина или его фрагментов, который специфично связывает и распознает антиген. Применение термина антитело подразумевает включение полных антител, поликлональных, моноклональных и рекомбинантных антител, их фрагментов, и, кроме того, включает одноцепочечные антитела, гуманизированные антитела; мышиные антитела; химерные, мышино-человеческие, мышино-приматные, приматно-человеческие моноклональные антитела, антиидиотипические антитела, фрагменты антител, такие как, например, фрагменты scFv, (scFv)2, Fab, Fab' и F(ab')2, F(ab1)2, Fv, dAb и Fd, диатела и родственные антителам полипептиды. Антитело включает биспецифичные антитела и мультиспецифичные антитела при условии, что они проявляют требуемую биологическую активность или функцию.
[00510] Термин "антигенсвязывающий фрагмент", применяемый в данном документе, относится к фрагментам интактного иммуноглобулина и любой части полипептида, включающего антигенсвязывающие области со способностью специфично связываться с антигеном. Например, антигенсвязывающий фрагмент может представлять собой фрагмент F(ab')2, фрагмент Fab', фрагмент Fab, фрагмент Fv или фрагмент scFv, но не ограничивается ими. Фрагмент Fab имеет один антигенсвязывающий участок и содержит вариабельные области легкой цепи и тяжелой цепи, константную область легкой цепи и первую константную область CH1 тяжелой цепи. Фрагмент Fab' отличается от фрагмента Fab тем, что фрагмент Fab' дополнительно включает шарнирную область тяжелой цепи, включающую по меньшей мере один цистеиновый остаток на C-конце области CH1 тяжелой цепи. Фрагмент F(ab')2 получают при условии, что цистеиновые остатки фрагмента Fab' соединяются дисульфидной связью в шарнирной области. Фрагмент Fv представляет собой минимальный фрагмент антитела, имеющий только вариабельные области тяжелой цепи и вариабельные области легкой цепи, и рекомбинантная методика получения фрагмента Fv хорошо известна из уровня техники. Двухцепочечные фрагменты Fv могут иметь структуру, при которой вариабельные области тяжелой цепи связаны с вариабельными областями легкой цепи посредством нековалентной связи. Одноцепочечные фрагменты Fv (scFv) обычно могут иметь димерную структуру, как и у двухцепочечных фрагментов Fv, в которых вариабельные области тяжелой цепи ковалентно связаны с вариабельными областями легкой цепи посредством пептидного линкера или вариабельные области тяжелой и легкой цепи напрямую связаны друг с другом на своих C-концах. Антигенсвязывающий фрагмент можно получить с применением протеазы (например, полное антитело расщепляется папаином с получением фрагментов Fab и расщепляется пепсином с получением фрагментов F(ab')2) и можно получить с помощью методики генетической рекомбинации. Фрагмент dAb состоит из VH-домена. Молекулы одноцепочечного антитела могут содержать полимер с рядом индивидуальных молекул, например, димер, тример или другие полимеры.
[00511] "Аппликатор" относится к любому устройству, применяемому для присоединения к субъекту. Он включает, например, иглы, канюли, катетеры и систему для внутривенных инфузий.
[00512] "Ассоциирован с", при применении для описания взаимоотношений между множественными соединениями и молекулами, охватывает, такие как, например, любое взаимодействие между экзогенным антигеном и мишенью или между экспрессирующей экзогенный антиген EHC и мишенью. Он включает ферментативное взаимодействие, ионное связывание, ковалентное связывание, нековалентное связывание, образование водородных связей, силы Лондона, вандерваальсовы силы, гидрофобное взаимодействие, липофильные взаимодействия, магнитные взаимодействия, электростатические взаимодействия и т.п.
[00513] "Ассоциирован с", при применении для описания взаимоотношений между мишенью, объектом, соединением, средством или молекулой и заболеванием, нарушением, состоянием, симптомом или фенотипом, представляет собой любую связь, которую можно обоснованно установить между ними, в том числе причинную связь или статистически значимую связь, эмпирически установленную связь, предполагаемую связь, независимо от того, вызывает ли она заболевание, нарушение, состояние, симптом или фенотип или нет.
[00514] "Аутоиммунные нарушения", как правило, представляют собой состояния, при которых иммунная система субъекта атакует собственные клетки организма, вызывая разрушение тканей. Аутоиммунные нарушения можно диагностировать с применением исследований крови, анализа спинномозговой жидкости, электромиограммы (измеряет функцию мышц) и магнитно-резонансной томографии головного мозга, однако исследование антител в крови в отношении аутореактивных антител (или аутоантител) является особенно применимым. Обычно с аутоиммунными заболеваниями ассоциирован класс антител IgG.
[00515] "Связывание" описывает взаимодействие между соединениями или молекулами, например, между экзогенным антигеном и мишенью или между экспрессирующей экзогенный антиген EHC и мишенью, которое происходит посредством ковалентного связывания или нековалентного связывания, в том числе ионного связывания, электростатических взаимодействий, образования водородных связей, сил Лондона, вандерваальсовых сил, гидрофобного взаимодействия, липофильных взаимодействий и подобного.
[00516] "Биологическая активность полипептида" относится к любой активности или фенотипу молекулы (как, например, связыванию, сигнальной трансдукции, катализу и т.д.), которые обусловлены полипептидом, таким как экзогенный полипептид антигена.
[00517] Применяемый в данном документе термин "биологический образец" относится к любому типу материала биологического происхождения, выделенному из субъекта, в том числе, например, к ДНК, РНК, липидам, углеводам и белку. Термин "биологический образец" включает ткани, клетки и биологические жидкости, выделенные из субъекта. Биологические образцы включают, например, но без ограничений, цельную кровь, плазму, сыворотку, сперму, слюну, слезы, мочу, фекальный материал, пот, буккальный эпителий, кожу, спинномозговую жидкость, костный мозг, желчь, волосы, биопсию мышц, ткань органов или другой материал биологического происхождения, известный специалистам в данной области. Биологические образцы можно получить, например, из биопсий внутренних органов или из злокачественных опухолей. Биологические образцы можно получить от субъекта для постановки диагноза или исследования или можно получить от здоровых субъектов в качестве контролей или для фундаментального исследования.
[00518] "Скорость клиренса", применяемую в данном документе, рассчитывают путем измерения количества или концентрации, например, экзогенного антигена, мишени-экзогенного антигена или экспрессирующих экзогенный антиген EHC-клеток, сохранившихся в кровеносной системе субъекта через некоторое время. Например, 1%, 2%, 3%, 4%, 5%, 10%, 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95% или 99% мишени, обнаруженной в первом образце, можут все еще обнаруживаться во втором образце, который взят через 1 час, 5 часов, 10 часов, 24 часа, 2 дня, 3 дня, 4 дня, 5 дней, 6 дней, 7 дней, 2 недели, 3 недели, 4 недели, 2 месяца, 3 месяца, 4 месяца, 5 месяцев, 6 месяцев, 7 месяцев, 8 месяцев, 9 месяцев, 10 месяцев, 11 месяцев, 12 месяцев, 2 года, 3 года, 4 года или 5 лет. В качестве альтернативы скорость клиренса может быть выражена как число объектов (например, мишень/экзогенный антиген) в единицу времени (например, в день). Повышением скорости клиренса является скорость, большая, чем проявляется у не подвергнутого лечению или здорового соответствующего контрольного субъекта. Снижением скорости клиренса является скорость, меньшая, чем проявляемая у не подвергнутого лечению или здорового соответствующего контрольного субъекта. Повышение или снижение может составлять 1%, 2%, 3%, 4%, 5%, 10%, 15%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 150%, 200%, 500%, 1000% или может составлять в 1,1 раза, 1,2 раза, 1,3 раза, 1,4 раза, 1,5 раза, 2 раза, 3 раза, 4 раза, 5 раз, 10 раз, 20 раз, 50 раз, 100 раз, 500 раз или 1000 раз.
[00519] "Расщепление", применяемое в данном документе, представляет собой процесс, который разрушает взаимодействие в виде связи, присутствующее в мишени, такой как полипептид или нуклеиновая кислота, например, с образованием двух или большего числа объектов, которые после расщепления могут отделяться друг от друга. Отделение может включать, например, разрушение ионной связи, ковалентной связи, полярной ковалентной связи, неполярной ковалентной связи или металлической связи. При применении расщепления в отношении полипептидных мишеней расщепление может включать распад одной или нескольких пептидных связей. При применении расщепления в отношении низкомолекулярных мишеней расщепление может включать распад одной или нескольких углеродных или сульфидных связей. При применении расщепления в отношении нуклеотидных последовательностей расщепление может включать распад одной или нескольких фосфодиэфирных связей. При применении расщепления в отношении микроорганизмов, таких как бактерии, грибы или вирусы, расщепление может включать лизис мембраны или капсидной структуры. Расщепление может осуществлять фермент, например, каталитически активный полипептид экзогенного антигена. Экзогенные антигены могут предусматривать, например, экзонуклеазную, эндонуклеазную или протеазную активность.
[00520] "Кровеносная система субъекта", применяемая в данном документе, охватывает пространство, занимаемое цельной кровью, и необязательно лимфатическую систему у человека, в том числе плазму и все циркулирующие в крови клетки и молекулы и распространенные по всем артериям, венам, капиллярам и лимфатическим сосудам всей тканей. "Концентрация в кровотоке" представляет собой концентрацию мишени, например, клетки, полипептида (такого как антитело, антиген патогена и т.д.), терапевтического средства, малой молекулы, метаболита или другого объекта, экзогенного антигена или экспрессирующей экзогенный антиген EHC в пространстве, определенном как кровеносная система. В определенных вариантах осуществления концентрация может определяться как число свободных (несвязанных) объектов в указанном объеме. В других вариантах осуществления концентрация может определяться как общее число объектов в указанном объеме.
[00521] Термин "участок, определяющий комплементарность (CDR)", применяемый в данном документе, относится к аминокислотной последовательности, обнаруженной в вариабельной области тяжелой цепи или легкой цепи иммуноглобулина. CDR определяют специфичность антитела и могут обеспечивать контактный остаток для связывания со специфичным эпитопом антигена. Тяжелая цепь и легкая цепь могут соответственно включать по три CDR (CDRH1, CDRH2, и CDRH3, и CDRL1, CDRL2 и CDRL3). Четыре каркасные области, которые имеют более высококонсервативные аминокислотные последовательности, чем CDR, разделяют CDR-участки в VH или VL.
[00522] "Комплекс", как применяется в данном документе, содержит ассоциацию двух или более объектов. Комплекс может содержать один или несколько полипептидов, нуклеиновых кислот, липидов, углеводов, неорганических соединений, органических соединений и т.п. Комплекс может быть функциональным (многокомпонентные полипептиды) или нефункциональным (например, агрегаты или преципитаты) и может обладать полезными или приносящими вред свойствами (например, иммунные комплексы). Комплексы могут быть встречающимися в природе или могут быть полученными человеком или синтетическими. Синтетические комплексы включают объекты высшего порядка, например, субклеточные структуры и клетки, если они содержат синтетическое вещество или молекулу.
[00523] Что касается аминокислотных последовательностей, специалист в данной области будет осознавать, что индивидуальные замены, делеции или добавления в последовательности нуклеиновой кислоты, пептидной, полипептидной или белковой последовательности, которые изменяют, добавляют или удаляют одну аминокислоту или небольшую процентную долю аминокислот в кодируемой последовательности, представляют собой "консервативно модифицированный вариант", когда изменение приводит к замене аминокислоты на химически подобную аминокислоту. Применяемый в данном документе термин "консервативная аминокислотная замена" иллюстрируется заменой аминокислот в пределах каждой из следующих групп: (1) глицин, аланин, валин, лейцин, и изолейцин, (2) фенилаланин, тирозин, и триптофан, (3) серин и треонин, (4) аспарагиновая кислота и глутаминовая кислота, (5) глутамин и аспарагин и (6) лизин, аргинин и гистидин.
[00524] "Снижение" в контексте симптома повергаемого лечению заболевания, нарушения или состояния относится к снижению измеряемых или ощущаемых параметров, ассоциированных с заболеванием или состоянием, которые проявляются как симптомы. Примерами измеряемых параметров является снижение температуры тела субъекта, снижение концентрации мишеней в образце, взятом у субъекта, снижение интенсивности воспаления или размера воспаленной области, снижение числа инфильтрующих клеток, снижение числа эпизодов, ассоциированных с заболеванием, нарушением или состоянием, увеличение/уменьшение размера органа, прибавка/потеря веса и т.д. Примерами ощущаемых параметров является, например, собственная оценка субъектом своего самочувствия и качества жизни. Например, в случае заболевания, опосредованного аутореактивными антителами, снижение можно оценить количественно в виде одного или комбинации следующих параметров: сниженное воспаление, сниженное обострение, сниженная утомляемость, сниженное свертывание крови, сниженная потливость, повышенная активность или увеличенный рост волос и т.д. Параметры, которые можно оценить количественно, представляют собой параметры, подходящие для оценки специфичного заболевания, нарушения или состояния, подлежащего лечению. Отсрочка, в контексте симптомов подвергаемого лечению заболевания, нарушения или состояния относится к значительному продлению управляемого состояния здоровья, которое в ином случае ухудшилось бы с применением лечения.
[00525] "Разрушение" определяется как процесс, при котором мишень либо напрямую, либо непрямым образом снижается, инактивируется, распадается, разлагается, лизируется, растворяется, разламывается, убавляется, нарушается, ослабляется, ухудшается, уменьшается или разделяется на части.
[00526] "Различное происхождение полипептидов" относится к организму или виду, от которого происходит генетическая последовательность, кодирующая пептид, полипептид или его часть. В определенных вариантах осуществления слияние, содержащее полипептиды с различным происхождением полипептидов, может включать полипептид экзогенного антигена, который кодируется генетической последовательностью для человеческой аденозиндезаминазы и генетической последовательностью для фенилаланингидроксилазы из Chromobacterium violaceum.
[00527] "Домен" представляет собой часть полипептида, такого как полипептид экзогенного антигена, которая обычно имеет 3-мерную структуру и может проявлять выраженную активность, функцию, такую как, например, каталитическая, ферментативная функция, структурная роль или функция связывания.
[00528] Под "обогащенная популяция клеток" подразумевают популяцию клеток, которая фактически состоит из конкретной клетки, представляющей интерес. В обогащенной популяции 50% или более клеток в популяции представляют собой клетки, представляющие интерес, например, 50%, 60%, 70%, обычно 80%, 85%, 90%, более обычно 92%, 95%, 96%, 97%, 98% или 99%, иногда вплоть до 100% клеток в популяции. Отделение клеток, представляющих интерес, из сложной смеси или гетерогенной культуры клеток можно осуществлять с помощью любых традиционных средств, известных из уровня техники, например, с помощью методик аффинного разделения, таких как магнитное разделение с применением магнитных гранул, покрытых аффинным реагентом, аффинная хроматография или "пэннинг" с аффинным реагентом, прикрепленным к твердой матрице, например, к планшету, или другая традиционная методика. Другие методики, обеспечивающие точное отделение включают устройства для сортировки флуоресцентно-активированных клеток, которые могут иметь различные степени усложнений, такие как множественные цветные каналы, каналы обнаружения с малым углом и каналы обнаружения со светорассеянием под тупыми углами, каналы сопротивления и т.д. Клетки можно отбирать в сравнении с погибшими клетками путем использования красителей, ассоциированных с погибшими клетками. Можно использовать любую технику, которая не является чрезмерно вредоносной для жизнеспособности требуемых клеток.
[00529] "Удаление ядра" представляет собой перевод клетки в нереплицируемое состояние, либо посредством инактивации, либо удаления ядра.
[00530] "Эпитоп" включает любой сегмент антигена, с которым связывается антитело, или другой лиганд, или связывающая молекула. Эпитоп может состоять из химически активных поверхностных группировок молекул, таких как аминокислоты или боковые цепи сахаров, и он обычно имеет специфичные особенности трехмерной структуры, а также специфичные особенности заряда. В некоторых вариантах осуществления экзогенные антигены содержат специфичные эпитопы. В некоторых вариантах осуществления мишени содержат специфичные эпитопы.
[00531] "Клетки эритроидного ряда", применяемые в данном документе, включают ядерные красные кровяные клетки, предшественники красных кровяных клеток, и безъядерные красные кровяные клетки, и перечисленные в таблице A1. Например, клетки эритроидного ряда представляют собой стволовую клетку пуповинной крови, CD34+ клетку, гемопоэтическую стволовую клетку (HSC), клетку колониеобразующей единицы селезенки (CFU-S), общую миелоидную клетку-предшественник (CMP), бластную колониеобразующую клетку, взрывообразующую единицу эритроидного ряда (BFU-E), эритро-мегакариоцитарную клетку-предшественника (MEP), колониеобразующую единицу эритроидного ряда (CFU-E), ретикулоцит, эритроцит, индуцированную плюрипотентную стволовую клетку (iPSC), мезенхимальную стволовую клетку (MSC), полихромный нормобласт, ортохромный нормобласт или их комбинацию. В некоторых вариантах осуществления клетки эритроидного ряда представляют собой бессмертные или иммортализованные клетки. Например, иммортализованные эритробластные клетки можно получать с помощью ретровирусной трансдукции CD34+ гемопоэтических прогениторных клеток для экспрессии Oct4, Sox2, Klf4, cMyc и супрессии TP53 (например, как описано в Huang et al., Mol Ther 2013, предварительная электронная публикация 3 сентября). Кроме того, клетки могут предполагаться для аутологичного применения или обеспечивать источник для аллогенного переливания. Клетки эритроидного ряда можно приводить в контакт с экзогенным антигеном с получением экспрессирующей экзогенный антиген EHC. Клетки эритроидного ряда, содержащие экзогенный антиген, представляют собой один пример экспрессирующей экзогенный антиген EHC. В некоторых вариантах осуществления клетки эритроидного ряда культивируют. В некоторых вариантах осуществления прогениторные клетки эритроидного ряда приводят в контакт с экзогенным антигеном с получением экспрессирующей экзогенный антиген EHC.
[00532] Применяемый в данном документе термин "клетка тромбоцитарного ряда" относится к клетке из линии дифференцировки стволовая клетка-мегакариоцит-тромбоцит, в том числе, например, к мегакариоцитам и тромбоцитам, или клеткам, которые индуцированы для дифференцировки с помощью тромбопоэтина, или клеткам, которые экспрессируют поверхностные маркеры, ассоциированные с этой линией дифференцировки, например CD41 (GP IIb/IIIa), CD42a (GPIX), CD42b (GPIb) и CD61 (avb3, витронектиновый рецептор), PAC-1 (активированный IIb/IIIa), CD62P (P-селектин), CD31 (PECAM) и CD63.
[00533] Применяемый в данном документе термин "безъядерная гемопоэтическая клетка" (EHC) относится к гемопоэтической клетке, человеческой или организма, не являющегося человеком, которая является безъядерной или которая стала безъядерной, как определено в данном документе. Данное определение охватывает как "клетки эритроидного ряда", так и "клетки тромбоцитарного ряда", как определено в данном документе.
[00534] Применяемый в данном документе термин "наполнитель" относится к инертному веществу, добавляемому в фармацевтическую композицию для дополнительного облегчения введения соединения. Примеры наполнителей включают без ограничений карбонат кальция, фосфат кальция, различные сахара и типы крахмалов, производные целлюлозы, желатин, растительные масла, антикоагулянты и полиэтиленгликоли.
[00535] Термин "экзогенный", как применяется в данном документе, означает клеточный компонент или функцию, выполняемую углеводом, полисахаридом, липидом, олигонуклеотидом или полипептидом, не обнаруживаемым естественным образом в клетке, или улучшение или манипуляцию с клеточным компонентом или функцией, которые являются эндогенными в отношении клетки, в том числе, например, белок слияния, содержащий экзогенный полипептидный антиген, и эндогенный белок или его функциональный фрагмент. Экзогенный антиген, в том числе полипептид экзогенного антигена, является "экзогенным" или "гетерологичным", так что он может не существовать естественным образом, так как представляет собой слияние или химеру, содержащую домены с различным происхождением полипептидов или домены, происходящие от различных видов, он может не встречаться естественным образом во встречающейся в природе клетке, такой как немодифицированный эритроцит или тромбоцит, он может не функционировать также, как функционировал бы встречающийся в природе полипептид, или он может не встречаться в природе в количестве, в котором встречается полипептид экзогенного антигена, например, в вариантах осуществления, в которых экзогенный антиген является сверхэкспрессированным по сравнению с экспрессией встречающегося в природе полипептида в немодифицированной клетке. В некоторых вариантах осуществления полипептид экзогенного антигена экспрессируется за счет экзогенной нуклеиновой кислоты. В некоторых вариантах осуществления экзогенный антиген является выделенным из источника и загруженным в экспрессирующую экзогенный антиген EHC или конъюгированным с ней. Термин "экзогенный", при применении в контексте нуклеиновой кислоты, включает трансген и рекомбинантные нуклеиновые кислоты.
[00536] Применяемый в данном документе термин "экспрессия" или "осуществление экспрессии" относится к процессу получения полипептида, такого как полипептид экзогенного антигена, в том числе транскрипции и трансляции. Экспрессия может представлять собой, например, повышение числа способов реализации, в том числе: повышение числа генов, кодирующих полипептид, повышение транскрипции гена (как, например, путем помещения гена под контроль конститутивного промотора), повышение трансляции гена, нокаут конкурирующего гена или комбинация этих и/или других способов реализации. Термин "экспрессия" или "осуществление экспрессии" также включает EHC-клетки, которые содержат экзогенный полипептид, который в одно время активно экспрессировался EHC, но после этого активная экспрессия (определяемая как транскрипция и трансляция) прекратилась. Например, полипептид экзогенного антигена активно экспрессировался (т.е. транскрибировался и транслировался) в EHC до события удаления ядра, и полипептид антигена сохраняется в EHC после удаления ядра, но более не подвергается активной экспрессии, например, из-за отсутствия кодирующей нуклеиновой кислоты. Например, EHC может содержать полипептид экзогенного антигена, кодируемый экзогенной нуклеиновой кислотой. Во время удаление ядра полипептид экзогенного антигена сохраняется в EHC, тогда как экзогенная нуклеиновая кислота не сохраняется, о такой EHC говорят, что она является "антиген-экспрессирующей" или "экспрессирующей антиген" даже в случае, если активная экспрессия (транскрипция и трансляция) полипептида антигена фактически завершена и/или EHC не содержит значительного количества реплицирующейся нуклеиновой кислоты.
[00537] "Функциональный" экзогенный антиген или экспрессирующая экзогенный антиген EHC проявляет требуемую или обозначенную активность или особенность, в том числе ферментативную, каталитическую или метаболическую активность, структурную целостность, иммуногенную комплементарность, связывание мишени и правильную локализацию, или способен стимулировать требуемый или обозначенный эффект или фенотип.
[00538] "Слияние или химера" представляет собой полипептидную последовательность или соответствующую кодирующую нуклеотидную последовательность, которая происходит из комбинации двух или более последовательностей, которые не встречаются вместе в природе. Она может представлять собой комбинацию отдельных последовательностей, происходящих из отдельных генов в пределах одного генома, или из гетерологичных генов, происходящих из геномов определенно отличающихся видов.
[00539] "Генетический материал" относится к молекулам нуклеиновой кислоты с нуклеотидными последовательностями из аденозина, тимина, урацила, цитозина и гуанина, способными кодировать ген.
[00540] Термин "тяжелая цепь", применяемый в данном документе, подразумевает включение полноразмерной тяжелой цепи, включающей вариабельную область (VH) с аминокислотными последовательностями, которые определяют специфичность в отношении антигенов, и константную область с тремя константными доменами (CH1, CH2 и CH3), и ее фрагментов. Кроме того, термин "легкая цепь", применяемый в данном документе, подразумевает включение полноразмерной легкой цепи, включающей вариабельную область (VL) с аминокислотными последовательностями, которые определяют специфичность в отношении антигенов, и константную область (CL), и ее фрагментов.
[00541] Термин "гомолог" обозначает полипептиды, в том числе полипептид экзогенного антигена, который имеет такие же или консервативные остатки в соответствующем положении в их первичной, вторичной или третичной структуре. Функциональные гомологи включают экзогенные антигены и другие полипептиды, которые проявляют подобную функцию и/или специфичность (например, в отношении конкретной мишени).
[00542] Встречающееся в природе интактное антитело или иммуноглобулин включает четыре полипептида: две полноразмерные легкие цепи и две полноразмерные тяжелые цепи, в котором каждая легкая цепь связана с тяжелой цепью дисульфидными связями. Каждая тяжелая цепь имеет константную область и вариабельную область. Аналогично, каждая легкая цепь имеет константную область и вариабельную область. Имеется пять классов (изотипов) тяжелых цепей: гамма (γ), мю (μ), альфа (α), дельта (δ) или эпсилон (ε), и дополнительно несколько подклассов гамма 1 (γ1), гамма 2 (γ2), гамма 3 (γ3), гамма 4 (γ4), альфа 1(α1) и альфа 2(α2). Константная область легкой цепи может иметь тип либо каппа (κ), либо лямба (λ). Вариабельные области отличаются последовательностью у разных антител и используются в связывании и определяют специфичность данного антитела в отношении его конкретного антигена.
[00543] Применяемые в данном документе термины "повышать", "усиливать", "стимулировать" и/или "индуцировать" (и подобные термины) в целом относятся к действию по увеличению или повышению, либо напрямую, либо опосредованно, концентрации, уровня, функции, активности или поведения по сравнению с природным, ожидаемым или средним или по сравнению с контрольным состоянием.
[00544] Применяемый в данном документе термин "ингибировать", "подавлять", "уменьшать", "препятствовать" и/или "снижать" (и подобные термины) в целом относится к действию по снижению, либо напрямую, либо опосредованно, концентрации, уровня, функции, активности или поведения по сравнению с природным, ожидаемым или средним или по сравнению с контрольным состоянием.
[00545] "Библиотека", применяемая в данном документе, включает набор молекул нуклеиновой кислоты (например, ДНК, РНК), имеющих различные последовательности нуклеиновой кислоты, набор генетически различных клонов, набор из различных полипептидов, набор из различных клеток, таких как EHC-клетки, и т.д.
[00546] Применяемый в данном документе "субъект-млекопитающее" включает всех млекопитающих, в том числе без ограничений, человека, домашних животных (например, собак, кошек и т.п.), сельскохозяйственных животных (например, коров, овец, свиней, лошадей и т.п.) и лабораторных животных (например, обезьян, крыс, мышей, кроликов, морских свинок и т.п.). Термины "индивидуум", "субъект", "хозяин" и "пациент" применяются в данном документе взаимозаменяемо и относятся к любому субъекту-млекопитающему, в отношении которого требуется постановка диагноза, лечение или терапия, в частности, к человеку. Способы, описанные в данном документе, применимы как для терапии человека, так и для применений в ветеринарии. В некоторых вариантах осуществления субъект представляет собой млекопитающее, и в других вариантах осуществления субъектом является человек.
[00547] "Медицинское устройство" относится к любому устройству, прибору или аппарату, применяемому для доставки дозы экспрессирующей экзогенный антиген EHC и/или терапевтического средства. Оно включает контейнеры, бутыли, флаконы, шприцы, пакеты, картриджи, кассеты, обоймы, цилиндры или канистры.
[00548] "Медицинский набор" относится к упакованной единице, которая включает медицинское устройство или аппликатор, соответствующую дозировку экспрессирующей экзогенный антиген EHC, необязательно включающей терапевтическое средство и соответственные этикетки и инструкции.
[00549] Применяемый в данном документе термин "модулировать", "осуществлять модулирование", "модифицировать" и/или "модулятор" в целом относится к способности изменять, путем повышения или уменьшения, например, напрямую или опосредованно активировать/стимулировать/повышающе регулировать или мешать/ингибировать/понижающе регулировать специфичную концентрацию, уровень, экспрессию, функцию или поведение, так, например, действовать как антагонист или агонист. В некоторых случаях модулятор может повышать и/или уменьшать определенную концентрацию, уровень, активность или функцию по сравнению с контролем, или по сравнению со средним уровнем активности, который в целом ожидался бы, или по сравнению с контрольным уровнем активности.
[00550] "Мембрана", как применяется в данном документе, представляет собой пограничный слой, который отделяет внутреннее пространство от наружного пространства, содержащий одно или несколько биологических соединений, как правило, липиды, и необязательно полипептиды. Мембраны могут представлять собой липидные бислои. В определенных вариантах осуществления мембраны содержат одно или несколько из фосфатидилхолина, сфингомиелина, лизофосфатидилхолина, фосфатидилэтаноламина, фосфатидилсерина, фосфатидилинозитола или фосфатидной кислоты. В некоторых вариантах осуществления мембраны содержат один или несколько полипептидов, таких как анкирин и кофермент Q10. В определение мембраны включаются клеточные мембраны, содержащие, например, фосфолипидный бислой и полипептиды, ассоциированные с клеточной мембраной.
[00551] Фраза "молекула нуклеиновой кислоты" относится к одно- или двухнитевому полимеру из дезоксирибонуклеотидных или рибонуклеоидных оснований. Она включает хромосомную ДНК и самореплицирующиеся плазмиды, векторы, мРНК, тРНК, siRNA и т.д., которые могут быть рекомбинантными и за счет которых могут экспрессироваться экзогенный полипептиды, когда нуклеиновая кислота введена в клетку.
[00552] Ортологи определяются как гены у различных видов, которые возникли от общего предкового гена в ходе видообразования.
[00553] Термин "фармацевтически приемлемый" и его грамматические варианты относится к композициям, носителям, разбавителям и реагентам, допускающим введение субъекту или нанесение на субъекта без выработки нежелательных физиологических эффектов в такой степени, которая воспрепятствовала бы введению композиции. Например, "фармацевтически приемлемый наполнитель" включает наполнитель, который применим в получении фармацевтической композиции, то есть является, в целом, безопасным, нетоксичным и желательным, и включает наполнители, которые подходят для применения в ветеринарии, а также для применения в фармацевтических препаратах для человека. Такие наполнители могут быть твердыми, жидкими, мягкими или, в случае аэрозольной композиции, газообразными.
[00554] Применяемый в данном документе термин "фармацевтически приемлемый носитель" включает любой из стандартных фармацевтических носителей, такой как фосфатно-солевой буфер, вода, эмульсии, такие как масло/вода или вода/масло, и различные типы смачивающих средств. Термин также охватывает любое из средств, одобренных регулирующим органом правительства США или перечисленных в фармакопее США для применения на животных, в том числе человеке, а также любой носитель или разбавитель, который не вызывает значительного раздражения у субъекта и не нейтрализует биологическую активность и свойства введенного соединения.
[00555] Некоторые средства можно вводить в виде "фармацевтически приемлемой соли", например, полученные с помощью неорганических и органических кислот. Неорганические кислоты для получения солей включают соляную кислоту, бромистоводородную кислоту, серную кислоту, азотную кислоту, фосфорную кислоту и т.п. Органические кислоты для получения солей включают уксусную кислоту, пропионовую кислоту, гликолевую кислоту, пировиноградную кислоту, щавелевую кислоту, яблочную кислоту, малоновую кислоту, янтарную кислоту, малеиновую кислоту, фумаровую кислоту, винную кислоту, лимонную кислоту, бензойную кислоту, коричную кислоту, миндальную кислоту, метансульфоновую кислоту, этансульфоновую кислоту, п-толуолсульфоновую кислоту, салициловую кислоту и т.п. Соли иногда также получают с помощью неорганических и органических оснований. Соли, полученные с помощью неорганических оснований, включают, исключительно для примера, соли натрия, калия, лития, аммония, кальция и магния. Соли, полученные с помощью органических оснований, включают без ограничений соли первичных, вторичных и третичных аминов. Любой рядовой специалист в данной области будет знать, как выбрать правильный фармацевтически приемлемый носитель, его фармацевтически приемлемую соль для реализации настоящего изобретения без излишнего проведения экспериментов.
[00556] Применяемый в данном документе термин "фармацевтическая композиция" относится к одному или нескольким соединениям, описанным в данном документе, таким как, например, экспрессирующая экзогенный антиген EHC, смешанная, или перемешанная с, или суспендированная в одном или нескольких других химических компонентах, таких как физиологически приемлемые носители и наполнители. Одним назначением фармацевтической композиции является облегчение введения соединения субъекту.
[00557] В определенных вариантах осуществления представлены различные полипептидные молекулы с последовательностями, ассоциированными с требуемой функцией или активностью, такие как полипептиды экзогенного антигена. Полипептид представляет собой термин, который относится к цепи из аминокислотных остатков, независимо от посттрансляционной модификации (например, фосфорилирования или гликозилирования) и/или образования комплексов с дополнительными полипептидами, синтеза в многокомпонентных комплексах с нуклеиновыми кислотами и/или углеводами, или другими молекулами. Протеогликаны, следовательно, также упоминаются в данном документе как полипептиды. В определенных вариантах осуществления экспрессирующая экзогенный антиген EHC содержит полипептидный экзогенный антиген. В определенных вариантах осуществления экспрессирующая экзогенный антиген EHC содержит один или несколько полипептидов, не относящихся к экзогенному антигену, которые необязательно ассоциированы с мембраной.
[00558] Термин "фармацевтически активное средство" или "фармацевтическое средство" определяется как любое соединение, например, низкомолекулярное лекарственное средство или биологическое средство (например, полипептидное лекарственное средство или лекарственное средство на основе нуклеиновой кислоты), которое при введении субъекту оказывает измеряемый или ощущаемый эффект в отношении субъекта, например, оно ослабляет или уменьшает симптом заболевания, нарушения или состояния. В некоторых вариантах осуществления фармацевтическое средство может вводиться перед, в комбинации с или после доставки экспрессирующей экзогенный антиген EHC. В некоторых вариантах осуществления фармацевтически активное средство оказывает синергетический лечебный эффект с экспрессирующей экзогенный антиген EHC. В некоторых вариантах осуществления фармацевтически активное средство оказывает аддитивный лечебный эффект с экспрессирующей экзогенный антиген EHC.
[00559] "Промотор" определяется как набор последовательностей контроля нуклеиновой кислоты, который управляет транскрипцией функционально связанной нуклеиновой кислоты. Промоторы включают необходимые последовательности нуклеиновой кислоты вблизи сайта инициации транскрипции. Промотор также необязательно включает дистальные энхансерные или репрессорные элементы. "Конститутивный" промотор представляет собой промотор, который является активным в большинстве условий окружения и состояний развития. "Индуцируемый" промотор представляет собой промотор, который активен при регуляции условиями окружения или состояниями развития. Термин "функционально связанный" относится к функциональной связи между последовательностью контроля экспрессии нуклеиновой кислоты (такой как промотор, или набор сайтов связывания факторов транскрипции) и второй последовательностью нуклеиновой кислоты, где последовательность контроля экспрессии управляет транскрипцией нуклеиновой кислоты, соответствующей второй последовательности.
[00560] "Экзогенный антиген", как применяется в данном документе, представляет собой объект, способный взаимодействовать с мишенью, например, ассоциироваться или связываться с мишенью. Экзогенный антиген может содержать или состоять фактически из полипептида. В некоторых вариантах осуществления экзогенный антиген содержит полипептид, углевод, нуклеиновую кислоту, липид, малую молекулу или их комбинацию. В вариантах осуществления, в которых экзогенный антиген представляет собой встречающееся в природе соединение или молекулу, антиген является "экзогенным" в том смысле, что он представляет собой экзогенное или гетерологичное соединение или молекулу относительно их присутствия в EHC. В других вариантах осуществления антиген является "экзогенным" в том смысле, что он представляет собой созданное человеком соединение или молекулу, такие как слияние или химера, не встречающийся в природе полипептид, углевод, нуклеиновая кислота, липид или их комбинация, или созданную человеком малую молекулу или другое терапевтическое средство. Например, экзогенный антиген может содержать слияние или химеру, содержащую один или несколько из S-домена, A-домена и U-домена. S-домен представляет собой поверхностный домен, подверженный воздействию окружающей среды вокруг EHC, такой как кровеносная система субъекта. A-домен представляет собой якорный домен, который прикрепляет S-домен к клеточной мембране EHC. U-домен обращен к не подвергающейся воздействию стороне EHC или расположен внутри ее (т.е. в ее внутриклеточном пространстве). Вне зависимости от какого-либо домена экзогенный антиген может быть расположен на поверхности экспрессирующей экзогенный антиген EHC или может быть расположен внутри EHC. Экзогенный антиген может быть ассоциирован с мембраной экспрессирующей экзогенный антиген EHC, например, экзогенный антиген заякорен в мембране, конъюгирован с ней или иным способом связан с мембраной. В некоторых вариантах осуществления экзогенный антиген может быть конъюгирован с мембраной экспрессирующей экзогенный антиген EHC с помощью химической или ферментативной конъюгации. В других вариантах осуществления экзогенный антиген не конъюгирован с мембраной. В некоторых вариантах осуществления экзогенный антиген не ассоциирован с мембраной экспрессирующей экзогенный антиген EHC и расположен внутри заключенного в мембрану внутриклеточного пространства EHC. В некоторых вариантах осуществления экзогенный антиген, расположенный внутри внутриклеточного пространства EHC, подвергаться незначительной диффузии из EHC и/или может не проникать через мембрану. В других вариантах осуществления экзогенный антиген может подвергаться значительной диффузии из EHC и/или может проникать через мембрану. В некоторых вариантах осуществления экзогенный антиген загружают, например, вводят внутрь или помещают на EHC. Экзогенный антиген, который загружают, не синтезируется биологическим путем в экспрессирующей экзогенный антиген EHC. Экзогенный антиген, пригодный для загрузки, например, может продуцироваться в клеточной системе экспрессии, быть выделенным из биологического образца, синтезированным химически или ферментативно и затем загруженным в EHC или на нее. В некоторых вариантах осуществления экзогенный антиген может быть дополнительно модифицирован экспрессирующей экзогенный антиген EHC после загрузки. В других вариантах осуществления экзогенный антиген не модифицируется после загрузки. В некоторых вариантах осуществления полипептид экзогенного антигена не загружают на EHC или в нее. В некоторых вариантах осуществления экзогенный антиген производится, например, синтезируется биологическим путем в экспрессирующей экзогенный антиген EHC. Как правило, полипептид экзогенного антигена экспрессируется в экспрессирующей экзогенный антиген EHC за счет экзогенной молекулы нуклеиновой кислоты (например, ДНК или мРНК), которая была введена в EHC. Экзогенный антиген может обладать биологической функцией, которая сохраняется, когда антиген экспрессируется на EHC. Экзогенный антиген может связываться с мишенью и/или изолировать ее. В качестве альтернативы или дополнительно экзогенный антиген может проявлять каталитическую активность в отношении мишени, например, экзогенный антиген может превращать или модифицировать мишень, или может разрушать мишень. Затем продукт необязательно может высвобождаться из экзогенного антигена.
[00561] "Пребывание" экспрессирующей экзогенный антиген EHC относится периоду времени, которое она проводит в физиологическом местонахождении. Специфическое местонахождение экспрессирующей экзогенный антиген EHC может меняться в ходе ее жизни и "пребывание" применимо к периоду времени, проведенному в различном окружении, в том числе в сосудистом русле, периферических тканях, капиллярах, пищеварительной системе, дыхательной системе, тканях носа, поверхности эпидермиса и интерстициальной ткани. В конкретных вариантах осуществления экспрессирующая экзогенный антиген EHC пребывает в кровеносной системы субъекта.
[00562] "Реплицирующаяся нуклеиновая кислота" относится дезоксирибонуклеиновой кислоте (ДНК), которая может подвергаться копированию под действием ферментов, предназначенных для увеличения числа копий ДНК. Обычно, репликация ДНК приводит к образованию двух идентичных копий из одной оригинальной молекулы ДНК. Репликация ДНК относится к поочередному включению нуклеотидов, совпадающих с нитью-матрицей, в растущую нить ДНК под действием ДНК-полимеразы посредством образования фосфодиэфирных связей.
[00563] "Изоляция" определяется как заточение, закрывание, отделение, разделение, скрывание, ограждение или выделение мишени и предотвращение ее свободного взаимодействия с окружающей средой.
[00564] "Специфичное связывание" или "специфичное взаимодействие", как применяется в данном документе, описывает любое взаимодействие между двумя объектами (например, мишень и экзогенный антиген, как, например, антитело и антиген, рецептор и лиганд, фермент и субстрат, биотин и авидин и т.д.), которое характеризуется насыщаемостью, зачастую обратимостью и, таким образом, является конкурентным, как эти термины понимают обычные специалисты в области химии и биохимии. Например, специфичное связывание с участием биологических молекул, таких как, например, белки, пептиды и нуклеиновая кислота, происходит, когда один элемент пары связывания имеет участок, имеющий такую форму и распределение заряженных, полярных или гидрофобных компонентов, что взаимодействие родственного лиганда с таким участком отличается благоприятными энергетическими характеристиками (т.е. отрицательной свободной энергией связывания). Специфичность взаимодействия можно измерить или выразить как константу связывания (Kd). Kd может варьировать от диапазона мМ до диапазона фМ, включая диапазон пМ, диапазоны мкМ и диапазоны нМ. Типичные значения Kd составляют ниже приблизительно 10-6 M, ниже приблизительно 10-7 M, ниже приблизительно 10-8 M и в некоторых вариантах осуществления ниже приблизительно 10-9 M.
[00565] Применяемый в данном документе термин "значительно" или "значительный" относится, например, к присутствию, уровню или концентрации объекта в конкретном пространстве, эффекту одного объекта в отношении другого объекта или эффекту лечения. Например, активность, уровень или концентрация объекта значительно повышены, если повышение составляет в 2 раза, 3 раза, 4 раза, 5 раз, 10 раз, 50 раз, 100 раз или 1000 раз по сравнению с исходным уровнем. Активность, уровень или концентрация объекта также значительно повышены, если повышение составляет 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 200% или 500% по сравнению с исходным уровнем. Объект может в значительной степени присутствовать в конкретном пространстве, если его можно обнаружить с помощью способов, известных из уровня техники. Объект может в незначительной степени присутствовать в конкретном пространстве, если он присутствует на уровне ниже предела обнаружения для анализов и способов, известных из уровня техники. В некоторых вариантах осуществления объект может не присутствовать в значительной степени в конкретном пространстве, если он едва обнаруживается, но только в нефункциональных количествах или минимальных количествах, так что он не обусловливает или не изменяет фенотип. В других вариантах осуществления объект может не присутствовать в значительной степени в конкретной популяции, если он присутствует и может быть обнаружен только на небольшой части элементов, составляющих популяцию, например, менее 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3% 2% или менее 1%, 0,5%, 0,1% элементов популяции. Например, экзогенная нуклеиновая кислота может не сохраняться после удаления ядра, клетка становится не реплицирующейся, и безъядерная клетка неспособна продолжать экспрессию полипептида экзогенного антигена, кодируемого экзогенной нуклеиновой кислотой. Утрата способности клетки к продолжению значительной степени трансляции экзогенного полипептида "фактически прекращает" экспрессию белка. В определенных вариантах осуществления экспрессирующая экзогенный антиген EHC в значительной степени неспособна к воспроизведению, например, репликации нуклеиновых кислот. Например, экспрессирующая экзогенный антиген EHC не может в значительной степени включать нуклеозид при контакте с меченным нуклеозидом, таким как тимидин, в анализе включения. В некоторых вариантах осуществления экспрессирующая экзогенный антиген EHC не содержит значительное количество самореплицирующихся нуклеиновых кислот. Термин "значительная идентичность" полинуклеотидных последовательностей или последовательностей нуклеиновой кислоты означает, что полинуклеотид содержит последовательность, которая характеризуется по меньшей мере 25% идентичностью последовательности. В качестве альтернативы, процент идентичности может составлять любое целое число от 25% до 100%. Более предпочтительные варианты осуществления включают по меньшей мере 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 99% в сравнении с эталонной последовательностью с применением программ, описанных в данном документе; преимущественно BLAST с применением стандартных параметров.
[00566] "Синтетический" относится к соединению или молекуле, которая является либо созданной человеком, либо не встречающейся в природе или, если она является встречающейся в природе, помещена в условия или местоположение, в которых она не существовала бы естественным образом, или, если она существует естественным образом в определенных условиях или местоположении, находящуюся в состоянии чистоты, или присутствующую в количестве, концентрации или числе, в которых она бы не присутствовала естественным образом в определенных условиях или местоположении. Синтетические объекты могут быть выделенными или очищенными соединениями, которые необязательно химически или ферментативно модифицированы в сравнении с их природным состоянием, экзогенными нуклеиновыми кислотами, экзогенными (гетерологичными) антигенами и т.п. Присутствие синтетического соединения или молекулы, как определено в данном документе, в любом объекте придает целому объекту характеристику "синтетический". Например, клетка, содержащая экзогенный антиген, является синтетической клеткой.
[00567] "Мишень", используемая в данном документе, представляет собой объект, способный взаимодействовать с экзогенным антигеном, например, ассоциироваться или связываться с экзогенным антигеном. "Мишень" включает без ограничений полипептид (например, антитело или родственный антителу полипептид, элемент комплемента, амилоидный белок, патоген, токсин, прион), молекулу (например, метаболит, стероид, гормон, углевод; олигосахарид; химическое вещество; полисахарид, ДНК; РНК; липид, аминокислоту, химический элемент, токсин или патоген), комплекс (например, иммунный комплекс) или клетку (например, раковая клетка, макрофаг, бактерия, гриб, вирус или паразит). Подразумевается, что мишень обнаруживается, диагностируется, ослабляется, разрушается или изменяется (например, функционально комплементируется) с помощью способов, представленных в данном документе. Специфичная мишень может быть несвязанной или ассоциированной с другими объектами в кровеносной системе субъекта.
[00568] "Аутореактивное антитело-мишень", применяемой в данном документе, представляет собой аутореактивное антитело, ассоциированное с аутоиммунным заболеванием. Такие аутореактивные антитела можно обнаруживать и анализировать с применением исследований связывания антитела, предусматривающих приведение антител субъекта в контакт с образцами собственной ткани субъекта, обычно ткани щитовидной железы, желудка, печени и почки. Антитела, связывающиеся с "собственной" тканью (содержащей собственные антигены), указывают на аутоиммунное нарушение.
[00569] "Трансген" или "экзогенная нуклеиновая кислота" относятся к чужеродной или нативной нуклеотидной последовательности, которая введена в EHC. Трансген и экзогенная нуклеиновая кислота применяются взаимозаменяемо в данном документе и охватывают рекомбинантные нуклеиновые кислоты.
[00570] Применяемые в данном документе "лечить", "осуществлять лечение" и/или "лечение" представляют собой подход для получения благоприятных или требуемых клинических результатов, фармакологического и/или физиологического эффекта, например, облегчение симптомов, предотвращение или устранение таких симптомов, и относятся как к терапевтическому лечению, так и к профилактическому или предупредительному лечению определенного заболевания, нарушения или состояния. Благоприятные или требуемые клинические результаты, фармакологический и/или физиологический эффект включают без ограничений предотвращение заболевания, нарушения или состояния от появления у субъекта, который может быть предрасположен к данному заболеванию, нарушению или состоянию, но который еще не испытал или у которого еще не проявились симптомы заболевания (профилактическое лечение), облегчение симптомов заболевания, нарушения или состояния, уменьшение выраженности заболевания, нарушения или состояния, стабилизация (т.е. отсутствие ухудшения) заболевания, нарушения или состояния, предотвращение распространения заболевания, нарушения или состояния, отсрочка или замедление прогрессирования заболевания, нарушения или состояние, уменьшение интенсивности или временное ослабление заболевания, нарушения или состояния и их комбинации, а также продление выживания по сравнению с ожидаемым выживанием в отсутствие лечения.
[00571] "Терапевтическое средство" или "терапевтическая молекула" включает соединение или молекулу, которая, если присутствует в эффективном количестве, приводит к требуемому терапевтическому эффекту, фармакологическому и/или физиологическому эффекту в отношении субъекта, нуждающегося в этом.
[00572] Термин "терапевтически эффективное количество" или "эффективное количество" представляет собой количество средства, вводимого субъекту, которое является достаточным для достижения благоприятных или требуемых клинических результатов, фармакологических и/или физиологических эффектов. Эффективное количество можно вводить за одно или несколько введений. Эффективное количество, как правило, является достаточным для временного ослабления, уменьшения интенсивности, стабилизации, обратного развития, замедления или отсрочки прогрессирования болезненного состояния. Таким образом, эффективное количество относится к количеству средства или частоте введения определенного количества средства, которые являются достаточными для обоснованного достижения требуемого терапевтического и/или профилактического эффекта. Например, оно может включать количество, которое приводит к предотвращению, лечению или снижению симптомов, ассоциированных с заболеванием или состоянием, которое подвергается лечению, например, заболеванием или медицинскими состояниями, ассоциированными с аутоиммунным ответом, сверхактивной активацией иммунной системы, или образованием ингибиторных антител, в отношении которых требуется иммунологическая толерантность, или заболеванием, или медицинскими состояниями, ассоциированными с полипептидом-мишенью. Количество терапевтической композиции, вводимой субъекту, будет зависеть от типа и тяжести заболевания и от характеристик индивидуума, таких как общее состояние здоровья, патологические состояния, диета, возраст, пол, масса тела и переносимость лекарственных средств. Оно будет также зависеть от степени, тяжести и типа заболевания. Кроме того, эффективное количество будет зависеть от применяемых способов составления и введения, например, времени введения, пути введения, скорости выведения и чувствительности реакции. Специалист в данной области будет способен определить подходящие дозировки в зависимости от этих и других факторов. Композиции также можно вводить в комбинации с одним или несколькими дополнительными терапевтическими соединениями. Требуемая дозировка фармацевтической композиции может находиться в диапазоне от приблизительно 0,001 до 100 мг/кг для взрослого организма. В одном примере внутривенное введение начинают при дозе, которая является минимально эффективной, и дозу повышают в течение предварительно установленного периода времени вплоть до наблюдения положительного эффекта. Впоследствии выполняют пошаговое повышение дозировки, ограничиваясь уровнями, которые приводят к соответствующему повышению эффекта, при этом учитывая любые побочные эффекты, которые могут возникнуть. Неограничивающие примеры соответствующих дозировок могут варьировать, например, от 1×1010 до 1×1014, от 1×1011 до 1×1013 или от 5×1011 до 5×1012 экспрессирующих экзогенный антиген EHC-клеток по настоящему изобретению. Конкретные примеры включают приблизительно 5×1010, 6×1010, 7×1010, 8×1010, 9×1010, 1×1011, 2×1011, 3×1011, 4×1011, 5×1011, 6×1011, 7×1011, 8×1011, 9×1011, 1×1012 или болееэкспрессирующих экзогенный антиген EHC-клеток по настоящему изобретению. Каждую дозу экспрессирующих экзогенный антиген EHC-клеток можно вводить с интервалами, такими как один раз в день, один раз в неделю, два раза в неделю, один раз в месяц или два раза в месяц.
[00573] "Несвязанный" относится к состоянию мишени, с которой способен взаимодействовать экзогенный антиген. Несвязанная мишень не ассоциирована с другим объектом или экзогенным антигеном. Несвязанный экзогенный антиген не ассоциирован с другим объектом или мишенью. Мишень считается "связанной", когда она ассоциирована с экзогенным антигеном или другим объектом. Несвязанные мишени включают растворимые формы мишени в кровотоке. Связанные мишени включают мишени, которые внедрены, ассоциированы, связаны или иным образом взаимодействуют с объектами в кровотоке или периферической ткани. Объекты, с которыми может взаимодействовать мишень, включают циркулирующие в крови клетки, периферический эндотелий, иммунные комплексы, гликолипиды, микроорганизмы, иммуноглобулины, сывороточный альбумин, факторы свертывания, липопротеины и электролиты.
[00574] "Вариант" представляет собой полипептид, который отличается от оригинального белка по одной или нескольким аминокислотным заменам, делециям, вставкам или другим модификациям. Эти модификации не приводят к значительному изменению биологической активности оригинального белка. Во многих случаях вариант сохраняет по меньшей мере 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95% или 100% биологической активности оригинального белка. Биологическая активность варианта также может быть выше активности оригинального белка. Вариант может быть встречающимся в природе, таким как произошедшим в результате аллельной вариации или полиморфизма, или быть полученным с помощью способов генной инженерии специально.
[00575] Аминокислотная последовательность варианта в значительной степени идентична последовательности оригинального белка. Во многих вариантах осуществления вариант разделяет по меньшей мере 50%, 60%, 70%, 80%, 85%, 90%, 95%, 99%, или более глобальной идентичности или сходства последовательности с оригинальным белком. Идентичность или сходство последовательности можно определять с применением различных способов, известных из уровня техники, таких как средство основного локального выравнивания (BLAST), анализ точечных матриц или способ динамического программирования. В одном примере идентичность или сходство последовательности определяют с применением GAP (алгоритм Нидлмана-Вунша) из программ Genetics Computer Group (GCG). Аминокислотные последовательности варианта и оригинального белка могут быть в значительной степени идентичными в одной или нескольких областях, но отличаться в других областях.
[00576] Применяемый в данном документе термин "вектор" представляет собой молекулу нуклеиновой кислоты, предпочтительно самореплицирующуюся, которая переносит и/или реплицирует вставленную молекулу нуклеиновой кислоты, такую как трансген или экзогенная нуклеиновая кислота в клетки-хозяева и/или между ними. Он включает плазмиду или вирусную хромосому, в геном которых вставлена рекомбинантная ДНК и которые применяются для введения рекомбинантной ДНК или трансгена в EHC.
Примеры
[00577] Следующие примеры представлены в целях иллюстрации, а не в целях ограничения.
Пример 1. Культура клеток эритроидного ряда с экспрессией гетерологичного гена
[00578] Клетки CD34 выделяли из периферической крови посредством отбора с супермагнитными микрогранулами с применением колонок Mini-MACS (Miltenyi Biotec; чистота 94% ± 3%). Клетки культивировали в среде для индукции эритроидной дифференцировки (EDM) на основе IMDM, дополненной стабилизированным глутамином, 330 мкг/мл человеческого голотрансферрина, 10 мкг/мл рекомбинантного человеческого инсулина, 2 МЕ/мл гепарина и 5% плазмы крови, вирусинактивированной с помощью растворителя/детергента. Процедура размножения клеток включала 3 стадии. На первой стадии (день 0 - день 7) 104/мл клеток CD34 культивировали в EDM в присутствии 1 мкM гидрокортизона, 100 нг/мл SCF, 5 нг/мл IL-3 и 3 МЕ/мл Epo. В день 4 1 объем культуры клеток разводили в 4 объемах свежей среды, содержащей SCF, IL-3, Epo и гидрокортизон. На второй стадии (день 7 - день 11) клетки ресуспендировали из расчета 10^5/мл в EDM, дополненной SCF и Epo. На третьей стадии (день 11 - день 18) клетки культивировали в EDM, дополненной только Epo. Количество клеток доводили до 7,5×105-1×106 и 5-10×106 клеток/мл в дни 11 и 15, соответственно. После дня 18 культуральную среду, содержащую Epo, обновляли дважды в неделю. Культуры поддерживали при 37°C с 5% CO2 в воздухе.
[00579] Кодирующую последовательность антигена, представляющего интерес, помещали под контроль промотора, специфичного для клеток эритроидного ряда, например, GATA-1, и в конец помещали поли(А)-хвост, см., например, Repik et al., Clin Exp Immunol 2005, 140:230. Данная последовательность закодирована в лентивирусном векторе (например, EF1, System Biosciences, Inc.). Вектор получали с помощью стандартных способов в клетках 293T. Лентивирусный вектор трансдуцировали в человеческую гемопоэтическую клетку-предшественник, например, CD34+ клетку, или иммортализованный эритробласт, или iPS-клетку, например, как описано в Chang et al., Nat Biotechnol 2006, 24:1017, в течение дней 1-4 культивирования. Последующие стадии размножения и дифференцировки выполняли, как описано выше.
Пример 2. Загрузка белка в клетку эритроидного ряда посредством циклов с гипотоническими/гипертоническими условиями
[00580] Антиген, в данном случае OVA, добавляли в суспензию RBC с гематокритом (Hct), составляющим 70%, при конечной концентрации 0,5 или 5 мг/мл. После процесса гипотонического диализа (50 мосмоль/кг) обеспечивали закрытие пор в RBC путем добавления гипертонического раствора (1900 мосмоль/кг) при 37°C течение 30 минут. OVA-загруженные RBC промывали четыре раза с помощью 0,9% NaCl+0,2% раствора глюкозы, центрифугировали при 1000×g в течение 10 мин при 4°C и доводили до Hct 50% плазмой крови или буфером.
Пример 3. Мечение клеточной поверхности с помощью гетеробифункционального сшивающего средства
[00581] Антиген c открытой восстановленной тиольной группой (антиген-SH) получали путем инкубации с 5 мМ TCEP. Восстанавливающее средство удаляли с помощью эксклюзионной хроматографии перед проведением конъюгации. Клетки инкубировали с малеимид-PEG-NHS-сшивающим средством, например, SM(PEG)x (Thermo Scientific) в течение 30 минут. Группа NHS вступала в реакцию со свободными аминогруппами на антигенах клеточной поверхности. Прореагировавшие клетки промывали в PBS для удаления избытка сшивающего средства. Раствор антиген-SH вводили в раствор клеток и сшивающего средства и оставляли для протекания реакции малеимид-SH на 30 минут. Клетки осаждали центрифугированием и промывали для удаления несвязанного антигена.
Пример 4. Мечение клеточной поверхности с помощью сортазы
[00582] Клетки, экспрессирующие поверхностные белки, которые содержат акцепторную последовательность для сортазы A, LPXTG, инкубировали со 100 мкM сортазы A в реакционном буфере, состоящем из 50 мМ Tris-Cl, pH 7,5, 150 мМ NaCl, 1 мМ CaCl2, в течение 4 ч при 37°C. Вводили 10x молярный избыток нуклеофила, GGG-антигена, и реакцию оставляли протекать при комнатной температуре в течение ночи. После завершения реакции избыток сортазы и непрореагировавший нуклеофил удаляли путем осаждения и промывания клеток.
Пример 5. Мечение клеточной поверхности с применением клик-химии
[00583] Осуществляли реакцию клеток со сложным эфиром алкин-NHS для конъюгации алкиновой группы с открытыми первичными аминами на клеточной поверхности в соответствии с инструкциями производителя (например, Glen Research). Избыток сложного эфира алкин-NHS удаляли путем осаждения и промывания клеток. Антиген вводили в реакцию со сложны эфиром азид-NHS для конъюгации азидной группы с открытыми первичными аминами на антигене в соответствии с инструкциями производителя (например, Thermo Scientific). Избыток сложного эфира азид-NHS удаляли с помощью эксклюзионной хроматографии. Проводили реакцию между клетками с алькиновыми группами и антигеном с азидной группой посредством циклоприсоединения, катализируемого медью, при комнатной температуре.
Пример 6. Измерение пролиферации Т-клеток у мыши
[00584] CD8+ Т-клетки из селезенок мышей OTI (CD45.2+) выделяли с применением набора для отрицательного отбора CD8 с магнитными микрогранулами (Miltenyi Biotec) в соответствии с инструкциями производителя. Свежевыделенные CD8+-клетки OTI ресуспендировали в PBS и метили с помощью 1 мкM CFSE (Invitrogen) в течение 6 мин. при комнатной температуре и реакцию гасили в течение 1 мин. с помощью равного объема модифицированной среды Дульбекко (IMDM) от Iscove с 10% (об./об.) FBS (Gibco). Клетки промывали, подсчитывали и ресуспендировали в чистой IMDM перед проведением инъекции. В общей сложности 3×106 CFSE-меченных CD8+-клеток OTI инъецировали i.v. в хвостовую вену мышей-реципиентов CD45.1+. Для кратковременных исследований пролиферации OVA-содержащие клетки эритроидного ряда инъецировали через 24 ч после адоптивного переноса. Спленоциты собирали через 5 дней после введения антигена и окрашивали для анализа с помощью проточной цитометрии.
Пример 7. Модель антигенной стимуляции мыши с помощью OVA
[00585] В общей сложности 3×105 CFSE-меченных CD8+ клеток OTI иньецировали мышам-реципиентам CD45.1+, как описано выше. В день 1 и 6 после адоптивного переноса мышам вводили i.v. в хвостовую вену 100 мкл OVA-содержащих клеток эритроидного ряда. В день 15 после адоптивного переноса мышей стимулировали с помощью 5 мкг OVA и 25 нг ультрачистого LPS E. сoli (InvivoGen) в 25 мкл физиологического раствора, инъецируемых интрадермально в подушечку каждой задней лапы (способ инъекции в пятку: общая доза 10 мкг OVA и 50 нг LPS). Количественное определение антиген-специфичных B- и Т-клеток и титра сывороточных антител описано ниже.
Пример 8. Количественное определение антиген-специфичных B- и Т-клеток у мыши
[00586] Мышей умерщвляли через 4 дня после стимуляции с помощью OVA, описанной выше, и клетки селезенки и дренирующих лимфатических узлов выделяли для повторной стимуляции. Для анализа внутриклеточных цитокинов с помощью проточной цитометрии клетки повторно стимулировали в присутствии 1 мг/мл OVA или 1 мкг/мл пептида SIINFEKL (Genscript) в течение 3 ч. Добавляли брефелдин-A (5 мкг/мл; Sigma) и повторную стимуляцию продолжали в течение дополнительных 3 ч перед проведением окрашивания и анализа с помощью проточной цитометрии. Для измерения секретируемых факторов с помощью ELISA клетки повторно стимулировали в присутствии 100 мкг/мл OVA или 1 мкг/мл пептида SIINFEKL в течение 4 дней. Клетки центрифугировали и среду собирали для анализа ELISA с применением наборов Ready-Set-Go для IFN-γ и IL-10 (eBioscience) в соответствии с инструкциями производителя.
Пример 9. Количественное определение титра циркулирующих в крови антител
[00587] OVA-специфичный IgG сыворотки крови обнаруживали путем инкубации мышиной сыворотки крови при различных разведениях на покрытых OVA планшетах с последующей конечной инкубацией с конъюгированными с HRP антителами козы к IgG мыши (Southern Biotech).
Пример 10. Индукция толерантности с применением внеклеточного SpyTag-SpyCatcher
[00588] Кассету экспрессии, содержащую кодирующую последовательность Kell и SpyTag, синтезировали и вставляли в лентивирусный вектор EF1. Популяцию CD34+ клеток трансформировали вектором. Экспрессию белка слияния Kell-SpyTag количественно определяли с помощью FACS. Клетки, которые экспрессировали Kell-SpyTag вне клетки, затем помещали в раствор пептидов Ara h (1-6), слитых с последовательностью SpyCatcher и меткой cMyc. После инкубации клетки сортировали с применением FACS для количественного определения ковалентной конъюгации с образованием изопептидной связи Kell-SpyTag и SpyCatcher-ArahX-cMyc. Также клетки лизировали в гипотонических условиях и присутствие Kell-SpyTag-SpyCatcher-ArahX-cMyc количественно определяли с помощью вестерн-блот-анализа.
Пример 11. Сборка гена
[00589] ДНК, кодирующую следующие гены: гликофорина A (ID в базе Uniprot P02724), Kell (ID в базе Uniprot P23276), scFv-антитела к поверхностному антигену гепатита B (Bose et al. 2003 Mol Immunol 40(9):617, ID в базе GenBank AJ549501.1), аденозиндезаминазы (ID в базе Uniprot P00813), фенилаланингидроксилазы из Chromobacterium violaceum (ID в базе GenBank AF146711.1), рецептора комплемента 1 (ID в базе Uniprot P17927), CD46 (GenBank: BAA12224.1), CD55 (ID в базе Uniprot P08174), CD59 (ID в базе Uniprot P13987), зеленого флуоресцентного белка (ID в базе Uniprot P42212), тимидинфосфорилазы (ID в базе Uniprot P19971), глюкоцереброзидазы (ID в базе Uniprot P04062), бета-2-гликопротеина 1 (ID в базе Uniprot P02749), рецептора фосфолипазы a2 (ID в базе Uniprot Q13018), коллагена альфа-3(IV) (ID в базе Uniprot Q01955), амилоида P сыворотки (ID в базе Uniprot P02743), липопротеинлипазы (ID в базе Uniprot P06858), аспарагиназы (ID в базе Uniprot P00805), фактора IX (ID в базе Uniprot F2RM35), ADAMTS13 (ID в базе Uniprot Q76LX8), приобретали в виде кДНК у Dharmacon (GE Life Sciences) или синтезировали de novo у DNA2.0 и Genscript.
1. Клонирование одного гена (CR1)
[00590] Гены собирали в векторах экспрессии с помощью стандартных способов молекулярной биологии, известных из уровня техники. Ген рецептора комплемента 1 (CR1) синтезировали у коммерческого поставщика (DNA2.0) и доставляли в стандартном векторе для клонирования (серия pJ). Ген амплифицировали из вектора pJ с помощью полимеразной цепной реакции (ПЦР) с применением олигонуклеотидов с негомологичными терминальными последовательностями для подготовки к вставке в вектор экспрессии для клетки млекопитающего (System Biosciences, серия pM): олигонуклеотид, гибридизирующийся против хода транскрипции, состоял из 25 нт, гомологичных сайту встраивания pM, расположенному против хода транскрипции, и 25 нт, гомологичных началу CR1; олигонуклеотид, гибридизирующийся по ходу транскрипции, состоял из 25 нт, гомологичных сайту встраивания pM, расположенному по ходу транскрипции, и 25 нт, гомологичных концу CR1. Продукт амплификации очищали с помощью гель-электрофореза (Qiagen). Вектор pM линеализировали с помощью ПЦР с олигонуклеотидами, соединенными "хвост к хвосту", гомологичными сайтам встраивания, расположенным по ходу и против хода транскрипции, и очищали путем очистки ПЦР-продуктов (Qiagen). Ампликон CR1 лигировали в линеаризованный вектор pM путем сборки по Гибсону, подробно описанной в Gibson 2011, Methods Enzymology Vol 498, p. 394. Последовательности подтверждали путем секвенирования по Сэнгеру.
2. Слияние двух генов (мембранный Kell-scFv)
[00591] Ген Kell приобретали в виде кДНК и доставляли в стандартном векторе для клонирования (серия pJ). Ген scFv-антитела, специфичного в отношении поверхностного антигена гепатита B (scFv, описанный в Bose 2003, Molecular Immunology 40:617), синтезировали у коммерческого поставщика (DNA2.0) и доставляли в стандартном векторе для клонирования (серия pJ). Гены амплифицировали из векторов pJ с помощью полимеразной цепной реакции (ПЦР) с применением олигонуклеотидов с негомологичными терминальными последовательностями для подготовки к вставке в вектор экспрессии для клетки млекопитающего (System Biosciences, серия pM). Kell амплифицировали с помощью олигонуклеотида, гибридизирующегося против хода транскрипции, состоящего из 25 нт, гомологичных сайту встраивания pM, расположенному против хода транскрипции, и 25 нт, гомологичных 5'-концу Kell, и олигонуклеотида, гибридизирующегося по ходу транскрипции, состоящего из 25 нт, гомологичных 5'-концу scFv, и 25 нт, гомологичных 3'-концу Kell. scFv амплифицировали с помощью олигонуклеотида, гибридизирующегося против хода транскрипции, состоящего из 25 нт, гомологичных 3'-концу сайта встраивания Kell, и 25 нт, гомологичных 5'-концу scFv, и олигонуклеотида, гибридизирующегося по ходу транскрипции, состоящего из 25 нт, гомологичных сайту встраивания pM, расположенному по ходу транскрипции, и 25 нт, гомологичных 3'-концу scFv. Продукты амплификации очищали с помощью гель-электрофореза (Qiagen). Вектор pM линеализировали с помощью ПЦР с олигонуклеотидами, соединенными "хвост к хвосту", гомологичными сайтам встраивания, расположенным по ходу и против хода транскрипции, и очищали путем очистки ПЦР-продуктов (Qiagen). Ампликоны Kell и scFv лигировали в линеаризованный вектор pM путем одностадийной сборки по Гибсону, подробно описанной в Gibson 2011, Methods Enzymology Vol 498, p. 394. Последовательности подтверждали путем секвенирования по Сэнгеру.
3. Сборка с линкером между генами (Kell-scfv)
[00592] Ген Kell приобретали в виде кДНК и доставляли в стандартном векторе для клонирования (серия pJ). Ген scFv-антитела, специфичного в отношении поверхностного антигена гепатита B (scFv, описанный в Bose 2003, Molecular Immunology 40:617), синтезировали у коммерческого поставщика и доставляли в стандартном векторе для клонирования (серия pJ). Гены амплифицировали из векторов pJ с помощью полимеразной цепной реакции (ПЦР) с применением олигонуклеотидов с негомологичными терминальными последовательностями для подготовки к вставке в вектор экспрессии для клетки млекопитающего (System Biosciences, серия pM). Kell амплифицировали с помощью олигонуклеотида, гибридизирующегося против хода транскрипции, состоящего из 25 нт, гомологичных сайту встраивания pM, расположенному против хода транскрипции, и 25 нт, гомологичных 5'-концу Kell; и олигонуклеотида, гибридизирующегося по ходу транскрипции, состоящего из 25 нт, гомологичных 5'-концу scFv, 24 нт, кодирующих спейсер (GlyGlyGlySer)x2, и 25 нт, гомологичных 3'-концу Kell. scFv амплифицировали с помощью олигонуклеотида, гибридизирующегося против хода транскрипции, состоящего из 25 нт, гомологичных 3'-концу сайта встраивания Kell, 24 нт, кодирующих спейсер (GlyGlyGlySer)x2, и 25 нт, гомологичных 5'-концу scFv; и олигонуклеотида, гибридизирующегося по ходу транскрипции, состоящего из 25 нт, гомологичных сайту встраивания pM, расположенному по ходу транскрипции, и 25 нт гомологичных 3'-концу scFv. Продукты амплификации очищали с помощью гель-электрофореза (Qiagen). Вектор pM линеализировали с помощью ПЦР с олигонуклеотидами, соединенными "хвост к хвосту", гомологичными сайтам встраивания, расположенным по ходу и против хода транскрипции, и очищали путем очистки ПЦР-продуктов (Qiagen). Ампликоны Kell и scFv лигировали в линеаризованный вектор pM путем одностадийной сборки по Гибсону, подробно описанной в Gibson 2011, Methods Enzymology Vol 498, p. 394. Последовательности подтверждали путем секвенирования по Сэнгеру.
4. Прикрепление эпитопной метки (Kell-scFv)
[00593] Ген Kell приобретали в виде кДНК и доставляли в стандартном векторе для клонирования (серия pJ). Ген scFv-антитела, специфичного в отношении поверхностного антигена гепатита B (scFv, описанный в Bose 2003, Molecular Immunology 40:617), синтезировали у коммерческого поставщика (DNA2.0) и доставляли в стандартном векторе для клонирования (серия pJ). Гены амплифицировали из векторов pJ с помощью полимеразной цепной реакции (ПЦР) с применением олигонуклеотидов с негомологичными терминальными последовательностями для подготовки к вставке в вектор экспрессии для клетки млекопитающего (System Biosciences, серия pM). Kell амплифицировали с помощью олигонуклеотида, гибридизирующегося против хода транскрипции, состоящего из 25 нт, гомологичных сайту встраивания pM, расположенному против хода транскрипции, и 25 нт, гомологичных 5'-концу Kell; и олигонуклеотида, гибридизирующегося по ходу транскрипции, состоящего из 25 нт, гомологичных 5'-концу scFv, 24 нт, кодирующих спейсер (GlyGlyGlySer)x2, и 25 нт, гомологичных 3'-концу Kell. scFv амплифицировали с помощью олигонуклеотида, гибридизирующегося против хода транскрипции, состоящего из 25 нт, гомологичных 3'-концу сайта встраивания Kell, 24 нт, кодирующих спейсер (GlyGlyGlySer)x2, и 25 нт, гомологичных 5'-концу scFv; и олигонуклеотида, гибридизирующегося по ходу транскрипции, состоящего из 25 нт, гомологичных сайту встраивания pM, расположенному по ходу транскрипции, и последовательности из 27 нт tacccctatgacgtgcccgactatgcc (Seq. ID No. 8), кодирующей HA-эпитопную метку, и 25 нт, гомологичных 3'-концу scFv. Продукты амплификации очищали с помощью гель-электрофореза (Qiagen). Вектор pM линеализировали с помощью ПЦР с олигонуклеотидами, соединенными "хвост к хвосту", гомологичными сайтам встраивания, расположенным по ходу и против хода транскрипции. Праймер, гибридизирующийся по ходу транскрипции, дополнительно содержал последовательность из 27 нт tacccctatgacgtgcccgactatgcc (Seq. ID No. 8), кодирующую HA-эпитопную метку. Линеаризованный вектор очищали путем очистки ПЦР-продуктов (Qiagen). Ампликоны Kell и scFv лигировали в линеаризованный вектор pM путем одностадийной сборки по Гибсону, подробно описанной в Gibson 2011, Methods Enzymology Vol 498, p. 394. Последовательности подтверждали путем секвенирования по Сэнгеру.
5. Слияние из двух генов (сборка с репортером) (GPA-HA)
Гены рецептора комплемента 1 (CR1) и зеленого флуоресцентного белка (GFP) синтезировали у коммерческого поставщика (DNA2.0) и доставляли в стандартных векторах для клонирования (серия pJ). Ген CR1 амплифицировали из вектора pJ с помощью полимеразной цепной реакции (ПЦР) с применением олигонуклеотидов с негомологичными терминальными последовательностями для подготовки к вставке в вектор экспрессии в клетке млекопитающего (System Biosciences, серия pM): олигонуклеотид, гибридизирующийся против хода транскрипции, состоял из 25 нт, гомологичных сайту встраивания pM, расположенному против хода транскрипции, и 25 нт, гомологичных началу CR1; олигонуклеотид, гибридизирующийся по ходу транскрипции, состоял из 54 нт, гомологичных вирусной последовательности T2A gagggcagaggaagtcttctaacatgcggtgacgtggaggsgsstcccggccct (Seq. ID No. 7). Ген GFP амплифицировали из вектора pJ с помощью полимеразной цепной реакции (ПЦР) с применением олигонуклеотидов с негомологичными терминальными последовательностями для подготовки к вставке в вектор экспрессии для клетки млекопитающего (System Biosciences, серии pM): олигонуклеотид, гибридизирующийся против хода транскрипции, состоял из 54 нт, гомологичных вирусной последовательности T2A gagggcagaggaagtcttctaacatgcggtgacgtggaggsgsstcccggccct (Seq. ID No. 7) и 25 нт, гомологичных началу GFP; олигонуклеотид, гибридизирующийся по ходу транскрипции, состоял из 25 нт, гомологичных сайту встраивания pM, расположенному по ходу транскрипции, и 25 нт, гомологичных концу GFP. Продукты амплификации очищали с помощью гель-электрофореза (Qiagen). Вектор pM линеализировали с помощью ПЦР с олигонуклеотидами, соединенными "хвост к хвосту", гомологичными сайтам встраивания, расположенным по ходу и против хода транскрипции, и очищали путем очистки ПЦР-продуктов (Qiagen). Ампликоны CR1 и GFP лигировали вместе и встраивали в линеаризованный вектор pM путем сборки по Гибсону, подробно описанной в Gibson 2011, Methods Enzymology Vol 498, p. 394. Последовательности подтверждали путем секвенирования по Сэнгеру.
Пример 12. Сборка мРНК
[00594] Ген, представляющий интерес, клонировали в сайт множественного клонирования вектора pSP64 (Promega) с применением стандартных способов молекулярной биологии. Вектор расщепляли с помощью EcoRI (NEB) для получения линеаризованного вектора из dsDNA, содержащего промотор SP6, ген, представляющий интерес, и поли(А)-хвост длиной 30 нуклеотидов. мРНК синтезировали с помощью реакции с РНК-полимеразой SP6 (Promega) в соответствии с инструкциями производителя, включая рекомендуемые концентрации аналога 5'-кэпа (ARCA) для синтеза кэпированного мРНК-транскрипта. Затем реакционную смесь обрабатывали ДНКазой для расщепления матрицы вектора (Riboprobe от Promega) и мРНК очищали с применением набора для очистки РНК EZNA MicroElute RNA Clean-Up (Omega).
Пример 13. Культура клеток
1. Человеческие красные кровяные клетки (RBC)
[00595] Клетки CD34 выделяли из периферической крови посредством отбора с супермагнитными микрогранулами с применением колонок Mini-MACS (Miltenyi Biotec; чистота 94% ± 3%). Клетки культивировали в среде для индукции эритроидной дифференцировки (EDM) на основе IMDM, дополненной стабилизированным глутамином, 330 мкг/мл человеческого голотрансферрина, 10 мкг/мл рекомбинантного человеческого инсулина, 2 МЕ/мл гепарина и 5% плазмы, вирусинактивированной с помощью растворителя/детергента. Процедура размножения клеток включала 3 стадии. На первой стадии (день 0 - день 7) 104/мл клеток CD34 культивировали в EDM в присутствии 1 мкM гидрокортизона, 100 нг/мл SCF, 5 нг/мл IL-3 и 3 МЕ/мл EPO. В день 4 1 объем культуры клеток разводили в 4 объемах свежей среды, содержащей SCF, IL-3, EPO и гидрокортизон. На второй стадии (день 7 - день 11) клетки ресуспендировали из расчета 105/мл в EDM, дополненной SCF и EPO. На третьей стадии (день 11 - день 18) клетки культивировали в EDM, дополненной только EPO. Количество клеток доводили до 7,5×105-1×106 и 5-10×106 клеток/мл в дни 11 и 15, соответственно. После дня 18 культуральную среду, содержащую EPO, обновляли дважды в неделю. Культуры поддерживали при 37°C с 5% CO2 в воздухе.
2. Мышиные красные кровяные клетки
[00596] Способы культивирования мышиных клеток эритроидного ряда из мышиных клеток-предшественников эритроидного ряда, полученных из фетальной печени, известны из уровня техники, см., например, Shi et al. 2014, PNAS 2014 111(28):10131.
[00597] Мышиные клетки-предшественники эритроидного ряда выделяли из фетальной печени. Фетальную печень приобретали у Charles River Labs. Печень помещали в 1 мл PBS на льду. Пипетировали вверх-вниз для получения раствора суспензии отдельных клеток и пропускали через фильтр с размером пор 70 мкм (BD Falcon 35-2235). Промывали сито с помощью 1 мл PBS. Объединяли фильтрат (1 мл на эмбрион). Осаждали клетки при 1,5 тыс. об/мин в течение 5 мин, повторно суспендировали в буфере для лизиса красных кровяных клеток (раствор хлорида аммония от Stemcell) и инкубировали на льду в течение 10 минут. Осаждали клетки при 1,5 тыс. об/мин в течение 5 мин, удаляли буфер для лизиса и повторно суспендировали в 10 мл PBS-2% FBS. Добавляли IgG крысы chromPure (Jackson ImmunoResearch, #012-000-003) из расчета 50 мкл/мышь и инкубировали при 4°C в течение 5 мин. Добавляли биотинилированное антитело к TER119 мыши (BD Pharmingen, #553672) из расчета (1 мкл/1*106 клеток) и инкубировали при 4°C в течение 15 мин. К клеткам добавляли Ms Lineage Panel (Fisher Scientific (Thermo Fisher Scientific) # BDB559971) из расчета (2 мкл/1*10^6 клеток) и инкубировали при 4°C в течение15 мин. Промывали один раз с помощью 10x объема PBS и центрифугировали клетки при 1,5 тыс. об/мин в течение 5 мин при 4 градусах. Добавляли Streptavidin Particles Plus - DM (магнитные гранулы) (BD Pharmigen, #557812) (5 мкл/1*106 клетки) и инкубировали при 4°C в течение 30 мин. Готовили 2-4 пробирки для FACS на магнитном держателе. Отмеряли 2 мл клеток в каждую пробирку (в общей сложности 4 мл), и осторожно извлекали клетки из пробирки и помещали в другую пробирку на противоположной стороне, избегая нарушения выстроенных столбиком магнитных микрогранул. Повторяли ту же процедуру и переносили Ter119-отрицательные и клетки, отрицательные по панели антител для линий дифференцировки, в новую пробирку. Концентрировали клетки и ресуспендировали клетки в 50-100 мкл PBS (2% FBS).
[00598] Очищенные клетки-предшественники эритроидного ряда культивировали в среде для индукции дифференцировки, содержащей (на 40 мл): IMDM: 29 мл, FBS (стволовые клетки): 6 мл (конечная концентрация 15%), 10% BSA в IMDM (стволовые клетки): 4 мл (конечная концентрация 1%), 10 мг/мл голотрансферрина: 2000 мкл (конечная концентрация: 500 мкг/мл), 100*L-глутамин: 400 мкл, 100* пенициллин-стрептомицин: 400 мкл, 10 Ед./мл Epo: 2 мкл (конечная концентрация: 0,5 Ед./мл), 10 мг/мл инсулина: 40 мкл (конечная концентрация: 10 мкг/мл). Культивировали 2*105 клеток/мл в среде для индукции дифференцировки в 24-луночном планшете при 37°C. После культивирования в общей сложности 44-48 часов выполняли анализы, например, с помощью проточной цитометрии, осуществляемой в данном документе. Безъядерные красные кровяные клетки гейтировали с применением (окраски с помощью Hoechst) для анализа профиля дифференцировки. Успешная культура будет давать 16 кратное повышение.
3. Тромбоциты
[00599] Донорские CD34+ клетки получали из Онкологического научного центра Фреда Хатчинсона. CD34+обогащенные клетки высевали в бессывороточную среду из расчета 2-4×104 клеток/мл и среду обновляли в день 4 путем добавления равного объема среды. В день 6 клетки подсчитывали и анализировали: 1,5×105 клеток промывали и помещали в 1 мл той же среды, дополненной смесью цитокинов, состоящей из 30 нг/мл TPO, 1 нг/мл SCF, 7,5 нг/мл интерлейкина (IL)-6 и 13,5 нг/мл IL-9 для индукции дифференцировки мегакариоцитов. В день 10 1/2-1/4 суспензионной культуры заменяли свежей средой. Все цитокины приобретали у Peprotech. Культуры инкубировали в увлажненной атмосфере (10% CO2) при 39°C в течение первых 6 дней культивирования и при 37°C в течение последних 8 дней. Жизнеспособные ядерные клетки подсчитывали с помощью гемоцитометра (0,4% трипанового синего; Invitrogen, Берлингтон, Онтарио, Канада).
[00600] Клоногенные прогениторные клетки (CPC) анализировали с применением MethoCult H4436 для миелоидных CPC и MegaCult-C для колониеобразующих единиц-мегакариоцитов (CFU-Mk) в соответствии с инструкциями производителя (StemCell Technologies, Ванкувер, Британская Колумбия, Канада). Для оценки дифференцировки клетки окрашивали антителами к CD61m, CD42b, CD41, CD61 и CD49b и анализировали с помощью проточной цитометрии с применением FACS-Calibur (Becton Dickinson). Для анализа клеточного цикла клетки промывали фосфатно-солевым буфером (PBS), фиксировали 2% формальдегидом (Sigma, Сент-Луис, Миссури, США) в течение 5 мин. и пермеабилизировали с помощью 0,1% Triton X-100 (Bio-Rad, Геркулес, Калифорния, США). Затем клетки метили с помощью mAb-Ki-67-FITC (BD Bioscience, Сан-Хосе, Калифорния, США), промывали и ресуспендировали в 0,5 мл PBS-1% фетальной бычьей сыворотки (FBS)- 0,01% азида 7-амино-актиномицина D (7-AAD), следуя инструкциям производителя (BD Biosciences).
Пример 14. Выделение клеток
1. Первичные RBC
[00601] Цельную кровь собирали с применением асептических методик в пробирки, содержащие низкомолекулярный гепарин, дальтепарин натрия (9 единиц/мл крови). Кровь центрифугировали при 5000 x g в течение 5 минут и после удаления плазмы крови и лейкоцитарной пленки (обе могут быть сохранены для последующего использования) эритроциты промывали дважды в холодном (4°C) фосфатно-солевом буфере (PBS) с центрифугированием. Полученную популяцию красных кровяных клеток хранили при 4°C в антикоагулянте CPDA-1 или растворе глицерина с целью длительного хранения.
2. Первичные тромбоциты
[00602] Цельную кровь (40 мл) собирали в 3,8% цитрат натрия (соотношение цитрата и крови 1:9 об./об.) от здоровых индивидуумов при использовании соответствующего протокола IRB. Кровь центрифугировали при 200g в течение 15 минут для отделения обогащенной тромбоцитами плазмы крови (PRP). Затем тромбоциты промывали в модифицированном буфере Тирода (содержащем 138 мМ NaCl, 5,5 мМ декстрозы, 12 мМ NaHCO3, 0,8 мМ CaCl2, 0,4 мМ MgCl2, 2,9 мМ KCl2, 0,36 мМ Na2HPO4 и 20 мМ Hepes, pH 7,4) в присутствии 1 мкM простагландина I2 и ресуспендировали в том же буфере.
Пример 15. Облучение первичных или культивируемых клеток
[00603] Облучение популяции экпрессирующих экзогенный антиген EHC-клеток можно выполнять, чтобы удостовериться в том, что они не способны к размножению. Такие протоколы подобны известным из уровня техники для облучения клеток, например, первичных красных кровяных клеток. Вкратце, брали одну дозу (350 мл) цельной крови и делили на две аликвоты по 175 мл каждая, 10 таких доз, таким образом, делили на 20 аликвот. Одну аликвоту (175 мл) из каждой дозы крови подвергали гамма-облучению 25 Гр, но не выше 50 Гр, с помощью автономного гамма-облучателя клеток (GammaCell 1000, Theratronics). Затем кровь хранили при 4°C при стандартных условиях заготовки крови. Отбор образцов производили из этих 10 облученных и 10 необлученных пакетов с кровью в дни 0, 7, 14 и 21 при помощи переходника для отбора образцов (Fenwal, США). Проводили тесты на пролиферацию клеток, в том числе анализ включения тимидина для количественного определения любого митотического потенциала. Супернатант анализировали на наличие свободного гемоглобина с помощью спектроскопии поглощения и на свободную лактатдегидрогеназу с помощью колометрического анализа (Pierce) для оценки уровней лизиса клеток.
Пример 16. Удаление ядер из клеток эритроидного ряда
[00604] Клетки эритроидного ряда выращивали до полуконфлюэнтного монослоя (1-4X104 клеток на см2) на покрытых коллагеном покровных стеклах диаметром 12 мм в IMDM, дополненной 100 единиц/мл пенициллина и 100 единиц/мл стрептомицина. Коллаген необходим для предотвращения опадания клеток с покровного стекла во время центрифугирования. Клетки выращивали до монослоев (5X104 клеток на см2) на покровных стеклах либо на той же среде, либо на среде Игла в модификации Дульбекко с 10% телячьей сыворотки. Покрытие покровных стекол с клетками коллагеном не является необходимым. Для удаления ядер из клеток покровные стекла переворачивали (стороной с клетками вниз) и помещали на дно центрифужных пробирок Corex на 15 мл, содержащих 2-5 мл среды с 10 г цитохалазина B на мл. Центрифужные пробирки с покровными стеклами сразу же помещали в центрифугу Sorvall RC-2, которую нагревали до 37°C путем вращения (SS 34) ротора с головкой в рабочем положении в течение приблизительно 1 ч при 10000 об/мин (при этом регулятор температуры установлен на 37-39°). Период времени и скорость центрифугирования являются важнейшими факторами для успешного удаление ядер. Клетки центрифугировали при 9000 об/мин в течение 1 ч при 37±20 и клетки центрифугировали при 6500 об/мин. В течение 50 мин при 37±- 20. После центрифугирования покровные стекла (Falcon) вынимали из центрифуги и помещали стороной с клетками вверх в чашки для культивирования ткани на 35 мм (Biolquest), содержащие 3 мл среды без цитохалазина B. Не позже чем через 30-60 мин. при 37°C клетки были морфологически нормальными и у 90-99% отсутствовали ядра. Безъядерные клетки снимали с покровных стекол путем обработки трипсином-EDTA (Grand Island Biological Co.) и клетки суспендировали в нормальной среде. Затем безъядерные клетки пересевали небольшими каплями на покровные стекла 22 мм2, которые находились в чашках для культивирования ткани на 35 мм, и помещали в термостат. Через определенные интервалы времени после пересева покровные стекла монтрировали на предметные стекла (12) и наблюдения за безъядерными клетками проводили с помощью фазового контраста Zeiss, поляризованного света и оптического прибора Nomarski.
Пример 17. Приведение клеток в контакт
1. Нуклеиновая кислота - трансфекция
[00605] Количество нуклеиновой кислоты, представляющей интерес, увеличивали с обеспечением приблизительно 5 мкг нуклеиновой кислоты на 105 EHC-клеток, предполагаемых для загрузки, например, клетки, такой как клетка эритроидного ряда, тромбоцит или гемопоэтическая клетка-предшественник. Нуклеиновую кислоту разводили в среде Opti-MEM (Life Technologies) при соотношении 1 мкг на 50 мкл среды. Разбавленную нуклеиновую кислоту затем объединяли с реагентом для трансфекции (Trans-IT для ДНК, Trans-IT mRNA для мРНК, Trans-IT siRNA для siRNA, Mirus Bio) при объемном соотношении 1:1 и обеспечивали возможность образовывать комплексы в течение 5 минут при комнатной температуре. Комплекс нуклеиновой кислоты добавляли в клеткам на 12-24 часа. Необязательно после этого периода времени среду можно заменить свежей средой так, чтобы реагенты для трансфекции больше не присутствовали.
2. Нуклеиновая кислота - вирусная трансдукция
[00606] Ген, представляющий интерес, клонировали в сайт множественного клонирования лентивирусного вектора pCDH с промоторной последовательностью MSCV от System Biosciences.
[00607] Лентивирус получали в клетках 293Т путем трансфекции клеток с помощью липофектамина. 5×106 клеток 293Т (линия клеток Lenti-X 293T, № в каталоге Clontech 632180) высевали в чашку Петри P10 за день перед трансфекцией. Конфлюэнтность монослоя клеток должна составлять приблизительно 70%. Одну чашку трансфицировали одной конструкцией. 20 мкл (10 мкг) смеси плазмиды pPACKH1 (System Biosciences)+2 мкг лентивирусной конструкции+20 мкл реагента Plus (LifeTechnologies, № по каталогу 11514-015) объединяли в 400 мкл Optimem и инкубировали 15 мин при RT. 30 мкл LF2000 (LifeTechnologies, № по каталогу 11668-019) разводили в 400 мкл Optimem, добавляли по каплям в смесь ДНК и инкубировали в течение 15 мин при RT. Смесь ДНК добавляли к клеткам (клетки находились в 9 мл Optimem). Клетки инкубировали в течение 6 часов и затем среду заменяли на DMEM/10% FBS. Супернатант, содержащий вирусы, собирали через 48 часов после трансфекции путем центрифугирования при 1500 об/мин в течение 5 минут. Супернатант собирали и замораживали в аликвотах по 1 мл при -80°C.
[00608] Клетки-мишени трансдуцировали в день 3-7 процесса культивирования, описанного в данном документе. 5×105 культивируемых клеток высевали в 500 мкл среды, содержащей 20 мкг/мл полибрена, в 24-луночный планшет. Для трансдукции каждым вируса предусматривали клетки в трех повторных лунках. Супернатант, содержащий вирусы, добавляли в другие 500 мкл среды и образец перемешивали пипетированием. Инфицирования обеспечивали с помощью инокуляции посредством центрифугирования, при этом центрифугируя планшет при 2000 об/мин в течение 90 минут при комнатной температуре. После инокуляции посредством центрифугирования клетки инкубировали при 37°C на протяжении ночи и на следующий день добавляли 1 мл свежей среды IMDM с соответствующими цитокинами.
3. Нуклеиновая кислота - катионный полимер
[00609] мРНК, кодирующую трансген, представляющий интерес, и содержащую последовательность промотора, расположенную против хода транскрипции, и поли(А)-хвост, расположенный по ходу транскрипции, можно приобрести у нескольких коммерческих поставщиков (например, IDT-DNA, Coralville IA). Трансфекции с помощью РНК проводили с применением средств доставки на основе катионных липидов RNAIMax (Invitrogen, Карлсбад, Калифорния) или TRANSIT-mRNA (Mims Bio, Мэдисон, Висконсин). РНК и реагент вначале разбавляли в минимальной среде Opti-MEM (Invitrogen, Карлсбад, Калифорния). 100 нг/мкл РНК разбавляли в 5× и 5 мкл RNAIMax на мкг РНК разводили в 10×. Разведенные компоненты объединяли и инкубировали 15 минут при комнатной температуре перед тем, как их вносили в культуральную среду. Для трансфекции с помощью TRANSIT-mRNA 100 нг/мкл РНК разбавляли в 10× в Opti-MEM и добавляли реагент BOOST (при концентрации 2 мкл на мкг РНК), добавляли TRANSIT-mRNA (при концентрации 2 мкл на мкг РНК) и затем комплексы РНК-липид доставляли в культуральную среду после 2-минутной инкубации при комнатной температуре. Трансфекцию с помощью РНК проводили в среде Nutristem xenofree hES (STEMGENT®, Кембридж, Массачусетс) или Opti-MEM с 2% FBS. Успешное введение мРНК-транскрипта в клетки-хозяева можно отслеживать с применением различных известных способов, таких как применение флуоресцентной метки или репортерного белка, такого как зеленый флуоресцентный белок (GFP). Успешную трансфекцию модифицированной мРНК можно также определять путем измерения уровня белковой экспрессии целевого полипептида с помощью, например, вестерн-блоттинга или иммуноцитохимии. Аналогичные способы можно осуществлять в случае больших объемов, увеличенных до формата культивирования в множестве литров (5-10000 л), исходя из соотношений комплекса РНК-липид.
4. Нуклеиновая кислота - электропорация
[00610] мРНК, кодирующую трансген, представляющий интерес, и содержащую последовательность промотора, расположенную против хода транскрипции, и расположенный по ходу транскрипции поли(А)-хвост, можно приобрести у нескольких коммерческих поставщиков (например, IDT-DNA, Coralville IA). Параметры электропорации оптимизировали путем трансфекции клеток эритроидной линии дифференцировки с помощью мРНК-транскриптов и измерения эффективности трансфекции с помощью количественной RT-PCR с праймерами, разработанными для специфичного обнаружения экзогенных транскриптов. В случае определенных препаратов клеток разрядка конденсатора емкостью 150 мкФ в 2,5×106 клеток, суспендированных в 50 мкл Opti-MEM (Invitrogen, Карлсбад, Калифорния), в стандартной кювете для электропорации с зазором 2 мм является достаточной для воспроизводимой доставки свыше 10000 копий модифицированных мРНК-транскриптов на клетку, как определено с применением способа калибровочной кривой, при этом с сохранением высокой жизнеспособности (>70%). Плотность клеток можно варьировать от 1×106 клеток/50 мкл до плотности 2,5×106 клеток/500, и требуется от 110 В дo 145 В для трансфекции клеток с аналогичной эффективностью, измеренной в копиях транскрипта на клетку. Электропорацию большого объема в множество литров (5-10000 л) можно выполнять аналогично стратегиям проточной электропорации большого объема, аналогичных описанным способам с описанными выше ограничениями (Li et al., 2002; Geng et al., 2010).
5. Полипептид - липосома
[00611] Клетки, в том числе первичные терминально дифференцированные клетки, например эритроциты, можно загружать экзогенным белком на их поверхности и в их цитоплазме. Загрузку белков можно выполнять с применением липосом.
[00612] Липиды (реагент Pro-Ject, Pierce) в органическом растворителе высушивали в атмосфере азота в тонкую пленку в стеклянном сцинтилляционном флаконе. Приблизительно по 2 мкл липидов применяли на 105 клеток. Поликлональный IgG мыши (Abcam) метили с помощью Dylight-650 (Pierce) в соответствии с инструкциями производителя. Раствор белка из расчета 0,1 мг/мл в PBS добавляли в смесь высушенных липидов. Раствор пипетировали несколько раз, инкубировали в течение 5 минут при комнатной температуре, затем энергично перемешивали на вортексе с получением инкапсулирующих липосом. Добавляли бессывороточную среду для доведения общего объема до 500 мкл на 105 клеток. Затем липосомальную смесь инкубировали с клетками в течение 3-4 часов при 37°C.
[00613] На фигуре 1 показана загрузка экзогенного белка, в данном случае флуоресцентно меченного IgG, в первичные эритроциты с помощью липосом. Загрузку измеряли с помощью проточной цитометрии. Загрузка является дозозависимой, поскольку 0,06% клеток являются флуоресцирующими в случае отсутствия липосом, ~60% клеток являются флуоресцирующими при низкой дозе липосом и ~85% клеток являются флуоресцирующими при высокой дозе липосом. Данные на фигуре 1 являются убедительным доказательством того, что экзогенные белки можно загружать в клетки эритроидного ряда с помощью липосом.
6. Полипептид - механическое разрушение
[00614] Клетки можно загружать с применением микрофлюидного устройства, содержащие каналы шириной 1 мкм, 2 мкм, 3 мкм, 4 мкм, 5 мкм, 10 мкм, которые позволяют образовать временные поры в клетках, что обеспечивает возможность проникновения загрузки, когда клетки продавливаются через систему.
[00615] Устройства на основе кремния производят в центре микротехнологий Массачусетского технологического института с применением методик фотолитографии и глубокого реактивного ионного травления. В данном способе 6" кремниевые полупроводниковые пластины толщиной 450 мкм обрабатывали гексаметилдисилазаном, наносили покрытие из фоторезиста (OCG934; FujiFilm) с помощью центрифугирования в течение 60 с при 3000 об/мин, подвергали действию УФ-излучения (EV1; EVG) через хромированный шаблон с конфигурации сужения канала и проявляли в растворе AZ405 (AZ Electronic Materials) в течение 100 с. После 20 мин запекания при 90°C полупроводникову пластину травили путем глубокого реактивного ионного травления (SPTS Technologies) до требуемой глубины (как правило, 15 мкм). Способ повторяли на противоположной стороне полупроводниковой пластины (т.e. стороне, не содержащей протравленные каналы) с применением другого шаблона, который содержал паттерны из смотровых отверстий, и с применением более толстого фоторезиста AZ9260 (AZ Electronic Materials). Затем применяли влажное оксидирование для выращивания 100-200 нм оксида кремния перед проведением анодного связывания полупроводниковой пластины с полупроводниковой пластиной Pyrex и разрезания на отдельные устройства. Перед каждым экспериментом устройства визуально осматривали и устанавливали на держатель с впускным и выпускным резервуарами (все разработано внутри лаборатории и произведено в Firstcut). Данные резервуары соединяли с устройством с помощью уплотнительных колец из бутадиенакрилонитрильного каучука (McMaster-Carr) для обеспечением надлежащей герметизации. Впускной резервуар соединяли с системой регуляции давления собственного производства с применением тефлоновой трубки для обеспечения необходимой движущей силы для проталкивания материала через устройство. Популяцию клеток эритроидного ряда сначала суспендировали в требуемом буфере для доставки [питательная среда, PBS или PBS, дополненный 3% FBS и 1% плюроников F-68 (Sigma)], смешивали с требуемым материалом для доставки и помещали во впускной резервуар устройства. Данный резервуар соединяли с линией подачи сжатого воздуха, контролируемой регулятором, и выбранное давление (0-70 фунтов/кв. дюйм) применяли для продвижения жидкости через устройство. Обработанные клетки затем собирали из выпускного резервуара. Клетки инкубировали при комнатной температуре в растворе для доставки в течение 5-20 мин после обработки, чтобы убедиться в закрытии отверстий, прежде чем подвергать клетки какой-либо дальнейшей обработке. Для доставки флуоресцентно меченной фенилаланин-аммоний-гидроксилазы (PAH) эксперименты осуществляли, как описано выше, так что буфер для доставки содержал 0,1-0,3 мг/мл PAH. Нокдаун GFP измеряли как процентную долю снижения средней интенсивности флуоресценции популяции клеток по сравнению с необработанными контролями.
7. Полипептид - конъюгация с поверхностью клетки
[00616] Клеточную поверхность обрабатывали реагентом Трота (2-иминотиолан HCl, Pierce) для тиолирования первичных аминов. Реагент Трота разводили в Tris-буфере, pH 8, с EDTA для предотвращения окисления сульфгидрилов. Приблизительно 1 пмоль реагента Трота применяли для обработки 106 клеток. Реагент Трота инкубировали с клетками в течение 1 часа при комнатной температуре. Избыточный или непрореагировавший реагент удаляли путем центрифугирования и промывания клеток. Количество доступных сульфгидрильных групп можно измерить с применением реагента Эллмана. Одновременно обрабатывали соответствующий полипептид экзогенного антигена средством для сшивки амина с сульфгидрилом, таким как SMCC (Pierce), в соответствии с инструкциями производителя. Избыток сшивающего реагента удаляли путем обессоливания. Затем малеимид-функционализированный белок инкубировали с тиолированными клетками в течение нескольких часов. Непрореагировавший белок отделяли от конъюгированного с клетками путем центрифугирования и промывки.
8. Полипептид - нековалентное присоединение к поверхности
[00617] Ген scFv-антитела против поверхностного антигена гепатита B (scFv, описанное в Bose 2003, Molecular Immunology 40:617) сливали с 6-гистидиновой аффинной меткой и с геном, кодирующим полипептидную последовательность, которая связывает мышиный гликофорин A, HWMVLPWLPGTLDGGSGCRG, в векторе экспрессии для клеток млекопитающего (Genlantis). Полный белок слияния получали путем временной трансфекции клеток HEK-293Т с применением стандартных способов и очищали на афинной смоле Ni-NTA (Pierce) в соответствии с инструкциями производителя. Очищенный белок слияния инкубировали с мышиными эритроцитами при концентрации >100 нM, что обеспечивает быстрое уравновешивание и связывание пептида с гликофорином A.
9. Полипептид - вставка липида в мембрану
[00618] Реагент Трота (Thermo Fisher) применяли для образования сульфгидрильных групп на амин-содержащей молекуле полипептида соответствующего экзогенного антигена, следуя протоколу производителя. Реакционную смесь инкубировали в течение 1 ч при комнатной температуре (RT) на встряхивателе и промывали посредством центрифужной колонки для обессоливания (Zeba, MWCO 7K, Thermo Scientific), следуя инструкциям производителя, для удаления непрореагировавшего реагента Трота. Образование сульфгидрильных групп на модифицированном полипептиде количественно оценивали с применением реагента Эллмана (Pierce) на основании протокола производителя.
[00619] DSPE-PEG3400-mal (1×10-3 M в PBS, 4 мкл, молярное соотношение липид:полипептид=1:1) (все липиды приобретали у Avanti Polar Lipids и хранили как раствор в хлороформе в атмосфере аргона при -20°C) добавляли в раствор обессоленного полипептида и инкубировали при RT на встряхивателе. Через 1 ч раствор образца фильтровали с применением устройства с центрифужным фильтром (Microcon, Millipore Co.) при 14 000 g в течение 15 мин при 4°C для удаления малых молекул и суспендировали в 600 мкл PBS (1 мг/мл полипептида).
[00620] 200 мкл цельной крови суспендировали в 1000 мкл PBS и центрифугировали при 1500 g в течение 30 с, повторяли четыре раза. Наконец, RBC суспендировали в 800 мкл PBS. Конъюгацию RBC/DSPE-PEG-полипептид проводили путем смешивания указанных выше суспензий RBC и различных количеств раствора DSPE-PEG-полипептид (1 мг на мл) с последующей инкубацией в течение 15-30 мин при 37°C. Смесь оставляли на 5 мин при комнатной температуре, затем промывали три раза в PBS и ресуспендировали до конечной концентрации RBC, составляющей 5×108 на мл. Автоматизированное устройство для подсчета клеток (Countess, Invitrogen) применяли для измерения концентрации клеток.
10. Полипептид - загрузка в гипотонических условиях
[00621] Полипептид соответствующего экзогенного антигена, в данном случае IgG мыши, приобретали у Abcam и добавляли из расчета 0,25 мг/мл в суспензию RBC в изотоническом растворе при гематокрите (Hct), составляющем 70%. Суспензию диализировали в 250 мл гипотонического раствора, содержащего 10 мМ фосфата натрия, pH 7,4, 10 мМ бикарбоната натрия и 20 мМ глюкозы, перемешивали при 15 об/мин в течение 1 часа при 4°C. Затем обеспечивали закрытие пор в клетках в изотоническом растворе путем добавления 1/10 объема раствора для закрытия, содержащего 5 мМ аденина, 100 мМ инозина, 100 мМ пирувата натрия, 100 мМ фосфата натрия, 100 мМ глюкозы, 12% (вес/об.) NaCl при pH 7,4. Затем клетки инкубировали при 37°C в течение 30 минут.
11. Полипептид - проникающий в клетку пептид
[00622] Производство конъюгированного с протамином полипептида известно из уровня техники, см., например, Kwon et al. 2009 J Contr Rel 139(3):182. 5 мг/мл низкомолекулярного протамина (LMWP) в 50 мМ буфера Hepes (pH 8) смешивали с гетеробифункциональным сшивающим средством, 3-(2-пиридилдитио)пропионовая кислота N-гидроксисукцинимид (SPDP, Sigma-Aldrich), при молярном соотношении 1:10 в DMSO и встряхивали в течение 1 ч при комнатной температуре. Затем реакционную смесь обрабатывали с помощью 50 мМ дитиотреитола (DTT, Sigma-Aldrich) и тиолированный LMWP очищали с помощью HPLC на колонке для аффинной хроматографии с гепарином. Продукт собирали с помощью ультрафильтрации, лиофилизировали и хранили при -20°C до дальнейшего использования.
[00623] Для конъюгации 5 мг/мл полипептида соответствующего экзогенного антигена смешивали с SPDP (40 мкл 0,1 M SPDP в этаноле на 1 мл раствора белка) в фосфатном буфере и перемешивали при комнатной температуре в течение 1 ч. Непрореагировавший SPDP удаляли путем быстрого обессоливания и замены буфера с помощью FPLC на 0,1 M фосфатный буфер (pH 7,4). Затем активированный полипептид конъюгировали с 10-кратным молярным избытком приготовленного выше LMWP-SH в течение 24 ч при 4°C. Конъюгаты LMWP-полипептид выделяли с помощью ионообменной хроматографии с применением колонки для аффинной хроматографии с гепарином с последующими пятью циклами фильтрования на центрифуге (отсечение по молекулярной массе: 5000 Дa). Объединенные конъюгаты LMWP-полипептид концентрировали и степень конъюгации определяли с помощью масс-спектроскопии MALDI-TOF.
[00624] Для экспериментов по поглощению свежие эритроциты овцы (MP Biomedicals, Солон, Огайо) суспендировали в сбалансированном солевом растворе Хенкса (HBSS) при плотности 5×108 клеток/мл и затем инкубировали с 0,5 мг/мл раствором конъюгатов LMWP-полипептид в течение 30 мин при комнатной температуре при осторожном встряхивании. Затем RBC промывали с помощью HBSS и хранили при 2-8°C.
12. Полипептид - химическая проницаемость
[00625] 3×108 RBC предварительно инкубировали в течение 30 мин. с хлорпромазином (Sigma Aldrich) из расчета 200 мкМ в растворе Рингера. Сразу после этого добавляли полипептид соответствующего экзогенного антигена в растворе Рингера (1-4 мкМ) до конечного объема 400 мкл и инкубировали в течение 30 мин. при комнатной температуре при легком перемешивании. После инкубации клетки дважды промывали, ресуспендировали в растворе Рингера и собирали для анализа.
13. Полипептид - ферментативная конъюгация
[00626] Виды ферментативной конъюгации с клеточной поверхностью с применением сортазы известны из уровня техники, см., например, Shi et al. PNAS 2014 111(28):10131. Для мечения N-конца GPA с помощью полипептида 30 мкл 500 мкM сортазы S. aureus и 1 мМ полипептида с LPETGG на C-конце предварительно инкубировали в 50 мМ Tris, pH 7,5, 150 мМ NaCl на льду в течение 15 минут и добавляли к 5×107 RBC в DMEM. Сортазу и смесь клеток инкубировали на льду в течение 30 мин. с периодическим осторожным перемешиванием, затем центрифугировали при 500 xg в течение 2 мин при 4°C для удаления буфера/DMEM, затем промывали три раза с помощью 1 мл ледяного PBS.
Пример 18. Оценка присутствия полипептида
1. Флуоресцентный трансген
[00627] Клетки эритроидного ряда культивировали, как описано в данном документе. Трансген, кодирующий гликофорин A с HA-меткой на С-конце, слитый с GFP через промежуточный вирусный пептид T2A, конструировали путем сборки по Гибсону, как описано в данном документе. Трансген вводили в клетки эритроидного ряда путем лентивирусной трансдукции, как описано в данном документе. Через два дня после трансдукции клетки собирали, промывали в буфере PBS и анализировали на проточном цитометре (Attune, Life Technologies). Эффективность трансдукции оценивали как процентную долю GFP-положительных клеток в популяции.
2. Белки клеточной поверхности
[00628] Что касается белков клеточной поверхности, уровень экспрессии белка можно определять уже через 2 дня после трансфекции с помощью проточной цитометрии с антителами, специфичными в отношении белка или в отношении совместно экспрессируемой эпитопной метки. Клетки эритроидного ряда культивировали, как описано в данном документе. Трансген, кодирующий гликофорин A с HA-меткой на N-конце, конструировали путем сборки по Гибсону, как описано в данном документе. Трансген вводили в клетки эритроидного ряда путем лентивирусной трансдукции, как описано в данном документе. Через два дня после трансдукции клетки собирали, промывали в буфере PBS и окрашивали с помощью 1:50 разведения мышиных антител к HA (Abcam) в течение 1 ч. Клетки промывали и затем окрашивали с помощью 1:100 разведения меченных alexa 488 вторичных антител козы к мыши (Life Technologies) в течение 30 минут на льду. Клетки промывали и анализировали на проточном цитометре (Attune, Life Technologies). Эффективность трансдукции оценивали как процентную долю alexa 488-положительных клеток в популяции.
3. Внутриклеточные белки
[00629] Что касается внутриклеточных белков, уровень экспрессии белка можно определять уже через 8-12 часов после трансфекции с помощью вестерн-блот-анализа. Клетки эритроидного ряда культивировали, как описано в данном документе. Трансген, кодирующий аденозиндезаминазу с HA-меткой на С-конце, конструировали путем сборки по Гибсону, как описано в данном документе. Трансген вводили в клетки эритроидного ряда путем лентивирусной трансдукции, как описано в данном документе. Через два дня после трансдукции клетки собирали, промывали в буфере PBS и лизировали в буфере RIPA для лизиса клеток (Pierce). Клеточный лизат денатурировали путем кипячения в 100 мМ DTT, затем загружали на готовый гель NuPage для SDS-PAGE. После электрофореза и переноса на нитроцеллюлозную мембрану полосы белка проявляли путем окрашивания с помощью 1:5000 разведения мышиных антител к HA (Abcam) с последующим окрашиванием с помощью 1:5000 разведения антител козы к иммуноглобулину мыши, конъюгированных с HRP (Pierce), и последующей обработкой с помощью субстрата для HRP (SuperSignal, Pierce). Изображения получали с применением визуализатора Amersham (GE healthcare).
Пример 19. Приведение клеток в контакт с химическим модифицирующим средством
[00630] Для повышения фагоцитоза антиген-презентирующими клетками и для содействия направленной доставке в печень композиции клеток, описанные в данном документе, обрабатывали в течение 30 мин. с помощью 0,15 микроM кальциевого ионофора A23187 (Sigma Aldrich, Сен-Кантен-Фаллавье, Франция) при 37°C или с помощью 5 мМ BS3 (Fischer Bioblock Scientific, Илькирш, Франция) при комнатной температуре (RT). После обработки конечные продукты хранили при 2-8°C в соответствующем буфере.
Пример 20. Оценка экспрессии и активности
[00631] Экспрессию экзогенных белков в культивируемых клетках и на них можно количественно оценивать с помощью проточной цитометрии (если белок экспрессируется на поверхности) или с помощью вестерн-блот-анализа (для белков, экспрессируемых в цитоплазме).
1. Количественная проточная цитометрия
[00632] Гранулы для количественной проточной цитометрии, связывающиеся с Fc антител мыши (Simply Cellular Calibration), приобретали у Bangs Labs. Флуоресцентно меченные антитела мыши против соответствующих рецепторов клеточной поверхности - гликофорина A, Ckit, и трансферринового рецептора - приобретали у BioLegend. Флуоресцентно меченные антитела мыши к HA-эпитопной метке приобретали у Life Technologies. Клетки эритроидного ряда культивировали, как описано в данном документе. Трансген, кодирующий гликофорин A с HA-меткой на N-конце, конструировали путем сборки по Гибсону, как описано в данном документе. Трансген вводили в клетки эритроидного ряда путем лентивирусной трансдукции, как описано в данном документе. По меньшей мере через два дня после трансдукции 2×105 клеток собирали, промывали в буфере PBS и окрашивали с помощью 1:100 разведения одного из перечисленных выше антител в течение 1 ч. Клетки промывали и анализировали на проточном цитометре (Attune, Life Technologies). Протокол повторяли для каждого из четырех антител, перечисленных выше. Количественное определение осуществляли в соответствии с инструкциями производителя. Вкратце, по одной капле каждого из пяти образцов с микрогранулами инкубировали с 1:100 разведением упомянутого выше антитела. Микрогранулы инкубировали в течение 1 ч, промывали в PBS и анализировали на проточном цитометре (Attune, Life Technologies). Протокол повторяли для каждого из четырех антител, перечисленных выше. Калибровочные кривые строили с применением предоставленных производителем таблиц в формате Excel, из которых выводили количественную оценку интенсивности флуоресценции для сигналов клеток.
2. Количественный вестерн-блот-анализ
[00633] Клетки эритроидного ряда культивировали, как описано в данном документе. Трансген, кодирующий аденозиндезаминазу с HA-меткой на С-конце, конструировали путем сборки по Гибсону, как описано в данном документе. Трансген вводили в клетки эритроидного ряда путем лентивирусной трансдукции, как описано в данном документе. Через два дня после трансдукции клетки собирали, промывали в буфере PBS и лизировали в буфере RIPA для лизиса клеток (Pierce).
[00634] Трансген вводили в клетки HEK293Т с помощью временной трансфекции с применением липофектамина 2000 (Life technologies). Клетки культивировали в течение одной недели и собирали супернатант. Рекомбинантный белок очищали на колонке для аффинной хроматографии для HA (Pierce) в соответствии с инструкциями производителя. Концентрацию белка оценивали по поглощению при длине волны 280 нм.
[00635] Вестерн-блоттинг выполняли, как описано в данном документе. В дополнение к образцам клеточного лизата на том же геле прогоняли известные количества рекомбинантной аденозиндезаминазы. После получения изображений значения интенсивности для полос рекомбинантного белка использовали для получения калибровочной кривой для оценки количества белка, присутствующего в образцах клеток.
[00636] Устойчивая экспрессия трансгенов на высоких уровнях играет важную роль в терапевтической активности конечной популяции клеток. На фигуре 2 приведена количественная оценка экспрессии трех поверхностных белков, являющихся показателями дифференцировки, и одного экзогенного трансгена с помощью количественной проточной цитометрии и показано, что трансген устойчиво экспрессируется на высоком уровне.
[00637] Клетки эритроидного ряда в культуре собирали в семь моментов времени во время четырехстадийного процесса дифференцировки in vitro. В первый момент времени ("Expand D6") клетки представляли собой ядерные гемопоэтические предшественники. К конечному моменту времени ("Diff 3 D8") клетки представляли собой преимущественно безъядерные клетки эритроидного ряда. GPA (треугольники, соединенные сплошной линией), канонический маркер клеток эритроидного ряда, сначала присутствует на низком уровне в клетках-предшественниках и быстро достигает >1×106 копий на клетку. CKIT (квадратики, соединенные пунктирной линией), рецептор фактора стволовых клеток, сначала присутствует на высоком уровне, затем снижается до <1×104 копий на клетку с наступлением дифференцировки. TR (ромбы, соединенные точечным пунктиром), необходимый для транспорта железа в клетки эритроидного ряда, сначала повышается, затем постепенно снижается до <1×105 копий на клетку. Трансген (незаштрихованные кружки) вводят в конце второй стадии дифференцировки ("Diff 1"), и он устойчиво экспрессируется на уровне приблизительно 1×105 копий на клетку на протяжении всей дифференцировки. Приведенные выше данные указывают на то, что трансгены устойчиво экспрессируются в культивируемых клетках.
[00638] Экспрессию экзогенных белков в культивируемых клетках и на них можно оценивать с помощью проточной цитометрии (если белок экспрессируется на поверхности), как описано в данном документе, или с помощью вестерн-блот-анализа (для белков, экспрессируемых в цитоплазме), как описано в данном документе. В случаях, когда экзогенный ген находится в конструкции для получения одного транскрипта, которая содержит последовательность флуоресцентного репортерного белка, расположенного по ходу транскрипции, флуоресценцию репортерного белка можно использовать как показатель экспрессии гена, расположенного против хода транскрипции, и оценивать с помощью проточной цитометрии, как описано в данном документе.
[00639] На фигурах 3 A-S показана экзогенная экспрессия поверхностных и цитоплазматических белков в безъядерных культивируемых клетках эритроидного ряда. Приведенные выше данные убедительно демонстрируют, что множество классов белков - в том числе цитоплазматические, поверхностные, интактные, слияния с мембранными белками I типа, слияния с мембранными белками II типа, слияния с GPI-связанными мембранными белками, внутриклеточные слияния, сверхэкспрессируемые и экспрессируемые de novo - можно экспрессировать на многих типах клеток, в том числе культивируемых безъядерных клетках эритроидного ряда, культивируемых ядерных клетках-предшественниках эритроидного ряда и эритролейкозных клетках K562.
[00640] На фигурах 3B и 3F показана одновременная экспрессия двух экзогенных белков в безъядерной культивируемой клетке.
[00641] Показанные на фигуре 3B клетки эритроидного ряда культивировали, как описано в данном документе. Трансгенную конструкцию, кодирующую сигнальную последовательность гликофорина A, HA-эпитопную метку, кодирующую последовательность гликофорина A, расщепляемую вирусную последовательность T2A и GFP, собирали путем сборки по Гибсону, как описано в данном документе. Трансген вводили в клетки эритроидного ряда путем лентивирусной трансдукции, как описано в данном документе. Клетки культивировали до терминальной дифференцировки, как описано в данном документе. Клетки анализировали с помощью проточной цитометрии, как описано в данном документе, с применением флуоресцентного антитела к HA и флуоресценции GFP для обнаружения экспрессии обоих трансгенов.
[00642] Показанные на фигуре 3F клетки эритроидного ряда культивировали, как описано в данном документе. Трансгенную конструкцию, кодирующую сигнальную последовательность гликофорина A, scFv-антитело, специфичное в отношении поверхностного антигена гепатита B, HA-эпитопную метку, кодирующую последовательность гликофорина A, расщепляемую вирусную последовательность T2A и GFP, собирали путем сборки по Гибсону, как описано в данном документе. Трансген вводили в клетки эритроидного ряда путем лентивирусной трансдукции, как описано в данном документе. Клетки культивировали до терминальной дифференцировки, как описано в данном документе. Клетки анализировали с помощью проточной цитометрии, как описано в данном документе, с применением флуоресцентного антитела к HA и флуоресценции GFP для обнаружения экспрессии обоих трансгенов.
Пример 21. Экспрессия белка за счет мРНК в тромбоцитах
[00643] Экспрессию в тромбоцитах экзогенных белков, транслируемых с экзогенной трансфицированной мРНК, измеряли с помощью проточной цитометрии. Вкратце, обогащенную тромбоцитами сыворотку крови центрифугировали при 190g в течение 15 минут для удаления эритроцитов и лейкоцитов. Затем супернатант центрифугировали в течение дополнительных 5 минут при 2500g для осаждения тромбоцитов. Тромбоциты ресуспендировали в 5 мл буфера Тирода с 1 мкM простагландина, промывали и ресуспендировали в 750 мкл буфера Тирода с 1 мкM простагландина. мРНК, кодирующую ген, представляющий интерес, в данном примере GFP, смешивали с липофектамином при соотношении 1:1 мг/мл. Смесь инкубировали в течение 5 минут, затем добавляли к популяции промытых тромбоцитов. Смесь инкубировали в течение 24 часов при комнатной температуре с медленным покачиванием. Экспрессию трансгена в тромбоцитах анализировали с помощью проточной цитометрии, измеряя флуоресценцию GFP. Поверхностные белки также можно анализировать с помощью проточной цитометрии. Цитоплазматические или другие экспрессируемые внутри клетки белки также можно анализировать с помощью вестерн-блот-анализа.
[00644] Терапевтическую значимость имеет введение экзогенных белков в тромбоциты или на них. Поскольку тромбоциты не имеют ядра или аппарата транскрипции РНК, трансфекция ДНК является неподходящим способом индукции экспрессии экзогенного белка в тромбоцитах. Однако трансфекция и трансляция мРНК представляет собой путь введения экзогенных белков в клетки. Полагают, что тромбоциты содержат аппарат трансляции мРНК, но в настоящее время неизвестно, способны ли они воспринимать экзогенную мРНК и транслировать ее в белок.
[00645] На фигуре 4 представлен ряд графиков результатов проточной цитометрии, которые демонстрируют трансляцию экзогенной мРНК, кодирующей трансген, в данном случае GFP, в тромбоцитах. GFP обнаруживали с помощью флуоресценции в канале FL1 после возбуждения лазером с длиной волны 488 нм. (4A) Нетрансфицированные тромбоциты (1,7% GFP+). (4B) Тромбоциты, трансфицированные с помощью 3 мкг мРНК GFP (8,6% GFP+). (4C) Тромбоциты, трансфицированные с помощью 6,8 мкг мРНК GFP (3,3% GFP+).
[00646] Данные впервые убедительно демонстрируют трансляцию экзогенной мРНК в экзогенный белок в тромбоцитах.
Пример 22. Активность ферментов
[00647] На фигуре 5 показана активность ферментов, содержащихся на клетках эритроидного ряда. Биохимическую активность цитоплазматических ферментов оценивали с помощью вестерн-блот-анализа в отношении сохранения белка в ходе дифференцировки. Биологическую активность цитоплазматических ферментов оценивали с помощью in vitro анализа ферментативной активности.
[00648] На фигуре 5 показана активность двух разных внутриклеточных ферментов, экспрессируемых в культивируемых клетках эритроидного ряда.
1. Аденозиндезаминаза
[00649] Трансген, кодирующий аденозиндезаминазу с HA-меткой на С-конце, конструировали путем сборки по Гибсону, как описано в данном документе. Трансген вводили в клетки HEK-293Т с помощью трансфекции с применением липофектамина (Life Technologies), как описано в данном документе. Ферментативную активность анализировали с применением протокола, взятого из Helenius 2012, Biochim Biophys Acta 1823(10):1967, в котором с помощью специфичной смеси ферментов преобразовывали пурины в мочевую кислоту и H2O2 с последующим флуориметрическим обнаружением образованной H2O2. Вкратце, через два дня после трансфекции клетки собирали, среду отсасывали и раствор фосфата глюкозы по Кребс-Рингеру (KRPG; содержащий: 145 мМ NaCl, 5,7 мМ фосфата натрия, 4,86 мМ KCl, 0,54 мМ CaCl2, 1,22 мМ MgSO4 и 5,5 мМ глюкозы; pH 7,35) добавляли в клетки из расчета 2×105 клеток/мл. Аденозин добавляли из расчета 50 мкM. После осуществления реакции в течение 6 часов супернатант собирали и инактивировали нагреванием в течение 5 минут при 60°C. Аликвоты супернатанта переносили в лунки белого 96-луночного микропланшета, содержащего 0,25 Ед./мл бактериальной пурин-нуклеозидфосфорилазы (PNP) и 0,15 Ед./мл микробной ксантиноксидазы (XO), обе от Sigma. После 20 мин. инкубации при RT 30 мкл смеси для обнаружения H2O2, содержащей HRP (конечная концентрация 1 Ед./мл, Sigma) и реагент Amplex Red (60 мкм, Invitrogen, Molecular Probes), добавляли в микролунки с последующим измерением интенсивности флуоресценции при длинах волны излучения и возбуждения, составляющих 545 и 590 нм, соответственно (Tecan Infinite M200).
2. Фенилаланингидроксилаза
[00650] Клетки эритроидного ряда культивировали, как описано в данном документе. Трансген, кодирующий фенилаланингидроксилазу с HA-меткой на С-конце, конструировали путем сборки по Гибсону, как описано в данном документе. Трансген вводили в клетки эритроидного ряда путем лентивирусной трансдукции, как описано в данном документе. Через два дня после трансдукции клетки собирали, промывали в буфере PBS и лизировали в буфере RIPA для лизиса клеток (Pierce). Клеточные лизаты (64 мкг общего белка) добавляли в 1 мл реакционного буфера, содержащего 100 мМ Tris-HCl, pH 7,5, 4 мМ DTT, 4 мМ фенилаланина, 33 мкг каталазы и 0,4 мМ DMPH4 (все от Sigma). Реакции проводили в течение ночи при 37°C. После инкубации образцы депротеинизировали путем фильтрования на центрифуге с центрифужным фильтром Amicon Ultra-4 10 кДа (Millipore UFC801024) с центрифугированием при 3700 об/мин в течение 10 мин. Образцы собирали и анализировали в отношении концентрации тирозина по поглощению при длине волны 540 нм.
[00651] Оба данных экзогенных белка сохранялись вплоть до конца терминальной дифференцировки, нетривиальный факт, с учетом того, что из уровня техники известно, что клетки эритроидного ряда подвергаются строгой программе элиминации белков, не являющихся необходимыми для основной функции (Liu J et al. (2010) Blood 115(10):2021-2027, Lodish HF et al. (1975) Developmental Biology 47(1):59). На фигуре 5A показано, что экзогенно сверхэкспрессированный белок аденозиндезаминазу обнаруживали с помощью вестерн-блот-анализа с антителом к HA в разные моменты времени в ходе дифференцировки от ядерных клеток-предшественников ("Diff I D5") вплоть до безъядерных клеток эритроидного ряда ("Diff III D8"). На фигуре 5C показано, что экзогенно экспрессированный микробный белок фенилаланингидроксилазу обнаруживали с помощью вестерн-блот-анализа с антителом к HA в разные моменты времени в ходе дифференцировки от ядерных клеток-предшественников ("Diff I D5") вплоть до безъядерных клеток эритроидного ряда ("Diff III D8").
[00652] Кроме того, оба данных фермента сохраняли свою способность ферментативно превращать субстрат в продукт. На фигуре 5B показано ферментативное превращение аденозина в инозин под действием интактных клеток 293Т, экспрессирующих аденозиндезаминазу. На фигуре 5D показано ферментативное превращение фенилаланина в тирозин под действием лизатов культивируемых безъядерных клеток эритроидного ряда, экспрессирующих фенилаланингидроксилазу.
[00653] Эти данные убедительно демонстрируют, что экзогенные ферменты сохраняются на клетках эритроидного ряда на протяжении всего процесса культивирования и что они являются ферментативно активными в клетках эритроидного ряда, что имеет огромное терапевтическое значение.
Пример 23. Активность CR1
[00654] На фигуре 6 показана как биохимическая, так и биологическая активность рецептора комплемента 1 (CR1), сверхэкспрессированного на поверхности культивируемых клеток эритроидного ряда. Биохимическую активность CR1 оценивали с помощью проточной цитометрии в отношении связывания с иммунным комплексом. Биологическую активность CR1 оценивали по переносу иммунных комплексов к макрофагам в анализе совместного культивирования.
1. Связывание иммунных комплексов клетками, экспрессирующими CR1
[00655] Клетки эритроидного ряда культивировали, как описано в данном документе. Трансгенную конструкцию, кодирующую рецептор комплемента 1 (CR1), конструировали путем сборки по Гибсону, как описано в данном документе. Трансген вводили в клетки эритроидного ряда путем лентивирусной трансдукции, как описано в данном документе. Уровни экспрессии трансгена оценивали с помощью проточной цитометрии, как описано в данном документе, с применением антитела к CR1 (Abcam). Клетки культивировали до терминальной дифференцировки, как описано в данном документе.
[00656] Альбумин бычьей сыворотки, меченный с помощью Dylight 650 (BSA-650), инкубировали с поликлональным антителом кролика к BSA (Abcam) при избытке антитела в течение 30 минут при комнатной температуре. Затем комплексы затем смешивали с человеческой сывороткой при объемном соотношении 1:1 в течение 30 минут при 37°C. Контрольные комплексы либо не смешивали с человеческой сывороткой, либо смешивали с инактивированной нагреванием человеческой сывороткой.
[00657] Комплексы инкубировали с клетками, экспрессирующими CR1, в течение 30 минут при 37°C. Клетки промывали и анализировали с помощью проточной цитометрии в отношении захвата иммунных комплексов путем обнаружения флуоресценции Dylight 650.
2. Перенос иммунных комплексов к макрофагам
[00658] Культивируемые моноциты U937 активировали путем инкубации со 100 нM форболмиристатацетата (PMA) в течение 24 часов при 37°C. Клетки, покрытые иммунными комплексами (см. выше), инкубировали с активированными макрофагами U937 в течение 30 минут при 37°C. Совместное культивирование анализировали с помощью проточной цитометрии. Макрофаги идентифицировали путем гейтирования FSC/SSC. Присутствие иммунного комплекса на макрофагах анализировали путем обнаружения флуоресценции Dylight 650 в популяции макрофагов.
[00659] На фигурe 6 показана биохимическая и биологическая активность рецептора комплемента 1 (CR1), экзогенно сверхэкспрессированного на культивируемых клетках эритроидного ряда.
[00660] На фигуре 6A показана биохимическая активность CR1, определяемая как захват иммунных комплексов in vitro. Черная гистограмма показывает захват иммунных комплексов на основе BSA с помощью CR1, сверхэкспрессированного на культивируемых клетках эритроидного ряда. Заштрихованная гистограмма показывает минимальное фоновое связывание с комплексами BSA и IgG, в которых отсутствует человеческий фактор комплемента, что показывает, что событие связывания является CR1-опосредованным.
[00661] На фигуре 6B показана биологическая активность CR1, определяемая как перенос иммунных комплексов от культивируемых клеток эритроидного ряда к макрофагам. Это является стандартным анализом в данной области, см.: Repik et al. 2005 Clin Exp Immunol. 140:230; Li et al. 2010 Infection Immunity 78(7):3129. Перенос оценивали с помощью проточной цитометрии и измеряли как интенсивность флуоресценции, происходящей от меченных иммунных комплексов на макрофагах. В данном анализе макрофаги, которые инкубировали в отсутствие иммунных комплексов (черные столбцы), не становились флуоресцентными. Макрофаги, которые инкубировали с комплексами BSA и IgG, в которых отсутствовал комплемент (и следовательно не связанные CR1), захватывали только небольшое количество иммунных комплексов (однотонные серые столбцы), независимо от присутствия культивируемых CR1-сверхэкспрессирующих клеток эритроидного ряда. Это поглощение, вероятно, связано с Fc-гамма рецепторами на клетках U937, взаимодействующих с Fc-участками молекул IgG. Макрофаги, которые инкубировали с иммунными комплексами (BSA+IgG+комплемент) в отсутствие клеток, сверхэкспрессирующих CR1 (заштрихованный столбец, слева), захватывали такое же количество иммунных комплексов, как и в отсутствие комплемента, вероятно при участии такого же Fc-гамма-опосредованного способа. Однако макрофаги, которые инкубировали с иммунными комплексами в присутствии клеток, сверхэкспрессирующих CR1(заштрихованный столбец, справа), захватывали приблизительно в два раза большее количество иммунных комплексов, как измерено с помощью флуоресценции.
[00662] Эти данные убедительно демонстрируют, что сверхэкспрессия CR1 на культивируемых клетках эритроидного ряда обеспечивает возможность захвата иммунных комплексов на указанных клетках эритроидного ряда, облегчает перенос иммунных комплексов от клеток эритроидного ряда на макрофаги и значительно повышает уровень и количество иммунных комплексов, захваченных макрофагами.
Пример 24. Активность scFv
[00663] На фигуре 7 показана биохимическая и биологическая активность scFv-антитела, экзогенно экспрессируемого на поверхности культивируемых клеток эритроидного ряда в виде слияния с трансмембранным белком GPA.
[00664] Клетки эритроидного ряда культивировали, как описано в данном документе. Трансгенную конструкцию, кодирующую лидерную последовательность гликофорина A, scFv-антитело, специфичное в отношении поверхностного антигена гепатита B (scFv, описанное в Bose 2003, Molecular Immunology 40:617), HA-эпитопную метку, гибкий линкер [Gly-3-Ser]2 и основную часть гликофорина A, собирали путем сборки по Гибсону, как описано в данном документе. Трансген вводили в клетки эритроидного ряда путем лентивирусной трансдукции, как описано в данном документе. Экспрессию трансгена оценивали с помощью проточной цитометрии, как описано в данном документе, с применением антитела к HA (Abcam). Клетки культивировали до терминальной дифференцировки, как описано в данном документе. Биохимическую активность scFv-антитела оценивали с помощью проточной цитометрии в отношении связывания с целевым белком, в данном случае поверхностным антигеном гепатита B (HBsAg). Рекомбинантный белок HBsAg (Abcam) метили флуорофором Dylight-650 (Pierce). Клетки, экспрессирующие scFv, инкубировали со 100 нM меченного белка, промывали в PBS и анализировали в отношении флуоресценции Dylight 650 с помощью проточной цитометрии, как описано в данном документе.
[00665] Биологическую активность антитела scFv оценивали с помощью in vivo захвата HBsAg, обнаруживаемого с помощью проточной цитометрии. Рекомбинантный белок HBsAg (Abcam) метили флуорофором Dylight-650 (Pierce). Клетки, экспрессирующие scFv, флуоресцентно метили CFSE (Sigma). Мышам NSG с ослабленным иммунитетом (Jackson labs) инъецировали ~400 пмоль меченного HBsAg в хвостовую вену. Через несколько минут тем самым мышам инъецировали 2×107 клеток, экспрессирующих scFv. Кровь собирали путем подчелюстной пункции через равные промежутки времени в пробирку, содержащую EDTA. Собранные кровяные клетки промывали и анализировали с помощью проточной цитометрии, как описано в данном документе. Человеческие клетки идентифицировали как клетки, которые были CFSE-положительными. Захват HBsAg обнаруживали как флуоресценцию Dylight 650 на человеческих клетках.
[00666] На фигурах 7A-7B показана биохимическая активность scFv-антитела, определяемая как связывание его соответственного антигена, поверхностного антигена гепатита B (HBsAg). На фигуре 7A показано, что клетки, которые экспрессировали (черный) или не экспрессировали (зашхрихованные серым) scFv-антитело, инкубировали с 450 нM HBsAg и окрашивали биотинилированным антителом к HBsAg и флуоресцентным стрептавидином. Клетки, которые экспрессировали scFv-антитело (~45% клеток в данной культуре), связывались с антигеном. На фигуре 7B показано, что клетки, которые экспрессировали scFv, инкубировали с различными концентрациями HBsAg и окрашивали, как описано выше, что показывает, что событие связывания является дозозависимым с афинностью, составляющей приблизительно 10 нM.
[00667] На фигурах 7C-7D показана биологическая активность scFv-антитела, определяемая как захват соответственного антигена HBsAg в кровотоке мыши. В данном эксперименте мышам NSG с ослабленным иммунитетом инъецировали ~400 пмоль флуоресцентно меченного HBsAg через хвостовую вену. Через пять минут через хвостовую вену инъецировали культивируемые безъядерные клетки эритроидного ряда (7C) или культивируемые безъядерные клетки эритроидного ряда, которые экспрессировали экзогенное scFv-антитело (7D). Перед инъекцией все культивируемые клетки метили флуоресцентным красителем CFSE. Кровь собирали через 6 часов, анализировали на проточном цитометре и гейтировали по CFSE+ клеткам человека. "Голые" культивируемые клетки не связывались с HBsAg (7C), тогда как клетки, экспрессирующие scFv-антитело, связывались с HBsAg (7D). Согласно эксперименту по биохимической активности приблизительно 45% клеток в данной культуре экспрессировали scFv-антитело.
[00668] Эти данные демонстрируют, что scFv-антитело является биохимически активным при экспрессии на поверхности культивируемых клеток эритроидного ряда и что scFv-антитело на клетке эритроидного ряда способно in vivo связываться с его мишенью, находящейся кровотоке. Это имеет огромное значение для терапевтических подходов, в которых требуется захват, секвестрация и клиренс вещества в кровотоке.
Пример 25. Активность - клиренс из кровотока
[00669] На фигуре 8 показаны как биохимическая, так и биологическая активности средств захвата поверхностных молекул, применяемых для клиренса мишени из кровотока.
[00670] Биохимическую активность средств захвата, в данном случае полипептида HA и биотина, оценивали с помощью проточной цитометрии в отношении связывания с целевым белком, в данном случае антитела к HA и антитела к биотину. Биологическую активность средств захвата оценивали с помощью in vivo захвата и клиренса целевого белка, которые обнаруживали с помощью проточной цитометрии и количественной оценки белка плазмы крови.
1. Захват антитела к биотину с помощью химически модифицированных клеток
[00671] Эритроциты от нормального донора-человека приобретали (Research Blood Components). Клетки метили с помощью CFSE (Sigma) в соответствии с инструкциями производителя в течение 20 минут при 37°C. Затем клетки биотинилировали с помощью NHS-биотина (Sigma) в соответствии с инструкциями производителя с применением 0,02 объема 2 мM исходного биотинового реагента в течение 30 минут при комнатной температуре. Антитело к биотину (Abcam) флуоресцентно метили с помощью Dylight 650 (Pierce). Эффективность мечения клеток оценивали с помощью проточной цитометрии с применением меченного антитела к биотину и флуоресценции CFSE в качестве маркеров обнаружения. 250 мкг меченного антитела инъецировали мыши NSG (Jackson Labs) внутривенно через хвостовую вену. Через четыре часа 1×108 биотинилированных клеток инъецировали внутривенно через хвостовую вену. Кровь собирали путем подчелюстной пункции через равные промежутки времени в пробирку, содержащую EDTA. Собранные кровяные клетки промывали и анализировали с помощью проточной цитометрии, как описано в данном документе. Человеческие клетки идентифицировали как клетки, которые были CFSE-положительными. Захват антитела к биотину обнаруживали как флуоресценцию Dylight 650 на человеческих клетках. Плазму крови из свежевзятой крови анализировали с помощью ELISA с применением покрытого биотином микропланшета (Pierce) в соответствии с инструкциями производителя для определения уровня антитела в кровотоке.
2. Захват антитела к HA трансгенными культивируемыми клетками
[00672] Клетки эритроидного ряда культивировали, как описано в данном документе. Трансгенную конструкцию, кодирующую сигнальную последовательность гликофорина A, HA-эпитопную метку, кодирующую последовательность гликофорина A, расщепляемую вирусную последовательность T2A и GFP, собирали путем сборки по Гибсону, как описано в данном документе. Трансген вводили в клетки эритроидного ряда путем лентивирусной трансдукции, как описано в данном документе. Клетки культивировали до терминальной дифференцировки, как описано в данном документе. Клетки анализировали с помощью проточной цитометрии, как описано в данном документе, с применением антитела к HA (Life Technologies), флуоресцентно меченного с помощью Dylight 650 (Pierce), и флуоресценции GFP для обнаружения экспрессия обоих трансгенов. 250 мкг меченного антитела к HA инъецировали мыши NSG (Jackson Labs) внутривенно через хвостовую вену. Через четыре часа 1×108 культивируемых клеток инъецировали внутривенно через хвостовую вену. Кровь собирали путем подчелюстной пункции через равные промежутки времени в пробирку, содержащую EDTA. Собранные кровяные клетки промывали и анализировали с помощью проточной цитометрии, как описано в данном документе. Человеческие клетки идентифицировали как клетки, которые были CFSE-положительными. Захват антитела к HA обнаруживали как флуоресценцию Dylight 650 на человеческих клетках. Плазму крови из свежевзятой крови анализировали с помощью ELISA с применением покрытого пептидом HA микропланшета (Pierce) в соответствии с инструкциями производителя для определения уровня антитела в кровотоке.
[00673] На фигуре 8 показана биохимическая и биологическая активность (8A-8B) полипептида HA, экспрессируемого на поверхности культивируемых клеток эритроидного ряда в виде слияния с GPA, и (8C-8D) биотина, химически конъюгированного с поверхностью первичных эритроцитов. Биохимическую активность определяли как захват целевого белка in vitro. Биологическую активность определяли как усиленный клиренс целевого белка in vitro.
[00674] На фигуре 8A показано, что полипептид HA, экспрессируемый как слияние с N-концом GPA, захватывает мышиное антитело к HA in vivo. Мышам NSG инъецировали флуоресцентно меченные мышиные антитела к HA с последующей инъекцией культивируемых человеческих клеток эритроидного ряда, которые либо не экспрессировали (верхняя панель), либо экспрессировали (нижняя панель) HA-эпитопную метку на их поверхности в виде слияния с GPA. Кровь собирали и клетки анализировали на проточном цитометре. По оси Х отложена флуоресценция CFSE. По оси Y отложена флуоресценция Dylight 650 антитела к HA. CFSE-положительные культивируемые человеческие эритроциты наблюдали в обоих образцах, но только клетки, экспрессирующие HA-эпитопную метку, были способны захватывать циркулирующие в крови антитела к HA.
[00675] На фигуре 8B показано, что мышам инъецировали антитело к HA, затем необязательно культивируемые человеческие клетки эритроидного ряда, которые либо не экспрессировали, либо экспрессировали пептид HA на своей поверхности в виде слияния с GPA. Плазму крови собирали в несколько моментов времени и уровень антитела к HA в плазме крови оценивали с помощью ELISA с применением планшета, покрытого пептидом HA, в качестве субстрата. Мыши, которым инъецировали только антитело к HA (незаштрихованные кружки, сплошная линия - эта мышь погибла через 120 минут по причинам, не связанным с обработкой) или антитело к HA с последующей инъекцией клеток, которые не экспрессировали пептид HA на своей поверхности (пунктирная линия), имели значительный уровень антител в кровотоке через 24 часа после инъекции клеток. Напротив, мыши, которым инъецировали антитело к HA с последующей инъекцией клеток, которые экспрессировали пептид HA на своей поверхности, становились истощенными по целевому антителу в пределах нескольких минут. Эти данные убедительно демонстрируют, что целевое антитело подвергается быстрому и специфичному клиренсу из кровотока с помощью культивируемых клеток эритроидного ряда, которые экспрессируют полипептид экзогенного антигена на своей поверхности.
[00676] На фигуре 8C показано, что молекула биотина, конъюгированная с поверхностью клеток эритроидного ряда путем химической функционализации аминной группы, захватывает мышиное антитело к биотину. В обоих этих случаях захват оценивали с помощью проточной цитометрии. Клетки, которые были CFSE-меченными и биотинилированными, обозначались как двойные положительные при окрашивании флуоресцентным антителом к биотину (нижняя точечная диаграмма), тогда как CFSE-меченные клетки, которые не были биотинилированы, обозначались как просто положительные (верхняя точечная диаграмма).
[00677] На фигуре 8D показано, что мышам инъецировали антитело к биотину, затем необязательно культивируемые человеческие клетки эритроидного ряда, которые либо не были конъюгированы, либо были конъюгированы с биотином на своей поверхности. Плазму крови собирали в несколько моментов времени и уровень антитела к биотину в плазме крови оценивали с помощью ELISA с применением планшета, покрытого биотином, в качестве субстрата. Мыши, которым инъецировали только антитело к биотину (незаштрихованные кружки, сплошная линия) или антитело к биотину с последующей инъекцией клеток, которые не были конъюгированы с биотином на своей поверхности (пунктирная линия), имели значительный уровень антитела в кровотоке через 24 часа после инъекции клеток. Напротив, мыши, которым инъецировали антитело к биотину с последующей инъекцией клеток, которые были конъюгированы с биотином на своей поверхности, становились истощенными по целевому антителу в пределах нескольких минут. Эти данные убедительно демонстрируют, что целевые антитела подвергаются быстрому и специфичному клиренсу из кровотока с помощью культивируемых клеток эритроидного ряда, которые содержат полипептид экзогенного антигена на своей поверхности.
[00678] Совместно данные убедительно демонстрируют, что соответствующие экзогенные антигены на экпрессирующих экзогенный антиген EHC-клетках способны связывать свои целевые молекулы in vivo и опосредовать быстрый клиренс циркулирующих в крови целевых молекул при нахождении в кровотоке, что имеет огромное терапевтическое значение.
Пример 26. Активность регуляторов комплемента
[00679] Активность регуляции комплемента у экспрессирующих экзогенный антиген EHC-клеток оценивали с помощью стандартных анализов CH50 и AH50, известных из уровня техники (см., например, Kabat et al. 1961 Exp Immunochem pp. 133-239 и Platts-Mills et al. 1974 J Immunol 113:348).
[00680] Вкратце, в анализе CH50 использовали эритроциты овцы (SRBC) в качестве клеток-мишеней. Вкратце, суспензию, содержащую 1×109 SRBC/мл, получали в буфере GVB(2+) (желатин/забуференный вероналом физиологический раствор с Ca2+ и Mg2+), pH 7,35. Гемолизин (антисыворотка кролика, содержащая антитела к иммуноглобулину овцы) титровали для определения оптимального разведения для сенсибилизирования SRBC. Разведенный гемолизин (1:800) смешивали с равным объемом SRBC (1×109 SRBC/мл) и полученную смесь инкубировали при 37°C в течение 15 минут. Это приводило к получению 5×108/мл покрытых антителами эритроцитов (EA). EA (100 мкл) инкубировали со 100 мкл пяти серийных двукратных разведений (1:20, 1:40, 1:80, 1:160 и 1:320) нормальной человеческой сыворотки (NHS) или подобным разведением смеси NHS и экспрессирующей экзогенный антиген EHC при 37°C в течение 1 часа. NHS, инкубируемую с буфером GVB2+, применяли в качестве контроля. Фоновый контроль получали путем инкубации EA только с буфером (сыворотку крови не добавляли), а полный лизис (100% гемолиз) определяли путем добавления к EA дистилированной воды. Реакцию останавливали с применением 1,2 мл ледяной 0,15 M NaCl, смесь центрифугировали для осаждения не подвергнутых лизису клеток и оптическую плотность супернатанта определяли спектрофотометрическим способом (412 нм). Процентную долю гемолиза определяли по отношению к контролю со 100% лизисом. Активность комплемента количественно оценивали путем определения разведения сыворотки крови, требуемого для лизиса 50% клеток в смеси для анализа. Результаты выражали как обратную величину данного разведения в CH50-единицы/мл сыворотки крови.
[00681] Вкратце, анализ AH50 зависит от лизиса несенсибилизированных эритроцитов кролика (Erab) с помощью человеческой сыворотки путем активации альтернативного пути. Активацию кальций-зависимого классического пути предотвращали путем добавления хелатора кальция этиленгликоль-тетрауксусной кислоты (EGTA) в буфер для анализа, и в буфер добавляли магний, необходимый для обоих путей. Вкратце, клеточную суспензию RBC кролика (2×108 клеток/мл) готовили в буфере GVB-Mg2+-EGTA. Серийное 1,5-кратное разведение (1:4, 1:6, 1:9, 1:13,5 и 1:20,25) нормальной человеческой сыворотки (NHS) или аналогичное разведение смеси NHS и экспрессирующей экзогенный антиген EHC получали в буфере GVB-Mg2+-EGTA и 100 мкл каждого разведения сыворотки крови добавляли в 50 мкл стандартизированных Erab. NHS, инкубируемую с буфером GVB-Mg2+-EGTA, применяли в качестве контроля. Затем смесь инкубировали в течение 60 минут при 37°C на водяной бане со встряхиванием для поддержания клеток в виде суспензии и 1,2 мл ледяного NaCl (0,15 M) применяли для остановки реакции. Пробирки центрифугировали при 1250 g при 4°C в течение 10 минут для осаждения клеток и оптическую плотность супернатанта определяли спектрофотометрическим способом (412 нм). Фоновый контроль содержал 100 мкл буфера GVB-Mg2+-EGTA и 50 мкл Erab и не превышал 10% полного лизиса. В пробирке для контроля полного лизиса 100 мкл дистилированной воды добавляли к 50 мкл суспензии Erab и процентную долю гемолиза определяли по отношению к контролю 100% лизиса. Подсчитывали результаты анализа и активность комплемента количественно оценивали путем определения разведения сыворотки крови, требуемого для лизиса 50% клеток в смеси для анализа. Результаты выражали как обратную величину данного разведения в AH50-единицы/мл сыворотки крови.
Пример 27. Активность тимидинфосфорилазы, загруженной в тромбоциты
[00682] Трансген, кодирующий тимидинфосфорилазу с HA-меткой на С-конце, конструировали путем сборки по Гибсону, как описано в данном документе. Тромбоциты культивировали из клеток-предшественников, как описано в данном документе. Трансген вводили в культивируемые клетки-предшественники тромбоцитов путем лентивирусной трансдукции, как описано в данном документе. Экспрессию тимидинфосфорилазы внутри культивируемых тромбоцитов оценивали с помощью вестерн-блоттинга с применением детекторного антитела к HA, как описано в данном документе.
[00683] Активность тимидинфосфорилазы определяли в образцах тромбоцитов путем количественной оценки скорости превращения тимидина в тимин. Предварительные эксперименты проводили для определения кинетики образования линейного метаболита в зависимости от времени и разведения фермента; показано, что способ является линейным вплоть до 16 мин, в диапазоне тиминфосфорилазы 4,0-719 нмоль/мин/мл (что соответствует диапазону разведения образца 10-9088). Лизаты предварительно подвергнутых диализу образцов культивируемых тромбоцитов и образцов контрольных тромбоцитов готовили путем разведения оттаявших образцов 1:710 с использованием 125 мМ Tris-HCl, pH 7,4. Двадцать пять мкл лизата тромбоцитов затем добавляли к 100 мкл буферного раствора на основе фосфата натрия (100 мM, pH 6,5) и 25 мкл стандарта тимидина (10 мM), смешивали и инкубировали при 37°C в течение 10 мин. Реакцию останавливали с помощью 25 мкл 40% TCA. Холостые пробы анализа готовили путем добавления TCA в смесь для инкубации с буферным раствором на основе фосфата натрия/тимидином перед добавлением лизата тромбоцитов. Образцы центрифугировали при 13400×g в течение 2 мин. и супернатант промывали дважды насыщенным водой диэтиловым эфиром в течение 2 мин на встряхивателе для экстракции TCA. Чтобы избежать противодействия эфира разделению с помощью HPLC, его эффективного удаления достигали путем оставления матрицы на воздухе в течение 5 мин для обеспечения возможности испарения эфира. Образец объемом 10 мкл инъецировали в HPLC.
[00684] Хроматографического разделения субстрата и продукта достигали с применением хроматографии с обращенной фазой с изократическим элюированием с применением системы Waters Alliance HPLC 2795. Предварительно упакованную C18-колонку (Spherisorb ODS 125 мм × 4,6 мм ID, размер частиц 5 мкм, Waters) применяли как стационарную стадию. Аналиты элюировали с применением подвижной фазы из ацетата аммония (40 мM) с агентом для образования пары ионов, тетрабутиламмоний гидросульфатом (5 мM), доведенной до pH 2,70 с помощью HCl, вводимой при скорости потока 1,0 мл/мин, со временем анализа, составляющим 8 мин. УФ-обнаружение проводили при 254 нм и 0,1 оптических единиц на всю шкалу. Метаболиты идентифицировали путем сравнения спектров с чистыми стандартами.
Пример 28. Активность антигена Гудпасчера, представленного на тромбоцитах
[00685] Трансген, кодирующий антиген NC1-домен коллагена альфа-3 (IV) (COL4A3), слитый с N-концом CD42b (GP1B, № в Genbank AAH27955.1), с промежуточной HA-меткой, конструировали путем сборки по Гибсону, как описано в данном документе. Тромбоциты культивировали из клеток-предшественников, как описано в данном документе. Трансген вводили в культивируемые клетки-предшественники тромбоцитов путем лентивирусной трансдукции, как описано в данном документе. Экспрессию экзогенного антигена на культивируемых тромбоцитах оценивали с помощью проточной цитометрии с применением детекторного антитела к HA, как описано в данном документе.
[00686] Сыворотку крови собирали от пациентов, страдающих от синдрома Гудпасчера, и сыворотку крови тестировали на наличие антител к COL4A3 с помощью коммерческого набора ELISA (набор ELISA MyBioSource COL4A3). Связывающую способность экпрессирующих антиген тромбоцитов оценивали с помощью проточной цитометрии, как описано в данном документе, с применением сыворотки крови к COL4A3 в качестве первичного детекторного антитела и флуоресцентного антитела к IgG человека в качестве вторичного детекторного антитела.
[00687] Облегченный тромбоцитами клиренс циркулирующего в крови антигена in vivo моделировали на мыши с применением экпрессирующих антиген тромбоцитов. Мышам NSG инъецировали 100 мкл мышиного антитела к COL4A3 человека (Creative BioMart), флуоресцентно меченного красителем Dylight 650. CFSE-меченные культивируемые тромбоциты (108 на мышь), которые экспрессировали экзогенный антиген, затем инъецировали через хвостовую вену. Кровь собирали из подчелюстного узла в моменты времени 10 мин, 30 мин, 2 ч, 12 ч и 24 ч. Кровь центрифугировали для сбора обогащенной тромбоцитами фракции, которую затем окрашивали и анализировали с помощью проточной цитометрии, как описано в данном документе. Захват антитела тромбоцитами определяли путем отслеживания двойной положительной в отношении CFSE-Dylight 650 популяции.
Пример 29. Активность in vivo (мышь)
[00688] Мышиные клетки эритроидного ряда культивировали, как описано в данном документе. Клетки-предшественники эритроидного ряда трансдуцировали соответствующим трансгеном полипептида экзогенного антигена, например, кодирующим рецептор комплемента 1 (CR1), с применением лентивируса, как описано в данном документе. Клетки культивировали до терминальной дифференцировки, как описано в данном документе. Присутствие экзогенного белка в клетках оценивали с помощью проточной цитометрии, как описано в данном документе. Клетки метили флуоресцентным красителем, например CFSE (Sigma Aldrich), в соответствии с инструкциями производителя для содействия их обнаружению. Клетки инъецировали в мышиную модель волчанки NZBWF1/J или другую пригодную модель заболевания или активности, соответствующую подходящему полипептиду экзогенного антигена, при этом приблизительно 1×108 клеток инъецировали через хвостовую вену. Кровь собирали в несколько моментов времени путем подчелюстной пункции. Уровни иммунных комплексов в плазме крови обнаруживали с помощью анализа клеток Раджи, см. например, Theofilopoulos et al. 1976, J Clin Invest 57(1):169. Фармакокинетику культивируемых клеток оценивали с помощью проточной цитометрии, как описано в данном документе, путем отслеживания процентной доли CFSE-флуоресцентных клеток в образце свежевзятой крови. Общее состояние здоровья мыши оценивали с помощью общей аутопсии, включающей гистологическое исследование ткани почки, для отслеживания снижения уровня отложения иммунных комплексов и опосредованного воспалением повреждения.
Пример 30. Быстрый скрининг
[00689] Линии клеток, например 293T и K562, имеют более короткие циклы экспрессии и культивирования (~1 день) по сравнению с культивируемыми клетками эритроидного ряда (дни - недели). Эти линии клеток можно применять для быстрой итерации по библиотеке генов, кодирующих полипептиды соответствующего экзогенного антигена, чтобы идентифицировать полипептид экзогенного антигена с наивысшей экспрессией или активностью.
[00690] Библиотеку трансгенов полипептида соответствующего экзогенного антигена, например, полноразмерных и более коротких вариантов рецептора комплемента 1 (CR1), конструировали с помощью полимеразной цепной реакции и сборки по Гибсону, как описано в данном документе. Библиотеку трансгенов трансфицировали в клетки HEK293Т параллельным способом в титрационном микропланшете с применением липофектамина, как описано в данном документе, и трансдуцировали в клетки K562 с применением лентивируса, как описано в данном документе. Экспрессию экзогенных антигенов оценивали с помощью проточной цитометрии, как описано в данном документе, через 24-48 часов. Активность каждого из экзогенных антигенов в библиотеке оценивали по захвату флуоресцентного иммунного комплекса, обнаруживаемому с помощью проточной цитометрии, как описано в данном документе, и по переносу флуоресцентных иммунных комплексов к культивируемым моноцитам, обнаруживаемому с помощью проточной цитометрии, как описано в данном документе. Экзогенные антигены из библиотеки, которые являются наиболее функциональными - например, экспрессируются на самом высоком уровне, захватывают наибольшее количество иммунных комплексов или лучше всего переносят иммунные комплексы к моноцитам - затем индивидуально трансдуцировали в параллельные культуры клеток эритроидного ряда, как описано в данном документе, с применением лентивируса, как описано в данном документе. Экспрессию каждого экзогенного антигена на культивируемых клетках эритроидного ряда оценивали с помощью проточной цитометрии, как описано в данном документе. Активность каждого экзогенного антигена на культивируемых клетках эритроидного ряда оценивали по захвату флуоресцентного иммунного комплекса, обнаруживаемому с помощью проточной цитометрии, как описано в данном документе, и по переносу флуоресцентных иммунных комплексов к культивируемым моноцитам, обнаруживаемому с помощью проточной цитометрии, как описано в данном документе.
Пример 31. Оценка скорости клиренса RBC in vivo
[00691] Скорость клиренса клеток эритроидного ряда оценивали in vivo на модели мыши с ослабленным иммунитетом. Мышей NSG обрабатывали в день -1 с помощью 100 мкл раствора клодронатных липосом (Clodrosomes.com) для селективного истощения макрофагов. Клетки метили флуоресцентной меткой CFSE и приблизительно 1×108 клеток инъецировали в каждую мышь через хвостовую вену. Через равные промежутки времени кровь собирали путем подчелюстной пункции и собирали кровяные клетки. Клетки совместно окрашивали с помощью антитела к человеческому GPA и анализировали с помощью проточной цитометрии. Человеческие клетки эритроидного ряда отличали от мышиных клеток эритроидного ряда по сигналу CFSE и по сигналу человеческого GPA.
[00692] Для терапевтических применений важно, чтобы культивируемые клетки эритроидного ряда и культивируемые клетки эритроидного ряда, содержащие экзогенный белок либо внутри клетки, либо на поверхности, нормально циркулировали в крови in vivo. Это показано на фигуре 9 с применением стандартной модели мыши с ослабленным иммунитетом. На фигуре 9A показано, что кровь, собранную у подвергнутой инъекции мыши, анализировали на проточном цитометре. Культивируемые человеческие клетки эритроидного ряда идентифицировали в верхнем правом квадранте графика как двойные положительные по CFSE и человеческому GPA. На фигуре 9B показано, что мышам инъецировали человеческие красные кровяные клетки (кружки, соединенные сплошной линией), культивируемые безъядерные клетки эритроидного ряда (ромбы, соединенные пунктиром), культивируемые безъядерные клетки эритроидного ряда, которые экспрессируют внутриклеточный экзогенный белок (квадратики, соединенные точечным пунктиром), и культивируемые безъядерные клетки эритроидного ряда, которые экспрессируют поверхностный экзогенный белок (незакрашенные треугольники). Скорость клиренса человеческих клеток измеряли как процентную долю CFSE+ клеток, сохраняющихся в течение некоторого времени, приведенную к доле в начальный момент времени (20 минут после инъекции). Среди четырех образцов не отмечено значительного отличия в скорости клиренса.
[00693] Эти данные четко показывают, что культивируемые безъядерные клетки эритроидного ряда характеризуются циркуляцией, в значительной степени аналогичной циркуляции нормальных человеческих красных кровяных клеток. Кроме того, экзогенные белки, экспрессируемые либо во внутриклеточном пространстве, либо на поверхности клеток, не оказывали значительного влияния на характер циркуляции этих клеток. Это является важным результатом для терапевтического применения данной технологии.
Пример 32. Оценка побочных эффектов в кровотоке
[00694] Частоту побочных эффектов, вызванных культивируемыми клетами эритроидного ряда в кровотоке, оценивали путем обнаружения продуктов распада фибриногена в крови и гистологического исследования животных, которым инъецировали культивируемые клетки эритроидного ряда.
[00695] Обнаружение продуктов распада фибриногена. Мышам инъецировали культивируемые клетки эритроидного ряда, как описано в данном документе. Кровь собирали у мышей путем подчелюстной пункции в пробирку, содержащую EDTA. Клетки разделяли путем центрифугирования и собирали плазму крови. Уровни продуктов распада фибриногена, фибринопептида A и фибринопептида B, измеряли в плазме крови мыши с помощью ELISA (MyBiosource) в соответствии с инструкциями производителя.
[00696] Гистологическое исследование. Образцы ткани от тех самых мышей собирали после аутопсии. Ткани иссекали, заливали в твердый парафин и делали срезы. Срезы ткани окрашивали с помощью окрашивания H&E и трихромного окрашивания. Изображения под микроскопом получали при увеличении 10× и 20×.
[00697] Для терапевтических применений важно, чтобы культивируемые клетки эритроидного ряда и культивируемые клетки эритроидного ряда, которые содержат экзогенные белки (либо внутри клетки, либо на поверхности), не индуцировали побочные эффекты, такие как активация системы свертывания крови и образование ткани тромба.
[00698] На фигурах 10A и 10B показаны уровни фибринопептидов A и B в плазме крови мыши у мышей, которым инъецировали (1) человеческие красные кровяные клетки, (2) культивируемые безъядерные клетки эритроидного ряда, (3) культивируемые безъядерные клетки эритроидного ряда, экспрессирующие внутриклеточный экзогенный белок, (4) культивируемые безъядерные клетки эритроидного ряда, экспрессирующие поверхностный экзогенный белок, и (5) только рекомбинантный белок. Уровни фибринопептидов A и B, маркеров распада фибриногена и активации системы свертывания крови, в значительной степени аналогичны для всех образцов.
[00699] На фигурах 10C и 10D показаны окрашенные гистологические срезы селезенки мышей, которым инъецировали культивируемые безъядерные клетки эритроидного ряда (10C) и рекомбинантный белок (10D). Значительная разница между тканями отсутствовала и не наблюдалось идентифицируемого повреждения ткани в селезенке, печени, легком, головном мозге, сердце и почке у какого-либо из образцов.
[00700] Эти данные убедительно демонстрируют, что культивируемые клетки эритроидного ряда с экзогенным белком или без него не индуцируют какие-либо побочные эффекты при нахождении в кровотоке мыши.
Пример 33. Оценка сохранения экзогенного белка в кровотоке
[00701] Сохранение экзогенных белков в культивируемых безъядерных клетках эритроидного ряда и на них оценивали с помощью проточной цитометрии и вестерн-блоттинга.
1. Сохранение экзогенного белка, оцениваемое с помощью проточной цитометрии
[00702] Клетки эритроидного ряда культивировали, как описано в данном документе. Трансгенную конструкцию, кодирующую сигнальную последовательность гликофорина A, scFv-антитело, специфичное в отношении поверхностного антигена гепатита B, HA-эпитопную метку и кодирующую последовательность гликофорина A, собирали путем сборки по Гибсону, как описано в данном документе. Трансген вводили в клетки эритроидного ряда путем лентивирусной трансдукции, как описано в данном документе. Клетки культивировали до терминальной дифференцировки, как описано в данном документе. Клетки флуоресцентно метили с помощью CFSE и инъецировали в мышь NSG с ослабленным иммунитетом (Jackson Labs) через хвостовую вену (1×10^8 клеток на мышь). Через равные промежутки времени кровь собирали путем подчелюстной пункции. Собранные клетки окрашивали флуоресцентным антителом к HA (Abcam) и анализировали с помощью проточной цитометрии. Человеческие клетки идентифицировали как CFSE+ клетки и сохранение экзогенного белка оценивали по фракции CFSE+ клеток, которые также окрашивали положительно в отношении эпитопной метки.
2. Сохранение экзогенного белка, оцениваемое с помощью вестерн-блот-анализа
[00703] Клетки эритроидного ряда культивировали, как описано в данном документе. Трансгенную конструкцию, кодирующую аденозиндезаминазу и HA-эпитопную метку, собирали путем сборки по Гибсону, как описано в данном документе. Трансген вводили в клетки эритроидного ряда путем лентивирусной трансдукции, как описано в данном документе. Клетки культивировали до терминальной дифференцировки, как описано в данном документе. Клетки флуоресцентно метили с помощью CFSE и инъецировали в мышь NSG с ослабленным иммунитетом (Jackson Labs) через хвостовую вену (1×108 клеток на мышь). Через равные промежутки времени кровь собирали путем подчелюстной пункции. Собранные клетки промывали, лизировали и анализировали с помощью вестерн-блот-анализа, как описано в данном документе, с использованием детекторного антитела к HA-эпитопной метке.
[00704] Для терапевтических применений важно, чтобы культивируемые клетки эритроидного ряда, которые содержат экзогенные белки либо внутри клетки, либо на поверхности, сохраняли эти трансгены при нахождении в кровотоке. Этот факт является нетривиальным, с учетом того, что в уровне техники часто выдвигались гипотезы о том, что клетки эритроидного ряда подвергаются строгой программе созревания и элиминации белков, не являющихся необходимыми для основной функции при нахождении в кровотоке по мере их созревания (Liu J et al. (2010) Blood 115(10):2021-2027, Lodish HF et al. (1975) Developmental Biology 47(1):59).
[00705] На фигуре 11 показано, что экзогенные белки, экспресируемые в культивируемых безъядерных клетках эритроидного ряда или на них, сохранялись в кровотоке. На фигуре 11A мышам инъецировали культивирумые безъядерные клетки эритроидного ряда, которые экспрессировали scFv-антитело на своей поверхности. Процентная доля клеток, положительных по scFv-антителу, начиналась со значения 50% и сохранялась устойчивой на этом уровне приблизительно в течение всего многодневного исследования в кровотоке. На фигуре 11B показано, что мышам инъецировали либо культивируемые безъядерные клетки эритроидного ряда, которые экспрессировали цитоплазматический фермент с HA-меткой, либо рекомбинантный фермент с HA меткой. При анализе с помощью вестерн-блот-анализа ясно видно, что фермент сохранялся внутри культивируемых клеток на всем протяжении эксперимента. Снижение интенсивности полос можно объяснить клиренсом клеток во время эксперимента, а не удалением экзогенного фермента из указанных клеток.
[00706] Эти данные четко показывают, что экзогенные белки, экспрессируемые в культивируемых безъядерных клетках эритроидного ряда или на них, сохраняются в клетках в кровотоке и на них, что имеет огромное и беспрецедентное значение для терапевтической актуальности.
Пример 34. Оценка продления периода полувыведения in vivo
[00707] Клетки эритроидного ряда культивировали, как описано в данном документе. Трансгенную конструкцию, кодирующую аденозиндезаминазу и HA-эпитопную метку, собирали путем сборки по Гибсону, как описано в данном документе. Трансген вводили в клетки эритроидного ряда путем лентивирусной трансдукции, как описано в данном документе. Клетки культивировали до терминальной дифференцировки, как описано в данном документе. Клетки флуоресцентно метили с помощью CFSE и инъецировали в мышь NSG с ослабленным иммунитетом (Jackson Labs) через хвостовую вену (1×10^8 клеток на мышь). Через равные промежутки времени кровь собирали путем подчелюстной пункции. Собранные клетки промывали, лизировали и анализировали с помощью вестерн-блот-анализа, как описано в данном документе, с использованием детекторного антитела к HA-эпитопной метке.
[00708] Трансген, кодирующий аденозиндезаминазу с HA-меткой на С-конце, конструировали путем сборки по Гибсону, как описано в данном документе. Трансген вводили в клетки HEK-293Т с помощью трансфекции с применением липофектамина (Life Technologies), как описано в данном документе. Белок очищали от супернатанта культуры клеток через 7 дней с применением афинной смолы к HA (Pierce) в соответствии с инструкциями производителя. Концентрацию белка оценивали по поглощению света при длине волны 280 нм. Белок (40 мкг) инъецировали в мышь NSG с ослабленным иммунитетом (Jackson Labs) через хвостовую вену. Через равные промежутки времени кровь собирали путем подчелюстной пункции. Плазму крови анализировали с помощью вестерн-блот-анализа, как описано в данном документе, с использованием детекторного антитела против HA-эпитопной метки.
[00709] На фигуре 11B показано, что мышам инъецировали либо культивируемые безъядерные клетки эритроидного ряда, которые экспрессировали цитоплазматический фермент с HA-меткой, либо рекомбинантный фермент с HA-меткой. При анализе с помощью вестерн-блот-анализа чсно видно, что период полувыведения циркулирующего в крови фермента значительно продлевается при экспрессии внутри циркулирующей в крови клетке по сравнению с инъекцией в растворимой форме.
Пример 35. Оценка скорости клиренса in vivo - тромбоциты
[00710] Популяцию тромбоцитов, экспрессирующих экзогенную тимидинфосфорилазу, культивировали с применением процедуры, подробно описанной в данном документе, и метили с помощью CFSE, и инъецировали в мышь NSG через хвостовую вену. Популяцию нативных тромбоцитов, полученных от человека, аналогичным образом метили с помощью CFSE и инъецировали в другую мышь. Проводили забор образцов у обеих мышей в моменты времени 10 мин, 1 ч, 4 ч, 8 ч, 24 ч и 48 ч и применяли проточную цитометрию для количественного оценки уровней тромбоцитов в кровотоке. Сравнивали периоды полувыведения природных и культивируемых тромбоцитов.
Пример 36. Оценка побочных эффектов в кровотоке - тромбоциты
[00711] Для терапевтических применений важно, чтобы культивируемые тромбоциты и культивируемые тромбоциты, которые содержат экзогенные белки (либо внутри клетки, либо на поверхности), не индуцировали побочные эффекты, такие как активация системы свертывания крови и образование ткани тромба. После инъекции культивируемых тромбоцитов в мышь NSG через хвостовую вену продукты распада фибриногена, фибринопептид A и фибринопептид B, определяли в плазме крови мыши с помощью ELISA в соответствии с протоколом производителя (MyBiosource). Образцы ткани от мышей NSG собирали после аутопсии. Ткани иссекали, заливали в твердый парафин и делали срезы. Срезы ткани окрашивали с помощью окрашивания H&E и трихромного окрашивания. Изображения под микроскопом получали при увеличении 10× и 20×, и их оценивал квалифицированный патологоанатом на наличие любых патогенных признаков.
Пример 37. Оценка сохранения экзогенного белка в кровотоке - тромбоциты
[00712] Сохранение экзогенных белков в культивируемых тромбоцитах и на них оценивали с помощью проточной цитометрии и вестерн-блоттинга.
[00713] CFSE-меченные тромбоциты, которые содержали внутриклеточный экзогенный белок, инъецировали в мышь через хвостовую вену. Через равные промежутки времени кровь собирали путем подчелюстной пункции. Кровь центрифугировали для отделения обогащенной тромбоцитами плазмы крови, которую затем лизировали и анализировали с помощью вестерн-блот-анализа с окрашиванием в отношении эпитопной метки, присутствующей на экзогенном белке.
Пример 38. Получение донорных клеток для производства
[00714] После получения информированного согласия здоровым донорам CD34+ стволовых клеток принимали rhG-CSF (Granocyte или Neupogen), 10 мкг/кг/день s.c., в течение 5 дней для мобилизации стволовых клеток в периферическую кровь и затем их подвергали аферезу в течение 2 последующих дней для сбора активированных CD34+ HSC. Мононуклеарные клетки (MNC) выделяли из мобилизованной периферической крови путем центрифугирования в градиенте плотности Ficoll и разделяли на две части. Одну часть использовали для очистки CD34+ клеток путем применения магнитных микрогранул, покрытых антителами к CD34 (Miltenyi Biotec, Inc., Германия), в соответствии с протоколом Miltenyi. Чистоту CD34+ фракций контролировали. Затем CD34+-обогащенные HSC применяли непосредственно в двухстадийном способе культивирования или замораживали до применения в одностадийном способе культивирования.
[00715] Применяемая полная среда (CM) представляла собой RPMI 1640 (Eurobio, Франция), дополненную 2 мМ L-глутамина и 100 МЕ/мл пенициллина-стрептомицина (Gibco, Гранд-Айленд, Нью-Йорк, США), а также 10% инактивированной нагреванием FBS (Gibco). IMDM (Gibco), дополненную 10% инактивированной нагреванием FBS, применяли для размножения. Рекомбинантный человеческий фактор стволовых клеток (rhSCF), тромбопоэтин (TPO), лиганд тирозинкиназы 3 фетальной печени (Flt-3L), GM-CSF и TNF-альфа приобретали у R&D Systems (Миннеаполис, Миннесота, США).
Пример 39. Увеличение объема для производства
[00716] Клетки эритроидного ряда последовательно увеличивали в объеме, поддерживая клетки при плотности, составляющей приблизительно от 1×105 до 2×106 клеток/мл в статической культуре. Посев для стадии размножения проводили при концентрации 105/мл, и она включала 3-7 переносов с увеличением объема; 100 мл, 500 мл, 1 л, 10 л, 50 л, 100 л, 100 л. Во время производства клеточная среда включала комбинацию IMDM, FBS, BSA, голотрансферрина, инсулина, глутамина, дексаметазона, бета-эстрадиола, IL-3, SCF и эритропоэтина. Когда клетки достигали объема, соответствующего для посева в производственный биореактор, их переносили в производственный биореактор для конечного увеличения объема и дифференцировки.
Пример 40. Культивирование клеток в биореакторе (Wave)
[00717] Систему 2/10 биореактора WAVE настраивали в соответствии с руководством по эксплуатации. Вкратце, Cellbag собирали на качающемся узле, который помещали на модуль перфузии. После нагнетания в мешок воздуха вес устанавливали на ноль. Затем мешок заполняли соответствующим количеством культуральной среды и инкубировали в течение по меньшей мере двух часов, позволяя среде достигнуть 37°C. Среду и клетки переносили в мешок посредством флакона для переноса, специально разработанного стеклянной бутыли DURAN с двумя отверстиями. В верхней части к отверстию присоединяли фильтр. Во втором отверстии, на дне флакона, монтировали трубку. Трубку на флаконе для переноса соединяли с патрубком для загрузки Cellbag. Флакон для переноса устанавливали в LAF-бокс для снижения риска загрязнения.
[00718] Перед началом перфузии систему трубок и контейнеры для сбора и загрузки присоединяли к Cellbag. Систему трубок готовили следующим образом: силиконовую трубку Saniflex ASTP-ELP (Gore/Saniflex AB) длиной 50 или 70 см с внутренним и внешним диаметром 3,2 и 6,4 мм, соответственно, оснащали присоединительными конусами луер-лок на обоих концах. Силиконовую трубку присоединяли к одному концу трубки C-Flex посредством канюли луер-лок. На другом конце трубки C-Flex монтировали конус луер-лок и затем систему трубок автоклавировали. Замки луер-лок удерживались на месте с помощью хомутиков-стяжек на всех трубках. Перед началом перфузии силиконовую часть присоединяли к Cellbag, а часть C-Flex к контейнеру на 5 л (Hyclone Labtainer) как для загрузки, так и для сбора. Все соединения выполняли в шкафу с ламинарным потоком воздуха.
[00719] Контроль факторов внешней среды и метаболических факторов может приводить к изменениям экспрессии или активности факторов транскрипции и регуляторных белков генов клеток эритроидного ряда в культуре, см. например, Csaszar et al., 2009 Biotechnol Bioeng 103(2):402; Csaszar et al. 2012 Cell Stem Cell 10(2):218. Для обеспечения контроля ввода и вывода из реактора создавали систему доставки микрообъемов, ключевым компонентом которой являлся капилляр из плавленого кварца длиной 60-80 см (#TSP100375, Polymicro Technologies) с внутренним диаметром 100 мкм. На входном конце к капилляру подводили шприц для исходной культуры с наконечником луер-лок (#309585 BD), соединенный посредством PEEK-луера с адаптером MicroTight (#P-662, Upchurch Scientific). Шприц для исходной культуры загружали на шприцевой насос Twin, модель 33 (#553333, Harvard Apparatus), храянщийся в холодильнике при 4°C. На выходном конце капилляр входил в биореактор: мешок для культуры клеток FEP с двумя отверстиями (#2PF-0002, VueLife), помещенный на орбитальный встряхиватель в термостат для культуры клеток при 37°C с 5% CO2. Капилляр проходил через самоуплотняющуюся резиновую мембрану (#B-IIS, InterLink) с помощью иголы в средину биореактора. Противоположный разъем на биореакторе замещали дополнительной самоуплотняющейся резиновой мембраной. Шприцы для исходной культуры и капилляры для доставки блокировали в течение ночи перед использованием раствором PBS с 10% фетальной бычьей сыворотки для предотвращения прилипания белка к стенкам шприца и капилляра.
[00720] LabVIEW 7.1 от National Instruments применяли для создания программы для контроля инъекций с помощью шприцевого насоса. Основная стратегия дозирования согласно программе представляет собой первоначальную инъекцию до концентрации L1 с последующим временем ожидания t1 и последующими инъекциями, каждая до концентрации L2, и последующим временем ожидания t2, повторяющимися n раз. Пользователь задает скорость потока, концентрацию исходной культуры, начальный объем культуры, требуемую концентрацию после инъекций, время между инъекциями и общее число инъекций.
Пример 41. Оценка иммуногенности и индукции толерантности
1. Индукция толерантности у мышей
[00721] У мышей толерантность можно индуцировать путем 3 последовательных внутривенных инъекций композиции клеток по настоящему изобретению, содержащей антигенный белок, в данном случае oвальбумин (OVA). Не подвергавшимся воздействию мышам инъецировали в дни -7, -3 и -1 либо свободный OVA, либо OVA, экспрессируемый внутри композиции клеток по настоящему изобретению (клеточный OVA). Затем мышей иммунизировали с помощью OVA путем двух инъекций антигена, смешанного с адъювантом поли(I:C) (Invivogen, Сан-Диего, Калифорния), для индукции сильного иммунного ответа.
2. Оценка титра антител
[00722] Уровни IgG в сыворотке крови мыши оценивали с помощью стандартного ELISA. Вкратце, сыворотку крови получали в разные моменты времени из образцов крови мышей, которым была инъецированы композиция клеток по настоящему изобретению, содержащая антигенный белок, например овальбумин (OVA), и мышей, которым был инъецирован свободный или рекомбинантный OVA. Применяли стандартный анализ ELISA с OVA в качестве антигена (1 мкг/мл в 50 мМ карбонатного буфера, pH 9,7), адсорбированного на аналитических планшетах. Образцы сыворотки крови серийно разводили в диапазоне 1:50-1:200 для сыворотки крови перед обработкой или без обработки и 1:400-1:500000 для сыворотки крови после обработки и тестировали в двух повторностях. Связывание антител к OVA в сыворотке крови с адсорбированным рекомбинантным OVA обнаруживали колориметрическим способом с помощью вторичного антитела к иммуноглобулину мыши, конъюгированного с пероксидазой хрена, с последующей обработкой хромогенным субстратом.
3. Анализ Т-клеточных иммунных ответов
[00723] Толерантность индуцировали к антигенному белку OVA, как описано в данном документе. Мышей умерщвляли через 7 дней после 2-го введения инъекции OVA фазы иммунизации и их селезенки собирали. Суспензии клеток селезенки получали путем процеживания органов через фильтр для клеток с размером пор 70 микрон и после лизиса RBC с помощью 0,8% раствора хлорида аммония (Stem-Cell Technologies, Гренобль, Франция). Все образцы инкубировали с антителом к Fc-рецептору (очищенное антитело к CD16/32, Ozyme, Сан-Диего, Калифорния) для предотвращения неспецифичного связывания перед инкубациями с антителами для анализа. Для окрашивания клеток селезенки использовали следующие моноклональные антитела (Ab). Конъюгат PC5-антитело к CD62L (MEL14) и конъюгат PC7-антитело к CD8 приобретали у Biolegend. Тетрамеры пептид OVA-MHC (тетрамеры PE-Kb-SIINFEKL) приобретали у Beckmann Coulter. OVA-специфичные Т-клетки идентифицировали путем проточной цитометрии как двойные положительные клетки при окрашивании антителом к CD8 и тетрамерами пептид OVA-MCH. Из данной популяции клеток процентную долю OVA-специфичных CD8 Т-клеток, которые были активированными, определяли по фракции клеток, которые были положительными в отношении окрашивания антителом к CD62L.
4. Анализ опосредованного Т-клетками лизиса in vivo
[00724] Не подвергавшиеся воздействию клетки селезенки активировали с помощью 10 микрограммов/мл пептида SIINFEKL (Genscript, Пискатауэй, Нью-Джерси) при 37°C в течение 1 часа и затем метили с помощью 0,4 микроM CFSE (низкая концентрация CFSE). Контрольную популяцию необработанных спленоцитов метили с помощью 4 микроM CFSE (высокая концентрация CFSE). Клетки с низкой концентрацией CFSE и высокой концентрацией CFSE объединяли в соотношении 1:1 и 1E7 клеток на мышь инъецировали i.v. путем мышам, у которых предварительно индуцировали толерантность к антигену OVA, как описано в данном документе, или мышам, которых иммунизировали антигеном OVA, как описано в данном документе. Через шестнадцать часов суспензии отдельных клеток селезенки готовили, как описано в данном документе, и анализировали с применением проточной цитометрии для определения соотношения клеток с низкой концентрацией CFSE/высокой концентрацией CFSE.
Пример 42. Оценка размножения и дифференцировки культивируемых клеток эритроидного ряда
[00725] Для оценки размножения, дифференцировки и удаления ядра in vitro дифференцированных клеток важно убедиться в том, что введение трансгена не влияет негативно на качество клеток в культуре. Размножение оценивали путем подсчета клеток. Дифференцировку оценивали с помощью проточной цитометрии, вестерн-блот-анализа и RT-PCR. Удаление ядра оценивали с помощью проточной цитометрии.
[00726] Оценка скорости размножения путем подсчета клеток. Клетки эритроидного ряда культивировали, как описано в данном документе. В разные моменты времени клетки собирали, промывали с помощью PBS и подсчитывали с применением автоматического устройства для подсчета клеток Countess (Life Technologies). Скорость размножения клеток определяли по росту количества клеток в зависимости от времени.
[00727] Оценка дифференцировки с помощью проточной цитометрии. Клетки эритроидного ряда культивировали, как описано в данном документе. В разные моменты времени клетки собирали, промывали с помощью PBS и окрашивали с помощью 1:100 разведений флуоресцентных антител к маркерам клеточной поверхности GPA (CD235a), CKIT (CD117) и TR (CD71), приобретенных у Life Technologies. Меченные клетки анализировали с помощью проточной цитометрии, как описано в данном документе.
[00728] Оценка дифференцировки с помощью вестерн-блот-анализа. Клетки эритроидного ряда культивировали, как описано в данном документе. В разные моменты времени клетки собирали, промывали с помощью PBS, лизировали с помощью буфера RIPA и анализировали с помощью вестерн-блот-анализа, как описано в данном документе, с применением антител к маркерам дифференцировки GATA1, GATA2, Band3, CD44 и актину (Abcam).
[00729] Оценка удаления ядра с помощью проточной цитометрии. Клетки эритроидного ряда культивировали, как описано в данном документе. В разные моменты времени клетки собирали, промывали с помощью PBS и окрашивали флуоресцентным антителом против гликофорина A (Life Technologies) и красителем для нуклеиновой кислоты DRAQ5 (Pierce) при рекомендуемых производителем разведениях и анализировали на проточном цитометре Attune, как описано в данном документе.
[00730] Оценка удаления ядра с помощью микроскопии (бензидин-краситель Гимза). Клетки эритроидного ряда культивировали, как описано в данном документе. В разные моменты времени клетки собирали, промывали с помощью PBS и центрифугировали на предметных стеклах с применением Cytospin (Thermo Scientific). Клетки фиксировали после центифугирования на Cytospin с помощью метанола с температурой -20°C в течение 2 мин при комнатной температуре, промывали водой и высушивали на воздухе. Таблетку бензидина (Sigma#D5905) растворяли в 10 мл PBS, к которому добавляли 10 мкл H2O2. Раствор фильтровали с помощью шприцевого фильтра с размером пор 0,22 мкм. Пятно с клетками на покровном стекле покрывали 300-500 мкл раствора бензидина, инкубировали при комнатной температуре в течение 1 ч, затем промывали водой. Краситель Гимза разводили водой (Sigma#GS500) в соотношении 1:20. Пятно с клетками на покровном стекле покрывали 300-500 мкл раствора Гимза, инкубировали при комнатной температуре в течение 40 минут, промывали водой и высушивали на воздухе. Стекла монтировали на предметные стекла и запечатывали перед визуализацией под микроскопом.
[00731] На фигуре 12A показана скорость размножения клеток эритроидного ряда в культуре в течение семидневного окна размножения и дифференцировки для клеток, которые содержат трансгены (пунктирная линия и точечная пунктирная линия), и клеток, которые не содержат трансген (сплошная линия). Следует отметить, что скорость размножения культивируемых клеток, которые содержат трансген, неотличима от скорости размножения клеток, которые не содержат трансген.
[00732] На фигуре 12B представлен ряд графиков результатов проточной цитометрии клеток, окрашенных с помощью антител против маркеров дифференцировки, расположенных на клеточной поверхности, GPA и CKIT. На данной конкретной стадии дифференцировки у культуры исчезала экспрессия CKIT и повышалась экспрессия GPA, поскольку клетки приближались к терминальной стадии созревания. Следует отметить, культивируемые клетки, которые содержат трансген, неотличимы от тех, которые не содержат трансген, по данному критерию дифференцировки.
[00733] На фигуре 12C представлен ряд графиков результатов проточной цитометрии клеток, которые окрашивали с помощью антитела против маркера клеточной поверхности GPA и флуоресцентного красителя ДНК. Бросаются в глаза три популяции клеток: (1) клетки с высоким уровнем GPA и низким уровнем ДНК, включающие безъядерные клетки эритроидного ряда; (2) клетки с высоким уровнем GPA и высоким уровнем ДНК, включающие клетки эритроидного ряда, в которых все еще содержится генетический материал; и (3) клетки с низким уровнем GPA и высоким уровнем ДНК, включающие пиреноциты или инкапсулированные в мембрану ядра, выброшенные из безъядерных клеток. Следует отметить, что культивируемые клетки, которые содержат трансген, неотличимы от тех, которые не содержат трансген, по данному критерию удаления ядра.
[00734] Введение трансгена в культуру клеток не влияет заметным образом на скорость размножения, дифференцировку или скорость удаления ядра из клетки в культуре.
Пример 43. Оценка содержания гемоглобина
1. Общий гемоглобин
[00735] Содержание гемоглобина в эритроцитах определяли с помощью реагента Драбкина (Sigma-Aldrich, продукт D5941) в соответствии с инструкциями производителя. Вкратце, кровяные клетки объединяли с реагентом в водном буфере, тщательно перемешивали и поглощение света при длине волны 540 нм измеряли с применением стандартного спектрофотометра. Калибровочную кривую растворимого гемоглобина применяли для количественного оценки содержания гемоглобина в клетках.
2. Типирование гемоглобина с помощью RT-PCR
[00736] Клетки лизировали и общую РНК собирали. Обратную транскрипцию осуществляли с применением системы синтеза первой нити SuperScript для RT-PCR (Life Technologies) в соответствии с протоколом производителя. Вкратце, общую РНК (5 мкг) инкубировали со 150 нг случайного гексамерного праймера и 10 нмоль смеси dNTP в 10 мкл H2O в течение пяти минут при 65°C, затем 1 минуту на льду. Исходную смесь для реакции готовили с помощью 2 мкл 10× RT-буфера, 4 мкл 25 мМ MgCl2, 2 мкл 0,1 M DTT и 1 мкл RNAseOUT. Реакционную смесь добавляли в смесь РНК/праймер, недолго перемешивали и затем оставляли при комнатной температуре на 2 мин. 1 мкл (50 единиц) SuperScript II RT добавляли в каждую пробирку, смешивали и инкубировали при 25°C в течение 10 мин. Реакционную смесь инкубировали при 42°C в течение 50 мин, инактивировали нагреванием при 70°C в течение 15 мин, затем хранили на льду. 1 мкл РНКазы H добавляли и инкубировали при 37°C в течение 20 мин. Этот продукт реакции, 1ую нить кДНК, затем хранили при -20°C до применения в RT-PCR.
[00737] Праймеры для амплификации различных генов гемоглобина и контрольные гены приобретали у IDT-DNA. Праймеры были следующими: hHBB_F - tcctgaggagaagtctgccgt (Seq. ID No. 9); hHBB_R - ggagtggacagatccccaaag (Seq. ID No. 10); hHBA_F1 - tctcctgccgacaagaccaa (Seq. ID No. 11); hHBA_R1 - gcagtggcttagcttgaagttg (Seq. ID No. 12); hHBA_F2 - caacttcaagctaagccactgc (Seq. ID No. 13); hHBA_R2 - cggtgctcacagaagccag (Seq. ID No. 14); hHBD_F - gactgctgtcaatgccctgt (Seq. ID No. 15); hHBD_R - aaaggcacctagcaccttctt (Seq. ID No. 16); hHBG2_F - cactggagctacagacaagaaggtg (Seq. ID No. 17); hHBG2_R - tctcccaccatagaagataccagg (Seq. ID No. 18); hHBE_F - aagagcctcaggatccagcac (Seq. ID No. 19); hHBE_R - tcagcagtgatggatggacac (Seq. ID No. 20); h18S-RNA-F - cgcagctaggaataatggaatagg (Seq. ID No. 21); h18S-RNA-R - catggcctcagttccgaaa (Seq. ID No. 22).
[00738] Реакционную смесь для RT PCR готовили с 25 мкл SYBR Green Mix (2x) (Applied Biosystems), 0,5 мкл 1ой нити кДНК, 2 мкл смеси пары прямой/обратный праймер (каждый праймер при концентрации 5 пмоль/мкл) в общем объеме 50 мкл H2O. Реакции прогоняли в приборе ABI Prism SDS 7000 (Applied Biosystems) с применением следующего цикла амплификации: 50°C - 2 мин, 1 цикл; 95°C - 10 мин, 1 цикл; 95°C - 15 с -> 60°C - 30 с -> 72°C - 30 с, 40 циклов; 72°C - 10 мин, 1 цикл. Анализа кривой диссоциации и результаты RT-PCR получали с применением прибора SDS 7000.
Пример 44. Оценка дифференцировки культивируемых тромбоцитов - FACS
[00739] Состояние дифференцировки тромбоцитов в культуре можно оценивать с помощью проточной цитометрии. Мегакариоциты (MK) представляют собой клетки с отличительной морфологией, которые предшествуют терминальной дифференцировке тромбоцитов. Для определения степени созревания в направлении MK 1×106 культивируемых клеток (LAMA-84 и CD34+ клетки) промывали и затем метили с помощью (a) конъюгата антитело к CD41-FITC (GpIIb/IIIa; BD Bioscience, Сан-Хосе, Калифорния, США) или конъюгата антитело к CD71-FITC, или (b) конъюгата антитело к CD33-FITC, конъюгата антитело к CD41-PE, конъюгата антитело к CD45-PerCp и CD34-APC (Beckman Coulter, Фуллертон, Калифорния,США) и анализировали на процентную долю образованных клеток CD41.
[00740] Для определения величины плоидности дифференцированные клетки LAMA-84 фиксировали в течение ночи в 75% этаноле при 4 °C, метили пропидиум иодидом (PI, 50 мкг/мл) и анализировали с применением FACScalibur (Becton Dickinson), при этом в день 14 дифференцированные CD34+ клетки количествено анализировали под микроскопом после окрашивания Майя-Грюнвальда/Гимза путем подсчета числа ядер на клетку и определения специфичной морфологии MK с помощью данного красителя. Анализировали только клетки с морфологией MK. Присутствие многоядерных клеток в препаратах после Cytospin указывает на присутствие полиплоидных MK. Дифференцированные CD34+ клетки анализировали на присутствие многоядерных зрелых MK по морфологии.
°C, метили пропидиум иодидом (PI, 50 мкг/мл) и анализировали с применением FACScalibur (Becton Dickinson), при этом в день 14 дифференцированные CD34+ клетки количествено анализировали под микроскопом после окрашивания Майя-Грюнвальда/Гимза путем подсчета числа ядер на клетку и определения специфичной морфологии MK с помощью данного красителя. Анализировали только клетки с морфологией MK. Присутствие многоядерных клеток в препаратах после Cytospin указывает на присутствие полиплоидных MK. Дифференцированные CD34+ клетки анализировали на присутствие многоядерных зрелых MK по морфологии.
Пример 45. Оценка дифференцировки культивируемых тромбоцитов - qPCR
[00741] Состояние дифференцировки тромбоцитов в культуре можно оценивать с помощью количественной ПЦР. РНК тромбоцитов экстрагировали для дополнительного определения характеристик культивируемых клеток. Общую РНК экстрагировали с применением реагента TRIzol (Invitrogen). Чистоту каждого препарата тромбоцитов оценивали с помощью ПЦР-анализа маркеров тромбоцитов (GPIIIa) и лейкоцитов (CD45). Целостность РНК тромбоцитов оценивали с помощью Bioanalyzer 2100 (Agilent) перед дальнейшими анализами.
[00742] Общую РНК собирали из клеточного лизата и библиотеку кДНК создавали с применением коммерческого набора для синтеза (Clontech). Меченные кДНК количественно оценивали с помощью набора Quant-iT PicoGreen dsDNA (Invitrogen) и разводили до 3 пМ для загрузки в одну полосу и секвенирования на анализаторе генома Illumina 1G (Solexa).
[00743] Необработанные последовательностей пропускали через серийные критерии контроля качества. Прежде всего, подтверждали присутствие по меньшей мере 6 нт из 3′-адаптера Solexa. Риды последовательностей, которые не соответствовали данному критерию, отбрасывали, тогда как остальные подрезали для удаления адаптерной последовательности, удерживаемой на 3′-конце. Остальные метки дополнительно фильровали с учетом их длины (>10 нт), количества копий (>4 ридов) и возможности считывания (<9 неидентифицированных нуклеотидов, аннотированный N). Риды, соответствующие всем данным критериям, впоследствии определяли как пригодные к использованию риды.
[00744] Все пригодные к использованию риды выравнивали с пре-микроРНК, полученными из базы данных miRBase. Метки последовательности, которые полностью соответствовали более чем одному предшественнику, равномерно распределяли между ними. Для учета дефектного расщепления с помощью Drosha и Dicer любую метку последовательности, которая полностью соответствовала пре-микроРНК в участке зрелой микроРНК, допускающей сдвиг до 4 нт по сравнению с положением в эталонной зрелой микроРНК, считали зрелой микроРНК. Уровень экспрессии микроРНК определяли как число ридов, соответствующих каждой зрелой микроРНК, нормированное к общему числу пригодных к использованию ридов, принимая во внимание, что общее количество малых РНК является постоянным. Относительное обилие каждой микроРНК определяли как число ридов, соответствующих каждой микроРНК, по сравнению с общим числом ридов, соответствующих зрелым микроРНК.
Пример 46. Очистка путем центрифугирования
[00745] Фракции культивируемых клеток можно очищать и отделять от ядер и загрязняющих и типов клеток с отличающейся плотностью посредством центрифугирования. Клетки центрифугировали при 200g в течение 15 минут для выделения фракций, богатых эритроцитами и ретикулоцитами. Супернатант собирали пипеткой и требуемую фракцию клеток затем промывали в модифицированном буфере Тирода (содержащем 138 мМ NaCl, 5,5 мМ декстрозы, 12 мМ NaHCO3, 0,8 мМ CaCl2, 0,4 мМ MgCl2, 2,9 мМ KCl2, 0,36 мМ Na2HPO4 и 20 мМ Hepes, pH 7,4) в присутствии 1 мкМ простагландина I2 и ресуспендировали в этом же буфере.
Пример 47. Очистка путем удаления ядра химическим способом
[00746] Удаление ядра культивируемых клеток можно стимулировать с помощью химических добавок в культуру, которые могут помочь увеличить безъядерную фракцию клеток перед очисткой. Клетки эритроидного ряда культивировали, как описано в данном документе. За 48 часов перед сбором клетки инкубировали с 210 мМ Me2SO. Затем клетки собирали с помощью центрифугирования при 350 X g в течение 5 мин при комнатной температуре, ресуспендировали на уровне 3 X 105 клеток на мл в свежей среде, содержащей 210 мМ Me2SO и 5 мкг/мл цитохалазина B (или другой молекулы, управляющей актином или ядром, т.e. p38 MAPK, псоралены) и инкубировали при 37°C. Долю клеток без ядер оценивали с помощью проточной цитометрии, как описано в данном документе, с применением DRAQ5 в качестве красителя для нуклеиновой кислоты и антител против гликофорина A в качестве маркера дифференцировки на поверхности клеток эритроидного ряда.
Пример 48. Очистка путем акустофореза
[00747] Для получения однородной популяции клеток можно использовать несколько систем механического разделения. Акустофорез со свободным потоком жидкости представляет собой один способ механического разделения (Petersson 2007, American Chemical Society). Суспензию в физиологическом растворе (0,9 мг/мл) с добавками питательных веществ, в том числе CsCl (0,22 г/мл), вносили в физиологический раствор. Образец суспензии, содержащей культивируемые клетки эритроидного ряда, обрабатывали с применением чипа для акустофореза (Cell-Care) с двумя активными выпускными отверстиями (скорость потока 0,10 мл/мн на выпускное отверстие).
[00748] Шприцевые насосы (WPI SP260P, World Precision Instruments Inc., Сарасота, Флорида) использовали для контроля скоростей потока в чипе. Все выпускные отверстия индивидуально соединяли с высокоточными стеклянными шприцами (1005 TLL и 1010 TLL, Hamilton Bonaduz AG, Бонадуц, Швейцария) посредством инжекторов с применением тефлоновой трубки, обеспечивая возможность независимого контроля скоростей выходного потока. Впускное отверстие для чистой жидкости соединяли со шприцевым насосом, а впускное отверстие для суспензии клеток с отрезком тефлоновой трубки длиной 50 мм (i.d. 0,3 мм), при этом ее другой конец погружали в лабораторный химический стакан, из которого тестируемая суспензия отсасывается при скорости, определяемой разницей между общим выходным потоком и потоком через впускное отверстие для чистой жидкости.
[00749] Ультразвук использовали для индукции стоячей волны между стенками канала для разделения, образованного с применением 20×20 мм пьезоэлектрической керамики (Pz26, Ferroperm Piezoceramics AS, Квистгард, Дания), прикрепленной к обратной стороне чипа. Ультразвуковой гель (Aquasonic Clear, Parker Laboratories Inc., Фейрфилд, Нью-Джерси) обеспечивал хорошую акустическую связь между этими двумя частями. Пьезоэлектрическую керамику приводили в действие посредством усилителя мощности (модель 75A250, Amplifier Research, Саудертон, Пенсильвания), соединенного с функциональным генератором (HP 3325A, Hewlett-Packard Inc., Пало-Альто, Калифорния). Хотя звуковые волны попадают в чип с обратной стороны, стоячую волну индуцировали между боковыми стенками канала для разделения как результат объединения механических колебаний по трем осям кристаллической структуры.
[00750] Процесс разделения отслеживали с применением стандартного микроскопа и ваттметра (ваттметр 43 Thruline, Bird Electronic Corp., Кливленд, Огайо). Процесс можно было в дальнейшем контролировать путем настройки частоты сигналов, силы возбуждения и скоростей потока.
[00751] Распределение клеток по размеру в образцах анализировали с применением счетчика Коултера (Multisizer 3, Beckman Coulter Inc., Фуллертон, Калифорния). Каждый образец смешивали с электролитом (Isoton II, Beckman Coulter Inc.) и анализировали с применением апертуры на 100 мкм. Уровень гемолиза, т.e. концентрацию свободного гемоглобина из поврежденных красных клеток, измеряли с применением фотометра (фотометр для определения низкого уровня HB в плазме крови, HemoCue AB, Ангелхольм, Швеция).
Пример 49. Очистка путем осуществления созревания ex vivo
[00752] Клетки эритроидного ряда, которые не являются полностью созревшими, можно довести до созревания с помощью инкубации ex vivo в системе, которая воспроизводит природные механизмы запуска созревания in vivo.
1. Совместное культивирование со стромальными клетками
[00753] На конечной стадии культивирования клетки эритроидного ряда культивировали на слое прикрепленных стромальных клеток в свежей среде без цитокинов. Культуры поддерживали при 37°C c 5% CO2 в воздухе. Слой прикрепленных клеток состоял либо из линии стромальных клеток MS-5, либо из мезенхимальных стромальных клеток (MSC), постоянных клеточных линий, происходящих из цельного костного мозга нормального взрослого организма (см. Prockop, DJ (1997) Science 276:71), в RPMI (Invitrogen), дополненной 10% фетальной телячьей сыворотки. Прикрепленные MSC размножали и очищали на протяжении по меньшей мере двух успешных пассажей перед применением в совместном культивировании.
2. Культивирование в планшетах, покрытых фибронектином
[00754] На конечной стадии культивирования клетки эритроидного ряда культивировали в планшетах с адсорбированным человеческим фибронектином. Для получения таких планшетов фибронектин (Sigma Aldrich) восстанавливали с помощью 1 мл стерильной H2O/мг белка и оставляли растворяться в течение по меньшей мере 30 минут при 37°C. Допускался остаток небольшого количества нерастворенного материала. Это не будет влиять на рабочие характеристики продукта Раствор фибронектина разбавляли 100x в стерильном сбалансированном солевом растворе и добавляли на поверхность культуры в минимальном объеме. Поверхности культуры позволяли высохнуть на воздухе в течение по меньшей мере 45 минут при комнатной температуре. Избыток фибронектина удаляли путем аспирации.
Пример 50. Очистка с помощью магнитофореза
[00755] Стратегии разделения, обогащения и/или очистки клеток эритроидного ряда с помощью магнитофореза известны из уровня техники, см. например, Zborowski et al., 2003, Biophys J 84(4) 2638 и Jin & Chalmers 2012, PLOS One 2012 7(8):e39491. Коммерческую систему магнитного разделения (сепаратор QuadroMACS™, объединяющий четыре сепарационные установки MidiMACS™ и LD-колонки, Miltenyi Biotec, Оберн, Калифорния) применяли для магнитного эритроцитного обогащения культур эритроцитов, происходящих из HSC. Клетки дезоксигенировали в надувной камере с перчатками Glove-Bag™ (Cole Parmer, Вернон-Хиллс, Иллинойс), заполненной азотом (азот от Medipure™, концентрация>99%, Praxair, Inc., Дэнбери, Коннектикут). Перед дезоксигенацией все материалы и оборудование, включая систему разделения, дегазированный стерильный буфер (PBS +2 мМ EDTA+0,5% BSA) и стерильные пробирки для сбора, помещали в камеру с перчатками, которую затем плотно закрывали. Дезоксигенированные культуры загружали непосредственно в LD-колонку MACS®, которую помещали в сепаратор QuadroMACS™, находящийся в бескислородных условиях внутри надувной камеры с перчатками, заполненной газом N2. Клетки, которые проходили через колонку, находящуюся внутри магнита, обозначали как отрицательную фракцию и, как ожидалось, они являются "немагнитными", включающими HSC-клетки и клетки эритроидного ряда до окончательного созревания. Клетки, оставшиеся в разделительной колонке, обозначали как положительную фракцию, которая является "магнитной" и состоит из созревающих RBC-подобных клеток, практически заполненных функциональным гемоглобином. Их элюировали из LD-колонки после удаления от магнита. По завершению разделения оксигенированные клетки обратимо восстанавливали путем воздействия воздуха на собранные клетки.
Пример 51. Очистка с помощью FACS
[00756] Популяцию культивируемых клеток эритроидного ряда сортировали с применением устройства для сортировки клеток Becton-Dickinson Aria IIu. Перед сортировкой клетки собирали, промывали с помощью PBS и окрашивали флуоресцентным антителом против гликофорина A (Life Technologies) и красителем для нуклеиновой кислоты DRAQ5 (Pierce) при рекомендуемых производителем разведениях. Применяли наконечник 100 мкм при частоте раскапывания 28000 капель/секунда. Пороговое значение образца составляло приблизительно 4000 объектов/секунда. Опцию контроля температуры применяли для поддержания образца и пробирок для сбора при 4°C на протяжении всего периода сортировки. Дополнительно устройство для перемешивания образца применяли при 200 об/мин для предотвращения осаждения образца на протяжении сортировки. Образец сортировали в аликвотах приблизительно 750 мкл, отмеряемых шприцем. В то же время, во время таких пауз пробирки для сбора поддерживали при 4°C, защищали от света и осторожно перемешивали до возобновления сортировки. Отсортированные образцы собирали в пробирку для сбора из боросиликатного стекла размером 12×75 мм, содержащую 250 мл DMEM, дополненной 10% FCS.
Пример 51. Очистка путем ферментативной обработки клеток
[00757] При использовании в качестве источников аллогенных эритроцитов можно извлечь пользу из удаления антигенов A и B для создания универсально совместимого продукта. Его можно проводить с помощью набора ферментов, способных селективно расщеплять группы галактозы, что делает клетки эритроидного ряда более подходящими по иммуногенным параметрам.
[00758] Два типа рекомбинантных белков эндо-ß-галактозидазы, которые первоначально были идентифицированы у Clostridium perfringens, получают в E. coli BL-21 с применением стандартных способов клонирования. ABазу получают для высвобождения A/B Ag, а эндо-β-галактозидазу C (EndoGalC) для высвобождения Galα1-3Galβ1-4GlcNAc (Gal Ag), который, как известно, обладает высокой иммуногенностью при ксенотрансплантации и имеет углеводную структуру, подобную A/B Ag. ABаза расщепляет связь Galβ1-4GlcNAc в антигене типа крови A [GalNAcα1-3(Fucα1-2) Galβ1-4GlcNAc] и в антигене типа крови B [Galα1-3(Fucα1-2) Galβ1-4GlcNAc].
[00759] Вкратце, после клонирования ABазы плазмиду экспрессии с C-терминальной His-меткой конструировали в eabC вектора pET-15b без сигнального пептида. Данный экзогенный ген трансформировали в клетки E. coli BL-21. Фермент, продуцируемый в клетках в виде фракции растворимого белка очищали на колонке, содержащей никель-нитрилотриуксусную кислоту (QIAGEN GmbH, Хильден, Германия). Наконец, 5 мл очищенной рекомбинантной ABазы получали в концентрации 3,6 мг/мл со специфической активностью 1500 Ед./мг. Одну единицу ферментативной активности определяли как количество фермента, требуемое для гидролиза 1 мкмоль субстрата в минуту.
[00760] Исследовали эффект обработки ABазой на присутствие Ag, связывание Ab и активацию комплемента. A/B RBC человека расщепляли с помощью ABазы и подвергали анализу проточной цитометрии после инкубации с перекрестнореагирующей сывороткой крови человека (содержащей антитело к A, или антитело к B, или антитела к A и B; типа B, типа A или типа O, соответственно). Среднюю интенсивность флуоресценции (MFI) применяли для количественной оценки уровня экспрессии антигенов типов крови A, B и Gal Ag. Уровень расщепления выражали как процентную долю Ag типов крови A или B, экспрессируемого на RBC после инкубации в отсутствие ABазы.
[00761] Свежую сыворотку крови типа O объединяли от трех здоровых людей-добровольцев и замораживали при -80°C для сохранения активности эндогенного комплемента до момента применения. Инактивированную нагреванием (в течение 30 мин при 56°C) сыворотку крови применяли для анализа связывания Ab. RBC после расщепления ферментом (ABаза) и без него инкубировали с 50% сывороткой крови типа O (100 мкл), разведенной фосфатно-солевым буфером, содержащим 0,2% бычьего сывороточного альбумина (PBS/BSA), в течение 30 мин при 37°C. После промывки проводили реакцию RBC с FITC-меченными антителами к человеческим IgG/IgM (DAKO, Глоструп, Дания) (×30, 100 мкл) в течение 30 мин при 4°C и затем подвергали анализу проточной цитометрии.
[00762] Ингибирующий эффект обработки ферментом на активацию комплемента также измеряли по изменению отложения C3d. После инкубации RBC с 50% сывороткой крови человека при наличии активности комплемента в течение 15 мин при 37°C, проводили реакцию RBC с FITC-меченным Ab кролика к C3d человека (DAKO, Глоструп, Дания) (×100, 100 мкл)в течение 30 мин. при 4°C и затем применяли для анализа проточной цитометрии. Процентную долю контрольного уровня (в отсутствие фермента) рассчитывали на основании MFI для оценки ингибирующего эффекта обработки ферментом на связывание Ab и отложение C3d.
Пример 52. Очистка тромбоцитов путем центрифугирования
[00763] Тромбоциты можно очищать из смешанных суспензий клеток путем центрифугирования. Примерно 40 мл цельной крови распределяли в пробирки для сбора образцов крови с цитратом натрия из расчета 3,2%, используемым в качестве антикоагулянта. Пробирки центрифугировали при 400×g в течение 10 мин. После этой стадии четко разграниченными были три слоя: плазма крови, красные кровяные клетки и промежуточная зона. Плазма крови с тромбоцитами находилась сверху, красные кровяные клетки находились на дне по причине их большей плотности, а тонкая беловатая промежуточную зона, называемая лейкоцитарной пленкой, состояла из более крупных тромбоцитов и лейкоцитов. С применением иголки Jelco 18G верхнюю часть плазмы крови с тромбоцитами сцеживали, а лейкоцитарную пленку помещали в две другие пробирки, в этот раз без добавок: одна пробирка для получения плазмы крови (P-пробирка) и вторая пробирка для получения тромбина (T-пробирка). Для получения тромбина использовали только 1,5 мл плазмы крови, в которую добавляли 0,5 мл глюконата кальция при концентрации 10%, в течение 15 мин. в двойном паровом котле при 37°C. Затем две пробирки снова центрифугировали, в этот раз при 800×g, в течение такого же промежутка времени (T=10 мин). После этого финального центрифугирования T-пробирка содержала богатую тромбином жидкость, в то время как P-пробирка содержала осажденные тромбоциты и некоторое количество красных кровяных клеток (эритроцитарно-тромбоцитарный сгусток). На данной стадии объем сокращали за счет удаления двух третей общего объема плазмы крови. Удаленная часть была обедненной тромбоцитами, в то время как оставшаяся часть с осажденными тромбоцитами (которые легко диспергируются посредством перемешивания) была обогащенной тромбоцитами.
Пример 53. Включение тимидина
[00764] Потенциал самовоспроизведения популяции клеток можно оценивать с применением анализа включения тимидина, известного из уровня техники, см., например, Harkonen et al. 1991 Exp Cell Res 186L288 и Tanaka et al. 1992 PNAS 89:8928.
[00765] Вкратце, однородно 13C- и 15N-обогащенный тимидин [U-13C, 15N-TdR] получали от Martek Biosciences (Коламбия, Мэриленд), а 3H-TdR (80 Ки/ммоль) приобретали у ICN Radiochemicals (Ирвин, Калифорния). Среды и буферы получали от Fisher Scientific (Питтсбург, Пенсильвания). Все ферменты за исключением фосфодиэстеразы были от Boehinger Mannheim (Индианаполис, Индиана). Фосфодиэстеразу II получали от Worthington Biochemical Corporation (Лейквуд, Нью-Джерси). Растворители для высокоэффективной жидкостной хроматографии (HPLC) были от EM Science (Гибстаун, Нью-Джерси) и содержали <0,1 ppm сухого остатка.
[00766] Клетки эритроидного ряда культивировали, как описано в данном документе. После культивирования клетки собирали для применения в анализе включения тимидина.
[00767] Клетки метили с помощью [U-13C, 15N]-TdR из расчета 1,6 мкг/мл в течение 18 ч с добавлением необогащенного тимидина для получения конечной концентрации тимидина 1 мкM. После промывки фосфатно-солевым буфером клетки культивировали в дополненной DMEM в течение еще 6 ч перед добавлением 3H-TdR при указанных концентрациях (0,1-10 мкКи/мл) для инкубации в течение дополнительных 18 часов. Немеченный тимидин добавляли к образцам для получения конечной концентрации тимидина 0,13 мкM, которая являлась эквивалентной концентрации 3H-TdR в образцах, получавших 10 мкКи радиоактивной метки/мл. После удаления 3H-TdR клетки инкубировали в дополненной DMEM в течение дополнительных 6-54 ч перед выделением ДНК.
[00768] ДНК экстрагировали с применением модифицированного набора для выделения ДНК Puregene (Gentra Systems, Миннеаполис, Миннесота). На основании числа клеток в образце для определения объемов добавляемых реагентов применяли процедуру пропорционального увеличения/уменьшения объема. Например, при использовании 1×107 клеток к 3 мл раствора для лизиса клеток добавляли 21 мкл раствора, содержащего 328 мкг протеиназы K. После перемешивания образец оставляли на протяжении ночи при комнатной температуре. На следующий день добавляли 10 мкг РНКазы и образец перемешивали и инкубировали в течение 2 ч при 37°C. Добавляли раствор для осаждения белка (1 мл) и образец инкубировали на льду в течение 5 мин. После центрифугирования в течение 10 мин при 2000g супернатант, содержащий ДНК, смешивали с 3 мл 100% 2-пропанола и осторожно переворачивали 50 раз или до тех пор, пока белые нити ДНК не становились видны. Затем образец центрифугировали при 2000g в течение 5 мин. Полученный осадок ДНК высушивали в течение 5 мин. перед промывкой в 3 мл 70% этанола и повторным центрифугированием в течение 5 мин при 2000g. Конечный осадок высушивали на воздухе и затем повторно гидратировали в деионизированной H2O и проводили количественную оценку путем измерения поглощения при 260 нм. Ту же самую процедуру применяли для CD34+ стволовых клеток в качестве контроля способности к воспроизведению.
[00769] Все ДНК денатурировали путем кипячения в течение 3 мин, затем быстро охлаждали на льду. Процедуру ферментативного гидролиза проводили с ДНК в концентрации 0,5 мг/мл. В следующем протоколе описан объем реагента, добавляемого на миллилитр раствора ДНК. ДНК гидролизовали с применением 10 мкл нуклеазы P1 (0,5 Ед./мкл) и 5 мкл ДНКазы I (4 Ед./мкл) в 10 мкл буфера, содержащего 200 мМ MgCl2, 100 мМ ZnCl2 и 1 M Tris, pH 7,2, в течение 2 ч при 45°C с последующим добавлением 20 мкл фосфодиэстеразы (4 мЕд./мкл) и последующей инкубацией в течение 2 ч при 37°C. Наконец, добавляли 5 мкл 10 M ацетата аммония (pH 9,0) и 10 мкл щелочной фосфатазы (1 Ед./мкл) и образцы инкубировали в течение дополнительных 2 ч при 37°C.
[00770] Расщепленный образец ДНК фильтровали с применением полиамидного фильтра с размером пор 0,22 мкм. Данный образец анализировали с помощью системы HPLC/CRI/IRMS с применением колонки для HPLC 4,6×250 мм Supelcosil LC-18-S (Supelco, Беллефонт, Пенсильвания). Такую же систему растворителей применяли при скорости 1 мл/мин и линейном градиенте от 5% до 25% B за 15 мин.
[00771] После разделения с помощью HPLC дезоксинуклеотиды анализировали с применением масс-спектрометрии с реакционно-химическим интерфейсом (CRIMS). В данном способе дезоксинуклеотиды вливают в систему распыления и десольватации, движимые потоком гелия, откуда они выходят в виде пучка сухих частиц. Количественное соотношение 13CO2/ 12CO2 из данного образованного в потоке CO2 определяли с помощью масс-спектрометра для определения соотношений изотопов Delta S Finnigan/MAT (ThermoFinnigan, Сан-Хосе, Калифорния) и его сопутствующей системы данных Isodat. 5-фтордезоксиуридин (Sigma) применяли в качестве внутреннего стандарта изотопного отношения.
[00772] Соотношения изотопов (IR в уравнении, которое следует далее) для трех нуклеотидов получали из каждого образца: T, dA и dG. Обогащение CO2, выделяющимся из каждого дезоксинуклеозида, происходящего из ДНК, вычисляли при помощи уравнения (13)CO2 (на тысячу)=1000 × (экспериментальное IR - стандартное IR)/стандартное IR Для поддержания наивысшего уровня внутренней совместимости и во избежание любого отклонения в разных экспериментах соотношение изотопов для dG вычитали во всех экспериментах из соотношения изотопов для T. Оценивали данные, полученные от конца периода мечения стабильным изотопом (день 0) до конца вымывания (день 3).
Пример 54. Количественное определение ядерного материала
[00773] Число ДНК-содержащих клеток в смешанной популяции оценивали с помощью проточной цитометрии с применением красителя для ДНК DRAQ5 (Pierce). Клетки инкубировали с красителем в соответствии с инструкциями производителя и анализировали на проточном цитометре, например, цитометре Attune (Life Technologies). Количественно оценивали процентную долю клеток с содержанием ядерного материала выше определенного порога.
Пример 55. Анализ канцерогенности in vitro
[00774] Для оценки потенциала воспроизведения клеток можно выполнять анализ образования колоний на мягком агаре. Вкратце, слой основного агара готовили путем приготовления 0,5% раствора агара+1x RPMI+10% FCS, при этом все компоненты нагревали до 40°C, и внесения 1,5 мл раствора в 35 мм чашку Петри. Агар оставляли застыть в течение 30 мин при комнатной температуре перед применением.
[00775] Верхний слой агарозы готовили путем расплавления 0,7% агарозы в микроволновой печи и охлаждения до 40°C. Раствор 2x RPMI+20% FCS нагревали до 40°C. Клетки подсчитывали и подготавливали к высеванию из расчета 5000 клеток на чашку при плотности 200000 клеток на мл. 0,1 мл суспензии клеток вносили в пробирки на 10 мл, с последующим добавлением 3 мл теплой 0,7% агарозы и 3 мл теплого раствора RPMI/FCS. Раствор осторожно перемешивали путем взбалтывания и вносили (1,5 мл) в каждую из трех или четырех параллельных чашек с основным агаром.
[00776] Чашки инкубировали при 37°C в увлаженном термостате в течение 10-30 дней. Клетки подпитывали 1-2 раза в неделю с помощью культуральной среды, 0,75 мл/чашка.
[00777] Для оценки образования колоний чашки окрашивали с помощью 0,5 мл 0,005% кристаллического фиолетового в течение >1 ч. Колонии подсчитывали с применением препаровального микроскопа.
Пример 56. Анализ канцерогенности in vivo
[00778] Терминально дифференцированные культивируемые клетки эритроидного ряда имплантировали в различные животные модели для оценки потенциала канцерогенности. Несколько тканей собирали от различных моделей и анализировали с помощью гистологического, иммунохимического и флуоресцентного анализов для количественной оценки канцерогенности.
[00779] Животным ежедневно вводили внутрибрюшинные инъекции CsA (10 мг/кг, Sandimmune, Novatis Pharma, Нюрнберг), начиная за два дня до имплантации. Для истощения NK-клеток некоторым крысам в дополнение к CsA вводили внутрибрюшинные инъекции моноклонального антитела (mAb), антитела к NKR-P1A (клон 10/78, мышиный IgG1, BD Biosciences, Гейдельберг, Германия) или соответствующего изотопического контроля (клон PPV-06, мышиный IgG1, Exbio, Прага, Чехия). mAb к NKR-P1A (клон 10/78) направлено против того же эпитопа, что и mAb (клон 3.2.3). Один мг соответствующих антител вводили за день до инъекции клеток эритроидного ряда с последующем введением 0,5 мг в день 4 после трансплантации клеток.
[00780] Образцы крови собирали перед началом этих экспериментов, в день 0 и через 4 дня после трансплантации клеток эритроидного ряда, а также при аутопсии (день 92), чтобы определить долю NK-клеток в крови с помощью проточной цитометрии. Для анализа роста подкожной опухоли клетки эритроидного ряда инъецировали в 100 мкл фосфатно-солевого буфера (PBS) в бок животных. Рост опухоли отслеживали через день путем пальпации и размер регистрировали с применением линейного штангенциркуля. Животных умерщвляли перед днем 100, если объем опухоли достигал 1 см3 у мышей и 5 см3 у крыс, если наблюдалась потеря веса более 10% или если наблюдались любые поведенческие признаки боли или страдания. Осуществляли аутопсию всех животных.
[00781] Ткани мыши возле места инъекции сразу же замораживали в жидком азоте или помещали в забуференный фосфатом 4% формалин на 16 ч и затем заливали в парафин. Селезенки и лимфатические узлы извлекали для последующих иммунологических анализов. Выполняли трансплантацию клеток эритроидного ряда в полосатое тело крыс с односторонним поражением под действием 6-OHDA. Этих животных умерщвляли через 6 недель после трансплантации.
[00782] Ткань животных анализировали с помощью проточной цитометрии. Соответствующие флуоресцентные и PE-конъюгированные антитела против установленных биомаркеров раковых клеток, CD133, CD3, CD, CD16, CD19, CD20, CD56, CD44, CD24 и CD133, добавляли к иссеченным образцам тканей и анализировали для количественного оценки канцерогенного потенциала.
Пример 57. Определение деформируемости с помощью EKTA
[00783] Клетки эритроидного ряда, культивируемые как описано в данном документе, оценивали в отношении характеристик деформируемости по сравнению с образцами природных эритроцитов посредством эктацитометрии.
[00784] Эктацитометр, состоящий из вискозиметра Куэтта, объединенного с гелий-неоновым лазером, применяли для получения дифракционного изображения красных клеток, суспендированных в вязкой жидкости между двумя цилиндрами. Когда вискозиметр вращается, нормальные красные клетки удлиняются в поле сдвига, в связи с чем дифракционное изображение становится эллиптическим. Эллиптичность изображения измеряли путем количественного оценки интенсивности света вдоль большой (A) и малой (B) осей дифракционной картины и выражали его как соотношение (A - B)/(A+B), индекс деформируемости (DI) или индекс удлинения (EI). Вязкость среды подбирали так, чтобы она превышала внутреннюю вязкость клеток эритроидного ряда с наибольшей плотностью. Раствор поливинилпирролидона (PVP) с концентрацией 31 г/литр, mw=360000, в 0,04 M фосфатном буфере, состоящем из K2HP04 и KH2P04 в дистиллированной воде, характеризовался вязкостью 0,20 пуаз при 25°C и 12 пуаз при 37°C.
[00785] Осмолярность регулировали с помощью NaCl до требуемого уровня и измеряли в осмометре Реблинга по точке замерзания Конечный pH варьировали с применением небольших добавок 1M растворов NaOH и HCl и измеряли в анализаторе газов крови Technicon BG I1. Азид натрия добавляли в качестве консерванта в исходные растворы с получением концентрации 0,4 г/л.
[00786] С помощью эктацитометра получали три главных показателя образцов клеток эритроидного ряда и сравнивали их с показателями нативных эритроцитов; минимальная осмоляльность (Omin), индекс деформируемости (Dimax) и осмоляльность, при которой DI достигает половины своего максимального значения (Ohyp).
[00787] Omin зависит от отношения площади поверхности к объему, и было обнаружено, что она равняется точке 50% гемолиза в классическом тесте осмотической хрупкости.
[00788] Dimax представляет собой максимальное значение индекса деформируемости, обычно достигаемое при 290 мосмоль (значение физиологической осмоляльности). Оно показывает максимальную деформируемость клетки при напряжении сдвига и зависит от ряда факторов, таких как площадь поверхности, объем, внутренняя вязкость и механические свойства клеточной мембраны.
[00789] Ohyp представляет собой осмоляльность, при которой DI достигает половины своего максимального значения. Он указывает на положение гипертонической части кривой, которая зависит от внутренней вязкости клетки, а также механических свойств мембраны, так как, например, как она будет искривляться под действием силы (жесткость).
[00790] Параметры, полученные для культивируемых клеток эритроидного ряда, сравнивали с такими же значениями первичных клеток эритроидного ряда.
Пример 58. Измерение деформируемости с помощью LORCA
[00791] Деформируемость для популяций очищенных cRBC измеряли с помощью методики лазерной дифракции (LORCA, лазерно-стимулированный оптический ротационный анализатор клеток, R&R Mechanotrics). Вкратце, сильно разведенную суспензию клеток подвергали деформации сдвига в системе Куэтта с зазором 0,3 мм между 2 цилиндрами, один из которых способен вращаться для индуцирования напряжений сдвига. Лазерный пучок пропускали через суспензию и дифракционную картину измеряли при 37°C. При низком напряжении сдвига клетки представляли собой круглые диски, тогда как при высоком напряжении сдвига клетки становились эллиптическими. Деформируемость клеток выражали в виде индекса удлинения (EI), который зависит от эллиптичности деформирующихся клеток. Аликвоты, содержащие 12,5 мкл осажденных центрифугированием осадков RBC, разводили в 5 мл раствора поливинилпирролидона (молекулярная масса 360000). Значения EI при 30 Па (называемые как EImax) и 3 Па выбирали как репрезентативные значения деформируемости для легкого сравнения образцов при разных напряжениях сдвига.
Пример 59. Оценка окклюзии сосуда - сосудистая система крысы ex vivo
[00792] Потенциал окклюзии сосуда клетками эритроидного ряда можно оценивать с помощью выделенной искусственно перфузируемой сосудистой системы крысы с применением способов, известных из уровня техники, см., например, Kaul et al. 1983, J Clin Invest 72:22. Вкратце, анестезированным (пентобарбитал натрия, 30 мг/кг) крысам линии Wistar, 120-150 г, правую подвздошно-ободочную артерию и вену канюлировали с помощью гепаринизированной (100 мкл/мл) силиконовой трубки в участке, расположенном на расстоянии 3 см от илеоцекального соединения. При стабильной перфузии раствором Рингера, содержащим 1% бычьего сывороточного альбумина, восходящую ободочную кишку и терминальный отдел подвздошной кишки (по 3 см каждой) разрезали между перевязками. После выполнения кровеостанавливающих перевязок всех сосудистых соединений ткань выделяли. Выделенную брыжеечку червеобразного отростка осторожно растягивали на оптически прозрачном блоке Lucite на предметном столике микроскопа. Препарат целиком покрывали полиэтиленовой пищевой пленкой за исключением выпускных отверстий канюль и объектива микроскопа.
[00793] Контрольные артериальное перфузионное давление (Ppa) и давления венозного оттока (Pv) поддерживали постоянными при 80 и 3,8 мм рт.ст., соответственно, и отслеживали с помощью датчиков давления Statham-Gould P-50 (Stathan Instruments Inc, Окснард, Калифорния). Скорость венозного оттока (Fv) отслеживали с применением фотоэлектрического счетчика капель и выражали в мл/мин. Временной интервал 10-12 мин предусмотрен для установления равновесия в ткани и стабилизации Fv. Применяли только препараты, демонстрирующие микрососудистую систему брыжеечки червеобразного отростка, не содержащую кровяных клеток хозяина, и со стабильной Fv, составляющей 4,6 +/- 0,5 (среднее +/- SD). Эксперименты проводили при 37°C.
[00794] Клетки эритроидного ряда выделяли, как описано в данном документе. После контрольных измерений Ppa и Fv клетки эритроидного ряда (0,2 мл, Hct 30%) осторожно вводили через устройство для ввода пробы на 15 см удаленное от места канюлирования артерии и изменения Ppa и Fv регистрировали на ленточной диаграмме полиграфа Грасса (Grass Instrument Co, Квинси, Массачусетс). Препараты ткани перфузировали в течение 10-15 мин перед инфузией в образцов с раствором Рингера, что обеспечивало стабилизацию ткани и освобождение сосудистой системы от оставшихся кровяных клеток животного-хозяина. Можно устранять непроходимость, возникающую в результате инфузии клеток, и восстанавливать поток с помощью кратковременного (2-3 мин.) перфузирования сосудистой системы с помощью полностью насыщенного кислородом раствора Рингера при высоком давлении (100 мм рт.ст.).
[00795] В конце каждого эксперимента препарат ткани целиком (без канюль и содержимого полостей) взвешивали. Единицы общего периферического сопротивления сосудов (PRU) рассчитывали и выражали как PRU=ΔP/Q=мм рт.ст./мл/(мин.-г), где ΔP (мм рт.ст.) представляет собой артериовенозную разность давления, а Q (мл/мин.-г) представляет собой скорость венозного оттока на грамм ткани.
[00796] В каждом эксперименте время восстановления пока под давлением (Tpf) определяли после болюсной инфузии образцов. Tpf обозначали как время (в секундах), требуемое для возвращения значений Ppa и Fv к их исходным уровням после введения указанного образца, и оно представляет собой общее время прохождения через сосудистую систему брыжеечки червеобразного отростка. Значения параметров, полученные для культивируемых клеток эритроидного ряда, сравнивали со значениями, полученными для первичных клеток эритроидного ряда.
Пример 60. Оценка окклюзии сосуда - проточная камера in vitro
[00797] Способы оценки окклюзии сосуда клетами эритроидного ряда с применением in vitro проточных камер с градиентом высоты известны из уровня техники, см. например, Zennadi et al 2004, Blood 104(12):3774.
[00798] Вкратце, проточные камеры с градуированной высотой применяли для количественной оценки адгезии клеток эритроидного ряда с эндотелиальными клетками (EC). Предметные стекла, покрытые EC, промывали сбалансированным физиологическим раствором Хенкса (HBSS) с 1,26 мМ Ca2+, 0,9 мМ Mg2+ (Gibco, Гранд-Айленд, Нью-Йорк), предварительно нагретым до 37°C, и затем помещали проточную камеру с регулируемой высотой. Проточную камеру устанавливали на предметный столик инвертированного фазово-контрастного микроскопа (Diaphot; Nikon, Мелвилл, Нью-Йорк), соединенный с Thermoplate (Tokai Hit, Фудзиномия-ши, Япония), установленным на 37°C. Клетки наблюдали с применением видеокамеры (RS Photometrics, Туксон, Аризона), прикрепленной к микроскопу и соединенной с компьютером Macintosh G4 (Apple, Купертино, Калифорния). Клетки эритроидного ряда культивировали, как описано в данном документе, и метили с помощью флуоресцентного красителя PKH 26 из набора красных флуоресцентных красителей для клетки (Sigma), следуя инструкциям производителя. Клетки (3 мл), суспендированные из расчета 0,2% (об./об.) в HBSS с Ca2+, Mg2+, вливали в проточную камеру и оставляли для прикрепления к предметному стеклу в течение 15 минут без применения потока. Перед воздействием потока как минимум 3 поля на каждом из 7 различных участков вдоль линии, ориентированной под прямым углом к будущему потоку, исследовали в отношении общего числа флуоресцирующих клеток. Затем запускали поток жидкости (HBSS с Ca2+, Mg2+) с применением калиброванного шприцевого насоса. После воздействия потока поля снова исследовали и подсчитывали число прикрепленных клеток. Долю прикрепленных клеток представляли следующим образом: Число клеток, прикрепленных после воздействию потока/клетки, присутствующие на поле перед воздействием потока. Напряжение сдвига на стенке рассчитывали следующим образом: τw=(6 мкQ)/(wH [x]2), где τw обозначает напряжение сдвига на стенке (дина/см2); Q, объемная скорость потока (см3/с); μ, вязкость среды; w, ширина проточного канала; и H(x), высота проточной камеры в зависимости от расположения по отношению к предметному стеклу микроскопа.
Пример 61. Оценка окклюзии сосуда - витальная микроскопия
[00799] Способы оценки окклюзии сосуда клетами эритроидного ряда с применением витальной микроскопии известны из уровня техники, см., например, Zennadi et al., 2007, Blood 110(7):2708.
[00800] Вкратце, общую анестезию подопытного животного обеспечивали путем внутрибрюшинной инъекции 100 мг/кг кетамина (Abbott Laboratory, Чикаго, Иллинойс) и 10 мг/кг ксилазина (Bayer, Шоуни Мишн, Канзас). Двухстороннюю титановую камеру с оптическим окном имплантировали хирургическим путем в спинную кожную складку в стерильных условиях с применением вытяжного шкафа с ламинарным потоком воздуха. Оперативное вмешательство включало осторожное удаление слоев эпидермиса и дермы с одной стороны спинной кожной складки, обнажение кровеносных сосудов подкожной клетчатки, прилегающих к поперечнополосатым мышцам противоположной стороне кожной складки, и затем прикрепление 2 сторон камеры к коже с применением винтов из нержавеющей стали и шовного материала. Стеклянное окно помещали в камеру для покрытия обнаженной ткани и прикрепляли с помощью прижимного кольца. Впоследствии животных содержали при 32°C - 34°C до проведения in vivo исследований через 3 дня после оперативного вмешательства.
[00801] Анестезированных животных с оптическим окном помещали на предметный столик микроскопа Axoplan (Carl Zeiss, Торнвуд, Нью-Йорк); температуру поддерживали на уровне 37°C с применением грелки-подушки с термостатической регулировкой. Все инфузии проводили через дорсальную часть хвостовой вены. Клетки эритроидного ряда культивировали, как описано в данном документе. Затем клетки метили красителями Dil или DiO (Molecular Probes, Юджин, Орегон) в соответствии с инструкциями производителя. Проводили инфузию меченных клеток (300 мкл; гематокрит [Hct] 0,50 [50%] в PBS с Ca2+ и Mg2+), и адгезию RBC и динамику потока крови наблюдали в подкожных сосудах в течение по меньшей мере 30 минут с применением объективов LD Achroplan 20×/0,40 Korr и Fluar 5×/0,25. Процессы в микроциркуляторном русле и адгезию клеток одновременно регистрировали с применением видеомонитора Trinitron Color (PVM-1353 MD; Sony, Токио, Япония) и кассетного видеомагнитофона JVC (BR-S3784; VCR King, Дарем, Северная Каролина), соединенного с цифровой видеокамерой C2400 (Hamamatsu Photonics KK, Хамамацу, Япония). Тридцать сегментов венул исследовали для каждого набора условий. Артериолы отличали от венул на основании (1) наблюдения расходящегося потока в противоположность к сходящемуся потоку; (2) внешнего вида стенок сосудов, обладающих свойствами двойного лучепреломления при применении трансиллюминации, что является характерной особенностью гладкой мускулатуры сосудистых стенок артериол; и (3) относительно прямой направленности сосуда без признаков извитости.
[00802] Измерение потока и адгезии красных клеток выполняли путем изучения видеолент с применением увеличения 20×. Прикрепление клеток количественно оценивали при рассматривании клеток, прикрепленных к стенкам сосудов и их неподвижности в течение 1 минуты. Процентную долю длины сосудов с диаметрами до 25 мкм или более 25 мкм, занятой SS RBC, количественно ооценивали следующим образом: % длины венул, занятых SS RBC=(длина стенки сосуда с прикрепленными клетками/общая длина анализируемых сегментов сосудов) × 100. Изменения потока RBC рассчитывали следующим образом: поток=число циркулирующих в крови флуоресцентных человеческих RBC, пересекающих одну точку, обозначенную на сосуде, имеющую менее 50 мкм в диаметре, за минуту.
Пример 62. Оценка окклюзии сосуда - тромбоциты
[00803] Способы оценки окклюзии сосуда тромбоцитами с применением человеческих клеток эндотелия сосудов (HUVEC) можно адаптировать из подобных способов для эритроцитов. Вкратце, суспензию объемом 2 мл с гематокритом 0,05% добавляли к конфлюэнтным HUVEC в чашке Петри для культивирования ткани. Аппарат типа "конус-пластина" собирали в пределах 1 мин после добавления тромбоцитов и помещали на инвентированный фазово-контрастный микроскоп Nikon Diaphot-TMD (Southern Micro Instruments, Атланта, Джорджия). Запускали двигатель для поворота конуса и прикрепление непрерывно отслеживали при напряжении сдвига 0,1 или 1 дина/см2 в течение 30 мин. Температуру поддерживали постоянной при 37°C с помощью термостата с воздушной завесой (Nicholson Precision Instruments, Inc., Бетезда, Мэриленд), продувающего на устройство для тестирования адгезии. Прикрепление тромбоцитов визуализировали и регистрировали каждые 5 мин. путем фокусировки на 8 разных полях зрения в течение 20 с на поле в каждый момент времени. Весь эксперимент рассматривали под общим увеличением 400× посредством автоматической фотокамеры CCD-72 (Dage-MTI, Inc., Мичиган-Сити, Индиана) и регистрировали на видеоленте с применением кассетного видеомагнитофона SVO 2000 (Sony Electronics, Сан-Хосе, Калифорния). Прикрепление количественно оценивали в автономном режиме в конце каждого эксперимента путем подсчета индивидуальных прикрепленных клеток во время ручного воспроизведения записанных видеоизображений. Количество клеток в 8 полях в каждый момент времени усредняли и нормировали к количеству прикрепленных красных клеток на квадратный миллиметр эндотелия.
Пример 63. Оценка массы/объема/плотности с помощью вибратора
[00804] Двойную суспензионную микроканальную вибрационную (SMR) систему применяли для характеристики массы, объема и плотности популяции терминально дифференцированных клеток эритроидного ряда на основании Bryan et al., LabChip, 2014. В начале измерения плотности клеток систему сначала промывали профильтрованной средой Percoll, которая служила в качестве жидкости с высокой плотностью. Затем обходной канал для образца заполняли разведенным образцом клеток, и наивысшие точки сосуда на впускном отверстии и выпускном отверстии для образца регулировали так, чтобы направлять поток жидкости в первую SMR. Давление на впускном отверстии для жидкости с высокой плотностью применяли для установки плотности жидкости 2, и давление на сливном отверстии контролировало общую скорость потока в устройстве. Для сведения к минимуму вероятности искажения размера за счет осаждения более тяжелых клеток на дне сосуда или трубки с образцом, свежий образец вводили через равные промежутки времени путем вливания в обходной канал для образца. Данные получали посредством LabVIEW и обрабатывали с помощью MATLAB.
[00805] Концентрацию клеток отслеживали с применением счетчика Коултера. Измерения клеток выполняли на культурах, выращиваемых до 5 × 105-1 × 106 клеток/мл. Жидкость с высокой плотностью, вводимую для измерения во второй SMR, составляли как раствор 50% (об./об.) Percoll (Sigma), 1,38% (вес./об.) порошкообразной среды L15 (Sigma), 0,4% (вес./об.) глюкозы, 100 МЕ пенициллина и 100 мкг/л- 1 стрептомицина. pH среды регулировали до 7,2. Эту среду Percoll хранили при 4°C и фильтровали непосредственно перед применением в двойной SMR.
Пример 64. Оценка содержания фосфатидилсерина с помощью аннексина V
[00806] Клетки эритроидного ряда культивировали, как описано в данном документе. 50 мкл суспензии клеток промывали в растворе Рингера, содержащем 5 мМ CaCl2, и затем окрашивали с помощью аннексина-V-FITC (разведение 1:200; ImmunoTools, Фризойте, Германия) в данном растворе при 37°C в течение 20 мин. с защитой от света. Клетки промывали и анализировали с помощью проточной цитометрии, как описано в данном документе, и интенсивность флуоресценции аннексина-V измеряли при длине волны возбуждения 488 нм и длине волны излучения 530 нм. Относительный выход фосфатидилсерина оценивали по флуоресценции аннексина-V.
Пример 65. Оценка содержания липидов с помощью хроматографии
[00807] Липиды экстрагировали из промытых экпрессирующих экзогенный антиген EHC-клеток путем трех экстракций с применением метанола-хлороформа в соотношении 1:1 при комнатной температуре в присутствии антиоксиданта BHT (Sigma Aldrich). Объединенные экстракты промывали с помощью 0,05 M KCl по способу из Folch, Lees and Sloane Stanley1957, J Biol Chem 226:497. Вкратце, для первой экстракции 15 мл метанола, содержащего 0,05 мг/мл BHT, добавляли к промытым комплексам в центрифужную пробирку и оставляли отстояться в течение 30 мин. с эпизодическим перемешиванием для разбиения осадка. Затем добавляли 15 мл хлороформа и смесь оставляли отстояться в течение 30 мин. с эпизодическим перемешиванием для разбиения сгустков. Пробирки центрифугировали в течение 5 минут при 1500g и фракции супернатанта сливали в делительные воронки, оснащенные тефлоновыми кранами. Вторую и третью экстракции выполняли аналогичным образом, при этом к остатку добавляли 15 мл метанола-BHT, а затем 15 мл хлороформа, за исключением того, что экстракты отстаивались только в течение 10 минут с эпизодическим перемешиванием после каждого добавления. После центрифугирования фракции супернатанта объединяли в делительной воронке, затем добавляли 48 мл хлороформа и 28 мл 0,05 M KCl и перемешивали. Смесь оставляли отстояться на протяжении ночи в темноте при 4°C для разделения фаз. После повторного нагревания до комнатной температуры самую нижнюю из двух прозрачных фаз собирали и выпаривали досуха в вакууме при 40°C в ротационном вакуумном испарителе. Липиды переносили количественно в мерную колбу на 10 мл с хлороформом и хранили при -22°C.
[00808] Концентрацию свободного холестерина в липидном экстракте определяли следующим образом. Липидный экстракт подвергали хроматографии на 0,5 мм слое силикагеля HR (Brinkmann Instruments, Inc., Вестбери, Нью-Йорк) c гексаном-диэтиловым эфиром-ледяной уксусной кислотой 80: 20: 1, пластину для TLC окрашивали путем распыления раствора 2,7-дихлорфлуоресцеина (см. ниже), пятно свободного холестерина соскабливали в коническую центрифужную пробирку и экстрагировали один раз с помощью 2,0 мл и трижды с помощью 1,0 мл хлороформа, экстракт выпаривали досуха в вакууме при 40°C в ротационном вакуумном испарителе, и количество холестерина оценивали с помощью способа с применением хлорида железа по Mann 1961 Clin Chem 7:275 без омыления. Стандарт свободного холестерина, приготовленный из коммерческого сертифицированного химически чистого материала путем выделения через дибромидное производное (см. например, Fieser J Amer Chem Soc 1953 75:5421), проводили через процедуру хроматографии и оценивали с применением каждого ряда определений. В каждом определении для значения свободного холестерина вносили поправку на восстановления стандарта, которое в среднем составляло 95%. TLC необходима для удаления BHT, который в противном случае препятствует способу с применением хлорида железа, приводя к образованию коричневого продукта, который поглощает при длине волны 560 нм.
[00809] Распределение фосфолипида определяли в трех повторностях с помощью TLC аликвот общего липидного экстракта при 4°C на силикагеле HR с толщиной 0,5 мм с хлороформметанолом-ледяной уксусной кислотой-водой 25: 15: 4: 2, к которому добавляли BHT при концентрации 50 мг/100 мл для предотвращения самоокисления во время хроматографии; пластины для TLC готовили с применением воды ("нейтральные" пластины). Применение «методики с клиновидным наконечником» для нанесения липидного образца на начало пластины (см., например, Stahl 1965 Thin-Layer Chromatography, Academic Press Inc.) приводит к превосходному разделению индивидуальных фосфолипидов. В частности, способ обеспечивает полное разделение фосфатидилэтаноламина (PE), фосфатидилсерина (PS), лецитина и сфингомиелина; отдельное пятно перемещается между PS и лецитином, которое идентифицируют как фосфатидилинозитол (PI). Пятна визуализировали под УФ-светом путем распыления раствора 2,7-дихлорфлуоресцеина (33,3 мг/100 мл водного 2 мМ раствора NaOH) и затем соскабливали непосредственно в пробирки Крамера-Гиттлемана, где фосфолипиды подвергали расщеплению при 190°C в течение 60 мин с помощью 1,0 мл 70% перхлорной кислоты. Остальную часть процедуры выполняли, как описано выше, за исключением того, что после проявления цвета силикагель удаляли путем центрифугирования при 3000g в течение 5 мин и поглощение определяли в прозрачном растворе супернатанта. Вводили поправки на поглощение соответствующих участков пустых полос.
[00810] Газово-жидкостную хроматографию выполняли на растворенных в гексане образцах с применением прибора Барбера-Колмана, модель 5000, оснащенного спаренными 8-футовыми колонками EGSS-X (сложный полиэфир этиленгликоля и сукцината, объединенный с силиконом) 8% на Gas-Chrom P, 100-120 меш (Applied Science Laboratories Inc.) и двойными пламенно-ионизационными детекторами. Скорость потока азота на входе составляла 50 мл/мин. Температуру колонки поддерживали на 165°C в течение 10 мин после инъекции образца, затем повышали на 2°C/мин до 200°C.
Пример 66. Оценка вязкости мембраны
[00811] Вязкость мембраны в популяции клеток можно оценивать с помощью анализа фотообесцвечивания флуоресценции. 0,5 мл образца клеток эритроидного ряда собирали и промывали один раз в забуференном HEPES физиологическом растворе (132 мМ NaCl, 4,7 мМ KCl, 2,0 мМ CaCl2, 1,2 мМ MgSO4, 20 мМ HEPES, доведенный до pH до 7,4). Затем клеточную массу промывали один раз в 145 мМ NaCl - 10 мМ NaHCO3, pH 9,5, и ресуспендировали в том же буфере с 1 мг/мл DTAF (получен от Research Organics, Кливленд, Огайо). Клетки инкубировали на льду в течение 1 ч, затем промывали дважды в 50 мМ глицина - 95 мМ NaCl - 10 мМ NaHCO3, pH 9,5, для удаления любого красителя, который не связан ковалентно с белком. Наконец, клетки промывали дважды и ресуспендировали до гематокрита -2% в забуференном HEPES физиологическом растворе с 1 мг/мл бычьего сывороточного альбумина. Ту же обработку применяли для контрольных нативных эритроцитов.
[00812] Проточную камеру устанавливали на предметный столик инвертированного микроскопа Leitz Diavert (Rockleigh, NJ), оснащенного для проведения флуоресцентной микроскопии с падающим светом. Дихроичное зеркало и фильтры возбуждения/эмиссии представляют собой стандартную комбинацию для применения с флуоресцеиновыми красителями (обозначение по Лейтцу 12), с длиной волны возбуждения в диапазоне 450-490 нм. Объектив принадлежал к типу для масляной иммерсии с увеличением 100x и числовой апертурой 1,25. Дуговая ртутная лампа высокого давления на 100 ватт (Осрам, Мюнхен) с соответствующим блоком питания и корпусом (Oriel, Стамфорд, Коннектикут) служила в качестве источника возбуждения флуоресценции.
[00813] Электронный затвор с компьютерным управлением (Vincent Associates, Рочестер, Нью-Йорк) ограничивал продолжительность облучения и был синхронизирован с электронной системой подсчета фотонов для измерения интенсивности флуоресценции. Диафрагму поля осветителя падающего света применяли для ограничения возбуждения до круглой областью диаметром 20-40 мкм. Через равные промежутки времени выходной импульс из компьютера вызывал открытие затвора в течение типичной продолжительности 20 мс. Свет от недолгого флуоресцентного изображения распадался с помощью серии призм, так что половина света направлялась на видеокамеру SIT с низким уровнем освещенности (модель 66-SIT, Dage-MTI, Мичиган-Сити, Индиана) и половина на фотоэлектронный умножитель (модель 8850, RCA, Харрисон, Нью-Джерси), заключенный в корпус с температурой окружающей среды. В течение времени, когда электронный затвор оставался открытым, процессор видеоизображений (модель 794, Hughes Aircraft, Карлсбад, Калифорния) приводился в действие для получения флуоресцентного изображения, путем создания моментального видеокадра, который можно было отслеживать, чтобы удостовериться в том, что субъект остается в фокусе и что посторонний объект не попал в поле зрения. Расстояния на видеоэкране измеряли с помощью штангенциркуля видеоизображения и калибровали путем сравнения с видеоизображением объект-микрометра. Также в течение времени, когда затвор оставался открытым, сигнал фотоэлектронного умножителя обрабатывался с помощью методики подсчета фотонов. Усилитель/дискриминатор (модель AD6, Pacific Instruments, Конкорд, Калифорния) вырабатывал числовые логические импульсы для каждого импульса сигнала выше заданной величины, и эти цифровые импульсы подсчитывали на счетчике, гейтированном на 100 МГц (модель 770, EG&G Ortec, Окридж, Теннесси). Микрокомпьютер контролировал выполнение гейтирования, возврата в начальное положение и регистрацию счета фотонов.
[00814] Типичный эксперимент состоял из ряда предварительных измерений флуоресценции, осуществляемых во время коротких (20 мс) импульсов света возбуждения, с последующим продолжительным периодом освещения (как правило, 30 с), во время которого образцы клеток обесцвечивались, с последующими дополнительными сериями коротких облучений, каждая по 15-30 с, пока не становилось очевидным окончание восстановления флуоресценции.
[00815] Время восстановления и другие параметры, полученные для культивируемых клеток эритроидного ряда, сравнивали с такими же значениями, полученными для первичных клеток эритроидного ряда.
Пример 67. Оценка среднего объема клетки с помощью гематологического анализатора Advia
[00816] Средний объем клетки (MCV) культивируемых клеток эритроидного ряда измеряли с применением полного электрического сопротивления в гематологическом анализаторе Advia 120 (Siemens Healthcare). Результаты сравнивали с результатами природных эритроцитов человека.
Пример 68. Испытание культивируемых клеток эритроидного ряда на патогены
[00817] RT-PCR применяли для количественного определения присутствия занесенного вируса в популяциях культивируемых клеток эритроидного ряда и подтверждения отсутствия контаминации (анализ № 003000.BSV, BioReliance). Испытание на стерильность необработанного и конечного балка, готовых флаконов, клеток перед помещение в банк и клеточных и вирусных банков выполняли путем инокуляции популяции клеток эритроидного ряда в 2 разных типа сред, которые поддерживают рост аэробных и анаэробных бактерий, соответственно. Образцы инкубировали в течение 14 дней с последующим испытанием на контаминацию микроорганизмами с применением протокола испытания на стерильность USP 71 от BioReliance.
Пример 69. Оценка осмотической хрупкости
[00818] Осмотическую хрупкость оценивали для измерения устойчивости клеток эритроидного ряда к лизису при воздействии гипотонических растворов. Растворы NaCl в воде готовили при концентрациях, охватывающих интервал от 0% дo 1%. Клетки инкубировали в каждом из солевых растворов в течение 15 минут. Образцы центрифугировали для осаждения интактных клеток. Супернатант анализировали на содержание гемоглобина по поглощению света при длине волны 540 нм с применением спектрофотометра. Точку, при которой происходит 50% гемолиз, рассчитывали и сравнивали со значением, полученным для первичных эритроцитов.
Пример 70. Оценка розеткообразования/иммуногенности
[00819] С помощью прямого антиглобулинового теста, также известного как тест Кумбса, оценивали агглютинацию или розеткообразование клеток эритроидного ряда, вызванные связыванием поликлональных антител из сыворотки крови с поверхностными антигенами на клетке. Его можно выполнять с применением объединенной сыворотки крови человека для оценки общей аллогенной иммуногенности или с сывороткой крови от предполагаемого реципиента для прогнозирования специфичной иммуногенности.
[00820] Вкратце, в реакционную пробирку вносили 1-2 капли клеток, хранимых в пробирке с EDTA. Трижды промывали эту пробирку изотоническим физиологическим раствором. После третьего промывки готовили 3% суспензию из промытых клеток. Помечали 2 пробирки A и B. Добавляли одну каплю 3% суспензии промытых клеток в каждую пробирку. Промывали эти пробирки еще один раз. При сливе размещали пробирку так, чтобы сгусток клеток находился вверху. Это будет предотвращать потерю слишком большого количества клеток в процессе промывки. Обеспечивали надлежащее стекание и промокали досуха с помощью салфетки для применения в биологических исследованиях. Сразу же добавляли одну каплю тестируемой человеческой сыворотки в обе пробирки и встряхивали для перемешивания. Оставляли пробирку B для инкубации при комнатной температуре в течение 5 минут. Центрифугировали пробирку A в течение времени, определенного для центрифугирования по тесту Кумбса, на Serofuge. Сразу же осторожно ресуспендировали и исследовали агглютинацию с применением освещаемого агглютиноскопа (Beckton Dickinson). Если пробирка А была положительной, не было необходимости считывать пробирку B, а также исследовать пробирку А при помощи микроскопа. Если пробирка А была отрицательной при исследовании с помощью освещаемого агглютиноскопа, агглютинацию исследовали под микроскопом. Если пробирка А была отрицательной при считывании с помощью микроскопа, центрифугировали пробирку В после ее инкубационного периода и повторяли этапы 2-4 с образцом в пробирке В. Если пробирка В также была отрицательной, в пробирку добавляли одну каплю контрольных клеток для теста Кумбса (проверочные клетки), покрытых IgG и центрифугировали. Исследовали агглютинацию. Агглютинация должна была присутствовать на данной стадии, либо результаты теста признавались недействительными.
[00821] Если агглютинация отсутствовала на любой из стадий до добавления проверочных клеток (ccc), результаты теста интерпретировали как отрицательные. Если агглютинация наблюдалась на любой из стадий до добавления проверочных клеток, результаты теста интерпретировали как положительные.
Пример 71. Оценка кислородсвязывающей способности
[00822] Равновесные кривые связывания кислорода при 37°C определяли в тонометре, связанном с кюветой с длиной пути 1 см. Спектрометрию выполняли с помощью спектрофотометра (Cary 50; Variant Inc) и температуру контролировали с помощью модуля Пелтье. Анализы выполняли в 50 мM буфере bis-Tris (pH 7,2), содержащем 140 мM NaCl и 2 мM глюкозы. После тщательной дезоксигенации в атмосфере азота суспензии красных клеток уравновешивали при разных парциальных давлениях кислорода путем инъекции известных объемов чистого кислорода в тонометр через резиновую крышку с помощью шприца Гамильтона. Фракционное насыщение оценивали путем моделирования спектров поглощения в видимой области и области Сорэ как линейной комбинации полностью дезоксигенированных и оксигенированных спектров суспензии RBC путем регрессии методом наименьших квадратов.
Пример 72. Оценка метаболического состояния клеток
[00823] Популяцию клеток эритроидного ряда можно проверять на метаболическую активность с применением множества различных анализов, основанных на ферментах, для количественной оценки важных конечных продуктов метаболизма. Активный гликолиз представляет собой главный метаболический путь для оценки, и его можно измерять с применением следующего анализа (набор для анализа гликолиза на основе клеток, Cayman Chemical, позиция 600450).
[00824] 450 мкл буфера для анализа отмеряли в пробирку с последующим добавлением 50 мкл стандартного раствора L-молочной кислоты и тщательно перемешивали. Кривую титрования строили с применением стандартных концентраций уксусной кислоты, начиная с разведения 1 мM.
[00825] Клетки добавляли в 96-луночный планшет и центрифугировали при 1000 об./мин. в течение 5 минут. 100 мкл стандартных растворов переносили в отдельный 96-луночный планшет. Затем в каждую лунку добавляли 90 мкл буфера для анализа. 10 мкл супернатанта из каждой лунки с клетками затем переносили в соответствующие новые лунки. В каждую лунку добавляли 100 мкл реакционного раствора с применением с пипеточного дозатора-репитера. Затем планшеты инкубировали на орбитальном встряхивателе в течение 30 минут при RT. Показатели поглощения считывали при 490 нм с помощью планшет-ридера. Результаты сравнивали с природными клетками для идентификации каких-либо отличий в метаболизме.
Пример 73. Оценка агрегации тромбоцитов
[00826] Можно отслеживать склонность к агрегации культивируемых тромбоцитов или первичных тромбоцитов, полученных из источников. Осуществляли анализ взбалтывания тромбоцитов посредством встряхивания их перед источником света, при этом результаты проявлялись как присутствие или отсутствие двойного лучепреломления. Порции тромбоцитарной массы, полученные в объеме 50-70 мл, оставляли постоять в течение одного часа и помещали в линейный встряхиватель (C-Mar®) при 70 об/мин при контролируемой температуре 22±2°C (71,6±3,6°F).
[00827] Тесты на тромбоцитарной массе (подсчет количества тромбоцитов, исследование агрегации тромбоцитов и pH) проводили в дни 1, 3 и 5 после обработки; подсчет количества лейкоцитов выполняли только в день 1, а микробиологический контроль выполняли только на 5-й день хранения. Для получения аликвот из образцов тромбоцитарной массы применяли стерильное соединительное устройство (Haemonetics®), которое обеспечивало целостность окружающей среды. Агрегацию тромбоцитов проводили с применением турбидиметрической методики агрегометрии с применением двухканального Chronolog (Crono-Log Corporation®) в пределах четырех часов от момента сбора крови. Для данной цели клетки сначала получали путем слабого центрифугирования при 1000 об/мин в течение пяти минут и затем центрифугировали при 3000 об/мин в течение пятнадцати минут (Eppendorf®). В образцах подсчитывали количество тромбоцитов в автоматическом устройстве для подсчета (Human Count®).
[00828] После доведения концентрации тромбоцитов агрегацию оценивали с применением различных концентраций индуцирующих агонистов: 2,0 мкг/мл коллагена и 7,0 мкг/мл АДФ (Crono-Log Corporation®). Для каждого теста применяли 400 мкл PRP и 400 мкл PPP, каждый в отдельной кювете, после того, как дожидались спонтанной агрегации. Кривую агрегации наблюдали через пять минут после стимуляции с помощью индуцирующих агонистов и вскоре после этого агрегацию измеряли и выражали как процентную долю в соответствии с кривыми, полученными в ходе данных тестов. Результат теста обычно выражали как процентную долю агрегации по количеству света, прошедшего через исследуемый раствор; агрегацию классифицировали как нормальную, низкую или высокую.
Пример 74. Процесс аутологичного культивирования
[00829] Культуру клеток эритроидного ряда с применением CD34+ клеток-предшественников, полученных из аутологичного источника, получали с целью оптимизации иммуносовместимости клеток для пациентов. CD34+ клетки из костного мозга мобилизировали в периферическую кровь у пациента с применением GM-CSF, как описано в данном документе. 106-108 CD34+ клеток собирали и культивировали с применением упомянутого выше 22-дневного протокола с применением указанных сред. В течение дня 4 клетки трансфицировали лентивирусным вектором, содержащим ген, который кодирует экспрессию терапевтического средства. После завершения протокола культивирования клетки очищали и оценивали по нескольким критериям контроля качества, в том числе физическим свойствам, которые коррелируют с жизнеспособностью в кровотоке, иммуногенностью, репликативным потенциалом, чистотой и терапевтической дозой. Затем клетки хранили в пригодном стабилизирующем растворе и составляли в шприце или пригодном средстве доставки. Затем клетки инфузировали тому же пациенту, который был донором исходных CD34+ клеток.
Пример 75. Процесс аутологичной загрузки
[00830] Что касается получения терапевтических клеток эритроидного ряда, загруженных подходящим экзогенным антигеном, эритроциты, полученные из аутологичного источника, можно использовать с целью оптимизации иммуносовместимости клеток для пациентов. Кровь собирали у пациента и центрифугировали при 5000g в течение 20 минут. Лейкоцитарную пленку удаляли и оставшиеся красные клетки ресуспендировали в антикоагулирующем буфере при плотности 108 клеток/мл с получением в общей сложности 1010 клеток. Клетки загружали терапевтическим экзогенным антигеном, представляющим интерес, с помощью одного из способов, описанных выше. После завершения протокола загрузки клетки очищали и оценивали по нескольким критериям контроля качества, в том числе физическим свойствам, которые коррелируют с жизнеспособностью в кровотоке, иммуногенностью, репликативным потенциалом, чистотой и терапевтической дозой. Затем клетки хранили в пригодном стабилизирующем растворе и составляли в шприце или пригодном средстве доставки. Клетки инфузировали тому же пациенту, который был донором исходных эритроцитов.
Пример 76. Процесс аллогенного культивирования
[00831] Для создания масштабируемого универсального терапевтического средства клетки эритроидного ряда можно культивировать из аллогенного источника. Культуру клеток эритроидного ряда с применением CD34+ клеток-предшественников, полученных из аллогенного источника, получали с целью упрощения способа и культивирования некоторого объема терапевтического средства, с помощью которого можно лечить пациентов в требуемом масштабе. У доноров определяли группу крови по основным антигенам крови, включая A, B, Rh, для идентификации универсальных доноров (например, O Rh- или Бомбей Rh-). CD34+ клетки из костного мозга мобилизировали в периферическую кровь у подходящего донора с применением GM-CSF, как описано в данном документе. 106-108 CD34+ клеток собирали и культивировали с применением упомянутого выше 22-дневного протокола с применением указанных сред. В течение дня 4 клетки трансфицировали лентивирусным вектором, содержащим ген, который кодирует экспрессию терапевтического средства. После завершения протокола культивирования клетки очищали и оценивали по нескольким критериям контроля качества, в том числе физическим свойствам, которые коррелируют с жизнеспособностью в кровотоке, иммуногенностью, репликативным потенциалом, чистотой и терапевтической дозой. Затем клетки хранили в пригодном стабилизирующем растворе и составляли в шприце или пригодном средстве доставки. Затем клетки инфузировали пациентам, независимо от их основных групп крови.
Пример 77. Процесс аллогенной загрузки
[00832] Культуру клеток эритроидного ряда с применением CD34+ клеток-предшественников, полученных из аллогенного источника, получали с целью упрощения способа получения больших объемов терапевтических клеток, с помощью которых можно лечить пациентов в требуемом масштабе. У доноров определяли группу крови по основным антигенам крови, включая A, B, Rh, для идентификации универсальных доноров (например, O Rh- или Бомбей Rh-). Клетки загружали терапевтическим экзогенным антигеном, представляющим интерес, с помощью одного из способов, описанных выше. После завершения протокола загрузки клетки очищали и оценивали по нескольким критериям контроля качества, в том числе физическим свойствам, которые коррелируют с жизнеспособностью в кровотоке, иммуногенностью, репликативным потенциалом, чистотой и терапевтической дозой. Затем клетки хранили в пригодном стабилизирующем растворе и составляли в шприце или пригодном средстве доставки. Затем клетки инфузировали пациентам, независимо от их основных групп крови.
Пример 78. Хранение
1. Хранение в буферном растворе в условиях холодильника
[00833] Стандартные протоколы для хранения красных кровяных клеток известны из уровня техники, см., например, Meryman and Hornblower 1986, Transfusion 26(6):500. Стандартный протокол для хранения красных кровяных клеток (в течение до 42 дней) представляет собой сбор крови в антикоагулирующие растворы (цитрат-декстроза-фосфат). Клетки эритроидного ряда культивировали, как описано в данном документе. Эритроцитарную массу готовили путем удаления плазмы крови с помощью центрифугирования. Клетки хранили при 4±2°C в слегка гипертоническом растворе с добавками, SAGM (натрий, аденин, глюкоза, маннит, 376 мосмоль/л).
2. Хранение в буферном растворе с замораживанием
[00834] Способы глицеролизации, замораживания и оттаивания клеток эритроидного ряда известны из уровня техники, см., например, Meryman and Hornblower 1977 Transfusion 17(5):4348. Человеческую кровь в цитрат-фосфат-декстрозе подвергали глицеролизации и замораживали в пределах 4 дней с момента сбора. Для получения обработанных глицерином RBC приблизительно 10 мл цельной крови сначала центрифугировали при 1400g в течение 10-15 мин и плазму крови удаляли. Затем полученную клеточную массу подвергали глицеролизации в две стадии с применением водного раствора глицерина следующего состава: 57,1 г глицерина, 0,03 г хлорида калия, 0,085 г хлорида магния гексагидрата, 0,08 г динатрия фосфата и 1,6 г лактата натрия в общем объеме 100 мл, доводенного до pH 6,8.42. На первом этапе 1,5 мл данного раствора глицерина добавляли по каплям к клеточной массе при осторожном перемешивании на протяжении 3 мин. Затем смеси давлаи уравновеситься, оставляя по меньшей мере на 5 мин. На второй стадии глицеролизации 5 мл раствора глицерина добавляли по каплям, в то же время осторожного перемешивали смесь на протяжении 3 мин, с получением конечной композиции глицерина ~40% вес./об. Весь процесс глицеролизации проводили при комнатной температуре. Затем обработанные глицерином RBC разделяли на аликвоты 0,6-1,1 мл в криогенных флаконах, помещали в контейнер для замораживания NalgeneVR Cryo "Mr. Frosty" (Thermo Scientific, Северная Каролина) и выдерживали в морозильной камере при -80°C в течение по меньшей мере 12 ч и на протяжении до 10 лет. Замороженные RBC оттаивали путем помещения криогенного флакона на водяную баню при 37°C на 1 мин. Все обработанные глицерином образцы крови применяли в экспериментах по деглицеролизации в пределах 2 ч от момента оттаивания.
3. Состав в виде шприца
[00835] Популяцию клеток можно вводить внутривенно с помощью шприца. Терапевтические клетки разводили до плотности 107 клеток/мл с применением стандартного солевого буфера при 37°C, при этом доставляли объем 100 мл или 109 клеток. Раствор клеток загружали в шприц на 150 см3, с 20 калибром иглы, и инъецировали пациенту через медиальную подкожную вену руки при скорости 5 см3/мин. Во время проведения инъекции отслеживали основные жизненные показатели пациента на предмет каких-либо иммуногенных реакций или реакций свертывания.
4. Состав в виде пакета
[00836] Популяцию клеток можно вводить внутривенно посредством шприца, соединенного с пакетом и капельницей (т.e. капельницей для внутривенного вливания). Терапевтические клетки разводили до плотности 107 клеток/мл с применением стандартного солевого буфера при 37°C, при этом доставляли объем 100 мл или 109 клеток. Раствор клеток загружали в полиэтиленовый пакет на 1 л, подсоединяли к катетеру и обеспечивали вытекание под действием сил гравитации в пациента через медиальную подкожную вену руки. Во время проведения инфузии отслеживали основные жизненные показатели пациента на предмет каких-либо иммуногенных реакций или реакций свертывания.
Пример 79. Лечение заболеваний
1. Гемофилия
[00837] Устанавливали диагноз пациенту, страдающему от гемофилии A. Композицию безъядерных гемопоэтических клеток, экспрессирующих экзогенный FVIII, готовили, как описано в данном документе. Пациенту внутривенно вводили 109 клеток. Скорость свертывания оценивали с помощью стандартного анализа времени свертывания крови in vitro, известного из уровня техники. Циркулирующие в крови антитела против FVIII обнаруживали в сыворотке крови, как описано в данном документе. Уровни циркулирующих в крови антител оценивали для отслеживания эффективности индукции иммунологической толерантности. Если активность системы свертывания крови была недостаточной для обеспечения нормальной коагуляции, одновременно внутривенно вводили рекомбинантный или выделенный FVIII с целью ослабления симптомов гемофилии A.
2. Атипичный гемолитико-уремический синдром
[00838] Устанавливали диагноз пациенту, страдающему от атипичного гемолитико-уремического синдрома (aHUS). Композицию безъядерных гемопоэтических клеток, экспрессирующих экзогенный CFH, готовили, как описано в данном документе. Пациенту внутривенно вводили 109 клеток. Скорость симптоматического гемолиза оценивали с помощью стандартного анализа гемолиза в моче, известного из уровня техники. Циркулирующие в крови антитела против CFH обнаруживали в сыворотке крови, как описано в данном документе. Уровни циркулирующих в крови антител оценивали для отслеживания эффективности индукции иммунологической толерантности. Пациенту вводили терапевтическое средство, пока не отмечалось ослабление симптомов с применением анализов, описанных в данном документе.
[00839] 3. Рассеянный склероз
[00840] Индивидууму с рассеянным склерозом (MS) вводили однократную инфузию 1×109 экспрессирующих антиген безъядерных гемопоэтических клеток, экспрессирующих антигенный полипептид основного белка миелина (MBP), полученных и составленных, как описано в данном документе. В день введения исследуемого лекарственного средства за пациентом наблюдали в стационарном отделении для проведения фазы 1 клинических исследований в течение 24 часов. Измерение первичных результатов выполняли в месяц 3 и дополнительное последующее наблюдение для оценки безопасности выполняли до месяца 6 с проведением последовательных клинических, MRI и общих физических осмотров, а также клинических и лабораторных анализов для оценки побочных эффектов и отслеживания активности заболевания MS. Процедуру повторяли до индукции толерантности, так что симптомы MS у индивидуума ослаблялись. См., например, Andreas Lutterotti et al. Sci Transl Med 5, 188ra75 (2013).
[00841] Встречаемость различных субпопуляций клеток анализировали в цельной крови (пробирки с EDTA) с помощью проточной цитометрии со следующими панелями антител: для субпопуляций иммунных клеток (гранулоциты, эозинофилы, моноциты и B-, T-, NK-, и NK Т-клетки)-антитело к CD45 (PE-Cy7, eBioscience), антитело к CD16 (APC-Cy7, BioLegend), антитело к CD19 [флуоресцеинизотиоцианат (FITC), BD], антитело к CD14 (V450, BD), антитело к CD3 [перидинин-хлорофилл-белок (PerCP), BD] и антитело к CD56 [фикоэритрин (PE), eBioscience]; для субпопуляций T-клеток, в том числе CD4+, FoxP3+ Treg, регуляторных CD8+CD57+ILT2+ и провоспалительные Т-клеток с высоким уровнем экспрессии CD8+CD161 -антитело к CD3 (PE-Cy7, eBioscience), антитело к CD4 (APC, eBioscience), антитело к CD8 [Pacific Blue (PB), Dako-Biozol], антитело к FoxP3 (PE, Miltenyi), антитело к CD25 (APC, eBioscience), антитело к CD57 (FITC, BD), антитело к ILT2 (PE, Beckman) и антитело к CD161 (APC, Miltenyi). Соответствующие изотопические контроли включали во все окрашивания. Клетки анализировали с помощью проточного цитометра LSR-II (BD) и программного обеспечения FACSDiva (BD).
[00842] Мононуклеарные клетки периферической крови (PBMC) выделяли путем центрифугирования в градиенте плотности Ficoll (PAA) и функциональный фенотип Т-клеток оценивали с помощью внутриклеточного окрашивания цитокинов следующим образом: 5×105 свежевыделенных PBMC инкубировали в течение ночи в 200 мл X-VIVO 15 (Lonza) в стерильной пробирке для FACS. На следующий день клетки стимулировали с помощью форбол-12-миристат-13-ацетата (50 нг/мл, Sigma) и иономицина (1 мг/мл, Sigma) в присутствии брефелдина A (10 мг/мл, eBioscience) в течение 5 часов. После промывания фосфатно-солевым буфером клетки окрашивали с помощью набора LiveDead (AmCyan, Invitrogen), фиксировали, пермеабилизировали и окрашивали различными антителами: антителом к IL-17 (Alexa Fluor 647; eBioscience), антителом к IL-4 (PE-Cy7, BioLegend), антителом к IFN-g (FITC, BioLegend), антителом к IL-10 (PE; BioLegend), антителом к CD3 (PE, DakoCytomation), антителом к CD4 (PB, DakoCytomation) и антителом к CD8 (PB, BioLegend), или с помощью соответствующих изотопических контролей.
[00843] Антиген-специфичные Т-клеточные ответы в отношении миелинового пептида, применяемого в исследовании, измеряли на свежевыделенных PBMC перед процедурой толеризации и через 3 месяца. Антиген-специфичные Т-клеточные ответы анализировали с помощью анализов пролиферации с включением тимидина. Вкратце, выделенные PBMC высевали в 96-луночные планшеты из расчета 1,5×105 PBMC на лунку в среду X-VIVO 15 (Lonza) с 1 мМ пептида. В каждом планшете сорок восемь лунок засевали для введения антигена и в шесть лунок вводили только среду в качестве отрицательного контроля. В качестве положительного контроля применяли TTx (5 мг/мл) (Novartis Behring). В день 7 планшеты инкубировали в течение 15 часов с 1 мКи [3H] тимидина (Hartmann Analytic). Планшеты с импульсами от [3H] тимидина анализировали с помощью сцинтилляционного b-счетчика (Wallac 1450, PerkinElmer). Измеряли сцинтилляционный счет (CPM) для каждой лунки. Лунки, демонстрирующие CPM выше среднего+3 SD для непростимулированных лунок, рассматривали как положительные.
[00844] Многие модификации и другие варианты осуществления изобретений, изложенные в данном документе, придут на ум специалисту в области, к которой принадлежат настоящие изобретения, с получением пользы от идей, представленных в вышеизложенном описании и ассоциированных графических материалах. Следовательно, следует понимать, что настоящие изобретения не следует ограничивать конкретными раскрытыми вариантами осуществления, и подразумевается, что модификации и другие варианты осуществления включены в объем прилагаемой формулы изобретения. Хотя в данном документе используются специальные термины, они применяются только в общем и описательном смысле, а не с целью ограничения.
[00845] Все публикации и патентные заявки, упомянутые в описании, свидетельствуют об уровне специалистов в области, к которой относится настоящее изобретение. Все публикации и патентные заявки включены в данный документ посредством ссылки в такой же полноте, как если бы каждая отдельная публикация или патентная заявка были бы специфично и индивидуально указаны как включенные посредством ссылки.
Таблицы A-J
Таблица A. Циркулирующие в крови клетки
Таблица A1. Клетки эритроидного ряда
Таблица B. Белки, ассоциированные с циркулирующими в крови клетками
Таблица C. Белки, ассоциированные с эритроцитами
Таблица C1. Трансмембранные белки эритроцитов
Таблица C2. Белки эритроцитов, присоединенные к GPI
Таблица C3. Внутриклеточные белки эритроцитов
Таблица D. Способы конъюгации
Таблица D1. Способы ферментативной конъюгации
Таблица E. Химия реактивных групп
Таблица F. Аутоиммунные заболевания и антигены
Таблица G. Воспалительные заболевания и антигены
Таблица H. Триггерные факторы аллергических заболеваний
Таблица I. Терапевтические белки для лечения заболеваний
Таблица J. Классы терапевтических белков для лечения заболеваний
Таблицы 1-8
Таблица 3. Последовательности рецептора комплемента 1
4A. Предшественник изоформы S CR1, эталонная последовательность Homo sapiens в NCBI № NP_000642.3
1 mgassprspe pvgppapglp fccggsllav vvllalpvaw gqcnapewlp farptnltde
61 fefpigtyln yecrpgysgr pfsiiclkns vwtgakdrcr rkscrnppdp vngmvhvikg
121 iqfgsqikys ctkgyrligs ssatciisgd tviwdnetpi cdripcglpp titngdfist
181 nrenfhygsv vtyrcnpgsg grkvfelvge psiyctsndd qvgiwsgpap qciipnkctp
241 pnvengilvs dnrslfslne vvefrcqpgf vmkgprrvkc qalnkwepel pscsrvcqpp
301 pdvlhaertq rdkdnfspgq evfyscepgy dlrgaasmrc tpqgdwspaa ptcevkscdd
361 fmgqllngrv lfpvnlqlga kvdfvcdegf qlkgssasyc vlagmeslwn ssvpvceqif
421 cpsppvipng rhtgkplevf pfgktvnytc dphpdrgtsf dligestirc tsdpqgngvw
481 sspaprcgil ghcqapdhfl faklktqtna sdfpigtslk yecrpeyygr pfsitcldnl
541 vwsspkdvck rkscktppdp vngmvhvitd iqvgsrinys cttghrligh ssaecilsgn
601 aahwstkppi cqripcglpp tiangdfist nrenfhygsv vtyrcnpgsg grkvfelvge
661 psiyctsndd qvgiwsgpap qciipnkctp pnvengilvs dnrslfslne vvefrcqpgf
721 vmkgprrvkc qalnkwepel pscsrvcqpp pdvlhaertq rdkdnfspgq evfyscepgy
781 dlrgaasmrc tpqgdwspaa ptcevkscdd fmgqllngrv lfpvnlqlga kvdfvcdegf
841 qlkgssasyc vlagmeslwn ssvpvceqif cpsppvipng rhtgkplevf pfgktvnytc
901 dphpdrgtsf dligestirc tsdpqgngvw sspaprcgil ghcqapdhfl faklktqtna
961 sdfpigtslk yecrpeyygr pfsitcldnl vwsspkdvck rkscktppdp vngmvhvitd
1021 iqvgsrinys cttghrligh ssaecilsgn aahwstkppi cqripcglpp tiangdfist
1081 nrenfhygsv vtyrcnpgsg grkvfelvge psiyctsndd qvgiwsgpap qciipnkctp
1141 pnvengilvs dnrslfslne vvefrcqpgf vmkgprrvkc qalnkwepel pscsrvcqpp
1201 pdvlhaertq rdkdnfspgq evfyscepgy dlrgaasmrc tpqgdwspaa ptcevkscdd
1261 fmgqllngrv lfpvnlqlga kvdfvcdegf qlkgssasyc vlagmeslwn ssvpvceqif
1321 cpsppvipng rhtgkplevf pfgkavnytc dphpdrgtsf dligestirc tsdpqgngvw
1381 sspaprcgil ghcqapdhfl faklktqtna sdfpigtslk yecrpeyygr pfsitcldnl
1441 vwsspkdvck rkscktppdp vngmvhvitd iqvgsrinys cttghrligh ssaecilsgn
1501 tahwstkppi cqripcglpp tiangdfist nrenfhygsv vtyrcnlgsr grkvfelvge
1561 psiyctsndd qvgiwsgpap qciipnkctp pnvengilvs dnrslfslne vvefrcqpgf
1621 vmkgprrvkc qalnkwepel pscsrvcqpp peilhgehtp shqdnfspgq evfyscepgy
1681 dlrgaaslhc tpqgdwspea prcavkscdd flgqlphgrv lfplnlqlga kvsfvcdegf
1741 rlkgssvshc vlvgmrslwn nsvpvcehif cpnppailng rhtgtpsgdi pygkeisytc
1801 dphpdrgmtf nligestirc tsdphgngvw sspaprcels vraghcktpe qfpfasptip
1861 indfefpvgt slnyecrpgy fgkmfsiscl enlvwssved ncrrkscgpp pepfngmvhi
1921 ntdtqfgstv nyscnegfrl igspsttclv sgnnvtwdkk apiceiisce ppptisngdf
1981 ysnnrtsfhn gtvvtyqcht gpdgeqlfel vgersiycts kddqvgvwss ppprcistnk
2041 ctapevenai rvpgnrsfft lteiirfrcq pgfvmvgsht vqcqtngrwg pklphcsrvc
2101 qpppeilhge htlshqdnfs pgqevfysce psydlrgaas lhctpqgdws peaprctvks
2161 cddflgqlph grvllplnlq lgakvsfvcd egfrlkgrsa shcvlagmka lwnssvpvce
2221 qifcpnppai lngrhtgtpf gdipygkeis yacdthpdrg mtfnligess irctsdpqgn
2281 gvwsspaprc elsvpaacph ppkiqnghyi gghvslylpg mtisyicdpg yllvgkgfif
2341 ctdqgiwsql dhyckevncs fplfmngisk elemkkvyhy gdyvtlkced gytlegspws
2401 qcqaddrwdp plakctsrth dalivgtlsg tiffilliif lswiilkhrk gnnahenpke
2461 vaihlhsqgg ssvhprtlqt neensrvlp (Seq. ID No. 1)
4B. Предшественник изоформы F CR1, эталонная последовательность Homo sapiens в NCBI № NP_000564.2
1 mgassprspe pvgppapglp fccggsllav vvllalpvaw gqcnapewlp farptnltde
61 fefpigtyln yecrpgysgr pfsiiclkns vwtgakdrcr rkscrnppdp vngmvhvikg
121 iqfgsqikys ctkgyrligs ssatciisgd tviwdnetpi cdripcglpp titngdfist
181 nrenfhygsv vtyrcnpgsg grkvfelvge psiyctsndd qvgiwsgpap qciipnkctp
241 pnvengilvs dnrslfslne vvefrcqpgf vmkgprrvkc qalnkwepel pscsrvcqpp
301 pdvlhaertq rdkdnfspgq evfyscepgy dlrgaasmrc tpqgdwspaa ptcevkscdd
361 fmgqllngrv lfpvnlqlga kvdfvcdegf qlkgssasyc vlagmeslwn ssvpvceqif
421 cpsppvipng rhtgkplevf pfgktvnytc dphpdrgtsf dligestirc tsdpqgngvw
481 sspaprcgil ghcqapdhfl faklktqtna sdfpigtslk yecrpeyygr pfsitcldnl
541 vwsspkdvck rkscktppdp vngmvhvitd iqvgsrinys cttghrligh ssaecilsgn
601 aahwstkppi cqripcglpp tiangdfist nrenfhygsv vtyrcnpgsg grkvfelvge
661 psiyctsndd qvgiwsgpap qciipnkctp pnvengilvs dnrslfslne vvefrcqpgf
721 vmkgprrvkc qalnkwepel pscsrvcqpp pdvlhaertq rdkdnfspgq evfyscepgy
781 dlrgaasmrc tpqgdwspaa ptcevkscdd fmgqllngrv lfpvnlqlga kvdfvcdegf
841 qlkgssasyc vlagmeslwn ssvpvceqif cpsppvipng rhtgkplevf pfgkavnytc
901 dphpdrgtsf dligestirc tsdpqgngvw sspaprcgil ghcqapdhfl faklktqtna
961 sdfpigtslk yecrpeyygr pfsitcldnl vwsspkdvck rkscktppdp vngmvhvitd
1021 iqvgsrinys cttghrligh ssaecilsgn tahwstkppi cqripcglpp tiangdfist
1081 nrenfhygsv vtyrcnlgsr grkvfelvge psiyctsndd qvgiwsgpap qciipnkctp
1141 pnvengilvs dnrslfslne vvefrcqpgf vmkgprrvkc qalnkwepel pscsrvcqpp
1201 peilhgehtp shqdnfspgq evfyscepgy dlrgaaslhc tpqgdwspea prcavkscdd
1261 flgqlphgrv lfplnlqlga kvsfvcdegf rlkgssvshc vlvgmrslwn nsvpvcehif
1321 cpnppailng rhtgtpsgdi pygkeisytc dphpdrgmtf nligestirc tsdphgngvw
1381 sspaprcels vraghcktpe qfpfasptip indfefpvgt slnyecrpgy fgkmfsiscl
1441 enlvwssved ncrrkscgpp pepfngmvhi ntdtqfgstv nyscnegfrl igspsttclv
1501 sgnnvtwdkk apiceiisce ppptisngdf ysnnrtsfhn gtvvtyqcht gpdgeqlfel
1561 vgersiycts kddqvgvwss ppprcistnk ctapevenai rvpgnrsfft lteiirfrcq
1621 pgfvmvgsht vqcqtngrwg pklphcsrvc qpppeilhge htlshqdnfs pgqevfysce
1681 psydlrgaas lhctpqgdws peaprctvks cddflgqlph grvllplnlq lgakvsfvcd
1741 egfrlkgrsa shcvlagmka lwnssvpvce qifcpnppai lngrhtgtpf gdipygkeis
1801 yacdthpdrg mtfnligess irctsdpqgn gvwsspaprc elsvpaacph ppkiqnghyi
1861 gghvslylpg mtisyicdpg yllvgkgfif ctdqgiwsql dhyckevncs fplfmngisk
1921 elemkkvyhy gdyvtlkced gytlegspws qcqaddrwdp plakctsrth dalivgtlsg
1981 tiffilliif lswiilkhrk gnnahenpke vaihlhsqgg ssvhprtlqt neensrvlp
(Seq. ID No. 2)
4C. Предсказанная изоформа X1 CR1, Homo sapiens, эталонная последовательность в NCBI № XP_005273121.1
1 mclgrmgass prspepvgpp apglpfccgg sllavvvlla lpvawgqcna pewlpfarpt
61 nltdefefpi gtylnyecrp gysgrpfsii clknsvwtga kdrcrrkscr nppdpvngmv
121 hvikgiqfgs qikysctkgy rligsssatc iisgdtviwd netpicdrip cglpptitng
181 dfistnrenf hygsvvtyrc npgsggrkvf elvgepsiyc tsnddqvgiw sgpapqciip
241 nkctppnven gilvsdnrsl fslnevvefr cqpgfvmkgp rrvkcqalnk wepelpscsr
301 vcqpppdvlh aertqrdkdn fspgqevfys cepgydlrga asmrctpqgd wspaaptcev
361 kscddfmgql lngrvlfpvn lqlgakvdfv cdegfqlkgs sasycvlagm eslwnssvpv
421 ceqifcpspp vipngrhtgk plevfpfgkt vnytcdphpd rgtsfdlige stirctsdpq
481 gngvwsspap rcgilghcqa pdhflfaklk tqtnasdfpi gtslkyecrp eyygrpfsit
541 cldnlvwssp kdvckrksck tppdpvngmv hvitdiqvgs rinyscttgh rlighssaec
601 ilsgnaahws tkppicqrip cglpptiang dfistnrenf hygsvvtyrc npgsggrkvf
661 elvgepsiyc tsnddqvgiw sgpapqciip nkctppnven gilvsdnrsl fslnevvefr
721 cqpgfvmkgp rrvkcqalnk wepelpscsr vcqpppdvlh aertqrdkdn fspgqevfys
781 cepgydlrga asmrctpqgd wspaaptcev kscddfmgql lngrvlfpvn lqlgakvdfv
841 cdegfqlkgs sasycvlagm eslwnssvpv ceqifcpspp vipngrhtgk plevfpfgkt
901 vnytcdphpd rgtsfdlige stirctsdpq gngvwsspap rcgilghcqa pdhflfaklk
961 tqtnasdfpi gtslkyecrp eyygrpfsit cldnlvwssp kdvckrksck tppdpvngmv
1021 hvitdiqvgs rinyscttgh rlighssaec ilsgnaahws tkppicqlcq pppdvlhaer
1081 tqrdkdnfsp gqevfyscep gydlrgaasm rctpqgdwsp aaptcevksc ddfmgqllng
1141 rvlfpvnlql gakvdfvcde gfqlkgssas ycvlagmesl wnssvpvceq ifcpsppvip
1201 ngrhtgkple vfpfgkavny tcdphpdrgt sfdligesti rctsdpqgng vwsspaprcg
1261 ilghcqapdh flfaklktqt nasdfpigts lkyecrpeyy grpfsitcld nlvwsspkdv
1321 ckrkscktpp dpvngmvhvi tdiqvgsrin yscttghrli ghssaecils gntahwstkp
1381 picqripcgl pptiangdfi stnrenfhyg svvtyrcnlg srgrkvfelv gepsiyctsn
1441 ddqvgiwsgp apqciipnkc tppnvengil vsdnrslfsl nevvefrcqp gfvmkgprrv
1501 kcqalnkwep elpscsrvcq pppeilhgeh tpshqdnfsp gqevfyscep gydlrgaasl
1561 hctpqgdwsp eaprcavksc ddflgqlphg rvlfplnlql gakvsfvcde gfrlkgssvs
1621 hcvlvgmrsl wnnsvpvceh ifcpnppail ngrhtgtpsg dipygkeisy tcdphpdrgm
1681 tfnligesti rctsdphgng vwsspaprce lsvraghckt peqfpfaspt ipindfefpv
1741 gtslnyecrp gyfgkmfsis clenlvwssv edncrrkscg pppepfngmv hintdtqfgs
1801 tvnyscnegf rligspsttc lvsgnnvtwd kkapiceiis ceppptisng dfysnnrtsf
1861 hngtvvtyqc htgpdgeqlf elvgersiyc tskddqvgvw ssppprcist nkctapeven
1921 airvpgnrsf ftlteiirfr cqpgfvmvgs htvqcqtngr wgpklphcsr vcqpppeilh
1981 gehtlshqdn fspgqevfys cepsydlrga aslhctpqgd wspeaprctv kscddflgql
2041 phgrvllpln lqlgakvsfv cdegfrlkgr sashcvlagm kalwnssvpv ceqifcpnpp
2101 ailngrhtgt pfgdipygke isyacdthpd rgmtfnlige ssirctsdpq gngvwsspap
2161 rcelsvpaac phppkiqngh yigghvslyl pgmtisyicd pgyllvgkgf ifctdqgiws
2221 qldhyckevn csfplfmngi skelemkkvy hygdyvtlkc edgytlegsp wsqcqaddrw
2281 dpplakctsr thdalivgtl sgtiffilli iflswiilkh rkgnnahenp kevaihlhsq
2341 ggssvhprtl qtneensrvl p (Seq. ID No. 3)
| название | год | авторы | номер документа |
|---|---|---|---|
| ФУНКЦИОНАЛИЗИРОВАННЫЕ ЭРИТРОИДНЫЕ КЛЕТКИ | 2018 |
|
RU2779315C2 |
| ИСКУССТВЕННЫЕ АНТИГЕНПРЕЗЕНТИРУЮЩИЕ КЛЕТКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2763798C1 |
| НОВЫЙ СПОСОБ ПОЛУЧЕНИЯ ДИФФЕРЕНЦИРОВАННЫХ КЛЕТОК | 2010 |
|
RU2661107C1 |
| НОВЫЙ СПОСОБ ПОЛУЧЕНИЯ ДИФФЕРЕНЦИРОВАННЫХ КЛЕТОК | 2010 |
|
RU2603094C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ИММУНОТЕРАПИИ | 2014 |
|
RU2680010C2 |
| МЕЧЕНЫЕ ХИМЕРНЫЕ ЭФФЕКТОРНЫЕ МОЛЕКУЛЫ И ИХ РЕЦЕПТОРЫ | 2014 |
|
RU2729463C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ БУСТИНГА ЭФФЕКТИВНОСТИ АДОПТИВНОЙ КЛЕТОЧНОЙ ИММУНОТЕРАПИИ | 2015 |
|
RU2716716C2 |
| ВНУТРИКЛЕТОЧНАЯ ДОСТАВКА БИОЛОГИЧЕСКИХ МОЛЕКУЛ ДЛЯ ИНДУЦИРОВАНИЯ ТОЛЕРАНТНОСТИ | 2017 |
|
RU2770492C2 |
| СПОСОБ ПОЛУЧЕНИЯ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР | 2013 |
|
RU2665548C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, ОБЛАДАЮЩАЯ ПРОТИВООПУХОЛЕВЫМ ЭФФЕКТОМ, И СПОСОБ СНИЖЕНИЯ ИНГИБИРУЮЩЕГО ЭФФЕКТА PD-L1 НА Т-КЛЕТКИ ЧЕЛОВЕКА | 2016 |
|
RU2688692C2 |
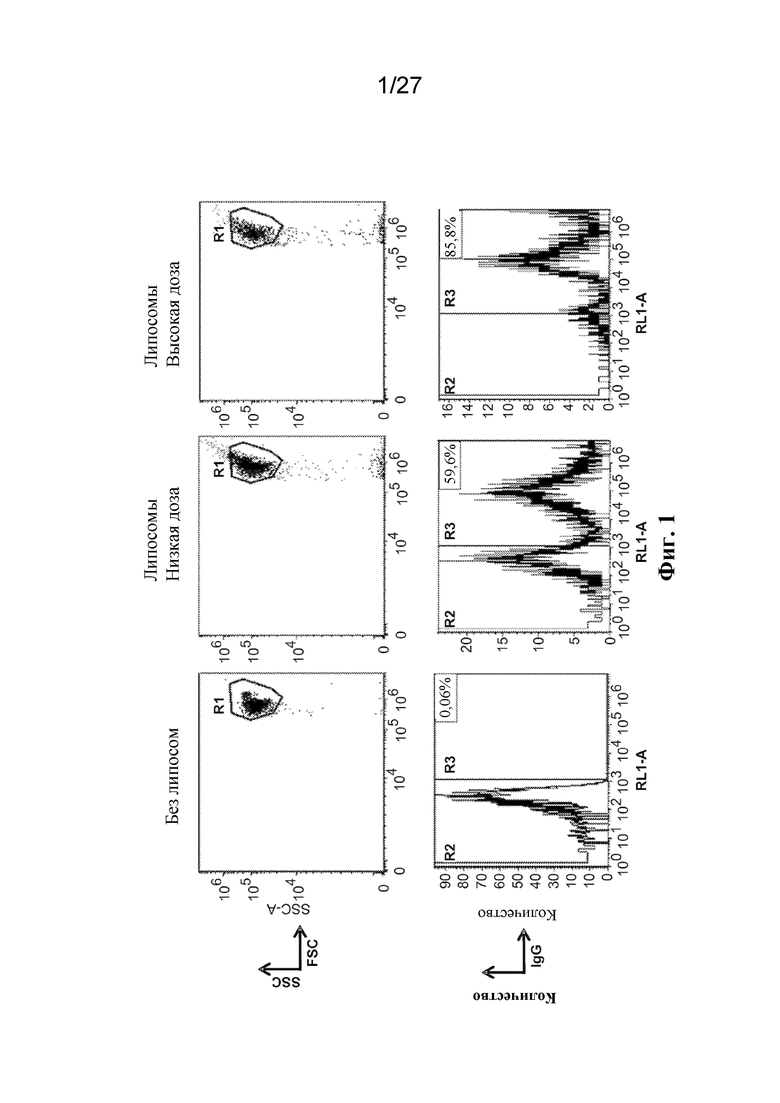
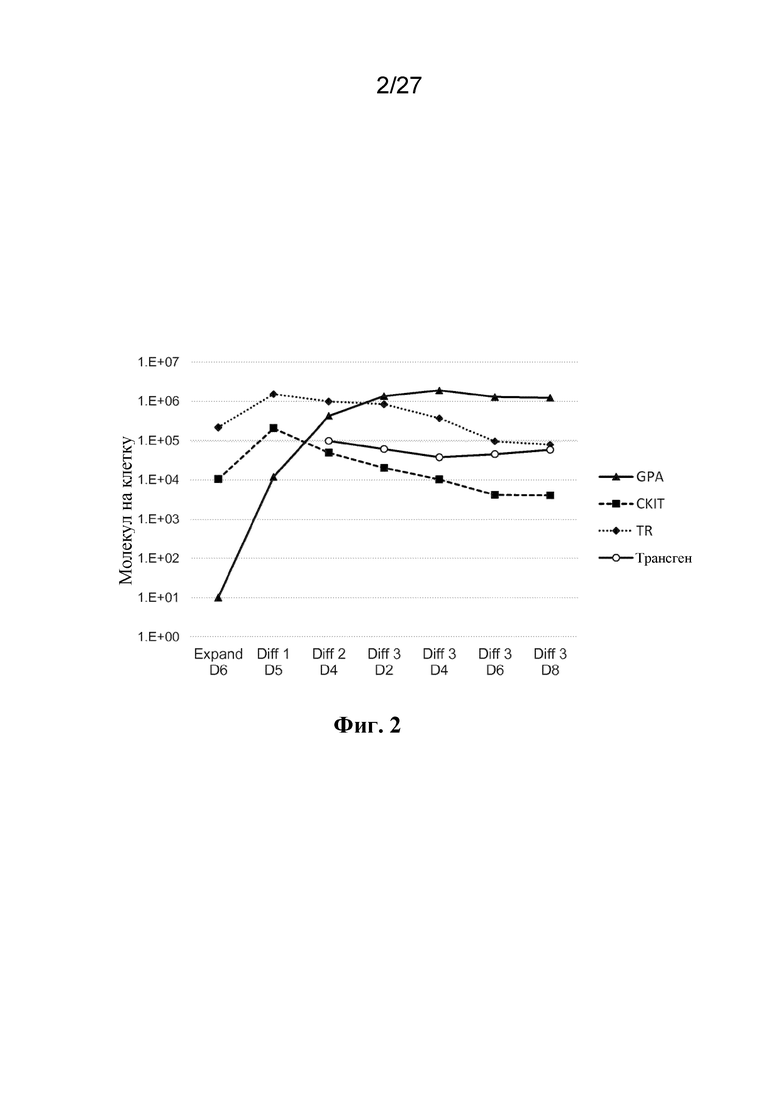
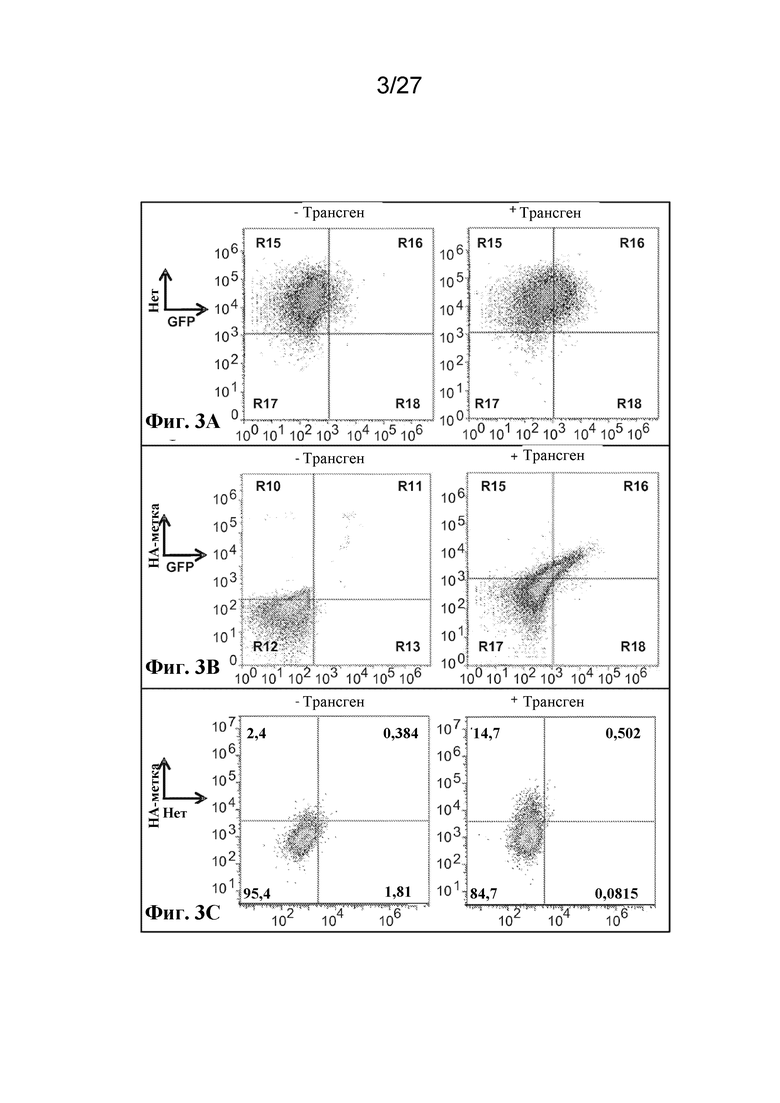
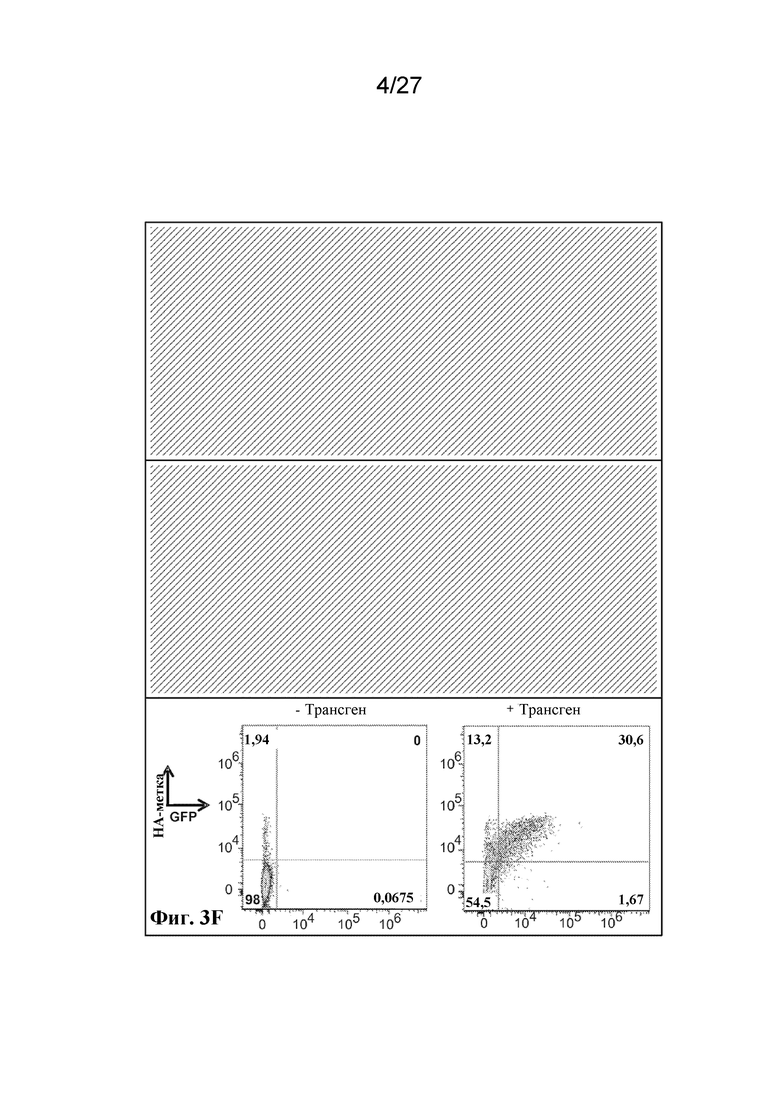


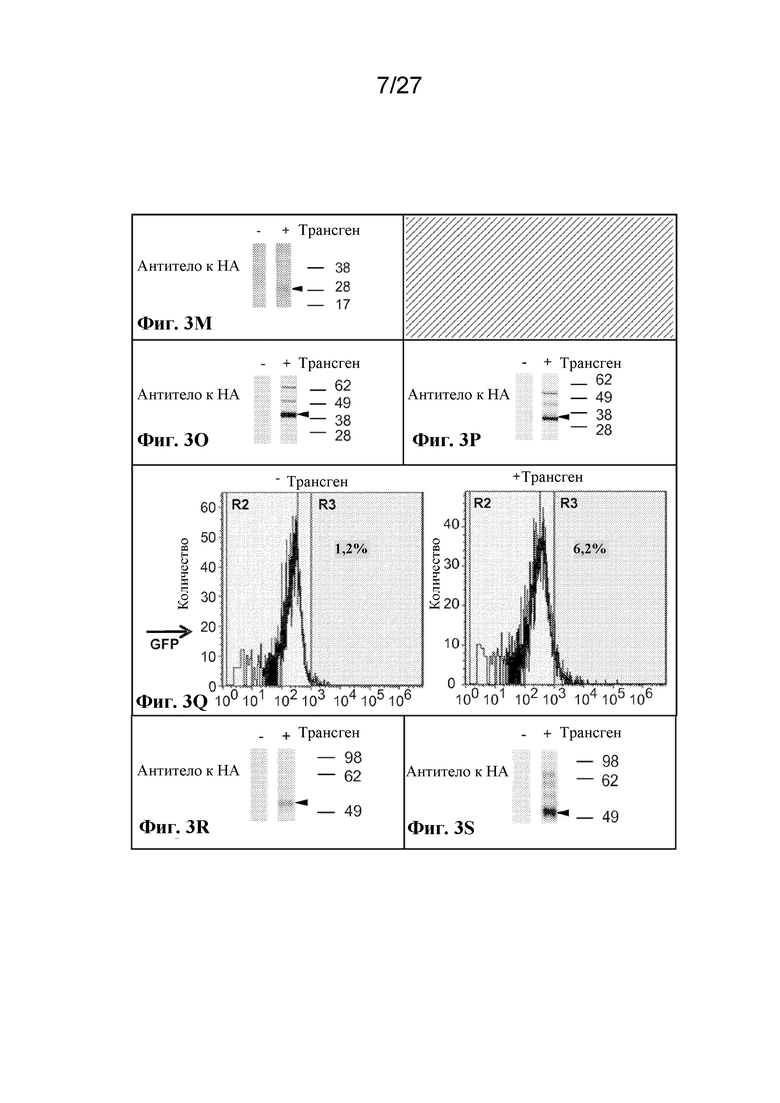
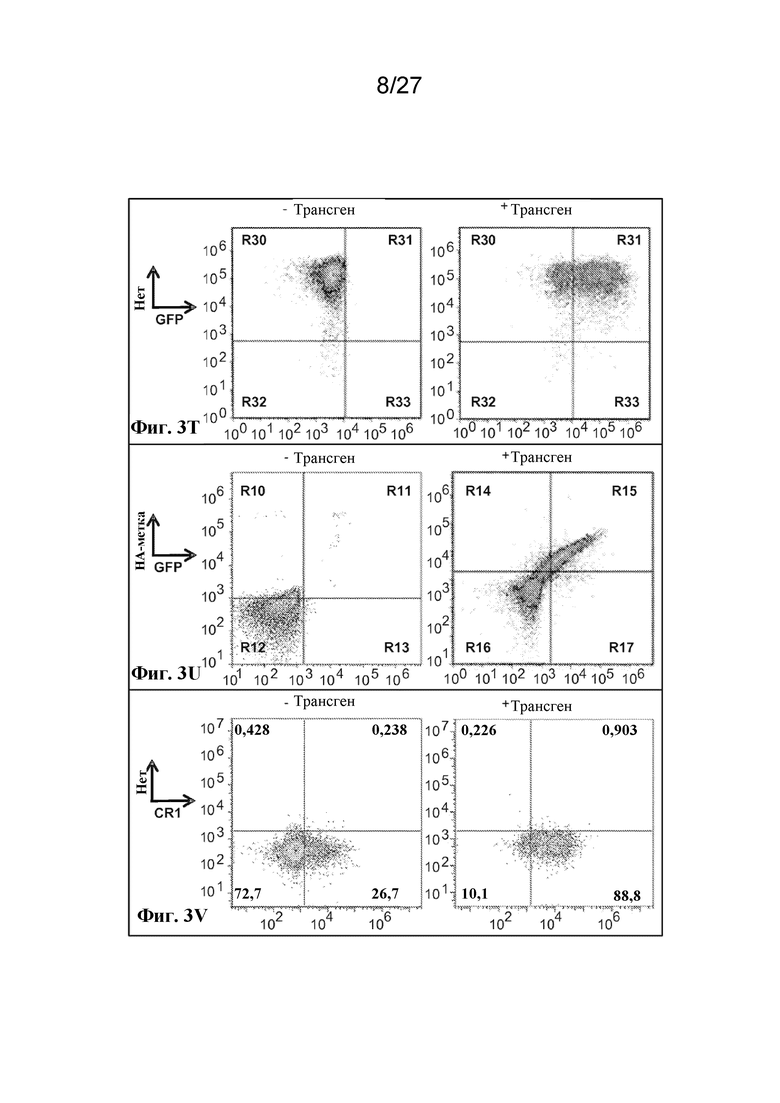
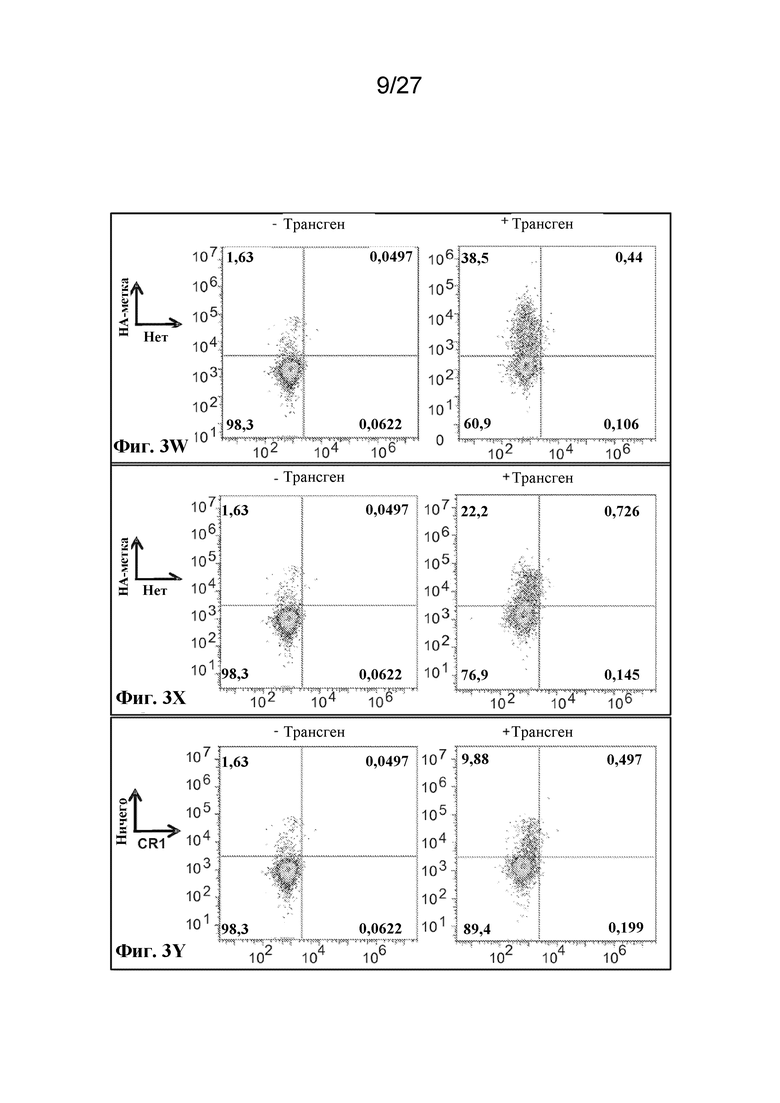
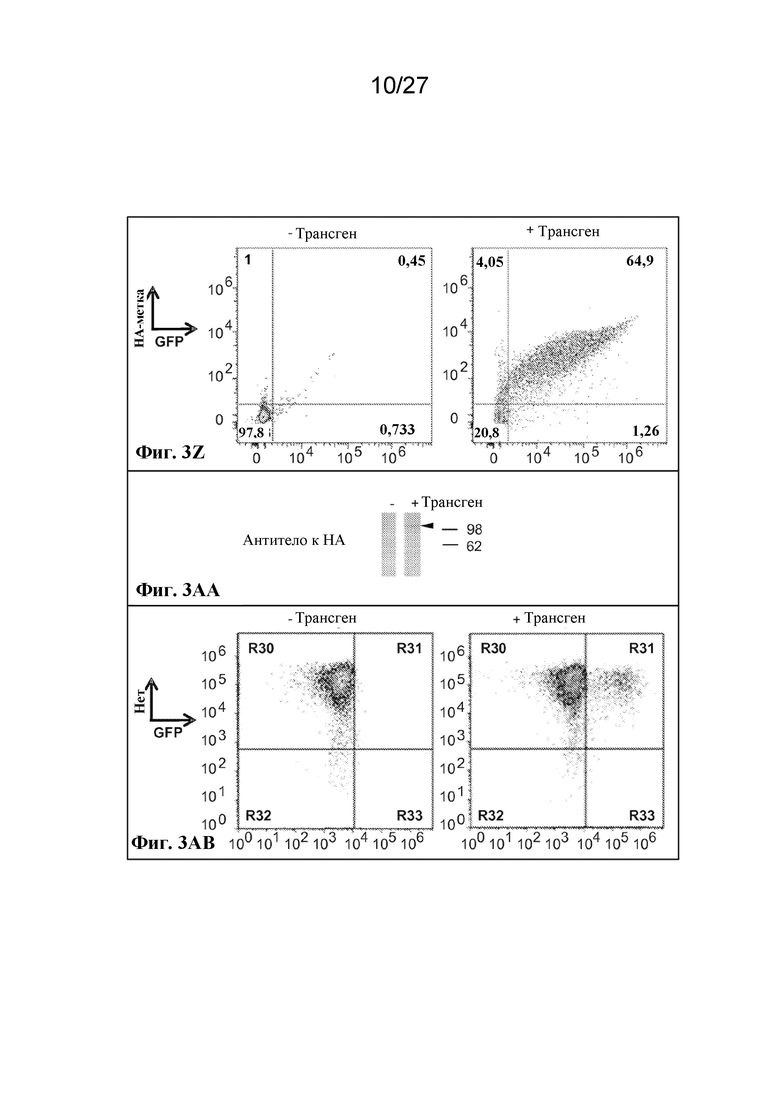

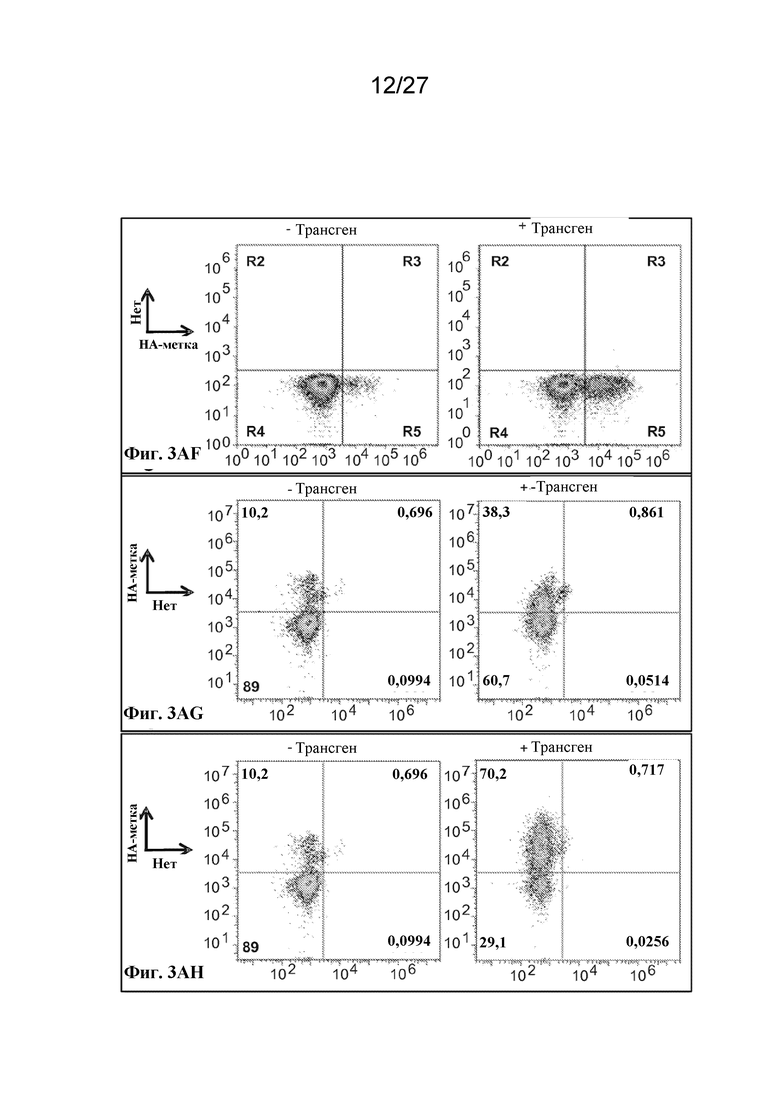
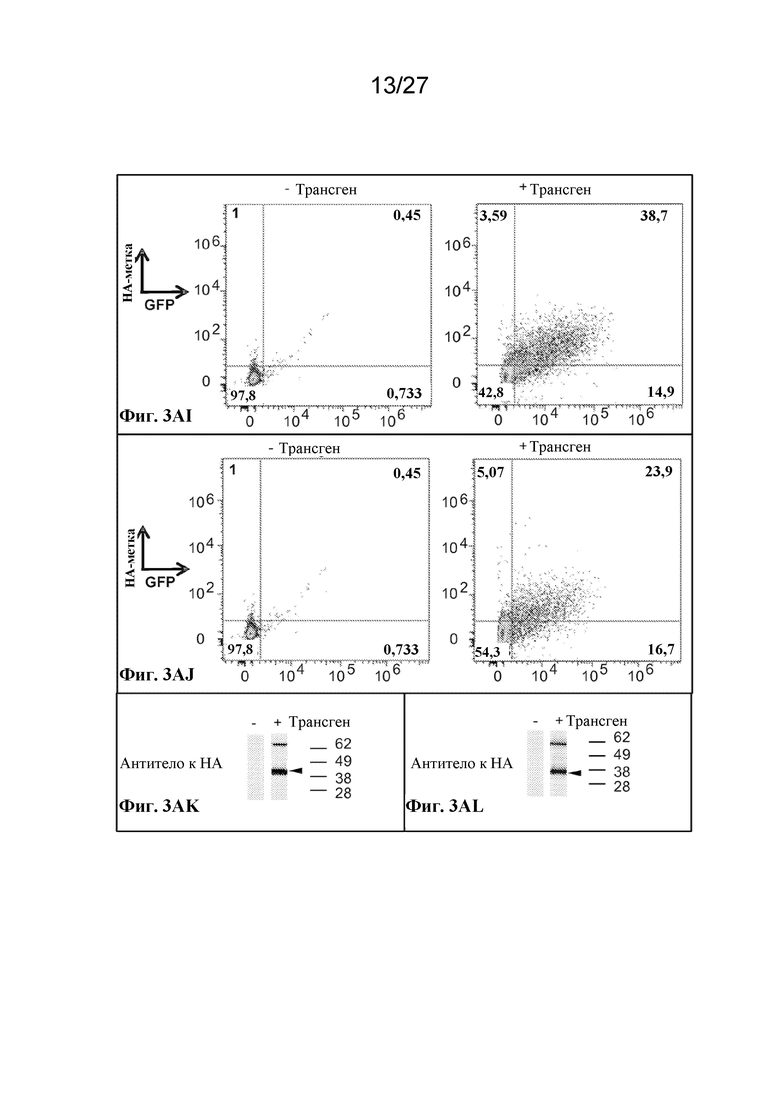

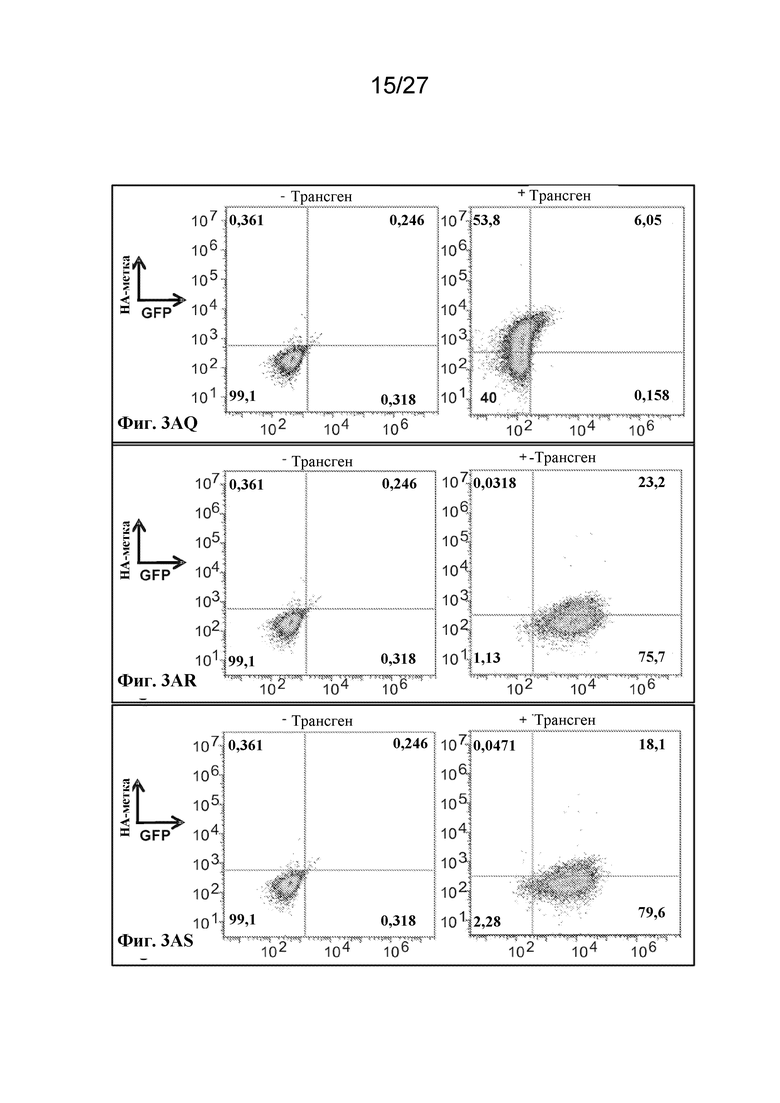


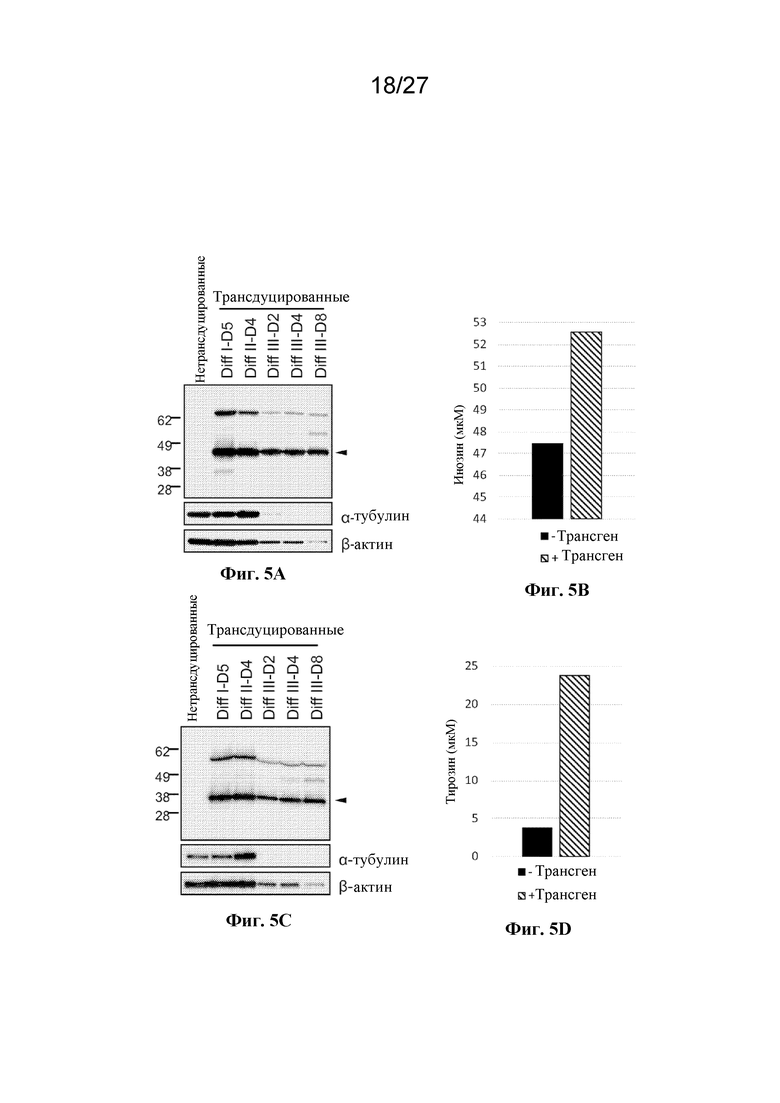

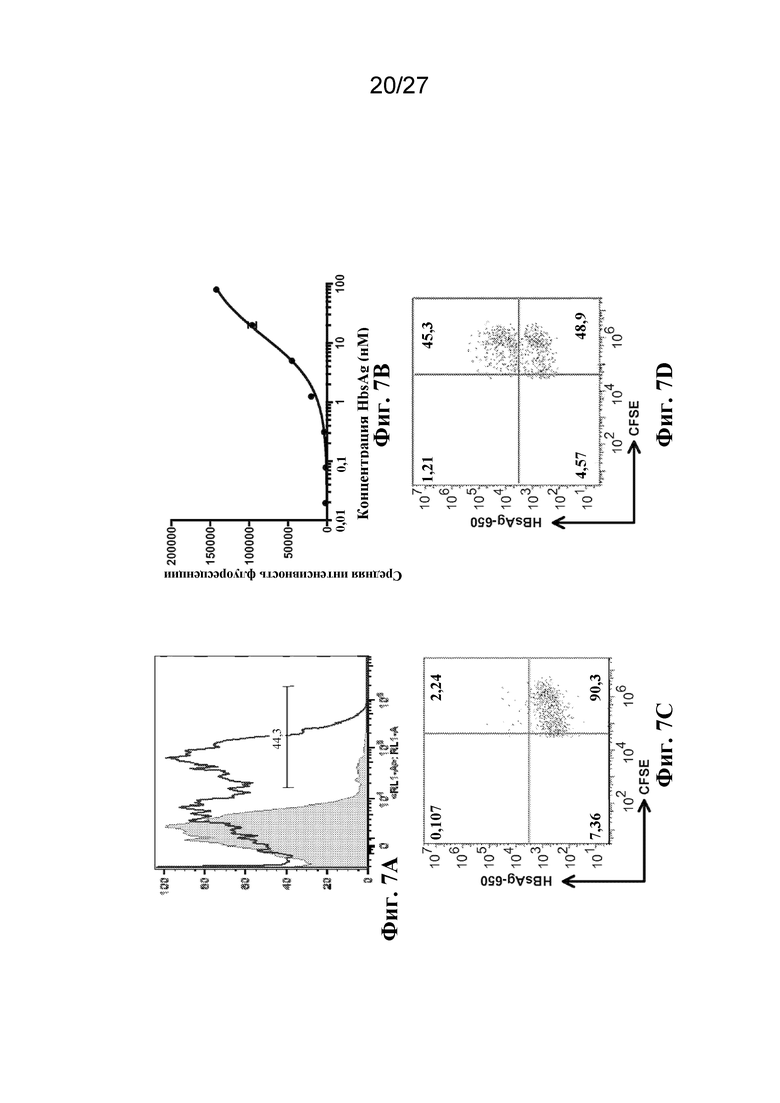

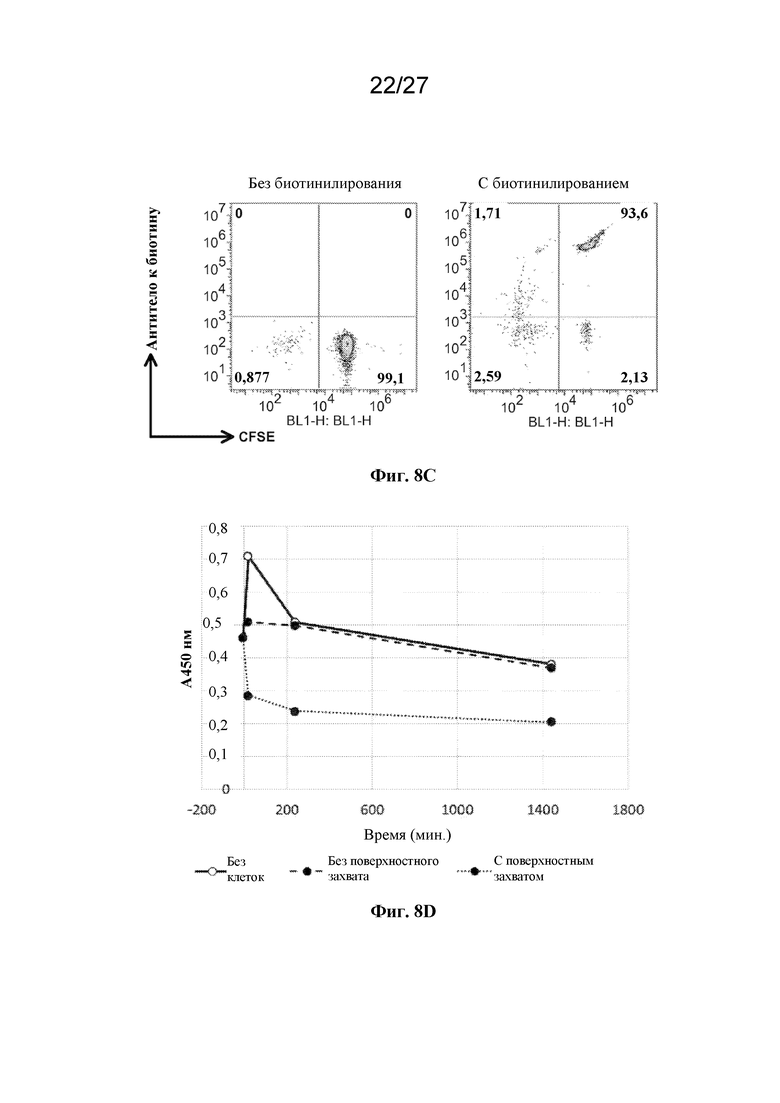
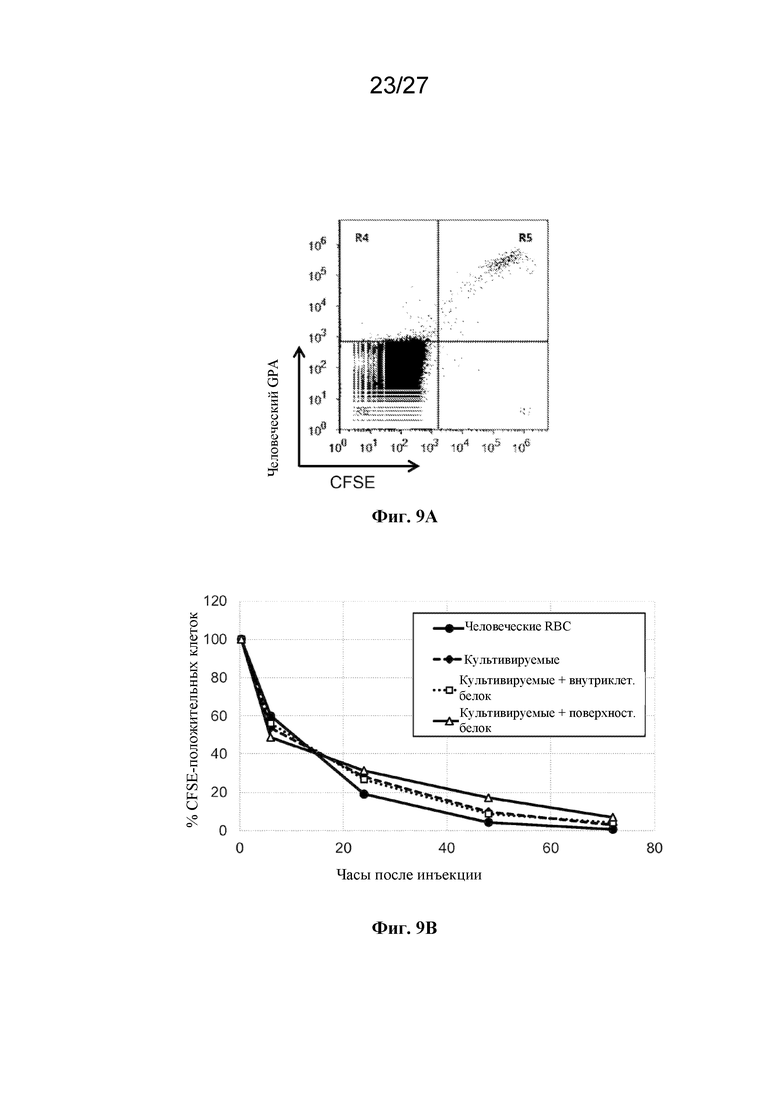

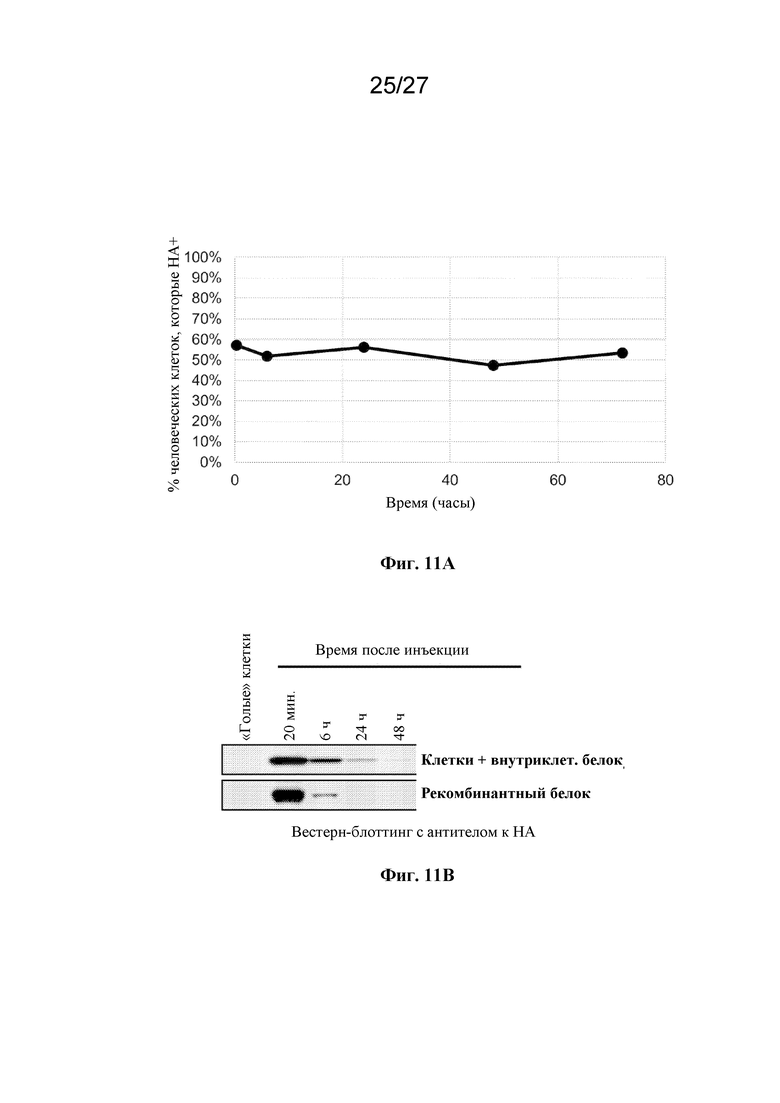
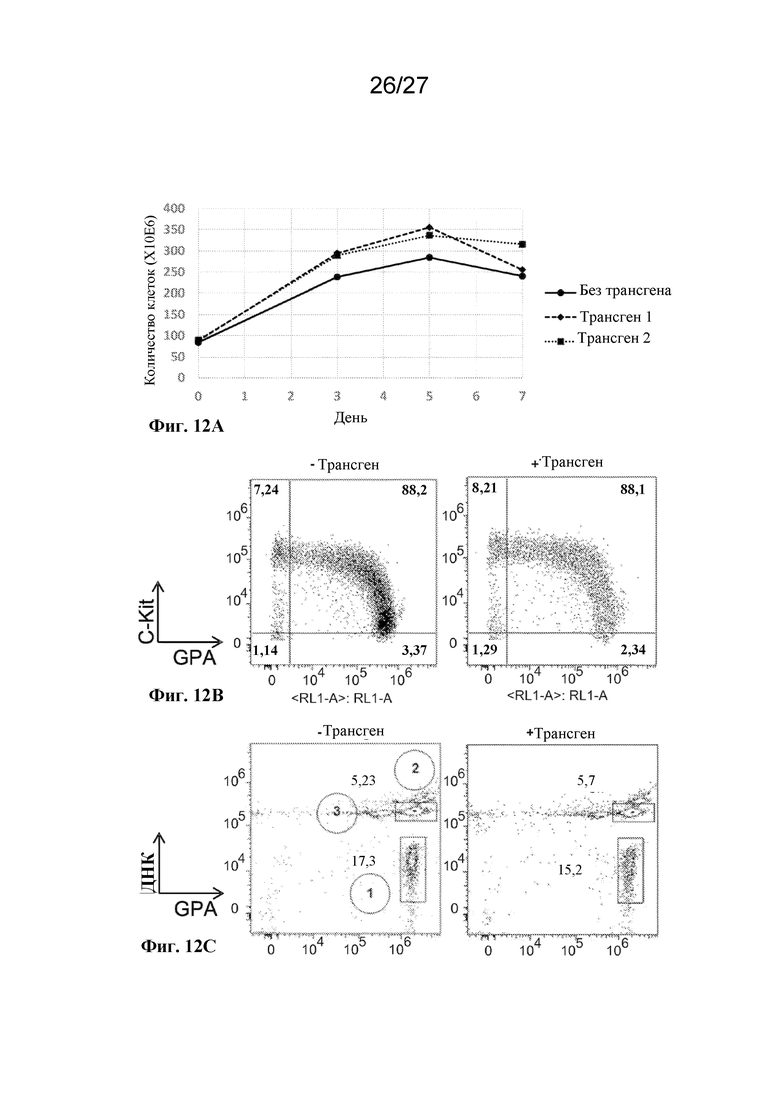
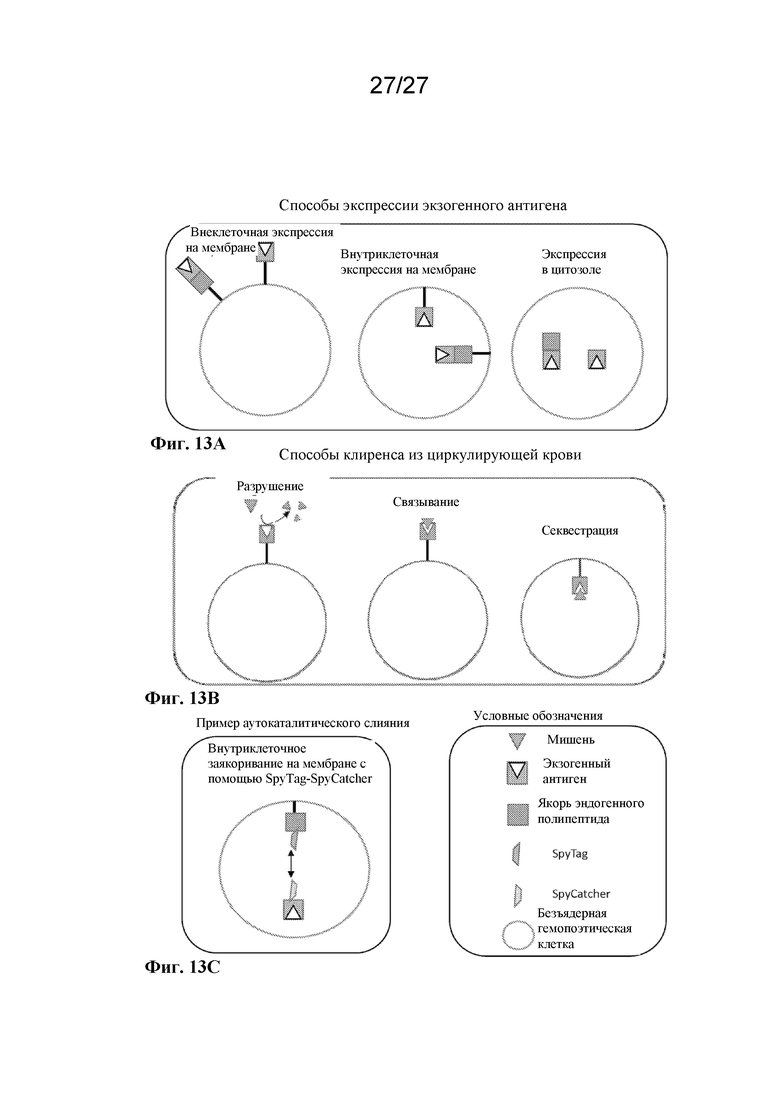
Группа изобретений относится к медицине, а именно к иммунологии, и может быть использована для индукции иммунной толерантности. Для этого фармацевтическая композиция для индукции иммунной толерантности содержит эффективное количество популяции безъядерных эритроидных клеток, где безъядерные эритроидные клетки содержат слитый полипептид, который содержит экзогенный антиген, выбранный из гликопротеина миелина олигодендроцитов и основного белка миелина, или его антигенный фрагмент, и фармацевтически приемлемый носитель. Группа изобретений относится также к способу получения данной композиции и применению ее для лечения рассеянного склероза у пациента. Использование данной группы изобретений позволяет применять заявленную композицию для лечения рассеянного склероза, обеспечивая иммунную толерантность, стимулируя иммуносупрессивный фенотип в антигенпрезентирующих клетках. 5 н. и 28 з.п. ф-лы, 79 пр., 23 табл., 13 ил.
1. Фармацевтическая композиция для индукции иммунной толерантности, содержащая эффективное количество популяции безъядерных эритроидных клеток, где безъядерные эритроидные клетки содержат слитый полипептид, который содержит экзогенный антиген, выбранный из гликопротеина миелина олигодендроцитов и основного белка миелина, или его антигенный фрагмент, и фармацевтически приемлемый носитель.
2. Фармацевтическая композиция для индукции иммунной толерантности, содержащая популяцию безъядерных эритроидных клеток, нагруженных в негипотонических условиях, содержащих по меньшей мере 1000 копий полипептида, который содержит экзогенный антиген, выбранный из гликопротеина миелина олигодендроцитов и основного белка миелина, или его антигенный фрагмент.
3. Фармацевтическая композиция по п.1 или 2, где экзогенный антиген связан с клеточной мембраной безъядерных эритроидных клеток.
4. Фармацевтическая композиция по любому из пп.1-3, где экзогенный антиген локализуется внеклеточно на поверхности безъядерных эритроидных клеток.
5. Фармацевтическая композиция по любому из пп.1-3, где экзогенный антиген является внутриклеточным.
6. Фармацевтическая композиция по п.1, где слитый полипептид содержит якорный домен и поверхностный домен, где поверхностный домен содержит экзогенный антиген.
7. Фармацевтическая композиция по п.6, где якорный домен содержит трансмембранную область.
8. Фармацевтическая композиция по любому из пп.1-7, где безъядерные эритроидные клетки содержит по крайней мере 10 копий, 100 копий, 1000 копий, 10000 копий, 25000 копий, 50000 копий, 100000 копий, 500000 копий, 1000000 копий или 2000000 копий экзогенного антигена.
9. Фармацевтическая композиция по любому из пп.1-8, где безъядерными эритроидными клетками являются эритроциты.
10. Фармацевтическая композиция по любому из пп.1-8, где безъядерными эритроидными клетками являются ретикулоциты.
11. Фармацевтическая композиция по любому из пп.1-10, где безъядерными эритроднными клетками являются клетки, имеющие человеческое происхождение.
12. Фармацевтическая композиция по п.1, полученная способом, включающим:
получение ядерных эритроидных клеток или их предшественников, содержащих экзогенную нуклеиновую кислоту, кодирующую слитый полипептид; и
культивирование ядерных эритроидных клеток или их предшественников, в условиях, походящих для разрушения ядра ядерных эритроидных клеток и получения слитого полипептида.
13. Фармацевтическая композиция по п.12, где способ получения дополнительно включает введение экзогенной нуклеиновой кислоты в ядерные эритроидные клетки или их предшественники.
14. Фармацевтическая композиция по п.1 или 2, где безъядерные клетки получены из ядерных клеток-предшественников способом, основанным на культивировании, которое приводит к разрушению ядра.
15. Фармацевтическая композиция по п.1 или 2, где безъядерные эритроидные клетки загружены в негипотонических условиях.
16. Фармацевтическая композиция по любому из пп.1-15, где безъядерная эритроидная клетка имеет по существу такую же осмотическую хрупкость мембраны, как и соответствующая выделенная, немодифицированная, некультивированная безъядерная клетка эритроидного ряда.
17. Фармацевтическая композиция по любому из пп.1-16, которая составлена в форме жидкой суспензии для внутривенного введения.
18. Фармацевтическая композиция по любому из пп.1-17, содержащая популяцию эритроидных клеток, которая более чем на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 99%, 99,5% или 99,9% является безъядерной.
19. Применение фармацевтической композиции по любому из пп.1-18 для лечения или профилактики рассеянного склероза у пациента.
20. Применение фармацевтической композиции по любому из пп.1-18 в способе индукции иммунной толерантности у человека, страдающего рассеянным склерозом или подверженного риску его развития.
21. Применение по п.19 или 20, где пациент страдает рассеянным склерозом.
22. Применение по любому из пп.19-21, где фармацевтическая композиция составлена для введения по меньшей мере два раза в период лечения так, чтобы происходило лечение рассеянного склероза или уменьшался его симптом.
23. Применение по любому из пп.19-22, где фармацевтическая композиция составлена для введения в достаточном количестве раз в течение периода лечения так, чтобы процент антиген-специфических иммунных клеток в организме человека существенно снижался в период лечения.
24. Способ получения фармацевтической композиции по п.1, включающий: получение ядерных эритроидных клеток или их предшественников, содержащих экзогенную нуклеиновую кислоту, кодирующую слитый полипептид;
культивирование ядерных эритроидных клеток или их предшественников, в условиях, подходящих для разрушения ядра ядерных эритроидных клеток и для получения слитого полипептида, таким образом, получая популяцию безъядерных эритроидных клеток, содержащих слитый полипептид; и
смешивание популяции безъядерных эритроидных клеток с фармацевтически приемлемым носителем, получая таким образом фармацевтическую композицию.
25. Способ по п.24, где экзогенный антиген локализуется внеклеточно на поверхности безъядерных эритроидных клеток.
26. Способ по п.24, где экзогенный антиген является внутриклеточным.
27. Способ по п.24, где слитый полипептид содержит якорный домен и поверхностный домен, где поверхностный домен содержит экзогенный антиген.
28. Способ по п.24, где якорный домен содержит трансмембранную область.
29. Способ по п.24, где безъядерные эритроидные клетки содержат по крайней мере 10 копий, 100 копий, 1000 копий, 10000 копий, 25000 копий, 50000 копий, 100000 копий, 500000 копий, 1000000 копий или 2000000 копий экзогенного антигена.
30. Способ по п.24, где безъядерные эритроидные клетки являются эритроцитами.
31. Способ по п.24, где безъядерные эритроидные клетки являются ретикулоцитами.
32. Способ по п.24, где безъядерные эритроидные клетки являются клетками человеческого происхождения.
33. Способ по п.24, дополнительно включающие введение экзогенной нуклеиновой кислоты в ядерные эритроидные клетки или их предшественники.
| RONCAROLO M.G | |||
| et al | |||
| Regulatory T-cell immunotherapy for tolerance to self antigens and alloantigens in humans // Nat Rev Immunol | |||
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| WO2005118780 A1, 15.12.2005 | |||
| Andreas Lutterotti et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

