Настоящее изобретение относится к выявлению болезни Лайма и других заболеваний, передаваемых клещами. Настоящее изобретение также относится к выявлению антител в биологическом образце. В частности, согласно настоящему изобретению предложена мультиплексная и многофункциональная платформа для выявления микроорганизмов, вызывающих заболевания, передаваемые клещами (tick-borne disease, TBD).
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Микроорганизмов, передаваемых клещами (tick-borne microbes, ТВМ), определяют как макроскопические вирулентные объекты, проникающие в организм хозяина при укусе клеща. Клещи являются исключительными переносчиками для передачи заболеваний и обитают почти на всех континентах, при этом число их видов превышает 850. Наиболее распространенным заболеванием, передаваемым клещами (TBD), как в Европе, так и в Северной Америке, является болезнь Лайма, вызываемая спирохетами Borrelia spp1,2. По всему миру болезнь Лайма эндемична для 80 стран, включая 27 стран ЕС и Центральной Азии3,4. Помимо боррелий, существует множество других бактерий и даже вирусов, которые также могут приводить к инфекциям, такие как Babesia, Rickettsia, Ehrlichia, Bartonella, вирус клещевого энцефалита и так далее5,6. Центры по контролю заболеваний (Center of Disease Control) в США и Европе ежегодно сообщают о 300000 и 85000 случаев TBD, соответственно. Тем не менее, по данным Всемирной организации здравоохранения, ежегодные показатели общего числа случаев TBD сильно занижены7.
Клиническая диагностика у пациента с заболеванием может быть затруднена, поскольку инфекции, вызываемые ТВМ, сначала проявляются как неспецифическое лихорадочное заболевание с поражением каких-либо конкретных систем органов, имитируя гриппоподобные симптомы2,5,8, или без него. Высокая частота вторичных инфекций, вызываемых микоплазмами, хламидиями, вирусом Эпштейна-Барр или другими вирусами, у этих пациентов дополнительно усложняет протоколы лечения6. Из-за недооценки, диагностических ошибок, сочетанных инфекций и вторичных инфекций, неправильное лечение может приводить к развитию тяжелых клинических состояний, таких как усталость, боль в мышцах/суставах, сердечно-сосудистые/когнитивные расстройства и так далее9. Из-за неправильной диагностики у пациентов развиваются тяжелые клинические состояния, а лечение приводит к снижению качества их жизни, увеличивая, как следствие, нагрузку на здравоохранение9,10. Поскольку клинические симптомы разнообразны и неспецифичны, для своевременного и правильного лечения пациентов крайне важны надежные методы диагностики4,6,11,12.
Сложность диагностики инфекций, передаваемых клещами, заключается в том, что применение методов прямого выявления, таких как культивирование и полимеразная цепная реакция (ПЦР), затруднено ввиду малого числа жизнеспособных патогенов, присутствующих в биоптатах пациентов. Это приводит к отрицательным результатам и не позволяет исключить наличие у пациента активных инфекций или различных стадий заболевания2,5,13. Непрямые методы, такие как твердофазный иммуноферментный анализ (ELISA), представляют собой анализы, ограниченные использованием антител, которые на ранних стадиях инфекции или заболевания могут присутствовать в недостаточном количестве или отсутствовать. При применении этих анализов, основанных на антителах, также велико число ложноположительных результатов, обусловленных перекрестной реактивностью в отношении других видов бактерий. При этом положительный специфический гуморальный иммунный ответ может сохраняться на протяжении нескольких месяцев или лет после успешного лечения инфекции. Эти методы, применяемые в настоящее время, не позволяют выявить первую стадию заболеваний, передаваемых клещами, почти у 80% пациентов и не позволяют отличать острые инфекции от хронических4,11. Кроме того, большинство диагностических наборов, основанных на ELISA, предназначены для применения у животных, а не у человека, и обычно позволяют проводить анализ на предмет одного, а не нескольких ТВМ3.
В диагностических инструментах, применяемых на данный момент, не учтены данные современных исследований. В последние годы новые научные данные, относящиеся к круглым тельцам боррелий (Borrelia Round Bodies)14, важности определения видов боррелий15,16, полимикробным инфекциям12 и IgM-иммунной дисфункции17 у пациентов с TBD, привели к изменению клинических представлений о TBD. Круглые тельца боррелий являются одной из плеоморфных структур спирохет боррелий14. В разные годы плеоморфные формы боррелий называли формами, лишенными клеточной стенки (cell-wall deficient, CWD), L-формами, сферопластами, протопластами, пропагулами или цистами5,8,18-20. Лишь недавно электронные микрофотографии, полученные  et al. (2015), позволили ликвидировать разногласия относительно плеоморфной морфологии боррелий и сделать вывод о том, что плеоморфные формы боррелий представляют собой круглые тельца (round body, RB). et al. (2015) обнаружили RB боррелий в человеческой сыворотке и продемонстрировали сферическое RB с интактной, но гибкой клеточной стенкой, которое было метаболически неактивно и обладало уникальными биохимическими характеристиками. Несмотря на неоднократные сообщения о клинических проявлениях, связанных с плеоморфными формами боррелий, их патогенетическая роль при TBD является объектом дискуссий и критики. Диагностические инструменты, применяемые на данный момент, не позволяют обследовать пациентов с TBD на предмет круглых телец боррелий8,21-25.
et al. (2015), позволили ликвидировать разногласия относительно плеоморфной морфологии боррелий и сделать вывод о том, что плеоморфные формы боррелий представляют собой круглые тельца (round body, RB). et al. (2015) обнаружили RB боррелий в человеческой сыворотке и продемонстрировали сферическое RB с интактной, но гибкой клеточной стенкой, которое было метаболически неактивно и обладало уникальными биохимическими характеристиками. Несмотря на неоднократные сообщения о клинических проявлениях, связанных с плеоморфными формами боррелий, их патогенетическая роль при TBD является объектом дискуссий и критики. Диагностические инструменты, применяемые на данный момент, не позволяют обследовать пациентов с TBD на предмет круглых телец боррелий8,21-25.
Диагностические инструменты, применяемые в настоящее время, позволяют проводить анализ на разные спирохеты боррелий, по отдельности или совместно, поскольку они приводят к разным клиническим проявлениям у индивидов16. В последнее время мультиплексные инструменты для диагностики TBD позволяют проводить анализ на различные рекомбинантные белки боррелий, однако уже известно, что TBD являются полимикробными инфекционными заболеваниями, а диагностические инструменты, применяемые на данный момент, не предназначены для диагностики у индивидов вторичных оппортунистических инфекций, сочетанных инфекций, а также аутоиммунных состояний, связанных с этими инфекциями5,13,22-25
Для устранения недостатков инструментов выявления TBD, применяемых на данный момент, согласно настоящему изобретению предложена новая твердая подложка, содержащая по меньшей мере один иммобилизованный антиген, полученный из группы, состоящей из плеоморфных круглых телец рода Borrelia; например, Borrelia burgdorferi, Borrelia afzelii и Borrelia garinii. Представленные результаты впервые демонстрируют, что иммунная система индивида может специфично отвечать только на круглые тельца боррелий и что этот иммунный ответ может быть связан с персистирующей стадией болезни Лайма.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Задача настоящего изобретения состоит в обеспечении новой платформы для выявления, позволяющей различать острые, прошедшие и, особенно, хронические или персистирующие стадии TBD у пациента. Кроме того, настоящее изобретение может также быть направлено на полимикробные аспекты и аспекты иммунной дисфункции, связанные с TBD.
Таким образом, в одном аспекте согласно настоящему изобретению предложена твердая подложка для выявления присутствия антител в биологическом образце, содержащая микробные антигены, иммобилизованные на указанной твердой подложке, включающие по меньшей мере один антиген, полученный из группы, состоящей из плеоморфных круглых телец вида рода Borrelia.
В другом аспекте согласно настоящему изобретению предложен способ выявления микроорганизма, передаваемого клещами, в биологическом образце, включающий:
(а) приведение биологического образца в контакт с твердой подложкой, содержащей микробные антигены, иммобилизованные на указанной твердой подложке, для образования комплекса, содержащего микробный антиген, иммобилизованный на указанной твердой подложке, и антитело из указанного биологического образца, связанное с указанным микробным антигеном, где указанные микробные антигены включают по меньшей мере один антиген, полученный из группы, состоящей из плеоморфных круглых телец вида рода Borrelia;
(с) выявление присутствия комплекса, полученного на стадии (а), где присутствие комплекса, содержащего антиген, полученный из плеоморфных круглых телец по меньшей мере одного вида рода Borrelia, указывает на присутствие микроорганизма, передаваемого клещами, в указанном биологическом образце.
В другом аспекте согласно изобретению предложена твердая подложка, как определено выше, для применения в диагностике болезни Лайма.
В другом аспекте согласно изобретению предложено применение твердой подложки, как определено выше, для изготовления диагностического теста для выявления микроорганизма, передаваемого клещами, в биологическом образце.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1. (А) Сводные данные по IgM-иммунным ответам на все антигены боррелий, (В) только на спирохеты боррелий и (С) только на круглые тельца боррелий. На 1А и 1В сокращения Вb, Ва и Bg означают Borrelia burgdorferi sensu stricto B31, Borrelia afzelii P12 и Borrelia garinii Fuji P1, соответственно.
Фиг. 2. (А) Сводные данные по IgG-иммунным ответам на все антигены боррелий, (В) только на спирохеты боррелий и (С) только на круглые тельца боррелий. На 2А и 2В сокращения Вb, Ва и Bg означают Borrelia burgdorferi sensu stricto B31, Borrelia afzelii P12 и Borrelia garinii Fuji P1, соответственно.
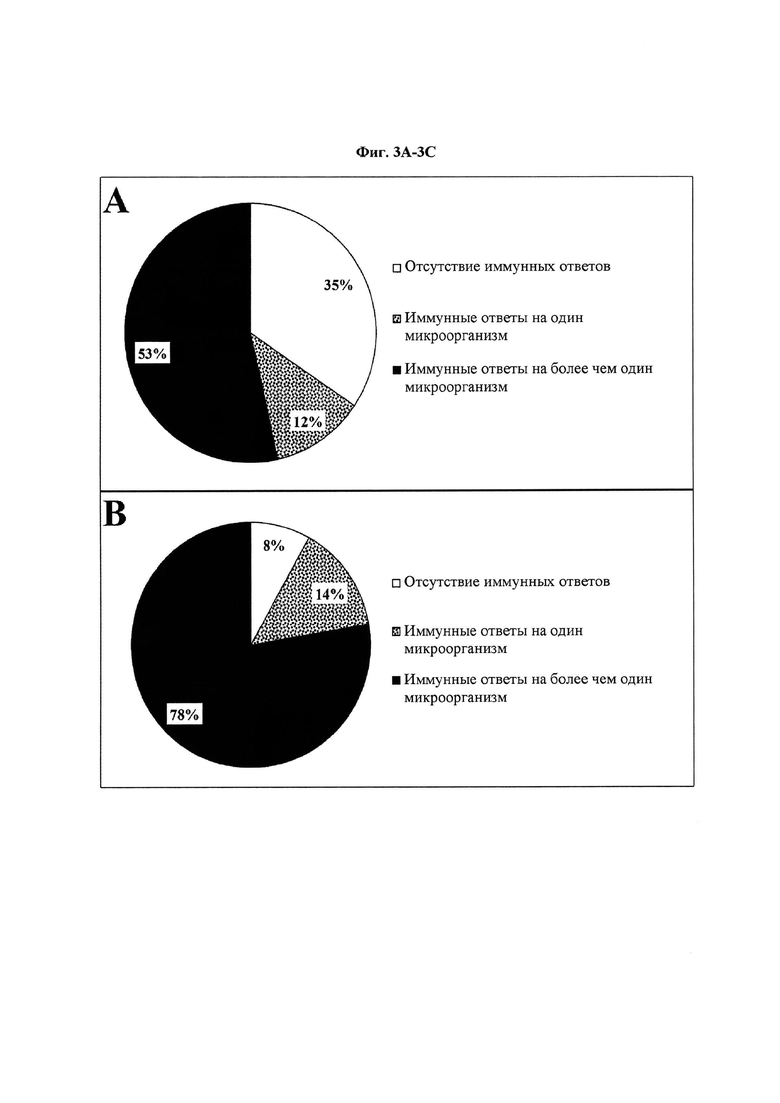
Фиг. 3. Оценка (A) IgM- и (В) IgG-иммунных ответов против одного или нескольких микробных антигенов. Для оценки того, отвечают ли индивиды только на один микробный антиген или на несколько микробных антигенов, была использована сыворотка 443 человек. В анализ также были включены индивиды без иммунного ответа на 20 антигенов.
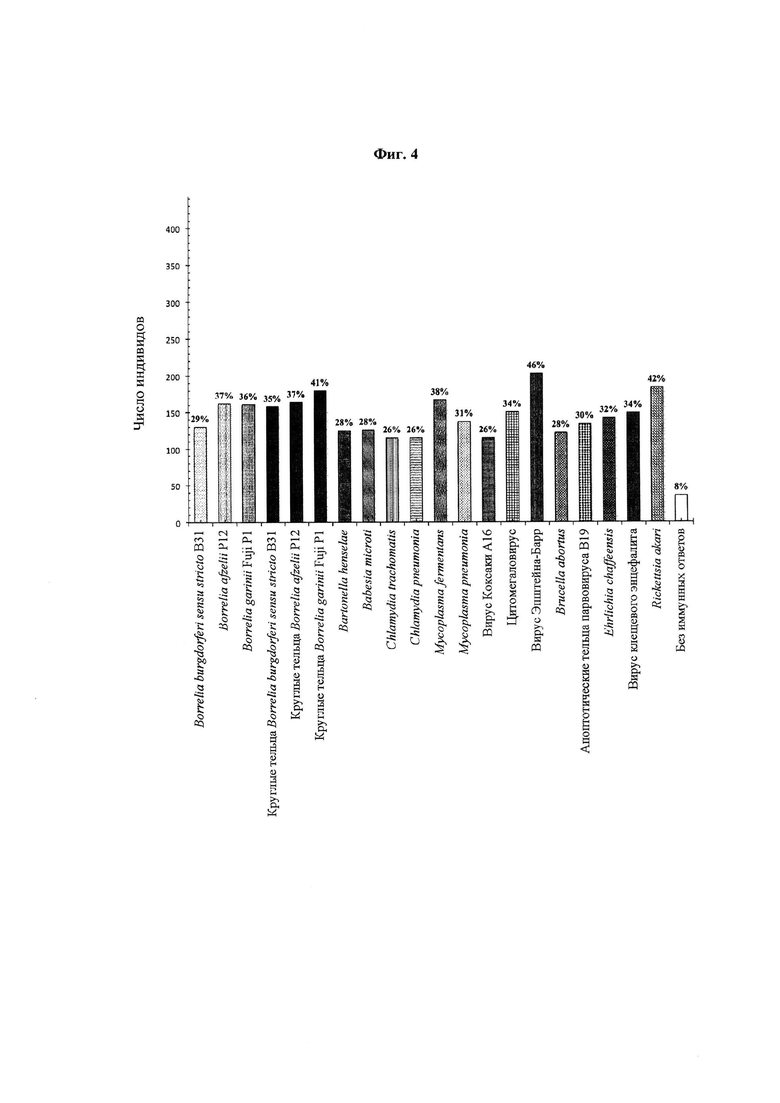
Фиг. 4. IgG-иммунные ответы на отдельные микробные антигены. Для оценки общего числа иммунных ответов на каждый микробный антиген, использованный в данном исследовании, была использована сыворотка 443 человек. В анализ также были включены индивиды без иммунного ответа на 20 антигенов.
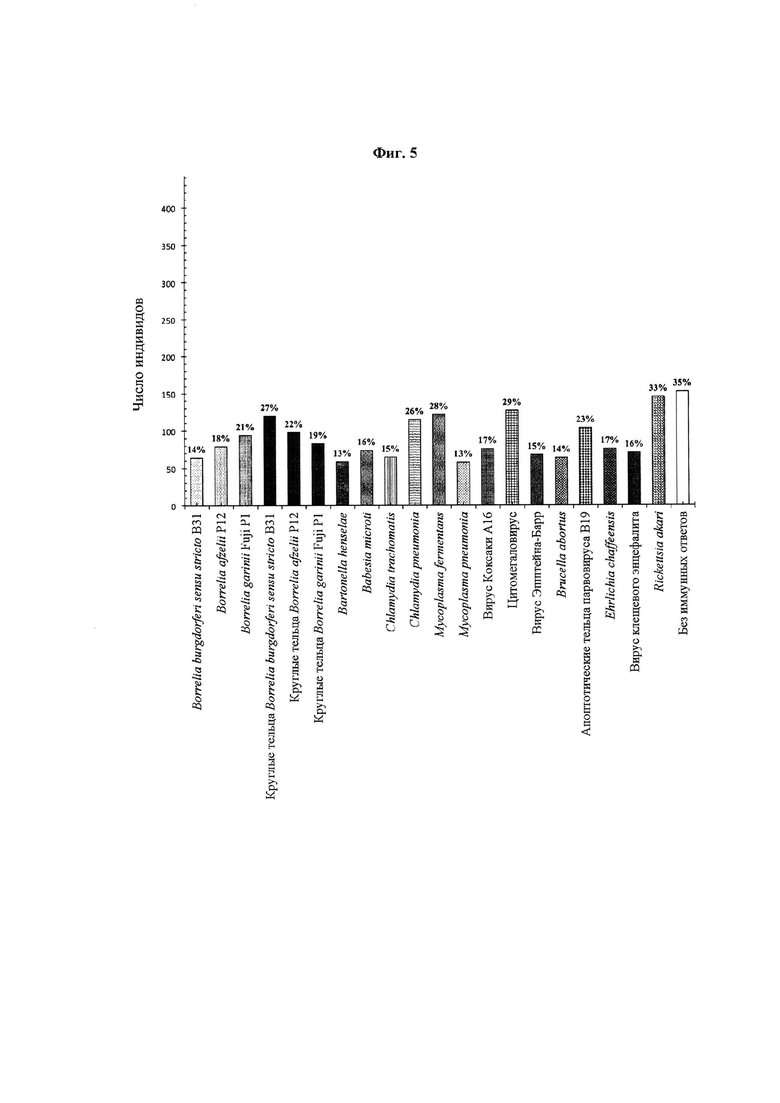
Фиг. 5. IgM-иммунные ответы на отдельные микробные антигены. Для оценки общего числа иммунных ответов на каждый микробный антиген, использованный в данном исследовании, была использована сыворотка 443 человек. В анализ также были включены индивиды без иммунного ответа на 20 антигенов.
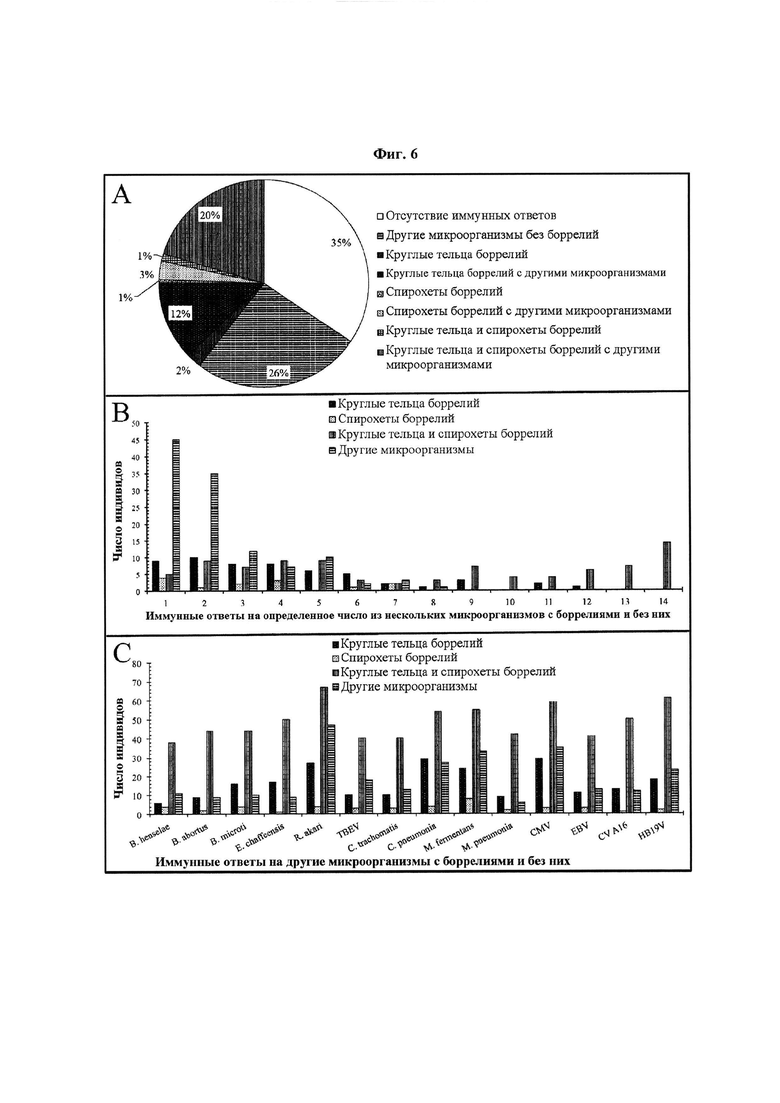
Фиг. 6. (А) Сводные данные по долям IgM-иммунных ответов индивидов на другие микроорганизмы с боррелиями и без них, (В) IgM-иммунных ответов индивидов на определенное число из нескольких других микроорганизмов с боррелиями и без них, и (С) IgM-иммунных ответов индивидов на конкретные другие микроорганизмы с боррелиями и без них. Для сравнения частоты IgM-иммунных ответов на несколько других микроорганизмов и их конкретные типы у индивидов, отвечавших только на спирохеты боррелий, только на круглые тельца боррелий или на комбинацию спирохет и круглых телец боррелий, была использована сыворотка 443 человек. Термин «другие микроорганизмы» включает сочетанные инфекции, вторичные и аутоиммунные антигены, такие как Bartonella henselae (В. henselae), Brucella abortus (В. abortus), Babesia microti (B. microti), Ehrlichia chaffeensis (E. chaffeensis), Rickettsia akari (R. akari), вирус клещевого энцефалита (tick borne encephaltis virus, TBEV), Chlamydia trachomatis (C. trachomatis), Chlamydia pneumonia (C. pneumonia), Mycoplasma fermentans (M fermentans), Mycoplasma pneumonia (M. pneumonia), цитомегаловирус (CMV), вирус Эпштейна-Барр (Epstein-barr virus, EBV), вирус Коксаки A16 (Coxsackie virus A16, CV A16) и парвовирус человека В19 (Human Parvovirus В19, HB19V).
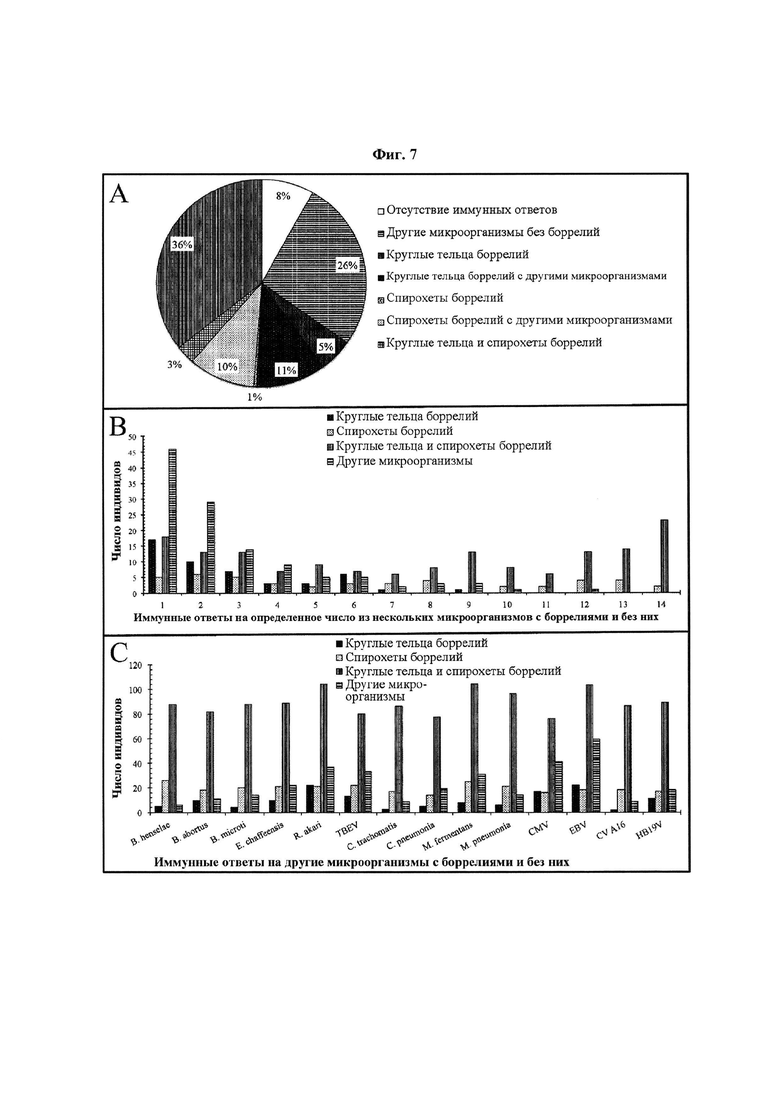
Фиг. 7. (А) Сводные данные по долям IgG-иммунных ответов индивидов на другие микроорганизмы с боррелиями и без них, (В) IgG-иммунных ответов индивидов на определенное число из нескольких других микроорганизмов с боррелиями и без них, и (С) IgG-иммунных ответов индивидов на конкретные другие микроорганизмы с боррелиями и без них. Для сравнения частоты IgG-иммунных ответов на несколько других микроорганизмов и их конкретные типы у индивидов, отвечавших только на спирохеты боррелий, только на круглые тельца боррелий или на комбинацию спирохет и круглых телец боррелий, была использована сыворотка 443 человек. Термин «другие микроорганизмы» включает сочетанные инфекции, вторичные и аутоиммунные антигены, такие как Bartonella henselae (В. henselae), Brucella abortus (В. abortus), Babesia microti (B. microti), Ehrlichia chaffeensis (E. chaffeensis), Rickettsia akari (R. akari), вирус клещевого энцефалита (TBEV), Chlamydia trachomatis (C. trachomatis), Chlamydia pneumonia (C. pneumonia), Mycoplasma fermentans (M. fermentans), Mycoplasma pneumonia (M pneumonia), цитомегаловирус (CMV), вирус Эпштейна-Барр (EBV), вирус Коксаки A16 (CV A16) и парвовирус человека В19 (HB19V).
ОПИСАНИЕ ВОПЛОЩЕНИЙ
На сегодняшний день существующие инструменты диагностики TBD основаны на скрининге одного иммунного ответа (IgG или IgM) для одного заболевания и, в случае положительного результата, часто требуют проведения повторного подтверждающего анализа. Согласно настоящему изобретению предложены средства и способы выявления хронических, латентных или персистирующих стадий болезни Лайма посредством выявления иммунного ответа против плеоморфных круглых телец вида рода Borrelia.
Известно, что возбудителями болезни Лайма или боррелиоза являются по меньшей мере 18 видов рода Borrelia, передаваемые клещами48. Основными патогенами, вызывающими болезнь Лайма, являются Borrelia burgdorferi, Borrelia afzelii и Borrelia garinii. Другими являются, например, Borrelia miyamotoi, Borrelia tanukii, Borrelia turdi, Borrelia valaisiana, Borrelia carolinensis, Borrelia americana, Borrelia lusitaniae, Borrelia japonica и Borrelia sinica.
Как мультиплексная и многофункциональная платформа настоящие аспекты могут быть использованы для диагностики у индивидов, направленной на выявление нескольких микроорганизмов и классов антител одновременно. Микробные антигены, используемые при диагностике первичной, персистирующей, вторичной, сочетанной инфекции и аутоиммунных состояний у индивидов с TBD, перечислены ниже в Таблице 1.
Настоящее изобретение направлено на твердую подложку для выявления присутствия антител в биологическом образце, содержащую микробные антигены, иммобилизованные на указанной твердой подложке, включающие по меньшей мере один антиген, полученный из группы, состоящей из плеоморфных круглых телец вида рода Borrelia, такого как Borrelia burgdorferi, Borrelia afzelii и Borrelia garinii.
Термин «плеоморфный» относится здесь к плеоморфизму, который в микробиологии определяют как способность некоторых бактерий менять свою форму или размер в ответ на условия внешней среды. Плеоморфные круглые тельца, как определено в настоящем изобретении, могут быть индуцированы, как раскрыто в  et al. (2015) или как раскрыто в «Экспериментальном разделе» ниже. Без ограничения теорией, причиной того, что цилиндрические спирохеты (то есть длинные штопорообразные клетки, средняя длина которых составляет 20 мкм) меняют свою форму, образуя плеоморфные круглые тельца (то есть сферические клетки, средний диаметр которых составляет 2,8 плюс/минус 0,46 мкм), состоит в физиологическом давлении на бактерии со стороны окружающей среды. Поэтому помимо изменений условий среды, в которой находятся бактерии, стрессовые условия, такие как осмотическое давление, также способствуют индукции круглых телец47.
et al. (2015) или как раскрыто в «Экспериментальном разделе» ниже. Без ограничения теорией, причиной того, что цилиндрические спирохеты (то есть длинные штопорообразные клетки, средняя длина которых составляет 20 мкм) меняют свою форму, образуя плеоморфные круглые тельца (то есть сферические клетки, средний диаметр которых составляет 2,8 плюс/минус 0,46 мкм), состоит в физиологическом давлении на бактерии со стороны окружающей среды. Поэтому помимо изменений условий среды, в которой находятся бактерии, стрессовые условия, такие как осмотическое давление, также способствуют индукции круглых телец47.
Раньше круглые тельца (RB) В. burgdorferi называли по-разному. Эти названия включают CWD- и L-формы, сферопласты, протопласты, пропагулы и даже цисты. Тем не менее, все эти обозначения описывают одни и те же сферические структуры14.
В одном воплощении по меньшей мере один антиген, полученный из группы, состоящей из плеоморфных круглых телец вида рода Borrelia, специфичен для плеоморфных круглых телец вида рода Borrelia.
В одном воплощении иммобилизованный антиген на твердой подложке представляет собой лизат или часть лизата культивированных плеоморфных круглых телец рода Borrelia; например, Borrelia burgdorferi, Borrelia afzelii или Borrelia garinii. Указанный иммобилизованный антиген может также представлять собой белковый или пептидный препарат указанных плеоморфных круглых телец. В данном изобретении могут также быть использованы другие известные препараты, содержащие антигены микробных клеток, полученные, например, с применением сдвига рН, человеческих сывороток, изменений концентрации соли.
Для одновременного выявления острых и хронических или персистирующих стадий болезни Лайма указанная твердая подложка может дополнительно содержать по меньшей мере один иммобилизованный антиген, полученный из группы, состоящей из рода Borrelia, например, Borrelia burgdorferi, Borrelia afzelii и Borrelia garinii, в форме нативных спирохет или их лизатов.
В одном воплощении по меньшей мере один иммобилизованный антиген, полученный из группы, состоящей из вида рода Borrelia в форме нативных спирохет, специфичен для этого вида рода Borrelia в форме нативных спирохет.
В одном воплощении анализ направлен на выявление одного определенного вида боррелий, например, где: (1) указанная твердая подложка содержит иммобилизованный антиген, полученный из плеоморфных круглых телец Borrelia burgdorferi, и иммобилизованный антиген, полученный из Borrelia burgdorferi в форме нативных спирохет; (2) указанная твердая подложка содержит иммобилизованный антиген, полученный из плеоморфных круглых телец Borrelia afzelii, и иммобилизованный антиген, полученный из Borrelia afzelii в форме нативных спирохет; или (3) указанная твердая подложка содержит иммобилизованный антиген, полученный из плеоморфных круглых телец Borrelia garinii, и иммобилизованный антиген, полученный из Borrelia garinii в форме нативных спирохет.
В одном воплощении иммобилизованный антиген, полученный из плеоморфных круглых телец Borrelia burgdorferi, специфичен для плеоморфных круглых телец Borrelia burgdorferi, и иммобилизованный антиген, полученный из Borrelia burgdorferi в форме нативных спирохет, специфичен для Borrelia burgdorferi в форме нативных спирохет.
В одном воплощении иммобилизованный антиген, полученный из плеоморфных круглых телец Borrelia afzelii, специфичен для плеоморфных круглых телец Borrelia afzelii, и иммобилизованный антиген, полученный из Borrelia afzelii в форме нативных спирохет, специфичен для Borrelia afzelii в форме нативных спирохет.
В одном воплощении иммобилизованный антиген, полученный из плеоморфных круглых телец Borrelia garinii, специфичен для плеоморфных круглых телец Borrelia garinii, и иммобилизованный антиген, полученный из Borrelia garinii в форме нативных спирохет, специфичен для Borrelia garinii в форме нативных спирохет.
В одном воплощении твердая подложка изготовлена для мультиплексного анализа и включает иммобилизованные антигены, полученные из плеоморфных круглых телец вида рода Borrelia, предпочтительно Borrelia burgdorferi, Borrelia afzelii и/или Borrelia garinii. В другом воплощении мультиплексный анализ включает также иммобилизованные антигены, полученные из вида рода Borrelia, такого как Borrelia burgdorferi, Borrelia afzelii и/или Borrelia garinii, в форме нативных спирохет.
В одном воплощении иммобилизованные антигены, полученные из плеоморфных круглых телец Borrelia burgdorferi, Borrelia afzelii и Borrelia garinii, специфичны для плеоморфных круглых телец Borrelia burgdorferi, Borrelia afzelii и Borrelia garinii, соответственно.
Мультиплексный анализ может также включать по меньшей мере один иммобилизованный антиген, полученный из группы, состоящей из Mycoplasma fermentans, Mycoplasma pneumonia, Bartonella henselae, Brucella abortus, Babesia microti, Chlamydia trachomatis, Chlamydia pneumonia, Ehrlichia chaffeensis, вируса Коксаки A16, вируса Эпштейна-Барр (EBV), цитомегаловируса (CMV), апоптотических телец парвовируса человека В19, вируса клещевого энцефалита (TBEV) и Rickettsia akari.
В одном воплощении по меньшей мере один иммобилизованный антиген, полученный из группы, состоящей из Mycoplasma fermentans, Mycoplasma pneumonia, Bartonella henselae, Brucella abortus, Babesia microti, Chlamydia trachomatis, Chlamydia pneumonia, Ehrlichia chaffeensis, вируса Коксаки A16, вируса Эпштейна-Барр, цитомегаловируса, апоптотических телец парвовируса человека В19, вируса клещевого энцефалита и Rickettsia akari, специфичен для Mycoplasma fermentans, Mycoplasma pneumonia, Bartonella henselae, Brucella abortus, Babesia microti, Chlamydia trachomatis, Chlamydia pneumonia, Ehrlichia chaffeensis, вируса Коксаки A16, вируса Эпштейна-Барр, цитомегаловируса, апоптотических телец парвовируса человека В19, вируса клещевого энцефалита и Rickettsia akari, соответственно.
Указанная твердая подложка может быть изготовлена из стекла или пластика, такого как полистирол или полипропилен. Примерами твердой подложки по настоящему изобретению являются биочип с антигенами или микролуночный планшет.Биочин с антигенами является формой белкового биочипа, также известного как белковый чип.Биочип представляет собой твердую подложку (обычно стекло), на которой тысячи разных белков (в данном случае антигенов) иммобилизованы в разных пространственных расположениях с получением точечной белковой матрицы высокой плотности. Микролуночный планшет представляет собой плоский планшет с множеством «лунок», каждую из которых используют для одного конкретного образца. Микролуночный планшет является стандартным инструментом, используемым в клинико-диагностических лабораториях. Его очень часто используют в твердофазном иммуноферментном анализе (ELISA).
В одном воплощении настоящее изобретение направлено на твердую подложку, как определено здесь, для применения в диагностике болезни Лайма, такой как хроническая/персистирующая болезнь Лайма.
В другом воплощении настоящее изобретение направлено на применение твердой подложки, как определено здесь, для изготовления диагностического теста для выявления микроорганизма, передаваемого клещами, в биологическом образце. В одном воплощении указанный диагностический тест предназначен для выявления болезни Лайма, такой как хроническая/персистирующая болезнь Лайма, у пациента.
«Пациент», «индивид» или «донор» может представлять собой субъекта-млекопитающего, такого как субъект-человек.
Настоящее изобретение также направлено на способ выявления микроорганизма, передаваемого клещами, в биологическом образце, включающий:
(a) приведение биологического образца в контакт с твердой подложкой, содержащей микробные антигены, иммобилизованные на указанной твердой подложке, для образования комплекса, содержащего микробный антиген, иммобилизованный на указанной твердой подложке, и антитело из указанного биологического образца, связанное с указанным микробным антигеном, где указанные микробные антигены включают по меньшей мере один антиген, полученный из группы, состоящей из плеоморфных круглых телец вида рода Borrelia; и
(b) выявление присутствия комплекса, полученного на стадии (а), где присутствие комплекса, содержащего антиген, полученный из плеоморфных круглых телец рода Borrelia, указывает на присутствие микроорганизма, передаваемого клещами, в указанном биологическом образце.
В одном воплощении присутствие комплекса, полученного на стадии (а), выявляют путем приведения указанной твердой подложки в контакт с реагентом против антитела для образования комплекса указанного микробного антигена, указанного антитела, связанного с указанным микробным антигеном, и указанного реагента против антитела.
Настоящее изобретение также обеспечивает возможность специфичного и чувствительного скрининга на предмет IgG- и IgM- или IgA-ответа у индивида против нескольких микроорганизмов с использованием одного набора. Соответственно, указанный реагент против антитела может представлять собой антитело против IgG, антитело против IgM или антитело против IgA. Например, указанный реагент против антитела может представлять собой антитело против человеческого IgG, антитело против человеческого IgM или антитело против человеческого IgA.
В одном воплощении указанный биологический образец представляет собой образец крови, сыворотки, мочи, слюны или слезы, образец спинномозговой жидкости или образец синовиальной жидкости, такой как образец сыворотки.
В одном воплощении настоящий способ включает предшествующую стадию культивирования вида рода Borrelia, такого как Borrelia burgdorferi, Borrelia afzelii или Borrelia garinii, в условиях, приводящих к образованию плеоморфных круглых телец, проведения лизиса культивированных клеток и покрытия или «распечатывания» твердой подложки полученным лизатом или его частью. Указанные условия получения плеоморфных круглых телец представляют собой условия, раскрытые в et al. (2015) или раскрытые в «Экспериментальном разделе» ниже, такие как инкубация клеток боррелий в форме спирохет в дистиллированной воде, или при изменяющихся концентрациях соли, или в присутствии человеческой сыворотки, или со сдвигом рН культуры в кислую сторону. После стадии культивирования в данном аспекте могут также быть применены другие известные методики получения антигенов из микробных клеток, отличные от лизиса клеток. Например, антигенные пептиды и белки для стадии покрытия или «распечатывания» могут быть получены из указанных плеоморфных круглых телец.
Теперь, после того, как изобретение описано в общих чертах, оно будет более понятным со ссылкой на последующий «Экспериментальный раздел», приведенный для наглядности и не являющийся ограничивающим. Несмотря на то, что в практическом применении или испытании настоящего изобретения могут быть применены методы и материалы, сходные с описанными здесь или эквивалентные им, подходящие методы и материалы описаны ниже.
ЭКСПЕРИМЕНТАЛЬНЫЙ РАЗДЕЛ
Материалы и методы
Разрешение этического комитета на сбор образцов сыворотки
Был осуществлен сбор в общей сложности 532 образцов человеческой сыворотки, проведенный в Аугсбургском центре боррелиозов (Borreliose Centrum Augsburg, ВСА), Германия; 10-й клинике ревматических болезней имени короля Кристиана (King Christian 10th Hospital for Rheumatic Diseases), Дания; и в нескольких европейских клиниках / специализированных лабораториях, разрешенный Федеральным институтом лекарственных средств и изделий медицинского назначения (Federal Institute for Drugs and Medical Devices), Германия (номер разрешения: 95.10-5661-7066); датским агентством по защите данных и региональным этическим комитетом Южной Дании (номер разрешения: S-20110029); и Западным экспертным советом (Western Institutional Review Board) (номер разрешения: USMA201441), соответственно. Из 532 образцов человеческой сыворотки 51 образец отрицательного контроля был использован для анализа IgG и еще 51 образец отрицательного контроля был использован для анализа IgM. Образцы отрицательного контроля были использованы для установления пороговых значений для качественного определения антител обоих классов.
Получение антигенов для ELISA
Все 532 образца человеческой сыворотки были проанализированы на предмет гуморальных иммунных ответов с IgM и IgG против 20 микробных антигенов. Все 20 антигенов приведены в Таблице 1. Спирохеты боррелий, круглые тельца боррелий и апоптотические тельца парвовируса человека В19 были культивированы и выделены без привлечения внешних специалистов. Апоптотические тельца парвовируса человека В19 были культивированы и выделены в соответствии с методикой, описанной в других публикациях26,27. Очищенные и дезактивированные лизаты Rickettsia akari были предоставлены Dr. Marco Quevendo Diaz (Акадения наук Словакии). Остальные 18 микроорганизмов были заказаны у GeneCust в форме лиофилизированных микробных пептидов. В случае Rickettsia akari и всех микробных пептидов для непосредственного использования в ELISA, были приготовлены исходные растворы в концентрации 1 мг/мл.
Культивирование и выделение видов боррелий в форме спирохет или в плеоморфной форме
Культуры боррелий были получены из Американской коллекции типовых культур (American Type Culture Collection, АТСС). Для выращивания культур всех трех боррелий была использована среда Барбора-Штенера-Келли (Barbour-Stoenner-Kelly, BSK). Среда BSK была приготовлена с соблюдением опубликованных ранее инструкций39. Для культивирования и выделения видов боррелий в форме нативных спирохет каждый штамм боррелий независимо выращивали в среде BSK при 37°С на протяжении 5-7 сут. После инкубации клетки боррелий выделяли центрифугированием культуральных пробирок при 5000 g на протяжении 10 мин. Супернатант удаляли и клеточный осадок хранили при -80°С до последующего использования14.
Для культивирования различных штаммов боррелий в форме круглых телец клеточные осадки соответствующих боррелий в форме спирохет ресуспендировали в 2 мл дистиллированной воды (ddH2O). Клетки боррелий в форме спирохет инкубировали в воде, или при изменяющихся концентрациях соли, или со сдвигом рН в кислую сторону, или в присутствии человеческой сыворотки при 37°С на протяжении 2 ч. После инкубации клетки боррелий центрифугировали при 5000 g на протяжении 10 мин. Супернатант удаляли и осадок круглых телец боррелий хранили при -80°С до последующего использования14.
Культивирование и выделение апоптотических телец парворируса человека В19
Kivovich et al. (2010) и Thammasri et al. (2013) сообщили об образовании апоптотических телец, индуцированных парвовирусом человека В19 (B19V), и выделении этих апоптотических телец, называемых здесь апоптотическими тельцами B19V (B19V Apobods). Кратко, неструктурный белок B19V (NS1) клонировали вместе с усиленным зеленым флуоресцентным белком (EGFP) в модифицированный вектор pFastBac1. Модифицированный вектор pFastBac1 использовали для получения рекомбинантного бакуловируса в вирусном векторе Autographa californica. Полученная структура была названа AcCMV-EGFP-NS1. Исходные рекомбинантные бакуловирусы получали с использованием бакуловирусной экспрессионной системы Bac-to-Bac® Baculovirus Expression System. Для амплификации исходных вирусов использовали монослойную культуру клеток насекомого Spodoptera frugiperda (клетки Sf9, АТСС CRL-1711, Манассас, штат Вирджиния, США). Исходные вирусы содержали рекомбинантную бакмидную ДНК. После инфицирования (PI) получали 3 поколения исходного вируса, каждое через 48 или 72 ч PI. После центрифугирования и фильтрации клеток определяли эффективность трансдукции выращиванием клеток HepG2 в течение ночи и трансдукцией рекомбинантными AcEGFP или AcEGFP-NS1. Для подтверждения 70%-й трансдукционной эффективности вирусов для их дальнейшего использования в индукции апоптотических телец использовали проточный цитометр BD FACSCALIBUR (Becton-Dickinson, штат Нью-Джерси, США). Затем клетки HepG2 трансдуцировали вирусами AcEGFP-NS1 третьего поколения с трансдукционной эффективностью 70%. В завершение, через 72 часа после трансдукции супернатант культуры центрифугировали, выделяли осадок и хранили его при -80°С до последующего использования.
Обработка выделенных микробных осадков для использования в ELISA
Осадки спирохет боррелий, круглых телец боррелий и апоптотических телец B19V размораживали на льду и ресуспендировали в 100 мкл забуференного фосфатом физиологического раствора (PBS, рН 7,4). Для диссоциации с получением лизатов и равномерного растворения содержимого в PBS все растворы попарно обрабатывали ультразвуком на протяжении 15 мин (Bransoni С220), нагревали при 99,9°С в течение 15 мин и еще раз обрабатывали ультразвуком на протяжении 15 мин. По завершении, все исходные растворы антигенов в концентрации 1 мг/мл хранили при +4°С.
Методика проведения ELISA
Исходные растворы антигенов (1 мг/мл) разводили 1:100 в 0,1 М карбонатном буфере (0,1 М Nа2СО3 + 0,1 М NаНСО3, рН 9,5). Для исходных растворов микроорганизмов с двумя пептидными последовательностями объем разведения делили поровну. В данном исследовании использовали два положительных контроля, человеческий IgG (Sigma) и человеческий IgM (Sigma). Кроме того, человеческий IgG (Sigma) и человеческий IgM (Sigma #18260) взаимозаменяемо использовали в качестве отрицательных контролей друг для друга. Исходные контрольные растворы (1 мг/мл) разводили 1:100 в 0,1 М карбонатном буфере. Положительные и отрицательные контроли использовали для поддержания одинаковых значений оптической плотности (optical density, OD) при 450 нм.
По 100 мкл антигенов и контролен сорбировали в двух повторах на плоскодонный 96-луночный полистироловый планшет для ELISA (Nunc) и инкубировали при +4°С в течение ночи. После инкубации планшеты промывали три раза с использованием 300 мкл PBS-Tween (PBS + 0,05% Tween 20) и затем покрывали их 100 мкл 2%-го BSA (Sigma #А7030) в PBS. После инкубации в течение ночи при +4°С 2%-й BSA удаляли. Затем добавляли 100 мкл сыворотки пациента, разведенной 1:200 в 1% BSA/PBS. Затем планшеты инкубировали в течение 2 ч при комнатной температуре (к.т.). После инкубации планшеты промывали пять раз с использованием 300 мкл PBS-Tween. В планшеты вносили пероксидазу хрена (Horse Radish Peroxidase, HRP), конъюгированную с мышиным антителом против человеческого IgG (Abcam) или кроличьим антителом против человеческого IgM (Antibodies Online) в количестве 100 мкл с кратностью разведения 1:10000 или 1:1000, соответственно. После 1,5 ч инкубации при к.т. планшеты промывали пять раз с использованием 300 мкл PBS-Tween и затем вносили 100 мкл 3, 3', 5, 5'-тетраметилбензидинового субстрата (ТМВ, 1-Step ultra TMB-ELISA substrate, Thermo-Piercenet #34028). Планшеты, в которые ранее была внесена HRP, конъюгированная с мышиным антителом против человеческого IgG или IgM, инкубировали при к.т. в течение 5 мин или 1 ч, соответственно. Взаимодействие между вторичными антителами и ТМВ-субстратом останавливали добавлением 100 мкл 2 М H2SO4. Затем с использованием многоканального аппарата для прочтения планшетов Victor™ X4 (Perkin Elmer 2030 Manger) измеряли значения OD при 450 нм за 0,1 с.
Обработка данных
В целях обеспечения качества проводили оценку каждого повтора, что убедиться, что они отличались друг от друга не более чем на 30%. Вместо анализа повторов на предмет их нахождения в пределах 30% от их среднего значения40, повторы анализировали на предмет их нахождения в пределах 30% друг от друга. Поскольку нахождение повторов в пределах 30% друг от друга не зависит от их среднего, разница между результатами измерений сильно ограничена в сравнении с нахождением повторов в пределах 30% от их среднего значения. Для установления пороговых значений для качественного определения антител к 20 антигенам использовали группу из 51 отрицательного контроля для IgG и еще одну группу из 51 отрицательного контроля для IgM. Пороговое значение для антигена устанавливали, добавляя среднее значение всех усредненных значений OD к утроенному стандартному отклонению всех усредненных значений OD41. После определения пороговых значений для 20 антигенов все средние значения OD делили на соответствующие пороговые значения антигенов для нормализации набора данных. Нормализуя все значения OD, получали набор данных по показателю оптической плотности (optical density index, ODI) для обоих типов антител. В завершение, значения ODI преобразовывали в бинарный набор данных со значениями «1» или «О» для обозначения положительных или отрицательных результатов, соответственно.
Вариабельность оценивали, рассчитывая внутри- и межаналитическую вариабельность42. Внутрианалитическую вариабельность определяли удвоением результатов измерений по одному образцу с высоким титром и одному образцу с низким титром на одном и том же планшете. Межаналитическую вариабельность определяли, используя результаты измерений по шести образцам с высоким титром и шести образцам с низким титром, полученные на разных планшетах в разные дни разными операторами.
Использованное оборудование
Для измерения концентрации белка в лизатах клеток при 280 нм использовали спектрофотометр ND 1000 (Finnzymes). Для измерения значений OD при 450 нм за 0,1 с использовали многоканальный аппарат для прочтения планшетов Victor™ X4 (Perkin Elmer 2030 Manger). Для промывки микропланшетов для ELISA использовали аппарат для промывки микропланшетов DNX-9620G (Nanjing Perlove Medical Equipment Co., Ltd).
Результаты
На Фиг. 1 и 2 продемонстрированы иммунные ответы 443 индивидов на комбинацию спирохет и круглых телец боррелий, только спирохеты боррелий и только круглые тельца боррелий. Общее число IgM- и IgG-иммунных ответов (Фиг. 1А и 2А) только на круглые тельца боррелий стабильно больше общего числа IgM- и IgG-иммунных ответов только на спирохеты боррелий. Кроме того, общее число IgM- и IgG-иммунных ответов (Фиг. 1А и 2А) на различные комбинации спирохет и круглых телец боррелий больше общего числа IgM- и IgG-иммунных ответов только на спирохеты боррелий и только на круглые тельца боррелий. Более того, на Фиг. 1В и 2В, спирохеты различных видов боррелий приводили к большему числу иммунных ответов по сравнению с общим числом иммунных ответов, отмеченным при различных комбинациях спирохет боррелий. Сходным образом, на Фиг. 1С и 2С, для круглых телец различных видов боррелий было отмечено большее число иммунных ответов, чем при различных комбинациях круглых телец боррелий. Фиг. 1 и 2 показывают, что, в дополнение к спирохетам различных видов боррелий, круглые тельца различных видов боррелий могут способствовать значительному повышению эффективности диагностических инструментов выявления инфекций, вызываемых боррелиями, у индивидов.
На Фиг. 1А, 95 (21%), 15 (3%) и 65 (15%) индивидов с IgM отвечали на спирохеты и круглые тельца боррелий, только спирохеты боррелий и только круглые тельца боррелий, соответственно. Общее число иммунных ответов только на круглые тельца боррелий было приблизительно в 5 раз больше общего числа иммунных ответов только на спирохеты боррелий. Остальные 268 (61%) индивидов не отвечали ни на какие антигены боррелий. Круглые тельца боррелий представляют собой дремлющую или латентную форму5,9,14 нативной спирохетной структуры боррелий. Более выраженный IgM-ответ пациента на круглые тельца боррелий, а не на их спирохетную структуру, указывает на IgM-иммунную дисфункцию17. Сходным образом, на Фиг. 2А, 171 (38%), 47 (11%) и 71 (16%) индивид с IgG отвечали на спирохеты и круглые тельца боррелий, только спирохеты боррелий и только круглые тельца боррелий, соответственно. Общее число иммунных ответов только на круглые тельца боррелий было приблизительно в 2 раза больше общего числа иммунных ответов только на спирохеты боррелий. Остальные 154 (35%) индивида не отвечали ни на какие антигены боррелий. Большее количество иммунных ответов на круглые тельца боррелий указывает на то, что диагностический набор только со спирохетами боррелий не может обеспечить индивидам полной и надежной диагностики инфекций, вызываемых боррелиями. Таким образом, данное исследование впервые демонстрирует необходимость использования круглых телец боррелий наряду со спирохетами боррелий для диагностики у пациентов с TBD.
Индивидам, инфицированным разными штаммами боррелий, необходимо разное лечение16. Таким образом, диагностика, проводимая у индивидов, должна включать выявление разных штаммов боррелий. Иммунные ответы только на спирохеты боррелий и только на круглые тельца боррелий (Фиг. 1А и 2А) были проанализированы более подробно (на Фиг. 1В, 1С, 2В и 2С) для оценки того, превышает ли общее число иммунных ответов на отдельные штаммы боррелий общее число иммунных ответов на различные комбинации штаммов боррелий. Общее число иммунных ответов на отдельные штаммы боррелий было стабильно больше общего числа иммунных ответов на различные комбинации штаммов боррелий (Фиг. 1В, 1С, 2В и 2С).
На Фиг. 1А, 15 (3%) индивидов, отвечавшие только на спирохеты боррелий, были проанализированы и изучены более подробно на Фиг. 1В. Из этих 15 (3%) индивидов, 1 (7%), 5 (33%) и 5 (33%) индивидов отвечали на спирохеты Borrelia burgdorferi (Bb), Borrelia afzeilii (Ва) и Borrelia garinii (Bg), соответственно. Кроме того, 3 (20%) и 1 (7%) индивид отвечали на комбинации спирохет Ba+Bg и Bb+Ba+Bg, соответственно. Из этих 15 индивидов, 4 (27%) индивида отвечали на комбинации различных штаммов боррелий, в то время как 11 (73%) индивидов отвечали на различные штаммы боррелий. Сходным образом, на Фиг. 2А, 47 (11%) индивидов, отвечавшие только на спирохеты боррелий, были проанализированы и изучены более подробно на Фиг. 2В. Из этих 47 (11%) индивидов, 3 (6%), 10 (21%) и 13 (28%) индивидов отвечали на спирохеты Вb, Ва и Bg, соответственно. Кроме того, 4 (9%), 7 (15%) и 10 (21%) индивидов отвечали на комбинации спирохет Bb+Bg, Ba+Bg и Bb+Ba+Bg, соответственно. Из этих 47 (11%) индивидов, 21 (45%) индивид отвечали на комбинации различных штаммов боррелий, в то время как 26 (55%) индивидов отвечали на различные штаммы боррелий. На комбинацию Bb+Ва не было отмечено ни IgM- (Фиг. 1В), ни IgG-иммунных ответов (Фиг. 2В). Кроме того, на Фиг. 1В не было отмечено иммунных ответов на комбинацию Bb+Bg.
На Фиг. 1А, 65 (15%) индивидов, отвечавшие только на круглые тельца боррелий, были проанализированы и изучены более подробно на Фиг. 1С. Из этих 65 (15%) индивидов, 16 (25%), 12 (18%) и 13 (20%) индивидов отвечали на круглые тельца Вb, Ва и Bg, соответственно. Кроме того, 9 (14%), 8 (12%) и 7 (11%) индивидов отвечали на комбинации круглых телец Bb+Ва, Bb+Bg и Bb+Ba+Bg, соответственно. Из этих 65 (15%) индивидов, 24 (37%) индивида отвечали на комбинации различных штаммов боррелий, в то время как 41 (63%) индивид отвечали на различные штаммы боррелий. Сходным образом, на Фиг. 2А, 71 (16%) индивид, отвечавшие только на круглые тельца боррелий, были проанализированы и изучены более подробно на Фиг. 2С. Из этих 71 индивида, 4 (6%), 5 (7%) и 30 (42%) индивидов отвечали на круглые тельца Вb, Ва и Bg, соответственно. Кроме того, 2 (3%), 16 (22%) и 12 (17%) индивидов отвечали на комбинации круглых телец Bb+Ва, Bb+Bg, Ba+Bg и Bb+Ba+Bg, соответственно. Из этих 71 индивида, 32 (45%) индивида отвечали на комбинации различных штаммов боррелий, в то время как 39 (55%) индивидов отвечали на различные штаммы боррелий. На комбинацию Ba+Bg не было отмечено ни IgM- (Фиг. 1С), ни IgG-иммунных ответов (Фиг. 2С). Очевидно, общее число иммунных ответов на отдельные штаммы боррелий превышает общее число иммунных ответов на комбинации штаммов боррелий (Фиг. 1В, 1С, 2В и 2С). Большее число иммунных ответов на отдельные штаммы боррелий указывает на распространенность этипопов, различающихся у разных штаммов боррелий43. Исключение различных штаммов боррелий из диагностического инструмента может ограничить его чувствительность44.
На Фиг. 3 представлены IgM- (3А) и IgG-иммунные ответы (3В) 443 индивидов на один или несколько микробных антигенов и оценена актуальность полимикробных состояний при TBD. В целом, медицинским сообществом и диагностической индустрией признано существование полимикробных инфекций при многих заболеваниях, таких как корь, туберкулез, гепатит, синдром приобретенного иммунодефицита (СПИД) и другие12,45. Тем не менее, применительно к полимикробным инфекциям, диагностика TBD пока не претерпела изменений46. На Фиг. 3А, 237 (53%) индивидов отвечали на несколько микробных антигенов, в то время как 53 (12%) индивида отвечали на какой-либо один микробный антиген. Сходным образом, на Фиг. 3В определено, что 344 (78%) индивида отвечали на несколько микробных антигенов, в то время как 63 (14%) индивида отвечали на какой-либо один микробный антиген. Экспериментальные данные о полимикробных инфекциях при TBD, представленные на Фиг. 3, указывают на необходимость изменения парадигмы в области диагностики TBD. У остальных 153 (35%) и 36 (8%) индивидов не было иммунного ответа на микробные антигены при анализе на IgM и IgG, соответственно. Индивидов с IgM-ответом на несколько микроорганизмов (Фиг. 3А) приблизительно в 5 раз больше, чем индивидов с ответом на один микроорганизм. Сходным образом, на Фиг. 3В, индивидов с ответом на несколько микроорганизмов приблизительно в 6 раз больше, чем индивидов с ответом на один микроорганизм. IgM-ответ на несколько антигенов (53%) (3А) указывает на то, что иммунная дисфункция может быть частым феноменом у индивидов с TBD17. Более того, Фиг. 3А и 3В показывают, что полимикробные инфекции могут быть более распространенным явлением, которое следует оценивать по IgG, а не по IgM.
На Фиг. 4 и 5 представлены IgM- и IgG-иммунные ответы на отдельные микробные антигены, соответственно. Общее число IgG-иммунных ответов на каждый отдельный антиген было стабильно больше общего числа соответствующих IgM-ответов. Иммунных ответов на круглые тельца боррелий было больше или примерно столько же, сколько и на спирохеты соответствующих штаммов. Эквивалентное число индивидов, иммунных к круглым тельцам боррелий, в сравнении со спирохетами боррелий указывает на то, что круглые тельца боррелий могут помочь максимизировать чувствительность диагностических инструментов выявления боррелий. 130 (29%) и 64 (14%) индивида отвечали на Borrelia burgdorferi sensu stricto B31 no IgG и IgM, соответственно; 162 (37%) и 79 (18%) индивидов отвечали на Borrelia afzelii Р12 по IgG и IgM, соответственно; 161 (36%) и 94 (21%) индивида отвечали на Borrelia garinii Fuji P1 no IgG и IgM, соответственно; 158 (35%) и 120 (27%) индивидов отвечали на круглые тельца Borrelia burgdorferi sensu stricto B31 no IgG и IgM, соответственно; 164 (37%) и 98 (22%) индивидов отвечали на круглые тельца Borrelia afzelii Р12 по IgG и IgM, соответственно; и 180 (41%) и 83 (19%) индивида отвечали на круглые тельца Borrelia garinii Fuji P1 no IgG и IgM, соответственно.
На Фиг. 4 и 5 иммунные ответы на антигены помимо спирохет / круглых телец боррелий указывают на необходимость проведения у индивидов анализа на вторичные и сочетанные инфекции и на аутоиммунные состояния. Такие IgG- и IgM-иммунные ответы обнаружены у следующего числа индивидов: 125 (28%) и 59 (13%) индивидов отвечали на Bartonella henselae, соответственно; 126 (28%) и 74 (16%) индивида отвечали на Babesia microti, соответственно; 115 (26%) и 65 (15%) индивидов отвечали на Chlamydia trachomatis, соответственно; 115 (26%) индивидов отвечали на Chlamydia pneumonia, соответственно; 167 (38%) и 122 (28%) индивида отвечали на Mycoplasma fermentans, соответственно; 137 (31%) и 58 (13%) индивидов отвечали на Mycoplasma pneumonia, соответственно; 115 (26%) и 76 (17%) индивидов отвечали на вирус Коксаки А16, соответственно; 150 (34%) и 127 (29%) индивидов отвечали на цитомегаловирус, соответственно; 203 (46%) и 68 (15%) индивидов отвечали на вирус Эпштейна-Барр, соответственно; 122 (28%) и 64 (14%) индивида отвечали на Brucella abortus, соответственно; 134 (30%) и 104 (23%) индивида отвечали на апоптотические тельца парвовируса В19, соответственно; 142 (32%) и 77 (17%) индивидов отвечали на Ehrlichia chaffeensis, соответственно; 149 (34%) и 71 (16%) индивид отвечали на вирус клещевого энцефалита, соответственно; 184 (47%) и 146 (33%) индивидов отвечали на Rickketsia akari, соответственно; и 36 (8%) и 153 (35%) не отвечали ни на один из 20 антигенов, соответственно.
На Фиг. 6 и 7 показаны различия иммунных ответов 443 индивидов на другие микроорганизмы со спирохетами боррелий, круглыми тельцами боррелий или комбинацией спирохет и круглых телец боррелий или без боррелий. По сути, на Фиг. 6 и 7 показаны различия частоты иммунного ответа на определенное число из нескольких других микроорганизмов и на каждый конкретный другой микроорганизм с и без круглых телец боррелий. Было отмечено, что индивиды, отвечающие на комбинацию спирохет и круглых телец боррелий, более склонны отвечать не только на определенное число из нескольких других микроорганизмов, но также на конкретный другой микроорганизм. Фиг. 6 и 7 указывают на то, что диагностический инструмент с антигенами спирохет боррелий, круглых телец боррелий, сочетанных и вторичных инфекций и аутоиммунными антигенами обеспечит индивидам полную и надежную диагностику TBD. Термин «другие микроорганизмы» включает сочетанные инфекции, вторичные и аутоиммунные антигены, такие как, без ограничения, Bartonella henselae (В. henselae), Brucella abortus (В. abortus), Babesia microti (В. microti), Ehrlichia chaffeensis (E. chaffeensis), Rickettsia akari (R. akari), вирус клещевого энцефалита (TBEV), Chlamydia trachomatis (C. trachomatis), Chlamydia pneumonia (C. pneumonia), Mycoplasma fermentans (M. fermentans), Mycoplasma pneumonia (M pneumonia), цитомегаловирус (CMV), вирус Эпштейна-Барр (EBV), вирус Коксаки A16 (CV A16) и парвовирус человека В19 (HB19V).
На Фиг. 6А и 7А, приблизительно четверть (26%) из 443 индивидов отвечали на другие микроорганизмы без боррелий. IgM- и IgG-иммунные ответы 115 (26%) и 118 (26%) индивидов на другие микроорганизмы без боррелий указывают на то, что у индивидов также необходимо проводить скрининг на другие микроорганизмы, отличные от боррелий. Кроме того, на Фиг. 6А и 7А представлены иммунные ответы индивидов только на боррелий и на другие микроорганизмы с боррелиями. Было отмечено, что число индивидов, отвечавших на другие микроорганизмы с боррелиями, было существенно больше числа индивидов, отвечавших только на антигены боррелий. На Фиг. 6А, из 443 индивидов 10 (2%), 2(1%) и 5(1%) индивидов отвечали на круглые тельца боррелий, спирохеты боррелий и комбинацию спирохет и круглых телец боррелий, соответственно. Тем не менее, из 443 индивидов 55 (12%), 13 (3%) и 90 (20%) индивидов отвечали на круглые тельца боррелий, спирохеты боррелий и комбинацию спирохет и круглых телец боррелий с другими микроорганизмами, соответственно. Сходным образом, на Фиг. 7А, из 443 индивидов 23 (5%), 2 (1%) и 13 (3%) индивидов отвечали на круглые тельца боррелий, спирохеты боррелий и комбинацию спирохет и круглых телец боррелий, соответственно. Однако, из 443 индивидов 48 (11%), 45 (10%) и 158 (36%) индивидов отвечали на круглые тельца боррелий, спирохеты боррелий и комбинацию спирохет и круглых телец боррелий с другими микроорганизмами, соответственно.
На Фиг. 6А и 7А, индивиды, отвечающие на круглые тельца боррелий, более склонны отвечать на другие микроорганизмы, чем индивиды, отвечающие на спирохеты боррелий. Тем не менее, индивиды, отвечающие на комбинацию спирохет и круглых телец боррелий, приблизительно в 3 раза более склонны отвечать на другие микробы, чем индивиды, отвечающие на круглые тельца боррелий или спирохеты боррелий. По IgM (Фиг. 6А), число индивидов, отвечающих на другие микроорганизмы с круглыми тельцами боррелий, приблизительно в 4 раза превышает число индивидов, отвечающих на другие микроорганизмы со спирохетами боррелий. Однако, по IgG (Фиг. 7А), число индивидов, отвечающих на другие микроорганизмы с круглыми тельцами боррелий, приблизительно соответствует числу индивидов, отвечающих на другие микроорганизмы со спирохетами боррелий. По IgM, из 443 индивидов 55 (12%) индивидов отвечали на другие микроорганизмы с круглыми тельцами боррелий, в то время как на другие микроорганизмы со спирохетами боррелий отвечали 13 (3%) индивидов (Фиг. 6А). Сходным образом, 48 (11%) индивидов отвечали на другие микроорганизмы с круглыми тельцами боррелий, а 45 (10%) индивидов отвечали на другие микроорганизмы со спирохетами боррелий.
На Фиг. 6В и 7В показаны различия в микробной нагрузке у индивидов, отвечавших на другие микроорганизмы с и без боррелий. Во-первых, индивиды, отвечавшие на другие микроорганизмы (Фиг. 6А и 7А), не отвечали более чем на восемь микроорганизмов по обоим классам антител (Фиг. 6В и 7В). Тем не менее, более 75% индивидов, отвечавших на другие микроорганизмы, не отвечали более чем на три микроорганизма. Из 115 (26%) индивидов с IgM-ответом на другие микроорганизмы (Фиг. 6А) 92 (80%) индивида не отвечали более чем на три микроорганизма. Сходным образом, из 118 (26%) индивидов с IgG-ответом на другие микроорганизмы (Фиг. 7А) 89 (75%) индивидов не отвечали более чем на три микроорганизма. Интересно, что индивиды, отвечавшие на боррелий, были более склонны отвечать на несколько других микроорганизмов, чем индивиды без ответа на боррелий (Фиг. 6В и 7В).
Индивиды с IgM-ответом на круглые тельца боррелий были более склонны к отвечать на несколько других микроорганизмов, чем индивиды, отвечавшие на спирохеты боррелий (Фиг. 6В) В противоположность этому, индивиды с IgG-ответом на спирохеты боррелий были более склонны к отвечать на несколько других микроорганизмов, чем индивиды, отвечавшие на круглые тельца боррелий (Фиг. 7В). Однако, индивиды, отвечавшие на комбинацию спирохет и круглых телец боррелий, стабильно более склонны отвечать на несколько микроорганизмов, чем индивиды, отвечавшие на круглые тельца боррелий или спирохеты боррелий. Более 50% индивидов, отвечавших на другие микроорганизмы с комбинацией спирохет и круглых телец боррелий, отвечали на 8-14 других микроорганизмов. Концентрация индивидов, отвечавших на другие микроорганизмы с комбинацией спирохет и круглых телец боррелий, максимальна при 14 микроорганизмах по обоим классам антител (Фиг. 6В и 7В). Из 90 (20%) индивидов с IgM-ответом на другие микроорганизмы с комбинацией спирохет и круглых телец боррелий (Фиг. 6А) 14 (16%) индивидов отвечали на 14 других микроорганизмов (Фиг. 6В). Сходным образом, из 158 (36%) индивидов с IgG-ответом на другие микроорганизмы с комбинацией спирохет и круглых телец боррелий (Фиг. 7А) 23 (15%) индивида отвечали на 14 других микроорганизмов (Фиг. 7В).
На Фиг. 6С и 7С показаны различия иммунных ответов 443 индивидов на отдельные другие микроорганизмы с и без боррелий. Антигены боррелий, продемонстрировавшие наибольшую микробную нагрузку на Фиг. 6В и 7В, также продемонстрировали наибольшую частоту иммунных ответов на отдельные другие микроорганизмы на Фиг. 6С и 7С. Исходя из Фиг. 6В и 7В, круглые тельца боррелий и спирохеты боррелий продемонстрировали наибольшую микробную нагрузку у индивидов по IgM и IgG, соответственно. Так, индивиды с IgM-ответом на круглые тельца боррелий отвечали на все другие микроорганизмы в среднем в 5 раз чаще, чем индивиды, отвечавшие на спирохеты боррелий (Фиг. 6С). Кроме того, индивиды с IgG-ответом на спирохеты боррелий отвечали на все другие микроорганизмы в среднем в 2 раза чаще, чем индивиды, отвечавшие на круглые тельца боррелий (Фиг. 7С). Тем не менее, наибольшую микробную нагрузку по обоим классам антител продемонстрировала комбинация спирохет и круглых телец боррелий (Фиг. 6В и 7В). Так, индивиды с IgM-ответом на комбинацию спирохет и круглых телец боррелий отвечали на все другие микроорганизмы приблизительно в 3 раза чаще, чем индивиды, отвечавшие на круглые тельца боррелий (Фиг. 6С). Кроме того, индивиды с IgG-ответом на комбинацию спирохет и круглых телец боррелий отвечали на все другие микроорганизмы приблизительно в 5 раз чаще, чем индивиды, отвечавшие на спирохеты боррелий (Фиг. 7С).
Внутри- и межаналитическая вариабельность
Расчетная внутри- и межаналитическая вариабельность настоящего метода составила 4,6% и 15,6%, соответственно. 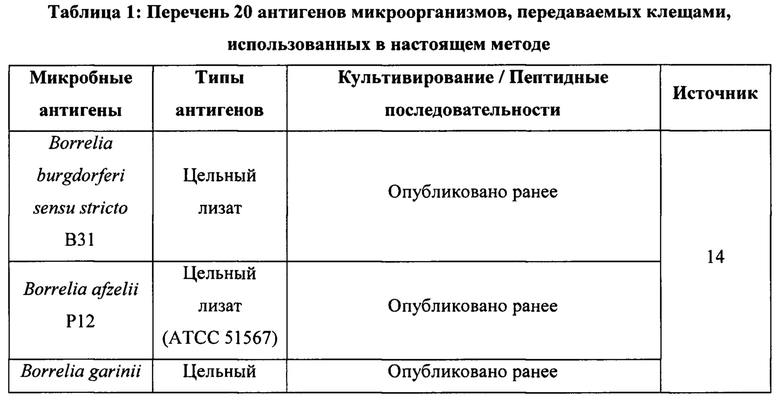
СПИСОК ЛИТЕРАТУРЫ
1. Steere AC, Coburn J, Glickstein L. The emergence of Lyme disease. J Clin Invest. 2004 Apr 4; 113(8):1093-101.
2. Steere AC. Lyme disease. N Engl J Med. 2001 Jul 4;345(2):115-25.
3. Chomel B. Lyme disease. Rev - Off Int Epizoot. 2015 Aug 6;34(2):569-76.
4. Mead PS. Epidemiology of Lyme disease. Infect Dis Clin North Am. 2015 Jun 1;29(2):187-210.
5. Strieker RB, Johnson L. Lyme disease: the next decade. Infect Drug Resist. 2011 Jan 6;4:1-9.
6. Berghoff W. Chronic Lyme Disease and Co-infections: Differential Diagnosis. Open Neurol J. 2012 Jan;6:158-78.
7. Lindgren E, Jaenson TGT. Lyme borreliosis in Europe: influences of climate and climate change, epidemiology, ecology and adaptation measures. WHO Regional Office for Europe. WHO Regional Office for Europe; 2006;EUR/04(/5046250):34.
8. Donta S. Issues in the Diagnosis and Treatment of Lyme Disease. Open Neurology J. bentham; 2012;6(1):140-5.
9. Johnson L, Wilcox S, Mankoff J, Stricker RB. Severity of chronic Lyme disease compared to other chronic conditions: a quality of life survey. PeerJ. 2014 Jan 3;2:e322.
10. Adrion ER, Aucott J, Lemke KW, Weiner JP. Health care costs, utilization and patterns of care following Lyme disease. PLoS ONE. 2015 Jan 4;10(2):e0116767.
11. Wilske B. Epidemiology and diagnosis of Lyme borreliosis. Ann Med. 2005 Jan 6;37(8):568-79.
12. Brogden KA, Guthmiller JM, Taylor CE. Human polymicrobial infections. Lancet. 2005 Jan 6;365(9455):253-5.
13. Aguero-Rosenfeld M, Wang G, Schwartz I, Wormser G. Diagnosis of Lyme Borreliosis. Clin Microbiol Rev. highwire; 2005;18(3):484-509.
14.  Herranen A, Schwarzbach A, Gilbert L. Morphological and biochemical features of Borrelia burgdorferi pleomorphic forms. Microbiology (Reading, Engl). 2015 Mar; 161 (Pt 3):516-27.
Herranen A, Schwarzbach A, Gilbert L. Morphological and biochemical features of Borrelia burgdorferi pleomorphic forms. Microbiology (Reading, Engl). 2015 Mar; 161 (Pt 3):516-27.
15. Seinost G, Golde WT, Berger BW, Dunn JJ, Qiu D, Dunkin DS, et al. Infection with multiple strains of Borrelia burgdorferi sensu stricto in patients with Lyme disease. Arch Dermatol. 1999 Nov 1;135(11):1329-33.
16.  Basse-Guerineau AL, Bachmeyer C, Christoforov B, Assous MV. [Lyme borreliosis: therapeutic aspects]. Presse Med. 1998 Dec 6;27(39):2043-7.
Basse-Guerineau AL, Bachmeyer C, Christoforov B, Assous MV. [Lyme borreliosis: therapeutic aspects]. Presse Med. 1998 Dec 6;27(39):2043-7.
17. Kalish, McHugh, Granquist, Shea, Ruthazer, Steere. Persistence of immunoglobulin M or immunoglobulin G antibody responses to Borrelia burgdorferi 10-20 years after active Lyme disease. Clin Infect Dis Official Publ Infect Dis Soc Am. highwire; 2001;33(6):780-5.
18. Mursic VP, Wanner G, Reinhardt S, Wilske B, Busch U, Marget W. Formation and cultivation of Borrelia burgdorferi spheroplast-L-form variants. Infection. 1996 Jan 1;24(3):218-26.
19. Domingue, Woody. Bacterial persistence and expression of disease. Clin Microbiol Rev. 1997;10(2):320-44.
20. Murgia R, Piazzetta C, Cinco M. Cystic forms of Borrelia burgdorferi sensu lato: induction, development, and the role of RpoS. Wien Klin Wochenschr. 2002 Jul 3;114(13-14):574-9.
21. Schenk J, Doebis C,  von Baehr V. Evaluation of a New Multiparametric Microspot Array for Serodiagnosis of Lyme Borreliosis. Clin Lab. 2015 Jan 4;61(11):1715-25.
von Baehr V. Evaluation of a New Multiparametric Microspot Array for Serodiagnosis of Lyme Borreliosis. Clin Lab. 2015 Jan 4;61(11):1715-25.
22. Lahey LJ, Panas MW, Mao R, Delanoy M, Flanagan JJ, Binder SR, et al. Development of a Multiantigen Panel for Improved Detection of Borrelia burgdorferi Infection in Early Lyme Disease. J Clin Microbiol. 2015 Dec 2;53(12):3834-41.
23. Embers ME, Hasenkampf NR, Barnes MB, Didier ES, Philipp MT, Tardo AC. A Five-Antigen Fluorescent Bead-based Assay for Diagnosis of Lyme Disease. Clin Vaccine Immunol. 2016 Feb 3.
24. Porwancher RB, Hagerty CG, Fan J, Landsberg L, Johnson BJ, Kopnitsky M, et al. Multiplex immunoassay for Lyme disease using VlsE1-IgG and рерС10-IgM antibodies: improving test performance through bioinformatics. Clin Vaccine Immunol. 2011 May;18(5):851-9.
25. Dessau RB,  Kolmos B, Henningsson AJ. Multiplex assay (Mikrogen recomBead) for detection of serum IgG and IgM antibodies to 13 recombinant antigens of Borrelia burgdorferi sensu lato in patients with neuroborreliosis: the more the better? J Med Microbiol. 2015 Mar;64(Pt 3):224-31.
Kolmos B, Henningsson AJ. Multiplex assay (Mikrogen recomBead) for detection of serum IgG and IgM antibodies to 13 recombinant antigens of Borrelia burgdorferi sensu lato in patients with neuroborreliosis: the more the better? J Med Microbiol. 2015 Mar;64(Pt 3):224-31.
26. Kivovich V, Gilbert L, Vuento M, Naides SJ. Parvovirus B19 genotype specific amino acid substitution in NS1 reduces the protein's cytotoxicity in culture. Int J Med Sci. 2010 Jan 5;7(3):110-9.
27. Thammasri K,  Wang L, Filippou A, Kivovich V,
Wang L, Filippou A, Kivovich V,  et al. Human parvovirus B19 induced apoptotic bodies contain altered self-antigens that are phagocytosed by antigen presenting cells. PLoS ONE. 2013 Jan 2;8(6):e67179.
et al. Human parvovirus B19 induced apoptotic bodies contain altered self-antigens that are phagocytosed by antigen presenting cells. PLoS ONE. 2013 Jan 2;8(6):e67179.
28. US 6699678 В1,Chlamydia trachomatis specific peptides and their use in diagnostic assays. United States Patent.
29. Mitchell WM, Stratton CW. Diagnosis and management of infection caused by chlamydia. United States Patent; US 6579854 B1, 1998.
30. Theiss P, Karpas A, Wise KS. Antigenic topology of the P29 surface lipoprotein of Mycoplasma fermentans: differential display of epitopes results in high-frequency phase variation. Infect Immun. 1996 May 3;64(5): 1800-9.
31. Jacobs E, Pilatschek A, Gerstenecker B, Oberle K, Bredt W. Immunodominant epitopes of the adhesin of Mycoplasma pneumoniae. J Clin Microbiol. 1990 Jun 5;28(6):1194-7.
32. Huang L, Hoey J, Adelson M, Mordechai E. Recombinant fragments and synthetic peptides of 17-kda polypeptide useful in detecting bartonella henselae. European Patent; EP 2326660 A2, 2011.
33. Zhang J, Guo F, Huang X, Chen C, Liu R, Zhang H, et al. A novel Omp25-binding peptide screened by phage display can inhibit Brucella abortus 2308 infection in vitro and in vivo. J Med Microbiol. 2014 Jun;63(Pt 6):780-7.
34. Flores O, Schwarzch A, Rredo B, Altieri GU. Biochip, antigen bouquet, optical reader and method for detecting and monitoring diseases. WIPO; WO 2014185803 A2, 2014.
35. Shi J, Huang X, Liu Q, Huang Z. Identification of conserved neutralizing linear epitopes within the VP1 protein of coxsackievirus A16. Vaccine. 2013 Apr 5;31(17):2130-6.
36. Middeldorp JM, van Grunsven WMJ. Peptides and nucleic acid sequences related to the Epstein Barr virus. United States Patent; US 7507804 B2, 2009.
37. Landini MP, Ripalti A, Sra K, Pouletty P. Human cytomegalovirus structural proteins: immune reaction against pp150 synthetic peptides. J Clin Microbiol. 1991 Sep;29(9): 1868-72.
38. Holzmann H, Utter G, Norrby E, Mandl CW, Kunz C, Heinz FX. Assessment of the antigenic structure of tick-borne encephalitis virus by the use of synthetic peptides. J Gen Virol. 1993 Sep3;74(Pt 9):2031-5.
39. Barbour AG, Hayes SF. Biology of Borrelia species. Microbiol Rev. 1986 Dec 1;50(4):381-400.
40. Dudal S, Baltrukonis D, Crisino R, Goyal MJ, Joyce A, Osterlund K, etal. Assay formats: Recommendation for best practices and harmonization from the global bioanalysis consortium harmonization team. AAPS J. 2014 Mar 6; 16(2): 194-205.
41. Puttaraksa K, Merilainen L, Capillo A, Schwarzbach A, garcia P, Gilbert L. Indirect ELISA diagnostic test for Lyme Disease.  2015.
2015.
42. Reed GF, Lynn F, Meade BD. Use of coefficient of variation in assessing variability of quantitative assays. Clin Diagn Lab Immunol. 2002 Nov 5;9(6): 1235-9.
43. Shoberg RJ, Jonsson M, Sadziene A,  Thomas DD. Identification of a highly cross-reactive outer surface protein В epitope among diverse geographic isolates of Borrelia spp.causing Lyme disease. J Clin Microbiol. 1994 Feb 2;32(2):489-500.
Thomas DD. Identification of a highly cross-reactive outer surface protein В epitope among diverse geographic isolates of Borrelia spp.causing Lyme disease. J Clin Microbiol. 1994 Feb 2;32(2):489-500.
44. Wormser GP, Liveris D,  Brisson D, Ludin S, Stracuzzi VJ, et al. Effect of Borrelia burgdorferi genotype on the sensitivity of C6 and 2-tier testing in North American patients with culture-confirmed Lyme disease. Clin Infect Dis. 2008 Oct 3;47(7):910-4.
Brisson D, Ludin S, Stracuzzi VJ, et al. Effect of Borrelia burgdorferi genotype on the sensitivity of C6 and 2-tier testing in North American patients with culture-confirmed Lyme disease. Clin Infect Dis. 2008 Oct 3;47(7):910-4.
45. O'Connor SMM, Taylor CE, Hughes JM. Emerging infectious determinants of chronic diseases. Emerging Infect Dis. 2006 Jul 6; 12(7): 1051-7.
46. Wormser GP, Dattwyler RJ, Shapiro ED, Halperin JJ, Steere AC, Klempner MS, et al. The clinical assessment, treatment, and prevention of lyme disease, human granulocytic anaplasmosis, and babesiosis: clinical practice guidelines by the Infectious Diseases Society of America. Clin Infect Dis. 2006 Nov 3;43(9):1089-134.
47. Miklossy, J., Kasas, S., Zurn, A., McCall, S., Yu, S., and McGeer. Persisting atypical and cystic forms of Borrelia burgdorferi and local inflammation in Lyme neuroborreliosis. J Neuroinflammation. Journal of Neuroinflammation, 2008, 5:40.
48. Cook, Michael J. Lyme borreliosis: a review of data on transmission time after tick attachment. International Journal of General Medicine, 2015, 8:1-8.






Группа изобретений относится к выявлению болезни Лайма и других заболеваний, передаваемых клещами. Раскрыта твердая подложка для выявления присутствия антител в биологическом образце, содержащая микробные антигены, иммобилизованные на указанной твердой подложке, включающие по меньшей мере один антиген, полученный из группы, состоящей из плеоморфных круглых телец вида рода Borrelia, где указанный вид рода Borrelia выбран из группы, состоящей из Borrelia afzelii и Borrelia garinii, и указанный антиген представляет собой лизат указанных плеоморфных круглых телец вида рода Borrelia. Также раскрыты способ выявления антител в биологическом образце и применение указанной твердой подложки для изготовления диагностического теста. Группа изобретений обеспечивает создание новой платформы, позволяющей различать острые, прошедшие, хронические и персистирующие стадии заболеваний, передаваемых клещами. 3 н. и 16 з.п. ф-лы, 7 ил., 1 табл., 1 пр.
1. Твердая подложка для выявления присутствия антител в биологическом образце, содержащая микробные антигены, иммобилизованные на указанной твердой подложке, включающие по меньшей мере один антиген, полученный из группы, состоящей из плеоморфных круглых телец вида рода Borrelia,
где указанный вид рода Borrelia выбран из группы, состоящей из Borrelia afzelii и Borrelia garinii, и
указанный антиген представляет собой лизат указанных плеоморфных круглых телец вида рода Borrelia.
2. Твердая подложка по п. 1, дополнительно содержащая антиген, представляющий собой лизат плеоморфных круглых телец Borrelia burgdorferi.
3. Твердая подложка по п. 1 или 2, представляющая собой микролуночный планшет или биочип с антигенами.
4. Твердая подложка по любому из пп. 1-3, дополнительно содержащая по меньшей мере один иммобилизованный антиген, полученный из группы, состоящей из вида рода Borrelia в форме нативных спирохет.
5. Твердая подложка по п. 4, где указанный вид рода Borrelia выбран из группы, состоящей из Borrelia burgdorferi, Borrelia afzelii и Borrelia garinii.
6. Твердая подложка по п. 4 или 5, содержащая иммобилизованный антиген, полученный из плеоморфных круглых телец Borrelia burgdorferi, и иммобилизованный антиген, полученный из Borrelia burgdorferi в форме нативных спирохет.
7. Твердая подложка по п. 4 или 5, содержащая иммобилизованный антиген, полученный из плеоморфных круглых телец Borrelia afzelii, и иммобилизованный антиген, полученный из Borrelia afzelii в форме нативных спирохет.
8. Твердая подложка по п. 4 или 5, содержащая иммобилизованный антиген, полученный из плеоморфных круглых телец Borrelia garinii, и иммобилизованный антиген, полученный из Borrelia garinii в форме нативных спирохет.
9. Твердая подложка по любому из пп. 2-5, содержащая иммобилизованные антигены, полученные из плеоморфных круглых телец Borrelia burgdorferi, Borrelia afzelii и Borrelia garinii.
10. Твердая подложка по любому из пп. 1-9, дополнительно содержащая по меньшей мере один иммобилизованный антиген, полученный из группы, состоящей из Mycoplasma fermentans, Mycoplasma pneumonia, Bartonella henselae, Brucella abortus, Babesia microti, Chlamydia trachomatis, Chlamydia pneumonia, Ehrlichia chaffeensis, вируса Коксаки A16, вируса Эпштейна-Барр, цитомегаловируса, апоптотических телец парвовируса человека B19, вируса клещевого энцефалита и Rickettsia akari.
11. Твердая подложка по любому из пп. 4-10, где указанные антигены, полученные из группы, состоящей из вида рода Borrelia, предпочтительно Borrelia burgdorferi, Borrelia afzelii и Borrelia garinii в форме нативных спирохет, представляют собой лизаты, белковые препараты или пептидные препараты указанных нативных спирохет.
12. Способ выявления антител в биологическом образце, включающий:
(a) приведение биологического образца в контакт с твердой подложкой, содержащей микробные антигены, иммобилизованные на указанной твердой подложке, для образования комплекса, содержащего микробный антиген, иммобилизованный на указанной твердой подложке, и антитело из указанного биологического образца, связанное с указанным микробным антигеном, где указанные микробные антигены включают по меньшей мере один антиген, полученный из группы, состоящей из плеоморфных круглых телец вида рода Borrelia, где указанная твердая подложка представляет собой твердую подложку по любому из пп. 1-11; и
(b) выявление присутствия комплекса, полученного на стадии (a), где присутствие комплекса, содержащего антиген, полученный из плеоморфных круглых телец рода Borrelia, указывает на присутствие антител в указанном биологическом образце.
13. Способ по п. 12, где присутствие комплекса, полученного на стадии (a), выявляют путем приведения указанной твердой подложки в контакт с реагентом против антитела для образования комплекса указанного микробного антигена, указанного антитела, связанного с указанным микробным антигеном, и указанного реагента против антитела.
14. Способ по п. 13, где указанный реагент против антитела представляет собой антитело против иммуноглобулина G (IgG), антитело против IgM или антитело против IgA.
15. Способ по п. 13 или 14, где указанный реагент против антитела представляет собой антитело против человеческого IgG, антитело против человеческого IgM или антитело против человеческого IgA.
16. Способ по любому из пп. 12-15, где указанный биологический образец представляет собой образец крови или сыворотки, образец слюны, образец спинномозговой жидкости, образец синовиальной жидкости или образец слезы.
17. Применение твердой подложки по любому из пп. 1-11 для изготовления диагностического теста для выявления антител в биологическом образце.
18. Применение по п. 17, где указанный диагностический тест предназначен для выявления болезни Лайма у пациента.
19. Применение по п. 18, где указанный диагностический тест предназначен для выявления хронической или персистирующей болезни Лайма у пациента.
| MERILAINEN L | |||
| Characterization and Immunological Aspects of Borrelia Burgdorferi Pleomorphic Round Bodies // Academic dissertation to be publicly discussed, by permission of the Faculty of Mathematics and Science of the University of Jyvaskyla, 2015 | |||
| US 20140274925 A1, 18.09.2014 | |||
| US 20130115634 A1, 09.05.2013 | |||
| КОНЕВА О.А | |||
| и др | |||
| Изучение уровня |

