Область изобретения
Настоящее изобретение относится к анти-PCSK9 антителу, его антигенсвязывающим фрагментам, химерным антителам и гуманизированным антителам, содержащим определяющие комплементарность области (CDR; от англ. complementarity determining regions) анти-PCSK9 антитела, а также к фармацевтической композиции, содержащей анти-PCSK9 антитело и его антигенсвязывающие фрагменты, а также к его применению в качестве лекарственного средства для снижения уровня липидов в крови.
Предшествующий уровень техники
Гиперхолестеринемия представляет собой заболевание с патологически измененным метаболизмом липидов, характеризующееся повышенным уровнем холестерина в сыворотке крови. Ее основным проявлением является повышенный уровень холестерина в сыворотке крови, который вызывает агрегацию холестерина в кровеносных сосудах и впоследствии приводит к образованию атеросклероза. На основании многочисленных результатов клинических и экспериментальных исследований доказано, что патологически измененный метаболизм липидов тесно связан с возникновением и развитием ишемической болезни сердца. Следовательно, снижение концентрации холестерина в крови становится основным средством лечения и профилактики атеросклероза.
С быстрым улучшением национального стандарта жизни в Китае дислипидемия становится основным фактором, представляющим угрозу безопасности для городского и сельского населения Китая. Согласно статистическим данным 2012 г. около 40% смертельных случаев в год в Китае было связано с сердечно-сосудистыми заболеваниями. Заболеваемость дислипидемией у взрослых в Китае составляет 18,6%, и в настоящее время по оценкам дислипидемией страдает 160 миллионов человек. Заболеваемость различными типами дислипидемии составляет: 2,9% для гиперхолестеринемии, 11,9% для гипертриглицеридемии, 7,4% для липопротеинемии низкой и высокой плотности и 3,9% для незначительного повышения уровня холестерина в крови. Было упомянуто, что в Китае гиперхолестеринемией страдает 33 миллиона человек, однако по данным Консенсуса экспертов Китая в области профилактики и контроля хронических заболеваний, Отделение профилактики и контроля хронических заболеваний Комитета по профилактике и контролю заболеваний Министерства здравоохранения, заболеваемость дислипидемией в местных областях значительно более серьезна по сравнению с описанными выше данными.
В настоящее время лекарственные средства, применяемые клинически для контроля уровней липидов, в основном сосредоточены на статинах. Липитор как наиболее широко применяемый и являющийся лидером продаж препарат, снижающий уровень холестерина, снижает продукцию холестерина посредством блокирования действия холестерин-продуцирующего фермента в печени и, следовательно, усиливает поглощение печенью холестерина из крови, за счет чего уменьшается концентрация холестерина в крови. Однако Липитор обладает рядом недостатков. Во-первых, на основании данных понятно, что применение препарата Липитор приводит к снижению концентрации липопротеина низкой плотности на 30%-40%, однако у многих пациентов достичь эффективного снижения уровня липидов в крови все же не удается (липопротеин низкой плотности менее 50 мг/дл). Во-вторых, среди пациентов существуют расовые различия по частоте ответа на препарат Липитор. По этим причинам пациенты нуждаются в более эффективном лекарственном средстве, снижающем уровень липидов в крови.
Семейная гиперхолестеринемия (СГХ) представляет собой аутосомно-доминантное моногенное наследственное заболевание, клиническими признаками которого являются статистически значимо повышенный уровень общего холестерина (ОХ) и холестерина липопротеинов низкой плотности (ЛПНП-х) в крови, ксантелазма, дуга роговицы и раннее сердечно-сосудистое заболевание. Мутация в гене рецептора липопротеинов низкой плотности (рецептор ЛПНП, англ. LDLR) приводит к дефициту или отсутствию LDLR, вследствие чего ЛПНП-х не транспортируется в печень для выведения, и, следовательно, повышается уровень ЛПНП-х в крови. В настоящее время идентифицировано 3 гена, коррелирующих с возникновением СГХ. Это ген LDLR, ген аполипопротеина В100 и ген пропротеинконвертазы субтилизин/кексин 9 типа (PCSK9) соответственно.
Пропротеинконвертаза субтилизин/кексин 9 типа (PCSK9) представляет собой пропротеинконвертазу, относящуюся к подсемейству протеазы К, принадлежащей к семейству секреторных белков Bacillus subtilis. Кодируемый белок синтезируется в виде растворимого профермента и претерпевает самокатализируемый внутримолекулярный процессинг в эндоплазматическом ретикулуме. Согласно экспериментальным данным PCSK9 стимулирует расщепление рецептора ЛПНП и, следовательно, увеличивает количество ЛПНП в плазме крови, поскольку рецептор ЛПНП опосредует процесс эндоцитоза ЛПНП в печени, являющийся основным путем метаболизма для удаления ЛПНП из системы кровообращения. Исследователи обнаружили, что у 12,5% пациентов с гиперхолестеринемией (ADH) идентифицированы мутации в гене PCSK9. Существуют различные типы мутаций PCSK9. В зависимости от различных типов влияния мутаций на регулируемый PCSK9 уровень ЛПНП-х эти мутации могут быть разделены на две группы - мутации типа утраты функции и мутации типа приобретения функции. Мутации типа утраты функции связаны с низким уровнем холестерина в крови и обладают эффектом предотвращения возникновения атеросклеротического заболевания сердца. Частоты мутаций PCSK9, связанных с низким уровнем холестерина, выше у населения негроидной расы, чем у других рас. Мутации PCSK9 типа приобретения функции приводят к повышению уровня холестерина в плазме крови за счет усиления функции PCSK9 и снижения экспрессии гена LDLR, что может вызвать серьезную гиперхолестеринемию и раннюю ишемическую болезнь сердца атеросклеротического происхождения. В настоящее время обнаружено, что мутации PCSK9 типа приобретения функции включают D374Y, S127R, F216L, N157K, R306S и т.д. По сравнению с PCSK9 дикого типа у мутанта D374Y количество рецепторов ЛПНП на поверхности клеток уменьшается на 36%, а у мутанта S127R оно уменьшается на 10%.
Ген PCSK9 в качестве потенциальной новой мишени стал важнейшим предметом исследований гиперхолестеринемии. Авторам изобретения важно лучше понять механизм метаболизма холестерина и найти новую терапевтическую стратегию. Многие международные фармацевтические компании разрабатывают моноклональные антитела к PCSK9, которые повышают концентрацию рецептора ЛПНП на поверхности клеток печени и снижают концентрацию ЛПНП в крови посредством нейтрализации PCSK9 в крови. К этому относятся патенты WO 2011111007, WO 2011072263, WO 2013170367, WO 2013169886, WO 2013148284, WO 2013091103, WO 2013039958, WO 2013039969, WO 2013016648, WO 2013008185, WO 2012170607, WO 2012168491, WO 2012154999, WO 2012109530, WO 2012101251, WO 2012088313, US 8829165B2, US 8563698B2, US 8859741 B2, US 8871913 B2, US 8871914 B2, US 8883983 B2, WO 2012058137 и WO 2012054438.
В настоящем изобретении предложены антитела к PCSK9, обладающие более высокой аффинностью, более высокой селективностью и более высокой биологической активностью.
Сущность изобретения
В настоящем изобретении предложено анти-PCSK9 антитело или его антигенсвязывающие фрагменты, содержащие одну или более CDR область, выбранную из следующих: HCDR, как показано в SEQ ID NO 12, SEQ ID NO 13 или SEQ ID NO 14, или HCDR, как показано в последовательности, обладающей по меньшей мере 95% идентичностью с SEQ ID NO 12, SEQ ID NO 13 или SEQ ID NO 14; и LCDR, как показано в SEQ ID NO 15, SEQ ID NO 16 или SEQ ID NO 17, или LCDR, как показано в последовательности, обладающей по меньшей мере 95% идентичностью с SEQ ID NO 15, SEQ ID NO 16 или SEQ ID NO 17.
В другом предпочтительном воплощении настоящего изобретения анти-PCSK9 антитело или его антигенсвязывающие фрагменты в соответствии с настоящим изобретением содержат HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO 12, SEQ ID NO 13 и SEQ ID NO 14 соответственно; или содержат HCDR1, HCDR2 и HCDR3, как показано в последовательности, обладающей по меньшей мере 95% идентичностью с SEQ ID NO 12, SEQ ID NO 13 и SEQ ID NO 14 соответственно.
В другом предпочтительном воплощении настоящего изобретения анти-PCSK9 антитело или его антигенсвязывающие фрагменты в соответствии с настоящим изобретением содержат LCDR1, LCDR2 и LCDR3, как показано в SEQ ID NO 15, SEQ ID NO 16 и SEQ ID NO 17 соответственно; или содержат LCDR1, LCDR2 и LCDR3, как показано в последовательности, обладающей по меньшей мере 95% идентичностью с SEQ ID NO 15, SEQ ID NO 16 и SEQ ID NO 17 соответственно.
Аминокислотная последовательность, обладающая 95% идентичностью, может быть получена путем индукции мутаций в CDR областях по настоящему изобретению с помощью стратегии созревания аффинности.
В другом предпочтительном воплощении настоящего изобретения анти-PCSK9 антитело или его антигенсвязывающие фрагменты в соответствии с настоящим изобретением представляют собой мышиное антитело или его фрагменты.
В другом предпочтительном воплощении анти-PCSK9 антитела или его антигенсвязывающих фрагментов в соответствии с настоящим изобретением вариабельная область легкой цепи анти-PCSK9 антитела дополнительно содержит каркасные (FR; от англ.. framework regions) области, имеющие происхождение от цепи к мыши или ее варианта, или FR области легкой цепи, имеющие происхождение от цепи λ мыши или ее варианта; вариабельная область тяжелой цепи анти-PCSK9 антитела дополнительно содержит FR области, имеющие происхождение от IgG1 мыши или его варианта, или FR области тяжелой цепи, имеющие происхождение от IgG2 мыши или его варианта, или FR области тяжелой цепи, имеющие происхождение от IgG3 мыши или его варианта.
В другом предпочтительном воплощении анти-PCSK9 антитела или его антигенсвязывающих фрагментов в соответствии с настоящим изобретением мышиное антитело содержит последовательность вариабельной области тяжелой цепи SEQ ID NO 10 и последовательность вариабельной области легкой цепи SEQ ID NO 11.
В другом предпочтительном воплощении анти-PCSK9 антитела или его антигенсвязывающих фрагментов в соответствии с настоящим изобретением легкая цепь анти-PCSK9 антитела дополнительно содержит константные области легкой цепи, имеющие происхождение от цепи к мыши или ее варианта, или константные области легкой цепи, имеющие происхождение от цепи λ мыши или ее варианта; вариабельная область тяжелой цепи анти-PCSK9 антитела дополнительно содержит константные области, имеющие происхождение от IgG1 мыши или его варианта, или константные области тяжелой цепи, имеющие происхождение от IgG2 мыши или его варианта, или константные области тяжелой цепи, имеющие происхождение от IgG3 мыши или его варианта.
В другом предпочтительном воплощении анти-PCSK9 антитела или его антигенсвязывающих фрагментов в соответствии с настоящим изобретением антитело или его антигенсвязывающие фрагменты представляют собой химерное антитело или его фрагменты.
В другом предпочтительном воплощении анти-PCSK9 антитела или его антигенсвязывающих фрагментов в соответствии с настоящим изобретением антитело или его антигенсвязывающие фрагменты представляют собой гуманизированное антитело или его фрагменты.
В другом предпочтительном воплощении анти-PCSK9 антитела или его антигенсвязывающих фрагментов в соответствии с настоящим изобретением последовательность FR тяжелой цепи вариабельной области тяжелой цепи гуманизированного антитела имеет происхождение от комбинированной последовательности тяжелых цепей зародышевой линии человека IGHV1-2*02 и hjh2 и ее мутантной последовательности; предпочтительно содержит FR1, FR2, FR3 тяжелой цепи зародышевой линии человека IGHV1-2*02 и FR4 hjh2 и ее мутантной последовательности или аминокислотную последовательность, обладающую по меньшей мере 95% идентичностью ее последовательностям.
В другом предпочтительном воплощении анти-PCSK9 антитела или его антигенсвязывающих фрагментов в соответствии с настоящим изобретением гуманизированное антитело содержит вариабельную область тяжелой цепи, как показано в SEQ ID NO 18, или вариабельную область тяжелой цепи, как показано в варианте SEQ ID NO 18; где вариант SEQ ID NO 18 представляет собой последовательность с 0-10 заменами аминокислот в вариабельной области тяжелой цепи, как показано в SEQ ID NO 18. Замена в аминокислотной последовательности может быть получена на основе существующей в данной области техники технологии для увеличения аффинности или периода полувыведения, например, модификации аминокислоты CDR с использованием метода созревания аффинности или модификации аминокислоты FR с использованием метода обратных мутаций.
В другом предпочтительном воплощении анти-PCSK9 антитела или его антигенсвязывающих фрагментов в соответствии с настоящим изобретением последовательность FR тяжелой цепи гуманизированного антитела имеет 0-10 обратных мутаций аминокислот, предпочтительно одну или более обратных мутаций, выбранных из группы, состоящей из T30N, R87T, R72A, T74K, M48I, V68A, M70L, R38K и R67K.
В другом предпочтительном воплощении анти-PCSK9 антитела или его антигенсвязывающих фрагментов в соответствии с настоящим изобретением гуманизированное антитело содержит вариабельную область тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO 19, SEQ ID NO 20, SEQ ID NO 21, SEQ ID NO 22 и SEQ ID NO 23.
В другом предпочтительном воплощении анти-PCSK9 антитела или его антигенсвязывающих фрагментов в соответствии с настоящим изобретением последовательность FR легкой цепи вариабельной области легкой цепи гуманизированного антитела имеет происхождение от комбинированной последовательности легких цепей зародышевой линии человека IGKV1-39*01 и hjk2.1 и ее мутантной последовательности; содержит FR1, FR2, FR3 IGKV1-39*01 и FR4 hjk2.1 и ее мутантной последовательности или аминокислотную последовательность, обладающую по меньшей мере 95% идентичностью ее последовательностям.
В другом предпочтительном воплощении анти-PCSK9 антитела или его антигенсвязывающих фрагментов в соответствии с настоящим изобретением гуманизированное антитело содержит вариабельную область легкой цепи, как показано в SEQ ID NO 24, или вариабельную область легкой цепи, как показано в варианте SEQ ID NO 24; вариант SEQ ID NO 24 имеет 1-10 замен аминокислот в вариабельной области легкой цепи, как показано в SEQ ID NO 24. Замены в аминокислотной последовательности могут быть получены на основании существующей в данной области техники технологии для увеличения аффинности или периода полувыведения, например, модификации аминокислоты CDR с использованием метода созревания аффинности или модификации аминокислоты FR с использованием метода обратных мутаций.
В другом предпочтительном воплощении анти-PCSK9 антитела или его антигенсвязывающих фрагментов в соответствии с настоящим изобретением вариант SEQ ID NO 24 имеет 0-10 обратных мутаций аминокислот в последовательности FR вариабельной области легкой цепи, как показано в SEQ ID NO 24; предпочтительно обратная мутация выбрана из группы, состоящей из T5S, S66D, Q3V и A49S; предпочтительно A49S.
В другом предпочтительном воплощении анти-PCSK9 антитела или его антигенсвязывающих фрагментов в соответствии с настоящим изобретением гуманизированное антитело содержит вариабельную область легкой цепи, выбранную из группы, состоящей из SEQ ID NO 25, SEQ ID NO 26 и SEQ ID NO 27.
В другом предпочтительном воплощении анти-PCSK9 антитела или его антигенсвязывающих фрагментов в соответствии с настоящим изобретением гуманизированное антитело содержит вариабельную область тяжелой цепи и/или вариабельную область легкой цепи, где вариабельная область тяжелой цепи выбрана из группы, состоящей из SEQ ID NO 19, SEQ ID NO 20, SEQ ID NO 21, SEQ ID NO 22 и SEQ ID NO 23, или вариабельная область тяжелой цепи выбрана из группы, состоящей из последовательностей, обладающих по меньшей мере 95% идентичностью с SEQ ID NO 19, SEQ ID NO 20, SEQ ID NO 21, SEQ ID NO 22 и SEQ ID NO 23; вариабельная область легкой цепи выбрана из группы, состоящей из SEQ ID NO 25, SEQ ID NO 26 и SEQ ID NO 27, или вариабельная область легкой цепи выбрана из группы, состоящей из последовательностей, обладающих по меньшей мере 95% идентичностью с SEQ ID NO 25, SEQ ID NO 26 и SEQ ID NO 27.
В другом предпочтительном воплощении анти-PCSK9 антитела или его антигенсвязывающих фрагментов в соответствии с настоящим изобретением анти-PCSK9 антитело содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, выбранные из группы, состоящей из:
1) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 18 и вариабельной области легкой цепи с последовательностью SEQ ID NO 25;
2) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 18 и вариабельной области легкой цепи с последовательностью SEQ ID NO 26;
3) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 18 и вариабельной области легкой цепи с последовательностью SEQ ID NO 27;
4) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 19 и вариабельной области легкой цепи с последовательностью SEQ ID NO 24;
5) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 19 и вариабельной области легкой цепи с последовательностью SEQ ID NO 25;
6) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 19 и вариабельной области легкой цепи с последовательностью SEQ ID NO 26;
7) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 19 и вариабельной области легкой цепи с последовательностью SEQ ID NO 27;
8) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 20 и вариабельной области легкой цепи с последовательностью SEQ ID NO 24;
9) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 20 и вариабельной области легкой цепи с последовательностью SEQ ID NO 25;
10) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 20 и вариабельной области легкой цепи с последовательностью SEQ ID NO 26;
11) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 20 и вариабельной области легкой цепи с последовательностью SEQ ID NO 27;
12) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 21 и вариабельной области легкой цепи с последовательностью SEQ ID NO 24;
13) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 21 и вариабельной области легкой цепи с последовательностью SEQ ID NO 25;
14) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 21 и вариабельной области легкой цепи с последовательностью SEQ ID NO 26;
15) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 21 и вариабельной области легкой цепи с последовательностью SEQ ID NO 27;
16) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 22 и вариабельной области легкой цепи с последовательностью SEQ ID NO 24;
17) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 22 и вариабельной области легкой цепи с последовательностью SEQ ID NO 25;
18) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 22 и вариабельной области легкой цепи с последовательностью SEQ ID NO 26;
19) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 22 и вариабельной области легкой цепи с последовательностью SEQ ID NO 27;
20) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 23 и вариабельной области легкой цепи с последовательностью SEQ ID NO 24;
21) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 23 и вариабельной области легкой цепи с последовательностью SEQ ID NO 25;
22) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 23 и вариабельной области легкой цепи с последовательностью SEQ ID NO 26;
23) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 23 и вариабельной области легкой цепи с последовательностью SEQ ID NO 27; и
24) вариабельной области тяжелой цепи с последовательностью SEQ ID NO 18 и вариабельной области легкой цепи с последовательностью SEQ ID NO 24.
В другом предпочтительном воплощении анти-PCSK9 антитела или его антигенсвязывающих фрагментов в соответствии с настоящим изобретением тяжелая цепь анти-PCSK9 антитела дополнительно содержит константные области тяжелой цепи, имеющие происхождение от IgG1, IgG2, IgG3 или IgG4 человека или его варианта, либо аминокислотной последовательности, обладающей 95% идентичностью с их последовательностями; предпочтительно содержит константные области тяжелой цепи, имеющие происхождение от IgG1, IgG2 или IgG4 человека или содержит константные области тяжелой цепи вариантов IgG1, IgG2 или IgG4, продлевающих период полувыведения антитела из сыворотки крови за счет мутации аминокислоты, наиболее предпочтительно содержит константные области тяжелой цепи IgG1, IgG2 или IgG4, в которые введена мутация YTE;
Легкая цепь анти-PCSK9 антитела дополнительно содержит константную область, имеющую происхождение от цепи к человека, цепи λ человека или аминокислотной последовательности, обладающей по меньшей мере 95% идентичностью их последовательностям.
В другом предпочтительном воплощении анти-PCSK9 антитела или его антигенсвязывающих фрагментов в соответствии с настоящим изобретением гуманизированное антитело содержит тяжелую цепь и легкую цепь, выбранные из группы, состоящей из:
1) тяжелой цепи с последовательностью SEQ ID NO 28 и легкой цепи с последовательностью SEQ ID NO 30; и
2) тяжелой цепи с последовательностью SEQ ID NO 32 и легкой цепи с последовательностью SEQ ID NO 30.
В настоящем изобретении дополнительно предложена фармацевтическая композиция, содержащая терапевтически эффективную дозу анти-PCSK9 антитела или его антигенсвязывающих фрагментов в соответствии с изобретением и одно или более из фармацевтически приемлемых носителей, разбавителей или вспомогательных веществ.
В настоящем изобретении дополнительно предложена молекула ДНК, кодирующая обсуждаемые выше анти-PCSK9 антитело или его антигенсвязывающие фрагменты.
В настоящем изобретении дополнительно предложен вектор, содержащий молекулу ДНК, как описано выше.
В настоящем изобретении дополнительно предложена клетка-хозяин, трансформированная экспрессионным вектором, как описано выше, которая выбрана из группы, состоящей из прокариотических клеток и эукариотических клеток, предпочтительно эукариотических клеток, более предпочтительно клеток млекопитающих.
В настоящем изобретении дополнительно предложено применение анти-PCSK9 антитела или его антигенсвязывающих фрагментов или фармацевтической композиции в соответствии с изобретением в получении лекарственного средства для лечения PCSK9-опосредованного заболевания или расстройства, где заболевание или расстройство предпочтительно представляет собой заболевание, связанные с повышенным уровнем холестерина (включая «заболевания, связанные с повышенным уровнем сывороточного холестерина»); более предпочтительно заболевание или расстройство выбрано из группы, состоящей из гиперхолестеринемии, заболевания сердца, метаболического синдрома, сахарного диабета, ишемической болезни сердца, инсультов, сердечно-сосудистого заболевания, болезни Альцгеймера и общей дислипидемии; наиболее предпочтительно гиперхолестеринемии, дислипидемии, атеросклероза, сердечнососудистого заболевания (ССЗ) или ишемической болезни сердца.
Примеры заболеваний, которые могут быть диагностированы с помощью антитела в соответствии с настоящим изобретением, включают заболевания, связанные с повышенным уровнем холестерина (включая «заболевания, связанные с повышенным уровнем сывороточного холестерина»), которые включают одно или более выбранных из гиперхолестеринемии, заболевания сердца, метаболического синдрома, сахарного диабета, ишемической болезни сердца, инсультов, сердечнососудистого заболевания, болезни Альцгеймера и общей дислипидемии (которая характеризуется повышенным уровнем общего сывороточного холестерина, повышенным уровнем ЛПНП, повышенным уровнем триглицеридов, повышенным уровнем липопротеина очень низкой плотности (ЛПОНП) и/или сниженным уровнем ЛПВП).
С одной стороны, в настоящем изобретении предложен способ лечения или профилактики гиперлипидемии и/или по меньшей мере одного симптома, выбранного из дислипидемии, атеросклероза, сердечно-сосудистого заболевания (ССЗ) и ишемической болезни сердца, где способ включает введение индивиду эффективного количества анти-PCSK9 антитела. В настоящем изобретении также предложено применение эффективного количества анти-PCSK9 антитела к внеклеточному или циркулирующему PCSK9 в получении лекарственного средства, которое предназначено для лечения или профилактики гиперхолестеринемии и/или по меньшей мере одного симптома, выбранного из дислипидемии, атеросклероза, ССЗ или ишемической болезни сердца.
Краткое описание графических материалов
Фиг. 1: Схема дизайна праймеров для конструирования векторов антител в соответствии с настоящим изобретением.
Фиг. 2: Схема конструирования векторов антител в соответствии с настоящим изобретением.
Фиг. 3: Изменение захвата ЛПНП клетками HepG2 при различных концентрациях антитела h001-4-YTE PCSK9. Эти результаты показывают, что анти-PCSK9 антитело может стимулировать захват ЛПНП клетками HepG2.
Фиг. 4: Изменение захвата ЛПНП клетками HepG2 при различных концентрациях антитела h001-4-WT PCSK9. Эти результаты показывают, что анти-PCSK9 антитело может стимулировать захват ЛПНП клетками HepG2.
Фиг. 5: Изменение сывороточных концентраций ЛПНП-х со временем у мышей после инъекции антитела h001-4-WT PCSK9 (*: р<0,05 по сравнению с IgG, **: р<0,01 по сравнению с IgG). Эти результаты показывают, что анти-PCSK9 антитело может вызывать снижение сывороточной концентрации ЛПНП-х у PCSK9-гиперэкспрессирующих мышей.
Фиг. 6: Изменение сывороточных концентраций ЛПНП-х у мышей, которым вводили путем инъекции антитело h001-4-WT PCSK9, относительно группы, которой вводили IgG. Эти результаты показывают, что по сравнению с группой, которой вводили IgG, анти-PCSK9 антитело может вызывать снижение сывороточной концентрации ЛПНП-х у PCSK9-гиперэкспрессирующих мышей.
Фиг. 7: Фармакодинамическое и фармакокинетическое тестирование антитела в соответствии с настоящим изобретением in vivo у яванских макак. На фигуре показано, что оба антитела h001-4-WT и h001-4-YTE могут вызывать статистически значимое снижение содержания ЛПВП у яванских макак, и продолжительность снижения, вызванного h001-4-YTE, лучше по сравнению с вызванным h001-4-WT.
Подробное описание изобретения
1. Термины
Чтобы обеспечить лучшее понимание изобретения, некоторые технические и научные термины конкретно определены ниже. Если в данном документе где-либо не определено иное, все остальные используемые в настоящем документе технические и научные термины имеют значение, обычно понимаемое специалистом средней квалификации в области техники, к которой принадлежит данное изобретение.
Используемые в настоящем документе однобуквенный код и трехбуквенный код для аминокислот являются такими, как описано в J. Biol. Chem, 243, (1968) p. 3558.
При использовании в настоящем документе «антитело» относится к иммуноглобулину, имеющему структуру из четырех пептидных цепей, соединенных вместе дисульфидными связями между двумя идентичными тяжелыми цепями и двумя идентичными легкими цепями. Константные области тяжелых цепей различных иммуноглобулинов проявляют различный аминокислотный состав и порядковое расположение, поэтому они обладают различными видами антигенности. Соответственно, иммуноглобулины можно разделить на пять категорий, или так называемых изотипов, а именно IgM, IgD, IgG, IgA и IgE, при этом соответствующие тяжелые цепи представляют собой μ цепь, δ цепь, γ цепь, α цепь и ε цепь соответственно. В соответствии с аминокислотным составом его шарнирной области и количеством и локализацией дисульфидных связей тяжелой цепи один и тот же тип Ig может быть разделен на различные подтипы, например, IgG может быть разделен на IgG1, IgG2, IgG3 и IgG4. Легкая цепь может быть разделена на цепи κ или λ в соответствии с различными константными областями. Каждый из пяти типов IgG может иметь κ или λ цепь.
В настоящем изобретении упомянутая в настоящем документе легкая цепь антитела дополнительно к вариабельной области содержит константную область легкой цепи, которая содержит κ, λ цепь человека или мыши или ее вариант.
В настоящем изобретении упомянутая в настоящем документе тяжелая цепь антитела дополнительно к вариабельной области содержит константную область тяжелой цепи, которая содержит IgG1, 2, 3, 4 человека или мыши или его вариант.
Вблизи N-конца тяжелой и легкой цепей около 110 аминокислот, известных как вариабельная область (Fv область), значительно изменяются; остальные аминокислоты вблизи С-конца, известные как константная область (С область), относительно стабильны. Вариабельная область содержит три гипервариабельные области (HVR) и четыре относительно консервативных каркасных области (FR). Эти три гипервариабельные области, также известные как определяющие комплементарность области (CDR), определяют специфичность антитела. Каждая вариабельная область легкой цепи (LCVR) и каждая вариабельная область тяжелой цепи (HCVR) состоит из трех CDR областей и четырех FR областей, которые располагаются в следующем последовательном порядке от амино-конца к карбокси-концу: FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Три CDR легкой цепи обозначают как LCDR1, LCDR2 и LCDR3; три CDR тяжелой цепи обозначают как HCDR1, HCDR2 и HCDR3. Количество и локализация аминокислотных остатков CDR областей в LCVR и HCVR областях антитела или его антигенсвязывающих фрагментов в данном случае удовлетворяют известным критериям номенклатуры Кэбота (LCDR1-3, HCDE2-3) или удовлетворяют критериям номенклатуры Кэбота и Чотиа (HCDR1).
Антитело по настоящему изобретению включает в себя мышиное антитело, химерное антитело и гуманизированное антитело, предпочтительно гуманизированное антитело.
Термин «мышиное антитело» в настоящем изобретении относится к моноклональному антителу к PCSK9 человека, которое получают в соответствии со знаниями и навыками в данной области техники. В процессе его получения в исследуемый объект вводили PCSK9 антиген, а затем отбирали гибридому, экспрессирующую антитело, которое обладает желаемыми последовательностями или функциональными характеристиками. В предпочтительном воплощении настоящего изобретения мышиное анти-PCSK9 антитело или его антигенсвязывающие фрагменты дополнительно содержат константную область легкой цепи κ, λ мыши или ее вариант или дополнительно содержат константную область тяжелой цепи IgG1, IgG2, IgG3 или IgG4 мыши или ее вариант.
Термин «химерное антитело» представляет собой антитело, образованное путем слияния вариабельной области мышиного антитела с константной областью человеческого антитела; химерное антитело может ослаблять индуцированный мышиным антителом иммунный ответ. Для создания химерного антитела сначала создают гибридому, секретирующую специфичное мышиное моноклональное антитело, клонируют ген вариабельной области из клеток мышиной гибридомы, затем по мере необходимости клонируют ген константной области человеческого антитела, ген мышиной вариабельной области лигируют с геном человеческой константной области с образованием химерного гена, который может быть встроен в человеческий вектор, и, наконец, молекулу химерного антитела экспрессируют в эукариотической или прокариотической системе производства. В предпочтительном воплощении настоящего изобретения легкая цепь химерного анти-PCSK9 антитела дополнительно содержит Fc области легкой цепи κ, λ человека или ее варианта. Тяжелая цепь химерного анти-PCSK9 антитела дополнительно содержит Fc области тяжелой цепи IgG1, IgG2, IgG3 или IgG4 человека или ее варианта, предпочтительно содержит константную область тяжелой цепи IgG1, IgG2, IgG3 или IgG4 человека или предпочтительно содержит константную область тяжелой цепи вариантов IgG1, IgG2 или IgG4 с мутациями аминокислот (например, с мутациями YTE) для продления периода полувыведения антитела из сыворотки.
Термин «гуманизированное антитело», также известное как CDR-трансплантированное антитело, относится к антителу, полученному путем трансплантации мышиных CDR последовательностей в каркас вариабельной области человеческого антитела, а именно в каркасную последовательность человеческого антитела зародышевой линии различного типа. Гуманизированное антитело позволяет преодолеть недостатки, связанные с сильным ответом антител, вызванным химерным антителом, которое несет большое количество компонентов мышиных белков. Такие каркасные последовательности могут быть получены из общедоступной базы данных ДНК, охватывающей последовательности генов антител зародышевой линии, или из источников опубликованных данных. Например, последовательности ДНК генов вариабельной области тяжелой и легкой цепи зародышевой линии человека можно найти в базе данных VBase последовательностей зародышевой линии человека (доступной на веб-сайте www.mrccpe.com.ac.uk/vbase), а также найти в кн. Kabat, ЕА, et al., 1991 Sequences of Proteins of Immunological Interest, 5-е изд. Во избежание снижения активности при снижении иммуногенности каркасные последовательности в вариабельной области человеческого антитела подвергают минимальным обратным мутациям, чтобы сохранить активность. Гуманизированное антитело по настоящему изобретению также включает в себя гуманизированное антитело, в котором созревание аффинности CDR осуществляют методом фагового дисплея. В предпочтительном воплощении настоящего изобретения мышиные CDR последовательности PCSK9 гуманизированных антител выбраны из группы, состоящей из SEQ ID NO 12, 13, 14, 15, 16 и 17; каркас вариабельной области человеческого антитела конструируют путем выбора FR последовательности вариабельной области легкой цепи антитела, имеющей происхождение от комбинированной последовательности легких цепей зародышевой линии человека IGKV1-39*01 и hjk2.1; при этом FR последовательность вариабельной области тяжелой цепи антитела имеет происхождение от комбинированной последовательности тяжелых цепей зародышевой линии человека IGHV1-2*02 и hjh2. Во избежание снижения активности, вызванного снижением иммуногенности, вариабельную область описанного в настоящем документе человеческого антитела можно подвергать минимальным обратным мутациям, чтобы сохранить активность антитела.
«Антигенсвязывающий фрагмент» в настоящем изобретении относится к Fab фрагменту, Fab' фрагменту, F(ab')2 фрагменту, обладающему антигенсвязывающей активностью, а также к Fv фрагменту scFv фрагмента, связывающемуся с PCSK9 человека; он содержит одну или более CDR областей антител, описанных в настоящем изобретении, выбранных из группы, состоящей из SEQ ID NO 12 - SEQ ID NO 17. Fv фрагмент содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи без константной области и представляет собой минимальный фрагмент антитела, обладающий всеми антигенсвязывающими участками. Как правило, Fv антитело дополнительно содержит полипептидный линкер между VH и VL доменами и способно к образованию структуры, необходимой для связывания антигена. Различные линкеры можно также использовать для соединения вариабельных областей двух антител с образованием полипептидной цепи, называемой одноцепочечным антителом или одноцепочечным Fv (scFv). Термин «связывание с PCSK9» в данном изобретении означает способность к взаимодействию с PCSK9 человека. Термин «антигенсвязывающие участки» в настоящем изобретении относится к трехмерным, не являющимся непрерывными участкам на антигене, которые распознаются антителом или антигенсвязывающим фрагментом по настоящему изобретению.
Термин «Fc область» в настоящем документе используют для определения С-концевого участка тяжелой цепи иммуноглобулина, который содержит по меньшей мере часть константной области. Этот термин включает Fc участки нативной последовательности и варианты участков Fc. В одном воплощении изобретения Fc область тяжелой цепи IgG человека проходит от Cys226 или от Pro230 до карбоксильного конца тяжелой цепи. Однако С-концевой лизин (Lys447) Fc области может присутствовать или отсутствовать. Если в настоящем документе не указано иное, номенклатура аминокислотных остатков в Fc области или константной области соответствует номенклатуре ЕС, также называемой индексом ЕС, как описано в Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991. Fc область существенна для эффекторных функций антител. Эффекторные функции включают инициацию комплементзависимой цитотоксичности (CDC, от англ. complement-dependent cytotoxicity), инициацию фагоцитоза и антителозависимой клеточно-опосредованной цитотоксичности (ADCC, от англ. antibody-dependent cell-mediated cytotoxicity) и перенесения антител через клеточные барьеры посредством трансцитоза. Кроме того, Fc область играет критическую роль для поддержания периода полувыведения антитела класса IgG из сыворотки крови (Ward and Ghetie, Ther. Immunol. 2: 77-94 (1995)). Исследователи обнаружили, что период полувыведения антитела IgG из сыворотки крови опосредован связыванием Fc с неонатальным рецептором Fc (FcRn). FcRn представляет собой гетеродимер, состоящий из трансмембранной цепи α и растворимой цепи β (β2-микроглобулин). В патенте США №6,165,745 раскрыт способ получения антитела со сниженным биологическим периодом полувыведения путем введения мутации в кодирующий антитело сегмент ДНК. Эта мутация включает замены аминокислот в положении 253, 310, 311, 433 или 434 Fc-шарнирного домена. В патенте США №6,277,375 BI раскрыта композиция, содержащая мутантную молекулу IgG, обладающую увеличенным периодом полувыведения из сыворотки крови по сравнению с IgG дикого типа, где мутантная молекула IgG содержит следующие замены аминокислот: треонин на лейцин в положении 252, треонин на серин в положении 254 или треонин на фенилаланин в положении 256 (M252Y, S254T и Т256Е). Также раскрыт мутант IgG с заменами аминокислот в положении 433, 435 или 436. В патенте США №6,528,624 раскрыт вариант антитела, содержащий Fc область IgG, где вариант включает в себя замены аминокислот в одном или более положений Fc области IgG человека (положения 270, 322, 326, 327, 329, 331, 333 и 334). В WO 02/060919 А2 раскрыт модифицированный IgG, содержащий константный домен IgG, включающий в себя одну или более модификаций аминокислот относительно константного домена IgG дикого типа, где модифицированный IgG обладает увеличенным периодом полувыведения по сравнению с периодом полувыведения IgG, имеющего константный домен IgG дикого типа, и где одна или более модификаций аминокислот в одном или более положений выбраны из группы, состоящей из положений 251, 253, 255, 285-290, 308-314, 385-389 и 428-435. Конкретно раскрытая в настоящем документе «мутация YTE или YET» относится к комбинации мутаций в Fc областях IgG I для стимуляции связывания между Fc областью и FcRn человека, приводящего к продлению периода полувыведения антитела у человека. Мутант YTE содержит комбинацию трех «мутаций YTE» M252Y, S254T и Т256Е. Номенклатура остатков основана на номенклатуре системы ЕС, которую также называют индексом ЕС, такой как номенклатура тяжелых цепей IgG по Кэботу и соавт. (см. патент США №7,658,921). По сравнению с антителами дикого типа мутанты YTE антител значительно продлевают период полувыведения антител из сыворотки крови, см., например, Dall'Acqua et al., J. Biol. Chem. 281: 23514-24 (2006) и патент США №7,083,784.
Способы получения и очистки антител и антигенсвязывающих фрагментов хорошо известны в данной области техники, и их можно найти, например, в Antibody Experimental Technology Guide of Cold Spring Harbor, главы 5-8 и 15. Например, мышей можно иммунизировать PCSK9 человека или его фрагментами, а затем полученные в результате антитела подвергать ренатурации, очистке и секвенированию, используя традиционные способы, хорошо известные в данной области техники. Антигенсвязывающие фрагменты могут быть также получены традиционными способами. Антитело или антигенсвязывающие фрагменты по настоящему изобретению конструируют методами генной инженерии с введением одной или более каркасных областей (FR) человека в CDR не человеческого происхождения. Последовательности FR зародышевой линии человека могут быть получены из баз данных ImMunoGeneTics (IMGT) посредством веб-сайта http://imgt.cines.fr или The Immunoglobulin FactsBook, 2001ISBN012441351.
Сконструированные антитело или антигенсвязывающие фрагменты по настоящему изобретению могут быть получены и очищены с использованием традиционных способов. Например, последовательности кДНК, кодирующие тяжелую цепь (SEQ ID NO 28) и легкую цепь (SEQ ID NO 30), можно клонировать и рекомбинировать в экспрессионном векторе GS. Впоследствии рекомбинированный экспрессионный вектор для иммуноглобулина можно стабильно трансфицировать в клетки яичника китайского хомячка (СНО). В результате использования экспрессионных систем млекопитающих как чаще всего рекомендуемого способа, хорошо известного в данной области техники, гликозилирование антител в характерном случае получают в высоко консервативном N-конце Fc области. Стабильные клоны могут быть получены посредством экспрессии антитела, специфично связывающегося с PCSK9 человека. Положительные клоны можно размножать в культуральной среде без сыворотки для продукции антител в биореакторах. Культуральную среду, в которой секретируется антитело, можно очищать традиционными методами. Например, удобно наносить среду на колонку с белком А или G или с FF-сефарозой, уравновешенную отрегулированным буферным раствором. Колонку промывают, чтобы удалить неспецифично связанные компоненты. Связанное антитело элюируют градиентом рН, и фрагменты антитела определяют с помощью электрофореза в полиакриламидном геле с додецилсульфатом натрия (ДСН-ПААГ) и впоследствии объединяют. Антитело можно фильтровать и концентрировать общепринятыми методами. Растворимые агрегаты и мультимеры можно эффективно удалять общепринятыми методами, включая эксклюзионные или ионообменные. Полученный продукт можно немедленно замораживать, например при -70°С, или лиофилизировать.
«Введение» и «воздействие» применительно к животному, человеку, экспериментальному объекту, клетке, ткани, органу или биологической текучей среде относится к приведению в контакт экзогенного фармацевтического, терапевтического, диагностического агента или композиции с животным, человеком, объектом, клеткой, тканью, органом или биологической текучей средой. «Введение» и «воздействие» могут относиться, например, к терапевтическим, фармакокинетическим, диагностическим, исследовательским и экспериментальным способам. Воздействие на клетку охватывает приведение реактива в контакт с клеткой, а также приведение реактива в контакт с биологической текучей средой, где текучая среда находится в контакте с клеткой. «Введение» и «воздействие» также означают воздействия in vitro и ex vivo, например воздействие на клетку реактива, диагностического, связывающего соединения или другой клетки. «Воздействие» применительно к человеку, ветеринарному или исследовательскому объекту относится к терапевтическому воздействию, профилактическим или превентивным мерам, к применению для исследования и диагностики.
«Лечить» означает «вводить терапевтический агент», такой как композиция, содержащая любое из связывающих соединений по настоящему изобретению, внутрь или снаружи, пациенту, пациенту, имеющему один или более симптомов, для которых этот агент обладает известной терапевтической активностью. В характерном случае агент вводят в количестве, эффективном для ослабления одного или более симптомов заболевания у подлежащего лечению пациента или популяции так, чтобы вызвать регрессию или ингибировать прогрессирование такого(-их) симптома(-ов) до какой-либо клинически измеримой степени. Количество терапевтического агента, эффективное для ослабления любого конкретного симптома заболевания (также называемое «терапевтически эффективным количеством»), может изменяться в соответствии с факторами, такими как болезненное состояние, возраст и масса тела пациента, и способностью лекарственного средства вызывать желаемый ответ у пациента. Наличие или отсутствие ослабления симптома заболевания можно оценивать с помощью любого клинического измерения, обычно применяемого врачами или другими квалифицированными медицинскими работниками для оценки тяжести или статуса прогрессирования этого симптома. Хотя воплощение настоящего изобретения (например, способ лечения или изделие производства) может не обладать эффективностью при ослаблении интересующего(-их) симптома(-ов) заболевания у каждого пациента, оно должно приводить к ослаблению интересующего(-их) симптома(-ов) заболевания у статистически значимого количества пациентов, которое определяют с помощью любого известного в данной области техники статистического критерия, такого как t-критерий Стьюдента, критерий хи-квадрат, U-критерий Манна и Уитни, критерий Краскела-Уоллиса (Н-критерий), критерий Джонкхиера-Терпстра и критерий Уилкоксона.
«Консервативные модификации» или «консервативное замещение или замена» относятся к заменам аминокислот в белке другими аминокислотами, обладающими сходными характеристиками (например, зарядом, размером боковой цепи, гидрофобностью/гидрофильностью, конформацией и жесткостью основной цепи и т.д.) так, что можно часто получать эти изменения без изменения биологической активности белка. Специалисты в данной области техники признают, что, как правило, отдельная замена аминокислоты в несущественных областях полипептида по существу не изменяет биологическую активность (см., например, Watson et al. (1987) Molecular Biology of the Gene, The Benjamin/Cummings Pub. Co., p. 224 (4.sup.th Ed.)). Кроме того, замены структурно или функционально сходных аминокислот с меньшей вероятностью нарушают биологическую активность.
«Эффективное количество» охватывает количество, достаточное для ослабления или предотвращения симптома или проявления медицинского состояния. Эффективное количество также означает количество, достаточное для обеспечения возможности или облегчения диагностики. Эффективное количество для конкретного пациента или ветеринарного объекта может изменяться в зависимости от факторов, таких как подлежащее лечению состояние, общее состояние здоровья пациента, путь, доза введения и тяжесть побочных эффектов. Эффективное количество может представлять собой максимальную дозу или протокол дозирования, позволяющие избежать побочных эффектов или токсических эффектов.
«Экзогенный» относится к веществам, которые продуцируются вне организма, клетки или человеческого тела в зависимости от контекста. «Эндогенный» относится к веществам, которые продуцируются внутри клетки, организма или человеческого тела в зависимости от контекста.
«Гомология» относится к подобию последовательности между двумя полинуклеотидными последовательностями или между двумя полипептидами. Когда положение в обоих из двух сравниваемых последовательностей занято одним и тем же основанием или мономерной аминокислотной субъединицей, например, если положение в каждой из двух молекул ДНК занято аденином, в данном положении молекулы гомологичны. Процент гомологии между двумя последовательностями представляет собой функцию количества совпадающих или гомологичных положений, общих для двух последовательностей, деленного на количество сравниваемых положений, затем умноженного на 100. Например, если 6 из 10 положений в двух последовательностях совпадают или гомологичны при оптимальном выравнивании последовательностей, две последовательности гомологичны на 60%. Как правило, сравнение проводят при выравнивании двух последовательностей с получением максимального процента гомологии.
Используемые в настоящем документе выражения «клетка», «линия клеток» и «культура клеток» используют взаимозаменяемо, и все такие обозначения включают в себя потомство. Таким образом, слова «трансформанты» и «трансформированные клетки» включают в себя первичную рассматриваемую клетку и полученные из нее культуры без учета числа пересевов. Также понятно, что все потомство может не быть точно идентичным по содержанию ДНК вследствие повреждений или случайных мутаций. Включено мутантное потомство, обладающее такой же функцией или биологической активностью, которые определены на скрининге первоначально трансформированных клеток. Если предусмотрены отдельные обозначения, это должно быть четко видно из контекста.
При использовании в настоящем документе «полимеразная цепная реакция» или «ПЦР» относится к методике или методу, в которых минимальные количества определенного фрагмента нуклеиновой кислоты, РНК и/или ДНК, амплифицируют, как описано, например, в патенте США №4,683,195. Как правило, чтобы можно было сконструировать олигонуклеотидные праймеры, должна быть доступна информация о концах интересующей области; эти праймеры будут идентичны или подобны по последовательности соответствующим нитям амплифицируемой матрицы. 5'-концевые нуклеотиды двух праймеров могут быть идентичны концам амплифицируемого материала. ПЦР можно использовать для амплификации определенных последовательностей РНК, определенных последовательностей ДНК из суммарной геномной ДНК и «ДНК, транскрибированной с суммарной клеточной РНК, последовательностей бактериофага или плазмиды и т.д. См. общую информацию в Mullis et al. (1987) Cold Spring Harbor Symp. Ouant. Biol. 51: 263; Erlich, ed., (1989) PCR TECHNOLOGY (Stockton Press, N.Y.). При использовании в настоящем документе ПЦР рассматривают как один, но не единственный пример метода полимеразной реакции нуклеиновой кислоты для амплификации нуклеиновой кислоты в исследуемом образце, включающий в себя использование известной нуклеиновой кислоты в качестве праймера и полимеразы нуклеиновой кислоты для амплификации или получения определенного фрагмента нуклеиновой кислоты.
«Необязательный» или «необязательно» означает, что следующие за ним событие или ситуация могут наступить, но не обязательно наступают, и описание включает в себя случаи, в которых это событие или обстоятельство наступает или не наступает. Например, «необязательно содержит 1-3 вариабельные области тяжелой цепи антитела» означает возможное, но необязательное присутствие вариабельной области тяжелой цепи антитела с определенной последовательностью.
«Фармацевтическая композиция» относится к композиции, содержащей смесь одного или более соединений в соответствии с настоящим изобретением или их физиологически/фармацевтически приемлемой соли или пролекарства с другими химическими компонентами, а также дополнительными компонентами, такими как физиологически/фармацевтически приемлемые носители и вспомогательные вещества. Фармацевтическая композиция нацелена на стимуляцию введения в организм, облегчение абсорбции активного ингредиента и, следовательно, проявление биологического эффекта.
Примеры и тесты
Далее настоящее изобретение дополнительно описано со ссылкой на примеры. Однако объем настоящего изобретения не ограничен ими. В примерах настоящего изобретения, где не указаны конкретные условия, эксперименты, как правило, проводят в традиционных условиях, как описано в Antibody Technology Laboratory Manual and Molecular Cloning Manual of Cold Spring Harbor, или в условиях, предложенных производителями материала или продукта. Если источник реактивов конкретно не указан, реактивы представляют собой традиционные имеющиеся в продаже реактивы.
Пример 1. Получение PCSK9 антигена и исследуемого белка
Конструирование и экспрессия белка
В качестве матрицы PCSK9 по изобретению для конструирования аминокислотных последовательностей антигена и исследуемого белка использовали пропротеинконвертазу субтилизин/кексин 9 типа из базы данных Uniprot (PCSK9 человека, номер Uniprot: Q8MBP7). Гибридные белки PCSK9 необязательно получали с различными метками, такими как гистидиновая (his) метка или пептид, стимулирующий иммунизацию, такой как пептид универсального HLA-DR-связывающего эпитопа (PADRE, от англ. Pan HLA-DR-binding Epitope), затем их клонировали в векторах рТТ5 (Biovector, № по каталогу 102762) или в векторах pTargeT (Promega, А1410) соответственно, транзиторно экспрессировали в клетках 293 или стабильно экспрессировали в клетках CHO-S и очищали. Наконец, получили антиген и исследуемый белок по изобретению.
PCSK9 с His-меткой: PCSK9-His6, используемый в качестве иммуногена для иммунизации мышей или используемый в качестве детектирующего реактива.

Примечание: Подчеркнутая последовательность представляет собой сигнальный пептид, а часть, выделенная курсивом, представляет собой последовательность His-метки (His6-метки).
PCSK9 с пептидом PADRE и His-меткой: PCSK9-PADRE-His6, используемый в качестве иммуногена, где содержащийся в нем пептид PADRE может стимулировать иммунизацию;

Примечание: Подчеркнутая последовательность представляет собой сигнальный пептид, дважды подчеркнутая последовательность представляет собой линкер, последовательность, подчеркнутая пунктирной линией, представляет собой пептид PADRE, а часть, выделенная курсивом, - His6-метку.
Гибридный белок PCSK9 с сайтом расщепления TEV и His-меткой: PCSK9-TEV-His6, N-PCSK9 (N-концевой домен PCSK9), используемый в качестве иммуногена, может быть получен с помощью фермента TEV;


Примечание: Подчеркнутая последовательность представляет собой сигнальный пептид, дважды подчеркнутая последовательность представляет собой сайт расщепления TEV, а часть, выделенная курсивом, представляет собой His6-метку.
Мутантный белок PCSK9-D374Y с His-меткой: PCSK9-D374Y-His6, используемый в качестве детектирующего реактива:

Примечание: Подчеркнутая последовательность представляет собой сигнальный пептид, а часть, выделенная курсивом, представляет собой His6-метку.
Белок PCSK9 со вставкой пептида-реципиента биотина ВР15 и His-метки: PCSK9-BP15-His6. В качестве детектирующего реактива биотиновая метка будет присоединена в положении пептида ВР15 в процессе экспрессии, что позволяет избежать мечения биотином in vitro и, следовательно, избежать возможных конформационных изменений.


Примечание: Подчеркнутая последовательность представляет собой сигнальный пептид, дважды подчеркнутая последовательность представляет собой пептид-реципиент биотина, а часть, выделенная курсивом, представляет собой His6-метку.
Мутантный белок PCSK9 D374Y со вставкой пептида-реципиента биотина ВР15 и His-метки: PCSK9-D374Y-BP15-His6, в качестве детектирующего белка:

Примечание: Подчеркнутая последовательность представляет собой сигнальный пептид, дважды подчеркнутая последовательность представляет собой пептид-реципиент биотина, а часть, выделенная курсивом, представляет собой His6-метку.
Внеклеточный домен белка-рецептора PCSK9 LDLR с меткой Flag и His-меткой: LDLR-ECD-Flag-His6 в качестве детектирующего реактива:

Примечание: Подчеркнутая последовательность представляет собой сигнальный пептид, дважды подчеркнутая последовательность представляет собой метку Flag, а часть, выделенная курсивом, представляет собой His6-метку.
LCDR-Fc, гибридный белок укороченного внеклеточного домена LDLR с hIgG1 Fc (с PCSK9-связывающей активностью): LDLR-sECD -Fc (hIgG1) в качестве детектирующего реактива:

Примечание: Подчеркнутая последовательность представляет собой сигнальный пептид, дважды подчеркнутая последовательность представляет собой укороченный внеклеточный домен LDLR с PCSK9-связывающей активностью (LDLR-sECD), а часть, выделенная курсивом, представляет собой hIgG1-Fc.
Гибридный белок более укороченного внеклеточного домена LDLR с hIgG1 Fc (с PCSK9-связывающей активностью): LDLR-ssECD-Fc (hIgG1) в качестве детектирующего реактива:

Примечание: Подчеркнутая последовательность представляет собой сигнальный пептид, дважды подчеркнутая последовательность представляет собой более укороченный внеклеточный домен LDLR с PCSK9-связывающей активностью (LDLR-ssECD), а часть, выделенная курсивом, представляет собой hIgG1-Fc.
Пример 2. Очищенный рекомбинантный белок PCSK9 и родственный LDLR рекомбинантный белок и очистка антитела гибридомы и рекомбинантного антитела
1. Стадии очистки рекомбинантных белков с His-меткой:
Образцы супернатантов экспрессирующих клеток центрифугировали методом высокоскоростного центрифугирования и удаляли примеси. Буферный раствор заменяли фосфатно-солевым буферным раствором (ФСБ) и добавляли имидазол до конечной концентрации 5 ммоль/л. Колонку с никелем уравновешивали раствором ФСБ, содержащим 5 ммоль/л имидазола, и промывали 2-5 объемами колонки. Образец супернатанта после замены буфера наносили на колонку IMAC. Колонку промывали раствором ФСБ, содержащим 5 ммоль/л имидазола, до тех пор, пока показание оптической плотности А280 не снижалось до исходного значения. Затем хроматографическую колонку промывали ФСБ с добавлением 10 ммоль/л имидазола для удаления неспецифично связанных белков и собирали поток, выходящий из колонки. Целевой белок элюировали раствором ФСБ, содержащим 300 ммоль/л имидазола, и собирали пик элюции. Собранный элюат концентрировали и дополнительно очищали гель-хроматографией (GE) Superdex 200, подвижная фаза которой представляла собой ФСБ. Пик мультимера удаляли и собирали пики элюции. Полученные белки идентифицировали методом электрофореза, пептидного картирования и жидкостной хроматографии с масс-спектрометрией (ЖХ-МС). PCSK9-His6 (SEQ ID NO 1), PCSK9-PADRE-His6 (SEQ ID NO 2), PCSK9-TEV-His6 (SEQ ID NO 3), PCSK9-D374Y-His6 (SEQ ID NO 4), PCSK9-BP15-His6 (SEQ ID NO 5) и PCSK9-D374Y-BP15-His6 (SEQ ID NO 6) были получены и использованы в качестве иммуногена и детектирующего реактива по изобретению. PCSK9-TEV-His6 очищали и расщепляли ферментом TEV, и фермент TEV, не полностью расщепленный PCSK9-TEV-His6 или фрагмент С-концевого домена с His-меткой удаляли из полученного продукта пропусканием через колонку IMAC. Выходящий продукт из колонки IMAC концентрировали, оставляя только N-концевой фрагмент домена PCSK9, и использовали его в качестве иммуногена для иммунизации мышей.
2. Стадии очистки рекомбинантного белка LDLR-ECD-Flag-His6 (SEQ ID NO 7) с His-меткой и Flag-меткой
Образцы центрифугировали методом высокоскоростного центрифугирования и удаляли примеси, а затем образцы концентрировали до надлежащего объема. Аффинную колонку Flag уравновешивали 0,5-кратным ФСБ и промывали 2-5 объемами колонки. После удаления примесей образцы супернатантов экспрессирующих клеток наносили на колонку. Колонку промывали 0,5-кратным раствором ФСБ до тех пор, пока показание оптической плотности А280 не снижалось до исходного значения. Колонку промывали ФСБ, содержащим 0,3 моль/л NaCl, и белки вымывали и собирали. Целевые белки элюировали раствором 0,1 моль/л уксусной кислоты (рН 3,5-4,0) и собирали, а затем значение рН доводили до нейтрального. Собранный элюат концентрировали и дополнительно очищали гель-хроматографией (GE) Superdex 200, подвижная фаза которой представляла собой ФСБ. Пик мультимера удаляли и собирали пики элюции. Полученные белки идентифицировали методом электрофореза, пептидного картирования и жидкостной хроматографии с масс-спектрометрией (ЖХ-МС). LDLR-ECD-Flag-His6 (SEQ ID NO 7) с метками FLAG/His6 были получены и использованы для экспериментального определения характеристик антитела по настоящему изобретению.
3. Стадии очистки гибридного белка LDLR Fc
Образцы супернатантов экспрессирующих клеток центрифугировали методом высокоскоростного центрифугирования и удаляли примеси, а затем образцы концентрировали до надлежащего объема и наносили на колонку с белком А. Колонку промывали ФСБ до тех пор, пока показание оптической плотности А280 не снижалось до исходного значения. Целевые белки элюировали раствором 100 ммоль/л ацетата натрия, рН 3,0, а затем нейтрализовали раствором 1 моль/л Трис-HCl. Элюированные образцы концентрировали надлежащим образом и дополнительно очищали гель-хроматографией на колонке Superdex 200 (GE), предварительно уравновешенной ФСБ. Собирали пики без мультимеров. Этот способ использовали для очистки LDLR-sECD-Fc (hIgG1) (SEQ ID NO 8) и LDLR-ssECD-Fc (hIgG1) (SEQ ID NO 9). Оба белка можно использовать для экспериментального определения характеристик антитела по настоящему изобретению.
Пример 3. Получение моноклональных антител к PCSK9 человека из гибридомы
1. Иммунизация
Моноклональное антитело к PCSK9 человека было получено путем иммунизации мышей. Самок экспериментальных белых мышей линии SJL в возрасте 6 недель (Beijing Weitong Lihua Experimental Animal Technology Co., Ltd., номер лицензии на разведение животных: SCXK (г. Пекин) 2012-0001). Условия окружающей среды для вскармливания: уровень «свободный от специфических патогенов» (SPF). После приобретения мышей животных содержали в лаборатории в течение 1 недели при цикле светлого/темного времени 12/12 часов, температуре 20-25°С, влажности 40-60%. Мышей, которые адаптировались к условиям окружающей среды, иммунизировали в соответствии с двумя описанными ниже схемами по 6-10 мышей на группу. Иммуноген представлял собой PCSK9-His6 (SEQ ID NO 1) человека с His-меткой, PCSK9-PADRE-His6 (SEQ ID NO 2) и N-PCSK9 (SEQ ID NO 3).
Схема А: получение эмульсии с адъювантом Фрейнда (Sigma, номер по каталогу: F5881/F5506): первая иммунизация с полным адъювантом Фрейнда (CFA, от англ. Complete Freund's adjuvant), бустер-иммунизация с неполным адъювантом Фрейнда (IFA, от англ. Incomplete Freund's adjuvant). Соотношение количеств антигена и адъюванта составляло 1:1, 100 мкг/мышь (для первой иммунизации), 50 мкг/мышь (для бустер-иммунизации). В день 0 мышам вводили путем интраперитонеальной (и/п) инъекции 100 мкг/мышь эмульсионных антигенов, после первой иммунизации один раз в две недели, в общей сложности в течение 6-8 недель.
Схема В: Мышей перекрестно иммунизировали с адъювантом Titermax (Sigma, номер по каталогу: Т2684) и квасцами (Alum) (Thremo, номер по каталогу: 77161). Соотношение количеств антигена и адъюванта (Titermax) составляло 1:1, и соотношение количеств антигена и адъюванта (Alum) составляло 3:1, 10-20 мкг/мышь (для первой иммунизации), 5 мкг/мышь (для бустер-иммунизации). В день 0 мышам вводили путем интраперитонеальной (и/п) инъекции 20/10 мкг/мышь эмульсионных антигенов, раз в неделю после первой иммунизации, поочередно используя Titermax и Alum, в общей сложности в течение 6-11 недель. Через четыре недели после иммунизации инъекцию антигена в область спины или интраперитонеальную инъекцию выбирали в зависимости от состояния отечности на спине и животе.
2. Слияние клеток
Для слияния спленоцитов выбирали мышей с высоким титром антител в сыворотке (см. тесты 1 и 2 в комбинации с твердофазным иммуноферментным анализом (ELISA, англ. enzyme-linked immunosorbent assay) для определения PCSK9) и с титром, склонным к выходу на плато. За 72 часа до слияния выбранных мышей иммунизировали 10 мкг/мышь PCSK9-His6 посредством интраперитонеальной инъекции. Проводили слияние лимфоцитов селезенки и клеток миеломы Sp2/0 (Американская коллекция типовых культур (АТСС®) CRL-8287™) с получением клеток гибридомы с помощью оптимизированной методики слияния, опосредованного полиэтиленгликолем (ПЭГ). Слитые клетки гибридомы ресуспендировали в полной гипоксантин-аминоптеринтимидиновой среде (ГАТ) (среда RPMI-1640, содержащая 20% фетальную бычью сыворотку (ФБС), 1-кратный раствор ГАТ и 1-кратный OPI), а затем добавляли в 96-луночный планшет для культур клеток (1×105/150 мкл/лунка) и инкубировали при 37°С и 5% CO2. В день 5 после слияния добавляли 50 мкл/лунка полной среды ГАТ, инкубировали при 37°С и 5% CO2. В день 7-день 8 после слияния в зависимости от роста клеток на основании их плотности среду полностью заменяли 200 мкг/лунка полной гипоксантин-тимидиновой (ГТ) среды (среда RPMI-1640, содержащая 20% ФБС, 1-кратный раствор ГТ и 1-кратный OPI) и инкубировали при 37°С и 5% CO2.
3. Скрининг клеток гибридомы
В день 10-день 11 после слияния в зависимости от роста клеток на основании их плотности проводили тесты ELISA по определению связывания PCSK9 или PCSK9-Y (см. тесты 1 и 2). Клетки, для которых был получен положительный результат в анализе связывания ELISA, использовали для тестирования блокирования связывания PCSK9 или PCSK9-Y с LDLR в анализе блокирования методом ELISA (см. тесты 3 и 4). Среду в положительных лунках заменяли, и клетки рассевали в 24-луночный планшет на основании плотности клеток. Линии клеток, пересеянные в 24-луночный планшет, консервировали, и после повторного тестирования проводили первое субклонирование. Клетки, для которых был получен положительный результат на скрининге после первого субклонирования (см. тесты 1 и 2), консервировали и подвергали второму субклонированию. Клетки, для которых был получен положительный результат на скрининге после второго субклонирования (см. тесты 1 и 2), консервировали для экспрессии белка. Клетки гибридомы, способные к блокированию связывания PCSK9 или PCSK9-Y с LDLR, были получены после многократного слияния.
Клон гибридомы mAb-001 был получен путем скрининга в соответствии с анализом блокирования и анализом связывания. Затем антитело было получено путем культивирования клеток в среде без сыворотки. Антитела очищали в соответствии с иллюстративными стадиями очистки и использовали в детектировании.
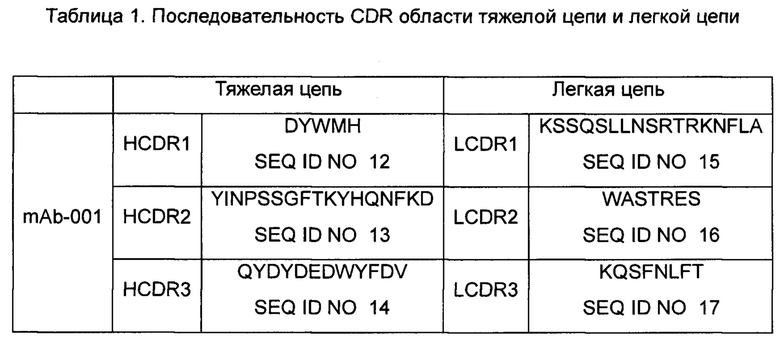
Последовательность вариабельной области мыши клона гибридомы mAb-001 приведена ниже:

Примечание: Порядок представляет собой FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4, часть, выделенная курсивом, представляет собой последовательность FR, а подчеркнутая часть - последовательность CDR.
Пример 4. Гуманизация моноклонального антитела к PCSK9 человека из гибридомы
1. Выбор гуманизированного каркаса для клона гибридомы mAb-001
Путем сравнения с генами вариабельной области тяжелой и легкой цепи зародышевой линии человека из базы данных IMGT с помощью программного обеспечения МОЕ были отобраны гены вариабельной области тяжелой и легкой цепи mAb-001 в качестве матриц. CDR этих двух мышиных антител соответственно трансплантировали на соответствующие матрицы человека с образованием последовательностей вариабельной области в следующем порядке: FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4. Аминокислотные остатки были пронумерованы и обозначены в соответствии с номенклатурой системы Кэбота.
Матрицами гуманизированной легкой цепи мышиного антитела mAb-001 были IGKV1-39*01 и hjk2.1, а матрицами гуманизированной тяжелой цепи были IGHV1-2*02 и hjh2. Последовательность вариабельной области гуманизированного антитела h001-1 после гуманизации показана ниже:


Примечание: Порядок представляет собой FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4, часть, выделенная курсивом, представляет собой последовательность FR, а подчеркнутая часть - последовательность CDR.
2. Выбор матрицы и дизайн обратных мутаций для клона гибридомы mAb-001 показаны в таблице 2. Комбинация гуманизированных последовательностей после обратной мутации гибридомы показана в таблице 2.
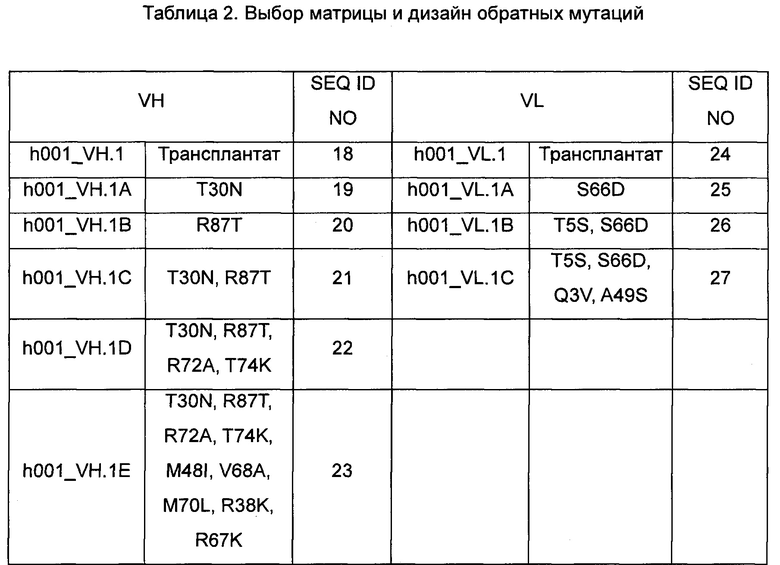
Примечание: Например, в соответствии с системой номенклатуры Кэбота S66D означает S в положении 66, подвергнутый обратной мутации с образованием D.
Трансплантаты представляют собой CDR мышиного антитела, которые были трансплантированы на последовательности каркасных областей FR человека, как показано в таблице 3.
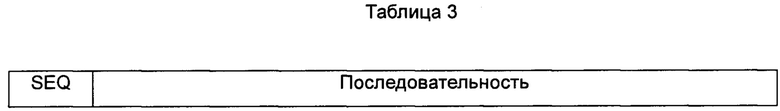

Примечание: Подчеркнутая часть представляет собой CDR область.


Примечание: В таблице показана комбинация вариабельных областей гуманизированного антитела, полученная путем комбинирования каждой последовательности и ее мутантной последовательности. Например, h001-1 показывает, что вариабельная область гуманизированного антитела h001-1 состоит из вариабельных областей легкой цепи h001_VL1 и тяжелой цепи h001_VH.1A. Другие получены аналогичным образом.
3. Приведенные выше гуманизированные последовательности комбинировали с образованием антитела, в котором константная область тяжелой цепи принадлежит IgG1 человека, а константная область легкой цепи - цепи каппа человека. Было приобретено соответствующее гуманизированное антитело и определено его связывание с PCSK9 методом ELISA (см. тест 1) и связывание с PCSK9-Y методом ELISA (см. тест 2). Клетки, для которых был получен положительный результат определения связывания описанным выше методом ELISA, подвергали дополнительному определению блокирования связывания PCSK9/LDLR в анализе блокирования методом ELISA (см. тест 4) и дополнительному определению блокирования связывания PCSK9-Y/LDLR в анализе блокирования методом ELISA (см. тест 3); результаты показаны в таблице 5-8.
Эти результаты показывают, что анти-PCSK9 антитела, полученные в изобретении, обладают высокой аффинностью связывания с PCSK9 и PCSK9-Y, и эти антитела могут также эффективно блокировать связывание PCSK9/PCSK9-Y с LDLR.
Пример 5. Конструирование и экспрессия гуманизированных антител к PCSK9 человека форматов IgG1 и IgG1-YTE
Способ конструирования и экспрессии гуманизированных антител к PCSK9 человека показан ниже:
1. Дизайн праймеров: Для синтеза VH/VK были сконструированы множественные праймеры, содержащие необходимые для рекомбинации фрагменты генов, с использованием программного обеспечения онлайн DNAWorks (v3.2.2, http://helixweb.nih.gov/dnaworks/): 5'-Сигнальный пептид 30 п.о. плюс VH/VK плюс CH1/CL 30 п.о.-3'. Принцип дизайна праймеров: если целевой ген 2 отличается от целевого гена 1 на 2 аминокислоты, конструировали следующий праймер, расположенный в участке мутации, как показано на Фиг. 1.
2. Сплайсинг фрагментов: в соответствии с методическими рекомендациями для ДНК полимеразы TakaRa Primer STAR GXL проводили двухстадийную амплификацию методом ПЦР с множественными праймерами, сконструированными, как описано выше, и получали содержащие VH/VK фрагменты генов, необходимые для рекомбинации.
3. Конструирование экспрессионного вектора pHr (с сигнальным пептидом и фрагментом гена константной области (CH1-FC/CL)) и расщепление ферментом рестрикции.
Экспрессионный вектор pHr (с сигнальным пептидом и фрагментом гена константной области (CH1-FC/CL)) был разработан и сконструирован с помощью некоторых специальных ферментов рестрикции, таких как BsmBI, которые распознают последовательность, отличающуюся от сайта расщепления ферментом, как показано на Фиг. 2. BsmBI использовали для разрезания вектора, фрагмент вырезали из геля и выделяли для использования.
4. Конструирование рекомбинантного экспрессионного вектора VH-CH1-FC-pHr/VK-CL-pHr.
Содержащие VH/VK фрагменты генов, необходимые для рекомбинации, и выделенный экспрессионный вектор pHr расщепляли ферментом BsmBI (с фрагментом гена сигнального пептида и константной области (CH1-FC/CL)), добавляли к компетентным клеткам DH5 альфа в соотношении 3:1, инкубировали на ледяной бане при 0°С в течение 30 мин, подвергали тепловому шоку в течение 90 секунд при 42°С и добавляли 5-кратный объем питательной среды Лурия-Бертани (LB), инкубировали при 37°С в течение 45 мин, высевали на чашку со средой LB с ампициллином (LB-Amp) и культивировали при 37°С в течение ночи. Отдельные клоны собирали и секвенировали.
Антитело по данному изобретению может быть сконструировано вышеописанным способом, но не ограниченным им. Например, дизайн антитела h001-4 и его варианта и их получение описаны ниже: 1) h001-4-WT: формат IgG h001-4, т.е. комбинация гуманизированной последовательности h001-4 с константной областью тяжелой цепи из IgG1 человека константной областью легкой цепи из цепи каппа человека; 2) h001-4-YTE: формат h001-4-IgG1-YTE, т.е. комбинация гуманизированной последовательности h001-4 с константной областью тяжелой цепи мутантного (мутация YTE) IgG1 человека и константной областью легкой цепи из цепи каппа человека. Мутантный IgG1 может иметь другие формы мутаций. Аффинность полученных антител и мутантных антител определяли с помощью анализа BIAcore (тест 6), и результаты показаны в таблице 9.
Последовательности сконструированных и экспрессированных гуманизированных антител к PCSK9 (его форматы IgG1 и IgG1-YTE) показаны ниже:
Формат Н001-4 IgG1, константная область тяжелой цепи получена из IgG1 человека, а константная область легкой цепи - из цепи каппа человека: Аминокислотная последовательность тяжелой цепи (IgG1 человека):

Последовательность ДНК тяжелой цепи:


h001-4-каппа
Аминокислотная последовательность легкой цепи:

Последовательность ДНК легкой цепи:

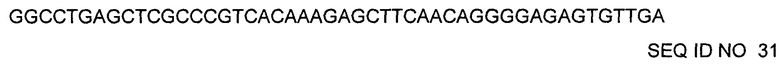
h001-4-IgG1-YTE (легкая цепь представляет собой h001-4-каппа: SEQ ID NO 30)
Аминокислотная последовательность тяжелой цепи: IgG1-YTE

Последовательность ДНК тяжелой цепи:

Примечание: Подчеркнутая часть представляет собой последовательность ДНК сигнального пептида.
Характеристики и преимущества настоящего изобретения подтверждают с помощью биохимических тестов, как показано ниже.
Тест 1. Анализ связывания анти-PCSK9 антител с белком PCSK9 дикого типа методом ELISA
Связывающую способность антител к PCSK9 по настоящему изобретению с PCSK9 определяли путем измерения количества антител, связывающихся с белком PCSK9 дикого типа (WT PCSK9, SEQ ID NO 5), фиксированном на планшете для ELISA.
Стрептавидин (Sigma, № по каталогу S4762) разводили до 2 мкг/мл ФСБ и наносили в виде покрытия в лунки 96-луночного планшета для ELISA при 4°С в течение ночи. Планшет промывали, а затем блокировали Трис-буфером (включающим 0,9 ммоль/л CaCl2, 0,05% Твин 20 и 5% обезжиренного молока) при 37°С в течение 2 часов. Затем планшет снова промывали и добавляли в лунки по 100 мкл/лунка меченого биотином PCSK9, полученного в лаборатории (bio-WT-PCSK9, разведенного Трис-буфером, содержащим 0,9 ммоль/л CaCl2, 0,05% Твин 20 и 1% обезжиренного молока), и инкубировали при 37°С в течение 1 часа. После промывания в лунки планшета вносили различные концентрации образцов анти-PCSK9 антитела и инкубировали при 37°С в течение 1 часа. Затем планшет снова промывали и добавляли козье античеловеческое антитело, меченое пероксидазой хрена (HRP) (H+L) (Jackson, № по каталогу 109-035-088) и инкубировали при 37°С в течение 1 часа. Затем планшет снова промывали и добавляли раствор тетраметилбензидина для проявления. Наконец, добавляли раствор для остановки реакции и измеряли показания OD450 на ридере для микропланшетов, а затем рассчитывали ЕС50.
Результаты анализа ELISA для связывающей способности химерных антител и антител, несущих обратные мутации, по настоящему изобретению с белком PCSK9 человека показаны в таблице 5.

Данные показывают, что гуманизированные антитела по настоящему изобретению обладают более высокой активностью связывания с белком PCSK9 человека.
Тест 2. ELISA анализ связывания анти-PCSK9 антител с белком PCSK9-Y
Связывающую способность антител к PCSK9 по настоящему изобретению с PCSK9-Y определяли путем измерения количества антител, связывающихся с белком PCSK9-Y (мутантный PCSK9, SEQ ID NO 6), фиксированном на планшете для ELISA.
Стрептавидин (Sigma № по каталогу S4762) разводили до 2 мкг/мл ФСБ и наносили в виде покрытия в лунки 96-луночного планшета для ELISA при 4°С в течение ночи. Планшет промывали, а затем блокировали Трис-буфером (включающим 0,9 ммоль/л CaCl2, 0,05% Твин 20 и 5% обезжиренного молока) при 37°С в течение 2 часов. Затем планшет снова промывали и добавляли в лунки по 100 мкл/лунка меченого биотином PCSK9-Y, полученного в лаборатории (bio-PCSK9-Y, разведенного Трис-буфером, содержащим 0,9 ммоль/л CaCl2, 0,05% Твин 20 и 1% обезжиренного молока), и инкубировали при 37°С в течение 1 часа. После промывания в лунки планшета вносили различные концентрации образцов анти-PCSK9 антитела и инкубировали при 37°С в течение 1 часа. Затем планшет снова промывали и добавляли козье античеловеческое антитело, меченое пероксидазой хрена (HRP) (H+L) (Jackson, № по каталогу 109-035-088) и инкубировали при 37°С в течение 1 часа. Затем планшет снова промывали и добавляли раствор тетраметилбензидина для проявления. Наконец, добавляли раствор для остановки реакции и измеряли показания OD450 на ридере для микропланшетов, а затем рассчитывали ЕС50.
Результаты анализа ELISA для связывающей способности химерных антител и антител, несущих обратные мутации, по настоящему изобретению с белком PCSK9 человека показаны в таблице 6.
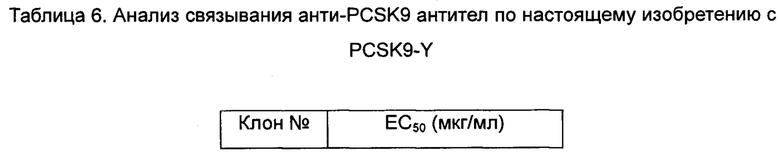
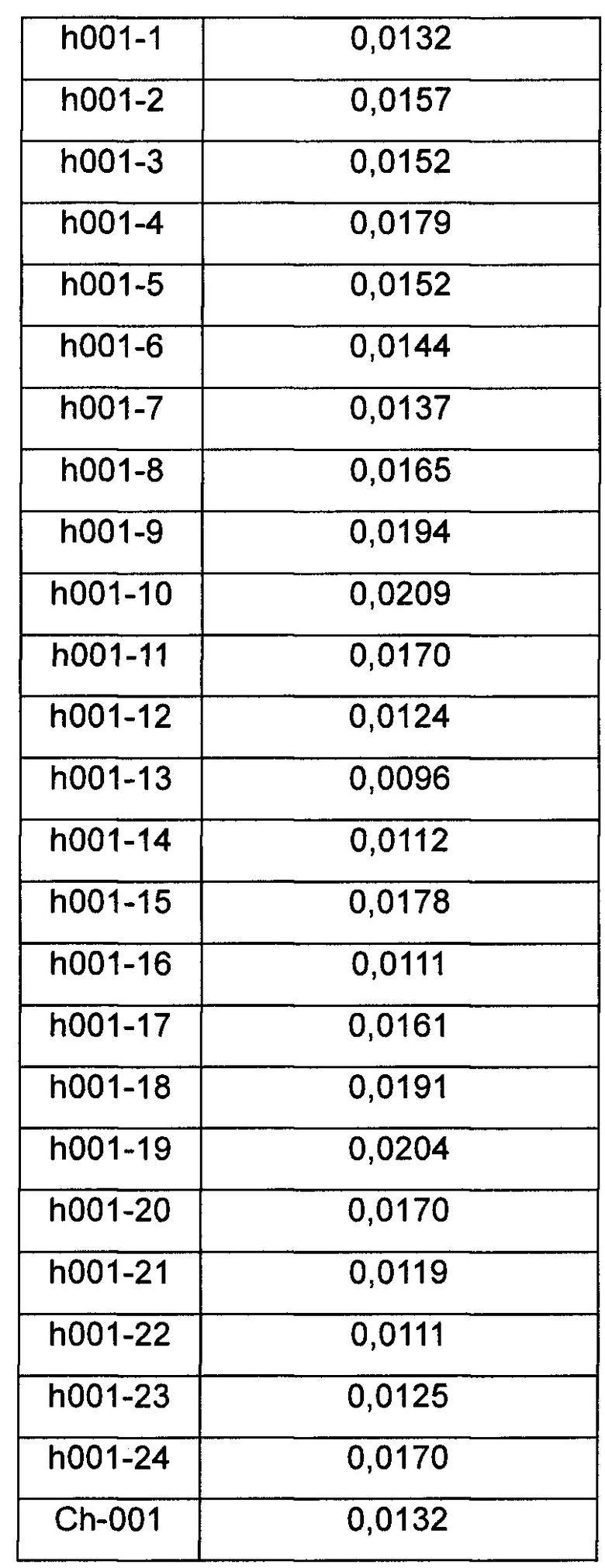
Данные показывают, что гуманизированные антитела по настоящему изобретению обладают более высокой активностью связывания с PCSK9-Y.
Тест 3. Антитела к PCSK9 блокируют связывание LDLR-FC/PCSK9-Y
Способности антител к PCSK9 к блокированию связывания LDLR-FC (SEQ ID NO 8) с PCSK9-Y (мутант PCSK9, SEQ ID NO 6) определяли путем измерения количества PCSK9-Y, связывающегося с LDLR в присутствии антител.
LDLR-FC разводили до 2 мкг/мл фосфатным буфером и наносили в виде покрытия в лунки 96-луночного планшета для ELISA (Costar, № по каталогу 3590), затем инкубировали при 4°С в течение ночи. Планшет промывали, а затем блокировали Трис-буфером (включающим 0,9 ммоль/л CaCl2, 0,05% Твин 20 и 5% обезжиренного молока) при 37°С в течение 2 часов. Затем планшет снова промывали и добавляли в лунки по 100 мкл/лунка смеси меченого биотином PCSK9-Y (био-PCSK9-Y, разведенного до конечной концентрации 1 мкг/мл Трис-буфером, содержащим 0,9 ммоль/л CaCl2, 0,05% Твин 20 и 1% обезжиренного молока) и образцов антитела (разведенного Трис-буфером, содержащим 0,9 ммоль/л CaCl2, 0,05% Твин 20 и 1% обезжиренного молока) и инкубировали при 37°С в течение 1 часа. Затем планшет снова промывали и добавляли меченый пероксидазой хрена стрептавидин (Sigma, № по каталогу S2438) и инкубировали при 37°С в течение 1 часа. Затем планшет промывали и добавляли раствор тетраметилбензидина для проявления. Наконец, добавляли раствор для остановки реакции и измеряли показания OD450 на ридере для микропланшетов, а затем рассчитывали IC50.
Результаты анализа блокирования для блокирующего действия химерных антител и антител, несущих обратные мутации, по настоящему изобретению на связывание LDLR-FC/ PCSK9-Y показаны в таблице 7.
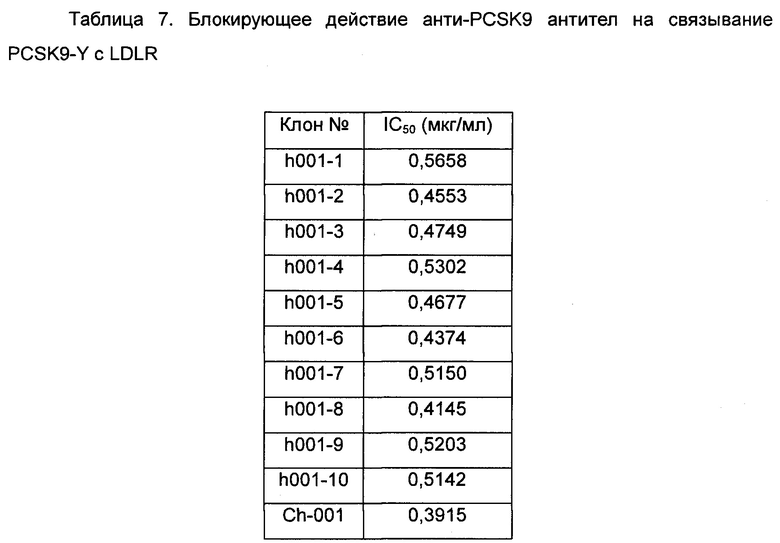
Данные показывают, что анти-PCSK9 антитела по настоящему изобретению могут эффективно блокировать связывание PCSK9-Y с LDLR.
Блокирующие действия анти-PCSK9 антител по настоящему изобретению на связывание других форматов LDLR-FC (полученного в лаборатории, последовательность показана в SEQ ID NO 7 или SEQ ID NO 9) с PCSK9-Y (SEQ ID NO 5) также исследовали описанными выше методами. Результаты показывают, что анти-PCSK9 антитела по настоящему изобретению могут эффективно блокировать связывание PCSK9-Y с укороченными LDLR.
Тест 4. Антитела к PCSK9 блокируют связывание LDLR-FC/PCSK9
Блокирующие действия анти-PCSK9 антител по настоящему изобретению на связывание LDLR-FC (полученного в лаборатории, последовательность показана в SEQ ID NO 8) с PCSK9 (SEQ ID NO 5) определяли путем измерения количества PCSK9, связывающегося с LDLR в присутствии антител.
LDLR-FC разводили до 5 мкг/мл фосфатным буфером и наносили в виде покрытия в лунки 96-луночного планшета для ELISA, затем инкубировали при 4°С в течение ночи. Планшет промывала а затем блокировали Трис-буфером (включающим 0,9 ммоль/л CaCl2, 0,05% Твин 20 и 5% обезжиренного молока) при 37°С в течение 2 часов. Затем планшет снова промывали и добавляли в лунки по 100 мкл/лунка смеси меченого биотином PCSK9 (био-WT-PCSK9, разведенного до конечной концентрации 2 мкг/мл Трис-буфером, содержащим 0,9 ммоль/л CaCl2, 0,05% Твин 20 и 1% обезжиренного молока) и образцов антитела (разведенного Трис-буфером, содержащим 0,9 ммоль/л CaCl2, 0,05% Твин 20 и 1% обезжиренного молока) и инкубировали при 37°С в течение 1 часа. Затем планшет снова промывали и добавляли меченый пероксидазой хрена стрептавидин (Sigma, № по каталогу S2438) и инкубировали при 37°С в течение 1 часа. Затем планшет промывали и добавляли раствор тетраметилбензидина для проявления. Наконец, добавляли раствор для остановки реакции и измеряли показания OD450 на ридере для микропланшетов, а затем рассчитывали IC50.
Результаты анализа блокирования для блокирующего действия химерных антител и антител, несущих обратные мутации, по настоящему изобретению на связывание LDLR-FC/ PCSK9 показаны в таблице 8.
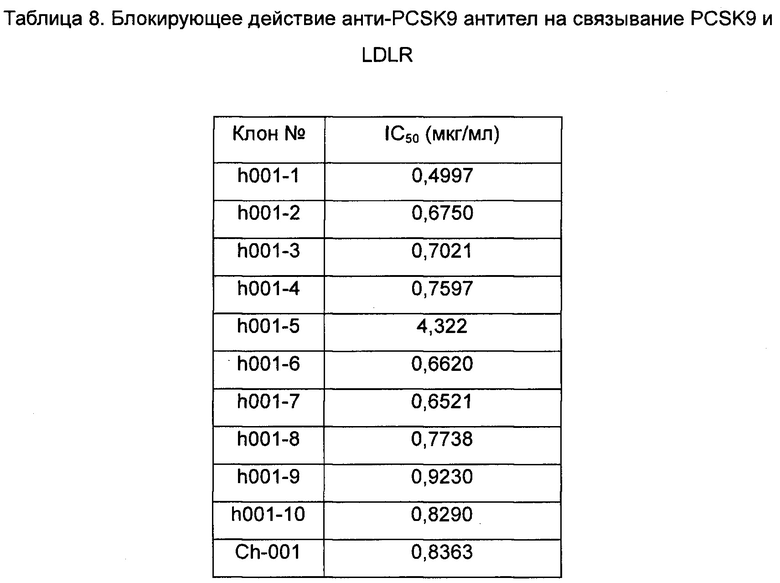
Данные показывают, что анти-PCSK9 антитела по настоящему изобретению могут эффективно блокировать связывание PCSK9 с LDLR.
Блокирующие действия анти-PCSK9 антител по настоящему изобретению на связывание других форматов LDLR-FC (полученного в лаборатории, последовательность показана в SEQ ID NO 7 или SEQ ID NO 9) и PCSK9 (SEQ ID NO 5) также исследовали описанными выше методами. Результаты показывают, что анти-PCSK9 антитела по настоящему изобретению могут эффективно блокировать связывание PCSK9 с укороченными LDLR.
Тест 5. Действия PCSK9 на захват ЛПНП
Клетки HepG2 (банк клеток Китайской Академии наук, № по каталогу TCHu72) культивировали в модифицированной Дульбекко среде Игла (DMEM) (Hyclone, № по каталогу SH30243.01B) (содержащей 10% ФБС, Gibco, № по каталогу 10099-141). Когда клетки покрывали 80-90% планшета, клетки расщепляли путем продувания и считали, 1,5*104 клеток/лунка высевали в 96-луночный планшет. Через 24 часа питательную среду заменяли средой DMEM и 10% сывороткой без липопротеина (Millipore, № по каталогу LP4). Через 48 часов планшет дважды промывали буфером ФСБ, затем в лунки добавляли смесь, предварительно проинкубированную при 4°С в течение 1 часа, содержащую PCSK9 (SEQ ID NO 1, конечная концентрация 10 мкг/мл) и образцы антитела (разведенные до различных концентраций питательной средой) и ЛПНП BODIPY-® в конечной концентрации 10 мкг/мл (Invitrogen, № по каталогу L3483), инкубировали при 37°С. Через 6 часов планшет дважды промывали буфером ФСБ. Показания флуоресценции считывали на ридере для микропланшетов (длина волны возбуждения 485 нм/длина волны испускания 535 нм), затем в лунки добавляли 50 мкл/лунка реактива для определения активности люминесценции клеток CellTiter-Glo® (Promega, G7571) и считывали показания хемилюминесценции. Результаты по захвату ЛПНП представлены на Фиг. 3 и 4 и показывают, что анти-PCSK9 антитела по настоящему изобретению могут стимулировать захват ЛПНП клетками HepG2.
Тест 6. Анализ аффинности антитела PCSK9 методом BIAcore
В соответствии со способом, описанным в наборе реактивов Human Fab Capture Kit (№ по каталогу 28-9583-25, GE) иммобилизующую молекулу Fab человека ковалентно связывали с биочипом СМ5 (« по каталогу BR-1000-12, GE) так, чтобы аффинно иммобилизовать исследуемые антитела. Таким образом, антиген PCSK9 человека (PCSK9 человека с His меткой: PCSK9- His6, SEQ ID NO 1) пропускали через поверхность биочипа, и сигнал реакции обнаруживали в реальном времени, используя прибор Biacore для получения кривых ассоциации и диссоциации. Наконец, были получены значения аффинности путем подбора и представлены в таблице 9 ниже. После окончания каждого цикла диссоциации в эксперименте биочип промывали и регенерировали раствором для регенерации в наборе реактивов Human Fab Capture kit (GE).
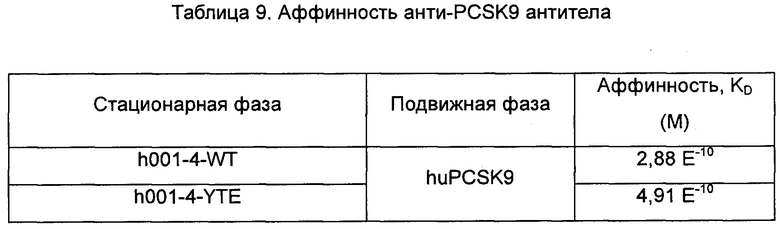
Этот результат демонстрирует, что анти-PCSK9 антитела по настоящему изобретению обладает сильной аффинностью к антигену PCSK9.
Тот же способ также использовали для определения аффинности антител по настоящему изобретению к PCSK9-Y (SEQ ID NO 4) и продемонстрировали результаты, показывающие, что анти-PCSK9 антитела по настоящему изобретению обладают сильной аффинностью к антигену PCSK9-Y.
Тест 7. Фармакодинамический тест анти-PCSK9 антител in vivo
Была создана модель мышей, гиперэкспрессирующих PCSK9 человека, и этим мышам вводили антитело PCSK9 путем инъекции в хвостовую вену. Была проведена оценка действия анти-PCSK9 антител в соответствии с настоящим изобретением на снижение уровня ЛПНП-х in vivo у мышей, гиперэкспрессирующих PCSK9 человека. В качестве стандартного образца использовали IgG человека (иммуноглобулин человека, очищенного из смешанной нормальной сыворотки крови человека традиционным методом аффинной хроматографии, например на белке А).
Мышей C57BI/6 (приобретенных в г. Шанхай, Sippr-BK Laboratory Animal Co., Ltd.) адаптировали в течение 5 дней к условиям лаборатории и вводили им 4×1011 геномных копий вируса AAV-PCSK9 (Benyuan Zhengyang Gene Technology Co., Ltd.) посредством инъекции в хвостовую вену. После инъекции вируса мыши находились в состоянии натощак в течение ночи. Затем на следующий день брали кровь из века и определяли ЛПНП-х с помощью набора реактивов для количественного определения ЛПВП и ЛПНП/ЛПОНП холестерина (HDL and LDL/VLDL Cholesterol Quantification Kit), (приобретенный у компании BioVision, номер по каталогу K613-100). Мышей случайным образом делили на группы (6 мышей/группа (n=6)) в соответствии с концентрацией ЛПНП-х и вводили антитела путем инъекции в хвостовую вену. IgG человека и антитело h001-4-WT, полученное в лаборатории, вводили в дозе 10 мг/кг (и IgG человека, и антитело h001-4-WT готовили в ФСБ в концентрации 1 мг/мл). Перед забором крови мыши находились в состоянии натощак в течение шести часов. Через 24 ч, 48 ч, 72 ч и 96 ч после введения брали кровь из века, хранили при 37°С в течение 1 часа, центрифугировали при 3500 об/мин в течение 10 минут, и сыворотку хранили при -80°С.
После последнего сбора сыворотки исследование всей замороженной сыворотки проводили в один и тот же день. Концентрацию ЛПНП-х в сыворотке крови определяли с помощью набора реактивов для количественного определения холестерина ЛПВП и ЛПНП/ЛПОНП в соответствии с инструкциями к набору.
Как показано на Фиг. 5, результаты показывают, что концентрация ЛПНП-х в сыворотке крови нормальной мыши составляет около 12 мг/дл. После инъекции вируса AAV8-PCSK9 концентрация ЛПНП-х в сыворотке крови в среднем составляла 40 мг/дл. Мышей делили на группы и вводили препараты. Через 24 ч после введения концентрация ЛПНП-х в группе, которой вводили h001-4-WT, снижалась на 50% по сравнению с группой, получавшей IgG; через 48 ч после введения концентрация ЛПНП-х в группе, которой вводили h001-4-WT, снижалась на 49%; через 72 ч после введения после введения концентрация ЛПНП-х в группе, которой вводили h001-4-WT, снижалась на 32%; через 96 ч после введения после введения концентрация ЛПНП-х в группе, которой вводили h001-4-WT, снижалась на 20%, как показано в таблице 10 и на Фиг. 6.
Таким образом, антитело h001-4-WT обладало способностью к снижению концентрации ЛПНП-х в сыворотке крови мышей, гиперэкспрессирующих PCSK9 человека, и этот эффект продолжается в течение 72 часов.
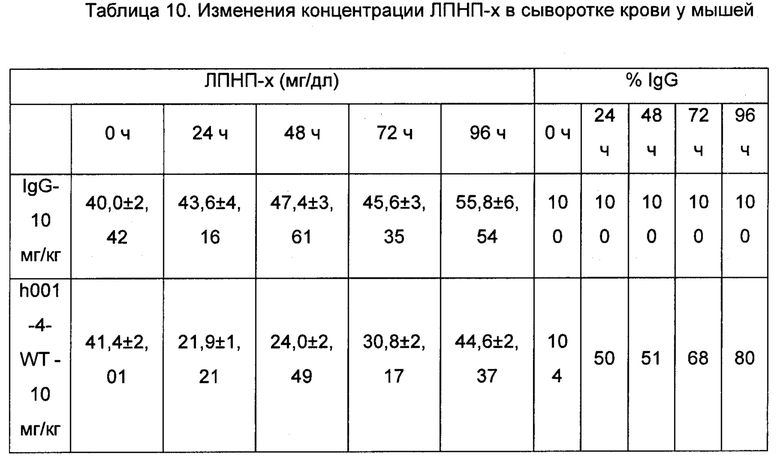
Тест 8. Сравнительный эксперимент
В сравнительном эксперименте ELISA планшет покрывали одним антителом в течение ночи. Затем совместно добавляли биотин-PCSK9-his и сравнительное антитело в концентрации, в 50 раз превышающей концентрацию покрывающего антитела. Покрывающее антитело должно конкурировать со сравнительным антителом за связывание антигена. Затем определяли сигнал антигена на планшете. Результаты показывают, что антитело h001-4 и 21В12 (US 8030457 B2) как таковое конкурирует за связывание с антигеном, однако явное конкурентное связывание между двумя антителами отсутствует, что позволяет предположить, что эпитопы антигена для этих двух антител различаются.
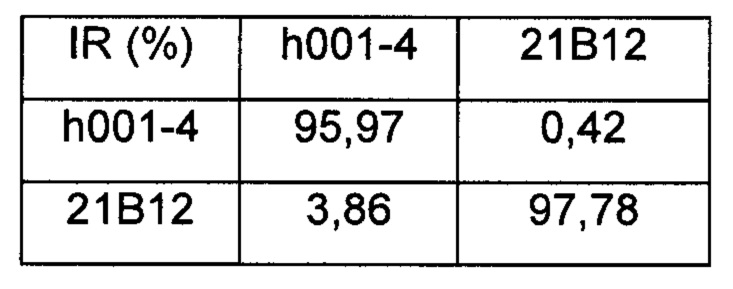
Тест 9. Исследование фармакодинамики и фармакокинетики у яванских макак in vivo
Для исследования действия и метаболизма антител по настоящему изобретению in vivo h001-4-WT и h001-4-YTE вводили in vivo яванским макакам соответственно. Вводимая доза составляла 3 мг/кг путем внутривенного введения, и в каждую группу включали 3 самцов яванских макак. Внутривенную инъекцию проводили со скоростью около 2-4 мл/мин. Образцы крови брали в различные моменты времени для определения концентрации липопротеина, в частности, липопротеина низкой плотности (ЛПНП), и концентрации антител в сыворотке. Моментами времени для определения липопротеина были момент перед введением препаратов и 1, 4, 8, 12, 16, 20, 24, 28 дни после введения препарата. Моменты времени забора образцов для ФК были момент перед введением препаратов и через 15 минут, 30 минут, 1 час, 3 часа, 8 часов, 12 часов, 24 часа, 48 часов, 72 часа, 96 часов, 120 часов, 144 часа, 168 часов, 336 часов, 504 часа, 672 часа после введения препарата.
Результаты (Фиг. 7) показывают, что оба антитела h001-4-WT и h001-4-YTE могут вызывать статистически значимое снижение содержания ЛПВП у яванских макак, и продолжительность снижения, вызванного h001-4-YTE, лучше по сравнению с h001-4-WT.
Содержание h001-4-WT и h001-4-YTE в образцах сыворотки крови, взятых для ФК, определяли методом ELISA. Способ описан в тесте 1, и результаты показывают, что период полувыведения h001-4-WT у яванских макак составляет 4 суток, при этом период полувыведения h001-4-YTE у яванских макак составляет 7,3 суток. Период полувыведения YTE in vivo статистически значимо увеличивается по сравнению с диким типом.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> JIANGSU HENGRUI MEDICINE CO.,LTD SHANGHAI HENGRUI PHARMACEUTICAL CO.,LTD
<120> АНТИ-PCSK9 АНТИТЕЛО, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ
<130> 760083CPCT
<160> 33
<170> PatentIn version 3.5
<210> 1
<211> 698
<212> ПРТ
<213> Искусственная
<220>
<223> PCSK9 с His-меткой: PCSK9-His6, используемый в качестве иммуногена для иммунизации мышей или в качестве детектирующего реактива
<400> 1
Met Gly Thr Val Ser Ser Arg Arg Ser Trp Trp Pro Leu Pro Leu Leu
1 5 10 15
Leu Leu Leu Leu Leu Leu Leu Gly Pro Ala Gly Ala Arg Ala Gln Glu
20 25 30
Asp Glu Asp Gly Asp Tyr Glu Glu Leu Val Leu Ala Leu Arg Ser Glu
35 40 45
Glu Asp Gly Leu Ala Glu Ala Pro Glu His Gly Thr Thr Ala Thr Phe
50 55 60
His Arg Cys Ala Lys Asp Pro Trp Arg Leu Pro Gly Thr Tyr Val Val
65 70 75 80
Val Leu Lys Glu Glu Thr His Leu Ser Gln Ser Glu Arg Thr Ala Arg
85 90 95
Arg Leu Gln Ala Gln Ala Ala Arg Arg Gly Tyr Leu Thr Lys Ile Leu
100 105 110
His Val Phe His Gly Leu Leu Pro Gly Phe Leu Val Lys Met Ser Gly
115 120 125
Asp Leu Leu Glu Leu Ala Leu Lys Leu Pro His Val Asp Tyr Ile Glu
130 135 140
Glu Asp Ser Ser Val Phe Ala Gln Ser Ile Pro Trp Asn Leu Glu Arg
145 150 155 160
Ile Thr Pro Pro Arg Tyr Arg Ala Asp Glu Tyr Gln Pro Pro Asp Gly
165 170 175
Gly Ser Leu Val Glu Val Tyr Leu Leu Asp Thr Ser Ile Gln Ser Asp
180 185 190
His Arg Glu Ile Glu Gly Arg Val Met Val Thr Asp Phe Glu Asn Val
195 200 205
Pro Glu Glu Asp Gly Thr Arg Phe His Arg Gln Ala Ser Lys Cys Asp
210 215 220
Ser His Gly Thr His Leu Ala Gly Val Val Ser Gly Arg Asp Ala Gly
225 230 235 240
Val Ala Lys Gly Ala Ser Met Arg Ser Leu Arg Val Leu Asn Cys Gln
245 250 255
Gly Lys Gly Thr Val Ser Gly Thr Leu Ile Gly Leu Glu Phe Ile Arg
260 265 270
Lys Ser Gln Leu Val Gln Pro Val Gly Pro Leu Val Val Leu Leu Pro
275 280 285
Leu Ala Gly Gly Tyr Ser Arg Val Leu Asn Ala Ala Cys Gln Arg Leu
290 295 300
Ala Arg Ala Gly Val Val Leu Val Thr Ala Ala Gly Asn Phe Arg Asp
305 310 315 320
Asp Ala Cys Leu Tyr Ser Pro Ala Ser Ala Pro Glu Val Ile Thr Val
325 330 335
Gly Ala Thr Asn Ala Gln Asp Gln Pro Val Thr Leu Gly Thr Leu Gly
340 345 350
Thr Asn Phe Gly Arg Cys Val Asp Leu Phe Ala Pro Gly Glu Asp Ile
355 360 365
Ile Gly Ala Ser Ser Asp Cys Ser Thr Cys Phe Val Ser Gln Ser Gly
370 375 380
Thr Ser Gln Ala Ala Ala His Val Ala Gly Ile Ala Ala Met Met Leu
385 390 395 400
Ser Ala Glu Pro Glu Leu Thr Leu Ala Glu Leu Arg Gln Arg Leu Ile
405 410 415
His Phe Ser Ala Lys Asp Val Ile Asn Glu Ala Trp Phe Pro Glu Asp
420 425 430
Gln Arg Val Leu Thr Pro Asn Leu Val Ala Ala Leu Pro Pro Ser Thr
435 440 445
His Gly Ala Gly Trp Gln Leu Phe Cys Arg Thr Val Trp Ser Ala His
450 455 460
Ser Gly Pro Thr Arg Met Ala Thr Ala Val Ala Arg Cys Ala Pro Asp
465 470 475 480
Glu Glu Leu Leu Ser Cys Ser Ser Phe Ser Arg Ser Gly Lys Arg Arg
485 490 495
Gly Glu Arg Met Glu Ala Gln Gly Gly Lys Leu Val Cys Arg Ala His
500 505 510
Asn Ala Phe Gly Gly Glu Gly Val Tyr Ala Ile Ala Arg Cys Cys Leu
515 520 525
Leu Pro Gln Ala Asn Cys Ser Val His Thr Ala Pro Pro Ala Glu Ala
530 535 540
Ser Met Gly Thr Arg Val His Cys His Gln Gln Gly His Val Leu Thr
545 550 555 560
Gly Cys Ser Ser His Trp Glu Val Glu Asp Leu Gly Thr His Lys Pro
565 570 575
Pro Val Leu Arg Pro Arg Gly Gln Pro Asn Gln Cys Val Gly His Arg
580 585 590
Glu Ala Ser Ile His Ala Ser Cys Cys His Ala Pro Gly Leu Glu Cys
595 600 605
Lys Val Lys Glu His Gly Ile Pro Ala Pro Gln Glu Gln Val Thr Val
610 615 620
Ala Cys Glu Glu Gly Trp Thr Leu Thr Gly Cys Ser Ala Leu Pro Gly
625 630 635 640
Thr Ser His Val Leu Gly Ala Tyr Ala Val Asp Asn Thr Cys Val Val
645 650 655
Arg Ser Arg Asp Val Ser Thr Thr Gly Ser Thr Ser Glu Gly Ala Val
660 665 670
Thr Ala Val Ala Ile Cys Cys Arg Ser Arg His Leu Ala Gln Ala Ser
675 680 685
Gln Glu Leu Gln His His His His His His
690 695
<210> 2
<211> 714
<212> ПРТ
<213> Искусственная
<220>
<223> PCSK9 с пептидом PADRE и His-меткой: PCSK9-PADRE-His6 в качестве иммуногена, где содержащийся в нем пептид PADRE может стимулировать иммунизацию
<400> 2
Met Gly Thr Val Ser Ser Arg Arg Ser Trp Trp Pro Leu Pro Leu Leu
1 5 10 15
Leu Leu Leu Leu Leu Leu Leu Gly Pro Ala Gly Ala Arg Ala Gln Glu
20 25 30
Asp Glu Asp Gly Asp Tyr Glu Glu Leu Val Leu Ala Leu Arg Ser Glu
35 40 45
Glu Asp Gly Leu Ala Glu Ala Pro Glu His Gly Thr Thr Ala Thr Phe
50 55 60
His Arg Cys Ala Lys Asp Pro Trp Arg Leu Pro Gly Thr Tyr Val Val
65 70 75 80
Val Leu Lys Glu Glu Thr His Leu Ser Gln Ser Glu Arg Thr Ala Arg
85 90 95
Arg Leu Gln Ala Gln Ala Ala Arg Arg Gly Tyr Leu Thr Lys Ile Leu
100 105 110
His Val Phe His Gly Leu Leu Pro Gly Phe Leu Val Lys Met Ser Gly
115 120 125
Asp Leu Leu Glu Leu Ala Leu Lys Leu Pro His Val Asp Tyr Ile Glu
130 135 140
Glu Asp Ser Ser Val Phe Ala Gln Ser Ile Pro Trp Asn Leu Glu Arg
145 150 155 160
Ile Thr Pro Pro Arg Tyr Arg Ala Asp Glu Tyr Gln Pro Pro Asp Gly
165 170 175
Gly Ser Leu Val Glu Val Tyr Leu Leu Asp Thr Ser Ile Gln Ser Asp
180 185 190
His Arg Glu Ile Glu Gly Arg Val Met Val Thr Asp Phe Glu Asn Val
195 200 205
Pro Glu Glu Asp Gly Thr Arg Phe His Arg Gln Ala Ser Lys Cys Asp
210 215 220
Ser His Gly Thr His Leu Ala Gly Val Val Ser Gly Arg Asp Ala Gly
225 230 235 240
Val Ala Lys Gly Ala Ser Met Arg Ser Leu Arg Val Leu Asn Cys Gln
245 250 255
Gly Lys Gly Thr Val Ser Gly Thr Leu Ile Gly Leu Glu Phe Ile Arg
260 265 270
Lys Ser Gln Leu Val Gln Pro Val Gly Pro Leu Val Val Leu Leu Pro
275 280 285
Leu Ala Gly Gly Tyr Ser Arg Val Leu Asn Ala Ala Cys Gln Arg Leu
290 295 300
Ala Arg Ala Gly Val Val Leu Val Thr Ala Ala Gly Asn Phe Arg Asp
305 310 315 320
Asp Ala Cys Leu Tyr Ser Pro Ala Ser Ala Pro Glu Val Ile Thr Val
325 330 335
Gly Ala Thr Asn Ala Gln Asp Gln Pro Val Thr Leu Gly Thr Leu Gly
340 345 350
Thr Asn Phe Gly Arg Cys Val Asp Leu Phe Ala Pro Gly Glu Asp Ile
355 360 365
Ile Gly Ala Ser Ser Asp Cys Ser Thr Cys Phe Val Ser Gln Ser Gly
370 375 380
Thr Ser Gln Ala Ala Ala His Val Ala Gly Ile Ala Ala Met Met Leu
385 390 395 400
Ser Ala Glu Pro Glu Leu Thr Leu Ala Glu Leu Arg Gln Arg Leu Ile
405 410 415
His Phe Ser Ala Lys Asp Val Ile Asn Glu Ala Trp Phe Pro Glu Asp
420 425 430
Gln Arg Val Leu Thr Pro Asn Leu Val Ala Ala Leu Pro Pro Ser Thr
435 440 445
His Gly Ala Gly Trp Gln Leu Phe Cys Arg Thr Val Trp Ser Ala His
450 455 460
Ser Gly Pro Thr Arg Met Ala Thr Ala Val Ala Arg Cys Ala Pro Asp
465 470 475 480
Glu Glu Leu Leu Ser Cys Ser Ser Phe Ser Arg Ser Gly Lys Arg Arg
485 490 495
Gly Glu Arg Met Glu Ala Gln Gly Gly Lys Leu Val Cys Arg Ala His
500 505 510
Asn Ala Phe Gly Gly Glu Gly Val Tyr Ala Ile Ala Arg Cys Cys Leu
515 520 525
Leu Pro Gln Ala Asn Cys Ser Val His Thr Ala Pro Pro Ala Glu Ala
530 535 540
Ser Met Gly Thr Arg Val His Cys His Gln Gln Gly His Val Leu Thr
545 550 555 560
Gly Cys Ser Ser His Trp Glu Val Glu Asp Leu Gly Thr His Lys Pro
565 570 575
Pro Val Leu Arg Pro Arg Gly Gln Pro Asn Gln Cys Val Gly His Arg
580 585 590
Glu Ala Ser Ile His Ala Ser Cys Cys His Ala Pro Gly Leu Glu Cys
595 600 605
Lys Val Lys Glu His Gly Ile Pro Ala Pro Gln Glu Gln Val Thr Val
610 615 620
Ala Cys Glu Glu Gly Trp Thr Leu Thr Gly Cys Ser Ala Leu Pro Gly
625 630 635 640
Thr Ser His Val Leu Gly Ala Tyr Ala Val Asp Asn Thr Cys Val Val
645 650 655
Arg Ser Arg Asp Val Ser Thr Thr Gly Ser Thr Ser Glu Gly Ala Val
660 665 670
Thr Ala Val Ala Ile Cys Cys Arg Ser Arg His Leu Ala Gln Ala Ser
675 680 685
Gln Glu Leu Gln Gly Ser Gly Ala Lys Phe Val Ala Ala Trp Thr Leu
690 695 700
Lys Ala Ala Ala His His His His His His
705 710
<210> 3
<211> 704
<212> ПРТ
<213> Искусственная
<220>
<223> Гибридный белок PCSK9 с сайтом расщепления TEV и His-меткой: PCSK9-TEV-His6, N-PCSK9 (N-концевой домен PCSK9), используемый в качестве иммуногена, может быть получен с помощью фермента TEV
<400> 3
Met Gly Thr Val Ser Ser Arg Arg Ser Trp Trp Pro Leu Pro Leu Leu
1 5 10 15
Leu Leu Leu Leu Leu Leu Leu Gly Pro Ala Gly Ala Arg Ala Gln Glu
20 25 30
Asp Glu Asp Gly Asp Tyr Glu Glu Leu Val Leu Ala Leu Arg Ser Glu
35 40 45
Glu Asp Gly Leu Ala Glu Ala Pro Glu His Gly Thr Thr Ala Thr Phe
50 55 60
His Arg Cys Ala Lys Asp Pro Trp Arg Leu Pro Gly Thr Tyr Val Val
65 70 75 80
Val Leu Lys Glu Glu Thr His Leu Ser Gln Ser Glu Arg Thr Ala Arg
85 90 95
Arg Leu Gln Ala Gln Ala Ala Arg Arg Gly Tyr Leu Thr Lys Ile Leu
100 105 110
His Val Phe His Gly Leu Leu Pro Gly Phe Leu Val Lys Met Ser Gly
115 120 125
Asp Leu Leu Glu Leu Ala Leu Lys Leu Pro His Val Asp Tyr Ile Glu
130 135 140
Glu Asp Ser Ser Val Phe Ala Gln Ser Ile Pro Trp Asn Leu Glu Arg
145 150 155 160
Ile Thr Pro Pro Arg Tyr Arg Ala Asp Glu Tyr Gln Pro Pro Asp Gly
165 170 175
Gly Ser Leu Val Glu Val Tyr Leu Leu Asp Thr Ser Ile Gln Ser Asp
180 185 190
His Arg Glu Ile Glu Gly Arg Val Met Val Thr Asp Phe Glu Asn Val
195 200 205
Pro Glu Glu Asp Gly Thr Arg Phe His Arg Gln Ala Ser Lys Cys Asp
210 215 220
Ser His Gly Thr His Leu Ala Gly Val Val Ser Gly Arg Asp Ala Gly
225 230 235 240
Val Ala Lys Gly Ala Ser Met Arg Ser Leu Arg Val Leu Asn Cys Gln
245 250 255
Gly Lys Gly Thr Val Ser Gly Thr Leu Ile Gly Leu Glu Phe Ile Arg
260 265 270
Lys Ser Gln Leu Val Gln Pro Val Gly Pro Leu Val Val Leu Leu Pro
275 280 285
Leu Ala Gly Gly Tyr Ser Arg Val Leu Asn Ala Ala Cys Gln Arg Leu
290 295 300
Ala Arg Ala Gly Val Val Leu Val Thr Ala Ala Gly Asn Phe Arg Asp
305 310 315 320
Asp Ala Cys Leu Tyr Ser Pro Ala Ser Ala Pro Glu Val Ile Thr Val
325 330 335
Gly Ala Thr Asn Ala Gln Asp Gln Pro Val Thr Leu Gly Thr Leu Gly
340 345 350
Thr Asn Phe Gly Arg Cys Val Asp Leu Phe Ala Pro Gly Glu Asp Ile
355 360 365
Ile Gly Ala Ser Ser Asp Cys Ser Thr Cys Phe Val Ser Gln Ser Gly
370 375 380
Thr Ser Gln Ala Ala Ala His Val Ala Gly Ile Ala Ala Met Met Leu
385 390 395 400
Ser Ala Glu Pro Glu Leu Thr Leu Ala Glu Leu Arg Gln Arg Leu Ile
405 410 415
His Phe Ser Ala Lys Asp Val Ile Asn Glu Ala Trp Phe Pro Glu Asp
420 425 430
Gln Arg Val Leu Thr Pro Asn Leu Val Ala Ala Leu Pro Pro Ser Thr
435 440 445
His Glu Asn Leu Tyr Phe Gln Gly Ala Gly Trp Gln Leu Phe Cys Arg
450 455 460
Thr Val Trp Ser Ala His Ser Gly Pro Thr Arg Met Ala Thr Ala Val
465 470 475 480
Ala Arg Cys Ala Pro Asp Glu Glu Leu Leu Ser Cys Ser Ser Phe Ser
485 490 495
Arg Ser Gly Lys Arg Arg Gly Glu Arg Met Glu Ala Gln Gly Gly Lys
500 505 510
Leu Val Cys Arg Ala His Asn Ala Phe Gly Gly Glu Gly Val Tyr Ala
515 520 525
Ile Ala Arg Cys Cys Leu Leu Pro Gln Ala Asn Cys Ser Val His Thr
530 535 540
Ala Pro Pro Ala Glu Ala Ser Met Gly Thr Arg Val His Cys His Gln
545 550 555 560
Gln Gly His Val Leu Thr Gly Cys Ser Ser His Trp Glu Val Glu Asp
565 570 575
Leu Gly Thr His Lys Pro Pro Val Leu Arg Pro Arg Gly Gln Pro Asn
580 585 590
Gln Cys Val Gly His Arg Glu Ala Ser Ile His Ala Ser Cys Cys His
595 600 605
Ala Pro Gly Leu Glu Cys Lys Val Lys Glu His Gly Ile Pro Ala Pro
610 615 620
Gln Glu Gln Val Thr Val Ala Cys Glu Glu Gly Trp Thr Leu Thr Gly
625 630 635 640
Cys Ser Ala Leu Pro Gly Thr Ser His Val Leu Gly Ala Tyr Ala Val
645 650 655
Asp Asn Thr Cys Val Val Arg Ser Arg Asp Val Ser Thr Thr Gly Ser
660 665 670
Thr Ser Glu Gly Ala Val Thr Ala Val Ala Ile Cys Cys Arg Ser Arg
675 680 685
His Leu Ala Gln Ala Ser Gln Glu Leu Gln His His His His His His
690 695 700
<210> 4
<211> 698
<212> ПРТ
<213> Искусственная
<220>
<223> Мутантный белок PCSK9-D374Y с His-меткой: PCSK9-D374Y-His6 в качестве детектирующего реактива
<400> 4
Met Gly Thr Val Ser Ser Arg Arg Ser Trp Trp Pro Leu Pro Leu Leu
1 5 10 15
Leu Leu Leu Leu Leu Leu Leu Gly Pro Ala Gly Ala Arg Ala Gln Glu
20 25 30
Asp Glu Asp Gly Asp Tyr Glu Glu Leu Val Leu Ala Leu Arg Ser Glu
35 40 45
Glu Asp Gly Leu Ala Glu Ala Pro Glu His Gly Thr Thr Ala Thr Phe
50 55 60
His Arg Cys Ala Lys Asp Pro Trp Arg Leu Pro Gly Thr Tyr Val Val
65 70 75 80
Val Leu Lys Glu Glu Thr His Leu Ser Gln Ser Glu Arg Thr Ala Arg
85 90 95
Arg Leu Gln Ala Gln Ala Ala Arg Arg Gly Tyr Leu Thr Lys Ile Leu
100 105 110
His Val Phe His Gly Leu Leu Pro Gly Phe Leu Val Lys Met Ser Gly
115 120 125
Asp Leu Leu Glu Leu Ala Leu Lys Leu Pro His Val Asp Tyr Ile Glu
130 135 140
Glu Asp Ser Ser Val Phe Ala Gln Ser Ile Pro Trp Asn Leu Glu Arg
145 150 155 160
Ile Thr Pro Pro Arg Tyr Arg Ala Asp Glu Tyr Gln Pro Pro Asp Gly
165 170 175
Gly Ser Leu Val Glu Val Tyr Leu Leu Asp Thr Ser Ile Gln Ser Asp
180 185 190
His Arg Glu Ile Glu Gly Arg Val Met Val Thr Asp Phe Glu Asn Val
195 200 205
Pro Glu Glu Asp Gly Thr Arg Phe His Arg Gln Ala Ser Lys Cys Asp
210 215 220
Ser His Gly Thr His Leu Ala Gly Val Val Ser Gly Arg Asp Ala Gly
225 230 235 240
Val Ala Lys Gly Ala Ser Met Arg Ser Leu Arg Val Leu Asn Cys Gln
245 250 255
Gly Lys Gly Thr Val Ser Gly Thr Leu Ile Gly Leu Glu Phe Ile Arg
260 265 270
Lys Ser Gln Leu Val Gln Pro Val Gly Pro Leu Val Val Leu Leu Pro
275 280 285
Leu Ala Gly Gly Tyr Ser Arg Val Leu Asn Ala Ala Cys Gln Arg Leu
290 295 300
Ala Arg Ala Gly Val Val Leu Val Thr Ala Ala Gly Asn Phe Arg Asp
305 310 315 320
Asp Ala Cys Leu Tyr Ser Pro Ala Ser Ala Pro Glu Val Ile Thr Val
325 330 335
Gly Ala Thr Asn Ala Gln Asp Gln Pro Val Thr Leu Gly Thr Leu Gly
340 345 350
Thr Asn Phe Gly Arg Cys Val Asp Leu Phe Ala Pro Gly Glu Asp Ile
355 360 365
Ile Gly Ala Ser Ser Tyr Cys Ser Thr Cys Phe Val Ser Gln Ser Gly
370 375 380
Thr Ser Gln Ala Ala Ala His Val Ala Gly Ile Ala Ala Met Met Leu
385 390 395 400
Ser Ala Glu Pro Glu Leu Thr Leu Ala Glu Leu Arg Gln Arg Leu Ile
405 410 415
His Phe Ser Ala Lys Asp Val Ile Asn Glu Ala Trp Phe Pro Glu Asp
420 425 430
Gln Arg Val Leu Thr Pro Asn Leu Val Ala Ala Leu Pro Pro Ser Thr
435 440 445
His Gly Ala Gly Trp Gln Leu Phe Cys Arg Thr Val Trp Ser Ala His
450 455 460
Ser Gly Pro Thr Arg Met Ala Thr Ala Val Ala Arg Cys Ala Pro Asp
465 470 475 480
Glu Glu Leu Leu Ser Cys Ser Ser Phe Ser Arg Ser Gly Lys Arg Arg
485 490 495
Gly Glu Arg Met Glu Ala Gln Gly Gly Lys Leu Val Cys Arg Ala His
500 505 510
Asn Ala Phe Gly Gly Glu Gly Val Tyr Ala Ile Ala Arg Cys Cys Leu
515 520 525
Leu Pro Gln Ala Asn Cys Ser Val His Thr Ala Pro Pro Ala Glu Ala
530 535 540
Ser Met Gly Thr Arg Val His Cys His Gln Gln Gly His Val Leu Thr
545 550 555 560
Gly Cys Ser Ser His Trp Glu Val Glu Asp Leu Gly Thr His Lys Pro
565 570 575
Pro Val Leu Arg Pro Arg Gly Gln Pro Asn Gln Cys Val Gly His Arg
580 585 590
Glu Ala Ser Ile His Ala Ser Cys Cys His Ala Pro Gly Leu Glu Cys
595 600 605
Lys Val Lys Glu His Gly Ile Pro Ala Pro Gln Glu Gln Val Thr Val
610 615 620
Ala Cys Glu Glu Gly Trp Thr Leu Thr Gly Cys Ser Ala Leu Pro Gly
625 630 635 640
Thr Ser His Val Leu Gly Ala Tyr Ala Val Asp Asn Thr Cys Val Val
645 650 655
Arg Ser Arg Asp Val Ser Thr Thr Gly Ser Thr Ser Glu Gly Ala Val
660 665 670
Thr Ala Val Ala Ile Cys Cys Arg Ser Arg His Leu Ala Gln Ala Ser
675 680 685
Gln Glu Leu Gln His His His His His His
690 695
<210> 5
<211> 719
<212> ПРТ
<213> Искусственная
<220>
<223> Белок PCSK9 со вставкой пептида-реципиента биотина BP15 и His-метки: PCSK9-BP15-His6. В качестве детектирующего реактива биотиновая метка будет присоединена в положении пептида BP15 в процессе экспрессии, что позволяет избежать мечения биотином in vitro и, следовательно, избежать возможных конформационных изменений.
<400> 5
Met Gly Thr Val Ser Ser Arg Arg Ser Trp Trp Pro Leu Pro Leu Leu
1 5 10 15
Leu Leu Leu Leu Leu Leu Leu Gly Pro Ala Gly Ala Arg Ala Gln Glu
20 25 30
Asp Glu Asp Gly Asp Tyr Glu Glu Leu Val Leu Ala Leu Arg Ser Glu
35 40 45
Glu Asp Gly Leu Ala Glu Ala Pro Glu His Gly Thr Thr Ala Thr Phe
50 55 60
His Arg Cys Ala Lys Asp Pro Trp Arg Leu Pro Gly Thr Tyr Val Val
65 70 75 80
Val Leu Lys Glu Glu Thr His Leu Ser Gln Ser Glu Arg Thr Ala Arg
85 90 95
Arg Leu Gln Ala Gln Ala Ala Arg Arg Gly Tyr Leu Thr Lys Ile Leu
100 105 110
His Val Phe His Gly Leu Leu Pro Gly Phe Leu Val Lys Met Ser Gly
115 120 125
Asp Leu Leu Glu Leu Ala Leu Lys Leu Pro His Val Asp Tyr Ile Glu
130 135 140
Glu Asp Ser Ser Val Phe Ala Gln Ser Ile Pro Trp Asn Leu Glu Arg
145 150 155 160
Ile Thr Pro Pro Arg Tyr Arg Ala Asp Glu Tyr Gln Pro Pro Asp Gly
165 170 175
Gly Ser Leu Val Glu Val Tyr Leu Leu Asp Thr Ser Ile Gln Ser Asp
180 185 190
His Arg Glu Ile Glu Gly Arg Val Met Val Thr Asp Phe Glu Asn Val
195 200 205
Pro Glu Glu Asp Gly Thr Arg Phe His Arg Gln Ala Ser Lys Cys Asp
210 215 220
Ser His Gly Thr His Leu Ala Gly Val Val Ser Gly Arg Asp Ala Gly
225 230 235 240
Val Ala Lys Gly Ala Ser Met Arg Ser Leu Arg Val Leu Asn Cys Gln
245 250 255
Gly Lys Gly Thr Val Ser Gly Thr Leu Ile Gly Leu Glu Phe Ile Arg
260 265 270
Lys Ser Gln Leu Val Gln Pro Val Gly Pro Leu Val Val Leu Leu Pro
275 280 285
Leu Ala Gly Gly Tyr Ser Arg Val Leu Asn Ala Ala Cys Gln Arg Leu
290 295 300
Ala Arg Ala Gly Val Val Leu Val Thr Ala Ala Gly Asn Phe Arg Asp
305 310 315 320
Asp Ala Cys Leu Tyr Ser Pro Ala Ser Ala Pro Glu Val Ile Thr Val
325 330 335
Gly Ala Thr Asn Ala Gln Asp Gln Pro Val Thr Leu Gly Thr Leu Gly
340 345 350
Thr Asn Phe Gly Arg Cys Val Asp Leu Phe Ala Pro Gly Glu Asp Ile
355 360 365
Ile Gly Ala Ser Ser Asp Cys Ser Thr Cys Phe Val Ser Gln Ser Gly
370 375 380
Thr Ser Gln Ala Ala Ala His Val Ala Gly Ile Ala Ala Met Met Leu
385 390 395 400
Ser Ala Glu Pro Glu Leu Thr Leu Ala Glu Leu Arg Gln Arg Leu Ile
405 410 415
His Phe Ser Ala Lys Asp Val Ile Asn Glu Ala Trp Phe Pro Glu Asp
420 425 430
Gln Arg Val Leu Thr Pro Asn Leu Val Ala Ala Leu Pro Pro Ser Thr
435 440 445
His Gly Ala Gly Trp Gln Leu Phe Cys Arg Thr Val Trp Ser Ala His
450 455 460
Ser Gly Pro Thr Arg Met Ala Thr Ala Val Ala Arg Cys Ala Pro Asp
465 470 475 480
Glu Glu Leu Leu Ser Cys Ser Ser Phe Ser Arg Ser Gly Lys Arg Arg
485 490 495
Gly Glu Arg Met Glu Ala Gln Gly Gly Lys Leu Val Cys Arg Ala His
500 505 510
Asn Ala Phe Gly Gly Glu Gly Val Tyr Ala Ile Ala Arg Cys Cys Leu
515 520 525
Leu Pro Gln Ala Asn Cys Ser Val His Thr Ala Pro Pro Ala Glu Ala
530 535 540
Ser Met Gly Thr Arg Val His Cys His Gln Gln Gly His Val Leu Thr
545 550 555 560
Gly Cys Ser Ser His Trp Glu Val Glu Asp Leu Gly Thr His Lys Pro
565 570 575
Pro Val Leu Arg Pro Arg Gly Gln Pro Asn Gln Cys Val Gly His Arg
580 585 590
Glu Ala Ser Ile His Ala Ser Cys Cys His Ala Pro Gly Leu Glu Cys
595 600 605
Lys Val Lys Glu His Gly Ile Pro Ala Pro Gln Glu Gln Val Thr Val
610 615 620
Ala Cys Glu Glu Gly Trp Thr Leu Thr Gly Cys Ser Ala Leu Pro Gly
625 630 635 640
Thr Ser His Val Leu Gly Ala Tyr Ala Val Asp Asn Thr Cys Val Val
645 650 655
Arg Ser Arg Asp Val Ser Thr Thr Gly Ser Thr Ser Glu Gly Ala Val
660 665 670
Thr Ala Val Ala Ile Cys Cys Arg Ser Arg His Leu Ala Gln Ala Ser
675 680 685
Gln Glu Leu Gln Gly Ser Thr Ser Gly Ser Gly Leu Asn Asp Ile Phe
690 695 700
Glu Ala Gln Lys Ile Glu Trp His Glu His His His His His His
705 710 715
<210> 6
<211> 719
<212> ПРТ
<213> Искусственная
<220>
<223> Мутантный белок PCSK9 D374Y со вставкой пептида-реципиента биотина BP15 и His-метки: PCSK9-D374Y-BP15-His6, в качестве детектирующего белка
<400> 6
Met Gly Thr Val Ser Ser Arg Arg Ser Trp Trp Pro Leu Pro Leu Leu
1 5 10 15
Leu Leu Leu Leu Leu Leu Leu Gly Pro Ala Gly Ala Arg Ala Gln Glu
20 25 30
Asp Glu Asp Gly Asp Tyr Glu Glu Leu Val Leu Ala Leu Arg Ser Glu
35 40 45
Glu Asp Gly Leu Ala Glu Ala Pro Glu His Gly Thr Thr Ala Thr Phe
50 55 60
His Arg Cys Ala Lys Asp Pro Trp Arg Leu Pro Gly Thr Tyr Val Val
65 70 75 80
Val Leu Lys Glu Glu Thr His Leu Ser Gln Ser Glu Arg Thr Ala Arg
85 90 95
Arg Leu Gln Ala Gln Ala Ala Arg Arg Gly Tyr Leu Thr Lys Ile Leu
100 105 110
His Val Phe His Gly Leu Leu Pro Gly Phe Leu Val Lys Met Ser Gly
115 120 125
Asp Leu Leu Glu Leu Ala Leu Lys Leu Pro His Val Asp Tyr Ile Glu
130 135 140
Glu Asp Ser Ser Val Phe Ala Gln Ser Ile Pro Trp Asn Leu Glu Arg
145 150 155 160
Ile Thr Pro Pro Arg Tyr Arg Ala Asp Glu Tyr Gln Pro Pro Asp Gly
165 170 175
Gly Ser Leu Val Glu Val Tyr Leu Leu Asp Thr Ser Ile Gln Ser Asp
180 185 190
His Arg Glu Ile Glu Gly Arg Val Met Val Thr Asp Phe Glu Asn Val
195 200 205
Pro Glu Glu Asp Gly Thr Arg Phe His Arg Gln Ala Ser Lys Cys Asp
210 215 220
Ser His Gly Thr His Leu Ala Gly Val Val Ser Gly Arg Asp Ala Gly
225 230 235 240
Val Ala Lys Gly Ala Ser Met Arg Ser Leu Arg Val Leu Asn Cys Gln
245 250 255
Gly Lys Gly Thr Val Ser Gly Thr Leu Ile Gly Leu Glu Phe Ile Arg
260 265 270
Lys Ser Gln Leu Val Gln Pro Val Gly Pro Leu Val Val Leu Leu Pro
275 280 285
Leu Ala Gly Gly Tyr Ser Arg Val Leu Asn Ala Ala Cys Gln Arg Leu
290 295 300
Ala Arg Ala Gly Val Val Leu Val Thr Ala Ala Gly Asn Phe Arg Asp
305 310 315 320
Asp Ala Cys Leu Tyr Ser Pro Ala Ser Ala Pro Glu Val Ile Thr Val
325 330 335
Gly Ala Thr Asn Ala Gln Asp Gln Pro Val Thr Leu Gly Thr Leu Gly
340 345 350
Thr Asn Phe Gly Arg Cys Val Asp Leu Phe Ala Pro Gly Glu Asp Ile
355 360 365
Ile Gly Ala Ser Ser Tyr Cys Ser Thr Cys Phe Val Ser Gln Ser Gly
370 375 380
Thr Ser Gln Ala Ala Ala His Val Ala Gly Ile Ala Ala Met Met Leu
385 390 395 400
Ser Ala Glu Pro Glu Leu Thr Leu Ala Glu Leu Arg Gln Arg Leu Ile
405 410 415
His Phe Ser Ala Lys Asp Val Ile Asn Glu Ala Trp Phe Pro Glu Asp
420 425 430
Gln Arg Val Leu Thr Pro Asn Leu Val Ala Ala Leu Pro Pro Ser Thr
435 440 445
His Gly Ala Gly Trp Gln Leu Phe Cys Arg Thr Val Trp Ser Ala His
450 455 460
Ser Gly Pro Thr Arg Met Ala Thr Ala Val Ala Arg Cys Ala Pro Asp
465 470 475 480
Glu Glu Leu Leu Ser Cys Ser Ser Phe Ser Arg Ser Gly Lys Arg Arg
485 490 495
Gly Glu Arg Met Glu Ala Gln Gly Gly Lys Leu Val Cys Arg Ala His
500 505 510
Asn Ala Phe Gly Gly Glu Gly Val Tyr Ala Ile Ala Arg Cys Cys Leu
515 520 525
Leu Pro Gln Ala Asn Cys Ser Val His Thr Ala Pro Pro Ala Glu Ala
530 535 540
Ser Met Gly Thr Arg Val His Cys His Gln Gln Gly His Val Leu Thr
545 550 555 560
Gly Cys Ser Ser His Trp Glu Val Glu Asp Leu Gly Thr His Lys Pro
565 570 575
Pro Val Leu Arg Pro Arg Gly Gln Pro Asn Gln Cys Val Gly His Arg
580 585 590
Glu Ala Ser Ile His Ala Ser Cys Cys His Ala Pro Gly Leu Glu Cys
595 600 605
Lys Val Lys Glu His Gly Ile Pro Ala Pro Gln Glu Gln Val Thr Val
610 615 620
Ala Cys Glu Glu Gly Trp Thr Leu Thr Gly Cys Ser Ala Leu Pro Gly
625 630 635 640
Thr Ser His Val Leu Gly Ala Tyr Ala Val Asp Asn Thr Cys Val Val
645 650 655
Arg Ser Arg Asp Val Ser Thr Thr Gly Ser Thr Ser Glu Gly Ala Val
660 665 670
Thr Ala Val Ala Ile Cys Cys Arg Ser Arg His Leu Ala Gln Ala Ser
675 680 685
Gln Glu Leu Gln Gly Ser Thr Ser Gly Ser Gly Leu Asn Asp Ile Phe
690 695 700
Glu Ala Gln Lys Ile Glu Trp His Glu His His His His His His
705 710 715
<210> 7
<211> 802
<212> ПРТ
<213> Искусственная
<220>
<223> Внеклеточный домен белка-рецептора PCSK9 LDLR с меткой Flag и His-меткой: LDLR-ECD-Flag-His6 в качестве детектирующего реактива
<400> 7
Met Gly Pro Trp Gly Trp Lys Leu Arg Trp Thr Val Ala Leu Leu Leu
1 5 10 15
Ala Ala Ala Gly Thr Ala Val Gly Asp Arg Cys Glu Arg Asn Glu Phe
20 25 30
Gln Cys Gln Asp Gly Lys Cys Ile Ser Tyr Lys Trp Val Cys Asp Gly
35 40 45
Ser Ala Glu Cys Gln Asp Gly Ser Asp Glu Ser Gln Glu Thr Cys Leu
50 55 60
Ser Val Thr Cys Lys Ser Gly Asp Phe Ser Cys Gly Gly Arg Val Asn
65 70 75 80
Arg Cys Ile Pro Gln Phe Trp Arg Cys Asp Gly Gln Val Asp Cys Asp
85 90 95
Asn Gly Ser Asp Glu Gln Gly Cys Pro Pro Lys Thr Cys Ser Gln Asp
100 105 110
Glu Phe Arg Cys His Asp Gly Lys Cys Ile Ser Arg Gln Phe Val Cys
115 120 125
Asp Ser Asp Arg Asp Cys Leu Asp Gly Ser Asp Glu Ala Ser Cys Pro
130 135 140
Val Leu Thr Cys Gly Pro Ala Ser Phe Gln Cys Asn Ser Ser Thr Cys
145 150 155 160
Ile Pro Gln Leu Trp Ala Cys Asp Asn Asp Pro Asp Cys Glu Asp Gly
165 170 175
Ser Asp Glu Trp Pro Gln Arg Cys Arg Gly Leu Tyr Val Phe Gln Gly
180 185 190
Asp Ser Ser Pro Cys Ser Ala Phe Glu Phe His Cys Leu Ser Gly Glu
195 200 205
Cys Ile His Ser Ser Trp Arg Cys Asp Gly Gly Pro Asp Cys Lys Asp
210 215 220
Lys Ser Asp Glu Glu Asn Cys Ala Val Ala Thr Cys Arg Pro Asp Glu
225 230 235 240
Phe Gln Cys Ser Asp Gly Asn Cys Ile His Gly Ser Arg Gln Cys Asp
245 250 255
Arg Glu Tyr Asp Cys Lys Asp Met Ser Asp Glu Val Gly Cys Val Asn
260 265 270
Val Thr Leu Cys Glu Gly Pro Asn Lys Phe Lys Cys His Ser Gly Glu
275 280 285
Cys Ile Thr Leu Asp Lys Val Cys Asn Met Ala Arg Asp Cys Arg Asp
290 295 300
Trp Ser Asp Glu Pro Ile Lys Glu Cys Gly Thr Asn Glu Cys Leu Asp
305 310 315 320
Asn Asn Gly Gly Cys Ser His Val Cys Asn Asp Leu Lys Ile Gly Tyr
325 330 335
Glu Cys Leu Cys Pro Asp Gly Phe Gln Leu Val Ala Gln Arg Arg Cys
340 345 350
Glu Asp Ile Asp Glu Cys Gln Asp Pro Asp Thr Cys Ser Gln Leu Cys
355 360 365
Val Asn Leu Glu Gly Gly Tyr Lys Cys Gln Cys Glu Glu Gly Phe Gln
370 375 380
Leu Asp Pro His Thr Lys Ala Cys Lys Ala Val Gly Ser Ile Ala Tyr
385 390 395 400
Leu Phe Phe Thr Asn Arg His Glu Val Arg Lys Met Thr Leu Asp Arg
405 410 415
Ser Glu Tyr Thr Ser Leu Ile Pro Asn Leu Arg Asn Val Val Ala Leu
420 425 430
Asp Thr Glu Val Ala Ser Asn Arg Ile Tyr Trp Ser Asp Leu Ser Gln
435 440 445
Arg Met Ile Cys Ser Thr Gln Leu Asp Arg Ala His Gly Val Ser Ser
450 455 460
Tyr Asp Thr Val Ile Ser Arg Asp Ile Gln Ala Pro Asp Gly Leu Ala
465 470 475 480
Val Asp Trp Ile His Ser Asn Ile Tyr Trp Thr Asp Ser Val Leu Gly
485 490 495
Thr Val Ser Val Ala Asp Thr Lys Gly Val Lys Arg Lys Thr Leu Phe
500 505 510
Arg Glu Asn Gly Ser Lys Pro Arg Ala Ile Val Val Asp Pro Val His
515 520 525
Gly Phe Met Tyr Trp Thr Asp Trp Gly Thr Pro Ala Lys Ile Lys Lys
530 535 540
Gly Gly Leu Asn Gly Val Asp Ile Tyr Ser Leu Val Thr Glu Asn Ile
545 550 555 560
Gln Trp Pro Asn Gly Ile Thr Leu Asp Leu Leu Ser Gly Arg Leu Tyr
565 570 575
Trp Val Asp Ser Lys Leu His Ser Ile Ser Ser Ile Asp Val Asn Gly
580 585 590
Gly Asn Arg Lys Thr Ile Leu Glu Asp Glu Lys Arg Leu Ala His Pro
595 600 605
Phe Ser Leu Ala Val Phe Glu Asp Lys Val Phe Trp Thr Asp Ile Ile
610 615 620
Asn Glu Ala Ile Phe Ser Ala Asn Arg Leu Thr Gly Ser Asp Val Asn
625 630 635 640
Leu Leu Ala Glu Asn Leu Leu Ser Pro Glu Asp Met Val Leu Phe His
645 650 655
Asn Leu Thr Gln Pro Arg Gly Val Asn Trp Cys Glu Arg Thr Thr Leu
660 665 670
Ser Asn Gly Gly Cys Gln Tyr Leu Cys Leu Pro Ala Pro Gln Ile Asn
675 680 685
Pro His Ser Pro Lys Phe Thr Cys Ala Cys Pro Asp Gly Met Leu Leu
690 695 700
Ala Arg Asp Met Arg Ser Cys Leu Thr Glu Ala Glu Ala Ala Val Ala
705 710 715 720
Thr Gln Glu Thr Ser Thr Val Arg Leu Lys Val Ser Ser Thr Ala Val
725 730 735
Arg Thr Gln His Thr Thr Thr Arg Pro Val Pro Asp Thr Ser Arg Leu
740 745 750
Pro Gly Ala Thr Pro Gly Leu Thr Thr Val Glu Ile Val Thr Met Ser
755 760 765
His Gln Ala Leu Gly Asp Val Ala Gly Arg Gly Asn Glu Lys Lys Pro
770 775 780
Ser Ser Val Arg Asp Tyr Lys Asp Asp Asp Asp Lys His His His His
785 790 795 800
His His
<210> 8
<211> 331
<212> ПРТ
<213> Искусственная
<220>
<223> Гибридный белок укороченного внеклеточного домена LDLR с hIgG1 Fc (с PCSK9-связывающей активностью): LDLR-sECD–Fc (hIgG1) в качестве детектирующего реактива
<400> 8
Met Glu Phe Gly Leu Ser Trp Leu Phe Leu Val Ala Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Gly Thr Asn Glu Cys Leu Asp Asn Asn Gly Gly Cys Ser
20 25 30
His Val Cys Asn Asp Leu Lys Ile Gly Tyr Glu Cys Leu Cys Pro Asp
35 40 45
Gly Phe Gln Leu Val Ala Gln Arg Arg Cys Glu Asp Ile Asp Glu Cys
50 55 60
Gln Asp Pro Asp Thr Cys Ser Gln Leu Cys Val Asn Leu Glu Gly Gly
65 70 75 80
Tyr Lys Cys Gln Cys Glu Glu Gly Phe Gln Leu Asp Pro His Thr Lys
85 90 95
Ala Cys Lys Glu Pro Lys Ser Ser Asp Lys Thr His Thr Cys Pro Pro
100 105 110
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
115 120 125
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
130 135 140
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
145 150 155 160
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
165 170 175
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
180 185 190
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
195 200 205
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
210 215 220
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
225 230 235 240
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
245 250 255
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
260 265 270
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
275 280 285
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
290 295 300
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
305 310 315 320
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 9
<211> 294
<212> ПРТ
<213> Искусственная
<220>
<223> Гибридный белок более укороченного внеклеточного домена LDLR с hIgG1 Fc (с PCSK9-связывающей активностью): LDLR-ssECD-Fc (hIgG1) в качестве детектирующего реактива
<400> 9
Met Glu Phe Gly Leu Ser Trp Leu Phe Leu Val Ala Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Gly Thr Asn Glu Cys Leu Asp Asn Asn Gly Gly Cys Ser
20 25 30
His Val Cys Asn Asp Leu Lys Ile Gly Tyr Glu Cys Leu Cys Pro Asp
35 40 45
Gly Phe Gln Leu Val Ala Gln Arg Arg Cys Glu Asp Ile Asp Glu Pro
50 55 60
Lys Ser Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
65 70 75 80
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
85 90 95
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
100 105 110
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
115 120 125
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
130 135 140
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
145 150 155 160
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
165 170 175
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
180 185 190
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
195 200 205
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
210 215 220
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
225 230 235 240
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
245 250 255
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
260 265 270
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
275 280 285
Ser Leu Ser Pro Gly Lys
290
<210> 10
<211> 121
<212> ПРТ
<213> Мышиная
<220>
<223> mAb-001 VH
<400> 10
Gln Val His Leu Gln Gln Ser Gly Ala Glu Leu Ala Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Asn Asp Tyr
20 25 30
Trp Met His Trp Val Lys Glu Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Phe Thr Lys Tyr His Gln Asn Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Tyr Asp Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Tyr Asp Tyr Asp Glu Asp Trp Tyr Phe Asp Val Trp Gly
100 105 110
Thr Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 11
<211> 112
<212> ПРТ
<213> Мышиная
<220>
<223> mAb-001VL
<400> 11
Asp Ile Val Met Ser Gln Ser Pro Ser Ser Leu Ala Val Ser Ala Gly
1 5 10 15
Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Phe Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Arg Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Lys Gln
85 90 95
Ser Phe Asn Leu Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 12
<211> 5
<212> ПРТ
<213> Мышиный
<220>
<223> mAb-001 HCDR1
<400> 12
Asp Tyr Trp Met His
1 5
<210> 13
<211> 17
<212> ПРТ
<213> Мышиная
<220>
<223> mAb-001 HCDR2
<400> 13
Tyr Ile Asn Pro Ser Ser Gly Phe Thr Lys Tyr His Gln Asn Phe Lys
1 5 10 15
Asp
<210> 14
<211> 12
<212> ПРТ
<213> Мышиная
<220>
<223> mAb-001 HCDR3
<400> 14
Gln Tyr Asp Tyr Asp Glu Asp Trp Tyr Phe Asp Val
1 5 10
<210> 15
<211> 17
<212> ПРТ
<213> Мышиная
<220>
<223> mAb-001 LCDR1
<400> 15
Lys Ser Ser Gln Ser Leu Leu Asn Ser Arg Thr Arg Lys Asn Phe Leu
1 5 10 15
Ala
<210> 16
<211> 7
<212> ПРТ
<213> Мышиная
<220>
<223> mAb-001 LCDR2
<400> 16
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 17
<211> 8
<212> ПРТ
<213> Мышиная
<220>
<223> mAb-001 LCDR3
<400> 17
Lys Gln Ser Phe Asn Leu Phe Thr
1 5
<210> 18
<211> 121
<212> ПРТ
<213> Искусственная
<220>
<223> h001-VH.1
<400> 18
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Phe Thr Lys Tyr His Gln Asn Phe
50 55 60
Lys Asp Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Tyr Asp Tyr Asp Glu Asp Trp Tyr Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 19
<211> 121
<212> ПРТ
<213> Искусственная
<220>
<223> h001-VH.1A
<400> 19
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Asn Asp Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Phe Thr Lys Tyr His Gln Asn Phe
50 55 60
Lys Asp Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Tyr Asp Tyr Asp Glu Asp Trp Tyr Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 20
<211> 121
<212> ПРТ
<213> Искусственная
<220>
<223> h001-VH.1B
<400> 20
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Phe Thr Lys Tyr His Gln Asn Phe
50 55 60
Lys Asp Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Thr Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Tyr Asp Tyr Asp Glu Asp Trp Tyr Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 21
<211> 121
<212> ПРТ
<213> Искусственная
<220>
<223> h001-VH.1C
<400> 21
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Asn Asp Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Phe Thr Lys Tyr His Gln Asn Phe
50 55 60
Lys Asp Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Thr Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Tyr Asp Tyr Asp Glu Asp Trp Tyr Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 22
<211> 121
<212> ПРТ
<213> Искусственная
<220>
<223> h001-VH.1D
<400> 22
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Asn Asp Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Phe Thr Lys Tyr His Gln Asn Phe
50 55 60
Lys Asp Arg Val Thr Met Thr Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Thr Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Tyr Asp Tyr Asp Glu Asp Trp Tyr Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 23
<211> 121
<212> ПРТ
<213> Искусственная
<220>
<223> h001-VH.1E
<400> 23
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Asn Asp Tyr
20 25 30
Trp Met His Trp Val Lys Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Phe Thr Lys Tyr His Gln Asn Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Thr Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Tyr Asp Tyr Asp Glu Asp Trp Tyr Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 24
<211> 112
<212> ПРТ
<213> Искусственная
<220>
<223> h001-VL.1 (трансплантация CDR)
<400> 24
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Phe Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln
85 90 95
Ser Phe Asn Leu Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 25
<211> 112
<212> ПРТ
<213> Искусственная
<220>
<223> h001-VL.1A
<400> 25
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Phe Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln
85 90 95
Ser Phe Asn Leu Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 26
<211> 112
<212> ПРТ
<213> Искусственная
<220>
<223> h001-VL.1B
<400> 26
Asp Ile Gln Met Ser Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Phe Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln
85 90 95
Ser Phe Asn Leu Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 27
<211> 112
<212> ПРТ
<213> Искусственная
<220>
<223> h001-VL.1C
<400> 27
Asp Ile Val Met Ser Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Phe Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln
85 90 95
Ser Phe Asn Leu Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 28
<211> 451
<212> ПРТ
<213> Искусственная
<220>
<223> h001-4-IgG1 аминокислотная последовательность тяжелой цепи
<400> 28
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Phe Thr Lys Tyr His Gln Asn Phe
50 55 60
Lys Asp Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Tyr Asp Tyr Asp Glu Asp Trp Tyr Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 29
<211> 1413
<212> ДНК
<213> Искусственная
<220>
<223> h001-4-IgG1 Последовательность ДНК тяжелой цепи
<400> 29
atggagtttg ggctgagctg gctttttctt gtcgcgattc ttaagggtgt ccagtgccag 60
gtgcagctgg tgcagagcgg cgctgaggtg aagaagcccg gagcgagcgt aaaggtgagc 120
tgcaaggcca gcggatacac cttcaccgac tactggatgc actgggtgag gcaggcccca 180
ggacagggcc tggagtggat gggctacatc aaccccagca gcggctttac caagtatcac 240
cagaacttca aagacagggt gaccatgacc agggacacca gcatcagcac cgcctacatg 300
gagctgagca ggctgaggag cgacgacacc gccgtgtact actgcgccag gcaatacgac 360
tacgacgagg actggtactt cgacgtgtgg ggccaaggaa ccaccgtgac tgtgagcagc 420
gcttcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 480
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 540
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 600
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 660
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 720
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 780
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 840
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 900
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 960
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 1020
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1080
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 1140
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1200
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1260
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 1320
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1380
cagaagagcc tctccctgtc tccgggtaaa tga 1413
<210> 30
<211> 219
<212> ПРТ
<213> Искусственная
<220>
<223> h001-4-каппа Аминокислотная последовательность легкой цепи
<400> 30
Asp Ile Val Met Ser Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Phe Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Lys Gln
85 90 95
Ser Phe Asn Leu Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 31
<211> 726
<212> ДНК
<213> Искусственная
<220>
<223> h001-4-каппа Последовательность ДНК легкой цепи
<400> 31
atggacatgc gcgtgcccgc ccagctgctg ggcctgctgc tgctgtggtt ccccggctcg 60
cgatgcgaca tcgtgatgtc tcagagccca tctagcctga gcgccagcgt gggcgacagg 120
gtaaccatca cctgcaagag cagccaaagc ctgctgaaca gcaggacccg caagaacttc 180
ctggcttggt atcagcagaa gcccggcaag tctcccaagt tgctgatcta ctgggccagc 240
accagggaga gcggcgtgcc cgacaggttc agcggctccg gcagcggcac cgacttcacc 300
ctgaccatct ctagtctgca gcccgaggac ttcgccacct actactgcaa gcagagcttc 360
aatctgttca ccttcggcca gggcaccaag ctggagatca agcgtacggt ggctgcacca 420
tctgtcttca tcttcccgcc atctgatgag cagttgaaat ctggaactgc ctctgttgtg 480
tgcctgctga ataacttcta tcccagagag gccaaagtac agtggaaggt ggataacgcc 540
ctccaatcgg gtaactccca ggagagtgtc acagagcagg acagcaagga cagcacctac 600
agcctcagca gcaccctgac gctgagcaaa gcagactacg agaaacacaa agtctacgcc 660
tgcgaagtca cccatcaggg cctgagctcg cccgtcacaa agagcttcaa caggggagag 720
tgttga 726
<210> 32
<211> 451
<212> ПРТ
<213> Искусственная
<220>
<223> h001-4-IgG1-YTE Аминокислотная последовательность тяжелой цепи
<400> 32
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Phe Thr Lys Tyr His Gln Asn Phe
50 55 60
Lys Asp Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Tyr Asp Tyr Asp Glu Asp Trp Tyr Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Tyr
245 250 255
Ile Thr Arg Glu Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 33
<211> 1413
<212> ДНК
<213> Искусственная
<220>
<223> h001-4-IgG1-YTE Последовательность ДНК тяжелой цепи
<400> 33
atggagtttg ggctgagctg gctttttctt gtcgcgattc ttaagggtgt ccagtgccag 60
gtgcagctgg tgcagagcgg cgctgaggtg aagaagcccg gagcgagcgt aaaggtgagc 120
tgcaaggcca gcggatacac cttcaccgac tactggatgc actgggtgag gcaggcccca 180
ggacagggcc tggagtggat gggctacatc aaccccagca gcggctttac caagtatcac 240
cagaacttca aagacagggt gaccatgacc agggacacca gcatcagcac cgcctacatg 300
gagctgagca ggctgaggag cgacgacacc gccgtgtact actgcgccag gcaatacgac 360
tacgacgagg actggtactt cgacgtgtgg ggccaaggaa ccaccgtgac tgtgagcagc 420
gcttcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 480
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 540
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 600
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 660
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 720
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 780
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tctacatcac ccgggagcct 840
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 900
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 960
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 1020
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1080
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag 1140
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1200
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1260
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 1320
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1380
cagaagagcc tctccctgtc tccgggtaaa tga 1413
<---
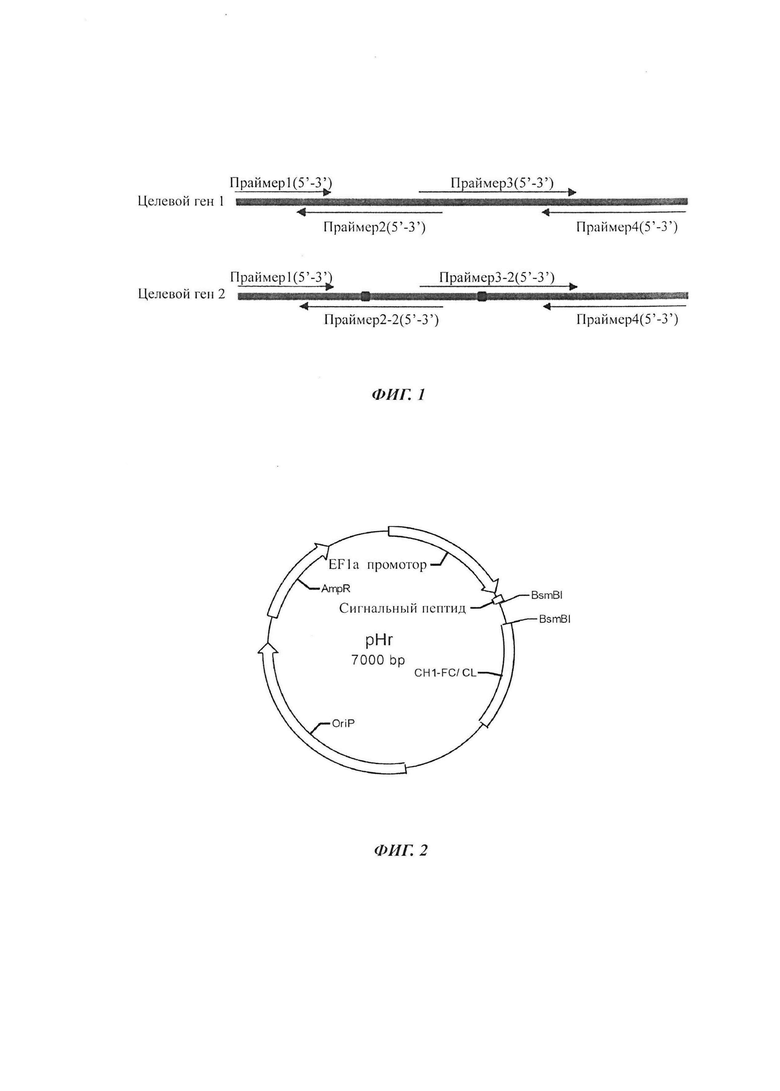
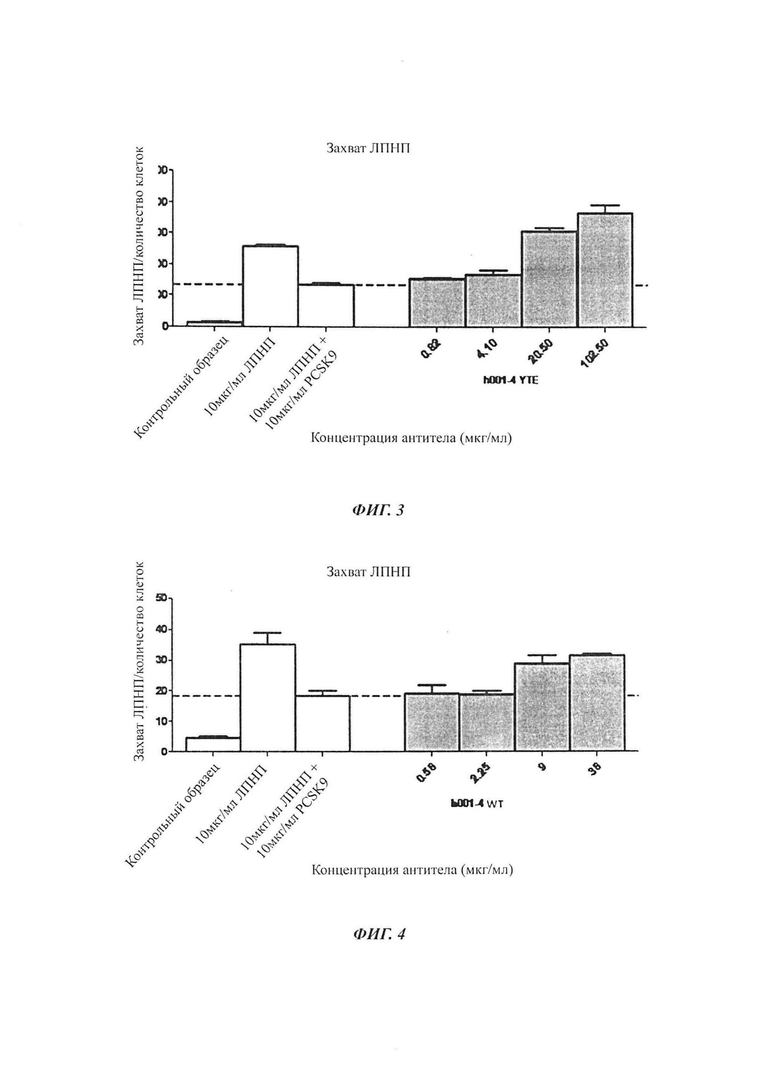
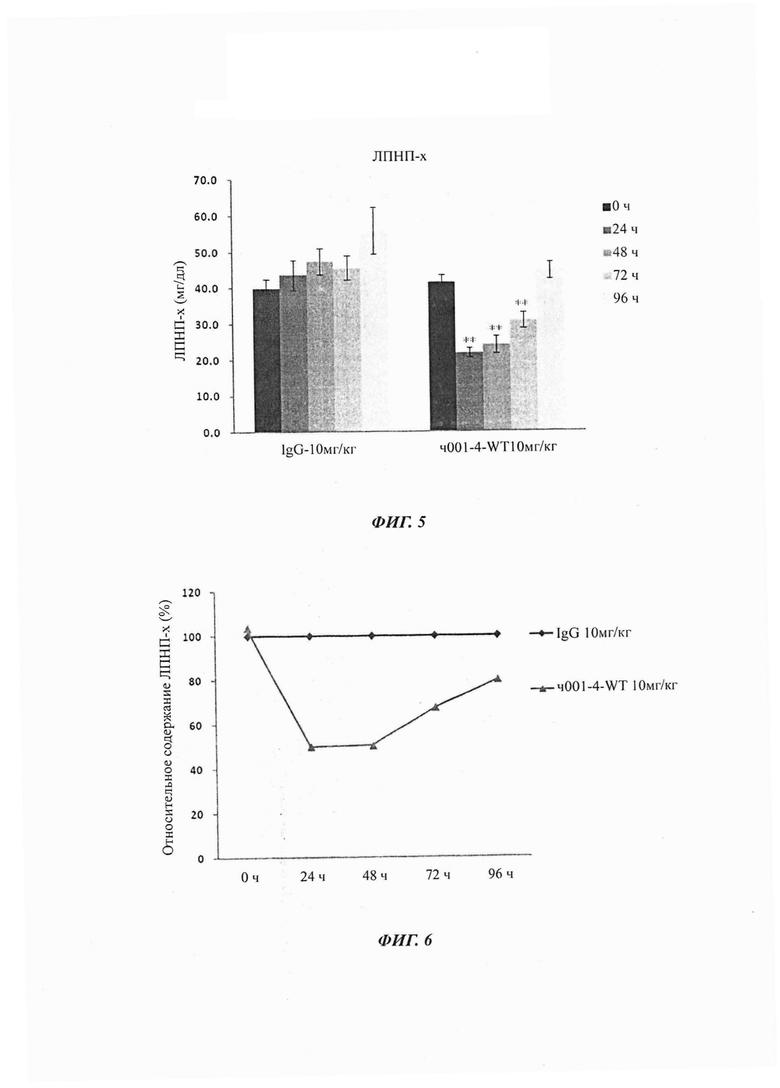
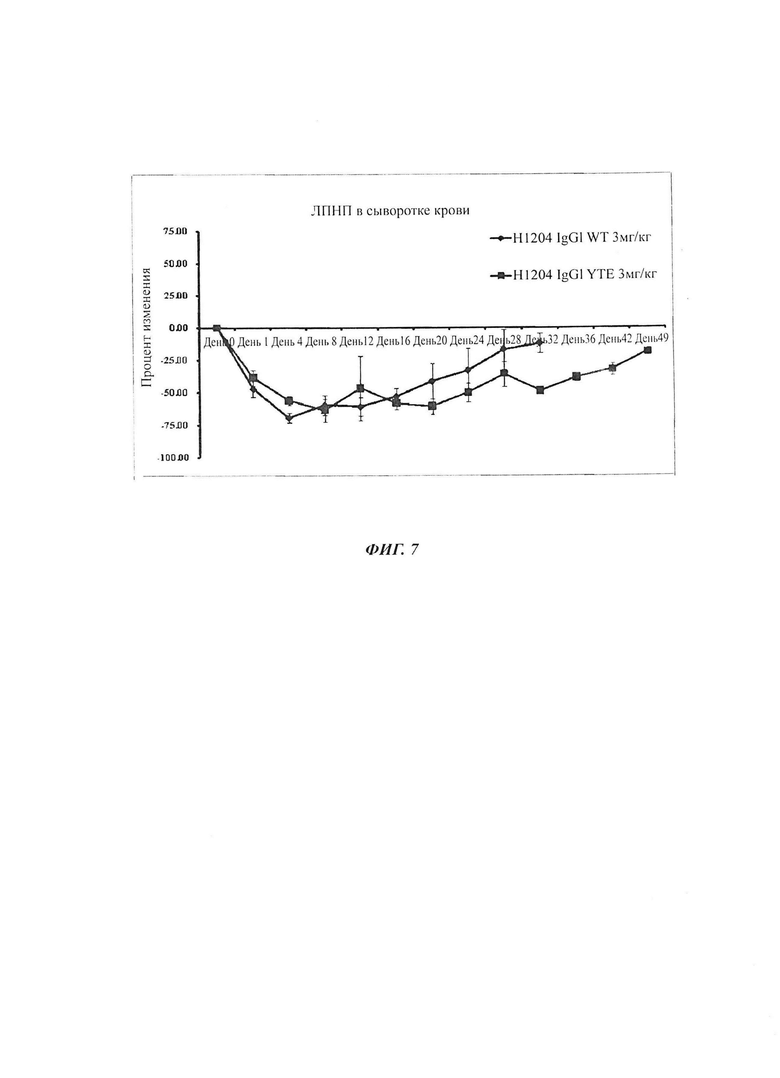
Изобретение относится к области биотехнологии. Предложено анти-PCSK9 антитело, его антигенсвязывающий фрагмент, кодирующие их нуклеиновые кислоты и экспрессионные векторы. Также изобретение относится к фармацевтической композиции, содержащей анти-PCSK9 антитело, и применению указанного антитела в качестве гиполипидемического средства. Предложенное антитело к PCSK9В обладает повышенной аффинностью, высокой селективностью и высокой биологической активностью, и может быть использовано для снижения уровня липидов в крови. 6 н. и 18 з.п. ф-лы, 10 табл., 5 пр., 7 ил.
1. Анти-PCSK9 антитело или его антигенсвязывающий фрагмент, содержащие: HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 12, SEQ ID NO: 13 и SEQ ID NO: 14 соответственно; и LCDR1, LCDR2 и LCDR3, как показано в SEQ ID NO: 15, SEQ ID NO: 16 и SEQ ID NO: 17 соответственно.
2. Анти-PCSK9 антитело или его антигенсвязывающий фрагмент по п. 1, где анти-PCSK9 антитело или его антигенсвязывающий фрагмент представляют собой мышиное антитело или его фрагмент.
3. Анти-PCSK9 антитело или его антигенсвязывающий фрагмент по п. 1 или 2, где вариабельная область легкой цепи анти-PCSK9 антитела дополнительно содержит каркасные (FR) области легкой цепи, имеющие происхождение от цепи κ мыши или ее варианта, или FR области легкой цепи, имеющие происхождение от цепи λ мыши или ее варианта; где вариабельная область тяжелой цепи анти-PCSK9 антитела дополнительно содержит FR области тяжелой цепи, имеющие происхождение от IgG1 мыши или его варианта, или FR области тяжелой цепи, имеющие происхождение от IgG2 мыши или его варианта, или FR области тяжелой цепи, имеющие происхождение от IgG3 мыши или его варианта.
4. Анти-PCSK9 антитело или его антигенсвязывающий фрагмент по п. 3, содержащие вариабельную область тяжелой цепи с последовательностью SEQ ID NO: 10 и вариабельную область легкой цепи с последовательностью SEQ ID NO: 11.
5. Анти-PCSK9 антитело или его антигенсвязывающий фрагмент по любому из пп. 1–4, где легкая цепь анти-PCSK9 антитела дополнительно содержит константные области легкой цепи, имеющие происхождение от цепи κ мыши или ее варианта, или константные области легкой цепи, имеющие происхождение от цепи λ мыши или ее варианта; где тяжелая цепь анти-PCSK9 антитела дополнительно содержит константные области тяжелой цепи, имеющие происхождение от IgG1 мыши или его варианта, или константные области тяжелой цепи, имеющие происхождение от IgG2 мыши или его варианта, или константные области тяжелой цепи, имеющие происхождение от IgG3 мыши или его варианта.
6. Анти-PCSK9 антитело или его антигенсвязывающий фрагмент по п. 1, где анти-PCSK9 антитело или его антигенсвязывающий фрагмент представляют собой химерное антитело или его фрагмент.
7. Анти-PCSK9 антитело или его антигенсвязывающий фрагмент по п. 1, где анти-PCSK9 антитело или его антигенсвязывающий фрагмент представляют собой гуманизированное антитело или его фрагмент.
8. Анти-PCSK9 антитело или его антигенсвязывающий фрагмент по п. 7, где последовательность FR тяжелой цепи вариабельной области тяжелой цепи гуманизированного антитела имеет происхождение от комбинированной последовательности тяжелых цепей зародышевой линии человека IGHV1-2*02 и hjh2 или ее мутантной последовательности; где гуманизированное антитело содержит FR1, FR2, FR3 тяжелой цепи зародышевой линии человека IGHV1-2*02 и FR4 hjh2 или ее мутантной последовательности.
9. Анти-PCSK9 антитело или его антигенсвязывающий фрагмент по п. 8, где гуманизированное антитело содержит вариабельную область тяжелой цепи с последовательностью SEQ ID NO: 18; или вариабельную область тяжелой цепи, как показано в варианте SEQ ID NO: 18; где вариант SEQ ID NO: 18 представляет собой последовательность с 1–10 заменами аминокислот в вариабельной области тяжелой цепи, как показано в SEQ ID NO: 18.
10. Анти-PCSK9 антитело или его антигенсвязывающий фрагмент по п. 9, где вариант SEQ ID NO: 18 имеет 1-10 обратных мутаций аминокислот в FR области вариабельной области тяжелой цепи, как показано в SEQ ID NO: 18; предпочтительно обратная мутация выбрана из группы, состоящей из T30N, R87T, R72А, T74K, M48I , V68A, M70L, R38K и R67K.
11. Анти-PCSK9 антитело или его антигенсвязывающий фрагмент по п. 8, где гуманизированное антитело содержит вариабельную область тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 19, SEQ ID NO: 20, SEQ ID NO: 21, SEQ ID NO: 22 и SEQ ID NO: 23.
12. Анти-PCSK9 антитело или его антигенсвязывающий фрагмент по п. 7, где последовательность FR легкой цепи вариабельной области легкой цепи гуманизированного антитела имеет происхождение от комбинированной последовательности легких цепей зародышевой линии человека IGKV1 -39*01 и hjk2.1 или ее мутантной последовательности; где гуманизированное антитело содержит FR1, FR2, FR3 зародышевой линии человека IGKV1-39*01 и FR4 hjk2.1 или ее мутантной последовательности.
13. Анти-PCSK9 антитело или его антигенсвязывающий фрагмент по п. 12, где гуманизированное антитело дополнительно содержит вариабельную область легкой цепи, как показано в SEQ ID NO: 24, или вариабельную область легкой цепи, как показано в варианте SEQ ID NO: 24; где вариант SEQ ID NO: 24 имеет 1-10 замен аминокислот в вариабельной области легкой цепи, как показано в SEQ ID NO: 24.
14. Анти-PCSK9 антитело или его антигенсвязывающий фрагмент по п. 13, где вариант SEQ ID NO: 24 имеет 1-10 обратных мутаций аминокислот в FR области вариабельной области легкой цепи, как показано в SEQ ID NO: 24; предпочтительно обратная мутация выбрана из группы, состоящей из T5S, S66D, Q3V и A49S, предпочтительно A49S.
15. Анти-PCSK9 антитело или его антигенсвязывающий фрагмент по п. 12, где гуманизированное антитело содержит вариабельную область легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 25, SEQ ID NO: 26 и SEQ ID NO: 27.
16. Анти-PCSK9 антитело или его антигенсвязывающий фрагмент по п. 7, где гуманизированное антитело содержит вариабельную область тяжелой цепи и/или вариабельную область легкой цепи, где вариабельная область тяжелой цепи выбрана из группы, состоящей из SEQ ID NO: 19, SEQ ID NO: 20, SEQ ID NO: 21, SEQ ID NO: 22 и SEQ ID NO: 23, или вариабельная область тяжелой цепи выбрана из группы, состоящей из последовательностей, обладающих по меньшей мере 95% идентичностью с SEQ ID NO: 19, SEQ ID NO: 20, SEQ ID NO: 21, SEQ ID NO: 22 и SEQ ID NO: 23; где вариабельная область легкой цепи выбрана из группы, состоящей из SEQ ID NO: 25, SEQ ID NO: 26 и SEQ ID NO: 27, или вариабельная область легкой цепи выбрана из группы, состоящей из последовательностей, обладающих по меньшей мере 95% идентичностью с SEQ ID NO: 25, SEQ ID NO: 26 и SEQ ID NO: 27.
17. Анти-PCSK9 антитело или его антигенсвязывающий фрагмент по п. 7, где анти-PCSK9 антитело содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, выбранные из группы, состоящей из:
1) последовательности вариабельной области тяжелой цепи SEQ ID NO: 18 и последовательности вариабельной области легкой цепи SEQ ID NO: 25;
2) последовательности вариабельной области тяжелой цепи SEQ ID NO: 18 и последовательности вариабельной области легкой цепи SEQ ID NO: 26;
3) последовательности вариабельной области тяжелой цепи SEQ ID NO: 18 и последовательности вариабельной области легкой цепи SEQ ID NO: 27;
4) последовательности вариабельной области тяжелой цепи SEQ ID NO: 19 и последовательности вариабельной области легкой цепи SEQ ID NO: 24;
5) последовательности вариабельной области тяжелой цепи SEQ ID NO: 19 и последовательности вариабельной области легкой цепи SEQ ID NO: 25;
6) последовательности вариабельной области тяжелой цепи SEQ ID NO: 19 и последовательности вариабельной области легкой цепи SEQ ID NO: 26;
7) последовательности вариабельной области тяжелой цепи SEQ ID NO: 19 и последовательности вариабельной области легкой цепи SEQ ID NO: 27;
8) последовательности вариабельной области тяжелой цепи SEQ ID NO: 20 и последовательности вариабельной области легкой цепи SEQ ID NO: 24;
9) последовательности вариабельной области тяжелой цепи SEQ ID NO: 20 и последовательности вариабельной области легкой цепи SEQ ID NO: 25;
10) последовательности вариабельной области тяжелой цепи SEQ ID NO: 20 и последовательности вариабельной области легкой цепи SEQ ID NO: 26;
11) последовательности вариабельной области тяжелой цепи SEQ ID NO: 20 и последовательности вариабельной области легкой цепи SEQ ID NO: 27;
12) последовательности вариабельной области тяжелой цепи SEQ ID NO: 21 и последовательности вариабельной области легкой цепи SEQ ID NO: 24;
13) последовательности вариабельной области тяжелой цепи SEQ ID NO: 21 и последовательности вариабельной области легкой цепи SEQ ID NO: 25;
14) последовательности вариабельной области тяжелой цепи SEQ ID NO: 21 и последовательности вариабельной области легкой цепи SEQ ID NO: 26;
15) последовательности вариабельной области тяжелой цепи SEQ ID NO: 21 и последовательности вариабельной области легкой цепи SEQ ID NO: 27;
16) последовательности вариабельной области тяжелой цепи SEQ ID NO: 22 и последовательности вариабельной области легкой цепи SEQ ID NO: 24;
17) последовательности вариабельной области тяжелой цепи SEQ ID NO: 22 и последовательности вариабельной области легкой цепи SEQ ID NO: 25;
18) последовательности вариабельной области тяжелой цепи SEQ ID NO: 22 и последовательности вариабельной области легкой цепи SEQ ID NO: 26;
19) последовательности вариабельной области тяжелой цепи SEQ ID NO: 22 и последовательности вариабельной области легкой цепи SEQ ID NO: 27;
20) последовательности вариабельной области тяжелой цепи SEQ ID NO: 23 и последовательности вариабельной области легкой цепи SEQ ID NO: 24;
21) последовательности вариабельной области тяжелой цепи SEQ ID NO: 23 и последовательности вариабельной области легкой цепи SEQ ID NO: 25;
22) последовательности вариабельной области тяжелой цепи SEQ ID NO: 23 и последовательности вариабельной области легкой цепи SEQ ID NO: 26;
23) последовательности вариабельной области тяжелой цепи SEQ ID NO: 23 и последовательности вариабельной области легкой цепи SEQ ID NO: 27; и
24) последовательности вариабельной области тяжелой цепи SEQ ID NO: 18 и последовательности вариабельной области легкой цепи SEQ ID NO: 24.
18. Анти-PCSK9 антитело или его антигенсвязывающий фрагмент по любому из пп. 6-17, где тяжелая цепь анти-PCSK9 антитела дополнительно содержит константные области тяжелой цепи, имеющие происхождение от IgG1, IgG2, IgG3 или IgG4 человека или его варианта, либо аминокислотные последовательности, обладающие по меньшей мере 95% идентичностью с их последовательностями; предпочтительно содержит константные области тяжелой цепи, имеющие происхождение от IgG1, IgG2 или IgG4 человека или содержит константные области тяжелой цепи вариантов IgG1, IgG2 или IgG4 человека, продлевающих период полувыведения антитела из сыворотки крови за счет мутации аминокислоты, наиболее предпочтительно содержит константные области тяжелой цепи IgG1, IgG2 или IgG4, в которые введена мутация YTE; где анти-PCSK9 антитело дополнительно содержит константную область легкой цепи, имеющую происхождение от цепи κ человека, цепи λ человека или ее варианта, либо аминокислотные последовательности, обладающие по меньшей мере 95% идентичностью их последовательностям.
19. Анти-PCSK9 антитело или его антигенсвязывающий фрагмент по п. 18, где гуманизированное антитело содержит тяжелую цепь и легкую цепь, выбранные из группы, состоящей из:
1) тяжелой цепи с последовательностью SEQ ID NO: 28 и легкой цепи с последовательностью SEQ ID NO: 30; и 2) тяжелой цепи с последовательностью SEQ ID NO: 32 и легкой цепи с последовательностью SEQ ID NO: 30.
20. Фармацевтическая композиция для лечения PCSK9-опосредованного заболевания или расстройства, содержащая терапевтически эффективное количество анти-PCSK9 антитела или его антигенсвязывающего фрагмента по любому из пп. 1-19 и одно или более из фармацевтически приемлемых носителей, разбавителей или вспомогательных веществ.
21. Молекула ДНК, кодирующая анти-PCSK9 антитело или его антигенсвязывающий фрагмент по любому из пп. 1–19.
22. Экспрессионный вектор, содержащий молекулу ДНК по п. 21.
23. Клетка-хозяин, трансформированная экспрессионным вектором по п. 22, для продуцирования анти-PCSK9 антитела или его антигенсвязывающего фрагмента по любому из пп. 1–19, которая выбрана из группы, состоящей из прокариотических клеток и эукариотических клеток, предпочтительно эукариотических клеток, более предпочтительно клеток млекопитающих.
24. Применение анти-PCSK9 антитела или его антигенсвязывающего фрагмента по любому из пп. 1–19 или фармацевтической композиции по п. 20 в получении лекарственного средства для лечения PCSK9-опосредованного заболевания или расстройства, где заболевание или расстройство предпочтительно представляет собой заболевание, связанные с повышенным уровнем холестерина; более предпочтительно выбрано из группы, состоящей из гиперхолестеринемии, заболевания сердца, метаболического синдрома, сахарного диабета, ишемической болезни сердца, инсультов, сердечно-сосудистого заболевания (ССЗ), болезни Альцгеймера и общей дислипидемии; наиболее предпочтительно гиперхолестеринемии, дислипидемии, атеросклероза, ССЗ или ишемической болезни сердца.
| US 2009142352 A1, 04.06.2009 | |||
| WO 2010029513 A2, 18.03.2010 | |||
| ВЫСОКОАФФИННЫЕ ЧЕЛОВЕЧЕСКИЕ АНТИТЕЛА К PCSK9 | 2009 |
|
RU2552169C2 |
| SUAREZ E | |||
| et al., Rearrangement of only one human IGHV gene is sufficient to generate a wide repertoire of antigen specific antibody responses in transgenic mice, Molecular Immunology, 2006, Volume 43, Issue 11, pp.1827-1835. | |||

