Данная заявка испрашивает приоритет по китайской патентной заявке №201910480169.4 (название: Антитело против фактора роста соединительной ткани и его применение, дата приоритета: 4 июня 2019 года).
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее раскрытие относится к области биомедицины и в частности относится к антителам, связывающимся с фактором роста соединительной ткани (CTGF - от англ. Connective Tissue Growth Factor), и их применениям.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Согласно описаниям в данном документе предложена только общая информация о настоящем раскрытии, и они не обязательно составляют предшествующий уровень техники.
Экспрессия CTGF индуцируется членами суперсемейства трансформирующих факторов роста бета (TGFβ - от англ. Transforming Growth Factor beta). Данное суперсемейство включает TGFβ-1, -2 и -3, костный морфогенетический белок (BMP - от англ. Bone Morphogenetic Protein)-2 и активин. Много регуляторов (включая дексаметазон, тромбин, фактор роста эндотелия сосудов (VEGF - от англ. Vascular Endothelial Growth Factor) и ангиотензин II) и напряжение, обусловленное воздействием окружающей среды (включая гипергликемию и гипертензию), также индуцируют экспрессию CTGF (см., например, Franklin (1997) Int J Biochem Cell Biol 29:79-89; Wunderlich (2000) Graefes Arch Clin Exp Ophthalmol 238: 910-915; Denton and Abraham (2001) Curr Opin Rheumatol 13: 505-511; и Riewald (2001) Blood 97: 3109-3116; Riser et al. (2000) J Am SocNephrol 11: 25-38; и International Publication WO 00/13706).
Стимулирующее действие TGFβ на экспрессию CTGF является быстрым и длительным, и нет необходимости в целенаправленном применении TGFβ (Igarashi et al. (1993) Mol BiolCell 4:637-645). TGFβ активирует транскрипцию через регуляторные элементы ДНK, находящиеся в промоторе CTGF, приводя к усиленной экспрессии CTGF (Grotendorst et al. (1996) Cell Growth Differ 7: 469-480; Grotendorst and Bradham, Патент США №6069006; Holmes et al. (2001) J Biol Chem 276: 10594-10601).
Экспрессия CTGF подвергается повышающей регуляции при гломерулонефрите, IgA-неф ропати и, интерфокальном и сегментарном гломерулосклерозе и диабетической нефропатии (см., например, Riser et al. (2000) J Am Soc Nephrol 11:25 -38). Увеличение числа клеток, экспрессирующих CTGF, также наблюдалось в участках хронического тубулоинтерстициального поражения, и уровень CTGF коррелирует со степенью поражения (Ito et al, (1998) Kidney Int 53:853-861). Кроме того, при нефропатии различного рода, ассоциированной с рубцеванием почечной паренхимы и склерозом, также повышается уровень экспрессии CTGF в клубочке или тубулоинтерстиции. Повышенные уровни CTGF также ассоциированы с фиброзом печени, инфарктом миокарда и фиброзом легких. Например, CTGF подвергается сильной повышающей регуляции в клетках из биопсий и жидкости бронхоальвеолярного лаважа у пациентов со спонтанным фиброзом легких (IPF) (Ujike et al, (2000) Biochem Biophys Res Commun 277:448-454; Abou -Shady et al. (2000) Liver 20: 296-304; Williams et al. (2000) J Hepatol 32: 754-761). Таким образом, CTGF представляет собой эффективную терапевтическую мишень при заболеваниях, описанных выше.
Однако, в настоящее время в клинической практике не используется эффективное средство на основе антитела, нацеленного на CTGF, и все еще существует необходимость в разработке новых безопасных и эффективных средств на основе антитела против CTGF.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В настоящем раскрытии предложено антитело против CTGF. Антитело против CTGF включает полноразмерные антитела против человеческого CTGF и его антигенсвязывающие фрагменты.
В некоторых воплощениях антитело против CTGF содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, где:
i) вариабельная область тяжелой цепи содержит такие же последовательности HCDR1, HCDR2 и HCDR3, как и последовательности HCDR1, HCDR2 и HCDR3 вариабельной области тяжелой цепи, показанной в SEQ ID NO: 6, и вариабельная область легкой цепи содержит такие же последовательности LCDR1, LCDR2 и LCDR3, как и последовательности LCDR1, LCDR2 и LCDR3 вариабельной области легкой цепи, показанной в SEQ ID NO: 7; или
ii) вариабельная область тяжелой цепи содержит такие же последовательности HCDR1, HCDR2 и HCDR3, как и последовательности HCDR1, HCDR2 и HCDR3 вариабельной области тяжелой цепи, показанной в SEQ ID NO: 8,
и вариабельная область легкой цепи содержит такие же последовательности LCDR1, LCDR2 и LCDR3, как и последовательности LCDR1, LCDR2 и LCDR3 вариабельной области легкой цепи, показанной в SEQ ID NO: 9.
В некоторых воплощениях антитело против CTGF содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, где:
ii-i) вариабельная область тяжелой цепи содержит такие же последовательности HCDR1, HCDR2 и HCDR3, как последовательности HCDR1, HCDR2 и HCDR3 вариабельной области тяжелой цепи, показанной в SEQ ID NO: 69, и вариабельная область легкой цепи содержит такие же последовательности LCDR1, LCDR2 и LCDR3, как и последовательности LCDR1, LCDR2 и LCDR3 вариабельной области легкой цепи, показанной в SEQ ID NO: 70; или
ii-ii) вариабельная область тяжелой цепи содержит такие же последовательности HCDR1, HCDR2 и HCDR3, как последовательности HCDR1, HCDR2 и HCDR3 вариабельной области тяжелой цепи, показанной в SEQ ID NO: 85, и вариабельная область легкой цепи содержит такие же последовательности LCDR1, LCDR2 и LCDR3, как и последовательности LCDR1, LCDR2 и LCDR3 вариабельной области легкой цепи, показанной в SEQ ID NO: 70.
В некоторых воплощениях антитело против CTGF содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, где:
iii) вариабельная область тяжелой цепи содержит HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 10, SEQ ID NO: 11 и SEQ ID NO: 12, соответственно, и вариабельная область легкой цепи содержит LCDR1, LCDR2 и LCDR3, как показано в SEQ ID NO: 13, SEQ ID NO: 14 и SEQ ID NO: 15, соответственно; или
iv) вариабельная область тяжелой цепи содержит HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 16, SEQ ID NO: 17 и SEQ ID NO: 18, соответственно, и вариабельная область легкой цепи содержит LCDR1, LCDR2 и LCDR3, как показано в SEQ ID NO: 19, SEQ ID NO: 20 и SEQ ID NO: 21, соответственно.
В некоторых воплощениях антитело против CTGF содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, где:
iv-i) вариабельная область тяжелой цепи содержит HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 71, SEQ ID NO: 72 и SEQ ID NO: 73, соответственно, и вариабельная область легкой цепи содержит LCDR1, LCDR2 и LCDR3, как показано в SEQ ID NO: 74, SEQ ID NO: 75 и SEQ ID NO: 76, соответственно; или
iv-ii) вариабельная область тяжелой цепи содержит HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 102, SEQ ID NO: 103 и SEQ ID NO: 104, соответственно, и вариабельная область легкой цепи содержит LCDR1, LCDR2 и LCDR3, как показано в SEQ ID NO: 74, SEQ ID NO: 75 и SEQ ID NO: 76, соответственно.
В некоторых воплощениях антитело против CTGF представляет собой мышиное антитело, химерное антитело или гуманизированное антитело.
В некоторых воплощениях антитело против CTGF содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, где:
(v-1) аминокислотная последовательность вариабельной области тяжелой цепи имеет по меньшей мере 90%-ную идентичность последовательностей с SEQ ID NO: 6, 27, 28 или 29; и аминокислотная последовательность вариабельной области легкой цепи имеет по меньшей мере 90%-ную идентичность последовательностей с SEQ ID NO: 7, 22, 23, 24, 25 или 26;
(vi-1) аминокислотная последовательность вариабельной области тяжелой цепи имеет по меньшей мере 90%-ную идентичность последовательностей с SEQ ID NO: 8, 33, 34, 35 или 36; и аминокислотная последовательность вариабельной области легкой цепи обладает по меньшей мере 90%-ной идентичностью последовательностей с SEQ ID NO: 9, 30, 31 или 32; или
(vi-i) аминокислотная последовательность вариабельной области тяжелой цепи имеет по меньшей мере 90%-ную идентичность последовательностей с SEQ ID NO: 69, 81, 82, 83, 84 или 85, и аминокислотная последовательность вариабельной области легкой цепи обладает по меньшей мере 90%-ной идентичностью последовательностей с SEQ ID NO: 70, 77, 78, 79 или 80.
В некоторых воплощениях антитело против CTGF содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, где:
(v) вариабельная область тяжелой цепи обладает по меньшей мере 90%-ной, 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%-ной, 96%-ной, 97%-ной, 98%-ной, 99%-ной или 100%-ной идентичностью с вариабельной областью тяжелой цепи, как показано в SEQ ID NO: 6, 27, 28 или 29, и вариабельная область легкой цепи обладает по меньшей мере 90%-ной, 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%-ной, 96%-ной, 97%-ной, 98%-ной, 99%-ной или 100%-ной идентичностью с вариабельной областью легкой цепи, как показано в SEQ ID NO: 7, 22, 23, 24, 25 или 26; или
(vi) вариабельная область тяжелой цепи обладает по меньшей мере 90%-ной, 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%-ной, 96%-ной, 97%-ной, 98%-ной, 99%-ной или 100%-ной идентичностью с вариабельной областью тяжелой цепи, как показано в SEQ ID NO: 8, 33, 34, 35 или 36, и вариабельная область легкой цепи обладает по меньшей мере 90%-ной, 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%-ной, 96%-ной, 97%-ной, 98%-ной, 99%-ной или 100%-ной идентичностью с вариабельной областью легкой цепи, как показано в SEQ ID NO: 9, 30, 31 или 32; или
вариабельная область тяжелой цепи обладает по меньшей мере 90%-ной, 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%-ной, 96%-ной, 97%-ной, 98%-ной, 99%-ной или 100%-ной идентичностью с вариабельной областью тяжелой цепи, как показано в SEQ ID NO: 6, и вариабельная область легкой цепи обладает по меньшей мере 90%-ной, 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%-ной, 96%-ной, 97%-ной, 98%-ной, 99%-ной или 100%-ной идентичностью с вариабельной областью легкой цепи, как показано в SEQ ID NO: 7;
вариабельная область тяжелой цепи обладает по меньшей мере 90%-ной, 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%-ной, 96%-ной, 97%-ной, 98%-ной, 99%-ной или 100%-ной идентичностью с вариабельной областью тяжелой цепи, как показано в SEQ ID NO: 27, 28 или 29, и вариабельная область легкой цепи обладает по меньшей мере 90%-ной, 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%м, 96%-ной, 97%-ной, 98%-ной, 99%-ной или 100%-ной идентичностью с вариабельной областью легкой цепи, как показано в SEQ ID NO: 22, 23, 24, 25 или 26;
вариабельная область тяжелой цепи обладает по меньшей мере 90%-ной, 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%-ной, 96%-ной, 97%-ной, 98%-ной, 99%-ной или 100%-ной идентичностью с вариабельной областью тяжелой цепи, как показано в SEQ ID NO: 8, и вариабельная область легкой цепи обладает по меньшей мере 90%-ной, 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%-ной, 96%-ной, 97%-ной, 98%-ной, 99%-ной или 100%-ной идентичностью с вариабельной областью легкой цепи, как показано в SEQ ID NO: 9;
вариабельная область тяжелой цепи обладает по меньшей мере 90%-ной, 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%-ной, 96%-ной, 97%-ной, 98%-ной, 99%-ной или 100%-ной идентичностью с вариабельной областью тяжелой цепи, как показано в SEQ ID NO: 33, 34, 35 или 36, и вариабельная область легкой цепи обладает по меньшей мере 90%-ной, 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%-ной, 96%-ной, 97%-ной, 98%-ной, 99%-ной или 100%-ной идентичностью с вариабельной областью легкой цепи, как показано в SEQ ID NO: 30, 31 или 32.
В некоторых воплощениях антитело против CTGF содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, где:
(vi-ii) вариабельная область тяжелой цепи обладает по меньшей мере 90%-ной, 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%-ной, 96%-ной, 97%-ной, 98%-ной, 99%-ной или 100%-ной идентичностью с вариабельной областью тяжелой цепи, как показано в SEQ ID NO: 69, 81, 82, 83, 84 или 85, вариабельная область легкой цепи обладает по меньшей мере 90%-ной, 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%-ной, 96%-ной, 97%-ной, 98%-ной, 99%-ной или 100%-ной идентичностью с вариабельной областью легкой цепи, как показано в SEQ ID NO: 70, 77, 78, 79 или 80.
В некоторых воплощениях антитела против CTGF, где антитело против CTGF представляет собой гуманизированное антитело, которое содержит каркасную область человеческого антитела или вариант каркасной области, и данный вариант каркасной области имеет вплоть до 10 обратных мутаций в каждой из каркасной области легкой цепи и/или каркасной области тяжелой цепи антитела человека;
В некоторых воплощениях антитело против CTGF содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, как описано ниже:
(a) вариабельная область легкой цепи содержит LCDR1, LCDR2 и LCDR3, как показано в SEQ ID NO: 13, SEQ ID NO: 14 и SEQ ID NO: 15, соответственно, и содержит одну или более обратных мутаций аминокислоты(от), выбранных из группы, состоящей из 4L, 36F, 43S, 45K, 47W, 58V или 71Y, и/или вариабельная область тяжелой цепи содержит HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 10, SEQ ID NO: 11 и SEQ ID NO: 12, соответственно, и содержит одну или более обратных мутаций аминокислоты(от), выбранных из группы, состоящей из 28S, 30N, 49А, 75Е, 76S, 93V, 94Е или 104D; или
(b) вариабельная область легкой цепи содержит LCDR1, LCDR2 и LCDR3, как показано в SEQ ID NO: 19, SEQ ID NO: 20 и SEQ ID NO: 21, соответственно, и содержит одну или более обратных мутаций, выбранных из группы, состоящей из 36V, 44F, 46G или 49G, и/или вариабельная область тяжелой цепи содержит HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 16, SEQ ID NO: 17 и SEQ ID NO: 18, соответственно, и содержит одну или более обратных мутаций, выбранных из группы, состоящей из 44G, 49G, 27F, 48L, 67L, 71 K, 78V или 80F.
В некоторых воплощениях антитело против CTGF содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, как описано ниже:
(b-i) вариабельная область легкой цепи содержит LCDR1, LCDR2 и LCDR3, как показано в SEQ ID NO: 74, SEQ ID NO: 75 и SEQ ID NO: 76, соответственно, и одну или более обратных мутаций, выбранных из группы, состоящей из 45Р, 46W, 48Y, 69S или 70Y, и вариабельная область тяжелой цепи содержит HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 71, SEQ ID NO: 72 и SEQ ID NO: 73, соответственно, и одну или более обратных мутаций, выбранных из группы, состоящей из 27F, 38K, 48I, 67K, 68А, 70L или 72F; или
(b-ii) вариабельная область легкой цепи содержит LCDR1, LCDR2 и LCDR3, как показано в SEQ ID NO: 74, SEQ ID NO: 75 и SEQ ID NO: 76, соответственно, и содержит одну или более обратных мутаций, выбранных из группы, состоящей из 45Р, 46W, 48Y, 69S или 70Y, и вариабельная область тяжелой цепи содержит HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 102, SEQ ID NO: 103 и SEQ ID NO: 104, соответственно, и содержит одну или более обратных мутаций, выбранных из группы, состоящей из 27F, 38K, 48I, 67K, 68А, 70L или 72F.
В некоторых воплощениях антитело против CTGF содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, как описано ниже:
(vii) последовательность вариабельной области тяжелой цепи представляет собой такую, как показано в SEQ ID NO: 6, и последовательность вариабельной области легкой цепи представляет собой такую, как показано в SEQ ID NO: 7;
(viii) последовательность вариабельной области тяжелой цепи представляет собой такую, как показано в SEQ ID NO: 27, 28 или 29, и последовательность вариабельной области легкой цепи представляет собой такую, как показано в SEQ ID NO: 22, 23, 24, 25 или 26;
(ix) последовательность вариабельной области тяжелой цепи представляет собой такую, как показано в SEQ ID NO: 8, и последовательность вариабельной области легкой цепи представляет собой такую, как показано в SEQ ID NO: 9; или
(x) последовательность вариабельной области тяжелой цепи представляет собой такую, как показано в SEQ ID NO: 33, 34, 35 или 36, и последовательность вариабельной области легкой цепи представляет собой такую, как показано в SEQ ID NO: 30, 31 или 32.
В некоторых воплощениях антитело против CTGF содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, как описано ниже:
(xi) аминокислотная последовательность вариабельной области тяжелой цепи представляет собой такую, как показано в SEQ ID NO: 69, аминокислотная последовательность вариабельной области легкой цепи представляет собой такую, как показано в SEQ ID NO: 70; или
(xii) аминокислотная последовательность вариабельной области тяжелой цепи представляет собой такую, как показано в SEQ ID NO: 81, 82, 83, 84 или 85, и аминокислотная последовательность вариабельной области легкой цепи представляет собой такую, как показано в EQ ID NO: 77, 78, 79 или 80.
В некоторых воплощениях антитело против CTGF содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, как показано ниже в Таблице а, Таблице b и Таблице с:

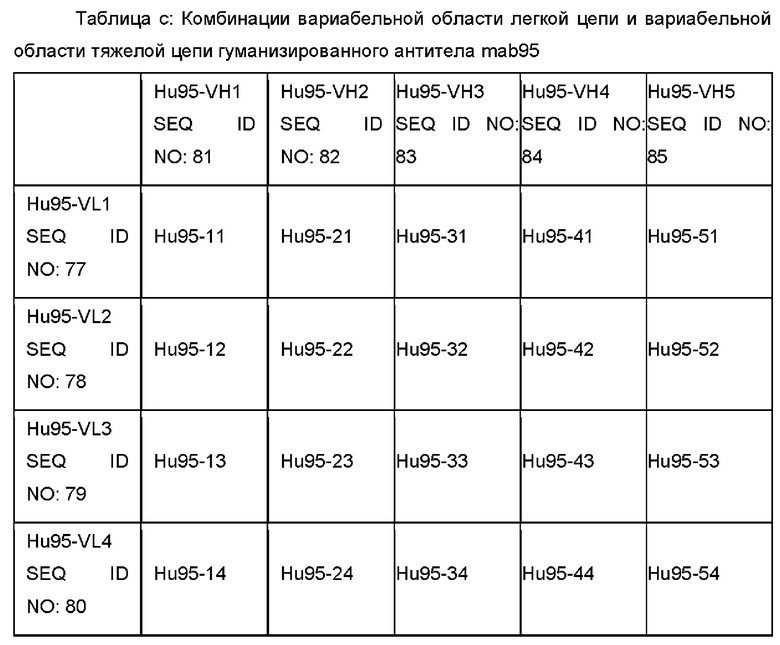
В некоторых воплощениях антитело против CTGF содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, как описано ниже:
(xiii) аминокислотная последовательность вариабельной области тяжелой цепи представляет собой такую, как показано в SEQ ID NO: 27, и аминокислотную последовательность вариабельной области легкой цепи представляет собой такую, как показано в SEQ ID NO: 22;
(xiv) аминокислотная последовательность вариабельной области тяжелой цепи представляет собой такую, как показано в SEQ ID NO: 34, и аминокислотную последовательность вариабельной области легкой цепи представляет собой такую, как показано в SEQ ID NO: 30; или
(xv) аминокислотная последовательность вариабельной области тяжелой цепи представляет собой такую, как показано в SEQ ID NO: 85, и аминокислотную последовательность вариабельной области легкой цепи представляет собой такую, как показано в SEQ ID NO: 77.
В некоторых воплощениях антитело против CTGF, где антитело дополнительно содержит константную область тяжелой цепи и константную область легкой цепи антитела; предпочтительно, константная область тяжелой цепи выбрана из группы, состоящей из константных областей человеческого IgG1, IgG2, IgG3 и IgG4 и их традиционных вариантов, и константная область легкой цепи выбрана из группы, состоящей из константных областей цепей к и А, человеческого антитела и их традиционных вариантов; более предпочтительно, антитело содержит константную область тяжелой цепи, как показано в SEQ ID NO: 37 или 38, и константную область легкой цепи, как показано в SEQ ID NO: 39 или 40. В некоторых воплощениях антитело против CTGF содержит:
(c) тяжелую цепь, обладающую по меньшей мере 85%-ной, 90%-ной, 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%-ной, 96%-ной, 97%-ной, 98%-ной, 99%-ной или 100%-ной идентичностью с тяжелой цепью, как показано в SEQ ID NO: 41, 43, 44, 45, 46, 47 или 48, и легкую цепь, обладающую по меньшей мере 85%-ной, 90%-ной, 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%-ной, 96%-ной, 97%-ной, 98%-ной, 99%-ной или 100%-ной идентичностью с легкой цепью, как показано в SEQ ID NO: 42, 49, 50, 51, 52 или 53;
(d) тяжелую цепь, обладающую по меньшей мере 85%-ной, 90%-ной, 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%-ной, 96%-ной, 97%-ной, 98%-ной, 99%-ной или 100%-ной идентичностью с тяжелой цепью, как показано в SEQ ID NO: 54, 56, 57, 58, 59, 60, 61, 62 или 63, и легкую цепь, обладающую по меньшей мере 85%-ной, 90%-ной, 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%-ной, 96%-ной, 97%-ной, 98%-ной, 99%-ной или 100%-ной идентичностью с легкой цепью, как показано в SEQ ID NO: 55, 64, 65 или 66;
(e) тяжелую цепь, как показано в SEQ ID NO: 41, 43, 44, 45, 46, 47 или 48, и легкую цепь, как показано в SEQ ID NO: 42, 49, 50, 51, 52 или 53; или
(f) тяжелую цепь, как показано в SEQ ID NO: 54, 56, 57, 58, 59, 60, 61, 62 или 63, и легкую цепь, как показано в SEQ ID NO: 55, 64, 65 или 66.
В некоторых воплощениях антитело против CTGF содержит: (д) тяжелую цепь, обладающую по меньшей мере 85%-ной, 90%-ной, 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%-ной, 96%-ной, 97%-ной, 98%-ной, 99%-ной или 100%-ной идентичностью последовательностей с SEQ ID NO: 86, 88, 89, 90, 91, 92, 93, 94, 95, 96 или 97, и легкую цепь, обладающую по меньшей мере 85%-ной, 90%-ной, 91%-ной, 92%-ной, 93%-ной, 94%-ной, 95%-ной, 96%-ной, 97%-ной, 98%-ной, 99%-ной или 100%-ной идентичностью последовательностей с SEQ ID NO: 87, 98, 99, 100 или 101; или
(h) тяжелую цепь, как показано в SEQ ID NO: 86, 88, 89, 90, 91, 92, 93, 94, 95, 96 или 97, и легкую цепь, как показано в SEQ ID NO: 87, 98, 99, 100 или 101.
В некоторых воплощениях антитело против CTGF содержит: (j) тяжелую цепь, как показано в SEQ ID NO: 46, и легкую цепь, как показано в SEQ ID NO: 49;
(k) тяжелую цепь, как показано в SEQ ID NO: 61, и легкую цепь, как показано в SEQ ID NO: 64; или
(l) тяжелую цепь, как показано в SEQ ID NO: 97, и легкую цепь, как показано в SEQ ID NO: 98.
В других аспектах настоящего раскрытия предложено антитело против CTGF, которое конкурентно с антителом против CTGF или его антигенсвязывающими фрагментами, как описано выше, за связывание с человеческим CTGF.
В других аспектах настоящего раскрытия предложена молекула нуклеиновой кислоты, кодирующая антитело против CTGF, как описано выше.
В других аспектах настоящего раскрытия предложена клетка-хозяин, содержащая молекулу нуклеиновой кислоты, как описано выше.
В других аспектах настоящего раскрытия предложена фармацевтическая композиция, содержащая терапевтически эффективное количество или профилактически эффективное количество антитела против CTGF, как описано выше, или молекулы нуклеиновой кислоты, как описано выше, и один или более фармацевтически приемлемых носителей, разбавителей, буферов или вспомогательных веществ.
В некоторых конкретных воплощениях терапевтически эффективное количество или профилактически эффективное количество означает, что лекарственная однократная доза композиции содержит 0,1-3000 мг или 1-1000 мг антитела против CTGF, как описано выше.
В других аспектах настоящего раскрытия предложен способ иммунологического анализа или определения CTGF, включающий стадию применения антитела против CTGF, как описано выше.
В других аспектах настоящего раскрытия предложен способ иммунологического анализа или определения CTGF, включающий стадию приведения антитела против CTGF, как описано выше, в контакт с субъектом или его образцом.
В других аспектах настоящего раскрытия предложен набор, содержащий антитело против CTGF, как описано выше.
В других аспектах настоящего раскрытия предложен способ лечения заболеваний, связанных с CTGF, включающий введение субъекту терапевтически эффективного количества антитела против CTGF, как описано выше, или молекулы нуклеиновой кислоты, как описано выше, или фармацевтической композиций, как описано выше, где заболевание предпочтительно представляет собой фиброзное заболевание (фиброзное заболевание предпочтительно представляет собой спонтанный фиброз легких, диабетическую нефропатию, диабетическую ретинопатию, остеоартрит, склеродерму, хроническую сердечную недостаточность, цирроз печени или почечный фиброз), гипертензию, диабет, инфаркт миокарда, артрит, клеточное пролиферативное заболевание, связанное с CTGF, атеросклероз, глаукому или рак (рак предпочтительно представляет собой острый лимфобластный лейкоз, дерматофиброму, рак молочной железы, ангиолипому, ангиолейомиому, рак, происходящий из соединительной ткани, рак предстательной железы, рак яичника, рак толстой и прямой кишки, рак предстательной железы, рак желудочно-кишечного тракта или рак печени).
В других аспектах настоящего раскрытия предложено применение антитела против CTGF, как описано выше, или молекулы нуклеиновой кислоты, как описано выше, или фармацевтической композиции, как описано выше, в получении лекарственного средства для лечения заболеваний, связанных с CTGF, заболевание, связанное с CTGF, включает фиброзное заболевание (фиброзное заболевание предпочтительно представляет собой спонтанный фиброз легких, диабетическую нефропатию, диабетическую ретинопатию, остеоартрит, склеродерму, хроническую сердечную недостаточность, цирроз печени или почечный фиброз), гипертензию, диабет, инфаркт миокарда, артрит, клеточное пролиферативное заболевание, связанное с CTGF, атеросклероз, глаукому или рак (рак предпочтительно представляет собой острый лимфобластный лейкоз, дерматофиброму, рак молочной железы, ангиолипому, ангиолейомиому, рак, происходящий из соединительной ткани, рак предстательной железы, рак яичника, рак толстой и прямой кишки, рак предстательной железы, рак желудочно-кишечного тракта или рак печени).
В других аспектах настоящего раскрытия предложено антитело против CTGF, как описано выше, или молекула нуклеиновой кислоты, кодирующая антитело против CTGF, как описано выше, или фармацевтическая композиция, как описано выше, для применения в лечении заболеваний, связанных с CTGF, заболевание, связанное с CTGF, включает фиброзное заболевание (фиброзное заболевание предпочтительно представляет собой спонтанный фиброз легких, диабетическую нефропатию, диабетическую ретинопатию, остеоартрит, склеродерму, хроническую сердечную недостаточность, цирроз печени или почечный фиброз), гипертензию, диабет, инфаркт миокарда, артрит, клеточное пролиферативное заболевание, связанное с CTGF, атеросклероз, глаукому или рак (рак предпочтительно представляет собой острый лимфобластный лейкоз, дерматофиброму, рак молочной железы, ангиолипому, ангиолейомиому, рак, происходящий из соединительной ткани, рак предстательной железы, рак яичника, рак толстой и прямой кишки, рак предстательной железы, рак желудочно-кишечного тракта или рак печени).
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1: Аффинность гуманизированного антитела, происходящего из mab164, в отношении человеческого CTGF, выявленная посредством ELISA (от англ. enzyme-linked immunosorbent assay - твердофазный иммуноферментный анализ).
Фиг. 2: Результаты эксперимента по гуманизированному антителу, происходящему из mab147 или mab164, в ингибировании ксенотрансплантатной опухоли мыши SU86.86.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Терминология
Для более легкого понимания настоящего раскрытия некоторые технические и научные термины конкретно определены ниже. Если в данном документе явным образом не определено иное, все другие технические и научные термины, используемые в данном документе, имеют значение, обычно подразумеваемое специалистами в области, к которой принадлежит данное раскрытие.
Трехбуквенные коды и однобуквенные коды для аминокислот, используемые в настоящем раскрытии, представляют собой такие, как описано в J. Biol. Chem, 243, р3558 (1968).
Фактор роста соединительной ткани (CTGF) представляет собой богатый цистеином гепарин-связывающий секретируемый гликопротеин, массой 36 кДа, который исходно выделен из эндотелиальных клеток пупочной вены человека (см., например, Bradham et al. (1991) J Cell BioM 14: 1285-1294; Grotendorst and Bradham, патент США №5408040). CTGF принадлежит к семейству белков CCN (CTGF, Cyr61, Nov) (секретируемые гликопротеины), и данное семейство включает индуцированный сывороткой продукт непосредственно раннего гена Cyr61, предполагаемый онкоген Nov, ЕСМ-связанный белок FISP-12, src-индуцируемый ген CEF-10, Wnt-индуцируемый секретируемый белок WISP-3 и антипролиферативный белок HICP/rCOP (Brigstock (1999) Endocr Rev 20: 189-206; О'Brian et al. (1990) Mol Cell Biol 10: 3569-3577; Joliot et al. (1992) Mol Cell Biol 12: 10-21; Ryseck et al. (1990) Cell Growth and Diff 2: 225-233; Simmons et al. (1989) Proc Natl Acad Sci USA 86: 1178-1182; Pennica ef at. (1998) Proc Natl Acad Sci USA, 95: 14717-14722; и Zhang et at. (1998) Mol Cell Biol 18: 6131-6141). Белок CCN характеризуется консервативными 38 остатками цистеина. 38 Остатков цистеина составляют более 10% общего содержания аминокислот и образуют модульную структуру с N-концевыми и С-концевыми доменами. Модульная структура CTGF включает консервативные мотивы для белка, связывающего инсулиноподобный фактор роста (IGF-BP - от англ. insulin-like growth factor binding protein), и фактора фон Виллебранда (VWC - от англ. von Willebrand factor) в N-концевом домене и тромбоспондина (TSP1 - от англ. thrombospondin), и цистеиновый мотив в С-концевом домене.
В том виде, в котором он используется в данном документе, термин «антитело» относится к иммуноглобулину, и полноразмерное антитело представляет собой структуру из четырех пептидных цепей, соединенных вместе межцепочечной(ыми) дисульфидной(ыми) связью(ями) между двумя идентичными тяжелыми цепями и двумя идентичными легкими цепями. Константные области тяжелой цепи иммуноглобулина демонстрируют разные составы и расположения аминокислот, следовательно, представляют разную антигенность. Соответственно, иммуноглобулины могут быть подразделены на пять типов, или называемых изотипами иммуноглобулинов, а именно IgM, IgD, IgG, IgA и IgE, и соответствующие тяжелые цепи представляют собой μ, δ, γ, α и ε, соответственно. В соответствии со своим аминокислотным составом шарнирной области, а также числом и локализацией дисульфидных связей тяжелой цепи, один и тот же тип Ig может быть подразделен на разные подтипы; например, IgG может быть подразделен на IgG1, IgG2, IgG3 и IgG4. Легкие цепи могут быть подразделены на цепь κ или λ, с учетом разных константных областей. Каждый из пяти типов IgG может иметь цепь каппа или цепь лямбда.
Последовательности, примерно из 110 аминокислот, рядом с N-концом тяжелой и легкой цепей антитела являются высоковариабельными, известны как вариабельные области (области Fv); остальные аминокислотные последовательности близко к С-концу являются относительно стабильными, известны как константные области. Вариабельная область включает 3 гипервариабельные области (HVR - от англ. hypervariable region) и 4 каркасные области (FR - от англ. framework region) с относительно консервативными последовательностями. Данные три гипервариабельные области, которые определяют специфичность антитела, также известны как области, определяющие комплементарность (CDR - от англ. complementarity determining region). Каждая вариабельная область легкой цепи (VL) и каждая вариабельная область тяжелой цепи (VH) состоит из трех областей CDR и четырех областей FR, расположенных от N-конца к С-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Данные три области CDR легкой цепи относятся к LCDR1, LCDR2 и LCDR3; и данные три области CDR тяжелой цепи относятся к HCDR1, HCDR2 и HCDR3.
Антитела по настоящему раскрытию включают мышиные антитела, химерные антитела и гуманизированные антитела.
В том виде, в котором он используется в данном документе, термин «мышиное антитело» относится к монокпональным антителам против человеческого CTGF, полученным в соответствии со знанием и умениями в данной области. Во время получения анализируемому субъекту инъецируют антиген CTGF, и затем выделяют гибридому, экспрессирующую антитело, которое имеет желаемую последовательность или функциональные характеристики. В предпочтительном воплощении настоящего раскрытия мышиное антитело против CTGF или его антигенсвязывающие фрагменты могут дополнительно сдержать константную область легкой цепи мышиной цепи каппа, лямбда или ее варианты или дополнительно содержат константную область тяжелой цепи мышиного IgG1, IgG2, IgG3 или ее варианты.
Термин «химерное антитело» представляет собой антитело, полученное в результате слияния вариабельной области мышиного антитела с константной областью человеческого антитела, и такое антитело может смягчать иммунный ответ, вызываемый мышиным антителом. Для создания химерного антитела сначала создают гибридому, секретирующую специфичное мышиное моноклональное антитело, и ген вариабельной области клонируют из мышиной гибридомы. Затем клонируют ген константной области из человеческого антитела при необходимости. Ген вариабельной области мыши соединяют с геном константной области человека с образованием химерного гена, который может быть впоследствии вставлен в экспрессионный вектор. Наконец, молекула химерного антитела будет экспрессироваться в эукариотической или прокариотической системе. В предпочтительном воплощении настоящего раскрытия легкая цепь антитела химерного антитела дополнительно содержит константную область легкой цепи капа, лямбда человека или ее варианты. Тяжелая цепь антитела химерного антитела против CTGF дополнительно содержит константную область тяжелой цепи человеческого IgG1, IgG2, IgG3, IgG4 или их вариантов, предпочтительно содержит константную область тяжелой цепи человеческого IgG1, IgG2 или IgG4 или варианта IgG1, IgG2 или IgG4 с мутацией (я ми) аминокислоты (как например, мутацией L234A и/или L235A, и/или мутацией S228P).
Термин «гуманизированное антитело», также известное как CDR-привитое антитело, относится к антителу, созданному посредством прививания мышиных последовательностей CDR в каркасы вариабельных областей человеческого антитела, а именно, антителу, полученному в разных типах последовательностей каркасов антитела зародышевой линии человека. Гуманизированное антитело может преодолевать гетерологичные ответы, вызываемые химерным антителом, которое несет большое число мышиных белковых компонентов. Такие последовательности каркасов могут быть получены из общедоступной базы данных ДНK, охватывающей последовательности генов антитела зародышевой линии, или опубликованные ссылки. Например, последовательности ДНК зародышевой линии генов вариабельной области тяжелой и легкой цепей человека могут быть найдены в базе данных последовательностей зародышевой линии человека «VBase» (доступной на www.mrccpe.com.ac.uk/vbase), а также в Kabat, ЕА, et al. 1991 Sequences of Proteins of Immunological Interest, 5th Ed. Для того, чтобы избежать снижения активности, вызываемого сниженной иммуногенностью, последовательности каркасов в вариабельной области человеческого антитела могут подвергаться минимальной(ым) обратной(ым) мутации(ям) или обратной(ым) мутации(ям) с сохранением или повышением активности. Гуманизированные антитела по настоящему раскрытию также включают гуманизированные антитела, на которых осуществляют созревание аффинности CDR посредством дрожжевого дисплея.
За счет остатков, контактирующих с антигенов, прививка CDR может приводить к снижению аффинности антитела или его антигенсвязывающего фрагмента в отношении антигена из-за изменения остатков каркаса, с которыми контактирует антиген. Такие взаимодействия могут быть результатом высокосоматических мутаций. Таким образом, может быть все еще необходимо прививать такие донорские аминокислоты каркаса на каркасы гуманизированного антитела. Аминокислотные остатки, участвующие в связывании антигена и происходящие из антитела, не являющегося человеческим, или его антигенсвязывающего фрагмента, могут быть идентифицированы посредством проверки последовательности и структуры вариабельной области моноклонального антитела животного. Аминокислотные остатки донорского CDR каркаса, которые отличаются от зародышевых линий, можно считать родственными. Если невозможно определить наиболее близкородственную зародышевую линию, последовательность можно сравнивать с общей последовательностью, общей у подтипов, или с последовательностью антитела животного с высоким процентом сходства. Считается, что редкие остатки в каркасе являются результатом высокой мутации в соматических клетках и играют важную роль в связывании.
В одном воплощении настоящего раскрытия антитело или его антигенсвязывающий фрагмент дополнительно содержит константную область легкой цепи, происходящую из человеческой или мышиной цепи к, А, или ее вариант, или дополнительно содержит константную область тяжелой цепи, происходящую из человеческих или мышиных IgG1, IgG2, IgG3, IgG4, или ее вариант; оно предпочтительно включает константную область тяжелой цепи, происходящую из человеческого IgG1, IgG2 или IgG4, или варианта IgG1, IgG2 или IgG4 с мутацией(ями) аминокислоты(от) (как например, мутацией L234A и/или L235A, и/или мутацией S228P).
В том виде, в котором они используется в данном документе, «общепринятые варианты» константной области тяжелой цепи человеческого антитела и константной области легкой цепи человеческого антитела относятся к вариантам константной области тяжелой и легкой цепей человека, раскрытым в предшествующем уровне техники, которые не меняют структуру и функцию вариабельных областей антитела. Типичные варианты включают варианты константных областей тяжелой цепи IgG1, IgG2, IgG3 или IgG4, полученные посредством сайт-направленной модификации и аминокислотных замен на константной области тяжелой цепи. Конкретные замены представляют собой, например, мутацию YTE, мутации L234A и/или L235A, мутацию S228P и/или мутации, приводящие к получению структуры «выступ-во-впадину» (что приводит к тому, что тяжелая цепь антитела имеет комбинацию выступ-Fc и впадина-Fc) и т.д. Оказалось, что данные мутации придают антителу новые свойства, не меняя функцию вариабельной области антитела.
Термины «человеческое антитело (НиМАb)», «антитело, происходящее из человека», «полноразмерное человеческое антитело» и «полное человеческое антитело» можно использовать взаимозаменяемо, и они могли бы представлять собой антитела, происходящие из человека, или антитела, полученные из генетически модифицированного организма, который «сконструирован» любым способом, известным в данной области, для получения специфичных человеческих антител в ответ на стимуляцию антигеном, и могут быть получены. В некоторых технологиях элементы локусов тяжелой и легкой цепей человека вводят в линии клеток организмов, происходящие из линий эмбриональных стволовых клеток, и эндогенные локусы тяжелой и легкой цепей в данных клеточныхлиниях являются мишенью и нарушаются. Эндогенные локусы тяжелой и легкой цепей - мишени, включенные в данные клеточные линии, нарушаются. Трансгенные организмы могут синтезировать человеческие антитела, специфичные к человеческим антигенам, и данные организмы могут быть использованы для получения гибридомы, которая секретирует человеческие антитела. Человеческое антитело может также представлять собой такое антитело, в котором тяжелая и легкая цепи кодируются нуклеотидными последовательностями, происходящими из одного или более источников ДНК человека. Полноразмерные человеческие антитела могут быть также сконструированы с помощью способов генной или хромосомной трансфекции и технологии фагового дисплея или сконструированы из В-клеток, активированных in vitro, все из которых известны в данной области.
Термины «полноразмерное антитело», «целое антитело» и «полное антитело» используются взаимозаменяемо в данном документе и относятся к антителу в по существу интактной форме, которое отличается от антигенсвязывающих фрагментов, определенных ниже. Термин конкретно относится к антителам, которые содержат константные области в легкой и тяжелой цепях.
«Антитело» по настоящему раскрытию включает «полноразмерные антитела» и их антигенсвязывающие фрагменты.
В некоторых воплощениях полноразмерное антитело по настоящему раскрытию включает полноразмерные антитела, образованные посредством связывания вариабельной области легкой цепи с константной областью легкой цепи, и связывания вариабельной области тяжелой цепи с константной областью тяжелой цепи, как показано в комбинациях легкой и тяжелой цепи, перечисленных ниже в Таблицах 1, 2 и 3. Специалисты в данной области могут выбрать константную область легкой цепи и константную область тяжелой цепи из разных источников антител в соответствии с фактической необходимостью, как например, происходящая из человеческого антитела константная область легкой цепи и константная область тяжелой цепи. В то же время, разные комбинации вариабельных областей легкой и тяжелой цепей, описанные в Таблицах 1, 2 и 3, могут образовывать одноцепочечное антитело (scFv - от англ. single-chain variable fragment - одноцепочечный вариабельный фрагмент), Fab или другие формы антигенсвязывающего фрагмента, содержащие scFv или Fab.


Термин «антигенсвязывающий фрагмент» или «функциональный фрагмент» относится к одному или более фрагментам антитела, которые сохраняют способность специфично связываться с антигеном (например, CTGF). Показано, что фрагменты полноразмерного антитела могут использоваться для достижения функции связывания с конкретным антигеном. Примеры связывающих фрагментов, включенных в термин «антигенсвязывающий фрагмент» антитела, включают: (i) фрагмент Fab, моновалентный фрагмент, состоящий из доменов VL, VH, CL и СН1; (ii) фрагмент F(ab')2, бивалентный фрагмент, содержащий два фрагмента Fab, соединенных дисульфидным(и) мостиком(ами) в шарнирной области, (iii) фрагмент Fd, состоящий из доменов VH и СН1; (iv) фрагмент Fv, состоящий из доменов VH и VL одного плеча антитела; (v) dsFv, стабильный антигенсвязывающий фрагмент, образованный VH и VL посредством межцепочечной(ых) дисульфидной(ых) связи(ей); и (vi) диатело, биспецифичное антитело и мультиспецифичное антитело, содержащее фрагменты, такие как scFv, dsFv и Fab. Кроме того, два домена, домен VL и VH, фрагмента Fv кодируются двумя отдельными генами, однако, они могут быть связаны синтетическим линкером посредством использования методов генной инженерии с образованием одной белковой цепи, в которой моновалентная молекула образована в результате объединения домена VL и VH (называемая одноцепочечным Fv (scFv); см., например, Bird et al. (1988) Science 242: 423-426; и Huston et al (1988) Proc. Natl. Acad. Sci USA85:5879-5883). Подразумевается, что такие одноцепочечные антитела также включены в термин «антигенсвязывающий фрагмент» антитела. Такие фрагменты антител получают, используя общепринятые методики, известные в данной области, и подвергаются скринингу в отношении функциональных фрагментов посредством использования того же способа, как способ для интактного антитела. Антигенсвязывающие участки могут быть получены посредством технологии генной инженерии или посредством ферментативного или химического разрушения интактного иммуноглобулина. Антитела могут находиться в виде разных изотипов, например, антитела IgG (например, подтип IgG 1, IgG2, IgG3 или IgG4), lgA1, lgA2, IgD, IgE или IgM.
Fab представляет собой фрагмент антитела, полученный посредством обработки молекулы антитела IgG папаином (который расщепляет аминокислотный остаток в положении 224 цепи Н),и фрагмент антитела имеет молекулярную массу примерно 50000 и обладает антигенсвязывающей активностью, в котором примерно половина N-концевой стороны цепи Н на вся цепь L связаны вместе дисульфидной(ыми) связью(ями).
«F(ab')2» представляет собой фрагмент антитела с молекулярной массой примерно 100000 и обладающий антигенсвязывающей активностью, и он получен в результате расщепления пепсином части, ниже двух дисульфидных связей в шарнирной области IgG. F(ab')2 содержит два Fab, соединенных в шарнирной области.
Fab' представляет собой фрагмент антитела, имеющий молекулярную массу примерно 50000 и обладающий антигенсвязывающей активностью, который получается в результате расщепления дисульфидной связи в шарнирной области упомянутого выше F(ab')2. Fab' по настоящему раскрытию может быть получен в результате обработки F(ab')2 по настоящему раскрытию, который специфично распознает CTGF и связывается с аминокислотными последовательностями внеклеточной области или ее трехмерной структурой, восстанавливающим средством, таким как дитиотреитол.
Кроме того, Fab' может быть получен посредством вставки ДНK, кодирующей Fab' антитела, в прокариотический экспрессионный вектор или эукариотический экспрессионный вектор и введения данного вектора в прокариота или эукариота для экспрессии Fab'.
Термин «одноцепочечное антитело», «одноцепочечный Fv» или «scFv» относится к молекуле, содержащей вариабельный домен тяжелой цепи антитела (или область; VH), соединенный с вариабельным доменом легкой цепи антитела (или областью; VL) посредством линкера. Такие молекулы scFv имеют общую структуру NH2-VL-линкер-\/Н-СООН или NH2-\/Н-линкер-VL-СООН. Подходящий линкер в предшествующем уровне технике состоит из повторяющейся аминокислотной последовательности GGGGS или ее варианта, например, варианта с 1-4 повторами (Holliger et al. (1993) Proc. Natl. Acad. Sci. USA 90: 6444-6448). Другие линкеры, которые могут быть использованы в настоящем раскрытии, описаны Alfthan et al. (1995), Protein Eng. 8:725-731; Choi etal. (2001), Eur. J. Immunol. 31:94-106; Hu et al. (1996), Cancer Res. 56:3055-3061; Kipriyanov et al. (1999), J. Mol. Biol. 293:41-56 и Roovers et al. (2001), Cancer Immunol.
Диатело представляет собой фрагмент антитела, в котором scFv или Fab димеризован, и оно представляет собой фрагмент антитела, обладающий двухвалентной антигенсвязывающей активностью. При двухвалентной антигенсвязывающей активности два антигена могут быть одинаковыми или разными.
Биспецифичное и мультиспецифичное антитело относятся к антителу, которое может одновременно связываться с двумя или более антигенами или антигенными детерминантами, включая фрагменты scFv или Fab, которые могут связывать CTGF.
Диатело по настоящему раскрытию может быть получено посредством следующих стадий: получение кДНK, кодирующих VH и VL моноклонального антитела по настоящему раскрытию, которое специфично распознает человеческий CTGF и связывается с внеклеточной областью или ее трехмерной структурой, конструирование ДНK, кодирующей scFv, таким образом, что длина пептида линкера составляет 8 или меньше аминокислотных остатков, вставка ДНК в прокариотический экспрессионный вектор или эукариотический экспрессионный вектор и затем введение экспрессионного вектора в прокариота или эукариота с экспрессией диатела.
dsFv получают посредством осуществления замены одного аминокислотного остатка в каждом из VH и VL остатком цистеина, и затем соединения полипептидов с заменой дисульфидной связью между двумя остатками цистеина. Аминокислотные остатки, подлежащие замене остатком цистеина, могут быть выбраны на основе предсказания трехмерной структуры антитела в соответствии с известными способами (например, Protein Engineering, 7, 697 (1994)).
Полноразмерные антитела или их антигенсвязывающий фрагменты по настоящему раскрытию могут быть получены посредством следующих стадий: получение кДНK, кодирующих антитело по настоящему раскрытию, которое специфично распознает человеческий CTGF и связывается с аминокислотной последовательностью внеклеточной области или ее трехмерной структурой, конструирование ДНK, кодирующей dsFv, вставка ДНК в прокариотический экспрессионный вектор или эукариотический экспрессионный вектор и затем введение экспрессионного вектора в прокариота или эукариота с экспрессией dsFv.
Термин «отличие по аминокислоте» или «мутация аминокислоты» относится к изменениям или мутациям аминокислоты в варианте белка или полипептида, по сравнению с исходным белком или полипептидом, включая 1, 2, 3 или более вставок, делеций или замен аминокислот, исходя из исходного белка или полипептида.
Термин «каркас антитела» или «область FR» относится к части вариабельного домена, или VL или VH, которая служит каркасом для антигенсвязывающих петель (CDR) данного вариабельного домена. По существу, это представляет собой вариабельный домен без CDR.
Термин «область, определяющая комплементарность», «CDR» или «гипервариабельная область» относится к одной из шести гипервариабельных областей, присутствующих в вариабельном домене антитела, которые главным образом содействуют связыванию антигена. Обычно, три CDR (HCDR1, HCDR2, HCDR3) находятся в каждой вариабельной области тяжелой цепи, и три CDR (LCDR1, LCDR2, LCDR3) - в каждой вариабельной области легкой цепи. Границы аминокислотных последовательностей CDR могут быть определены с помощью любой из множества хорошо известных схем, включая критерии нумерации «Kabat» (см. Kabat et al. (1991), «Sequences of Proteins of Immunological Interest», 5th edition, Public Health Service, National Institutes of Health, Bethesda, MD), критерии нумерации «Chothia» (см. Al-Lazikani et al., (1997) JMB 273:927-948) и критерии нумерации ImMunoGenTics (IMGT) (Lefranc MP, Immunologist, 7, 132- 136 (1999); Lefranc, MP, etc., Dev. Сотр. Immunol., 27, 55-77 (2003)) и т.п.Например, в случае классического формата, следуя критериям Kabat, аминокислотные остатки CDR в вариабельном домене тяжелой цепи (VH) пронумерованы как 31-35 (HCDR1), 50-65 (HCDR2) и 95-102 (HCDR3); и аминокислотные остатки CDR в вариабельном домене легкой цепи (VL) пронумерованы как 24-34 (LCDR1), 50-56 (LCDR2) и 89-97 (LCDR3). В соответствии с критериями Chothia аминокислотные остатки CDR в VH пронумерованы как 26-32 (HCDR1), 52-56 (HCDR2) и 95-102 (HCDR3); и аминокислотные остатки в VL пронумерованы как 26-32 (LCDR1), 50-52 (LCDR2) и 91-96 (LCDR3). Объединяя как Kabat, так и Chothia для определения CDR, CDR состоят из аминокислотных остатков 26-35 (HCDR1), 50-65 (HCDR2) и 95-102 (HCDR3) в VH человека и аминокислотных остатков 24-34 (LCDR1), 50-56 (LCDR2) и 89-97 (LCDR3) в VL человека. В соответствии с критериями IMGT аминокислотные остатки CDR в VH приблизительно пронумерованы как 26-35 (CDR1), 51-57 (CDR2) и 93-102 (CDR3), и аминокислотные остатки CDR в VL приблизительно пронумерованы как 27-32 (CDR1), 50-52 (CDR2) и 89-97 (CDR3). В соответствии с критериями IMGT области CDR антитела могут быть определены, используя программу IMGT/DomainGapAlign. Если не указано иное, вариабельные области антитела и последовательности CDR, включенные в воплощения настоящего раскрытия, применимы для критериев нумерации «Kabat».
Термин «эпитоп» или «антигенная детерминанта» относится к сайту на антигене, с которым специфично связывается иммуноглобулин или антитело (например, конкретный сайт на молекуле CTGF). Эпитопы обычно включают по меньшей мере 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 последовательных или непоследовательных аминокислот в уникальной третичной конформации. См., например, Epitope Mapping Protocols in Methods in Molecular Biology, Vol.66, ed. G. E. Morris, Ed. (1996).
Термин «специфично связываются с», «селективно связываются с», «селективно связывается с» или «специфично связывается с» относится к связыванию антитела с заданным эпитопом на антигене. Обычно антитело связывается с аффинностью (KD) меньше чем примерно 10-8 М, например, меньше чем примерно 10-9 М, 10-10 М, 10-11 М, 10-12 М или даже меньше.
Термин «KD» относится к константе равновесия диссоциации для конкретного взаимодействия антитело-антиген. Обычно антитело по настоящему раскрытию связывается с CTGF с константой равновесия диссоциации (KD) меньше чем примерно 10-7 М, например, меньше чем примерно 10-8 М или 10-9 М, например, аффинность антитела в настоящем раскрытии в отношении антигена клеточной поверхности определяют посредством измерения значения KD способом FACS (от англ. fluorescence-activated cell sorting - сортировка клеток с активированной флуоресценцией).
Когда термин «конкурирование» используется в контексте антигенсвязывающих белков (например, белков, нейтрализующих связывание антигена, или нейтрализующих антител), которые конкурируют за один и тот же эпитоп, он означает, что между антигенсвязывающими белками происходит конкуренция, которая определяется анализами, в которых антигенсвязывающий белок, подлежащий тестированию (например, антитело или его иммунологически функциональный фрагмент), предотвращает или ингибирует (например, уменьшает) специфичное связывание референсного антигенсвязывающего белка (например, лиганда или референсного антитела) с общим антигеном (например, антигеном CTGF или его фрагментами). Множество типов анализов конкурентного связывания доступно для определения того, конкурирует ли один антигенсвязывающий белок с другим. Данные анализы представляют собой, например, твердофазный прямой или непрямой радиоиммунологический анализ (RIA - от англ. radioimmunoassay), твердофазный прямой или непрямой иммуноферментный анализ (EIA - от англ. enzyme immunoassay), конкурентный «сэндвич»-анализ (см., например, Stahli et al, 1983, Methods in Enzymology 9: 242-253); твердофазный прямой EIA с биотином-авидином (см., например, Kirkland et al, 1986, J. Immunol. 137: 3614-3619); твердофазный прямой анализ на основе мечения, твердофазный прямой «сэндвич»-анализ на основе мечения (см., например, Harlow and Lane, 1988, Antibodies, A Laboratory Manual, Cold Spring Harbor Press); твердофазный прямой RIA на основе мечения меткой Г125 (см., например, Morel et al, 1988, Molec. Immunol. 25: 7-15); твердофазный прямой EIA на основе мечения с биотином-авидином (см., например, Cheung, et al, 1990, Virology 176: 546-552); и прямой RIA на основе мечения (Moldenhauer et al, 1990, Scand. J. Immunol. 32: 77-82). Обычно анализ включает применение очищенного антигена, способного связываться с твердой поверхностью или клеткой, которую загружают как немеченым тестируемым антигенсвязывающим белком, так и меченым референсным антигенсвязывающим белком. Конкурентное ингибирование определяется измерением количества метки, связанной с твердой поверхностью или с клеткой, в присутствии тестируемого антигенсвязывающего белка. Обычно, тестируемый антигенсвязывающий белок находится в избытке. Антигенсвязывающие белки, идентифицируемые посредством конкурентного анализа (конкуренция с антигенсвязывающим белком), включают: антигенсвязывающие белки, которые связываются с тем же эпитопом, что и референсный антигенсвязывающий белок; и антигенсвязывающие белки, которые связываются с эпитопом, который достаточно близок к эпитопу, с которым связывается референсный антигенсвязывающий белок, где данные два эпитопа пространственно мешают друг другу, препятствуя связыванию. Дополнительные подробности относительно способов определения конкурентного связывания предложены в данном документе в разделе Примеры. Обычно, когда конкурирующий антигенсвязывающий белок находится в избытке, он будет ингибировать (например, уменьшать) по меньшей мере на 40-45%, 45-50%, 50-55%, 55-60%, 60-65%, 65-70%, 70-75% или 75% или даже больше специфичное связывание референсного антигенсвязывающего белка с общим антигеном. В некоторых случаях связывание ингибируется по меньшей мере на 80-85%, 85-90%, 90-95%, 95-97% или 97% или даже больше.
Термин «молекула нуклеиновой кислоты», в том виде, в котором он используется в данном документе, относится к молекулам ДНК и молекулам РНК. Молекула нуклеиновой кислоты может быть одноцепочечной или двухцепочечной, и предпочтительно является двухцепочечной ДНК или одноцепочечной мРНК или модифицированной мРНК. Нуклеиновая кислота является «функционально связанной», когда она находится в функциональной взаимосвязи с другой последовательностью нуклеиновой кислоты. Например, промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию данной последовательности.
Термин «идентичность» аминокислотных последовательностей относится к проценту аминокислотных остатков, которые идентичны у первой и второй последовательности при выравнивании данных аминокислотных последовательностей (введение гэпов при необходимости) для достижения максимального процента идентичности последовательностей, и любая консервативная замена не считается частью идентичности последовательностей. С целью определения процента идентичности аминокислотных последовательностей, выравнивание может быть достигнуто множеством путей в объеме данной области, например, используя имеющееся в открытом доступе программное обеспечение, такое как программное обеспечение BLAST, BLAST-2, ALIGN, ALIGN-2 или Megalign (DNASTAR). Специалисты в данной области могут определять параметры, подходящие для измерения выравнивания, включая любой алгоритм, требуемый для достижения оптимального выравнивания по всей длине сравниваемых последовательностей.
Термин «экспрессионный вектор» относится к молекуле нуклеиновой кислоты, способной транспортировать другую нуклеиновую кислоту, с которой она связана. В одном воплощении вектор представляет собой «плазмиду», которая относится к петле кольцевой двухцепочечной ДНK, в которую могут быть лигированы дополнительные сегменты ДНК. В еще одном воплощении вектор представляет собой вирусный вектор, в котором дополнительные сегменты ДНК могут быть лигированы в вирусный геном. Векторы, раскрытые в данном документе, способны к саморепликации в клетке-хозяине, в которую их вводят (например, бактериальные векторы, имеющие бактериальный репликатор, и эписомальные векторы млекопитающего), или могли бы быть интегрированы в геном клетки-хозяина при введении в клетку-хозяина и, таким образом, реплицируются вместе с геномом хозяина (например, векторы млекопитающего, не являющиеся эписомальными).
Способы получения и очистки антител и их антигенсвязывающих фрагментов хорошо известны в данной области, например, A Laboratory Manual for Antibodies, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, chapters 5-8 and 15. Например, мыши могут быть иммунизированы человеческим CTGF или его фрагментом, и полученные антитела могут быть затем ренатурированы, очищены и секвенированы в отношении аминокислотных последовательностей посредством использования традиционных способов, хорошо известных в данной области. Антигенсвязывающие фрагменты также могут быть получены традиционными способами. Антитела или их антигенсвязывающие фрагменты по настоящему раскрытию конструируют, включая одну или более каркасных областей человека в области CDR, происходящие из антитела, не являющегося человеческим. Последовательности FR зародышевой линии человека могут быть получены из ImMunoGeneTics (IMGT) посредством их веб-сайта http://imgt.cines.fr или из The Immunoglobulin Facts Book, 2001, ISBN 012441351, посредством выравнивания против последовательностей базы данных генов вариабельных областей антител зародышевой линии человека, используя программное обеспечение МОЕ.
Термин «клетка-хозяин» относится к клетке, в которую был введен экспрессионный вектор. Клетки-хозяева могут включать клетки бактерий, микроорганизмов, растений или животных. Бактерии, которые легко трансформируются, включают члены Enterobacteriaceae, такие как штаммы Escherichia coil или Salmonella; Baciiiaceae, как например Bacillus subtilis; Pneumococcus; Streptococcus и Haemophilus influenzae. Подходящие микроорганизмы включают Saccharomyces cerevisiae и Pichia pastoris. Подходящие линии животных клеток-хозяев включают СНО (от англ. Chinese Hamster Ovary - яичник китайского хомячка), клетки 293 и клетки NS0.
Сконструированные антитела или антигенсвязывающие фрагменты по настоящему раскрытию могут быть получены и очищены общепринятыми способами. Например, последовательности кДНK, кодирующие тяжелую и легкую цепи, можно клонировать и рекомбинировать в экспрессионный вектор GS. Затем, векторами, экспрессирующими рекомбинантный иммуноглобулин, можно стабильно трансфицировать клетки СНО. В качестве более рекомендованного способа, хорошо известного в данной области, экспрессионные системы млекопитающих могут приводить к гликозилированию антител, особенно на высоко консервативных N-концевых участках в области Fc. Стабильные клоны могут быть получены посредством экспрессии антитела, специфично связывающегося с человеческим CTGF. Позитивные клоны могут быть размножены в бессывороточной культуральной среде в биореакторах для получения антител. Культуральную среду, в которую было секретировано антитело, можно очищать традиционными методиками. Например, очистку можно проводить на колонке с сефарозой FF с белком А или G, которая была модифицирована буфером. Компоненты неспецифичного связывания вымывают.Связанное антитело элюируют с помощью градиента рН, и фрагменты антитела выявляют посредством SDS-PAGE (от англ. sodium dodecyl sulphate-polyacrylamide gel electrophoresis - электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия) и затем объединяют.Антитела можно фильтровать и концентрировать, используя обычные методики. Растворимые смеси и мультимеры можно эффективно удалять обычными методиками, такими как гель-фильтрация или ионный обмен. Затем полученный продукт нужно сразу же заморозить, например, при -70°С, или лиофилизировать.
Термин «введение», «осуществление дозирования» и «обработка» при применении к животному, человеку, субъекту эксперимента, клетке, ткани, органу или биологической жидкости, относится к приведению экзогенного фармацевтического, терапевтического, диагностического средства или композиции в контакт с животным, человеком, субъектом, клеткой, тканью, органом или биологической жидкостью. Термины «введение», «осуществление дозирования» или «обработка» могут относиться, например, к терапевтическим, фармакокинетическим, диагностическим, исследовательским и экспериментальным способам. Обработка клетки охватывает приведение реагента в контакт с клеткой, а также приведение реагента в контакт с жидкостью, где данная жидкость находится в контакте с клеткой. Термин «введение», «осуществление дозирования» или «обработка» также означает обработки in vitro или ex vivo, например клетки, реагентом, диагностическим, связывающим соединением или другой клеткой. Термин «лечение», при применении к субъекту-человеку, субъекту ветеринарии или субъекту исследования, относится к терапевтическому лечению, профилактическим или превентивным мерам, к исследовательским и диагностическим применениям.
Термин «лечение» означает введение терапевтического средства, такого как композиция, содержащая любое из связывающих соединений по настоящему раскрытию, внутрь или наружно пациенту, имеющему один или более симптомов заболевания, в отношении которого данное средств обладает известной терапевтической активностью. Типично средство вводят в количестве, эффективном для облегчения одного или более симптомов заболевания у пациента или популяции, подлежащих лечению, вызывая регрессию или ингибируя прогрессирование такого(их) симптома(ов) до какой-либо клинически измеряемой степени. Количество терапевтического средства, которое является эффективным для облегчения какого-либо конкретного симптома заболевания (также называемое «терапевтически эффективным количеством»), может варьировать в зависимости от разных факторов, таких как патологическое состояние, возраст и масса тела пациента, и способности лекарственного средства вызывать желаемый ответ у пациента. Был ли облегчен симптом заболевания можно оценить посредством любого клинического измерения, обычно используемого лечащими врачами или другими квалифицированными медицинскими работниками, для оценки тяжести или статуса прогрессирования того симптома. В то время как одно воплощение настоящего раскрытия (например, способ лечения или продукт изготовления) может не быть эффективным в облегчении целевого(ых) симптома(ов) заболевания у каждого пациента, оно должно облегчать целевой(ые) симптом(ы) заболевания у статистически значимого числа пациентов, как определено любым статистическим критерием, известным в данной области, таким как t-критерий Стьюдента, критерий хи-квадрат, U-критерий Манна и Уитни, критерий Краскела - Уоллиса (Н-критерий), критерий Джонкхира-Терпстры и критерий Уилкоксона.
Термин «консервативная модификация» или «консервативная замена или замещение» относится к заменам аминокислот в белке другими аминокислотами, имеющими похожие характеристики (например, заряд, размер боковых цепей, гидрофобность/гидрофильность, конформация и жесткость остова и т.д.), таким образом, что данные изменения могут часто происходить без изменения биологической активности белка. Специалисты в данной области осознают, что в общем одна единственная аминокислотная замена в заменимой области полипептида существенно не изменяет биологическую активность (см., например, Watson et al. (1987) Molecular Biology of the Gene, The Benjamin/Cummings Pub. Co., page 224, (4th edition)). Кроме того, замены структурно или функционально похожими аминокислотами с меньшей вероятностью нарушат биологическую активность. Типичные консервативные замены изложены ниже.

Термин «эффективное количество» или «эффективная доза» относится к количеству лекарственного средства, соединения или фармацевтической композиции, необходимому для получения какого-либо одного или более полезных или желательных результатов. Для профилактических применений полезные или желательные результаты включают исключение или снижение риска, ослабление тяжести или задержку начала заболевания, включая биологические, гистологические и поведенческие проявления патологического состояния, его осложнений и промежуточных патологических фенотипов на протяжении развития данного состояния. Для терапевтических применений полезные или желательные результаты включают клинические результаты, такие как уменьшение частоты возникновения разных состояний, ассоциированных с антигеном-мишенью по настоящему раскрытию, или улучшение одного или более симптомов состояния, уменьшение дозировки/дозы других средств, требующихся для лечения патологического состояния, усиление эффективности другого средства и/или задержка прогрессирования патологического состояния, ассоциированного с антигеном-мишенью по настоящему раскрытию, у пациентов.
Термин «экзогенный» относится к веществам, которые образуются вне организмов, клеток или человека, в зависимости от обстоятельств. Термин «эндогенный» относится к веществам, которые образуются в клетках, организмах или организме человека, в зависимости от обстоятельств.
Термин «гомология» относится к сходству последовательностей между двумя полинуклеотидными последовательностями или между двумя полипептидными последовательностями. Когда положение в обеих из данных двух последовательностей, подлежащих сравнению, занято одной и той же мономерной субъединицей - основанием или аминокислотой - например, когда положение в каждой из двух молекул ДНК занято аденином, тогда данные молекулы являются гомологичными в данном положении. Процент гомологии двух последовательностей является функцией числа совпадающих или гомологичных положений, являющихся общими у данных двух последовательностей, деленного на число сравниваемых положений и затем умноженного на 100. Например, при оптимальном выравнивании двух последовательностей, если 6 из 10 положений в данных двух последовательностях совпадают или являются гомологичными, тогда данные две последовательности обладают 60%-ной гомологией; если 95 из 100 положений в двух последовательностях совпадают или являются гомологичными, тогда данные две последовательности обладают 95%-ной гомологией. Обычно, когда две последовательности подвергают выравниванию, проводят сравнение с получением максимального процента гомологии. Например, сравнение можно проводить посредством алгоритма BLAST, в котором параметры алгоритма выбраны для предоставления максимального совпадения между каждой последовательностью по всей длине каждой референсной последовательности. Следующие ссылки относятся к алгоритму BLAST, часто используемому для анализа последовательностей: алгоритм BLAST (BLAST ALGORITHMS): Altschul, SF et al., (1990) J. Mol. Biol. 215:403-410; Gish, W. et al., (1993) Nature Genet. 3:266-272; Madden, TL etal., (1996) Meth. Enzymol. 266:131-141; Altschul, SFetal., (1997) Nucleic Acids Res. 25:3389-3402; Zhang, J. et al. (1997) Genome Res. 7:649-656. Другие общепринятые алгоритмы BLAST, такие как алгоритмы, имеющиеся у NCBI BLAST, также хорошо известны специалистам в данной области.
В том виде, в котором они используются в данном документе, выражения «клетка», «клеточная линия» и «клеточная культура» используются взаимозаменяемо, и все такие обозначения включают потомство. Таким образом, термины «трансформант» и «трансформированная клетка» включают первичные исследуемые клетки и полученные из них культуры без учета числа переносов. Также следует понимать, что все потомство не может быть с точностью идентичным по содержанию ДНК вследствие целенаправленных или случайных мутаций. Также включено потомство с мутациями, которое имеет такую же функцию или обладает такой же биологической активностью, как показано в исходно трансформированных клетках. Когда предполагаются отличающиеся обозначения, это будет явно понятно из контекста.
В том виде, в которой она используется в данном документе, фраза «полимеразная цепная реакция» или «ПЦР» относится к способу или методике, в которой малые количества конкретного фрагмента нуклеиновой кислоты, РНК и/или ДНK, амплифицируют, как описано, например, в патенте США №4683195. Обычно существует необходимость в наличии доступной информации о последовательности на концах или за пределами исследуемой области, таким образом, чтобы можно было сконструировать олигонуклеотидные праймеры; данные праймеры будут идентичными или похожими по последовательности с комплементарными цепями матрицы, подлежащей амплификации. 5'-Концевые нуклеотиды данных двух праймеров могут соответствовать концам амплифицируемого материала. ПЦР можно использовать для амплификации конкретных последовательностей РНK, конкретных последовательностей ДНК из тотальной геномной ДНК и кДНK, транскрибируемой с тотальной РНК клетки, последовательностей бактериофагов или плазмид и т.д. Обычно см. Mullis et al. (1987) Cold Spring Harbor Symp.Ouant.Biol. 51:263; Erlich, ed., (1989) PCR TECHNOLOGY (Stockton Press, N.Y.). ПЦР, используемая в настоящем раскрытии, считается одним, но не единственным, примером способа амплификации образца нуклеиновой кислоты, подлежащего анализу, на основе полимеразной реакции. Способ включает применение последовательностей нуклеиновой кислоты, известных как праймеры, вместе с полимеразой нуклеиновой кислоты для амплификации или образования конкретного фрагмента нуклеиновой кислоты.
Термин «выделенный» относится к очищенному состоянию, в котором обозначенная молекула по существу не содержит других биологических молекул, таких как нуклеиновые кислоты, белки, липиды, углеводы или другие вещества, такие как обломки клеток и среда для выращивания. В общем, термин «выделенный» не предназначен для обозначения полного отсутствия данных веществ или отсутствия воды, буферов или солей, если они не находятся в количестве, которое значимо мешает экспериментальному или терапевтическому применению соединения, как описано в данном документе.
Термин «возможный» или «возможно» означает, что событие или ситуация, которая следует, может происходить, но не обязательно происходит, и описание включает примеры, в которых событие или ситуация происходит или не происходит.
Термин «фармацевтическая композиция» относится к смеси, содержащей одно или более соединений согласно настоящему раскрытию или их физиологически/фармацевтически приемлемую соль или пролекарство и другие химические компоненты, такие как физиологически/фармацевтически приемлемые носители и вспомогательные вещества. Целью фармацевтической композиции является содействие введению в организм, облегчение поглощения активного вещества и оказание, таким образом, биологического действия.
Термин «фармацевтически приемлемый носитель» относится к любому неактивному веществу, подходящему для применения в композиции для доставки антител или антигенсвязывающих фрагментов. Носитель может представлять собой антиадгезивное средство, связывающее вещество, покрывающий агент, разрыхлитель, наполнитель или разбавитель, консервант (такой как антиоксидант, антибактериальное или противогрибковое средство), подсластитель, средство, препятствующее поглощению, увлажнитель, эмульгатор, буфер и т.п. Примеры подходящих фармацевтически приемлемых носителей включают воду, этанол, полиол (такой как глицерин, пропиленгликоль, полиэтиленгликоль и т.п.), декстрозу, растительное масло (такое как оливковое масло), физиологический раствор, буфер, забуференный физиологический раствор и изотонический агент (как например, сахара, полиол, сорбит и хлорид натрия).
Кроме того, настоящее раскрытие включает средство для лечения заболевания, ассоциированного с клетками, позитивными в отношении антигена-мишени (таким как CTGF), причем данное средство содержит антитело против CTGF или его фрагмент по настоящему раскрытию в качестве активного ингредиента. Активный ингредиент вводят субъекту в терапевтически эффективном количестве, и он способен лечить заболевание, ассоциированное с CTGF-позитивными клетками у субъекта. Терапевтически эффективное количество означает, что однократная доза композиции содержит 0,1-3000 мг полноразмерного антитела или его антигенсвязывающего фрагмента, которое специфично связывается с человеческим CTGF, как описано выше.
CTGF-связанное заболевание по настоящему раскрытию не ограничено, при условии, что это заболевание, связанное с CTGF. Например, терапевтический ответ, индуцируемый молекулой по настоящему раскрытию, может играть роль за счет связывания с человеческим CTGF и затем предотвращения связывания CTGF со своим рецептором/лигандом или посредством уничтожения опухолевых клеток, сверхэкспрессирующих CTGF. Таким образом, молекулы по настоящему раскрытию, когда находятся в препарате или композиции, подходящей для терапевтических применений, очень полезны для таких людей, страдающих от опухоли или рака, возможно включая фиброз легких, фиброз почек, развитие и рост опухоли, глаукому, клеточное пролиферативное заболевание, катаракту, хориоидальную неоваскуляризацию, отслойку сетчатки, пролиферативную витреоретинопатию, макулодистрофию, диабетическую ретинопатию, рубцевание роговицы, помутнение роговицы, кисту, уменьшенную кальцификацию сосудов, протоковую аденокарциному поджелудочной железы, рак поджелудочной железы, меланому, постлучевой фиброз (RIF - от англ. radiation-induced fibrosis), идиопатический легочный фиброз, заболевание на основе ремоделирования легких, выбранное из группы, состоящей из астмы, хронического бронхита, хронической обструктивной болезни легких (ХОБЛ), кистозного фиброза, эмфиземы и т.д., предпочтительно рак поджелудочной железы, фиброз легких и фиброз почек.
Кроме того, настоящее раскрытие относится к способам иммунологического анализа или определения антигенов-мишеней (например, CTGF), реагентам для иммунологического анализа или определения антигенов-мишеней (например, CTGF), способам иммунологического анализа или определения клеток, экспрессирующих антигены-мишени (например, CTGF), и диагностическим средствам для диагностирования заболеваний, ассоциированных с антигеном-мишенью, например, CTGF-позитивных клеток, содержащим антитело или фрагмент антитела по настоящему раскрытию (в качестве активного ингредиента), которое специфично распознает антигены-мишени (например, человеческий CTGF) и связывается с аминокислотными последовательностями внеклеточной области или с ее третичной структурой.
В настоящем раскрытии способ выявления или измерения количества антигена-мишени (например, CTGF) может представлять собой любой известный способ. Например, он включает способы иммунологического анализа или определения.
Способ иммунологического анализа или определения представляет собой способ выявления или измерения количества антитела или антигена с использованием меченого антигена или антитела. Примеры способов иммунологического анализа или определения включают иммунологический способ с использованием антитела, меченного радиоактивным веществом (RIA), иммуноферментный анализ (EIA или ELISA), флуоресцентный иммунологический анализ (FIA - от англ. fluorescence immunoassay), иммунолюминесцентный анализ, вестерн-блоттинг, физико-химический способ и т.д.
Упомянутые выше заболевания, ассоциированные с CTGF-позитивными клетками, могут быть диагностированы посредством выявления или измерения CTGF-экспрессирующих клеток с использованием антител или фрагментов антител по настоящему раскрытию.
Клетки, экспрессирующие полипептид, можно выявлять известными способами иммунодетекции, предпочтительно посредством иммунопреципитации, окрашивания клеток флуоресцентными красителями, иммуногистохимического окрашивания и т.п. Кроме того, можно использовать способ, такой как способ окрашивания на основе флуоресцирующих антител с использованием системы FMAT8100HTS (Applied Biosystem).
В настоящем раскрытии образцы, подлежащие выявлению или измерению в отношении антигена-мишени (например, CTGF), конкретным образом не ограничены, в том случае, если они могут содержать клетки, экспрессирующие антиген-мишень (например, CTGF), как например, ткани, клетки, кровь, плазма, сыворотка, секреция поджелудочной железы, моча, кал, тканевая жидкость или культуральная среда.
В зависимости от требуемого способа диагностики, диагностическое средство, содержащее моноклональное антитело или фрагмент антитела по настоящему раскрытию, может также содержать реагенты для проведения реакции антиген-антитело или реагенты для выявления данной реакции. Реагенты для проведения реакции антиген-антитело включают буферы, соли и т.п. Реагенты, используемые для выявления, включают средства, обычно используемые в способах иммунологического анализа или иммунодетекции, например, меченое вторичное антитело, которое распознает моноклональное антитело, фрагмент антитела или его конъюгат, и субстрат, соответствующий метке.
Подробности одного или более воплощений настоящего раскрытия изложены в приведенном выше описании изобретения. Предпочтительные методы и материалы описаны ниже, несмотря на то, что любой метод или материал, похожий или идентичный методу или материалу, описанному в данном документе, можно использовать на практике или в анализе настоящего раскрытия. На примере описания изобретения и формулы изобретения, другие признаки, цели и преимущества настоящего раскрытия станут очевидными. В описании изобретения и формуле изобретения формы единственного числа предназначены для того, чтобы включать формы множественного числа, если контекстом явным образом не продиктовано иное. Если в данном документе явным образом не определено иное, все технические и научные термины, используемые в данном документе, имеют значение, обычно подразумеваемое специалистами в области, к которой принадлежит данное раскрытие. Все патенты и публикации, приведенные в описании изобретения, включены посредством ссылки. Следующие примеры представлены для более полной иллюстрации предпочтительных воплощений настоящего раскрытия. Данные примеры не следует рассматривать как ограничивающие каким-либо образом объем настоящего раскрытия, и объем настоящего раскрытия определяется формулой изобретения.
ПРИМЕРЫ
Пример 1. Получение антигена CTGF и антитела 1. Конструирование и экспрессия антигена
Человеческий белок CTGF (номер Genbank: NM_001901.2) использовали в качестве матрицы для конструирования аминокислотной последовательности антигена, и данный белок использовали для выявления. Возможно, белок CTGF или его фрагмент сливали с разными метками, и отдельно клонировали в вектор рТТ5 или вектор рХС, и подвергали временной экспрессии в клетках 293 или стабильной экспрессии в клетках СНО LONZA с получением антигенов и белков, используемых для выявления настоящего раскрытия. Следующие антигены CTGF относятся к человеческому CTGF, если конкретно не указано иное.
Человеческий полноразмерный белок CTGF (для иммунизации), SEQ ID NO: 1
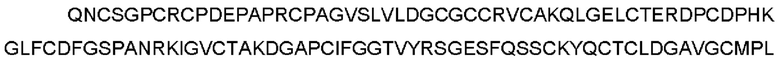

CTGF-his (слитый белок человеческого зрелого белка CTGF с his- меткой) может быть использован для выявления, SEQ ID NO: 2

(Примечание: одинарное подчеркивание представляет сигнальный пептид, выделение курсивом представляет внеклеточную область hCTGF и двойное подчеркивание представляет his-метку.)
mCTGF-mFc (слитый белок кодирующей области CTGF мыши с мышиной Fc-меткой, который может быть использован для выявления) SEQ ID NO: 3

(Примечание: одинарное подчеркивание представляет сигнальный пептид, выделение курсивом представляет кодирующую область CTGF мыши и двойное подчеркивание представляет mFc-метку.)
Мышиный белок CTGF-His-метка, SEQ ID NO: 4


(Примечание: одинарное подчеркивание представляет сигнальный пептид, выделение курсивом представляет кодирующую область мышиного CTGF, и двойное подчеркивание представляет his-метку).
CTGF-Fc яванского макака (слитый белок кодирующей области CTGF яванского макака с Fc, который можно использовать для выявления), SEQ ID NO: 5

(Примечание: одинарное подчеркивание представляет сигнальный пептид, выделение курсивом представляет кодирующую область CTGF яванского макака и двойное подчеркивание представляет Fc-метку.)
II. Очистка CTGF-связанного рекомбинантного белка и очистка антитела гибридомы против человеческого CTGF и рекомбинантного антитела
1. Выделение и очистка супернатанта гибридомы посредством аффинной хроматографии на основе белка G
Для очистки супернатанта мышиной гибридомы аффинная хроматография, проводимая с помощью белка G, была предпочтительной. Полученную гибридому после культивирования центрифугировали, и вынимали супернатант, и, исходя из объема супернатанта, 10-15% объем 1 М Tris-HCl (рН 8,0-8,5) добавляли для доведения рН супернатанта до нейтрального. Колонку с белком G промывали 3-5х объемом колонки 6 М гуанидин гидрохлорида и затем промывали 3-5х объемом колонки чистой воды; колонку уравновешивали 3-5х объемом колонки 1×PBS (от англ. Phosphate buffered saline - фосфатно-солевой буферный раствор) (рН 7,4); супернатант клеток загружали при низкой скорости потока для связывания, и скорость потока контролировали таким образом, что время удерживания составляло примерно 1 мин или дольше; колонку промывали 3-5х объемом колонки 1×PBS (рН 7,4) до тех пор, пока поглощение УФ не падало до исходного уровня; образцы элюировали 0,1 М буфером уксусная кислота/ацетат натрия (рН 3,0), пики элюирования объединяли в соответствии с УФ-детектированием. рН элюированного продукта быстро доводили до рН 5-6 посредством 1 М Tris-HCl (рН 8,0). Элюируемый продукт может подвергаться замене раствора в соответствии со способами, хорошо известными специалистам в данной области, например, замене раствора желательной буферной системой посредством ультрафильтрации-концентрирования с помощью ультрафильтрационной трубки, или замене раствора желательной буферной системой посредством использования молекулярного исключения, как например, обессоливание G-25, или удалению агрегатов из элюированного продукта для улучшения чистоты образца посредством использования молекулярной эксклюзионной колонки, такой как Superdex 200.
2. Очистка антитела посредством аффинной хроматографии на основе белка А
Сначала, супернатант клеточной культуры, экспрессирующей антитело, центрифугировали при высокой скорости для сбора супернатанта. Колонку для аффинной хроматографии с белком А промывали 3-5х объемом колонки 6 М гуанидин гидрохлорида и затем промывали 3-5х объемом колонки чистой воды. Колонку уравновешивали 3-5х объемом колонки, например, 1×PBS (рН 7,4). Клеточный супернатант загружали при низкой скорости потока для связывания, и скорость потока контролировали таким образом, что время удерживания составляло примерно 1 мин или дольше; сразу после окончания связывания, колонку промывали 3-5х объемом колонки 1×PBS (рН 7,4) до тех пор, пока поглощение УФ не падало до исходного уровня. Образцы элюировали 0,1 М буфером уксусная кислота/ацетат натрия (рН 3,0-3,5), и пики элюирования объединяли в соответствии с УФ-выявлением. рН элюируемого продукта быстро подводили с помощью 1 М Tris-HCl (рН 8,0) до рН 5-6. Элюируемый продукт можно подвергать замене растворителя в соответствии со способами, хорошо известными специалистам в данной области, например, замене раствора желательной буферной системой посредством ультрафильтрации-концентрирования с помощью ультрафильтрационной трубки, или замене раствора желательной буферной системой посредством использования молекулярного исключения, такого как, обессоливание G-25, или удалению агрегатов из элюированного продукта для улучшения чистоты образца посредством использования молекулярной эксклюзионной колонки высокого разрешения, такой как Superdex 200.
3. Очистка CTGF-связанных рекомбинантных белков (таки как человеческий CTGF-his) посредством никелевой колонки
Образец супернатанта клеточной экспрессии центрифугировали при высокой скорости для удаления примесей, буфер заменяли PBS, и добавляли имидазол до конечной концентрации 5 мМ. Никелевую колонку уравновешивали раствором PBS, содержащим 5 мМ имидазол, и промывали 2-5х объемом колонки. Образец супернатанта после замены загружали на колонку для связывания, и никелевые колонки, имеющиеся у разных компаний, могут быть выбраны в качестве сред. Колонку промывали раствором PBS, содержащим 5 мМ имидазол, до тех пор, пока измеренное значение при А280 не падало до исходного уровня. Хроматографическую колонку промывали PBS плюс 10 мМ имидазол для удаления неспецифично связанных белковых примесей, и собирали эффлюент.Целевой белок элюировали раствором PBS, содержащим 300 мМ имидазол, и собирали пик элюирования.
Собранный продукт элюирования концентрировали, и затем он может быть дополнительно очищен посредством колонки для гель-хроматографии Superdex200 (GE) с помощью PBS в качестве подвижной фазы для удаления агрегатов и пиков белковой примеси и для сбора пика элюирования целевого продукта. Полученный белок идентифицировали посредством электрофореза, составления пептидных карт и ЖХ-МС (жидкостная хроматография-масс-спектрометрия), и правильные белки аликвотировали для применения.
4. Очистка CTGF-связанных рекомбинантных белков (таких как человеческий CTGF-Fc, CTGF-Fc яванского макака) посредством аффинной хроматографии с белком А
Сначала, супернатант культуры центрифугировали при высокой скорости для сбора супернатанта. Колонку для аффинной хроматографии с белком А промывали 3-5х объемом колонки 6 М гуанидин гидрохлорида и затем промывали 3-5х объемом колонки чистой воды. Колонку уравновешивали 3-5х объемом колонки, например, 1×PBS (рН 7,4). Супернатант клеточной культуры загружали при низкой скорости потока для связывания, и скорость потока контролировали таким образом, что время удерживания составляло примерно 1 мин или дольше; сразу после окончания связывания, колонку промывали 3-5х объемом колонки 1 xPBS (рН 7,4) до тех пор, пока поглощение УФ не падало до исходного уровня. Образцы элюировали 0,1 М буфером уксусная кислота/ацетат натрия (рН 3,0-3,5), и пики элюирования объединяли в соответствии с УФ-выявлением.
Собранные продукт элюирования концентрировали, и затем он может быть дополнительно очищен посредством гель-хроматографии Superdex200 (GE) с помощью PBS в качестве подвижной фазы для удаления агрегатов и пиков белковых примесей, и для сбора пика элюирования целевого продукта. Полученный белок идентифицировал посредством электрофореза, составления пептидных карт и ЖХ-МС, и правильные белки аликвотировали для применения.
Пример 2. Получение моноклонального антитела против человеческого
CTGF
I. Иммунизация
Моноклональные антитела против человеческого CTGF получали посредством иммунизации мышей. Для эксперимента использовали самок белых мышей SJL в возрасте 6-8 недель (Beijing Charles River Experimental Animal Technology Co., Ltd., номер лицензии на производство животных: SCXK (Пекин) 2012-0001). Среда кормления: уровень SPF (от англ. specific pathogen free -свободный от специфической патогенной микрофлоры). После приобретения мышей их держали в лабораторных условиях в течение 1 недели, с 12/12-часовым циклом свет/темнота, при температуре 20-25°С; влажности 40-60%. Затем мышей, которые были адаптированы к окружающей среде, иммунизировали в соответствии со следующими схемами. CTGF (R&D) и mCTGF-mFc использовали в качестве антигена для иммунизации.
Схема иммунизации: мышей иммунизировали белком CTGF, 25 микрограмм/мышь/время, посредством внутрибрюшинной инъекции. 100 мкл/мышь инъецировали внутрибрюшинно (в.б.) в сутки 0, и затем проводили повторную иммунизацию один раз в 14 суток. Образы крови собирали в сутки 21, 35, 49 и 63, титр антител в мышиной сыворотке определяли посредством способа ELISA. После 7-9 иммунизаций мышей с высоким титром антител в сыворотке, достигающим плато, отбирали для слияния со спленоцитами. За трое суток до слияния со спленоцитами раствор белкового антигена CTGF, полученный в физиологическом растворе, инъецировали внутрибрюшинно (в.б.), 25 мкг/мышь, для повторной иммунизации.
II. Слияние со спленоцитами
Клетки гибридомы получали посредством слияния лимфоцитов селезенки с клетками миеломы Sp2/0 (АТСС® CRL-8287™) посредством использования способа слияния, опосредованного ПЭГ (полиэтиленгликоль). Клетки слитой гибридомы ресуспенд провал и в полной среде (среда IMDM, содержащая 20% FBS (от англ. Fetal Bovine Serum - фетальная телячья сыворотка), 1×HAT и 1×ОР1) при плотности 0,5×106 - 1×106/мл, засевали в 96-луночный планшет в количестве 100 мкл/лунка, инкубировали при 37°С и 5% СO2 в течение 3-4 суток, дополняли 100 мкл/лунка полной среды HAT и продолжали культивировать на протяжении 3-4 суток до образования точно установленных клонов. Супернатант удаляли, добавляли в него 200 мкл/лунка полной среды НТ (среда IMDM, содержащая 20% FBS, 1×НТ и 1×ОРI), инкубировали при 37°С, 5% СO2 в течение 3 суток и затем подвергали ELISA-выявлению.
III. Скрининг клеток гибридомы
В соответствии с плотностью роста клеток гибридомы, супернатант культуры гибридомы выявляли способом ELISA связывания. Клеточный супернатант в позитивных лунках выявляли анализом ELISA в отношении связывания с белком CTGF обезьяны/ CTGF мыши/ CTGF человека (конкретные способы представляли собой такие же, как способы Примера 3). Клетки в лунках, позитивных в случае анализа ELISA связывания белка, своевременно размножали и криоконсервировали; и субклонировали 2 - 3 раза до получения клона одиночной клетки.
Клетки после каждого субклонирования также анализировали посредством ELISA связывания CTGF. Клоны гибридомы получали посредством осуществления скрининга с помощью указанного выше анализа. Антитела дополнительно получали способом на основе клеточной культуры, не содержащей сыворотку. Антитела очищали в соответствии с примером очистки для использования в примерах анализа.
IV. Секвенирование позитивных клонов гибридомы
Способы клонирования последовательностей из позитивной гибридомы выглядели следующим образом. Клетки гибридомы собирали в логарифмическую фазу роста. РНК выделяли посредством Тризола (Invitrogen, кат. №15596-018) в соответствии с инструкцией к набору и подвергали обратной транскрипции с использованием набора для обратной транскрипции PrimeScript™ (Takara, кат. №2680А). кДНK, полученные в результате обратной транскрипции, амплифицировали посредством ПЦР с использованием набора праймеров к мышиному Ig (Novagen, ТВ326 Rev. В 0503), и амплифицированные продукты подвергали секвенированию. После секвенирования получали мышиные антитела против CTGF maM47, maM64 и mab95. Аминокислотные последовательности вариабельных областей выглядят следующим образом:
Аминокислотная последовательность VH mab147 представляет собой такую, как показано в SEQ ID NO: 6,

Аминокислотная последовательность VL mab147 представляет собой такую, как показано в EQ ID NO: 7,
Аминокислотная последовательность VH mab164 представляет собой такую, как показано в SEQ ID NO: 8,

Аминокислотная последовательность VL mab164 представляет собой такую, как показано в SEQ ID NO: 9,

Аминокислотная последовательность VH mab95 представляет собой такую, как показано в SEQ ID NO: 69,

Аминокислотная последовательность VL mab95 представляет собой такую, как показано в SEQ ID NO: 70,

В упомянутых выше последовательностях вариабельных областей антитела mab147, maM64 и mab95, выделение курсивом представляет FR-последовательность, выделение курсивом с подчеркиванием представляет последовательности CDR, и порядок последовательности выглядит следующим образом: FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4.
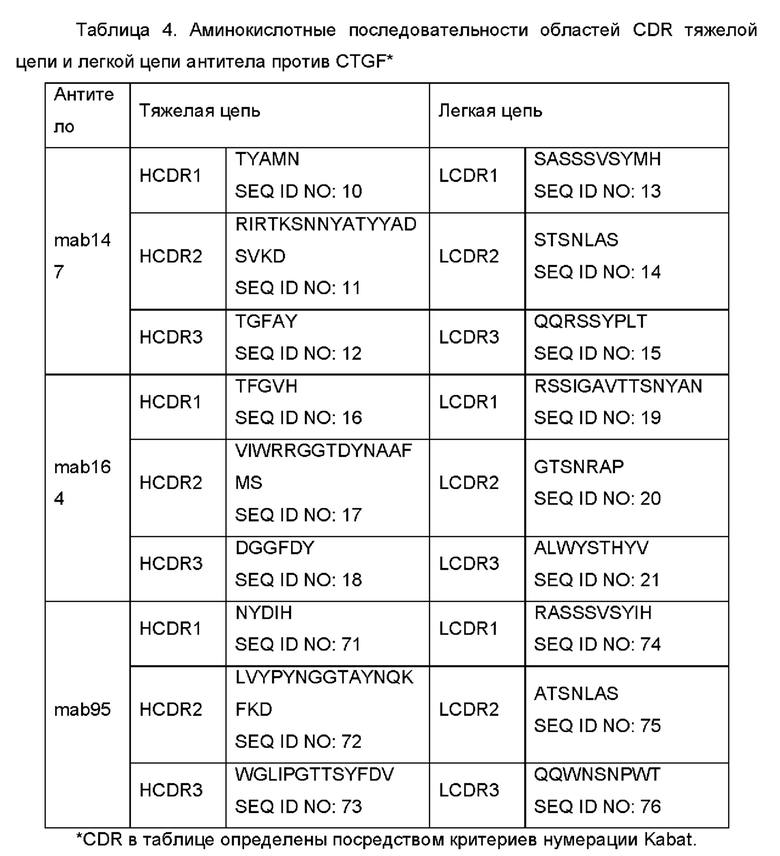
V. Получение химерного антитела IgG1 человека
Молекулы-кандидаты mab147, mab164 и mab95 амплифицировали и секвенировали с получением последовательностей гена, кодирующих вариабельные области. Прямые и обратные праймеры конструировали на основе последовательностей, полученных посредством секвенирования, ген, подлежащий секвенированию, использовали в качестве матрицы для конструирования фрагмента гена VH/VK для каждого антитела посредством ПЦР и затем вставляли в экспрессионный вектор рНr (с сигнальным пептидом и фрагментом гена константной области hIgG1/hkappa/hlambda (CH1-Fc/CL)) посредством гомологичной рекомбинации для конструирования экспрессионной плазмиды для полноразмерного рекомбинантного химерного антитела VH-CH1-Fc-pHr/VL-CL-pHr, что приводило к получению трех химерных антител Сh147, Сh164 и Сh95.
VI. Гуманизация мышиного антитела против человеческого CTGF Гены вариабельных областей тяжелой цепи и легкой цепи зародышевой линии, обладающие высокой гомологией с мышиными антителами, отбирали в качестве матриц посредством выравнивания против базы данных генов вариабельных областей тяжелой и легкой цепи антител зародышевой линии человека IMGT (http://imgt.cines.fr) посредством использования программного обеспечения МОЕ (Molecular Operating Environment). CDR антитела мыши прививали на соответствующие матрицы человека с образованием последовательностей вариабельных областей в следующем порядке FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4.
1. Гуманизация mab147
Для мышиного антитела mab147 матрицы легкой цепи для гуманизации представляли собой IGKV3-1T01 и IGKJ2*01, или IGKV1-39*01 и IGKJ2*01, и матрицы тяжелой цепи для гуманизации представляли собой IGHV3-72*01 и IGHJ1×01. Каждую из CDR мышиного антитела mab147 прививали на его человеческую матрицу, и затем аминокислоты FR гуманизированного антитела подвергали обратной мутации. Среди них, обратная(ые) мутация(и) в вариабельной области легкой цепи вкпючает(ют) одну или более, выбранных из группы, состоящей из 4L, 36F, 43S, 45K, 47VV, 58V или 71Y, и обратная(ые) мутация(и) в вариабельной области тяжелой цепи включает(ют) одну или более, выбранных из группы, состоящей из 28S, 30N, 49А, 75Е, 76S, 93V, 94Е или 104D (положения обратной мутации в вариабельных областях легкой и тяжелой цепей определяли в соответствии с критериями нумерации Kabat), что приводило к получению вариабельных областей легкой цепи и вариабельных областей тяжелой цепи с разными последовательностями. Получали гуманизированные антитела, содержащие CDR mab147, и последовательности их вариабельных областей выглядят следующим образом:
Специфичные последовательности вариабельных областей гуманизированных антител тАМ47 выглядят следующим образом:
Аминокислотная последовательность Hu147-VL1 представляет собой такую, как показано в SEQ ID NO: 22

Аминокислотная последовательность Hu147-VL2 представляет собой такую, как показано в SEQ ID NO: 23

Аминокислотная последовательность Hu147-VL3 представляет собой такую, как показано в SEQ ID NO: 24

Аминокислотная последовательность Hu147-VL4 представляет собой такую, как показано в SEQ ID NO: 25

Аминокислотная последовательность Hu147-VL5 представляет собой такую, как показано в SEQ ID NO: 26

Аминокислотная последовательность Hu147-VH1 представляет собой такую, как показано в SEQ ID NO: 27

Аминокислотная последовательность Hu147-VH2 представляет собой такую, как показано в SEQ ID NO: 28

Аминокислотная последовательность Hu147-VH3 представляет собой такую, как показано в SEQ ID NO: 29

Примечание: Последовательности CDR, определенные в соответствии с критериями Kabat, подчеркнуты, и данные последовательности расположены в следующем порядке: FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4.
2. Гуманизация mab164
Для мышиного антитела mab164 матрицы легкой цепи для гуманизации состояли из IGLV7-43*01 и IGLJ2*01, IGLV8-61×01 и IGLJ2*01, или IGLV1-40*02 и IGLJ2*01, соответственно, и матрицы тяжелой цепи для гуманизации состояли из IGHV2-26*01 и IGHJ6*01 или IGHV4-31×02 и IGHJ6*01, соответственно. Каждую из CDR мышиного антитела mab164 прививали на его человеческую матрицу, и затем аминокислоты FR гуманизированного антитела подвергали обратной мутации. Среди них, обратная(ые) мутация(и) в вариабельной области легкой цепи включает(ют) одну или более, выбранных из группы, состоящей из 36V, 44F, 46G или 49G, и обратная(ые) мутация(и) в вариабельной области тяжелой цепи включает(ют) одну или более, выбранных из группы, состоящей из 44G, 49G, 27F, 48L, 67L, 71 K, 78V или 80F (положения обратной мутации в вариабельных областях легкой и тяжелой цепей определяли в соответствии с критериями нумерации Kabat), что приводило к получению гуманизированных тяжелой и легкой цепи mab164 с последовательностями вариабельных областей, которые выглядят следующим образом:
Специфичные последовательности вариабельных областей гуманизированных антител тАМ64 выглядят следующим образом:
Аминокислотная последовательность Hu164-VL7 представляет собой такую, как показано в SEQ ID NO: 30

Аминокислотная последовательность Hu164-VL8 представляет собой такую, как показано в SEQ ID NO: 31

Аминокислотная последовательность Hu164-VL9 представляет собой такую, как показано в SEQ ID NO: 32

Аминокислотная последовательность Hu164-VH5 представляет собой такую, как показано в SEQ ID NO: 33

Аминокислотная последовательность Hu164-VH6 представляет собой такую, как показано в SEQ ID NO: 34

Аминокислотная последовательность Hu164-VH7 представляет собой такую, как показано в SEQ ID NO: 35

Аминокислотная последовательность Hu164-VH8 представляет собой такую, как показано в SEQ ID NO: 36

Примечание: В упомянутых выше последовательностях антитела Ни164 подчеркивание представляет последовательность CDR, и порядок последовательности представляет собой FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4, где последовательности CDR определены в соответствии с критериями Kabat.
3. Гуманизация mab95
Для мышиного антитела mab95 матрицы легкой цепи для гуманизации представляли собой IGKV3-20*02 и IGKV1-40*01, и матрица тяжелой цепи для гуманизации представляла собой IGHV1-3*01. Каждую из CDR мышиного антитела mab95 прививали на его человеческую матрицу, и затем аминокислоты FR гуманизированного антитела подвергали обратной мутации. Среди них, обратная(ые) мутация(и) в вариабельной области легкой цепи включает(ют) одну или более, выбранных из группы, состоящей из 45Р, 46W, 48Y, 69S или 70Y, и обратная(ые) мутация(и) в вариабельной области тяжелой цепи включает(ют) одну или более, выбранных из группы, состоящей из 27F, 38K, 48I, 67K, 68А, 70L или 72F (положения обратной мутации в вариабельных областях легкой и тяжелой цепей определяли в соответствии с критериями нумерации Kabat), что приводило к получению вариабельных областей легкой цепи и вариабельных областей тяжелой цепи с разными последовательностями. Получали гуманизированные антитела, содержащие CDR mab95, и последовательности их вариабельных областей выглядят следующим образом:
Специфичные последовательности вариабельных областей гуманизированных антител тАЬ95 выглядят следующим образом:
Аминокислотная последовательность Hu95-VL1 представляет собой такую, как показано в SEQ ID NO: 77
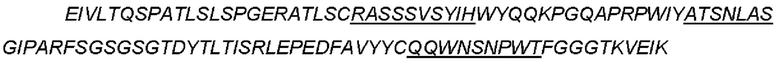
Аминокислотная последовательность Hu95-VL2 представляет собой такую, как показано в SEQ ID NO: 78

Аминокислотная последовательность Hu95-VL3 представляет собой такую, как показано в SEQ ID NO: 79

Аминокислотная последовательность Hu95-VL4 представляет собой такую, как показано в SEQ ID NO: 80

Аминокислотная последовательность Hu95-VH1 представляет собой такую, как показано в SEQ ID NO: 81

Аминокислотная последовательность Hu95-VH2 представляет собой такую, как показано в SEQ ID NO: 82

Аминокислотная последовательность Hu95-VH3 представляет собой такую, как показано в SEQ ID NO: 83
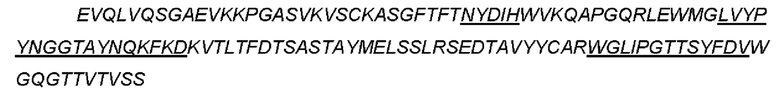
Аминокислотная последовательность Hu95-VH4 представляет собой такую, как показано в SEQ ID NO: 84 
Аминокислотная последовательность Hu95-VH5 представляет собой такую, как показано в SEQ ID NO: 85

Примечание: Последовательности CDR, определенные в соответствии с критериями Kabat, подчеркнуты, и последовательности расположены в следующем порядке FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4.
Среди них, области CDR Hu95-VH5 дополнительно модифицированы, где HCDR1 представляет собой такую, как показано в SEQ ID NO: 102 (NYAIH), HCDR2 представляет собой такую, как показано в SEQ ID NO: 103 (LVYPYTGGTAYNQKFKD), и HCDR3 представляет собой такую, как показано в SEQ ID NO: 104 (WGMIPGTNSYFDV).
4. Конструирование и экспрессия гуманизированного антитела против
CTGF
Конструировали праймеры, фрагмент гена VH/VL каждого гуманизированного антитела конструировали посредством ПЦР и затем вставляли в экспрессионный вектор рНг (с сигнальным пептидом и фрагментом гена константной области (CH/CL)) посредством гомологичной рекомбинации для конструирования экспрессионного вектора для полноразмерного антитела VH-CH -pHr/VL-CL-pHr. Для гуманизированного антитела константная область может быть выбрана из группы, состоящей из константной области легкой цепи к, X человека, и может быть выбрана из группы, состоящей из константной области тяжелой цепи IgG1, IgG2, IgG3 или IgG4 или ее варианта. Неограничивающие примеры включают оптимизацию константной области человеческого IgG1, IgG2 или IgG4 для улучшения функции антитела. Например, период полувыведения антитела может быть продлен посредством точечных мутаций в константной области, таких как мутации в сайте M252Y/S254T/T256E (YTE) и мутации, такие как S228P, F234A и L235A в константной области.
Иллюстративная последовательности константной области гуманизированного антитела против CTGF выглядит следующим образом:
Аминокислотная последовательность константной области тяжелой цепи представляет собой такую, как показано в SEQ ID NO: 37 (тяжелую цепь полноразмерного антитела называют с суффиксом: w):

Аминокислотная последовательность константной области тяжелой цепи сконструированного антитела YTE представляет собой такую, как показано в SEQ ID NO: 38 (тяжелую цепь полноразмерного антитела называют с суффиксом: у):

Аминокислотная последовательность константной области легкой цепи каппа гуманизированного антитела против CTGF представляет собой такую, как показано в SEQ ID NO: 39: (тяжелую цепь полноразмерного антитела называют с суффиксом: k):

Аминокислотная последовательность константной области легкой цепи лямбда гуманизированного антитела против CTGF представляет собой такую, как показано в SEQ ID NO: 40: (тяжелую цепь полноразмерного антитела называют с суффиксом: l):
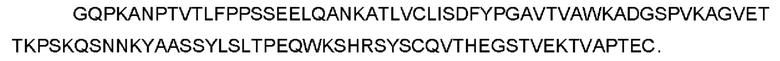
Вариабельная область тяжелой цепи гуманизированного антитела была связана с константной областью тяжелой цепи, как показано в SEQ ID NO: 37, и вариабельная область легкой цепи гуманизированного антитела была связана с константной областью легкой цепи каппа, как показано в SEQ ID NO: 39, что приводило к получению полноразмерного гуманизированного антитела, происходящего из mab147, включая:


Вариабельная область тяжелой цепи гуманизированного антитела была связана с YTE вариантом константной области тяжелой цепи, как показано в SEQ ID NO: 38, и вариабельная область легкой цепи гуманизированного антитела была связана с константной областью легкой цепи каппа, как показано в SEQ ID NO: 39, что приводило к получению полноразмерного гуманизированного антитела, происходящего из mab147, включая:
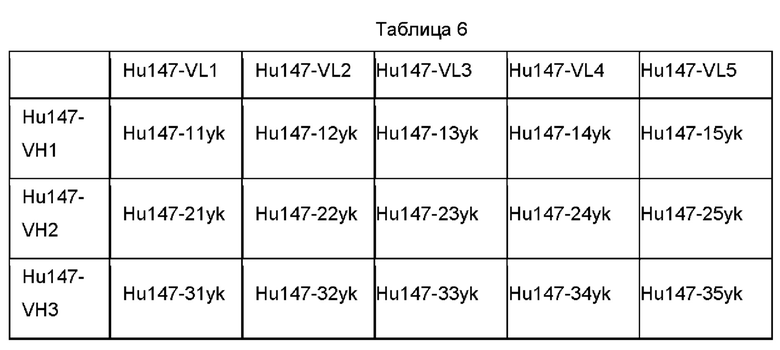
Вариабельная область тяжелой цепи гуманизированного антитела была связана с константной областью тяжелой цепи, как показано в SEQ ID NO: 37, и вариабельная область легкой цепи гуманизированного антитела была связана с константной областью легкой цепи лямбда, как показано в SEQ ID NO: 40, что приводило к получению полноразмерного гуманизированного антитела, происходящего из mab164, включая:
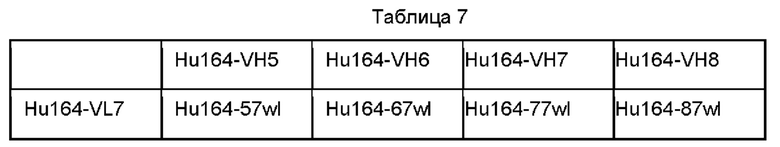

Вариабельная область тяжелой цепи гуманизированного антитела была связана с YTE вариантом константной области тяжелой цепи, как показано в SEQ ID NO: 38, и вариабельная область легкой цепи гуманизированного антитела была связана с константной областью легкой цепи лямбда, как показано в SEQ ID NO: 40, что приводило к получению полноразмерного гуманизированного антитела, происходящего из mab164, включая:

Вариабельная область тяжелой цепи гуманизированного антитела была связана с константной областью тяжелой цепи, как показано в SEQ ID NO: 37, и вариабельная область легкой цепи гуманизированного антитела была связана с константной областью легкой цепи каппа, как показано в SEQ ID NO: 39, что приводило к получению полноразмерного гуманизированного антитела, происходящего из mab95, включая:

Вариабельная область тяжелой цепи гуманизированного антитела была связана с вариантом YTE константной области тяжелой цепи, как показано в SEQ ID NO: 38, и вариабельная область легкой цепи гуманизированного антитела была связана с константной областью легкой цепи каппа, как показано в SEQ ID NO: 39, что приводило к получению полноразмерного гуманизированного антитела, происходящего из mab95, включая:

Иллюстративные аминокислотные последовательности полноразмерного антитела против CTGF выглядят следующим образом:














Последовательности антитела против CTGF положительного контроля гпАМ (из CN1829740 В, с названием антитела памревлумаб, также известного как FG-3019), выглядят следующим образом:
Тяжелая цепь: (SEQ ID NO: 67)


Легкая цепь: (SEQ ID NO: 68)

Пример 3. Выявление активности связывания антитела против CTGF I. Анализ ELISA связывания антитела против CTGF с человеческим белком CTGF
Активность связывания антитела против человеческого CTGF с человеческим белком CTGF анализировали посредством анализа ELISA и Biacore. 96-Луночный планшет для микротитрования непосредственно покрывали рекомбинантным белком CTGF. Интенсивность сигнала при добавлении антитела использовали для определения активности связывания антитела с CTGF. Конкретные способы экспериментов выглядели следующим образом:
Белок CTGF (R&D, кат. №9190-СС) разводили буфером PBS, рН 7,4 (Shanghai Yuanpei, кат. №В320), до концентрации 0,2 мкг/мл,, из которых 50 мкл/лунка добавляли в 96-луночный планшет для микротитрования (Corning, кат. №CLS3590-100EA), и инкубировали в инкубаторе при 37°С в течение 2 часов или при 4°С в течение ночи (16-18 часов). Жидкость удаляли, добавляли 250 мкл/лунка блокирующего раствора (5%-ное обезжиренное молоко (обезжиренное молоко BD, кат. №232100), разведенное в PBS) и инкубировали в инкубаторе при 37°С в течение 3 часов или при 4°С в течение ночи (16-18 часов) для блокирования. После окончания блокирования блокирующий раствор удаляли, планшет промывали 5 раз буфером PBST (PBS, содержащим 0,1% Tween-20, рН 7,4), добавляли 50 мкл/лунка разных концентраций каждого из тестируемых антител (антител, очищенных из гибридомы, или гуманизированных антител), разведенных раствором для разведения образца, и инкубировали в инкубаторе при 37°С в течение 1 часа. После окончания инкубации планшет промывали 3 раза PBST, добавляли 50 мкл/лунка вторичного антитела козы против мышиного антитела, меченного HRP (от англ. horseradish peroxidase - пероксидаза хрена) (Jackson Immuno Research, кат. №109-035-003), или вторичного антитела козы против человеческого антитела (Jackson Immuno Research, кат. №109-035-003), разведенного раствором для разведения образца, и инкубировали при 37°С в течение 1 часа. Планшет промывали 3 раза PBST, 50 мкл/лунка хромогенного субстрата ТМВ (от англ. tetramethylbenzidine -тетраметилбензидин) (KPL, кат. №52-00-03) добавляли, инкубировали при комнатной температуре в течение 5-10 мин и 50 мкл/лунка 1 М H2S04 добавляли для остановки реакции. Значение поглощения считывали посредством микропланшет-ридера (Molecular Devices, VERSA max) при длине волны 450 нм, и данные анализировали посредством GraphPad Prism 5. Связывание антитела против CTGF с человеческим белком CTGF рассчитывали в виде значения ЕС50 (от англ. half maximal effective concentration - полумаксимальная эффективная концентрация). Результаты эксперимента показаны на Фиг. 1 и в Таблице 12.

Результаты эксперимента показывают, что все гуманизированные полноразмерные антитела против CTGF по настоящему раскрытию оказывают превосходное связывающее действие в отношении человеческого белка CTGF.
II. Анализ ELISA связывания антитела против CTGF с белком CTGF обезьяны - яванского макака
Активность связывания антитела против CTGF обезьяны с белком CTGF обезьяны анализировали посредством анализа ELISA. 96-луночный планшет для микротитрования непосредственно покрывали белком против mFc, и затем белок cyno-CTGF с Fc-меткой связывали с планшетом, и затем добавляли антитело. Интенсивность сигнала при добавлении антитела использовали для определения активности связывания антитела с cyno-CTGF. Конкретные способы эксперимента выглядели следующим образом:
Белок против mFc (Sigma, кат. №М4280) разводили буфером PBS, рН7,4 (Shanghai Yuanpei, кат. №В320) до концентрации 2 мкг/мл, который добавляли в 96-луночный планшет для микротитрования (Corning, кат. №CLS3590-100EA) в количестве 50 мкл/лунка, и инкубировали в инкубаторе при 37°С в течение 2 часов. Жидкость удаляли, добавляли 250 мкл/лунка блокирующего раствора (5%-ное обезжиренное молоко (обезжиренное молоко BD, кат. №232100), разведенное в PBS) и инкубировали в инкубаторе при 37°С в течение 3 часов или при 4°С в течение ночи (16-18 часов) для блокирования. После окончания блокирования блокирующий раствор удаляли, планшет 3 раза промывали буфером PBST (PBS, содержащим 0,1% Tween-20, рН 7,4), в каждую лунку добавляли 50 мкп 1 мкг/мл белка cyno-CTGF-mFc, инкубировали в инкубаторе при 37°С в течение 1 часа, и затем планшет 3 раза промывали буфером PBST (PBS, содержащим 0,1% Tween-20, рН 7,4). Добавляли 50 мкл/лунка разных концентраций каждого из анализируемых антител (антител, очищенных из гибридомы, или гуманизированных антител), разведенных раствором для разведения образца, и инкубировали в инкубаторе при 37°С в течение 1 часа. После окончания инкубации планшет 3 раза промывали PBST, добавляли 50 мкл/лунка вторичного антитела козы против мышиного антитела, меченного HRP (Jackson Immuno Research, кат. №109-035-003), или вторичного антитела козы против человеческого антитела (Jackson Immuno Research, кат. №109-035-003), разведенного раствором для разведения образца, и инкубировали при 37°С в течение 1 часа. Планшет 5 раз промывали PBST, добавляли 50 мкл/лунка хромогенного субстрата ТМВ (KPL, кат. №52-00-03), инкубировали при комнатной температуре в течение 5-10 мин, и 50 мкл/лунка 1 М H2SO4 добавляли для остановки реакции. Значение поглощения считывали посредством микропланшет-ридера (Molecular Devices, VERSA max) при длине волны 450 нм, и данные анализировали посредством GraphPad Prism 5. Связывание антитела против CTGF с белком cyno-CTGF рассчитывали в виде значения ЕС50. Результаты эксперимента показаны в Таблице 13.

Данные эксперимента показывают, что все гуманизированные антитела против CTGF по настоящему раскрытию оказывают превосходное связывающее действие в отношении белка CTGF обезьяны.
III. Анализ ELISA связывания антитела против CTGF с мышиным белком
CTGF
Активность связывания антитела против мышиного CTGF с мышиным белком CTGF анализировали посредством анализа ELISA. 96-луночный планшет для микротитрования непосредственно покрывали слитым белком CTGF мыши. Интенсивность сигнала при добавлении антитела использовали для определения активности связывания антитела с мышиным CTGF. Конкретные способы эксперимента выглядели следующим образом:
Белок мышиный CTGF-his разводили буфером PBS, рН 7,4 (Shanghai Yuanpei, кат. №В320), до концентрации 0,5 мкг/мл, добавляли в 96-луночный планшет для микротитрования (Corning, кат. №CLS3590-100EA) в количестве 50 мкл/лунка и инкубировали в инкубаторе при 37°С в течение 2 часов. Жидкость удаляли, добавляли 250 мкл/лунка блокирующего раствора (5%-ное обезжиренное молоко (обезжиренное молоко BD, кат. №232100), разведенное в PBS) и инкубировали в инкубаторе при 37°С в течение 3 часов или при 4°С в течение ночи (16-18 часов) для блокирования. После окончания блокирования блокирующий раствор удаляли, планшет 3 раза промывали буфером PBST (PBS, содержащим 0,1% Tween-20, рН 7,4), добавляли 50 мкл/лунка разных концентраций каждого из анализируемых антител (антител, очищенных из гибридомы, или гуманизированных антител), разведенных раствором для разведения образца, и инкубировали в инкубаторе при 37°С в течение 1 часа. После окончания инкубации планшет 3 раза промывали PBST, добавляли 50 мкл/лунка вторичного антитела козы против мышиного антитела, меченного HRP (Jackson Immuno Research, кат. №109-035-003), или вторичного антитела козы против человеческого антитела (Jackson Immuno Research, кат. №109-035-003), разведенного раствором для разведения образца, и инкубировали при 37°С в течение 1 часа. Планшет 5 раз промывали PBST, добавляли 50 мкл/лунка хромогенного субстрата ТМВ (KPL, кат. №52-00-03), инкубировали при комнатной температуре в течение 5-10 мин и 50 мкл/лунка 1 М H2SO4 добавляли для остановки реакции. Значение поглощения считывали посредством микропланшет-ридера (Molecular Devices, VERSA max) при длине волны 450 нм, и данные анализировали посредством GraphPad Prism 5. Связывание антитела против CTGF с мышиным белком CTGF рассчитывали в виде значения ЕС50. Результаты эксперимента показаны в Таблице 14.

Результаты эксперимента показывают, что все гуманизированные полноразмерные антитела против CTGF по настоящему раскрытию оказывают превосходное связывающее действие в отношении мышиного белка CTGF.
Пример 4. Анализ функции-блокирования антитела против CTGF
В данном анализе прибор Biacore использовали для выявления активности гуманизированного антитела против CTGF, подлежащего анализу, в отношении блокирования функции человеческого TGF-(31.
Сначала, антиген человеческий CTGF иммобилизировали на биосенсорном чипе СМ5 (кат. №BR-1005-30, GE) посредством связывания аминогрупп с уровнем иммобилизации 1500 RU, и затем высокой концентрации антитела против CTGF (150 мкг/мл) давали протекать через поверхность чипа на протяжении 150 с; 50 нМ белка TGF-01 инъецировали на протяжении 120 с, и сигналы взаимодействия выявляли в реальном времени с использованием прибора Biacore Т200 с получением кривой связывания и диссоциации. После завершения диссоциации в каждом из экспериментальных циклов биосенсорный чип промывали и регенерировали буфером для регенерации.
Полученные экспериментальные данные статистически анализировали посредством программного обеспечения GE Biacore Т200, ознакомительная версия 3.0.
Эффективность блокирования=[1-(значение отклика образца/значение отклика холостого контроля)]* 100%. Конкретные результаты показаны в Таблице 15.

Результаты эксперимента показывают, что все гуманизированные антитела против CTGF по настоящему раскрытию обладают высокой активностью функции-блокирования.
Определение аффинности антитела против CTGF посредством Biacore В данном эксперименте прибор Biacore использовали для определения аффинности гуманизированных антител против CTGF, подлежащих анализу, посредством человеческого CTGF и мышиного CTGF.
Определенное количество каждого из анализируемых антител подвергалось аффинному захвату с помощью биосенсорного чипа с белком А (кат. №29127556, GE), и затем определенной концентрации человеческого или мышиного CTGF давали протекать через поверхность чипа. Сигналы взаимодействия выявляли в реальном времени посредством прибора Biacore с получением кривой связывания и диссоциации. После завершения диссоциации в каждом цикле биосенсорный чип промывали и регенерировали регенерирующим раствором глицина-HCl, рН 1,5 (кат. №BR-1003-54, GE). Подвижный буфер для анализа представлял собой буферный раствор 1×HBS-EP (кат. №BR-1001-88, GE).
Полученные данные аппроксимировали в соответствии с (1:1) моделью Ленгмюра с использованием ознакомительной версии 3.0 программного обеспечения GE Biacore Т200, и получали значения аффинности, как более подробно показано в Таблице 16 и 17. 


Результаты экспериментов показывают, что все химерные антитела СМ 47 и СМ64 и гуманизированные антитела против CTGF, происходящие из мышиных антител тАМ47 и тАМ64, могут связываться с человеческим и мышиным CTGF с высокой аффинностью. Ни замены, ни обратная(ые) мутация(ии), находящаяся(иеся) в областях FR антитела зародышевой линии человека, не препятствуют аффинности гуманизированного антитела, и некоторые модификации могут даже усиливать аффинность антитела в отношении антигена.
Пример 5. Анализ антитела против CTGF в отношении in vitro ингибирования миграции клеток С2С12, индуцируемой TGFβ
Клетки С2С12 (мышиные миобласты, Банк клеток Китайской академии наук, #GNM26) расщепляли 0,25%-ным трипсином (Gibco, #25200-072), центрифугировали и ресуспендировали в бессывороточной среде DMEM (Gibco, #10564-029). Смесь клетки-антитела и смесь TGFβl-антитела готовили с бессывороточной средой DMEM, в которой плотность клеток составляла 2 * 105/мл, концентрация рекомбинантного человеческого TGFβl (Cell signaling Technology, #8915LC) составляла 10 нг/мл, и концентрация антитела, подлежащего анализу, составляла 30 мкг/мл.
Крышку и фильтрующую мембрану ChemoTx ® Disposable Chemotaxis System (Neuro Probe, #106-8) открывали, и смесь TGFβ-антитела добавляли в нижнюю камеру, 30 мкл/лунка, 4 пары лунок для каждой группы. Фильтрующую мембрану покрывали, и смесь клетки-антитела добавляли на мембрану, 50 мкл/лунка, 4 пары лунок для каждой группы. Крышку закрывали и помещали в инкубатор (37°С, 5% СO2).
Через 48 часов после инкубации, жидкость на фильтрующей мембране удаляли посредством чистого бумажного полотенца, фильтрационную мембрану открывали, 10 мкп предварительно охлажденного 0,25%-ного трипсина добавляли к каждой лунке в нижнюю камеру, и фильтрующую мембрану покрывали. Через 5 минут после расщепления образцы центрифугировали при 1000 об./мин. в течение 1 минуты. Фильтрующую мембрану открывали, 20 мкп раствора Cell Titer-Glo (Promega, #G7573) добавляли в каждую лунку в нижнюю камеру и инкубировали при комнатной температуре в течение 10 минут. Жидкость переносили в 384-луночный планшет с белым дном (Thermo Scientific, #267462), и осуществляли считывание показаний с планшета посредством микропланшет-ридера (BMG, #PheraStar) по хемилюминесценции. Данные анализировали и обрабатывали посредством Graphpad Prism 6.

Результаты эксперимента показаны в Таблице 18.

Результаты показывают, что гуманизированные антитела против CTGF по настоящему раскрытию могут ингибировать способность к миграции клеток С2С12 in vitro, индуцируемую TGFβl, и демонстрируют более сильное ингибирующее действие на способность к миграции клеток С2С12, по сравнению со способностью к миграции клеток С2С12 антитела положительного контроля тАМ.
Пример 6. Анализ, демонстрирующий ингибирующее действие in vitro антитела против CTGF на миграцию клеток PANC1, индуцируемую TGFβ
Клетки PANC-1 (раковые клетки поджелудочной железы человека, АТСС, #CRL-1469) расщепляли 0,25%-ным трипсином (Gibco, #25200-072), центрифугировали и ресуспендировали посредством среды DMEM (Gibco, #10564-029), содержащей 0,1% фетальную телячью сыворотку (Gibco, #10099-141). Смесь клетки-антитела и смесь TGFβ-антитела готовили посредством среды DMEM, содержащей 0,1% фетальную телячью сыворотку, в которой плотность клеток составляла 4×105/мл, концентрация рекомбинантного человеческого TGFβ (Cell signaling Technology, #8915LC) составляла 10 нг/мл, и концентрация антитела, подлежащего анализу, составляла 30 мкг/мл.
Крышку и фильтрующую мембрану ChemoТх ® Disposable Chemotaxis System (Neuro Probe, #106-8) открывали, и смесь TGFβ-антитела добавляли в нижнюю камеру, 30 мкл/лунка, 4 пары лунок для каждой группы. Фильтрующую мембрану покрывали, и смесь клетки-антитела добавляли на мембрану, 50 мкл/лунка, 4 пары лунок для каждой группы. Крышку закрывали и помещали в инкубатор (37°С, 5% С02).
Через 48 часов после инкубации, жидкость на фильтрующей мембране удаляли посредством чистого бумажного полотенца, фильтрующую мембрану открывали, 10 мкп предварительно охлажденного 0,25%-ного трипсина добавляли к каждой лунке в нижнюю камеру, и фильтрующую мембрану покрывали. Через 5 минут после расщепления образцы центрифугировали при 1000 об./мин. в течение 1 минуты. Клетки, которые прилипали ко дну планшета, подсчитывали под инвертированным микроскопом (Leica, #DMIL LED). Данные анализировали и обрабатывали посредством Graphpad Prism 6. Степень ингибирования=[(среднее значение TGFβ - среднее значение выборки)/(среднее значение группы TGFβ -среднее значение необработанной группы)]×100%
Результаты эксперимента показаны в Таблице 19.

Результаты показывают, что все гуманизированные антитела против CTGF по настоящему раскрытию могут ингибировать способность к миграции клеток PANC1 in vitro, индуцируемую TGFβl.
Пример 7. Анализ антитела против CTGF в отношении ингибирующего действия in vitro на пролиферацию PANC1 на мягком агаре
Деионизированную воду добавляли в агар (BD, #214220) и нагревали с получением 1,4%-ого раствора геля. 2*DMEM среду (Thermo, #12100046) смешивали с раствором геля в соотношении 1:1 по объему, 500 мкп добавляли в каждую лунку 24-луночного планшета (Costar, #3524) и размещали при 4 градусах для отверждения.
Клетки PANC-1 (раковые клетки поджелудочной железы человека, АТСС, #CRL-1469) расщепляли 0,25%-ным трипсином (Gibco, #25200-072), центрифугировали и доводили до уровня средой DMEM (GE, #SH30243.01), содержащей 4% фетальную телячью сыворотку (Gibco, #10099-141) для получения плотности клеток 5x104 клеток/мл. 2 мг/мл антитела получали посредством среды 2xDMEM. Клетки, антитело и 1,4% раствор геля смешивали в соотношении 2:1:1 по объему, и 200 мкп добавляли в каждую лунку 24-луночного планшета, покрытого нижним слоем геля. После отверждения верхнего геля, 200 мкп среды DMEM, содержащей 2% фетальную телячью сыворотку, добавляли в 24-луночный планшет.Через 28 суток после инкубации в инкубаторе (37°С, 5% С02), систему анализа высокого содержания (Molecular Devices, ImageXpress) использовали для развития визуализации, и анализировали площадь образованного клона клеток.
Степень ингибирования = (1 - среднее значение выборки/среднее значение необработанной группы)×100%. Результаты эксперимента показаны в Таблице 20.

Результаты показывают, что гуманизированные антитела, происходящие из клонов тАМ47 и тАМ64, обладают значимым ингибирующим действием на пролиферацию раковых клеток поджелудочной железы человека (клетки PANC-1) in vitro.
Пример 8: Биологическая активность in vivo антитела против CTGF у животных
I. Анализ антитела против CTGF в отношении его ингибирующего действия на фиброз легких мыши, вызываемый блеомицином
Протокол эксперимента: 40 мышей случайным образом разделяли на следующие 4 группы в соответствии с массой тела: группа нормального контроля (PBS, в.б., один раз в два дня), группа гпАМ (10 мг/кг, в.б., один раз в два дня), группа Hu164-67wl (10 мг/кг, в.б., один раз в два дня) и группа Hu164-67yl (мг/кг, в.б., один раз в два дня). В каждой группе находится 10 животных, и конкретное разделение на группы показано ниже в Таблице 21. В сутки 1 эксперимента каждый из растворителей и антител инъецировали внутрибрюшинно в соответствии с массой тела (10 мл/кг, нормальный контроль и эталонные группы обеспечивали PBS); спустя 2 часа, использовали аэрозольный блеомицин (распыление 50 мг/8 мл в течение 30 минут и выдерживание в течение 5 минут) для создания моделей. Затем, растворитель или антитело инъецировали внутрибрюшинно через день в соответствии с указанными выше группами. Период эксперимента составлял 21 сутки, в общей сложности с 11 внутрибрюшинными введениями. В сутки 22 эксперимента, мышам давали наркоз посредством внутрибрюшинной инъекции 1% пентобарбитала натрия (10 мл/кг), и затем фиксировали на операционном столе в лежачем положении. Делали кожный разрез 1 см вдоль срединной линии шеи, и нижний конец разреза достигал вода в грудную клетку. Пинцет использовали для прямого разделения подкожных соединительных тканей и мышц с обнажением трахеи. Соединительные ткани с обеих сторон трахеи и между трахеей и пищеводом отделяли для выделения свободной трахеи. Вставляли вживляемую в вену иглу и фиксировали посредством швов с помощью связывания в месте, где канюля поступала в трахею. Шприц на 1 мл, наполненный 0,8 мл PBS, соединяли с впускным концом вживленной в вену иглы, PBS медленно инъецировали в трахею, оставляли на 30 секунд и затем PBS медленно извлекали обратно. Во время изъятия могли наблюдать молочную пенящуюся жидкость. Промывание повторяли три раза, промывную жидкость переносили в ЕР пробирки, и затем 0,5 мл PBS отбирали, повторяли указанные выше стадии. Две промывные жидкости смешивали и центрифугировали при 1500 об./мин. в течение 5 мин, и супернатант хранили при -20°С для анализа. Супернатант BALF еще раз центрифугировали при 10000 об./мин. в течение 10 мин, и супернатант отбирали для выявления растворимого коллагена (набор: Biocolor, №партии АА883).

Использовали программное обеспечение для статистического анализа Excel: среднее значение рассчитывали как avg (Среднее); значение SD (Стандартное отклонение) рассчитывали как STDEV (стандартное отклонение); значение SEM (стандартная ошибка среднего; стандартное отклонение выборочного среднего) рассчитывали как STDEV/SQRT (квадратный корень) (число животных в каждой группе); программное обеспечение GraphPad Prism использовали для построения графика, двухфакторный ANOVA (двухфакторный дисперсионный анализ) или однофакторный ANOVA (однофакторный дисперсионный анализ) использовали для статистического анализа данных.
Результаты эксперимента показаны в Таблице 22.

Результаты показывают, что Hu164-67wl или Hu164-67yl демонстрируют сильное ингибирующее действие на фиброз легких у мышей.
II. Анализ антитела против CTGF в отношении ингибирования подкожно трансплантированной опухоли раковых клеток поджелудочной железы человека Su86.86
Протокол эксперимента: 200 мкп клеток SU86.86 (АТСС, CRL-1837, 3,0×106 клеток) инокулировали подкожно в правые ребра бестимусных мышей Nu/Nu. Через 11 суток после инокуляции, когда объем опухоли составлял примерно 140 мм3, мышей с слишком большой или маленькой массой тельца или объемом опухоли исключали. Мышей случайным образом делили на 5 групп в соответствии с объемом опухоли, 10 мышей/группа, и введение начинали в те же сутки. Антитело инъецировали внутрибрюшинно, два раза в неделю, на протяжении в общей сложности 18 суток. Объем опухоли и массу тела измеряли 1-2 раза в неделю, и данные записывали. Деление на группы и схема дозирования показаны в Таблице 23.

Использовали программное обеспечение для статистического анализа Excel: среднее значение рассчитывали как avg; значение SD рассчитывали как STDEV; значение SEM рассчитывали как STDEV/SQRT (число животных в каждой группе); программное обеспечение GraphPad Prism использовали для построения графика, двухфакторный ANOVA или однофакторный ANOVA использовали для статистического анализа данных.
Объем опухоли (V) рассчитывали в соответствии со следующей формулой:
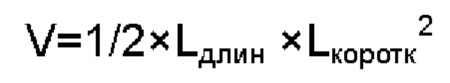
Относительная степень пролиферации опухоли Т/С (%)=(Tt-T0)/(Ct-С0)×100, где Tt и Ct представляют собой объемы опухоли группы обработки и контрольной группы в конце эксперимента; Т0 и С0 представляют собой объем опухоли в начале эксперимента.

Результаты эксперимента:
С D1 по D18, подкожную инъекцию проводили два раза в неделю, и выявляли воздействие антител против CTGF на ингибирование роста опухоли SU86.86. Результаты эксперимента показаны в Таблице 24 и на Фиг. 2. По сравнению с холостой контрольной группой того же изотипа человеческого IgG, степени ингибирования опухоли группы тАМ-40 мг/кг, Hu164-67yl-40 мг/кг, Hu164-67yl-20 мг/кг и Hu147-33wk (40 мг/кг) составляли 45%, 53%, 47% и 54%, соответственно. Группы тАМ-40 мг/кг, Hu164-67yl -40 мг/кг, HM64-67yl-20 мг/кг и Hu147-33wk (40 мг/кг) могут значимо ингибировать рост опухолей Su86.86 (р<0,001), и ингибирующее действие на опухоль SU86.86 является дозозависимым. Кроме того, масса тела мышей в каждой группе значимо не менялась.
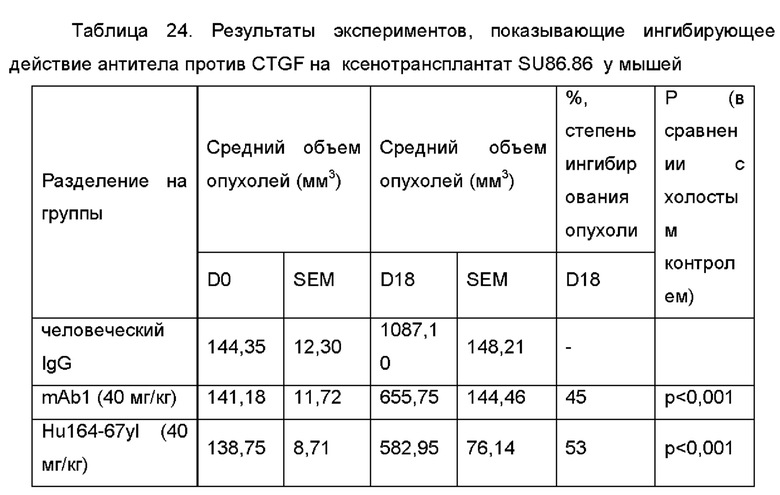

--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> JIANGSU HENGRUI MEDICINE CO., LTD.;
SHANGHAI HENGRUI PHARMACEUTICAL CO., LTD.
<120> АНТИТЕЛО ПРОТИВ ФАКТОРА РОСТА СОЕДИНИТЕЛЬНОЙ ТКАНИ И ЕГО ПРИМЕНЕНИЕ
<160> 104
<170> SIPOSequenceListing 1.0
<210> 1
<211> 323
<212> ПРТ
<213> Homo sapiens
<220>
<221> ПЕПТИД
<223> полноразмерный белок CTGF человека
<400> 1
Gln Asn Cys Ser Gly Pro Cys Arg Cys Pro Asp Glu Pro Ala Pro Arg
1 5 10 15
Cys Pro Ala Gly Val Ser Leu Val Leu Asp Gly Cys Gly Cys Cys Arg
20 25 30
Val Cys Ala Lys Gln Leu Gly Glu Leu Cys Thr Glu Arg Asp Pro Cys
35 40 45
Asp Pro His Lys Gly Leu Phe Cys Asp Phe Gly Ser Pro Ala Asn Arg
50 55 60
Lys Ile Gly Val Cys Thr Ala Lys Asp Gly Ala Pro Cys Ile Phe Gly
65 70 75 80
Gly Thr Val Tyr Arg Ser Gly Glu Ser Phe Gln Ser Ser Cys Lys Tyr
85 90 95
Gln Cys Thr Cys Leu Asp Gly Ala Val Gly Cys Met Pro Leu Cys Ser
100 105 110
Met Asp Val Arg Leu Pro Ser Pro Asp Cys Pro Phe Pro Arg Arg Val
115 120 125
Lys Leu Pro Gly Lys Cys Cys Glu Glu Trp Val Cys Asp Glu Pro Lys
130 135 140
Asp Gln Thr Val Val Gly Pro Ala Leu Ala Ala Tyr Arg Leu Glu Asp
145 150 155 160
Thr Phe Gly Pro Asp Pro Thr Met Ile Arg Ala Asn Cys Leu Val Gln
165 170 175
Thr Thr Glu Trp Ser Ala Cys Ser Lys Thr Cys Gly Met Gly Ile Ser
180 185 190
Thr Arg Val Thr Asn Asp Asn Ala Ser Cys Arg Leu Glu Lys Gln Ser
195 200 205
Arg Leu Cys Met Val Arg Pro Cys Glu Ala Asp Leu Glu Glu Asn Ile
210 215 220
Lys Lys Gly Lys Lys Cys Ile Arg Thr Pro Lys Ile Ser Lys Pro Ile
225 230 235 240
Lys Phe Glu Leu Ser Gly Cys Thr Ser Met Lys Thr Tyr Arg Ala Lys
245 250 255
Phe Cys Gly Val Cys Thr Asp Gly Arg Cys Cys Thr Pro His Arg Thr
260 265 270
Thr Thr Leu Pro Val Glu Phe Lys Cys Pro Asp Gly Glu Val Met Lys
275 280 285
Lys Asn Met Met Phe Ile Lys Thr Cys Ala Cys His Tyr Asn Cys Pro
290 295 300
Gly Asp Asn Asp Ile Phe Glu Ser Leu Tyr Tyr Arg Lys Met Tyr Gly
305 310 315 320
Asp Met Ala
<210> 2
<211> 348
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> CTGF-his
<400> 2
Met Glu Phe Gly Leu Ser Trp Leu Phe Leu Val Ala Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Gln Asn Cys Ser Gly Pro Cys Arg Cys Pro Asp Glu Pro
20 25 30
Ala Pro Arg Cys Pro Ala Gly Val Ser Leu Val Leu Asp Gly Cys Gly
35 40 45
Cys Cys Arg Val Cys Ala Lys Gln Leu Gly Glu Leu Cys Thr Glu Arg
50 55 60
Asp Pro Cys Asp Pro His Lys Gly Leu Phe Cys Asp Phe Gly Ser Pro
65 70 75 80
Ala Asn Arg Lys Ile Gly Val Cys Thr Ala Lys Asp Gly Ala Pro Cys
85 90 95
Ile Phe Gly Gly Thr Val Tyr Arg Ser Gly Glu Ser Phe Gln Ser Ser
100 105 110
Cys Lys Tyr Gln Cys Thr Cys Leu Asp Gly Ala Val Gly Cys Met Pro
115 120 125
Leu Cys Ser Met Asp Val Arg Leu Pro Ser Pro Asp Cys Pro Phe Pro
130 135 140
Arg Arg Val Lys Leu Pro Gly Lys Cys Cys Glu Glu Trp Val Cys Asp
145 150 155 160
Glu Pro Lys Asp Gln Thr Val Val Gly Pro Ala Leu Ala Ala Tyr Arg
165 170 175
Leu Glu Asp Thr Phe Gly Pro Asp Pro Thr Met Ile Arg Ala Asn Cys
180 185 190
Leu Val Gln Thr Thr Glu Trp Ser Ala Cys Ser Lys Thr Cys Gly Met
195 200 205
Gly Ile Ser Thr Arg Val Thr Asn Asp Asn Ala Ser Cys Arg Leu Glu
210 215 220
Lys Gln Ser Arg Leu Cys Met Val Arg Pro Cys Glu Ala Asp Leu Glu
225 230 235 240
Glu Asn Ile Lys Lys Gly Lys Lys Cys Ile Arg Thr Pro Lys Ile Ser
245 250 255
Lys Pro Ile Lys Phe Glu Leu Ser Gly Cys Thr Ser Met Lys Thr Tyr
260 265 270
Arg Ala Lys Phe Cys Gly Val Cys Thr Asp Gly Arg Cys Cys Thr Pro
275 280 285
His Arg Thr Thr Thr Leu Pro Val Glu Phe Lys Cys Pro Asp Gly Glu
290 295 300
Val Met Lys Lys Asn Met Met Phe Ile Lys Thr Cys Ala Cys His Tyr
305 310 315 320
Asn Cys Pro Gly Asp Asn Asp Ile Phe Glu Ser Leu Tyr Tyr Arg Lys
325 330 335
Met Tyr Gly Asp Met Ala His His His His His His
340 345
<210> 3
<211> 575
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> mCTGF-Fc
<400> 3
Met Glu Phe Gly Leu Ser Trp Leu Phe Leu Val Ala Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Gln Asp Cys Ser Ala Gln Cys Gln Cys Ala Ala Glu Ala
20 25 30
Ala Pro His Cys Pro Ala Gly Val Ser Leu Val Leu Asp Gly Cys Gly
35 40 45
Cys Cys Arg Val Cys Ala Lys Gln Leu Gly Glu Leu Cys Thr Glu Arg
50 55 60
Asp Pro Cys Asp Pro His Lys Gly Leu Phe Cys Asp Phe Gly Ser Pro
65 70 75 80
Ala Asn Arg Lys Ile Gly Val Cys Thr Ala Lys Asp Gly Ala Pro Cys
85 90 95
Val Phe Gly Gly Ser Val Tyr Arg Ser Gly Glu Ser Phe Gln Ser Ser
100 105 110
Cys Lys Tyr Gln Cys Thr Cys Leu Asp Gly Ala Val Gly Cys Val Pro
115 120 125
Leu Cys Ser Met Asp Val Arg Leu Pro Ser Pro Asp Cys Pro Phe Pro
130 135 140
Arg Arg Val Lys Leu Pro Gly Lys Cys Cys Glu Glu Trp Val Cys Asp
145 150 155 160
Glu Pro Lys Asp Arg Thr Ala Val Gly Pro Ala Leu Ala Ala Tyr Arg
165 170 175
Leu Glu Asp Thr Phe Gly Pro Asp Pro Thr Met Met Arg Ala Asn Cys
180 185 190
Leu Val Gln Thr Thr Glu Trp Ser Ala Cys Ser Lys Thr Cys Gly Met
195 200 205
Gly Ile Ser Thr Arg Val Thr Asn Asp Asn Thr Phe Cys Arg Leu Glu
210 215 220
Lys Gln Ser Arg Leu Cys Met Val Arg Pro Cys Glu Ala Asp Leu Glu
225 230 235 240
Glu Asn Ile Lys Lys Gly Lys Lys Cys Ile Arg Thr Pro Lys Ile Ala
245 250 255
Lys Pro Val Lys Phe Glu Leu Ser Gly Cys Thr Ser Val Lys Thr Tyr
260 265 270
Arg Ala Lys Phe Cys Gly Val Cys Thr Asp Gly Arg Cys Cys Thr Pro
275 280 285
His Arg Thr Thr Thr Leu Pro Val Glu Phe Lys Cys Pro Asp Gly Glu
290 295 300
Ile Met Lys Lys Asn Met Met Phe Ile Lys Thr Cys Ala Cys His Tyr
305 310 315 320
Asn Cys Pro Gly Asp Asn Asp Ile Phe Glu Ser Leu Tyr Tyr Arg Lys
325 330 335
Met Tyr Gly Asp Met Ala Glu Pro Arg Gly Pro Thr Ile Lys Pro Cys
340 345 350
Pro Pro Cys Lys Cys Pro Ala Pro Asn Leu Leu Gly Gly Pro Ser Val
355 360 365
Phe Ile Phe Pro Pro Lys Ile Lys Asp Val Leu Met Ile Ser Leu Ser
370 375 380
Pro Ile Val Thr Cys Val Val Val Asp Val Ser Glu Asp Asp Pro Asp
385 390 395 400
Val Gln Ile Ser Trp Phe Val Asn Asn Val Glu Val His Thr Ala Gln
405 410 415
Thr Gln Thr His Arg Glu Asp Tyr Asn Ser Thr Leu Arg Val Val Ser
420 425 430
Ala Leu Pro Ile Gln His Gln Asp Trp Met Ser Gly Lys Glu Phe Lys
435 440 445
Cys Lys Val Asn Asn Lys Asp Leu Pro Ala Pro Ile Glu Arg Thr Ile
450 455 460
Ser Lys Pro Lys Gly Ser Val Arg Ala Pro Gln Val Tyr Val Leu Pro
465 470 475 480
Pro Pro Glu Glu Glu Met Thr Lys Lys Gln Val Thr Leu Thr Cys Met
485 490 495
Val Thr Asp Phe Met Pro Glu Asp Ile Tyr Val Glu Trp Thr Asn Asn
500 505 510
Gly Lys Thr Glu Leu Asn Tyr Lys Asn Thr Glu Pro Val Leu Asp Ser
515 520 525
Asp Gly Ser Tyr Phe Met Tyr Ser Lys Leu Arg Val Glu Lys Lys Asn
530 535 540
Trp Val Glu Arg Asn Ser Tyr Ser Cys Ser Val Val His Glu Gly Leu
545 550 555 560
His Asn His His Thr Thr Lys Ser Phe Ser Arg Thr Pro Gly Lys
565 570 575
<210> 4
<211> 348
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> белок-His метка CTGF мыши
<400> 4
Met Glu Phe Gly Leu Ser Trp Leu Phe Leu Val Ala Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Gln Asp Cys Ser Ala Gln Cys Gln Cys Ala Ala Glu Ala
20 25 30
Ala Pro His Cys Pro Ala Gly Val Ser Leu Val Leu Asp Gly Cys Gly
35 40 45
Cys Cys Arg Val Cys Ala Lys Gln Leu Gly Glu Leu Cys Thr Glu Arg
50 55 60
Asp Pro Cys Asp Pro His Lys Gly Leu Phe Cys Asp Phe Gly Ser Pro
65 70 75 80
Ala Asn Arg Lys Ile Gly Val Cys Thr Ala Lys Asp Gly Ala Pro Cys
85 90 95
Val Phe Gly Gly Ser Val Tyr Arg Ser Gly Glu Ser Phe Gln Ser Ser
100 105 110
Cys Lys Tyr Gln Cys Thr Cys Leu Asp Gly Ala Val Gly Cys Val Pro
115 120 125
Leu Cys Ser Met Asp Val Arg Leu Pro Ser Pro Asp Cys Pro Phe Pro
130 135 140
Arg Arg Val Lys Leu Pro Gly Lys Cys Cys Glu Glu Trp Val Cys Asp
145 150 155 160
Glu Pro Lys Asp Arg Thr Ala Val Gly Pro Ala Leu Ala Ala Tyr Arg
165 170 175
Leu Glu Asp Thr Phe Gly Pro Asp Pro Thr Met Met Arg Ala Asn Cys
180 185 190
Leu Val Gln Thr Thr Glu Trp Ser Ala Cys Ser Lys Thr Cys Gly Met
195 200 205
Gly Ile Ser Thr Arg Val Thr Asn Asp Asn Thr Phe Cys Arg Leu Glu
210 215 220
Lys Gln Ser Arg Leu Cys Met Val Arg Pro Cys Glu Ala Asp Leu Glu
225 230 235 240
Glu Asn Ile Lys Lys Gly Lys Lys Cys Ile Arg Thr Pro Lys Ile Ala
245 250 255
Lys Pro Val Lys Phe Glu Leu Ser Gly Cys Thr Ser Val Lys Thr Tyr
260 265 270
Arg Ala Lys Phe Cys Gly Val Cys Thr Asp Gly Arg Cys Cys Thr Pro
275 280 285
His Arg Thr Thr Thr Leu Pro Val Glu Phe Lys Cys Pro Asp Gly Glu
290 295 300
Ile Met Lys Lys Asn Met Met Phe Ile Lys Thr Cys Ala Cys His Tyr
305 310 315 320
Asn Cys Pro Gly Asp Asn Asp Ile Phe Glu Ser Leu Tyr Tyr Arg Lys
325 330 335
Met Tyr Gly Asp Met Ala His His His His His His
340 345
<210> 5
<211> 574
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> CTGF-Fc яванского макака
<400> 5
Met Glu Phe Gly Leu Ser Trp Leu Phe Leu Val Ala Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Gln Asn Cys Ser Gly Pro Cys Arg Cys Pro Ala Glu Gln
20 25 30
Ala Pro Arg Cys Pro Ala Gly Val Ser Leu Val Leu Asp Gly Cys Gly
35 40 45
Cys Cys Arg Val Cys Ala Lys Gln Leu Gly Glu Leu Cys Thr Glu Arg
50 55 60
Asp Pro Cys Asp Pro His Lys Gly Leu Phe Cys Asp Phe Gly Ser Pro
65 70 75 80
Ala Asn Arg Lys Ile Gly Val Cys Thr Ala Lys Asp Gly Ala Pro Cys
85 90 95
Ile Phe Gly Gly Thr Val Tyr Arg Ser Gly Glu Ser Phe Gln Ser Ser
100 105 110
Cys Lys Tyr Gln Cys Thr Cys Leu Asp Gly Ala Val Gly Cys Met Pro
115 120 125
Leu Cys Ser Met Asp Val Arg Leu Pro Ser Pro Asp Cys Pro Phe Pro
130 135 140
Arg Arg Val Lys Leu Pro Gly Lys Cys Cys Glu Glu Trp Val Cys Asp
145 150 155 160
Glu Pro Lys Asp Gln Thr Val Val Gly Pro Ala Leu Ala Ala Tyr Arg
165 170 175
Leu Glu Asp Thr Phe Gly Pro Asp Pro Thr Met Ile Arg Ala Asn Cys
180 185 190
Leu Val Gln Thr Thr Glu Trp Ser Ala Cys Ser Lys Thr Cys Gly Met
195 200 205
Gly Ile Ser Thr Arg Val Thr Asn Asp Asn Ala Ser Cys Arg Leu Glu
210 215 220
Lys Gln Ser Arg Leu Cys Met Val Arg Pro Cys Glu Ala Asp Leu Glu
225 230 235 240
Glu Asn Ile Lys Lys Gly Lys Lys Cys Ile Arg Thr Pro Lys Ile Ser
245 250 255
Lys Pro Ile Lys Phe Glu Leu Ser Gly Cys Thr Ser Val Lys Thr Tyr
260 265 270
Arg Ala Lys Phe Cys Gly Val Cys Thr Asp Gly Arg Cys Cys Thr Pro
275 280 285
His Arg Thr Thr Thr Leu Pro Val Glu Phe Lys Cys Pro Asp Gly Glu
290 295 300
Val Met Lys Lys Asn Met Met Phe Ile Lys Thr Cys Ala Cys His Tyr
305 310 315 320
Asn Cys Pro Gly Asp Asn Asp Ile Phe Glu Ser Leu Tyr Tyr Arg Lys
325 330 335
Met Tyr Gly Asp Met Ala Glu Pro Lys Ser Ser Asp Lys Thr His Thr
340 345 350
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
355 360 365
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
370 375 380
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
385 390 395 400
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
405 410 415
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
420 425 430
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
435 440 445
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
450 455 460
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
465 470 475 480
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
485 490 495
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
500 505 510
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
515 520 525
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
530 535 540
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
545 550 555 560
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
565 570
<210> 6
<211> 116
<212> ПРТ
<213> Mus musculus
<220>
<221> ДОМЕН
<223> mab147 VH
<400> 6
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Glu Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Thr Lys Ser Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Glu Ser Met
65 70 75 80
Leu Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Met Tyr
85 90 95
Tyr Cys Val Glu Thr Gly Phe Ala Tyr Trp Asp Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ala
115
<210> 7
<211> 106
<212> ПРТ
<213> Mus musculus
<220>
<221> ДОМЕН
<223> mab147 VL
<400> 7
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Trp Ile Tyr
35 40 45
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Ser Tyr Pro Leu Thr
85 90 95
Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 8
<211> 114
<212> ПРТ
<213> Mus musculus
<220>
<221> ДОМЕН
<223> mab164 VH
<400> 8
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Thr Phe
20 25 30
Gly Val His Trp Ile Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Arg Gly Gly Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Ser Ile Thr Lys Asp Asn Ser Arg Ser His Val Phe Phe
65 70 75 80
Lys Met Thr Ser Leu Gln Thr Asp Asp Ser Ala Ile Tyr Tyr Cys Ala
85 90 95
Arg Asp Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val
100 105 110
Ser Ser
<210> 9
<211> 109
<212> ПРТ
<213> Mus musculus
<220>
<221> ДОМЕН
<223> mab164 VL
<400> 9
Gln Ala Val Val Thr Gln Glu Ser Ala Leu Thr Thr Ser Pro Gly Gly
1 5 10 15
Thr Val Ile Leu Thr Cys Arg Ser Ser Ile Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Glu Lys Pro Asp His Leu Phe Thr Gly
35 40 45
Leu Ile Gly Gly Thr Ser Asn Arg Ala Pro Gly Val Pro Val Arg Phe
50 55 60
Ser Gly Ser Leu Ile Gly Asp Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Glu Asp Asp Ala Met Tyr Phe Cys Ala Leu Trp Tyr Ser Thr
85 90 95
His Tyr Val Phe Gly Gly Gly Thr Lys Val Thr Val Leu
100 105
<210> 10
<211> 5
<212> ПРТ
<213> Mus musculus
<220>
<221> ДОМЕН
<223> mab147 HCDR1
<400> 10
Thr Tyr Ala Met Asn
1 5
<210> 11
<211> 19
<212> ПРТ
<213> Mus musculus
<220>
<221> ДОМЕН
<223> mab147 HCDR2
<400> 11
Arg Ile Arg Thr Lys Ser Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser
1 5 10 15
Val Lys Asp
<210> 12
<211> 5
<212> ПРТ
<213> Mus musculus
<220>
<221> ДОМЕН
<223> mab147 HCDR3
<400> 12
Thr Gly Phe Ala Tyr
1 5
<210> 13
<211> 10
<212> ПРТ
<213> Mus musculus
<220>
<221> ДОМЕН
<223> mab147 LCDR1
<400> 13
Ser Ala Ser Ser Ser Val Ser Tyr Met His
1 5 10
<210> 14
<211> 7
<212> ПРТ
<213> Mus musculus
<220>
<221> ДОМЕН
<223> mab147 LCDR2
<400> 14
Ser Thr Ser Asn Leu Ala Ser
1 5
<210> 15
<211> 9
<212> ПРТ
<213> Mus musculus
<220>
<221> ДОМЕН
<223> mab147 LCDR3
<400> 15
Gln Gln Arg Ser Ser Tyr Pro Leu Thr
1 5
<210> 16
<211> 5
<212> ПРТ
<213> Mus musculus
<220>
<221> ДОМЕН
<223> mab164 HCDR1
<400> 16
Thr Phe Gly Val His
1 5
<210> 17
<211> 16
<212> ПРТ
<213> Mus musculus
<220>
<221> ДОМЕН
<223> mab164 HCDR2
<400> 17
Val Ile Trp Arg Arg Gly Gly Thr Asp Tyr Asn Ala Ala Phe Met Ser
1 5 10 15
<210> 18
<211> 6
<212> ПРТ
<213> Mus musculus
<220>
<221> ДОМЕН
<223> mab164 HCDR3
<400> 18
Asp Gly Gly Phe Asp Tyr
1 5
<210> 19
<211> 14
<212> ПРТ
<213> Mus musculus
<220>
<221> ДОМЕН
<223> mab164 LCDR1
<400> 19
Arg Ser Ser Ile Gly Ala Val Thr Thr Ser Asn Tyr Ala Asn
1 5 10
<210> 20
<211> 7
<212> ПРТ
<213> Mus musculus
<220>
<221> ДОМЕН
<223> mab164 LCDR2
<400> 20
Gly Thr Ser Asn Arg Ala Pro
1 5
<210> 21
<211> 9
<212> ПРТ
<213> Mus musculus
<220>
<221> ДОМЕН
<223> mab164 LCDR3
<400> 21
Ala Leu Trp Tyr Ser Thr His Tyr Val
1 5
<210> 22
<211> 106
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> Hu147-VL1
<400> 22
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Gln Ser Pro Arg Leu Leu Ile Tyr
35 40 45
Ser Thr Ser Asn Leu Ala Ser Gly Ile Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Ser Tyr Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 23
<211> 106
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> Hu147-VL2
<400> 23
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile Tyr
35 40 45
Ser Thr Ser Asn Leu Ala Ser Gly Ile Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Ser Tyr Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 24
<211> 106
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> Hu147-VL3
<400> 24
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Trp Ile Tyr
35 40 45
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Ser Tyr Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 25
<211> 106
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> Hu147-VL4
<400> 25
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile Tyr
35 40 45
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Ser Tyr Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 26
<211> 106
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> Hu147-VL5
<400> 26
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Trp Ile Tyr
35 40 45
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Ser Tyr Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 27
<211> 116
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> Hu147-VH1
<400> 27
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Arg Thr Lys Ser Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Glu Thr Gly Phe Ala Tyr Trp Asp Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 28
<211> 116
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> Hu147-VH2
<400> 28
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Thr Lys Ser Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Glu Thr Gly Phe Ala Tyr Trp Asp Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 29
<211> 116
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> Hu147-VH3
<400> 29
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Thr Lys Ser Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Glu Ser Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Glu Thr Gly Phe Ala Tyr Trp Asp Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 30
<211> 109
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> Hu164-VL7
<400> 30
Glu Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Ile Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Phe Arg Gly
35 40 45
Leu Ile Gly Gly Thr Ser Asn Arg Ala Pro Trp Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Ser Thr
85 90 95
His Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 31
<211> 109
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> Hu164-VL8
<400> 31
Glu Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Ile Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Thr Pro Gly Gln Ala Phe Arg Gly
35 40 45
Leu Ile Gly Gly Thr Ser Asn Arg Ala Pro Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Leu Trp Tyr Ser Thr
85 90 95
His Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 32
<211> 109
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> Hu164-VL9
<400> 32
Glu Ser Val Val Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Arg Ser Ser Ile Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Leu Pro Gly Thr Ala Phe Lys Gly
35 40 45
Leu Ile Gly Gly Thr Ser Asn Arg Ala Pro Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Leu Trp Tyr Ser Thr
85 90 95
His Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 33
<211> 114
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> Hu164-VH5
<400> 33
Glu Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Thr Phe
20 25 30
Gly Val His Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu
35 40 45
Ala Val Ile Trp Arg Arg Gly Gly Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Asp Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 34
<211> 114
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> Hu164-VH6
<400> 34
Glu Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Thr Phe
20 25 30
Gly Val His Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Arg Gly Gly Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Phe
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Asp Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 35
<211> 114
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> Hu164-VH7
<400> 35
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Ile Ser Thr Phe
20 25 30
Gly Val His Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Val Ile Trp Arg Arg Gly Gly Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Val Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 36
<211> 114
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> Hu164-VH8
<400> 36
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Ile Ser Thr Phe
20 25 30
Gly Val His Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Arg Gly Gly Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Ser Phe
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 37
<211> 330
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> вариабельный домен тяжелой цепи w
<400> 37
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 38
<211> 330
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> вариабельный домен тяжелой цепи y
<400> 38
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Tyr Ile Thr Arg Glu Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 39
<211> 107
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> константная область легкой цепи каппа гуманизированного антитела
<400> 39
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 40
<211> 105
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> константная область легкой цепи лямбда гуманизированного антитела
<400> 40
Gly Gln Pro Lys Ala Asn Pro Thr Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Gly Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys
100 105
<210> 41
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> тяжелая цепь Ch147
<400> 41
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Glu Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Thr Lys Ser Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Glu Ser Met
65 70 75 80
Leu Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Met Tyr
85 90 95
Tyr Cys Val Glu Thr Gly Phe Ala Tyr Trp Asp Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 42
<211> 213
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Ch147 легкая цепь
<400> 42
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Trp Ile Tyr
35 40 45
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Ser Tyr Pro Leu Thr
85 90 95
Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 43
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu147-11wk тяжелая цепь
<400> 43
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Arg Thr Lys Ser Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Glu Thr Gly Phe Ala Tyr Trp Asp Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 44
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu147-21wk тяжелая цепь
<400> 44
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Thr Lys Ser Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Glu Thr Gly Phe Ala Tyr Trp Asp Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 45
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu147-31wk тяжелая цепь
<400> 45
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Thr Lys Ser Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Glu Ser Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Glu Thr Gly Phe Ala Tyr Trp Asp Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 46
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu147-11yk тяжелая цепь
<400> 46
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Arg Thr Lys Ser Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Glu Thr Gly Phe Ala Tyr Trp Asp Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Tyr Ile Thr Arg Glu Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 47
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu147-21yk тяжелая цепь
<400> 47
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Thr Lys Ser Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Glu Thr Gly Phe Ala Tyr Trp Asp Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Tyr Ile Thr Arg Glu Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 48
<211> 446
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu147-31yk тяжелая цепь
<400> 48
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Asn Thr Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Thr Lys Ser Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Glu Ser Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Glu Thr Gly Phe Ala Tyr Trp Asp Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Tyr Ile Thr Arg Glu Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 49
<211> 213
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu147-11wk легкая цепь
<400> 49
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Gln Ser Pro Arg Leu Leu Ile Tyr
35 40 45
Ser Thr Ser Asn Leu Ala Ser Gly Ile Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Ser Tyr Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 50
<211> 213
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu147-12wk легкая цепь
<400> 50
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile Tyr
35 40 45
Ser Thr Ser Asn Leu Ala Ser Gly Ile Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Ser Tyr Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 51
<211> 213
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu147-13wk легкая цепь
<400> 51
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Trp Ile Tyr
35 40 45
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Ser Tyr Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 52
<211> 213
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu147-14wk легкая цепь
<400> 52
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile Tyr
35 40 45
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Ser Tyr Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 53
<211> 213
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu147-15wk легкая цепь
<400> 53
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Trp Ile Tyr
35 40 45
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Ser Tyr Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 54
<211> 444
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Ch164 тяжелая цепь
<400> 54
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Thr Phe
20 25 30
Gly Val His Trp Ile Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Arg Gly Gly Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Ser Ile Thr Lys Asp Asn Ser Arg Ser His Val Phe Phe
65 70 75 80
Lys Met Thr Ser Leu Gln Thr Asp Asp Ser Ala Ile Tyr Tyr Cys Ala
85 90 95
Arg Asp Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 55
<211> 214
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Ch164 легкая цепь
<400> 55
Gln Ala Val Val Thr Gln Glu Ser Ala Leu Thr Thr Ser Pro Gly Gly
1 5 10 15
Thr Val Ile Leu Thr Cys Arg Ser Ser Ile Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Glu Lys Pro Asp His Leu Phe Thr Gly
35 40 45
Leu Ile Gly Gly Thr Ser Asn Arg Ala Pro Gly Val Pro Val Arg Phe
50 55 60
Ser Gly Ser Leu Ile Gly Asp Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Glu Asp Asp Ala Met Tyr Phe Cys Ala Leu Trp Tyr Ser Thr
85 90 95
His Tyr Val Phe Gly Gly Gly Thr Lys Val Thr Val Leu Gly Gln Pro
100 105 110
Lys Ala Asn Pro Thr Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu
115 120 125
Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro
130 135 140
Gly Ala Val Thr Val Ala Trp Lys Ala Asp Gly Ser Pro Val Lys Ala
145 150 155 160
Gly Val Glu Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala
165 170 175
Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg
180 185 190
Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr
195 200 205
Val Ala Pro Thr Glu Cys
210
<210> 56
<211> 444
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu164-57wl тяжелая цепь
<400> 56
Glu Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Thr Phe
20 25 30
Gly Val His Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu
35 40 45
Ala Val Ile Trp Arg Arg Gly Gly Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Asp Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 57
<211> 444
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu164-67wl тяжелая цепь
<400> 57
Glu Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Thr Phe
20 25 30
Gly Val His Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Arg Gly Gly Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Phe
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Asp Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 58
<211> 444
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu164-77wl тяжелая цепь
<400> 58
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Ile Ser Thr Phe
20 25 30
Gly Val His Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Val Ile Trp Arg Arg Gly Gly Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Val Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 59
<211> 444
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu164-87wl тяжелая цепь
<400> 59
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Ile Ser Thr Phe
20 25 30
Gly Val His Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Arg Gly Gly Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Ser Phe
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 60
<211> 444
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu164-57yl тяжелая цепь
<400> 60
Glu Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Thr Phe
20 25 30
Gly Val His Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu
35 40 45
Ala Val Ile Trp Arg Arg Gly Gly Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Leu
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Asp Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Tyr Ile Thr Arg Glu Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 61
<211> 444
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu164-67yl тяжелая цепь
<400> 61
Glu Val Thr Leu Lys Glu Ser Gly Pro Val Leu Val Lys Pro Thr Glu
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Thr Phe
20 25 30
Gly Val His Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Arg Gly Gly Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Ser Gln Val Val Phe
65 70 75 80
Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Arg Asp Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Tyr Ile Thr Arg Glu Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 62
<211> 444
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu164-77yl тяжелая цепь
<400> 62
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Ile Ser Thr Phe
20 25 30
Gly Val His Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Val Ile Trp Arg Arg Gly Gly Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Val Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Tyr Ile Thr Arg Glu Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 63
<211> 444
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu164-87yl тяжелая цепь
<400> 63
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Ile Ser Thr Phe
20 25 30
Gly Val His Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Arg Gly Gly Thr Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Thr Ile Ser Lys Asp Thr Ser Lys Asn Gln Val Ser Phe
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Tyr Ile Thr Arg Glu Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 64
<211> 214
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu164-57wl легкая цепь
<400> 64
Glu Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Ile Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Phe Arg Gly
35 40 45
Leu Ile Gly Gly Thr Ser Asn Arg Ala Pro Trp Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Ala Leu Trp Tyr Ser Thr
85 90 95
His Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro
100 105 110
Lys Ala Asn Pro Thr Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu
115 120 125
Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro
130 135 140
Gly Ala Val Thr Val Ala Trp Lys Ala Asp Gly Ser Pro Val Lys Ala
145 150 155 160
Gly Val Glu Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala
165 170 175
Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg
180 185 190
Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr
195 200 205
Val Ala Pro Thr Glu Cys
210
<210> 65
<211> 214
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu164-58wl легкая цепь
<400> 65
Glu Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Ile Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Thr Pro Gly Gln Ala Phe Arg Gly
35 40 45
Leu Ile Gly Gly Thr Ser Asn Arg Ala Pro Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Leu Trp Tyr Ser Thr
85 90 95
His Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro
100 105 110
Lys Ala Asn Pro Thr Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu
115 120 125
Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro
130 135 140
Gly Ala Val Thr Val Ala Trp Lys Ala Asp Gly Ser Pro Val Lys Ala
145 150 155 160
Gly Val Glu Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala
165 170 175
Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg
180 185 190
Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr
195 200 205
Val Ala Pro Thr Glu Cys
210
<210> 66
<211> 214
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu164-59wl легкая цепь
<400> 66
Glu Ser Val Val Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Arg Ser Ser Ile Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Leu Pro Gly Thr Ala Phe Lys Gly
35 40 45
Leu Ile Gly Gly Thr Ser Asn Arg Ala Pro Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Leu Trp Tyr Ser Thr
85 90 95
His Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro
100 105 110
Lys Ala Asn Pro Thr Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu
115 120 125
Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro
130 135 140
Gly Ala Val Thr Val Ala Trp Lys Ala Asp Gly Ser Pro Val Lys Ala
145 150 155 160
Gly Val Glu Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala
165 170 175
Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg
180 185 190
Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr
195 200 205
Val Ala Pro Thr Glu Cys
210
<210> 67
<211> 450
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> тяжелая цепь mAb1
<400> 67
Glu Gly Gln Leu Val Gln Ser Gly Gly Gly Leu Val His Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Gly Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Gly Thr Gly Gly Gly Thr Tyr Ser Thr Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Met Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Asp Tyr Tyr Gly Ser Gly Ser Phe Phe Asp Cys Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 68
<211> 214
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> mAb1 легкая цепь
<400> 68
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Glu Lys Ala Pro Lys Ser Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 69
<211> 122
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> mab95 VH
<400> 69
Glu Val Leu Leu Gln Gln Ser Gly Pro Val Leu Val Lys Pro Gly Pro
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Asn Tyr
20 25 30
Asp Ile His Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Leu Val Tyr Pro Tyr Asn Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Phe Asp Thr Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Leu Ile Pro Gly Thr Thr Ser Tyr Phe Asp Val Trp
100 105 110
Gly Thr Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 70
<211> 106
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> mab95 VL
<400> 70
Gln Ile Val Leu Ser Gln Ser Pro Pro Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Arg Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Asn Ser Asn Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Arg Leu Glu Ile Lys
100 105
<210> 71
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> mab95 HCDR1
<400> 71
Asn Tyr Asp Ile His
1 5
<210> 72
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> mab95 HCDR2
<400> 72
Leu Val Tyr Pro Tyr Asn Gly Gly Thr Ala Tyr Asn Gln Lys Phe Lys
1 5 10 15
Asp
<210> 73
<211> 13
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> mab95 HCDR3
<400> 73
Trp Gly Leu Ile Pro Gly Thr Thr Ser Tyr Phe Asp Val
1 5 10
<210> 74
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> mab95 LCDR1
<400> 74
Arg Ala Ser Ser Ser Val Ser Tyr Ile His
1 5 10
<210> 75
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> mab95 LCDR2
<400> 75
Ala Thr Ser Asn Leu Ala Ser
1 5
<210> 76
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> mab95 LCDR3
<400> 76
Gln Gln Trp Asn Ser Asn Pro Trp Thr
1 5
<210> 77
<211> 106
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-VL1
<400> 77
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Ser Ser Val Ser Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Pro Trp Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Ile Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp Asn Ser Asn Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 78
<211> 106
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-VL2
<400> 78
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Ser Ser Val Ser Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Arg Pro Trp Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp Asn Ser Asn Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 79
<211> 105
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-VL3
<400> 79
Glu Ser Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Arg Ala Ser Ser Ser Val Ser Tyr Ile His
20 25 30
Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Pro Trp Ile Tyr Ala
35 40 45
Thr Ser Asn Leu Ala Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Lys
50 55 60
Ser Gly Thr Ser Tyr Ser Leu Ala Ile Thr Gly Leu Gln Ala Glu Asp
65 70 75 80
Glu Ala Asp Tyr Tyr Cys Gln Gln Trp Asn Ser Asn Pro Trp Thr Phe
85 90 95
Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 80
<211> 105
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-VL4
<400> 80
Glu Ile Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Arg Ala Ser Ser Ser Val Ser Tyr Ile His
20 25 30
Trp Tyr Gln Gln Leu Pro Gly Thr Ser Pro Lys Pro Trp Ile Tyr Ala
35 40 45
Thr Ser Asn Leu Ala Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Lys
50 55 60
Ser Gly Thr Ser Tyr Ser Leu Ala Ile Thr Gly Leu Gln Ala Glu Asp
65 70 75 80
Glu Ala Asp Tyr Tyr Cys Gln Gln Trp Asn Ser Asn Pro Trp Thr Phe
85 90 95
Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 81
<211> 122
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-VH1
<400> 81
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Asn Tyr
20 25 30
Asp Ile His Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Leu Val Tyr Pro Tyr Asn Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Val Thr Leu Thr Phe Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Leu Ile Pro Gly Thr Thr Ser Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 82
<211> 122
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-VH2
<400> 82
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Asn Tyr
20 25 30
Asp Ile His Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Leu Val Tyr Pro Tyr Asn Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Ala Thr Leu Thr Phe Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Leu Ile Pro Gly Thr Thr Ser Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 83
<211> 122
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-VH3
<400> 83
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Asn Tyr
20 25 30
Asp Ile His Trp Val Lys Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Leu Val Tyr Pro Tyr Asn Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Val Thr Leu Thr Phe Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Leu Ile Pro Gly Thr Thr Ser Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 84
<211> 122
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-VH4
<400> 84
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Asn Tyr
20 25 30
Asp Ile His Trp Val Lys Gln Ala Pro Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Leu Val Tyr Pro Tyr Asn Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Phe Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Leu Ile Pro Gly Thr Thr Ser Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 85
<211> 122
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-VH5
<400> 85
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Arg Ala Ser Gly Phe Thr Phe Thr Asn Tyr
20 25 30
Ala Ile His Trp Val Lys Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Leu Val Tyr Pro Tyr Thr Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Val Thr Leu Thr Phe Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Met Ile Pro Gly Thr Asn Ser Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 86
<211> 452
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Ch95 тяжелая цепь
<400> 86
Glu Val Leu Leu Gln Gln Ser Gly Pro Val Leu Val Lys Pro Gly Pro
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Asn Tyr
20 25 30
Asp Ile His Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Leu Val Tyr Pro Tyr Asn Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Phe Asp Thr Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Leu Ile Pro Gly Thr Thr Ser Tyr Phe Asp Val Trp
100 105 110
Gly Thr Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 87
<211> 213
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Ch95 легкая цепь
<400> 87
Gln Ile Val Leu Ser Gln Ser Pro Pro Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Arg Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Asn Ser Asn Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Arg Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 88
<211> 452
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-11wk тяжелая цепь
<400> 88
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Asn Tyr
20 25 30
Asp Ile His Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Leu Val Tyr Pro Tyr Asn Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Val Thr Leu Thr Phe Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Leu Ile Pro Gly Thr Thr Ser Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 89
<211> 452
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-21wk тяжелая цепь
<400> 89
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Asn Tyr
20 25 30
Asp Ile His Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Leu Val Tyr Pro Tyr Asn Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Ala Thr Leu Thr Phe Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Leu Ile Pro Gly Thr Thr Ser Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 90
<211> 452
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-31wk тяжелая цепь
<400> 90
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Asn Tyr
20 25 30
Asp Ile His Trp Val Lys Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Leu Val Tyr Pro Tyr Asn Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Val Thr Leu Thr Phe Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Leu Ile Pro Gly Thr Thr Ser Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 91
<211> 452
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-41wk тяжелая цепь
<400> 91
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Asn Tyr
20 25 30
Asp Ile His Trp Val Lys Gln Ala Pro Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Leu Val Tyr Pro Tyr Asn Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Phe Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Leu Ile Pro Gly Thr Thr Ser Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 92
<211> 452
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-51wk тяжелая цепь
<400> 92
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Arg Ala Ser Gly Phe Thr Phe Thr Asn Tyr
20 25 30
Ala Ile His Trp Val Lys Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Leu Val Tyr Pro Tyr Thr Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Val Thr Leu Thr Phe Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Met Ile Pro Gly Thr Asn Ser Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 93
<211> 452
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-11yk тяжелая цепь
<400> 93
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Asn Tyr
20 25 30
Asp Ile His Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Leu Val Tyr Pro Tyr Asn Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Val Thr Leu Thr Phe Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Leu Ile Pro Gly Thr Thr Ser Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Tyr Ile Thr Arg Glu Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 94
<211> 452
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-21yk тяжелая цепь
<400> 94
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Asn Tyr
20 25 30
Asp Ile His Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Leu Val Tyr Pro Tyr Asn Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Arg Ala Thr Leu Thr Phe Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Leu Ile Pro Gly Thr Thr Ser Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Tyr Ile Thr Arg Glu Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 95
<211> 452
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-31yk
<400> 95
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Asn Tyr
20 25 30
Asp Ile His Trp Val Lys Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Leu Val Tyr Pro Tyr Asn Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Val Thr Leu Thr Phe Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Leu Ile Pro Gly Thr Thr Ser Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Tyr Ile Thr Arg Glu Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 96
<211> 452
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-41yk тяжелая цепь
<400> 96
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Thr Asn Tyr
20 25 30
Asp Ile His Trp Val Lys Gln Ala Pro Gly Gln Arg Leu Glu Trp Ile
35 40 45
Gly Leu Val Tyr Pro Tyr Asn Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Phe Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Leu Ile Pro Gly Thr Thr Ser Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Tyr Ile Thr Arg Glu Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 97
<211> 452
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-51yk тяжелая цепь
<400> 97
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Arg Ala Ser Gly Phe Thr Phe Thr Asn Tyr
20 25 30
Ala Ile His Trp Val Lys Gln Ala Pro Gly Gln Arg Leu Glu Trp Met
35 40 45
Gly Leu Val Tyr Pro Tyr Thr Gly Gly Thr Ala Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Val Thr Leu Thr Phe Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Gly Met Ile Pro Gly Thr Asn Ser Tyr Phe Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Tyr Ile Thr Arg Glu Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 98
<211> 213
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-11wk легкая цепь
<400> 98
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Ser Ser Val Ser Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Pro Trp Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Ile Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp Asn Ser Asn Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 99
<211> 213
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-12wk легкая цепь
<400> 99
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Ser Ser Val Ser Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Arg Pro Trp Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu
65 70 75 80
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp Asn Ser Asn Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 100
<211> 212
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-13wk легкая цепь
<400> 100
Glu Ser Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Arg Ala Ser Ser Ser Val Ser Tyr Ile His
20 25 30
Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Pro Trp Ile Tyr Ala
35 40 45
Thr Ser Asn Leu Ala Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Lys
50 55 60
Ser Gly Thr Ser Tyr Ser Leu Ala Ile Thr Gly Leu Gln Ala Glu Asp
65 70 75 80
Glu Ala Asp Tyr Tyr Cys Gln Gln Trp Asn Ser Asn Pro Trp Thr Phe
85 90 95
Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro Ser
100 105 110
Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala
115 120 125
Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val
130 135 140
Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser
145 150 155 160
Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr
165 170 175
Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys
180 185 190
Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn
195 200 205
Arg Gly Glu Cys
210
<210> 101
<211> 212
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> Hu95-14wk легкая цепь
<400> 101
Glu Ile Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Arg Ala Ser Ser Ser Val Ser Tyr Ile His
20 25 30
Trp Tyr Gln Gln Leu Pro Gly Thr Ser Pro Lys Pro Trp Ile Tyr Ala
35 40 45
Thr Ser Asn Leu Ala Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Lys
50 55 60
Ser Gly Thr Ser Tyr Ser Leu Ala Ile Thr Gly Leu Gln Ala Glu Asp
65 70 75 80
Glu Ala Asp Tyr Tyr Cys Gln Gln Trp Asn Ser Asn Pro Trp Thr Phe
85 90 95
Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro Ser
100 105 110
Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala
115 120 125
Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val
130 135 140
Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser
145 150 155 160
Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr
165 170 175
Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys
180 185 190
Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn
195 200 205
Arg Gly Glu Cys
210
<210> 102
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> Hu95-VH5 HCDR1
<400> 102
Asn Tyr Ala Ile His
1 5
<210> 103
<211> 17
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> Hu95-VH5 HCDR2
<400> 103
Leu Val Tyr Pro Tyr Thr Gly Gly Thr Ala Tyr Asn Gln Lys Phe Lys
1 5 10 15
Asp
<210> 104
<211> 13
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> Hu95-VH5 HCDR3
<400> 104
Trp Gly Met Ile Pro Gly Thr Asn Ser Tyr Phe Asp Val
1 5 10
<---


Изобретение относится к области биотехнологии. Предложены антитело против CTGF, молекула нуклеиновой кислоты, кодирующая антитело, клетка-хозяин для получения антитела, фармацевтическая композиция. Также изобретение относится к способу определения CTGF и способу лечения или предупреждения заболевания, связанного с CTGF. Изобретение может найти применение в терапии заболеваний и состояний, связанных с CTGF, таких, как фиброзные заболевания, гипертензия, диабет, инфаркт миокарда, артрит, клеточное пролиферативное заболевание, связанное с CTGF, атеросклероз, глаукома или рак. 6 н. и 12 з.п. ф-лы, 2 ил., 27 табл., 8 пр.
1. Антитело против фактора роста соединительной ткани (CTGF), содержащее вариабельную область тяжелой цепи и вариабельную область легкой цепи, где:
i) вариабельная область тяжелой цепи содержит такие же последовательности HCDR1, HCDR2 и HCDR3, как и последовательности HCDR1, HCDR2 и HCDR3 вариабельной области тяжелой цепи, показанной в SEQ ID NO: 6, и вариабельная область легкой цепи содержит такие же последовательности LCDR1, LCDR2 и LCDR3, как и последовательности LCDR1, LCDR2 и LCDR3 вариабельной области легкой цепи, показанной в SEQ ID NO: 7;
ii) вариабельная область тяжелой цепи содержит такие же последовательности HCDR1, HCDR2 и HCDR3, как и последовательности HCDR1, HCDR2 и HCDR3 вариабельной области тяжелой цепи, показанной в SEQ ID NO: 8, и вариабельная область легкой цепи содержит такие же последовательности LCDR1, LCDR2 и LCDR3, как и последовательности LCDR1, LCDR2 и LCDR3 вариабельной области легкой цепи, показанной в SEQ ID NO: 9;
ii-i) вариабельная область тяжелой цепи содержит такие же последовательности HCDR1, HCDR2 и HCDR3, как и последовательности HCDR1, HCDR2 и HCDR3 вариабельной области тяжелой цепи, показанной в SEQ ID NO: 69, и вариабельная область легкой цепи содержит такие же последовательности LCDR1, LCDR2 и LCDR3, как и последовательности LCDR1, LCDR2 и LCDR3 вариабельной области легкой цепи, показанной в SEQ ID NO: 70; или
ii-ii) вариабельная область тяжелой цепи содержит такие же последовательности HCDR1, HCDR2 и HCDR3, как последовательности HCDR1, HCDR2 и HCDR3 вариабельной области тяжелой цепи, показанной в SEQ ID NO: 85, и вариабельная область легкой цепи содержит такие же последовательности LCDR1, LCDR2 и LCDR3, как и последовательности LCDR1, LCDR2 и LCDR3 вариабельной области легкой цепи, показанной в SEQ ID NO: 70.
2. Антитело против CTGF по п. 1, содержащее вариабельную область тяжелой цепи и вариабельную область легкой цепи, где:
iii) вариабельная область тяжелой цепи содержит HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 10, SEQ ID NO: 11 и SEQ ID NO: 12, соответственно, и вариабельная область легкой цепи содержит LCDR1, LCDR2 и LCDR3, как показано в SEQ ID NO: 13, SEQ ID NO: 14 и SEQ ID NO: 15, соответственно;
iv) вариабельная область тяжелой цепи содержит HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 16, SEQ ID NO: 17 и SEQ ID NO: 18, соответственно, и вариабельная область легкой цепи содержит LCDR1, LCDR2 и LCDR3, как показано в SEQ ID NO: 19, SEQ ID NO: 20 и SEQ ID NO: 21, соответственно;
iv-i) вариабельная область тяжелой цепи содержит HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 71, SEQ ID NO: 72 и SEQ ID NO: 73, соответственно, и вариабельная область легкой цепи содержит LCDR1, LCDR2 и LCDR3, как показано в SEQ ID NO: 74, SEQ ID NO: 75 и SEQ ID NO: 76, соответственно; или
iv-ii) вариабельная область тяжелой цепи содержит HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 102, SEQ ID NO: 103 и SEQ ID NO: 104, соответственно, и вариабельная область легкой цепи содержит LCDR1, LCDR2 и LCDR3, как показано в SEQ ID NO: 74, SEQ ID NO: 75 и SEQ ID NO: 76, соответственно.
3. Антитело против CTGF по п. 1 или 2, где указанное антитело против CTGF представляет собой мышиное антитело, химерное антитело или гуманизированное антитело.
4. Антитело против CTGF по любому из пп. 1-3, содержащее вариабельную область тяжелой цепи и вариабельную область легкой цепи, где:
(v) аминокислотная последовательность вариабельной области тяжелой цепи имеет по меньшей мере 90%-ную идентичность последовательностей с SEQ ID NO: 6, 27, 28 или 29, и аминокислотная последовательность вариабельной области легкой цепи имеет по меньшей мере 90%-ную идентичность последовательностей с SEQ ID NO: 7, 22, 23, 24, 25 или 26;
(vi) аминокислотная последовательность вариабельной области тяжелой цепи имеет по меньшей мере 90%-ную идентичность последовательностей с SEQ ID NO: 8, 33, 34, 35 или 36, и аминокислотная последовательность вариабельной области легкой цепи имеет по меньшей мере 90%-ную идентичность последовательностей с SEQ ID NO: 9, 30, 31 или 32; или
(vi-i) аминокислотная последовательность вариабельной области тяжелой цепи имеет по меньшей мере 90%-ную идентичность последовательностей с SEQ ID NO: 69, 81, 82, 83, 84 или 85, и аминокислотная последовательность вариабельной области легкой цепи имеет по меньшей мере 90%-ную идентичность последовательностей с SEQ ID NO: 70, 77, 78, 79 или 80.
5. Антитело против CTGF по п. 2, где указанное антитело против CTGF представляет собой гуманизированное антитело, причем указанное гуманизированное антитело содержит каркасную область человеческого антитела или вариант его каркасной области, причем указанный вариант каркасной области имеет вплоть до 10 обратных мутаций на каждой из каркасной области легкой цепи и/или каркасной области тяжелой цепи антитела человека.
6. Антитело против CTGF по п. 5, где указанное гуманизированное антитело содержит вариабельную область легкой цепи и вариабельную область тяжелой цепи, выбранные из группы, состоящей из следующих (a), (b) или (b-i):
(a) вариабельная область легкой цепи, содержащая:
LCDR1, LCDR2 и LCDR3, как показано в SEQ ID NO: 13, SEQ ID NO: 14 и SEQ ID NO: 15, соответственно, и одну или более обратных мутаций, выбранных из группы, состоящей из 4L, 36F, 43S, 45K, 47W, 58V и 71Y, и
вариабельная область тяжелой цепи, содержащая:
HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 10, SEQ ID NO: 11 и SEQ ID NO: 12, соответственно, и одну или более обратных мутаций, выбранных из группы, состоящей из 28S, 30N, 49A, 75E, 76S, 93V, 94E и 104D;
(b) вариабельная область легкой цепи, содержащая:
LCDR1, LCDR2 и LCDR3, как показано в SEQ ID NO: 19, SEQ ID NO: 20 и SEQ ID NO: 21, соответственно, и одну или более обратных мутаций, выбранных из группы, состоящей из 36V, 44F, 46G и 49G, и
вариабельная область тяжелой цепи, содержащая:
HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 16, SEQ ID NO: 17 и SEQ ID NO: 18, соответственно, и одну или более обратных мутаций, выбранных из группы, состоящей из 44G, 49G, 27F, 48L, 67L, 71K, 78V и 80F;
(b-i) вариабельная область легкой цепи, содержащая:
LCDR1, LCDR2 и LCDR3, как показано в SEQ ID NO: 74, SEQ ID NO: 75 и SEQ ID NO: 76, соответственно, и одну или более обратных мутаций, выбранных из группы, состоящей из 45P, 46W, 48Y, 69S и 70Y, и
вариабельная область тяжелой цепи, содержащая:
HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 71, SEQ ID NO: 72 и SEQ ID NO: 73, соответственно, и одну или более обратных мутаций, выбранных из группы, состоящей из 27F, 38K, 48I, 67K, 68A, 70L и 72F; или
(b-ii) вариабельная область легкой цепи, содержащая:
LCDR1, LCDR2 и LCDR3, как показано в SEQ ID NO: 74, SEQ ID NO: 75 и SEQ ID NO: 76, соответственно, и одну или более обратных мутаций, выбранных из группы, состоящей из 45P, 46W, 48Y, 69S и 70Y, и
вариабельная область тяжелой цепи, содержащая:
HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 102, SEQ ID NO: 103 и SEQ ID NO: 104, соответственно, и одну или более обратных мутаций, выбранных из группы, состоящей из 27F, 38K, 48I, 67K, 68A, 70L и 72F.
7. Антитело против CTGF по п. 6, содержащее вариабельную область тяжелой цепи и вариабельную область легкой цепи, в которых:
(vii) аминокислотная последовательность вариабельной области тяжелой цепи представляет собой такую, как показано в SEQ ID NO: 6, и аминокислотная последовательность вариабельной области легкой цепи представляет собой такую, как показано в SEQ ID NO: 7;
(viii) аминокислотная последовательность вариабельной области тяжелой цепи представляет собой такую, как показано в SEQ ID NO: 27, 28 или 29, и аминокислотная последовательность вариабельной области легкой цепи представляет собой такую, как показано в SEQ ID NO: 22, 23, 24, 25 или 26;
(ix) аминокислотная последовательность вариабельной области тяжелой цепи представляет собой такую, как показано в SEQ ID NO: 8, и аминокислотная последовательность вариабельной области легкой цепи представляет собой такую, как показано в SEQ ID NO: 9;
(x) аминокислотная последовательность вариабельной области тяжелой цепи представляет собой такую, как показано в SEQ ID NO: 33, 34, 35 или 36, и аминокислотная последовательность вариабельной области легкой цепи представляет собой такую, как показано в SEQ ID NO: 30, 31 или 32;
(xi) аминокислотная последовательность вариабельной области тяжелой цепи представляет собой такую, как показано в SEQ ID NO: 69, и аминокислотная последовательность вариабельной области легкой цепи представляет собой такую, как показано в SEQ ID NO: 70; или
(xii) аминокислотная последовательность вариабельной области тяжелой цепи представляет собой такую, как показано в SEQ ID NO: 81, 82, 83, 84 или 85, и аминокислотная последовательность вариабельной области легкой цепи представляет собой такую, как показано в SEQ ID NO: 77, 78, 79 или 80.
8. Антитело против CTGF по п. 7, которое содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, как показано ниже:
(xiii) аминокислотная последовательность вариабельной области тяжелой цепи представляет собой такую, как показано в SEQ ID NO: 27, и аминокислотная последовательность вариабельной области легкой цепи представляет собой такую, как показано в SEQ ID NO: 22;
(xiv) аминокислотная последовательность вариабельной области тяжелой цепи представляет собой такую, как показано в SEQ ID NO: 34, и аминокислотная последовательность вариабельной области легкой цепи представляет собой такую, как показано в SEQ ID NO: 30; или
(xv) аминокислотная последовательность вариабельной области тяжелой цепи представляет собой такую, как показано в SEQ ID NO: 85, и аминокислотная последовательность вариабельной области легкой цепи представляет собой такую, как показано в SEQ ID NO: 77.
9. Антитело против CTGF по любому из пп. 1-8, где указанное антитело дополнительно содержит константную область тяжелой цепи и константную область легкой цепи антитела; причем константная область тяжелой цепи выбрана из группы, состоящей из константных областей человеческого IgG1, IgG2, IgG3 и IgG4 и их вариантов, и константная область легкой цепи выбрана из группы, состоящей из константных областей цепей κ и λ человеческого антитела и их вариантов.
10. Антитело против CTGF по п. 9, где антитело содержит константную область тяжелой цепи, как показано в SEQ ID NO: 37 или 38, и константную область легкой цепи, как показано в SEQ ID NO: 39 или 40.
11. Антитело против CTGF по любому из пп. 1-10, содержащее:
(c) тяжелую цепь, имеющую по меньшей мере 85%-ную идентичность последовательностей с SEQ ID NO: 41, 43, 44, 45, 46, 47 или 48, и легкую цепь, имеющую по меньшей мере 85%-ную идентичность последовательностей с SEQ ID NO: 42, 49, 50, 51, 52 или 53;
(d) тяжелую цепь, имеющую по меньшей мере 85%-ную идентичность последовательностей с SEQ ID NO: 54, 56, 57, 58, 59, 60, 61, 62 или 63, и легкую цепь, имеющую по меньшей мере 85%-ную идентичность последовательностей с SEQ ID NO: 55, 64, 65 или 66;
(e) тяжелую цепь, как показано в SEQ ID NO: 41, 43, 44, 45, 46, 47 или 48, и легкую цепь, как показано в SEQ ID NO: 42, 49, 50, 51, 52 или 53;
(f) тяжелую цепь, как показано в SEQ ID NO: 54, 56, 57, 58, 59, 60, 61, 62 или 63, и легкую цепь, как показано в SEQ ID NO: 55, 64, 65 или 66;
(g) тяжелую цепь, имеющую по меньшей мере 85%-ную идентичность последовательностей с SEQ ID NO: 86, 88, 89, 90, 91, 92, 93, 94, 95, 96 или 97, и легкую цепь, имеющую по меньшей мере 85%-ную идентичность последовательностей с SEQ ID NO: 87, 98, 99, 100 или 101; или
(h) тяжелую цепь, как показано в SEQ ID NO: 86, 88, 89, 90, 91, 92, 93, 94, 95, 96 или 97, и легкую цепь, как показано в SEQ ID NO: 87, 98, 99, 100 или 101.
12. Антитело против CTGF по п. 11, содержащее:
(j) тяжелую цепь, как показано в SEQ ID NO: 46, и легкую цепь, как показано в SEQ ID NO: 49;
(k) тяжелую цепь, как показано в SEQ ID NO: 61, и легкую цепь, как показано в SEQ ID NO: 64; или
(l) тяжелую цепь, как показано в SEQ ID NO: 97, и легкую цепь, как показано в SEQ ID NO: 98.
13. Молекула нуклеиновой кислоты, кодирующая антитело против CTGF по любому из пп. 1-12.
14. Клетка-хозяин для получения антитела против CTGF, содержащая молекулу нуклеиновой кислоты по п. 13.
15. Фармацевтическая композиция для лечения или предупреждения заболевания, связанного с CTGF, содержащая терапевтически эффективное количество антитела против CTGF по любому из пп. 1-12 и один или более фармацевтически приемлемых носителей, разбавителей, буферов или вспомогательных веществ.
16. Способ определения CTGF, включающий стадию приведения антитела против CTGF по любому из пп. 1-12 в контакт с субъектом или его образцом.
17. Способ лечения или предупреждения заболевания, связанного с CTGF, включающий введение субъекту терапевтически эффективного количества или профилактически эффективного количества антитела против CTGF по любому из пп. 1-12 или фармацевтической композиции по п. 15, где заболевание, связанное с CTGF, представляет собой фиброзные заболевания, гипертензию, диабет, инфаркт миокарда, артрит, клеточное пролиферативное заболевание, связанное с CTGF, атеросклероз, глаукому или рак.
18. Способ по п. 17, где фиброзное заболевание выбрано из группы, состоящей из: спонтанного фиброза легких, диабетической нефропатии, диабетической ретинопатии, остеоартрита, склеродермы, хронической сердечной недостаточности, цирроза печени или фиброза почек;
рак выбран из группы, состоящей из: острого лимфобластного лейкоза, дерматофибромы, рака молочной железы, ангиолипомы, ангиолейомиомы, рака, происходящего из соединительной ткани, рака предстательной железы, рака яичника, колоректального рака, рака поджелудочной железы, рака желудочно-кишечного тракта или рака печени.
| WO9933878 A1, 08.07.1999 | |||
| US 2010291098 A1, 18.11.2010 | |||
| АНТИТЕЛА К РОСТОВОМУ ФАКТОРУ СОЕДИНИТЕЛЬНОЙ ТКАНИ | 2004 |
|
RU2330861C2 |
| OHARA Y | |||
| et al | |||
| Connective tissue growth factor-specific monoclonal antibody inhibits growth of malignant mesothelioma in an orthotopic mouse model, Oncotarget, 2018, vol.9(26), pp.18494-18509 | |||
| MAKINO K | |||
| et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |

