ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к антителам к CD3, таким как гуманизированные антитела к CD3, и к мультиспецифическим антителам, одновременно связывающимся с CD3 и опухолеассоциированным (-и) антиген (-ами).
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
В описании предшествующего уровня техники представлена только общая информация о настоящем изобретении, которая необязательно составляет предшествующий уровень техники.
CD3 представляет собой T-клеточный корецептор, состоящий из четырех различных цепей (Wucherpfennig, KW et al. (2010) Structural Biology of The T cell Receptor: Insights Into Receptor Assembly, Ligand Recognition, and Initiation of Signaling, Cold Spring Harb. Perspect. Biol. 2(4):A005140; pages 1-14; Chetty, R. et al. (1994) CD3:Structure, Function, And Role Of Immunostaining In Clinical Practice, J. Pathol. 173(4):303-307; Guy, C.S. et al. (2009) Organization of Proximal Signal Initiation at the TCR:CD3 Complex, Immunol.Rev.232(1):7-21).
У млекопитающих комплекс, образованный мульти-субъединицами CD3, ассоциирован с молекулой T-клеточного рецептора (TCR), в результате чего он генерирует сигналы активации в T-лимфоцитах (Smith-Garvin, JE et al. (2009) T Cell Activation, Annu. Rev. Immunol. 27:591-619). В отсутствие CD3 TCR либо не может осуществить правильную сборку, либо происходит его деструкция (Thomas, S. et al. (2010) Molecular Immunology Lessons From Therapeutic T cell Receptor Gene Transfer, Immunology 129(2): 170-177). В исследованиях было обнаружено, что CD3 связывается с мембранами всех зрелых T-клеток и прочно связывается с другими типами клеток (Janeway, C.A. et al. (2005): Immunobiology: The Immune System in Health and Disease, 6th Edition, Garland Science Publishing, NY, pp.214-216; Sun, Z.J. et al. (2001) Mechanisms Contributing to T Cell Receptor Signaling and Assembly Revealed by the Solution Structure of an Ectodomain Fragment of the CD3ε: γ Heterodimer, Cell 105 (7): 913-923; Kuhns, M.S. et al. (2006) Deconstructing the Form and Function of the TCR/CD3 Complex, Immunity. 2006 Feb,24(2): 133-139).
Постоянный сигнальный компонент CD3ε комплекса T-клеточного рецептора (TCR) на T-клетках был использован в качестве мишени для стимуляции образования иммунологических синапсов между T-клетками и опухолевыми клетками. Одновременное взаимодействие CD3 и опухолевого(-ых) антигена(-ов) активирует T-клетки, что приводит в результате к лизису опухолевых клеток, экспрессирующих опухолевый(-ые) антиген(-ы) (Baeuerle et al. (2011) Bispecific T Cell Engager for Cancer Therapy, In: Bispecific Antibodies, Kontermann, R.E. (Ed.) Springer-Verlag; 2011:273-287). Данный способ, широко применяемый к ряду опухолевых антигенов клеточной поверхности, позволяет биспецифическим антителам полностью взаимодействовать с T-клеточными компартментами со столь же высокой специфичностью, как и с опухолевыми клетками.
B7H3 является представителем семейства B7 и относится к трансмембранным белкам I типа. Он содержит сигнальный пептид на амино-конце, внеклеточную иммуноглобулин-подобную вариабельную область (IgV), константную область (IgC), трансмембранную область и область цитоплазматического «хвоста», содержащую 45 аминокислот (Tissue Antigens. 2007 Aug; 70 (2): 96-104). Как известно в настоящее время, B7H3 главным образом имеет две формы сплайсинга B7H3a и B7H3b. Внеклеточный сегмент B7H3a, также известный как 2IgB7H3, состоит из двух иммуноглобулиновых доменов IgV-IgC, тогда как внеклеточный сегмент B7H3b, также известный как 4IgB7H3, состоит из четырех иммуноглобулиновых доменов IgV-IgC-IgV-IgC.
Белок B7H3 отсутствует в нормальных тканях и клетках или экспрессируется в нормальных тканях и клетках на крайне низком уровне; тем не менее, его экспрессия в опухолевых тканях высока и тесно связана с прогрессированием опухоли, выживанием пациента и прогнозом. Недавно согласно клиническим данным сообщали, что сверхэкспрессия B7H3 происходит в ряде злокачественных опухолевых тканей, в частности при немелкоклеточном раке легких, раке почки, эпительальном раке мочеполовых путей, колоректальном раке, раке предстательной железы, мультиформной глиобластоме, раке яичника и раке поджелудочной железы. (Lung Cancer. 2009 Nov; 66(2): 245-249; Clin Cancer Res. 2008 Aug 15; 14(16): 5150-5157). Кроме того, согласно литературным данным также сообщали о наличии положительной корреляции интенсивности экспрессии B7H3 с клиническими патологическими злокачественными свойствами при раке предстательной железы, такими как объем опухоли, инвазией вне предстательной железы и баллом по шкале Глисона, а также корреляции с прогрессированием рака (Cancer Res. 2007 Aug 15; 67 (16): 7893-7900). Аналогичным образом, существует отрицательная корреляция B7H3 с выживаемостью без событий при мультиформной глиобластоме и корреляция экспрессии B7H3 с метастазами в лимфатические узлы и патологическим прогрессированием при раке поджелудочной железы. Таким образом, B7H3 рассматривают как новый опухолевый маркер и потенциальную терапевтическую мишень.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В описании настоящего изобретения предложены антитела или их антигенсвязывающие фрагменты, которые могут специфично связываться с человеческим CD3.
В одном аспекте в описании настоящего изобретения предложено антитело или его антигенсвязывающий фрагмент, специфично связывающиеся с человеческим CD3, причем антитело или его антигенсвязывающий фрагмент содержит вариабельную область тяжелой цепи и вариабельную область легкой цепи, при этом вариабельная область легкой цепи содержит LCDR1 (определяющая комплементарнсоть область 1 легкой цепи), LCDR2 и LCDR3, как показано в SEQ ID NO: 48, 49 и 50 соответственно, а вариабельная область тяжелой цепи представляет собой любое выбранное из группы, состоящей из следующих i)-v):
i) вариабельной области тяжелой цепи, содержащей HCDR1 (определяющая комплемантарность область 1 тяжелой цепи), HCDR2 и HCDR3, как показано в SEQ ID NO: 37, 38 и 39 соответственно;
ii) вариабельной области тяжелой цепи, содержащей HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 37, 40 и 41 соответственно;
iii) вариабельной области тяжелой цепи, содержащей HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 37, 40 и 42 соответственно;
iv) вариабельной области тяжелой цепи, содержащей HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 37, 40 и 43 соответственно; и
v) вариабельной области тяжелой цепи, содержащей HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 37, 47 и 45 соответственно.
В некоторых воплощениях изобретения антитело или его антигенсвязывающий фрагмент, специфично связывающиеся с человеческим CD3, представляет собой гуманизированное антитело или его антигенсвязывающий фрагмент.
В некоторых воплощениях изобретения антитело или его антигенсвязывающий фрагмент, специфично связывающиеся с человеческим CD3, содержит вариабельную область легкой цепи SEQ ID NO: 36 и/или вариабельную область тяжелой цепи, как показано в любой из последовательностей, выбранной из группы, состоящей из SEQ ID NO: 29, 30, 31, 32 и 35.
В некоторых воплощениях изобретения антитело или его антигенсвязывающий фрагмент, специфично связывающиеся с человеческим CD3, дополнительно содержит константную область легкой цепи антитела и/или константную область тяжелой цепи антитела; необязательно, константная область легкой цепи представляет собой константную область легкой цепи человеческой цепи каппа, лямбда или ее вариант, а константная область тяжелой цепи представляет собой константную область тяжелой цепи человеческого IgG1, IgG2, IgG3, IgG4 или ее вариант.
В некоторых воплощениях изобретения антигенсвязывающий фрагмент выбран из Fab, Fab', F(ab')2, димеризованной области V (диатело) и области V, стабилизированной дисульфидными связями (dsFv).
В другом аспекте в описании настоящего изобретения предложено одноцепочечное антитело, содержащее вариабельную область легкой цепи и вариабельную область тяжелой цепи антитела или его антигенсвязывающего фрагмента, специфично связывающихся с человеческим CD3, как описано выше.
В некоторых воплощениях изобретения последовательность одноцепочечного антитела является такой, как показано в SEQ ID NO: 55, 56, 57, 58, 61, 62, 63, 64, 65 или 68.
В другом аспекте в описании настоящего изобретения предложено мультиспецифическое антитело, специфично связывающееся с человеческим CD3 и опухолеассоциированным(-и) антигеном(-ами) (tumor-associated antigen(s), TAA), причем мультипецифическое антитело включает в себя одноцепочечное антитело, либо антитело или его антигенсвязывающий фрагмент, специфично связывающиеся с человеческим CD3, как описано выше.
В некоторых воплощениях мультиспецифического антитела опухолеассоциированный антиген выбран из группы, состоящей из AFP, ALK, B7H3, белка BAGE, BCMA, BIRC5 (сурвивин), BIRC7, β-катенина, brc-ab1, BRCA1, BORIS, CA9, CA125, карбоангидразы IX, каспазы-8, CALR, CCR5, CD19, CD20(MS4A1), CD22, CD30, CD33, CD38, CD40, CD123, CD133, CD138, CDK4, CEA, клаудина 18.2, циклина-B1, CYP1B1, EGFR, EGFRvIII, ErbB2/Her2, ErbB3, ErbB4, ETV6-AML, EpCAM, EphA2, Fra-1, FOLR1, белка GAGE (такого как GAGE-1, -2), GD2, GD3, GloboH, глипикана-3, GM3, gp100, Her2, HLA/B-raf, HLA/k-ras, HLA/MAGE-A3, hTERT, IL13Rα2, LMP2, κ-Light, LeY, белка MAGE (такого как MAGE-1, -2, -3, -4, -6 и -12), MART-1, мезотелина, ML-IAP, MOv-γ, Muc1, Muc2, Muc3, Muc4, Muc5, Muc16(CA-125, MUM1, NA17, NKG2D, NY-BR1, NY-BR62, NY-BR85, NY-ESO1, OX40, p15, p53, PAP, PAX3, PAX5, PCTA-1, PLAC1, PRLR, PRAME, PSMA (FOLH1), белка RAGE, Ras, RGS5, Rho, ROR1, SART-1, SART-3, STEAP1, STEAP2, TAG-72, TGF-β, TMPRSS2, антигена Thompson-nouvelle (Tn), TRP-1, TRP-2, тирозиназы, уроплакина-3 и 5T4 (трофобластного гликопротеина). Предпочтительно опухолеассоциированный антиген выбран из группы, состоящей из B7H3, BCMA, CEA, CD19, CD20, CD38, CD138, клаудина 18.2, PSMA и мезотелина.
В другом аспекте в описании настоящего изобретения предложена фармацевтическая композиция, содержащая терапевтически эффективное количество антитела или его антигенсвязывающего фрагмента, специфично связывающихся с человеческим CD3, одноцепочечного антитела или мультиспецифического антитела, как описано выше, и один или более фармацевтически приемлемых носителей, разбавителей, буферных растворов или эксципиентов. В некоторых воплощениях терапевтически эффективное количество означает, что композиция содержит единичную дозу 0,1-3000 мг (более предпочтительно 1-1000 мг) антитела или его антигенсвязывающего фрагмента, специфично связывающихся с человеческим CD3, одноцепочечного антитела или мультиспецифического антитела, как описано выше.
В другом аспекте в описании настоящего изобретения предложена выделенная молекула нуклеиновой кислоты, кодирующая антитело или его антигенсвязывающий фрагмент, специфично связывающиеся с человеческим CD3, кодирующая одноцепочечное антитело или кодирующая мультиспецифическое антитело, как описано выше.
В другом аспекте в описании настоящего изобретения предложен рекомбинантный вектор, содержащий выделенную молекулу нуклеиновой кислоты, как описано выше.
В другом аспекте в описании настоящего изобретения предложена клетка-хозяин, трансформированная вышеуказанным рекомбинантным вектором, причем клетка-хозяин выбрана из прокариотической клетки и эукариотической клетки, предпочтительно эукариотической клетки, более предпочтительно клетки млекопитающего или клетки насекомого.
В другом аспекте в описании настоящего изобретения предложен способ получения антитела или его антигенсвязывающего фрагмента, специфично связывающихся с человеческим CD3, одноцепочечного антитела или мультиспецифического антитела, как описано выше, причем способ включает культивирование клетки-хозяина, как описано выше, в питательной среде для образования и накопления антитела или его антигенсвязывающего фрагмента, специфично связывающихся с человеческим CD3, одноцепочечного антитела или мультиспецифического антитела, как описано выше, и выделение антитела или его антигенсвязывающего фрагмента, специфично связывающихся с человеческим CD3, одноцепочечного антитела или мультиспецифического антитела из культуры.
В другом аспекте в описании настоящего изобретения предложено антитело или его антигенсвязывающий фрагмент, специфично связывающиеся с человеческим CD3, одноцепочечное антитело, мультиспецифическое антитело, фармацевтическая композиция или выделенная молекула нуклеиновой кислоты, как описано выше, в качестве лекарственного средства. В некоторых воплощениях изобретения лекарственное средство представляет собой лекарственное средство для активации T-клеток; в некоторых воплощениях изобретения лекарственное средство представляет собой лекарственное средство для лечения рака, аутоиммунного заболевания или воспалительного заболевания.
В другом аспекте в описании настоящего изобретения предложено применение антитела или его антигенсвязывающего фрагмента, специфично связывающихся с человеческим CD3, одноцепочечного антитела, мультиспецифического антитела, фармацевтической композиции или выделенной молекулы нуклеиновой кислоты, как описано выше, в получении лекарственного средства для активации T-клеток.
В другом аспекте в описании настоящего изобретения предложен способ активации T-клеток, включающий введение субъекту терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента, специфично связывающихся с человеческим CD3, одноцепочечного антитела, мультиспецифического антитела, фармацевтической композиции или выделенной молекулы нуклеиновой кислоты, как описано выше. В некоторых воплощениях изобретения этот способ включает введение субъекту композиции, содержащей единичную дозу 0,1-3000 мг (более предпочтительно 1-1000 мг) мультиспецифического антитела, как описано выше, или фармацевтической композиции, или выделенной молекулы нуклеиновой кислоты, как описано выше.
В другом аспекте в описании настоящего изобретения предложен способ лечения рака, аутоиммунного заболевания или воспалительного заболевания, включающий введение субъекту терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента, специфично связывающихся с человеческим CD3, одноцепочечного антитела, мультиспецифического антитела, фармацевтической композиции или выделенной молекулы нуклеиновой кислоты, как описано выше. В некоторых воплощениях изобретения этот способ включает введение субъекту композиции, содержащей единичную дозу 0,1-3000 мг (более предпочтительно 1-1000 мг) мультиспецифического антитела, как описано выше, или фармацевтической композиции, или выделеннлой молекулы нуклеиновой кислоты, как описано выше.
В другом аспекте в описании настоящего изобретения предложено применение антитела или его антигенсвязывающего фрагмента, специфично связывающихся с человеческим CD3, одноцепочечного антитела, мультиспецифического антитела, фармацевтической композиции или выделенной молекулы нуклеиновой кислоты, как описано выше, в получении лекарственного средства для лечения рака, аутоиммунного заболевания или воспалительного заболевания.
В некоторых воплощениях изобретения описанный выше рак представляет собой любой выбранный из группы, состоящей из карциномы, лимфомы, бластомы, саркомы, лейкоза или лимфоидных злокачественных новообразований. Более конкретные примеры рака включают плоскоклеточный рак, миелому, мелкоклеточный рак легких, немелкоклеточный рак легких (НМКРЛ), плоскоклеточный рак головы и шеи (ПКРГШ), глиому, лимфому Ходжкина, неходжкинскую лимфому, диффузную крупноклеточную B-клеточную лимфому (DLBCL), фолликулярную лимфому, острый лимфобластный лейкоз (ОЛЛ), острый миелоидный лейкоз (ОМЛ), хронический лимфоцитарный лейкоз (ХЛЛ), хронический миелоидный лейкоз (ХМЛ), первичную крупноклеточную B-клеточную лимфому средостения, лимфому мантийных клеток (ЛМК), мелкоклеточную лимфоцитарную лимфому (МЛЛ), обогащенную T-клетками/гистиоцитами крупноклеточную B-клеточную лимфому, множественную миелому, миелоидный лейкоз с экспрессией белка 1 (Mcl-1), миелодиспластический синдром (МДС), рак желудочно-кишечного тракта, рак почки, рак яичника, рак печени, лимфобластный лейкоз, лимфоцитарный лейкоз, колоректальный рак, рак эндометрия, рак почки, рак предстательной железы, рак щитовидной железы, меланому, хондросаркому, нейробластому, рак поджелудочной железы, мультиформную глиобластому, рак желудка, рак кости, саркому Юинга, рак шейки матки, рак головного мозга, рак желудка, рак мочевого пузыря, печеночно-клеточную опухоль, рак молочной железы, рак ободочной кишки, печеночно-клеточный рак (ПКР), светлоклеточную почечно-клеточную карциному (СПК), рак головы и шеи, фаринго-ларингеальный рак, рак печени и желчевыводящих путей, рак центральной нервной системы, рак пищевода, злокачественную плевральную мезотелиому, системный амилоидоз легкой цепи, лимфоплазмацитарную лимфому, миелодиспластический синдром, миелопролиферативную опухоль, нейроэндокринную опухоль, рак из клеток Меркеля, рак яичка и рак кожи. В некоторых воплощениях изобретения рак представляет собой рак из B7-H3-положительных клеток; предпочтительно рак молочной железы, рак яичника, рак предстательной железы, рак поджелудочной железы, рак почки, рак легких, рак печени, рак желудка, рак ободочной кишки, рак мочевого пузыря, рак пищевода, рак шейки матки, рак желчного пузыря, глиобластому или меланому.
В некоторых воплощениях изобретения описанное выше аутоиммунное заболевание или воспалительное заболевание представляет собой любое выбранное из группы, состоящей из ревматоидного артрита, псориаза, болезни Крона, анкилозирующего спондилита, рассеянного склероза, сахарного диабета I типа, гепатита, миокардита, синдрома Шегрена, аутоиммунной гемолитической анемии после отторжения трансплантата, везикулезного пемфигоида, болезни Грейвса, тиреоидита Хасимото, системной красной волчанки (СКВ), миастении гравис, пузырчатки и пернициозной анемии.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг.1A-1B: Фиг.1A представляет собой схематическую диаграмму бивалентного биспецифического антитела, а Фиг.1B представляет собой схематическую диаграмму моновалентного биспецифического антитела.
Фиг.2A-2D: Определение активности связывания антител с клетками при наличии или в отсутствие экспрессии соответствующего антигена методом проточной цитометрии. На Фиг.2A показано определение активности связывания различных антител с клетками A498, экспрессирующими человеческий B7H3; на Фиг.2B показано определение активности связывания различных антител с клетками CT26 со сверхэкспрессией человеческого B7H3; на Фиг.2C показано определение активности связывания различных антител с клетками CT26, которые не экспрессируют человеческий B7H3, и результаты показывают, что ни одно из антител не связывается с клетками CT26, которые не экспрессируют человеческий B7H3; на Фиг.2D показано определение активности связывания различных антител с рекомбинантными клетками линии Jurkat, экспрессирующими CD3. Вертикальная ось на Фиг.2A-2D представляет собой среднее геометрическое сигнала флуоресценции.
Фиг.3A-3B: Определение цитолитической активности биспецифических антител, содержащих различные scFv CD3, для клеток A498. На Фиг.3A показана цитолитическая активность моновалентного биспецифического антитела к B7H3. На Фиг.3B показана цитолитическая активность бивалентного биспецифического антитела к B7H3. Все биспецифические антитела демонстрируют очевидную цитолитическую активность независимо от моновалентности или бивалентности антитела к B7H3, за исключением 155, 156, 185 и 186, которые обладают более слабой цитолитической активностью для клеток A498.
Фиг.4A-4B: Сравнение цитолитической активности для клеток A498 моно- и бивалентных биспецифических антител к B7H3, содержащих один и тот же scFv CD3. На Фиг.4A показано сравнение цитолитической активности моновалентных (181) и бивалентных (131) биспецифических антител к B7H3, содержащих HRH1. На Фиг.4B показано сравнение цитолитической активности моновалентных (187) и бивалентных (177) биспецифических антител к B7H3, содержащих HRH7. Все экспериментальные результаты показывают, что бивалентные биспецифические антитела к B7H3 обладают более очевидной цитолитической активностью для клеток A498 по сравнению с моновалентными биспецифическими антителами к B7H3. В то же время бивалентные биспецифические антитела к B7H3 обладают значимо более выраженной цитолитической активностью, чем моновалентные биспецифические антитела к B7H3.
Фиг.5A-5C: Определение цитолитической активности для клеток A498 бивалентных биспецифических антител к B7H3, содержащих одинаковую вариабельную область тяжелой цепи CD3, но с расположением в другом порядке. На Фиг.5A показано сравнение цитолитической активности бивалентных биспецифических антител, содержащих первую полипептидную цепь, содержащую HRH2, где первая полипептидная цепь располагается в другом порядке (AFF1, AFF2, AFF3, AFF4). На Фиг.5B показано сравнение цитолитической активности бивалентных биспецифических антител, содержащих вторую полипептидную цепь, содержащую HRH2, где вторая полипептидная цепь расположена в другом порядке (AFF3, AFF3-B). Результаты показывают, что все бивалентные биспецифические антитела с одинаковой последовательностью, но с различным порядком расположения VH и VL, обладают значимой цитолитической активностью для клеток A498, и молекулы с расположением в другом порядке обладают одинаковой цитолитической активностью. На Фиг.5C показано сравнение цитолитической активности между биспецифическими антителами, содержащими одинаковые scFv B7H3 и scFv CD3, но с различными структурами. Все три исследуемых биспецифических антитела 127, 201 и 202 обладают способностью к цитолизу опухолевых клеток A498; из них биспецифическое антитело 127 проявляет цитолитическую активность, превосходящую активность 201 и 202.
Фиг.6A-6B: Определение активации рекомбинантных клеток Jurkat различными антителами. На Фиг.6A показана антителоопосредованная B7H3 мишень-специфическая активация рекомбинантных клеток Jurkat в присутствии клеток A498; на Фиг.6B показана антителоопосредованная, неспецифичная к мишени B7H3 активация рекомбинантных клеток Jurkat в отсутствие клеток A498. Для Фиг.6A-6B обозначения антител одинаковы.
Фиг.7A-7B: Определение активации рекомбинантных клеток Jurkat биспецифическими антителами, содержащими одинаковые CD3scFv, но с различными валентностями. На Фиг.7A показана антителоопосредованная B7H3 мишень-специфическая активация рекомбинантных клеток Jurkat в присутствии клеток A498; на Фиг.7B показана антителоопосредованная, неспецифичная к мишени B7H3 активация рекомбинантных клеток Jurkat моно/бивалентными биспецифическими антителами в отсутствие клеток A498.
Фиг.8A-8C: Исследовали стимуляцию мононуклеарных клеток периферической крови (МКПК) на выработку секреции B7H3-мишень-специфических цитокинов различными антителами в присутствии клеток A498. На Фиг.8A показано сравнение уровней секреции IFNγ (интерферон γ) из МКПК, стимулированных различными антителами; на Фиг.8B показано сравнение уровней секреции TNFα (фактор некроза опухоли α) из МКПК, стимулированных различными антителами; и на Фиг 8C показано сравнение уровней секреции IL-2 (иммуноглобулин-2) из МКПК, стимулированных различными антителами. На Фиг.8A-8C показано, что антитела 118, 127 и 132 могут значимо стимулировать МКПК на выработку секреции B7H3-мишень-специфических цитокинов. Для Фиг.8A-8C обозначения антител одинаковы.
Фиг.9A-9C: Исследовали стимуляцию мононуклеарных клеток периферической крови (МКПК) на выработку секреции неспецифичных к мишени B7H3 цитокинов различными антителами в присутствии клеток CHOK1, не экспрессирующих B7H3. На Фиг.9A показано сравнение уровней IFNγ, секретируемого из МКПК, стимулированных различными антителами; на Фиг.9B показано сравнение уровней TNFα, секретируемого из МКПК, стимулированных различными антителами; и на Фиг 9C показано сравнение уровней IL-2, секретируемого из МКПК, стимулированных различными антителами. На Фиг.9A-9C показано, что антитела 118, 127 и 132 могут значимо стимулировать МКПК на выработку секреции неспецифичных к мишени B7H3 цитокинов. Для Фиг.9A-9C обозначения антител одинаковы.
Фиг.10A-10E: Определение противоопухолевой эффективности биспецифических антител в мышиной модели A498, реконструированной с человеческими МКПК. На Фиг.10A показано определение противоопухолевой активности низкой дозы бивалентных биспецифических антител к B7H3. Оба антитела 118 и 119 в низкой дозе все же проявляют некоторую противоопухолевую активность и проявляют некоторую степень зависимости от дозы. На Фиг.10B показано определение противоопухолевой активности бивалентных биспецифических антител к B7H3 в дозах 0,3 мг/кг и 0,6 мг/кг. Антитело 113 проявляет дозозависимую активность ингибирования опухоли in vivo. На Фиг.10C показано определение противоопухолевой активности бивалентных биспецифических антител к B7H3 в дозах 0,12 мг/кг и 0,36 мг/кг. Антитело 118 в обеих дозах проявляет значимую противоопухолевую активность. На Фиг.10D показано определение противоопухолевой активности бивалентных биспецифических антител к B7H3 в дозе 0,36 мг/кг. Все антитела 126, 127 и 128 проявляют значимую противоопухолевую активность. На Фиг.10E показана противоопухолевая активность антитела 127 в различных дозах и при различной периодичности дозирования. На Фиг.10A-10E «раствор-носитель» представляет собой группу отрицательного контроля, которой вводили фосфатно-солевой буферный раствор (ФСБ).
Фиг.11A-11B: Противоопухолевая эффективность биспецифических антител в мышиной модели hCD3 KI. На Фиг.11A и Фиг.11B показано противоопухолевое действие антител 118 и 132 в мышиной модели hCD3 KI соответственно.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Терминология (определения)
Трехбуквенные и однобуквенные коды аминокислот, используемые в настоящем описании, являются такими, как описано в J. Biol. Chem, 243, p3558 (1968).
Термин «мультиспецифическая молекула белка» относится к молекуле белка, способной специфично связываться с двумя или более антигенов-мишеней или эпитопов антигена-мишени. Молекулу белка, которая может специфично связываться с двумя антигенами-мишенями или эпитопами антигена-мишени, называют биспецифической молекулой белка, включая антитела или антигенсвязывающие фрагменты антител (такие как одноцепочечные антитела). Термин «биспецифическая молекула белка» в настоящем документе используется взаимозаменяемо с термином «биспецифическое антитело».
Термин «связывающая область» для антигена относится к области или участку в мультиспецифической молекуле белка или молекуле антитела, которые могут специфично связываться с антигеном. Антигенсвязывающая область может представлять собой лиганд-связывающий домен, который может непосредственно связываться с антигеном, или может представлять собой домен, содержащий вариабельную область антитела, которая может непосредственно связываться с антигеном.
Термин «антитело (Ат)» включает любую антигенсвязывающую молекулу или молекулярный комплекс, который включает в себя по меньшей мере одну область, определяющую комплементарность (CDR), которая специфично связывается или взаимодействует со специфичным антигеном (например, CD3). Термин «антитело» включает в себя: четыре полипептидных цепи, соединенных друг с другом посредством дисульфидной(-ых) связи(-ей), молекулы иммуноглобулинов, содержащие две тяжелых (H) цепи и две легких (L) цепи, и их мультимеры (например, IgM). Каждая тяжелая цепь включает в себя вариабельную область тяжелой цепи (далее сокращенно обозначенную как HCVR или VH) и константную область тяжелой цепи. Эта константная область тяжелой цепи содержит три области (домена): CH1, CH2 и CH3. Каждая легкая цепь включает в себя вариабельную область легкой цепи (далее сокращенно обозначенную как LCVR или VL) и константную область легкой цепи. Константная область легкой цепи содержит одну область (домен, CL1). Области VH и VL можно дополнительно подразделить на гипервариабельные области, называемые областями, определяющими комплементарность (CDR), между которыми распределены более консервативные области, называемые каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенные в следующем порядке от амино-конца к карбокси-концу: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. В различных воплощениях описания настоящего изобретения FR антител к CD3 (или их антигенсвязывающих участков), антител к B7H3 (или их антигенсвязывающих участков) или антител к другим целевым антигенам могут иметь такую же последовательность, как последовательность зародышевой линии человека, или могут быть естественно или искусственно модифицированными. Антитела могут представлять собой антитела различных подклассов, например, антитело IgG (например, подкласса IgG1, IgG2, IgG3 или IgG4), IgA1, IgA2, IgD, IgE или IgM.
Термин «антитело» также охватывает антигенсвязывающие фрагменты или полноразмерные молекулы антитела. Используемый в настоящем документе термин «антигенсвязывающий участок», «антигенсвязывающий домен», «антигенсвязывающий фрагмент» и т.д. антитела включает любой встречающийся в природе, полученный ферментативным путем, синтетический или сконструированный методами генной инженерии полипептид или гликопротеин, который специфично связывается с антигеном с образованием комплекса. Антигенсвязывающие фрагменты антитела могут быть получены, например, из полноразмерной молекулы антитела путем использования любой подходящей стандартной методики, такой как протеолитическое расщепление или рекомбинантные методы генной инженерии, в которых задействованы манипуляции с ДНК, кодирующими вариабельные области и, необязательно, константные области антитела, и ее экспрессия. Эта ДНК известна и/или может быть легко получена, например, из коммерческих источников, баз данных ДНК (включая, например, базы данных фагового дисплея антител), или синтезирована. ДНК можно секвенировать и проводить манипуляции с ней химическим путем или с использованием молекулярной биотехнологии, например, располагая одну или более вариабельных и/или константных областей в подходящей конфигурации, или путем введения кодонов, образующих остатки цистеина, модификации, добавления или делеции аминокислот и т.д.
Не имеющие ограничительного характера примеры антигенсвязывающих фрагментов включают: (i) Fab фрагмент; (ii) F(ab')2 фрагмент; (iii) Fd фрагмент; (iv) Fv фрагмент; (v) одноцепочечную молекулу Fv (scFv); (vi) dAb фрагмент.В используемый в настоящем документе термин также включены другие молекулы, сконструированные методами генной инженерии, такие как область-специфичные антитела, однодоменные антитела, антитела с делетированной областью, химерные антитела, CDR-привитые антитела, диатела, тритела, тетратела, минитела, нанотела (например, моновалентные антитела, бивалентные антитела и т.д.), иммунофармацевтическое средство на основе модульного белка малого размера (SMIP) и вариабельные области IgNAR акулы.
Антигенсвязывающий фрагмент антитела будет, как правило, содержать по меньшей мере одну вариабельную область. Вариабельная область может представлять собой область любого размера или аминокислотного состава и по существу будет содержать один или более CDR, примыкающих к каркасным последовательностям или находящихся внутри них. В антигенсвязывающем фрагменте, имеющим VH область, ассоциированную с VL областью, области VH и VL могут быть расположены друг напротив друга в любом подходящем порядке. Например, вариабельная область может быть димеризована и содержать димеры VH-VL или VL-VH.
В некоторых воплощениях изобретения антигенсвязывающий фрагмент антитела имеет любую конфигурацию вариабельной области и константной области, причем вариабельная область и константная область могут быть соединены непосредственно в любом порядке или могут быть соединены посредством полноразмерной или частичной шарнирной или линкерной области. Шарнирная область может состоять из по меньшей мере 2 (например, 5, 10, 15, 20, 40, 60 или более) аминокислот, так чтобы получить гибкое или полужесткое соединение между примыкающими вариабельными и/или константными областями в одной молекуле полипептида. Кроме того, антигенсвязывающие фрагменты антител по настоящему изобретению могут включать гомодимеры или гетеродимеры (или другие мультимеры) в любой конфигурации вариабельной области и константной области, как показано выше, при этом вариабельная область и константная область могут быть нековалентно соединены друг с другом и/или соединены с одной или более мономерных областей VH или VL (например, посредством дисульфидной(-ых) связи(-ей)).
Используемое в настоящем документе «мышиное антитело» относится к моноклональным антителам мышиного происхождения, полученным в соответствии со знаниями и навыками специалистов в данной области техники. В процессе получения испытуемым субъектам вводят антиген путем инъекции, а затем выделяют гибридому, экспрессирующую антитело, которое обладает требуемыми последовательностями или функциональными характеристиками. Если испытуемые субъекты представляют собой мышей, полученное в результате антитело будет представлять собой мышиное антитело.
«Химерное антитело» представляет собой антитело, полученное путем слияния вариабельной области мышиного антитела вместе с константной областью человеческого антитела, и такое антитело может ослаблять иммунный ответ, вызванный мышиным антителом. Чтобы создать химерное антитело, сначала создают гибридому, секретирующую специфичное мышиное моноклональное антитело, и клонируют ген вариабельной области из мышиной гибридомы. Ген константной области клонируют из человеческого антитела в соответствии с потребностями. Ген мышиной вариабельной области соединяют с геном человеческой константной области с образованием химерного гена, который может быть впоследствии встроен в экспрессионный вектор. Наконец, молекула химерного антитела будет экспрессироваться в эукариотической или прокариотической системе. В предпочтительном воплощении описания настоящего изобретения легкая цепь химерного антитела дополнительно содержит константную область человеческой легкой цепи каппа, лямбда или ее вариант. Тяжелая цепь химерного антитела дополнительно содержит константную область тяжелой цепи человеческого IgG1, IgG2, IgG3, IgG4 или ее вариант, предпочтительно содержит константную область тяжелой цепи человеческого IgG1, IgG2 или IgG4, или содержит вариант константной области тяжелой цепи человеческого IgG1, IgG2 или IgG4 с мутациями аминокислот (такими как мутация YTE или обратная мутация, мутация L234A и/или L235A, либо мутация S228P).
Термин «гуманизированное антитело», включая CDR-привитое антитело, относится к антителу, полученному путем прививания последовательностей CDR антитела животного происхождения, например мышиного антитела, в каркасные участки вариабельной области человеческого антитела (т.е. каркасные области). Гуманизированные антитела позволяют преодолевать гетерологичные ответы, индуцируемые химерными антителами, несущими большое количество компонентов гетерологичных белков. Такие каркасные последовательности могут быть получены из общедоступной базы данных ДНК, охватывающей последовательности генов антител зародышевой линии, или из опубликованных источников. Например, последовательности ДНК человеческих генов вариабельной области тяжелой и легкой цепи зародышевой линии можно найти в базе данных последовательностей человеческой зародышевой линии VBase (доступна на сайте http://www.vbase2.org/), а также в работе Kabat, EA, et al. 1991 Sequences of Proteins of Immunological Interest, 5th Ed. Во избежание снижения активности, вызванного снижением иммуногенности, каркасные последовательности вариабельной области человеческого антитела можно подвергать минимальным реверсивным или обратным мутациям для сохранения его активности. Гуманизированное антитело по описанию настоящего изобретения также включает гуманизированное антитело, на котором созревание аффинности CDR выполняют методом фагового дисплея.
В связи с остатками, контактирующими с антигеном, прививание CDR может привести в результате к сниженной аффинности антитела или его антигенсвязывающего фрагмента к антигену из-за остатков каркасной области, контактирующих с антигеном. Такие взаимодействия могут возникать в результате большого числа соматических мутаций. Поэтому все же может быть необходимым прививать аминокислоты донорского каркаса на каркас гуманизированного антитела. Аминокислотные остатки, вовлеченные в связывание антигена и полученные из нечеловеческого антитела или его антигенсвязывающего фрагмента, можно идентифицировать путем проверки последовательности и структуры вариабельной области моноклонального антитела животного. Аминокислотные остатки каркаса донорского CDR, отличающиеся от зародышевых линий, можно рассматривать как родственные. Если невозможно определить наиболее близкородственную зародышевую линию, последовательность можно сравнивать с консенсус-последовательностью, общей для последовательностей подтипов антитела животного, с высоким процентом подобия. Редкие каркасные остатки считаются результатом высокой частоты мутаций в соматических клетках и играют важную роль в связывании.
В одном воплощении настоящего изобретения антитело или его антигенсвязывающий фрагмент могут дополнительно содержать константную область легкой цепи человеческой или мышиной цепи κ, λ или ее вариант, либо дополнительно содержать каркасную область тяжелой цепи человеческого или мышиного IgG1, IgG2, IgG3, IgG4 или ее вариант.
«Человеческое антитело» и «антитело человеческого происхождения» могут использоваться взаимозаменяемо и могут представлять собой антитела, имеющие происхождение от человека, или антитела, полученные из генетически модифицированного организма, который «сконструирован» или получен любым способом, известным в данной области техники для получения специфичных человеческих антител в ответ на стимуляцию антигеном. В таких методиках в клеточные линии организмов, полученных из эмбриональных стволовых клеточных линий, вводят элементы локусов человеческой тяжелой и легкой цепи, которые нацеливаются на эндогенные локусы тяжелой и легкой цепи и разрушают их. Нацеливаемые эндогенные локусы тяжелой и легкой цепи, включенные в эти клеточные линии, разрушаются. Трансгенные организмы могут синтезировать человеческие антитела, специфичные к человеческим антигенам, и можно использовать эти организмы для получения гибридом, секретирующих человеческие антитела. Человеческое антитело может также представлять собой такое антитело, в котором тяжелая и легкая цепи кодируются нуклеотидными последовательностями, имеющими происхождение от одного или более источников человеческой ДНК. Полностью человеческие антитела можно также конструировать с помощью методов трансфекции генов или хромосом и технологии фагового дисплея, либо конструировать из активированных in vitro B-клеток; все эти методы известны в данной области техники.
«Моноклональное антитело» относится к антителу, полученному из по существу однородной популяции антител, то есть отдельные антитела, составляющие эту популяцию, являются идентичными и/или связываются с одним и тем же эпитопом, за исключением возможных вариантов антител (например, вариантов, содержащих мутации, возникающие естественным путем, или мутации, полученные в ходе производства препаратов антител, и эти мутации обычно присутствуют в минимальных количествах). В отличие от препаратов поликлональных антител, которые обычно содержат различные антитела, направленные на различные детерминанты (эпитопы), каждое моноклональное антитело препарата (лекарственной формы) моноклональных антител направлено на единственную детерминанту антигена. Таким образом, определение «моноклональный» указывает на характеристики антитела, полученного из по существу однородной популяции антител, и не должно рассматриваться как требующее какого-либо определенного метода для производства антитела. Например, моноклональные антитела, используемые в соответствии с настоящим изобретением, могут быть получены с помощью различных методик, включающих без ограничений гибридомные методы, методы рекомбинантных ДНК, методы фагового дисплея и методы использования трансгенных животных, полностью или частично содержащих локусы человеческого иммуноглобулина. Такие методы и другие иллюстративные методы получения моноклональных антител описаны в настоящем документе.
В настоящем документе термины «полноразмерное антитело», «полное антитело», «целое антитело» и «полноценное антитело» используются взаимозаменяемо и относятся к антителу в по существу полноценной форме в отличие от описанных ниже антигенсвязывающих фрагментов. Этот термин конкретно относится к антителу, тяжелая цепь которого содержит Fc область.
Кроме того, домен VL и домен VH фрагмента Fv кодируются двумя отдельными генами, тем не менее они могут быть связаны посредством синтетического линкера с использованием рекомбинантных методов с образованием единой белковой цепи, в которой моновалентная молекула образуется в результате спаривания доменов VL и VH (называется одноцепочечным Fv (scFv); см., например, Bird et al. (1988): 423-426; Science 242, и Huston et al. (1988) Proc. Natl. Acad. Sci USA85:5879-5883). Подразумевают, что такие одноцепочечные антитела также включены в термин «антигенсвязывающий фрагмент» антитела. Такие фрагменты антител получают с использованием общепринятых методик, известных в данной области техники, и подвергают скринингу на функциональные фрагменты, используя такой же метод, как для интактного антитела. Антигенсвязывающие участки могут быть получены с помощью технологии рекомбинантных ДНК, либо путем ферментативного или химического разрушения интактного иммуноглобулина.
Антигенсвязывающие фрагменты могут быть также включены в одноцепочечную молекулу, содержащую пару тандемных Fv фрагментов (VH-CH1-VH-CH1), и эта пара тандемных Fv фрагментов образует пару антигенсвязывающих областей вместе с комплементарными полипептидами легкой цепи (Zapata et al., 1995 Protein Eng. 8(10): 1057-1062; и патент США №5,641,870).
Fab представляет собой фрагмент антитела, полученный путем обработки молекулы антитела IgG папаином, который расщепляет ее в аминокислотным остатке 224 H-цепи), и этот фрагмент антитела, в котором около половины N-концевой стороны H-цепи и полноразмерная L-цепь связаны вместе посредством дисульфидной (-ых) связи (-ей), имеет молекулярную массу около 50 ООО Да и обладает антигенсвязывающей активностью.
F(ab')2 представляет собой фрагмент антитела, имеющий молекулярную массу около 100 ООО Да и обладающий антигенсвязывающей активностью, а также содержащий две Fab-области, которые связаны в положении шарнирной области; он может быть получен путем отщепления пепсином части вниз от двух дисульфидных связей шарнирной области IgG.
Fab' представляет собой фрагмент антитела, имеющий молекулярную массу около 50 ООО Да и обладающий антигенсвязывающей активностью, который получают путем расщепления дисульфидных связей в шарнирной области указанного выше F(ab')2. Fab' может быть получен путем обработки F(ab')2, который специфично распознает антиген и связывается с ним, восстанавливающим агентом, таким как дитиотрейтол.
Дополнительно Fab’ может быть получен путем встраивания ДНК, кодирующей Fab’ антитела, в прокариотический экспрессионный вектор или эукариотический экспрессионный вектор и введения этого вектора в прокариоту или эукариоту для экспрессии Fab’.
Термин «одноцепочечное антитело», «одноцепочечный Fv» или «scFv» относится к молекуле, содержащей вариабельный домен тяжелой цепи антитела (или область VH), соединенный с вариабельным доменом легкой цепи антитела (или область VL) посредством линкера. Такие молекулы scFv имеют общую структуру NH2-VL-линкер-VH-COOH или NH2-VH-линкер-VL-COOH. Подходящие линкеры предшествующего уровня техники состоят из повторяющейся аминокислотной последовательности GGGGS или ее варианта, например варианта с 1-4 (включая 1, 2, 3 или 4) повторами (Holliger et al. (1993), Proc Natl Acad Sci USA, 90: 6444-6448). Другие линкеры, полезные для описания настоящего изобретения, описаны Alfthan et al. (1995), Protein Eng. 8:725-731, Choi et al. (2001), Eur J Immuno.31:94-106, Hu et al. (1996), Cancer Res.56:3055-3061, Kipriyanov et al. (1999), J Mol Biol. 293:41-56 и Roovers et al. (2001), Cancer Immunol Immunother. 50:51-59.
«Мультиспецифическое антитело» относится к антителу, которое содержит два или более антигенсвязывающих домена и способно к связыванию двух или более различных эпитопов (например, двух, трех, четырех или более различных эпитопов), и эпитоп может присутствовать в одном и том же или в разных антигенах. Примеры мультиспецифических антител включают «биспецифические антитела», которые связываются с двумя различными эпитопами.
Термин «бивалентное биспецифическое антитело» к опухолеассоциированному антигену относится к биспецифическому антителу, в котором на опухолеассоциированный антиген-мишень направлены две антигенсвязывающие области. Например, бивалентное биспецифическое антитело к B7H3 относится к биспецифическому антителу, содержащему две антигенсвязывающие области, нацеленные на B7H3. Термин «моновалентное биспецифическое антитело» относится к биспецифическому антителу, в котором только одна антигенсвязывающая область направлена на определенную мишень. Например, моновалентное биспецифическое антитело к B7H3 относится к биспецифическому антителу, содержащему одну антигенсвязывающую область, нацеленную на B7H3.
«Линкер» или «линкерный фрагмент» относится к L1, локализованному между двумя доменами белка, для соединения двух доменов, а также относится к соединительной пептидной последовательности, используемой для соединения двух доменов белка. Обычно он обладает некоторой степенью гибкости, и использование линкеров не вызовет утрату исходных функций домена белка.
Диатело представляет собой фрагмент антитела, в котором scFv димеризован, и представляет собой фрагмент антитела, обладающий бивалентной антигенсвязывающей активностью. При бивалентной антигенсвязывающей активности два антигена могут быть одинаковыми или разными.
dsFv получают путем замены одного аминокислотного остатка в каждой из VH и VL остатком цистеина, а затем соединения замещенных полипептидов посредством дисульфидной связи между двумя остатками цистеина. Аминокислотные остатки, подлежащие замене остатком цистеина, могут быть выбраны на основании прогнозирования трехмерной структуры антитела в соответствии с известными способами (Protein Engineering, 7, 697 (1994)).
В некоторых воплощениях описания настоящего изобретения антигенсвязывающий фрагмент может быть получен путем следующих стадий: получения кДНК, кодирующей VH и/или VL моноклонального антитела по описанию настоящего изобретения, которое специфично распознает антиген и связывается с ним, и кДНК, кодирующей по мере необходимости другие домены; конструирование ДНК, кодирующей антигенсвязывающий фрагмент; встраивание ДНК в прокариотический или эукариотический экспрессионный вектор с последующим встраиванием этого экспрессионного вектора в прокариоту или эукариоту для экспрессии антигенсвязывающего фрагмента.
«Fc область» может представлять собой природную последовательность или вариант Fc области. Границы Fc области тяжелой цепи иммуноглобулина изменчивы; тем не менее Fc область тяжелой цепи человеческого иммуноглобулина IgG обычно определяют как область, проходящую от аминокислотного остатка в положении Cys226 или от Pro230 до карбокси-конца. Нумерация остатков в области Fc соответствует индексу нумерации ЕС по Кэботу, Kabat et al., Sequences of Proteins of Immunological Interest, 5th Edition Public Health Service, National Institutes of Health, Bethesda, Md., 1991. Область Fc иммуноглобулина обычно имеет два константных домена CH2 и CH3. В данном случае «первую Fc» также называют Fc1, а вторую Fc также называют Fc2.
В Va1-L1-Vb1-L2-Vc2-L2-Vd2-L4-Fc1 и Ve3-L5-Vf3-L6-Fc2 Va1, Vb1, Vc2, Vd2, Ve3 и Vf3 представляют собой вариабельную область легкой цепи или вариабельную область тяжелой цепи антитела, при этом Va1 и Vb1 связываются с первым эпитопом антигена, Vc2 и Vd2 связываются со вторым эпитопом антигена, а Ve3 и Vf3 связываются с третьим эпитопом. Первый эпитоп, второй эпитоп и третий эпитоп могут быть одинаковыми или неодинаковыми.
Как и VHTAA-L1-VLTAA-L2-VHCD3-L3-VLCD3-L4-Fc1, VHTAA и VLTAA представляют собой вариабельную область антитела, связывающуюся с эпитопом опухолеассоциированного антигена, а VHCD3 и VLCD3 представляют собой вариабельную область антитела, связывающуюся с эпитопом CD3.
В описании настоящего изобретения «knob-Fc» относится к подобной выступу пространственной структуре, образованной в результате включения точечной мутации T366W в область Fc антитела. Соответственно, «hole-Fc» относится к подобной впадине пространственной структуре, образованной в результате включения точечных мутаций T366S, L368A и Y407V в область Fc антитела. Knob-Fc и hole-Fc с наибольшей вероятностью образуют гетеродимеры за счет стерического затруднения. Чтобы дополнительно стимулировать образование гетеродимеров, в участки knob-Fc и hole-Fc соответственно можно ввести точечные мутации S354C и Y349C, которые дополнительно стимулируют образование гетеродимеров посредством дисульфидных связей. В то же время для устранения или ослабления влияния АЗКЦ (антителозавсисимая клеточная цитотоксичность), вызванного антителом Fc, в Fc можно также ввести мутации типа замен 234A и 235A. Предпочтительные knob-Fc и hole-Fc по описанию настоящего изобретения показаны, например, в SEQ ID NO: 69 и 70 соответственно. В биспецифическом антителе knob-Fc или hole-Fc можно использовать либо в качестве Fc области первой полипептидной цепи, либо в качестве Fc области второй полипептидной цепи. В одном биспецифическом антителе Fc области первой и второй полипептидной цепи могут не представлять собой обе knob-Fc или hole-Fc.
Термин «различие аминокислот» или «мутация аминокислоты» относится к изменениям или мутациям аминокислот в варианте белка или полипептида по сравнению с исходным белком или полипептидом, в которых задействованы инсерция, делеция или замена одной или более аминокислот на основе исходного белка или полипептида.
«Вариабельная область» антитела относится к вариабельной области легкой цепи антитела (VL) или к вариабельной области тяжелой цепи антитела (VH), отдельно или в комбинации. Как известно в данной области техники, каждая из вариабельных областей тяжелой и легкой цепи состоит из трех областей, определяющих комплементарность (CDR) (также называемых гипервариабельными областями), соединенных с четырьмя каркасными областями (FR). CDR в каждой цепи прочно удерживаются вместе посредством FR и участвуют в формировании антигенсвязывающего участка антитела вместе с CDR от другой цепи. Существует по меньшей мере две методики определения CDR: (1) метод, основанный на межвидовой вариабельности последовательностей (т.е. Kabat et al. Sequences of Proteins of Immunological Interest, (5th edition, 1991, National Institutes of Health, Bethesda MD)); и (2) метод, основанный на кристаллографических исследованиях комплексов антиген-антитело (Al-Lazikani et al., J. Molec. Biol. 273:927-948 (1997)). Используемые в настоящем документе CDR могут относиться к тем, которые определены любым из этих методов или комбинацией двух методов.
Термин «каркасная область антитела» или «FR область» относится к участку вариабельного домена, либо VL, либо VH, который выступает в качестве стержневой структуры для антигенсвязывающих петель (CDR) этого вариабельного домена. По существу она представляет собой вариабельный домен без CDR.
Термин «CDR» относится к одной из шести гипервариабельных областей, присутствующих в вариабельном домене антитела, которые вносят основной вклад в связывание антигена. Одно из наиболее часто используемых определений 6 CDR предложено Kabat E.A. et al. ((1991) Sequences of Proteins of Immunological Interest. NIH Publication 91-3242). При использовании в некоторых воплощениях в настоящем документе CDR могут быть определены в соответствии с критериями Кэбота (Kabat et al. Sequences of Proteins of Immunological Interest (5th edition, 1991, National Institutes of Health, Bethesda MD)) для определения CDR1, CDR2 и CDR3 вариабельных доменов легких цепей (LCDR1, LCDR2 и LCDR3) и CDR1, CDR2 и CDR3 вариабельных доменов тяжелых цепей (HCDR1, HCDR2 и HCDR3), например, для определения CDR антитела к CD3 по описанию настоящего изобретения. В других воплощениях изобретения CDR могут быть также определены в соответствии с критериями базы данных иммуногенетики IMGT и т.п. Например, CDR антитела B7H3 определяют в соответствии с критериями IMGT.
Термин «опухолевый антиген» относится к веществу, вырабатываемому опухолевыми клетками, необязательно представляющему собой белок, включая «опухолеассоциированный антиген» или TAA (который относится к белку, вырабатываемому опухолевыми клетками и дифференциально экспрессирующемуся в злокачественных новообразованиях по сравнению с соответствующими нормальными тканями) и «опухолеспецифический антиген» или TSA (который относится к опухолевому антигену, вырабатываемому опухолевыми клетками и специфично экспрессирующемуся или аномально экспрессирующемуся в злокачественных новообразованиях по сравнению с соответствующими нормальными тканями).
Не имеющие ограничительного характера примеры «опухолеассоциированного антигена» включают, например, AFP, ALK, B7H3, белок BAGE, BCMA, BIRC5 (сурвивин), BIRC7, β-катенин, brc-ab1, BRCA1, BORIS, CA9, CA125, карбоангидразу IX, каспазу-8, CALR, CCR5, CD19, CD20(MS4A1), CD22, CD30, CD33, CD38, CD40, CD123, CD133, CD138, CDK4, CEA, клаудин 18.2, циклин-B1, CYP1B1, EGFR, EGFRvIII, ErbB2/Her2, ErbB3, ErbB4, ETV6-AML, EpCAM, EphA2, Fra-1, FOLR1, белок GAGE (такой как GAGE-1, -2), GD2, GD3, GloboH, глипикан-3, GM3, gp100, Her2, HLA/B-raf, HLA/k-ras, HLA/MAGE-A3, hTERT, IL13Rα2, LMP2, κ-Light, LeY, белок MAGE (такой как MAGE-1, -2, -3, -4, -6 и -12), MART-1, мезотелин, ML-IAP, MOv-γ, Muc1, Muc2, Muc3, Muc4, Muc5, Muc16(CA-125, MUM1, NA17, NKG2D, NY-BR1, NY-BR62, NY-BR85, NY-ESO1, OX40, p15, p53, PAP, PAX3, PAX5, PCTA-1, PLAC1, PRLR, PRAME, PSMA (FOLH1), белка RAGE, Ras, RGS5, Rho, ROR1, SART-1, SART-3, STEAP1, STEAP2, TAG-72, TGF-β, TMPRSS2, антиген Thompson-nouvelle (Tn), TRP-1, TRP-2, тирозиназу, уроплакин-3 и 5T4 (трофобластный гликопротеин).
«CD3» относится к антигену, экспрессирующемуся на T-клетках в составе мультимолекулярного T-клеточного рецептора (TCR) и представляет собой гомодимер или гетеродимер, образованный двумя из следующих четырех цепей рецептора: CD3-ε, CD3-δ, CD3-ζ и CD3-γ. Человеческий CD3-ε (hCD3ε) содержит аминокислотную последовательность, описанную в базе данных UniProtKB/Swiss-Prot: P07766.2. Человеческий CD3-δ (hCD3δ) содержит аминокислотную последовательность, описанную в базе данных UniProtKB/Swiss-Prot: P04234.1. Таким образом, термин «CD3» относится к человеческому CD3, если специально не указано, что он имеет происхождение от вида, отличающегося от человека, например, «мышиный CD3», «обезьяний CD3» и т.д.
«Эпитоп» или «антиегнная детерминанта» относится к участку на антигене, с которым специфично связывается иммуноглобулин или антитело. Эпитопы обычно включают в себя по меньшей мере 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 последовательных или непоследовательных аминокислот в уникальной пространственной конформации. См, например, протоколы картирования эпитопов в кн. Methods in Molecular Biology, Vol.66, G.E. Morris, Ed. (1996).
Термин «специфично связывается с», «селективно связывается с», «селективное связывание» или «специфичное связывание» относится к связыванию антитела с предопределенным эпитопом или антигеном. Как правило, антитело связывается с аффинностью (константой диссоциации, KD) менее чем около 10-8 M, например, менее чем около 10-9 M, 10-10 M или 10-11 M или даже менее.
Термин «аффинность» относится к силе взаимодействия между антителом и антигеном или отдельным эпитопом. В пределах каждого антигенного участка вариабельная область «плеча» антитела взаимодействует с антигеном в нескольких аминокислотных участках посредством слабых нековалентных сил; чем больше это взаимодействие, тем сильнее аффинность. При использовании в настоящем документе термин «высокая аффинность» антитела или его антигенсвязывающего фрагмента (например, Fab фрагмента) по существу относится к антителу или антигенсвязывающему фрагменту с KD 1E-9 M или менее (например, KD 1E-10M или менее, KD 1E-11 M или менее, KD 1E-12 M или менее, KD 1E-13 M или менее, KD 1E-14 M или менее и т.д.).
Термин «KD» или «KD» относится к константе равновесной диссоциации для конкретного взаимодействия антитело-антиген. Как правило, антитело связывается с антигеном с константой равновесной диссоциации (KD) менее чем около 1E-8 M, например, менее чем около 1E-9 M, 1E-10 M или 1E-11 M или даже менее, определенной, например, с помощью технологии поверхностного плазмонного резонанса (SPR) на приборе Biacore. Чем меньше значение KD, тем больше аффинность.
Термин «молекула нуклеиновой кислоты» относится к молекулам ДНК и к молекулам РНК. Молекула нуклеиновой кислоты может быть однонитевой или двунитевой, но предпочтительно представляет собой двунитевую ДНК. Нуклеиновая кислота «функционально связана», если она находится в функциональной взаимосвязи с другой нуклеиново-кислотной последовательностью. Например, промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию этой последовательности.
Термин «вектор» означает конструкцию, способную доставлять один или более целевых генов или последовательностей и предпочтительно экспрессировать их в клетке-хозяине. Примеры векторов включают без ограничений вирусные векторы, депротеинизированные экспрессионные ДНК- или РНК-векторы, плазмиды, космиды или фаговые векторы, экспрессионные ДНК- или РНК-векторы, ассоциированные с катионными коагулянтами, экспрессионные ДНК- или РНК-векторы, инкапсулированные в липосомы, и некоторые эукариотические клетки, такие как клетки-продуценты.
Методы получения и очистки антител и антигенсвязывающих фрагментов хорошо известны в данной области техники, например, Antibodies: A Laboratory Manual, Cold Spring Harbor, New York, главы 5-8 и 15. Например, можно иммунизировать мышей антигеном или его фрагментом, а затем полученные в результате антитела можно ренатурировать, очищать и секвенировать для определения аминокислотных последовательностей, используя общепринятые методы, хорошо известные в данной области техники. Антигенсвязывающие фрагменты также можно получать общепринятыми методами. Антитела или антигенсвязывающие фрагменты по описанию настоящего изобретения конструируют методами генной инженерии так, что в них одна или более человеческих каркасных областей встроена на CDR области, выделенные из не человеческого антитела. Человеческие последовательности FR зародышевой линии можно получить на вебсайте http://imgt.cines.fr или из кн. The Immunoglobulin Facts Book, 2001, ISBN 012441351 путем их выравнивания в сравнении с базой данных генов вариабельных областей человеческих антител зародышевой линии IMGT с помощью программного обеспечения MOE.
Термин «клетка-хозяин» относится к клетке, в которую вводят экспрессионный вектор. Клетки-хозяева могут включать клетки бактерий, микроорганизмов, растений или животных. Бактерии, которые легко трансформируются, включают представителей семейства Enterobacteriaceae, таких как штаммы Escherichia coli или Salmonella; Bacillaceae, таких как Bacillus subtilis; Pneumococcus; Streptococcus и Haemophilus influenzae. Подходящие микроорганизмы включают Saccharomyces cerevisiae и Pichia pastoris. Подходящие линии клеток-хозяев животных включают CHO (линия клеток яичника китайского хомячка), клетки HEK293 (не имеющие ограничительного характера примеры, такие как клетки HEK293E) и клетки NS0.
Сконструированные антитела или антигенсвязывающие фрагменты могут быть получены и очищены общепринятыми методами. Например, последовательности кДНК, кодирующие тяжелую и легкую цепи, можно клонировать и рекомбинировать с экспрессионным вектором GS. Рекомбинантным экспрессионным вектором иммуноглобулина можно стабильно трансфицировать клетки CHO. В качестве альтернативы предшествующему уровню техники экспрессионные системы млекопитающих могут приводить к гликозилированию антител, особенно в высоко консервативных N-концевых участках Fc области. Стабильные клоны были получены путем экспрессии антитела, специфично связывающегося с антителом. Положительные клоны можно размножать в питательной среде без сыворотки в биореакторах для продукции антител. Питательную среду, в которую секретируется антитело, можно очищать с помощью общепринятых методик. Например, очистку можно выполнять на колонке с сефарозой FF с белком A или с белком G. Компоненты неспецифического связывания вымывают. Связанное антитело элюируют с помощью градиента pH и определяют фрагменты антитела методом электрофореза в полиакриламидном геле с додецилсульфатом натрия (ДСН-ПААГ-электрофореза), а затем объединяют. Антитела можно фильтровать и концентрировать, используя обычные методики. Растворимые смеси и мультимеры можно эффективно удалять с помощью обычных методик, например эксклюзионной или ионообменной хроматографии. Полученный в результате продукт необходимо сразу замораживать, например, при минус 70°C, или лиофилизировать.
«Введение» или «лечение» применительно к животному, человеку, экспериментальному субъекту, клетке, ткани, органу или биологической жидкости относится к приведению в контакт экзогенного фармацевтического, терапевтического, диагностического агента или композиции с животным, человеком, экспериментальным субъектом, клеткой, тканью, органом или биологической жидкостью. «Введение» и «лечение» могут относиться, например, к терапевтическим, фармакокинетическим, диагностическим, научно-исследовательским и экспериментальным методам. Обработка клетки охватывает приведение в контакт реагента с клеткой, а также приведение в контакт реагента с жидкостью, которая находится в контакте с клеткой. «Введение» или «лечение» также означают обработки in vitro или ex vivo, например, клетки, реагентом, диагностическим, связывающим соединением или другой клеткой. «Лечение» применительно к человеку, ветеринарному или исследуемому субъекту относится к терапевтическому лечению, профилактическим или превентивным мерам, научно-исследовательским и диагностическим видам применения.
«Лечить» означает вводить терапевтический агент, такой как композиция, содержащая любое из соединений по описанию настоящего изобретения, внутрь или снаружи, пациенту, имеющему один или более симптомов, в отношении которых этот агент обладает известной терапевтической активностью. Как правило, этот агент вводят в количестве, эффективном для ослабления одного или более симптомов заболевания у пациента или в популяции, подлежащих лечению, путем индукции регресса или ингибирования прогрессирования такого(-их) симптома(-ов) до любой клинически измеримой степени. Количество терапевтического агента, эффективное для ослабления какого-либо конкретного симптома заболевания (также называемое «терапевтически эффективным количеством») может изменяться в соответствии с различными факторами, такими как болезненное состояние, возраст и масса тела пациента, и способностью лекарственного препарата вызывать желаемый ответ у пациента. Оценивать, ослаблен ли симптом заболевания, можно с помощью любого клинического измерения, обычно используемого врачами или другими квалифицированными медицинскими работниками для оценки степени тяжести или статуса прогрессирования симптома. Хотя воплощение описания настоящего изобретения (например, способ лечения или изделие производства) может быть эффективным в ослаблении целевого(-ых) симптома(-ов) заболевания не у каждого пациента, оно должно ослаблять целевой(-ые) симптом(-ы) заболевания у статистически значимого количества пациентов, определяемого с помощью любого известного в данной области техники статистического критерия, такого как t-критерий Стьюдента, критерий хи-квадрат, U-критерий Манна и Уитни, критерий Краскела-Уоллиса (H-критерий), критерий Джонкхира-Терпстра и критерий Уилкоксона.
«Консервативная модификация аминокислоты» или «консервативная замена аминокислоты» означает, что аминокислоты в белке заменены другими аминокислотами с подобными характеристиками (такими как заряд, размер боковой цепи, гидрофобность/гидрофильность, конформация и жесткость структуры и т.д.) так, чтобы эти изменения можно было вводить часто без изменения биологической активности или других необходимых характеристик (таких как аффинность и/или специфичность к антигену) белка или полипептида. Специалистам в данной области техники понятно, что, как правило, замена одной аминокислоты в несущественных областях полипептида не приводит к существенному изменению биологической активности (см., например, Watson et al. (1987) Molecular Biology of the Gene, The Benjamin/Cummings Pub. Co., p.224 (4th Ed.)). Кроме того, замены структурно или функционально подобными аминокислотами с меньшей вероятностью нарушают биологическую активность. Примеры консервативных замен представлены в таблице ниже, «Иллюстративные консервативные замены аминокислот».
Иллюстративные консервативные замены аминокислот
«Эффективное количество» или «эффективная доза» относится к количеству лекарственного средства, соединения или фармацевтической композиции, необходимому для получения любого одного или более полезных или желательных результатов. Для профилактического применения полезные или желательные результаты включают устранение или снижение риска, уменьшение тяжести или задержку начала заболевания, включая биохимические, гистологические и поведенческие манифестации состояния, его осложнения и промежуточные патологические фенотипы в процессе развития состояния. Для терапевтического применения полезные или желательные результаты включают клинические результаты, такие как снижение частоты новых случаев различных состояний, связанных с целевым антигеном по описанию настоящего изобретения, или улучшение одного или более симптомов состояния, уменьшение дозировки других агентов, необходимых для лечения состояния, усиление эффективности другого агента и/или задержку прогрессирования состояния, связанного с целевым антигеном по описанию настоящего изобретения у пациентов.
«Экзогенный» относится к веществам, полученным вне организмов, клеток или людей в зависимости от обстоятельств. «Эндогенный» относится к веществам, полученным в клетках, организмах или в человеческом организме в зависимости от обстоятельств.
В настоящем документе «гомология» и «идентичность» взаимозаменяемы и относятся к подобию последовательностей между двумя полинуклеотидными последовательностями или между двумя полипептидными последовательностями. Если положение в обоих из двух сравниваемых последовательностей занято одним и тем же азотистым основанием или мономерной аминокислотной субъединицей, например, если положение в каждой из двух молекул ДНК занято аденином, то указанные молекулы гомологичны в этом положении. Процент гомологии между двумя последовательностями представляет собой функцию результата деления числа совпадений или гомологичных положений, общих для двух последовательностей, на число сравниваемых положений, умноженного на 100. Например, если при оптимальном выравнивании двух последовательностей 6 из 10 положений в двух последовательностях совпадают или гомологичны, то эти две последовательности гомологичны на 60%, если 95 из 100 положений в двух последовательностях совпадают или гомологичны, то эти две последовательности гомологичны на 95%. Как правило, при выравнивании двух последовательностей проводят сравнение до получения максимального процента гомологии. Например, сравнение можно выполнять в соответствии с алгоритмом BLAST, в котором параметры алгоритма выбраны так, чтобы получить максимальное совпадение между каждой последовательностью по всей длине каждой сравниваемой последовательности.
Приведенные ниже ссылки относятся к алгоритму BLAST, часто используемому для анализа последовательностей: Алгоритм BLAST (BLAST ALGORITHMS): Altschul, SF et al., (1990) J. Mol. Biol. 215:403-410; Gish, W. et al., (1993) Nature Genet. 3:266-272; Madden, TL et al., (1996) Meth. Enzymol. 266:131-141; Altschul, SF et al., (1997) Nucleic Acids Res. 25:3389-3402; Zhang, J. et al. (1997) Genome Res. 7:649-656. Другие алгоритмы BLAST, например, доступные на сайте Национального центра биотехнологической информации (NCBI) BLAST, также хорошо известны специалистам в данной области техники.
«Выделенный» относится к очищенному состоянию, в котором обозначенная молекула по существу не содержит других биологических молекул, таких как нуклеиновые кислоты, белки, липиды, углеводы или другие материалы, такие как клеточный дебрис и ростовая среда. Как правило, термин «Выделенный» не подразумевает полное отсутствие этих материалов или отсутствие воды, буферных растворов или солей, если они не присутствуют в количестве, которое значимо влияет на экспериментальное или терапевтическое применение соединения, как описано в настоящем документе.
«Необязательный» или «необязательно» означает, что событие или обстоятельство, которое следует за этим, может произойти, но необязательно произойдет, и описание включает случаи, в которых это событие или обстоятельство происходит или не происходит. Например, «необязательно содержит 1-3 вариабельных области тяжелой цепи антитела» означает, что вариабельная область с указанной последовательностью может присутствовать, но необязательно присутствует.
«Фармацевтическая композиция» относится к смеси, содержащей одно или более соединений в соответствии с описанием настоящего изобретения или их физиологически приемлемую соль или пролекарство и другие химические компоненты, такие как физиологически/фармацевтически приемлемые носители и эксципиенты. Фармацевтическая композиция нацелена на то, чтобы способствовать введению в организм, облегчить всасывание активного ингредиента и тем самым вызвать биологический эффект.
Термин «фармацевтически приемлемый носитель» относится к любому неактивному веществу, подходящему для применения в лекарственной форме для доставки антител или антигенсвязывающих фрагментов. Носитель может представлять собой антиадгезионный агент, адгезионный агент, покрывающий агент, разрыхлитель, наполнитель или разбавитель, консервант (такой как антиоксидант, антибактериальный или противогрибковый агент), подсластитель, агент, замедляющий всасывание, смачивающий агент, эмульгатор, буферный раствор и т.п. Примеры подходящих фармацевтически приемлемых носителей включают воду, этанол, полиолы (такие как глицерин, пропиленгликоль, полиэтиленгликоль и т.п.), декстрозу, растительное масло (такое как оливковое масло), физиологический раствор, буферный раствор, буферный физиологический раствор и изотонический агент, такой как сахара, полиолы, сорбит и хлорид натрия.
Термин «рак», «раковый» или «злокачественный» относится к физиологическому состоянию у млекопитающих или описывает его, которое по существу характеризуется нерегулируемым ростом клеток. Примеры рака включают без ограничений карциному, лимфому, бластому, саркому и лейкоз или лимфоидные злокачественные новообразования. Более конкретные примеры рака включают плоскоклеточный рак, миелому, мелкоклеточный рак легких, немелкоклеточный рак легких (НМКРЛ), плоскоклеточный рак головы и шеи (ПКРГШ), глиому, лимфому Ходжкина, неходжкинскую лимфому, диффузную крупноклеточную B-клеточную лимфому (DLBCL), фолликулярную лимфому, острый лимфобластный лейкоз (ОЛЛ), острый миелоидный лейкоз (ОМЛ), хронический лимфоцитарный лейкоз (ХЛЛ), хронический миелоидный лейкоз (ХМЛ), первичную крупноклеточную B-клеточную лимфому средостения, лимфому мантийных клеток (ЛМК), мелкоклеточную лимфоцитарную лимфому (МЛЛ), обогащенную T-клетками/гистиоцитами крупноклеточную B-клеточную лимфому, множественную миелому, миелоидный лейкоз с экспрессией белка 1 (Mcl-1), миелодиспластический синдром (МДС), рак желудочно-кишечного тракта, рак почки, рак яичника, рак печени, лимфобластный лейкоз, лимфоцитарный лейкоз, колоректальный рак, рак эндометрия, рак почки, рак предстательной железы, рак щитовидной железы, меланому, хондросаркому, нейробластому, рак поджелудочной железы, мультиформную глиобластому, рак желудка, рак кости, саркому Юинга, рак шейки матки, рак головного мозга, рак желудка, рак мочевого пузыря, печеночно-клеточную опухоль, рак молочной железы, рак ободочной кишки, печеночно-клеточный рак (ПКР), светлоклеточную почечно-клеточную карциному (СПК), рак головы и шеи, рак печени и желчевыводящих путей, рак центральной нервной системы, рак пищевода, злокачественную плевральную мезотелиому, системный амилоидоз легкой цепи, лимфоплазмацитарную лимфому, миелодиспластический синдром, миелопролиферативную опухоль, нейроэндокринную опухоль, рак из клеток Меркеля, рак яичка и рак кожи.
«Воспалительное расстройство» относится к любому заболеванию, расстройству или синдрому, в котором избыточный или нерегулируемый воспалительный ответ приводит в результате к избыточным воспалительным симптомам, повреждению ткани хозяина или утрате функции ткани. «Воспалительное заболевание» также относится к патологическому состоянию, опосредованному хемотаксическим объединением лейкоцитов или нейтрофилов.
«Воспаление» относится к локальному защитному ответу, вызванному повреждением или деструкцией ткани, и полезному для деструкции, ослабления или удаления (выделения) вредных веществ и поврежденных тканей. Воспаление значимо связано с хемотаксическим объединением лейкоцитов или нейтрофилов. Воспаление может быть вызвано патогенными организмами и вирусами, а также неинфекционными причинами, такими как травма, реперфузия после инфаркта миокарда, инсульт, иммунный ответ на чужеродные антигены и аутоиммунный ответ.
«Аутоиммунное заболевание» относится к любой группе заболеваний, при которых повреждение ткани обусловлено ответами на собственные компоненты организма, опосредованными гуморальным или клеточным иммунитетом. Не имеющие ограничительного характера примеры аутоиммунных заболеваний включают ревматоидный артрит, псориаз, болезнь Крона, анкилозирующий спондилит, рассеянный склероз, сахарный диабет I типа, гепатит, миокардит, синдром Шегрена, аутоиммунную гемолитическую анемию вследствие отторжения трансплантата, везикулезный пемфигоид, болезнь Грейвса, тиреоидит Хасимото, системную красную волчанку (СКВ), миастению гравис, пузырчатку, пернициозную анемию и т.д.
Кроме того, другой аспект описания настоящего изобретения относится к способам иммунологического анализа или определения целевых антигенов, реагентам для иммунологического анализа или определения целевых антигенов, способам иммунологического анализа или определения клеток, экспрессирующих целевые антигены, и к диагностическим агентам для диагностики заболеваний, связанных с положительными по целевому антигену клетками, включающим в качестве активного ингредиента моноклональные антитела или фрагменты антител по описанию настоящего изобретения, которые специфично распознают целевой антиген и связываются с ним.
В описании настоящего изобретения способ выявления или измерения количества целевого антигена может представлять собой любой известный способ. Например, он включает иммуноанализ или способ иммунологического анализа.
Способ иммунопробы или способ иммунологического анализа представляет собой способ выявления или измерения количества антитела или антигена с помощью меченого антигена или антитела. Примеры способов иммунопробы или иммунологического анализа включают способ определения антитела, меченого радиоактивным веществом (RIA), ферментный иммуноанализ (EIA или ELISA), флуоресцентный иммуноанализ (FIA), люминесцентный иммуноанализ, вестерн-блоттинг, физико-химический метод и т.п.
Вышеупомянутые заболевания, связанные с положительными по целевому антигену клетками, можно диагностировать путем выявления или измерения количества клеток, экспрессирующих целевой антиген, используя антитела или фрагменты антител по описанию настоящего изобретения.
Клетки, экспрессирующие антиген, можно выявлять известными методами иммунодетектирования, предпочтительно методом иммунопреципитации, флуоресцентного окрашивания клеток, иммунологического окрашивания тканей и т.п. Кроме того, можно использовать метод, такой как метод флуоресцентного окрашивания антител с помощью системы FMAT8100HTS (Applied Biosystem).
В описании настоящего изобретения образцы для выявления или количественного измерения целевого антигена конкретно не ограничены, если возможно, что они содержат клетки, экспрессирующие целевой антиген, такие как клетки тканей, кровь, плазма крови, сыворотка крови, панкреатический сок, моча, кал, тканевая жидкость или питательная среда.
В зависимости от требуемого метода диагностики диагностический агент, содержащий моноклональное антитело или его фрагмент антитела по описанию настоящего изобретения может также содержать реагенты для выполнения реакции антиген-антитело или реагенты для детектирования этой реакции. Реагенты для выполнения реакции антиген-антитело включают буферные растворы, соли и т.п. Реагенты для детектирования включают агенты, обычно используемые в методах иммунологического анализа или иммунологического детектирования, например, меченое вторичное антитело, которое распознает моноклональное антитело, фрагмент антитела или его конъюгат и субстрат в соответствии с маркировкой.
Подробные описания одного или более воплощений описания настоящего изобретения изложены в приведенном выше описании. Предпочтительные способы и материалы описаны ниже, несмотря на то, что на практике или при тестировании описания настоящего изобретения можно использовать любой метод или материал, подобный или идентичный описанному в настоящем документе. Другие признаки, цели и преимущества описания настоящего изобретения станут очевидными из всего текста описания и формулы изобретения. В описании и формуле изобретения формы единственного числа включают в себя аспекты множественного числа, если контекст явным образом не требует иного. Если в настоящем документе явным образом не определено иное, все технические и научные термины, используемые в настоящем описании изобретения, в общем понятны специалистам в данной области техники, к которой относится данное описание изобретения. Все патенты и публикации, цитируемые в описании, включены путем отсылки. Следующие примеры приведены для более полной иллюстрации предпочтительных воплощений описания настоящего изобретения. Эти примеры не должны быть никоим образом истолкованы как ограничивающие объем описания настоящего изобретения, определенный формулой изобретения.
Примеры
Получение и скрининг антител
Методы получения моноклональных антител известны в данной области техники. Один из методов, которые можно использовать, представляет собой метод, описанный в работе Kohler, G. et al. (1975) Continuous Cultures Of Fused Cells Secreting Antibody Of Predefined Specificity, Nature 256:495-497, или в его модифицированной форме. Как правило, моноклональные антитела образуются в биологических видах, не являющихся человеком, таких как мыши. Как правило, для иммунизации используют мышей или крыс, но также можно использовать других животных, таких как кролики или альпака. Антитела получают путем иммунизации мышей иммуногенными количествами клеток, клеточных экстрактов или препаратов белка, содержащих человеческий CD3 или другие целевые антигены (такие как B7H3). Иммуноген может представлять собой без ограничений первичные клетки, культивируемые клеточные линии, раковые клетки, нуклеиновые кислоты или ткани.
В одном воплощении изобретения моноклональные антитела, связывающиеся с целевым антигеном, получают путем использования в качестве иммуногена клеток-хозяев, в которых осуществляется сверхэкспрессия целевого антигена. Такие клетки включают, например, но без ограничений, человеческие T-клетки, клетки, в которых происходит сверхэкспрессия человеческого B7H3.
Для мониторинга ответа антител можно получить небольшое количество образца (например, крови) от животного и тестировать титр антител к иммуногену. Селезенку или другие крупные лимфатические узлы можно извлекать и диссоциировать на отдельные клетки. При желании клетки селезенки могут быть выбраны путем нанесения клеточной суспензии на покрытый антигеном планшет или лунку (после удаления прикрепившихся неспецифичных клеток). B-клетки, экспрессирующие мембраносвязанные антиген-специфические иммуноглобулины, будут связаны с планшетом и не будут смываться остальной суспензией. Впоследствии все полученные в результате B-клетки или все диссоциированные клетки селезенки могут быть слиты с клетками миеломы (например, X63-Ag8.653 и клетки, доступные от Института биологических исследований Саика, Центр распределения клеток, Сан-Диего, штат калифорния, США). Для слияния клеток селезенки или лимфоцитов с клетками миеломы с образованием гибридом можно использовать полиэтиленгликоль (ПЭГ). Затем гибридомы культивируют в селективной среде (например, в среде с гипоксантином, аминоптерином, тимидином, иначе называемой средой HAT). Впоследствии полученные в результате гибридомы высевают в планшет методом ограничивающего разведения и анализируют продукцию антител, которые специфично связываются с иммуногеном, используя, например, метод FACS (сортировка флуорецентно-активированных клеток) или иммуногистохимическое окрашивание (ИГХ). Впоследствии отобранные гибридомы, секретирующие антитела, культивируют in vitro (например, о флаконе для клеточных культур или в реакторе с холофайбером) или in vivo (например, в виде асцитов у мышей).
В качестве другой альтернативы технологии клеточного слияния для получения моноклональных антител по описанию настоящего изобретения можно использовать B-клетки, иммортализованные вирусом Эпштейна-Барре (EBV). При необходимости гибридому размножают и субклонируют, и антииммуногенную активность супернатанта анализируют традиционными методами анализа (например, FACS, ИГХ, радиоиммунологический анализ, ферментный иммуноанализ, флуоесцентный иммуноанализ и т.д.).
В другой альтернативе моноклональные антитела к целевым антигенам (таким как CD3, B7H3) и другие эквивалентные антитела можно секвенировать и получать рекомбинантным путем любым известным в данной области техники способом (например, гуманизация, получение полностью человеческих антител с использованием трансгенных мышей, технология фагового дисплея и т.д.). В одном воплощении изобретения моноклональные антитела к целевым антигенам (например, CD3, B7H3) секвенируют, а затем клонируют полинуклеотидные последовательности в векторе для экспрессии или размножения. Последовательность, кодирующую интересующее антитело, можно поддерживать в векторе в клетке-хозяине, а затем размножать клетку-хозяина и замораживать для последующего использования.
Полинуклеотидные последовательности моноклональных антител к CD3 и к любому другому эквивалентному антигену, используемые для генетических манипуляций в целях получения гуманизированных антител для улучшения аффинности или других характеристик антител. Общий принцип гуманизированных антител включает сохранение основной последовательности антигенсвязывающего участка антитела, при этом не человеческий участок антитела заменен последовательностью человеческого антитела. Для гуманизации моноклональных антител обычно используют четыре стадии. Эти стадии состоят в следующем: (1) определение нуклеотидных последовательностей и предполагаемых аминокислотных последовательностей вариабельных доменов легкой и тяжелой цепей исходного антитела; (2) конструирование гуманизированного антитела, то есть определение, какая каркасная область будет использоваться в процессе гуманизации; (3) актуальные методы/способы гуманизации и (4) трансфекция и экспрессия гуманизированных антител. См., например, патенты США №№US4816567, US5807715, US5866692 и US6331415.
1. Получение и скрининг антител к B7H3
B-клетки выделяли, используя человеческие мононуклеарные клетки периферической крови (МКПК), селезенку и ткани лимфоузлов, и экстрагировали РНК для получения библиотеки природных однонитевых фаговых антител. Сконструированную библиотеку однонитевых фаговых антител упаковывали с образованием фаговых частиц, которые подвергали скринингу путем пэннинга, используя жидкофазный метод. Фаг был ассоциирован с биотинилированной жидкой фазой B7H3, а затем его отделяли с помощью магнитных гранул, покрытых стрептавидином. Чтобы получить положительную последовательность, которая связывается с человеческим B7H3, для пэннинга использовали биотинилированный человеческий B7H3. Несколько моноклональных колоний собирали и упаковывали в фаговые однонитевые антитела для тестирования методом твердофазного иммуносорбентного ферментного анализа (ELISA). Моноклональные фаги тестировали на их способность к связыванию человеческого B7H3 и мышиного B7H3 соответственно, и после скрининга получили антитела к B7H3.
Антиген B7H3, используемый для детектирования, представлен ниже:
Человеческий антиген B7H3 для детектирования
Имеющийся в продаже продукт (SinoBiological, номер по каталогу 11188-H08H)
Последовательность представлена ниже:

SEQ ID NO: 1
Примечание: Подчеркнутый участок представляет собой внеклеточную область B7H3; курсивом представлена His-метка.
Обезьяний антиген B7H3 для детектирования
Имеющийся в продаже продукт (SinoBiological, номер по каталогу 90806-C08H)
Последовательность представлена ниже:

SEQ ID NO: 2
Примечание: Подчеркнутый участок представляет собой внеклеточную область B7H3; курсивом представлена His-метка.
Мышиный антиген B7H3 для детектирования
Имеющийся в продаже продукт (SinoBiological, номер по каталогу 50973-M08H)
Последовательность представлена ниже:

SEQ ID NO: 3
Примечание: Подчеркнутый участок представляет собой внеклеточную область B7H3; курсивом представлена His-метка.
Полноразмерная аминокислотная последовательность человеческого B7H3

SEQ ID NO: 4
Примечание: Участок с двойным подчеркиванием представляет собой сигнальный пептид (сигнальный пептид: 1-28); Подчеркнутый участок представляет собой внеклеточную область B7H3 (внеклеточный домен: 29-466), где 29-139 относится к домену Ig-подобного V-типа 1, 145-238 относится к домену Ig-подобного C2-типа 1; 243-357 относится к домену Ig-подобного V-типа 2 и 363-456 относится к домену Ig-подобного C2-типа 2; пунктирной линией представлена трансмембранная область (трансмембранный домен: 467-487); курсивом представлена внутриклеточная область (цитоплазматический домен: 488-534).
Полноразмерная аминокислотная последовательность обезьяньего B7H3

SEQ ID NO: 5
Примечание:
Участок с двойным подчеркиванием представляет собой сигнальный пептид (сигнальный пептид: 1-28); подчеркнутый участок представляет собой внеклеточную область B7H3 (внеклеточный домен: 29-466), где 29-139 относится к домену Ig-подобного V-типа 1, 145-238 относится к домену Ig-подобного C2-типа 1; 243-357 относится к домену Ig-подобного V-типа 2 и 363-456 относится к домену Ig-подобного C2-типа 2; пунктирной линией представлена трансмембранная область (трансмембранный домен: 467-487); курсивом представлена внутриклеточная область (цитоплазматический домен: 488-534).
Полноразмерная аминокислотная последовательность мышиного B7H3

SEQ ID NO: 6
Примечание:
Участок с двойным подчеркиванием представляет собой сигнальный пептид (сигнальный пептид: 1-28); Подчеркнутый участок представляет собой внеклеточную область B7H3 (внеклеточный домен: 29-248); пунктирной линией представлена трансмембранная область (трансмембранный домен: 249-269); курсивом представлена внутриклеточная область (цитоплазматический домен: 270-316).
Для антитела к B7H3 h1702, полученного путем скрининга, последовательности и CDR последовательности, определенные критериями нумерации IMGT, представлены ниже:
>h1702 VH

SEQ ID NO: 7
>h1702 VL
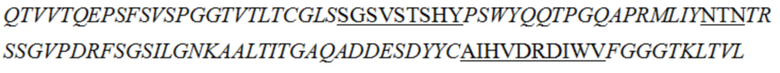
SEQ ID NO: 8
Примечание: Расположенные в порядке FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4, последовательности курсивом представляют собой FR, а подчеркнутые последовательности представляют собой CDR.
Таблица 1. Последовательности CDR легкой и тяжелой цепей антитела B7H3 h1702
SEQ ID NO: 9
SEQ ID NO: 12
SEQ ID NO: 10
SEQ ID NO: 13
SEQ ID NO: 11
SEQ ID NO: 14
Чтобы дополнительно улучшить функциональную способность биспецифических антител, мутации замены цистеином были введены в VH и VL антитела B7H3 h1702. Мутация G103C (нумерация соответствует природной аминокислотной последовательности, положение 103 SEQ ID NO: 16) была введена в вариабельную область легкой цепи, а мутация G44C (нумерация соответствует природной аминокислотной последовательности, положение 44 SEQ ID NO: 15) была введена в вариабельную область тяжелой цепи, так чтобы образовалась пара дисульфидных мостиков. Вариабельные области тяжелой и легкой цепей одноцепочечного антитела к B7H3 после мутации приведены ниже:
B7H3 VH44C:
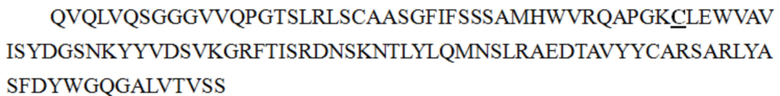
SEQ ID NO: 15
B7H3 VL103C:
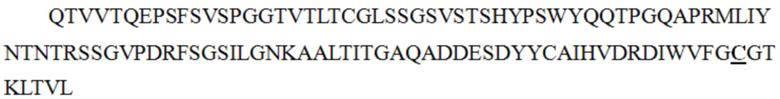
SEQ ID NO: 16.
2. Получение и скрининг антител к CD3
Гуманизированные антитела к CD3 могут быть получены на основе мышиных антител к CD3 такими методами, как мутация, конструирование библиотеки, гуманизация методами генной инженерии и скрининг.
Информация о последовательности, относящаяся к антигену CD3, приведена ниже
Человеческий антиген CD3 для детектирования
Имеющийся в продаже продукт (SinoBiological номер по каталогу CT038-H2508H)
Последовательности представлены ниже:
Человеческий CD3ε (Человеческий CD3ε)
 SEQ ID NO: 17
SEQ ID NO: 17
Примечание:
Подчеркнутый участок представляет собой внеклеточную область CD3ε (внеклеточный домен: 23-126); курсивом представлена His-метка.
Человеческий CD3δ

SEQ ID NO: 18
Примечание:
Подчеркнутый участок представляет собой внеклеточную область CD3δ (внеклеточный домен: 22-105); курсивом представлена Flag-метка.
Обезьяний антиген CD3 для детектирования
Имеющийся в продаже продукт (Acro Biosystem номер по каталогу CDD-C52W4-100 мкг)
Последовательности представлены ниже:
Обезьяний CD3ε

SEQ ID NO: 19
Примечание:
Подчеркнутый участок представляет собой внеклеточную область CD3ε (внеклеточный домен: 22-117); курсивом представлена His-метка.
Обезьяний CD3δ

SEQ ID NO: 20
Примечание:
Подчеркнутый участок представляет собой внеклеточную область CD3δ (внеклеточный домен: 22-105); курсивом представлена Flag-метка.
Мышиный антиген CD3 для детектирования
Имеющийся в продаже продукт (SinoBiological номер по каталогу CT033-M2508H). Последовательности представлены ниже:
Мышиный CD3ε

SEQ ID NO: 21
Примечание:
Подчеркнутый участок представляет собой внеклеточную область CD3ε (внеклеточный домен: 22-108); курсивом представлена His-метка.
Мышиный CD3δ
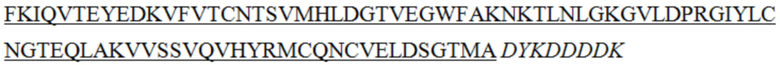
SEQ ID NO: 22
Примечание:
Подчеркнутый участок представляет собой внеклеточную область CD3δ (внеклеточный домен: 22-105); курсивом представлена Flag-метка.
Полноразмерная аминокислотная последовательность человеческого CD3ε

SEQ ID NO: 23
Примечание:
Участок с двойным подчеркиванием представляет собой сигнальный пептид (сигнальный пептид: 1-28); подчеркнутый участок представляет собой внеклеточную область CD3ε (внеклеточный домен: 23-126), где 32-112 относится к Ig-подобному домену; пунктирной линией представлена трансмембранная область (трансмембранный домен: 127-152); курсивом представлена внутриклеточная область (цитоплазматический домен: 153-207).
Полноразмерная аминокислотная последовательность человеческого CD3δ

SEQ ID NO: 24
Примечание:
Участок с двойным подчеркиванием представляет собой сигнальный пептид (сигнальный пептид: 1-21); подчеркнутый участок представляет собой внеклеточную область CD3δ (внеклеточный домен: 22-105); пунктирной линией представлена трансмембранная область (трансмембранный домен: 106-126); курсивом представлена внутриклеточная область (цитоплазматический домен: 127-171).
Полноразмерная аминокислотная последовательность обезьяньего CD3ε

SEQ ID NO: 25
Примечание:
Участок с двойным подчеркиванием представляет собой сигнальный пептид (сигнальный пептид: 1-21); подчеркнутый участок представляет собой внеклеточную область CD3δ (внеклеточный домен: 22-117); пунктирной линией представлена трансмембранная область (трансмембранный домен: 118-138); курсивом представлена внутриклеточная область (цитоплазматический домен: 139-198).
Полноразмерная аминокислотная последовательность обезьяньего CD3δ

SEQ ID NO: 26
Примечание:
Участок с двойным подчеркиванием представляет собой сигнальный пептид (сигнальный пептид: 1-21); подчеркнутый участок представляет собой внеклеточную область CD3δ (внеклеточный домен: 22-105); пунктирной линией представлена трансмембранная область (трансмембранный домен: 106-126); курсивом представлена внутриклеточная область (цитоплазматический домен: 127-171).
Полноразмерная аминокислотная последовательность мышиного CD3ε

SEQ ID NO: 27
Примечание:
Участок с двойным подчеркиванием представляет собой сигнальный пептид (сигнальный пептид: 1-21); подчеркнутый участок представляет собой внеклеточную область CD3δ (внеклеточный домен: 22-108); пунктирной линией представлена трансмембранная область (трансмембранный домен: 109-134); курсивом представлена внутриклеточная область (цитоплазматический домен: 135-189).
Полноразмерная аминокислотная последовательность мышиного CD3δ

SEQ ID NO: 28
Примечание:
Участок с двойным подчеркиванием представляет собой сигнальный пептид (сигнальный пептид: 1-21); подчеркнутый участок представляет собой внеклеточную область CD3δ (внеклеточный домен: 22-105); пунктирной линией представлена трансмембранная область (трансмембранный домен: 106-126); курсивом представлена внутриклеточная область (цитоплазматический домен: 127-173).
После повторного анализа и оптимизации была получена серия последовательностей гуманизированного антитела к CD3. Последовательности вариабельной области тяжелой цепи представлены ниже:
Таблица 2. Последовательность вариабельной области тяжелой цепи гуманизированных антител к CD3

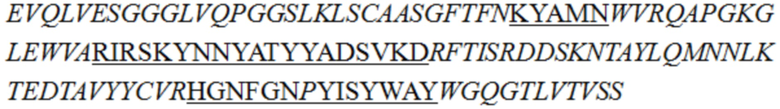
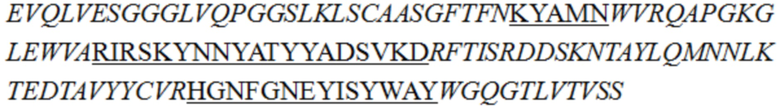
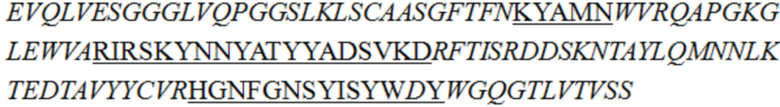
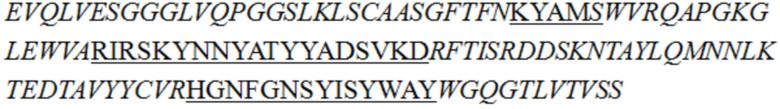
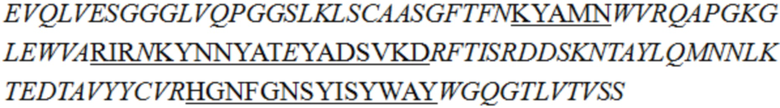
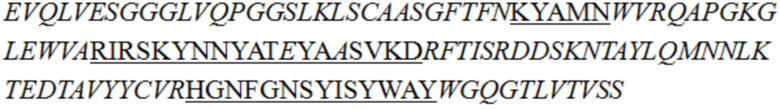
Последовательности вариабельной области легкой цепи представлены ниже:
>HRL

SEQ ID NO: 36
Примечание: Расположенные в порядке FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4, последовательности курсивом представляют собой FR, а подчеркнутые последовательности представляют собой CDR. Число и положение CDR вариабельной области легкой и тяжелой цепи (LCDR1-LCDR3 и HCDR1-HCDR3) гуманизированных антител к CD3, перечисленных здесь и в таблице 3 ниже, соответствуют хорошо известным критериям нумерации по Кэботу.
Таблица 3. Последовательности CDR антител к CD3
SEQ ID NO: 37
SEQ ID NO: 48
SEQ ID NO: 38
SEQ ID NO: 49
SEQ ID NO: 39
SEQ ID NO: 50
SEQ ID NO: 37
SEQ ID NO: 40
SEQ ID NO: 41
SEQ ID NO: 37
SEQ ID NO: 40
SEQ ID NO: 42
SEQ ID NO: 37
SEQ ID NO: 40
SEQ ID NO: 43
SEQ ID NO: 44
SEQ ID NO: 40
SEQ ID NO: 45
SEQ ID NO: 37
SEQ ID NO: 46
SEQ ID NO: 45
SEQ ID NO: 37
SEQ ID NO: 47
SEQ ID NO: 45
Конструирование и получение одноцепочечных антител
scFv к B7H3 и scFv к CD3 были получены путем соединения вариабельных областей легкой и тяжелой цепи, полученных из указанных выше антител к B7H3, и путем соединения вариабельных областей легкой и тяжелой цепи, полученных из антитела к CD3 соответственно, где линкер может быть выбран из хорошо известных в данной области техники. Иллюстративный линкер может быть выбран из: (GGGGS)n или (GGGGS)n GGG, где n может быть равно 1, 2, 3 или 4.
Иллюстративные scFv антитела к B7H3 представлены ниже:
Таблица 4. Перечень последовательностей различных одноцепочечных антител (scFv) к B7H3
(VLB7H3-линкер-VHB7H3)
(SEQ ID NO: 51)
(SEQ ID NO: 52)
(SEQ ID NO: 53)
(SEQ ID NO: 54)
Иллюстративные scFv антитела к CD3 представлены ниже:
Таблица 5. Перечень последовательностей одноцепочечных антител (scFv) к CD3
(VHCD3-линкер-VLCD3)
(SEQ ID NO: 55)
(VHCD3-линкер-VLCD3)
(SEQ ID NO: 56)
(VHCD3-линкер-VLCD3)
(SEQ ID NO: 57)
(VHCD3-линкер-VLCD3)
(SEQ ID NO: 58)
(VHCD3-линкер-VLCD3)
(SEQ ID NO: 59)
(VHCD3-линкер-VLCD3)
(SEQ ID NO: 60)
(VHCD3-линкер-VLCD3)
(SEQ ID NO: 61)
(VLCD3-линкер-VHCD3)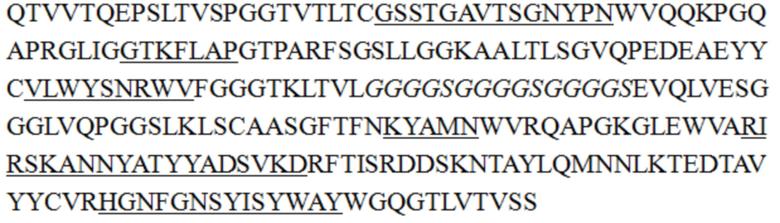
(SEQ ID NO: 62)
(VLCD3-линкер-VHCD3)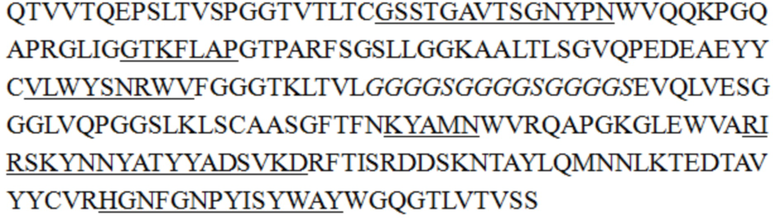
(SEQ ID NO: 63)
(VLCD3-линкер-VHCD3)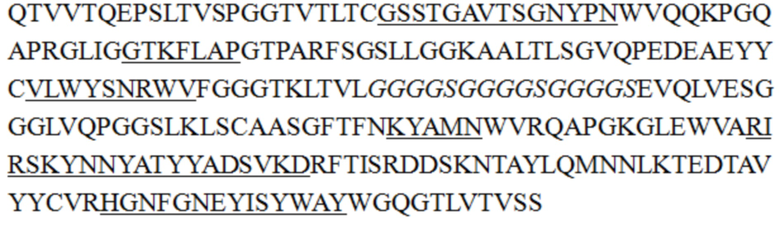
(SEQ ID NO: 64)
(VLCD3-линкер-VHCD3)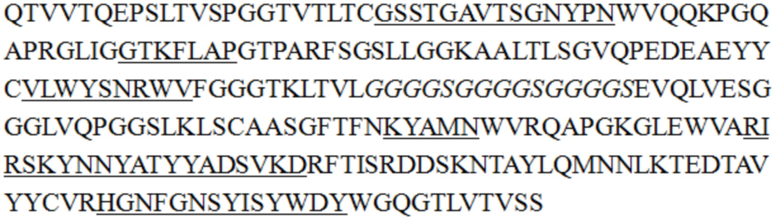
(SEQ ID NO: 65)
(VLCD3-линкер-VHCD3)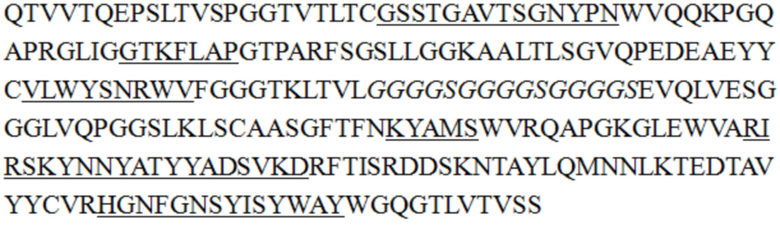
(SEQ ID NO: 66)
(VLCD3-линкер-VHCD3)
(SEQ ID NO: 67)
(VLCD3-линкер-VHCD3)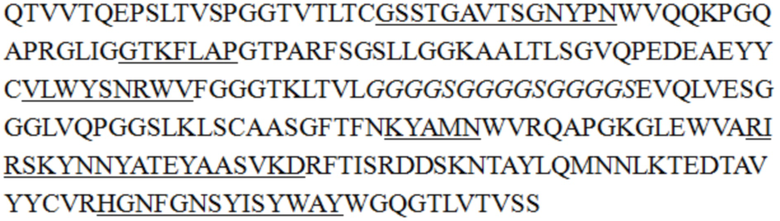
(SEQ ID NO: 68)
Конструирование и получение биспецифических антител
Бивалентные биспецифические антитела к B7H3 и моновалентные биспецифические антитела к B7H3
В некоторых воплощениях описания настоящего изобретения структура бивалентных биспецифических антител к B7H3 показана на Фиг.1A, где C-конец антитела может быть немеченым или меченным His-меткой. Два антигенсвязывающих домена B7H3 и один антигенсвязывающий домен CD3 организованы в сконструированную асимметрическую структуру из Fc-содержащих цепей, в которой каждый антигенсвязывающий домен B7H3 находится на каждой из этих двух цепей соответственно, и все антигенсвязывающие домены находятся в форме scFv. Fc-область может обеспечивать возможность сохранения нормального периода полувыведения и благоприятной стабильности антитела. Конструкция из двух цепей значительно уменьшает вероятность несоответствий и улучшает однородность образца и выход целевого антитела. Специфичная молекулярная структура (формат) биспецифических антител показана ниже в таблице 6. Кроме того, молекулярная структура моновалентных биспецифических антител к B7H3, используемая в некоторых воплощениях описания настоящего изобретения, имеет один домен Fc во второй пептидной цепи, не содержащей антигенсвязывающий домен, и такая структура показана на Фиг.1B.
Таблица 6. Структурное представление биспецифических антител
Примечание: В этой таблице карбоксильный конец первой или второй полипептидной цепи может быть немеченым или меченным His-меткой. L1, L2, L3, L4, L5 и L6 представляют собой линкеры для соединения каждого антигенсвязывающий домен с областью Fc.
Таблица 7. Выбор линкерной последовательности
Где n представляет собой из 1, 2, 3 или 4; предпочтительно n в L1 представляет собой 2 или 3, более предпочтительно 3; n в L2 представляет собой 1 или 2, более предпочтительно 1; n в L3 или L5 представляет собой 3. Необязательно, линкер, используемый для соединения антигенсвязывающего домена и области Fc, может быть выбран из любых других линкеров, которые можно использовать для соединения функциональных доменов антитела, и не ограничен линкерами, определенными приведенными выше последовательностями.
Fc1 и Fc2, указанные в таблице 6 выше, могут представлять собой Fc с одной и той же последовательностью, либо могут представлять собой knob-Fc и hole-Fc соответственно, либо hole-Fc и knob-Fc соответственно. В некоторых воплощениях настоящего изобретения последовательности knob-Fc и hole-Fc предпочтительно являются такими, как показано в таблице 8:
Таблица 8. Перечень последовательностей различных Fc


Для вышеупомянутых вариабельных областей легкой и тяжелой цепей, одноцепочечных антител и биспецифических антител, ДНК, кодирующие вышеупомянутые полипептиды или их антигенсвязывающие фрагменты, можно конструировать на основе кДНК, кодирующих VH и/или VL и другие необходимые домены, и встраивать ДНК в прокариотический(-ие) экспрессионный(-ые) вектор(-ы) или эукариотический(-ие) экспрессионный(ые) вектор(-ы), и впоследствии экспрессионный(ые) вектор(-ы) вводят в прокариотический или эукариотический организм для экспрессии полипептидов или антигенсвязывающих фрагментов.
Пример 1. Получение молекул биспецифического антитела, положительных контрольных молекул и отрицательных контрольных молекул
В соответствии со способом конструирования молекул биспецифического антитела по описанию настоящего изобретения были сконструированы и получены молекулы специфичного биспецифического антитела. Примеры аминокислотных последовательностей приведены в таблице 9 ниже:
Таблица 9. Перечень последовательностей биспецифических антител

(SEQ ID NO: 72)
 (SEQ ID NO: 71)
(SEQ ID NO: 71)
(SEQ ID NO: 73)
(SEQ ID NO: 74)
(SEQ ID NO: 75)
(SEQ ID NO: 76)
(SEQ ID NO: 77) (SEQ ID NO: 78)
(SEQ ID NO: 78)
(SEQ ID NO: 79)
(SEQ ID NO: 80) (SEQ ID NO: 81)
(SEQ ID NO: 81)
(SEQ ID NO: 82)
(SEQ ID NO: 83)
(SEQ ID NO: 84)
(SEQ ID NO: 85)
(SEQ ID NO: 86)
(SEQ ID NO: 87)
(SEQ ID NO: 76)


(SEQ ID NO: 88)
(SEQ ID NO: 72)
Примечание: Вторые полипептидные цепи молекул бивалентных биспецифических антител к B7H3 113, 118, 119, 126, 127, 128, 131, 132, 154, 155, 156, 161, 162, 171, 172 и 177, указанные в таблице выше, представляют собой VL B7H3-L5-VHB7H3-L6-hole-Fc, как показано в SEQ ID NO: 71; а вторые полипептидные цепи молекул моновалентных биспецифических антител к B7H3 181-187 представляют собой hole-Fc, как показано в SEQ ID NO: 70.
Аминокислотные последовательности отрицательных контрольных (NC1, NC2, NC3) и положительных контрольных (MGD009) биспецифических антител, используемых в данном описании изобретения, являются следующими:
NC1: B7H3-связывающий домен заменен неродственным антителом (антитело к флуоресцеину, антифлуоресцеин), но CD3-связывающий домен сохранен. Справочная литература по его аминокислотной последовательности: The anti-fluorescein antibody used to form the control DART diabody was antibody 4-4-20 (Gruber. M.et al.(1994)).
Цепь 1 (VHCD3-VLCD3-VLctrl-VHctrl-knob-Fc)
SEQ ID NO: 89
Цепь 2 (VLctrl-VH ctrl-hole-Fc)
DIQMTQSPSSVSASVGDRVTITCRASQDIANYLSWYQQKPGKSPKLLIYGTSNLEVGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCLQDKEFPRTFGGGTKVEIKGGGGSGGGGSGGGGSEVQLVESGGGLVQPGGSLRLSCAASGFTFSHYYMAWVRQAPGKGLEWVTSISYEGDITYYGDSVKGRFTISRDNSKNTLYLQMNSLRAEDTATYYCASQTLRESFDYWGQGTLVTVSSGGGDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVCTLPPSREEMTKNQVSLSCAVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLVSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
SEQ ID NO: 90
NC2: B7H3-связывающий домен сохранен, только CD3-связывающий домен замещен неродственным антителом антифлуоресцеином.
Цепь 1 (VHctrl-VLctrl-VLB7H3-VHB7H3-knob-Fc)

SEQ ID NO: 91
Цепь 2

SEQ ID NO: 92
Примечание: Порядок расположения: VLB7H3-линкер-VHB7H3-линкер-Fc. Подчеркнутые последовательности представляют собой последовательности антитела к B7H3, а курсивом представлена последовательность hole-Fc.
NC3
Цепь 1 (VHctrl-VLctrl-VLCD3-VHCD3-knob-Fc-His метка)

SEQ ID NO: 93
Цепь 2 (VLctrl-VH ctrl-hole-Fc)
DIQMTQSPSSVSASVGDRVTITCRASQDIANYLSWYQQKPGKSPKLLIYGTSNLEVGVPSRFSGSRSGTDFTLTISSLQPEDFATYYCLQDKEFPRTFGGGTKVEIKGGGGSGGGGSGGGGSEVQLVESGGGLVQPGGSLRLSCAASGFTFSHYYMAWVRQAPGKGLEWVTSISYEGDITYYGDSVKGRFTISRDNSKNTLYLQMNSLRAEDTATYYCASQTLRESFDYWGQGTLVTVSSGGGDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVCTLPPSREEMTKNQVSLSCAVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLVSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
SEQ ID NO: 94
Положительный контроль MGD009 содержит три цепи, и его получение и аминокислотные последовательности можно найти в опубликованной заявке на патент WO2017030926A1. Его аминокислотные последовательности приведены ниже:
Цепь 1 (B7H3VL-CD3VH-Fc)
DIQLTQSPSFLSASVGDRVTITCKASQNVDTNVAWYQQKPGKAPKALIYSASYRYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYNNYPFTFGQGTKLEIKGGGSGGGGEVQLVESGGGLVQPGGSLRLSCAASGFTFSTYAMNWVRQAPGKGLEWVGRIRSKYNNYATYYADSVKDRFTISRDDSKNSLYLQMNSLKTEDTAVYYCVRHGNFGNSYVSWFAYWGQGTLVTVSSGGCGGGEVAALEKEVAALEKEVAALEKEVAALEKGGGDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLWCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
SEQ ID NO: 95
Цепь 2 (CD3VL-B7H3VH)
QAVVTQEPSLTVSPGGTVTLTCRSSTGAVTTSNYANWVQQKPGQAPRGLIGGTNKRAPWTPARFSGSLLGGKAALTITGAQAEDEADYYCALWYSNLWVFGGGTKLTVLGGGGSGGGGEVQLVESGGGLVQPGGSLRLSCAASGFTFSSFGMHWVRQAPGKGLEWVAYISSDSSAIYYADTVKGRFTISRDNAKNSLYLQMNSLRDEDTAVYYCGRGRENIYYGSRLDYWGQGTTVTVSSGGCGGGKVAALKEKVAALKEKVAALKEKVAALKE
SEQ ID NO: 96
Цепь 3 (Fc)
DKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLSCAVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLVSKLTVDKSRWQQGNVFSCSVMHEALHNRYTQKSLSLSPGK
SEQ ID NO: 97
201 (DART-Fc трехцепочечная структура) 201 цепь 1 (B7H3VL-CD3VH-E-Fc)
QTVVTQEPSFSVSPGGTVTLTCGLSSGSVSTSHYPSWYQQTPGQAPRMLIYNTNTRSSGVPDRFSGSILGNKAALTITGAQADDESDYYCAIHVDRDIWVFGGGTKLTVLGGGSGGGGEVQLVESGGGLVQPGGSLKLSCAASGFTFNKYAMNWVRQAPGKGLEWVARIRSKYNNYATYYADSVKDRFTISRDDSKNTAYLQMNNLKTEDTAVYYCVRHGNFGNEYISYWAYWGQGTLVTVSSGGCGGGEVAALEKEVAALEKEVAALEKEVAALEKGGGDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLWCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALH NHYTQKSLSLSPGKHHHHHH
SEQ ID NO: 99
201 цепь 2 (CD3VL-B7H3VH-K)
QTVVTQEPSLTVSPGGTVTLTCGSSTGAVTSGNYPNWVQQKPGQAPRGLIGGTKFLAPGTPARFSGSLLGGKAALTLSGVQPEDEAEYYCVLWYSNRWVFGGGTKLTVLGGGGSGGGGQVQLVQSGGGVVQPGTSLRLSCAASGFIFSSSAMHWVRQAPGKGLEWVAVISYDGSNKYYVDSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARSARLYASFDYWGQGALVTVSSGGCGGGKVAALKEKVAALKEKVAALKEKVAALKE
SEQ ID NO: 100
201 Цепь 3
DKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLSCAVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLVSKLTVDKSRWQQGNVFSCSVMHEALHNRYTQKSLSLSPGK
SEQ ID NO: 97
202 (четырехцепочечная структура, где массовое соотношение четырех цепей составляет: цепь 1:цепь 2:цепь 3:цепь 4 представляет собой 1:2:1:1)
202 цепь 1 (B7H3VH-CH1 Fc)
QVQLVQSGGGVVQPGTSLRLSCAASGFIFSSSAMHWVRQAPGKGLEWVAVISYDGSNKYYVDSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARSARLYASFDYWGQGALVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPCREEMTKNQVSLWCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKHHHHHH
SEQ ID NO: 101
202 цепь 2 (B7H3VL-CL)
QTVVTQEPSFSVSPGGTVTLTCGLSSGSVSTSHYPSWYQQTPGQAPRMLIYNTNTRSSGVPDRFSGSILGNKAALTITGAQADDESDYYCAIHVDRDIWVFGGGTKLTVLRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
SEQ ID NO: 102
202 цепь 3 (B7H3 VH-CH1-CD3VH-CL)
QVQLVQSGGGVVQPGTSLRLSCAASGFIFSSSAMHWVRQAPGKGLEWVAVISYDGSNKYYVDSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARSARLYASFDYWGQGALVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDGGGGSGGGGSEVQLVESGGGLVQPGGSLKLSCAASGFTFNKYAMNWVRQAPGKGLEWVARIRSKYNNYATYYADSVKDRFTISRDDSKNTAYLQMNNLKTEDTAVYYCVRHGNFGNEYISYWAYWGQGTLVTVSSASVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGECDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVCTLPPSREEMTKNQVSLSCAVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLVSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
SEQ ID NO: 103
202 цепь 4 (CD3VL-CH1)
QTVVTQEPSLTVSPGGTVTLTCGSSTGAVTSGNYPNWVQQKPGQAPRGLIGGTKFLAPGTPARFSGSLLGGKAALTLSGVQPEDEAEYYCVLWYSNRWVFGGGTKLTVLSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSC
SEQ ID NO: 104.
Пример 2. Экспрессия и очистка биспецифических антител CD3-B7H3
Клетки HEK293E трансфицировали плазмидой, экспрессирующей биспецифическое антитело (цепь 1:цепь 2 представялет собой 1:1), супернатант после экспрессии собирали спустя 6 дней и удаляли примеси высокоскоростным центрифугированием. Осветленный супернатант очищали на колонке Ni Sepharose excel (GE Healthcare). Колонку промывали фосфатно-солевым буферным раствором (ФСБ) до тех пор, пока показания при A280 не снижались до исходного уровня, а затем колонку промывали ФСБ с 10 ммоль/л имидазола, чтобы удалить примеси неспецифично связанных белков, и собирали элюат.Наконец, целевой белок элюировали раствором ФСБ, содержащим 300 ммоль/л имидазола, и собирали пики элюирования. Образцы элюата концентрировали соответствующим образом и впоследствии дополнительно очищали гель-хроматографией на колонке Superdex200 (GE), предварительно уравновешенной буферным раствором 550 (10 ммоль/л уксусной кислоты, pH 5,5, 135 ммоль/л NaCl). Целевой пик собирали. Образец уравновешивали против буферного раствора 559 (10 ммоль/л уксусной кислоты, pH 5,5, 9% сахарозы) на обессоливающей колонке или в центрифужной пробирке для ультрафильтрации, делили на аликвоты и хранили при минус 80°C.
Тестовый пример 1. Аффинность биспецифических антител к B7H3 и CD3, определенная в анализе BIAcore
Определение аффинности антител к B7H3 и CD3 выполняли в форме иммобилизованного антитела. БсАт иммобилизовали на биосенсорном чипе CM5 (№по каталогу BR-1005-30, GE) или связывали биосенсорный чип с белком A (№по каталогу 29127556, GE) с антителом к IgG человека (№по каталогу BR-1008-39, №партии 10260416, GE), а затем каждый антиген пропускали через поверхность чипа. Сигналы взаимодействия детектировали в реальном времени с помощью прибора Biacore T200 с получением кривых связывания и диссоциации. После завершения диссоциации в каждом экспериментальном цикле чип промывали и регенерировали с помощью буферного раствора для регенерации Glycine1.5 (№ по каталогу BR100354, GE) или 3 ммоль/л MgCl2 (из набора человеческих иммобилизованных антител, №по каталогу BR100839, GE). Данные согласовывали с моделью Ленгмюра (1:1), используя программное обеспечение GE Biacore T200 Evaluation версии 3.0, и получали значения аффинности.
При неизменном порядке расположения аффинность биспецифических антител к CD3 незначительно изменялась при варьировании последовательностей VH антитела к CD3. При использовании последовательностей HRH-6 и HRH-5 аффинность антитела к CD3 была самой слабой, и связывание CD3 не детектировалось прибором Biacore.
Таблица 10. Результаты анализа Biacore антигенсвязывающей аффинности биспецифических антител со структурой AFF3
В качестве примера для анализа были отобраны антитела, содержащие HRH3 в качестве антигенсвязывающего домена CD3 вариабельной области тяжелой цепи. Среди отобранных антител исследуемые антитела 118, 127 и 132 обладали аффинностью к человеческому B7H3 и человеческому CD3 на уровне 10-9 и 10-8 моль/л соответственно, что сопоставимо со значениями для MGD009. Все эти антитела обладали сильной перекрестной связывающей способностью как с обезьяньим (cyno) B7H3, так и с человеческим CD3.
Таблица 11. Результаты анализа Biacore антигенсвязывающей аффинности биспецифических антител, содержащих HRH3, расположенные в различном порядке
Тестовый пример 2. Определение связывающей способности на клеточном уровне
Способность биспецифических антител к связыванию с антигенами клеточной поверхности определяли методом сортировки флуоресцентно-активированных клеток (FACS). Рекомбинантные линии клеток A498 (ATCC, HTB-44), CT26/hB7H3 (рекомбинантная линия клеток с сверхэкспрессией человеческого B7H3 в мышиных клетках CT26, сконструирована внутри лаборатории, CT26 была получена из банка клеток Академии Наук Китая, TCM37) и Jurkat (клетки Jurkat были получены из Американской коллекции типовых культур (ATCC), PTS-TIB-152; рекомбинантная линия клеток была получена на основе клеток Jurkat путем создания сверхэкспрессии гена люциферазы и встраивания элемента ответа NFAT выше гена) использовали по отдельности для связывания с антигенами B7H3 и CD3 на клеточной поверхности.
Буферный раствор FACS (98% ФСБ, 2% фетальной бычьей сыворотки (ФБС)) вносили в 96-луночный планшет с U-образными лунками (Corning, 3795) для ресуспендирования клеток, добавляли серийно разведенные антитела, инкубировали при 4°C в течение 1 часа и дважды промывали планшет буферным раствором FACS. Впоследствии в каждую лунку добавляли антитело APC к Fc IgG человека (Biolegend, №по каталогу 409306, разведение 1:50), инкубировали при 4°C в течение 30 минут, дважды промывали, ресуспендировали клетки в буферном растворе FACS и, наконец, считывали значения сигналов флуоресценции на устройстве FACS CantoII (BD).
Результаты показали, что бивалентные биспецифические антитела к B7H3 118, 127 и 132 и отрицательное контрольное антитело NC2 (в котором был сохранен B7H3-связывающий домен, а CD3-связывающий домен был замещен неродственным антителом) способны к связыванию линии клеток A498 (с высокой экспрессией B7H3) (см. Фиг.2A), проявляющему градиент-зависимый эффект с более сильной связывающей способностью по сравнению с MGD009, и специфичны к мишени B7H3. Отрицательное контрольное антитело NC1 (в котором B7H3-связывающий домен был замещен неродственным антителом, но CD3-связывающий домен был сохранен) не связывается с A498. Аналогичным образом, биспецифические антитела 118, 127 и 132, MGD009 и NC2 активно связывались с линией клеток CT26/hB7H3 (см. Фиг.2B), но не связывались с линией клеток CT26, не экспрессирующей B7H3 (см. Фиг.2C), что также полностью демонстрирует, что исследуемые биспецифические антитела специфично связываются с мишенью B7H3 на поверхности клеточной мембраны. Антитела 118, 127 и 132 проявляют связывающую способность, отличающуюся от MGD009; такая разница в связывающей способности гораздо более значима на линии клеток CT26/hB7H3 с сверхэкспрессией B7H3, чем на линии клеток A498, что свидетельствует о более значимом преимуществе бивалентных биспецифических антител B7H3 для связывания с клетками с более высокой экспрессией B7H3 и их лучшем окне безопасности по сравнению с моновалентным биспецифическим антителом к B7H3, MGD009.
Биспецифические антитела 118, 127 и 132 и отрицательное контрольное антитело NC1 могут связываться с линией рекомбинантных клеток Jurkat (см. Фиг.2D), проявляя градиент-зависимый эффект.Среди этих антител 118 и NC1 обладают связывающей способностью по отношению к линии рекомбинантных клеток Jurkat, эквивалентной MGD009, при этом 127 и 132 проявляют значительно более слабую связывающую способность. Это может быть потому, что CD3-связывающий домен локализован между B7H3-связывающим доменом и FC, и некоторое стерическое затруднение может влиять на связывание с рекомбинантными клетками Jurkat. Отрицательное контрольное антитело NC2 без CD3-связывающего домена не связывается с рекомбинантными клетками Jurkat, что свидетельствует о специфичности связывания биспецифических антител с Jurkat к мишени CD3.
Тестовый пример 3. Анализ цитолитической активности МКПК in vitro
Опосредованная биспецифическими антителами цитолитическая активность МКПК в отношении опухолевых клеток была достигнута на основании количественного определения пролиферации клеток. Содержание АТФ, являющееся индикатором метаболизма живых клеток и прямо пропорциональная количеству клеток в культуре, определяли с помощью тест-системы Cell Titer-glo на клетках.
Использовали четыре различных линии клеток-мишеней (T), включающие три линии опухолевых клеток с различными уровнями экспрессии B7H3 (A498, U87 (Банк клеток Академии наук Китая, TCHu138), Detroit562 (ATCC, CCL-138)), и одну отрицательную контрольную линию клеток CHOK1, не экспрессирующую B7H3 (ATCC, CCL-61). Эффекторные клетки (E) представляли собой МКПК, полученные от здоровых добровольцев. Клетки-мишени высевали в 96-луночный планшет, культивировали в течение ночи, и на следующий день в каждую лунку добавляли равные количества только что выделенных МКПК и серийно разведенных исследуемых биспецифических антител (самая высокая конечная концентрация составляла 300 нмоль/л, разведение 1:3) или ФСБ (контроль с эффекторными клетками и клетками-мишенями без антитела). Ставили холостые контрольные пробы (холостая проба: только среда, среда без клеток или без антител). Соотношения E:T составляли 10:1, 5:1, 5:1 и 5:1 соответственно для клеток A498, U87, Detroit562 и CHOK1. Клетки инкубировали в течение 48 часов и детектировали с помощью тест-системы Cell Titer-glo (см. Инструкцию по применению). Значения сигналов считывали на ридере для микропланшетов и, наконец, преобразовывали в Степень ингибирования. Обработку и анализ данных выполняли с помощью программного обеспечения Graphpad Prism 5.
Степень ингибирования, % (% ингибирования)=100% - (значение сигналаобразец - значение сигналахолостая проба)/(значение сигналаконтроль - значение сигналахолостая проба).
3.1 Сравнение антител, содержащих различные антиген-связывающие домены CD3 с варьирующей аффинностью
Для конструирования различных биспецифических антител, которые проявляют различные цитолитические эффекты in vitro (см. Фиг.3A и Фиг.3B), использовали scFv к CD3 с различными значениями аффинности. Биспецифические антитела 155, 156, 185 и 186, содержащие HRH5 и HRH6 соответственно, проявляют самый слабый цитолитический эффект, что согласуется с результатами анализа аффинности Biacore.
3.2 Сравнение моновалентных и бивалентных биспецифических антител к B7H3
Сравнение структуры AFF3 (131 и 177 использовали в качестве примеров антител с этой структурой) и AF3 (181 и 187 использовали в качестве примеров антител с этой структурой) было выполнено в качестве иллюстративного примера биспецифических антител, сконструированных из scFv, содержащих различные вариабельные области тяжелой цепи антитела к CD3 (см. Фиг.4A и Фиг.4B). Бивалентные биспецифические антитела к B7H3 со структурой AFF3 CD3-B7H3 значимо усиливали цитолитическую активность in vitro по сравнению с моновалентными биспецифическими антителами к B7H3 со структурой AF3. Это применимо ко всем биспецифическим антителам, содержащим различные VH к CD3.
Таблица 12. Порядок расположения участков в антителе
3.3 Влияние различных структур бивалентных биспецифических антител к B7H3 на противоопухолевую цитолитическую активность
Молекулы бивалентных биспецифических антител к B7H3 161, 162, 113 и 126 (см. Фиг.5A) и 113 и 143 (см. Фиг.5B), который имеют одинаковые общие компоненты антигенсвязывающего домена, но различный порядок их расположения, параллельно тестировали на противоопухолевую цитолитическую активность. Все вышеуказанные молекулы имеют HRH2 в качестве антигенсвязывающего домена вариабельной области тяжелой цепи антитела к CD3. Результаты показывают, что все молекулы бивалентного биспецифического антитела к B7H3 с различным порядком расположения участков обладают значимыми цитолитическими эффектами в отношении клеток A498. Среди этих антител 161, 162, 113 и 126 обладали цитолитической активностью, эквивалентной или несколько повышенной активностью по сравнению с MGD009. Порядок расположения различных структур оказывал незначительное влияние на противоопухолевую цитолитическую активность бивалентных биспецифических антител к B7H3.
Таблица 13. Сравнение структуры различных исследуемых антител

3.4 Биспецифические антитела обладают цитолитическими эффектами в отношении линий опухолевых клеток с различными уровнями экспрессии B7H3
Три исследуемых биспецифических антитела 118, 127 и 132 тестировали на цитолитические эффекты in vitro на линиях опухолевых клеток A498, U87 и Detroit562. Цитолитический эффект проявлял положительную корреляцию с уровнем экспрессии B7H3. Например, 118 характеризовалось EC50 0,34, 2,4 и 14,5 нмоль/л в отношении A498, U87 и Detroit562 соответственно. Эту тенденцию проявляли все три молекулы антитела. Ни одно из биспецифических антител не обладало цитолитическим эффектом на B7H3-отрицательной контрольной линии клеток CHOK1, а отрицательное контрольное биспецифическое антитело NC1 не обладало цитолитическим эффектом ни на одной линии клеток-мишеней. Эти два аспекта в совокупности указывают на то, что цитолитическое действие на клетки специфично по отношению к мишени, что требует перенаправления эффекторных клеток на B7H3-положительные клетки-мишени под действием биспецифических антител.
Таблица 14. Перенаправление МКПК, опосредованное исследуемыми биспецифическими антителами, на цитолитическое действие на различные линии клеток-мишеней
3.5 Сравнение цитолитических эффектов биспецифических антител с различными структурами на клетках A498
Три исследуемых биспецифических антитела 127, 201 и 202 тестировали на цитолитические эффекты in vitro на линии опухолевых клеток A498. Результаты показывают (см. Фиг.5C), что все биспецифические антитела с тремя структурами обладают противоопухолевой цитолитической активностью, среди которых биспецифическое антитело 127 обладает превосходящей цитолитической активностью по сравнению с 201 или 202.
Тестовый пример 4. Анализ активации T-клеток in vitro
Чтобы определить активирующую T-клетки функцию биспецифических антител, измеряли экспрессию NFAT-управляемого гена-репортера люциферазы после активации Jurkat, используя линию рекомбинантных клеток Jurkat в присутствии или в отсутствие линии опухолевых клеток A498.
Клетки A498 высевали в 96-луночный планшет для клеточных культур (1 на 105/мл, 100 мкл/лунка) и помещали в инкубатор на 37°C, 5% CO2 на 20-24 ч. На следующий день после удаления супернатанта клеточной культуры в каждую лунку добавляли 90 мкл суспензии рекомбинантных клеток Jurkat (5,5 на 10 5/мл) и 10 мкл серийно разведенного исследуемого биспецифического антитела (самой высокой конечной концентрации 500 нмоль/л, градиентное разведение 1:3) и ставили отрицательный контроль (который представлял собой клетки A498 и рекомбинантные клетки Jurkat без антител) и холостой контроль (который представлял собой среду без клеток или без антител) и инкубировали в инкубаторе на 37°C, 5% CO2 в течение 5-6 часов. Для определения специфической активации неопухолевых рекомбинантных клеток Jurkat в холостую пробу 96-луночного планшета для клеточных культур непосредственно добавляли рекомбинантные клетки Jurkat и исследуемые антитела. После совместного культивирования в каждую лунку добавляли 100 мкл реагента Bright-Glo (Bright-Glo™ Luciferase Assay System, Promega, № по каталогу: E2620), помещали при комнатной температуре 5-10 минут и считывали сигналы хемилюминесценции на многофункциональном микропланшете. Кратность усиления флуоресценции рассчитывали по формуле:
Кратность усиления=(Сигналобразец - сигналхолостая проба)/Сигналконтроль - сигналхолостая проба).
4.1 Все бивалентные молекулы антител к B7H3 с различным порядком расположения участков могут эффективно активировать T-клетки
Бивалентные биспецифические антитела к B7H3 118, 127 и 132 тестировали на активацию рекомбинантных клеток Jurkat в присутствии или в отсутствие A498 для верификации эффектов специфичной и неспецифичной активации T-клеток биспецифическими антителами. Результаты показывают, что молекулы биспецифических антител к B7H3 118, 127 и 132 с различным порядком расположения участков могут эффективно активировать линию рекомбинантных клеток Jurkat и значимо индуцировать экспрессию люциферазы в присутствии линии опухолевых клеток A498 (см. Фиг.6A). Это демонстрирует, что активация рекомбинантных клеток Jurkat специфична к мишени B7H3, поскольку отрицательное контрольное антитело NC1 не может индуцировать экспрессию люциферазы. Для активации рекомбинантных клеток Jurkat посредством биспецифических антител требуется одновременное рекрутирование рекомбинантных клеток Jurkat, экспрессирующих CD3, и опухолевых клеток, экспрессирующих B7H3. В том случае, если рекомбинантные клетки Jurkat присутствуют отдельно в отсутствие клеток A498 (см. Фиг.6B), экспрессия люциферазы очень низка, и только несколько слабых сигналов можно детектировать в нескольких точках самой высокой концентрации антител.
Таблица 15. Порядок расположения участков в антителе
4.2 Сравнение моновалентных и бивалентных биспецифических антител к B7H3
Бивалентные биспецифические антитела к CD3-B7H3 обладают значимо усиленной мишень-специфической активацией T-клеток по сравнению с моновалентными антителами к B7H3, что согласуется с усиленной цитолитической способностью бивалентных молекул к B7H3 in vitro по сравнению с моновалентными молекулами к B7H3, как показано в тестовом примере 3. В то же время, неспецифичная к мишени активация T-клеток остается без изменений. Таким образом, бивалентная молекула к B7H3 (131) обладает более сильной эффективностью, чем моновалентная молекула B7H3 (181) (см. Фиг.7A), при этом побочные эффекты, вызываемые неспецифической активацией T-клеток, не усиливаются (см. Фиг.7B).
Таблица 16. Структура антитела
Тестовый пример 5. Анализ секреции цитокинов in vitro
Эффекторные клетки перенаправляются на клетки-мишени при посредстве биспецифического антитела и высвобождают цитокины при цитолизе клеток-мишеней. Секрецию цитокинов анализировали путем количественного определения содержания цитокинов (включая IL2, IFNγ и TNFα) в супернатанте клеточной культуры методом твердофазного иммуноферментного анализа (ELISA).
Дизайн эксперимента и используемые антитела были такими же, как описаны в тестовом примере 4. Супернатант клеточной культуры собирали в конце анализа на цитолиз in vitro и вносили в лунки 96-луночного планшета (Corning №3795), и хранили при минус 20°C для последующего использования. Для анализа ELISA доставали замороженный супернатант культуры, размораживали при комнатной температуре, центрифугировали при 3500 об/мин в течение 10 минут и собирали супернатант для анализа ELISA. Методики ELISA следовали инструкциям, поставляемым с набором (набор реагентов Human IL-2 ELISA, набор реагентов Human IFN-γ ELISA, набор реагентов Human TNF-α ELISA, Neobioscience, №№по каталогу EHC003.96, EHC102g.96, EHC103a.96).
Результаты показывают, что исследуемые биспецифические антитела могут эффективно индуцировать секрецию IL2, IFNγ и TNFα МКПК в присутствии и МКПК, и B7H3-положительных клеток-мишеней A498 (см. Фиг.8A-8C); среди этих антител MGD009 и 118 индуцируют самый высокий уровень секреции цитокинов, следующий уровень показали 127 и 132, а отрицательное контрольное антитело NC1 индуцирует секрецию цитокинов на уровне за пределами диапазона чувствительности обнаружения. MGD009 может значимо индуцировать высвобождение IFNγ и TNFα в трех точках самой высокой концентрации в присутствии и МКПК, и B7H3-отрицательных клеток CHOK1 (см. Фиг.9A-9C), при этом три исследуемых биспецифических антитела 118, 127 и 132 не могут индуцировать высвобождение IFNγ и TNFα, что указывает на лучшую безопасность трех исследуемых биспецифических антител по сравнению с MGD009 в плане секреции неспецифичных к мишени цитокинов.
Тестовый пример 6. Фармакокинетический тест в мышиной модели A498, реконструированной человеческими МКПК
В этом тестовом примере противоопухолевую эффективность исследуемых биспецифических антител к CD3-B7H3 по настоящему изобретению у мышей оценивали путем использования модели A498 (ATCC) на мышах NOG (Beijing Charles River Experimental Animal Co., Ltd.), реконструированной человеческими МКПК.
5×106 клеток/мышь/100 мкл (содержащих 50% матригеля) клеток A498 инокулировали подкожно в правый бок мышей NOG. Когда объем опухоли у несущих опухоли мышей достигал около 130-150 мм3, мышей случайным образом распределяли в группы по 5-6 животных на группу, и день распределения в группы определяли как день 0 эксперимента. В день 0 или в день 1 МКПК, выделенные в необработанном виде у двух добровольцев, смешивали в соотношении к 1:1 и вводили мышам NOG интраперитонеально 5 на 106 клеток/100 мкл; каждое антитело вводили интраперитонеально два раза в неделю, в общей сложности 6 доз. Мониторинг объемов опухоли и массы тела животных проводили два раза в неделю и регистрировали данные. Раствор-носитель означает отрицательную контрольную группу, которой вместо антитела вводили буферный раствор ФСБ.
Антитела 118 и 119 показали некоторую противоопухолевую эффективность в низких дозах (Фиг.10A) и проявляли дозозависимый эффект.Антитело 118 характеризовалось степенью ингибирования опухоли (tumor inhibition rate; TGI) 22,17% и 60,39% при дозах 0,01 мг/кг и 0,03 мг/кг соответственно по окончании эксперимента (день 20).
Антитело 113 показало некоторый противоопухолевый эффект в день 14, а степени ингибирования опухоли в группах, получавших дозы 0,6 мг/кг и 0,3 мг/кг, достигали 70,05% (p менее 0,05) и 60,78% (p<0,05) соответственно (Фиг.10B). На 20-й день противоопухолевые эффекты продолжали возрастать дозозависимым образом, и противоопухолевые коэффициенты составляют более 100% (p менее 0,001) и 77,92% (p менее 0,05) соответственно.
В условиях дозирования 0,12 мг/кг и 0,36 мг/кг (Фиг.10C) антитело 118 показало степень ингибирования опухоли 39,18% и 57,44% (p<0,001) при дозах,12 мг/кг и 0,36 мг/кг соответственно в день 12, а в день 21 степени ингибирования опухоли достигали 81,72% (p<0,01) и более 100% (p<0,001) соответственно. Среди этих результатов при дозе 0,36 мг/кг у одной мыши проявлялась даже полная регрессия опухоли (1/6).
В дозе 0,36 мг/кг (Фиг.10D) антитело 126 обладало степенью ингибирования опухоли 47,78% в день 21 (p<0,01). Антитело 128 показало значимый противоопухолевый эффект в день 19 (TGI=56,37%). К 21-му дню степень ингибирования опухоли повысился до 69,28% (p<0,001). Антитело 127 проявляло степень ингибирования опухоли 76,20% в день 12 (p<0,001), и эффект ингибирования опухоли продолжал возрастать к дню 21, а степень ингибирования опухоли составляет более 100% (p<0,001). У 3 из 5 животных объемы опухоли регрессировали по сравнению с днем распределения в группы, а объемы опухоли у 2 других животных полностью регрессировали.
Противоопухолевая активность антитела 127 повторилась в другом эксперименте (Фиг.10E). Степень ингибирования опухоли достигал 90,6% в день 14 (p<0,001) и возрастал до 95,80% в день 17 (p<0,001). 127 был еще эффективен в более низкой дозе (0,12 мг/кг) и при более низкой частоте введения (один раз в неделю, 127-0,36 мг/кг - 1 р/нед.), при этом степень ингибирования опухоли в день 17 достигали 51,37% (p<0,001) и 96,20% (p<0,001) соответственно.
Тестовый пример 7. Фармакодинамический тест в модели мышей hCD3 KI
В этом эксперименте мышам Balb/c-hCD3 инокулировали подкожно линию опухолевых клеток CT26-hB7H3 (клетки CT26 были получены из Банка клеток Академии наук Китая, TCM37, а клетки CT26-hB7H3 были получены путем экспрессии hB7H3) для оценки ингибирующего действия биспецифических антител CD3-B7H3 по настоящему изобретению на рост опухоли у мышей.
Самок трансгенных мышей hCD3E Balb/c приобретали в Центре исследования моделей на животных Нанкинского университета (номер сертификата 201801374/5/6, лицензия SCXK (Цзянсу) 2015-0001).
Клетки CT26-hB7H3 (8 на 105 клеток/мышь/100 мкл) инокулировали подкожно в правый бок мышей hCD3. Когда объем опухоли у несущих опухоли мышей достигал около 80-120 мм3, мышей случайным образом делили на различные группы по 7 мышей на группу. День распределения в группы определяли как день 0 эксперимента и выполняли интраперитонеальное введение каждого антитела два раза в неделю, в общей сложности 5 доз. Мониторинг объемов опухоли и массы тела животных проводили два раза в неделю и регистрировали данные. Раствор-носитель означает отрицательную контрольную группу, которой вместо антитела вводили буферный раствор ФСБ.
Результаты показывают, что антитело 118 проявляет выраженную эффективность после начального введения в дозе 1 мг/кг (Фиг.11A), и степень ингибирования опухоли достигал 38,34% в день 13 (p менее 0,05).
Антитело 132 проявляло тенденцию к ингибированию роста опухоли в дозе 3,6 мг/кг (Фиг.11B), и степень ингибирования опухоли достигал 26,35% в день 13.
Тестовый пример 8. ФК эксперимент на крысах
В этом эксперименте для оценки метаболизма биспецифических антител к CD3-B7H3 у крыс линии SD (Спрег-Доули) вводили биспецифические антитела к CD3-B7H3 в хвостовую вену крыс линии SD и определяли концентрации антитела в сыворотке крови крыс в различных точках контроля.
Исследуемые лекарственные препараты вводили в хвостовую вену крыс в дозе 3 мг/кг, а объем введения составлял 5 мл/кг. Кровь собирали в различных точках контроля, то есть перед введением и через 5 мин, 8 ч, 1 д, 2 д, 4 д, 7 д, 10 д, 14 д, 21 д, 28 д после введения. Концентрации антитела в сыворотке крови определяли методом ELISA. Использовали два различных метода ELISA, где антиген B7H3 (1 мкг/мл) или антиген CD3 (1 мкг/мл) наносили на планшет, а антитело к человеческому Fc-HRP (Abcam, ab98624) использовали в качестве вторичного антитела. Параметры фармакокинетики исследуемых лекарственных препаратов рассчитывали с помощью программного обеспечения Winnolin, и полученные в результате параметры фармакокинетики показаны в таблице 17.
Антитела 118, 127 и 132 обладали периодом полувыведения 4,9-8,1 дня в отношении антигенсвязывающей области для B7H3, несколько дольше, чем MGD009, и достигающим уровня обычных антител IgG; и периодом полувыведения 3,2-5,6 дней в отношении антигенсвязывающей области для CD3. При этом кинетические параметры двух различных антигенсвязывающих участков для B7H3 и CD3 в антителе 118 различаются незначительно, что свидетельствует о благоприятной целостности молекулы in vivo, и их периоды полувыведения составляют соответственно 4,9 и 4,4 дня. Антитело 127 имеет периоды полувыведения 4,9 и 3,2 дня соответственно в отношении двух различных антигенсвязывающих областей для B7H3 и CD3. Различия в количественной экспозиции и скорости клиренса очевидны, при этом показатели для участка CD3 хуже, чем для участка B7H3. Вероятнее, это вызвано ослаблением связывающей способности CD3, чем распадом молекулы, поскольку участок CD3 находится внутри молекулярной структуры. Антитело 132 было получено путем включения пары дисульфидных связей в scFv к B7H3 на основе молекулярной последовательности антитела 127. Эта модификация значительно увеличивает период полувыведения молекулы (65-75%), а также значительно улучшает количество экспозиции и скорость клиренса.
Таблица 17. Основные фармакокинетические параметры у крыс
Покрытый
Покрытый
Покрытый
Покрытый
Покрытый
Покрытый
Покрытый
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЦЗЯНСУ ХЭНЖУЙ МЕДСИН КО., ЛТД.,
ШАНХАЙ ХЭНЖУЙ ФАРМАСЬЮТИКАЛ КО., ЛТД.
<120> АНТИТЕЛО К CD3 И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ
<150> 201811491781.3
<151> 2018-12-7
<160> 104
<170> SIPOSequenceListing 1.0
<210> 1
<211> 439
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> человеческий антиген B7H3 для обнаружения
<400> 1
Leu Glu Val Gln Val Pro Glu Asp Pro Val Val Ala Leu Val Gly Thr
1 5 10 15
Asp Ala Thr Leu Cys Cys Ser Phe Ser Pro Glu Pro Gly Phe Ser Leu
20 25 30
Ala Gln Leu Asn Leu Ile Trp Gln Leu Thr Asp Thr Lys Gln Leu Val
35 40 45
His Ser Phe Ala Glu Gly Gln Asp Gln Gly Ser Ala Tyr Ala Asn Arg
50 55 60
Thr Ala Leu Phe Pro Asp Leu Leu Ala Gln Gly Asn Ala Ser Leu Arg
65 70 75 80
Leu Gln Arg Val Arg Val Ala Asp Glu Gly Ser Phe Thr Cys Phe Val
85 90 95
Ser Ile Arg Asp Phe Gly Ser Ala Ala Val Ser Leu Gln Val Ala Ala
100 105 110
Pro Tyr Ser Lys Pro Ser Met Thr Leu Glu Pro Asn Lys Asp Leu Arg
115 120 125
Pro Gly Asp Thr Val Thr Ile Thr Cys Ser Ser Tyr Gln Gly Tyr Pro
130 135 140
Glu Ala Glu Val Phe Trp Gln Asp Gly Gln Gly Val Pro Leu Thr Gly
145 150 155 160
Asn Val Thr Thr Ser Gln Met Ala Asn Glu Gln Gly Leu Phe Asp Val
165 170 175
His Ser Ile Leu Arg Val Val Leu Gly Ala Asn Gly Thr Tyr Ser Cys
180 185 190
Leu Val Arg Asn Pro Val Leu Gln Gln Asp Ala His Ser Ser Val Thr
195 200 205
Ile Thr Pro Gln Arg Ser Pro Thr Gly Ala Val Glu Val Gln Val Pro
210 215 220
Glu Asp Pro Val Val Ala Leu Val Gly Thr Asp Ala Thr Leu Arg Cys
225 230 235 240
Ser Phe Ser Pro Glu Pro Gly Phe Ser Leu Ala Gln Leu Asn Leu Ile
245 250 255
Trp Gln Leu Thr Asp Thr Lys Gln Leu Val His Ser Phe Thr Glu Gly
260 265 270
Arg Asp Gln Gly Ser Ala Tyr Ala Asn Arg Thr Ala Leu Phe Pro Asp
275 280 285
Leu Leu Ala Gln Gly Asn Ala Ser Leu Arg Leu Gln Arg Val Arg Val
290 295 300
Ala Asp Glu Gly Ser Phe Thr Cys Phe Val Ser Ile Arg Asp Phe Gly
305 310 315 320
Ser Ala Ala Val Ser Leu Gln Val Ala Ala Pro Tyr Ser Lys Pro Ser
325 330 335
Met Thr Leu Glu Pro Asn Lys Asp Leu Arg Pro Gly Asp Thr Val Thr
340 345 350
Ile Thr Cys Ser Ser Tyr Arg Gly Tyr Pro Glu Ala Glu Val Phe Trp
355 360 365
Gln Asp Gly Gln Gly Val Pro Leu Thr Gly Asn Val Thr Thr Ser Gln
370 375 380
Met Ala Asn Glu Gln Gly Leu Phe Asp Val His Ser Val Leu Arg Val
385 390 395 400
Val Leu Gly Ala Asn Gly Thr Tyr Ser Cys Leu Val Arg Asn Pro Val
405 410 415
Leu Gln Gln Asp Ala His Gly Ser Val Thr Ile Thr Gly Gln Pro Met
420 425 430
Thr His His His His His His
435
<210> 2
<211> 443
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> обезьяний антиген B7H3 для обнаружения
<400> 2
Leu Glu Val Gln Val Pro Glu Asp Pro Val Val Ala Leu Val Gly Thr
1 5 10 15
Asp Ala Thr Leu Arg Cys Ser Phe Ser Pro Glu Pro Gly Phe Ser Leu
20 25 30
Ala Gln Leu Asn Leu Ile Trp Gln Leu Thr Asp Thr Lys Gln Leu Val
35 40 45
His Ser Phe Thr Glu Gly Arg Asp Gln Gly Ser Ala Tyr Ala Asn Arg
50 55 60
Thr Ala Leu Phe Leu Asp Leu Leu Ala Gln Gly Asn Ala Ser Leu Arg
65 70 75 80
Leu Gln Arg Val Arg Val Ala Asp Glu Gly Ser Phe Thr Cys Phe Val
85 90 95
Ser Ile Arg Asp Phe Gly Ser Ala Ala Val Ser Leu Gln Val Ala Ala
100 105 110
Pro Tyr Ser Lys Pro Ser Met Thr Leu Glu Pro Asn Lys Asp Leu Arg
115 120 125
Pro Gly Asp Thr Val Thr Ile Thr Cys Ser Ser Tyr Arg Gly Tyr Pro
130 135 140
Glu Ala Glu Val Phe Trp Gln Asp Gly Gln Gly Ala Pro Leu Thr Gly
145 150 155 160
Asn Val Thr Thr Ser Gln Met Ala Asn Glu Gln Gly Leu Phe Asp Val
165 170 175
His Ser Val Leu Arg Val Val Leu Gly Ala Asn Gly Thr Tyr Ser Cys
180 185 190
Leu Val Arg Asn Pro Val Leu Gln Gln Asp Ala His Gly Ser Ile Thr
195 200 205
Ile Thr Pro Gln Arg Ser Pro Thr Gly Ala Val Glu Val Gln Val Pro
210 215 220
Glu Asp Pro Val Val Ala Leu Val Gly Thr Asp Ala Thr Leu Arg Cys
225 230 235 240
Ser Phe Ser Pro Glu Pro Gly Phe Ser Leu Ala Gln Leu Asn Leu Ile
245 250 255
Trp Gln Leu Thr Asp Thr Lys Gln Leu Val His Ser Phe Thr Glu Gly
260 265 270
Arg Asp Gln Gly Ser Ala Tyr Ala Asn Arg Thr Ala Leu Phe Leu Asp
275 280 285
Leu Leu Ala Gln Gly Asn Ala Ser Leu Arg Leu Gln Arg Val Arg Val
290 295 300
Ala Asp Glu Gly Ser Phe Thr Cys Phe Val Ser Ile Arg Asp Phe Gly
305 310 315 320
Ser Ala Ala Val Ser Leu Gln Val Ala Ala Pro Tyr Ser Lys Pro Ser
325 330 335
Met Thr Leu Glu Pro Asn Lys Asp Leu Arg Pro Gly Asp Thr Val Thr
340 345 350
Ile Thr Cys Ser Ser Tyr Arg Gly Tyr Pro Glu Ala Glu Val Phe Trp
355 360 365
Gln Asp Gly Gln Gly Ala Pro Leu Thr Gly Asn Val Thr Thr Ser Gln
370 375 380
Met Ala Asn Glu Gln Gly Leu Phe Asp Val His Ser Val Leu Arg Val
385 390 395 400
Val Leu Gly Ala Asn Gly Thr Tyr Ser Cys Leu Val Arg Asn Pro Val
405 410 415
Leu Gln Gln Asp Ala His Gly Ser Val Thr Ile Thr Gly Gln Pro Met
420 425 430
Thr Phe Pro Pro Glu His His His His His His
435 440
<210> 3
<211> 222
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> мышиный антиген B7H3 для обнаружения
<400> 3
Val Glu Val Gln Val Ser Glu Asp Pro Val Val Ala Leu Val Asp Thr
1 5 10 15
Asp Ala Thr Leu Arg Cys Ser Phe Ser Pro Glu Pro Gly Phe Ser Leu
20 25 30
Ala Gln Leu Asn Leu Ile Trp Gln Leu Thr Asp Thr Lys Gln Leu Val
35 40 45
His Ser Phe Thr Glu Gly Arg Asp Gln Gly Ser Ala Tyr Ser Asn Arg
50 55 60
Thr Ala Leu Phe Pro Asp Leu Leu Val Gln Gly Asn Ala Ser Leu Arg
65 70 75 80
Leu Gln Arg Val Arg Val Thr Asp Glu Gly Ser Tyr Thr Cys Phe Val
85 90 95
Ser Ile Gln Asp Phe Asp Ser Ala Ala Val Ser Leu Gln Val Ala Ala
100 105 110
Pro Tyr Ser Lys Pro Ser Met Thr Leu Glu Pro Asn Lys Asp Leu Arg
115 120 125
Pro Gly Asn Met Val Thr Ile Thr Cys Ser Ser Tyr Gln Gly Tyr Pro
130 135 140
Glu Ala Glu Val Phe Trp Lys Asp Gly Gln Gly Val Pro Leu Thr Gly
145 150 155 160
Asn Val Thr Thr Ser Gln Met Ala Asn Glu Arg Gly Leu Phe Asp Val
165 170 175
His Ser Val Leu Arg Val Val Leu Gly Ala Asn Gly Thr Tyr Ser Cys
180 185 190
Leu Val Arg Asn Pro Val Leu Gln Gln Asp Ala His Gly Ser Val Thr
195 200 205
Ile Thr Gly Gln Pro Leu Thr Phe His His His His His His
210 215 220
<210> 4
<211> 534
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> полноразмерная аминокислотная последовательность человеческого
B7H3
<400> 4
Met Leu Arg Arg Arg Gly Ser Pro Gly Met Gly Val His Val Gly Ala
1 5 10 15
Ala Leu Gly Ala Leu Trp Phe Cys Leu Thr Gly Ala Leu Glu Val Gln
20 25 30
Val Pro Glu Asp Pro Val Val Ala Leu Val Gly Thr Asp Ala Thr Leu
35 40 45
Cys Cys Ser Phe Ser Pro Glu Pro Gly Phe Ser Leu Ala Gln Leu Asn
50 55 60
Leu Ile Trp Gln Leu Thr Asp Thr Lys Gln Leu Val His Ser Phe Ala
65 70 75 80
Glu Gly Gln Asp Gln Gly Ser Ala Tyr Ala Asn Arg Thr Ala Leu Phe
85 90 95
Pro Asp Leu Leu Ala Gln Gly Asn Ala Ser Leu Arg Leu Gln Arg Val
100 105 110
Arg Val Ala Asp Glu Gly Ser Phe Thr Cys Phe Val Ser Ile Arg Asp
115 120 125
Phe Gly Ser Ala Ala Val Ser Leu Gln Val Ala Ala Pro Tyr Ser Lys
130 135 140
Pro Ser Met Thr Leu Glu Pro Asn Lys Asp Leu Arg Pro Gly Asp Thr
145 150 155 160
Val Thr Ile Thr Cys Ser Ser Tyr Gln Gly Tyr Pro Glu Ala Glu Val
165 170 175
Phe Trp Gln Asp Gly Gln Gly Val Pro Leu Thr Gly Asn Val Thr Thr
180 185 190
Ser Gln Met Ala Asn Glu Gln Gly Leu Phe Asp Val His Ser Ile Leu
195 200 205
Arg Val Val Leu Gly Ala Asn Gly Thr Tyr Ser Cys Leu Val Arg Asn
210 215 220
Pro Val Leu Gln Gln Asp Ala His Ser Ser Val Thr Ile Thr Pro Gln
225 230 235 240
Arg Ser Pro Thr Gly Ala Val Glu Val Gln Val Pro Glu Asp Pro Val
245 250 255
Val Ala Leu Val Gly Thr Asp Ala Thr Leu Arg Cys Ser Phe Ser Pro
260 265 270
Glu Pro Gly Phe Ser Leu Ala Gln Leu Asn Leu Ile Trp Gln Leu Thr
275 280 285
Asp Thr Lys Gln Leu Val His Ser Phe Thr Glu Gly Arg Asp Gln Gly
290 295 300
Ser Ala Tyr Ala Asn Arg Thr Ala Leu Phe Pro Asp Leu Leu Ala Gln
305 310 315 320
Gly Asn Ala Ser Leu Arg Leu Gln Arg Val Arg Val Ala Asp Glu Gly
325 330 335
Ser Phe Thr Cys Phe Val Ser Ile Arg Asp Phe Gly Ser Ala Ala Val
340 345 350
Ser Leu Gln Val Ala Ala Pro Tyr Ser Lys Pro Ser Met Thr Leu Glu
355 360 365
Pro Asn Lys Asp Leu Arg Pro Gly Asp Thr Val Thr Ile Thr Cys Ser
370 375 380
Ser Tyr Arg Gly Tyr Pro Glu Ala Glu Val Phe Trp Gln Asp Gly Gln
385 390 395 400
Gly Val Pro Leu Thr Gly Asn Val Thr Thr Ser Gln Met Ala Asn Glu
405 410 415
Gln Gly Leu Phe Asp Val His Ser Val Leu Arg Val Val Leu Gly Ala
420 425 430
Asn Gly Thr Tyr Ser Cys Leu Val Arg Asn Pro Val Leu Gln Gln Asp
435 440 445
Ala His Gly Ser Val Thr Ile Thr Gly Gln Pro Met Thr Phe Pro Pro
450 455 460
Glu Ala Leu Trp Val Thr Val Gly Leu Ser Val Cys Leu Ile Ala Leu
465 470 475 480
Leu Val Ala Leu Ala Phe Val Cys Trp Arg Lys Ile Lys Gln Ser Cys
485 490 495
Glu Glu Glu Asn Ala Gly Ala Glu Asp Gln Asp Gly Glu Gly Glu Gly
500 505 510
Ser Lys Thr Ala Leu Gln Pro Leu Lys His Ser Asp Ser Lys Glu Asp
515 520 525
Asp Gly Gln Glu Ile Ala
530
<210> 5
<211> 534
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> полноразмерная аминокислотная последовательность обезьянего
B7H3
<400> 5
Met Leu His Arg Arg Gly Ser Pro Gly Met Gly Val His Val Gly Ala
1 5 10 15
Ala Leu Gly Ala Leu Trp Phe Cys Leu Thr Gly Ala Leu Glu Val Gln
20 25 30
Val Pro Glu Asp Pro Val Val Ala Leu Val Gly Thr Asp Ala Thr Leu
35 40 45
Arg Cys Ser Phe Ser Pro Glu Pro Gly Phe Ser Leu Ala Gln Leu Asn
50 55 60
Leu Ile Trp Gln Leu Thr Asp Thr Lys Gln Leu Val His Ser Phe Thr
65 70 75 80
Glu Gly Arg Asp Gln Gly Ser Ala Tyr Ala Asn Arg Thr Ala Leu Phe
85 90 95
Leu Asp Leu Leu Ala Gln Gly Asn Ala Ser Leu Arg Leu Gln Arg Val
100 105 110
Arg Val Ala Asp Glu Gly Ser Phe Thr Cys Phe Val Ser Ile Arg Asp
115 120 125
Phe Gly Ser Ala Ala Val Ser Leu Gln Val Ala Ala Pro Tyr Ser Lys
130 135 140
Pro Ser Met Thr Leu Glu Pro Asn Lys Asp Leu Arg Pro Gly Asp Thr
145 150 155 160
Val Thr Ile Thr Cys Ser Ser Tyr Arg Gly Tyr Pro Glu Ala Glu Val
165 170 175
Phe Trp Gln Asp Gly Gln Gly Ala Pro Leu Thr Gly Asn Val Thr Thr
180 185 190
Ser Gln Met Ala Asn Glu Gln Gly Leu Phe Asp Val His Ser Val Leu
195 200 205
Arg Val Val Leu Gly Ala Asn Gly Thr Tyr Ser Cys Leu Val Arg Asn
210 215 220
Pro Val Leu Gln Gln Asp Ala His Gly Ser Ile Thr Ile Thr Pro Gln
225 230 235 240
Arg Ser Pro Thr Gly Ala Val Glu Val Gln Val Pro Glu Asp Pro Val
245 250 255
Val Ala Leu Val Gly Thr Asp Ala Thr Leu Arg Cys Ser Phe Ser Pro
260 265 270
Glu Pro Gly Phe Ser Leu Ala Gln Leu Asn Leu Ile Trp Gln Leu Thr
275 280 285
Asp Thr Lys Gln Leu Val His Ser Phe Thr Glu Gly Arg Asp Gln Gly
290 295 300
Ser Ala Tyr Ala Asn Arg Thr Ala Leu Phe Leu Asp Leu Leu Ala Gln
305 310 315 320
Gly Asn Ala Ser Leu Arg Leu Gln Arg Val Arg Val Ala Asp Glu Gly
325 330 335
Ser Phe Thr Cys Phe Val Ser Ile Arg Asp Phe Gly Ser Ala Ala Val
340 345 350
Ser Leu Gln Val Ala Ala Pro Tyr Ser Lys Pro Ser Met Thr Leu Glu
355 360 365
Pro Asn Lys Asp Leu Arg Pro Gly Asp Thr Val Thr Ile Thr Cys Ser
370 375 380
Ser Tyr Arg Gly Tyr Pro Glu Ala Glu Val Phe Trp Gln Asp Gly Gln
385 390 395 400
Gly Ala Pro Leu Thr Gly Asn Val Thr Thr Ser Gln Met Ala Asn Glu
405 410 415
Gln Gly Leu Phe Asp Val His Ser Val Leu Arg Val Val Leu Gly Ala
420 425 430
Asn Gly Thr Tyr Ser Cys Leu Val Arg Asn Pro Val Leu Gln Gln Asp
435 440 445
Ala His Gly Ser Val Thr Ile Thr Gly Gln Pro Met Thr Phe Pro Pro
450 455 460
Glu Ala Leu Trp Val Thr Val Gly Leu Ser Val Cys Leu Val Ala Leu
465 470 475 480
Leu Val Ala Leu Ala Phe Val Cys Trp Arg Lys Ile Lys Gln Ser Cys
485 490 495
Glu Glu Glu Asn Ala Gly Ala Glu Asp Gln Asp Gly Glu Gly Glu Gly
500 505 510
Ser Lys Thr Ala Leu Gln Pro Leu Lys His Ser Asp Ser Lys Glu Asp
515 520 525
Asp Gly Gln Glu Leu Ala
530
<210> 6
<211> 316
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> полноразмерная аминокислотная последовательность мышиного
B7H3
<400> 6
Met Leu Arg Gly Trp Gly Gly Pro Ser Val Gly Val Cys Val Arg Thr
1 5 10 15
Ala Leu Gly Val Leu Cys Leu Cys Leu Thr Gly Ala Val Glu Val Gln
20 25 30
Val Ser Glu Asp Pro Val Val Ala Leu Val Asp Thr Asp Ala Thr Leu
35 40 45
Arg Cys Ser Phe Ser Pro Glu Pro Gly Phe Ser Leu Ala Gln Leu Asn
50 55 60
Leu Ile Trp Gln Leu Thr Asp Thr Lys Gln Leu Val His Ser Phe Thr
65 70 75 80
Glu Gly Arg Asp Gln Gly Ser Ala Tyr Ser Asn Arg Thr Ala Leu Phe
85 90 95
Pro Asp Leu Leu Val Gln Gly Asn Ala Ser Leu Arg Leu Gln Arg Val
100 105 110
Arg Val Thr Asp Glu Gly Ser Tyr Thr Cys Phe Val Ser Ile Gln Asp
115 120 125
Phe Asp Ser Ala Ala Val Ser Leu Gln Val Ala Ala Pro Tyr Ser Lys
130 135 140
Pro Ser Met Thr Leu Glu Pro Asn Lys Asp Leu Arg Pro Gly Asn Met
145 150 155 160
Val Thr Ile Thr Cys Ser Ser Tyr Gln Gly Tyr Pro Glu Ala Glu Val
165 170 175
Phe Trp Lys Asp Gly Gln Gly Val Pro Leu Thr Gly Asn Val Thr Thr
180 185 190
Ser Gln Met Ala Asn Glu Arg Gly Leu Phe Asp Val His Ser Val Leu
195 200 205
Arg Val Val Leu Gly Ala Asn Gly Thr Tyr Ser Cys Leu Val Arg Asn
210 215 220
Pro Val Leu Gln Gln Asp Ala His Gly Ser Val Thr Ile Thr Gly Gln
225 230 235 240
Pro Leu Thr Phe Pro Pro Glu Ala Leu Trp Val Thr Val Gly Leu Ser
245 250 255
Val Cys Leu Val Val Leu Leu Val Ala Leu Ala Phe Val Cys Trp Arg
260 265 270
Lys Ile Lys Gln Ser Cys Glu Glu Glu Asn Ala Gly Ala Glu Asp Gln
275 280 285
Asp Gly Asp Gly Glu Gly Ser Lys Thr Ala Leu Arg Pro Leu Lys Pro
290 295 300
Ser Glu Asn Lys Glu Asp Asp Gly Gln Glu Ile Ala
305 310 315
<210> 7
<211> 119
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> h1702 VH
<400> 7
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Ala Leu Val Thr Val Ser Ser
115
<210> 8
<211> 110
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> h1702 VL
<400> 8
Gln Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser
20 25 30
His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Met
35 40 45
Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Ile His Val Asp Arg
85 90 95
Asp Ile Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 9
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> h1702 HCDR1
<400> 9
Gly Phe Ile Phe Ser Ser Ser Ala
1 5
<210> 10
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> h1702 HCDR2
<400> 10
Ile Ser Tyr Asp Gly Ser Asn Lys
1 5
<210> 11
<211> 12
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> h1702 HCDR3
<400> 11
Ala Arg Ser Ala Arg Leu Tyr Ala Ser Phe Asp Tyr
1 5 10
<210> 12
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> h1702 LCDR1
<400> 12
Ser Gly Ser Val Ser Thr Ser His Tyr
1 5
<210> 13
<211> 3
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> h1702 LCDR2
<400> 13
Asn Thr Asn
1
<210> 14
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> h1702 LCDR3
<400> 14
Ala Ile His Val Asp Arg Asp Ile Trp Val
1 5 10
<210> 15
<211> 119
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> B7H3 VH44C
<400> 15
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Ala Leu Val Thr Val Ser Ser
115
<210> 16
<211> 110
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> B7H3 VL103C
<400> 16
Gln Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser
20 25 30
His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Met
35 40 45
Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Ile His Val Asp Arg
85 90 95
Asp Ile Trp Val Phe Gly Cys Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 17
<211> 110
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> Человеческий эпсилон CD3
<400> 17
Asp Gly Asn Glu Glu Met Gly Gly Ile Thr Gln Thr Pro Tyr Lys Val
1 5 10 15
Ser Ile Ser Gly Thr Thr Val Ile Leu Thr Cys Pro Gln Tyr Pro Gly
20 25 30
Ser Glu Ile Leu Trp Gln His Asn Asp Lys Asn Ile Gly Gly Asp Glu
35 40 45
Asp Asp Lys Asn Ile Gly Ser Asp Glu Asp His Leu Ser Leu Lys Glu
50 55 60
Phe Ser Glu Leu Glu Gln Ser Gly Tyr Tyr Val Cys Tyr Pro Arg Gly
65 70 75 80
Ser Lys Pro Glu Asp Ala Asn Phe Tyr Leu Tyr Leu Arg Ala Arg Val
85 90 95
Cys Glu Asn Cys Met Glu Met Asp His His His His His His
100 105 110
<210> 18
<211> 92
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> Человеческий дельта CD3
<400> 18
Phe Lys Ile Pro Ile Glu Glu Leu Glu Asp Arg Val Phe Val Asn Cys
1 5 10 15
Asn Thr Ser Ile Thr Trp Val Glu Gly Thr Val Gly Thr Leu Leu Ser
20 25 30
Asp Ile Thr Arg Leu Asp Leu Gly Lys Arg Ile Leu Asp Pro Arg Gly
35 40 45
Ile Tyr Arg Cys Asn Gly Thr Asp Ile Tyr Lys Asp Lys Glu Ser Thr
50 55 60
Val Gln Val His Tyr Arg Met Cys Gln Ser Cys Val Glu Leu Asp Pro
65 70 75 80
Ala Thr Val Ala Asp Tyr Lys Asp Asp Asp Asp Lys
85 90
<210> 19
<211> 102
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> Человеческий эпсилон CD3
<400> 19
Gln Asp Gly Asn Glu Glu Met Gly Ser Ile Thr Gln Thr Pro Tyr Gln
1 5 10 15
Val Ser Ile Ser Gly Thr Thr Val Ile Leu Thr Cys Ser Gln His Leu
20 25 30
Gly Ser Glu Ala Gln Trp Gln His Asn Gly Lys Asn Lys Glu Asp Ser
35 40 45
Gly Asp Arg Leu Phe Leu Pro Glu Phe Ser Glu Met Glu Gln Ser Gly
50 55 60
Tyr Tyr Val Cys Tyr Pro Arg Gly Ser Asn Pro Glu Asp Ala Ser His
65 70 75 80
His Leu Tyr Leu Lys Ala Arg Val Cys Glu Asn Cys Met Glu Met Asp
85 90 95
His His His His His His
100
<210> 20
<211> 92
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> Человеческий дельта CD3
<400> 20
Phe Lys Ile Pro Val Glu Glu Leu Glu Asp Arg Val Phe Val Lys Cys
1 5 10 15
Asn Thr Ser Val Thr Trp Val Glu Gly Thr Val Gly Thr Leu Leu Thr
20 25 30
Asn Asn Thr Arg Leu Asp Leu Gly Lys Arg Ile Leu Asp Pro Arg Gly
35 40 45
Ile Tyr Arg Cys Asn Gly Thr Asp Ile Tyr Lys Asp Lys Glu Ser Ala
50 55 60
Val Gln Val His Tyr Arg Met Cys Gln Asn Cys Val Glu Leu Asp Pro
65 70 75 80
Ala Thr Leu Ala Asp Tyr Lys Asp Asp Asp Asp Lys
85 90
<210> 21
<211> 93
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> Эпсилон CD3 мыши
<400> 21
Asp Asp Ala Glu Asn Ile Glu Tyr Lys Val Ser Ile Ser Gly Thr Ser
1 5 10 15
Val Glu Leu Thr Cys Pro Leu Asp Ser Asp Glu Asn Leu Lys Trp Glu
20 25 30
Lys Asn Gly Gln Glu Leu Pro Gln Lys His Asp Lys His Leu Val Leu
35 40 45
Gln Asp Phe Ser Glu Val Glu Asp Ser Gly Tyr Tyr Val Cys Tyr Thr
50 55 60
Pro Ala Ser Asn Lys Asn Thr Tyr Leu Tyr Leu Lys Ala Arg Val Cys
65 70 75 80
Glu Tyr Cys Val Glu Val Asp His His His His His His
85 90
<210> 22
<211> 92
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> Дельта CD3 мыши
<400> 22
Phe Lys Ile Gln Val Thr Glu Tyr Glu Asp Lys Val Phe Val Thr Cys
1 5 10 15
Asn Thr Ser Val Met His Leu Asp Gly Thr Val Glu Gly Trp Phe Ala
20 25 30
Lys Asn Lys Thr Leu Asn Leu Gly Lys Gly Val Leu Asp Pro Arg Gly
35 40 45
Ile Tyr Leu Cys Asn Gly Thr Glu Gln Leu Ala Lys Val Val Ser Ser
50 55 60
Val Gln Val His Tyr Arg Met Cys Gln Asn Cys Val Glu Leu Asp Ser
65 70 75 80
Gly Thr Met Ala Asp Tyr Lys Asp Asp Asp Asp Lys
85 90
<210> 23
<211> 207
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> полноразмерная аминокислотная последовательность человеческого
эпсилон CD3
<400> 23
Met Gln Ser Gly Thr His Trp Arg Val Leu Gly Leu Cys Leu Leu Ser
1 5 10 15
Val Gly Val Trp Gly Gln Asp Gly Asn Glu Glu Met Gly Gly Ile Thr
20 25 30
Gln Thr Pro Tyr Lys Val Ser Ile Ser Gly Thr Thr Val Ile Leu Thr
35 40 45
Cys Pro Gln Tyr Pro Gly Ser Glu Ile Leu Trp Gln His Asn Asp Lys
50 55 60
Asn Ile Gly Gly Asp Glu Asp Asp Lys Asn Ile Gly Ser Asp Glu Asp
65 70 75 80
His Leu Ser Leu Lys Glu Phe Ser Glu Leu Glu Gln Ser Gly Tyr Tyr
85 90 95
Val Cys Tyr Pro Arg Gly Ser Lys Pro Glu Asp Ala Asn Phe Tyr Leu
100 105 110
Tyr Leu Arg Ala Arg Val Cys Glu Asn Cys Met Glu Met Asp Val Met
115 120 125
Ser Val Ala Thr Ile Val Ile Val Asp Ile Cys Ile Thr Gly Gly Leu
130 135 140
Leu Leu Leu Val Tyr Tyr Trp Ser Lys Asn Arg Lys Ala Lys Ala Lys
145 150 155 160
Pro Val Thr Arg Gly Ala Gly Ala Gly Gly Arg Gln Arg Gly Gln Asn
165 170 175
Lys Glu Arg Pro Pro Pro Val Pro Asn Pro Asp Tyr Glu Pro Ile Arg
180 185 190
Lys Gly Gln Arg Asp Leu Tyr Ser Gly Leu Asn Gln Arg Arg Ile
195 200 205
<210> 24
<211> 171
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> полноразмерная аминокислотная последовательность человеческого
дельта CD3
<400> 24
Met Glu His Ser Thr Phe Leu Ser Gly Leu Val Leu Ala Thr Leu Leu
1 5 10 15
Ser Gln Val Ser Pro Phe Lys Ile Pro Ile Glu Glu Leu Glu Asp Arg
20 25 30
Val Phe Val Asn Cys Asn Thr Ser Ile Thr Trp Val Glu Gly Thr Val
35 40 45
Gly Thr Leu Leu Ser Asp Ile Thr Arg Leu Asp Leu Gly Lys Arg Ile
50 55 60
Leu Asp Pro Arg Gly Ile Tyr Arg Cys Asn Gly Thr Asp Ile Tyr Lys
65 70 75 80
Asp Lys Glu Ser Thr Val Gln Val His Tyr Arg Met Cys Gln Ser Cys
85 90 95
Val Glu Leu Asp Pro Ala Thr Val Ala Gly Ile Ile Val Thr Asp Val
100 105 110
Ile Ala Thr Leu Leu Leu Ala Leu Gly Val Phe Cys Phe Ala Gly His
115 120 125
Glu Thr Gly Arg Leu Ser Gly Ala Ala Asp Thr Gln Ala Leu Leu Arg
130 135 140
Asn Asp Gln Val Tyr Gln Pro Leu Arg Asp Arg Asp Asp Ala Gln Tyr
145 150 155 160
Ser His Leu Gly Gly Asn Trp Ala Arg Asn Lys
165 170
<210> 25
<211> 198
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> полноразмерная аминокислотная последовательность обезьяньего
эпсилон CD3
<400> 25
Met Gln Ser Gly Thr Arg Trp Arg Val Leu Gly Leu Cys Leu Leu Ser
1 5 10 15
Ile Gly Val Trp Gly Gln Asp Gly Asn Glu Glu Met Gly Ser Ile Thr
20 25 30
Gln Thr Pro Tyr Gln Val Ser Ile Ser Gly Thr Thr Val Ile Leu Thr
35 40 45
Cys Ser Gln His Leu Gly Ser Glu Ala Gln Trp Gln His Asn Gly Lys
50 55 60
Asn Lys Glu Asp Ser Gly Asp Arg Leu Phe Leu Pro Glu Phe Ser Glu
65 70 75 80
Met Glu Gln Ser Gly Tyr Tyr Val Cys Tyr Pro Arg Gly Ser Asn Pro
85 90 95
Glu Asp Ala Ser His His Leu Tyr Leu Lys Ala Arg Val Cys Glu Asn
100 105 110
Cys Met Glu Met Asp Val Met Ala Val Ala Thr Ile Val Ile Val Asp
115 120 125
Ile Cys Ile Thr Leu Gly Leu Leu Leu Leu Val Tyr Tyr Trp Ser Lys
130 135 140
Asn Arg Lys Ala Lys Ala Lys Pro Val Thr Arg Gly Ala Gly Ala Gly
145 150 155 160
Gly Arg Gln Arg Gly Gln Asn Lys Glu Arg Pro Pro Pro Val Pro Asn
165 170 175
Pro Asp Tyr Glu Pro Ile Arg Lys Gly Gln Gln Asp Leu Tyr Ser Gly
180 185 190
Leu Asn Gln Arg Arg Ile
195
<210> 26
<211> 171
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> полноразмерная аминокислотная последовательность обезьяньего
дельта CD3
<400> 26
Met Glu His Ser Thr Phe Leu Ser Gly Leu Val Leu Ala Thr Leu Leu
1 5 10 15
Ser Gln Val Ser Pro Phe Lys Ile Pro Val Glu Glu Leu Glu Asp Arg
20 25 30
Val Phe Val Lys Cys Asn Thr Ser Val Thr Trp Val Glu Gly Thr Val
35 40 45
Gly Thr Leu Leu Thr Asn Asn Thr Arg Leu Asp Leu Gly Lys Arg Ile
50 55 60
Leu Asp Pro Arg Gly Ile Tyr Arg Cys Asn Gly Thr Asp Ile Tyr Lys
65 70 75 80
Asp Lys Glu Ser Ala Val Gln Val His Tyr Arg Met Cys Gln Asn Cys
85 90 95
Val Glu Leu Asp Pro Ala Thr Leu Ala Gly Ile Ile Val Thr Asp Val
100 105 110
Ile Ala Thr Leu Leu Leu Ala Leu Gly Val Phe Cys Phe Ala Gly His
115 120 125
Glu Thr Gly Arg Leu Ser Gly Ala Ala Asp Thr Gln Ala Leu Leu Arg
130 135 140
Asn Asp Gln Val Tyr Gln Pro Leu Arg Asp Arg Asp Asp Ala Gln Tyr
145 150 155 160
Ser Arg Leu Gly Gly Asn Trp Ala Arg Asn Lys
165 170
<210> 27
<211> 189
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> полноразмерная аминокислотная последовательность мышиного
эпсилон CD3
<400> 27
Met Arg Trp Asn Thr Phe Trp Gly Ile Leu Cys Leu Ser Leu Leu Ala
1 5 10 15
Val Gly Thr Cys Gln Asp Asp Ala Glu Asn Ile Glu Tyr Lys Val Ser
20 25 30
Ile Ser Gly Thr Ser Val Glu Leu Thr Cys Pro Leu Asp Ser Asp Glu
35 40 45
Asn Leu Lys Trp Glu Lys Asn Gly Gln Glu Leu Pro Gln Lys His Asp
50 55 60
Lys His Leu Val Leu Gln Asp Phe Ser Glu Val Glu Asp Ser Gly Tyr
65 70 75 80
Tyr Val Cys Tyr Thr Pro Ala Ser Asn Lys Asn Thr Tyr Leu Tyr Leu
85 90 95
Lys Ala Arg Val Cys Glu Tyr Cys Val Glu Val Asp Leu Thr Ala Val
100 105 110
Ala Ile Ile Ile Ile Val Asp Ile Cys Ile Thr Leu Gly Leu Leu Met
115 120 125
Val Ile Tyr Tyr Trp Ser Lys Asn Arg Lys Ala Lys Ala Lys Pro Val
130 135 140
Thr Arg Gly Thr Gly Ala Gly Ser Arg Pro Arg Gly Gln Asn Lys Glu
145 150 155 160
Arg Pro Pro Pro Val Pro Asn Pro Asp Tyr Glu Pro Ile Arg Lys Gly
165 170 175
Gln Arg Asp Leu Tyr Ser Gly Leu Asn Gln Arg Ala Val
180 185
<210> 28
<211> 173
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> полноразмерная аминокислотная последовательность мышиного
дельта CD3
<400> 28
Met Glu His Ser Gly Ile Leu Ala Ser Leu Ile Leu Ile Ala Val Leu
1 5 10 15
Pro Gln Gly Ser Pro Phe Lys Ile Gln Val Thr Glu Tyr Glu Asp Lys
20 25 30
Val Phe Val Thr Cys Asn Thr Ser Val Met His Leu Asp Gly Thr Val
35 40 45
Glu Gly Trp Phe Ala Lys Asn Lys Thr Leu Asn Leu Gly Lys Gly Val
50 55 60
Leu Asp Pro Arg Gly Ile Tyr Leu Cys Asn Gly Thr Glu Gln Leu Ala
65 70 75 80
Lys Val Val Ser Ser Val Gln Val His Tyr Arg Met Cys Gln Asn Cys
85 90 95
Val Glu Leu Asp Ser Gly Thr Met Ala Gly Val Ile Phe Ile Asp Leu
100 105 110
Ile Ala Thr Leu Leu Leu Ala Leu Gly Val Tyr Cys Phe Ala Gly His
115 120 125
Glu Thr Gly Arg Pro Ser Gly Ala Ala Glu Val Gln Ala Leu Leu Lys
130 135 140
Asn Glu Gln Leu Tyr Gln Pro Leu Arg Asp Arg Glu Asp Thr Gln Tyr
145 150 155 160
Ser Arg Leu Gly Gly Asn Trp Pro Arg Asn Lys Lys Ser
165 170
<210> 29
<211> 125
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRH-1
<400> 29
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Ala Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 30
<211> 125
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRH-2
<400> 30
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Pro Tyr Ile Ser Tyr Trp
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 31
<211> 125
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRH-3
<400> 31
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Glu Tyr Ile Ser Tyr Trp
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 32
<211> 125
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRH-4
<400> 32
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp
100 105 110
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 33
<211> 125
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRH-5
<400> 33
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 34
<211> 125
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRH-6
<400> 34
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Asn Lys Tyr Asn Asn Tyr Ala Thr Glu Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 35
<211> 125
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRH-7
<400> 35
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Glu Tyr Ala Ala
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 36
<211> 109
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRL
<400> 36
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly
20 25 30
Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn
85 90 95
Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 37
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRH-1 HCDR1
<400> 37
Lys Tyr Ala Met Asn
1 5
<210> 38
<211> 19
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRH-1 HCDR2
<400> 38
Arg Ile Arg Ser Lys Ala Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser
1 5 10 15
Val Lys Asp
<210> 39
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRH-1 HCDR3
<400> 39
His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp Ala Tyr
1 5 10
<210> 40
<211> 19
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRH-2 HCDR2
<400> 40
Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser
1 5 10 15
Val Lys Asp
<210> 41
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRH-2 HCDR3
<400> 41
His Gly Asn Phe Gly Asn Pro Tyr Ile Ser Tyr Trp Ala Tyr
1 5 10
<210> 42
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRH-3 HCDR3
<400> 42
His Gly Asn Phe Gly Asn Glu Tyr Ile Ser Tyr Trp Ala Tyr
1 5 10
<210> 43
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRH-4 HCDR3
<400> 43
His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp Asp Tyr
1 5 10
<210> 44
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRH-5 HCDR1
<400> 44
Lys Tyr Ala Met Ser
1 5
<210> 45
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRH-5 HCDR3
<400> 45
His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp Ala Tyr
1 5 10
<210> 46
<211> 19
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRH-6 HCDR2
<400> 46
Arg Ile Arg Asn Lys Tyr Asn Asn Tyr Ala Thr Glu Tyr Ala Asp Ser
1 5 10 15
Val Lys Asp
<210> 47
<211> 19
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRH-7 HCDR2
<400> 47
Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Glu Tyr Ala Ala Ser
1 5 10 15
Val Lys Asp
<210> 48
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRL LCDR1
<400> 48
Gly Ser Ser Thr Gly Ala Val Thr Ser Gly Asn Tyr Pro Asn
1 5 10
<210> 49
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRL LCDR2
<400> 49
Gly Thr Lys Phe Leu Ala Pro
1 5
<210> 50
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ДОМЕН
<223> HRL LCDR3
<400> 50
Val Leu Trp Tyr Ser Asn Arg Trp Val
1 5
<210> 51
<211> 244
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> B7H3-scFv1
<400> 51
Gln Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser
20 25 30
His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Met
35 40 45
Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Ile His Val Asp Arg
85 90 95
Asp Ile Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
115 120 125
Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser Ala Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Val Ile
165 170 175
Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val Lys Gly Arg
180 185 190
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Ser
210 215 220
Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly Ala Leu Val
225 230 235 240
Thr Val Ser Ser
<210> 52
<211> 244
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> B7H3-scFv2
<400> 52
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Ala Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln Glu Pro Ser Phe
130 135 140
Ser Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly Leu Ser Ser
145 150 155 160
Gly Ser Val Ser Thr Ser His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro
165 170 175
Gly Gln Ala Pro Arg Met Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser
180 185 190
Gly Val Pro Asp Arg Phe Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala
195 200 205
Leu Thr Ile Thr Gly Ala Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys
210 215 220
Ala Ile His Val Asp Arg Asp Ile Trp Val Phe Gly Gly Gly Thr Lys
225 230 235 240
Leu Thr Val Leu
<210> 53
<211> 244
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> B7H3-scFv3
<400> 53
Gln Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser
20 25 30
His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Met
35 40 45
Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Ile His Val Asp Arg
85 90 95
Asp Ile Trp Val Phe Gly Cys Gly Thr Lys Leu Thr Val Leu Gly Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
115 120 125
Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser Ala Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val Ala Val Ile
165 170 175
Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val Lys Gly Arg
180 185 190
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Ser
210 215 220
Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly Ala Leu Val
225 230 235 240
Thr Val Ser Ser
<210> 54
<211> 244
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> B7H3-scFv4
<400> 54
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Ala Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln Glu Pro Ser Phe
130 135 140
Ser Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly Leu Ser Ser
145 150 155 160
Gly Ser Val Ser Thr Ser His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro
165 170 175
Gly Gln Ala Pro Arg Met Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser
180 185 190
Gly Val Pro Asp Arg Phe Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala
195 200 205
Leu Thr Ile Thr Gly Ala Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys
210 215 220
Ala Ile His Val Asp Arg Asp Ile Trp Val Phe Gly Cys Gly Thr Lys
225 230 235 240
Leu Thr Val Leu
<210> 55
<211> 249
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> CD3-scFv1H
<400> 55
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Ala Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
115 120 125
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Thr Val Val
130 135 140
Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly Thr Val Thr Leu
145 150 155 160
Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly Asn Tyr Pro Asn
165 170 175
Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly Leu Ile Gly Gly
180 185 190
Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe Ser Gly Ser Leu
195 200 205
Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val Gln Pro Glu Asp
210 215 220
Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn Arg Trp Val Phe
225 230 235 240
Gly Gly Gly Thr Lys Leu Thr Val Leu
245
<210> 56
<211> 249
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> CD3-scFv2H
<400> 56
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Pro Tyr Ile Ser Tyr Trp
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
115 120 125
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Thr Val Val
130 135 140
Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly Thr Val Thr Leu
145 150 155 160
Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly Asn Tyr Pro Asn
165 170 175
Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly Leu Ile Gly Gly
180 185 190
Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe Ser Gly Ser Leu
195 200 205
Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val Gln Pro Glu Asp
210 215 220
Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn Arg Trp Val Phe
225 230 235 240
Gly Gly Gly Thr Lys Leu Thr Val Leu
245
<210> 57
<211> 249
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> CD3-scFv3H
<400> 57
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Glu Tyr Ile Ser Tyr Trp
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
115 120 125
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Thr Val Val
130 135 140
Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly Thr Val Thr Leu
145 150 155 160
Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly Asn Tyr Pro Asn
165 170 175
Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly Leu Ile Gly Gly
180 185 190
Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe Ser Gly Ser Leu
195 200 205
Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val Gln Pro Glu Asp
210 215 220
Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn Arg Trp Val Phe
225 230 235 240
Gly Gly Gly Thr Lys Leu Thr Val Leu
245
<210> 58
<211> 249
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> CD3-scFv4H
<400> 58
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp
100 105 110
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
115 120 125
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Thr Val Val
130 135 140
Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly Thr Val Thr Leu
145 150 155 160
Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly Asn Tyr Pro Asn
165 170 175
Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly Leu Ile Gly Gly
180 185 190
Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe Ser Gly Ser Leu
195 200 205
Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val Gln Pro Glu Asp
210 215 220
Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn Arg Trp Val Phe
225 230 235 240
Gly Gly Gly Thr Lys Leu Thr Val Leu
245
<210> 59
<211> 249
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> CD3-scFv5H
<400> 59
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
115 120 125
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Thr Val Val
130 135 140
Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly Thr Val Thr Leu
145 150 155 160
Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly Asn Tyr Pro Asn
165 170 175
Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly Leu Ile Gly Gly
180 185 190
Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe Ser Gly Ser Leu
195 200 205
Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val Gln Pro Glu Asp
210 215 220
Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn Arg Trp Val Phe
225 230 235 240
Gly Gly Gly Thr Lys Leu Thr Val Leu
245
<210> 60
<211> 249
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> CD3-scFv6H
<400> 60
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Asn Lys Tyr Asn Asn Tyr Ala Thr Glu Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
115 120 125
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Thr Val Val
130 135 140
Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly Thr Val Thr Leu
145 150 155 160
Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly Asn Tyr Pro Asn
165 170 175
Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly Leu Ile Gly Gly
180 185 190
Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe Ser Gly Ser Leu
195 200 205
Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val Gln Pro Glu Asp
210 215 220
Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn Arg Trp Val Phe
225 230 235 240
Gly Gly Gly Thr Lys Leu Thr Val Leu
245
<210> 61
<211> 249
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> CD3-scFv7H
<400> 61
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Glu Tyr Ala Ala
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
115 120 125
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Thr Val Val
130 135 140
Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly Thr Val Thr Leu
145 150 155 160
Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly Asn Tyr Pro Asn
165 170 175
Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly Leu Ile Gly Gly
180 185 190
Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe Ser Gly Ser Leu
195 200 205
Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val Gln Pro Glu Asp
210 215 220
Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn Arg Trp Val Phe
225 230 235 240
Gly Gly Gly Thr Lys Leu Thr Val Leu
245
<210> 62
<211> 249
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> CD3-scFv1L
<400> 62
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly
20 25 30
Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn
85 90 95
Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu
115 120 125
Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Lys Leu
130 135 140
Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr Ala Met Asn Trp
145 150 155 160
Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Arg Ile Arg
165 170 175
Ser Lys Ala Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Asp
180 185 190
Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Ala Tyr Leu Gln
195 200 205
Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg
210 215 220
His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp Ala Tyr Trp Gly
225 230 235 240
Gln Gly Thr Leu Val Thr Val Ser Ser
245
<210> 63
<211> 249
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> CD3-scFv2L
<400> 63
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly
20 25 30
Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn
85 90 95
Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu
115 120 125
Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Lys Leu
130 135 140
Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr Ala Met Asn Trp
145 150 155 160
Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Arg Ile Arg
165 170 175
Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Asp
180 185 190
Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Ala Tyr Leu Gln
195 200 205
Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg
210 215 220
His Gly Asn Phe Gly Asn Pro Tyr Ile Ser Tyr Trp Ala Tyr Trp Gly
225 230 235 240
Gln Gly Thr Leu Val Thr Val Ser Ser
245
<210> 64
<211> 249
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> CD3-scFv3L
<400> 64
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly
20 25 30
Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn
85 90 95
Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu
115 120 125
Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Lys Leu
130 135 140
Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr Ala Met Asn Trp
145 150 155 160
Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Arg Ile Arg
165 170 175
Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Asp
180 185 190
Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Ala Tyr Leu Gln
195 200 205
Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg
210 215 220
His Gly Asn Phe Gly Asn Glu Tyr Ile Ser Tyr Trp Ala Tyr Trp Gly
225 230 235 240
Gln Gly Thr Leu Val Thr Val Ser Ser
245
<210> 65
<211> 249
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> CD3-scFv4L
<400> 65
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly
20 25 30
Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn
85 90 95
Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu
115 120 125
Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Lys Leu
130 135 140
Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr Ala Met Asn Trp
145 150 155 160
Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Arg Ile Arg
165 170 175
Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Asp
180 185 190
Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Ala Tyr Leu Gln
195 200 205
Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg
210 215 220
His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp Asp Tyr Trp Gly
225 230 235 240
Gln Gly Thr Leu Val Thr Val Ser Ser
245
<210> 66
<211> 249
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> CD3-scFv5L
<400> 66
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly
20 25 30
Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn
85 90 95
Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu
115 120 125
Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Lys Leu
130 135 140
Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr Ala Met Ser Trp
145 150 155 160
Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Arg Ile Arg
165 170 175
Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Asp
180 185 190
Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Ala Tyr Leu Gln
195 200 205
Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg
210 215 220
His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp Ala Tyr Trp Gly
225 230 235 240
Gln Gly Thr Leu Val Thr Val Ser Ser
245
<210> 67
<211> 249
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> CD3-scFv6L
<400> 67
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly
20 25 30
Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn
85 90 95
Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu
115 120 125
Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Lys Leu
130 135 140
Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr Ala Met Asn Trp
145 150 155 160
Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Arg Ile Arg
165 170 175
Asn Lys Tyr Asn Asn Tyr Ala Thr Glu Tyr Ala Asp Ser Val Lys Asp
180 185 190
Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Ala Tyr Leu Gln
195 200 205
Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg
210 215 220
His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp Ala Tyr Trp Gly
225 230 235 240
Gln Gly Thr Leu Val Thr Val Ser Ser
245
<210> 68
<211> 249
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> CD3-scFv7L
<400> 68
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly
20 25 30
Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn
85 90 95
Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu
115 120 125
Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Lys Leu
130 135 140
Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr Ala Met Asn Trp
145 150 155 160
Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Arg Ile Arg
165 170 175
Ser Lys Tyr Asn Asn Tyr Ala Thr Glu Tyr Ala Ala Ser Val Lys Asp
180 185 190
Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Ala Tyr Leu Gln
195 200 205
Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg
210 215 220
His Gly Asn Phe Gly Asn Ser Tyr Ile Ser Tyr Trp Ala Tyr Trp Gly
225 230 235 240
Gln Gly Thr Leu Val Thr Val Ser Ser
245
<210> 69
<211> 217
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> knob-Fc
<400> 69
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215
<210> 70
<211> 217
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> hole-Fc
<400> 70
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215
<210> 71
<211> 474
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> вторая полипептидная цепь 113
<400> 71
Gln Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser
20 25 30
His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Met
35 40 45
Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Ile His Val Asp Arg
85 90 95
Asp Ile Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
115 120 125
Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser Ala Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Val Ile
165 170 175
Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val Lys Gly Arg
180 185 190
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Ser
210 215 220
Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly Ala Leu Val
225 230 235 240
Thr Val Ser Ser Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys
245 250 255
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
260 265 270
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
275 280 285
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
290 295 300
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
305 310 315 320
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
325 330 335
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
340 345 350
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
355 360 365
Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Glu Glu
370 375 380
Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr
385 390 395 400
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
405 410 415
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
420 425 430
Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
435 440 445
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
450 455 460
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
465 470
<210> 72
<211> 734
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> первая полипептидная цепь113
<400> 72
Gln Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser
20 25 30
His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Met
35 40 45
Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Ile His Val Asp Arg
85 90 95
Asp Ile Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
115 120 125
Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser Ala Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Val Ile
165 170 175
Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val Lys Gly Arg
180 185 190
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Ser
210 215 220
Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly Ala Leu Val
225 230 235 240
Thr Val Ser Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser
245 250 255
Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala
260 265 270
Ala Ser Gly Phe Thr Phe Asn Lys Tyr Ala Met Asn Trp Val Arg Gln
275 280 285
Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Arg Ile Arg Ser Lys Tyr
290 295 300
Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Asp Arg Phe Thr
305 310 315 320
Ile Ser Arg Asp Asp Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn Asn
325 330 335
Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg His Gly Asn
340 345 350
Phe Gly Asn Pro Tyr Ile Ser Tyr Trp Ala Tyr Trp Gly Gln Gly Thr
355 360 365
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
370 375 380
Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr
385 390 395 400
Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly
405 410 415
Ala Val Thr Ser Gly Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly
420 425 430
Gln Ala Pro Arg Gly Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly
435 440 445
Thr Pro Ala Arg Phe Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu
450 455 460
Thr Leu Ser Gly Val Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val
465 470 475 480
Leu Trp Tyr Ser Asn Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr
485 490 495
Val Leu Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
500 505 510
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
515 520 525
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
530 535 540
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
545 550 555 560
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
565 570 575
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
580 585 590
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
595 600 605
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
610 615 620
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr
625 630 635 640
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
645 650 655
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
660 665 670
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
675 680 685
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
690 695 700
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
705 710 715 720
Ser Leu Ser Leu Ser Pro Gly Lys His His His His His His
725 730
<210> 73
<211> 734
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> первая полипептидная цепь118
<400> 73
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Glu Tyr Ile Ser Tyr Trp
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
115 120 125
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Thr Val Val
130 135 140
Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly Thr Val Thr Leu
145 150 155 160
Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly Asn Tyr Pro Asn
165 170 175
Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly Leu Ile Gly Gly
180 185 190
Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe Ser Gly Ser Leu
195 200 205
Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val Gln Pro Glu Asp
210 215 220
Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn Arg Trp Val Phe
225 230 235 240
Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Gly Gly Ser Gln Thr
245 250 255
Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly Thr Val
260 265 270
Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser His Tyr
275 280 285
Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Met Leu Ile
290 295 300
Tyr Asn Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe Ser Gly
305 310 315 320
Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala Gln Ala
325 330 335
Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Ile His Val Asp Arg Asp Ile
340 345 350
Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Gly Gly
355 360 365
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Val
370 375 380
Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr Ser Leu Arg Leu Ser
385 390 395 400
Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser Ala Met His Trp Val
405 410 415
Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Val Ile Ser Tyr
420 425 430
Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val Lys Gly Arg Phe Thr
435 440 445
Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser
450 455 460
Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Ser Ala Arg
465 470 475 480
Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly Ala Leu Val Thr Val
485 490 495
Ser Ser Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
500 505 510
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
515 520 525
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
530 535 540
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
545 550 555 560
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
565 570 575
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
580 585 590
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
595 600 605
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
610 615 620
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr
625 630 635 640
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
645 650 655
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
660 665 670
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
675 680 685
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
690 695 700
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
705 710 715 720
Ser Leu Ser Leu Ser Pro Gly Lys His His His His His His
725 730
<210> 74
<211> 732
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> первая полипептидная цепь 119
<400> 74
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Glu Tyr Ile Ser Tyr Trp
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
115 120 125
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gln Thr Val Val Thr Gln
130 135 140
Glu Pro Ser Leu Thr Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys
145 150 155 160
Gly Ser Ser Thr Gly Ala Val Thr Ser Gly Asn Tyr Pro Asn Trp Val
165 170 175
Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly Leu Ile Gly Gly Thr Lys
180 185 190
Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe Ser Gly Ser Leu Leu Gly
195 200 205
Gly Lys Ala Ala Leu Thr Leu Ser Gly Val Gln Pro Glu Asp Glu Ala
210 215 220
Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn Arg Trp Val Phe Gly Gly
225 230 235 240
Gly Thr Lys Leu Thr Val Leu Gly Gly Gly Gly Ser Gln Thr Val Val
245 250 255
Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly Thr Val Thr Leu
260 265 270
Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser His Tyr Pro Ser
275 280 285
Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Met Leu Ile Tyr Asn
290 295 300
Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Ile
305 310 315 320
Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala Gln Ala Asp Asp
325 330 335
Glu Ser Asp Tyr Tyr Cys Ala Ile His Val Asp Arg Asp Ile Trp Val
340 345 350
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Gly Gly Ser Gly
355 360 365
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Val Gln Ser
370 375 380
Gly Gly Gly Val Val Gln Pro Gly Thr Ser Leu Arg Leu Ser Cys Ala
385 390 395 400
Ala Ser Gly Phe Ile Phe Ser Ser Ser Ala Met His Trp Val Arg Gln
405 410 415
Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Val Ile Ser Tyr Asp Gly
420 425 430
Ser Asn Lys Tyr Tyr Val Asp Ser Val Lys Gly Arg Phe Thr Ile Ser
435 440 445
Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg
450 455 460
Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Ser Ala Arg Leu Tyr
465 470 475 480
Ala Ser Phe Asp Tyr Trp Gly Gln Gly Ala Leu Val Thr Val Ser Ser
485 490 495
Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
500 505 510
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
515 520 525
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
530 535 540
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
545 550 555 560
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
565 570 575
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
580 585 590
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
595 600 605
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
610 615 620
Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn
625 630 635 640
Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
645 650 655
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
660 665 670
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
675 680 685
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
690 695 700
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
705 710 715 720
Ser Leu Ser Pro Gly Lys His His His His His His
725 730
<210> 75
<211> 734
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> первый полипептид 126
<400> 75
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Pro Tyr Ile Ser Tyr Trp
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
115 120 125
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Thr Val Val
130 135 140
Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly Thr Val Thr Leu
145 150 155 160
Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly Asn Tyr Pro Asn
165 170 175
Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly Leu Ile Gly Gly
180 185 190
Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe Ser Gly Ser Leu
195 200 205
Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val Gln Pro Glu Asp
210 215 220
Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn Arg Trp Val Phe
225 230 235 240
Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Gly Gly Ser Gln Thr
245 250 255
Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly Thr Val
260 265 270
Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser His Tyr
275 280 285
Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Met Leu Ile
290 295 300
Tyr Asn Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe Ser Gly
305 310 315 320
Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala Gln Ala
325 330 335
Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Ile His Val Asp Arg Asp Ile
340 345 350
Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Gly Gly
355 360 365
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Val
370 375 380
Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr Ser Leu Arg Leu Ser
385 390 395 400
Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser Ala Met His Trp Val
405 410 415
Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Val Ile Ser Tyr
420 425 430
Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val Lys Gly Arg Phe Thr
435 440 445
Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser
450 455 460
Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Ser Ala Arg
465 470 475 480
Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly Ala Leu Val Thr Val
485 490 495
Ser Ser Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
500 505 510
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
515 520 525
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
530 535 540
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
545 550 555 560
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
565 570 575
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
580 585 590
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
595 600 605
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
610 615 620
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr
625 630 635 640
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
645 650 655
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
660 665 670
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
675 680 685
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
690 695 700
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
705 710 715 720
Ser Leu Ser Leu Ser Pro Gly Lys His His His His His His
725 730
<210> 76
<211> 734
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> первый полипептид 127
<400> 76
Gln Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser
20 25 30
His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Met
35 40 45
Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Ile His Val Asp Arg
85 90 95
Asp Ile Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
115 120 125
Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser Ala Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Val Ile
165 170 175
Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val Lys Gly Arg
180 185 190
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Ser
210 215 220
Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly Ala Leu Val
225 230 235 240
Thr Val Ser Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser
245 250 255
Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala
260 265 270
Ala Ser Gly Phe Thr Phe Asn Lys Tyr Ala Met Asn Trp Val Arg Gln
275 280 285
Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Arg Ile Arg Ser Lys Tyr
290 295 300
Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Asp Arg Phe Thr
305 310 315 320
Ile Ser Arg Asp Asp Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn Asn
325 330 335
Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg His Gly Asn
340 345 350
Phe Gly Asn Glu Tyr Ile Ser Tyr Trp Ala Tyr Trp Gly Gln Gly Thr
355 360 365
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
370 375 380
Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr
385 390 395 400
Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly
405 410 415
Ala Val Thr Ser Gly Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly
420 425 430
Gln Ala Pro Arg Gly Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly
435 440 445
Thr Pro Ala Arg Phe Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu
450 455 460
Thr Leu Ser Gly Val Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val
465 470 475 480
Leu Trp Tyr Ser Asn Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr
485 490 495
Val Leu Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
500 505 510
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
515 520 525
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
530 535 540
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
545 550 555 560
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
565 570 575
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
580 585 590
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
595 600 605
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
610 615 620
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr
625 630 635 640
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
645 650 655
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
660 665 670
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
675 680 685
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
690 695 700
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
705 710 715 720
Ser Leu Ser Leu Ser Pro Gly Lys His His His His His His
725 730
<210> 77
<211> 739
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> первый полипептид 128
<400> 77
Gln Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser
20 25 30
His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Met
35 40 45
Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Ile His Val Asp Arg
85 90 95
Asp Ile Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
115 120 125
Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser Ala Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Val Ile
165 170 175
Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val Lys Gly Arg
180 185 190
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Ser
210 215 220
Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly Ala Leu Val
225 230 235 240
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val
245 250 255
Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu
260 265 270
Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr Ala Met
275 280 285
Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Arg
290 295 300
Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser Val
305 310 315 320
Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr Ala Tyr
325 330 335
Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys
340 345 350
Val Arg His Gly Asn Phe Gly Asn Pro Tyr Ile Ser Tyr Trp Ala Tyr
355 360 365
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser
370 375 380
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln
385 390 395 400
Glu Pro Ser Leu Thr Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys
405 410 415
Gly Ser Ser Thr Gly Ala Val Thr Ser Gly Asn Tyr Pro Asn Trp Val
420 425 430
Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly Leu Ile Gly Gly Thr Lys
435 440 445
Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe Ser Gly Ser Leu Leu Gly
450 455 460
Gly Lys Ala Ala Leu Thr Leu Ser Gly Val Gln Pro Glu Asp Glu Ala
465 470 475 480
Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn Arg Trp Val Phe Gly Gly
485 490 495
Gly Thr Lys Leu Thr Val Leu Gly Gly Gly Asp Lys Thr His Thr Cys
500 505 510
Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu
515 520 525
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
530 535 540
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
545 550 555 560
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
565 570 575
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
580 585 590
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
595 600 605
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
610 615 620
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys
625 630 635 640
Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys
645 650 655
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
660 665 670
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
675 680 685
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
690 695 700
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
705 710 715 720
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys His His His
725 730 735
His His His
<210> 78
<211> 734
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> первый полипептид 132
<400> 78
Gln Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser
20 25 30
His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Met
35 40 45
Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Ile His Val Asp Arg
85 90 95
Asp Ile Trp Val Phe Gly Cys Gly Thr Lys Leu Thr Val Leu Gly Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
115 120 125
Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser Ala Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val Ala Val Ile
165 170 175
Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val Lys Gly Arg
180 185 190
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Ser
210 215 220
Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly Ala Leu Val
225 230 235 240
Thr Val Ser Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser
245 250 255
Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala
260 265 270
Ala Ser Gly Phe Thr Phe Asn Lys Tyr Ala Met Asn Trp Val Arg Gln
275 280 285
Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Arg Ile Arg Ser Lys Tyr
290 295 300
Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Asp Arg Phe Thr
305 310 315 320
Ile Ser Arg Asp Asp Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn Asn
325 330 335
Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg His Gly Asn
340 345 350
Phe Gly Asn Glu Tyr Ile Ser Tyr Trp Ala Tyr Trp Gly Gln Gly Thr
355 360 365
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
370 375 380
Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr
385 390 395 400
Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly
405 410 415
Ala Val Thr Ser Gly Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly
420 425 430
Gln Ala Pro Arg Gly Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly
435 440 445
Thr Pro Ala Arg Phe Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu
450 455 460
Thr Leu Ser Gly Val Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val
465 470 475 480
Leu Trp Tyr Ser Asn Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr
485 490 495
Val Leu Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
500 505 510
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
515 520 525
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
530 535 540
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
545 550 555 560
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
565 570 575
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
580 585 590
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
595 600 605
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
610 615 620
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr
625 630 635 640
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
645 650 655
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
660 665 670
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
675 680 685
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
690 695 700
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
705 710 715 720
Ser Leu Ser Leu Ser Pro Gly Lys His His His His His His
725 730
<210> 79
<211> 734
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> первый полипептид 131
<400> 79
Gln Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser
20 25 30
His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Met
35 40 45
Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Ile His Val Asp Arg
85 90 95
Asp Ile Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
115 120 125
Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser Ala Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Val Ile
165 170 175
Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val Lys Gly Arg
180 185 190
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Ser
210 215 220
Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly Ala Leu Val
225 230 235 240
Thr Val Ser Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser
245 250 255
Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala
260 265 270
Ala Ser Gly Phe Thr Phe Asn Lys Tyr Ala Met Asn Trp Val Arg Gln
275 280 285
Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Arg Ile Arg Ser Lys Ala
290 295 300
Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Asp Arg Phe Thr
305 310 315 320
Ile Ser Arg Asp Asp Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn Asn
325 330 335
Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg His Gly Asn
340 345 350
Phe Gly Asn Ser Tyr Ile Ser Tyr Trp Ala Tyr Trp Gly Gln Gly Thr
355 360 365
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
370 375 380
Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr
385 390 395 400
Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly
405 410 415
Ala Val Thr Ser Gly Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly
420 425 430
Gln Ala Pro Arg Gly Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly
435 440 445
Thr Pro Ala Arg Phe Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu
450 455 460
Thr Leu Ser Gly Val Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val
465 470 475 480
Leu Trp Tyr Ser Asn Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr
485 490 495
Val Leu Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
500 505 510
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
515 520 525
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
530 535 540
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
545 550 555 560
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
565 570 575
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
580 585 590
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
595 600 605
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
610 615 620
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr
625 630 635 640
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
645 650 655
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
660 665 670
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
675 680 685
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
690 695 700
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
705 710 715 720
Ser Leu Ser Leu Ser Pro Gly Lys His His His His His His
725 730
<210> 80
<211> 734
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> первый полипептид 154
<400> 80
Gln Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser
20 25 30
His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Met
35 40 45
Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Ile His Val Asp Arg
85 90 95
Asp Ile Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
115 120 125
Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser Ala Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Val Ile
165 170 175
Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val Lys Gly Arg
180 185 190
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Ser
210 215 220
Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly Ala Leu Val
225 230 235 240
Thr Val Ser Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser
245 250 255
Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala
260 265 270
Ala Ser Gly Phe Thr Phe Asn Lys Tyr Ala Met Asn Trp Val Arg Gln
275 280 285
Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Arg Ile Arg Ser Lys Tyr
290 295 300
Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Asp Arg Phe Thr
305 310 315 320
Ile Ser Arg Asp Asp Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn Asn
325 330 335
Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg His Gly Asn
340 345 350
Phe Gly Asn Ser Tyr Ile Ser Tyr Trp Asp Tyr Trp Gly Gln Gly Thr
355 360 365
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
370 375 380
Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr
385 390 395 400
Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly
405 410 415
Ala Val Thr Ser Gly Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly
420 425 430
Gln Ala Pro Arg Gly Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly
435 440 445
Thr Pro Ala Arg Phe Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu
450 455 460
Thr Leu Ser Gly Val Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val
465 470 475 480
Leu Trp Tyr Ser Asn Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr
485 490 495
Val Leu Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
500 505 510
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
515 520 525
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
530 535 540
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
545 550 555 560
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
565 570 575
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
580 585 590
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
595 600 605
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
610 615 620
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr
625 630 635 640
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
645 650 655
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
660 665 670
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
675 680 685
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
690 695 700
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
705 710 715 720
Ser Leu Ser Leu Ser Pro Gly Lys His His His His His His
725 730
<210> 81
<211> 734
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> первый полипептид 156
<400> 81
Gln Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser
20 25 30
His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Met
35 40 45
Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Ile His Val Asp Arg
85 90 95
Asp Ile Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
115 120 125
Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser Ala Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Val Ile
165 170 175
Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val Lys Gly Arg
180 185 190
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Ser
210 215 220
Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly Ala Leu Val
225 230 235 240
Thr Val Ser Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser
245 250 255
Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala
260 265 270
Ala Ser Gly Phe Thr Phe Asn Lys Tyr Ala Met Asn Trp Val Arg Gln
275 280 285
Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Arg Ile Arg Asn Lys Tyr
290 295 300
Asn Asn Tyr Ala Thr Glu Tyr Ala Asp Ser Val Lys Asp Arg Phe Thr
305 310 315 320
Ile Ser Arg Asp Asp Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn Asn
325 330 335
Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg His Gly Asn
340 345 350
Phe Gly Asn Ser Tyr Ile Ser Tyr Trp Ala Tyr Trp Gly Gln Gly Thr
355 360 365
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
370 375 380
Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr
385 390 395 400
Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly
405 410 415
Ala Val Thr Ser Gly Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly
420 425 430
Gln Ala Pro Arg Gly Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly
435 440 445
Thr Pro Ala Arg Phe Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu
450 455 460
Thr Leu Ser Gly Val Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val
465 470 475 480
Leu Trp Tyr Ser Asn Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr
485 490 495
Val Leu Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
500 505 510
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
515 520 525
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
530 535 540
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
545 550 555 560
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
565 570 575
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
580 585 590
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
595 600 605
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
610 615 620
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr
625 630 635 640
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
645 650 655
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
660 665 670
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
675 680 685
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
690 695 700
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
705 710 715 720
Ser Leu Ser Leu Ser Pro Gly Lys His His His His His His
725 730
<210> 82
<211> 734
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> первый полипептид 155
<400> 82
Gln Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser
20 25 30
His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Met
35 40 45
Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Ile His Val Asp Arg
85 90 95
Asp Ile Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
115 120 125
Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser Ala Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Val Ile
165 170 175
Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val Lys Gly Arg
180 185 190
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Ser
210 215 220
Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly Ala Leu Val
225 230 235 240
Thr Val Ser Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser
245 250 255
Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala
260 265 270
Ala Ser Gly Phe Thr Phe Asn Lys Tyr Ala Met Ser Trp Val Arg Gln
275 280 285
Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Arg Ile Arg Ser Lys Tyr
290 295 300
Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Asp Arg Phe Thr
305 310 315 320
Ile Ser Arg Asp Asp Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn Asn
325 330 335
Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg His Gly Asn
340 345 350
Phe Gly Asn Ser Tyr Ile Ser Tyr Trp Ala Tyr Trp Gly Gln Gly Thr
355 360 365
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
370 375 380
Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr
385 390 395 400
Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly
405 410 415
Ala Val Thr Ser Gly Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly
420 425 430
Gln Ala Pro Arg Gly Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly
435 440 445
Thr Pro Ala Arg Phe Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu
450 455 460
Thr Leu Ser Gly Val Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val
465 470 475 480
Leu Trp Tyr Ser Asn Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr
485 490 495
Val Leu Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
500 505 510
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
515 520 525
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
530 535 540
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
545 550 555 560
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
565 570 575
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
580 585 590
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
595 600 605
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
610 615 620
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr
625 630 635 640
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
645 650 655
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
660 665 670
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
675 680 685
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
690 695 700
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
705 710 715 720
Ser Leu Ser Leu Ser Pro Gly Lys His His His His His His
725 730
<210> 83
<211> 734
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> первый полипептид 177
<400> 83
Gln Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser
20 25 30
His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Met
35 40 45
Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Ile His Val Asp Arg
85 90 95
Asp Ile Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
115 120 125
Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser Ala Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Val Ile
165 170 175
Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val Lys Gly Arg
180 185 190
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Ser
210 215 220
Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly Ala Leu Val
225 230 235 240
Thr Val Ser Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser
245 250 255
Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala
260 265 270
Ala Ser Gly Phe Thr Phe Asn Lys Tyr Ala Met Asn Trp Val Arg Gln
275 280 285
Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Arg Ile Arg Ser Lys Tyr
290 295 300
Asn Asn Tyr Ala Thr Glu Tyr Ala Ala Ser Val Lys Asp Arg Phe Thr
305 310 315 320
Ile Ser Arg Asp Asp Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn Asn
325 330 335
Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg His Gly Asn
340 345 350
Phe Gly Asn Ser Tyr Ile Ser Tyr Trp Ala Tyr Trp Gly Gln Gly Thr
355 360 365
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
370 375 380
Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr
385 390 395 400
Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly
405 410 415
Ala Val Thr Ser Gly Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly
420 425 430
Gln Ala Pro Arg Gly Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly
435 440 445
Thr Pro Ala Arg Phe Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu
450 455 460
Thr Leu Ser Gly Val Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val
465 470 475 480
Leu Trp Tyr Ser Asn Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr
485 490 495
Val Leu Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
500 505 510
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
515 520 525
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
530 535 540
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
545 550 555 560
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
565 570 575
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
580 585 590
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
595 600 605
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
610 615 620
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr
625 630 635 640
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
645 650 655
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
660 665 670
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
675 680 685
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
690 695 700
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
705 710 715 720
Ser Leu Ser Leu Ser Pro Gly Lys His His His His His His
725 730
<210> 84
<211> 734
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> первый полипептид 172
<400> 84
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Ala Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln Glu Pro Ser Phe
130 135 140
Ser Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly Leu Ser Ser
145 150 155 160
Gly Ser Val Ser Thr Ser His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro
165 170 175
Gly Gln Ala Pro Arg Met Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser
180 185 190
Gly Val Pro Asp Arg Phe Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala
195 200 205
Leu Thr Ile Thr Gly Ala Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys
210 215 220
Ala Ile His Val Asp Arg Asp Ile Trp Val Phe Gly Gly Gly Thr Lys
225 230 235 240
Leu Thr Val Leu Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln Glu
245 250 255
Pro Ser Leu Thr Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly
260 265 270
Ser Ser Thr Gly Ala Val Thr Ser Gly Asn Tyr Pro Asn Trp Val Gln
275 280 285
Gln Lys Pro Gly Gln Ala Pro Arg Gly Leu Ile Gly Gly Thr Lys Phe
290 295 300
Leu Ala Pro Gly Thr Pro Ala Arg Phe Ser Gly Ser Leu Leu Gly Gly
305 310 315 320
Lys Ala Ala Leu Thr Leu Ser Gly Val Gln Pro Glu Asp Glu Ala Glu
325 330 335
Tyr Tyr Cys Val Leu Trp Tyr Ser Asn Arg Trp Val Phe Gly Gly Gly
340 345 350
Thr Lys Leu Thr Val Leu Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
355 360 365
Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
370 375 380
Val Gln Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe
385 390 395 400
Thr Phe Asn Lys Tyr Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys
405 410 415
Gly Leu Glu Trp Val Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala
420 425 430
Thr Tyr Tyr Ala Asp Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp
435 440 445
Asp Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu
450 455 460
Asp Thr Ala Val Tyr Tyr Cys Val Arg His Gly Asn Phe Gly Asn Glu
465 470 475 480
Tyr Ile Ser Tyr Trp Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
485 490 495
Ser Ser Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
500 505 510
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
515 520 525
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
530 535 540
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
545 550 555 560
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
565 570 575
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
580 585 590
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
595 600 605
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
610 615 620
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr
625 630 635 640
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
645 650 655
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
660 665 670
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
675 680 685
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
690 695 700
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
705 710 715 720
Ser Leu Ser Leu Ser Pro Gly Lys His His His His His His
725 730
<210> 85
<211> 734
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> первый полипептид 171
<400> 85
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Ala Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln Glu Pro Ser Phe
130 135 140
Ser Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly Leu Ser Ser
145 150 155 160
Gly Ser Val Ser Thr Ser His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro
165 170 175
Gly Gln Ala Pro Arg Met Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser
180 185 190
Gly Val Pro Asp Arg Phe Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala
195 200 205
Leu Thr Ile Thr Gly Ala Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys
210 215 220
Ala Ile His Val Asp Arg Asp Ile Trp Val Phe Gly Gly Gly Thr Lys
225 230 235 240
Leu Thr Val Leu Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser
245 250 255
Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala
260 265 270
Ala Ser Gly Phe Thr Phe Asn Lys Tyr Ala Met Asn Trp Val Arg Gln
275 280 285
Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Arg Ile Arg Ser Lys Tyr
290 295 300
Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Asp Arg Phe Thr
305 310 315 320
Ile Ser Arg Asp Asp Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn Asn
325 330 335
Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg His Gly Asn
340 345 350
Phe Gly Asn Glu Tyr Ile Ser Tyr Trp Ala Tyr Trp Gly Gln Gly Thr
355 360 365
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
370 375 380
Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr
385 390 395 400
Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly
405 410 415
Ala Val Thr Ser Gly Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly
420 425 430
Gln Ala Pro Arg Gly Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly
435 440 445
Thr Pro Ala Arg Phe Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu
450 455 460
Thr Leu Ser Gly Val Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val
465 470 475 480
Leu Trp Tyr Ser Asn Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr
485 490 495
Val Leu Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
500 505 510
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
515 520 525
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
530 535 540
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
545 550 555 560
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
565 570 575
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
580 585 590
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
595 600 605
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
610 615 620
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr
625 630 635 640
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
645 650 655
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
660 665 670
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
675 680 685
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
690 695 700
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
705 710 715 720
Ser Leu Ser Leu Ser Pro Gly Lys His His His His His His
725 730
<210> 86
<211> 734
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> первый полипептид 161
<400> 86
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Ala Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln Glu Pro Ser Phe
130 135 140
Ser Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly Leu Ser Ser
145 150 155 160
Gly Ser Val Ser Thr Ser His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro
165 170 175
Gly Gln Ala Pro Arg Met Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser
180 185 190
Gly Val Pro Asp Arg Phe Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala
195 200 205
Leu Thr Ile Thr Gly Ala Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys
210 215 220
Ala Ile His Val Asp Arg Asp Ile Trp Val Phe Gly Gly Gly Thr Lys
225 230 235 240
Leu Thr Val Leu Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser
245 250 255
Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala
260 265 270
Ala Ser Gly Phe Thr Phe Asn Lys Tyr Ala Met Asn Trp Val Arg Gln
275 280 285
Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Arg Ile Arg Ser Lys Tyr
290 295 300
Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Asp Arg Phe Thr
305 310 315 320
Ile Ser Arg Asp Asp Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn Asn
325 330 335
Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg His Gly Asn
340 345 350
Phe Gly Asn Pro Tyr Ile Ser Tyr Trp Ala Tyr Trp Gly Gln Gly Thr
355 360 365
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
370 375 380
Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr
385 390 395 400
Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly
405 410 415
Ala Val Thr Ser Gly Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly
420 425 430
Gln Ala Pro Arg Gly Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly
435 440 445
Thr Pro Ala Arg Phe Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu
450 455 460
Thr Leu Ser Gly Val Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val
465 470 475 480
Leu Trp Tyr Ser Asn Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr
485 490 495
Val Leu Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
500 505 510
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
515 520 525
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
530 535 540
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
545 550 555 560
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
565 570 575
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
580 585 590
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
595 600 605
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
610 615 620
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr
625 630 635 640
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
645 650 655
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
660 665 670
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
675 680 685
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
690 695 700
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
705 710 715 720
Ser Leu Ser Leu Ser Pro Gly Lys His His His His His His
725 730
<210> 87
<211> 734
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> первый полипептид 162
<400> 87
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Ala Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln Glu Pro Ser Phe
130 135 140
Ser Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly Leu Ser Ser
145 150 155 160
Gly Ser Val Ser Thr Ser His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro
165 170 175
Gly Gln Ala Pro Arg Met Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser
180 185 190
Gly Val Pro Asp Arg Phe Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala
195 200 205
Leu Thr Ile Thr Gly Ala Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys
210 215 220
Ala Ile His Val Asp Arg Asp Ile Trp Val Phe Gly Gly Gly Thr Lys
225 230 235 240
Leu Thr Val Leu Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln Glu
245 250 255
Pro Ser Leu Thr Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly
260 265 270
Ser Ser Thr Gly Ala Val Thr Ser Gly Asn Tyr Pro Asn Trp Val Gln
275 280 285
Gln Lys Pro Gly Gln Ala Pro Arg Gly Leu Ile Gly Gly Thr Lys Phe
290 295 300
Leu Ala Pro Gly Thr Pro Ala Arg Phe Ser Gly Ser Leu Leu Gly Gly
305 310 315 320
Lys Ala Ala Leu Thr Leu Ser Gly Val Gln Pro Glu Asp Glu Ala Glu
325 330 335
Tyr Tyr Cys Val Leu Trp Tyr Ser Asn Arg Trp Val Phe Gly Gly Gly
340 345 350
Thr Lys Leu Thr Val Leu Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
355 360 365
Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
370 375 380
Val Gln Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe
385 390 395 400
Thr Phe Asn Lys Tyr Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys
405 410 415
Gly Leu Glu Trp Val Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala
420 425 430
Thr Tyr Tyr Ala Asp Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp
435 440 445
Asp Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu
450 455 460
Asp Thr Ala Val Tyr Tyr Cys Val Arg His Gly Asn Phe Gly Asn Pro
465 470 475 480
Tyr Ile Ser Tyr Trp Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
485 490 495
Ser Ser Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
500 505 510
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
515 520 525
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
530 535 540
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
545 550 555 560
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
565 570 575
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
580 585 590
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
595 600 605
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
610 615 620
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr
625 630 635 640
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
645 650 655
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
660 665 670
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
675 680 685
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
690 695 700
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
705 710 715 720
Ser Leu Ser Leu Ser Pro Gly Lys His His His His His His
725 730
<210> 88
<211> 474
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> второй полипептид 142
<400> 88
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Ala Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln Glu Pro Ser Phe
130 135 140
Ser Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly Leu Ser Ser
145 150 155 160
Gly Ser Val Ser Thr Ser His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro
165 170 175
Gly Gln Ala Pro Arg Met Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser
180 185 190
Gly Val Pro Asp Arg Phe Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala
195 200 205
Leu Thr Ile Thr Gly Ala Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys
210 215 220
Ala Ile His Val Asp Arg Asp Ile Trp Val Phe Gly Gly Gly Thr Lys
225 230 235 240
Leu Thr Val Leu Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys
245 250 255
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
260 265 270
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
275 280 285
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
290 295 300
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
305 310 315 320
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
325 330 335
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
340 345 350
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
355 360 365
Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Glu Glu
370 375 380
Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr
385 390 395 400
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
405 410 415
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
420 425 430
Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
435 440 445
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
450 455 460
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
465 470
<210> 89
<211> 730
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> цепь 1 NC1
<400> 89
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Lys Tyr
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg His Gly Asn Phe Gly Asn Glu Tyr Ile Ser Tyr Trp
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
115 120 125
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Thr Val Val
130 135 140
Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly Thr Val Thr Leu
145 150 155 160
Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly Asn Tyr Pro Asn
165 170 175
Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly Leu Ile Gly Gly
180 185 190
Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe Ser Gly Ser Leu
195 200 205
Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val Gln Pro Glu Asp
210 215 220
Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn Arg Trp Val Phe
225 230 235 240
Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Gly Gly Ser Asp Ile
245 250 255
Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly Asp Arg
260 265 270
Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ala Asn Tyr Leu Ser
275 280 285
Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile Tyr Gly
290 295 300
Thr Ser Asn Leu Glu Val Gly Val Pro Ser Arg Phe Ser Gly Ser Arg
305 310 315 320
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp
325 330 335
Phe Ala Thr Tyr Tyr Cys Leu Gln Asp Lys Glu Phe Pro Arg Thr Phe
340 345 350
Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly
355 360 365
Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly
370 375 380
Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala
385 390 395 400
Ser Gly Phe Thr Phe Ser His Tyr Tyr Met Ala Trp Val Arg Gln Ala
405 410 415
Pro Gly Lys Gly Leu Glu Trp Val Thr Ser Ile Ser Tyr Glu Gly Asp
420 425 430
Ile Thr Tyr Tyr Gly Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg
435 440 445
Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala
450 455 460
Glu Asp Thr Ala Thr Tyr Tyr Cys Ala Ser Gln Thr Leu Arg Glu Ser
465 470 475 480
Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly
485 490 495
Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
500 505 510
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
515 520 525
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
530 535 540
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
545 550 555 560
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
565 570 575
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
580 585 590
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
595 600 605
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
610 615 620
Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val
625 630 635 640
Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
645 650 655
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
660 665 670
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
675 680 685
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
690 695 700
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
705 710 715 720
Ser Pro Gly Lys His His His His His His
725 730
<210> 90
<211> 470
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> цепь 2 NC1
<400> 90
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ala Asn Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Thr Ser Asn Leu Glu Val Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Asp Lys Glu Phe Pro Arg
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu
115 120 125
Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys
130 135 140
Ala Ala Ser Gly Phe Thr Phe Ser His Tyr Tyr Met Ala Trp Val Arg
145 150 155 160
Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Thr Ser Ile Ser Tyr Glu
165 170 175
Gly Asp Ile Thr Tyr Tyr Gly Asp Ser Val Lys Gly Arg Phe Thr Ile
180 185 190
Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu
195 200 205
Arg Ala Glu Asp Thr Ala Thr Tyr Tyr Cys Ala Ser Gln Thr Leu Arg
210 215 220
Glu Ser Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
225 230 235 240
Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
245 250 255
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
260 265 270
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
275 280 285
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
290 295 300
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
305 310 315 320
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
325 330 335
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
340 345 350
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
355 360 365
Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
370 375 380
Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile
385 390 395 400
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
405 410 415
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys
420 425 430
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
435 440 445
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
450 455 460
Ser Leu Ser Pro Gly Lys
465 470
<210> 91
<211> 725
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> цепь 1 NC2
<400> 91
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Tyr
20 25 30
Tyr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Thr Ser Ile Ser Tyr Glu Gly Asp Ile Thr Tyr Tyr Gly Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Ser Gln Thr Leu Arg Glu Ser Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
115 120 125
Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val
130 135 140
Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln
145 150 155 160
Asp Ile Ala Asn Tyr Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ser
165 170 175
Pro Lys Leu Leu Ile Tyr Gly Thr Ser Asn Leu Glu Val Gly Val Pro
180 185 190
Ser Arg Phe Ser Gly Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile
195 200 205
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Asp
210 215 220
Lys Glu Phe Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
225 230 235 240
Gly Gly Gly Gly Ser Gln Thr Val Val Thr Gln Glu Pro Ser Phe Ser
245 250 255
Val Ser Pro Gly Gly Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly
260 265 270
Ser Val Ser Thr Ser His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro Gly
275 280 285
Gln Ala Pro Arg Met Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser Gly
290 295 300
Val Pro Asp Arg Phe Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu
305 310 315 320
Thr Ile Thr Gly Ala Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala
325 330 335
Ile His Val Asp Arg Asp Ile Trp Val Phe Gly Gly Gly Thr Lys Leu
340 345 350
Thr Val Leu Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
355 360 365
Gly Ser Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro
370 375 380
Gly Thr Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser
385 390 395 400
Ser Ser Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu
405 410 415
Trp Val Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp
420 425 430
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr
435 440 445
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
450 455 460
Tyr Cys Ala Arg Ser Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly
465 470 475 480
Gln Gly Ala Leu Val Thr Val Ser Ser Gly Gly Gly Asp Lys Thr His
485 490 495
Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
500 505 510
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
515 520 525
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
530 535 540
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
545 550 555 560
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
565 570 575
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
580 585 590
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
595 600 605
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
610 615 620
Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu
625 630 635 640
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
645 650 655
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
660 665 670
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
675 680 685
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
690 695 700
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys His
705 710 715 720
His His His His His
725
<210> 92
<211> 474
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> цепь 2 NC2
<400> 92
Gln Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser
20 25 30
His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Met
35 40 45
Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Ile His Val Asp Arg
85 90 95
Asp Ile Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
100 105 110
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
115 120 125
Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr Ser Leu Arg
130 135 140
Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser Ala Met His
145 150 155 160
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Val Ile
165 170 175
Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val Lys Gly Arg
180 185 190
Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met
195 200 205
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Ser
210 215 220
Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly Ala Leu Val
225 230 235 240
Thr Val Ser Ser Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys
245 250 255
Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
260 265 270
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
275 280 285
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
290 295 300
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
305 310 315 320
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
325 330 335
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
340 345 350
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
355 360 365
Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Glu Glu
370 375 380
Met Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr
385 390 395 400
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
405 410 415
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
420 425 430
Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
435 440 445
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
450 455 460
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
465 470
<210> 93
<211> 730
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> цепь 1 NC3
<400> 93
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ala Asn Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Thr Ser Asn Leu Glu Val Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Asp Lys Glu Phe Pro Arg
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu
115 120 125
Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys
130 135 140
Ala Ala Ser Gly Phe Thr Phe Ser His Tyr Tyr Met Ala Trp Val Arg
145 150 155 160
Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Thr Ser Ile Ser Tyr Glu
165 170 175
Gly Asp Ile Thr Tyr Tyr Gly Asp Ser Val Lys Gly Arg Phe Thr Ile
180 185 190
Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu
195 200 205
Arg Ala Glu Asp Thr Ala Thr Tyr Tyr Cys Ala Ser Gln Thr Leu Arg
210 215 220
Glu Ser Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
225 230 235 240
Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
245 250 255
Val Gln Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe
260 265 270
Thr Phe Asn Lys Tyr Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys
275 280 285
Gly Leu Glu Trp Val Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala
290 295 300
Thr Tyr Tyr Ala Asp Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp
305 310 315 320
Asp Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr Glu
325 330 335
Asp Thr Ala Val Tyr Tyr Cys Val Arg His Gly Asn Phe Gly Asn Glu
340 345 350
Tyr Ile Ser Tyr Trp Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val
355 360 365
Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
370 375 380
Ser Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly
385 390 395 400
Gly Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser
405 410 415
Gly Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg
420 425 430
Gly Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg
435 440 445
Phe Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly
450 455 460
Val Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser
465 470 475 480
Asn Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
485 490 495
Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
500 505 510
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
515 520 525
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
530 535 540
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
545 550 555 560
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
565 570 575
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
580 585 590
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
595 600 605
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
610 615 620
Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val
625 630 635 640
Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
645 650 655
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
660 665 670
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
675 680 685
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
690 695 700
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
705 710 715 720
Ser Pro Gly Lys His His His His His His
725 730
<210> 94
<211> 470
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> цепь 2 NC3
<400> 94
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ala Asn Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Thr Ser Asn Leu Glu Val Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Asp Lys Glu Phe Pro Arg
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu
115 120 125
Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys
130 135 140
Ala Ala Ser Gly Phe Thr Phe Ser His Tyr Tyr Met Ala Trp Val Arg
145 150 155 160
Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Thr Ser Ile Ser Tyr Glu
165 170 175
Gly Asp Ile Thr Tyr Tyr Gly Asp Ser Val Lys Gly Arg Phe Thr Ile
180 185 190
Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu
195 200 205
Arg Ala Glu Asp Thr Ala Thr Tyr Tyr Cys Ala Ser Gln Thr Leu Arg
210 215 220
Glu Ser Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
225 230 235 240
Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
245 250 255
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
260 265 270
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
275 280 285
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
290 295 300
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
305 310 315 320
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
325 330 335
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
340 345 350
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
355 360 365
Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
370 375 380
Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile
385 390 395 400
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
405 410 415
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys
420 425 430
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
435 440 445
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
450 455 460
Ser Leu Ser Pro Gly Lys
465 470
<210> 95
<211> 504
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> цепь 1 MGD009
<400> 95
Asp Ile Gln Leu Thr Gln Ser Pro Ser Phe Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asn Val Asp Thr Asn
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Ala Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Asn Tyr Pro Phe
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Gly Gly Gly Ser Gly
100 105 110
Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
115 120 125
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
130 135 140
Ser Thr Tyr Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
145 150 155 160
Glu Trp Val Gly Arg Ile Arg Ser Lys Tyr Asn Asn Tyr Ala Thr Tyr
165 170 175
Tyr Ala Asp Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser
180 185 190
Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr
195 200 205
Ala Val Tyr Tyr Cys Val Arg His Gly Asn Phe Gly Asn Ser Tyr Val
210 215 220
Ser Trp Phe Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
225 230 235 240
Gly Gly Cys Gly Gly Gly Glu Val Ala Ala Leu Glu Lys Glu Val Ala
245 250 255
Ala Leu Glu Lys Glu Val Ala Ala Leu Glu Lys Glu Val Ala Ala Leu
260 265 270
Glu Lys Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
275 280 285
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
290 295 300
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
305 310 315 320
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
325 330 335
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
340 345 350
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
355 360 365
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
370 375 380
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
385 390 395 400
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
405 410 415
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
420 425 430
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
435 440 445
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
450 455 460
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
465 470 475 480
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
485 490 495
Ser Leu Ser Leu Ser Pro Gly Lys
500
<210> 96
<211> 274
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> цепь 2 MGD009
<400> 96
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Arg Ser Ser Thr Gly Ala Val Thr Thr Ser
20 25 30
Asn Tyr Ala Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Asn Lys Arg Ala Pro Trp Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Leu Trp Tyr Ser Asn
85 90 95
Leu Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
115 120 125
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
130 135 140
Phe Thr Phe Ser Ser Phe Gly Met His Trp Val Arg Gln Ala Pro Gly
145 150 155 160
Lys Gly Leu Glu Trp Val Ala Tyr Ile Ser Ser Asp Ser Ser Ala Ile
165 170 175
Tyr Tyr Ala Asp Thr Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn
180 185 190
Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Asp Glu Asp
195 200 205
Thr Ala Val Tyr Tyr Cys Gly Arg Gly Arg Glu Asn Ile Tyr Tyr Gly
210 215 220
Ser Arg Leu Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
225 230 235 240
Gly Gly Cys Gly Gly Gly Lys Val Ala Ala Leu Lys Glu Lys Val Ala
245 250 255
Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu Lys Val Ala Ala Leu
260 265 270
Lys Glu
<210> 97
<211> 227
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> цепь 3 MGD009
<400> 97
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn Arg Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 98
<211> 13
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ПЕПТИД
<223> линкер
<400> 98
Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10
<210> 99
<211> 513
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> цепь 1 201
<400> 99
Gln Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser
20 25 30
His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Met
35 40 45
Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Ile His Val Asp Arg
85 90 95
Asp Ile Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly
115 120 125
Leu Val Gln Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly
130 135 140
Phe Thr Phe Asn Lys Tyr Ala Met Asn Trp Val Arg Gln Ala Pro Gly
145 150 155 160
Lys Gly Leu Glu Trp Val Ala Arg Ile Arg Ser Lys Tyr Asn Asn Tyr
165 170 175
Ala Thr Tyr Tyr Ala Asp Ser Val Lys Asp Arg Phe Thr Ile Ser Arg
180 185 190
Asp Asp Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn Asn Leu Lys Thr
195 200 205
Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg His Gly Asn Phe Gly Asn
210 215 220
Glu Tyr Ile Ser Tyr Trp Ala Tyr Trp Gly Gln Gly Thr Leu Val Thr
225 230 235 240
Val Ser Ser Gly Gly Cys Gly Gly Gly Glu Val Ala Ala Leu Glu Lys
245 250 255
Glu Val Ala Ala Leu Glu Lys Glu Val Ala Ala Leu Glu Lys Glu Val
260 265 270
Ala Ala Leu Glu Lys Gly Gly Gly Asp Lys Thr His Thr Cys Pro Pro
275 280 285
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
290 295 300
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
305 310 315 320
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
325 330 335
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
340 345 350
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
355 360 365
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
370 375 380
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
385 390 395 400
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
405 410 415
Glu Met Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe
420 425 430
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
435 440 445
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
450 455 460
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
465 470 475 480
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
485 490 495
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys His His His His His
500 505 510
His
<210> 100
<211> 271
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> цепь 2 201
<400> 100
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly
20 25 30
Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn
85 90 95
Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Gln Val Gln Leu Val Gln Ser Gly Gly Gly
115 120 125
Val Val Gln Pro Gly Thr Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
130 135 140
Phe Ile Phe Ser Ser Ser Ala Met His Trp Val Arg Gln Ala Pro Gly
145 150 155 160
Lys Gly Leu Glu Trp Val Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys
165 170 175
Tyr Tyr Val Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn
180 185 190
Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp
195 200 205
Thr Ala Val Tyr Tyr Cys Ala Arg Ser Ala Arg Leu Tyr Ala Ser Phe
210 215 220
Asp Tyr Trp Gly Gln Gly Ala Leu Val Thr Val Ser Ser Gly Gly Cys
225 230 235 240
Gly Gly Gly Lys Val Ala Ala Leu Lys Glu Lys Val Ala Ala Leu Lys
245 250 255
Glu Lys Val Ala Ala Leu Lys Glu Lys Val Ala Ala Leu Lys Glu
260 265 270
<210> 101
<211> 455
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> цепь 1 202
<400> 101
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Ala Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Trp
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys His His His His His His
450 455
<210> 102
<211> 217
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> цепь 2 202
<400> 102
Gln Thr Val Val Thr Gln Glu Pro Ser Phe Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Ser Thr Ser
20 25 30
His Tyr Pro Ser Trp Tyr Gln Gln Thr Pro Gly Gln Ala Pro Arg Met
35 40 45
Leu Ile Tyr Asn Thr Asn Thr Arg Ser Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Ile Leu Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Ala Asp Asp Glu Ser Asp Tyr Tyr Cys Ala Ile His Val Asp Arg
85 90 95
Asp Ile Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Arg Thr
100 105 110
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
115 120 125
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
130 135 140
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
145 150 155 160
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
165 170 175
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
180 185 190
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
195 200 205
Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 103
<211> 692
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> цепь 3 202
<400> 103
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Thr
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ile Phe Ser Ser Ser
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Ala Arg Leu Tyr Ala Ser Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Ala Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Gly
210 215 220
Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser
225 230 235 240
Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Lys Leu Ser Cys Ala
245 250 255
Ala Ser Gly Phe Thr Phe Asn Lys Tyr Ala Met Asn Trp Val Arg Gln
260 265 270
Ala Pro Gly Lys Gly Leu Glu Trp Val Ala Arg Ile Arg Ser Lys Tyr
275 280 285
Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp Ser Val Lys Asp Arg Phe Thr
290 295 300
Ile Ser Arg Asp Asp Ser Lys Asn Thr Ala Tyr Leu Gln Met Asn Asn
305 310 315 320
Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys Val Arg His Gly Asn
325 330 335
Phe Gly Asn Glu Tyr Ile Ser Tyr Trp Ala Tyr Trp Gly Gln Gly Thr
340 345 350
Leu Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile
355 360 365
Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val
370 375 380
Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys
385 390 395 400
Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu
405 410 415
Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu
420 425 430
Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr
435 440 445
His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu
450 455 460
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
465 470 475 480
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
485 490 495
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
500 505 510
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
515 520 525
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
530 535 540
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
545 550 555 560
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
565 570 575
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
580 585 590
Val Cys Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
595 600 605
Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
610 615 620
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
625 630 635 640
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr
645 650 655
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
660 665 670
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
675 680 685
Ser Pro Gly Lys
690
<210> 104
<211> 214
<212> ПРТ
<213> Искусственная последовательность
<220>
<221> ЦЕПЬ
<223> цепь 4 202
<400> 104
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Thr Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Ser Ser Thr Gly Ala Val Thr Ser Gly
20 25 30
Asn Tyr Pro Asn Trp Val Gln Gln Lys Pro Gly Gln Ala Pro Arg Gly
35 40 45
Leu Ile Gly Gly Thr Lys Phe Leu Ala Pro Gly Thr Pro Ala Arg Phe
50 55 60
Ser Gly Ser Leu Leu Gly Gly Lys Ala Ala Leu Thr Leu Ser Gly Val
65 70 75 80
Gln Pro Glu Asp Glu Ala Glu Tyr Tyr Cys Val Leu Trp Tyr Ser Asn
85 90 95
Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Ser Ser Ala
100 105 110
Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser
115 120 125
Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe
130 135 140
Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly
145 150 155 160
Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu
165 170 175
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr
180 185 190
Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys
195 200 205
Val Glu Pro Lys Ser Cys
210
<---


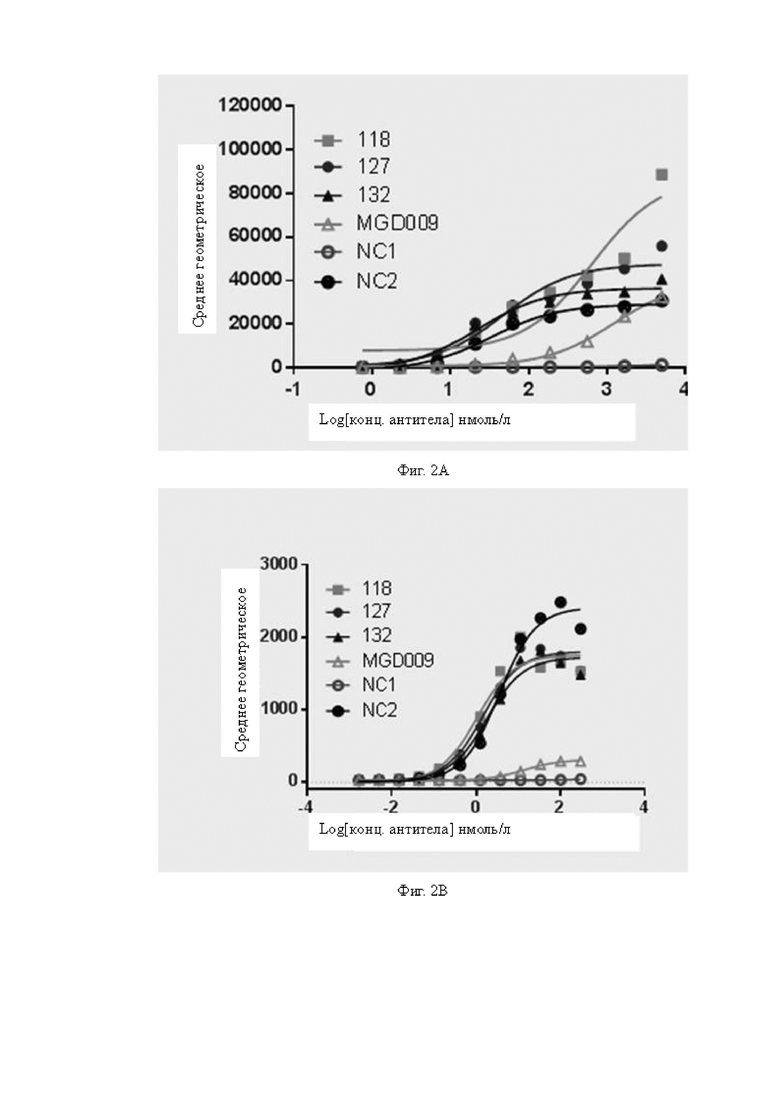
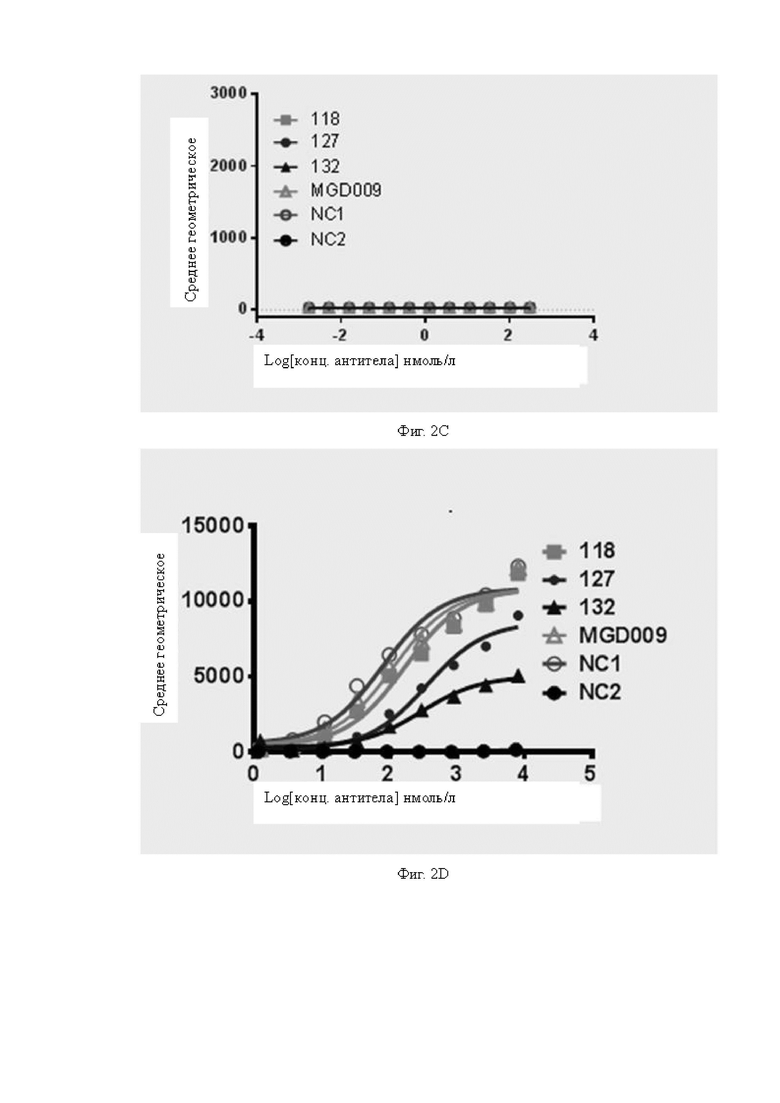
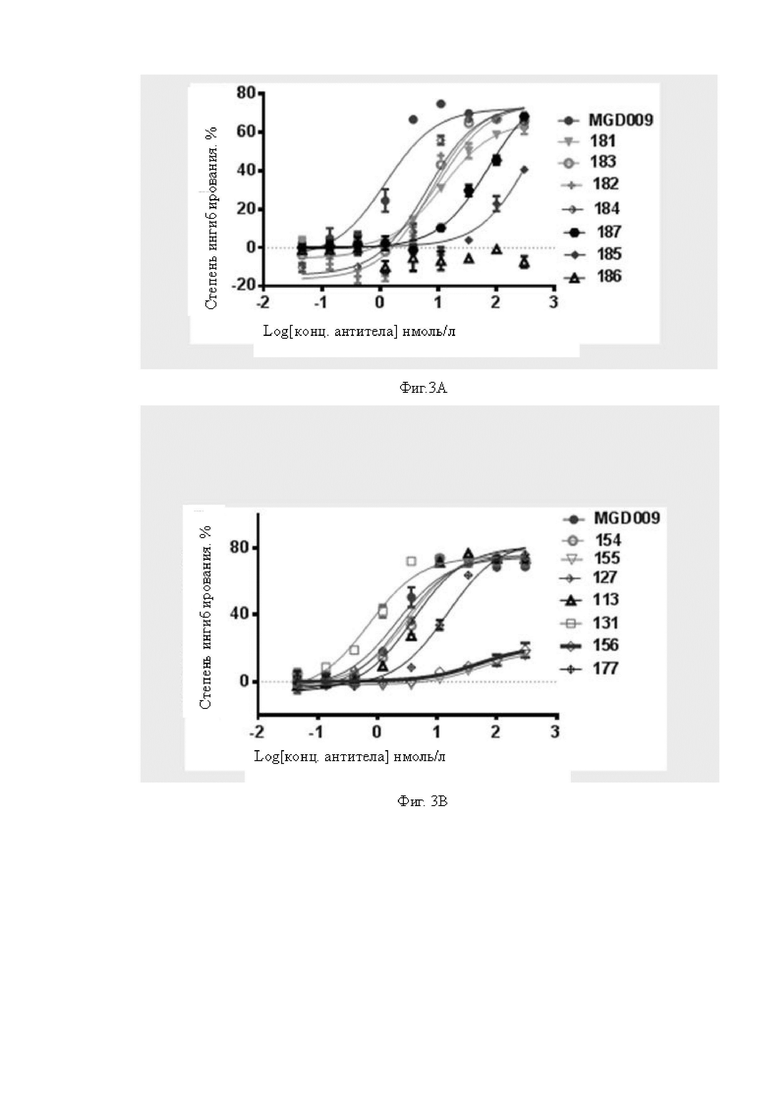
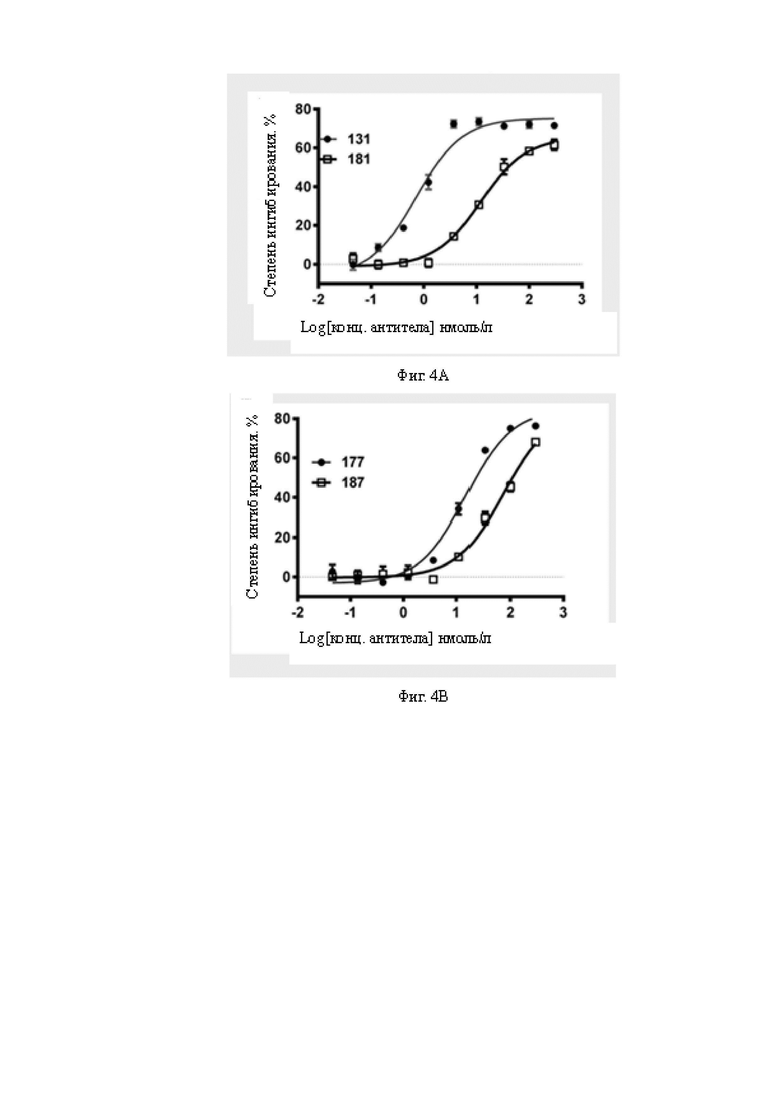
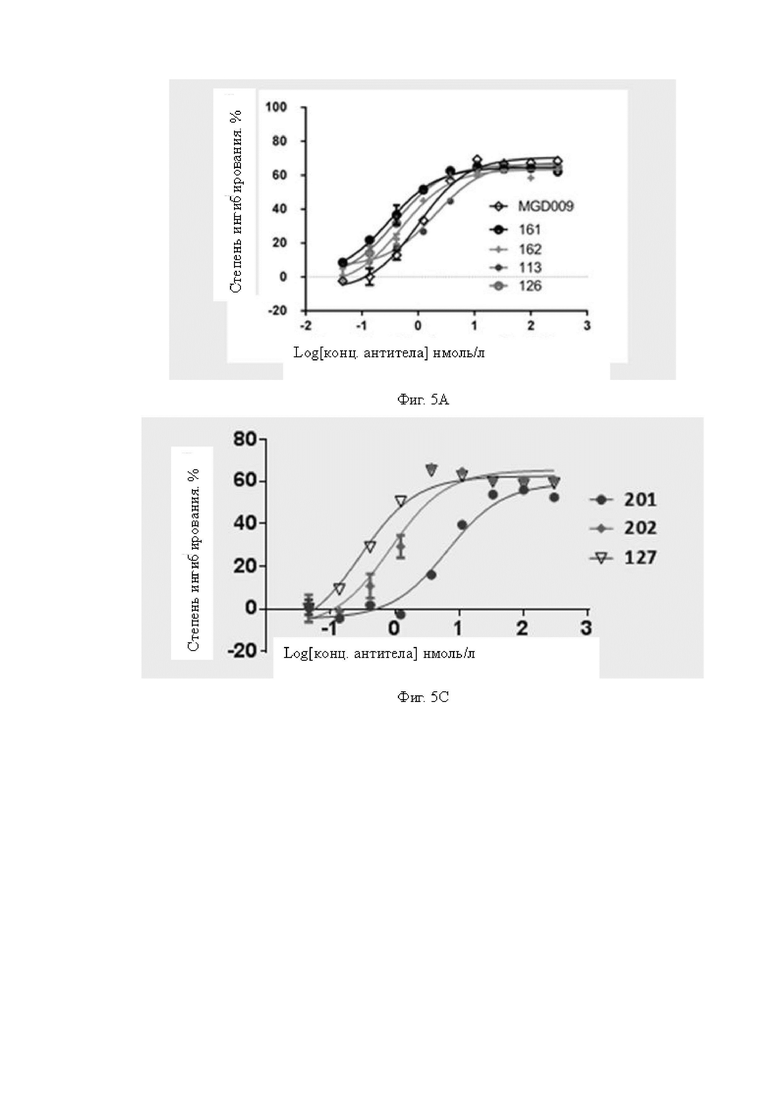
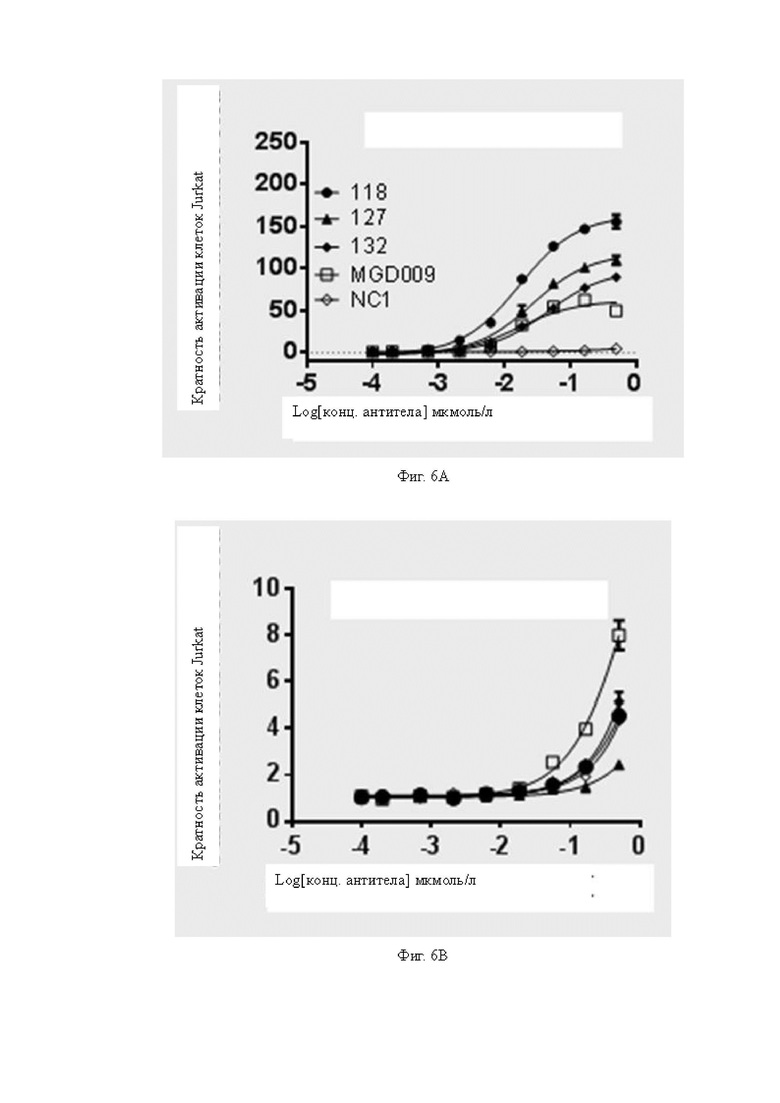
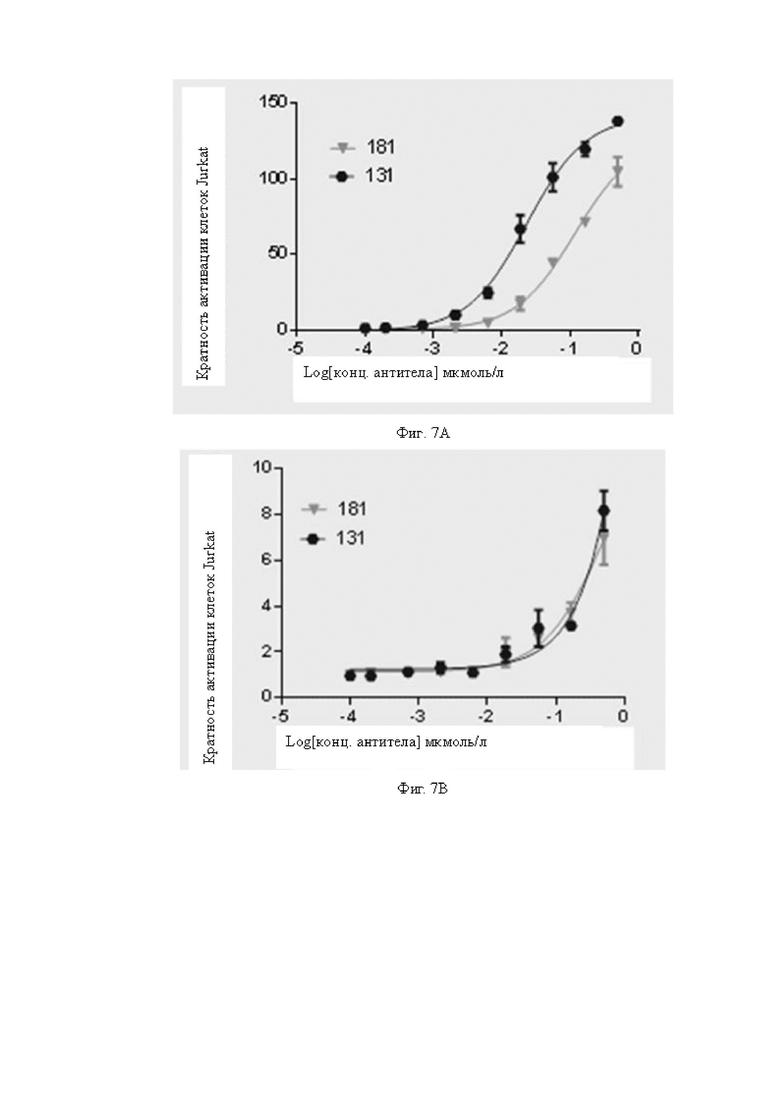
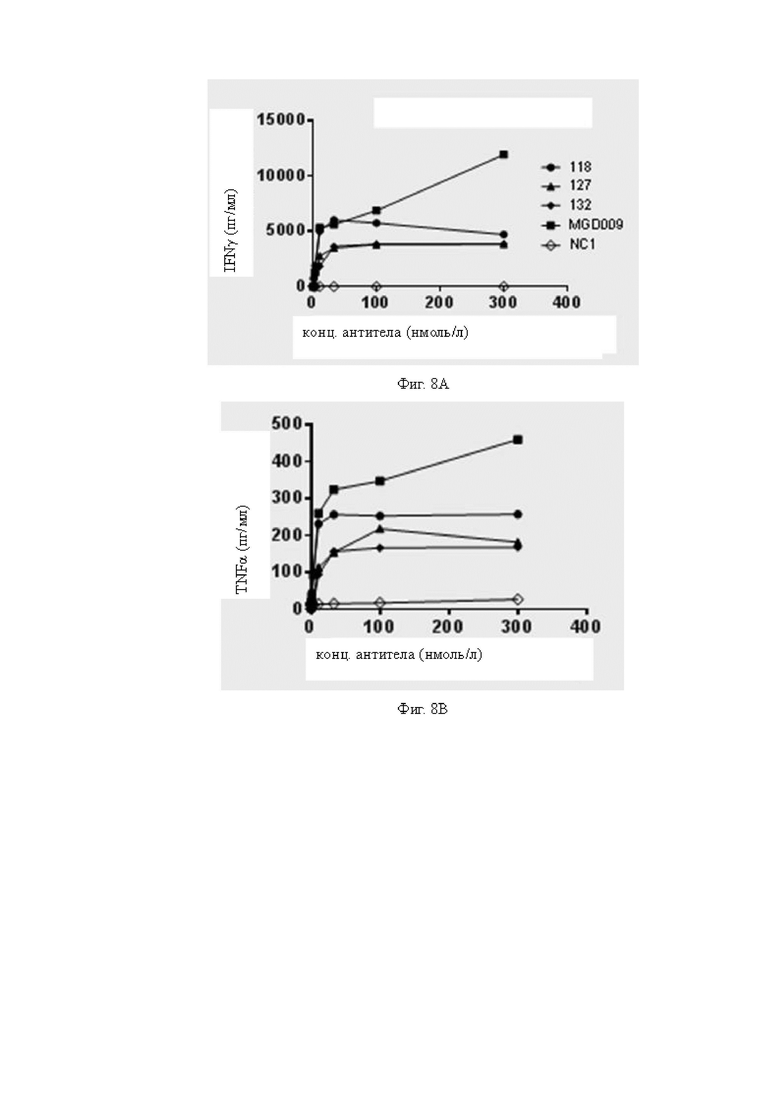
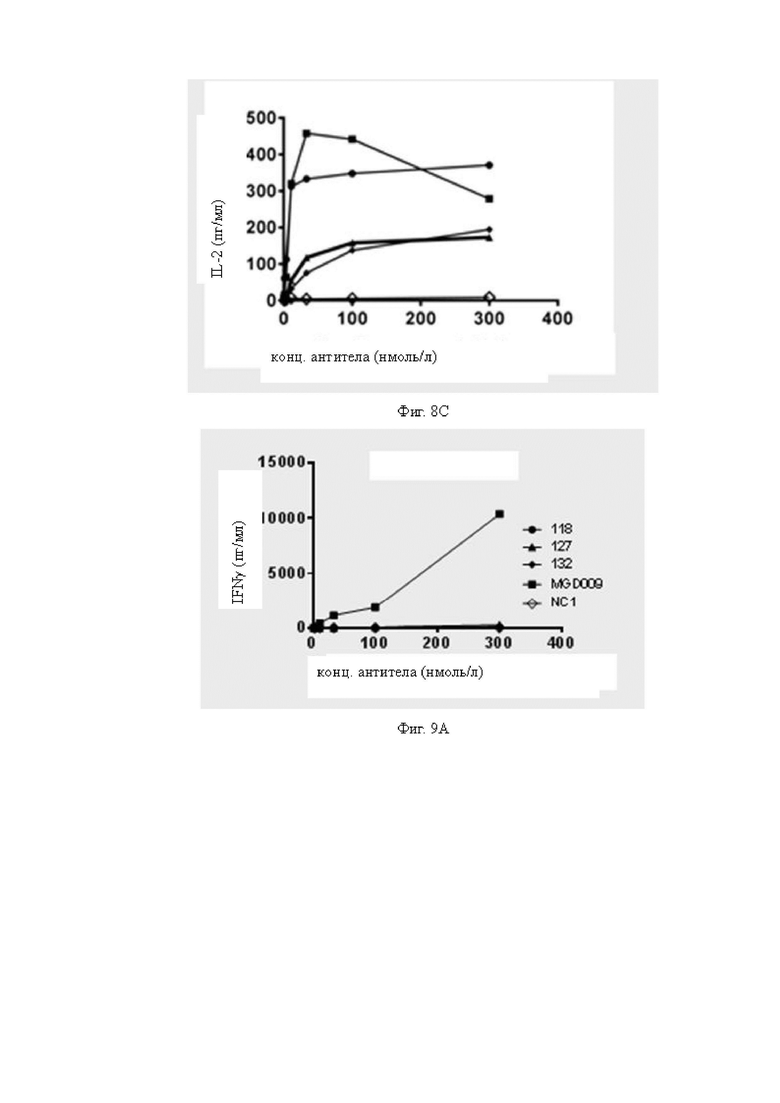
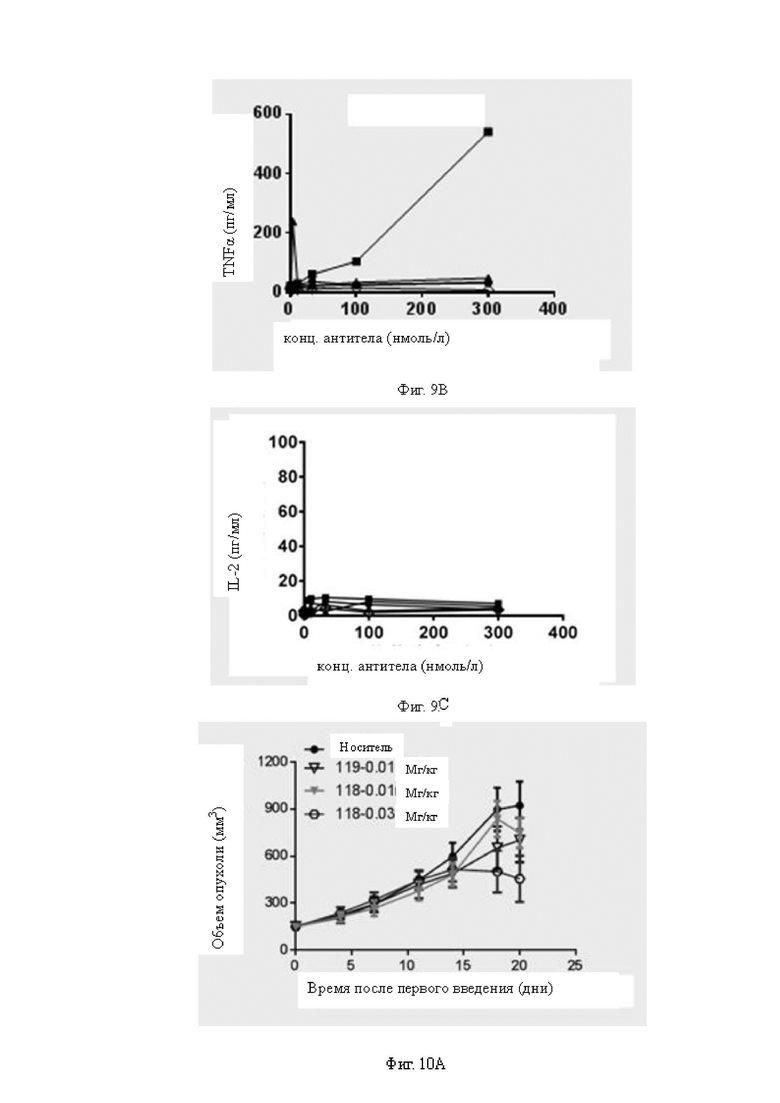
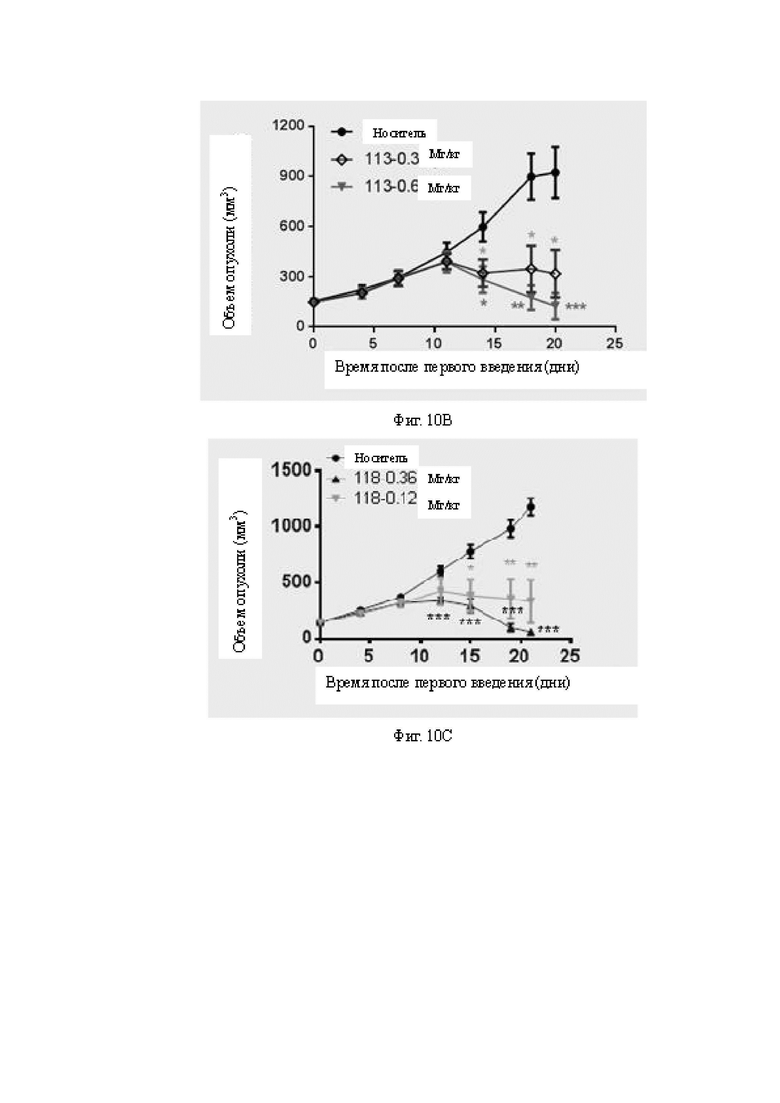
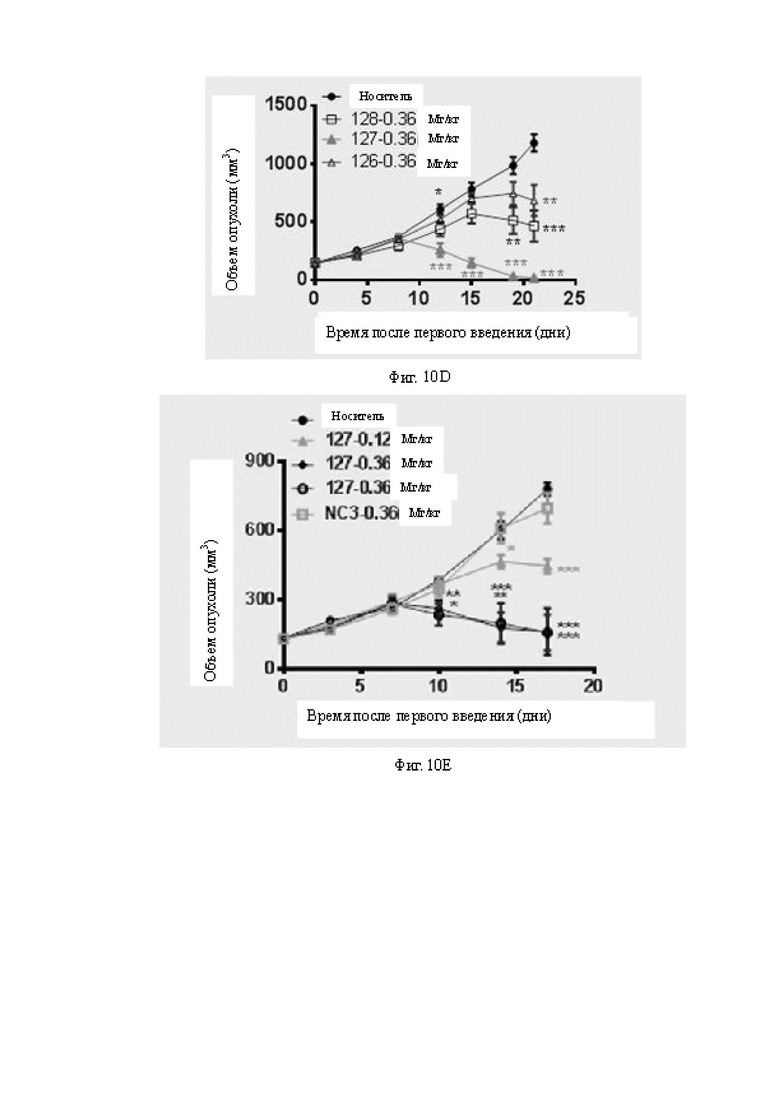
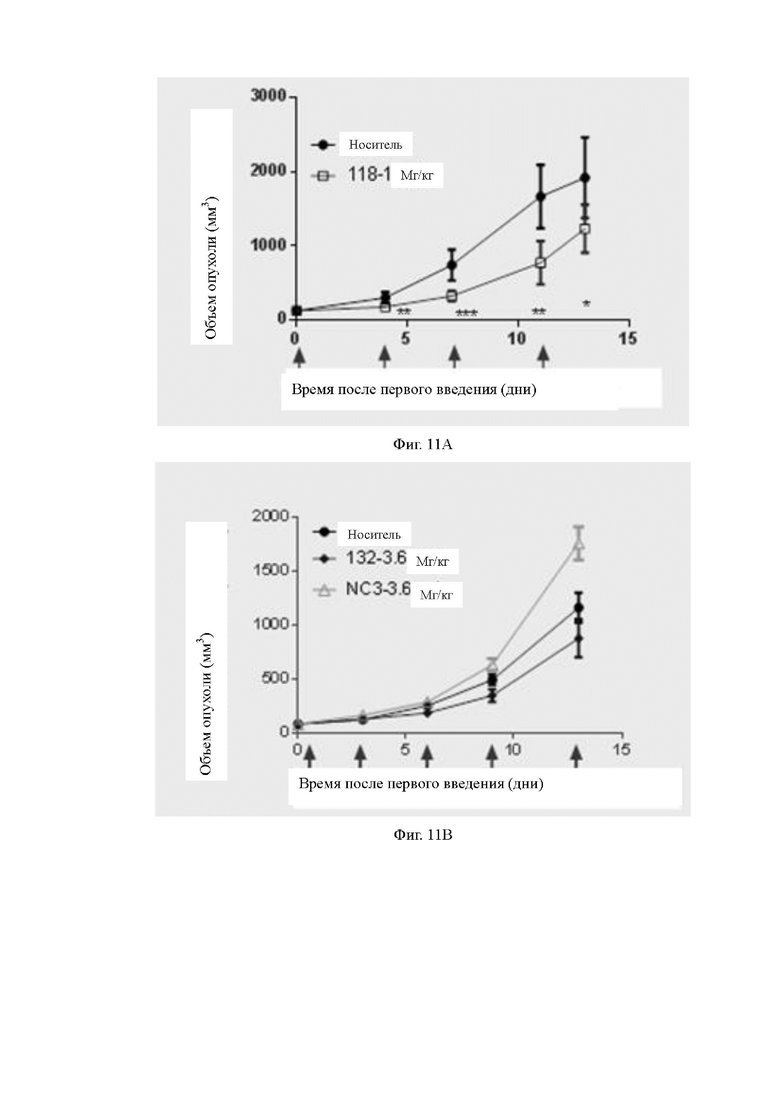
Изобретение относится к области биохимии, в частности к антителу или его антигенсвязывающему фрагменту, специфично связывающимся с человеческим CD3. Также раскрыты одноцепочечное антитело, специфично связывающееся с человеческим CD3; биспецифическое антитело, специфично связывающееся с человеческим CD3; фармацевтическая композиция, содержащая указанные антитела; выделенная молекула нуклеиновой кислоты, кодирующая указанное антитело; рекомбинантный вектор и клетка-хозяин, содержащая указанную молекулу нуклеиновой кислоты. Раскрыт способ получения указанного антитела. Изобретение позволяет эффективно лечить заболевания, ассоциированные с CD3. 8 н. и 6 з.п. ф-лы, 8 пр., 17 табл.,11 ил.
1. Антитело или его антигенсвязывающий фрагмент, специфично связывающиеся с человеческим CD3, причем антитело или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи и вариабельную область легкой цепи, где:
вариабельная область легкой цепи содержит LCDR1 (определяющая комплементарность область 1 легкой цепи), LCDR2 и LCDR3, как показано в SEQ ID NO: 48, SEQ ID NO: 49 и SEQ ID NO: 50 соответственно, и
вариабельная область тяжелой цепи представляет собой любую выбранную из группы, состоящей из следующих i)-v):
i) вариабельной области тяжелой цепи, содержащей HCDR1 (определяющая комплементарность область 1 тяжелой цепи), HCDR2 и HCDR3, как показано в SEQ ID NO: 37, SEQ ID NO: 38 и SEQ ID NO: 39 соответственно;
ii) вариабельной области тяжелой цепи, содержащей HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 37, SEQ ID NO: 40 и SEQ ID NO: 41 соответственно;
iii) вариабельной области тяжелой цепи, содержащей HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 37, SEQ ID NO: 40 и SEQ ID NO: 42 соответственно;
iv) вариабельной области тяжелой цепи, содержащей HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 37, SEQ ID NO: 40 и SEQ ID NO: 43 соответственно; и
v) вариабельной области тяжелой цепи, содержащей HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 37, SEQ ID NO: 47 и SEQ ID NO: 45 соответственно.
2. Антитело или его антигенсвязывающий фрагмент, специфично связывающиеся с человеческим CD3, по п. 1, где антитело или его антигенсвязывающий фрагмент представляет собой гуманизированное антитело или его антигенсвязывающий фрагмент.
3. Антитело или его антигенсвязывающий фрагмент, специфично связывающиеся с человеческим CD3, по п. 2, содержащие вариабельную область легкой цепи, как показано в SEQ ID NO: 36, и/или
вариабельную область тяжелой цепи, как показано в любой из последовательностей, выбранной из группы, состоящей из SEQ ID NO: 29, SEQ ID NO: 30, SEQ ID NO: 31, SEQ ID NO: 32 и SEQ ID NO: 35.
4. Антитело или его антигенсвязывающий фрагмент, специфично связывающиеся с человеческим CD3, по п. 3, дополнительно содержащие константную область легкой цепи и/или константную область тяжелой цепи антитела;
необязательно, константная область легкой цепи представляет собой константную область человеческой легкой цепи каппа, цепи лямбда или ее вариант, и константная область тяжелой цепи представляет собой константную область тяжелой цепи человеческого IgG1, IgG2, IgG3, IgG4 или ее вариант.
5. Антитело или его антигенсвязывающий фрагмент, специфично связывающиеся с человеческим CD3, по любому из пп. 1-4, где антигенсвязывающий фрагмент выбран из Fab, Fab', F(ab')2, димеризованной области V и стабилизированной дисульфидом области V (dsFv).
6. Одноцепочечное антитело, специфично связывающееся с человеческим CD3, содержащее вариабельную область легкой цепи и вариабельную область тяжелой цепи антитела или его антигенсвязывающего фрагмента, где:
вариабельная область легкой цепи содержит LCDR1, LCDR2 и LCDR3, как показано в SEQ ID NO: 48, SEQ ID NO: 49 и SEQ ID NO: 50, соответственно, и
вариабельная область тяжелой цепи представляет собой любую выбранную из группы, состоящей из следующих i)-v):
i) вариабельной области тяжелой цепи, содержащей HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 37, SEQ ID NO: 38 и SEQ ID NO: 39, соответственно;
ii) вариабельной области тяжелой цепи, содержащей HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 37, SEQ ID NO: 40 и SEQ ID NO: 41, соответственно;
iii) вариабельной области тяжелой цепи, содержащей HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 37, SEQ ID NO: 40 и SEQ ID NO: 42, соответственно;
iv) вариабельной области тяжелой цепи, содержащей HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 37, SEQ ID NO: 40 и SEQ ID NO: 43, соответственно; и
v) вариабельной области тяжелой цепи, содержащей HCDR1, HCDR2 и HCDR3, как показано в SEQ ID NO: 37, SEQ ID NO: 47 и SEQ ID NO: 45, соответственно.
7. Одноцепочечное антитело по п. 6, где последовательность одноцепочечного антитела является такой, как показано в SEQ ID NO: 55, 56, 57, 58, 61, 62, 63, 64, 65 или 68.
8. Биспецифическое антитело, специфично связывающееся с человеческим CD3 и опухолеассоциированным(-ыми) антигеном(-ами), причем биспецифическое антитело включает одноцепочечное антитело по п. 6 или 7 или антитело или его антигенсвязывающий фрагмент, специфично связывающиеся с человеческим CD3, по любому из пп. 1-5, где опухолеассоциированный антиген выбран из группы, состоящей из B7H3, BCMA, CD20, CEA.
9. Фармацевтическая композиция для лечения рака или аутоиммунного заболевания, содержащая терапевтически эффективное количество антитела или его антигенсвязывающего фрагмента, специфично связывающихся с человеческим CD3, по любому из пп. 1-5, одноцепочечного антитела по п. 6 или 7 или биспецифического антитела по п. 8 и один или более фармацевтически приемлемых носителей, разбавителей, буферных растворов или эксципиентов.
10. Выделенная молекула нуклеиновой кислоты, кодирующая антитело или его антигенсвязывающий фрагмент, специфично связывающиеся с человеческим CD3, по любому из пп. 1-5, кодирующая одноцепочечное антитело по п. 6 или 7 или кодирующая биспецифическое антитело по п. 8.
11. Рекомбинантный вектор для экспрессии антитела или его антигенсвязывающего фрагмента, специфично связывающихся с человеческим CD3, по любому из пп. 1-5, содержащий выделенную молекулу нуклеиновой кислоты по п. 10.
12. Клетка-хозяин для экспрессии антитела или его антигенсвязывающего фрагмента, специфично связывающихся с человеческим CD3, по любому из пп. 1-5, или одноцепочечного антитела по п. 6 или 7, или биспецифического антитела по п. 8, где клетка-хозяин трансформирована рекомбинантным вектором по п. 11, причем указанная клетка-хозяин выбрана из прокариотической клетки и эукариотической клетки, предпочтительно эукариотической клетки, более предпочтительно клетки млекопитающего или клетки насекомого.
13. Способ получения антитела или его антигенсвязывающего фрагмента, специфично связывающихся с человеческим CD3, по любому из пп. 1-5, одноцепочечного антитела по п. 6 или 7 или биспецифического антитела по п. 8, причем способ включает культивирование клетки-хозяина по п. 12 в питательной среде для образования и накопления антитела или его антигенсвязывающего фрагмента, специфично связывающихся с человеческим CD3, по любому из пп. 1-5, одноцепочечного антитела по п. 6 или 7 или биспецифического антитела по п. 8, и выделение антитела или его антигенсвязывающего фрагмента, одноцепочечного антитела или биспецифического антитела из культуры.
14. Антитело или его антигенсвязывающий фрагмент, специфично связывающиеся с человеческим CD3, по любому из пп. 1-5, одноцепочечное антитело по п. 6 или 7, биспецифическое антитело по п. 8, фармацевтическая композиция по п. 9 или выделенная молекула нуклеиновой кислоты по п. 10 в качестве лекарственного средства для лечения рака или аутоиммунного заболевания.
| Способ изготовления цементирующей массы | 1931 |
|
SU30437A1 |
| ТЕРАПЕВТИЧЕСКИЕ DLL4-СВЯЗЫВАЮЩИЕ БЕЛКИ | 2010 |
|
RU2570639C2 |
| WO 2011070088 A1, 16.06.2011 | |||
| WO 2011117329 A1, 29.09.2011. | |||
