Настоящее изобретение относится к генам, придающим устойчивость к Podosphaera leucotricha, растениям, частям растений и семенам, содержащим рассматриваемые гены, обеспечивающие устойчивость, и их применению для селекции растений, устойчивых к Podosphaera leucotricha.
Мучнистая роса (powdery mildew, PM) представляет собой основное грибковое заболевание для тысяч видов растений, включая культивированные розоцветные (Rosaceae), такие как яблоня домашняя (Malus domestica), абрикос (Prunus armeniaca), персик (Prunus persica) и клубника (Fragaria x ananassa). Мучнистая роса встречается во всех основных регионах выращивания розоцветных, приводя к высоким потерям.
Мучнистая роса яблони, вызванная грибом P. leucotricha, является одной из наиболее экономически актуальных болезней яблонь в мире. Симптомами являются белые пятна на молодых зеленых тканях, особенно листьях в первые дни после открытия, при этом зрелые листья проявляют некоторую устойчивость. Зараженные листья сморщиваются, скручиваются и преждевременно опадают. Цветки и фрукты не являются главными мишенями гриба PM, но возможны инфекции этих тканей. Споры мучнистой росы зимуют в почках, а затем весной в ходе вегетативного возобновления они начинают новую инфекцию. Мучнистая роса яблонь появляется во всех регионах мира, занимающихся выращиванием яблок. Заболевание причиняет экономический ущерб, уменьшая силу деревьев, образование цветочных бутонов и качество плодов. Мучнистая роса яблонь проявляет симптомы на молодых побегах, листьях, цветках и плодах. Как правило, симптомы наиболее заметны на листьях и плодах.
P. leucotricha представляет собой сумчатый гриб (аскомицет) семейства Erysiphaceae. В течение вегетационного периода этот облигатный биотроф непрерывно вырабатывает споры бесполого размножения (конидии) на специальных коротких ветвях, называемых конидиеносцами, и споры полового размножения (аскоспоры) в сумкоподобных асках, заключенных в плодовые тела (аскокарпы). Конидии являются гиалиновыми (прозрачными, бесцветными) и содержат фиброзиновые тельца самой различной формы, тогда как каждый аскокарп (черный) содержит один аск с восьмью эллиптическим аскоспорами, плотно сгруппированными вместе. Фиброзиновые тельца гиалина представляют собой рефрактивные тельца включения, которые имеют разнообразную форму, включая стержни и конусы. Аскокарпы не играют никакой известной роли в инициировании новых эпидемий, так как аскоспоры не могут легко прорастать. Конидии разносятся ветром и не требуют свободной влаги для прорастания. При попадании на восприимчивую ткань они инициируют заражение и продуцируют мицелий.
В коммерческих садах фунгициды почти всегда используют для контроля мучнистой росы, а также других болезней яблонь. Однако P. leucotricha проявил способность к развитию устойчивости к этим фунгицидам при повторном применении. Бензимидазолы обладают активностью против P. leucotricha, но их применение для контроля заболеваний у яблонь в настоящее время ограничено из-за широкого развития устойчивости к грибу Venturia inaequalis (парша яблони). Фунгициды обычно применяют с интервалом 7-10 дней от фазы обособления бутонов до завершения роста терминальных побегов для того, чтобы обеспечить совпадение применения с быстрым развитием листьев и периодом конца цветения, и чтобы новый рост не оставался незащищенным на протяжении длительного времени. Кроме того, интенсивное применение фунгицидов имеет несколько недостатков. Прежде всего, доказано воздействие на окружающую среду. Во-вторых, затраты на химические вещества и их применение могут доходить до 20% от всех затрат на выращивание яблонь в некоторых областях. В-третьих, развитие устойчивых популяций патогена уже было доказано Baudoin et al. (2008) и Dufour et al. (2011), сильно снижая эффективность химических обработок. Таким образом, существует растущий интерес к разработке новых альтернативных методов контроля PM.
Создание устойчивых к PM сортов является одним из лучших способов сделать устойчивое выращивание яблонь реальным, сохраняя при этом доходы производителей. К самым эффективным средствам предотвращения РМ относится использование менее восприимчивых сортов яблонь. Сорта яблонь, включая Jonafree, Prima и Enterprise, демонстрируют природную устойчивость к PM, но широко не выращиваются. На выбор сорта в большей степени влияет коммерческая привлекательность, качества плода, товарные качества и характеристики опыления, чем устойчивость к болезням. Яблочные сорта, такие как Golden Delicious, Idared и Granny Smith, широко выращиваются, но являются высоко восприимчивыми к PM и требуют химического контроля болезни.
Наиболее распространенная стратегия создания устойчивых растений нацелена на интрогрессию генов устойчивости (R-генов). R-гены кодируют белки, которые распознают эффекторы патогена и запускают защитный ответ, опосредуемый сигнальной сетью, в которой гормоны растений играют важную роль (Pavan et al., 2010). Устойчивость проявляется в виде локализованного гиперчувствительного ответа в месте заражения (Robert-Seilaniantz et al., 2007; Bruce and Pickett, 2007; Bari and Jones, 2009). R-гены являются недостаточно надежными, поскольку мутации эффекторов патогена, позволяет преодолеть устойчивость (Parlevliet et al., 1993). Кроме того, R-гены часто происходят из диких родственников культивированных видов, и поэтому их интрогрессии могут препятствовать барьеры межвидовой совместимости. Более того, в случае успешного скрещивания с R-геном вводится несколько нежелательных качеств, что требует обширного обратного скрещивания, являющегося затратным по времени у древесных видов, таких как яблоня.
Альтернативный подход основан на инактивации генов восприимчивости (S-генов), определенных как гены, потеря функции которых приводит к рецессивно унаследованной устойчивости (Pavan et al., 2010). Некоторые патогены способны подавлять защитную систему растений путем активации белков растений, функцией которых является негативная регуляция иммунной системы растений. Гены, кодирующие эти белки растений, известны как гены восприимчивости (S-гены), а их нокаут ослабляет супрессию защитной системы растений и приводит к устойчивости (Pavan et al., 2010). Недостатком S-генов являются плейотропные фенотипы, иногда связанные с их нокаутом (Pavan et al., 2011). Гены Mildew Locus O (MLO) являются типичным примером S-генов РМ.
Устойчивость вследствие нокаута гена MLO (устойчивость mlo) была обнаружена в ячмене в 1992 году (Jørgensen, 1992) и долгое время считалась уникальной формой устойчивости. Однако последующие исследования выявили, что гены MLO являются в значительной степени консервативными в растительном мире, и потеря их функции приводит к устойчивости у ряда видов, таких как Arabidopsis (Consonni et al., 2006), горох (Pavan et al., 2011), томат (Bai et al., 2008) и перец (Zheng et al., 2013). Не все гены MLO являются S-генами, и члены семейства MLO подразделяются на семь клад (Acevedo-Garcia et al., 2014; Pessina et al., 2014). Только две клады содержат S-гены: клада IV содержит все S-гены однодольных растений (Panstruga et al., 2005; Reinstädler et al., 2010); и клада V содержит все S-гены двудольных растений (Consonni et al., 2006; Bai et al., 2008; Feechan et al., 2008; Winterhagen et al., 2008). Не все члены клад IV и V являются S-генами.
Учитывая экономическое влияние инфекции P. leucotricha на производство яблок, имеется постоянная потребность в данной области в генах, обеспечивающих устойчивость к P. leucotricha.
Целью настоящего изобретения, помимо других целей, является удовлетворение этой потребности в данной области.
В соответствии с настоящим изобретением вышеупомянутая цель, помимо других целей, достигается путем обеспечения нарушенных генов, обеспечивающих устойчивость к P. leucotricha, как указано в прилагаемой формуле изобретения.
В частности, вышеупомянутая цель, помимо других целей, достигается в соответствии с первым аспектом настоящего изобретения путем обеспечения генов, придающих устойчивость к P. leucotricha, при этом аминокислотная последовательность, кодируемая геном, придающим устойчивость, представляет собой первичную аминокислотную последовательность, представленную SEQ ID NO: 1, или первичную аминокислотную последовательность, обладающую более чем 70%-ной идентичностью, предпочтительно более чем 80%-ной идентичностью, более предпочтительно более чем 90%-ной идентичностью и наиболее предпочтительно более чем 95%-ной идентичностью с последовательностью SEQ ID NO: 1, при условии, что рассматриваемые гены, придающие устойчивость, являются нарушенными.
Идентичность последовательности, используемая в настоящем документе, определяется как число идентичных, следующих друг за другом выровненных нуклеотидов или аминокислот по всей длине рассматриваемых последовательностей, деленое на число нуклеотидов или аминокислот рассматриваемых последовательностей полной длины и умноженное на 100%. Например, последовательность, обладающая 80%-ной идентичностью с последовательностью SEQ ID NO: 1, содержит по всей длине 589 аминокислот последовательности SEQ ID NO: 1, 471 или 472 идентичные выравненные аминокислоты, то есть 471 или 472/589*100% = 80%.
Нарушенный ген, придающий устойчивость, в соответствии с настоящим изобретением означает ген, обеспечивающий сниженную или даже отсутствующую восприимчивость к Podosphaera leucotricha, как видно по порошкоподобным пятнам на листьях и стеблях.
Нарушенные гены, придающие устойчивость, в соответствии с настоящим изобретением представляют собой мутировавшие гены. Мутация или мутации в рассматриваемых генах могут приводить к нарушению посредством различных механизмов. Например, одна или более мутаций в последовательностях ДНК, кодирующих белок, могут привести к образованию мутантных, усеченных или нефункциональных белков. Одна или более мутаций в некодирующих последовательностях ДНК могут вызывать альтернативный сплайсинг, трансляцию или миграцию белков. В качестве альтернативы, одна или более мутаций, приводящих к измененной транскрипционной активности гена, который определяет количество мРНК, доступное для трансляции в белок, может привести к устойчивости благодаря низкому уровню или полному отсутствию кодированных белков. Кроме того, нарушение рассматриваемых генов может быть вызвано после трансляции, т.е. на белковом уровне.
Термин «нарушенный» также обозначает в настоящем документе кодирование нефункционального гена или белка. Хотя функция рассматриваемых генов еще не идентифицирована, нефункциональный ген или белок может быть легко определен по установлению устойчивости к Podosphaera leucotricha (нефункциональный) или восприимчивости к Podosphaera leucotricha (функциональный) у растения. На устойчивость растения к Podosphaera leucotricha (нефункциональный) указывает наличие гена, который является мутированным на белковом уровне по сравнению с SEQ ID NO: 1, или наблюдается отсутствие или пониженные уровни мРНК, содержащей SEQ ID NO: 2.
Функциональные и нефункциональные гены или белки могут быть также определены с помощью экспериментов на комплементацию. Например, трансформация растения Malus Domestica, устойчивого к Podosphaera leucotricha, копией рассматриваемых генов под контролем конститутивного промотора приводит к получению растения Malus domestica, восприимчивого к Podosphaera leucotricha.
В соответствии с настоящим изобретением рассматриваемые гены, придающие устойчивость к Podosphaera leucotricha, обеспечивают устойчивость к Podosphaera leucotricha в случае их нарушения. На нарушение в соответствии с настоящим изобретением может указывать отсутствие или уменьшение содержания белка, идентифицированного в настоящем документе последовательностью SEQ ID NO: 1. В данной области известно множество механизмов, приводящих к нарушению гена на транскрипционном, трансляционном или белковом уровне.
Например, нарушение на уровне транскрипции может являться результатом одной или более мутаций в последовательностях, регулирующих транскрипцию, таких как последовательности промоторов, энхансеров, инициации, терминации или сплайсинга интронов. Эти последовательности обычно расположены на 5'-конце, 3'-конце или в пределах кодирующих последовательностей, представленных SEQ ID NO: 2. Нарушение может быть также обеспечено путем делеции, перестановки или вставки в рассматриваемых генах.
Нарушение на уровне трансляции может быть обеспечено преждевременными стоп-кодонами или другой РНК для механизмов контроля белков или посттрансляционных модификаций, влияющих, например, на сворачивание белка или клеточную миграцию.
Нарушение на белковом уровне может быть обеспечено усеченными, неправильно упакованными белками или нарушенными белок-белковыми взаимодействиями.
Независимо от лежащего в основе механизма, на нарушение в соответствии с настоящим изобретением указывает снижение содержания или отсутствие функционального белка в соответствии с SEQ ID NO: 1.
С учетом вышесказанного, в соответствии с вариантом осуществления первого аспекта настоящего изобретения нарушение в соответствии с настоящим изобретением представляет собой одну или более мутаций в рассматриваемых генах, что приводит к отсутствию продукта экспрессии белка с первичной аминокислотной последовательностью, представленной SEQ ID NO: 1, или мРНК, содержащей SEQ ID NO: 2.
В соответствии с другим вариантом осуществления первого аспекта настоящего изобретения рассматриваемое нарушение представляет собой одну или более мутаций в рассматриваемых генах, приводящих к образованию нефункционального продукта экспрессии белка.
В соответствии с еще одним вариантом осуществления этого первого аспекта настоящего изобретения рассматриваемое нарушение представляет собой пониженный уровень транскрипции, что приводит к пониженному уровню мРНК, соответствующей SEQ ID NO: 2.
В соответствии с еще одним вариантом осуществления этого первого аспекта настоящего изобретения рассматриваемое нарушение представляет собой пониженный уровень трансляции мРНК, соответствующей SEQ ID NO: 2.
В соответствии с особенно предпочтительным вариантом осуществления изобретения рассматриваемый ген, придающий устойчивость к Podosphaera leucotricha, получен из Malus domestica.
В соответствии со вторым аспектом настоящее изобретение относится к растениям Malus Domestica, содержащим в своем геноме нарушенный ген, придающий устойчивость к Podosphaera leucotricha, как описано выше, при этом нарушение обеспечивает устойчивость к Podosphaera leucotricha.
В соответствии с предпочтительным вариантом осуществления этого второго аспекта настоящего изобретения рассматриваемые растения Malus domestica демонстрируют экспрессию или транскрипцию рассматриваемых генов, придающих устойчивость к Podosphaera leucotricha, которая снижена по меньшей мере на 10% по сравнению с растением Malus domestica, восприимчивым к Podosphaera leucotricha, предпочтительно при этом экспрессия или транскрипция снижена по меньшей мере на 20% по сравнению с растением Malus domestica, восприимчивым к Podosphaera leucotricha, предпочтительно по меньшей мере на 30%, более предпочтительно по меньшей мере на 50%, еще более предпочтительно по меньшей мере на 70% и наиболее предпочтительно по меньшей мере на 80%, то есть на 25%, 35%, 40%, 45%, 55%, 60%, 65% или 75%.
В соответствии с другим предпочтительным вариантом осуществления этого второго аспекта настоящего изобретения рассматриваемые растения Malus domestica проявляют отсутствие экспрессии или трансляции рассматриваемых генов, придающих устойчивость к Podosphaera leucotricha.
В соответствии с особенно предпочтительным вариантом осуществления этого второго аспекта настоящего изобретения рассматриваемые растения Malus domestica содержат в своем геноме нарушенный ген, придающий устойчивость к Podosphaera leucotricha, кодирующий белок с первичной аминокислотной последовательностью SEQ ID NO: 1, или первичной аминокислотной последовательностью, которая обладает более чем 70%-ной идентичностью, предпочтительно более чем 80%-ной идентичностью, более предпочтительно более чем 90%-ной идентичностью, и наиболее предпочтительно более 95%-ной идентичностью с SEQ ID NO: 1; иными словами, настоящее изобретение в соответствии с особенно предпочтительным вариантом осуществления относится к растениям Malus domestica, содержащим нарушенный ген Mdmlo19 или содержащим нарушенный ген MdMLO19.
В соответствии с третьим аспектом настоящее изобретение относится к семенам, частям растений или материалу для размножения рассматриваемых растений, устойчивых к Podosphaera leucotricha, содержащих в своем геноме рассматриваемый нарушенный ген, придающий устойчивость к Podosphaera leucotricha, обеспечивающий устойчивость к Podosphaera leucotricha.
В соответствии с четвертым аспектом настоящее изобретение относится к выделенной нуклеотидной последовательности, представленной SEQ ID NO: 2, или нуклеотидным последовательностям, которые обладают более чем 70%-ной идентичностью, предпочтительно более чем 80%-ной идентичностью, более предпочтительно более чем 90%-ной идентичностью, и наиболее предпочтительно более чем 95%-ной идентичностью с SEQ ID NO: 2.
В соответствии с пятым аспектом настоящее изобретение относится к выделенным аминокислотным последовательностям, представленным SEQ ID NO: 1, или аминокислотным последовательностям, которые обладают более чем 70%-ной идентичностью, предпочтительно более чем 80%-ной идентичности, более предпочтительно более чем 90%-ной идентичностью, и наиболее предпочтительно более чем 95%-ной идентичностью с SEQ ID NO: 1.
В соответствии с шестым аспектом настоящее изобретение относится к применению рассматриваемых генов, придающих устойчивость к Podosphaera leucotricha, рассматриваемой выделенной нуклеотидной последовательности или рассматриваемой выделенной аминокислотной последовательности для селекции растений Malus domestica, устойчивых к Podosphaera leucotricha, с использованием, например, рассматриваемой последовательности для разработки молекулярных маркеров.
Настоящее изобретение будет описано более подробно в приведенном ниже примере. В этом примере делается ссылка на фигуры, среди которых:
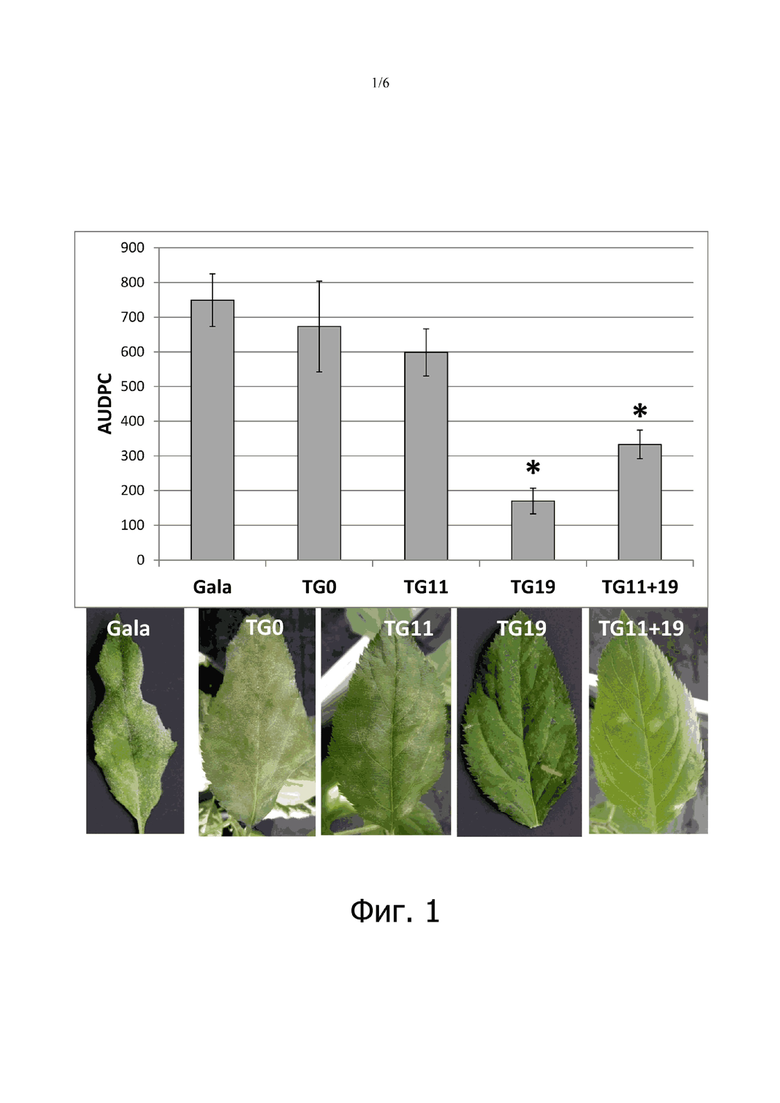
на фигуре 1: показана площадь под кривой, описывающей динамику развития болезни (AUDPC) у четырех линий яблонь mlo и контрольной «Gala», инокулированных Podosphaera leucotricha. Среднее значение AUDPC рассчитано из 15-24 биологических повторов, рассмотренных в четырех экспериментах. Планки погрешностей показывают стандартные ошибки среднего. Статистически значимые различия при сравнении с «Gala» в соответствии с (post-hoc)-тестами Тьюки и Геймса-Ховелла (P = 0,05) обозначены звездочками;
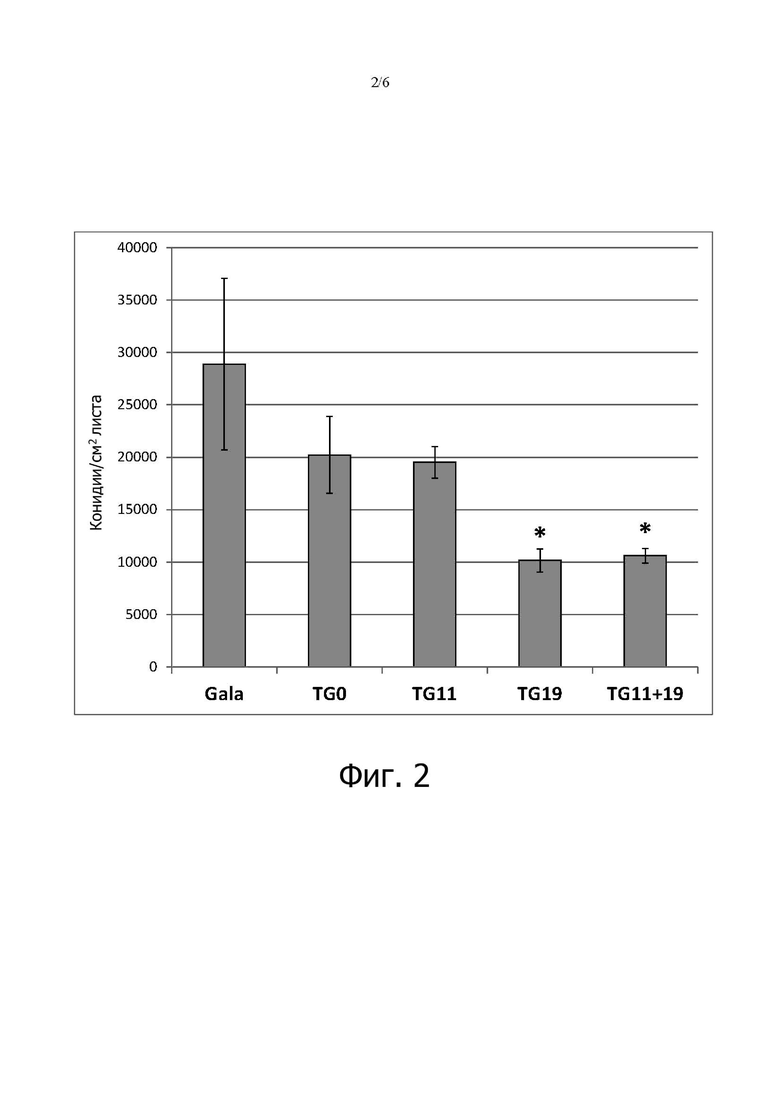
на фигуре 2: показано число конидий на см2 поверхности листа «Gala» и линий mlo TG0, TG11, TG19 и TG11+19, инокулированных P. leucotricha, через 21 день после инокуляции (dpi). Столбцы обозначают среднее число конидий, измеренное в двух экспериментах. Планки погрешностей показывают стандартные ошибки среднего. Звездочки обозначают статистически значимые различия по сравнению с «Gala» в соответствии с (post-hoc)-тестом Тьюки (P = 0,01);
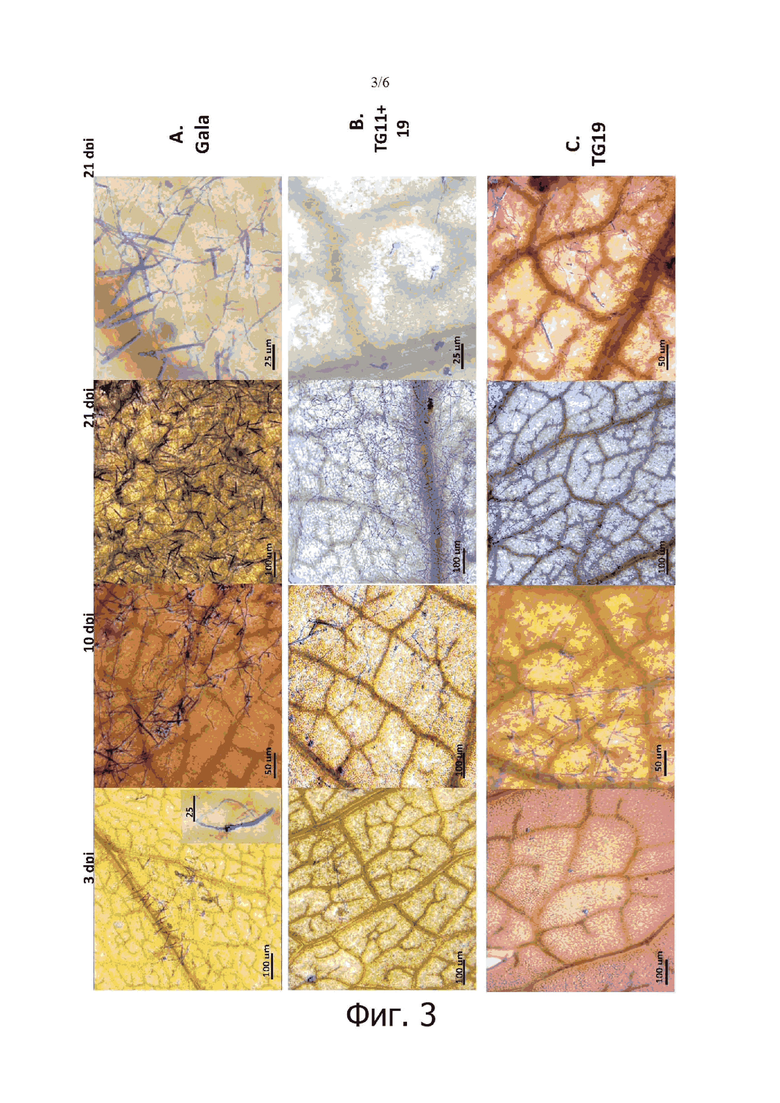
на фигуре 3: показаны изображения светлопольной микроскопии инфицированных листьев «Gala» и линий TG11+19 и TG19, полученные через 3, 10 и 21 день после инокуляции (dpi). Для Gala через 3 дня после инокуляции при более высоком увеличении показано прорастание спор P. leucotricha;
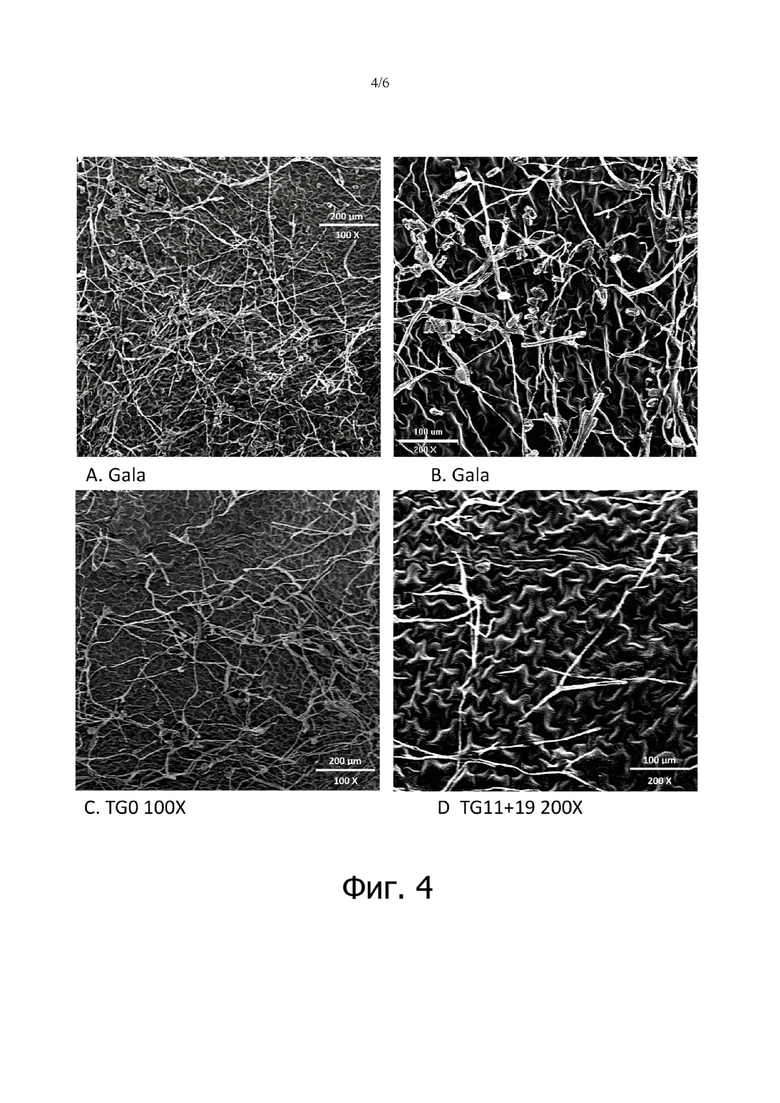
на фигуре 4: показаны изображения сканирующего электронного микроскопа (SEM) инфицированных листьев «Gala», восприимчивой линии TG0 и устойчивой линии TG11+19. Изображения получали через 21 день после инокуляции;
на фигуре 5: показано формирование через 3 дня после инокуляции папиллом в инфицированных листьях «Gala» (A, B) и в устойчивых линиях TG11+19 (C, D) и TG19 (E). Изображения слева получены с помощью светлопольного микроскопа, справа - с помощью флуоресцентного микроскопа. Для линии TG19 показано только изображение, полученное с помощью флуоресцентного микроскопа;
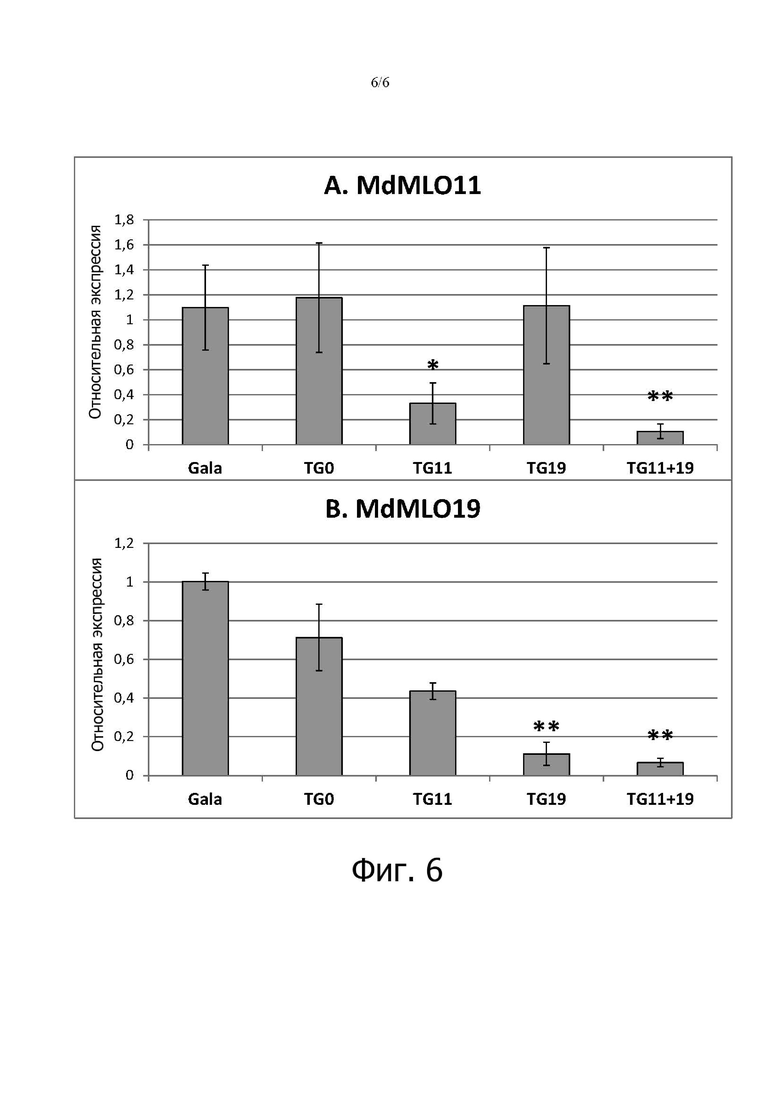
на фигуре 6: показана экспрессия двух генов яблонь MLO в пяти линиях mlo в отсутствие инфекции P. leucotricha. Каждый столбец представляет усредненную по линиям относительную экспрессию, оцененную у трех-пяти растений. Планки погрешностей показывают стандартные ошибки среднего. Звездочки обозначают значительные различия при сравнении линий mlo с «Gala» на основе (post-hoc)-тестов Тьюки и Геймса-Ховелла (P = 0,05).
Пример
Malus - это род, состоящий из множества широко распространенных и разнообразных таксонов, но почти все мировое коммерческое производство яблок сосредоточено в пределах одного вида, Malus domestica, который является родным для Азии. M. Domestica высоко восприимчив к PM, с относительно незначительными различиями между сортами и селекциями.
Контроль PM в коммерческих садах обычно требует значительного использования фунгицидов, что имеет экономическое воздействие для производителей яблок. Многочисленные органические и неорганические фунгициды применяют для контроля P. leucotricha. Однако P. leucotricha имеет склонность к развитию устойчивости к этим веществам в случае их неоднократного применения, а также эти синтетические продукты не являются экологически безопасными.
Для повышения устойчивости яблонь к этой болезни специалисты в области селекции начали вводить генетическую устойчивость от дикого Malus spp. в M. domestica с получением множества гибридов Malus (таких как White Angel).
Три бинарных вектора RNAi разработали для сайленсинга 2 генов MLO. Инокуляция трансгенных «Gala», мутированных по генам MLO, показала, что сайленсинг гена MdMLO19 повышает устойчивость M. domestica к P. leucotricha. Снижение до 75% восприимчивости к этой болезни наблюдалось в мутированной линии «Gala» с уровнем сайленсинга MLO около 90%.
При скрининге данных ресеквенирования 63 сортов яблонь была обнаружена естественная мутация с потерей функции в MdMLO19. Подобных мутаций не было обнаружено у других четырех рассматриваемых генов MLO (MdMLO5, 7, 11 и 18). Когда эта мутация присутствует в обеих аллелях, сорт яблони является устойчивым.
Материалы и методы
Конструкции для нокдауна MdMLO11 и MdMLO19 у яблонь
Фрагменты генов для RNAi амплифицировали из MdMLO11 и MdMLO19 (номера доступа в таблице 1) с использованием праймеров, указанных в таблице 2, и клонировали в вектор Gateway pENTR/SD-TOPO (Thermo Fisher Scientific, Waltham, USA). Кроме того, разработали химерную конструкцию, объединяющую фрагменты RNAi, которые должны осуществлять одновременный сайленсинг MdMLO11 и MdMLO19 (Abbott et al., 2002). Для этого сайт рестрикции для EcoRI добавляли на 3'-конце фрагмента RNAi MdMLO11 и на 5'-конце фрагмента RNAi MdMLO19. Оба фрагмента рестрицировали с помощью EcoR1 и соединяли ДНК-лигазой T4 (New England Biolabs, Ipswich, USA). Полученную конструкцию клонировали в вектор pENTR. После секвенирования все фрагменты клонировали в целевой вектор pHELLSGATE 12 (Thermo Fisher Scientific, Waltham, USA). Конечные конструкции проверяли путем секвенирования и вставляли в штамм A. tumefaciens AGL0 посредством электропорации. Трансформированные клетки A. tumefaciens тестировали с помощью PCR на присутствие конструкций, используя специфические праймеры, предназначенные для отжига на векторных и MLO последовательностях.
Таблица 1. Праймеры для qPCR
a Доступен по адресу http://bioinformatics.psb.ugent.be/plaza
b Доступен по адресу http://www.rosaceae.org/gb/gbrowse/malus_x_domestica/
Таблица 2. Праймеры для RNAi
GCAC
GGAC
a Доступен по адресу://www.rosaceae.org/gb/gbrowse/malus_x_domestica/.
Создание саженцев яблони с применением РНК-интерференции (RNAi)
RNAi-конструкции переносили в яблоню, как описано Joshi et al. (2011). Экспланты из четырех верхних листьев 4-недельных размноженных in vitro побегов яблони сорта Gala выдерживали на среде с канамицином (Joshi et al., 2011) и выращивали в ростовой камере с использованием цикла 16 часов света/8 часов темноты при 24°C. Для подтверждения наличия конструкций с помощью PCR геномную ДНК из регенерированных ростков экстрагировали с помощью набора Illustra Nucleon Phytopure kit (GE Healthcare). Прямой праймер, гибридизованный на промоторе CaMV 35S (5’- CGCACAATCCCACTATCCTT - 3’), и обратные праймеры были специфическими в отношении фрагментов RNAi (таблица 2). Полимеразную цепную реакцию (PCR) проводили при помощи GoTaq® Green Master Mix (Promega, Fitchburg, USA). Растения, положительные в отношении конструкции, перемещали в среду для размножения побегов (Shoot Propagation Medium, SPM): 4,4 г/л среды Мурасиге-Скуга с витаминами, 30 г/л сахарозы, 0,7 мг/л BAP, 96 мг/л FeEDDHA, pH 5,8. Для стимуляции укоренения растения переносили на среду, содержащую IBA. После формирования корней растения постепенно акклиматизировали к условиям теплицы (25°C, цикл 16-часов света/8-часов темноты, относительная влажность 70±5%) в 125 мл горшках, покрытых полиэтиленовыми пакетами и содержащих влажный стерилизованный в автоклаве торф (“Terriccio Vegetal Radic” - Tercomposti Spa, Brescia, Italy). Каждые 5-7 дней в течение трех недель влажность воздуха понижали, чтобы способствовать формированию кутикулы листьев. Затем полиэтиленовые пакеты удаляли и растения переносили в горшки объемом 1 литр. Контроль (нетрансформированные in vitro выращенные «Gala») акклиматизировали, как описано выше.
Инокуляция P. leucotricha и оценка тяжести болезни у яблони
Для получения инокулята PM местные штаммы Podosphaera leucotricha выделяли из инфицированных листьев сада, расположенного в провинции Трентино (Италия). Гриб поддерживали путем серийных инокуляций на саженцах M. domestica в тепличных условиях. Растения подвергали сухой инокуляции путем обработки адаксиального эпидермиса листьями зараженных саженцев. Для содействия проникновению гриба растения инкубировали в теплице при 25°C с относительной влажностью 100% в течение 6 часов. Затем растения выдерживали при 25°С и относительной влажности 80±10% до завершения оценки.
Четыре эксперимента по инокуляции проводили в разные периоды года. В каждом тесте было рассмотрено от трех до восьми биологических повторов каждой трансгенной линии. Линии тестировали по меньшей мере в трех из четырех экспериментов, и общее количество повторов варьировалось от 15 до 24. Тяжесть болезни визуально оценивали на всех инокулированных листьях через 7, 14 и 21 дней после инокуляции. Тяжесть болезни выражали в процентах (интервалы в 5%) площади адаксиальной стороны листа, покрытой мицелием PM, и рассчитывали среднее значение для одного растения. Уменьшение тяжести болезни у трансформированных растений выражали как [(тяжести у контролей - тяжесть у трансгенных растений)/тяжесть у контролей] × 100%. Для рассмотрения вместе всех временных точек рассчитывали площадь под кривой, описывающей динамику развития болезни (AUDPC), отражая интенсивность болезни во времени (Campbell and Madden, 1990; Madden et al., 2007). Количество конидий P. leucotricha, присутствующих на инфицированных листьях, оценивали, как в Angeli et al. (2012), но с незначительными изменениями: три листа собирали от каждого повтора через 21 день после инокуляции и четыре диска диаметром 0,8 см для каждого листа разрезали в общей сложности на 12 частей на повтор. Диски листьев переносили в 50 мл пробирки, содержащие 5 мл дистиллированной воды с 0,01% Tween 20 (Sigma-Aldrich, Sant Louis, USA). Пробирки встряхивали на вортексе в течение одной минуты, и концентрацию конидий на мл определяли путем подсчета с помощью гемоцитометра под световым микроскопом. Количество конидий выражали в виде количества на квадратный сантиметр (см2) листа.
Гистологический анализ инокулированных листьев яблони
Два инокулированных листа для каждого повтора собирали через 3, 10 и 21 день после инокуляции для наблюдений с помощью светлопольной микроскопии. Для визуализации гифов грибов листья очищали в смеси этанол:уксусная кислота (3:1 объем/объем) до удаления хлорофилла (приблизительно 48 часов). Образцы окрашивали в течение 15 минут трипановым синим при концентрации 250 мкг/мл в смеси молочной кислоты, глицерина и воды (1:1:1). После промывки и заключения в среду, как описано у Vogel и Somerville (2000), гифы визуализировали при освещении по методу светлого поля на микроскопе Leica LMD7000 (Wetzlar, Germany). Листья, предназначенные для оценки методом сканирующей электронной микроскопии (Hitachi S-2300, Tokyo, Japan), фиксировали в 3% глутаровом альдегиде на 0,1 M фосфатном буфере Соренсена, pH 7. Через 24 часа листья промывали в буфере Соренсена без глутарового альдегида в течение двух часов при умеренном перемешивании (80-100 об/мин). После этого образцы последовательно обезвоживали путем четырех промывок этанолом при концентрациях от 40 до 100%, сушили и хранили в пробирках фирмы Falcon до наблюдения. Перед наблюдением фрагменты листьев металлизировали золотом. Изображения обрабатывали с помощью программного обеспечения ImageJ (http://imagej.nih.gov/ij/).
Для детекции папиллом листья очищали в смеси этанол:уксусная кислота (3:1, об./об.) до удаления хлорофилла и уравновешивали в течение ночи в растворе молочной кислоты, глицерина и воды (1:1:1). Папилломы визуализировали с использованием фильтра LMD (BP-фильтр возбуждения 380-420 нм, 415 дихроическое зеркало и BP эмиссии 445-485 нм) микроскопа Leica LMD6500 (Leica Microsystem, Wetzlar, Germany).
Анализ экспрессии генов
Для идентификации линий, демонстрирующих эффекты сайленсинга, в первом исследовании экспрессии генов использовали выращенные in vitro трансгенные растения, реплицированные три раза. Во втором исследовании, касающемся акклиматизированных трансгенных растений, образцы листьев собирали непосредственно перед инокуляцией PM, через 24 часа после инокуляции (hpi) и через 10 дней после инокуляции (dpi). Для каждой линии в каждый момент времени образцы листьев собирали от пяти разных растений. Образцы замораживали в жидком азоте и хранили при 80°C. Тотальную РНК выделяли с помощью набора Spectrumtm Plant Total RNA kit (Sigma-Aldrich), обрабатывали DNAse I (Sigma-Aldrich) и обратно транскрибировали с использованием обратной транскриптазы SuperScript III (Invitrogen, Life Technologies, Waltham, USA). Количественный PCR-анализ (qPCR) проводили с использованием супермикса SsoAdvanced Universal SYBR Green Supermix (Bio-Rad, Hercules, USA) в реакционном объеме 15 мкл с использованием системы PCR в режиме реального времени с детекцией CFX96 Touchtm (Bio-Rad, Hercules, USA) и программным обеспечением CFX Manager. Образцы испытывали в двух технических повторах в соответствии со следующими параметрами термического цикла: при 95°C в течение 3 мин, при 95°C в течение 10 сек, при 55°C в течение 30 сек (повтор 40 раз), при 95°C в течение 10 сек. Для анализа MdMLO19 использовали пары праймеров, рассмотренных в предыдущей работе (таблица 1; Pessina et al., 2014). Для MdMLO11 и для экспрессии 17 генов, вовлеченных во взаимодействие между яблоней и P. leucotricha, были разработаны новые пары праймеров с помощью инструмента NCBI Primer Designing Tool (http://www.ncbi.nlm.nih.gov/tools/primer-blast/) (таблица 1). Серийные разведения кДНК (1/10, 1/100, 1/1000 и 1/10000) позволили рассчитать эффективность пар праймеров; ожидаемые размеры продуктов подтверждали с использованием электрофореза в агарозном геле. Кривую специфической конечной диссоциации определяли после каждого цикла qPCR с постепенным повышением температуры от 65 до 95°C (каждый шаг 0,5°C, 5 сек). В качестве референсных генов использовали фактор элонгации 1, убиквитин и 8283 (таблица 1). Все они, как известно, являются стабильными референсными генами для яблони (Botton et al., 2011; Pessina et al., 2014). Анализ с помощью программы geNorm (medgen.ugent.be/~jvdesomp/genorm) показал значения М ниже 1 для всех трех референсных генов, при этом значения М ниже 1,5 считаются адекватными (Ling and Salvaterra, 2011). Значения порогового цикла (Ct) были преобразованы в относительные уровни экспрессии, как в Hellemans et al. (2007), используя в качестве входных данных среднее значение Ct двух технических повторов. В качестве референсного значения Ct было принято среднее значение Ct для «Gala» дикого типа непосредственно после инокуляции (0 hpi).
Статистика
Тяжесть болезни
Данные по тяжести болезни анализировали с помощью статистического пакета SPSS (IBM, Armonk, USA). Для яблони и A. thaliana данные по тяжести болезни для листьев от одного и того же растения усредняли перед последующими анализами. Были рассмотрены данные по тяжести болезни для яблони от восьми более молодых листьев растения, а для A. thaliana - от всех листьев. Перед любым анализом данные, как было показано, имели нормальное распределение (тесты Колмогорова-Смирнова и Шапиро-Уилка, P>0,05) и имели однородные дисперсии (тест Левена, P> 0,05). Однофакторный дисперсионный анализ (ANOVA) с (post-hoc)-тестом Тьюки был выбран для детекции значительных различий (P <0,05) в каждый момент времени. Данные были преобразованы в соответствии с y=arcsin(x) для удовлетворения условий ANOVA. В случае неоднородных дисперсий применяли (post-hoc)-тест Геймса-Ховелла. Перед объединением данных из независимых экспериментов тестировали эффект одиночных экспериментов: значительного эффекта экспериментов не наблюдалось. Объединенные данные анализировали независимо в моменты времени 14 и 21 dpi. Данные AUDPC обрабатывали, как описано выше для данных по тяжести болезни. Данные по количеству конидий анализировали с помощью однофакторного дисперсионного анализа (ANOVA), применяя (post-hoc)-тест Тьюки (P <0,05).
Анализ данных qPCR
Для оценки генной экспрессии значения относительной экспрессии логарифмически преобразовывали в соответствии с Y=ln(x) (Pessina et al., 2014) для удовлетворения нормальным распределениям и однородностям дисперсий, которые оценивались соответственно с помощью теста Шапиро-Уилка (P ≤ 0,05) и Левена (P ≤ 0,05). Парное сравнение гомоскедастических данных проводили с помощью теста Тьюки (P <0,05), тогда как негомоскедастические данные анализировали с помощью теста Геймса-Ховелла (P <0,05) с использованием статистического пакета SPSS (IBM). Для детекции значительных различий в экспрессии применяли однофакторный дисперсионный анализ (ANOVA) с (post-hoc)-тестом Тьюки (P <0,05) к данным, полученным от образцов непосредственно после инокуляции (0 hpi). Анализ экспрессии связанных с защитой генов тестировали с помощью (post-hoc)-теста Фишера.
Корреляции
Двухсторонний корреляционный тест Пирсона был выбран для исследования корреляций между AUDPC и относительной экспрессией генов MLO через 10 dpi, и между степенью тяжести болезни и числом конидий через 21 dpi. Все данные были преобразованы согласно y=arcsin(x) для достижения нормального распределения.
Метаболиты
Данные, полученные от фенольных метаболитов, подвергали однофакторному дисперсионному анализу (ANOVA) с (post-hoc)-тестом Фишера. В случае негомоскедастических данных применяли (post-hoc)-тест Геймса-Ховелла, и непараметрический тест Крускала-Уоллиса для данных, имеющих ненормальное распределение.
Результаты
Создание саженцев яблони с использованием RNAi
Были созданы три конструкции RNAi, две из которых нацелены на индивидуальный нокдаун генов MdMLO11 и MdMLO19 (i = KD-MdMLO11, ii = KD-MdMLO19), третья нацелена на одновременный нокдаун генов MdMLO11 и MdMLO19 (iii = KD-MdMLO11+19). Было получено восемьдесят регенерированных линий, из которых 48 несли вставку RNAi, как описано в разделе «Материалы и методы» (таблица 3). Для оценки уровня экспрессии генов MLO тестировали 48 трансгенных линий с помощью qPCR, но значительный нокдаун наблюдался только в трех из них (таблица 3). В этих трех линиях нецелевые нокдауны не были обнаружены для двух других генов клады V яблони (MdMLO5 и 7). Три линии нокдауна, названные TG11 (Transgenic Gala MdMLO11), TG19 и TG11+19, акклиматизировали к условиям теплицы, а также контрольные дикого типа «Gala» и TG0, линия, несущая конструкцию RNAi для MdMLO19, но не демонстрирующая значительного нокдауна генов MLO. TG0, TG11, TG19 и TG11+19 будут указаны как трансгенные линии, и только TG11, TG19 и TG11+19 как линии mlo.
Показатель выживаемости растений после процедуры акклиматизации составил более 90%. В тепличных условиях линии mlo показали нормальный рост по сравнению с «Gala».
Таблица 3. Обобщенные результаты переноса генов
Сниженная восприимчивость к P. leucotricha у RNAi-растений яблони
Четыре трансгенные линии и контроль тестировали на их восприимчивость к PM в четырех независимых экспериментах. TG0, линия, не проявляющая какого-либо нокдауна генов MLO, показала уровень восприимчивости к P. leucotricha, сравнимый с уровнем восприимчивости у контроля; то же самое наблюдалось для TG11, тогда как TG11+19 и TG19 имели очевидное снижение тяжести болезни (фигура 1). Хотя листья растений TG11+19 и TG19 были частично инфицированы (фигура 1), участок площади на адаксиальной поверхности листа, покрытый спорами, был заметно меньше по сравнению с контролем (фигура 1). В таблице 4 показаны обобщенные результаты уменьшения тяжести болезни.
У всех трансгенных линий уменьшилось число конидий, присутствующих на листьях (фигура 2), но уменьшение было статистически значимым (P <0,05) только для TG11+19 и TG19. Это хорошо согласуется с оценкой тяжести болезни, представленной на фигуре 1: по сравнению с «Gala» TG11+19 показала уменьшение числа конидий на 63,3%, а TG19 - на 64,8%. Значимая (P = 0,01), но умеренная положительная корреляция (коэффициент Пирсона 0,525) была обнаружена между тяжестью болезни через 21 dpi и числом конидий через 21 dpi.
Линии TG11+19 и TG19 вместе с «Gala» дополнительно анализировали с помощью светлопольной микроскопии и сканирующей электронной микроскопии (SEM) для отслеживания развития инфекции P. leucotricha. В «Gala» хорошо развитая инфекция на листьях наблюдалась уже через 3 dpi (фигура 3А), тогда как развитие грибов все еще было ограничено у TG11+19 и TG19 (фигура 3В и 3С). Через 10 dpi конидиофоры наблюдались на листьях всех рассматриваемых линий, но их число было выше в «Gala» (фигура 3). Через 21 dpi листья «Gala» были полностью покрыты спорами, и визуально наблюдалось большое количество конидиофор (фигура 3А). Поверхность листьев TG11+19 и TG19 была частично колонизирована спорообразующим мицелием, но также наблюдались изолированные споры, неспособные к развитию, а также меньшее количество конидиофор по сравнению с ситуацией, отмеченной для «Gala» (фигура 3В и 3С). SEM-изображения показали уменьшенный рост мицелия на TG11+19 по сравнению с TG0 и «Gala» (фигура 4).
Формирование папиллом наблюдалось через 3 dpi во всех линиях, как устойчивых, так и восприимчивых (фигура 6). По сравнению с TG11+19 и TG19 папилломы у «Gala» (фигура 6А, В) были меньше, форма более четкая и излучаемая флуоресценция более интенсивная (фигура 6С, D, Е).
Таблица 4. Уменьшение тяжести заболевания мучнистой росой (%) в линиях, трансформированных RNAi-конструкциями MLO.
* Статистически значимое различие по сравнению с контролем в соответствии с (post-hoc)-тестом Тьюки (P = 0,05).
# Gala использовали в качестве контроля (19 растений) и предположили, что имеется 0% уменьшения заболевания.
° Линия TG11 показала более высокий уровень инфекции по сравнению с Gala через 21 dpi.
Экспрессия генов MLO в линиях яблонь mlo
Анализ генной экспрессии ранее выбранных линий mlo повторяли на растениях, акклиматизированных к теплице. Ген MdMLO11 значительно меньше экспрессировался в TG11+19 (P=0,01) и TG11 (P=0,05) (фигура 6А), в то время как экспрессия MdMLO19 была сниженной в TG11+19 (P=0.01) и TG19 (P=0,01) (фигура 6B). MdMLO5 и MdMLO7, два других члена клады V яблони, также были подвергнуты тестированию, но значительного снижения не наблюдалось ни в одной трансгенной линии, подтверждая отсутствие нецелевого сайленсинга (данные не показаны).
Корреляция между экспрессией MdMLO19 и AUDPC - показателем тяжести болезни - была статистически значимой (P = 0,05), хотя и умеренной (коэффициент Пирсона = 0,515). Напротив, значительной корреляции не было обнаружено между AUDPC и экспрессией MdMLO11.
Обсуждение
Природные и искусственные мутации с потерей функции S-генов MLO снижают восприимчивость к патогенам PM, как описано, в ячмене (Büschges et al., 1997), A. thaliana (Consonni et al., 2006), горохе (Pavan et al., 2011), томате (Bai et al., 2008) и перце (Zheng et al., 2013). В двудольных растениях все гены восприимчивости к PM принадлежат кладе V (Consonni et al., 2006; Bai et al., 2008; Feechan et al., 2008; Winterhagen et al., 2008). В предыдущей работе авторы идентифицировали три гена MLO M. domestica, которые имели повышенную экспрессию на ранних стадий инфекции РМ (Pessina et al., 2014). Два из них, MdMLO11 и MdMLO19, принадлежат двудольному растению клады V, и MdMLO18 принадлежит кладе VII. Так как гены MLO вне клады V, действующие как S-гены, являются неизвестными, только MdMLO11 и MdMLO19 рассматривались в качестве приемлемых кандидатов, подлежащих нокдауну в яблоне.
Гены MdMLO11 и MdMLO19 подвергали нокдауну для оценки их роли в поддержании восприимчивости яблонь к РМ. RNAi выбирали для снижения экспрессии этих двух генов MLO и, несмотря на большое число созданных трансгенных линий «Gala» (48), только для трех из них было детектировано значительное снижение экспрессии генов-мишеней. Отчасти это ожидалось, поскольку короткие фрагменты RNAi размером менее 150 п.о. (пар оснований), как те, которые использовали в экспериментах, известны своей ограниченной эффективностью нокдауна. С другой стороны, они обладают преимуществом, состоящим в более высокой специфичности, избегая тем самым генерации нецелевого сайленсинга других генов MLO клады V, как было обнаружено в экспериментах.
У некоторых видов нокаут или нокдаун генов MLO вызывает плейотропные фенотипы, такие как формирование некротических пятен на листьях и снижение урожая зерна у ячменя (Jørgensen, 1992), медленный рост у A. thaliana (Consonni et al., 2006) и уменьшенный размер растения у перца (Zheng et al., 2013). Эти или другие неожиданные плейотропные фенотипы не наблюдались в тепличных условиях, указанных в разделе «Материалы и методы».
Инокуляция в теплице трансгенных линий яблонь привела к статистически значимому снижению тяжести болезни в линиях TG11+19 и TG19. Можно предположить, что благодаря нокдауну в обеих устойчивых линиях, ген MdMLO19 являлся самым эффективным геном, ответственным за снижение восприимчивости к РМ. Нокдаун MdMLO11 не привел к значительному снижению восприимчивости, и даже его нокдаун в комбинации с MdMLO19 не привел к какому-либо дополнительному снижению восприимчивости. Вывод состоит в том, что из двух генов клады V, индуцированных PM в яблоне, только MdMLO19 является функциональным S-геном. Также, MdMLO18, ген клады VII, индуцируемый инокуляцией P. Leucotricha, не следует рассматривать как S-ген PM. Линия TG0 рассматривалась для оценки эффекта вставки «неэффективной в отношении мишени» конструкции RNAi на восприимчивость к PM. TG0 получали в результате переноса, нацеленного на нокдаун MdMLO19. В этой линии отмечалось уменьшение экспрессии MdMLO19, хотя и незначительное, а также умеренное незначительное снижение восприимчивости к РМ. Сделан вывод о том, что вставка «неэффективной» конструкции RNAi может иметь функциональную значимость, но это не может быть статистически доказано.
Точный механизм, посредством которого потеря функции S-генов MLO снижает восприимчивость к патогенам PM, до сих пор полностью не ясен. Тем не менее, известно, что устойчивость mlo связана с миграцией секреторных пузырьков (Miklis et al., 2007; Feechan et al., 2011) и формированием аппозиций клеточных стенок, называемых папиллами (Consonni et al., 2006). Папилла состоит из каллозной матрицы, обогащенной белками и аутофлуорогенными фенольными смолами (Vanacker et al. 2000), образование которых зависит от актин-зависимого эндомембранного транспорта (Hückelhoven, 2014). Линии «Gala», TG11+19 и TG19 характеризовались наличием папилл через 3 dpi, но форма и размеры различались в устойчивых и восприимчивых линиях. Быстрое формирование папилл (Lyngkjaer et al., 2000), увеличенный размер папилл на участках попытки проникновения (Stolzenburg et al., 1984) и различный биохимический состав (Chowdhury et al., 2014) могут дать объяснение указанным различиям между эффективной и неэффективной папиллой. В линиях mlo, особенно TG19, размер папилл был больше, чем в контроле, что подтверждает гипотезу о том, что более крупные размеры повышают эффективность папилл. В работе Chowdhury et al. (2014) показано, что различие между эффективными и неэффективными папиллами состоит в более высокой концентрации каллозы, целлюлозы и арабиноксилана у эффективных папилл. Это, возможно, отражает наблюдаемые различия во флуоресценции между папиллами устойчивых и восприимчивых линий. На самом деле, белки MLO считаются отрицательными регуляторами связанных с везикулами и актин-зависимых путей защиты (Panstruga, 2005), которые, будучи под контролем гриба, индуцируют актиновые нити на снабжение растущих гиф питательными веществами (Miklis et al., 2007). Представленные здесь данные подтверждают мнение о том, что в яблонях диких типов после проникновения патоген контролирует транспорт материала к клеточной стенке, изменяя состав папилл и делая их неэффективными. Аналогичная работа, проведенная на культурном винограде (Pessina et al., не опубликовано), поддерживает эту интерпретацию: по сравнению с контролем, линии винограда mlo показали более крупные и менее четкие папиллы, аналогично наблюдаемым у яблонь mlo.
Ссылки
Abbott JC, Barakate A, Pinçon G, Legrand M, Lapierre C, Mila I, Schuch W, Halpin C: Simultaneous suppression of multiple genes by single transgenes. Down-regulation of three unrelated lignin biosynthetic genes in tobacco. Plant Physiol 2002, 128(3): 844-853
Acevedo-Garcia J, Kusch S, Panstruga R: Magical mystery tour: MLO proteins in plant immunity and beyond. New Phytol 2014, 204(2):273-81
Angeli D, Puopolo G, Maurhofer M, Gessler C, Pertot I: Is the mycoparasitic activity of Ampelomyces quisqualis biocontrol strains related to phylogeny and hydrolytic enzyme production? Biological Control 2012, 63; 348-358
Bai Y, Pavan S, Zheng Z, Zappel N, Reinstädler A, Lotti C, De Giovanni C, Ricciardi L, Lindhout P, Visser RGF, Theres K, Panstruga R: Naturally occurring broad-spectrum powdery mildew resistance in a Central American tomato accession is caused by loss of MLO function. MPMI 2008, 21: 30-39
Bari R, Jones JDG: Role of plant hormones in plant defense responses. Plant Mol Biol, 2009, 69:473-488.
Botton A, Eccher G, Forcato C, Ferrarini A, Begheldo M, Zermiani M, Moscatello S, Battistelli A, Velasco R, Ruperti B, Ramina A: Signaling pathways mediating the induction of apple fruitlet abscission. Plant Physiol 2011, 155(1):185-208
Büschges R, Hollricher K, Panstruga R, Simons G, Wolter M, Frijters A, van Daelen R, van der Lee T, Diergaarde P, Groenendijk J, Töpsch S, Vos P, Salamini F, Schulze-Lefert P: The barley Mlo gene: a novel control element of plant pathogen resistance. Cell 1997, 88(5):695-705
Campbell CL, Madden LV: Introduction to plant disease epiderniology. John Wiley and Sons 1990, New York. 532pp.
Chen Z, Noir S, Kwaaitaal M, Hartmann A, Wu MJ, Mudgil Y, Sukumar P, Muday G, Panstruga R, Jones AM: Two seven-transmembrane domain MILDEW RESISTANCE LOCUS O proteins cofunction in Arabidopsis root thigmomorphogenesis. Plant Cell 2009, 21:1972-1991.
Chowdhury J, Henderson M, Schweizer P, Burton RA, Fincher GB, Little A: Differential accumulation of callose, arabinoxylan and cellulose in nonpenetrated versus penetrated papillae on leaves of barley infected with Blumeria graminis f. sp. Hordei. New Phytol 2014, 204: 650-660
Consonni C, Humphry ME, Hartmann HA, Livaja M, Durner J, Westphal L, Vogel J, Lipka V, Kemmerling B, Schulze-Lefert P, Somerville SC, Panstruga R: Conserved requirement for a plant host cell protein in powdery mildew pathogenesis. Nature Genetics 2006, 38(6):716-720.
Dufour MC, Fontaine S, Montarry J, Corio-Costet MF: Assessment of fungicide resistance and pathogen diversity in Erysiphe necator using quantitative real-time PCR assays. Pest Manag. Sci. 2011, 67, 60-69.
Feechan A, Jermakow AM,Torregrosa L, Panstruga R, Dry IB: Identification of grapevine MLO gene candidates involved in susceptibility to powdery mildew. Funct Plant Biol 2008, 35:1255-1266
Feechan A, Kabbara s, Dry IB: Mechanisms of powdery mildew resistance in the Vitaceae family. Mol Plant Pathology 2011, 12(3):263-274
Hellemans J, Mortier G, De Paepe A, Speleman F and Vandesompele J: qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol. 2007, 8:R19
Hückelhoven R: The effective papilla hypothesis. New Phytol 2014, 204: 438-440
Joshi SG, Schaart JG, Groenwold R, Jacobsen E, Schouten HJ, Krens FA: Functional analysis and expression profiling of HcrVf1 and HcrVf2 for development of scab resistant cisgenic and intragenic apples. Plant Mol Biol 2011, 75:579-591
Jørgensen JH: Discovery, characterization and exploitation of Mlo powdery mildew resistance in barley. Euphytica 1992, 63:141-152.
Ling D, Salvaterra PM: Robust RT-qPCR data normalization: validation and selection of internal reference genes during post-experimental data analysis. PLoS One 2011, 6:3.
Lyngkjaer M F, Newton AC, Atzema JL, Baker SJ: The Barley mlo-gene: an important powdery mildew source. Agronomie 2000, 20 745-756.
Madden LV, Hughes G, Van Den Bosch F: The Study of plant disease epidemics. APS press, St. Paul, 2007.
Miklis M, Consonni C, Bhat RA, Lipka V, Schulze-Lefert P, Panstruga R: Barley MLO modulates actin-dependent and actin-independent antifungal defense pathways at the cell periphery. Plant Physiol 2007, 144:1132-1143
Panstruga R: Serpentine plant MLO proteins as entry portals for powdery mildew fungi. Biochem Soc Transact 2005, 33(Pt 2):389-392
Parlevliet JE: What is durable resistance, a general outline. In Durability of Disease Resistance. Edited by Jacobs TH, Parlevliet JE. Dordrecht: Kluwer; 1993:23-29.
Pavan S, Jacobsen E, Visser RGF, Bai Y: Loss of susceptibility as a novel breeding strategy for durable and broad-spectrum resistance. Mol Breed 2010, 25:1-12.
Pavan S, Schiavulli A, Appiano M, Marcotrigiano AR, Cillo F, Visser RGF, Bai Y, Lotti C, Ricciardi L: Pea powdery mildew er1 resistance is associated to loss-of-function mutations at a MLO homologous locus. Theor Appl Gen 2011, 123:1425-1431
Pessina S, Pavan S, Catalano D, Gallotta A, Visser RGF, Bai Y, Malnoy M, Schouten HJ: Characterization of the MLO gene family in Rosaceae and gene expression analysis in Malus domestica. BMC genomics 2014, 15:618
Pessina S, Lenzi L, Perrazzolli M, Campa M, Dalla Costa L, Urso S, Valè G, Velasco R, Salamini F, Malnoy M: MLO genes Knock-down reduces susceptibility to powdery mildew in grapevine. Unpublished
Reinstädler A, Müller J, Czembor JH, Piffanelli P, Panstruga R: Novel induced mlo mutant alleles in combination with site-directed mutagenesis reveal functionally important domains in the heptahelical barley Mlo protein. BMC Plant Biol 2010, 10:31.
Stolzenburg M, Aist JR, Israel HW: The role of papillae in resistance to powdery mildew conditioned by the ml-o gene in barley. I Correlative evidence. Physiological Plant Pathology 1984, 25, 337-346
Turechek WW, Carroll JE, Rosenberger DA: Powdery mildew of apple. NY State Integrated Pest Management Program: Cornell University 2004 (www.nysipm.cornell.edu/factsheets/treefruit/diseases/pm/apple_pm.pdf).
Vanacker H, Carver TLW, Foyer CH: Early H2O2 accumulation in mesophyll cells leads to induction of glutathione during hypersensitive response in the barley-powdery mildew interaction. Plant Physiology 2000, 123: 1289-1300
Wightwick A, Walters R, Allinson G, Reichman S, Menzies N: Environmental risks of fungicides used in horticultural production systems. Fungicides 2010, Odile Carisse (Ed.), ISBN: 978-953-307-266-1
Winterhagen P, Howard SF, Qiu W, Kovács LG: Transcriptional up-regulation of grapevine MLO genes in response to powdery mildew infection. Am J Enol Vitic 2008, 59:2
Zheng Z, Nonomura T, Appiano M, Pavan S, Matsuda Y, Toyoda H, Wolters AA, Visser RGF1, Bai Y: loss of function in Mlo orthologs reduces susceptibility of pepper and tomato to powdery mildew disease caused by Leveillula taurica. PLoS One 2013, 8(7):e70723
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> FONDAZIONE EDMUND MACH
<120> PODOSPHAERA LEUCOTRICHA RESISTENCE PROVIDING GENES IN MALUS DOMESTICA
<130> 4/2UD42/2P
<150> PCT/EP2015/073135
<151> 2015-10-07
<160> 2
<170> BiSSAP 1.3
<210> 1
<211> 589
<212> PRT
<213> Malus sp.
<400> 1
Met Ala Gly Gly Lys Lys Gly Arg Ser Leu Glu Gln Thr Pro Thr Trp
1 5 10 15
Ala Val Ala Val Val Cys Phe Val Leu Val Leu Ile Ser Ile Leu Ile
20 25 30
Glu Tyr Phe Ile His Leu Ile Gly Lys Trp Leu Lys Lys Arg Asn Lys
35 40 45
Arg Ala Leu Tyr Glu Ala Leu Glu Lys Ile Lys Ser Glu Leu Met Leu
50 55 60
Leu Gly Phe Leu Ser Leu Leu Leu Thr Val Gly Gln Gly Pro Ile Ser
65 70 75 80
Asn Ile Cys Ile Ser Lys Ala Val Gly Ala Thr Trp His Pro Cys Ser
85 90 95
Lys Lys Gln Glu Val Lys Ser Asp Lys Asn Glu Asp Lys Ser Ser Val
100 105 110
Ser Asp Asp Asn Ala Arg Arg Arg Leu Leu Ser Ala Leu Asp Ser Ser
115 120 125
Gly Gly Gly Arg Arg Val Leu Ala Ala Ala Gly Tyr Asp Lys Cys Ala
130 135 140
Ala Lys Asn Lys Val Pro Phe Val Ser Tyr Tyr Gly Ile His Gln Leu
145 150 155 160
His Ile Leu Ile Phe Val Leu Ala Val Phe His Val Leu Tyr Cys Ile
165 170 175
Thr Thr Leu Val Leu Gly Arg Ala Lys Met Arg Lys Trp Lys Thr Trp
180 185 190
Glu Leu Glu Thr Lys Thr Ala Ala Tyr Gln Phe Ser His Asp Pro Glu
195 200 205
Arg Phe Arg Phe Ala Arg Asp Thr Ser Phe Gly Arg Arg His Leu Ser
210 215 220
Phe Trp Ser Arg Ser Pro Ile Ser Leu Trp Ile Val Cys Phe Phe Arg
225 230 235 240
Gln Phe Val Arg Ser Val Pro Lys Val Asp Tyr Leu Thr Leu Arg His
245 250 255
Gly Phe Ile Ala Ala His Leu Ala Pro Gln Ser Gln Thr Lys Phe Asp
260 265 270
Phe Gln Lys Tyr Ile Asn Arg Ser Leu Glu Glu Asp Phe Lys Val Val
275 280 285
Val Gly Ile Ser Pro Thr Ile Trp Leu Phe Ala Val Leu Ile Leu Met
290 295 300
Ser Asn Thr His Gly Ser Arg Ser Tyr Leu Trp Leu Pro Phe Val Pro
305 310 315 320
Leu Val Met Ile Leu Met Val Gly Thr Lys Leu Gln Val Ile Ile Thr
325 330 335
Lys Met Gly Leu Lys Leu Ser Glu Arg Gly Glu Val Val Arg Gly Thr
340 345 350
Pro Leu Val Glu Pro Gly Asp His Leu Phe Trp Phe Asn Asn Pro Arg
355 360 365
Leu Leu Leu Tyr Ile Ile His Phe Val Leu Phe Gln Asn Ala Phe Ala
370 375 380
Leu Ala Phe Phe Ala Trp Thr Trp Ser Gly Lys Gly Gly Leu His Phe
385 390 395 400
Thr Leu Leu Gln Tyr Glu Phe Gly Leu Lys Ser Cys Phe His Glu Lys
405 410 415
Leu Glu Asp Val Val Leu Arg Ile Ser Met Gly Val Ile Ile Gln Ile
420 425 430
Leu Cys Ser Tyr Val Thr Leu Pro Leu Tyr Ala Leu Val Thr Gln Met
435 440 445
Gly Ser Thr Met Lys Pro Val Ile Phe Asn Asp Arg Val Ala Thr Ala
450 455 460
Leu Lys Lys Trp His Ile Ala Ala Lys Lys His Val Lys His Lys Asn
465 470 475 480
Ala Ser Pro Ala Ser Ala Pro Gly Thr Pro Leu His Ser Met Ser Pro
485 490 495
Val His Leu Leu Arg Asn Tyr Lys Tyr Glu Gln Asp Ile Asp Ser Ile
500 505 510
Gln Thr Ser Pro Arg Met Pro Tyr Phe Asp Asn Glu Gly Ser Asp Ser
515 520 525
Pro Phe His His Gln Asp Asn Leu Thr Trp Ser Gln Gln Gly Thr Asn
530 535 540
Met Glu Gly Gln Lys Glu Glu Ile Ser Ala His Gly Pro Asn Ala Glu
545 550 555 560
Ser Asn Ala Leu Gly Ala Tyr Gly Ser Ile Ile Gln His Glu Ile Gln
565 570 575
Ile His Ser Ala Ala Leu Thr Phe Glu Lys Thr Glu Arg
580 585
<210> 2
<211> 1733
<212> DNA
<213> Malus sp.
<400> 2
atggagcaac ttggcatcca tgcagtaaga agcaagaggt caaatcagac aagaacgaag 60
ataagagtag tgtttcagat gacaacgccc gcagaaggct tctctcagcc ttggattcca 120
gtgggggtgg ccgacgtgtt ttagcagctg ctggatatga caaatgtgct gccaaggtaa 180
tttgtgtcat attatgggat ccaccagctt cacatactca tctttgtgct agccgtcttc 240
cacgtgcttt actgcataac aaccctagtt ttgggcagag caaaggtctc tctctcgaaa 300
acatgggaac tggaaacaaa aacagctgct accaattctc acatggcagg aggaaaaaaa 360
ggaagatctt tggagcaaac accaacttgg gcagttgccg tgtttgtttc gttttggttt 420
tatttcaatt ctcatcgaat atttcattca tttaattgga aaggtgttga aaaagagaaa 480
caaaagagct ctctatgaag cacttgagaa gatcaagtcg agcttatgct attagggttc 540
ctatccttgc ttctaacagt aggacaaggc cccatttcaa atatttgtat atcgaaggct 600
gttggtatgc tacctcagat ttgccagaga cacatccttt gggagaagac acttgagctt 660
ttggagccgt tctcccatta gcttatggat tgtaagagtt gaaaattgta agatctgtgc 720
ccaaagttga ttacttgact ctgcgacatg ggtttattgc gtaagtctct tactaaattt 780
aaactaagtt tgacttccag aagtacatta acaggtcact tgaagaggat ttcaaggttg 840
tcgtagggat caggtgggtc actagtctag gcatactcat gtcaaacacc cacggtaagc 900
ttccctttac ttgctaacct gtttttcccc tagttgtaag cgccaacccc accgttcaat 960
cctttttaat aacaaagatg gggctcaaat tatcagaaag aggtgaagtg gttaggggaa 1020
ccccactggt tgagcctggt gaccatctct tctggttcaa caacccccga ctcctgcttt 1080
atatcatcca ctttgttctc ttcaggtaat tcactttctt tccaattgga tttagtgatt 1140
gtaagtaaaa ctgtaaaccc atatttactt tcacggatga atttggcttg aagtcttgct 1200
tccatgagaa gttggaagat gtcgttttaa gaatatcaat ggggtaactc aaattatttt 1260
tacatttttt ttgttcactc ctctttatgc attggtaaca caggtaatta agtggccatg 1320
atgaaaatta gatgctaaaa agtggcgact gcacttaaaa aatggcacat tgcagcgaag 1380
aagcacgtaa aacacaagaa tgcaagtcca gccagtgcac caggcactcc tttgcactcc 1440
atgtcccctg ttcatctact acgcaactat aaatatgaac aagacattga cagcattcag 1500
acatcaccaa gaatgcctta ttttgataac gaaggttcag actcaccgtt tcatcaccaa 1560
gacaatttga cttggtctca gcaaggtaca aacatggagg gtcagaagga ggagattagt 1620
gctcatggac ctaacgcaga gagcaatgct ttaggcgctt atggttcgat aattcaacat 1680
gagattcaaa ttcactcggc ggcactcaca tttgagaaaa cagaaagagc tga 1733
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ИНСЕКТИЦИДНЫЕ ПОЛИПЕПТИДЫ С ШИРОКИМ СПЕКТРОМ АКТИВНОСТИ И ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2750459C2 |
| СКОНСТРУИРОВАННЫЕ ПЕСТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2017 |
|
RU2816526C2 |
| СКОНСТРУИРОВАННЫЕ ПЕСТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2017 |
|
RU2817591C2 |
| КОНТРОЛЬ УСТОЙЧИВОСТИ НАСЕКОМЫХ С ПОМОЩЬЮ КОМБИНАЦИИ БЕЛКОВ Cry1Be И Cry1F | 2010 |
|
RU2575611C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2740312C2 |
| КОМПОЗИЦИИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2015 |
|
RU2745322C2 |
| КОМПОЗИЦИИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2015 |
|
RU2741833C2 |
| НОВЫЙ ШТАММ PAENIBACILLUS, ПРОТИВОГРИБКОВЫЕ СОЕДИНЕНИЯ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2763542C2 |
| ЭКСПРЕССИЯ ПОЛИПЕПТИДОВ НИТРОГЕНАЗЫ В РАСТИТЕЛЬНЫХ КЛЕТКАХ | 2020 |
|
RU2833880C2 |
| РАСТЕНИЯ С МОДИФИЦИРОВАННЫМИ ПРИЗНАКАМИ | 2017 |
|
RU2809117C2 |

Изобретение относится к области биохимии, в частности к гену, придающему устойчивость к Podosphaera leucotricha. Также раскрыты растение Malus Domestica, устойчивое к Podosphaera leucotricha, содержащее указанный ген, семя для выращивания устойчивого к Podosphaera leucotricha указанного растения Malus Domestica. Изобретение позволяет эффективно бороться с Podosphaera leucotricha. 3 н. и 9 з.п. ф-лы, 6 ил., 4 табл., 1 пр.
1. Ген, придающий устойчивость к Podosphaera leucotricha, при этом аминокислотная последовательность, кодируемая указанным геном, придающим устойчивость, представляет собой первичную аминокислотную последовательность, представленную SEQ ID NO: 1, и при этом указанный ген, придающий устойчивость, является нарушенным.
2. Ген, придающий устойчивость к Podosphaera leucotricha, по п. 1, отличающийся тем, что указанное нарушение представляет собой одну или более мутаций в указанном гене, приводящих к отсутствию продукта экспрессии белка с первичной аминокислотной последовательностью SEQ ID NO: 1 или мРНК, содержащей SEQ ID NO: 2.
3. Ген, придающий устойчивость к Podosphaera leucotricha, по п. 1, отличающийся тем, что указанное нарушение представляет собой одну или более мутаций в указанном гене, приводящих к образованию нефункционального продукта экспрессии белка.
4. Ген, придающий устойчивость к Podosphaera leucotricha, по п. 1, отличающийся тем, что указанное нарушение представляет собой сниженный уровень транскрипции, что приводит к снижению уровня мРНК, содержащей SEQ ID NO: 2.
5. Ген, придающий устойчивость к Podosphaera leucotricha, по п. 1, отличающийся тем, что указанное нарушение представляет собой сниженный уровень трансляции.
6. Ген, придающий устойчивость к Podosphaera leucotricha, по любому из пп. 1-5, отличающийся тем, что указанный ген получен из Malus Domestica.
7. Растение Malus Domestica, устойчивое к Podosphaera leucotricha, содержащее в своем геноме нарушенный ген, придающий устойчивость к Podosphaera leucotricha, по любому из пп. 1-6, отличающееся тем, что указанное нарушение обеспечивает устойчивость к Podosphaera leucotricha.
8. Растение Malus domestica по п. 7, отличающееся тем, что экспрессия или транскрипция указанного гена, придающего устойчивость к Podosphaera leucotricha, снижена по меньшей мере на 10% по сравнению с растением Malus domestica, восприимчивым к Podosphaera leucotricha.
9. Растение Malus domestica по п. 8, отличающееся тем, что экспрессия или транскрипция указанного гена, придающего устойчивость к Podosphaera leucotricha, снижена по меньшей мере на 20% по сравнению с растением Malus domestica, восприимчивым к Podosphaera leucotricha, предпочтительно по меньшей мере на 30%, более предпочтительно по меньшей мере на 50%, даже более предпочтительно по меньшей мере на 70% и наиболее предпочтительно по меньшей мере на 80%.
10. Растение Malus domestica по любому из пп. 7-9, отличающееся тем, что экспрессия или транскрипция указанного гена, придающего устойчивость к Podosphaera leucotricha, отсутствует.
11. Растение Malus domestica по любому из пп. 7-10, содержащее в своем геноме нарушенный ген, придающий устойчивость к Podosphaera leucotricha, кодирующий белок с первичной аминокислотной последовательностью SEQ ID NO: 1.
12. Семя для выращивания устойчивого к Podosphaera leucotricha растения Malus Domestica, где семя из растения по любому из пп. 7-11, содержащее в своем геноме нарушенные гены, придающие устойчивость к Podosphaera leucotricha, как определено по любому из пп. 1-6, при этом указанное нарушение обеспечивает устойчивость к Podosphaera leucotricha.
| СПОСОБ ДИАГНОСТИРОВАНИЯ АДАПТИВНОГО ПОТЕНЦИАЛА СОРТОВ ПЛОДОВЫХ КУЛЬТУР | 2005 |
|
RU2305930C2 |
| Состав для вспышки | 1929 |
|
SU19042A1 |
| СПОСОБ И ВЕЩЕСТВО ДЛЯ СТИМУЛЯЦИИ РОСТА РАСТЕНИЙ | 2007 |
|
RU2438312C2 |

