Изобретение относится к медицине, а именно к фтизиатрии, и может быть использовано для лабораторного определения микобактерий туберкулеза, относящихся к основным филогенетическим сублиниям генотипа Beijing.
В настоящее время при снижении заболеваемости туберкулезом в России имеет место нарастание доли множественно/широко-лекарственно-устойчивых штаммов (MDR/XDR ТВ - multi/extensively drug resistant ТВ). Устойчивые к антибиотикам и одновременно способные к быстрому распространению штаммы Mycobacterium tuberculosis угрожают борьбе с глобальной эпидемией туберкулеза.
М. tuberculosis является строго клональным видом, подразделяемым на 8 филогенетических линий, которые включают генетические семейства и клональные комплексы. Такие различные геногруппы имеют и различия в клинической значимости и патогенетическом потенциале как результат различий адаптивной коэволюции с человеком. Восточно-Азиатская линия является одной из наиболее распространенных линий M. tuberculosis, а генетическое семейство Beijing - наиболее известным и основным ее представителем. Впервые открытые в Китае в 1995 году (van Soolingen et al., 1995), сейчас штаммы Beijing глобально распространены, часто, но не всегда ассоциированы с лекарственной устойчивостью и определяются как гипервирулентные.
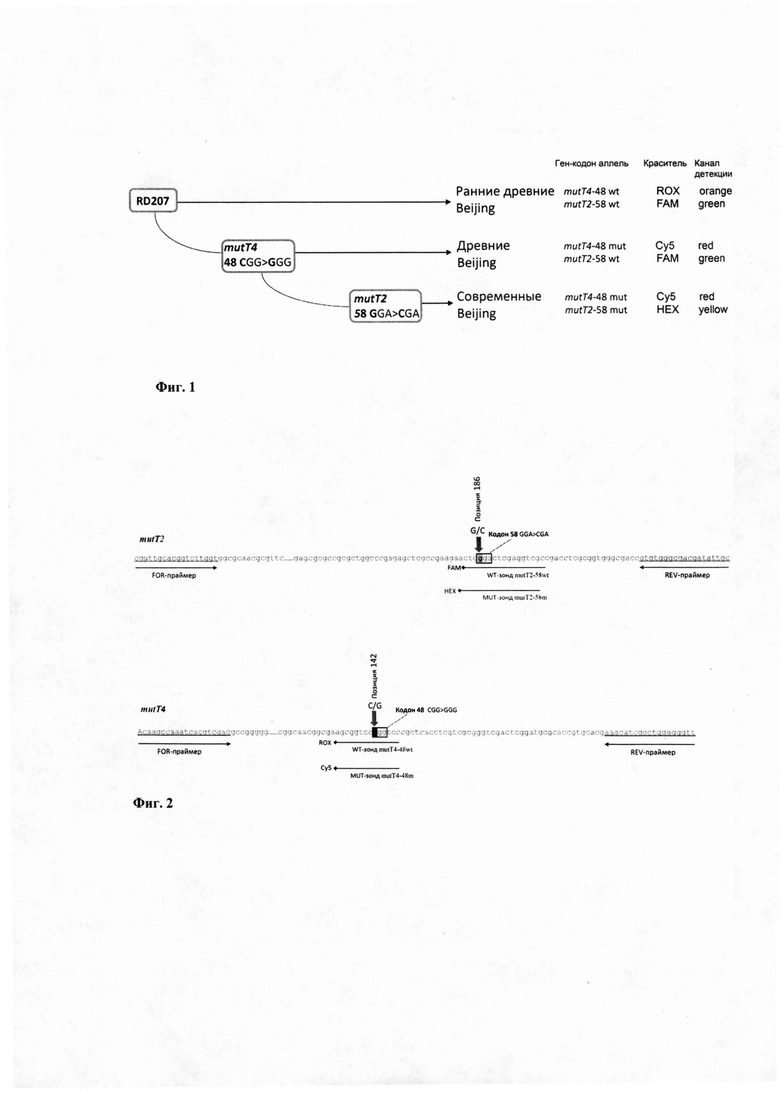
Генетическое семейство Beijing разделяют на крупные филогенетические сублинии древних и современных штаммов (Mokrousov et al., 2002, 2006), которые выделяют на основе ряда маркеров генома, а именно делеции RD181, полиморфизмы в генах mutT2 и mutT4, и полиморфизма в локусе NTF (наличие/отсутствие IS6110) (Фиг. 1). Биоинформационный и филогенетический анализ генетических данных штаммов М. tuberculosis различных генотипов, включая штаммы семейства Beijing, позволил выявить однонуклеотидные полиморфизмы (мутация mutT2-58 GGA>CGA, что соответствует позиции 1286766 в геноме референтного штамма H37Rv NC_000962.3 и мутация mutT4-48 CGG>GGG что соответствует позиции 4393590 в геноме референтного штамма H37Rv NC_000962.3), позволяющие разграничить ранние древние, древние и современные штаммы генотипа Beijing (Ebrahimi-Rad et al., 2003; Wada et al., 2009; Yin et al., 2015).
В то время как современные штаммы Beijing циркулируют по всему миру, древние сублинии преобладают в Корее (Shamputa et al., 2010) и Японии (Wada et al., 2009), реже встречаются в Китае (Mokrousov et al., 2005; Luo et al., 2015) и Вьетнаме (Maeda et al., 2014) и редко в других странах. Например, в России, где генотип Beijing составляет от 35 до 65% от общей популяции М. tuberculosis, древние штаммы были обнаружены в 5-14% популяций Beijing на северо-западе России и в Западной Сибири (Mokrousov et al., 2002, 2019). Следует отметить, что различия в ассоциации устойчивости к определенным препаратам были отмечены еще в первых работах в Китае (Mokrousov et al., 2006) и Японии (Iwamoto et al., 2008). Во Вьетнаме устойчивость к изониазиду и стрептомицину чаще наблюдали у древних штаммов Beijing (Maeda et al. 2014). С другой стороны, нарастание циркуляции современных штаммов было отмечено в Японии среди молодого населения (Iwamoto et al., 2009) и увеличенная кластеризация, как прокси-маркер трансмиссивности была описана как среди древних, так и современных штаммов Beijing по сравнению со штаммами других, не-Beijing генотипов во Вьетнаме (Maeda et al., 2014). Недавно был описан клональный кластер относящийся к ранней древней сублинии генотипа Beijing в Западной Сибири в относительно высокой доле (14,3% популяции Beijing) и ассоциированный с множественной и широкой лекарственной устойчивостью (Mokrousov et al., 2019).
Таким образом, разработка способа быстрой детекции крупных филогенетических сублиний внутри генотипа Beijing имеет и практическую значимость.
Известно определение штаммов древних и современных сублиний Beijing секвенированием фрагментов генов mutT2 и mutT4 (Ebrahimi Rad 2003, Kremer 2004) или с использованием мультиплексной ПЦР локуса NTF (Nakanishi et al., 2013). Также описана дифференциация штаммов древних и современных сублиний Beijing на основе анализа длины гена Rv3135, наличия вставок IS6110 в локусе IS61547 (Mokrousov et al., 2002), анализа структуры локуса NTF на наличие и количество вставок IS6110 (Mokrousov et al., 2005).
Недостатками данных подходов является трудоемкость анализа, требующего секвенирование генов mutT2 и mutT4, нестабильность локуса NTF, приводящая к неспецифическим продуктам ПЦР, отсутствие валидации методов на основе анализа генов Rv315 и IS1547.
Таким образом, разработка быстрого и простого способа детекции трех крупных филогенетических сублиний М. tuberculosis Beijing является актуальной задачей эпидемиологического мониторинга возбудителя туберкулеза.
Задачей предлагаемого изобретения является разработка способа быстрой и надежной детекции сублиний генотипа Beijing микобактерий туберкулеза (ранние древние, древние и современные штаммы) на основе анализа кодона 58 в гене mutT2 и кодона 48 в гене mutT4.
Задача реализуется за счет того, что одновременно определяют наличие нуклеотидных замен в данных кодонах с помощью ПЦР в формате реального времени с использованием олигонуклеотидных праймеров (5'-CGGTTGCACGGTCTTGGT, 5'-GCAATATCGTCGCCCACAC, 5'-ACAAGCCAAATCACGTCGAC, 5'-AACCCTCCAGCCGATGTTT), и флуоресцентно-меченых зондов (FAM-5'-CTGGGACTCGAGGTCGC-RTQ1, НЕХ-5'-ACTGCGACTCGAGGTCG-BHQ2, ROX-5'-AGCGGTCCCGGTCC-BHQ2, Cy5-5'-AGCGGTCCGGGTCC-BHQ2), с оценкой результатов ПЦР между 15-35 циклами и при регистрации экспоненциального роста сигнала флуоресценции по НЕХ-каналу с длиной волны 555 нм и Cy5 каналу с длиной волны 670 нм судят о принадлежности штамма к современной сублинии генотипа Beijing М. tuberculosis, при регистрации экспоненциального роста по FAM-каналу с длиной волны 510 нм и Cy5 каналу с длиной волны 670 нм судят о принадлежности штамма к древней сублинии генотипа Beijing М. tuberculosis, а при регистрации экспоненциального роста по FAM-каналу с длиной волны 510 нм и ROX каналу с длиной волны 600 нм судят о принадлежности штамма к ранней древней сублинии генотипа Beijing М. tuberculosis.
Преимущества предлагаемого способа: 1. быстрота (менее одного дня от момента выделения ДНК); 2. простая и однозначная интерпретация результатов; 3. возможность быстрого анализа больших коллекций штаммов М. tuberculosis для оценки их принадлежности к сублиниям генотипа Beijing М. Tuberculosis; 4. возможность выявления контаминации и микс-инфекции.
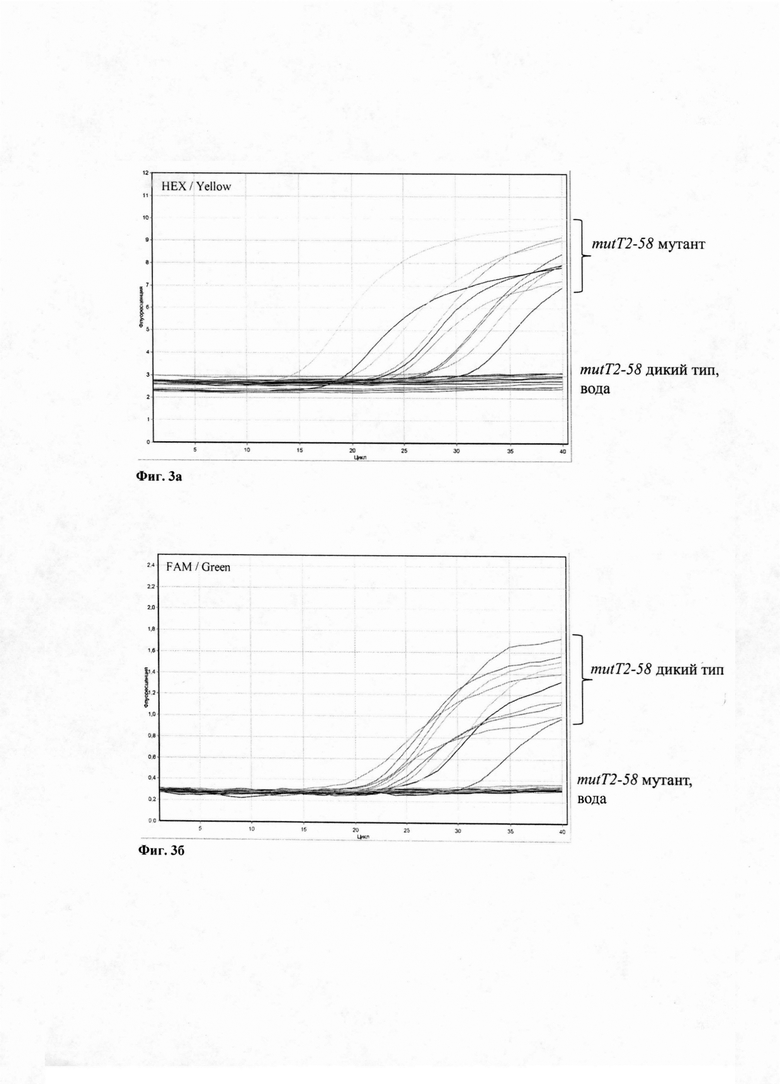
Аллель-специфическая ПЦР основана на использовании сконструированных нами 4 праймеров и 4 флуоресцентно-меченых зондов для одновременного выявления аллелей mutT2-58 GGA>CGA и muT4-48 CGG) (Фиг. 2).
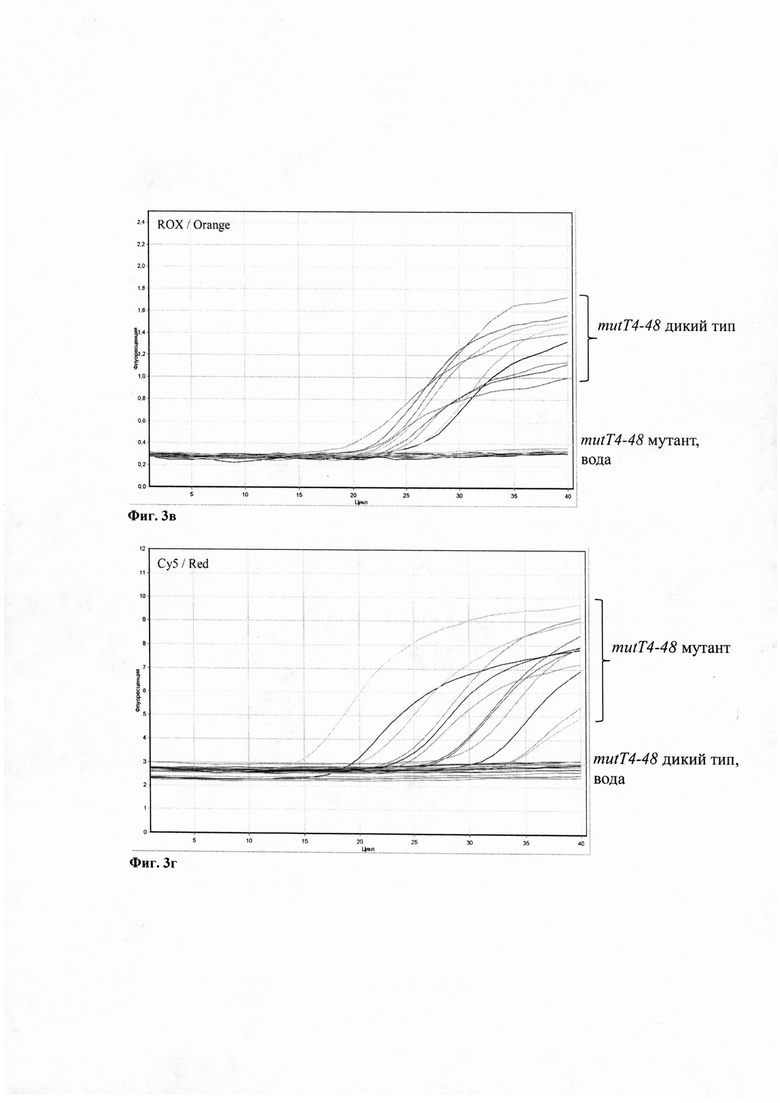
Изобретение поясняется чертежами, где Фиг. 1 - Схема эволюции генотипа Beijing с указанием ключевых эволюционных событий, разделяющих три сублинии. Фиг. 2 - Схема ПЦР (не в масштабе) участков двух генов mutT2 и mutT4, где тонкие стрелки - позиции праймеров и зондов в геноме референс-штамма H37Rv; а широкая стрелка обозначает позиции маркерных нуклеотидных замен в генах mutT2-58 и mutT4-48. Фиг. 3 - накопление сигнала флуоресценции: (а) по HEX каналу детекции, на современную сублинию М. tuberculosis Beijing, (б) по FAM каналу детекции, на штамм ранней древней или древней сублинии генотипа Beijing М. tuberculosis. (в) по ROX каналу детекции, на раннюю древнюю сублинию генотипа Beijing М. tuberculosis. и (г) Cy5 каналу детекции, на штамм древней или современной сублинии генотипа Beijing М. tuberculosis. Вода служит отрицательным контрольным образцом.
Способ был отработан и условия были оптимизированы на 37 штаммах М. tuberculosis различных генотипов по которым ранее были получены данные полногеномного секвенирования (депонированы в NCBI Sequence Read Archive как PRJNA305488 и PRJNA489691) и соответственно были известны их аллели генов mutT2-58 и mutTE4-48.
Способ осуществляется следующим образом.
(1) Выделение ДНК из культуры М. tuberculosis, выращенной на среде Левенштейна-Йенсена, проводят по van Embden et al. [1993]: суспендируют 1 стандартную бактериологическую петлю культуры в 400 мкл буфера ТЕ ×1 и инкубируют 20 мин при 85°С.Дальнейшую обработку проводят с использованием лизоцима, протеиназы К, додецилсульфата натрия и цетилтриметиламмонийбромида. Полученный клеточный лизат обрабатывают смесью фенол-хлороформ-изоамиловый спирт (25:24:1), центрифугируют, осаждают изопропанолом, промывают 70% этанолом, осадок высушивают и растворяют в 30-50 мкл ТЕ ×l.
(2) Выявление генотипа Beijing любым удобным способом, например, методом сполготипирования (Kamerbeek et al., 1997). Дальнейший анализ проводят только для штаммов генотипа Beijing.
(3) Реализация разработанного нами способа детекции сублиний генотипа Beijing.
Были использованы следующие сконструированные нами праймеры и зонды для ПЦР в реальном времени для проведения реакции ПЦР и детекции флуоресценции по 4 каналам в одной пробирке одновременно: праймеры mutT2-58F mutT2-58R, mutT4-48F, mutT4-48R, и зонды mutT2-58wt FAM, mutT2-58m HEX, mutT4-48wt ROX, mutT4-48m Cy5 (Фиг. 2).
Для детекции штамма генотипа Beijing современной сублинии служили зонды mutT2-58m HEX и mutT4-48m Су5 (Фиг. 2), детекцию сигнала проводили по HEX и Су5 каналам детекции (Фиг. 3а). Для детекции штамма генотипа Beijing древней сублинии служили зонды mutT2-58wt FAM и mutT4-48m Су5 (Фиг. 2), детекцию сигнала проводили по FAM и Су5 каналам детекции (Фиг. 3а). Для детекции штамма генотипа Beijing ранней древней сублинии служили зонды mutT2-58wt FAM и mutT4-48wt ROX (Фиг. 2), детекцию сигнала проводили по FAM и ROX каналам детекции (Фиг. 3а).
Очищенная ДНК (0.01 мкл ДНК разведенной 1 к 10) добавляется к смеси ПЦР (конечный объем 30 мкл) содержащей 1,5 mM MgCh, 1 U Taq ДНК полимеразы для ПЦР в реальном времени в режиме «горячего старта», 200 μМ каждого из дНТФ, праймеры и зонды (по 5 пмоль каждый кроме зонда mutT2-58m-HEX). ПЦР проводили в термоциклере RotorGene6000 (Corbette Research) в следующих условиях: 95°С, 3 мин; далее 40 циклов 95°С, 15 с; 63°С, 40 с. Считывание сигнала флуоресценции - при 63°С по каналам с длиной волны 510, 555, 600, 670 нм.
Оценка результатов
Полученные данные - кривые накопления флуоресцентного сигнала по 4 каналам - анализируют с помощью программного обеспечения прибора, используемого для проведения ПЦР в формате реального времени. Результаты интерпретируют по экспоненциальному накоплению сигнала флуоресценции по тому или иному каналу детекции между 15-35 циклами ПЦР. По каналам HEX и Су5 регистрируется накопление фрагментов ДНК, специфических для современной сублинии генотипа М. tuberculosis Beijing (Фиг. 3аг), по каналам FAM и Су5 - фрагментов ДНК, специфических для древней сублинии генотипа М. tuberculosis Beijing (Фиг. 3бг), по каналам FAM и ROX - фрагментов ДНК, специфических для ранней древней сублинии генотипа М. tuberculosis Beijing (Фиг. 3бв).
При отсутствии сигнала флуоресценции по всем каналам делается вывод о недостаточном количестве и/или качестве ДНК или ингибировании реакции ПЦР и невозможности какого-либо вывода о наличии или отсутствии ДНК штамма Beijing той или иной сублини в образце.
Способ был апробирован на коллекциях ДНК клинических изолятов М. tuberculosis из коллекции лаборатории молекулярной микробиологии ФБУН НИИЭМ им. Пастера, представляющих страны с различными популяциями возбудителя и различной долей штаммов Beijing древних и современных сублиний, что показано на примерах ниже.
Примеры
Пример 1
Анализ ДНК изолятов М. tuberculosis из Северо-Запада России, региона с незначительной долей ранних древних штаммов Beijing (5% от всех Beijing) (Mokrousov et al., 2002).
Всего было проверено 126 образцов ДНК изолятов М. tuberculosis генотипа Beijing, выделенных от больных туберкулезом легких в 2017-2018 годах в Республике Коми (73 изолята) и Вологодской области (53 изолята). Все образцы ДНК М. tuberculosis были охарактеризованы методом 24-MIRU-VNTR типирования (Supply et al., 2006) с последующим сравнением с базой данных MIRU-VNTRplus.org. 2 изолята имели профили 1071-32, который как было показано ранее, включает штаммы ранней древний сублинии Beijing (Mokrousov et al., 2019). В результате применения разработанного способа ПЦР в формате реального времени нами было установлено, что данные 2 изолята имели аллели дикого типа в кодонах mutT4-48 и mutT2-58 и, таким образом, относились к ранней древней сублинии, а остальные 124 изолята имели мутантные аллели в кодонах mutT4-48 и mutT2-58 и, таким образом, относились к современной филогенетической сублинии генотипа Beijing, что соответствовало результатам 24-MIRU-VNTR анализа.
Пример 2. Анализ ДНК изолятов М. tuberculosis выделенных во Вьетнаме.
Изучено 35 изолятов М. tuberculosis Beijing выделенных от больных туберкулезом легких из Вьетнама. Все образцы ДНК М. tuberculosis были охарактеризованы методом сполиготипирования (Kamerbeek et al., 1997) и 24-MIRU-VNTR типирования (Supply et al., 2006) с последующим сравнением с базой данных MIRU-VNTRplus.org. Все штаммы были проанализированы с помощью мультиплексной ПЦР локуса NTF, позволяющей разделить штаммы Beijing на древние и современные сублинии по наличию или отсутствию вставки IS6110 (Mokrousov et al., 2005).
Далее ДНК всех штаммов была протестирована разработанным нами способом ПЦР в формате реального времени для определения филогенетических сублиний генотипа Beijing микобактерий туберкулеза (ранние древние, древние и современные штаммы). В результате проведения ПЦР было выявлено 2 изолята ранней древней сублинии (дикие аллели mutT4-48 и mutT2-58), 15 изолятов древней сублинии (дикий аллель mutT4-48 и мутантный аллель mutT2-58) и 18 изолятов современной сублинии Beijing (мутантные аллели в кодонах mutT4-48 и mutT2-58). Эти результаты соответствовали результатам типирования локуса NTF.
Пример 3. Анализ ДНК изолятов М. tuberculosis выделенных в Китае.
Изучено 25 изолятов М. tuberculosis генотипа Beijing, выделенных от больных туберкулезом легких из северного Китая (Пекин и соседние провинции).
Все образцы ДНК М. tuberculosis были охарактеризованы методом сполиготипирования (Kamerbeek et al., 1997), 24-MIRU-VNTR типирования (Supply et al., 2006) и мультиплексной ПЦР локуса NTF позволяющему разделить штаммы Beijing на древние и современные сублинии по наличию или отсутствию вставки IS6110 (Mokrousov et al., 2005).
Далее ДНК всех штаммов была протестирована предлагаемым способом ПЦР в формате реального времени для определения филогенетических сублиний генотипа Beijing микобактерий туберкулеза (ранние древние, древние и современные штаммы). В результате проведения ПЦР, было выявлено 12 изолятов ранней древней сублинии Beijing, 12 изолятов древней сублинии Beijing и один изолят определен как современный Beijing, что полностью соответствовало результатам 24-MIRU-VNTR типирования и мультиплексной ПЦР локуса NTF.
Работа выполнена при поддержке Российского фонда фундаментальных исследований (грант 19-04-00263).
СПИСОК ЛИТЕРАТУРЫ
Ebrahimi-Rad М, Bifani Р, Martin С, Kremer K, Samper S, Rauzier J, et al. Mutations in putative mutator genes of Mycobacterium tuberculosis strains of the W-Beijing family. Emerg Infect Dis. 2003; 9:838-45.
Iwamoto T, Yoshida S, Suzuki K, Wada T. Population structure analysis of the Mycobacterium tuberculosis Beijing family indicates an association between certain sublineages and multidrug resistance. Antimicrob Agents Chemother. 2008; 52:3805-9.
Iwamoto T, Fujiyama R, Yoshida S, Wada T, Shirai C, Kawakami Y. Population structure dynamics of Mycobacterium tuberculosis Beijing strains during past decades in Japan. J Clin Microbiol. 2009; 47:3340-3.
Kamerbeek J, Schouls L, Kolk A, van Agterveld M, van Soolingen D, Kuijper S, et al. Simultaneous detection and strain differentiation of Mycobacterium tuberculosis for diagnosis and epidemiology. J Clin Microbiol. 1997; 35:907-14.
Kremer K, Glynn JR, Lillebaek T, Niemann S, Kurepina NE, Kreiswirth BN, et al. Definition of the Beijing/W lineage of Mycobacterium tuberculosis on the basis of genetic markers. J Clin Microbiol. 2004; 42:4040-9.
Luo T, Comas I, Luo D, Lu B, Wu J, Wei L, et al. Southern East Asian origin and coexpansion of Mycobacterium tuberculosis Beijing family with Han Chinese. Proc Natl Acad Sci USA. 2015; 112:8136-41.
Maeda S, Hang NT, Lien LT, Thuong PH, Hung NV, Hoang NP, et al. Mycobacterium tuberculosis strains spreading in Hanoi, Vietnam: Beijing sublineages, genotypes, drug susceptibility patterns, and host factors. Tuberculosis (Edinb). 2014; 94:649-56.
Mokrousov I, Vyazovaya A, Pasechnik O, Gerasimova A, Dymova M, Chernyaeva E, et al. Early ancient sublineages of Mycobacterium tuberculosis Beijing genotype: unexpected clues from phylogenomics of the pathogen and human history. Clin Microbiol Infect. 2019; 25:1039.el-1039.e6.
Mokrousov I, Ly HM, Often T, Lan NN, Vyshnevskyi B, Hoffner S, et al. Origin and primary dispersal of the Mycobacterium tuberculosis Beijing genotype: clues from human phylogeography. Genome Res. 2005; 15:1357-64.
Mokrousov I, Narvskaya O, Often T, Vyazovaya A, Limeschenko E, Steklova L, et al. Phylogenetic reconstruction within Mycobacterium tuberculosis Beijing genotype in northwestern Russia. Res. Microbiol. 2002; 153: 629-37
Mokrousov I, Jiao WW, Sun GZ, Liu JW, Valcheva V, Li M, Narvskaya O, Shen AD. Evolution of drug resistance in different sublineages of Mycobacterium tuberculosis Beijing genotype. Antimicrob Agents Chemother. 2006 Aug; 50(8):2820-3.
Nakanishi N, Wada T, Arikawa K, Millet J, Rastogi N, Iwamoto T. Evolutionary robust SNPs reveal the misclassification of Mycobacterium tuberculosis Beijing family strains into sublineages. Infect Genet Evol. 2013 Jun; 16:174-7. doi: 10.1016/j.meegid.2013.02.007.
Shamputa IC, Lee J, Allix-Béguec C, Cho EJ, Lee JI, Rajan V, et al. Genetic diversity of Mycobacterium tuberculosis isolates from a tertiary care tuberculosis hospital in South Korea. J Clin Microbiol. 2010; 48:387-94.
Supply P, Allix C, Lesjean S, Cardoso-Oelemann M, Rüsch-Gerdes S, Willery E, et al. Proposal for standardization of optimized mycobacterial interspersed repetitive unit-variable-number tandem repeat typing of Mycobacterium tuberculosis. J Clin Microbiol. 2006; 44:4498-510.
van Embden J, Cave M, Crawford J, Dale JW, Eisenach KD, Gicquel B, et al. Strain identification on Mycobacterium tuberculosis by DNA fingerprinting: recommendations for a standardized methodology. J. Clin. Microbiol. 1993; 31:406-409.
van Soolingen D, Qian L, de Haas PE, Douglas JT, Traore H, Portaels F, Qing HZ, Enkhsaikan D, Nymadawa P, van Embden JD. Predominance of a single genotype of Mycobacterium tuberculosis in countries of east Asia. J Clin Microbiol. 1995 Dec; 33(12):3234-8.
Wada Т., Iwamoto Т., Maeda S. FEMS Microbiol Lett, 2009, 291: 35-43
Yin QQ, Liu HC, Jiao WW, Li QJ, Han R, Tian JL, et al. Evolutionary History and Ongoing Transmission of Phylogenetic Sublineages of Mycobacterium tuberculosis Beijing Genotype in China. SciRep. 2016; 6:34353.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ детекции генотипа Mycobacterium tuberculosis Beijing 1071-32-кластер в формате реального времени | 2021 |
|
RU2768021C1 |
| Способ детекции генотипа Mycobacterium tuberculosis Beijing 14717-15-кластер методом полимеразной цепной реакции в формате реального времени | 2023 |
|
RU2839699C2 |
| Способ детекции изолятов Mycobacterium tuberculosis Beijing 94-32-кластера в формате реального времени | 2017 |
|
RU2689800C1 |
| Способ детекции генотипа Mycobacterium tuberculosis Beijing 14717-15-кластер | 2022 |
|
RU2812351C1 |
| Способ выявления микобактерий туберкулеза Центрально-Азиатского эпидемического кластера генотипа Beijing | 2019 |
|
RU2735415C1 |
| Способ детекции S-генотипа Mycobacterium tuberculosis | 2024 |
|
RU2832725C1 |
| СПОСОБ ВЫЯВЛЕНИЯ МИКОБАКТЕРИЙ ТУБЕРКУЛЕЗА ГЕНОТИПА ВЕIJING В РЕЖИМЕ РЕАЛЬНОГО ВРЕМЕНИ | 2011 |
|
RU2528866C2 |
| Способ выявления микобактерий туберкулеза генотипа Beijing В0-кластер в формате реального времени | 2017 |
|
RU2684314C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ СУБЛИНИЙ ВОЗБУДИТЕЛЯ ТУБЕРКУЛЕЗА ЛИНИИ L2 Beijing НА БИОЛОГИЧЕСКИХ МИКРОЧИПАХ | 2022 |
|
RU2790296C1 |
| СПОСОБ ВЫЯВЛЕНИЯ МИКОБАКТЕРИЙ ТУБЕРКУЛЕЗА ГЕНОТИПА BEIJING | 2008 |
|
RU2405836C2 |
Изобретение относится к биотехнологии, а именно к способу определения филогенетических сублиний генотипа Beijing M. tuberculosis. Проводят одновременный анализ кодона 58 в гене mutT2 и кодона 48 в гене mutT4 с помощью ПЦР в формате реального времени. Используют олигонуклеотидные праймеры: 5'-CGGTTGCACGGTCTTGGT, 5'-GCAATATCGTCGCCCACAC, 5'-ACAAGCCAAATCACGTCGAC, 5'-AACCCTCCAGCCGATGTTT, и флуоресцентно-меченые зонды: FAM-5'-CTGGGACTCGAGGTCGC-RTQ1, НЕХ-5'-ACTGCGACTCGAGGTCG-BHQ2, ROX-5'-AGCGGTCCCGGTCC-BHQ2, Су5-5'-AGCGGTCCGGGTCC-BHQ2. Результаты ПЦР оценивают между 15-35 циклами. При регистрации экспоненциального роста сигнала флуоресценции по НЕХ-каналу с длиной волны 555 нм и Су5-каналу с длиной волны 670 нм судят о принадлежности штамма к современной сублинии генотипа Beijing М. tuberculosis, по FAM-каналу с длиной волны 510 нм и Су5-каналу с длиной волны 670 нм судят о принадлежности штамма к древней сублинии, по FAM-каналу с длиной волны 510 нм и ROX-каналу с длиной волны 600 нм судят о принадлежности штамма к ранней древней сублинии. Способ позволяет быстро и надежно детектировать сублинии генотипа Beijing М. tuberculosis, проводить анализ больших коллекций штаммов М. tuberculosis. 3 ил., 3 пр.
Способ детекции филогенетических сублиний генотипа Beijing Mycobacterium tuberculosis в формате реального времени путем анализа кодона 58 в гене mutT2 и кодона 48 в гене mutT4, отличающийся тем, что одновременно определяют наличие нуклеотидных замен в данных кодонах с помощью ПЦР в формате реального времени с использованием олигонуклеотидных праймеров 5'-CGGTTGCACGGTCTTGGT, 5'-GCAATATCGTCGCCCACAC, 5'-ACAAGCCAAATCACGTCGAC, 5'-AACCCTCCAGCCGATGTTT, и флуоресцентно-меченых зондов FAM-5'-CTGGGACTCGAGGTCGC-RTQ1, HEX-5'-ACTGCGACTCGAGGTCG-BHQ2, ROX-5'-AGCGGTCCCGGTCC-BHQ2, Cy5-5'-AGCGGTCCGGGTCC-BHQ2, с оценкой результатов ПЦР между 15-35 циклами и при регистрации экспоненциального роста сигнала флуоресценции по НЕХ-каналу с длиной волны 555 нм и Су5 каналу с длиной волны 670 нм судят о принадлежности штамма к современной сублинии генотипа Beijing М. tuberculosis, при регистрации экспоненциального роста по FAM-каналу с длиной волны 510 нм и Су5 каналу с длиной волны 670 нм судят о принадлежности штамма к древней сублинии генотипа Beijing М. tuberculosis, а при регистрации экспоненциального роста по FAM-каналу с длиной волны 510 нм и ROX каналу с длиной волны 600 нм судят о принадлежности штамма к ранней древней сублинии генотипа Beijing М. tuberculosis.
| СПОСОБ ВЫЯВЛЕНИЯ МИКОБАКТЕРИЙ ТУБЕРКУЛЕЗА ГЕНОТИПА BEIJING | 2008 |
|
RU2405836C2 |
| СПОСОБ ВЫЯВЛЕНИЯ МИКОБАКТЕРИЙ ТУБЕРКУЛЕЗА ГЕНОТИПА ВЕIJING В РЕЖИМЕ РЕАЛЬНОГО ВРЕМЕНИ | 2011 |
|
RU2528866C2 |
| Способ выявления микобактерий туберкулеза генотипа Beijing В0-кластер в формате реального времени | 2017 |
|
RU2684314C2 |
| Способ детекции изолятов Mycobacterium tuberculosis Beijing 94-32-кластера в формате реального времени | 2017 |
|
RU2689800C1 |
| WO 2003016562 A2, 27.02.2003. | |||

