Настоящее изобретение относится к медицине, касается способа снижения алкогольной мотивации и может быть использовано с целью купирования патологических симптомов при алкоголизме.
Алкоголизм представляет собой глобальную медико-социальную проблему современного общества. В течение последних десятилетий во всем мире сохраняется тенденция к увеличению потребления психоактивных веществ (ПАВ), в том числе и алкоголя, что приводит к значительному увеличению числа больных алкоголизмом и росту частоты сопутствующих соматических заболеваний.
Известно много способов снижения алкогольной мотивации и купирования симптомов алкогольной интоксикации путем перорального, парентерального и других способов введения химических препаратов, как искусственного, так и естественного происхождения в различных лекарственных формах, в том числе гомеопатических препаратов, однократного и по заданным схемам (RU 1806743, A61K 31/485, 1993; RU 2090190, A61K 31/485, 1997; RU 2252756, A61K 33/00, 2995; RU 2393868, A61K 33/38, 2010; RU 2665629, А61М 21/00, 2018).
Основными недостатками данных способов, основанных на фармакотерапевтическом подходе, являются сложности в стандартизации многокомпонентных и гомеопатических препаратов естественного происхождения; низкая избирательность действия при системном введении и необходимость введения больших доз для создания эффективных действующих концентраций в средах организма. Кроме того, фармпрепараты при алкоголизме увеличивают токсическую нагрузку на организм и обладают выраженными побочными эффектами, в том числе, ко многим из них развивается привыкание в процессе терапии. Немаловажным фактом является также увеличение числа больных с терапевтической резистентностью и толерантностью к психофармакотерапии.
Известен ряд способов снижения алкогольной мотивации путем физиотерапевтических процедур и различных психокоррекционных интервенций, таких как гипноз, нейролингвистическое программирование, бихевиоральная терапия, и другие (RU 1128954 A61K 31/00, 1984; RU 2234344, А61М 21/00, 2004; RU 2277943, А61М 21/00, 2006; RU 2405581, А61М 21/00, 2010; RU 2474440, А61М 21/00, 2011).
Основными недостатками данной группы методов, основанных на физиотерапевтических и психокоррекционных воздействиях, исключающих применение фармпрепаратов, являются сложности в стандартизации физиотерапевтических и психокоррекционных процедур; существенное влияние на процесс психокорреции как индивидуальных особенностей личности терапевта, так и индивидуальных особенностей личности пациента; существенное влияние на процесс психокоррекции микросоциального окружения личностей, вовлеченных в процесс психокорреции.
Известны способы снижения алкогольной мотивации, сочетающие применение препаратов и физиотерапевтических и психокоррекционных воздействий (RU 2159636, А61М 21/00, 2000; RU 2250115, А61М 21/00, 2005; RU 2402974, А61В 5/00, 2010). Данные способы сочетают в себе недостатки обеих групп вышеописанных способов.
Существенным звеном в патогенезе алкоголизма является изменение функциональной активности иммунной системы и ее клеточных элементов. Потребление 10%-го этанола приводит к дизрегуляции пролиферации Т-клеток (Lau А.Н., Abe М., Thomson A.W. Ethanol affects the generation, cosignaling molecule expression, and function of plasmacytoid and myeloid dendritic cell subsets in vitro and in vivo // Journal of leukocyte biology. - 2006. - T. 79. - №. 5. - C. 941-953; Sureshchandra S., Raus A., Jankeel A., Ligh B.J.K., Walter N.A.R., Newman N., Grant K.A., Messaoudi I. Dose-dependent effects of chronic alcohol drinking on peripheral immune responses // Sci Rep., - 2019 - Т.9 - №1-С. 7847; Lawrimore C.J., Coleman L.G., Zou J., Crews F.T. Ethanol Induction of Innate Immune Signals Across BV2 Microglia and SH-SY5Y Neuroblastoma Involves Induction of IL-4 and IL-13 // Brain Sciences. - 2019. - T.9. - №9. - C. 228). Длительное употребление алкоголя снижает эффективность клеточно-опосредованного и гуморального иммунного ответа на инфекцию и вакцинацию, что может быть связано с конверсией фенотипа наивных Т-лимфоцитов, с сокращением числа CD4 и CD8 субпопуляций лимфоцитов и модуляцией их функциональной активности (Gheorghiu М. et al. Ethanol-induced dysfunction of hepatocytes and leukocytes in patients without liver failure // Roumanian archives of microbiology and immunology. - 2004. - T. 63. - №. 1-2. - C. 5-33; Percival S.S., Sims C.A. Wine modifies the effects of alcohol on immune cells of mice // The Journal of nutrition. - 2000. - T. 130. - №. 5. - C. 1091-1094; Zhang H., Meadows G.G. Chronic alcohol consumption in mice increases the proportion of peripheral memory T cells by homeostatic proliferation // Journal of leukocyte biology. - 2005. - Т. 78. - №. 5. - С.1070-1080; Boyd A., Almeida J.R., Darrah Р.А., Sauce D., Seder R.A., et al. Pathogen-specific T cell polyfunctionality is a correlate of T cell efficacy and immune protection //PloS one. - 2015. - T. 10. - №. 6. - e0128714).
ГАМКА-рецепторы (ГАМКА-Р), представляют основную ингибиторную нейротрансмиттерную систему в ЦНС и играют центральную роль в опосредовании эффектов этанола (Trantham-Davidson Н. et al. Chronic alcohol disrupts dopamine receptor activity and the cognitive function of the medial prefrontal cortex // Journal of Neuroscience. - 2014. - T. 34. - №. 10. - C. 3706-3718; Follesa P. et al. Chronic intermittent ethanol regulates hippocampal GABA (A) receptor delta subunit gene expression // Frontiers in cellular neuroscience. - 2015. - T. 9. - C. 445). Описано также наличие функциональных ГАМКА-Р на поверхности Т-лимфоцитов (Geigerseder С. et al. Evidence for a GABAergic system in rodent and human testis: local GABA production and GABA receptors // Neuroendocrinology. - 2003. - T. 77. - №. 5. - C. 314-323). Изменение активности ГАМКА-Р, аналогично эффектам на нейрональных клетках, вызывает модуляцию функциональной активности клеток иммунной системы, изменяя их пролиферативную активность в периферической крови (Bandyopadhyay В.С., Poddar М.K. Hypothalamic GABA-ergic activity and T-cell proliferation in aged mammal: effect of dietary protein // Neurochemistry international. - 1998. - T. 32. - №. 2. - C. 191-196) и в костном мозге (Idova G.V., Alperina E.L, Cheido M.A. Contribution of brain dopamine, serotonin and opioid receptors in the mechanisms of neuroimmunomodulation: evidence from pharmacological analysis // International immunopharmacology. - 2012. - T. 12. - №. 4. - C. 618-625). Посредством ГАМКА-Р показана также модуляция клеточно-опосредованного иммунного ответа (Tian J. et al. GABAA receptors mediate inhibition of T cell responses // Journal of neuroimmunology. - 1999. - T. 96. - №. 1. - C. 21-28).
Известна способность иммунокомпетентных клеток с определенной функциональной активностью в экспериментальных условиях после трансплантации направленно изменять особенности поведения реципиентов (Маркова Е.В., Абрамов В.В., Козлов В.А. Регуляция ориентировочно-исследовательского поведения у животных путем трансплантации иммунокомпетентных клеток // Успехи современной биологии. - 2009. Т. - 129. - №4. - С. 348-354; Маркова Е.В. Иммунная система и высшая нервная деятельность. Механизмы нейроиммунных взаимодействий в реализации и регуляции поведенческих реакций. Saarbruken. - 2012; Маркова Е.В., Княжева М.А., Козлов В.А. Клеточные механизмы нейроиммунных взаимодействий в регуляции ориентировочно-исследовательского поведения // Сибирский вестник психиатрии и наркологии. - 2013. - №1 (76). - С. 49-52; Markova E.V., Anikeeva О.S. Immune cells influence on the behavioral stereotypes formation in ontogenesis // In the World of Scientific Discoveries, Series В. - 2015. - Т. 3. - №1. - С. 53-61).
Однонаправленное влияние большинства психоактивных препаратов на ЦНС и иммунную систему позволяет рассматривать иммунные клетки в качестве модельных объектов для воздействия на межсистемную функциональную взаимосвязь. Так, в частности, ранее нами была показана возможность и механизмы стимуляции стресс-индуцированных депрессивно-подобного и агрессивного поведения у экспериментальных животных трансплантацией спленоцитов, с модулированной in vitro ПАВ функциональной активностью (Markova Е., Knyazheva М., Shushpanova Т. Neuroleptic effect in aggressive mice after the transplantation of immune cells treated in vitro with chlorpromazine // European Psychiatry. - 2016. - T. 33(S) - C. S263; Маркова E.B., Княжева M.A., Савкин И.В., Тихонова М.А., Амстиславская Т.Г. Купирование поведенческих паттернов депрессивно-подобного состояния у экспериментальных животных трансплантацией модулированных психоактивным веществом иммунных клеток // Медицинская иммунология. - 2017. - Т. 19. - №S. - С. 63; Markova E.V., Knyazheva М.А., Savkin I.V., Amstislavskaya T.G. Psychoneuroimmunomodulating effect of immune cells treated with psychoactive drug in depressive - like animals // European Psychiatry. - 2018. - T. 48 (S) - C. S293; Маркова E.B., Княжева M.A. Эффект трансплантации модулированных нейролептиком иммунокомпетентных клеток у экспериментальных животных в состоянии агрессии // Медицинская иммунология. - 2015. - Т. 17 (S) - С. 44). Показан также был позитивный эффект трансплантации иммунокомпетентных клеток при опиатной зависимости (Маркова Е.В., Абрамов В.В., Старостина М.В., Козлов В.А. Влияние трансплантации иммунокомпетентных клеток на поведенческие и иммунологические параметры у животных с синдромом хронической морфиновой зависимости // Наркология. - 2006. - Т. 5. - №5 (53) - С. 27-31; Чухрова М.Г., Дресвянников В.Л., Маркова Е.В. Наркотическая зависимость: современные стратегии исследования // Saint-Louis, Missouri, USA. - 2015; Markova E., Kozlov V. Perspectives of the cells therapy in the treatment of drug abuse // European Psychiatry. - 2015. - T. 30(S). - C. S1567). Это обуславливает возможность и перспективность иммунотерапии аддиктивноподобного состояния аутологичными иммуноцитами с модулированной экстракорпорально психоактивными веществами функциональной активностью.
Принимая во внимание тот факт, что одним из иммунологических механизмов хронической алкогольной интоксикации является изменение функциональной активности иммунных клеток и приобретение лимфоцитами способности усиливать алкогольную мотивацию, методологически оправданной является иммунотерапия аутологичными иммунными клетками с экстракорпорально модулированной функциональной активностью.
Известны два синтетических лиганда ГАМКА-Р комплекса, усиливающие аффинность связывания ГАМК, с выявленными иммуномодулирующими свойствами: мета-хлорбензгидрилмочевина (РФ 2691143, A61K 31/17, 2019) и 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты (РФ 2702114, A61K 31/515, 2019). Для указанных соединений показана способность восстанавливать измененные вследствие хронического токсического влияния этанола функциональные свойства клеток иммунной системы в условиях in vitro ГАМКА-Р опосредованным образом, а также in vivo модулировать активность нервной и иммунной систем при экспериментальном алкоголизме (Маркова Е.В., Савкин И.В., Княжева М.А., Шушпанова Т.В., Новожеева Т.П. Психонейроиммуномодулирующее влияние оригинального антиконвульсанта при экспериментальном алкоголизме // Российский иммунологический журнал. - 2019. - Т. 13 (22). - №3. - С. 1217-1223; Markova Е., Savkin I., Anikeeva О., Shushpanova Т. Immunomodulatory effect of original anticonvulsant meta-chloro-benzhydryl-urea in mice with experimental alcoholism // European Psychiatry. - 2019. -V. 56S. -. S. 662-663; Маркова E.B., Савкин И.В. Влияние оригинального антиконвульсанта на показатели функциональной активности нервной и иммунной систем при алкоголизме // Журнал научных статей Здоровье и образование в XXI веке. - 2018. - Т. 20. - №10. - С. 54-57).
Известен способ реабилитации больных алкоголизмом при абстинентном синдроме, состоящий в том, что при проведении фармакотерапии дополнительно вводят лиганд ГАМКА-Р, а именно - галодиф (мета-хлорбензгидрилмочевину) в дозе 100 мг 3 раза вдень курсом 21 день (РФ 2436573, A61K 31/17, 2011).
К недостатку метода стоит отнести необходимость использования высоких доз препарата для достижения в организме его действующей концентрации; вероятность побочных эффектов со стороны ЖКТ, печени, почек и других систем, и органов, возникающих при системном (пероральном) введении препарата и ограничивающих его применение; а также необходимость изготовления таблетированной формы препарата, применяемой для перорального введения.
Известен способ снижения алкогольной мотивации у мышей, включающий забор спленоцитов у алкоголизированных мышей, их инкубацию in vitro, последующую отмывку спленоцитов и внутривенное введение полученных спленоцитов сингенным алкоголизированным мышам. Алкоголизацию мышей проводят в течение 10 дней. Инкубацию in vitro производят поликлональными кроличьими антителами против серотонина в течение 24 часов. При использовании данного способа происходит изменение субпопуляционного состава инкубированных спленоцитов в части увеличения количества спленоцитов, специфичных к антителам против серотонина, пригодных для использования в качестве антиидиотипической вакцины (Давыдова Т.В., Евсеев В.А., Фомина В.Г. Подавление антителами к серотонину усиленного потребления алкоголя, вызванного адаптивным переносом спленоцитов от животных с абстинентным синдромом //Бюллетень экспериментальной биологии и медицины. - 1998. - Т. 126. - №. 9. - С. 328-330).
Указанный способ подтверждает существенную роль клеток иммунной системы в патогенезе алкоголизма. Недостатком данного способа является необходимость применения поликлональных антител к серотонину, сложных в получении и очистке. Кроме того, существенным ограничением применения поликлональных антител является высокая вероятность возникновения вторичного иммунного ответа на структурные компоненты антител при длительном и даже при повторном применении.
Задачей изобретения является снижение алкогольной мотивации у мышей путем экстракорпорального опосредованного ГАМКА-Р воздействия иммуномодуляторами на лимфоциты в составе спленоцитов с целью восстановления функциональной активности спленоцитов, измененной вследствие длительного токсического воздействия этанола, и последующего введения данных спленоцитов в кровоток.
Указанная задача решается тем, что в способе снижения алкогольной мотивации у мышей, включающем забор спленоцитов у алкоголизированных мышей, их инкубацию in vitro с помощью иммуномодулятора, последующую отмывку спленоцитов и внутривенное введение полученных спленоцитов сингенным алкоголизированным мышам при проведении инкубации спленоцитов в течение 10-25 мин в качестве иммуномодулятора используют мета-хлорбензгидрилмочевину или 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты в действующей концентрации 10 мкг/мл на 15×106 клеток на мышь.
В качестве иммуномодулятора используют мета-хлорбензгидрилмочевину (РФ 2691143, A61K 31/17, 2019) либо 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты (РФ 2702114, A61K 31/515, 2019).
Предложенное изобретение, на наш взгляд, является новым. Существенные признаки изобретения направлены на достижение указанной цели. Внутривенное введение длительно алкоголизированным мышам прекультивированных определенным способом с мета-хлорбензгидрилмочевиной или 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты спленоцитов способствует у алкоголизированных мышей-реципиентов снижению потребления алкоголя. Также наблюдалась стимуляции локомоторной активности и других поведенческих и иммунологических показателей, свидетельствующих о позитивном патогенетическом эффекте модулированных мета-хлорбензгидрилмочевиной или 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты спленоцитов при экспериментальном алкоголизме. Существенным отличием предложенного нами способа является то, что в известном методе -прототипе эффект достигается за счет изменения субпопуляционного состава лимфоцитов, содержащихся в сингенных спленоцитах, что по сути является вакцинацией, а в предложенном нами способе эффект достигается за счет изменения функциональной активности спленоцитов под действием иммуномодуляторов in vitro, что является неспецифической иммунотерапией, существенно отличающейся от вакцинального подхода.
Способ осуществляется следующим образом. Предварительно мыши-гибриды (СВА х C57BI/6)F1, самцы, были подвергнуты длительной алкоголизации методом принудительного спаивания 10% раствора этанола в течении 6 месяцев (алкоголизированные млекопитающие).
Учитывая наличие в популяции самцов мышей (CBA x C57BI/6)F1 с активным и пассивным типами поведения, представители которых характеризуются определенными структурно-функциональными характеристиками нервной и иммунной систем и различной психофизиологической реакцией на прием ПАВ (Маркова Е.В. Иммунная система и высшая нервная деятельность. Механизмы нейроиммунных взаимодействий в реализации и регуляции поведенческих реакций.- Saarbruken. 2012; Чухрова М.Г., Дресвянников В.П., Маркова Е.В. Наркотическая зависимость: современные стратегии исследования. // Saint-Louis, Missouri, USA: Publishing House Science and Innovation Center, 2015; Гольдина И.А., Маркова E.B., Гольдин Б.Г., Княжева М.А., Гайдуль К.В. Протекторные свойства экстракта куркумы при этанол-индуцированных нарушениях поведения Саратовский научно-медицинский журнал. 2017. - Т. 13. - №. - 1. - С. 131-135; Маркова Е.В., Савкин И.В., Княжева М.А., Шушпанова Т.В., Новожеева Т.П. Психонейроиммуномодулирующее влияние оригинального антиконвульсанта при экспериментальном алкоголизме // Российский иммунологический журнал. - 2019. - Т. 13 (22). - №3. - С. 1217-1223), с целью формирования однородных экспериментальных групп животных, все мыши были предварительно протестированы в «открытом поле» и в исследование были включены только особи со средним уровнем поведения. Эти мыши были разделены на 2 группы: мыши первой группы подвергались принудительной алкоголизации (10% раствор этанола в качестве единственного источника жидкости) в течение 6 месяцев; мыши второй группы содержались в обычных условиях и составили контрольную группу интактных млекопитающих.
У длительно алкоголизированых мышей (в данном случае мыши-гибриды (СВА х C57BI/6)F1) производили забор зрелых периферических иммунных клеток селезенки (спленоцитов) в стерильных условиях путем перфузии селезенки. После чего выделенные спленоциты подвергали in vitro инкубации с иммуномодулятором (мета-хлорбензгидрилмочевина или 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты), в концентрации 10 мкг/мл на 15×106 клеток на мышь в течение 10-25 минут. Затем производили трехкратную отмывку спленоцитов от иммуномодулятора в физиологическом растворе и вводили полученные клетки внутривенно сингенным длительно алкоголизированным мышам-реципиентам. Алкогольная мотивация длительно алкоголизированных мышей -реципиентов оценивалась по потреблению 10% раствора этанола в условиях свободного выбора с водой (двух-бутылочный тест) индивидуально, каждой мышью-реципиентом в течение 10 дней.
Оценка эффективности способа осуществлялась путем сравнения трех групп млекопитающих по потреблению 10% раствора этанола в условиях свободного выбора с водой. Группу длительно алкоголизированных мышей-реципиентов, получивших инъекцию спленоцитов, инкубированных in vitro с иммуномодулятором, сравнивали с группой длительно алкоголизированных мышей и с группой интактных мышей соответствующего возраста.
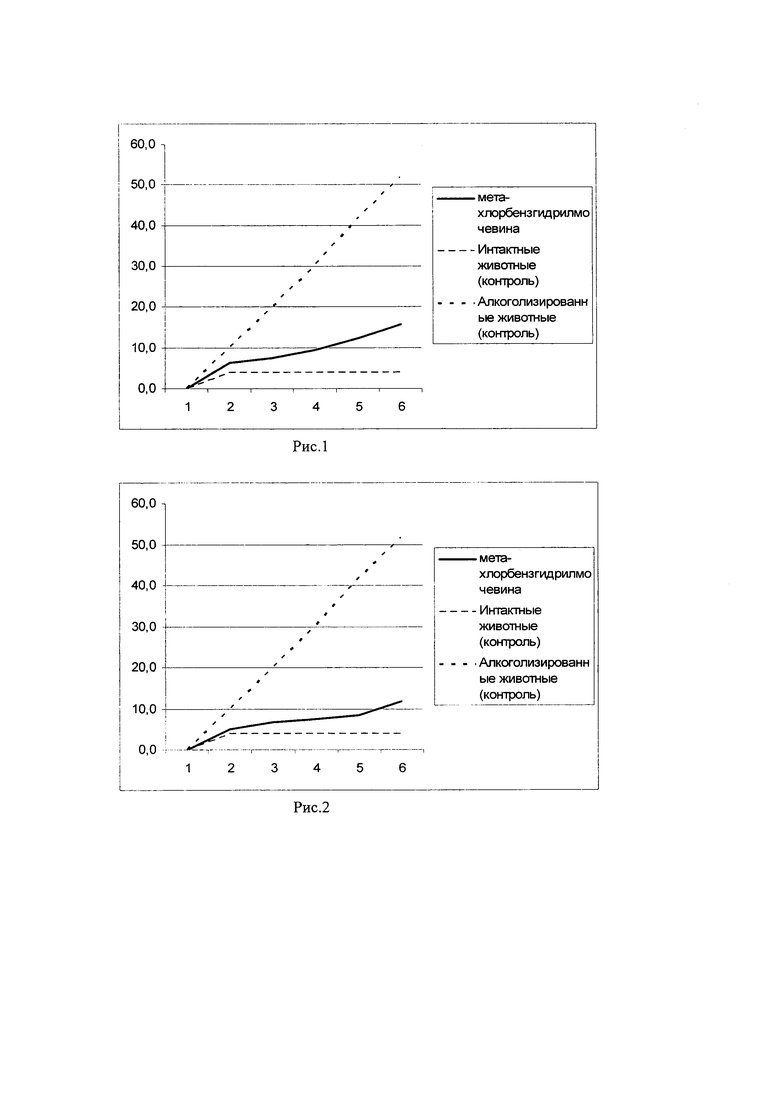
Пример 1. У длительно алкоголизированных мышей (мыши-гибриды (СВА х C57BI/6)F1) производили забор спленоцитов. Затем выделенные клетки подвергали in vitro инкубации с мета-хлорбензгидрилмочевиной, в концентрации 10 мкг/мл на 15×106 клеток на мышь в течение 10 минут. Затем производили трехкратную отмывку спленоцитов от мета-хлорбензгидрилмочевины в физиологическом растворе и вводили полученные клетки внутривенно сингенным длительно алкоголизированным мышам-реципиентам. Алкогольная мотивация длительно алкоголизированных мышей-реципиентов оценивалась по потреблению 10% раствора этанола в условиях свободного выбора с водой в сравнении с длительно алкоголизированными и интактыми мышами соответствующего возраста (результаты представлены на рис. 1).
На рис. 1 показано накопительное потребление этанола длительно алкоголизированными мышами после внутривенного введения сингенных спленоцитов с функциональной активностью, модулированной in vitro мета-хлорбензгидрилмочевиной. Представлено среднесуточное потребление этанола (мл/мышь), начиная с 1-го дня после введения спленоцитов в условиях свободного выбора 10% раствора этанола с водой (двух-бутылочный тест). Как видно из рис. 1, внутривенное введение сингенных спленоцитов с функциональной активностью, модулированной in vitro мета-хлорбензгидрилмочевиной, снижает потребление этанола в группе длительно алкоголизированными мышей-реципиентов и, следовательно, снижает алкогольную мотивацию.
Пример 2. У длительно алкоголизированных мышей (мыши-гибриды (СВА х C57BI/6)F1) производили забор спленоцитов. Затем выделенные клетки подвергали in vitro инкубации с мета-хлорбензгидрилмочевиной, в концентрации 10 мкг/мл на 15×106 клеток на мышь в течение 18 мин. Затем производили трехкратную отмывку спленоцитов от мета-хлорбензгидрилмочевины в физиологическом растворе и вводили полученные клетки внутривенно сингенным длительно алкоголизированным мышам-реципиентам. Алкогольная мотивация длительно алкоголизированных мышей-реципиентов оценивалась по потреблению 10% раствора этанола в условиях свободного выбора с водой в сравнении с длительно алкоголизированными и интактыми мышами соответствующего возраста (результаты представлены на рис. 2).
На рис. 2 показано накопительное потребление этанола длительно алкоголизированными мышами после внутривенного введения сингенных спленоцитов с функциональной активностью, модулированной in vitro мета-хлорбензгидрилмочевиной. Представлено среднесуточное потребление этанола (мл/мышь), начиная с 1-го дня после введения спленоцитов в условиях свободного выбора. Как видно из рис. 2, внутривенное введение сингенных спленоцитов с функциональной активностью, модулированной in vitro мета-хлорбензгидрилмочевиной, снижает потребление этанола в группе длительно алкоголизированных мышей-реципиентов и, следовательно, снижает алкогольную мотивацию.
Пример 3. У длительно алкоголизированных мышей (мыши-гибриды (СВА х C57BI/6)F1) производили забор спленоцитов. Затем выделенные клетки подвергали in vitro инкубации с мета-хлорбензгидрилмочевиной, в концентрации 10 мкг/мл на 15×106 клеток на мышь в течение 25 мин. Затем производили трехкратную отмывку спленоцитов от мета-хлорбензгидрилмочевины в физиологическом растворе и вводили полученные клетки внутривенно сингенным длительно алкоголизированным мышам-реципиентам. Алкогольная мотивация длительно алкоголизированных мышей-реципиентов оценивалась по потреблению 10% раствора этанола в условиях свободного выбора с водой в сравнении с длительно алкоголизированными и интактыми мышами соответствующего возраста (результаты представлены на рис. 3).
На рис. 3 показано накопительное потребление этанола длительно алкоголизированными мышами после внутривенного введения сингенных спленоцитов с функциональной активностью, модулированной in vitro мета-хлорбензгидрилмочевиной. Представлено среднесуточное потребление этанола (мл/мышь), начиная с 1-го дня после введения спленоцитов в условиях свободного выбора 10% раствора этанола с водой (двух-бутылочный тест). Как видно из рис. 3, внутривенное введение сингенных спленоцитов с функциональной активностью, модулированной in vitro мета-хлорбензгидрилмочевиной, снижает потребление этанола в группе длительно алкоголизированных мышей-реципиентов и, следовательно, снижает алкогольную мотивацию.
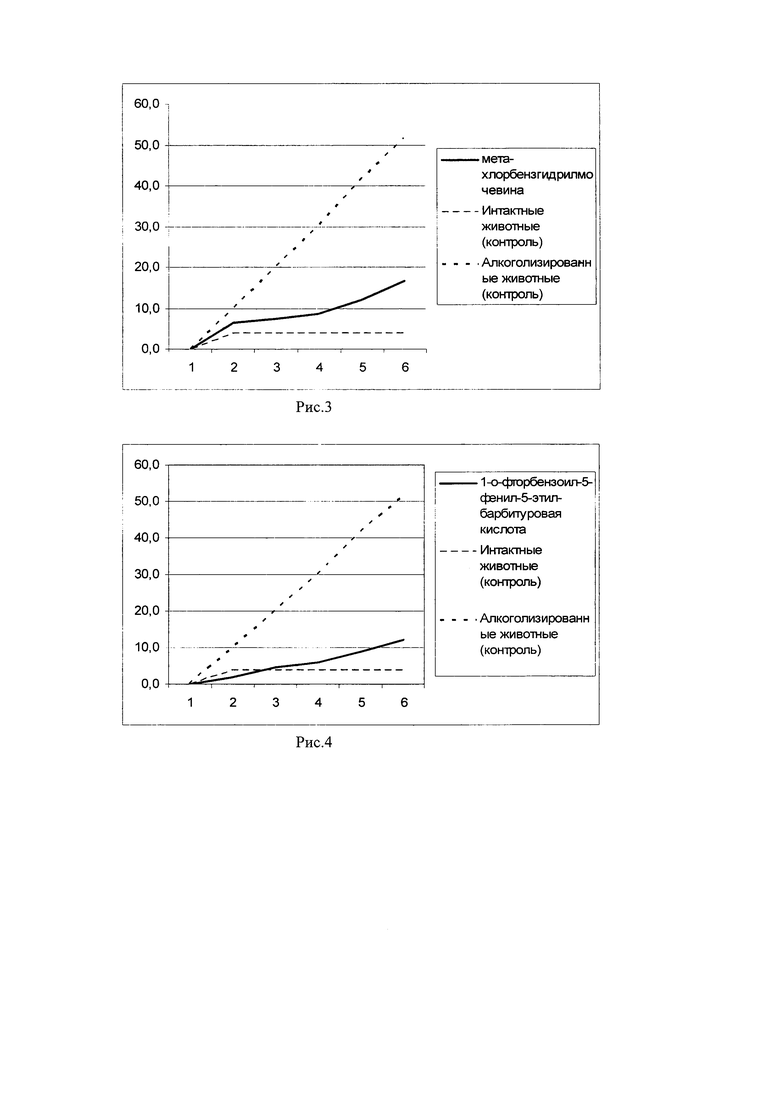
Пример 4. У длительно алкоголизированных мышей (мыши-гибриды (СВА х C57BI/6)F1) производили забор спленоцитов. После чего выделенные клетки подвергали in vitro инкубации с 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты, в концентрации 10 мкг/мл на 15×106 клеток на мышь в течение 10 минут. Затем производили трехкратную отмывку спленоцитов от 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты в физиологическом растворе и вводили полученные клетки в кровоток сингенным длительно алкоголизированным мышам-реципиентам. Алкогольная мотивация длительно алкоголизированных мышей-реципиентов оценивалась по потреблению 10% раствора этанола в условиях свободного выбора с водой в сравнении с длительно алкоголизированными и интактыми мышами соответствующего возраста (результаты представлены на рис. 4).
На рис. 4 показано накопительное потребление этанола длительно алкоголизированными мышами после внутривенного введения сингенных спленоцитов с функциональной активностью, модулированной in vitro 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислот. Представлено среднесуточное потребление этанола (мл/мышь), начиная с 1-го дня после введения спленоцитов в условиях свободного выбора. Как видно из рис. 4, внутривенное введение сингенных спленоцитов с функциональной активностью, модулированной in vitro 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты, снижает потребление этанола в группе длительно алкоголизированных мышей-реципиентов и, следовательно, снижает алкогольную мотивацию.
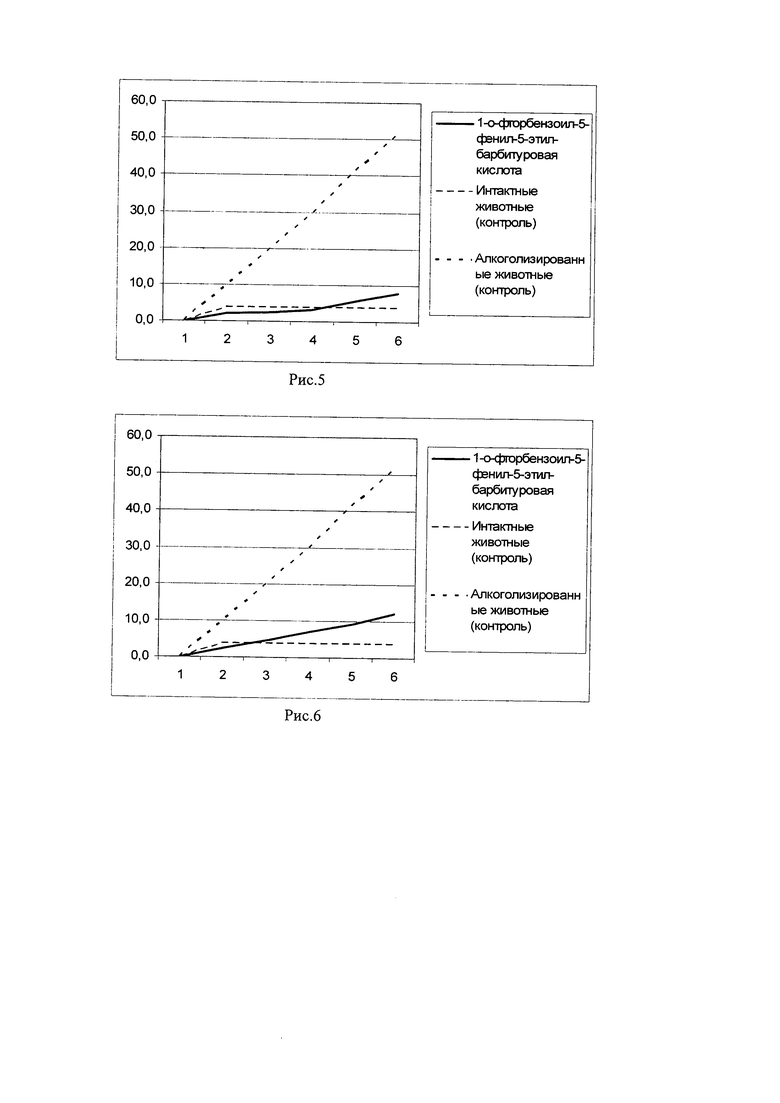
Пример 5. У длительно алкоголизированных мышей (мыши-гибриды (СВА х C57BI/6)F1) производили забор спленоцитов. После чего выделенные клетки подвергали in vitro инкубации с 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты, в концентрации 10 мкг/мл на 15×106 клеток на мышь в течение 18 мин. Затем производили трехкратную отмывку спленоцитов от 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты в физиологическом растворе и вводили полученные клетки в кровоток сингенным длительно алкоголизированным мышам-реципиентам. Алкогольная мотивация длительно алкоголизированных мышей-реципиентов оценивалась по потреблению 10% раствора этанола в условиях свободного выбора с водой в сравнении с длительно алкоголизированными и интактыми мышами соответствующего возраста (результаты представлены на рис. 5).
На рис. 5 показано накопительное потребление этанола длительно алкоголизированными мышами после внутривенного введения сингенных спленоцитов с функциональной активностью, модулированной in vitro 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты. Представлено среднесуточное потребление этанола (мл/мышь), начиная с 1-го дня после введения спленоцитов в условиях свободного выбора 10% раствора этанола с водой (двух-бутылочный тест). Как видно из рис. 5, внутривенное введение сингенных спленоцитов с функциональной активностью, модулированной in vitro 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты, снижает потребление этанола в группе длительно алкоголизированных мышей-реципиентов и, следовательно, снижает алкогольную мотивацию.
Пример 6. У длительно алкоголизированных мышей (мыши-гибриды (СВА х C57BI/6)F1) производили забор спленоцитов. После чего выделенные клетки подвергали in vitro инкубации с 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты, в концентрации 10 мкг/мл на 15×106 клеток на мышь в течение 25 минут. Затем производили трехкратную отмывку спленоцитов от 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты в физиологическом растворе и вводили полученные клетки в кровоток сингенным длительно алкоголизированным мышам-реципиентам. Алкогольная мотивация длительно алкоголизированных мышей-реципиентов оценивалась по потреблению 10% раствора этанола в условиях свободного выбора с водой в сравнении с длительно алкоголизированными и интактыми мышами соответствующего возраста (результаты представлены на рис. 6).
На рис. 6 показано накопительное потребление этанола длительно алкоголизированными мышами после внутривенного введения сингенных спленоцитов с функциональной активностью, модулированной in vitro 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты. Представлено среднесуточное потребление этанола (мл/мышь), начиная с 1-го дня после введения спленоцитов в условиях свободного выбора 10% раствора этанола с водой (двух-бутылочный тест). Как видно из рис. 6, внутривенное введение сингенных спленоцитов с функциональной активностью, модулированной in vitro 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты, снижает потребление этанола в группе длительно алкоголизированных мышей-реципиентов и, следовательно, снижает алкогольную мотивацию.
Стоит отметить, что во всех примерах время инкубации с иммуномодулятором значительно не влияло на потребление этанола длительно алкоголизированными мышами-реципиентами. Однако, при инкубации меньше 10 минут воздействие иммуномодулятора на спленоциты не проявлялось в полной мере и эффект от введения клеток снижался (данные не приводятся). При инкубации больше 25 мин жизнеспособность спленоцитов начинала существенно снижаться, и, соответственно, эффект от введения клеток также снижался (данные не приводятся).
Изобретение позволяет снизить алкогольную мотивацию у мышей за счет восстановления нарушенной длительным токсическим воздействием этанола функциональной активности зрелых периферических клеток иммунной системы (спленоцитов). Изобретение позволяет провести направленное воздействие на функциональную активность периферических зрелых иммунных клеток селезенки (спленоцитов) in vitro мета-хлорбензгидрилмочевиной или 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты, не подвергая негативному избыточному токсическому стрессу органы и системы организма, а затем воздействовать модулированными мета-хлорбензгидрилмочевиной или 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты сингенными спленоцитами после их внутривенного введения на паттерны поведения длительно алкоголизированных мышей, в частности на алкогольную мотивацию.
| название | год | авторы | номер документа |
|---|---|---|---|
| ИММУНОМОДУЛЯТОР | 2018 |
|
RU2691143C1 |
| ИММУНОМОДУЛЯТОР | 2019 |
|
RU2702114C1 |
| ИММУНОМОДУЛЯТОР | 2020 |
|
RU2747986C1 |
| СПОСОБ СТИМУЛЯЦИИ НЕЙРОГЕНЕЗА В ГИППОКАМПЕ | 2017 |
|
RU2675111C2 |
| Способ индукции иммунологической толерантности на трансплантационные антигены у млекопитающих | 2018 |
|
RU2717011C1 |
| Способ регенерации клеток головного мозга с восстановлением его функций | 2023 |
|
RU2816789C1 |
| СРЕДСТВО, СНИЖАЮЩЕЕ АЛКОГОЛЬНУЮ МОТИВАЦИЮ | 1999 |
|
RU2155034C1 |
| ВАКЦИНА НА ОСНОВЕ В-КЛЕТОК, НАГРУЖЕННЫХ ЛИГАНДОМ Т-КЛЕТОК-ПРИРОДНЫХ КИЛЛЕРОВ И АНТИГЕНОМ | 2006 |
|
RU2420311C2 |
| СПОСОБ ПОДАВЛЕНИЯ ИММУННЫХ РЕАКЦИЙ | 2008 |
|
RU2393481C1 |
| СПОСОБ ПОЛУЧЕНИЯ ПОЛЯРИЗОВАННЫХ ЛИМФОЦИТОВ ДЛЯ МОДЕЛИРОВАНИЯ Th2-ИНДУЦИРОВАННОГО ОТЕКА | 2006 |
|
RU2318525C1 |
Изобретение относится к медицине. Предложен способ снижения алкогольной мотивации у мышей, включающий забор спленоцитов у алкоголизированных мышей, их инкубацию in vitro в течение 10-25 мин с иммуномодулятором мета-хлорбензгидрилмочевины либо 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты в действующей концентрации 10 мкг/мл, последующую отмывку спленоцитов и внутривенное введение полученных спленоцитов сингенным алкоголизированным мышам в количестве 15×106 клеток на мышь. Изобретение обеспечивает восстановление функциональной активности спленоцитов, измененной вследствие длительного токсического воздействия этанола и может быть использовано с целью купирования патологических симптомов при алкогольной зависимости. 6 пр., 6 ил.
Способ снижения алкогольной мотивации у мышей, включающий забор спленоцитов у алкоголизированных мышей, их инкубацию in vitro с помощью иммуномодулятора, последующую отмывку спленоцитов и внутривенное введение полученных спленоцитов сингенным алкоголизированным мышам, отличающийся тем, что при проведении инкубации спленоцитов в течение 10-25 мин в качестве иммуномодулятора используют мета-хлорбензгидрилмочевину либо 1-о-фторбензоил-5-фенил-5-этил-барбитуровой кислоты в действующей концентрации 10 мкг/мл на 15×106 клеток на мышь.
| ДАВЫДОВА Т.В | |||
| и др | |||
| "Подавление антителами к серотонину усиленного потребления алкоголя, вызванного адаптивным переносом спленоцитов от животных с абстинентным синдромом"//Бюллетень экспериментальной биологии и медицины, 1998, т.26, N 9, с.328-330 | |||
| МАРКОВА Е.В | |||
| и др | |||
| "Влияние оригинального антиконвульсанта на показатели функциональной активности |

