ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к выделенным пептидам, их вариантам и фрагментам, которые специфически связывают тромбин, который вызывает коагулопатии у человека. Также предусматривается использование таких пептидов в методах диагностики и лечения заболеваний, связанных с тромбозом, и для покрытия медицинских изделий.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Гемостаз представляет собой физиологический процесс, который минимизирует экстравазацию крови после повреждения сосудов. Коагуляция крови, часть этого процесса, включает активацию циркулирующих зимогенов ограниченным протеолизом последовательным образом и завершается образованием фибринового сгустка. Тромбин (FIIa) играет ключевую роль в гемостазе (Stubbs M. T. and Bode W. Throm Res 69, 1-58 (1993)). В своей прогемостатической роли: (а) тромбин расщепляет растворимый фибриноген до мономеров фибрина, которые полимеризуются с образованием зарождающегося сгустка (Versteeg H. H., et al., Physiol Rev 93, 327–358 (2013)); (b) активирует трансглутаминазу (FXIII), которая ковалентно сшивает мономеры фибрина для стабилизации сгустка; (c) активирует неферментативные кофакторы (FV и FVIII), необходимые для его собственного усиления; (d) активирует FXI, который, в свою очередь, активирует собственный путь (Versteeg H. H., et al., Physiol Rev 93, 327–358 (2013)); и (e) активирует тромбоциты путем расщепления рецепторов, активированных протеазой, что приводит к изменению их формы, дегрануляции и агрегации (Monroe D. M., et al., Arterioscler Thromb Vasc Biol 22, 1381-1389 (2002)). В то же время тромбин также играет важную роль в качестве антикоагулянта, снижая прогрессирование и усиление процесса коагуляции крови: при связывании с тромбомодулином он активирует белок С, который, в свою очередь, инактивирует как кофакторы FVa, так и FVIIIa для смягчения дальнейшей генерации тромбина (Di Cera E. Mol Aspects Med 29(4), 203–254 (2008)). Эти парадоксальные прокоагулянтные и антикоагулянтные роли тромбина поддерживают баланс между неконтролируемым кровотечением и образованием обструктивных тромбов с достаточным образованием тромба, когда это необходимо.
Сердечно-сосудистые заболевания являются первой причиной смертности во всем мире и являются мощным источником бремени неинфекционных заболеваний (Chaudhari K., et al., Nat. Rev. Drug. Discov 13, 571-572 (2014)). Ишемическая болезнь сердца и инсульт, оба из которых являются патологическими проявлениями тромбоза, являются наиболее распространенными примерами сердечно-сосудистых заболеваний и являются причиной вплоть до одной из четырех смертей во всем мире (Raskob G. Thromb Haem 112(5), 843-943 (2014)). Антикоагулянты, такие как прямые ингибиторы тромбина (DTI), прямые ингибиторы фактора Xa (FXa) и антагонисты витамина К (VKA), составляют значительную долю существующих терапевтических возможностей антитромботических препаратов. Некоторыми примерами DTI, используемыми в качестве терапевтических вариантов, являются бивалирудин, синтетический аналог гирудина, который является двухвалентным ингибитором, связывающимся с активным центром тромбина и экзосайтом-I; аргатробан и дабигатран, маломолекулярные одновалентные DTI, которые связываются только с активным центром; и гепарины с низкой молекулярной массой (LMWH), которые ингибируют тромбин антитромбинозависимым образом (Michiel Coppens, et al., Circ Res 112, 920-931 (2012)). Несмотря на то, что они являются популярными вариантами антикоагуляционной терапии, эти классы связаны ограничениями, такими как узкое терапевтическое окно, индивидуальное дозирование, высокий риск кровотечений, плохая биологическая доступность и высокие взаимодействия препаратов с пищей (Bauer K. A. Haem 464-470 (2013)). Поэтому ищут новые лучшие антикоагулянты с большими преимуществами.
Животные-гематофаги приспособились к кормовому рациону из крови и разработали ассортимент молекул, которые контролируют гемостаз хозяина, чтобы обеспечить непрерывный кровоток для успешного питания. Среди антикоагулянтов ингибиторы тромбина занимают центральную ступень у этих кровососущих паразитов (Koh C. Y. and Kini R. M. Expert Rev. Haematol 1(2), 135-139 (2008)). Гирудин, гемадин, триабин, орнитодорин и роднин являются одними из наиболее широко изученных примеров конкретных семейств ингибиторов тромбина у животных-гематофагов (Huntington J. A. Thromb Haemost 111, 583–589 (2014)). Мы предварительно выделили и охарактеризовали новый ингибитор тромбина, который мы назвали вариегин (PCT/IB2008/002109), пептид из 32 остатков, который является быстрым, плотно связывающим и конкурентным ингибитором тромбина из экстрактов слюнной железы твердого клеща - Amblyomma variegatum (Koh C. Y., J Biol Chem 282 (40), 29101–29113 (2007)).
Необходимо производить более эффективные пептиды в качестве терапевтических агентов для лечения сердечно-сосудистых и цереброваскулярных заболеваний. Примеры использования включают в себя лечение и профилактику артериального и венозного тромбоза, вызывающего сердечные приступы, инсульты и эмболию; для антикоагуляции при нестабильной стенокардии, коронарной ангиопластике, чрескожном коронарном вмешательстве и операциях на сердце. Кроме того, эти пептиды также могут быть разработаны в качестве реагентов, в качестве противосвертывающих агентов в пробирках для взятия крови и в качестве материалов для поверхностного покрытия для медицинских устройств, таких как стенты, катетеры и трубки для интубации.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение направлено на решение или улучшение описанных выше проблем и получение новых пептидов и их вариантов с улучшенной аффинностью к тромбину. Хотя вновь идентифицированные пептиды «аватрин» и «ультравариегин» демонстрируют ограниченную идентичность последовательности с вариегином, они избирательно ингибируют тромбин в аналогичном быстром конкурентном режиме с жестким связыванием с Ki 545 пМ и 4,4 пМ соответственно. Эти аффинности примерно в 5 и 650 раз выше, соответственно, для мишени (тромбина), чем у аналогичного ингибитора тромбина на основе пептида, используемого в клинике (BivalirudinTM), который страдает от отсутствия четкой эффективности и необходимости непрерывной инфузии. Мы определили их важные функциональные центры с участием трехмерных структур с высоким разрешением и структурно-функциональных соотношений ряда вариантов этих ингибиторов тромбина. Разработали и синтезировали новые варианты этих пептидов с улучшенной аффинностью к тромбину. Мы успешно продемонстрировали эффективность этих пептидов in vivo с помощью модели тромбоза мышиных артерий. Также идентифицированы и изучены другие новые пептиды из иксодовых клещей.
Первый аспект изобретения предусматривает ингибиторы тромбина, содержащие аминокислотную последовательность, выбранную из группы, включающей SGGHQTAVPKISKQGLGGDFEEIPSDEIIE (SEQ ID NO: 1), ее вариант или фрагмент, SDEAVRAIPKMYSTAPPGDFEEIPDDAIEE (SEQ ID NO: 2), ее вариант или фрагмент, SEQ ID NO: ID NO: 22, ее вариант или фрагмент, SEQ ID NO: 23, ее вариант или его фрагмент, SEQ ID NO: 24, ее вариант или фрагмент и SEQ ID NO: 25, ее вариант или фрагмент.
В предпочтительном варианте осуществления аминокислотную последовательность выбирают из группы, включающей:
QTAVPKISKQGLGGDFEEIPSDEIIE (SEQ ID NO: 3);
ISKQGLGGDFEEIPSDEIIE (SEQ ID NO: 4);
SGGHQTAVPKIAKQGLGGDFEEIPSDEIIE (SEQ ID NO: 5);
SGGHQTAVPKIHKQGLGGDFEEIPSDEIIE (SEQ ID NO: 6);
SGGHQTAVPRISKQGLGGDFEEIPSDEIIE (SEQ ID NO: 7);
SGGHQTAVPXISKQGLGGDFEEIPSDEIIE (SEQ ID NO: 8), где X представляет собой β-гомоаргинин;
SDEAVRAIPXMYSTAPPGDFEEIPDDAIEE (SEQ ID NO: 9), где X представляет собой β-гомоаргинин;
SDQGDVAIPKMYSTAPPGDFEEIPDDAIEE (SEQ ID NO: 10);
SDEAVRAEPKMHKTAPPGDFEEIPDDAIEE (SEQ ID NO: 11);
SDEAVRAIPKMYSTAPPGDFEEIPEEYLDDES (SEQ ID NO: 12);
MYSTAPPGDFEEIPDDAIEE (SEQ ID NO: 13);
SDEAVRAIPKMYSTAPPGDFEEIPDDEIEE (SEQ ID NO: 14);
SDEAVRAIPKMYSQAPPGDFEEIPDDAIEE (SEQ ID NO: 15);
SDQGDVAEPKMYSTAPPFDFEAIPEEYLDDES (SEQ ID NO: 16);
SDQGDVAEPXMHSTAPPFDFEAIPEEYLDDES (SEQ ID NO: 17) вариант SEQ ID NO: 2, где X представляет собой β-гомоаргинин;
CDEAVRAIPKMYSTAPPGDFEEIPDDAIEE (SEQ ID NO: 18) вариант SEQ ID NO: 2;
SDEAVRAIPKMYSTAPPGDFEEIPDDAIEECA (SEQ ID NO: 19) вариант SEQ ID NO: 2;
MYSTAPPGDFEEIPDDAIEEGCCC (SEQ ID NO: 20) вариант SEQ ID NO: 2;
SDEAVRAIPKMYSTAPPGDFEEIPDDAIEEGCCC (SEQ ID NO: 21) вариант SEQ ID NO: 2;
SGEDHTAVPKMSRKGLGGDFEDIPPEAYERALEAR (SEQ ID NO: 22);
ELESGDEDSEGGDSQSSPTESAAPRLHQREGGGGDFENVEYDQDQK (SEQ ID NO: 23);
SDVAPADYESDEGDNDGGHDGSEVAKPKMPRGNGGGGDFEEIPEVE (SEQ ID NO: 24); и
TGSDDDDEYDMYESDGDSNEGNDNDEFETAVPRLPNPNSGRDSEHIPMPVN (SEQ ID NO: 25).
Другим аспектом изобретения является выделенный ингибитор тромбина, содержащий аминокислотную последовательность в соответствии с любым аспектом изобретения для профилактики или лечения заболевания, связанного с тромбом.
Другим аспектом изобретения является способ ингибирования активности тромбина, где способ включает контактирование тромбина с по меньшей мере одним ингибитором тромбина в соответствии с любым аспектом изобретения.
В соответствии с другим аспектом изобретения предложено применение ингибитора тромбина в соответствии с любым аспектом изобретения для приготовления лекарственного средства для профилактики и/или лечения заболевания, связанного с тромбом.
В соответствии с другим аспектом изобретения предложен способ профилактики и/или лечения заболевания, связанного с тромбом, включающий введение субъекту, нуждающемуся в такой профилактике и/или обработке, эффективного количества ингибитора тромбина в соответствии с любым аспектом настоящего изобретения.
В соответствии с другим аспектом изобретения предложен способ обнаружения накопления тромбина у субъекта, включающий введение по меньшей мере одного ингибитора в соответствии с любым аспектом изобретения субъекту или в образец ткани, выделенный у субъекта, и обнаружение присутствия указанного по меньшей мере одного ингибитора тромбина, связанного с тромбином.
В соответствии с другим аспектом изобретения предложена фармацевтическая композиция, содержащая эффективное количество по меньшей мере одного ингибитора тромбина в соответствии с любым аспектом изобретения.
В соответствии с другим аспектом изобретения предложена выделенная молекула нуклеиновой кислоты, кодирующая ингибитор тромбина в соответствии с любым аспектом изобретения. В предпочтительном варианте осуществления последовательность нуклеиновой кислоты кодирует SEQ ID NO: 1 и представлена формулой
TCGGGTGGCCATCAGACTGCTGTTCCGAAGATATCTAAGCAAGGCTTGGGGGAGACTTTGAAGAAATTCCAAGTGATGAAATAATCGAG (SEQ ID NO: 26).
В соответствии с другим аспектом изобретения предложен набор, содержащий по меньшей мере один ингибитор тромбина, как здесь определено.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг. 1 показана локализация мРНК, кодирующей белковый белок-предшественник BAD29729 подобный вариегину в слюнных железах Amblyomma variegatum. Гибридизация in situ по всему объему выявила экспрессию BAD29729 в ацинусах II типа, расположенных вблизи основного слюнного канала, но не в других типах ацинусов (ацинусы III типа обозначены звездочками). Стрелки указывают центры экспрессии. А-С. Слюнные железы самки через 5 дней питания (A) и самца через 12 дней питания (B) демонстрируют сильную экспрессию BAD29729 в 2-4 больших клетках (C). D-H. Экспрессия BAD29729 у нимф в процессе питания кровью. Некормленные (D) нимфы, нимфы через 2 дня питания (E) и через 4 дня питания (F), демонстрирующие окрашивание в базальных участках ацинусов II типа. Это окрашивание отсутствовало у особей через 6 дней питания (G) и было очень слабым у полностью напитавшихся кровью и отсоединившихся (H) нимф. I. Закрый ацинус II типа у нимф через 2 дня питания, демонстрирующий окрашивание в двух базальных клетках. Масштабная черта 100 мкм (A-B), 50 мкм (D-H) и 25 мкм (C, I). J. Выравненные последовательности вариегина и аватрина (активный ингибирующий тромбин пептид из белка-предшественника).
На Фиг. 2 показан синтез и очистка аватрина. Аватрин синтезировали с использованием твердофазного пептидного синтеза и очищали с использованием хроматографии с обращенной фазой. A. Очистка аватрина на колонке с обращенной фазой Jupiter Proteo (5 мкм, 250 мм × 10 мм, 90 Å) с использованием градиента ацетонитрила от 10 % растворителя B до 70 % растворителя B (Растворитель A: 0 % ацетонитрила, 99,9 % воды качества Milli-Q и 0,1 % трифторуксусной кислоты, растворитель B: 0 % ацетонитрила, 99,9 % воды качества Milli-Q и 0,1 % трифторуксусной кислоты). B. Дальние УФ-спектры CD (260-190 нм) аватрина и его усеченного варианта - IS20, растворенного в 10 мМ натрий-фосфатном буфере (рН 7,4). Оба спектра были типичными для случайной спирали. C. Чистоту и массу аватрина определяли с помощью масс-спектрометрии с электрораспылением. Аватрин ионизировался в трех разных состояниях заряда и показывал значения m/z 785,84 а.е.м., 1047,55 а.е.м. и 1570,58 а.е.м. соответствующие +4, +3 и +2, соответственно. D. Деконволюционный масс-спектр аватрина показал, что наблюдаемая масса аватрина соответствует ожидаемой массе 3139,65 ± 0,83 Да (табл. 1).
Фиг. 3 показывает ингибирующую активность тромбина аватрина. A. Влияние различных концентраций аватрина на амидолитическую активность тромбина (0,81 нМ) измеряли с использованием S2238 (100 мкМ) с (10 мин) или без предварительной инкубации. Аватрин показал ингибирование амидолитической активности тромбина дозозависимым образом. IC50 и коэффициент Хилла ингибирования составляют 6,95 ± 0,42 нМ и 0,92 ± 0,01 при 0 мин; и 4,86 ± 0,36 нМ и 0,94 ± 0,02 нМ при 10 мин соответственно. Каждая точка данных представляет собой среднее значение ± стандартное отклонение (S.D.) по меньшей мере трех экспериментов. B. Проводили амидолитический анализ тромбина (0,81 нМ) с использованием S2238 (100 мкМ) в присутствии различных концентраций аватрина и получали линейные кривые прогрессирования ингибирования тромбина в присутствии аватрина - характеристика ингибитора быстрого связывания. C. Остаточную тромбиновую амидолитическую активность при наличии различных концентраций аватрина измеряли при различных концентрациях S2238 и определяли Ki’ (видимая Ki). Реакции начинались с добавления тромбина (0,81 нМ). Данные были привязаны к уравнению жесткой связи по Morrison с использованием программного обеспечения GraphPad Prizm. Каждая точка данных представляет собой среднее значение ± S.D. по меньшей мере трех экспериментов. D. График концентрации Ki’ по отношению к S2238 увеличивался линейно, что указывает на то, что аватрин является конкурентным ингибитором. Ингибирующую константу Ki определили как равную 545,3 ± 3,1 пМ.
На Фиг. 4 показано ингибирование фибриногенолитической активности тромбина, в результате чего аватрин, IS20, GL16 и ультравариегин удлиняют время свертывания фибриногена дозозависимым образом.
На Фиг. 5 показана селективность сериновой протеазы аватрина. Аватрин подвергали скринингу на селективность по отношению к 13 сериновым протеазам, включая прокоагулянтные, антикоагулянтные, фибринолитические и классические сериновые протеазы (тромбин, трипсин, fIXa, fXIa, fXa, химотрипсин, tPA, fVIIa, плазмин, APC, калликреин, урокиназа и fXIIa). Конечные концентрации протеаз и субстратов, используемых для амидолитических анализов, приведены в скобках в нM и мкМ соответственно, если не указано иное: α-тромбин/S2238 (0,81/100), трипсин/S2222 (0,87/100), fIXa // Spectrozyme® fIXa (333/0,4), fXIa/S2366 (0,125/1000), fXa/S2765 (0,24/650), химотрипсин/S2586 (1,2/0,67), tPA/S2288 (36,9/1000), fVIIa/S2288 (460/1200), плазмин/S2251 (3,61/1200), APC/S2366 (2,74/600), калликреин/S2302 (0,93/1100), урокиназа/S2444 (32 ед/мл/650), fXIIa/S2302 (20/1000). Активность тромбина тестировали при более низких концентрациях аватрина (1000 нМ, 100 нМ и 10 нМ), в то время как другие протеазы тестировали при гораздо более высоких концентрациях аватрина (100 мкМ, 10 мкМ и 1 мкМ). Каждая точка данных представляет собой среднее значение ± S.D. как минимум трех независимых экспериментов.
На Фиг. 6 показано расщепление аватрина тромбином. A. Оценка расщепления аватрина тромбином с помощью ВЭЖХ-хроматограмм. Аватрин (150 мкМ) инкубировали с тромбином (5 мкМ) в разное время и реакционные смеси разделяли с использованием высоко эффективной жидкостной хроматографии с обращенной фазой (ОФ-ВЭЖХ); массы продуктов расщепления анализировали с помощью масс-спектрометрии с электрораспылением. Через 0 мин (верхняя панель) был идентифицирован один пик, соответствующий полной длине аватрина (масса 3139 Да). Через 120 мин (средняя панель) в дополнение к пику аватрина были идентифицированы два новых пика, соответствующие N-концевому продукту расщепления (SGGHQTAVPK, масса 982 Да) и C-концевому продукту расщепления (ISKQGGGGDFEEIPSDEIIE, масса 2176 Да). Через 600 мин (нижняя панель) наблюдали два пика, соответствующие N- и С-терминальным продуктам расщепления, в то время как пик аватрина не наблюдали, что указывало на полное расщепление. B. Зависимое от времени расщепление аватрина тромбином. Относительные количества в процентах аватрина, его N- и С-концевых продуктов расщепления определяли путем расчета площадей под кривой. Каждая точка данных представляет собой среднее значение ± S.D. по меньшей мере трех независимых экспериментов. C. Влияние расщепления тромбина на ингибирующие свойства аватрина. Аватрин инкубировали с тромбином (0,81 нМ) в течение 36 часов и анализировали в разные моменты времени его способность ингибировать амидолитическую активность тромбина на хромогенном субстрате S2238. При 25 нМ аватрина ингибитор присутствовал при приблизительно 30-кратном избытке тромбина (0,81 нм), и эти соотношения были аналогичны тем, которые использовали при ВЭЖХ-анализе продуктов расщепления. Через 24 часа продукты расщепления сохраняли более 50 % от первоначальной ингибирующей активности, хотя полный аватрин был полностью расщеплен через примерно 10 ч. Таким образом, продукты расщепления, в частности С-концевой продукт расщепления аватрина, по-видимому, остаются связанными с тромбином и продолжают ингибировать тромбин. Каждая точка данных представляет собой среднее значение ± S.D. по меньшей мере трех независимых экспериментов.
На Фиг. 7 показаны эффекты влияния усеченных мутантов аватрина на амидолитическую активность тромбина. A. Измеряли влияние QT26 и IS20 на амидолитическую активность тромбина (0,81 нМ). Оба пептида ингибируют тромбин дозозависимым образом. IC50 и коэффициент Хилла пептида QT26 составляли 8,94 ± 0,64 нМ и 0,88 ± 0,03 при 0 мин; и 13,54 ± 0,81 и 0,92 ± 0,03 при 10 мин соответственно. IC50 и коэффициент Хилла пептида IS20 составляли 12,38 ± 0,32 нМ и 0,86 ± 0,02 при 0 мин; и 22,70 ± 0,94 нМ и 0,87 ± 0,01 при 10 мин соответственно. Каждая точка данных представляет собой среднее значение ± S.D. по меньшей мере трех независимых экспериментов. B. GL16 (3, 10, 30, 100 и 300 мкМ) неспособны ингибировать амидолитическую активность тромбина; вместо этого при высоких концентрациях они несколько усиливали гидролиз S2238 тромбином. Каждая точка данных представляет собой среднее значение ± S.D. по меньшей мере трех независимых экспериментов. C и D. Измеряли остаточную амидолитическую активность тромбина в присутствии QT26 и IS20 и определяли Ki’ (видимую Ki). Оба QT26 и IS20 были ингибиторами сильного связывания. Каждая точка данных представляет собой среднее значение ± S.D. по меньшей мере трех экспериментов. E. График концентрации Ki’ по отношению к S2238 увеличивался линейно, что указывает на то, что QT26 является конкурентным ингибитором. Установлено, что ингибирующая константа Ki составляет 760,32 ± 0,91 пМ. F. График концентрации Ki’ по отношению к S2238 уменьшался криволинейно, что указывает на то, что IS20 является неконкурентным ингибитором (с α менее 1). Установлено, что ингибирующая константа Ki составляет 5760 ± 230 пМ.
На Фиг. 8 показано влияние вариантов аватрина на амидолитическую активность тромбина по сравнению с той же активностью аватрина. Все варианты ингибируют активность тромбина дозозависимым образом. A. IC50 и коэффициент Хилла S12A составляли 101,20 ± 1,32 нМ и 0,62 ± 0,01 соответственно. IC50 S12A был в 20 раз ниже, чем аватрина, что подтверждает важность Ser12 в аватрине для ингибирования тромбина. IC50 и коэффициент Хилла S12H составляли 18,51 ± 0,32 нМ и 0,88 ± 0,02 соответственно. Каждая точка данных представляет собой среднее значение ± S.D. по меньшей мере трех экспериментов. B. IC50 и коэффициент Хилла L16P, G17P составляли 181,32 ± 3,76 нМ и 0,54 ± 0,02 соответственно. IC50 и коэффициент Хилла G2D, Q5D составляли 12,98 ± 1,23 нМ и 0,71 ± 0,03 соответственно. Каждая точка данных представляет собой среднее значение ± S.D. по меньшей мере трех независимых экспериментов. C. IC50 и коэффициент Хилла β-аватрина составляли 332,16 ± 1,32 нМ и 0,62 ± 0,01 соответственно. Хотя β-аватрин не расщепляли до 72 ч, имелось сильное снижение его активности. Каждая точка данных представляет собой среднее значение ± S.D. по меньшей мере трех независимых экспериментов. D. IC50 и коэффициент Хилла K10R составляли 1,15 ± 0,45 нМ и 1,10 ± 0,01 соответственно. K10R имел 5-кратный прирост активности, но был расщеплен со скоростью намного быстрее, чем аватрин (3 часа). Каждая точка данных представляет собой среднее значение ± S.D. по меньшей мере трех независимых экспериментов. E. IC50 и коэффициент Хилла β-вариегина составляли 117,90 ± 1,16 нМ и 0,93 ± 0,05 соответственно. Каждая точка данных представляет собой среднее значение ± S.D. по меньшей мере трех независимых экспериментов.
На Фиг. 9 показаны ВЭЖХ-хроматограммы вариантов расщепления аватрина K10R (A) и β-аватрина (B) тромбином. Пептиды инкубировали с тромбином в разное время и реакционные смеси разделяли с использованием ОФ-ВЭЖХ, и массы продуктов расщепления анализировали с использованием масс-спектрометрии с электрораспылением. В то время как K10R-аватрин полностью расщепляется через 3 часа (со скоростью намного быстрее, чем аватрин), β-аватрин не был расщеплен даже через 72 часа.
На Фиг. 10 показаны параметры кинетики ингибирования вариантов аватрина. Остаточные амидолитические активности тромбина в присутствии A. S12H, B. G2D, Q5D и C. K10R измеряли при различных концентрациях S2238 и определяли Ki’ (видимая Ki). Реакции начинались с добавления тромбина (0,81 нМ). S12H, G2D, Q5D и K10R были ингибиторами сильного связывания тромбина с величинами Ki 1,23 ± 0,04 нМ, 0,93 ± 0,01 нМ и 0,17 ± 0,00 нМ соответственно. Каждая точка данных представляет собой среднее значение ± S.D. по меньшей мере трех независимых экспериментов.
На Фиг. 11 показаны графики Лайнуивера-Берка ингибирования тромбина β-аватрином и S12A. Остаточную амидолитическую активность тромбина в присутствии β-аватрина (300 нМ, А) и S12A (30 нМ, В) измеряли при различных концентрациях S2238 (сплошная линия - без ингибитора, пунктирная линия - с ингибитором). И β-аватрин, и S12A не являются ингибиторами сильного связывания тромбина, поскольку они не способны ингибировать тромбин при эквимолярных концентрациях. Двойные взаимно обратные графики показывают β-аватрин и S12A имел значения Ki 32,04 ± 0,36 нМ и 6,07 ± 0,18 нМ соответственно. Каждая точка данных представляет собой среднее значение ± S.D. по меньшей мере трех независимых экспериментов.
На Фиг. 12 показана модель кристаллической структуры комплекса аватрин-тромбин. A. Обзор комплекса тромбин-аватрин. Аватрин показан как модель из палочек (углерод-желтый, азот-синий, кислород-красный), а также поверхностная презентация тромбина показана светло-голубым. Шесть остатков A(5QTAVPK10) и 10 остатков A(19DFEEIPSDEI28) были идентифицированы в непосредственной близости от активного центра тромбина и в углублении экзосайта-1. B. По аналогии с (A) показаны карты электронной плотности, рассчитанные для аватрина. Карта 2Fo-Fc (серый, очерченный на уровне 1,0 сигма) и карта Fo-FC (зеленый для позитивной карты, очерченной на 3,0 сигма, красный для негативной карты, очерченной на 3,0 сигма). C. Остатки тромбина (светло-голубая линия) в радиусе 4,5 Å N-концевого пептида аватрина от неустойчивой химической связи (желтые палочки) показаны так, что изображают взаимодействие между ними. D. Тромбиновые остатки (светло-голубая линия) в радиусе 4,5 Å пептида аватрина (желтые палочки), связанного с экзозитом-I, показаны так, что изображают взаимодействие между ними. E. По аналогии с (C) показаны карты электронной плотности, рассчитанные для аватрина. Карта 2Fo-Fc (серый, очерченный на уровне 1,0 сигма) и карта Fo-FC (зеленый для позитивной карты, очерченный на 3,0 сигма, красный для негативной карты, очерченный на 3,0 сигма). F. По аналогии с (D) показаны карты электронной плотности, рассчитанные для аватрина. Карта 2Fo-Fc (серый, очерченный на уровне 1,0 сигма) и карта Fo-FC (зеленый для позитивной карты, очерченный на 3,0 сигма, красный для негативной карты, очерченный на 3,0 сигма).
На Фиг. 13 показано ингибирование связанного сгустком тромбина с помощью аватрина. Эффект влияния аватрина на амидолитическую активность связанного сгустком тромбина (0,81 нМ) измеряли с использованием S2238 (100 мкМ). Аватрин ингибирует связанный сгустком тромбин дозозависимым образом с IC50, составляющим 1,74 ± 0,35 мкМ.
На Фиг. 14 показано влияние аватрина на время окклюзии в мышиной модели тромбоза сонной артерии, вызванной хлоридом железа, по сравнению с гирулог-1. Две дозы (3 мг/кг и 10 мг/кг) аватрина или гирулога-1, растворенные в 100 мкл физиологического раствора (0,9 %), вводили внутривенно через хвостовую вену мышей (n = 6). Тромбоз индуцировали путем помещения фильтровальной бумаги (2 мм × 2 мм), насыщенной 10 % FeCl3 на сонную артерию, и времени на окклюзию (ТТО) после измерения FeCl3-тромбоза. Контрольным животным вводили внутривенно или перитонеально 100 мкл физиологического раствора (n равно 6). От 7,24 ± 1,46 мин у контрольных животных ТТО увеличивалось до 15,03 ± 3,23 мин и 22,51 ± 4,19 мин у животных, которым вводили соответственно 3 мг/кг и 10 мг/кг аватрина. У мышей, которым вводили 3 мг/кг и 10 мг/кг гирулога-1, ТТО составляли 9,76 ± 3,15 мин и 15,23 ± 3,39 мин соответственно. Значительные различия между двумя группами были рассчитаны с использованием t-теста (* P менее 0,001). Аватрин демонстрирует лучшую эффективность по сравнению с гирулогом-1 в этой модели.
На Фиг. 15 показано ингибирование тромбинового амидолитического анализа пептидами семейства вариегина. Выбранные пептиды в разных концентрациях (0,0003-3000 нМ) испытывали на их способность ингибировать тромбин. Ультравариегин, пептид, идентифицированный из сиаломы Amblyomma variegatum, был наиболее мощным пептидом с IC50, составляющим 0,26 ± 0,008 нМ. Все другие пептиды имели IC50 в диапазоне от 14 до 130 нМ.
На Фиг. 16 показано ингибирование тромбина вариантами ультравариегина UV003 и UV004. А и С. Различные концентрации (от 0,03 нМ до 100 нМ) испытывали на их способность ингибировать амидолитическую активность тромбина и сравнивали с ультравариегином. UV003 и UV004 ингибировали тромбин с значениями IC50 0,60 ± 0,20 нМ и 0,46 ± 0,08 нМ соответственно. B и D. Остаточные тромбиновые амидолитические активности в присутствии UV003 и UV004 измеряли при различных концентрациях S2238 и определяли их Ki. Было обнаружено, что Ki UV003 и UV004 составляют 4,21 ± 0,96 пМ и 4,55 ± 0,37 пМ соответственно.
На Фиг. 17 показано ингибирование тромбина по вариантам ультравариегина UV005 и UV011. A и C. Различные концентрации (от 0,03 нМ до 100 нМ) тестировали на их способность ингибировать амидолитическую активность тромбина, сравнивали с ультравариегином. UV005 и UV011 ингибировали тромбин с значениями IC50 0,91 ± 0,47 пМ и 1,66 ± 0,77 нМ. B и D. Остаточные тромбиновые амидолитические активности в присутствии UV005 и UV011 измеряли при различных концентрациях S2238 и определяли их Ki. Было установлено, что Ki UV005 и UV011 составляют 16,0 ± 3,05 и 1387 ± 230 пМ.
На Фиг. 18 показано ингибирование тромбина вариантами UV012, UV013, Вариегин YS, UV007, UV008, UV014 и UV015. A, C и E. Различные концентрации (от 0,003 нМ до 3000 нМ) испытывали на их способность ингибировать амидолитическую активность тромбина, сравнивали с ультравариегином. Значения IC50 различных вариантов приведены в табл. 5. B и D. Остаточные амидолитические активности тромбина в присутствии UV012, UV013, Вариегин YS, UV007, UV008, UV014 и UV015 для определения их значений Ki путем подбора данных в уравнениях ингибиторов сильного связывания. Значения Ki соответствующих вариантов приведены в табл. 4.
На Фиг. 19 показана ингибирующая константа Ki ультравариегина. Ki ультравариегина, наиболее мощного члена семейства вариегинов, показана в качестве представителя. Ультравариегин является сильно связывающим ингибитором тромбина. Различные концентрации ультравариегина смешивали с различными концентрациями S2238 (50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ, 250 мкМ, 300 мкМ, 350 мкМ и 400 мкМ и определяли Ki’. Реакции начинались с добавлением тромбина (0,81 нМ). Данные были установлены на уравнение Моррисона с использованием программного обеспечения GraphPad prizm (n равно 3, значения погрешности представляют собой S.D.). (B) График Ki’ по отношению к концентрации субстрата увеличивался линейно, что указывает на то, что ультравариегин конкурентно ингибировал амидолитическую активность тромбина и константу ингибирования Ki определяли как 4,4 ± 0,35 пМ (значения погрешности представляют собой S.D.).
На Фиг. 20 показано расщепление ультравариегина тромбином. Анализ временного курса ультравариегина, инкубированного с тромбином, показал, что ультравариегин действительно расщепляется тромбином. Отделение ОФ-ВЭЖХ реакционной смеси, инкубированной в разные моменты времени, проводили для разделения и количественного определения продуктов расщепления. Количественное определение продуктов расщепления ультравариегина проводили путем пиковой интеграции и расчета площади под кривой. С увеличением времени инкубации с тромбином уменьшалось количество ультравариегина полной длины и наблюдали количество продуктов расщепления, соответствующее расщеплению связи Lys10-Met11. Через 18 ч ультравариегин был полностью расщеплен и наблюдались только пики, соответствующие продуктам расщепления.
На Фиг. 21 показана селективность сериновой протеазы ультравариегина. Ультравариегин анализировали по отношению к 13 сериновым протеазам: фибринолитические сериновые протеазы (плазмин и ТРА), антикоагулянтная сериновая протеаза (урокиназа), прокоагулянтные сериновые протеазы (FXIIa, FXIa, FXa, FIXa, FVIIa, калликреин и тромбин) и классические сериновые протеазы (трипсин и химотрипсин). Тромбин испытывали по отношению к трем концентрациям ультравариегина: 100, 10 и 1 нМ. Для других протеаз использовали гораздо более высокие концентрации аватрина: 100, 10 и 1 мкМ.
На Фиг. 22 показана ингибирующая константа Ki пептида из Rhipicephalus sanguineus. Ki’ пептида Rhipicephalus sanguineus определяли путем смешивания различных концентраций пептида с различными концентрациями S2238 (50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ, 250 мкМ, 300 мкМ, 350 мкМ и 400 мкМ). Реакции начинались с добавления тромбина (0,81 нМ). Данные подставляли в уравнения Моррисона с использованием программного обеспечения GraphPad prizm (n равно 3, значения погрешности представляют собой S.D.). (B) График Ki’ по отношению к концентрации субстрата увеличивался линейно, указывая на то, что пептид конкурентно ингибировал амидолитическую активность тромбина и определяли, что ингибирующая константа Ki составляет 8,79 ± 0,65 нМ (значения погрешности представляют собой S.D.).
На Фиг. 23 показана селективность сериновых протеаз пептида Rhipicephalus sanguineus, проанализированного по отношению к 13 сериновым протеазам: фибринолитические сериновые протеазы (плазмин и ТРА), антикоагулянтная сериновая протеаза (урокиназа), прокоагулянтные сериновые протеазы (FXIIa, FXIa, FXa, FIXa, FVIIa, калликреин и тромбин) и классические сериновые протеазы (трипсин и химотрипсин). Тромбин тестировали на трех концентрациях пептида: 1000, 100 и 10 нМ. Для других протеаз использовали гораздо более высокие концентрации пептида: 100, 10 и 1 мкМ.
На Фиг. 24 показана ингибирующая константа Ki пептидов Amblyomma americanum и Hyalomma marginatum rufipes. Ki’ обоих пептидов определяли путем смешивания различных концентраций пептида с различными концентрациями S2238 (50 мкМ, 100 мкМ, 150 мкМ, 200 мкМ, 250 мкМ, 300 мкМ, 350 мкМ и 400 мкМ). Реакции начинались с добавления тромбина (0,81 нМ). Данные подставляли в уравнения Моррисона с использованием программного обеспечения GraphPad prizm (n равно 3, значения погрешности представляют собой S.D.). (B) График Ki’ по отношению к концентрации субстрата увеличивался линейно, указывая на то, что оба пептида конкурентно ингибируют амидолитическую активность тромбина (значения погрешности представляют собой S.D.). Было определено, что Ki пептидов из Amblyomma americanum и Hyalomma marginatum rufipes составляют 1,63 ± 0,61 нМ и 6,135 ± 0,39 нМ соответственно.
Фиг. 25 показывает селективность сериновых протеаз на пептидах (A) Amblyomma americanum и (B) Hyalomma marginatum rufipes. Оба пептида подвергали анализу по отношению к фибринолитическим сериновым протеазам (плазмин и ТРА), антикоагулянтной сериновой протеазе (урокиназы), прокоагулянтным сериновым протеазам (FXIIa, FXIa, FXa, FIXa, FVIIa, калликреина и тромбина) и классическим сериновым протеазам (трипсин и химотрипсин). Тромбин тестировали по отношению к трем концентрациям пептида: 1000, 100 и 10 нМ. Для других протеаз использовали гораздо более высокие концентрации пептида: 100, 10 и 1 мкМ.
На Фиг. 26 показана ответная реакция на АДФ-индуцированную агрегацию тромбоцитов в крови, под действием различных антикоагуляционных ингибиторов тромбина, гирудина, ультравариегина, аватрина, вариегина и цитрата, измеренная в разные моменты времени после взятия крови.
На Фиг. 27 показаны взаимодействия аватрина и других ингибиторов с экзосайтом тромбина. Поверхностное представление экзосайта тромбина показано светло-голубым; ингибиторы тромбина показаны на моделях из палочек. С-концевые сегменты аватрина (желтый), вариегин (пурпурный) и гирулог-1 (зеленый) перекрываются.
На Фиг. 28 показано выравнивание последовательности аватрина с вариегином и родственными пептидами, идентифицированными из транскриптомов слюнных желез Amblyomma variegatum, Amblyomma americanum и Amblyomma cajannense. Каждый транскрипт кодирует белок-предшественник, который посттрансляционно расщепляется на три-пять зрелых пептидов. Показана только одна репрезентативная пептидная последовательность из каждого транскрипта.
Библиографические ссылки, упомянутые в настоящем описании, для удобства перечислены в виде списка ссылок и добавлены в конце примеров. Все содержание данных библиографических ссылок включено в данную заявку в качестве ссылки.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Определения
Для удобства здесь собраны некоторые термины, используемые в описании, примерах и прилагаемой формуле изобретения.
Термин «содержащий» здесь определяется как то, где различные компоненты, ингредиенты или стадии могут быть совместно использованы при осуществлении настоящего изобретения. Соответственно, термин «содержащий» охватывает более ограничивающие термины «состоящие в основном из» и «состоящие из».
Термин «выделенный» здесь определяется как биологический компонент (такой как нуклеиновая кислота, пептид или белок), который был по существу отделен, получен отдельно или очищен от других биологических компонентов в клетке организма, в котором компонент происходит естественным образом, т. е. другая хромосомная и внехромосомная ДНК и РНК и белки. Таким образом, нуклеиновые кислоты, пептиды и белки, которые были выделены, включают нуклеиновые кислоты и белки, очищенные стандартными методами очистки. Термин также охватывает нуклеиновые кислоты, пептиды и белки, полученные рекомбинантной экспрессией в клетке-хозяине, а также химически синтезированные нуклеиновые кислоты.
Термин «вариант» здесь обозначен как тот, который имеет по меньшей мере одну нуклеотидную последовательность, которая получается из эталонной последовательности путем замещения, делеции или добавления по меньшей мере одной нуклеиновой кислоты, но кодирует аминокислотную последовательность, которая сохраняет способность распознавать, связывать и ингибировать тромбин. Термин «вариант» также относится к аминокислотной последовательности, которая получается из по меньшей мере одной эталонной последовательности путем замещения, делеции или добавления по меньшей мере одной аминокислоты, но сохраняет способность распознавать, связывать и ингибировать тромбин. В частности, варианты могут быть естественными или могут быть рекомбинантными или синтетически произведенными. Более конкретно, вариант может иметь по меньшей мере 30 %, по меньшей мере 40 %, по меньшей мере 50 %, по меньшей мере 60 %, по меньшей мере 70 %, по меньшей мере 80 %, по меньшей мере 90 % или по меньшей мере 95 % последовательности идентичной с эталонными последовательностями. Например, аминокислотную последовательность Аватрин K10R, представленную в SEQ ID NO: 7, создали путем замены аминокислот в аминокислотной последовательности Аватрина, указанной в SEQ ID NO: 1, и ее можно рассматривать как вариант SEQ ID NO: 1.
Термин «вариант» может включать пептидные ингибиторы тромбина, которые имеют «консервативные» изменения, причем замещенная аминокислота имеет сходные структурные или химические свойства (например, замещение лейцина изолейцином). Реже вариант может иметь «неконсервативные» изменения (например, замена глицина триптофаном). Аналогичные незначительные вариации могут также включать делеции или вставки аминокислот, или и то, и другое. Рекомендации по определению, какие аминокислотные остатки могут быть замещены, вставлены или удалены без отмены биологической активности, могут быть найдены с использованием компьютерных программ, хорошо известных в данной области, например, программного обеспечения DNASTAR® (DNASTAR, Inc. Madison, Висконсин, США). Другая форма варианта включает замещение аминокислоты модифицированной аминокислотой, такой как бета-гомоаргинин в β-ультравариегин (SDEAVRAIPMYSTAPPGDFEEIPDDAIEE), обозначенной как «X» в SEQ ID NO: 9. Для целей перечня последовательностей, бета-гомоаргинин будет представлен как «X» или «Xaa».
Используемый здесь термин «фрагмент» относится к аминокислотной последовательности, которая имеет изменения одной или нескольких аминокислот, но сохраняет способность распознавать и связывать один и тот же конформационный эпитоп на тромбине, что и эталонная последовательность. Например, аминокислотная последовательность QTAVPKISKQGGGDFEEIPSDEIIE (QT26), представленная в SEQ ID NO: 3, короче, чем последовательность Аватрина, представленная в SEQ ID NO: 1 (SGGHQTAVPKISKQGLGGDFEEIPSDEIIE), и может рассматриваться как фрагмент SEQ ID NO: 1, поскольку она сохраняет способность распознавать, связывать и ингибировать тромбин. Аналогично, аминокислотная последовательность IS20 (ISKQGLGGDFEEIPSDEIIE), представленная в SEQ ID NO: 4, короче, чем последовательность Аватрина, представленная в SEQ ID NO: 1 (SGGHQTAVPKISKQGLGGDFEEIPSDEIIE), и может рассматриваться как фрагмент SEQ ID NO: 1, поскольку она сохраняет способность распознавать, связывать и ингибировать тромбин.
Используемый здесь термин «образец» используется в самом широком смысле. Биологический образец, предположительно содержащий тромбин, может содержать физиологическую жидкость или ткань.
Термин «лечение», используемый в контексте изобретения, относится к профилактическому, улучшающему, терапевтическому воздействию или радикальному лечению.
Термин «субъект» здесь определяется как позвоночное, в особенности млекопитающее, более конкретно человек. Для целей исследования субъектом может быть, в частности, по меньшей мере одна модель животного, например мышь, крыса и тому подобное. В частности, для лечения тромба и/или заболеваний, связанных с тромбом, субъект может быть человеком.
Настоящее изобретение в одном аспекте описывает выделенный ингибитор тромбина, содержащий аминокислотную последовательность, выбранную из группы, включающей SGGHQTAVPKISKQGLGGDFEEIPSDEIIE (SEQ ID NO: 1), ее вариант или фрагмент, SDEAVRAIPKMYSTAPPGDFEEIPDDAIEE (SEQ ID NO: 2), ее вариант или фрагмент, SEQ ID NO: 22, ее вариант или фрагмент, SEQ ID NO: 23, ее вариант или фрагмент, SEQ ID NO: 24, ее вариант или фрагмент и SEQ ID NO: 25, ее вариант или фрагмент. Понятно, что эти последовательности, описанные здесь, могут быть изменены и по-прежнему сохранять способность ингибировать активность тромбина.
Например, было обнаружено, что остатки 7 (A), 9 (P), 10 (K), 19-21 (DFE), 23 (I) и 24 (P) SEQ ID NO: 1 и SEQ ID NO: 2 важны для ингибирования активности тромбина, поскольку они являются наименее терпимыми к изменениям. Если большинство из этих ключевых остатков сохраняются, мы продуцировали пептидные варианты с минимальной достоверностью 30 %, которая сохраняет способность ингибировать активность тромбина.
В предпочтительном варианте осуществления выделенный ингибитор тромбина содержит аминокислотную последовательность, имеющую по меньшей мере 30 %, по меньшей мере 40 %, по меньшей мере 50 %, по меньшей мере 60 %, по меньшей мере 70 %, по меньшей мере 80 %, по меньшей мере 90 % или более предпочтительно по меньшей мере 95 % идентичности последовательности с SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24 или SEQ ID NO: 25. Более предпочтительно, выделенный ингибитор тромбина содержит аминокислотную последовательность, имеющую по меньшей мере 60 %, по меньшей мере 70 %, по меньшей мере 80 %, по меньшей мере 90 % или более предпочтительно по меньшей мере 95 % идентичности последовательности с SEQ ID NO: 1 или SEQ ID NO: 2 ,
В предпочтительном варианте осуществления выделенная аминокислотная последовательность ингибитора тромбина выбрана из группы, включающей:
QTAVPKISKQGLGGDFEEIPSDEIIE (SEQ ID NO: 3) фрагмент SEQ ID NO: 1;
ISKQGLGGDFEEIPSDEIIE (SEQ ID NO: 4) фрагмент SEQ ID NO: 1;
SGGHQTAVPKIAKQGLGGDFEEIPSDEIIE (SEQ ID NO: 5) S12A вариант SEQ ID NO: 1;
SGGHQTAVPKIHKQGLGGDFEEIPSDEIIE (SEQ ID NO: 6) S12H вариант SEQ ID NO: 1;
SGGHQTAVPRISKQGLGGDFEEIPSDEIIE (SEQ ID NO: 7) K10R вариант SEQ ID NO: 1;
SGGHQTAVPXISKQGLGGDFEEIPSDEIIE (SEQ ID NO: 8) β-гомоаргинин вариант SEQ ID NO: 1;
SDEAVRAIPXMYSTAPPGDFEEIPDDAIEE (SEQ ID NO: 9) β-гомоаргинин вариант SEQ ID NO: 2.
SDQGDVAIPKMYSTAPPGDFEEIPDDAIEE (SEQ ID NO: 10) вариант SEQ ID NO: 2;
SDEAVRAEPKMHKTAPPGDFEEIPDDAIEE (SEQ ID NO: 11) вариант SEQ ID NO: 2;
SDEAVRAIPKMYSTAPPGDFEEIPEEYLDDES (SEQ ID NO: 12) вариант SEQ ID NO: 2;
MYSTAPPGDFEEIPDDAIEE (SEQ ID NO: 13) вариант SEQ ID NO: 2;
SDEAVRAIPKMYSTAPPGDFEEIPDDEIEE (SEQ ID NO: 14) вариант SEQ ID NO: 2;
SDEAVRAIPKMYSQAPPGDFEEIPDDAIEE (SEQ ID NO: 15) вариант SEQ ID NO: 2;
SDQGDVAEPKMYSTAPPFDFEAIPEEYLDDES (SEQ ID NO: 16) вариант SEQ ID NO: 2;
SDQGDVAEPXMHSTAPPFDFEAIPEEYLDDES (SEQ ID NO: 17) вариант SEQ ID NO: 2;
CDEAVRAIPKMYSTAPPGDFEEIPDDAIEE (SEQ ID NO: 18) вариант SEQ ID NO: 2;
SDEAVRAIPKMYSTAPPGDFEEIPDDAIEECA (SEQ ID NO: 19) вариант SEQ ID NO: 2;
MYSTAPPGDFEEIPDDAIEEGCCC (SEQ ID NO: 20) a variant of SEQ ID NO: 2;
SDEAVRAIPKMYSTAPPGDFEEIPDDAIEEGCCC (SEQ ID NO: 21) a variant of SEQ ID NO: 2;
SGEDHTAVPKMSRKGLGGDFEDIPPEAYERALEAR (SEQ ID NO: 22);
ELESGDEDSEGGDSQSSPTESAAPRLHQREGGGGDFENVEYDQDQK (SEQ ID NO: 23);
SDVAPADYESDEGDNDGGHDGSEVAKPKMPRGNGGGGDFEEIPEVE (SEQ ID NO: 24); и
TGSDDDDEYDMYESDGDSNEGNDNDEFETAVPRLPNPNSGRDSEHIPMPVN (SEQ ID NO: 25);
Предпочтительно указанный ингибитор имеет аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 2, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20 и SEQ ID NO: 21.
В другом предпочтительном варианте осуществления указанный ингибитор тромбина ингибирует фибриногенолитическую активность тромбина и/или ингибирует амидолитическую активность тромбина.
В другом предпочтительном варианте осуществления IC50 ингибитора тромбина составляет менее 400 нМ, предпочтительно менее 300 нМ, менее 200 нМ, менее 100 Нм, менее 50 нм, менее 10 нМ, предпочтительно менее 9 нМ, менее 8 нМ, менее 7 нМ, менее 6 нМ, менее 5 нМ, менее 4 нМ, менее 3 нМ, менее 2 нМ или менее 1 нМ при оценке амидолитическим анализом. Более предпочтительно IC50 составляет менее 2 нМ. Предполагается, что значения IC50 между теми, которые перечислены конкретно, входят в объем изобретения.
В другом предпочтительном варианте осуществления Ki ингибитора тромбина составляет менее 6000 нМ, предпочтительно менее 2000, менее 500, менее 400 нМ, менее 300 нМ, менее 200 нМ, менее 100 Нм, менее 50 нм, менее 10 нМ, предпочтительно менее 9 нМ, менее 8 нМ, менее 7 нМ, менее 6 нМ, менее 5 нМ, менее 4 нМ, менее 3 нМ, менее 2 нМ или менее 1 нM при оценке амидолитическим анализом. Более предпочтительно Ki составляет менее 2 нМ. Предполагается, что значения Ki между теми, которые перечислены конкретно, находятся в пределах объема изобретения.
Другим аспектом изобретения является выделенный ингибитор тромбина, содержащий аминокислотную последовательность в соответствии с любым аспектом изобретения для профилактики или лечения заболевания, связанного с активностью тромбина.
Другим аспектом изобретения является способ ингибирования активности тромбина, где способ включает контактирование тромбина с по меньшей мере одним ингибитором тромбина в соответствии с любым аспектом изобретения. В предпочтительном варианте осуществления по меньшей мере один ингибитор тромбина присутствует в качестве противосвертыващего агента в пробирках для взятия крови или в качестве материала для поверхностного покрытия на медицинских устройствах, таких как стенты, катетеры и другие трубки для интубации.
В соответствии с другим аспектом изобретения предложено применение ингибитора тромбина в соответствии с любым аспектом изобретения для приготовления лекарственного средства для профилактики и/или лечения заболевания, связанного с тромбом. В предпочтительном варианте осуществления заболевание, связанное с тромбом, выбрано из артериального и венозного тромбоза, вызывающего сердечные приступы, инсульт и эмболию; для антикоагуляции при нестабильной стенокардии, коронарной ангиопластике, чрескожном коронарном вмешательстве и операциях на сердце.
Подходящие способы введения терапевтической композиции в соответствии со способами настоящего изобретения включают (но не ограничиваются ими) системное введение, парентеральное введение (включая внутрисосудистое, внутримышечное, внутриартериальное введение), пероральное введение, местное введение, буккальное введение, ректальное введение, вагинальное введение, подкожное введение, внутрибрюшинное введение, хирургическая имплантация, местная инъекция и гипертоническая инъекция/бомбардировка. Там, где это применимо, непрерывная инфузия может усиливать накопление лекарственного средства в целевом центре (см., например, патент U.S. № 6, 180, 082).
Независимо от способа введения пептиды по настоящему изобретению обычно вводят в количестве, эффективном для достижения желаемой ответной реакции. Используемые здесь термины «эффективное количество» и «терапевтически эффективное количество» относятся к количеству терапевтической композиции (например, композиции, содержащей полипептид ингибитора тромбина, и фармацевтический наполнитель, носитель или эксципиент), достаточные для получения измеримой биологической ответной реакции (например, уменьшение количества тромба или заболевания, связанного с тромбом). Фактические уровни дозировки активных ингредиентов в терапевтической композиции по настоящему изобретению могут варьироваться, чтобы вводить количество активного полипептида(-ов), которое является эффективным для достижения желаемой терапевтической ответной реакции для конкретного субъекта и/или применения. Разумеется, эффективное количество в любом конкретном случае будет зависеть от множества факторов, включая активность терапевтической композиции, состав, пути введения, комбинации с другими лекарственными средствами или лечения, тяжесть состояния, подлежащего лечению, и физическое состояние и предшествующая история болезни подлежащего лечению субъекта. Предпочтительно вводить минимальную дозу, и дозу эскалируют при отсутствии токсичности, ограничивающей дозу, до минимально эффективного количества. Определение и корректировка терапевтически эффективной дозы, а также оценка того, когда и как сделать такие корректировки, известны специалистам в данной области.
Дополнительные рекомендации относительно состава и дозы см. в патенты США №№ 5 326 902 и 5 234 933; международная публикация PCT № WO 93/25521; Berkow, et al., (1997) The Merck Manual of Medical Information, Home ed. Merck Research Laboratories, Whitehouse Station, N.J.; Goodman, et al., (2006) Goodman & Gilman's the Pharmacological Basis of Therapeutics, 11th ed. McGraw-Hill Health Professions Division, Нью-Йорк.
В соответствии с другим аспектом изобретения предложен способ профилактики и/или лечения заболевания, связанного с активностью тромбина, включающий введение субъекту, нуждающемуся в такой профилактике и/или лечении, эффективного количества ингибитора тромбина в соответствии с любым аспектом изобретения. В предпочтительном варианте осуществления заболевание, связанное с тромбом, выбрано из артериального и венозного тромбоза, вызывающего сердечные приступы, инсульт и эмболию; для антикоагуляции при нестабильной стенокардии, коронарной ангиопластике, чрескожном коронарном вмешательстве и операциях на сердце.
Ингибиторы тромбина по изобретению также могут быть использованы местно для лечения, например, синяков с гематомой или без нее. Пептидные ингибиторы по изобретению могут быть введены в крем таким же образом, как и ранее опробовано для гирудина, где 280 UI/100 г применяли к односторонним острым травмам опорно-двигательного аппарата (синяками) в 3-4 ежедневных применений в течение 5 дней [Stamenova PK., et al, Eur Rev Med Pharmacol Sci. 5(2):37-42 (2001)]. Также была разработана рецептура ингибитора тромбина для местного применения (r-гирудин 1120 IU/40 г, MINAPHARM Pharmaceuticals, Каир, Египет) для лечения тромбоза артериовенозного шунта, ушибов головного мозга, растяжений, разрывов мышц, травматических гематом, отека, эритемы, варикозного расширения, перифлебита и анального перифлебита с геморроем, особенно связанных с тромбоэмболическими осложнениями [см. Thrombexx® at minapharm.com]. Молекулы по настоящему изобретению составляют менее половины размера гирудина, что должно увеличивать проникновение активного ингредиента (т. е. ингибитора тромбина) через барьер кожи при местном применении.
В соответствии с другим аспектом изобретения предложен способ обнаружения накопления тромбина у субъекта, включающий введение по меньшей мере одного ингибитора в соответствии с любым аспектом изобретения субъекту или в образец ткани, выделенный у субъекта, и обнаружение присутствия указанного по меньшей мере одного ингибитора тромбина, связанного с тромбином.
В предпочтительном варианте осуществления предложен способ обнаружения накопления тромбина у субъекта, причем указанный способ включает:
а. получение образца ткани у пациента; а также
б. обнаружение того, накопился ли тромбин в образце путем контактирования образца с по меньшей мере одним ингибитором в соответствии с любым аспектом изобретения и обнаружение связывания между тромбином и по меньшей мере одним ингибитором тромбина.
Предпочтительно ингибитор тромбина включает аминокислотную последовательность, выбранную из группы, включающей SGGHQTAVPKISKQGLGGDFEEIPSDEIIE (SEQ ID NO: 1), ее вариант или фрагмент и SDEAVRAIPKMYSTAPPGDFEEIPDDAIEE (SEQ ID NO: 2), ее вариант или его фрагмент.
В другом предпочтительном варианте осуществления настоящего изобретения ингибитор тромбина включает аминокислотную последовательность, выбранную из группы, включающей SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20, SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24 и SEQ ID NO: 25.
Предпочтительно указанный ингибитор имеет аминокислотную последовательность, выбранную из группы, состоящей из of SEQ ID NO: 2, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15 и SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20 и SEQ ID NO: 21.
В соответствии с другим аспектом изобретения предложен способ диагностики заболевания или состояния, связанного с тромбом, включающий введение по меньшей мере одного ингибитора в соответствии с любым аспектом изобретения субъекту или в образец ткани, выделенный у указанного субъекта, и обнаружение присутствия указанного ингибитора тромбина, связанного с тромбином, причем обнаружение повышенного уровня указанного ингибитора, связанного с тромбином, по сравнению с уровнем ингибитора, связанным с нормальным уровнем тромбина, указывает на указанное заболевание или состояние.
В другом предпочтительном варианте осуществления способа ингибитор тромбина включает аминокислотную последовательность, выбранную из группы, включающей SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20, SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24 и SEQ ID NO: 25.
Предпочтительно указанный ингибитор имеет аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 2, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20 и SEQ ID NO: 21.
В соответствии с другим аспектом изобретения предложена фармацевтическая композиция, содержащая эффективное количество по меньшей мере одного ингибитора тромбина в соответствии с любым аспектом изобретения.
В соответствии с другим аспектом изобретения предложена выделенная молекула нуклеиновой кислоты, кодирующая ингибитор тромбина в соответствии с любым аспектом изобретения. В предпочтительном варианте осуществления последовательность нуклеиновой кислоты кодирует SEQ ID NO: 1 и представлена:
TCGGGTGGCCATCAGACTGCTGTTCCGAAGATATCTAAGCAAGGCTTGGGTGGAGACTTTGAAGAAATTCCAAGTGATGAAATAATCGAG (SEQ ID NO: 26).
В другом предпочтительном варианте осуществления выделенная молекула нуклеиновой кислоты кодирует ингибитор тромбина, содержащий аминокислотную последовательность, выбранную из группы, включающей SEQ ID NO: 1 и SEQ ID NO: 2, их вариантов или фрагментов.
В предпочтительном варианте осуществления выделенная нуклеиновая молекула кодирует SEQ ID NO: 2, ее фрагмент или вариант и может быть получена путем модификации последовательности нуклеиновой кислоты, представленной;
TCAGACGAAGCTGTCAGGGCGATTCCCAAGATGTACTCGACTGCCCCACCGGGAGATTTCGAAACAATCCCTGACGACGCTATTGAGGAG (SEQ ID NO: 27), ее фрагмент или вариант. Предпочтительно, чтобы нуклеотидная последовательность SEQ ID NO: 27 изменялась путем замены кодона, кодирующего Thr22 на подходящий кодон для кодирования Glu22, с получением пептида SEQ ID NO: 2. Было бы понятно, что нативная нуклеотидная последовательность (SEQ ID NO: 27) кодирует подходящий ингибитор тромбина в соответствии с изобретением.
В предпочтительном варианте осуществления нуклеиновая кислота имеет последовательность, представленную SEQ ID NO: 26 или SEQ ID NO: 27, их вариантами или производными.
В соответствии с другим аспектом изобретения предложен вектор, содержащий молекулу нуклеиновой кислоты по изобретению, как описано здесь.
Согласно другому аспекту изобретения предложена клетка-хозяин, содержащая молекулу нуклеиновой кислоты или вектор в соответствии с любым аспектом изобретения. Клетка-хозяин может быть прокариотической или эукариотической, но предпочтительно эукариотической.
В соответствии с другим аспектом изобретения предложен набор для профилактики или лечения заболевания или состояния, связанного с тромбом, содержащий по меньшей мере один ингибитор тромбина или лекарственное средство в соответствии с любым аспектом изобретения. Набор может содержать медицинское устройство, такое как стент, катетер или трубку другой формы, покрытую ингибитором тромбина по изобретению.
Теперь изобретение в целом описано, и то же самое будет более легким для понимания при помощи ссылок на следующие примеры, которые приведены в качестве иллюстрации и не предназначены для ограничения настоящего изобретения. Способы, методики и химические вещества описаны в приведенных ссылках или в протоколах стандартных учебников по биотехнологии и молекулярной биологии.
ПРИМЕРЫ
Материалы и способы
Калликрейн, фибриноген человека и бычий трипсин приобретали у Merck. Chemicals Ltd. (Ноттингем, Великобритания). Бычий химотрипсин, гексагидрат хлорида железа и бычий сывороточный альбумин приобретали у Sigma-Aldrich (Сент-Луис, штат Миссури, США). Все другие сериновые протеазы приобретали у Hematologic Technologies, Inc. (Эссекс Джанкшен, штат Вермонт, США). Рекомбинантный тромбин был подарен Хемо-серо-терапевтическим исследовательским институтом (Какецукен, Япония). Хромогенные субстраты приобретали у Chromogenix, а Spectrozyme FIXa у American Diagnostica. Все другие химические вещества и реагенты были аналитического качества.
Синтез и очистка пептидов
Все пептиды синтезировали с использованием твердофазного пептидного синтеза на пептидном синтезаторе Intavis MultiPep RSi. (Intavis Bioanalytical Instruments, Кельн, Германия) и отщепляли от смолы, как описано ранее [Koh CY, et al., J Biol Chem 282: 29101–13 (2007)]. Неочищенные пептиды очищали с использованием ВЭЖХ с обращенной фазой (ОФ-ВЭЖХ) на очистителе AKTA от GE Healthcare (Уппсала, Швеция) с колонкой Jupiter Proteo (5 мкм, 250 мм × 10 мм, 90 Å). Чистоту и массу всех пептидов определяли с помощью масс-спектрометрии с электрораспылением с использованием LCQ Fleet Ion Trap MS от Thermo Fisher Scientific (Уолтем, штат Массачусетс, США).
CD-спектроскопия
Дальние УФ-спектры CD (260-190 нм) аватрина, QT26 и IS20, растворенные в 10 мМ натрий-фосфатном буфере (рН 7,4), измеряли с использованием спектрополяриметра Jasco™ J-810 (Истон, штат Мэриленд). Все измерения проводили при комнатной температуре с использованием кюветки длины пути 0,1 см со скоростью сканирования 50 нм/мин, шириной полосы 2 нм и разрешением 0,2 нм.
Ингибирование амидолитической и фибриногенолитической активности тромбина и протеазной активности.
Анализы тромбиновой амидолитической активности для всех пептидов проводили в 96-луночных планшетах для микротитрования в 50 мМ Трис-буфере (рН 7,4), содержащем 100 мМ NaCl и 1 мг/мл бычьего сывороточного альбумина. Как правило, 100 мкл пептида и 100 мкл тромбина предварительно инкубировали в течение различных временных интервалов до того, как в реакционные лунки добавляли 100 мкл S2238. За уровнями образования п-нитроанилина следовало измерение поглощения при 405 нм в течение 10 мин с помощью микропланшетного прибора Tecan InfinitePro M200 фирмы Tecan (Меннедорф, Швейцария). Кривые доза-ответ установливали с использованием программного обеспечения GraphPad Prizm (Сан-Диего, Калифорния, США) для расчета значений IC50 и коэффициентов Хилла. Для измерения констант ингибирования использовали различные концентрации S2238, а остаточные скорости определяли с использованием уравнения сильной связи Моррисона. Ингибирование фибриногенолитической активности тромбина пептидов тестировали путем измерения поглощения при 650 нм с использованием считывателя микропланшета Sunrise от Tecan (Меннедорф, Швейцария), как описано ранее [Koh CY, et al., J Biol Chem 282: 29101–13 (2007)]. Селективность аватрина исследовали в присутствии 13 сериновых протеаз (Фиг. 5). Эффекты влияния аватрина на амидолитическую активность этих сериновых протеаз определяли с использованием соответствующих хромогенных субстратов. Расщепление аватрина тромбином. Аватрин (150 мкМ) инкубировали с тромбином (5 мкМ) в 50 мМ Трис-буфере (рН 7,4), содержащем 150 мМ NaCl и 1 мг/мл бычьего сывороточного альбумина (BSA). После различного времени инкубации реакции гасили 1 % трифторуксусной кислотой (pH 1,8) и загружали в колонку Jupiter Proteo (4 мкм, 90Å, 100 × 1,0 мм), прикрепленную к системе нано-ВЭЖХ Dionex от Thermo Fisher Scientific (Уолтем, штат Массачусетс, США) и элюировали с использованием градиента ацетонитрила с 0,05 % трифторуксусной кислоты и 99,95 % воды качества Milli-Q в качестве растворителя A и 0,05 % трифторуксусной кислоты, 19,95 % воды качества Milli-Q и 80 % ацетонитрила в качестве растворителя B. Массы всех пиков измеряли для идентификации продуктов расщепления, которые впоследствии количественно определяли путем интегрирования пиков и расчета площади под кривыми. Чтобы измерить влияние различных периодов предварительной инкубации на ингибирующую активность тромбина, проводили амидолитические анализы для различных периодов инкубации (до 36 часов).
Рентгеновская кристаллография
Рекомбинантный α-тромбин (в 150 мМ NaCl) обессоливали с использованием 3000 центробежных фильтров с номинальным отсечением по молекулярной массе в 20 мМ NH4HCO3 и лиофилизировали перед кристаллизацией. Также использовали и оптимизировали условия кристаллизации для других комплексов ингибиторов, тромбин с вариегином, гиругеном и гирулогом [Koh CY, et al., PLoS One 2011; 6.; Skrzypczak-Jankun E, et al., J Mol Biol 221: 1379–93 (1991)]. Аватрин (81,7 мкМ) растворяли в 50 мМ буфере HEPES (рН 7,4), содержащем 375 мМ NaCl. Тромбин растворяли в растворе аватрина до конечной концентрации 54,5 мкМ. Кристаллизации достигали с использованием метода диффузии оседающих паров. Как правило, 1 мкл смеси, содержащей аватрин и тромбин, смешивали с 1 мкл осадителя (100 мМ HEPES, pH 7,4, от 20 до 25 % (масса/объем) PEG 8000) и оставляли при 4 °C. Кристаллы появились примерно через шесть недель. Кристаллы пропитывали раствором криопротектора, содержащего маточный раствор, дополненный 25 % (об./об.) глицерином, и охлаждали в вакууме при 100 К в потоке холодного газообразного азота (Oxford Cryosystem, Оксфорд, Великобритания). Собрали набор данных из 180 кадров (осцилляция 180°) с использованием ПЗС, смонтированного на вращающемся аноде рентгеновского генератора Rigaku. Набор данных обрабатывали и масштабировали с использованием Mosflm [Battye TGG, et al., Acta Crystallogr Sect D Biol Crystallogr 67: 271–81 (2011)] и AIMLESS [Evans PR, and Murshudov GN. Acta Crystallogr Sect D Biol Crystallogr 69: 1204–14 (2013)] соответственно [Leslie AGW, and Powell HR. Evolving methods for macromolecular Crystallography. 2007]. Структуру комплекса определяли заменой молекул с использованием в качестве шаблона программу Phaser и кристаллической структуры тромбина-вариегина (PDB:3B23) [McCoy AJ. Acta Crystallogr Sect D Biol Crystallogr International Union of Crystallography 63: 32–41 (2006)]. Построение и уточнение модели выполняли с использованием COOT (автономный пакет для молекулярного моделирования) [Emsley P, and Cowtan K. Acta Crystallogr Sect D Biol Crystallogr International Union of Crystallography 60: 2126–32 (2004)].
Ингибирование связанного сгустком тромбина
Активность связанного сгустком тромбина тестировали с использованием S2238. Вкратце, фибриновые сгустки получали путем инкубации 100 мкл фибриногена 2 мг/мл (в 50 мМ буфера HEPES, pH 7,5, 150 мМ NaCl, 10 мг/мл CaCl2) с помощью 100 мкл 30 нМ тромбина. Через 2 часа при 37 °С сгустки интенсивно промывали тем же буфером. Эту промывку повторяли каждые три часа в течение 24 часов. Затем к сгусткам добавляли различные концентрации аватрина и инкубировали в течение 60 мин. Затем добавляли хромогенный субстрат S2238 (конечная концентрация 200 мкМ) и реакционную смесь инкубировали в течение 90 мин при 37 °С. Аликвоты брали, и гидролиз субстрата оценивали по показателям конечной точки при 405 нм, используя считыватель микропланшет Tecan Infinite®Pro. Эксперименты проводили четыре раза, и процентное ингибирование наносили на график.
Модель тромбоза сонной артерии, вызванного хлоридом железа
Все эксперименты на животных проводили в соответствии с протоколом 041/12, одобренным Институтом по уходу и использованию животных Национального университета Сингапура. Модель тромбоза сонной артерии, индуцированная хлоридом железа, выполняли, как описано ранее [Eckly A, et al., J Thromb Haemost 9: 779-89 (2011)] с небольшими изменениями. Как правило, самцов мышей C57BL/6 (9-11 недель, 24,5-27,5 г) анестезировали внутрибрюшинной инъекцией кетамина (75 мг/кг) и медетомидина (1 мг/кг). 100 мкл различных доз аватрина вводили мышам через хвостовую вену. Правую сонную артерию расчленили с использованием тупого рассечения и вызвали сосудистую травму с применением фильтровальной бумаги толщиной 2 мм × 2 мм, насыщенной FeCl3 поверх сонной артерии. Через 3 мин фильтровальную бумагу удаляли и сосуд промывали стерильным физиологическим раствором. Чтобы определить время окклюзии, вокруг сонной артерии помещали миниатюрный доплеровский проточный зонд (Transonic Systems Inc., Ithaca, Нью-Йорк, США), и кровоток регистрировали с использованием расходомера Transonic® от ADInstruments (Данедин, Новая Зеландия). Максимальное время мониторинга кровотока после травмы составляло 30 мин. Мышей подвергали эвтаназии дислокацией шейки матки сразу после завершения эксперимента и до завершения действия анестезии.
Взятие крови в пробирки для взятия крови с пептидами
Индивидуальные пептиды растворяют в забуференном фосфатом физиологическом растворе для получения 10× пептидных растворов (т.е. концентрации пептидов в 10 раз выше, чем конечные концентрации, при которых они должны быть проверены). 0,3 мл 10× пептидных растворов помещают в пробирки без каких-либо добавок. Кровь берут у здоровых добровольцев с помощью шприцев и сразу же переносят в коническую центрифужную пробирку объемом 50 мл (пробирка Falcon) перед отмериванием пипеткой 2,7 мл крови в пробирки для взятия крови с пептидными растворами. Конечные концентрации пептидов приведены в табл. 5 и 6.
Антикоагуляционное действие пептидов в пробирках для взятия крови
Пробирки для взятия крови с различными пептидами в разных концентрациях оставляли стоять при комнатной температуре до тех пор, пока их не проверили в назначенное время. В разные моменты времени пробирки несколько раз переворачивали и визуально проверяли на нерастворимые материалы в качестве признаков образования сгустка.
Сохранение функции тромбоцитов в пробирках для взятия крови, содержащих пептиды
Пробирки для взятия крови с различными пептидами в разных концентрациях оставляли стоять при комнатной температуре до тех пор, пока их не проверили в назначенное время. В разные моменты времени пробирки несколько раз переворачивали, и образцы крови вынимали для тестов агрегации тромбоцитов с использованием агрегометра Multiplate® Platelet с аденозиндифосфатом (АДФ) в качестве агониста в соответствии с рекомендованными изготовителем протоколами.
Результаты
Обнаружение вариегин-подобных транскриптов
в слюнных железах Amblyomma variegatum
Вырожденные праймеры на основе последовательности вариегина амплифицировали транскрипт (AB183707), кодирующий белок-предшественник из остатка 219 (BAD29729) из кДНК слюнных желез самок Amblyomma variegatum после 9 дней питания. Этот предшественник содержит предполагаемый сигнал секреции и пять идентичных повторов из 30 остатков с предполагаемыми центрами расщепления между ними, что позволяет посттрансляционное расщепление предшественника до пяти активных пептидов. Экспрессия этого белка-предшественника в слюнных железах была подтверждена гибридизацией in situ. Зонд гибридизуется в цитоплазме крупных базальных гранулярных клеток слюнной железы типа II. Локализация транскриптов в слюнных железах нимф и взрослых самок и самцов клещей продемонстрировала различия в выраженности во время кормления, а также индивидуальную изменчивость в начале экспрессии. Наиболее сильное выражение было обнаружено у нимф 2-4 дней питания, самок 5 дней питания и самцов 12 дней питания, то есть в то время, когда самки были напитаны кровью и начали отделяться (Фиг. 1A-I). Мы рассматривали только небольшой участок последовательности в пределах 219 аминокислотной последовательности, достаточной для активности, поэтому синтезировали SEQ ID NO: 1 аватрин (30 аминокислот). Нуклеотидная последовательность, кодирующая SEQ ID NO: 1 представлена SEQ ID NO: 21.
Активные пептиды, кодируемые этим транскриптом, показали приблизительно 40 % идентичности последовательности с вариегином. Эти пептиды также показали несколько отличий в ключевых функциональных остатках по сравнению с вариегином:
(i) У вариегина есть кислотный N-конец, который постулируется как важный для быстрой кинетики связывания [Koh CY, et al., J Biol Chem 282: 29101–13 (2007)]. Напротив, кислотные остатки, как правило, отсутствуют в этих подобных вариегину пептидах;
(ii) Вариегин ингибирует тромбин, нарушая систему ретрансляции заряда каталитической триады активного центра его возможной водородной связью His12, возможно, с Ser195 тромбина [Koh CY, et al., PLoS One 2011; 6]. В этих подобных вариегину пептидах этот функциональный гистидин заменен серином;
(iii) Thr14 в нативном вариегине гликозилирован, и он показал 14-кратно более высокую аффинность, чем синтетический вариегин [Koh CY, et al., J Biol Chem 282: 29101–13 (2007)]. В подобных вариегину пептидах глутамин, который не может быть гликозилирован, присутствует в этом положении;
(iv) Pro16 и Pro17 в вариегине вводят перегибы скелета, возможно, ограничивая конформационную гибкость в связи между активным центром и сегментом связывания экзосайта-I [Koh CY, et al., PLoS One 2011; 6]. Подобная область в подобных вариегину пептидах содержит три глицина, что придает пептиду гораздо большую гибкость.
Все эти различия дали толчок для оценки структурно-функциональных соотношений этих подобных вариегину пептидов. Следовательно, мы продолжили синтез активного пептида из белка-предшественника (BAD29729), чтобы дополнительно охарактеризовать его ингибирующее действие на α-тромбин человека.
Аватрин является мощным и селективным ингибитором тромбина
Активный пептид, названный аватрином (ингибитор тромбина Amblyomma variegatum), синтезировали твердофазным пептидным fmoc-синтезом и очищали до гомогенности (Фиг. 2). Аватрин ингибировал амидолитическую активность тромбина на небольшом пептидил хромогенном субстрате S2238 дозозависимым образом с IC50 и коэффициентом Хилла, составляющими 6,95 ± 0,42 нМ и 0,92 ± 0,01 соответственно. Значительное ингибирование (14,33 ± 1,39 %) наблюдали для эквимолярных концентраций тромбина и аватрина (0,81 нМ тромбина, 1 нМ аватрина), соответствующих описаниям типичных ингибиторов сильного связывания [Copeland R a. Enzymes: A practical Introduction to Structure, mechanism, and data analysis. 2000] (Фиг. 3А). Кривые хода реакции показали, что устойчивое равновесие было достигнуто при перемешивании, что указывает на режим быстрого связывания (Фиг. 3B). Реакционные скорости тромбина определяли в присутствии различных концентраций аватрина для получения видимой константы ингибирования Ki’. График Ki’ линейно возрастал с увеличением концентрации S2238, что указывает на то, что аватрин является конкурентным ингибитором тромбина по отношению к S2238 (Ki 545,3 ± 3,1 пМ) (Фиг. 3C, 3D). Таким образом, аватрин является быстрым, конкурентным ингибитором тромбина сильного связывания. Аватрин также продлевал время свертывания фибриногена дозозависимым образом, что указывает на то, что он также ингибирует фибриногенолитическую активность тромбина (Фиг. 4).
Селективность аватрина исследовали путем скрининга его в присутствии 13 сериновых протеаз. При 10 нМ аватрин ингибировал 65 % активности тромбина. Однако даже при 100 мкМ ингибирование других протеаз составляло менее 30 % (Фиг. 5), что указывает на то, что аватрин является высокоселективным ингибитором с по меньшей мере на четыре порядка большей избирательностью для тромбина.
Аватрин демонстрирует длительное ингибирование тромбина
Конкурентоспособное ингибирование пептидного субстрата тромбина S2238 указывает на то, что аватрин связывается с активным центром и, следовательно, он может быть восприимчивым к протеолитическому расщеплению тромбином, подобно всем другим макромолекулярным субстратам или ингибиторам, таким как вариегин и бивалирудин. Мы исследовали расщепление аватрина тромбином путем инкубации его с ферментом (отношение 30:1) для увеличения количества времени и проанализировали реакцию с помощью хроматографии с обращенной фазой и масс-спектрометрии с электрораспылением. При инкубациях были идентифицированы два новых пика, соответствующих SGGHQTAVPK (981,3 Да) и ISKQGLGGDFEEIPSDEIIE (2176,3 Да), что указывает на расщепление по связи Lys10-Ile11 (Фиг. 6A). Расщепление количественно определяли путем расчета площадей под пиками на хроматограммах (Фиг. 6В). С увеличением продолжительности времени количество расщепленных продуктов увеличивалось, а количество пептида полной длины уменьшалось, а аватрин полностью расщеплялся через приблизительно 10 ч. Эффект влияния расщепления на ингибирующую активность оценивали (Фиг. 6C). При том же соотношении аватрин:тромбин (1:30), используемом для экспериментов по расщеплению, более 45 % амидолитической активности тромбина ингибировалось через 24 часа, что указывало на длительное ингибирование тромбина даже после полного расщепления аватрина. Вариегин проявлял подобное поведение, так как его расщепленный по связи C-концевой пептид продолжает ингибировать тромбин после расщепления. Чтобы проверить, что продукт расщепления аватрина также ингибирует тромбин, соответствующий пептид (IS20) очищают и испытывают на ингибирование. IS20 ингибирует амидолитическую активность тромбина с IC50 12,38 ± 0,32 нМ (Фиг. 7A). Ki’ уменьшается криволинейно с увеличением концентраций S2238, что указывает на то, что IS20 является неконкурентным ингибитором относительно небольшого пептидного субстрата с общим Ki 5,76 ± 0,23 нМ (Фиг. 7B и 7C). IS20 также ингибирует фибриногенолитическую активность тромбина дозозависимым образом (Фиг. 4). Таким образом, аватрин проявлял длительное ингибирование через свой С-концевой пептид, который сохранял аффинность сильного связывания к тромбину.
Тромбин-связывающие сегменты на аватрине
Мы синтезировали два дополнительных усеченных варианта аватрина, а именно QT26 и GL16, для локализации сегментов связывания тромбина на аватрине. Четыре и 15 N-концевых остатков аватрина удаляли соответственно в QT26 и GL16. Оба пептида испытывали на их способность ингибировать амидолитическую и фибриногенолитическую активность тромбина. QT26 ингибирует амидолитическую активность тромбина (IC50 = 8,94 ± 0,64 нМ и Ki = 760,32 ± 0,91 пМ) (Фиг. 7A, 7D и 7E) и фибриногенолитическую активность (Фиг. 4). По сравнению с полноразмерным аватрином потеря активности из-за удаления 4 N-концевых остатков в QT26 является минимальной. GL16, напротив, не ингибировал амидолитическую активность даже при 300 мкМ и вместо этого проявлял небольшую активацию (5-10 %) (Фиг. 7F). Однако он ингибировал фибриногенолитическую активность (Фиг. 4). В совокупности это указывает на то, что QT26 содержит как активные центры, так и последовательности связывания экзосайта-I, но GL16 содержит только последовательности связывания экзосайта-I. Поскольку расщепляемая связь находится между Lys10-Ile11, участок связывания активного центра аватрина находится в пределах последовательности 5QTAVPKISKQ14.
Структурно-функциональные соотношения взаимодействия тромбина и аватрина
Несмотря на низкую общую идентичность последовательности вариегина и аватрина и изменения в нескольких ключевых функциональных остатках, как описано выше, функционально аватрин показал высокую степень сходства с вариегином в его ингибирующей активности в отношении тромбина. Для дальнейшего изучения значимости различий в двух последовательностях мы оценили серию аватрин замещенных мутантов, будучи проинформированными о предыдущих исследованиях структурной функции вариегина [Koh CY, et al., PLoS One 2011; 6]:
(i) Ключевым функциональным остатком является VH12, который наиболее вероятно разрушает каталитическую триаду тромбина, и заменяется ASer12 в аватрине. Мы синтезировали два мутанта с заменами ASer12 на Ala (S12A) или His (S12H). S12A проявлял сходное снижение активности (более 10-кратного) до наблюдаемого у аналогичного мутанта вариегина (Фиг. 8A), что указывает на важность ASer12 для ингибирующего действия аватрина. S12H ингибировал тромбин с IC50 18,51 ± 0,32 нМ (Фиг. 8А) и был в 2 раза менее сильным, чем аватрин, что указывает на то, что серин делает аватрин более сильным ингибитором тромбина, чем если бы он имел гистидин в этом положении.
(ii) Вариегин содержит два остатка Glu на своем N-конце, и подозревали, что эти два кислотных остатка направляли вариегин в сторону экзосайта-II тромбина и обеспечивали путь ингибирования быстрого связывания [Koh CY, et al., J Biol Chem 282: 29101 -13 (2007)]. Хотя аватрин демонстрировал быструю кинетику связывания без кислотного N-конца, нам было интересно исследовать роль кислотного N-конца в активности аватрина при ингибировании тромбина. В одном двойном мутантном пептиде G2D, Q5D кислые остатки вводили в N-конец аватрина, чтобы имитировать возможную роль электростатического направляющего действия в придании быстрой кинетики связывания вариегину [Koh CY, et al., J Biol Chem 282: 29101–13 (2007)]. Этот мутант демонстрировал ингибирование слабее (менее 2 раз), чем аватрин и QT26 (Фиг. 8В). Таким образом, присутствие кислотного N-конца вряд ли будет важно для быстрой кинетики связывания, как предполагалось ранее [Koh CY, et al., J Biol Chem 282: 29101–13 (2007)].
(iii) Чтобы проверить гипотезу о том, что более жесткий, богатый пролином линкер между активным центром и связывающими сегментами экзосайта-I в вариегине (15APPF18) полезен по сравнению с гибким обогащенным глицином линкером в аватрине (15GLGG18), пептид двойной мутант L16P, G17P синтезировали и испытывали. Этот пептид регистрировал падение активности более 25-кратного (IC50, 181,32 ± 3,76 нМ) по сравнению с аватрином (рис. 8B), что указывает на то, что гибкость в линкере необходима для аватрина.
(iv) Известно, что в тромбине предпочтителен остаток аргинина в P1 [Berliner LJ. Journal of Chemical Information and Modeling. (1992)], и замещение в P1 Arg на Lys вызывает десятикратный скачок активности [Gallwitz M, et al., PLoS One 2012; 7]. Как вариегин, так и аватрин имеют лизин в P1, а мутация в P1 Lys-Arg в вариегине приводит к небольшому усилению активности (менее 3 раз) [Koh CY, et al., PLoS One 2011; 6]. Поэтому мы заменили в аватрине в P1 Lys на Arg (K10R) и наблюдали аналогичный 3-4-кратный прирост активности (IC50, 1,15 ± 0,45 нМ) (Фиг. 8C). Однако расщепление этого пептида тромбином также протекает быстрее, что приводит к полному расщеплению в течение 3 ч (Фиг. 9А).
(v) Затем мы синтезировали вариант с названием β-аватрин, в котором расщепляемая пептидная связь (VLys10-VIle11) замещена протеолитически стабильной связью (β-гомоArg10-Ile11). β-аватрин был более чем в 100 раз менее сильным чем аватрин, и ингибировал тромбин с IC50 332 ± 1,32 нМ (Фиг. 8D). Хотя он не был расщеплен тромбином до 72 ч (Фиг. 9В), сдвиг, возникающий из-за дополнительного атома углерода вдоль основной цепи пептида, по-видимому, крайне отрицательно влияет на активность. Аналогичное падение активности также наблюдали с вариантами гирулога, в которых были заменены расщепляемые связи, чтобы сделать пептид более устойчивым к расщеплению resistant [Bourdon P. Biochem Biophys Res Commun 177: 1049–55 (1991)]. Значения Ki этих пептидов показаны на Фиг. 10 и 11. Результаты по аватрину и пептидам вариантам аватрина сведены в табл. 1 и 2.
Таблица 1. Ожидаемые и наблюдаемые массы аватрина и его вариантов
Таблица 2. Параметры кинетики ингибирования тромбина аватрином и его вариантами
* И.О.- Ингибирование отсутствует; ** Н.О.- Не определено
Кристаллическая структура комплекса тромбин-аватрин
Комплекс тромбин-аватрин кристаллизовали в С2-пространственной группе и доводили до 2,09 Å (табл. 3). Плотность электронов для большинства остатков тромбина хорошо определена, за исключением остатков на концах легкой цепи; и в петле автолиза и С-конце тяжелой цепи. К сожалению, не все остатки в связанном аватрине имеют плотность, достаточную для однозначного построения модели. Не наблюдали плотности для первых четырех (1SGGH4) и двух последних остатков (29IE30). Плотность электронов для аватрина также является прерывистой от С-конца до расщепляемой связи между AIle11 и AGly18. Таким образом, мы построили два сегмента пептида, представляющие N-конец активного центра связывания расщепляемой связи (5QTAVPK10) и сегмента связывания экзосайта-I (19DFEEIPSDEI28) (Фиг. 12A и B). Мы выдвигаем гипотезу о том, что при кристаллизации происходит расщепление аватрина. После расщепления остатки между AIle11 и AGly18 не имели упорядоченной связанной структуры в комплексе с тромбином. Из-за малой плотности боковые цепи AGln5, AGlu22 и ASer25 также не могут быть построены.
Таблица 3. Статистика сбора и уточнения кристаллографических данных
*Значения в скобках для оболочки с высоким разрешением
# Статистика карты Рамачандрана сообщается сервером Molprobity
Ингибирование тромбина аватрином, по-видимому, происходит путем блокирования активного центра, как ожидается, из амидолитических анализов, демонстрирующих конкурентное ингибирование с небольшими пептидильными субстратами. Модель показала состояние активного центра после расщепления, в котором, по-видимому, находится система регенерации тромбина. Oγ TSer195 составляет 2,7 Å от Nε THis57, а Nδ THis57, в свою очередь, в 2,7 Å от Oδ TAsp102. Кажется, что Cα ALys10 (P1) смещается дальше от нуклеофила (Oγ TSer195) при 3,2 Å, так как происходит расщепление. Карбонильный кислород ALys10 все еще стабилизирован в оксианионном отверстии, расположенном на расстоянии 3,1 Å от основного азота TGly193. P1 ALys10 связывается на субцентре S1, как ожидалось, со своим амином боковой цепи, образующим водородную связь с TAsp189 на дне кармана специфичности. P2 APro9 пирролидиновое кольцо, по-видимому, взаимодействует с ароматическими боковыми цепями THis57 и TTyr60A перпендикулярно, что напоминает типичные pi взаимодействия конец-к-началу. Боковая цепь P3 AVal8 подвержена действию растворителя и не имеет особых взаимодействий. Метильная группа боковой цепи P4 AAla7, напротив, полностью закрыта в гидрофобный карман, образованный TAsn98, TLeu99, TIle174 и TTrp216. P5 AThr6 и P6 AGln5 одновременно подвержены действию растворителя, поскольку пептид приближается к экзосайту-II, однако существует недостаток электронной плотности в аватрине за этой областью, так что невозможно определить, протягивается ли пептид к экзосайту II (Фиг. 12C).
Остатки, расположенные непосредственно от C-конца расщепляемой связи, не демонстрировали хорошей электронной плотности вплоть до окружения экзосайта-I. В целом, эта часть аватрина связывается в канале экзосайта-I, как гируген, гирулог-1 и вариегин [Koh CY, et al., PLoS One 2011; 6; Skrzypczak-Jankun E, et al., J Mol Biol 221: 1379–93 (1991); Qiu X, et al., Biochemistry 31: 11689–97 (1992)]. Представляется, что как электростатическое, так и гидрофобное взаимодействия важны для этого связывания. Наблюдаются три электростатических взаимодействия (AAsp19-TArg73, AGlu21-TArg75 и AGlu27-TArg77A) между C-концом аватрина и экзосайтом-I (Фиг. 12D). Три гидрофобные боковые цепи (APhe20, AIle23 и AIle28) погружены в зону взаимодействия аватрин-тромбин. AIle23 и AIle28 погружены внутрь большого гидрофобного кармана, который образован боковыми цепями TPhe34, TLeu65, TArg67, TTyr76 и TIle82. Кроме того, частично обнаженный APro24, по-видимому, имеет некоторые благоприятные контакты с фенольным кольцом TTyr76. Остатки AGlu22 и ASer25 являются растворимыми и имеют недостаток плотности боковой цепи, поэтому их взаимодействие с тромбином не является интерпретируемым. Кроме того, после AIle28 наблюдается очень низкая плотность электронов, что не позволяет разместить AIle29 и AGlu30.
Ингибирование связанного сгустком тромбина
Гемостатические фибриновые сгустки удерживают активный тромбин, ограничивая его циркуляцию [Francischetti IMB, et al., Biochemistry 38: 16678–85 (1999)]. Этот связанный сгустком тромбин защищен от ингибирования комплексом гепарин-антитромбин III и действует как резервуар активного тромбина, который, как считается, играет важную роль при повторном тромбозе [Francischetti IMB, et al., Biochemistry 38: 16678–85 (1999); Bridge KI, et al.,Thromb Haemost 112: 1–8 (2014)]. Таким образом, ингибирование связанного сгустком тромбина может предотвратить повторный тромбоз. Поэтому мы оценили способность аватрина ингибировать связанный сгустком тромбин. Аватрин ингибировал связанный сгустком тромбин дозозависимым образом с IC50 1,74 ± 0,35 мкМ (Фиг. 13), которая выше, чем у тромбина в растворе. Ингибиторы активного центра, такие как аргатробан, быстро и обратимо ингибируют связанный сгустком тромбин с зависимым от концентрации образом с IC50 2,7 мкМ [Berry C, et al., Thromb Haemost 72: 381–6 (1994)].
Модель FeCl3-индуцированного тромбоза сонной артерии
Чтобы оценить антитромботическую эффективность аватрина in vivo, мы использовали модель индуцированного FeCl3 тромбоза сонной артерии у мышей [Wan C, et al., J Thromb Haemost 13: 248–61 (2015); Eckly a, et al., J Thromb Haemost 2011; 9: 779–89]. Среднее время окклюзии (TTO) увеличивалось дозозависимым образом у мышей, которым внутривенно вводили аватрин. У контрольных животных ТТО увеличилось с 7,24 ± 1,46 мин до 15,03 ± 3,23 и 22,51 ± 4,19 мин у животных, которым вводили 3 и 10 мг/кг соответственно. Эффективность аватрина сравнивали с гирулогом-1 в качестве препарата сравнения. TTO у мышей, инъецированных 3 и 10 мг/кг гирулог-1, составлял 9,70 ± 3,15 и 15,22 ± 3,39 мин соответственно. Таким образом, аватрин показал лучшую антитромботическую эффективность по сравнению с гирулогом-1 (Фиг. 14).
Идентификация пептидных последовательностей из транскриптомов иксодовых клещей
Пептидные последовательности, которые были похожи на вариегин и аватрин, идентифицировали путем выполнения автономного анализа BLAST опубликованных транскриптомов Amblyomma variegatum, Rhipicephalus pulchellus, Amblyomma americanum, Amblyomma cajenesse, Amblyomma maculatum и Hyalomma marginatum rufipes. Эти последовательности вручную выравнивали с вариегином и аватрином, и один пептид из каждого клеща выбирали для дальнейшего анализа.
Ингибирование амидолитической активности и селективность других пептидов
Еще несколько белковых последовательностей со сходством с транскриптом аватрина A. variagatum и других твердых клещей можно найти в базе данных NCBI. Некоторые из этих последовательностей синтезировали и тестировали на их ингибирующую активность в отношении тромбина (Фиг. 15 и табл. 4). Эти последовательности имеют номера доступа DAA34688.1, DAA34160.1 и DAA34258.1. Подобно транскриптам аватрина, они содержат несколько повторов, которые могут быть обработаны посредством посттрансляционных модификаций в короткие, похожие на вариегин зрелые пептиды. Репрезентативную последовательность синтезировали, тестировали на ингибирование тромбина и назвали ультравариегином. Ультравариегин на 50 % идентичен вариегину.
Таблица 4. Молекулярные массы, значения IC50 и Ki членов семейства вариегина
Ультравариегин представляет собой пептид из 30 остатков, представленный SEQ ID NO: 2, и основанный на отрезке из 30 остатков 212 аминокислот последовательности белка, полученной из транскрипта из базы данных. Однако мы заменили один аминокислотный остаток в отрезке последовательности ультравариегина из 30 остатков, обнаруженной в последовательности белка из 212 аминокислот (Thr22Glu, чтобы получить SEQ ID NO: 2). Было обнаружено, что пептид SEQ ID NO: 2 ингибирует тромбин с Ki 4,4 пМ. По сравнению с вариегином, Ki которого составляет 342 пМ, ультравариегин более чем в 70 раз сильнее. Для дальнейшего понимания структурно-функциональных соотношений ультравариегина в ингибировании тромбина синтезировали еще несколько вариантов ультравариегина следующим образом. UV003, UV004 и UV005 представляют собой гибридные пептиды ультравариегин-вариегин. На основании последовательности ультравариегина первые 7 остатков на N-конце заменялись последовательностью вариегина в UV003. Последующие 7 остатков заменяли таким же образом в UV004. В UV005 последние 6 остатков в ультравариегине заменяли на последние 8 остатков вариегина (у вариегина есть два дополнительных остатка). Было обнаружено, что Ki UV003 и UV004 аналогичны этим же величинам ультравариегина, показывая, что замена последовательностей ультравариегина на последовательности вариегина в первых 14 остатках на N-конце в значительной степени несущественна для его активности (Фиг. 16). Тем не менее, UV005 показал примерно 3,6-кратное снижение аффиности к тромбину. Поэтому основное различие в ингибирующей активности между вариегином и ультравариегином происходило от их С-концов. Последовательности в С-конце ультравариегина делают его более мощным ингибитором, чем вариегин (Фиг. 17А и С).
Для выяснения роли С-конца ультравариегина С-концевой продукт расщепления ультравариегина тромбином синтезировали и испытывали как UV011 (Фиг. 17 В и D). Этот пептид имел Ki 1,39 нМ, что примерно в 10 раз лучше, чем у C-концевого продукта расщепления вариегина (MH22, Ki = 14,1 нМ) и примерно в 4 раза лучше, чем у C-концевого продукта расщепления аватрина (IS20, Ki = 5,76 нМ). Мутация Ala27 на Glu в UV012 также вызывала 5-кратное снижение аффинности по сравнению с ультравариегином. Мутация Thr14-Gln не меняла активности ультравариегина (Фиг. 18 A и B). Мутации последовательности вариегина от VHis12-VLys13 до Tyr-Ser, тогда как ультравариегин имеют UTyr12-USer13 в этом положении, не значительно улучшали активность вариегина (Фиг. 18 C и D). Результаты всех пептидов ультравариегина приведены в табл. 5.
Четыре пептида с остатком(-ами) цистеина либо на N- или С-конце синтезировали и тестировали их активность. Варианты пептидов с остатком(-ами) цистеина обеспечивают среднюю ковалентную иммобилизацию пептидов на поверхностях для покрытия. До сих пор добавление цистеинов на концах пептидов обычно вызывало небольшое, но приемлемое снижение активности по сравнению с аналогичными последовательностями без цистеина(-ов). Значения IC50 и Ki пептидов приведены в табл. 5.
Таблица 5. Последовательность, IC50 и Ki ультравариегина и его вариантов.
Тестировали ингибирование амидолитической активности тромбина и селективность пептидов других клещей, о которых говорилось выше. Было обнаружено, что все пептиды избирательно ингибируют тромбин. Кинетика и селективность этих пептидов подробно показаны на Фиг. 19-25.
Ингибиторы тромбина в качестве стабилизирующего агента в устройствах для взятия крови
Мы протестировали вариегин, ультравариегин и аватрин на антикоагуляционный эффект в пробирках для взятия крови при комнатной температуре в трех концентрациях: 75 мкМ, 150 мкМ и 300 мкМ. Вариегин, но не аватрин и ультравариегин, ранее включали в качестве добавки в пробирки для взятия крови в патентной заявке (WO2012075407A2). Аватрин (Ki=545 пМ) имеет схожую аффинность к вариегину (Ki=318 пМ), и он показал эффект антикоагуляции, сопоставимый с эффектом вариегина (табл. 6).
Таблица 6. Антикоагулянтный эффект вариегина и связанных с ним пептидов в крови
Время образования сгустка, которое наблюдали, одинаково как для аватрина, так и для вариегина при той же концентрации (например, 52-66 ч при 150 мкМ). Как и ожидалось, ультравариегин превосходил по своим техническим характеристикам аватрин и вариегин (табл. 3). Это согласуется с более 200-кратной более сильной аффинностью ультравариегина (Ki равно 1,5 пМ) по сравнению с двумя последними пептидами. Даже при самой низкой концентрации, антикоагуляционный эффект ультравариегина (91-101 ч при 75 мкМ) длился дольше, чем при самой высокой концентрации протестированных пектина/аватрина (52-66 ч при 300 мкМ). Напротив, в другом варианте (β-вариегин) Ki составляет более 30 нМ (т.е. более чем в 100 раз слабее, чем вариегин) и способен только предотвратить образование сгустков в течение 3-4 часов. Данные показали, что аффинность пептидов к тромбину хорошо коррелирует с их антикоагуляционным эффектом в пробирках для взятия крови.
Расширенная стабильность крови для теста функции тромбоцитов
Мы также тестировали ответную реакцию крови на агрегацию тромбоцитов под антикоагуляционным действием ультравариегина, аватрина и вариегина при 150 мкМ и 300 мкМ с использованием анализатора Multiplate®. Мы использовали АДФ (6,5 мкМ) в качестве агониста. Мы использовали цитрат и гирудин в качестве двух контролей. Полученные результаты показаны в табл. 7 и на Фиг. 26.
Таблица 7. Сводная таблица ответных реакций на агрегацию тромбоцитов (в виде области под кривой, U) крови (АДФ в качестве агониста) в пробирках для взятия крови, содержащих различные стабилизирующие агенты.
** Считывания не были получены из-за образования сгустка
* Концентрация гирудина является такой, как определено в информации для продукта пробирки гирудина Vacuette®
Поскольку в литературе нет консенсуса по нижнему пределу ответной реакции, который можно считать надежным показателем, мы решили использовать отсечку 32 U (Фиг. 26, пунктирная линия). Это ответная реакция, получаемая в содержащей цитрат крови в наших экспериментах через 4 часа, что является максимальным временем анализа, рекомендованным в стандарте тестирования функции тромбоцитов CLSI Guideline H58-P.
Пробирки для взятия крови с ультравариегином, аватрином и вариегином все показали по меньшей мере 6-кратное увеличение окна стабилизации по сравнению с золотым стандартом - цитратом. В первые 24 ч кровь с гирудином (Фиг. 26, оранжевый) проявляла свойства аналогичные в сравнении с ультравариегином (Фиг. 26, серый), аватрином (Фиг. 26, синий) и вариегином (Фиг. 26, желтый), что может быть неудивительно, поскольку гирудин также является мощным прямым ингибитором тромбина [Warkentin TE. Best Pract Res Clin Haematol 17: 105–25 (2004)]. Относительно более сильная аффинность ультравариегина и гирудина к тромбину приводила к более высоким ответным реакциям на агрегацию через 48 ч и 72 ч по сравнению с двумя другими пептидами. Тем не менее, эти показатели немного ниже установленной отсечки 32 U (Фиг. 26).
Обсуждение
Несмотря на низкое общее сходство последовательностей аватрина и вариегина (40 %) и вариации ключевых функциональных остатков, как описано выше, аватрин, по-видимому, функционирует аналогично вариегину. Оба ингибируют тромбин в качестве быстрых двухвалентных ингибиторов сильной связи, нацеленных на активный центр и экзосайт-I с похожими константами ингибирования (Ki вариегина = 342 пM, Ki аватрина = 545 пM). Как вариегин, так и аватрин канонически связываются с тромбином и, следовательно, расщепляются тромбином при связывании. Расщепление вариегина (4 часа до завершения) тромбином происходит быстрее, чем аватрин (10 ч до завершения). Более медленное расщепление аватрина может быть связано с общей гибкостью, обеспечиваемой AGly15, AGly17 и AGly18. Продукты расщепления обоих пептидов ингибировали тромбин, не будучи конкурентоспособными по отношению к малым пептидным субстратам, причем аффинность продукта расщепления вариегина (MH22) приблизительно в 2 раза ниже, чем у продукта расщепления аватрина (IS20) [Koh CY, et al., ChemBioChem 10: 2155–8 (2009)]. Более медленная скорость расщепления аватрина, вероятно, может быть связана с более высокой аффинностью IS20, что снижает доступность свободного тромбина для расщепления пептида в равновесии. Мы также показали, что серин в положении 12, по-видимому, обеспечивает немного лучшую аффинность, чем гистидин, тогда как кислотные остатки на N-конце не определяют кинетику быстрого связывания. Сравнение последовательностей вариегина и аватрина позволило провести эти целенаправленные структурно-функциональные исследования и помогло понять взаимодействия ингибиторов тромбина.
Структуру комплекса тромбин-аватрин лучше сравнивать с комплексом тромбин-гирулог-1 (RMSD 0,41 Å для 279 остатков), так как оба являются ингибиторами пептидил-бивалентного тромбина, кристаллизованными как расщепленные пептиды [Skrzypczak-Jankun E, et al., J Mol Biol 221: 1379–93 (1991); Bourdon P, et al., FEBS Lett 294: 163–6 (1991)]. Гирулог-1 представляет собой более короткий пептид по сравнению с аватрином, но два пептида проявляют значительную степень идентичности в активном центре их тромбина и последовательностях связывания экзосайта-I. После расщепления, только сегмент N-конца до расщепляемой связи и сегмента в экзосайте-I могут быть построены в их соответствующих структурах. Оба кристалла имеют одинаковые размеры элементарной ячейки (C2; a/b/c приблизительно составляет 70/72/72 Å; β приблизительно составляет 100°). Напротив, комплекс тромбин-вариегин кристаллизован в другой кристаллической форме (C2; a/b/c составляет 125/51/62 Å; β составляет 99°) и, по-видимому, связан только с расщепленным C-концевым пептидом.
Как аватрин, так и гирулог-1 имеют свои остатки P1, связывающиеся с одним и тем же карманом S1 на тромбине. Хотя P1 Lys гирулога-1 связывается с TAsp189 на дне кармана S1 через молекулу воды [Bode W. Blood Cells, Mol Dis 36: 122–30 (2006)], мы наблюдали прямое взаимодействие между ALys10 и TAsp189. В том же исследовании с гирулогом-1 показано уменьшение аффинности в 10 раз, когда Arg заменен на Lys, но мы видим меньшее изменение (приблизительно трехкратное) с ALys10 до мутации Arg, что согласуется с прямым взаимодействием, наблюдаемым в структуре. Гирулог-1 и аватрин имеют одну и ту же P2-аминокислоту (Pro) и, следовательно, взаимодействуют подобным образом с сайтом S2. Гирулог-1 P3 представляет собой D-Phe, занимая тот же самый гидрофобный карман, который занимает аватрин P4 AAla7 [Skrzypczak-Jankun E, et al., J Mol Biol 221: 1379–93 (1991)]. Хромогенный субстрат S2238, используемый во всех наших ферментативных анализах, почти идентичен гирулогу-1 при P3-P1 (D-Phe-Пипеколиновая кислота-Arg). Учитывая, что аватрин и гирулог-1 связываются с одними и теми же центрами на тромбине в этих положениях, конкурентный механизм ингибирования согласуется со структурой
Первые шесть остатков экзосайта-1 связующих сегментов аватрина (DFEEIPSDEIIE), гирулога-1 (DFEEIPEEYL) и вариегина (DFEAIPEEYLDDES) почти идентичны (подчеркнуты). Эти остатки также хорошо выровнены в кристаллических структурах (Фиг. 27). Взаимодействия в трех пептидах с тромбином в основном сохраняются во всех структурах. Единственный неконсервативный остаток в этом сегменте (AGlu22 по отношению к HGlu57 по отношению к VAla22) является гидрофильным во всех трех структурах без специфических взаимодействий. Вариегин и гирулог-1 используют следующие 4 остатка, EEYL, которые соответствуют последовательности SDEI в аватрине. Эти остатки наблюдаются в комплексах аватрин и вариегин-тромбин, но не в комплексе гирулог-1-тромбина. Среди соответствующих 4 остатков аватрина (SDEI), ASer25 и AAsp26 являются гидрофильными. AGlu27 и VGlu26 являются структурно эквивалентными и имеют электростатическое взаимодействие с TArg77A. Анализ последующих остатков затруднен из-за отсутствия четкой плотности.
Несмотря на использование последовательности вариегина для разработки праймеров для амплификации, нам не удалось амплифицировать ген вариегина, что свидетельствует о высокой степени вариабельности пептидов, продуцируемых множеством генов в этих клещах в разные моменты питания. Более удивительно, что как аватрин, так и вариегин, как представляется, синтезируются в виде более крупных белков-предшественников, содержащих множественные повторы, и процессируются в более короткие активные пептиды, которые обладают активностью ингибирования тромбина. Несмотря на низкую общую идентичность последовательности и вариации в некоторых ключевых функциональных остатках, оба пептида имеют сходные ингибирующие механизмы и эффекты на тромбин. Поиск в базе данных выявил более похожие последовательности таких белков-предшественников, содержащих повторы подобных вариегину пептидов Amblyomma variegatum (BM291228: три пептида, неполная транскрипция, BM293052: пять пептидов, BM289492: пять пептидов), Amblyomma americanum (ACG76173: пять пептидов, неполная транскрипция) и Amblyomma cajennense (ACAJ0085C_1: четыре пептида) [Nene V, et al., Int J Parasitol 32: 1447–56 (2002); Batista IFC, et al., Toxicon 51: 823–34 (2008)] (Фиг. 28). Имеются также свидетельства наличия подобных пептидов в слюнных железах других твердых клещей, таких как Rhipicephalus sanguineus и Hyalomma marginatum rufipes. Большинство предшественников можно процессировать в три-пять пептидов с образованием нового семейства близкородственных ингибиторов пептидил-тромбина. Мы называем это семейство ингибиторов тромбина иксотрины (ингибиторы тромбина Ixodidae). Иксотрины являются небольшими ингибиторами тромбина без каких-либо дисульфидных связей и встречаются в семействе Ixodidae (твердых клещей). Они связываются с экзосайтом-1 и активным центром тромбина. Несколько копий иксотринов производятся в одном предшественнике. Каждый предшественник имеет сигнальный пептид, и посттрансляционная обработка происходит либо в эндоплазматическом ретикулуме, либо в слюне в случае зрелых иксотринов. По-видимому, такой многопродукционный подход при производстве ингибиторов пептидил-тромбина довольно распространен среди твердых клещей. Множественность компонентов слюны для нацеливания на молекулу коагуляции одного хозяина не является чем-то необычным [Francischetti IMB, et al., J Proteomics 71: 493–512 (2008); Fontaine A, et al., Parasit Vectors BioMed Central Ltd; 4: 187 (2011)], хотя мы считаем, что это один из первых примеров, когда множественность также встроена в единственный белок-предшественник.
В заключение мы продемонстрировали, что твердые клещи инактивируют тромбин, который является решающим ферментом при коагуляции, используя множество разнообразных последовательностей, сохраняя в значительной степени аналогичный общий скаффолд и функцию. Мы продемонстрировали, что аватрин и несколько других пептидов предотвратили тромбоз лучше, чем гирулог-1, несмотря на сходство в последовательностях, в модели тромбоза сонной артерии, индуцированного FeCl3. Это семейство молекул может производить антикоагулянты, полезные для нескольких клинических показаний и сердечно-сосудистых процедур, таких как профилактика артериального тромбоза и реокклюзии во время инвазивных процедур, профилактика венозного тромбоза после ортопедической хирургии и лечение инфаркта миокарда после детальной оценки профилей безопасности и эффективности [Bauer K A. Hematology Am Soc Hematology Educ Program 2013: 464-70 (2013)].
Ссылки
Batista IFC, Chudzinski-Tavassi AM, Faria F, Simons SM, Barros-Batestti DM, Labruna MB, Leo LI, Ho PL, Junqueira-de-Azevedo ILM. Expressed sequence tags (ESTs) from the salivary glands of the tick Amblyomma cajennense (Acari: Ixodidae). Toxicon 2008; 51: 823–34.
Battye TGG, Kontogiannis L, Johnson O, Powell HR, Leslie AGW. iMOSFLM: A new graphical interface for diffraction-image processing with MOSFLM. Acta Crystallogr Sect D Biol Crystallogr 2011; 67: 271–81.
Bauer K. A. Pros and cons of new oral anticoagulants. Haem. 2013. 464-470.
Berliner LJ. Thrombin Structure and Function. Journal of Chemical Information and Modeling. 1992.
Bourdon P. Hirulog peptides with sciccile bond replacements resistant to thrombin cleavage. Biochem Biophys Res Commun 1991; 177: 1049–55.
Bourdon P, Jablonski J a., Chao BH, Maraganore JM. Structure-function relationships of hirulog peptide interactions with thrombin. FEBS Lett 1991; 294: 163–6.
Bridge KI, Philippou H, Ariëns R a S. Clot properties and cardiovascular disease. Thromb Haemost 2014; 112: 1–8.
Chaudhari K., Hamad B. and Syed B.A. Antithrombotic drugs market. Nat. Rev. Drug. Discov. 2014, 13, 571-572.
Copeland R a. Enzymes: A Practical Introduction to Structure, mechanism and Data analysis. Enzymes: A practical Introduction to Structure, mechanism, and data analysis. 2000.
Di Cera E. Thrombin. Mol Aspects Med. 2008, 29(4), 203–254.
Eckly a, Hechler B, Freund M, Zerr M, Cazenave J-P, Lanza F, Mangin PH, Gachet C. Mechanisms underlying FeCl3-induced arterial thrombosis. J Thromb Haemost 2011; 9: 779–89.
Emsley P, Cowtan K. Coot: Model-building tools for molecular graphics. Acta Crystallogr Sect D Biol Crystallogr International Union of Crystallography; 2004; 60: 2126–32.
Evans PR, Murshudov GN. How good are my data and what is the resolution? Acta Crystallogr Sect D Biol Crystallogr 2013; 69: 1204–14.
Francischetti IMB, Meng Z, Mans BJ, Gudderra N, Hall M, Veenstra TD, Pham VM, Kotsyfakis M, Ribeiro JMC. An insight into the salivary transcriptome and proteome of the soft tick and vector of epizootic bovine abortion, Ornithodoros coriaceus. J Proteomics 2008, 71: 493–512.
Fontaine A, Diouf I, Bakkali N, Missé D, Pagès F, Fusai T, Rogier C, Almeras L. Implication of haematophagous arthropod salivary proteins in host-vector interactions. Parasit Vectors BioMed Central Ltd; 2011, 4: 187.
Gallwitz M, Enoksson M, Thorpe M, Hellman L. The extended cleavage specificity of human thrombin. PLoS One 2012; 7.
Huntington J. A. Natural inhibitors of thrombin. Thromb Haemost. 2014, 111, 583–589.
Koh C. Y., Kazimirova M., Trimnell A., Takac P., Labuda M., Nuttall P. A. and Kini R. M., Variegin, a Novel Fast and Tight Binding Thrombin Inhibitor from the Tropical Bont Tick, 2007, 282 (40), 29101–29113.
Koh C. Y. and Kini R. M. Pros and cons of new oral anticoagulants. Expert Rev. Haematol. 2008, 1(2), 135-139.
Koh CY, Kazimirova M, Nuttall P a., Kini RM. Noncompetitive inhibitor of thrombin. ChemBioChem 2009; 10: 2155–8.
Koh CY, Kumar S, Kazimirova M, Nuttall P a., Radhakrishnan UP, Kim S, Jagadeeswaran P, Imamura T, Mizuguchi J, Iwanaga S, Swaminathan K, Kini RM. Crystal structure of thrombin in complex with s-variegin: Insights of a novel mechanism of inhibition and design of tunable thrombin inhibitors. PLoS One 2011; 6.
Leslie AGW, Powell HR. Processing diffraction data with MOSFLM. Evolving methods for macromolecular Crystallography. 2007.
McCoy AJ. Solving structures of protein complexes by molecular replacement with Phaser. Acta Crystallogr Sect D Biol Crystallogr International Union of Crystallography; 2006; 63: 32–41.
Michiel Coppens, John W. Eikelboom, David Gustafsson, Jeffrey I. Weitz, Jack Hirsh. Development of Direct Thrombin Inhibitors, Circ Res. 2012, 112, 920-931.
Monroe D. M., Hoffman M. and Roberts H. R. Platelets and Thrombin Generation. Arterioscler Thromb Vasc Biol., 2002, 22, 1381-1389.
Nene V, Lee D, Quackenbush J, Skilton R, Mwaura S, Gardner MJ, Bishop R. AvGI, an index of genes transcribed in the salivary glands of the ixodid tick Amblyomma variegatum. Int J Parasitol 2002; 32: 1447–56.
Qiu X, Padmanabhan KP, Carperos VE, Tulinsky a, Kline T, Maraganore JM, Fenton JW. Structure of the hirulog 3-thrombin complex and nature of the S’ subsites of substrates and inhibitors. Biochemistry 1992; 31: 11689–97.
Raskob G. Thrombosis: A major contributor to global disease burden. Thromb Haem. 2014, 112(5), 843-943.
Skrzypczak-Jankun E, Carperos VE, Ravichandran KG, Tulinsky a, Westbrook M, Maraganore JM. Structure of the hirugen and hirulog 1 complexes of alpha-thrombin. J Mol Biol 1991; 221: 1379–93.
Stamenova PK, Marchetti T, Simeonov I. Efficacy and safety of topical hirudin (Hirudex): a double-blind, placebo-controlled study. Eur Rev Med Pharmacol Sci. 2001 Mar-Apr; 5(2):37-42.
Stubbs M. T. and Bode W. A player of many parts: the spotlight falls on thrombin’s structure. Throm Res.1993, 69, 1-58.
Versteeg H. H., Heemskerk J. W. M., Levi M., and. Reitsma P. H. New fundamentals in hemostasis. Physiol Rev. 2013, 93, 327–358.
Wan C, Carvalho LPD, Chan MY, Kini RM, Kang TS. Fasxiator, a novel factor XIa inhibitor from snake venom, and its site-specific mutagenesis to improve potency and selectivity. J Thromb Haemost 2015; 13: 248–61.
Warkentin TE. Bivalent direct thrombin inhibitors: Hirudin and bivalirudin. Best Pract Res Clin Haematol 2004; 17: 105–25.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> National University of Singapore
Institute of Zoology, Slovak Academy of Sciences
<120> НОВЫЕ ИНГИБИТОРЫ ТРОМБИНА
<130> FP8143
<150> US 62/230,923
<151> 2015-06-18
<160> 27
<170> PatentIn version 3.5
<210> 1
<211> 30
<212> PRT
<213> Amblyomma variegatum
<400> 1
Ser Gly Gly His Gln Thr Ala Val Pro Lys Ile Ser Lys Gln Gly Leu
1 5 10 15
Gly Gly Asp Phe Glu Glu Ile Pro Ser Asp Glu Ile Ile Glu
20 25 30
<210> 2
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> Thr22Glu substitution of native Amblyomma variegatum sequence
<400> 2
Ser Asp Glu Ala Val Arg Ala Ile Pro Lys Met Tyr Ser Thr Ala Pro
1 5 10 15
Pro Gly Asp Phe Glu Glu Ile Pro Asp Asp Ala Ile Glu Glu
20 25 30
<210> 3
<211> 26
<212> PRT
<213> Amblyomma variegatum
<400> 3
Gln Thr Ala Val Pro Lys Ile Ser Lys Gln Gly Leu Gly Gly Asp Phe
1 5 10 15
Glu Glu Ile Pro Ser Asp Glu Ile Ile Glu
20 25
<210> 4
<211> 20
<212> PRT
<213> Amblyomma variegatum
<400> 4
Ile Ser Lys Gln Gly Leu Gly Gly Asp Phe Glu Glu Ile Pro Ser Asp
1 5 10 15
Glu Ile Ile Glu
20
<210> 5
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> Ser12Ala substitution
<400> 5
Ser Gly Gly His Gln Thr Ala Val Pro Lys Ile Ala Lys Gln Gly Leu
1 5 10 15
Gly Gly Asp Phe Glu Glu Ile Pro Ser Asp Glu Ile Ile Glu
20 25 30
<210> 6
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> Ser12His substitution
<400> 6
Ser Gly Gly His Gln Thr Ala Val Pro Lys Ile His Lys Gln Gly Leu
1 5 10 15
Gly Gly Asp Phe Glu Glu Ile Pro Ser Asp Glu Ile Ile Glu
20 25 30
<210> 7
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> Lys10Arg substitution
<400> 7
Ser Gly Gly His Gln Thr Ala Val Pro Arg Ile Ser Lys Gln Gly Leu
1 5 10 15
Gly Gly Asp Phe Glu Glu Ile Pro Ser Asp Glu Ile Ile Glu
20 25 30
<210> 8
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> Lys10beta homoarginine substitution
<220>
<221> MOD_RES
<222> (10)..(10)
<223> beta-homoarginine
<400> 8
Ser Gly Gly His Gln Thr Ala Val Pro Xaa Ile Ser Lys Gln Gly Leu
1 5 10 15
Gly Gly Asp Phe Glu Glu Ile Pro Ser Asp Glu Ile Ile Glu
20 25 30
<210> 9
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> Lys10beta homoarginine substitution
<220>
<221> MOD_RES
<222> (10)..(10)
<223> beta-homoarginine
<400> 9
Ser Asp Glu Ala Val Arg Ala Ile Pro Xaa Met Tyr Ser Thr Ala Pro
1 5 10 15
Pro Gly Asp Phe Glu Glu Ile Pro Asp Asp Ala Ile Glu Glu
20 25 30
<210> 10
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> ultravariegin-variegin hybrid
<400> 10
Ser Asp Gln Gly Asp Val Ala Ile Pro Lys Met Tyr Ser Thr Ala Pro
1 5 10 15
Pro Gly Asp Phe Glu Glu Ile Pro Asp Asp Ala Ile Glu Glu
20 25 30
<210> 11
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> ultravariegin-variegin hybrid
<400> 11
Ser Asp Glu Ala Val Arg Ala Glu Pro Lys Met His Lys Thr Ala Pro
1 5 10 15
Pro Gly Asp Phe Glu Glu Ile Pro Asp Asp Ala Ile Glu Glu
20 25 30
<210> 12
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> ultravariegin-variegin hybrid
<400> 12
Ser Asp Glu Ala Val Arg Ala Ile Pro Lys Met Tyr Ser Thr Ala Pro
1 5 10 15
Pro Gly Asp Phe Glu Glu Ile Pro Glu Glu Tyr Leu Asp Asp Glu Ser
20 25 30
<210> 13
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> ultravariegin C-terminal thrombin cleavage product
<400> 13
Met Tyr Ser Thr Ala Pro Pro Gly Asp Phe Glu Glu Ile Pro Asp Asp
1 5 10 15
Ala Ile Glu Glu
20
<210> 14
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> ultravariegin Ala27Glu substitution
<400> 14
Ser Asp Glu Ala Val Arg Ala Ile Pro Lys Met Tyr Ser Thr Ala Pro
1 5 10 15
Pro Gly Asp Phe Glu Glu Ile Pro Asp Asp Glu Ile Glu Glu
20 25 30
<210> 15
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> ultravariegin Thr14Gln substitution
<400> 15
Ser Asp Glu Ala Val Arg Ala Ile Pro Lys Met Tyr Ser Gln Ala Pro
1 5 10 15
Pro Gly Asp Phe Glu Glu Ile Pro Asp Asp Ala Ile Glu Glu
20 25 30
<210> 16
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> variegin variant
<400> 16
Ser Asp Gln Gly Asp Val Ala Glu Pro Lys Met Tyr Ser Thr Ala Pro
1 5 10 15
Pro Phe Asp Phe Glu Ala Ile Pro Glu Glu Tyr Leu Asp Asp Glu Ser
20 25 30
<210> 17
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> beta homoarginine variegin variant
<220>
<221> MOD_RES
<222> (10)..(10)
<223> beta homoarginine
<400> 17
Ser Asp Gln Gly Asp Val Ala Glu Pro Xaa Met His Ser Thr Ala Pro
1 5 10 15
Pro Phe Asp Phe Glu Ala Ile Pro Glu Glu Tyr Leu Asp Asp Glu Ser
20 25 30
<210> 18
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<223> ultravariegin N-terminal Cys
<400> 18
Cys Asp Glu Ala Val Arg Ala Ile Pro Lys Met Tyr Ser Thr Ala Pro
1 5 10 15
Pro Gly Asp Phe Glu Glu Ile Pro Asp Asp Ala Ile Glu Glu
20 25 30
<210> 19
<211> 32
<212> PRT
<213> Artificial Sequence
<220>
<223> ultravariegin C-terminal Cys
<400> 19
Ser Asp Glu Ala Val Arg Ala Ile Pro Lys Met Tyr Ser Thr Ala Pro
1 5 10 15
Pro Gly Asp Phe Glu Glu Ile Pro Asp Asp Ala Ile Glu Glu Cys Ala
20 25 30
<210> 20
<211> 24
<212> PRT
<213> Artificial Sequence
<220>
<223> ultravariegin fragment c-terminal Cys CysCys
<400> 20
Met Tyr Ser Thr Ala Pro Pro Gly Asp Phe Glu Glu Ile Pro Asp Asp
1 5 10 15
Ala Ile Glu Glu Gly Cys Cys Cys
20
<210> 21
<211> 34
<212> PRT
<213> Artificial Sequence
<220>
<223> ultravariegin C-terminal CysCysCys
<400> 21
Ser Asp Glu Ala Val Arg Ala Ile Pro Lys Met Tyr Ser Thr Ala Pro
1 5 10 15
Pro Gly Asp Phe Glu Glu Ile Pro Asp Asp Ala Ile Glu Glu Gly Cys
20 25 30
Cys Cys
<210> 22
<211> 35
<212> PRT
<213> Amblyomma americanum
<400> 22
Ser Gly Glu Asp His Thr Ala Val Pro Lys Met Ser Arg Lys Gly Leu
1 5 10 15
Gly Gly Asp Phe Glu Asp Ile Pro Pro Glu Ala Tyr Glu Arg Ala Leu
20 25 30
Glu Ala Arg
35
<210> 23
<211> 46
<212> PRT
<213> Amblyomma maculatum
<400> 23
Glu Leu Glu Ser Gly Asp Glu Asp Ser Glu Gly Gly Asp Ser Gln Ser
1 5 10 15
Ser Pro Thr Glu Ser Ala Ala Pro Arg Leu His Gln Arg Glu Gly Gly
20 25 30
Gly Gly Asp Phe Glu Asn Val Glu Tyr Asp Gln Asp Gln Lys
35 40 45
<210> 24
<211> 46
<212> PRT
<213> Rhipicephalus sanguineus
<400> 24
Ser Asp Val Ala Pro Ala Asp Tyr Glu Ser Asp Glu Gly Asp Asn Asp
1 5 10 15
Gly Gly His Asp Gly Ser Glu Val Ala Lys Pro Lys Met Pro Arg Gly
20 25 30
Asn Gly Gly Gly Gly Asp Phe Glu Glu Ile Pro Glu Val Glu
35 40 45
<210> 25
<211> 51
<212> PRT
<213> Hyalomma marginatum
<400> 25
Thr Gly Ser Asp Asp Asp Asp Glu Tyr Asp Met Tyr Glu Ser Asp Gly
1 5 10 15
Asp Ser Asn Glu Gly Asn Asp Asn Asp Glu Phe Glu Thr Ala Val Pro
20 25 30
Arg Leu Pro Asn Pro Asn Ser Gly Arg Asp Ser Glu His Ile Pro Met
35 40 45
Pro Val Asn
50
<210> 26
<211> 90
<212> DNA
<213> Artificial Sequence
<220>
<223> Avathrin
<400> 26
tcgggtggcc atcagactgc tgttccgaag atatctaagc aaggcttggg tggagacttt 60
gaagaaattc caagtgatga aataatcgag 90
<210> 27
<211> 90
<212> DNA
<213> Artificial Sequence
<220>
<223> ultravariegin native
<400> 27
tcagacgaag ctgtcagggc gattcccaag atgtactcga ctgccccacc gggagatttc 60
gaaacaatcc ctgacgacgc tattgaggag 90
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ ЛЕЧЕНИЯ СОСТОЯНИЙ, АССОЦИИРОВАННЫХ С MASP-2-ЗАВИСИМОЙ АКТИВАЦИЕЙ КОМПЛЕМЕНТА | 2014 |
|
RU2718850C2 |
| СПОСОБЫ ЛЕЧЕНИЯ СОСТОЯНИЙ, АССОЦИИРОВАННЫХ С MASP-2-ЗАВИСИМОЙ АКТИВАЦИЕЙ КОМПЛЕМЕНТА | 2014 |
|
RU2830289C2 |
| СПОСОБЫ ЛЕЧЕНИЯ СОСТОЯНИЙ, СВЯЗАННЫХ С MASP-2 ЗАВИСИМОЙ АКТИВАЦИЕЙ КОМПЛЕМЕНТА | 2012 |
|
RU2743409C2 |
| АНТИТЕЛА К ФАКТОРУ XI И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2739946C2 |
| ХИМЕРНЫЕ РЕЦЕПТОРЫ АНТИГЕНА ПРОТИВ МЕЗОТЕЛИНА ЧЕЛОВЕКА И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2714902C2 |
| KLK5-ИНГИБИРУЮЩИЙ ПЕПТИД | 2019 |
|
RU2826422C2 |
| НАЦЕЛЕННЫЕ НА uPARAP КОНЪЮГАТЫ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО | 2017 |
|
RU2740311C2 |
| ПЕПТИД ДЛЯ ЛЕЧЕНИЯ ВОЗРАСТНОЙ МАКУЛЯРНОЙ ДЕГЕНЕРАЦИИ | 2017 |
|
RU2788083C2 |
| АНТИ-αvβ8 АНТИТЕЛА И КОМПОЗИЦИИ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2812478C2 |
| КОНСТРУКЦИИ ДНК-АНТИТЕЛ ДЛЯ ПРИМЕНЕНИЯ ПРОТИВ БОЛЕЗНИ ЛАЙМА | 2017 |
|
RU2813829C2 |
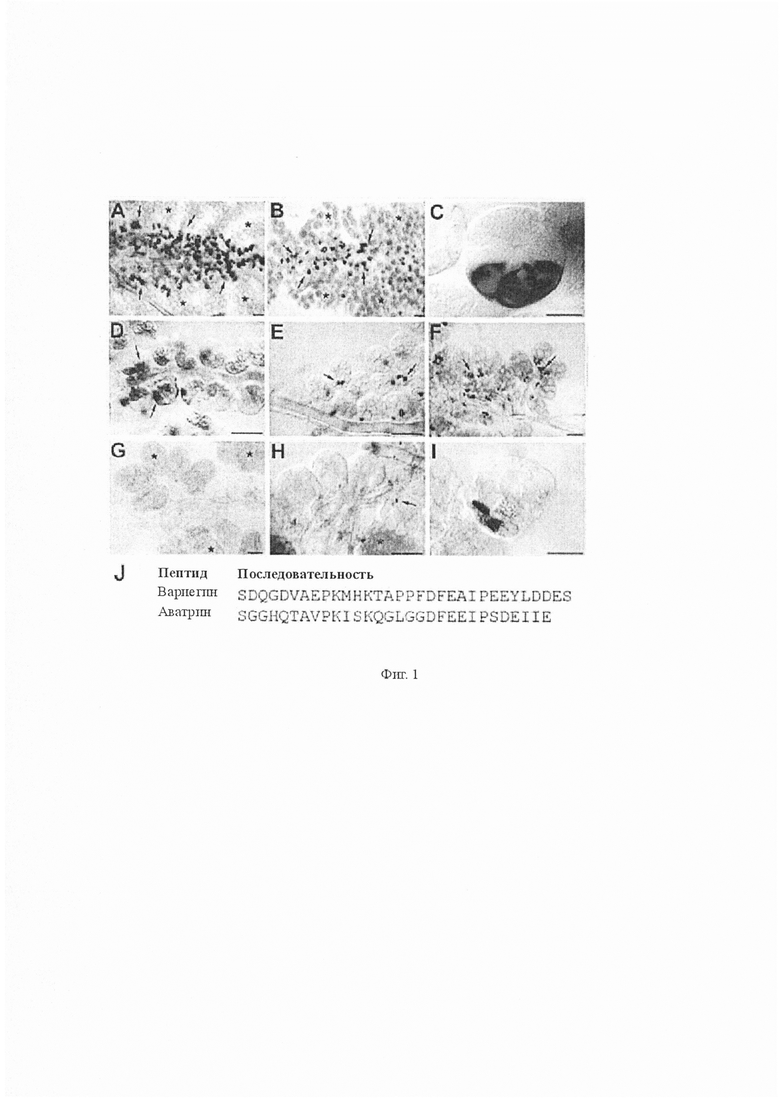
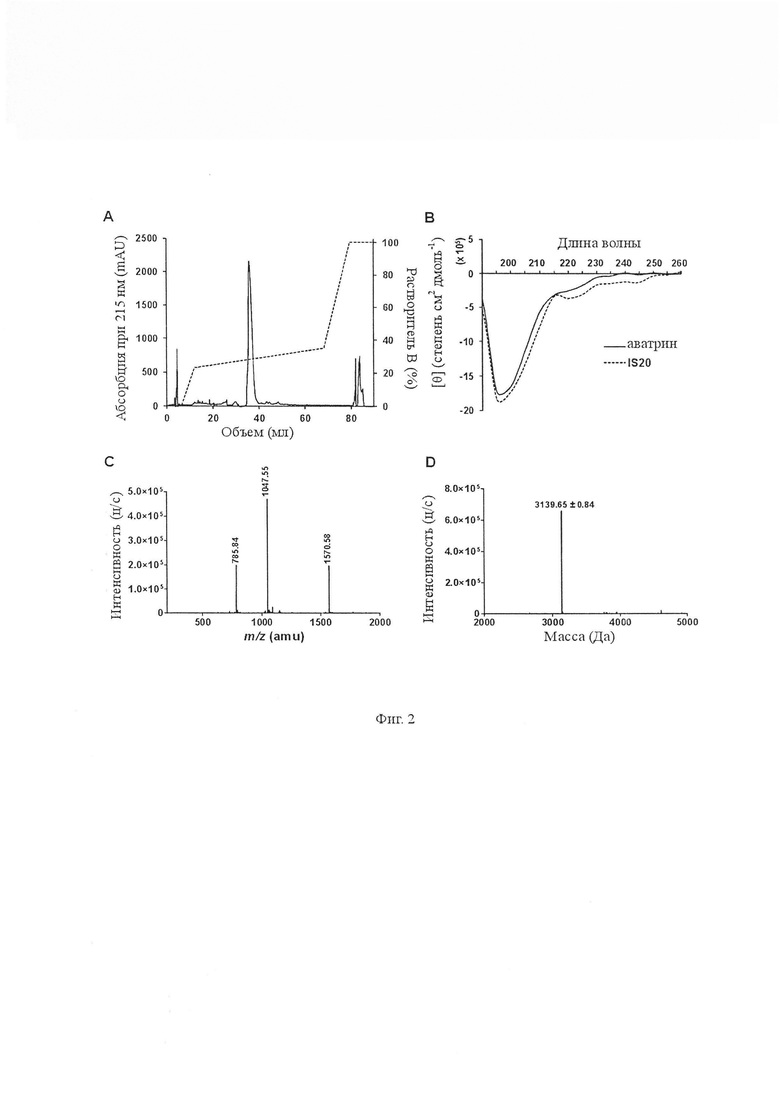
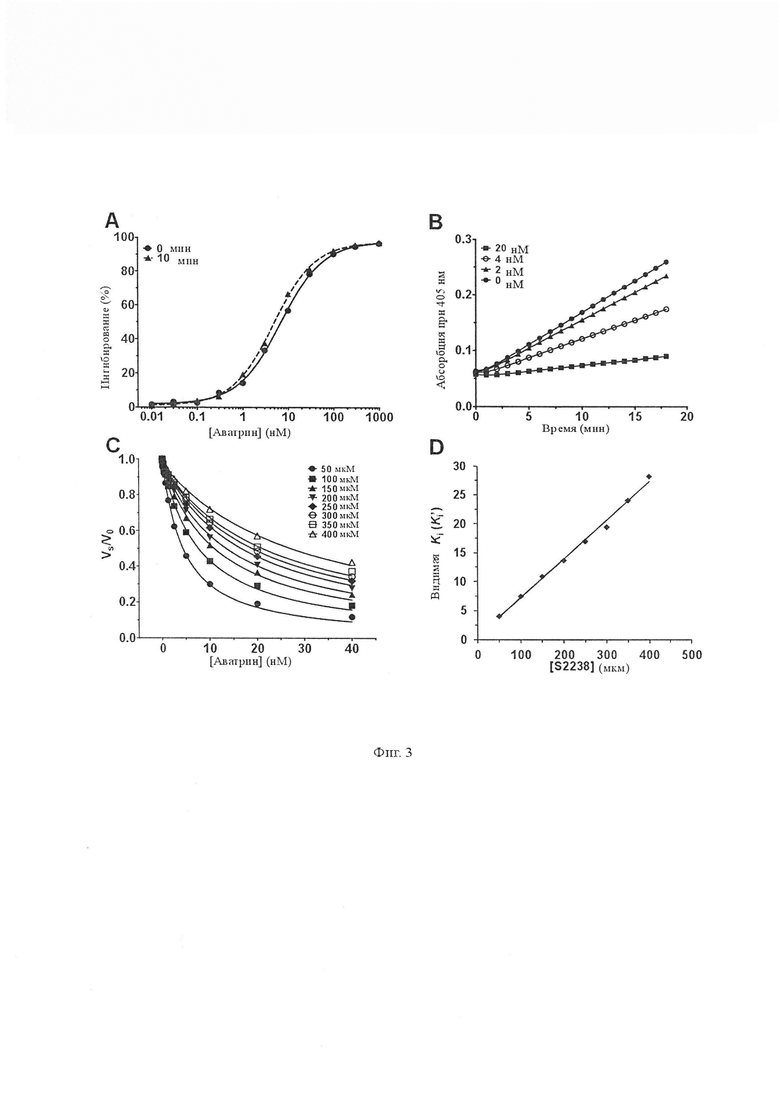
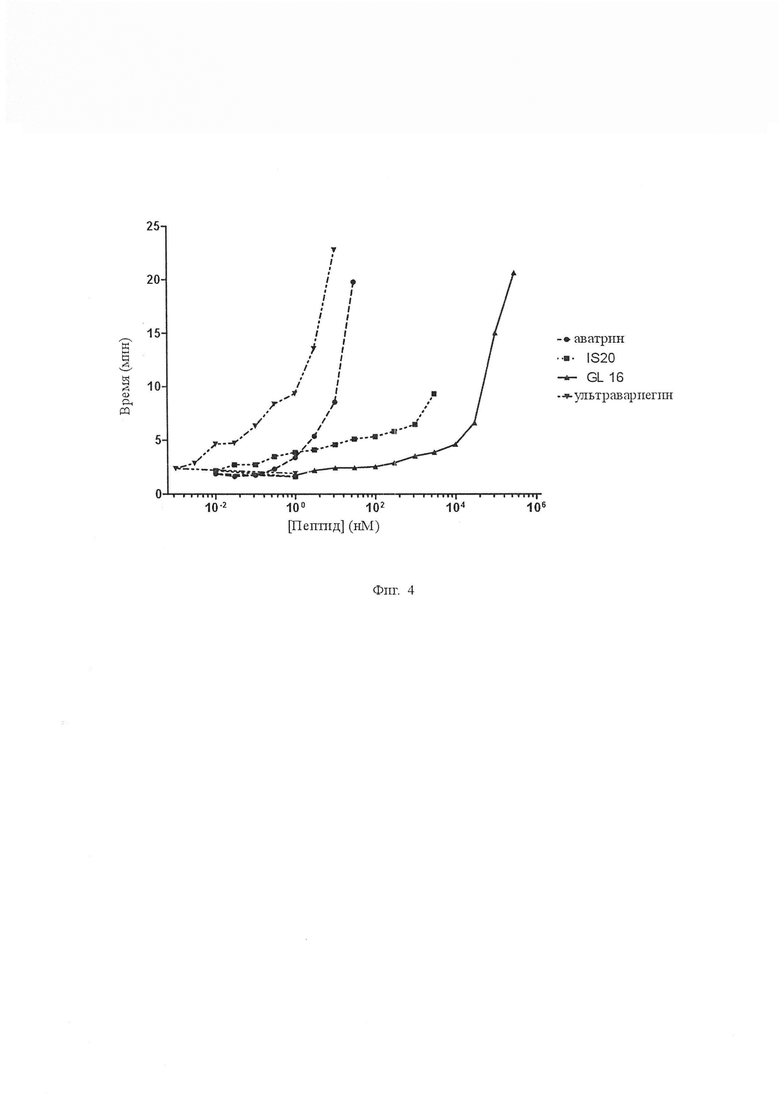
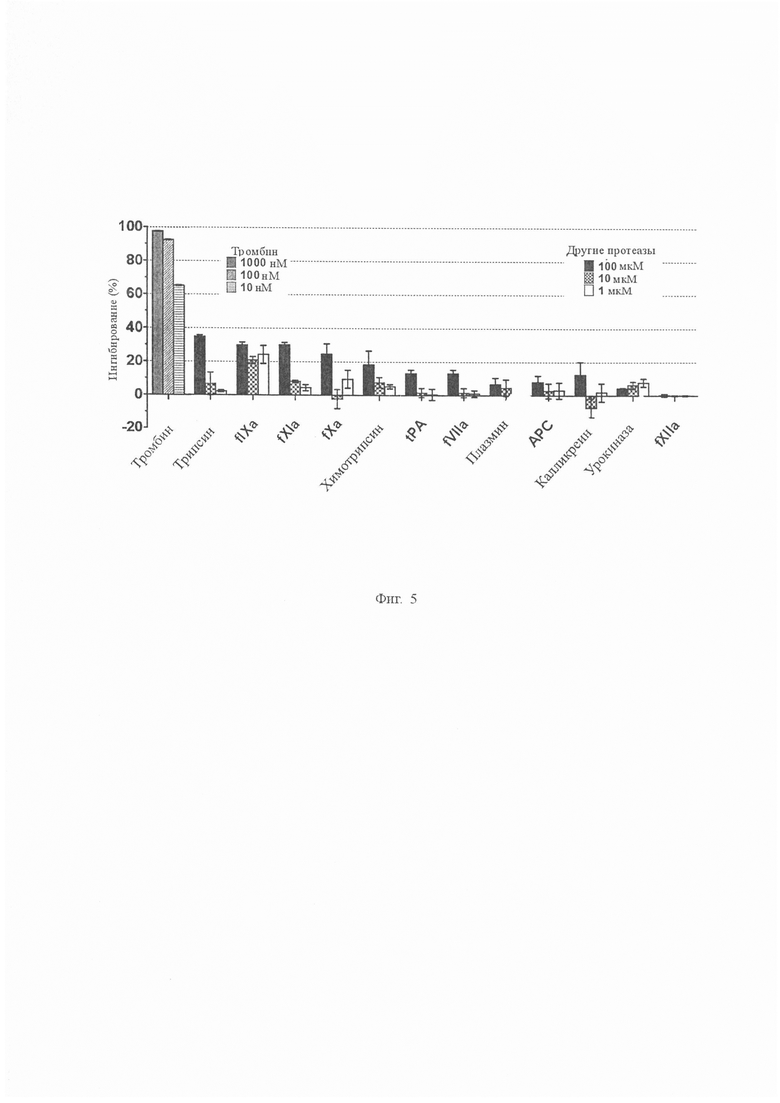
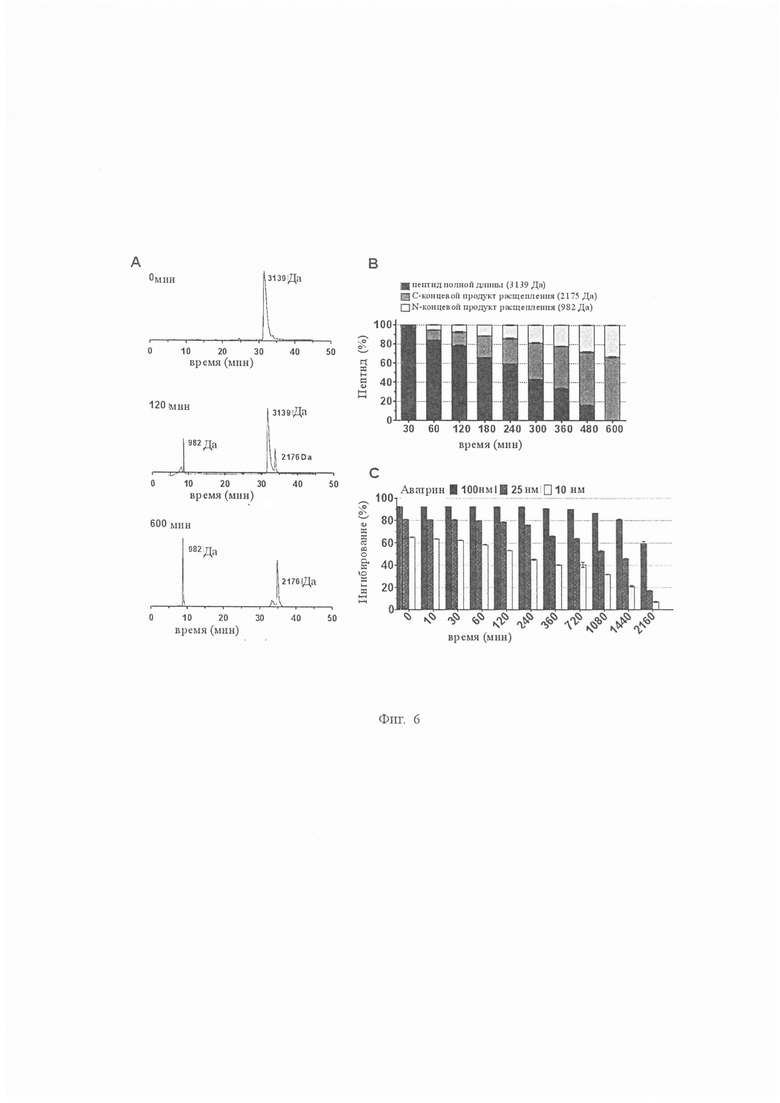
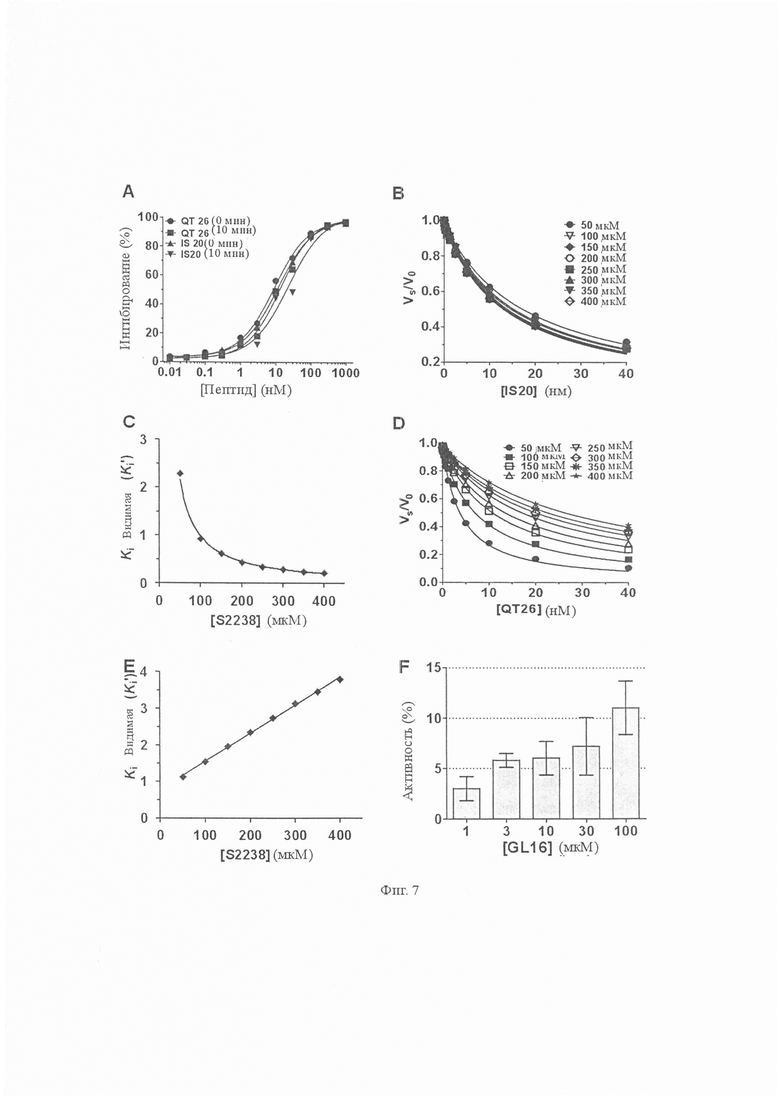
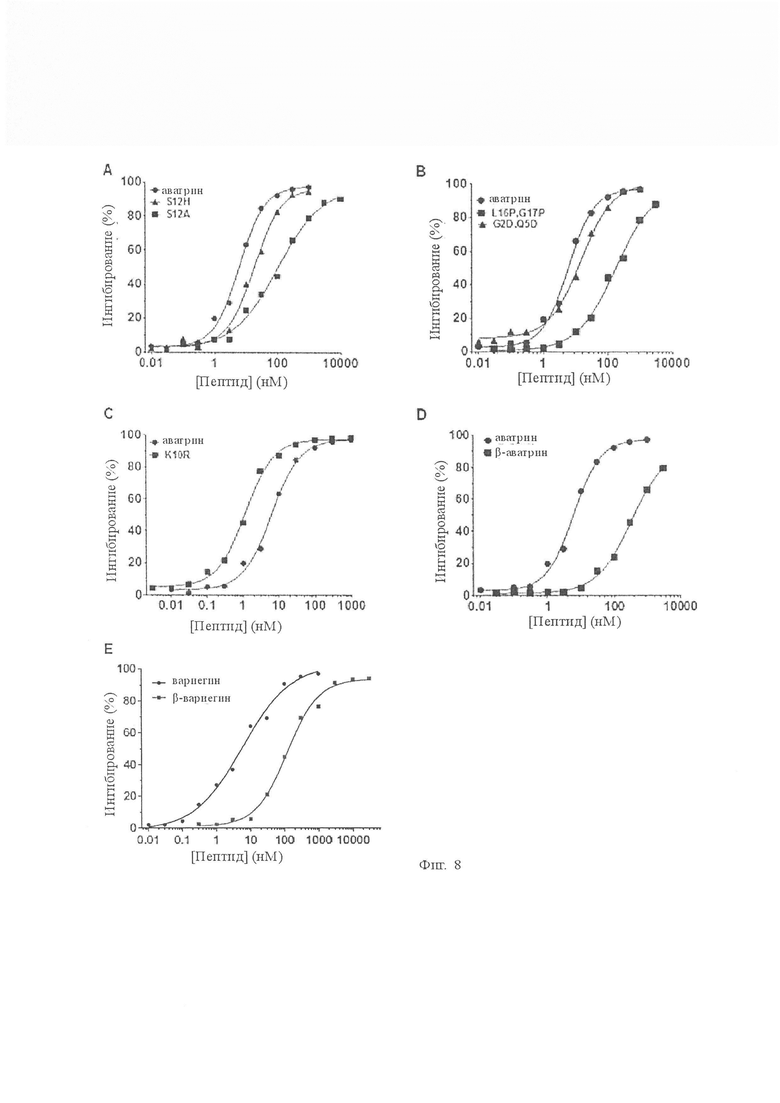


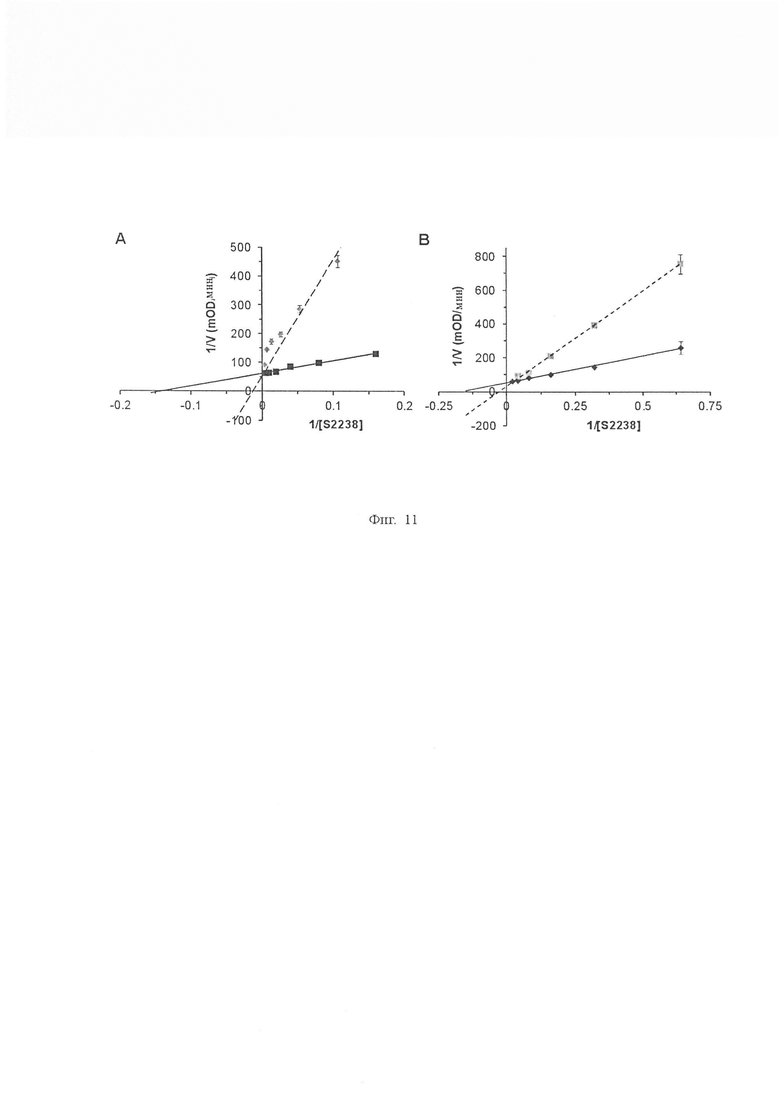
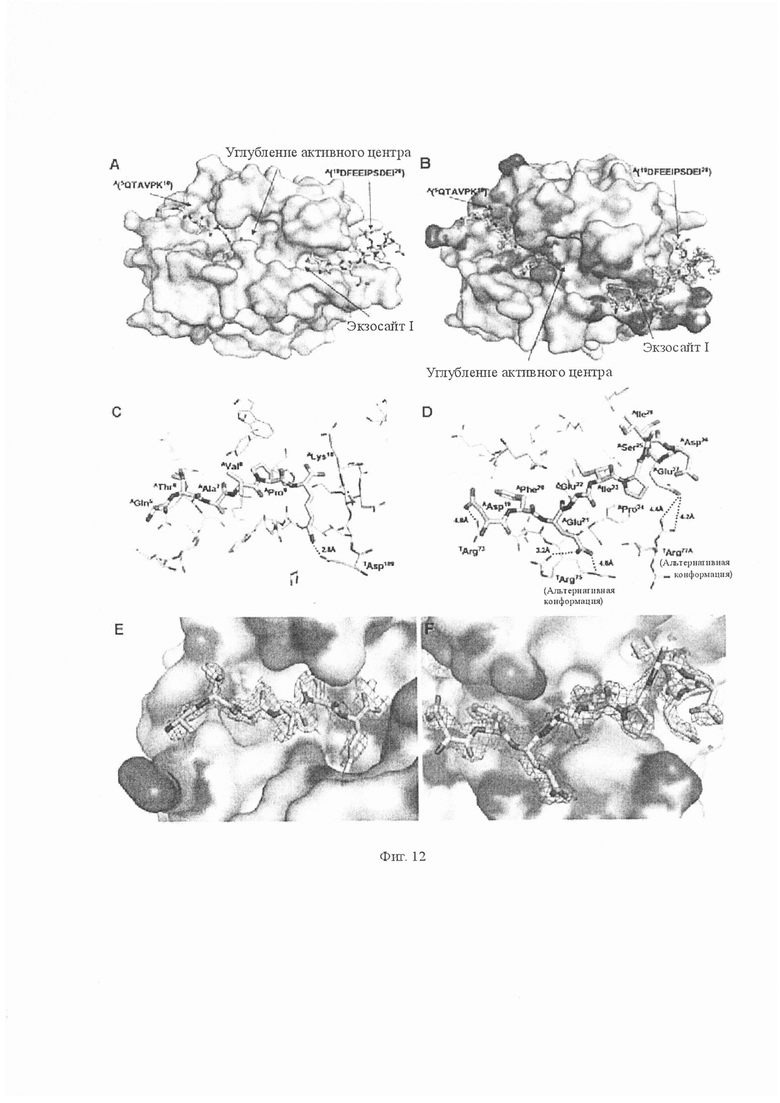

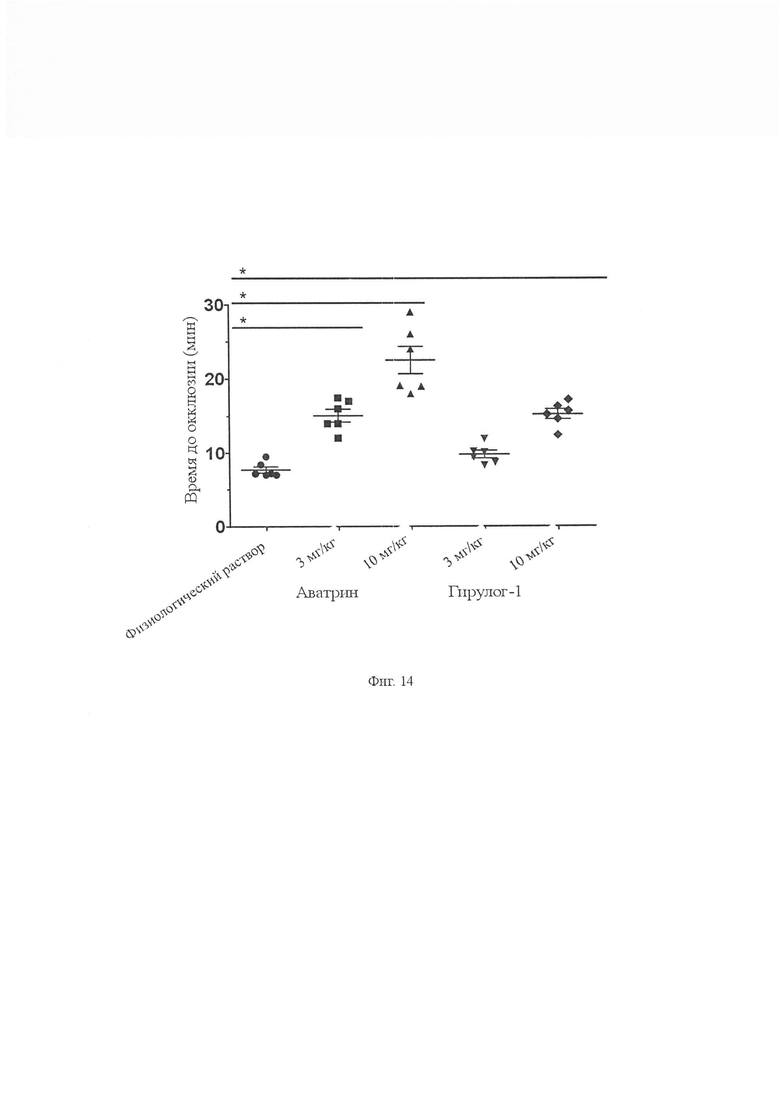
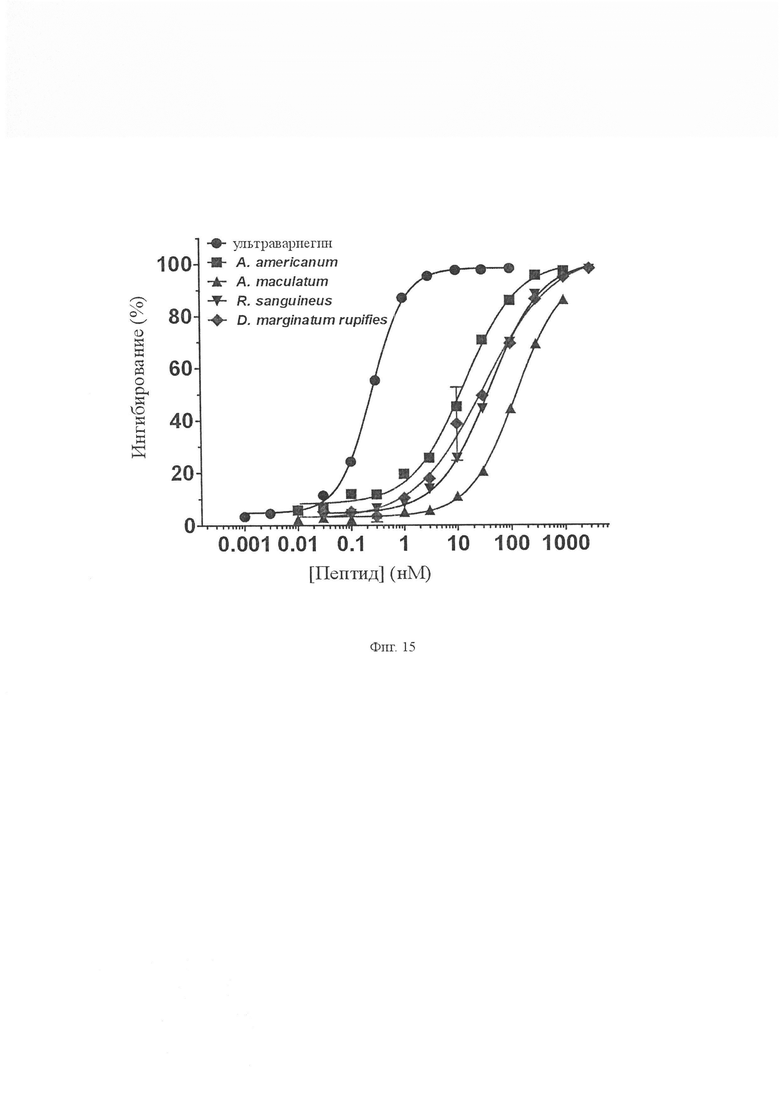
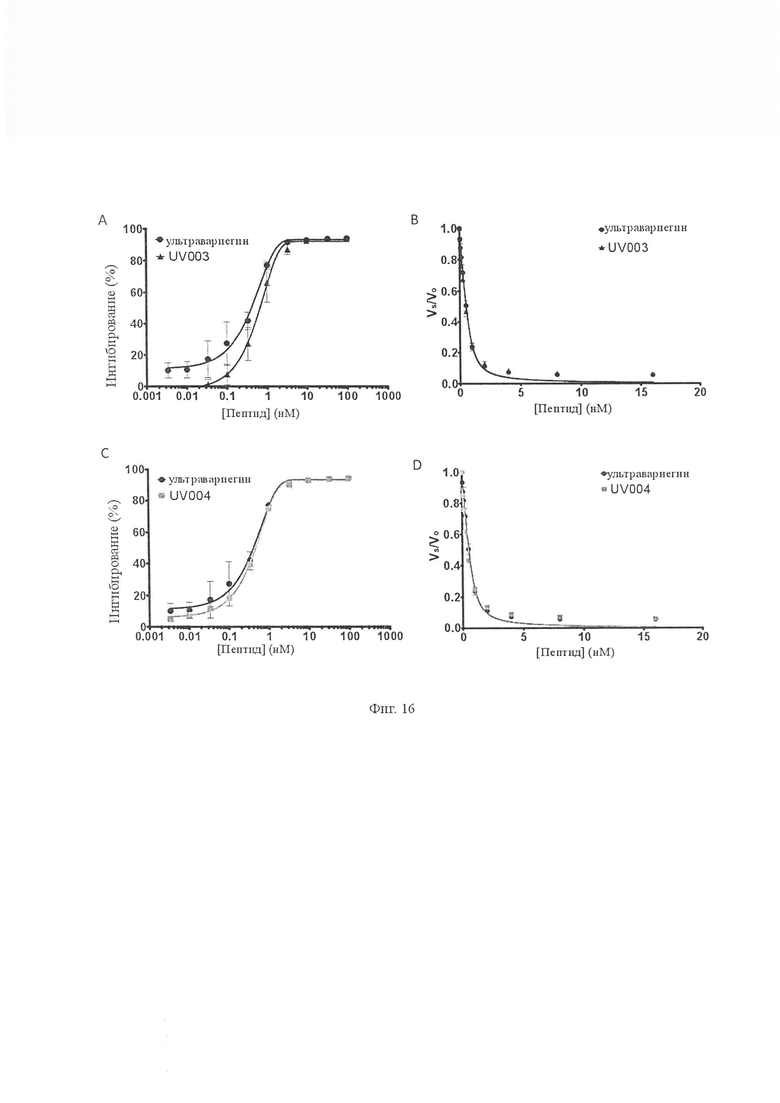

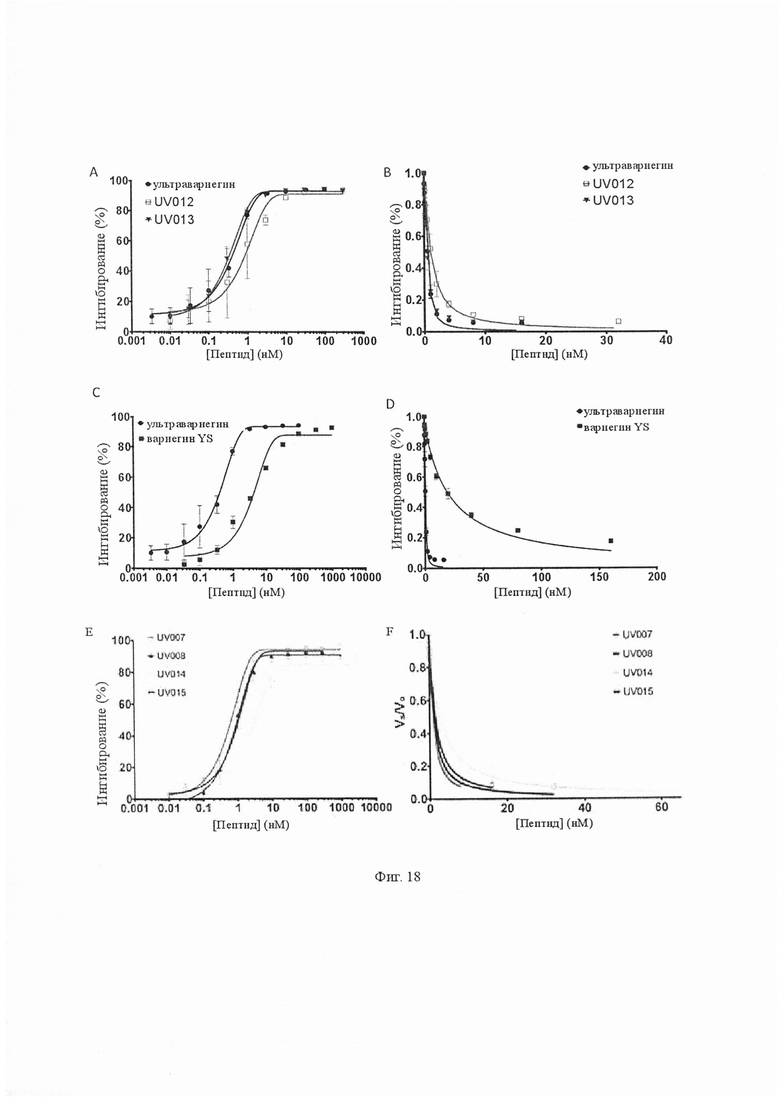
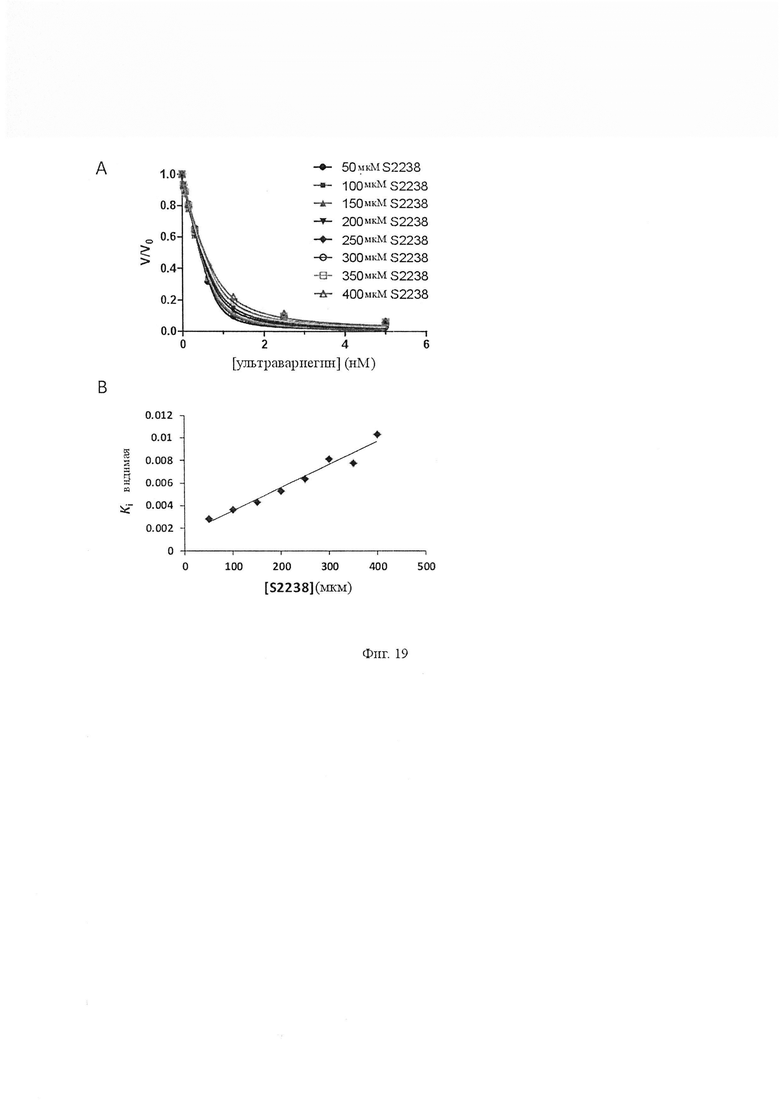
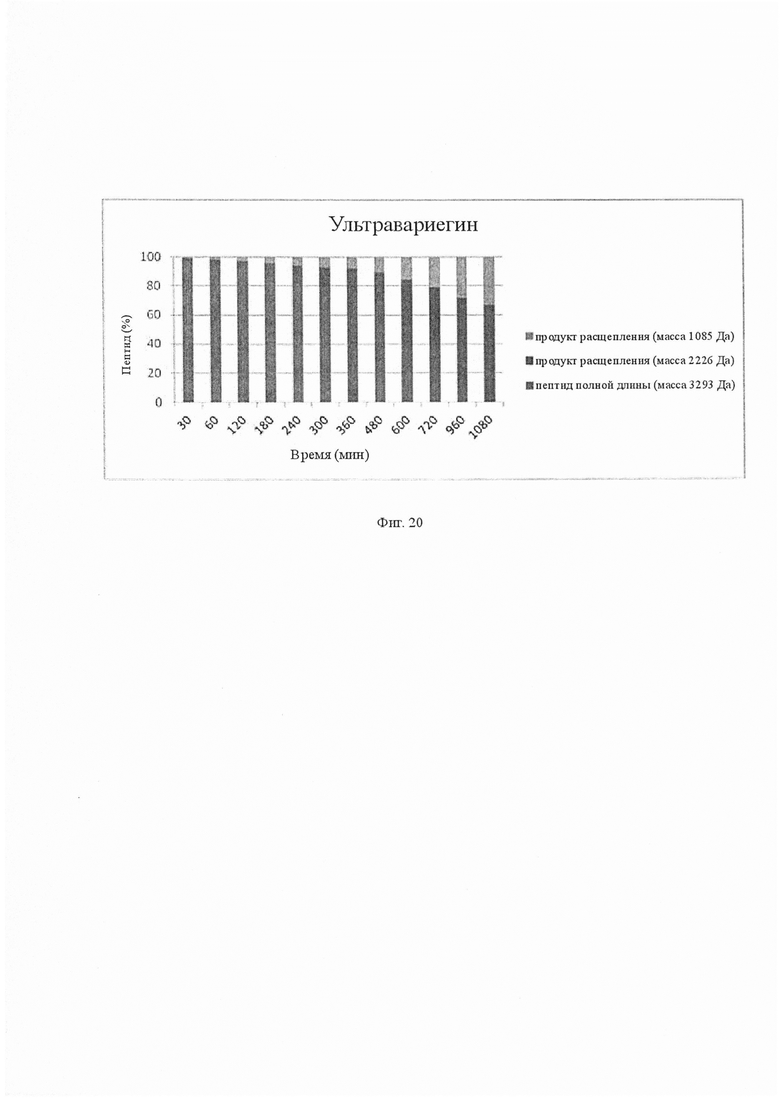

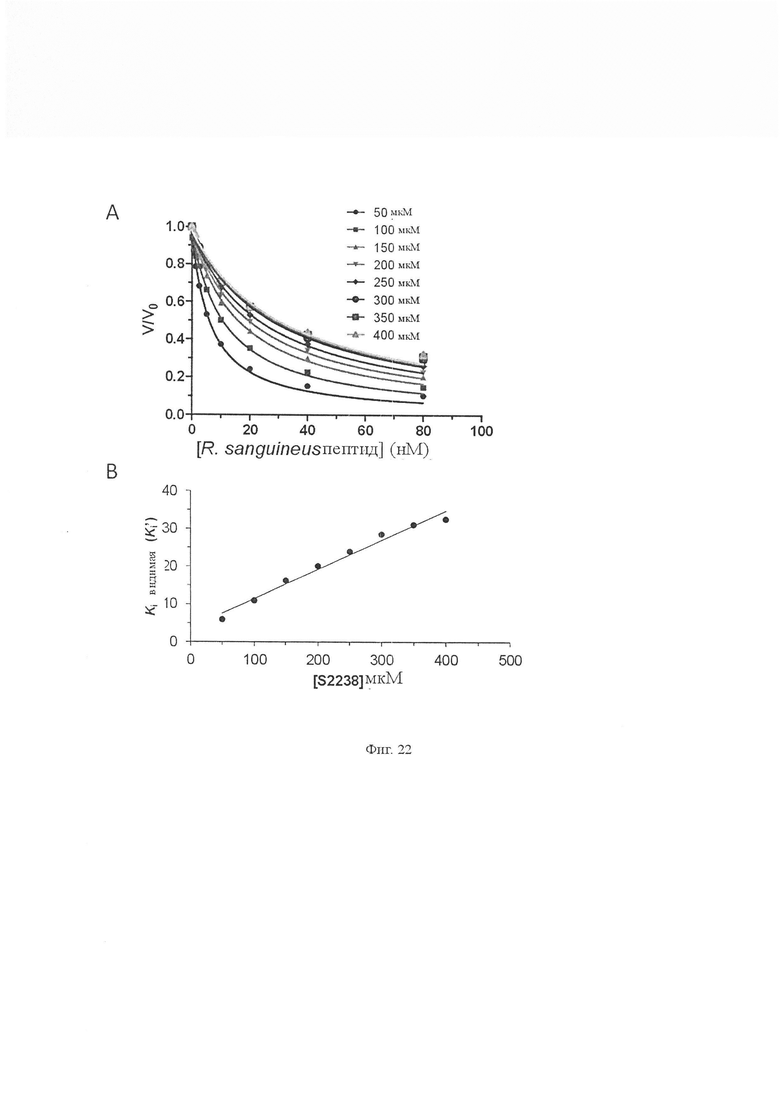

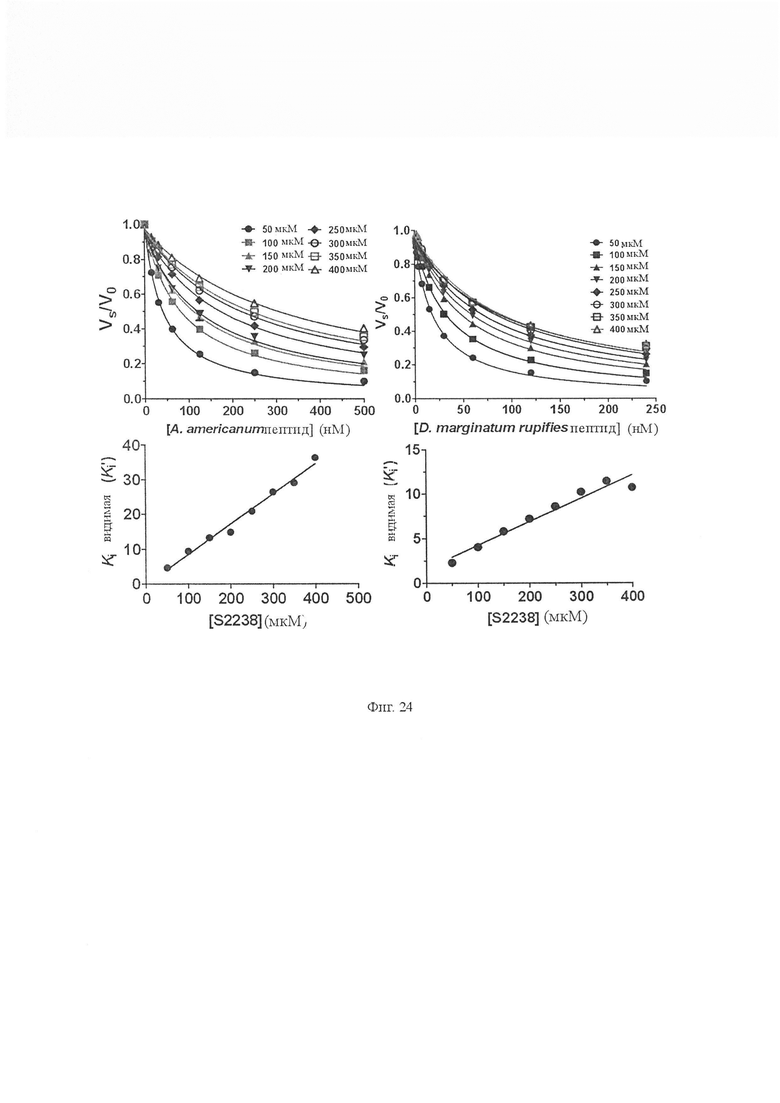
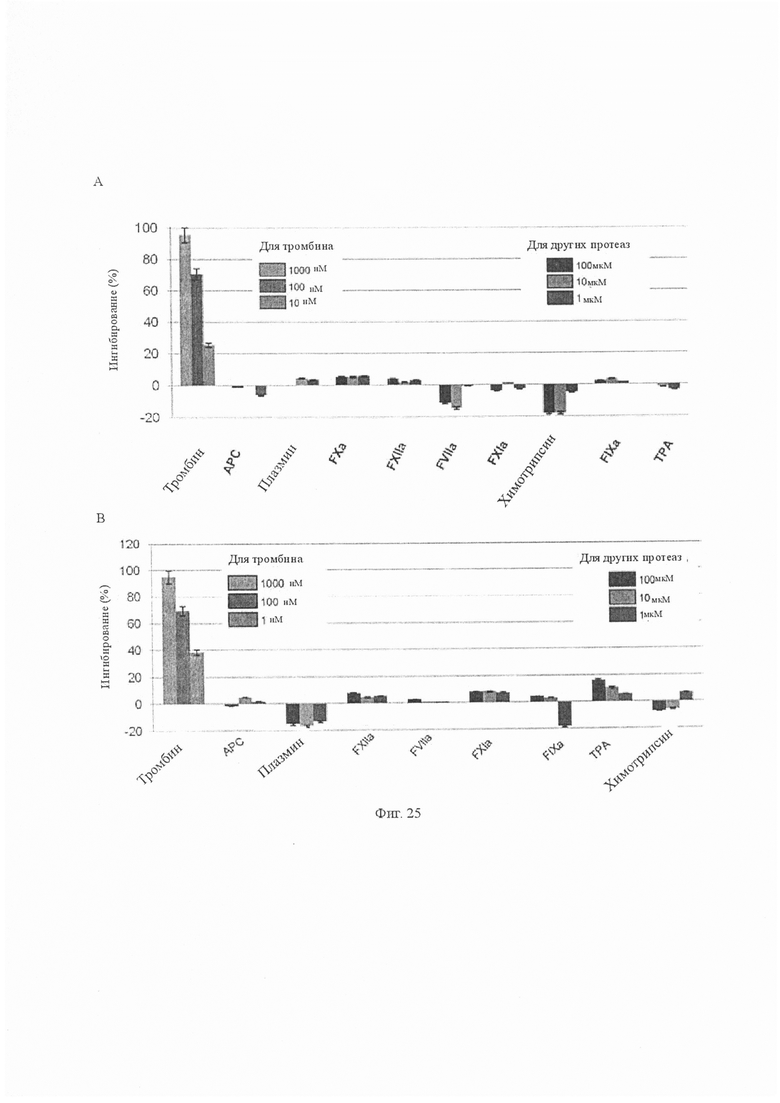


Настоящее изобретение относится к области биотехнологии, конкретно к пептидным ингибиторам тромбина, и может быть использовано в медицине для профилактики или лечения заболевания, связанного с тромбозом, а также для обнаружения накопления тромбина у субъекта. Изобретение обеспечивает получение пептидов, которые способны связываться с высоким уровнем специфичности с тромбином и ингибировать его активность. 13 н. и 6 з.п. ф-лы, 28 ил., 7 табл., 1 пр.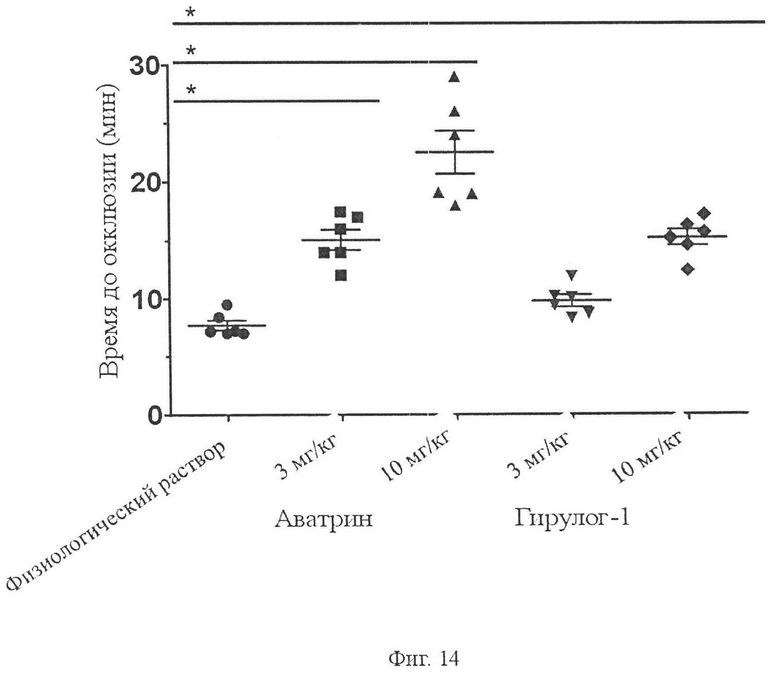
1. Выделенный пептидный ингибитор тромбина, состоящий из аминокислотной последовательности, выбранной из группы, содержащей SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20, SEQ ID NO: 21, SEQ ID NO: 22, SEQ ID NO: 23, SEQ ID NO: 24 и SEQ ID NO: 25.
2. Выделенный пептидный ингибитор тромбина по п. 1, где указанный ингибитор ингибирует фибриногенолитическую активность тромбина и/или ингибирует амидолитическую активность тромбина.
3. Выделенный пептидный ингибитор тромбина по п. 2, где IC50 ингибитора тромбина составляет менее 400 нМ, предпочтительно менее 300 нМ, менее 200 нМ, менее 100 нМ, менее 50 нМ, менее 10 нМ, предпочтительно менее 9 нМ, менее 8 нМ, менее 7 нМ, менее 6 нМ, менее 5 нМ, менее 4 нМ, менее 3 нМ, менее 2 нМ или менее 1 нМ при оценке в амидолитическом анализе.
4. Выделенный пептидный ингибитор тромбина по п. 3, где Ki ингибитора тромбина составляет менее 6000 нМ, предпочтительно менее 2000 нМ, менее 500 нМ, менее 400 нМ, менее 300 нМ, менее 200 нМ, менее 100 нМ, менее 50 нМ, менее 10 нМ, предпочтительно менее 9 нМ, менее 8 нМ, менее 7 нМ, менее 6 нМ, менее 5 нМ, менее 4 нМ, менее 3 нМ, менее 2 нМ или менее 1 нМ при оценке в амидолитическом анализе.
5. Применение выделенного пептидного ингибитора тромбина по любому из пп. 1-4 для профилактики или лечения заболевания, связанного с тромбозом, где связанное заболевание выбрано из артериального и венозного тромбоза, вызывающего сердечные приступы, инсульт и эмболию; тромбоза глубоких вен; тромбофлебита; легочной эмболии; эмболических и микроэмболических приступов, где источником могут быть сердце, атеросклеротическая бляшка, клапанные или сосудистые протезы или неизвестный источник; диссеминированной внутрисосудистой коагуляции.
6. Применение выделенного пептидного ингибитора тромбина по любому из пп. 1-4 для профилактики или лечения заболевания, связанного с тромбозом, где пептидный ингибитор тромбина предназначен для антикоагуляции при нестабильной стенокардии, коронарной ангиопластике, чрескожном коронарном вмешательстве и операции на сердце.
7. Способ ингибирования активности тромбина, в котором способ включает контактирование тромбина с пептидным ингибитором тромбина по любому из пп. 1-4.
8. Способ по п. 7, в котором пептидный ингибитор тромбина присутствует в качестве:
а) противосвертывающего агента в пробирках для взятия крови; или
б) материала поверхностного покрытия на медицинских устройствах, таких как стенты, катетеры и другие трубки для интубации.
9. Применение пептидного ингибитора тромбина по любому из пп. 1-4 для получения лекарственного средства для профилактики или лечения заболевания, связанного с повышенной активностью тромбина.
10. Применение по п. 9, в котором ассоциированное заболевание выбрано из артериального и венозного тромбоза, вызывающего сердечные приступы, инсульт и эмболию; тромбоза глубоких вен; тромбофлебита; легочной эмболии; эмболических и микроэмболических приступов, где источником могут быть сердце, атеросклеротическая бляшка, клапанные или сосудистые протезы или неизвестный источник; диссеминированной внутрисосудистой коагуляции.
11. Применение по п. 9, в котором лекарственное средство предназначено для местного применения для профилактики или лечения тромбоза артериовенозного шунта, ушибов головного мозга, растяжений, разрывов мыщц, травматических гематом, отеков, эритемы, варикозов, перифлебита и анального перифлебита с геморроем, особенно связанных с тромбоэмболическими осложнениями.
12. Способ профилактики или лечения заболевания, связанного с тромбозом, где связанное заболевание выбрано из артериального и венозного тромбоза, вызывающего сердечные приступы, инсульт и эмболию; тромбоза глубоких вен; тромбофлебита; легочной эмболии; эмболических и микроэмболических приступов, где источником могут быть сердце, атеросклеротическая бляшка, клапанные или сосудистые протезы или неизвестный источник; диссеминированной внутрисосудистой коагуляции, включающий введение субъекту, нуждающемуся в такой профилактике или лечении, эффективного количества пептидного ингибитора тромбина по любому из пп. 1-4.
13. Способ профилактики или лечения заболевания, связанного с тромбозом, включающий введение субъекту, нуждающемуся в антикоагуляции при нестабильной стенокардии, коронарной ангиопластике, чрескожном коронарном вмешательстве и операции на сердце, эффективного количества пептидного ингибитора тромбина по любому из пп. 1-4.
14. Способ диагностики заболевания или состояния, связанного с тромбозом, где связанное заболевание выбрано из артериального и венозного тромбоза, вызывающего сердечные приступы, инсульт и эмболию; тромбоза глубоких вен; тромбофлебита; легочной эмболии; эмболических и микроэмболических приступов, где источником могут быть сердце, атеросклеротическая бляшка, клапанные или сосудистые протезы или неизвестный источник; диссеминированной внутрисосудистой коагуляции, включающий введение пептидного ингибитора по любому из пп. 1-4 в образец ткани, выделенный у указанного субъекта, и обнаружение присутствия указанного ингибитора тромбина, связанного с тромбином, где обнаружение повышенного уровня указанного ингибитора, связанного с тромбином, по сравнению с уровнем ингибитора, связанного с нормальным уровнем тромбина, указывает на указанное заболевание или состояние.
15. Способ обнаружения накопления тромбина у субъекта, включающий введение пептидного ингибитора по любому из пп. 1-4 в образец ткани, выделенный у субъекта, обнаружение присутствия указанного ингибитора тромбина, связанного с тромбином, и сравнение уровня указанного ингибитора, связанного с тромбином, в указанном образце ткани с уровнем ингибитора, связанного с нормальным уровнем тромбина, где обнаружение повышенного уровня указанного ингибитора, связанного с тромбином, по сравнению с уровнем ингибитора, связанного с нормальным уровнем тромбина, указывает на накопление тромбина.
16. Фармацевтическая композиция для профилактики или лечения тромбоза, содержащая эффективное количество пептидного ингибитора тромбина по любому из пп. 1-4 и фармацевтически приемлемый наполнитель, носитель или эксципиент.
17. Выделенная молекула нуклеиновой кислоты, кодирующая пептидный ингибитор по п. 1.
18. Вектор экспрессии, содержащий молекулу нуклеиновой кислоты по п. 17.
19. Клетка-хозяин для получения пептидных ингибиторов тромбина, содержащая молекулу нуклеиновой кислоты по п. 17, оперативно связанную с промотором, или вектор экспрессии по п. 18, где клетка-хозяин не является эмбриональной или зародышевой клеткой человека.
| KOH C.Y | |||
| ET AL | |||
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |

