Область техники
Изобретение относится к ДНК-диагностике патологичных состояний, вызванных изменением числа макросателлитных повторов и может быть использовано для определения числа макросателлитных повторов D4Z4 на каждом из аллелей хромосомы 4 для диагностики миодистрофии Ландузи-Дежерина 1-го типа (FSHD1, MIM: 158900).
Предшествующий уровень техники
Макросателлитные повторы - нуклеотидные последовательности протяженностью от 100 пар оснований (п.о.) до нескольких тысяч п.о., зачастую тандемно сгруппированные и широко распространенные в геноме человека. Наиболее хорошо изученным являются макросателлитные повторы D4Z4 хромосомы 4, длина каждого повтора D4Z4 составляет 3300 п.о. Изменение их количества является пусковым механизмом в развитии миодистрофии Ландузи-Дежерина 1-го типа. Так в норме на каждом из аллелей насчитывается от 11 до 100 единиц D4Z4, в то время как у пациентов с миодистрофией Ландузи-Дежерина 1-го типа хотя бы один из аллелей хромосомы 4 имеет частичную делецию массива повторов D4Z4 до 1-10 единиц.
Известна методика молекулярного комбинга для определения числа повторов D4Z4 в отдельности на каждом из аллелей хромосомы 4 и 10 [1]. В данной методике используются флуоресцентно окрашенные ДНК-зонды для in-situ гибридизации с единичными молекулами ДНК, линейно растянутыми и фиксированными на покровном стекле. Линейное растяжение молекул ДНК достигается за счет погружения покровного стекла в раствор с ДНК и вытягиванием его из раствора с определенной постоянной скоростью специализированным оборудованием. Детекция флуоресцентного сигнала производится с использованием специализированного аппаратно-программного комплекса для FISH-анализа.
К недостаткам данной методики следует отнести то, что она требует использования дорогостоящего узкоспециализированного оборудования, кроме того ее использование для определения количества макросателлитных повторов на сегодня ограничивается только повторами D4Z4 хромосом 4 и 10.
Также известно применение метода оптического картирования для подсчета количества макросателлитных повторов [2]. В данной методике используется вставка флуоресцентных нуклеотидов в сайты никирования определенных эндонуклеаз. Полученные таким образом молекулы ДНК разделяются на единичные продольно вытянутые молекулы в специализированных микрочипах. Далее флуоресцентный сигнал считывается специализированным аппаратно-программным комплексом с каждой молекулы ДНК и делается вывод количестве тех или иных макросателлитных последовательностей. Кроме того, полученные данные могут служить для анализа структурных перестроек и de-novo сборки геномов.
В этой методике лимитирующим фактором для использования данной методики в стандартных молекулярно-генетических лабораториях являются высокая стоимость расходных материалов и необходимость использования дорогостоящего оборудования также является.
Существует также метод нанопорового секвенирования для определения числа макросателлитных повторов. При этом способе секвенирования, ДНК протягивается через нанопоры и каждый проходящий нуклеотид вызывает определенное изменение силы тока. По этим изменениям делается вывод о нуклеотидном составе и длине молекулы ДНК. На экспериментальных образцах была показана возможность получения непрерывных прочтений нуклеотидной последовательности тандема макросателлитных повторов D4Z4 длиной 25 тысяч пар оснований (6 повторов D4Z4) [3].
Однако, в большинстве случаев в диагностических целях необходимо получение прочтений большей длинны. Поэтому, на сегодняшний день, нанопоровое секвенирование имеет технические ограничения, что не позволяет использовать его для диагностики.
Существуют способы определения числа макросателлитных повторов, каждого из интересующих аллелей, на основе методики гибридизации по Саузерну с использованием радиоактивно меченных ДНК-зондов [4], а также модификации данной методики с использованием нерадиоактивного мечения ДНК-зондов биотином и дигоксигенином. При данном способе массивы тандемно расположенных макросателлитных повторов вырезаются из хромосомы эндонуклеазами рестрикции по фланкирующим сайтам. Полученные образцы разделяются в агарозном геле методом пульс-электрофореза, после чего ДНК-фрагменты переносятся из агарозного геля на нейлоновую мембрану и далее гибридизуются со специфичными ДНК-зондами к исследуемым последовательностям и ДНК-зондом к маркеру молекулярного веса. Определение количества интересующих макросателлитных фрагментов осуществляется соотнесением специфичного сигнала с сигналами маркера молекулярного веса.
Недостатком методики является то, что трудо- и ресурсоемкость процесса гибридизации и - в некоторых случаях - необходимость использования радиоактивных ДНК-зондов накладывают существенные ограничения на использование гибридизации по Саузерну в стандартных молекулярно-генетических лабораториях.
Раскрытие сущности изобретения
Задачей, на решение которой направлено заявленное изобретение, является создание способа определения числа макросателлитных тандемных повторов D4Z4 на каждом аллеле хромосомы 4.
Технический результат, получаемый при решении поставленной задачи, выражается в получении способа определения числа макросателлитных тандемных повторов D4Z4 на каждом аллеле хромосомы 4 с применением количественной ПЦР с фрагментами агарозного геля после пульс-электрофореза, исключающего недостатки известных способов и позволяющего проводить достоверное определение числа макросателлитных тандемных повторов на каждом аллеле в исследуемом образце ДНК.
Для решения поставленной задачи способ определения числа макросателлитных тандемных повторов D4Z4 на каждом аллеле хромосомы 4 включает следующие стадии:
А) выделение высокомолекулярной ДНК, с длинной более 300 тысяч пар оснований, в агарозных блоках и обработка полученной ДНК эндонуклеазами рестрикции с сайтами разрезания, фланкирующими область тандемных макросателлитных повторов;
Б) проведение пульс-электрофореза с образцами ДНК после обработки их эндонуклеазами рестрикции в трис-боратном электрофорезном буфере без добавления ЭДТА, но с добавлением дитиотрейтола, при этом тандемы макросателлитных повторов, разделяются в агарозном геле в зависимости от количества повторов в каждом тандеме;
В) окраска бромистым этидием и фрагментирование дорожек агарозного геля, содержащих рестрицированную ДНК, поперечными разрезами в соответствии с маркером молекулярного веса таким образом, чтобы соседние фрагменты различались на одну-две единицы макросателлитных повторов;
Г) фотодокуметирование полученной схемы фрагментации с точным соотнесением положения фрагментов дорожки агарозного геля с маркером молекулярного веса;
Д) постановка количественной ПЦР с использование фрагментов дорожек агарозного геля в качестве матриц и с использованием пары праймеров: прямой 5`- GTGCTTGCGCCACCCACGT -3` и обратный 5’- GCCGCGCGGAGGCGGAG -3`, а также флуоресцентным «TaqMan» ДНК-зондом 5’- FAM AGTCCGTGGTGGGGCTGGGG BHQ1 -3`.
Интерпретация результатов производится путем сравнения пороговых циклов полученных при амплификации матриц фрагментов дорожек агарозного геля с пороговым циклом отрицательного контроля - фрагментом геля из зоны вне дорожек агарозного геля, демонстрирующего общую контаминацию агарозного геля фрагментами таргетной ДНК.
Осуществление изобретения
Работоспособность заявляемого изобретения подтверждается следующими примерами конкретного выполнения.
ПРИМЕР 1.
Пациент №120, согласно результатам гибридизации по Саузерну имеет два аллеля массива повторов D4Z4 хромосомы 4: 6 повторов на одном аллеле, более 11 повторов на втором.
Пробоподготовка. Из 10 мл венозной крови выделялись лимфоциты стандартной методикой центрифугирования в градиенте плотности фиколла (ρ = 1,077 г/мл). Лимфоцитарная масса разводилась в растворе жидкой низкоплавкой агарозы (CleanCut™ agarose, BioRad) до концентрации 1×106 клеток в 100 мкл. Полученная суспензия разносилась по лункам пластикового молдинга, объём каждой лунки равен 100 мкл.
После полимеризации агарозы в лунках молдинга, полученные агарозные блоки помещались в 10 мл раствора 1% лауроилсаркозин-N натриевой соли и протеиназы К (конечная концентрация 0,6 мг/мл) и оставлялись на двое суток при +37°С. Далее агарозные блоки дважды промывались в 15 мл дистилированной воды и помещались в 10 мл раствора EDTA (0,5 М, pH 8.0) для хранения. Для обработки ДНК эндонуклеазой рестрикции ½ агарозного блока отмывалась от EDTA в 800 мкл стандартного 1xTE-буфера дважды по 1,5 часа на качалке (MR-1, Biosan).
Затем агарозный блок переносился в 250 мкл рестрикционного буфера (2x Tango (Thermo Fisher Scientific); 1 mM DTT; 3.3 mM spermidine (Sigma Aldrich)) и инкубировался при комнатной температуре в течение 2 часов. Затем агарозный блок переносился в свежий рестрикционный буфер с добавлением 30 единиц активности эндонуклеазы рестрикции EcoRI (Thermo Fisher Scientific) и инкубировался в течение 16 часов при +37°С. Таким образом геномная ДНК рестрицировалсь непосредственно в агарозном блоке.
После обработки эндонуклеазой рестрикции проводился пульс-электрофорез на комплекте оборудования: электрофорезная камера с гексогональным электродом и охлаждающим контуром «Gene Navigator» (Pharmacia Biotech, Sweden), блок питания «2301 MacroDrive 1 Power Supply» (LKB Bromma, Stockholm, Sweden), блок управления пульсацией «2015 Pulsaphor Plus Control Unit» (LKB Bromma, Stockholm, Sweden). Температура в электрофорезной камере поддерживалась на уровне 5°С при помощи водяной бани с внешней циркуляцией «2219-001 Multitemp II Thermostatic Circulator» (LKB Bromma, Stockholm, Sweden). Агарозный гель для электрофореза имел размеры 17×17×0,5 см.
Агарозные блоки после обработки эндонуклеазой рестрикции помещались в лунки агарозного геля для электрофореза. Использовались следующие маркеры молекулярного веса: «Lambda Mix Marker, 19» (Fermentas, Warsaw, Poland), «ДНК-маркер 1 Kb» (СибЭнзим, Новосибирск, Россия). Электрофорез проводился при вольтаже 450V; интервал пульсации 0,5 сек; продолжительность электрофореза 3,5 часа, температура электрофорезного буфера +4°С.
После проведения пульс-электрофореза агарозный гель окрашивался в электрофорезном буфере с добавлением бромистого этидия (конечная концентрация 0,5 мкг/мл). Отмывка агарозного геля производилась в дистилированной воде, 3 раза по 20 мин. Фотодокументирование агарозных гелей производилась прибором «ChemiDoc XRS+» (Bio-Rad Laboratories, Veenendaal, The Netherlands).
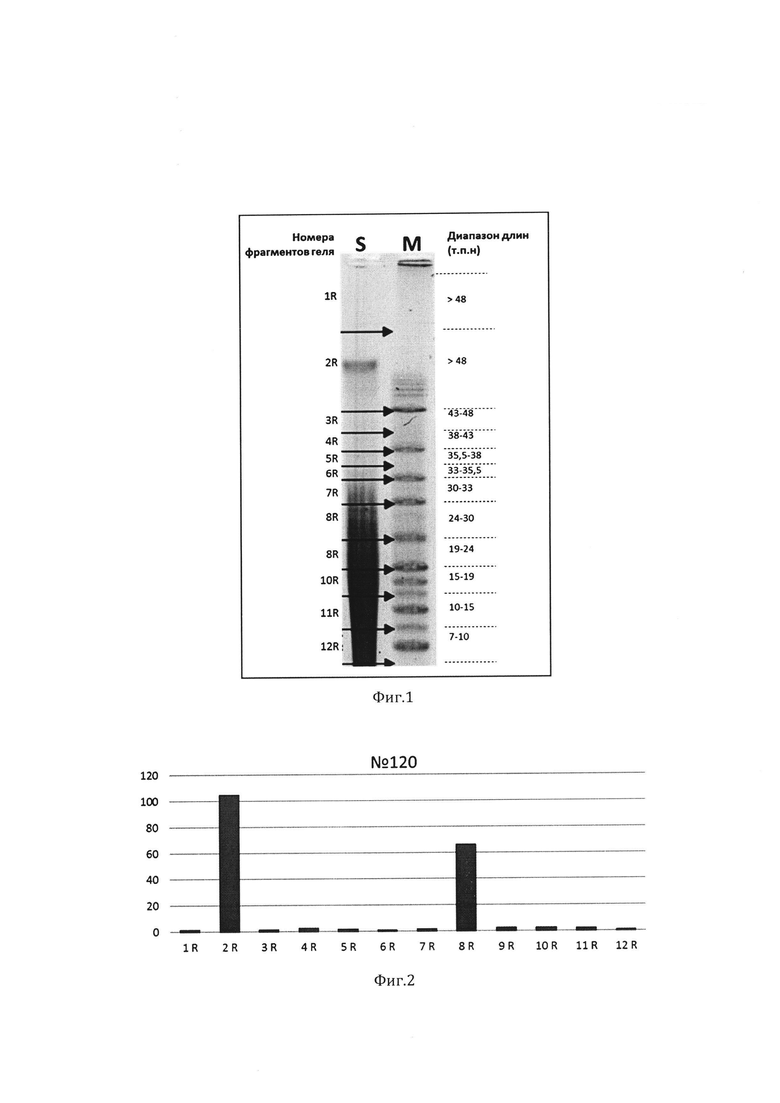
Изображения обрабатывались программой «Image Lab v4.1» (Bio-Rad Laboratories, Veenendaal, The Netherlands). Дорожки агарозного геля, содержащие ДНК, фрагментировались вручную в соответствии с длинами маркеров молекулярного веса, схема фрагментирования геля документировалась на изображении (Фиг.1) в программе «Image Lab v4.1» и в дальнейшем использовалась при анализе данных количественной ПЦР. 10 мкл каждого фрагмента агарозного геля разводились в «MQ» воде и использовались в качестве матрицы для количественной ПЦР.
Количественная ПЦР. Для количественной ПЦР. использовались 0,25 единиц активности ДНК-плимераза «SmarNG-Taq» (Диалат ЛТД, Москва, Россия), 1х Taq-буфер (Диалат ЛТД, Москва, Россия), бетаина 1,2 М (Tokyo Chemical Industry Co., Ltd., Tokyo, Japan), раствор MgCl2 2,5 мМ (Диалат ЛТД, Москва, Россия), дезоксинуклеотидтрифосфаты 0,2 mM (Диалат ЛТД, Москва, Россия).
Праймеры для специфичной амплификации повторов D4Z4 хромосомы 4: Rep1.5F 5’-GTGCTTGCGCCACCCACGT-3’ 0,4 мМ, Rep1R 5’-GCCGCGCGGAGGCGGAG-3’ 0,4 мМ. «TaqMan»-зонд: zond2Rep1 10 мМ 5’FAM-AGTCCGTGGTGGGGCTGGGG-BHQ1 3’.
Амплификацию проводили на приборе «StepOne Plus» (Applied Biosystems) по следующей схеме: горячий старт при 95°С - 3 минуты, далее 35 циклов: 95°С - 30 сек., 64°С - 8 сек., 72°С - 4 сек. ПЦР проводилась в двух повторностях для каждого фрагмента геля.
Средние значения пороговых циклов каждого фрагмента геля обрабатывались в программе Microsoft Excel. Обсчет данных ПЦР осуществлялся методом 2-ΔΔCt. При расчетах вычислялась разница между двумя типами фрагментов геля: фрагменты вне зоны дорожек и фрагменты дорожки геля. Внутренним контролем служил агарозный гель из зоны вне дорожек, т.к. он отражает контаминированность агарозного геля таргетными нуклеотидными последовательностями. Фрагменты геля с максимальными значениями относительной концентрации D4Z4 указывают на наличие в них таргетных нуклеотидных последовательностей.
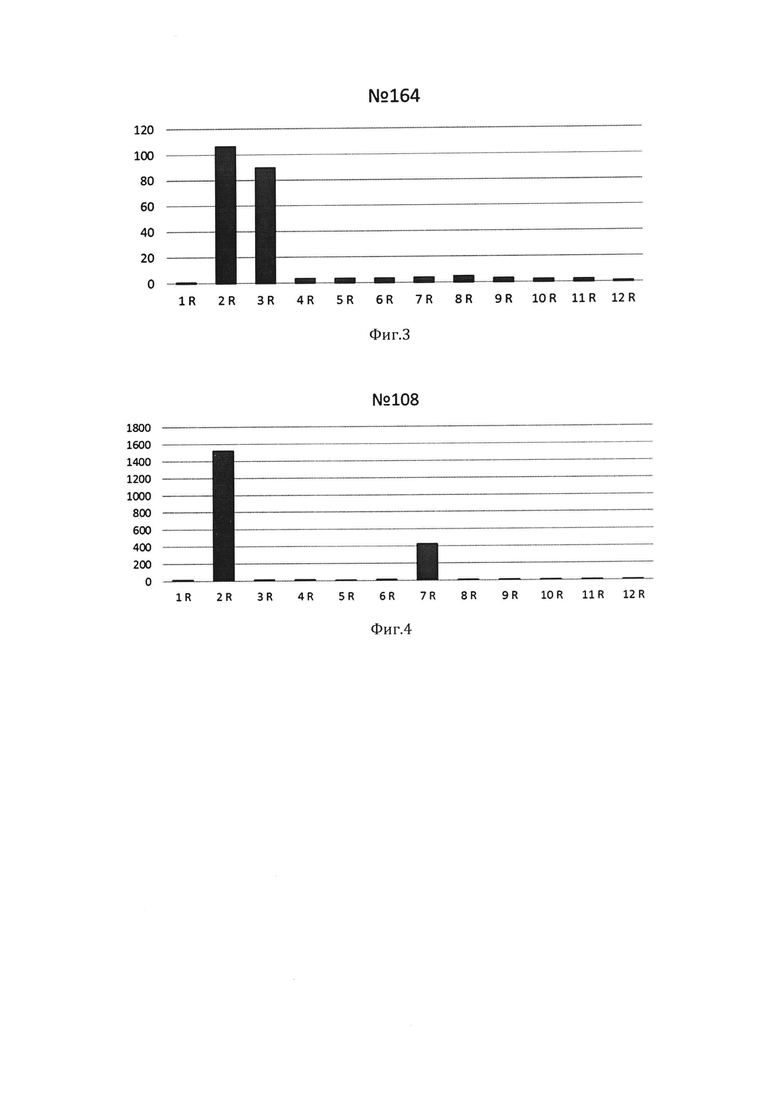
Сопоставив положение данных фрагментов геля относительно маркера молекулярного веса, определяли, что пациент имеет два аллеля массивов повторов D4Z4 хромосомы 4: один аллель имел 6 повторов, второй более 11 (Фиг.2).
ПРИМЕР 2
Пациент №164, согласно результатам гибридизации по Саузерну имеет два аллеля массива повторов D4Z4 хромосомы 4 с числом повторов более 11 для обоих аллелей.
Этапы пробоподготовки и количественной ПЦР аналогичны описанным в Примере 1.
Сопоставление фрагментов геля с максимальными значениями относительной концентрации D4Z4 относительно маркера молекулярного веса, указывало на то, что пациент имеет два аллеля массивов повторов D4Z4 хромосомы 4, с числом повторов более 11 для обоих аллелей (Фиг.3).
ПРИМЕР 3
Пациент №108, согласно результатам гибридизации по Саузерну имеет два аллеля массива повторов D4Z4 хромосомы 4 с числом повторов равном 7 на одном аллеле и более 11 на другом.
Этапы пробоподготовки и количественной ПЦР аналогичны описанным в Примере 1.
Сопоставление фрагментов геля с максимальными значениями относительной концентрации D4Z4 относительно маркера молекулярного веса, указывало на то, что пациент имеет два аллеля массивов повторов D4Z4 хромосомы 4, с числом повторов более 11 на одном аллеле и 7 на другом (Фиг.4).
Преимущества заявленного способа состоят в следующем:
Отсутствие необходимости использовать малораспространенные и дорогостоящие методики гибридизации по Саузерну и молекулярного комбинга, что делает возможным определения числа тандемных макросателлитных повторов в условиях стандартных молекулярно-биологических лабораторий.
Экономится до двух рабочих дней за счет использования количественной ПЦР вместо гибридизации с ДНК зондами в методиках гибридизации по Саузерну и молекулярного комбинга.
В частности, для диагностики FSHD1, отсутствует необходимость повторной гибридизации для определения гаплотипа патогенного массива повторов D4Z4, данный шаг заменяется аллельспецифичной ПЦР.
Возможность использования ДНК из фрагментов геля после пульс-электрофореза в целях секвенирования, определения уровня метилирования, клонирования в генно-инженерные конструкции.
Краткое описание чертежей
Фигура 1. Схема фрагментации агарозного геля после пульс-электрофореза образа геномной ДНК обработанного эндонуклеазой рестрикции EcoRI. Дорожка S - горизонтальными черными стрелками указаны границы фрагментов геля. Дорожка М - маркер молекулярного веса «Lambda Mix Marker, 19». Слева от фото геля указаны номера фрагментов геля по порядку, справа - диапазоны длин, в которых расположен фрагмент геля (т.п.н.).
Фигура 2. Результаты количественно ПЦР с фрагментами геля после пульс-электрофореза образца ДНК пациента №120. По оси абсцисс - номера фрагментов геля. По оси ординат - относительные значения концентрации таргетных нуклеотидных последовательностей. Максимальные значения относительной концентрации во фрагментах геля 2R и 8R указывают на наличие в них аллелей массивов D4Z4. По результатам сделан вывод о наличии двух аллелей: один более 11 повторов D4Z4, второй 6 повторов D4Z4.
Фигура 3. Результаты количественно ПЦР с фрагментами геля после пульс-электрофореза образца ДНК пациента №120. По оси абсцисс - номера фрагментов геля. По оси ординат - относительные значения концентрации таргетных нуклеотидных последовательностей. Максимальные значения относительной концентрации во фрагментах геля 2R и 3R указывают на наличие в них аллелей массивов D4Z4. По результатам сделан вывод о наличии двух аллелей, оба более 11 повторов D4Z4.
Фигура 4. Результаты количественно ПЦР с фрагментами геля после пульс-электрофореза образца ДНК пациента №120. По оси абсцисс - номера фрагментов геля. По оси ординат - относительные значения концентрации таргетных нуклеотидных последовательностей. Максимальные значения относительной концентрации во фрагментах геля 2R и 8R указывают на наличие в них аллелей массивов D4Z4. По результатам сделан вывод о наличии двух аллелей: один более 11 повторов D4Z4, второй 7 повторов D4Z4.
Список литературы
1. Vasale J, Jocson M, Sulcova V, Chan P, Liaquat K, Hoffman C, et al. Molecular combing compared to Southern blot for measuring D4Z4 contractions in FSHD. 2015. doi:10.1016/j.nmd.2015.08.008.
2. Young E, Abid HZ, Kwok P-Y, Riethman H, Xiao M. Comprehensive Analysis of Human Subtelomeres by Whole Genome Mapping. PLOS Genet. 2020;16:e1008347. doi:10.1371/journal.pgen.1008347.
3. Mitsuhashi S, Mitsuhashi H, Ueda M, Nakagawa S, Imanishi T. Nanopore-based single molecule sequencing for D4Z4 array responsible for facioscapulohumeral muscular dystrophy. J Neurol Sci. 2017;381:457-8.
4. Schaap M, Lemmers RJLF, Maassen R, van der Vliet PJ, Hoogerheide LF, van Dijk HK, et al. Genome-wide analysis of macrosatellite repeat copy number variation in worldwide populations: Evidence for differences and commonalities in size distributions and size restrictions. BMC Genomics. 2013;14:143. doi:10.1186/1471-2164-14-143.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ ДЛЯ ПОЛУЧЕНИЯ СТАБИЛЬНЫХ ТРАНСФОРМАНТОВ ГРИБОВ РОДА Rhizopus | 2009 |
|
RU2439157C2 |
| Способ преимплантационного генетического тестирования анемии Фанкони | 2022 |
|
RU2792147C1 |
| Способ преимплантационного генетического тестирования семейной гипертрофической кардиомиопатии | 2021 |
|
RU2772938C1 |
| Способ преимплантационного генетического тестирования Гемофилии А | 2022 |
|
RU2795796C1 |
| Способ преимплантационного генетического тестирования наследственных множественных остеохондром типа 1 | 2022 |
|
RU2795824C1 |
| Способ преимплантационного генетического тестирования лице-лопаточно-плечевой мышечной дистрофии | 2023 |
|
RU2808833C1 |
| Способ преимплантационного генетического тестирования семейной фокальной эпилепсии 1 типа | 2021 |
|
RU2777084C1 |
| Способ преимплантационного генетического тестирования синдрома Альпорта | 2022 |
|
RU2795481C1 |
| Способ преимплантационного генетического тестирования наследственной зонулярной катаракты | 2022 |
|
RU2799541C1 |
| Способ преимплантационного генетического тестирования Синдрома Марфана | 2023 |
|
RU2837158C1 |
Изобретение относится к области биотехнологии. Предложен способ определения количества макросателлитных повторов на каждом из аллелей в геноме, включающий выделение геномной ДНК в агарозных блоках, обработку эндонуклеазами рестрикции образцов геномной ДНК в агарозных блоках, разделение рестрицированной геномной ДНК и маркера молекулярного веса методом горизонтального пульс-электрофореза в агарозном геле, окраску, фотодокументирование агарозного геля после пульс-элетрофореза, фрагментацию дорожек агарозного геля, содержащих образцы рестрицированной геномной ДНК, полученных после пульс-электрофореза, в соответствии с длинами маркера молекулярного веса, постановку количественной ПЦР со специфичной парой праймеров с фрагментами агарозного геля в качестве ПЦР-матриц, анализ данных количественной ПЦР для обнаружения фрагментов агарозного геля, содержащих максимальное количество таргетных последовательностей, соотнесение фрагментов агарозного геля с максимальным количеством таргетных последовательностей с длинами маркера молекулярного веса по фото, полученным после фотодокументации, для определения числа таргетных повторяющихся последовательностей. Изобретение обеспечивает получение более простого и экономичного способа определения количества макросателлитных повторяющихся последовательностей генома, в частности связанных с развитием наследственной патологии. 7 з.п. ф-лы, 4 ил., 3 пр.
1. Способ обнаружения и определения числа тандемных макросателлитных повторяющихся последовательностей D4Z4 для каждого аллеля хромосомы 4:
выделение геномной ДНК в агарозных блоках из лимфоцитов венозной крови;
обработка эндонуклеазами рестрикции образцов геномной ДНК в агарозных блоках, полученных на стадии а);
разделение фрагментов геномной ДНК, полученных после проведения стадии b), и маркера молекулярного веса методом горизонтального пульс-электрофореза в агарозном геле
окраска, фотодокументирование агарозного геля после стадии c);
фрагментация дорожек агарозного геля, содержащих образцы рестрицированной геномной ДНК, полученных после стадии c), в соответствии с длинами маркера молекулярного веса;
постановка количественной ПЦР со специфичными к повторам D4Z4 хромосомы 4 парой праймеров: прямой 5`-GTGCTTGCGCCACCCACGT-3` и обратный 5’-GCCGCGCGGAGGCGGAG-3`, а также флуоресцентным «TaqMan» ДНК-зондом 5’-FAM AGTCCGTGGTGGGGCTGGGG BHQ1-3`;
постановка количественной ПЦР с фрагментами агарозного геля, полученными на стадии e), в качестве ПЦР-матриц;
анализ данных количественной ПЦР для обнаружения фрагментов агарозного геля, содержащих максимальное количество таргетных последовательностей;
соотнесение фрагментов агарозного геля с максимальным количеством таргетных последовательностей с длинами маркера молекулярного веса по фото, полученным на стадии d), для определения числа таргетных повторяющихся последовательностей.
2. Способ по п.1, характеризующийся тем, что выделение геномной ДНК производится в агарозных блоках, осуществляют следующим образом:
живые клетки суспензируют в расплавленной агарозе; полученный раствор разливают по пластиковым формам по 100 микролитров каждая и оставляют до полной полимеризации агарозы;
полученные агарозные блоки извлекаются из агарозных форм и помещаются в депротеинизирующий раствор (1% раствор лауриил-саркозила + 6 миллиграммов протеиназы К) на двое суток, после чего депротеинизирующий раствор сливается, а агарозные блоки помещаются в 10 миллилитров раствора 0,5 M EDTA.
3. Способ по п.1, характеризующийся тем, что обработка геномной ДНК эндонуклеазами рестрикции проводится следующим образом:
агарозные блоки с геномной ДНК извлекаются из раствора EDTA и отмываются в TE-буфере 2 раза по 1,5 часа;
после отмывки агарозные блоки уравновешиваются в буфере для соответствующей эндонуклеазы рестрикции и подвергаются рестрикции в течение 14-16 часов.
4. Способ по п.1, характеризующийся тем, что разделение рестрицированной ДНК в агарозном геле методом электрофореза проводится следующим образом:
подготавливается агарозный гель в 0,5Х TB буфере с добавлением DTT до 10 mM;
после обработки эндонуклеазами рестрикции агарозные блоки загружаются в лунки агарозного геля;
в свободные лунки загружается маркер молекулярного веса; горизонтальный пульс-электрофорез проводят в 0,5Х TB буфере с добавлением DTT до 10 mM при температуре буфера +5°С, при постоянной пульсации с временем пульса 0,5 с, в течение 3,5 часов.
5. Способ по п.1, характеризующийся тем, что окраска агарозного геля проводится следующим образом:
после горизонтального пульс-электрофореза агарозный гель окрашивается в электрофорезном буфере бромистым этидием с конечной концентрацией 0,5 мкг/мл в течение 5 минут;
на следующем этапе агарозный гель отмывается в дистиллированной воде 3 раза по 20 минут.
6. Способ по п.1, характеризующийся тем, что фрагментирование агарозного геля проводится следующим образом:
отмытый гель фотографируется с помощью гель-документирующей системы;
дорожки геля фрагментируются вручную соответственно маркеру молекулярного веса, и каждый фрагмент помещается в отдельную пробирку; также в отдельную пробирку помещается гель из зоны вне дорожек для контроля общей контаминации агарозного геля молекулами ДНК.
7. Способ по п.1, характеризующийся тем, что количественная ПЦР с фрагментами геля в качестве матрицы проводится следующим образом:
фрагменты геля плавятся при 95°С в течение 10 минут;
10 мкл раствора фрагментов расплавленного геля разводятся в 10 раз водой; полученный раствор используется для постановки количественной ПЦР;
для количественной ПЦР используются пара праймеров: прямой 5`-GTGCTTGCGCCACCCACGT-3` и обратный 5’-GCCGCGCGGAGGCGGAG-3` и «TaqMan» зонд 5’-FAM AGTCCGTGGTGGGGCTGGGG BHQ1-3`, специфичные к повторам D4Z4 хромосомы 4;
в качестве контролей используются разведенный в 10 раз гель из зоны вне дорожек с добавлением и без геномной ДНК; положительный контроль с добавлением геномной ДНК; отрицательный контроль с добавлением воды для ПЦР.
8. Способ по п.1, характеризующийся тем, что анализ данных количественной ПЦР с фрагментами геля в качестве матрицы проводится следующим образом:
полученные средние значения пороговых циклов каждого фрагмента геля обрабатываются в программе Microsoft Excel; обсчет данных ПЦР осуществляется методом 2-ΔΔCt,
при расчетах вычисляется разница между двумя типами фрагментов геля: фрагменты вне зоны дорожек и фрагменты дорожки геля; внутренним контролем служит агарозный гель из зоны вне дорожек, т.к. он отражает контаминированность агарозного геля таргетными молекулами, таким образом, изменения в типе образцов вне зоны дорожек геля сводится к нулю, в результате уравнение для обсчета данных сводится к виду: N(i)=2 - (Ct(i) - Ct(g)), где N(i) - относительное значение концентрации последовательностей D4Z4 в исследуемом фрагменте геля; Ct(g) - среднее значение порогового цикла в образце контрольного геля «G-»; Ct(i) - среднее значение порогового цикла в исследуемом фрагменте геля, по полученным данным строится гистограмма;
фрагменты геля с максимальными значениями относительной концентрации таргетных молекул указывают на наличие в них таргетных аллелей, сопоставив положение данных фрагментов геля относительно маркера молекулярного веса, делается вывод о количестве макросателлитных повторов D4Z4 хромосомы 4 в обнаруженных аллелях.
| VASALE J | |||
| et al., Molecular combing compared to Southern blot for measuring D4Z4 contractions in FSHD | |||
| Устройство для закрепления лыж на раме мотоциклов и велосипедов взамен переднего колеса | 1924 |
|
SU2015A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| YOUNG E | |||
| et al, Comprehensive Analysis of Human Subtelomeres by Whole Genome Mapping | |||
| PLOS Genet | |||
| Способ восстановления спиралей из вольфрамовой проволоки для электрических ламп накаливания, наполненных газом | 1924 |
|
SU2020A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| MITSUHASHI S | |||
| et al, Nanopore-based single molecule | |||

