ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к созданию синтетического белкового каркаса, который проявляет определенные функциональные и биофизические характеристики, полезные при использовании синтетического белка для доставки и презентации антигенов иммунной системе хозяина даже тогда, когда указанные антигены слабо иммуногенны или неиммуногенны в организме хозяина, вследствие чего у хозяина индуцируется выработка специфического гуморального иммунного ответа на антигены. Описывается синтетический белок, который является иммуногенным для иммунных систем млекопитающих и который может собираться в стабильные определенные мультимеры. Кроме того, описывается способ применения указанного белка для обеспечения иммуногенности и индуцирования образования специфических антител против слабо иммуногенных или неиммуногенных пептидов.
ПЕРЕКРЕСТНЫЕ ССЫЛКИ НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка заявляет приоритет заявки на патент США №61/791268, поданной 15 марта 2013 года, под названием «Синтетические самособирающиеся иммуногенные белки», которая включена в данный документ посредством ссылки во всей своей полноте.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Введение чужеродного (не своего) вещества, то есть антигена, в иммунную систему позвоночного животного обычно приводит к индукции иммунного ответа хозяина, направленного против этого антигена. Обычно это вызывает стимуляцию В- и/или Т-лимфоцитов и продукцию молекул иммуноглобулинов (антител), которые узнают и связывают этот антиген. Существует очень много факторов, которые влияют на степень индуцирования веществом иммунного ответа в организме хозяина. Степень чужеродности важна, поскольку иммунная система возникла в процессе эволюции и развилась с целью быть нечувствительной к «своему». Размер также является важным фактором, так как молекулы большего размера обычно являются более иммуногенными, чем молекулы меньшего размера. Молекулы, имеющие молекулярную массу менее ~1000 Да (относятся к категории гаптенов), слишком малы, чтобы быть замеченными иммунной системой в изолированном виде, и поэтому они являются неиммуногенными, хотя они все еще могут быть антигенными.
Молекулы большего размера будут более сложноорганизованными и поэтому более вероятно, что они будут содержать многочисленные иммуногенные эпитопы, а также легче будут поглощаться и перерабатываться антигенпрезентирующими клетками (АРС). Состав вещества также является важным, причем белки, бесспорно, являются наиболее иммуногенными. Полисахариды гораздо менее иммуногенны (в изолированном виде), а нуклеиновые кислоты и липиды в основном неиммуногенны. Аналогично, корпускулярные или денатурированные антигены являются более иммуногенными, чем растворимые и природные молекулы. Путь поступления и биологические активности чужеродных веществ могут также значительно влиять на природу и степень любого иммунного ответа хозяина. Например, парентеральная инъекция вещества, которое взаимодействует с компонентами или клетками иммунной системы, будет приводить к намного более сильному ответу на относительно инертное или неактивное вещество, чем поступление через слизистую оболочку (прием внутрь/ингаляция).
Т-клетки и В-клетки узнают чужеродные антигены и реагируют на них по-разному. Специализированные антигенпрезентирующие клетки или АРС (макрофаги, дендритные клетки и В-клетки) непрерывно исследуют свою среду, захватывая молекулы из внеклеточного пространства, в том числе макромолекулы и целые микроорганизмы, и перерабатывая их белковый состав. Экзогенные белки перевариваются группой протеолитических ферментов в эндосомах, и результирующие пептиды презентируются на поверхности клеток в канавке молекул главного комплекса гистосовместимости (МНС) класса II. Они, в свою очередь, узнаются специализированными рецепторами на поверхности Т-клеток (TCR). Процесс развития Т-клеток обеспечивает, что Т-клетки, несущие рецепторы, которые реагируют на МНС II, содержащий аутологичные пептиды, удаляются, и только те клетки, которые распознают чужеродные последовательности, созревают успешно. Пептиды, которые узнаются Т-клетками (Т-клеточные эпитопы), являются без исключения линейными, но они не всегда экспонированы или доступны на нативном свернутом белке, из которого они были получены.
Напротив, поверхностные рецепторы В-клеток или иммуноглобулины (BCR) узнают и взаимодействуют в основном с растворимыми белками (включая конформационные и денатурированные эпитопы), гаптенами, полисахаридами и в меньшей степени некоторыми липидами и нуклеиновыми кислотами. Специфичность BCR идентична тому антителу, которое В-клетка может секретировать. После связывания с когнатным антигеном BCR интернализуется, и связанный с ним антиген процессируется. Только когда антиген является белком или когда он связан с белковым компонентом, антиген будет презентироваться на клеточной поверхности как часть комплекса МНС II. При этих условиях В-клетка может быть доступна для активации при помощи клетки Т-хелпера, которая содержит TCR, распознающий презентированный пептид. В случае большого или сложного бежа В-клетка может быть активирована многими различными Т-клетками, ни одна из которых не должна обязательно узнавать такой же антигенный эпитоп, как BCR, но все из которых будут узнавать пептидный компонент того же самого белка. Именно эта способность иммунных систем позвоночных позволяет им вырабатывать антитела против антигенных детерминант, которые не иммуногенны сами по себе.
Чтобы разработать эффективные вакцины, необходимо презентовать антигенные эпитопы иммунной системе хозяина таким способом, чтобы стимулировать сильный иммунный ответ, вовлекающий Т- и В-лимфоциты. Иммунные ответы, которые не включают активацию эффекторных (хелперных) Т-клеток и последующую стимуляцию ими В-клеток, являются обычно непродолжительными и не приводят к антигенной памяти, т.е. не приводят к более агрессивному и более быстрому гуморальному иммунному ответу, когда хозяин подвергается этому иммуногену во второй раз.
Также часто требуется, чтобы вакцины активировали антитела, способные ингибировать, блокировать или иным способом нейтрализовать функциональную активность мишени, и тем самым обеспечивали защиту хозяина. Это может представлять сложную задачу по многим причинам. Часто те эпитопы, которые должны выбираться в качестве мишени для гуморального иммунного ответа, не были идентифицированы из-за нехватки структурно-функциональных данных, касающихся этой мишени. Даже когда детальная информация о мишени и ее взаимодействиях установлена, идентифицированный(е) эпитоп(ы) может(могут) не быть иммунно-доминантным(и) и поэтому может(могут) не вызывать ответы, искомые для большинства пациентов. В других случаях ключевые защитные антигенные детерминанты могут быть не белком, а, например, полисахаридами в патогенных гликопротеинах, которые обычно неиммунногенны (Т-зависимые) в изолированном виде.
Преобладающее большинство вакцин вводят парентеральными путями; однако, есть много преимуществ для доставки через слизистую оболочку, таких как соблюдение пациентом режима, самостоятельное введение, сниженный риск инфекции и возможность стимулирования как иммунных свойств слизистых оболочек, так и системного иммунитета. Существует также много препятствий, которые нужно преодолеть, таких как разбавление вакцины, присутствие микрофлоры, потребность противостоять низкому pH при пероральном введении, мембранная проницаемость и потребность в сильных адъювантах (Vajdy et al. 2004). Кроме того, введение через слизистую оболочку может привести к толерантности В-клеток, а не к иммунному ответу. Дозировка также может оказывать большой эффект на иммунные ответы. Если иммуноген практически не воспринимается иммунной системой или если система загружена слишком высокой дозой, то может индуцироваться толерантность. Наоборот, слишком маленькая доза может также привести к толерантности или может быть просто не способной стимулировать иммунные клетки в достаточной мере.
Были разработаны многочисленные подходы, чтобы способствовать преодолению этих трудностей. В большинстве случаев вакцины вводят вместе с некоторой формой адъюванта. По существу адъювантами является любой состав, который при совместном введении с иммуногеном вызывает одно или несколько из следующих явлений: стойкость иммунногена в месте инъекции, усиление костимулирующих сигналов, неспецифическая стимуляция пролиферации лимфоцитов или формирование гранулемы. Существует множество форм адъювантов, например, полный адъювант Фрейнда содержит инактивированные Mycobacterium, тогда как другие содержат эмульсию масла (например, сквалена) в воде. Такие адъюванты обычно используются для животных, поскольку они могут вызвать неблагоприятные реакции в месте инъекции. Некоторые органические адъюванты используются в вакцинах для человека, таких как Montanide (основана на минеральном масле с добавкой растительных компонентов), хотя в большинстве случаев они являются неорганическими, как, например, соли алюминия.
Среди самых распространенных и широко используемых методов для преодоления низкой иммуногенности может применяться объединение известного или желаемого антигена или антигенной детерминанты с высоко иммуногенным носителем. Таким является белок или полипептид, полученный из различных организмов, например бычий сывороточный альбумин (BSA) и гемоцианин фиссуреллы (KLH), которые часто используются в качестве носителей для химически конъюгированных гаптенов и небольших пептидов для того, чтобы индуцировать антитела у животных (Berzofsky and Berzofsky, 1993). Носитель презентует гаптены на достаточно большой молекуле для узнавания и процессирования иммунной системой хозяина, а также стимулирует иммунный ответ хозяина, будучи по своей сути иммунногенным.
В общем, чем дальше филогенетически отдалены источники белков-носителей от реципиента, тем лучше. Тогда, вероятно, носитель будет сильно отличаться от белков хозяина, и, следовательно, будет более чужеродным. Еще одной важной особенностью при выборе белка-носителя является возможность того, что если этот белок является гомологом белка хозяина и, следовательно, имеет значительную гомологию, то вызванный иммунный ответ может также реагировать на белки хозяина и приводить к неблагоприятным побочным эффектам. Небелковые антигены могут быть соединены только химически, что может ограничить контроль над тем, где на носителе они прикреплены и как они презентуются. Небольшие пептиды могут быть соединены химически или на генетическом уровне. В других отраслях научных исследований современные разработки в биоинформатике привели к развитию рационального дизайна иммуногенов, и, в частности, пептидов.
Концепция пептидных вакцин основана на идентификации и химическом синтезе эпитопов В-клеток и Т-клеток, которые являются иммунно-доминантными и могут индуцировать специфические иммунные ответы, например, связывание В-клеточного эпитопа целевой молекулы с хорошо известным Т-клеточным эпитопом для обеспечения его иммуногенности (Naz R.K. and Dabir P. 2007). Считается, что пептиды относительно легко производить по сравнению с большими и более сложными белковыми антигенами. Они также могут обладать необходимой химической стабильностью и отсутствием онкогенного или инфекционного потенциала, что делает их привлекательными вакцинными кандидатами. Однако несколько препятствий ограничивают широкое распространение использования пептидных вакцин, включая их часто слабую собственную иммуногенность и потребность в сильных адъювантах и носителях. Другие исследования предложили, что можно получать более иммуногенные рекомбинантные химерные белки, если эпитопы Т-хелперов соединять в виде тандемных повторов (Kjerrulf М., et al. 1997).
Другим популярным классом белка-носителя являются бактериальные токсоиды. В случае вакцин против бактериальной инфекции, когда симптомы инфекции вызваны действием токсинов, тогда эти токсины могут использоваться в качестве вакцин. Конечно, необходимо сделать их неактивными или химически, или при помощи нетоксичного компонента. Такие аттенуированые токсины, например, вакцины против дифтерии и столбняка, которые были разработаны в XX столетии, называются токсоидами. Полисахаридно-белковые конъюгированные вакцины, которые используются или находятся на заключительной стадии разработки компаниями, такими как Wyeth (Pfizer), Aventis Pasteur, GSK, Merck и другими, используют столбнячный, дифтерийный или другие токсоиды.
Субъединица В холерного токсина или теплоустойчивый энтеротоксин (LT) Е. coli были предложены многими как полезные белковые носители для различных применений вакцин (Nemchinov, L.G et al. 2000, George-Chandy, A. et al. 2001, патент США №6153203). Они высоко иммуногенны, а в отсутствии СТ-А субъединицы являются нетоксичными. При формировании основы широкого использования противохолерной вакцины она характеризуется подтвержденным профилем безопасности при систематическом применении. Относительно небольшая (~12 кДа), она может собираться в стабильные пентамеры, которые придают ей намного более высокую молекулярная массу.
Для многих исследователей особый интерес представляет использование аффинности СТВ и пентамеров энтеротоксина к GMI-ганглиозиду, разветвленному пента-сахариду, который был обнаружен на поверхности ядросодержащих клеток. Во время инфекции холеры именно это связывание облегчает перенос голотоксина через кишечный эпителий. В научной литературе было много сообщений о том, что вакцины, основанные на СТВ-соединениях, химически или генетически, могут быть эффективными при стимулировании иммунных свойств слизистых оболочек (George-Chandy, A. et al. 2001, Flouhui Song et al. 2003, Shenghua Li et al. 2009, Harakuni, A. et al. 2005), если они применяются перорально или интраназально. Для сохранения способности взаимодействовать с GMI-ганглиозидами, которые присоединяются в полости, сформированной между смежными субъединицами СТВ, важно, чтобы целевой антиген не блокировал доступ к сайту связывания GMI и не предотвращал сборку СТВ-мультимеров.
Было продемонстрировано, что слияния на генетическом уровне могут осуществляться с СТВ, которая успешно сохраняет GMI-связывание, хотя есть также и ограничения. Liljeqvist, S. и др. (1997) показали, что связывающий домен сывороточного альбумина стрептококкового белка G может быть слит на генетическом уровне или с N-, или с С-концом СТВ, или с обеими концами одновременно при сохранении GMI-связывания. Тем не менее, было отмечено, что N-концевой слитый белок и двойной слитый белок были значительно менее эффективными при формировании устойчивых пентамеров и менее эффективными при связывании с G GMI-Подобным образом было продемонстрировано, что большие слияния на генетическом уровне не способны формировать пентамеры, если гетерогенная смесь СТВ и химерного, и дикого типа не присутствует (Harakuni, A. et al. 2005).
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Согласно настоящему раскрытию высоко иммуногенная природа синтетического носителя способна увеличивать иммунный ответ хозяина на включенные вариабельные последовательности благодаря его изначально иммуностимуляторным и адъювант-подобным свойствам. Дополнительно раскрывается синтетический носитель, который согласно изобретению будет вызывать у хозяина выработку гуморального иммунного ответа на «свои» антигенные детерминанты, которые кодируются, по меньшей мере от части, вариабельными последовательностями.
В иллюстративном варианте осуществления рекомбинантный синтетический белок может собираться в стабильные гомопентамеры, где каждый мономер включает одну или несколько антигенных детерминант, полученных из целевых белков.
В другом иллюстративном варианте осуществления рекомбинантный синтетический белок может собираться в стабильные гетеропентамеры, где мономеры, которые экспрессируют различные антигенные детерминанты, собраны вместе.
В дополнительном иллюстративном варианте осуществления изобретения рекомбинантный синтетический белок в значительной степени сходен со следующей последовательностью:
И в еще одном дополнительном иллюстративном варианте осуществления рекомбинантный синтетический белок включает линкерную или спейсерную последовательности, посредством которых вариабельные последовательности, размещенные на одном или обоих концах синтетического носителя, отделяются от синтетического носителя таким способом, чтобы дать возможность связаться синтетическим элементам носителя из нескольких рекомбинантных белков. В одном иллюстративном варианте осуществления рекомбинантный синтетический белок, содержащий линкерную последовательность, которая присоединяется к фактору роста, в значительной степени сходен со следующей последовательностью:

В другом иллюстративном варианте осуществления рекомбинантный синтетический белок будет собираться в стабильные пентамеры в форме кольца. Другие мультимерные сборки, например, без ограничения димеры, тримеры, тетрамеры и большие мультимеры, также предусмотрены в пределах объема настоящего раскрытия. Линкерные последовательности могут быть различными, но, как минимум, они должны иметь достаточную длину, чтобы предотвращать стерическое ингибирование сборки мультимеров различных последовательностей антигенных детерминант при связывании доменов синтетического носителя. В пределах объема настоящего раскрытия предусмотрено, что линкеры или спейсеры могут быть гибкими, позволяя формировать стабильные пентамеры.
В другом варианте осуществления пентамерная структура мультимера стабилизируется введением по меньшей мере одной стабилизирующей молекулы. Стабилизирующая молекула может быть коэкспрессированна в тех же самых клетках, что и синтетическая молекула, или альтернативно может быть добавлена экзогенно.
В одном варианте осуществления изобретения стабилизирующая молекула включает последовательности в значительной степени схожие с доменом СТ-А2 холерного токсина.
В другом варианте осуществления изобретения стабилизирующая молекула содержит последовательности, которые взаимодействуют с синтетическим белком для того, чтобы стабилизировать мультимеры. Дополнительно предусматривается, что эта стабилизация может приводить к большей пропорции пентамерных форм с использованием общепризнанных методик производства, известных в данной области техники.
В дополнительном иллюстративном варианте осуществления стабилизирующая молекула может служить для прикрепления антигенного домена к мультимерной форме синтетического белка.
В еще одном дополнительном варианте осуществления прерывающиеся повторы стабилизирующей молекулы могут служить для связывания двух или нескольких мультимерных форм синтетического белка.
В иллюстративном варианте осуществления линкерные или спейсерные последовательности будут в основном гидрофильными и будут гибкими, таким образом, они не будут включать определенной вторичной структуры. В другом иллюстративном варианте осуществления изобретения линкерные последовательности будут такими, которые прямо или косвенно не влияют на сворачивание домена синтетического носителя или различных доменов антигенных детерминант.
В дополнительном иллюстративном варианте осуществления изобретения линкерные последовательности будут устойчивыми к протеолизу внеклеточными протеазами.
В другом иллюстративном варианте осуществления изобретения линкерные или спейсерные последовательности включают без ограничения следующие:
В одном иллюстративном варианте осуществления линкерные последовательности будут включать последовательности, которые или отдельно, или совместно с фланкирующими последовательностями из различных антигенных детерминант или синтетического носителя будут формировать Т-клеточные эпитопы. Предпочтительно Т-клеточные эпитопы будут Т-клеточными эпитопами, которые распознаются Т-хелперами человека, когда они связаны с белками МНС (главного комплекса гистосовместимости) класса II. Предпочтительно Т-клеточные эпитопы будут связаны с молекулами МНС II HLA (лейкоцитарного антигена человека) подкласса DR.
В другом иллюстративном варианте осуществления изобретения синтетический рекомбинантный белок является структурно гомологичным В-субъединице группы А1В5 бактериальных голотоксинов.
В дополнительном иллюстративном варианте осуществления мультимеры рекомбинантного синтетического белка способны связываться с GМI-ганглиозидом.
В иллюстративном варианте осуществления изобретения синтетический рекомбинантный белок является иммуногенным белком, который индуцирует иммунный ответ в организмах хозяев млекопитающих.
В другом иллюстративном варианте осуществления изобретения синтетический рекомбинантный белок является иммуногенным белком, включающим одну или несколько вариабельных последовательностей, которые презентуют антигенные детерминанты, на которые желательно вызвать иммунный ответ у пациентов. Вариабельные последовательности могут быть расположены на N- и/или С- конце последовательности, кодирующей синтетический носитель, или могут быть включены в последовательность, кодирующую синтетический носитель, таким образом, что они презентуются надлежащим образом клеткам иммунной системы. В предпочтительном варианте осуществления изобретения вариабельные последовательности представлены таким образом, чтобы воспроизводить конформацию, которую эти последовательности проявляют в природных молекулах, из которых они получены, и быть доступными для иммуноглобулиновых поверхностных рецепторов В-клеток.
В другом иллюстративном варианте осуществления изобретения различные антигенные детерминанты получают из сигнальных молекул, включая без ограничения факторы роста, такие как адреномедуллин (AM), ангиопоэтин (Ang), костные морфогенетические белки, эпидермальный фактор роста (EGF), фактор роста фибробластов (FGF), фактор роста гепатоцитов (HGF), инсулиноподобный фактор роста (IGF), фактор роста нервов (NGF) и другие нейротрофины, фактор роста тромбоцитов (PDGF), трансформирующий фактор роста альфа (TGF-α), трансформирующий фактор роста бета (TGF-β), фактор некроза опухоли альфа (TNF-α) и фактор роста эндотелия сосудов (VEGF).
В другом варианте осуществления вариабельные антигенные детерминанты получают из других молекул лигандов, которые вовлечены в развитие или прогрессирование болезни, таких как без ограничения PDL1.
В другом варианте осуществления изобретения различные антигенные детерминанты получают из рецепторов, найденных на поверхности клеток и вовлеченных в сигнальные события, которые регулируют рост клеток, таких как природные рецепторы факторов роста.
В дополнительном варианте осуществления вариабельные антигенные детерминанты получают из опухолевых антигенов.
В еще одном варианте осуществления различные антигенные детерминанты получают из бактериальных, вирусных, грибковых или других патогенов.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Варианты осуществления, описанные в настоящем раскрытии, проиллюстрированы на фигурах прилагаемых графических материалов, которые предназначены для иллюстративных целей, а не для ограничения, в которых аналогичные ссылки предназначены для обозначения аналогичных или соответствующих частей, в которых:
на фигуре 1 показаны сигналы ELISA, произведенные группой растворимых мутантных клонов, связывающихся с иммобилизованной молекулой, содержащей галактозу (GM1), после 3-х циклов отбора;
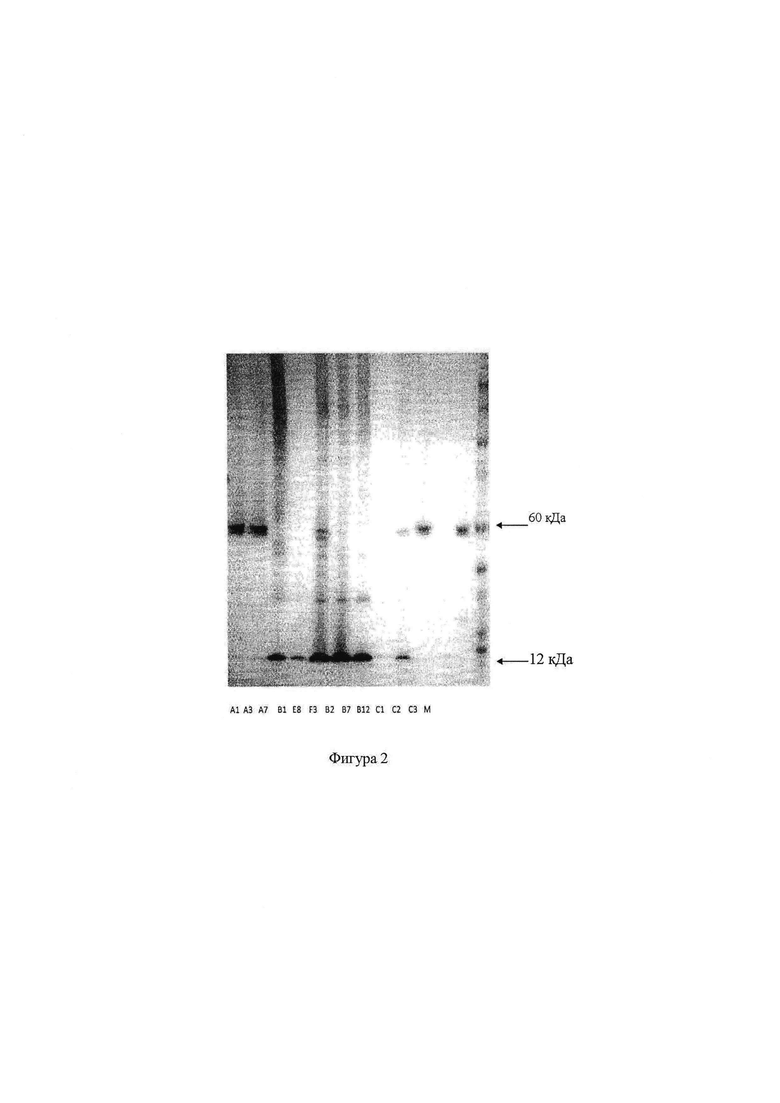
на фигуре 2 показан Вестерн-блоттинг растворимых мутантных белков одной из мутантных библиотек, которые подвергали электрофорезу в геле с SDS (додецилсульфатом натрия), некоторые из которых сохраняют способность формировать пентамеры, и других, которые не сохраняют эту способность;
на фигуре 3 показан отбор мутаций, которые были идентифицированы после отбора и скрининга нескольких мутантных библиотек последовательностей при отборе мутантов на фигуре 3(a), каждая из которых выделена из одной из 7 отдельных библиотек мутантов и иллюстрирует распределения мутантных остатков, и последовательностей синтетических белков на фигуре 3(b), которые были получены из отобранных мутаций, полученных из нескольких различных мутантных клонов;
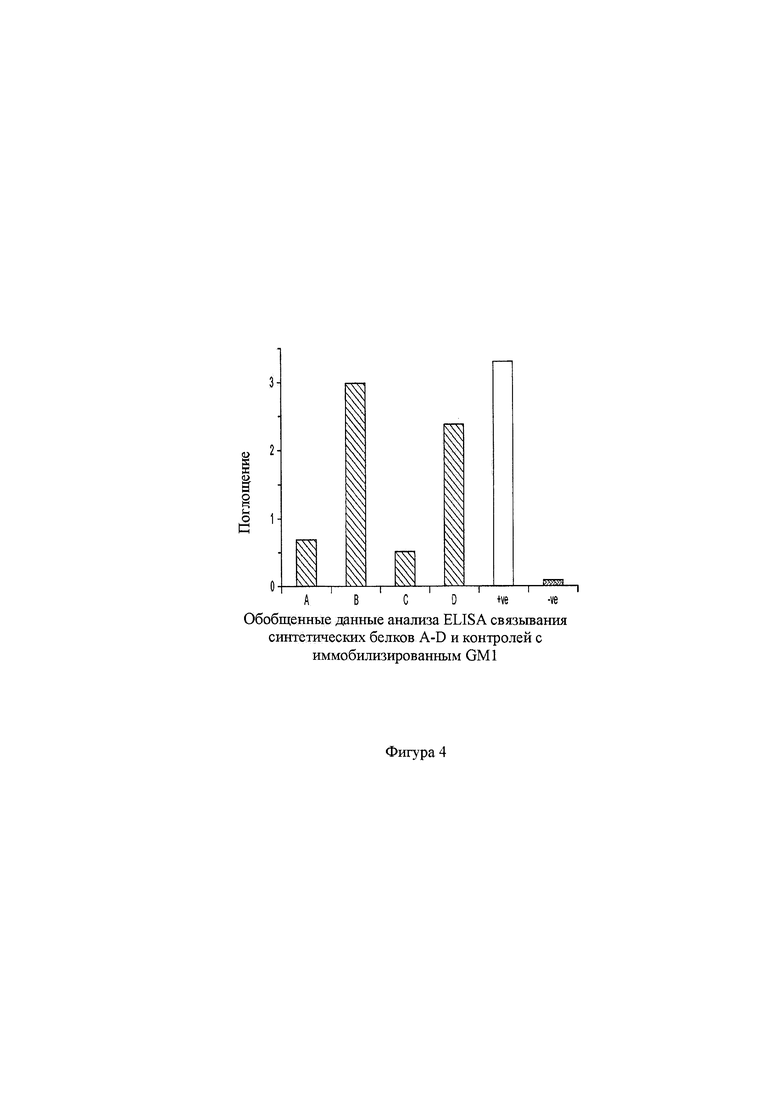
на фигуре 4 показаны результаты анализа ELISA GM1-связывания, полученные от четырех отобранных мутантов, демонстрирующие, что два клона сохраняли галактозо-связывающую активность и два не сохраняли;
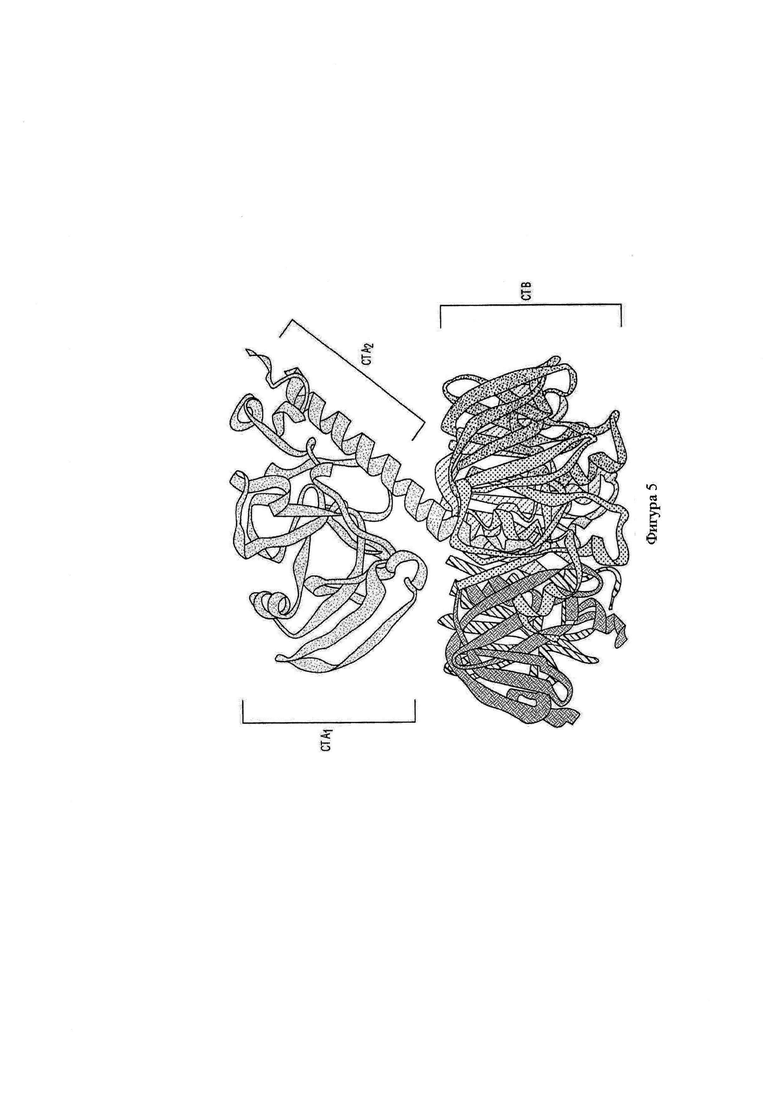
на фигуре 5 показана структура целой молекулы холерного голотоксина и
на фигуре 6 показаны последовательности синтетического белка согласно настоящему раскрытию (Sequence ID: 1) и дополнительные последовательности синтетического белка согласно настоящему раскрытию включающие последовательности, кодирующие человеческий TGF-бета 1 (жирный шрифт), гибкий линкер (выделено курсивом) и синтетический белок носитель (Sequence ID: 2).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В данном документе раскрыты подробные варианты осуществления данных рекомбинантных белков или вакцин, однако, нужно понимать, что раскрытые варианты осуществления изобретения являются только иллюстративными и могут быть осуществлены в различных формах. Поэтому, специфические функциональные детали, раскрытые в данном документе, не должны интерпретироваться как ограничивающие, а исключительно как основа для формулы и как примерная основа для обучения специалистов в данной области в целях различного применения рекомбинантного белка, раскрытого в данном документе.
Настоящее раскрытие предлагает синтетический рекомбинантный белок для улучшения презентирования максимального количества эпитопов фактора роста, эпитопов опухолевых антигенов и/или рецептор-связывающих участков как элементов иммуногенного рекомбинантного белка.
В одном иллюстративном варианте осуществления описан синтетический рекомбинантный белок, показанный в Sequence ID: 2, содержащий по меньшей мере один фактор роста, включая без ограничения трансформирующий фактор роста человека (TGF), опухолевый антиген и/или рецептор. В альтернативных иллюстративных вариантах осуществления белок может представлять собой другие иммуногенные рекомбинантные белки, которые моделируют, основываясь на известных иммуногенных белках. В пределах объема настоящего раскрытия предполагается, что такие рекомбинантные белки будут представлять собой полипептиды, которые являются высоко иммуногенными для человеческой иммунной системы. Предпочтительно рекомбинантные белки придадут дополнительные свойства химерному белку, например, высокий экспрессионный выход и простоту изготовления, стабильность при оральном применении и способность проникать из пищеварительного тракта в кровоток и/или существующее безопасное применение у человека.
Определенные проиллюстрированные варианты осуществления изобретения, представленные в данном документе, включают рекомбинантные белки согласно настоящему раскрытию в композициях вакцин и в композициях иммунологических адъювантов, включая фармацевтические составы, которые содержат, в дополнение к рекомбинантным белкам, по меньшей мере один адъювант, который относится к компоненту таких композиций, которые имеют адъювантную активность.
Адъювант, обладающий такой адъювантной активностью, включает композицию, которая, при введении субъекту, такому как человек (например, больной человек), примат за исключением человека, млекопитающее или другие высшие эукариотические организмы, который имеет изученную иммунную систему, способен к изменению (то есть, увеличению или уменьшению статистически значимым образом и в определенных предпочтительных вариантах осуществления к усилению или увеличению) силы и/или длительности иммунного ответа. В определенных проиллюстрированных вариантах осуществления, раскрытых в данном документе, желаемый антиген и/или антигены содержатся в белке-носителе, и необязательно один или несколько адъювантов могут таким образом изменять, например, вызывать или увеличить, иммунный ответ, который направлен против желаемого антигена и/или антигенов, которые можно вводить одновременно или которые могут быть разделены во времени и/или пространстве (например, в разных частях тела) при их введении, но определенные проиллюстрированные варианты осуществления изобретения не предназначены для ограничения таким образом, и поэтому также предусматривают введение рекомбинантного белка в композиции, которая не включает указанный антиген, но которая может также включать без ограничения один или несколько коадъювантов, имидазохинолиновый иммуномодулятор.
Соответственно и как отмечено выше, адъюванты включают композиции, которые характеризуются адъювантными эффектами, такие как сапонины и миметики сапонина, включая миметики QS21 и QS21 (см., например, патент США. №5057540; ЕР 0362279 B1; WO95/17210), квасцы, растительные алкалоиды, такие как томатин, детергенты, такие как (без ограничения) сапонин, полисорбат 80, Span 85 и стерил-тирозин, один или несколько цитокинов (например, GM-CSF, IL-2, IL-7, IL-12, TNF-альфа, IFN-гамма), имидазохинолиновый иммуномодулятор и иммуномодулятор со структурой типа «двухцепочечный стебель-петля» (dSLIM, например, Weeratna et al. 2005 Vaccine 23: 5263).
Детергенты, включающие сапонины, описаны, например, в патенте США №6544518; Facaille-Dubois, М. and Wagner FI. (1996 Phytomedicine 2: 363-386), патенте США №5057540, Kensil, Crit. Rev Ther Drug Carrier Syst, 1996,12 (1-2): 1-55, и ЕР 0362279 B1. Корпускулярные структуры, которые называются иммуностимулирующими комплексами (ISCOMS), которые включают фракции Quil А (сапонин), являются гемолитическими и ранее использовались при изготовлении вакцин (Morein, В., ЕР 0109942 В1). Сообщалось, что эти структуры имеют адъювантную активность (ЕР 0109942 B1; WO 96/11711). Гемолитические сапонины QS21 и QS17 (очищенные с помощью ВЭЖХ фракции Quil А) были описаны как мощные системные адъюванты, и метод их производства описан в патенте США №5057540 и ЕР 0362279 В1. В этих ссылках также дано описание использования QS7 (не гемолитической фракции Quil-A), которая действует как сильный адъювант для системных вакцин. Использование QS21 дополнительно описано в Kensil et al., 1991. J. Immunology 146: 431-437).
Также известны комбинации QS21 и полисорбата или циклодекстрина (WO 99/10008). Корпускулярные адъювантные системы, включающие фракции QuilA, такие как QS21 и QS7, описаны в WO 96/33739 и WO 96/11711. Другие сапонины, которые использовались в исследованиях системной вакцинации, охватывают сапонины, полученные из других видов растений, таких как Gypsophila и Saponaria (Bomford et al., Vaccine, 10 (9): 572-577, 1992). [0203] Эсцин является другим детергентом, родственным сапонинам, для использования в адъювантных композициях в вариантах осуществления, раскрытых в данном документе. Эсцин описан в Merck Index (12-е дополненное издание, статья 3737) как смесь сапонинов, получаемых из семян дерева конского каштана, Aesculus hippocastanum. Для его выделения описана хроматография и очистка (Fiedler, Arzneimittel-Forsch. 4, 213 (1953)), и ионообменные носители (Erbring et al., Патент США №3238190). Фракции эсцина (также известного как аэсцин) были очищены и продемонстрированы как обладающие биологической активностью (Yoshikawa М., et al. (Chem Pharm Bull (Токио) 1996 August; 44 (8): 1454-1464)). Дигитонин является другим детергентом, также описанным в Merck index (12-е издание, статья 3204) как сапонин, получаемый из семян Digitalis purpurea и очищенный согласно процедуре, описанной в Gisvold et al., J. Am. Pharm. Assoc., 1934, 23, 664; и Rubenstroth-Bauer, Physiol. Chem., 1955, 301, 621.
Другие адъюванты или коадъюванты для использования согласно раскрытым в данном документе вариантами осуществления изобретения включают блок-сополимер или биоразлагаемый полимер, которые относятся к классу полимерных соединений, с которыми будут знакомы специалисты в данной области. Примеры блок-сополимера или биоразлагаемого полимера, которые могут быть включены в вакцинных композиций или иммунологического адъюванта, включают Pluronic. RTM. L121 (BASF Corp., Маунт Олив, Нью-Джерси; см., например, Yeh et al., 1996 Pharm. Res. 13: 1693).
Некоторые дополнительные варианты осуществления изобретения предусматривают иммунологические адъюванты, которые включают без ограничения масло, которое в некоторых таких вариантах осуществления изобретения может вносить вклад в адъювантную активность, а в других таких вариантах осуществления могут дополнительно или альтернативно обеспечивать фармацевтически приемлемый носитель или наполнитель. Любое число соответствующих масел известно и может быть подобрано для включения в состав вакцин и иммунологических адъювантных составов, основываясь на настоящем изобретении. Примеры таких масел для иллюстрации, а не для ограничения, включают сквален, сквалан, минеральное масло, оливковое масло, холестерин и маннид моноолеат.
Иммуномодуляторы, такие как имидазохинолин, также известны в данной области техники и также могут быть включены как адъюванты или коадъюванты в определенных раскрытых в данном документе вариантах осуществления. Также, как отмечено выше, одним из типов адъюванта или коадъюванта для использования в вакцинной композиции в соответствии с раскрытием, описанным в данном документе, могут быть алюминиевые коадъюванты, которые обычно называются "квасцы". Квасцовые коадъюванты основаны на следующих: оксигидроксиде алюминия, фосфате алюминия или различных патентованных солях. Квасцовые коадъюванты обладают полезными свойствами, потому что они имеют хорошие показатели безопасности, увеличивают гуморальные иммунные ответы, стабилизируют антигены и относительно просты для крупномасштабного производства (Edelman 2002 Mol. Biotechnol. 21:129-148; Edelman, R. 1980 Rev. Infect. Dis. 2: 370-383).
Фармацевтические композиции
В определенных иллюстративных вариантах осуществления фармацевтическая композиция представляет собой вакцинную композицию, которая включает рекомбинантный белок согласно данному раскрытию и может кроме того включать один или несколько компонентов, представленных в данном документе, которые выбраны из TLR-агониста, коадъюванта (включая, например, цитокин, имидазохинолиновый иммуномодулятор и/или dSLIM) и т.п., и/или конструкцию для рекомбинантной экспрессии в комбинации с фармацевтически приемлемым носителем, наполнителем или разбавителем.
Иллюстративные носители будут нетоксичны для реципиентов при используемых дозировках и концентрациях. Для вакцин, содержащих рекомбинантный белок, будут осуществляться введения от приблизительно 0,01 мкг/кг до приблизительно 100 мг/кг массы тела, как правило, внутрикожным, подкожным, внутримышечным, или внутривенным путем, или другими путями. Для специалистов в данной области будет очевидным, что количество и частота введения будет зависеть от ответа организма хозяина. «Фармацевтически приемлемые носители» для терапевтических средств хорошо известны в области фармации и описаны, например, в Remington’s Pharmaceutical Sciences, Mack Publishing Co. (A.R. Gennaro edit. 1985). Например, при физиологическом значении рН могут использоваться стерильный солевой раствор и фосфатный буферный раствор. Консерванты, стабилизаторы, красители и даже ароматизирующие средства могут быть представлены в фармацевтической композиции. Например, бензоат натрия, аскорбиновая кислота и сложные эфиры п-гидроксибензойной кислоты могут быть добавлены как консерванты. Кроме того, могут использоваться антиоксиданты и суспендирующие агенты.
Фармацевтические композиции могут быть представлены в любой форме, которая позволяет вводить композицию пациенту. Например, композиция может быть в твердой, жидкой или газообразной (аэрозоль) форме. Типичные пути введения включают без ограничения пероральный, местный, парентеральный (например, сублингвальный или трансбуккальный), сублингвальный, ректальный, вагинальный и интраназальный (например, в виде спрея). Термин парентеральный, используемый в данном документе, включает ионтофоретическую доставку, сонофоретическое, пассивное трансдермальное введение, введение при помощи микроиглы, а также подкожные инъекции, внутривенную, внутримышечную, внутригрудинную, интракавернозную, внутриоболочечную, внутримуральную, внутриуретральную инъекцию или методики инфузии. В определенном варианте осуществления изобретения композиция, описанная в данном документе, (включая вакцинные и фармацевтические композиции) вводится интрадермально при использовании техники, выбранной из ионтофореза, микрокавитации, сонофореза или микро иглоукалывания.
Фармацевтический состав формулируется так, чтобы дать возможность активным ингредиентам, содержащемся в составе, быть биодоступными после введения композиции пациенту. Составы, которые будут вводиться пациенту, могут быть в форме одной или нескольких единиц дозировки, где, например, таблетка может быть одной единицей дозировки, а контейнер, содержащий одно или несколько соединений этого изобретения в форме аэрозоля, может содержать несколько единиц дозировки.
Для перорального введения может присутствовать наполнитель и/или связующее вещество. Примерами являются сахароза, каолин, глицерин, крахмальные декстрины, альгинат натрия, карбоксиметилцеллюлоза и этилцеллюлоза. Могут присутствовать красящие и/или ароматизирующие средства. Может использоваться покрывающая оболочка. Композиция может быть в форме жидкости, например, эликсира, сиропа, раствора, эмульсии или суспензии. Жидкость может использоваться для перорального введения или для доставки путем инъекции, например. В случае предназначения для перорального введения предпочтительные композиции содержат один или несколько подсластителей, консерванты, краситель/пигмент и усилитель вкуса. В композицию, предназначенную для введения путем инъекции, может быть включено одно или несколько поверхностно-активных веществ, консервант, смачивающее средство, диспергирующее средство, суспендирующее средство, буфер, стабилизатор и изотоническое средство.
Жидкая фармацевтическая композиция, используемая в данном документе или в форме раствора, суспензии, или в другой подобной форме, может включать один или несколько из следующих носителей или наполнителей: стерильные разбавители, такие как вода, для инъекций, солевой раствор, предпочтительно физиологический солевой раствор, раствор Рингера, изотонический раствор хлорида натрия, нелетучие масла, такие как сквален, сквалан, минеральное масло, маннид моноолеат, холестерин и/или синтетические моно- или ди-глицериды, которые могут служить в качестве растворителя или суспендирующей среды, полиэтиленгликоли, глицерин, пропиленгликоль или другие растворители; бактерицидные средства, такие как бензиловый спирт или метилпарабен; антиоксиданты, такие как аскорбиновая кислота или бисульфит натрия; хелирующие средства, такие как этилендиаминтетрауксусная кислота; буферы, такие как ацетаты, цитраты или фосфаты, и средства для регулирования тоничности, такие как хлорид натрия или декстроза. Парентеральный препарат может быть заключен в ампулы, шприцы для одноразового применения или в многодозовые, сделанные из стекла или пластика флаконы. Инъецируемая фармацевтическая композиция обязательно является стерильной.
В определенном варианте осуществления фармацевтическая или вакцинная композиция по настоящему изобретению содержит стабильную водную суспензию, менее чем 0,2 мкм, и дополнительно содержит, по меньшей мере один компонент, отобранный из группы, состоящей из фосфолипидов, жирных кислот, поверхностно-активных веществ, детергентов, сапонинов, фторированных липидов и т.п.
Также желательно включать другие компоненты в вакцинную или фармацевтическую композицию, такие как средства доставки, которые включают без ограничения алюминиевые соли, эмульсии «вода-в-масле», биоразлагаемые масляные среды, эмульсии «масло-в-воде», биоразлагаемые микрокапсулы и липосомы. Примеры дополнительных иммунностимуляторных веществ (коадъювантов) для использования в таких средах также описаны выше и могут включать N-ацетилмурамил-L-аланин-D-изоглутамин (MDP), глюкан, IL-12, GM-CSF, гамма-интерферон и IL-12.
Хотя любой подходящий носитель, известный специалистам в данной области техники, может быть применен в фармацевтических композициях по этому изобретению, тип носителя может меняться в зависимости от способа введения и от того, требуется ли замедленное освобождение. Для парентерального введения, такого как подкожная инъекция, носитель предпочтительно включает воду, солевой раствор, спирт, жир, воск или буфер. Для перорального введения можно применять любой из вышеупомянутых носителей или твердых носителей, таких как маннит, лактоза, крахмал, стеарат магния, сахарин натрия, тальк, целлюлоза, глюкоза, сахароза и карбонат магния. Биоразлагаемые микросферы (например, полимолочный галактид) также можно применять в качестве носителей для фармацевтических составов по настоящему изобретению.
Фармацевтические составы могут также содержать разбавители, такие как буферы, антиоксиданты, такие как аскорбиновая кислота, низкомолекулярные (меньше чем приблизительно 10 остатков) полипептиды, белки, аминокислоты, углеводы, включая глюкозу, сахарозу или декстрины, хелирующие средства, такие как ЭДТА, глутатион и другие стабилизаторы и наполнители. Нейтральный забуференный физиологический раствор или солевой раствор, смешанный с неспецифическим сывороточным альбумином, являются иллюстративными пригодными разбавителями. Предпочтительно продукт может быть составлен как лиофилизат с применением пригодных растворов наполнителей (например, сахарозы) в качестве разбавителей.
В иллюстративном варианте настоящего изобретения эпитоп или рецептор, несущий домен рекомбинантного белка, полученный либо из естественной, либо из синтетической полипептидной последовательности, должен обладать способностю самособираться в олигомерные мультимеры при соответствующих химических/окружающих условиях или редуцироваться до мономеров при альтернативных условиях. В идеале, домены, способные к мультимеризации, будут собираться в стабильные мультимеры с определенным количеством субъединиц, например, димеров, тримеров, тетрамеров, пентамеров, и т.д., так, что образуется продукт однородного размера. Без связи с каким-либо конкретным теоретическим объяснением считается, что рекомбинантный синтетический белок, указанный в Sequence ID: 1, будет делать возможной сборку в стабильные мультимеры с небольшим количеством субъединиц. Примеры природных полипептидов включают без ограничения белковые домены «лейциновые молнии», белок лактозный репрессор, стрептавидин/авидин, В-субъединицу холерного токсина, тримеризационный домен Pseudomonas и белки вирусного капсида.
Согласно настоящему раскрытию рекомбинантные белки, факторы роста или их части, клеточные рецепторы или их части, опухолеве антигены или их части, относятся к широкому спектру либо клеточных путей, вовлеченных в хроническое заболевание, либо к раковым заболеваниям относительно факторов роста и рецепторов и к максимально широкому спектру солидных опухолей для применения опухолевых антигенов в составе вышеуказанных синтетических белков. Эти белки находятся в форме рекомбинантного белка и могут применяться в лечении хронических заболеваний, например, раковых заболеваний груди, легких, мочевого пузыря, яичников, наружных женских половых органов, толстой кишки, легочной артерии, мозга, прямой кишки, кишечника, головы, шеи и пищевода. Поскольку в вышеупомянутых заболеваниях могут экспрессироваться различные опухолевые антигены, и могут сверхэкспрессироваться многие клеточные рецепторы и факторы роста, белки, описанные ниже, могут содержать один или несколько различных опухолевых антигенов, один или несколько различных рецепторов или факторов роста одного или многих клеточных путей, связанных с заболеванием. Эти белки называются «мультивалентными».
В контексте настоящего раскрытия «нейтрализующий домен» определен как область или области одного члена или обоих членов специфической связывающейся пары, например, фактора роста и его родственного рецептора, где связывание третьей молекулы, которая не является членом специфической связывающейся пары с вышеупомянутой(ыми) областью(ями), будет предотвращать последующее связывание двух членов специфической связывающейся пары. Третья молекула может быть другой молекулой белка, включая без ограничения антитело, или может быть небольшой молекулой небелковой природы и может быть либо природной, либо синтетической по происхождению. Нейтрализующий домен будет обычно включать те области членов специфической связывающейся пары, которые находятся в прямом контакте во время связывания, и будет также включать области за пределами вышеописанных областей, где после связывания третья молекула вводит достаточное стерическое препятствие для предотвращения непосредственного связывания членов специфической связывающейся пары.
В данной области хорошо известно, что специфическое распознавание лиганда его когнатным рецептором определяется взаимодействием между сайтом связывания рецептора и особым молекулярным «портретом» (эпитопом) лиганда. Таким образом, антитело, которое либо связывается с сайтом связывания, либо иным способом блокирует сайт связывания рецептора, либо связывается с эпитопом, либо иным способом блокирует распознавание эпитопа лиганда, будет предотвращать взаимодействие рецептора и лиганда. Такие антитела описаны как "нейтрализующие". В контексте настоящего раскрытия желательно, чтобы нейтрализующие антитела вырабатывались организмом хозяина на введение рекомбинантного белка, и таким образом последовательность белка может экспрессировать или включать одну или несколько из общего количества подходящих последовательностей, полученных из фактора роста или опухолевого антигена, таким образом, что эпитопы, требуемые для связывания с рецептором, будут представлены в функциональной (природной) конформации.
В дополнение к экспрессии многочисленных копий одного опухолевого антигена, рецептора и/или фактора роста, представленного как одиночный опухолевый антиген, рецептора и/или фактора роста или как его части из расчета на естественный сайт, и/или как цепочки повторяющегося опухолевого антигена, рецептора и/или последовательностей фактора роста (например, n=1 или больше); белок согласно настоящему раскрытию, может также включать экспрессию одного или нескольких эпитопов или сайтов связывания двух или нескольких различных опухолевых антигенов, рецепторов и/или факторов роста, представленных в одиночном виде или в виде цепочек в различных позициях в пределах последовательности рекомбинантного белка.
В иллюстративном варианте осуществления раскрыт белок, состоящий из гомогенного рекомбинантного белка, представляющего собой один или несколько нейтрализующих доменов эпидермального фактора роста (EGF). Этот белок представлен в форме рекомбинантного белка и может применяться в лечении хронических заболеваний, например раковых заболеваний груди, легких, мочевого пузыря, яичников, наружных женских половых органов, толстой кишки, легочной артерии, мозга, прямой кишки, кишечника, головы, шеи и пищевода. В иллюстративном варианте осуществления белок является рекомбинантным белком, представляющим собой или включающим последовательности EGF и последовательности синтетического полипептида согласно настоящему раскрытию. В одном иллюстративном варианте осуществления последовательность синтетического полипептида в значительной степени сходна с Sequence ID: 1.
В другом иллюстративном варианте осуществления раскрыт белок, состоящий из гомогенного рекомбинантного белка, представляющего собой один фактор роста фибробластов (FGF).
В дополнительном иллюстративном варианте осуществления белок является рекомбинантным белком, представляющим собой или включающим последовательности FGF и последовательности синтетического полипептида согласно настоящему раскрытию.
В еще одном иллюстративном варианте осуществления раскрыт белок, состоящий из гомогенного рекомбинантного белка, представляющего собой один трансформирующий фактора роста бета-1 (TGF-β1). В иллюстративном варианте осуществления белок является рекомбинантным белком, представляющим собой или включающим последовательности TGF-β1 и последовательности синтетического полипептида согласно настоящему раскрытию.
И еще в одном иллюстративном варианте осуществления раскрыт белок, включающий гомогенный рекомбинантный белок, экспрессирующий один трансформирующий фактор роста бета-1 (TGF-β1). В иллюстративном варианте осуществления белок является рекомбинантным белком, представляющим собой или включающим последовательности TGF-pi и последовательности синтетического полипептида, в значительной степени сходные с Sequence ID: 2 согласно настоящему раскрытию.
В другом иллюстративном варианте осуществления описывается один экспрессирующийся инсулиноподобный фактор роста 1 (IGF-1). В иллюстративном варианте осуществления белок является рекомбинантным белком, представляющим собой или включающим последовательности IGF-1 и последовательности синтетического полипептида, белком, включающим, гомогенный рекомбинантный белок, представляющий собой один фактор роста гепатоцитов (HGF).
В дополнительном иллюстративном варианте осуществления белок является рекомбинантным белком, экспрессирующим или включающим последовательности HGF и последовательности синтетического полипептида согласно данному раскрытию.
В дополнительном иллюстративном варианте осуществления настоящего изобретения раскрыт белок, состоящий из гомогенного рекомбинантного белка, представляющего собой один инсулиноподобный фактор роста-1 (IGF-1) и один инсулиноподобный фактор роста-2. В иллюстративном варианте осуществления белок является рекомбинантным белком, представляющим собой или включающим последовательности IGF-1, последовательности IGF-2 и последовательности синтетического полипептида согласно настоящему раскрытию.
В еще одном иллюстративном варианте осуществления раскрыт белок, состоящий из гомогенного рекомбинантного белка, представляющего собой один фактор роста эндотелия сосудов-А (VEGF-A) и один фактор роста эндотелия сосудов-C (VEGF-C). В иллюстративном варианте осуществления белок является рекомбинантным белком, представляющим собой или включающим последовательности нейтрализующего домена VEGF-A, последовательности VEGF-C и последовательности синтетического полипептида согласно настоящему раскрытию.
ПРИМЕРЫ
Аспекты настоящего раскрытия дополнительно подробно описаны в следующих примерах. Однако следующие примеры не предназначены для ограничения объема настоящего раскрытия с целью уточнения деталей методики или разработки, излагаемых ниже. Практические и иллюстративные варианты осуществления изобретения проиллюстрированы и описаны в следующих примерах. Тем не менее, специалистам в данной области будет понятно, что можно осуществлять модификации и усовершенствования в пределах сути и объема настоящего изобретения.
ПРИМЕР 1. Разработка селектируемой углевод-связывающей дисплейной системы
Требовалась система, при помощи которой библиотеки мутантов могли создаваться и скринироваться на наличие мутантов с желаемыми характеристиками, которая включала бы физическую связь между отбираемым фенотипом и кодирующим его генотипом для разнообразных популяций клонов. Такую систему изготавливали при использовании признанных технологий, используемых с большим успехом в инженерии антител в частности.
Ген, кодирующий выбранный углеводсвязывающий домен, клонировали внутри рамки и против хода транскрипции от гена минорного белка оболочки (ген III) бактериофага М13 как в полностью независимом функциональном 'фаговом' векторе М13 с подходящим селектируемым маркером ('фаг' М13-К07), так и в 'фагмидном' векторе, который включает только область упаковки F1 и ген, кодирующий минорный белок оболочки вируса. Вышеупомянутый вектор при введении в подходящего бактериального хозяина, такого как Е. coli, и размноженный соответственно, будет образовывать вирусные частицы, которые представят пять копий выбранного углеводсвязывающего домена на одном конце нитевидной вирусной частицы в виде N-концевых слияний минорного белка оболочки Р3 (кодируемого геном III).
Последний, благодаря природе размножения вируса, при использовании фаговой системы помощника/фагмиды, известной специалисту в данной области, будет производить вирусные частицы, в которых только небольшая часть популяции будет представлять, как правило, одну или меньше копий одинакового углеводсвязывающего домена Р3 слитого белка.
Клоны, полученные из каждого вектора, размножали при соответствующих условиях, которые хорошо известны специалистам в данной области, и супернатант культуры, содержащий вирусные частицы, скринировали при помощи ELISA для анализа связывающей активности с главным углеводом, который узнается углеводсвязывающим доменом. В этом примере использовали комплексную молекулу, которая включала дистальную галактозную группу и которую подвергали иммобилизации путем адсорбции на обычном 96-луночном иммунологическом планшете. Для связывания с углеводной группой необходимо, что бы два или несколько углеводсвязывающих домена ассоциировали в комплекс, поскольку углеводсвязывающий карман образуется между смежными доменами.
В предыдущем 'фаговом' примере карман может потенциально сформироваться между смежными слитыми белками Р3 на одиночном вирионе, при условии, что приемлемая ориентация ограничивается слитым белком или между отдельными вирусными частицами. При использовании фагмидной системы такие ассоциации будут, наиболее вероятно, формироваться между двумя или несколькими отдельными вирусными частицами, поскольку пропорциональная часть вирусов, включающих два или несколько слитых белков на одиночном вирионе, как ожидается, будет очень маленькая.
В результате скрининга вышеупомянутых клонов значительно более сильный сигнал ELISA получали от 'одновалентного' полученного от фагмиды клона, чем от пентавалентного фагового клона, последний также производил на много меньший титр инфекционных вирусных частиц. Низкая производительность фаговой системы в этом примере не ограничивает использование этой системы в описанном процессе, но более вероятно, как полагают, является функцией или стерических/ориентационных ограничений, налагаемых на систему специфической генетической связью, используемой в данном случае, и/или ограничений инфекционности, налагаемых слитым белком (минорный белок оболочки Р3 является медиатором вирусной инфекции). Поэтому система, основанная на фагмиде, была выбрана для того, чтобы создавать и отбирать различные мутантные белки.
ПРИМЕР 2. Создание библиотек мутантных клонов
Библиотеки мутантных полипептидкодирующих клонов создавали из генетической матрицы, полученной от углеводсвязывающего самособирающего белкового домена. В настоящем примере белковый домен выбирали из группы А1В5 бактериальных голотоксинов. Структурную и функциональную информацию, полученную из опубликованных баз данных, таких как Protein Database, и из научной литературы, использовали для того, чтобы идентифицировать области и/или определенные остатки среди потенциально или доказано вовлеченных в способность домена белка матрицы формировать стабильные гомопентамеры или взаимодействовать со специфическими углеводсодержащими частями, найденными на поверхности многих типов клеток млекопитающих. Эти области/остатки исключали из мутагенеза и последующих раундов отбора мутантов.
Области или остатки, определенные выше как являющиеся потенциально поддающимися мутации и не вовлеченными в требуемые характеристики, отбирали и делали мишенью для рационального или случайного мутагенеза, считаемого целесообразным. Чтобы максимизировать шансы создания и отбора углеводсвязывающих вариантов, которые являлись отличными от домена матрицы, мутации ограничивали небольшим числом остатков в непосредственной близости от каждого последовательного раунда скрининга разработки библиотеки.
В зависимости от региона или выбранного в качестве мишени остатка мутация была либо случайной (т.е. включающей потенциально все 20 аминокислот), либо функционально ограниченной до возможных пределов теми остатками, которые проявляют похожие биофизические и/или химические свойства с теми, которые найдены в домене матрицы.
Характеристики, которые считались значащими в таких случаях, включали без ограничения размер боковой цепи, заряд, полярность, гидрофобность/гидрофильность и способностью участвовать в формировании специфической вторичной структуры, такой как α-спирали. Где возможно/допустимо, что аминокислотный остаток, кодируемый матрицей в любом данном положении, исключали из библиотек мутантов во избежание отбора немутантов.
Библиотеки создавали с помощью конструирования олигонуклеотидных праймеров, которые включали и области последовательности, которые были гомологичными с геном матрицы, таким образом, что праймер (и особенно его 3'-конец) отжигался бы с ДНК матрицы при соответствующих условиях, и дополнительные области, в которых соответствующее вырождение предусматривали, например, с целью кодирования многообразия аминокислот в одном или нескольких положениях.
В данном примере такие олигонуклеотиды производили при использовании заданного вырождения, известного специалистам в данной области, где основание ДНК в любой различной позиции включает равные количества из 2-4 оснований (G, А, Т или С) в соответствии с требованием. Альтернативно, более контролируемое многообразие предусматривается, когда олигонуклеотиды получают из позиционных смесей тринуклеотид-фосфорамидитов, использованных так, что специфические остатки (аминокислоты) и их относительные пропорции точно контролируются в каждом различном положении.
Когда мутации нужно было вводить в одном из концов, одиночную ПЦР-реакцию выполняли (и повторяли, если необходимо) при использовании одного или двух вырожденных праймеров для того, чтобы ввести необходимое генетическое разнообразие. Полученный продукт, включающий (полученные от праймера) фланкирующие сайты рестрикции, клонировали в фагмидный вектор, описанный в примере 1 выше, таким образом, что библиотека различных белков была закодирована как слияния в рамке считывания с минорным белок-кодирующим геном Р3 (ген III) бактериофага М13.
В том случае, если требовалось ввести разнообразие в область(и) отдаленную(ые) к концам матричного гена, две пары праймеров использовали таким образом, что каждая пара включала один терминальный (5'- или 3'-) праймер, который точно соответствовал последовательности матрицы, и один вырожденный праймер, который кодировал некоторые, все или ни одной из требуемого разнообразия. 3'-концы вырожденных праймеров конструировали таким образом, что i) каждый точно соответствовал последовательности матрицы и ii) каждый был точно комплементарным вырожденному праймеру второй пары праймеров.
Таким образом, получали два ПЦР-продукта и затем отжигали и связали друг с другом посредством комплементарной перекрывающейся последовательности, полученной с использованием вырожденных праймеров. Полученный продукт затем клонировали в фагмидный вектор как слияние в рамке считывания с геном III, геном минорного белка оболочки. Последующие библиотеки строили либо на основании продуктов скрининга и отбора одного или нескольких клонов из предыдущей библиотеки, либо строили и скринировали в параллельных реакциях из матрицы или одного или нескольких отобранных мутантных клонов.
Все библиотеки генетических конструкций вводили в клетки TG1 хозяина Е. coli при помощи электропорации с использованием стандартных методологий и трансформанты высевали на селективную среду. После соответствующей инкубации колонии бактерий собирали из чашек и хранили и/или скринировали в соответствии с требованиями.
ПРИМЕР 3. Скрининг мутантных библиотек
Для проведения отборов мутантов из библиотек культуры (библиотеки) инокулировали в жидкие среды (2xTY, 100 мкг/мл ампициллина, 1% глюкозы) таким образом, чтобы достаточное количество клеток включали 100-1000-кратную копийность ожидаемого разнообразия и достаточный объем с целью обеспечения нахождения значения OD600 в области 0,1. Клеточные культуры затем инкубировали (с перемешиванием) при приблизительно 37°С до того, как значение OD600 достигало 0,4-0,6 (т.е. лог-фазы роста). Культуры затем инфицировали добавлением фага-помощника М13-K07 в соотношении ~20 фагов-помощников на одну бактериальную клетку (OD60o со значением 1,0 принимали равной ~8,0×108 клеток/мл). После приблизительно 30 минут статической инкубации клетки инкубировали в течение приблизительно 30 дополнительных минут с перемешиванием. Добавляли до ~50 мкг/мл канамицина и культуру инкубировали на протяжении ночи при ~30°C с перемешиванием.
На следующее утро культуру центрифугировали в течение приблизительно 25-30 минут при ~8000 g для осаждения клеток и супернатант удаляли и сохраняли. Клеточный осадок отбрасывали. 20% объем 200 мМ NaCl, 20% PEG 6000 добавляли к культуральному супернатанту и инкубировали на льду приблизительно 1 час для осаждения фаговых частиц. Фаг осаждали центрифугированием при ~8000 g в течение приблизительно 25-30 минут и ресуспендировали в 20 процентах от исходного объема PBS (натрий-фосфатного буфера). Повторно добавляли полученный 20% объем 200 мМ NaCl, 20% PEG 6000 и фаг инкубировали на льду в течение приблизительно 25-30 минут. Фаг снова осаждали центрифугированием при ~8000 g в течение 25-30 минут и осадок ресуспендировали в ~2 мл натрий-фосфатного буфера (PBS). Полученную суспензию переносили в пробирку(ки) типа эппендорф и осаждали центрифугированием на максимальной скорости в течение 5 минут для того, чтобы осадить любые оставшиеся бактериальные клетки/дебрис.Фаговую суспензию затем использовали для отбора.
Для проведений отбора иммунологическую пробирку покрывали подходящим антигеном, таким как способное к иммобилизации производное галактозы или природный лиганд В-субъединицы, в концентрации 1-10 мкг/мл (как правило 5 мл) и оставляли в течение ночи при 4-8°С или при комнатной температуре в течение 1 часа. После промывки 3-5 раз с PBS (просто вливая в иммунологическую пробирку и снова выливая), пробирку блокировали добавлением MPBS (PBS, содержащим 2% сухое молоко) в течение 1-2 часов при 37°С. Промывали с MPBS, как описано выше, и добавляли от приблизительно 1×1011 до 1×1012 или больше фаговых частиц.
Объем доводили до ~5 мл и иммунологическую пробирку запечатывали, например, парафиновой пленкой. Затем пробирку 'перемешивали переворачиванием' (с донышка на крышку) в течение приблизительно 30 минут, затем инкубировали в вертикальном положении в течение приблизительно 90 минут, с целью позволить тем фагам, которые представляют белок из библиотеки мутантов, способный узнавать иммобилизованный лиганд, связаться с ним. Фаг, который не связывался или только слабо связывался, удаляли с помощью промывки. Строгость отбора изменяли в соответствии с требованиями при помощи количества применяемых промывок, использованием PBST (PBS, содержащий 0,1% Tween 20) для промывок или изменением концентрации лиганда при покрытии.
Связанные частицы фага элюировали из иммунологической пробирки с добавлением 1 мл 100 мМ триэтиламина (TEA) и премешивали переворачиванием максимум в течение 10 минут (более длинная инкубация оказывает негативное влияние на жизнеспособность фага), и затем сразу же выливали в 0,5 мл 1 М Tris-HCl (рН 7,4) для нейтрализации. Приблизительно 0,75 мл элюированного фага добавляли к ~10 мл находящейся в лог-фазе клеточной культуры подходящего штамма Е. coli, такого как TG1 (Agilent). Культуру инкубировали при приблизительно 37°С без перемешивания для обеспечения инфицирования. Серийные разведения образцов инфицированных клеток наносили на маленькие TYE-чашки, содержащие 100 мкг/мл ампициллина и 1% глюкозу. Остальные клетки наносили на большие чашки для биологического анализа с той же самой средой. Все инкубировали в течение ночи при приблизительно 30°С. Другими 0,75 мл фага, -75 мкл инфицировали, как описано выше, ~1 мл клеток НВ2151 Е. coli, находящихся в лог-фазе, и серийные разведения осаждали и инкубировали, как описано выше. Остальную часть фага хранили при приблизительно -80°С в виде глицериновой смеси (~15% глицерина).
На следующий день большие чашки для биологического анализа соскабливали, чтобы собрать клетки, которые затем хранили в виде глицериновых смесей или выращивали и 'освобождали' с помощью фагового помощника М13 К07, как описано ранее, для получения обогащенной популяции фага для следующего раунда отбора. Обычно проводили два или три последовательных раунда отбора. После завершения отбора отдельные колонии из маленьких чашек, содержащих серийные разведения клеток НВ2151, выбирали с помещением на 96-луночные культуральные планшеты, содержащие ~100 мкл/лунку 2 х TY с 100 мкг/мл ампициллина, и инкубировали с перемешиванием в течение ночи при 37°С. На следующий день приблизительно 5 мкл культуры из каждой лунки иннокулировали на новый планшет с ~150 мкл среды/на лунку и инкубировали при приблизительно 37°С с перемешиванием в течение приблизительно 2 часов. Добавляли дополнительную среду, содержащую 1 М IPTG, с обеспечением конечной концентрации 1 мМ IPTG и чашку инкубировали с перемешиванием в течение ночи при 30°С. В клетках НВ2151 индукция IPTG приводила к экспрессии растворимых мутантных белков, которые включали детектируемую пептидную метку с-myc, а не белки, слитые с компонентом вирусной частицы, как в случае TG1 клеток Е. coli. Исходный 96-луночный культуральный планшет хранили при приблизительно -80°С.
На следующий день культуральные супернатанты из индуцированного 96-луночного планшета анализировали в отношении связывания с иммобилизованными углеводами при использовании стандартных протоколов ELISA, и связывание было обнаружено с использованием легкодоступного HRP-меченного анти-с-Мус антитела (фиг. 1). Клоны, которые были очень активными при связывании с GM1, показаны в затененных ячейках. Положительные и отрицательные контроли взяты в рамку.
Культуральные супернатанты клонов, которые были очень активными при связывании в анализе ELISA, дополнительно анализировали при помощи SDS PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия) и Вестерн-блоттинга, чтобы проанализировать присутствие пентамеров и других мультимерных состояний, проиллюстрированых на фигуре 2. Образцы не гидрировали или не подвергали кипячению перед электрофорезом и белок обнаруживали при помощи HRP-меченных анти-с-Мус антител. В показанном примере клоны С1 и С3 являются позитивными контролями, которые быстро собираются в пентамеры ~60 кДа. Клон С2 является негативным контролем, который двигается в виде 12 кДа мономера при используемых условиях. Можно видеть, что клоны А1 и A3 двигаются, главным образом, как пентамеры, тогда как значительная часть клона В12 формирует мономеры. Остальные клоны формируют либо мономер (В1, В7), либо формируют много мультимерных форм.
ПРИМЕР 4. Обобщение данных мутаций
С целью оценки способности максимального количества аминокислотных остатков вмещать мутации производили и скринировали несколько различных библиотек, каждая из которых ориентирована на различные остатки или области матрицы. В результате идентифицировали большое количество клонов, которые проявляли желаемые критерии отбора по связыванию с иммобилизованными производными углевода и формированию преимущественно пентамеров, однако каждый из которых включал много уникальных мутаций/различий, как проиллюстрировано на фигуре 3А. Чтобы создать клоны с потенциально улучшенными характеристиками, необходимо было объединить эти различные последовательности в меньшем количестве клонов.
Поскольку в процессе получения и отбора мутантов было идентифицировано несколько положений, где могли размещаться три или несколько различных остатков, в общей сложности разработали четыре комбинации мутантов (A-D), как проиллюстрировано на фигуре 3В. Экспрессировали четыре клона, содержащие комбинированные мутации, и синтетические белки оценивали по их связыванию в сравнении с соответствующими контролями при использовании ELISA, как проиллюстрировано на фигуре 4. Было обнаружено, что два клона (А и С) обладали только слабым связыванием, однако другие два (В и D) были способны связываться с GM1 очень прочно. Это указывает на то, что мутации, полученные из отдельных клонов, успешно объединяться, однако, усиление или даже сохранение селективных характеристик связывания не гарантировано.
ПРИМЕР 5. Стабилизация гомопентамеров
В их природной полностью функциональной форме бактериальные голотоксины, описанные под названием группы 'А1В5', состоят из пяти В-субъединиц и одной А-субъединицы. В-субъединицы собираются в пентамерное кольцо с центральной 'порой'. Единственная А-субъединица находится на верхушке этого кольца и выступает вниз в свободное место, как проиллюстрировано на фигуре 5. Собственно А-субъединица состоит из 2 отдельных доменов. Домен А1, который освобождается, когда голотоксин транслоцируется в клетку, характеризуется ферментативной активностью, ассоциированной с токсичностью белкового комплекса. Он удерживается как часть голотоксина доменом А2. Последний имеет альфа-спиральную структуру, в которой домен А1 находится HaN-конце, и С-конец выступает в пору, сформированную с помощью 5 В-субъединиц. Некоторые остатки С-конца А2 формируют взаимодействия с остатками В-субъединиц. Поскольку есть подтвержденные взаимодействия между определенными остатками В-субъединиц и домена А2 (Zhang R.G., et al. (1995)), логично предположить, что присутствие А-субъединицы, и особенно домена А2, способствует стабилизации пентамерного кольца В-субъединицы. Вследствие структурных и функциональных различий между двумя доменами А-субъединицы, не будучи подтвержденным конкретными доказательствами, считается, что взятий отдельно домен А2 будет придавать этот эффект стабилизации В-субъединичному пентамеру. Подобным образом, поскольку известно, что В-субъединицы способны формировать пентамерные кольца в полном отсутствии А-субъединицы, вполне вероятно, что домен А2 может связываться с ним и следовательно оказывать стабилизирующий эффект на пентамеры В-субъединицы, либо будучи коэкспрессированным с В-субъединицами, либо будучи добавленным экзогенно.
В связи с этим предлагается, что если синтетический или неприродный полипептид является таким, который имеет достаточно похожую конформацию с В-субъединицами голотоксина, который способен собраться в мультимеры, включая пентамерные кольца, и в котором остатки, соответствующие тем, которые взаимодействуют с А2 в В-субъединицах голотоксинов, являются теми же или имеют схожие биофизические свойства, тогда такой пентамер синтетического белка будет испытывать подобный стабилизирующий эффект.Кроме того предлагается, что в отсутствие природного домена А2 синтетический полипептид может быть изолирован из подходящей библиотеки полипептидов с использованием технологий, известных специалистам в данной области, таких как 'фаговый дисплей', который будет связываться, по меньшей мере частично, с подвергающимися воздействию поверхностями двух или нескольких природных или синтетических мономеров, когда они собраны в пентамеры, и таким образом стабилизировать структуру.
Поскольку природные домены А2 поддерживают структурно независимый домен А1, который сам по себе не взаимодействует ни с каким другим компонентом голотоксина, тогда вероятно, что домен А2 или функциональный синтетический эквивалент могут обеспечить средство для закрепления другого домена полипептида. Не будучи связанным с какой-либо конкретной теорией, считается, что неприродный полипептид, который имеет достаточно похожую конформацию с полипептидом, описанном в Sequence ID: 1, сам способствует тому, чтобы формировать пентамерные кольца, которые представляют стабилизированную пентамерную кольцевую структуру, которая позволяет прикрепление эпитопов, описанных в настоящем раскрытии.
Хотя устройства, системы и способы были описаны и проиллюстрированы в связи с определенными вариантами осуществления настоящего изобретения, много вариантов и модификаций будут очевидны для специалистов в данной области и могут быть осуществлены без отступления от сути и объема настоящего раскрытия. Таким образом, настоящее раскрытие не должно ограничиваться точными деталями методологии или конструкционным набором, изложенным выше, поскольку такие варианты и модификации предназначены для включения в объем настоящего раскрытия.
Термины и выражения, которые были применены, используются как термины для описания, а не для ограничения, и нет никакого намерения, чтобы при использовании таких терминов и выражений исключать любые равнозначные показанные и описанные свойства или их части, но очевидно, что различные модификации возможны в пределах объема настоящего изобретения. Таким образом, нужно понимать, что хотя настоящее изобретение было специально раскрыто с помощью предпочтительных вариантов осуществления и дополнительных свойств, специалисты в данной области могут прибегнуть к модификациям и вариациям понятий, раскрытых в данном документе, и что такие модификации и варианты рассматриваются в рамках настоящего изобретения, как определено прилагаемой формулой изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| САМОСОБИРАЮЩИЕСЯ СИНТЕТИЧЕСКИЕ БЕЛКИ | 2014 |
|
RU2801230C2 |
| РЕКОМБИНАНТНЫЕ БЕЛКИ И ИХ ПРИМЕНЕНИЯ В ТЕРАПЕВТИЧЕСКИХ ЦЕЛЯХ | 2012 |
|
RU2650574C2 |
| РЕКОМБИНАНТНЫЕ БЕЛКИ И ИХ ПРИМЕНЕНИЯ В ТЕРАПЕВТИЧЕСКИХ ЦЕЛЯХ | 2018 |
|
RU2766595C2 |
| ИММУНОГЕННЫЕ КОМПОЗИЦИИ И СПОСОБЫ | 2008 |
|
RU2468034C2 |
| САПОВИРУСНЫЕ ВАКЦИНЫ | 2022 |
|
RU2839769C2 |
| ИММУНОГЕННАЯ КОМПОЗИЦИЯ ПРЕЗЕНТАЦИИ МНОЖЕСТВЕННЫХ АНТИГЕНОВ, ОТНОСЯЩИЕСЯ К НЕЙ СПОСОБЫ И ПРИМЕНЕНИЯ | 2012 |
|
RU2619176C2 |
| МОДИФИЦИРОВАННЫЙ БИОТИН-СВЯЗЫВАЮЩИЙ БЕЛОК, СЛИТЫЕ БЕЛКИ НА ЕГО ОСНОВЕ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2632651C2 |
| ЛЕЧЕНИЕ ИЛИ ПРОФИЛАКТИКА ИНФЕКЦИИ | 2011 |
|
RU2617399C2 |
| ВАКЦИНЫ ПРОТИВ FMDV НА ОСНОВЕ РЕКОМБИНАНТНОГО АДЕНОВИРУСНОГО ВЕКТОРА И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2725495C2 |
| ИММУНОСТИМУЛИРУЮЩИЕ И ВАКЦИННЫЕ КОМПОЗИЦИИ | 2011 |
|
RU2578420C2 |



Изобретение относится к области биотехнологии, конкретно к рекомбинантным терапевтическим белкам, и может быть использовано в медицине для лечения раковых заболеваний груди, легких, мочевого пузыря, яичников, наружных женских половых органов, толстой кишки, легочной артерии, мозга, прямой кишки, кишечника, головы, шеи и пищевода. Предложен синтетический белок на основе мутантной B-субъединицы группы A1B5 бактериальных голотоксинов, полученной из B-субъединицы холерного токсина (СТВ) и соединенной через пептидный спейсер с полипептидом, включающим нейтрализующий домен трансформирующего фактора роста (TGF) человека. Изобретение обеспечивает получение стабильных гомопентамерных конструкций без потери функции TGF - индукции иммунного ответа в организмах хозяев млекопитающих с образованием нейтрализующих антител против TGF. 2 н. и 5 з.п. ф-лы, 6 ил., 5 пр.
1. Рекомбинантный синтетический белок для лечения хронических заболеваний, выбранных из группы, состоящей из раковых заболеваний груди, легких, мочевого пузыря, яичников, наружных женских половых органов, толстой кишки, легочной артерии, мозга, прямой кишки, кишечника, головы, шеи и пищевода, включающий
мономерную полипептидную последовательность, способную собираться в стабильные гомопентамеры, где каждая мономерная последовательность включает
синтетическую В-субъединицу группы A1B5 бактериальных голотоксинов, полученную из B-субъединицы холерного токсина (СТВ), где а) B-субъединица включает миссенс-мутации T1F, Р2Т, Q3D, N4I, M37I, A38I, I39L, I40V, T41N, T78S, E79N, A80S, A95S, A102V, N103R, L8I, A10G, F25L, A32V, N44G и V82I и характеризуется последовательностью SEQ ID NO: 1 или b) вариант B-субъединицы, изложенный в подпункте а), в котором миссенс-мутации L8I, A10G, F25L, A32V, N44G и V82I отсутствуют;
пептидный спейсер; и
полипептид, включающий нейтрализующий домен трансформирующего фактора роста (TGF) человека, где полипептид отделен от синтетической B-субъединицы пептидным спейсером.
2. Рекомбинантный белок по п. 1, где B-субъединица не включает миссенс-мутацию, выбранную из группы, состоящей из L8I, A10G, F25L, A32V, N44G и V82I.
3. Рекомбинантный белок по п. 1, где полипептид включает полную длину или нейтрализующий домен фактора роста человека как одиночный домен или как два или более многократных повторов.
4. Рекомбинантный белок по п. 3, где полипептидный спейсер выбран из группы, состоящей из SSG, SSGGG, SGG, GGSGG, GGGGS, SSGGGSGGSSG, GGSGGTSGGGSG, SGGTSGGGGSGG, GGSGGTSGGGGSGG, SSGGGSGGSSG, SSGGGGSGGGSSG, SSGGGSGGSSGGG и SSGGGGSGGGSSGGG.
5. Рекомбинантный белок по п. 4, где полипептидный спейсер включает аминокислотную последовательность фактора роста или его нейтрализующего домена.
6. Рекомбинантный белок по п. 5, где полипептидный спейсер включает один или несколько Т-клеточных эпитопов хозяина.
7. Способ получения мультивалентной молекулы, включающий смешивание нескольких рекомбинантных синтетических белков по п. 1 в необходимых условиях для образования стабильных гомопентамеров.
| WO 2010080538 A1, 15.07.2010 | |||
| LI S | |||
| ET AL., Pentabody-mediated antigen delivery induces antigen-specific mucosal immune response, MOLECULAR IMMUNOLOGY, 2009, v.46, n | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| LEBENS M ET AL., A mucosally administered recombinant fusion protein vaccine against schistosomiasis protecting against immunopathology and infection, VACCINE, | |||

