Область техники, к которой относится изобретение
Настоящее изобретение охватывает РНК-репликон, который может быть реплицирован происходящей от альфавируса репликазой. РНК-репликон содержит элементы последовательности, необходимые для репликации репликазой, но эти элементы последовательности не кодируют ни один белок или его фрагмент, такой как альфавирусный неструктурный белок или его фрагмент. Таким образом, в РНК-репликоне в соответствии с настоящим изобретением элементы последовательности, необходимые для репликации с помощью репликазы, и кодирующие белок области являются несвязанными. В соответствии с настоящим изобретением разъединение достигается удалением по меньшей мере одного инициирующего кодона по сравнению с нативной альфавирусной геномной РНК. РНК-репликон может содержать ген, кодирующий представляющий интерес белок, такой как фармацевтически активный белок. Репликаза может кодироваться РНК-репликоном или отдельной молекулой нуклеиновой кислоты.
Предшествующий уровень техники изобретения
Молекулы нуклеиновой кислоты, содержащие чужую генетическую информацию, кодирующую один или несколько полипептидов для профилактических и терапевтических целей, изучались в биомедицинских исследованиях на протяжении многих лет. Подходы предшествующего уровня техники характеризуются доставкой молекулы нуклеиновой кислоты в целевую клетку или организм, но различаются по типу молекулы нуклеиновой кислоты и/или системы доставки: под влиянием опасений за безопасность, связанных с применением молекул дезоксирибонуклеиновой кислоты (ДНК), молекулы рибонуклеиновой кислоты (РНК) в последние годы получили повышенное внимание. Были предложены различные подходы, включающие в себя введение одноцепочечной или двухцепочечной РНК в форме голой РНК или в связанной в комплекс или упакованной форме, например, в невирусных или вирусных средствах доставки. В вирусах и в вирусных средствах доставки генетическая информация, как правило, инкапсулируется белками и/или липидами (вирусная частица). Например, в качестве средства доставки для обработки растений (публикация международной заявки WO 2000/053780 A2) или для вакцинации млекопитающих были предложены сконструированные частицы РНК-вирусов, полученные из РНК-вирусов (Tubulekas et al., 1997, Gene, vol. 190, pp. 191-195).
В общем, РНК-вирусы представляют собой разнообразную группу инфекционных частиц с геномом РНК. РНК-вирусы могут быть сгруппированы в вирусы с одноцепочечной РНК (оцРНК) и двухцепочечной РНК (дцРНК), а вирусы с оцРНК могут быть дополнительно разделены на вирусы с положительной нитью ((+)-нить) и/или отрицательной нитью [(-)-нить]. РНК-вирусы с положительной нитью являются, на первый взгляд, привлекательными в качестве системы доставки в биомедицине, потому что их РНК может служить непосредственно в качестве матрицы для трансляции в клетке-хозяине.
Альфавирусы являются типичными представителями вирусов с положительной нитью РНК. Хозяева альфавирусов включают в себя широкий спектр организмов, охватывающих насекомых, рыб и млекопитающих, таких как домашние животные и люди. Альфавирусы реплицируются в цитоплазме инфицированных клеток (для обзора жизненного цикла альфавируса смотрите публикацию José et al., Future Microbiol., 2009, vol. 4, pp. 837-856). Общая длина генома многих альфавирусов, как правило, составляет от 11000 до 12000 нуклеотидов, и геномная РНК, как правило, содержит 5'-кэп и 3'-поли(А)-хвост. Геном альфавирусов кодирует неструктурные белки (вовлеченные в транскрипцию, модификацию и репликацию вирусной РНК и в модификацию белка) и структурные белки (образующие вирусную частицу). В геноме, как правило, имеются две открытые рамки считывания (ORF). Четыре неструктурных белка (nsP1-nsP4), как правило, кодируются вместе первой ORF, начинающейся возле 5′-конца генома, тогда как структурные белки альфавирусов вместе кодируются второй ORF, которая находится ниже по ходу транскрипции от первой ORF и продолжается вблизи 3′-конца генома. Как правило, первая ORF больше, чем вторая ORF, причем отношение составляет приблизительно 2:1.
В инфицированных альфавирусом клетках из геномной РНК транслируется только последовательность нуклеиновой кислоты, кодирующая неструктурные белки, тогда как генетическая информация, кодирующая структурные белки является транслируемой из субгеномного транскрипта, который представляет собой молекулу РНК, характеризующуюся сходством с эукариотической матричной РНК (мРНК, Gould et al., 2010, Antiviral Res., vol. 87, pp. 111-124). После инфицирования, то есть на ранних стадиях жизненного цикла вируса, (+)-нитевая геномная РНК непосредственно действует в качестве матричной РНК для трансляции открытой рамки считывания, кодирующей неструктурный полипротеин (nsP1234). В некоторых альфавирусах существует опал стоп-кодон между кодирующими последовательностями nsP3 и nsP4: полипротеин P123, содержащий nsP1, nsP2 и nsP3, производится тогда, когда трансляция заканчивается в опал стоп-кодоне, а полипротеин P1234, содержащий дополнительно nsP4, производится при считывания через этот опал-кодон (Strauss & Strauss, Microbiol. Rev., 1994, vol. 58, pp. 491-562; Rupp et al., 2015, J. Gen. Virology, vol. 96, pp. 2483-2500). nsP1234 аутопротеолитически расщепляется на фрагменты nsP123 и nsP4. Полипептиды nsP123 и nsP4 соединяются с образованием репликазного комплекса с (-)-нитью, который транскрибирует (-)-нитевую РНК, используя (+)-нитевую геномную РНК в качестве матрицы. Как правило, на более поздних стадиях фрагмент nsP123 полностью расщепляется на отдельные белки nsP1, nsP2 и nsP3 (Shirako & Strauss, 1994, J. Virol. vol. 68, pp. 1874-1885). Все четыре белка образуют (+)-нитевый репликазный комплекс, который синтезирует новые (+)-нитевые геномы с использованием (-)-нитевого комплемента геномной РНК в качестве матрицы (Kim et al., 2004, Virology, vol. 323, pp. 153-163, Vasiljeva et al., 2003, J. Biol. Chem. Vol. 278, pp. 41636-41645).
В инфицированных клетках субгеномная РНК, а также новая геномная РНК обеспечивается 5'-кэпом посредством nsP1 (Pettersson et al. 1980, Eur. J. Biochem. 105, 435-443; Rozanov et al., 1992, J. Gen. Virology, vol. 73, pp. 2129-2134) и обеспечивается полиаденилатным (поли(А)) хвостом посредством nsP4 (Rubach et al., Virology, 2009, vol. 384, pp. 201-208). Таким образом, как субгеномная РНК, так и геномная РНК являются похожими на матричную РНК (мРНК).
Альфавирусные структурные белки (белок C сердцевины нуклеокапсида, белок E2 оболочки и белок E1 оболочки, все составляющие вирусной частицы), как правило, кодируются одной единственной открытой рамкой считывания под контролем субгеномного промотора (Strauss & Strauss, Microbiol. Rev., 1994, vol. 58, pp. 491-562). Субгеномный промотор распознается альфавирусными неструктурными белками, действующими in cis. В частности, альфавирусная репликаза синтезирует (+)-нитевый субгеномный транскрипт с использованием (-)-нитевого комплемента геномной РНК в качестве матрицы. (+)-нитевый субгеномный транскрипт кодирует структурные белки альфавируса (Kim et al., 2004, Virology, vol. 323, pp. 153-163, Vasiljeva et al., 2003, J. Biol. Chem. vol. 278, pp. 41636-41645). Субгеномный транскрипт РНК служит в качестве матрицы для трансляции открытой рамки считывания, кодирующей структурные белки, в виде одного полипротеина, а полипротеин расщепляется с образованием структурных белков. На поздней стадии альфавирусной инфекции в клетке-хозяине сигнал упаковки, который находится в кодирующей последовательности nsP2, обеспечивает селективную упаковку геномной РНК в почкующиеся вирионы, упакованные посредством структурных белков (White et al., 1998, J. Virol., vol. 72, pp. 4320-4326).
В инфицированных клетках синтез (-)-нити РНК, как правило, наблюдается только в первые 3-4 часа после инфицирования и на поздних стадиях она не обнаруживается, при этом наблюдается синтез только (+)-нити РНК (как геномной, так и субгеномной). Согласно публикации Frolov et al., 2001, RNA, vol. 7, pp. 1638-1651, преобладающая модель регуляции синтеза РНК предполагает зависимость от процессинга неструктурного полипротеина: начальное расщепление неструктурного полипротеина nsP1234 приводит к получению nsP123 и nsP4; nsP4 действует как РНК-зависимая РНК-полимераза (RdRp), которая активна при синтезе (-)-нити, но неэффективна для производства (+)-нитей РНК. Дальнейший процессинг полипротеина nsP123, включая расщепление в месте соединения nsP2/nsP3, изменяет специфичность матрицы репликазы для увеличения синтеза (+)-нити РНК и для уменьшения или прекращения синтеза (-)-нити РНК.
Синтез альфавирусной РНК также регулируется цис-действующими РНК-элементами, включающими в себя четыре консервативных элемента последовательности (CSE, Strauss & Strauss, Microbiol. Rev., 1994, vol. 58, pp. 491-562 и Frolov, 2001, RNA, vol. 7, pp. 1638-1651).
В общем, 5' репликационная последовательность распознавания генома альфавируса характеризуется низкой общей гомологией между различными альфавирусами, но имеет консервативную прогнозируемую вторичную структуру. 5' репликационная последовательность распознавания генома альфавируса участвует не только в инициации трансляции, но также содержит 5' репликационную последовательность распознавания, состоящую из двух консервативных элементов последовательности, участвующих в синтезе вирусной РНК, CSE 1 и CSE 2. Считается, что для функции CSE 1 и 2, вторичная структура важнее линейной последовательности (Strauss & Strauss, Microbiol. Rev., 1994, vol. 58, pp. 491-562).
Напротив, 3'-концевая последовательность генома альфавируса, то есть последовательность, расположенная непосредственно выше против хода транскрипции от поли(А)-последовательности, характеризуется консервативной первичной структурой, в частности, посредством консервативного элемента последовательности 4 (CSE 4), также называемого «консервативной последовательностью из 19 нуклеотидов», что важно для инициации синтеза (-)-нитей.
CSE 3, также называемый «последовательностью места соединения», представляет собой консервативный элемент последовательности на (+)-нити альфавирусной геномной РНК, а комплемент CSE 3 на (-)-нити действует как промотор для транскрипции субгеномной РНК (Strauss & Strauss, Microbiol. Rev., 1994, vol. 58, pp. 491-562; Frolov et al., 2001, RNA, vol. 7, pp. 1638-1651). CSE 3, как правило, перекрывается с областью, кодирующей C-концевой фрагмент nsP4.
В дополнение к альфавирусным белкам, также факторы клетки-хозяина, предположительно белки, могут связываться с консервативными элементами последовательности (Strauss & Strauss, смотрите выше).
Были предложены полученные из альфавирусов векторы для доставки чужой генетической информации в целевые клетки или целевые организмы. В простых подходах открытая рамка считывания, кодирующая альфавирусные структурные белки, заменяется открытой рамкой считывания, кодирующей представляющий интерес белок. Системы транс-репликации на основе альфавирусов основаны на элементах нуклеотидной последовательности альфавируса на двух отдельных молекулах нуклеиновой кислоты: одна молекула нуклеиновой кислоты кодирует вирусную репликазу (как правило, в виде полипротеина nsP1234), а другая молекула нуклеиновой кислоты способна реплицироваться указанной репликазой in trans (отсюда обозначение система транс-репликации). Транс-репликация требует наличия обеих этих молекул нуклеиновой кислоты в данной клетке-хозяине. Молекула нуклеиновой кислоты, способная реплицироваться репликазой in trans, должна содержать определенные элементы альфавирусной последовательности, которые позволяют распознавать и синтезировать РНК с помощью альфавирусной репликазы. Соответствующий репликон иллюстрируется как «РНК-матрица WT-RRS» на фиг. 1. Такой репликон связан с преимуществом, позволяющим амплифицировать представляющий интерес ген под контролем субгеномного промотора; однако более универсальные векторы трудно разработать, поскольку открытая рамка считывания, кодирующая nsP1234, перекрывается с 5' репликационной последовательностью распознавания генома альфавируса (кодирующей последовательностью для nsP1) и, как правило, также с субгеномным промотором, содержащим CSE 3 (кодирующей последовательностью для nsP4),
Например, Michel с соавт. (2007, Virology, vol. 362, pp. 475-487) описывают, что введение 95 молчащих мутаций (т.е. мутаций, которые не влияют на закодированную последовательность белка) в кодирующую область nsP1 вируса венесуэльского лошадиного энцефалита (VEEV) альфавируса полностью отменяло способность VEEV реплицироваться в клетках, по-видимому, потому что молчащие мутации разрушали вторичную структуру РНК. В публикации международной заявки WO 2008/156829 A2 и в публикации Kamrud et al. (2010, J. Gen. Virol., vol. 91, pp. 1723-1727) описана РНК-помощник (т.е. транс-реплицирующая РНК, экспрессирующая капсид и оболочку VEEV), которая была модифицирована таким образом, что специфический триплет оснований AUG, который служит в качестве стартового кодона для nsP1 в обнаруженном в природе VEEV, можно удалить (путем преобразования в стоп-кодон) для создания модифицированных конструкций. Согласно публикации Kamrud et al., преобразование стартового кодона nsP1 в стоп-кодон сохраняло репликативный потенциал РНК, и эти модифицированные РНК-помощники приводили к получению частиц VEEV только слегка уменьшенного титра. Однако авторы заметили, что преобразование всех AUG, обнаруженных в области CSE1/2 в стоп-кодоны, приводит к тому, что РНК-помощник плохо реплицируется в присутствии альфавирусной репликазы. Авторы приписывают плохую репликацию предполагаемому нарушению вторичной структуры РНК. Нарушенная вторичная структура является весьма вероятным объяснением, так как авторы не упомянули, что они контролировали правильную укладку их модифицированной РНК-помощника.
Тот факт, что 5' репликационная последовательность распознавания, требуемая для репликации РНК, содержит стартовый кодон AUG для nsP1 и, таким образом, перекрывается с кодирующей последовательностью для N-концевого фрагмента альфавирусного неструктурного белка, представляет собой серьезную помеху для конструирования основанных на альфавирусе векторов, потому что репликон, содержащий 5' репликационную последовательность распознавания, как правило, кодирует (по меньшей мере) часть неструктурного белка альфавируса, как правило, N-концевой фрагмент nsP1. Это невыгодно в нескольких аспектах:
В случае цис-репликонов это перекрывание ограничивает, например, адаптацию использования кодонов ORF репликазы к различным целевым клеткам млекопитающих (человека, мыши, сельскохозяйственных животных). Возможно, что вторичная структура 5' репликационной последовательности распознавания, как она обнаружена в вирусах, не является оптимальной в каждой целевой клетке. Однако вторичная структура не может быть свободно изменена, так как возможные полученные изменения аминокислот в ORF репликазы необходимо рассматривать и проверять на предмет влияния на функцию репликазы. Также невозможно обменять полную ORF репликазы на репликазы гетерологичного происхождения, так как это может привести к нарушению структуры 5' репликационной последовательности распознавания.
В случае транс-репликонов это перекрытие приводит к синтезу фрагмента белка nsP1, так как 5' репликационная последовательность распознавания должна сохраняться в транс-репликонах. Фрагмент nsP1, как правило, не требуется и не желателен: нежелательная трансляция накладывает ненужную нагрузку на клетку-хозяина, а РНК-репликоны, предназначенные для терапевтических применений, которые кодируют в дополнение к фармацевтически активному белку фрагмент nsP1, могут сталкиваться с регуляторными проблемами. Например, необходимо будет продемонстрировать, что усеченный nsP1 не создает нежелательных побочных эффектов. Кроме того, наличие стартового кодона AUG для nsP1 в 5' репликационной последовательности распознавания предотвращает разработку транс-репликонов, кодирующих представляющий интерес гетерологичный ген таким образом, при котором стартовый кодон для трансляции представляющего интерес гена находится в наиболее 5' положении, доступном для инициации рибосомной трансляции. В свою очередь, 5'-кэп-зависимая трансляция трансгенов из транс-репликона РНК предшествующего уровня техники является сложной, если только он не клонируется как слитый белок в рамке считывания со стартовым кодоном nsP1 (такие конструкции слияния описаны, например, Michel et al., 2007, Virology, vol. 362, pp. 475-487). Такие конструкции слияния приводят к той же ненужной трансляции упомянутого выше фрагмента nsP1, что поднимает те же проблемы, что и выше. Более того, слитые белки вызывают дополнительные проблемы, поскольку они могут изменять функцию или активность представляющего интерес слитого трансгена или когда он используется в качестве вектора вакцины, пептиды, охватывающие область слияния, могут изменять иммуногенность слитого антигена.
Необходимо преодолеть эти недостатки. Например, необходимо обеспечить улучшенные репликоны для экспрессии нуклеиновой кислоты, кодирующей представляющий интерес белок, такой как фармацевтически активный белок, безопасным и эффективным способом. Как описано в настоящем документе, аспекты и варианты осуществления настоящего изобретения направлены на эту потребность.
Краткое раскрытие изобретения
Иммунотерапевтические стратегии предоставляют перспективные варианты профилактики и лечения, например, инфекционных заболеваний и онкологических заболеваний. Идентификация растущего числа связанных с патогенами и опухолями антигенов привела к широкому набору подходящих мишеней для иммунотерапии. Настоящее изобретение охватывает улучшенные средства и способы, подходящие для эффективной экспрессии антигенов, подходящих для иммунотерапевтического лечения для профилактики и лечения заболеваний.
Согласно первому аспекту в настоящем изобретении предусмотрен РНК-репликон, содержащий 5' репликационную последовательность распознавания, причем 5' репликационная последовательность распознавания характеризуется тем, что в ней имеется удаление по меньшей мере одного инициирующего кодона по сравнению с нативной альфавирусной 5' репликационной последовательностью распознавания.
Согласно одному варианту осуществления РНК-репликон по настоящему изобретению содержит (модифицированную) 5' репликационную последовательность распознавания и первую открытую рамку считывания, кодирующую представляющий интерес белок, например, функциональный альфавирусный неструктурный белок или трансген, который предпочтительно не происходит от альфавируса, в частности неструктурного белка альфавируса, расположенную ниже по ходу транскрипции от 5' репликационной последовательности распознавания, причем 5' репликационная последовательность распознавания и первая открытая рамка считывания. кодирующая представляющий интерес белок, не перекрываются и предпочтительно, чтобы 5' репликационная последовательность распознавания не перекрывалась с какой-либо открытой рамкой считывания РНК-репликона, например, 5' репликационная последовательность распознавания не содержала бы функциональный инициирующий кодон и предпочтительно не содержала бы никаких инициирующих кодонов. Наиболее предпочтительно, чтобы инициирующий кодон первой открытой рамки считывания, кодирующей представляющий интерес белок, находился бы в направлении 5'→3' от первого функционального инициирующего кодона РНК-репликона, предпочтительно первого инициирующего кодона. Согласно одному варианту осуществления первая открытая рамка считывания, кодирующая представляющий интерес белок, кодирует функциональный альфавирусный неструктурный белок. Согласно одному варианту осуществления первая открытая рамка считывания, кодирующая представляющий интерес белок и предпочтительно весь РНК-репликон, не экспрессирует нефункциональный альфавирусный неструктурный белок, такой как фрагмент неструктурного белка альфавируса, в частности фрагмент nsP1 и/или nsP4. Согласно одному варианту осуществления функциональный альфавирусный неструктурный белок является гетерологичным для 5' репликационной последовательности распознавания. Согласно одному варианту осуществления первая открытая рамка считывания, кодирующая представляющий интерес белок, не находится под контролем субгеномного промотора. Согласно одному варианту осуществления РНК-репликон содержит по меньшей мере одну дополнительную открытую рамку считывания, кодирующую представляющий интерес белок, который находится под контролем субгеномного промотора. Согласно одному варианту осуществления субгеномный промотор и первая открытая рамка считывания, кодирующая представляющий интерес белок, не перекрываются.
Согласно одному варианту осуществления 5' репликационная последовательность распознавания РНК-репликона, которая характеризуется удалением по меньшей мере одного инициирующего кодона, содержит последовательность, гомологичную приблизительно 250 нуклеотидам на 5'-конце альфавируса. Согласно предпочтительному варианту осуществления она содержит последовательность, гомологичную приблизительно 300-500 нуклеотидам на 5'-конце альфавируса. Согласно предпочтительному варианту осуществления она содержит 5'-концевую последовательность, необходимую для эффективной репликации специфических форм альфавирусов, которые являются источниками для векторной системы.
Согласно одному варианту осуществления 5' репликационная последовательность распознавания РНК-репликона содержит последовательности, гомологичные консервативному элементу последовательности 1 (CSE 1) и консервативному элементу последовательности 2 (CSE 2) альфавируса.
Согласно предпочтительному варианту осуществления РНК-репликон содержит CSE 2 и дополнительно характеризуется тем, что он содержит фрагмент открытой рамки считывания неструктурного белка из альфавируса. Согласно более предпочтительному варианту осуществления указанный фрагмент открытой рамки считывания неструктурного белка не содержит никакого инициирующего кодона.
Согласно одному варианту осуществления 5' репликационная последовательность распознавания содержит последовательность, гомологичную открытой рамке считывания неструктурного белка или его фрагмента из альфавируса, причем последовательность, гомологичная открытой рамке считывания неструктурного белка или его фрагмента из альфавируса отличается тем, что она характеризуется удалением по меньшей мере одного инициирующего кодона по сравнению с нативной альфавирусной последовательностью.
Согласно предпочтительному варианту осуществления последовательность, гомологичная открытой рамке считывания неструктурного белка или его фрагмента из альфавируса, отличается тем, что она характеризуется удалением по меньшей мере нативного стартового кодона открытой рамки считывания неструктурного белка.
Согласно предпочтительному варианту осуществления последовательность, гомологичная открытой рамке считывания неструктурного белка или его фрагмента из альфавируса, характеризуется тем, что в ней имеется удаление одного или нескольких инициирующих кодонов, отличных от нативного стартового кодона открытой рамки считывания неструктурного белка. Согласно более предпочтительному варианту осуществления указанная последовательность нуклеиновой кислоты дополнительно характеризуется удалением нативного стартового кодона открытой рамки считывания неструктурного белка, предпочтительно nsP1.
Согласно предпочтительному варианту осуществления 5' репликационная последовательность распознавания содержит одну или несколько структур «петля-на-стебле», обеспечивающих функциональность 5' репликационной последовательности распознавания в отношении репликации РНК. Согласно предпочтительному варианту осуществления одна или несколько структур «петля-на-стебле» 5' репликационной последовательности распознавания не удаляются или не нарушаются. Более предпочтительно, чтобы одна или несколько структур «петля-на-стебле» 1, 3 и 5, предпочтительно все структуры «петля-на-стебле» 1, 3 и 4 или структуры «петля-на-стебле» 3 и 4 не были удалены или нарушены. Более предпочтительно, чтобы ни одна из структур «петля-на-стебле» 5' репликационной последовательности распознавания не удалялась или не нарушалась.
Согласно предпочтительному варианту осуществления РНК-репликон содержит одно или несколько нуклеотидных изменений, компенсирующих нарушения спаривания нуклеотидов в пределах одной или нескольких структур «петля-на-стебле», введенных посредством удаления по меньшей мере одного инициирующего кодона.
Согласно одному варианту осуществления РНК-репликон не содержит открытую рамку считывания, кодирующую усеченный альфавирусный неструктурный белок.
Согласно одному варианту осуществления РНК-репликон содержит 3' репликационную последовательность распознавания.
Согласно одному варианту осуществления РНК-репликон содержит первую открытую рамку считывания, кодирующую представляющий интерес белок.
Согласно одному варианту осуществления РНК-репликон характеризуется тем, что представляющий интерес белок, кодируемый первой открытой рамкой считывания, может экспрессироваться из РНК-репликона в качестве матрицы.
Согласно одному варианту осуществления РНК-репликон характеризуется тем, что он содержит субгеномный промотор. Как правило, субгеномный промотор контролирует производство субгеномной РНК, содержащей открытую рамку считывания, кодирующую представляющий интерес белок.
Согласно предпочтительному варианту осуществления представляющий интерес белок, кодируемый первой открытой рамкой считывания, может быть экспрессирован из РНК-репликона в качестве матрицы. Согласно более предпочтительному варианту осуществления представляющий интерес белок, кодируемый первой открытой рамкой считывания, может дополнительно быть экспрессирован из субгеномной РНК.
Согласно предпочтительному варианту осуществления РНК-репликон дополнительно характеризуется тем, что он содержит субгеномный промотор, контролирующий производство субгеномной РНК, содержащей вторую открытую рамку считывания, кодирующую представляющий интерес белок. Представляющий интерес белок может представлять собой второй белок, который идентичен представляющему интерес белку, кодируемому первой открытой рамкой считывания, или отличаться от него.
Согласно более предпочтительному варианту осуществления субгеномный промотор и вторая открытая рамка считывания, кодирующая представляющий интерес белок, расположены ниже по ходу транскрипции от первой открытой рамки считывания, кодирующей представляющий интерес белок.
Согласно одному варианту осуществления представляющий интерес белок, кодируемый первой и/или второй открытой рамкой считывания, представляет собой функциональный альфавирусный неструктурный белок.
Согласно одному варианту осуществления РНК-репликон содержит открытую рамку считывания, кодирующую функциональный альфавирусный неструктурный белок.
Согласно одному варианту осуществления открытая рамка считывания, кодирующая функциональный альфавирусный неструктурный белок, не перекрывается с 5' репликационной последовательностью распознавания.
Согласно одному варианту осуществления РНК-репликон, который кодирует функциональный альфавирусный неструктурный белок, может быть реплицирован функциональным альфавирусным неструктурным белком.
Согласно одному варианту осуществления РНК-репликон не содержит открытую рамку считывания, кодирующую функциональный альфавирусный неструктурный белок. Согласно этому варианту осуществления функциональный альфавирусный неструктурный белок для репликации репликона может быть предусмотрен in trans, как описано в настоящем документе.
Согласно второму аспекту в настоящем изобретении предусмотрена система, содержащая:
конструкцию РНК для экспрессии функционального альфавирусного неструктурного белка,
РНК-репликон в соответствии с первым аспектом настоящего изобретения, который может быть реплицирован функциональным альфавирусным неструктурным белком in trans. Предпочтительно РНК-репликон дополнительно характеризуется тем, что он не кодирует функциональный альфавирусный неструктурный белок.
Согласно одному варианту осуществления РНК-репликон в соответствии с первым аспектом или система в соответствии со вторым аспектом характеризуется тем, что альфавирус представляет собой вирус леса Семлики.
Согласно третьему аспекту в настоящем изобретении предусмотрена ДНК, содержащая последовательность нуклеиновой кислоты, кодирующую РНК-репликон в соответствии с первым аспектом настоящего изобретения.
Согласно четвертому аспекту в настоящем изобретении предусмотрен способ получения представляющего интерес белка в клетке, предусматривающий следующие стадии:
(а) получение РНК-репликона в соответствии с первым аспектом настоящего изобретения, который содержит открытую рамку считывания, кодирующую функциональный альфавирусный неструктурный белок, который может быть реплицирован функциональным альфавирусным неструктурным белком, и который дополнительно содержит открытую рамку считывания, кодирующую представляющий интерес белок, и
(b) инокуляция РНК-репликона в клетку.
Согласно различным вариантам осуществления способа РНК-репликон является таким, как определено выше для репликона по настоящему изобретению.
Согласно пятому аспекту в настоящем изобретении предусмотрен способ получения представляющего интерес белка в клетке, предусматривающий следующие стадии:
(а) получение конструкции РНК для экспрессии функционального альфавирусного неструктурного белка,
(b) получение РНК-репликона в соответствии с первым аспектом настоящего изобретения, который может быть реплицирован функциональным альфавирусным неструктурным белком в соответствии с (а) in trans и который содержит открытую рамку считывания, кодирующую представляющий интерес белок, и
(c) инокуляция смеси РНК-конструкции для экспрессии функционального альфавирусного неструктурного белка и РНК-репликона в клетку.
Согласно различным вариантам осуществления способа конструкция РНК для экспрессии функционального альфавирусного неструктурного белка и/или РНК-репликон являются такими, как определено выше для системы по настоящему изобретению. Согласно пятому аспекту, РНК-репликон, как правило, сам по себе не кодирует функциональный альфавирусный неструктурный белок.
Согласно шестому аспекту в настоящем изобретение предусмотрена клетка, содержащая репликон первого аспекта или систему второго аспекта. Согласно одному варианту осуществления клетку инокулируют в соответствии со способом четвертого аспекта или в соответствии со способом пятого аспекта настоящего изобретения. Согласно одному варианту осуществления клетка может быть получена способом четвертого аспекта или способом пятого аспекта настоящего изобретения. Согласно одному варианту осуществления клетка представляет собой часть организма.
Согласно седьмому аспекту в настоящем изобретении предусмотрен способ получения представляющего интерес белка у субъекта, предусматривающий следующие стадии:
(а) получение РНК-репликона в соответствии с первым аспектом настоящего изобретения, который содержит открытую рамку считывания, кодирующую функциональный альфавирусный неструктурный белок, который может быть реплицирован функциональным альфавирусным неструктурным белком, и который дополнительно содержит открытую рамку считывания, кодирующую представляющий интерес белок, и
(b) введение РНК-репликона субъекту.
Согласно различным вариантам осуществления способа РНК-репликон является таким, как определено выше для репликона по настоящему изобретению.
Согласно восьмому аспекту в настоящем изобретении предусмотрен способ получения представляющего интерес белка у субъекта, предусматривающий следующие стадии:
(а) получение конструкции РНК для экспрессии функционального альфавирусного неструктурного белка,
(b) получение РНК-репликона в соответствии с первым аспектом настоящего изобретения, который может быть реплицирован функциональным альфавирусным неструктурным белком согласно (а) in trans и который содержит открытую рамку считывания, кодирующую представляющий интерес белок, и
(c) введение конструкции РНК для экспрессии функционального альфавирусного неструктурного белка и РНК-репликона субъекту.
Согласно различным вариантам осуществления способа конструкция РНК для экспрессии функционального альфавирусного неструктурного белка и/или РНК-репликон являются такими, как определено выше для системы по настоящему изобретению. Согласно восьмому аспекту РНК-репликон, как правило, сам по себе не кодирует функциональный альфавирусный неструктурный белок.
Краткое описание графических материалов
Фиг.1: Схематическое представление РНК-репликонов, содержащих немодифицированную или модифицированную 5' репликационную последовательность распознавания
Сокращения: AAAA = поли(A)-хвост; ATG = стартовый кодон/инициирующий кодон (ATG на уровне ДНК, AUG на уровне РНК); 5x ATG = последовательность нуклеиновой кислоты, содержащая все стартовые кодоны в последовательности нуклеиновой кислоты, кодирующей nsP1* (в случае последовательности нуклеиновой кислоты, кодирующей nsP1*, из вируса леса Семлики 5x ATG соответствует пяти специфическим стартовым кодонам, смотрите пример 1); Δ5ATG = последовательность нуклеиновой кислоты, соответствующая последовательности нуклеиновой кислоты, кодирующей nsP1*; однако не содержащая никаких стартовых кодонов последовательности нуклеиновой кислоты, кодирующей nsP1* в альфавирусе, обнаруженном в природе (в случае nsP1*, происходящем от вируса леса Семлики, «Δ5ATG» соответствует удалению пяти специфических стартовых кодонов по сравнению с вирусом леса Семлики, обнаруженным в природе, смотрите пример 1); EcoRV = сайт рестрикции EcoRV; nsP = последовательность нуклеиновой кислоты, кодирующая альфавирусный неструктурный белок (например, nsP1, nsP2, nsP3, nsP4); nsP1* = последовательность нуклеиновой кислоты, кодирующая фрагмент nsP1, причем фрагмент не содержит C-концевой фрагмент nsP1; *nsP4 = последовательность нуклеиновой кислоты, кодирующая фрагмент nsP4, причем фрагмент не содержит N-концевой фрагмент nsP4; RRS = 5' репликационная последовательность распознавания; SalI = сайт рестрикции SalI; SGP = субгеномный промотор; SL = структура «петля-на-стебле» (например, SL1, SL2, SL3, SL4); положения SL1-4 графически проиллюстрированы; UTR = нетранслируемая область (например, 5'-UTR, 3'-UTR); WT = дикий тип.
Цис-репликон WT-RRS: РНК-репликон, по существу соответствующий геному альфавируса, за исключением того, что последовательность нуклеиновой кислоты, кодирующая структурные белки альфавируса, была заменена открытой рамкой считывания, кодирующей представляющий интерес ген («трансген»). Когда в клетку вводится «репликон WT-RRS», продукт трансляции открытой рамки считывания, кодирующей репликазу (nsP1234 или ее фрагмент), может управлять репликацией РНК-репликона in cis и управлять синтезом последовательности нуклеиновой кислоты (субгеномного транскрипта) ниже по ходу транскрипции субгеномного промотора (SGP).
Транс-репликон или РНК-матрица WT-RRS: РНК-репликон, по существу соответствующий «репликону WT-RRS», за исключением того, что большая часть последовательности нуклеиновой кислоты, кодирующей альфавирусные неструктурные белки nsP1-4, была удалена. Более конкретно, последовательность нуклеиновой кислоты, кодирующая nsP2 и nsP3, была полностью удалена; последовательность нуклеиновой кислоты, кодирующая nsP1, была усечена так, что «РНК-матрица WT-RRS» кодирует фрагмент nsP1, причем этот фрагмент не содержит C-концевой фрагмент nsP1 (но он содержит N-концевой фрагмент nsP1; nsP1*); последовательность нуклеиновой кислоты, кодирующая nsP4, была усечена так, что «РНК-матрица WT-RRS» кодирует фрагмент nsP4, причем этот фрагмент не содержит N-концевой фрагмент nsP4 (но он содержит C-концевой фрагмент nsP4; *nsP4). Эта усеченная последовательность nsP4 частично перекрывается с полностью активным субгеномным промотором. Последовательность нуклеиновой кислоты, кодирующая nsP1*, содержит все инициирующие кодоны последовательности нуклеиновой кислоты, которая кодирует nsP1* в альфавирусе, обнаруженном в природе (в случае nsP1* из вируса леса Семлики, пять специфических инициирующих кодонов).
Δ5ATG-RRS: РНК-репликон, по существу соответствующий «РНК-матрице WT-RRS», за исключением того, что она не содержит никаких инициирующих кодонов последовательности нуклеиновой кислоты, которая кодирует nsP1* в альфавирусе, обнаруженном в природе (в случае вируса леса Семлики, «Δ5ATG-RRS» соответствует удалению пяти специфических инициирующих кодонов по сравнению с вирусом леса Семлики, обнаруженным в природе). Все изменения нуклеотидов, введенные для удаления стартовых кодонов, компенсировались дополнительными нуклеотидными изменениями для сохранения предсказанной вторичной структуры РНК.
Δ5ATG-RRSΔSGP: РНК-репликон, по существу соответствующий «Δ5ATG-RRS», за исключением того, что он не содержит субгеномный промотор (SGP) и не содержит последовательность нуклеиновой кислоты, которая кодирует *nsP4. «Трансген 1» = представляющий интерес ген.
Δ5ATG-RRS-бицистронный: РНК-репликон, по существу соответствующий «Δ5ATG-RRS», за исключением того, что он содержит первую открытую рамку считывания, кодирующую первый представляющий интерес ген («трансген 1») выше против хода транскрипции от субгеномного промотора, и вторую открытую рамку считывания, кодирующую второй представляющий интерес ген («трансген 2») ниже по ходу транскрипции от субгеномного промотора. Локализация второй открытой рамки считывания соответствует локализации представляющего интерес гена («трансгена») в РНК-репликоне «Δ5ATG-RRS».
Цис-репликон Δ5ATG-RRS: РНК-репликон, по существу соответствующий «Δ5ATG-RRS-бицистронный», за исключением того, что открытая рамка считывания, кодирующая первый представляющий интерес ген, кодирует функциональный альфавирусный неструктурный белок (как правило, одну открытую рамку считывания, кодирующую полипротеин nsP1-nsP2-nsP3-nsP4, т.е. nsP1234). «Трансген» в «цис-репликоне Δ5ATG-RRS» соответствует «трансгену 2» в «Δ5ATG-RRS-бицистронном». Функциональный альфавирусный неструктурный белок способен распознавать субгеномный промотор и синтезировать субгеномные транскрипты, содержащие последовательность нуклеиновой кислоты, кодирующую представляющий интерес ген («трансген»). «Цис-репликон Δ5ATG-RRS» кодирует функциональный альфавирусный неструктурный белок in cis, как и «цис-репликон WT-RRS»; однако не требуется, чтобы кодирующая последовательность для nsP1, кодируемая «цис-репликоном Δ5ATG-RRS», содержала точную последовательность нуклеиновой кислоты «цис-репликона WT-RRS», включая в себя все структуры «петля на стебле».
Фиг. 2: Удаление стартовых кодонов в 5' репликационной последовательности распознавания не влияет на репликацию РНК транс-репликонов. A: Иллюстрация молекул нуклеиновой кислоты, используемых в примере 2. B: Измеренная экспрессия люциферазы подвергнутых электропорации клеток BHK21. Подробнее смотрите пример 2. Показано среднее ± SD из 2 независимых экспериментов с 2 независимо производимыми партиями РНК-репликона («матрица»); (N = 4).
Фиг. 3: Удаление стартовых кодонов в 5' репликационной последовательности распознавания обеспечивает возможность зависящей от кэпа трансляции. Слева: иллюстрация молекул нуклеиновой кислоты (РНК-репликоны), используемых в примере 3. Справа: измеренная экспрессия люциферазы подвергнутых электропорации фибробластов крайней плоти человека. Подробнее смотрите пример 3. Показано среднее ± SD одного эксперимента, выполненного в трех повторениях.
Фиг. 4: Сразу после трансфекции зависящая от кэпа трансляция транс-репликона, характеризующаяся удалением стартовых кодонов в 5' репликационной последовательности распознавания, сильнее, чем трансляция из субгеномного транскрипта. Слева: иллюстрация молекул нуклеиновых кислот. РНК-репликоны, используемые в примере 4, показаны как «Δ5ATG-RRS» и «ΔATAT-RRSΔSGP». Справа: измеренная экспрессия люциферазы подвергнутых электропорации клеток BHK21. Подробнее смотрите пример 4. Показано среднее из одного эксперимента, выполненного в трех повторениях.
Фиг. 5: Кэпированный транс-репликон, характеризующийся удалением исходных кодонов в 5' репликационной последовательности распознавания, делает возможной экспрессию трансгена на ранних стадиях. Слева: иллюстрация молекулы нуклеиновой кислоты «Δ5ATG-RRSΔSGP», используемой в примере 5. Справа: измеренная экспрессия люциферазы подвергнутых электропорации клеток BHK21. Подробнее смотрите пример 5. Показано среднее ± SD одного эксперимента, выполненного в трех повторениях.
Фиг. 6. Структуры динуклеотидов кэпа. Вверху: природный динуклеотид кэпа, m7GpppG. Внизу: фосфоротиоатный аналог кэпа динуклеотид бета-S-ARCA. Существует два диастереомера бета-S-ARCA из-за стереогенного P-центра, которые обозначены D1 и D2 в соответствии с их характеристиками элюирования в ВЭЖХ с обращенной фазой.
Фиг. 7. Реконструированные цис-репликоны с удаленным ATG RRS, являются функциональными. ORF репликазы SFV вставляли в «Δ5ATG-RRS», кодирующую люциферазу светлячков ниже по ходу транскрипции от субгеномного промотора (SGP). Внутри вставленной репликазы области, соответствующие CSE2 и SGP сердцевины, были разрушены нуклеотидными обменами (заштрихованые прямоугольники), чтобы избежать дублирования этих регуляторных областей. Это привело к реконструированному цис-репликону. Клетки BHK21 подвергали совместной электропорации с 2,5 мкг либо «цис-репликона WT-RRS», либо «цис-репликона Δ5ATG-RRS». Через 24 часа после электропорации измеряли экспрессию люциферазы. Среднее ± SD одного эксперимента в трех повторах.
Фиг. 8. Бицистронные транс-репликоны экспрессируют оба трансгена. Секретируемую нано-люциферазу (SNL) клонировали ниже по ходу транскрипции от субгеномного промотора (SGP) транс-репликона WT-RSS. Положение выше по ходу транскрипции от SGP не кодирует трансген (-)SGP(SNP). В нижней конструкции SNL клонировали ниже по ходу транскрипции от ΔATG-RSS и люциферазу светлячков (Luc) вставляли ниже по ходу транскрипции от SGP (SNL)SGP(Luc). Клетки BHK21 подвергали совместной электропорации с 0,9 мкг транс-реплицирующей РНК и 5 мкг кодирующей SFV-репликазу мРНК, через 48 часов после электропорации измеряли экспрессию SNL и Luc. Данные одного эксперимента.
Фиг. 9. Транс-репликоны вируса Синдбис, не содержащие стартовые кодоны в репликационной последовательности распознавания, эффективно реплицируются. Транс-репликоны конструировали из генома вируса Синдбис путем синтеза генов, и GFP вставляли ниже по ходу транскрипции от субгеномного промотора (SGP). В дополнение к этому транс-репликону с немодифицированной репликационной последовательностью распознавания (WT-RSS) создавали два его варианта. В ΔATG-RRS исходный стартовый кодон плюс 4 дополнительных ATG удаляли из WT-RRS. При необходимости также вводили компенсаторные нуклеотидные изменения для сохранения вторичной структуры РНК. Для получения ΔATG-RRSΔSGP область, соответствующую субгеномному промотору, удаляли из ΔATG-RRS, получая в результате вектор с GFP непосредственно ниже по ходу транскрипции от 5'RRS с удалением ATG. Клетки BHK21 подвергали совместной электропорации с 0,1 мкг транс-реплицирующей РНК и 2,4 мкг кодирующей SFV-репликазу мРНК. Через 24 часа после электропорации оценивали экспрессию GFP (скорость трансфекции [%] и среднюю интенсивность флуоресценции (MFI)). Данные одного эксперимента.
Подробное описание изобретения
Хотя настоящее изобретение подробно описано ниже, следует понимать, что настоящее изобретение не ограничено конкретными способами, протоколами и реагентами, описанными в настоящем документе, поскольку они могут различаться. Следует также понимать, что используемая в настоящем документе терминология предназначена только для описания конкретных вариантов осуществления и не предназначена для ограничения объема настоящего изобретения, который будет ограничен только прилагаемой формулой настоящего изобретения. Если не указано иное, все технические и научные термины, используемые в настоящем документе, имеют те же значения, которые, как правило, понимаются специалистом в настоящей области техники.
Предпочтительно используемые в настоящем документе термины определяются, как описано в «Многоязычном словаре биотехнологических терминов: (Рекомендации IUPAC)», H.G.W. Leuenberger, B. Nagel и H. Kölbl, Eds., Helvetica Chimica Acta, CH-4010 Basel, Switzerland, (1995).
В практике настоящего изобретения будут использованы, если не указано иное, традиционные способы химии, биохимии, клеточной биологии, иммунологии и рекомбинантной ДНК, которые объясняются в литературе в этой области (смотрите, например, Molecular Cloning: A Laboratory Manual, 2nd Edition, J. Sambrook et al. eds., Cold Spring Harbor Laboratory Press, Cold Spring Harbor 1989).
Ниже будут описаны элементы настоящего изобретения. Эти элементы перечислены в конкретных вариантах осуществления, однако следует понимать, что они могут быть объединены любым способом и в любом числе для создания дополнительных вариантов осуществления. Различные описанные примеры и предпочтительные варианты осуществления не должны толковаться как ограничивающие настоящее изобретение только явно описанными вариантами осуществления. Это описание следует понимать как раскрытие и объединение в себе вариантов осуществления, которые объединяют явно описанные варианты осуществления с любым количеством раскрытых и/или предпочтительных элементов. Кроме того, любые перестановки и комбинации всех описанных элементов в этом приложении следует рассматривать как раскрытые этим описанием, если контекст не указывает иное.
Термин «приблизительно» означает около или примерно, и в контексте численного значения или диапазона, изложенного в настоящем документе, предпочтительно означает +/- 10% от указанного или заявленного численного значения или диапазона.
Неопределенные и определенные артикли и аналогичные ссылки, используемые в контексте описания настоящего изобретения (особенно в контексте формулы изобретения), должны толковаться как охватывающие как единственное, так и множественное число, если иное не указано в настоящем документе или явно не противоречит контексту. Перечисление диапазонов значений в настоящем документе просто предназначено для использования в качестве сокращенного способа индивидуальной ссылки на каждое отдельное значение, входящее в диапазон. Если не указано иное, каждое индивидуальное значение включается в спецификацию, как если бы оно было отдельно представлено в настоящем документе. Все описанные в настоящем документе способы могут быть выполнены в любом подходящем порядке, если иное не указано в настоящем документе или иначе явно не противоречит контексту. Использование любых и всех примеров или иллюстративного языка (например, «такой как»), приведенного в настоящем документе, предназначено просто для лучшей иллюстрации настоящего изобретения и не представляет собой ограничение объема настоящего изобретения, заявленного иным образом. Никакой язык в спецификации не следует истолковывать как указание на какой-либо незаявленный элемент, необходимый для практики настоящего изобретения.
Если прямо не указано иное, термин «содержащий» используется в контексте настоящего документа, чтобы указать на то, что дополнительные представители могут необязательно присутствовать в дополнение к представителям перечня, представленного посредством термина «содержащий». Однако в качестве конкретного варианта осуществления настоящего изобретения предполагается, что термин «содержащий» охватывает возможность отсутствия каких-либо дополнительных представителей, то есть для целей этого варианта осуществления «содержащий» следует понимать как имеющий значение «состоящий из».
Указания относительных количеств компонента, характеризуемого общим термином, означают общее количество всех конкретных вариантов или представителей, охватываемых указанным общим термином. Если определенный компонент, определяемый общим термином, указывается как присутствующий в определенном относительном количестве, и если этот компонент дополнительно характеризуется как специфический вариант или представитель, охватываемый общим термином, подразумевается, что никакие другие варианты или представители, охватываемые общим термином, дополнительно не присутствуют таким образом, что общее относительное количество компонентов, охватываемых общим термином, превышает указанную относительную величину; более предпочтительно нет других вариантов или представителей, охватываемых общим термином.
В тексте данного описания настоящего изобретения приводится несколько документов. Каждый из приведенных в настоящем документе документов (включая все патенты, заявки на патент, научные публикации, описания производителя, инструкции и т.д.), независимо от того, представлены ли они выше или ниже, полностью включены в настоящий документ посредством ссылки. Ничто в настоящем документе не должно толковаться как признание того, что настоящее изобретение не имеет права датировать более ранним числом такое раскрытие.
Используемые в настоящем документе такие термины, как «уменьшить» или «ингибировать», означают способность вызывать общее снижение, предпочтительно на 5% или более, 10% или более, 20% или более, более предпочтительно 50% или более и наиболее предпочтительно 75% или более. Термин «ингибировать» или подобные фразы включают в себя полное или практически полное ингибирование, то есть снижение до нуля или по существу до нуля.
Такие термины, как «увеличение» или «повышение» предпочтительно относятся к увеличению или повышению приблизительно на 10%, предпочтительно по меньшей мере на 20%, предпочтительно по меньшей мере на 30%, более предпочтительно по меньшей мере на 40%, более предпочтительно по меньшей мере на 50%, еще более предпочтительно по меньшей мере на 80% и наиболее предпочтительно по меньшей мере на 100%.
Термин «суммарный заряд» относится к заряду на целом объекте, таком как соединение или частица.
Ион, имеющий общий суммарный положительный заряд, представляет собой катион, тогда как ион, имеющий общий суммарный отрицательный заряд, представляет собой анион. Таким образом, согласно настоящему изобретению анион представляет собой ион с большим количеством электронов, чем протоны, что дает ему суммарный отрицательный заряд; а катион представляет собой ион с меньшим количеством электронов, чем протоны, что дает ему суммарный положительный заряд.
Такие термины как «заряженные», «суммарный заряд», «отрицательно заряженные» или «положительно заряженные» применительно к данному соединению или частице относятся к электрическому суммарному заряду данного соединения или частицы при растворении или суспендировании в воде при рН 7,0.
Термин «нуклеиновая кислота» согласно настоящему изобретению также предусматривает химическое получение производных нуклеиновой кислоты на нуклеотидном основании, на сахаре или на фосфате, и нуклеиновые кислоты, содержащие неприродные нуклеотиды и нуклеотидные аналоги. Согласно некоторым вариантам осуществления нуклеиновая кислота представляет собой дезоксирибонуклеиновую кислоту (ДНК) или рибонуклеиновую кислоту (РНК). В общем, молекула нуклеиновой кислоты или последовательность нуклеиновой кислоты относится к нуклеиновой кислоте, которая предпочтительно представляет собой дезоксирибонуклеиновую кислоту (ДНК) или рибонуклеиновую кислоту (РНК). В соответствии с настоящим изобретением нуклеиновые кислоты включают в себя геномную ДНК, кДНК, мРНК, вирусную РНК, рекомбинантно полученные и химически синтезированные молекулы. В соответствии с настоящим изобретением нуклеиновая кислота может быть в форме одноцепочечной или двухцепочечной и линейной или ковалентно замкнутой кольцевой молекулы.
В соответствии с настоящим изобретением «последовательность нуклеиновой кислоты» относится к последовательности нуклеотидов в нуклеиновой кислоте, например, рибонуклеиновой кислоте (РНК) или дезоксирибонуклеиновой кислоте (ДНК). Этот термин может относиться ко всей молекуле нуклеиновой кислоты (такой как единственная нить всей молекулы нуклеиновой кислоты) или к части (например, ее фрагменту).
В соответствии с настоящим изобретением термин «РНК» или «молекула РНК» относится к молекуле, которая содержит рибонуклеотидные остатки и которая предпочтительно полностью или по существу состоит из рибонуклеотидных остатков. Термин «рибонуклеотид» относится к нуклеотиду с гидроксильной группой в 2'-положении β-D-рибофуранозильной группы. Термин «РНК» включает в себя двухцепочечную РНК, одноцепочечную РНК, выделенную РНК, такую как частично или полностью очищенная РНК, по существу чистую РНК, синтетическую РНК и рекомбинантно произведенную РНК, такую как модифицированная РНК, которая отличается от природной РНК добавлением, делецией, заменой и/или изменением одного или нескольких нуклеотидов. Такие изменения могут включать в себя добавление ненуклеотидного материала, например, на конце (концах) РНК или внутри, например, к одному или нескольким нуклеотидам РНК. Нуклеотиды в молекулах РНК могут также содержать нестандартные нуклеотиды, такие как не встречающиеся в природе нуклеотиды или химически синтезированные нуклеотиды или дезоксинуклеотиды. Эти измененные РНК можно назвать аналогами, особенно аналогами встречающихся в природе РНК.
Согласно настоящему изобретению РНК может быть одноцепочечной или двухцепочечной. Согласно некоторым вариантам осуществления настоящего изобретения одноцепочечная РНК является предпочтительной. Термин «одноцепочечная РНК», как правило, относится к молекуле РНК, с которой не связана ни одна комплементарная молекула нуклеиновой кислоты (как правило, ни одна комплементарная молекула РНК). Одноцепочечная РНК может содержать самокомплементарные последовательности, которые позволяют частям РНК конъюгировать «в себе» и образовывать мотивы вторичной структуры, включая в себя, без ограничения, пары оснований, структуры «стебель», структуры «петля-на-стебле» и структуры «петли». Одноцепочечная РНК может существовать как минус-нить [(-)-нить] или как плюс-нить [(+)-нить]. (+)-нить представляет собой нить, которая содержит или кодирует генетическую информацию. Генетическая информация может представлять собой, например, полинуклеотидную последовательность, кодирующую белок. Когда (+)-нитевая РНК кодирует белок, (+)-нить может служить непосредственно в качестве матрицы для трансляции (синтеза белка). (-)-нить является комплементарной к (+)-нити. В случае двухцепочечной РНК (+)-нить и (-)-нить представляют собой две отдельные молекулы РНК, и обе эти молекулы РНК связываются друг с другом с образованием двухцепочечной РНК («дуплексная РНК»).
Термин «стабильность» РНК относится к «периоду полужизни» РНК. «Период полужизни» относится к периоду времени, который необходим для устранения половины активности, количества или числа молекул. В контексте настоящего изобретения период полужизни РНК является показателем стабильности указанной РНК. Период полужизни РНК может влиять на «продолжительность экспрессии» РНК. Можно ожидать, что РНК, характеризующаяся длительным периодом полужизни, будет экспрессироваться в течение длительного периода времени.
Термин «эффективность трансляции» относится к количеству продукта трансляции, предоставленного молекулой РНК в течение определенного периода времени.
«Фрагмент» по отношению к последовательности нуклеиновой кислоты относится к части последовательности нуклеиновой кислоты, то есть последовательности, которая представляет собой последовательность нуклеиновой кислоты, укороченную на 5'- и/или 3'-конце(-ах). Предпочтительно фрагмент последовательности нуклеиновой кислоты содержит по меньшей мере 80%, предпочтительно по меньшей мере 90%, 95%, 96%, 97%, 98% или 99% нуклеотидных остатков от указанной последовательности нуклеиновой кислоты. В настоящем изобретении предпочтительными являются те фрагменты молекул РНК, которые сохраняют стабильность РНК и/или эффективность трансляции.
«Фрагмент» по отношению к аминокислотной последовательности (пептиду или белку) относится к части аминокислотной последовательности, то есть последовательности, которая представляет собой аминокислотную последовательность, укороченную на N-конце и/или С-конце. Фрагмент, укороченный на С-конце (N-концевой фрагмент), можно получить, например, путем трансляции усеченной открытой рамки считывания, которая не содержит 3'-конца открытой рамки считывания. Фрагмент, укороченный на N-конце (С-концевой фрагмент), можно получить, например, путем трансляции усеченной открытой рамки считывания, которая не содержит 5'-конца открытой рамки считывания, при условии, что усеченная открытая рамка считывания содержит стартовый кодон, который служит для инициации трансляции. Фрагмент аминокислотной последовательности содержит, например, по меньшей мере 1%, по меньшей мере 2%, по меньшей мере 3%, по меньшей мере 4%, по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% аминокислотных остатков из аминокислотной последовательности.
Термин «вариант» в отношении, например, последовательностей нуклеиновых кислот и аминокислот согласно настоящему изобретению включает в себя любые варианты, в частности мутанты, варианты вирусных штаммов, варианты сращивания, конформации, изоформы, аллельные варианты, видовые варианты и видовые гомологи, в частности те, которые присутствуют в природе. Аллельный вариант относится к изменению нормальной последовательности гена, значение которого часто неясно. Полное секвенирование генов часто идентифицирует многочисленные аллельные варианты для данного гена. Что касается молекул нуклеиновой кислоты, термин «вариант» включает в себя вырожденные последовательности нуклеиновой кислоты, причем вырожденная нуклеиновая кислота согласно настоящему изобретению представляет собой нуклеиновую кислоту, которая отличается от эталонной нуклеиновой кислоты в последовательности кодонов из-за вырожденности генетического кода. Видовой гомолог представляет собой нуклеиновую кислоту или аминокислотную последовательность, происходящую от другого вида, чем данная последовательность нуклеиновой кислоты или аминокислотная последовательность. Гомолог вируса представляет собой нуклеиновую кислоту или аминокислотную последовательность, происходящую от другого вируса, чем данная последовательность нуклеиновой кислоты или аминокислотная последовательность.
В соответствии с настоящим изобретением варианты нуклеиновой кислоты включают в себя одиночные или множественные нуклеотидные делеции, добавления, мутации, замены и/или вставки по сравнению с эталонной нуклеиновой кислотой. Делеции включают в себя удаление одного или нескольких нуклеотидов из эталонной нуклеиновой кислоты. Варианты добавлений предусматривают 5'- и/или 3'-концевые слияния одного или нескольких нуклеотидов, например, 1, 2, 3, 5, 10, 20, 30, 50 или более нуклеотидов. В случае замен по меньшей мере один нуклеотид в последовательности удаляется и на его место вставляется по меньшей мере один другой нуклеотид (например, трансверсии и транзиции). Мутации включают в себя участки с удаленными азотистыми основаниями, сшитые сайты и химически измененные или модифицированные основания. Вставки включают в себя добавление по меньшей мере одного нуклеотида в эталонную нуклеиновую кислоту.
В соответствии с настоящим изобретением «нуклеотидное изменение» может относиться к одиночным или множественным нуклеотидным делециям, добавлениям, мутациям, заменам и/или вставкам по сравнению с эталонной нуклеиновой кислотой. Согласно некоторым вариантам осуществления «нуклеотидное изменение» выбирают из группы, состоящей из делеции одного нуклеотида, добавления одного нуклеотида, мутации одного нуклеотида, замены одного нуклеотида и/или вставки одного нуклеотида, по сравнению с эталонной нуклеиновой кислотой. В соответствии с настоящим изобретением вариант нуклеиновой кислоты может содержать одно или несколько нуклеотидных изменений по сравнению с эталонной нуклеиновой кислотой.
Варианты конкретных последовательностей нуклеиновых кислот предпочтительно характеризуются по меньшей мере одним функциональным свойством указанных конкретных последовательностей и предпочтительно являются функционально эквивалентными указанным конкретным последовательностям, например, последовательностям нуклеиновых кислот, обладающим свойствами, идентичными или сходными таковым конкретных последовательностей нуклеиновых кислот.
Как описано ниже, некоторые варианты осуществления настоящего изобретения характеризуются, среди прочего, последовательностями нуклеиновой кислоты, которые являются гомологичными последовательностям нуклеиновых кислот альфавируса, такого как обнаруженный в природе альфавирус. Эти гомологичные последовательности представляют собой варианты последовательностей нуклеиновых кислот альфавируса, такого как обнаруженный в природе альфавирус.
Предпочтительно степень идентичности между данной последовательностью нуклеиновой кислоты и последовательностью нуклеиновой кислоты, которая представляет собой вариант указанной данной последовательности нуклеиновой кислоты, будет составлять по меньшей мере 70%, предпочтительно по меньшей мере 75%, предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90% или наиболее предпочтительно по меньшей мере 95%, 96%, 97%, 98% или 99%. Степень идентичности предпочтительно определяется для области, составляющей по меньшей мере приблизительно 30, по меньшей мере приблизительно 50, по меньшей мере приблизительно 70, по меньшей мере приблизительно 90, по меньшей мере приблизительно 100, по меньшей мере приблизительно 150, по меньшей мере приблизительно 200, по меньшей мере приблизительно 250, по меньшей мере приблизительно 300 или по меньшей мере приблизительно 400 нуклеотидов. Согласно предпочтительным вариантам осуществления степень идентичности определяется для всей длины эталонной последовательности нуклеиновой кислоты.
«Сходство последовательностей» указывает на процент аминокислот, которые либо идентичны, либо представляют собой консервативные аминокислотные замены. «Идентичность последовательностей» между двумя последовательностями полипептида или нуклеиновой кислоты показывает процент аминокислот или нуклеотидов, которые идентичны между последовательностями.
Термин «% идентичности» предназначен, в частности, для обозначения процента нуклеотидов, которые идентичны при оптимальном выравнивании между двумя подлежащими сравнению последовательностями, причем указанный процент является чисто статистическим, а различия между этими двумя последовательностями могут быть случайно распределены по всей длине последовательности, и подлежащая сравнению последовательность может содержать добавления или удаления по сравнению с эталонной последовательностью, чтобы получить оптимальное выравнивание между двумя последовательностями. Сравнение двух последовательностей, как правило, выполняется путем сравнения указанных последовательностей после оптимального выравнивания относительно сегмента или «окна сравнения» для идентификации локальных областей соответствующих последовательностей. Оптимальное выравнивание для сравнения может быть выполнено вручную или с помощью алгоритма поиска локальной гомологии от Smith and Waterman, 1981, Ads App. Math. 2, 482, с помощью алгоритма поиска локальной гомологии от Needleman and Wunsch, 1970, J. Mol. Biol. 48, 443 и с помощью алгоритма поиска подобия от Pearson and Lipman, 1988, Proc. Natl Acad. Sci. USA 85, 2444 или с помощью компьютерных программ с использованием указанных алгоритмов (GAP, BESTFIT, FASTA, BLAST P, BLAST N и TFASTA в пакете программного обеспечения Wisconsin Genetics, Genetics Computer Group, 575 Science Drive, Madison, Wis.).
Процент идентичности получают путем определения количества идентичных положений, в которых соответствуют подлежащие сравнению последовательности, деля их на количество сравниваемых положений и умножая этот результат на 100.
Например, может быть использована программа BLAST «последовательности BLAST 2», которая доступна на веб-сайте http://www.ncbi.nlm.nih.gov/blast/bl2seq/wblast2.cgi.
Нуклеиновая кислота «способна гибридизоваться» или «гибридизуется» с другой нуклеиновой кислотой, если две последовательности являются комплементарными друг другу. Нуклеиновая кислота является «комплементарной» по отношению к другой нуклеиновой кислоте, если две последовательности способны образовывать стабильный дуплекс друг с другом. Согласно настоящему изобретению гибридизацию предпочтительно проводят в условиях, которые позволяют специфическую гибридизацию между полинуклеотидами (строгие условия). Строгие условия описаны, например, в публикации Molecular Cloning: A Laboratory Manual, J. Sambrook et al., Editors, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989 или Current Protocols in Molecular Biology, F.M. Ausubel et al., Editors, John Wiley & Sons, Inc., New York и, например, относятся к гибридизации при температуре 65°C в буфере для гибридизации (3,5 × SSC, фиколл в концентрации 0,02%, поливинилпирролидон в концентрации 0,02%, бычий сывороточный альбумин в концентрации 0,02%, NaH2PO4 в концентрации 2,5 мМ (pH 7), ДСН в концентрации 0,5%, ЭДТА в концентрации 2 мМ). SSC представляет сбой 0,15 М хлорида натрия/0,15 М цитрата натрия, рН 7. После гибридизации мембрану, на которую переносили ДНК, промывают, например, в 2 × SSC при комнатной температуре, а затем в 0,1-0,5 × SSC/0,1 × SDS при температурах до 68°C.
Процент комплементарности указывает на процент смежных остатков в молекуле нуклеиновой кислоты, которая может образовывать водородные связи (например, спаривание оснований Уотсона-Крика) со второй последовательностью нуклеиновой кислоты (например, 5, 6, 7, 8, 9, 10 из 10 комплементарны на 50%, 60%, 70%, 80%, 90% и 100%). «Совершенно комплементарный» или «полностью комплементарный» означает, что все смежные остатки последовательности нуклеиновой кислоты будут связаны посредством водородной связи с таким же количеством смежных остатков во второй последовательности нуклеиновой кислоты. Предпочтительно, степень комплементарности согласно настоящему изобретению составляет по меньшей мере 70%, предпочтительно по меньшей мере 75%, предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90% или наиболее предпочтительно по меньшей мере 95% 96%, 97%, 98% или 99%. Наиболее предпочтительно, степень комплементарности согласно настоящему изобретению составляет 100%.
Термин «производное» включает в себя любое получение нуклеиновой кислоты химическим способом на нуклеотидном основании, на сахаре или на фосфате. Термин «производное» также включает в себя нуклеиновые кислоты, которые содержат нуклеотиды и нуклеотидные аналоги, не встречающиеся в природе. Предпочтительно, получение нуклеиновой кислоты химическим способом повышает ее стабильность.
В соответствии с настоящим изобретением «последовательность нуклеиновой кислоты, которая получена из последовательности нуклеиновой кислоты» относится к нуклеиновой кислоте, которая представляет собой вариант нуклеиновой кислоты, из которой она получена. Предпочтительно последовательность, которая представляет собой вариант по отношению к определенной последовательности, когда она заменяет конкретную последовательность в молекуле РНК, сохраняет стабильность РНК и/или эффективность трансляции.
«nt» представляет собой сокращение для нуклеотида или для нуклеотидов, предпочтительно последовательных нуклеотидов в молекуле нуклеиновой кислоты.
В соответствии с настоящим изобретением термин «кодон» относится к триплету оснований в кодирующей нуклеиновой кислоте, которая указывает, какая аминокислота будет добавляться далее во время синтеза белка на рибосоме.
Термины «транскрипция» и «транскрибирование» относятся к процессу, в ходе которого молекула нуклеиновой кислоты с определенной последовательностью нуклеиновой кислоты («матрица нуклеиновой кислоты») считывается РНК-полимеразой, так что РНК-полимераза производит одноцепочечную молекулу РНК. Во время транскрипции генетическая информация в матрице нуклеиновой кислоты транскрибируется. Матрица нуклеиновой кислоты может представлять собой ДНК; однако, например, в случае транскрипции из матрицы альфавирусной нуклеиновой кислоты матрица, как правило, представляет собой РНК. Впоследствии транскрибируемая РНК может быть транслирована в белок. В соответствии с настоящим изобретением термин «транскрипция» включает в себя «транскрипцию in vitro», причем термин «транскрипция in vitro» относится к процессу, при котором РНК, в частности мРНК, синтезируется in vitro в бесклеточной системе. Предпочтительно векторы клонирования применяются для производства транскриптов. Эти векторы клонирования, как правило, обозначаются как векторы транскрипции и согласно настоящему изобретению охватываются термином «вектор». Векторы клонирования предпочтительно представляют собой плазмиды. В соответствии с настоящим изобретением РНК предпочтительно представляет собой транскрибированную in vitro РНК (IVT-РНК) и может быть получена посредством транскрипции in vitro соответствующей матрицы ДНК. Промотором для контроля транскрипции может быть любой промотор для любой РНК-полимеразы. Матрица ДНК для транскрипции in vitro может быть получена путем клонирования нуклеиновой кислоты, в частности кДНК, и введения ее в соответствующий вектор для транскрипции in vitro. кДНК может быть получена путем обратной транскрипции РНК.
Одноцепочечная молекула нуклеиновой кислоты, полученная во время транскрипции, как правило, содержит последовательность нуклеиновой кислоты, которая представляет собой комплементарную последовательность по отношению к матрице.
В соответствии с настоящим изобретению термины «матрица» или «матрица нуклеиновой кислоты» или «матричная нуклеиновая кислота», как правило, относятся к последовательности нуклеиновой кислоты, которая может быть реплицирована или транскрибирована.
«Последовательность нуклеиновой кислоты, транскрибируемая из последовательности нуклеиновой кислоты» и сходные термины относятся к последовательности нуклеиновой кислоты, где это применимо, в виде части полной молекулы РНК, которая представляет собой продукт транскрипции последовательности-матрицы нуклеиновой кислоты. Как правило, транскрибированная последовательность нуклеиновой кислоты представляет собой одноцепочечную молекулу РНК.
«3'-конец нуклеиновой кислоты» относится в соответствии с настоящим изобретением к тому концу, который содержит свободную гидроксигруппу. В схематическом представлении двухцепочечных нуклеиновых кислот, в частности ДНК, 3'-конец всегда находится с правой стороны. «5'-конец нуклеиновой кислоты» относится в соответствии с настоящим изобретением к тому концу, который содержит свободную фосфатную группу. В схематическом представлении двухцепочечных нуклеиновых кислот, в частности ДНК, 5'-конец всегда находится с левой стороны.
5'-конец 5'--P-NNNNNNN-OH-3' 3'-конец
3'-HO-NNNNNNN-P--5'
«Выше против хода транскрипции» описывает относительное позиционирование первого элемента молекулы нуклеиновой кислоты по отношению ко второму элементу этой молекулы нуклеиновой кислоты, причем оба элемента содержатся в одной и той же молекуле нуклеиновой кислоты, причем первый элемент расположен ближе к 5'-концу молекулы нуклеиновой кислоты, чем второй элемент этой молекулы нуклеиновой кислоты. Второй элемент потому называется как находящийся «ниже по ходу транскрипции» от первого элемента этой молекулы нуклеиновой кислоты. Элемент, который расположен «выше против хода транскрипции» от второго элемента, может быть синонимично обозначен как расположенный «5'» от этого второго элемента. Для двухцепочечной молекулы нуклеиновой кислоты указания относительно «выше против хода транскрипции» и «ниже по ходу транскрипции» даются относительно (+)-нити.
В соответствии с настоящим изобретением «функциональная связь» или «функционально связанный» относится к соединению в рамках функциональной связи. Нуклеиновая кислота «функционально связана», если между ней и другой последовательностью нуклеиновой кислоты установлена функциональная связь. Например, промотор функционально связан с кодирующей последовательностью, если он влияет на транскрипцию указанной кодирующей последовательности. Функционально связанные нуклеиновые кислоты, как правило, примыкают друг к другу, где соответствующие нуклеиновые кислоты отделены друг от друга дополнительными последовательностями нуклеиновых кислот и, согласно конкретным вариантам осуществления, транскрибируются РНК-полимеразой с получением одной молекулы РНК (общий транскрипт).
Согласно конкретным вариантам осуществления нуклеиновая кислота функционально связана в соответствии с настоящим изобретением с последовательностями контроля экспрессии, которые могут быть гомологичными или гетерологичными по отношению к нуклеиновой кислоте.
Термин «последовательность контроля экспрессии» включает в себя согласно настоящему изобретению промоторы, связывающие рибосомы последовательности и другие контрольные элементы, которые контролируют транскрипцию гена или трансляцию полученной РНК. Согласно конкретным вариантам осуществления настоящего изобретения последовательности контроля экспрессии можно регулировать. Точная структура последовательностей контроля экспрессии может варьировать в зависимости от вида или типа клеток, но, как правило, включает в себя 5'-нетранскрибируемые и 5'- и 3'-нетранслируемые последовательности, участвующие в инициации транскрипции и трансляции, соответственно. Более конкретно, 5'-нетранскрибируемые последовательности контроля экспрессии включают в себя промоторную область, которая охватывает последовательность промотора для контроля транскрипции функционально связанного гена. Последовательности контроля экспрессии могут также включать в себя энхансерные последовательности или последовательности активаторов выше против хода транскрипции. Последовательность контроля экспрессии молекулы ДНК, как правило, включает в себя 5'-нетранскрибируемые и 5'- и 3'-нетранслируемые последовательности, такие как TATA-бокс, кэпирующую последовательность, последовательность CAAT и тому подобное. Последовательность контроля экспрессии альфавирусной РНК может включать в себя субгеномный промотор и/или один или несколько элементов консервативной последовательности. Конкретная последовательность контроля экспрессии в соответствии с настоящим изобретением представляет собой субгеномный промотор альфавируса, как описано в настоящем документе.
Указанные в настоящем документе последовательности нуклеиновой кислоты, в частности транскрибируемые и кодирующие последовательности нуклеиновой кислоты, могут быть объединены с любыми последовательностями контроля экспрессии, в частности промоторами, которые могут быть гомологичными или гетерологичными по отношению к указанным последовательностям нуклеиновых кислот, причем термин «гомологичный» относится к факту того, что последовательность нуклеиновой кислоты также функционально связана природным образом с последовательностью контроля экспрессии, а термин «гетерологичный» относится к тому факту, что последовательность нуклеиновой кислоты функционально не связана природным образом с последовательностью контроля экспрессии.
Транскрибируемая последовательность нуклеиновой кислоты, в частности последовательность нуклеиновой кислоты, кодирующая пептид или белок, и последовательность контроля экспрессии «функционально» связаны друг с другом, если они ковалентно связаны друг с другом таким образом, что транскрипция или экспрессия транскрибируемой и, в частности, кодирующей последовательности нуклеиновой кислоты находится под контролем или под влиянием последовательности контроля экспрессии. Если последовательность нуклеиновой кислоты подлежит трансляции в функциональный пептид или белок, индукция последовательности контроля экспрессии, функционально связанной с кодирующей последовательностью, приводит к транскрипции указанной кодирующей последовательности, не вызывая сдвига рамки в кодирующей последовательности, или кодирующая последовательность не может быть транслирована в желаемый пептид или белок.
Термин «промотор» или «область промотора» относится к последовательности нуклеиновой кислоты, которая контролирует синтез транскрипта, например, транскрипта, содержащего кодирующую последовательность, путем предоставления сайта распознавания и связывания для РНК-полимеразы. Область промотора может включать в себя дополнительные сайты распознавания или связывания для дополнительных факторов, связанных с регуляцией транскрипции указанного гена. Промотор может контролировать транскрипцию прокариотического или эукариотического гена. Промотор может быть «индуцируемым» и инициировать транскрипцию в ответ на индуктор или быть «конститутивным», если транскрипция не контролируется индуктором. Индуцируемый промотор экспрессируется только в очень малой степени или вообще не экспрессируется, если индуктор отсутствует. В присутствии индуктора ген «включен» или уровень транскрипции увеличивается. Это, как правило, опосредуется связыванием специфического фактора транскрипции. Специфический промотор в соответствии с настоящим изобретением представляет собой субгеномный промотор альфавируса, как описано в настоящем документе. Другими специфическими промоторами являются геномные плюс-нитевые или минус-нитевые промоторы альфавируса.
Термин «основной промотор» относится к последовательности нуклеиновой кислоты, которая состоит из промотора. Основной промотор, как правило, представляет собой минимальную часть промотора, необходимую для правильной инициации транскрипции. Основной промотор, как правило, включает в себя сайт начала транскрипции и сайт связывания для РНК-полимеразы.
«Полимераза», как правило, относится к молекулярному объекту, способному катализировать синтез полимерной молекулы из мономерных структурных блоков. «РНК-полимераза» представляет собой молекулярный объект, способный катализировать синтез молекулы РНК из рибонуклеотидных структурных блоков. «ДНК-полимераза» представляет собой молекулярный объект, способный катализировать синтез молекулы ДНК из дезоксирибонуклеотидных структурных блоков. В случае ДНК-полимераз и РНК-полимераз молекулярный объект, как правило, представляет собой белок или сборку или комплекс из нескольких белков. Как правило, ДНК-полимераза синтезирует молекулу ДНК на основе матричной нуклеиновой кислоты, которая, как правило, представляет собой молекулу ДНК. Как правило, РНК-полимераза синтезирует молекулу РНК на основе матричной нуклеиновой кислоты, которая представляет собой либо молекулу ДНК (в этом случае РНК-полимераза представляет собой ДНК-зависимую РНК-полимеразу, DdRP), либо представляет собой молекулу РНК (в этом случае РНК полимераза представляет собой РНК-зависимую РНК-полимеразу, RdRP).
«РНК-зависимая РНК-полимераза» или «RdRP» представляет собой фермент, который катализирует транскрипцию РНК из матрицы РНК. В случае альфавирусной РНК-зависимой РНК-полимеразы последовательный синтез (-)-нитевого комплемента геномной РНК и (+)-нитевой геномной РНК приводит к репликации РНК. Таким образом, альфавирусная РНК-зависимая РНК-полимераза синонимично называется «РНК-репликазой». В природе РНК-зависимые РНК-полимеразы, как правило, кодируются всеми РНК-вирусами, кроме ретровирусов. Типичными представителями вирусов, кодирующих РНК-зависимую РНК-полимеразу, являются альфавирусы.
В соответствии с настоящим изобретением «репликация РНК», как правило, относится к молекуле РНК, синтезированной на основе нуклеотидной последовательности данной молекулы РНК (молекула-матрица РНК). Молекула РНК, которая синтезируется, может быть, например, идентичной или комплементарной молекуле-матрице РНК. В общем случае репликация РНК может происходить посредством синтеза промежуточного соединения ДНК или может происходить непосредственно с помощью РНК-зависимой репликации РНК, опосредованной РНК-зависимой РНК-полимеразой (RdRP). В случае альфавирусов репликация РНК не происходит через промежуточное звено ДНК, но опосредуется РНК-зависимой РНК-полимеразой (RdRP): матричная цепь РНК (первая цепь РНК) - или ее часть - служит в качестве матрицы для синтеза второй цепи РНК, которая является комплементарным первой цепи РНК или ее части. Вторая цепь РНК - или ее часть - может, в свою очередь, необязательно служить в качестве матрицы для синтеза третьей цепи РНК, которая комплементарна второй цепи РНК или ее части. Таким образом, третья цепь РНК идентична первой цепи РНК или ее части. Таким образом, РНК-зависимая РНК-полимераза способна непосредственно синтезировать комплементарную цепь РНК-матрицы и косвенно синтезировать идентичную цепь РНК (через комплементарную промежуточную цепь).
В соответствии с настоящим изобретением термин «РНК-матрица» относится к РНК, которая может быть транскрибирована или реплицирована РНК-зависимой РНК-полимеразой.
Согласно настоящему изобретению термин «ген» относится к определенной последовательности нуклеиновой кислоты, которая отвечает за производство одного или нескольких клеточных продуктов и/или за достижение одной или нескольких межклеточных или внутриклеточных функций. Более конкретно, указанный термин относится к сектору нуклеиновой кислоты (как правило, ДНК, но РНК в случае РНК-вирусов), который содержит нуклеиновую кислоту, кодирующую специфический белок или функциональную или структурную молекулу РНК.
Используемая в настоящем документе «выделенная молекула» предназначена для обозначения молекулы, которая по существу не содержит других молекул, таких как другой клеточный материал. Термин «выделенная нуклеиновая кислота» означает в соответствии с настоящим изобретением, что нуклеиновая кислота была (i) амплифицирована in vitro, например, посредством полимеразной цепной реакции (ПЦР), (ii) рекомбинантно произведена клонированием, (iii) очищена, например путем расщепления и гель-электрофоретического фракционирования, или (iv) синтезирована, например, путем химического синтеза. Выделенная нуклеиновая кислота представляет собой нуклеиновую кислоту, доступную для манипуляции рекомбинантными способами.
Термин «вектор» используется в настоящем документе в его самом общем значении и включает в себя любые промежуточные носители для нуклеиновой кислоты, которые, например, позволяют вводить указанную нуклеиновую кислоту в прокариотические и/или эукариотические клетки-хозяева и, при необходимости, интегрироваться в геном. Такие векторы предпочтительно реплицируются и/или экспрессируются в клетке. Векторы содержат плазмиды, фагемиды, вирусные геномы и их фракции.
Термин «рекомбинантный» в контексте настоящего изобретения означает «сделанный с помощью генной инженерии». Предпочтительно, «рекомбинантный объект», такой как рекомбинантная клетка в контексте настоящего изобретения, является неприродным.
Используемый в настоящем документе термин «природный» относится к тому факту, что объект может быть обнаружен в природе. Например, пептид или нуклеиновая кислота, которая присутствует в организме (включая в себя вирусы) и может быть выделена из источника в природе и которая не была намеренно модифицирована человеком в лаборатории является природной. Термин «обнаруженный в природе» означает «присутствующий в природе» и включает в себя известные объекты, а также объекты, которые еще не были обнаружены в природе и/или выделены из природного источника, но которые могут быть обнаружены и/или выделены в будущем из природного источника.
В соответствии с настоящим изобретением термин «экспрессия» используется в его наиболее общем значении и включает в себя получение РНК или РНК и белка. Он также включает в себя частичную экспрессию нуклеиновых кислот. Кроме того, экспрессия может быть временной или стабильной. Что касается РНК, термин «экспрессия» или «трансляция» относится к процессу в рибосомах клетки, с помощью которой нить кодирующей РНК (например, матричная информационной РНК) направляет сборку последовательности аминокислот для получения пептида или белка.
В соответствии с настоящим изобретением термин «мРНК» означает «матричную РНК» и относится к транскрипту, который, как правило, производится с использованием ДНК-матрицы и кодирует пептид или белок. Как правило, мРНК содержит 5'-UTR, область кодирования белка, 3'-UTR и поли(А)-последовательность. мРНК может быть получена путем транскрипции in vitro из ДНК-матрицы. Специалисту в настоящей области техники известен способ транскрипции in vitro. Например, существует множество коммерчески доступных наборов транскрипции in vitro. В соответствии с настоящим изобретением мРНК может быть модифицирована путем стабилизации модификаций и кэпирования.
В соответствии с настоящим изобретением термины «поли(А)-последовательность» или «поли(А)-хвост» относятся к непрерывной или прерывающейся последовательности остатков аденилата, которая, как правило, расположена на 3'-конце молекулы РНК. Непрерывная последовательность характеризуется последовательными остатками аденилата. В природе характерна непрерывная поли(А)-последовательность. Хотя поли(А)-последовательность, как правило, не кодируется в эукариотической ДНК, но прикрепляется при транскрипции эукариот в ядре клетки к свободному 3'-концу РНК с помощью независимой от матрицы РНК-полимеразы после транскрипции, настоящее изобретение охватывает поли(А)-последовательности, кодируемые ДНК.
В соответствии с настоящим изобретением термин «первичная структура» применительно к молекуле нуклеиновой кислоты относится к линейной последовательности нуклеотидных мономеров.
В соответствии с настоящим изобретением термин «вторичная структура» применительно к молекуле нуклеиновой кислоты относится к двумерному представлению молекулы нуклеиновой кислоты, которая отражает спаривание оснований; например, в случае однонитевой молекулы РНК, в частности, спаривание внутримолекулярных оснований. Хотя каждая молекула РНК содержит только одну полинуклеотидную цепь, молекула, как правило, характеризуется областями (внутримолекулярными) пар оснований. В соответствии с настоящим изобретением термин «вторичная структура» предусматривает структурные мотивы, включающие в себя, без ограничения, пары оснований, структуры «стебли», структуры «петля-на-стебле», выпуклости, петли, такие как внутренние петли и петли с многократным ветвлением. Вторичная структура молекулы нуклеиновой кислоты может быть представлена двумерным рисунком (плоским графом) с показанными спариваниями оснований (для получения дополнительной информации о вторичной структуре молекул РНК смотрите публикацию Auber et al., J. Graph Algorithms Appl., 2006, vol. 10, pp. 329-351). Как описано в настоящем документе, вторичная структура некоторых молекул РНК имеет значение в контексте настоящего изобретения.
В соответствии с настоящим изобретением вторичная структура молекулы нуклеиновой кислоты, в частности одноцепочечной молекулы РНК, определяется путем прогнозирования с использованием веб-сервера для прогнозирования вторичной структуры РНК (http://rna.urmc.rochester.edu/RNAstructureWeb/Servers/Predict1/Predict1.html). Предпочтительно, в соответствии с настоящим изобретением «вторичная структура», применительно к молекуле нуклеиновой кислоты, конкретно относится ко вторичной структуре, определенной указанным прогнозированием. Прогнозирование также может быть выполнено или подтверждено с использованием прогнозирования структуры MFOLD (http://unafold.rna.albany.edu/?q=mfold).
В соответствии с настоящим изобретением «пара оснований» представляет собой структурный мотив вторичной структуры, в котором два нуклеотидных основания связываются друг с другом через водородные связи между донорными и акцепторными сайтами на основаниях. Комплементарные основания A:U и G:C образуют устойчивые пары оснований через водородные связи между донорными и акцепторными сайтами на основаниях; пары оснований A:U и G:C называются Уотсон-Криковскими парами оснований. Более слабая пара оснований (так называемая неоднозначная пара оснований) образована основаниями G и U (G:U). Пары оснований A:U и G:C называются каноническими парами оснований. Другие пары оснований, такие как G:U (которые довольно часто встречаются в РНК) и другие редкие пары оснований (например, A:C; U:U) называются неканоническими парами оснований.
В соответствии с настоящим изобретением «нуклеотидное спаривание» относится к двум нуклеотидам, которые связываются друг с другом, так что их основания образуют пару оснований (каноническую или неканоническую пару оснований, предпочтительно каноническую пару оснований, наиболее предпочтительно Уотсон-Криковскую пару оснований).
В соответствии с настоящим изобретением термины «петля-на-стебле» или «шпилька» или «петля-шпилька» применительно к молекуле нуклеиновой кислоты, все взаимозаменяемо относятся к конкретной вторичной структуре молекулы нуклеиновой кислоты, как правило, одноцепочечной молекулы нуклеиновой кислоты, такой как одноцепочечная РНК. Конкретная вторичная структура, представленная структурой «петля-на-стебле», состоит из последовательной последовательности нуклеиновой кислоты, содержащей стебель и (концевую) петлю, также называемую петлей-шпилькой, причем стебель образован двумя соседними полностью или частично комплементарными элементами последовательности; которые разделены короткой последовательностью (например, 3-10 нуклеотидами), которая образует петлю структуры «петля-на-стебле». Две соседние полностью или частично комплементарные последовательности могут быть определены как, например, элементы структуры «петля-на-стебле» стебель 1 и стебель 2. Структура «петля-на-стебле» образуется, когда эти две соседние полностью или частично обратные комплементарные последовательности, например, элементы структуры «петля-на-стебле» стебель 1 и стебель 2 образуют пары оснований друг с другом, что приводит к двухцепочечной последовательности нуклеиновой кислоты, содержащей неспаренную петлю на ее конце, образованную короткой последовательностью, расположенной между элементами структуры «петля-на-стебле» стебель 1 и стебель 2. Таким образом, структура «петля-на-стебле» состоит из двух стеблей (стебель 1 и стебель 2), которые на уровне вторичной структуры молекулы нуклеиновой кислоты образуют пары оснований друг с другом и которые - на уровне первичной структуры молекулы нуклеиновой кислоты - разделены короткой последовательностью, которая не является частью стебля 1 или стебля 2. Для иллюстрации двумерное представление структуры «петля-на-стебле» напоминает структуру в форме леденца. Образование структуры «петля-на-стебле» требует наличия последовательности, которая может укладываться сама по себе, чтобы образовать спаренную двойную нить; спаренная двойная нить образуется стеблем 1 и стеблем 2. Стабильность спаренных элементов структуры «петля-на-стебле», как правило, определяется длиной, количеством нуклеотидов стебля 1, которые способны образовывать пары оснований (предпочтительно канонические пары оснований, более предпочтительно Уотсон-Криковские пары оснований) с нуклеотидами стебля 2, по сравнению с количеством нуклеотидов стебля 1, которые не способны образовывать такие пары оснований с нуклеотидами стебля 2 (несоответствия или выпуклости). В соответствии с настоящим изобретением оптимальная длина петли составляет 3-10 нуклеотидов, более предпочтительно 4-7 нуклеотидов, например, 4 нуклеотида, 5 нуклеотидов, 6 нуклеотидов или 7 нуклеотидов. Если заданная последовательность нуклеиновой кислоты характеризуется структурой «петля-на-стебле», соответствующая комплементарная последовательность нуклеиновой кислоты, как правило, также характеризуется структурой «петля-на-стебле». Структура «петля-на-стебле», как правило, образуется одноцепочечными молекулами РНК. Например, несколько структур «петля-на-стебле» присутствуют в 5' репликационной последовательности распознавания альфавирусной геномной РНК (показано на фиг. 1).
В соответствии с настоящим изобретением «нарушение» или «нарушать» применительно к конкретной вторичной структуре молекулы нуклеиновой кислоты (например, структуре «петля-на-стебле») означает, что конкретная вторичная структура отсутствует или изменена. Как правило, вторичная структура может быть нарушена вследствие изменения по меньшей мере одного нуклеотида, который представляет собой часть вторичной структуры. Например, структура «петля-на-стебле»может быть нарушена посредством изменения одного или нескольких нуклеотидов, которые образуют стебель, так что нуклеотидное спаривание невозможно.
В соответствии с настоящим изобретением «компенсирует нарушение вторичной структуры» или «компенсация нарушения вторичной структуры» относится к одному или нескольким нуклеотидным изменениям в последовательности нуклеиновой кислоты; более типично это относится к одному или нескольким нуклеотидным изменениям в последовательности нуклеиновой кислоты, причем последовательность нуклеиновой кислоты также содержит одно или несколько первых нуклеотидных изменений, характеризующихся следующим образом: в то время как одно или несколько первых нуклеотидных изменений, в отсутствие одного или нескольких вторых нуклеотидных изменений, вызывают нарушение вторичной структуры последовательности нуклеиновой кислоты, совместное присутствие одного или нескольких первых нуклеотидных изменений и одного или несколько вторых нуклеотидных изменений не вызывает нарушения вторичной структуры нуклеиновой кислоты. Совместное присутствие означает наличие как одного или нескольких первых нуклеотидных изменений, так и одного или нескольких вторых нуклеотидных изменений. Как правило, одно или несколько первых нуклеотидных изменений и одно или несколько вторых нуклеотидных изменений присутствуют вместе в одной и той же молекуле нуклеиновой кислоты. Согласно конкретному варианту осуществления одно или несколько нуклеотидных изменений, которые компенсируют нарушение вторичной структуры, представляют собой одно или несколько нуклеотидных изменений, которые компенсируют одно или несколько нарушений спаривания нуклеотидов. Таким образом, согласно одному варианту осуществления «компенсация нарушения вторичной структуры» означает «компенсацию нарушений спаривания нуклеотидов», то есть одно или несколько нарушений спариваний нуклеотидов, например, одно или несколько нарушений спариваний нуклеотидов в пределах одной или нескольких структур «петля-на-стебле». Одно или несколько нарушений спаривания нуклеотидов, возможно, были введены путем удаления по меньшей мере одного инициирующего кодона. Каждое из одного или нескольких нуклеотидных изменений, которое компенсирует нарушение вторичной структуры, представляет собой нуклеотидное изменение, которое может быть независимо выбрано из делеции, добавления, замены и/или вставки одного или нескольких нуклеотидов. В иллюстративном примере, когда нуклеотидное спаривание A:U было нарушено заменой A на C (C и U, как правило, не подходят для образования пары нуклеотидов); то нуклеотидное изменение, которое компенсирует нарушение спаривания нуклеотидов, может представлять собой замену U на G, что позволяет образовать нуклеотидное спаривание C:G. Таким образом, замена U на G компенсирует нарушение спаривания нуклеотидов. В альтернативном примере, когда нуклеотидное спаривание A:U было нарушено заменой А на С; то нуклеотидное изменение, которое компенсирует нарушение нуклеотидного спаривания, может представлять собой замену C на A, тем самым восстанавливая образования исходного нуклеотидного спаривания A:U. В общем, согласно настоящему изобретению предпочтительными являются те нуклеотидные изменения, компенсирующие нарушение вторичной структуры, которые не восстанавливают исходную последовательность нуклеиновой кислоты и не создают новые триплеты AUG. В приведенном выше наборе примеров замена U на G является предпочтительной по сравнению с заменой C на A.
В соответствии с настоящим изобретением термин «третичная структура» применительно к молекуле нуклеиновой кислоты относится к трехмерной структуре молекулы нуклеиновой кислоты, определенной по атомным координатам.
В соответствии с настоящим изобретением такая нуклеиновая кислота, как РНК, например, мРНК, может кодировать пептид или белок. Соответственно, транскрибируемая последовательность нуклеиновой кислоты или ее транскрипт может содержать открытую рамку считывания (ORF), кодирующую пептид или белок.
В соответствии с настоящим изобретением термин «нуклеиновая кислота, кодирующая пептид или белок», означает, что нуклеиновая кислота, если она присутствует в подходящей окружающей среде, предпочтительно внутри клетки, может направлять сборку аминокислот для получения пептида или белка во время процесса трансляции. Предпочтительно, кодирующая РНК согласно настоящему изобретению способна взаимодействовать с механизмом трансляции клеток, обеспечивая трансляцию кодирующей РНК с получением пептида или белка.
В соответствии с настоящим изобретением термин «пептид» включает в себя олиго- и полипептиды и относится к веществам, которые содержат два или более, предпочтительно 3 или более, предпочтительно 4 или более, предпочтительно 6 или более, предпочтительно 8 или более, предпочтительно 10 или более, предпочтительно 13 или более, предпочтительно 16 или более, предпочтительно 20 или более и предпочтительно до 50, предпочтительно до 100 или предпочтительно до 150 последовательных аминокислот, связанных друг с другом посредством пептидных связей. Термин «белок» относится к большим пептидам, предпочтительно пептидам, содержащим по меньшей мере 151 аминокислоту, но термины «пептид» и «белок», как правило, используются в настоящем документе как синонимы.
Термины «пептид» и «белок» включают в себя в соответствии с настоящим изобретением вещества, которые содержат не только аминокислотные компоненты, но и неаминокислотные компоненты, такие как сахара и фосфатные структуры, а также содержат вещества, содержащие связи, такие как сложноэфирная, тиоэфирная или дисульфидная связи.
В соответствии с настоящим изобретением термины «инициирующий кодон» и «стартовый кодон» синонимически относятся к кодону (триплету оснований) молекулы РНК, который потенциально представляет собой первый кодон, который транслируется рибосомой. Такой кодон, как правило, кодирует аминокислоту метионин у эукариот и модифицированный метионин у прокариот. Наиболее распространенным инициирующим кодоном у эукариот и прокариот является AUG. Если в настоящем документе специально не указано, что подразумевается отличный от AUG инициирующий кодон, термины «инициирующий кодон» и «стартовый кодон» применительно к молекуле РНК относятся к кодону AUG. В соответствии с настоящим изобретением термины «инициирующий кодон» и «стартовый кодон» также используются для обозначения соответствующего триплета оснований дезоксирибонуклеиновой кислоты, а именно триплета оснований, кодирующего инициирующий кодон РНК. Если инициирующим кодоном матричной РНК является AUG, триплет оснований, кодирующий AUG, представляет собой ATG. В соответствии с настоящим изобретением термины «инициирующий кодон» и «стартовый кодон» предпочтительно относятся к функциональному инициирующему кодону или стартовому кодону, то есть к инициирующему кодону или стартовому кодону, который используется или будет использоваться в качестве кодона рибосомой для начала трансляции. Могут существовать кодоны AUG в молекуле РНК, которые не используются в качестве кодонов рибосомой для начала трансляции, например, из-за короткого расстояния между кодоном и кэпом. Эти кодоны не охватываются терминами функционального инициирующего кодона или стартового кодона.
В соответствии с настоящим изобретением термины «стартовый кодон открытой рамки считывания» или «инициирующий кодон открытой рамки считывания» относятся к триплету оснований, который служит в качестве инициирующего кодона для синтеза белка в кодирующей последовательности, например, в кодирующей последовательности обнаруженной в природе молекулы нуклеиновой кислоты. В молекуле РНК стартовому кодону открытой рамки считывания часто предшествует 5'-нетранслируемая область (5'-UTR), хотя строго это не требуется.
В соответствии с настоящим изобретением термины «нативный стартовый кодон открытой рамки считывания» или «нативный инициирующий кодон открытой рамки считывания» относятся к триплету оснований, который служит в качестве инициирующего кодона для синтеза белка в нативной кодирующей последовательности. Нативная кодирующая последовательность может представлять собой, например, кодирующую последовательность обнаруженной в природе молекулы нуклеиновой кислоты. Согласно некоторым вариантам осуществления в настоящем изобретении предусмотрены варианты обнаруженных в природе молекул нуклеиновой кислоты, которые характеризуются тем, что нативный стартовый кодон (который присутствует в нативной кодирующей последовательности) был удален (так, что он отсутствует в вариантной молекуле нуклеиновой кислоты).
В соответствии с настоящим изобретением «первый AUG» означает находящийся выше против хода транскрипции относительно всех триплетов оснований AUG молекулы матричной РНК, предпочтительно находящийся выше против хода транскрипции относительно всех триплетов оснований AUG молекулы матричной РНК, который используется или будет использоваться в качестве кодона рибосомой для начала трансляции. Соответственно, «первый ATG» относится к триплету оснований ATG кодирующей последовательности ДНК, которая кодирует первый AUG. В некоторых случаях первый AUG молекулы мРНК представляет собой стартовый кодон открытой рамки считывания, то есть кодон, который используется в качестве стартового кодона во время рибосомного синтеза белка.
В соответствии с настоящим изобретением термины «имеется удаление» или «характеризуется удалением» и подобные термины применительно к определенному элементу варианта нуклеиновой кислоты означают, что указанный определенный элемент не является функциональным или не присутствует в нуклеиновой кислоте по сравнению с исходной молекулой нуклеиновой кислоты. Без ограничения, удаление может представлять собой удаление всего или части определенного элемента, замену всего или части определенного элемента или изменения функциональных или структурных свойств определенного элемента. Удаление функционального элемента последовательности нуклеиновой кислоты требует, чтобы функция не проявлялась в положении варианта нуклеиновой кислоты, содержащем удаление. Например, вариант РНК, характеризующийся удалением определенного инициирующего кодона, требует, чтобы рибосомный синтез белка не инициировался в положении варианта РНК, характеризующегося удалением. Удаление структурного элемента последовательности нуклеиновой кислоты требует, чтобы структурный элемент не присутствовал в положении варианта нуклеиновой кислоты, содержащем удаление. Например, вариант РНК, характеризующийся удалением определенного триплета оснований AUG, то есть триплета оснований AUG в определенном положении, может быть охарактеризован, например, путем удаления части или всего определенного триплета оснований AUG (например, ΔAUG) или путем замены одного или нескольких нуклеотидов (A, U, G) определенного триплета оснований AUG любым одним или несколькими различными нуклеотидами, так что в результате нуклеотидная последовательность варианта не содержит указанного триплета оснований AUG. Подходящими заменами одного нуклеотида являются те, которые преобразуют триплет оснований AUG в триплет оснований GUG, CUG или UUG, или в триплет оснований AAG, ACG или AGG, или в триплет оснований AUA, AUC или AUU. Могут быть выбраны, соответственно, подходящие замены большего количества нуклеотидов.
В соответствии с настоящим изобретением термин «альфавирус» следует понимать в широком смысле, и он включает в себя любую вирусную частицу, которая обладает характеристиками альфавирусов. Характеристики альфавируса включают в себя наличие (+)-нитевой РНК, которая кодирует генетическую информацию, подходящую для репликации в клетке-хозяине, включая в себя активность РНК-полимеразы. Дополнительные характеристики многих альфавирусов описаны, например, в публикации Strauss & Strauss, Microbiol. Rev., 1994, vol. 58, pp. 491-562. Термин «альфавирус» включает в себя альфавирус, обнаруженный в природе, а также любой его вариант или производное. Согласно некоторым вариантам осуществления вариант или производное не обнаружены в природе.
Согласно одному варианту осуществления альфавирус представляет собой альфавирус, обнаруженный в природе. Как правило, обнаруженный в природе альфавирус является инфекционным для любого одного или нескольких эукариотических организмов, таких как животное (включая в себя позвоночных, таких как человек, и членистоногих, таких как насекомое).
Альфавирус, обнаруженный в природе, предпочтительно выбирают из группы, состоящей из следующих вирусов: комплекс вирусов леса Барма (содержащий вирус леса Барма); комплекс восточного лошадиного энцефалита (содержащий семь антигенных типов вируса восточного лошадиного энцефалита); комплекс вирусов Мидделбурга (содержащий вирус Мидделбурга); комплекс вирусов Ндуму (содержащий вирус Ндуму); комплекс вирусов леса Семлики (содержащий вирус Бебару, вирус Чикунгунья, вирус Майаро и его подтип вирус Уна, вирус О'Нйонг Нйонг и его подтип вирус Игбо-Ора, вирус Росс-ривер и его подтипы вирус Бебару, вирус Гета, вирус Сагияма, вирус леса Семлики и его подтип вирус Me Tri); комплекс венесуэльского лошадиного энцефалита (содержащий вирус Кабассоу, вирус Эверглейдс, вирус Mosso das Pedras, вирус Мукамбо, вирус Paramana, вирус Пиксуна, вирус Рио-Негро, вирус Трокара и его подтип вирус Бижу-Бридж, вирус венесуэльского лошадиного энцефалита); комплекс западного лошадиного энцефалита (содержащий вирус Аура, вирус Бабанки, вирус Кызылагач, вирус Синдбис, вирус Окелбо, вирус Ватароа, вирус Багги-Крик, вирус Форт-Морган, вирус Хайлендс Джи, вирус западного лошадиного энцефалита); и некоторые неклассифицированные вирусы, включающие в себя вирус панкреатической болезни лосося; вирус нарколептической болезни; вирус южного морского слона; вирус Tonate. Более предпочтительно, чтобы альфавирус был выбран из группы, состоящей из комплекса вирусов леса Семлики (содержащего указанные выше типы вирусов, включая в себя вирус леса Семлики), комплекса западного лошадиного энцефалита (содержащего указанные выше типы вирусов, включая в себя вирус Синдбис), комплекса восточного лошадиного энцефалита (содержащего указанные выше типы вирусов), комплекса венесуэльского лошадиного энцефалита (содержащего указанные выше типы вирусов, включая вирус венесуэльского лошадиного энцефалита).
Согласно дополнительному предпочтительному варианту осуществления альфавирус представляет собой вирус леса Семлики. Согласно альтернативному дополнительному предпочтительному варианту осуществления альфавирус представляет собой вирус Синдбис. Согласно альтернативному дополнительному предпочтительному варианту осуществления альфавирус представляет собой вирус венесуэльского лошадиного энцефалита.
Согласно некоторым вариантам осуществления настоящего изобретения альфавирус представляет собой не обнаруженный в природе альфавирус. Как правило, не обнаруженный в природе альфавирус представляет собой вариант или производное обнаруженного в природе альфавируса, который отличается от обнаруженного в природе альфавируса по меньшей мере одной мутацией в нуклеотидной последовательности, то есть геномной РНК. Мутация в нуклеотидной последовательности может быть выбрана из вставки, замены или делеции одного или нескольких нуклеотидов по сравнению с обнаруженным в природе альфавирусом. Мутация в нуклеотидной последовательности может быть связана или не связана с мутацией в полипептиде или белке, кодируемом нуклеотидной последовательностью. Например, не обнаруженный в природе альфавирус может представлять собой аттенуированный альфавирус. Не обнаруженный в природе аттенуированный альфавирус представляет собой альфавирус, который, как правило, содержит по меньшей мере одну мутацию в своей нуклеотидной последовательности, благодаря которой он отличается от обнаруженного в природе альфавируса, и который либо не является инфекционным вообще, либо является инфекционным, но характеризуется более низкой болезнетворной способностью или вообще не характеризуется болезнетворной способностью. В качестве иллюстративного примера TC83 представляет собой аттенуированный альфавирус, который отличается от обнаруженного в природе вируса венесуэльского лошадиного энцефалита (VEEV) (McKinney et al., 1963, Am. J. Trop. Med. Hyg., 1963, vol. 12; pp. 597-603).
Представители рода альфавирусов также могут быть классифицированы на основе их относительных клинических особенностей у людей: альфавирусы, связанные прежде всего с энцефалитом, и альфавирусы, связанные прежде всего с лихорадкой, сыпью и полиартритом.
Термин «альфавирусный» означает обнаруженный в альфавирусе или происходящий от альфавируса, или полученный из альфавируса, например, посредством генной инженерии.
В соответствии с настоящим изобретением «SFV» означает вирус леса Семлики. В соответствии с настоящим изобретением «SIN» или «SINV» означает вирус Синдбис. В соответствии с настоящим изобретением «VEE» или «VEEV» означает вирус венесуэльского лошадиного энцефалита.
В соответствии с настоящим изобретением термин «альфавирус» относится к сущности происхождения из альфавируса. Для иллюстрации белок альфавируса может относиться к белку, который обнаружен в альфавирусе и/или к белку, который кодируется альфавирусом; и последовательность нуклеиновой кислоты альфавируса может относиться к последовательности нуклеиновой кислоты, которая обнаружена в альфавирусе и/или к последовательности нуклеиновой кислоты, которая кодируется альфавирусом. Предпочтительно, чтобы последовательность нуклеиновой кислоты «альфавируса» относилась к последовательности нуклеиновой кислоты «генома альфавируса» и/или «геномной РНК альфавируса».
В соответствии с настоящим изобретением термин «альфавирусная РНК» относится к любой одной или нескольким из альфавирусной геномной РНК (т.е. (+)-нити), комплементу альфавирусной геномной РНК (т.е. (-)-нити) и субгеномному транскрипту (т.е. (+)-нити) или фрагменту любого из них.
В соответствии с настоящим изобретением «геном альфавируса» относится к геномной (+)-нитевой РНК альфавируса.
В соответствии с настоящим изобретением термин «нативная последовательность альфавируса» и подобные термины, как правило, относятся к последовательности (например, нуклеиновой кислоты) встречающегося в природе альфавируса (обнаруженного в природе альфавируса). Согласно некоторым вариантам осуществления термин «нативная последовательность альфавируса» также включает в себя последовательность аттенуированного альфавируса.
В соответствии с настоящим изобретением термин «5' репликационная последовательность распознавания» предпочтительно относится к непрерывной последовательности нуклеиновой кислоты, предпочтительно к последовательности рибонуклеиновой кислоты, которая идентична или гомологична 5'-фрагменту генома альфавируса. «5' репликационная последовательность распознавания» представляет собой последовательность нуклеиновой кислоты, которая может быть распознана альфавирусной репликазой. Термин 5' репликационная последовательность распознавания включает в себя нативные 5' репликационные последовательности распознавания, а также их функциональные эквиваленты, такие как, например, функциональные варианты 5' репликационной последовательности распознавания обнаруженной в природе альфавируса. В соответствии с настоящим изобретением функциональные эквиваленты включают в себя производные 5' репликационных последовательностей распознавания, характеризующиеся удалением по меньшей мере одного инициирующего кодона, как описано в настоящем документе. 5' репликационная последовательность распознавания необходима для синтеза (-)-нитевого комплемента альфавирусной геномной РНК и необходима для синтеза (+)-нити вирусной геномной РНК на основе (-)-нитевой матрицы. Нативная 5' репликационная последовательность распознавания, как правило, кодирует по меньшей мере N-концевой фрагмент nsP1; но не содержит всю открытую рамку считывания, кодирующую nsP1234. Ввиду того факта, что нативная 5' репликационная последовательность распознавания, как правило, кодирует по меньшей мере N-концевой фрагмент nsP1, нативная 5' репликационная последовательность распознавания, как правило, содержит по меньшей мере один инициирующий кодон, как правило, AUG. Согласно одному варианту осуществления 5' репликационная последовательность распознавания содержит консервативный элемент последовательности 1 (CSE 1) генома альфавируса или его вариант и консервативный элемент последовательности 2 (CSE 2) генома альфавируса или его вариант. 5' репликационная последовательность распознавания, как правило, способна образовывать четыре структуры «петля-на-стебле» (SL), то есть SL1, SL2, SL3, SL4. Нумерация этих структур «петля-на-стебле» начинается с 5'-конца 5' репликационной последовательности распознавания.
В соответствии с настоящим изобретением термин «на 5'-конце альфавируса» относится к 5'-концу генома альфавируса. Последовательность нуклеиновой кислоты на 5'-конце альфавируса охватывает нуклеотид, расположенный на 5'-конце геномной РНК альфавируса, плюс необязательно последовательную последовательность дополнительных нуклеотидов. Согласно одному варианту осуществления последовательность нуклеиновой кислоты на 5'-конце альфавируса идентична последовательности 5' репликационной последовательности распознавания генома альфавируса.
Термин «консервативный элемент последовательности» или «CSE» относится к нуклеотидной последовательности, обнаруженной в альфавирусной РНК. Эти элементы последовательности называются «консервативными», поскольку ортологи присутствуют в геноме разных альфавирусов, а ортологические CSE разных альфавирусов предпочтительно характеризуются высоким процентом идентичности последовательности и/или аналогичной вторичной или третичной структурой. Термин CSE включает в себя CSE 1, CSE 2, CSE 3 и CSE 4.
В соответствии с настоящим изобретением «CSE 1» или «CSE размером 44 нуклеотида» синонимично относятся к нуклеотидной последовательности, которая требуется для синтеза (+)-нити из (-)-нити. Термин «CSE 1» относится к последовательности на (+)-нити, а комплементарная последовательность CSE 1 (на (-)-нити) функционирует как промотор для синтеза (+)-нити. Предпочтительно, термин CSE 1 включает в себя самый 5' нуклеотид генома альфавируса. CSE 1, как правило, образует консервативную структуру «петля-на-стебле». Без ограничения конкретной теорией считается, что для CSE 1 вторичная структура важнее первичной структуры, то есть линейной последовательности. В геномной РНК модельного относящегося к альфавирусам вируса Синдбис CSE 1 состоит из последовательной последовательности из 44 нуклеотидов, которая образована расположенными наиболее 5' 44 нуклеотидами геномной РНК (Strauss & Strauss, Microbiol. Rev., 1994, vol. 58 , pp. 491-562).
В соответствии с настоящим изобретением термины «CSE 2» или «CSE размером 51 нуклеотид» синонимично относятся к нуклеотидной последовательности, которая требуется для синтеза (-)-нити из (+)-нитевой матрицы. (+)-нитевая матрица представляет собой, как правило, геномную РНК альфавируса или РНК-репликон (следует обратить внимание, что субгеномный РНК-транскрипт, который не содержит CSE 2, не функционирует как матрица для синтеза (-)-нити). В альфавирусной геномной РНК CSE 2, как правило, локализуется в кодирующей последовательности для nsP1. В геномной РНК модельного относящегося к альфавирусам вируса Синдбис CSE из 51 нуклеотида находится в нуклеотидных положениях 155-205 геномной РНК (Frolov et al., 2001, RNA, vol. 7, pp. 1638-1651). CSE 2, как правило, представляет собой две консервативные структуры «петля-на-стебле». Эти структуры «петля-на-стебле»обозначаются как «петля-на-стебле» 3 (SL3) и «петля-на-стебле» 4 (SL4), потому что они являются третьей и четвертой консервативными структурами «петля-на-стебле», соответственно, альфавирусной геномной РНК, отсчитываемой от 5'-конца геномной РНК альфавируса. Без ограничения какой-либо конкретной теорией считается, что для CSE 2 вторичная структура важнее первичной структуры, то есть линейной последовательности.
Согласно настоящему изобретению термины «CSE 3» или «последовательность места соединения» синонимически относятся к нуклеотидной последовательности, которая происходит из альфавирусной геномной РНК и которая содержит стартовый сайт субгеномной РНК. Комплемент этой последовательности в (-)-нити действует для содействия субгеномной транскрипции РНК. В альфавирусной геномной РНК CSE 3, как правило, перекрывается с областью, кодирующей C-концевой фрагмент nsP4, и продолжается до короткой некодирующей области, расположенной выше по ходу транскрипции от открытой рамки считывания, кодирующей структурные белки. Согласно публикации Strauss & Strauss (Microbiol. Rev., 1994, vol. 58, pp. 491-562), CSE 3 характеризуется консенсусной последовательностью
ACCUCUACGGCGGUCCUAAAUAGG (SEQ ID NO: 1, консенсусная последовательность места соединения (консенсус CSE 3), подчеркнутые нуклеотиды представляют собой первые пять нуклеотидов субгеномного транскрипта)
Согласно одному варианту осуществления настоящего изобретения CSE 3 состоит из SEQ ID NO: 1 или его варианта или содержит их, причем вариант предпочтительно характеризуется степенью идентичности последовательности по отношению к SEQ ID NO: 1, составляющей 80% или более, 85% или более, 90% или более, 95% или более, 96% или более, 97% или более, 98% или более или 99% или более.
В соответствии с настоящим изобретением термины «CSE 4» или «консервативная последовательность размером 19 нуклеотидов», или «CSE из 19 нуклеотидов» синонимически относятся к нуклеотидной последовательности из альфавирусной геномной РНК, непосредственно перед поли(А)-последовательностью в 3'нетранслируемой области генома альфавируса. CSE 4, как правило, состоит из 19 последовательных нуклеотидов. Без ограничения конкретной теорией CSE 4 понимается как функционирующий в качестве основного промотора для инициации синтеза (-)-нитей (José et al., Future Microbiol., 2009, vol. 4, pp. 837-856) и/или CSE 4 и поли(А)-хвост геномной РНК альфавируса понимаются как функционирующие совместно для эффективного синтеза (-)-нитей (Hardy & Rice, J. Virol., 2005, vol. 79, pp. 4630-4639 ).
Согласно Strauss & Strauss, CSE 4 характеризуется консервативной последовательностью AUUUUGUUUUUAAUAUUUC (SEQ ID NO: 2; консервативная последовательность из 19 нуклеотидов)
Согласно одному варианту осуществления настоящего изобретения CSE 4 состоит из SEQ ID NO: 2 или его варианта или содержит их, причем вариант предпочтительно характеризуется степенью идентичности последовательности по отношению к SEQ ID NO: 2, составляющей 80% или более, 85% или более, 90% или более, 95% или более, 96% или более, 97% или более, 98% или более, или 99% или более.
В соответствии с настоящим изобретением термин «субгеномный промотор» или «SGP» относится к последовательности нуклеиновой кислоты выше против хода транскрипции (5') от последовательности нуклеиновой кислоты (например, кодирующей последовательности), которая контролирует транскрипцию указанной последовательности нуклеиновой кислоты путем обеспечения сайта распознавания и связывания для РНК-полимеразы, как правило, РНК-зависимой РНК-полимеразы, в частности функционального альфавирусного неструктурного белка. SGP может включать в себя дополнительные сайты распознавания или связывания для дополнительных факторов. Субгеномный промотор, как правило, представляет собой генетический элемент вируса РНК с положительной нитью, такого как альфавирус. Субгеномный промотор альфавируса представляет собой последовательность нуклеиновой кислоты, содержащуюся в вирусной геномной РНК. Субгеномный промотор, как правило, характеризуется тем, что он позволяет инициировать транскрипцию (синтез РНК) в присутствии РНК-зависимой РНК-полимеразы, например, функционального альфавирусного неструктурного белка. (-)-нить РНК, то есть комплемент альфавирусной геномной РНК, служит в качестве матрицы для синтеза (+)-нити субгеномного транскрипта, а синтез (+)-нитевого субгеномного транскрипта, как правило, инициируется в субгеномном промоторе или вблизи него. Используемый в настоящем документе термин «субгеномный промотор» не ограничивается какой-либо конкретной локализацией в нуклеиновой кислоте, содержащей такой субгеномный промотор. Согласно некоторым вариантам осуществления SGP идентичен CSE 3 или перекрывается с CSE 3 или содержит CSE 3.
Термины «субгеномный транскрипт» или «субгеномная РНК» синонимично относятся к молекуле РНК, которая может быть получена в результате транскрипции с использованием молекулы РНК в качестве матрицы («РНК-матрица»), причем РНК-матрица содержит субгеномный промотор, который контролирует транскрипцию субгеномного транскрипта. Субгеномный транскрипт можно получить в присутствии РНК-зависимой РНК-полимеразы, в частности функционального альфавирусного неструктурного белка. Например, термин «субгеномный транскрипт» может относиться к транскрипту РНК, который получают в клетке, инфицированной альфавирусом, с использованием (-)-нитевого комплемента альфавирусной геномной РНК в качестве матрицы. Однако используемый в настоящем документе термин «субгеномный транскрипт» не ограничивается этим, а также включает в себя транскрипты, полученные с использованием гетерологичной РНК в качестве матрицы. Например, субгеномные транскрипты также могут быть получены с использованием (-)-нитевого комплемента SGP-содержащих репликонов в соответствии с настоящим изобретением в качестве матрицы. Таким образом, термин «субгеномный транскрипт» может относиться к молекуле РНК, которая может быть получена путем транскрибирования фрагмента геномной РНК альфавируса, а также к молекуле РНК, которая может быть получена путем транскрибирования фрагмента репликона в соответствии с настоящим изобретением.
Термин «аутологичный» используется для описания всего, что получают от одного и того же субъекта. Например, «аутологичная клетка» относится к клетке, полученной от одного и того же субъекта. Введение аутологичных клеток в субъект выгодно, поскольку эти клетки преодолевают иммунологический барьер, который в противном случае приводит к отторжению.
Термин «аллогенный» используется для описания всего, что получают от разных особей одного и того же вида. Два или более индивидуумов считаются аллогенными друг по отношению к другу, когда гены в одном или нескольких локусах не идентичны.
Термин «сингенный» используется для описания всего, что получают от индивидуумов или из тканей, имеющих идентичные генотипы, то есть идентичных близнецов или животных одного и того же инбредного штамма, или их тканей или клеток.
Термин «гетерологичный» используется для описания того, что состоит из нескольких разных элементов. В качестве примера, введение клетки одного индивидуума другим индивидуумам представляет собой гетерологичную трансплантацию. Гетерологичный ген представляет собой ген, полученный из источника, отличного от субъекта.
Ниже приводятся конкретные и/или предпочтительные варианты отдельных особенностей настоящего изобретения. Настоящее изобретение также рассматривает в качестве особенно предпочтительных вариантов осуществления те варианты осуществления, которые получают путем объединения двух или более конкретных и/или предпочтительных вариантов осуществления, описанных для двух или более особенностей настоящего изобретения.
РНК-репликон
Согласно первому аспекту в настоящем изобретении предусмотрен репликон, содержащий 5' репликационную последовательность распознавания, причем 5' репликационная последовательность распознавания характеризуется тем, что в ней имеется удаление по меньшей мере одного инициирующего кодона по сравнению с нативной альфавирусной 5' репликационной последовательностью распознавания. Репликон предпочтительно представляет собой РНК-репликон.
5' репликационная последовательность распознавания, которая характеризуется тем, что в ней имеется удаление по меньшей мере одного инициирующего кодона по сравнению с нативной альфавирусной 5' репликационной последовательностью распознавания в соответствии с настоящим изобретением, может упоминаться в настоящем документе как «модифицированная 5' репликационная последовательность распознавания» или «5' репликационная последовательность распознавания в соответствии с настоящим изобретением». Как описано ниже, 5' репликационная последовательность распознавания в соответствии с настоящим изобретением может быть необязательно охарактеризована наличием одного или нескольких дополнительных нуклеотидных изменений.
Конструкция нуклеиновой кислоты, которая может быть реплицирована репликазой, предпочтительно альфавирусной репликазой, называется репликоном. Согласно настоящему изобретению термин «репликон» определяет молекулу РНК, которая может быть реплицирована РНК-зависимой РНК-полимеразой, получая, без промежуточной ДНК, одну или несколько идентичных или по существу идентичных копий репликона РНК. «Без промежуточной ДНК» означает, что в процессе образования копий РНК-репликона не образуется копия или комплемент дезоксирибонуклеиновой кислоты (ДНК) репликона и/или что никакая молекула дезоксирибонуклеиновой кислоты (ДНК) не используется в качестве матрицы в процессе образования копий РНК-репликона или их комплемента. Функция репликазы, как правило, предоставляется функциональным альфавирусным неструктурным белком.
В соответствии с настоящим изобретением термины «может быть реплицирована» и «способна к репликации», как правило, описывают, что можно получить одну или несколько идентичных или по существу идентичных копий нуклеиновой кислоты. При использовании вместе с термином «репликаза», например, «способна к репликации репликазой», термины «может быть реплицирована» и «способна к репликации» описывают функциональные характеристики молекулы нуклеиновой кислоты, например, РНК-репликона, по отношению к репликазе. Эти функциональные характеристики предусматривают по меньшей мере одно из следующего: (i) репликаза способна распознавать репликон и (ii) репликаза способна действовать как РНК-зависимая РНК-полимераза (RdRP). Предпочтительно, репликаза способна как (i) распознавать репликон, так и (ii) действовать как РНК-зависимая РНК-полимераза.
Выражение «способна распознавать» описывает, что репликаза способна физически связываться с репликоном, и предпочтительно, что репликаза способна связываться с репликоном, как правило, нековалентно. Термин «связывание» может означать, что репликаза обладает способностью связываться с любым одним или несколькими из следующего: консервативный элемент последовательности 1 (CSE 1) или его комплементарная последовательность (если составлена репликоном), консервативный элемент последовательности 2 (CSE 2) или его комплементарная последовательность (если составлена репликоном), консервативный элемент последовательности 3 (CSE 3) или его комплементарная последовательность (если составлена репликоном), консервативный элемент последовательности 4 (CSE 4) или его комплементарная последовательность (если составлена репликоном). Предпочтительно репликаза способна связываться с CSE 2 [т.е. с (+)-нитью] и/или с CSE 4 [т.е. с (+)-нитью] или связываться с комплементом CSE 1 [т.е. с (-)-нитью] и/или с комплементом CSE 3 [т.е. с (-)-нитью].
Выражение «способное действовать как RdRP» означает, что репликаза способна катализировать синтез (-)-нитевого комплемента альфавирусной геномной (+)-нитевой РНК, причем (+)-нитевая РНК характеризуется функцией матрицы, и/или что репликаза способна катализировать синтез (+)-нитевой альфавирусной геномной РНК, причем (-)-нитевая РНК характеризуется функцией матрицы. В общем, выражение «способна действовать как RdRP» также может включать в себя то, что репликаза способна катализировать синтез (+)-нитевого субгеномного транскрипта, причем (-)-нитевая РНК характеризуется функцией матрицы и причем синтез (+)-нитевого субгеномного транскрипта, как правило, инициируется в субгеномном промоторе альфавируса.
Выражения «способны связываться» и «способны действовать как RdRP» относятся к способности в нормальных физиологических условиях. В частности, они относятся к условиям внутри клетки, которая экспрессирует функциональный альфавирусный неструктурный белок или которая была трансфицирована нуклеиновой кислотой, которая кодирует функциональный альфавирусный неструктурный белок. Клетка предпочтительно представляет собой эукариотическую клетку. Способность связываться и/или способность действовать как RdRP может быть экспериментально испытана, например, в бесклеточной системе in vitro или в эукариотической клетке. Необязательно, указанная эукариотическая клетка представляет собой клетку от вида, к которому конкретный альфавирус, от которого происходит репликаза, характеризуется инфекционностью. Например, когда используется альфавирусная репликаза из конкретного альфавируса, который является инфекционным для человека, нормальные физиологические условия представляют собой условия в клетке человека. Более предпочтительно, чтобы эукариотическая клетка (в одном примере человеческая клетка) происходила бы из той же ткани или органа, к которой конкретный альфавирус, от которого происходит репликаза, характеризуется инфекционностью.
В соответствии с настоящим изобретением «по сравнению с нативной альфавирусной последовательностью» и подобные термины относятся к последовательности, которая представляет собой вариант нативной альфавирусной последовательности. Вариант, как правило, сам не представляет собой нативную альфавирусную последовательность.
Согласно одному варианту осуществления РНК-репликон содержит 3' репликационную последовательность распознавания. 3' репликационная последовательность распознавания представляет собой последовательность нуклеиновой кислоты, которая может быть распознана функциональным альфавирусным неструктурным белком. Другими словами, функциональный альфавирусный неструктурный белок способен распознавать 3' репликационную последовательность распознавания. Предпочтительно, 3' репликационная последовательность распознавания расположена на 3'-конце репликона (если репликон не содержит поли(A)-хвост) или непосредственно выше против хода транскрипции от поли(A)-хвоста (если репликон содержит поли(А)-хвост). Согласно одному варианту осуществления 3' репликационная последовательность распознавания состоит из CSE 4 или содержит ее.
Согласно одному варианту осуществления 5' репликационная последовательность распознавания и 3' репликационная последовательность распознавания способны направлять репликацию РНК-репликона в соответствии с настоящим изобретением в присутствии функционального альфавирусного неструктурного белка. Таким образом, когда они присутствуют отдельно или, предпочтительно, вместе, эти последовательности распознавания направляют репликацию РНК-репликона в присутствии функционального альфавирусного неструктурного белка.
Предпочтительно, чтобы функциональный альфавирусный неструктурный белок был предусмотрен in cis (кодируемый как представляющий интерес белок посредством открытой рамки считывания на репликоне) или in trans (кодируемый как представляющий интерес белок посредством открытой рамки считывания на отдельной репликазной конструкции, как описано во втором аспекте), который способен распознавать как модифицированную 5' репликационную последовательность распознавания, так и 3' репликационную последовательность распознавания. Согласно одному варианту осуществления это достигается, когда 3' репликационная последовательность распознавания является нативной для альфавируса, из которой происходит функциональный альфавирусный неструктурный белок, и когда модифицированная 5' репликационная последовательность распознавания представляет собой вариант 5' репликационной последовательности распознавания, который является нативным для альфавируса, из которого получен функциональный альфавирусный неструктурный белок. Нативный означает, что природное происхождение этих последовательностей представлено одним и тем же альфавирусом. Согласно альтернативному варианту осуществления модифицированная 5' репликационная последовательность распознавания и/или 3' репликационная последовательность распознавания не являются нативными для альфавируса, из которого получен функциональный альфавирусный неструктурный белок, при условии, что функциональный альфавирусный неструктурный белок способен распознавать как модифицированную 5' репликационную последовательность распознавания, так и 3' репликационную последовательность распознавания репликона. Другими словами, функциональный альфавирусный неструктурный белок совместим с модифицированной 5' репликационной последовательностью распознавания и 3' репликационной последовательностью распознавания. Когда ненативный функциональный альфавирусный неструктурный белок способен распознавать соответствующую последовательность или элемент последовательности, функциональный альфавирусный неструктурный белок считается совместимым (межвирусная совместимость). Любая комбинация (3'/5') репликационных последовательностей распознавания и CSE, соответственно, с функциональным альфавирусным неструктурным белком возможна до тех пор, пока существует межвирусная совместимость. Межвирусная совместимость может быть легко исследована квалифицированным специалистом, работающим в соответствии с настоящим изобретением, путем инкубации подлежащего исследованию функционального альфавирусного неструктурного белка вместе с РНК, причем РНК содержит подлежащие исследованию 3'- и (необязательно модифицированные) 5'- репликационные последовательности распознавания, в условиях, подходящих для репликации РНК, например, в подходящей клетке-хозяине. Если происходит репликация, (3'/5') репликационные последовательности распознавания и функциональный альфавирусный неструктурный белок определяются как совместимые.
Удаление по меньшей мере одного инициирующего кодона обеспечивает несколько преимуществ по сравнению с известными транс-репликонами (такими как, например, представленные посредством «РНК-матрицы WT-RRS» на фиг. 1). Отсутствие инициирующего кодона в последовательности нуклеиновой кислоты, кодирующей nsP1*, как правило, будет приводить к тому, что nsP1* (N-концевой фрагмент nsP1) не будет транслироваться. Кроме того, поскольку nsP1* не транслируется, открытая рамка считывания, кодирующая представляющий интерес белок («трансген»), представляет собой расположенную выше всех против хода транскрипции открытую рамку считывания, доступную для рибосомы; таким образом, когда репликон присутствует в клетке, трансляция начинается на первом AUG открытой рамки считывания (РНК), кодирующей представляющий интерес ген. Это представляет собой преимущество перед транс-репликонами предшествующего уровня техники, такими как описанные в публикации Spuul et al. (J. Virol., 2011, vol. 85, pp. 4739-4751): репликоны согласно Spuul с соавт. направляют экспрессию N-концевой части nsP1, пептида из 74 аминокислот. Из предшествующего уровня техники известно также, что построение РНК-репликонов из полноразмерных вирусных геномов не является тривиальным, поскольку определенные мутации могут сделать РНК неспособной к репликации (публикация международной заявки WO 2000/053780 A2) и удаление некоторых частей структуры 5', которая важна для репликации альфавируса, влияет на эффективность репликации (Kamrud et al., 2010, J. Gen. Virol., vol. 91, pp. 1723-1727).
Преимущество перед цис-репликонами предшествующего уровня техники состоит в том, что удаление по меньшей мере одного инициирующего кодона разъединяет кодирующую область для альфавирусного неструктурного белка и 5' репликационную последовательность распознавания. Это позволяет дополнительно сконструировать цис-репликоны, например, путем замены нативной 5' репликационной последовательности распознавания на искусственную последовательность, мутированную последовательность или гетерологичную последовательность, взятую из другого РНК-вируса. Подобная манипуляция последовательностями в цис-репликонах предшествующего уровня техники ограничена аминокислотной последовательностью nsP1. Любая точечная мутация или кластеры точечных мутаций потребовала бы экспериментальной оценки влияния на репликацию, и небольшие вставки или делеции, приводящие к мутациям сдвига рамки считывания, невозможны из-за их пагубного воздействия на белок.
Удаление по меньшей мере одного инициирующего кодона согласно настоящему изобретению может быть достигнуто любым подходящим способом, известным в настоящей области техники. Например, подходящая молекула ДНК, кодирующая репликон в соответствии с настоящим изобретением, то есть характеризующаяся удалением инициирующего кодона, может быть разработана in silico и впоследствии синтезирована in vitro (синтез гена); альтернативно, подходящая молекула ДНК может быть получена путем сайт-направленного мутагенеза последовательности ДНК, кодирующей репликон. В любом случае соответствующая молекула ДНК может служить в качестве матрицы для транскрипции in vitro, тем самым обеспечивая репликон в соответствии с настоящим изобретением.
Удаление по меньшей мере одного инициирующего кодона по сравнению с нативной альфавирусной 5' репликационной последовательностью распознавания конкретно не ограничено и может быть выбрано из любой нуклеотидной модификации, включающей в себя замену одного или нескольких нуклеотидов (включая в себя, на уровне ДНК, замену А и/или T и/или G инициирующего кодона); удаление одного или нескольких нуклеотидов (включая в себя, на уровне ДНК, делецию A и/или T и/или G инициирующего кодона) и вставку одного или нескольких нуклеотидов (включая в себя, на уровне ДНК, вставку одного или нескольких нуклеотидов между A и T и/или между T и G инициирующего кодона). Независимо от того, представляет ли собой нуклеотидная модификация замену, вставку или делецию, нуклеотидная модификация не должна приводить к образованию нового инициирующего кодона (в качестве иллюстративного примера: вставка, на уровне ДНК, не должна представлять собой вставку ATG).
5' репликационная последовательность распознавания РНК-репликона, которая характеризуется удалением по меньшей мере одного инициирующего кодона (то есть модифицированная 5' репликационная последовательность распознавания в соответствии с настоящим изобретением), предпочтительно представляет собой вариант 5' репликационной последовательности распознавания генома обнаруженного в природе альфавируса. Согласно одному варианту осуществления модифицированная 5' репликационная последовательность распознавания в соответствии с настоящим изобретением предпочтительно характеризуется степенью идентичности последовательности, составляющей 80% или более, предпочтительно 85% или более, более предпочтительно 90% или более, еще более предпочтительно 95% или более, по отношению к 5' репликационной последовательности распознавания генома по меньшей мере одного обнаруженного в природе альфавируса.
Согласно одному варианту осуществления 5' репликационная последовательность распознавания РНК-репликона, которая характеризуется удалением по меньшей мере одного инициирующего кодона, содержит последовательность, гомологичную приблизительно 250 нуклеотидам на 5'-конце альфавируса, то есть на 5'-конце альфавирусного генома. Согласно предпочтительному варианту осуществления она содержит последовательность, гомологичную приблизительно 250-500, предпочтительно приблизительно 300-500 нуклеотидам на 5'-конце альфавируса, то есть на 5'-конце альфавирусного генома. «На 5'-конце альфавирусного генома» означает последовательность нуклеиновой кислоты, начинающуюся на нуклеотиде альфавирусного генома, расположенном выше всех против хода транскрипции, и включающую в себя его. Другими словами, находящийся выше всех против хода транскрипции нуклеотид альфавирусного генома обозначен нуклеотидом № 1 и, например, «250 нуклеотидов на 5'-конце альфавирусного генома» означают нуклеотиды от 1 до 250 альфавирусного генома. Согласно одному варианту осуществления 5'-репликационная последовательность распознавания РНК-репликона, которая характеризуется удалением по меньшей мере одного инициирующего кодона, характеризуется степенью идентичности последовательности, составляющей 80% или более, предпочтительно 85% или более, более предпочтительно 90% или более, еще более предпочтительно 95% или более, по отношению по меньшей мере к 250 нуклеотидам на 5'-конце генома по меньшей мере одного обнаруженного в природе альфавируса. По меньшей мере 250 нуклеотидов включают в себя, например, 250 нуклеотидов, 300 нуклеотидов, 400 нуклеотидов, 500 нуклеотидов.
5' репликационная последовательность распознавания обнаруженного в природе альфавируса, как правило, характеризуется по меньшей мере одним инициирующим кодоном и/или консервативными вторичными структурными мотивами. Например, нативная 5' репликационная последовательность распознавания вируса леса Семлики (SFV) содержит пять специфических триплетов оснований AUG, соответствующих триплетам оснований ATG в ДНК SEQ ID NO: 4 (смотрите пример 1). В соответствии с анализом Frolov et al. (2001, RNA, vol. 7, pp. 1638-1651) с помощью MFOLD показал, что нативная 5' репликационная последовательность распознавания вируса леса Семлики предположительно образует четыре структуры «петля-на-стебле» (SL), называемых структуры «петля-на-стебле» 1-4 (SL1, SL2, SL3, SL4). Согласно Frolov с соавт. анализ с помощью MFOLD показал, что также нативная 5' репликационная последовательность распознавания другого альфавируса, вируса Синдбис, предположительно будет образовывать четыре структуры «петля-на-стебле»: SL1, SL2, SL3, SL4. В примере 1 настоящего изобретения предсказанное присутствие четырех структур «петля-на-стебле»в нативной 5' репликационной последовательности распознавания вируса леса Семлики было подтверждено двумя веб-серверами для прогнозирования вторичной структуры РНК (http://rna.urmc.rochester.edu /RNAstructureWeb/Servers/Predict1/Predict1.html)
(http://unafold.rna.albany.edu/?q=mfold)
Известно, что 5'-конец альфавирусного генома содержит элементы последовательности, которые позволяют реплицировать альфавирусный геном функциональным альфавирусным неструктурным белком. Согласно одному варианту осуществления настоящего изобретения 5' репликационная последовательность распознавания РНК-репликона содержит последовательность, гомологичную консервативному элементу последовательности 1 (CSE 1), и/или последовательность, гомологичную консервативному элементу последовательности 2 (CSE 2) альфавируса.
Консервативный элемент последовательности 2 (CSE 2) геномной РНК альфавируса, как правило, представлен SL3 и SL4, которому предшествует SL2, содержащий по меньшей мере нативный инициирующий кодон, который кодирует первый аминокислотный остаток альфавирусного неструктурного белка nsP1. В этом описании, однако, согласно некоторым вариантам осуществления консервативный элемент последовательности 2 (CSE 2) геномной РНК альфавируса относится к области, охватывающей от SL2 до SL4 и содержащей нативный инициирующий кодон, который кодирует первый аминокислотный остаток альфавирусного неструктурного белка nsP1. Согласно предпочтительному варианту осуществления РНК-репликон содержит CSE 2 или последовательность, гомологичную CSE 2. Согласно одному варианту осуществления РНК-репликон содержит последовательность, гомологичную CSE 2, которая предпочтительно характеризуется степенью идентичности последовательности, составляющей 80% или более, предпочтительно 85% или более, более предпочтительно 90% или более, еще более предпочтительно 95% или более, по отношению к последовательности CSE 2 по меньшей мере одного обнаруженного в природе альфавируса.
Согласно предпочтительному варианту осуществления 5' репликационная последовательность распознавания содержит последовательность, которая гомологична CSE 2 альфавируса. CSE 2 альфавируса может содержать фрагмент открытой рамки считывания неструктурного белка из альфавируса.
Таким образом, согласно предпочтительному варианту осуществления РНК-репликон характеризуется тем, что он содержит последовательность, гомологичную открытой рамке считывания неструктурного белка или его фрагмента из альфавируса. Последовательность, гомологичная открытой рамке считывания неструктурного белка или его фрагмента, как правило, представляет собой вариант открытой рамки считывания неструктурного белка или его фрагмента обнаруженного в природе альфавируса. Согласно одному варианту осуществления последовательность, гомологичная открытой рамке считывания неструктурного белка или его фрагмента, предпочтительно характеризуется степенью идентичности последовательности, составляющей 80% или более, предпочтительно 85% или более, более предпочтительно 90% или более, еще более предпочтительно 95% или более, по отношению к открытой рамке считывания неструктурного белка или его фрагмента по меньшей мере одного обнаруженного в природе альфавируса.
Согласно более предпочтительному варианту осуществления последовательность, гомологичная открытой рамке считывания неструктурного белка, которая состоит из репликона по настоящему изобретению, не содержит нативный инициирующий кодон неструктурного белка и более предпочтительно не содержит любой инициирующий кодон неструктурного белка. Согласно предпочтительному варианту осуществления последовательность, гомологичная CSE 2, характеризуется удалением всех инициирующих кодонов по сравнению с последовательностью CSE 2 нативного альфавируса. Таким образом, последовательность, гомологичная CSE 2, предпочтительно не содержит никакого инициирующего кодона.
Когда последовательность, гомологичная открытой рамке считывания, не содержит какого-либо инициирующего кодона, последовательность, гомологичная открытой рамке считывания, сама по себе не является открытой рамкой считывания, поскольку она не служит в качестве матрицы для трансляции.
Согласно одному варианту осуществления 5' репликационная последовательность распознавания содержит последовательность, гомологичную открытой рамке считывания неструктурного белка или его фрагмента из альфавируса, причем последовательность, гомологичная открытой рамке считывания неструктурного белка или его фрагмента из альфавируса характеризуется тем, что в ней имеется удаление по меньшей мере одного инициирующего кодона по сравнению с нативной альфавирусной последовательностью.
Согласно предпочтительному варианту осуществления последовательность, гомологичная открытой рамке считывания неструктурного белка или его фрагмента из альфавируса, характеризуется тем, что в ней имеется удаление по меньшей мере нативного стартового кодона открытой рамки считывания неструктурного белка. Предпочтительно она характеризуется тем, что в ней имеется удаление по меньшей мере нативного стартового кодона открытой рамки считывания, кодирующей nsP1.
Нативный стартовый кодон представляет собой триплет оснований AUG, в котором трансляция на рибосомах в клетке-хозяине начинается, когда РНК присутствует в клетке-хозяине. Другими словами, нативный стартовый кодон представляет собой первый триплет оснований, который транслируется во время рибосомного синтеза белка, например, в клетке-хозяине, в которую была инокулирована РНК, содержащая нативный стартовый кодон. Согласно одному варианту осуществления клетка-хозяин представляет собой эукариотическую клетку, которая представляет собой природного хозяина конкретного альфавируса, который содержит нативную альфавирусную 5' репликационную последовательность распознавания. Согласно предпочтительному варианту осуществления клетка-хозяин представляет собой клетку BHK21 из клеточной линии «BHK21 [C13] (ATCC® CCL10™)», доступную из Американской коллекции типовых культур, Манассас, штат Вирджиния, США.
Геномы многих альфавирусов были полностью секвенированы и являются общедоступными, а также последовательности неструктурных белков, кодируемые этими геномами, общедоступны. Такая информация о последовательности позволяет определить нативный стартовый кодон in silico.
Для иллюстрации в примере 1 описан триплет оснований ДНК, соответствующий нативному стартовому кодону nsP1 SFV.
Согласно одному варианту осуществления нативный стартовый кодон состоит из последовательности Козак или функционально эквивалентной последовательности. Последовательность Козак представляет собой последовательность, первоначально описанную Kozak (1987, Nucleic Acids Res., vol. 15, pp. 8125-8148). Последовательность Козак на молекуле мРНК распознается рибосомой как сайт начала трансляции. Согласно этой ссылке последовательность Козак содержит стартовый кодон AUG, за которым сразу следует высококонсервативный нуклеотид G: AUGG (смотрите также инициирующий кодон в последовательности ДНК в соответствии с SEQ ID NO: 4 (пример 1)). В частности, по этой ссылке было описано, что последовательность Козак может быть идентифицирована посредством (gcc)gccRccAUGG (SEQ ID NO: 6) следующим образом: (i) строчные буквы обозначают наиболее распространенное основание в положении, где основание тем не менее может меняться; (ii) прописные буквы указывают на высококонсервативные основания (например, «AUGG»); (iii) «R» обозначает пурин (аденин или гуанин); (iv) последовательность в скобках ((gcc)) имеет неопределенное значение; (v) подчеркнутый триплет оснований AUG представляет собой стартовый кодон. Согласно одному варианту осуществления настоящего изобретения последовательность, гомологичная открытой рамке считывания неструктурного белка или его фрагмента из альфавируса, характеризуется тем, что она содержит удаление инициирующего кодона, который представляет собой часть последовательности Козак.
Согласно одному варианту осуществления настоящего изобретения 5' репликационная последовательность распознавания репликона характеризуется удалением по меньшей мере всех тех инициирующих кодонов, которые, на уровне РНК, представляют собой часть последовательности AUGG.
Согласно предпочтительному варианту осуществления последовательность, гомологичная открытой рамке считывания неструктурного белка или его фрагмента из альфавируса, характеризуется тем, что она содержит удаление одного или нескольких инициирующих кодонов, отличных от нативного стартового кодона открытой рамки считывания неструктурного белка. Согласно более предпочтительному варианту осуществления указанная последовательность нуклеиновой кислоты дополнительно характеризуется удалением нативного стартового кодона. Например, помимо удаления нативного стартового кодона, один или два, или три, или четыре, или более четырех (например, пять) инициирующих кодонов могут быть удалены.
Если репликон характеризуется удалением нативного стартового кодона и, необязательно, удалением одного или нескольких инициирующих кодонов, отличных от нативного стартового кодона, открытой рамки считывания неструктурного белка, последовательность, гомологичная открытой рамка считывания сама по себе не является открытой рамкой считывания, поскольку она не служит в качестве матрицы для трансляции.
Один или несколько инициирующих кодонов, отличных от нативного стартового кодона, который удаляется, предпочтительно в дополнение к удалению нативного стартового кодона, предпочтительно выбирают из триплета оснований AUG, который может инициировать трансляцию. Триплет оснований AUG, который характеризуется потенциалом к инициации трансляции, может упоминаться как «потенциальный инициирующий кодон». Может ли данный триплет оснований AUG характеризоваться потенциалом к инициации трансляции, можно определить in silico или в анализе in vitro на основе клеток.
Согласно одному варианту осуществления in silico определяется, может ли данный триплет оснований AUG характеризоваться потенциалом к инициации трансляции: согласно этому варианту осуществления исследуется нуклеотидная последовательность, и определяется, что триплет оснований AUG характеризуется потенциалом к инициации трансляции, если он представляет собой часть последовательности AUGG, предпочтительно часть последовательности Козак (SEQ ID NO: 6).
Согласно одному варианту осуществления в анализе in vitro на основе клеток определяют, характеризуется ли данный триплет оснований AUG потенциалом инициировать трансляцию: РНК-репликон, характеризующийся удалением нативного стартового кодона и содержащий данный триплет оснований AUG ниже по ходу транскрипции от положения удаления нативного стартового кодона вводится в клетку-хозяина. Согласно одному варианту осуществления клетка-хозяин представляет собой эукариотическую клетку, которая является природным хозяином конкретного альфавируса, который содержит нативную альфавирусную 5' репликационную последовательность распознавания. Согласно предпочтительному варианту осуществления клетка-хозяин представляет собой клетку BHK21 из клеточной линии «BHK21 [C13] (ATCC® CCL10™)», доступную из Американской коллекции типовых культур, Манассас, штат Вирджиния, США. Предпочтительно, чтобы никакой другой триплет оснований AUG не присутствовал между положением удаления нативного стартового кодона и данным триплетом оснований AUG. Если после переноса РНК-репликона, характеризующегося удалением нативного стартового кодона и содержащего данный триплет оснований AUG, в клетку-хозяина, трансляция инициируется в данном триплете оснований AUG, данный триплет оснований AUG определяется как характеризующийся потенциалом к инициации трансляции. Независимо от инициации трансляции, может быть определен любой подходящий способ, известный в настоящей области техники. Например, репликон может кодировать, ниже по ходу транскрипции от данного триплета оснований AUG и внутри рамки с данным триплетом оснований AUG, метку, которая облегчает обнаружение продукта трансляции (если он есть), например, myc-метку или HA-метку; независимо от того, может ли быть определен продукт экспрессии, содержащий закодированную метку, например, посредством Вестерн-блоттинга. Согласно этому варианту осуществления предпочтительно, чтобы не было никакого дополнительного триплета оснований AUG между данным триплетом оснований AUG и последовательностью нуклеиновой кислоты, кодирующей метку. Анализ на основе клеток in vitro может выполняться индивидуально для более чем одного данного триплета оснований AUG: в каждом случае предпочтительно, чтобы отсутствовал дополнительный триплет оснований AUG между положением удаления нативного стартового кодона и данного триплета оснований AUG. Этого можно добиться, удалив все триплеты оснований AUG (если есть) между положением удаления нативного стартового кодона и данным триплетом оснований AUG. Таким образом, данный триплет оснований AUG представляет собой первый триплет оснований AUG ниже по ходу транскрипции от положения удаления нативного стартового кодона.
Предпочтительно, репликон в соответствии с настоящим изобретением характеризуется удалением всех потенциальных инициирующих кодонов, которые находятся ниже по ходу транскрипции от положения удаления нативного стартового кодона и которые находятся в открытой рамке считывания альфавирусного неструктурного белка или его фрагмента. Таким образом, в соответствии с настоящим изобретением 5' репликационная последовательность распознавания предпочтительно не содержит открытую рамку считывания, которая может быть транслирована в белок.
Согласно предпочтительному варианту осуществления 5' репликационная последовательность распознавания РНК-репликона в соответствии с настоящим изобретением характеризуется вторичной структурой, которая эквивалентна вторичной структуре 5' репликационной последовательности распознавания альфавирусной геномной РНК. Согласно предпочтительному варианту осуществления 5' репликационная последовательность распознавания РНК-репликона в соответствии с настоящим изобретением характеризуется предсказанной вторичной структурой, которая эквивалентна предсказанной вторичной структуре 5' репликационной последовательности распознавания альфавирусной геномной РНК. В соответствии с настоящим изобретением вторичная структура молекулы РНК предпочтительно предсказывается веб-сервером для прогнозирования вторичной структуры РНК http://rna.urmc.rochester.edu/RNAstructureWeb/Servers/Predict1/Predict1.html.
Сравнивая вторичную структуру или предсказанную вторичную структуру 5' репликационной последовательности распознавания РНК-репликона, характеризующегося удалением по меньшей мере одного инициирующего кодона по сравнению с нативной альфавирусной 5' репликационной последовательностью распознавания, может быть идентифицировано наличие или отсутствие нарушения спаривания нуклеотидов. Например, по меньшей мере одна пара оснований может отсутствовать в данном положении по сравнению с нативной альфавирусной 5' репликационной последовательностью распознавания, например, пару оснований в структуре «петля-на-стебле», в частности, стебле структуры «петля-на-стебле».
Согласно предпочтительному варианту осуществления одна или несколько структур «петля-на-стебле» 5' репликационной последовательности распознавания не удаляются или не нарушаются. Более предпочтительно, чтобы структуры «петля-на-стебле» 3 и 4 не удалялись и не нарушались. Более предпочтительно, чтобы ни одна из структур «петля-на-стебле» 5' репликационной последовательности распознавания не удалялась или не нарушалась.
Согласно одному варианту осуществления удаление по меньшей мере одного инициирующего кодона не нарушает вторичную структуру 5' репликационной последовательности распознавания. Согласно альтернативному варианту осуществления удаление по меньшей мере одного инициирующего кодона нарушает вторичную структуру 5' репликационной последовательности распознавания. Согласно этому варианту осуществления удаление по меньшей мере одного инициирующего кодона может быть причиной отсутствия по меньшей мере одной пары оснований в данном положении, например, пары оснований в структуре «петля-на-стебле» по сравнению с нативной альфавирусной 5' репликационной последовательностью распознавания. Если в структуре «петля-на-стебле» отсутствует пара оснований, по сравнению с нативной альфавирусной 5' репликационной последовательностью распознавания, определяют удаление по меньшей мере одного инициирующего кодона, чтобы ввести нарушение спаривания нуклеотидов в структуре «петля-на-стебле». Пара оснований в структуре «петля-на-стебле», как правило, представляет собой пару оснований в стебле структуры «петля-на-стебле».
Согласно предпочтительному варианту осуществления РНК-репликон содержит одно или несколько нуклеотидных изменений, компенсирующих нарушения спаривания нуклеотидов в пределах одной или нескольких структур «петля-на-стебле», введенных удалением по меньшей мере одного инициирующего кодона.
Если удаление по меньшей мере одного инициирующего кодона вводит нарушение спаривания нуклеотидов в пределах структуры «петля-на-стебле» по сравнению с нативной альфавирусной 5' репликационной последовательностью распознавания, может быть введено одно или несколько нуклеотидных изменений, которые, как ожидается, будут компенсировать нарушение спаривания нуклеотидов, и полученная таким образом вторичная структура или предсказанная вторичная структура могут быть сравнены с нативной альфавирусной 5' репликационной последовательностью распознавания.
На основе общих знаний и на раскрытии настоящего документа специалист в настоящей области техники может ожидать, что определенные нуклеотидные изменения компенсируют нарушения спариваний нуклеотидов. Например, если пара оснований нарушена в заданном положении вторичной структуры или предсказанной вторичной структуры данной 5' репликационной последовательности распознавания РНК-репликона, характеризующегося удалением по меньшей мере одного инициирующего кодона, по сравнению с нативной альфавирусной 5' репликационной последовательностью распознавания, нуклеотидное изменение, которое восстанавливает пару оснований в этом положении, предпочтительно без повторного введения инициирующего кодона, как ожидается, компенсирует нарушение спаривания нуклеотидов. В одном примере 5' репликационная последовательность распознавания репликона, содержащая одно или несколько нуклеотидных изменений, компенсирующих нарушения спаривания нуклеотидов в пределах одной или нескольких структур «петля-на-стебле», введенных удалением по меньшей мере одного инициирующего кодона, кодируется последовательностью ДНК, представленной SEQ ID NO: 5 (пример 1).
Согласно предпочтительному варианту осуществления 5' репликационная последовательность распознавания репликона не перекрывается или не включает в себя транслируемую последовательность нуклеиновой кислоты, то есть транслируемую в пептид или белок, в частности nsP, в частности nsP1, или фрагмент любого из них. Для того чтобы нуклеотидная последовательность была «транслируемой», она требует наличия инициирующего кодона; инициирующий кодон кодирует большинство N-концевых аминокислотных остатков пептида или белка. Согласно одному варианту осуществления 5' репликационная последовательность распознавания репликона не перекрывается с транслируемой последовательностью нуклеиновой кислоты, кодирующей N-концевой фрагмент nsP1, или не включает ее.
В некоторых сценариях, которые подробно описаны ниже, РНК-репликон содержит по меньшей мере один субгеномный промотор. Согласно предпочтительному варианту осуществления субгеномный промотор репликона не перекрывается с транслируемой последовательностью нуклеиновой кислоты или не включает в себя ее, т.е. транслируемой в пептид или белок, в частности nsP, в частности nsP4, или фрагмент любого из них. Согласно одному варианту осуществления субгеномный промотор репликона не перекрывается с транслируемой последовательностью нуклеиновой кислоты, которая кодирует С-концевой фрагмент в nsP4, или не содержит ее. РНК-репликон, содержащий субгеномный промотор, который не перекрывается с транслируемой последовательностью нуклеиновой кислоты или не содержит ее, например, может быть получен путем удаления части кодирующей последовательности для nsP4 (как правило, части, кодирующей N-концевую часть nsP4), и/или путем удаления триплетов оснований AUG в части кодирующей последовательность для nsP4, которая не была удалена. Если триплеты оснований AUG в кодирующей последовательности для nsP4 или ее части удалены, триплеты оснований AUG, которые удалены, предпочтительно представляют собой потенциальные инициирующие кодоны. Альтернативно, если субгеномный промотор не перекрывается с последовательностью нуклеиновой кислоты, которая кодирует nsP4, вся нуклеотидная последовательность, кодирующая nsP4, может быть удалена.
Согласно одному варианту осуществления РНК-репликон не содержит открытую рамку считывания, кодирующую усеченный альфавирусный неструктурный белок. В контексте этого варианта осуществления особенно предпочтительно, чтобы РНК-репликон не содержал открытую рамку считывания, кодирующую N-концевой фрагмент nsP1, и, необязательно, не содержал открытую рамку считывания, кодирующую C-концевой фрагмент nsP4. N-концевой фрагмент nsP1 представляет собой усеченный альфавирусный белок; С-концевой фрагмент nsP4 также представляет собой усеченный альфавирусный белок.
Согласно некоторым вариантам осуществления репликон в соответствии с настоящим изобретением не содержит структуру «петля-на-стебле» 2 (SL2) 5'-конца генома альфавируса. Согласно Frolov с соавт., смотрите выше, структура «петля-на-стебле» 2 представляет собой консервативную вторичную структуру, обнаруженную на 5'-конце генома альфавируса, выше против хода транскрипции от CSE 2, но недоступную для репликации.
Согласно одному варианту осуществления 5' репликационная последовательность распознавания репликона не перекрывается с последовательностью нуклеиновой кислоты, которая кодирует альфавирусный неструктурный белок или его фрагмент. Таким образом, настоящее изобретение охватывает репликоны, которые характеризуются по сравнению с геномной альфавирусной РНК удалением по меньшей мере одного инициирующего кодона, как описано в настоящем документе, необязательно в сочетании с делецией кодирующей области для одного или нескольких альфавирусных неструктурных белков или их части. Например, кодирующая область для nsP2 и nsP3 может быть удалена, или кодирующая область для nsP2 и nsP3 может быть удалена вместе с удалением кодирующей области для C-концевого фрагмента nsP1 и/или кодирующей области для N-концевого фрагмента nsP4, и один или несколько оставшихся инициирующих кодонов, то есть оставшихся после указанного удаления, могут быть удалены, как описано в настоящем документе.
Удаление кодирующей области для одного или нескольких альфавирусных неструктурных белков может быть достигнуто стандартными способами, например, на уровне ДНК, удаление с помощью рестрикционных ферментов, предпочтительно рестрикционных ферментов, которые распознают уникальные сайты рестрикции в открытой рамке считывания (в качестве иллюстрации: смотрите пример 1). Необязательно, уникальные сайты рестрикции могут быть введены в открытую рамку считывания посредством мутагенеза, например, сайт-направленного мутагенеза. Соответствующую ДНК можно использовать в качестве матрицы для транскрипции in vitro.
Сайт рестрикции представляет собой последовательность нуклеиновой кислоты, например, ДНК, которая необходима и достаточна для прямой рестрикции (расщепления) молекулы нуклеиновой кислоты, например, молекулы ДНК, в которой содержится сайт рестрикции, с помощью специфического фермента рестрикции. Сайт рестрикции уникален для данной молекулы нуклеиновой кислоты, если одна копия сайта рестрикции присутствует в молекуле нуклеиновой кислоты.
Фермент рестрикции представляет собой эндонуклеазу, которая разрезает молекулу нуклеиновой кислоты, например, молекулу ДНК, в участке рестрикции или вблизи него.
Альтернативно, последовательность нуклеиновой кислоты, характеризующаяся делецией части или всей открытой рамки считывания, может быть получена синтетическими способами.
РНК-репликон в соответствии с настоящим изобретением предпочтительно представляет собой одноцепочечную молекулу РНК. РНК-репликон в соответствии с настоящим изобретением, как правило, представляет собой (+)-нитевую молекулу РНК. Согласно одному варианту осуществления РНК-репликон по настоящему изобретению представляет собой выделенную молекулу нуклеиновой кислоты.
По меньшей мере одна открытая рамка считывания, содержащаяся в репликоне
Согласно одному варианту осуществления РНК-репликон в соответствии с настоящим изобретением содержит по меньшей мере одну открытую рамку считывания, кодирующую представляющий интерес пептид или представляющий интерес белок. Предпочтительно представляющий интерес белок кодируется гетерологичной последовательностью нуклеиновой кислоты. Ген, кодирующий представляющий интерес пептид или белок, синонимично называется «представляющим интерес геном» или «трансгеном». Согласно различным вариантам осуществления представляющий интерес пептид или белок кодируется гетерологичной последовательностью нуклеиновой кислоты. В соответствии с настоящим изобретением термин «гетерологичный» относится к тому факту, что последовательность нуклеиновой кислоты не является функционально или структурно связанной с последовательностью нуклеиновой кислоты альфавируса.
Репликон в соответствии с настоящим изобретением может кодировать один полипептид или множество полипептидов. Множество полипептидов можно кодировать в виде одного полипептида (слитого полипептида) или в виде отдельных полипептидов. Согласно некоторым вариантам осуществления репликон в соответствии с настоящим изобретением может содержать более чем одну открытую рамку считывания, каждая из которых может быть независимо выбрана, чтобы быть под контролем субгеномного промотора или нет. Альтернативно, полипротеин или слитый полипептид содержит отдельные полипептиды, разделенные необязательно аутокаталитическим сайтом расщепления протеазой (например, белок 2A вируса ящура) или интеин.
Представляющие интерес белки могут, например, быть выбраны из группы, состоящей из репортерных белков, фармацевтически активных пептидов или белков, ингибиторов внутриклеточной передачи сигналов интерферонов (IFN) и функционального альфавирусного неструктурного белка.
Репортерный белок
Согласно одному варианту осуществления открытая рамка считывания кодирует репортерный белок. Согласно этому варианту осуществления открытая рамка считывания содержит репортерный ген. Определенные гены могут быть выбраны в качестве репортеров, потому что характеристики, которые они придают клеткам или организмам, экспрессирующим их, могут быть легко идентифицированы и измерены, или потому что они являются селектируемыми маркерами. Репортерные гены часто используются в качестве индикатора того, был ли поглощен определенный ген или экспрессирован в популяции клеток или организмов. Предпочтительно, продукт экспрессии репортерного гена визуально обнаруживается. Обычные визуально обнаруживаемые репортерные белки, как правило, содержат флуоресцентные или люминесцирующие белки. Примеры конкретных репортерных генов включают в себя ген, который кодирует зеленый флуоресцентный белок медузы (GFP), который заставляет клетки, которые его экспрессируют, светить зеленым под синим светом, фермент люциферазу, который катализирует реакцию с люциферином для получения света, и красный флуоресцентный белок (RFP). Возможны варианты любого из этих специфических репортерных генов, если варианты обладают визуально обнаруживаемыми свойствами. Например, eGFP представляет собой вариант GFP с точечной мутацией. Вариант осуществления репортерного белка особенно подходит для исследования экспрессии, смотрите, например, примеры 2-5.
Фармацевтически активный пептид или белок
В соответствии с настоящим изобретением согласно одному варианту осуществления РНК репликона содержит или состоит из фармацевтически активной РНК. «Фармацевтически активная РНК» может представлять собой РНК, которая кодирует фармацевтически активный пептид или белок. Предпочтительно, чтобы РНК-репликон в соответствии с настоящим изобретением кодировал фармацевтически активный пептид или белок. Предпочтительно, чтобы открытая рамка считывания кодировала фармацевтически активный пептид или белок. Предпочтительно, чтобы РНК-репликон содержал открытую рамку считывания, которая кодирует фармацевтически активный пептид или белок, предпочтительно, под контролем субгеномного промотора.
«Фармацевтически активный пептид или белок» оказывает положительное или благоприятное действие на состояние или заболевание субъекта при введении субъекту в терапевтически эффективном количестве. Предпочтительно, фармацевтически активный пептид или белок обладает лечебными или паллиативными свойствами и может вводиться для улучшения, ослабления, облегчения, обращения, задержки начала или уменьшения тяжести одного или нескольких симптомов заболевания или нарушения. Фармацевтически активный пептид или белок может характеризоваться профилактическими свойствами и может быть использован для замедления начала заболевания или уменьшения тяжести такого заболевания или патологического состояния. Термин «фармацевтически активный пептид или белок» включает в себя целые белки или полипептиды и также может относиться к их фармацевтически активным фрагментам. Он также может включать в себя фармацевтически активные аналоги пептида или белка. Термин «фармацевтически активный пептид или белок» включает в себя пептиды и белки, которые представляют собой антигены, то есть пептид или белок вызывает иммунный ответ у субъекта, который может быть терапевтическим или частично или полностью защитным.
Согласно одному варианту осуществления фармацевтически активный пептид или белок представляет собой или содержит иммунологически активное соединение или антиген или эпитоп.
Согласно настоящему изобретению термин «иммунологически активное соединение» относится к любому соединению, изменяющему иммунный ответ, предпочтительно путем индуцирования и/или подавления созревания иммунных клеток, индуцирования и/или подавления биосинтеза цитокинов и/или изменения гуморального иммунитета с помощью стимулирования производства антител В-клетками. Согласно одному варианту осуществления иммунный ответ включает в себя стимуляцию ответа антитела (как правило, включающего в себя иммуноглобулин G (IgG)). Иммунологически активные соединения обладают мощной иммуностимулирующей активностью, включая в себя, без ограничения, противовирусную и противоопухолевую активность, а также могут понижающе регулировать другие аспекты иммунного ответа, например, сдвигать иммунный ответ от иммунного ответа TH2, что применимо для лечения широкого спектра заболеваний, опосредованных TH2.
В соответствии с настоящим изобретением термин «антиген» или «иммуноген» охватывает любое вещество, которое будет вызывать иммунный ответ. В частности, «антиген» относится к любому веществу, которое специфически реагирует с антителами или Т-лимфоцитами (Т-клетками). В соответствии с настоящим изобретением термин «антиген» включает в себя любую молекулу, которая содержит по меньшей мере один эпитоп. Предпочтительно, антиген в контексте настоящего изобретения представляет собой молекулу, которая, необязательно после процессинга, индуцирует иммунную реакцию, которая предпочтительно является специфической для антигена. В соответствии с настоящим изобретением может быть использован любой подходящий антиген, который является кандидатом для иммунной реакции, причем иммунная реакция может представлять собой как гуморальную, так и клеточную иммунную реакцию. В контексте вариантов осуществления настоящего изобретения антиген предпочтительно представлен клеткой, предпочтительно антигенпрезентирующей клеткой, в контексте молекул МНС, что приводит к иммунной реакции против антигена. Антиген предпочтительно представляет собой продукт, который соответствует природному антигену или происходит из него. Такие встречающиеся в природе антигены могут включать в себя или могут происходить из аллергенов, вирусов, бактерий, грибов, паразитов и других инфекционных агентов и патогенов или антиген также может представлять собой опухолевый антиген. В соответствии с настоящим изобретением антиген может соответствовать встречающемуся в природе продукту, например, вирусному белку или его части. Согласно предпочтительным вариантам осуществления антиген представляет собой поверхностный полипептид, то есть полипептид, естественно отображаемый на поверхности клетки, патоген, бактерию, вирус, гриб, паразит, аллерген или опухоль. Антиген может вызывать иммунный ответ против клетки, патогена, бактерии, вируса, гриба, паразита, аллергена или опухоли.
Термин «патоген» относится к патогенному биологическому материалу, способному вызывать заболевание в организме, предпочтительно в организме позвоночных. Патогены включают в себя микроорганизмы, такие как бактерии, одноклеточные эукариотические организмы (простейшие), грибы, а также вирусы.
Термины «эпитоп», «антигенный пептид», «антигенный эпитоп», «иммуногенный пептид» и «связывающий МНС пептид» используются в настоящем документе взаимозаменяемо и относятся к антигенной детерминанте в молекуле, такой как антиген, то есть к части или фрагменту иммунологически активного соединения, которое распознается иммунной системой, например, которое распознается Т-клеткой, в частности, когда оно представлено в контексте молекул МНС. Эпитоп белка предпочтительно содержит непрерывную или прерывистую часть указанного белка и предпочтительно составляет от 5 до 100, предпочтительно от 5 до 50, более предпочтительно от 8 до 30, наиболее предпочтительно от 10 до 25 аминокислот в длину, например, эпитоп может составлять предпочтительно 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 аминокислот в длину. В соответствии с настоящим изобретением эпитоп может связываться с молекулами MHC, такими как молекулы MHC на поверхности клетки и, таким образом, может представлять собой «связывающий MHC пептид» или «антигенный пептид». Термин «основной комплекс гистосовместимости» и сокращение «MHC» включает в себя молекулы MHC класса I и MHC класса II и относится к комплексу генов, который присутствует у всех позвоночных. Белки или молекулы MHC важны для передачи сигналов между лимфоцитами и антигенпрезентирующими клетками или пораженными клетками в иммунных реакциях, причем белки или молекулы MHC связывают пептиды и презентируют их для распознавания Т-клеточными рецепторами. Белки, кодируемые MHC, экспрессируются на поверхности клеток и отображают как аутоантигены (пептидные фрагменты из самой клетки), так и неаутоантигены (например, фрагменты вторгающихся микроорганизмов) для Т-клетки. Предпочтительные такие иммуногенные части связываются с молекулой МНС класса I или класса II. Как используется в настоящем документе, иммуногенная часть называется частью, которая «связывается с» молекулой МНС класса I или класса II, если такое связывание обнаруживается с использованием любого анализа, известного в настоящей области техники. Термин «связывающий MHC пептид» относится к пептиду, который связывается с молекулой MHC класса I и/или MHC класса II. В случае комплексов MHC класса I с пептидом связывающие пептиды, как правило, составляют 8-10 аминокислот в длину, хотя могут быть эффективными и более длинные или более короткие пептиды. В случае комплексов MHC класса II с пептидом связывающие пептиды, как правило, составляют в длину 10-25 аминокислот и, в частности, составляют 13-18 аминокислот, тогда как могут быть эффективными и более длинные и более короткие пептиды.
Согласно одному варианту осуществления представляющий интерес белок в соответствии с настоящим изобретением содержит эпитоп, подходящий для вакцинации целевого организма. Специалисту в настоящей области техники известно, что один из принципов иммунобиологии и вакцинации основан на том факте, что иммунозащитная реакция на заболевание вырабатывается путем иммунизации организма антигеном, который является иммунологически релевантным по отношению к подлежащему лечению заболеванию. В соответствии с настоящим изобретением антиген выбирают из группы, содержащей аутоантиген и неаутоантиген. Неаутоантиген предпочтительно представляет собой бактериальный антиген, вирусный антиген, грибковый антиген, аллерген или паразитный антиген. Предпочтительно, чтобы антиген содержал эпитоп, который способен вызывать иммунный ответ в целевом организме. Например, эпитоп может вызывать иммунный ответ против бактерии, вируса, гриба, паразита, аллергена или опухоли.
Согласно некоторым вариантам осуществления неаутоантиген представляет собой бактериальный антиген. Согласно некоторым вариантам осуществления антиген вызывает иммунный ответ против бактерии, которая заражает животных, включая в себя птиц, рыб и млекопитающих, включая в себя домашних животных. Предпочтительно, бактерия, против которой вызывается иммунный ответ, представляет собой патогенную бактерию.
Согласно некоторым вариантам осуществления неаутоантиген представляет собой вирусный антиген. Вирусный антиген может представлять собой, например, пептид из поверхностного белка вируса, например, капсидный полипептид или полипептид шипа. Согласно некоторым вариантам осуществления антиген вызывает иммунный ответ против вируса, который заражает животных, включая в себя птиц, рыб и млекопитающих, включая в себя домашних животных. Предпочтительно, вирус, против которого вызывается иммунный ответ, представляет собой патогенный вирус.
Согласно некоторым вариантам осуществления неаутоантиген представляет собой полипептид или белок из гриба. Согласно некоторым вариантам осуществления антиген вызывает иммунный ответ против гриба, который заражает животных, включая в себя птиц, рыб и млекопитающих, включая в себя домашних животных. Предпочтительно, гриб, против которого вызывается иммунный ответ, представляет собой патогенный гриб.
Согласно некоторым вариантам осуществления неаутоантиген представляет собой полипептид или белок из одноклеточного эукариотического паразита. Согласно некоторым вариантам осуществления антиген вызывает иммунный ответ против одноклеточного эукариотического паразита, предпочтительно патогенного одноклеточного эукариотического паразита. Патогенные одноклеточные эукариотические паразиты могут быть, например, из рода Plasmodium, например, P. falciparum, P. vivax, P. malariae или P. ovale, из рода Leishmania или из рода Trypanosoma, например, T. cruzi или T. brucei.
Согласно некоторым вариантам осуществления неаутоантиген представляет собой аллергенный полипептид или аллергенный белок. Аллергенный белок или аллергенный полипептид подходит для иммунотерапии аллергеном, также известной как гипосенсибилизация.
Согласно некоторым вариантам осуществления антиген представляет собой аутоантиген, в частности опухолевый антиген. Опухолевые антигены и их определение известны специалисту в настоящей области техники.
В контексте настоящего изобретения термин «опухолевый антиген» или «опухолеассоциированный антиген» относится к белкам, которые в нормальных условиях специфически экспрессируются в ограниченном числе тканей и/или органов или на конкретных стадиях развития, например, опухолевый антиген может в нормальных условиях специфически экспрессироваться в ткани желудка, предпочтительно в слизистой оболочке желудка, в репродуктивных органах, например, в яичках, в трофобластической ткани, например, в плаценте или в клетках зародышевой линии, и экспрессируются или аберрантно экспрессируются в одной или нескольких опухолевых или злокачественных тканях. В этом контексте «ограниченное число» предпочтительно означает не более 3, более предпочтительно не более 2. Опухолевые антигены в контексте настоящего изобретения включают в себя, например, антигены дифференцировки, предпочтительно специфические к клеточному типу антигены дифференцировки, т.е. белки, которые в нормальных условиях специфически экспрессируются в определенном типе клеток на определенной стадии дифференциации, антигены злокачественной опухоли/яичка, то есть белки, которые в нормальных условиях специфически экспрессируются в яичках, а иногда и в плаценте, и специфические к зародышевой линии антигены. В контексте настоящего изобретения опухолевый антиген предпочтительно ассоциирован с клеточной поверхностью злокачественной клетки и предпочтительно не экспрессируется или только редко экспрессируется в нормальных тканях. Предпочтительно, опухолевый антиген или аберрантная экспрессия опухолевого антигена идентифицирует злокачественные клетки. В контексте настоящего изобретения опухолевый антиген, который экспрессируется злокачественной клеткой у субъекта, например, страдающего от злокачественного заболевания пациента, предпочтительно представляет собой собственный белок у указанного субъекта. Согласно предпочтительным вариантам осуществления опухолевый антиген в контексте настоящего изобретения экспрессируется в нормальных условиях, особенно в ткани или органе, который является несущественным, т.е. ткани или органы, которые при повреждении иммунной системой не приводят к гибели субъекта, или в органах или структурах организма, которые недоступны или едва ли доступны иммунной системе. Предпочтительно, аминокислотная последовательность опухолевого антигена идентична между опухолевым антигеном, который экспрессируется в нормальных тканях, и опухолевым антигеном, который экспрессируется в злокачественных тканях.
Примерами опухолевых антигенов, которые могут быть использованы в настоящем изобретении, являются p53, ART-4, BAGE, β-катенин/m, Bcr-abL CAMEL, CAP-1, CASP-8, CDC27/m, CDK4/m, CEA, белки клеточной поверхности семейства клаудин, такие как CLAUDIN-6, CLAUDIN-18.2 и CLAUDIN-12, c-MYC, CT, Cyp-B, DAM, ELF2M, ETV6-AML1, G250, GAGE, GnT-V, Gap100 , HAGE, HER-2/neu, HPV-E7, HPV-E6, HAST-2, hTERT (или hTRT), LAGE, LDLR/FUT, MAGE-A, предпочтительно, MAGE-A1, MAGE-A2, MAGE-A3, MAGE-A, MAGE-A5, MAGE-A6, MAGE-A7, MAGE-A8, MAGE-A9, MAGE-A10, MAGE-A11 или MAGE-A12, MAGE-B, MAGE-C, MART-1/мелан-A, MC1R, миозин/m, MUC1, MUM-1, -2, -3, NA88-A, NF1, NY-ESO-1, NY-BR-1, p190 минорный BCR-abL, Pm1/RARa, PRAME , протеиназа 3, PSA, PSM, RAGE, RU1 или RU2, SAGE, SART-1 или SART-3, SCGB3A2, SCP1, SCP2, SCP3, SSX, SURVIVIN, TEL/AML1, TPI/m, TRP-1, TRP- 2, TRP-2/INT2, TPTE и WT. Особенно предпочтительные опухолевые антигены включают в себя CLAUDIN-18.2 (CLDN18.2) и CLAUDIN-6 (CLDN6).
Согласно некоторым вариантам осуществления не требуется, чтобы фармацевтически активный пептид или белок был антигеном, который вызывает иммунный ответ. Подходящие фармацевтически активные пептиды или белки могут быть выбраны из группы, состоящей из цитокинов и белков иммунной системы, таких как иммунологически активные соединения (например, интерлейкины, колониестимулирующий фактор (CSF), гранулоцитарный колониестимулирующий фактор (G-CSF), гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF), эритропоэтин, фактор некроза опухоли (TNF), интерфероны, интегрины, адрессины, селектины, «хоминг»-рецепторы, Т-клеточные рецепторы, иммуноглобулины), гормоны (инсулин, гормон щитовидной железы, катехоламины, гонадотрофины, трофические гормоны, пролактин, окситоцин, дофамин, бычий соматотропин, лептины и тому подобное), гормоны роста (например, гормон роста человека), факторы роста (например, эпидермальный фактор роста, фактор роста нервов, инсулиноподобный фактор роста и тому подобное), рецепторы фактора роста , ферменты (тканевый активатор плазминогена, стрептокиназа, биосинтеза или деградации холестерина, стероидогенные ферменты, киназы, фосфодиэстеразы, метилазы, деметилазы, дегидразы, целлюлазы, протеазы, липазы, фосфолипазы, ароматазы, цитохромы, аденилат- или гуанилатциклазы, нейрамидазы и тому подобное), рецепторы (рецепторы стероидных гормонов, пептидные рецепторы), связывающие белки (связывающие гормон роста или связывающие фактор роста белки и т.п.), факторы транскрипции и трансляции, подавляющие опухолевый рост белки (например, белки, которые ингибируют ангиогенез), структурные белки (такие как коллаген, фибрин, фибриноген, эластин, тубулин, актин и миозин), белки крови (тромбин, сывороточный альбумин, фактор VII, фактор VIII, инсулин, фактор IX, фактор X, активатор тканевого плазминогена, белок C, фактор фон Виллебранда, антитромбин III, глюкоцереброзидаза, эритропоэтин, гранулоцитарный колониестимулирующий фактор (GCSF) или модифицированный фактор VIII, антикоагулянты и т.п. Согласно одному варианту осуществления фармацевтически активный белок согласно настоящему изобретению представляет собой цитокин, который участвует в регуляции лимфоидного гомеостаза, предпочтительно, цитокин, который участвует и предпочтительно индуцирует или усиливает развитие, праймирование, размножение, дифференцировку и/или выживание Т-клеток. Согласно одному варианту осуществления цитокин представляет собой интерлейкин, т.е. IL-2, IL-7, IL-12, IL-15 или IL-21.
Ингибитор передачи сигналов интерферона (IFN)
Дополнительный подходящий белок, кодируемый открытой рамкой считывания, представляет собой ингибитор передачи сигналов интерферона (IFN). Хотя сообщалось, что жизнеспособность клеток, в которые была введена РНК для экспрессии, может быть уменьшена, в частности, если клетки трансфицируют РНК несколько раз, ингибирующие IFN средства, как было установлено, повышают жизнеспособность клеток, в которых РНК должна быть экспрессирована (публикация международной заявки WO 2014/071963 A1). Предпочтительно ингибитор представляет собой ингибитор передачи сигналов IFN типа I. Предотвращение вовлечения рецептора IFN с помощью внеклеточного IFN и ингибирование внутриклеточной передачи сигналов IFN в клетках делает возможной стабильную экспрессию РНК в клетках. Альтернативно или дополнительно, предотвращение вовлечения рецептора IFN с помощью внеклеточного IFN и ингибирование внутриклеточной передачи сигналов IFN повышает выживаемость клеток, в частности, если клетки повторно трансфицируют с РНК. Без ограничения теорией предполагается, что внутриклеточная передача сигналов IFN может привести к ингибированию трансляции и/или деградации РНК. Это может быть устранено путем ингибирования одного или нескольких индуцируемых IFN обладающих антивирусной активностью эффекторных белков. Индуцируемый IFN обладающий антивирусной активностью эффекторный белок может быть выбран из группы, состоящей из РНК-зависимой протеинкиназы (PKR), 2',5'-олигоаденилатсинтетазы (OAS) и RNaseL. Ингибирование внутриклеточной передачи сигналов IFN может предусматривать ингибирование PKR-зависимого пути и/или OAS-зависимого пути. Подходящим представляющим интерес белком является белок, который способен ингибировать PKR-зависимый путь и/или OAS-зависимый путь. Ингибирование PKR-зависимого пути может предусматривать ингибирование фосфорилирования eIF2-alpha. Ингибирование PKR может предусматривать обработку клетки по меньшей мере одним ингибитором PKR. Ингибитор PKR может представлять собой вирусный ингибитор PKR. Предпочтительный вирусный ингибитор PKR представляет собой вирус осповакцины E3. Если пептид или белок (например, E3, K3) предназначен для ингибирования внутриклеточной передачи сигналов IFN, предпочтительна внутриклеточная экспрессия пептида или белка. Вирус осповакцины E3 представляет собой дцРНК-связывающий белок размером 25 кДа (кодируемый геном E3L), который связывается и отделяет дцРНК, чтобы предотвратить активацию PKR и OAS. E3 может связываться непосредственно с PKR и ингибировать его активность, что приводит к снижению фосфорилирования eIF2-альфа. Другими подходящими ингибиторами передачи сигналов IFN являются вирус ICP34.5 простого герпеса, вирус NSS тосканской лихорадки, вирус ядерного полиэдроза PK2 Bombyx mori и HCV NS34A.
Ингибитор передачи сигналов IFN может быть предоставлен клетке в виде последовательности нуклеиновой кислоты (например, РНК), кодирующей ингибитор передачи сигналов IFN.
Согласно одному варианту осуществления ингибитор внутриклеточной или внеклеточной передачи сигналов IFN кодируется молекулой мРНК. Эта молекула мРНК может содержать модификацию, изменяющую неполипептид-кодирующую последовательность, как описано в настоящем документе, например, кэп, 5'-UTR, 3'-UTR, поли(A)-последовательность, адаптацию использования кодонов. Соответствующие варианты осуществления проиллюстрированы, например, в примере 3.
Согласно альтернативному варианту осуществления ингибитор внутриклеточной или внеклеточной передачи сигналов IFN кодируется репликоном, предпочтительно транс-репликоном или описанным в настоящем документе транс-репликоном. Репликон содержит элементы последовательности нуклеиновой кислоты, которые позволяют реплицировать альфавирусную репликазу, как правило, CSE 1, CSE 2 и CSE 4; и предпочтительно также элементы последовательности нуклеиновой кислоты, которые позволяют производить субгеномный транскрипт, то есть субгеномный промотор, как правило, содержащий CSE 3. Репликон может дополнительно содержать одну или несколько модификаций, изменяющих неполипептид-кодирующие последовательности, как описано в настоящем документе, например, кэп, поли(A)-последовательность, адаптация использования кодонов. Если в репликоне присутствует множество открытых рамок считывания, то ингибитор внутриклеточной передачи сигналов IFN может кодироваться любой из них, необязательно под контролем субгеномного промотора или без него. Согласно предпочтительному варианту осуществления ингибитор внутриклеточной передачи сигналов IFN кодируется расположенной выше всех против хода транскрипции открытой рамкой считывания РНК-репликона. Когда ингибитор внутриклеточной передачи сигналов IFN кодируется расположенной выше всех против хода транскрипции открытой рамкой считывания РНК-репликона, генетическая информация, кодирующая ингибитор внутриклеточной передачи сигналов IFN, будет транслироваться вскоре после введения РНК-репликона в клетку-хозяина, и полученный белок может впоследствии ингибировать внутриклеточную передачу сигналов IFN.
Функциональный альфавирусный неструктурный белок
Еще один подходящий белок, кодируемый открытой рамкой считывания, представляет собой функциональный альфавирусный неструктурный белок. Термин «альфавирусный неструктурный белок» включает в себя каждую котрансляционно или посттрансляционно модифицированную форму, включая в себя модифицированные углеводами (такие как гликозилированные) и липид-модифицированные формы альфавирусного неструктурного белка.
Согласно некоторым вариантам осуществления термин «альфавирусный неструктурный белок» относится к любому одному или нескольким отдельным происходящим от альфавируса неструктурным белкам (nsP1, nsP2, nsP3, nsP4) или к полипротеину, содержащему полипептидную последовательность более чем одного происходящего от альфавируса неструктурного белка. Согласно некоторым вариантам осуществления «альфавирусный неструктурный белок» относится к nsP123 и/или к nsP4. Согласно другим вариантам осуществления «альфавирусный неструктурный белок» относится к nsP1234. Согласно одному варианту осуществления представляющий интерес белок, кодируемый открытой рамкой считывания, состоит из всех nsP1, nsP2, nsP3 и nsP4 в виде одного единственного, необязательно расщепляемого полипротеина: nsP1234. Согласно одному варианту осуществления представляющий интерес белок, кодируемый открытой рамкой считывания, состоит из nsP1, nsP2 и nsP3 в виде одного единственного, необязательно расщепляемого полипротеина: nsP123. Согласно этому варианту осуществления nsP4 может представлять собой дополнительный представляющий интерес белок и может быть кодирован другой открытой рамкой считывания.
Согласно некоторым вариантам осуществления альфавирусный неструктурный белок способен образовывать комплекс или объединение, например, в клетке-хозяине. Согласно некоторым вариантам осуществления «альфавирусный неструктурный белок» относится к комплексу или объединению nsP123 (синонимично P123) и nsP4. Согласно некоторым вариантам осуществления «альфавирусный неструктурный белок» относится к комплексу или объединению nsP1, nsP2 и nsP3. Согласно некоторым вариантам осуществления «альфавирусный неструктурный белок» относится к комплексу или объединению nsP1, nsP2, nsP3 и nsP4. Согласно некоторым вариантам осуществления «альфавирусный неструктурный белок» относится к комплексу или объединению любого из одного или нескольких выбранных из группы, состоящей из nsP1, nsP2, nsP3 и nsP4. Согласно некоторым вариантам осуществления альфавирусный неструктурный белок содержит по меньшей мере nsP4.
Термины «комплекс» или «объединение» относятся к двум или более одинаковым или различным молекулам белка, которые находятся в пространственной близости. Белки комплекса предпочтительно находятся в прямом или косвенном физическом или физико-химическом контакте друг с другом. Комплекс или объединение может состоять из нескольких различных белков (гетеромультимер) и/или нескольких копий одного конкретного белка (гомомультимер). В контексте альфавирусного неструктурного белка термин «комплекс или объединение» описывает множество по меньшей мере из двух молекул белка, из которых по меньшей мере один представляет собой альфавирусный неструктурный белок. Комплекс или объединение могут состоять из нескольких копий одного конкретного белка (гомомультимер) и/или нескольких разных белков (гетеромультимер). В контексте мультимера «мульти» означает более одного, например, два, три, четыре, пять, шесть, семь, восемь, девять, десять или более десяти.
Термин «функциональный альфавирусный неструктурный белок» включает в себя альфавирусный неструктурный белок, который характеризуется функцией репликазы. Таким образом, «функциональный альфавирусный неструктурный белок» включает в себя альфавирусную репликазу. «Функция репликазы» предусматривает функцию РНК-зависимой РНК-полимеразы (RdRP), то есть фермента, который способен катализировать синтез (-)-нити РНК на основе (+)-нити РНК-матрицы и/или который способен катализировать синтеза (+)-нити РНК на основе (-)-нити РНК-матрицы. Таким образом, термин «функциональный альфавирусный неструктурный белок» может относиться к белку или комплексу, который синтезирует (-)-нитевую РНК с использованием (+)-нитевой (например, геномной) РНК в качестве матрицы к белку или комплексу, который синтезирует новые (+)-нитевые РНК с использованием (-)-нитевого комплемента геномной РНК в качестве матрицы, и/или к белку или комплексу, который синтезирует субгеномный транскрипт, используя фрагмент (-)-нитевого комплемента геномной РНК в качестве матрицы. Функциональный альфавирусный неструктурный белок может дополнительно характеризоваться одной или несколькими дополнительными функциями, такими как, например, функции протеиназы (для ауторасщепления), геликазы, концевой аденилаттрансферазы (для добавления поли(А)-хвоста), метилтрансферазы и гуанилтрансферазы (для получения нуклеиновой кислоты с 5'-кэпом), сайтов ядерной локализации, трифосфатазы (Gould et al., 2010, Antiviral Res., vol. 87, pp. 111-124; Rupp et al., 2015, J. Gen. Virol., vol. 96, pp. 2483-500).
В соответствии с настоящим изобретением термин «альфавирусная репликаза» относится к альфавирусной РНК-зависимой РНК-полимеразе, включая в себя РНК-зависимую РНК-полимеразу из встречающегося в природе альфавируса (альфавируса, обнаруженного в природе) и РНК-зависимую РНК-полимеразу из варианта или производного альфавируса, например, из аттенуированного альфавируса. В контексте настоящего изобретения термины «репликаза» и «альфавирусная репликаза» используются взаимозаменяемо, если только контекст не указывает, что какая-либо конкретная репликаза не представляет собой альфавирусную репликазу.
Термин «репликаза» предусматривает все варианты, в частности посттрансляционно модифицированные варианты, конформации, изоформы и гомологи альфавирусной репликазы, которые экспрессируются инфицированными альфавирусами клетками или которые экспрессируются клетками, которые были трансфицированы нуклеиновой кислотой, которая кодирует альфавирусную репликазу. Более того, термин «репликаза» включает в себя все формы репликазы, которые были получены и могут быть получены рекомбинантными способами. Например, репликаза, содержащая метку, которая облегчает обнаружение и/или очистку репликазы в лаборатории, например, myc-метка, HA-метка или олигогистидиновая метка (His-метка), может быть получена рекомбинантными способами.
Необязательно, альфавирусная репликаза дополнительно функционально определяется способностью связываться с любым одним или несколькими из консервативного элемента последовательности альфавируса 1 (CSE 1) или его комплементарной последовательности, консервативного элемента последовательности 2 альфавируса (CSE 2) или его комплементарной последовательности, консервативного элемента последовательности альфавируса 3 (CSE 3) или его комплементарной последовательности, консервативного элемента последовательности 4 альфавируса (CSE 4) или его комплементарной последовательности. Предпочтительно репликаза способна связываться с CSE 2 [т.е. с (+)-нитью] и/или с CSE 4 [т.е. с (+)-нитью] или связываться с комплементом CSE 1 [т.е. с (-)-нитью] и/или с комплементом CSE 3 [т.е. с (-)-нитью].
Происхождение репликазы не ограничивается каким-либо конкретным альфавирусом. Согласно предпочтительному варианту осуществления альфавирусная репликаза содержит неструктурный белок из вируса леса Семлики, включая в себя встречающийся в природе вируса леса Семлики и вариант или производное вируса леса Семлики, например, аттенуированный вирус леса Семлики. Согласно альтернативному предпочтительному варианту осуществления альфавирусная репликаза содержит неструктурный белок из вируса Синдбис, включая в себя природный вирус Синдбис и вариант или производное вируса Синдбис, такое как аттенуированный вирус Синдбис. Согласно альтернативному предпочтительному варианту осуществления альфавирусная репликаза содержит неструктурный белок из вируса венесуэльского лошадиного энцефалита (VEEV), включая в себя природный VEEV и вариант или производное VEEV, такое как аттенуированный VEEV.
Репликаза может также содержать неструктурные белки более чем из одного альфавируса. Таким образом, гетерологичные комплексы или объединения, содержащие альфавирусный неструктурный белок и характеризующиеся функцией репликазы, одинаково включены в настоящее изобретение. Для иллюстративных целей репликаза может содержать один или несколько неструктурных белков (например, nsP1, nsP2) из первого альфавируса и один или несколько неструктурных белков (nsP3, nsP4) из второго альфавируса. Неструктурные белки более чем из одного другого альфавируса могут кодироваться отдельными открытыми рамками считывания или могут кодироваться единственной открытой рамкой считывания в виде полипротеина, например, nsP1234.
Согласно некоторым вариантам осуществления функциональный альфавирусный неструктурный белок способен образовывать мембранные комплексы репликации и/или вакуоли в клетках, в которых экспрессируется функциональный альфавирусный неструктурный белок.
Если функциональный альфавирусный неструктурный белок, т.е. альфавирусный неструктурный белок с функцией репликазы, кодируется молекулой нуклеиновой кислоты в соответствии с настоящим изобретением, предпочтительно, чтобы субгеномный промотор репликона, если он присутствует, был совместим с указанной репликазой. Совместимость в этом контексте означает, что альфавирусная репликаза способна распознавать субгеномный промотор, если он присутствует. Согласно одному варианту осуществления это достигается, когда субгеномный промотор является нативным для альфавируса, из которого получена репликаза, т.е. естественное происхождение этих последовательностей представляет собой один и тот же альфавирус. Согласно альтернативному варианту осуществления субгеномный промотор не является нативным для альфавируса, из которого получена альфавирусная репликаза, при условии, что альфавирусная репликаза способна распознавать субгеномный промотор. Другими словами, репликаза совместима с субгеномным промотором (межвирусная совместимость). Примеры межвирусной совместимости в отношении субгеномного промотора и репликазы, происходящих из разных альфавирусов, известны в настоящей области техники. Любая комбинация субгеномного промотора и репликазы возможна, если существует межвирусная совместимость. Межвирусная совместимость может быть легко исследована специалистом, работающим по настоящему изобретению, путем инкубирования исследуемой репликазы вместе с РНК, причем РНК содержит субгеномный промотор, подлежащий исследованию, в условиях, подходящих для синтеза РНК из субгеномного промотора. Если получают субгеномный транскрипт, субгеномный промотор и репликаза определяются как совместимые. Известны различные примеры межвирусной совместимости (обзор представлен в публикации Strauss & Strauss, Microbiol. Rev., 1994, vol. 58, pp. 491-562).
Согласно одному варианту осуществления альфавирусный неструктурный белок не кодируется как слитый белок с гетерологичным белком, например, убиквитин.
В настоящем изобретении открытая рамка считывания, кодирующая функциональный альфавирусный неструктурный белок, может быть предусмотрена на РНК-репликоне или, альтернативно, может быть предусмотрена в виде отдельной молекулы нуклеиновой кислоты, например, мРНК. Отдельная молекула мРНК может необязательно содержать, например, кэп, 5'-UTR, 3'-UTR, поли(A)-последовательность и/или адаптацию использования кодонов. Отдельная молекула мРНК может быть предусмотрена in trans, как описано в настоящем документе для системы по настоящему изобретению.
Когда в РНК-репликоне предусмотрена открытая рамка считывания, кодирующая функциональный альфавирусный неструктурный белок, репликон предпочтительно может быть реплицирован функциональным альфавирусным неструктурным белком. В частности, РНК-репликон, который кодирует функциональный альфавирусный неструктурный белок, может быть реплицирован функциональным альфавирусным неструктурным белком, кодируемым репликоном. Этот вариант осуществления является сильно предпочтительным, когда никакая нуклеиновая кислота, кодирующая функциональный альфавирусный неструктурный белок, не предусмотрена in trans. Согласно этому варианту осуществления на это нацелена цис-репликация репликона. Согласно предпочтительному варианту осуществления РНК-репликон содержит открытую рамку считывания, кодирующую функциональный альфавирусный неструктурный белок, а также дополнительную открытую рамку считывания, кодирующую представляющий интерес белок, и может быть реплицирован функциональным альфавирусным неструктурным белком. Этот вариант осуществления особенно подходит для некоторых способов получения представляющего интерес белка в соответствии с настоящим изобретением. Пример соответствующего репликона показан на фиг. 1 («цис-репликон Δ5ATG-RRS»).
Если репликон содержит открытую рамку считывания, кодирующую функциональный альфавирусный неструктурный белок, предпочтительно, чтобы открытая рамка считывания, кодирующая функциональный альфавирусный неструктурный белок, не перекрывалась с 5' репликационной последовательностью распознавания. Согласно одному варианту осуществления открытая рамка считывания, кодирующая функциональный альфавирусный неструктурный белок, не перекрывается с субгеномным промотором, если он присутствует. Пример соответствующего репликона показан на фиг. 1 («цис-репликон Δ5ATG-RRS»).
Если на репликоне присутствует множество открытых рамок считывания, то функциональный альфавирусный неструктурный белок может кодироваться любой из них, необязательно под контролем субгеномного промотора или без него, предпочтительно не под контролем субгеномного промотора. Согласно предпочтительному варианту осуществления функциональный альфавирусный неструктурный белок кодируется расположенной выше всех против хода транскрипции открытой рамкой считывания РНК-репликона. Когда функциональный альфавирусный неструктурный белок кодируется расположенной выше всех против хода транскрипции открытой рамкой считывания РНК-репликона, генетическая информация, кодирующая функциональный альфавирусный неструктурный белок, будет транслирована сразу после введения РНК-репликона в клетку-хозяина, и в результате белок может впоследствии управлять репликацией и, возможно, производить субгеномный транскрипт в клетке-хозяине. Пример соответствующего репликона показан на фиг. 1 («цис-репликон Δ5ATG-RRS»).
Наличие открытой рамки считывания, кодирующей функциональный альфавирусный неструктурный белок, либо содержащейся в репликоне, либо содержащейся в отдельной молекуле нуклеиновой кислоты, которая предусмотрена in trans, позволяет реплицировать репликон, и, следовательно, что представляющий интерес ген, кодируемый репликоном, необязательно под контролем субгеномного промотора, экспрессируется на высоком уровне. Это связано с преимуществом в затратах по сравнению с другими трансгенными экспрессионными системами. Например, в случае вакцинации животных стоимость вакцины является ключом к ее успеху в ветеринарии и сельскохозяйственном сообществе. Поскольку репликон по настоящему изобретению может быть реплицирован в присутствии функционального альфавирусного неструктурного белка, например, в клетке вакцинированного животного могут быть достигнуты высокие уровни экспрессии представляющего интерес гена, даже если вводятся относительно небольшие количества РНК-репликона. Низкие количества РНК-репликона положительно влияют на стоимость вакцины на одного субъекта.
Положение по меньшей мере одной открытой рамки считывания в репликоне РНК
РНК-репликон подходит для экспрессии одного или нескольких генов, кодирующих представляющий интерес пептид или представляющий интерес белок, необязательно под контролем субгеномного промотора. Возможны различные варианты осуществления. В РНК-репликоне может присутствовать одна или несколько открытых рамок считывания, каждая из которых кодирует представляющий интерес пептид или представляющий интерес белок. Расположенная выше всех против хода транскрипции открытая рамка считывания РНК-репликона называется «первой открытой рамкой считывания». Согласно некоторым вариантам осуществления «первая открытая рамка считывания» представляет собой единственную открытую рамку считывания РНК-репликона. Необязательно, одна или несколько дополнительных открытых рамок считывания могут присутствовать ниже по ходу транскрипции от открытой рамки считывания. Одна или несколько дополнительных открытых рамок считывания ниже по ходу транскрипции от первой открытой рамки считывания могут упоминаться как «вторая открытая рамка считывания», «третья открытая рамка считывания» и т.д. в порядке (от 5' к 3'), в котором они находятся ниже по ходу транскрипции от первой открытой рамки считывания. Предпочтительно каждая открытая рамка считывания содержит стартовый кодон (триплет оснований), как правило, AUG (в молекуле РНК), соответствующий ATG (в соответствующей молекуле ДНК).
Если репликон содержит 3' репликационную последовательность распознавания, предпочтительно, чтобы все открытые рамки считывания были локализованы выше против хода транскрипции от 3' репликационной последовательности распознавания.
Когда РНК-репликон, содержащий одну или несколько открытых рамок считывания, вводится в клетку-хозяина, трансляция предпочтительно не инициируется в любом положении выше против хода транскрипции от первой открытой рамки считывания из-за удаления по меньшей мере одного инициирующего кодона из 5' репликационной последовательности распознавания. Следовательно, репликон может служить непосредственно в качестве матрицы для трансляции первой открытой рамки считывания. Предпочтительно репликон содержит 5'-кэп. Это полезно для экспрессии гена, кодируемого первой открытой рамкой считывания, непосредственно из репликона.
Согласно некоторым вариантам осуществления по меньшей мере одна открытая рамка считывания репликона находится под контролем субгеномного промотора, предпочтительно субгеномного промотора альфавируса. Субгеномный промотор альфавируса очень эффективен и поэтому подходит для гетерологичной экспрессии генов на высоких уровнях. Предпочтительно субгеномный промотор представляет собой промотор для субгеномной транскрипции в альфавирусе. Это означает, что субгеномный промотор является нативным по отношению к альфавирусу и предпочтительно контролирует транскрипцию открытой рамки считывания, кодирующей один или несколько структурных белков в указанном альфавирусе. Альтернативно, субгеномный промотор представляет собой вариант субгеномного промотора альфавируса; любой вариант, который функционирует как промотор для субгеномной транскрипции РНК в клетке-хозяине, является подходящим. Если репликон содержит субгеномный промотор, предпочтительно, чтобы репликон содержал консервативный элемент последовательности 3 (CSE 3) или его вариант.
Предпочтительно, чтобы по меньшей мере одна открытая рамка считывания под контролем субгеномного промотора была локализована ниже по ходу транскрипции от субгеномного промотора. Предпочтительно, субгеномный промотор контролирует производство субгеномной РНК, содержащей транскрипт открытой рамки считывания
Согласно некоторым вариантам осуществления первая открытая рамка считывания находится под контролем субгеномного промотора. Когда первая открытая рамка считывания находится под контролем субгеномного промотора, ее локализация напоминает локализацию открытой рамки считывания, кодирующей структурные белки в геноме альфавируса. Когда первая открытая рамка считывания находится под контролем субгеномного промотора, ген, кодируемый первой открытой рамкой считывания, может экспрессироваться как из репликона, так и из его субгеномного транскрипта (последний в присутствии функционального альфавирусного неструктурного белка). Соответствующий вариант осуществления иллюстрируется репликоном «Δ5ATG-RRS» на фиг. 1. Предпочтительно «Δ5ATG-RRS» не содержит никакого инициирующего кодона в последовательности нуклеиновой кислоты, кодирующей C-концевой фрагмент nsP4 (*nsP4). Одна или несколько дополнительных открытых рамок считывания, каждая под контролем субгеномного промотора, может присутствовать ниже по ходу транскрипции от первой открытой рамки считывания, которая находится под контролем субгеномного промотора (не показана на фиг. 1). Гены, кодируемые одной или несколькими дополнительными открытыми рамками считывания, например, посредством второй открытой рамки считывания, могут быть транслированы из одного или нескольких субгеномных транскриптов, каждый из которых находится под контролем субгеномного промотора. Например, РНК-репликон может содержать субгеномный промотор, контролирующий производство транскрипта, который кодирует второй представляющий интерес белок.
Согласно другим вариантам осуществления первая открытая рамка считывания не находится под контролем субгеномного промотора. Когда первая открытая рамка считывания не находится под контролем субгеномного промотора, ген, кодируемый первой открытой рамкой считывания, может экспрессироваться из репликона. Соответствующий вариант осуществления иллюстрируется репликоном «Δ5ATG-RRSΔSGP» на фиг. 1. Одна или несколько дополнительных открытых рамок считывания, каждая под контролем субгеномного промотора, может присутствовать ниже по ходу транскрипции от первой открытой рамки считывания (для иллюстрации двух иллюстративных вариантов осуществления смотрите «Δ5ATG-RRS-бицистронный» и «цис-репликон Δ5ATG-RRS» на фиг. 1). Гены, кодируемые одной или несколькими дополнительными открытыми рамками считывания, могут экспрессироваться из субгеномных транскриптов.
В клетке, которая содержит репликон в соответствии с настоящим изобретением, репликон может быть амплифицирован функциональным альфавирусным неструктурным белком. Кроме того, если репликон содержит одну или несколько открытых рамок считывания под контролем субгеномного промотора, ожидается, что один или несколько субгеномных транскриптов будут получены функциональным альфавирусным неструктурным белком. Функциональный альфавирусный неструктурный белок может быть предусмотрен in trans или может кодироваться открытой рамкой считывания репликона.
Если репликон содержит более одной открытой рамки считывания, кодирующей представляющий интерес белок, предпочтительно, чтобы каждая открытая рамка считывания кодировала другой белок. Например, белок, кодируемый второй открытой рамкой считывания, отличается от белка, кодируемого первой открытой рамкой считывания.
Согласно некоторым вариантам осуществления представляющий интерес белок, кодируемый первой и/или дальнейшей открытой рамкой считывания, предпочтительно первой открытой рамкой считывания, представляет собой функциональный альфавирусный неструктурный белок или ингибитор передачи сигналов IFN, например, E3. Согласно некоторым вариантам осуществления представляющий интерес белок, кодируемый первой и/или дальнейшей открытой рамкой считывания, например, второй открытой рамкой считывания, представляет собой фармацевтически активный пептид или белок или репортерный белок.
Согласно одному варианту осуществления представляющий интерес белок, кодируемый первой открытой рамкой считывания, представляет собой функциональный альфавирусный неструктурный белок. Согласно этому варианту осуществления репликон предпочтительно содержит 5'-кэп. В частности, когда представляющий интерес белок, кодируемый первой открытой рамкой считывания, представляет собой функциональный альфавирусный неструктурный белок, и, предпочтительно, когда репликон содержит 5'-кэп, последовательность нуклеиновой кислоты, кодирующая функциональный альфавирусный неструктурный белок, может быть эффективно транслирована из репликона, и полученный белок может впоследствии управлять репликацией репликона и управлять синтезом субгеномного транскрипта(ов). Этот вариант осуществления может быть предпочтительным, когда никакая дополнительная молекула нуклеиновой кислоты, кодирующая функциональный альфавирусный неструктурный белок, не используется или не присутствует вместе с репликоном. Согласно этому варианту осуществления цис-репликация нацелена на репликон.
Один вариант осуществления, при котором первая открытая рамка считывания кодирует функциональный альфавирусный неструктурный белок, иллюстрируется посредством «цис-репликона Δ5ATG-RRS» на фиг. 1. После трансляции последовательности нуклеиновой кислоты, кодирующей nsP1234, продукт трансляции (nsP1234 или его фрагмент(ы)) может выступать в качестве репликазы и управлять синтезом РНК, т.е. репликацией репликона и синтезом субгеномного транскрипта, содержащего вторую открытую рамку считывания («трансген» на фиг. 1).
Система транс-репликации
Согласно второму аспекту в настоящем изобретение предусмотрена система, содержащая:
конструкцию РНК для экспрессии функционального альфавируса неструктурного белка,
РНК-репликон в соответствии с первым аспектом настоящего изобретения, который может быть реплицирован функциональным альфавирусным неструктурным белком in trans.
Согласно второму аспекту предпочтительно, чтобы РНК-репликон не содержал открытую рамку считывания, кодирующую функциональный альфавирусный неструктурный белок.
Таким образом, в настоящем изобретении предусмотрена система, содержащая две молекулы нуклеиновой кислоты: первую конструкцию РНК для экспрессии функционального альфавирусного неструктурного белка (то есть кодирующую функциональный альфавирусный неструктурный белок); и вторую молекулу РНК, РНК-репликон. Конструкция РНК для экспрессии функционального альфавирусного неструктурного белка синонимично называется в настоящем документе как «конструкция РНК для экспрессии функционального альфавирусного неструктурного белка» или как «репликазная конструкция».
Функциональный альфавирусный неструктурный белок является таким, как определено выше, и, как правило, кодируется открытой рамкой считывания, содержащейся в репликазной конструкции. Функциональный альфавирусный неструктурный белок, кодируемый репликазной конструкцией, может представлять собой любой функциональный альфавирусный неструктурный белок, который способен реплицировать репликон.
Когда система по настоящему изобретению вводится в клетку, предпочтительно эукариотическую клетку, может быть транслирована открытая рамка считывания, кодирующая функциональный альфавирусный неструктурный белок. После трансляции функциональный альфавирусный неструктурный белок способен реплицировать отдельную молекулу РНК (РНК-репликон) in trans. Таким образом, в настоящем изобретении предусмотрена система для репликации РНК in trans. Следовательно, система по настоящему изобретению представляет собой систему транс-репликации. Согласно второму аспекту, репликон представляет собой транс-репликон.
В настоящем документе транс (например, в контексте транс-действующего, транс-регуляторного), в общем, означает «действие из другой молекулы» (то есть межмолекулярное). Это противоположно цис (например, в контексте цис-действующего, цис-регуляторного), что, в общем, означает «действующий из той же молекулы» (то есть внутримолекулярный). В контексте синтеза РНК (включая в себя транскрипцию и репликацию РНК) транс-действующий элемент включает в себя последовательность нуклеиновой кислоты, которая содержит ген, кодирующий фермент, способный синтезировать РНК (РНК-полимераза). РНК-полимераза использует вторую молекулу нуклеиновой кислоты, то есть молекулу нуклеиновой кислоты, отличную от той, которой она кодируется, в качестве матрицы для синтеза РНК. Говорят, что как РНК-полимераза, так и последовательность нуклеиновой кислоты, которая содержит ген, кодирующий РНК-полимеразу, «действуют in trans» на вторую молекулу нуклеиновой кислоты. В контексте настоящего изобретения РНК-полимераза, кодируемая транс-действующей РНК, представляет собой функциональный альфавирусный неструктурный белок. Функциональный альфавирусный неструктурный белок способен использовать вторую молекулу нуклеиновой кислоты, которая представляет собой РНК-репликон, в качестве матрицы для синтеза РНК, включая в себя репликацию РНК-репликона. РНК-репликон, который может быть реплицирован репликазой in trans в соответствии с настоящим изобретением, синонимично упоминается в настоящем документе как «транс-репликон».
В системе настоящего изобретения роль функционального альфавирусного неструктурного белка заключается в амплификации репликона и получении субгеномного транскрипта, если на репликоне присутствует субгеномный промотор. Если репликон кодирует представляющий интерес ген для экспрессии, уровни экспрессии представляющего интерес гена и/или продолжительность экспрессии могут регулироваться in trans путем модификации уровней функционального альфавирусного неструктурного белка.
Тот факт, что альфавирусная репликаза, как правило, способна распознавать и реплицировать РНК-матрицу in trans, была первоначально обнаружена в 1980-х годах, но потенциал транс-репликации для биомедицинских применений не был признан, в частности, поскольку транс-реплицированная РНК считалась ингибирующей эффективную репликацию: она была обнаружена в случае дефектной интерферирующей (DI) РНК, которая совместно реплицирует альфавирусные геномы в инфицированных клетках (Barrett et al., 1984, J. Gen. Virol., vol. 65 ( Pt 8), pp. 1273-1283; Lehtovaara et al., 1981, Proc. Natl. Acad. Sci. U. S. A, vol. 78, pp. 5353-5357; Pettersson, 1981, Proc. Natl. Acad. Sci. U. S. A, vol. 78, pp. 115-119). DI РНК представляют собой транс-репликоны, которые могут происходить квази-естественным образом во время инфекций клеточных линий с высокой вирусной нагрузкой. DI-элементы кореплицируются настолько эффективно, что они уменьшают вирулентность родительского вируса и, таким образом, действуют как ингибирующая паразитная РНК (Barrett et al., 1984, J. Gen. Virol., Vol. 65 (Pt 11), pp. 1909-1920). Хотя потенциал для биомедицинских применений не был распознан, феномен транс-репликации использовался в нескольких базовых исследованиях, направленных на выяснение механизмов репликации без необходимости экспрессировать репликазу из той же молекулы in cis; кроме того, разделение репликазы и репликона также позволяет проводить функциональные исследования с участием мутантов вирусных белков, даже если соответствующие мутанты представляют собой мутанты с потерями функциональности (Lemm et al., 1994, EMBO J., vol. 13, pp. 2925-2934). Эти исследования потери функции и DI РНК не предполагали, что системы транс-активации, основанные на альфавирусных элементах, могут в конечном итоге стать доступными для терапевтических целей.
Система по настоящему изобретению содержит по меньшей мере две молекулы нуклеиновой кислоты. Таким образом, она может содержать две или более, три или более, четыре или более, пять или более, шесть или более, семь или более, восемь или более, девять или более или десять или более молекул нуклеиновой кислоты, которые предпочтительно представляют собой молекулы РНК. Согласно предпочтительному варианту осуществления система состоит ровно из двух молекул РНК, репликона и репликазной конструкции. Согласно альтернативным предпочтительным вариантам осуществления система содержит более одного репликона, каждый из которых предпочтительно кодирует по меньшей мере один представляющий интерес белок, а также содержит репликазную конструкцию. Согласно этим вариантам осуществления функциональный альфавирусный неструктурный белок, кодируемый репликазной конструкцией, может действовать на каждый репликон для управления репликацией и производства субгеномных транскриптов, соответственно. Например, каждый репликон может кодировать фармацевтически активный пептид или белок. Это выгодно, например, если требуется вакцинация субъекта против нескольких разных антигенов.
Предпочтительно, для репликазной конструкции не хватает по меньшей мере одного консервативного элемента последовательности (CSE), который необходим для синтеза (-)-нити на основе матрицы (+)-нити и/или для синтеза (+)-нити на основе матрицы (-)-нити. Более предпочтительно, репликазная конструкция не содержит каких-либо альфавирусных консервативных элементов последовательности (CSE). В частности, среди четырех CSE альфавирусов (Strauss & Strauss, Microbiol. Rev., 1994, vol. 58, pp. 491-562; José et al., Future Microbiol., 2009, vol. 4, pp. 837-856), любая одна или несколько из следующих CSE предпочтительно не присутствуют на репликазной конструкции: CSE 1; CSE 2; CSE 3; CSE 4. В частности, в отсутствие какой-либо одной или нескольких альфавирусных CSE репликазная конструкция по настоящему изобретению напоминает типичную эукариотическую мРНК намного больше, чем она напоминает альфавирусную геномную РНК.
Репликазная конструкция по настоящему изобретению отличается от альфавирусной геномной РНК по меньшей мере тем, что она не способна к саморепликации и/или что она не содержит открытую рамку считывания под контролем субгеномного промотора. Когда она не может самореплицироваться, репликазная конструкция может также называться «суицидная конструкция».
Система транс-репликации связана со следующими преимуществами:
Прежде всего, универсальность системы транс-репликации позволяет разрабатывать и/или получать репликон и репликазную конструкцию в разное время и/или в разных местах . Согласно одному варианту осуществления репликазную конструкцию получают в первый момент времени, а репликон получают в более поздний момент времени. Например, после его получения репликазная конструкция может быть сохранена для использования в более поздний момент времени. В настоящем изобретении предусмотрена повышенная гибкость по сравнению с цис-репликонами: при появлении нового патогена система по настоящему изобретению может быть разработана для вакцинации путем клонирования в репликон нуклеиновой кислоты, кодирующей полипептид, который вызывает иммунный ответ против нового патогена. Предварительно полученную репликазную конструкцию можно восстановить из хранилища. Таким образом, не требуется, чтобы во время разработки и получения репликазной конструкции была известна природа конкретного патогена или антигена(ов) конкретного патогена. Следовательно, не требуется, чтобы во время разработки и получения репликазной конструкции, был доступен репликон, кодирующий полипептид, который вызывает иммунный ответ против конкретного нового патогена. Другими словами, репликазная конструкция может быть разработана и получена независимо от любого конкретного репликона. Это позволяет быстро реагировать на появление новых патогенов или на патогены, характеризующиеся экспрессией по меньшей мере одного нового антигена, поскольку получение репликона, лишенного репликазы, требует меньше усилий и ресурсов, чем получение цис-репликонов. История говорит, что необходима система, позволяющая быстро реагировать на патогены: это иллюстрируется, например, появлением в последние годы патогенов, вызывающих тяжелый острый респираторный синдром (SARS), Эбола и различные подтипы вируса гриппа.
Во-вторых, транс-репликон в соответствии с настоящим изобретением, как правило, представляет собой более короткую молекулу нуклеиновой кислоты, чем типичный цис-репликон. Это обеспечивает более быстрое клонирование репликона, кодирующего представляющий интерес белок, например, иммуногенный полипептид, и обеспечивает высокие выходы представляющего интерес белка.
Дополнительные преимущества системы по настоящему изобретению включают в себя независимость от ядерной транскрипции и наличия ключевой генетической информации о двух отдельных молекулах РНК, что обеспечивает беспрецедентную свободу дизайна. Ввиду своих универсальных элементов, которые сочетаются друг с другом, настоящее изобретение позволяет оптимизировать экспрессию репликазы для желаемого уровня амплификации РНК, для желаемого целевого организма, для желаемого уровня производства представляющего интерес белка и т.д. Система в соответствии с настоящим изобретением позволяет совместно трансфицировать различные количества или соотношения репликона и репликазной конструкции для любого данного типа клеток - покоящегося или циклирующего, in vitro или in vivo. Система транс-репликации по настоящему изобретению подходит для инокуляции клетки-хозяина и для экспрессии представляющего интерес гена в клетке-хозяине (смотрите, например, примеры 2, 3 и 5).
Репликазная конструкция в соответствии с настоящим изобретением предпочтительно представляет собой одноцепочечную молекулу РНК. Репликазная конструкция в соответствии с настоящим изобретением, как правило, представляет собой (+)-нитевую молекулу РНК. Согласно одному варианту осуществления репликазная конструкция по настоящему изобретению представляет собой выделенную молекулу нуклеиновой кислоты.
Предпочтительные особенности молекул РНК в соответствии с настоящим изобретением
Молекулы РНК в соответствии с настоящим изобретением могут быть необязательно охарактеризованы дополнительными особенностями, например, структурами 5'-кэп, 5'-UTR, 3'-UTR, поли(A)-последовательность и/или адаптацией использования кодонов. Подробности описаны ниже.
Кэп
Согласно некоторым вариантам осуществления репликон в соответствии с настоящим изобретением содержит 5'-кэп.
Согласно некоторым вариантам осуществления репликазная конструкция в соответствии с настоящим изобретением содержит 5'-кэп.
Термины «5'-кэп», «кэп», «структура 5'-кэп», «структура кэп» используются синонимично для обозначения динуклеотида, который находится на 5'-конце некоторых первичных транскриптов эукариот, таких как предшественник матричной РНК. 5'-кэп представляет собой структуру, в которой (необязательно модифицированный) гуанозин связан с первым нуклеотидом молекулы мРНК через 5'-5'-трифосфатную связь (или модифицированную трифосфатную связь в случае некоторых аналогов кэпа). Термины могут относиться к обычному кэпу или к аналогу кэпа. Для иллюстрации некоторые конкретные динуклеотиды кэпа (включая в себя аналоги динуклеотидов кэпа) показаны на фиг. 6.
Термин «РНК, которая содержит 5'-кэп» или «РНК, которая предусмотрена с 5'-кэпом», или «РНК, которая модифицирована 5'-кэпом», или «кэпированная РНК», относится к РНК, которая содержит 5'-кэп. Например, обеспечение РНК 5'-кэпом может быть достигнуто путем транскрипции in vitro матрицы ДНК в присутствии указанного 5'-кэпа, причем указанный 5'-кэп котранскрипционно встраивается в произведенную цепь РНК или РНК может быть получена, например, путем транскрипции in vitro, и 5'-кэп может быть присоединен к РНК после транскрипции с использованием кэпирующих ферментов, например, кэпирующих ферментов вируса осповакцины. В кэпированной РНК 3'-положение первого основания (кэпированной) молекулы РНК связано с 5'-положением последующего основания молекулы РНК («второе основание») через фосфодиэфирную связь.
Присутствие кэпа на молекуле РНК является очень предпочтительным, если желательна трансляция последовательности нуклеиновой кислоты, кодирующей белок на ранних стадиях после введения соответствующей РНК в клетки-хозяева или в организм-хозяин. Например, как показано в примере 4, наличие кэпа делает возможным, чтобы представляющий интерес ген, закодированный РНК-репликоном, эффективно транслировался на ранних стадиях после введения соответствующей РНК в клетки-хозяева. Термин «ранние стадии», как правило, означает в течение первого часа или в течение первых двух часов, или в течение первых трех часов после введения РНК.
Присутствие кэпа на молекуле РНК также является предпочтительным, если желательно, чтобы трансляция происходила в отсутствие функциональной репликазы или когда в клетке-хозяине присутствуют только незначительные уровни репликазы. Например, даже если молекула нуклеиновой кислоты, кодирующая репликазу, вводится в клетку-хозяина, на ранних стадиях после введения уровни репликазы, как правило, будут незначительными.
В системе в соответствии с настоящим изобретением предпочтительно, чтобы конструкция РНК для экспрессии функционального альфавирусного неструктурного белка содержала 5'-кэп.
В частности, когда РНК-репликон в соответствии с настоящим изобретением не используется или не предусмотрен вместе со второй молекулой нуклеиновой кислоты (например, мРНК), которая кодирует функциональный альфавирусный неструктурный белок, предпочтительно, чтобы РНК-репликон содержал 5'-кэп. Независимо, РНК-репликон может также содержать 5'-кэп, даже если он используется или предусмотрен вместе со второй молекулой нуклеиновой кислоты, которая кодирует функциональный альфавирусный неструктурный белок.
Термин «обычный 5'-кэп» относится к встречающемуся в природе 5'-кэпу, предпочтительно к 7-метилгуанозиновому кэпу. В 7-метилгуанозиновом кэпе гуанозин кэпа представляет собой модифицированный гуанозин, в котором модификация состоит из метилирования в 7-положении (вверху на фиг. 6).
В контексте настоящего изобретения термин «аналог 5'-кэпа» относится к молекулярной структуре, которая напоминает обычный 5'-кэп, но модифицирована, чтобы обладать способностью стабилизировать РНК, если она присоединена к ней, предпочтительно in vivo и/или в клетке. Аналог кэпа не представляет собой обычный 5'-кэп.
В случае эукариотической мРНК 5'-кэп также, как правило, был описан как включенный в эффективную трансляцию мРНК: в целом у эукариот трансляция инициируется только на 5'-конце молекулы матричной РНК (мРНК), если IRES отсутствует. Эукариотические клетки способны обеспечить РНК 5'-кэпом во время транскрипции в ядре: вновь синтезированные мРНК, как правило, модифицируют с помощью структуры 5'-кэпа, например, когда транскрипт достигает длины от 20 до 30 нуклеотидов. Сначала 5'-концевой нуклеотид pppN (ppp, представляющий собой трифосфат, N, представляющий собой любой нуклеозид), превращается в клетке в 5' GpppN с помощью кэпирующего фермента, характеризующегося 5'-трифосфатазной и гуанилилтрансферазной активностью РНК. Затем GpppN может быть метилирован в клетке вторым ферментом с (гуанин-7)-метилтрансферазной активностью с образованием монометилированного кэпа m7GpppN. Согласно одному варианту осуществления 5'-кэп, используемый в настоящем изобретении, представляет собой природный 5'-кэп.
В настоящем изобретении природный 5'-кэп-динуклеотид, как правило, выбирают из группы, состоящей из неметилированного кэпа-динуклеотида (G(5')ppp(5')N, также называемого GpppN) и метилированного кэпа-динуклеотида ((m7G(5')ppp(5')N, также называемого m7GpppN). m7GpppN (где N представляет собой G) представлен следующей формулой:
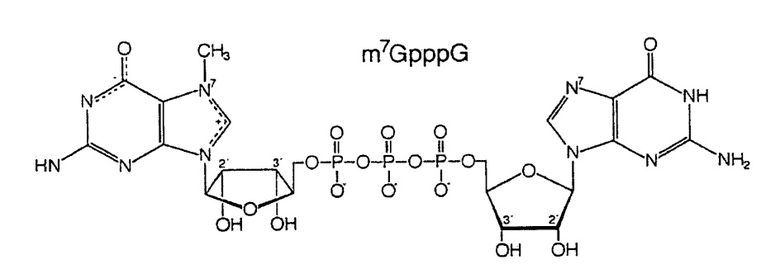
Кэпированная РНК по настоящему изобретению может быть получена in vitro и, следовательно, не зависит от инструмента кэпирования в клетке-хозяине. Наиболее часто используемый способ получения кэпированных РНК in vitro состоит в том, чтобы транскрибировать ДНК-матрицу либо с помощью бактериальной, либо с помощью бактериофаговой РНК-полимеразы в присутствии всех четырех рибонуклеозидтрифосфатов и кэпа-динуклеотида, такого как m7G(5')ppp(5')G (также называемого m7GpppG). РНК-полимераза инициирует транскрипцию с нуклеофильной атакой 3'-OH гуанозинового фрагмента m7GpppG на α-фосфате следующего матричного нуклеозидтрифосфата (pppN), в результате чего получают промежуточное m7GpppGpN (где N является вторым основанием молекулы РНК). Образование конкурирующего инициированного GTP продукта pppGpN подавляется путем установления молярного отношения кэпа к GTP от 5 до 10 при транскрипции in vitro.
Согласно предпочтительным вариантам осуществления настоящего изобретения 5'-кэп (если присутствует) представляет собой аналог 5'-кэпа. Эти варианты осуществления особенно подходят, если РНК получена путем транскрипции in vitro, например, представляет собой транскрибированную in vitro РНК (IVT-РНК). Аналоги кэпа изначально описаны для облегчения крупномасштабного синтеза транскриптов РНК посредством транскрипции in vitro.
Для матричной РНК некоторые известные аналоги кэпа (синтетические структуры кэп), как правило, описаны на сегодняшний день, и все они могут использоваться в контексте настоящего изобретения. В идеале выбирают аналог кэпа, который связан с более высокой эффективностью трансляции и/или повышенной устойчивостью к деградации in vivo и/или повышенной устойчивостью к деградации in vitro.
Предпочтительно используют аналог кэпа, который может быть включен только в цепь РНК в одной ориентации. В публикации Pasquinelli et al. (1995, RNA J., vol., 1, pp. 957-967) показано, что во время транскрипции in vitro бактериофаговые РНК-полимеразы используют блок 7-метилгуанозина для инициирования транскрипции, при этом приблизительно 40-50% транскриптов с кэпом обладают кэпом-динуклеотидом в обратной ориентации (т.е. исходный продукт реакции представляет собой Gpppm7GpN). По сравнению с РНК с правильным кэпом, РНК с обратным кэпом не являются функциональными по отношению к трансляции последовательности нуклеиновой кислоты в белок. Таким образом, желательно включить кэп в правильной ориентации, то есть в результате получить РНК со структурой, по существу соответствующей m7GpppGpN и т.д. Было показано, что обратное интегрирование кэпа-динуклеотида ингибируется замещением либо 2'-, либо 3'-OH группы метилированного гуанозина (Stepinski et al., 2001; RNA J., vol. 7, pp. 1486-1495; Peng et al., 2002; Org. Lett., vol. 24, pp. 161-164). РНК, которые синтезируются в присутствии таких «аналогов кэп-структур с правильной ориентацией», транслируются более эффективно, чем РНК, которые транскрибируются in vitro в присутствии обычного 5'-кэпа m7GpppG. С этой целью один аналог кэпа, в котором группа 3'-OH метилированного гуанозина замещена OCH3, описан в публикации Holtkamp et al., 2006, Blood, vol. 108, pp. 4009-4017 (7-метил(3'-O-метил) GpppG, аналог кэп-структуры с правильной ориентацией (ARCA)). ARCA является подходящим кэпом-динуклеотидом в соответствии с настоящим изобретением.
Согласно предпочтительному варианту осуществления настоящего изобретения РНК по настоящему изобретению по существу не подвержена декэпированию. Это важно, потому что в целом количество белка, полученного из синтетических мРНК, вводимых в культивируемые клетки млекопитающих, ограничено естественной деградацией мРНК. Один из путей in vivo для деградации мРНК начинается с удаления кэпа мРНК. Это удаление катализируется гетеродимерной пирофосфатазой, которая содержит регуляторную субъединицу (Dcp1) и каталитическую субъединицу (Dcp2). Каталитическая субъединица расщепляется между α- и β-фосфатными группами трифосфатного мостика. В настоящем изобретении может быть выбран или может присутствовать аналог кэпа, который не восприимчив или менее восприимчив к этому типу расщепления. Соответствующий аналог кэпа для этой цели может быть выбран из кэпа-динуклеотида согласно формуле (I):
где R1 выбран из группы, содержащей необязательно замещенный алкил, необязательно замещенный алкенил, необязательно замещенный алкинил, необязательно замещенный циклоалкил, необязательно замещенный гетероциклил, необязательно замещенный арил и необязательно замещенный гетероарил,
R2 и R3 независимо выбраны из группы, состоящей из H, галогена, ОН и необязательно замещенного алкокси, или R2 и R3 вместе образуют O-X-O, где X выбран из группы, состоящей из необязательно замещенных CH2, CH2CH2, CH2CH2CH2, CH2CH(CH3) и C(CH3)2, или R2 объединяют с атомом водорода в положении 4' кольца, к которому присоединен R2, с образованием -O-CH2- или -CH2-O-,
R5 выбран из группы, состоящей из S, Se и BH3,
R4 и R6 независимо выбраны из группы, состоящей из O, S, Se и BH3,
n равно 1, 2 или 3.
Предпочтительные варианты осуществления для R1, R2, R3, R4, R5, R6 раскрыты в публикации международной заявки WO 2011/015347 A1 и могут быть соответственно выбраны в настоящем изобретении.
Например, согласно предпочтительному варианту осуществления настоящего изобретения РНК по настоящему изобретению содержит фосфоротиоатный аналог кэпа. Фосфоротиоатные аналоги кэпа представляют собой специфические аналоги кэпа, в которых один из трех немостиковых атомов O в трифосфатной цепи замещен атомом S, то есть один из R4, R5 или R6 в формуле (I) представляет собой S. Фосфоротиоатные аналоги кэпа были описаны в публикации J. Kowalska et al., 2008, RNA, vol. 14, pp. 1119-1131, в качестве решения нежелательного процесса декэпирования и, таким образом, для повышения стабильности РНК in vivo. В частности, замена атома кислорода атомом серы в бета-фосфатной группе 5'-кэпа приводит к стабилизации против Dcp2. Согласно этому варианту осуществления, который является предпочтительным в настоящем изобретении, R5 в формуле (I) представляет собой S, а R4 и R6 представляют собой O.
Согласно еще одному предпочтительному варианту осуществления настоящего изобретения РНК по настоящему изобретению содержит фосфоротиоатный аналог кэпа, в котором фосфоротиоатная модификация 5'-кэпа РНК сочетается с модификацией в виде «аналога кэп-структуры с правильной ориентацией» (ARCA). Соответствующие фосфоротиоатные аналоги ARCA-кэпа описаны в публикации международной заявки WO 2008/157688 A2, и все они могут быть использованы в РНК по настоящему изобретению. Согласно этому варианту осуществления по меньшей мере один из R2 или R3 в формуле (I) не представляет собой OH, предпочтительно один из R2 и R3 представляет собой метокси (OCH3), а другой из R2 и R3 предпочтительно представляет собой ОН. Согласно предпочтительному варианту осуществления атом кислорода заменен атомом серы в бета-фосфатной группе (так что R5 в формуле (I) представляет собой S, а R4 и R6 представляют собой O). Считается, что фосфоротиоатная модификация ARCA гарантирует, что α, β и γ фосфоротиоатные группы точно расположены в активных сайтах связывающих кэп белков в обоих трансляционных и декэпирующих инструментах. По меньшей мере некоторые из этих аналогов по существу устойчивы к пирофосфатазе Dcp1/Dcp2. Было описано, что модифицированные фосфоротиоатом ARCA имеют гораздо более высокое сродство к eIF4E, чем соответствующие ARCA, лишенные фосфоротиоатной группы.
Соответствующий аналог кэпа, который является особенно предпочтительным в настоящем изобретении, т.е. m2’7,2’-OGppspG, называется бета-S-ARCA (публикация международной заявки WO 2008/157688 A2; Kuhn et al., Gene Ther., 2010, vol. 17, pp. 961-971). Таким образом, согласно одному варианту осуществления настоящего изобретения РНК по настоящему изобретению модифицирована бета-S-ARCA. Бета-S-ARCA представлен следующей структурой:
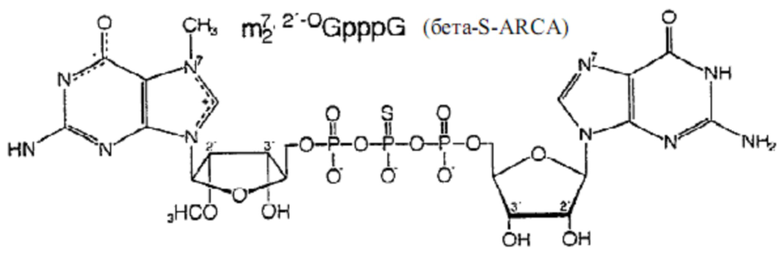
В общем случае замещение атома кислорода атомом серы в мостиковом фосфате приводит к образованию фосфоротиоатных диастереомеров, обозначенных D1 и D2, в соответствии с их профилем элюирования в ВЭЖХ. Вкратце, диастереомер D1 «бета-S-ARCA» или «бета-S-ARCA(D1)» представляет собой диастереомер бета-S-ARCA, который элюирует первым на колонке ВЭЖХ по сравнению с диастереомером D2 бета-S-ARCA (бета-S-ARCA(D2)) и, таким образом, проявляет более короткое время удерживания. Определение стереохимической конфигурации с помощью ВЭЖХ описано в публикации международной заявки WO 2011/015347 A1.
Согласно первому особенно предпочтительному варианту осуществления настоящего изобретения РНК по настоящему изобретению модифицирована диастереомером бета-S-ARCA(D2). Два диастереомера бета-S-ARCA отличаются чувствительностью к нуклеазам. Было показано, что РНК, несущая диастереомер D2 бета-S-ARCA, практически полностью устойчива к расщеплению Dcp2 (всего лишь 6% расщепления по сравнению с РНК, которая была синтезирована в присутствии немодифицированного 5'-кэпа ARCA), тогда как РНК с 5'-кэпом бета-S-ARCA(D1) обладает промежуточной чувствительностью к расщеплению Dcp2 (расщепление 71%). Дополнительно было показано, что повышенная устойчивость к расщеплению Dcp2 коррелирует с повышенной экспрессией белка в клетках млекопитающих. В частности, было показано, что РНК, несущие кэп бета-S-ARCA(D2), более эффективно транслируются в клетках млекопитающих, чем РНК, несущие кэп бета-S-ARCA(D1). Поэтому согласно одному варианту осуществления настоящего изобретения РНК по настоящему изобретению модифицирована с помощью аналога кэпа согласно формуле (I), характеризующегося стереохимической конфигурацией на атоме Р, содержащего заместитель R5 в формуле (I), который соответствует таковому у атома Pβ диастереомера D2 бета-S-ARCA. Согласно этому варианту осуществления R5 в формуле (I) представляет собой S, а и R4 и R6 представляют собой O. Кроме того, по меньшей мере один из R2 или R3 в формуле (I) предпочтительно не является OH, предпочтительно один из R2 и R3 представляет собой метокси (OCH3), а другой из R2 и R3 предпочтительно представляет собой ОН.
Согласно второму особенно предпочтительному варианту осуществления РНК по настоящему изобретению модифицирована диастереомером бета-S-ARCA(D1). Этот вариант осуществления особенно подходит для переноса кэпированной РНК в незрелые антигенпрезентирующие клетки, например, в целях вакцинации. Было продемонстрировано, что диастереомер бета-S-ARCA(D1) при переносе соответственно кэпированной РНК в незрелые антигенпрезентирующие клетки особенно подходит для повышения стабильности РНК, повышения эффективности трансляции РНК, продления трансляции РНК, повышения общей белковой экспрессии РНК и/или повышения иммунного ответа против антигена или антигенного пептида, кодируемого указанной РНК (Kuhn et al., 2010, Gene Ther., Vol. 17, pp. 961-971). Следовательно, согласно альтернативному варианту осуществления настоящего изобретения РНК по настоящему изобретению модифицирована с помощью аналога кэпа согласно формуле (I), характеризующегося стереохимической конфигурацией у атома Р, содержащего заместитель R5 в формуле (I), который соответствует таковому у атома Pβ диастереомера D1 бета-S-ARCA. Соответствующие аналоги и варианты их осуществления описаны в публикации международной заявки WO 2011/015347 A1 и в публикации Kuhn et al., 2010, Gene Ther., vol. 17, pp. 961-971. Любой аналог кэпа, описанный в публикации международной заявки WO 2011/015347 A1, в котором стереохимическая конфигурация у атома P, включающая заместитель R5, соответствует таковому у атома Pβ диастереомера D1 бета-S-ARCA, может быть использован в настоящем изобретении. Предпочтительно R5 в формуле (I) представляет собой S, а R4 и R6 представляют собой O. Кроме того, по меньшей мере один из R2 или R3 в формуле (I) предпочтительно не представляет собой OH, предпочтительно один из R2 и R3 представляет собой метокси (OCH3), а другой из R2 и R3 предпочтительно представляет собой ОН.
Согласно одному варианту осуществления РНК по настоящему изобретению модифицирована структурой 5'-кэп в соответствии с формулой (I), причем любая фосфатная группа замещена боранофосфатной группой или фосфороселеноатной группой. Такие структуры кэп повышают стабильность как in vitro, так и in vivo. Необязательно, соответствующее соединение содержит 2'-O- или 3'-O-алкильную группу (где алкил предпочтительно представляет собой метил); соответствующие аналоги структуры кэп называются BH3-ARCA или Se-ARCA. Соединения, которые особенно подходят для кэпирования мРНК, включают в себя β-BH3-ARCA и β-Se-ARCA, как описано в публикации международной заявки WO 2009/149253 A2. Для этих соединений предпочтительной является стереохимическая конфигурация у атома Р, содержащая заместитель R5 в формуле (I), который соответствует таковому у атома Pβ диастереомера D1 бета-S-ARCA.
UTR
Термин «нетранслируемая область» или «UTR» относится к области в молекуле ДНК, которая транскрибируется, но не транслируется в аминокислотную последовательность, или к соответствующей области в молекуле РНК, такой как молекула мРНК. Нетранслируемая область (UTR) может присутствовать 5' (выше по ходу транскрипции) от открытой рамки считывания (5'-UTR) и/или 3' (ниже по ходу транскрипции) от открытой рамки считывания (3'-UTR).
3'-UTR, если она присутствует, расположена на 3'-конце гена ниже по ходу транскрипции от кодона терминации кодирующей белок области, но термин «3'-UTR» предпочтительно не включает в себя поли(А)-хвост. Таким образом, 3'-UTR находится выше против хода транскрипции от поли(А)-хвоста (если он присутствует), например, непосредственно примыкая к поли(А)-хвосту.
5'-UTR, если присутствует, расположена на 5'-конце гена, выше против хода транскрипции от стартового кодона кодирующей белок области. 5'-UTR находится ниже по ходу транскрипции от 5'-кэпа (если он присутствует), например, непосредственно примыкая к 5'-кэпу.
5'- и/или 3'-нетранслируемые области могут, в соответствии с настоящим изобретением, быть функционально связаны с открытой рамкой считывания для того, чтобы эти области были связаны с открытой рамкой считывания таким образом, чтобы увеличивать стабильность и/или эффективность трансляции РНК, содержащей указанную открытую рамку считывания.
Согласно некоторым варианта осуществления репликазная конструкция в соответствии с настоящим изобретением содержит 5'-UTR и/или 3'-UTR.
Согласно предпочтительному варианту осуществления репликазная конструкция в соответствии с настоящим изобретением содержит
(1) 5'-UTR,
(2) открытую рамку считывания и
(3) 3'-UTR.
UTR участвуют в стабильности и эффективности трансляции РНК. И то и то может быть улучшено, помимо структурных модификаций, касающихся 5'-кэпа и/или 3' поли(А)-хвоста, как описано в настоящем документе, путем выбора специфических 5' и/или 3' нетранслируемых областей (UTR). Как правило, элементы последовательности в UTR влияют на эффективность трансляции (в основном, 5'-UTR) и стабильность РНК (в основном 3'-UTR). Предпочтительно, чтобы присутствовала 5'-UTR, которая активна для повышения эффективности трансляции и/или стабильности репликазной конструкции. Независимо или дополнительно предпочтительно, чтобы присутствовала 3'-UTR, которая активна для повышения эффективности трансляции и/или стабильности репликазной конструкции.
Термины «активна для повышения эффективности трансляции» и/или «активна для повышения стабильности» применительно к первой последовательности нуклеиновой кислоты (например, UTR), означает, что первая последовательность нуклеиновой кислоты способна к модификации в общем транскрипте со второй последовательностью нуклеиновой кислоты, влияя на эффективность трансляции и/или стабильность указанной второй последовательности нуклеиновой кислоты таким образом, что эффективность трансляции и/или стабильность трансляции увеличивается по сравнению с эффективностью трансляции и/или стабильностью указанной второй последовательности нуклеиновой кислоты в отсутствие указанной первой последовательности нуклеиновой кислоты.
Согласно одному варианту осуществления репликазная конструкция в соответствии с настоящим изобретением содержит 5'-UTR и/или 3'-UTR, которая является гетерологичной или ненативной для альфавируса, из которого получен функциональный альфавирусный неструктурный белок. Это позволяет создавать нетранслируемые области в соответствии с требуемой эффективностью трансляции и стабильностью РНК. Таким образом, гетерологичные или ненативные UTR обеспечивают высокую степень гибкости, и эта гибкость является предпочтительной по сравнению с нативными альфавирусными UTR. В частности, хотя известно, что альфавирусная (нативная) РНК также содержит 5'-UTR и/или 3'-UTR, альфавирусные UTR выполняют двойную функцию, то есть (i) для управления репликацией РНК, а также (ii) для управления трансляцией. В то время как сообщалось, что альфавирусные UTR были неэффективны для трансляции (Berben-Bloemheuvel et al., 1992, Eur. J. Biochem., vol. 208, pp. 581-587), они, как правило, не могут быть легко заменены более эффективными UTR из-за их двойной функции. Однако в настоящем изобретении 5'-UTR и/или 3'-UTR, содержащиеся в репликазной конструкции, для репликации in trans могут быть выбраны независимо от их потенциального влияния на репликацию РНК.
Предпочтительно, репликазная конструкция в соответствии с настоящим изобретением содержит 5'-UTR и/или 3'-UTR, которая происходит не от вирусов; в особенности не от альфавирусов. Согласно одному варианту осуществления репликазная конструкция содержит 5'-UTR, полученную из эукариотической 5'-UTR, и/или 3'-UTR, полученную из эукариотической 3'-UTR.
5'-UTR в соответствии с настоящим изобретением может содержать любую комбинацию из более чем одной последовательности нуклеиновой кислоты, необязательно разделенной линкером. 3'-UTR в соответствии с настоящим изобретением может содержать любую комбинацию из более чем одной последовательности нуклеиновой кислоты, необязательно разделенной линкером.
Термин «линкер» в соответствии с настоящим изобретением относится к последовательности нуклеиновой кислоты, добавляемой между двумя последовательностями нуклеиновой кислоты для соединения указанных двух последовательностей нуклеиновой кислоты. Нет никаких особых ограничений относительно последовательности линкера.
3'-UTR, как правило, составляет в длину от 200 до 2000 нуклеотидов, например, от 500 до 1500 нуклеотидов. 3'-нетранслируемые области мРНК иммуноглобулина относительно короткие (менее 300 нуклеотидов), тогда как 3'-нетранслируемые области других генов относительно длинные. Например, 3'-нетранслируемая область tPA составляет в длину приблизительно 800 нуклеотидов, а фактор VIII составляет в длину приблизительно 1800 нуклеотидов, а длина эритропоэтина составляет приблизительно 560 нуклеотидов. 3'-нетранслируемые области мРНК млекопитающих, как правило, содержат область гомологии, известную как гексануклеотидная последовательность AAUAAA. Эта последовательность, по-видимому, представляет собой сигнал присоединения поли(A) и часто находится на расстоянии от 10 до 30 оснований выше против хода транскрипции от сайта связывания поли(A). 3'-нетранслируемые области могут содержать один или несколько инвертированных повторов, которые могут складываться, чтобы создавать структуры «петля-на-стебле», которые действуют как барьеры для экзорибонуклеаз или взаимодействуют с белками, известными, как повышающие стабильность РНК (например, РНК-связывающими белками).
3'-UTR бета-глобина человека, особенно две последовательные идентичные копии 3'-UTR бета-глобина человека, способствует высокой стабильности транскрипции и эффективности трансляции (Holtkamp et al., 2006, Blood, vol. 108, pp. 4009-4017). Таким образом, согласно одному варианту осуществления репликазная конструкция в соответствии с настоящим изобретением содержит две последовательные идентичные копии 3'-UTR бета-глобина человека. Таким образом, она содержит в направлении 5'→3': (а) необязательно 5'-UTR; (b) открытую рамку считывания; (c) 3'-UTR; указанная 3'-UTR содержит две последовательные идентичные копии 3'-UTR бета-глобина человека, ее фрагмент или вариант 3'-UTR бета-глобина человека или ее фрагмент.
Согласно одному варианту осуществления репликазная конструкция в соответствии с настоящим изобретением содержит 3'-UTR, которая активна для повышения эффективности трансляции и/или стабильности, но которая не представляет собой 3'-UTR бета-глобина человека, ее фрагмент или вариант 3'-UTR бета-глобина человека или ее фрагмента.
Согласно одному варианту осуществления репликазная конструкция в соответствии с настоящим изобретением содержит 5'-UTR, которая активна для повышения эффективности трансляции и/или стабильности.
UTR-содержащую репликазную конструкцию в соответствии с настоящим изобретением можно получить, например, посредством транскрипции in vitro. Это может быть достигнуто путем генетической модификации молекулы-матрицы нуклеиновой кислоты (например, ДНК) таким образом, что она позволяет транскрипцию РНК с 5'-UTR и/или 3'-UTR.
Как показано на фиг. 1, также репликон может быть охарактеризован посредством 5'-UTR и/или 3'-UTR. UTR репликона, как правило, представляют собой альфавирусные UTR или их варианты.
Поли(A)-последовательность
Согласно некоторым вариантам осуществления репликон в соответствии с настоящим изобретением содержит 3' поли(А)-последовательность. Если репликон содержит консервативный элемент последовательности 4 (CSE 4), 3' поли(А)-последовательность репликона предпочтительно присутствует ниже по ходу транскрипции от CSE 4, наиболее предпочтительно непосредственно примыкает к CSE 4.
Согласно некоторым вариантам осуществления репликазная конструкция в соответствии с настоящим изобретением содержит 3' поли(А)-последовательность.
В соответствии с настоящим изобретением согласно одному варианту осуществления поли(А)-последовательность содержит или по существу состоит, или состоит по меньшей мере из 20, предпочтительно по меньшей мере из 26, предпочтительно по меньшей мере из 40, предпочтительно по меньшей мере из 80, предпочтительно по меньшей мере из 100 и содержит предпочтительно до 500, предпочтительно до 400, предпочтительно до 300, предпочтительно до 200 и, в частности, до 150, нуклеотидов А и, в частности, приблизительно 120 нуклеотидов А. В этом контексте «по существу состоит из» означает, что большинство нуклеотидов в поли(А)-последовательности, как правило, по меньшей мере на 50% и предпочтительно по меньшей мере на 75% по количеству нуклеотидов в «поли(А)-последовательности» представляют собой нуклеотиды А, но позволено, чтобы оставшиеся нуклеотиды были нуклеотидами, отличными от нуклеотидов А, такими как нуклеотиды U (уридилат), нуклеотиды G (гуанилат), нуклеотиды C (цитидилат). В этом контексте «состоит из» означает, что все нуклеотиды в поли(А)-последовательности, то есть 100% по количеству нуклеотидов в поли(А)-последовательности, являются нуклеотидами А. Термин «нуклеотид А» или «А» относится к аденилату.
Действительно, было продемонстрировано, что 3' поли(А)-последовательность приблизительно из 120 нуклеотидов А оказывает благотворное влияние на уровни РНК в трансфицированных эукариотических клетках, а также на уровни белка, который транслируется из открытой рамки считывания (5') 3' поли(А)-последовательности (Holtkamp et al., 2006, Blood, vol. 108, pp. 4009-4017).
В альфавирусах считается, что 3' поли(А)-последовательность по меньшей мере из 11 последовательных аденилатных остатков или по меньшей мере из 25 последовательных аденилатных остатков важна для эффективного синтеза минусовой нити. В частности, понятно, что в альфавирусах 3' поли(А)-последовательность по меньшей мере из 25 последовательных аденилатных остатков функционирует вместе с консервативным элементом последовательности 4 (CSE 4) для содействия синтезу (-)-нити (Hardy & Rice, J. Virol., 2005, vol. 79, pp. 4630-4639).
В настоящем изобретении предусмотрена 3' поли(А)-последовательность, которая должна быть присоединена во время транскрипции РНК, то есть во время получения транскрибируемой in vitro РНК на основе ДНК-матрицы, содержащей повторяющиеся нуклеотиды dT (дезокситимидилат) в цепи, комплементарной кодирующей цепи. Последовательность ДНК, кодирующая поли(А)-последовательность (кодирующая цепь), называется поли(А)-кассетой.
Согласно предпочтительному варианту осуществления настоящего изобретения 3' поли(А)-кассета, присутствующая в кодирующей цепи ДНК, по существу состоит из нуклеотидов dA, но прерывается случайной последовательностью, имеющей равное распределение четырех нуклеотидов (dA, dC, dG, dT). Такая случайная последовательность может составлять от 5 до 50, предпочтительно от 10 до 30, более предпочтительно от 10 до 20 нуклеотидов. Такая кассета раскрыта в публикации международной заявки WO 2016/005004 A1. В настоящем изобретении может быть использована любая поли(А)-кассета, раскрытая в публикации международной заявки WO 2016/005004 A1. Поли(А)-кассета, которая по существу состоит из нуклеотидов dA, но прерывается случайной последовательностью, характеризующейся равным распределением четырех нуклеотидов (dA, dC, dG, dT) и составляющая в длину, например, от 5 до 50 нуклеотидов на уровне ДНК, показывает, на уровне ДНК, постоянное воспроизведение плазмидной ДНК в E.coli и по-прежнему связана, на уровне РНК, с полезными свойствами в отношении поддержания стабильности РНК и эффективности трансляции.
Следовательно, согласно предпочтительному варианту осуществления настоящего изобретения 3' поли(А)-последовательность, содержащаяся в молекуле РНК, описанной в настоящем документе, по существу состоит из нуклеотидов А, но прерывается случайной последовательностью, характеризующейся равным распределением четырех нуклеотидов (А, C, G, U). Такая случайная последовательность может составлять от 5 до 50, предпочтительно от 10 до 30, более предпочтительно от 10 до 20 нуклеотидов.
Использование кодонов
В общем, вырожденность генетического кода сделает возможным замену некоторых кодонов (триплеты оснований, кодирующие аминокислоту), которые присутствуют в последовательности РНК другими кодонами (триплетами оснований), сохраняя при этом ту же самую способность кодирования (так что замещающий кодон кодирует ту же самую аминокислоту, что и замещенный кодон). Согласно некоторым вариантам осуществления настоящего изобретения по меньшей мере один кодон открытой рамки считывания, состоящий из молекулы РНК, отличается от соответствующего кодона в соответствующей открытой рамке считывания у видов, из которых происходит открытая рамка считывания. Согласно этому варианту осуществления кодирующая последовательность открытой рамки считывания называется «адаптированной» или «модифицированной». Кодирующая последовательность открытой рамки считывания, содержащаяся в репликоне, может быть адаптирована. Альтернативно или дополнительно, кодирующая последовательность для функционального альфавирусного неструктурного белка, содержащаяся в репликазной конструкции, может быть адаптирована.
Например, когда кодирующая последовательность открытой рамки считывания адаптирована, могут быть выбраны часто используемые кодоны: в публикации международной заявки WO 2009/024567 A1 описана адаптация кодирующей последовательности молекулы нуклеиновой кислоты с заменой редких кодонов более часто используемыми кодонами. Поскольку частота использования кодонов зависит от клетки-хозяина или организма-хозяина, этот тип адаптации подходит для подгонки последовательности нуклеиновой кислоты для экспрессии в конкретной клетке-хозяине или организме-хозяине. В общем случае, чаще используемые кодоны, как правило, более эффективно транслируются в клетке-хозяине или в организме-хозяине, хотя адаптация всех кодонов открытой рамки считывания не всегда требуется.
Например, когда кодирующая последовательность открытой рамки считывания адаптирована, содержание остатков G (гуанилата) и остатков C (цитидилата) может быть изменено путем выбора кодонов с наибольшим богатым на GC содержанием для каждой аминокислоты. Сообщалось, что молекулы РНК с богатыми на GC открытыми рамками считывания характеризуются потенциалом для снижения иммунной активации и улучшения трансляции и полужизни РНК (Thess et al., 2015, Mol. Ther., 23, 1457-1465).
Когда репликон в соответствии с настоящим изобретением кодирует альфавирусный неструктурный белок, кодирующая последовательность альфавирусного неструктурного белка может быть адаптирована по желанию. Эта свобода возможна, поскольку открытая рамка считывания, кодирующая альфавирусный неструктурный белок, не перекрывается с 5' репликационной последовательностью распознавания репликона.
Особенности безопасности вариантов осуществления настоящего изобретения
Следующие особенности предпочтительны в настоящем изобретении отдельно или в любой подходящей комбинации:
Предпочтительно репликон или система по настоящему изобретению не являются образующими частицы. Это означает, что после инокуляции клетки-хозяина репликоном или системой по настоящему изобретению клетка-хозяин не производит вирусные частицы, такие как вирусные частицы следующего поколения. Согласно одному варианту осуществления все молекулы РНК в соответствии с настоящим изобретением полностью не содержат генетической информации, кодирующей любой структурный белок альфавируса, такой как белок C нуклеокапсида сердцевины, белок оболочки P62 и/или белок оболочки E1. Этот аспект настоящего изобретения обеспечивает дополнительную ценность с точки зрения безопасности по сравнению с известными системами, в которых структурные белки кодируются транс-реплицирующей РНК-помощником (например, Bredenbeek et al., J. Virol, 1993, vol. 67, pp. 6439- 6446).
Предпочтительно, система по настоящему изобретению не содержит какого-либо структурного белка альфавируса, такого как белок C нуклеокапсида сердцевины, белок P62 оболочки и/или белок E1 оболочки.
Предпочтительно, репликон и репликазная конструкция системы по настоящему изобретения не идентичны друг другу. Согласно одному варианту осуществления репликон не кодирует функциональный альфавирусный неструктурный белок. Согласно одному варианту осуществления репликазная конструкция не содержит по меньшей мере один элемент последовательности (предпочтительно, по меньшей мере один CSE), который требуется для синтеза (-)-нити на основе (+)-нитевой матрицы и/или для синтеза (+)-нити на основе (-)-нитевой матрицы. Согласно одному варианту осуществления репликазная конструкция не содержит CSE 1 и/или CSE 4.
Предпочтительно, чтобы ни репликон в соответствии с настоящим изобретением, ни репликазная конструкция в соответствии с настоящим изобретением не содержали сигнал упаковки альфавируса. Например, сигнал упаковки альфавируса, содержащийся в кодирующей области nsP2 в SFV (White et al. 1998, J. Virol., vol. 72, pp. 4320-4326), может быть удален, например, путем удаления или мутации. Подходящий способ удаления сигнала упаковки альфавируса предусматривает адаптацию использования кодонов в кодирующей области nsP2. Вырожденность генетического кода может позволить удалить функцию сигнала упаковки, не затрагивая аминокислотную последовательность закодированного nsP2.
Согласно одному варианту осуществления система по настоящему изобретению представляет собой выделенную систему. Согласно этому варианту осуществления система отсутствует внутри клетки, например, внутри клетки млекопитающего, или отсутствует внутри капсида вируса, например, внутри слоя, содержащего структурные белки альфавируса. Согласно одному варианту осуществления система по настоящему изобретению присутствует in vitro.
ДНК
Согласно третьему аспекту в настоящем изобретении предусмотрена ДНК, содержащая последовательность нуклеиновой кислоты, кодирующую РНК-репликон в соответствии с первым аспектом настоящего изобретения или и то и другое.
Предпочтительно, ДНК является двухцепочечной.
Согласно предпочтительному варианту осуществления ДНК в соответствии с третьим аспектом настоящего изобретения представляет собой плазмиду. Используемый в настоящем документе термин «плазмида», как правило, относится к конструкции внехромосомного генетического материала, как правило, к кольцевому дуплексу ДНК, который может реплицироваться независимо от хромосомной ДНК.
ДНК по настоящему изобретению может содержать промотор, который может быть распознан ДНК-зависимой РНК-полимеразой. Это позволяет транскрибировать закодированную РНК in vivo или in vitro, например, РНК по настоящему изобретению. IVT-векторы могут использоваться стандартным образом в качестве матрицы для транскрипции in vitro. Примерами промоторов, предпочтительных в соответствии с настоящим изобретением, являются промоторы для полимеразы SP6, T3 или T7.
Согласно одному варианту осуществления ДНК по настоящему изобретению представляет собой выделенную молекулу нуклеиновой кислоты.
Способы получения РНК
Любая молекула РНК в соответствии с настоящим изобретением, будь она частью системы по настоящему изобретению или нет, может быть получена путем транскрипции in vitro. Транскрибируемая in vitro РНК (IVT-РНК) представляет особый интерес в настоящем изобретении. IVT-РНК получают путем транскрипции из молекулы нуклеиновой кислоты (в частности, молекулы ДНК). Молекулы ДНК согласно третьему аспекту настоящего изобретения пригодны для таких целей, особенно если они содержат промотор, который может быть распознан ДНК-зависимой РНК-полимеразой.
РНК в соответствии с настоящим изобретением может быть синтезирована in vitro. Это позволяет добавлять аналоги кэпа в реакцию транскрипции in vitro. Как правило, поли(A)-хвост кодируется поли(dT)-последовательностью на матрице ДНК. Альтернативно, кэпирование и добавление поли(А)-хвоста могут быть достигнуты ферментативно после транскрипции.
Специалисту в настоящей области техники известен способ транскрипции in vitro. Например, как упоминалось в публикации международной заявки WO 2011/015347 A1, множество наборов транскрипции in vitro являются коммерчески доступными.
Набор
В настоящем изобретении также предусмотрен набор, содержащий РНК-репликон в соответствии с первым аспектом настоящего изобретения или систему в соответствии со вторым аспектом настоящего изобретения.
Согласно одному варианту осуществления составляющие набора присутствуют в виде отдельных объектов. Например, одна молекула нуклеиновой кислоты набора может присутствовать в одном объекте, а другая нуклеиновая кислота набора может присутствовать в отдельном объекте. Например, открытый или закрытый контейнер является подходящим объектом. Предпочтительным является закрытый контейнер. Используемый контейнер предпочтительно должен быть без РНКазы или по существу без РНКазы.
Согласно одному варианту осуществления набор по настоящему изобретению содержит РНК для инокуляции клетки и/или для введения человеку или животному.
Набор в соответствии с настоящим изобретением необязательно содержит этикетку или другую форму информационного элемента, например, электронный носитель данных. Этикетка или информационный элемент предпочтительно содержит инструкции, например, напечатанные инструкции или инструкции в электронной форме, которые могут быть распечатаны. Инструкции могут касаться по меньшей мере одного подходящего возможного применения набора.
Фармацевтическая композиция
Описанная в настоящем документе репликазная конструкция и/или репликон может присутствовать в форме фармацевтической композиции. Фармацевтическая композиция в соответствии с настоящим изобретением может содержать по меньшей мере одну молекулу нуклеиновой кислоты в соответствии с настоящим изобретением. Фармацевтическая композиция в соответствии с настоящим изобретением содержит фармацевтически приемлемый разбавитель и/или фармацевтически приемлемое вспомогательное вещество, и/или фармацевтически приемлемый носитель, и/или фармацевтически приемлемый наполнитель. Выбор фармацевтически приемлемого носителя, наполнителя, вспомогательного вещества или разбавителя конкретно не ограничен. Можно использовать любой подходящий фармацевтически приемлемый носитель, наполнитель, вспомогательное вещество или разбавитель, известный в настоящей области техники.
Согласно одному варианту осуществления настоящего изобретения фармацевтическая композиция может дополнительно содержать такой растворитель, как водный растворитель или любой растворитель, который позволяет сохранить целостность РНК. Согласно предпочтительному варианту осуществления фармацевтическая композиция представляет собой водный раствор, содержащий РНК. Водный раствор может необязательно содержать растворенные вещества, например, соли.
Согласно одному варианту осуществления настоящего изобретения фармацевтическая композиция находится в форме лиофилизированной композиции. Лиофилизированную композицию можно получить путем лиофилизации соответствующей водной композиции.
Согласно одному варианту осуществления фармацевтическая композиция содержит по меньшей мере один катионный объект. Как правило, катионные липиды, катионные полимеры и другие вещества с положительными зарядами могут образовывать комплексы с отрицательно заряженными нуклеиновыми кислотами. Можно стабилизировать РНК в соответствии с настоящим изобретением путем комплексообразования с катионными соединениями, предпочтительно поликатионными соединениями, такими как, например, катионный или поликатионный пептид или белок. Согласно одному варианту осуществления фармацевтическая композиция в соответствии с настоящим изобретением содержит по меньшей мере одну катионную молекулу, выбранную из группы, содержащей протамин, полиэтиленимин, поли-L-лизин, поли-L-аргинин, гистон или катионный липид.
В соответствии с настоящим изобретением катионный липид представляет собой катионную амфифильную молекулу, например, молекулу, которая содержит по меньшей мере один гидрофильный и липофильный фрагменты. Катионный липид может быть монокатионным или поликатионным. Катионные липиды, как правило, содержат липофильный фрагмент, такой как стерол, ацильную или диацильную цепь, и характеризуются общим положительным зарядом. Головная группа липида, как правило, несет положительный заряд. Катионный липид предпочтительно имеет положительный заряд от 1 до 10 валентностей, более предпочтительно положительный заряд составляет от 1 до 3 валентностей и более предпочтительно положительный заряд составляет 1 валентность. Примеры катионных липидов включают в себя, без ограничения, 1,2-ди-O-октадеценил-3-триметиламмонийпропан (DOTMA); диметилдиоктадециламмоний (DDAB); 1,2-диолеил-3-триметиламмонийпропан (DOTAP); 1,2-диолеил-3-диметиламмонийпропан (DODAP); 1,2-диацилокси-3-диметиламмонийпропаны; 1,2-диалкилокси-3-диметиламмонийпропаны; хлорид диоктадецилдиметиламмония (DODAC), 1,2-димиристоилоксипропил-1,3-диметилгидроксиэтиламмоний (DMRIE) и трифторацетат 2,3-диолеилокси-N-[2(сперминкарбоксамид)этил]-N,N-диметил-1-пропанамида (DOSPA). Катионные липиды также включают в себя липиды с третичной аминогруппой, включая в себя 1,2-дилинолилокси-N,N-диметил-3-аминопропан (DLinDMA). Катионные липиды подходят для получения составов РНК в липидных составах, как описано в настоящем документе, таких как липосомы, эмульсии и липоплексы. Как правило, положительные заряды вносятся по меньшей мере одним катионным липидом, а отрицательные заряды вносят РНК. Согласно одному варианту осуществления фармацевтическая композиция содержит по меньшей мере один вспомогательный липид, в дополнение к катионному липиду. Вспомогательный липид может быть нейтральным или анионным липидом. Вспомогательный липид может представлять собой природный липид, такой как фосфолипид, или аналог природного липида, или полностью синтетический липид, или липидоподобную молекулу, не имеющую сходства с природными липидами. В случае, когда фармацевтическая композиция содержит как катионный липид, так и вспомогательный липид, молярное отношение катионного липида к нейтральному липиду может быть соответствующим образом определено с учетом стабильности состава и тому подобного.
Согласно одному варианту осуществления фармацевтическая композиция в соответствии с настоящим изобретением содержит протамин. В соответствии с настоящим изобретением протамин применим в качестве катионного средства-носителя. Термин «протамин» относится к любому из различных сильноосновных белков с относительно низкой молекулярной массой, которые богаты аргинином и встречаются, в частности, с ДНК вместо соматических гистонов в клетках спермы животных, таких как рыба. В частности, термин «протамин» относится к белкам, обнаруженным в сперме рыб, которые являются сильноосновными, растворимы в воде, не коагулируют при нагревании и содержат несколько мономеров аргинина. В соответствии с настоящим изобретением используемый в настоящем документе термин «протамин» подразумевает, что он содержит любую аминокислотную последовательность протаминов, полученную или происходящую из природных или биологических источников, включая в себя их фрагменты и мультимерные формы указанной аминокислотной последовательности или ее фрагмента. Кроме того, этот термин охватывает (синтезированные) полипептиды, которые являются искусственными и специально разработаны для конкретных целей и не могут быть выделены из природных или биологических источников.
Согласно некоторым вариантам осуществления композиции по настоящему изобретению могут содержать один или несколько адъювантов. Адъюванты могут быть добавлены к вакцинам для стимуляции ответа иммунной системы; адъюванты, как правило, не обеспечивают иммунитет сами. Иллюстративные адъюванты включают в себя, без ограничения, следующие: неорганические соединения (например, квасцы, гидроксид алюминия, фосфат алюминия, гидроксид фосфата кальция); минеральное масло (например, парафиновое масло), цитокины (например, IL-1, IL-2, IL-12); иммуностимулирующий полинуклеотид (такой как РНК или ДНК, например, CpG-содержащие олигонуклеотиды); сапонины (например, сапонины растений из килайи, сои, Polygala senega); масляные эмульсии или липосомы; полиоксиэтиленовый эфир и полиоксиэтиленовые сложные эфиры; полифосфазен (PCPP); мурамиловые пептиды; соединения имидазохинолинов; тиосемикарбазоновые соединения; лиганд Flt3 (публикация международной заявки WO 2010/066418 A1) или любой другой адъювант, который известен специалисту в настоящей области техники. Предпочтительным адъювантом для введения РНК в соответствии с настоящим изобретением является лиганд Flt3 (публикация международной заявки WO 2010/066418 A1). Когда лиганд Flt3 вводят вместе с РНК, которая кодирует антиген, может наблюдаться сильное увеличение антигенспецифических CD8+ Т-клеток.
Фармацевтической композиции согласно изобретению можно придать буферные свойства (например, ацетатным буфером, цитратным буфером, сукцинатным буфером, трис-буфером, фосфатным буфером).
РНК-содержащие частицы
Согласно некоторым вариантам осуществления из-за неустойчивости незащищенной РНК предпочтительно предоставлять молекулы РНК по настоящему изобретению в виде комплексов или в инкапсулированной форме. Соответствующие фармацевтические композиции предусмотрены в настоящем изобретении. В частности, согласно некоторым вариантам осуществления фармацевтическая композиция по настоящему изобретению содержит содержащие нуклеиновую кислоту частицы, предпочтительно содержащие РНК частицы. Соответствующие фармацевтические композиции называются состоящими из частиц составами. В состоящих из частиц составах в соответствии с настоящим изобретением частица содержит нуклеиновую кислоту в соответствии с настоящим изобретением и фармацевтически приемлемый носитель или фармацевтически приемлемый наполнитель, который подходит для доставки нуклеиновой кислоты. Содержащие нуклеиновую кислоту частицы могут быть, например, в форме белковых частиц или в виде липидсодержащих частиц. Подходящие белки или липиды называются образующими частицы средствами. Белковые частицы и липидсодержащие частицы были описаны ранее, как пригодные для доставки альфавирусной РНК в форме частиц (например, публикация Strauss & Strauss, Microbiol. Rev., 1994, vol. 58, pp. 491-562). В частности, альфавирусные структурные белки (предоставленные, например, вирусом-помощником) являются подходящим носителем для доставки РНК в форме белковых частиц.
Когда система в соответствии с настоящим изобретением составлена в виде состоящего из частиц состава, возможно, что каждый вид РНК (например, репликон, репликазная конструкция и необязательные дополнительные виды РНК, такие как РНК, кодирующая белок, подходящий для ингибирования IFN), отдельно составляется в виде отдельного состоящего из частиц состава. В этом случае каждый отдельный состоящий из частиц состав будет содержать один вид РНК. Отдельные состоящие из частиц составы могут присутствовать в виде отдельных объектов, например, в отдельных контейнерах. Такие составы можно получить, предоставляя каждый вид РНК отдельно (обычно каждый в форме РНК-содержащего раствора) вместе с образующим частицы средством, тем самым позволяя образовывать частицы. Соответствующие частицы будут содержать исключительно специфические виды РНК, которые предоставляются, когда образуются частицы (отдельные состоящие из частиц составы).
Согласно одному варианту осуществления фармацевтическая композиция в соответствии с настоящим изобретением содержит более одного отдельного состоящего из частиц состава. Соответствующие фармацевтические композиции называются смешанными состоящими из частиц составами. Смешанные состоящие из частиц составы согласно настоящему изобретению можно получить, образуя, раздельно, отдельные состоящие из частиц составы, как описано выше, с последующей стадией смешивания отдельных состоящих из частиц составов. На стадии смешивания можно получить один состав, содержащий смешанную популяцию РНК-содержащих частиц (для иллюстрации: например, первая популяция частиц может содержать репликон в соответствии с настоящим изобретением, а второй состав частиц может содержать репликазную конструкцию в соответствии с настоящим изобретением). Отдельные состоящие из частиц популяции могут быть вместе в одном контейнере, состоящем из смешанной популяции отдельных состоящих из частиц составов.
Альтернативно, возможно, что все виды РНК фармацевтической композиции (например, репликон, репликазная конструкция и необязательные дополнительные виды, такие как РНК, кодирующая белок, подходящий для ингибирования IFN), составляют вместе в виде комбинированного состоящего из частиц состава. Такие составы можно получить путем обеспечения комбинированного состава (как правило, комбинированного раствора) всех видов РНК вместе с образующим частицы средством, тем самым позволяя образовывать частицы. В отличие от смешанного состоящего из частиц состава комбинированный состоящий из частиц состав, как правило, содержит частицы, которые содержат более одного вида РНК. В комбинированном состоящем из частиц составе различные виды РНК, как правило, присутствуют вместе в одной частице.
Согласно одному варианту осуществления состоящий из частиц состав по настоящему изобретению представляет собой состоящий из наночастиц состав. Согласно этому варианту осуществления композиция в соответствии с настоящим изобретением содержит нуклеиновую кислоту в соответствии с настоящим изобретением в форме наночастиц. Состоящие из наночастиц составы могут быть получены посредством различных протоколов и с различными комплексообразующими соединениями. Липиды, полимеры, олигомеры или амфифилы являются типичными составляющими состоящих из наночастиц составов.
Используемый в настоящем документе термин «наночастица» относится к любой частице, имеющей диаметр, который делает частицу подходящей для системного, в частности парентерального, введения, в частности нуклеиновые кислоты, как правило, диаметром 1000 нанометров (нм) или менее. Согласно одному варианту осуществления наночастицы характеризуются средним диаметром в диапазоне от приблизительно 50 нм до приблизительно 1000 нм, предпочтительно от приблизительно 50 нм до приблизительно 400 нм, предпочтительно от приблизительно 100 нм до приблизительно 300 нм, например, от приблизительно 150 нм до приблизительно 200 нм. Согласно одному варианту осуществления наночастицы характеризуются диаметром в диапазоне от приблизительно 200 до приблизительно 700 нм, от приблизительно 200 до приблизительно 600 нм, предпочтительно от приблизительно 250 до приблизительно 550 нм, в частности от приблизительно 300 до приблизительно 500 нм или от приблизительно 200 до приблизительно 400 нм.
Согласно одному варианту осуществления показатель полидисперсности (PI) описанных в настоящем документе наночастиц, измеренный динамическим рассеянием света, составляет 0,5 или менее, предпочтительно 0,4 или менее или даже более предпочтительно 0,3 или менее. «Индекс полидисперсности» (PI) представляет собой измерение однородного или гетерогенного распределения по размеру отдельных частиц (таких как липосомы) в смеси частиц и указывает ширину распределения частиц в смеси. PI можно определить, например, как описано в публикации международной заявки WO 2013/143555 A1.
Используемый в настоящем документе термин «состоящий из наночастиц состав» или аналогичные термины относятся к любому состоящему из частиц составу, содержащему по меньшей мере одну наночастицу. Согласно некоторым вариантам осуществления состоящая из наночастиц композиция представляет собой однородный набор наночастиц. Согласно некоторым вариантам осуществления состоящая из наночастиц композиция представляет собой липидсодержащий фармацевтический состав, такой как липосомный состав или эмульсия.
Липидсодержащие фармацевтические композиции
Согласно одному варианту осуществления фармацевтическая композиция по настоящему изобретению содержит по меньшей мере один липид. Предпочтительно, по меньшей мере один липид представляет собой катионный липид. Указанная липидсодержащая фармацевтическая композиция содержит нуклеиновую кислоту в соответствии с настоящим изобретением. Согласно одному варианту осуществления фармацевтическая композиция в соответствии с настоящим изобретением содержит РНК, инкапсулированную в везикуле, например, в липосоме. Согласно одному варианту осуществления фармацевтическая композиция в соответствии с настоящим изобретением содержит РНК в форме эмульсии. Согласно одному варианту осуществления фармацевтическая композиция в соответствии с настоящим изобретением содержит РНК в комплексе с катионным соединением, образуя, например, так называемые липоплексы или полиплексы. Инкапсуляция РНК в везикулах, таких как липосомы, отличается, например, от комплексов липид/РНК. Комплексы липид/РНК могут быть получены, например, когда РНК, например, смешивают с предварительно образованными липосомами.
Согласно одному варианту осуществления фармацевтическая композиция в соответствии с настоящим изобретением содержит РНК, инкапсулированную в везикулу. Такой состав представляет собой конкретный состоящий из частиц состав в соответствии с настоящим изобретением. Везикула представляет собой липидный бислой, свернутый в сферическую оболочку, вмещающий небольшое пространство и отделяющий это пространство от пространства вне везикулы. Как правило, пространство внутри везикулы представляет собой водное пространство, т.е. содержит воду. Как правило, пространство вне везикулы представляет собой водное пространство, т.е. содержит воду. Липидный бислой образован одним или несколькими липидами (образующими везикулы липидами). Мембрана, вмещающая везикулу, представляет собой ламеллярную фазу, аналогичную таковой плазматической мембраны. Везикула в соответствии с настоящим изобретением может представлять собой многослойную везикулу, однослойную везикулу или их смесь. При инкапсулировании в везикулу РНК, как правило, отделена от любой внешней среды. Таким образом, она присутствует в защищенной форме, функционально эквивалентной защищенной форме в природном альфавирусе. Подходящими везикулами являются частицы, особенно наночастицы, как описано в настоящем документе.
Например, РНК может быть инкапсулирована в липосому. Согласно этому варианту осуществления фармацевтическая композиция представляет собой или содержит липосомный состав. Инкапсуляция внутри липосомы, как правило, будет защищать РНК от расщепления РНКазой. Возможно, что липосомы включают в себя некоторую внешнюю РНК (например, на своей поверхности), но по меньшей мере половина РНК (а в идеале вся она) инкапсулируется в сердцевине липосомы.
Липосомы представляют собой микроскопические липидные везикулы, часто содержащие один или несколько бислоев образующего везикулы липида, такого как фосфолипид, и способны инкапсулировать лекарственное средство, например, РНК. В контексте настоящего изобретения могут быть использованы различные типы липосом, включая в себя, без ограничения, многослойные везикулы (MLV), небольшие однослойные везикулы (SUV), большие однослойные везикулы (LUV), стерически стабилизированные липосомы (SSL), мультивезикулярные везикулы (MV) и большие мультивезикулярные везикулы (LMV), а также другие двуслойные формы, известные в настоящей области техники. Размер и слойность липосомы будут зависеть от способа получения. Существует несколько других форм супрамолекулярной организации, в которых липиды могут присутствовать в водной среде, включая в себя ламеллярные фазы, гексагональную и обратную гексагональные фазы, кубические фазы, мицеллы, обратные мицеллы, состоящие из монослоев. Эти фазы также могут быть получены в комбинации с ДНК или РНК, а взаимодействие с РНК и ДНК может существенно влиять на фазовое состояние. Такие фазы могут присутствовать в состоящих из наночастиц РНК составах по настоящему изобретению.
Липосомы могут быть образованы с использованием стандартных способов, известных специалисту в настоящей области техники. Соответствующие способы предусматривают способ обратного испарения, способ инъекции этанола, способ регидратации дегидратации, ультразвук или другие подходящие способы. После образования липосом липосомы могут быть отсортированы по размеру для получения популяции липосом, характеризующихся по существу однородным диапазоном размеров.
Согласно предпочтительному варианту осуществления настоящего изобретения РНК присутствует в липосоме, которая содержит по меньшей мере один катионный липид. Соответствующие липосомы могут быть образованы из одного липида или из смеси липидов при условии использования по меньшей мере одного катионного липида. Предпочтительные катионные липиды содержат атом азота, который способен к протонированию; предпочтительно такие катионные липиды представляют собой липиды с третичной аминогруппой. Особенно подходящим липидом с третичной аминогруппой является 1,2-дилинолеилокси-N,N-диметил-3-аминопропан (DLinDMA). Согласно одному варианту осуществления РНК в соответствии с настоящим изобретением присутствует в составе липосом, как описано в публикации международной заявки WO 2012/006378 A1: липосома, содержащая липидный бислой, инкапсулирующий водную сердцевину, содержащую РНК, причем липидный бислой содержит липид с pKa в диапазоне от 5,0 до 7,6, который предпочтительно содержит третичную аминогруппу. Предпочтительные катионные липиды с третичной аминогруппой содержат DLinDMA (pKa 5.8) и, как правило, описаны в публикации международной заявки WO 2012/031046 A2. Согласно публикации международной заявки WO 2012/031046 A2 липосомы, содержащие соответствующее соединение, особенно подходят для инкапсуляции РНК и, следовательно, для липосомной доставки РНК. Согласно одному варианту осуществления РНК в соответствии с настоящим изобретением присутствует в составе липосом, причем липосома содержит по меньшей мере один катионный липид, головная группа которого содержит по меньшей мере один атом азота (N), который способен к протонированию, причем липосома и РНК имеет отношение N:P от 1:1 до 20:1. В соответствии с настоящим изобретением «отношение N:P» относится к молярному соотношению атомов азота (N) в катионном липиде к атомам фосфата (Р) в РНК, содержащейся в липидсодержащей частице (например, липосоме), как описано в публикации международной заявки WO 2013/006825 A1. Отношение N:P от 1:1 до 20:1 связано с суммарным зарядом липосомы и эффективностью доставки РНК в клетку позвоночных.
Согласно одному варианту осуществления РНК в соответствии с настоящим изобретением присутствует в липосомном составе, который содержит по меньшей мере один липид, который включает в себя фрагмент полиэтиленгликоля (ПЭГ), причем РНК инкапсулируется в пегилированную липосому, так что фрагмент ПЭГ присутствует на внешней стороне липосомы, как описано в публикациях международных заявок WO 2012/031043 A1 и WO 2013/033563 A1.
Согласно одному варианту осуществления РНК в соответствии с настоящим изобретением присутствует в составе липосом, причем липосома характеризуется диаметром в диапазоне 60-180 нм, как описано в публикации международной заявки WO 2012/030901 A1.
Согласно одному варианту осуществления РНК в соответствии с настоящим изобретением присутствует в составе липосом, причем РНК-содержащие липосомы характеризуются суммарным зарядом, близким к нулю или отрицательным, как описано в публикации международной заявки WO 2013/143555 A1.
Согласно другим вариантам осуществления РНК в соответствии с настоящим изобретением присутствует в форме эмульсии. Ранее были описаны эмульсии для доставки молекул нуклеиновых кислот, таких как молекулы РНК, в клетки. Предпочтительными в настоящем документе являются эмульсии масло-в-воде. Соответствующие частицы эмульсии содержат сердцевину-масло и катионный липид. Более предпочтительными являются катионные эмульсии масло-в-воде, в которых РНК в соответствии с настоящим изобретением образует комплексы с частицами эмульсии. Частицы эмульсии содержат сердцевину-масло и катионный липид. Катионный липид может взаимодействовать с отрицательно заряженной РНК, тем самым привязывая РНК к частицам эмульсии. В эмульсии масло-в-воде частицы эмульсии диспергируют в водной дисперсионной фазе. Например, средний диаметр частиц эмульсии, как правило, может составлять от приблизительно 80 до 180 нм. Согласно одному варианту осуществления фармацевтическая композиция по настоящему изобретению представляет собой катионную эмульсию масло-в-воде, причем частицы эмульсии содержат сердцевину-масло и катионный липид, как описано в публикации международной заявки WO 2012/006380 A2. РНК в соответствии с настоящим изобретением может присутствовать в форме эмульсии, содержащей катионный липид, причем отношение N:P эмульсии составляет по меньшей мере 4:1, как описано в публикации международной заявки WO 2013/006834 A1. РНК в соответствии с настоящим изобретением может присутствовать в форме катионной липидной эмульсии, как описано в публикации международной заявки WO 2013/006837 A1. В частности, композиция может содержать РНК, образовавшую комплекс с частицей катионной эмульсии масло-в-воде, причем отношение масло/липид составляет по меньшей мере приблизительно 8:1 (моль:моль).
Согласно другим вариантам осуществления фармацевтическая композиция в соответствии с настоящим изобретением содержит РНК в формате липоплекса. Термин «липоплекс» или «РНК-липоплекс» относится к комплексу липидов и нуклеиновых кислот, таких как РНК. Липоплексы могут образовываться из катионных (положительно заряженных) липосом и анионной (отрицательно заряженной) нуклеиновой кислоты. Катионные липосомы могут также содержать нейтральный липид-«помощник». В простейшем случае липоплексы образуются спонтанно путем смешивания нуклеиновой кислоты с липосомами с определенным протоколом смешивания, однако могут применяться различные другие протоколы. Понятно, что электростатические взаимодействия между положительно заряженными липосомами и отрицательно заряженной нуклеиновой кислотой являются движущей силой образования липоплекса (публикация международной заявки WO 2013/143555 A1). Согласно одному варианту осуществления настоящего изобретения суммарный заряд частиц липоплекса РНК близок к нулю или отрицателен. Известно, что электро-нейтральные или отрицательно заряженные липоплексы РНК и липосом приводят к значительной экспрессии РНК в дендритных клетках селезенки (DC) после системного введения и не связаны с повышенной токсичностью, о которой сообщалось для положительно заряженных липосом и липоплексов (смотрите публикацию международной заявки WO 2013/143555 A1). Поэтому согласно одному варианту осуществления настоящего изобретения фармацевтическая композиция в соответствии с настоящим изобретением содержит РНК в формате наночастиц, предпочтительно наночастиц-липоплексов, в которых (i) число положительных зарядов в наночастицах не превышает число отрицательных зарядов в наночастицах и/или (ii) наночастицы характеризуются нейтральным или суммарным отрицательным зарядом, и/или (iii) отношение зарядов положительных зарядов к отрицательным зарядам в наночастицах составляет 1,4:1 или менее, и/или (iv) зета-потенциал наночастиц составляет 0 или менее. Как описано в публикации международной заявки WO 2013/143555 A1, зета-потенциал представляет собой научный термин для электрокинетического потенциала в коллоидных системах. В настоящем изобретении (а) зета-потенциал и (b) отношение заряда катионного липида к РНК в наночастицах может быть рассчитано как описано в публикации международной заявки WO 2013/143555 A1. Таким образом, фармацевтические композиции, представляющие собой состоящие из наночастиц-липоплексов составы с определенным размером частиц, в которых суммарный заряд частиц близок к нулю или отрицателен, как описано в публикации международной заявки WO 2013/143555 A1, представляют собой предпочтительные фармацевтические композиции в контексте настоящего изобретения.
Способы получения белка
Согласно четвертому аспекту в настоящем изобретении предусмотрен способ получения представляющего интерес белка в клетке, предусматривающий следующие стадии:
(а) получение РНК-репликона в соответствии с первым аспектом настоящего изобретения, который содержит открытую рамку считывания, кодирующую функциональный альфавирусный неструктурный белок, который может быть реплицирован функциональным альфавирусным неструктурным белком, и который дополнительно содержит открытую рамку считывания, кодирующую представляющий интерес белок и
(b) инокуляция репликона РНК в клетку.
Согласно различным вариантам осуществления способа РНК-репликон является таким, как определено выше для РНК-репликона по настоящему изобретению, при условии, что РНК-репликон содержит открытую рамку считывания, кодирующую функциональный альфавирусный неструктурный белок, и открытую рамку считывания, кодирующую представляющий интерес белок, и может быть реплицирован посредством функционального альфавирусного неструктурного белка.
Согласно пятому аспекту в настоящем изобретении предусмотрен способ получения представляющего интерес белка в клетке, предусматривающий следующие стадии:
(а) получение конструкции РНК для экспрессии функционального альфавирусного неструктурного белка,
(b) получение РНК-репликона в соответствии с первым аспектом настоящего изобретения, который может быть реплицирован посредством функционального альфавирусного неструктурного белка в соответствии с (а) in trans и который содержит открытую рамку считывания, кодирующую представляющий интерес белок, и
(c) совместная инокуляция конструкции РНК для экспрессии функционального альфавирусного неструктурного белка и РНК-репликона в клетку.
Согласно различным вариантам осуществления способа конструкция РНК для экспрессии функционального альфавирусного неструктурного белка и/или РНК-репликон являются такими, как определено выше для системы по настоящему изобретению, при условии, что РНК-репликон может быть реплицирован посредством функционального альфавирусного неструктурного белка in trans и содержит открытую рамку считывания, кодирующую представляющий интерес белок. Конструкция РНК для экспрессии функционального альфавирусного неструктурного белка и РНК-репликон могут быть либо инокулированы в один и тот же момент времени, либо могут быть альтернативно инокулированы в разные моменты времени. Во втором случае конструкцию РНК для экспрессии функционального альфавирусного неструктурного белка, как правило, инокулируют в первый момент времени, а репликон, как правило, инокулируют во второй, более поздний момент времени. В этом случае предполагается, что репликон будет немедленно реплицирован, поскольку репликаза уже будет синтезирована в клетке. Второй момент времени, как правило, происходит вскоре после первого момента времени, например, через от 1 минуты до 24 часов после первого момента времени.
Клетка, в которую можно инокулировать одну или несколько молекул нуклеиновой кислоты, можно назвать «клеткой-хозяином». В соответствии с настоящим изобретением термин «клетка-хозяин» относится к любой клетке, которая может быть трансформирована или трансфицирована экзогенной молекулой нуклеиновой кислоты. Термин «клетка» предпочтительно представляет собой интактную клетку, т.е. клетку с неповрежденной мембраной, которая не высвобождает свои нормальные внутриклеточные компоненты, такие как ферменты, органеллы или генетический материал. Интактная клетка предпочтительно представляет собой жизнеспособную клетку, т.е. живую клетку, способную выполнять свои нормальные метаболические функции. Термин «клетка-хозяин» включает в себя, в соответствии с настоящим изобретением, прокариотические (например, E.coli) или эукариотические клетки (например, клетки человека и животных, растительные клетки, дрожжевые клетки и клетки насекомых). Особое предпочтение отдается клеткам млекопитающих, таким как клетки людей, мышей, хомяков, свиней, домашних животных, включая в себя лошадей, коров, овец и коз, а также приматов. Клетки могут быть получены из множества типов тканей и содержать первичные клетки и клеточные линии. Конкретные примеры включают в себя кератиноциты, лейкоциты периферической крови, стволовые клетки костного мозга и эмбриональные стволовые клетки. Согласно другим вариантам осуществления клетка-хозяин представляет собой антигенпрезентирующую клетку, в частности дендритную клетку, моноцит или макрофаг. Нуклеиновая кислота может присутствовать в клетке-хозяине в одной или нескольких копиях и согласно одному варианту осуществления экспрессируется в клетке-хозяине.
Клетка может представлять собой прокариотическую клетку или эукариотическую клетку. Прокариотические клетки пригодны в настоящем документе, например, для увеличения количества ДНК в соответствии с настоящим изобретением, а эукариотические клетки пригодны в настоящем документе, например, для экспрессии открытой рамки считывания репликона.
В способе по настоящему изобретению можно использовать любое из системы в соответствии с настоящим изобретением или набора в соответствии с настоящим изобретением, или фармацевтической композиции в соответствии с настоящим изобретением. РНК может быть использована в форме фармацевтической композиции или в виде голой РНК, например, для электропорации.
Согласно способу настоящего изобретения может быть достигнута эффективная экспрессия представляющего интерес гена в клетке-хозяине (смотрите, например, примеры 2-5).
Согласно одному варианту осуществления дополнительная молекула РНК, предпочтительно молекула мРНК, может быть введена в клетку. Необязательно, дополнительная молекула РНК кодирует белок, подходящий для ингибирования IFN, такой как E3, как описано в настоящем документе. Необязательно, дополнительная молекула РНК может быть введена до введения репликона или репликазной конструкции или системы согласно настоящему изобретению.
В способе получения белка в клетке в соответствии с настоящим изобретением клетка может представлять собой антигенпрезентирующую клетку, и этот способ может быть использован для экспрессии РНК, кодирующей антиген. С этой целью настоящее изобретение может предусматривать введение антигена, кодирующего РНК, в антигенпрезентирующие клетки, такие как дендритные клетки. Для трансфекции антигенпрезентирующих клеток, таких как дендритные клетки, может быть использована фармацевтическая композиция, содержащая кодирующую антиген РНК.
Согласно одному варианту осуществления способ получения белка в клетке представляет собой способ in vitro. Согласно одному варианту осуществления способ получения белка в клетке не предусматривает удаление клетки из субъекта-человека или субъекта-животного хирургическим путем или терапией.
Согласно этому варианту осуществления клетка, инокулированная в соответствии с четвертым аспектом настоящего изобретения, может вводиться субъекту, чтобы производить белок у субъекта и обеспечивать субъекта белком. Клетка может быть аутологичной, сингенной, аллогенной или гетерологичной по отношению к субъекту.
Согласно другим вариантам осуществления клетка в способе получения белка в клетке может присутствовать у субъекта, такого как пациент. Согласно этим вариантам осуществления способ получения белка в клетке представляет собой способ in vivo, который предусматривает введение молекулы РНК субъекту.
В этом отношении в настоящем изобретении также предусмотрен способ получения представляющего интерес белка у субъекта, предусматривающий следующие стадии:
(а) получение РНК-репликона в соответствии с первым аспектом настоящего изобретения, который содержит открытую рамку считывания, кодирующую функциональный альфавирусный неструктурный белок, который может быть реплицирован функциональным альфавирусным неструктурным белком, и который дополнительно содержит открытую рамку считывания, кодирующую представляющий интерес белок, и
(b) введение РНК-репликона субъекту.
Согласно различным вариантам осуществления способа РНК-репликон является таким, как определено выше для РНК-репликона по настоящему изобретению, при условии, что РНК-репликон содержит открытую рамку считывания, кодирующую функциональный альфавирусный неструктурный белок, и открытую рамку считывания, кодирующую представляющий интерес белок, и может быть реплицирован посредством функционального альфавирусного неструктурного белка.
В настоящем изобретении дополнительно предусмотрен способ получения представляющего интерес белка у субъекта, предусматривающий следующие стадии:
(а) получение конструкции РНК для экспрессии функционального альфавирусного неструктурного белка,
(b) получение РНК-репликона в соответствии с первым аспектом настоящего изобретения, который может быть реплицирован функциональным альфавирусным неструктурным белком согласно (а) in trans и который содержит открытую рамку считывания, кодирующую представляющий интерес белок, и
(c) введение конструкции РНК для экспрессии функционального альфавирусного неструктурного белка и РНК-репликона субъекту.
Согласно различным вариантам осуществления способа конструкция РНК для экспрессии функционального альфавирусного неструктурного белка и/или РНК-репликон являются такими, как определено выше для системы по настоящему изобретению, при условии, что РНК-репликон может быть реплицирован посредством функционального альфавирусного неструктурного белка in trans и содержит открытую рамку считывания, кодирующую представляющий интерес белок. Конструкция РНК для экспрессии функционального альфавирусного неструктурного белка и РНК-репликон может либо вводиться в один и тот же момент времени, либо может альтернативно вводиться в разные моменты времени. Во втором случае конструкцию РНК для экспрессии функционального альфавирусного неструктурного белка, как правило, вводят в первый момент времени, а РНК-репликон, как правило, вводят во второй, более поздний момент времени. В этом случае предполагается, что репликон будет немедленно реплицирован, поскольку репликаза уже будет синтезирована в клетке. Второй момент времени, как правило, происходит вскоре после первого момента времени, например, через от 1 минуты до 24 часов после первого момента времени. Предпочтительно введение РНК-репликона проводят в одном и том же месте и посредством одного и того же пути введения, что и введение конструкции РНК для экспрессии функционального альфавирусного неструктурного белка, с тем чтобы увеличить перспективы того, что РНК-репликон и конструкция РНК для экспрессии функционального альфавирусного неструктурного белка достигают одной и той же целевой ткани или клетки. «Место» относится к части тела субъекта. Подходящими местами являются, например, левая рука, правая рука и т.д.
Согласно одному варианту осуществления субъекту может быть введена дополнительная молекула РНК, предпочтительно молекула мРНК. Необязательно, дополнительная молекула РНК кодирует белок, подходящий для ингибирования IFN, такой как E3, как описано в настоящем документе. Необязательно, дополнительную молекулу РНК можно вводить перед введением репликона или репликазной конструкции или системы согласно настоящему изобретению.
Что угодно из РНК-репликона в соответствии с настоящим изобретением, системы в соответствии с настоящим изобретением или набора в соответствии с настоящим изобретением, или фармацевтической композиции в соответствии с настоящим изобретением может быть использовано в способе получения белка у субъекта в соответствии с настоящим изобретением. Например, в способе по настоящему изобретению РНК может быть использована в форме фармацевтической композиции, например, как описано в настоящем документе, или в виде голой РНК.
Принимая во внимание потенциальную возможность, подлежащую введению субъекту, каждый из РНК-репликона в соответствии с настоящим изобретением, системы в соответствии с настоящим изобретением или набора в соответствии с настоящим изобретением, или фармацевтической композиции в соответствии с настоящим изобретением могут упоминаться как «лекарственное средство» или тому подобное. Настоящее изобретение предусматривает, что РНК-репликон, система, набор, фармацевтическая композиция по настоящему изобретению предназначены для применения в качестве лекарственного средства. Лекарственное средство можно использовать для лечения субъекта. Под «лечением» подразумевается введение соединения или композиции или другого объекта, как описано в настоящем документе, субъекту. Термин включает в себя способы лечения организма человека или животного путем терапии.
Вышеописанное лекарственное средство, как правило, не содержит ДНК и, таким образом, связано с дополнительными особенностями безопасности по сравнению с ДНК-вакцинами, описанными в предшествующем уровне техники (например, в публикации международной заявки WO 2008/119827 A1).
Альтернативное медицинское применение в соответствии с настоящим изобретением предусматривает способ получения белка в клетке в соответствии с четвертым аспектом настоящего изобретения, причем клетка может представлять собой антигенпрезентирующую клетку, такую как дендритная клетка, с последующим введением указанной клетки субъекту. Например, РНК, кодирующая фармацевтически активный белок, такой как антиген, может быть введена (трансфицирована) в антигенпрезентирующие клетки ex vivo, например, антигенпрезентирующие клетки, взятые у субъекта, и антигенпрезентирующие клетки, необязательно клонированно размноженные ex vivo, могут быть повторно введены одному и тому же или другому субъекту. Трансфицированные клетки могут быть повторно введены субъекту с использованием любых средств, известных в настоящей области техники.
Лекарственное средство в соответствии с настоящим изобретением может вводиться нуждающемуся в этом субъекту. Лекарственное средство по настоящему изобретению может быть использовано как в профилактических, так и в терапевтических способах лечения субъекта.
Лекарственное средство в соответствии с настоящим изобретением вводят в эффективном количестве. «Эффективное количество» относится к количеству, которого достаточно, отдельно или вместе с другими дозами, чтобы вызвать реакцию или желаемый эффект. В случае лечения определенного заболевания или определенного состояния у субъекта желаемым эффектом является ингибирование прогрессирования заболевания. Это включает в себя замедление прогрессирования заболевания, в частности прерывание прогрессирования заболевания. Желаемый эффект при лечении заболевания или состояния может также представлять собой задержку вспышки заболевания или ингибирование вспышки заболевания.
Эффективное количество будет зависеть от подвергаемого лечению состояния, тяжести заболевания, индивидуальных параметров пациента, включающих в себя возраст, физиологическое состояние, размер и массу, продолжительности лечения, типа сопровождающей терапии (если таковая имеется), конкретного способа введения и других факторов.
Вакцинация
Термин «иммунизация» или «вакцинация», как правило, относится к процессу лечения субъекта по терапевтическим или профилактическим причинам. Лечение, в частности профилактическое лечение, представляет собой или предусматривает, предпочтительно, лечение, направленное на индуцирование или усиление иммунного ответа субъекта, например, против одного или нескольких антигенов. Если в соответствии с настоящим изобретением желательно индуцировать или усиливать иммунный ответ с использованием РНК, как описано в настоящем документе, иммунный ответ может быть вызван или усилен посредством РНК. Согласно одному варианту осуществления в настоящем изобретении предусмотрено профилактическое лечение, которое представляет собой или предусматривает предпочтительно вакцинацию субъекта. Вариант осуществления настоящего изобретения, при котором репликон кодирует в качестве представляющего интерес белка фармацевтически активный пептид или белок, который является иммунологически активным соединением или антигеном, особенно применим для вакцинации.
Ранее была описана РНК для вакцинации против чужеродных патогенов, включающих в себя патогены или злокачественную опухоль (недавно рассмотренная в публикации Ulmer et al., 2012, Vaccine, vol. 30, pp. 4414-4418). В отличие от общих подходов в предшествующем уровне техники репликон в соответствии с настоящим изобретением является особенно подходящим элементом для эффективной вакцинации из-за способности реплицироваться посредством описанного в настоящем изобретении функционального альфавирусного неструктурного белка. Вакцинация в соответствии с настоящим изобретением может быть использована, например, для индукции иммунного ответа на слабо иммуногенные белки. В случае РНК-вакцин согласно настоящему изобретению белковый антиген никогда не подвергается воздействию сывороточных антител, а производится самими трансфицированными клетками после трансляции РНК. Поэтому анафилаксия не должна быть проблемой. Таким образом, настоящее изобретение позволяет повторную иммунизацию пациента без риска аллергических реакций.
В способах, предусматривающих вакцинацию в соответствии с настоящим изобретением, лекарственное средство по настоящему изобретению вводится субъекту, в частности, если желательно лечить субъекта, имеющего заболевание с участием антигена или подверженного риску заболевания с участием антигена.
В способах, предусматривающих вакцинацию в соответствии с настоящим изобретением, представляющий интерес белок, кодируемый репликоном в соответствии с настоящим изобретением, кодирует, например, бактериальный антиген, против которого должен быть направлен иммунный ответ, или вирусный антиген, против которого должен быть направлен иммунный ответ, или антиген злокачественной опухоли, против которой должен быть направлен иммунный ответ, или антиген одноклеточного организма, против которого должен быть направлен иммунный ответ. Эффективность вакцинации может быть оценена известными стандартными способами, такими как измерение антигенспецифических IgG-антител из организма. В способах, предусматривающих аллергенспецифическую иммунотерапию в соответствии с настоящим изобретением, представляющий интерес белок, кодируемый репликоном в соответствии с настоящим изобретением, кодирует антиген, имеющий отношение к аллергии. Аллергенспецифическая иммунотерапия (также известная как гипосенсибилизация) определяется как введение предпочтительно увеличенной дозы аллергенной вакцины в организм с одной или несколькими аллергиями для достижения состояния, при котором симптомы, связанные с последующим воздействием возбудительного аллергена облегчаются. Эффективность аллергенспецифической иммунотерапии может быть оценена известными стандартными способами, такими как измерение аллергенспецифических антител IgG и IgE в организме.
Лекарственное средство по настоящему изобретению может вводиться субъекту, например, для лечения субъекта, включая вакцинацию субъекта.
Термин «субъект» относится к позвоночным, особенно млекопитающим. Например, млекопитающими в контексте настоящего изобретения являются люди, отличные от человека приматы, одомашненные млекопитающие, такие как собаки, кошки, овцы, крупный рогатый скот, козы, свиньи, лошади и т.д., лабораторные животные, такие как мыши, крысы, кролики, морские свинки и т. д., а также животные в неволе, такие как животные зоопарков. Термин «субъект» также относится к немлекопитающим позвоночным, таким как птицы (особенно одомашненные птицы, такие как куры, утки, гуси, индюки), и к рыбе (особенно выращиваемая рыба, например, лосось или сом). Используемый в настоящем документе термин «животное» также включает в себя людей.
Введение одомашненным животным, таким как собаки, кошки, кролики, морские свинки, хомяки, овцы, крупный рогатый скот, козы, свиньи, лошади, куры, утки, гуси, индюки, или диким животным, например, лисицам, предпочтительнее согласно некоторым вариантам осуществления. Например, профилактическая вакцинация в соответствии с настоящим изобретением может быть пригодна для вакцинации популяции животных, например, в сельском хозяйстве или в популяции диких животных. Могут быть вакцинированы другие популяции животных в неволе, такие как домашние животные или животные зоопарков.
При введении субъекту репликон и/или репликазная конструкция, используемые в качестве лекарственного средства, предпочтительно не содержат последовательностей из типа альфавируса, который является инфекционным для вида или рода, к которому относится подвергаемый лечению субъект. Предпочтительно, в этом случае репликон и/или репликазная конструкция не содержат никакой нуклеотидной последовательности из альфавируса, который может инфицировать соответствующие виды или род. Этот вариант осуществления имеет то преимущество, что рекомбинация с инфекционным (например, полностью функциональным или диким) альфавирусом невозможна, даже если субъект, которому вводится РНК, (например, случайно) поражен инфекционным альфавирусом. В качестве иллюстративного примера, для лечения свиней, используемый репликон и/или репликазная конструкция не содержит никакой нуклеотидной последовательности из альфавируса, который может инфицировать свиней.
Способ введения
Лекарственное средство в соответствии с настоящим изобретением может быть применено к субъекту посредством любого подходящего пути введения.
Например, лекарственное средство можно вводить системно, например, внутривенно (i.v.), подкожно (s.c.), внутрикожно (i.d.) или путем ингаляции.
Согласно одному варианту осуществления лекарственное средство в соответствии с настоящим изобретением вводят в мышечную ткань, такую как скелетная мышца или кожа, например, подкожно. Как правило, понимается, что перенос РНК в кожу или мышцы приводит к высокой и устойчивой локальной экспрессии, параллельно с сильной индукцией гуморальных и клеточных иммунных реакций (Johansson et al., 2012, PLoS. One., 7, e29732; Geall et al., 2012, Proc. Natl. Acad. Sci. US A, vol. 109, pp. 14604-14609).
Альтернативы введению в мышечную ткань или кожу включают в себя, без ограничения: внутрикожное, интраназальное, внутриглазное, внутрибрюшинное, внутривенное, интерстициальное, трансбуккальное, трансдермальное или подъязычное введение. Внутрикожное и внутримышечное введение представляют собой два предпочтительных пути.
Введение может быть достигнуто различными способами. Согласно одному варианту осуществления лекарственное средство в соответствии с настоящим изобретением вводят путем инъекции. Согласно предпочтительному варианту осуществления инъекцию осуществляют через иглу. В качестве альтернативы можно использовать инъекцию без иглы.
Настоящее изобретение описано подробно и иллюстрируется графическими материалами и примерами, которые используются только для иллюстративных целей и не предназначены для ограничения. Благодаря описанию и примерам, дополнительные варианты осуществления, которые также включены в настоящее изобретение, доступны для квалифицированного работника.
Примеры
Материал и способы:
Следующие материалы и способы были использованы в примерах, которые описаны ниже.
Клонирование плазмид, транскрипция in vitro, очистка РНК
Плазмиды клонировали с использованием стандартной технологии. Детали клонирования отдельных плазмид, используемых в примерах по настоящему изобретению, описаны в примере 1. Транскрипцию in vitro с использованием плазмид, описанных в примере 1, и РНК-полимеразы Т7, и очистку РНК проводили, как описано ранее (Holtkamp et al., 2006, Blood, vol. 108, pp. 4009-4017, Kuhn et al., 2010, Gene Ther., vol.17, pp. 961-971).
Качество очищенной РНК оценивали с помощью спектрофотометрии и анализа на BioAnalyzer 2100 (Agilent, Santa Clara, USA). Все РНК, трансфицированные в клетки в примерах, представляли собой транскрибированные in vitro IVT-РНК.
Трансфекция РНК:
Для электропорации РНК электропорировали в клетки млекопитающих при комнатной температуре с использованием устройства для электропорации с квадратной волной (BTX ECM 830, Harvard Apparatus, Holliston, MA, USA), используя следующие настройки: для клеток BHK21: 750 В/см, 1 импульс 16 мс; для фибробластов крайней плоти человека: 500 В/см, 1 импульс 24 мс. Смеси различных видов РНК готовили в пробирках без РНК и оставляли на льду до трансфекции. Для электропорации РНК или смеси РНК ресуспендировали в конечном объеме 62,5 мкл/мм зазора формующей щели кюветы.
Для липофекции клетки высевали в количестве приблизительно 20000 клеток/см2 площади роста и трансфицировали общим количеством 260 нг/см2 РНК и 1 мкл/см2 реагента MessengerMax, следуя инструкциям производителя (Life Technologies, Darmstadt, Germany).
Клеточная культура: все среды для роста, телячья фетальная сыворотка (FCS), антибиотики и другие добавки были предоставлены Life Technologies/Gibco, за исключением случаев, когда указано иное. Фибробласты крайней плоти человека, полученные из System Bioscience (HFF, неонатальный) или ATCC (CCD-1079Sk), культивировали в минимальных поддерживающих средах (MEM), содержащих FCS в концентрации 15%, незаменимые аминокислоты в концентрации 1%, пируват натрия в концентрации 1 мМ при температуре 37°С. Клетки выращивали при температуре 37°С в увлажненной атмосфере, уравновешенной до 5% СО2. Клетки BHK21 из клеточной линии «BHK21 [C13] (ATCC® CCL10™)», доступные из Американской коллекции типовых культур, Манассас, Вирджиния, США, выращивали в минимальной поддерживающей среде Игла, дополненной 10% FCS.
Проточная цитометрия:
Экспрессию РНК, кодирующей GFP, измеряли с помощью проточной цитометрии с использованием проточного цитометра FACS Canto II (BD Bioscience, Heidelberg, Germany), а полученные данные анализировали с помощью соответствующего программного обеспечения Diva или программного обеспечения FlowJo (Tree Star Inc., Ashland, OR, USA).
Анализ люциферазы:
Для оценки экспрессии люциферазы светлячков трансфицированные клетки высевали в 96-луночные черные микропланшеты, а супернатанты из трансфицированных Nanoluc клеток переносили на 96-луночные черные микропланшеты (Nunc, Langenselbold, Germany). Экспрессию люциферазы светлячков измеряли с использованием системы анализа люциферазы Bright-Glo, экспрессию Nanoluc измеряли с использованием системы анализа люциферазы Nano-Glo (оба Promega, Madison, WI, USA) в соответствии с инструкциями производителя. Биолюминесценцию измеряли с использованием микропланшетного ридера люминесценции Infinite M200 (Tecan Group, Männedorf, Switzerland). Данные представлены в относительных световых единицах [RLU], отрицательные по люциферазе клетки использовали для вычитания фонового сигнала.
Пример 1. Клонирование плазмид
A. Плазмиды, кодирующие репликон на основе вируса леса Семлики (SFV), получали с использованием способов бесшовного клонирования на основе ПЦР. Бесшовное клонирование означает способы клонирования, основанные на рекомбинации производимого ПЦР фрагмента в линеаризованные векторы с использованием гомологичных фрагментов секвенирования. Таким образом, последовательность ДНК, кодирующая репликон, который соответствует геному SFV, за исключением отсутствия открытой рамки считывания, кодирующей вирусные структурные гены, переносили из pSFV-ген-GFP (Ehrengruber & Lundstrom, 1999, Proc. Natl. Acad. Sci. U. S. A, vol. 96, pp. 7041-7046; Lundstrom, 2001, Histochem. Cell Biol., vol. 115, pp. 83-91) в остов плазмиды pST1 (Holtkamp et al., 2006, Blood, vol. 108, pp. 4009-4017) сразу ниже по ходу транскрипции от промотора РНК-полимеразы T7-фага. Кодируемую плазмидой поли(А)-кассету либо из 120 остатков аденилата (Holtkamp et al., смотрите выше), либо модифицированную поли(А)-кассету, состоящую из 30 и 70 остатков аденилата, разделенных 10-нуклеотидной случайной последовательностью (публикация международной заявки WO 2016/005004 A1), добавляли сразу же ниже по ходу транскрипции от самого последнего нуклеотида SFV 3'-UTR. Сайт рестрикции SapI помещали сразу же ниже по ходу транскрипции от поли(А)-кассеты или модифицированной поли(А)-кассеты. Кроме того, кодирующую последовательность, кодирующую myc-метку, вставляли в сайт XhoI, который находится в области кодирования для вариабельной области nsP3 SFV. Вставки в вариабельную область nsP3 не влияют на активность репликазного полипротеина (Spuul et al., 2010, J. Virol, vol. 85, pp. 7543-7557). Полученная в результате плазмида содержит последовательность ДНК, кодирующую 5' репликационную последовательность распознавания SFV под контролем промотора для полимеразы T7. Последовательность ДНК, кодирующая 5' репликационную последовательность распознавания SFV, представлена посредством SEQ ID NO: 4:
ATGGCGGATGTGTGACATACACGACGCCAAAAGATTTTGTTCCAGCTCCTGCCACCTCCG
CTACGCGAGAGATTAACCACCCACGATGGCCGCCAAAGTGCATGTTGATATTGAGGCTGA
CAGCCCATTCATCAAGTCTTTGCAGAAGGCATTTCCGTCGTTCGAGGTGGAGTCATTGCA
GGTCACACCAAATGACCATGCAAATGCCAGAGCATTTTCGCACCTGGCTACCAAATTGA (SED ID NO:4)
В приведенном выше представлении SEQ ID NO: 4 первый подчеркнутый ATG служит в качестве инициирующего кодона для синтеза N-концевого фрагмента nsP1; основания, транслированные в белок, представлены жирным шрифтом. Дальнейшие ATG в области кодирования nsP1 также подчеркнуты. Во время транскрипции in vitro плазмиды, содержащей 5' репликационную последовательность распознавания SFV (представленную посредством SEQ ID NO: 4) с помощью полимеразы Т7, получают транскрипт, содержащий последовательность РНК, соответствующую SEQ ID NO: 3: 5'-концевой G соответствует первому нуклеотиду, который транскрибируется посредством полимеразы Т7. Этот G предшествует альфавирусной последовательности и необходим для эффективной транскрипции с помощью полимеразы T7.
GATGGCGGATGTGTGACATACACGACGCCAAAAGATTTTGTTCCAGCTCCTGCCACCTCCGCTACGCGAGAGATTAACCACCCACGATGGCCGCCAAAGTGCATGTTGATATTGAGGCTGACAGCCCATTCATCAAGTCTTTGCAGAAGGCATTTCCGTCGTTCGAGGTGGAGTCATTGCAGGTCACACCAAATGACCATGCAAATGCCAGAGCATTTTCGCACCTGGCTACCAAATTGATCGAGCAGGAGACTGACAAAGACACACTCATCTTGGATATC (SED ID NO:3)
В приведенном выше представлении SEQ ID NO: 3 подчеркнуты пять специфических триплетов ATG. Уникальный сайт рестрикции EcoRV в кодирующей области для репликазы SFV (GATATC) выделен жирным шрифтом.
В результате получали плазмиду А. Плазмида А содержит открытую рамку считывания для функционального альфавирусного неструктурного белка.
B. Плазмиду, кодирующую транс-репликон с немодифицированной 5' репликационной последовательностью распознавания, получали путем удаления кодирующей неструктурный белок последовательности от EcoRV до SalI в плазмиде A (смотрите выше). Таким образом, основная часть открытой рамки считывания nsP1234, то есть последовательность, проходящая от уникального сайта рестрикции EcoRV до уникального сайта рестрикции SalI, была удалена. Для иллюстрации: GATATC (наиболее удаленные к 3'-концу шесть нуклеотидов SEQ ID NO: 3, полужирный шрифт в приведенном выше представлении) соответствует уникальному сайту рестрикции EcoRV кодирующей последовательности репликазы SFV. После удаления основной части открытой рамки считывания, кодирующей nsP1234, 5' репликационная последовательность распознавания и субгеномный промотор все еще присутствуют. Из-за удаления основной части открытой рамки считывания, кодирующей nsP1234, РНК, закодированная соответствующей плазмидой, если она присутствует в клетке-хозяине, не способна реплицировать in cis, но для репликации требуется наличие функционального альфавирусного неструктурного белка.
Открытую рамку считывания, кодирующую люциферазу светлячков (трансген), вставляли ниже по ходу транскрипции от субгеномного промотора (SGP). Таким образом получали плазмиду В. РНК-репликон, кодируемый соответствующей плазмидой, представлен как «РНК-матрица WT-RRS» на фиг. 1.
С использованием веб-сервера для прогнозирования вторичной структуры РНК (http://rna.urmc.rochester.edu/RNAstructureWeb/Servers/Predict1/Predict1.html) было подтверждено что предсказанная немодифицированная 5' репликационная последовательность распознавания (РНК) укладывается в четыре структуры «петля-на-стебле», неотличимые от родительской РНК и в соответствии с литературой (Frolov, 2001, RNA, vol. 7, pp. 1638-1651).
С-1. Начиная с плазмиды В, плазмиду, кодирующую транс-репликон с модификацией 5' репликационной последовательности распознавания (содержащей удаление триплета оснований ATG, который кодирует первый аминокислотный остаток nsP1, а также удаление четырех дополнительных триплетов оснований ATG в 5' репликационной последовательности распознавания), производили однонуклеотидными изменениями, то есть заменой одного основания соответствующего триплета оснований ATG (т.е. заменой A или T или G) на другое основание. Другими словами, те триплеты оснований ATG в SEQ ID NO: 3, которые были подчеркнуты выше, удаляли, каждый путем изменения одного нуклеотида (замены), соответственно.
Укладка РНК транс-репликона, кодируемой плазмидой, была предсказана с использованием веб-сервера для прогнозирования вторичной структуры РНК (http://rna.urmc.rochester.edu/RNAstructureWeb/Servers/Predict1/Predict1.html) и Mfold (http://unafold.rna.albany.edu/?q=mfold).
Предсказанную укладку модифицированной 5' репликационной последовательности распознавания, характеризующейся удалением ATG, сравнивали с предсказанной укладкой соответствующей немодифицированной 5' репликационной последовательности распознавания. В случае, если предсказание показывало, что вторичная структура модифицированной 5' репликационной последовательности распознавания отличалась от вторичной структуры немодифицированной 5' репликационной последовательности распознавания, выполняли одно или несколько дополнительных нуклеотидных изменений (замен) способом проб и ошибок до тех пор, пока не будет предсказано, что укладка РНК 5' репликационной последовательности распознавания без ATG (Δ5ATG-RRS) будет идентичной укладке РНК немодифицированной 5' репликационной последовательности распознавания, кодируемой плазмидой B. В результате получали последовательность ДНК в соответствии с SEQ ID NO: 5:
GATGGCGGATGTGTGACATACACGACGCCAAAAGATTTTGTTCCAGCTCCTGCCACCTCCGCTACGCGAGAGATTAACCACCCACGACGGCCGCCAAAGTGCTTGTTGATATTGAGGCTGACAGCCCATTCATCAAGTCTTAGCAGAAGGCATTTCCGTCGTTCGAGGTGGAGTCATTGGAGGTGACACCAAATCACCATCCAAATCCCAGAGCATTTTCGCACCTGGGTACCAAATTGATCGAGCAGGAGACTGACAAAGACACACTCATCTTGGATATC (SED ID NO:5)
Плазмида, кодирующая модифицированную 5' репликационную последовательности распознавания, характеризующуюся предсказанной идентичной укладкой, то есть плазмида, содержащая SEQ ID NO: 5, обозначается в настоящем документе C-1.
РНК-репликон, кодируемый соответствующей плазмидой, показан на фиг. 1, обозначенный как «Δ5ATG-RRS».
Понятно, что удаление пяти специфических ATG, как показано в SEQ ID NO: 5, предотвратит синтез nsP1 или его фрагмента. Понятно, что из-за удаления нативного инициирующего кодона для nsP1 синтез белка инициируется в первом инициирующем кодоне ниже по ходу транскрипции от субгеномного промотора (SGP), что приводит к транскрипции открытой рамки считывания, кодирующей люциферазу светлячков (трансген).
С-2. Начиная с С-1, SGP удаляли путем расщепления с EcoRV и SmaI и повторного лигирования. Соответствующий РНК-репликон схематически изображен на фиг. 1, обозначенный как «Δ5ATG-RRSΔSGP». Если соответствующий РНК-репликон содержит 5'-кэп, кодирующий люциферазу трансген может быть помещен под прямой трансляционный контроль 5'-кэпа.
С-3. Начиная с C-1, открытую рамку считывания, кодирующую репликазу SFV, вставляли с использованием сайтов рестрикции EcoRV и SalI. Таким образом, получали плазмиду, кодирующую РНК, способную к саморепликации, причем ORF репликазы не перекрывается ни с 5' репликационной последовательности распознавания, ни с субгеномным промотором. Соответствующий РНК-репликон схематически изображен на фиг. 1, обозначенный как «цис-репликон Δ5ATG-RRS».
D. Чтобы обеспечить возможность трансляции последовательности нуклеиновой кислоты, кодирующей репликазу, из нереплицирующей мРНК, открытую рамку считывания, кодирующую репликазу SFV, клонировали в плазмиду, содержащую 5'-UTR альфа-глобина человека, синтетическую 3'-UTR и кодированный плазмидой поли(А)-хвост из 30 плюс 70 аденилатных остатков, разделенных стабилизирующим линкером из 10 нуклеотидов (публикация международной заявки WO 2016/005324 A1). Открытую рамку считывания, кодирующую репликазу SFV, клонировали ниже по ходу транскрипции от 5'-UTR альфа-глобина человека. Плазмида содержала промотор T7 для транскрипции открытой рамки считывания, кодирующей репликазу SFV.
E. Чтобы обеспечить возможность трансляции последовательности нуклеиновой кислоты, кодирующей ингибитор Е3 протеинкиназы вируса осповакцины, из нереплицирующей мРНК, открытую рамку считывания, кодирующую ингибитор Е3 протеинкиназы R вируса осповакцины («Е3»), клонировали в плазмиду, содержащую 5'-UTR альфа-глобина человека, синтетическую 3'-UTR и кодированный плазмидой поли(А)-хвост из 30 плюс 70 аденилатных остатков, разделенных стабилизирующим линкером из 10 нуклеотидов (публикация международной заявки WO 2016/005324 A1). Кодирующую E3 открытую рамку считывания клонировали ниже по ходу транскрипции от 5'-UTR альфа-глобина человека. Плазмида содержала промотор T7 для транскрипции открытой рамки считывания, кодирующей E3.
Пример 2. Удаление стартовых кодонов в 5' репликационной последовательности распознавания не влияет на репликацию РНК транс-репликонов.
Клетки BHK21 подвергали совместной электропорации с (i) 5 мкг мРНК, кодирующей репликазу SFV (кодируемую плазмидой D примера 1), и (ii) различными количествами РНК транс-репликонов (кодируемой плазмидами B или C-1 примера 1, в фиг. 2: «матрица»). Транс-репликон кодирует люциферазу светлячков; транс-репликон содержит либо 5' репликационную последовательность распознавания дикого типа (фиг. 2: «WT-RRS», кодируемую плазмидой B); либо 5' репликационную последовательность распознавания, характеризующуюся удалением всех инициирующих кодонов (фиг. 2: «Δ5ATG-RRS», кодируемую плазмидой C-1). РНК транс-репликона была некэпирована (без кэпа). 5000 подвергнутых электропорации клеток BHK21 высевали в каждую лунку 96-луночных планшетов для измерения экспрессии люциферазы через 24 часа после электропорации. Результаты показаны на фиг. 2B.
Пример 3. Удаление стартовых кодонов в 5' репликационной последовательности распознавания позволяет осуществлять трансляцию трансгена под контролем кэпа
Использовали транс-репликон, характеризующийся удалением всех инициирующих кодонов и содержащий субгеномный промотор (фиг. 3: «Δ5ATG-RRS», кодируемый плазмидой C-1 примера 1), транс-репликон, характеризующийся удалением всех инициирующих кодонов, но не содержащий субгеномный промотор (фиг. 3: «Δ5ATG-RRSΔSGP», кодируемый плазмидой C-2 примера 1), и транс-репликон, имеющий все инициирующие кодоны и содержащий субгеномный промотор (фиг. 3 «РНК-матрица WT-RRS»). Фибробласты крайней плоти человека подвергали совместной электропорации с (i) 0,45 мкг соответствующей РНК транс-репликона, как показано на фиг. 3, и либо (ii-a) 2,5 мкг мРНК, кодирующей белок E3 вируса осповакцины (кодируемой плазмидой E примера 1, на фиг. 3: «E3»), либо (ii-b) 2,5 мкг мРНК, кодирующей репликазу (кодируемой плазмидой D примера 1 на фиг. 3: «репликаза») плюс 2,5 мкг мРНК, кодирующей белок E3 вируса осповакцины (кодируемой плазмидой Е примера 1). мРНК, кодирующую белок E3 вируса осповакцины, добавляли для ингибирования активации протеинкиназы R и тем самым стимулировали экспрессию люциферазы и репликазы. РНК транс-репликона была либо некэпирована (фиг. 3: «без кэпа»), либо котранскрипционно кэпирована аналогом кэпа бета-S-ARCA (D2) (фиг. 3: «D2-кэп»). Экспрессию люциферазы оценивали через 24 часа. Результаты показаны на фиг. 3 (правая панель).
Этот пример демонстрирует, что удаление инициирующих кодонов из 5' репликационной последовательности распознавания позволяет трансгену эффективно транслироваться непосредственно из кэпированной РНК транс-репликона.
Пример 4. На ранних стадиях после трансфекции зависимая от кэпа трансляция из транс-репликона, характеризующегося удалением стартовых кодонов в 5' репликационной последовательности распознавания, является более сильной, чем из субгеномной РНК.
Использовали транс-репликон, характеризующийся удалением всех инициирующих кодонов и содержащий субгеномный промотор (фиг. 4: «Δ5ATG-RRS», кодируемый плазмидой C-1 примера 1), и транс-репликон, характеризующийся удалением всех инициирующих кодонов, но не содержащий субгеномный промотор (на фиг. 4: «ΔATAT-RRSΔSGP», кодируемый плазмидой C-2 примера 1). Клетки BHK21 подвергали совместной электропорации с (i) 0,45 мкг соответствующей РНК транс-репликона и с (ii) 2,5 мкг мРНК, кодирующей репликазу (кодируемой плазмидой D примера 1). РНК транс-репликона либо не показана (фиг. 4: «без кэпа»), либо котранскрипционно кэпирована аналогом кэпа бета-S-ARCA(D2) (фиг. 4: «D2-кэп»). Экспрессию люциферазы оценивали с течением времени. Результаты показаны на фиг. 4 (правая панель).
Пример 5: Зависимая от кэпа трансляции из транс-репликона, характеризующаяся удалением стартовых кодонов в 5' репликационной последовательности распознавания, позволяет экспрессировать трансген на ранних стадиях, не завися от предшествующей экспрессии репликазы.
Использовали транс-репликон, характеризующийся удалением всех инициирующих кодонов, но не содержащий субгеномный промотор (фиг. 5: «ΔATAT-RRSΔSGP», кодируемый плазмидой C-2 примера 1). РНК транс-репликона либо некэпирована (фиг. 5: «без кэпа»), либо котранскрипционно кэпирована аналогом кэпа бета-S-ARCA (D2) (фиг. 5: «D2-кэп»). Клетки BHK21 подвергали электропорации с (i) 0,45 мкг соответствующей РНК транс-репликона и, если это указано («репликаза» на фиг. 5), дополнительно с (ii) 2,5 мкг кодирующей репликазу мРНК (кодируемой плазмидой D примера 1). Экспрессию люциферазы оценивали с течением времени. Результаты показаны на фиг. 5 (правая панель).
Пример 6. Реконструированные цис-репликоны с репликационной последовательностью распознавания с удалением ATG являются функциональными. ORF репликазы SFV вставляли в «Δ5ATG-RRS» (фиг. 7A: «Δ5ATG-RRS», кодированная плазмидой C-1 примера 1), что приводило в результате к получению плазмиды C-3 примера 1 «цис-репликон Δ5ATG-RRS» и кодированию люциферазы светлячков ниже по ходу транскрипции от субгеномного промотора (SGP). Внутри вставленной репликазы области, соответствующие CSE2 и SGP сердцевины, разрушали нуклеотидными обменами (заштрихованные прямоугольники), чтобы избежать дупликации этих регуляторных областей. Это привело к реконструированному цис-репликону. Клетки BHK21 подвергали совместной электропорации с 2,5 мкг либо «цис-репликона WT-RRS», либо «цис-репликона Δ5ATG-RRS». Через 24 ч после электропорации измеряли экспрессию люциферазы и продемонстрировали, что этот реконструированный цис-репликон является функциональным (фиг. 7В).
Пример 7. Бицистронные транс-репликоны экспрессируют оба трансгена. Секретируемую нано-люциферазу (SNL) клонировали ниже по ходу транскрипции от субгеномного промотора (SGP) транс-репликона WT-RSS (плазмида B примера 1). Положение выше против хода транскрипции от SGP не кодирует трансген, поскольку он недоступен для трансляции (фиг. 8A). Во второй конструкции SNL клонировали ниже по ходу транскрипции от ΔATG-RSS (плазмида C-1 примера 1) и люциферазу светлячков (Luc) вставляли ниже по ходу транскрипции от SGP. Клетки BHK21 подвергали совместной электропорации с 0,9 мкг транс-реплицирующей РНК и 5 мкг кодирующей SFV-репликазу мРНК. Через 48 часов после электропорации измеряли экспрессию SNL и Luc (фиг. 8B). Этот эксперимент свидетельствует о том, что трансгены экспрессируются из обоих положений в «Δ5ATG-RRS-бицистронных» транс-репликонах.
Пример 8: Транс-репликоны вируса Синдбис, не содержащие стартовые кодоны в репликационной последовательности распознавания, эффективно реплицируются. Транс-репликоны конструировали из генома вируса Синдбис путем синтеза генов аналогично конструкциям, описанным в примере 1 для SFV. Помимо транс-репликона с немодифицированной репликационной последовательностью распознавания (WT-RSS) производили два варианта. Первый (ΔATG-RRS) содержит делеции исходного стартового кодона плюс 4 дополнительных ATG и соответствующие компенсаторные нуклеотидные изменения для сохранения вторичной структуры РНК. На следующей стадии область, соответствующую субгеномному промотору, дополнительно удаляли для получения ΔATG-RRSΔSGP. GFP вставляли в РНК транс-репликона непосредственно ниже по ходу транскрипции от 5'RRS с удаленными ATG в векторы Δ5ATG-RRSΔSGP или ниже по ходу транскрипции от субгеномного промотора (SGP) в векторы Δ5ATG-RRS и WT-RRS (фиг. 9A). Клетки BHK21 подвергали совместной электропорации с 0,1 мкг транс-реплицирующей РНК и 2,4 мкг кодирующей SFV-репликазу мРНК, и через 24 ч оценивали экспрессию GFP (частоту трансфекции [%] и среднюю интенсивность флуоресценции (MFI) (фиг. 9В)). Этот эксперимент показывает, что тот же принцип модификации последовательности, который применялся к сконструированным репликонам SFV, может быть применен к сконструированным репликонам вируса Синдбис.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> БИОНТЭК РНА ФАРМАСЬЮТИКАЛЗ
ТРОН - ТРАНСЛАЦИОНАЛЕ ОНКОЛОГИ АН ДЕР УНИФЕРЗИТЕТСМЕДИЦИН ДЕР ИОГАНН
ГУТЕНБЕРГ-УНИФЕРЗИТЕТ МАЙНЦ ГЕМАЙННЮТЦИГЕ ГМБХ
<120> РНК-РЕПЛИКОН ДЛЯ УНИВЕРСАЛЬНОЙ И ЭФФЕКТИВНОЙ ГЕННОЙ ЭКСПРЕССИИ
<130> 674-173 PCT2
<150> PCT/EP2016/056165
<151> 2016-03-21
<160> 6
<170> PatentIn version 3.5
<210> 1
<211> 24
<212> RNA
<213> Artificial Sequence
<220>
<223> conserved sequence element 3
<400> 1
accucuacgg cgguccuaaa uagg 24
<210> 2
<211> 19
<212> RNA
<213> Artificial Sequence
<220>
<223> conserved sequence element 4
<400> 2
auuuuguuuu uaauauuuc 19
<210> 3
<211> 281
<212> DNA
<213> Artificial Sequence
<220>
<223> 5' replication recognition sequence
<400> 3
gatggcggat gtgtgacata cacgacgcca aaagattttg ttccagctcc tgccacctcc 60
gctacgcgag agattaacca cccacgatgg ccgccaaagt gcatgttgat attgaggctg 120
acagcccatt catcaagtct ttgcagaagg catttccgtc gttcgaggtg gagtcattgc 180
aggtcacacc aaatgaccat gcaaatgcca gagcattttc gcacctggct accaaattga 240
tcgagcagga gactgacaaa gacacactca tcttggatat c 281
<210> 4
<211> 239
<212> DNA
<213> Artificial Sequence
<220>
<223> 5' replication recognition sequence
<400> 4
atggcggatg tgtgacatac acgacgccaa aagattttgt tccagctcct gccacctccg 60
ctacgcgaga gattaaccac ccacgatggc cgccaaagtg catgttgata ttgaggctga 120
cagcccattc atcaagtctt tgcagaaggc atttccgtcg ttcgaggtgg agtcattgca 180
ggtcacacca aatgaccatg caaatgccag agcattttcg cacctggcta ccaaattga 239
<210> 5
<211> 281
<212> DNA
<213> Artificial Sequence
<220>
<223> 5' replication recognition sequence
<400> 5
gatggcggat gtgtgacata cacgacgcca aaagattttg ttccagctcc tgccacctcc 60
gctacgcgag agattaacca cccacgacgg ccgccaaagt gcttgttgat attgaggctg 120
acagcccatt catcaagtct tagcagaagg catttccgtc gttcgaggtg gagtcattgg 180
aggtgacacc aaatcaccat ccaaatccca gagcattttc gcacctgggt accaaattga 240
tcgagcagga gactgacaaa gacacactca tcttggatat c 281
<210> 6
<211> 13
<212> RNA
<213> Artificial Sequence
<220>
<223> Kozak sequence
<220>
<221> misc_feature
<222> (7)..(7)
<223> purine
<400> 6
gccgccncca ugg 13
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ТРАНС-РЕПЛИЦИРУЮЩАЯ РНК | 2017 |
|
RU2752580C2 |
| СОСТАВ ДЛЯ ВВЕДЕНИЯ РНК | 2017 |
|
RU2746118C2 |
| РЕЦЕПТУРА ДЛЯ ВВЕДЕНИЯ РНК | 2019 |
|
RU2797147C2 |
| СПОСОБ ПОВЫШЕНИЯ ЭКСПРЕССИИ РНК В КЛЕТКЕ | 2018 |
|
RU2784654C2 |
| ПЛАТФОРМЫ ДОСТАВКИ АНТИГЕНОВ | 2011 |
|
RU2597974C2 |
| ЭМУЛЬСИИ ТИПА "МАСЛО В ВОДЕ", КОТОРЫЕ СОДЕРЖАТ НУКЛЕИНОВЫЕ КИСЛОТЫ | 2012 |
|
RU2606846C2 |
| МИКРОЧАСТИЦЫ ДЛЯ ДОСТАВКИ ГЕТЕРОЛОГИЧНЫХ НУКЛЕИНОВЫХ КИСЛОТ | 2001 |
|
RU2295954C2 |
| ПОСЛЕДОВАТЕЛЬНОСТИ 3'-UTR ДЛЯ СТАБИЛИЗАЦИИ РНК | 2016 |
|
RU2720934C2 |
| МАЛЕНЬКИЕ ЛИПОСОМЫ ДЛЯ ДОСТАВКИ КОДИРУЮЩЕЙ ИММУНОГЕН РНК | 2011 |
|
RU2671482C2 |
| ВАКЦИНА ПРОТИВ БОЛЕЗНИ ЛАЙМА У СОБАК | 2018 |
|
RU2785620C2 |
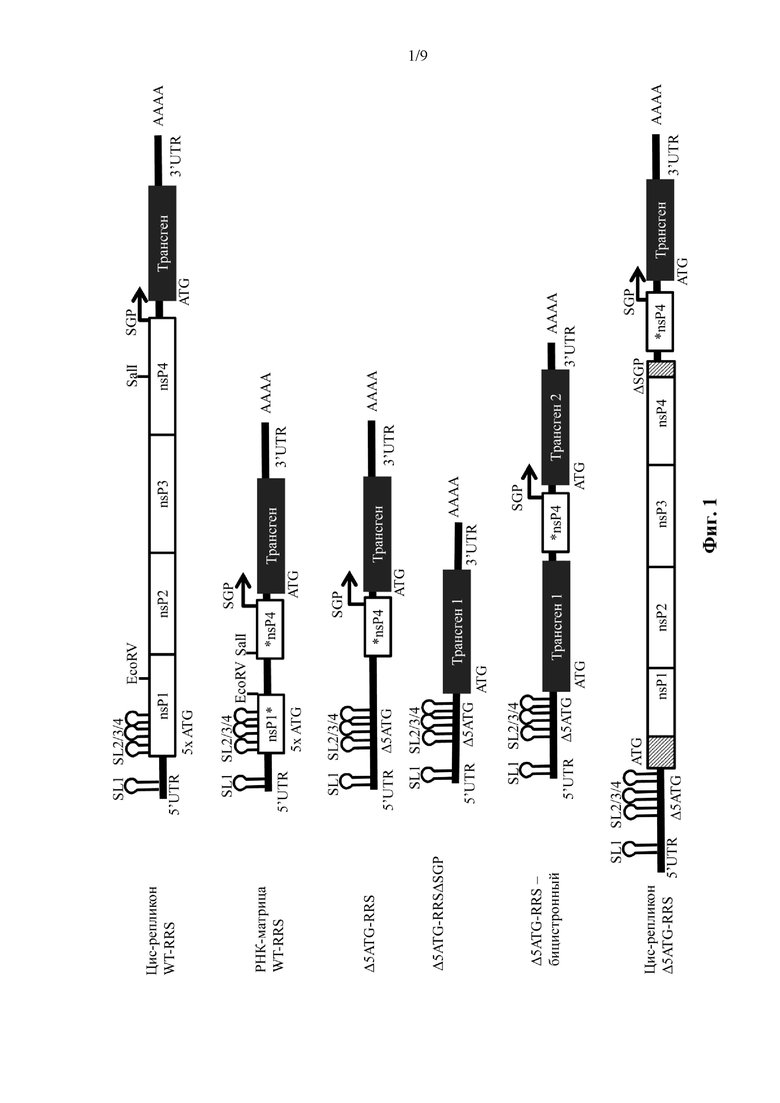
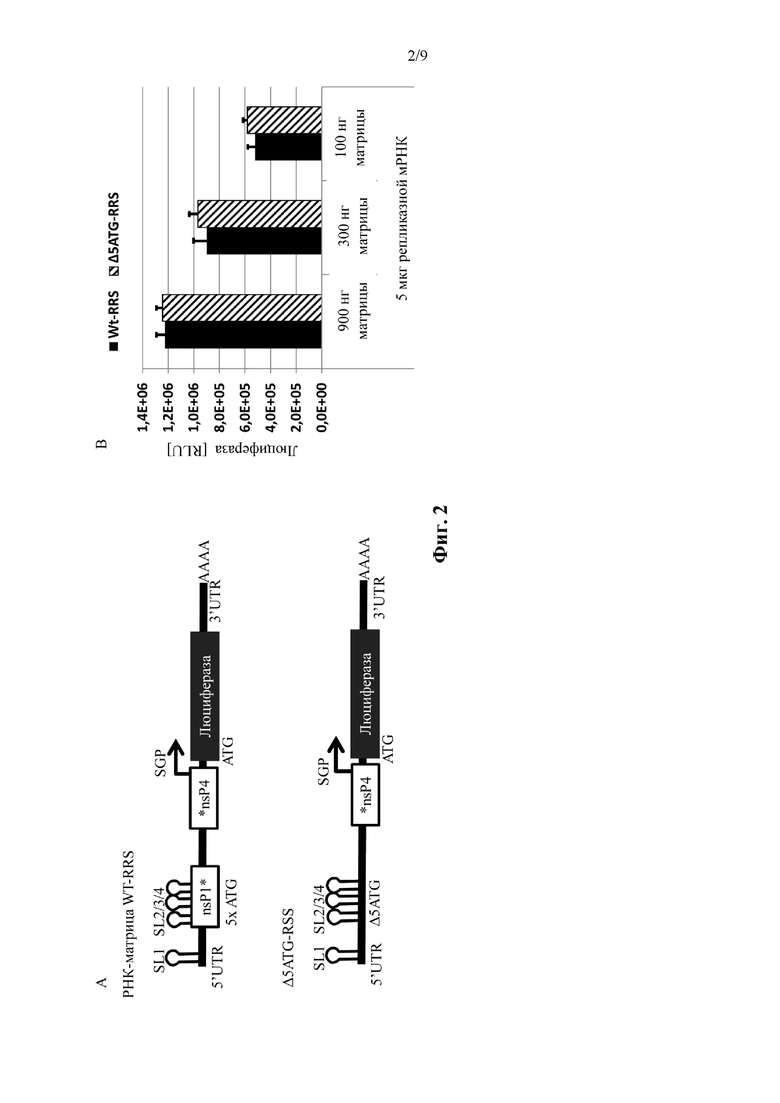
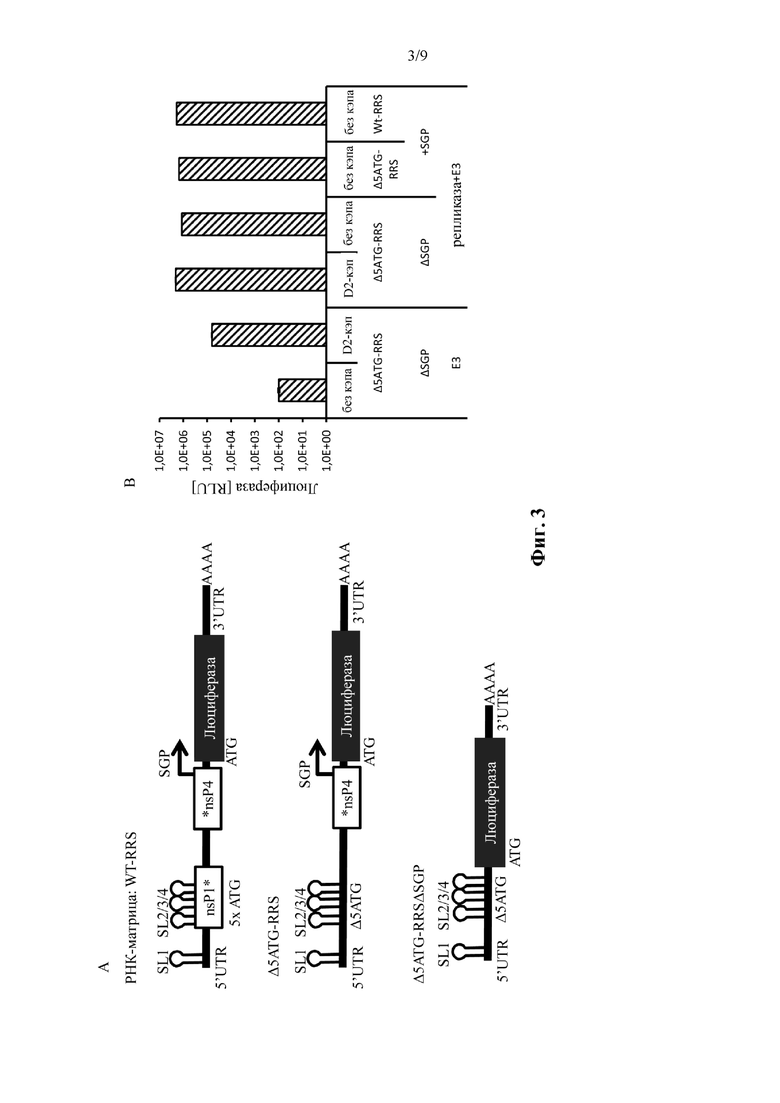
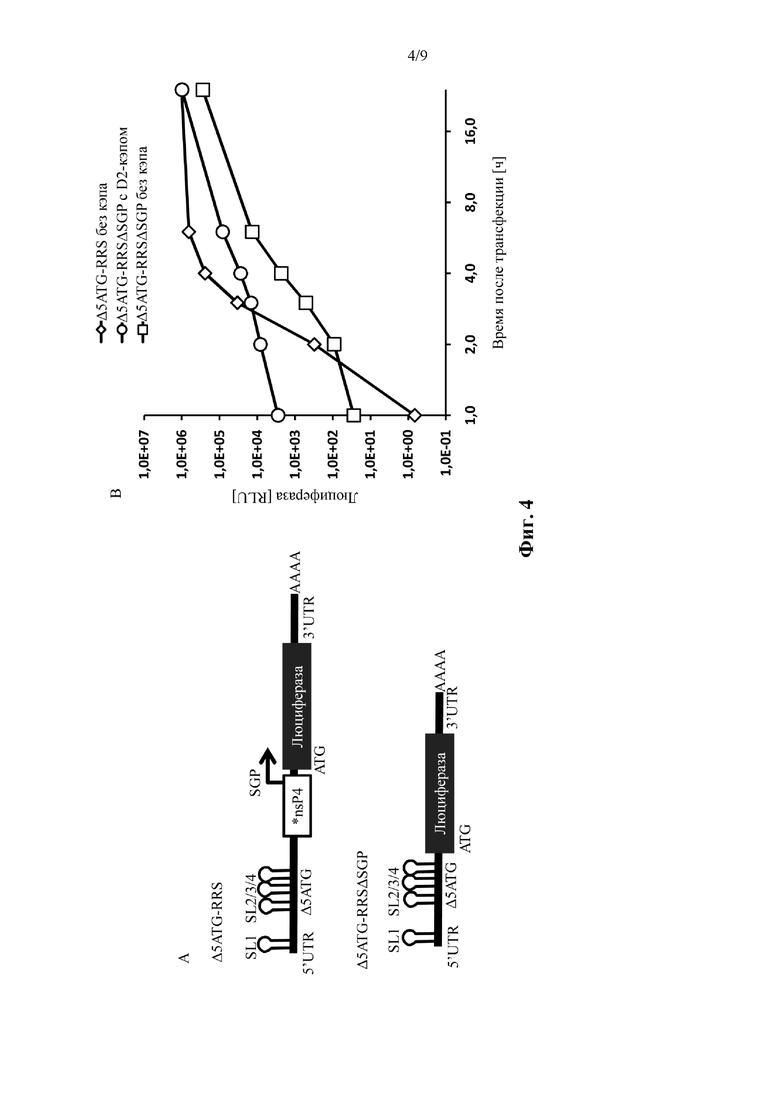
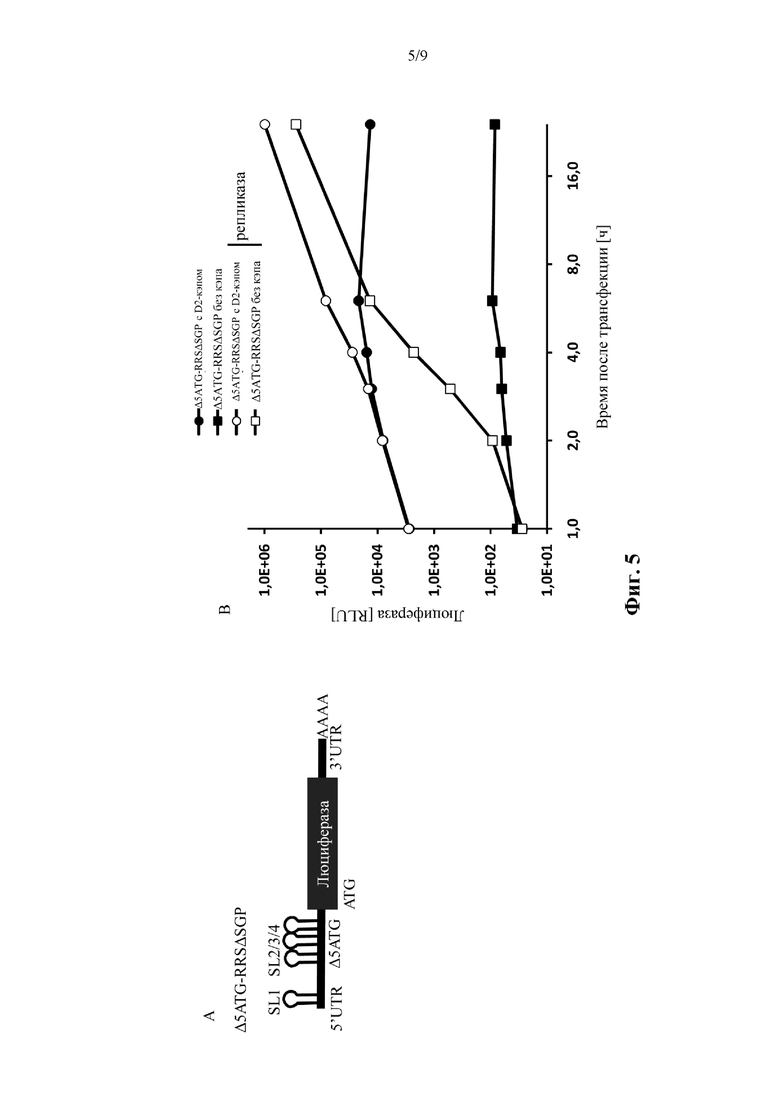
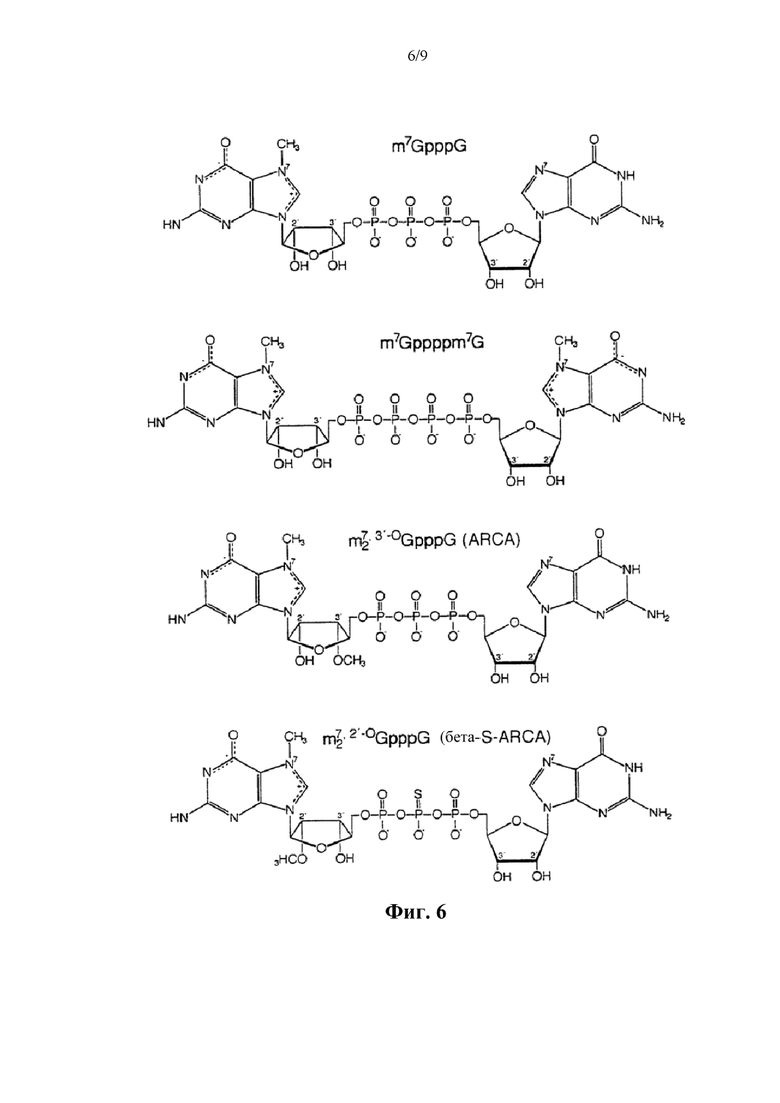
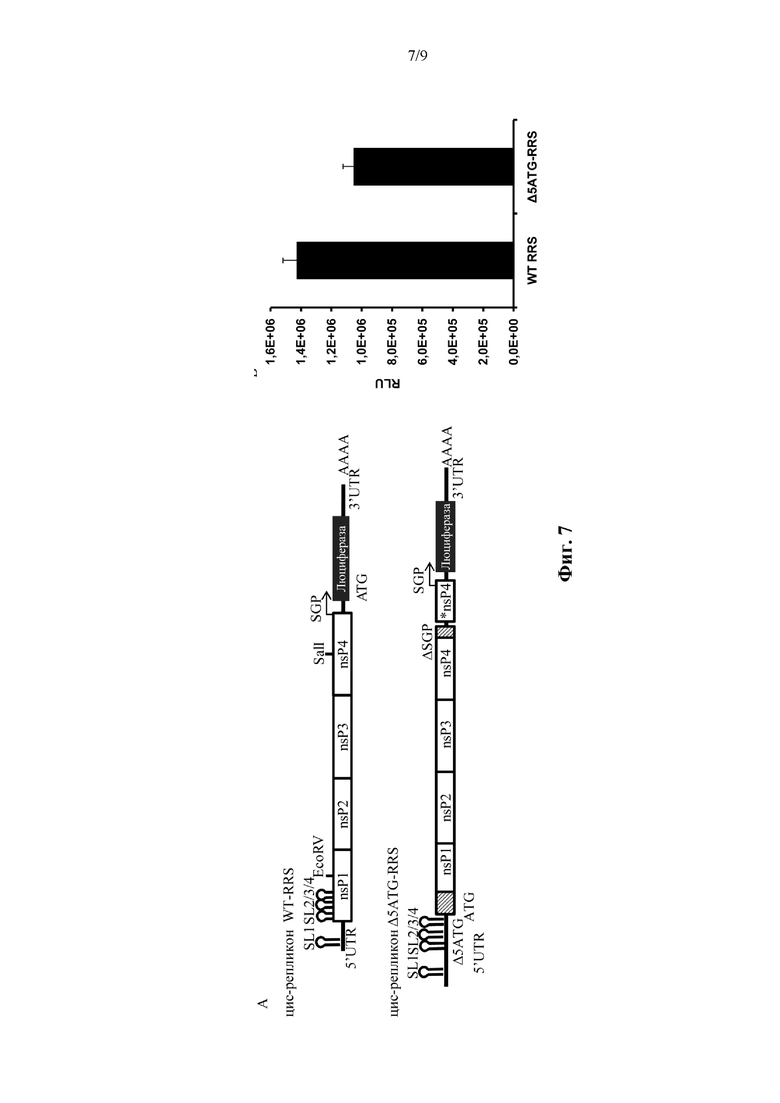
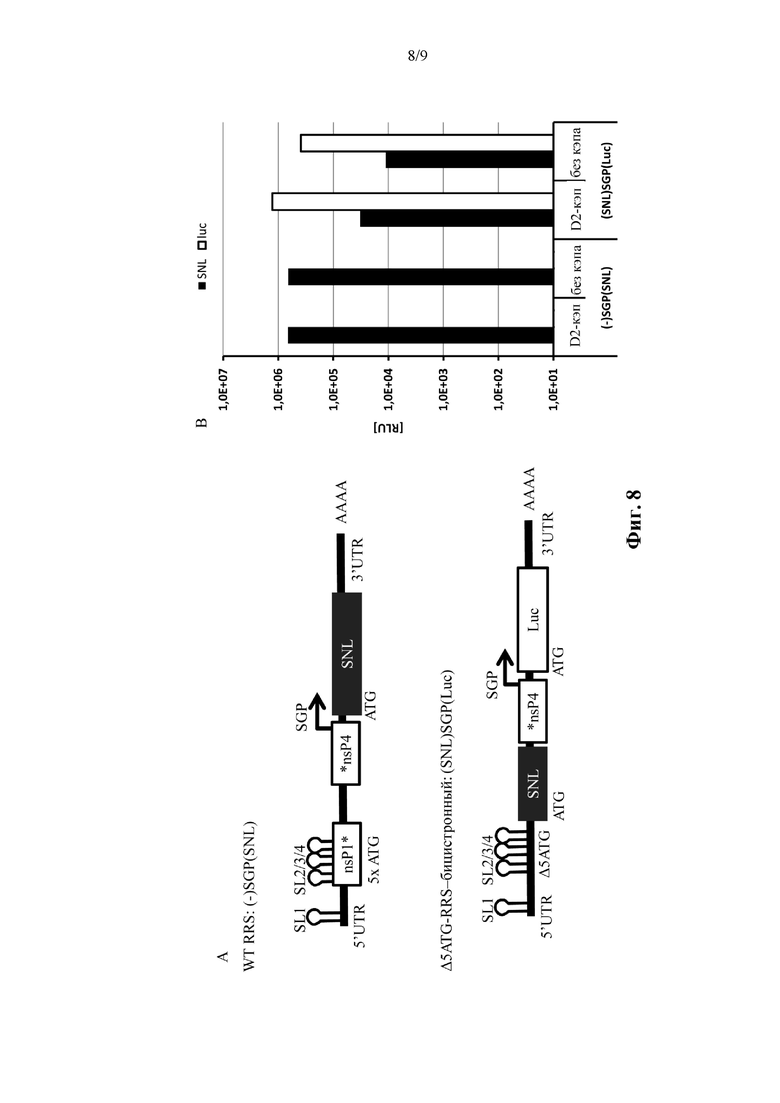
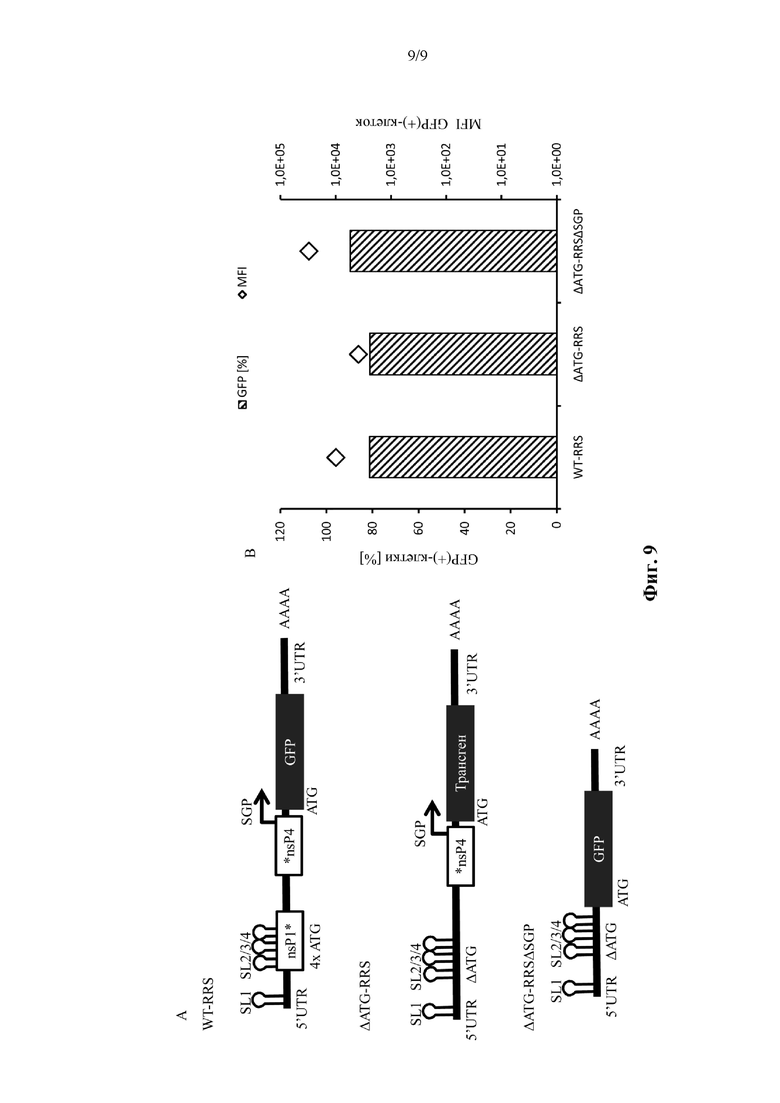
Изобретение относится к области биотехнологии. Описана группа изобретений, включающая РНК-репликон для экспрессии белка, систему транс-репликации для экспрессии белка, включающую вышеуказанный РНК-репликон, ДНК для использования в качестве матрицы для транскрипции in vivo или in vitro, способ получения представляющего интерес белка в клетке (варианты), клетку-хозяин для получения белка, способ получения представляющего интерес белка у субъекта (варианты). В одном из вариантов реализации РНК-репликон содержит модифицированную 5' репликационную последовательность распознавания альфавируса, причем 5' репликационная последовательность распознавания модифицирована посредством удаления всех инициирующих трансляцию кодонов по сравнению с нативной альфавирусной 5' репликационной последовательностью распознавания, где альфавирус представляет собой вирус леса Семлики или вирус Синбис. Изобретение расширяет арсенал средств для репликации белка. 8 н. и 15 з.п. ф-лы, 9 ил., 8 пр.
1. РНК-репликон для экспрессии белка, где репликон содержит модифицированную 5' репликационную последовательность распознавания альфавируса, причем 5' репликационная последовательность распознавания модифицирована посредством удаления всех инициирующих трансляцию кодонов по сравнению с нативной альфавирусной 5' репликационной последовательностью распознавания, причем альфавирус представляет собой вирус леса Семлики или вирус Синбис.
2. РНК-репликон по п. 1, который содержит одно или несколько изменений нуклеотидов, компенсирующих нарушения спаривания нуклеотидов в одной или нескольких структурах «петля-на-стебле», полученных в результате удаления по меньшей мере одного инициирующего кодона.
3. РНК-репликон по п. 1 или 2, который не содержит открытую рамку считывания, кодирующую усеченный альфавирусный неструктурный белок.
4. РНК-репликон по любому из пп. 1-3, который содержит 3' репликационную последовательность распознавания.
5. РНК-репликон по любому из пп. 1-4, который содержит первую открытую рамку считывания, кодирующую представляющий интерес белок.
6. РНК-репликон по п. 5, в котором представляющий интерес белок, кодируемый первой открытой рамкой считывания, может быть экспрессирован из РНК-репликона в качестве матрицы.
7. РНК-репликон по п. 5 или 6, который содержит субгеномный промотор, контролирующий производство субгеномной РНК, содержащей первую открытую рамку считывания, кодирующую представляющий интерес белок.
8. РНК-репликон по п. 7, причем представляющий интерес белок, кодируемый первой открытой рамкой считывания, может быть экспрессирован из РНК-репликона и субгеномной РНК.
9. РНК-репликон по п. 5 или 6, который содержит субгеномный промотор, контролирующий производство субгеномной РНК, содержащей вторую открытую рамку считывания, кодирующую представляющий интерес белок.
10. РНК-репликон по п. 9, в котором субгеномный промотор и вторая открытая рамка считывания, кодирующая представляющий интерес белок, расположены ниже по ходу транскрипции от первой открытой рамки считывания, кодирующей представляющий интерес белок.
11. РНК-репликон по любому из пп. 5-10, в котором представляющий интерес белок, кодируемый первой и/или второй открытой рамкой считывания, представляет собой функциональный альфавирусный неструктурный белок.
12. РНК-репликон по любому из пп. 1-11, который содержит открытую рамку считывания, кодирующую функциональный альфавирусный неструктурный белок.
13. РНК-репликон по п. 11 или 12, в котором открытая рамка считывания, кодирующая функциональный альфавирусный неструктурный белок, не перекрывается с 5' репликационной последовательностью распознавания.
14. РНК-репликон по любому из пп. 11-13, который может быть реплицирован посредством функционального альфавирусного неструктурного белка.
15. РНК-репликон по любому из пп. 1-10, который не содержит открытую рамку считывания, кодирующую функциональный альфавирусный неструктурный белок.
16. Система транс-репликации для экспрессии белка, где система транс-репликации содержит:
конструкцию РНК для экспрессии функционального альфавирусного неструктурного белка,
РНК-репликон по любому из пп. 1-10 и 15, который может быть реплицирован посредством функционального альфавирусного неструктурного белка in trans.
17. РНК-репликон по любому из пп. 1-15 или система по п. 16, в которой альфавирус представляет собой вирус леса Семлики.
18. ДНК для использования в качестве матрицы для транскрипции in vivo или in vitro, содержащая последовательность нуклеиновой кислоты, кодирующую РНК-репликон по любому из пп. 1-15 и 17.
19. Способ получения представляющего интерес белка в клетке, предусматривающий стадии:
(а) получения РНК-репликона по любому из пп. 1-14 и 17, который содержит открытую рамку считывания, кодирующую функциональный альфавирусный неструктурный белок, может быть реплицирован функциональным альфавирусным неструктурным белком и содержит открытую рамку считывания, кодирующую представляющий интерес белок, и
(b) инокуляции РНК-репликона в клетку.
20. Способ получения представляющего интерес белка в клетке, предусматривающий стадии:
(а) получения конструкции РНК для экспрессии функционального альфавирусного неструктурного белка,
(b) получения РНК-репликона по любому из пп. 1-10, 15 и 17, который может быть реплицирован функциональным альфавирусным неструктурным белком in trans и содержит открытую рамку считывания, кодирующую представляющий интерес белок, и
(c) инокуляции смеси конструкции РНК для экспрессии функционального альфавирусного неструктурного белка и РНК-репликона в клетку.
21. Клетка-хозяин для получения белка, где клетка-хозяин инокулирована в соответствии со способом по п. 19 или 20.
22. Способ получения представляющего интерес белка у субъекта, предусматривающий стадии:
(а) получения РНК-репликона по любому из пп. 1-14 и 17, который содержит открытую рамку считывания, кодирующую функциональный альфавирусный неструктурный белок, может быть реплицирован функциональным альфавирусным неструктурным белком и содержит открытую рамку считывания, кодирующую представляющий интерес белок, и
(b) введения РНК-репликона субъекту.
23. Способ получения представляющего интерес белка у субъекта, предусматривающий стадии:
(а) получения РНК, кодирующей функциональный альфавирусный неструктурный белок,
(b) получения РНК-репликона по любому из пп. 1-10, 15 и 17, который может быть реплицирован функциональным альфавирусным неструктурным белком in trans и содержит открытую рамку считывания, кодирующую представляющий интерес белок, и
(c) введения конструкции РНК для экспрессии функционального альфавирусного неструктурного белка и РНК-репликона субъекту.
| WO 2008156829 A2, 24.12.2008 | |||
| Духовой шкаф | 1991 |
|
SU1791678A1 |
| US 7425337 B2, 16.09.2008 | |||
| ПЛАТФОРМЫ ДОСТАВКИ АНТИГЕНОВ | 2011 |
|
RU2597974C2 |
| Gregory J | |||
| Atkins, Marina N | |||
| Fleeton and Brian J | |||
| Sheahan "Therapeutic and prophylactic applications of alphavirus vectors", Expert reviews in molecular medicine, 2008, 10, e33, DOI 10.1017/S1462399408000859. | |||

