Область техники, к которой относится изобретение
Настоящее изобретение касается композиций, содержащих полиплексные составы для доставки РНК в целевой орган или целевые клетки после парентерального введения, в частности, после внутримышечного введения. Более точно, настоящее изобретение касается составов для введения РНК типа самореплицирующейся РНК, в частности, путем внутримышечного введения. Более конкретно, полиплексные частицы содержат одноцепочечную РНК, предпочтительно самореплицирующуюся или самоамплифицирующуюся РНК, и полиалкиленимин. РНК может кодировать представляющий интерес белок типа фармацевтически активного белка. РНК поглощается клеткой и затем предпочтительно транслируется в пептид или белок, который может проявлять свою физиологическую активность. Композиции по изобретению применимы для запуска или усиления иммунного ответа. Они также применимы при профилактическом и/или терапевтическом лечении заболеваний, связанных с антигенами типа белков. Кроме того, настоящее изобретение касается способов получения стабильных композиций, включающих композиции РНК-полиплекс, причем данные составы РНК-полиплекс содержат одноцепочечную РНК и полиалкиленимин. Композиции частиц РНК-полиплекс, описанные здесь, можно замораживать и оттаивать либо обезвоживать и регидратировать без потери качества продукта, в частности, без существенной потери активности РНК. В частности, составы частиц РНК-полиплекс, описанные здесь, можно замораживать и оттаивать либо обезвоживать и регидратировать методами лиофилизации, распылительной сушки или родственными методами, позволяющими достигать увеличения срока хранения продуктов в отношении хранения жидкости. Кроме того, композиции частиц РНК-полиплекс, описанные здесь, могут соответствовать требованиям к фармацевтическим продуктам, более конкретно, требованиям по правилам организации производства и контроля качества лекарственных средств и требованиям по качеству фармацевтических продуктов для парентерального применения. Описанные здесь рецептуры РНК-полиплексов особенно применимы для вакцинации людей или животных, напр., против инфекционных заболеваний.
Уровень техники
Введение чужеродных нуклеиновых кислот, кодирующих один или несколько полипептидов для профилактических и терапевтических целей, было целью биомедицинских исследований в течение многих лет. Для подходов предшествующего уровня техники характерна доставка молекул нуклеиновой кислоты в целевые клетки или организмы, но эти подходя различаются в зависимости от типа молекул нуклеиновой кислоты и/или систем доставки: под влиянием проблем с безопасностью, связанных с применением молекул дезоксирибонуклеиновой кислоты (ДНК), в последние годы все большее внимание привлекают молекулы рибонуклеиновой кислоты (РНК). Предлагались различные подходы, включая введение одноцепочечной или двухцепочечной РНК в виде голой РНК или в виде комплекса либо в упакованном виде, напр., в безвирусных или вирусных средствах доставки. В вирусах и в вирусных средствах доставки нуклеиновая кислота обычно инкапсулирована белками и/или липидами (вирусная частица). Например, сконструированные частицы РНК вируса, полученные из РНК-вирусов, предлагались в качестве средства доставки для обработки растений (WO 2000/053780 A2) или для вакцинации млекопитающих (Tubulekas et al., 1997, Gene, vol. 190, pp. 191-195). Ввиду проблем с безопасностью медицинское и ветеринарное сообщество противится введению частиц РНК-вируса людям или животным. Невирусные средства доставки, которые могли быть применимы к РНК, проходили тщательные исследования для разработки терапевтических средств на основе доставки генов. Однако по разным причинам внедрение безвирусных подходов к доставке генов в клиническую практику было не очень успешным. Причины могут быть связаны с неудовлетворительным уровнем экспрессии генов, технологическими и нормативными проблемами в связи с фармацевтической разработкой таких сложных продуктов и соображениями безопасности.
Таким образом, существует потребность в фармацевтических продуктах для безопасной и эффективной доставки РНК, кодирующей белок с терапевтической ценностью, типа вакцины, у пациентов и животных. Как описано здесь, аспекты и воплощения настоящего изобретения удовлетворяют эту потребность.
Cущность изобретения
иммунотерапевтические стратегии представляют многообещающие варианты для профилактики и терапии, напр., инфекционных заболеваний и раковых заболеваний. Выявление растущего числа связанных с патогенами и опухолями антигенов привело к широкому набору подходящих мишеней для иммунотерапии. Настоящее изобретение охватывает улучшенные средства и способы, подходящие для эффективной экспрессии антигенов, подходящие для иммунотерапевтического лечения для профилактики и терапии заболеваний.
В одном аспекте изобретения предусмотрены фармацевтические композиции, содержащие:
(a) одноцепочечную РНК; и
(b) полиалкиленимин.
В другом аспекте изобретения предусмотрены композиции, содержащие:
(a) одноцепочечную РНК; и
(b) полиалкиленимин;
для применения в качестве лекарственного средства.
В одном воплощении всех аспектов изобретения молярное отношение числа атомов азота (N) в полиалкиленимине к числу атомов фосфора (P) в одноцепочечной РНК (соотношение N:P) составляет от 1,0 до 30, предпочтительно от 2,0 до 15,0, более предпочтительно от 6,0 до 12,0.
В следующем аспекте изобретения предусмотрены композиции, содержащие:
(a) одноцепочечную РНК; и
(b) полиалкиленимин;
причем молярное отношение числа атомов азота (N) в полиалкиленимине к числу атомов фосфора (P) в одноцепочечной РНК (соотношение N:P) составляет от 1,0 до 30,0, предпочтительно от 2,0 до 15,0, более предпочтительно от 6,0 до 12,0.
В одном воплощении всех аспектов изобретения ионная сила композиции составляет 50 мМ или меньше, при этом предпочтительно концентрация положительно заряженных одновалентных ионов составляет 25 мМ или меньше, а концентрация свободных положительно заряженных двухвалентных катионных ионов составляет 20 мкМ или меньше.
В следующем аспекте изобретения предусмотрены композиции, содержащие:
(a) одноцепочечную РНК; и
(b) полиалкиленимин;
причем ионная сила составляет 50 мМ или меньше.
В одном воплощении концентрация положительно заряженных одновалентных ионов составляет 25 мМ или меньше, а концентрация положительно заряженных двухвалентных катионных ионов составляет 20 мкМ или меньше.
В одном воплощении всех аспектов изобретения композиция предназначена для внутримышечного введения типа внутримышечной инъекции.
В одном воплощении всех аспектов изобретения одноцепочечная РНК и полиалкиленимин находятся в полиплексных частицах.
В одном воплощении всех аспектов изобретения полиалкиленимин имеет следующую общую формулу (I):
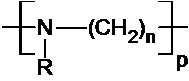 ,
,
где R означает H, ацил или группу, имеющую следующую общую формулу (II):
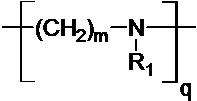 ,
,
где R1 означает Н или группу, имеющую следующую общую формулу (III):
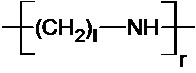 ,
,
n, m и l выбраны независимо из целых чисел от 2 до 10; а
p, q и r – целые числа, причем сумма p, q и r такова, что средняя молекулярная масса полимера составляет от 1,5⋅102 до 107 Да, предпочтительно от 5000 до 105 Да, более предпочтительно от 10000 до 40000 Да, более предпочтительно от 15000 до 30000 Да, еще более предпочтительно от 20000 до 25000 Да.
В одном воплощении n, m и l выбраны независимо из 2, 3, 4 и 5, предпочтительно из 2 и 3. В одном воплощении R1 означает H. В одном воплощении R означает H или ацил.
В одном воплощении всех аспектов изобретения полиалкиленимин включает полиэтиленимин и/или полипропиленимин, предпочтительно полиэтиленимин.
В одном воплощении всех аспектов изобретения по меньшей мере 92% атомов N в полиалкиленимине являются протонируемыми.
В одном воплощении всех аспектов изобретения композиции по изобретению содержат одну или несколько добавок. В одном воплощении одну или несколько добавок выбирают из группы, состоящей из буферных веществ, сахаридов, стабилизаторов, криопротекторов, лиопротекторов и хелатообразующих веществ. В одном воплощении всех аспектов изобретения композиции по изобретению содержат один или несколько полимеров. В одном воплощении буферные вещества включают по меньшей мере одно, выбранное из группы, состоящей из 4-(2-гидроксиэтил)-1-пиперазинэтансульфокислоты (HEPES), 2-(N-морфолино)этансульфокислоты (MES), 3-морфолино-2-гидроксипропансульфокислоты (MOPSO), буферных систем на основе уксусной кислоты и аналогов, буферных систем на основе фосфорной кислоты или на основе лимонной кислоты. В одном воплощении всех аспектов изобретения композиции по изобретению содержат буферы для забуферивания в диапазоне от рН 4 до 8, предпочтительно от 5 до 7,5. Примерами таких буферных систем являются ацетатные буферы или буферы HEPES или фосфатные буферы или буферы на основе уксусной кислоты. В одном воплощении сахариды включают по меньшей мере один, выбранный из группы, состоящей из моносахаридов, дисахаридов, трисахаридов, олигосахаридов и полисахаридов, предпочтительно из глюкозы, трегалозы, сахарозы и декстрана. В одном воплощении добавка представляет собой декстран со средней молекулярной массой от 1 кДа до 100 кДа. В одном воплощении криопротекторы включают по меньшей мере один, выбранный из группы, состоящей из таких гликолей, как этиленгликоль, пропиленгликоль и глицерин. В одном воплощении хелатообразующие агенты включают ЭДТА. В одном воплощении липиды включают по меньшей мере один, выбранный из группы, состоящей из катионных липидов, нейтральных липидов и анионных липидов. В одном воплощении композиции по изобретению содержат один или несколько блок-сополимеров, включающих строительные блоки из этиленоксида и пропиленоксида. В одном воплощении композиции по изобретению содержат сополимеры, включающие этилендиаминовые группы. В одном воплощении композиции по изобретению содержат амфифильный блок-сополимер, предпочтительно включающий строительные блоки из этиленоксида и пропиленоксида, необязательно также включающий этилендиаминовые группы.
В одном воплощении всех аспектов изобретения композиции содержат забуференную HEPES глюкозу (HBG или HBG×1), забуференную MES глюкозу (MBG или MBG×1), забуференную ацетатом глюкозу (ABG) или забуференную HEPES трегалозу (HBT или HBT×1). В одном воплощении всех аспектов изобретения композиции содержат глюкозу или трегалозу или сахарозу в буфере на основе уксусной кислоты при концентрации в пределах от 0,1 мМ до 10 мМ. В одном воплощении всех аспектов изобретения композиции содержат глюкозу или трегалозу или сахарозу в фосфатном буфере при концентрации в пределах от 0,1 мМ до 10 мМ.
В одном воплощении всех аспектов изобретения z-средний размер частиц составляет менее 200 нм, предпочтительно менее 150 нм и более предпочтительно менее 100 нм. В одном воплощении z-средний размер частиц составляет от 50 нм до 200 нм. В одном воплощении всех аспектов изобретения дзета-потенциал частиц составляет 20 мВ или больше, предпочтительно от 25 до 40 мВ. В одном воплощении всех аспектов изобретения электрофоретическая подвижность (µ) частиц составляет от 1 до 1,6 мкм×см/В×См. В одном воплощении всех аспектов изобретения z-средний размер частиц и/или дзета-потециал и/или электрофоретическая подвижность определяются в суспензии, содержащей полиплексные частицы и забуференную HEPES глюкозу (HBG) или забуференную HEPES трегалозу (HBT). В одном воплощении HBG содержит 5% глюкозы (вес./об.) и 10 мМ HEPES, pH 7,1, а HBT содержит 10% трегалозы (вес./об.) и 10 мМ HEPES, pH 7,1. В одном воплощении z-средний размер частиц определяется методом динамического рассеяния света с анализом данных по кумулянтному алгоритму. В одном воплощении измеряется коэффициент поступательной диффузии методом динамического светорассеяния. Затем применяется уравнение Стока-Эйнштейна для вычисления Z-оценки. В одном воплощении электрофоретическая подвижность измеряется методом лазерно-доплеровского электрофореза. Затем для расчета ζ-потенциала применяется уравнение Генри или уравнение Смолуховского.
В одном воплощении всех аспектов изобретения частицы являются нейтральными или положительно заряженными, предпочтительно при физиологическом pH или при pH от 4,5 до 7,5.
В одном воплощении всех аспектов изобретения одноцепочечная РНК представляет собой молекулу от 6000 до 15000 нуклеотидов, предпочтительно от 9000 до 12000 нуклеотидов. В одном воплощении всех аспектов изобретения одноцепочечная РНК кодирует по меньшей мере один представляющий интерес белок. В одном воплощении всех аспектов изобретения одноцепочечная РНК представляет собой репликон, предпочтительно самореплицирующуюся или самоамплифицирующуюся РНК. В одном воплощении репликон может реплицироваться репликазой из альфавируса, причем репликон предпочтительно содержит 5′-последовательность распознавания репликации из альфавируса или её вариант и 3′-последовательность распознавания репликации из альфавируса или её вариант. В одном воплощении всех аспектов изобретения одноцепочечная РНК содержит открытую рамку считывания, кодирующую представляющий интерес пептид или белок типа фармацевтически активного пептида или белка.
В одном воплощении всех аспектов изобретения описанные здесь композиции предназначены для применения в терапии. В одном воплощении всех аспектов изобретения описанные здесь композиции представляют собой вакцинные композиции.
В следующем аспекте изобретения предусмотрено применение описанных здесь композиций для введения РНК в клетки, в частности, для экспрессии РНК в клетках. В одном воплощении клетки представлены мышечными клетками.
В следующем аспекте изобретения предусмотрено применение описанных здесь композиций для внутримышечного введения РНК.
В следующем аспекте изобретения предусмотрен способ внутримышечного введения РНК, включающий стадию внутримышечного введения описанных здесь композиций.
В следующем аспекте изобретения предусмотрены замороженные, лиофилизованные или высушенные распылением композиции, содержащие:
(a) одноцепочечную РНК; и
(b) полиалкиленимин;
причем композиции содержат криопротекторы и/или лиопротекторы, предпочтительно дисахариды типа трегалозы или полисахариды типа декстрана.
В одном воплощении композиции также содержат хелатообразующее вещество типа ЭДТА.
В одном воплощении композиции получают из водных композиций, содержащих дисахарид при 5-20% (вес./об.) и необязательно хелатообразующее вещество от 20 мкМ до 10 мМ, как-то от 80 мкМ до 5 мМ. В одном воплощении водные композиции содержат трегалозу, HEPES и EDTA типа 10% трегалозы (вес./об.), 2,8 мМ HEPES, 80 мкМ EDTA, pH 7,1.
В следующем аспекте изобретения предусмотрены водные композиции, получаемые путем оттаивания описанных здесь замороженных композиций или восстановления описанных здесь лиофилизованных или высушенных распылением композиций.
В следующем аспекте изобретения предусмотрен способ получения замороженных, лиофилизованных или высушенных распылением композиций, включающий:
(i) приготовление водной композиции, содержащей одноцепочечную РНК, полиалкиленимин и криопротектор и/или лиопротектор, предпочтительно дисахарид типа трегалозы или полисахарид типа декстрана, и
(ii) замораживание, лиофилизацию или распылительную сушку композиции.
В одном воплощении водные композиции также содержат хелатообразующее вещество типа ЭДТА. В одном воплощении водные композиции содержат дисахарид при 5-20% (вес./об.) и необязательно хелатообразующее вещество от 20 мкМ до 10 мМ, как-то от 80 мкМ до 5 мМ. В одном воплощении водные композиции содержат трегалозу, HEPES и EDTA типа 10% трегалозы (вес./об.), 2,8 мМ HEPES, 80 мкМ EDTA, pH 7,1.
В следующем аспекте изобретения предусмотрено применение криопротекторов и/или лиопротекторов, предпочтительно дисахаридов типа трегалозы или полисахаридов типа декстрана для получения замороженных, лиофилизованных или высушенных распылением композиций, содержащих:
(a) одноцепочечную РНК; и
(b) полиалкиленимин.
В одном воплощении дисахариды применяются в сочетании с хелатообразующим веществом типа ЭДТА.
Замороженные или лиофилизованные или высушенные распылением композиции либо водные композиции для приготовления замороженных или лиофилизованных или высушенных распылением композиций могут содержать одно или несколько из следующего:
(i) неводные растворители, такие как этиленгликоль, глицерин, диметилсульфоксид и диметилформамид;
(ii) поверхностно-активные вещества, такие как Tween 80, Brij 35, Brij 30, Lubrol PX, Triton X-100, Pluronic F127 (сополимер полиоксиэтилена-полиоксипропилена), также известный как полоксамер, полоксамин и додецилсульфат натрия;
(iii) дисахариды, такие как трегалоза, сахароза, лактоза и мальтоза;
(iv) полимеры (которые могут иметь различные молекулярные массы), такие как полиэтиленгликоль, декстран, поливиниловый спирт, гидроксипропилметилцеллюлоза, желатин, поливинилпирролидон, гидроксиэтилцеллюлоза, фиколл и альбумин;
(v) аминокислоты, такие как глицин, пролин, 4-гидроксипролин, L-серин, глутамат, аланин, лизин, саркозин и гамма-аминомасляная кислота.
В следующем аспекте изобретения предусмотрен способ непрерывного производства полиплексных рецептур РНК при помощи насосов непрерывного действия и смесительного устройства, в котором две водные жидкости смешиваются по каналам миллиметрового или микрометрового размера.
Краткое описание фигур
Фиг. 1. A. Токсичность свободного чистого PEI на клетках HEK-293 in vitro. IC50 = 77 мкМ азота (свободного). B. Токсичность полиплексов PEI/РНК репликона на клетках HEK-293 in vitro. IC50 = 542 мкМ азота (полиплексной композиции).
Фиг. 2. Относительная люминесценция от мышечных клеток C2C12 после инкубации с полиплексами PEI/РНК репликона при N/P = 11,6 при различных условиях хранения после 1-недельного хранения.
Фиг. 3. Относительная целостность РНК в полиплексах PEI/РНК репликона при N/P = 11,6 при различных условиях хранения после 2-недельного хранения.
Фиг. 4. Поли(2-этил-2-оксазолин) получают путем изомеризационной полимеризации с размыканием цикла 2-этил-2-оксазолина в присутствии инициаторов.
Фиг 5. Синтез полностью деацилированных линейных PEI22, PEI87 и PEI217 путем кислотного гидролиза PEOZ. Условия: (i) 24% (вес./об.) HCl, 110°C, 96 ч; n = 504 для PEOZ в 50 кДа, 2018 для PEOZ в 200 кДа и 5044 для PEOZ в 500 кДа.
Фиг. 6. Кинетика агрегации полиплексов с IVT-РНК (A) и РНК репликона (B) при возрастающих концентрациях соли.
Фиг 7. Физико-химические параметры полиплексов до (исходно) и после (фильтр.) фильтрования. A и B. Диаметр и полидисперсность полиплексов измеряли методом DLS. C. Из полиплексов высвобождали РНК гепарином и измеряли по УФ-поглощению при 260 нм. D. Концентрацию PEI измеряли методом с CuSO4.
Фиг. 8. Сравнение химической структуры PEI высокой чистоты и PEI обычной чистоты. n = 58 для PEI в 25 кДа. Среднее количество мономеров -CH2CH2NH- у PEI в 25 кДа составляет 581, что также является длиной непрерывного отрезка потенциально протонируемых атомов азота. Предполагая равномерное распределение N-пропиониловых фрагментов в обычном PEI25, непрерывный отрезок протонируемых азотов у него составляет лишь 64.
Фиг. 9. Трансфекция мышечных клеток C2C12 in vitro полиплексами с РНК репликона, которые получали с использованием PEI различного уровня чистоты при различных соотношениях N/P.
Фиг. 10. Полиплексы с РНК репликона получали при соотношениях N/P = 1 (-) и 11,6 (+) с PEI высокой чистоты (jetPEI) и PEI обычной чистоты (25 кДа). В качестве контроля использовали свободную РНК. Препараты вводили внутримышечно (в/м) в задние конечности мышей (n = 3). Регистрировали сигналы люминесценции из мышц мышей.
Фиг. 11. Полиплексы РНК репликона получали при соотношениях N/P = 7,7 и 11,6 с PEI высокой чистоты: jetPEI (фирмы Polyplus), PEI-Max 40000 (фирмы Polyscience) и Exgen 500 (фирмы Euromedex). Все препараты готовили в буфере HBGx1, за исключением лиофилизированного препарата, который готовили в буфере HBTx1. Препараты вводили внутримышечно (в/м) в задние конечности мышей (n = 3). Регистрировали сигналы люминесценции из мышц мышей.
Фиг 12. Лиофилизованные комки полиплексов JetPEI/РНК репликона готовили при N/P = 11,6 с использованием различных буферов.
Фиг 13. Мышечные клетки C2C12 трансфицировали in vitro с помощью IVT-РНК, кодирующей люциферазу. РНК образовывала комплексы с JetPEI при различных соотношениях N/P в буфере HBGx1. Через 24 ч после трансфекции измеряли сигналы люминесценции.
Фиг. 14. Получали полиплексы с IVT-РНК при соотношениях N/P = 5,8 и 11,6 с очищенным PEI в буфере HBGx1. В качестве контроля использовали свободную IVT-РНК в буфере HBGx1. Препараты вводили внутримышечно (в/м) в задние конечности мышей (n = 3) в дозах РНК по 2-8 мкг на инъекцию. Через 6 ч после введения регистрировали сигналы люминесценции из мышц мышей.
Фиг. 15. Получали полиплексы РНК-репликона и jetPEI при различных концентрациях РНК при соотношении N/P = 11,6 в буфере HBGx1. Для измерения размеров методом DLS полиплексы разбавляли до концентрации РНК 10 мг/л.
Фиг 16. Мышечные клетки C2C12 трансфицировали in vitro полиплексами из фиг. 16. Через 24 ч после трансфекции измеряли сигналы люминесценции.
Фиг. 17. Получали полиплексы РНК-репликона при соотношении N/P = 11,6 с чистым PEI в буфере HBGx1 при различных концентрациях РНК, как на фиг. 16. Препараты вводили внутримышечно (в/м) в задние конечности мышей (n = 3) в дозах РНК по 2-8 мкг на инъекцию. Регистрировали сигналы люминесценции из мышц мышей.
Фиг. 18. Исследования in vitro с полиплексами PEI/РНК репликона на дендритных клетках человека (DC) и мышечных клетках мыши (C2C12) A. Токсичность (выражена в % жизнеспособных клеток после обработки полиплексами). B. Трансфекция (выражена в виде люминесцентного излучения после обработки полиплексами). Результаты трансфекции представлены только для клеток C2C12.
Фиг. 19. Получали полиплексы РНК-репликона при соотношении N/P = 11,6 или 15,8 с PEI фирмы Polyplus или Polytheragene в буфере HBGx1 или 10 мМ HEPES. Перед введением мышам полиплексы разбавляли в буфере HBGx1 или Opti-MEM. Препараты вводили внутримышечно (в/м) в задние конечности мышей (n = 3) в дозах РНК по 2 мкг на инъекцию. Регистрировали сигналы люминесценции из мышц мышей.
Фиг. 20. A. Через 4, 7 и 11 дней после внутримышечного (в/м) введения в обе мышцы tibialis posterior у мышей Balb/с по 2 мкг не составленной (буферный раствор) или составленной РНК репликона, кодирующей люциферазу, животных подвергали неинвазивной визуализации по биолюминесценции in vivo. Регистрировали исходящие из белка люциферазы фотоны в течение одной минуты, которые показаны в виде наложения на фотографию визуализируемых мышей. B. Графическое отображение измеряемых фотонов в секунду (p/s) на месте инъекции.
Фиг. 21. A. Через 7 дней после интрадермального (и/д) введения по 2 мкг не составленной (буферный раствор) или составленной РНК репликона, кодирующей люциферазу, в два места инъекции на дорсальной коже мышей Balb/c, животных подвергали неинвазивной визуализации по биолюминесценции in vivo. Регистрировали исходящие из белка люциферазы фотоны в течение одной минуты, которые показаны в виде наложения на фотографию визуализируемых мышей. Черными стрелками указаны места инъекции. B. Графическое отображение измеряемых фотонов в секунду (p/s) в месте инъекции.
Фиг. 22. Благотворное влияние препарата РНК репликона в качестве вакцины.
Фиг. 23. Благотворное влияние препарата РНК репликона в качестве вакцины.
Фиг. 24. Результаты распылительной сушки РНК репликона, составленной с PEI в 10% (вес./об.) трегалозе.
Фиг. 25. Нормализованная люминесценция от мышечных клеток C2C12 после инкубации с полиплексами PEI/РНК репликона при различных соотношениях N/P, до (исходно) и после (стерилизован) стерилизации фильтрованием.
Фиг. 26. Влияние комбинации коротких и длинных PEI на эффективность трансфекции in vitro согласно примеру 16. Фиг. 26A: эффективность трансфекции полиплексов с короткими линейными PEI и длинными PEI при 250 нг РНК на лунку. Фиг. 26B: эффективность трансфекции полиплексов с короткими разветвленными PEI и длинными PEI при 250 нг РНК на лунку. По сравнению с эталоном in vivo JetPEI и при таком же общем соотношении N/P достигались более высокие уровни экспрессии в различных временных интервалах с комбинациями коротких PEI (фиг. 26A: линейный; фиг. 26B: разветвленный) и длинных PEI (напр., in vivo jetPEI).
Фиг. 27. Эффективность трансфекции полиплексов РНК репликона с длинными JetPEI + короткими PEI в сравнении с эталоном (т.е. in vivo JetPEI NP12) согласно примеру 17: короткие PEI (разветвленные в 1,8 кДа) и длинные PEI в различных комбинациях (NP4 + NP8 или NP1.15 + NP11) при общем N/P = 12.
Фиг. 28. Влияние вариаций соли (напр., NaCl) на эффективность трансфекции РНК репликона (saRNA) in vivo согласно примеру 18. Регистрировали сигналы биолюминесценции на 3-й (фиг. 28A), 6-й (фиг. 28B), 9-й (фиг. 28C) и 13-й день (фиг. 28D). Уровень сигналов сравнивали на фиг. 28E. Наиболее интенсивный сигнал в области мышц у мышей выявляется на 6-й день после внутримышечного (в/м) введения у мышей, получавших полиплексы PEI/РНК репликона (напр., с длинным PEI при N/P 12) с добавлением низких концентраций соли (от 5 до 10 мМ).
Фиг. 29. Влияние доведения pH на эффективность трансфекции препаратов РНК репликона (saRNA) согласно примеру 18. Хорошие результаты получали с препаратами полиплексов saRNA-PEI со значениями pH между 6,5 и 7,1. Наиболее интенсивный сигнал выявляется с препаратом saRNA-длинный PEI NP12, доведенным до pH 6,5. В качестве эталона использовали полиплексы saRNA-JetPEI NP12 без доведения pH (BM) или в HBG (20 мМ HEPES, pH 7,4, 5% мас. глюкозы).
Фиг. 30. Электрофоретическая подвижность полиплексов in vivo jetPEI/РНК репликона (N/P = 4) при доведении до различных значений pH согласно примеру 19.
Фиг. 31. Нормализованная люминесценция от мышечных клеток C2C12 после инкубации с различными дозами полиплексов in vivo jetPEI/РНК репликона при соотношении N/P = 4 и различных значениях pH (pH 6,5 – pH 8,5) согласно примеру 19.
Фиг. 32. Экспрессия люциферазы после трансфекции различными количествами PEI с избытком положительных зарядов в препаратах PEI согласно примеру 20.
Фиг. 33. Оптимизация трансфекции полиплексов посредством 2-стадийного комплексообразования согласно примеру 21.
Фиг. 34. Эксперимент по иммунизации согласно примеру 22, демонстрирующий превосходный эффект полиплексов saRNA, составленных с забуференной MES глюкозой (MBG), в сравнении с забуференной HEPES глюкозой (HBG).
Фиг. 35. У животных развивается иммунный ответ нейтрализующих антител после внутримышечной (в/м) иммунизации с помощью составленной с PEI самоамплифицирующейся РНК (saRNA), кодирующей HA вируса A/California/7/2009 (H1N1) (HA H1N1/Cf7).
(A) Мышей BALB/c в день 0 иммунизировали один раз буфером или 1/25 дозой человеческой вакцины или 0,1 мкг составленной с PEI saRNA VEEV или saRNA SFV, кодирующей HA H1N1/Cf7, при соотношении N/P = 12:1. Через 28 и 48 дней у животных брали кровь и проводили анализ сыворотки на антитела против HA методом нейтрализации вируса (VNT; n = 4).
(B) Домашних поросят в день 0 иммунизировали один раз буфером или 1 дозой человеческой вакцины или 90 мкг составленной с PEI saRNA VEEV или saRNA SFV, кодирующей HA H1N1/Cf7, при соотношении N/P = 12/:1. У поросят брали кровь на 14, 21, 28 и 35-й день после иммунизации и проводили анализ иммунного ответа нейтрализующих антител против HA методом VNT (n = 8; группа буфера: n = 4).
У группы животных, получавших составленную с saRNA VEEV вакцину, развивался такой же иммунный ответ, как у животных, которым вводили положительный контроль. saRNA SFV также приводила к развитию иммунного ответа нейтрализующих антител, но с более низким титром, чем после иммунизации saRNA VEEV. На графике представлены средние значения ± стандартная погрешность средней величины (SEM).
Фиг. 36. У животных развивается иммунный ответ антител после внутримышечной (в/м) иммунизации с помощью составленной с PEI самоамплифицирующейся РНК (saRNA), кодирующей белок cap_EU свиного цирковируса-2 (PCV2).
Мышей BALB/c иммунизировали дважды в день 0 и день 35 буфером или 1 мкг составленной с PEI saRNA SFV или saRNA VEEV, кодирующей белок cap_EU PCV2, при соотношении N/P = 12:1. На 14, 34 и 56-й день у животных брали кровь и проводили анализ сыворотки на антитела против PCV2-cap при помощи коммерчески доступного набора для ELISA (INgezim Circo IgG, Ingenasa; n = 4).
Группы животных, получавших составленные с saRNA SFV или VEEV вакцины, проявляли сходные ответы антител против белка cap_EU PCV2. Иммунный ответ антител после однократной вакцинации saRNA SFV был несколько выше, чем для saRNA VEEV. После двух иммунизаций ответ антител был почти одинаковым для обоих типов вакцин с saRNA. На графике представлены средние значения ± SEM.
Фиг. 37. Составляли комплексы с РНК репликона (saRNA) при N/P = 12 в различных буферных системах и условиях рН. Оба типа буферов, ацетатный или MES, содержали буферное вещество в конечной концентрации 10 мМ и D-глюкозу в конечной концентрации 5% вес./об.. Полиплексы saRNA/PEI хранили в соответствующем буфере при 4°C в течение различного времени (1, 2, 4 и 8 дней после образования комплексов). Сразу же после образования комплексов в различных препаратах измеряли целостность РНК (t = 0). Целостность РНК измеряли методом капиллярного электрофореза. saRNA, образующая комплекс в полиплексах, может высвобождаться после 20-минутной инкубации при комнатной температуре с сильным избытком полианиона, который вызывает электростатическое взаимодействие с полимером, высвобождая РНК, заключенную в полиплексах. Для анализа методом капиллярного электрофореза использовали по 200 мкг высвобожденной РНК строго в соответствии с протоколом, прилагаемым к соответствующему набору (набор DF471 для анализа РНК стандартной чувствительности). Для каждой временной точки использовали контрольную saRNA для определения целостности saRNA.
Более высокие значения pH в буфере для препарата приводят к значительному усилению деградации saRNA. Наименьшая потеря целостности saRNA при образовании комплексов достигалась при pH 4 с ацетатным буфером.
Фиг. 38. Составленная с PEI saRNA VEEV, кодирующая HA вируса A/California/7/2009 (H1N1; Cf7/HA), индуцирует сильный и более продолжительный ответ антител по сравнению с коммерческой вакциной, но дополнительно индуцирует сильный T-клеточный ответ, который белковые вакцины не вызывают.
Мышей BALB/c иммунизировали в/м дважды в день 0 и день 35 (на графиках они обозначены стрелками) буфером (черные символы) или 20 мкл человеческой лицензированной вакцины против сезонных штаммов вируса гриппа (Begripal 2016/2017; hLIC; серые символы) или 0,5 мкг составленной с PEI вакцины на основе saRNA VEEV, кодирующей Cf7/HA (темно-серые символы). В разные моменты времени мышей забивали и A) отбирали спленоциты для проведения Cf7/HA-специфичных анализов ELISpot на суспензиях отдельных клеток. Для анализа ELISpot использовали различные пулы CF7/HA-специфичных пептидов для стимуляции ответа T-клеток CD8+ (слева) или T-клеток CD4+ (справа), измеряемого по секреции IFN-γ. Кроме того, отбирали образцы сыворотки для проведения B) анализа Cf7/HA-специфичной нейтрализации вируса для сывороточных антител на их функциональность в ингибировании вирусной инфекции клеток. Отметим, что для серологического анализа использовали вирус A/California/4/2009 (H1N1; Cf4); а данные показывают средние значения ± SEM (группа буфера: n=3; группы вакцины: n=4).
Раскрытие сущности изобретения
Хотя настоящее изобретение подробно описано ниже, следует понимать, что это изобретение не ограничивается конкретными методиками, протоколами и реагентами, описанными здесь, поскольку они могут варьироваться. Также следует понимать, что используемая здесь терминология предназначена только для описания конкретных воплощений и не должна ограничивать объем настоящего изобретения, который должен ограничиваться только прилагаемой формулой изобретения. Если не указано иначе, все технические и научные термины, используемые здесь, имеют те же значения, которые обычно понимаются рядовыми специалистами в данной области.
Предпочтительно используемые здесь термины определяются, как описано в “A multilingual glossary of biotechnological terms: (IUPAC Recommendations)”, H.G.W. Leuenberger, B. Nagel, and H. Kölbl, Eds., Helvetica Chimica Acta, CH-4010 Basel, Switzerland, (1995).
В практике настоящего изобретения должны применяться, если не указано иначе, стандартные методы химии, биохимии, клеточной биологии, иммунологии и методы рекомбинантной ДНК, которые изложены в литературе в данной области (напр., см. Molecular Cloning: A Laboratory Manual, 2nd Edition, J. Sambrook et al., eds., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1989).
Далее будут описаны элементы настоящего изобретения. Эти элементы приводятся с конкретными воплощениями, однако следует понимать, что их можно комбинировать любым образом и в любом количестве для создания дополнительных воплощений. Описанные по-разному примеры и предпочтительные воплощения не должны рассматриваться как ограничивающие настоящее изобретение только явно описанными воплощениями. Следует иметь в виду, что данное описание раскрывает и охватывает воплощения, которые объединяют прямо описанные воплощения с любым числом раскрытых и/или предпочтительных элементов. Кроме того, любые перестановки и комбинации всех описанных элементов в данной заявке должны рассматриваться как раскрытые этим описанием, если из контекста не следует иное.
Термин “примерно” означает приблизительно или почти, а в контексте приведенных здесь числовых значений или диапазонов предпочтительно означает ±10% от указанного или заявленного числового значения или диапазона.
Приведенные в единственном числе термины и аналогичные ссылки, используемые в контексте описания изобретения (особенно в контексте формулы изобретения), следует рассматривать как охватывающие и единственное, и множественное число, если не указано здесь иначе или явно не противоречит контексту. Приведение диапазонов значений здесь просто служит кратким способом индивидуального указания каждого отдельного значения, попадающего в диапазон. Если не указано здесь иначе, каждое отдельное значение включается в описание, как если бы оно было отдельно указано здесь. Все описанные здесь способы могут выполняться в любом подходящем порядке, если не указано здесь иначе или явно не противоречит контексту. Использование всяческих примеров или примерных формулировок (напр., “таких как”), приведенных здесь, служит просто для лучшей иллюстрации изобретения и не налагает никаких ограничений на объем заявленного изобретения. Никакие выражения в описании не должны рассматриваться как указывающие на какой-нибудь не заявленный элемент, существенный для практики изобретения.
Если прямо не указано иначе, термин “включающий” применяется в контексте настоящего документа для того, чтобы указать, что могут необязательно присутствовать дополнительные элементы в дополнение к элементам списка, введенным через “включающий”. Однако в качестве особого воплощения настоящего изобретения предусматривается, что термин “включающий” охватывает и возможность отсутствия дополнительных элементов, то есть в целях этого воплощения “включающий” следует понимать как имеющий значение “состоящий из”.
По всему тексту настоящего описания цитируются некоторые документы. Каждый из процитированных здесь документов (включая все патенты, патентные заявки, научные публикации, спецификации изготовителя, инструкции и т.д.), будь то выше или ниже, настоящим включены сюда путем ссылки во всей полноте. При этом ничто не должно истолковываться как признание того, будто настоящее изобретение не было способно предшествовать такому раскрытию.
Далее будут представлены определения, которые применимы ко всем аспектам настоящего изобретения.
Термины типа “снижать” или “ингибировать” в настоящем изобретении означают способность вызывать общее снижение уровня, предпочтительно на 5% или больше, 10% или больше, 20% или больше, более предпочтительно на 50% или больше и наиболее предпочтительно на 75% или больше. Термин “ингибировать” и подобные выражения включают в себя полное или практически полное ингибирование, то есть снижение до нуля или практически до нуля.
Термины типа “повышать” или “усиливать” предпочтительно означают повышение или усиление по меньшей мере на 10%, предпочтительно по меньшей мере на 20%, предпочтительно по меньшей мере на 30%, более предпочтительно по меньшей мере на 40%, более предпочтительно по меньшей мере на 50%, еще более предпочтительно по меньшей мере на 80% и наиболее предпочтительно по меньшей мере на 100%.
“Фрагмент” в отношении последовательности нуклеиновой кислоты означает часть последовательности нуклеиновой кислоты, то есть последовательность, которая представляет последовательность этой нуклеиновой кислоты, укороченную на 5′- и/или 3′-конце. Предпочтительно фрагмент последовательности нуклеиновой кислоты включает по меньшей мере 80%, предпочтительно по меньшей мере 90%, 95%, 96%, 97%, 98% или 99% нуклеотидных остатков из данной последовательности нуклеиновой кислоты. В настоящем изобретении предпочтительными являются те фрагменты молекул РНК, которые сохраняют стабильность РНК и/или эффективность трансляции.
“Фрагмент” в отношении аминокислотной последовательности (пептида или белка) означает часть аминокислотной последовательности, то есть последовательность, которая представляет эту аминокислотную последовательность, укороченную на N-конце и/или С-конце. Фрагмент, укороченный на С-конце (N-концевой фрагмент), можно получить, напр., путем трансляции усеченной открытой рамки считывания, лишенной 3′-конца открытой рамки считывания. Фрагмент, укороченный на N-конце (С-концевой фрагмент), можно получить, напр., путем трансляции усеченной открытой рамки считывания, лишенной 5′-конца открытой рамки считывания, если только усеченная открытая рамка считывания содержит старт-кодон, который служит для инициации трансляции. Фрагмент аминокислотной последовательности включает, напр., по меньшей мере 1%, по меньшей мере 2%, по меньшей мере 3%, по меньшей мере 4%, по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% аминокислотных остатков из аминокислотной последовательности.
Термин “ионная сила” относится к математической зависимости между количеством различных видов ионных частиц в определенном растворе и соответствующими им зарядами. Таким образом, ионная сила I математически выражается формулой:
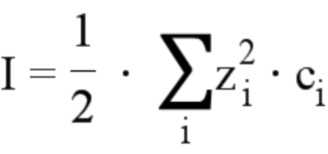
где с – молярная концентрация определенного ионного компонента, а z – абсолютная величина его заряда. Сумма Σ берется по всем различным видам ионов (i) в растворе.
Предпочтительно ионная сила описанных здесь композиций составляет 50 мМ или меньше, предпочтительно 25 мМ или меньше, предпочтительно 20 мМ или меньше, 19 мМ или меньше, 18 мМ или меньше, 17 мМ или меньше, 16 мМ или меньше, 15 мМ или меньше, 10 мМ или меньше, или 5 мМ или меньше. Предпочтительно ионная сила описанных здесь композиций является достаточно низкой, чтобы предотвратить агрегацию полиплексных частиц.
Согласно изобретению, термин “ионная сила” предпочтительно относится к присутствию одновалентных ионов. Что касается присутствия двухвалентных ионов, в частности, двухвалентных катионов, то их концентрация или эффективная концентрация (наличие свободных ионов) вследствие присутствия хелатообразующих веществ предпочтительно будет достаточно низкой с тем, чтобы предотвратить деградацию РНК. В одном особенно предпочтительном воплощении концентрация или эффективная концентрация двухвалентных ионов будет ниже каталитического уровня для гидролиза фосфодиэфирных связей между нуклеотидами РНК. В одном особенно предпочтительном воплощении концентрация свободных двухвалентных ионов составляет 20 мкМ или меньше, предпочтительно свободных двухвалентных ионов нет или практически нет.
Предпочтительно значение рН описанных здесь композиций составляет от 4 до 8; более предпочтительно от 5,5 до 8, как-то от 6 до 7,5, напр., от 6,5 до 7,1, от 6,5 до 7 или от 6,5 до 6,9.
Термин “дисахарид” относится к углеводам, состоящим из двух моносахаридных остатков, соединенных гликозидными связями. Типичные примеры дисахаридов включают трегалозу, мальтозу, сахарозу, лактозу, лактулозу, целлобиозу, изомальтозу, гентибиозу, дисахарид ламинарин (ламинарабиозу), хитобиозу, ксилобиозу, дисахарид инулин и сахарид маннобиозу. Предпочтительное содержание дисахарида в описанных здесь композициях составляет 5-20% (вес./об.), как-то 5-15% (вес./об.), 7-15% (вес./об.) или 8-12% (вес./об.). Согласно изобретению, предпочтительными являются дисахариды, имеющие высокие температуры стеклования.
Термин “хелатообразующий агент” означает соединение, которое образует хелаты с ионами металлов, предпочтительно с двухвалентными или многовалентными ионами металлов. Хелатообразующий агент содержит несколько групп, напр., -OH, -COOH, способных образовывать кольцевые структуры с ионами металлов. Примерами хелатообразующих веществ являются: этилендиаминтетрауксусная кислота (EDTA), диэтилентриаминпентауксусная кислота (DTPA), транс-1,2-диаминоциклогексантетрауксусная кислота моногидрат, N-гидроксиэтилэтилендиаминтриуксусная кислота (HEDTA), лимонная кислота и хелатообразующие вещества на основе фосфорной кислоты (напр., Dequest 2000). Согласно изобретению, предпочтительной является этилендиаминтетрауксусная кислота (EDTA). Предпочтительно хелатообразующий агент присутствует в описанных здесь композициях в концентрации по меньшей мере 20 мкМ, по меньшей мере 40 мкМ, по меньшей мере 60 мкМ или по меньшей мере 80 мкМ. Предпочтительно хелатообразующий агент присутствует в описанных здесь композициях в концентрации вплоть до 10 мМ, до 5 мМ, до 2 мМ, до 1 мМ, до 0,5 мМ, до 0,2 мМ или до 0,1 мм.
Термин “замораживание” означает затвердевание жидкости, обычно с отводом тепла.
Термин “лиофилизировать” или “лиофилизация” означает высушивание вещества вымораживанием путем его замораживания, а затем снижения окружающего давления, что позволяет замороженной среде в веществе сублимироваться прямо из твердой фазы в газовую фазу.
Термин “распылительная сушка” означает высушивание вещества распылением путем смешивания (нагретого) газа с жидкостью, которая пульверизируется (распыляется) в сосуде (распылительной сушилке), где из образовавшихся капель испаряется растворитель и образуется сухой порошок.
Термин “криопротектор” означает вещество, которое добавляют к препарату для защиты активных ингредиентов на стадиях замораживания.
Термин “лиопротектор” означает вещество, которое добавляют к препарату для защиты активных ингредиентов на стадиях сушки.
Термин “восстанавливать” означает добавление растворителя типа воды к высушенному продукту, чтобы вернуть его в жидкое состояние типа его исходного жидкого состояния.
Термин “аутологичный” применяется для описания всего, что происходит от одного и того же субъекта. Например, “аутологичные клетки” означает клетки, полученные от одного и того же субъекта. Введение аутологичных клеток субъекту выгодно, так как эти клетки преодолевают иммунологический барьер, который в противном случае приводит к отторжению.
Термин “аллогенный” применяется для описания всего, что происходит от разных особей одного и того же вида. Говорят, что два или несколько индивидов аллогенны друг другу, когда гены в одном или нескольких локусах не идентичны.
Термин “сингенный” применяется для описания всего, что происходит от индивидов или тканей, имеющих идентичные генотипы, то есть идентичных близнецов или животных одной и той же инбредной линии либо их тканей или клеток.
Термин “гетерологичный” применяется для описания чего-либо, состоящего из множества различных элементов. Например, введение клеток одного индивида другому индивиду составляет гетерологичный трансплантат. Гетерологичный ген – это ген, происходящий из другого источника, чем субъект.
В соответствии с изобретением, из-за нестабильности незащищенной РНК выгодно представлять молекулы РНК в закомплексованном виде. В частности, в некоторых воплощениях композиции по настоящему изобретению включают частицы, содержащие РНК и полиалкиленимин.
Когда система по настоящему изобретению составлена в виде препарата частиц, каждый вид РНК (напр., репликон, репликазная конструкция и необязательно другие виды РНК типа РНК, кодирующей белок, подходящий для ингибирования IFN) может быть отдельно составлен в виде индивидуального препарата частиц. В этом случае каждый индивидуальный препарат частиц будет содержать один вид РНК. Индивидуальные препараты частиц могут присутствовать в виде отдельных объектов, напр., в отдельных контейнерах. Такие препараты получают путем предоставления каждого вида РНК отдельно (обычно каждый в виде РНК-содержащего раствора) вместе с образующим частицы реагентом, тем самым обеспечивая образование частиц. Соответствующие частицы будут содержать исключительно тот конкретный вид РНК, который предоставлялся при образовании частиц (индивидуальный препарат частиц).
В одном воплощении композиции по изобретению содержат более чем один индивидуальный препарат частиц. Соответствующие композиции называются смешанными препаратами частиц. Смешанные препараты частиц по изобретению получают путем получения индивидуальных препаратов частиц по отдельности, как описано выше, с последующей стадией смешивания индивидуальных препаратов частиц. На стадии смешивания получают препарат, содержащий смешанную популяцию РНК-содержащих частиц (для иллюстрации: напр., первая популяция частиц может содержать репликон, а вторая популяция частиц может содержать репликазную конструкцию). Индивидуальные популяции частиц могут находиться вместе в одном контейнере, содержащем смешанную популяцию индивидуальных препаратов частиц.
С другой стороны, все виды РНК в композиции (напр., репликон, репликазная конструкция и необязательно другие виды типа РНК, кодирующей белок, подходящий для ингибирования IFN) могут быть составлены вместе в виде комбинированного препарата частиц. Такие препараты получают путем предоставления объединенного состава (обычно объединенного раствора) всех видов РНК вместе с образующим частицы реагентом, тем самым обеспечивая образование частиц. В отличие от смешанного препарата частиц, комбинированный препарат частиц обычно будет содержать частицы, которые содержат более одного вида РНК. В комбинированной композиции частиц различные виды РНК обычно присутствуют вместе в одной частице.
В одном воплощении препараты частиц по настоящему изобретению представляют собой препараты из наночастиц. В этом воплощении композиции по настоящему изобретению содержат РНК в виде наночастиц.
По общему определению, термин “наночастицы” относится к любым частицам, имеющим диаметр от 1 нм до 1000 нанометров (нм).
В контексте настоящего изобретения термин “частицы” относится к структурированным объектам, образованным молекулами или комплексами молекул. В одном воплощении термин “частицы” относится к микро- или наноразмерным структурам типа микро- или наноразмерных компактных структур.
Термины “In vivo-jetPEI™”, “in vivo jetPEI™”, “in vivo jetPEI”, “jetPEI”, “jet PEI” и “JetPEI” все относятся к коммерчески доступному реагенту In vivo-jetPEI™, кат. № 201-50G фирмы Polyplus-Transfection SA (Illkirch, Франция).
Термин “полиплекс” в настоящем изобретении относится к комплексам полимера и нуклеиновой кислоты типа РНК, образованным посредством электростатических взаимодействий. В тех случаях, когда полиплекс содержит РНК, его также можно называть “РНК-комплексом” или “РНК-полиплексом”.
Настоящим изобретением предусмотрены полиплексные частицы, образованные из по меньшей мере одной одноцепочечной РНК и по меньшей мере одного полиалкиленимина.
В одном воплощении описанные здесь частицы имеют средний диаметр менее 200 нм, предпочтительно менее 150 нм и более предпочтительно менее 100 нм. В одном воплощении описанные здесь частицы имеют средний диаметр по меньшей мере 30 нм, по меньшей мере 40 нм, по меньшей мере 50 нм, по меньшей мере 60 нм, по меньшей мере 70 нм, по меньшей мере 80 нм, по меньшей мере 90 нм или по меньшей мере 100 нм.
Термин “средний диаметр” означает средний гидродинамический диаметр частиц при измерении методом динамического рассеяния света с анализом данных по так называемому кумулянтному алгоритму, который в качестве результата дает так называемый Zaverage с размерностью длины и индекс полидисперсности (PI), который является безразмерным (Koppel D., J. Chem. Phys. 57, 1972, pp. 4814-4820, ISO 13321). При этом “средний диаметр”, “диаметр” или “размер” частиц применяются синонимично с этим значением Zaverage.
Термин “результирующий заряд” относится к общей сумме зарядов типа положительных и отрицательных зарядов. Например, если частица содержит большее количество отрицательных зарядов, чем положительных зарядов, то результирующий заряд частицы будет отрицательным. Если частица содержит большее количество положительных зарядов, чем отрицательных зарядов, то результирующий заряд частицы будет положительным. Если частица содержит одинаковое количество положительных и отрицательных зарядов, то результирующий заряд частицы будет нейтральным, а именно электронейтральным. Таким образом, результирующий заряд частиц по настоящему изобретению может быть отрицательным, положительным или нейтральным. В одном воплощении результирующий заряд частиц положителен. В одном воплощении результирующий заряд частиц отрицателен.
Термины “заряженный”, “результирующий заряд”, “отрицательно заряженный” или “положительно заряженный” относятся к результирующему электрическому заряду данного соединения или частиц при растворении или суспендировании в водном буфере при соответствующем рН (напр., 7.1).
Согласно настоящему изобретению, термины “соотношение N/P”, “отношение NP”, “соотношение N:P”, “N/P” и “NP” относятся к молярному соотношению атомов азота (N) в полиэтиленимине к атомам фосфора (P) в РНК.
Согласно изобретению, молярное соотношение числа атомов азота (N) в полиалкиленимине к числу атомов фосфора (P) в РНК (соотношение N/P) предпочтительно составляет от 2,0 до 15,0, предпочтительно от 8,0 до 12,0, от 6,0 до 14,0 или от 6,0 до 12,0.
Согласно изобретению, описанные здесь композиции предпочтительно доводят до конечного соотношения N/P более чем за 1 стадию, как-то за 2, 3, 4 или больше стадий. Например, композицию можно довести до первого соотношения N/P, которое ниже, чем конечное соотношение N/P, на первой стадии, к примеру, используя длинный полиалкиленимин. При дальнейшем добавлении полиалкиленимина, к примеру, короткого полиалкиленимина или длинного полиалкиленимина типа длинного полиалкиленимина, использовавшегося на первой стадии, соотношение N/P можно довести до конечного соотношения N/P. В одном воплощении окончательное соотношение N/P составляет от 8 до 16, как-то от 9 до 14, напр., между 10 и 12. В одном воплощении соотношение N/P, получаемое на первой стадии, составляет от 1 до 6, как-то от 2 до 5 типа 3 или 4.
Полиалкиленимины
Используемые здесь полиалкиленимины предпочтительно имеют следующую общую формулу (I):
,
где R означает H, ацил или группу, имеющую следующую общую формулу (II):
,
где R1 означает Н или группу, имеющую следующую общую формулу (III):
,
n, m и l выбраны независимо из целых чисел от 2 до 10; а
p, q и r – целые числа, причем сумма p, q и r такова, что средняя молекулярная масса полимера составляет от 1,5⋅102 до 107 Да, предпочтительно от 5000 до 105 Да, более предпочтительно от 10000 до 40000 Да, более предпочтительно от 15000 до 30000 Да, еще более предпочтительно от 20000 до 25000 Да.
В одном воплощении n, m и l выбраны независимо из 2, 3, 4 и 5, предпочтительно из 2 и 3 и более предпочтительно равны 2. В одном воплощении R1 означает H. В одном воплощении R означает H или ацил.
В одном воплощении полиалкиленимин включает полиэтиленимин и/или полипропиленимин, предпочтительно полиэтиленимин. Предпочтительным полиалкиленимином является полиэтиленимин (PEI). Средняя молекулярная масса PEI предпочтительно составляет от 1,5⋅102 Да до 107 Да, предпочтительно от 5000 до 105 Да, более предпочтительно от 10000 до 40000 Да, более предпочтительно от 15000 до 30000 Да, еще более предпочтительно от 20000 до 25000 Да.
Предпочтительным согласно изобретению является линейный полиалкиленимин типа линейного полиэтиленимина (PEI). В одном воплощении линейный PEI получают путем изомеризационной полимеризации с размыканием цикла 2-этил-2-оксазолина с получением поли(2-этил-2-оксазолина) (PEOX; N-пропионил-PEI), который затем подвергают гидролизу кислотой с отщеплением N-пропионильных групп, получая PEI.
Согласно изобретению, линейный PEI предпочтительно получают путем полного или практически полного деацилирования PEOX. Например, PEOX с молекулярной массой 50 кДа дает линейный PEI с молекулярной массой 22 кДа. Предпочтительно по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или практически 100% заместителей у атомов азота в полиалкиленимине типа полиэтиленимина представлены водородом (т.е. R в приведенной выше формуле означает H). При этом предпочтительно по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или практически 100% атомов азота в полиалкиленимине типа полиэтиленимина являются протонируемыми.
Предпочтительным полиалкиленимином по изобретению является полиэтиленимин (PEI), в частности, линейный полиэтиленимин. Такой линейный полиэтиленимин предпочтительно имеет молярную массу от 15 кДа до 30 кДа и предпочтительно применяется в сочетании с самореплицирующейся или самоамплифицирующейся РНК, причем соотношение N/P предпочтительно составляет от 6 до 15, а линейный полиэтиленимин и самореплицирующаяся или самоамплифицирующаяся РНК предпочтительно находятся в полиплексных частицах размером менее 200 нм, предпочтительно менее 150 нм и еще более предпочтительно менее 100 нм.
В одном воплощении изобретения полиалкиленимин представляет собой комбинацию из короткого полиалкиленимина типа короткого полиэтиленимина, от 0,6 до 11 кДа, предпочтительно от 1 до 6 кДа или от 1 до 4 кДа, как-то от 1 до 3 кДа (линейного и/или разветвленного) и длинного полиалкиленимина типа длинного полиэтиленимина, от 20 до 40 кДа (линейного и/или разветвленного), причем общее соотношение N/P предпочтительно составляет от 8 до 16, как-то от 9 до 14, напр., от 10 до 12. В одном воплощении соотношение N/P между длинным полиалкиленимином и РНК составляет от 1 до 6, как-то от 2 до 5 типа 3 или 4.
Полиэтиленимин (PEI) представляет собой органическую макромолекулу с высокой плотностью катионных зарядов. PEI может компактировать нуклеиновые кислоты в положительно заряженные частицы, способные взаимодействовать с анионными протеогликанами на поверхности клетки и облегчать проникновение частиц посредством эндоцитоза.
Существует несколько способов получения PEI. Согласно изобретению, линейный полиэтиленимин предпочтительно синтезируют и получают способом, включающим стадии, исходя из определенного количества мономера 2-этил-2-оксазолина с чистотой свыше 99%, тщательного высушивания данного количества мономера и полимеризации данного количества мономера для получения поли(2-этил-2-оксазолина) (PEOX) путем:
– после тщательного высушивания заданного количества ацетонитрила, используя данный ацетонитрил в качестве растворителя в указанном количестве высушенного мономера, в то же время добавления заданного количество тщательно высушенного инициатора реакции полимеризации и смешивания их вместе,
– очистки полученного PEOX выпариванием для удаления данного растворителя при одновременном выполнении по меньшей мере трех последовательных операций промывки/осаждения метанолом и диэтиловым эфиром и соответствующих фильтрований,
причем данные операции высушивания, полимеризации и очистки организованы так, чтобы получить (i) при проведении тестов 1H-ЯМР – правильную идентификацию данного полимера PEOX, подтверждение отсутствия мономера на уровне <1,0% и подтверждение отсутствия растворителя на уровне <5,0%, а (ii) при проведении гель-проникающей хроматографии – среднюю молекулярную массу (Mw) >23000 Да и полидисперсность (Mw/Mn) данного PEOX <1,5,
– гидролиза данного PEOX соляной кислотой для получения указанного PEI настолько эффективно, чтобы при проведении тестов 1H-ЯМР он имел остаточное содержание боковых цепей или пропионовой кислоты <5% и идентифицировался как PEI в виде единственного пика.
Под тщательным высушиванием определенного количества мономера, ацетонитрила или инициатора следует понимать достижение, непосредственно перед использованием, снижения влажности ниже 10 ppm воды, которого можно достичь путем сушки на гидриде кальция в течение 48 ч, а затем дистилляции и отбора мономера при температуре выше 129°C.
Одна или несколько из следующих характеристик являются предпочтительными согласно изобретению:
(i) средняя молекулярная масса (Mw) PEOX составляет 40000 Да <Mw <60000 Да;
(ii) соотношение мономер/инициатор составляет около 500 (под около следует понимать ±5%);
(iii) соотношение мономер/инициатор равно 480;
(iv) мономер имеет чистоту свыше 99,95%;
(v) инициатор смешивают с ацетонитрилом перед добавлением к мономеру;
(vi) полимеризация проводится в течение более 20 часов при температуре свыше 85°C;
(vii) температура при полимеризации превышает или равна 105°C;
(viii) после первого фильтрования остаток щедро промывают растворителем типа MeOH, а после добавления диэтилового эфира поли(2-этил-2-оксазолин) отделяется от раствора естественным образом в виде масла, причем общий растворитель декантируют и стадии промывки и отделения повторяют по меньшей мере четыре раза перед сушкой in vacuo;
(ix) стадия гидролиза включает удаление из реакционной смеси отработанной пропионовой кислоты, получаемой при азеотропной перегонке регулярно и в течение по меньшей мере одного дня, в то же время отслеживая процесс реакции методом 1Н-ЯМР-спектроскопии;
(x) остаток, полученный в конце процесса реакции, разбавляют водой и упаривают по меньшей мере три раза для удаления следов пропионовой кислоты, а затем остаток опять растворяют в воде и фильтруют перед лиофилизацией;
(xi) фильтрование проводится через стерильную мембрану с размером ячеек от 0,20 до 0,25 мкм, предпочтительно через стерильную пористую ацетатную мембрану.
Предпочтительно линейный PEI для применения по изобретению отличается тем, что промежуточный PEOX имеет молекулярную массу Mw типа 40000 <Mw <60000 Да.
Степень полимеризации контролируется по соотношению мономер/инициатор и по выходу продукта. Определение молекулярной массы может проводиться методом гель-проникающей хроматографией (GPC).
Термин “нуклеиновая кислота” согласно изобретению включает дезоксирибонуклеиновую кислоту (ДНК), рибонуклеиновую кислоту (РНК) и блокированную нуклеиновую кислоту (LNA). Согласно изобретению, нуклеиновая кислота включает геномную ДНК, кДНК, мРНК, вирусную РНК, полученные рекомбинантно и химически синтезированные молекулы. Согласно изобретению, нуклеиновая кислота может быть в виде одноцепочечной или двухцепочечной и линейной или ковалентно замкнутой кольцевой молекулы. Термин “нуклеиновая кислота” согласно изобретению также включает химические производные нуклеиновой кислоты по основаниям нуклеотидов, по сахарам или по фосфатам, и нуклеиновые кислоты, содержащие неприродные нуклеотиды и аналоги нуклеотидов. Описанные нуклеиновые кислоты могут представлять собой выделенные и/или рекомбинантные нуклеиновые кислоты.
Термин “выделенные” в настоящем изобретении служит для обозначения таких молекул, которые практически не содержат других молекул типа другого клеточного материала. Термин “выделенная нуклеиновая кислота” согласно изобретению означает, что нуклеиновая кислота была (i) амплифицирована in vitro, к примеру, методом полимеразной цепной реакции (ПЦР), (ii) получена рекомбинантно путем клонирования, (iii) очищена, к примеру, путем расщепления и фракционирования методом гель-электрофореза, или (iv) синтезирована, к примеру, путем химического синтеза. Выделенная нуклеиновая кислота представляет собой нуклеиновую кислоту, доступную для манипулирования рекомбинантными методами.
Термин “рекомбинантные” в контексте настоящего изобретения означает “полученные при помощи генной инженерии”. Предпочтительно “рекомбинантный объект” в контексте настоящего изобретения не встречается в природе.
Термин “встречающийся в природе” в настоящем изобретении означает то, что объект может встречаться в природе. Например, пептид или нуклеиновая кислота, которые присутствуют в организме (включая вирусы) и могут быть выделены из источника в природе и не были преднамеренно модифицированы человеком в лаборатории, являются природными. Термин “встречается в природе” означает “присутствующий в природе” и включает известные объекты, а также объекты, которые еще не были открыты и/или выделены из природы, но которые могут быть открыты и/или выделены в будущем из природного источника.
Согласно изобретению “последовательность нуклеиновой кислоты” означает последовательность нуклеотидов в нуклеиновой кислоте, напр., рибонуклеиновой кислоте (РНК) или дезоксирибонуклеиновой кислоте (ДНК). Термин может относиться ко всей молекуле нуклеиновой кислоты (типа одной нити всей молекулы нуклеиновой кислоты) или к её части (напр., фрагменту).
“3′-конец нуклеиновой кислоты” согласно изобретению означает тот конец, который содержит свободную гидроксигруппу. При схематическом представлении двухцепочечных нуклеиновых кислот, в частности ДНК, 3′-конец всегда находится с правой стороны. “5′-конец нуклеиновой кислоты” согласно изобретению означает тот конец, который содержит свободную фосфатную группу. При схематическом представлении двухцепочечных нуклеиновых кислот, в частности ДНК, 5′-конец всегда находится с левой стороны.
5'-конец 5'--P-NNNNNNN-OH-3' 3'-конец
3'-HO-NNNNNNN-P--5'
“Впереди от” описывает относительное положение первого элемента молекулы нуклеиновой кислоты по отношению ко второму элементу этой молекулы нуклеиновой кислоты, причем оба элемента содержатся в одной и той же молекуле нуклеиновой кислоты, при этом первый элемент расположен ближе к 5′-концу молекулы нуклеиновой кислоты, чем второй элемент этой молекулы нуклеиновой кислоты. Тогда говорят, что второй элемент находится “позади” от первого элемента этой молекулы нуклеиновой кислоты. Элемент, который располагается “впереди” от второго элемента, может синонимически называться расположенным “5′” от этого второго элемента. Для двухцепочечной молекулы нуклеиновой кислоты указания типа “впереди” и “позади” приводятся относительно (+)-нити.
Согласно изобретению, термин “ген” относится к определенной последовательности нуклеиновой кислоты, которая отвечает за вырабатывание одного или нескольких клеточных продуктов и/или за осуществление одной или нескольких межклеточных или внутриклеточных функций. Более конкретно, данный термин относится к такому отрезку нуклеиновой кислоты (ДНК или РНК), который содержит нуклеиновую кислоту, кодирующую определенный белок либо функциональную или структурную молекулу РНК.
Термин “вектор” применяется здесь в самом общем значении и включает любые промежуточные носители для нуклеиновой кислоты, которые, к примеру, способствуют введению данной нуклеиновой кислоты в прокариотические и/или эукариотические клетки хозяина и, если потребуется, встраиванию в геном. Такие векторы предпочтительно реплицируются и/или экспрессируются в клетке. Векторы включают плазмиды, фагемиды, вирусные геномы и их части.
В контексте настоящего изобретения термин “РНК” относится к молекулам, которые содержат рибонуклеотидные остатки, а предпочтительно полностью или практически полностью состоят из рибонуклеотидных остатков, и включает все типы РНК, описанные здесь. Термин “рибонуклеотид” относится к нуклеотидам с гидроксильной группой в 2′-положении β-D-рибофуранозильной группы. Термин “РНК” включает двухцепочечную РНК, одноцепочечную РНК, выделенную РНК типа частично или полностью очищенной РНК, практически чистую РНК, синтетическую РНК и полученную рекомбинантно РНК типа модифицированной РНК, которая отличается от природной РНК добавлением, делецией, заменой и/или изменением одного или нескольких нуклеотидов. Такие изменения могут включать добавление безнуклеотидного материала типа по концам РНК или внутри, к примеру, по одному или нескольким нуклеотидам РНК. Нуклеотиды в молекулах РНК также могут содержать нестандартные нуклеотиды, как-то не встречающиеся в природе нуклеотиды или химически синтезированные нуклеотиды или дезоксинуклеотиды. Такие измененные РНК могут называться аналогами, в частности, аналогами встречающихся в природе РНК. РНК по настоящему изобретению может иметь известный состав или же состав РНК может быть частично или полностью не известен.
Термин “стабильность” РНК относится к “периоду полураспада” РНК. “Период полураспада” означает такой период времени, который необходим для устранения половины активности, содержания или количества молекул. В контексте настоящего изобретения период полужизни РНК является показателем стабильности данной РНК. Период полураспада РНК может влиять на “продолжительность экспрессии” РНК. Можно ожидать, что РНК с длительным периодом полураспада будет экспрессироваться в течение продолжительного времени.
Термин “эффективность трансляции” относится к количеству продукта трансляции, выдаваемому молекулой РНК за определенный период времени.
Согласно изобретению, “двухцепочечная РНК” или “дцРНК” означает РНК с двумя частично или полностью комплементарными нитями.
Согласно изобретению, РНК предпочтительно представлена одноцепочечной РНК (оцРНК). Термин “одноцепочечная РНК” обычно означает такую молекулу РНК, с которой не связана никакая комплементарная молекула нуклеиновой кислоты (обычно никакая комплементарная молекула РНК). Одноцепочечная РНК может содержать самокомплементарные последовательности, которые позволяют частям РНК сворачиваться и образовывать вторичные структурные мотивы, в том числе, без ограничения, пары оснований, стебли, шпильки и выступы. Одноцепочечная РНК может существовать в виде минус-нити [(-)-нити] или плюс-нити [(+)-нити]. (+)-Нить – это нить, которая содержит или кодирует генетическую информацию. Генетическая информация может представлять собой, к примеру, последовательность полинуклеотида, кодирующего белок. Когда (+)-нить РНК кодирует белок, то (+)-нить может непосредственно служить матрицей для трансляции (синтеза белка). (-)-Нить комплементарна (+)-нити. В случае двухцепочечной РНК (+)-нить и (-)-нить представляют собой две отдельные молекулы РНК, и обе эти молекулы РНК связываются друг с другом, образуя двухцепочечную РНК (“дуплекс-РНК”).
Особенно предпочтительной одноцепочечной РНК по изобретению является мРНК и РНК репликона типа самореплицирующейся РНК. Согласно настоящему изобретению, РНК может представлять собой кодирующую РНК, то есть РНК, кодирующую пептид или белок. Предпочтительно РНК является фармацевтически активной РНК.
“Фармацевтически активная РНК” есть такая РНК, которая кодирует фармацевтически активный пептид или белок типа антигена или иммунологически активное соединение (которое не кодирует антиген) либо является фармацевтически активной сама по себе, напр., обладает одной или несколькими фармацевтическими активностями типа тех, что описаны для фармацевтически активных белков.
Согласно изобретению, термин “РНК, кодирующая пептид или белок” означает, что РНК, если она находится в соответствующем окружении, предпочтительно в клетке, может направлять сборку аминокислот с образованием пептида или белка в процессе трансляции. Предпочтительно кодирующая РНК по изобретению способна взаимодействовать с клеточным механизмом трансляции, а при трансляции кодирующей РНК образуется пептид или белок.
Согласно изобретению, термин “мРНК” означает “матричная РНК” и относится к транскриптам, которые обычно образуются с использованием ДНК-матрицы и кодируют пептид или белок. Как правило, мРНК содержит 5′-UTR, кодирующую белок область, 3′-UTR и последовательность поли(A). мРНК может быть получена путем транскрипции in vitro из ДНК-матрицы. Методология транскрипции in vitro известна специалистам. Например, имеются различные коммерчески доступные наборы для транскрипции in vitro. Согласно изобретению, мРНК может быть модифицирована посредством стабилизирующих модификаций и кэппинга.
Термин “нетранслируемая область” или “UTR” относится к такой области в молекуле ДНК, которая транскрибируется, но не транслируется в аминокислотную последовательность, или к соответствующей области в молекуле РНК типа молекулы мРНК. Нетранслируемая область (UTR) может находиться 5′ (впереди) от открытой рамки считывания (5′-UTR) и/или 3′ (позади) от открытой рамки считывания (3′-UTR).
3′-UTR, если она есть, располагается на 3′-конце гена, позади от стоп-кодона кодирующей белок области, но термин “3′-UTR” предпочтительно не включает поли(А)-хвост. Так, 3′-UTR находится впереди от поли(А)-хвоста (если он есть), напр., непосредственно примыкает к поли(А)-хвосту. 5′-UTR, если она есть, располагается на 5′-конце гена, перед старт-кодоном кодирующей белок области. 5′-UTR находится позади от 5′-кэпа (если он есть), напр., непосредственно примыкает к 5′-кэпу. 5′- и/или 3′-нетранслируемые области, согласно изобретению, могут быть функционально связаны с открытой рамкой считывания, причем эти области связываются с открытой рамкой считывания таким образом, что повышается стабильность и/или эффективность трансляции РНК, содержащей данную открытую рамку считывания.
Согласно изобретению, термины “последовательность поли(A)” или “поли(A)-хвост” относятся к непрерывной или прерывистой последовательности аденилатных остатков, которая обычно располагается на 3′-конце молекулы РНК. Непрерывная последовательность характеризуется последовательными аденилатными остатками. В природе типичной является непрерывная последовательность поли(A). Хотя последовательность поли(A) обычно не кодируется в эукариотической ДНК, но присоединяется при эукариотической транскрипции в ядре клетки к свободному 3′-концу РНК с помощью независимой от матрицы РНК-полимеразы после транскрипции, однако настоящее изобретение охватывает последовательности поли(A), кодируемые ДНК.
Термины типа “5′-кэп”, “кэп”, “структура 5′-кэпа” или “структура кэпа” применяются синонимически для обозначения динуклеотида, который находится на 5′-конце некоторых эукариотических первичных транскриптов типа предшественников матричной РНК. 5′-кэп представляет собой структуру, в которой (необязательно модифицированный) гуанозин образует связь с первым нуклеотидом молекулы мРНК через 5′-5′-трифосфатную связь (или модифицированную трифосфатную связь в случае некоторых аналогов кэпа). Эти термины могут относиться к обычным кэпам или к аналогам кэпов.
Молекулы РНК по изобретению могут характеризоваться 5′-кэпом, 5′-UTR, 3′-UTR, последовательностью поли(A) и/или адаптацией по использованию кодонов.
Молекулы РНК для применения по изобретению предпочтительно имеют размеры более 2000 нуклеотидов, предпочтительно более 3000 нуклеотидов, более 4000 нуклеотидов, более 5000 нуклеотидов, более 6000 нуклеотидов, более 7000 нуклеотидов, более 8000 нуклеотидов, более 9000 нуклеотидов или более 10000 нуклеотидов. Молекулы РНК для применения по изобретению предпочтительно имеют размеры от 6000 до 20000 нуклеотидов, предпочтительно от 6000 до 15000 нуклеотидов, предпочтительно от 9000 до 12000 нуклеотидов.
Согласно изобретению, термин “экспрессия” применяется в самом общем значении и включает вырабатывание РНК и/или белка. Он также включает частичную экспрессию нуклеиновых кислот. Кроме того, экспрессия может быть краткосрочной или стабильной. Что касается РНК, то термин “экспрессия” или “трансляция” относится к такому процессу в рибосомах клетки, посредством которого цепь кодирующей РНК (напр., матричной РНК) направляет сборку последовательности аминокислот для получения пептида или белка.
Термины “транскрипция” и “транскрибирование” относятся к процессу, при котором молекула нуклеиновой кислоты с определенной последовательностью нуклеотидов (“матрица из нуклеиновой кислоты”) считывается РНК-полимеразой так, что РНК-полимераза вырабатывает одноцепочечную молекулу РНК. При транскрипции генетическая информация в матрице нуклеиновой кислоты транскрибируется. Матрица нуклеиновой кислоты может представлять собой ДНК; однако, напр., в случае транскрипции с матрицы альфавирусной нуклеиновой кислоты эта матрица обычно представляет собой РНК. Впоследствии транскрибированная РНК может транслироваться в белок. Согласно настоящему изобретению, термин “транскрипция” включает “транскрипцию in vitro”, причем термин “транскрипция in vitro” относится к процессу, при котором РНК, в частности мРНК, синтезируется in vitro в бесклеточной системе. Предпочтительно для получения транскриптов применяются клонирующие векторы. Эти клонирующие векторы обычно называют векторами транскрипции и они по настоящему изобретению охватываются термином “вектор”. Клонирующими векторами предпочтительно являются плазмиды. Согласно настоящему изобретению, РНК предпочтительно представляет собой транскрибированную in vitro РНК (IVT-РНК) и может быть получена путем транскрипции in vitro соответствующей ДНК-матрицы. Промотором для контроля транскрипции может быть любой промотор для любой РНК-полимеразы. ДНК-матрица для транскрипции in vitro может быть получена путем клонирования нуклеиновой кислоты, в частности кДНК, и введения её в подходящий вектор для транскрипции in vitro. кДНК может быть получена путем обратной транскрипции РНК.
Молекулы одноцепочечной нуклеиновой кислоты, вырабатываемые при транскрипции, обычно имеют последовательности нуклеотидов, которые комплементарны последовательности матрицы.
Согласно изобретению, термины “матрица” или “матрица из нуклеиновой кислоты” или “нуклеиновая кислота матрицы” обычно обозначают последовательность нуклеиновой кислоты, которая может реплицироваться или транскрибироваться.
Термин “последовательность контроля экспрессии” согласно изобретению включает промоторы, связывающиеся с рибосомами последовательности и другие контрольные элементы, которые контролируют транскрипцию гена или трансляцию полученной РНК. В определенных воплощениях изобретения последовательности контроля экспрессии могут регулироваться. Точная структура последовательностей контроля экспрессию может варьироваться в зависимости от вида или типа клетки, но обычно включает 5′-нетранскрибируемые и 5′- и 3′-нетранслируемые последовательности, участвующие в инициации транскрипции и трансляции, соответственно. В частности, 5′-нетранскрибируемые последовательности контроля экспрессии область промотора, которая включает последовательность промотора для контроля транскрипции функционально связанного гена. Последовательности контроля экспрессии также могут включать последовательности энхансеров или последовательности вышележащих активаторов. Последовательности контроля экспрессии молекул ДНК обычно включают 5′-нетранскрибируемые и 5′- и 3′-нетранслируемые последовательности типа TATA-бокс, кэппирующие последовательности, последовательность CAAT и др. Последовательность контроля экспрессии альфавирусной РНК может включать субгеномный промотор и/или один или несколько консервативных элементов последовательности. Специфической последовательностью контроля экспрессии по настоящему изобретению является субгеномный промотор альфавируса, как описано здесь.
Термин “промотор” или “область промотора” относится к такой последовательности нуклеиновой кислоты, которая контролирует синтез транскрипта, напр., транскрипта, содержащего кодирующую последовательность, обеспечивая сайт узнавания и связывания для РНК-полимеразы. Промоторная область может включать и другие сайты узнавания или связывания для других факторов, участвующих в регуляции транскрипции данного гена. Промотор может контролировать транскрипцию прокариотического или эукариотического гена. Промотор может быть “индуцибельным” и инициировать транскрипцию в ответ на индуктор или же “конститутивным”, если транскрипция не контролируется индуктором. Индуцибельный промотор экспрессируется лишь в очень небольшой степени или вообще не экспрессируется, если отсутствует индуктор. В присутствии индуктора ген “включается” или повышается уровень транскрипции. Обычно это опосредуется связыванием специфического фактора транскрипции. Специфическим промотором по настоящему изобретению является субгеномный промотор альфавируса, как описано здесь. Другими специфическими промоторами являются геномные промоторы плюс-нити или минус-нити альфавируса.
Термин “основа промотора” относится к последовательности нуклеиновой кислоты, которая содержится в промоторе. Основа промотора обычно составляет минимальную часть промотора, необходимую для правильной инициации транскрипции. Основа промотора обычно включает в себя сайт запуска транскрипции и сайт связывания для РНК-полимеразы.
Приведенные здесь последовательности нуклеиновых кислот, в частности, транскрибируемые и кодирующие последовательности нуклеиновых кислот, могут комбинироваться с любыми последовательностями контроля экспрессии, которые могут быть гомологичными или гетерологичными данным последовательностям нуклеиновых кислот, причем термин “гомологичная” означает то, что последовательность нуклеиновой кислоты также по природе функционально связана с последовательностью контроля экспрессии, а термин “гетерологичная” означает то, что последовательность нуклеиновой кислоты по природе функционально не связана с последовательностью контроля экспрессии.
Последовательность нуклеиновой кислоты, в частности, последовательность нуклеиновой кислоты, кодирующая пептид или белок, и последовательность контроля экспрессии “функционально” связаны друг с другом, если они ковалентно связаны друг с другом таким образом, что транскрипция или экспрессия транскрибируемой и/или кодирующей последовательности нуклеиновой кислоты находится под контролем или под влиянием последовательности контроля экспрессии.
Согласно изобретению, “функциональная связь” или “функционально связанные” означает связи в пределах функциональных отношений. Нуклеиновая кислота является “функционально связанной”, если она функционально связана с другой последовательностью нуклеиновой кислоты. Например, промотор функционально связан с кодирующей последовательностью, если он влияет на транскрипцию данной кодирующей последовательности. Функционально связанные нуклеиновые кислоты обычно примыкают друг к другу, а если нужно, то разделяются другими последовательностями нуклеиновых кислот.
В определенных воплощениях нуклеиновая кислота по изобретению функционально связана с последовательностями контроля экспрессии, которые могут быть гомологичными или гетерологичными по отношению к нуклеиновой кислоте.
“Полимераза” обычно означает молекулярный объект, способный катализировать синтез полимерной молекулы из мономерных строительных блоков. “РНК-полимераза” –молекулярный объект, способный катализировать синтез молекул РНК из строительных блоков-рибонуклеотидов. “ДНК-полимераза” – молекулярный объект, способный катализировать синтез молекул ДНК из строительных блоков-дезоксирибонуклеотидов. В случае ДНК-полимераз и РНК-полимераз молекулярный объект обычно представляет собой белок или совокупность или комплекс из нескольких белков. Как правило, ДНК-полимераза синтезирует молекулы ДНК на основе матрицы из нуклеиновой кислоты, которой обычно является молекула ДНК. Как правило, РНК-полимераза синтезирует молекулы РНК на основе матрицы из нуклеиновой кислоты, которой является либо молекула ДНК (в этом случае РНК-полимераза представляет собой ДНК-зависимую РНК-полимеразу, DdRP), либо молекула РНК (в этом случае РНК-полимераза представляет собой РНК-зависимую РНК-полимеразу (RdRP).
“РНК-зависимая РНК-полимераза” или “RdRP” – фермент, который катализирует транскрипцию РНК с РНК-матрицы. В случае альфавирусной РНК-зависимой РНК-полимеразы последовательный синтез комплементарной (-)-нити геномной РНК и (+)-нити геномной РНК приводит к репликации РНК. Поэтому альфавирусная РНК-зависимая РНК-полимераза синонимически называется “РНК-репликазой”. В природе РНК-зависимые РНК-полимеразы обычно кодируются всеми РНК-вирусами, кроме ретровирусов. Типичными представителями вирусов, кодирующих РНК-зависимую РНК-полимеразу, являются альфавирусы.
Согласно настоящему изобретению, “репликация РНК” обычно означает синтез молекул РНК на основе нуклеотидной последовательности данной молекулы РНК (матричной молекулы РНК). Синтезируемые молекулы РНК могут быть, напр., идентичными или комплементарными матричной молекуле РНК. Как правило, репликация РНК может происходить посредством синтеза промежуточной ДНК или же непосредственно путем РНК-зависимой репликации РНК при помощи РНК-зависимой РНК-полимеразы (RdRP). В случае альфавирусов репликация РНК происходит не через промежуточную ДНК, а с помощью РНК-зависимой РНК-полимеразы (RdRP): матричная нить РНК (первая нить РНК) или её часть служит матрицей для синтеза второй нити РНК, комплементарной первой нити РНК или её части. Вторая нить РНК или её часть, в свою очередь, может необязательно служить матрицей для синтеза третьей цепи РНК, комплементарной второй нити РНК или её части. При этом третья нить РНК будет идентична первой нити РНК или её части. Таким образом, РНК-зависимая РНК-полимераза способна прямо синтезировать комплементарную матрице нить РНК и косвенно синтезировать идентичную нить РНК (через комплементарную промежуточную нить).
Согласно изобретению, термин “РНК-матрица” означает такую РНК, которая может транскрибироваться или реплицироваться РНК-зависимой РНК-полимеразой.
В предпочтительном воплощении изобретения РНК, используемая по изобретению, представляет собой РНК репликона или просто “репликон”, в частности, самореплицирующуюся РНК. В одном особенно предпочтительном воплощении репликон или самореплицирующаяся РНК происходит из или содержит элементы, происходящие из вируса с оцРНК, в частности, вируса с положительной нитью оцРНК типа альфавируса.
В общем, РНК-вирусы представляют собой разнообразную группу инфекционных частиц с РНК-геномом. РНК-вирусы можно подразделить на вирусы с одноцепочечной РНК (оцRNA) и двухцепочечной РНК (дцRNA), а вирусы с оцRNA в целом еще подразделить на вирусы с положительной нитью [(+)-нитью] и/или с отрицательной нитью [(-)-нитью]. Вирусы с положительной нитью РНК прежде всего привлекательны в качестве системы доставки в биомедицине, так как их РНК может служить непосредственно в качестве матрицы для трансляции в клетках хозяина.
Типичными представителями вирусов с положительной нитью РНК являются альфавирусы. Хозяевами альфавирусов является широкий спектр организмов, включая насекомых, рыб и млекопитающих типа домашних животных и людей. Альфавирусы реплицируются в цитоплазме инфицированных клеток (см. обзор жизненного цикла альфавируса в José et al., Future Microbiol., 2009, v. 4, pp. 837-856). Общая длина генома многих альфавирусов обычно составляет от 11000 до 12000 нуклеотидов, а геномная РНК обычно имеет 5′-кэп и 3′-поли(A)-хвост. Геном альфавирусов кодирует неструктурные белки (участвующие в транскрипции, модификации и репликации вирусной РНК и в модификации белков) и структурные белки (образующие вирусные частицы). Обычно в геноме есть две открытые рамки считывания (ORF). Четыре неструктурных белка (nsP1–nsP4) обычно кодируются вместе первой ORF, начинающейся возле 5′-конца генома, а структурные белки альфавируса кодируются вместе второй ORF, которая находится позади от первой ORF и доходит почти до 3′-конца генома. Как правило, первая ORF больше, чем вторая ORF, а их соотношение составляет примерно 2:1.
В инфицированных альфавирусом клетках из геномной РНК транслируется лишь последовательность нуклеиновой кислоты, кодирующая неструктурные белки, тогда как генетическая информация, кодирующая структурные белки, транслируется из субгеномного транскрипта, который представляет собой молекулу РНК наподобие эукариотической матричной РНК (мРНК; Gould et al., 2010, Antiviral Res., vol. 87 pp. 111-124). После инфицирования, то есть на ранних стадиях жизненного цикла вируса, (+)-нить геномной РНК непосредственно действует в качестве матричной РНК для трансляции открытой рамки считывания, кодирующей неструктурный полибелок (nsP1234). У некоторых альфавирусов имеется стоп-кодон opal между кодирующими последовательностями nsP3 и nsP4: когда трансляция заканчивается на стоп-кодоне opal, то вырабатывается полибелок P123, содержащий nsP1, nsP2 и nsP3, а если считывание переходит дальше за этот кодон opal, то вырабатывается полибелок P1234, дополнительно содержащий nsP4 (Strauss & Strauss, Microbiol. Rev., 1994, vol. 58, pp. 491-562; Rupp et al., 2015, J. Gen. Virology, vol. 96, pp. 2483-2500). nsP1234 аутопротеолитически расщепляется на фрагменты nsP123 и nsP4. Полипептиды nsP123 и nsP4 связываются, образуя репликазный комплекс (-)-нити, который транскрибирует (-)-нить РНК, используя (+)-нить геномной РНК в качестве матрицы. Обычно на более поздних стадиях фрагмент nsP123 полностью расщепляется на отдельные белки nsP1, nsP2 и nsP3 (Shirako & Strauss, 1994, J. Virol., vol. 68, pp. 1874-1885). Все четыре белка образуют репликазный комплекс (+)-нити, который синтезирует новые (+)-нити генома, используя комплементарную (-)-нить геномной РНК в качестве матрицы (Kim et al., 2004, Virology, vol. 323, pp. 153-163; Vasiljeva et al., 2003, J. Biol. Chem., vol. 278, pp. 41636-41645).
В инфицированных клетках и субгеномная РНК, и новая геномная РНК снабжается 5′-кэпом под действием nsP1 (Pettersson et al., 1980, Eur. J. Biochem. 105, 435-443; Rozanov et al., 1992, J. Gen. Virology, vol. 73, pp. 2129-2134) и полиаденилатным [поли(A)] хвостом под действием nsP4 (Rubach et al., Virology, 2009, vol. 384, pp. 201-208). Таким образом, и субгеномная РНК, и геномная РНК похожа на матричную РНК (мРНК).
Структурные белки альфавируса обычно кодируются единой открытой рамкой считывания под контролем субгеномного промотора (Strauss & Strauss, Microbiol. Rev., 1994, vol. 58, pp. 491-562). Субгеномный промотор распознается неструктурными белками альфавируса, действующими в цис-положении. В частности, репликаза альфавируса синтезирует (+)-нить субгеномного транскрипта, используя комплементарную (-)-нить геномной РНК в качестве матрицы. (+)-Нить субгеномного транскрипта кодирует структурные белки альфавируса (Kim et al., 2004, Virology, vol. 323, pp. 153-163; Vasiljeva et al., 2003, J. Biol. Chem. vol. 278, pp. 41636-41645). Субгеномный РНК-транскрипт служит матрицей для трансляции открытой рамки считывания, кодирующей структурные белки в виде одного полибелка, и этот полибелок расщепляется с образованием структурных белков. На поздней стадии альфавирусной инфекции в клетках хозяина сигнал упаковки, который находится внутри кодирующей последовательности nsP2, обеспечивает селективную упаковку геномной РНК в почкующиеся вирионы, упакованные структурными белками (White et al., 1998, J. Virol., vol. 72, pp. 4320-4326).
В инфицированных клетках синтез (-)-нити РНК обычно наблюдается только в первые 3-4 ч после инфицирования и не обнаруживается на поздних стадиях, на которых наблюдается только синтез (+)-нитей РНК (как геномной, так и субгеномной). Согласно Frolov et al., 2001, RNA, vol. 7, pp. 1638-1651, преобладающая модель для регуляции синтеза РНК предполагает зависимость от процессинга неструктурного полибелка: первоначальное расщепление неструктурного полибелка nsP1234 дает nsP123 и nsP4; а nsP4 действует как РНК-зависимая РНК-полимераза (RdRp), которая активна при синтезе (-)-нити, но неэффективна для создания (+)-нитей РНК. Дальнейший процессинг полибелка nsP123, включая расщепление на стыке nsP2/nsP3, изменяет матричную специфичность репликазы на усиление синтеза (+)-нити РНК и на уменьшение или прекращение синтеза (-)-нити РНК.
Синтез альфавирусной РНК также регулируется цис-действующими элементами РНК, включая четыре консервативных элемента последовательности (CSEs; Strauss & Strauss, Microbiol. Rev., 1994, vol. 58, pp. 491-562; и Frolov, 2001, RNA, vol. 7, pp. 1638-1651).
В целом, 5′-последовательность распознавания репликации в геноме альфавируса характеризуется низкой общей гомологией между различными альфавирусами, но имеет консервативную предсказанную вторичную структуру. 5′-последовательность распознавания репликации в геноме альфавируса не только участвует в инициации трансляции, но также включает 5′-последовательность распознавания репликации, содержащую два консервативных элемента последовательности, участвующих в синтезе вирусной РНК, CSE 1 и CSE 2. Для функции CSE 1 и 2 вторичная структура считается более важной, чем линейная последовательность (Strauss & Strauss, Microbiol. Rev., 1994, vol. 58, pp. 491-562).
Напротив, 3′-концевая последовательность генома альфавируса, то есть последовательность непосредственно перед последовательностью поли(A), характеризуется консервативной первичной структурой, в частности, консервативным элементом последовательности 4 (CSE 4), также именуемым “консервативной последовательностью в 19 нт.”, которая важна для инициации синтеза (-)-нити.
CSE 3, также именуемый “стыковочной последовательностью”, является консервативным элементом последовательности на (+)-нити альфавирусной геномной РНК, а комплементарный CSE 3 на (-)-нити действует в качестве промотора для транскрипции субгеномной РНК (Strauss & Strauss, Microbiol Rev., 1994, vol. 58, pp. 491-562; Frolov et al., 2001, RNA, vol. 7, pp. 1638-1651). CSE 3 обычно перекрывается с участком, кодирующим C-концевой фрагмент nsP4.
Наряду с белками альфавируса, с консервативными элементами последовательности могут связываться клеточные факторы хозяина, предположительно белки (Strauss & Strauss, выше).
Для доставки чужеродной генетической информации в целевые клетки или целевые организмы были предложены происходящие из альфавируса векторы. В простых подходах открытая рамка считывания, кодирующая структурные белки альфавируса, заменяется открытой рамкой считывания, кодирующей представляющий интерес белок. Системы транс-репликации на основе альфавируса базируются на элементах нуклеотидной последовательности альфавируса на двух отдельных молекулах нуклеиновой кислоты: одна молекула нуклеиновой кислоты кодирует вирусную репликазу (обычно в виде полибелка nsP1234), а другая молекула нуклеиновой кислоты способна реплицироваться данной репликазой как транс (отсюда и обозначение системы транс-репликации). транс-Репликация требует наличия обеих этих молекул нуклеиновой кислоты в данной клетке хозяина. Молекула нуклеиновой кислоты, способная реплицироваться репликазой как транс, должна содержать определенные элементы последовательности альфавируса, чтобы обеспечить распознавание и синтез РНК альфавирусной репликазой.
Согласно изобретению, термин “альфавирус” следует понимать в широком смысле, и он включает любые вирусные частицы, которые обладают характеристиками альфавирусов. Характеристики альфавирусов включают наличие (+)-нити РНК, которая кодирует генетическую информацию, подходящую для репликации в клетках хозяина, включая активность РНК-полимеразы. Другие характеристики многих альфавирусов описаны, напр., в Strauss & Strauss, Microbiol. Rev., 1994, vol. 58, с. 491-562. Термин “альфавирус” включает альфавирусы, встречающиеся в природе, а также любые их варианты или производные. В некоторых воплощениях варианты или производные не встречаются в природе.
В одном воплощении альфавирус представлен альфавирусом, встречающимся в природе. Как правило, встречающийся в природе альфавирус является инфекционным для одного или нескольких эукариотических организмов типа животных (включая позвоночных типа человека и членистоногих типа насекомых).
Встречающийся в природе альфавирус предпочтительно выбирают из группы, состоящей из следующего: комплекс вирусов Barmah Forest (включающего вирус Barmah Forest); комплекса восточного энцефалита лошадей (включающего 7 антигенных типов вируса восточного энцефалита лошадей); комплекса вирусов Middelburg (включающего вирус Middelburg); комплекса вирусов Ndumu (включающего вирус Ndumu); комплекса вирусов Semliki Forest (включающего вирус Bebaru, вирус Chikungunya, вирус Mayaro и его подтип вирус Una, вирус O’Nyong Nyong и его подтип вирус Igbo-Ora, вирус Ross River и его подтипы вирус Bebaru, вирус Getah, вирус Sagiyama, вирус Semliki Forest и его подтип вирус Me Tri); комплекса венесуэльского энцефалита лошадей (включающего вирус Cabassou, вирус Everglades, вирус Mosso das Pedras, вирус Mucambo, вирус Paramana, вирус Pixuna, вирус Rio Negro, вирус Trocara и его подтип вирус Bijou Bridge, вирус венесуэльского энцефалита лошадей); комплекса западного энцефалита лошадей (включающего вирус Aura, вирус Babanki, вирус Kyzylagach, вирус Sindbis, вирус Ockelbo, вирус Whataroa, вирус Buggy Creek, вирус Fort Morgan, вирус Highlands J, вирус западного энцефалита лошадей); и некоторые неклассифицированные вирусы, включая вирус панкреатической болезни лосося; вирус сонной болезни; вирус южного морского слона; вирус Tonate. Более предпочтительно альфавирус выбирают из группы, состоящей из комплекса вирусов Semliki Forest (включающего типы вирусов, указанные выше, включая вирус Semliki Forest), комплекса западного энцефалита лошадей (включающего типы вирусов, указанные выше, включая вирус Sindbis), вируса восточного энцефалита лошадей (включающего типы вирусов, указанные выше), комплекса венесуэльского энцефалита лошадей (включающего типы вирусов, указанные выше, включая вирус венесуэльского энцефалита лошадей).
В другом предпочтительном воплощении альфавирус представлен вирусом Semliki Forest. В альтернативном другом предпочтительном воплощении альфавирус представлен вирусом Sindbis. В альтернативном другом предпочтительном воплощении альфавирус представлен вирусом венесуэльского энцефалита лошадей.
В некоторых воплощениях настоящего изобретения альфавирус не является встречающимся в природе альфавирусом. Как правило, не встречающийся в природе альфавирус является вариантом или производным альфавируса, встречающегося в природе, который отличается от альфавируса, встречающегося в природе, по меньшей мере одной мутацией в последовательности нуклеотидов, т.е. геномной РНК. Мутация в последовательности нуклеотидов может быть из числа вставок, замен или делеций одного или нескольких нуклеотидов по сравнению со встречающимся в природе альфавирусом. Мутация в последовательности нуклеотидов может быть или не быть связана с мутацией
в полипептиде или белке, кодируемом последовательностью нуклеотидов. Например, не встречающийся в природе альфавирус может быть аттенюированным альфавирусом. Аттенюированный альфавирус, не встречающийся в природе, это такой альфавирус, который обычно имеет по меньшей мере одну мутацию в своей нуклеотидной последовательности, по которой он отличается от встречающегося в природе альфавируса, и который либо вообще не является инфекционным, либо является инфекционным, но обладает меньшей болезнетворной способностью или не обладает ею вообще. В качестве иллюстративного примера: TC83 является аттенуированным альфавирусом, который отличается от вируса венесуэльского энцефалита лошадей (VEEV), встречающегося в природе (McKinney et al., 1963, Am. J. Trop. Med. Hyg., 1963, vol. 12, pp. 597-603).
Представителей рода альфавирусов также можно классифицировать на основании их относительных клинических особенностей у людей: альфавирусы, связанные главным образом с энцефалитом, и альфавирусы, связанные главным образом с лихорадкой, сыпью и полиартритом.
Термин “альфавирусный” означает встречающийся у альфавируса либо происходящий из альфавируса или полученный из альфавируса, напр., посредством генной инженерии.
Согласно изобретению, “SFV” означает вирус Semliki Forest. Согласно изобретению, “SIN” или “SINV” означает вирус Sindbis. Согласно изобретению, “VEE” или “VEEV” означает вирус венесуэльского энцефалита лошадей.
Согласно изобретению, термин “из альфавируса” или “получен из альфавируса” относится к объекту, происходящему из альфавируса. Для иллюстрации: белок альфавируса может означать белок, который находится в альфавирусе, и/или белок, который кодируется альфавирусом; а последовательность нуклеиновой кислоты альфавируса может означать последовательность нуклеиновой кислоты, которая находится в альфавирусе, и/или последовательность нуклеиновой кислоты, которая кодируется альфавирусом. Предпочтительно последовательность нуклеиновой кислоты “из альфавируса” означает последовательность нуклеиновой кислоты “из генома альфавируса” и/или “из геномной РНК альфавируса”.
Согласно изобретению, термин “альфавирусная РНК” означает какую-либо одну или несколько из альфавирусных геномных РНК, а именно: (+)-нить, комплементарную нить альфавирусной геномной РНК, т.е. (-)-нить, и субгеномный транскрипт, т.е. (+)-нить, или фрагмент любой из них.
Согласно изобретению, “геном альфавируса” относится к геномной (+)-нити РНК альфавируса.
Согласно изобретению, термин “нативная последовательность альфавируса” и подобные термины обычно относятся к последовательности (напр., нуклеиновой кислоты) природного альфавируса (альфавируса, встречающегося в природе). В некоторых воплощениях термин “нативная последовательность альфавируса” также включает последовательность аттенюированного альфавируса.
Согласно изобретению, термин “5′-последовательность распознавания репликации” предпочтительно относится к непрерывной последовательности нуклеиновой кислоты, предпочтительно последовательности рибонуклеиновой кислоты, которая идентична или гомологична 5′-фрагменту генома альфавируса. “5′-последовательность распознавания репликации” представляет собой последовательность нуклеиновой кислоты, которая может распознаваться альфавирусной репликазой. Термин 5′-последовательность распознавания репликации включает нативные 5′-последовательности распознавания репликации, а также их функциональные эквиваленты, такие, напр., как функциональные варианты 5′-последовательности распознавания репликации альфавирусов, встречающихся в природе. 5′-последовательность распознавания репликации необходима для синтеза комплементарной (-)-нити геномной РНК альфавируса, а также для синтеза (+)-нити геномной РНК вируса на основе матрицы из (-)-нити. Нативная 5′-последовательность распознавания репликации обычно кодирует как минимум N-концевой фрагмент nsP1; но содержит не всю открытую рамку считывания, кодирующую nsP1234. Ввиду того, что нативная 5′-последовательность распознавания репликации обычно кодирует по меньшей мере N-концевой фрагмент nsP1, то нативная 5′-последовательность распознавания репликации обычно содержит как минимум один инициирующий кодон, обычно AUG. В одном воплощении 5′-последовательность распознавания репликации содержит консервативный элемент 1 последовательности генома альфавируса (CSE 1) или его вариант и консервативный элемент 2 последовательности генома альфавируса (CSE 2) или его вариант. 5′-последовательность распознавания репликации обычно способна образовывать 4 петли на стеблях (SL), то есть SL1, SL2, SL3, SL4. Нумерация этих петель начинается с 5′-конца 5′-последовательности распознавания репликации.
Согласно изобретению, термин “3′-последовательность распознавания репликации” предпочтительно относится к непрерывной последовательности нуклеиновой кислоты, предпочтительно последовательности рибонуклеиновой кислоты, которая идентична или гомологична 3′-фрагменту генома альфавируса. “3′-последовательность распознавания репликации” представляет собой последовательность нуклеиновой кислоты, которая может распознаваться альфавирусной репликазой. Термин 3′-последовательность распознавания репликации включает нативные 3′-последовательности распознавания репликации, а также их функциональные эквиваленты, такие, напр., как функциональные варианты 3′-последовательности распознавания репликации альфавирусов, встречающихся в природе. 5′-последовательность распознавания репликации необходима для синтеза комплементарной (-)-нити геномной РНК альфавируса. В одном воплощении 3′-последовательность распознавания репликации содержит консервативный элемент 4 последовательности генома альфавируса (CSE 4) или его вариант и необязательно поли(A)-хвост генома альфавируса.
Термин “консервативный элемент последовательности” или “CSE” относится к последовательностям нуклеотидов, встречающимся в альфавирусной РНК. Эти элементы последовательности называются “консервативными”, так как их ортологи присутствуют в геноме различных альфавирусов, а ортологичные CSE различных альфавирусов предпочтительно имеют высокий процент идентичности последовательности и/или близкую вторичную или третичную структуру. Термин CSE включает в себя CSE 1, CSE 2, CSE 3 и CSE 4.
Согласно изобретению, термины “CSE 1” или “CSE в 44 нт.” синонимически относятся к последовательности нуклеотидая, которая необходима для синтеза (+)-нити из матрицы (-)-нити. Термин “CSE 1” относится к последовательности на (+)-нити, а комплементарная последовательность CSE 1 на (-)-нити действует в качестве промотора для синтеза (+)-нити. Предпочтительно термин CSE 1 включает большинство 5′-нуклеотидов генома альфавируса. CSE 1 обычно образует консервативную структуру петли на стебле. Не придерживаясь какой-либо определенной теории, полагают, что для CSE 1 более важна вторичная структура, чем первичная структура, то есть линейная последовательность. CSE 1 в геномной РНК модельного альфавируса – вируса Sindbis состоит из последовательности в 44 последовательных нуклеотида, которая образована большинством из 44 5′-нуклеотидов геномной РНК (Strauss & Strauss, Microbiol. Rev., 1994, vol. 58, pp. 491-562).
Согласно изобретению, термины “CSE 2” или “CSE в 51 нт.” синонимически относятся к последовательности нуклеотидов, которая необходима для синтеза (-)-нити из матрицы (+)-нити. Матрица из (+)-нити обычно представляет собой геномную РНК альфавируса или РНК-репликон (отметим, что субгеномный РНК-транскрипт, который не содержит CSE 2, не функционирует в качестве матрицы для синтеза (-)-нити). В геномной РНК альфавируса CSE 2 обычно локализуется в кодирующей последовательности для nsP1. CSE в 51 нт. в геномной РНК модельного альфавируса – вируса Sindbis находится в положении нуклеотидов 155-205 геномной РНК (Frolov et al., 2001, RNA, vol. 7, pp. 1638-1651). CSE 2 обычно образует две консервативные структуры типа петель на стеблях. Эти структуры петель на стеблях обозначаются как петля 3 (SL3) и петля 4 (SL4), так как это третья и четвертая консервативная петля на стебле, соответственно, в геномной РНК альфавируса, считая от 5′-конца геномной РНК альфавируса. Не придерживаясь какой-либо определенной теории, полагают, что для CSE 2 более важна вторичная структура, чем первичная структура, то есть линейная последовательность.
Согласно изобретению, термины “CSE 3” или “стыковочная последовательность” синонимически относятся к последовательности нуклеотидов, которая происходит из альфавирусной геномной РНК и содержит начальный сайт субгеномной РНК. Комплементарная ей последовательность в (-)-нити стимулирует транскрипцию субгеномной РНК. В геномной РНК альфавируса CSE 3 обычно перекрывается с участком, кодирующим C-концевой фрагмент nsP4, и доходит до короткой некодирующей области, расположенной перед открытой рамкой считывания, кодирующей структурные белки.
Согласно изобретению, термины “CSE 4” или “консервативная последовательность в 19 нт.” или “CSE в 19 нт.” синонимически относятся к последовательности нуклеотидов из альфавирусной геномной РНК непосредственно перед последовательностью поли(А) в 3′-нетранслируемой области генома альфавируса. CSE 4 обычно состоит из 19 последовательных нуклеотидов. Не придерживаясь какой-либо определенной теории, полагают, что CSE 4 действует в качестве базового промотора для инициации синтеза (-)-нити (José et al., Future Microbiol., 2009, vol. 4, pp. 837-856) и/или что CSE 4 и поли(A)-хвост геномной РНК альфавируса действуют вместе для эффективного синтеза (-)-нити (Hardy & Rice, J. Virol., 2005, vol. 79, pp. 4630-4639).
Согласно изобретению термин “субгеномный промотор” или “SGP” относится к последовательности нуклеиновой кислоты впереди (5′) от последовательности нуклеиновой кислоты (напр., кодирующей последовательности), которая контролирует транскрипцию данной последовательности нуклеиновой кислоты, обеспечивая сайт распознавания и связывания для РНК-полимеразы, обычно РНК-зависимой РНК-полимеразы, в частности, для функционального неструктурного белка альфавируса. SGP может содержать и другие сайты распознавания или связывания для других факторов. Субгеномный промотор, как правило, представляет собой генетический элемент РНК-вируса с положительной нитью типа альфавируса. Субгеномный промотор альфавируса представляет собой последовательность нуклеиновой кислоты, содержащейся в вирусной геномной РНК. Субгеномный промотор обычно характеризуется тем, что он обеспечивает инициацию транскрипции (синтеза РНК) в присутствии РНК-зависимой РНК-полимеразы, напр., функционального неструктурного белка альфавируса. (-)-Нить РНК, комплементарная геномной РНК альфавируса, служит матрицей для синтеза (+)-нити субгеномного транскрипта, а синтез (+)-нити субгеномного транскрипта обычно инициируется на или рядом с субгеномным промотором. Термин “субгеномный промотор” в настоящем изобретении не ограничивается какой-либо конкретной локализацией в нуклеиновой кислоте, содержащей такой субгеномный промотор. В некоторых воплощениях SGP идентичен CSE 3 или перекрывается с CSE 3 или включает CSE 3.
Термины “субгеномный транскрипт” или “субгеномная РНК” синонимически относятся к молекуле РНК, которая получается в результате транскрипции с использованием молекулы РНК в качестве матрицы (“РНК-матрицы”), причем РНК-матрица содержит субгеномный промотор, который контролирует транскрипцию субгеномного транскрипта. Субгеномный транскрипт можно получить в присутствии РНК-зависимой РНК-полимеразы, в частности, функционального неструктурного белка альфавируса. Например, термин “субгеномный транскрипт” может относиться к РНК-транскриптам, которые получают в клетках, инфицированных альфавирусом, используя комплементарную (-)-нить геномной РНК альфавируса в качестве матрицы. Однако термин “субгеномный транскрипт” в настоящем изобретении не ограничивается этим и также включает транскрипты, получаемые с использованием гетерологичной РНК в качестве матрицы. Например, субгеномные транскрипты также можно получить, используя в качестве матрицы комплементарную (-)-нить содержащих SGP репликонов по настоящему изобретению. Таким образом, термин “субгеномный транскрипт” может относиться к молекулам РНК, получаемым при транскрипции фрагментов геномной РНК альфавируса, а также к молекулам РНК, получаемым при транскрипции фрагментов репликона по настоящему изобретению.
Согласно изобретению, конструкция из нуклеиновой кислоты, которая способна реплицироваться репликазой, предпочтительно альфавирусной репликазой, называется репликоном. Согласно изобретению, термин “репликон” определяет молекулу РНК, которая может реплицироваться РНК-зависимой РНК-полимеразой, давая, без промежуточной ДНК, одну или несколько идентичных или практически идентичных копий РНК-репликона. “Без промежуточной ДНК” означает то, что в процессе образования копий РНК-репликона не образуется ни одна копия дезоксирибонуклеиновой кислоты (ДНК) или комплементарного репликона и/или что в процесс формирования копий РНК-репликона или комплементарного репликона не используются молекулы дезоксирибонуклеиновой кислоты (ДНК) в качестве матрицы. Функция репликазы обычно обеспечивается функциональным неструктурным белком альфавируса.
Согласно изобретению, термины “может реплицироваться” и “способна к репликации” обычно означают то, что можно получить одну или несколько идентичных или практически идентичных копий нуклеиновой кислоты. При использовании вместе с термином “репликаза” типа “способна реплицироваться репликазой” термины “может реплицироваться” и “способна к репликации” описывают функциональные характеристики молекулы нуклеиновой кислоты, напр., РНК-репликона по отношению к репликазе. Эти функциональные характеристики включают по меньшей мере одно из: (i) репликаза способна распознавать репликон и (ii) репликаза способна действовать в качестве РНК-зависимой РНК-полимеразы (RdRP). Предпочтительно репликаза способна и (i) распознавать репликон, и (ii) действовать в качестве РНК-зависимой РНК-полимеразы.
Выражение “способна распознавать” означает, что репликаза способна физически ассоциироваться с репликоном, предпочтительно способна связываться с репликоном, как правило, нековалентно. Термин “связываться” может означать, что репликаза обладает способностью связываться с одним или несколькими из числа консервативного элемента последовательности 1 (CSE 1) или комплементарной ему последовательности (если она содержится в репликоне), консервативного элемента последовательности 2 (CSE 2) или комплементарной ему последовательности (если она содержится в репликоне), консервативного элемента последовательности 3 (CSE 3) или комплементарной ему последовательности (если она содержится в репликоне), консервативного элемента последовательности 4 (CSE 4) или комплементарной ему последовательности (если она содержится в репликоне). Предпочтительно репликаза способна связываться с CSE 2 [т.е. с (+)-нитью] и/или с CSE 4 [т.е. с (+)-нитью] или же с комплементарным CSE 1 [т.е. с (-)-нитью] и/или с комплементарным CSE 3 [т.е. с (-)-нитью].
Выражение “способна действовать в качестве RdRP” означает то, что репликаза способна катализировать синтез (-)-нити, комплементарной альфавирусной геномной (+)-нити РНК, причем (+)-нить РНК обладает функцией матрицы, и/или что репликаза способна катализировать синтез (+)-нити альфавирусной геномной РНК, при этом (-)-нить РНК обладает функцией матрицы. В общем, выражение “способна действовать в качестве RdRP” также может означать то, что репликаза способна катализировать синтез (+)-нити субгеномного транскрипта, при этом (-)-нить РНК обладает функцией матрицы, причем синтез (+)-нити субгеномного транскрипта обычно инициируется субгеномным промотором альфавируса.
Выражения “способна связываться” и “способна действовать в качестве RdRP” относятся к способности в нормальных физиологических условиях. В частности, они относятся к условиям внутри клеток, в которых экспрессируется функциональный неструктурный белок альфавируса или которые были трансфицированы нуклеиновой кислотой, кодирующей функциональный неструктурный белок альфавируса. Клетки предпочтительно являются эукариотическими клетками. Способность связываться и/или способность действовать в качестве RdRP можно проверить экспериментально, напр., в бесклеточной системе in vitro или в эукариотических клетках. Необязательно данные эукариотические клетки представляют собой клетки из того вида, для которого конкретный альфавирус, представляющий происхождение репликазы, является инфекционным. Например, при использовании альфавирусной репликазы из определенного альфавируса, который является инфекционным для человека, нормальными физиологическими условиями являются условия в клетках человека. Более предпочтительно эукариотические клетки (в одном примере человеческие клетки) происходят из той же ткани или органа, для которых конкретный альфавирус, представляющий происхождение репликазы, является инфекционным.
Согласно изобретению, “по сравнению с нативной последовательностью альфавируса” и подобные термины относятся к последовательности, которая является вариантом нативной последовательности альфавируса. Вариант, как правило, сам по себе не является природной последовательностью альфавируса.
В одном воплощении РНК-репликон содержит последовательность распознавания репликации, как-то 5′-последовательность распознавания репликации и 3′-последовательность распознавания репликации. Последовательность распознавания репликации представляет собой последовательность нуклеиновой кислоты, которая может распознаваться функциональным неструктурным белком альфавируса. Иными словами, функциональный неструктурный белок альфавируса способен распознавать последовательность распознавания репликации. Предпочтительно 5′-последовательность распознавания репликации располагается на 5′-конце репликона. В одном воплощении 5′-последовательность распознавания репликации состоит из или содержит CSE 1 и 2. Предпочтительно 3′-последовательность распознавания репликации располагается на 3′-конце репликона (если репликон не содержит поли(А)-хвоста) или непосредственно перед поли(А)-хвостом (если репликон содержит поли(А)-хвост). В одном воплощении 3′-последовательность распознавания состоит из или содержит CSE 4.
В одном воплощении 5′-последовательность распознавания репликации и 3′-последовательность распознавания репликации способны направлять репликацию РНК-репликона в присутствии функционального неструктурного белка альфавируса. Таким образом, когда они присутствуют по отдельности или предпочтительно вместе, эти распознающие последовательности направляют репликацию РНК-репликона в присутствии функционального неструктурного белка альфавируса.
Предпочтительно функциональный неструктурный белок альфавируса представлен как цис (кодируется как данный белок открытой рамкой считывания на репликоне) или как транс (кодируется как данный белок открытой рамкой считывания на отдельной репликазной конструкции, которая способна распознавать и 5′-последовательность распознавания репликации, и 3′-последовательность распознавания репликации репликона. В одном воплощении это достигается, когда 5′- и 3′-последовательности распознавания репликации являются нативными для того альфавируса, из которого происходит функциональный неструктурный белок альфавируса. Нативный означает то, что природным источником этих последовательностей является один и тот же альфавирус. В альтернативном воплощении 5′-последовательность распознавания репликации и/или 3′-последовательность распознавания репликации не являются нативными для того альфавируса, из которого происходит функциональный неструктурный белок альфавируса, при условии, что функциональный неструктурный белок альфавируса способен распознавать и 5′-последовательность распознавания репликации, и 3′-последовательность распознавания репликации репликона. Иными словами, функциональный неструктурный белок альфавируса совместим и с 5′-последовательностью распознавания репликации, и с 3′-последовательностью распознавания репликации. Если ненативный функциональный неструктурный белок альфавируса способен распознавать соответствующую последовательность или элемент последовательности, то говорят, что функциональный неструктурный белок альфавируса является совместимым (межвирусная совместимость). Возможны любые комбинации (3′/5′)-последовательностей распознавания репликации и CSE, соответственно, с функциональным неструктурным белком альфавируса, если существует межвирусная совместимость. Межвирусная совместимость может быть легко проверена специалистами, работающими с настоящим изобретением, путем инкубации подлежащего проверке функционального неструктурного белка альфавируса вместе с РНК, причем РНК содержит 3′- и 5′-последовательности распознавания репликации для тестирования, в условиях, подходящих для репликации РНК, напр., в подходящих клетках хозяина. Если происходит репликация, то (3′/5′)-последовательности распознавания репликации и функциональный неструктурный белок альфавируса определяются как совместимые.
В одном воплощении изобретения репликон является частью системы транс-репликации, поэтому репликон является транс-репликоном. В этом воплощении предпочтительно РНК-репликон не содержит открытой рамки считывания, кодирующей функциональный неструктурный белок альфавируса. Так, в этом воплощении настоящего изобретения предусмотрена система, содержащая две молекулы нуклеиновой кислоты: первую РНК-конструкцию для экспрессии функционального неструктурного белка альфавируса (то есть кодирующую функциональный неструктурный белок альфавируса); и вторую молекулу РНК – РНК-репликон. РНК-конструкция для экспрессии функционального неструктурного белка альфавируса синонимически именуется здесь как “РНК-конструкция для экспрессии функционального неструктурного белка альфавируса” или как “репликазная конструкция”. Функциональный неструктурный белок альфавируса является таким, как определено выше, и обычно кодируется открытой рамкой считывания, содержащейся в репликазной конструкции. Функциональный неструктурный белок альфавируса, кодируемый репликазной конструкцией, может представлять собой любой функциональный неструктурный белок альфавируса, способный реплицировать репликон. Согласно изобретению, репликазная конструкция может находиться с репликоном в одной и той же композиции, напр., в виде смешанного препарата частиц или комбинированного препарата частиц или же в отдельных композициях, напр., в виде индивидуальных препаратов частиц. Когда система по настоящему изобретению вводится в клетки, предпочтительно в эукариотические клетки, то открытая рамка считывания, кодирующая функциональный неструктурный белок альфавируса, может транслироваться. После трансляции функциональный неструктурный белок альфавируса способен реплицировать отдельные молекулы РНК (РНК-репликоны) как транс.
При этом транс (напр., в контексте транс-действия, транс-регуляции), в общем, означает “действовать через другую молекулу” (т.е. межмолекулярно). Это противоположно цис (напр., в контексте цис-действия, цис-регуляци), что, в общем, означает “действующий из той же самой молекулы” (т.е. внутримолекулярно). В контексте синтеза РНК (включая транскрипцию и репликацию РНК) транс-действующий элемент включает последовательность нуклеиновой кислоты, которая содержит ген, кодирующий фермент, способный к синтезу РНК (РНК-полимеразу). РНК-полимераза использует вторую молекулу нуклеиновой кислоты, то есть молекулу нуклеиновой кислоты, отличную от той, которой она кодируется, в качестве матрицы для синтеза РНК. Говорят, что и РНК-полимераза, и последовательность нуклеиновой кислоты, которая содержит ген, кодирующий РНК-полимеразу, “действует как транс” на вторую молекулу нуклеиновой кислоты. В контексте настоящего изобретения РНК-полимераза, кодируемая транс-действующей РНК, может представлять собой функциональный неструктурный белок альфавируса. Функциональный неструктурный белок альфавируса может использовать вторую молекулу нуклеиновой кислоты, которая представляет собой РНК-репликон, в качестве матрицы для синтеза РНК, включая репликацию РНК-репликона. РНК-репликон, который может реплицироваться репликазой как транс согласно настоящему изобретению, синонимически именуется здесь как “транс-репликон”.
Согласно настоящему изобретению, роль функционального неструктурного белка альфавируса заключается в амплификации репликона и получении субгеномного транскрипта, если на репликоне присутствует субгеномный промотор. Если репликон кодирует представляющий интерес ген для экспрессии, то уровни экспрессии данного гена и/или продолжительность экспрессии можно регулировать как транс путем модификации уровня функционального неструктурного белка альфавируса.
Система транс-репликации по настоящему изобретению содержит по меньшей мере две молекулы нуклеиновой кислоты. В предпочтительном воплощении система состоит ровно из двух молекул РНК, репликона и репликазной конструкции. В альтернативных предпочтительных воплощениях система содержит более одного репликона, каждый из которых предпочтительно кодирует по меньшей мере один представляющий интерес белок, а также содержит репликазную конструкцию. В этих воплощениях функциональный неструктурный белок альфавируса, кодируемый репликазной конструкцией, может воздействовать на каждый репликон, вызывая репликацию и необязательно продукцию субгеномных транскриптов, соответственно. Например, каждый репликон может кодировать фармацевтически активный пептид или белок. Это выгодно, напр., если желательна вакцинация субъекта против нескольких различных антигенов.
Предпочтительно репликазная конструкция лишена по меньшей мере одного консервативного элемента последовательности (CSE), необходимого для синтеза (-)-нити на основе (+)-нити матрицы и/или для синтеза (+)-нити на основе (-)-нити матрицы. Более предпочтительно репликазная конструкция не содержит никаких консервативных элементов последовательности (CSE) альфавируса. В частности, среди четырех CSE альфавируса (Strauss & Strauss, Microbiol. Rev., 1994, vol. 58, pp. 491-562; José et al., Future Microbiol., 2009, vol. 4, pp. 837-856) в репликазной конструкции предпочтительно отсутствует один или несколько из следующих CSE: CSE 1; CSE 2; CSE 3; CSE 4. В частности, в отсутствие одного или нескольких альфавирусных CSE репликазная конструкция по настоящему изобретению гораздо больше похожа на типичную эукариотическую мРНК, чем на геномную РНК альфавируса.
Репликазная конструкция по настоящему изобретению предпочтительно отличается от альфавирусной геномной РНК по крайней мере тем, что она не способна к саморепликации и/или не содержит открытой рамки считывания под контролем субгеномного промотора. Если репликазная конструкция не способна к саморепликации, то её также можно назвать “суицидной конструкцией”.
Репликазная конструкция по настоящему изобретению представляет собой предпочтительно одноцепочечную молекулу РНК. Репликазная конструкция по настоящему изобретению обычно представляет собой (+)-нить молекулы РНК. В одном воплощении репликазная конструкция по настоящему изобретению представляет собой выделенную молекулу нуклеиновой кислоты.
В одном воплощении РНК типа репликона по настоящему изобретению содержит по меньшей мере одну открытую рамку считывания, кодирующую представляющий интерес пептид или белок. В различных воплощениях представляющий интерес пептид или белок кодируется гетерологичной последовательностью нуклеиновой кислоты. Согласно настоящему изобретению, термин “гетерологичная” означает то, что последовательность нуклеиновой кислоты не является по природе функционально или структурно связанной с последовательностью нуклеиновой кислоты типа последовательности нуклеиновой кислоты альфавируса.
РНК по настоящему изобретению может кодировать один полипептид или несколько полипептидов. Несколько полипептидов могут кодироваться в виде единого полипептида (слитого полипептида) или в виде отдельных полипептидов. В некоторых воплощениях РНК по настоящему изобретению может содержать более одной открытой рамки считывания, каждая из которых в случае репликона может независимо быть под контролем субгеномного промотора или нет. В качестве альтернативы полибелок или слитый полипептид содержит индивидуальные полипептиды, разделенные необязательно аутокаталитическим сайтом расщепления протеазой (напр., белком 2A вируса ящура) или интеином.
Представляющие интерес белки могут, напр., быть выбраны из группы, состоящей из репортерных белков, фармацевтически активных пептидов или белков, ингибиторов сигнализации внутриклеточного интерферона (IFN) и функционального неструктурного белка альфавируса.
Согласно изобретению, термин “пептид” включает олиго- и полипептиды и относится к веществам, которые содержат две или больше, предпочтительно 3 или больше, предпочтительно 4 или больше, предпочтительно 6 или больше, предпочтительно 8 или больше, предпочтительно 10 или больше, предпочтительно 13 или больше, предпочтительно 16 или больше, предпочтительно 20 или больше и вплоть до предпочтительно 50, предпочтительно 100 или предпочтительно 150 последовательных аминокислот, связанных друг с другом пептидными связями. Термин “белок” относится к крупным пептидам, предпочтительно пептидам, содержащим по меньшей мере 151 аминокислоту, но термины “пептид” и “белок” применяются здесь обычно в качестве синонимов.
Термины “пептид” и “белок” включают, согласно изобретению, вещества, которые содержат не только аминокислотные компоненты, но также и не аминокислотные компоненты, как-то сахара и фосфатные структуры, а также включают вещества, содержащие такие связи, как сложноэфирные, тиоэфирные или дисульфидные связи.
Термин “вариант” в отношении, к примеру, последовательностей нуклеиновых кислот и аминокислотных последовательностей по изобретению включает любые варианты, в частности мутанты, варианты вирусных штаммов, сплайс-варианты, конформации, изоформы, аллельные варианты, видовые варианты и видовые гомологи, в частности те, что присутствуют в природе. Аллельные варианты относятся к изменениям нормальной последовательности гена, значение которых зачастую неясно. Полное секвенирование генов часто выявляет многочисленные аллельные варианты для данного гена. В отношении молекул нуклеиновых кислот термин “вариант” включает вырожденные последовательности нуклеиновых кислот, причем вырожденная нуклеиновая кислота по изобретению представляет собой нуклеиновую кислоту, которая отличается от стандартной нуклеиновой кислоты по последовательности кодонов из-за вырожденности генетического кода (напр., вследствие адаптации употребления кодонов). Видовые гомологи представляют собой такие последовательности нуклеиновых кислот или аминокислотные последовательности, которые происходят из другого вида, чем данная последовательность нуклеиновой кислоты или аминокислотная последовательность. Вирусные гомологи представляют собой такие последовательности нуклеиновых кислот или аминокислотные последовательности, которые происходят из другого вируса, чем данная последовательность нуклеиновой кислоты или аминокислотная последовательность.
Согласно изобретению, варианты нуклеиновых кислот включают одиночные или множественные делеции, добавления, мутации, замены и/или вставки нуклеотидов по сравнению с эталонной нуклеиновой кислотой. Делеции включают удаление одного или нескольких нуклеотидов из эталонной нуклеиновой кислоты. Варианты с добавлением включают 5′- и/или 3′-концевые слияния одного или нескольких нуклеотидов типа 1, 2, 3, 5, 10, 20, 30, 50 или большее нуклеотидов. В случае замен по меньшей мере один нуклеотид удаляется из последовательности, а на его место вставляется по меньшей мере один другой нуклеотид (типа трансверзии и транзиции). Мутации включают абазические сайты, перекрестносшитые сайты и химически измененные или модифицированные основания. Вставки включают добавление по меньшей мере одного нуклеотида в эталонную нуклеиновую кислоту.
Согласно изобретению, “нуклеотидные изменения” могут означать единичные или множественные делеции, добавления, мутации, замены и/или вставки нуклеотидов по сравнению с эталонной нуклеиновой кислотой. В некоторых воплощениях “нуклеотидные изменения” выбирают из группы, состоящей из делеции одного нуклеотида, добавления одного нуклеотида, мутации одного нуклеотида, замены одного нуклеотида и/или вставки одного нуклеотида по сравнению с эталонной нуклеиновой кислотой. Согласно изобретению, вариант нуклеиновой кислоты может содержать одно или несколько нуклеотидных изменений по сравнению с эталонной нуклеиновой кислотой.
Варианты определенных последовательностей нуклеиновых кислот предпочтительно обладают по меньшей мере одним функциональным свойством данных конкретных последовательностей и предпочтительно функционально эквивалентны данным конкретным последовательностям, напр., последовательности нуклеиновых кислот, проявляющие свойства, идентичные или близкие свойствам данных конкретных последовательностей нуклеиновых кислот.
Предпочтительно степень идентичности между данной последовательностью нуклеиновой кислоты и последовательностью нуклеиновой кислоты, которая является вариантом данной последовательности нуклеиновой кислоты, должна составлять по меньшей мере 70%, предпочтительно по меньшей мере 75%, предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95%, 96%, 97%, 98% или 99%. Степень идентичности предпочтительно приводится для участка по меньшей мере в 30, по меньшей мере 50, по меньшей мере 70, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 300 или по меньшей мере около 400 нуклеотидов. В предпочтительных воплощениях степень идентичности приводится для всей длины эталонной последовательности нуклеиновой кислоты.
“Сходство последовательностей” означает процент аминокислот, которые либо идентичны, либо представляют консервативные замены аминокислот. “Идентичность последовательностей” между двумя последовательностями полипептидов или нуклеиновых кислот означает процент аминокислот или нуклеотидов, которые идентичны между этими последовательностями.
Термин “% идентичны” служит для обозначения, в частности, процента таких нуклеотидов, которые идентичны при оптимальном совмещении между двумя сравниваемыми последовательностями, причем данный процент является чисто статистическим и различия между этими двумя последовательностями могут иметь случайное распределение по всей длине последовательности, а сравниваемая последовательность может включать добавления или делеции в сравнении с эталонной последовательностью с тем, чтобы получить оптимальное совмещение между двумя последовательностями. Сравнение двух последовательностей обычно проводится путем сравнения данных последовательностей после оптимального совмещения в отношении сегмента или “окна сравнения” с тем, чтобы идентифицировать локальные участки соответствия последовательностей. Оптимальное совмещение для сравнения может проводиться вручную или с помощью алгоритма локальной гомологии Smith and Waterman, 1981, Ads App. Math. 2, 482, с помощью алгоритма локальной гомологии Needleman and Wunsch, 1970, J. Mol. Biol. 48, 443, с помощью алгоритма поиска сходства Pearson and Lipman, 1988, Proc. Natl Acad. Sci. USA 85, 2444, или же с помощью компьютерных программ, использующих данные алгоритмы (GAP, BESTFIT, FASTA, BLAST P, BLAST N и TFASTA in Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Drive, Madison, Wis.).
Процент идентичности получается путем определения количества идентичных положений, в которых сравниваемые последовательности совпадают, деления этого числа на количество сравниваемых положений и умножения этого результата на 100.
Например, можно использовать BLAST-программу “BLAST 2 sequences”, которая доступна на веб-сайте http://www.ncbi.nlm.nih.gov/blast/bl2seq/wblast2.cgi.
Нуклеиновая кислота “способна гибридизироваться” или “гибридизуется” с другой нуклеиновой кислотой, если две последовательности комплементарны друг другу. Нуклеиновая кислота “комплементарна” другой нуклеиновой кислоте, если две последовательности способны образовывать стабильный дуплекс друг с другом. Согласно изобретению, гибридизация предпочтительно проводится в условиях, обеспечивающих специфическую гибридизацию между полинуклеотидами (строгих условиях). Строгие условия описаны, к примеру, в Molecular Cloning: A Laboratory Manual, J. Sambrook et al., Editors, 2nd Edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989; или в Current Protocols in Molecular Biology, F.M. Ausubel et al., Editors, John Wiley & Sons, Inc., New York; и означают, к примеру, гибридизацию при 65°C в буфере для гибридизации (3,5×SSC, 0,02% фиколла, 0,02% поливинилпирролидона, 0,02% бычьего сывороточного альбумина, 2,5 мМ NaH2PO4 (рН 7), 0,5% SDS, 2 мМ EDTA). SSC означает 0,15 М хлорида натрия/0,15 М цитрата натрия, рН 7. После гибридизации мембрану, на которую была перенесена ДНК, промывают, к примеру, в 2×SSC при комнатной температуре, а затем в 0,1-0,5×SSC/0,1×SDS при температуре вплоть до 68°C.
Степень комплементарности означает процент смежных остатков в молекуле нуклеиновой кислоты, которые могут образовывать водородные связи (напр., пары оснований Уотсона-Крика) со второй последовательностью нуклеиновой кислоты (напр., 5, 6, 7, 8, 9, 10 из 10 означает комплементарность на 50%, 60%, 70%, 80%, 90% и 100%). “Совершенно комплементарны” или “полностью комплементарны” означает, что все смежные остатки последовательности нуклеиновой кислоты будут образовывать водородные связи с таким же числом смежных остатков во второй последовательности нуклеиновой кислоты. Предпочтительно степень комплементарности согласно изобретению составляет по меньшей мере 70%, предпочтительно по меньшей мере 75%, предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95%, 96%, 97%, 98% или 99%. Наиболее предпочтительно степень комплементарности согласно изобретению составляет 100%.
Термин “производная” включает любую химическую дериватизацию нуклеиновой кислоты по основаниям нуклеотидов, по сахарам или по фосфатам. Термин “производная” также включает нуклеиновые кислоты, которые содержат нуклеотиды и аналоги нуклеотидов, не встречающиеся в природе. Предпочтительно дериватизация нуклеиновой кислоты повышает её стабильность.
Согласно изобретению, “последовательность нуклеиновой кислоты, которая произведена из последовательности нуклеиновой кислоты” означает, что нуклеиновая кислота может быть вариантом той нуклеиновой кислоты, из которой она получена.
В одном воплощении открытая рамка считывания кодирует репортерный белок. В этом воплощении открытая рамка считывания содержит репортерный ген. Некоторые гены могут быть выбраны в качестве репортеров потому, что они придают экспрессирующим их клеткам или организмам характеристики, которые можно легко идентифицировать и измерить, или потому, что они являются отборочными маркерами. Репортерные гены часто применяются как показатели того, что определенный ген был захвачен или же экспрессируется в популяции клеток или организмов. Предпочтительно продукт экспрессии репортерного гена обнаруживается визуально. Распространенные визуально детектируемые репортерные белки обычно обладают флуоресцентными или люминесцентными свойствами. Примеры конкретных репортерных генов включают ген, кодирующий зеленый флуоресцентный белок (GFP) медузы, который заставляет экспрессирующие его клетки светиться зеленым при голубом свете, фермент люциферазу, которая катализирует реакцию с люциферином с испусканием света, и красный флуоресцентный белок (RFP). Возможны варианты любого из этих специфических репортерных генов, если только эти варианты обладают визуально детектируемыми свойствами. Например, eGFP является вариантом GFP типа точечного мутанта.
Согласно изобретению, в одном воплощении РНК содержит или состоит из фармацевтически активной РНК. “Фармацевтически активная РНК” может представлять собой РНК, кодирующую фармацевтически активный пептид или белок. Предпочтительно РНК по настоящему изобретению кодирует фармацевтически активный пептид или белок. Предпочтительно открытая рамка считывания кодирует фармацевтически активный пептид или белок. Предпочтительно РНК содержит открытую рамку считывания, которая кодирует фармацевтически активный пептид или белок, в случае РНК-репликона необязательно под контролем субгеномного промотора.
“Фармацевтически активный пептид или белок” оказывает положительное или благоприятное действие на состояние или заболевание у субъекта при введении ему в терапевтически эффективном количестве. Предпочтительно фармацевтически активный пептид или белок обладает лечебными или паллиативными свойствами и может вводиться для улучшения, ослабления, облегчения, регрессии, замедления возникновения или уменьшения тяжести одного или нескольких симптомов заболевания или расстройства. Фармацевтически активный пептид или белок может обладать профилактическими свойствами и может применяться для замедления возникновения заболевания или для уменьшения тяжести такого заболевания или патологического состояния. Термин “фармацевтически активный пептид или белок” охватывает целые белки или полипептиды, а также их фармацевтически активные фрагменты. Он также может включать и фармацевтически активные аналоги пептида или белка. Термин “фармацевтически активный пептид или белок” охватывает пептиды и белки, которые являются антигенами, то есть пептид или белок вызывают иммунный ответ у субъекта, который может быть терапевтическим либо частично или полностью защитным.
В одном воплощении фармацевтически активный пептид или белок представляет собой или включает иммунологически активное соединение либо антиген или эпитоп.
Согласно изобретению, термин “иммунологически активное соединение” относится к любым соединениям, изменяющим иммунный ответ, предпочтительно путем индукции и/или подавления созревания иммунных клеток, индукции и/или подавления биосинтеза цитокинов и/или изменения гуморального иммунитета путем стимуляции вырабатывания антител B-клетками. В одном воплощении иммунный ответ включает стимуляцию антительного ответа (обычно включающего иммуноглобулин G (IgG)) и/или клеточного ответа типа T-клеточного ответа. Иммунологически активные соединения могут обладать сильным иммуностимулирующим действием, включая, без ограничения, противовирусное и противоопухолевое действие, а также могут подавлять другие аспекты иммунного ответа, к примеру, смещать иммунный ответ прочь от иммунного ответа TH2, что полезно для лечения широкого спектра опосредованных TH2 заболеваний.
Согласно изобретению, термин “антиген” или “иммуноген” охватывает любые вещества, которые вызывают иммунный ответ. В частности, “антиген” относится к любым веществам, которые специфически реагируют с антителами или T-лимфоцитами (T-клетками). Согласно настоящему изобретению, термин “антиген” включает любые молекулы, которые содержат по меньшей мере один эпитоп. Предпочтительно антиген в контексте настоящего изобретения представляет собой молекулу, которая, необязательно после процессинга, индуцирует иммунную реакцию, которая предпочтительно специфична для антигена. Согласно настоящему изобретению, можно использовать любой подходящий антиген, который является кандидатом для иммунной реакции, причем иммунная реакция может означать как гуморальный, так и клеточный иммунитет. В контексте воплощений настоящего изобретения антиген предпочтительно презентируется клетками, предпочтительно антигенпрезентирующими клетками, в контексте молекул MHC, что приводит к иммунной реакции против антигена. Антигеном предпочтительно является продукт, который соответствует природному антигену или происходит из него. Такие природные антигены могут включать или происходить из аллергенов, вирусов, бактерий, грибков, паразитов и других возбудителей инфекций и патогенов, или же антиген также может быть опухолевым антигеном. Согласно настоящему изобретению, антиген может соответствовать природному продукту, к примеру, вирусному белку или его части. В предпочтительных воплощениях антиген представляет собой поверхностный полипептид, то есть полипептид, проявляющийся в природе на поверхности клеток, патогенов, бактерий, вирусов, грибков, паразитов, аллергенов или опухолей. Антиген может вызывать иммунный ответ против клеток, патогенов, бактерий, вирусов, грибков, паразитов, аллергенов или опухолей.
Термин “связанный с заболеванием антиген” применяется в самом широком смысле для обозначения любых антигенов, связанных с заболеванием. Связанный с заболеванием антиген представляет собой молекулу, которая содержит эпитопы, которые будут стимулировать иммунную систему хозяина на выработку клеточного антигенспецифичного иммунного ответа и/или гуморального антительного ответа против заболевания. Поэтому связанные с заболеванием антигены можно использовать в терапевтических целях. Связанные с заболеванием антигены предпочтительно связаны с микробными инфекциями, обычно это микробные антигены, или же связаны с раком, обычно с опухолями.
Термин “патоген” относится к патогенным биологическим материалам, способным вызывать заболевания в организме, предпочтительно в организме позвоночных. Патогены включают микроорганизмы типа бактерий, одноклеточные эукариотические организмы (простейшие), грибки, а также вирусы.
Термины “эпитоп”, “антигенный пептид”, “антигенный эпитоп”, “иммуногенный пептид” и “MHC-связывающий пептид” применяются здесь взаимозаменяемо и относятся к антигенной детерминанте в молекулах типа антигена, то есть к части или фрагменту иммунологически активного соединения, которое распознается иммунной системой, к примеру, распознается T-клетками, в частности, когда оно презентировано в контексте молекул MHC. Эпитоп белка предпочтительно составляет непрерывную или прерывистую часть данного белка и предпочтительно имеет длину от 5 до 100, предпочтительно от 5 до 50, более предпочтительно от 8 до 30, наиболее предпочтительно от 10 до 25 аминокислот, к примеру, эпитоп предпочтительно может иметь длину в 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 аминокислот. Согласно изобретению, эпитоп может связываться с молекулами MHC типа молекул MHC на поверхности клетки, поэтому это может быть “MHC-связывающий пептид” или “антигенный пептид”. Термин “главный комплекс гистосовместимости” и сокращенно “MHC” включает молекулы MHC класса I и MHC класса II и относится к комплексу генов, которые есть у всех позвоночных. Белки или молекулы MHC важны для сигнализации между лимфоцитами и антигенпрезентирующими клетками или больными клетками при иммунных реакциях, в которых белки или молекулы MHC связывают пептиды и презентируют их для распознавания T-клеточными рецепторами. Кодируемые MHC белки экспрессируются на поверхности клеток и предъявляют T-клеткам как аутоантигены (пептидные фрагменты из самой клетки), так и несобственные антигены (напр., фрагменты вторгающихся микроорганизмов). Предпочтительно такие иммуногенные части связываются с молекулами MHC класса I или класса II. В настоящем изобретении считается, что иммуногенная часть “связывается” с молекулой MHC класса I или класса II, если такое связывание обнаруживается любым методом, известным в данной области. Термин “MHC-связывающий пептид” относится к пептидам, которые связываются с молекулами MHC класса I и/или MHC класса II. В случае комплексов MHC класса I/пептид связывающие пептиды обычно имеют длину в 8-10 аминокислот, хотя и более длинные или короткие пептиды могут быть эффективными. В случае комплексов MHC класса II/пептид связывающие пептиды обычно имеют длину в 10-25 аминокислот, в частности, 13-18 аминокислот, хотя и более длинные или короткие пептиды могут быть эффективными.
В одном воплощении представляющий интерес белок по настоящему изобретению содержит эпитоп, подходящий для вакцинации целевого организма. Специалистам в данной области должно быть известно, что один из принципов иммунобиологии и вакци нации состоит в том, что иммунопротективная реакция на заболевание вырабатывается при иммунизации организма антигеном, который иммунологически релевантен в отношении заболевания, подлежащего лечению. Согласно настоящему изобретению, антиген выбирают из группы, включающей аутоантигены и несобственные антигены. Несобственные антигены предпочтительно представляет собой бактериальные антигены, вирусные антигены, грибковые антигены, аллергены или антигены паразитов. Предпочтительно антиген содержит эпитоп, который способен вызвать иммунный ответ в целевом организме. Например, эпитоп может вызывать иммунный ответ против бактерий, вирусов, грибков, паразитов, аллергенов или опухолей.
В некоторых воплощениях несобственный антиген представляет собой бактериальный антиген. В некоторых воплощениях антиген вызывает иммунный ответ против бактерий, которые инфицируют животных, включая птиц, рыб и млекопитающих, в том числе домашних животных. Предпочтительно бактерии, против которых вырабатывается иммунный ответ, являются патогенными бактериями.
В некоторых воплощениях несобственный антиген представляет собой вирусный антиген. Вирусный антиген, к примеру, может представлять собой белок, полипептид или пептид из поверхностного белка вируса, напр., мембраносвязанный гликопротеин, белок или полипептид капсида либо белок или полипептид пепломеров. В некоторых воплощениях антиген вызывает иммунный ответ против вируса, который инфицирует животных, включая птиц, рыб и млекопитающих, в том числе домашних животных. Предпочтительно вирус, против которого вырабатывается иммунный ответ, является патогенным вирусом.
В некоторых воплощениях несобственный антиген представляет собой полипептид или белок из грибка. В некоторых воплощениях антиген вызывает иммунный ответ против грибка, который инфицирует животных, включая птиц, рыб и млекопитающих, в том числе домашних животных. Предпочтительно грибок, против которого вырабатывается иммунный ответ, является патогенным грибком.
В некоторых воплощениях несобственный антиген представляет собой полипептид или белок из одноклеточного эукариотического паразита. В некоторых воплощениях антиген вызывает иммунный ответ против одноклеточного эукариотического паразита, предпочтительно патогенного одноклеточного эукариотического паразита. Патогенные одноклеточные эукариотические паразиты могут быть, напр., из рода Plasmodium, напр., P. falciparum, P. vivax, P. malariae или P. ovale, из рода Leishmania или из рода Trypanosoma, напр., T. cruzi или T. brucei.
В некоторых воплощениях несобственный антиген представляет собой аллергенный полипептид или аллергенный белок. Аллергенный белок или аллергенный полипептид подходит для иммунотерапии аллергеном, также известной как гипосенсибилизация.
В некоторых воплощениях антиген представляет собой аутоантиген, в частности, опухолевый антиген. Опухолевые антигены и их определение известны специалистам.
В контексте настоящего изобретения термин “опухолевый антиген” или “связанный с опухолью антиген” относится к белкам, которые в нормальных условиях специфически экспрессируются в ограниченном числе тканей и/или органов или на определенных стадиях развития, к примеру, опухолевый антиген в нормальных условиях может специфически экспрессироваться в ткани желудка, предпочтительно в слизистой желудка, в репродуктивных органах, напр., в яичках, в трофобластической ткани, напр., в плаценте или в клетках зародышевой линии, и экспрессироваться или аномально экспрессироваться в одной или нескольких опухолевых или раковых тканях. В этом контексте “ограниченное число” предпочтительно означает не более 3, более предпочтительно не более 2. Опухолевые антигены в контексте настоящего изобретения включают, к примеру, антигены дифференцировки, предпочтительно антигены дифференцировки, специфичные для типа клеток, т.е. белки, которые в нормальных условиях специфически экспрессируются в определенном типе клеток на определенной стадии дифференцировки, раковые/яичковые антигены, т.е. белки, которые в нормальных условиях специфически экспрессируются в яичках и иногда в плаценте, и специфические антигены зародышевой линии. В контексте настоящего изобретения опухолевые антигены предпочтительно связаны с клеточной поверхностью раковых клеток и предпочтительно не экспрессируются или только редко экспрессируются в нормальных тканях. Предпочтительно опухолевые антигены или аберрантная экспрессия опухолевых антигенов идентифицирует раковые клетки. В контексте настоящего изобретения опухолевый антиген, который экспрессируется раковыми клетками у субъекта, напр., пациента, страдающего раковым заболеванием, предпочтительно является собственным белком у данного субъекта. В предпочтительных воплощениях опухолевый антиген в контексте настоящего изобретения специфически экспрессируется в нормальных условиях в ткани или органе, который не является незаменимым, то есть в тканях или органах, которые при повреждении их иммунной системой не приводят к гибели субъекта, либо в органах или структурах организма, которые недоступны или почти недоступны для иммунной системы. Предпочтительно аминокислотная последовательность опухолевого антигена идентична между опухолевым антигеном, который экспрессируется в нормальных тканях, и опухолевым антигеном, который экспрессируется в раковых тканях.
Примеры опухолевых антигенов, которые могут быть полезными в настоящем изобретении: p53, ART-4, BAGE, β-катенин/m, Bcr-Abl, CAMEL, CAP-1, CASP-8, CDC27/m, CDK4/m, CEA, белки клеточной поверхности из семейства клаудина, как-то CLAUDIN-6, CLAUDIN-18.2 и CLAUDIN-12, c-MYC, CT, Cyp-B, DAM, ELF2M, ETV6-AML1, G250, GAGE, GnT-V, Gap100, HAGE, HER-2/neu, HPV-E7, HPV-E6, HAST-2, hTERT (или hTRT), LAGE, LDLR/FUT, MAGE-A, предпочтительно MAGE-A1, MAGE-A2, MAGE-A3, MAGE-A4, MAGE-A5, MAGE-A6, MAGE-A7, MAGE-A8, MAGE-A9, MAGE-A10, MAGE-A11 или MAGE-A12, MAGE-B, MAGE-C, MART-1/Melan-A, MC1R, миозин/m, MUC1, MUM-1, -2, -3, NA88-A, NF1, NY-ESO-1, NY-BR-1, p190 minor BCR-Abl, Pm1/RARa, PRAME, протеиназа 3, PSA, PSM, RAGE, RU1 или RU2, SAGE, SART-1 или SART-3, SCGB3A2, SCP1, SCP2, SCP3, SSX, SURVIVIN, TEL/AML1, TPI/m, TRP-1, TRP-2, TRP-2/INT2, TPTE и WT. Особенно предпочтительные опухолевые антигены: CLAUDIN-18.2 (CLDN18.2) и CLAUDIN-6 (CLDN6).
Согласно изобретению, было отмечено, что эффективный иммунный ответ можно вызвать иммунизацией с помощью составленной с PEI самореплицирующейся (самоамплифицирующейся) РНК для доставки антигена. Составленная с PEI самореплицирующаяся РНК, полученная из вируса венесуэльского энцефалита лошадей (VEEV), была особенно эффективной для доставки тех антигенов, которые являются мембранными белками.
Согласно изобретению, термин “мембранный белок” относится к белкам, которые ассоциированы или связаны с клеточной мембраной. Они включают интегральные мембранные белки, которые заякорены на постоянной основе или являются частью мембраны, и периферические мембранные белки, которые только временно прикреплены к липидному бислою или к другим интегральным белкам. Интегральные мембранные белки классифицируются как трансмембранные белки, которые проходят через мембрану, и монотопные интегральные белки, которые прикрепляются только к одной стороне мембраны. Например, мембранные белки могут относиться к различным общим типам:
1) Мембранные белки I-го типа: эти белки имеют один трансмембранный домен в зрелом белке. N-конец внеклеточный, а C-конец – цитоплазматический. Характерно, что N-концевая часть белков содержит классическую последовательность сигнального пептида, которая направляет белок для импорта в ER. Эти белки подразделяются на тип Ia (содержащие отщепляемую сигнальную последовательность) и тип 1b (без отщепляемой сигнальной последовательности). Примеры мембранных белков I-го типа включают, без ограничения: HA вируса гриппа, инсулиновый рецептор, гликофорин, рецептор LDL и вирусные G-белки.
2) Мембранные белки II-го типа: у этих белков с одним мембранным доменом C-конец внеклеточный, а N-конец – цитоплазматический. N-конец может содержать последовательность сигнального якоря. Примеры белков этого типа включают, без ограничения: нейраминидазу вируса гриппа, галактозилтрансферазу Гольджи, сиалилтрансферазу Гольджи, предшественник сукразы-изомальтазы, рецептор асиалогликопротеина и рецептор трансферрина.
3) Многопролетные трансмембранные белки: у мембранных белков типа I и II полипептид пересекает липидный бислой один раз, тогда как у многопролетных мембранных белков полипептид пересекает мембрану несколько раз. Многопролетные трансмембранные белки также подразделяются на типы IIIa и IIIb. Белки типа IIIa имеют отщепляемые сигнальные последовательности. У белков типа IIIb N-концы выходят на наружную поверхность мембраны, но они не имеют отщепляемой сигнальной последовательности. Белки типа IIIa включают, без ограничения, пептиды M и L центров фотореакции. Белки типа IIIb включают, без ограничения, цитохром P450 и лидерную пептидазу E. coli. Другие примеры многопролетных трансмембранных белков – мембранные переносчики типа переносчиков сахаров (глюкозы, ксилозы) и переносчиков ионов.
4) Заякоренные липидной цепью мембранные белки: эти белки связаны с бислойной мембраной посредством одной или нескольких ковалентно связанных цепей жирных кислот или липидных цепей другого типа, называемых пренильными группами.
5) Заякоренные GPI мембранные белки: эти белки заякорены в мембране посредством гликозилфосфатидилинозитола (GPI).
6) Периферические мембранные белки: эти белки связаны с мембраной косвенно посредством нековалентных взаимодействий с другими мембранными белками.
Термин “мембранный белок” в настоящем изобретении включает клеточные мембранные белки человеческих или нечеловеческих клеток, а также белки вирусной оболочки. Воплощением мембранного белка является гемагглютинин вируса гриппа (HA) – гликопротеин, который находится на поверхности вирусов гриппа. Он отвечает за связывание вируса с клетками через сиаловые кислоты на мембранах типа клеток верхних дыхательных путей или эритроцитов. Он также отвечает за слияние вирусной оболочки с мембраной эндосом после снижения pH. Другие мембранные белки вируса гриппа – белок M2, который экспрессируется в избытке на клеточной поверхности, и нейраминидаза (NA).
Соответственно, в одном аспекте изобретения предусмотрены фармацевтические композиции, содержащие:
(a) одноцепочечную самореплицирующуюся РНК, кодирующую пептид или белок, содержащий антиген или эпитоп; и
(b) полиалкиленимин.
В другом аспекте изобретения предусмотрены композиции, содержащие:
(a) одноцепочечную самореплицирующуюся РНК, кодирующую пептид или белок, содержащий антиген или эпитоп; и
(b) полиалкиленимин;
для применения в качестве лекарственного средства.
В одном воплощении всех аспектов изобретения молярное отношение числа атомов азота (N) в полиалкиленимине к числу атомов фосфора (P) в одноцепочечной РНК (соотношение N:P) составляет от 1,0 до 30, предпочтительно от 2,0 до 15,0, более предпочтительно от 6,0 до 12,0.
В следующем аспекте изобретения предусмотрены композиции, содержащие:
(a) одноцепочечную самореплицирующуюся РНК, кодирующую пептид или белок, содержащий антиген или эпитоп; и
(b) полиалкиленимин;
причем молярное отношение числа атомов азота (N) в полиалкиленимине к числу атомов фосфора (P) в одноцепочечной РНК (соотношение N:P) составляет от 1,0 до 30,0, предпочтительно от 2,0 до 15,0, более предпочтительно от 6,0 до 12,0.
В одном воплощении всех аспектов изобретения ионная сила композиции составляет 50 мМ или меньше, при этом предпочтительно концентрация положительно заряженных одновалентных ионов составляет 25 мМ или меньше, а концентрация свободных положительно заряженных двухвалентных катионных ионов составляет 20 мкМ или меньше.
В следующем аспекте изобретения предусмотрены композиции, содержащие:
(a) одноцепочечную самореплицирующуюся РНК, кодирующую пептид или белок, содержащий антиген или эпитоп; и
(b) полиалкиленимин;
причем ионная сила составляет 50 мМ или меньше.
В одном воплощении концентрация положительно заряженных одновалентных ионов составляет 25 мМ или меньше, а концентрация положительно заряженных двухвалентных катионных ионов составляет 20 мкМ или меньше.
В одном воплощении всех аспектов изобретения одноцепочечная самореплицирующаяся РНК представляет собой цис-репликон.
В одном воплощении одноцепочечная самореплицирующаяся РНК происходит из вируса венесуэльского энцефалита лошадей (VEEV). В одном воплощении одноцепочечная самореплицирующаяся РНК соответствует или в основном соответствует геномной РНК VEEV или его аттенюированной формы, причем открытая рамка считывания, кодирующая структурные белки, заменена на открытую рамку считывания, кодирующую пептид или белок, содержащий антиген или эпитоп. В одном воплощении антиген либо пептид или белок, содержащий антиген или эпитоп, является мембранным белком типа белка оболочки вируса. В одном воплощении антигеном является гемагглютинин вируса гриппа.
В одном воплощении одноцепочечная самореплицирующаяся РНК происходит из вируса Semliki Forest (SFV). В одном воплощении одноцепочечная самореплицирующаяся РНК соответствует или в основном соответствует геномной РНК SFV или его аттенюированной формы, причем открытая рамка считывания, кодирующая структурные белки, заменена на открытую рамку считывания, кодирующую пептид или белок, содержащий антиген или эпитоп. В одном воплощении антиген или пептид либо белок, содержащий антиген или эпитоп, не является мембранным белком. В одном воплощении антигеном является капсидный белок вируса.
В одном воплощении всех аспектов изобретения композиция предназначена для внутримышечного введения типа внутримышечной инъекции.
В одном воплощении всех аспектов изобретения одноцепочечная РНК и полиалкиленимин находятся в полиплексных частицах.
В одном воплощении всех аспектов изобретения полиалкиленимин имеет следующую общую формулу (I):
,
где R означает H, ацил или группу, имеющую следующую общую формулу (II):
,
где R1 означает Н или группу, имеющую следующую общую формулу (III):
,
n, m и l выбраны независимо из целых чисел от 2 до 10; а
p, q и r – целые числа, причем сумма p, q и r такова, что средняя молекулярная масса полимера составляет от 1,5⋅102 до 107 Да, предпочтительно от 5000 до 105 Да, более предпочтительно от 10000 до 40000 Да, более предпочтительно от 15000 до 30000 Да, еще более предпочтительно от 20000 до 25000 Да.
В одном воплощении n, m и l выбраны независимо из 2, 3, 4 и 5, предпочтительно из 2 и 3. В одном воплощении R1 означает H. В одном воплощении R означает H или ацильную группу.
В одном воплощении всех аспектов изобретения полиалкиленимин включает полиэтиленимин и/или полипропиленимин, предпочтительно полиэтиленимин.
В одном воплощении всех аспектов изобретения по меньшей мере 92% атомов N в полиалкиленимине являются протонируемыми.
В одном воплощении всех аспектов изобретения композиции по изобретению содержат одну или несколько добавок. В одном воплощении всех аспектов изобретения композиции по изобретению содержат одну или несколько добавок. В одном воплощении одну или несколько добавок выбирают из группы, состоящей из буферных веществ, сахаридов, стабилизаторов, криопротекторов, лиопротекторов и хелатообразующих веществ. В одном воплощении всех аспектов изобретения композиции по изобретению содержат один или несколько полимеров. В одном воплощении буферные вещества включают по меньшей мере одно, выбранное из группы, состоящей из 4-(2-гидроксиэтил)-1-пиперазинэтансульфоновой кислоты (HEPES), 2-(N-морфолино)этансульфоновой кислоты (MES), 3-морфолино-2-гидроксипропансульфоновой кислоты (MOPSO), буферных систем на основе уксусной кислоты и аналогов, буферных систем на основе фосфорной кислоты или на основе лимонной кислоты. В одном воплощении всех аспектов изобретения композиции по изобретению содержат буферы для забуферивания в диапазоне от рН 4 до 8, предпочтительно от 5 до 7,5. Примерами таких буферных систем являются ацетатные буферы или буферы HEPES или фосфатные буферы или буферы на основе уксусной кислоты. В одном воплощении сахариды включают по меньшей мере один, выбранный из группы, состоящей из моносахаридов, дисахаридов, трисахаридов, олигосахаридов и полисахаридов, предпочтительно из глюкозы, трегалозы, сахарозы и декстрана. В одном воплощении добавка представляет собой декстран со средней молекулярной массой от 1 кДа до 100 кДа. В одном воплощении криопротекторы включают по меньшей мере один, выбранный из группы, состоящей из таких гликолей, как этиленгликоль, пропиленгликоль и глицерин. В одном воплощении хелатообразующие агенты включают EDTA. В одном воплощении липиды включают по меньшей мере один, выбранный из группы, состоящей из катионных липидов, нейтральных липидов и анионных липидов. В одном воплощении композиции по изобретению включают один или несколько блок-сополимеров, содержащих строительные блоки из этиленоксида и пропиленоксида. В одном воплощении композиции по изобретению включают сополимеры, содержащие этилендиаминовые группы. В одном воплощении композиции по изобретению включают амфифильный блок-сополимер, предпочтительно содержащий строительные блоки из этиленоксида и пропиленоксида, необязательно также содержащий этилендиаминовые группы.
В одном воплощении всех аспектов изобретения композиции содержат забуференную HEPES глюкозу (HBG или HBG×1), забуференную MES глюкозу (MBG или MBG×1), забуференную ацетатом глюкозу (ABG) или забуференную HEPES трегалозу (HBT или HBT×1). В одном воплощении всех аспектов изобретения композиции содержат глюкозу или трегалозу или сахарозу в буфере на основе уксусной кислоты при концентрации в пределах от 0,1 мМ до 10 мМ. В одном воплощении всех аспектов изобретения композиции содержат глюкозу или трегалозу или сахарозу в фосфатном буфере при концентрации в пределах от 0,1 мМ до 10 мМ.
В одном воплощении всех аспектов изобретения z-средний размер частиц составляет менее 200 нм, предпочтительно менее 150 нм и более предпочтительно менее 100 нм. В одном воплощении z-средний размер частиц составляет от 50 нм до 200 нм. В одном воплощении всех аспектов изобретения дзета-потенциал частиц составляет 20 мВ или больше, предпочтительно от 25 до 40 мВ. В одном воплощении всех аспектов изобретения электрофоретическая подвижность (µ) частиц составляет от 1 до 1,6 мкм×см/В×См. В одном воплощении всех аспектов изобретения z-средний размер частиц и/или дзета-потециал и/или электрофоретическая подвижность определяются в суспензии, содержащей полиплексные частицы и забуференную HEPES глюкозу (HBG) или забуференную HEPES трегалозу (HBT). В одном воплощении HBG содержит 5% глюкозы (вес./об.) и 10 мМ HEPES, pH 7,1, а HBT содержит 10% трегалозы (вес./об.) и 10 мМ HEPES, pH 7,1. В одном воплощении z-средний размер частиц определяется методом динамического рассеяния света с анализом данных по кумулянтному алгоритму. В одном воплощении измеряется коэффициент поступательной диффузии методом динамического светорассеяния. Затем применяется уравнение Стока-Эйнштейна для вычисления Z-оценки. В одном воплощении электрофоретическая подвижность измеряется методом лазерно-доплеровского электрофореза. Затем для расчета дзета-потециала применяется уравнение Генри или уравнение Смолуховского.
В одном варианте осуществления MBG содержит 5% глюкозы (вес / объем) и 10 мМ MES. В одном варианте осуществления ацетатно-буферная глюкоза содержит 5% глюкозы (вес / объем) и 10 мМ ацетата.
В одном воплощении всех аспектов изобретения частицы являются нейтральными или положительно заряженными, предпочтительно при физиологическом pH или при pH от 4,5 до 7,5.
В одном воплощении всех аспектов изобретения одноцепочечная РНК представляет собой молекулу от 6000 до 15000 нуклеотидов, предпочтительно от 9000 до 12000 нуклеотидов.
В одном воплощении всех аспектов изобретения описанные здесь композиции предназначены для применения в терапии. В одном воплощении всех аспектов изобретения описанные здесь композиции представляют собой вакцинные композиции.
В следующем аспекте изобретения предусмотрены описанные здесь композиции для индукции иммунного ответа. В одном воплощении композиции вводятся внутримышечно. В одном воплощении иммунный ответ направлен против антигена или эпитопа.
В следующем аспекте изобретения предусмотрен способ индукции иммунного ответа, включающий стадию введения описанных здесь композиций. В одном воплощении композиции вводятся внутримышечно. В одном воплощении иммунный ответ направлен против антигена или эпитопа.
В некоторых воплощениях фармацевтически активный пептид или белок не обязательно является антигеном, вызывающим иммунный ответ. Подходящие фармацевтически активные белки или пептиды могут быть выбраны из группы, состоящей из цитокинов и белков иммунной системы типа иммунологически активных соединений (напр., интерлейкинов, колониестимулирующего фактора (CSF), колониестимулирующего фактора гранулоцитов (G-CSF), колониестимулирующего фактора гранулоцитов-макрофагов (GM-CSF), эритропоэтина, фактора некроза опухолей (TNF), интерферонов, интегринов, адрессинов, селетинов, хоминг-рецепторов, T-клеточных рецепторов, иммуноглобулинов), гормонов (инсулина, тиреоидного гормона, катехоламинов, гонадотропинов, трофических гормонов, пролактина, окситоцина, дофамина, бычьего соматотропина, лептинов и т.п.), гормонов роста (напр., гормона роста человека), факторов роста (напр., эпидермального фактора роста, фактора роста нервов, инсулиноподобного фактора роста и т.п.), рецепторов факторов роста, ферментов (тканевого активатора плазминогена, стрептокиназы, ферментов биосинтеза или деградации холестерина, стериодогенных ферментов, киназ, фосфодиэстераз, метилаз, деметилаз, дегидрогеназ, целлюлаз, протеаз, липаз, фосфолипаз, ароматаз, цитохромов, аденилат- или гуанилатциклаз, нейраминидаз и т.п.), связывающих белков (белков, связывающих гормоны роста или факторы роста и т.п.), факторов транскрипции и трансляции, белков, подавляющих рост опухолей (напр., белков, подавляющих ангиогенез), структурных белков (типа коллагена, фиброина, фибриногена, эластина, тубулина, актина и миозина), белков крови (тромбина, сывороточного альбумина, фактора VII, фактора VIII, инсулина, фактора IX, фактора X, тканевого активатора плазминогена, белка C, фактора фон Виллебранда, антитромбина III, глюкоцереброзидазы, эритропоэтина, колониестимулирующего фактора гранулоцитов (GCSF) или модифицированного фактора VIII, антикоагулянтов и т.п. В одном воплощении фармацевтически активным белком по изобретению является цитокин, который участвует в регуляции лимфоидного гомеостаза, предпочтительно цитокин, который участвует в и предпочтительно индуцирует или усиливает развитие, прайминг, размножение, дифференцировку и/или выживание T-клеток. В одном воплощении цитокин представляет собой интерлейкин, напр., IL-2, IL-7, IL-12, IL-15 или IL-21.
Другие подходящие и представляющие интерес белки, кодируемые открытой рамкой считывания – это ингибиторы сигнализации интерферона (IFN). Хотя сообщалось, что жизнеспособность клеток, в которые вводилась РНК для экспрессии, может снижаться, в частности, если клетки многократно трансфецируются РНК, однако оказалось, что ингибирующие IFN агенты повышают жизнеспособность клеток, в которых должна экспрессироваться РНК (WO 2014/071963 A1). Предпочтительно ингибитор представляет собой ингибитор сигнализации IFN типа I. Предотвращение перекрытия рецептора IFN внеклеточным IFN и ингибирование внутриклеточной сигнализации IFN в клетках обеспечивает стабильную экспрессию РНК в клетках. С другой стороны, предотвращение перекрытия рецептора IFN внеклеточным IFN и ингибирование внутриклеточной сигнализации IFN повышает выживаемость клеток, в частности, если клетки неоднократно трансфецируются РНК. Не придерживаясь какой-либо теории, предполагается, что внутриклеточная сигнализация IFN может приводить к ингибированию трансляции и/или к деградации РНК. Это можно решить путем ингибирования одного или нескольких индуцируемых IFN активных антивирусных эффекторных белков. Индуцируемый IFN активный антивирусный эффекторный белок может быть выбран из группы, состоящей из РНК-зависимой протеинкиназы (PKR), 2′,5′-олигоаденилатсинтетазы (OAS) и РНКазы L. Ингибирование внутриклеточной сигнализации IFN может включать ингибирование PKR-зависимого пути и/или OAS-зависимого пути. Подходящим для этого белком является белок, способный ингибировать PKR-зависимый путь и/или OAS-зависимый путь. Ингибирование PKR-зависимого пути может включать ингибирование фосфорилирования eIF2-α. Ингибирование PKR может включать обработку клеток по меньшей мере одним ингибитором PKR. Ингибитор PKR может быть вирусным ингибитором PKR. Предпочтительным вирусным ингибитором PKR является E3 вируса коровьей оспы. Если пептид или белок (напр., E3, K3) должен ингибировать внутриклеточную сигнализацию IFN, то предпочтительна внутриклеточная экспрессия пептида или белка. Белок E3 вируса коровьей оспы представляет собой связывающий дцРНК белок в 25 кДа (кодируемый геном E3L), который связывает и секвестрирует дцРНК для предотвращения активации PKR и OAS. E3 может напрямую связываться с PKR и ингибировать её активность, что приводит к снижению фосфорилирования eIF2-α. Другие подходящие ингибиторы сигнализации IFN: ICP34.5 вируса простого герпеса, NSs вируса Toscana, нуклеополиэдровирус PK2 Bombyx mori и NS34A HCV.
В одном воплощении ингибитор внутриклеточной или внеклеточной сигнализации IFN кодируется репликоном. Репликон содержит элементы последовательности нуклеиновой кислоты, которые обеспечивают репликацию альфавирусной репликазой, обычно это CSE 1, CSE 2 и CSE 4; и предпочтительно также элементы последовательности нуклеиновой кислоты, которые обеспечивают продукцию субгеномного транскрипта, т.е. субгеномный промотор, обычно включающий CSE 3. Репликон может дополнительно включать одну или несколько модификаций, не модифицирующих полипептидную последовательность, как описано здесь, напр., кэп, последовательность поли(A), адаптацию употребления кодонов. Если на репликоне находится несколько открытых рамок считывания, то ингибитор внутриклеточной сигнализации IFN может кодироваться любым из них, необязательно под контролем субгеномного промотора или нет. В предпочтительном воплощении ингибитор внутриклеточной сигнализации IFN кодируется самой передней открытой рамкой считывания РНК-репликона. Когда ингибитор внутриклеточной сигнализации IFN кодируется самой передней открытой рамкой считывания РНК-репликона, генетическая информация, кодирующая ингибитор внутриклеточной сигнализации IFN, будет транслироваться сразу же после введения РНК-репликона в клетки хозяина, а полученный белок может впоследствии ингибировать внутриклеточную сигнализацию IFN.
Другой подходящий и представляющий интерес белок, кодируемый открытой рамкой считывания – функциональный неструктурный белок альфавируса. Термин “неструктурный белок альфавируса” включает всевозможные ко- или посттрансляционно модифицированные формы, в том числе модифицированные углеводами (как-то гликозилированные) и модифицированные липидами формы неструктурного белка альфавируса.
В некоторых воплощениях термин “неструктурный белок альфавируса” относится к одному или нескольким индивидуальным неструктурным белкам альфавирусного происхождения (nsP1, nsP2, nsP3, nsP4) либо к полибелку, содержащему полипептидную последовательность из более чем одного неструктурного белка альфавирусного происхождения. В некоторых воплощениях “неструктурный белок альфавируса” означает nsP123 и/или nsP4. В других воплощениях “неструктурный белок альфавируса” означает nsP1234. В одном воплощении данный белок, кодируемый открытой рамкой считывания, состоит из всех nsP1, nsP2, nsP3 и nsP4 в виде единого, необязательно расщепляемого полибелка: nsP1234. В одном воплощении данный белок, кодируемый открытой рамкой считывания, состоит из nsP1, nsP2 и nsP3 в виде единого, необязательно расщепляемого полибелка: nsP123. В этом воплощении nsP4 может быть дополнительным представляющим интерес белком и может кодироваться дополнительной открытой рамкой считывания.
В некоторых воплощениях неструктурный белок альфавируса способен образовывать комплекс или ассоциацию, напр., в клетках хозяина. В некоторых воплощениях “неструктурный белок альфавируса” означает комплекс или ассоциацию nsP1, nsP2 и nsP3. В некоторых воплощениях “неструктурный белок альфавируса” означает комплекс или ассоциацию nsP1, nsP2, nsP3 и nsP4. В некоторых воплощениях “неструктурный белок альфавируса” означает комплекс или ассоциацию одного или нескольких из группы, состоящей из nsP1, nsP2, nsP3 и nsP4. В некоторых воплощениях неструктурный белок альфавируса включает по меньшей мере nsP4.
Термины “комплекс” или “ассоциация” относятся к двум или нескольким одинаковым или разным белковым молекулам, находящимся в пространственной близости. Белки комплекса предпочтительно находятся в прямом или непрямом физическом или физико-химическом контакте друг с другом. Комплекс или ассоциация может состоять из нескольких разных белков (гетеромультимер) и/или нескольких копий одного определенного белка (гомомультимер). В контексте неструктурного белка альфавируса термин “комплекс или ассоциация” означает множество из по меньшей мере двух молекул белка, из которых по меньшей мере одна представляет собой неструктурный белок альфавируса. Комплекс или ассоциация может состоять из нескольких копий одного определенного белка (гомомультимер) и/или нескольких разных белков (гетеромультимер). В контексте мультимера “мульти” означает более одного, как-то два, три, четыре, пять, шесть, семь, восемь, девять, десять или более десяти.
Термин “функциональный неструктурный белок альфавируса” включает такие неструктурные белки альфавируса, которые обладают функцией репликазы. Так, “функциональный неструктурный белок альфавируса” включает репликазу альфавируса. Функция репликазы включает функцию РНК-зависимой РНК-полимеразы (RdRP), т.е. фермента, который способен катализировать синтез (-)-нити РНК на основе (+)-нити РНК-матрицы и/или способен катализировать синтез (+)-нити РНК на основе (-)-нити РНК-матрицы. Таким образом, термин “функциональный неструктурный белок альфавируса” может относиться к белку или комплексу, который синтезирует (-)-нить РНК, используя (+)-нить (напр., геномную) РНК в качестве матрицы, к белку или комплексу, который синтезирует новую (+)-нить РНК, используя комплементарную (-)-нить геномной РНК в качестве матрицы, и/или к белку или комплексу, который синтезирует субгеномный транскрипт, используя фрагмент комплементарной (-)-нити геномной РНК в качестве матрицы. Функциональный неструктурный белок альфавируса еще может иметь одну или несколько дополнительных функций, таких, напр., как протеаза (для ауторасщепления), геликаза, терминальная аденилилтрансфераза (для добавления поли(A)-хвоста), метилтрансфераза и гуанилилтрансфераза (для обеспечения нуклеиновой кислоты 5′-кэпом), сайты ядерной локализации, трифосфатаза (Gould et al., 2010, Antiviral Res., vol. 87 pp. 111-124; Rupp et al., 2015, J. Gen. Virol., vol. 96, pp. 2483-500).
Согласно изобретению, термин “репликаза альфавируса” относится к альфавирусной РНК-зависимой РНК-полимеразе, включая РНК-зависимую РНК-полимеразу из природного альфавируса (встречающегося в природе альфавируса) и РНК-зависимую РНК-полимеразу из варианта или производного альфавируса типа аттенюированного альфавируса. В контексте настоящего изобретения термины “репликаза” и “репликаза альфавируса” применяются взаимозаменяемо, если только контекст не предписывает, что какая-то конкретная репликаза не является альфавирусной репликазой.
Термин “репликаза” включает все варианты, в частности, посттрансляционно модифицированные варианты, конформации, изоформы и гомологи репликазы альфавируса, которые экспрессируются клетками, инфицированными альфавирусом, или клетками, которые были трансфицированы нуклеиновой кислотой, кодирующей репликазу альфавируса. Кроме того, термин “репликаза” включает все формы репликазы, которые были получены и могут быть получены рекомбинантными методами. Например, рекомбинантными методами может быть получена репликаза, содержащая метку, которая облегчает обнаружение и/или очистку репликазы в лаборатории, напр., myc-метку, HA-метку или олигогистидиновую метку (His-метку).
Необязательно репликаза альфавируса также функционально определяется способностью к связыванию с одним или несколькими из числа консервативного элемента последовательности 1 (CSE 1) альфавируса или комплементарной ему последовательности, консервативного элемента последовательности 2 (CSE 2) или комплементарной ему последовательности, консервативного элемента последовательности 3 (CSE 3) или комплементарной ему последовательности, консервативного элемента последовательности 4 (CSE 4) или комплементарной ему последовательности. Предпочтительно репликаза способна связываться с CSE 2 [т.е. с (+)-нитью] и/или с CSE 4 [т.е. с (+)-нитью] или же с комплементарным CSE 1 [т.е. с (-)-нитью] и/или с комплементарным CSE 3 [т.е. с (-)-нитью].
Происхождение репликазы не ограничивается каким-либо конкретным альфавирусом. В предпочтительном воплощении альфавирусная репликаза содержит неструктурный белок из вируса Semliki Forest, включая природный вирус Semliki Forest и варианты или производные вируса Semliki Forest типа аттенюированного вируса Semliki Forest. В другом предпочтительном воплощении альфавирусная репликаза содержит неструктурный белок из вируса Sindbis, включая природный вирус Sindbis и варианты или производные вируса Sindbis типа аттенюированного вируса Sindbis. В альтернативном предпочтительном воплощении альфавирусная репликаза содержит неструктурный белок из вируса венесуэльского энцефалита лошадей (VEEV), включая природный вирус VEEV и варианты или производные VEEV типа аттенюированного VEEV. В альтернативном предпочтительном воплощении альфавирусная репликаза содержит неструктурный белок из вируса чикунгунья (CHIKV), включая природный вирус CHIKV и варианты или производные CHIKV типа аттенюированного CHIKV.
Репликаза также может содержать неструктурные белки из более чем одного альфавируса. Так, настоящим изобретением предусмотрены и гетерологичные комплексы или ассоциации, включающие неструктурный белок альфавируса и обладающие функцией репликазы. Только в целях иллюстрации: репликаза может содержать один или несколько неструктурных белков (напр., nsP1, nsP2) из первого альфавируса и один или несколько неструктурных белков (nsP3, nsP4) из второго альфавируса. Неструктурные белки из более чем одного другого альфавируса могут кодироваться отдельными открытыми рамками считывания или же одной открытой рамкой считывания в виде полибелка, напр., nsP1234.
В некоторых воплощениях функциональный неструктурный белок альфавируса способен образовывать мембранные комплексы репликации и/или вакуоли в клетках, в которых экспрессируется функциональный неструктурный белок альфавируса.
Если функциональный неструктурный белок альфавируса, то есть неструктурный белок альфавируса с функцией репликазы, кодируется молекулой нуклеиновой кислоты по настоящему изобретению, то предпочтительно, чтобы субгеномный промотор репликона, если он есть, был совместимым с данной репликазой. Совместимый в этом контексте означает то, что репликаза альфавируса способна распознавать субгеномный промотор, если он есть. В одном воплощении это достигается, когда субгеномный промотор является нативным по отношению к альфавирусу, из которого происходит репликаза, то есть природным источником этих последовательностей является один и тот же альфавирус. В альтернативном воплощении субгеномный промотор не является нативным для альфавируса, из которого происходит альфавирусная репликаза, при условии, что альфавирусная репликаза способна распознавать субгеномный промотор. Иными словами, репликаза совместима с субгеномным промотором (межвирусная совместимость). Примеры межвирусной совместимости в отношении субгеномного промотора и репликазы, происходящих из разных альфавирусов, известны в данной области. Возможны любые комбинации субгеномного промотора и репликазы, если только существует межвирусная совместимость. Межвирусная совместимость может быть легко проверена специалистами, работающими с настоящим изобретением, путем инкубации подлежащей проверке репликазы вместе с РНК, причем РНК содержит подлежащий проверке субгеномный промотор, в условиях, подходящих для синтеза РНК из субгеномного промотора. Если получается субгеномный транскрипт, то субгеномный промотор и репликаза определяются как совместимые. Известны различные примеры межвирусной совместимости (см. Strauss & Strauss, Microbiol. Rev., 1994, v. 58, pp. 491-562).
В настоящем изобретении открытая рамка считывания, кодирующая функциональный неструктурный белок альфавируса, может быть представлена на РНК-репликоне или же в виде отдельной молекулы нуклеиновой кислоты, напр., молекулы мРНК. Отдельная молекула мРНК необязательно может включать, напр., кэп, 5′-UTR, 3′-UTR, последовательность поли(A) и/или адаптацию употребления кодонов. Отдельная молекула мРНК может быть представлена как транс, как описано здесь.
Когда на РНК-репликоне представлена открытая рамка считывания, кодирующая функциональный неструктурный белок альфавируса, то репликон предпочтительно может реплицироваться функциональным неструктурным белком альфавируса. В частности, РНК-репликон, кодирующий функциональный неструктурный белок альфавируса, может реплицироваться функциональным неструктурным белком альфавируса, кодируемым репликоном. Это воплощение особо предпочтительно, когда молекула нуклеиновой кислоты, кодирующая функциональный неструктурный белок альфавируса, не представлена как транс. В этом воплощении предусмотрена цис-репликация репликона. В предпочтительном воплощении РНК-репликон содержит открытую рамку считывания, кодирующую функциональный неструктурный белок альфавируса, а также по меньшей мере еще одну открытую рамку считывания, кодирующую представляющий интерес белок, и может реплицироваться функциональным неструктурным белком альфавируса.
Если на репликоне представлено несколько открытых рамок считывания, то функциональный неструктурный белок альфавируса может кодироваться любой из них, необязательно под контролем субгеномного промотора или нет, предпочтительно не под контролем субгеномного промотора. В предпочтительном воплощении функциональный неструктурный белок альфавируса кодируется самой передней открытой рамкой считывания РНК-репликона. Когда функциональный неструктурный белок альфавируса кодируется самой передней открытой рамкой считывания РНК-репликона, то генетическая информация, кодирующая функциональный неструктурный белок альфавируса, будет транслироваться сразу после введения РНК-репликона в клетки хозяина, а полученный белок может впоследствии управлять репликацией и, необязательно, продукцией субгеномного транскрипта в клетках хозяина.
Наличие открытой рамки считывания, кодирующей функциональный неструктурный белок альфавируса, содержащейся в репликоне либо в отдельной молекуле нуклеиновой кислоты, представленной как транс, обеспечивает то, что репликон реплицируется, и тем самым и ген, кодируемый репликоном, необязательно под контролем субгеномного промотора, экспрессируется на высоком уровне.
РНК-репликон подходит для экспрессии одного или нескольких генов, кодирующих представляющий интерес пептид или белок, необязательно под контролем субгеномного промотора. Возможны различные воплощения. На РНК-репликоне может находиться одна или несколько открытых рамок считывания, каждая из которых кодирует представляющий интерес пептид или белок. Самая передняя открытая рамка считывания РНК-репликона именуется как “первая открытая рамка считывания”. В некоторых воплощениях “первая открытая рамка считывания” является единственной открытой рамкой считывания РНК-репликона. Необязательно после первой открытой рамки считывания может находиться одна или несколько других открытых рамок считывания. Одна или несколько других открытых рамок считывания после первой открытой рамки считывания могут упоминаться как “вторая открытая рамка считывания”, “третья открытая рамка считывания” и т.д. в том порядке (от 5′ к 3′), в котором они следуют за первой открытой рамкой считывания. Предпочтительно каждая открытая рамка считывания содержит старт-кодон (триплет оснований), обычно AUG (в молекуле РНК), что соответствует ATG (в соответствующей молекуле ДНК).
Если репликон содержит 3′-последовательность распознавания репликации, то предпочтительно все открытые рамки считывания располагаются перед 3′-последовательностью распознавания репликации.
Когда РНК-репликон, содержащий одну или несколько открытых рамок считывания, вводят в клетку-хозяина, то репликон может непосредственно служить матрицей для трансляции первой открытой рамки считывания. Предпочтительно репликон содержит 5′-кэп. Это полезно для экспрессии гена, кодируемого первой открытой рамкой считывания, непосредственно с репликона.
В некоторых воплощениях по меньшей мере одна открытая рамка считывания репликона находится под контролем субгеномного промотора, предпочтительно субгеномного промотора альфавируса. Субгеномный промотор альфавируса очень эффективен и поэтому подходит для экспрессии гетерологичных генов на высоком уровне. Предпочтительно субгеномный промотор является промотором для субгеномного транскрипта у альфавируса. Это значит, что субгеномный промотор является таким, который естественен для альфавируса и предпочтительно контролирует транскрипцию открытой рамки считывания, кодирующей один или несколько структурных белков у данного альфавируса. В качестве альтернативы субгеномный промотор является вариантом субгеномного промотора альфавируса; подходит любой вариант, который функционирует в качестве промотора для транскрипции субгеномной РНК в клетках хозяина. Если репликон содержит субгеномный промотор, то предпочтительно он содержит и консервативный элемент 3 последовательности (CSE 3) или его вариант.
Предпочтительно по меньшей мере одна открытая рамка считывания под контролем субгеномного промотора располагается позади от субгеномного промотора. Предпочтительно субгеномный промотор контролирует продукцию субгеномной РНК, содержащей транскрипт открытой рамки считывания.
В некоторых воплощениях первая открытая рамка считывания находится под контролем субгеномного промотора. Когда первая открытая рамка считывания находится под контролем субгеномного промотора, её локализация похожа на локализацию открытой рамки считывания, кодирующей структурные белки в геноме альфавируса. Когда первая открытая рамка считывания находится под контролем субгеномного промотора, то предпочтительно, чтобы ген, кодируемый первой открытой рамкой считывания, мог экспрессироваться как из репликона, так и из его субгеномного транскрипта (последнее в присутствии функционального неструктурного белка альфавируса). Позади от первой открытой рамки считывания, которая находится под контролем субгеномного промотора, может находиться одна или несколько других открытых рамок считывания, каждая под контролем субгеномного промотора. Гены, кодируемые одной или несколькими другими открытыми рамками считывания, напр., второй открытой рамкой считывания, могут транслироваться с одного или нескольких субгеномных транскриптов, каждый под контролем субгеномного промотора. Например, РНК-репликон может содержать субгеномный промотор, контролирующий продукцию транскрипта, кодирующего второй представляющий интерес белок.
В других воплощениях первая открытая рамка считывания не находится под контролем субгеномного промотора. Когда первая открытая рамка считывания не находится под контролем субгеномного промотора, ген, кодируемый первой открытой рамкой считывания, может экспрессироваться из репликона. Следом за первой открытой рамкой считывания может находиться одна или несколько других открытых рамок считывания, каждая под контролем субгеномного промотора. Гены, кодируемые одной или несколькими другими открытыми рамками считывания, могут экспрессироваться из субгеномных транскриптов.
В клетках, содержащих репликон по настоящему изобретению, репликон может амплифицироваться функциональным неструктурным белком альфавируса. Кроме того, если репликон содержит одну или несколько открытых рамок считывания под контролем субгеномного промотора, то ожидается, что будет вырабатываться один или несколько субгеномных транскриптов функциональным неструктурным белком альфавируса. Функциональный неструктурный белок альфавируса может быть представлен как транс или же кодироваться открытой рамкой считывания репликона.
Если репликон содержит более одной открытой рамки считывания, кодирующей представляющий интерес белок, то предпочтительно, чтобы каждая открытая рамка считывания кодировала отдельный белок, напр., другой фармацевтически активный пептид или белок. Например, белок, кодируемый второй открытой рамкой считывания, должен отличаться от белка, кодируемого первой открытой рамкой считывания.
В некоторых воплощениях представляющий интерес белок, кодируемый первой и/или другой открытой рамкой считывания, предпочтительно первой открытой рамкой считывания, представляет собой функциональный неструктурный белок альфавируса или же ингибитор сигнализации IFN, напр., E3. В некоторых воплощениях представляющий интерес белок, кодируемый первой и/или другой открытой рамкой считывания, напр., второй открытой рамкой считывания, представляет собой фармацевтически активный пептид или белок либо репортерный белок.
В одном воплощении представляющий интерес белок, кодируемый первой открытой рамкой считывания, представляет собой функциональный неструктурный белок альфавируса. В этом воплощении репликон предпочтительно содержит 5′-кэп. В частности, когда представляющий интерес белок, кодируемый первой открытой рамкой считывания, представляет собой функциональный неструктурный белок альфавируса, а репликон предпочтительно содержит 5′-кэп, то последовательность нуклеиновой кислоты, кодирующая функциональный неструктурный белок альфавируса, может эффективно транслироваться из репликона, а полученный белок может впоследствии управлять репликацией репликона и управлять синтезом субгеномного транскрипта. Это воплощение может быть предпочтительным, когда не используется или не присутствует вместе с репликоном никакая дополнительная молекула нуклеиновой кислоты, кодирующая функциональный неструктурный белок альфавируса. В этом воплощении предусмотрена цис-репликация репликона.
Описанные здесь композиции могут вводиться для лечения заболеваний типа описанных здесь, напр., заболеваний, связанных с антигеном, кодируемым вводимой РНК.
Термин “заболевание” относится к аномальным состояниям, которые поражают организм индивида. Заболевания часто понимаются как заболевания, связанные с конкретными симптомами и признаками. Заболевания могут быть вызваны факторами, исходящими из внешнего источника, типа инфекционных заболеваний, или же могут быть вызваны внутренними дисфункциями, типа аутоиммунных заболеваний. У людей “заболевание” применяется в более широком смысле для обозначения любых состояний, которые вызывают боль, дисфункцию, дистресс, социальные проблемы или смерть у пораженного индивида или аналогичные проблемы у тех, кто находится в контакте с ним. В этом более широком смысле оно иногда включает травмы, инвалидность, расстройства, синдромы, инфекции, отдельные симптомы, отклонения в поведении и нетипичные изменения структуры и функции, тогда как в других контекстах и для других целей их можно рассматривать как отличимые категории. Заболевания обычно поражают людей не только физически, но и эмоционально, так как появление многих болезней и жизнь с ними может изменить взгляды на жизнь и личность человека.
Термин “заболевание, связанное с антигеном” или “заболевание с участием антигена” относится к любым заболеваниям, которые связаны с антигенами, напр., к заболеваниям, которые характеризуются присутствием антигена. Заболевание с участием антигена может представлять собой инфекционное заболевание, аутоиммунное заболевание или раковое заболевание или же просто рак. Как указано выше, антиген может быть связанным с заболеванием антигеном типа связанного с опухолью антигена, вирусного антигена или бактериального антигена.
Термин “инфекционное заболевание” относится к любым заболеваниям, которые могут передаваться от человека к человеку или от организма к организму и вызываются микробными агентом (напр., простуда). Инфекционные заболевания известны в данной области и включают, к примеру, вирусные заболевания, бактериальные заболевания или паразитарные заболевания, причем эти заболевания вызываются вирусами, бактериями и паразитами, соответственно. В этом отношении инфекционным заболеванием может быть, к примеру, гепатит, заболевания, передающиеся половым путем (напр., хламидиоз или гонорея), туберкулез, ВИЧ/синдром приобретенного иммунодефицита (СПИД), дифтерия, гепатит B, гепатит C, холера, тяжелый острый респираторный синдром (SARS), птичий грипп, грипп, такие заболевания животных, как ящур, чума мелких жвачных (peste des petits ruminants), вирус репродуктивного и респираторного синдрома свиней или паразитарные заболевания типа болезни Чагаса, малярии и др.
Термин “аутоиммунное заболевание” относится к любым заболеваниям, при которых организм вырабатывает иммуногенный (т.е. иммунной системы) ответ на какой-то компонент своей собственной ткани. Иными словами, иммунная система теряет способность распознавать какую-то ткань или систему в организме как собственную и целится и атакует её, как будто она чужеродная. Аутоиммунные заболевания можно подразделить на те, при которых поражается преимущественно один орган (напр., гемолитическая анемия и аутоиммунный тиреоидит), и те, при которых процесс аутоиммунного заболевания распространяется по многим тканям (напр., системная красная волчанка). Например, считается, что рассеянный склероз вызван тем, что T-клетки атакуют оболочки, окружающие нервные волокна головного и спинного мозга. Это приводит к потере координации, слабости и нечеткости зрения. Аутоиммунные заболевания известны в данной области и включают, к примеру, тиреоидит Хашимото, болезнь Грейвса, волчанку, рассеянный склероз, ревматический артрит, гемолитическую анемию, аутоиммунный тиреоидит, системную красную волчанку, целиакию, болезнь Крона, колит, диабет, склеродермию, псориаз и др.
Термины “раковое заболевание” или “рак” относятся или описывают физиологическое состояние индивида, которое обычно характеризуется нерегулируемым ростом клеток. Примеры раковых заболеваний включают, без ограничения, карциному, лимфому, бластому, саркому и лейкемию. Более конкретно, примеры таких видов рака включают рак костей, рак крови, рак легких, рак печени, рак поджелудочной железы, рак кожи, рак головы или шеи, кожную или внутриглазную меланому, рак матки, рак яичников, рак прямой кишки, рак анального канала, рак желудка, рак толстой кишки, рак молочной железы, рак простаты, рак матки, карциномы половых и репродуктивных органов, болезнь Ходжкина, рак пищевода, рак тонкой кишки, рак эндокринной системы, рак щитовидной железы, рак околощитовидной железы, рак надпочечников, саркомы мягких тканей, рак мочевого пузыря, рак почек, почечно-клеточный рак, рак почечной лоханки, неоплазмы центральной нервной системы (ЦНС), нейроэктодермальный рак, опухоли позвоночника, глиому, менингиому и аденому гипофиза. Термин “рак” согласно изобретению также включает и метастазы рака.
Термин “иммунный ответ” относится к реакциям иммунной системы типа на иммуногенные организмы, такие как бактерии или вирусы, клетки или вещества. Термин “иммунный ответ” включает в себя врожденный иммунитет и адаптивный иммунитет. Предпочтительно иммунный ответ связан с активацией иммунных клеток, индукцией биосинтеза цитокинов и/или выработки антител.
Предпочтительно иммунный ответ, индуцируемый композициями по настоящему изобретению, включает стадии активации антигенпрезентирующих клеток типа дендритных клеток и/или макрофагов, презентации антигена или его фрагмента данными антиген-презентирующими клетками и активации цитотоксических T-клеток вследствие этой презентации.
Термин “иммунные клетки” относится к клеткам иммунной системы, участвующим в защите организма человека. Термин “иммунные клетки” охватывает конкретные типы иммунных клеток и их предшественников, включая лейкоциты, в том числе макрофаги, моноциты (предшественники макрофагов), гранулоциты типа нейтрофилов, эозинофилов и базофилов, дендритные клетки, тучные клетки и лимфоциты типа B-клеток, T-клеток и клеток природных киллеров (NK). Макрофаги, моноциты (предшественники макрофагов), нейтрофилы, дендритные клетки и тучные клетки являются фагоцитарными клетками.
Термин “иммунотерапия” относится к лечению заболеваний или состояний путем запуска, усиления или подавления иммунного ответа. Иммунотерапия, которая должна вызывать или усиливать иммунный ответ, классифицируется как активационная иммунотерапия, тогда как иммунотерапия, которая уменьшает или подавляет иммунный ответ, классифицируется как супрессивная иммунотерапия. Термин “иммунотерапия” включает антигенную иммунизацию или вакцинацию либо противоопухолевую иммунизацию или вакцинацию. Термин “иммунотерапия” также относится к манипулированию иммунными реакциями с тем, чтобы модулировать неадекватные иммунные реакции в более подходящие в контексте таких аутоиммунных заболеваний, как ревматический артрит, аллергия, диабет или рассеянный склероз.
Термины “иммунизация” или “вакцинация” описывают процесс введения антигена индивиду с целью индуцирования иммунного ответа, к примеру, по терапевтическим или профилактическим причинам.
Термин “терапевтическое лечение” или просто “лечение” означает такое лечение, которое улучшает состояние здоровья и/или продлевает (повышает) продолжительность жизни индивида. Такое лечение может устранять заболевание у индивида, останавливать или замедлять развитие заболевания у индивида, ингибировать или замедлять развитие заболевания у индивида, уменьшать частоту или тяжесть симптомов у индивида и/или снижать рецидивы у индивида, который в настоящее время имеет или у кого ранее было заболевание.
Термин “профилактическое лечение” или “превентивное лечение” означает такое лечение, которое должно предотвратить возникновение заболевания у индивида. Термины “профилактическое лечение” или “превентивное лечение” применяются здесь взаимозаменяемым образом.
Термины “защищать”, “предотвращать”, “профилактическое”, “превентивное” или “защитное” относятся к предупреждению и/или лечению возникновения и/или распространения заболевания, напр., опухоли, у индивида. Например, профилактическое проведение иммунотерапии, напр., путем введения композиции по настоящему изобретению, может защитить получающего индивида от возникновения опухоли. Например, терапевтическое проведение иммунотерапии, напр., путем введения композиции по настоящему изобретению, может остановить развитие заболевания, напр., привести к торможению прогрессирования/роста опухоли. Это включает и замедление прогрессирования/роста опухоли, в частности, нарушение прогрессирования опухоли, которое предпочтительно приводит к устранению опухоли. Терапевтическое проведение иммунотерапии может защитить индивида, к примеру, от распространения или метастазирования существующих опухолей.
Термин “индивид” или “субъект” относится к позвоночным, в особенности к млекопитающим. Например, млекопитающими в контексте настоящего изобретения являются люди, другие приматы, такие домашние млекопитающие, как собаки, кошки, овцы, крупный рогатый скот, козы, свиньи, лошади и др., такие лабораторные животные, как мыши, крысы, кролики, морские свинки и др., а также животные в неволе типа животных в зоопарке. Термин “субъект” также относится к позвоночным, не являющимся млекопитающими, как-то птицам (особенно к таким домашним птицам, как куры, утки, гуси, индейки) и рыбам (особенно к культивируемым рыбам, напр., лососям или сомам). Термин “животные” в настоящем изобретении также включает людей.
Такие средства, как описанные здесь полиплексные частицы, можно вводить в виде любой подходящей фармацевтической композиции. Термин “фармацевтическая композиция” относится к препаратам, содержащим терапевтически эффективное средство или его соль, предпочтительно вместе с такими фармацевтическими наполнителями, как буферы, консерванты и модификаторы тоничности. Такие фармацевтические композиции применимы для лечения, профилактики или уменьшения тяжести заболевания или расстройства путем введения данной фармацевтической композиции индивидам. Фармацевтические композиции также известны в данной области как фармацевтические препараты. Фармацевтические композиции можно вводить локально или системно. В контексте настоящего изобретения фармацевтические композиции содержат описанные здесь частицы.
Термин “системное введение” означает введение терапевтически эффективного средства так, чтобы оно широко распространялось в организме индивида в значительном количестве и проявляло биологический эффект. Согласно настоящему изобретению, введение предпочтительно осуществляется путем парентерального введения.
Термин “парентеральное введение” означает введение терапевтически эффективного средства так, чтобы оно не проходило через кишечник. Термин “парентеральное введение” включает внутривенное введение, подкожное введение, внутрикожное введение или внутриартериальное введение, но не ограничивается этим.
В одном особенно предпочтительном воплощении композиции по настоящему изобретению вводятся в мышечную ткань типа скелетных мышц. Таким образом, внутримышечное введение типа внутримышечной инъекции является предпочтительным способом введения.
Введение может осуществляться различными способами. В одном воплощении композиции по настоящему изобретению вводятся путем инъекции. В предпочтительном воплощении инъекции проводятся через иглу. В качестве альтернативы можно применять безыгольные инъекции.
Фармацевтические композиции по настоящему изобретению могут содержать по меньшей мере один адъювант. Термин “адъювант” относится к соединениям, которые при введении индивиду в комбинации с антигеном или антигенным пептидом пролонгируют или усиливают или ускоряют иммунный ответ. Предполагается, что адъюванты проявляют свою биологическую активность по одному или нескольким механизмам, включая увеличение поверхности антигена, продление удержания антигена в организме, замедление высвобождения антигена, наведение антигена в макрофаги, усиление поглощения антигена, усиление процессинга антигена, стимуляцию высвобождения цитокинов, стимуляцию и активацию иммунных клеток типа B-клеток, макрофагов, дендритных клеток, T-клеток и неспецифическую активацию иммунных клеток. Адъюванты составляют гетерогенную группу соединений типа масляных эмульсий (напр., адъюванты Фрейнда), минеральных соединений (типа квасцов), бактериальных продуктов (типа токсина Bordetella pertussis) или иммуностимулирующих комплексов. Примеры адъювантов включают сапонины, неполные адъюванты Фрейнда, полные адъюванты Фрейнда, токоферол или квасцы, но не ограничиваются этим.
Фармацевтические композиции по настоящему изобретению обычно применяется
в “фармацевтически эффективном количестве” и в виде “фармацевтически приемлемых препаратов”.
Термин “фармацевтически эффективное количество” означает такое количество, которое дает требуемую реакцию или требуемый эффект само по себе или вместе с дополнительными дозами. В случае лечения конкретного заболевания требуемая реакция предпочтительно означает торможение течения заболевания. Это включает замедление течения заболевания, в частности, прерывание или обращение течения заболевания. Требуемая реакция при лечении заболевания также может представлять собой замедление возникновения или предотвращение возникновения данного заболевания или состояния. Эффективное количество описанных здесь композиций будет зависеть от подлежащего лечению заболевания, степени тяжести заболевания, индивидуальных параметров пациента, включая возраст, физиологическое состояние, размер и вес, продолжительности лечения, типа сопутствующей терапии (если она есть), конкретного способа введения и подобных факторов. Соответственно, вводимые дозы описанных здесь композиций могут зависеть от различных таких параметров. В том случае, когда реакция у пациента недостаточна при начальной дозе, можно применять большие дозы (или эффективно большие дозы, достигаемые при другом, более локализованном способе введения).
Термин “фармацевтически приемлемый” относится к нетоксичности материала, который не нарушает действие активного компонента фармацевтической композиции.
Фармацевтические композиции по настоящему изобретению могут содержать соли, буферы, консерванты, носители и необязательно другие терапевтические средства. Предпочтительно фармацевтические композиции по настоящему изобретению содержат один или несколько фармацевтически приемлемых носителей, разбавителей и/или наполнителей.
Термин “наполнитель” служит для обозначения всех веществ в фармацевтической композиции, которые не являются активными ингредиентами, как-то связующих, смазывающих веществ, загустителей, поверхностно-активных веществ, консервантов, эмульгаторов, буферов, ароматизаторов или красителей.
Термин “разбавитель” относится к разбавителям и/или разжижающим веществам. Кроме того, термин “разбавитель” включает любые текучие, жидкие или твердые суспендирующие и/или смесительные среды.
Термин “носитель” относится к таким совместимым твердым или жидким заполнителям или разбавителям, которые подходят для введения людям. Термин “носитель” относится к таким природным или синтетическим органическим или неорганическим компонентам, которые объединяют с активным компонентом для того, чтобы облегчить применение активного компонента. Предпочтительно компонентами носителя являются стерильные жидкости типа воды или масла, включая те, что происходят из минерального масла, животных или растений, как-то арахисовое масло, соевое масло, кунжутное масло, подсолнечное масло и др. В качестве водных соединений-носителей также можно использовать солевые растворы и водные растворы декстрозы и глицерина.
Фармацевтически приемлемые носители или разбавители для терапевтического применения хорошо известны в области фармацевтики и описаны, к примеру, в Remington’s Pharmaceutical Sciences, Mack Publishing Co. (A.R Gennaro, edit. 1985). Примеры подходящих носителей включают, к примеру, карбонат магния, стеарат магния, тальк, сахар, лактозу, пектин, декстрин, крахмал, желатин, трагакант, метилцеллюлозу, натриевую карбоксиметилцеллюлозу, легкоплавкий воск, масло какао и т.п. Примеры подходящих разбавителей включают этанол, глицерин и воду.
Фармацевтические носители, наполнители или разбавители можно выбирать с учетом предполагаемого способа введения и стандартной фармацевтической практики. Фармацевтические композиции по настоящему изобретению могут включать в качестве или в дополнение к носителям, эксципиентам или разбавителям любые подходящие связующие, смазывающие, суспендирующие вещества, покровные вещества и/или солюбилизирующие вещества. Примеры подходящих связующих включают крахмал, желатин, такие природные сахара, как глюкоза, безводная лактоза, сыпучая лактоза, бета-лактоза, кукурузные подсластители, такие натуральные и синтетические камеди, как гуммиарабик, трагакант или альгинат натрия, карбоксиметилцеллюлоза и полиэтиленгликоль. Примеры подходящих смазывающих веществ включают олеат натрия, стеарат натрия, стеарат магния, бензоат натрия, ацетат натрия, хлорид натрия и т.п. В фармацевтических композициях могут находиться консерванты, стабилизаторы, красители и даже ароматизаторы. Примеры консервантов включают бензоат натрия, сорбиновую кислоту и сложные эфиры п-гидроксибензойной кислоты. Также можно использовать антиоксиданты и суспендирующие вещества.
В одном воплощении композиции представляют собой водные композиции. Водные композиция могут необязательно содержать растворенные вещества, напр., соли. В одном воплощении композиции имеют вид лиофилизованных композиций. Лиофилизованные композиции могут быть получены путем лиофилизации соответствующей водной композиции.
Приведенные здесь средства и композиции могут применяться по отдельности или в сочетании с другими схемами лечения, такими как хирургия, облучение, химиотерапия и/или трансплантация костного мозга (аутологичная, сингенная, аллогенная или неродственная).
Настоящее изобретение описано подробно и проиллюстрировано на фигурах и примерах, которые приводятся только в целях иллюстрации и не должны быть ограничительными. Благодаря описанию и примерам, специалистам доступны дополнительные воплощения, которые также включены в изобретение.
ПРИМЕРЫ
Пример 1. Токсичность полиплексов in vitro
Материалы и методы
Реагент in vivo-jetPEI™, кат. № 201-50G, приобретали у Polyplus-Transfection (Illkirch, Франция). In vivo-jetPEI™ поставляется при 150 мМ (выражается по концентрации остатков азота) в стерильной апирогенной воде. JetPEI разбавляли в буфере 10 мМ HEPES, pH 7,1, 5% глюкозы (HBGx1) до требуемой концентрации (выраженной по концентрации остатков азота).
Анализ цитотоксичности in vitro
Клетки HEK-293 высеивали в 96-луночный планшет (плоскодонный) в концентрации 2×104 клеток на лунку. Клетки поддерживали при 37°C и 7,5% CO2. Через 24 часа супернатанты отбрасывали и заменяли на 50 мкл среды DMEM (+ 10% FCS). PEI разбавляли (1:5) в среде RPMI с 10% FCS и преинкубировали в течение ~15 мин. Затем к клеткам добавляли 50 мкл раствора PEI до конечного объема среды 100 мкл. Еще через 18 ч проводили XTT-анализ (набор для жизнеспособности клеток XTT #9095, New England Biolabs GmbH, Frankfurt, Германия) в соответствии с приведенными инструкциями. Данные по гибели клеток в зависимости от концентрации PEI аппроксимировали по сигмоидальному уравнению:
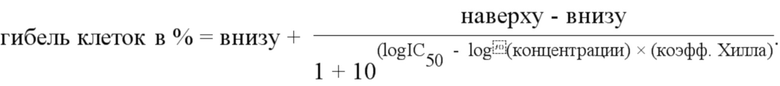
На фиг. 1A представлена токсичность свободного чистого PEI на клетках HEK-293 in vitro. IC50 = 77 мкМ азота (свободного). На фиг. 1B представлена токсичность полиплексов PEI/РНК-репликон на клетках HEK-293 in vitro. IC50 = 542 мкМ азота (препарата полиплексов).
Результаты и выводы
Свободный PEI приводит к гибели клеток при концентрации азота более 18 мкМ (фиг. 1A). Конечная концентрация свободного PEI в клеточной среде должна быть ниже вышеуказанного предела, чтобы избежать проблем с токсичностью.
Полиплексы вызывают меньшую гибель клеток, чем свободный PEI, но их токсичность тоже повышается с повышением концентрации PEI (фиг. 1B). Гибель клеток после добавления полиплексов можно рассчитать по уравнению:
гибель клеток в % = 21,13 + 78,59/(1+10((-0,27-log(концентрации))×3,13)).
Для полиплексов с N/P = 11,6 (концентрация РНК 10 мг/л, PEI = 348 мкМ) гибель клеток составляет 36,8%, тогда как для полиплексов с N/P = 15,8 гибель клеток составляет 52,3%.
Пример 2. Исследование стабильности полиплексов
Материалы и методы
Использовали реагент in vivo-jetPEI™ из примера 1. РНК, кодирующая люциферазу, репликон Construct D1-824, ID R076 1, была предоставлена Отделом биохимии РНК (BioNTech RNA Pharmaceuticals GmbH, Mainz, Германия).
Получение полиплексов
Перед получением уравновешивали in vivo-jetPEI™ и раствор сахара при комнатной температуре. Получение комплексов in vivo-jetPEI™/РНК проводили в ламинарном шкафу, используя стерильные растворы сахара (табл. 1). Конечная концентрация сахара в препаратах составляла 5-10% вес./об..
Все препараты готовили при концентрации РНК 250 мг/л и соотношении N/P 11,6.
Стадии получения:
1. Разбавляли РНК концентрированным сахарным буфером (HBGx2, MBGx2 или HBTx2), получая раствор в ½ конечного объема. Плавно перемешивали на вибромешалке.
2. Разбавляли реагент in vivo-jetPEI™ тем же сахарным буфером и стерильной водой, получая раствор в 3/5 конечного объема. Плавно перемешивали на вибромешалке.
3. В разбавленную РНК быстро добавляли половину объема разбавленного in vivo jetPEI™ всю сразу и плавно перемешивали на вибромешалке.
4. Инкубировали полиплексы в течение 15-20 мин при комнатной температуре, а затем переводили в соответствующие условия хранения.
Таблица 1. Среды для получения и хранения полиплексов
Лиофилизация полиплексов
Препараты помещали в коллекторы настольной сублимационной сушки Epsilon 2-4 LSCplus (Martin Christ Gefriertrocknungsanlagen GmbH, Osterode, Германия).
Замораживали образцы до -40°C (~2°C/мин) при 1 атм. Это занимало 60-90 мин.
Снижали давление до 0,2 атм. в течение 10 мин при -40°C.
Сушили образцы в течение 4 ч при 0,2 атм. и -40°C.
Нагревали образцы до -16°C при 0,2 атм. в течение 2 ч (полого).
Продолжали сушить образцы в течение 4 ч при -16°C и 0,2 атм.
Снижали давление до 0,01 атм. в течение 10 мин при -16°C.
Продолжали сушить образцы в течение 4 ч при -16°C и 0,01 атм.
Нагревали образцы до 20°C при 0,01 атм. в течение 2 ч (полого).
Продолжали сушить образцы в течение 8 ч при 20°C и 0,01 атм.
Высвобождение РНК из полиплексов
Готовили 12 пробирок по 90 мкл образцов в соответствии с табл. 2.
Таблица 2. Подготовка образцов для измерения целостности и концентрации РНК
В каждую пробирку добавляли 10 мкл гепарина 50 г/л в 500 мМ NaCl. Смесь инкубировали 20 мин при 30°C на качалке со скоростью 300 об/мин.
Целостность РНК
Использовали 5 мкл высвобожденной РНК для измерения на биоанализаторе. РНК смешивали с 5 мкл формамида. Определение целостности РНК проводили на приборе Agilent 2100 Bioanalyzer. Уравновешивали набор Agilent RNA 6000 Nano Kit при комнатной температуре в течение 30 мин. Центрифугировали 400 мкл “матрикса геля Nano Gel” при 1500 g в течение 10 мин. Смешивали 65 мкл супернатанта с 1 мкл хорошо перемешанного “концентрата красителя Nano Dye” и центрифугировали при 15000 g в течение 10 мин, получая “смесь гель-краситель”. Подготовленные образцы денатурировали нагреванием 10 минут при 70°C, а также нагревали ступеньку “Nano Ladder” 2 минуты при такой же температуре. Заполняли чип с помощью блока “Priming Station”, добавляя 9 мкл “смеси гель-краситель” в отмеченное положение G и создавая давление на чип в течение 30 сек. Затем добавляли 9 мкл “смеси гель-краситель” в оба других положения G. Добавляли 7,5 мкл “Маркера” в положение ступеньки и по 5 мкл в каждое положение для образцов. В положение ступеньки добавляли денатурированную “Nano Ladder” (1,5 мкл), а затем по 1 мкл образца вносили во все 12 лунок для образцов (в неиспользуемые лунки добавляли 1 мкл H2O). Чип помещали на установку IKA Vortex и запускали вибромешалку на 1 мин при 2000 об/мин. Этот чип измеряли на приборе, и пик РНК репликона проявлялся через 47-57 сек. Целостность РНК рассчитывали по программе Expert 2100 методом анализа пятен, выделяя область РНК-репликона.
Относительную целостность РНК в % рассчитывали по следующему уравнению:

Анализ трансфекции in vitro
Клетки C2C12 высеивали в 96-луночный планшет (плоскодонный) в концентрации 2×104 клеток на лунку. Клетки поддерживали при 37°C и 7,5% CO2. Через 24 часа супернатанты отбрасывали и заменяли на 50 мкл среды DMEM (+ 10% FCS). Полиплексы разбавляли (1:5) в среде RPMI с 10% FCS и преинкубировали в течение ~15 мин. Затем к клеткам добавляли 50 мкл раствора полиплекса до конечного объема среды 100 мкл. Еще через 48 ч проводили анализ на люциферазу Bright-Glo™ (кат. № E2610, Promega GmbH, Mannheim, Германия) в соответствии с приведенными инструкциями.
Относительную люминесценцию в % рассчитывали по следующему уравнению:
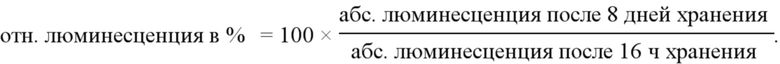
На фиг. 2 представлена относительная люминесценция от мышечных клеток C2C12 после инкубации с полиплексами PEI/РНК репликона при N/P = 11,6 при различных условиях хранения после 1 недели хранения.
На фиг. 3 представлена относительная целостность РНК в полиплексах PEI/РНК репликона при N/P = 11,6 при различных условиях хранения после 2 недель хранения.
Результаты и выводы
Полиплексы обладают слабой стабильностью при хранении в жидком состоянии
(4 и 25°C). Стабильность полиплексов значительно лучше в твердом состоянии (замороженном или лиофилизованном), чем в жидком состоянии. Стабильность полиплексов при хранении в буфере HBT+EDTA в твердом состоянии значительно лучше, чем при хранении в буфере HBGx1.
Для полиплексов в HBT+EDTA возможна стабильность при длительном хранении в твердом состоянии, тогда как для полиплексов в HBGx1 стабильность при длительном хранении в жидком состоянии очень маловероятна.
Препараты полиплексов с РНК репликона можно стабилизировать добавлением 5-20% (вес./об.) трегалозы и от 80 мкМ до 5 мМ EDTA.
Пример 3. Описание расчета м.в. линейных PEI
Линейные PEI синтезируют из 2-этил-2-оксазолина в две стадии. Сначала получают поли(2-этил-2-оксазолин) путем изомеризационной полимеризации с размыканием цикла 2-этил-2-оксазолина в присутствии инициаторов (фиг. 4). Затем гидролизуют PEOX (N-пропионил-PEI) кислотой с отщеплением N-пропионильных групп, получая PEI (фиг. 5).
Полное деацилирование PEOX (N-пропионил-PEI) с молекулярной массой 50 кДа дает линейный PEI с молекулярной массой 22 кДа. Определение молекулярной массы промежуточного продукта (PEOX) проводили методом гель-проникающей хроматографии с детектором показателя преломления или многоугольным детектором рассеяния света. Полные технические подробности описаны в: Adib, Abdennaji, Fabrice Stock, and Patrick Erbacher. “Method for Manufacturing Linear Polyethylenimine (PEI) for Transfection Purpose and Linear PEI Obtained with Such Method.” U.S. Patent Application No. 12/671.312.
Согласно изобретению, PEI, синтезированные из PEOX в диапазоне молекулярных масс 40-60 кДа, являются сильными реагентами для трансфекции РНК репликона.
PEI для применения по изобретению можно приобрести: in vivo JetPEI у Polyplus-Transfection SA (Illkirch-Graffenstaden, Франция), PEI MAX 40000 у Polysciences Europe GmbH (Eppelheim, Германия) и Exgen 500 у Euromedex (Souffelweyersheim, Франция).
Пример 4. Кинетика агрегации полиплексов
Материалы и методы
Полиплексы получали в HBGx1 при концентрации РНК в 200 мг/л, как описано ранее в Примере 2. Их разбавляли в HBGx1 и фосфатно-солевом буфере pH 7,4 (PBS) до концентрации РНК в 10 мг/л. Для повышения ионной силы раствора использовали PBS. 96-луночный планшет очищали фильтрованным воздухом перед внесением разбавленных полиплексов на планшет. Размеры измеряли на считывающем устройстве DynaPro II фирмы Wyatt Technology GmbH (Dernbach, Германия). Для расчета размеров мономодальных образцов использовали кумулянтную аппроксимацию, а для мультимодальных – метод регуляризации.
На фиг. 6 представлена кинетика агрегации полиплексов JetPEI с IVT-РНК (A) и РНК репликона (B) при возрастающих концентрациях соли.
Результаты и выводы
Повышение ионной силы приводит к увеличению размера полиплексов (фиг. 6). Ионная сила в препаратах полиплексов должна составлять ≤20 мМ, чтобы предотвратить агрегацию полиплексов.
Пример 5. Стерилизация полиплексов фильтрованием
Материалы и методы
Полиплексы готовили при концентрации РНК 100 мг/л и соотношении N/P = 11,5, 13,5 и 15,5, как описано ранее в разделе «Исследование стабильности полиплексов».
Концентрация и целостность РНК
РНК высвобождали из полиплексов путем инкубации с гепарином. Смешивали 90 мкл свободной РНК (100 мг/л) или полиплексов с 10 мкл гепарина 20 г/л в 10 мМ HEPES, pH 7,4, 1 мМ EDTA. Смесь инкубировали 20 мин при 30°C на вибромешалке. Пять мкл этой смеси смешивали с 5 мкл формамида. Затем измеряли целостность РНК на биоанализаторе с пико-чипами. Целостность РНК рассчитывали, как описано в примере 2. Концентрацию РНК измеряли методом Ribogreen с помощью набора “Quant-iT RiboGreen RNA Reagent and Kit” (кат. № R11490, Thermo Fischer Scientific) в соответствии с инструкциями производителя для метода с высокой чувствительностью. Вкратце, смесь полиплексов с гепарином инкубировали с флуорофором Ribogreen в буфере 10 мМ трис, рН 7,5, 1 мМ EDTA. Флуоресценцию Ribogreen измеряли при длине волны возбуждения 485 нм и излучения 535 нм.
Концентрация PEI
Готовили раствор 23 мг CuSO4 (безводного) в 100 мл 0,1 М Na-ацетата, pH 5,4 (реагент CSS). 600 мкл реагента CSS смешивали с полиплексами и инкубировали 5 мин при окружающей температуре. Измеряли поглощение каждого раствора при 285 нм на УФ-спектрофотометре в кювете на 1 см против холостой пробы. Составляли калибровочную кривую по известным концентрациям PEI (0-1,55 мМ) и использовали её для расчета концентрации PEI в неизвестных образцах. Из всех образцов вычитали фоновое поглощение свободной РНК.
Измерение электрофоретической подвижности (µ)
Полиплексы разбавляли в HBGx1 до концентрации РНК 20 мг/л в 3,1 мл. Затем их центрифугировали при 600 g в течение 2 мин. Для каждого препарата готовили три образца по 1,05 мл в пластиковых кюветах. Электрофоретическую подвижность полиплексов измеряли методом лазерно-доплеровского электрофореза на приборе ζ-Wallis (Corduan Technologies, Франция). Проводили измерения со средним разрешением по 1 серии из 10 прогонов для каждого образца. Измерения с низким отношением сигнал/шум или с экстремальными значениями µ (> 3 или < -3 мкм×см/В×См) исключали из окончательного анализа. Все препараты измеряли в трех повторах.
Фильтрование полиплексов
Готовили 3 пробирки по 2,68 мл полиплексов при трех различных соотношениях N/P. Полиплексы (1,34 мл) фильтровали через стерильные шприц-фильтры Millex-GP Med с порами на 220 нм (кат. № SLMPL25SS, Merck Millipore). Измеряли физико-химические свойства до и после фильтрования.
На фиг. 7 представлены физико-химические параметры полиплексов до (исходно) и после (фильтр.) фильтрования. A и B. Из полиплексов высвобождали РНК гепарином. Затем измеряли целостность РНК-репликона (А) методом капиллярного электрофореза на биоанализаторе. А концентрацию РНК-репликона (B) измеряли по флуоресценции Ribogreen. C. Концентрацию PEI измеряли методом с CuSO4. D. Электрофоретическую подвижность (µ) измеряли методом лазерно-доплеровского электрофореза.
Результаты и выводы
Шприц-фильтровые элементы являются подходящим методом для стерилизации полиплексов. Для стерилизации фильтрованием полиплексы должны быть небольшими <120 нм. При соотношении N/P 11,6 физико-химические свойства полиплексов не изменяются при фильтровании. А при повышении соотношения N/P свыше 11,6 происходит потеря РНК во время фильтрования.
Пример 6. Влияние чистоты PEI на трансфекцию препаратов РНК репликона/PEI
Материалы и методы
Приобретали следующие PEI высокой чистоты: in vivo JetPEI у фирмы Polyplus-Transfection SA (Illkirch-Graffenstaden, Франция) и PEI MAX 40000 у фирмы Polysciences Europe GmbH (Eppelheim, Германия).
Приобретали следующий PEI обычной чистоты: PEI 25000 у фирмы Polysciences Europe GmbH (Eppelheim, Германия).
Полиплексы готовили в HBGx1 при концентрации РНК 100 мг/л, как описано ранее в Примере 2.
Трансфекция in vivo
Мышей анестезировали изофлураном, брили заднюю сторону задних лап и дезинфицировали 70% раствором EtOH. В мышцу tibialis posterior вводили 20 мкл исследуемого препарата инсулиновым шприцем, снабженным канюлей размером 30G. Мышей наблюдали до прихода в сознание на признаки боли, страдания и дистресса.
В день измерения мышам вводили внутрибрюшинно раствор люциферина. После этого мышей анестезировали изофлураном и помещали на нагревательный мат (37°C) внутри камеры для визуализации IVIS® Spectrum (Perkin Elmer) с постоянной подачей изофлурана/кислорода через индивидуальные маски для анестезии. Через 5 мин после введения люциферина проводили детектирование биолюминесценции на протяжении 1 мин с помощью камеры. Полученные изображения анализировали с помощью программы “LivingImage” (Perkin Elmer).
На фиг. 8 представлено сравнение химической структуры PEI высокой чистоты и PEI обычной чистоты. n = 58 для PEI в 25 кДа. Среднее число мономеров -CH2CH2NH- у PEI в 25 кДа составляет 581, что также является длиной непрерывного отрезка потенциально протонируемых атомов азота. Предполагая равномерное распределение N-пропиониловых групп в обычном PEI25, непрерывный отрезок протонируемых азотов у него составляет всего лишь 64.
На фиг. 9 представлена трансфекция мышечных клеток C2C12 in vitro полиплексами с РНК репликона, которые получали с использованием PEI различного уровня чистоты при различных соотношениях N/P.
Согласно фиг. 10, получали полиплексы с РНК репликона при соотношениях N/P = 1 (-) и 11,6 (+) с PEI высокой чистоты (jetPEI) и PEI обычной чистоты (25 кДа). В качестве контроля использовали свободную РНК. Препараты вводили внутримышечно (в/м) в задние конечности мышей (n = 3). Регистрировали сигналы люминесценции из мышц мышей.
Результаты и выводы
Полиплексы, которые получали с использованием PEI высокой чистоты, трансфецируют мышечные клетки in vitro лучше, чем PEI обычной чистоты. Полиплексы проявляют наибольшую эффективность трансфекции in vivo при N/P 11,6 после в/м введения.
Катионные полиплексы с PEI высокой чистоты при N/P 11,6 могут трансфицировать мышечную ткань in vivo значительно лучше (разница в 4-5 раз), чем свободная РНК репликона.
Анионные полиплексы с PEI высокой чистоты при N/P = 1, полиплексы с PEI обычной чистоты при N/P = 1 и катионные полиплексы с PEI обычной чистоты при N/P 11,6 трансфецируют мышечную ткань хуже, чем свободная РНК репликона.
Пример 7. Высокая эффективность трансфекции РНК-репликона очищенными PEI от разных поставщиков и лиофилизованными полиплексами
Материалы и методы
Приобретали следующие PEI высокой чистоты: in vivo JetPEI у фирмы Polyplus-Transfection SA (Illkirch-Graffenstaden, Франция), Exgen 500 у фирмы Euromedex (Souffelweyersheim, Франция) и PEI MAX 40000 у фирмы Polysciences Europe GmbH (Eppelheim, Германия).
Полиплексы получали в HBGx1 при концентрации РНК в 100 мг/л, как описано ранее в Примере 2. Все препараты готовили в буфере HBGx1, за исключением лиофилизованного препарата, который готовили в буфере HBTx1. Лиофилизацию полиплексов в буфере HBTx1 проводили, как описано ранее в Примере 2. Препараты вводили в/м в задние конечности мышей (n = 3) и регистрировали сигналы люменесценции из мышц мышей, как описано ранее в Примере 6.
Для эксперимента на фиг. 11, полиплексы с РНК репликона получали при соотношениях N/P 7,7 и 11,6 с PEI высокой чистоты: jetPEI (фирмы Polyplus), PEI-Max 40000 (фирмы Polyscience) и Exgen 500 (фирмы Euromedex).
Таблица 3. Критерий Манна-Уитни, двусторонний, для статистической значимости (значения p) отличий между препаратами на фиг. 11.
На фиг 12 представлены лиофилизованные комки полиплексов JetPEI/РНК репликона, полученные при N/P = 11,6 с различными буферами.
Результаты и выводы
Полиплексы с РНК репликона эффективно трансфецируют мышечную ткань после в/м введения (фиг. 11). Для получения полиплексов можно использовать PEI высокой чистоты: jetPEI (фирмы Polyplus), PEI-Max 40000 (фирмы Polyscience) и Exgen 500 (фирмы Euromedex). Полиплексы с Exgen 500 лучше трансфецируют при N/P 7,7, чем при 11,6.
Лиофилизаты полиплексов в трегалозе имеют комковатую морфологию, тогда как в глюкозе они спадаются и с трудом растворяются в воде (фиг. 12).
Трегалоза предпочтительнее глюкозы для лиофилизации полиплексов. Лиофилизованные полиплексы действуют in vivo аналогично свежеприготовленным жидким полиплексам.
Пример 8. Трансфекция IVT-РНК в мышечные клетки полиплексами с очищенным PEI
Материалы и методы
Реагент in vivo-jetPEI™, кат. № 201-50G, приобретали у Polyplus-Transfection (Illkirch, Франция). Транскрибированная in vitro (IVT) мРНК, кодирующая люциферазу, Construct pST1-475, была предоставлена Отделом биохимии РНК (BioNTech RNA Pharmaceuticals, Mainz, Германия).
Полиплексы c IVT-РНК и PEI получали, как описано ранее в Примере 2. Различные соотношения N/P получали, поддерживая постоянную концентрацию РНК и увеличивая концентрацию PEI.
Трансфекцию мышечных клеток in vitro проводили, как описано ранее в Примере 2.
Исследования по трансфекции in vivo проводили, как описано ранее в Примере 6.
Согласно фиг. 13, мышечные клетки C2C12 трансфицировали in vitro с помощью IVT-РНК, кодирующей люциферазу. РНК образовывала комплексы с JetPEI при различных соотношениях N/P в буфере HBGx1. Через 24 ч после трансфекции измеряли сигналы люминесценции.
Согласно фиг. 14, получали полиплексы с IVT-РНК при соотношениях N/P = 5,8 и 11,6 с очищенным PEI в буфере HBGx1. В качестве контроля использовали свободную IVT-РНК в буфере HBGx1. Препараты вводили внутримышечно (в/м) в задние конечности мышей (n = 3) в дозах РНК по 2-8 мкг на инъекцию. Через 6 ч после введения регистрировали сигналы люминесценции из мышц мышей.
Результаты и выводы
Полиплексы IVT-РНК с PEI высокой чистоты могут эффективно трансфицировать мышечные клетки in vitro. Существует положительная корреляция между эффективностью трансфекции и соотношением N/P (фиг. 13). Свободная IVT-РНК не трансфецирует клетки in vitro.
Полиплексы с PEI высокой чистоты при соотношениях N/P = 5,8 и 11,6 трансфецируют мышечную ткань in vivo значительно хуже, чем свободная IVT-РНК (фиг. 14).
Пример 9. Влияние размера частиц и условий получения на трансфекцию полиплексами с РНК репликона
Материалы и методы
Полиплексы РНК-репликона и PEI получали, как описано ранее в Примере 2. Полиплексы получали при 5 различных концентрациях РНК: 100, 250, 500, 750, 1000 мг/л. Концентрацию PEI повышали соответствующим образом, чтобы поддерживать соотношение N/P на уровне 11,6 для всех препаратов.
Размер полиплексов измеряли, как описано ранее в Примере 4.
Трансфекцию мышечных клеток in vitro проводили, как описано ранее в Примере 2.
Исследования трансфекции in vivo проводили, как описано ранее в Примере 6.
Согласно фиг. 15, получали полиплексы РНК-репликона и jetPEI при различных концентрациях РНК при соотношении N/P = 11,6 в буфере HBGx1. Для измерения размеров методом DLS полиплексы разбавляли до концентрации РНК 10 мг/л.
Согласно фиг. 16, мышечные клетки C2C12 трансфицировали in vitro полиплексами из фиг. 16. Через 24 ч после трансфекции измеряли сигналы люминесценции.
Согласно фиг. 17, получали полиплексы РНК-репликона при соотношении N/P = 11,6 с очищенным PEI в буфере HBGx1 при различных концентрациях РНК, как на фиг. 16. Препараты вводили внутримышечно (в/м) в задние конечности мышей (n = 3) в дозах РНК по 2-8 мкг на инъекцию. Регистрировали сигналы люминесценции из мышц мышей.
Результаты и выводы
Полиплексы являются хорошими средствами для трансфекции РНК-репликона. На биоактивность полиплексов с точки зрения эффективности трансляции РНК в мышечной ткани не влияет концентрация РНК (0,1-1 г/л) во время образования полиплексов.
Размеры полиплексов в диапазоне 60-200 нм не влияют на эффективность трансфекции полиплексов in vitro и in vivo (внутримышечное введение).
Пример 10. Различные полиплексы работают по-разному после подкожного или внутримышечного введения мышам
Материалы и методы
Реагент in vivo-jetPEI™, кат. № 201-50G, приобретали у Polyplus-Transfection (Illkirch, Франция). Линейный PEI в 22 кДа был предоставлен профессором Cheradame (Polytheragene, EVRY cedex, Франция). РНК, кодирующая люциферазу, Construct pST1-475, была предоставлена Отделом биохимии РНК (BioNTech RNA Pharmaceuticals, Mainz, Германия).
Полиплексы с JetPEI получали в HBGx1 при концентрации РНК в 500 мг/л, как описано ранее в Примере 2.
Полиплексы с PEI от Polytheragene получали аналогично процедуре с JetPEI, но с двумя модификациями:
1) для получения полиплексов использовали буфер 10 мМ HEPES, pH 7,4 вместо HBGx1;
2) использовали соотношение N/P = 15,8 вместо 11,6.
Получение полиплексов с PEI от Polytheragene проводили в соответствии со следующей статьей: Démoulins Thomas et al. “Polyethylenimine-based polyplex delivery of self-replicating RNA vaccines.” Nanomedicine: Nanotechnology, Biology and Medicine (2015).
Полиплексы разбавляли в буфере HBGx1 или Opti-MEM (кат. № 31985062, Thermo Fisher Scientific, Schwerte, Германия) до конечной концентрации РНК 5 мг/л для исследований in vitro и 100 мг/л для исследований in vivo.
Исследования in vitro проводили, как описано ранее в Примерах 1 и 2.
Исследования in vivo проводили, как описано ранее в Примере 6.
Результаты и выводы
На фиг. 18 представлены исследования in vitro с полиплексами PEI/РНК репликона на дендритных клетках человека (DC) и мышечных клетках мыши (C2C12) A. Токсичность (выражена в % жизнеспособных клеток после обработки полиплексами). B. Трансфекция (выражена в виде люминесцентного излучения после обработки полиплексами). Результаты трансфекции представлены только для клеток C2C12.
Для результатов на фиг. 19 получали полиплексы РНК-репликона при соотношении N/P = 11,6 или 15,8 с PEI фирмы Polyplus или Polytheragene в буфере HBGx1 или 10 мМ HEPES. Перед введением мышам полиплексы разбавляли в буфере HBGx1 или Opti-MEM. Препараты вводили внутримышечно (в/м) в задние конечности мышей (n = 3) в дозах РНК по 2 мкг на инъекцию. Регистрировали сигналы люминесценции из мышц мышей.
Результаты и выводы
Таблица 4. Сравнение размеров у различных полиплексов
Ионная сила Opti-MEM составляет более 20 мМ. Полиплексы в Opti-MEM быстро агрегируют (табл. 4), поэтому этот состав не подходит для фармацевтической разработки.
Описанные здесь полиплексы (PEI от Polyplus, N/P 11,6, HBGx1) имеют совсем другие характеристики, чем ранее описанные полиплексы (PEI от Polytheragene, N/P 15,8, Opti-MEM). Полиплексы в Opti-MEM имеют диаметр >1000 нм и мультимодальное распределение по размерам. Описанные здесь полиплексы имеют диаметр ~100 нм и низкую полидисперсность.
In vitro все полиплексы более токсичны для дендритных клеток, чем для мышечных клеток. Вероятно, дендритные клетки после подкожного введения поглощают полиплексы, тогда как мышечные клетки после внутримышечного введения трансфецируются полиплексами.
Трансфекция мышечных клеток in vitro описанными здесь полипеплексами аналогична ранее описанным полиплексам.
Трансфекция in vivo описанными здесь полипеплексами отличается от ранее описанных полиплексов и зависит от способа введения. Описанные здесь полиплексы хорошо трансфецируются после внутримышечного введения и хуже после подкожного введения. Ранее описанные полиплексы вообще не трансфецируются после внутримышечного введения, хотя после подкожного введения и наблюдается слабый сигнал.
Пример 11. Введение мышам РНК-репликона внутримышечно в сравнении с интрадермальным введением
РНК-репликон, кодирующий фермент люциферазу, растворяли в буфере HBGx1 или получали комплекс в полиплексах, как описано в примере 2. Эти препараты вводили внутримышечно в мышцы tibialis posterior мышей Balb/c в дозах по 2 мкг РНК. Через 4, 7 и 10 дней после введения мышей анастезировали изофлураном. Затем им вводили субстрат люциферин и регистрировали излучение люминесценции из мышц CCD-камерой.
Регистрировали исходящие из белка люциферазы фотоны в течение одной минуты, что показано в виде наложения на фотографию визуализируемых мышей (фиг. 20A). На фиг. 20B представлено графическое отображение измеряемых фотонов за секунду (p/s) на месте инъекции.
Через 7 дней после интрадермального (и/д) введения по 2 мкг не составленной (HBGx1) или составленной РНК репликона, кодирующей люциферазу, в два места инъекции на дорсальной коже мышей Balb/c, животных подвергали неинвазивной визуализации по биолюминесценции in vivo. Регистрировали исходящие из белка люциферазы фотоны в течение одной минуты, что показано в виде наложения на фотографию визуализируемых мышей. Черными стрелками указано место инъекции (фиг. 21A). На фиг. 21B представлено графическое отображение измеряемых фотонов за секунду (p/s) в месте инъекции.
Пример 12. Благотворное влияние препарата РНК в качестве вакцины
Мышей иммунизировали дважды в дни испытания 0 и 21 композицией одноцепочечной РНК репликона, кодирующей гемагглютинин (HA) вируса гриппа H1N1 штамма A/PuertoRico/8/1934 (H1N1/PR8), составленной с PEI при соотношении N/P 11,6, либо не составленной одноцепочечной РНК. Все животные получали дозу в 1,25 мкг РНК. Третья группа получала только физраствор в качестве группы контроля на буфер. Как видно из фиг. 22A, через 19 дней после первой иммунизации и незадолго до второй иммунизации у всех животных, получавших составленную РНК, развивался иммунный ответ против HA при анализе методом нейтрализации вируса (VNT, предел обнаружения 1280). Напротив, только у 5 из 8 животных, получавших несоставленную РНК, произошла сероконверсия против HA. Как видно из фиг. 22B, через 35 дней после первой иммунизации все получавшие РНК животные были положительными на HA-специфические антитела, а титр антител против HA повышался. Титры у животных в группе составленной РНК, но не в группе с несоставленной РНК, значительно повышались по сравнению с группой контроля на физраствор. Как видно из фиг. 22C, через 54 дня после иммунизации у мышей, получавших 1,25 мкг составленной РНК, кодирующей HA H1N1/PR8, развивался значительно более высокий титр антител по сравнению с животными, получавшими 1,25 мкг несоставленной РНК, кодирующей HA H1N1/PR8 (значимость рассчитывали односторонним методом ANOVA; * p <0,001).
Мышей иммунизировали дважды в дни испытания 0 и 21 композицией одноцепочечной РНК репликона, кодирующей гемагглютинин (HA) вируса гриппа H1N1 штамма A/PuertoRico/8/1934 (H1N1/PR8), составленной с PEI при соотношении N/P 11,6, либо не составленной одноцепочечной РНК. Все животные получали 0,25 мкг РНК. Третья группа получала только физраствор в качестве группы контроля на буфер. Как видно из фиг. 23A, через 54 дня после иммунизации все получавшие РНК животные были положительными на HA-специфические антитела, а титры антител в группе составленной РНК значительно повышались по сравнению с группой контроля на физраствор и с животными, получавшими 0,25 мкг несоставленной РНК, кодирующей HA H1N1/PR8 (значимость рассчитывали односторонним методом ANOVA; ** р <0,001; * р <0,05). Через 55 дней после иммунизации всех мышей инфицировали 10-кратной средней летальной дозой (MLD50) H1N1/PR8. Наблюдали за выживанием. Как видно из фиг. 23B, все контрольные животные погибли в пределах 8 дней. После контрольного заражения выжили 4 из 5 животных, получавших несоставленную РНК, кодирующую HA H1N1/PR8. Напротив, все мыши, получавшие составленную РНК, кодирующую HA H1N1/PR8, выжили после контрольного заражения, что свидетельствует о благотворном эффекте композиции РНК/полиалкиленимин.
Пример 13. Распылительная сушка РНК репликона в составе с PEI
Получали препарат с соотношением N:P = 12, дважды следуя схеме пипетирования, приведенной в табл. 1, с последующим объединением полученного материала. При этом получили 5,0 мл полиплексов РНК репликона с концентрацией РНК в 0,1 мг/мл.
Высушивали распылением 3,5 мл этого препарата и получили 533 мг материала (выход: 76,1%). Размер частиц свежеприготовленных полиплексов РНК репликона не определяли, так как материал был выгружен в демонстрационной лаборатории Büchi. После восстановления водой (смесь: 20 мг высушенных распылением полиплексов и 200 мкл воды для инъекций (wfi), что дает 10 мМ трегалозы и концентрацию РНК в 0,1 мг/мл) размер частиц (z-среднее) составлял 289 нм, а PDI составлял 0,238 (Nicomp, 15 мин). На фиг. 24A представлено исследование in vitro экспрессии люциферазы из полиплексов saRNA до и после распылительной сушки. Люциферазная активность saRNA не теряется
в процессе распылительной сушки. Различия в абсолютной высоте сигналов обусловлены колебаниями метода.
В дополнительном эксперименте некомплексованную мРНК в 10% (вес./об.) трегалозе высушивали распылением и исследовали целостность мРНК после распылительной сушки методом капиллярного электрофореза.
Смешивали 2,50 мл мРНК (R36-05.2-DP; c(РНК) = 0,5 мг/мл; 10 мМ HEPES; 0,1 мМ EDTA; pH 7,0) с 2,50 мл 20% (вес./об.) раствора трегалозы (соотношение объемов 1:1), получая следующий состав: c(РНК) = 0,25 мг/мл; 10% (вес./об.) трегалозы; 5 мМ HEPES; 0,05 мМ EDTA. Во время распылительной сушки материал образца держали на льду. В целом 2,5 мл этого раствора сушили распылением. Во время распылительной сушки температура на выходе повышалась с 33 до 37°C за 10 минут. Чтобы уменьшить повышение температуры, расход газа повышали с 98 до 101 л/мин. Во время распылительной сушки температура еще повышалась до 41°C за следующие 60 минут. После полного распыления было получено 165 мг высушенного материала (выход: 66,0%). Для анализа целостности РНК материал растворяли в воде wfi (смесь: 20 мг высушенной распылением РНК и 200 мкл воды wfi, что дает содержание трегалозы в 10% и концентрацию РНК в 0,25 мг/мл). Растворенную РНК анализировали на биоанализаторе Agilent 2100. Результаты этих анализов представлены в виде электрофоретограммы высушенной распылением РНК на фиг. 24B.
Было показано, что сохраняется целостность некомплексованной РНК, а распылительная сушка не приводит к измеримой деградации РНК.
Эксперименты по сушке распылением проводили следующим образом.
Для распылительной сушки содержащих РНК составов использовали Nano Spray Dryer B-90 фирмы Büchi. Для приготовления этих составов использовали следующие соединения:
• мРНК (R36-05.2-DP; c(РНК) = 0,5 мг/мл; 10 мМ HEPES; 0,1 мМ EDTA; pH 7,0)
• РНК репликона, кодирующая люциферазу (D2 RNA-A1310 29-01 pST1-SFV4-TRON-I2m2-A30L70, wfi, c(РНК) = 1 мг/мл)
• вода для инъекций (wfi)
• NaCl (1,5 М в воде wfi)
• трегалоза 20% (вес./об.) в воде wfi (Pfanstiehl, № партии: 35261A)
• забуференная HEPES 2×трегалоза, 20% (вес./об.) в воде wfi (Pfanstiehl, № партии: 35261A, 20 мМ HEPES)
• JetPEI (Polyplus; № партии: 13081A1S; 150 мМ азота).
Для распылительной сушки были выбраны следующие параметры процесса:
• форсунка: 4 мкм
• температура на входе: 80°C
• температура на выходе: 30°C
• расход газа: 98 мл/мин
• подаваемый ток: 15000 В; 350 мкА.
Перед всеми экспериментами установку очищали с помощью РНКазы Zapp и протирали этанолом, а крышку очищали в ультразвуковой ванне.
Пример 14. Микрофлюидика для производства полиплексов
Использовали NanoAssemblr™ (Precision Nanosystems, BC, Vancouver, Канада) с микрожидкостным чипом, предоставленным производителем (1029-036). В качестве РНК использовали РНК репликона, произведенную на месте (партия № R071_1_2). Полиэтиленимин (Max PEI 40) был от Polysciences (Eppelheim, Германия). В качестве шприцов использовали BD Plastipak на 1 мл; 1508006, BD Biosciences (Heidelberg, Германия). Два компонента смешивали в соотношении 1:1 при скорости подачи в 12 мл/мин. Образцы готовили в двух разных концентрациях, а именно 50 мг/л и 250 мг/л.
Для измерения размера частиц методом динамического рассеяния света использовали субмикронный анализатор частиц/дзета-потенциала 380 ZLS фирмы Nicomp (PSS Nicomp, Santa Barbara, CA).
Для каждого условия готовили 1,5 мл. Для измерения размеров образцы разбавляли буфером HBG.
Схема пипетирования была следующей:
Эксперименты по микрожидкостному перемешиванию проводили с помощью устройства, содержащего микрожидкостный смеситель Y-типа, где смешиваются два компонента, приведенные в стандартных шприцах. Два компонента смешивали в соотношении 1:1 при постоянной скорости потока 12 мл/мин. Образцы готовили в двух разных концентрациях, а именно 50 мг/л и 250 мг/л, и измеряли размеры частиц полученных полиплексов.
Полиплексы получали с помощью микрожидкостного устройства, что свидетельствует о возможности поточного производства для расширения и производства по GMP. Частицы формировались без проблем и не наблюдалось признаков образования агрегатов или закупоривания. Размеры частиц измеряли методом динамического рассеяния света. Для полиплексов, производимых при 0,05 мг/л, размер составлял около 123 нм, а для частиц, производимых при 0,25 мг/л, размер составлял около 314 нм. Полученные результаты подробно приведены в следующей таблице. Все образцы измеряли при концентрации РНК 0,02 мг/мл (разбавленной HBGx1).
Была показана возможность получения полиплексов при помощи микрофлюидики. Изготовление проводилось с помощью простого смесителя Y-типа и не требовалось никаких особых процедур типа гидродинамической фокусировки для обеспечения бесперебойного производства. При подходящих условиях можно получать частицы размером значительно меньше 200 нм, что позволяет проводить терминальную стерилизацию фильтрованием с помощью общепринятых и соответствующих GMP стерильных фильтров. Поскольку не отмечалось признаков агрегации или закупоривания, то делается вывод о том, что масштабирование до более крупных производственных партий будет возможным без проблем. Другие варианты масштабирования включают запараллеливание нескольких идентичных устройств. В целом, результаты можно принимать как свидетельство об общей осуществимости совместимого с GMP микрофлюидного производства полиплексов PEI/РНК.
Пример 15. Стерилизация полиплексов фильтрованием
Получали полиплексы при концентрации РНК репликона 100 мг/л и соотношении N/P = 11,5, 13,5 и 15,5, как описано ранее в разделе «Исследование стабильности полиплексов».
Готовили три пробирки по 2,68 мл полиплексов при трех различных соотношениях N/P. Полиплексы (1,34 мл) фильтровали через стерильные шприц-фильтры Millex-GP Med с порами на 220 нм (кат. № SLMPL25SS, Merck Millipore).
Полиплексы разбавляли до концентрации РНК 10 мг/л в HBGx1. Затем их разбавляли до концентрации РНК 5 мг/л в 0,9% NaCl за 30 мин до добавления к клеткам.
Клетки C2C12 высеивали в 96-луночный планшет (плоскодонный) в концентрации 2×104 клеток на лунку. Клетки поддерживали при 37°C и 7,5% CO2. Через 24 часа супернатанты отбрасывали и заменяли на 50 мкл среды DMEM (+ 10% FCS). Полиплексы разбавляли (1:5) в среде RPMI с 10% FCS и преинкубировали в течение ~15 мин. Затем к клеткам добавляли 50 мкл раствора полиплекса до конечного объема среды 100 мкл. Еще через 48 ч проводили анализ на люциферазу Bright-Glo™ (кат. № E2610, Promega GmbH, Mannheim, Германия) в соответствии с приведенными инструкциями.
Параллельно с трансфекцией также измеряли количество клеток с помощью набора для пролиферации клеток XTT II (Roche, кат. № 11465015001). При обоих анализах каждый образец полиплексов тестировали в трех биологических повторах. В качестве отрицательного контроля высеивали клетки без обработки (“необработанные”). Для XTT-анализа также высеивали среду без клеток, эквивалентную фону (BG), тоже в трех повторах. Наконец, измеряли люминесценцию (Bright-Glo™) и поглощение (XTT) на считывающем устройстве “Infinite 200pro” (Tecan). Рассчитывали нормализованную люминесценцию путем деления сигнала люминесценции на поглощение (пропорциональное числу клеток).
На фиг. 25 представлена нормализованная люминесценция от мышечных клеток C2C12 после инкубации с полиплексами PEI/РНК репликона при различных соотношениях N/P, до (исходно) и после (стерил.) стерилизации фильтрованием.
Можно сделать вывод, что шприц-фильтровые элементы подходят для стерилизации полиплексов. Эффективность трансфекции полиплексов не изменяется из-за процесса стерилизации.
Пример 16. Оптимизация трансфекции полиплексов путем комбинирования коротких и длинных PEI
Получали комплексы РНК-репликона, кодирующего люциферазу, с различными комбинациями коротких PEI между 0,6 и 11 кДа (напр., линейный короткий PEI в 2,5 кДа или разветвленный короткий PEI в 1,8 кДа) и длинных PEI между 20 и 40 кДа (напр., in vivo-JetPEI в 22 кДа) в различных сочетаниях при общем N/P = 10 или 12 в буфере MBG (конечная концентрация 5% вес./об.. глюкозы, 10 мМ MES, pH 6,1). В качестве эталона использовали in vivo-JetPEI NP12 сам по себе. Комплексообразование полиплексов с короткими и длинными PEI происходило в две стадии. На первой стадии проводили комплексообразование РНК-репликона с in vivo-JetPEI при требующемся N/P. На второй стадии в состав добавляли избыток короткого PEI для достижения требуемого общего соотношения N/P; т.е. первое число означает NP длинного PEI, а второе число – короткого PEI (напр., NP4+8 = NP4 длинного PEI и NP8 короткого PEI). Анализ на люциферазу проводили, как описано ранее в Примере 15.
Результаты представлены на фиг. 26. На фиг. 26A представлена эффективность трансфекции полиплексов с коротким линейным PEI и длинным in vivo-JetPEI при 250 нг РНК на лунку. На фиг. 26B представлена эффективность трансфекции полиплексов с коротким разветвленными PEI и длинным in vivo-JetPEI при 250 нг РНК на лунку.
Вывод: по сравнению с результатами эталона только с длинным PEI (напр., in vivo JetPEI) эффективность трансфекции может значительно улучшиться при использовании комбинаций длинных PEI и коротких PEI.
Интересно, что в отношении комбинаций с линейным коротким PEI сигнал экспрессии люциферазы достигал пика в первые 24 часа после трансфекции, тогда как в отношении комбинаций с разветвленным коротким PEI сигнал экспрессии люциферазы достигал пика в первые 48 часов после трансфекции. Таким образом, в ситуациях, когда требуется экспрессия в определенном временном интервале, то будет разумным тщательный выбор правильной комбинации короткого PEI и длинного PEI.
Пример 17. Оптимизация трансфекции полиплексов за счет снижения количества длинного PEI
Получали комплексы РНК-репликона, кодирующего секретируемую люциферазу, с различными комбинациями коротких PEI (напр., разветвленного в 1,8 кДа) и длинных PEI (напр., in vivo-JetPEI) в различных сочетаниях при общем N/P = 12 в буфере MBG (конечная концентрация 5% вес./об.. глюкозы, 10 мМ MES, pH 6,1). В качестве эталона использовали in vivo-JetPEI NP12. Комплексообразование полиплексов с короткими + длинными PEI происходило в две стадии. На первой стадии проводили комплексообразование РНК-репликона с in vivo-JetPEI при требующемся N/P. На второй стадии в состав добавляли избыток короткого PEI для достижения требуемого общего соотношения N/P; т.е. первое число означает NP длинного PEI, а второе число – короткого PEI (напр., NP4+8 = NP4 длинного PEI и NP8 короткого PEI). Секретируемую люциферазу измеряли в соответствии с протоколом производителя (Nano-GLO, Promega, США) при 125 нг РНК на лунку. Анализ жизнеспособности клеток проводили, как описано ранее в Примере 15.
Результаты представлены на фиг. 27. По сравнению с эталоном и при таком же общем соотношении N/P, более высокие уровни экспрессии достигались с комбинациями, напр., NP4+8 и NP1.15+11. Таким образом, эффективность трансфекции можно значительно повысить путем снижения концентрации длинного PEI в составе и повышения концентрации менее токсичного короткого PEI (напр., разветвленного в 1,8 кДа).
Пример 18. Влияние вариаций соли и/или рН на эффективность трансфекции in vivo
Мышей BALB/c приобретали у Janvier Laboratories и использовали в эксперименте в 8-недельном возрасте. Перед введением указанных тест-препаратов мышей подвергали изофлурановой анестезии и удаляли шерсть с задних лап электрической бритвой. После этого в обе мышцы tibialis posterior вводили полиплексы РНК репликона (saRNA) с PEI (напр., полиплексы saRNA + in vivo JetPEI NP12) в дозах по 2 мкг РНК в общем объеме 20 мкл, по три мыши на группу. Препараты варьировались в отношении pH и/или концентрации соли (напр., NaCl). В указанные моменты времени мышам внутрибрюшинно вводили раствор D-люциферина (100 мг/кг массы тела) и регистрировали биолюминесценцию неинвазивно у анестезированных изофлураном мышей в течение 1 мин на приборе IVIS® Spectrum (Perkin Elmer). На графиках представлены сигналы биолюминесценции в фотонах за секунду [p/s] при определении с помощью программы Living Image® (Perkin Elmer) в задаваемых вручную участках (ROI) мышц мышей (места инъекции) на этих снимках, в общей сложности по 6 значений на группу.
Влияние вариаций соли (напр., NaCl) на эффективность трансфекции представлено на фиг. 28. Внутримышечное введение полиплексов PEI/РНК репликона при различных соотношениях N/P приводило к продолжительным сигналам биолюминесценции в области мышц у мышей при измерении на 3, 6, 9 и 13-й день. Сила детектируемого сигнала возрастала с 3-го до 6-го дня (пик) после введения, но проявлялась даже на 13-й день. Наиболее интенсивный сигнал в области мышц у мышей обнаруживался на 6-й день после в/м введения у мышей, получавших полиплексы РНК репликона с PEI (напр., длинным PEI N/P 12) при добавлении низких концентраций соли (5-10 мМ).
Влияние вариаций pH на эффективность трансфекции представлено на фиг. 29. Хорошие результаты получали с препаратами полиплексов saRNA-PEI со значениями pH между 6,5 и 7,1, предпочтительно от 6,5 до 6,9. Наиболее интенсивный сигнал проявлялся с препаратом РНК репликона с длинным PEI NP12, доведенным до pH 6,5. В качестве эталона использовали полиплексы РНК репликона с JetPEI NP12 без доведения pH (BM) или в HBG (20 мМ HEPES, pH 7,4, 5% мас. глюкозы).
Пример 19. Зависимые от рН эффекты на электрофоретическую подвижность и эффективность трансфекции полиплексов с PEI in vitro
Получали комплексы РНК-репликона, кодирующего люциферазу, с длинным PEI (напр., струйным in vivo-jetPEI) при соотношении N/P = 4 в буфере HBG (конечная концентрация 4,5% вес./об.. глюкозы, 10 мМ HEPES, pH 7,1) при комнатной температуре в течение 15 минут и разделяли на 11 образцов. Доводили pH образцов добавлением HCl или NaOH в зависимости от конечного значения pH при тестировании. Проводили измерения электрофоретической подвижности (µ), как описано в Примере 5. Трансфекцию in vitro мышечных клеток C2C12 мыши, анализы люциферазы и жизнеспособности клеток проводили, как описано ранее в Примере 15.
Зависимые от рН эффекты на электрофоретическую подвижность и эффективность трансфекции полиплексов с PEI in vitro представлены на фиг. 30 и 31. На фиг. 30 представлена электрофоретическая подвижность полипеплексов in vivo jetPEI/РНК-репликона (N/P 4) при доведении до различных значений pH. На фиг. 31 представлена нормализованная люминесценция мышечных клеток C2C12 после инкубации с различными дозами полиплексов in vivo jetPEI/РНК-репликона при соотношении N/P = 4 и различных значениях pH (pH 6,5 – pH 8,5).
Электрофоретическая подвижность полиплексов отрицательно коррелирует с рН. Нейтральные полиплексы получаются примерно при рН 8,9. Повышение плотности положительных зарядов на полиплексах PEI путем снижения общего pH полиплексов, предпочтительно до значений pH от 6,5 до 7,1, приводило к большей эффективности трансфекции клеток C2C12 без влияния на жизнеспособность клеток. Полимеры PEI содержат первичные и вторичные амины, состояние протонирования которых зависит от общего рН. Таким образом, результаты предполагают эффективный способ контроля заряда полиплексов и эффективности их трансфекции путем доведения и оптимизации общего рН.
Пример 20. Влияние избытка положительных зарядов в составах с длинным PEI
Получали комплексы РНК-репликона, кодирующего секретируемую люциферазу, при различных соотношениях N/P от 2 до 12 в буфере MBG (конечная концентрация 5% вес./об.. глюкозы, 10 мМ MES, pH 6,1). Секретируемую люциферазу измеряли в соответствии с протоколом производителя (Nano-GLO, Promega, США) при 125 нг РНК на лунку. Избыток положительных зарядов рассчитывали, исходя из соотношения N/P у РНК-репликона с длинным PEI (т.е. in vivo-JetPEI) и точного соотношения N/P, при котором происходит полное комплексообразование этого РНК-репликона с in vivo-JetPEI. По разности между используемым соотношением N/P и известным соотношением N/P для полного комплексообразования можно рассчитать избыток положительных зарядов в составе.
В качестве примера на фиг. 32 представлены результаты по трансфекции с помощью полиплексов in vivo-JetPEI при 250 нг РНК. В общем, повышение концентрации положительных зарядов в составе экспоненциально пропорционально уровню экспрессии люциферазы. Избыток положительных зарядов вплоть до 30 нМ за счет повышения количества PEI оказался полезным.
Пример 21. Оптимизация трансфекции полиплексов с однокомпонентным PEI посредством 2-стадийного комплексообразования
При использовании только одного варианта PEI (как-то, напр., длинного PEI) эффективность трансфекции можно улучшить посредством 2-стадийного метода комплексообразования. Получали комплексы РНК-репликона, кодирующего секретируемую люциферазу, в две стадии при общем N/P = 12 в буфере MBG (конечная концентрация 5% вес./об.. глюкозы, 10 мМ MES, pH 6,1). В качестве эталона использовали 1-стадийное комплексообразование с in vivo-JetPEI NP12. Например, комплексообразование в 2 стадии с использованием in vivo-JetPEI происходило следующим образом. На первой стадии проводили комплексообразование РНК-репликона при требующемся исходном N/P. На второй стадии в состав добавляли избыток in vivo-JetPEI для достижения требуемого общего соотношения N/P; т.е. первое число означает исходное NP PEI, а второе число – избыток данного in vivo-JetPEI на второй стадии (напр., NP4+8 = NP4 JetPEI на первой стадии и NP8 JetPEI на второй стадии). Секретируемую люциферазу измеряли в соответствии с протоколом производителя (Nano-GLO, Promega, США) при 125 нг РНК на лунку. Анализ жизнеспособности клеток проводили, как описано ранее в Примере 15.
Результаты по 2-стадийному комплексообразованию представлены на фиг. 33. По сравнению с эталоном 1-стадийного комплексообразования с in vivo-JetPEI NP12 и при таком же общем соотношении NP, при двухстадийном комплексообразовании достигались более высокие уровни экспрессии.
Пример 22. Влияние буфера в составе полиплексов на эффективность иммунизации
Составляли полиплексы saRNA с использованием in vivo-jetPEI. Полиплексы получали с использованием забуференной HEPES глюкозы (HBG) или забуференной MES глюкозы (MBG) (5% D-глюкозы, 10 мМ MES, pH 6,1). Мышей иммунизированы в день 0 по однократной схеме. В день 45 после иммунизации получали сыворотку и определяли содержание специфичных к Cf07-HA антител в сыворотке HA-специфическим методом ELISA. Проводили титрование разведений сыворотки до конечной точки и определяли площадь под кривой (AUC). Представлено увеличение площади под кривой у полученных в MBG полиплексов в процентах по сравнению с полученными в HBG полиплексами, которые принимали за 100%.
Результаты представлены на фиг. 34. По сравнению с полиплексами, составленными с HBG, полиплексы, полученные с использованием забуференной MES глюкозы, дают намного больший сигнал ELISA после однократной вакцинации мышей.
Пример 23. У животных развивается иммунный ответ нейтрализующих антител после внутримышечной (в/м) иммунизации с помощью составленной с PEI самоамплифицирующейся РНК (saRNA), кодирующей HA вируса A/California/7/2009 (H1N1) (H1N1/Cf7-HA)
Мышей BALB/c в день 0 иммунизировали один раз буфером или 1/25 дозой человеческой вакцины или 0,1 мкг составленной с PEI saRNA VEEV или saRNA SFV, кодирующей HA H1N1/Cf7, при соотношении N/P = 12:1. Через 28 и 48 дней у животных брали кровь и проводили анализ сыворотки на антитела против HA методом нейтрализации вируса (VNT; n = 4). Результаты представлены на фиг. 35(A).
Домашних поросят в день 0 иммунизировали один раз буфером или 1 дозой человеческой вакцины или 90 мкг составленной с PEI saRNA VEEV или saRNA SFV, кодирующей HA H1N1/Cf7, при соотношении N/P = 12^1. У поросят брали кровь на 14, 21, 28 и 35-й день после иммунизации и проводили анализ иммунного ответа нейтрализующих антител против HA методом VNT (n = 8; группа буфера: n = 4). Результаты представлены на фиг. 35(B).
У группы животных, получавших составленную с saRNA VEEV вакцину, развивался такой же иммунный ответ, как у животных, которым вводили положительный контроль. saRNA SFV также приводила к развитию иммунного ответа нейтрализующих антител, но с более низким титром, чем после иммунизации saRNA VEEV. На графике представлены средние значения ± SEM.
Пример 24. У животных развивается иммунный ответ антител после внутримышечной (в/м) иммунизации с помощью составленной с PEI самоамплифицирующейся РНК (saRNA), кодирующей белок cap_EU свиного цирковируса-2 (PCV2)
Мышей BALB/c иммунизировали дважды в день 0 и день 35 буфером или 1 мкг составленной с PEI saRNA SFV или saRNA VEEV, кодирующей белок cap_EU PCV2, при соотношении N/P = 12:1. На 14, 34 и 56-й день у животных брали кровь и проводили анализ сыворотки на антитела против PCV2-cap при помощи коммерчески доступного набора для ELISA (INgezim Circo IgG, Ingenasa; n = 4).
Как видно из фиг. 36, группы животных, получавших составленные с saRNA SFV или VEEV вакцины, проявляли сходные ответы антител против белка cap_EU PCV2. Иммунный ответ антител после однократной вакцинации saRNA SFV был несколько выше, чем для saRNA VEEV. После двух иммунизаций ответ антител был почти одинаковым для обоих типов вакцин с saRNA. На графике представлены средние значения ± SEM.
Пример 25. Влияние рН на стабильность самоамплифицирующейся РНК (saRNA)
Составляли комплексы с РНК репликона (saRNA) при N/P = 12 в различных буферных системах и условиях рН. Оба типа буферов, ацетатный или MES, содержали буферное вещество в конечной концентрации 10 мМ и D-глюкозу в конечной концентрации 5% вес./об.. Полиплексы saRNA/PEI хранили в соответствующем буфере при 4°C в течение различного времени (1, 2, 4 и 8 дней после образования комплексов). Сразу же после образования комплексов в различных препаратах измеряли целостность РНК (t = 0). Целостность РНК измеряли методом капиллярного электрофореза. saRNA, образующая комплекс в полиплексах, может высвобождаться после 20-минутной инкубации при комнатной температуре с сильным избытком полианиона, который вызывает электростатическое взаимодействие с полимером, высвобождая РНК, заключенную в полиплексах. Для анализа методом капиллярного электрофореза использовали по 200 мкг высвобожденной РНК строго в соответствии с протоколом, прилагаемым к соответствующему набору (набор DF471 для анализа РНК стандартной чувствительности). Для каждой временной точки использовали эталонную saRNA для определения целостности saRNA.
Как видно из фиг. 37, более высокие значения pH в буфере для препарата приводят к значительному усилению деградации saRNA. Наименьшая потеря целостности saRNA при образовании комплексов достигалась при pH 4 с ацетатным буфером.
Пример 26. Составленная с PEI saRNA VEEV, кодирующая HA вируса A/California/7/2009 (H1N1; Cf7/HA), индуцирует сильный и более продолжительный ответ антител по сравнению с коммерческой вакциной, но дополнительно индуцирует сильный T-клеточный ответ, который белковые вакцины не вызывают
На фиг. 38 мышей BALB/c иммунизировали в/м дважды в день 0 и день 35 (на графиках они обозначены стрелками) буфером (черные символы) или 20 мкл человеческой лицензированной вакцины против сезонных штаммов вируса гриппа (Begripal 2016/2017; hLIC; серые символы) или 0,5 мкг составленной с PEI вакцины на основе saRNA VEEV, кодирующей Cf7/HA (темно-серые символы). В различные моменты времени мышей забивали и A) отбирали спленоциты для проведения Cf7/HA-специфичных анализов ELISpot на суспензиях отдельных клеток. Для анализа ELISpot использовали различные пулы CF7/HA-специфичных пептидов для стимуляции ответа T-клеток CD8+ (слева) или T-клеток CD4+ (справа), измеряемого по секреции IFN-γ. Кроме того, отбирали образцы сыворотки для проведения B) анализа Cf7/HA-специфичной нейтрализации вируса для сывороточных антител на их функциональность в ингибировании вирусной инфекции клеток. Отметим, что для серологического анализа использовали вирус A/California/4/2009 (H1N1; Cf4); а данные показывают средние значения ± SEM (группа буфера: n = 3; группы вакцины: n = 4).
СПИСОК СОКРАЩЕНИЙ И ОПРЕДЕЛЕНИЙ
АТМ: атмосферное давление
C: концентрация
CSS: раствор 23 мг CuSO4 в 100 мл 0,1M Na-ацетата, pH 5,4
DLS: динамическое рассеяние света
EDTA: этилендиаминтетрауксусная кислота
FCS: фетальная телячья сыворотка
h: ч
HBGx1: забуференная 10 мМ HEPES (pH 7,1) 5% глюкоза
HBGx2: забуференная 20 мМ HEPES (pH 7,1) 10% глюкоза
HBTx1: забуференная 10 мМ HEPES (pH 7,1) 10% трегалоза
HBTx2: забуференная 20 мМ HEPES (pH 7,1) 20% трегалоза
HBTx1+EDTA: забуференная 8 мМ HEPES (pH 7,1) 10% трегалоза с 80 мкМ EDTA
HEPES: 4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота
IVI: Институт вирусологии и иммунологии, Mittelhäusern, Швейцария
IVT: транскрибированная in vitro мРНК
kDa: 1000 дальтон
Lyo: лиофилизация
мин: минуты
MBGx1: 5% D-глюкоза, 10 мM MES, pH 6,1
MES: 2-(N-морфолино)этансульфоновая кислота
N/P: соотношение между числом аминогрупп в PEI и фосфатных групп в РНК
PEI: полиэтиленимин
RNA: рибонуклеиновая кислота
UV: ультрафиолет.
| название | год | авторы | номер документа |
|---|---|---|---|
| СОСТАВ ДЛЯ ВВЕДЕНИЯ РНК | 2017 |
|
RU2746118C2 |
| РНК-РЕПЛИКОН ДЛЯ УНИВЕРСАЛЬНОЙ И ЭФФЕКТИВНОЙ ГЕННОЙ ЭКСПРЕССИИ | 2017 |
|
RU2748892C2 |
| ТРАНС-РЕПЛИЦИРУЮЩАЯ РНК | 2017 |
|
RU2752580C2 |
| СПОСОБ ПОВЫШЕНИЯ ЭКСПРЕССИИ РНК В КЛЕТКЕ | 2018 |
|
RU2784654C2 |
| ПЛАТФОРМЫ ДОСТАВКИ АНТИГЕНОВ | 2011 |
|
RU2597974C2 |
| ЧАСТИЦА РЕПЛИКОНА АЛЬФАВИРУСА | 2018 |
|
RU2795596C2 |
| МИКРОЧАСТИЦЫ ДЛЯ ДОСТАВКИ ГЕТЕРОЛОГИЧНЫХ НУКЛЕИНОВЫХ КИСЛОТ | 2001 |
|
RU2295954C2 |
| ВАКЦИНА ПРОТИВ БОЛЕЗНИ ЛАЙМА У СОБАК | 2018 |
|
RU2785620C2 |
| ЭМУЛЬСИИ ТИПА "МАСЛО В ВОДЕ", КОТОРЫЕ СОДЕРЖАТ НУКЛЕИНОВЫЕ КИСЛОТЫ | 2012 |
|
RU2606846C2 |
| ВАКЦИНА ПРОТИВ ВИРУСА СВИНОГО ГРИППА А, СОДЕРЖАЩАЯ ДВЕ РАЗНЫЕ ЧАСТИЦЫ РНК-РЕПЛИКОНА | 2021 |
|
RU2839757C1 |
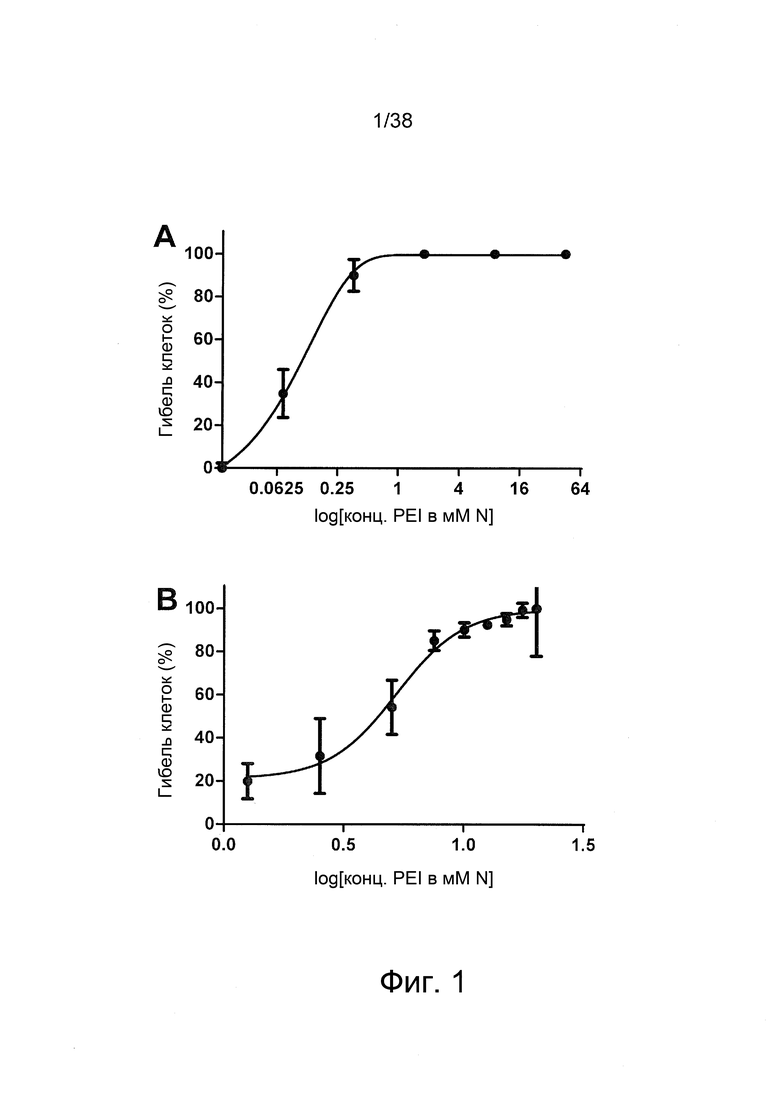
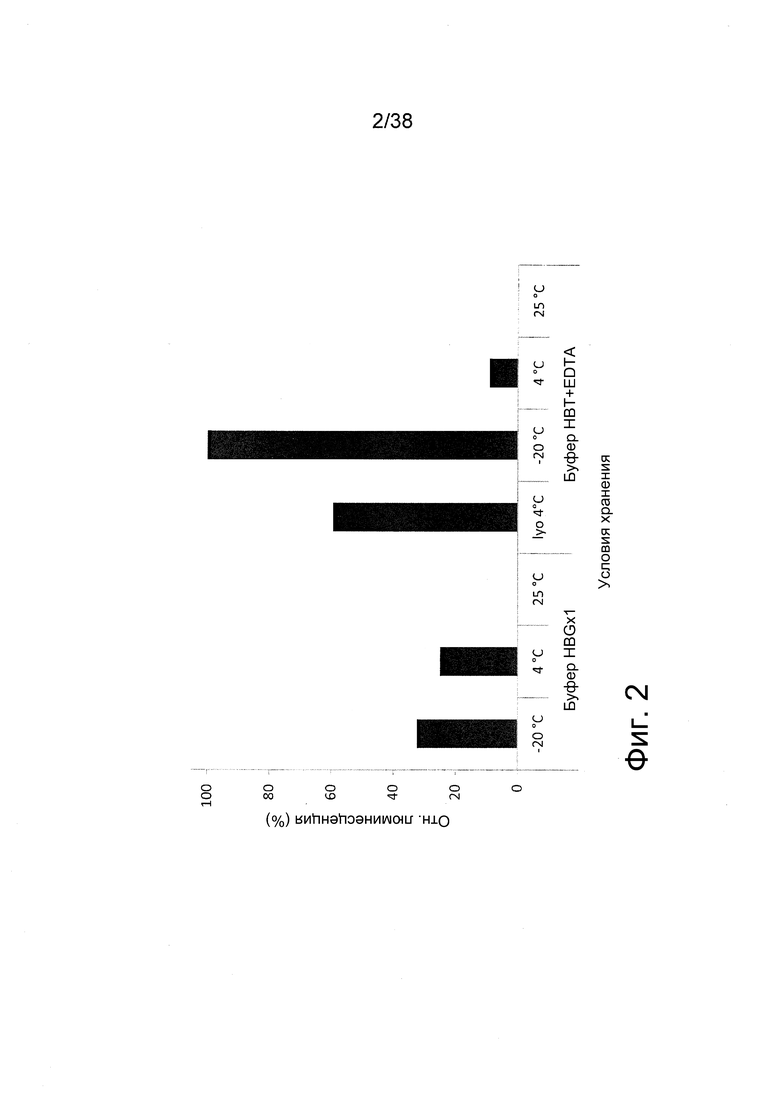

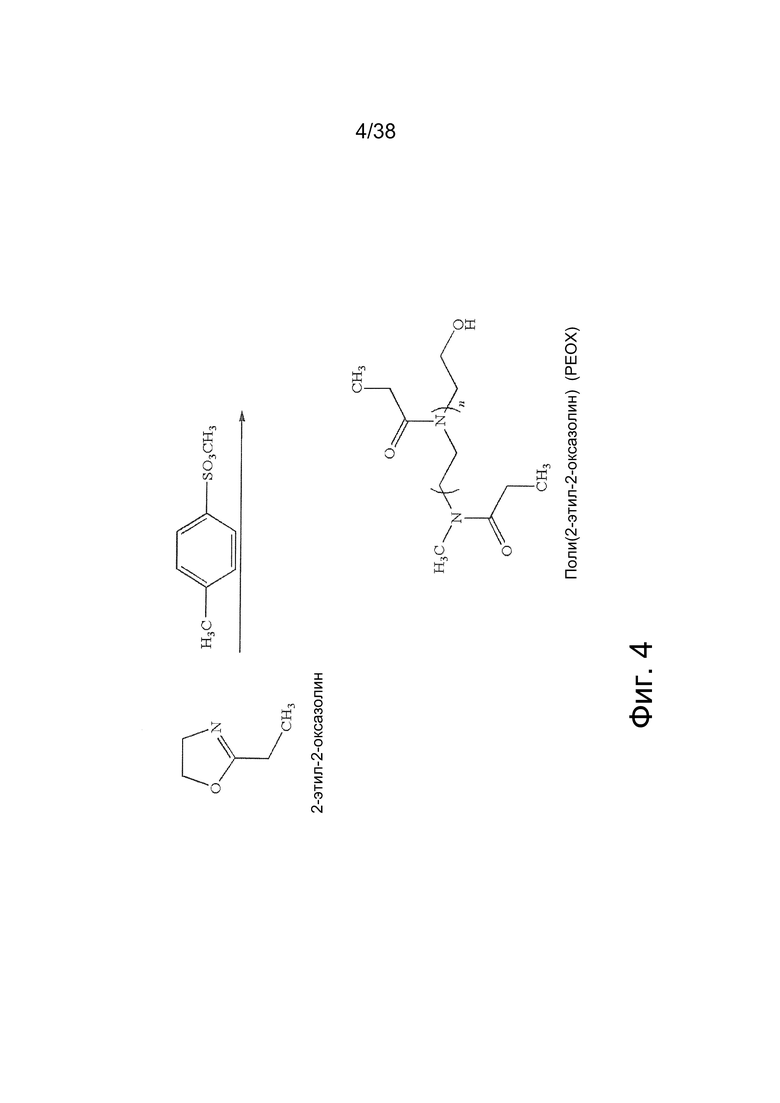

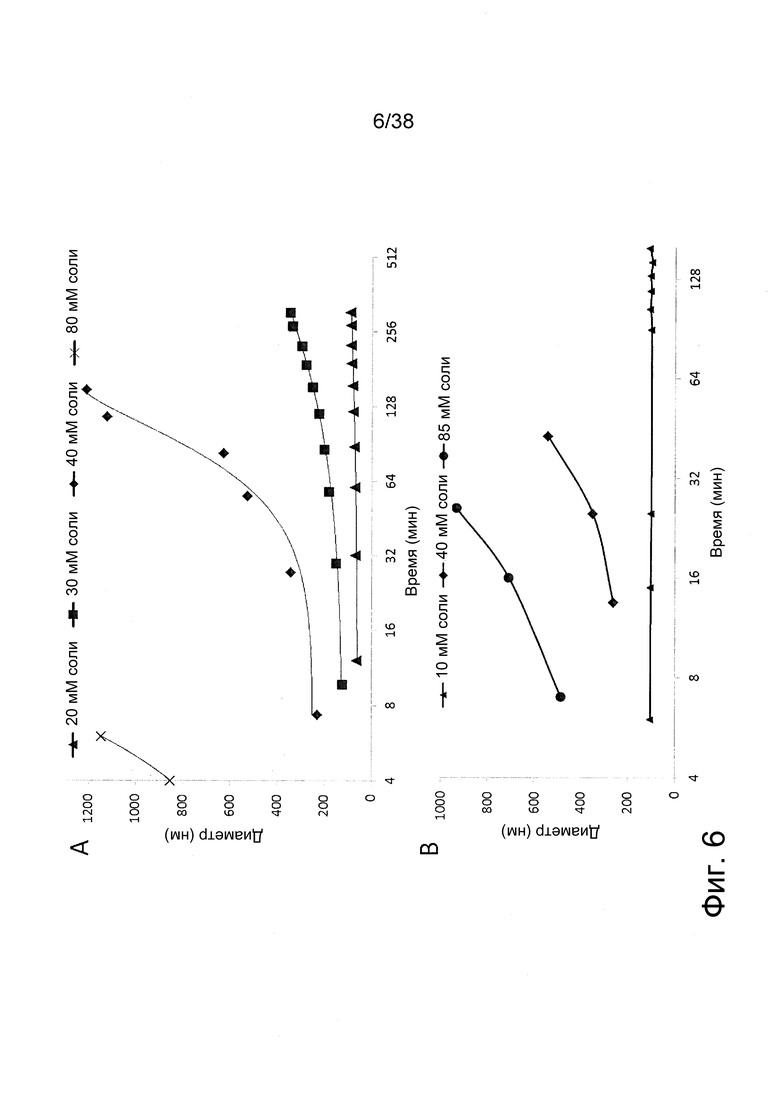


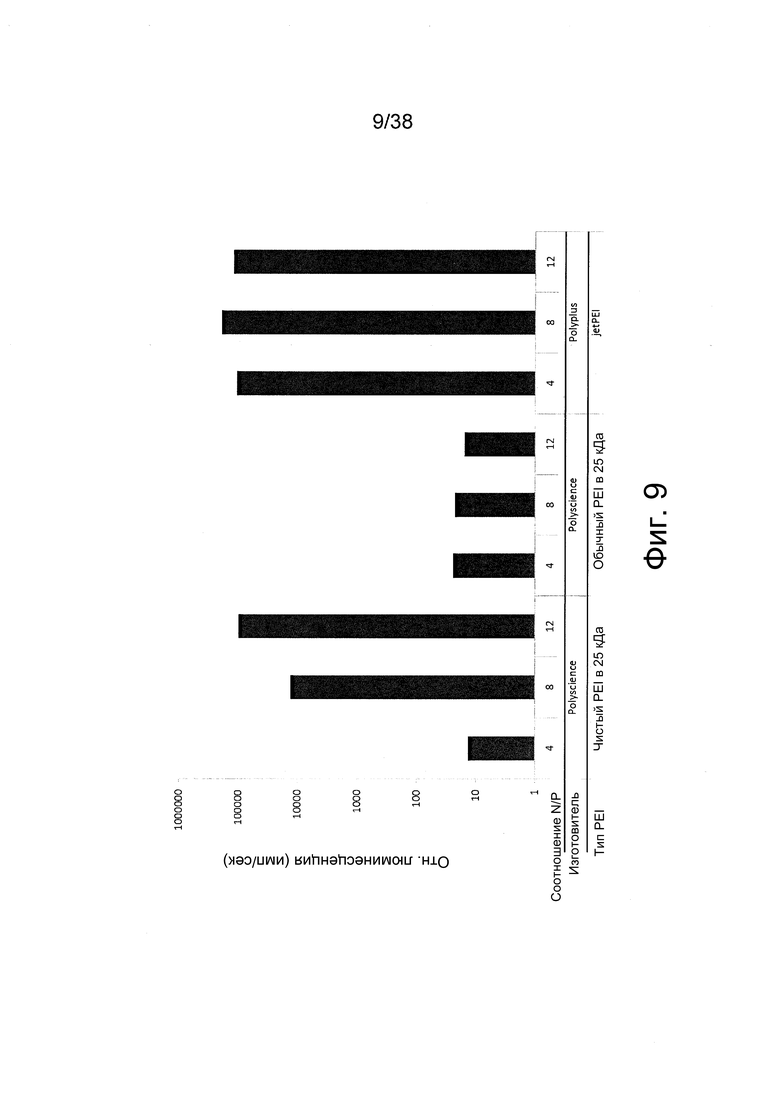
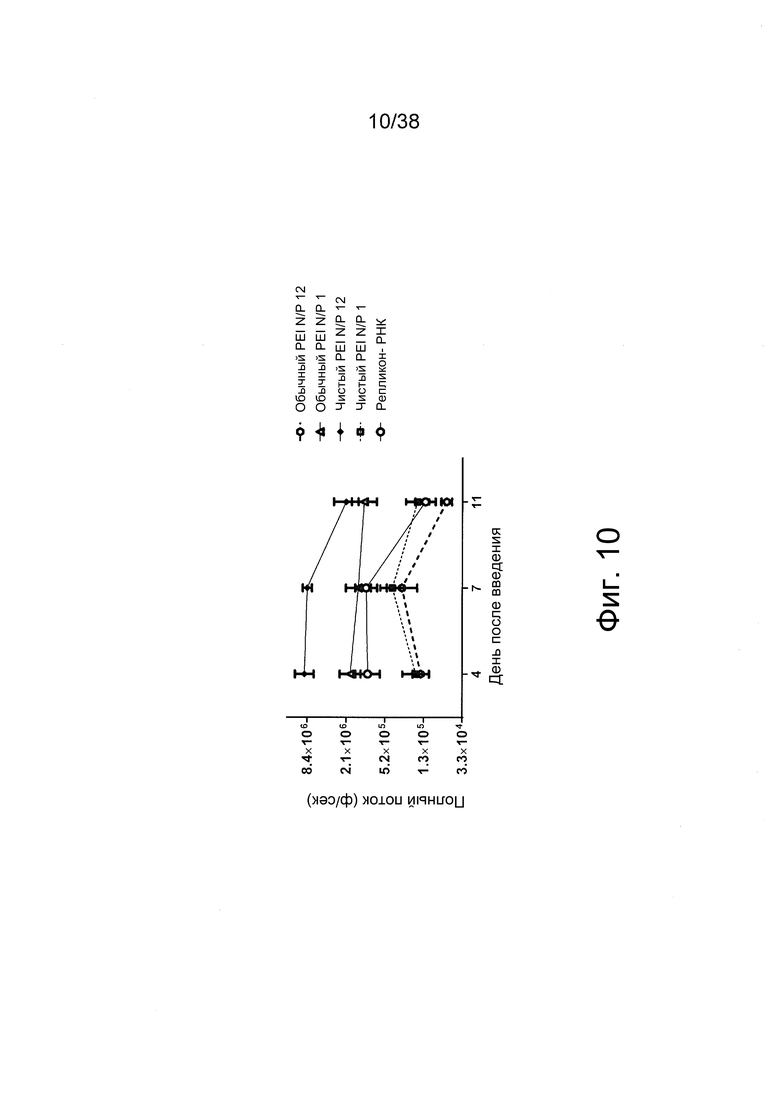
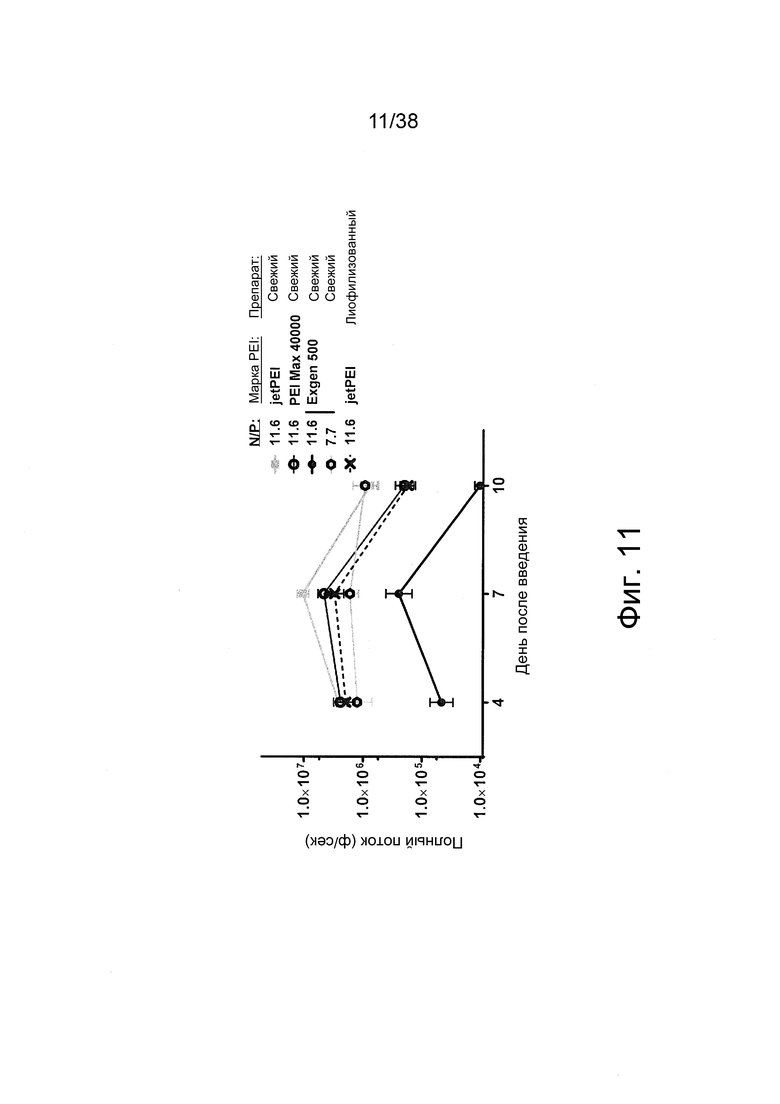

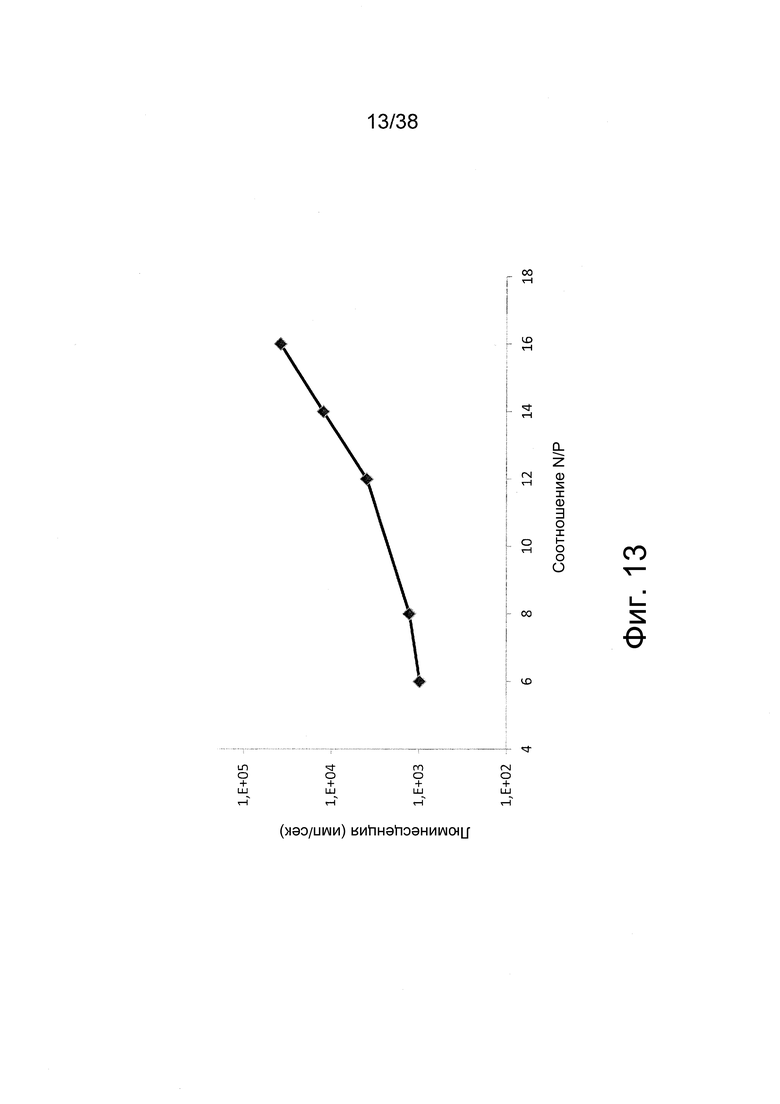
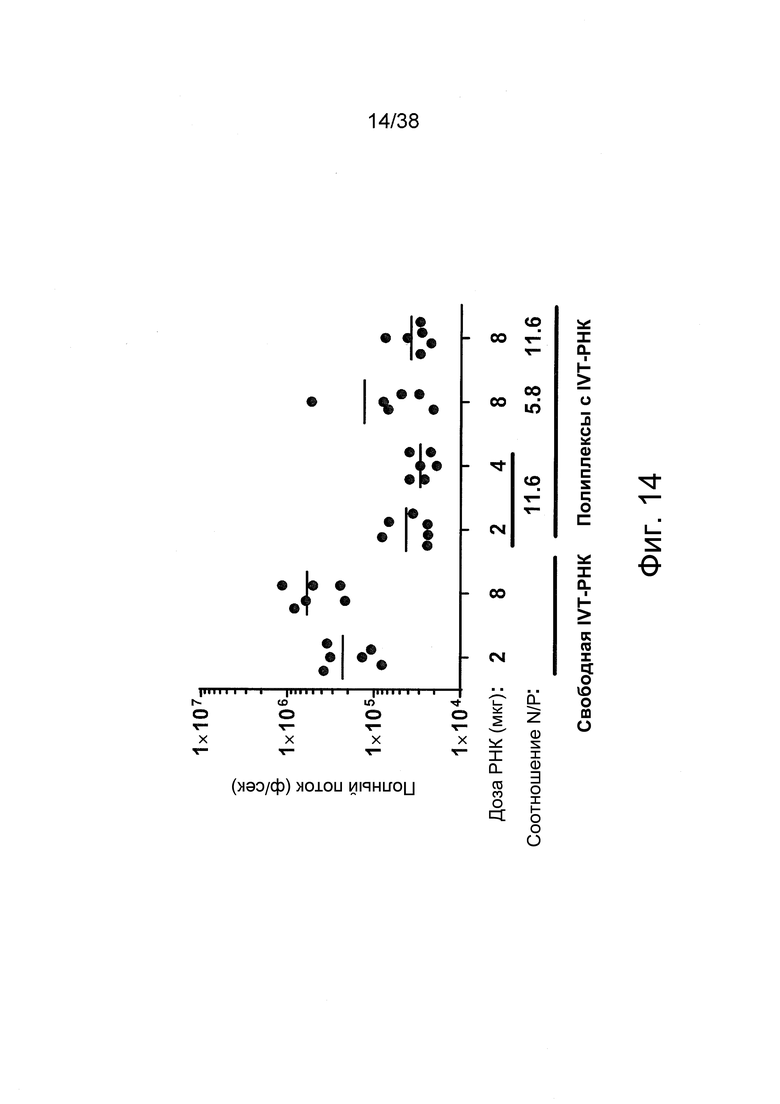
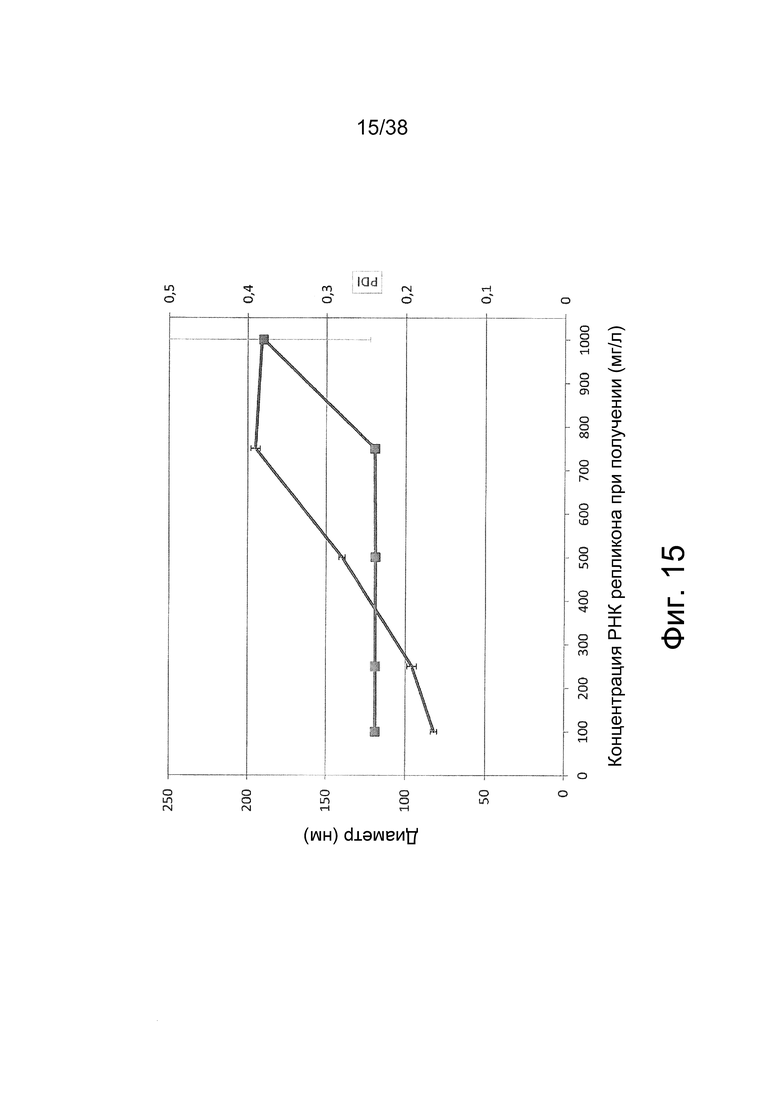
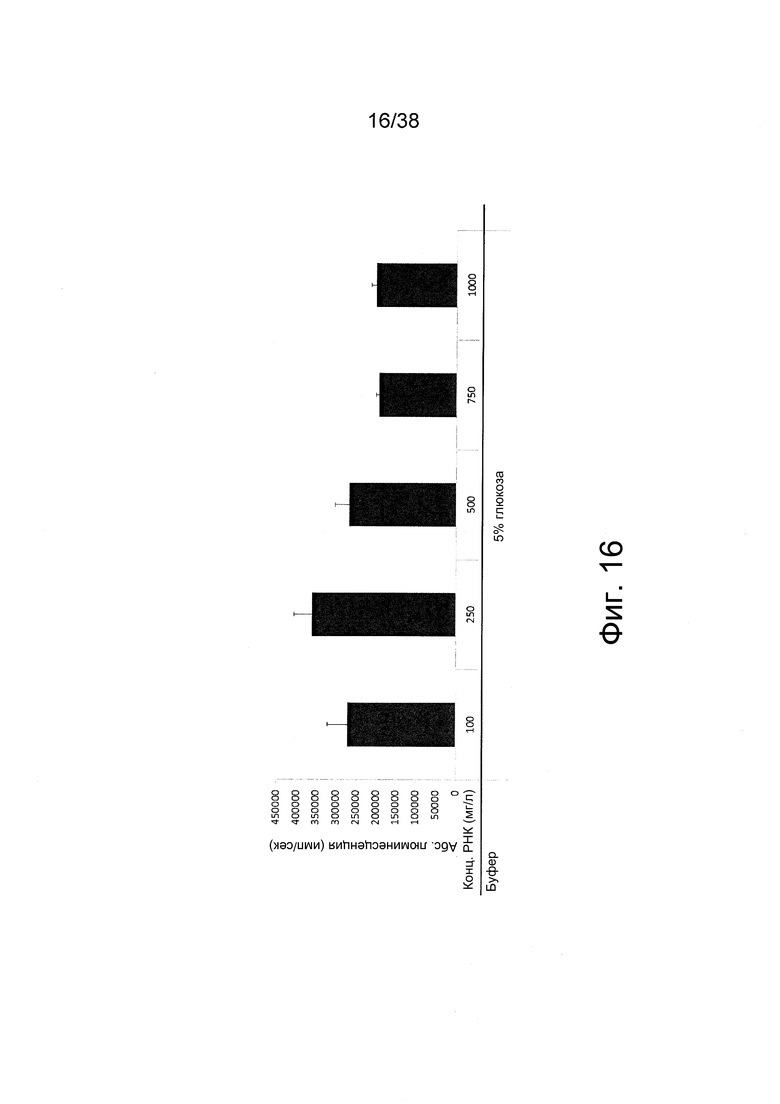
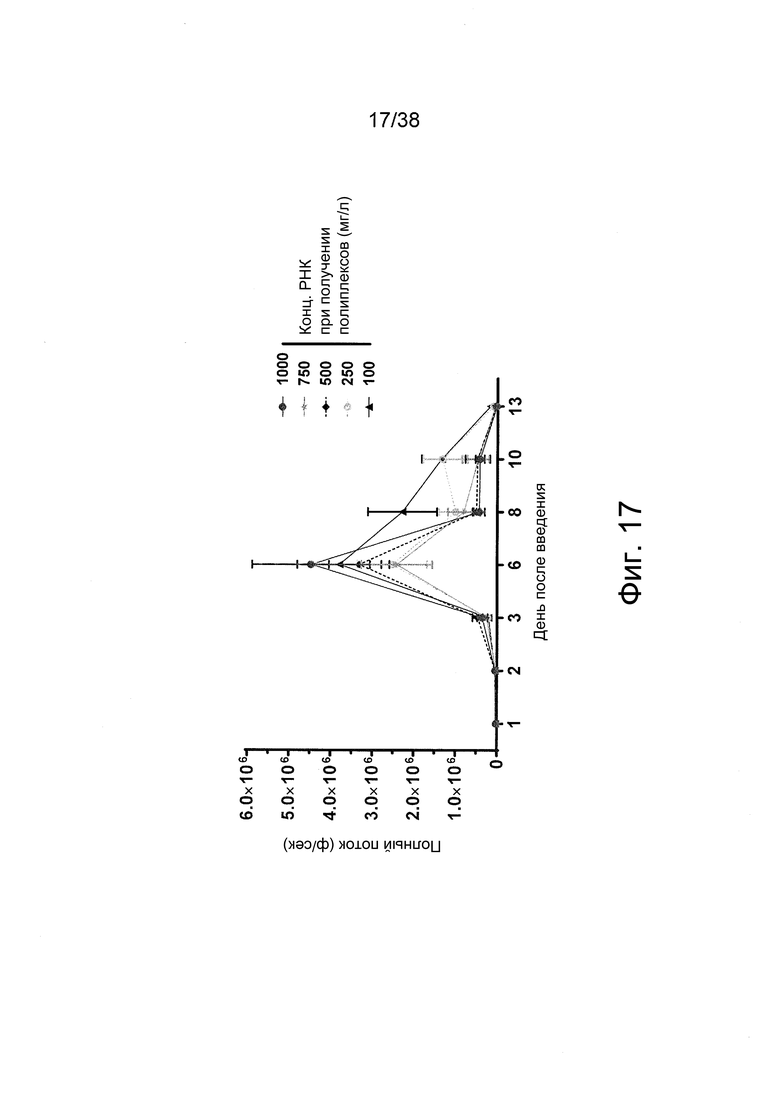
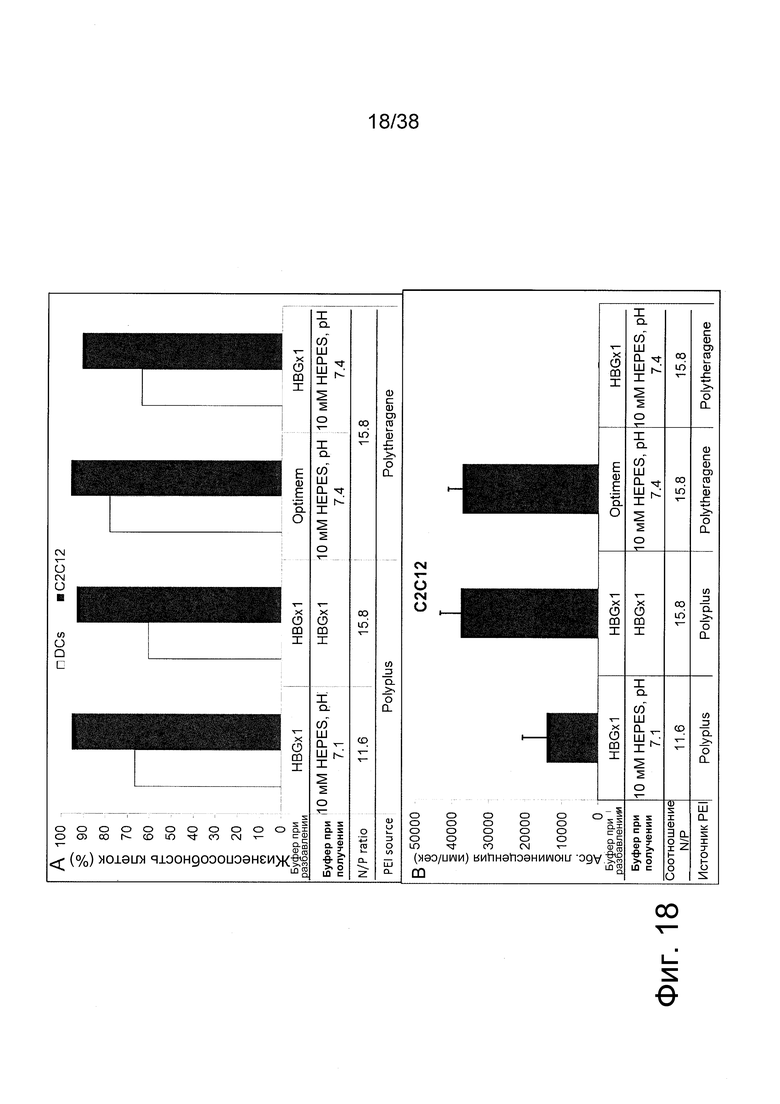
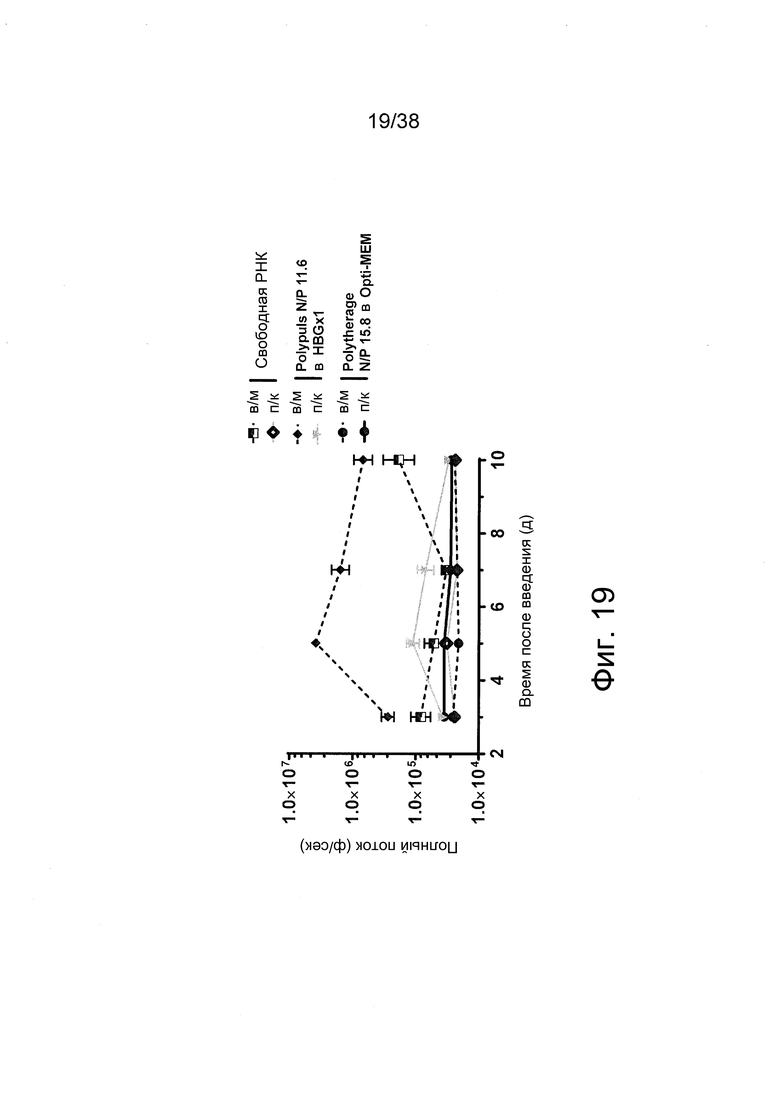
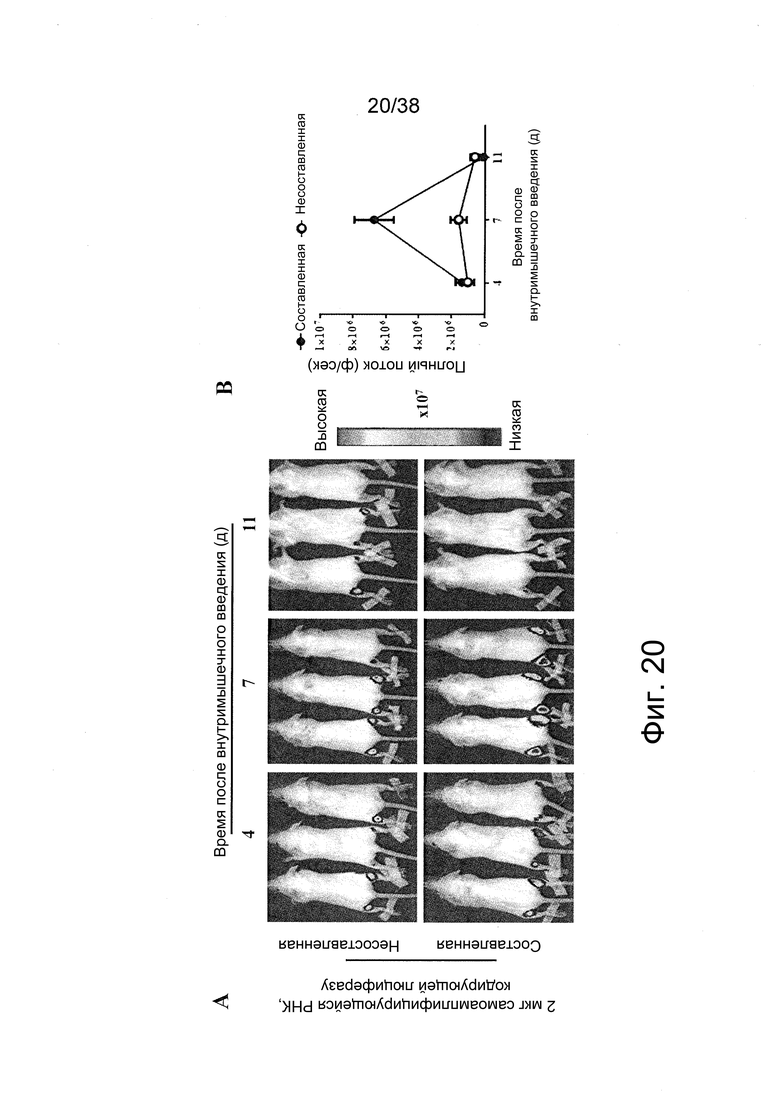
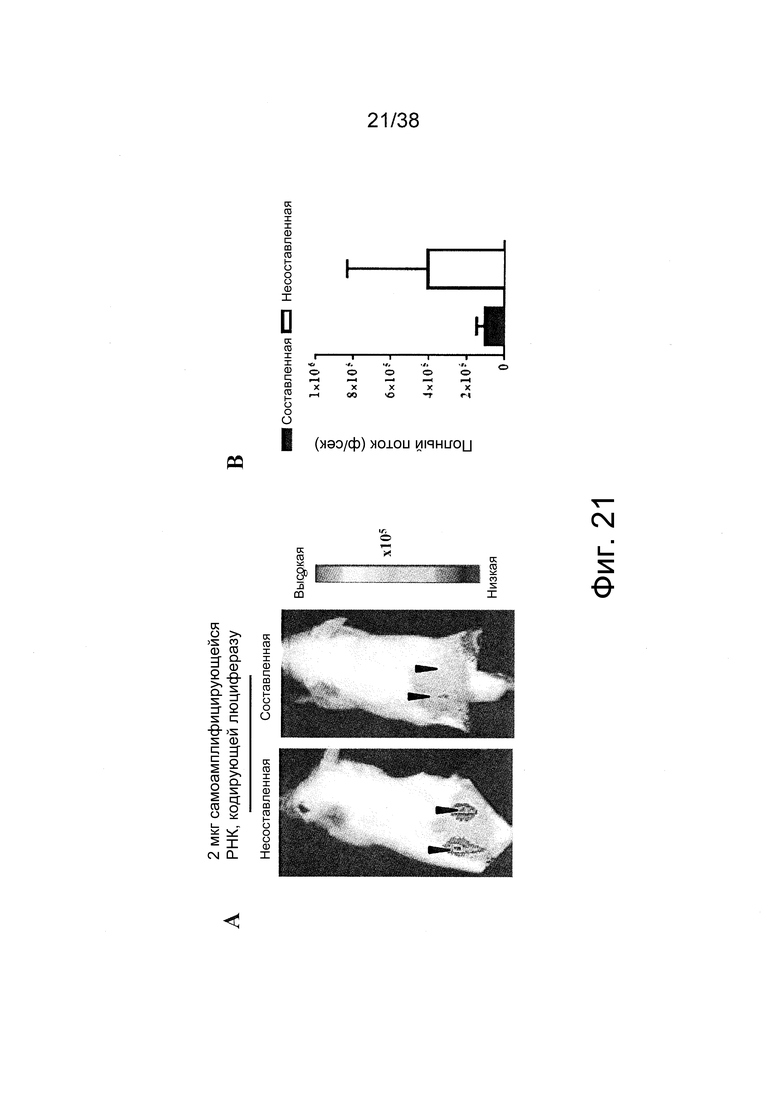




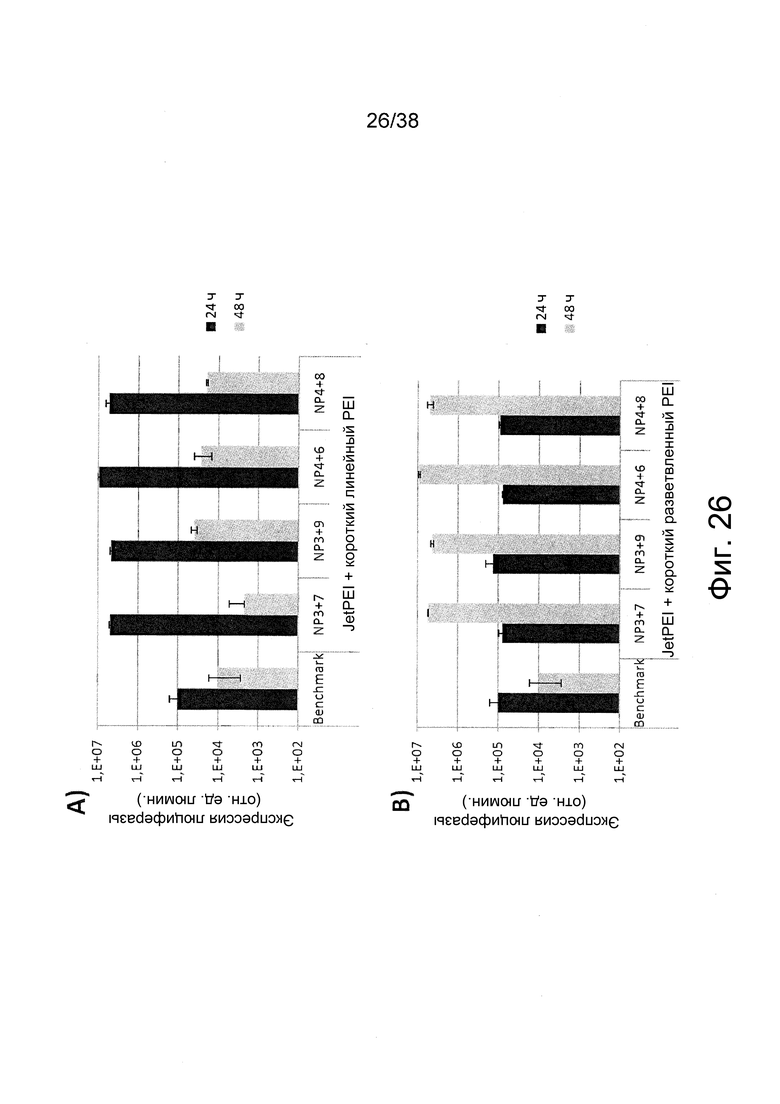

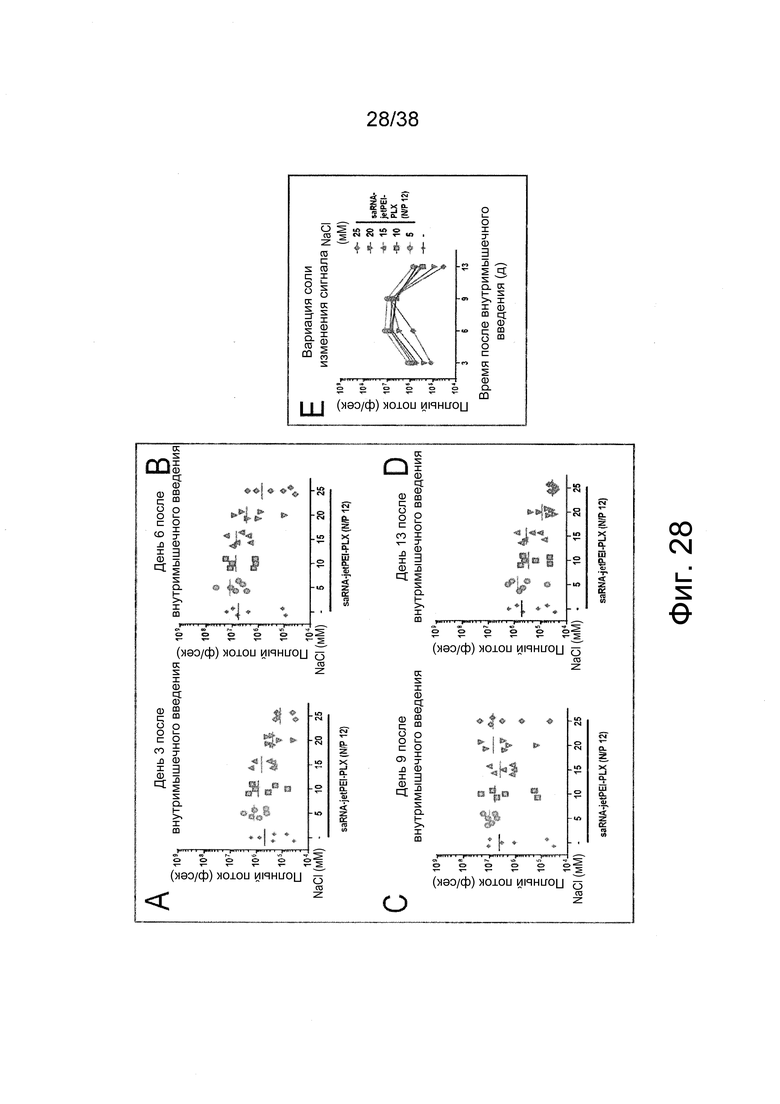
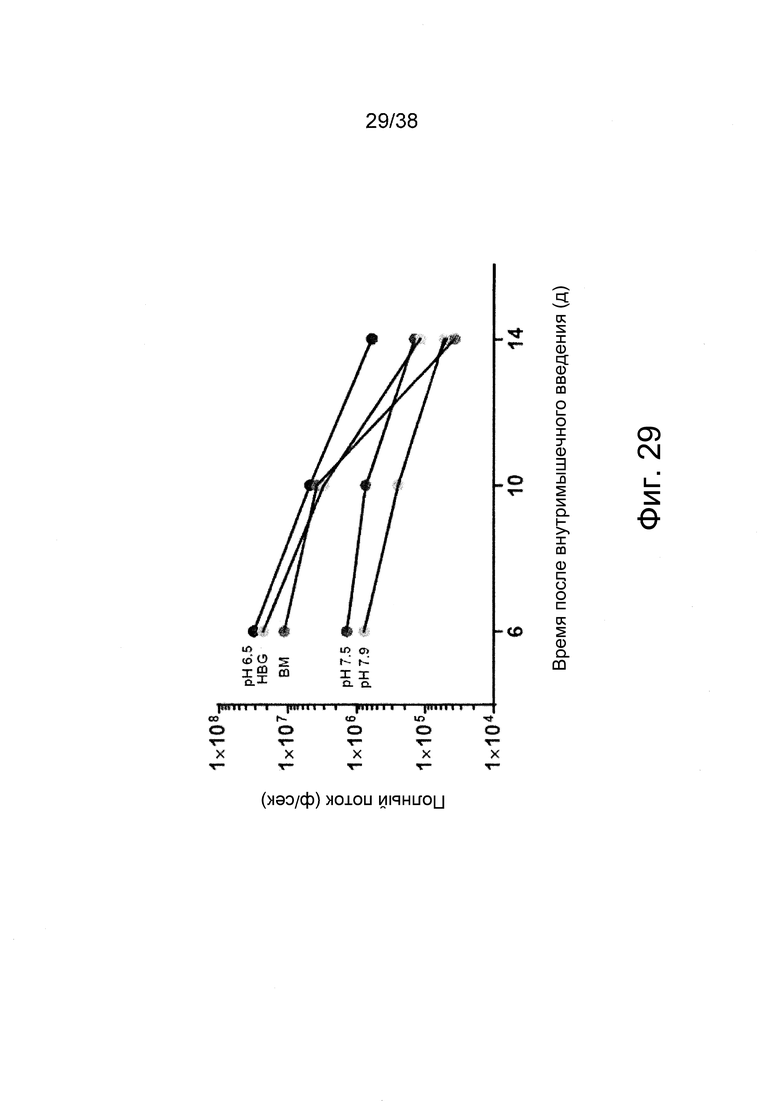
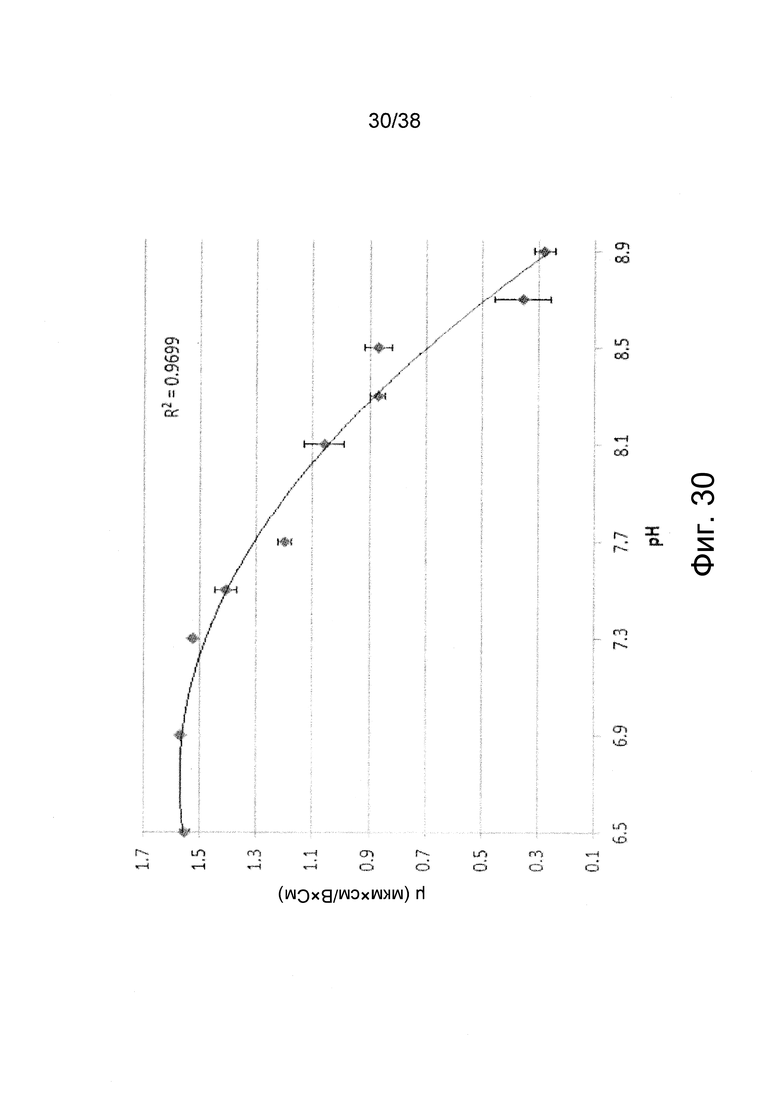
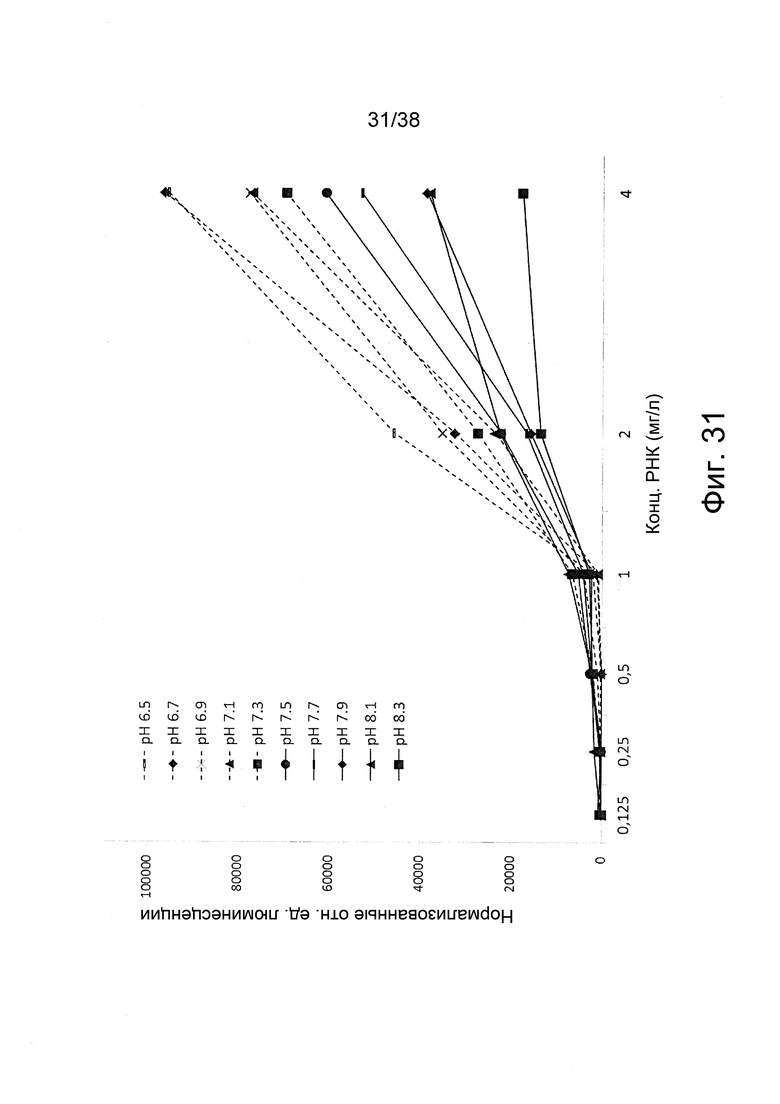

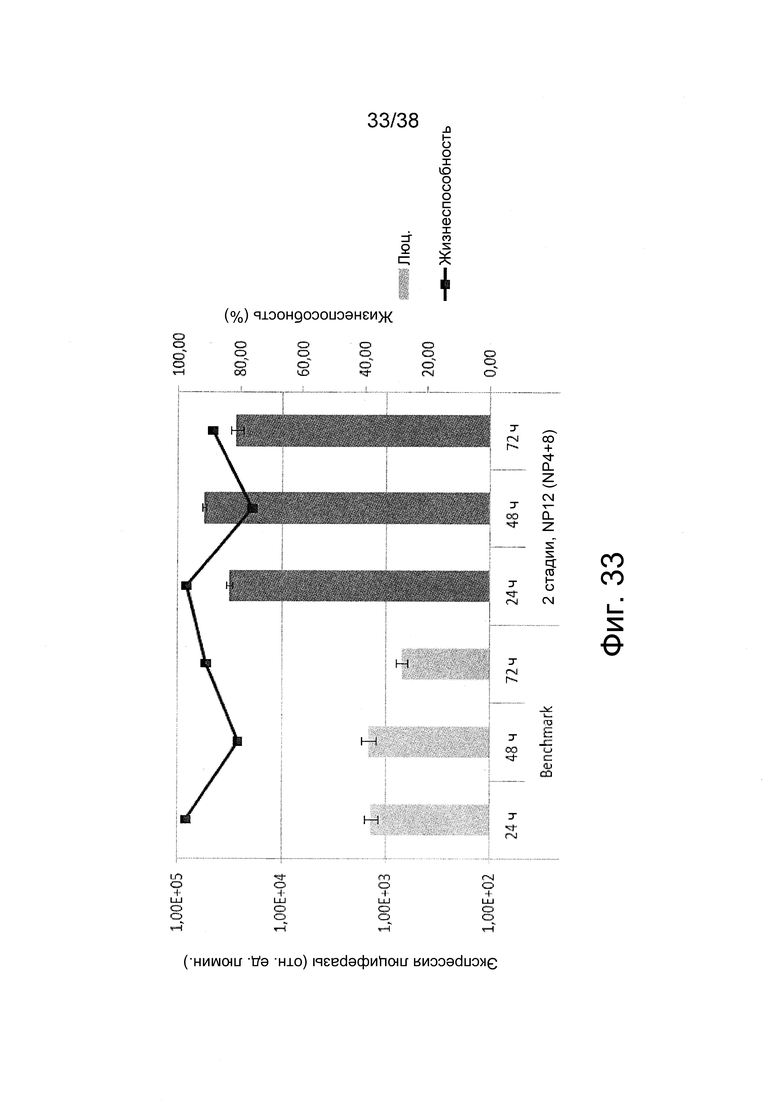
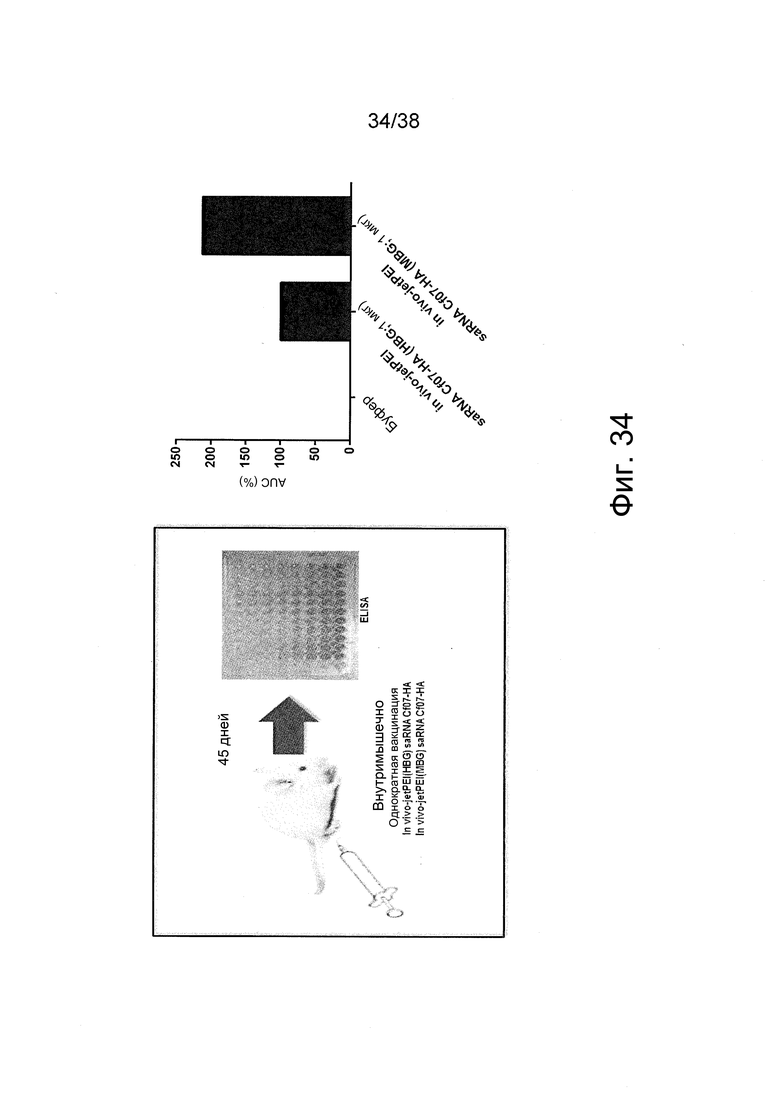
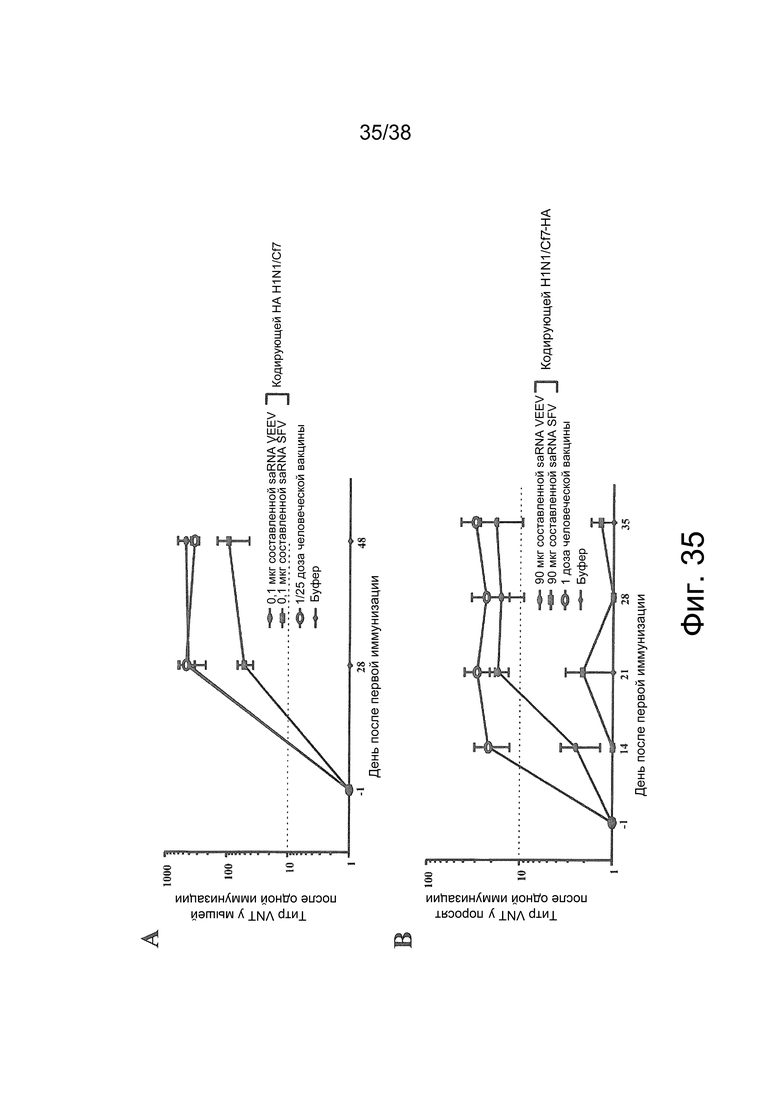
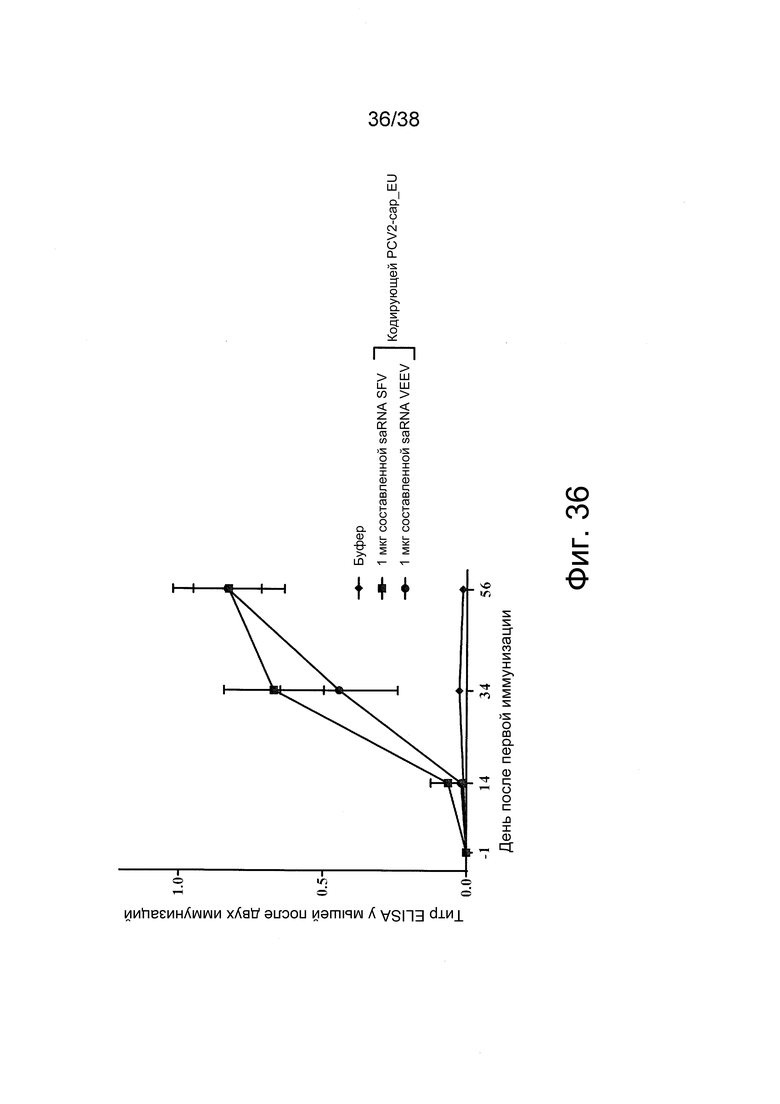
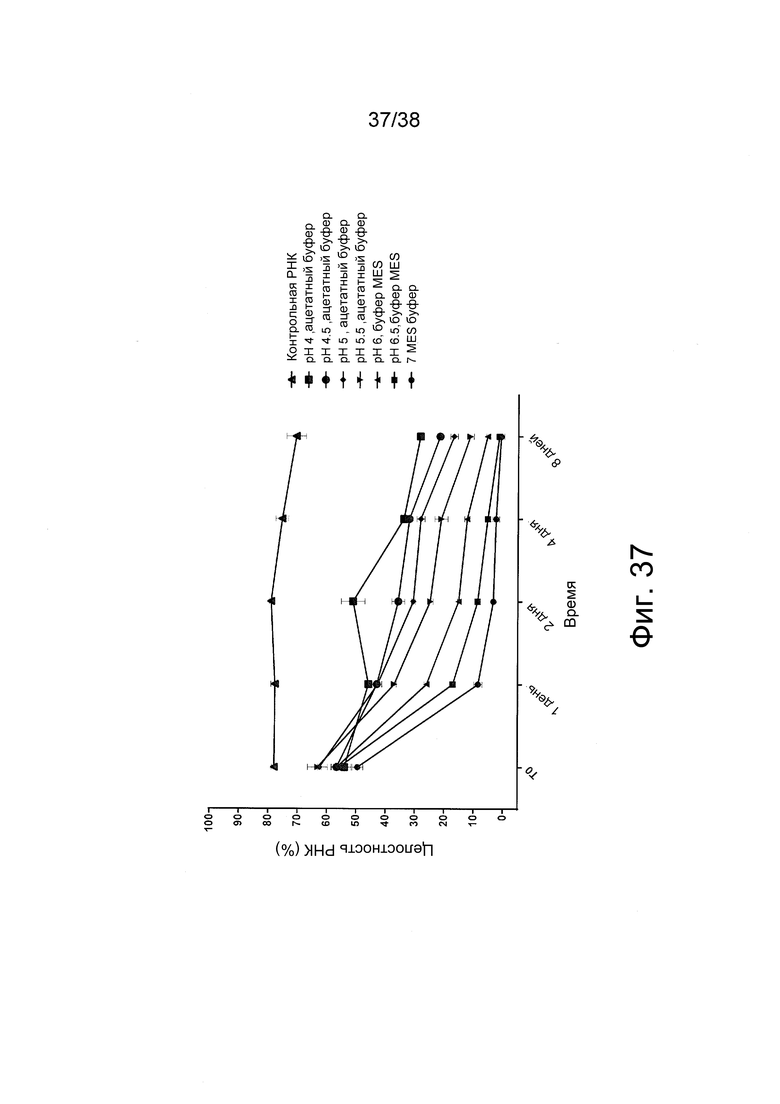
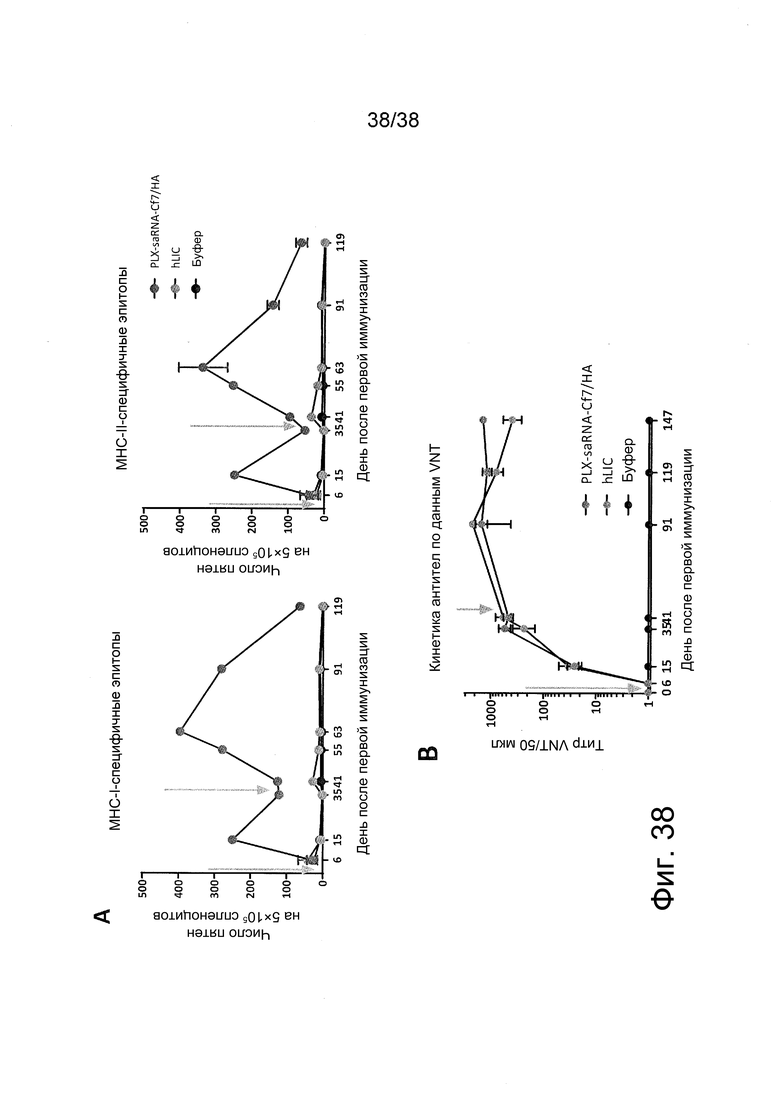
Группа изобретений относится к области фармацевтики и медицины. 1 объект представляет собой фармацевтическую композицию для внутримышечной инъекции, включающую активный ингредиент, который представляет собой одноцепочечную самореплицирующуюся РНК, кодирующую пептид или белок, содержащий антиген или эпитоп, где указанная самореплицирующаяся РНК получена из вируса венесуэльского энцефалита лошадей (VEEV) или из вируса Semliki Forest (SFV), и полиэтиленимин, причем указанная самореплицирующаяся РНК и полиэтиленимин находятся в полиплексных частицах, и молярное отношение числа атомов азота (N) в полиэтиленимине к числу атомов фосфора (P) в одноцепочечной РНК (соотношение N:P) составляет от 2,0 до 15,0. 2 объект – способ индуцирования иммунного ответа, включающий стадию введения композиции. Технический результат заключается в индуцировании иммунного ответа при внутримышечном введении с применением полиплексов РНК и полиэтиленимина, содержащих одноцепочечную самореплицирующуюся РНК, полученную из VEEV или из SFV. 2 н. и 34 з.п. ф-лы, 38 ил., 7 табл., 26 пр.
1. Фармацевтическая композиция для внутримышечной инъекции, включающая активный ингредиент, отличающаяся тем, что активный ингредиент представляет собой одноцепочечную самореплицирующуюся РНК, кодирующую пептид или белок, содержащий антиген или эпитоп, где указанная самореплицирующаяся РНК получена из вируса венесуэльского энцефалита лошадей (VEEV) или из вируса Semliki Forest (SFV); и указанная композиция дополнительно содержит полиэтиленимин;
где указанная самореплицирующаяся РНК и полиэтиленимин находятся в полиплексных частицах, и
где молярное отношение числа атомов азота (N) в полиэтиленимине к числу атомов фосфора (P) в одноцепочечной РНК (соотношение N:P) составляет от 2,0 до 15,0.
2. Композиция по п. 1, при этом молярное отношение числа атомов азота (N) в полиэтиленимине к числу атомов фосфора (P) в одноцепочечной РНК (соотношение N:P) составляет от 6,0 до 12,0.
3. Композиция по п. 1 или 2, при этом ионная сила составляет 50 мМ или меньше, при этом предпочтительно концентрация одновалентных катионных ионов составляет 25 мМ или меньше, а концентрация двухвалентных катионных ионов составляет 20 мкМ или меньше.
4. Композиция по любому из пп. 1-3, при этом одноцепочечная самореплицирующаяся РНК представляет собой цис-репликон.
5. Композиция по любому из пп. 1-4, при этом одноцепочечная самореплицирующаяся РНК соответствует или в основном соответствует геномной РНК VEEV или его аттенюированной формы, причем открытая рамка считывания, кодирующая структурные белки, заменена на открытую рамку считывания, кодирующую пептид или белок, содержащий антиген или эпитоп.
6. Композиция по п. 5, при этом антиген либо пептид или белок, содержащий антиген или эпитоп, является мембранным белком типа белка оболочки вируса.
7. Композиция по п. 5 или 6, при этом антигеном является гемагглютинин вируса гриппа.
8. Композиция по любому из пп. 1-7, при этом одноцепочечная самореплицирующаяся РНК соответствует или в основном соответствует геномной РНК SFV или его аттенюированной формы, причем открытая рамка считывания, кодирующая структурные белки, заменена на открытую рамку считывания, кодирующую пептид или белок, содержащий антиген или эпитоп.
9. Композиция по п. 8, при этом антиген либо пептид или белок, содержащий антиген или эпитоп, не является мембранным белком.
10. Композиция по п. 8 или 9, при этом антигеном является капсидный белок вируса.
11. Композиция по любому из пп. 1-10, при этом полиэтиленимин имеет следующую общую формулу (I):
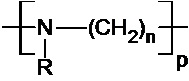 ,
,
где R означает H, ацил или группу, имеющую следующую общую формулу (II):
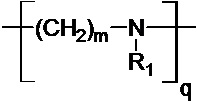 ,
,
где R1 означает Н или группу, имеющую следующую общую формулу (III):
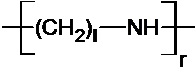 ,
,
n равно двум,
m и l выбраны независимо из целых чисел от 2 до 10; а
p, q и r – целые числа, причем сумма p, q и r такова, что средняя молекулярная масса полимера составляет от 1,5⋅102 до 107 Да, предпочтительно от 5000 до 105 Да, более предпочтительно от 10000 до 40000 Да, более предпочтительно от 15000 до 30000 Да, еще более предпочтительно от 20000 до 25000 Да.
12. Композиция по п. 11, при этом m и l выбраны независимо из 2, 3, 4 и 5, предпочтительно из 2 и 3.
13. Композиция по п. 11 или 12, при этом R1 означает H.
14. Композиция по любому из пп. 11-13, при этом R означает H или ацил.
15. Композиция по любому из пп. 1-14, при этом по меньшей мере 92% атомов N в полиэтиленимине являются протонируемыми.
16. Композиция по любому из пп. 1-15, дополнительно содержащая одну или несколько добавок.
17. Композиция по п. 16, при этом одна или несколько добавок выбраны из группы, состоящей из буферных веществ, сахаридов, стабилизаторов, криопротекторов, лиопротекторов и хелатообразующих веществ.
18. Композиция по п. 17, при этом буферные вещества включают по меньшей мере одно, выбранное из группы, состоящей из 4-(2-гидроксиэтил)-1-пиперазинэтансульфоновой кислоты (HEPES), 2-(N-морфолино)этансульфоновой кислоты (MES), 3-морфолино-2-гидроксипропансульфоновой кислоты (MOPSO), буферов на основе уксусной кислоты и ацетата и их аналогов, буферов на основе фосфорной кислоты и фосфата и буферов на основе лимонной кислоты и цитрата.
19. Композиция по п. 17 или 18, при этом сахариды включают по меньшей мере один, выбранный из группы, состоящей из моносахаридов, дисахаридов, трисахаридов, олигосахаридов и полисахаридов, предпочтительно из глюкозы, трегалозы и сахарозы.
20. Композиция по любому из пп. 17-19, при этом криопротекторы включают по меньшей мере один, выбранный из группы, состоящей из таких гликолей, как этиленгликоль, пропиленгликоль и глицерин.
21. Композиция по любому из пп. 17-20, при этом хелатообразующее вещество включает EDTA.
22. Композиция по любому из пп. 1-21, при этом композиция содержит забуференную HEPES глюкозу (HBG), забуференную MES глюкозу (MBG), забуференную ацетатом глюкозу или забуференную HEPES трегалозу (HBT).
23. Композиция по любому из пп. 1-22, при этом z-средний размер частиц по данным динамического рассеяния света составляет менее 200 нм, предпочтительно менее 150 нм и более предпочтительно менее 100 нм, и/или индекс полидисперсности частиц по данным динамического рассеяния света составляет менее 0,5, предпочтительно менее 0,3 и более предпочтительно менее 0,2.
24. Композиция по любому из пп. 1-23, при этом дзета-потенциал частиц составляет 20 мВ или больше, предпочтительно от 25 до 40 мВ.
25. Композиция по любому из пп. 22-24, при этом HBG содержит 5% глюкозы (вес./об.) и 10 мМ HEPES, pH 7,1.
26. Композиция по любому из пп. 22-24, где MBG содержит 5% глюкозы (вес./об.) и 10 мМ MES, pH 6,1.
27. Композиция по любому из пп. 22-24, при этом HBT содержит 10% трегалозы (вес./об.) и 10 мМ HEPES, pH 7,1.
28. Композиция по любому из пп. 1-27, при этом частицы являются нейтральными или положительно заряженными при физиологическом pH.
29. Композиция по любому из пп. 1-28, при этом одноцепочечная РНК представляет собой молекулу из от 6000 до 15000 нуклеотидов, предпочтительно от 9000 до 12000 нуклеотидов.
30. Композиция по любому из пп. 1-29 для применения в терапии.
31. Композиция по любому из пп. 1-30, которая является вакцинной композицией.
32. Композиция по любому из пп. 1-31 для индуцирования иммунного ответа.
33. Композиция по п. 32, которая вводится внутримышечной инъекцией.
34. Способ индуцирования иммунного ответа, включающий стадию введения композиции по любому из пп. 1-31.
35. Способ по п. 34, при этом композицию вводят внутримышечной инъекцией.
36. Композиция по п. 32 или 33 либо способ по п. 34 или 35, при этом иммунный ответ направлен против антигена или эпитопа.
| WO 2016165825 A1, 20.10.2016 | |||
| US 2007044929 A1, 01.03.2007 | |||
| US 2007219365 A1, 20.09.2007 | |||
| WO 2017162266 A1, 28.09.2017 | |||
| US 2016074506 A1, 17.03.2016 | |||
| US 2012237565 A1, 20.09.2012 | |||
| US 2016128944 A1, 12.05.2016 | |||
| US 2010285111 A1, 11.11.2010 | |||
| WO 2014145839 A2, 18.09.2014 | |||
| ROSENKRANZ A.A., SOBOLEV A.S | |||
| Polyethylenimine-based polyplex nanoparticles |
