Иастоящее изобретение относится к полифункциональной, например трифункциональной, антигенсвязывающей молекуле и ее терапевтическому применению, например, в иммунотерапии. Данная молекула представляет собой Fv-производное антитела. В определенных вариантах реализации изобретение относится к мультимерным, например, димерным, антигенсвязывающим молекулам.
Биспецифичные, то есть бифункциональные, антитела можно применять для поражения двух разных терапевтических мишеней или для выполнения двух различных функций. Подобные антитела можно применять, например, для привлечения иммуноэффекторной клетки, например, Т-клетки или NK-клетки, к конкретной клетке-мишени. Множество различных молекул на основе фрагментов антител известны и исследуются, например, в целях терапии рака.
Бифункциональные и димерные антитела можно сконструировать с использованием только вариабельных доменов антител. Например, последовательность линкера между доменами VH и VL можно укоротить до такой степени, что они не смогут складываться и связываться друг с другом внутримолекулярным способом. Подобные короткие линкеры, например, из 2-12 остатков, препятствуют указанному фолдингу молекулы одноцепочечного Fv (scFv) и благоприятствуют созданию межмолекулярных пар VH-VL между комплементарными вариабельными доменами различных полипептидных цепей, образующих димерное «диатело» (Holliger et al., 1993, Proc. Natl. Acad. Sci. USA 90, 6444-6448). Подобное диатело можно использовать для конструирования бифункциональных антител, которые получают путем нековалентной ассоциации двух одноцепочечных полипептидных продуктов гибридизации, каждый из которых состоит из домена VH от одного антитела, соединенного коротким линкером с доменом VL другого антитела.
В WO 03/025018 раскрыта биспецифичная и мультимерная антигенсвязывающая молекула, структура которой образована идентичными одноцепочечными полипептидами с по меньшей мере четырьмя связывающими доменами. Домен VH и домен VL в концевой части каждой полипептидной цепи связаны коротким линкером и ассоциируют межмолекулярно с соответствующими доменами VH и VL другой полипептидной цепи, тогда как другие домены VH и VL каждой полипептидной цепи связаны внутримолекулярно друг с другом в пределах одной и той же цепи, в результате чего образуется антигенсвязывающая scFv-единица. Подобные конструкции представляют собой гомодимеры, то есть они состоят из идентичных одноцепочечных полипептидов, ассоциированных друг с другом.
В рамках настоящей заявки предложены полифункциональные антигенсвязывающие молекулы, которые являются по меньшей мере трифункциональными. В некоторых вариантах реализации трифункциональная антигенсвязывающая молекула является по меньшей мере триспецифичной, то есть обладает специфичностью к по меньшей мере трем разным эпитопам антигена.
Антигенсвязывающая молекула согласно настоящему изобретению представляет собой Fv-производное, которое содержит только вариабельные (Fv) домены антитела, но лишено константных доменов антитела. Вариабельные (Fv) домены антитела антигенсвязывающий молекулы соединены друг с другом пептидным линкером или пептидной связью. Антигенсвязывающая молекула согласно настоящему изобретению может представлять собой мономер одной полипептидной цепи или мультимер многоцепочечного полипептида. Мультимерная антигенсвязывающая молекула может представлять собой, например, димер, имеющий две полипептидные цепи, тример, имеющий три полипептидные цепи, или тетрамер, имеющий четыре полипептидные цепи.
В некоторых вариантах реализации триспецифичная антигенсвязывающая молекула является по меньшей мере четырехвалентной. «Четырехвалентный» означает, что антигенсвязывающая молекула включает четыре антигенсвязывающих сайта, при этом каждый из антигенсвязывающих сайтов включает пару VH/VL, имеющую вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL) со специфичностью к одному и тому же эпитопу антигена, ассоциированные друг с другом. Таким образом, подобная четырехвалентная антигенсвязывающая молекула включает по меньшей мере восемь вариабельных доменов антитела, а именно четыре вариабельных домена тяжелой цепи (VH) и четыре вариабельных домена легкой цепи (VL). Триспецифичная и четырехвалентная антигенсвязывающая молекула включает антигенсвязывающий сайт, обладающий специфичностью против первого эпитопа антигена, антигенсвязывающий сайт, обладающий специфичностью против второго эпитопа антигена, и два антигенсвязывающих сайта, обладающих специфичностью против третьего эпитопа антигена. Таким образом, данная триспецифичная и четырехвалентная антигенсвязывающая молекула обладает разными специфичностями для трех разных эпитопов антигена. Например, подобная антигенсвязывающая молекула включает первый антигенсвязывающий сайт, обладающий специфичностью против первого эпитопа антигена, второй антигенсвязывающий сайт, обладающий специфичностью против второго эпитопа антигена, третий и четвертый антигенсвязывающие сайты, обладающие специфичностью против третьего эпитопа антигена. В некоторых вариантах реализации, где триспецифичная и четырехвалентная антигенсвязывающая молекула представляет собой мультимер, антигенсвязывающая молекула является гетеродимерной, то есть включает по меньшей мере две разные полипептидные цепи, при этом эти две полипептидные цепи отличаются по меньшей мере по одному вариабельному домену, например, одна полипептидная цепи включает только VH-домен, а другая включает только соответствующий VL-домен с той же специфичностью к эпитопу антигена.
Поскольку четырехвалентная антигенсвязывающая молекула включает восемь вариабельных доменов антитела, ее молекулярная масса составляет свыше 100 кДа, что приводит к более продолжительному периоду полувыведения подобной молекулы по сравнению с трехвалентными и триспецифичными молекулами одноцепочечного Fv-производного.
Кроме того, каждая триспецифичная и четырехвалентная антигенсвязывающая молекула включает два антигенсвязывающих сайта, обладающих специфичностью к одному и тому же эпитопу антигена. Таким образом повышена авидность, то есть сила взаимодействия между эпитопом антигена и антигенсвязывающей молекулой. Преимуществами более высокой авидности являются увеличенные стабильность взаимодействия и время удержания на мишени. Например, если мишенью является цитотоксичная иммуноэффекторная клетка, такая как Т-клетка или NK-клетка, более высокая авидность может привести к повышенной цитолитической активности антигенсвязывающей молекулы. В другом примере, если мишенью является опухолевая клетка, более высокая авидность улучшает время удержания на мишени и снижает показатели диссоциации с мишенью. В определенном варианте реализации настоящего изобретения триспецифичная и четырехвалентная антигенсвязывающая молекула включает первый и второй антигенсвязывающие сайты, специфичные к двум разным эпитопам антигена одного и того же вида опухолевой клетки, и третий и четвертый антигенсвязывающие сайты, специфичные к эпитопу антигена на иммуноэффекторной клетке, такой как Т-клетка или NK-клетка. Подобная антигенсвязывающая молекула приводит к повышенной специфичности, а также авидности, в отношении конкретного вида опухолевой клетки и к повышенной авидности в отношении активации рецептора на иммуноэффекторной клетке, что в результате дает преимущественно повышенную специфичную цитолитическую активность антигенсвязывающей молекулы. Связывание с двумя различными опухолевыми эпитопами антигена приводит к увеличению направленной специфичности и расширению терапевтического окна путем снижения побочных токсических явлений. Важно, что, несмотря на сложность структуры, подобная триспецифичная и четырехвалентная антигенсвязывающая молекула согласно настоящему изобретению является стабильной.
Следовательно, антигенсвязывающую молекулу согласно настоящему изобретению можно применять различными способами для перенаправления цитотоксической активности иммуноэффекторных клеток на разрушение опухолевых клеток или инфекционных агентов. В некоторых вариантах реализации триспецифичная антигенсвязывающая молекула может связываться с двумя разными эпитопами антигена на мишени. Например, два разных эпитопа могут быть на одном и том же антигене во избежание «ускользающих» мутантов или для усиления эффективности или два эпитопа могут быть на двух разных антигенах мишени. В других вариантах реализации триспецифичная антигенсвязывающая молекула может связываться с двумя разными эпитопами антигена на иммуноэффекторных клетках. Например, первый антигенсвязывающий сайт обладает специфичностью к активирующему рецептору, например, CD16A или CD3, и второй антигенсвязывающий сайт обладает специфичностью к костимулирующему рецептору, например, CD137 или CD28. В другом примере первый антигенсвязывающий сайт обладает специфичностью к CD16A и второй антигенсвязывающий сайт обладает специфичностью к другому активирующему рецептору на NK-клетках, например, NKG2D, DNAM, рецепторам NCR).
В другом варианте реализации триспецифичная антигенсвязывающая молекула имеет первый антигенсвязывающий сайт, обладающий специфичностью к эпитопу антигена на опухолевой клетке, второй антигенсвязывающий сайт, обладающий специфичностью к антигенсвязывающему эпитопу на иммуноэффекторной клетке, и третий антигенсвязывающий сайт, обладающий специфичностью к эпитопу антигена на растворимом белке, выбранном из группы, состоящей из факторов роста, цитокинов, хемокинов, митогенов и альбуминов. Примерами подобного растворимого белка являются интерлейкин-6 (ИЛ-6), фактор активации В-клеток (BAFF), индуцирующий пролиферацию лиганд (APRIL), трансформирующий фактор роста-бета (TGF-beta, ТФР-бета), ИЛ-10, сосудистый эндотелиальный фактор роста A (VEGF-A), гепарин-связывающий фактор роста, подобный эпидермальному фактору роста (HB-EGF), ангиопоетин-2 и сывороточный альбумин человека (САЧ).
В одном альтернативном варианте реализации антигенсвязывающая молекула имеет один антигенсвязывающий сайт, обладающий специфичностью к эпитопу антигена, презентированному на одном типе клетки, и три антигенсвязывающих сайта, обладающих специфичностью к эпитопам антигена на одном или нескольких других типах клеток.
«Эффекторные клетки» представляют собой клетки иммунной системы, которые могут стимулировать или запускать цитотоксичность, фагоцитоз, презентацию антигена, высвобождение цитокинов. Подобными эффекторными клетками являются, например, Т-клетки, клетки-естественные киллеры (NK-клетки), гранулоциты, моноциты, макрофаги, дендритные клетки и антиген-презентующие клетки, но не ограничиваются только ими. Примеры подходящих специфичностей к эффекторным клеткам включают (но не ограничиваются) CD2, CD3 и субъединицы CD3, такие как CD3ε, CD5, CD28 и другие компоненты Т-клеточного рецептора (TCR) в случае Т-клеток; CD16, CD16A, CD25, CD38, CD44, CD56, CD69, CD94, CD335 (NKp46), CD336 (NKp44), CD337 (NKp30), NKp80, NKG2C и NKG2D, DNAM, рецепторы NCR в случае NK-клеток; CD18, CD64 и CD89 в случае гранулоцитов; CD18, CD32, CD64, CD89 и маннозный рецептор в случае моноцитов и макрофагов; CD64 и маннозный рецептор в случае дендритных клеток; а также CD35. В определенных вариантах реализации настоящего изобретения данные специфичности, то есть молекулы клеточной поверхности, эффекторных клеток подходят для опосредования уничтожения клеток при связывании триспецифичной антигенсвязывающей молекулы с подобной молекулой клеточной поверхности и, таким образом, индуцирования цитолиза или апоптоза.
Антиген CD3 ассоциирован с комплексом Т-клеточного рецептора на Т-клетках. В случае, когда специфичность к эффекторной клетке представляет собой CD3, связывание антигенсвязывающей молекулы согласно настоящему изобретению с CD3 запускает цитотоксическую активность Т-клеток. Посредством связывания антигенсвязывающей молекулы с CD3 и с клеткой-мишенью, например, опухолевой клеткой, может быть индуцирован клеточный лизис клетки-мишени.
Антиген CD16A (FcγIIIA) представляет собой рецептор, экспрессируемый на поверхности NK-клеток. NK-клетки обладают врожденной цитолитической активностью, и посредством связывания антигенсвязывающей молекулы согласно настоящему изобретению с CD16 или CD16A может быть запущена цитотоксическая активность NK-клеток по отношению к мишени.
«Мишень» представляет собой сайт, на котором расположен эпитоп антигена и с которым должна связываться антигенсвязывающая молекула. Примерами мишеней являются клетки, инфекционные агенты, такие как вирусные или бактериальные патогены, например, вирус денге, простой вирус герпеса, вирус гриппа, вирус иммунодефицита человека (ВИЧ), вирус гепатита С, или клетки, несущие аутоиммунные мишени, такие как ИЛ-2/рецептор ИЛ-2 (IL2R), аутоиммунный маркер или аутоиммунный антиген, или опухолевые клетки. В вариантах реализации, где по меньшей мере один из антигенсвязывающих сайтов обладает специфичностью к эффекторной клетке, мишенью может быть опухолевая клетка, к которой должна быть перенаправлена эффекторная клетка для индуцирования или запуска соответствующего биологического, например, иммунного, ответа.
Подходящими специфичностями к опухолевым клеткам могут быть опухолевые антигены и антигены клеточной поверхности на соответствующей опухолевой клетке, например, специфичные опухолевые маркеры. Термин «опухолевый антиген», используемый в настоящей заявке, включает опухолеассоциированный антиген (ОАА) и опухолеспецифичный антиген (ОСА). Термин «опухолеассоциированный антиген» (ОАА), используемый в настоящей заявке, относится к белку, который присутствует на опухолевых клетках, и на нормальных клетках в ходе эмбрионального периода (раково-эмбриональные антигены), и после рождения в отдельных органах, но в значительно меньшей концентрации, чем на опухолевых клетках. ОАА также может присутствовать в строме вблизи опухолевой клетки, но экспрессироваться в меньших количествах в строме в другом месте в организме. Напротив, термин «опухолеспецифичный антиген» (ОСА) относится к белку, экспрессируемому опухолевыми клетками. Термин «антиген клеточной поверхности» относится к молекуле любого антигена или его фрагмента, который может быть распознан антителом на поверхности клетки. Примеры специфичностей к опухолевым клеткам включают CD19, CD20, CD26, CD29, CD30, CD33, CD52, CD200, CD267, рецептор эпидермального фактора роста (EGFR), EGFR2, EGFR3, EGFRvIII, HER2, HER3, рецептор инсулиноподобного фактора роста (IGFR), IGF-1R, молекулу адгезии эпителиальных клеток (Ер-САМ), плацентарную щелочную фосфатазу (PLAP), антиген Томсена-Фриденрайха (TF), член надсемейства рецепторов факторов некроза опухоли 17 (TNFRSF17), gpA33, MUC-1 (муцин), IGFR, CD5, альфа-рецептор ИЛ-4 (IL4-R альфа), рецептор ИЛ-13 (IL13-R), высокоаффинный рецептор IgE I типа (FcεRI), комплексы главного комплекса гистосовместимости класса I (MHCI) с пептидом и IgE, но не ограничиваются ими.
Антигенсвязывающие молекулы согласно настоящему изобретению, где специфичность к опухоли направлена в отношении антигена CD 19, можно применять для иммунотерапии В-клеточных опухолей, потому что антиген CD 19 экспрессируется на практически всех опухолях В-клеточного происхождения от острого лимфобластного лейкоза (ОЛЛ) до неходжкинской лимфомы (НХЛ).
Антигенсвязывающие молекулы согласно настоящему изобретению, где специфичность к опухоли направлена в отношении CD30, могут быть особенно полезными в терапии болезни Ходжкина и Т-клеточных лимфом.
Для увеличения периода полувыведения из сыворотки антигенсвязывающей молекулы согласно настоящему изобретению в организме антигенсвязывающая молекула при желании может быть присоединена к альбумину, например, САЧ, или пегилирована, сиалирована или гликозилирована (см., например, Stork et al., 2008, J. Biol. Chem., 283:7804-7812).
В некоторых вариантах реализации триспецифичная антигенсвязывающая молекула включает по меньшей мере один антигенсвязывающий сайт, при этом домены VH и VL пары VH/VL антигенсвязывающего сайта нековалентно связаны друг с другом, то есть домены VH и VL этой пары VH/VL не соединены пептидным линкером или пептидной связью. В определенных вариантах реализации данные нековалентно связанные домены VH и VL расположены на разных, то есть на первой и на второй, полипептидных цепях мультимерной антигенсвязывающей молекулы. В других вариантах реализации данные нековалентно связанные домены VH и VL расположены на одной и той же полипептидной цепи мономерной антигенсвязывающей молекулы, при этом по меньшей мере другой вариабельный домен размещен на мономере между этими нековалентно связанными доменами VH и VL. В некоторых вариантах реализации каждый из данных нековалентно связанных доменов VH и VL этого антигенсвязывающего сайта связан пептидным линкером или пептидной связью с доменом VH или VL второй пары VH/VL смежного антигенсвязывающего сайта. Предпочтительно, чтобы подобный пептидный линкер к домену VH или VL пары VH/VL смежного антигенсвязывающего сайта был коротким, чтобы предотвратить внутримолекулярный фолдинг между смежными доменами и подтолкнуть ассоциацию двух нековалентно связанных доменов VH и VL друг с другом. Например, пептидный линкер включает 12 или менее аминокислотных остатков, предпочтительно от 3 до 9 аминокислотных остатков. Подобное формирование по меньшей мере одного антигенсвязывающего сайта двумя нековалентно связанными доменами VH и VL дает преимущество в отношении стабильности антигенсвязывающей молекулы, потому что приводит к получению более компактной антигенсвязывающей молекулы.
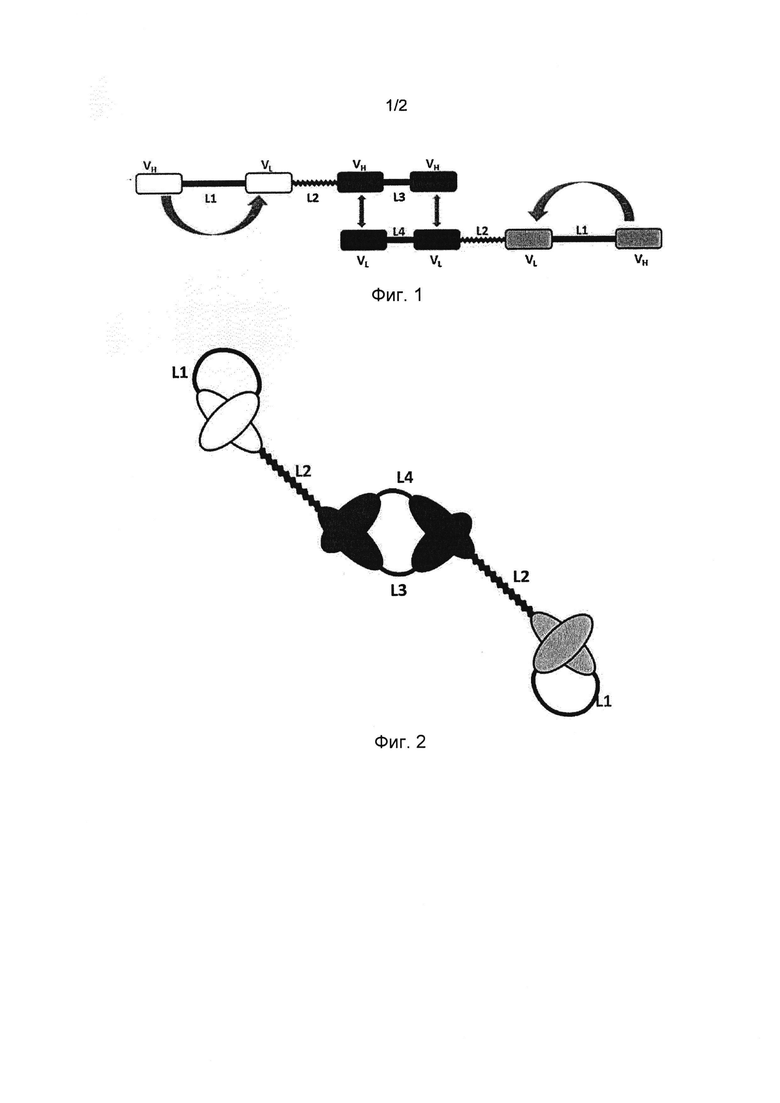
Например, на Фиг. 1 и 2 показана триспецифичная антигенсвязывающая молекула, где домены VH и VL центральных пар VH/VL (показаны черным цветом) нековалентно связаны друг с другом. В этом примере нековалентно связанные домены VH и VL расположены на разных полипептидных цепях. Каждый из данных нековалентно связанных доменов VH и VL этого антигенсвязывающего сайта связан пептидным линкером L3 или L4 с доменом VH или VL второй пары VH/VL смежного антигенсвязывающего сайта.
В дополнительных вариантах реализации триспецифичная антигенсвязывающая молекула включает по меньшей мере один первый антигенсвязывающий сайт, при этом домены VH и VL пары VH/VL данного первого антигенсвязывающего сайта нековалентно связаны друг с другом, то есть домены VH и VL этой пары VH/ VL не соединены пептидным линкером или пептидной связью, и нековалентно связанный домен VH данного первого антигенсвязывающего сайта связан пептидным линкером с доменом VH пары VH/VL второго антигенсвязывающего сайта, расположенного смежно с первым антигенсвязывающим сайтом, и нековалентно связанный домен VL первого антигенсвязывающего сайта связан пептидным линкером с доменом VL пары VH/VL второго антигенсвязывающего сайта, расположенного смежно с первым антигенсвязывающим сайтом. В вариантах реализации, где антигенсвязывающая молекула является одноцепочечным, то есть мономерным, полипептидом, домены VH и VL размещены на одной и той же полипептидной цепи. В вариантах реализации, где антигенсвязывающая молекула является мультимерным, то есть многоцепочечным, полипептидом, домен VH первого антигенсвязывающего сайта, связанный пептидным линкером или пептидной связью с доменом VH второго антигенсвязывающего сайта, расположен на первом полипептиде, и домен VL первого антигенсвязывающего сайта, связанный пептидным линкером или пептидной связью с доменом VL второго антигенсвязывающего сайта, расположен на втором полипептиде. Предпочтительно, чтобы пептидный линкер был коротким, например, менее 12 аминокислотных остатков, предпочтительно от 3 до 9 аминокислотных остатков, чтобы предотвратить внутримолекулярный фолдинг между смежными доменами VH-VH и смежными доменами VL-VL соответственно и подтолкнуть ассоциацию доменов VH-VH с доменами VL-VL для образования первого и второго антигенсвязывающих сайтов. Данное расположение доменов VH-VH и VL-VL облегчает правильный фолдинг триспецифичной антигенсвязывающей молекулы.
Термин «антигенсвязывающая молекула» относится к молекуле производного иммуноглобулина с мультивалентными антигенсвязывающими свойствами, предпочтительно имеющей по меньшей мере четыре антигенсвязывающих сайта. Антигенсвязывающая молекула может быть одноцепочечным, то есть мономерным, полипептидом или многоцепочечным, то есть мультимерным, полипептидом. Каждый полипептид антигенсвязывающей молекулы включает вариабельные (Fv) домены антитела, соединенные друг с другом пептидным линкером или пептидной связью. Каждый антигенсвязывающий сайт образован присущим антителам, то есть иммуноглобулиновым, вариабельным доменом тяжелой цепи (VH) и вариабельным доменом антитела легкой цепи (VL), которые связываются с одним и тем же эпитопом антигена. Эпитоп антигена может быть на одном и том же или на разных антигенах. Предпочтительно, чтобы антигенсвязывающая молекула согласно настоящему изобретению была лишена константных доменов иммуноглобулина или их фрагментов.
Термин «полипептид» относится к полимеру из аминокислотных остатков, соединенных амидными связями. Полипептид является предпочтительно одноцепочечным гибридным белком, который не имеет разветвлений. В полипептиде вариабельные (Fv) домены антитела соединены друг за другом. Полипептид может иметь последовательные аминокислотные остатки в дополнении к N-концевым и/или С-концевым. Например, полипептид может содержать маркерную последовательность (Tag-последовательность), предпочтительно на С-конце, которая может быть полезна для очистки полипептида. Примерами маркерных последовательностей являются His-маркер (гистидиновый маркер), например, His-маркер, состоящий из шести остатков гистидина, маркер FLAG, например, октапептид DYKDDDDK (SEQ ID NO: 5), или маркер STREP® II, например, октапептид WSHPQFEK (SEQ ID NO: 6). В случае мультимерной антигенсвязывающей молекулы предпочтительно, чтобы для разных полипептидов использовали разные маркерные последовательности.
Касательно аминокислотного состава пептидных линкеров, выбирают пептиды, которые не препятствуют ассоциации доменов, а также не препятствуют мультимеризации, например, димеризации, мультимерных молекул. Например, линкеры, включающие остатки глицина и серина главным образом обеспечивают устойчивость к действию протеаз. Аминокислотную последовательность линкеров можно оптимизировать, например, с помощью способов с использованием фаговых дисплеев для улучшения связывания с антигенами и конечного выхода продукта антигенсвязывающей молекулы. В одном варианте реализации применяют пептидные линкеры (G2S)x.
В некоторых вариантах реализации настоящего изобретения по меньшей мере один, предпочтительно все, вариабельные домены антитела являются полностью человеческими, гуманизированными или химерными доменами. Гуманизированные антитела можно получить хорошо разработанными способами, такими как, например, пересадка определяющих комплементарность участков (CDR) (см., например, Antibody engineering: methods and protocols / edited by Benny K.C. Lo; Benny K.C. II Series: Methods in molecular biology (Totowa, N.J.). Таким образом, специалист способен легко создать гуманизированную или полностью человеческую версию антигенсвязывающей молекулы и вариабельных доменов из нечеловеческих, например, мышиных или не относящихся к приматам, источников происхождения с помощью стандартных способов молекулярной биологии, известных в данной области техники, для снижения иммуногенности и улучшения эффективности действия антигенсвязывающей молекулы в иммунной системе человека. В одном предпочтительном варианте реализации настоящего изобретения все вариабельные домены антитела являются гуманизированными или полностью человеческими; наиболее предпочтительно, антигенсвязывающая молекула согласно настоящему изобретению является гуманизированной или полностью человеческой. Термин «полностью человеческий», используемый в настоящей заявке, означает, что аминокислотные последовательности вариабельных доменов и пептидов, соединяющих вариабельные домены в полипептиде, имеют человеческое происхождение или могут быть обнаружены у человека. В определенных вариантах реализации настоящего изобретения вариабельные домены могут быть человеческими или гуманизированными, но не пептиды, соединяющие вариабельные домены антитела.
В некоторых вариантах реализации согласно настоящему изобретению предложен полифункциональный антигенсвязывающий полипептидный мультимер.
В некоторых вариантах реализации согласно настоящему изобретению предложена антигенсвязывающая молекула трифункционального антигенсвязывающего полипептидного мультимера, разработанная для направленного воздействия на три разных антигена или эпитопа. Подобный мультимер включает первый полипептид и второй полипептид. Каждый из двух полипептидов является одноцепочечным гибридным пептидом, имеющим по меньшей мере четыре вариабельных домена антитела, соединенные друг за другом от N-конца к С-концу каждого полипептида. Каждый из полипептидов включает два вариабельных домена антитела, соединенных коротким линкером для предотвращения внутримолекулярного спаривания в пределах одного и того же полипептида, и одноцепочечную Fv-единицу, имеющую пару вариабельных доменов антитела из двух других вариабельных доменов, способных внутримолекулярно образовывать антигенсвязывающий сайт парой вариабельных доменов в пределах одного и того же полипептида. Мультимер образуется путем нековалентной ассоциацией между двумя полипептидами, тогда как два вариабельных домена антитела, соединенные коротким линкером одного полипептида, ассоциируют с двумя соответствующими вариабельными доменами антитела другого полипептида, тем самым формируя два дополнительных антигенсвязывающих сайта. Таким образом, этот мультимер включает по меньшей мере четыре антигенсвязывающих сайта и является по меньшей мере четырехвалентным. В конкретном аспекте настоящего изобретения мультимер представляет собой димер, то есть состоит из двух полипептидных цепей.
Для создания подобного триспецифичного и четырехвалентного антигенсвязывающего димера два полипептида должны иметь разные составы вариабельных доменов антитела, потому что по отношению к по меньшей мере одной из трех специфичностей соответствующие вариабельный домен легкой цепи антитела и вариабельный домен тяжелой цепи антитела должны быть вставлены в разные полипептиды, так что один из полипептидов содержит только вариабельный домен тяжелой цепи, а другой полипептид содержит только вариабельный домен легкой цепи для этой специфичности. Таким образом, подобный димер согласно настоящему изобретению является гетеродимерным, потому что он состоит из двух разных полипептидов.
Можно предпринять особые меры для обеспечения возможности правильной ассоциации двух разных полипептидов, включающих вариабельные домены антитела для трех разных специфичностей, и для предотвращения неправильной гомодимеризации между двумя идентичными полипептидами. Например, авторы настоящего изобретения достигали правильной гетеродимеризации между двумя разными триспецифичными полипептидами путем вставки двух вариабельных доменов тяжелой цепи антитела, соединенных коротким линкером, в один полипептид и вставки двух соответствующих вариабельных доменов легкой цепи антитела, соединенных коротким линкером, в другой полипептид. Удивительно, образовывались только гетеродимерные виды триспецифичных антигенсвязывающих полипептидных димеров.
Таким образом, в одном варианте реализации настоящего изобретения предложена
триспецифичная антигенсвязывающая молекула, при этом указанная молекула является триспецифичным антигенсвязывающим полипептидным димером, включающим первый полипептид и второй полипептид, каждый полипептид, имеющий по меньшей мере четыре вариабельных домена антитела, соединенных друг за другом, и
(а) указанный первый полипептид включает первый и второй вариабельные домены тяжелой цепи антитела (VH), соединенные друг с другом первым линкером, препятствующим внутримолекулярному спариванию в пределах одного и того же полипептида, например, из приблизительно 12 или менее аминокислотных остатков, и антигенсвязывающую одноцепочечную Fv-единицу, имеющую третий вариабельный домен тяжелой цепи антитела (VH), соединенный вторым пептидным линкером с вариабельным доменом легкой цепи антитела (VL), указанный третий вариабельный домен тяжелой цепи антитела (VH) и вариабельный домен легкой цепи антитела (VL) способны ассоциировать с образованием первого антигенсвязывающего сайта, при этом второй вариабельный домен тяжелой цепи антитела (VH) соединен с антигенсвязывающей одноцепочечной Fv-единицей третьим пептидным линкером; и
(b) указанный второй полипептид включает первый и второй вариабельные домены легкой цепи антитела (VL), соединенные друг с другом вторым пептидным линкером, препятствующим внутримолекулярному спариванию в пределах одного и того же полипептида, например, из приблизительно 12 или менее аминокислотных остатков, и антигенсвязывающую одноцепочечную Fv-единицу, имеющую третий вариабельный домен легкой цепи антитела (VL), соединенный вторым пептидным линкером с вариабельным доменом тяжелой цепи антитела (VH), указанный третий вариабельный домен легкой цепи антитела (VL) и вариабельный домен тяжелой цепи антитела (VH) способны ассоциировать с образованием второго антигенсвязывающего сайта, при этом второй вариабельный домен легкой цепи антитела (VL) соединен с антигенсвязывающей одноцепочечной Fv-единицей третьим пептидным линкером; и
(c) первый и второй вариабельные домены тяжелой цепи антитела (VH) первого полипептида ассоциируют с первым и вторым вариабельными доменами легкой цепи антитела (VL) второго полимера с образованием двух дополнительных, то есть третьего и четвертого, антигенсвязывающих сайтов, тогда как в одном предпочтительном варианте реализации первый вариабельный домен тяжелой цепи антитела (VH) первого полипептида ассоциирует со второй областью легкой цепи антитела (VL) второго полипептида с образованием третьего антигенсвязывающего сайта, и второй вариабельный домен тяжелой цепи антитела (VH) первого полипептида ассоциирует с первым вариабельным доменом легкой цепи антитела (VL) второго полипептида с образованием четвертого антигенсвязывающего сайта.
Триспецифичный антигенсвязывающий полипептидный димер образуется, когда два из указанных четырех антигенсвязывающих сайта специфичны к одному и тому же антигену.
Подобный триспецифичный димер распознает три разных специфичности и может направленно воздействовать, например, на два разных антигена или эпитопа на клетке-мишени и с учетом третьей функциональности, то есть специфичности, связываться, например, с иммуноэффекторной клеткой, такой как, например, Т-клетка или NK-клетка.
Триспецифичный димер согласно настоящему изобретению можно применять различными способами.
Например, вариабельные домены антитела могут быть размещены в полипептиде так, что два вариабельных домена антитела, ассоциированные с двумя соответствующими вариабельными доменами антитела другого полипептида, могут быть расположены, например, на N-конце или С-конце полипептида. Эти два вариабельных домена антитела могут иметь одну и ту же специфичность или отличающиеся специфичности. Например, оба могут быть специфичны к одной и той же иммуноэффекторной клетке или иметь различные специфичности к двум антигенам на опухолевой клетке.
Дополнительно, два вариабельных домена, образующих одноцепочечную Fv-единицу, могут располагаться, например, в последовательности VH-VL или VL-VH в направлении от N-конца к С-концу полипептида. Одноцепочечные Fv-единицы двух димеризованных полипептидов могут иметь одинаковые или разные специфичности. Например, если два вариабельных домена антитела, ассоциирующих с двумя соответствующими вариабельными доменами антитела другого полипептида, обладают одинаковой специфичностью, одноцепочечные Fv-единицы двух полипептидов обладают разными специфичностями, чтобы получить триспецифичный димер.
Таким образом, по меньшей мере четыре вариабельных домена антитела могут быть размещены, например, так, что два вариабельных домена антитела, ассоциирующих с двумя соответствующими вариабельными доменами антитела другого полипептида, являются специфичными к иммуноэффекторной клетке и одноцепочечные Fv-единицы двух полипептидов обладают специфичностями к двум различным опухолевым антигенам или два вариабельных домена антитела, ассоциирующих с двумя соответствующими вариабельными доменами антитела другого полипептида, являются специфичными к разным опухолевым антигенам и обе одноцепочечные Fv-единицы двух полипептидов обладают одинаковой специфичностью к иммуноэффекторной клетке.
Антигенсвязывающий полипептид представляет собой «димер», термин, который относится к комплексу из первого и второго полипептидного мономера. В одном из аспектов антигенсвязывающий полипептидный димер представляет собой «гетеродимер», термин, который означает, что антигенсвязывающий полипептид состоит из двух разных полипептидных мономеров, которые кодируются двумя различными полинуклеотидами.
Предпочтительно, чтобы в антигенсвязывающем димере первый и второй полипептиды были нековалентно ассоциированы друг с другом, в частности, с условием, что между первым и вторым полипептидом не существует ковалентной связи. Однако при желании два полипептида могут быть дополнительно стабилизированы с помощью по меньшей мере одной ковалентной связи, например, с помощью дисульфидного мостика между остатками цистеина у разных полипептидов.
Длина линкеров влияет на гибкость антигенсвязывающего полипептидного димера. Требуемая гибкость антигенсвязывающего полипептидного димера зависит от плотности антигена-мишени и доступности антигена-мишени, то есть эпитопов. Более длинные линкеры обеспечивают получение более гибкого антигенсвязывающего полипептидного димера с более подвижными антигенсвязывающими сайтами. Влияние длины линкера на формирование димерных антигенсвязывающих полипептидов описано, например, в Todorovska et al., 2001 Journal of Immunological Methods 248:47-66; Perisic et al., 1994 Structure 2:1217-1226; Le Gall et al., 2004, Protein Engineering 17:357-366 и WO 94/13804.
Согласно настоящему изобретению предпочтительно, чтобы длина первого пептидного линкера первого и второго вариабельных доменов тяжелой цепи антитела первого полипептида и первого и второго вариабельных доменов легкой цепи антитела второго полипептида была такой, что домены первого полипептида могли межмолекулярно ассоциировать с доменами второго полипептида с образованием димерного антигенсвязывающего полипептида. Подобные линкеры являются «короткими», то есть состоят из 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или приблизительно 12 аминокислотных остатков. В случае 0 аминокислотных остатков линкер представляет собой пептидную связь. Подобные короткие линкеры способствуют правильной димеризации первого полипептида со вторым путем связывания и образования антигенсвязывающих сайтов между вариабельными доменами легкой цепи антитела и доменами тяжелой цепи антитела разных полипептидов. Укорачивание линкера до приблизительно 12 или менее аминокислотных остатков главным образом препятствует взаимодействию соседних доменов одной и той же полипептидной цепи друг с другом. В одном варианте реализации настоящего изобретения данные линкеры состоят из приблизительно 3 до приблизительно 10, например, 7 последовательных аминокислотных остатков. Кроме того, в принципе возможно, что два полипептида, имеющих линкер с более 12 аминокислотными остатками между вариабельными доменами антитела, правильно димеризуются друг с другом (см., например, Le Gall et al., 2004, Protein Engineering 17:357-366).
В случае одноцепочечных Fv-единиц полипептидов второй пептидный линкер является длинным и гибким (в основном состоит из приблизительно 12 или более аминокислотных остатков) для фолдинга внутримолекулярно по типу «голова к хвосту» и образования антигенсвязывающей одноцепочечной (scFv) единицы. Дополнительные аминокислотные остатки обеспечивают дополнительную гибкость. Например, данный линкер между доменами VH и VL или VL и VH одноцепочечной Fv-единицы в полипептиде может состоять из приблизительно 12 до приблизительно 35, в частности, от 15 до 25 последовательных аминокислотных остатков.
Третий пептидный линкер полипептида для соединения одноцепочечной Fv-единицы с двумя другими вариабельными доменами антитела, которые ассоциируют с соответствующими вариабельными доменами другого полипептида, может быть длиною, например, от 5 до 30, предпочтительно по меньшей мере 6, 7, 8, 9, 10, 11 или 12 последовательных аминокислотных остатков.
В одном варианте реализации настоящего изобретения триспецифичный антигенсвязывающий полипептидный димер является биспецифичным к двум различным антигенам на опухолевой клетке и дополнительно специфичным к эффекторной клетке, в частности, Т-клетке или NK-клетке. Подходящими специфичностями к опухолевым клеткам могут быть опухолевые антигены и антигены клеточной поверхности на соответствующей опухолевой клетке, например, специфичные опухолевые маркеры. Подобный триспецифичный антигенсвязывающий димер связывается бифункционально с опухолевой клеткой и с иммуноэффекторной клеткой, тем самым запуская цитотоксический ответ, индуцированный Т-клеткой или NK-клеткой.
Антигенсвязывающая молекула согласно любому из вариантов реализации, описанных в настоящей заявке ранее, можно получить путем экспрессии полинуклеотидов, кодирующих отдельные полипептидные цепи, которые образуют антигенсвязывающую молекулу. Следовательно, дополнительный вариант реализации настоящего изобретения представляет собой полинуклеотиды, например, ДНК или РНК, кодирующие полипептидные цепи антигенсвязывающей молекулы, как описано в настоящей заявке выше.
Полинуклеотиды можно сконструировать способами, известными специалисту, например, путем объединения генов, кодирующих вариабельные домены антитела, либо разделенные пептидными линкерами, либо напрямую соединенными пептидной связью полипептидов, в генетической конструкции, функционально связанной с подходящим промотором и необязательно подходящим терминатором транскрипции, и экспрессии конструкции в бактериальной или другой подходящей экспрессионной системе, такой как, например, клетки яичника китайского хомячка (клетки СНО). В зависимости от векторной системы и используемого хозяина можно использовать любое число подходящих транскрипционных и трансляционных элементов, включая конститутивные и индуцируемые промоторы. Промотор выбирают такой, чтобы он запускал экспрессию полинуклеотидов в соответствующей клетке-хозяине.
Полинуклеотиды можно вставлять в вектора, предпочтительно экспрессионные вектора, что представляет собой дополнительный вариант реализации настоящего изобретения. Данные рекомбинантные вектора можно сконструировать согласно способам, хорошо известным специалисту в данной области техники.
Можно применить множество различных систем экспрессионный вектор-хозяин для включения в состав и экспрессии полинуклеотидов, кодирующих полипептидные цепи согласно настоящему изобретению. Примерами экспрессионных векторов для экспрессии в E.coli является pSKK (LeGall et al., J Immunol Methods. (2004) 285(1):111-27) или pcDNA5 (Invitrogen) для экспрессии в клетках млекопитающих.
Таким образом, антигенсвязывающую молекулу, как описано в настоящей заявке, можно получить посредством введения вектора, кодирующего полипептидные цепи, как описано выше, в клетку-хозяина и культивирования указанной клетки-хозяина в условиях, при которых полипептидные цепи экспрессируют, могут выделять и необязательно дополнительно очищать.
В дополнительном варианте реализации настоящего изобретения предложены композиции, включающие антигенсвязывающую молекулу или полинуклеотиды, как описано в настоящей заявке выше, и по меньшей мере один дополнительный компонент.
Согласно настоящему изобретению дополнительно предложен способ, где антигенсвязывающую молекулу, как описано в настоящей заявке выше, вводят в эффективной дозе субъекту, например, пациенту, для лечения рака (например, неходжкинской лимфомы; хронического лимфоцитарного лейкоза). Антигенсвязывающую молекулу можно применять в качестве лекарственного средства.
Специалист легко способен без лишних трудностей сконструировать и получить антигенсвязывающую молекулу, описанную в настоящей заявке, используя разработанные техники и стандартные способы, известные в данной области техники, см., например, Sambrook, Molecular Cloning A Laboratory Manual, Cold Spring Harbor Laboratory (1989) N.Y.; The Protein Protocols Handbook, edited by John M. Walker, Humana Press Inc. (2002); или Antibody engineering: methods and protocols / edited by Benny K.C. Lo; Benny K.C. II Series: Methods in molecular biology (Totowa, N.J.)).
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На Фигуре 1 показан первый и второй полипептиды для образования трифункционального, то есть триспецифичного, антигенсвязывающего полипептидного димера согласно настоящему изобретению. Первый полипептид имеет четыре вариабельных домена антитела VH, VL, VH, VH, соединенных друг за другом. Первый и второй вариабельные домены антитела VH (черные) обладают одинаковой первой специфичностью и соединены коротким линкером L3 для предотвращения внутримолекулярного спаривания в пределах одного и того же полипептида, и одноцепочечная Fv-единица, имеющая пару вариабельных доменов антитела из других третьего вариабельного домена антитела VL и четвертого вариабельного домена антитела VH (белые), соединенных вторым линкером L1, способная внутримолекулярно образовывать антигенсвязывающий сайт со второй специфичностью с помощью пары вариабельных доменов в пределах одного и того же полипептида. Второй вариабельный домен антитела VH и третий вариабельный домен антитела VL с разными специфичностями соединены третьим линкером L2.
Второй полипептид имеет четыре вариабельных домена антитела VL, VL, VL, VH, соединенных друг за другом. Первый и второй вариабельные домены антитела VL (черные) имеют одинаковую первую специфичность и соединены коротким линкером L4 для предотвращения внутримолекулярного спаривания в пределах одного и того же полипептида, и одноцепочечная Fv-единица, имеющая пару вариабельных доменов антитела из других третьего вариабельного домена антитела VL и четвертого вариабельного домена антитела VH, имеющих третью специфичность (серые) и соединенных вторым линкером L1, способная внутримолекулярно образовывать антигенсвязывающий сайт с помощью пары вариабельных доменов в пределах одного и того же полипептида. Второй вариабельный домен антитела VL и третий вариабельный домен антитела VL с разными специфичностями соединены третьим линкером L2.
На Фигуре 2 показан антигенсвязывающий полипептидный димер, образованный нековалентной ассоциацией между двумя полипептидами Фигуры 1, тогда как два вариабельных домена антитела VH, соединенных коротким линкером первого полипептида, ассоциируют с двумя соответствующими вариабельными доменами антитела VL второго полипептида, тем самым образуя два антигенсвязывающих сайта, имеющих одинаковую специфичность (черные), тогда как вторая специфичность обеспечена одноцепочечной Fv-единицей первого полипептида (белый цвет) и третья специфичность обеспечена одноцепочечной Fv-единицей второго полипептида (серый цвет).
На Фигуре 3 показана трифункциональная антигенсвязывающая молекула, в частности, трифункциональный антигенсвязывающий полипептид, который согласно настоящему изобретению является триспецифичным антителом для двойного направленного воздействия на опухолевые клетки. Данное антитело, то есть антигенсвязывающий полипептид, разработан для направленного воздействия на две разные мишени/эпитопа на опухолевой клетке и с учетом третьей функциональности для связывания с высокой аффинностью с эффекторной клеткой. Антигенсвязывающий полипептид состоит из четырех антигенсвязывающих сайтов, при этом два центральных антигенсвязывающих сайта связываются с двумя разными антигенами на опухолевой клетке и два периферийных антигенсвязывающих сайта связываются с эффекторной клеткой.
Нижеследующие примеры дополнительно иллюстрируют настоящее изобретения без ограничения объема настоящего изобретения.
Пример 1
ДНК-конструкции:
Плазмидную ДНК, кодирующую полипептидные цепи, получают с помощью генной инженерии ДНК или синтеза генов и секвенирования. Экспрессионные конструкции для транзиентной или стабильной трансфекции клеток млекопитающих основаны на эукариотическом экспрессионном векторе pCDNA5/FRT (Life Technologies) и включают интересующий продукт гена под контролем вирусного или убиквитарного промотора, а также кассету устойчивости к гигромицину в качестве селективного маркера. Для очистки и анализа полипептидные цепи продукта экспрессируют с His-маркером, FLAG-маркером или StrepII-маркером.
Клеточные линии и культивирование клеток:
Клетки Flp-In СНО (Life Technologies), производные от клеток яичника китайского хомячка СНО-К1 (АТСС, CCL-61) (Као and Puck, 1968), культивируют в питательной смеси Хэма F-12, в которую добавили L-глутамин, 10% эмбриональную телячью сыворотку (ЭТС) и 100 мкг/мл зеоцина. Адгезивные клетки отделяли от поверхности 0,25% раствором трипсин-ЭДТА и пересеивают согласно стандартным протоколам культивирования клеток.
Для адаптации к росту в суспензии клетки отделяли от поверхности флаконов для культивирования тканей и помещают в бессывороточную среду для последующей инкубации во встряхиваемых колбах (Corning) при 37°С, 5% СО2 и скорости 120 оборотов/мин. Стандартной средой для культивирования адаптированных к росту в суспензии клеток Flp-In СНО является среда HyClone CDM4 СНО (Thermo Scientific), в которую добавили L-глутамин (Life Technologies), гипоксантин и тимидин - НТ Supplement (Life Technologies), пенициллин/стрептомицин (Life Technologies) и 100 мкг/мл зеоцина (Life Technologies). Адаптированные к росту в суспензии клетки пересеивают каждые 2-3 дня с плотностями посева, составляющими от 2×106 до 3×106 клеток/мл. Концентрацию клеток и жизнеспособность определяют у всех культур с помощью теста на вытеснения трипанового синего. Клетки криоконсервируют в среде с 10% диметилсульфоксидом (ДМСО) и проверяют на отсутствие микоплазмы с использованием набора MycoAlert Mycoplasma detection Kit (Lonza).
Получение стабильно трансфицированных популяций клеток:
Рекомбинантные линии клеток Flp-In СНО, стабильно экспрессирующие триспецифичные потенциальные антитела, получают трансфекцией адаптированных к росту в суспензии клеток. Для этого клетки помещают в стандартную среду без зеоцина за день до котрансфекции экспрессионными плазмидами (2,5 мкг), кодирующими интересующий белок (pcDNA5/FRT) и Flp-рекомбиназу (pOG44, Life Technologies), с использованием полиэтиленимина (ПЭИ). Вкратце, векторную ДНК и реагент для трансфекции смешивают с массовым соотношении ДНК:ПЭИ 1:3 в общем объеме 100 мкл среды OptiMEM I (Life Technologies) и инкубируют в течение 10 минут перед добавлением 2×106 клеток Flp-In СНО, суспендированных в 1 мл среды CHO-S-SFMII (Life Technologies). После инкубации в течение 24 часов начинают селекцию стабильно трансфицированных клеток путем добавления 500 мкг/мл гигромицина Б вслед за разбавлением культур до плотности 0,1×106 жизнеспособных клеток/мл в среде CHO-S-SFMII и посевом в культуральные флаконы Т75. Flp-рекомбиназа опосредует вставку Flp-In-экспрессионной конструкции в геном на месте интегрированного FRT-сайта посредством сайт-специфичной ДНК-рекомбинации (О' Gorman et al 1991). В процессе селекции плотности жизнеспособных клеток измеряют два раза в неделю, и клетки центрифугируют и ресуспендируют в свежей среде для селекции с максимальной плотностью 0,1×106 жизнеспособных клеток/мл. Популяции клеток, стабильно экспрессирующие продукты рекомбинантные белковые продукты, выделяют после приблизительно 3 недель селекции, в этот момент клетки переносят на стандартную культуральную среду во встряхиваемых колбах. Экспрессию рекомбинантных секретируемых белков подтверждают белковым гель-электрофорезом супернатантов клеточных культур с использованием технологии Criterion Stain-Free (Bio-Rad). Стабильные популяции клеток криоконсервируют в среде, содержащей 50% ProFreeze (Lonza) и 7,5% ДМСО.
Получение рекомбинантного белка в суспензионных культурах клеток СНО с подпиткой:
Рекомбинантные белки получают в 10-дневных подпитываемых культурах стабильно трансфицированных линий клеток СНО путем секреции в культуральный супернатант. Для этого производят посев популяций клеток, стабильно экспрессирующих интересующий продукт, с начальными плотностями 6×105 клеток/мл в стандартной культуральной среде в поликарбонатных колбах Эрленмейера с газопроницаемыми крышками (Corning) и инкубируют при 37°С и 5% CO2 со скоростью встряхивания 140 оборотов/мин. В процессе культивирования с подпиткой в среду добавляют 40 мл/л ActiCHO Feed А (РАА) и 4 мл/л ActiCHO Feed В (РАА) в 0 день (начальный день) и двойные количества на 3, 5 и 7 дни. Собирают супернатанты клеточных культур после 10 дней при жизнеспособности культур, составляющей, как правило, более 75%. Отбирают образцы из производственных культур через день перед подпиткой и оценивают плотность и жизнеспособность клеток. В день сбора супернатанты клеточных культур очищают центрифугированием и вакуумной фильтрацией (0,22 мкм) с использованием мембранных фильтров Millipore Express PLUS Membrane Filters (Millipore) перед дальнейшим применением.
Определение титра экспрессии:
Титры экспрессии белка и целостность продукта в культуральных супернатантах анализируют электрофорезом в полиакриламидном геле с додецилсульфатом натрия (ДСН-ПААГ-электрофорез) с использованием системы визуализации гель-электрофореза Criterion Stain-Free (Bio-Rad) на 7 и 10 дни (перед и после фильтрации на фильтре с размерами пор 0,22 мкм). Титры продукта определяют полуколичественно путем сравнения с референтным белком известной концентрации.
Очистка триспецифичных антигенсвязывающих полипептидов:
Продукты с His-маркером очищают из супернатантов культур клеток СНО двуэтапной процедурой, включающей Ni-NTA-хроматографию (хроматографию на сорбенте с никель-нитрилотриацетатной кислотой) и препаративную гель-хроматографию. Во-первых, супернатанты очищают вакуумной фильтрацией (0,22 мкм) и подводят до 5 мМ имидазола перед загрузкой на хроматографическую колонку HisTrap FF (GE Healthcare), уравновешенную в буфере А для аффинной хроматографии на иммобилизованных ионах металла (IMAC), с линейной скоростью потока 5 мл/мин. После этого колонки промывают 5 объемами буфера А для IMAC и 10 объемами смеси буфера А для IMAC и буфера В для IMAC (7%). Продукты с His-маркером затем элюируют последовательным промыванием 10 объемами 30% буфера В для IMAC и 5 объемами 100% буфера В для IMAC при той же линейной скорости потока. Собирают 2,5 мл фракции элюата и оценивают содержание и чистоту белка, подвергая каждую фракцию одномерному ДСН-ПААГ-электрофорезу с последующей визуализацией белка с использованием технологии Criterion Stain-Free (Bio-Rad). Объединяют фракции, содержащие продукты, и концентрируют ультрафильтрацией. Далее сконцентрированные образцы очищают гель-фильтрацией с использованием колонки HiLoad 26/600 Superdex 200 pg (GE Healthcare) и элюируют в буфере для гель-хроматографии (20 мМ Трис-HCl, 100 мМ NaCl, рН 7,5) со скоростью 2,5 мл/мин. Фракции, содержащие очищенный продукт, как определено путем сравнения объемов элюирования с задержкой в колонке маркерных белков с различной молекулярной массой (GE Healthcare), собирают и объединяют. После последней замены буфера (10 мМ ацетат натрия, рН 5,0) с использованием высаливающей колонки PD-10 (GE Healthcare) образцы концентрируют до концентрации 1,0-1,5 мг/мл ультрафильтрацией, как описано выше. Чистоту и гомогенность (как правило, более 90%) конечных образцов оценивают системой визуализации гель-электрофореза белков Criterion Stain-Free после ДСН-ПААГ в восстанавливающих и в невосстанавливающих условиях, как описано выше, в отдельных случаях с последующим иммуноблоттингом со специфичными антителами и аналитической гель-хроматографией соответственно. Очищенные белки хранят в виде аликвот при -80°С до дальнейшего применения.
Пример 2. Триспецифичные молекулы CD3xCD19xCD30
Антигенсвязывающие полипептидные димеры, содержащие CD3-, CD 19- и CD30-вариабельные связывающие домены антитела, которые происходят от антител ОКТ3, HD37 и HRS3 соответственно, получают согласно Примеру 1:
Триспецифичное антитело 1:
VH(CD3)-(G2S)2- VH(CD3)-(G2S)3-VH(CD30)-(G2S)5-VL(CD30)-His6 (SEQ ID NO: 1)
VL(CD3)-(G2S)2-VL(CD3)-(G2S)3-VH(CD19)-(G2S)5VL(CD19)-FLAG (SEQ ID NO: 2)
Триспецифичное антитело 2:
VH(CD30)-(G2S)2-VH(CD19)-(G2S)2-VH(CD3)-(G2S)5-VL(CD3)-His6 (SEQ ID NO: 3)
VL(CD19)-(G2S)2-VL(CD30)-(G2S)2-VH(CD3)-(G2S)5-VL(CD3)-FLAG (SEQ ID NO: 4)
Линкер 1 = (G2S)2, Линкер 2 = (G2S)5, Линкер 3 = (G2S)3
Иммунопреципитация Триспецифичного антитела 1 и Триспецифичного антитела 2 показывает, что обнаруживаются только гетеродимерные виды антигенсвязывающего полипептидного димера. Триспецифичное антитело 1 и Триспецифичное антитело 2 демонстрируют превосходную стабильность при 40°С после 7 дней и при рН 3,5 после 1 часа.
Пример: Оценка цитотоксической активности, опосредованной триспецифичными антителами.
Процедуры исследования
Выделение мононуклеарных клеток периферической крови (МКПК) и обогащение Т-клеток:
МКПК выделяют из лейкоцитарных пленок центрифугированием в градиенте плотности. Т-клетки обогащают из популяции МКПК с использованием набора EasySep™ Human Т Cell Enrichment Kit для иммуномагнитного выделения нетронутых Т-клеток человека и магнита Big Easy EasySep™ Magnet в соответствии с инструкциями производителя.
Анализ цитотоксичности на основе сортировки флуоресцентно-активированных клеток (FACS):
Т-клеткам, которые применяют в качестве эффекторных клеток, дают характеристику на основе поточной цитометрии, как описано.
Клетки-мишени (МЕС-1: номер в каталоге DSMZ: АСС 497; NALM-6: номер в каталоге DSMZ: АСС 128) культивируют в стандартных условиях, как описано ниже. Для анализа цитотоксичности клетки-мишени собирают, промывают два раза средой RPMI 1640 без ЭТС и ресуспендируют в растворителе С, предоставленном в наборе РКН67 Green Fluorescent Cell Linker Mini Kit, до плотности 2×107/мл. Суспензию клеток затем смешивают с равным объемом сконцентрированного в два раза раствора для мечения красителем РКН67 (например, 1 мкл РКН67 в 250 мкл растворителя С) и инкубируют согласно инструкциям производителя. Останавливают реакцию окрашивания. После промывки меченых клеток-мишеней полной средой RPMI клетки подсчитывают и ресуспендируют до плотности 2×105/мл в полной среде RPMI.
Проводят посев 2×104 клеток-мишеней вместе с Т-клетками и указанными антителами в отдельные лунки. Определяют спонтанную смерть клеток и уничтожение мишеней эффекторами в отсутствии антител.
После инкубации культуры промывают один раз буфером для FACS, а затем ресуспендируют в 150 мкл буфера для FACS, в который добавили 2 мкг/мл пропидия иодида. Абсолютное количество живых клеток-мишеней, для которых характерно положительное зеленое окрашивание красителем РКН67, но они являются отрицательными в отношении окрашивания пропидия иодидом, измеряют с использованием проточного цитометра Beckman-Coulter FC500 MPL (Beckman-Coulter) или проточного цитометра Millipore Guava EasyCyte (Merck Millipore).
Основываясь на измеренном количестве оставшихся живых клеток, рассчитывают процент специфичного лизиса клеток в соответствии со следующей формулой: [1-(число живых клеток-мишеней (образец)) / (число живых клеток-мишеней (спонтанно))]×100%. Сигмоидальные кривые зависимости доза-эффект и значения 50% эффективной концентрации (ЭК50) рассчитывают с помощью нелинейной регрессии/логистического подбора по 4 параметрам с использованием программного обеспечения GraphPad Prism (GraphPad Prism, версия 6.00 для Windows, GraphPad Software, Ла-Хойя, Калифорния, США).
Статистический анализ
Значения величин лизиса, полученные для заданной концентрации антитела, определяют и анализируют с помощью анализа сигмоидальной кривой зависимости доза-эффект/логистического подбора по 4 параметрам с использованием программного обеспечения Prism (GraphPad Prism, версия 6.00 для Windows, GraphPad Software, Ла-Хойя, Калифорния, США) и используют для расчета значений ЭК50, среднего значения и среднеквадратичного отклонения параллельных измерений процента лизиса.
Результаты:
Триспецифичное антитело 1 и Триспецифичное антитело 2 демонстрируют более высокую цитотоксическую активность в отношении линий двойных положительных клеток (CD19+ и CD30+) при сравнении с соответствующими линиями клеток, положительными по одному маркеру.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> AFFIMED GMBH
<120> Трифункциональная антигенсвязывающая молекула
<130> A 3264PCT
<150> EP14164523.4
<151> 2014-04-13
<160> 6
<170> PatentIn версия 3.5
<210> 1
<211> 509
<212> PRT
<213> искусственная последовательность
<220>
<223> полипептидная цепь
<400> 1
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Asp Asp His Tyr Cys Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Leu Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gln Val Gln
115 120 125
Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys
130 135 140
Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His
145 150 155 160
Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile
165 170 175
Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys
180 185 190
Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu
195 200 205
Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr
210 215 220
Tyr Asp Asp His Tyr Cys Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu
225 230 235 240
Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gln Val Gln
245 250 255
Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys
260 265 270
Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr Thr Ile His
275 280 285
Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile Gly Tyr Ile
290 295 300
Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe Lys Gly Lys
305 310 315 320
Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr Met Gln Leu
325 330 335
Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Arg
340 345 350
Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr Trp Gly Gln
355 360 365
Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly Ser Gly Gly
370 375 380
Ser Gly Gly Ser Gly Gly Ser Asp Ile Val Met Thr Gln Ser Pro Lys
385 390 395 400
Phe Met Ser Thr Ser Val Gly Asp Arg Val Thr Val Thr Cys Lys Ala
405 410 415
Ser Gln Asn Val Gly Thr Asn Val Ala Trp Phe Gln Gln Lys Pro Gly
420 425 430
Gln Ser Pro Lys Val Leu Ile Tyr Ser Ala Ser Tyr Arg Tyr Ser Gly
435 440 445
Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
450 455 460
Thr Ile Ser Asn Val Gln Ser Glu Asp Leu Ala Glu Tyr Phe Cys Gln
465 470 475 480
Gln Tyr His Thr Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu
485 490 495
Ile Asn Ala Ala Ala Gly Ser His His His His His His
500 505
<210> 2
<211> 490
<212> PRT
<213> искусственная последовательность
<220>
<223> полипептидная цепь
<400> 2
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala His Phe Arg Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Gly Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr
85 90 95
Phe Gly Ser Gly Thr Lys Leu Glu Ile Asn Gly Gly Ser Gly Gly Ser
100 105 110
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
115 120 125
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
130 135 140
Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr
145 150 155 160
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala His Phe Arg Gly Ser
165 170 175
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Gly Met Glu Ala Glu
180 185 190
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr
195 200 205
Phe Gly Ser Gly Thr Lys Leu Glu Ile Asn Gly Gly Ser Gly Gly Ser
210 215 220
Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg
225 230 235 240
Pro Gly Ser Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe
245 250 255
Ser Ser Tyr Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
260 265 270
Glu Trp Ile Gly Gln Ile Trp Pro Gly Asp Gly Asp Thr Asn Tyr Asn
275 280 285
Gly Lys Phe Lys Gly Lys Ala Thr Leu Thr Ala Asp Glu Ser Ser Ser
290 295 300
Thr Ala Tyr Met Gln Leu Ser Ser Leu Ala Ser Glu Asp Ser Ala Val
305 310 315 320
Tyr Phe Cys Ala Arg Arg Glu Thr Thr Thr Val Gly Arg Tyr Tyr Tyr
325 330 335
Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Gly
340 345 350
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Asp Ile
355 360 365
Leu Leu Thr Gln Thr Pro Ala Ser Leu Ala Val Ser Leu Gly Gln Arg
370 375 380
Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Tyr Asp Gly Asp
385 390 395 400
Ser Tyr Leu Asn Trp Tyr Gln Gln Ile Pro Gly Gln Pro Pro Lys Leu
405 410 415
Leu Ile Tyr Asp Ala Ser Asn Leu Val Ser Gly Ile Pro Pro Arg Phe
420 425 430
Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Asn Ile His Pro Val
435 440 445
Glu Lys Val Asp Ala Ala Thr Tyr His Cys Gln Gln Ser Thr Glu Asp
450 455 460
Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Asn Ala Ala Ala
465 470 475 480
Gly Ser Asp Tyr Lys Asp Asp Asp Asp Lys
485 490
<210> 3
<211> 513
<212> PRT
<213> искусственная последовательность
<220>
<223> полипептидная цепь
<400> 3
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Thr Ile His Trp Val Arg Gln Arg Pro Gly His Asp Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Ser Asp Tyr Asn Gln Asn Phe
50 55 60
Lys Gly Lys Thr Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Asp Tyr Gly Asn Tyr Glu Tyr Thr Trp Phe Ala Tyr
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Ser Gly Gly
115 120 125
Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Pro Gly
130 135 140
Ser Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe Ser Ser
145 150 155 160
Tyr Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp
165 170 175
Ile Gly Gln Ile Trp Pro Gly Asp Gly Asp Thr Asn Tyr Asn Gly Lys
180 185 190
Phe Lys Gly Lys Ala Thr Leu Thr Ala Asp Glu Ser Ser Ser Thr Ala
195 200 205
Tyr Met Gln Leu Ser Ser Leu Ala Ser Glu Asp Ser Ala Val Tyr Phe
210 215 220
Cys Ala Arg Arg Glu Thr Thr Thr Val Gly Arg Tyr Tyr Tyr Ala Met
225 230 235 240
Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Gly Gly Ser
245 250 255
Gly Gly Ser Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu
260 265 270
Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Ala Ser Gly
275 280 285
Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro Gly
290 295 300
Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr
305 310 315 320
Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys
325 330 335
Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp
340 345 350
Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Cys Leu
355 360 365
Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Ser
370 375 380
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gln Ile Val Leu
385 390 395 400
Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val Thr
405 410 415
Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln
420 425 430
Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys
435 440 445
Leu Ala Ser Gly Val Pro Ala His Phe Arg Gly Ser Gly Ser Gly Thr
450 455 460
Ser Tyr Ser Leu Thr Ile Ser Gly Met Glu Ala Glu Asp Ala Ala Thr
465 470 475 480
Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr Phe Gly Ser Gly
485 490 495
Thr Lys Leu Glu Ile Asn Ala Ala Ala Gly Ser His His His His His
500 505 510
His
<210> 4
<211> 486
<212> PRT
<213> искусственная последовательность
<220>
<223> полипептидная цепь
<400> 4
Asp Ile Leu Leu Thr Gln Thr Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Tyr Asp
20 25 30
Gly Asp Ser Tyr Leu Asn Trp Tyr Gln Gln Ile Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Asp Ala Ser Asn Leu Val Ser Gly Ile Pro Pro
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Asn Ile His
65 70 75 80
Pro Val Glu Lys Val Asp Ala Ala Thr Tyr His Cys Gln Gln Ser Thr
85 90 95
Glu Asp Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Asn Gly
100 105 110
Gly Ser Gly Gly Ser Asp Ile Val Met Thr Gln Ser Pro Lys Phe Met
115 120 125
Ser Thr Ser Val Gly Asp Arg Val Thr Val Thr Cys Lys Ala Ser Gln
130 135 140
Asn Val Gly Thr Asn Val Ala Trp Phe Gln Gln Lys Pro Gly Gln Ser
145 150 155 160
Pro Lys Val Leu Ile Tyr Ser Ala Ser Tyr Arg Tyr Ser Gly Val Pro
165 170 175
Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
180 185 190
Ser Asn Val Gln Ser Glu Asp Leu Ala Glu Tyr Phe Cys Gln Gln Tyr
195 200 205
His Thr Tyr Pro Leu Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Asn
210 215 220
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gln Val Gln Leu Gln Gln Ser
225 230 235 240
Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys
245 250 255
Ala Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln
260 265 270
Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg
275 280 285
Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr
290 295 300
Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr
305 310 315 320
Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His
325 330 335
Tyr Cys Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
340 345 350
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gln
355 360 365
Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu
370 375 380
Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met Asn
385 390 395 400
Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp
405 410 415
Thr Ser Lys Leu Ala Ser Gly Val Pro Ala His Phe Arg Gly Ser Gly
420 425 430
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Gly Met Glu Ala Glu Asp
435 440 445
Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr Phe
450 455 460
Gly Ser Gly Thr Lys Leu Glu Ile Asn Ala Ala Ala Gly Ser Trp Ser
465 470 475 480
His Pro Gln Phe Glu Lys
485
<210> 5
<211> 8
<212> PRT
<213> искусственная последовательность
<220>
<223> Маркерная последовательность
<400> 5
Asp Tyr Lys Asp Asp Asp Asp Lys
1 5
<210> 6
<211> 8
<212> PRT
<213> искусственная последовательность
<220>
<223> Маркерная последовательность
<400> 6
Trp Ser His Pro Gln Phe Glu Lys
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МУЛЬТИВАЛЕНТНЫЕ FV-АНТИТЕЛА | 2016 |
|
RU2785766C2 |
| CD3-СВЯЗЫВАЮЩИЙ ДОМЕН | 2015 |
|
RU2742691C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2810924C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2832669C2 |
| БИСПЕЦИФИЧЕСКИЙ АНТИГЕНСВЯЗЫВАЮЩИЙ БЕЛОК EGFR/CD16 | 2019 |
|
RU2792240C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2019 |
|
RU2797268C2 |
| СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, ВЫЗЫВАЮЩИЕ КЛЕТОЧНУЮ АКТИВАЦИЮ | 2019 |
|
RU2770620C1 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2020 |
|
RU2841523C2 |
| ПОЛИПЕПТИД, СОДЕРЖАЩИЙ АНТИГЕНСВЯЗЫВАЮЩИЙ ДОМЕН И ТРАНСПОРТИРУЮЩИЙ СЕГМЕНТ | 2017 |
|
RU2827545C2 |
| БИПАРАТОПНЫЕ FR-АЛЬФА АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ | 2020 |
|
RU2832083C2 |

Настоящее изобретение относится к биотехнологии и генной инженерии, в частности к триспецифичной антигенсвязывающей молекуле, при этом указанная антигенсвязывающая молекула является по меньшей мере четырехвалентной и содержит антигенсвязывающий сайт, обладающий специфичностью против первого эпитопа антигена, антигенсвязывающий сайт, обладающий специфичностью против второго эпитопа антигена, и два антигенсвязывающих сайта, обладающих специфичностью против третьего эпитопа антигена. Изобретение позволяет эффективно применять такую молекулу в качестве лекарственного средства для терапии опухолей. 4 н. и 11 з.п. ф-лы, 3 ил., 2 пр.
1. Тетравалентная молекула, которая представляет собой антигенсвязывающую молекулу, способную связывать три различных антигена, где указанная молекула представляет собой полипептидный димер, включающий первый полипептид и второй полипептид, при этом каждый полипептид содержит четыре вариабельных домена, которые соединены последовательно, причём:
(а) указанный первый полипептид содержит первый и второй вариабельные домены тяжелой цепи антитела (VH), соединенные друг с другом линкером из приблизительно 12 или менее аминокислотных остатков, и антигенсвязывающую одноцепочечную Fv-единицу, имеющую третий вариабельный домен тяжелой цепи антитела (VH), соединенный с вариабельным доменом легкой цепи антитела (VL), указанные третий вариабельный домен тяжелой цепи антитела (VH) и вариабельный домен легкой цепи антитела (VL) способны ассоциировать с образованием первого антигенсвязывающего сайта, при этом указанный первый или второй вариабельный домен тяжелой цепи антитела (VH) соединен с указанной антигенсвязывающей одноцепочечной Fv-единицей пептидным линкером;
(b) указанный второй полипептид содержит первый и второй вариабельные домены легкой цепи антитела (VL), соединенные друг с другом линкером из приблизительно 12 или менее аминокислотных остатков, и антигенсвязывающую одноцепочечную Fv-единицу, имеющую третий вариабельный домен легкой цепи антитела (VL), соединенный с вариабельным доменом тяжелой цепи антитела (VH), причем указанные третий вариабельный домен легкой цепи антитела (VL) и вариабельный домен тяжелой цепи антитела (VH) способны ассоциировать с образованием второго антигенсвязывающего сайта, при этом первый или второй вариабельный домен легкой цепи антитела (VL) соединен с антигенсвязывающей одноцепочечной Fv-единицей пептидным линкером;
(с) указанный первый вариабельный домен тяжелой цепи антитела (VH) первого полипептида ассоциирует со вторым вариабельным доменом легкой цепи антитела (VL) второго полипептида с образованием третьего антигенсвязывающего сайта; и
(d) указанный второй вариабельный домен тяжелой цепи антитела (VH) первого полипептида ассоциирует с первым вариабельным доменом легкой цепи антитела (VL) второго полипептида с образованием четвертого антигенсвязывающего сайта.
2. Тетравалентная молекула по п. 1, отличающаяся тем, что указанные первый и второй вариабельные домены первого полипептида и указанные первый и второй вариабельные домены второго полипептида соединены линкером, имеющим от 3 до 9 аминокислотных остатков.
3. Тетравалентная молекула по п. 1 или 2, отличающаяся тем, что указанные второй вариабельный домен и одноцепочечная Fv-единица первого полипептида и указанные второй вариабельный домен и одноцепочечная Fv-единица второго полипептида соединены линкером, имеющим от 2 до 35 аминокислотных остатков.
4. Тетравалентная молекула по любому из пп. 1-3, отличающаяся тем, что указанные вариабельный домен тяжелой цепи и вариабельный домен легкой цепи одноцепочечной Fv-единицы первого полипептида и указанные вариабельный домен легкой цепи и вариабельный домен тяжелой цепи одноцепочечной Fv-единицы второго полипептида соединены линкером, имеющим 12 или более аминокислотных остатков.
5. Тетравалентная молекула по любому из пп. 1-4, отличающаяся тем, что указанные первый и второй антигенсвязывающие сайты двух одноцепочечных Fv-единиц специфичны к одному и тому же антигену.
6. Тетравалентная молекула по любому из пп. 1-5, отличающаяся тем, что указанные третий и четвертый антигенсвязывающие сайты, образованные посредством ассоциации между первым и вторым полипептидом, специфичны к одному и тому же антигену.
7. Тетравалентная молекула по любому из пп. 1-6, отличающаяся тем, что два указанных антигенсвязывающих сайта, специфичных к одному и тому же антигену, специфичны к антигену, презентированному на Т-клетке или клетке-естественном киллере (NK-клетке).
8. Тетравалентная молекула по п. 7, отличающаяся тем, что указанный антиген представляет собой CD3, CD16 или CD16A.
9. Тетравалентная молекула по п. 7 или 8, отличающаяся тем, что два других связывающих сайта специфичны к двум разным антигенам на одной и той же клетке.
10. Тетравалентная молекула по п. 9, отличающаяся тем, что указанная клетка представляет собой клетку опухоли.
11. Тетравалентная молекула по п. 10, отличающаяся тем, что два разных антигена выбраны из группы, состоящей из CD19, CD20, CD26, CD29, CD30, CD33, CD200, CD267, рецептора эпидермального фактора роста (EGFR), EGFRvIII, HER2, HER3, рецептора инсулиноподобного фактора роста (IGFR), IGF-1R, молекулы адгезии эпителиальных клеток (Ep-CAM), плацентарной щелочной фосфатазы (PLAP), антигена Томсена-Фриденрайха (TF), MUC-1 (муцина), CD5, альфа-рецептора ИЛ-4 (IL4-R альфа), рецептора ИЛ-13 (IL13-R), высокоаффинного рецептора IgE I типа (FcεRI) и IgE, gpA33, комплексов главного комплекса гистосовместимости класса I (MHCI) c пептидом.
12. Тетравалентная молекула по любому из пп. 1-5, отличающаяся тем, что первый антигенсвязывающий сайт и второй антигенсвязывающий сайт являются специфичными к двум разным эпитопам антигенов, презентированным на клетках-естественных киллерах (NK-клетках).
13. Вектор для экспрессии тетравалентной молекулы по любому из пп. 1-12, содержащий полинуклеотиды, кодирующие полипептидные цепи антигенсвязывающей молекулы по любому из пп. 1-12.
14. Клетка-хозяин для получения тетравалентной молекулы по любому из пп. 1-12, трансформированная вектором по п. 13.
15. Применение тетравалентной молекулы по п. 10 или 11 в терапии опухолей.
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| БИСПЕЦИФИЧЕСКАЯ МОЛЕКУЛА АНТИТЕЛА ДЛЯ ЛИЗИСА ОПУХОЛЕВЫХ КЛЕТОК, СПОСОБ ПОЛУЧЕНИЯ F(AB') 2 ФРАГМЕНТА БИСПЕЦИФИЧЕСКОЙ МОЛЕКУЛЫ АНТИТЕЛА, МОНОКЛОНАЛЬНОЕ АНТИТЕЛО (ВАРИАНТЫ), ФАРМАЦЕВТИЧЕСКИЙ ПРЕПАРАТ, ФАРМАЦЕВТИЧЕСКИЙ НАБОР ДЛЯ ЛИЗИСА ОПУХОЛЕВЫХ КЛЕТОК (ВАРИАНТЫ), СПОСОБ ЛИЗИСА ОПУХОЛЕВЫХ КЛЕТОК EX VIVO ПРИ АУТОГЕННОЙ ТРАНСПЛАНТАЦИИ КОСТНОГО МОЗГА | 1994 |
|
RU2203319C2 |

