Область техники, к которой относится изобретение
Настоящее изобретение относится к новому способу лечения рака, в котором применяют биспецифическое антитело.
Предпосылки создания изобретения
Рак является одной из основных причин смертности в мире. За исключением некоторых карцином, рак часто является неоперабельным в момент его обнаружения, и исход лечения, которое основано на применении химиотерапевтических агентов, что представляет собой основной терапевтический метод, не обязательно является благоприятным. Гетерогенность раковых клеток сама по себе является не единственным фактором, который затрудняет лечение рака, и предполагается, что основную роль играет микроокружение опухоли (незапатентованный документ 1). В последние годы было выдвинуто предположение о том, что нерезектабельную злокачественную меланому можно излечивать с помощью антитела к CTLA-4, которое ослабляет супрессорные Т-клетки (незапатентованный документ 2). Это позволяет предположить, что иммуностимуляция опухоли может служить основой для разработки новых стратегий лечения рака.
Известно, что Т-клетки, которые играют важные роли в опухолевом иммунитете, активируются двумя сигналами: 1) связыванием Т-клеточного рецептора (TCR) с антигенным пептидом, который презентуется молекулами главного комплекса гистосовместимости (ГКГС), и активацией TCR; и 2) связыванием костимуляторной молекулы на поверхности Т-клеток с лигандами на антигенпрезентирующих клетках и активацией костимулятора. Кроме того, описано, что для Т-клеточной активации является важной активация молекул, принадлежащих к суперсемейству фактора некроза опухоли (TNF) и суперсемейству TNF-рецептора, таких как CD137(4-1BB), на поверхности Т-клеток (незапатентованный документ 3).
В суперсемейство TNF и суперсемейство TNF-рецептора входят такие молекулы, как CD137, CD137L, CD40, CD40L, ОХ40, OX40L, CD27, CD70, HVEM, LIGHT, RANK, RANKL, CD30, CD153, GITR и GITRL. Описано, что CD137 экспрессируется не только на поверхности Т-клеток, но также на поверхности других иммунных клеток, таких как дендритные клетки (ДК), В-клетки, NK-клетки и нейтрофилы (незапатентованный документ 4).
Уже известно, что агонистические антитела к CD137 обладают противоопухолевыми действиями, и это, как продемонстрировано экспериментальным путем, главным образом является следствием активации CD8-позитивных Т-клеток и NK-клеток (незапатентованный документ 5). Однако побочные действия, связанные с неспецифической гепатотоксичностью агонистических антител к CD137, оказались клинической и неклинической проблемой, и вследствие этого разработка фармацевтических агентов не расширялась (незапатентованные документы 6 и 7). Предполагается, что основная причина побочных действий обусловлена связыванием с Fcγ-рецептором через константную область антитела (незапатентованный документ 8). Кроме того, для агонистических антител, мишенью которых являются рецепторы, принадлежащие к суперсемейству TNF, установлено, что для проявления их агонистической активности in vivo необходимо, чтобы антитело перекрестно связывалось через экспрессирующие Fcγ-рецептор клетки (экспрессирующие FcγRII клетки) (незапатентованный документ 9). Более конкретно, медицинские действия агонистических антител к CD137, представляющие собой противоопухолевые действия, и побочные действия, включающие гепатотоксичность, оба включают связывание антител с Fcγ-рецепторами. Таким образом, если связывание антител с Fcγ-рецепторами повышают, то, как ожидается, улучшаются медицинские действия, но при этом также должны возрастать гепатотоксические побочные действия, а, если связывание антител с Fcγ-рецепторами понижают, то побочные действия должны уменьшаться, но при этом медицинские действия также могут снижаться, и до настоящего времени не описаны агонистические антитела к CD137, у которых медицинские действия отделены от побочных действий. Кроме того, противоопухолевые действия самих агонистических антител к CD137 в целом не являются сильными, и поэтому желательно избегать токсичности и одновременно дополнительно повышать их медицинское действие.
Биспецифические антитела отличаются наличием по меньшей мере двух связывающих доменов, и их молекулярная морфология уже хорошо известна специалистам в данной области. Среди прочего, сконструированы молекулы, в которых один из двух связывающих доменов специфически связывается с раковым поверхностным антигеном, а второй связывающий домен связывается с антигеном Т-клеточной поверхности CD3 (незапатентованный документ 10). Установлено, что такие биспецифические одноцепочечные антитела проявляют противоопухолевое действие путем активации Т-клеток с помощью зависимого от ракового антигена пути.
Глипикан 3 (GPC3) представляет собой белок, который принадлежит к семейству глипиканов, т.е. группе гепарансульфатпротеогликанов, связанных с клеточной поверхностью через гликозилфосфатидилинозитол (незапатентованный документ 11). Глипиканы играют важную роль в пролиферации, дифференцировке и миграции клеток. GPC3 экспрессируется в 70% или большем количестве тканей гепатом, полученных с помощью хирургического иссечения или бипсии, и редко экспрессируется или вообще не экспрессируется в соседних ненеопластических повреждениях печени и в большинстве тканей взрослых (незапатентованные документы 12 и 13). Кроме того, установлено, что пациенты с высоким уровнем экспрессии GPC3 в ткани гепатом имеют плохой прогноз (незапатентованный документ 14), и GPC3 рассматривается в качестве перспективной молекулы-мишени для гепатомы.
Документы, характеризующие известный уровень техники
Незапатентованные документы:
[незапатентованный документ 1] Hanahan, Cell, 144, 2011, сс. 646-674;
[незапатентованный документ 2] Prieto, Clin Cancer Res. 18, 2012, сс. 2039-2047;
[незапатентованный документ 3] Summers, Nat. Rev. Immunol., 12, 2012, сс. 339-351;
[незапатентованный документ 4] Vinay, Cell Biol Int., 33, 2009, cc. 453-465;
[незапатентованный документ 5] Houot, Blood, 114, 2009, cc. 3431-3438;
[незапатентованный документ 6] Ascierto, Semin Oncol., 37, 2010, cc. 508-516;
[незапатентованный документ 7] Dubrot, Cancer Immunol. Immunother., 59, 2010, cc. 1223-1233;
[незапатентованный документ 8] Schabowsky, Vaccine, 28, 2009, cc. 512-522;
[незапатентованный документ 9] Li, Proc Natl Acad Sci USA. 110(48), 2013, cc. 19501-19506;
[незапатентованный документ 10] Brandl, Cancer Immunol. Immunother., 56, 2007, cc. 1551-1563;
[незапатентованный документ 11] Filmus, J. Clin. Invest., 108, 2001, cc. 497-501;
[незапатентованный документ 12] Zhu-Zu-W, Gut, 48, 2001, cc. 558-564;
[незапатентованный документ 13] Yamauchi, Mod. Pathol., 18, 2005, cc. 1591-1598;
[незапатентованный документ 14] Yorita, Liver Int., 1, 2010, cc. 120-131.
Краткое изложение сущности изобретения
Техническая задача, положенная в основу настоящего изобретения
Настоящее изобретение было создано с учетом вышеуказанных обстоятельств. В основу настоящего изобретения была положена задача разработать антигенсвязывающие молекулы, которые обладают агонистической активностью в отношении представителей суперсемейства TNF или суперсемейства TNF-рецептора, которые не обладают токсичностью, активируя при этом иммунные клетки, и обладают очень высоким противоопухолевым действием. Другой задачей настоящего изобретения является создание фармацевтических композиций, содержащих антигенсвязывающую молекулу в качестве действующего вещества, или способов лечения рака с помощью фармацевтической композиции.
Средства решения указанных задач
При создании настоящего изобретения установлено, что даже, несмотря на то, что антигенсвязывающие молекулы, имеющие только один домен, связывающийся с представителем суперсемейства TNF, или только один домен, связывающийся с представителем суперсемейства TNF-рецептора, не обладают способностью активировать иммунные клетки, антигенсвязывающие молекулы, которые имеют домен, связывающийся со специфическим для рака антигеном, и домен, связывающийся с представителем суперсемейства TNF, или домен, связывающийся со специфическим для рака антигеном, и домен, связывающийся с представителем суперсемейства TNF-рецептора, активируют иммунные клетки посредством агонистической активности в отношении факторов, принадлежащих к суперсемейству TNF или суперсемейству TNF-рецептора, только в присутствии клеток, экспрессирующих специфический для рака антиген, и не обладают побочными действиями, такими как гепатотоксичность, сохраняя при этом противоопухолевую активность. Кроме того, при создании настоящего изобретения установлено, что путем применения антигенсвязывающих молекул в сочетании с антигенсвязывающими молекулами, которые имеет домен, связывающийся со специфическим для рака антигеном, и домен, связывающийся с комплексом Т-клеточного рецептора, можно избегать побочных действий и повышать противоопухолевую активность, и тем самым решать задачу, положенную в основу настоящего изобретения.
Более конкретно, в настоящем изобретении предложены:
[1] антигенсвязывающая молекула, содержащая:
(1) домен, связывающийся со специфическим для рака антигеном; и
(2) домен, связывающийся с представителем суперсемейства фактора некроза опухоли (TNF), или домен, связывающийся с представителем суперсемейства рецептора фактора некроза опухоли (TNF);
[2] антигенсвязывающая молекула по п. [1], дополнительно содержащая FcRn-связывающий домен;
[3] антигенсвязывающая молекула по п. [2], в которой FcRn-связывающий домен представляет собой Fc-область антитела с пониженной активностью связывания с Fcγ-рецептором;
[4] антигенсвязывающая молекула по одному из п.п. [1]-[3], в которой домен, связывающийся с представителем суперсемейства TNF, или домен, связывающийся с представителем суперсемейства TNF-рецептора, представляет собой CD137-связывающий домен;
[5] антигенсвязывающая молекула по одному из п.п. [1]-[4], которая представляет собой биспецифическое антитело;
[6] фармацевтическая композиция, содержащая в качестве действующего вещества антигенсвязывающую молекулу по одному из п.п. [1]-[5];
[7] фармацевтическая композиция по п. [6], которая представляет собой индуцирующую цитотоксичность композицию;
[8] фармацевтическая композиция по п. [6], которая представляет собой композицию, предназначенную для применения для лечения рака;
[9] фармацевтическая композиция, содержащая комбинацию первой антигенсвязывающей молекулы по одному из п.п. [1]-[5] и второй антигенсвязывающей молекулы, которая содержит:
(1) домен, связывающийся со специфическим для рака антигеном; и
(2) домен, связывающийся с комплексом Т-клеточного рецептора;
[10] фармацевтическая композиция по п. [9], в которой вторая антигенсвязывающая молекула представляет собой антигенсвязывающую молекулу, которая дополнительно содержит FcRn-связывающий домен;
[11] фармацевтическая композиция по п. [10], в которой FcRn-связывающий домен представляет собой Fc-область антитела с пониженной активностью связывания с Fcγ-рецептором;
[12] фармацевтическая композиция по одному из п.п. [9]-[11], в которой домен, связывающийся с комплексом Т-клеточного рецептора, представляет собой домен, связывающийся с Т-клеточным рецептором;
[13] фармацевтическая композиция по одному из п.п. [9]-[11], в которой домен, связывающийся с комплексом Т-клеточного рецептора, представляет собой CD3-связывающий домен;
[14] фармацевтическая композиция по одному из п.п. [9]-[13], в которой вторая антигенсвязывающая молекула представляет собой биспецифическое антитело;
[15] фармацевтическая композиция по одному из п.п. [9]-[14], в которой первая антигенсвязывающая молекула и вторая антигенсвязывающая молекула смешаны;
[16] фармацевтическая композиция по одному из п.п. [9]-[14], в которой первую антигенсвязывающую молекулу и вторую антигенсвязывающую молекулу применяют совместно;
[17] фармацевтическая композиция по одному из п.п. [9]-[14], в которой первую антигенсвязывающую молекулу и вторую антигенсвязывающую молекулу применяют одновременно;
[18] фармацевтическая композиция по одному из п.п. [9]-[14], в которой первую антигенсвязывающую молекулу и вторую антигенсвязывающую молекулу применяют раздельно;
[19] фармацевтическая композиция по одному из п.п. [9]-[18], которая представляет собой индуцирующую цитотоксичность композицию;
[20] фармацевтическая композиция по одному из п.п. [9]-[18], которая представляет собой композицию, предназначенную для применения для лечения рака;
[21] фармацевтическая композиция, содержащая в качестве действующего вещества первую антигенсвязывающую молекулу, которая содержит:
(1) домен, связывающийся со специфическим для рака антигеном; и
(2) домен, связывающийся с представителем суперсемейства фактора некроза опухоли (TNF), или домен, связывающийся с представителем суперсемейства рецептора фактора некроза опухоли (TNF),
предназначенная для совместного применения со второй антигенсвязывающей молекулой, которая содержит:
(1) домен, связывающийся со специфическим для рака антигеном; и
(2) домен, связывающийся с комплексом Т-клеточного рецептора;
[22] фармацевтическая композиция по п. [21], которая представляет собой индуцирующую цитотоксичность композицию;
[23] фармацевтическая композиция по п. [21], которая представляет собой композицию, предназначенную для применения для лечения рака;
[24] фармацевтическая композиция по одному из п.п. [21]-[23], в которой первая антигенсвязывающая молекула и/или вторая антигенсвязывающая молекула представляет собой антигенсвязывающую молекулу, которая дополнительно содержит FcRn-связывающий домен;
[25] фармацевтическая композиция по п. [24], в которой FcRn-связывающий домен представляет собой Fc-область антитела с пониженной активностью связывания с Fcγ-рецептором;
[26] фармацевтическая композиция по одному из п.п. [21]-[25], в которой домен, связывающийся с представителем суперсемейства TNF, или домен, связывающийся с представителем суперсемейства рецептора TNF, представляет собой CD137-связывающий домен или CD40-связывающий домен;
[27] фармацевтическая композиция по одному из п.п. [21]-[26], в которой домен, связывающийся с комплексом Т-клеточного рецептора, представляет собой домен, связывающийся с Т-клеточным рецептором;
[28] фармацевтическая композиция по одному из п.п. [21]-[26], в которой домен, связывающийся с комплексом Т-клеточного рецептора, представляет собой CD3-связывающий домен;
[29] фармацевтическая композиция по одному из п.п. [21]-[28], в которой первая антигенсвязывающая молекула и/или вторая антигенсвязывающая молекула представляет собой биспецифическое антитело;
[30] фармацевтическая композиция по одному из п.п. [21]-[29], которую применяют одновременно со второй антигенсвязывающей молекулой;
[31] фармацевтическая композиция по одному из п.п. [21]-[29], которую применяют отдельно от второй антигенсвязывающей молекулы;
[32] фармацевтическая композиция, содержащая в качестве действующего вещества вторую антигенсвязывающую молекулу, которая содержит:
(1) домен, связывающийся со специфическим раковым антигеном; и
(2) домен, связывающийся с комплексом Т-клеточного рецептора,
предназначенная для совместного применения с первой антигенсвязывающей молекулой, которая содержит:
(1) домен, связывающийся со специфическим для рака антигеном; и
(2) домен, связывающийся с представителем суперсемейства фактора некроза опухоли (TNF), или домен, связывающийся с представителем суперсемейства рецептора фактора некроза опухоли (TNF);
[33] фармацевтическая композиция по п. [32], которая представляет собой индуцирующую цитотоксичность композицию;
[34] фармацевтическая композиция по п. [32], которая представляет собой композицию, предназначенную для применения для лечения рака;
[35] фармацевтическая композиция по одному из п.п. [32]-[34], в которой первая антигенсвязывающая молекула и/или вторая антигенсвязывающая молекула представляет собой антигенсвязывающую молекулу, которая дополнительно содержит FcRn-связывающий домен;
[36] фармацевтическая композиция по п. [35], в которой FcRn-связывающий домен представляет собой Fc-область антитела с пониженной активностью связывания с Fcγ-рецептором;
[37] фармацевтическая композиция по одному из п.п. [32]-[36], в которой домен, связывающийся с комплексом Т-клеточного рецептора, представляет собой домен, связывающийся с Т-клеточным рецептором;
[38] фармацевтическая композиция по одному из п.п. [32]-[36], в которой домен, связывающийся с комплексом Т-клеточного рецептора, представляет собой CD3-связывающий домен;
[39] фармацевтическая композиция по одному из п.п. [32]-[38], в которой домен, связывающийся с представителем суперсемейства TNF, или домен, связывающийся с представителем суперсемейства TNF-рецептора, представляет собой CD137-связывающий домен или CD40-связывающий домен;
[40] фармацевтическая композиция по одному из п.п. [32]-[39], в которой первая антигенсвязывающая молекула и/или вторая антигенсвязывающая молекула представляет собой биспецифическое антитело;
[41] фармацевтическая композиция по одному из п.п. [32]-[40], которую применяют одновременно с первой антигенсвязывающей молекулой;
[42] фармацевтическая композиция по одному из п.п. [32]-[40], которую применяют отдельно от первой антигенсвязывающей молекулы;
[43] способ индукции цитотоксичности, подавления клеточной пролиферации, активирования иммунитета против раковой клетки или содержащей раковую клетку опухолевой ткани, или лечения или предупреждения рака, который включает стадию, на которых применяют антигенсвязывающую молекулу по одному из п.п. [1]-[5] или фармацевтическую композицию по одному из п.п. [6]-[42];
[44] антигенсвязывающая молекула по одному из п.п. [1]-[5] или фармацевтическая композиция по одному из п.п. [6]-[42], предназначенная для применения для индукции цитотоксичности, подавления клеточной пролиферации, активирования иммунитета против раковой клетки или содержащей раковую клетку опухолевой ткани, или лечения или предупреждения рака;
[45] применение антигенсвязывающей молекулы по одному из п.п. [1]-[5] для приготовления фармацевтической композиции по одному из п.п. [6]-[42]; и
[46] способ приготовления фармацевтической композиции по одному из п.п. [6]-[42], который включает стадию, на которой применяют антигенсвязывающую молекулу по одному из п.п. [1]-[5].
Кроме того, настоящее изобретение относится к способам лечения или предупреждения рака, заключающимся в том, что вводят антигенсвязывающую молекулу, предлагаемую в настоящем изобретении, или фармацевтическую композицию, предлагаемую в настоящем изобретении, пациенту, который нуждается в лечении. Настоящее изобретение относится также к набору, предназначенному для применения в способе, предлагаемом в настоящем изобретении, который содержит антигенсвязывающую молекулу, предлагаемую в настоящем изобретении. Настоящее изобретение относится также к применению антигенсвязывающей молекулы, предлагаемой в настоящем изобретении, для получения фармацевтической композиции, предназначенной для индукции цитотоксичности (например, фармацевтической композиции, предназначенной для лечения или предупреждения рака). Кроме того, настоящее изобретение относится к антигенсвязывающим молекулам, предлагаемым в настоящем изобретении, или фармацевтическим композициям, предлагаемым в настоящем изобретении, предназначенным для применения в способах, предлагаемых в настоящем изобретении.
Краткое описание чертежей
На чертежах показано:
на фиг 1 - график, иллюстрирующий результаты оценки воздействия антител к мышиному CD137 на активацию Т-клеток, полученные с помощью IFN-γ-ELISA. Ctrl mIgG1 обозначает применяемое в качестве отрицательного контроля мышиное антитело IgG1-изотипа;
на фиг. 2 - диаграмма, на которой продемонстрирована концепция активирующего воздействия на Т-клетки антитела к мышиному CD137 в различных молекулярных формах;
на фиг. 3 - диаграмма, на которой продемонстрирована концепция зависящего от антигена GPC3 активирующего воздействия на Т-клетки биспецифического антитела к человеческому GPC3/к мышиному CD137;
на фиг. 4 - график, иллюстрирующий результат оценки зависящего от антигена GPC3 активирующего воздействия на Т-клетки биспецифического антитела к человеческому GPC3/к мышиному CD137, полученные с помощью IFN-γ-ELISA;
на фиг. 5 - график, иллюстрирующий результат оценки влияния изменений в константных областях биспецифического антитела к человеческому GPC3/к мышиному CD137 на зависящее от антигена GPC3 активирующее воздействие на Т-клетки, полученный с помощью IFN-γ-ELISA;
на фиг. 6 - график, иллюстрирующий результат оценки повышающего Т-клеточную активацию воздействия, полученного в результате применения смеси биспецифического антитела к человеческому GPC3/к мышиному CD137 и биспецифического антитела к человеческому GPC3/к мышиному CD3, полученный с помощью IFN-γ-ELISA. Ctrl hIgG1 обозначает применяемое в качестве отрицательного контроля человеческое антитело IgG1-изотипа (фирма Alexis Corporation);
на фиг. 7 - график, иллюстрирующий противоопухолевое действие биспецифического антитела к человеческому GPC3/к мышиному CD137 на созданной на мышах с помощью сингенной трансплантации модели опухоли линии СТ26. Стрелками обозначено время введения антител;
на фиг. 8 - график, иллюстрирующий влияние биспецифического антитела к человеческому GPC3/к мышиному CD137 и антитела к мышиному CD137 на аспартатаминотрансферазу (AST) в крови при оценке на созданной на мышах с помощью сингенной трансплантации модели опухоли линии СТ26;
на фиг. 9 - график, иллюстрирующий влияние биспецифического антитела к человеческому GPC3/к мышиному CD137 и антитела к мышиному CD137 на аланинаминотрансферазу (ALT) в крови при оценке на созданной на мышах с помощью сингенной трансплантации модели опухоли линии СТ26;
на фиг. 10 - график, иллюстрирующий влияние биспецифического антитела к человеческому GPC3/к мышиному CD137 и антитела к мышиному CD137 на общий билирубин в крови при оценке на созданной на мышах с помощью сингенной трансплантации модели опухоли линии СТ26;
на фиг. 11 - фотографии, полученные при проведении гистопатологических исследований печени на созданной на мышах с помощью сингенной трансплантации модели опухоли линии СТ26 после обработки биспецифическим антителом к человеческому GPC3/к мышиному CD137 и антителом к мышиному CD137. На фотографиях представлены окрашенные гематоксилином-эозином изображения срезов печени репрезентативной мыши, при этом на фотографиях а) и г) представлены результаты, полученные после введения растворителя, на фотографиях б) и д) представлены результаты, полученные после введения 1D8-МВ492, и на фотографиях в) и е) представлены результаты, полученные после введения GPC3 ERY22-3-1D8. Наконечниками стрелок обозначены расщепленные или некротизированные клетки, а * обозначены области воспаления;
на фиг. 12 - график, демонстрирующий противоопухолевое действие совместного применения биспецифического антитела к человеческому GPC3/к мышиному CD137 и биспецифического антитела к человеческому GPC3/к мышиному CD3 на созданной на мышах с помощью сингенной трансплантации модели опухоли линии LLC. Стрелками обозначено время введения антител;
на фиг. 13 - взаимосвязь между аминокислотными остатками, образующими Fc-области IgG1, IgG2, IgG3 и IgG4, и EU-нумерацией по Кэботу (в контексте настоящего описания обозначена также как EU-индекс);
на фиг. 14-1 - результаты, полученные с помощью ELISA, оценки связывания антител к человеческому CD137 с фрагментированными слитыми белками человеческий CD137-Fc. На чертеже «нет» обозначает уровень проявления цвета при осуществлении ELISA в лунках, в которых не был иммобилизован антиген (несенсибилизированные лунки);
на фиг. 14-2 - величины (отношения к уровню в несенсибилизированных лунках), полученные путем деления уровней проявления цвета при осуществлении ELISA для каждого образца, представленного на фиг. 14-1, на уровень проявления цвета при осуществлении ELISA в несенсибилизированных лунках (связывание с лунками, в которых не был иммобилизован антиген);
на фиг. 15 - график, иллюстрирующий IFNγ-индуцирующую активность антител к человеческому CD137;
на фиг. 16 - данные об активирующем Т-клетки действии и профиле связывания антител к человеческому CD137;
на фиг. 17 - график, иллюстрирующий результаты оценки повышающего активацию Т-клеток действия, полученного в результате применения смеси биспецифического антитела к человеческому GPC3/к мышиному CD40 и биспецифического антитела к человеческому GPC3/к мышиному CD3, полученные с помощью IFN-γ-ELISA. Ctrl hIgG1 обозначает применяемое в качестве отрицательного контроля человеческое антитело IgG1-изотипа;
на фиг. 18 - график, иллюстрирующий результаты оценки повышающего активацию Т-клеток действия, полученного в результате применения биспецифического антитела к человеческому GPC3/к человеческому CD137 GPC3 FAE-BMS, полученные с помощью IFN-γ-ELISA. Ctrl hIgG1 обозначает применяемое в качестве отрицательного контроля человеческое антитело IgG1-изотипа;
на фиг. 19 - график, иллюстрирующий результаты оценки опосредуемой CD137 агонистической активности различных биспецифических антител к человеческому GPC3/к человеческому CD137, по уровню производства IL-6, который активирует В-клетки. Ctrl hIgG1 обозначает применяемое в качестве отрицательного контроля человеческое антитело IgG1-изотипа.
Варианты осуществления изобретения
Представленные ниже определения даны с целью облегчения понимания настоящего изобретения.
Антигенсвязывающие молекулы
В настоящем изобретении не существует ограничения касательно конкретных «антигенсвязывающих молекул», если в них присутствует «связывающий домен», предлагаемый в настоящем изобретении, и они могут содержать также пептид или белок, состоящий примерно из пяти или большего количества аминокислот. Пептид и белок не ограничены субстанциями, полученными из живого организма, и они, например, могут представлять собой полипептид, полученный из искусственно созданной последовательности. Они могут представлять собой также любой встречающийся в естественных условиях полипептид, синтетический полипептид, рекомбинантный полипептид и т.п.
Предпочтительным примером антигенсвязывающей молекулы, предлагаемой в настоящем изобретении, является антигенсвязывающая молекула, которая содержит FcRn-связывающий домен, входящий в Fc-область антитела. Метод удлинения времени полужизни в крови белка, введенного в живой организм, метод добавления FcRn-связывающего домена антитела к представляющему интерес белку и использования функции опосредуемого FcRn рециклинга хорошо известны.
В настоящем изобретении не существует ограничения касательно конкретного «FcRn-связывающего домена», если он обладает способностью связываться с FcRn, и его примеры включают вариабельные области антитела, Fab и Fc-области антитела, антигенами которых являются FcRn, и их фрагменты. Предпочтительным вариантом осуществления настоящего изобретения являются Fc-области антител или фрагменты, содержащие FcRn-связывающую область Fc-области. Так, например, в качестве «Fc-области» можно использовать Fc-область, полученную из встречающегося в естественных условиях IgG. Встречающийся в естественных условиях IgG означает полипептид, который содержит такую же аминокислотную последовательность, что и IgG, встречающийся в природе, и принадлежит к классу антител, которые в основном кодируются генами иммуноглобулина гамма. Встречающийся в естественных условиях человеческий IgG означает, например, встречающийся в естественных условиях человеческий IgG1, встречающийся в естественных условиях человеческий IgG2, встречающийся в естественных условиях человеческий IgG3 или встречающийся в естественных условиях человеческий IgG4. Встречающиеся в естественных условиях IgG включают также мутанты и субстанции, полученные из них в естественных условиях. Множество последовательностей аллотипов, образовавшихся в результате генетического полиморфизма, описано в Sequences of Proteins of Immunological Interest, NIH Publication №91-3242 для константной области антитела, такого как человеческий IgG1, человеческий IgG2, человеческий IgG3 и человеческий IgG4, и любую из последовательностей можно применять согласно настоящему изобретению. В частности, аминокислотная последовательность, простирающаяся от положения 356 до положения 358 согласно EU-нумерации, может представлять собой DEL или ЕЕМ в случае последовательности человеческого IgG1.
Известные в настоящее время Fc-области антител представляют собой, например, Fc-области IgA1-, IgA2-, IgD-, IgE-, IgG1-, IgG2-, IgG3-, IgG4- и IgM-типа. Например, Fc-область, полученную из встречающегося в естественных условиях антитела в виде человеческого IgG, можно применять в качестве Fc-области антитела согласно настоящему изобретению. Fc-области, полученные из константной области встречающегося в естественных условиях IgG, или более конкретно константной области, полученной из встречающегося в естественных условиях человеческого IgG1 (SEQ ID NO: 1), константной области, полученной из встречающегося в естественных условиях человеческого IgG2 (SEQ ID NO: 2), константной области, полученной из встречающегося в естественных условиях человеческого IgG3 (SEQ ID NO: 3), и константной области, полученной из встречающегося в естественных условиях человеческого IgG4 (SEQ ID NO: 4), можно применять в качестве Fc-области согласно настоящему изобретению. Встречающиеся в естественных условиях константные области IgG включают также мутанты и субстанции, полученные из них в естественных условиях.
Указанные Fc-области антител можно получать соответствующим методом, например, путем частичного расщепления антител, таких как моноклональные антитела, с помощью протеазы, такой как пепсин, с последующей адсорбцией образовавшихся фрагментов на колонке с белком А или колонке с белком G и последующей элюцией с использованием соответствующего буфера для элюции и т.п. Не существует ограничения касательно конкретной протеазы, если она обладает способностью избирательно расщеплять антитело, такое как моноклональное антитела, в соответствующих для данного фермента реакционных условиях, таких как значение рН, и ее примерами являются пепсин и фицин.
Изотип антитела определяют на основе структуры константной области. Константную область изотипов IgG1, IgG2, IgG3 и IgG4 обозначают как Cγ1, Сγ2, Сγ3 и Сγ4 соответственно. Аминокислотные последовательности полипептидов, образующих Fc-области человеческих Cγ1, Сγ2, Сγ3 и Сγ4, в качестве примера представлены в SEQ ID NO: 5, 6, 7 и 8. Взаимосвязь между аминокислотными остатками, образующими каждую из указанных аминокислотных последовательностей, и EU-нумерацией по Кэботу (обозначенной в контексте настоящего описания также как EU-индекс) представлена на фиг. 13.
Fc-область обозначает область, которая не включает F(ab')2-фрагмент, который состоит из двух легких цепей и двух тяжелых цепей, содержащих часть константной области между СН1-доменом и СН2-доменом, в результате между двумя тяжелыми цепями образуются межцепочечные дисульфидные связи. Fc-области, которые образуют антигенсвязывающие молекулы, представленные в настоящем описании, можно соответственно получать путем частичного расщепления моноклональных антител в виде IgG1, IgG2, IgG3 или IgG4 или т.п. с помощью протеазы, такой как пепсин, и последующей повторной элюции фракций, адсорбированных на колонке с белком А. Не существует ограничения касательно конкретной протеазы, если она обладает способностью избирательно расщеплять полноразмерное антитело в соответствующих для данного фермента реакционных условиях, таких как значение рН, с получением F(ab')2-фрагмента. Указанные протеазы представляют собой, например, пепсин и фицин.
Наиболее предпочтительным в качестве FcRn-связывающего домена, предлагаемого в настоящем изобретении, является домен с пониженной активностью связывания с Fcγ-рецептором. В контексте настоящего описания понятие Fcγ-рецептор (который обозначают в контексте настоящего описания также как Fcγ-рецептор, FcγR или FcgR) относится к рецептору, который может связываться с Fc-областью IgG1, IgG2, IgG3 или IgG4 и включает всех представителей, принадлежащих к семейству белков, кодируемых в основном генами Fcγ-рецептора. У человека указанное семейство включает (но, не ограничиваясь только ими) FcγRI (CD64), включая изоформы FcγRIa, FcγRIb и FcγRIc; FcγRII (CD32), включая изоформы FcγRIIa (включая аллотипы Н131 (тип Н) и R131 (тип R), FcγRIIb (включая FcγRIIb-1 и FcγRIIb-2) и FcγRIIc; и FcγRIII (CD16), включая изоформы FcγRIIIa (включая аллотипы V158 и F158) и FcγRIIIb (включая аллотипы FcγRIIIb-NA1 и FcγRIIIb-NA2); а также любые пока не открытые человеческие FcγR и изоформы или аллотипы FcγR. FcγR включают (но не ограничиваясь только ими) рецепторы человека, мышей, крыс, кроликов и обезьян, и могут иметь происхождение из любого организма. Мышиные FcγR включают (но, не ограничиваясь только ими) FcγRI (CD64), FcγRII (CD32), FcγRIII (CD16) и FcγRIII-2 (CD16-2), а также пока не открытые мышиные FcγR и изоформы или аллотипы FcγR. Приемлемые примеры указанных Fcγ-рецепторов включают человеческий FcγRI (CD64), FcγRIIa (CD32), FcγRIIb (CD32), FcγRIIIa (CD16) и/или FcγRIIIb (CD16).
Среди FcγR присутствуют активирующие рецепторы, которые несут активирующий мотив на основе тирозина иммунорецептора (ITAM), и ингибирующие рецепторы, которые несут ингибирующий мотив на основе тирозина иммунорецептора (ITIM). FcγR подразделяют на активирующие FcγR: FcγRI, FcγRIIa R, FcγRIIa H, FcγRIIIa и FcγRIIIb, и ингибирующий FcγR: FcγRIIb.
Полинуклеотидная последовательность и аминокислотная последовательность FcγRI представлены в NM_000566.3 и NP_000557.1 соответственно; полинуклеотидная последовательность и аминокислотная последовательность FcγRIIa представлены в ВС020823.1 и ААН20823.1 соответственно; полинуклеотидная последовательность и аминокислотная последовательность FcγRIIb представлены в ВС146678.1 и AAI46679.1 соответственно; полинуклеотидная последовательность и аминокислотная последовательность FcγRIIIa представлены в ВС033678.1 и ААН33678.1 соответственно и полинуклеотидная последовательность и аминокислотная последовательность FcγIIIb представлены в ВС128562.1 и AAI28563.1 соответственно (регистрационный номер RefSeq). Известно два типа генных полиморфизмов FcγRIIa, в результате которых аминокислота в положении 131 FcγRIIa заменена на гистидин (Н-тип) или аргинин (R-тип) (J. Exp. Med, 172, 1990, сс. 19-25). Кроме того, известно два типа генных полиморфизмов FcγRIIb, в результате которых аминокислота в положении 232 FcγRIIb заменена на изолейцин (1-тип) или треонин (Т-тип) (Arthritis. Rheum. 46, 2002, сс. 1242-1254). Кроме того, известно два типа генных полиморфизмов FcγRIIIa, в результате которых аминокислота в положении 158 FcγRIIIa заменена на валин (V-тип) или фенилаланин (F-тип) (J. Clin. Invest. 100(5), 1997, сс. 1059-1070). Известно также два типа генных полиморфизмов FcγRIIIb, которые представляют собой NA1-тип и NA2-тип (J. Clin. Invest. 85, 1990, сс. 1287-1295).
Снижение активности связывания с Fcγ-рецептором можно подтверждать с помощью хорошо известных методов, таких как FACS, формат ELISA, скрининг с использованием гомогенного анализа усиленной за счет эффекта близости люминесценции (ALPHA), BIACORE-метода на основе резонанса поверхностного плазмона (SPR) и др. (Proc. Natl. Acad. Sci. USA 103(11), 2006, cc. 4005-4010).
ALPHA-скрининг осуществляют на основе технологии ALPHA, которая основана на описанном ниже принципе, с использованием двух типов гранул, а именно, донорских и акцепторных гранулы. Люминесцентные сигналы поддаются обнаружению только тогда, когда происходит биологическое взаимодействие молекул, связанных с гранулами-донорами, с молекулами, связанными с гранулами-акцепторами, и когда обе гранулы находятся в непосредственной близости друг от друга. Возбужденный лазерным пучком фотосенсибилизатор в гранулах-донорах превращает кислород окружающей среды в возбужденный синглетный кислород. Когда синглетный кислород диффундирует из гранул-доноров и достигает гранул-акцепторов, локализованных в непосредственной близости, то индуцируется хемилюминесцентная реакция в гранулах-акцепторах, что в итоге приводит к испусканию света. Если молекулы, связанные с гранулами-донорами, не взаимодействуют с гранулами-акцепторами, то хемилюминесцентной реакции не происходит, поскольку синглетный кислород, который продуцируется гранулами-донорами, не достигает гранул-акцепторов.
Например, когда антигенсвязывающая молекула содержит Fc-область в качестве FcRn-связывающего домена, то получают антигенсвязывающую молекулу, которая имеет Fc-область дикого типа, и антигенсвязывающую молекулу, которая имеет мутантную Fc-область, полученную путем добавления аминокислотных мутаций для изменения связывания с Fcγ-рецептором, биотинилированную антигенсвязывающую молекулу связывают с гранулами-донорами, а меченный глутатион-S-трансферазой (GST) Fcγ-рецептор связывают с гранулами-акцепторами. В присутствии антигенсвязывающей молекулы, имеющей мутантную Fc-область, антигенсвязывающая молекула, имеющая Fc-область дикого типа, взаимодействует с Fcγ-рецептором и образует сигналы с длиной волны от 520 до 620 нм. Когда антигенсвязывающая молекула, имеющая мутантную Fc-область, является немеченой, то она конкурирует с антигенсвязывающей молекулой, имеющей Fc-область дикого типа, за взаимодействие с Fcγ-рецептором. Относительную аффинность связывания можно оценивать, определяя количественно снижение флуоресценции в результате конкуренции. Методы биотинилирования полипептидных комплексов, таких как антитела, с помощью сульфо-NHS-биотина или подобных агентов являются известными. Приемлемые методы введения GST-метки в Fcγ-рецептор включают метод, при котором экспрессируют Fcγ-рецептор и GST в клетке, несущий вектор, который обеспечивает экспрессию слитого гена, полученного путем слияния полинуклеотида, который кодирует Fcγ-рецептор, в рамке считывания с полинуклеотидом, который кодирует GST, и затем осуществляют очистку с помощью содержащей глутатион колонки, которую можно адаптировать соответствующим образом. Индуцированные сигналы можно анализировать, например, посредством подгонки к односайтовой модели конкуренции на основе нелинейного регрессионного анализа с использованием такой программы, как GRAPHPAD PRISM (фирма GraphPad; Сан-Диего).
Одну из субстанций (лиганд), предназначенных для исследования взаимодействия, иммобилизуют на тонком слое (пленке) золота сенсорного чипа и путем проникновения света на заднюю поверхность сенсорного чипа так, что имеет место полное отражение на границе раздела между тонким слоем золота и стеклом, интенсивность отраженного света в определенном сайте частично снижается (SPR-сигнал). Подготавливают другую субстанцию (аналит), предназначенную для исследования взаимодействия, для инъекции на поверхность сенсорного чипа; и когда лиганд связывается с аналитом, масса иммобилизованной молекулы-лиганда возрастает, и показатель преломления растворителя на поверхности сенсорного чипа изменяется. В результате указанного изменения показателя преломления положение SPR-сигнала сдвигается (и наоборот, положение сигнала возвращается в исходное, если происходит диссоциация указанного связывания). С помощью Biacore-системы определяют уровень описанного выше сдвига, или более конкретно изменение массы в зависимости от времени, откладывая изменение массы на поверхности сенсорного чипа по вертикальной оси, и таким образом получают количественные данные (сенсограмма). Кинетические параметры, такие как константа скорости ассоциации (ka) и константа скорости диссоциации (kd), определяют из представленных в виде кривых сенсограмм, и определяют аффинность (KD) как отношение указанных констант.BIACORE-метод можно применять также в качестве метода для анализа ингибирования. Примеры такого метода для анализа ингибирования описаны в Proc. Natl. Acad. Sci. USA 103(11), 2006, cc. 4005-4010.
В контексте настоящего описания «пониженная активность связывания с Fcγ-рецептором» означает, например, что при использовании описанного выше метода анализа активность связывания тестируемой антигенсвязывающей молекулы составляет 50% или менее, предпочтительно 45% или менее, 40% или менее, 35% или менее, 30% или менее, 20% или менее или 15% или менее или наиболее предпочтительно 10% или менее, 9% или менее, 8% или менее, 7% или менее, 6% или менее, 5% или менее, 4% или менее, 3% или менее, 2% или менее, или 1% или менее, по сравнению с активностью связывания контрольной антигенсвязывающей молекулы, содержащей Fc-область.
В качестве приемлемой контрольной антигенсвязывающей молекулы можно применять антигенсвязывающие молекулы, которые имеют, например, домен, содержащий Fc-область моноклонального антитела в виде IgG1, IgG2, IgG3 или IgG4. Структуры Fc-областей представлены в SEQ ID NO: 1 (А добавлен к N-концу последовательности, представленной в RefSeq под регистрационным номером ААС82527.1), SEQ ID NO: 2 (А добавлен к N-концу последовательности, представленной в RefSeq под регистрационным номером ААВ59393.1), SEQ ID NO: 3 (А добавлен к N-концу последовательности, представленной в RefSeq под регистрационным номером САА27268.1) и SEQ ID NO: 4 (А добавлен к N-концу последовательности, представленной в RefSeq под регистрационным номером ААВ59394.1). Кроме того, когда в качестве тестируемой субстанции используют антигенсвязывающую молекулу, содержащую мутант Fc-области антитела конкретного изотипа, то воздействие мутации мутанта на активность связывания с Fcγ-рецептором оценивают с использованием в качестве контроля антигенсвязывающей молекулы, которая имеет Fc-область антитела такого же конкретного изотипа. Таким путем можно получать антигенсвязывающие молекулы, содержащие мутантную Fc-область с установленной пониженной активностью связывания с Fcγ-рецептором.
Примеры таких мутантов включают мутанты с делецией аминокислот 231A-238S (WO 2009/011941) или мутанты C226S, C229S, P238S, (C220S) (J. Rheumatol 34, 2007, с. 11), C226S и C229S (Hum. Antibod. Hybridomas 1(1), 1990, сс. 47-54); C226S, C229S, Е233Р, L234V, и L235A (Blood 109, 2007, сс. 1185-1192), в которых нумерация аминокислоты соответствует EU-нумерации.
Таким образом, приемлемые примеры включают антигенсвязывающие молекулы, имеющие Fc-область с заменой любой из аминокислот в положениях 220, 226, 229, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 264, 265, 266, 267, 269, 270, 295, 296, 297, 298, 299, 300, 325, 327, 328, 329, 330, 331 и 332 согласно EU-нумерации в аминокислотной последовательности, образующей Fc-область антитела конкретного изотипа. Изобретение не ограничено конкретным изотипом антитела, из которого имеет происхождение Fc-область, и можно использовать соответствующую Fc-область, полученную из моноклонального антитела в виде IgG1, IgG2, IgG3 или IgG4, и можно применять Fc-область, полученную из встречающегося в естественных условиях человеческого антитела в виде IgG1.
Например, предпочтительно можно применять антигенсвязывающую молекулу, которая имеет Fc-область, содержащую любую из указанных ниже замен, положение которой определяется согласно EU-нумерации, аминокислот, образующих Fc-область антитела в виде IgG1 (каждый номер обозначает положение аминокислотного остатка согласно EU-нумерации; и однобуквенный код аминокислоты, который находится перед номером, обозначает аминокислотный остаток до его замены, а однобуквенный код аминокислоты, расположенный за номером, обозначает аминокислотный остаток после замены):
(a) L234F, L235E, P331S,
(б) C226S, C229S, P238S,
(в) C226S, C229S,
(г) C226S, C229S, Е233Р, L234V, L235A;
или Fc-область с делецией аминокислот в положениях 231-238 из аминокислот, образующих Fc-область антитела в виде IgG1.
Кроме того, предпочтительно можно применять антигенсвязывающую молекулу, которая имеет Fc-область, содержащую любую из указанных ниже замен, положение которой определяется согласно EU-нумерации, аминокислот, образующих Fc-область антитела в виде IgG2 (каждый номер обозначает положение аминокислотного остатка согласно EU-нумерации; и однобуквенный код аминокислоты, который находится перед номером, обозначает аминокислотный остаток до его замены, а однобуквенный код аминокислоты, расположенный за номером, обозначает аминокислотный остаток после замены)
(д) H268Q, V309L, A330S и P331S,
(е) V234A,
(ж) G237A,
(з) V234A и G237A,
(и) А235Е и G237A,
(к) V234A, А235Е, G237A.
Кроме того, предпочтительно можно применять антигенсвязывающую молекулу, которая имеет Fc-область, содержащую любую из указанных ниже замен, положение которой определяется согласно EU-нумерации, аминокислот, образующих Fc-область антитела в виде IgG3 (каждый номер обозначает положение аминокислотного остатка согласно EU-нумерации; и однобуквенный код аминокислоты, который находится перед номером, обозначает аминокислотный остаток до его замены, а однобуквенный код аминокислоты, расположенный за номером, обозначает аминокислотный остаток после замены)
(л) F241A,
(м) D265A,
(н) V264A.
Кроме того, предпочтительно можно применять антигенсвязывающую молекулу, которая имеет Fc-область, содержащую любую из указанных ниже замен, положение которой определяется согласно EU-нумерации, аминокислот, образующих Fc-область антитела в виде IgG4 (каждый номер обозначает положение аминокислотного остатка согласно EU-нумерации; и однобуквенный код аминокислоты, который находится перед номером, обозначает аминокислотный остаток до его замены, а однобуквенный код аминокислоты, расположенный за номером, обозначает аминокислотный остаток после замены):
(о) L235A, G237A, Е318А,
(п) L235E,
(р) F234A, L235A.
Другие предпочтительные примеры включают антигенсвязывающие молекулы, имеющие Fc-область, в которой любая из аминокислот в положениях 233, 234, 235, 236, 237, 327, 330 и 331 согласно EU-нумерации аминокислот, образующих Fc-область встречающегося в естественных условиях человеческого антитела в виде IgG1, заменена на аминокислоты, которые имеют соответствующую EU-нумерацию в соответствующем IgG2 или IgG4.
Другие предпочтительные примеры могут включать антигенсвязывающие молекулы, имеющие Fc-область, в которой одна или несколько аминокислот в положениях 234, 235 и 297 согласно EU-нумерации аминокислот, образующих Fc-область встречающегося в естественных условиях человеческого антитела в виде IgG1, заменена(ы) на другие аминокислоты. Настоящее изобретение не ограничено конкретным типом аминокислоты после замены, однако наиболее предпочтительной является антигенсвязывающая молекула, имеющая Fc-область, в которой одна или несколько аминокислот в положениях 234, 235 и 297 заменена(ы) на аланин.
Другие предпочтительные примеры могут включать антигенсвязывающие молекулы, имеющие Fc-область, в которой аминокислота в положении 265 согласно EU-нумерации аминокислот, образующих Fc-область антитела в виде IgG1, заменена на другую аминокислоту. Настоящее изобретение не ограничено конкретным типом аминокислоты после замены, однако наиболее предпочтительной является антигенсвязывающая молекула, имеющая Fc-область, в которой аминокислота в положении 265 заменена на аланин.
Понятия «домен, связывающийся со специфическим для рака антигеном», «домен, связывающийся с представителем суперсемейства фактора некроза опухоли (TNF)», «домен, связывающийся с представителем суперсемейства рецептора фактора некроза опухоли (TNF)» и «домен, связывающийся с комплексом Т-клеточного рецептора» (далее в контексте настоящего описания четыре связывающих домена вместе обозначают как антигенсвязывающие домены), включенные в антигенсвязывающие молекулы, предлагаемые в настоящем изобретении, относятся к областям, которые специфически связываются с полными антигенами или с их частями, которые представляют собой специфические для рака антигены, факторы, принадлежащие к суперсемейству TNF, факторы, принадлежащие к суперсемейству TNF-рецептора, или комплекс Т-клеточного рецептора; и примером связывающего домена является область, которая содержит антигенсвязывающую область антитела. Когда молекулярная масса антигена является большой, то антигенсвязывающая область антитела может связываться только с конкретным участком антигена. Указанный конкретный участок называют «эпитопом». Антигенсвязывающий домен может состоять из одного или нескольких вариабельных доменов антитела. Предпочтительно антигенсвязывающий домен содержит вариабельную область легкой цепи (VL) антитела и вариабельную область тяжелой цепи (VH) антитела. Указанные предпочтительные антигенсвязывающие домены включают, например, «одноцепочечный Fv (scFv)», «одноцепочечное антитело», «Fv», «одноцепочечный Fv2 (scFv2)», «Fab» и «F(ab')2» и т.п.
В контексте настоящего описания понятие «специфический для рака антиген» относится к антигену, экспрессируемому раковыми клетками, который обусловливает возможность различать раковые клетки и здоровые клетки; и, например, он включает антигены, которые экспрессируются на клетках, которые экспрессируются при приобретении клетками признака злокачественности, или содержат аномальные сахарные цепи, которые появляются на белковых молекулах или на клеточной поверхности, когда клетки становятся раковыми. Конкретные примеры включают ALK-рецептор (рецептор плейотрофина); плейотрофин; антиген карциномы поджелудочной железы KS 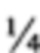 , антиген карциномы яичника (СА125); простатический кислый фосфат; простатсецифический антиген (PSA); ассоциированный с меланомой антиген р97; меланомный антиген gp75; высокомолекулярный меланомный антиген (HMW-MAA); простатспецифический мембранный антиген; карциноэмбриональный антиген (СЕА); антиген, представляющий собой полиморфный эпителиальный муцин; человеческий антиген жировых глобул молока; антигены, ассоциированные с колоректальным раком, такие как СЕА, TAG-72, СО17-1A, GICA 19-9, СТА-1 и LEA; антиген лимфомы Беркитта-38.13; CD19; антиген человеческой В-лимфомы CD20; CD33; специфические для меланомы антигены, такие как ганглиозид GD2, ганглиозид GD3, ганглиозид GM2 и ганглиозид GM3; опухольспецифический трансплантационный антиген клеточной поверхности (TSTA); индуцируемые вирусами опухолевые антигены, включая Т-антиген и оболочечные антигены опухолей, индуцированных ДНКовыми вирусами и РНКовыми вирусами; СЕА ободочной кишки; онкоэмбриональные антигены, такие как онкоэмбриональный трофобластный гликопротеин 5Т4, онкоэмбриональный антиген опухоли мочевого пузыря; α-фетопротеин; дифференцированные антигены, такие как антигены человеческой карциномы легкого L6 и L20; антигены фибросаркомы; антиген человеческого Т-клеточного лейкоза Gp37; неогликопротеин; сфинголипиды; антигены рака молочной железы, такие как EGFR (рецептор эпидермального фактора роста); NY-BR-16; NY-BR-16 и антиген HER2 (p185HER2); полиморфный эпителиальный муцин (РЕМ); антиген злокачественных человеческих лимфоцитов АРО-1; дифференцированные антигены, такие как антиген I, обнаруженный в фетальных эритроцитах; антиген I первичной эндодермы, обнаруженный в эритроцитах взрослых; эмбрионы до имплантации; I(Ма), обнаруженный при раке желудка; M18 и М39, обнаруженные в эпителии молочной железы; SSEA-1, VEP8, VEP9, Myl и VIM-D5, обнаруженные в миелодных клетках; D156-22, обнаруженный при колоректальном раке; TRA-1-85 (антиген группы крови Н); SCP-1, обнаруженный при раке яичек и яичников; С14, обнаруженный при колоректальном раке; F3, обнаруженный при раке легкого; АН6, обнаруженный при раке желудка; Y-гаптен; Ley (антиген группы крови Y Льюиса), обнаруженный в клетках эмбриональной карциномы; TL5 (антиген группы крови А); EGF-рецептор, обнаруженный в А431-клетках; антиген из Е1-серий (антиген группы крови В), обнаруженный в поджелудочной железе; FC10.2, обнаруженный в клетках эмбриональной карциномы; антиген рака желудка; СО-514 (антиген группы крови Lea), обнаруженный при аденокарциномах; NS-10, обнаруженный при аденокарциномах; СО-43 (антиген группы крови Leb); G49, обнаруженный в EGF-рецепторе А431-клеток; аМН2 (антиген группы крови ALeb/Ley), обнаруженный при раке ободочной кишки; 19.9, обнаруженный при раке ободочной кишки; муцины, характерные для рака желудка; Т5А7, обнаруженный в миелоидный клетках; R24, обнаруженный при меланоме; 4.2, GD3, D1.1, OFA-1, GM2, OFA-2, GD2 и М1:22:25:8, обнаруженные в клетках эмбриональной карциномы, а также SSEA-3 и SSEA-4, обнаруженные у эмбрионов на стадии 4-8 клеток; антиген подкожной Т-клеточной лимфомы; антиген MART-1; сиалилированный Tn-антиген (STn); антиген рака ободочной кишки NY-CO-45; антиген рака легкого NY-LU-12, вариант А; антиген аденокарциномы ART1; ассоциированный с паранеопластическим синдромом антиген рака головного мозга-яичек (онконевральный антиген МА2; параненеопластичесакий невральный антиген; невроонкологический вентральный антиген 2 (NOVA2); антиген карциномы гемоцитов, связанной с геном 520; опухольассоциированный антиген СО-029; опухольассоциированные антигены MAGE-C1 (раково-тестикуларный антиген СТ7), MAGE-B1 (антиген MAGE-XP), MAGE-B2 (DAM6), MAGE-2, MAGE-4a, MAGE-4b и MAGE-X2; раково-тестикуларный антиген (NY-EOS-1); YKL-40, фрагменты любого из вышеуказанных полипептидов или структуры, образовавшиеся в результате их модификации (например, упомянутая выше модифицированная фосфатная группа или сахарная цепь); ЕрСАМ; EREG; СА19-9; СА15-3; сиалилированный SSEA-1(SLX); HER2; PSMA; СЕА и CLEC12A. Специфические для рака антигены, которые становятся мишенями для доменов, которые связываются со специфическими для рака антигенами, предлагаемыми в настоящем изобретении, предпочтительно представляют собой, в частности, антигены, которые экспрессируются на клеточной поверхности, и примеры указанных специфических для рака антигенов включают CD19, CD20, EGFR, HER2, ЕрСАМ и EREG.
, антиген карциномы яичника (СА125); простатический кислый фосфат; простатсецифический антиген (PSA); ассоциированный с меланомой антиген р97; меланомный антиген gp75; высокомолекулярный меланомный антиген (HMW-MAA); простатспецифический мембранный антиген; карциноэмбриональный антиген (СЕА); антиген, представляющий собой полиморфный эпителиальный муцин; человеческий антиген жировых глобул молока; антигены, ассоциированные с колоректальным раком, такие как СЕА, TAG-72, СО17-1A, GICA 19-9, СТА-1 и LEA; антиген лимфомы Беркитта-38.13; CD19; антиген человеческой В-лимфомы CD20; CD33; специфические для меланомы антигены, такие как ганглиозид GD2, ганглиозид GD3, ганглиозид GM2 и ганглиозид GM3; опухольспецифический трансплантационный антиген клеточной поверхности (TSTA); индуцируемые вирусами опухолевые антигены, включая Т-антиген и оболочечные антигены опухолей, индуцированных ДНКовыми вирусами и РНКовыми вирусами; СЕА ободочной кишки; онкоэмбриональные антигены, такие как онкоэмбриональный трофобластный гликопротеин 5Т4, онкоэмбриональный антиген опухоли мочевого пузыря; α-фетопротеин; дифференцированные антигены, такие как антигены человеческой карциномы легкого L6 и L20; антигены фибросаркомы; антиген человеческого Т-клеточного лейкоза Gp37; неогликопротеин; сфинголипиды; антигены рака молочной железы, такие как EGFR (рецептор эпидермального фактора роста); NY-BR-16; NY-BR-16 и антиген HER2 (p185HER2); полиморфный эпителиальный муцин (РЕМ); антиген злокачественных человеческих лимфоцитов АРО-1; дифференцированные антигены, такие как антиген I, обнаруженный в фетальных эритроцитах; антиген I первичной эндодермы, обнаруженный в эритроцитах взрослых; эмбрионы до имплантации; I(Ма), обнаруженный при раке желудка; M18 и М39, обнаруженные в эпителии молочной железы; SSEA-1, VEP8, VEP9, Myl и VIM-D5, обнаруженные в миелодных клетках; D156-22, обнаруженный при колоректальном раке; TRA-1-85 (антиген группы крови Н); SCP-1, обнаруженный при раке яичек и яичников; С14, обнаруженный при колоректальном раке; F3, обнаруженный при раке легкого; АН6, обнаруженный при раке желудка; Y-гаптен; Ley (антиген группы крови Y Льюиса), обнаруженный в клетках эмбриональной карциномы; TL5 (антиген группы крови А); EGF-рецептор, обнаруженный в А431-клетках; антиген из Е1-серий (антиген группы крови В), обнаруженный в поджелудочной железе; FC10.2, обнаруженный в клетках эмбриональной карциномы; антиген рака желудка; СО-514 (антиген группы крови Lea), обнаруженный при аденокарциномах; NS-10, обнаруженный при аденокарциномах; СО-43 (антиген группы крови Leb); G49, обнаруженный в EGF-рецепторе А431-клеток; аМН2 (антиген группы крови ALeb/Ley), обнаруженный при раке ободочной кишки; 19.9, обнаруженный при раке ободочной кишки; муцины, характерные для рака желудка; Т5А7, обнаруженный в миелоидный клетках; R24, обнаруженный при меланоме; 4.2, GD3, D1.1, OFA-1, GM2, OFA-2, GD2 и М1:22:25:8, обнаруженные в клетках эмбриональной карциномы, а также SSEA-3 и SSEA-4, обнаруженные у эмбрионов на стадии 4-8 клеток; антиген подкожной Т-клеточной лимфомы; антиген MART-1; сиалилированный Tn-антиген (STn); антиген рака ободочной кишки NY-CO-45; антиген рака легкого NY-LU-12, вариант А; антиген аденокарциномы ART1; ассоциированный с паранеопластическим синдромом антиген рака головного мозга-яичек (онконевральный антиген МА2; параненеопластичесакий невральный антиген; невроонкологический вентральный антиген 2 (NOVA2); антиген карциномы гемоцитов, связанной с геном 520; опухольассоциированный антиген СО-029; опухольассоциированные антигены MAGE-C1 (раково-тестикуларный антиген СТ7), MAGE-B1 (антиген MAGE-XP), MAGE-B2 (DAM6), MAGE-2, MAGE-4a, MAGE-4b и MAGE-X2; раково-тестикуларный антиген (NY-EOS-1); YKL-40, фрагменты любого из вышеуказанных полипептидов или структуры, образовавшиеся в результате их модификации (например, упомянутая выше модифицированная фосфатная группа или сахарная цепь); ЕрСАМ; EREG; СА19-9; СА15-3; сиалилированный SSEA-1(SLX); HER2; PSMA; СЕА и CLEC12A. Специфические для рака антигены, которые становятся мишенями для доменов, которые связываются со специфическими для рака антигенами, предлагаемыми в настоящем изобретении, предпочтительно представляют собой, в частности, антигены, которые экспрессируются на клеточной поверхности, и примеры указанных специфических для рака антигенов включают CD19, CD20, EGFR, HER2, ЕрСАМ и EREG.
Кроме того, в качестве факторов, принадлежащих к «суперсемейству TNF» или к «суперсемейству TNF-рецептора», известны лиганды, имеющие трехмерную структуру, и рецепторы с трехмерной структурой, с которыми связываются лиганды, которые принимают участие в активации различных иммунных клеток (Nat. Rev. Immunol., 12, 2012, сс. 339-351). Примеры факторов, принадлежащих к суперсемейству TNF или суперсемейству TNF-рецептора, включают CD137, CD137L, CD40, CD40L, ОХ40, OX40L, CD27, CD70, HVEM, LIGHT, RANK, RANKL, CD30, CD153, GITR и GITRL. Предпочтительными факторами являются, например, CD137 и CD40. Наиболее предпочтительным фактором является, например, CD137.
Кроме того, «комплекс Т-клеточного рецептора» может представлять собой сам Т-клеточный рецептор или молекулу-адаптер, входящую в комплекс Т-клеточного рецептора вместе с Т-клеточным рецептором. Пригодной молекулой-адаптером является CD3.
Для Т-клеточного рецептора эпитопом, с которым связывается домен, связывающийся с Т-клеточным рецептором, может быть вариабельная область или константная область, но предпочтительным является эпитоп, присутствующий в константной области. Примеры последовательностей константной области включают α-цепь Т-клеточного рецептора, которая имеет регистрационный № RefSeq САА26636.1 (SEQ ID NO: 9), β-цепь Т-клеточного рецептора, которая имеет регистрационный № RefSeq С25777 (SEQ ID NO: 10), γ1-цепь Т-клеточного рецептора, которая имеет регистрационный № RefSeq А26659 (SEQ ID NO: 11), γ2-цепь Т-клеточного рецептора, которая имеет регистрационный № RefSeq ААВ63312.1 (SEQ ID NO: 12) и δ-цепь Т-клеточного рецептора, которая имеет регистрационный № RefSeq ААА61033.1 (SEQ ID NO: 13).
В настоящем изобретении, когда «CD3-связывающий домен» применяют в качестве домена, связывающегося с комплексом Т-клеточного рецептора, то CD3-связывающий домен может представлять собой один или несколько вариабельных доменов антитела. Предпочтительно CD3-связывающий домен включает вариабельную область легкой цепи (VL) и вариабельную область тяжелой цепи (VH) антитела к CD3. Примеры приемлемых указанных CD3-связывающих доменов включают «одноцепочечный Fv (scFv)», «одноцепочечное антитело», «Fv», «одноцепочечный Fv2 (scFv2)», «Fab», «F(ab')2» и т.п.
CD3-связывающий домен, предлагаемый в настоящем изобретении, может включать домены, которые связываются с любым эпитопом, если он в качестве эпитопа присутствует в γ-цепи, δ-цепи или ε-цепи, образующей человеческий CD3. В настоящем изобретении предпочтительно применять CD3-связывающий домен, который содержит вариабельную область легкой цепи (VL) антитела к CD3 и вариабельную область тяжелой цепи (VH) антитела к CD3 и который связывается с эпитопом, присутствующим во внеклеточной области ε-цепи человеческого комплекса CD3. В качестве указанного CD3-связывающего домена можно применять CD3-связывающий домен, который содержит вариабельную область легкой цепи (VL) и вариабельную область тяжелой цепи (VH) антитела OKT3 (Proc. Natl. Acad. Sci. USA 77, 1980, cc. 4914-4917) или различных известных антител к CD3. Предпочтительно можно применять CD3-связывающий домен, полученный из антитела к CD3, которое обладает требуемыми свойствами и которое получают путем иммунизации требуемого животного γ-цепью, δ-цепью или ε-цепью, образующей человеческий CD3, с использованием описанного выше метода. Предпочтительно можно применять человеческие и соответственно гуманизированные антитела, описанные ниже, в качестве антитела к CD3, которое служит источником CD3-связывающего домена. Структуры γ-цепи, δ-цепи или ε-цепи, которые образуют CD3, представлены в виде полинуклеотидных последовательностей SEQ ID NO: 14 (NM_000073.2), SEQ ID NO: 16 (NM_000732.4) и SEQ ID NO: 18 (NM_000733.3) соответственно и в виде полипептидных последовательностей SEQ ID NO: 15 (NP_000064.1), SEQ ID NO: 17 (NP_000723.1) и SEQ ID NO: 19 (NP_000724.1) соответственно (в скобках представлены регистрационные номера RefSeq).
В предпочтительном варианте осуществления изобретения «антигенсвязывающая молекула», предлагаемая в настоящем изобретении, включает антитело, которое содержит вариабельную область антитела, предлагаемого в настоящем изобретении.
Примерами антител, предлагаемых в настоящем изобретении, являются следующие антитела:
[1] антитело, содержащее аминокислотную последовательность SEQ ID NO: 66 в качестве вариабельной области тяжелой цепи и аминокислотную последовательность SEQ ID NO: 85 в качестве вариабельной области легкой цепи;
[2] антитело, содержащее аминокислотную последовательность SEQ ID NO: 67 в качестве вариабельной области тяжелой цепи и аминокислотную последовательность SEQ ID NO: 86 в качестве вариабельной области легкой цепи;
[3] антитело, содержащее аминокислотную последовательность SEQ ID NO: 70 в качестве вариабельной области тяжелой цепи и аминокислотную последовательность SEQ ID NO: 89 в качестве вариабельной области легкой цепи;
[4] а антитело, содержащее аминокислотную последовательность SEQ ID NO: 76 в качестве вариабельной области тяжелой цепи и аминокислотную последовательность SEQ ID NO: 95 в качестве вариабельной области легкой цепи;
[5] антитело, содержащее аминокислотную последовательность SEQ ID NO: 77 в качестве вариабельной области тяжелой цепи и аминокислотную последовательность SEQ ID NO: 96 в качестве вариабельной области легкой цепи;
[6] антитело, содержащее аминокислотную последовательность SEQ ID NO: 78 в качестве вариабельной области тяжелой цепи и аминокислотную последовательность SEQ ID NO: 97 в качестве вариабельной области легкой цепи;
[7] антитело по одному из п.п. [1]-[6], содержащее аминокислотную последовательность SEQ ID NO: 99 в качестве константной области тяжелой цепи и аминокислотную последовательность SEQ ID NO: 59 или аминокислотную последовательность SEQ ID NO: 60 в качестве константной области легкой цепи;
[8] антитело, обладающее активностью, эквивалентной активности антитела по одному из п.п. [1]-[7], и
[9] антитело, которое связывается с таким же эпитопом, что эпитоп, с которым связывается антитело по одному из п.п. [1]-[7].
Касательно антитела по п. [8] понятие «эквивалентная активность» относится к агонистической активности в отношении CD137, которая составляет 70% или более, предпочтительно 80% или более и более предпочтительно 90% или более от активности связывания антитела по одному из п.п. [1]-[7]. В настоящем изобретении предложено также антитело по п. [9], которое связывается с таким же эпитопом, что эпитоп, с которым связывается антитело к CD137, описанное в настоящем изобретении. Указанное антитело можно получать, например, с помощью описанного ниже метода.
Имеет ли тестируемое антитело общий эпитоп с определенным антителом, можно оценивать на основе конкуренции между двумя антителами за один и тот же эпитоп. Конкуренцию между антителами можно определять с помощью анализа перекрестной блокады или т.п. Например, анализ ELISA в условиях конкуренции представляет собой предпочтительный анализ перекрестной блокады. В частности, в анализе перекрестной блокады белок CD137, применяемый для сенсибилизации лунок титрационного микропланшета, предварительно инкубируют в присутствии потенциально конкурирующего антитела или без него, а затем добавляют антитело к CD137, предлагаемое в настоящем изобретении. Количество антитела к CD137, предлагаемого в настоящем изобретении, связанного с белком CD137 в лунках, косвенно коррелирует со способностью к связыванию потенциально конкурирующего антитела (тестируемого антитела), которое конкурирует за связывание с тем же самым эпитопом. Это означает, что чем выше аффинность тестируемого антитела к тому же самому эпитопу, тем меньше количество антитела к CD137, предлагаемого в настоящем изобретении, которое связано с сенсибилизированными белком CD137 лунками, и тем больше количество тестируемого антитела, которое связано с сенсибилизированными белком CD137 лунками.
Количество связанного с лунками антитела можно легко определять путем предварительного мечения антитела. Например, меченное биотином антитело можно количественно измерять, используя конъюгат авидин/пероксидаза и соответствующий субстрат. В частности, анализ перекрестной блокады, в котором применяют ферментную метку, такую как пероксидаза, обозначают как «конкурентный ELISA-анализ». Антитело можно метить другими предназначенными для мечения субстанциями, которые можно обнаруживать или количественно измерять. В частности, известны радиоактивные метки, флуоресцентные метки и т.п.
Кроме того, когда тестируемое антитело имеет константную область, полученную из вида, отличного от того, из которого получено антитело к CD137, предлагаемое в настоящем изобретении, то количество антитела, связанного с лунками, можно оценивать количественно, используя меченое антитело, которое распознает константную область этого антитела. Альтернативно этому, если антитела получают из одного и того же вида, но они принадлежат к различным классам, то количество антител, связанных с лунками, можно количественно оценивать, используя антитела, которые позволяют различать индивидуальные классы.
Если антитело-кандидат может блокировать связывание антитела к CD137 по меньшей мере на 20%, предпочтительно по меньшей мере на 20-50% и еще более предпочтительно по меньшей мере на 50% по сравнению с активностью связывания, полученной в контрольном эксперименте, который осуществляют в отсутствии потенциально конкурирующего антитела, то потенциально конкурирующее антитело представляет собой либо антитело, которое связывается практически с тем же самым эпитопом, либо антитело, которое конкурирует за связыванием с тем же самым эпитопом, что и антитело к CD137, предлагаемое в настоящем изобретении.
Предпочтительным примером антитела, которое связывается с тем же самым эпитопом, что и эпитоп, с которым связывается антитело по одному из п.п. [1]-[7], является, например, антитело, которое распознает область, содержащую последовательность
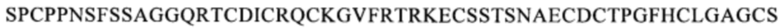
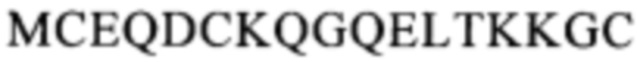 (SEQ ID NO: 113) в белке CD137. Другой пример включает антитело, которое распознает область, содержащую последовательность
(SEQ ID NO: 113) в белке CD137. Другой пример включает антитело, которое распознает область, содержащую последовательность 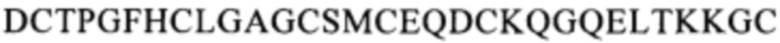 (SEQ ID NO: 108) в белке CD137.
(SEQ ID NO: 108) в белке CD137.
Биспецифическое антитело к противораковому антигену /к человеческому CD137, которое обладает требуемыми противоопухолевыми видами активности, можно получать путем модификации вышеописанного антитела к человеческому CD137 с помощью антитела к специфическому для рака антигену (например, антитела к человеческому GPC3) с получением биспецифического антитела, и оценки его зависящего от специфического для рака антигена агонистического действия в отношении CD137.
Одним из вариантов осуществления настоящего изобретения является (но, не ограничиваясь только им) биспецифическое антитело, которое содержит домен, связывающийся со специфическим для рака антигеном, и домен, связывающийся с человеческим CD137.
Примерами биспецифического антитела, предлагаемого в настоящем изобретении, являются следующие антитела:
[I] биспецифическое антитело, содержащее аминокислотную последовательность SEQ ID NO: 122 (вариабельная область тяжелой цепи) и аминокислотную последовательность SEQ ID NO: 123 (вариабельная область легкой цепи) в качестве связывающего человеческий CD137 домена;
[II] биспецифическое антитело, содержащее аминокислотную последовательность SEQ ID NO: 124 (вариабельная область тяжелой цепи) и аминокислотную последовательность SEQ ID NO: 82 (вариабельная область легкой цепи) в качестве связывающего человеческий CD137 домена;
[III] биспецифическое антитело, содержащее аминокислотную последовательность SEQ ID NO: 125 (вариабельная область тяжелой цепи) и аминокислотную последовательность SEQ ID NO: 84 (вариабельная область легкой цепи) в качестве связывающего человеческий CD137 домена;
[IV] антитело, которое связывается с тем же эпитопом, что и эпитоп, с которым связывается биспецифическое антитело по одному из п.п. [I]-[III].
В зависимости от ракового антигена-мишени специалисты в данной области легко могут выбирать последовательность вариабельной области тяжелой цепи и последовательность вариабельной области легкой цепи, которые связываются с раковым антигеном, в качестве вариабельной области тяжелой цепи и вариабельной области легкой цепи, которые можно включать в домен, связывающийся со специфическим для рака антигеном.
В настоящем изобретении предложено также биспецифическое антитело по п. [IV], которое связывается с тем же эпитопом, что и эпитоп, с которым связывается биспецифическое антитело к специфическому для рака антигену /к человеческому CD137, указанное в настоящем изобретении. Указанное антитело можно получать, например, с помощью описанного ниже метода.
Имеет ли тестируемое антитело общий эпитоп с определенным антителом, можно оценивать на основе конкуренции между двумя антителами за один и тот же эпитоп. Конкуренцию между антителами можно определять с помощью анализа перекрестной блокады или т.п. Например, анализ ELISA в условиях конкуренции представляет собой предпочтительный анализ перекрестной блокады. В частности, в анализе перекрестной блокады белок CD137, применяемый для сенсибилизации лунок титрационного микропланшета, предварительно инкубируют в присутствии потенциально конкурирующего антитела или без него, а затем добавляют антитело к CD137, предлагаемое в настоящем изобретении. Количество антитела к CD137, предлагаемого в настоящем изобретении, связанного с белком CD137 в лунках, косвенно коррелирует со способностью к связыванию потенциально конкурирующего антитела (тестируемого антитела), которое конкурирует за связывание с тем же самым эпитопом. Это означает, что чем выше аффинность тестируемого антитела к тому же самому эпитопу, тем меньше количества антитела к CD137, предлагаемого в настоящем изобретении, связано с сенсибилизированными белком CD137 лунками и тем больше количества тестируемого антитела связано с сенсибилизированными белком CD137 лунками.
Количество связанного с лунками антитела можно легко определять путем предварительного мечения антитела. Например, меченное биотином антитело можно количественно оценивать, используя конъюгат авидин/пероксидаза и соответствующий субстрат. В частности, анализ перекрестной блокады, в котором применяют ферментную метку, такую как пероксидаза, обозначают как «конкурентный ELISA-анализ». Антитело можно метить другими предназначенными для мечения субстанциями, которые можно обнаружить или количественно оценивать. В частности, известны радиоактивные метки, флуоресцентные метки и т.п.
Кроме того, когда тестируемое антитело имеет константную область, полученную из вида, отличного от того, из которого получено антитело к CD137, предлагаемое в настоящем изобретении, то количество антитела, связанного с лунками, можно измерять, используя меченое антитело, которое распознает константную область этого антитела. Альтернативно этому, если антитела получают из одного и того же вида, но они принадлежат к различным классам, то количество антител, связанных с лунками, можно измерять, используя антитела, которые позволяют различать индивидуальные классы.
Если антитело-кандидат может блокировать связывание антитела к CD137 по меньшей мере на 20%, предпочтительно по меньшей мере на 20-50% и еще более предпочтительно по меньшей мере на 50% по сравнению с активностью связывания, полученной в контрольном эксперименте, который осуществляют в отсутствии потенциально конкурирующего антитела, то потенциально конкурирующее антитело представляет собой либо антитело, которое связывается практически с тем же самым эпитопом, либо антитело, которое конкурирует за связыванием с тем же самым эпитопом, что и антитело к CD137, предлагаемое в настоящем изобретении.
В другом варианте осуществления изобретения способность тестируемого антитела конкурентно или перекрестно конкурентно связываться с другим антителом, специалисты в данной области могут легко определять с использованием стандартного анализа связывания, такого как BIAcore-анализ или анализ методом проточной цитометрии, которые известны в данной области.
Методы определения пространственной конформации эпитопов включают, например, рентгеновскую кристаллографию и двухмерный ядерный магнитный резонанс (см., например, Epitope Mapping Protocols in Methods in Molecular Biology, под ред. Morris, т. 66, 1996).
Предпочтительными примерами биспецифического антитела, которое связывается с тем же самым эпитопом, что и эпитоп человеческого CD137, с которым связывается биспецифическое антитело по одному из п.п. [I]-[III], являются биспецифические антитела, которое распознает область, содержащую последовательность 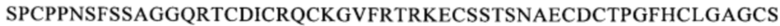
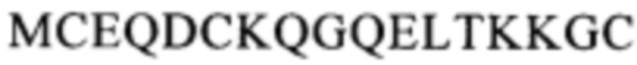 (SEQ ID NO: 113), область, содержащую последовательность
(SEQ ID NO: 113), область, содержащую последовательность 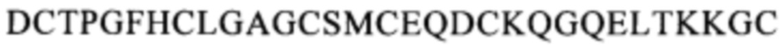 (SEQ ID NO: 108), область, содержащую последовательность
(SEQ ID NO: 108), область, содержащую последовательность 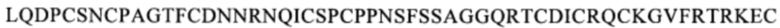
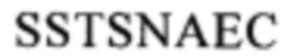 (SEQ ID NO: 111), или область, содержащую последовательность
(SEQ ID NO: 111), или область, содержащую последовательность 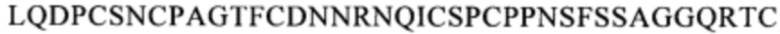 (SEQ ID NO: 106), в человеческом белке CD137. Более предпочтительные примеры включают биспецифические антитела, которые распознают область, содержащую последовательность
(SEQ ID NO: 106), в человеческом белке CD137. Более предпочтительные примеры включают биспецифические антитела, которые распознают область, содержащую последовательность 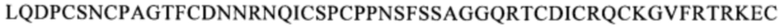
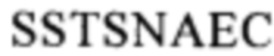 (SEQ ID NO: 111) или область, содержащую последовательность
(SEQ ID NO: 111) или область, содержащую последовательность 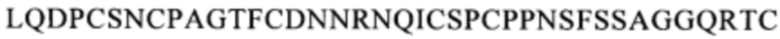 (SEQ ID NO: 106) в человеческом белке CD137.
(SEQ ID NO: 106) в человеческом белке CD137.
Вариантом осуществления настоящего изобретения является (но, не ограничиваясь только им) биспецифическое антитело, которое содержит домен, связывающийся со специфическим для рака антигеном, и домен, связывающийся с человеческим CD40.
В зависимости от ракового антигена-мишени специалисты в данной области легко могут выбирать последовательность вариабельной области тяжелой цепи и последовательность вариабельной области легкой цепи, которые связываются с раковым антигеном, в качестве вариабельной области тяжелой цепи и вариабельной области легкой цепи, которые можно включать в домен, связывающийся со специфическим для рака антигеном.
Связывающая активность антител
Антигенсвязывающую активность антитела можно оценивать с помощью известных методов (Antibodies: A Laboratory Manual (под ред. Harlow, David Lane, изд-во Cold Spring Harbor Laboratory, 1988). Например, можно применять твердофазный иммуноферментный анализ (ELISA), ферментный иммуноанализ (EIA), радиоиммуноанализ (РИА), метод разделения клеток на основе возбуждения флуоресценции (FACS), скрининг с использованием гомогенного анализа усиленной за счет эффекта близости люминесценции (ALPHA), BIACORE-метод на основе поверхностного плазмонного резонанса (SPR) или флуороиммуноанализ. Методы оценки связывающей активности антитела с антигеном, экспрессируемым клеткой, включают, например, методы, описанные на сс. 359-420 в «Antibodies: A Laboratory Manual».
В частности, методы, в которых используют проточный цитометр, можно успешно применять в качестве метода измерения связывания между антигеном, экспресируемым на поверхности клеток, суспендированных в буфере или т.п., и антителом против антигена. Проточные цитометры, которые можно использовать, включают, например, FACSCanto™ II, FACSAria™, FACSArray™, FACSVantage™ SE, и FACSCalibur™ (все вышеуказанные устройства фирмы BD Biosciences); и EPICS ALTRA HyPerSort Cytomics FC 500, EPICS XL-MCL ADC EPICS XL ADC и Cell Lab Quanta/Cell Lab Quanta SC (все вышеуказанные устройства фирмы Beckman Coulter).
Примеры приемлемого метода измерения связывающей активности тестируемого антитела к CD137 с антигеном включают метод, основанный на взаимодействии экспрессирующих CD137 клеток с тестируемым антителом и последующем окрашивании меченным с помощью ФИТЦ вторичным антителом, которое распознает тестируемое антитело, и последующее получение данных измерений с использованием FACSCalibur (фирма BD), и анализа полученной интенсивности флуоресценции с использованием программы CELL QUEST (фирма BD).
Антитело
В контексте настоящего описания «антитело» относится к встречающемуся в естественных условиях иммуноглобулину или иммуноглобулину, полученному полностью или частично путем синтеза. Антитела можно выделять из встречающихся в естественных условиях источников, таких как встречающиеся в естественных условиях плазма и сыворотка, или из супернатантов культур продуцирующих антитела клеток гибридом. Альтернативно этому, антитела можно частично или полностью синтезировать с использованием таких методик, как генетическая рекомбинация. Пригодными примерами антител являются, например, антитела, принадлежащие к какому-либо изотипу иммуноглобулинов или его подклассу. Известные человеческие иммуноглобулины включают антитела следующих девяти классов (изотипов): IgG1, IgG2, IgG3, IgG4, IgA1, IgA2, IgD, IgE и IgM. Из этих изотипов к антителам, предлагаемым в изобретении, относятся IgG1, IgG2, IgG3 и IgG4.
Методы получения антитела с требуемой активностью связывания известны специалистам в данной области, и антитела можно получать в виде поликлональных или моноклональных антител. В качестве антител, предлагаемых в настоящем изобретении, можно получать моноклональные антитела, происходящие из организма млекопитающих. Указанные происходящие из организма млекопитающих моноклональные антитела включают антитела, полученные с помощью гибридом, и антитела, полученные из клеток-хозяев, трансформированных экспрессионным вектором, который несет ген антитела, созданный с помощью методов генетической инженерии.
Отсутствует какое-либо конкретное ограничение, касающееся млекопитающих, подлежащих иммунизации с целью получения антител. Предпочтительно выбирать млекопитающих с учетом их совместимости с родительскими клетками, применяемыми для клеточного слияния для получения с помощью гибридом. В целом, предпочтительно применяют кроликов, обезьян и грызунов, таких как мыши, крысы и хомяки.
Вышеуказанных животных иммунизируют сенсибилизирующим антигеном с помощью известных методов. Общепринятым методом иммунизации является, например, введение млекопитающим сенсибилизирующего антигена путем внутрибрюшинной или подкожной инъекции. В частности, сенсибилизирующий антиген можно соответствующим образом разводить в ЗФР (забуференный фосфатом физиологический раствор), в физиологическом соляном растворе или т.п. При необходимости с антигеном смешивают общепринятый адъювант, такой как полный адъювант Фрейнда, и смесь эмульгируют. Затем сенсибилизирующий антиген вводят млекопитающему несколько раз с 4-21-дневными интервалами. При иммунизации сенсибилизирующим антигеном можно использовать соответствующие носители. В частности, когда в качестве сенсибилизирующего антигена используют низкомолекулярный неполный пептид, то иногда для иммунизации требуется сочетать пептид, представляющий собой сенсибилизирующий антиген, с белком-носителем, таким как альбумин или гемоцианин лимфы улитки.
Альтернативно этому, можно получать продуцирующие требуемое антитело гибридомы с помощью описанной ниже ДНК-иммунизации. ДНК-иммунизация представляет собой метод иммунизации, который обеспечивает иммуностимуляцию посредством экспрессии сенсибилизирующего антигена в организме иммунизированного животного в результате введения ДНК-вектора, сконструированного таким образом, чтобы он обеспечивал экспрессию гена, кодирующего антигенный белок, в организме животного. По сравнению с общепринятыми методами иммунизации, при которых животным, подлежащим иммунизации, вводят белковый антиген, ДНК-иммунизация, по-видимому, имеет следующие преимущества:
- можно осуществлять иммуностимуляцию, сохраняя при этом структуру мембранного белка; и
- отсутствует необходимость в очистке антигена для иммунизации.
Для получения моноклонального антитела, предлагаемого в настоящем изобретении, с использованием ДНК-иммунизации сначала вводят животному, подлежащему иммунизации, ДНК, экспрессирующую антигенный белок. ДНК, кодирующую антигенный белок, можно синтезировать с помощью известных методов, таких как ПЦР. Полученную ДНК встраивают в соответствующий экспрессионный вектор и затем его вводят животному, подлежащему иммунизации. Предпочтительно применяемые для этой цели экспрессионные векторы включают, например, поступающие в продажу экспрессионные векторы, такие как pcDNA3.1. Векторы можно вводить в организм с помощью общепринятых методов. Например, ДНК-иммунизацию осуществляют с использованием генной пушки для интродукции золотых частиц, покрытых экспрессионным вектором, в клетки тела животного, подлежащего иммунизации.
После описанной выше иммунизации млекопитающего у него подтверждают в сыворотке повышенный титр антигенсвязывающего антитела. После этого получают из организма млекопитающего иммунные клетки и затем используют их для клеточного слияния. В частности, в качестве иммунных клеток предпочтительно применяют спленоциты.
Клетку миеломы млекопитающих применяют в качестве клетки, подлежащей слиянию с вышеуказанными иммунными клетками. Клетки миеломы предпочтительно содержат приемлемый маркер селекции для скрининга. Маркер селекции придает клеткам характеристики, обеспечивающие их выживание (или гибель) в специфических условиях культивирования. В качестве маркера селекции известны дефицит гипоксантин-гуанин-фосфорибозилтрансферазы (сокращенно обозначенный далее в контексте настоящего описания как дефицит HGPRT) и дефицит тимидинкиназы (сокращенно обозначенный далее в контексте настоящего описания как дефицит TK). Клетки с дефицитом HGPRT или TK обладают чувствительностью к гипоксантин-аминоптерин-тимидину (далее в контексте настоящего описания сокращенно обозначена как ГАТ-чувствительность). Клетки с ГАТ-чувствительностью не могут синтезировать ДНК в селекционной ГАТ-среде и в результате погибают. Однако, когда клетки сливают со здоровыми клетками, они могут продолжать синтез ДНК с использованием «реутилизационного» пути здоровых клеток, и в результате они могут расти даже в селекционной ГАТ-среде.
Клетки с HGPRT-дефицитом и TK-дефицитом можно отбирать в среде, содержащей 6-тиогуанин, 8-азагуанин (далее в контексте настоящего описания сокращенно обозначенный как 8AG) или 5'-бромдезоксиуридин соответственно. Здоровые клетки уничтожаются, поскольку они включают эти пиримидиновые аналоги в их ДНК. При этом клетки с дефицитом этих ферментов могут выживать в селекционной среде, поскольку они не могут включать указанные пиримидиновые аналоги. Кроме того, к маркеру селекции относится устойчивость к G418, которая обеспечивается геном устойчивости к неомицину, придающим устойчивость к 2-дезоксистрептаминовым антибиотикам (аналоги гентамицина). Известны различные типы клеток миелом, которые можно применять для клеточного слияния.
Например, в качестве клеток миеломы предпочтительно можно применять следующие клетки:
P3(P3x63Ag8.653) (J. Immunol. 123 (4), 1979, сс. 1548-1550);
P3x63Ag8U.1 (Current Topics in Microbiology and Immunology 81, 1978, cc. 1-7);
NS-1 (C. Eur. J. Immunol. 6 (7), 1976, cc. 511-519);
MPC-11 (Cell 8 (3), 1976, cc. 405-415);
SP2/0 (Nature 276 (5685), 1978, cc. 269-270);
FO (J. Immunol. Methods 35 (1-2), 1980, cc. 1-21);
S194/5.XX0.BU.1 (J. Exp. Med. 148 (1), 1978, cc. 313-323);
R210 (Nature 277 (5692), 1979, cc. 131-133) и т.д.
Клеточное слияние иммуноцитов и клеток миеломы, как правило, осуществляют с помощью известных методов, например, метода, описанного у Kohler и Milstein и др. (Methods Enzymol. 73, 1981, сс. 3-46).
Более конкретно, клеточное слияние можно осуществлять, например, в общепринятой культуральной среде в присутствии усиливающего клеточное слияние агента. Усиливающие слияние агенты включают, например, полиэтиленгликоль (ПЭГ) и вирус Сендай (гемагглютинирующий японский вирус мышей) (HVJ). При необходимости для повышения эффективности слияния добавляют также вспомогательную субстанцию, такую как диметилсульфоксид.
Соотношение иммуноцитов и клеток миеломы можно определять по усмотрению исследователя, предпочтительно, например, одна клетка миеломы на каждые 1-10 иммуноцитов. Культуральные среды, применяемые для клеточных слияний, включают, например, среды, пригодные для выращивания клеточных линий миелом, такие как среда RPMI1640 и среда MEM, а также другая общепринятая культуральная среда, применяемая для данного типа клеточной культуры. Кроме того, в культуральную среду предпочтительно можно вносить добавки в виде сыворотки, такой как фетальная телячья сыворотка (FCS).
Для клеточного слияния указанные выше иммунных клетки и клетки миеломы, взятые в предварительно определенных количествах, хорошо перемешивают в указанной выше культуральной среде. Затем к ней добавляют предварительно нагретый до температуры примерно 37°С раствор ПЭГ (например, средняя молекулярная масса которого составляет примерно от 1000 до 6000) в концентрации, составляющей, как правило, от 30 до 60% (мас./об.). Смесь осторожно перемешивают до получения требуемых слитых клеток (гибридомы). Затем указанную выше культуральную среду постепенно добавляют к клеткам и повторно центрифугируют для удаления супернатанта. Таким путем можно удалять агенты для клеточного слияния, которые являются нежелательными для роста гибридом.
Полученные таким образом гибридомы можно отбирать путем культивирования, используя общепринятую селективную среду, например, ГАТ-среду (культуральная среда, содержащая гипоксантин, аминоптерин и тимидин). Культивирование в вышеуказанной ГАТ-среде продолжают в течение периода времени, достаточного для уничтожения клеток, отличных от требуемых гибридом (неслитые клетки). Как правило, период составляет от нескольких дней до нескольких недель. Затем осуществляют скрининг гибридом, продуцирующих требуемое антитело, и по отдельности клонируют с помощью общепринятых методов серийных разведений.
Полученные таким образом гибридромы можно отбирать, используя селекционную среду на основе маркера селекции, который несут клетки миеломы, применяемые для клеточного слияния. Например, клетки с HGPRT-или TK-дефицитом можно отбирать путем культивирования с использованием ГАТ-среды (культуральная среда, содержащая гипоксантин, аминоптерин и тимидин). В частности, когда для клеточного слияния используют чувствительные к ГАТ клетки миеломы, то клетки, для которых характерно успешное слияние со здоровыми клетками, могут избирательно размножаться в ГАТ-среде. Культивирование в вышеуказанной ГАТ-среде продолжают в течение периода времени, достаточного для уничтожения клеток, отличных от требуемых гибридом (неслитые клетки). В частности, требуемые гибридомы можно отбирать путем культивирования, как правило, в течение периода времени, составляющего от нескольких дней до нескольких недель. Затем осуществляют скрининг гибридом, продуцирующих требуемое антитело, и по отдельности клонируют с помощью общепринятых методов серийных разведений.
Требуемые антитела предпочтительно можно отбирать и по отдельности клонировать с помощью методов скрининга, основанных на известной реакции антиген/антитело. Например, можно осуществлять скрининг указанных моноклональных антител с помощью метода разделения клеток на основе возбуждения флуоресценции (FACS). FACS представляет собой систему, которая позволяет оценивать связывание антитела с клеточной поверхностью посредством анализа клеток, контактирующих с флуоресцентным антителом, с использованием лазерного пучка, и путем оценки флуоресценции, испускаемой индивидуальными клетками.
Для скрининга с использованием FACS в отношении гибридом, которые продуцируют моноклональное антитело, предлагаемое в настоящем изобретении, прежде всего, получают клетки, которые экспрессируют антиген, связанный с продуцируемым антителом. Клетки, которые предпочтительно используют для скрининга, представляют собой клетки млекопитающих, в которых происходит принудительная экспрессия антигена. В качестве контроля можно избирательно определять с использованием нетрансформированных клеток млекопитающих в качестве клеток-хозяев способность антитела связываться с расположенным на клеточной поверхности антигеном. В частности, гибридомы, продуцирующие моноклональное антитело, можно выделять путем отбора гибридом, которые продуцируют антитело, связывающееся с клетками, принудительно экспрессирующими антиген, но не с клетками-хозяевами.
Альтернативно этому, клетки, экспрессирующие представляющий интерес антиген, иммобилизуют, и способность антитела связываться с экспрессирующими антиген клетками можно оценивать на основе принципа ELISA. Например, экспрессирующие антиген клетки иммобилизуют на лунках планшета для ELISA. Супернатанты культур гибридом приводят в контакт с иммобилизованными клетками в лунках и выявляют антитела, которые связываются с иммобилизованными клетками. Когда моноклональные антитела имеют мышиное происхождение, то антитела, связанные с клетками, можно выявлять с помощью антитела к мышиному иммуноглобулину. Гибридомы, продуцирующие требуемое антитело, которое обладает антигенсвязывающей способностью, отбирают путем описанного выше скрининга, и их можно клонировать методом серийных разведений или сходным методом, или т.п.
Полученные таким путем гибридомы, продуцирующие моноклональные антитела, можно пересевать в общепринятую культуральную среду и хранить в жидком азоте в течение длительного периода времени.
Указанные выше гибридомы культивируют с помощью общепринятого метода и требуемые моноклональные антитела можно получать из супернатантов культур. Альтернативно этому, гибридомы интродуцируют и выращивают в пригодных для этой цели млекопитающих и моноклональные антитела можно получать из асцитов. Первый метод пригоден для получения антител с высокой степенью чистоты.
Предпочтительно можно применять также антитела, кодируемые генами антител, которые клонированы из продуцирующих антитела клеток, таких как указанные выше гибридомы. Клонированный ген антитела встраивают в соответствующий вектор и его интродуцируют в хозяина для экспрессии кодируемого геном антитела. Методы выделения генов антител, встраивания генов в векторы и трансформации клеток-хозяев разработаны ранее (см., например, Vandamme и др., Eur. J. Biochem. 192(3), 1990, сс. 767-775). Методы получения рекомбинантных антител также известны и описаны ниже.
Как правило, для получения кДНК, кодирующей вариабельную область (V-область) антитела, сначала общую РНК экстрагируют из гибридом. Например, следующие методы можно применять для экстракции мРНК из клеток:
- метод ультрацентрифугирования в присутствии гуанидина (Biochemistry 18(24), 1979, сс. 5294-5299) и
- AGPC (метод выделения РНК путем гуанидинтиоцинат-фенол-хлороформной экстракции) (Anal. Biochem. 162(1), 1987, сс. 156-159).
Экстрагированные мРНК можно очищать с помощью набора для очистки мРНК (фирма GE Healthcare Bioscience) или аналогичного набора. В качестве альтернативы можно применять также поступающие в продажу наборы для экстракции мРНК непосредственно из клеток, такие как набор для очистки мРНК QuickPrep (фирма GE Healthcare Bioscience). мРНК можно получать из гибридом с использованием таких наборов. Кодирующие V-область антитела кДНК можно синтезировать из полученных мРНК с помощью обратной транскриптазы. кДНК можно синтезировать с помощью набора для синтеза первой цепи кДНК, содержащего обратную транскриптазу AMV (Reverse Transcriptase First-strand cDNA Synthesis Kit) (фирма Seikagaku Co.) или аналогичного набора. Кроме того, для синтеза и амплификации кДНК можно применять набор для амплификации кДНК SMART RACE (фирма Clontech) и метод, основанный на ПЦР 5'-RACE (быстрая амплификация концов кДНК) (Proc. Natl. Acad. Sci. USA 85(23), 1988, cc. 8998-9002; Nucleic Acids Res. 17(8), 1989, cc. 2919-2932). При таком процессе синтеза кДНК соответствующие описанные ниже сайты, распознаваемые рестриктазами, можно интродуцировать на оба конца кДНК.
Представляющий интерес фрагмент кДНК очищают из полученного ПЦР-продукта и затем его встраивают путем лигирования в ДНК-вектор. Таким путем создают рекомбинантный вектор и интродуцируют в Е.coli или подобного хозяина. После селекции колоний требуемый рекомбинантный вектор можно получать из колониеобразующих Е.coli. Затем с использованием известного метода, такого как метод терминации нуклеотидной цепи (дидезокси-метод), определяют, имеет ли рекомбинантный вектор представляющую интерес нуклеотидную последовательность кДНК.
5'-RACE-метод, в котором используют праймеры для амплификации гена вариабельной области, как правило, применяют для выделения гена, кодирующего вариабельную область. Сначала конструируют библиотеку кДНК, применяемую для 5'-RACE (5'-RACE-библиотека кДНК) с использованием РНК, экстрагированных из клеток гибридомы, в качестве матрицы. Для синтеза 5'-RACE-библиотеки кДНК можно использовать поступающий в продажу набор, такой как набор для амплификации кДНК SMART RACE.
Ген антитела амплифицируют с помощью ПЦР, используя полученную 5'-RACE-библиотеку кДНК в качестве матрицы. Праймеры для амплификации гена мышиного антитела можно создавать на основе известных генных последовательностей антител. Нуклеотидные последовательности праймеров варьируются в зависимости от подкласса иммуноглобулина. Таким образом, предпочтительно предварительно определять подкласс с помощью доступного набора, такого как набор для изотипирования мышиных моноклональных антител Iso Strip (Iso Strip mouse monoclonal antibody isotyping kit) (фирма Roche Diagnostics).
В частности, например, для выделения генов, кодирующих мышиный IgG, применяют праймеры, которые обеспечивают амплификацию генов, кодирующих тяжелые цепи γ1, γ2а, γ2b и γ3 и легкие цепи κ и λ. В целом, праймер, сайт гибридизации («отжига») которого с константной областью расположен вблизи вариабельной области, применяют в качестве 3'-концевого праймера для амплификации гена вариабельной области IgG. При этом праймер, присоединенный к набору конструкций 5' RACE-библиотеки кДНК, применяют в качестве 5'-концевого праймера.
Амплифицированные таким образом ПЦР-продукты применяют для реконструирования иммуноглобулинов, состоящих из комбинации тяжелых и легких цепей. Требуемое антитело можно отбирать, используя в качестве показателя антигенсвязывающую активность реконструированного иммуноглобулина. Можно осуществлять скрининг, например, с использованием следующих стадий, на которых:
(1) приводят в контакт экспрессирующую требуемый антиген клетку с антителом, содержащим V-область, которая кодируется кДНК, выделенной из гибридомы;
(2) определяют связывание антитела с экспрессирующей антиген клеткой; и
(3) отбирают антитело, которое связывается с экспрессирующей антиген клеткой.
Методы определения связывания антитела с экспрессирующими антиген клетками, являются известными. В частности, связывание антитела с экспрессирующими антиген клетками можно определять с помощью описанных выше методик, таких как FACS. Иммобилизованные образцы экспрессирующих антиген клеток можно применять для оценки связывающей активности антитела.
Предпочтительные методы скрининга антител, в которых используют связывающую активность в качестве показателя, включают также методы пэннинга, основанные на использовании фаговых векторов. Методы скрининга с использованием фаговых векторов имеют преимущество, когда гены антитела выделяют из библиотек подкласса тяжелой цепи и легкой цепи из популяции клеток, экспрессирующих поликлональные антитела. Гены, кодирующие вариабельные области тяжелой цепи и легкой цепи, можно сшивать с помощью приемлемой линкерной последовательности с образованием одноцепочечного Fv (scFv). Фаги, экспрессирующие на своей поверхности scFv, можно получать путем встраивания гена, кодирующего scFv, в фаговый вектор. Фаги приводят в контакт с представляющим интерес антигеном. Затем ДНК, кодирующую scFv, который обладает представляющей интерес связывающей активностью, можно выделять путем сбора фагов, связанных с антигеном. Указанный процесс можно повторять при необходимости для обогащения scFv, которые обладают представляющей интерес связывающей активностью.
После выделения кДНК, кодирующей V-область представляющего интерес антитела, кДНК расщепляют рестриктазами, которые распознают сайты рестрикции, интродуцированные в оба конца кДНК. Предпочтительные рестриктазы распознают и расщепляют нуклеотидную последовательность, которая встречается с низкой частотой в нуклеотидной последовательности гена антитела. Кроме того, для встраивания однокопийного расщепленного фрагмента в правильной ориентации предпочтительно интродуцируют в представляющий интерес вектор сайт рестрикции для фермента, который образует «липкий» конец. Кодирующую V-область антитела кДНК расщепляют согласно описанному выше методу и встраивают в приемлемый экспрессионный вектор для создания экспрессионного вектора антитела. В том случае, когда ген, кодирующий константную область (С-область) антитела, и ген, кодирующий указанную выше V-область, сливают в рамке считывания, то получают химерное антитело. В контексте настоящего описания «химерное антитело» означает, что константная область и вариабельная область отличаются по своему происхождению. Так, помимо мышиных/человеческих гетерохимерных антител к химерным антителам, предлагаемым в настоящем изобретении, относятся также человеческие/человеческие аллохимерные антитела. Экспрессионный вектор химерного антитела можно создавать путем встраивания указанного выше гена V-области в экспрессионный вектор, который уже содержит константную область. В частности, например, последовательность, распознаваемую рестриктазой, которая вырезает указанный выше ген V-области, предпочтительно следует помещать в 5'-область экспрессионного вектора, несущего ДНК, которая кодирует константную область (С-область) требуемого антитела. Экспрессионный вектор химерного антитела создают путем слияния в рамке считывания двух генов, расщепленных одной и той же комбинацией рестриктаз.
Для получения моноклонального антитела гены антитела встраивают в экспрессионный вектор таким образом, чтобы экспрессия генов происходила под контролем регулирующей экспрессию области. Регулирующая экспрессию область, предназначенная для экспрессии антител, включают, например, энхансеры и промоторы. Кроме того, соответствующую сигнальную последовательность можно присоединять к аминоконцу таким образом, чтобы экспрессируемое антитело секретировалось из клеток наружу. Сигнальная последовательность отщепляется от карбоксильного конца экспрессируемого полипептида, и образовавшееся антитело может секретироваться из клеток наружу. Затем приемлемые клетки-хозяева трансформируют экспрессионным вектором и таким образом можно получать рекомбинантные клетки, экспрессирующие ДНК, которая кодирует антитело.
ДНК, которые кодируют тяжелую цепь антитела (Н-цепь) и легкую цепь антитела (L-цепь), встраивают по отдельности в различные экспрессионные векторы для экспрессии гена антитела. Молекулу антитела, имеющую Н- и L-цепи, можно экспрессировать, осуществляя для этой цели контрансфекцию одной и той же клетки-хозяина векторами, в которые встроены соответственно гены Н-цепи и L-цепи. В качестве альтернативы, клетки-хозяева можно трансформировать одним экспрессионным вектором, в который встроены ДНК, кодирующие Н- и L-цепи (см. WO 94/11523).
Известны различные комбинации клеток-хозяев/экспрессионных векторов для получения антитела путем интродукции выделенных генов антител в соответствующих хозяев. Все эти системы экспрессии можно применять для выделения доменов, связывающихся со специфическими для рака антигенами, предлагаемых в настоящем изобретении, доменов, связывающихся с представителями суперсемейства рецептора фактора некроза опухоли (TNFRSF) и комплексом Т-клеточного рецептора.
Приемлемыми эукариотическими клетками, которые применяют в качестве клеток-хозяев, являются клетки животных, клетки растений и клетки грибов. В частности, клетки животных представляют собой, например, следующие клетки:
(1) клетки млекопитающих: СНО, COS, миеломы, почки детеныша хомяка (BHK), HeLa, Vero или т.п.;
(2) клетки амфибий: ооциты шпорцевой лягушки (Xenopus) или т.п. и
(3) клетки насекомых: sf9, sf21, Tn5 или т.п.
Кроме того, в качестве растительной клетки известна система экспрессии генов антитела, в которой используют клетки, полученные из представителей рода Nicotiana, например, Nicotiana tabacum. Для трансформации растительных клеток можно применять культивированные клетки каллюса.
Кроме того, следующие клетки можно применять в качестве грибных клеток:
дрожжи: рода Saccharomyces, например, Saccharomyces cerevisiae, и рода Pichia, например, Pichia pastoris, и
нитчатые грибы: рода Aspergillus, например, Aspergillus niger.
Кроме того, известны системы экспрессии генов антитела, в которых используют прокариотические клетки. Например, согласно настоящему изобретению можно применять бактериальные клетки, т.е. клетки Е.coli, клетки Bacillus subtilis и т.п. Экспрессионные векторы, которые несут представляющие интерес гены антитела, интродуцируют в эти клетки путем трансфекции. Трансфектированные клетки культивируют in vitro, и требуемое антитело можно получать из культуры трансформированных клеток.
Помимо указанных выше клеток-хозяев для получения рекомбинантного антитела можно применять также трансгенных животных. Это означает, что антитело можно получать из животного, в организм которого интродуцирован ген, кодирующий представляющее интерес антитело. Например, ген антитела можно создавать в виде слитого гена посредством встраивания в рамке считывания в ген, кодирующий белок, который специфически образуется в молоке. В качестве белка, секретируемого в молоко, можно применять, например, козий β-казеин. ДНК-фрагменты, содержащие слитый ген, в который входит ген антитела, инъецируют в эмбрион козы и затем этот эмбрион интродуцируют в самку козы. Требуемые антитела можно получать в виде белка, слитого с молочным белком из молока трансгенной козы, родившейся от козы, являющейся реципиентом эмбриона (или ее потомства). Кроме того, для увеличения объема молока, содержащего требуемое антитело, которое продуцируется трансгенной козой, трансгенной козе можно при необходимости вводить гормоны (Bio/Technology 12 (7), 1994, сс. 699-702).
Когда антигенсвязывающую молекулу, представленную в настоящем описании, вводят человеку, то в качестве различных связывающих доменов в молекуле можно применять антигенсвязывающий домен, происходящий из антитела, полученного путем генетической рекомбинации, которое искусственно модифицировано для снижения гетерологичной антигенности в отношении человека и других животных, когда применяют домены, содержащие вариабельную область антитела. Указанные антитела, полученные путем генетической рекомбинации, включают, например, гуманизированные антитела. Эти модифицированные антитела можно получать с помощью известных методов.
Вариабельная область антитела, применяемая для получения различных связывающих доменов, представленных в настоящем описании, как правило, состоит из трех гипервариабельных участков (CDR), разделенных четырьмя каркасными участками (FR). CDR представляет собой область, которая в значительной степени определяет специфичность связывания антитела. Для аминокислотных последовательностей CDR характерна высокая степень вариабельности. С другой стороны, образующие FR аминокислотные последовательности часто обладают высокой идентичностью даже среди антител с различными специфичностями связывания. Таким образом, как правило, путем трансплантации CDR специфичность связывания конкретного антитела можно интродуцировать в другое антитело.
Гуманизированное антитело называют также реконструированным человеческим антителом. В частности, известны гуманизированные антитела, полученные путем трансплантации CDR антитела животного кроме человека, такого как мышиное антитело, в человеческое антитело и т.п. Известны также общепринятые методики генной инженерии, предназначенные для получения гуманизированных антител. В частности, например, ПЦР с перекрывающимися праймерами представляет собой известный метод трансплантации CDR мышиного антитела в человеческий FR. При осуществлении ПЦР с перекрывающимися праймерами нуклеотидную последовательность, которая кодирует CDR мышиного антитела, подлежащий трансплантации, добавляют к праймерам, предназначенным для синтеза FR человеческого антитела. Получают праймеры для каждого из четырех FR. Принято считать, что когда осуществляют трансплантацию мышиного CDR в человеческий FR, то для поддержания функции CDR целесообразно выбирать человеческий FR, обладающий высоким уровнем идентичности с мышиным FR. Таким образом, как правило, является предпочтительным применять человеческий FR, который содержит аминокислотную последовательность, обладающую высоким уровнем идентичности с аминокислотной последовательностью FR, который примыкает к подлежащему трансплантации мышиному CDR.
Нуклеотидные последовательности, подлежащие лигированию, создают таким образом, чтобы они были соединены друг с другом в рамке считывания. Человеческие FR синтезируют индивидуально с использованием соответствующих праймеров. В результате получают продукты, в которых ДНК, кодирующая мышиный CDR, присоединена к индивидуальным ДНК, кодирующим FR. Нуклеотидные последовательности, кодирующие мышиный CDR каждого продукта, создают таким образом, чтобы они перекрывались друг с другом. Затем осуществляют реакцию синтеза комплементарной цепи для «отжига» перекрывающихся CDR-участков продуктов, синтезированных с использованием гена человеческого антитела в качестве матрицы. С помощью этой реакции человеческие FR встраивают путем лигирования через мышиные CDR-последовательности.
Полноразмерный ген V-области, в которую, в конце концов, лигированы три CDR и четыре FR, амплифицируют с использованием праймеров, гибридизующихся с 5'- или 3'-концом, которые добавляют с последовательностями, распознаваемыми приемлемыми рестриктазами. Экспрессионный вектор для гуманизированного антитела можно получать путем встраивания полученной согласно описанному выше методу ДНК и ДНК, которая кодирует С-область человеческого антитела, в экспрессионный вектор таким образом, чтобы лигировать их в рамке считывания. После трансфекции хозяина рекомбинантным вектором для создания рекомбинантных клеток рекомбинантные клетки культивируют и ДНК, кодирующую гуманизированное антитело, экспрессируют с получением гуманизированного антитела в культуре клеток (см. публикацию европейского патента ЕР 239400 и публикацию международной заявки на патент WO 1996/002576).
Путем качественной или количественной оценки и измерения антигенсвязывающей активности гуманизированного антитела, полученного согласно описанному выше методу, можно отбирать FR человеческого антитела, которые позволяют CDR образовывать предпочтительный антигенсвязывающий центр при лигировании посредством CDR. Аминокислотные остатки в FR при необходимости можно заменять так, чтобы CDR реконструированного человеческого антитела образовывали приемлемый антигенсвязывающий центр. Например, мутации аминокислотной последовательности можно интродуцировать в FR, используя ПЦР-метод, применяемый для трансплантации мышиного CDR в человеческий FR. Более конкретно, мутации неполной нуклеотидной последовательности можно интродуцировать в праймеры, гибридизующиеся с FR. Мутации нуклеотидной последовательности интродуцируют в FR, синтезированные с использованием указанных праймеров. Мутантные последовательности FR, имеющие требуемые характеристики, можно отбирать путем измерения и оценки активности мутантного антитела с аминокислотной заменой в отношении связывания с антигеном с помощью упомянутого выше метода (Sato K. и др., Cancer Res. 53, 1993, сс. 851-856).
В качестве альтернативы, требуемые человеческие антитела можно получать путем иммунизации трансгенных животных, имеющих полный спектр генов человеческого антитела (см. WO 1993/012227; WO 1992/003918; WO 1994/002602; WO 1994/025585; WO 1996/034096; WO 1996/033735), с использованием ДНК-иммунизации.
Кроме того, известны методики получения человеческих антител путем пэннинга с использованием библиотек человеческих антител. Например, V-область человеческого антитела экспрессируют в виде одноцепочечного антитела (scFv) на поверхности фага с использованием метода фагового дисплея. Можно отбирать фаги, экспрессирующие scFv, который связывается с антигеном. Последовательность ДНК, кодирующую V-область человеческого антитела, которая связывается с антигеном, можно определять путем анализа генов отобранных фагов. Определяют последовательность ДНК scFv, который связывается с антигеном. Экспрессионный вектор получают путем слияния последовательности V-области в рамке считывания с последовательностью С-области требуемого человеческого антитела и последующего встраивания в приемлемый экспрессионный вектор. Экспрессионный вектор интродуцируют в клетку, пригодную для указанной выше экспрессии. Человеческое антитело можно получать путем экспрессии гена, кодирующего человеческое антитело, в клетках. Такие методы уже описаны (см. WO 1992/001047; WO 1992/020791; WO 1993/006213; WO 1993/011236; WO 1993/019172; WO 1995/001438; WO 1995/015388).
Помимо метода фагового дисплея методики, в которых используют бесклеточную систему трансляции, методики дисплея антигенсвязывающих молекул на поверхности вирусов или клеток и методики, в которых используют эмульсии, также известны в качестве методик, предназначенных для получения человеческих антител путем пэннинга с использованием библиотек человеческих антител. Например, метод рибосомного дисплея, при осуществлении которого формируют комплекс между транслируемым белком и мРНК посредством рибосомы путем удаления стоп-кодона и т.п. метод кДНК-дисплея или метод мРНК-дисплея, при осуществлении которых генную последовательность и транслируемый белок ковалентно связывают с использованием такого соединения, как пуромиин, метод CIS-дисплея, при осуществлении которого формируют комплекс между геном и транслируемым белком с использованием связывающего нуклеиновую кислоту белка, или т.п., можно применять в качестве методик для бесклеточной системы трансляции. В методике презентации антигенсвязывающих молекул на поверхности клеток или вирусов помимо метода фагового дисплея можно применять метод дисплея на Е.coli, метод дисплея на грамотрицательных бактериях, метод дрожжевого дисплея, метод дисплея на клетках млекопитающих, метод вирусного дисплея и т.п. В качестве методики, в которой применяют эмульсии, можно использовать метод вирусного дисплея in vitro, который заключается в том, что вносят гены и связанные с трансляцией молекулы в эмульсию и т.п. Эти методы уже известны научной общественности (Nat Biotechnol. 18(12), декабрь 2000 г., сс. 1287-1292; Nucleic Acids Res. 34(19), 2006, e127; Proc Natl Acad Sci USA. 101(9), 2 марта 2004 г., cc. 2806-2810; Proc Natl Acad Sci USA. 101(25), 22 июня 2004 г., cc. 9193-9198; Protein Eng Des Sel. 21(4), апреля 2008 г., cc. 247-255; Proc Natl Acad Sci USA. 97(20), 26 сентября 2000 г., cc. 10701-10705; MAbs. 2(5), сентябрь-октябрь 2010 г., cc. 508-518 и Methods Mol Biol. 911, 2012, cc. 183-198).
В настоящем изобретении «специфическое» означает состояние, при котором одна из молекул, участвующих в специфическом связывании, не обладает никаким специфическим связыванием с молекулами, отличными от одной или нескольких молекул, являющихся партнерами по связыванию Кроме того, понятие «специфический» используют также тогда, когда антигенсвязывающий домен является специфическим в отношении конкретного эпитопа из нескольких эпитопов, входящих в антиген. Когда эпитоп, с которым связывается антигенсвязывающий домен, входит в несколько различных антигенов, то антигенсвязывающая молекула, содержащая антигенсвязывающий домен, может связываться с различными антигенами, которые содержат эпитоп.
«Эпитоп» означает антигенную детерминанту в антигене и относится к антигенному сайту, с которым связывается антигенсвязывающая молекула, представленная в настоящем описании. Так, например, эпитоп можно характеризовать на основе его структуры. Альтернативному этому, эпитоп можно характеризовать на основе антигенсвязывающей активности антигенсвязывающей молекулы, которая распознает эпитоп. Когда антиген представляет собой пептид или полипептид, то эпитоп можно определять по аминокислотным остаткам, образующим эпитоп. Альтернативно этому, когда эпитоп представляет собой сахарную цепь, то эпитоп можно определять по специфической для него структуре сахарной цепи.
Линейный эпитоп представляет собой эпитоп, который содержит эпитоп, у которого распознается первичная аминокислотная последовательность. Указанный линейный эпитоп, как правило, содержит по меньшей мере три и наиболее часто по меньшей мере пять, например, примерно от 8 до 10 или от 6 до 20 аминокислот в определенной последовательности.
В отличие от линейного эпитопа «конформационный эпитоп» представляет собой эпитоп, в котором первичная аминокислотная последовательность, составляющая эпитоп, не является единственной детерминантой распознаваемого эпитопа (например, не является обязательным, чтобы первичная аминокислотная последовательность конформационного эпитопа распознавалась специфическим в отношении эпитопа антителом). Конформационные эпитопы могут содержать большее количество аминокислотных остатков по сравнению с линейными эпитопами. Распознающее конформационный эпитоп антитело распознает трехмерную структуру пептида или белка. Например, когда белковая молекула уложена и образует трехмерную структуру, аминокислоты и/или полипептидные основные цепи, которые образуют конформационный эпитоп, выравниваются, и эпитоп становится распознаваемым для антитела. Методы определения конформаций эпитопов включают (но не ограничиваясь только ими), например, рентгеновскую кристаллографию, двухмерную ядерную магнитно-резонансную спектроскопию, сайтспецифическое спиновое мечение и электронную парамагнитно-резонансную спектроскопию (см., например, Epitope Mapping Protocols in Methods in Molecular Biology, под ред. Morris, т. 66, 1996).
Примеры методов оценки связывания эпитопа в специфическом для рака антигене тестируемой антигенсвязывающей молекулой, описаны ниже. Можно применять также описанные ниже в качестве примеров методы оценки связывания эпитопа в антигене-мишени другим связывающим доменом.
Например, для подтверждения того, что тестируемая антигенсвязывающая молекула, которая содержит антигенсвязывающий домен для специфического для рака антигена, распознает линейный эпитоп в молекуле антигена, можно применять описанный ниже метод. Например, для указанной выше цели синтезируют линейный пептид, содержащий аминокислотную последовательность, образующую внеклеточный домен специфического для рака антигена. Пептид можно синтезировать химически или получать с помощью методов генной инженерии, используя область в кДНК специфического для рака антигена, которая кодирует аминокислотную последовательность, соответствующую внеклеточному домену. Затем тестируемую антигенсвязывающую молекулу, содержащую антигенсвязывающий домен для специфического для рака антигена, оценивают в отношении ее активности связывания с линейным пептидом, который содержит аминокислотную последовательность, образующую внеклеточный домен. Например, иммобилизованный линейный пептид можно применять в качестве антигена при осуществлении ELISA для оценки активности в отношении связывания антигенсвязывающей молекулы с пептидом. Альтернативно этому, активность связывания с линейным пептидом можно оценивать по уровню ингибирования линейным пептидом связывания антигенсвязывающей молекулы с клетками, экспрессирующими специфический для рака антиген. Эти анализы могут демонстрировать активность связывания антигенсвязывающей молекулы с линейным пептидом.
Для подтверждения того, что указанная выше антигенсвязывающая молекула, которая содержит антигенсвязывающий домен для антигена, распознает конформационный эпитоп, можно применять описанный ниже метод. Например, считается, что антигенсвязывающая молекула, которая содержит антигенсвязывающий домен для специфического для рака антигена, распознает конформационный эпитоп, если она при контакте отличается сильным связыванием с клетками, экспрессирующими специфический для рака антиген, но незначительно связывается с иммобилизованным линейным пептидом, который содержит аминокислотную последовательность, образующую внеклеточный домен специфического для рака антигена. В контексте настоящего описания «незначительно связывается» означает, что активность связывания составляет 80% или менее, как правило, 50% или менее, предпочтительно 30% или менее и наиболее предпочтительно 15% или менее по сравнению с активностью связывания с клетками, экспрессирующими антиген.
Методы оценки активности связывания тестируемой антигенсвязывающей молекулы, которая содержит антигенсвязывающий домен, с экспрессирующими антиген клетками включают, например, методы, описанные в: Antibodies: А Laboratory Manual (под ред. Harlow, David Lane, изд-во Cold Spring Harbor Laboratory, 1988, cc. 359-420). В частности, оценку можно осуществлять на основе принципа ELISA или метода разделения клеток на основе возбуждения флуоресценции (FACS) с использованием в качестве антигена экспрессирующие антиген клетки.
В формате ELISA активность связывания тестируемой антигенсвязывающей молекулы, которая содержит антигенсвязывающий домен, с экспрессирующими антиген клетками можно оценивать количественно путем сравнения уровней сигналов, возникающих в процессе ферментативной реакции. В частности, тестируемую антигенсвязывающую молекулу добавляют в планшет для ELISA, на котором иммобилизованы экспрессирующие антиген клетки. Затем тестируемую антигенсвязывающую молекулу, связанную с клетками, выявляют с помощью меченного ферментом антитела, которое распознает тестируемую антигенсвязывающую молекулу. Альтернативно этому, когда применяют FACS, приготавливают серийные разведения тестируемой антигенсвязывающей молекулы и титр антитела, связывающегося с экспрессирующими антиген клетками, можно определять путем сравнения активности связывания тестируемой антигенсвязывающей молекулы с экспрессирующими антиген клетками.
Связывание тестируемой антигенсвязывающей молекулы с антигеном, который экспрессируется на поверхности клеток, суспендированных в буфере или в сходной среде, можно выявлять с помощью проточного цитометра. Известными проточными цитометрами являются, например, следующие устройства:
FACSCanto™ II,
FACSAria™,
FACSArray™,
FACSVantage™ SE
FACSCalibur™ (все товарные знаки принадлежат фирме BD Biosciences),
EPICS ALTRA HyPerSort,
Cytomics FC 500,
EPICS XL-MCL ADC EPICS XL ADC,
Cell Lab Quanta/Cell Lab Quanta SC (все товарные знаки принадлежат фирме Beckman Coulter).
Приемлемыми методами оценки активности связывания указанной выше тестируемой антигенсвязывающей молекулы, которая содержит антигенсвязывающий домен, с антигеном является, например, следующий метод. Сначала экспрессирующие антиген клетки подвергают взаимодействию с тестируемой антигенсвязывающей молекулой, а затем осуществляют окрашивание меченным с помощью ФИТЦ вторичным антителом, которое распознает антигенсвязывающую молекулу. Тестируемую антигенсвязывающую молекулу соответствующим образом разводят приемлемым буфером с получением антигенсвязывающей молекулы в требуемой концентрации. Например, молекулу можно применять в концентрации, составляющей от 10 мкг/мл до 10 нг/мл. Затем определяют интенсивность флуоресценции и количество клеток с помощью FACSCalibur (фирма BD). Интенсивность флуоресценции, определенная с помощью анализов на основе программы CELL QUEST (фирма BD), а именно, выраженная в виде средних геометрических значений, отражает уровень связывания антитела с клетками. Это означает, что активность связывания тестируемой антигенсвязывающей молекулы, которая характеризуется количеством связанной тестируемой антигенсвязывающей молекулы, можно оценивать, определяя среднее геометрическое значение.
Имеет ли тестируемая антигенсвязывающая молекула, которая содержит антигенсвязывающий домен, предлагаемый в настоящем изобретении, общий эпитоп с другой антигенсвязывающей молекулой, можно оценивать по конкуренции между двумя молекулами за один и тот же эпитоп. Конкуренцию между антигенсвязывающими молекулами можно определять путем анализа перекрестной блокады или с помощью сходного анализа. Например, предпочтительным анализом перекрестной блокады является конкурентный ELISA-анализ.
В частности, при осуществлении анализа перекрестной блокады антиген, иммобилизованный в лунках титрационного микропланшета, предварительно инкубируют в присутствии антигенсвязывающей молекулы, рассматриваемой в качестве конкурента-кандидата, или без нее, а затем вносят тестируемую антигенсвязывающую молекулу. Количество связанной с антигеном тестируемой антигенсвязывающей молекулы в лунках косвенно коррелирует с активностью связывания антигенсвязывающей молекулы, рассматриваемой в качестве конкурента-кандидата, которая конкурирует за связывание с тем же эпитопом. Это означает, что чем выше аффинность антигенсвязывающей молекулы, рассматриваемой в качестве конкурента-кандидата, к тому же эпитопу, тем ниже активность связывания тестируемой антигенсвязывающей молекулы с сенсибилизированными антигеном лунками.
Количество тестируемой антигенсвязывающей молекулы, связанной через антиген с лунками, легко определять путем предварительного мечения антигенсвязывающей молекулы. Например, меченную биотином антигенсвязывающую молекулу оценивают с использованием конъюгата авидин/пероксидаза и соответствующего субстрата. В частности, анализ перекрестной блокады, в котором применяют в качестве меток ферменты, такие как пероксидаза, называют «ELISA-анализом в конкурентных условиях (конкурентный анализ)». Антигенсвязывающую молекулу можно метить также с помощью других предназначенных для мечения субстанций, которые можно выявлять или оценивать. В частности, известны радиоактивные метки, флуоресцентные метки и т.п.
Когда антигенсвязывающая молекула, рассматриваемая в качестве конкурента-кандидата, может блокировать связывание тестируемой антигенсвязывающей молекулы, которая содержит антигенсвязывающий домен, по меньшей мере на 20%, предпочтительно по меньшей мере на 20-50% и предпочтительно по меньшей мере на 50% по сравнению с активностью связывания, установленной в контрольном эксперименте, который проводят в отсутствии антигенсвязывающей молекулы, рассматриваемой в качестве конкурента-кандидата, то считается, что для тестируемой антигенсвязывающей молекулы характерна выраженная способность к связыванию с тем же эпитопом, с которым связывается антигенсвязывающая молекула, рассматриваемая в качестве конкурента-кандидата, или она конкурирует за связывание с тем же самым эпитопом.
Если структура эпитопа, связанного с тестируемой антигенсвязывающей молекулой, которая содержит антигенсвязывающий домен, предлагаемый в настоящем изобретении, уже идентифицирована, то для решения вопроса о том, имеют ли тестируемая и контрольная антигенсвязывающие молекулы общий эпитоп, можно сравнивать активности связывания двух антигенсвязывающих молекул с пептидом, полученным путем интродукции аминокислотных мутаций в пептид, образующий эпитоп.
Для оценки указанных выше активностей связывания, например, сравнивают активности связывания тестируемой и контрольной антигенсвязывающих молекул с линейным пептидом, в который интродуцирована мутация, с помощью указанного выше формата ELISA. Помимо метода ELISA активность связывания в отношении мутантного пептида, связанного с колонкой, можно определять, пропуская через колонку тестируемую и контрольную антигенсвязывающие молекулы и затем оценивая количество антигенсвязывающей молекулы, элюированной в раствор для элюции. Известны методы адсорбции мутантного пептида на колонке, например, в форме слитого с GST пептида.
Альтернативно этому, когда идентифицированный эпитоп представляет собой конформационный эпитоп, то для решения вопроса о том, имеют ли тестируемая и контрольная антигенсвязывающие молекулы общий эпитоп, можно применять следующий метод. Сначала получают клетки, экспрессирующие антиген, являющийся мишенью для антигенсвязывающей молекулы, и клетки, экспрессирующие антиген с мутацией, интродуцированной в эпитоп. Тестируемую и контрольную антигенсвязывающие молекулы добавляют в клеточную суспензию, полученную путем суспендирования этих клеток в приемлемом буфере, таком как ЗФР. Затем клеточные суспензии соответствующим образом промывают буфером и добавляют меченное с помощью ФИТЦ антитело, которое распознает тестируемую и контрольную антигенсвязывающие молекулы. Определяют интенсивность флуоресценции и количество клеток, окрашенных меченым антителом, используя FACSCalibur (фирма BD). Тестируемую и контрольную антигенсвязывающие молекулы соответствующим образом разводят приемлемым буфером и применяют в требуемых концентрациях. Например, их можно применять в концентрации от 10 мкг/мл до 10 нг/мл. Интенсивность флуоресценции определяют с помощью анализа, для оценки результатов которого применяют программу CELL QUEST (фирма BD), а именно, определяют среднее геометрическое значение, которое отражает количество меченого антитела, связанного с клетками. Это означает, что активности связывания тестируемой и контрольной антигенсвязывающих молекул, которые характеризуются количеством связанного меченого антитела, можно определять, оценивая среднее геометрическое значение.
«Антигенсвязывающая молекула», предлагаемая в настоящем изобретении, содержит как тяжелые, так и легкие цепи, которые образуют «вариабельную область антитела», предлагаемого в изобретении, в одной полипептидной цепи; однако она может представлять собой фрагмент антитела, лишенный константной области. Примеры указанных фрагментов антител включают димерное антитело (диабоди, Db), scFv, одноцепочечное антитело, sc(Fv)2 и sc(Fab')2.
Db представляет собой димер, состоящий из двух полипептидных цепей (Holliger Р. и др., Proc. Natl. Acad. Sci. USA 90, 1993, cc. 6444-6448; ЕР 404097; WO 93/11161). В каждой полипептидной цепи вариабельная область L-цепи (VL) и вариабельная область Н-цепи (VH) сцеплены с помощью линкера, который является достаточно коротким, чтобы не давать вступать в ассоциацию друг с другом указанным двум областям на одной и той же цепи, например, линкера, который состоит примерно из 5 остатков.
Поскольку линкер между VL и VH является слишком коротким, для того, чтобы происходило образование одноцепочечного фрагмента вариабельных областей, то VL и VH, кодируемые на одной и той же полипептидной цепи, димеризуются с образованием двух антигенсвязывающих центров.
Кроме того, в контексте настоящего описания понятия «scFv», «одноцепочечное антитело» и «sc(Fv)2» все относятся к фрагменту антитела в виде одной полипептидной цепи, которая содержит вариабельные области, происходящие из тяжелой и легкой цепей, но не содержит константную область. Как правило, одноцепочечное антитело содержит также полипептидный линкер между VH- и VL-доменами, позволяющий образовывать требуемую структуру, которая, вероятно, обеспечивает связывание антигена. Одноцепочечное антитело подробно описано у Pluckthun в: «The Pharmacology of Monoclonal Antibodies», т. 113, под ред. Rosenburg и Moore, изд-во Springer-Verlag, New York, 1994, cc. 269-315» (см. также публикацию международной заявки на патент WO 1988/001649; US №№4946778 и 5260203). В конкретном варианте осуществления изобретения одноцепочечное антитело может быть биспецфическим и/или гуманизированным.
scFv представляет собой антигенсвязывающий домен, в котором VH и VL, образующие Fv, сцеплены друг с другом с помощью пептидного линкера (Proc. Natl. Acad. Sci. U.S.A. 85(16), 1988, cc. 5879-5883). VH и VL могут удерживаться в непосредственной близости с помощью пептидного линкера.
sc(Fv)2 представляет собой одноцепочечное антитело, в котором четыре вариабельные области двух VL и двух VH сцеплены линкерами, такими как пептидные линкеры, с образованием одной цепи (J Immunol. Methods 231(1-2), 1999, сс. 177-189). Две VH и две VL можно получать из различных моноклональных антител. Указанные sc(Fv)2 предпочтительно включают, например, биспецифический sc(Fv)2, который распознает два типа эпитопов, присутствующих в одном антигене, что описано в Journal of Immunology 152(11), 1994, сс. 5368-5374. sc(Fv)2 можно получать методами, известными специалистам в данной области. Например, sc(Fv)2 можно получать путем связывания scFv с помощью линкера, такого как пептидный линкер.
Согласно настоящему описанию к форме антигенсвязывающего домена, образующего sc(Fv)2, относится антитело, в котором две единицы VH и две единицы VL организованы в следующем порядке: VH, VL, VH и VL ([VH]-линкер-[VL]-линкер-[VH]-линкер-[VL]), начиная с N-конца одноцепочечного полипептида. Порядок расположения двух единиц VH и двух единиц VL не ограничен указанной выше формой, и их можно организовывать в любом порядке. Пример порядка расположения в различных формах приведен ниже.
[VL]-линкер-[VH]-линкер-[VH]-линкер-[VL]
[VH]-линкер-[VL]-линкер-[VL]-линкер-[VH]
[VH]-линкер-[VH]-линкер-[VL]-линкер-[VL]
[VL]-линкер-[VL]-линкер-[VH]-линкер-[VH]
[VL]-линкер-[VH]-линкер-[VL]-линкер-[VH].
Молекулярная форма sc(Fv)2 подробно описана также в WO 2006/132352. Таким образом, согласно указанным описаниям специалисты в данной области могут получать требуемый sc(Fv)2, предназначенный для создания антигенсвязывающих молекул, представленных в настоящем описании.
В контексте настоящего описания понятие «вариабельный фрагмент (Fv)» относится к минимальной единице происходящего из антитела антигенсвязывающего домена, который состоит из пары, включающей вариабельную область легкой цепи антитела (VL) и вариабельную область тяжелой цепи антитела (VH). В 1988 г. Skerra и Pluckthun обнаружили, что гомогенные и активные антитела можно получать из периплазматической фракции Е.coli путем встраивания гена антитела по ходу транскрипции относительно бактериальной сигнальной последовательности и индукции экспрессии гена в Е.coli (Science 240(4855), 1988, сс. 1038-1041). В Fv, полученном из периплазматической фракции, VH ассоциируется с VL таким образом, чтобы связываться с антигеном.
Кроме того, антигенсвязывающую молекулу, предлагаемую в настоящем изобретении, можно конъюгировать с полимером-носителем, таким как ПЭГ, или органическим соединением, таким как противораковое средство. Альтернативно этому, можно встраивать последовательность (сайт) гликозилирования для соответствующего добавления последовательности сахарной цепи для того, чтобы сахарная цепь обеспечивала требуемое действие.
Линкеры, предназначенные для связывания вариабельных областей антитела, представляют собой произвольные пептидные линкеры, которые можно интродуцировать с помощью генной инженерии, синтетические линкеры и линкеры, описанные, например, в Protein Engineering, 9(3), 1996, сс. 299-305. Однако для целей настоящего изобретения наиболее предпочтительными являются пептидные линкеры. Длина пептидных линкеров специально не ограничена, и специалисты в данной области могут выбирать ее в зависимости от поставленной задачи. Предпочтительная длина составляет пять или большее количество аминокислот (при этом, верхний предел составляет (но не ограничиваясь только указанным), как правило, вплоть до 30 аминокислот или менее, предпочтительно 20 аминокислот или менее), и наиболее предпочтительно 15 аминокислот. Когда sc(Fv)2 содержит три пептидных линкера, то их длина может быть одинаковой или различной.
Например, указанные пептидные линкеры включают:
Ser,
Gly-Ser,
Gly-Gly-Ser,
Ser-Gly-Gly,
Gly-Gly-Gly-Ser (SEQ ID NO: 20),
Ser-Gly-Gly-Gly (SEQ ID NO: 21),
Gly-Gly-Gly-Gly-Ser (SEQ ID NO: 22),
Ser-Gly-Gly-Gly-Gly (SEQ ID NO: 23),
Gly-Gly-Gly-Gly-Gly-Ser (SEQ ID NO: 24),
Ser-Gly-Gly-Gly-Gly-Gly (SEQ ID NO: 25),
Gly-Gly-Gly-Gly-Gly-Gly-Ser (SEQ ID NO: 26),
Ser-Gly-Gly-Gly-Gly-Gly-Gly (SEQ ID NO: 27),
(Gly-Gly-Gly-Gly-Ser (SEQ ID NO: 22))n,
(Ser-Gly-Gly-Gly-Gly (SEQ ID NO: 23))n,
где n обозначает целое число, равное 1 или более высокому значению. Специалисты в данной области могут выбирать длину или последовательности пептидных линкеров в зависимости от поставленной задачи.
Как правило, для перекрестного сшивания используют синтетические линкеры (химические перекрестносшивающие агенты), они представляют собой, например:
N-гидроксисукцинимид (NHS),
дисукцинимидилсуберат (DSS),
бис(сульфосукцинимидил)суберат (BS3),
дитиобис(сукцинимидилпропионат) (DSP),
дитиобис(сульфосукцинимидилпропионат) (DTSSP),
этиленгликольбис(сукцинимидилсукцинат) (EGS),
этиленгликольбис(сульфосукцинимидилсукцинат) (сульфо-EGS),
дисукцинимидилтартрат (DST),
дисульфосукцинимидилтартрат (сульфо-DST),
бис[2-(сукцинимидоксикарбонилокси)этил]сульфон (BSOCOES) и
и бис[2-(сульфосукцинимидоксикарбонилокси)этил]сульфон (сульфо-BSOCOES). Эти перекрестносшивающие агенты поступают в продажу.
Как правило, требуется три линкера для соединения вместе четырех вариабельных областей антитела. Применяемые линкеры могут быть одного типа или различных типов.
«Fab» состоит из одной легкой цепи и СН1-домена и вариабельной области из одной тяжелой цепи. Тяжелая цепь молекулы Fab не может образовывать дисульфидные мостки с тяжелой цепью другой молекулы.
«F(ab')2» или «Fab» получают обработкой иммуноглобулина (моноклональное антитело) протеазой, такой как пепсин и папаин, и понятия относятся к фрагменту антитела, получаемого расщеплением иммуноглобулина (моноклональное антитело) вблизи дисульфидных мостиков, присутствующих между шарнирными областями в каждой из двух Н-цепей. Например, папаин расщепляет IgG перед дисульфидными мостиками, присутствующими между шарнирными областями в каждой из двух Н-цепей, с образованием двух гомологичных фрагментов антитела, в которых L-цепь, содержащая VL (вариабельная область L-цепи), и CL (константная область L-цепи), сцеплены с фрагментом Н-цепи, содержащим VH (вариабельный домен Н-цепи) и CHγ1 (γ1-область в константной области Н-цепи), через дисульфидный мостик в их С-концевых областях. Каждый из указанных двух гомологичных фрагментов антител обозначают как Fab'.
«F(ab')2» состоит из двух легких цепей и двух тяжелых цепей, содержащих в константной области СН1-домен и часть СН2-доменов, в результате между двумя тяжелыми цепями образуются дисульфидные мостики. F(ab')2, образующий антигенсвязывающую молекулу, представленную в настоящем описании, предпочтительно можно получать следующим образом. Полное моноклональное антитело или аналогичное антитело, содержащее требуемый антигенсвязывающий домен, частично расщепляют протеазой, такой как пепсин; и Fc-фрагменты удаляют путем адсорбции на колонке с белком А. Не существует ограничения касательно конкретной протеазы, если она обладает способностью избирательно расщеплять полное антитело с образованием F(ab')2 в соответствующих для данного фермента реакционных условиях, таких как значение рН. Указанные протеазы представляют собой, например, пепсин и фицин.
В предпочтительном варианте осуществления изобретения «антигенсвязывающая молекула», предлагаемая в настоящем изобретении, включает мультиспецифическое антитело. При применении Fc-области с пониженной активностью связывания с Fcγ-рецептором в качестве Fc-области мультиспецифического антитела, можно применять соответственно Fc-область, происходящую из мультиспецифического антитела. Из мультиспецифических антител, предлагаемых в настоящем изобретении, предпочтительными являются, в частности, биспецифические антитела.
Для ассоциации мультиспецифических антител можно применять интродукцию отталкивающего заряда на поверхность раздела второй константной области Н-цепи антитела (СН2) или третьей константной области Н-цепи (СН3) для подавления нежелательных ассоциаций между Н-цепями (WO 2006/106905).
При осуществлении методики подавления нежелательной ассоциации между Н-цепями путем интродукции отталкивающего заряда на поверхность раздела СН2 или СН3 примеры аминокислотных остатков, которые контактируют на поверхности раздела других константных областей Н-цепи, включают остатки из области, содержащей остаток в положении 356 (EU-нумерация), остаток в положении 439 (EU-нумерация), остаток в положении 357 (EU-нумерация), остаток в положении 370 (EU-нумерация), остаток в положении 399 (EU-нумерация) и остаток в положении 409 (EU-нумерация) в СН3-области.
Более конкретно, например, в случае антитела, которое содержит два типа СН3-областей Н-цепей, антитело можно создавать так, чтобы 1-3 пары аминокислотных остатков, выбранных из пар аминокислотных остатков, указанных ниже в подпунктах (1)-(3), в СН3-области первой Н-цепи имели одинаковый заряд: (1) аминокислотные остатки в положениях 356 и 439 (EU-нумерация), которые представляют собой аминокислотные остатки, входящие в СН3-область Н-цепи; (2) аминокислотные остатки в положениях 357 и 370 (EU-нумерация), которые представляют собой аминокислотные остатки, входящие в СН3-область Н-цепи; и (3) аминокислотные остатки в положениях 399 и 409 (EU-нумерация), которые представляют собой аминокислотные остатки, входящие в СН3-область Н-цепи.
Кроме того, антитело можно создавать так, чтобы 1-3 пары аминокислотных остатков, соответствующих парам аминокислотных остатков, которые указаны выше в подпунктах (1)-(3), имеющих одинаковый тип заряда, в СН3-области первой Н-цепи, которые представляют собой пары аминокислотных остатков, выбранные из пар аминокислотных остатков, которые указаны выше в подпунктах (1)-(3), в СН3-области второй Н-цепи, которая отличается от СН3-области первой Н-цепи, имели заряд, противоположный заряду соответствующих аминокислотных остатков в вышеуказанной СН3-области первой Н-цепи.
Соответствующие аминокислотные остатки, указанные выше в подпунктах (1)-(3), при ассоциации располагаются близко друг к другу. Специалисты в данной области могут установить для требуемой СН3-области Н-цепи или константной области Н-цепи сайты, которые соответствуют вышеуказанным остаткам, указанным в подпунктах (1)-(3), посредством моделирования гомологии и т.п., используя поступающую в продажу программу, и аминокислотные остатки этих сайтов можно подвергать при необходимости модификациям.
В указанных выше антителах «имеющие заряд аминокислотные остатки» предпочтительно выбирают, например, из аминокислотных остатков, которые входят в любую одну из указанных ниже групп (а) и (б):
(а) глутаминовая кислота (Е) и аспарагиновая кислота (D) и
(б) лизин (K), аргинин (R) и гистидин (Н).
Касательно указанных выше антител то, что они «имеют одинаковый тип заряда» означает, например, что два или большее количество аминокислотных остатков все представляют собой аминокислотные остатки, включенные в любую одну из вышеуказанных групп (а) и (б). Понятие «имеют противоположный заряд» означает, например, что, когда по меньшей мере один из двух или большего количества аминокислотных остатков представляет собой аминокислотный остаток, включенный в любую одну из вышеуказанных групп (а) и (б), оставшийся(иеся) остаток(ки) должен(ны) представлять собой аминокислотный остаток, включенный в другую группу.
В предпочтительном варианте вышеуказанного антитела СН3-область первой Н-цепи и СН3-область второй Н-цепи могут быть перекрестно сшиты дисульфидным мостиком.
Согласно настоящему изобретению аминокислотный остаток, подлежащий изменению, не ограничен аминокислотным остатком константной области или вариабельной области описанного выше антитела. Касательно полипептидных мутантов или гетеромультимеров специалисты в данной области могут установить аминокислотные остатки, которые образуют поверхность раздела, посредством моделирования гомологии и т.п., используя поступающую в продажу программу, и аминокислотные остатки этих сайтов можно подвергать при необходимости изменениям для того, чтобы регулировать ассоциацию.
Другие известные методики можно применять также для ассоциации мультиспецифических антител, предлагаемых в настоящем изобретении. Полипептиды с различными аминокислотами в Fc-области можно эффективно ассоциировать друг с другом путем замены боковой цепи аминокислоты, присутствующей в одной из вариабельных областей Н-цепи антитела, на более крупную боковую цепь («выступ») и замены боковой цепи аминокислоты в соответствующей вариабельной области другой Н-цепи на меньшую боковую цепь («впадина») так, что «выступ» помещается во «впадину» (WO 1996/027011; Ridgway J.В. и др., Protein Engineering 9, 1996, сс. 617-621; Merchant A.M. и др., Nature Biotechnology 16, 1998, сс. 677-681 и US 2013/0336973).
Кроме того, можно применять также другие известные методики для ассоциации мультиспецифических антител, предлагаемых в настоящем изобретении. Ассоциацию полипептидов, имеющих различные последовательности, можно эффективно индуцировать путем комплементарной ассоциации СН3-областей, используя сконструированный СН3-домен с обменом цепей, полученный путем изменения части СН3 в одной из Н-цепей антитела на соответствующую полученную из IgA последовательность и интродукции в комплементарную часть СН3 в другой Н-цепи соответствующей полученной из IgA последовательности (Protein Engineering Design & Selection, 23, 2010, cc. 195-202). Указанную известную методику можно применять также для эффективного получения представляющих интерес мультиспецифических антител.
Кроме того, для получения мультиспецифических антител можно применять следующие методики: методики получения антител на основе ассоциации СН1 и CL антитела и ассоциации VH и VL, которые описаны в WO 2011/028952, WO 2014/018572 и в Nat Biotechnol. 32(2), февраль 2014 г., сс. 191-198; методики получения биспецифических антител с использованием в комбинации полученных по отдельности моноклональных антител (обмен плечей Fab, (Fab arm exchange, FAE), описанные в WO 2008/119353 и WO 2011/131746; методики регуляции ассоциации между СН3-доменами тяжелых цепей антител, описанные в WO 2012/058768 и WO 2013/063702; методики получения биспецифических антител, состоящих из двух типов легких цепей и одного типа тяжелой цепи, описанные в WO 2012/023053; методики получения биспецифических антител с использованием двух штаммов бактериальных клеток, которые индивидуально экспрессируют одну из цепей антитела, содержащего одну Н-цепь и одну L-цепь, описанные Christoph и др., Nature Biotechnology т. 31, 2013, сс. 753-758.
Одним из вариантов путей получения мультиспецифического антитела являются методы получения биспецифических антител путем смешения двух типов моноклональных антител в присутствии восстановителя для расщепления дисульфидных связей в коровой шарнирной области с последующей повторной ассоциацией для гетеродимеризации (FAE), описанной выше. При этом интродукция электростатических взаимодействий в поверхность взаимодействия СН3-области (WO 2006/106905) может индуцировать еще более эффективную гетеродимеризация в процессе повторной ассоциации (WO 2015/046467). При получении FAE с использованием встречающегося в естественных условиях IgG повторная ассоциация может происходить случайно; и поэтому теоретически биспецифические антитела можно получать лишь с 50%-ной эффективностью; однако при применении этого метода биспецифические антитела можно получать с высоким выходом.
Альтернативно этому, даже, если представляющее интерес мультиспецифическое антитело нельзя получать эффективно, мультиспецифическое антитело, предлагаемое в настоящем изобретении, можно получать путем отделения представляющего интерес мультиспецифического антитела от полученных антител и его очистки. Например, описан метод, позволяющий осуществлять очистку двух типов гомологичных форм и представляющего интерес гетерологичного антитела с помощью ионообменной хроматографии, путем установления различия в изоэлектрических точках в результате интродукции аминокислотных замен в вариабельные области двух типов Н-цепей (WO 2007/114325). К настоящему времени в качестве метода для очистки гетерологичных форм описан метод, основанный на применении белка А для очистки гетеродимеризованного антитела, содержащего Н-цепь мышиного IgG2a, которая связывается с белком А, и Н-цепь крысиного IgG2b, которая не связывается с белком A (WO 98/050431 и WO 95/033844). Кроме того, гетеродимеризованное антитело само можно эффективно очищать с использованием колонки с белком А путем изменения взаимодействия между каждой из Н-цепей и белком А, путем применения Н-цепей, в которых аминокислотные остатки в сайте связывания IgG-белка А в положениях 435 и 436 (EU-нумерация) заменены на аминокислотные остатки, которые обеспечивают другую силу связывания с белком А, такие как Tyr, His или т.п.
Альтернативно этому, можно получать общую L-цепь, которая может обеспечивать способность связываться с множеством различных Н-цепей, и применять в качестве общей L-цепи мультиспецифического антитела. Для достижения эффективной экспрессии мультиспецифического IgG можно интродуцировать в клетки гены указанной общей L-цепи и множества различных Н-цепей и экспрессировать IgG (Nature Biotechnology, 16, 1998, сс. 677-681). Метод селекции общей L-цепи, для которой характерна выраженная способностью к связыванию с любыми различными Н-цепями, можно применять также для селекции общей Н-цепи (WO 2004/065611).
Кроме того, Fc-область, отличающуюся улучшенной С-концевой гетерогенностью, можно применять в качестве Fc-области, предлагаемой в настоящем изобретении. Более конкретно, предложены Fc-области, в которых отсутствуют глицин в положении 446 и лизин в положении 447 (EU-нумерация) в аминокислотных последовательностях двух полипептидов, образующих Fc-область, которая происходит из IgG1, IgG2, IgG3 или IgG4.
Несколько, например, две или большее количество, указанных методик можно применять в сочетании друг с другом. Кроме того, эти методики можно соответствующим образом и раздельно применять к двум Н-цепям, подлежащим ассоциации. Кроме того, указанные методики можно применять в комбинации с описанной выше Fc-областью, у которой снижена активность связывания с Fcγ-рецептором. Кроме того, антигенсвязывающая молекула, предлагаемая в настоящем изобретении, может представлять собой молекулу, полученную отдельно на основе антигенсвязывающей молекулы, подвергнутой таким описанным выше модификациям, что она имеет такую же аминокислотную последовательность.
Антигенсвязывающая молекула (первая антигенсвязывающая молекула), предлагаемая в настоящем изобретении, может содержать (1) указанный выше домен, связывающийся со специфическим для рака антигеном, и (2) домен, связывающийся с представителем суперсемейства фактора некроза опухоли (TNF), или домен, связывающийся с представителем суперсемейства рецептора фактора некроза опухоли (TNF), и ее структура не ограничена указанной. В результате присутствия двух указанных связывающих доменов первая антигенсвязывающая молекула специфически активирует клетки, которые экспрессируют молекулу, принадлежащую к суперсемейству TNF или суперсемейству рецептора TNF, и которая экспрессирует специфический для рака антиген, или клетки, которые присутствуют в опухолевых тканях, содержащих эти клетки, и индуцирует очень высокие (специфические) цитотоксические воздействия в отношении указанных клеток, экспрессирующих специфический для рака антиген, или опухолевых тканей, содержащих указанные клетки. Домен, связывающийся со специфическим для рака антигеном, домен, связывающийся с представителем суперсемейства TNF и домен, связывающийся с представителем суперсемейства рецептора TNF, можно соответственно отбирать, применяя специфический для рака антиген или антиген, принадлежащий к суперсемейству TNF или суперсемейству рецептора TNF, которые описаны выше, соответственно. Указанные связывающие домены могут быть соединены непосредственно с помощью пептидных линкеров или связаны через линкеры.
Антигенсвязывающие молекулы, предлагаемые в настоящем изобретении, могут содержать также FcRn-связывающий домен. При применении описанной выше Fc-области антитела в качестве FcRn-связывающего домена, предпочтительной является Fc-область с пониженной активностью связывания с Fcγ-рецептором. Снижение активности связывания с Fcγ-рецептором позволяет подавлять побочные действия, являющиеся результатом иммуностимуляции, такие как высвобождение цитокинов, вызываемое перекрестным сшиванием между клетками, которые экспрессируют Fcγ-рецептор, и клетками, которые экспрессируют факторы, принадлежащие к суперсемейству рецептора TNF.
Антигенсвязывающие молекулы, предлагаемые в настоящем изобретении, можно получать с помощью известных описанных выше методов. Например, когда применяют (1) F(ab')2 в качестве домена, связывающегося со специфическим для рака антигеном, (2) F(ab')2 в качестве домена, связывающегося с представителем суперсемейства TNF или представителем суперсемейства рецептора TNF, и (3) домен, который содержит Fc-область с пониженной активностью связывания с Fcγ-рецептором в качестве FcRn-связывающего домена, и когда аентигенсвязывающие домены, описанные в подпунктах (1) и (2), и содержащий Fc-область домен, описанный в подпункте (3), непосредственно сшивают с помощью пептидных связей, то связанные полипептиды могут образовывать структуру антитела. Указанные антитела можно получать путем очистки из описанной выше среды для культивирования гибридом, а также путем очистки антител из культуральной среды требуемых клеток-хозяев, которые стабильно несут полинуклеотиды, кодирующие полипептиды, из которых состоит антитело.
Помимо линкеров, примеры которых приведены выше, можно применять также линкеры с пептидными метками, такими как His-метка, НА-метка, myc-метка и FLAG-метка, в качестве линкеров, которые следует использовать, когда требуется соединение каждого из доменов через линкеры. Кроме того, предпочтительно можно использовать способность к взаимному связыванию на основе водородной связи, дисульфидной связи, ковалентной связи, ионного взаимодействия или свойство взаимного связывания в результате их комбинации. Например, можно использовать аффинность между СН1 и CL антитела и Fc-областями, происходящие из описанных выше мультиспецифических антител можно применять также для гетерологичной ассоциации Fc-области.
В настоящем изобретении первую антигенсвязывающую молекулу можно применять в сочетании со второй антигенсвязывающей молекулой.
Также как и в случае первой антигенсвязывающей молекулы, структура второй антигенсвязывающей молекулы не ограничена указанной, и она может содержать:
(1) домен, связывающийся со специфическим для рака антигеном, и
(2) домен, связывающийся с комплексом Т-клеточного рецептора;
и ее можно получать с помощью методов, аналогичных тем, которые описаны для первой антигенсвязывающей молекулы. Кроме того, поскольку вторая антигенсвязывающая молекула содержит домен, связывающийся со специфическим для рака антигеном, и домен, связывающийся с комплексом Т-клеточного рецептора, то ее структура не является такой же, что и у первой антигенсвязывающей молекулы. Специфический для рака антиген, связывающийся с доменом, который связывается со специфическим для рака антигеном первой антигенсвязывающей молекулы, и специфический для рака антиген, связывающийся с доменом, который связывается со специфическим для рака антигеном второй антигенсвязывающей молекулы, могут быть одинаковыми или различными, но предпочтительно они представляют собой один и тот же специфический для рака антиген. Когда специфические для рака антигены являются одинаковыми, то эпитопы, с которыми связываются первая и вторая антигенсвязывающие молекулы, могут быть одинаковыми или различными. Применение указанных первой и второй антигенсвязывающих молекул в сочетании друг с другом позволяет получать очень высокую цитотоксическую активность. Домен, связывающийся со специфическим для рака антигеном, и домен, связывающийся с комплексом Т-клеточного рецептора во второй антигенсвязывающей молекуле соответственно можно выбирать из указанных выше специфических для рака антигенов или антигенов, принадлежащих к комплексам Т-клеточного рецептора.
Аналогично первой антигенсвязывающей молекуле, вторая антигенсвязывающая молекула, предлагаемая в настоящем изобретении, может содержать также FcRn-связывающий домен. При применении описанной выше Fc-области антитела в качестве FcRn-связывающего домена, предпочтительной, также как и в случае первой антигенсвязывающей молекулы, является Fc-область с пониженной активностью связывания с Fcγ-рецептором. Снижение активности связывания с Fcγ-рецептором позволяет подавлять побочные действия, являющиеся результатом иммуностимуляции, такие как высвобождение цитокинов, вызываемое перекрестным сшиванием между клетками, которые экспрессируют Fcγ-рецептор, и клетками, которые экспрессируют факторы, принадлежащие к суперсемейству рецептора TNF.
Кроме того, настоящее изобретение относится к полинуклеотидам, кодирующим антигенсвязывающие молекулы, предлагаемые в настоящем изобретении, и их можно встраивать в любые экспрессионные векторы. Для получения клеток, экспрессирующих антигенсвязывающие молекулы, соответствующего хозяина можно трансформировать экспрессионным вектором. Антигенсвязывающие молекулы, кодируемые полинуклеотидом, можно получать путем культивирования клеток, экспрессирующих антигенсвязывающие молекулы, и сбора продукта экспрессии из супернатанта культуры. В частности, настоящее изобретение относится к векторам, несущим полинуклеотид, который кодирует антигенсвязывающую молекулу, предлагаемую в настоящем изобретении, клеткам, которые содержат векторы, и способам получения антигенсвязывающих молекул, заключающимся в том, что культивируют клетки и собирают антигенсвязывающие молекулы из супернатанта культуры. Для описанных выше целей можно использовать такие же методики, которые описаны выше для рекомбинантных антител.
Фармацевтические композиции
Другим объектом настоящего изобретения являются фармацевтические композиции, которые содержат в качестве действующего вещества описанную выше первую антигенсвязывающую молекулу. Кроме того, настоящее изобретение относится к фармацевтическим композициям, которые индуцируют цитотоксичность (терапевтические агенты для индукции цитотоксичности), ингибиторам клеточной пролиферации и противораковым агентам, которые содержат в качестве действующего вещества антигенсвязывающую молекулу. Фармацевтические композиции, предлагаемые в настоящем изобретении, можно применять в качестве средств для терапии или средств для профилактики рака. Согласно настоящему изобретению терапевтические агенты для индукции клеточной цитотоксичности, ингибиторы клеточной пролиферации и противораковые агенты, предлагаемые в настоящем изобретении, предпочтительно вводят индивидуумам, страдающим раком или имеющим вероятность рецидива рака.
Кроме того, согласно настоящему изобретению терапевтические агенты для индукции клеточной цитотоксичности, ингибиторы клеточной пролиферации и противораковые агенты, которые содержат в качестве действующего вещества первую антигенсвязывающую молекулу, можно применять в способах индукции цитотоксичности, способах подавления клеточной пролиферации, способах активизации иммунитета против раковых клеток или опухолевых тканей, содержащих раковые клетки, или способах предупреждения или лечения рака, заключающихся в том, что осуществляют стадию, на которой вводят антигенсвязывающую молекулу индивидууму; или в изобретении предложено применение антигенсвязывающей молекулы для получения фармацевтических композиций, предназначенных для индукции цитотоксичности, ингибиторов клеточной пролиферации и противораковых агентов. Альтернативно этому, предложены антигенсвязывающие молекулы для применения для индукции цитотоксичности, подавления клеточной пролиферации, активизации иммунитета против раковых клеток или опухолевых тканей, содержащих раковые клетки, или лечения или предупреждения рака.
В контексте настоящего описания понятие «содержат в качестве действующего вещества антигенсвязывающую молекулу» означает присутствие антигенсвязывающей молекулы в качестве основного действующего вещества, однако содержание антигенсвязывающей молекулы не ограничено конкретными значениями.
Кроме того, фармацевтические композиции или фармацевтические композиции, предназначенные для индукции цитотоксичности, ингибиторы клеточной пролиферации и противораковые агенты, предлагаемые в настоящем изобретении (далее в контексте настоящего описания обозначены как фармацевтические композиции или т.п.), можно применять в комбинации с вышеуказанными вторыми антигенсвязывающими молекулами. Применение второй антигенсвязывающей молекулы в комбинации с фармацевтической композицией или т.п., содержащей первую антигенсвязывающую молекулу, может усиливать цитотоксические действия в отношении экспрессирующих антиген клеток. В контексте настоящего описания «применение второй антигенсвязывающей молекулы в комбинации» может относиться к варианту, согласно которому смешивают вторую антигенсвязывающую молекулу с фармацевтической композицией или т.п., содержащей первую антигенсвязывающую молекулу, или варианту, согласно которому вторую антигенсвязывающую молекулу включают в фармацевтическую композицию или т.п., которая отличается от фармацевтической композиции или т.п., содержащей первую антигенсвязывающую молекулу. Их лекарственные формы могут быть одинаковыми или различными. Кроме того, когда первую антигенсвязывающую молекулу и вторую антигенсвязывающую молекулу включают в различные фармацевтические композиции или т.п., эти фармацевтические композиции или т.п. можно вводить индивидууму одновременно или по отдельности. Кроме того, эти фармацевтические композиции или т.п. можно включать в наборы.
В настоящем изобретении первую антигенсвязывающую молекулу или фармацевтическую композицию, которая содержит в качестве действующего вещества первую антигенсвязывающую молекулу, можно применять в качестве фармацевтической композиции, предназначенной для усиления цитотоксической активности или повышения индукции цитотоксической активности путем совместного ее применения с второй антигенсвязывающей молекулой или фармацевтической композицией или т.п., которая содержит в качестве действующего вещества вторую антигенсвязывающую молекулу. Кроме того, вторую антигенсвязывающую молекулу или фармацевтическую композицию, которая содержит в качестве действующего вещества вторую антигенсвязывающую молекулу, можно применять в качестве фармацевтической композиции, предназначенной для усиления цитотоксической активности или повышения индукции цитотоксической активности путем совместного ее применения с первой антигенсвязывающей молекулой или фармацевтической композицией или т.п., которая содержит в качестве действующего вещества первую антигенсвязывающую молекулу.
В контексте настоящего описания понятие «совместное применение» включает вариант, согласно которому фармацевтическую композицию или т.п., которая содержит в качестве действующего вещества первую антигенсвязывающую молекулу, и фармацевтическую композицию или т.п., которая содержит в качестве действующего вещества вторую антигенсвязывающую молекулу, вводят одновременно индивидууму, и вариант, согласно которому их вводят по отдельности индивидууму. Их лекарственные формы могут быть одинаковыми или различными. Кроме того, эти фармацевтические композиции или т.п. можно включать в наборы.
Кроме того, настоящее изобретение относится к способу, в котором используются действия, возникающие в результате совместного применения первой антигенсвязывающей молекулы, описанной выше, или фармацевтической композиции или т.п., которая содержит в качестве действующего вещества первую антигенсвязывающую молекулу, и второй антигенсвязывающей молекулы или фармацевтической композиции или т.п., которая содержит в качестве действующего вещества вторую антигенсвязывающую молекулу, для повышения цитотоксической активности или противоопухолевого действия второй антигенсвязывающей молекулы или фармацевтической композиции или т.п., которая содержит в качестве действующего вещества вторую антигенсвязывающую молекулу, с помощью первой антигенсвязывающей молекулы или фармацевтической композиции или т.п., которая содержит в качестве действующего вещества первую антигенсвязывающую молекулу. Кроме того, в настоящем изобретении предложен способ усиления цитотоксической активности или противоопухолевого действия первой антигенсвязывающей молекулы или фармацевтической композиции или т.п., которая содержит в качестве действующего вещества первую антигенсвязывающую молекулу, с помощью второй антигенсвязывающей молекулы или фармацевтической композиции или т.п., которая содержит в качестве действующего вещества вторую антигенсвязывающую молекулу.
Кроме того, фармацевтические композиции или т.п., предлагаемые в настоящем изобретении, можно применять путем объединения при необходимости нескольких типов первой антигенсвязывающей молекулы и/или второй антигенсвязывающей молекулы. Например, путем применения «коктейля» из нескольких антигенсвязывающих молекул, предлагаемых в изобретении, которые связываются с одним и тем же антигеном, можно повышать цитотоксическую активность в отношении клеток, экспрессирующих антиген.
При необходимости антигенсвязывающие молекулы, предлагаемые в настоящем изобретении, можно капсулировать в микрокапсулы (микрокапсулы, изготовленные из гидроксиметилцеллюлозы, желатина, поли(метилметакрилата) и т.п.), и можно включать в компоненты коллоидных систем, предназначенных для введения лекарственных веществ (липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) (например, см. «Remington's Pharmaceutical Science 16-ое изд.», под ред. Oslo, 1980). Кроме того, известны методы приготовления агентов в виде средств с замедленным высвобождением, и их можно применять для антигенсвязывающих молекул, предлагаемых в настоящем изобретении (J. Biomed. Mater. Res. 15, 1981, cc. 267-277; Chemtech. 12, 1982, cc. 98-105; US №3773719; опубликованные европейские патенты (ЕР) ЕР 58481 и ЕР 133988; Biopolymers 22, 1983, сс. 547-556).
Фармацевтические композиции, агенты, подавляющие клеточную пролиферацию, или противораковые агенты, предлагаемые в настоящем изобретении, можно вводить пациенту либо орально, либо парентерально. Предпочтительным является парентеральное введение. В частности, указанные методы введения включают инъекцию, введение в нос, транспульмонарное введение и чрескожное введение. Инъекции включают, например, внутривенные инъекции, внутримышечные инъекции, внутрибрюшинные инъекции и подкожные инъекции. Например, фармацевтические композиции, терапевтические агенты для индукции клеточной цитотоксичности, агенты, подавляющие клеточную пролиферацию, или противораковые агенты, предлагаемые в настоящем изобретении, можно применять местно или системно путем инъекции. Кроме того, приемлемые методы введения можно выбирать в зависимости от возраста пациента и симптомов. Вводимую дозу можно выбирать, например, из следующего диапазона: от 0,0001 до 1000 мг на кг веса тела на каждое введение. Альтернативно этому, дозу можно выбирать, например, из следующего диапазона: от 0,001 до 100000 мг/пациента. Однако доза фармацевтической композиции, предлагаемой в настоящем изобретении, не ограничена указанными дозами.
Фармацевтические композиции, предлагаемые в настоящем изобретении, можно приготавливать с помощью общепринятых методов (например, см. Remington's Pharmaceutical Science, последнее изд., изд-во Mark Publishing Company, Easton, U.S.A.), и они могут содержать также фармацевтически приемлемые носители и добавки. Их примерами являются (но не ограничиваясь только ими) поверхностно-активные вещества, эксципиенты, красители, ароматизаторы, консерванты, стабилизаторы, буферы, суспендирующие агенты, придающие изотоничность агенты, связующие вещества, разрыхлители, замасливатели, повышающие текучесть агенты и корригенты, но можно применять и другие общепринятые носители. Конкретными примерами носителей являются легкая безводная кремниевая кислота, лактоза, кристаллическая целлюлоза, маннит, крахмал, кармеллоза кальция, кармеллоза натрия, гидроксипропилцеллюлоза, гидроксипропилметилцеллюлоза, поливинилацетальдиэтиламиноацетат, поливинилпирролидон, желатин, триглицерид со средней длиной цепи, полиоксиэтиленированное гидрогенизированное касторовое масло 60, сахароза, карбоксиметилцеллюлоза, кукурузный крахмал, неорганическая соль и т.п.
В настоящем изобретении предложены также способы индукции повреждения клеток, экспрессирующих специфический для рака антиген, или опухолевых тканей, которые содержат клетки, экспрессирующие специфический для рака антиген, и способ подавления клеточной пролиферации указанных клеток или указанных опухолевых тканей, заключающиеся в том, что приводят в контакт клетки, экспрессирующие специфический для рака антиген, с первой антигенсвязывающей молекулой или с первой антигенсвязывающей молекулой, а также второй антигенсвязывающей молекулой, предлагаемой в настоящем изобретении, которая связывается со специфическим для рака антигеном. Изобретение не ограничено клетками, с которыми связывается антигенсвязывающая молекула, предлагаемая в настоящем изобретении, которая связывается со специфическим для рака антигеном, если они представляют собой клетки, которые экспрессируют специфический для рака антиген. В частности, в настоящем изобретении предпочтительными экспрессирующими раковый антиген клетками являются клетки рака яичника, клетки рака предстательной железы, клетки рака молочной железы, клетки рака матки, клетки рака печени, клетки рака легкого, клетки рака поджелудочной железы, клетки рака желудка, клетки рака мочевого пузыря и клетки рака ободочной кишки.
Согласно настоящему изобретению «приведение в контакт» можно осуществлять, например, добавляя антигенсвязывающую молекулу, предлагаемую в настоящем изобретении, которая связывается с раковым антигеном, в раствор к клеткам, экспрессирующим раковый антиген, которые культивируют in vitro. В этом случае добавляемую антигенсвязывающую молекулу можно применять в пригодной для этой цели форме, такой как раствор или твердое вещество, полученное путем сушки вымораживанием, и т.п.Когда осуществляют добавление в виде водного раствора, то можно применять водный раствор, содержащий только антигенсвязывающую молекулу, предлагаемую в изобретении, или можно применять раствор, содержащий поверхностно-активные вещества, эксципиенты, красители, ароматизаторы, консерванты, стабилизаторы, буферы, суспендирующие агенты, придающие изотоничность агенты, связующие вещества, разрыхлители, замасливатели, повышающие текучесть агенты и корргиенты, описанные выше. Концентрация добавляемой молекулы не имеет решающего значения; однако конечная концентрация в культуральном растворе предпочтительно составляет от 1 пг/мл до 1 г/мл, более предпочтительно от 1 нг/мл до 1 мг/мл и еще более предпочтительно от 1 мкг/мл до 1 мг/мл.
Согласно другому варианту осуществления настоящего изобретения «приведение в контакт» можно осуществлять также путем введения антигенсвязывающей молекулы, предлагаемой в настоящем изобретении, которая связывается с раковым антигеном, животным кроме человека, которым трансплантированы клетки, экспрессирующие специфический для рака антиген, и животным, у которых эндогенно происходит экспрессия специфических для рака антигенов. Метод введения может быть оральным или парентеральным, и предпочтительным является парентеральное введение. В частности, указанные методы введения включают инъекцию, введение в нос, транспульмонарное введение и чрескожное введение. Инъекции включают, например, внутривенные инъекции, внутримышечные инъекции, внутрибрюшинные инъекции и подкожные инъекции. Фармацевтическую композицию, предлагаемую в настоящем изобретении, или фармацевтическую композицию, предназначенную для индукции цитотоксичности, ингибитор клеточной пролиферации и противораковый агент можно применять местно или системно путем инъекции. Приемлемые методы введения можно выбирать в зависимости от возраста подопытного животного и симптомов. Когда осуществляют добавление в виде водного раствора, то можно применять водный раствор, содержащий только антигенсвязывающую молекулу, предлагаемую в изобретении, или можно применять раствор, содержащий поверхностно-активные вещества, эксципиенты, красители, ароматизаторы, консерванты, стабилизаторы, буферы, суспендирующие агенты, придающие изотоничность агенты, связующие вещества, разрыхлители, замасливатели, повышающие текучесть агенты и корргиенты, описанные выше. Дозу можно выбирать, например, из следующего диапазона: от 0,0001 до 1000 мг на кг веса тела на каждое введение. Альтернативно этому, дозу можно выбирать, например, из следующего диапазона: от 0,001 до 100000 мг/пациента. Однако доза антигенсвязывающей молекулы, предлагаемой в настоящем изобретении, не ограничена указанными дозами.
Описанный ниже метод предпочтительно используют в качестве метода оценки или количественного измерения цитотоксичности, индуцируемой в клетках, экспрессирующих специфический для рака антиген, которые связаны с доменом, связывающимся со специфическим для рака антигеном, который входит в антигенсвязывающую молекулу, предлагаемую в настоящем изобретении, в результате контакта антигенсвязывающей молекулы с клетками. Методы оценки или количественного измерения цитотоксической активности in vitro включают методы определения активности цитотоксических Т-клеток или т.п.Обладает ли антигенсвязывающая молекула, предлагаемая в настоящем изобретении, способностью индуцировать опосредуемую Т-клетками цитотоксичность, можно определять с помощью известных методов (см., например, Current protocols in Immunology, глава 7. Immunologic studies in humans, под ред. John E, Coligan и др., изд-во, John Wiley & Sons, Inc., 1993 и т.п.). При осуществлении анализа активности антигенсвязывающую молекулу, антигенсвязывающий домен которой связывается с антигеном, отличным от связывающегося молекулой антигена, предлагаемого в настоящем изобретении, и представляет собой антиген, который не экспрессируется на клетках, которые используют для анализа, можно применять в качестве контроля и анализировать таким же образом, что и антигенсвязывающую молекулу, предлагаемую в настоящем изобретении, и активность можно оценивать, определяя, обладает ли антигенсвязывающая молекула, предлагаемая в настоящем изобретении, более выраженной цитотоксической активностью, чем применяемая в качестве контроля антигенсвязывающая молекула.
Для оценки или количественного измерения цитотоксической активности in vivo, например, клетки, экспрессирующие антиген, который связывается доменом, связывающимся со специфическим для рака антигеном, который входит в антигенсвязывающую молекулу, предлагаемую в настоящем изобретении, трансплантируют внутрикожно или подкожно подопытному животному кроме человека, а затем, начиная с дня трансплантации или после него, вводят тестируемую антигенсвязывающую молекулу внутривенно или внутрибрюшинно каждый день или с интервалами, составляющими несколько дней. Размер опухоли определяют ежедневно и различие в размере опухоли можно рассматривать как цитотоксическую активность. Также как и в анализе in vitro, вводят контрольную антигенсвязывающую молекулу, и принимают, что можно считать, что антигенсвязывающая молекула, предлагаемая в настоящем изобретении, обладает цитотоксической активностью, когда размер опухолей является существенно меньшим в группе, которую обрабатывали антигенсвязывающей молекулой, предлагаемой в настоящем изобретении, чем в группе, которую обрабатывали контрольной антигенсвязывающей молекулой.
В качестве метода оценки или количественного измерения подавляющего воздействия на пролиферацию клеток, экспрессирующих антиген, который связывается доменом, связывающимся со специфическим для рака антигеном, который входит в антигенсвязывающую молекулу, предлагаемую в настоящем изобретении, путем контакта с антигенсвязывающей молекулой, предпочтительно можно применять метод количественного измерения поглощения меченного с помощью изотопа тимидина клетками или МТТ-метод. В качестве метода оценки или количественного измерения подавляющей клеточную пролиферацию активности in vivo предпочтительно можно применять такой же метод, который описан выше для оценки или количественного измерения цитотоксической активности in vivo.
В настоящем изобретении предложены также наборы, которые можно применять в способе, предлагаемом в настоящем изобретении, которые содержат антигенсвязывающую молекулу, предлагаемую в настоящем изобретении, или антигенсвязывающую молекулу, полученную способом, предлагаемым в настоящем изобретении. Кроме того, наборы могут быть упакованы вместе с дополнительным фармацевтически приемлемым носителем, растворителем и с инструкциями с описанием метода применения.
Кроме того, настоящее изобретение относится к антигенсвязывающей молекуле, предлагаемой в настоящем изобретении, или антигенсвязывающей молекуле, полученной способом, предлагаемым в настоящем изобретении, для применения в способе, предлагаемом в настоящем изобретении.
Специалистам в данной области должно быть очевидно, что оптимальные комбинации одного или нескольких вариантов осуществления изобретения, представленных в настоящем описании, включены в настоящее изобретение, если отсутствуют технические ограничения, определяемые общепринятыми техническими сведениями, которые известны специалистам в данной области.
Все процитированные в настоящем описании документы, характеризующие известный уровень техники, включены в настоящее описание в качестве ссылки.
Примеры
Ниже настоящее изобретение описано со ссылкой на конкретные примеры, которые не ограничивают объем изобретения.
Приведенный для справки пример 1. Конструирование экспрессионных векторов для антител и экспрессия и очистка антител
Синтез полноразмерных генов, кодирующих нуклеотидные последовательности вариабельных областей Н-цепи и L-цепи антитела, осуществляли согласно методам получения, известным специалистам в данной области, используя сборку с помощью ПЦР и т.п. Интродукцию аминокислотных замен осуществляли согласно методам получения, известным специалистам в данной области, используя ПЦР или т.п.Полученный фрагмент плазмиды встраивали в экспрессионный вектор для клеток животных и получения экспрессионного вектора для Н-цепи и экспрессионного вектора для L-цепи. Нуклеотидную последовательность полученных экспрессионных векторов определяли методами, известными специалистам в данной области. Полученные плазмиды кратковременно интродуцировали в клеточную линию HEK293H, полученную из раковых клеток почки человеческого эмбриона (фирма Invitrogen) или в клетки FreeStyle293 (фирма Invitrogen) для экспрессии антитела. Полученный супернатант культуры собирали и затем пропускали через фильтр MILLEX(R)-GV с размером отверстий 0,22 мкм (фирма Millipore) или через фильтр MILLEX(R)-GV с размером отверстий 0,45 мкм (фирма Millipore) для получения супернатанта культуры. Антитела очищали из полученного супернатанта культуры с помощью методов, известных специалистам в данной области, применяя систему rProtein A Sepharose™ Fast Flow (фирма GE Healthcare) или Protein G Sepharose™ 4 Fast Flow (фирма GE Healthcare). Для оценки концентрации очищенных антител измеряли абсорбцию при длине волны 280 нм с помощью спектрофотометра. На основе полученных данных рассчитывали концентрацию антитела на основе коэффициента экстинкции, определенного с помощью таких методов, как РАСЕ (Protein Science 4, 1995, сс. 2411-2423).
Приведенный для справки пример 2. Метод получения мышиного Fcγ-рецептора (mFcγR) и метод анализа взаимодействия между модифицированным антителом и mFcγR
Внеклеточные домены мышиных FcγR получали следующим методом. Сначала гены внеклеточных доменов FcγR синтезировали с помощью метода, хорошо известного специалистам в данной области. Для этого получали последовательность каждого FcγR на основе зарегистрированной в NCBI информации. В частности, mFcγRI получали на основе последовательности референс-последовательности NCBI: NP_034316.1; mFcγRII получали на основе последовательности референс-последовательности NCBI: NP_034317.1; mFcγRIII получали на основе последовательности референс-последовательности NCBI: NP_034318.2; и mFcγRIV получали на основе последовательности референс-последовательности NCBI: NP_653142.2. К С-концу указанных последовательностей присоединяли His-метку. Каждый из полученных генных фрагментов встраивали в экспрессионный вектор для клеток животных для создания экспрессионных векторов. Сконструированный экспрессионный вектор кратковременно интродуцировали в полученные из раковых клеток клетки почки человеческого эмбриона FreeStyle293 (фирма Invitrogen) для экспрессии представляющих интерес белков. Полученный супернатант культуры собирали и затем пропускали через фильтр MILLEX(R)-GV с размером отверстий 0,22 мкм с получением супернатанта культуры. Полученные супернатанты культур очищали согласно технологии, включающей 4 следующие стадии: стадия 1 - ионообменная колоночная хроматография, стадия 2 - аффинная колоночная хроматография для His-метки (HisTrap HP), стадия 3 - колоночная гель-фильтрация (Супердекс200) и стадия 4 - фильтрация в асептических условиях. Ионообменную колоночную хроматографию на стадии 1 осуществляли, используя Q Сефарозу HP для mFcγRI, используя SP Сефарозу FF для mFcγRII и mFcγRIV, и используя SP Сефарозу HP для mFcγRIII. Когда на стадии 3 применяли растворитель и последующая стадия включала D-ЗФР(-), то для mFcγRIII применяли D-ЗФР(-), содержащий 0,1M аргинин. Для очищенных белков измеряли абсорбцию при длине волны 280 нм с помощью спектрофотометра. На основе полученных данных рассчитывали концентрацию очищенных белков на основе коэффициента экстинкции, определенного с помощью таких методов, как РАСЕ (Protein Science 4, 1995, сс. 2411-2423). Взаимодействие между каждым модифицированным антителом и Fcγ-рецептором, полученным согласно описанному выше методу, анализировали с помощью Biacore Т100 (фирма GE Healthcare), Biacore Т200 (фирма GE Healthcare), Biacore A100 и Biacore 4000. Применяемый подвижный буфер представлял собой HBS-EP+ (фирма GE Healthcare) и температуру, при которой осуществляли измерения, устанавливали на 25°С. Применяемый чип представлял собой: сенсорный чип серий S СМ5 (фирма GE Healthcare) или сенсорный чип серий S СМ4 (фирма GE Healthcare), на котором иммобилизовали белок L (фирма ACTIGEN или фирма BioVision) с помощью метода аминного сочетания. Представляющие интерес антитела «захватывали» на указанных сенсорных чипах и mFcγR, разбавленному подвижным буфером, давали взаимодействовать с ним. Измеряли уровень связывания антителами и сравнивали между антителами. Однако, поскольку количество связанного mFcγR зависело от количества «захваченного» антитела, сравнение осуществляли с использованием скорректированных значений, полученных путем деления количества связанного mFcγR на количество каждого иммобилизованного антитела. Кроме того, 10 мМ глицин-HCl, рН 1,5 применяли для отмывки «захваченного» антитела от сенсорных чипов и сенсорный чип регенерировали и использовали повторно. Кинетический анализ для расчета величин KD каждого измененного антитела в отношении FcγR осуществляли с помощью описанного ниже метода. Сначала представляющие интерес антитела «захватывали» на вышеуказанных сенсорных чипах и mFcγR, разведенному подвижным буфером, давали взаимодействовать с ними. Для получения сенсограмм результаты измерений обрабатывали методом глобальной аппроксимации согласно модели связывания 1:1 Лэнгмюра, применяя программу Biacore Evaluation для расчета константы скорости ассоциации ka (л/моль/с) и константы скорости диссоциации kd (1/с). Их этих величин определяли константу диссоциации KD (моль/л).
Приведенный для справки пример 3. Подопытные животные и клеточные линии
Применяемые подопытные животные представляли собой самок мышей линии C57BL/6 (фирма Charles River Laboratories Japan, Inc.) или самок мышей линии Balb/c (фирма Charles River Laboratories Japan, Inc.). Их выращивали к виварии при постоянных условиях окружающей среды (температура: 20-26°С; освещение: 12-часовой цикл света-темнотые) со свободным доступом к корму и воде. Человеческий ген GPC3 интегрировали в хромосому мышиной клеточной линии рака легкого LLC (АТСС № CRL-1642) с помощью метода, хорошо известного специалистам в данной области, получая клеточную линию LLC-GPC3, которая, характеризуется высоким уровнем экспрессии человеческого GPC3. Уровень экспрессии человеческого GPC3 (2,3×105/клетку) определяли с помощью набора QIFI (фирма Dako) согласно рекомендованному производителем методу. Аналогично этому, человеческий ген GPC3 интегрировали в хромосому мышиной клеточной линии колоректального рака СТ-26 (АТСС №о. CRL-2638), получая клеточную линию CT26-GPC3, характеризующуюся высоким уровнем экспрессии (уровень экспрессии: 3,1×105/клетку). Для поддержания человеческого гена GPC3 указанные рекомбинантные линии клеток культивировали в рекомендованных АТСС средах, добавляя генетицин (фирма GIBCO) в дозе 400 мкг/мл для LLC-GPC3 и 200 мкг/мл для CT26-GPC3. После культивирования указанные клетки отделяли, применяя 2,5 г/л трипсина-1 мМ ЭДТК (фирма Nacalai Tesque), и затем применяли в каждом из экспериментов.
Пример 1. Получение мышиных антител к CD137 и оценка их агонистической активности
1-1. Получение мышиного антитела к мышиному CD137 и оценка способности связываться с mFcγR
1D8VH (SEQ ID NO: 28), вариабельную область антитела к мышиному CD137, описанному в WO 2005/017148, которую применяли в качестве вариабельной области Н-цепи антитела, и конструкцию 1D8VH-mIgG1 (SEQ ID NO: 29), имеющую константную область Н-цепи встречающегося в естественных условиях мышиного IgG1, которую применяли в качестве константной области Н-цепи антитела, получали согласно методу, описанному в приведенном для справки примере 1. 1D8VH-mF18 (SEQ ID NO: 30) получали путем интродукции в 1D8VH-mIgG1 модификации, приводящей к замене на Lys Pro в положении 235 (EU-нумерация), и модификации, приводящей к замене на Lys Ser в положении 239 (EU-нумерация), которые являются модификациями, которые элиминируют связывание с FcγR, что описано в WO 2012/133782. Кроме того, получали 1D8VH-MB492 (SEQ ID NO: 31) путем интродукции в 1D8VH-mIgG1 модификаций (Т230Е, V231P, P232N, S238E, S239D, N324D), которые повышают связывание mFcgRII. Конструкцию 1D8VL, описанную в WO 2005/017148, применяли в качестве вариабельной области L-цепи антитела, и конструкцию 1D8VL-mk0 (SEQ ID NO: 32), которая имеет константную область мышиной κ-цепи, применяли в качестве константной области L-цепи. Их экспрессировали и очищали согласно методу, описанному в приведенном для справки примере 1, с получением 1D8VH-mIgG1/1D8VL-mk0, 1D8VH-mF18/1D8VL-mk0 и 1D8VH-MB492/1D8VL-mk0. Ниже в контексте настоящего описания указанные антитела обозначали для простоты как 1D8-mIgG1, 1D8-mF18 и 1D8-MB492.
Кроме того, для измерения связывания с mFcγR каждой константной области получали H237-mIgG1 (SEQ ID NO: 34) и H237-MB492 (SEQ ID NO: 35), которые имеют вариабельную область Н237 антитела к человеческому рецептору интерлейкина 6 (SEQ ID NO: 33), описанного в WO 2009/125825, в качестве вариабельной области Н-цепи. MRAL-k0 (SEQ ID NO: 36), представляющую собой L-цепь тоцилизумаба, применяли в качестве L-цепи антитела. Экспрессию и очистку осуществляли согласно методу, описанному в приведенном для справки примере 1, с получением H237-mIgG1/MRAL-k0 и H237-MB492/MRAL-k0. Аналогичным образом получали mPM1H-mIgG1 (SEQ ID NO: 37) и mPM1H-mF18 (SEQ ID NO: 38), которые имели вариабельную область (mPM1H) мышиного РМ-1, т.е. мышиного антитела, которое связывается с человеческим IL6R (Sato, Cancer Res., 53, 1993, сс. 851-856), в качестве вариабельной области Н-цепи антитела. MRAL-k0 применяли в качестве L-цепи антитела. Экспрессию и очистку осуществляли согласно методу, описанному в приведенном для справки примере 1, с получением mPM1H-mIgG1/MRAL-k0 и mPM1H-mF18/MRAL-k0.
Способность mPM1H-mIgG1/MRAL-k0 и mPM1H-mF18/MRAL-k0 связываться с mFcγRII и mFcγRIII оценивали согласно методу, описанному в приведенном для справки примере 2. Встречающийся в естественных условиях мышиный IgG1 (mIgG1) не связывается с mFcγRI или mFcγRIV, а из четырех типов мышиного FcγR связывается только с mFcγRII и mFcγRIII (Nimmerjahn, Science, 310, 2005, сс. 1510-1512). Таким образом, можно ожидать, что интродукция модификаций, которые снижают связывание с mFcγR, во встречающийся в естественных условиях mIgG1 может приводить к получению вариантов, обладающих пониженной способностью связываться с mFcγRII и mFcγRIII, и таким образом к пониженному связыванию со всеми mFcγR. Результаты представлены в таблице 1.

Указанные выше результаты продемонстрировали, что константная область mF18 представляет собой вариант со значительно сниженной способностью связываться с mFcγR.
Аналогично этому, в таблице 2 представлены результаты оценки связывания H237-mIgG1/MRAL-k0 и H237-MB492/MRAL-k0 с mFcγRII и mFcγRIII.

«Относительная активность связывания» в таблице означает активность связывания МВ492, когда активность связывания встречающегося в естественных условиях mIgG1 с каждым из mFcγR принимают за 1. Указанные выше результаты продемонстрировали, что МВ492 представляет собой вариант, у которого в 621,5 раза увеличилось связывание с mFcγRII и в 10,9 раза увеличилось связывание с mFcγRIII по сравнению с mIgG1.
1-2. Оценка агонистического действия in vitro в отношении CD137 антител к мышиному CD137
Выделяли селезенки из наивных самок мышей линии C57BL/6. Клетки суспендировали в содержащей 10% FBS среде RPMI1640, дополненной 0,5 мкг/мл иономицина и 10 нг/ мл форбол-12 миристат-13-ацетата (ФМА), и высевали в 96-луночный планшет с плотностью 2×105 клеток/100 мкл/лунку. Антитела к мышиному CD137 добавляли в указанные лунки в концентрации 3 мкг/мл и клетки культивировали в следующих условиях: 37°С и 5% СО2 в течение 3 дней. Собирали супернатант культуры и определяли концентрацию мышиного IFN-γ в супернатанте с помощью ELISA для оценки активации селезеночных Т-клеток. ELISA осуществляли согласно инструкциям производителя, прилагаемым к набору (фирма PeproTech).
Установлено (фиг. 1), что среди полученных мышиных антител в виде IgG1 к мышиному CD137 антитело (1D8-mF18), которое обладало очень значительно пониженной способностью связывать FcγR, не обладало активностью, а антитело (1D8-mIgG1), которое имело Fc дикого типа, активировало Т-клетки. Кроме того, специфическая активность антитела (1D8-MB492), которое обладало повышенной способностью связываться с FcγRIIB, повышалась примерно в 8 раз по сравнению с активностью антитела, которое имело Fc дикого типа.
Это свидетельствует о том, что аналогично агонистическим антителам к другим TNFRSF, которые описаны в Proc Natl Acad Sci USA, 110(48), 2013, cc. 19501-19506, для того, чтобы антитела к CD137 обладали агонистической активностью, антитела должны связываться с FcγRII, и антитела к CD137, связывающиеся с экспрессирующими CD137 Т-клетками, должны перекрестно связываться с экспрессирующими FcγRII клетками (фиг. 2). FcγRII экспрессируется на многих иммунных клетках и фагоцитах, таких как В-клетки. Таким образом, агонистическая активность антител к CD137 может проявляться системно и в результате приводить к побочным действиям.
Пример 2. Получение биспецифических антител к человеческому GPC3/мышиному CD137 и оценка их агонистической активности
2-1. Концепция разработки зависимых от ракового антигена агонистического антитела на основе биспецифических антител к раковому антигену и CD137
Согласно данным, представленным в примере 1, поскольку агонистическая активность обычных антител к CD137 проявляется системно, то, вероятно, противоопухолевые действия и побочные действия в здоровой ткани (такие как Т-клеточная активация) не поддаются разделению. Таким образом, при создании настоящего изобретения было высказано предположение о том, что применение биспецифических антител против ракового антигена и CD137 может обеспечивать проявление агонистической активности антителом к CD137 только в раковых тканях, в которых присутствует раковый антиген, посредством перекрестного сшивания экспрессирующих CD137 Т-клеток и экспрессирующих раковый антиген клеток (таких как раковые клетки) через биспецифические антитела (фиг. 3).
2-1. Получение биспецифических антител к человеческому GPC3/мышиному CD137 (GPC3 ERY22-1D8, GPC3 ERY22-G2-1D8 и GPC3 ERY22-G4-1D81
Получали три типа биспецифических антител к человеческому GPC3/к мышиному CD137, которые имели константную область человеческого IgG1, IgG2 или IgG4 соответственно. Для создания этих молекул применяли технологию получения CrossMab, описанную у Schaefer с соавторами (Schaefer, Proc. Natl. Acad. Sci., 108, 2011, cc. 11187-11192), для регуляции ассоциации между Н- и L-цепями и эффективного получения биспецифических антител. Более конкретно, указанные молекулы получали путем обмена VH- и VL-доменов Fab к человеческому GPC3, согласно методу, описанному в WO 2012/073985. Для усиления гетерологичной ассоциации применяли технологию «Knobs-into-Holes» («выступы-во-впадины») для константной области Н-цепи антитела. Технология «Knobs-into-Holes» представляет собой метод, позволяющий получать представляющие интерес гетеродимеризованные антитела посредством усиления гетеродимеризации Н-цепей путем замены боковой цепи аминокислоты, присутствующей в СН3-области одной из Н-цепей, на более крупную боковую цепи («выступ») и замены боковой цепи аминокислоты в СН3-области другой Н-цепи на меньшую боковую цепь («впадина») так, чтобы «выступ» помещался во «впадину» (Burmeister, Nature, 372, 1994, сс. 379-383). Далее в контексте настоящего описания константную область, в которую интродуцирована модификация, приводящая к образованию «выступа», обозначают как Kn, а константную область, в которую интродуцирована модификация, приводящая к образованию «впадины», обозначают Hl. Кроме того, модификации, описанные в WO 2011/108714, применяли для снижения связывания с Fcγ. В частности, в IgG1-тип и IgG4-тип интродуцировали модификации, приводящие к замене на Ala аминокислот в положениях 234, 235 и 297 (EU-нумерация). В IgG2-тип интродуцировали модификации, приводящие к замене на Ala аминокислот в положении 234, 237 и 297. Gly в положении 446 и Lys в положении 447 (EU-нумерация) удаляли из С-концов Н-цепей антитела. Для дополнительного облегчения очистки после экспрессии антитела добавляли гистидиновую метку к С-концу Н-цепи антитела к человеческому GPC3, a FLAG-метку добавляли к С-концу Н-цепи антитела к мышиному CD137. Н-цепи антитела к человеческому GPC3, полученные путем интродукции указанных выше модификаций, представляли собой GC33(2)H-G1dKnHS (SEQ ID NO: 39), GC33(2)H-G2dmKnHS (SEQ ID NO: 40) и GC33(2)H-G4dKnHS (SEQ ID NO: 41). Полученные Н-цепи антитела к мышиному CD137 представляли собой 1D8VH-G1dH1FS (SEQ ID NO: 42), 1D8VH-G2dmH1FS (SEQ ID NO: 43) и 1D8VH-G4dH1FS (SEQ ID NO: 44). В GC33(2)H-G2dmKnHS и 1D8VH-G2dmH1FS, которые имеют константную область IgG2-типа, только СН1-домен и первая половина шарнирной области относились к IgG1-типу. В частности, они содержали, по сравнению с последовательностью СН1 встречающегося в естественных условиях IgG2, Ser в положении 131, Lys в положении 133 и Gly в положениях 137 и 138; а шарнирная область содержала Ser в положении 219 (EU-нумерация). L-цепи антител GC33(2)L-k0 (SEQ ID NO: 45) и 1D8VL-k0 (SEQ ID NO: 46), как правило, применяли в плече, представляющем собой антитело к человеческому GPC3, и в плече, представляющем собой антитело к мышиному CD137, соответственно. Антитела, имеющие комбинации, представленные в таблице 3, экспрессировали с получением представляющих интерес биспецифических антител. Указанные антитела экспрессировали путем кратковременной экспрессии в клетках FreeStyle293 (фирма Invitrogen) согласно методу, описанному в «1-1». Полученный супернатант культуры вносили на колонку с антителом к FLAG М2 (фирма Sigma) и колонку промывали, после чего элюировали с использованием FLAG-пептида (фирма Sigma) в концентрации 0,1 мг/мл. Содержащую антитело фракцию вносили на колонку HisTrap HP (фирма GE Healthcare) и колонку промывали, после чего элюировали с помощью концентрационного градиента имидазола. Содержащую антитело фракцию концентрировали с помощью ультрафильтрационной мембраны, а затем концентрированный раствор вносили на колонку с Супердекс 200 (фирма GE Healthcare). Собирали только мономерные антитела в элюате с получением очищенных антител.
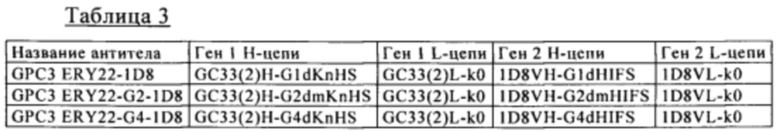
2-2. Оценка in vitro GPC3-зависимого агонистического действия в отношении CD137 биспецифических антител к человеческому GPC3/к мышиному CD137
Мышиную Т-клеточную линию CTLL-2 (АТСС каталожный №. TIB-214) суспендировали в содержащей 10% FBS среде RPMI1640, дополненной 0,5 мкг/мл иономицина и 10 нг/мл ФМА, и клетки высевали в 96-луночный планшет с плотностью 2×104 клеток/100 мкл/лунку. Экспрессирующие человеческий GPC3 клетки мышиного рака легкого линии LLC-GPC3 (приведенный для справки пример 3) суспендировали в такой же среде и высевали в такой же 96-луночный планшет с плотностью 2×104 клеток/100 мкл/лунку. Кроме того, получали суспензии каждой клеточной линии, содержащие одинаковое количество клеток CTLL-2 или LLC-GPC3, и затем клетки высевали в 96-луночный планшет с плотностью 4×104 клеток/100 мкл/лунку. В лунки добавляли биспецифическое антитело к человеческому GPC3/к мышиному CD137 в виде человеческого антитела IgG1-типа с чрезвычайно низкой способностью связываться с FcγR (GPC3 ERY22-1D8) или моноспецифическое антитело к человеческому GPC3 в виде человеческого антитела IgG-типа (GC33(2)-hG1S, содержащего GC33(2)H2-G1dS nGC33(2)L2-k0), в концентрации 5 мкг/мл и клетки культивировали в следующих условиях: 37°С и 5% СО2 в течение 24 ч. Собирали супернатант культуры и определяли концентрацию мышиного IFN-γ в супернатанте с помощью ELISA для оценки активации CTLL-2. ELISA осуществляли согласно инструкциям производителя, прилагаемым к набору (фирма PeproTech).
В результате установлено, что высокие уровни накопления мышиного IFN-γ обнаружены только в условиях, в которых присутствовали и клетки линии LLC-GPC3, и клетки линии CTLL-2 (фиг. 4). На основе этого результата можно предположить, что активация Т-клеток происходила в зависимости от ассоциации с CD137 на Т-клетках, обусловленной несколькими различными биспецифическими антителами, связывающимися с экспрессирующими GPC3 клетками (фиг. 3).
Кроме того, на фиг. 5 продемонстрировано наличие активности у биспецифических антител, Fc-область которых заменена на область человеческого IgG2-типа (GPC3 ERY22-G2-1D8) или человеческого IgG4-типа (GPC3 ERY22-G4-1D8), с чрезвычайно низкой способностью связываться с FcγR. Изменение подкласса антитела не приводило к каким-либо значительным изменениям в агонистической активности в отношении CD137.
Эти результаты подтверждают, что биспецифические антитела против ракового антигена (GPC3 в настоящих примерах) и CD137 с пониженной способностью связываться с FcγR обладают агонистической активностью после ассоциации с CD137 на Т-клетках только, когда присутствуют экспрессирующие раковый антиген клетки (раковые клетки и т.п.). Более конкретно, Т-клетки не активируются в здоровых тканях, в которых раковый антиген отсутствует, и это позволяет снижать или элиминировать побочные действия.
Пример 3. Повышающее действие в отношении Т-клеточной активации смеси биспецифического антитела к человеческому GPC3/к мышиному CD137 и биспецифического антитела к человеческому GPC3/к мышиному CD3
3-1. Концепция
Хотя агонистические антитела к CD137, как известно, обладают противоопухолевым действием в результате активации Т-клеток, это действие, как известно, является невысоким, когда агонистические антитела к CD137 применяют в качестве единственного агента. Таким образом, для повышения способности биспецифических антител к раковому антигену/к CD137 активировать Т-клетки и, как следствие, обладать более сильным противоопухолевым действием, оценивали совместное применение агентов со сходной способностью активировать Т-клетки. Биспецифические антитела к раковому антигену/к CD3 могут перенаправлять Т-клетки к раковому антигену и вызывать опосредуемую Т-клетками цитотоксическую активность в отношении раковых клеток. Однако противоопухолевое действие биспецифических антител к раковому антигену/к CD3 также не является достаточно высоким при их применении в качестве единственных агентов. Таким образом, изучали совместное применение биспецифического антитела к раковому антигену /к CD137 и биспецифического антитела к раковому антигену/к CD33 для того, чтобы продемонстрировать их синергетическое действие в отношении способности активировать Т-клетки и в отношении противоопухолевой активности.
3-2. Получение GPC3 ERY22-3-1D8 и GPC3 ERY22-3-2C11
Получали биспецифическое антитело к человеческому GPC3/к мышиному CD137, т.е. GPC3 ERY22-3-1D8, и биспецифическое антитело к человеческому GPC3/к мышиному CD3, т.е. GPC3 ERY22-3-2C11. GPC3 ERY22-3-1D8 получали, добавляя известные специалистам в данной области модификации в константной области биспецифического антитела GPC3 ERY22-1D8, полученного согласно методу, описанному в примере 2-1, для дополнительного упрощения очистки. В частности, GC33(2)H-G1dKnHSG3 (SEQ ID NO: 48) получали путем добавления известной специалистам в данной области модификации H435R к гену константной области Н-цепи антитела к человеческому GPC3 GC33(2)H-G1dKnHS для упрощения очистки. Параллельно получали 1D8VH-G1dH1S (SEQ ID NO: 47) путем удаления FLAG-метки из гена константной области Н-цепи антитела к мышиному CD137 1D8VH-G1dH1FS. Кроме того, получали 2C11VH-G1dH1S (SEQ ID NO: 50), используя последовательность 2C11VH (SEQ ID NO: 49) в качестве вариабельной области Н-цепи антитела к мышиному CD3. L-цепи GC33(2)L-k0, 1D8VL-k0 и 2C11VL-k0 (SEQ ID NO: 51) антител применяли для плеча, представляющего собой антитело к человеческому GPC3, представляющего собой антитело к мышиному CD137 и представляющего собой антитело к мышиному CD3, соответственно. Антитела, имеющие комбинации, представленные в таблице 4, экспрессировали с получением представляющих интерес биспецифических антител. Указанные антитела кратковременно экспрессировали в клетках FreeStyle293 согласно приведенному для справки примеру 1. Полученный супернатант культур вносили на колонку MabSelect SuRe (фирма GE Healthcare) и колонку промывали, после чего элюировали 50 мМ уксусной кислотой. Содержащую антитело фракцию вносили на колонку HisTrap HP (фирма GE Healthcare) или колонку Ni Сефароза FF (фирма GE Healthcare) и колонку промывали, после чего элюировали имидазолом. Содержащую антитело фракцию концентрировали с помощью ультрафильтрационной мембраны. Затем концентрированный раствор вносили на колонку с Супердекс 200 (фирма GE Healthcare). Собирали только мономерные антитела в элюате с получением очищенных антител.

Кроме того, в качестве применяемого для сравнения контроля получали также антитело GC33(2)-G1dS, которое обладает пониженной способностью связываться с FcγR, а также антитело к человеческому GPC3. GC33(2)-G1dS представляет собой встречающееся в естественных условиях антитело к человеческому GPC3, полученное без применения технологии CrossMab, и оно имеет константную область с пониженной способностью связываться с FcγR. В частности, получали конструкцию GC33(2)H2-G1dS (SEQ ID NO: 53), которая имеет GC33(2)H2 (SEQ ID NO: 52), соответствующую вариабельной области Н-цепи антитела, и имеет G1d, с интродуцированными мутациями L234A, L235A и N297A, в качестве константной области Н-цепи антитела. GC33(2)L2-k0 (SEQ ID NO: 54) применяли в качестве L-цепи антитела. Экспрессию и очистку осуществляли согласно методу, описанному в приведенном для справки примере 1, с получением GC33(2)H2-G1dS/GC33(2)L2-k0. Далее в контексте настоящего описания антитело для простоты обозначено как GC33(2)-G1dS.
3-3. Оценка in vitro повышающего действия в отношении Т-клеточной активации смеси биспецифического антитела к человеческому GPC3/к мышиному CD137 и биспецифического антитела к человеческому GPC3/к мышиному CD3
Выделяли селезенки из наивных самок мышей линии C57BL/6. Клетки суспендировали в содержащей 10% FBS среде RPMI1640, дополненной 10 нг/мл мышиного IL2, с плотностью 4×106 клеток/мл. Кроме того, клетки мышиного колоректального рака, экспрессирующие человеческий GPC3, линии CT26-GPC3 (приведенный для справки пример 3) суспендировали в такой же среде с плотностью 4×105 клеток/мл. Смешивали равные количества каждой клеточной суспензии и смесь высевали в 96-луночный планшет из расчета 100 мкл/лунку. В некоторые лунки дополнительно вносили 0,5 мкг/мл иономицина и 10 нг/мл ФМА. В лунки добавляли биспецифическое антитело к человеческому GPC3/к мышиному CD137 с чрезвычайно низкой способностью связываться с FcγR (GPC3 ERY22-1D8) и биспецифическое антитело к человеческому GPC3/к мышиному CD3 с чрезвычайно низкой способностью связываться с FcγR (GPC3 ERY22-2C11 : GPC3 ERY22-3-2C11, в котором модификацию H435R возвращали к ее исходному состоянию) в концентрации 3 мкг/мл, и клетки культивировали в следующих условиях: 37°С и 5% СО2 в течение 24 ч. Собирали супернатант культуры и определяли концентрацию мышиного IFN-γ в супернатанте с помощью ELISA для оценки активации селезеночных Т-клеток. ELISA осуществляли согласно инструкциям производителя, прилагаемым к набору (фирма PeproTech).
В результате установлено (фиг. 6), что 1D8-MB492 и GPC3 ERY22-1D8 обладали индуцирующей IFN-γ активностью при добавлении с иономицином и ФМА. Вероятно, это являлось результатом индукции CD137 в селезеночных Т-клетках в результате стимуляции митогеном и т.п. Кроме того, установлен высокий уровень накопления IFN-γ при применении смеси, содержащей GPC3 ERY22-1D8 и GPC3 ERY22-2C11. Это позволяет предположить, что одновременная стимуляция CD3 и CD137 вызывает сильную Т-клеточную активацию.
Пример 4. Противоопухолевая активность биспецифических антител к человеческому GPC3/к мышиному CD137 и их роль в снижении токсичности для печени
4-1. Сравнение эффективности в качестве лекарственных средств биспецифических антител к человеческому GPC3/к мышиному CD137 и антител к мышиному CD137
Клетки рекомбинантной линии мышиного колоректального рака СТ26-GPC3, которые экспрессируют человеческий GPC3 (приведенный для справки пример 3), помещали в сбалансированный соляной раствор Хэнкса (HBSS) из расчета 5×106 клеток/мл, и 200 мкл указанного раствора (1×106 клеток) трансплантировали подкожно в брюшную область мышей линии BALB/c (самки 7-нельного возраста, фирма Charles River Laboratories Japan Inc.). Животных произвольно разделяли на 5 групп по 5 особей в каждой, а затем с помощью внутривенной инъекции вводили в хвостовую вену антитела через 3 дня, 7 дней, 10 дней и 17 дней после трансплантации. Приготавливали препараты биспецифического антитела к человеческому GPC3/к мышиному CD137 (GPC3 ERY22-3-1D8) с концентрацией 0,75 и 0,15 мг/мл в растворителе (водный раствор, содержащий 150 мМ NaCl и 20 мМ His-HCl (рН 6,0) который пропускали через фильтр с размером отверстий 0,22 мкм), и вводили из расчета 10 мл/кг (7,5 и 1,5 мг/кг соответственно). Приготавливали препараты антитела к мышиному CD137 (1D8-MB492) с концентрацией 1,5 и 0,3 мг/мл в растворителе и вводили из расчета 10 мл/кг (15 и 3 мг/кг соответственно). Процент ингибирования роста опухоли (%) определяли на основе объема опухоли, рассчитанного с помощью приведенного ниже уравнения.
Объем опухоли (мм3) = максимальная ось (мм) × минимальная ось (мм) × минимальная ось (мм) / 2
Процент ингибирования роста опухоли (%) = [1-(Т-Т0)/(С-С0)] × 100
Т: Средний объем опухоли в каждой группе в каждый момент анализа.
Т0: Средний объем опухоли в каждой группе в первый день введения.
С: Средний объем опухоли в контрольной группе в каждый момент анализа
С0: Средний объем опухоли в контрольной группе в первый день введения.
Как продемонстрировано на фиг. 7, во всех группах, которые обрабатывали антителом, обнаружено сильное противоопухолевое действие с ингибированием роста опухоли на 95% или более. Более конкретно, установлено, что биспецифические антитела к человеческому GPC3/к мышиному CD137 обладали сильным противоопухолевым действием, сходным с действием антител к мышиному CD137, а также обладали сильным противоопухолевым действием при активации CD137 зависимым от ракового антигена образом.
4-2. Снижение повреждения печени при применении биспецифических антител к человеческому GPC3/к мышиному CD137. выявленное на модели, созданной путем подкожной трансплантации клеток CT26-GPC3
По окончании тестов по изучению эффективности при введении применяемых в качестве лекарственных средств антител животных умерщвляли путем обескровливания под анестезией и выделяли плазму. Плазму применяли для измерения аспартатаминотрансферазы (AST; метод JSCC Transferable), аланинаминотрансферазые (ALT; метод JSCC Transferable) и общего билирубина (TBIL; ферментативный метод) с помощью автоматического анализатора ТВА-120FR (фирма Toshiba Medical Systems Corporation). Печень получали в процессе аутопсии, фиксировали в 10%-ном забуференном до нейтрального значения растворе формалина с получением с помощью общепринятых методов тонких срезов залитых в парафин препаратов ткани (гематоксилин-эозин (НЕ)), и осуществляли гистопатологическую оценку с помощью светового микроскопа. Статистический анализ осуществляли с использованием непараметрического критерия Дуннетта для множественного сравнения с контрольной группой.
В результате установлено, что, как продемонстрировано на фиг. 8-11, в группе, которую обрабатывали антителом к мышиному CD137 (1D8-MB492), уровень AST, ALT и TBIL повышался или имел тенденцию к повышению при всех изученных дозах; и во всех случаях при гистопатологической оценке обнаружено повреждение печени, от слабого до умеренного, такое как дегенерация/некроз и воспаление клеток печени. С другой стороны, в группе, которую обрабатывали биспецифическим антителом к человеческому GPC3/к мышиному CD137 (GPC3 ERY22-3-1D8), не удалось обнаружить изменения уровней AST, ALT и TBIL в крови, которое могло быть связано с повреждением печени. Слабые с точки зрения гистопатологии дегенерацию/некроз или воспаление клеток печени были обнаружены в двух-трех случаях из пяти в каждой обработанной дозой группе, при этом печеночное нарушение было уменьшено. В одном случае в группе, которую обрабатывали этим же антителом в дозе 3 мг/кг, обнаружено существенное повышение уровней AST и ALT в крови, но при этом не обнаружено изменения уровней TBIL в крови. Поскольку при гистопатологической оценке печени не были обнаружены проявления, позволяющие заподозрить повреждение печени, источник ферментов не рассматривается как принимающий участие в повреждении печени.
Из приведенных выше результатов следует, что биспецифическое антитело к человеческому GPC3/к мышиному CD137 GPC3 ERY22-3-1D8 обладает сильной противоопухолевой активностью, не индуцируя серьезное повреждение печени, которое характерно для описанных к настоящему времени обычных агонистических антител к CD137. Более конкретно, биспецифическое антитело к раковому антигену и CD137 с пониженной способностью связываться с FcγR, и, как предлагается, обладающее зависимой от ракового антигена агонистической активностью в отношении CD137, и обладающее способностью активировать Т-клетки только в опухолях без активации Т-клеток в здоровых тканях, обладает избирательной цитотоксической активностью в отношении раковых клеток, но без побочных действий, таких как цитотоксичность и высвобождение цитокинов.
Пример 5. Противоопухолевое действие при совместном применении биспецифического антитела к человеческому GPC3/к мышиному CD137 и биспецифического антитела к человеческому GPC3/к мышиному CD3
Клетки мышиного рака легкого линии LLC-GPC3, которые экспрессируют человеческий GPC3 (приведенный для справки пример 3), суспендировали в HBSS из расчета 5×106 клеток/мл и 200 мкл суспензии (1×106 клеток) трансплантировали подкожно в брюшную область мышей линии C57BL/6N (самки, 6-недельного возраста, фирма Charles River Laboratories Japan Inc.). Через 10 дней после трансплантации животных разделяли на основе данных об объеме опухолей и веса тела на пять групп по пять особей в каждой, а затем вводили антитела с помощью внутривенной инъекции в хвостовую вену через 10 дней, 14 дней и 17 дней после трансплантации. Приготавливали препарат биспецифического антитела к человеческому GPC3/к мышиному CD137 (GPC3 ERY22-3-1D8) с концентрацией 0,5 мг/мл в растворителе (водный раствор, содержащий 150 мМ NaCl и 20 мМ His-HCl (рН 6,0) который пропускали через фильтр с размером отверстий 0,22 мкм), и вводили из расчета 10 мл/кг (5 мг/кг). Приготавливали препарат биспецифического антитела к человеческому GPC3/к мышиному CD3 (GPC3 ERY22-3-2C11) с концентрацией 0,45 мг/мл в растворителе и вводили из расчета 10 мл/кг (4,5 мг/кг). Кроме того, создавали группу, которой совместно вводили два типа антител. Процент ингибирования роста опухоли (%) определяли на основе объема опухоли, рассчитанного с помощью приведенного ниже уравнения.
Объем опухоли (мм3) = максимальная ось (мм) × минимальная ось (мм) × минимальная ось (мм) / 2
Процент ингибирования роста опухоли (%) = [1-(Т-Т0)/(С-С0)] × 100
Т: Средний объем опухоли в каждой группе в каждый момент анализа.
Т0: Средний объем опухоли в каждой группе в первый день введения.
С: Средний объем опухоли в контрольной группе в каждый момент анализа
С0: Средний объем опухоли в контрольной группе в первый день введения.
Как продемонстрировано на фиг. 12, процент ингибирования роста опухолей через 23 дня после трансплантации составлял 36% в группе, которой вводили только биспецифическое антитело к человеческому GPC3/к мышиному CD137, и 29% в группе, которой вводили только биспецифическое антитело к человеческому GPC3/к мышиному CD3, но в группе, которой совместно вводили указанные два антитела, обнаружено 100%-ное ингибирование и четко проявлялось синергетическое действие в результате совместного применения.
По окончании тестов по изучению эффективности применяемых в качестве лекарственных средств антител осуществляли анализ параметров функции печени (AST, ALT и TBIL) в плазме и гистопатологический анализ срезов ткани печени с помощью НЕ-окрашивания с применением методов, аналогичных описанным в примере «4-2». Изменения, позволяющие заподозрить повреждение печени, не обнаружены ни в одной из подвергающихся обработке групп.
Таким образом, установлено, что совместное применение биспецифического антитела против ракового антигена и CD137 и биспецифического антитела против ракового антигена и CD3 приводит к одновременной специфической и локальной ассоциации CD137 и CD3 на опухоли и вызывает сильную способность активировать Т-клетки, чего не удается достигать при индивидуальном введении каждого из антител по данным экспериментов in vitro, и при этом достигается противоопухолевое действие, которым также не обладают индивидуальные агенты in vivo.
Пример 6. Получение человеческих CD137-связывающих антител из библиотеки человеческих антител с помощью метода фагового дисплея
6-1. Получение фаговой дисплейной библиотеки наивных человеческих антител
Согласно методам, известным специалистам в данной области, полиА-хвост РНК, полученной из человеческих РВМС, и полиА-хвост поступающих в продажу РНК и т.п. применяли в качестве матрицы для создания фаговой дисплейной библиотеки человеческих антител, экспонирующих Fab-домены последовательностей человеческих антител, которые отличаются друг от друга.
6-2. Получение человеческих CD137-связывающих антител из библиотеки наивных человеческих антител с помощью пэннинга с использованием гранул
Антитела, у которых обнаружена антигенсвязывающая активность, отбирали путем скрининга из фаговой дисплейной библиотеки наивных человеческих антител, созданной согласно методу, описанному в примере 6-1. Более конкретно, собирали фаги, презентующие антитела, которые обладали активностью связывания с антигенами, иммобилизованными на гранулах. В качестве антигена применяли биотинилированный человеческий CD137. В частности, пэннинг осуществляли с использованием антигена, фиксированного на магнитных гранулах. В качестве магнитных гранул применяли сенсибилизированные нейтравидином гранулы (сенсибилизированные Sera-Mag SpeedBeads NeutrAvidin) или сенсибилизированные стрептавидином гранулы (Dynabeads М-280 Streptavidin).
Сначала фаги, полученные из Escherichia coli, несущие сконструированные фагмиды для фагового дисплея, очищали с помощью общепринятого метода. Затем получали суспензию фаговой библиотеки, которую подвергали диализу в противотоке TBS. Затем добавляли к суспензии фаговой библиотеки БСА до получения конечной концентрации 4%.
Затем добавляли к образовавшейся суспензии фаговой библиотеки 250 пмолей биотинилированного человеческого CD137, давая суспензии фаговой библиотеки контактировать с человеческим CD137 в течение 60 мин при комнатной температуре. Затем к суспензии фаговой библиотеки добавляли блокированные БСА магнитные гранулы и комплексам человеческий CD137-фаг давали связываться с магнитными гранулами в течение 15 мин при комнатной температуре. Гранулы однократно промывали TBS. Затем к гранулам добавляли 0,5 мл раствора трипсина с концентрацией 1 мг/мл, гранулы суспендировали при комнатной температуре в течение 15 мин и гранулы немедленно отделяли, используя магнитный стенд для сбора суспензии фагов. Собранную суспензию фагов добавляли к 10 мл штамма Е.coli ER2738 на логарифмической фазе роста (ОП600 от 0,4 до 0,7). Е.coli осторожно перемешивали и инкубировали при 37°С в течение 1 ч, давая фагам инфицировать Е.coli. Инфицированные Е.coli высевали в планшет (225 мм × 225 мм). Затем фаги собирали из культуральной среды, засеянной Е.coli, с получением суспензии фаговой библиотеки.
Во втором раунде пэннинга осуществляли обогащение фагов, обладающих способностью связываться с человеческим CD137. К полученной суспензии фаговой библиотеки добавляли 100 пмолей биотинилированного человеческого CD137 и давали суспензии фаговой библиотеки контактировать с человеческим CD137 в течение 60 мин при комнатной температуре. Затем к суспензии фаговой библиотеки добавляли блокированные БСА магнитные гранулы и комплексам человеческий CD137-фаг давали связываться с магнитными гранулами в течение 15 мин при комнатной температуре. Гранулы трижды промывали TBST (TBS, содержащий 0,1% Твин20). Затем к гранулам добавляли 0,5 мл раствора трипсина с концентрацией 1 мг/мл, гранулы суспендировали при комнатной температуре в течение 15 мин и гранулы немедленно отделяли, используя магнитный стенд для сбора суспензии фагов. Собранную суспензию фагов добавляли к 10 мл штамма Е.coli ER2738 на логарифмической фазе роста (ОП600 от 0,4 до 0,7). Е.coli осторожно перемешивали и инкубировали при 37°С в течение 1 ч, давая фагам инфицировать Е.coli. Инфицированные Е.coli высевали в планшет (225 мм × 225 мм). Затем фаги собирали из культуральной среды, засеянной Е.coli, с получением суспензии фаговой библиотеки.
Пэннинг, предназначенный для получения антител, которые обладают способностью связываться с человеческим CD137, повторяли трижды, применяя такую же процедуру. Четвертый пэннинг осуществляли, используя 40 пмолей биотинилированного человеческого CD137.
6-3. Конструирование фаговой дисплейной библиотеки синтетических человеческих антител
Фаговую дисплейную библиотеку синтетических человеческих антител конструировали с помощью метода, известного специалистам в данной области, применяя 10 типов последовательностей зародышевой линии тяжелых цепей и 7 типов последовательностей зародышевой линии легких цепей. Частоту встречаемости в популяции человеческих В-клеток и физико-химические свойства семейства вариабельных областей применяли в качестве индикаторов для отбора VH1-2, VH1-69, VH3-23, VH3-66, VH3-72, VH4-59, VH4-61, VH4-b, VH5-51, VH6-1, Vκ1-39, Vκ2-28, Vκ3-20, Vλ1-40, Vλ1-44, Vλ2-14 и Vλ3-21 для применения в качестве последовательной зародышевых линий. Антигенраспознающие сайты в библиотеке синтетических антител диверсифицировали, имитируя популяции человеческих антител, синтезируемых В-клетками.
6-4. Получение человеческих CD137-связывающих антител из библиотеки синтетических человеческих антитела с помощью пэннинга с использованием гранул
Антитела, у которых обнаружена антигенсвязывающая активность, отбирали путем скрининга из фаговой дисплейной библиотеки синтетических человеческих антител, созданной согласно методу, описанному в примере 6-3. Более конкретно, собирали фаги, презентующие антитела, обладающие активностью связывания с антигенами, иммобилизованными на гранулах. В качестве антигена применяли биотинилированный человеческий CD137.
Фаги, полученные из Escherichia coli, несущие сконструированные фагмиды для фагового дисплея, очищали с помощью общепринятого метода. Популяцию фагов осаждали из культуральной среды Е.coli, применяемой для получения фагов, путем добавления 2,5М NaCl/10% ПЭГ. Затем осадок разводили TBS с получением суспензии фаговой библиотеки. Затем добавляли к суспензии фаговой библиотеки БСА до получения конечной концентрации 4%.
Пэннинг осуществляли с использованием антигена, иммобилизованного на магнитных гранулах. В качестве магнитных гранул применяли сенсибилизированные нейтравидином гранулы (сенсибилизированные Sera-Mag SpeedBeads NeutrAvidin) или сенсибилизированные стрептавидином гранулы (Dynabeads М-280 Streptavidin).
Затем добавляли к образовавшейся суспензии фаговой библиотеки 250 пмолей биотинилированного человеческого CD137, давая суспензии фаговой библиотеки контактировать с человеческим CD137 в течение 60 мин при комнатной температуре. Затем к суспензии фаговой библиотеки добавляли блокированные БСА магнитные гранулы и комплексам человеческий CD137-фаг давали связываться с магнитными гранулами в течение 15 мин при комнатной температуре. Гранулы однократно промывали TBS. Затем к гранулам добавляли 0,5 мл раствора трипсина с концентрацией 1 мг/мл, гранулы суспендировали при комнатной температуре в течение 15 мин и гранулы немедленно отделяли, используя магнитный стенд для сбора суспензии фагов. Собранную суспензию фагов добавляли к 10 мл штамма Е.coli ER2738 на логарифмической фазе роста (ОП600 от 0,4 до 0,7). Е.coli перемешивали и инкубировали при 37°С в течение 1 ч, давая фагам инфицировать Е.coli. Инфицированные Е.coli высевали в планшет (225 мм × 225 мм). Затем фаги собирали из культуральной среды, засеянной Е.coli, с получением суспензии фаговой библиотеки.
На втором раунде пэннинга осуществляли обогащение фагов, обладающих способностью связываться с человеческим CD137. К полученной суспензии фаговой библиотеки добавляли 100 пмолей биотинилированного человеческого CD137 и давали суспензии фаговой библиотеки контактировать с человеческим CD137 в течение 60 мин при комнатной температуре. Затем к суспензии фаговой библиотеки добавляли блокированные БСА магнитные гранулы и комплексам человеческий CD137-фаг давали связываться с магнитными гранулами в течение 15 мин при комнатной температуре. Гранулы трижды промывали TBST и дважды TBS. Затем к гранулам добавляли 0,5 мл раствора трипсина с концентрацией 1 мг/мл, гранулы суспендировали при комнатной температуре в течение 15 мин и гранулы немедленно отделяли, используя магнитный стенд для сбора суспензии фагов. Собранную суспензию фагов добавляли к 10 мл штамма Е.coli ER2738 на логарифмической фазе роста (ОП600 от 0,4 до 0,7). Е.coli осторожно перемешивали и инкубировали при 37°С в течение 1 ч, давая фагам инфицировать Е.coli. Инфицированные Е.coli высевали в планшет (225 мм × 225 мм). Затем фаги собирали из культуральной среды, засеянной Е.coli, с получением суспензии фаговой библиотеки.
Пэннинг, предназначенный для получения антител, которые обладают способностью связываться с человеческим CD137, повторяли трижды, применяя такую же процедуру. Четвертый пэннинг осуществляли, используя 40 пмолей биотинилированного человеческого CD137.
6-5. Оценка способности связываться с человеческим CD137 с помощью ELISA, предназначенного для анализа фагов
Из единичных колоний Е.coli, полученных с помощью метода пэннинга, который описан в приведенных выше примерах, собирали содержащие фаги супернатанты культуры с помощью общепринятого метода (Methods Mol. Biol. 178, 2002, сс. 133-145).
Фаги с добавленным к ним TBS подвергали ELISA, осуществляя описанную ниже процедуру. Титрационные 96-луночные микропланшеты StreptaWell (покрытие стрептавидином) (фирма Roche) сенсибилизировали, используя 100 мкл содержащего TBS меченного биотином антигена (биотинилированный человеческий CD137), при комнатной температуре в течение 1 ч. Затем каждую лунку планшета промывали TBST (TBS, содержащий 0,1% Твин20) для удаления антигена, не связанного с планшетом, лунки блокировали с помощью 250 мкл 2% обезжиренного молока-TBS в течение 1 ч или более. 2% обезжиренное молоко-TBS удаляли, а затем в каждую лунку добавляли полученные фаги. Планшетам давали выстояться при комнатной температуре в течение 1 ч для обеспечения связывания экспонирующих антитело фагов с антигеном в каждой лунке. Затем каждую лунку промывали TBST, в лунки добавляли конъюгированное с HRP антитело к М13 (фирма Amersham Pharmacia Biotech), разведенное TBS, и планшеты инкубировали в течение 1 ч. После промывок с использованием TBST в каждую лунку добавляли только раствор ТМВ (фирма ZYMED). Хромогенную реакцию в растворе в каждой лунке прекращали, добавляя серную кислоту. Затем оценивали изменение цвета, измеряя абсорбцию при 450 нм.
Из 192 клонов, подвергнутых ELISA, предназначенного для анализа фагов, идентифицировали множество антител, обладающих активностью в отношении связывания с человеческим CD137. Результаты, полученные с помощью ELISA, предназначенного для анализа фагов, представлены в таблице 5.

6-6. Анализ последовательностей антител, которые связываются с биотинилированным человеческим CD137
Из клонов, для которых установлено наличие специфической активности связывания с человеческим CD137 с помощью ELISA, предназначенного для анализа фагов, описанного в примере 6-5, анализировали нуклеотидные последовательности генов, амплифицированные с использованием пар специфических праймеров (SEQ ID NO: 55 и 56 для библиотек наивных человеческих антител и SEQ ID NO: 57 и 56 для библиотек синтетических человеческих антител). Результат анализа подтвердил присутствие нескольких типов последовательностей антител, обладающих активностью в отношении связывания с человеческим CD137.
6-7. Получение антител, связывающихся с человеческим CD137
Из клонов полученных согласно методу, описанному в примере 6-6, для которых установлено наличие активности связывания с меченным биотином человеческим CD137, последовательности вариабельных областей тяжелых цепей и легких цепей 5 клонов, полученных из библиотеки наивных человеческих антител (R1-R5), и 14 клонов, полученных из библиотеки синтетических человеческих антител (R6-R19), связывали с константной областью тяжелой цепи антитела (SEQ ID NO: 58, которая представляет собой последовательность, полученную путем модификации константной области человеческого IgG1) или с последовательностью константной области легкой каппа-цепи (SEQ ID NO: 59), или с последовательностью константной области легкой лямбда-цепи (SEQ ID NO: 60), и затем каждую из них встраивали в плазмиды для экспрессии в животных. Последовательности вариабельных областей тяжелых цепей и легких цепей каждого из клонов представлены в таблице 6.

Каждое из антител экспрессировали и очищали с помощью метода, описанного в приведенном для справки примере 1. Кроме того, для повышения активирующего Т-клетки действия in vitro антител к человеческому CD137, гены, VH-область которых представлена в таблице 6, связывали с константной областью (SEQ ID NO: 99), которая обладала повышенной способностью связываться с человеческим FcγRIIB, гены встраивали в плазмидный вектор для экспрессии в клетках животных, и антитела экспрессировали и очищали аналогичным методом с получением комбинаций их вариабельных областей в виде комбинаций, представленных в таблице 6.
Пример 7. Анализ эпитопов антител к человеческому CD137
7-1. Получение слитых белков, содержащих фрагментированный человеческий CD137-Fc, и получение антител
Для анализа эпитопа полученных антител к человеческому CD137 получали слитые белки, содержащие фрагментированный человеческий CD137 и Fc-область антитела, в которых фрагментированный человеческий CD137 разделяли на домены на основе структуры, общей со структурой TNFRSF, и структур, образованных с помощью Cys-Cys, которые обозначены как CRD в J Exp Med., 211(7), 30 июня 2014 г., сс. 1433-1448 (таблица 7). Слитый белок фрагментированный человеческий CD137-Fc встраивали в плазмидный вектор для экспрессии в клетках животных с помощью метода, известного специалистам в данной области, получая каждый генный фрагмент с помощью ПЦР из полинуклеотида, кодирующего слитый белок полноразмерный человеческий CD137-Fc (SEQ ID NO: 100), так, чтобы он содержал аминокислотную последовательность, представленную в таблице 7. Слитый белок фрагментированный человеческий CD137-Fc очищали таким же образом, что и антитела, с помощью метода, описанного в приведенном для справки примере 1. Кроме того, в качестве контроля для ELISA получали антитела с помощью метода, описанного в приведенном для справки примере 1, путем включения в плазмидный вектор для экспрессии в клетках животных генов, кодирующих антитело (SEQ ID NO: 101 Н-цепи и SEQ ID NO: 102 L-цепи), полученное путем замены константной области Н-цепи антитела к человеческому CD137, которое описано в WO 2005/035584 А1 (сокращенно обозначено как В), на константную область с удаленными С-концевыми Gly и Lys в константной области Н-цепи человеческого IgG1, и кодирующих антитело (SEQ ID NO: 103 Н-цепи и SEQ ID NO: 104 L-цепи), полученное путем замены константной области антитела к человеческому CD137, которое описано в WO2012/145183A3 (сокращенно обозначено как М), на константную область с повышенной способностью связываться с человеческим FcγRIIB.

7-2-1. Анализ эпитопов с использованием слитых белков фрагментированный человеческий CD137-Fc
Слитые белки фрагментированный человеческий CD137-Fc, полученные согласно методу, описанному в примере 7-1, применяли для оценки связывания с помощью ELISA для выявления тех сайтов в человеческом CD137, которые связываются антителами (в которых применяют SEQ ID NO: 99 в качестве константной области тяжелой цепи), которые получали согласно методу, описанному выше в примере 6. Например, в случае антитела, которое связывается с доменом 1, можно предположить, что такое антитело должно связываться с содержащими домен 1 слитыми белками фрагментированный человеческий CD137-Fc, но не связываться со слитыми белками фрагментированный человеческий CD137-Fc, которые не содержат домен 1.
7-2-2. Метод ELISA
Слитые белки фрагментированный человеческий CD137-Fc разводили до концентрации 2 мкг/мл в водном растворе карбоната натрия, значение рН которого доводили до 9,6. Добавляли индивидуально в каждую лунку плоскодонного 96-луночного планшета Nunc MaxiSorp (фирма Nunc) по 50 мкл разведенного слитого белка фрагментированный человеческий CD137-Fc. Давали выстояться при 4°С в течение ночи или в течение более продолжительного периода времени, а затем планшету давали выстояться при комнатной температуре в течение 1 ч, чтобы температура планшета полностью сравнялась с комнатной температурой. Раствор, содержащий слитый белок фрагментированный человеческий CD137-Fc, удаляли путем опрокидывания и каждую лунку промывали трижды, используя 300 мкл промывочного буфера (TBS, содержащий 0,1% Твин20, фирма TaKaRa). Затем в каждую лунку добавляли по 150 мкл блокирующего буфера (TBS, содержащий 2% БСА) и давали выстояться в течение 1 ч или в течение более продолжительного периода времени. Блокирующий буфер удаляли путем опрокидывания и каждую лунку промывали трижды промывочным буфером, аналогично методу, описанному на предыдущей стадии. Затем в каждую лунку добавляли по 50 мкл раствора антитела, заранее приготовленного путем разведения с помощью TBS до концентрации 10 или 5 мкг/мл. Планшет центрифугировали при скорости 600 об/мин или близкой скорости в течение 1 ч при комнатной температуре для связывания антитела с иммобилизованным антигеном. После удаления раствора антитела путем опрокидывания каждую лунку промывали трижды промывочным буфером, аналогично методу, описанному на предыдущей стадии. В каждую лунку добавляли по 100 мкл раствора вторичного антитела, полученного путем 1000-краткого разведения TBS, содержащим 0,1% Твин20. В качестве вторичного антитела применяли «ANTIBODY ALKALINE PHOSPHATASE CONJUGATE HUMAN IMMUNOGLOBULIN ABSORBED», т.е. козье антитело к человеческой каппа-цепи, конъюгированное с щелочной фосфатазой, фирмы BIOSOURCE в случае антител, несущих каппа-цепь, и применяли антитело к человеческой легкой цепи, т.е. козье антитело к человеческой легкой лямбда-цепи, конъюгированное с щелочной фосфатазой, фирмы BETHYL LABORATORIES INC. в случае антител, несущих лямбда-цепь. После осуществления реакции в течение 1 ч путем инкубации при комнатной температуре раствор антитела удаляли путем опрокидывания и каждую лунку промывали трижды промывочным буфером, аналогично методу, описанному на предыдущей стадии. Для осуществления цветной реакции использовали набор BluePhos Microwell фирмы KPL. После завершения хромогенной реакции ее прекращали, используя стоп-раствор АР фирмы KPL, абсорбцию измеряли при 620 нм с помощью абсорциометра. Результаты представлены на фиг. 14. На фиг. 14 продемонстрировано, что каждое антитело характеризовалось разной степенью проявления окрашивания в присутствии соответствующего ему слитого белка фрагментированный человеческий CD137-Fc и связывалось с различным фрагментом слитого белка человеческий CD137-FC. Кроме того, установлено, что полученные антитела отличались от известных антител В и М.
Пример 8. Оценка способности антител к человеческому CD137 активировать Т-клетки in vitro
Т-клетки размножали культивированием из поступающих в продажу РВМС (фирма AllCells), применяя гранулы Dynabead с активатором человеческих Т-клеток CD3/CD28 (фирма Gibco, 11132D). Человеческие Т-клетки суспендировали с плотностью 4×105 клеток/мл в среде RPMI1640, содержащей 10% FBS, 60 ед./мл человеческого IL2, 0,5 мкг/мл иономицина, 10 нг/мл ФМА и пенициллин и стрептомицин в соответствующей концентрации. Кроме того, клетки человеческой В-клеточной лимфомы линии Raji суспендировали в такой же среде с плотностью 4×105 клеток/мл. Указанные клеточные суспензии смешивали в равных количествах и высевали в 96-луночный планшет из расчета 100 мкл/лунку. Добавляли связывающие человеческий CD137 антитела, полученные согласно методу, описанному в примере 6 (R1-R19; применяли те же антитела, которые использовали для осуществления анализа методом ELISA, описанным в примере 7) в концентрации 5 мкг/мл и клетки культивировали в следующих условиях: 37°С и 5% СО2 в течение 3 дней. Собирали супернатант культуры и концентрацию человеческого IFN-γ в супернатанте измеряли с помощью ELISA для оценки активации человеческих Т-клеток. ELISA осуществляли согласно инструкциям, прилагаемым производителем к набору для ELISA (фирма PeproTech).
В результате установлено (фиг. 15), что по сравнению с контрольным человеческим IgG (фирма Allexis, 804-133-С100: hIgG на фиг. 15) клоны, кроме R7 и R15, все обладали индуцирующей IFN-γ активностью. Определяли являются ли указанные антитела, обладающие IFN-γ-индуцирующей активностью, агонистическими антителами в отношении CD137.
Характеристики полученных антител обобщены на фиг. 16. Получали много антител, которые распознают эпитопы, отличные от эпитопов антител В и М к человеческому CD137, указанных в описанных выше примерах. Указанные антитела к человеческому CD137 модифицировали с получением биспецифических антител с использованием антитела к GC33 (антитело к человеческому GPC3), и оценивали зависимую от их ракового антигена (GPC3) агонистическую активность в отношении CD137. Таким путем можно получать биспецифические антитела к человеческому GPC3/к человеческому CD137, которые обладают требуемыми противоопухолевыми действиями.
Пример 9. Получение биспецифического антитела к человеческому GPC3/к мышиному CD40 (GPC3 FAE-FGK45)
Биспецифическое антитело к человеческому GPC3/к мышиному CD40, т.е. GPC3 FAE-FGK45, несущее константные области человеческого IgG1, получали с помощью описанной ниже процедуры. Для плеча, представляющего собой антитело к мышиному CD40, применяли FGK45VH6 (SEQ ID NO: 120) в качестве вариабельной области тяжелой цепи и применяли FGK45VL4 (SEQ ID NO: 121) в качестве вариабельной области легкой цепи. В этом случае F760nG3P17 (SEQ ID NO: 119) и k0 (SEQ ID NO: 118) применяли в качестве константных областей тяжелой и легкой цепей соответственно. Плечо, представляющее собой антитело к GPC3, как правило, несло вариабельную область тяжелой цепи Н0000 (SEQ ID NO: 115) и вариабельную область GL4 (SEQ ID NO: 116). В этом случае в качестве константных областей применяли константную область тяжелой цепи F760nN17 (SEQ ID NO: 117), модифицированную так, что в ней присутствовала гетерологическая ассоциация между двумя тяжелыми цепями, и она отличалась пониженной способностью связываться с Fcγ-рецептором, и константную область легкой цепи k0 (SEQ ID NO: 118). Указанные антитела экспрессировали следующим методом. Клетки почки человеческого эмбриона, полученные из линии FreeStyle 293-F (фирма Invitrogen), суспендировали в среде для экспрессии FreeStyle 293 (фирма Invitrogen) и высевали с плотностью 1,33×106 клеток/мл. Полученные плазмиды интродуцировали в клетки методом липофекции. Клетки культивировали в течение 4 дней в CO2-инкубаторе (37°С, 8% CO2, 90 об/мин) и антитела очищали из супернатантов культур с помощью rProtein A Sepharose™ Fast Flow (фирма Amersham Biosciences) или Protein G Sepharose 4 Fast Flow (фирма GE HEALTHCARE) с использованием метода, известного специалистам в данной области. Измеряли абсорбцию растворов очищенных антител при 280 нм с использованием спектрофотометра. Концентрации очищенных антител рассчитывали на основе полученных значений, используя коэффициент молярной экстинкции, рассчитанный с помощью метода РАСЕ (Protein Science 4, 1995, сс. 2411-2423). Каждую из очищенных гомологичных форм смешивали, используя комбинации, представленные в таблице 8, для получения представляющих интерес биспецифических антител с использованием методик, известных специалистам в данной области (WO 2015/046467).

Пример 10. Оценка усиливающего действия в отношении активации спленоцитов in vitro с помощью смеси биспецифического антитела к человеческому GPC3/к мышиному CD40 и биспецифичекого антитела к человеческому GPC3/к мышиному CD3
Выделяли селезенку у наивных самок мышей линии Balb/c и ее клетки суспендировали с плотностью 4×106 клеток/мл в среде, приготовленной путем добавления мышиного IL2 в концентрации 10 нг/мл к среде RPMI1640, содержащей 10% FBS, 0,5 мкг/мл иономицина и 10 нг/мл ФМА. Клетки мышиного колоректального рака линии CT26-GPC3, которые экспрессируют человеческий GPC3 (приведенный для справки пример 3), суспендировали также в такой же среде с плотностью 4×105 клеток/мл. Эти две клеточные суспензии смешивали в равных количествах и высевали в 96-луночный планшет из расчета 100 мкл/лунку. Биспецифическое антитело к человеческому GPC3/к мышиному CD40 с чрезвычайно низкой способностью связываться с FcγR (GPC3 ERY22-FGK45) добавляли в концентрации 3 мкг/мл, а биспецифическое антитело к человеческому GPC3/к мышиному CD3 с чрезвычайно низкой способностью связываться с FcγR (GPC3 ERY22-2C11) добавляли в концентрации 1 мкг/мл, и клетки культивировали в следующих условиях: 37°С и 5% СО2 в течение 72 ч. Собирали супернатанты культуры и определяли концентрацию IFN-γ в супернатанте с помощью ELISA для оценки активации Т-клеток, присутствующих в спленоцитах. ELISA осуществляли согласно инструкциям, прилагаемым производителем к набору для ELISA (фирма PeproTech).
В результате установлено (фиг. 17), что хотя антитело GPC3 ERY22-2C11 обладало IFN-γ-индуцирующей активностью при применении в качестве индивидуального агента, для антитела GPC3 ERY22-FGK45 при его применении в качестве индивидуального агента убедительно доказано отсутствие какой-либо активности. Однако при применении смеси GPC3 ERY22-FGK45 и GPC3 ERY22-2С11 установлен высокий уровень накопления IFN-γ. Это позволяет предположить, что одновременное применение CD3 и CD40 стимулирует различные смеси иммунных клеток, приводя к сильной активации Т-клеток.
Пример 11. Получение биспецифических антител к человеческому GPC3/к человеческому CD137 и оценка их агонистической активности
11-1. Получение биспецифических антител к человеческому GPC3/к человеческому CD137
Биспецифические антитела к человеческому GPC3/к человеческому CD137, несущие константные области человеческого IgG1, получали с помощью следующей процедуры. Последовательности (R3 и R5), для которых в примере 7 подтверждена способность связываться с человеческим CD137, модифицировали, используя праймеры, созданные для интродукции произвольных изменений в аминокислотах CDR3 тяжелой цепи. Последовательности вариабельных областей представлены в таблице 9. В этом случае, когда осуществляли модификацию R3 и R5, то последовательность, полученную путем добавления Gly-Lys (обозначают также как «GK») к С-концу последовательности F760nG3P17, сконструированной согласно методу, описанному в примере 9, и последовательность константной области лямбда-цепи (SEQ ID NO: 60) применяли в качестве константной области тяжелой цепи и константной области легкой цепи соответственно. Плечо, представляющее собой антитело к GPC3, как правило, несло вариабельную область тяжелой цепи Н0000 (SEQ ID NO: 115) и вариабельную область GL4 (SEQ ID NO: 116). В этом случае в качестве константных областей применяли константную область тяжелой цепи F760nN17 (SEQ ID NO: 117), модифицированную так, что в ней присутствовала гетерологическая ассоциация между двумя тяжелыми цепями, и она отличалась пониженной способностью связываться с Fcγ-рецептором, и константную область легкой цепи k0 (SEQ ID NO: 118). Указанные антитела экспрессировали следующим методом. Клетки почки человеческого эмбриона клетки, полученные из линии FreeStyle 293-F (фирма Invitrogen), суспендировали в среде для экспрессии FreeStyle 293 (фирма Invitrogen) и высевали с плотностью 1,33×106 клеток/мл. Полученные плазмиды интродуцировали в клетки методом липофекции. Клетки культивировали в течение 4 дней в СО2-инкубаторе (37°С, 8% СО2, 90 об/мин) и антитела очищали из супернатантов культур с помощью rProtein A Sepharose™ Fast Flow (фирма Amersham Biosciences) или Protein G Sepharose 4 Fast Flow (фирма GE HEALTHCARE) с использованием метода, известного специалистам в данной области. Измеряли абсорбцию растворов очищенных антител при 280 нм с использованием спектрофотометра. Концентрации очищенных антител рассчитывали на основе полученных значений, используя коэффициент молярной экстинкции, рассчитанный с помощью метода РАСЕ (Protein Science 4, 1995, сс. 2411-2423). Для антител к человеческому CD137 (полученных на основе R3 и R5) расчеты осуществляли с использованием Е1% = 14. Как продемонстрировано в таблице 9, антитело к человеческому GPC и гомологичные формы каждого из антител к человеческому CD137, для очистки которых применяли метод, описанный в примере 9. смешивали с получением представляющих интерес биспецифических антител с использованием методик, известных специалистам в данной области (WO 2015/046467).
11-2. Оценка in vitro GPC3-зависимого агонистического действия в отношении CD137 биспецифического антитела к человеческому GPC3/к человеческому CD137
Т-клетки размножали культивированием из поступающих в продажу РВМС (фирма AllCells), применяя гранулы Dynabead с активатором человеческих Т-клеток CD3/CD28 (фирма Gibco, 11132D). Человеческие Т-клетки суспендировали с плотностью 4×105 клеток/мл в среде RPMI1640, содержащей 10% FBS, 60 ед./мл человеческого IL2, 0,5 мкг/мл иономицина, 10 нг/мл ФМА и пенициллин и стрептомицин в соответствующей концентрации. Кроме того, клетки мышиного колоректального рака линии CT26-GPC3, которые экспрессируют человеческий GPC3 (приведенный для справки пример 3), суспендировали в такой же среде с плотностью 4×105 клеток /мл. Указанные две клеточные суспензии смешивали в равных количествах и высевали в 96-луночный планшет из расчета 100 мкл/лунку. Добавляли контрольный человеческий IgG (фирма Allexis, 804-133-С100: Ctrl hIgGl на фиг. 18) или антитело GPC3 FAE-BMS, полученное согласно методу, описанную в представленном выше примере 11-1 (биспецифическое антитело к человеческому GPC3/к человеческому CD 137 с чрезвычайной низкой способностью связываться с FcγR), в концентрации 10 мкг/мл и клетки культивировали в следующих условиях: 37°С и 5% CO2 в течение 3 дней. Собирали супернатант культуры и концентрацию человеческого IFN-γ в супернатанте измеряли с помощью ELISA для оценки активации Т-клеток. ELISA осуществляли согласно инструкциям, прилагаемым производителем к набору для ELISA (фирма PeproTech).
В результате установлено (фиг. 18), что биспецифическое антитело к человеческому GPC3/к человеческому CD137 обладает LFN-y-индуцирующей активностью. Это позволяет предположить, что и в человеческих Т-клетках стимуляция CD137 приводит к сильной активации Т-клеток, аналогично тому, что установлено при применении мышиных Т-клеток, которые использовали в примере 2.
11-3. Оценка in vitro GPC3-зависимого агонистического действия в отношении CD137 биспецифического антитела к человеческому GPC3/к человеческому CD137
Человеческий CD137 экспрессируется также в В-клеточной линии HDML-2, и агонистическую активность в отношении CD137 можно оценивать также с использованием HDML-2. Клетки человеческого В-клеточного рака линии HDLM-2 суспендировали с плотностью 8×105 клеток/мл в среде RPMI1640, содержащей 20% FBS и пенициллин-стрептомицин в соответствующей концентрации. Кроме того, клетки мышиного колоректального рака линии СТ26-GPC3, которые экспрессируют человеческий GPC3 (приведенный для справки пример 3) суспендировали в такой же среде с плотностью 4×105 клеток /мл. Указанные две клеточные суспензии смешивали в равных количествах и высевали в 96-луночный планшет из расчета 100 мкл/лунку. Добавляли контрольный человеческий IgG (фирма Allexis, 804-133-С100: Ctrl hIgG1 на фиг. 19) или биспецифическое антитело к человеческому GPC3/к человеческому CD137 с чрезвычайной низкой способностью связываться с FcγR, полученное согласно методу, описанному в представленном выше примере 11-1, в концентрации 10 мкг/мл и клетки культивировали в следующих условиях: 37°С и 5% СО2 в течение 3 дней. Собирали супернатант культуры и концентрацию человеческого IL-6 в супернатанте измеряли с помощью ELISA для оценки активации В-клеток. ELISA осуществляли согласно инструкциям, прилагаемым производителем к набору для ELISA (фирма PeproTech).
В результате установлено (фиг. 19), что биспецифическое антитело к человеческому GPC3/к человеческому CD137 обладает IL-6-индуцирующей активностью. Это свидетельствует о том, что стимуляцию CD137 можно оценивать также с использованием человеческих В-клеток таким же методом, который применяли для мышиных Т-клеток в примере 2 и человеческих Т-клеток в примере 11-2.
В примерах 11-2 и 11-3 продемонстрировано, что, аналогично результатам, полученным в примерах 2-5, в которых использовали мышиный CD137, биспецифические антитела обладают агонистической активностью в отношении человеческого CD137, и что, вероятно, человеческий CD137 может обладать такими же действиями, что и мышиный CD137.
Промышленная применимость
В настоящем изобретении предложены новые антигенсвязывающие молекулы или фармацевтические композиции, обладающие высокой безопасностью, очень высокой противоопухолевой активностью и не обладающие токсичностью, обусловленной повреждением здоровых тканей или цитокиновым «штормом», независимым от ракового антигена образом. Фармацевтические композиции, содержащие антигенсвязывающую молекулу, предлагаемую в настоящем изобретении, в качестве действующего вещества, активируют иммунные клетки зависимым от ракового антигена образом и осуществляют цитотоксические действия, направленные на различные клетки, включая раковые клетки. Это обеспечивает возможность лечения или предупреждения различных видов рака. Настоящее изобретение может не только обеспечивать малоопасное лечение, но также снижать физическую нагрузку и повышать удобство лечения, что является желательным для пациентов.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЧУГАИ СЕИЯКУ КАБУШИКИ КАИША
<120> Иммуноактивирующая антигенсвязывающая молекула
<130> C1-A1401Y1P
<140> PCT/JP2015/060794
<141> 2015-04-07
<150> JP 2014-078457
<151> 2014-04-07
<150> JP 2014-264589
<151> 2014-12-26
<160> 125
<170> PatentIn, верся 3.5
<210> 1
<211> 330
<212> PRT
<213> Homo sapiens
<400> 1
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 2
<211> 326
<212> PRT
<213> Homo sapiens
<400> 2
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 3
<211> 377
<212> PRT
<213> Homo sapiens
<400> 3
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr Cys Pro
100 105 110
Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg
115 120 125
Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys
130 135 140
Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
145 150 155 160
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
165 170 175
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
180 185 190
Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys Trp Tyr
195 200 205
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
210 215 220
Gln Tyr Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu His
225 230 235 240
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
245 250 255
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln
260 265 270
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
275 280 285
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
290 295 300
Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu Asn Asn
305 310 315 320
Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu
325 330 335
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Ile
340 345 350
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Phe Thr Gln
355 360 365
Lys Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 4
<211> 327
<212> PRT
<213> Homo sapiens
<400> 4
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 5
<211> 330
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 5
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 6
<211> 326
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 6
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 7
<211> 377
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 7
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr Cys Pro
100 105 110
Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg
115 120 125
Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys
130 135 140
Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
145 150 155 160
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
165 170 175
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
180 185 190
Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Lys Trp Tyr
195 200 205
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
210 215 220
Gln Tyr Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Leu His
225 230 235 240
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
245 250 255
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln
260 265 270
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
275 280 285
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
290 295 300
Ser Asp Ile Ala Val Glu Trp Glu Ser Ser Gly Gln Pro Glu Asn Asn
305 310 315 320
Tyr Asn Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu
325 330 335
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Ile
340 345 350
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Phe Thr Gln
355 360 365
Lys Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 8
<211> 327
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 8
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 9
<211> 141
<212> PRT
<213> Homo sapiens
<400> 9
Asp Ile Gln Asn Pro Asp Pro Ala Val Tyr Gln Leu Arg Asp Ser Lys
1 5 10 15
Ser Ser Asp Lys Ser Val Cys Leu Phe Thr Asp Phe Asp Ser Gln Thr
20 25 30
Asn Val Ser Gln Ser Lys Asp Ser Asp Val Tyr Ile Thr Asp Lys Thr
35 40 45
Val Leu Asp Met Arg Ser Met Asp Phe Lys Ser Asn Ser Ala Val Ala
50 55 60
Trp Ser Asn Lys Ser Asp Phe Ala Cys Ala Asn Ala Phe Asn Asn Ser
65 70 75 80
Ile Ile Pro Glu Asp Thr Phe Phe Pro Ser Pro Glu Ser Ser Cys Asp
85 90 95
Val Lys Leu Val Glu Lys Ser Phe Glu Thr Asp Thr Asn Leu Asn Phe
100 105 110
Gln Asn Leu Ser Val Ile Gly Phe Arg Ile Leu Leu Leu Lys Val Ala
115 120 125
Gly Phe Asn Leu Leu Met Thr Leu Arg Leu Trp Ser Ser
130 135 140
<210> 10
<211> 179
<212> PRT
<213> Homo sapiens
<400> 10
Glu Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe Glu Pro
1 5 10 15
Ser Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys Leu
20 25 30
Ala Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser Trp Trp Val Asn
35 40 45
Gly Lys Glu Val His Ser Gly Val Ser Thr Asp Pro Gln Pro Leu Lys
50 55 60
Glu Gln Pro Ala Leu Asn Asp Ser Arg Tyr Cys Leu Ser Ser Arg Leu
65 70 75 80
Arg Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe Arg Cys
85 90 95
Gln Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln Asp
100 105 110
Arg Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly Arg
115 120 125
Ala Asp Cys Gly Phe Thr Ser Glu Ser Tyr Gln Gln Gly Val Leu Ser
130 135 140
Ala Thr Ile Leu Tyr Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr Ala
145 150 155 160
Val Leu Val Ser Ala Leu Val Leu Met Ala Met Val Lys Arg Lys Asp
165 170 175
Ser Arg Gly
<210> 11
<211> 173
<212> PRT
<213> Homo sapiens
<400> 11
Asp Lys Gln Leu Asp Ala Asp Val Ser Pro Lys Pro Thr Ile Phe Leu
1 5 10 15
Pro Ser Ile Ala Glu Thr Lys Leu Gln Lys Ala Gly Thr Tyr Leu Cys
20 25 30
Leu Leu Glu Lys Phe Phe Pro Asp Val Ile Lys Ile His Trp Gln Glu
35 40 45
Lys Lys Ser Asn Thr Ile Leu Gly Ser Gln Glu Gly Asn Thr Met Lys
50 55 60
Thr Asn Asp Thr Tyr Met Lys Phe Ser Trp Leu Thr Val Pro Glu Lys
65 70 75 80
Ser Leu Asp Lys Glu His Arg Cys Ile Val Arg His Glu Asn Asn Lys
85 90 95
Asn Gly Val Asp Gln Glu Ile Ile Phe Pro Pro Ile Lys Thr Asp Val
100 105 110
Ile Thr Met Asp Pro Lys Asp Asn Cys Ser Lys Asp Ala Asn Asp Thr
115 120 125
Leu Leu Leu Gln Leu Thr Asn Thr Ser Ala Tyr Tyr Met Tyr Leu Leu
130 135 140
Leu Leu Leu Lys Ser Val Val Tyr Phe Ala Ile Ile Thr Cys Cys Leu
145 150 155 160
Leu Arg Arg Thr Ala Phe Cys Cys Asn Gly Glu Lys Ser
165 170
<210> 12
<211> 204
<212> PRT
<213> Homo sapiens
<400> 12
Lys Gln Leu Asp Ala Asp Val Ser Pro Lys Pro Thr Ile Phe Leu Pro
1 5 10 15
Ser Ile Ala Glu Thr Lys Leu Gln Lys Ala Gly Thr Tyr Leu Cys Leu
20 25 30
Leu Glu Lys Phe Phe Pro Asp Ile Ile Lys Ile His Trp Gln Glu Lys
35 40 45
Lys Ser Asn Thr Ile Leu Gly Ser Gln Glu Gly Asn Thr Met Lys Thr
50 55 60
Asn Asp Thr Tyr Met Lys Phe Ser Trp Leu Thr Val Pro Glu Glu Ser
65 70 75 80
Leu Asp Lys Glu His Arg Cys Ile Val Arg His Glu Asn Asn Lys Asn
85 90 95
Gly Ile Asp Gln Glu Ile Ile Phe Pro Pro Ile Lys Thr Asp Val Thr
100 105 110
Thr Val Asp Pro Lys Asp Ser Tyr Ser Lys Asp Ala Asn Asp Val Thr
115 120 125
Thr Val Asp Pro Lys Tyr Asn Tyr Ser Lys Asp Ala Asn Asp Val Ile
130 135 140
Thr Met Asp Pro Lys Asp Asn Trp Ser Lys Asp Ala Asn Asp Thr Leu
145 150 155 160
Leu Leu Gln Leu Thr Asn Thr Ser Ala Tyr Tyr Met Tyr Leu Leu Leu
165 170 175
Leu Leu Lys Ser Val Val Tyr Phe Ala Ile Ile Thr Cys Cys Leu Leu
180 185 190
Gly Arg Thr Ala Phe Cys Cys Asn Gly Glu Lys Ser
195 200
<210> 13
<211> 177
<212> PRT
<213> Homo sapiens
<400> 13
Pro Ser Tyr Thr Gly Gly Tyr Ala Asp Lys Leu Ile Phe Gly Lys Gly
1 5 10 15
Thr Arg Val Thr Val Glu Pro Arg Ser Gln Pro His Thr Lys Pro Ser
20 25 30
Val Phe Val Met Lys Asn Gly Thr Asn Val Ala Cys Leu Val Lys Glu
35 40 45
Phe Tyr Pro Lys Asp Ile Arg Ile Asn Leu Val Ser Ser Lys Lys Ile
50 55 60
Thr Glu Phe Asp Pro Ala Ile Val Ile Ser Pro Ser Gly Lys Tyr Asn
65 70 75 80
Ala Val Lys Leu Gly Lys Tyr Glu Asp Ser Asn Ser Val Thr Cys Ser
85 90 95
Val Gln His Asp Asn Lys Thr Val His Ser Thr Asp Phe Glu Val Lys
100 105 110
Thr Asp Ser Thr Asp His Val Lys Pro Lys Glu Thr Glu Asn Thr Lys
115 120 125
Gln Pro Ser Lys Ser Cys His Lys Pro Lys Ala Ile Val His Thr Glu
130 135 140
Lys Val Asn Met Met Ser Leu Thr Val Leu Gly Leu Arg Met Leu Phe
145 150 155 160
Ala Lys Thr Val Ala Val Asn Phe Leu Leu Thr Ala Lys Leu Phe Phe
165 170 175
Leu
<210> 14
<211> 549
<212> ДНК
<213> Homo sapiens
<400> 14
atggaacagg ggaagggcct ggctgtcctc atcctggcta tcattcttct tcaaggtact 60
ttggcccagt caatcaaagg aaaccacttg gttaaggtgt atgactatca agaagatggt 120
tcggtacttc tgacttgtga tgcagaagcc aaaaatatca catggtttaa agatgggaag 180
atgatcggct tcctaactga agataaaaaa aaatggaatc tgggaagtaa tgccaaggac 240
cctcgaggga tgtatcagtg taaaggatca cagaacaagt caaaaccact ccaagtgtat 300
tacagaatgt gtcagaactg cattgaacta aatgcagcca ccatatctgg ctttctcttt 360
gctgaaatcg tcagcatttt cgtccttgct gttggggtct acttcattgc tggacaggat 420
ggagttcgcc agtcgagagc ttcagacaag cagactctgt tgcccaatga ccagctctac 480
cagcccctca aggatcgaga agatgaccag tacagccacc ttcaaggaaa ccagttgagg 540
aggaattga 549
<210> 15
<211> 182
<212> PRT
<213> Homo sapiens
<400> 15
Met Glu Gln Gly Lys Gly Leu Ala Val Leu Ile Leu Ala Ile Ile Leu
1 5 10 15
Leu Gln Gly Thr Leu Ala Gln Ser Ile Lys Gly Asn His Leu Val Lys
20 25 30
Val Tyr Asp Tyr Gln Glu Asp Gly Ser Val Leu Leu Thr Cys Asp Ala
35 40 45
Glu Ala Lys Asn Ile Thr Trp Phe Lys Asp Gly Lys Met Ile Gly Phe
50 55 60
Leu Thr Glu Asp Lys Lys Lys Trp Asn Leu Gly Ser Asn Ala Lys Asp
65 70 75 80
Pro Arg Gly Met Tyr Gln Cys Lys Gly Ser Gln Asn Lys Ser Lys Pro
85 90 95
Leu Gln Val Tyr Tyr Arg Met Cys Gln Asn Cys Ile Glu Leu Asn Ala
100 105 110
Ala Thr Ile Ser Gly Phe Leu Phe Ala Glu Ile Val Ser Ile Phe Val
115 120 125
Leu Ala Val Gly Val Tyr Phe Ile Ala Gly Gln Asp Gly Val Arg Gln
130 135 140
Ser Arg Ala Ser Asp Lys Gln Thr Leu Leu Pro Asn Asp Gln Leu Tyr
145 150 155 160
Gln Pro Leu Lys Asp Arg Glu Asp Asp Gln Tyr Ser His Leu Gln Gly
165 170 175
Asn Gln Leu Arg Arg Asn
180
<210> 16
<211> 516
<212> ДНК
<213> Homo sapiens
<400> 16
atggaacata gcacgtttct ctctggcctg gtactggcta cccttctctc gcaagtgagc 60
cccttcaaga tacctataga ggaacttgag gacagagtgt ttgtgaattg caataccagc 120
atcacatggg tagagggaac ggtgggaaca ctgctctcag acattacaag actggacctg 180
ggaaaacgca tcctggaccc acgaggaata tataggtgta atgggacaga tatatacaag 240
gacaaagaat ctaccgtgca agttcattat cgaatgtgcc agagctgtgt ggagctggat 300
ccagccaccg tggctggcat cattgtcact gatgtcattg ccactctgct ccttgctttg 360
ggagtcttct gctttgctgg acatgagact ggaaggctgt ctggggctgc cgacacacaa 420
gctctgttga ggaatgacca ggtctatcag cccctccgag atcgagatga tgctcagtac 480
agccaccttg gaggaaactg ggctcggaac aagtga 516
<210> 17
<211> 171
<212> PRT
<213> Homo sapiens
<400> 17
Met Glu His Ser Thr Phe Leu Ser Gly Leu Val Leu Ala Thr Leu Leu
1 5 10 15
Ser Gln Val Ser Pro Phe Lys Ile Pro Ile Glu Glu Leu Glu Asp Arg
20 25 30
Val Phe Val Asn Cys Asn Thr Ser Ile Thr Trp Val Glu Gly Thr Val
35 40 45
Gly Thr Leu Leu Ser Asp Ile Thr Arg Leu Asp Leu Gly Lys Arg Ile
50 55 60
Leu Asp Pro Arg Gly Ile Tyr Arg Cys Asn Gly Thr Asp Ile Tyr Lys
65 70 75 80
Asp Lys Glu Ser Thr Val Gln Val His Tyr Arg Met Cys Gln Ser Cys
85 90 95
Val Glu Leu Asp Pro Ala Thr Val Ala Gly Ile Ile Val Thr Asp Val
100 105 110
Ile Ala Thr Leu Leu Leu Ala Leu Gly Val Phe Cys Phe Ala Gly His
115 120 125
Glu Thr Gly Arg Leu Ser Gly Ala Ala Asp Thr Gln Ala Leu Leu Arg
130 135 140
Asn Asp Gln Val Tyr Gln Pro Leu Arg Asp Arg Asp Asp Ala Gln Tyr
145 150 155 160
Ser His Leu Gly Gly Asn Trp Ala Arg Asn Lys
165 170
<210> 18
<211> 624
<212> ДНК
<213> Homo sapiens
<400> 18
atgcagtcgg gcactcactg gagagttctg ggcctctgcc tcttatcagt tggcgtttgg 60
gggcaagatg gtaatgaaga aatgggtggt attacacaga caccatataa agtctccatc 120
tctggaacca cagtaatatt gacatgccct cagtatcctg gatctgaaat actatggcaa 180
cacaatgata aaaacatagg cggtgatgag gatgataaaa acataggcag tgatgaggat 240
cacctgtcac tgaaggaatt ttcagaattg gagcaaagtg gttattatgt ctgctacccc 300
agaggaagca aaccagaaga tgcgaacttt tatctctacc tgagggcaag agtgtgtgag 360
aactgcatgg agatggatgt gatgtcggtg gccacaattg tcatagtgga catctgcatc 420
actgggggct tgctgctgct ggtttactac tggagcaaga atagaaaggc caaggccaag 480
cctgtgacac gaggagcggg tgctggcggc aggcaaaggg gacaaaacaa ggagaggcca 540
ccacctgttc ccaacccaga ctatgagccc atccggaaag gccagcggga cctgtattct 600
ggcctgaatc agagacgcat ctga 624
<210> 19
<211> 207
<212> PRT
<213> Homo sapiens
<400> 19
Met Gln Ser Gly Thr His Trp Arg Val Leu Gly Leu Cys Leu Leu Ser
1 5 10 15
Val Gly Val Trp Gly Gln Asp Gly Asn Glu Glu Met Gly Gly Ile Thr
20 25 30
Gln Thr Pro Tyr Lys Val Ser Ile Ser Gly Thr Thr Val Ile Leu Thr
35 40 45
Cys Pro Gln Tyr Pro Gly Ser Glu Ile Leu Trp Gln His Asn Asp Lys
50 55 60
Asn Ile Gly Gly Asp Glu Asp Asp Lys Asn Ile Gly Ser Asp Glu Asp
65 70 75 80
His Leu Ser Leu Lys Glu Phe Ser Glu Leu Glu Gln Ser Gly Tyr Tyr
85 90 95
Val Cys Tyr Pro Arg Gly Ser Lys Pro Glu Asp Ala Asn Phe Tyr Leu
100 105 110
Tyr Leu Arg Ala Arg Val Cys Glu Asn Cys Met Glu Met Asp Val Met
115 120 125
Ser Val Ala Thr Ile Val Ile Val Asp Ile Cys Ile Thr Gly Gly Leu
130 135 140
Leu Leu Leu Val Tyr Tyr Trp Ser Lys Asn Arg Lys Ala Lys Ala Lys
145 150 155 160
Pro Val Thr Arg Gly Ala Gly Ala Gly Gly Arg Gln Arg Gly Gln Asn
165 170 175
Lys Glu Arg Pro Pro Pro Val Pro Asn Pro Asp Tyr Glu Pro Ile Arg
180 185 190
Lys Gly Gln Arg Asp Leu Tyr Ser Gly Leu Asn Gln Arg Arg Ile
195 200 205
<210> 20
<211> 4
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 20
Gly Gly Gly Ser
1
<210> 21
<211> 4
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 21
Ser Gly Gly Gly
1
<210> 22
<211> 5
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 22
Gly Gly Gly Gly Ser
1 5
<210> 23
<211> 5
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 23
Ser Gly Gly Gly Gly
1 5
<210> 24
<211> 6
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 24
Gly Gly Gly Gly Gly Ser
1 5
<210> 25
<211> 6
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 25
Ser Gly Gly Gly Gly Gly
1 5
<210> 26
<211> 7
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 26
Gly Gly Gly Gly Gly Gly Ser
1 5
<210> 27
<211> 7
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 27
Ser Gly Gly Gly Gly Gly Gly
1 5
<210> 28
<211> 113
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 28
Gln Val Gln Leu Lys Glu Ala Gly Pro Gly Leu Val Gln Pro Thr Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Ser Asp
20 25 30
Gly Val His Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Tyr Tyr Asp Gly Gly Thr Asp Tyr Asn Ser Ala Ile Lys
50 55 60
Ser Arg Leu Ser Ile Ser Arg Asp Thr Ser Lys Ser Gln Val Phe Leu
65 70 75 80
Lys Ile Asn Ser Leu Gln Thr Asp Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Arg Ile His Phe Asp Tyr Trp Gly Gln Gly Val Met Val Thr Val Ser
100 105 110
Ser
<210> 29
<211> 437
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 29
Gln Val Gln Leu Lys Glu Ala Gly Pro Gly Leu Val Gln Pro Thr Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Ser Asp
20 25 30
Gly Val His Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Tyr Tyr Asp Gly Gly Thr Asp Tyr Asn Ser Ala Ile Lys
50 55 60
Ser Arg Leu Ser Ile Ser Arg Asp Thr Ser Lys Ser Gln Val Phe Leu
65 70 75 80
Lys Ile Asn Ser Leu Gln Thr Asp Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Arg Ile His Phe Asp Tyr Trp Gly Gln Gly Val Met Val Thr Val Ser
100 105 110
Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro Gly Ser
115 120 125
Ala Ala Gln Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val Lys Gly
130 135 140
Tyr Phe Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser Leu Ser
145 150 155 160
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr
165 170 175
Leu Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp Pro Ser Glu Thr
180 185 190
Val Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys
195 200 205
Lys Ile Val Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys Thr Val
210 215 220
Pro Glu Val Ser Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Val
225 230 235 240
Leu Thr Ile Thr Leu Thr Pro Lys Val Thr Cys Val Val Val Asp Ile
245 250 255
Ser Lys Asp Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val
260 265 270
Glu Val His Thr Ala Gln Thr Gln Pro Arg Glu Glu Gln Phe Asn Ser
275 280 285
Thr Phe Arg Ser Val Ser Glu Leu Pro Ile Met His Gln Asp Trp Leu
290 295 300
Asn Gly Lys Glu Phe Lys Cys Arg Val Asn Ser Ala Ala Phe Pro Ala
305 310 315 320
Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys Ala Pro
325 330 335
Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys Asp Lys
340 345 350
Val Ser Leu Thr Cys Met Ile Thr Asp Phe Phe Pro Glu Asp Ile Thr
355 360 365
Val Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys Asn Thr
370 375 380
Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe Val Tyr Ser Lys Leu
385 390 395 400
Asn Val Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr Cys Ser
405 410 415
Val Leu His Glu Gly Leu His Asn His His Thr Glu Lys Ser Leu Ser
420 425 430
His Ser Pro Gly Lys
435
<210> 30
<211> 437
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 30
Gln Val Gln Leu Lys Glu Ala Gly Pro Gly Leu Val Gln Pro Thr Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Ser Asp
20 25 30
Gly Val His Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Tyr Tyr Asp Gly Gly Thr Asp Tyr Asn Ser Ala Ile Lys
50 55 60
Ser Arg Leu Ser Ile Ser Arg Asp Thr Ser Lys Ser Gln Val Phe Leu
65 70 75 80
Lys Ile Asn Ser Leu Gln Thr Asp Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Arg Ile His Phe Asp Tyr Trp Gly Gln Gly Val Met Val Thr Val Ser
100 105 110
Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro Gly Ser
115 120 125
Ala Ala Gln Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val Lys Gly
130 135 140
Tyr Phe Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser Leu Ser
145 150 155 160
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr
165 170 175
Leu Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp Pro Ser Glu Thr
180 185 190
Val Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys
195 200 205
Lys Ile Val Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys Thr Val
210 215 220
Lys Glu Val Ser Lys Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Val
225 230 235 240
Leu Thr Ile Thr Leu Thr Pro Lys Val Thr Cys Val Val Val Asp Ile
245 250 255
Ser Lys Asp Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val
260 265 270
Glu Val His Thr Ala Gln Thr Gln Pro Arg Glu Glu Gln Phe Asn Ser
275 280 285
Thr Phe Arg Ser Val Ser Glu Leu Pro Ile Met His Gln Asp Trp Leu
290 295 300
Asn Gly Lys Glu Phe Lys Cys Arg Val Asn Ser Ala Ala Phe Pro Ala
305 310 315 320
Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys Ala Pro
325 330 335
Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys Asp Lys
340 345 350
Val Ser Leu Thr Cys Met Ile Thr Asp Phe Phe Pro Glu Asp Ile Thr
355 360 365
Val Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys Asn Thr
370 375 380
Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe Val Tyr Ser Lys Leu
385 390 395 400
Asn Val Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr Cys Ser
405 410 415
Val Leu His Glu Gly Leu His Asn His His Thr Glu Lys Ser Leu Ser
420 425 430
His Ser Pro Gly Lys
435
<210> 31
<211> 437
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 31
Gln Val Gln Leu Lys Glu Ala Gly Pro Gly Leu Val Gln Pro Thr Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Ser Asp
20 25 30
Gly Val His Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Tyr Tyr Asp Gly Gly Thr Asp Tyr Asn Ser Ala Ile Lys
50 55 60
Ser Arg Leu Ser Ile Ser Arg Asp Thr Ser Lys Ser Gln Val Phe Leu
65 70 75 80
Lys Ile Asn Ser Leu Gln Thr Asp Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Arg Ile His Phe Asp Tyr Trp Gly Gln Gly Val Met Val Thr Val Ser
100 105 110
Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro Gly Ser
115 120 125
Ala Ala Gln Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val Lys Gly
130 135 140
Tyr Phe Pro Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser Leu Ser
145 150 155 160
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr
165 170 175
Leu Ser Ser Ser Val Thr Val Pro Ser Ser Thr Trp Pro Ser Glu Thr
180 185 190
Val Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys
195 200 205
Lys Ile Val Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys Glu Pro
210 215 220
Asn Glu Val Glu Asp Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Val
225 230 235 240
Leu Thr Ile Thr Leu Thr Pro Lys Val Thr Cys Val Val Val Asp Ile
245 250 255
Ser Lys Asp Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val
260 265 270
Glu Val His Thr Ala Gln Thr Gln Pro Arg Glu Glu Gln Phe Asn Ser
275 280 285
Thr Phe Arg Ser Val Ser Glu Leu Pro Ile Met His Gln Asp Trp Leu
290 295 300
Asn Gly Lys Glu Phe Lys Cys Arg Val Asp Ser Ala Ala Phe Pro Ala
305 310 315 320
Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys Ala Pro
325 330 335
Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys Asp Lys
340 345 350
Val Ser Leu Thr Cys Met Ile Thr Asp Phe Phe Pro Glu Asp Ile Thr
355 360 365
Val Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys Asn Thr
370 375 380
Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr Phe Val Tyr Ser Lys Leu
385 390 395 400
Asn Val Gln Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr Cys Ser
405 410 415
Val Leu His Glu Gly Leu His Asn His His Thr Glu Lys Ser Leu Ser
420 425 430
His Ser Pro Gly Lys
435
<210> 32
<211> 213
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 32
Asp Ile Val Leu Thr Gln Ser Pro Thr Thr Ile Ala Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Ile Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Tyr Trp Tyr Gln Gln Lys Ser Gly Ala Ser Pro Lys Leu Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Asn Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Ala Ile Asn Thr Met Glu Thr Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Thr Pro Leu Thr
85 90 95
Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Arg Ala Asp Ala Ala Pro
100 105 110
Thr Val Ser Ile Phe Pro Pro Ser Ser Glu Gln Leu Thr Ser Gly Gly
115 120 125
Ala Ser Val Val Cys Phe Leu Asn Asn Phe Tyr Pro Lys Asp Ile Asn
130 135 140
Val Lys Trp Lys Ile Asp Gly Ser Glu Arg Gln Asn Gly Val Leu Asn
145 150 155 160
Ser Trp Thr Asp Gln Asp Ser Lys Asp Ser Thr Tyr Ser Met Ser Ser
165 170 175
Thr Leu Thr Leu Thr Lys Asp Glu Tyr Glu Arg His Asn Ser Tyr Thr
180 185 190
Cys Glu Ala Thr His Lys Thr Ser Thr Ser Pro Ile Val Lys Ser Phe
195 200 205
Asn Arg Asn Glu Cys
210
<210> 33
<211> 119
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 33
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 34
<211> 443
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 34
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr
115 120 125
Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met Val Thr Leu
130 135 140
Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Val Thr Trp
145 150 155 160
Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Pro Ser Ser
180 185 190
Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val Ala His Pro Ala Ser
195 200 205
Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys Gly Cys Lys
210 215 220
Pro Cys Ile Cys Thr Val Pro Glu Val Ser Ser Val Phe Ile Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro Lys Val Thr
245 250 255
Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val Gln Phe Ser
260 265 270
Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr Gln Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu Leu Pro Ile
290 295 300
Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg Val Asn
305 310 315 320
Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys
325 330 335
Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu
340 345 350
Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile Thr Asp Phe
355 360 365
Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly Gln Pro Ala
370 375 380
Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr
385 390 395 400
Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp Glu Ala Gly
405 410 415
Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu His Asn His His
420 425 430
Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
435 440
<210> 35
<211> 443
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 35
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly His Ser Ile Ser His Asp
20 25 30
His Ala Trp Ser Trp Val Arg Gln Pro Pro Gly Glu Gly Leu Glu Trp
35 40 45
Ile Gly Phe Ile Ser Tyr Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu
50 55 60
Gln Gly Arg Val Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Glu Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr
115 120 125
Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met Val Thr Leu
130 135 140
Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Val Thr Trp
145 150 155 160
Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Pro Ser Ser
180 185 190
Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val Ala His Pro Ala Ser
195 200 205
Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys Gly Cys Lys
210 215 220
Pro Cys Ile Cys Glu Pro Asn Glu Val Glu Asp Val Phe Ile Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro Lys Val Thr
245 250 255
Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val Gln Phe Ser
260 265 270
Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr Gln Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu Leu Pro Ile
290 295 300
Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg Val Asp
305 310 315 320
Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys
325 330 335
Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu
340 345 350
Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile Thr Asp Phe
355 360 365
Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly Gln Pro Ala
370 375 380
Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr
385 390 395 400
Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp Glu Ala Gly
405 410 415
Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu His Asn His His
420 425 430
Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
435 440
<210> 36
<211> 214
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 36
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 37
<211> 443
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 37
Asp Val Gln Leu Gln Glu Ser Gly Pro Val Leu Val Lys Pro Ser Gln
1 5 10 15
Ser Leu Ser Leu Thr Cys Thr Val Thr Gly Tyr Ser Ile Thr Ser Asp
20 25 30
His Ala Trp Ser Trp Ile Arg Gln Phe Pro Gly Asn Lys Leu Glu Trp
35 40 45
Met Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Thr Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Ile Ser Ile Thr Arg Asp Thr Ser Lys Asn Gln Phe Phe
65 70 75 80
Leu Gln Leu Asn Ser Val Thr Thr Gly Asp Thr Ser Thr Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Ser Val Thr Val Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr
115 120 125
Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met Val Thr Leu
130 135 140
Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Val Thr Trp
145 150 155 160
Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Pro Ser Ser
180 185 190
Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val Ala His Pro Ala Ser
195 200 205
Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys Gly Cys Lys
210 215 220
Pro Cys Ile Cys Thr Val Pro Glu Val Ser Ser Val Phe Ile Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro Lys Val Thr
245 250 255
Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val Gln Phe Ser
260 265 270
Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr Gln Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu Leu Pro Ile
290 295 300
Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg Val Asn
305 310 315 320
Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys
325 330 335
Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu
340 345 350
Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile Thr Asp Phe
355 360 365
Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly Gln Pro Ala
370 375 380
Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr
385 390 395 400
Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp Glu Ala Gly
405 410 415
Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu His Asn His His
420 425 430
Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
435 440
<210> 38
<211> 443
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 38
Asp Val Gln Leu Gln Glu Ser Gly Pro Val Leu Val Lys Pro Ser Gln
1 5 10 15
Ser Leu Ser Leu Thr Cys Thr Val Thr Gly Tyr Ser Ile Thr Ser Asp
20 25 30
His Ala Trp Ser Trp Ile Arg Gln Phe Pro Gly Asn Lys Leu Glu Trp
35 40 45
Met Gly Tyr Ile Ser Tyr Ser Gly Ile Thr Thr Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Ile Ser Ile Thr Arg Asp Thr Ser Lys Asn Gln Phe Phe
65 70 75 80
Leu Gln Leu Asn Ser Val Thr Thr Gly Asp Thr Ser Thr Tyr Tyr Cys
85 90 95
Ala Arg Ser Leu Ala Arg Thr Thr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Ser Val Thr Val Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr
115 120 125
Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met Val Thr Leu
130 135 140
Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Val Thr Trp
145 150 155 160
Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Pro Ser Ser
180 185 190
Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val Ala His Pro Ala Ser
195 200 205
Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys Gly Cys Lys
210 215 220
Pro Cys Ile Cys Thr Val Lys Glu Val Ser Lys Val Phe Ile Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro Lys Val Thr
245 250 255
Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val Gln Phe Ser
260 265 270
Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr Gln Pro Arg
275 280 285
Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu Leu Pro Ile
290 295 300
Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg Val Asn
305 310 315 320
Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys
325 330 335
Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu
340 345 350
Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile Thr Asp Phe
355 360 365
Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly Gln Pro Ala
370 375 380
Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp Gly Ser Tyr
385 390 395 400
Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp Glu Ala Gly
405 410 415
Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu His Asn His His
420 425 430
Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
435 440
<210> 39
<211> 450
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 39
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Arg Asn Thr Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala
35 40 45
Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Asn
85 90 95
Thr His Val Pro Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro His His His His His His
435 440 445
His His
450
<210> 40
<211> 446
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 40
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Arg Asn Thr Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala
35 40 45
Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Asn
85 90 95
Thr His Val Pro Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr
180 185 190
Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Thr Val Glu Arg Lys Ser Cys Val Glu Cys Pro Pro Cys Pro
210 215 220
Ala Pro Pro Ala Ala Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
225 230 235 240
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
245 250 255
Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val
260 265 270
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
275 280 285
Phe Ala Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln
290 295 300
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly
305 310 315 320
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro
325 330 335
Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
340 345 350
Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser
355 360 365
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
370 375 380
Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
385 390 395 400
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
405 410 415
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
420 425 430
Ser Leu Ser Leu Ser Pro His His His His His His His His
435 440 445
<210> 41
<211> 447
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 41
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Arg Asn Thr Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala
35 40 45
Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Asn
85 90 95
Thr His Val Pro Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys
115 120 125
Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
210 215 220
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
225 230 235 240
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
245 250 255
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
260 265 270
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
275 280 285
Gln Phe Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
290 295 300
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
305 310 315 320
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
325 330 335
Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Gln Glu Glu Met
340 345 350
Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro
355 360 365
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
370 375 380
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
385 390 395 400
Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val
405 410 415
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
420 425 430
Lys Ser Leu Ser Leu Ser Leu His His His His His His His His
435 440 445
<210> 42
<211> 449
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 42
Gln Val Gln Leu Lys Glu Ala Gly Pro Gly Leu Val Gln Pro Thr Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Ser Asp
20 25 30
Gly Val His Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Tyr Tyr Asp Gly Gly Thr Asp Tyr Asn Ser Ala Ile Lys
50 55 60
Ser Arg Leu Ser Ile Ser Arg Asp Thr Ser Lys Ser Gln Val Phe Leu
65 70 75 80
Lys Ile Asn Ser Leu Gln Thr Asp Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Arg Ile His Phe Asp Tyr Trp Gly Gln Gly Val Met Val Thr Val Ser
100 105 110
Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser
115 120 125
Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
130 135 140
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
145 150 155 160
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
165 170 175
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln
180 185 190
Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp
195 200 205
Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Cys
340 345 350
Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Asp Tyr Lys Asp Asp Asp Asp
435 440 445
Lys
<210> 43
<211> 445
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 43
Gln Val Gln Leu Lys Glu Ala Gly Pro Gly Leu Val Gln Pro Thr Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Ser Asp
20 25 30
Gly Val His Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Tyr Tyr Asp Gly Gly Thr Asp Tyr Asn Ser Ala Ile Lys
50 55 60
Ser Arg Leu Ser Ile Ser Arg Asp Thr Ser Lys Ser Gln Val Phe Leu
65 70 75 80
Lys Ile Asn Ser Leu Gln Thr Asp Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Arg Ile His Phe Asp Tyr Trp Gly Gln Gly Val Met Val Thr Val Ser
100 105 110
Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser
115 120 125
Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
130 135 140
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
145 150 155 160
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
165 170 175
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln
180 185 190
Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp
195 200 205
Lys Thr Val Glu Arg Lys Ser Cys Val Glu Cys Pro Pro Cys Pro Ala
210 215 220
Pro Pro Ala Ala Ala Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
225 230 235 240
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
245 250 255
Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
260 265 270
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
275 280 285
Ala Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp
290 295 300
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
305 310 315 320
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg
325 330 335
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Cys Glu Met Thr Lys
340 345 350
Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp
355 360 365
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
370 375 380
Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser
385 390 395 400
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
405 410 415
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
420 425 430
Leu Ser Leu Ser Pro Asp Tyr Lys Asp Asp Asp Asp Lys
435 440 445
<210> 44
<211> 446
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 44
Gln Val Gln Leu Lys Glu Ala Gly Pro Gly Leu Val Gln Pro Thr Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Ser Asp
20 25 30
Gly Val His Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Tyr Tyr Asp Gly Gly Thr Asp Tyr Asn Ser Ala Ile Lys
50 55 60
Ser Arg Leu Ser Ile Ser Arg Asp Thr Ser Lys Ser Gln Val Phe Leu
65 70 75 80
Lys Ile Asn Ser Leu Gln Thr Asp Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Arg Ile His Phe Asp Tyr Trp Gly Gln Gly Val Met Val Thr Val Ser
100 105 110
Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser
115 120 125
Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
130 135 140
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
145 150 155 160
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
165 170 175
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys
180 185 190
Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp
195 200 205
Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala
210 215 220
Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
225 230 235 240
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
245 250 255
Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val
260 265 270
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
275 280 285
Phe Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
290 295 300
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly
305 310 315 320
Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
325 330 335
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Cys Glu Met Thr
340 345 350
Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser
355 360 365
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
370 375 380
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val
385 390 395 400
Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe
405 410 415
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
420 425 430
Ser Leu Ser Leu Ser Leu Asp Tyr Lys Asp Asp Asp Asp Lys
435 440 445
<210> 45
<211> 222
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 45
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Ile Arg Gln Pro Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
115 120 125
Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
130 135 140
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
145 150 155 160
Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser
165 170 175
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala
180 185 190
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
195 200 205
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<210> 46
<211> 213
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 46
Asp Ile Val Leu Thr Gln Ser Pro Thr Thr Ile Ala Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Ile Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Tyr Trp Tyr Gln Gln Lys Ser Gly Ala Ser Pro Lys Leu Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Asn Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Ala Ile Asn Thr Met Glu Thr Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Thr Pro Leu Thr
85 90 95
Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 47
<211> 441
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 47
Gln Val Gln Leu Lys Glu Ala Gly Pro Gly Leu Val Gln Pro Thr Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Ser Asp
20 25 30
Gly Val His Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Tyr Tyr Asp Gly Gly Thr Asp Tyr Asn Ser Ala Ile Lys
50 55 60
Ser Arg Leu Ser Ile Ser Arg Asp Thr Ser Lys Ser Gln Val Phe Leu
65 70 75 80
Lys Ile Asn Ser Leu Gln Thr Asp Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Arg Ile His Phe Asp Tyr Trp Gly Gln Gly Val Met Val Thr Val Ser
100 105 110
Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser
115 120 125
Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
130 135 140
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
145 150 155 160
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
165 170 175
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln
180 185 190
Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp
195 200 205
Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Cys
340 345 350
Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440
<210> 48
<211> 450
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 48
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Arg Asn Thr Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala
35 40 45
Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Asn
85 90 95
Thr His Val Pro Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro His His His His His His
435 440 445
His His
450
<210> 49
<211> 116
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 49
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Lys
1 5 10 15
Ser Leu Lys Leu Ser Cys Glu Ala Ser Gly Phe Thr Phe Ser Gly Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Arg Gly Leu Glu Ser Val
35 40 45
Ala Tyr Ile Thr Ser Ser Ser Ile Asn Ile Lys Tyr Ala Asp Ala Val
50 55 60
Lys Gly Arg Phe Thr Val Ser Arg Asp Asn Ala Lys Asn Leu Leu Phe
65 70 75 80
Leu Gln Met Asn Ile Leu Lys Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Phe Asp Trp Asp Lys Asn Tyr Trp Gly Gln Gly Thr Met Val
100 105 110
Thr Val Ser Ser
115
<210> 50
<211> 444
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 50
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Lys
1 5 10 15
Ser Leu Lys Leu Ser Cys Glu Ala Ser Gly Phe Thr Phe Ser Gly Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Arg Gly Leu Glu Ser Val
35 40 45
Ala Tyr Ile Thr Ser Ser Ser Ile Asn Ile Lys Tyr Ala Asp Ala Val
50 55 60
Lys Gly Arg Phe Thr Val Ser Arg Asp Asn Ala Lys Asn Leu Leu Phe
65 70 75 80
Leu Gln Met Asn Ile Leu Lys Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Phe Asp Trp Asp Lys Asn Tyr Trp Gly Gln Gly Thr Met Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Cys Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440
<210> 51
<211> 214
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 51
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Pro Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Asn Cys Gln Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Asn Lys Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Arg Asp Ser Ser Phe Thr Ile Ser Ser Leu Glu Ser
65 70 75 80
Glu Asp Ile Gly Ser Tyr Tyr Cys Gln Gln Tyr Tyr Asn Tyr Pro Trp
85 90 95
Thr Phe Gly Pro Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 52
<211> 115
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 52
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Ile Arg Gln Pro Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 53
<211> 443
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 53
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Ile Arg Gln Pro Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
130 135 140
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
180 185 190
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
195 200 205
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440
<210> 54
<211> 219
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 54
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Arg Asn Thr Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala
35 40 45
Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Asn
85 90 95
Thr His Val Pro Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 55
<211> 20
<212> ДНК
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 55
cgcaacgcaa ttaatgtgag 20
<210> 56
<211> 21
<212> ДНК
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 56
tgagttccac gacaccgtca c 21
<210> 57
<211> 18
<212> ДНК
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 57
gcgtcacact ttgctatg 18
<210> 58
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 58
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Arg Gly Gly Pro Lys Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Lys Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Tyr Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 59
<211> 107
<212> PRT
<213> Homo sapiens
<400> 59
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 60
<211> 106
<212> PRT
<213> Homo sapiens
<400> 60
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 61
<211> 120
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 61
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Thr His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ala Pro Glu Tyr Ser Ser Ser Ser Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 62
<211> 124
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 62
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Ser
20 25 30
Ser Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile His Tyr Thr Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Leu Asn Pro Ser Ile Ala Ala Arg Pro Gly Ala Phe Asp
100 105 110
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 63
<211> 120
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 63
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Thr His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ala Pro Glu Tyr Ser Ser Ser Ser Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 64
<211> 120
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 64
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Ser Gly Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Asn Ser Gly Asp Thr Asn Tyr Ala Gln Asn Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Thr Gly Tyr Ser Ser Ser Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Pro Val Thr Val Ser Ser
115 120
<210> 65
<211> 117
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 65
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Asn Ser Gly Gly Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Thr Gly Asp Leu Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 66
<211> 119
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 66
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Tyr Ser Tyr Met Leu Arg Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 67
<211> 120
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 67
Gln Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Arg Ala Met Tyr His Pro Ser Phe Asn Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 68
<211> 124
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 68
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser Ser Gly
20 25 30
Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Ser Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln His Ser Phe Thr Arg Tyr Tyr Gly Tyr Tyr Tyr Phe Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 69
<211> 120
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 69
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Lys Trp Tyr Tyr Tyr Pro Ala Leu Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 70
<211> 118
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 70
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Asn Ser Gly Gly Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Asp Tyr Trp Met Leu Pro Asp Val Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 71
<211> 120
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 71
Gln Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Lys Trp Tyr Tyr Tyr Pro Ala Leu Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 72
<211> 119
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 72
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Leu Ser Tyr Trp Met Lys Trp Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 73
<211> 123
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 73
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Asp Ile Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Pro Ser Gly Gln Ser Thr Tyr Tyr Arg Arg Glu Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Gly Arg Glu Tyr Gly Gly Gly Trp Tyr Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 74
<211> 120
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 74
Gln Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Lys Trp Tyr Tyr Tyr Pro Ala Leu Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 75
<211> 119
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 75
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Tyr Tyr Tyr Tyr Tyr Lys Met Asp Val Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 76
<211> 119
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 76
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Pro Ser Tyr Tyr Pro Trp Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 77
<211> 118
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 77
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Asn Ser Gly Gly Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Asn Tyr Tyr Met Phe Met Asp Leu Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Tyr Ser
115
<210> 78
<211> 119
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 78
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Arg Tyr Gly Leu His Met Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 79
<211> 127
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 79
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Thr Lys Ser Ser Lys Ser Arg Gly Lys Arg Tyr Tyr Arg
100 105 110
Tyr Met Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 80
<211> 111
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 80
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Val Ser Asn Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ser
85 90 95
Ser Thr Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 81
<211> 110
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 81
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Ala Ala Pro Gly Gln
1 5 10 15
Lys Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Ala Asn Asn
20 25 30
Phe Val Ser Trp Phe Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asn Lys Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Thr Leu Gly Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Arg Ser Leu
85 90 95
Ser Ala Gly Val Phe Gly Gly Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 82
<211> 111
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 82
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Phe Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Val Ser Lys Arg Pro Ser Gly Val Ser Thr Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Asn Ser Tyr Thr Thr Ser
85 90 95
Ser Thr Arg Trp Met Phe Gly Gly Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 83
<211> 111
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 83
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Val Ser Asn Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ser
85 90 95
Ser Thr Arg Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 84
<211> 111
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 84
Gln Ser Ala Leu Thr Gln Pro Arg Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Val Tyr
20 25 30
Asp Tyr Val Ser Trp Tyr Gln Gln Phe Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Val Asn Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Cys Ser Tyr Ser Gly Thr
85 90 95
Asp Ile Arg Trp Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 85
<211> 107
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 85
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Phe His Ser Phe Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 86
<211> 107
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 86
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Phe Asn Ser Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 87
<211> 107
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 87
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Ser Asp Ser Ile Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 88
<211> 107
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 88
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Phe Tyr Ser Phe Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 89
<211> 107
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 89
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Phe Asp Asn Phe Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 90
<211> 107
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 90
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Phe Ser Arg Phe Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 91
<211> 107
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 91
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Phe Asn Gly Phe Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 92
<211> 107
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 92
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Phe Ser Arg Phe Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 93
<211> 107
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 93
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Val Tyr Arg Ser Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 94
<211> 107
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 94
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Phe Val Arg His Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 95
<211> 107
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 95
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Phe Asn Asn Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 96
<211> 107
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 96
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Phe Thr Ser Phe Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 97
<211> 107
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 97
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Phe Asn Ser Phe Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 98
<211> 107
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 98
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Val Tyr Ser Ser Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 99
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 99
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Glu His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 100
<211> 399
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 100
Leu Gln Asp Pro Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Asn Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser
20 25 30
Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly Val
35 40 45
Phe Arg Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp
50 55 60
Cys Thr Pro Gly Phe His Cys Leu Gly Ala Gly Cys Ser Met Cys Glu
65 70 75 80
Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys Asp
85 90 95
Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg Pro
100 105 110
Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly Thr
115 120 125
Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser Pro
130 135 140
Gly Ala Ser Ser Val Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly His
145 150 155 160
Ser Pro Gln Asp Ile Glu Gly Arg Met Asp Pro Lys Ser Cys Asp Lys
165 170 175
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
180 185 190
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
195 200 205
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
210 215 220
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
225 230 235 240
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
245 250 255
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
260 265 270
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
275 280 285
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
290 295 300
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
305 310 315 320
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
325 330 335
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
340 345 350
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
355 360 365
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
370 375 380
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
385 390 395
<210> 101
<211> 449
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 101
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Gly Gly Tyr Val Thr Tyr Asn Pro Ser Leu Glu
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro
<210> 102
<211> 216
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 102
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Ala Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 103
<211> 444
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 103
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Ser Thr Tyr
20 25 30
Trp Ile Ser Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Lys Ile Tyr Pro Gly Asp Ser Tyr Thr Asn Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Tyr Gly Ile Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Glu His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440
<210> 104
<211> 214
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 104
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Asn Ile Gly Asp Gln Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Lys Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Tyr Thr Gly Phe Gly Ser Leu
85 90 95
Ala Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gln Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 105
<211> 163
<212> PRT
<213> Homo sapiens
<400> 105
Leu Gln Asp Pro Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Asn Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser
20 25 30
Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly Val
35 40 45
Phe Arg Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp
50 55 60
Cys Thr Pro Gly Phe His Cys Leu Gly Ala Gly Cys Ser Met Cys Glu
65 70 75 80
Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys Asp
85 90 95
Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg Pro
100 105 110
Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly Thr
115 120 125
Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser Pro
130 135 140
Gly Ala Ser Ser Val Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly His
145 150 155 160
Ser Pro Gln
<210> 106
<211> 39
<212> PRT
<213> Homo sapiens
<400> 106
Leu Gln Asp Pro Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Asn Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser
20 25 30
Ala Gly Gly Gln Arg Thr Cys
35
<210> 107
<211> 41
<212> PRT
<213> Homo sapiens
<400> 107
Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser Ala Gly Gly Gln Arg Thr
1 5 10 15
Cys Asp Ile Cys Arg Gln Cys Lys Gly Val Phe Arg Thr Arg Lys Glu
20 25 30
Cys Ser Ser Thr Ser Asn Ala Glu Cys
35 40
<210> 108
<211> 31
<212> PRT
<213> Homo sapiens
<400> 108
Asp Cys Thr Pro Gly Phe His Cys Leu Gly Ala Gly Cys Ser Met Cys
1 5 10 15
Glu Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys
20 25 30
<210> 109
<211> 69
<212> PRT
<213> Homo sapiens
<400> 109
Lys Asp Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys
1 5 10 15
Arg Pro Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn
20 25 30
Gly Thr Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu
35 40 45
Ser Pro Gly Ala Ser Ser Val Thr Pro Pro Ala Pro Ala Arg Glu Pro
50 55 60
Gly His Ser Pro Gln
65
<210> 110
<211> 94
<212> PRT
<213> Homo sapiens
<400> 110
Leu Gln Asp Pro Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Asn Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser
20 25 30
Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly Val
35 40 45
Phe Arg Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp
50 55 60
Cys Thr Pro Gly Phe His Cys Leu Gly Ala Gly Cys Ser Met Cys Glu
65 70 75 80
Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys
85 90
<210> 111
<211> 63
<212> PRT
<213> Homo sapiens
<400> 111
Leu Gln Asp Pro Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Asn Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser
20 25 30
Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly Val
35 40 45
Phe Arg Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys
50 55 60
<210> 112
<211> 141
<212> PRT
<213> Homo sapiens
<400> 112
Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser Ala Gly Gly Gln Arg Thr
1 5 10 15
Cys Asp Ile Cys Arg Gln Cys Lys Gly Val Phe Arg Thr Arg Lys Glu
20 25 30
Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp Cys Thr Pro Gly Phe His
35 40 45
Cys Leu Gly Ala Gly Cys Ser Met Cys Glu Gln Asp Cys Lys Gln Gly
50 55 60
Gln Glu Leu Thr Lys Lys Gly Cys Lys Asp Cys Cys Phe Gly Thr Phe
65 70 75 80
Asn Asp Gln Lys Arg Gly Ile Cys Arg Pro Trp Thr Asn Cys Ser Leu
85 90 95
Asp Gly Lys Ser Val Leu Val Asn Gly Thr Lys Glu Arg Asp Val Val
100 105 110
Cys Gly Pro Ser Pro Ala Asp Leu Ser Pro Gly Ala Ser Ser Val Thr
115 120 125
Pro Pro Ala Pro Ala Arg Glu Pro Gly His Ser Pro Gln
130 135 140
<210> 113
<211> 72
<212> PRT
<213> Homo sapiens
<400> 113
Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser Ala Gly Gly Gln Arg Thr
1 5 10 15
Cys Asp Ile Cys Arg Gln Cys Lys Gly Val Phe Arg Thr Arg Lys Glu
20 25 30
Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp Cys Thr Pro Gly Phe His
35 40 45
Cys Leu Gly Ala Gly Cys Ser Met Cys Glu Gln Asp Cys Lys Gln Gly
50 55 60
Gln Glu Leu Thr Lys Lys Gly Cys
65 70
<210> 114
<211> 100
<212> PRT
<213> Homo sapiens
<400> 114
Asp Cys Thr Pro Gly Phe His Cys Leu Gly Ala Gly Cys Ser Met Cys
1 5 10 15
Glu Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys
20 25 30
Asp Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg
35 40 45
Pro Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly
50 55 60
Thr Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser
65 70 75 80
Pro Gly Ala Ser Ser Val Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly
85 90 95
His Ser Pro Gln
100
<210> 115
<211> 115
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 115
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Glu Met His Trp Ile Arg Gln Pro Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Asp Pro Lys Thr Gly Asp Thr Ala Tyr Ser Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Tyr Ser Tyr Thr Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 116
<211> 112
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 116
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Arg Asn Thr Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala
35 40 45
Pro Arg Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Asn
85 90 95
Thr His Val Pro Pro Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 117
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 117
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Arg Gly Gly Pro Lys Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Tyr Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Glu Ser Leu Ser Leu Ser Pro
325
<210> 118
<211> 107
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 118
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 119
<211> 328
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 119
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Arg Gly Gly Pro Lys Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Lys Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Tyr Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Arg Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro
325
<210> 120
<211> 116
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 120
Glu Val Gln Val Val Glu Ser Asp Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Lys Leu Pro Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ala Trp Val Arg Gln Ala Pro Thr Lys Gly Leu Glu Trp Val
35 40 45
Ala Ser Ile Ser Tyr Asp Gly Ser Ser Thr Tyr Tyr Arg Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Gly Arg His Ser Ser Tyr Phe Asp Tyr Trp Gly Gln Gly Val Met Val
100 105 110
Thr Val Ser Ser
115
<210> 121
<211> 106
<212> PRT
<213> Искусственная
<220>
<223> искусственно синтезированная последовательность
<400> 121
Asp Ile Glu Leu Thr Gln Ser Pro Ala Leu Ala Val Ser Pro Gly Glu
1 5 10 15
Arg Val Thr Ile Ser Cys Arg Ala Ser Asp Ser Val Ser Thr Leu Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Gln Pro Lys Leu Leu Ile Tyr
35 40 45
Leu Ala Ser His Leu Glu Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asp Pro Val Glu Ala Asp
65 70 75 80
Asp Thr Ala Thr Tyr Tyr Cys Gln Gln Ser Trp Asn Asp Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 122
<211> 121
<212> PRT
<213> Искусственная последовательность
<220>
<223> искусственно синтезированная последовательность
<400> 122
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Gly Gly Tyr Val Thr Tyr Asn Pro Ser Leu Glu
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 123
<211> 109
<212> PRT
<213> Искусственная последовательность
<220>
<223> искусственно синтезированная последовательность
<400> 123
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Ala Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 124
<211> 120
<212> PRT
<213> Искусственная последовательность
<220>
<223> искусственно синтезированная последовательность
<400> 124
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Thr His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ala Pro Glu Tyr Ser Ser Ser Ser Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 125
<211> 117
<212> PRT
<213> Искусственная последовательность
<220>
<223> искусственно синтезированная последовательность
<400> 125
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Asn Ser Gly Gly Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Ile Gly Asn His Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЕ СЛИТЫЕ БЕЛКИ, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ CD137 И GPC3 | 2020 |
|
RU2814653C2 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2022 |
|
RU2824285C2 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2015 |
|
RU2743464C2 |
| СЛИТЫЙ ПОЛИПЕПТИД С ПРОТИВОРАКОВОЙ АКТИВНОСТЬЮ | 2016 |
|
RU2754466C2 |
| АНТИ-CD137 АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2829536C2 |
| БЕЛОК, СВЯЗЫВАЮЩИЙСЯ С NKG2D, CD16 И С ОПУХОЛЕСПЕЦИФИЧЕСКИМ АНТИГЕНОМ | 2018 |
|
RU2788531C2 |
| НОВЫЙ СЛИТЫЙ БЕЛОК, СПЕЦИФИЧНЫЙ ДЛЯ CD137 И PD-L1 | 2019 |
|
RU2818349C2 |
| АНТИТЕЛО ПРОТИВ ГЛИПИКАНА-3 И ЕГО ПРИМЕНЕНИЕ | 2016 |
|
RU2744245C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПОЛИПЕПТИДНОГО ГЕТЕРОМУЛЬТИМЕРА | 2014 |
|
RU2758952C1 |
| СПОСОБ ПОЛУЧЕНИЯ ПОЛИПЕПТИДНОГО ГЕТЕРОМУЛЬТИМЕРА | 2014 |
|
RU2730594C2 |
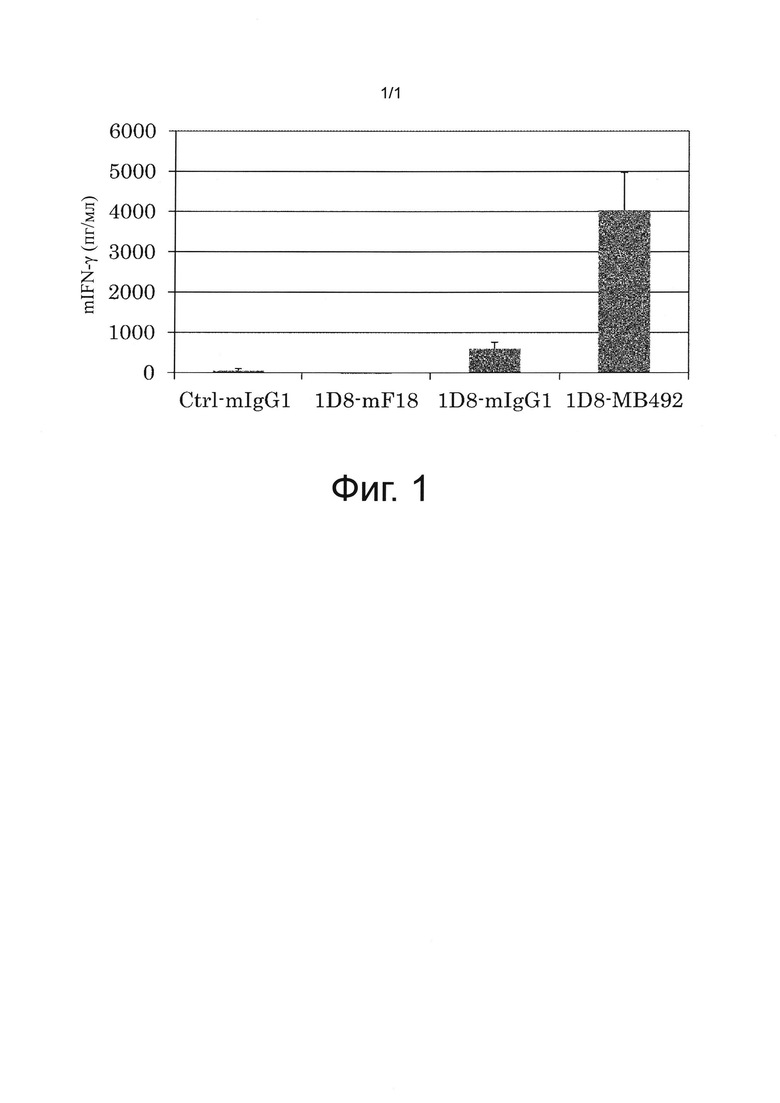
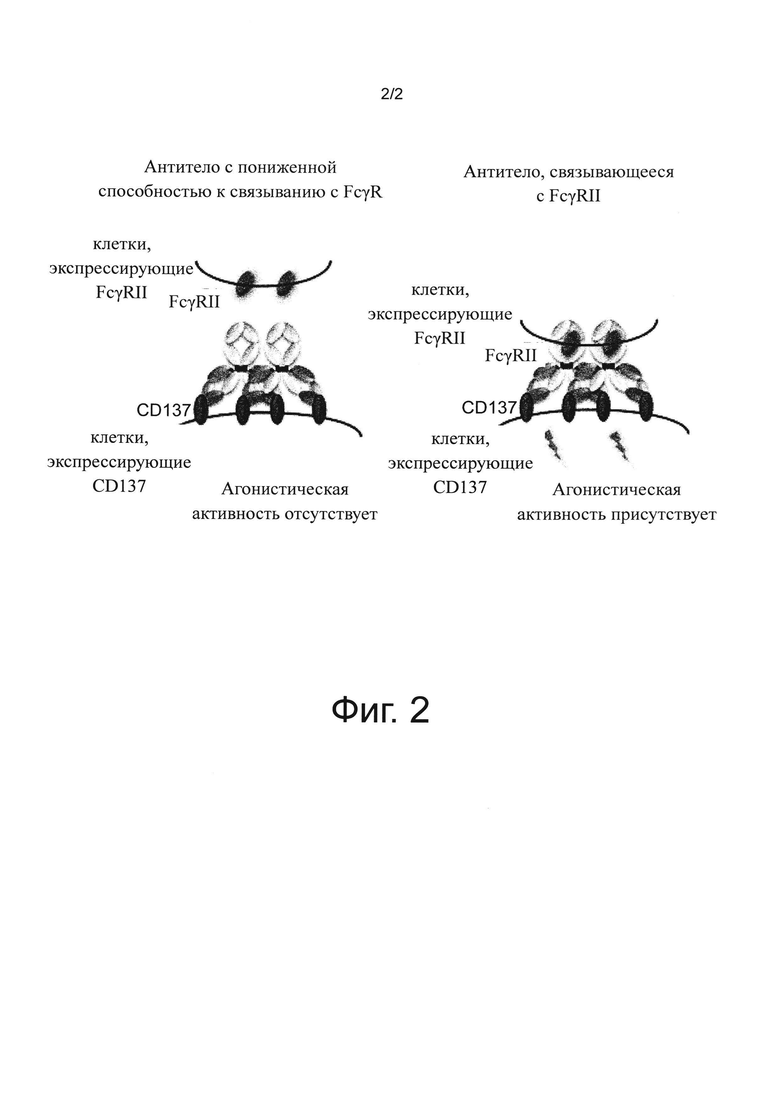
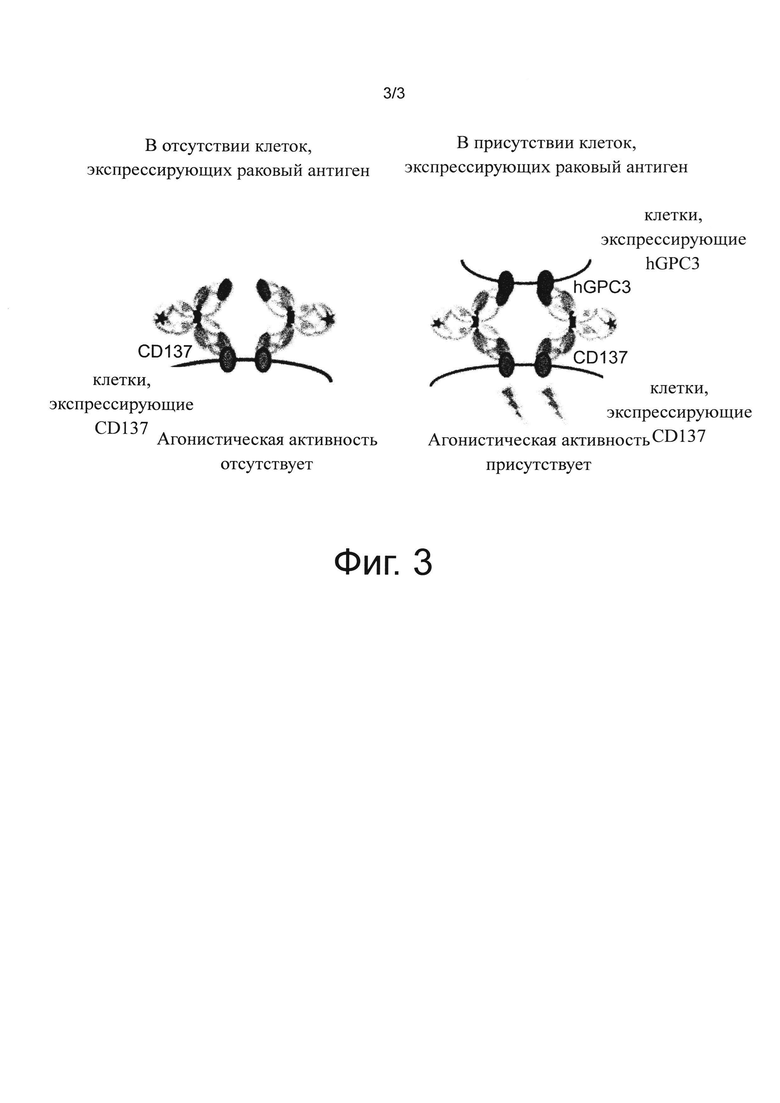
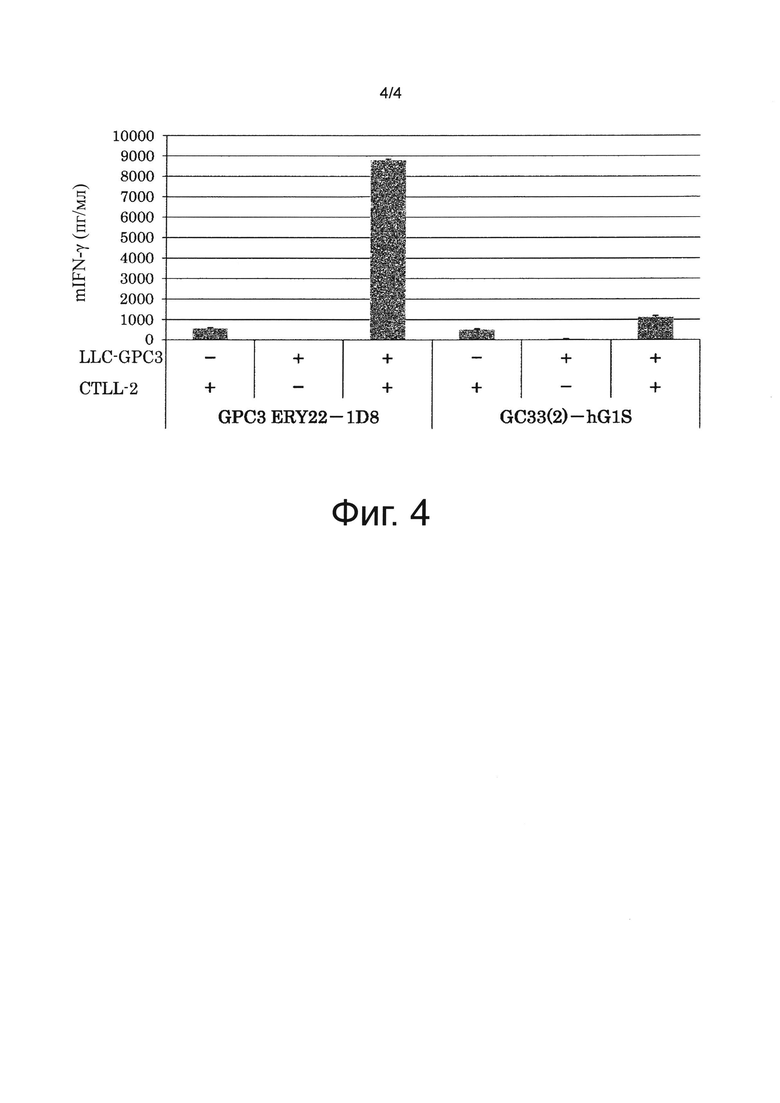
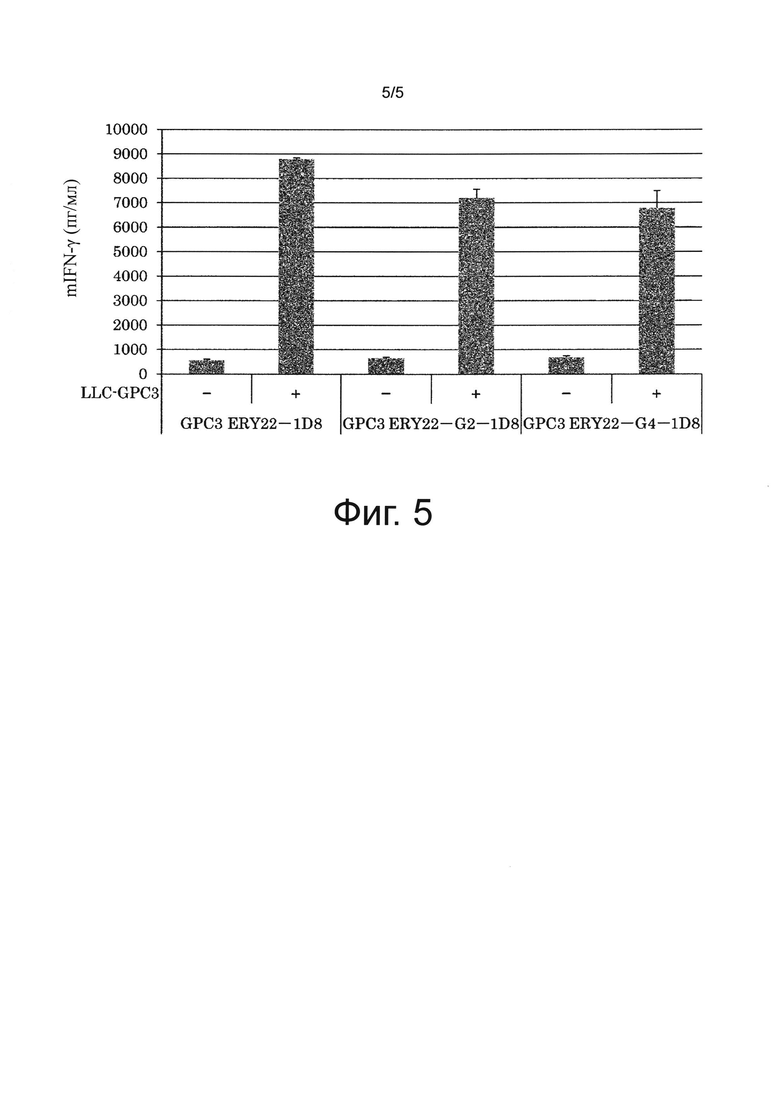
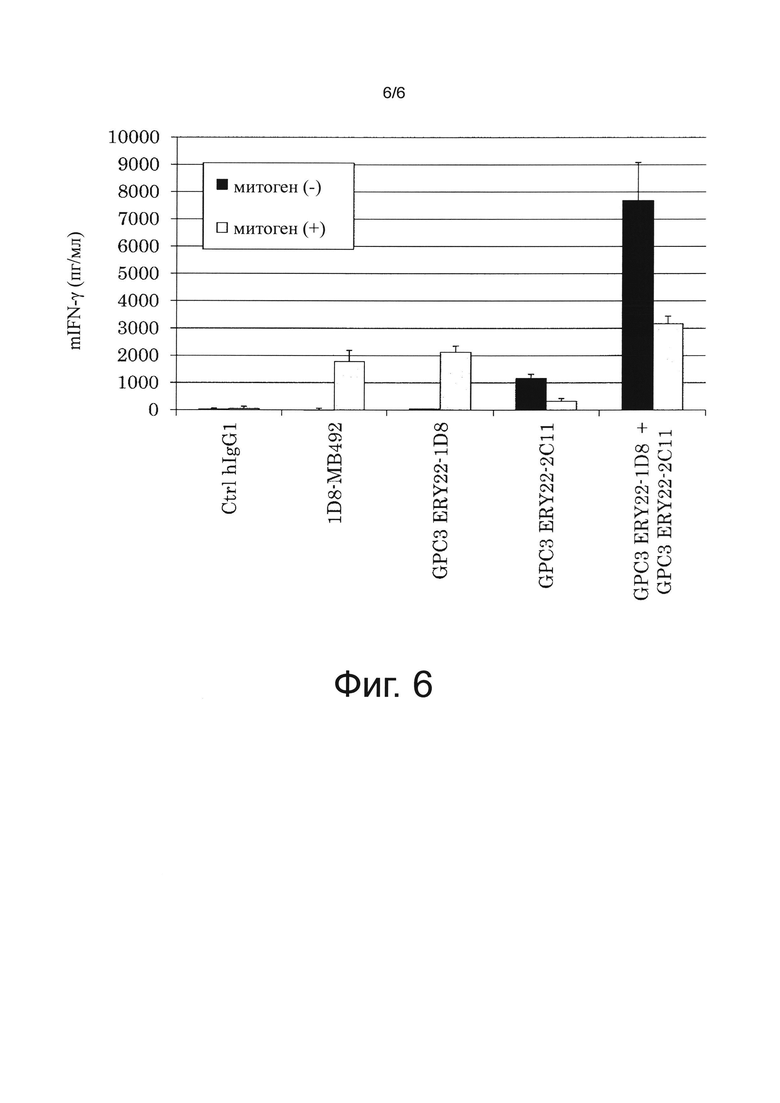
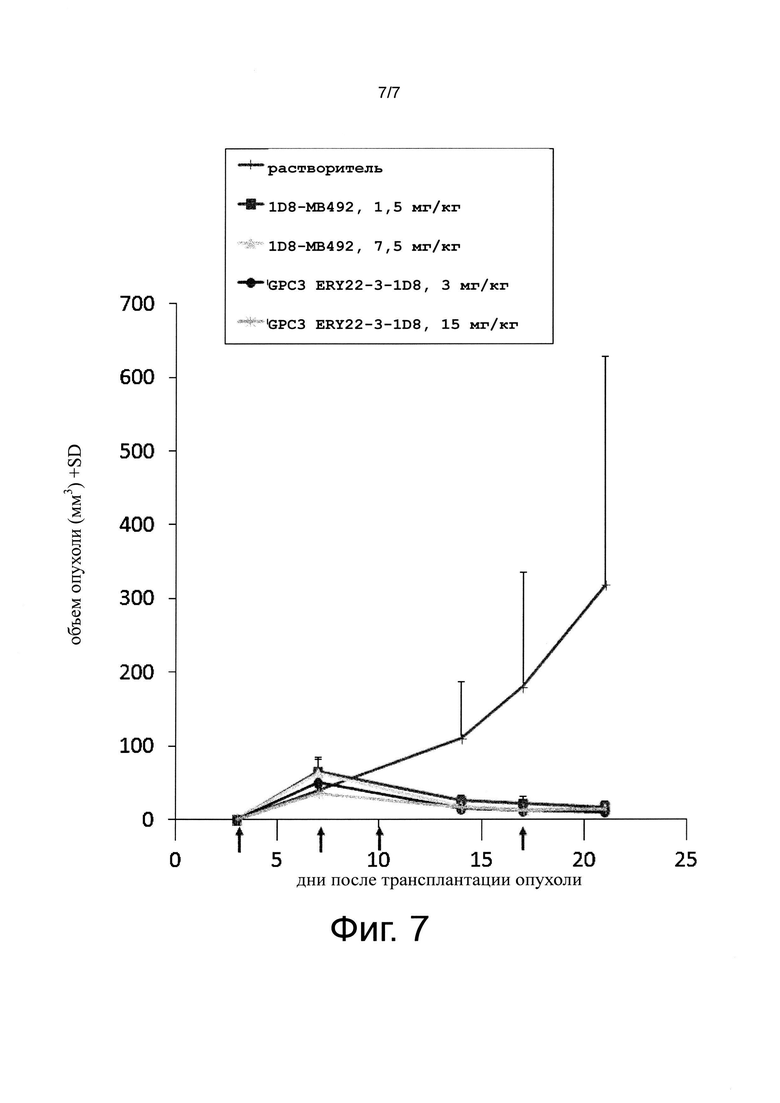
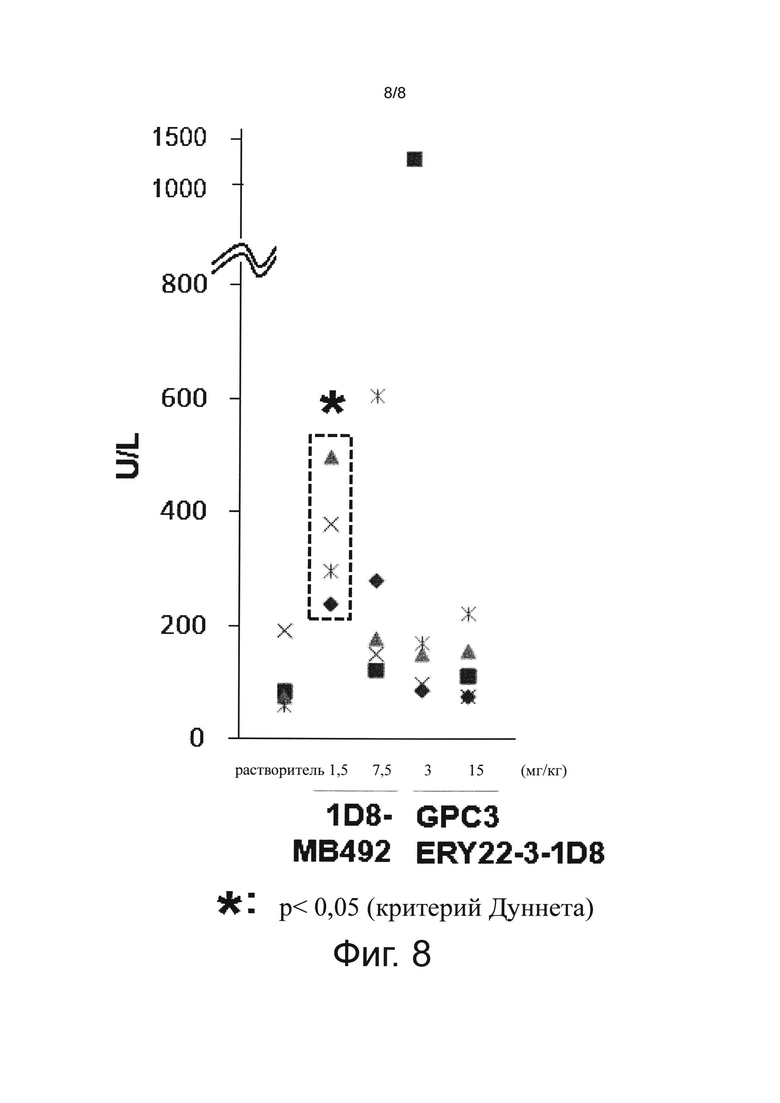
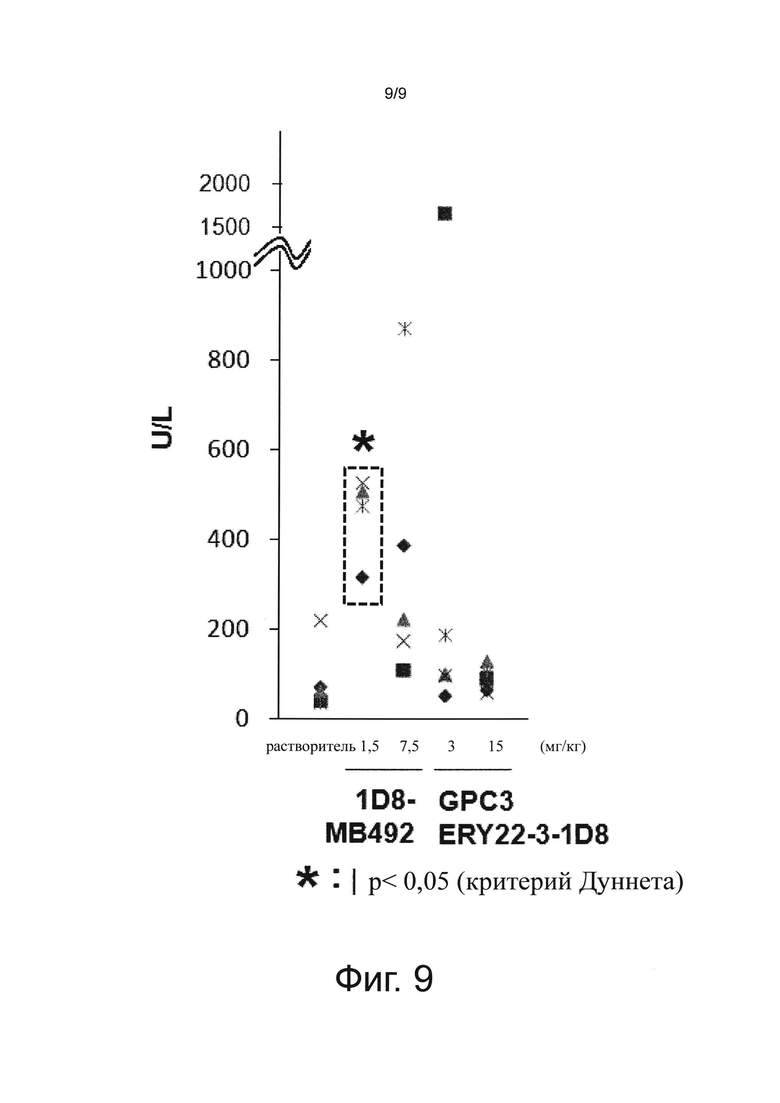
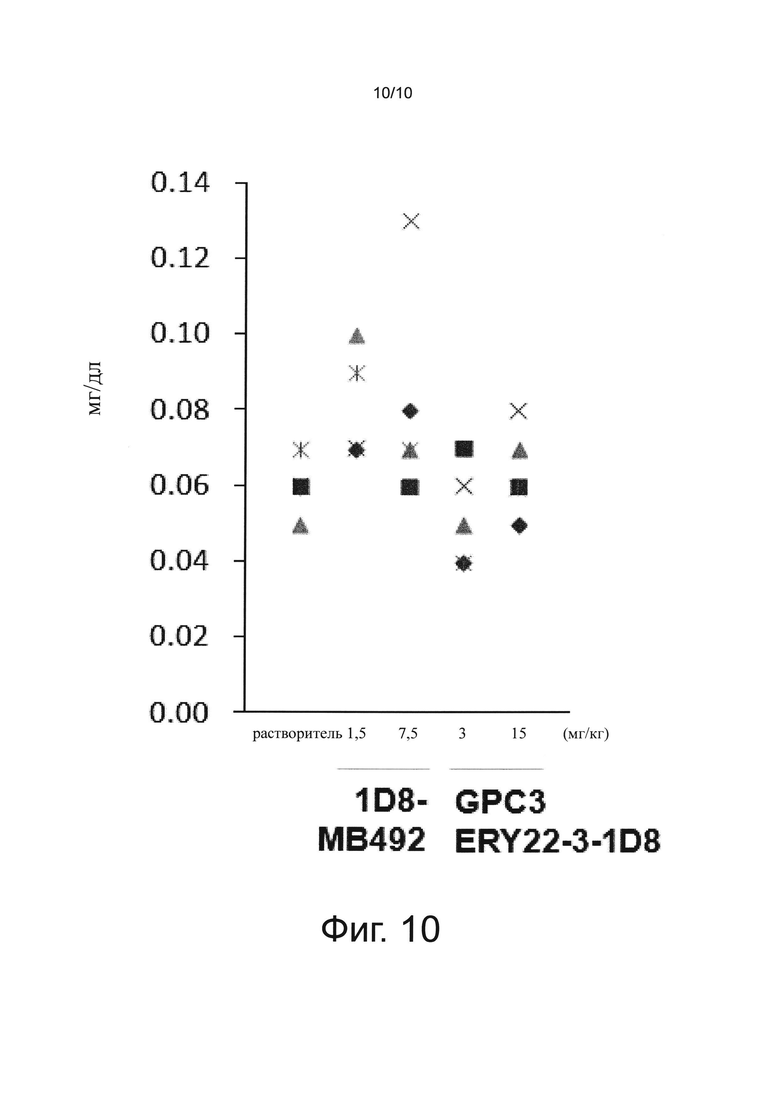
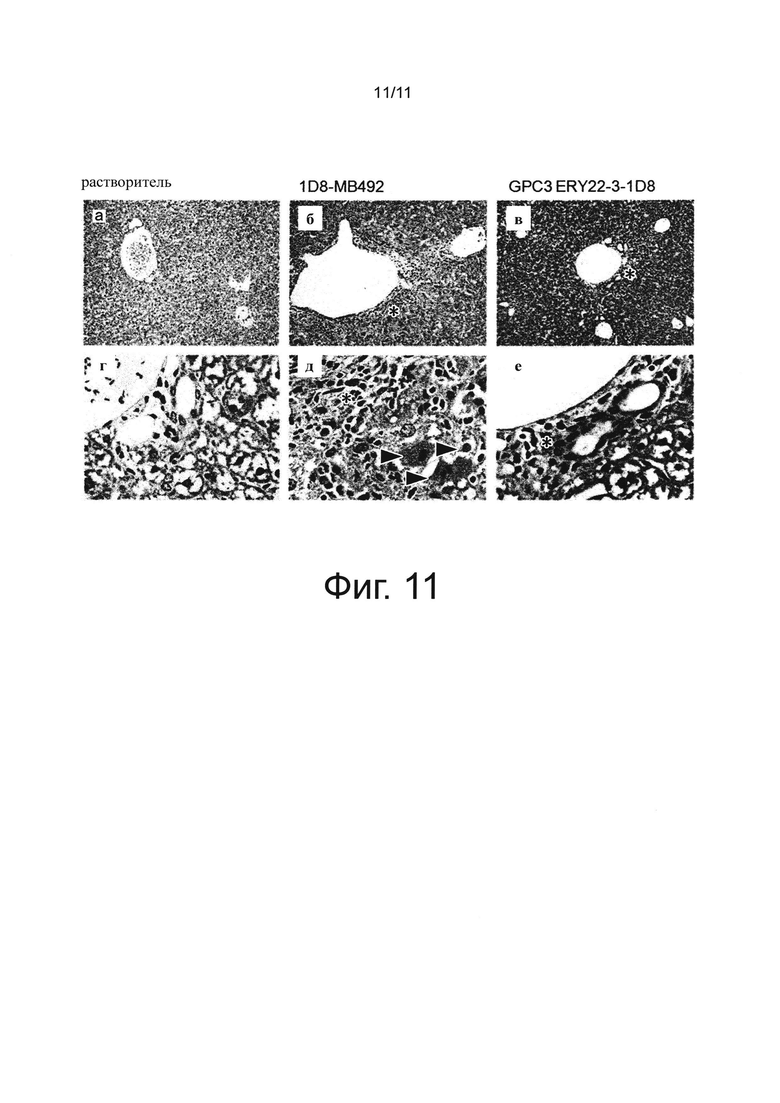
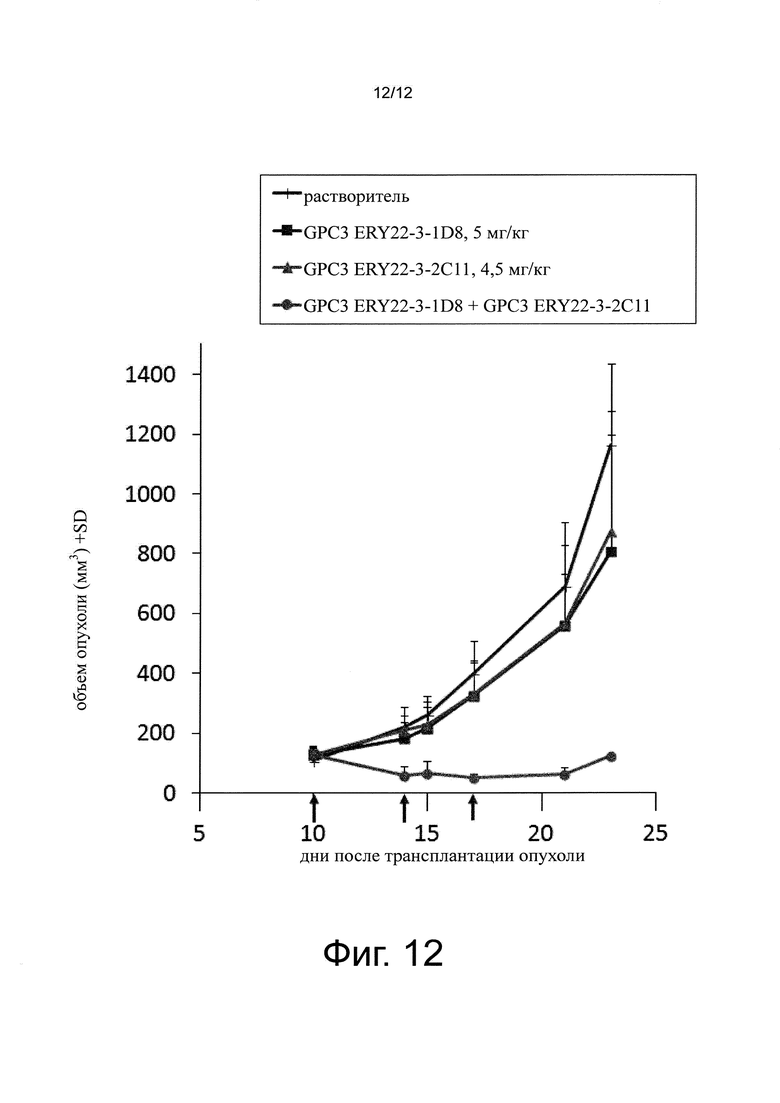
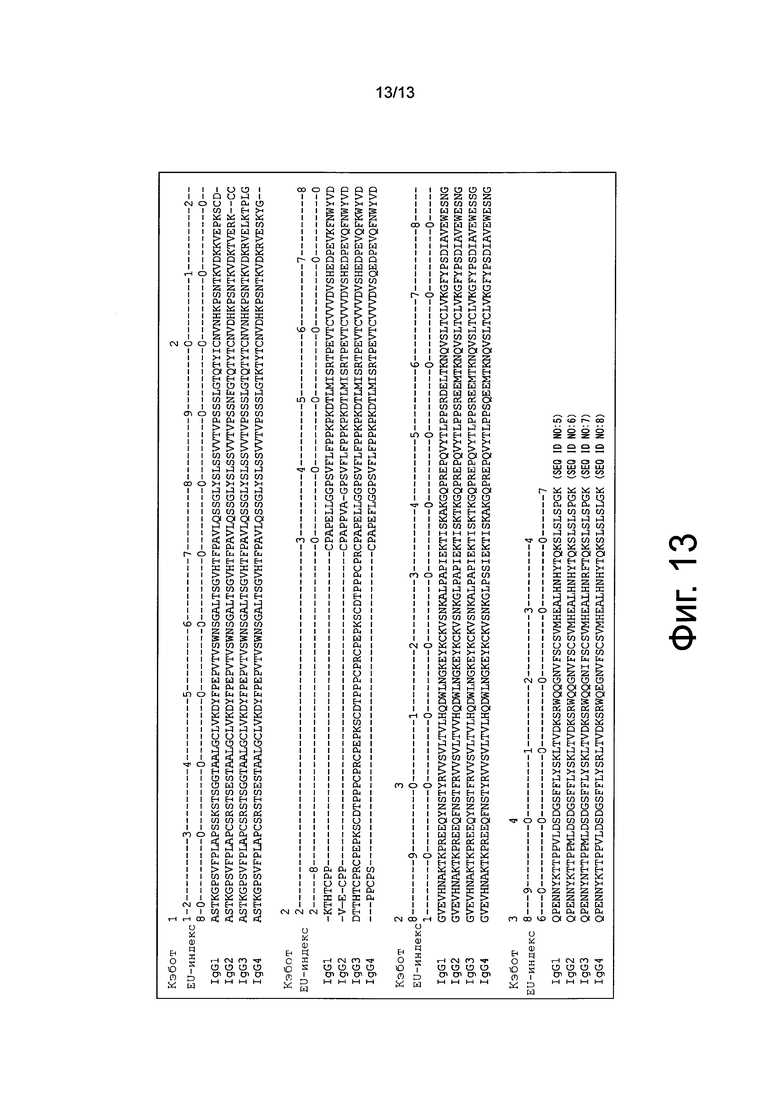
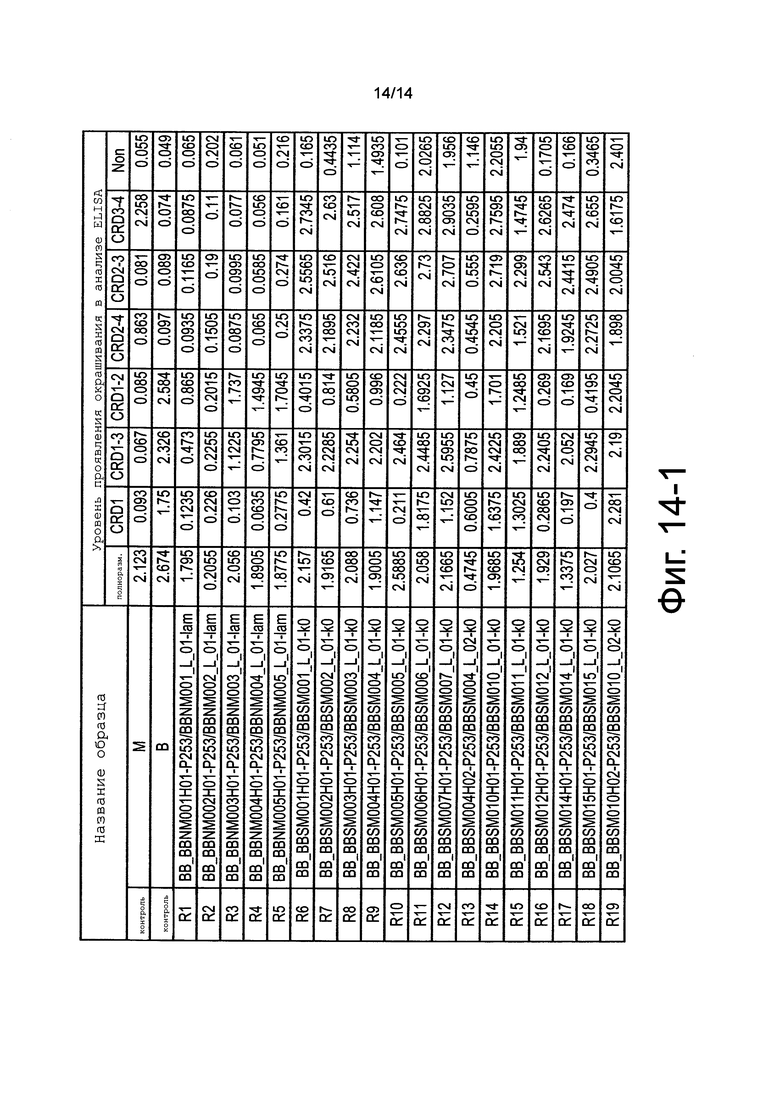
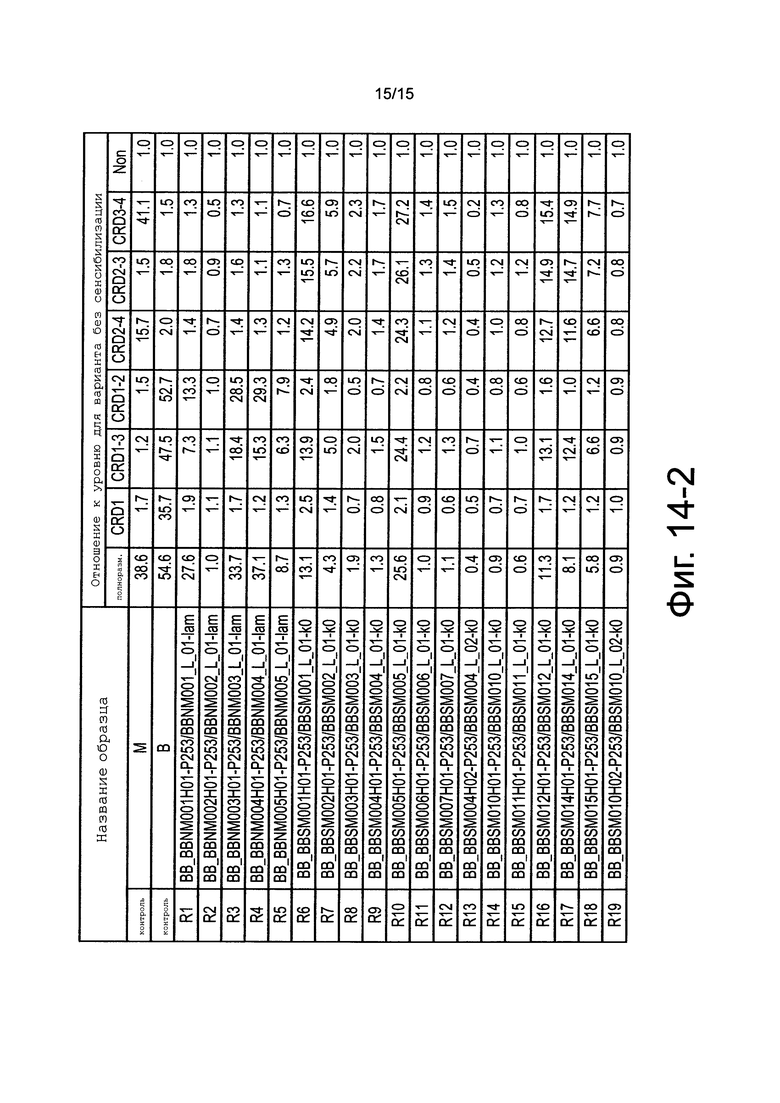
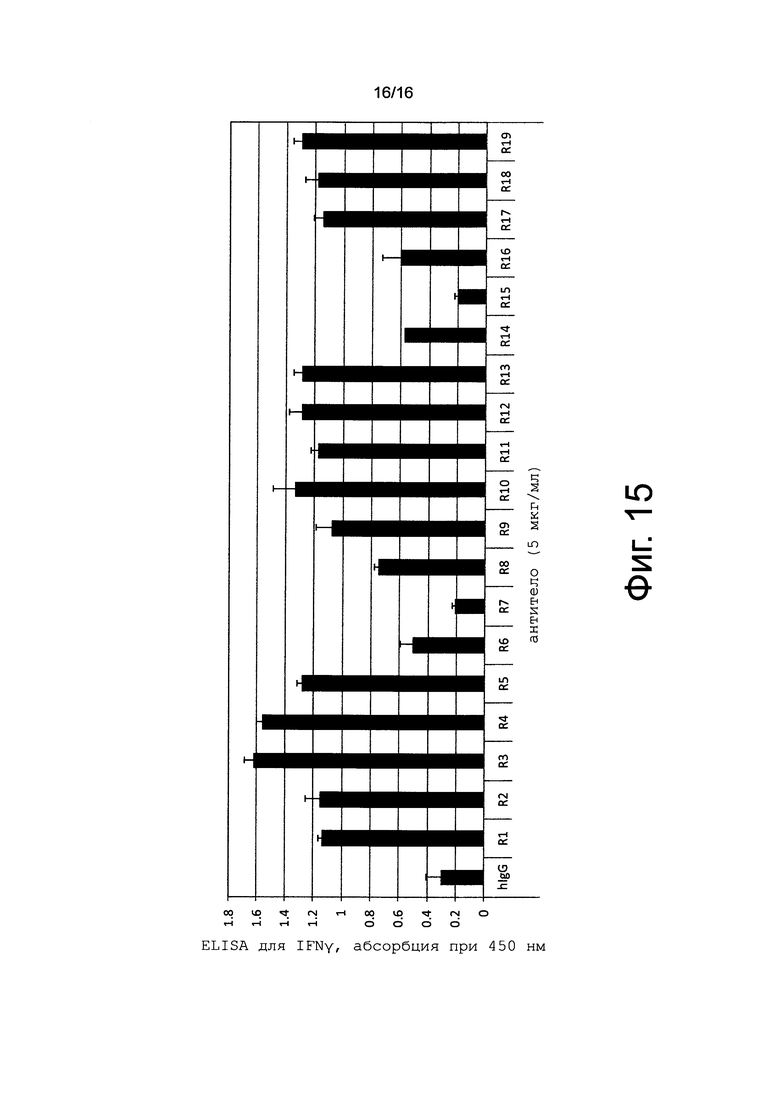
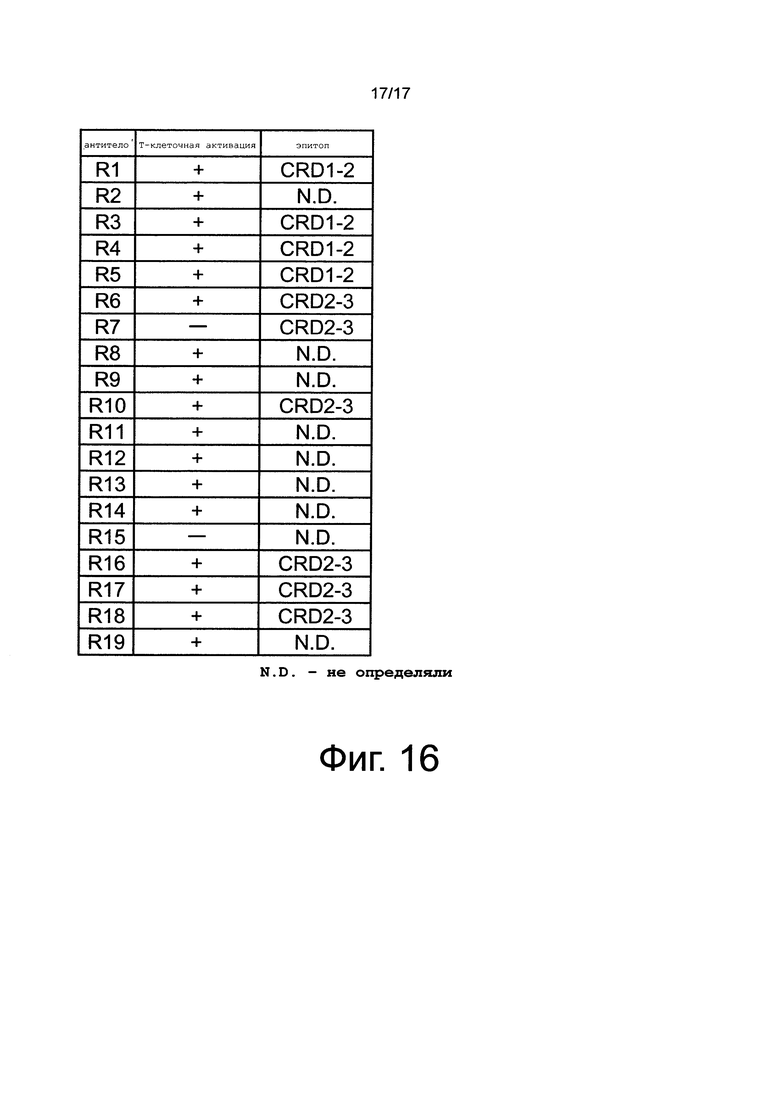
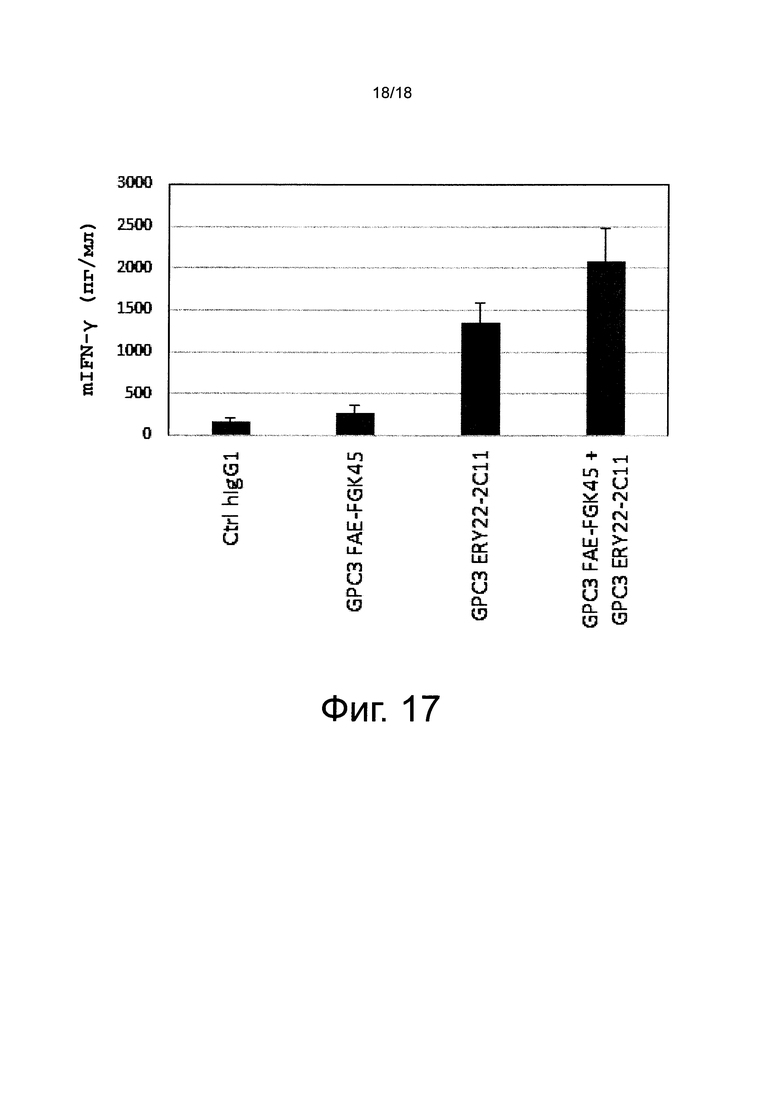

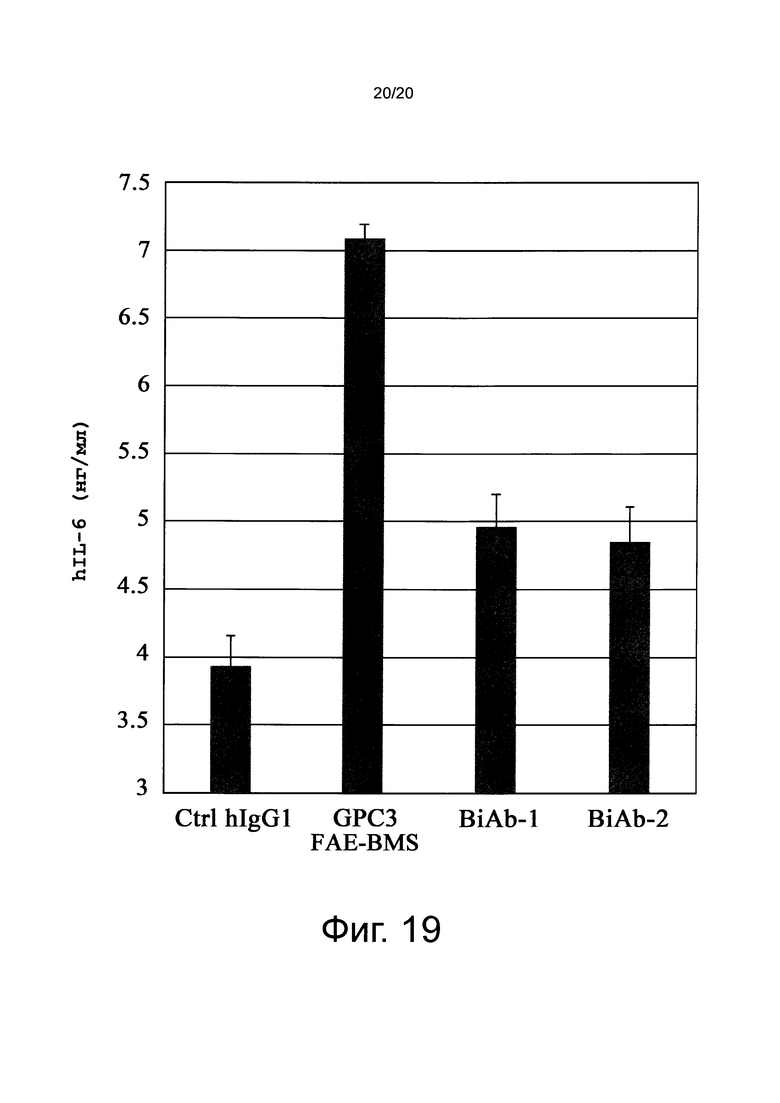
Изобретение относится биотехнологии. Описано антитело для лечения рака, содержащее: (1) специфический для рака антигенсвязывающий Fab домен; (2) CD137-связывающий Fab домен и (3) FcRn-связывающий домен, в котором FcRn-связывающий домен представляет собой Fc-область антитела с пониженной активностью связывания с Fcγ-рецептором. Также описана фармацевтическая композиция, содержащая указанное антитело. Изобретение предназначено для рака. Изобретение приводит к активации иммунных клеток и тем самым приводит к противоопухолевой активности при отсутствии побочных действий, таких как гепатотоксичность. 5 н. и 10 з.п. ф-лы, 19 ил., 9 табл., 11 пр.
1. Антитело для лечения рака, содержащее:
(1) специфический для рака антигенсвязывающий Fab домен;
(2) CD137-связывающий Fab домен и
(3) FcRn-связывающий домен, в котором FcRn-связывающий домен представляет собой Fc-область антитела с пониженной активностью связывания с Fcγ-рецептором.
2. Фармацевтическая композиция для лечения рака, содержащая в качестве действующего вещества антитело по п. 1 и фармацевтически приемлемые носители и/или добавки.
3. Фармацевтическая композиция по п. 2, которая представляет собой индуцирующую цитотоксичность композицию.
4. Фармацевтическая комбинация для лечения рака, содержащая первое антитело по п. 1 и второе антитело, которое содержит:
(1) домен, связывающийся со специфическим для рака антигеном;
(2) CD3-связывающий домен и
(3) FcRn-связывающий домен, который представляет собой Fc-область антитела с пониженной активностью связывания с Fcγ-рецептором.
5. Фармацевтическая комбинация по п. 4, в которой первое антитело и второе антитело присутствуют в смеси.
6. Фармацевтическая комбинация по п. 4, в которой первое антитело и второе антитело предназначены для совместного применения.
7. Фармацевтическая комбинация по п. 4, в которой первое антитело и второе антитело предназначены для одновременного применения.
8. Фармацевтическая комбинация по п. 4, в которой первое антитело и второе антитело предназначены для раздельного применения.
9. Фармацевтическая комбинация по любому из пп. 4-8, которая представляет собой индуцирующую цитотоксичность комбинацию.
10. Фармацевтическая композиция для лечения рака, содержащая в качестве действующего вещества антитело по п. 1 и антитело, которое содержит:
(1) домен, связывающийся со специфическим для рака антигеном;
(2) CD3-связывающий домен и
(3) FcRn-связывающий домен, который представляет собой Fc-область антитела с пониженной активностью связывания с Fcγ-рецептором, а также фармацевтически приемлемые носители.
11. Фармацевтическая композиция по п. 10, которая представляет собой индуцирующую цитотоксичность композицию.
12. Применение антитела по п. 1 и антитела, которое содержит:
(1) домен, связывающийся со специфическим для рака антигеном;
(2) CD3-связывающий домен и
(3) FcRn-связывающий домен, который представляет собой Fc-область антитела с пониженной активностью связывания с Fcγ-рецептором, в комбинированной терапии рака.
13. Применение по п. 12, при котором антитела вводят совместно.
14. Применение по п. 12, при котором антитела вводят одновременно.
15. Применение по п. 12, при котором антитела вводят раздельно.
| WO 2013070468 A1, 16.05.2013 | |||
| US 20120076727 A1, 29.03.2012 | |||
| WO 2009126920 A2, 15.10.2009 | |||
| US 20110059076 A1, 10.03.2011 | |||
| US 20090274649 A1, 05.11.2009 | |||
| US 8080250 B1, 20.12.2011 | |||
| ОПТИМИЗИРОВАННЫЕ Fc-ВАРИАНТЫ, ИМЕЮЩИЕ ИЗМЕНЕННОЕ СВЯЗЫВАНИЕ С FcγR, И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2004 |
|
RU2337107C2 |
| WO 2008051424 A2, 02.05.2008 | |||
| WO 2010037395 A2, 08.04.2010. | |||

