Настоящее изобретение относится к вакцинам для животных, более конкретно, относится к экзосомам, выделенным из животного, у которого нет патогена, вызывающего заболевание, к способу их получения, к фармацевтической или ветеринарной композиции, содержащей такую вакцину, и к применению для предотвращения или профилактики инфекционных заболеваний, например, инфекционного заболевания, вызванного вирусом репродуктивно-респираторного синдрома свиней (PRRSV).
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
В настоящее время существует обширная область исследований, направленная на поиск альтернативных стратегий разработки новых вакцин, которые могут решать многие существующие медицинские проблемы. Новым подходом в этой области являются вакцины на основе экзосом (Chaput & Théry. Exosomes: immune properties and potential clinical implementations. 2011. Semi. Immunopathol., 33:419-440).
Экзосомы представляют собой везикулы размером менее 300 нм, которые образуются путем слияния цитоплазматических мультивезикулярных тел с клеточной мембраной и последующего их высвобождения во внеклеточную среду в виде внеклеточных везикул. Первоначально экзосомы были описаны в тестах, проведенных с ретикулоцитами, клетками-предшественниками эритроцитов. В этих пионерских работах исследовался их биогенез, и было сделано предложение, чтобы экзосомы являются селективными молекулярными отходами плазматической мембраны, полученными в процессе дифференцировки ретикулоцитов в зрелые эритроциты. Однако, десять лет спустя было показано, что экзосомы не являются уникальными для ретикулоцитов, что их можно получать из других типов клеток и что они могут играть модулирующую роль в иммунном ответе (Raposo G. et al., ʺB lymphocytes secrete antigen-presenting vesiclesʺ, J. Exp. Med., 1996, том 183, стр.1161-1172). Так, Raposo и его коллеги продемонстрировали в своих работах с В-лимфоцитами, что эти клетки также продуцируют экзосомы с теми же физическими и химическими свойствами, как и у экзосом, полученных из ретикулоцитов. В дополнение, анализ их молекулярного состава показал, что экзосомы из В-лимфоцитов содержат белки комплекса гистосовместимости II (MHCII) и что если антигены связаны с ними, то они могут быть презентированы Т-лимфоцитам, вызывая антигенспецифические иммунные ответы. С тех пор в различных исследованиях было показано, что экзосомы могут быть секретированы практически всеми типами клеток, включая клетки иммунной системы, такие как дендритные клетки, макрофаги, а также Т- и В-лимфоциты. (Théry C. et al., ʺMembrane vesicles as conveyors of immune responsesʺ, Nat. Rev. Immunol., 2009, том 9, стр.581-593). Благодаря этому обнаружению стало возможным использовать эти везикулы в качестве новой терапевтической стратегии различных заболеваний, таких как злокачественные новообразования. Таким образом, после того как было обнаружено, что В-лимфоциты способны продуцировать экзосомы вместе с молекулами класса МНС-II и что они способны презентировать антиген Т-лимфоцитам, та же группа исследователей три года спустя показала, что дендритные клетки (DC), отвечающие за индукцию специфического иммунитета, секретируют экзосомы вместе с молекулами МНС класса I, связанными с пептидами, которые способны вызывать CD8 Т-цитотоксические противоопухолевые иммунные ответы (Zitvogel L. et al., ʺDendritic cells or their exosomes are effective biotherapies of cancerʺ, European journal of cancer, 1999, том 35, Supl 3:S36-38). Эти пионерские исследования установили молекулярную основу, согласно которой экзосомы участвуют в межклеточной коммуникации в иммунной системе и дали возможность использовать экзосомы в качестве новых противоопухолевых вакцин (Chaput & Théry. Exosomes: immune properties and potential clinical implementations. 2011. Semi. Immunopathol., 33:419-440). Фактически, в настоящее время проводятся клинические испытания фазы I у пациентов с метастатической меланомой, раком легкого и колоректальным раком, как упоминается в этом обзоре. Кроме того, недавно было проведено клиническое исследование фазы II с использованием экзосом, полученных из дендритных клеток, у пациентов с немелкоклеточным раком легкого (Besse et al., ʺDendritic cell-derived exosomes as maintenance immunotherapy after first line chemotherapy in NSCLCʺ, Oncoimmunology, 5(4): e1071008).
Как и клетки высших организмов, паразиты и бактерии также секретируют экзосомы. Аналогично, паразиты и внутриклеточные патогены (включая вирус) вызывают секрецию экзосом клеткой-хозяином. Фактически, в последние несколько лет было показано, что такие экзосомы содержат в своем молекулярном составе «характерный набор» патогена (Marcilla A. et al., ʺExtracellular vesicles in parasitic diseasesʺ, 2014, J Extracell. Vesicles 3, 25040, and Schorey J. S. & Bhatnagar S., ʺExosome function: from tumor immunology to pathogen biologyʺ, 2008, Traffic, 9:871-81).
Ввиду вышеизложенного, за последние несколько лет ведутся активные исследования возможного применения экзосом в качестве вакцин. Основными преимуществами применения вакцин на основе экзосом против патогенов являются: (i) экзосомы, полученные из патогенов, содержат несколько природных белков возбудителя с характерной укладкой; (ii) экзосомы имеют широкое биологическое распределение в организме, поскольку циркулируют в разных биологических жидкостях, достигая отдаленных органов, так как защищены от деградации и способны преодолевать сложные барьеры эндотелиальных клеток, например, гематоэнцефалический барьер; (iii) они имеют более эффективную связь с антиген-презентирующими клетками в виду наличия адгезинов наряду с другими молекулами.
Однако некоторые концептуальные и практические аспекты в области применения экзосом в качестве профилактических вакцин нуждаются в оценке. Один из них заключается в том, что во всех описанных до сих пор случаях экзосомы, используемые в качестве терапевтических агентов, были выделены из образцов пациентов, животных или клеточных культур с активной инфекцией. Это препятствует их применению в качестве вакцины, поскольку трудно показать, что такие выделенные экзосомы не содержат патогена, вызывающего заболевание. Для некоторых патогенов (особенно патогенов вирусных инфекций) это требование трудно выполнить из-за сходных физико-химических особенностей (размера и/или плотности) продуцируемой экзосомы и некоторых патогенов, вызывающих заболевание, например, вируса репродуктивно-респираторного синдрома свиней (PRRSV), поэтому требуются конкретные протоколы, гарантирующие получение препарата экзосом без патогенов. Это не всегда возможно, и, если возможно, то создание таких конкретных протоколов отрицательно влияет как на стоимость, так и на время, необходимые для производства вакцины.
Ввиду вышеизложенного необходимо найти стратегии для применения экзосом для профилактики и/или лечения различных патологий, альтернативные доступным стратегиям.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения обнаружили, что животное, перенесшее заболевание, вызванное патогеном, и у которого больше нет следов патогена, способно продуцировать экзосомы с определенным характерным молекулярным набором патогена. Как показано в примере 5, анализ с жидкостной хроматографией в комбинации с масс-спектрометрией, показал, что образцы сыворотки, экстрагированные из не зараженных вирусом животных, уже перенесших болезнь, содержали экзосомы с вирусными белками на поверхностях.
Этот результат является неожиданным, учитывая тот факт, что до сих пор считали, что для получения экзосом из организма, включая белки и других связанных с патогенами биомолекул, необходимо активное присутствие патогена.
Эти результаты являются важным шагом вперед в области вакцин. Тот факт, что организм, перенесший конкретную патологию, вызванную патогеном, способен продуцировать экзосомы с потенциальной иммуногенной активностью (благодаря тому, что на его поверхности экспрессируется белковый профиль патогена) без активного присутствия патогена, может в значительной степени облегчить производство вакцин, не содержащих патоген. Следовательно, нежелательные побочные эффекты, связанные с существующими в настоящее время вакцинами, основанными на аттенуированных патогенах, минимизируются.
Кроме того, изобретатели обнаружили, что экзосомы, продуцирующиеся у животного во время инфекции и у животного, перенесшего эту инфекцию, различаются. Как показано на фиг.2 и 3, у животного (свиньи), которое перенесло заболевание (репродуктивно-респираторный синдром «PRRS») и у которого отсутствуют следы патогенов в плазме, как диагностировано с помощью чувствительного и специфического способа, была обнаружена цистеинпротеаза c1ab, сходная с папаином, неописанный предполагаемый белок PRRSV, полипротеин PRRSV, NSP2, GP2b и ORF2a. Кроме белков, указанных на фиг.2 и 3, также было обнаружено, что экспрессия белка оболочки, нуклеокапсидного белка и белков ORF1a (пример 10 ниже) различается. Эти белки не обнаруживаются в экзосомах, продуцируемых свиньями во время инфекции.
Следовательно, в первом аспекте настоящее изобретение относится к экзосоме, выделенной у животного, где животное (а) перенесло заболевание, вызванное патогеном, и (b) у него отсутствует патоген, вызывающий это заболевание.
В настоящем изобретении под выражением «перенесло заболевание» понимают, что животное не имеет каких-либо клинических симптомов, характерных для заболевания, и которые использовали в момент диагностики заболевания. В одном варианте осуществления победа над заболеванием происходит естественным путем, то есть под воздействием на патоген иммунной системы хозяина, при условии, что лечение иммуногеном или вакциной не проводили. Альтернативно, в другом варианте осуществления победа над заболеванием происходит в результате проведения лечения фармакологическими препаратами.
В настоящем изобретении выражение «отсутствует патоген» и «нет следов патогена» используются взаимозаменяемо, и оно означает, что в образце, взятом у животного, не обнаружено никакой патогенной нагрузки. В одном варианте осуществления патогенная нагрузка определяется в образце жидкости организма (например, в плазме или сыворотке). Протоколы, раскрытые в уровне техники для диагностики заболеваний у животных, основанные на количественном определении патогена, также могут использоваться в контексте настоящего изобретения для оценки того, является ли животное животным, у которого «отсутствует патоген». С другой стороны, удобный протокол(ы) для определения патогенной нагрузки для конкретного патогена хорошо известен в уровне техники (Schimdt B. et al., ʺDiagnostic tools for animal diseasesʺ, Rev. Sci Tech. Off. Int. Epiz., 2005, том 24 (1), стр. 243-250).
В настоящем изобретении под «патогеном» понимают любой агент, способный вызвать заболевание или поражение в определенный момент времени у хозяина. В одном варианте осуществления патоген представляет собой микроорганизм. В другом варианте патоген представляет собой бактерию, гриб, вирус или паразит.
В варианте осуществления первого аспекта изобретения животное перенесло вирусное заболевание и у него отсутствует вирус, вызывающий это заболевание.
В еще одном варианте осуществления первого аспекта изобретения животное перенесло паразитическое заболевание и у него отсутствует паразит, вызывающий это заболевание.
В другом варианте осуществления первого аспекта изобретения животное перенесло бактериальное заболевание и у него отсутствует бактерия, вызывающая это заболевание.
В одном варианте осуществления животное является млекопитающим. В другом варианте осуществления животное является сельскохозяйственным животным. В качестве неограничивающего примера, термин «сельскохозяйственное животное» включает, в том числе, свиней, овец и лошадей. В одном из вариантов осуществления животное представляет собой свинью. Альтернативно, в другом варианте осуществления животное является человеком.
Иллюстративные неограничивающие примеры вирусов, бактерий и паразитов, поражающих сельскохозяйственных животных, перечислены в документе организации ООН по продовольствию и сельскому хозяйству (FAO) ʺManual on meat inspection for developing countriesʺ, 1994, повторная публикация 2000, части 3-8 (документ доступен онлайн на странице http://www.fao.org/docrep/003/t0756e/T0756E00.htm#TOC).
В другом варианте осуществления первого аспекта изобретения животное представляет собой млекопитающее, которое перенесло вирусное заболевание и у которого отсутствует вирус, вызывающий заболевание (т.е. является животным, «не зараженным вирусом»).
В еще одном варианте осуществления первого аспекта изобретения животное является сельскохозяйственным животным, которое перенесло вирусное заболевание и у которого отсутствует вирус, вызывающий это заболевание.
В еще одном варианте осуществления первого аспекта изобретения животное является свиньей, которая перенесла вирусное заболевание и у которой отсутствует вирус, вызывающий это заболевание.
В еще одном варианте осуществления первого аспекта изобретения животное является свиньей, которая перенесла репродуктивно-респираторный синдром свиней (PRSS) и у которой отсутствует вирус, вызывающий это заболевание.
Репродуктивно-респираторный синдром свиней характеризуется нарушениями в расплоде свиноматок и респираторным кризисом у поросят и растущих свиней, что является важной причиной значительных экономических потерь. Этот синдром вызывается небольшим оболочечным одноцепочечным положительным РНК-вирусом (+ssРНК), классифицированным в порядке Nidovirales, семейство Arteriviridae и род Arterivirus.
До сих пор было разработано два типа вакцин против PRRSV, один тип - содержащий мертвый вирус, а другой тип, разработанный в последнее время - содержащие аттенуированный вирус с адъювантом или без него. Следует подчеркнуть, что подавляющее большинство таких вакцин распространены только в странах, где они производятся, и все они нуждаются в существенном усовершенствовании с точки зрения эффективности и биобезопасности, особенно в отношении получения вакцины, в которой полностью отсутствуют следы патогена (Charerntantanakul W., ʺPorcine reproductive and respiratory syndrome virus Vaccines: immunogenicity, efficacy and safety aspectsʺ, 2012, World Journal of Virology, том 1(1), стр.23-30). Как уже упоминалось выше, с экзосомами по изобретению, которые выделены из животных, которые перенесли заболевание и у которых отсутствует патоген, получают вакцину, не содержащую остаточного патогена, что улучшает биологическую безопасность вакцин.
Настоящее изобретение вносит большой вклад в область иммунологических вакцин, поскольку впервые описано, что животные, у которых отсутствует патоген и которые уже перенесли заболевание, способны продуцировать экзосомы, которые являются иммуногенными, как показано ниже.
Как показано в примере 7 и на фиг. 4, белки, идентифицированные в выделенных экзосомах, распознаются иммунными сыворотками животных, которые перенесли инфекцию, что указывает на иммуногенный эффект, который могут вызывать экзосомы по настоящему изобретению.
В еще одном варианте осуществления первого аспекта изобретения животное является млекопитающим, которое перенесло паразитическое заболевание и у которого отсутствует паразит, вызывающий это заболевание.
В еще одном варианте осуществления первого аспекта изобретения животное является сельскохозяйственным животным, которое перенесло паразитическое заболевание и у которого отсутствует паразит, вызывающий это заболевание.
В еще одном варианте осуществления первого аспекта изобретения животное является коровой, которая перенесла паразитическое заболевание и у которой отсутствует паразит, вызывающий это заболевание.
В еще одном варианте осуществления первого аспекта изобретения животное является коровой, которая перенесла паразитическое заболевание тейлериоз и у которой отсутствует паразит, вызывающий это заболевание.
В другом варианте осуществления первого аспекта изобретения животное является млекопитающим, которое перенесло бактериальное заболевание и у которого отсутствует бактерия, вызывающая это заболевание.
В другом варианте осуществления первого аспекта изобретения животное является сельскохозяйственным животным, которое перенесло бактериальное заболевание и у которого отсутствует бактерия, вызывающая это заболевание.
В другом варианте осуществления первого аспекта изобретения животное является свиньей, которая перенесла бактериальное заболевание и у которой отсутствует бактерия, вызывающая это заболевание.
В другом варианте осуществления первого аспекта изобретения животное является свиньей, которая перенесла бактериальное заболевание микоплазмоз и у которой отсутствует бактерия, вызывающая это заболевание.
В последующих работах изобретатели по настоящему изобретению провели протеомический анализ экзосом, выделенных из образцов плазмы животных, инфицированных либо PRRSV, либо микоплазмозом, либо тейлериозом, и животных, которые перенесли заболевание и у которых отсутствуют следы патогена. На основании этого анализа были идентифицированы пептидные последовательности SEQ ID NO:1 - SEQ ID NO:15:
SEQ ID NO:1 VEVEGHLMTSK
SEQ ID NO:2 QAKKHEVAGANK
SEQ ID NO:3 AGKKQSQK
SEQ ID NO:4 NIAPMGNGQSVNQLCQLLGTMMK
SEQ ID NO:5 MAGRNQRQK
SEQ ID NO:6: LEELFK
SEQ ID NO:7: KGSIVDIENQK
SEQ ID NO:8: MQIFVK
SEQ ID NO:9: TITLEVEPSDTIENVK
SEQ ID NO:10: IENLSDTFLSNNGKPEYKR
SEQ ID NO:11: AGFAGDDAPR
SEQ ID NO:12: IWHHTFYNELR
SEQ ID NO:13: YPIEHGIVTNWEDMEK
SEQ ID NO:14: STELLIRK
SEQ ID NO:15: EGDGVCTITAKMPKDEQK
и было обнаружено, что они содержатся в различном количестве в экзосомах из группы свиней, которые перенесли заболевание, и отсутствуют у свиней с инфекцией.
В варианте осуществления первого аспекта изобретения, необязательно в комбинации с любым из вариантов осуществления, представленных выше или ниже, экзосома содержит пептид, содержащий последовательность, выбранную из группы, состоящей из: SEQ ID NO:1 - SEQ ID NO:15, и пептидную последовательность, которая по меньшей мере на 85% идентична любой из последовательностей SEQ ID NO:1-15. В другом варианте осуществления первого аспекта изобретения, необязательно в комбинации с любым из вариантов осуществления, представленных выше или ниже, экзосома содержит пептид, содержащий последовательность SEQ ID NO:1 или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:1, и пептид, содержащий последовательность SEQ ID NO:2 или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:2. В другом варианте осуществления первого аспекта изобретения, необязательно в комбинации с любым из вариантов осуществления, представленных выше или ниже, экзосома содержит пептид, содержащий последовательность SEQ ID NO:1 или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:1, и пептид, содержащий последовательность SEQ ID NO:3, 4 или 5 или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:3, 4 или 5, соответственно. В еще одном варианте осуществления первого аспекта изобретения, необязательно в комбинации с любым из вариантов осуществления, представленных выше или ниже, экзосома содержит пептид, содержащий последовательность SEQ ID NO:2 или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:2, и пептид, содержащий последовательность SEQ ID NO:3, 4 или 5 или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:3, 4 или 5. В еще одном варианте осуществления первого аспекта изобретения, необязательно в комбинации с любым из вариантов осуществления, представленных выше или ниже, экзосома содержит пептид, содержащий последовательность SEQ ID NO:1 или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:1, пептид, содержащий последовательность SEQ ID NO:2 или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:2, и пептид, содержащий последовательность SEQ ID NO:3, SEQ ID NO:4 или SEQ ID NO:5 или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:3, 4 или 5, соответственно. В другом варианте осуществления первого аспекта изобретения, необязательно в комбинации с любым из вариантов осуществления, представленных выше или ниже, экзосома содержит последовательности SEQ ID NO:1-5. В другом варианте осуществления первого аспекта изобретения, необязательно в комбинации с любым из вариантов осуществления, представленных выше или ниже, экзосома содержит пептид, содержащий последовательность SEQ ID NO:6 или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:6, и пептид, содержащий последовательность SEQ ID NO:7 или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:7. В другом варианте осуществления первого аспекта изобретения, необязательно в комбинации с любым из вариантов осуществления, представленных выше или ниже, экзосома содержит пептид, содержащий последовательность SEQ ID NO:6, и пептид, содержащий последовательность SEQ ID NO:7. В другом варианте осуществления первого аспекта изобретения экзосома содержит один или несколько пептидов, содержащих последовательность, выбранную из SEQ ID NO:8 - SEQ ID NO:15, или последовательность, которая по меньшей мере на 85% идентична любой из последовательностей SEQ ID NO:8-15. В другом варианте осуществления первого аспекта изобретения экзосома содержит пептид, содержащий последовательность SEQ ID NO:8 или последовательность, которая по меньшей мере на 85% идентична последовательности SEQ ID NO:8, пептид, содержащий последовательность SEQ ID NO:9 или последовательность, которая по меньшей мере на 85% идентична последовательности SEQ ID NO:9, пептид, содержащий последовательность SEQ ID NO:10 или последовательность, которая по меньшей мере на 85% идентична последовательности SEQ ID NO:10, пептид, содержащий последовательность SEQ ID NO:11 или последовательность, которая по меньшей мере на 85% идентична последовательности SEQ ID NO:11, пептид, содержащий последовательность SEQ ID NO:12 или последовательность, которая по меньшей мере на 85% идентична последовательности SEQ ID NO:12, пептид, содержащий последовательность SEQ ID NO:13 или последовательность, которая по меньшей мере на 85% идентична последовательности SEQ ID NO:13, пептид, содержащий последовательность SEQ ID NO:14 или последовательность, которая по меньшей мере на 85% идентична последовательности SEQ ID NO:14, и пептид, содержащий последовательность SEQ ID NO:15 или последовательность, которая по меньшей мере на 85% идентична последовательности SEQ ID NO:15. В другом варианте осуществления первого аспекта изобретения экзосома содержит пептид, содержащий последовательность SEQ ID NO:8, пептид, содержащий последовательность SEQ ID NO:9, пептид, содержащий последовательность SEQ ID NO:10, пептид, содержащий последовательность SEQ ID NO:11, пептид, содержащий последовательность SEQ ID NO:12, пептид, содержащий последовательность SEQ ID NO:13, пептид, содержащий последовательность SEQ ID NO:14, и пептид, содержащий последовательность SEQ ID NO: 15. Экзосомы этих вариантов осуществления, которые включают пептиды, содержащие один или несколько пептидов последовательности SEQ ID NO:1-15, могут альтернативно быть получены общеизвестными способами.
Таким образом, в следующем аспекте настоящее изобретение относится к везикуле, содержащей пептид, содержащий последовательность, выбранную из группы, состоящей из SEQ ID NO:1 и SEQ ID NO:15, и пептидную последовательность, которая по меньшей мере на 85% идентична любой из последовательностей SEQ ID NO:1-15. В варианте осуществления этого аспекта, необязательно в комбинации с любым из вариантов осуществления, представленных выше или ниже, везикула содержит пептид, содержащий последовательность SEQ ID NO:1 или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:1, и пептид, содержащий последовательность SEQ ID NO:2 или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:2. В другом варианте осуществления этого аспекта изобретения, необязательно в комбинации с любым из вариантов осуществления, представленных выше или ниже, везикула содержит пептид, содержащий последовательность SEQ ID NO:1, или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:1, и пептид, содержащий последовательность SEQ ID NO:3, 4 или 5, или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:3, 4 или 5, соответственно. В другом варианте осуществления этого аспекта изобретения, необязательно в комбинации с любым из вариантов осуществления, представленных выше или ниже, везикула содержит пептид, содержащий последовательность SEQ ID NO:2 или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:2, и пептид, содержащий последовательность SEQ ID NO:3, 4 или 5, или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:3, 4 или 5. В другом варианте осуществления этого аспекта, необязательно в комбинации с любым из вариантов осуществления, представленных выше или ниже, везикула содержит пептид, содержащий последовательность SEQ ID NO:1, или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:1, пептид, содержащий последовательность SEQ ID NO:2, или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:2, и пептид, содержащий последовательность SEQ ID NO:3, 4 или 5, или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:3, 4 или 5, соответственно. В другом варианте осуществления этого аспекта изобретения, необязательно в комбинации с любым из вариантов осуществления, представленных выше или ниже, везикула содержит последовательности SEQ ID NO:1-5. В другом варианте осуществления этого аспекта, необязательно в комбинации с любым из вариантов осуществления, представленных выше или ниже, везикула содержит пептид, содержащий последовательность SEQ ID NO:6 или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:6, и пептид, содержащий последовательность SEQ ID NO:7 или последовательность, которая по меньшей мере на 85% идентична SEQ ID NO:7. В другом варианте осуществления этого аспекта изобретения, необязательно в комбинации с любым из вариантов осуществления, представленных выше или ниже, везикула содержит пептид, содержащий последовательность SEQ ID NO:6, и пептид, содержащий последовательность SEQ ID NO:7. В другом варианте осуществления этого аспекта изобретения везикула содержит один или несколько пептидов, содержащих последовательность, выбранную из SEQ ID NO:8 - SEQ ID NO:15, или последовательность, которая по меньшей мере на 85% идентична любой из последовательностей SEQ ID NO:8-15. В другом варианте осуществления этого аспекта изобретения везикула содержит пептид, содержащий последовательность SEQ ID NO:8 или последовательность, которая по меньшей мере на 85% идентична последовательности SEQ ID NO:8, пептид, содержащий последовательность SEQ ID NO:9 или последовательность, которая по меньшей мере на 85% идентична последовательности SEQ ID NO:9, пептид, содержащий последовательность SEQ ID NO:10 или последовательность, которая по меньшей мере на 85% идентична последовательности SEQ ID NO:10, пептид, содержащий последовательность SEQ ID NO:11 или последовательность, которая по меньшей мере на 85% идентична последовательности SEQ ID NO:11, пептид, содержащий последовательность SEQ ID NO:12 или последовательность, которая по меньшей мере на 85% идентична последовательности SEQ ID NO:12, пептид, содержащий последовательность SEQ ID NO:13 или последовательность, которая по меньшей мере на 85% идентична последовательности SEQ ID NO:13, пептид, содержащий последовательность SEQ ID NO:14 или последовательность, которая по меньшей мере на 85% идентична последовательности SEQ ID NO:14, и пептид, содержащий последовательность SEQ ID NO:15 или последовательность, которая по меньшей мере на 85% идентична последовательности SEQ ID NO:15. В другом варианте осуществления этого аспекта изобретения везикула содержит пептид, содержащий последовательность SEQ ID NO:8, пептид, содержащий последовательность SEQ ID NO:9, пептид, содержащий последовательность SEQ ID NO:10, пептид, содержащий последовательность SEQ ID NO:11, пептид, содержащий последовательность SEQ ID NO:12, пептид, содержащий последовательность SEQ ID NO:13, пептид, содержащий последовательность SEQ ID NO:14, и пептид, содержащий последовательность SEQ ID NO:15.
Во втором аспекте настоящее изобретение относится к пептиду с длиной последовательности, равной или меньше 100 аминокислот, и содержащую последовательность, выбранную из SEQ ID NO:1-15, или пептидную последовательность, которая по меньшей мере на 85% идентична последовательности SEQ ID NO:1-15.
В настоящем изобретении термин «идентичность» относится к проценту остатков или оснований, которые идентичны в двух последовательностях при оптимальном выравнивании последовательностей. Если при оптимальном выравнивании положение в первой последовательности занято тем же аминокислотным остатком или нуклеотидом, что и в соответствующем положении второй последовательности, то последовательности идентичны по этому положению. Уровень идентичности двух последовательностей (или «процентная идентичность последовательностей») измеряется как отношение числа идентичных положений в последовательностях по отношению к размеру последовательностей (то есть, процент идентичности последовательностей=(число идентичных положений/общее число положений) × 100).
Ряд математических алгоритмов для быстрого получения оптимального выравнивания и вычисления идентичности двух или более последовательностей известен и включен в ряд доступных программ. Примеры таких программ включают, среди прочего, программы MATCH-BOX, MULTAIN, GCG, FASTA и ROBUST для анализа аминокислотной последовательности. Предпочтительное программное обеспечение для анализа включает программы ALIGN, CLUSTAL W и BLAST (например, BLAST 2.1, BL2SEQ и их более поздние версии).
Для анализа аминокислотной последовательности для определения идентичности используется матрица сравнения, такая как матрица BLOSUM (например, матрицы BLOSUM45, BLOSUM50, BLOSUM62 и BLOSUM80), матрица Gonnet или матрица PAM (например, матрицы PAM30, PAM70, PAM120, PAM160, PAM250 и PAM350).
Программы BLAST проводят анализ по меньшей мере двух аминокислотных последовательностей либо путем выравнивания выбранной последовательности с несколькими последовательностями из базы данных (например, GenSeq), либо с помощью BL2SEQ, путем выравнивания двух выбранных последовательностей. Программы BLAST предпочтительно модифицируют программами фильтрации с низкой степенью сложности, такими как программы DUST или SEG, которые предпочтительно интегрируют в операторы программы BLAST. Если используется штраф за наличие пропуска (или баллы за пропуск), то штраф за наличие пропуска предпочтительно составляет от приблизительно -5 до -15. Аналогичные параметры пропуска можно использовать в других подходящих программах. Программы BLAST и принципы, лежащие в их основе, дополнительно описаны, например, в Altschul et al., ʺBasic local alignment search toolʺ, 1990, J. Mol. Biol, том 215, стр.403-410.
Для анализа нескольких последовательностей можно использовать программу CULSTAL W. Желательно, запускать программу CLUSTAL W, используя «динамические» настройки (в противоположность «быстрым»). Аминокислотные последовательности оценивают с помощью переменного набора матриц BLOSUM в зависимости от уровня идентичности последовательностей. Программа CLUSTAL W и принципы, лежащие в основе ее работы, дополнительно описаны, например, в Higgins et al., ʺCLUSTAL V: improved software for multiple sequence alignmentʺ, 1992, CABIOS, 8(2), стр.189-191.
В одном варианте осуществления второго аспекта изобретения пептид имеет длину последовательности, равную или меньше 50 аминокислот. В другом варианте осуществления второго аспекта изобретения пептид представляет собой последовательность SEQ ID NO:16, 17, 18 или последовательность, которая по меньшей мере на 85% идентична любой из последовательностей 6-8:
SEQ ID NO:16 LDAKGRLYRWRSPVIIEKGGKVEVEGHLMTSKELC (которая содержит последовательность SEQ ID NO:1)
SEQ ID NO:17 QAKKHEVAGANKAEHLKHYSPPAEGNCGWHCISAI (которая содержит последовательность SEQ ID NO:2)
SEQ ID NO:18 MAGRNQSQKKKKNIAPMGNGQSVNQLCQLLGTMMK (которая содержит последовательности SEQ ID NO:3-5)
Неожиданно, как показано ниже, пептиды последовательности SEQ ID NO:16-18 обладают иммуногенными свойствами.
На основании результатов, приведенных ниже в примере 10, можно сделать вывод, что внеклеточная везикула по изобретению может быть как таковая использована в качестве иммуногена, а также в качестве источника для идентификации пептидов, являющихся кандидатами в иммуногены конкретного заболевания. И общая методика является простой. Иллюстративным способом идентификации иммуногенных кандидатов является сравнение протеомического анализа внеклеточной везикулы, выделенной у особи, страдающей заболеванием, и везикулы, выделенной у особи, которая успешно перенесла заболевание и у которой отсутствуют следы патогена, и те пептиды, наличие которых различается, могут быть выбраны в качестве потенциальных кандидатов в иммуногены. В частности, те пептиды, которые присутствуют во внеклеточной везикуле по изобретению, а не во внеклеточной везикуле, секретируемой во время инфекции, являются хорошими кандидатами в иммуногенные пептиды.
В других вариантах осуществления, как только кандидат был идентифицирован, он может быть протестирован для определения его эффекта в иммунной системе. Но, в качестве альтернативы, для специалиста в данной области может быть предпочтительным спрогнозировать последовательность эпитопа, используя бесплатные коммерческие программы, такие как программы, перечисленные в IEDB Analysis Resource (www.tools.iedb.org), из них можно использовать Bepipred. Как только информация об эпитопе получена, этот эпитоп может быть синтезирован с использованием, например, методов твердофазного синтеза (например, Fmoc) и тестирован для подтверждения эффекта на иммунную систему.
В третьем аспекте настоящее изобретение относится к способу получения внеклеточной везикулы по первому аспекту изобретения, который включает отделение внеклеточной везикулы от тестируемого образца, выбранного из биологической жидкости или образца ткани, взятого у животного, (а) которое перенесло заболевание, вызванное патогеном, и (b) у которого отсутствует патоген, вызывающий это заболевание.
Биологической жидкостью могут быть, например, сыворотка, плазма, пот, слезы, молоко или семенная жидкость.
Специалисту известны несколько методов, которые могут быть использованы для отделения экзосомы от других компонентов, содержащихся в плазме. Иллюстративные не ограничивающие примеры включают ультрацентрифугирование, фильтрацию и эксклюзионную хроматографию. Такая стадия отделения, указанная в способе по третьему аспекту изобретения, может включать использование одной или нескольких методик, например, можно провести отделение ультрацентрифугированием с последующей эксклюзионной хроматографией, или, альтернативно, можно определить, что одного метода достаточно. В варианте осуществления третьего аспекта изобретения отделение проводят с помощью эксклюзионной хроматографии. В другом варианте осуществления третьего аспекта изобретения внеклеточная везикула представляет собой экзосому, и отделение проводят с помощью эксклюзионной хроматографии через матрицу из сефарозы.
В варианте осуществления третьего аспекта изобретения способ включает стадию экзосомного обогащения образца, например, путем осаждения полигликолем, перед стадией отделения. В другом варианте осуществления третьего аспекта изобретения способ включает: (а) осаждение выделенного образца биологической жидкости полигликолем, таким как полиэтиленгликоль Mn6000 (8,5 масс.%/объем) при конечной концентрации хлорида натрия 0,4 М; (b) ресуспендирование осадка, полученного на стадии смешивания (а) с буфером; и (c) отделение экзосомы от суспензии. Изобретатели обнаружили, что можно увеличить масштаб этого процесса за счет проведения стадии осаждения полиолом перед стадией отделения, таким образом получая значительное количество экзосом. Стадию (а) можно проводить с любым полигликолем, хотя в одном варианте осуществления полигликолем является полиэтиленгликоль. Стадию (b) можно проводить с любым буфером, таким как забуференный фосфатом физиологический раствор. На стадии (с) можно использовать любую методику, указанную ранее для отделения экзосом, или их комбинацию (фильтрация, ультрацентрифугирование или эксклюзионная хроматография).
Альтернативно, в четвертом аспекте настоящее изобретение относится к способу получения экзосом по первому аспекту изобретения, который включает выделение и культивирование ретикулоцитов из образца крови, взятого у животного, и получения фракции экзосом из ретикулоцитов. Выделенную из ретикулоцитов фракцию экзосом можно отделить ультрацентрифугированием, эксклюзионной хроматографией или фильтрацией. В варианте осуществления четвертого аспекта способ включает, прежде всего, отделение экзосомы, стадию экзосомного обогащения, например, осаждение полигликолем. В другом варианте осуществления четвертого аспекта изобретения способ включает: (а) выделение и культивирование ретикулоцитов из образца крови, взятого у животного, (b) осаждение выделенного образца биологической жидкости полигликолем, таким как полиэтиленгликоль (8,5 масс.%/объем) при конечной концентрации хлорида натрия 0,4 М; (c) ресуспендирование осадка, полученного на предыдущей стадии (b) буфером; и (d) отделение экзосом в суспензии. Стадию (b) можно проводить с любым полигликолем, хотя в одном варианте осуществления полигликолем является полиэтиленгликоль. Стадию (c) можно проводить с любым буфером, таким как забуференный фосфатом физиологический раствор. На стадии (d) можно использовать любую указанную ранее методику выделения экзосом, либо самостоятельно, либо в комбинации (фильтрация, ультрацентрифугирование или эксклюзионная хроматография).
В пятом аспекте настоящее изобретение относится к внеклеточной везикуле, полученной способом по второму или третьему аспекту изобретения.
Термин «внеклеточная везикула», например, «экзосома», получаемая этим способом, используется в настоящем документе для определения внеклеточной везикулы посредством способа получения и относится к продукту, который может быть получен способом получения, который включает указанные стадии, как определено в настоящем документе. Для целей изобретения выражения «получаемая», «полученная» и аналогичные эквивалентные выражения используются взаимозаменяемо, и в любом случае выражение «получаемая» включает в себя выражение «полученная».
На основании данных, приведенных в примере 7 и примере 10, можно сделать вывод, что и экзосомы, и пептиды по изобретению имеют иммуногенный профиль, подходящий для терапии.
Таким образом, в шестом аспекте настоящее изобретение относится к фармацевтической или ветеринарной композиции, содержащей внеклеточную везикулу по первому или пятому аспекту, или пептид по второму аспекту изобретения, и один или несколько фармацевтически или ветеринарно приемлемых эксципиентов или носителей.
Термин «фармацевтически или ветеринарно приемлемый» относится к эксципиентам или носителям для их применения в фармацевтической или ветеринарной области, с целью получения композиций для медицинского применения у людей или животных. Каждый компонент должен быть фармацевтически или ветеринарно приемлемым в смысле совместимости с другими ингредиентами фармацевтической или ветеринарной композиции, соответственно. Он также должен быть пригодным при использовании в контакте с тканью или органом человека и животных, не вызывая чрезмерную токсичность, раздражение, аллергическую реакцию, иммуногенность или другие проблемы или осложнения, соизмеримо с разумным соотношением польза/риск.
В одном варианте осуществления шестого аспекта настоящее изобретение относится к композиции, содержащей терапевтически эффективное количество внеклеточной везикулы по первому или пятому аспекту изобретения или пептид по второму аспекту изобретения, и один или больше фармацевтически или ветеринарно приемлемых эксципиентов или носителей.
Выражение «терапевтически эффективное количество», как используется в настоящем документе, относится к количеству экзосомы или пептида, которое при введении является достаточным для предотвращения развития или для облегчения до определенной степени одного или нескольких симптомов заболевания. Конкретная доза экзосом, вводимая в соответствии с изобретением, будет определяться для конкретных обстоятельств, таких как, например, путь введения и заболевание, подлежащее лечению.
Композиции по настоящему изобретению могут быть получены с использованием известных в уровне техники способов. Специалист в данной области техники может определить эксципиенты и/или носители и подходящие используемые количества в зависимости от полученной рецептуры.
В варианте осуществления шестого аспекта фармацевтическая или ветеринарная композиция является вакциной.
Эксципиенты и адъюванты, которые могут быть введены в состав вакцины, хорошо известны специалисту в данной области и выбраны таким образом, чтобы они не оказывали отрицательного влияния на иммунологическую активность экзосомы.
Как указано выше, экзосомы, выделенные у свиней, которые перенесли синдром PRRSV, имеют подходящий иммуногенный профиль, что предполагает их применение в качестве вакцины. Тот же вывод можно сделать на основании экспериментальных данных, представленных для пептидов, выделенных из экзосом по изобретению (см. пример 10). Таким образом, анализ ELISA показал, что экзосомы, полученные у не зараженных вирусом животных, используемые в качестве покрывающих антигенов, специфически распознавались сыворотками животных, ранее инфицированных вирусом PRRSV, но не распознавались сыворотками животных, никогда не инфицированных. Кроме того, иммунизация наивных животных экзосомами, полученными у не зараженных вирусом животных, или пептидами по изобретению, как ранее было идентифицировано с помощью масс-спектрометрии, показала, что животные были способны индуцировать специфические гуморальные и клеточные иммунные ответы. Примечательно, что животные, которые были примированы экзосомами не зараженных вирусом животных и стимулированы синтетическими пептидами, также дают такие ответы, что однозначно демонстрирует, что такие экзосомы содержат и презентируют такие белки иммунной системе свиней. Наконец, поскольку у животных, иммунизированных экзосомами не зараженных вирусом животных, получавших 1 миллиграмм экзосом, никогда не развивалась инфекция PRRSV, то такой подход вакцинации не содержит вируса.
Таким образом, в вакцине по настоящему изобретению, содержащей внеклеточные везикулы или пептиды по предыдущим аспектам, отсутствуют вирусные частицы (что не допускает развития клинических симптомов и возможного распространения патогенов из-за реверсии вирулентности), и она может непосредственно влиять на эпидемиологическую ситуацию, что предпочтительно. В иллюстративном, но не ограничительном примере вакцина может влиять на эпидемиологическую ситуацию на ферме и может быть распознана сывороткой, полученной на другой ферме с другой эпидемиологической ситуацией (фиг. 4). Таким образом, вакцина способна действовать непосредственно на вирусный штамм, который вызывает инфекцию в разных местах и в то или иное время.
В дополнение к вышесказанному, в примере 10 показано, что иммунный ответ, вызванный введением экзосомы или пептида по настоящему изобретению, запускает продукцию белковых маркеров, отличных от маркеров, продукция которых запускается во время инфекционного процесса.
Вакцинация животных является наиболее важным инструментом в борьбе с болезнью и ее уничтожения. Однако существующие в настоящее время вакцины, основанные на аттенуированном вирусе или мертвом вирусе, стимулируют гуморальный ответ против антигена, крайне сходный с тем ответом, который возникает во время инфекции. Поскольку установлено, что самая точная диагностика основана на детекции антител, то невозможно безошибочно отличить вакцинированных животных от инфицированных.
Экзосомы и пептиды по изобретению решают эту задачу, представляя вакцину, которая наделяет вакцинированное животное «меткой», так, что врач или ветеринар, в зависимости от особи, может легко определить, присутствует ли у особи инфекционный процесс или же особь вакцинирована.
Этот отличительный белковый набор может помочь отличить вакцинированные животные от инфицированных животных.
Таким образом, в седьмом аспекте настоящее изобретение относится к применению внеклеточной везикулы по первому или пятому аспекту изобретения, для проведения различия между животными, вакцинированными везикулой по первому или пятому аспекту изобретения от животных, инфицированных тем же патогеном, на который ссылается первый аспект изобретения.
Кроме вышеуказанного, в примере 10 показано, что иммунный ответ, индуцированный в ответ на патоген и в ответ на экзосому по изобретению, различается.
Как указано выше, экзосомы по изобретению презентируют иммуногенный профиль, что делает их эффективными для лечения заболеваний.
Следовательно, в восьмом аспекте настоящее изобретение относится к применению экзосомы или пептида по любому из предыдущих аспектов в качестве иммуногена.
В девятом аспекте настоящее изобретение относится к применению экзосомы или пептида по любому из предыдущих аспектов в качестве лекарственного средства.
В десятом аспекте изобретение относится к внеклеточной везикуле или пептиду по любому из предыдущих аспектов для применения в профилактике или лечении вирусного, бактериального или паразитарного заболевания. Этот аспект также может быть сформулирован как применение внеклеточной везикулы или пептида по любому из предыдущих аспектов для получения лекарственного средства для профилактики или лечения вирусного, бактериального или паразитарного заболевания. Этот аспект можно альтернативно сформулировать как способ профилактики или лечения вирусного, бактериального или паразитарного заболевания, где способ включает стадию введения нуждающейся особи, терапевтически эффективного количества внеклеточной везикулы или пептида по любому из аспектов или вариантов осуществления настоящего изобретения.
В одном варианте осуществления десятого аспекта изобретения внеклеточная везикула или пептид используются для лечения или профилактики заболевания, выбранного из заболевания PRRSV, микоплазмоза и тейлериоза.
В одиннадцатом аспекте настоящее изобретение относится к способу идентификации пептида, являющегося кандидатом в иммуноген, где способ включает стадию анализа белковой композиции внеклеточной везикулы по любому из вышеуказанных аспектов.
Анализ белковой композиции экзосомы (количественная протеомика) может быть осуществлен любой из рутинных методик, таких как жидкостная хроматография и масс-спектрометрия (de Menezes-Neto et al., ʺSize exclusion chromatography as a stand-alone methodology identifies novel markers in mass spectrometry analyses of plasma-derived vesicles from healthy indivuals, J. Extracell. Ves., 2015, 4: 27378).
В одном варианте осуществления одиннадцатого аспекта настоящее изобретение относится к способу идентификации пептида, являющегося кандидатом в иммуногены, где способ включает:
(а) анализ белковых компонентов внеклеточной везикулы, такой как экзосома, по первому или пятому аспекту изобретения; и
(b) сравнение белкового профиля, полученного на стадии (а), с белковым профилем внеклеточной везикулы, такой как экзосома, выделенной у животного, страдающего тем же заболеванием, что и животное, из которого выделена экзосома, используемая на стадии (а);
причем, если пептид включен в везикулу по изобретению, а не включен в везикулу инфицированного животного, то это указывает на то, что пептид является кандидатом в иммуноген.
В описании и формуле изобретения слово «содержит» и варианты этого слова не предназначены для исключения других технических признаков, добавок, компонентов или стадий. Кроме того, слово «содержит» включает в себя случай «состоящий из». Дополнительные объекты, преимущества и признаки изобретения станут очевидными для специалистов в данной области после изучения описания или могут быть изучены при осуществлении изобретения. Следующие примеры и чертежи приведены в качестве иллюстрации, и они не предназначены для ограничения настоящего изобретения. Ссылочные знаки, относящиеся к чертежам и помещенные в скобки в формуле изобретения, предназначены исключительно для ясности формулы изобретения и не должны пониматься как ограничивающие объем формулы изобретения. Кроме того, настоящее изобретение охватывает все возможные комбинации конкретных и предпочтительных вариантов осуществления, описанных в настоящем документе.
КРАТКОЕ ОПИСАНИЕ ФИГУР
ФИГ. 1. Анализ профиля элюции белка с помощью анализ Брэдфорда и анализа маркеров CD63 (b) и CD81 (c), используя проточную цитометрию. На фигуре приведены наборы данных Брэдфорда (мг/мл) и медианной флуоресценции (MFI) для каждого из испытуемых образцов. Примечательны высокие значения флуоресценции перед элюированием растворимого белка в процессе эксклюзионной хроматографии фракции. Эти значения MFI позволяют отличить фракции, обогащенные везикулами, от фракций, которые содержат большое количество растворимого белка (не связанного с везикулами). А и В соответствуют образцам зараженных вирусом животных (201406-2PS и 201406-4PS); C и D соответствуют образцам не зараженных вирусом свиней (201406-6PS и 201406-10PS).
Фиг. 2. Электрофорез белков на полиакриламидном геле в восстанавливающих условиях. (M) Маркер молекулярного веса указан в килодальтонах. Набор белков получали для разных фракций четырех проанализированных образцов. Две зараженный вирусом свиньи (2PS и 4PS) и две свиньи, которые перенесли заболевание (6PS и 10PS).
ФИГ. 3 Диаграмма Венна для сравнения белкового разнообразия вируса респираторно-репродуктивного синдрома свиней (PRRSV) в образцах зараженных вирусом животных (201406-2PS/201406-4PS) и в образцах животных, которые перенесли вирусную инфекцию (201406-6PS/201406-10PS).
Фиг. 4. Оптическая плотность (OD) при 450 нм, полученная в анализе ELISA сэндвич-типа (CAPTURE ELISA), в котором оценивали сыворотки животных, перенесших инфекцию. (а) Не зараженные вирусом свиньи+невиремические сыворотки, (b) Наивные свиньи+невиремические сыворотки и (с) Экзосомы из ретикулоцитов человека+невиремические сыворотки.
На фиг. 5 представлена схема и хронограмма исследования на ферме, проведенного в примере 10.
Фиг. 6. Распознавание сыворотки свиньи (42-ой день) против вакцины Porcilis PRRSV. Ту же сыворотку 42-ого дня исследования иммунизации оценивали против аттенуированной вирусной частицы Porcilis PRRSV (используется в качестве вакцины против заболевания). На графике образцы выше пунктирной линии являются положительными на антитела против этого вируса (порогом является среднее значение отрицательного контроля плюс трехкратное стандартное отклонение, представляя статистическую значимость p <0,05).
Фиг. 7. Распознавание сыворотки свиньи (42 день) против синтетических пептидов. Ту же сыворотку 42-го дня исследования иммунизации оценивали против трех пептидных смесей (синтезировали, как показано в настоящем документе). На графике образцы выше пунктирной линии являются положительными на антитела против этого вируса (порогом является среднее значение отрицательного контроля плюс трехкратное стандартное отклонение, представляющее статистическую значимость p <0,05).
ФИГ. 8. Характеристика экзосом, полученных из сывороток свиней (животных, излечившихся от инфекции микоплазмой). В образцах смесь фракций, обогащенных экзосомами (фракции 6-9), отбирали в качестве фракций с наибольшей концентрацией экзосом и в дальнейшем анализировали. CD63 и CD81 использовали в качестве признака для отбора фракций для проведения протеомических анализов.
ФИГ.9. Характеристика экзосом, полученных из сывороток не содержащих паразитов коров. В обоих случаях фракция 8 представляет собой наиболее обогащенную фракцию молекулярных маркеров, связанных с экзосомами. CD63 и CD81 использовали в качестве признака для отбора фракций для проведения протеомических анализов.
ПРИМЕРЫ
1. Сбор образцов
Собирали образцы сыворотки животных с ферм, где была вспышка PRRSV. Для определения того, какие образцы принадлежат животным, которые перенесли заболевание, группой исследователей г.Льейда - the Group Porcine Sanitation (GSP - Grup de Sanejament Porcí) -был проведен слепой анализ этих образцов с помощью методик: (i) ОТ-ПЦР, которая обнаруживает активную вирусную РНК, что указывает на то, что сыворотка принадлежит зараженному вирусом животному (реагенты и контроли Taqman PRRSV) и (ii) ELISA для определения титров антител, используя коммерческий набор (тест IDEXX PRRS X3 Ab). Следует отметить, что группа GSP аккредитована в Каталонии для диагностики этого патогена (http://www.gsplleida.net/ca/content/laboratori).
Параллельно образцы сыворотки животных собирали на фермах города Льейда, на которых до сегодняшнего дня не были зарегистрированы вспышки PRRSV. Эти сыворотки были оценены группой GSP, используя те же тесты. В этой группе образцы, которые были отрицательными как по результатам ОТ-ПЦР (что означает отсутствие активного вируса), так и по данным ELISA (что означает отсутствие протекающей инфекции), считались отрицательными контролями.
На основании полученных результатов были отобраны два образца зараженных вирусом животных (далее обозначены 201406-2PS и 201406-4PS) и два образца не зараженных вирусом животных (201406-6PS и 201406-10PS).
ПРИМЕР 2. ВЫДЕЛЕНИЕ ЭКЗОСОМ СЫВОРОТКИ
Экзосомы имеют характерный размер частиц 30-100 нм. Поэтому, для сбора этих везикул из разных образцов использовали процесс разделения с помощью эксклюзионной хроматографии, используя в качестве разделяющей матрицы сефарозу CL2B (de Menezes-Neto et al., ʺSize exclusion chromatography as a stand-alone methodology identifies novel markers in mass spectrometry analyses of plasma-derived vesicles from healthy indivuals, J. Extracell. Ves., 2015, 4: 27378). Хотя существуют другие методики получения экзосом, способы с сефарозой лучше очищают экзосомы. Кратко, замороженные 3 мл аликвоты разных образцов сыворотки оттаивали на льду и центрифугировали при 500 g в течение 10 минут при комнатной температуре для того, чтобы избавиться от клеточного дебриса. Параллельно, 12 мл-овые шприцы наполняли сефарозой CL-2B (Sigma-Aldrich, St. Louis, MO, USA) до конечного объема 10 мл и уравновешивали фосфатно-солевым буферным раствором (PBS) 0,32% цитрата натрия (масс./об.). Затем, 2 мкл аликвот каждого образца добавляли на отдельные колонки с сефарозой CL-2B и для каждого образца собирали 18-20 фракций по 0,5 мл аликвот.
ПРИМЕР 3. МОЛЕКУЛЯРНАЯ ХАРАКТЕРИСТИКА ЭКЗОСОМ
После получения различных фракций экзосом наличие везикул подтверждали в анализе концентрации белка, используя анализ Брэдфорда, и в анализе с молекулярными маркерами, проводимым с помощью проточной цитометрии.
3.1. Анализ Брэдфорда
Концентрацию белка получали с помощью колориметрического анализа, используя метода Брэдфорда (Bradford M. M. ʺA rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye bindingʺ, 1976, Analytical biochemistry, том 7(72), стр. 248-254).
3.2. Проточная цитометрия
Параллельно, фракции также анализировали с помощью проточной цитометрии на наличие антигенов CD9, CD63 или CD81, двух тетраспанинов, которые являются специфическими маркерами экзосом (Raposo G. et al., ʺExtracellular vesicles: exosomes, microvesicles, and friendsʺ, 2013, The Journal of cell biology, том 18(200), стр.373-383). Каждую аликвоту обрабатывали по следующему протоколу: к каждой аликвоте добавляли латексные бусы, размером 4 микрон (альдегид сульфат) (Invitrogen, номер в каталоге A37304), и смесь оставляли в покое на 15 минут, а затем добавляли 1 мл буфера BCB (PBS ×1, 0,1% бычьего сывороточного альбумина, 0,01% азида натрия). Полученную смесь инкубировали в течение ночи при комнатной температуре при вращении, а затем инкубировали с первыми антителами (анти-CD63 и анти-CD81, любезно предоставлены доктором Francisco Sánchez-Madrid) в течение 30 минут при 4°C. Оба антитела использовали в разведении 1:10. После двух этапов промывки с помощью 150 мкл буфера PBS-BSA (фосфатно-солевой буфер/бычий сывороточный альбумин 0,1%) и центрифугирования при 2000 g в течение 10 минут добавляли вторые антитела, конъюгированные с FITC (разведение 1:100) или alexa 488 (разведение 1:1000) (Southern Biotec, номер в каталоге 1032-02), и смесь инкубировали в течение 30 минут при 4°C. После двух дополнительных стадий промывки с помощью 150 мкл буфера PBS-BSA при 0,1% при 2000 g в течение 10 минут латексные бусы ресуспендировали в 100 мкл 0,1% PBS-BSA.
Полученные образцы анализировали с помощью проточной цитометрии, используя проточный цитометр LRSFortessa (BD Biosciences) и корректировали порог подсчета при 10000 событиях. Используя программное обеспечение для анализа FlowJo, файлы FCS, соответствующие каждому обрабатываемому образцу, добавляли в рабочий список, отбирали область (прямой и боковой разброс), в которой была сосредоточена популяция латексных бус, и измеряли флуоресценцию для FITC, связанную с этой областью. Получали таблицу с данными средней интенсивности флуоресценции (MFI) и количеством бус, полученным в закрытой области для каждого проанализированного образца. 20000 отдельных латексных бус исследовали в каждом образце, и использовали MFI для сравнения фракций.
Следуя обоим протоколам, как показано на фиг.1, можно идентифицировать содержащие экзосомы фракции во фракциях, содержащих растворимый белок, подтверждая, дополнительно, что образцы с вирусом и без вируса имеют аналогичный характер элюирования: было обнаружено увеличение сигнала флуоресценции для молекулярных маркеров CD63 и CD81 непосредственно перед тем, как анализ Брэдфорде стал обнаруживать растворимый белок в анализируемых фракциях.
ПРИМЕР 4. АНАЛИЗ БЕЛКОВОГО ПРОФИЛЯ ЭКЗОСОМ
4.1. Анализ распределения и размера экзосом, используя анализ отслеживания наночастиц (NTA)
Использование NTA для количественной оценки, распределения и размера экзосом стал одним из наиболее часто используемых методов в области внеклеточных везикул (http://www.malvern.com/en/products/technology/nanoparticle-tracking-analysis/). Поэтому после подтверждения наличия маркеров, связанных с этих везикулами во фракциях, авторы решили определить количество и размер популяции микровезикул, присутствующих в анализируемых образцах. Для этого каждый анализируемый образец разводили в PBS до тех пор, пока камера NTA (Malvern Instruments Ltd) не зафиксировала значение от 20 до 100 частиц в поле. Достигнув этой идеальной концентрации и разведения, в камеру NTA вводили путем инъекции 400 мкл, и вручную регулировали область фиксации микроскопом. Активировали цифровой термометр и начинали фокусировку частиц с меньшим преломлением, используя микрометр микроскопа в области, ближайшей к периметру лазерного луча. Запускали автоматическую запись видео, и проводили анализ полученного видео, используя программное обеспечение, разработанное поставщиком оборудования. Таким образом, подтверждали, что большинство образцов имели концентрацию везикул в порядке 1010 частиц на миллилитр. Кроме того, измеряли диаметр модового пятна, и он составил от 40 до 150 нм.
4.2. Белковый электрофорез
Для обнаружения наличие белков экзосом и элюированных белков во фракциях, обогащенных экзосомами, и для определения диапазона их молекулярного веса, все выбранные фракции анализировали, используя электрофорез в восстанавливающих условиях и окрашивание серебром. Для этого брали из каждой фракции, собранной в примере 1, брали две аликвоты по 10 мкл, и к каждой из них добавляли 10 мкл буфера для расщепления (Bio-Rad). Затем каждый образец нагружали на предварительно подготовленный 10% SDS-PAGE гель (Bio-Rad). На фигуре 2 показаны полученные результаты. На основании результатов электрофореза можно сделать вывод, что белковая композиция в образцах с вирусом и без вируса отличается. Например, образцы без вируса давали полосу белка приблизительно 15 кДа, которая отсутствовала в образцах с вирусом. Тем не менее, чтобы однозначно показать, что молекулярный состав экзосом, выделенных из животных, которые перенесли заболевание, отличался от состава экзосом, полученных от зараженных вирусом животных, авторы провели протеомическую характеристику, используя жидкостную хроматографию-масс-спектрометрию.
ПРИМЕР 5. ПРОТЕОМИЧЕСКИЙ АНАЛИЗ С ИСПОЛЬЗОВАНИЕМ ЖИДКОСТНОЙ ХРОМАТОГРАФИИ И МАСС-СПЕКТРОМЕТРИИ
Жидкостную хроматографию (nanoLCULTRA-EKSIGENT) с последующей масс-спектрометрией (LC-MS/MS) проводили на оборудовании LTQ Orbitrap Velos (Thermo Fisher). Образцы экзосом в PBS восстанавливали 10 мМ DTT (дитиотреитол), алкилировали 55 мМ йодацетамида и осаждали 10% трихлоруксусной кислотой (TCA), промывали 100% ацетоном и восстанавливали в 2 мл 8 М мочевины. Перед ночной ферментной обработкой трипсином образцы ресуспендировали в 1,6 М растворе мочевины. Реакцию останавливали 1% муравьиной кислотой (об./об.), и трипсинизированные образцы пропускали через предварительную колонку (C18PepMap-100-Thermoscientific-5мм-ID300мкм-5мкм-100A), а затем вводили в аналитическую колонку (AcclaimPepMap100-Thermoscientific-15см-ID75мкм-3мкм-100A-C18). Образцы элюировали со скоростью 400 нл/мин с помощью подвижного фазового градиента: 0-40% растворителя В в растворителе А в течение первых 80-90 минут, а затем 40-100% растворителя В в растворителе А до окончания эксперимента через 100-110 минут (А: 3% ацетонитрил, 0,1% муравьиная кислота в воде, В: 97% ацетонитрил, 0,1% муравьиная кислота в воде). Наносили элюат на источник нанораспыления масс-спектрометра Orbitrap и анализировали полученное на Orbitrap полное сканирование масс-спектра в пределах диапазона масс 400-1500 m/z с разрешением 30000 и максимальным временем впрыска 500 мс. MS/MS проводили в LTQ и 20 более интенсивных пептидов выделяли и фрагментировали, используя 35% CID с низкой энергией соударения. Программное обеспечение Maxquant v1.5 использовали для анализа необработанных данных, используя режим количественного определения без метки (LFQ). Кроме того, для окончательной идентификации авторы использовали поисковую систему для последовательностей, Andromeda (модуль, включенный в программное обеспечение Maxquant v1.5), добавляя набор данных о последовательностях, созданный на основе последовательностей, полученных с сайта UniProtKB, включая все белковые последовательности PRRSV, которые были секвенированы до этого момента (приблизительно 14000 последовательностей).
На основании этого анализа во всех образцах идентифицировали множество вирусных белков. Неожиданно, как показано на диаграмме Венна на фиг.3, авторы настоящего изобретения обнаружили, что молекулярный состав экзосом, продуцирующихся у животного во время или после окончания инфекции, различен. Таким образом, можно сделать вывод о том, что экзосомы, продуцирующиеся у животного (свиньи), которое перенесло болезнь (вирус репродуктивно-респираторного синдрома «PRRSV»), и у которого отсутствуют следы патогена в плазме (201406-6PS и 201406- 10PS), экспрессируют белки: полипротеин репликазы 1ab/сходная с папаином цистеинпротеаза, неописанный предполагаемый белок PRRSV, полипротеин PRRSV, NSP2, GP2b, ORF2a, которые отсутствуют в экзосомах, продуцирующихся у животных во время острой инфекции (201406-4PS и 201406-2PS). Важно отметить, что все белки, идентифицированные Maxquant v1.5, имеют связанную вероятность, известную как PEP или вероятность последующей ошибки, что указывает на вероятность ошибочного определения одного белка при сравнении. Все белки, идентифицированные в этом анализе, имели PEP <0,0001, что подтвердило обоснованность этих результатов.
ПРИМЕР 6. ВЫДЕЛЕНИЕ ЭКЗОСОМ ИЗ КЛЕТОК РЕТИКУЛОЦИТОВ ЧЕЛОВЕКА
Для выделения ретикулоцитов использовали антикоагулированную кровь, взятую у здоровых доноров людей в объеме, достаточном, чтобы дать высокий выход в этом методе. Кровь переносили в 50 мл пробирки и центрифугировали 15 минут при 1000 g, ACC 8/DEC 3, для отделения плазмы (жидкости) от клеток (твердой части). Плазму удаляли, и глобулярную часть (клетки) разводили при 50% гематокрите средой RPMI 1640 и центрифугировали в течение 10 минут при 600 g, ACC 8/DEC 3, супернатант удаляли, и клетки снова ресуспендировали в RPMI 1640 для достижения 50% гематокрита. Для оценки начальной концентрации клеток в образце брали аликвоту (ретикулоциты составляют приблизительно 2% от общего количества клеток).
Чтобы удалить популяцию белых клеток (лейкоцитов) из разведения, глобулярную часть при 50% гематокрите пропускали через колонку CF11 (Whatman, 4021050), так что лейкоциты оставались в матрице из-за своего размера, а эритроциты (группа, в которой могут быть найдены ретикулоцитоы), пропускали через матрицу и собирали в 15 мл-ые пробирки Falcon.
Собранный объем (элюированный из колонки CF11), включая все эритроциты, исследовали визуально для обнаружения сгустков крови, и, если сгустки были найдены, то образец пропускали через нейлоновый фильтр. Затем образец центрифугировали 10 минут при 600 g, ACC 8/DEC 3, супернатант удаляли, и осадок ресуспендировали до 50% гематокрита.
Для обогащения ретикулоцитами использовали разделение в Percoll. Для этого, в 15 мкл-овые пробирки Falcon добавляли Percoll (GE Healthcare, 17-0891-02) (основной раствор Percoll: 9 частей чистого Percoll и 1 часть 1,5 М хлорида натрия - раствор для разделения в Percoll: 70% основного раствора Percoll и 30% хлорида натрия 0,15 М), а затем на стенки пробирки выливали образец, так что ретикулоциты находились в верхней части пробирки. Другую стадию центрифугирования проводили при 1200 g/15 мин/ACC 4/DEC 0, таким образом, ретикулоциты отделяли по плотности от зрелых эритроцитов, и они образовывали кольцо в середине пробирке. Наконец, ретикулоцитарные кольца собирали непосредственно из пробирок с градиентом Percoll, переносили в 15 мл-овые пробирки Falon и промывали два-три раза, добавляя среду RPMI 1640 до максимального объема (15 мл) для удаления следов Percoll при центрифугировании в течение 7 минут при 500 g между каждой стадией промывки средой. Мазки крови готовили как с бриллиантовым крезиловым синим (Sigma), так и с красителем Гимза (Sigma) из образцов, полученных с помощью этого метода в начальной точке (прямой образец), после уменьшения количества лейкоцитов с CF11 (Sigma) и после обогащения с помощью Percoll. Это гарантировало правильную работу всех процессов и тот факт, что критерии воспроизводимости следовали за контролем качества.
Кроме того, суспензию клеток ретикулоцитов культивировали в колбах для культивирования клеток с культуральной средой RPMI 1640 без добавления в течение 36 часов. Из супернатанта клеточной культуры выделяли экзосомы в соответствии с протоколами, описанными в примере 2.
ПРИМЕР 7. ИММУНОГЕННОСТЬ ВИРУСНЫХ БЕЛКОВ, СВЯЗАННЫХ С ЭКЗОСОМАМИ
7.1. Протокол ELISA для титрования антител
Протокол непрямого анализа ELISA использовали для титрования сывороток, полученных у свиней, которые перенесли инфекцию PRRSV (ПЦР (-), Ab (+), «NV»), свиней, у которых обнаружено виремическое состояние (ПЦР (+); Ab (+/-), «V») и свиней, которые никогда не контактировали с вирусом (ПЦР (-), Anb (-), «CN»). Общий протокол непрямого анализа ELISA описан компанией Abcam. Кратко, планшет из поливинилхлорида (ПВХ) с плоским дном покрывали интересующим антигеном, соответствующим аттенуированной вакцине PRRSV, доступной на рынке (Porcilis PRRS Vaccine «intervet», серия A200ED03). Разведение основного раствора антигена осуществляли с помощью карбонатно-бикарбонатного буфера (Na2CO3 0,015M/NaHCO3 0,035M) при pH 9,6 до достижения нормальной концентрации дозы для вакцинации свиней. В каждую лунку добавляли 50 мкл разведенного антигена и инкубировали в течение ночи при 4°C с пластиковым покрытием, чтобы избежать испарения. В конце стадии инкубирования антигена оставшийся объем в лунках удаляли, переворачивая планшет, и планшет 4 раза промывали 200 мкл PBS 1×/0,2% Tween 20. После того как планшет был промыт, проводили стадию блокирования, чтобы заполнить пустые места, где антиген не связался, чтобы избежать неспецифических результатов анализа, добавляя PBS 1×/5% обезжиренное сухое молоко. После блокирования планшет промывали четыре раза PBS 1×/0,2% Tween 20, а сыворотку различных групп инкубировали (NV, V и CN, разводили от 1/5 до 1/5000) в течение 1 часа при комнатной температуре, после чего четыре раза промывали PBS 1×/0,2% Tween 20. Наконец, планшет инкубировали со вторым антителом козел против свиньи (Fc): HRP (AbSerotec, AAI41P) в разведениях от 1/100 до 1/100000 в течение 1 часа при комнатной температуре и защищая от света. Затем проводили четыре промывки PBS 1×/0,2 мкм Tween 20.
Развитие реакции ELISA стимулировали субстратом TMB (3,3,5,5-тетраметилбензидин) от Abcam (Ab142042) в соответствии с инструкциями производителя (15-20 минут развития), и реакцию останавливали, добавляя 2M серную кислоту. Результаты ELISA считывали на Varioskan от Termmo-Scientific при 450 нм. Тому же протоколу следовали в последующих примерах для определения оптической плотности фракции в каждой лунке.
Как видно из результатов, полученных для сывороток разных групп и антител в вышеуказанном анализе, был сделан вывод, что лучшие разведения сывороток для того, чтобы увидеть различия между экспериментальными группами (NV, V и CN), составили от 1/50 до 1/100 в комбинации с разведением второго антитела 1/10000. Эта комбинация факторов была принята в качестве стандарта для последующих анализов, в которых необходимо было оценить способность сывороток разных экспериментальных групп распознавать антиген.
7.2. Распознавание вирусных белков, связанных с экзосомами, полученными у свиней, которые перенесли инфекцию
Одну часть микротитровального PVC планшета с плоским дном покрывали интересующим антигеном, соответствующим аттенуированной вакцине PRRSV, доступной на рынке (Porcilis PRRS Vaccine «intervet», лот A200ED03), а другую часть планшета покрывали 1/10 разведенным антителом мыши против CD63-клона TEA 3/18 человека (любезно предоставлен доктором Francisco Sanchez-Madrid). Основной раствор антигена разводили карбонатно-бикарбонатным буфером (Na2CO3 0,015M/NaHCO3 0,035M) рН 9,6 до достижения дозы вакцинации, а иммобилизованное антитело - 1/10. В каждую лунку планшета ELISA добавляли 50 мкл разведенного антигена/иммобилизованного антитела и инкубировали в течение ночи при температуре 4°С, накрывая липкой пластиковой крышкой, чтобы избежать испарения. В конце стадии инкубации (покрытия) антигена оставшийся объем в лунках удаляли, переворачивая планшет, и планшет 4 раза промывали 200 мкл PBS 1×/0,2% Tween 20. После того как планшет был промыт, проводили стадию блокирования, чтобы блокировать пустые места, где антиген не связался, чтобы избежать неспецифических результатов анализа, добавляя 100 мкл PBS 1×/5% обезжиренное сухое молоко. После блокирования планшет промывали четыре раза PBS 1×/Tween 20 0,2%, а лунки с иммобилизированным антителом инкубировали в течение 90 минут при 37°C с 100 мкл внеклеточных везикул, полученных у животных, перенесших инфекцию PRRSV (выделены полиэтиленгликолем (Sigma- Aldrich, номер в каталоге 81260-1KG)) и, в качестве контроля специфичности, с экзосомами, полученными из ретикулоцитов человека, поскольку PRRS является патогеном, специфичным для свиней; они были получены в соответствии с примером 6. Все экзосомы были получены с помощью эксклюзионной хроматографии, используя матрицу из сефарозы CL-2B (Sigma-Aldrich, номер в каталоге CL2B300-100ML).
Те лунки, в которые была добавлена коммерческая вакцина (использовалась в качестве положительного контроля), закрывали 100 мкл PBS 1× для поддержания минимального объема в лунке, чтобы избежать повреждения белка в результате высыхания. Затем каждую лунку промывали четыре раза 200 мкл PBS 1×/0,2% Tween 20. Затем планшет инкубировали с различными сыворотками (NV, V и CN, разведенными от 1/25 до 1/100) в течение 1 часа при комнатной температуре, затем четыре раза промывали PBS 1×/0,2% Tween 20 (об./об.). Наконец, планшет инкубировали со вторым антителом козел против свиньи (Fc): HRP (AbSerotec, AAI41P) при разведении 1/10000 в течение 1 часа при комнатной температуре и защищая от света, а затем четыре раза промывали PBS 1×/0,2% Tween 20. Развитие реакции ELISA стимулировали субстратом TMB (3,3,5,5-тетраметилбензидин) от Abcam (Ab142042) в соответствии с инструкциями производителя (время развития 15-20 минут), и реакцию останавливали, добавляя 2M серную кислоту (H2SO4). Планшет считывали на Varioskan (Thermo-Scientific) при 450 нм.
Результаты, полученные в сэндвич-анализе ELISA, показали, что оптические плотности везикул, полученных у животных, которые перенесли инфекцию PRRSV, были значительно (p=0,02) выше, чем у везикул, полученных у животных, которые не контактировали с вирусом (отрицательный контроль), и у везикул, полученных из ретикулоцитов человека (контроль специфичности).
Сравнивая эти два контроля с экспериментальной группой, можно наблюдать изменение специфичности распознавания антиген-антитело, сравнивая сыворотку Nv и Pigex Nv, которые отсутствуют у двух контролей (NC, отрицательный контроль и REX, специфичность).
ПРИМЕР 8. МАСШТАБИРУЕМОСТЬ СПОСОБА И ПРОДУКЦИЯ ЭКЗОСОМ, ПОЛУЧЕННЫХ У СВИНЕЙ, ПЕРЕНЕСШИХ ИНФЕКЦИЮ PRRSV
Сначала измеряли общий объем сыворотки, полученной путем разделения при центрифугировании общих образцов крови, собранных непосредственно на ферме у животных, которые перенесли инфекцию PRRSV (как указано в примере 1). После количественного определения общего объема сыворотки концентрацию доводили до конечной концентрации 0,4 М, добавляя полиэтиленгликоль «ПЭГ» (Sigma-Aldrich, номер в каталоге 81260) при 8,5% (масс./об.) и хлорид натрия (Sigma-Aldrich, номер в каталоге S5150-1L). Смесь инкубировали в течение ночи в холодной камере (4°C), перемешивая. После такой инкубации смесь центрифугировали на высокоскоростной центрифуге при 7000 g в течение 10 минут и 4°C. Супернатант удаляли, и осадок ресуспендировали в PBS 1×.
Из полученной суспензии брали аликвоты в 1,5 мл-овые пробирки Eppendorf и хранили при -80°C перед использованием. Аликвоту размером 2 мл использовали для продолжения разделения экзосомы методом эксклюзионной хроматографии (SEC).
Колонку для разделения по вышеуказанному методу готовили, используя шприц объемом 10 мл (одноразовый шприц BD™, номер в каталоге BD302188), в котором наконечник закрывали нейлоновой крышкой, чтобы избежать вытекания матрицы из сефарозы при уплотнении, а также клапан открытие/закрытие для контроля потока элюата, фильтруемого через колонку. После того, как была подготовлена система открытие/закрытие, колонку загружали матрицей из сефарозы CL-2B (Sigma-aldrich CL2B300-100ML) до 10 мл уплотненной матрицы из сефарозы. После завершения подготовки колонки, на колонку воздействовали ультрафиолетовым излучением в течение 10 минут в ламинарном вытяжном шкафу. Параллельно, готовили 1,5 мл-овые пробирки Eppendorf для сбора фракций и их маркировали кодом образца, датой исследования и номером фракции. Затем на колонку наносили образец объемом 2 мкл, полученный путем концентрирования в ПЭГ, и проводили разделение, собирая фракции объемом приблизительно 0,5 мл и добавляя в колонку PBS 1×/0,32% цитрат натрия, чтобы избежать высушивания матрицы. После сбора 20-и фракций объемом 0,5 мл клапан закрывали и фракции анализировали анализом Брэдфорда для количественной оценки концентрации белка в каждой фракции.
После количественного анализа белка по Брэдфорду был получен профиль концентрации белка. Этот профиль имел обнаруживаемый и количественно определяемый белковый пик во фракциях, обогащенных везикулами (небольшое увеличение во фракциях 6-10), уменьшенный после этих фракций и увеличенный во втором пике, соответствующем элюированным фракциям растворимого белка. Растворимые белки меньше, чем везикулы, и, таким образом, у них происходит задержка времени элюирования при эксклюзионной хроматографии. Этот профиль Брэдфорда позволяет определить дозу вакцины, которая может быть использована в клинических испытаниях, и выразить ее в количественных единицах Брэдфорда (мг/мл).
ПРИМЕР 9 МАСШТАБИРУЕМОСТЬ СПОСОБА (ИСПОЛЬЗУЯ РАЗЛИЧНОЕ НАСЫЩЕНИЕ ПРИ ЦЕНТРИФУГИРОВАНИИ) И ПОЛУЧЕНИЕ ЭКЗОСОМ, ПОЛУЧЕННЫХ У СВИНЕЙ, КОТОРЫЕ САМОИЗЛЕЧИЛИСЬ ОТ PRRSV
Сначала образцы крови, полученные у животных, которые, самоизлечились от инфекции PRRSV, центрифугировали при 1800 об/мин (15 минут) (центрифуга Beckman Coulter Allegra X-12R), сыворотки собирали (как указано в примере 1) и измеряли объем. Внеклеточные везикулы отделяли от клеточного дебриса путем центрифугирования при 500 g в течение 15 минут в 50 мл-овых пробирках Falcon. Супернатант сохраняли, а осадок удяляли. Второе центрифугирование в Beckman Coulter Optima XL-100K Ultracentrige (ротор SW-28, класс CDFGH, серия 5E 1427) для удаления микровезикул и апоптотических тел при 15000 g в течение 45 минут проводили в полипропиленовых пробирках (Beckman Coulter Ref.326823 - серия Z51002SCA). Супернатант сохраняли, а осадок удаляли. Наконец, для осаждения внеклеточных везикул и растворимого белка была необходима стадия ультрацентрифугирования (Beckman Coulter Optima XL-100K Ultracentrige и Rotor SW-28, класс CDFGH, серия 5E 1427) при 100000 g в течение 2 часов в полипропиленовых пробирках, в этом случае супернатант удаляли, а осадок ресуспендировали в PBS 1×.
Из полученной суспензии брали аликвоты в 1,5 мл-овые пробирки Eppendorf и хранили при -80°C перед использованием. Для разделения эксклюзионной хроматографией 2 мкл предыдущей суспензии оттаивали и помещали в разделительную колонку. Колонку эксклюзионной хроматографии готовили, используя шприц объемом 10 мл (одноразовый шприц BD™, номер в каталоге BD302188), в котором наконечник закрывали нейлоновой крышкой, чтобы избежать вытекания матрицы при уплотнении, а также клапан открытие/закрытие для контроля потока элюата через колонку. После того, как была подготовлена система открытие/закрытие, в колонку добавляли матрицу из сефарозы CL-2B (Sigma-Aldrich CL2B300-100ML) до 10 мл уплотненной матрицы из сефарозы. Когда процесс упаковки был завершен, колонку перемещали в вытяжной шкаф с ламинарным потоком и подвергали воздействию ультрафиолетового излучения в течение 10 минут, параллельно готовили 1,5 мл-овые пробирки Eppendorf для сбора фракций и их маркировали кодом образца, датой исследования и номером фракции. Затем в колонку вводили 2 мл концентрированного образца, полученного методом дифференциального центрифугирования, и начинали процедуру разделения, собирая фракции, объемом приблизительно 0,5 мл, и добавляя PBS 1×/0,32% цитрат натрия, чтобы избежать высушивания матрицы. После сбора 20-и фракций, клапан открытие/закрытие закрывали, и фракции анализировали с помощью анализа Брэдфорда для количественной оценки концентрации белка в каждой фракции.
После количественного анализа белка по Брэдфорду был получен профиль концентрации белка для каждого образца. Этот профиль имел обнаруживаемый и количественно определяемый белковый пик во фракциях, обогащенных везикулами (небольшое увеличение во фракциях 6-10), уменьшенный после этих фракций и увеличенный во втором пике, соответствующем элюированным фракциям растворимого белка. Растворимые белки меньше, чем везикулы, и, таким образом, у них происходит задержка времени элюирования при эксклюзионной хроматографии. Этот профиль Брэдфорда позволяет определить дозу вакцины, которая может быть использована в клинических испытаниях, и выразить ее в количественных единицах Брэдфорда (мг/мл).
ПРИМЕР 10
10.1. ЖИВОТНЫЕ, ИСПОЛЬЗОВАННЫЕ ДЛЯ ВЫДЕЛЕНИЯ ЭКЗОСОМ, ИХ ХАРАКТЕРИСТИКИ И ПРОДУКЦИИ ВАКЦИНЫ
Образцы сывороток для выделения, молекулярной характеристики и получения вакцин были взяты у животных, которые перенесли инфекцию PRRSV, что было подтверждено (i) ОТ-ПЦР (реагенты и контроль TaqMan® NA и EU PRRSV) для обнаружения циркулирующих вирусных частиц в крови и (ii) путем обнаружения постинфекционных антител (тест IDEXX PRRS X3 Ab). Независимая диагностическая лаборатория по заболеваниям свиней в г.Льейда, Grup de Sanejament Porci of Lleida (GSP), провела эти анализы в соответствии со своими стандартными рабочими методиками. Среди всех животных, оцененных и зарегистрированных по результатам теста UP1570743, три были отобраны для дальнейшего анализа и производства вакцины (таблица 1).
номер
номер фермы
10.2 ПРОТЕОМИЧЕСКИЙ АНАЛИЗ С ИСПОЛЬЗОВАНИЕМ ЖИДКОСТНОЙ ХРОМАТОГРАФИИ И МАСС-СПЕКТРОМЕТРИИ
Жидкостную хроматографию (nanoLCULTRA-EKSIGENT) с последующей масс-спектрометрией (LC-MS/MS) проводили на оборудовании LTQ Orbitrap Velos (Thermo Fisher). Образцы экзосом в PBS 1× восстанавливали 10 мМ DTT (дитиотреитол), алкилировали 55 мМ йодацетамида и осаждали 10% трихлоруксусной кислотой (TCA), промывали 100% ацетоном и восстанавливали в 2 мл 8 М мочевины. Перед ночной ферментной обработкой трипсином образцы ресуспендировали в 1,6 М растворе мочевины. После действия ферментов в течение ночи, реакцию останавливали 1% муравьиной кислотой (об./об.) и трипсинированные образцы пропускали через предварительную колонку (C18PepMap-100-Thermoscientific-5мм-ID300мкм-5мкм-100A), а затем из вводили в аналитическую колонку (AcclaimPepMap100-Thermoscientific-15см-ID75мкм-3мкм-100A-C18). Образцы элюировали со скоростью 400 нл/мин с помощью подвижного фазового градиента: 0-40% растворителя В в растворителе А в течение первых 80-90 минут, а затем 40-100% растворителя В в растворителе А до окончания эксперимента через 100-110 минут (А: 3% ацетонитрил, 0,1% муравьиная кислота в воде, В: 97% ацетонитрил, 0,1% муравьиная кислота в воде). Элюат наносили на источник нанораспыления массспектрометра Orbitrap и анализировали полученное на Orbitrap полное сканирование масс-спектра в пределах диапазона масс 400-1500 m/z с разрешением 30000 и максимальным временем впрыска 500 мс.
MS/MS проводили в LTQ и 20 более интенсивных пептидов выделяли и фрагментировали, используя 35% CID с низкой энергией соударения. Программное обеспечение Maxquant v1.5 использовали для анализа необработанных данных, используя режим количественного определения без метки (LFQ). Кроме того, для окончательной идентификации авторы использовали поисковую систему для последовательностей, Andromeda (модуль, включенный в программное обеспечение Maxquant v1.5), добавляя набор данных о последовательностях, созданный на основе последовательностей, полученных с сайта UniProtKB, включая все белковые последовательности PRRSV, которые были секвенированы до этого момента (приблизительно 14000 последовательностей).
Примеси отфильтровывали, и пептиды считали истинно положительными, если они отвечали следующим критериям: (i) доля ложно-положительных результатов (FDR)=1%, (ii) в независимых друг от друга образцах идентифицировали более двух пептидов одного и того же белка или был идентифицирован уникальный пептид, (iii) пептид должен присутствовать в экзосомах по меньшей мере двух разных животных.
оболочки
10.3. ПРОГНОЗИРОВАНИЕ ЭПИТОПА И КАРТИРОВАНИЕ
Белковые последовательности оценивали с использованием Bepipred 2.0 (http://www.iedb.org/). Bepipred 2.0 прогнозирует расположение линейных B-клеточных эпитопов, используя комбинации скрытой модели Маркова и метода подбора по показателю предрасположенности. Предполагается, что остатки с баллами выше порогового (по умолчанию 0,35) являются частью эпитопа и они окрашены в серый цвет на графике (где Y-оси изображают баллы остатков, а X-ось - положения остатков в последовательности). На значение баллов не влияет выбранное пороговое значение. Пептиды, идентифицированные MS/MS (таблица 2), затем были локализованы для определения того, картированы ли они в прогнозированных областях B-клеточных эпитопов.
10.4. СИНТЕТИЧЕСКИЕ ПЕПТИДЫ
После подписания соглашения о конфиденциальности с ʺPeptide synthesis facility of the Department of Experimental and Health Sciencesʺ при Университете Помпеу Фабра (Барселона, Испания) пептиды синтезировали твердофазным синтезом пептидов Fmoc, Merrifield RB Solid Phase. I. The synthesis of a tetrapeptide, J. Am Chem Soc., 1963, том 85, стр.2149-2154). Таким образом, пептидные цепи собирались поэтапно, по одной аминокислоте за один раз, при этом присоединяясь к нерастворимому полимерному носителю. Это позволило удалить побочные продукты реакции на каждой стадии простой промывкой. Аминокислоты были защищены на своем амино-конце с помощью группы Fmoc (9-флуоренилметоксикарбонил) и присоединены к растущей цепи после активации конца с группой карбоновой кислоты. Группу Fmoc удаляли обработкой пиперидином, и процесс повторяли. После того, как пептид был собран, его снимали с полимера путем обработки трифторуксусной кислотой (TFA). В то же время защитные группы на боковых цепях аминокислот также удаляли с получением неочищенного линейного пептида. Одностадийной очистки с помощью ВЭЖХ с обращенной фазой достаточно для получения пептида с чистотой >95%.
Характеристику каждого синтетического пептида проводили с использованием A-ВЭЖХ с колонкой Luna C18 (4,6 × 50 мм, 3 мкм; Phenomenex), градиент: линейный B (0,036% TFA в MeCN) в A (0,045% TFA в H2O) в течение 15 минут со скоростью потока 1 мл/мин и детекции при 220 нм. Все пептиды ресуспендировали в сверхчистой H2O (вода MiliQ), брали аликвоты и хранили при -20°C перед использованием.
10.5. ИССЛЕДОВАНИЕ С ВАКЦИНАЦИЕЙ НА ФЕРМЕ
Для определения иммуногенности этих пептидов, а также их содержащих экзосом, проводили исследование с вакцинацией свиней на экспериментальном фермерском хозяйстве Университета г. Льейда (фиг. 5). Эти исследования были одобрены этическим комитетом Университета г. Льейда (Испания) и их проводили в соответствии с рекомендациями по уходу за животными этого комитета (DAAM7684). Контрольные сыворотки определяли как образец, собранный в день ноль у каждого животного (пре-иммунные сыворотки - наивные животные).
В таблице 4 показаны экспериментальные группы, тестированные в ходе исследования с вакцинацией. Все животные, используемые в исследовании с вакцинацией, находились в одних и тех же окружающих условиях.
Адъювантом, используемым для проведения вакцинации, был Montanide ISA 206 VG (SEPPIC - серия 36022E/U42131). MONTANIDETM ISA 206 VG представляет собой адъювант на основе минерального масла, который был разработан для получения эмульсий "вода-в-масле-в-воде" (W/O/W). Он содержит высококачественное способное к инъекции минеральное масло и высокоочищенный эмульгатор, полученный из маннита и очищенной олеиновой кислоты растительного происхождения. MONTANIDETM ISA 206 VG не содержит ингредиентов животного происхождения. Препараты вакцины с ним вызывают кратковременный и долгосрочный иммунитет.
По сравнению с обычными эмульсиями, эмульсии MONTANIDETM ISA 206 VG стабильны, имеют низкую вязкость и легко вводятся инъекцией. Было продемонстрировано, что этот адьювант отлично стимулирует гуморальные и клеточные реакции. Этот продукт рекомендован для бактериальных, микоплазменных, вирусных или паразитных антигенов. Committee for Veterinary Medical Products (CVMP) считает адъюванты Montanide™ и их компоненты безопасными для использования в иммунологических продуктах и они включены в качестве разрешенных веществ в приложение к European Council Regulation № 470/2009 (ранее 2377/90/EC), которые не нуждаются в дополнительных исследованиях MRL или уже включены в зарегистрированные ветеринарные коммерческие продукты. Рекомендуемое соотношение для дозы вакцины составляет 1:1 монтанид/антиген вакцины (масс./масс.).
10.6. ИММУНОГЕННОСТЬ ПЕПТИДОВ И ЭКЗОСОМ
Непрямой анализ ELISA использовали для определения иммуногенности различных вакцинных составов. Кратко, на планшеты из поливинилхлорида (ПВХ) с плоским дном наносили 50 мкл аттенуированной вакцины PRRSV (Porcilis PRRS Vaccine «intervet», серия A200ED03) или пептидов, конечную концентрацию 5 мкг/мл получали в карбонатно-бикарбонатном буфере (Na2CO3 0,015M/NaHCO3 0,035M), в качестве покрывающих антигенов. Образцы вакцин разбавляли в карбонатно-бикарбонатном буфере до приблизительной концентрации, соответствующей нормальной дозе вакцины, указанной в протоколе получения вакцины (от 104 до 106 TCID50 для Porcilis PRRSV).
Разведение основных растворов антигенов проводили карбонатно-бикарбонатным буфером (Na2CO3 0,015M/NaHCO3 0,035M) при pH 9,6. В каждую лунку добавляли 50 мкл либо разведенного Porcilis, либо разведенных пептидов, и инкубировали в течение ночи при 4°C с пластиковым покрытием, чтобы избежать испарения. Затем растворы для покрытия удаляли, и планшеты промывали 4 раза 200 мкл PBS 1×/0,2% Tween 20. После промывки, проводили необходимую стадию блокирования с PBS 1×/5% обезжиренное сухое молоко для закрытия всех пустых мест без антигена в лунках, чтобы избежать неспецифического связывания. Проводили четыре стадии промывки PBS 1×/0,2% Tween 20 и инкубацию с сывороткой различных групп, разведенной 1/50 в PBS 1×/0,2% Tween 20 (из анализа иммунизации) в течение 1 часа при комнатной температуре, а затем проводили стадии промывки PBS 1×/0,2% Tween 20. Второе антитело козла против свиньи (Fc): HRP (AbSerotec, AAI41P) использовали в качестве детектирующего антитела при разведении 1/10000 в течение 1 часа при комнатной температуре и защищая от света, после чего проводили четыре стадии промывки PBS 1×/0,2% Tween 20. Развитие реакции ELISA стимулировали субстратом TMB (3,3,5,5-тетраметилбензидин) от Abcam (Ab142042) в соответствии с инструкциями производителя (15-20 минут развития), и реакцию останавливали, добавляя 2M серную кислоту (H2SO4). Планшет считывали на Varioskan (Thermo-Scientific) при 450 нм.
Порог для положительного результата определяли как среднее плюс трехкратное стандартное отклонение (доверие 95%) отрицательного контроля. Порог для положительного результата планшетов, покрытых Porcilis, составлял 0,3, а покрытых пептидами - 0,13 (фиг.6 и 7). Следует отметить, что сыворотки животных 81, 86, 87, 88 и 89, которые были вакцинированы пептидами или экзосомами по изобретению, продуцировали антитела, способные специфически взаимодействовать с антигенами Porcilis, что подтверждало способность как вакцин, содержащих пептидов, так и вакцин, содержащих экзосомы, индуцировать гуморальный иммунитет против антигенов Porcilis.
В настоящее время усиливается потребность в местной ликвидации PRRSV, но для этого требуются надежные вакцины, то есть вакцины, которые не могут снова стать вирулентными и распространиться на не вакцинированных животных и которые могут сохраняться в стадах свиней в долгое время. В идеале, следующее поколение вакцин PRRSV должно также включать маркеры отличия инфицированных животных от и вакцинированных (DIVA). Примечательно, что после вакцинации не было обнаружено антител в тесте IDEXX PRRS X3 Ab при рутинном анализе, проведенном в GSP ни у одного из животных этого исследования. Эти данные показывают, что с помощью подхода с вакцинами на основе экзосом можно отличить инфицированных животных от вакцинированных (DIVA).
10.7. ПРОДУКЦИЯ ГАММА ИНТЕРФЕРОНА ПОСЛЕ ВАКЦИНАЦИИ
Стерильные белые планшеты для анализа ELISPOT (Millipore, номер в каталоге S2EM004M99, серия R4Ma77120) активировали, используя 20 мкл 35% этанол в течение 30 секунд. После удаления этанола планшеты дважды промывали PBS 1× и покрывали 100 мкл анти-IFNg (BD bioscience - BD559961) при разведении 1/100 и инкубировали в течение ночи при 4°C. Растворы для покрытия удаляли, и планшеты промывали 200 мкл блокирующего буфера (BB: RPMI 1640 (Lonza, номер в каталоге BE12-167F)/10% фетальная бычья сыворотка (Life technologies, номер в каталоге 10270106)/1% L-глутамин (Cultek, номер в каталоге H3BE17-605E/1% пенициллин/стрептомицин (Cultek, номер в каталоге H3DE17-603E). После удаления растворов для покрытия планшеты инкубировали с ВВ в течение 2 часов при комнатной температуре. Для получения стимулирующего вещества аттеруированные вирусные частицы (Pyrsvac-183, SYVA) ресуспендировали в полной среде (CM: RPMI 1640 10% фетальная бычья сыворотка «FBS»/1% L-глутамин/1% пенициллин/стрептомицин) до концентрации нормальной дозы вакцины у свиней, одну часть помещали на планшет в нативной конформации, а другую денатурировали нагреванием при 90°C в течение 10 минут. Для пептидов использовали концентрацию 1 мкг каждого пептида на лунку (5 мкг/мл), разведенного в СМ. Положительным контролем был PHA-M в разведении 1/100, а отрицательным контролем был чистый CM. 100 мкл каждого стимулирующего вещества добавляли в планшет, дублируя, и инкубировали перед добавлением клеток.
Выделение мононуклеарных клеток периферической крови (РВМС), сбор цельной крови проводили в пробирках ЭДТА (объем приблизительно 10 мл - BD Bioscience, номер в каталоге 366643). Кровь разводили 1:1 PBS 1× (конечный объем 20 мл) и помещали в количестве свыше 15 мл Ficoll Histopaque®-1077 Hybri-Max™ (Sigma, h8889-500ML) в 50 мл-овые пробирки Falcon, центрифугировали при 1800 об/мин при комнатной температуре AC=9 Dac=3. С помощью пипеток Пастера собирали в 15 мл-овых конических пробирках кольца PBMC в центре градиента Ficoll, и клетки дважды промывали максимальным объемом PBS1× и центрифугировали при 400 g/5 минут при комнатной температуре. Количество клеток и их жизнеспособность оценивали с помощью проточного цитометра и анализа жизнеспособности от BD Pharmigen (PE Annexin V Apoptosis Detection Kit I, номер в каталоге 559763), следуя его протоколу. В цитометрические пробирки добавляли 15 мкл клеточной суспензии, 2 мкл 7AAD и 15 мкл калибровочных бус для количественной оценки клеток. Живые клетки подсчитывали в проточном цитометре (система BD FACSCanto™ II) и определяли концентрацию, используя следующие расчеты.
Клетки/мкл=количество подсчитанных клеток/количество подсчитанных бус (приблизительно 2000)
Общее количество клеток определяли следующим образом:
Общее количество клеток=клетки/мкл * 2000 мкл.
Готовили суспензию клеток для загрузки 500000 клеток/лунка и инкубировали при 37°С и 5% CO2 в течение 48 часов. После завершения инкубации со стимулирующим веществом клетки удаляли, а планшет промывали 200 мкл воды MiliQ. Раствор удаляли, и три раза промывали PBS 1×/0,05% Tween 20 (PBST). После этого 100 мкл моноклонального антитела против IFNg-биотин (BD bioscience - BD559958) разводили 1/250 в PBS 1×/10% FBS (PBSS) и инкубировали в течение 2 часов при комнатной температуре. Раствор для обнаружения антител удаляли, и планшет трижды промывали PBST. 100 мкл конъюгированного стрептавидина-HRPO, разведенного 1/100 в PBSS, помещали в планшет и инкубировали в течение 1 часа при комнатной температуре. Раствор конъюгированного стрептавидина-HRPO удаляли, и планшет четыре раза промывали PBST. Детекцию проводили, добавляя 100 мкл из набора BD ELISPOT AEC для субстрата (BD Bioscience номер в каталоге 551951, серия 4314987) в соответствии с инструкциями производителя. Стадию обнаружения останавливали дистиллированной водой, а затем подсчитывали пятна (таблица 5).
Метод иммуноферментных пятен (ELISPOT) позволяет при соответствующих условиях визуализировать секреторные продукты отдельных активированных или реагирующих клеток. Каждое пятно, которое образуется в анализе, представляет собой одну реактивную клетку. Таким образом, анализ ELISPOT дает как качественную (в отношении конкретного цитокина или другого секретируемой иммунной молекулы), так и количественную (частоту отвечающих клеток в пределах тестируемой популяции) информацию.
В этом конкретном случае были получены данные количественного анализа клеток, продуцирующих интерферон гамма (IFN-g), животных, вакцинированных пептидами или экзосомами (таблица 4), после стимуляции антигенами или пептидами вакцины, связанными с PRRSV.
Важность IFN-гамма в иммунной системе частично связана с его способностью ингибировать репликацию вируса напрямую, и, наиболее важно, с его иммуностимулирующими и иммуномодулирующими эффектами. IFN-гамма продуцируется преимущественно природными киллерами (NK) и природными киллерами Т-клетками (NKT) как часть врожденного иммунитета, а также CD4 Th1 и CD8-цитотоксическими Т-лимфоцитами (CTL), являющимися эффекторными Т-клетками, если развивается антиген-специфический иммунитет.
Как видно из результатов (табл. 5), РВМС, полученные у животных 81, 85 и 86, вакцинированных пептидами по изобретению, способны продуцировать гамма-интерферон, если эти клетки в дальнейшем стимулируют этими антигенами (пептидами). Кроме того, РВМС, полученные у животных 85 (вакцинированы пептидом по изобретению), продуцировали гамма-интерферон в присутствии аттенуированной вирусной частицы PRRSV (вакцина Pyrsvac-183), которая была другим антигеном, чем тот, который использовался для его вакцинации (см. таблицу 4). Наиболее важно, если животные, вакцинированные экзосомами не зараженных вирусом животных (87,90), получали одну стимуляцию пептидами, то эти животные продуцировали интерфон-гамма; таким образом, убедительно подтверждая, что экзосомы содержали и презентировали эти пептиды иммунной системе свиней. Эти данные показывают, что пептидная вакцина способна индуцировать клеточный иммунитет против антигенов PRRSV.
ПРИМЕР 11
11.1. Сбор образцов
Собирали образцы сывороток животных (свиней) с ферм, где была вспышка микоплазмоза, вызванного Mycoplasma suis. На этой ферме наблюдаются ежегодные вспышки инфекции микоплазмой. Чтобы определить, были ли эти животные заражены указанными бактериями, образцы крови анализировали рутинными лабораторными тестами (Zootecnia, Salamanca - Spain), и они дали положительные результаты на Mycoplasma suis в анализе ПЦР и микроскопии (род Mycoplasma) при их проведении в соответствии с их собственными стандартными рабочими процедурами.
Все эти образцы группировали как «излеченные животные» в виду того факта, что образцы, полученные в Innovex Therapeutics, были собраны после того, как болезнь и клинические симптомы исчезли в популяции.
11.2 Выделение экзосом сыворотки
Экзосомы имеют характерный размер частиц 30-100 нм. Следовательно, для сбора этих везикул из разных образцов был использован процесс разделения с использованием эксклюзионной хроматографии с сефарозой CL2B в качестве разделительной матрицы (Boing A. N. et al., «Single-step isolation of extracellular vesicles by size-exclusion chromatographyʺ, 2014, J. Extracell Vesicles, 3). Хотя существуют другие методики получения экзосом, способы с сефарозой лучше очищают экзосомы. Коротко, замороженные 3 мл аликвоты образцов различных сывороток оттаивали на льду и центрифугировали при 500g в течение 10 минут при комнатной температуре для удаления клеточного дебриса. Параллельно, 12 мл шприцы наполняли сефарозой CL-2B (Sigma-Aldrich, St. Louis, MO, USA) до конечного объема 10 мл и уравновешивали фосфатно-солевым буферным раствором (PBS) 0,32% цитрата натрия (масс./об.). Затем, 2 мкл аликвот каждого образца добавляли на отдельные колонки с сефарозой CL-2B и для каждого образца собирали 18-20 фракций по 0,5 мл аликвот.
11.3. Молекулярная характеристика экзосом
После получения различных фракций экзосом наличие везикул подтверждали в анализе концентрации белка, используя анализ Брэдфорда, и в анализе с молекулярными маркерами, проводимым проточной цитометрией.
11.3.1. Анализ Брэдфорда
Концентрацию белка получали с помощью колориметрического анализа, используя метода Брэдфорда (Bradford M. M. ʺA rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye bindingʺ, 1976, Analytical biochemistry, том 7(72), стр. 248-254).
11.3.2. Проточная цитометрия
Параллельно, фракции также анализировали с помощью проточной цитометрии на наличие антигенов CD9, CD63 или CD81, трех тетраспанинов, которые являются специфическими маркерами экзосом (Raposo G. et al., ʺExtracellular vesicles: exosomes, microvesicles, and friendsʺ, 2013, The Journal of cell biology, том 18(200), стр.373-383). Каждую аликвоту обрабатывали по следующему протоколу: к каждой аликвоте добавляли латексные бусы, размером 4 микрон (альдегид сульфат) (Invitrogen, номер в каталоге A37304), и смесь оставляли в покое на 15 минут, а затем добавляли 1 мл буфера BCB (PBS ×1, 0,1% бычьего сывороточного альбумина, 0,01% азида натрия). Полученную смесь инкубировали в течение ночи при комнатной температуре при вращении, а затем инкубировали с первыми антителами (анти-CD63, анти-CD9 и анти-CD81, любезно предоставлены доктором Francisco Sánchez-Madrid) в течение 30 минут при 4°C. Оба антитела использовали в разведении 1:10. После двух этапов промывки с помощью 150 мкл буфера PBS-BSA (фосфатно-солевой буфер/бычий сывороточный альбумин 0,1%) и центрифугирования при 2000 g в течение 10 минут добавляли вторые антитела, конъюгированные с FITC (разведение 1:100) или alexa 488 (разведение 1:1000) (Southern Biotec, номер в каталоге 1032-02), и смесь инкубировали в течение 30 минут при 4°C. После двух дополнительных стадий промывки с помощью 150 мкл буфера PBS-BSA при 0,1% при 2000 g в течение 10 минут латексные бусы ресуспендировали в 100 мкл 0,1% PBS-BSA.
Полученные образцы анализировали с помощью проточной цитометрии, используя проточный цитометр LRSFortessa (BD Biosciences) и корректировали порог подсчета при 10000 событиях. Используя программное обеспечение анализа FlowJo, файлы FCS, соответствующие каждому обрабатываемому образцу, были добавлены в рабочий список, выбирали область (прямой и боковой разброс), в которой была сосредоточена популяция латексных бус, и измеряли флуоресценцию для FITC этой области. Получали таблицу с данными средней интенсивности флуоресценции (MFI) и количеством бус, полученным в закрытой области для каждого проанализированного образца. 20000 отдельных латексных бус исследовали в каждом образце, и использовали MFI для сравнения фракций.
Следуя обоим протоколам, как показано на фиг.8, можно идентифицировать содержащие экзосомы фракции во фракциях, которые содержат растворимый белок, подтверждая, дополнительно, что образцы с вирусом и без вируса имеют аналогичный характер элюирования: было обнаружено увеличение сигнала флуоресценции для молекулярных маркеров CD63 и CD81 непосредственно перед тем, как в анализе Брэдфорде стал обнаруживаться растворимый белок в анализируемых фракциях.
11.4. АНАЛИЗ БЕЛКОВОГО ПРОФИЛЯ ЭКЗОСОМ
11.4.1. Анализ распределения и размера экзосом, используя анализ отслеживания наночастиц (NTA)
Использование NTA для количественной оценки, распределения и размера микровезикул, в частности, экзосом, стал одним из наиболее часто используемых методов в области внеклеточных везикул (http://www.malvern.com/en/products/technology/nanoparticle-tracking-analysis/). Поэтому после подтверждения наличия маркеров, связанных с этими везикулами во фракциях, авторы решили определить количество и размер популяции микровезикул, присутствующих в анализируемых образцах. Для этого каждый анализируемый образец разводили PBS до тех пор, пока камера NTA (Malvern Instruments Ltd) не фиксировала значение от 20 до 100 частиц в поле. Достигнув этой идеальной концентрации и разбавления, в камеру NTA вводили путем инъекции 400 мкл, и вручную регулировали область фиксации микроскопом. Активировали цифровой термометр и начинали фокусировку частиц с меньшим преломлением, используя микрометр микроскопа в области, ближайшей к периметру лазерного луча. Запускали автоматическую запись видео, и проводили анализ полученного видео, используя программное обеспечение, разработанное поставщиком оборудования. Таким образом, подтверждали, что большинство образцов имели концентрацию везикул в порядке 1010 частиц на миллилитр. Кроме того, измеряли диаметр модового пятна, и он составил от 40 до 150 нм.
11.5. ПРОТЕОМИЧЕСКИЙ АНАЛИЗ С ИСПОЛЬЗОВАНИЕМ ЖИДКОСТНОЙ ХРОМАТОГРАФИИ И МАСС-СПЕКТРОМЕТРИИ
Жидкостную хроматографию (nanoLCULTRA-EKSIGENT) с последующей масс-спектрометрией (LC-MS/MS) проводили на оборудовании LTQ Orbitrap Velos (Thermo Fisher). Образцы экзосом в PBS восстанавливали 10 мМ DTT (дитиотреитол), алкилировали 55 мМ йодацетамида и осаждали 10% трихлоруксусной кислотой (TCA), промывали 100% ацетоном и восстанавливали в 2 мл 8 М мочевины. Перед ночной ферментной обработкой трипсином образцы ресуспендировали в 1,6 М растворе мочевины. Реакцию останавливали 1% муравьиной кислотой (об./об.) и трипсинированные образцы пропускали через предварительную колонку (C18PepMap-100-Thermoscientific-5мм-ID300мкм-5мкм-100A), а затем вводили инъекцией в аналитическую колонку (AcclaimPepMap100-Thermoscientific-15см-ID75мкм-3мкм-100A-C18). Образцы элюировали со скоростью 400 нл/мин с помощью подвижного фазового градиента: 0-40% растворителя В в растворителе А в течение первых 80-90 минут, а затем 40-100% растворителя В в растворителе А до окончания эксперимента через 100-110 минут (А: 3% ацетонитрил, 0,1% муравьиная кислота в воде, В: 97% ацетонитрил, 0,1% муравьиная кислота в воде). Наносили элюат на источник нанораспыления массспектрометра Orbitrap и анализировали полученное на Orbitrap полное сканирование масс-спектра в пределах диапазона масс 400-1500 m/z с разрешением 30000 и максимальным временем впрыска 500 мс. MS/MS проводили в LTQ и 20 более интенсивных пептидов выделяли и фрагментировали, используя 35% CID с низкой энергией соударения. Программное обеспечение Maxquant v1.5 использовали для анализа необработанных данных, используя режим количественного определения без метки (LFQ). Кроме того, для окончательной идентификации авторы использовали поисковую систему для последовательностей, Andromeda (модуль, включенный в программное обеспечение Maxquant v1.5), добавляя набор данных о последовательностях, созданный на основе последовательностей, полученных с сайта UniProtKB, включая все белковые последовательности Mycoplasma suis, которые были секвенированы до этого момента (приблизительно 4566 последовательностей).
На основании этого анализа было идентифицировано два бактериальных белка и два из них с более чем двумя уникальными пептидами. Таким образом, можно сделать вывод, что экзосомы, продуцируемые у животного (свинья), которое перенесло болезнь (микоплазмоз), и у которого не обнаруживается следов патогена, экспрессируют белки, показанные в таблице ниже. Важно отметить, что все белки, идентифицированные Maxquant v1.5, имеют связанную вероятность, известную как PEP или вероятность последующей ошибки, что указывает на вероятность ошибочного определения одного белка при сравнении. Все белки, идентифицированные в этом анализе, имели PEP <0,0001, что подтвердило обоснованность этих результатов.
ПРИМЕР 12
12.1 Сбор образцов
Собирали образцы сывороток у животных (коровы породы Frisona) с ферм, на которых была вспышка тейлериоза. На этой ферме наблюдаются ежегодные вспышки инфекции тейлериоза (эндемическая вспышка на острове Менорка). Вспышка была между октябрем и декабрем 2015 года. Чтобы определить, были ли эти животные инфицированы указанным паразитом, образцы крови анализировали рутинными лабораторными тестами (Menorca) и они дали положительные (от умеренных до высоких) результаты на Theileria sp. Большинство испытуемых животных умерли после лабораторного диагноза и имели такие симптомы, как высокая температура, желтуха, анорексия, прострация и потеря молока у кормящих коров.
Все эти образцы были сгруппированы как «излеченные животные» в виду того факта, что они выздоровели после лечения.
декабрь
декабрь
12.2 ВЫДЕЛЕНИЕ ЭКЗОСОМ СЫВОРОТКИ
Экзосомы имеют характерный размер частиц 30-100 нм. Следовательно, для сбора этих везикул из разных образцов был использован процесс разделения с использованием эксклюзионной хроматографии с сефарозой CL2B в качестве разделительной матрицы (Boing A. N. et al., ʺSingle-step isolation of extracellular vesicles by size-exclusion chromatographyʺ, 2014, J. Extracell Vesicles, 3). Хотя существуют другие методики получения экзосом, способы с сефарозой лучше очищают экзосомы. Кратко, замороженные 3 мл аликвоты разных образцов сыворотки оттаивали на льду и центрифугировали при 500 g в течение 10 минут при комнатной температуре, чтобы избавиться от клеточного дебриса. Параллельно, 12 мл шприцы наполняли CL-2B (Sigma-Aldrich, St. Louis, MO, USA) до конечного объема 10 мл и уравновешивали фосфатно-солевым буферным раствором (PBS) 0,32% цитрата натрия (масс./об.). Затем, 2 мкл аликвоты каждого образца добавляли на отдельные колонки с сефарозой CL-2B и для каждого образца собирали 18-20 фракций по 0,5 мл аликвот.
12.3 МОЛЕКУЛЯРНАЯ ХАРАКТЕРИСТИКА ЭКЗОСОМ
После получения различных фракций экзосом наличие везикул подтверждали в анализе концентрации белка, используя анализ Брэдфорда, и в анализе с молекулярными маркерами, проводимым проточной цитометрией.
12.3.1. Анализ Брэдфорда
Концентрацию белка получали с помощью колориметрического анализа, используя метода Брэдфорда (Bradford M. M. ʺA rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye bindingʺ, 1976, Analytical biochemistry, том 7(72), стр.248-254).
12.3.2. Проточная цитометрия
Параллельно, фракции также анализировали с помощью проточной цитометрии на наличие антигенов CD63 или CD81, двух тетрасанинов, которые являются специфическими маркерами экзосом (Raposo G. et al., ʺExtracellular vesicles: exosomes, microvesicles, and friendsʺ, 2013, The Journal of cell biology, vol. 18(200), стр.373-383). Каждую аликвоту подвергали следующему протоколу: к каждой аликвоте добавляли латексные бусы размером 4 микрона (альдегид сульфат) (Invitrogen, номер в каталоге A37304), и смесь оставляли на 15 минут в условиях покоя, а затем добавляли 1 мл буфера BCB (PBS 1×, 0,1% бычьего сывороточного альбумина, 0,01% азида натрия). Полученную смесь инкубировали в течение ночи при комнатной температуре при вращении перед инкубацией с первичными антителами (анти-CD63 и анти-CD81, любезно предоставлены доктором Francisco Sánchez-Madrid) в течение 30 минут при 4°C. Оба антитела использовали в разведении 1:10. После двух этапов промывки 150 мкл буфера PBS-BSA (фосфатно-солевой буфер/бычий сывороточный альбумин 0,1%) и центрифугирования при 2000 g в течение 10 минут добавляли вторые антитела, конъюгированные с FITC (разведение 1:100) или alexa 488 (разведение 1:1000) (Southern Biotec, номер в каталоге 1032-02), и смесь инкубировали в течение 30 минут при 4°C. После двух дополнительных стадий промывки 150 мкл буфера PBS-BSA при 0,1% при 2000 g в течение 10 минут латексные бусы ресуспендировали в 100 мкл 0,1% PBS-BSA.
Полученные образцы анализировали с помощью проточной цитометрии, используя проточный цитометр LRSFortessa (BD Biosciences) и корректировали порог подсчета при 10000 событиях. Используя программное обеспечение для анализа FlowJo, файлы FCS, соответствующие каждому обрабатываемому образцу, добавляли в рабочий список, отбирали область (прямой и боковой разброс), где была сосредоточена популяция латексных бус, была измерена флуоресценция для FITC, связанная с этой областью. Получали таблицу с данными средней интенсивности флуоресценции (MFI) и количеством бус, полученным в закрытой области для каждого проанализированного образца. 20000 отдельных латексных бус исследовали в каждом образце, и использовали MFI для сравнения фракций.
Следуя обоим протоколам, как показано на фиг.9, можно идентифицировать содержащие экзосомы фракции во фракциях, которые содержат растворимый белок, подтверждая, дополнительно, что образцы с вирусом и без вируса имеют аналогичный характер элюирования: было обнаружено увеличение сигнала флуоресценции для молекулярных маркеров CD63 и CD81 непосредственно перед тем, как в анализе Брэдфорде стал обнаруживаться растворимый белок в анализируемых фракциях.
12.4. АНАЛИЗ БЕЛКОВОГО ПРОФИЛЯ ЭКЗОСОМ
12.4.1. Анализ распределения и размера экзосом, используя анализ отслеживания наночастиц (NTA)
Использование NTA для количественной оценки, распределения и размера микровезикул, в частности, экзосом, стал одним из наиболее часто используемых методов в области внеклеточных везикул (http://www.malvern.com/en/products/technology/nanoparticle-tracking-analysis/). Поэтому после подтверждения наличия маркеров, связанных с этими везикулами во фракциях, авторы решили определить количество и размер популяции микровезикул, присутствующих в анализируемых образцах. Для этого каждый анализируемый образец разбавляли в PBS до тех пор, пока камера NTA (Malvern Instruments Ltd) не фиксировала значение от 20 до 100 частиц в поле. Достигнув этой идеальной концентрации и разбавления, в камеру NTA вводили путем инъекции 400 мкл (!!!), и вручную регулировали область фиксации микроскопом. Активировали цифровой термометр и начинали фокусировку частиц с меньшим преломлением, используя микрометр микроскопа в области, ближайшей к периметру лазерного луча. Запускали автоматическую запись видео, и проводили анализ полученного видео, используя программное обеспечение, разработанное поставщиком оборудования. Таким образом, подтверждали, что большинство образцов имели концентрацию везикул в порядке 1010 частиц на миллилитр. Кроме того, измеряли диаметр модового пятна, и он составил от 40 до 150 нм.
12.5. ПРОТЕОМИЧЕСКИЙ АНАЛИЗ С ИСПОЛЬЗОВАНИЕМ ЖИДКОСТНОЙ ХРОМАТОГРАФИИ И МАСС-СПЕКТРОМЕТРИИ
Жидкостную хроматографию (nanoLCULTRA-EKSIGENT) с последующей масс-спектрометрией (LC-MS/MS) проводили на оборудовании LTQ Orbitrap Velos (Thermo Fisher). Образцы экзосом в PBS восстанавливали 10 мМ DTT (дитиотреитол), алкилировали 55 мМ йодацетамидом и осаждали 10% трихлоруксусной кислотой (TCA), промывали 100% ацетоном и восстанавливали в 2 мл 8 М мочевины. Перед ночной ферментной обработкой трипсином образцы ресуспендировали в 1,6 М растворе мочевины. Реакцию останавливали 1% муравьиной кислотой (об./об.), и трипсинированные образцы пропускали через предварительную колонку (C18PepMap-100-Thermoscientific-5мм-ID300мкм-5мкм-100A), а затем вводили в аналитическую колонку (AcclaimPepMap100-Thermoscientific-15см-ID75мкм-3мкм-100A-C18). Образцы элюировали со скоростью 400 нл/мин с помощью подвижного фазового градиента: 0-40% растворителя В в растворителе А в течение первых 80-90 минут, а затем 40-100% растворителя В в растворителе А до окончания эксперимента через 100-110 минут (А: 3% ацетонитрил, 0,1% муравьиная кислота в воде, В: 97% ацетонитрил, 0,1% муравьиная кислота в воде). Наносили элюат на источник нанораспыления массспектрометра Orbitrap и анализировали полученное на Orbitrap полное сканирование масс-спектра в пределах диапазона масс 400-1500 m/z с разрешением 30000 и максимальным временем впрыска 500 мс. MS/MS проводили в LTQ и 20 более интенсивных пептидов выделяли и фрагментировали, используя 35% CID с низкой энергией соударения. Программное обеспечение Maxquant v1.5 использовали для анализа необработанных данных, используя режим количественного определения без метки (LFQ). Кроме того, для окончательной идентификации авторы использовали поисковую систему для последовательностей, Andromeda (модуль, включенный в программное обеспечение Maxquant v1.5), добавляя набор данных о последовательностях, созданный на основе последовательностей, полученных с сайта UniProtKB, включая все белковые последовательности Theileria sp., которые были секвенированы до этого момента (приблизительно 18779 последовательностей).
На основании этого анализа было идентифицировано пять белков паразита и два из них с более чем двумя уникальными пептидами. Таким образом, можно сделать вывод, что экзосомы, продуцируемые у животного (корова), которое перенесло заболевание (тейлериоз), и у которого отсутствуют следы патогена, экспрессируют белки, показанные в таблице ниже. Важно отметить, что все белки, идентифицированные Maxquant v1.5, имеют связанную вероятность, известную как PEP или вероятность последующей ошибки, что указывает на вероятность ошибочного определения одного белка при сравнении. Все белки, идентифицированные в этом анализе, имели PEP <0,0001, что подтвердило обоснованность этих результатов.
| название | год | авторы | номер документа |
|---|---|---|---|
| СЛИТЫЕ БЕЛКИ ДЛЯ ПРИМЕНЕНИЯ В КАЧЕСТВЕ ИММУНОГЕННЫХ УСИЛИВАЮЩИХ АГЕНТОВ ДЛЯ ИНДУЦИРОВАНИЯ АНТИГЕНСПЕЦИФИЧЕСКОГО Т-КЛЕТОЧНОГО ОТВЕТА | 2013 |
|
RU2619187C2 |
| ВЕЗИКУЛЫ, СОДЕРЖАЩИЕ ИНГИБИТОР PTEN, И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2800729C2 |
| АДЕНОВИРУС, СОДЕРЖАЩИЙ АЛЬБУМИН-СВЯЗЫВАЮЩИЙ УЧАСТОК | 2015 |
|
RU2711371C2 |
| ВИРУСОПОДОБНЫЕ ЧАСТИЦЫ С ВЫСОКОПЛОТНЫМ ПОКРЫТИЕМ ДЛЯ ИНДУКЦИИ ЭКСПРЕССИИ АНТИТЕЛ | 2017 |
|
RU2813282C2 |
| СЛИТЫЕ БЕЛКИ ДЛЯ ПРИМЕНЕНИЯ В КАЧЕСТВЕ ИММУНОГЕННЫХ УСИЛИВАЮЩИХ АГЕНТОВ ДЛЯ ИНДУЦИРОВАНИЯ АНТИГЕНСПЕЦИФИЧЕСКОГО Т-КЛЕТОЧНОГО ОТВЕТА | 2013 |
|
RU2631002C2 |
| НОВЫЙ ПТИЧИЙ АСТРОВИРУС | 2009 |
|
RU2547589C2 |
| ИММУНОГЕНЫ ДЛЯ ВАКЦИНАЦИИ ПРОТИВ ВИЧ | 2013 |
|
RU2721274C2 |
| СЛИТЫЕ ПОЛИПЕПТИДЫ И ВАКЦИНЫ | 2014 |
|
RU2699006C2 |
| ИММУНОГЕНЫ ДЛЯ ВАКЦИНАЦИИ ПРОТИВ ВИЧ | 2013 |
|
RU2648791C2 |
| СПОСОБ БЫСТРОГО ОТБОРА ВАРИАНТОВ GP-120 ВИЧ | 2012 |
|
RU2603732C2 |




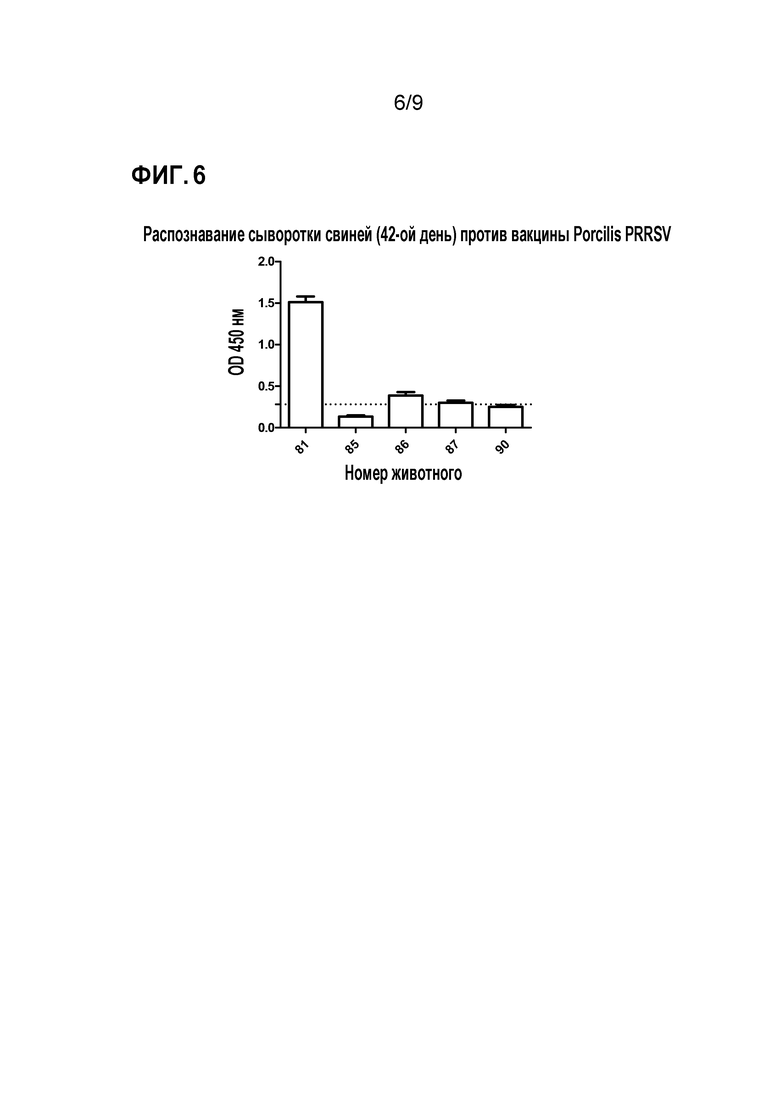
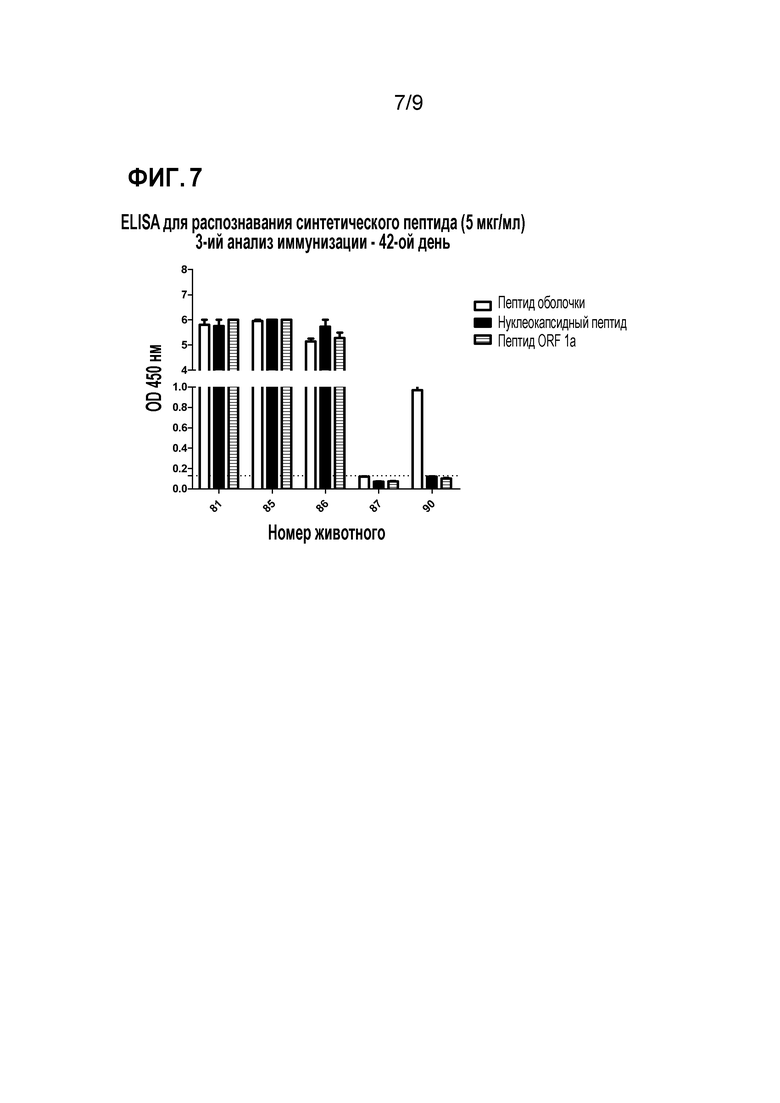
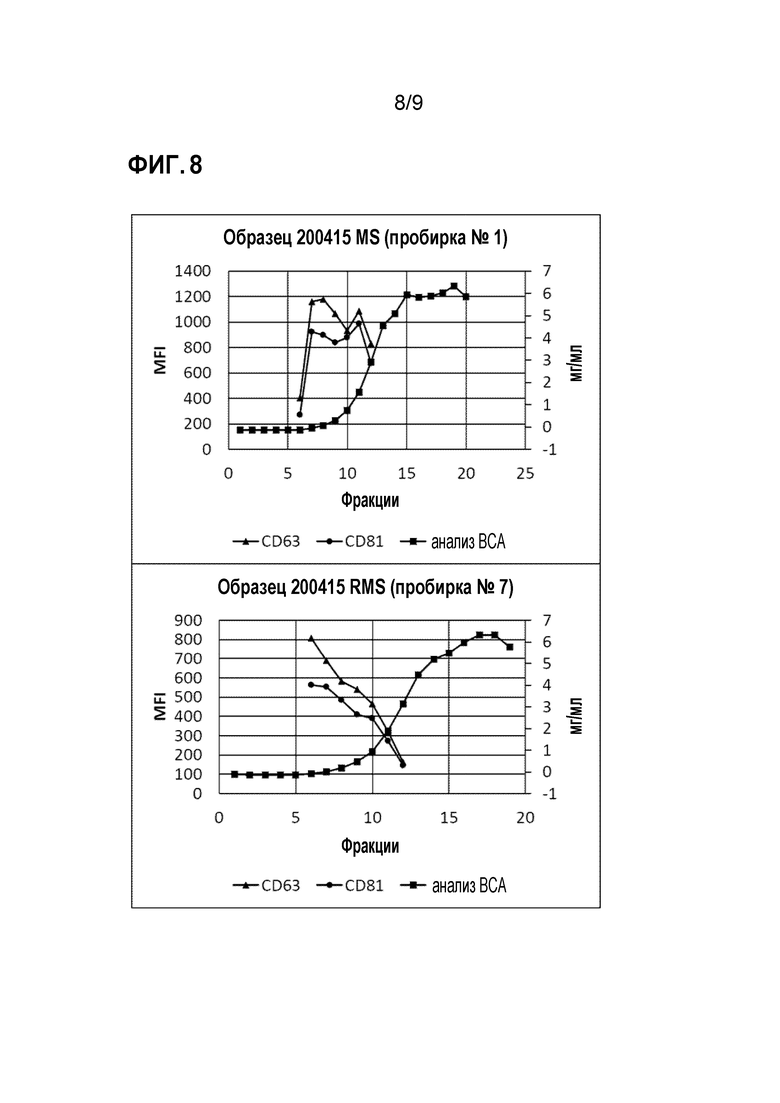
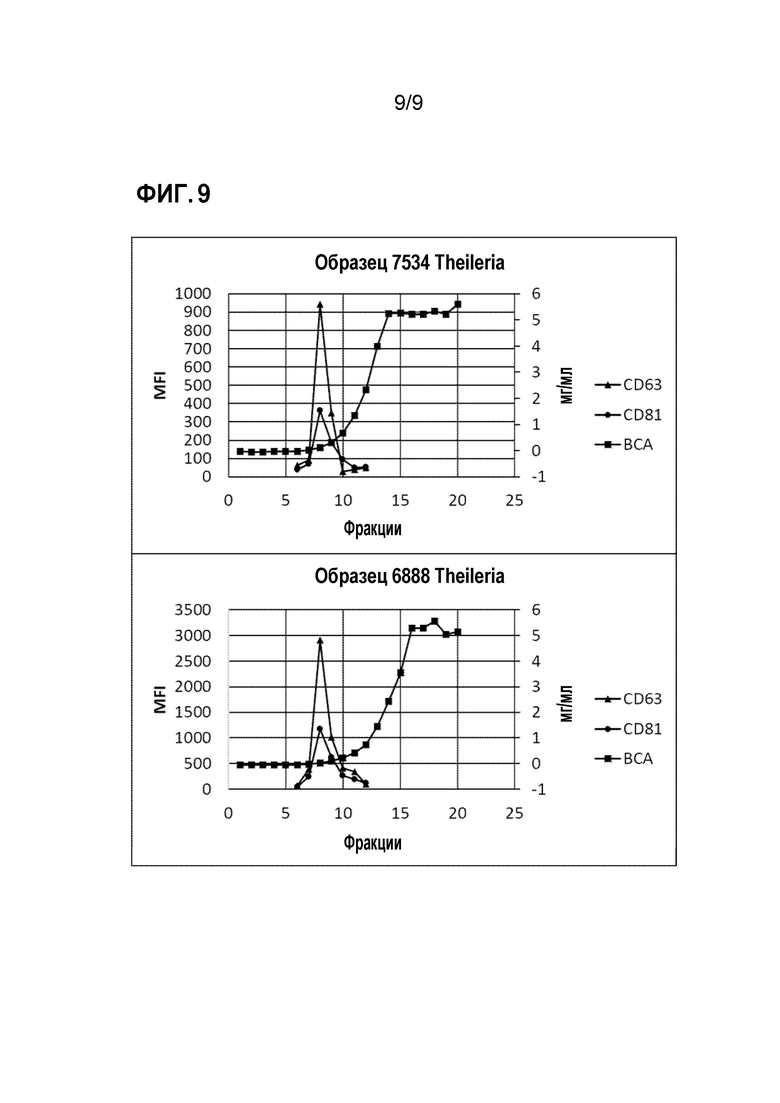
Изобретение относится к области биотехнологии, конкретно к получению экзосом с иммуногенными пептидами от свиней, перенесших респираторное заболевание, вызванное вирусом репродуктивно-респираторного синдрома свиней (PRRSV), и может быть использовано в ветеринарии для лечения или профилактики инфекционного заболевания, вызванного PRRSV. Из образца плазмы или сыворотки, в котором отсутствует PRRSV, получают свободный от PRRSV иммуногенный обогащенный препарат иммуногенных экзосом, которые содержат иммунногенный пептид PRRSV с SEQ ID NO:18. Указанный образец взят у свиньи, не имеющей вируса, которая (а) перенесла респираторное заболевание, вызванное вирусом PRRSV, и (b) у которой отсутствует PRRSV, вызывающий указанное заболевание; где признаки (а) и (b) определяют в указанном образце свиньи, осуществляя ОТ-ПЦР и выявляя антитела против PRRSV. При этом животное перенесло указанное заболевание и свободно от патогена, если ОТ-ПЦР является отрицательной, но определяются антитела против PRRSV. Изобретение обеспечивает индукцию иммунного ответа против PRRSV свиней. 6 н. и 8 з.п. ф-лы, 9 ил., 9 табл., 12 пр.
1. Свободный от вируса репродуктивно-респираторного синдрома свиней (PRRSV) иммуногенный обогащенный препарат иммуногенных экзосом для индукции иммунного ответа против PRRSV свиней,
где экзосомы содержат иммунногенный пептид PRRSV с последовательностью SEQ ID NO:18;
где экзосомы выделены из образца плазмы или сыворотки, в котором отсутствует PRRSV, где образец взят у свиньи, не имеющей вируса, и которая (а) перенесла респираторное заболевание, вызванное вирусом PRRSV, и (b) у которой отсутствует PRRSV, вызывающий указанное заболевание; где
признаки (а) и (b) определяют в указанном образце свиньи, осуществляя ОТ-ПЦР и выявляя антитела против PRRSV,
животное перенесло указанное заболевание и свободно от патогена, если ОТ-ПЦР является отрицательной, но определяются антитела против PRRSV.
2. Препарат по п.1, где свинья является сельскохозяйственным животным.
3. Препарат по п.1, где экзосома дополнительно содержит цистеинпротеазу c1ab, сходную с папаином, полипротеин PRRSV, белки NSP2, GP2b и ORF2a.
4. Способ получения препарата по любому из пп.1-3, где способ включает выделение экзосом из образца плазмы или сыворотки, в котором отсутствует PRRSV, где образец взят у свиньи, не имеющей вируса, которая (а) перенесла респираторное заболевание, вызванное вирусом PRRSV, и (b) у которой отсутствует PRRSV, вызывающий указанное заболевание,
где экзосомы содержат иммунногенный пептид PRRSV с последовательностью SEQ ID NO:18;
где экзосомы выделены с помощью эксклюзионной хроматографии, ультрацентрифугирования или фильтрации;
признаки (а) и (b) определяют в указанном образце свиньи, осуществляя ОТ-ПЦР и выявляя антитела против PRRSV;
животное перенесло указанное заболевание и свободно от патогена, если ОТ-ПЦР является отрицательной, но определяются антитела против PRRSV.
5. Способ по п.4, где выделенным образцом является образец сыворотки.
6. Способ по п.4 или п.5, где экзосомы выделены с помощью эксклюзионной хроматографии.
7. Ветеринарная композиция для лечения или профилактики инфекционного заболевания, вызванного PRRSV, содержащая эффективное количество препарата по любому из пп.1-4, и один или несколько приемлемых в ветеринарии эксципиентов.
8. Композиция по п.7, которая является вакциной.
9. Препарат по любому из пп.1-3 для применения в качестве иммуногена.
10. Препарат по любому из пп.1-3 для применения в качестве лекарственного средства.
11. Препарат по любому из пп.1-3 для применения в лечении или профилактики заболевания, вызываемого PRRSV.
12. Способ идентификации пептида, кандидата в иммуногены для заболевания, вызванного PRRSV, где способ включает:
(a) анализ препарата по любому из пп.1-3 на белковые компоненты экзосомы; и
(b) сравнение белкового профиля, полученного на стадии (a) с белковым профилем экзосомы, выделенной у животного, страдающего тем же заболеванием, вызванным PRRSV, что и заболевание, вызванное PRRSV, которое перенесла свинья, не имеющая вируса, из которой выделена экзосома на стадии (a);
где, если пептид включен в экзосому указанного препарата, а не в экзосому инфицированного животного, то это указывает на то, что пептид является кандидатом в иммуноген для заболевания, вызванного PRRSV.
13. Применение препарата по любому из пп.1-3 для проведения различия между животными, вакцинированными препаратом по любому из пп.1-3, и животными, инфицированными тем же патогеном, который вызывает заболевание PRRSV.
14. Способ повышения иммуногенности против PRRSV, где способ включает введение терапевтически эффективного количества препарата по любому из пп.1-3 пациенту.
| HINA KALRA et al | |||
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| Emerging Roles of Exosomes in Normal and Pathological Conditions: New Insights for Diagnosis and Therapeutic | |||

