ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[001] По настоящей заявке испрашивается приоритет предварительным заявкам на патент США № 62/301538, поданной 29 февраля 2016 года, и 62/415430, поданной 31 октября 2016 года, содержание каждой из которых включено в настоящее описание в качестве ссылки в полном объеме.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
[002] Изобретение относится к новым способам, композициям и устройствам для хранения и извлечения информации с использованием устройств с нанопорами для синтеза и секвенирования полимеров, например, нуклеиновых кислот.
УРОВЕНЬ ТЕХНИКИ
[003] Существует постоянная необходимость в хранении все большего количества данных на или в физических носителях, причем устройства для хранения становятся все в большей степени малыми по размеру, а их вместимость возрастает. Согласно имеющейся информации, количество хранимых данных удваивается каждые два года и, согласно одному из исследований, к 2020 году количество данных, которые создаются и копируются ежегодно, достигнут 44 зетабайт или 44 триллиона гигабайт. Более того, существующие носители для хранения данных, такие как жесткие диски, оптические носители и магнитные ленты, являются относительно нестабильными и повреждаются в результате длительного хранения.
[004] Существует острая необходимость в альтернативных подходах для хранения больших объемов данных в течение длительных периодов времени, например, десятилетий или веков.
[005] Было предложено использовать ДНК для хранения данных. ДНК чрезвычайно стабильна и в теории может кодировать огромные количества данных и хранить данных в течение очень длительных периодов времени. См., например, Bancroft, C., et al., Long-Термин Storage of Information in DNA, Science (2001) 293: 1763-1765. Кроме того, ДНК в качестве носителя для хранения информации не подвержена рискам недостаточного обеспечения безопасности, присущим традиционным цифровым носителям для хранения информации. Однако отсутствует практический подход для осуществления этой идеи.
[006] В WO 2014/014991, например, описан способ хранения данных на ДНК-олигонуклеотидах, где информация кодируется в двоичном формате, по одному биту на нуклеотид, с блоком данных из 96 бит (96 нуклеотидов), адресной последовательностью из 19 нуклеотидов и фланкирующими последовательностями для амплификации и секвенирования. Затем код считывается путем амплификации последовательностей с использованием ПЦР и секвенирования с использованием высокоскоростного секвенатора, такого как устройство Illumina HiSeq. Затем последовательности блоков данных организуются в правильном порядке с использованием адресных меток, адресные и фланкирующие последовательности отфильтровываются и данные последовательностей транслируются в двоичный код. Такой подход имеет значительные ограничения. Например, 96-битный блок данных может кодировать только 12 букв (с использованием общепринятого одного байта или 8 бит на букву или пробел). Соотношение хранимой полезной информации и информации "домашнего хозяйства" является низким - приблизительно 40% информации последовательности занимают адресная и фланкирующая ДНК. В описании описано кодирование книги с использованием 54898 олигонуклеотидов. Распечатанные на струйном принтере высокоточные ДНК-микрочипы, используемые для синтеза олигонуклеотидов, ограничивали размер олигонуклеотидов (описанные 159-меры были верхним пределом). Более того, считывание олигонуклеотидов требует амплификации и выделения, что приводит к дополнительному потенциалу ошибки. Также см. WO 2004/088585A2; WO 03/025123 A2; C. BANCROFT: "Long-Term Storage of Information in DNA", Science (2001) 293 (5536): 1763c-1765; COX J P L: "Long-термин data storage in DNA", Trends in Biotechnology (2001)19(7): 247-250.
[007] В то время как потенциальная плотность информации и стабильность ДНК делает ее привлекательным носителем для хранения данных, как признают на протяжении двадцати пяти лет, все еще отсутствует практический подход для записывания и считывания больших количеств данных в этой форме.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[008] Авторы настоящего изобретения разработали новый подход для хранения нуклеиновых кислот с использованием нанофлюидных систем для синтеза последовательностей нуклеиновых кислот и устройств считывания с нанопорами для считывания последовательностей. Подход, разработанный авторами изобретения, позволяет синтез, хранение и считывание цепей ДНК длиной сотни, тысячи и даже миллионы оснований. Поскольку последовательности являются длинными, только относительно небольшая часть последовательности отводится для идентификации информации, так что плотность информации является значительно более высокой, чем в описанном выше подходе. Более того, в некоторых вариантах осуществления синтезированная нуклеиновая кислота имеет конкретное положение на наночипе, так что последовательность может быть идентифицирована даже без идентифицирующей информации. Секвенирование, проводимое в нанокамерах, является очень быстрым, и считывание последовательности через нанопору может быть чрезвычайно быстрым, составляя порядка вплоть до одного миллиона оснований в секунду. Поскольку требуется только два типа оснований, секвенирование может быть более быстрым и более точным, чем методики секвенирования, которые должны различить четыре типа нуклеотидных оснований (аденин, тимин, цитозин, гуанин). В конкретных вариантах осуществления два основания не образуют пару друг с другом и не образуют вторичные структуры, и также имеют различные размеры. Например, аденин и цитозин будут лучшими для этой цели, чем аденин и тимин, которые имеют тенденцию к гибридизации, или аденин и гуанин, которые имеют сходный размер.
[009] В некоторых вариантах осуществления эту систему можно использовать для синтеза длинных полимеров, кодирующих данные, которые можно амплифицировать и/или извлекать, а затем секвенировать на различных секвенаторах. В других вариантах осуществления систему можно использовать для предоставления специализированных последовательностей ДНК. В других вариантах осуществления систему можно использовать для считывания последовательностей ДНК.
[011] Таким образом, изобретение относится, среди прочего, к следующим вариантам осуществления:
- наночип для синтеза электрически заряженного полимера, например ДНК, содержащего по меньшей мере два различных мономера, причем наночип содержит две или более реакционных камеры, разделенных одной или более нанопорами, где каждая реакционная камера содержит электролитическую жидкость, один или более электродов для втягивания электрически заряженного полимера в камеру и один или более реагентов для облегчения присоединения мономеров или олигомеров к полимеру. Наночип необязательно может быть оборудован функциональными элементами для направления, перемещения и/или контроля ДНК, необязательно он может быть покрыт или изготовлен с материалами, позволяющими плавное течение ДНК или прикрепление ДНК, и он может содержать элементы наноконтура для обеспечения и контроля электродов вблизи нанопор. Например, каждая из одной или более нанопор может быть сопряжена с электродами, которые могут контролировать прохождение полимера через нанопору и/или детектировать изменения электрического потенциала, тока, сопротивления или емкости на поверхности контакта нанопоры и полимера, тем самым определяя последовательность полимера по мере его прохождения через одну или более нанопор. В конкретных вариантах осуществления олигомеры синтезируются использованием полимераз или сайт-специфических рекомбиназ. В некоторых вариантах осуществления полимер секвенируется в ходе синтеза, чтобы обеспечить детектирование и необязательно коррекцию ошибок. В некоторых вариантах осуществления полимер, полученный таким образом, хранится на наночипе и может секвенироваться, когда желателен доступ к информации, закодированной в последовательности полимера.
- Способ синтеза полимера, например ДНК, с использованием наночипа, как описано.
- Одноцепочечная молекула ДНК, где последовательность по существу состоит из негибридизующихся нуклеотидов, например, аденина и цитозина (A и C), которые организованы в виде последовательности, соответствующей двоичному коду, например, для применения в способе хранения данных.
- Двухцепочечная ДНК, содержащая серию нуклеотидных последовательностей, соответствующих двоичному коду, где двухцепочечная ДНК дополнительно содержит
- Способ считывания двоичного кода, закодированного в ДНК, включающий использование секвенатора с нанопорами.
- Способ хранения данных и устройство для этого с использованием описанного выше наночипа для получения электрически заряженного полимера, например ДНК, содержащей по меньшей мере два различных мономера, где мономеры организованы в виде последовательности, соответствующей двоичному коду.
[012] Следующие аспекты и области применения настоящего изобретения станут очевидными из подробного описания, предоставленного далее. Следует понимать, что подробное описание и конкретные примеры, хотя и указывают на предпочтительные варианты осуществления изобретения, предназначены только для иллюстрации и не предназначены для ограничения объема изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[013] Настоящее изобретение станет более понятным из подробного описания и прилагаемых чертежей, где:
[014] На фиг.1 представлена диаграмма простой конструкции двухкамерного наночипа с разделяющей мембраной, перфорированной нанопорой и электродами на каждой стороне мембраны.
[015] На фиг.2 и 3 показано, как заряженный полимер, например ДНК, движется в направлении анода.
[016] На фиг.4 и 5 показано, что полимер может перемещаться обратно путем изменения полярности электродов.
[017] На фиг.6 показана конструкция двухкамерного наночипа для ДНК-систем, в котором фермент полимераза расположена в одной камере, деблокирующий фермент находится в другой камере, и ни один из них не проходит через нанопору.
[018] На фиг.7 представлено присоединение аденинового нуклеотида, когда 3'-блокированный dATP (A) проходит через левую камеру и устанавливается "прямой" ток для перемещения ДНК в эту камеру.
[019] На фиг.8 представлено удаление защитной группы из олигонуклеотида, чтобы мог быть присоединен дополнительный нуклеотид. Например, удаление защитной группы происходит после перемещения ДНК в камеру путем установки тока на "обратный".
[020] На фиг.9 представлено присоединение 3'-блокированного dCTP (C). В определенных вариантах осуществления для замены содержимого этой камеры используют поток жидкости, например, как изображено, ранее в этой камере был "А".
[021] На фиг.10 показано, как может быть предоставлено множество отдельных удерживающих камер, в то время как проточная камера становится единой линией для предоставления реагентов.
[022] На фиг.11 показан подход для удержания ДНК, ассоциированной с ее камерой, путем связывания с камерой (верхний фрагмент ДНК на фиг.) или путем присоединения к объемной группе, которая не может пройти через нанопору (нижний фрагмент ДНК на фиг.). В этой системе конец ДНК все еще может проходить в проточную камеру и получать дополнительные нуклеотиды, однако другой конец остается в удерживающей камере.
[023] На фиг.12 представлена конфигурация, где ДНК связана со стенкой камеры и контролируется множеством электродов.
[024] На фиг.13 показано, как ДНК при желании может удерживаться в камере просто путем контроля полярности электродов.
[025] На фиг.14 показана матрица со свободным потоком реагентов в обе стороны, где ДНК связана с поверхностью камеры.
[026] На фиг.15 представлена альтернативная конструкция с электродами на сторонах, соседних с разделяющей мембраной, что позволяет менее дорогостоящее производство.
[027] На фиг.16 представлена система из трех отделений, где ДНК может перемещаться из отделения в отделение посредством электродов. Эта система не требует значительного потока реагентов в ходе синтеза.
[028] На фиг.17 представлен пример того, как реагенты могут быть организованы в системе из трех отделений.
[029] На фиг.18 представлен олигонуклеотид, связанный рядом с нанопорой, где нанопора имеет электродные элементы с каждой стороны от мембраны.
[030] На фиг.19 представлена серия молекул ДНК, связанных вдоль мембраны, содержащей нанопоры, и каждая из которых контролируется электродами рядом с нанопорой, с линией потока с каждой стороны мембраны. Например, как представлено, левая линия потока обеспечивает поток промывочного буфера/3'-блокированного dATP (A)/промывочного буфера/3'-блокированного dCTP (C)/промывочного буфера, где молекулы ДНК перемещаются в проточную камеру, только когда присутствует желаемый. Правая линия обеспечивает деблокирующий агент(ы) для удаления защитной группы с 3'-конца нуклеотида и обеспечения присоединения другого нуклеотида. В одном варианте осуществления поток деблокирующего агента(ов) происходит, когда левая линия промывается буфером. В других вариантах осуществления агент(ы) для удаления защитной группы является слишком объемным, чтобы проходить в левую линию через нанопоры.
[031] На фиг.20-22 схематично представлены эксперименты для подтверждения правильности концепции, где биты, используемые для кодирования данных, представляют собой короткие олигомеры, присоединенные с использованием топоизомеразы.
[032] На фиг.23 представлен формат секвенатора с нанопорами, где последовательность полимера считывается с использованием емкостных колебаний. В этой схеме считывания данных емкости электроды образуют верхнюю и нижнюю пластины конденсатора, разделенные мембраной, содержащей нанопору. Конденсатор является частью колебательного контура, где пульсирующий постоянный ток может проводить заряженный полимер через нанопору. Это изменение емкости измеряют по мере того, как полимер, например ДНК, проходит через нанопору, с использованием высокочастотной импеданс-спектроскопии. Значительным преимуществом этого подхода, в частности, с ДНК, является то, что частота измерения может быть очень высокой (фактически, измерение для каждого цикла, так что частота 100 МГц соответствует 100 миллионам измерений в секунду), и значительно более высокой, чем скорость прохождения мономеров через нанопору (ДНК, например, если только не ограничена каким-либо образом, проходит через нанопору в ответ на электрический ток со скоростью порядка 1 миллиона нуклеотидов в секунду).
[033] На фиг.24 представлена конструкция с двумя камерами для присоединения, пригодная для присоединения двух различных типов мономеров или олигомеров, например, для 2-битного или двоичного кодирования. На верхней части фиг. представлен вид сверху. На нижней части представлено поперечное сечение вида сбоку. Полное устройство в этом варианте осуществления может быть собрано из вплоть до 3 независимо изготовленных слоев и объединено посредством сращивания пластин, или может быть сформировано путем гравировки единой подложки. Чип содержит электрический контрольный слой (1), флюидный слой (2), который содержит две камеры для присоединения над резервной камерой с заряженным полимером (например, ДНК), заякоренным между входами в нанопору в первой и второй камерах для присоединения, и слой заземления (3).
[034] На фиг.25 представлено функционирование конструкции с двумя камерами для присоединения, представленной на фиг.24. Можно наблюдать, что в основании каждой камеры для присоединения находится нанопора (4). Нанопора получена, например, путем просверливания посредством FIB, TEM, влажной или сухой гравировки или посредством диэлектрического пробоя. Мембрана (5), содержащая нанопоры, имеет толщину, например, от 1 атомного слоя до 10-ов нм. Она изготовлена, например, из SiN, BN, SiOx, графена, дихалькогенидов переходных металлов, например WS2 или MoS2. Под мембраной нанопор (5) находится резервная или деблокирующая камера (6), которая содержит реагенты для удаления защитной группы из полимера после присоединения мономера или олигомера в одной из камер для присоединения (для напоминания, мономеры или олигомеры присоединяются в форме с защищенными концами, так что за раз присоединяется только один мономер или олигомер). Полимер (7) может быть проведен в или из камер для присоединения путем изменения полярности электродов в слое электрорегулирования (1).
[035] На фиг.26 представлен вид сверху конструкции, сходной с конструкциями фиг.24 и 25, однако в данном случае присутствует четыре камеры для присоединения, которые имеют общую резервную или деблокирующую камеру, и полимер связан в положении (9) с доступом в каждую из четырех камер. Поперечное сечение этой конструкции может быть таким, как представлено на фиг.24 и 25, и заряженный полимер может перемещаться в каждую из четырех камер для присоединения под контролем электродов в слое электрорегулирования (1 на фиг.24).
[036] На фиг.27 представлен вид сверху чипа с нанопорами, имеющего множество наборов двойных камер для присоединения, как представлено на фиг.24 и 25, позволяющих синтез множества полимеров параллельно. Мономеры (в данном случае нуклеотиды dATP и dGTP, обозначаемые как as A и G) загружают в каждую камеру посредством последовательных линий потока. Одна или более общих деблокирующих проточных ячеек позволяют удаление защитной группы из полимера после присоединения мономера или олигомера в одной из камер для присоединения. Это также позволяет открепление полимеров при необходимости (например, с использованием фермента рестрикции в случае ДНК, или химическое открепление от поверхности рядом с нанопорой) и накопление снаружи. В этом конкретном варианте осуществления деблокирующие проточные ячейки перпендикулярны каналам загрузки, используемым для заполнения камер для присоединения.
[037] На фиг.28 представлены следующие детали электропроводки для схем камер с двойным присоединением. Слой электрорегулирования (1) включает электропроводку из металла или поликремния. Плотность электропроводки возрастает посредством 3D-укладки, причем электрическая изоляция обеспечивается покрытием диэлектриком (например, через PECVD, распыление, ALD и т.д.). Контакт (11) с верхней частью электрода в камере для присоединения в одном варианте осуществления осуществляется с использованием переходных отверстий в кремнии (TSV) посредством глубокой гравировки реактивными ионами (DRIE) (крио-процесс или процесс BOSCH). Индивидуальный контроль напряжения (12) позволяет обращаться к каждой камере для присоединения индивидуально, обеспечивая точный контроль последовательности множества полимеров параллельно. На правой стороне фиг. представлен вид сверху, иллюстрирующий электропроводку для множества ячеек для присоединения. Слой заземления (3) может быть общим (как показано) или разделенным для снижения взаимного влияния между ячейками.
[038] На фиг.29 представлена альтернативная конфигурация, где контрольные электроды (13) камер для присоединения могут быть расположены на стенке камеры в виде обертки вместо положения вверху камеры.
[039] На фиг.30 представлен гель SDS-PAGE, подтверждающий, что протокол присоединения посредством топоизомеразы, как описано в примере 3, работает, причем отчетливо видны полосы, соответствующие ожидаемым продуктам A5 и B5.
[040] На фиг.31 представлен агарозный гель, подтверждающий, что продукт ПЦР примера 5 имеет правильный размер. Дорожка 0 представляет собой маркер молекулярной массы из 25 пар оснований; дорожка 1 представляет собой продукт эксперимента - линию, соответствующую ожидаемой молекулярной массе; дорожка 2 представляет собой отрицательный контроль #1; дорожка 3 представляет собой отрицательный контроль #2; дорожка 4 представляет собой отрицательный контроль #4.
[041] На фиг.32 представлен агарозный гель, подтверждающий, что фермент рестрикции, как описано в примере 5, продуцирует ожидаемый продукт. Маркер молекулярной массы слева представляет собой маркер из 100 пар оснований. Дорожка 1 соответствует нерасщепленному NAT1/NAT9c, дорожка 2 соответствует расщепленному NAT1/NAT9c. Дорожка 3 соответствует нерасщепленному NAT1/NAT9cI, дорожка 4 соответствует расщепленному NAT1/NAT9cI.
[042] На фиг.33 представлена иммобилизация ДНК вблизи нанопоры. На панели (1) представлена ДНК со структурой оригами на одном конце в левой камере (в реальном наночипе в левой камере изначально присутствует множество таких структур оригами). На панели (2) проиллюстрирована система с анодом справа, которая движет ДНК к нанопоре. В то время как цепь ДНК способна проходить через нанопору, структура оригами является слишком большой, чтобы проходить, так что ДНК "застревает". Выключение тока (панель 3) позволяет ДНК диффундировать. С использованием подходящей химии конец цепи ДНК способен связываться, когда он контактирует с поверхностью вблизи нанопоры. На панели (4) добавлен фермент рестрикции, который отщепляет структуру оригами от ДНК. Камеру промывают для удаления фермента и остаточной ДНК. Конечным результатом является одна молекула ДНК, присоединенная вблизи нанопоры, способная к перемещению в одну и в другую сторону через нанопору.
[043] На фиг.34 представлена основа функционирования нанопоры. На каждой панели ось y представляет собой ток (нА) и ось x представляет собой время (с). На левой панели "скрининг шума RF" иллюстрирует пригодность клетки Фарадея. Чип без нанопоры помещают в проточную ячейку и применяют 300 мВ. Когда крышка клетки Фарадея закрыта (первая стрелка), можно наблюдать снижение шума. Небольшой импульс возникает, когда защелка закрыта (вторая стрелка). Следует отметить, что ток составляет ~0 нМ. После формирования поры (средняя панель), применение 300 мВ (стрелка) приводит к току ~3,5 нМ. Когда ДНК помещают в камеру заземления и применяют +300 мВ можно наблюдать перемещение ДНК (правая панель) в качестве временного снижения тока (следует отметить, что в этом случае используют буфер TS: 50 мМ Tris, pH 8, 1 M NaCl). Для этого эксперимента по перемещению ДНК используют лямбда-ДНК.
[044] На фиг.35 представлена упрощенная картина, иллюстрирующая основные признаки структуры ДНК-оригами: крупная одноцепочечная область, кубическая структура оригами и присутствие 2 участков рестрикции (SwaI и AlwN1) вблизи структуры оригами.
[045] На фиг.36 представлено изображение, полученное с использованием электронного микроскопа, изготовленной структуры ДНК-оригами, и продемонстрирована ожидаемая топология. Оригами получают в 5-мМ основе Tris, 1 мМ EDTA, 5 мМ NaCl, 5 мМ MgCl2. Для сохранения структуры оригами предпочтительно, чтобы концентрации Mg++ составляли ~5 мМ или концентрации Na+/K+ составляли 1 M. Структуру оригами хранят при 4°C в концентрации 500 нМ.
[046] На фиг.37 представлено рестрикционное расщепление ДНК-оригами для подтверждения правильной сборки и функции. На крайней дорожке слева представлены стандарты ММ. Участки рестрикции исследуют путем расщепления оригами посредством AlwN1 и Swa1. Четыре исследуемых дорожки содержат следующие реагенты (единицы представляют собой микролитры):
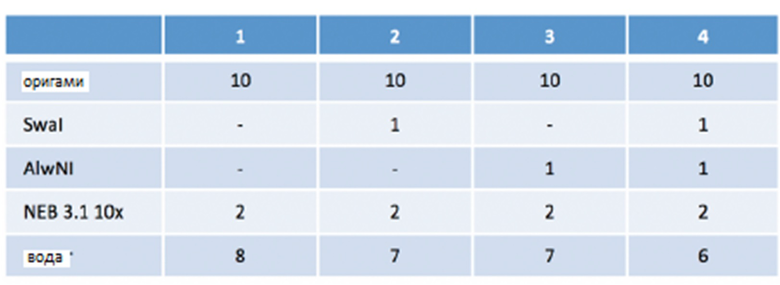
[047] Исследуемая дорожка (1) представляет собой отрицательный контроль; (2) представляет собой продукт расщепления посредством Swa1; (3) представляет собой продукт расщепления посредством AlwN1; (4) представляет собой продукт двойного расщепления посредством Swa1/AlwN1. Расщепление проводят при комнатной температуре в течение 60 минут, а затем при 37°C в течение 90 минут. Агарозный гель 1/2x TBE-Mg (1/2x TBE c 5 мМ MgCl2) визуализируют окрашиванием бромидом этидия. Индивидуальное расщепление каждым ферментом не демонстрирует эффекта подвижности в геле, однако расщепление обоими ферментами вместе (дорожка 4) приводит к двум фрагментом различной длины, как и ожидалось.
[048] На фиг.38 представлено связывание меченных биотином олигонуклеотидов с покрытыми стрептавидином гранулами против связывания с контрольными покрытыми BSA гранулами. По оси y приведены единицы флуоресценции, "до связывания" представляет собой флуоресценцию олигонуклеотида в исследуемом растворе до связывания с гранулами, (-) контроли представляют собой флуоресценцию, наблюдаемую после связывания с двумя различными партиями конъюгированных с BSA гранул, SA-1 и SA-2 представляют собой флуоресценцию, наблюдаемую после связывания с 2 различными партиями конъюгированных со стрептавидином гранул. В случае конъюгированных с BSA гранул наблюдают небольшой заметный уровень связывания, однако значительно большее связывание наблюдают в случае конъюгированных со стрептавидином гранул.
[049] На фиг.39 представлено связывание меченных биотином олигонуклеотидов с покрытыми стрептавидином гранулами против связывания с контрольными покрытыми BSA гранулами в различных буферных системах: буфере MPBS и HK. Левый столбик "отрицательный контроль" соответствует флуоресценции олигонуклеотида в исследуемом растворе до связывания с гранулами. В среднем столбце представлена флуоресценция "гранул BSA" и в правом столбце "гранул SA" после связывания с гранулами с BSA или стрептавидином, соответственно. В обеих буферных системах флуоресценция снижена в случае стрептавидиновых гранул относительно контролей, что указывает на то, что меченные биотином олигонуклеотиды хорошо связываются с покрытыми стрептавидином гранулами в различных буферных системах.
[050] На фиг.40 представлено функционирование SiO2-нанопоры с конъюгацией, где поверхность покрыта стрептавидном с одной стороны и покрыта BSA с другой. По оси x представлено время и по оси y представлен ток. Точка демонстрирует место, где происходит изменение тока на обратное. При изменении тока происходит кратковременный скачок, затем ток опускается приблизительно до той же абсолютной величины. Нанопора демонстрирует ток ~+3 нА при 200 мВ и ~-3 нА при -200 мВ.
[051] На фиг.41 изображена структура ДНК-оригами, помещенная в нанопору.
[052] На фиг.42 показано присоединение одноцепочечной ДНК к покрытой стрептавидином поверхности рядом с нанопорой.
[053] На фиг.43 представлены результаты эксперимента для ДНК-оригами, связанной с поверхностью вблизи нанопоры. Ток составляет+или - ~2,5 нА в обоих направлениях, что меньше исходного тока +/-~3нМ, что отражает частичное закупоривание структурой оригами. По оси x представлено время (с), по оси y представлен ток (нА), круги соответствуют точкам переключения напряжения.
[054] На фиг.44 представлено вхождение ДНК-оригами, что приводит к небольшому снижению тока. Оригами сразу выходи из нанопоры, как только ток исчезает. По оси x приведено время (с), по оси y приведен ток (нМ), круги соответствуют точка переключения напряжения.
[055] На фиг.45 представлено контролируемое движение цепи ДНК в одну и в другую сторону через нанопору посредством применения тока. С левой стороны ДНК находится в поре, так что наблюдаемый ток является более низким, чем в случае отсутствия ДНК в поре. Когда ток изменяют на обратный (правая сторона), ДНК в поре отсутствует, так что ток не изменяется.
[056] На фиг.46 представлены результаты эксперимента, подтверждающие это представление. Когда применяют положительное напряжение, ток составляет ~3 нА, что сравнимо с током, обычно наблюдаемым, когда пора открыта. Когда напряжение меняется на обратное, ток составляет ~-2,5 нА. Это ниже, чем ток, обычно наблюдаемый, когда пора открыта, и соответствует току, обычно наблюдаемому, когда пора блокирована цепью ДНК. несколько последовательных переключений напряжения демонстрирует стабильные результаты, указывая на то, что конфигурация ДНК изменяется, как представлено на фиг.45.
[057] На фиг.47 представлена другая химия конъюгации для связывания ДНК с поверхностью рядом с нанопорой.
ПОДРОБНОЕ ОПИСАНИЕ
[058] Приведенное ниже описание предпочтительного варианта(ов) осуществления является только иллюстративным и никоим образом не предназначено для ограничения изобретения, его назначения или применений.
[059] Как используют в настоящем описании, диапазоны используются в качестве сокращенного обозначения для описания каждой величины, входящей в данный диапазон. Любая величина в диапазоне может быть выбрана в качестве предела диапазона. Кроме того, все ссылки, цитированные в настоящем описании, включены в настоящее описание в качестве ссылок в полном объеме. В случае противоречия между определением в настоящем описании и определением в цитированной ссылке следует руководствоваться настоящим описанием.
[060] Если нет иных указаний, все проценты и количества, приведенные в данном разделе и в других разделах описания, следует понимать как относящиеся к процентам по массе. Приведенные количества основаны на активной массе материала.
[061] "Наночип", как используют в рамках изобретения, относится к нанофлюидному устройству, содержащему множество камер, содержащих жидкость, и необязательно каналы, обеспечивающие ток жидкости, где критические размеры элементов наночипа, например, ширина элементов, отделяющие камеры друг от друга, составляют от одного атома до 10 микрометров, например, менее одного микрометра, например, 0,01-1 микрометр. Поток материалов в наночипе может регулироваться электродами. Например, поскольку ДНК и РНК являются отрицательно заряженными, они будут двигаться к положительно заряженному электроду. См., например, Gershow, M, et al., Recapturing and Trapping Single Molecules with a Solid State Nanopore, Nat Nanotechnol. (2007) 2(12): 775-779, включенную в настоящее описание в качестве ссылки. В некоторых случаях поток жидкости также может регулироваться ограничительными элементами и путем прокачивания, инжектирования и/или выкачивания жидкостей в наночип и из него. Система способна к точному мультиплексному анализу нуклеиновых кислот (ДНК/РНК). В определенных вариантах осуществления наночип может быть изготовлен из кремниевого материала, например, диоксида кремния или нитрида кремния. Нитрид кремния (например, Si3N4) является особенно желательным для этой цели, поскольку он является химически относительно инертным и обеспечивает эффективный барьер против диффузии воды и ионов, даже при толщине несколько нм. Диоксид кремния (как используется в примерах настоящего описания) также является пригодным, поскольку он является подходящей поверхностью для химической модификации. Альтернативно в определенных вариантах осуществления наночип может быть изготовлен целиком или частично из материалов, которые могут образовывать листы толщиной всего в одну молекулу (иногда называемые однослойными материалами), например, графена, например, как описано в Heerema, SJ, et al., Graphene nanodevices for DNA sequencing, Nature Nanotechnology (2016) 11: 127-136; Garaj S et al., Graphene as a subnanometre trans-electrode membrane, Nature (2010) 467 (7312), 190-193, содержание каждой из которых включено в настоящее описание в качестве ссылки, или дихалькогенида переходного металла, например, дисульфида молибдена (MoS2), как описано в Feng, et al., Identification of single nucleotides in MoS2 nanopores, Nat Nanotechnol. (2015) 10(12):1070-1076, содержание которых включено в настоящее описание в качестве ссылки, или нитрид бора, как описано в Gilbert, et al. Fabrication of Atomically Precise Nanopores in Hexagonal Boron Nitride, eprint arXiv:1702,01220 (2017).
[062] В некоторых вариантах осуществления наночип содержит такой однослойный материал, который является относительно жестким и инертным, например, по меньшей мере настолько инертным и жестким, как графен, например MoS2. Однослойные материалы можно использовать, например, в качестве всей или части мембраны, содержащей нанопору. Наночип может быть частично покрыт металлом, например, на его стенки может иметь слои (например, металл-нитрид кремния-металл), а затем металл может быть организован так, чтобы в нем была предоставлена контролируемая пара электродов вблизи нанопоры, так чтобы нуклеиновая кислота могла двигаться в одну и в другую сторону через нанопору под действием электродвижущей силы, и также могла быть отсеквеирована путем измерения изменения электрического потенциала по мере прохождения нуклеиновой кислоты через нанопору.
[063] Нанофлуидные устройства в виде наночипов для секвенирования ДНК являются общеизвестными, например, как описано в Li, J., et al., Solid-state nanopore for detecting individual biopolymers, Methods Mol Biol. (2009)544:81-93; Smeets RM, et al. Noise in solid-state nanopores, PNAS (2008)105(2):417-21; Venta K, et al., Differentiation of short, single-stranded DNA homopolymers in solid-state nanopores, ACS Nano. (2013)7(5):4629-36; Briggs K, et al. Automated fabrication of 2-nm solid-state nanopores for nucleic acid analysis, Small (2014)10(10):2077-86; и Chen Z, DNA translocation through an array of kinked nanopores, Nat Mater. (2010)9(8):667-75; полное содержание каждой из которых включено в настоящее описание в качестве ссылки, например, в отношении их идей по конструкции и производству наночипов, содержащих нанопоры.
[064] "Нанопора", как используют в рамках изобретения, представляет собой пору диаметром менее 1 микрометра, например, диаметром 2-20 нм, например, порядка 2-5 нм. Одноцепочечная ДНК может проходить через 2-нм нанопору; одноцепочечная или двухцепочечная ДНК может проходить через 4-нм нанопору. В случае очень малой нанопоры, например, 2-5 нм, ДНК может проходить через нее, однако более крупные белковые фрагменты не могут, тем самым позволяя контролируемый синтез ДНК (или другого заряженного полимера). Когда используют более крупные нанопоры (или меньшие белковые ферменты), белковый фермент может быть конъюгирован с подложкой, что будет препятствовать его прохождению через нанопору, например, с более крупной молекулой, такой как более крупный белок, с гранулой или с поверхностью камеры. Известны различные типы нанопор. Например, биологические нанопоры образуются сборкой порообразующего белка в мембране, такой как липидный бислой. Например, α-гемолизин и сходные белковые поры встречаются в природе в клеточных мембранах, где они выступают в качестве каналов для ионов или молекул, подлежащих транспортировке в и наружу клеток, и такие белки могут быть приспособлены в качестве наноканалов. Твердофазные нанопоры образованы синтетическими материалами, такими как нитрид кремния или графен, например, путем внесения отверстий в синтетическую мембрану, например, с использованием контролируемого по принципу обратной связи моделирования низкоэнергетическим ионным пучком (IBS) или освещения высокоэнергетическим электронным пучком. Гибридные нанопоры могут быть изготовлены заключением порообразующего белка в синтетический материал. Когда на любом из концов или на любой стороне от нанопоры присутствует металлическая поверхность или электрод, может быть обеспечен электрический ток через нанопору в электролитной среде. Электроды могут быть изготовлены из любого проводящего материала, например, серебра, золота, платины, меди, диоксида титана, например, серебра, покрытого хлоридом серебра.
[065] Способы формирования твердофазной мембраны нанопор, например, из нитрида кремния, известны. В одном подходе кремниевую подложку покрывают материалом мембраны, например, нитридом кремния, и общую конфигурацию мембраны формируют с использованием фотолитографии и влажной химической гравировки, с получением мембран из нитрида кремния желаемого размера для включения в наночип, например, приблизительно 25×25 микрометров. Первоначальные отверстия или полости диаметром 0,1 микрометров пробивают в мембране из нитрида кремния с использованием фокусированного ионного пучка (FIB). Моделирование ионным пучком может сформировать нанопору путем сокращения поры большего размера, например, посредством индуцированного ионным пучком латерального переноса массы на поверхности мембраны, либо путем удаления материала мембраны ионным пучком, снимающим слой за слоем с плоской стороны мембраны, содержащей полость с противоположных сторон, так что, когда в конечном итоге формируется полость, нанопора имеет острые края. Когда ионный пучок угасает, тогда ток ионов, передаваемый через пору, является пригодным для желаемого размера пор. См., например. Li, J., et al., Solid-state nanopore for detecting individual biopolymers, Methods Mol Biol. (2009)544:81-93. Альтернативно нанопоры могут быть сформированы с использованием освещения высокоэнергетическим (200-300 кэВ) электронным пучком в TEM. С использованием полупроводниковых способов обработки, литографии e-пучком, реактивной ионной гравировки шаблонных слоев SiO2, и анизотропной KOH-гравировки Si, получают пирамидальные поры размером 20×20 нм и более в мембране толщиной 40 нм. Электронный пучок в TEM используют для уменьшения размера более крупных пор размером 20 нм до меньшего размера. TEM позволяет наблюдение процесса уменьшения размера в реальном времени. С использованием более тонкой мембраны (например, толщиной <10 нм) нанопоры могут быть просверлены посредством высокоэнергетичнеского сфокусированного электронного пучка в TEM. См., главным образом, Storm AJ, et al. Fabrication of solid-state nanopores with single-nanometre precision. Nature Materials (2003) 2:537-540; Storm AJ, et al. Translocation of double-stranded DNA through a silicon oxide nanopore. Phys. Rev. E (2005)71:051903; Heng JB, et al. Sizing DNA Using a Nanometer-Diameter Pore. Biophys. J (2004) 87(4):2905-11; содержание каждой из которых включено в настоящее описание в качестве ссылки.
[066] В других вариантах осуществления нанопоры получают с использованием диэлектрического пробоя с использованием относительно высокого потенциала через мембрану, где потенциал возрастает до тех пор, пока не будет обнаружен ток, например, как описано в Kwok, et al., "Nanopore Fabrication by Controlled Dielectric Breakdown", PLOS ONE (2014) 9(3): e92880, содержание которой включено в настоящее описание в качестве ссылки.
[067] С использованием этих способов и в зависимости от конкретного используемого способа, и толщины и состава мембраны, конечная форма нанопоры в твердом материале, таком как нитрид кремния, может быть в общих чертах похожа на две воронки, вершины которых соприкасаются в самой узкой точке, т.е. истинную нанопору. Такая форма в виде двух конусов позволяет проводить полимер через нанопору и обратно. Для подтверждения и измерения размера, положения и конфигурации наномембран, отверстий или полостей FIB и конечных нанопор можно использовать способы визуализации, например, атомно-силовую микроскопию (AFM) или трансмиссионную электронную микроскопию (TEM), в частности TEM.
[068] В некоторых вариантах осуществления один конец полимера, например ДНК, связан вблизи нанопоры или на внутренней стенке воронки, ведущей в нанопору. Если один конец полимера связан вблизи нанопоры, то, поскольку полимер первоначально приближается к нанопоре посредством диффузии, а затем движется под действием электрического градиента, обеспечиваемое градиентом движение максимизируется и диффузионное движение минимизируется, и, таким образом, скорость и эффективность повышается. См., например Wanunu M, Electrostatic focusing of unlabelled DNA into nanoscale pores using a salt gradient, Nat Nanotechnol. (2010) 5(2):160-5; Gershow M., Recapturing and trapping single molecules with a solid-state nanopore. Nat Nanotechnol. (2007) 2(12):775-9; Gershow, M., Recapturing and Trapping Single Molecules with a Solid State Nanopore. Nat Nanotechnol. (2007) 2(12): 775-779.
[069] В одном варианте осуществления один конец полимера, например ДНК, связан с гранулой, и полимер проходит через пору. Связывание с гранулой остановит движение полимера через нанопору на противоположном конце разделяющей мембраны в соседней камере. Затем ток выключается, и полимер, например ДНК, связывается с поверхностью рядом с нанопорой, в камере с другой стороны разделяющей мембраны. Например, в одном варианте осуществления один конец оцДНК ковалентно связан с гранулой размером 50 нм, а другой конец биотинилирован. Стрептавидин связан с областью в желаемой точке прикрепления в камере с другой стороны от разделяющей мембраны. ДНК втягивается через нанопору под действием электрического потенциала, и биотин связывается со стрептавидином. Связывание с гранулой и/или поверхностью рядом с нанопорой может осуществляться либо через ковалентные связи, либо через сильные нековалентные связи (такие как связь биотин-стрептавидин). Затем гранула отрезается ферментом и вымывается. В некоторых вариантах осуществления одноцепочечная ДНК расщепляется ферментом рестрикции, который расщепляет одноцепочечную ДНК, например, как описано в K. Nishigaki, Type II restriction endonuclease cleave single-stranded DNAs in general. Nucleic Acid Res. (1985) 13(16): 5747-5760, включенной в качестве ссылки. В других вариантах осуществления предоставляется комплементарный олигонуклеотид для обеспечения двухцепочечного участка рестрикции, который затем может расщепляться соответствующим ферментом рестрикции.
[070] По мере того, как полимер проходит через нанопору, изменение электрического потенциала или тока через нанопору, вызванное частичной блокадой нанопоры по мере прохождения полимера, может быть определено и может использоваться для идентификации последовательности мономеров в полимере, поскольку различные мономеры могут различаться их размерами и электрстатическими потенциалами.
[071] Использование наночипов, содержащих нанопоры, в способе изготовления ДНК, как описано в настоящем описании, не описано на уровне техники, однако такие чипы хорошо известны и коммерчески доступны для быстрого секвенирования ДНК. Например, MinION (Oxford Nanopore Technologies, Oxford, UK) является небольшим и может быть подключен к переносному компьютеру. Поскольку единственная цепь ДНК проходит через белковую нанопору со скоростью 30 оснований в секунду, MinION измеряет электрический ток. Цепи ДНК в поре нарушают ток ионов, что приводит к изменениям тока, соответствующим нуклеотидам в последовательности. Mikheyev, AS, et al. A first look at the Oxford Nanopore MinION sequencer, Mol. Ecol. Resour. (2014)14, 1097-1102. В то время как точность MinION является низкой, требуя повторного секвенирования, скорость и точность секвенирования с использованием наночипов по настоящему изобретению может быть значительно улучшена, если считываемая ДНК содержит только два легко различимых основания, например, A и C.
[072] В некоторых вариантах осуществления мембрана, содержащая нанопоры, может иметь трехслойную конфигурацию с металлической поверхностью на каждой стороне, изолирующей материал сердцевины, например, мембрану из нитрида кремния. В этом варианте осуществления металлические поверхности сформированы, например, способами литографии, для обеспечения микроконтура с парными электродами, по одному на каждом конце нанопоры, например, так чтобы между электродами мог быть обеспечен электрический ток через нанопору в электролитной среде, и этот ток мог проводить полимер через нанопору, а при изменении полярности мог проводить его обратно. По мере прохождения полимера через нанопору электроды могут измерять изменение электрического потенциала через нанопору, идентифицируя последовательность мономеров в полимере.
[073] В некоторых вариантах осуществления последовательность полимер организована для хранения данных. В некоторых вариантах осуществления данные хранятся в двоичном коде (1-ы и 0-и). В некоторых вариантах осуществления каждое основание соответствует 1 или 0. В других вариантах осуществления легко распознаваемая последовательность из двух или более оснований соответствует 1, а другая легко распознаваемая последовательность из двух или более оснований соответствует 0. В других вариантах осуществления данные могут храниться в третичном, четвертичном или другом коде. В конкретном варианте осуществления полимер представляет собой ДНК, например, одноцепочечную ДНК, где ДНК содержит только два типа оснований и не содержит никакие основания, способные к самогибридизации, например, где ДНК содержит остатки аденина и гуанина, аденина и цитозина, тимидина и гуанина, или тимидина и цитозина. В некоторых вариантах осуществления между двумя основаниями может быть вставлено одно или более дополнительных оснований, например, A и C могут содержать T в качестве "знака препинания" в последовательности, например, указывающего разрыв в кодирующей последовательности, с частотой, которая не приводит к значительной самогибридизации. В других вариантах осуществления, например, когда нуклеиновая кислота является двухцепочечной, можно использовать некоторые или все доступные основания.
[074] Нуклеотидные основания могут быть природными или в некоторых вариантах осуществления могут состоять из или включать неприродные основания, например, как описано в Malyshev, D. et al. "A semi-synthetic organism with an expanded genetic alphabet", Nature (2014) 509: 385-388, включенной в настоящее описание в качестве ссылки.
[075] В одном варианте осуществления данные хранят путем добавления единичных мономеров, например, единичных нуклеотидов в случае ДНК, к полимеру. В одном варианте осуществления полимер представляет собой ДНК и мономеры представляют собой остатки аденина (A) и цитозина (C). Остатки A и C имеют преимущество, поскольку (i) A и C имеют большее различие в размере, таким образом их различение в нанопоре должно упрощаться, (ii) A и C не образуют пары друг с другом, так что не формируют выраженную вторичную структуру, которая могла бы осложнить интерпретацию сигнала из нанопоры, и (iii) по той же причине G является менее предпочтительным, поскольку известно, что он образует гуаниновые тетрады. Нуклеотиды добавляются концевой трансферазой (или полинуклеотидфосфорилазой), однако нуклеотиды являются 3'-блокированными, так что за один раз добавляется только один нуклеотид. Блок удаляется перед присоединением следующего нуклеотида.
[076] В некоторых вариантах осуществления ДНК остается в наночипе. В других вариантах осуществления она извлекается и необязательно конвертируется в двухцепочечную ДНК и/или необязательно конвертируется в кристаллическую форму, например, для повышения долговременной стабильности. В других вариантах осуществления ДНК может амплифицироваться, и амплифицированная ДНК может извлекаться для долговременного хранения, в то время как исходная ДНК-матрица, например, ДНК связанная со стенкой камеры в наночипе, может оставаться в наночипе, где она может считываться и/или использоваться в качестве матрицы для создания дополнительной ДНК.
[077] В некоторых вариантах осуществления ДНК или другой полимер заякоривается на поверхности вблизи нанопоры в ходе синтеза. Например, в одном варианте осуществления каждая одноцепочечная молекула ДНК связана на 5'-конце с поверхностью вблизи нанопоры, где электрический ток в каждой нанопоре независимо может регулироваться электродами у этой нанопоры, так что 3'-конец молекулы ДНК может втягиваться через нанопору из удерживающей камеры в проточную камеру, содержащую поток 3'-защищенных dNTP вместе с полимеразой или концевой трансферазой для присоединения 3'-защищенных dNTP, или удерживаться в удерживающей камере, где нанопора не пропускает фермент, так что dNTP не присоединяется. См., например, изображения на фиг.12-16 и также на фиг.18 и 19. В других вариантах осуществления одноцепочечную ДНК конструируют присоединение к 5'-концу (с присоединенным 3'-концом) с использованием топоизомеразы, как более подробно описано ниже. Посредством контроля того, участвует ли каждая молекула ДНК в каждом цикле, последовательность каждой молекулы ДНК можно точно контролировать, например, следующим образом:
Поток A=3'-защищенный dATP
Поток C=3'-защищенный dCTP
Нанопора 1 и нанопора 2 на этой схеме ассоциированы с различными цепями ДНК, положения которых (в или вне проточных камер) контролируются по отдельности. Защитную группу из ДНК можно удалять либо посредством специфического фермента в удерживающей камере или путем изменения потока в проточной камере для обеспечения удаления защитной группы ферментативными, химическими, катализируемыми светами или другими средствами. В одном варианте осуществления поток деблокирующего агента(ов) между циклами потока A и потока C, например, когда проточную камеру промывают буфером, так что деблокирующий агент не удаляет защитную группу из нуклеотидных строительных блоков. В других вариантах осуществления агент(ы) удаления защитной группы является слишком объемным для прохождения в проточную камеру через нанопоры.
[078] Конечным результатом в вышеуказанном примере является присоединение A и C к ДНК в нанопоре 1, и присоединение C и C к ДНК в нанопоре 2.
[079] В другом варианте осуществления конфигурация камеры является сходной, но с двухцепочечной ДНК, заякоренной на поверхности вблизи нанопоры и каждый из олигонуклеотидных фрагментов, например, двух или более типов, соответствующих двоичному коду, добавляется последовательно, например, с использованием сайт-специфических рекомбиназ, т.е. ферментов, которые самопроизвольно распознают и расщепляют по меньшей мере одну цепь двойной цепи нуклеиновых кислот с сегментом последовательности, известным как последовательность сайт-специфической рекомбинации, например, с использованием нагруженных топоизомеразой олигонуклеотидов, как описано ниже.
[080] В определенных вариантах осуществления может быть желательным сохранение электрически заряженного полимера, например ДНК, в конденсированном состоянии после синтеза. Для этого несколько причин:
- полимер должен быть более стабильным в этой форме,
- конденсация полимера не допустит скучивание и позволит использование более длинных полимеров в малых объемах,
- упорядоченная конденсация может уменьшить возможность того, что полимер образует узлы или клубки,
- если какие-либо из камер взаимосоиденены, это поможет предотвратить чрезмерное удлинение полимера и прохождение его в пору, отличную от предполагаемой, когда применяют электрической ток,
- конденсация поможет удержать полимер вдали электродов, где электрохимия может повредить полимер.
Клетка человека имеет размер приблизительно 10 микрометров, но содержит 8 миллиардов пар оснований ДНК. В вытянутом состоянии они растянутся на метр. ДНК вмещается в клетку, поскольку она обвита вокруг гистонных белков. В определенных вариантах осуществления гистоны или сходные белки обеспечивают сходную функцию в наночипах по изобретению. В некоторых вариантах осуществления внутренние поверхности наночипов являются слабоположительно заряженными, так что электрически заряженный полимер, например ДНК, имеет тенденцию слабо прикрепляться к ним.
[081] В определенных вариантах осуществления заряженный полимер, например, одноцепочечная или двухцепочечная ДНК, связан с поверхностью вблизи нанопоры. Это можно осуществлять различными способами. Как правило, полимер локализуют вблизи нанопоры путем связывания полимера с относительно объемной структурой (например, гранулой, белком или структурой ДНК-оригами (описанной ниже), имеющей диаметр, слишком большой для вхождения в нанопору, например, >10 нм, например, приблизительно 20-50 нм), втягивания заряженного полимера через нанопору с использованием электрического тока, заякоривания конца полимера, дистального относительно объемной структуры, на поверхности рядом с нанопорой и отщепления объемной структуры.
[082] Стадию заякоривания конца полимера, дистального для объемной структуры, на поверхности, соседней с нанопорой, можно проводить различными путями. В одном варианте осуществления полимер представляет собой одноцепочечную ДНК и существуют предварительно связанные цепи ДНК (приблизительно 50 п.н.), которые комплементарны части одноцепочечной ДНК, так что одноцепочечная ДНК и предварительно связанные цепи ДНК могут объединяться посредством спаривания оснований. Если спаривание является достаточно сильным, этого будет достаточно для удержания ДНК заякоренной даже в ходе манипулирования. Преимущество этого способа присоединения состоит в том, что позволяет удаление ДНК из чипа с нанопорами при желании для длительного хранения ДНК. Альтернативно цепь связывают с поверхностью ковалентно, либо с использованием химии конъюгации, например, конъюгации стрептавидин-биотин, как описано в примере 1 ниже, либо клик-химии (см. Kolb, et al. Angew. Chem. Int. Ed. (2001)40: 2004-2021, включенную в настоящее описание в качестве ссылки) и/либо с использованием ферментативного присоединения, например, путем предварительного ковалентно связывания олигонуклеотидов с дистальной поверхностью, а затем с использованием ДНК-лигаза для их соединения.
[083] После связывания дистального конца цепи с поверхностью рядом с нанопорой объемная структура отщепляется, например, с использованием эндонуклеазы рестрикции, которая расщепляет в участке рестрикции вблизи объемной структуры.
[084] Объемная структура может представлять собой гранулу, объемную молекулу, например, белок, который обратимо связывается с цепью ДНК, или структуру ДНК-оригами. ДНК-оригами вовлекает применение спаривания оснований для формирования трехмерных ДНК-структур. Технологии ДНК-оригами в общих чертах описаны в Bell, et al, Nano Lett. (2012)12: 512-517, включенной в настоящее описание в качестве ссылки. Например, в рамках настоящего изобретения ДНК-оригами можно использовать для связывания одной молекулы ДНК с поверхностью рядом с нанопорой. В одном варианте осуществления эта структура представляет собой "блок в виде соты", например, с размером каждой стороны приблизительно 20 нм. Это частично препятствует прохождению этой части ДНК через нанопору (подобно тому, как в указанной статье). Со структурой оригами связана длинная цепь ДНК (одноцепочечная или двухцепочечная). Цепь ДНК проходит через нанопору до тех пор, пока блок оригами не натолкнется на нанопору и не заблокируют дальнейший прогресс. Затем электрический ток выключается и цепь связывается с поверхностью рядом с нанопорой.
[085] В другом варианте осуществления электрически заряженный полимер, например ДНК, со структурой оригами находится в средней камере трехкамерной конфигурации. Оригами удерживает ДНК от вхождения целиком в другие 2 камеры (или в другую камеру в случае 2-камерного примера). Таким образом, в этом примере полимер не должен быть заякорен на поверхности. Это снижает риск запутывания полимера и устраняет необходимость в стадии связывания одного конца полимера с поверхностью и отщепления объемной части на другом конце. Объем камеры с оригами должен быть минимальным, чтобы быть практичным, так чтобы полимер оставался относительно вблизи поры, что поможет обеспечить его быстрое перемещение при применении электрического тока. Следует отметить, что в то время как средняя камера, содержащая оригами-часть полимера, не может быть соединена с другими средними камерами (или иначе различные полимеры смешаются), другие камеры (или группы камер в примере с 3 камерами) могут быть соединены. Эти другие камеры могут иметь больший объем при желании, поскольку полимер обязательно будет находиться вблизи поры (в действительности, часть его будет находиться в поре), когда ДНК перемещается в эту камеру.
[086] В некоторых вариантах осуществления устройство содержит три камеры в линии, где камеры для присоединения являются последовательными, чтобы позволить поток и имеют общие электроды, в то время как камеры "для удаления защитной группы" являются изолированными в отношении жидкостей, за исключением потока через нанопору и имеют уникальные электроды.
[087] В других вариантах осуществления ДНК или другой заряженный полимер не являются заякоренными, но могут перемещаться между камерой(ами) для синтеза и камерой(ами) для удаления защитной группы под контролем электродов в камерах, в то время как движение полимеразы и агентов, удаляющие защитную группу, между камерами ограничено, поскольку они являются слишком объемными, чтобы проходить через нанопоры, соединяющие камеры, и/или заякорены на поверхности камеры. См., например, фиг.1-9 и 16-17.
[088] Электрический ток, требуемый для перемещения заряженного полимера через нанопору, зависит, например, от природы полимера, размера нанопоры, материала мембраны, содержащей нанопору, и концентрации соли и, таким образом, при необходимости он оптимизируется для конкретной системы. В случае ДНК, как используют в примерах настоящего описания, примерами напряжения и электрического тока являются, например, 50-500 мВ, как правило, 100-200 мВ, и 1-10 нА, например, приблизительно 4 нА, с концентрациями соли порядка от 100 мМ до 1 M.
[089] Движение заряженного полимера, например ДНК, через нанопору обычно является очень быстрым, например, от 1 до 5 мкс на основание, таким образом, порядка миллиона оснований в секунду (1 МГц согласно номенклатуре частоты), что затрудняет отличение правильного считывания от помех в системе. С использованием способов по настоящему изобретению, либо (i) нуклеотид должен быть повторен последовательно, например, последовательно 100 раз, для обеспечения поддающегося измерению характерного изменения, либо (ii) должны использоваться белковые поры, такие как на основе альфа-гемолизина (αHL) или порина A Mycobacterium smegmatis (MspA), которые обеспечивают относительно длинную пору с возможностью множественного считывания по мере продвижения основания через полимер, и в некоторых случаях, они могут быть адаптированы для обеспечения контролируемой подачи ДНК через пору по одному основанию за раз, в некоторых случаях с использованием экзонуклеазы для расщепления каждого основания по мере его прохождения. Возможны различные подходы, например,
- замедление скорости полимера от приблизительно 1 МГц до приблизительно 100-200 Гц, например с использованием среды, содержащей электрореологическую жидкость, которая становится более вязкой при применении напряжения, тем самым замедляя скорость прохождения полимера через нанопору, или плазмонную жидкостную систему, где вязкость среды может контролироваться светом; или молекулярный двигатель или защелку;
- предоставление последовательности в полимере, например, в одноцепочечной ДНК, которая образует объемную вторичную структуру, например, конфигурацию "шпильки", "головки молотка" или "гантели", которая должна линеаризоваться для прохождения через нанопору, таким образом, делая информацию менее плотной и предоставляя сигнал, имеющий более высокую длительность;
- обеспечения множества считываний одной и той же последовательности, например, с использованием быстро меняющегося тока, позволяя множество считываний для одной рамки последовательности, и в комбинации с кратковременными импульсами прямого тока для втягивания молекулы до следующей рамки последовательности, путем считывания всей последовательности множество раз или путем считывания множества идентичных последовательностей параллельно, в каждом случае сопоставляя данные считывания для предоставления консенсусной считанной последовательности, которая усиливает сигнал;
- измерение изменения импеданса высокочастотного сигнала, индуцированного изменением емкости по мере прохождения мономеров (например, нуклеотидов) через нанопору, вместо измерения изменений непосредственно электрического тока или сопротивления;
- усиление различий в токе, сопротивлении или емкости между различными основаниями, например, с использованием неприродных оснований, которые имеют большее отличие размера или в ином случае модифицировано для генерирования различных сигналов, или путем формирования более крупных вторичных структур в ДНК, таких как конфигурация "шпильки", "головки молотка" или "гантели", которые обеспечивают усиленный сигнал вследствие их большего размера;
- использование оптической считывающей системы, например с использованием встроенной оптической антенны рядом с нанопорой, которая действует в качестве оптического преобразователя (или оптического усилителя сигнала), дополняя или заменяя стандартное измерение тока ионов, например, как описано в Nam, et al., "Graphene Nanopore with a Self-Integrated Optical Antenna", Nano Lett. (2014)14: 5584-5589, содержание которой включено в настоящее описание в качестве ссылки. В некоторых вариантах осуществления мономеры, например, нуклеотиды ДНК, являются меченными флуоресцентными красителями, так что каждый отдельный мономер флуоресцирует с определенной интенсивностью по мере прохождения через место соединения нанопоры и ее оптической антенны. В некоторых вариантах осуществления флуоресцентные метки счищаются с твердофазной нанопоры, что приводит к серии поддающихся детектированию фотонных импульсов по мере прохождения полимера через нанопору с высокой скоростью, например, как описано в McNally et al., "Optical recognition of converted DNA nucleotides for single molecule DNA sequencing using nanopore arrays", Nano Lett. (2010)10(6): 2237-2244, и Meller A., "Towards Optical DNA Sequencing Using Nanopore Arrays", J Biomol Tech. (2011) 22(Suppl): S8-S9, содержание каждой из которых включено в настоящее описание в качестве ссылки.
[090] В одном варианте осуществления, заряженный полимер представляет собой нуклеиновую кислоту, например, одноцепочечную ДНК, где последовательности обеспечивают вторичную структуру. В Bell, et al., Nat Nanotechnol.(2016)11(7):645-51, включенной в настоящее описание в качестве ссылки, описано использование относительно короткой последовательности конфигураций гантели, поддающихся детектированию в формате твердофазной нанопоры, для мечения антигенов в иммуноанализе. Нанопоры, использованные Bell, et al., были относительно крупными, так что вся структура гантели могла проходить через пору, но при использовании нанопор, меньших чем диаметр конфигурации гантели, ДНК "распаковывается" и становится линеаризованной. Можно использовать более сложные конфигурации, например, где каждый бит соответствует последовательности, сходной с тРНК (см., например, Henley, et al. Nano Lett. (2016)16: 138-144, включенную в настоящее описание в качестве ссылки). Таким образом изобретение относится к заряженным полимерам, например, одноцепочечной ДНК, имеющим по меньшей мере два типа вторичной структуры, где вторичная структура кодирует данные (например, двоичные данные, где один тип вторичной структуры представляет собой 1, а второй 0). В других вариантах осуществления вторичные структуры используются для замедления прохождения ДНК через нанопору или для внесения разрывов в последовательность, облегчая считывание последовательности.
[091] В другом варианте осуществления в рамках изобретения используется молекула ДНК, содержащая серию по меньшей мере из двух различных мотивов ДНК, где каждый мотив специфически связывается с конкретным лигандом, например, регулирующим ген белком для двухцепочечной ДНК или тРНК для одноцепочечной ДНК, где по меньшей мере два различных мотива ДНК кодируют информацию, например, в двоичном коде, где один мотив представляет собой 1 и второй представляет собой 0, например, где лиганд усиливает различие сигнала (например, изменение тока или емкости) через нанопору по мере прохождения ДНК через нанопору.
[092] Как описано выше, когда различные мономеры проходят через нанопору, они влияют на ток через нанопору, в основном посредством физического блокирования нанопоры и изменения проводимости через нанопору. В существующих системах нанопор это изменение тока измеряют прямо. Проблема современных считывающих систем состоит в том, что в системе существует значительный шум и в случае ДНК, например, при определении флуктуаций тока по мере прохождении различных нуклеотидных элементов через нанопору, требуется относительно длительное время интеграции, порядка одной сотой секунды, для точного детектирования различий между различными мономерами, например, между различными основаниями. Недавно бело показано, что изменения импеданса и емкости могут быть пригодными для исследования клеток и биологических систем, несмотря на возможность сложных взаимодействий с солями и биологическими молекулами. Например, Laborde, et al. Nat Nano. (2015)10(9):791-5 (включена в настоящее описание в качестве ссылки) демонстрируют, что высокочастотную импедансную спектроскопию можно использовать для детектирования небольших изменений емкости в физиологических условиях солей и для визуализирующих микрочастиц и живых клеток за границами предела Дебая.
[093] В одном варианте осуществления изобретения, таким образом, проводят измерение изменения емкости, а не измерение изменения непосредственно тока, например, где последовательность заряженного полимера идентифицируют путем измерения изменения фазы радиочастотного сигнала, индуцированного изменением емкости по мере прохождения мономеров (например, нуклеотидов) через нанопору.
[094] Проще говоря, емкость существует в любом контуре, где присутствует щель между одним электрическим проводником и другим. В то время как ток прямо изменяется в зависимости от изменения емкости, он не изменяется одновременно с емкостью. Например, если нанести на график ток и напряжение с течением времени в емкостную цепь с переменным электрическим током, можно видеть, что, хотя как ток, так и напряжение образуют синусоидальную волну, волны находятся не в фазе. Когда происходит изменение тока, происходит изменение емкости, которое отражается изменением фазы сигнала. Радиочастотный переменный ток обеспечивает сигнал с фиксированной частотой и амплитудой, в то время как фаза сигнала варьируется в зависимости от емкости контура. В системе, разработанной авторами изобретения, они используют пульсирующий прямой ток вместо переменного тока (т.е. напряжение изменяется между двумя величинами, однако напряжение не пересекает линию "нуля", так что полярность сохраняется и один электрод остается положительным, а другой отрицательным), так что заряженный полимер может быть втянут через нанопору (в направлении положительного электрода в случае ДНК). Когда в нанопоре ничего нет, емкость имеет одну величину, которая изменяется по мере прохождения различных мономеров полимера через нанопору. Подходящие диапазоны частоты находятся в радиочастотном диапазоне, например от 1 МГц до 1 ГГц, например 50-200 МГц, например приблизительно 100 МГц, например ниже более высоких микроволновых частот, которые могут вызывать значительное диэлектрическое нагревание среды. Для уменьшения возможности интерференции можно использовать различные частоты для различных нанопор, так что можно проводить измерение множества нанопор одновременно с одной радиочастотной входной линией.
[095] Измерение изменений импеданса (вследствие, например, изменений емкости) при высоких частотах повышает соотношение сигнала и шума, доступное в определенный промежуток времени, поскольку это снижает эффекты шума 1/f или "розового" шума, свойственного электронным измерительным контурам. Использование высокочастотного сигнала повышает соотношение сигнала и шуму, поскольку множество измерений проводится за данный промежуток времени, обеспечивая более стабильный сигнал, который хорошо отличим от изменений импеданса вследствие внешних или обуславливаемых устройством изменений и флуктуаций.
[096] С использованием этих принципов для настоящего изобретения, в одном варианте осуществления изобретение относится к способу измерения изменения импеданса высокочастотного сигнала, индуцированного изменением емкости по мере прохождения мономеров (например, нуклеотидов) через нанопору, например, к способу считывания последовательности мономеров заряженного полимера, содержащего по меньшей мере два различных типа мономеров, например молекулы ДНК, включающему применение радиочастотного пульсирующего прямого тока, например, с частотой от 1 МГц до 1 ГГц, например 50-200 МГц, например, приблизительно 100 МГц, через нанопору, где пульсирующий прямой ток втягивает заряженный полимер через нанопору и последовательность мономеров считывается путем измерения изменения емкости через нанопору по мере прохождения заряженного полимера через нанопору.
[097] В некоторых вариантах осуществления изобретение относится к наночипу для секвенирования электрически заряженного полимера, например ДНК, содержащего по меньшей мере два различных мономера, причем наночип содержит по меньшей мере первую и вторую реакционные камеры, каждая из которых содержит электролитную среду и разделена мембраной, содержащей одну или более нанопор, где пара электродов (например в форме противостоящих пластин), подсоединенных к контуру, находится с каждой стороны мембраны, содержащей одну или более нанопор, причем электроды разделены расстоянием 1-30 микрометров, например, приблизительно 10 микрометров, так что щель между электродами имеет емкость при применении к электродам радиочастотного импульсного прямого тока, например от 1 МГц до 1 ГГц, так что электрически заряженный полимер втягивается через нанопору, например, из одной камеры в следующую, и так что фаза импульсного прямого радиочастотного тока изменяется при изменении емкости по мере прохождения электрически заряженного полимера через нанопору, тем самым позволяя детектирование последовательность мономеров электрически заряженного полимера. В определенных вариантах осуществления наночип содержит множество наборов реакционных камер, где реакционные камеры в наборе разделены мембраной, имеющей одну или более нанопор, и группы реакционных камер разделены экранирующим слоем для минимизации электрической интерференции и возможности их параллельного секвенирования.
[098] Например, в одном варианте осуществления электроды образуют верхнюю и нижнюю пластины конденсатора, являющегося частью колебательного контура, и изменение емкости измеряется по мере прохождения ДНК через пору между пластинами.
[099] В определенных вариантах осуществления наночип дополнительно содержит реагенты для синтеза полимера, например ДНК, например, согласно наночипу 1 и т.д., ниже.
[0100] В одном варианте осуществления, изобретение, таким образом, относится к способу (Способ 1) синтеза заряженного полимера [например, нуклеиновой кислоты (например, ДНК или РНК)], содержащей по меньшей мере два различных мономера, в наночипе, причем наночип содержит
одну или более камер для присоединения, содержащих реагенты для присоединения одного или более мономеров [например, нуклеотидов] или олигомеров [например, олигонуклеотидов] к заряженному полимеру, в буферном растворе в форме с защищенными концами, так что только один мономер или олигомер может присоединяться за один цикл реакции; и
одну или более резервных камер, содержащих буферный раствор, но не все реагенты, необходимые для присоединения одного или более мономеров или олигомеров,
где камеры разделены одной или более мембранами, содержащими одну или более нанопор, и
где заряженный полимер может проходить через нанопору, но по меньшей мере один из реагентов для присоединения одного или более мономеров не могут,
причем способ включает
a) перемещение первого конца заряженного полимера, имеющего первый конец и второй конец, под действием электрического притяжения в камеру для присоединения, причем мономеры и олигомеры присоединяются к указанному первому концу в блокированной форме,
b) перемещение первого конца заряженного полимера с добавленным мономером или олигомером в блокированной форме в резервную камеру,
c) деблокирование присоединенного мономера или олигомера, и
d) повторение стадий a-c, где мономеры или олигомеры, присоединенные на стадии a), являются такими же или отличаются, до тех пор, пока не получат желаемую последовательность полимера.
[0101] Например, изобретение относится к следующему
1.1. Способ 1, где полимер представляет собой нуклеиновую кислоту, например, где полимер представляет собой ДНК или РНК, например, где он представляет собой ДНК, например, дцДНК или оцДНК.
1.2. Любой из вышеуказанных способов, где второй конец полимера, например нуклеиновой кислоты, либо защищен, либо связан с подложкой рядом с нанопорой.
1.3. Любой из вышеуказанных способов, где электрическое притяжение обеспечивают путем применения электрического потенциала между электродами в каждой камере, где полярность и ток между электродами можно контролировать, например, так что нуклеиновая кислота притягивается к положительному электроду.
1.4. Любой из вышеуказанных способов, где полимер представляет собой нуклеиновую кислоту и
(i) указанный первый конец нуклеиновой кислоты представляет собой 3'-конец, присоединение нуклеотидов происходит в направлении от 5' к 3' и катализируется полимеразой, например, где затруднено прохождение полимеразы (например, вследствие ее размера или вследствие связывания с подложкой в первой камере) через нанопору, нуклеотиды являются 3'-защищенными при присоединении и после присоединения 3'-защищенного нуклеотида к 3'-концу нуклеиновой кислоты, 3'-защитная группа на нуклеиновой кислоте удаляется, например, в резервной камере; или
(ii) указанный первый конец нуклеиновой кислоты представляет собой 5'-конец, присоединение нуклеотидов происходит в направлении от 3' к 5', нуклеотиды являются 5'-защищенными при присоединении и после присоединения 5'-защищенного нуклеотида к 5'-концу нуклеиновой кислоты 5'-защитная группа удаляется, например, во второй камере; (например, где фосфат на 5'-защищенном нуклеотиде представляет собой нуклеозидфосфорамидит, связанный через 5'-защитную группу с объемной группой, которая не может проходить через нанопору, так что после связывания с нуклеиновой кислотой не вступившие в реакцию нуклеотиды вымываются, объемная 5'-защитная группа отщепляется от нуклеиновой кислоты и вымывается, и 5'-конец нуклеиновой кислоты может переместиться в резервную камеру);
где присоединение нуклеотидов к нуклеиновой кислоте контролируется перемещением первого конца нуклеиновой кислоты в и из одной или более камер для присоединения, и цикл продолжается до тех пор, пока не будет получена желаемая последовательность.
1.5. Любой из вышеуказанных способов, где последовательность мономеров или олигомеров в полимере [например, последовательность нуклеотидов в нуклеиновой кислоте], синтезированном таким образом, соответствует двоичному коду.
1.6. Любой из вышеуказанных способов, где полимер, синтезированный таким образом, представляет собой одноцепочечную ДНК.
1.7. Любой из вышеуказанных способов, где последовательность полимера [например, нуклеиновой кислоты] проверяется в процессе синтеза секвенированием мономеров или олигомеров [например, нуклеотидных оснований] по мере их прохождения через нанопору для идентификации ошибок в секвенировании.
1.8. Любой из вышеуказанных способов, где полимер, синтезированный таким образом, представляет собой одноцепочечную ДНК, где по меньшей мере 95%, например, по меньшей мере 99%, например, по существу все основания в последовательности выбраны из двух оснований, которые не гибридизуются с другими основаниями в цепи, например, оснований, выбранных из аденина и цитозина.
1.9. Любой из вышеуказанных способов, где множество полимеров [например, олигонуклеотидов] синтезируется независимо параллельно, так что получают полимеры [олигонуклеотиды], имеющие различные последовательности, путем контроля того, присутствуют ли они в одной или более камерах для присоединения или одной или более резервных камерах.
1.10. Любой из вышеуказанных способов, где присутствует по меньшей мере две камеры для присоединения, которые содержат реагенты, пригодные для присоединения различных мономеров или олигомеров, например, различных нуклеотидов, например, где существует одна или более камер для присоединения, содержащих реагенты, пригодные для присоединения первого мономера или олигомера, и одна или более камер для присоединения, содержащих реагенты, пригодные для присоединения второго отличающегося мономера или олигомера, например, где существует одна или более камер для присоединения, содержащих реагенты, пригодные для присоединения адениновых нуклеотидов, и одна или более камер для присоединения, содержащих реагенты, пригодные для присоединения цитозиновых нуклеотидов.
1.11. Любой из вышеуказанных способов, где по меньшей мере одна камера для присоединения представляет собой проточную камеру, обеспечивающую проточный цикл, включающий (i) подачу в проточную камеру реагентов, пригодных для присоединения первого мономера или олигомера, (ii) промывание, (iii) подачу в проточную камеру реагентов, пригодных для присоединения второго отличающегося мономера или олигомера, и (iv) промывание, и повторение цикла до завершения синтеза, где последовательность мономеров или олигомеров в полимере контролируется путем входа или выхода первого конца полимера из проточной камеры на стадии (i) или (iii) каждого цикла;
1.12. Любой из вышеуказанных способов, где полимер представляет собой ДНК и по меньшей мере одна камера для присоединения представляет собой проточную камеру, обеспечивающую проточный цикл, включающий (i) подачу в проточную камеру реагентов, пригодных для присоединения первого типа нуклеотида, (ii) промывание, (iii) подачу в проточную камеру реагентов, пригодных для присоединения второго типа нуклеотида, и (iv) промывание, и повторение цикла до завершения синтеза, где последовательность мономеров или олигомеров в полимере контролируется присутствием или отсутствием первого конца ДНК (например, 3'-конца) в проточной камере.
1.13. Любой из вышеуказанных способов, где полимер представляет собой ДНК и по меньшей мере одна камера для присоединения представляет собой проточную камеру, обеспечивающую проточный цикл, включающий (i) подачу в проточную камеру реагентов, пригодных для присоединения первого типа нуклеотида, (ii) промывание, (iii) подачу в проточную камеру реагентов, пригодных для присоединения второго типа нуклеотида, и (iv) промывание, (i) подачу в проточную камеру реагентов, пригодных для присоединения третьего типа нуклеотида, (ii) промывание, (iii) подачу в проточную камеру реагентов, пригодных для присоединения четвертого типа нуклеотида, и (iv) промывание, и повторение цикла до завершения синтеза, где последовательность мономеров или олигомеров в полимере контролируется присутствием или отсутствием первого конца ДНК (например, 3'-конца) в проточной камере, когда реагенты, пригодные для присоединения различных типов нуклеотидов, присутствуют.
1.14. Любой из вышеуказанных способов, где полимер представляет собой ДНК и наночип содержит две камеры для присоединения, которые представляют собой проточные камеры: (a) первую проточную камеру, обеспечивающую проточный цикл, включающий (i) подачу в проточную камеру реагентов, пригодных для присоединения первого типа нуклеотида, (ii) промывание, (iii) подачу в проточную камеру реагентов, пригодных для присоединения второго типа нуклеотида, и (iv) промывание, и повторение цикла до завершения синтеза, и (b) вторую проточную камеру, обеспечивающую проточный цикл, включающий (i) подачу в проточную камеру реагентов, пригодных для присоединения третьего типа нуклеотида, (ii) промывание, (iii) подачу в проточную камеру реагентов, пригодных для присоединения четвертого типа нуклеотида, и (iv) промывание, и повторение цикла до завершения синтеза, где нуклеотиды выбраны из dATP, dTTP, dCTP и dGTP и где последовательность контролируется путем направления первого конца ДНК (например, 3'-конца) в проточную камеру, где предоставляется следующий желаемый нуклеотид.
1.15. Любой из вышеуказанных способов, где полимер представляет собой ДНК и чип с нанопорами содержит одну или более камер для присоединения dATP, одну или более камер для присоединения dTTP, одну или более камер для присоединения dCTP, и одну или более камер для присоединения dGTP.
1.16. Любой из вышеуказанных способов, где каждый синтезированный полимер [например, нуклеиновая кислота] связан через его второй конец с поверхностью вблизи нанопоры.
1.17. Любой из вышеуказанных способов, где последовательность полимера [например, нуклеиновой кислоты] определяют после каждого цикла путем детектирования изменения электрического потенциала, тока, сопротивления, емкости и/или импеданса по мере прохождения полимера через нанопору.
1.18. Любой из вышеуказанных способов, где полимер представляет собой нуклеиновую кислоту и синтез нуклеиновой кислоты происходит в буферном растворе, например, растворе, содержащем буфер с pH 7-8,5, например приблизительно pH 8, например, буфер, содержащий трис(гидроксиметил)аминометан (Tris), подходящую кислоту и необязательно хелатор, например, этилендиаминтетрауксусную кислоту (EDTA), например, буфер TAE, содержащий смесь основания Tris, уксусной кислоты и EDTA, или буфер TBE, содержащий смесь оснований Tris, борной кислоты и EDTA; например, раствор, содержащий 10 мМ Tris pH 8, 1 мМ EDTA, 150 мМ KCl, или, например, 50 мМ ацетат калия, 20 мМ Tris-ацетат, 10 мМ ацетат магния, pH 7,9, при 25°C.
1.19. Любой из вышеуказанных способов, где полимер представляет собой одноцепочечную ДНК, дополнительно включающий конвертирование синтезированной одноцепочечной ДНК в двухцепочечную ДНК.
1.20. Любой из вышеуказанных способов, дополнительно включающий удаление полимера [например, нуклеиновой кислоты] из наночипа после завершения синтеза.
1.21. Любой из вышеуказанных способов, где полимер представляет собой нуклеиновую кислоту, дополнительно включающий амплификацию и извлечение копий синтезированной нуклеиновой кислоты с использованием подходящего праймера и полимеразы (например, Phi29).
1.22. Любой из вышеуказанных способов, где полимер представляет собой нуклеиновую кислоту, дополнительно включающий расщепление синтезированной нуклеиновой кислоты ферментом рестрикции и удаление нуклеиновой кислоты из наночипа.
1.23. Любой из вышеуказанных способов, где полимер представляет собой нуклеиновую кислоту, дополнительно включающий амплификацию синтезированной таим образом нуклеиновой кислоты.
1.24. Любой из вышеуказанных способов, дополнительно включающий удаления полимера [например, нуклеиновой кислоты] из наночипа и кристаллизацию полимера.
1.25. Любой из вышеуказанных способов, где полимер представляет собой нуклеиновую кислоту, дополнительно включающий стабилизацию нуклеиновой кислоты, например, путем сушки раствора, содержащего нуклеиновую кислоту, с одним или более из буфера (например, боратного буфера), антиоксиданта, увлажнителя, например, полиола, и необязательно хелатора, например, как описано в US 8283165 B2, включенной в настоящее описание в качестве ссылки; или путем формирования матрикса между нуклеиновой кислотой и полимером, таким как блок-сополимер полиэтиленгликоль-поли(l-лизин) (PEG-PLL) типа AB; или путем присоединения комплементарной цепи нуклеиновой кислоты или белка, который связывает ДНК.
1.26. Любой из вышеуказанных способов, включающий:
(i) реакцию нуклеиновой кислоты с 3'-защищенным нуклеотидом в камере для присоединения в присутствии полимеразы, которая катализирует присоединение 3'-защищенного нуклеотида к 3'-концу нуклеиновой кислоты;
(ii) втягивание по меньшей мере 3'-конца 3'-защищенной нуклеиновой кислоты, полученной таким образом, из камеры для присоединения через по меньшей мере одну нанопору в резервную камеру, где прохождение полимеразы через нанопору затруднено (например, вследствие ее размера или вследствие связывания с подложкой в первой камере);
(iii) удаление защитной группы из 3'-защищенной нуклеиновой кислоты, например, химически или ферментативно; и
(iv) если желательно присоединение к олигонуклеотиду дополнительного 3'-защищенного dNTP, втягивание 3'-конца олигонуклеотида в ту же или отличающуюся камеру для присоединения для повторения стадий (i)-(iii), или если это не является желательным, обеспечение нахождения 3'-конца нуклеиновой кислоты в резервной камере до следующего цикла, где в камеру для присоединения поступает желаемый 3'-защищенный dNTP; и
(v) повторение цикла стадий (i)- (iv) до тех пор, пока не будет получена желаемая последовательность нуклеиновой кислоты.
1.27. Любой из вышеуказанных способов, где полимер представляет собой одноцепочечную ДНК (оцДНК) и одна или более нанопор имеют диаметр, позволяющий проходить оцРНК, но двухцепочечной ДНК (дцДНК), например, диаметр приблизительно 2 нм.
1.28. Любой из вышеуказанных способов, где мономер представляет собой 3'-защищенный нуклеотид, например, дезоксинуклеотидтрифосфат (dNTP), например, выбранный из дезоксиаденозинтрифосфата (dATP), дезоксигуанозинтрифосфата (dGTP), дезоксицитидинтрифосфата (dCTP), дезокситимидинтрифосфата (dTTP), например, dATP или dCTP.
1.29. Любой из вышеуказанных способов, где полимер представляет собой нуклеиновую кислоту и присоединение нуклеотида к нуклеиновой кислоте катализируется полимеразой, например, независимой от матрицы полимеразой, например, концевой дезоксинуклеотидилтрансферазой (TdT), или полинуклеотидфосфорилазой, например, где полимераза катализирует включение дезоксинуклеотида на 3'-гидроксильный конец ДНК.
1.30. Любой из вышеуказанных способов, где мембрана содержит множество нанопор и множество полимеров, каждый из которых связан с поверхностью вблизи нанопоры, например, множество нуклеиновых кислот, каждая из которых связана через ее 5'-конец с поверхностью вблизи нанопоры.
1.31. Любой из вышеуказанных способов, где множество полимеров, каждый из которых связан с поверхностью вблизи нанопоры, например, множество нуклеиновых кислот, каждая из которых связана 5'-концом с поверхностью вблизи нанопоры, синтезируются независимо, где каждая нанопора имеет ассоциированную с ней пару электродов, где один электрод в паре расположен вблизи одного конца нанопоры, а другой электрод расположен вблизи другого конца нанопоры, так что каждый полимер может независимо перемещаться между первой и второй камерами под действием электрического тока, обеспечиваемого парой электродов.
1.32. Любой из вышеуказанных способов, где полимер представляет собой 3'-защищенную нуклеиновую кислоту, связанную 5'-концом с поверхностью вблизи нанопоры, и 3'-конец 3'-защищенной нуклеиновой кислоты втягивается через нанопору с использованием электрической силы, например, с использованием электрической силы, применяемой через электрод, в соседнюю камеру.
1.33. Способ 1.20, где новый 3'-защищенный dNTP является таким же или отличается от первого 3'-защищенного dNTP.
1.34. Способ 1.20, где 3'-защищенный dNTP, используемый на стадии (i) цикла, чередуется каждый цикл между 3'-защищенным dATP и 3'-защищенным dCTP.
1.35. Любой из вышеуказанных способов, где полимер представляет собой нуклеиновую кислоту и удаление защитной группы из нуклеиновой кислоты осуществляется посредством фермента, который удаляет 3'-защитную группу на оцДНК, но не на 3'-защищенном dNTP.
[0101] Например, изобретение относится к способу синтеза нуклеиновой кислоты в наночипе, содержащем по меньшей мере первую камеру и вторую камеру, разделенные мембраной, содержащей по меньшей мере одну нанопору, причем синтез осуществляется в буферном растворе посредством цикла присоединения нуклеотидов к первому концу нуклеиновой кислоты, имеющей первый конец и второй конец, где первый конец нуклеиновой кислоты перемещается под действием электрического притяжения между одной или более камерами для присоединения (которые содержат реагенты, способные присоединять нуклеотиды) и одной или более резервными камерами (которые не содержат реагенты, необходимыми для присоединения нуклеотидов), причем камеры разделены одной или более мембранами, каждая из которых содержит одну или более нанопор, где нанопора является достаточно крупной, чтобы обеспечить прохождение нуклеиновой кислоты, но слишком мелкой, чтобы обеспечить прохождение по меньшей мере одного реагента, необходимого для присоединения нуклеотида, например, где способ соответствует любому из способа 1 и т.д.
[0102] В определенных вариантах осуществления последовательность полимера соответствует двоичному коду, например, где полимер представляет собой нуклеиновую кислоту и последовательность соответствует двоичному коду, где каждый бит (0 или 1) соответствуют основанию, например, A или C.
[0103] В определенных вариантах осуществления полимер представляет собой ДНК.
[0104] В некоторых других вариантах осуществления каждому биту соответствует короткая последовательность мономеров, а не один мономер. Например, в одном таком варианте осуществления синтезируются блоки ДНК, где каждый блок генерирует уникальный сигнал через нанопору и соответствует нулю или единице. Этот вариант осуществления имеет определенные преимущества, состоящие в том, что единичные нуклеотиды труднее детектировать в нанопорах, особенно в твердофазных нанопорах, так что использование блоков менее подвержено ошибкам считывания, хотя плотность информации в полимере соответственно снижается.
[0105] Например, блоки (двухцепочечных) нуклеотидов можно присоединять с использованием сайт-специфических рекомбиназ, т.е. ферментов, которые самопроизвольно распознают и расщепляют по меньшей мере одну цепь двойной цепи нуклеиновых кислот в сегменте последовательности, известном как последовательность сайт-специфической рекомбинации. В одном таком варианте осуществления сайт-специфическая рекомбиназа представляет собой топоизомеразу, используемую для лигирования блока сопряженного с топоизомеразой дцДНК-олигонуклеотида с последовательностью. Эти олигонуклеотиды сами по себе не имеют структуру, совместимую с дальнейшим лигированием, если они не расщеплены ферментом рестрикции. Топоизмераза I вируса коровьей оспы специфически распознает последовательность ДНК 5´-(C/T)CCTT-3'. Топоизомераза связывается с двухцепочечной ДНК и расщепляет ее в участке расщепления 5´-(C/T)CCTT-3'. Следует отметить, что расщепление является неполным, поскольку топоизомераза расщепляет только одну цепь ДНК (хотя наличие вблизи разрыва одной цепи вызывает некий двухцепочечный разрыв), и при расщеплении топоизомераза ковалентно связывается с 3'-фосфатом 3'-нуклеотида. Затем фермент остается ковалентно связанным с 3'-концом ДНК и может либо религировать ковалентно удерживаемую цепь по той же связи, которая была первоначально расщеплена (как происходит в ходе релаксации ДНК), либо он может религировать с гетерологичной акцепторной ДНК, имеющей совместимые выступающие концы, создавая рекомбинантную молекулу. В этом варианте осуществления создаются донорные олигонуклеотиды на основе дц-ДНК (например, содержащие одну из по меньшей мере двух различных последовательностей, одной для "0", а другой для "1"), фланкированных участком рекомбинации топоизомеразой и участком рестрикции, который создает участок лигирования топоизомеразы. Кассеты являются заряженными топоизомеразой; т.е. они ковалентно связаны с топоизомеразой, которая будет связывать их с участком лигирования топоизомеразы на принимающем олигонуклеотиде. Когда растущая цепь принимающей ДНК расщепляется ферментом рестрикции, она становится способной к лигированию с нагруженной топоизомеразой кассетой. Так что необходимо только последовательно осуществлять цикл для растущей ДНК от фермента рестрикции до нагруженной топоизомеразы кассеты, причем на каждом цикле присоединяется другой донорный олигонуклеотид. Сходный подход описан для клонирования, см., например, Shuman S., Novel approach to molecular cloning and polynucleotide synthesis using vaccinia DNA topoisomerase. J Biol Chem. (1994); 269(51):32678-84, содержание которой включено в качестве ссылки.
[0106] Единичные основания можно присоединять с использованием сходной стратегии. В присутствии подходящей одноцепочечной "акцепторной" ДНК "с удаленной защитной группой", нагруженная топоизомеразой ДНК ферментативно и ковалентно лигируется ("присоединяется") с акцептором топоизомеразой, которая в этом процессе удаляется из ДНК. Затем фермент рестрикции типа IIS может расщеплять все присоединяемые ДНК за исключением одного основания (основания, которое "добавляется"). Этот процесс удаления защитной группы-присоединения можно повторять для присоединения дополнительных оснований (бит). Как продемонстрировано в примерах настоящего описания, является возможным использовать комбинацию топоизомераза/фермент рестрикции типа IIS для присоединения одного нуклеотида к 5'-концу одноцепочечной ДНК-мишени. Использование фермента рестрикции типа IIS позволяет расщепление ДНК в положении, отличающемся от положения последовательности распознавания (другие ферменты рестрикции типа IIS могут быть найдены на https://www.neb.com/tools-and-resources/selectioncharts/type-iis-restriction-enzymes). Использование остатков инозина (который выступает в качестве "универсального основания" и образует пару с любым другим основанием) в этой системе позволяет протекание этой реакции без каких-либо конкретных требований к последовательности ДНК-мишени. Нуклеотид, присоединяемый к одноцепочечной ДНК-мишени, представляет собой 3'-нуклеотид, с которым топоизомераза вируса коровьей оспы конъюгирована через 3'-фосфат. Поскольку последовательность распознавания топоизомеразы вируса коровьей оспы представляет собой (C/T)CCTT, авторы изобретения использовали эту систему для присоединения "T" к ДНК-мишени. Существует родственная топоизомераза, SVF, которую можно использовать для распознавания последовательности CCCTG (https://www.ncbi.nlm.nih.gov/pubmed/8661446). Таким образом SVF можно использовать для присоединения "G" вместо "T". В паре с топоизомеразой вируса коровьей оспы двоичные данные могут быть закодированы посредством T и G.
[0107] В другом подходе для присоединения одного основания на 5'-фосфате предоставлена блокирующая группа для обеспечения добавления одного основания в направлении от 3' к 5'. Реакция нагрузки обеспечивает нагрузку топоизомеразы T (или G, или другим нуклеотидом при необходимости), имеющим 5'-фосфатную группа. Когда нагруженная топоизомераза "видит" свободную 5'-неблокированную (нефосфорилированную) одноцепочечную цепь ДНК, она присоединяет T к этой цепи, обеспечивая ДНК с T, добавленным к 5'-концу. Это присоединение облегчается присутствием адаптерной ДНК, имеющей последовательности, с которыми может связываться топоизомераза и одноцепочечная акцепторная ДНК (следует отметить, что адаптерная ДНК является каталитической - ее можно вновь использовать в качестве матрицы в повторных реакциях). Присоединенный нуклеотид имеет 5'-фосфат на нем, так что он не будет субстратом для дальнейшего присоединения пока не подвергнется воздействию фосфатазы, которая удаляет 5'-фосфат. Этот процесс повторяется с использованием топоизомеразы вируса коровьей оспы для присоединения одного "T" к 5'-концу одноцепочечной ДНК-мишени и топоизомеразы SVF для присоединения одного "G", таким образом, позволяя конструирование последовательности, кодирующей двоичную информацию посредством T и G. Другие топоизомеразы можно использовать для присоединения A или C, хотя эта реакция является менее эффективной.
[0108] Одним преимуществом использования опосредуемой топоизомеразой стратегии является то, что мономер ковалентно связан с топоизомеразой, и, таким образом, не может "вырваться" для препятствования другим реакциям. Когда используют полимеразу мономеры могут диффундировать, так что полимеразы и/или деблокирующие агенты должны быть специфическими (например, селективными в отношении A против C, например) или альтернативно мономеры предоставляются посредством потока, так что они не имеют возможности смешаться.
[0109] В одном аспекте изобретение относится к топоизомеразе, нагруженной одним нуклеотидом, т.е. к топоизомеразе, конъюгированной с одним нуклеотидом, например, где топоизомераза конъюгирована через 3'-фосфат нуклеотида, и нуклеотид является защищенным, например, фосфорилированным, в 5'-положении.
[0110] В другом аспекте изобретение относится к способу (Способ A) синтеза молекулы ДНК с использованием опосредуемого топоизомеразой лигирования путем присоединения единичных нуклеотидов или олигомеров к цепи ДНК в направлении от 3' к 5', включающему (i) реакцию молекулы ДНК с топоизомеразой, нагруженной желаемым нуклеотидом или олигомером, где нуклеотид или олигомер блокирован от дальнейшего присоединения на 5'-конце, затем (ii) деблокирование 5'-конца ДНК, полученной таким образом, и повторение стадий (i) и (ii) до тех пор, пока не будет получена желаемая нуклеотидная последовательность, например:
A1.1. Способ A, который представляет собой способ синтеза молекулы ДНК путем присоединения единичных нуклеотидов в направлении от 3' к 5', включающий (i) реакцию молекулы ДНК с топоизомеразой, нагруженной желаемым нуклеотидом в 5'-защищенной форме, например, 5'-фосфорилированной форме, так что желаемый нуклеотид в 5'-защищенной форме присоединяется к 5'-концу ДНК, затем (ii) удаление защитной группы из 5'-конца ДНК, образовавшейся таким образом, с использованием фермента фосфатазы и повторение стадий (i) и (ii) до тех пор, пока не будет получена желаемая нуклеотидная последовательность; или
A1.2. Способ A, который представляет собой способ синтеза молекулы ДНК путем добавления олигомеров в направлении от 3' к 5', включающий (i) реакцию молекулы ДНК с топоизомеразой, нагруженной желаемым олигомером, тем самым лигируя олигомер с молекулой ДНК, затем (ii) использование фермента рестрикции для предоставления 5'-участка для опосредуемого топоизомеразой лигирования с другим олигомером и повторение стадий (i) и (ii) до тех пор, пока не будет получена желаемая последовательность олигомера.
A1.3. Любой из вышеуказанных способов, включающий предоставление лигазы и ATP для устранения разрывов в ДНК [примечание: лигирование посредством топоизомеразы лигирует только одну цепь].
A1.4. Любой из вышеуказанных способов, где нагруженный топоизомеразой донорный олигонуклеотид содержит 5'-выступающий конец на одной цепи, комплементарный цепи, имеющей топоизомеразу, содержащий последовательность полиинозина [примечание: остатки инозина выступают в качестве "универсальных оснований" и образуют пару с любым другим основанием].
A1.5. Любой из вышеуказанных способов, где фермент рестрикции представляет собой фермент рестрикции типа IIS, который может отщеплять всю присоединенную ДНК за исключением одного основания (основания, которое "присоединяется").
A1.6. Любой из вышеуказанных способов, где топоизомераза выбрана из топоизомеразы вируса коровьей оспы и топоизомеразы I SVF.
A1.7. Любой из вышеуказанных способов, где топоизомераза вируса коровьей оспы (которая распознает (C/T)CCTT) используется для присоединения нуклеотидов dTTP, и топоизомераза I SVF (которая распознает CCCTG) используется для присоединения нуклеотидов dGTP, например, для обеспечения двоичного кода.
A1.8. Любой из вышеуказанных способов, где ДНК является двухцепочечной и резервная камера дополнительно содержит лигазу и ATP для репарации цепи ДНК, не связанной с топоизомеразой.
A1.9. Любой из вышеуказанных способов, включающий использование ингибитора топоизомеразы для подавления связывания и активности свободной топоизомеразы в отношении ДНК-олигомера, например, где ингибиторы выбраны из новобиоцина и коумермицина.
A1.10. Любой из вышеуказанных способов, где цепь ДНК, предоставленная таким образом, имеет последовательность, содержащую тимидиновые (T) нуклеозиды и дезоксигуаниновые (G) нуклеозиды.
A1.11. Любой из вышеуказанных способов, где топоизомераза присоединяет одно основание, однако фермент рестрикции осуществляет расщепление в положении, которое находится на расстоянии одного нуклеотида в 5'-направлении от основания, присоединенного топоизомеразой.
A1.12. Любой из вышеуказанных способов, где цепь ДНК, предоставленная таким образом, имеет последовательность, содержащую последовательность динуклеотидов "TT" и "TG".
A1.13. Любой из вышеуказанных способов, где ДНК является одноцепочечной.
A1.14. Любой из вышеуказанных способов, где ДНК является двухцепочечной.
A1.15. Любой из вышеуказанных способов, где ДНК находится на подложке или магнитной грануле, где она при необходимости может селективно подвергаться воздействию или удаляться от реагентов для предоставления желаемой последовательности.
A1.16. Любой из вышеуказанных способов, где некоторые или все реагенты для присоединения или деблокирования ДНК предоставляются посредством потока и удаляются путем промывания.
A1.17. Любой из вышеуказанных способов, где присоединение единственных нуклеотидов или олигомеров к одноцепочечной ДНК облегчается присутствием адаптерной ДНК, имеющей последовательности, с которыми может связываться топоизомераза и одноцепочечная акцепторная ДНК.
A1.18. Любой из вышеуказанных способов, проводимый в системе, где нанопора отделяет камеру, содержащую топоизомеразу, от камеры, содержащей фосфатазу или фермент рестрикции, где нанопора позволяет движение ДНК под действием электрического притяжения, но не позволяет движение ферментов, например, как описано для любого из способа 2 и т.д.
[0111] Одной возможной проблемой является то, что последовательности поли-G могут образовывать вторичные структуры G-кватета. Путем смещения фермента рестрикции назад на одно основание (в 5'-направлении от последовательности топоизомеразы) и следуя сходной стратегии топоизомераза/IIS можно присоединять "TT" или "TG", каждый из которых может соответствовать отдельному биту. Хотя это требует 2 основания для кодирования бита, это имеет преимущество избегания последовательностей поли-G. В других вариантах осуществления другие основания на 3'-конце последовательности распознавания топоизомеразой - хотя и менее эффективные, чем (C/T)CCTT, могут позволить конъюгацию с использованием топоизомеразы поксвируса с (C/T)CCTA, (C/T)CCTC и (C/T)CCTG (https://www.ncbi.nlm.nih.gov/pubmed/17462694). Для повышения эффективности этих реакций также можно использовать способы белковой инженерии/селекции и для присоединения неканонических оснований можно использовать сходные подходы.
[0112] В определенных вариантах осуществления способ синтеза ДНК с использованием этого способа включает обработку ДНК лигазой и ATP. Топоизомераза соединяет только одну сторону ДНК (другая по существу разорвана). Лигаза репарирует разрыв и гарантирует, что сама по себе топоизомераза не разрежет продукт реакции вновь и не расщепит его.
[0113] В определенных вариантах осуществления способ включает использование ингибитора топоизомеразы для подавления связывания и активности свободной топоизомеразы в отношении ДНК-олигомера. Подходящие ингибиторы включают новобиоцин и коумермицин. Следует отметить, что полное ингибирование нежелательно, поскольку низкий уровень активности топоизомеразы может помочь "релаксировать" скрученную ДНК, что является особенно полезным при синтезе длинных цепей ДНК.
[0114] Таким образом, в другом варианте осуществления изобретение относится к способу (Способ 2) синтеза ДНК в наночипе, содержащем одну или более камер для присоединения, содержащих нагруженный топоизомеразой олигонуклеотид (т.е. олигонуклеотид, связанный 3'-концом с топоизомеразой), и одну или более резервных камер, содержащих фермент рестрикции и деблокирующий агент, например, фосфатазу, причем указанные камеры также содержат совместимый буферный раствор и отделены мембраной, содержащей по меньшей мере одну нанопору, где прохождение топоизомеразы и фермента рестрикции затруднено (например, поскольку они являются слишком большими и/или поскольку они связаны с субстратом в первой и второй камерах, соответственно), синтез проводится за один цикл присоединения единичных нуклеотидов или коротких олигонуклеотидных блоков к первому концу нуклеиновой кислоты, имеющей первый конец и второй конец, где первый конец нуклеиновой кислоты двигается под действием электрического притяжения между камерами для присоединения и резервной камерой, например в одном варианте осуществления следующим образом:
(i) движение 5'-конца принимающей ДНК (например, двухцепочечной ДНК) в первую камеру для присоединения под действием электрической силы,
(ii) предоставление в первой камере для присоединения нагруженного топоизомеразой донорного олигонуклеотида, где донорный олигонуклеотид содержит участок связывания топоизомеразы, информационную последовательность (например, выбранную из по меньшей мере двух различных нуклеотидов или последовательностей, например, где одна последовательность соответствует "0", а другая соответствует "1" в двоичном коде) и участок рестрикции, который при расщеплении ферментом рестрикции предоставляет участок лигирования топоизомеразы;
(iii) предоставление достаточного количества времени для лигирования донорного олигонуклеотида и, тем самым, продления приемной ДНК;
(iv) продвижение 5'-конца приемной ДНК, удлиненной таким образом, в резервную камеру, под действием электрической силы, например так, что фермент рестрикции расщепляет приемную ДНК, предоставляя участок лигирования топоизомеразы, или в случае присоединения единичного нуклеотида, деблокирующий агент, например фосфатаза, образует 5'-неблокированный нуклеотид на одноцепочечной ДНК; и
(v) повторение стадий (i)-(iv) цикла, присоединяя олигонуклеотиды, имеющие ту же или отличающуюся информационную последовательность до тех пор, пока не получат желаемую последовательность или последовательности ДНК.
[0115] Например, изобретение относится к следующему
2.1. Способ 2, где 3'-конец приемной ДНК связан вблизи нанопоры и 5'-конец приемного олигонуклеотида содержит участок лигирования топоизомеразы, и включающий стадию после стадии (iv), состоящую в присоединении дополнительного олигонуклеотида к 5'-концу приемной ДНК путем промывания первой камеры для присоединения и подачи нового нагруженного топоизомеразой донорного олигонуклеотида в первую камеру для присоединения, где новый донорный олигонуклеотид имеет информационную последовательность, отличающуюся от предшествующего донорного олигонуклеотида; и, если желательно, чтобы новый донорный олигонуклеотид присоединился к приемной ДНК, втягивание 5'-конца приемной нуклеиновой кислоты обратно в первую камеру и повторение стадий (i)-(iii), или, если не желательно, обеспечения возможности приемной ДНК оставаться во второй камере до тех пор, пока в первую камеру не поступит желаемый донорный олигонуклеотид.
2.2. Любой из вышеуказанных способов, где множество приемных молекул ДНК синтезируется независимо параллельно, так что молекулы ДНК, имеющие различные последовательности, получают путем контроля по отдельности, присутствуют ли они в первой камере.
2.3. Любой из вышеуказанных способов, где множество приемных молекул ДНК, каждая из которых связана на 3'-конце с поверхностью вблизи нанопоры, синтезируется независимо, где каждая нанопора имеет ассоциированную с ней пару электродов, где один электрод в паре расположен вблизи одного конца нанопоры, а другой электрод расположен вблизи другого конца нанопоры, так что каждая приемная молекула ДНК независимо может перемещаться между первой и второй камерой под действием электрического тока, обеспечиваемого парой электродов.
2.4. Любой из вышеуказанных способов, где донорные олигонуклеотиды, используемые на стадии (i) цикла, чередуются в каждом цикле между донорными олигонуклеотидами, содержащими первую информационную последовательность, и донорными олигонуклеотидами, содержащими вторую информационную последовательность.
2.5. Способ 2, включающий стадию присоединения дополнительного олигонуклеотида на 5'-конец принимающей ДНК путем возвращения 5'-конца принимающей ДНК в первую камеру для присоединения для добавления олигонуклеотида, имеющего ту же информационную последовательность, или перемещения 5'-конца принимающей ДНК во вторую камеру для присоединения, имеющую донорный олигонуклеотид, связанный на 3'-конце с топоизомеразой, где донорной олигонуклеотид во второй камере для присоединения имеет информационную последовательность, отличную от донорного олигонуклеотида в первой камере для присоединения.
2.6. Любой из вышеуказанных способов, где донорный олигонуклеотид содержит следующую структуру:
5' CGAAGGG <информационная последовательность A или B> GTCGACNNNNN
3' GCTTCCC <---------комплементарная последовательность----------> CAGCTGNNNNN
где N относится к любому нуклеотиду и фермент рестрикции представляет собой Acc1, который может разрезать ДНК (например, GTCGAC в вышеуказанной последовательности), обеспечивая подходящий выступающий конец.
2.7. Любой из вышеуказанных способов, где донорный олигонуклеотид имеет шпилечную структуру, например, 2.6, где группы NNNNN на верхней и нижней цепях соединены.
2.8. Любой из вышеуказанных способов, где по меньшей мере один из нагруженных топоизомеразой олигонуклеотидов имеет следующую структуру:
5' CGAAGGG <информационная последовательность A или B> GTCGACNNNNN
3' *TTCCC <---------комплементарная последовательность----------> CAGCTGNNNNN
(*=топоизомераза)
2.9. Любой из вышеуказанных способов, где по меньшей мере один из нагруженных топоизомеразой олигонуклеотидов имеет следующую структуру:
5' pCACGTCAGGCGTATCCATCCCTT*
3' GTGCAGTCCGCATAGGTAGGGAAGCGC
2.10. Предшествующий способ, где нагруженный топоизомеразой олигонуклеотид
2.11. Любой из вышеуказанных способов, где последовательность синтезированной ДНК определяется после каждого цикла путем детектирования изменения электрического потенциала, тока, сопротивления, емкости и/или импеданса по мере прохождения олигонуклеотида через нанопору.
2.12. Любой из вышеуказанных способов, где синтез ДНК происходит в буферном растворе, например, растворе, содержащем буфер с pH 7-8,5, например приблизительно pH 8, например, буфер, содержащий трис(гидроксиметил)аминометан (Tris), подходящую кислоту и необязательно хелатор, например, этилендиаминтетрауксусную кислоту (EDTA), например, буфер TAE, содержащий смесь основания Tris, уксусной кислоты и EDTA, или буфер TBE, содержащий смесь основания Tris, борной кислоты и EDTA; например, раствор, содержащий 10 мМ Tris, pH 8, 1 мМ EDTA, 150 мМ KCl, или например, 50 мМ ацетат калия, 20 мМ Tris-ацетат, 10 мМ ацетат магния, pH 7,9 при 25°C.
2.13. Любой из вышеуказанных способов, дополнительно включающий удаление ДНК из наночипа.
2.14. Любой из вышеуказанных способов, включающий амплификацию ДНК, синтезированной таким образом.
2.15. Любой из вышеуказанных способов, дополнительно включающий удаление ДНК из наночипа и кристаллизацию ДНК.
2.16. Любой из вышеуказанных способов, дополнительно включающий стабилизацию ДНК, например, путем высушивания раствора, содержащего ДНК вместе с одним или более из буфера (например, боратный буфер), антиоксиданта, увлажнителя, например полиола, и необязательно хелатора, например, как описано в US 8283165 B2, включенной в настоящее описание в качестве ссылки, или путем формирования матрикса между нуклеиновой кислотой и полимером, таким как блок-сополимер полиэтиленгликоль-поли(l-лизин) (PEG-PLL) типа AB.
2.17. Любой из вышеуказанных способов, включающий предоставление лигазы и ATP для устранения разрывов в ДНК [примечание: лигирование топоизомеразой лигирует только одну цепь].
2.18. Любой из вышеуказанных способов, где нагруженный топоизомеразой донорный олигонуклеотид содержит 5'-выступающий конец на цепи, комплементарной цепи, имеющей топоизомеразу, содержащий последовательность полиинозина [примечание: остатки инозина выступают в качестве "универсальных оснований" и образуют пару с любым другим основанием].
2.19. Любой из вышеуказанных способов, где фермент рестрикции представляет собой фермент рестрикции типа IIS, который может отщеплять всю присоединенную ДНК за исключением одного основания (основания, которое "присоединяется").
2.20. Любой из вышеуказанных способов, где топоизомераза выбрана из топоизомеразы вируса коровьей оспы и топоизомеразы I SVF.
2.21. Любой из вышеуказанных способов, где топоизомераза вируса коровьей оспы (которая распознает (C/T)CCTT) используется для присоединения нуклеотидов dTTP, и топоизомераза I SVF (которая распознает CCCTG) используется для присоединения нуклеотидов dGTP, например, для обеспечения двоичного кода.
2.22. Любой из вышеуказанных способов, где резервная камера дополнительно содержит лигазу и ATP для репарации цепи ДНК, не связанной с топоизомеразой.
2.23. Любой из вышеуказанных способов, включающий применение ингибитора топоизомеразы ингибитор для подавления связывания и активности свободной топоизомеразы в отношении ДНК-олигомера, например, где ингибиторы выбраны из новобиоцина и коумермицина.
2.24. Любой из вышеуказанных способов, где цепь ДНК, предоставленная таким образом, имеет последовательность, содержащую тимидиновые (T) нуклеозиды и дезоксигуаниновые (G) нуклеозиды.
2.25. Любой из вышеуказанных способов, где топоизомераза присоединяет одно основание, однако фермент рестрикции осуществляет расщепление в положении, которое находится на расстоянии одного нуклеотида в 5'-направлении от основания, присоединенного топоизомеразой.
2.26. Любой из вышеуказанных способов, где цепь ДНК, предоставленная таким образом, имеет последовательность, содержащую последовательность динуклеотидов "TT" и "TG".
2.27. Любой из вышеуказанных способов, который представляет собой способ синтеза молекулы ДНК путем присоединения единичных нуклеотидов в направлении от 3' к 5', включающий (i) реакцию молекулы ДНК c топоизомеразой, нагруженной желаемым нуклеотидом в 5'-защищенной форме, например, 5'-фосфорилированной форме, так что желаемый нуклеотид в 5'-защищенной форме присоединяется к 5'-концу ДНК, затем (ii) удаление защитной группы из 5'-конца ДНК, образованной таким образом, с использованием фермента фосфатазы, и повторение стадий (i) и (ii) до тех пор, пока не будет получена желаемая нуклеотидная последовательность.
2.28. Любой из вышеуказанных способов, который представляет собой способ синтеза молекулы ДНК путем присоединения олигомеров в направлении от 3' к 5', включающий (i) реакцию молекулы ДНК с топоизомеразой, нагруженной желаемым олигомером, тем самым лигируя олигомер с молекулой ДНК, затем (ii) использование фермента рестрикции для предоставления 5'-участка для опосредуемого топоизомеразой лигирования другого олигомера, и повторение стадий (i) и (ii) до тех пор, пока не будет получена желаемая нуклеотидная последовательность.
2.29. Любой из вышеуказанных способов, который представляет собой способ согласно любому из способа A и т.д.
[0116] Продукт реакций синтеза можно детектировать, изучать для контроля качества и считывать для извлечения данных, закодированных в полимере. Например, ДНК можно амплифицировать и секвенировать общепринятыми способами для подтверждения того, что секвенирование в нанопоре является надежным.
[0117] В другом варианте осуществления изобретение относится к олигонуклеотиду, содержащему участок связывания топоизомеразы, информационную последовательность (например, выбранную по меньшей мере из двух различных последовательностей, например, где одна последовательность соответствует "0", а другая соответствует "1" в двоичном коде), и участок рестрикции, который при расщеплении ферментом рестрикции обеспечивает участок лигирования топоизомеразой, например, содержащий следующую последовательность:
5' CGAAGGG <информационная последовательность A или B> GTCGAC
3' GCTTCCC <---------комплементарная последовательность----------> CAGCTG
где информационная последовательность A или B представляет собой последовательность из 3-12, например, приблизительно 8 нуклеотидов.
[0118] В другом варианте осуществления изобретение относится к нагруженному топоизомеразой олигонуклеотиду, где олигонуклеотид содержит участок связывания топоизомеразы, информационную последовательность (например, выбранную по меньшей мере из двух различных последовательностей, например, где одна последовательность соответствует "0", а другая соответствует "1" в двоичном коде), и участок рестрикции, который при расщеплении ферментом рестрикции обеспечивает участок лигирования топоизомеразой; например, к нагруженному топоизомеразой олигонуклеотиду, имеющему следующую структуру:
5' CGAAGGG <информационная последовательность A или B> GTCGACNNNNN
3' *TTCCC <---------комплементарная последовательность----------> CAGCTGNNNNN
где информационная последовательность A или B представляет собой последовательность из 3-12, например, приблизительно 8 нуклеотидов и * представляет собой топоизомеразу, ковалентно связанную с олигонуклеотидом; например, где топоизомераза представляет собой топоизомеразу I вируса коровьей оспы.
[0119] В другом варианте осуществления изобретение относится к одноцепочечной или двухцепочечной молекуле ДНК, как описано выше, где единичная цепь или кодирующая последовательность по существу состоит из негибридизующихся оснований, например, остатков аденина и цитозина (A и C), которые организованы в виде последовательности, соответствующей двоичному коду, например, для применения в способе хранения данных. Например, изобретение относится к ДНК (ДНК 1), где ДНК является одноцепочечной или двухцепочечной, имеющей длину по меньшей мере 1000 нуклеотидов, например, 1000-1000000 нуклеотидов или, например, от 5000 до 20000 нуклеотидов, где последовательность нуклеотидов соответствует двоичному коду; например,
1.1. ДНК 1, где ДНК является одноцепочечной.
1.2. ДНК 1, где ДНК является двухцепочечной.
1.3. Любая из вышеуказанных ДНК, где нуклеотиды в единичной цепи или в кодирующей цепи выбраны из адениновых, тиминовых и цитозиновых нуклеотидов, например, выбраны из адениновых и цитозиновых нуклеотидов или тиминовых и цитозиновых нуклеотидов.
1.4. Любая из вышеуказанных ДНК, состоящая в основном из негибридизующихся нуклеотидов, так что она не образует существенные вторичные структуры в форме единичной цепи.
1.5. Любая из вышеуказанных ДНК, где нуклеотиды по меньшей мере на 95%, например на 99%, например, на 100% представляют собой адениновые и цитозиновые нуклеотиды.
1.6. Любая из вышеуказанных ДНК, содержащая нуклеотид или последовательность нуклеотидов, добавляемую для разделения или прерывания нуклеотидов, соответствующих двоичному коду, например, для разделения 1-ц и 0-ей или групп 1-ц и 0-ей, так чтобы последовательные 1-цы или 0-и было легче считывать.
1.7. Любая из вышеуказанных ДНК, где (a) каждый бит в двоичном коде соответствует одному нуклеотиду, например, каждый из 1 и 0 соответствует A или C; или (b) каждый бит в двоичном коде соответствует серии из более чем 1 нуклеотида, например, из 2, 3 или 4 нуклеотидов, например, AAA или CCC.
1.8. Любая из вышеуказанных ДНК, которая является кристаллизованной.
1.9. Любая из вышеуказанных ДНК, которая предоставлена в сухой форме вместе с одним или более из буферной соли (например, боратный буфер), антиоксиданта, увлажнителя, например, полиола, и необязательно хелатора, например, как описано в US 8283165 B2, включенной в настоящее описание в качестве ссылки; и/или в матриксе между нуклеиновой кислотой и полимером, таким как блок-сополимер полиэтиленгликоль-поли(l-лизин) (PEG-PLL) типа AB; и/или вместе с комплементарной цепью нуклеиновой кислоты или белком, который связывает ДНК.
1.10. Любая из вышеуказанных ДНК, полученная любым из способа 1 и т.д., или способа 2 и т.д, или способа A и т.д.
[0120] Наночипы можно изготавливать, например, как изображено на фиг.23-29. Например, в одном формате каждая цепь полимера ассоциирована с двумя или четырьмя камерами для присоединения, где формат с двумя камерами для присоединения пригоден для кодирования двоичного кода в полимере и формат с четырьмя камерами для присоединения является особенно пригодным для получения специализированных последовательностей ДНК. Каждая камера для присоединения содержит отдельно контролируемый электрод. Камеры для присоединения содержат реагенты для присоединения мономеров к полимеру в буфере. Камеры для присоединения разделены мембраной, содержащей одну или более нанопор, от резервной камеры, которая может быть общей для нескольких камер для присоединения и которая содержит реагенты для удаления защитной группы и буфер, для удаления защитной группы из защищенных мономеров или олигомеров, присоединенных в камерах для присоединения. Наночипы содержат множество групп камер для присоединения, чтобы обеспечить параллельный синтез множества полимеров.
[0121] Для разрешения, соответствующего одному основанию ДНК, важен высокоскоростной и малошумный сенсор с нанопорами и электроника для детектирования. В определенных вариантах осуществления наночип электрически связан с дополняющим полупроводниковым металл-оксидным чипом (CMOS). Твердофазные нанопоры могут быть интегрированы в платформу CMOS вблизи электродов смещения и специализированной усиливающей электроники, например, как описано в Uddin, et al., ʺIntegration of solid-state nanopores in a 0,5 μm cmos foundry processʺ, Nanotechnology (2013) 24(15): 155501, содержание которых включено в настоящее описание в качестве ссылки.
[0122] В другом варианте осуществления изобретение относится к наночипу (наночип 1) для синтеза и/или секвенирования электрически заряженного полимера, например ДНК, содержащего по меньшей мере два различных мономера, причем наночип содержит по меньшей мере первую и вторую реакционные камеры, разделенные мембраной, содержащей одну или более нанопор, где каждая реакционная камера содержит один или более электродов для втягивания электрически заряженного полимера в камеру и, кроме того, содержит электролитную среду и необязательно реагенты для присоединения мономеров к полимеру, например,
1.1. Наночип 1, где нанопора имеет диаметр 2-20 нм, например 2-10 нм, например 2-5 нм.
1.2. Любой из вышеуказанных наночипов, где некоторые или все из стенок реакционных камер наночипа содержат кремниевый материал, например, кремний, диоксид кремния, нитрид кремния или их комбинации, например, нитрид кремния.
1.3. Любой из вышеуказанных наночипов, где некоторые или все из стенок реакционных камер наночипа содержат кремниевый материал, например, кремния, диоксид кремния, нитрид кремния или их комбинации, например, нитрид кремния, и некоторые или все из нанопор изготовлены посредством бомбардировки ионами.
1.4. Любой из вышеуказанных наночипов, где некоторые или все из нанопор состоят из порообразующего белка, α-гемолизина, в мембране, например, липидном бислое.
1.5. Любой из вышеуказанных наночипов, где некоторые или все из стенок реакционных камер являются покрытыми для минимизации взаимодействий с реагентами, например, покрыты полимером, таким как полиэтиленгликоль, или белком, таким как бычий сывороточный альбумин.
1.6. Любой из вышеуказанных наночипов, содержащий электролитную среду.
1.7. Любой из вышеуказанных наночипов, содержащий электролитную среду, содержащую буфер, например, буфер с pH 7-8,5, например, приблизительно pH 8, например, буфер, содержащий трис(гидроксиметил)аминометан (Tris), подходящую кислоту и необязательно хелатор, например, этилендиаминтетрауксусную кислоту (EDTA), например, буфер TAE, содержащий смесь основания Tris, уксусной кислоты и EDTA, или буфер TBE, содержащий смесь оснований Tris, борной кислоты и EDTA; например, раствор, содержащий 10 мМ Tris pH 8, 1 мМ EDTA, 150 мМ KCl, или, например, 50 мМ ацетат калия, 20 мМ Tris-ацетат, 10 мМ ацетат магния, pH 7,9, при 25°C.
1.8. Любой из вышеуказанных наночипов, содержащий реагенты для присоединения мономеров к полимеру.
1.9. Любой из вышеуказанных наночипов, способный как синтезировать (ʺзаписыватьʺ, например, посредством последовательного присоединения мономеров или групп мономеров к полимеру) и секвенирования (ʺсчитыванияʺ, например, путем измерения изменений тока и/или индуктивности по мере прохождения мономеров через нанопору) полимера.
1.10. Любой из вышеуказанных наночипов, где мембрана, содержащая одну или более нанопор, содержит металлическую поверхность на обеих сторонах, причем металлическая поверхность разделена изолирующим материалом, например, мембраной из нитрида кремния, причем поверхности металла организованы, например, способами литографии, для предоставления электродов на каждой стороне каждой нанопоры, например, так что в электролитной среде может быть сгенерирован электрический ток через нанопору, например, так что ток может втянуть полимер через нанопору и по мере прохождения полимера через нанопору изменение электрического потенциала через нанопору может быть измерено и использовано для идентификации последовательности мономеров в полимере.
1.11. Любой из вышеуказанных наночипов, содержащий электрически заряженный полимер, который представляет собой ДНК.
1.12. Любой из вышеуказанных наночипов, содержащий электрически заряженный полимер, который представляет собой одноцепочечную ДНК (оцДНК).
1.13. Любой из вышеуказанных наночипов, содержащий электрически заряженный полимер, который представляет собой ДНК, содержащую определенный участок рестрикции.
1.14. Любой из вышеуказанных наночипов, содержащий электрически заряженный полимер, который представляет собой ДНК, где ДНК представляет собой ДНК, как описано для любой из ДНК 1 и т.д., выше.
1.15. Любой из вышеуказанных наночипов, содержащий электрически заряженный полимер, который представляет собой ДНК, где ДНК по меньшей мере на 95%, например на 99%, например, на 100% состоит из остатков аденина и цитозина.
1.16. Любой из вышеуказанных наночипов, содержащий электрически заряженный полимер, который представляет собой ДНК, где ДНК содержит только остатки аденина и цитозина.
1.17. Любой из вышеуказанных наночипов, содержащий одно или более отверстий для возможности подачи и слива буфера и реагентов.
1.18. Любой из вышеуказанных наночипов, содержащий буферный раствор, например, раствор, содержащий буфер для pH 7-8,5, например, приблизительно pH 8, например, буфер, содержащий трис(гидроксиметил)аминометан (Tris), подходящую кислоту и необязательно хелатор, например, этилендиаминтетрауксусную кислоту (EDTA), например, буфер TAE, содержащий смесь основания Tris, уксусной кислоты и EDTA, или буфер TBE, содержащий смесь оснований Tris, борной кислоты и EDTA; например, раствор, содержащий 10 мМ Tris pH 8, 1 мМ EDTA, 150 мМ KCl, или, например, 50 мМ ацетат калия, 20 мМ Tris-ацетат, 10 мМ ацетат магния, pH 7,9, при 25°C.
1.19. Любой из вышеуказанных наночипов, который представляет собой или способен к лиофилизации для хранения, а затем регидратации, например, структура наночипа содержит гидратируемый или водопроницаемый полимер.
1.20. Любой из вышеуказанных наночипов, который синтезирован в сухой форме, например, где структура наночипа содержит гидратируемый или водопроницаемый полимер, с последующей гидратацией перед применением, необязательно с последующей лиофилизацией для длительного хранения после завершения процесса записывания.
1.21. Любой из вышеуказанных наночипов, где электрически заряженный полимер, например, ДНК, стабилизирован гистоном.
1.22. Любой из вышеуказанных наночипов, где внутренняя поверхность положительно заряжена.
1.23. Любой из вышеуказанных наночипов, где электроды функционально соединены в емкостной цепи, способной обеспечивать радиочастотный пульсирующий прямой ток, например, с частотой от 1 МГц до 1 ГГц, например 50-200 МГц, например приблизительно 100 МГц, через нанопору, например, где пульсирующий прямой ток может втягивать заряженный полимер через нанопору и последовательность мономеров может быть определена путем измерения изменения емкости через нанопору по мере прохождения заряженного полимера через нанопору.
1.24. Любой из вышеуказанных наночипов, содержащий резервную или деблокирующую камеру, которая содержит реагенты для удаления защитной группы из полимера после присоединения мономера или олигомера в одной из камер для присоединения.
1.25. Любой из вышеуказанных наночипов, содержащий множество пар камер для присоединения.
1.26. Любой из вышеуказанных наночипов, содержащий слой электрорегулирования, флюидный слой и слой заземления, например, как представлено на фиг.24, соединенные сращиванием.
1.27. Любой из вышеуказанных наночипов, где нанопора выполнена просверливанием посредством FIB, TEM, влажной или сухой гравировки.
1.28. Любой из вышеуказанных наночипов, где мембрана, содержащая нанопоры, имеет толщину от 1 атомного слоя до 30 нм.
1.29. Любой из вышеуказанных наночипов, где мембрана, содержащая нанопоры, изготовлена из SiN, BN, SiOx, графена, дихалькогенидов переходных металлов, например, WS2 или MoS2.
1.30. Любой из вышеуказанных наночипов, содержащий проводку, изготовленную из металла или поликремния.
1.31. Любой из вышеуказанных наночипов, где плотность проводки увеличена путем 3D-укладки, причем электрическая изоляция обеспечивается покрытием диэлектриком (например, через PECVD, распыление, ALD и т.д.).
1.32. Любой из вышеуказанных наночипов, где контакт с электродом в камере для присоединения осуществляется с использованием переходных отверстий в кремнии (TSV) посредством глубокой гравировки реактивными ионами (DRIE), например. с использованием крио-процесса или процесса BOSC, или посредством влажной кремниевой гравировки.
1.33. Любой из вышеуказанных наночипов, где индивидуальный контроль напряжения для электрода в каждой камере для присоединения позволяет индивидуальный контроль и мониторинг электрода в каждой камере для присоединения.
1.34. Любой из вышеуказанных наночипов, где каждый полимер ассоциирован с первой камерой для присоединения, второй камерой для присоединения и деблокирующей камерой.
1.35. Любой из вышеуказанных наночипов, где одна или более камер имеют поток жидкости.
1.36. Любой из вышеуказанных наночипов, где одна или более камер изолированы в отношении жидкостей.
1.37. Любой из вышеуказанных наночипов, где деблокирующая камера имеет поток жидкости.
1.38. Любой из вышеуказанных наночипов, где камеры для присоединения имеют общий поток жидкости.
1.39. Любой из вышеуказанных наночипов, где проводка между камерами является общей для камер сходного типа (например, для первых камер для присоединения, для вторых камер для присоединения и для деблокирующих камер).
1.40. Любой из вышеуказанных наночипов, где камеры для присоединения имеют индвидуальный контроль напряжения и деблокирующие камеры имеют общее заземление.
1.41. Любой из вышеуказанных наночипов, где деблокирующие камеры имеют индивидуальный контроль напряжения, первые камеры для присоединения имеют общее заземление, и вторые камеры для присоединения имеют общее заземление.
1.42. Любой из вышеуказанных наночипов, где наночип изготовлен путем сращивания слоев, и камеры предварительно заполняют желаемыми реагентами перед сращиванием.
1.43. Любой из вышеуказанных наночипов, где одна или более внутренних поверхностей являются силанизированными.
1.44. Любой из вышеуказанных наночипов, который имеет одно или более отверстий для подачи или извлечения жидкости.
1.45. Любой из вышеуказанных наночипов, где контакт электродов в камерах с полимером ограничен, например, где электрод помещен слишком далеко от нанопоры, чтобы его достиг заряженный полимер, связанный с поверхностью рядом с нанопорой, или где электрод защищен материалом, который позволяет прохождение воды и одноатомных ионов (например, ионов Na+, K+ и Cl-), но не прохождение полимера, или мономерных или олигомерных реагентов, подлежащих связыванию с полимером.
1.46. Любой из вышеуказанных наночипов, который электрически связан с дополняющим полупроводниковым металл-оксидным чипом (CMOS).
[0123] Например, в одном варианте осуществления изобретение относится к наночипу, например, согласно любому из наночипа 1 и т.д., для секвенирования электрически заряженного полимера, например ДНК, содержащего по меньшей мере два различных мономера, причем наночип содержит по меньшей мере первую и вторую реакционные камеры, содержащие электролитную среду и разделенные мембраной, содержащей одну или более нанопор, где каждая реакционная камера содержит по меньшей мере одну пару электродов, размещенную на противоположных сторонах мембраны, где электроды функционально соединены в емкостном контуре, способном генерировать радиочастотный пульсирующий прямой ток, например, с частотой от 1 МГц до 1 ГГц, например 50-200 МГц, например, приблизительно 100 МГц, через нанопору, например, где пульсирующий прямой ток может втягивать заряженный полимер через нанопору и последовательность мономеров может быть определена путем измерения изменения емкости через нанопору по мере прохождения заряженного полимера через нанопору.
[0124] В другом варианте осуществления, изобретение относится к способу считывания последовательности мономеров заряженного полимера, содержащего по меньшей мере два различных типа мономеров, например, молекулы ДНК, включающему применение радиочастотного пульсирующего прямого тока, например, с частотой от 1 МГц до 1 ГГц, например 50-200 МГц, например, приблизительно 100 МГц, через нанопору, где пульсирующий прямой ток втягивает заряженный полимер через нанопору и последовательность мономеров считывается путем измерения изменения емкости через нанопору по мере прохождения полимера через нанопору.
[0125] В другом варианте осуществления, изобретение относится к применению любой из ДНК 1 и т.д. в способе хранения информации.
[0126] В другом варианте осуществления изобретение относится к применению одноцепочечной ДНК в способе хранения информации, например, где последовательность по существу не является самогибридизующейся.
[0127] В другом варианте осуществления, изобретение относится к способу хранения данных и к устройству с использованием наночипа, например, любого из наночипа 1 и т.д., для получения электрически заряженного полимера, например ДНК, содержащего по меньшей мере два различных мономера или олигомера, где мономеры или олигомеры организованы в виде последовательности, соответствующей двоичному коду, например, в соответствии с любым из вышеуказанных способов 1 и/или 2 и т.д.
[0128] Например, в одном варианте осуществления наночип, содержащий полимер, синтезированный таким образом, обеспечивает устройство хранения данных, поскольку наночип может быть активирован и последовательность полимера может быть определена путем пропускания его через нанопору в любой момент времени. В других вариантах осуществления полимер извлекается из наночипа, или амплифицируется и амплифицированный полимер извлекается из наночипа, хранится до тех пор, пока не потребуется, а затем считывается с использованием общепринятого секвенатора, например, общепринятого секвенирующего устройства с нанопорами.
[0129] В другом варианте осуществления, изобретение относится к способу хранения информации, включающему синтез любой из ДНК 1 и т.д., например, в соответствии с любым из способов 1 и т.д. или способов 2 и т.д.
[0130] В другом варианте осуществления, изобретение относится к способу считывания двоичного кода, например, кодируемого любой из ДНК 1 и т.д., с использованием секвенатора с нанопорами, например с использованием наночипа 1 и т.д., как описано в настоящем описании.
[0131] Любой из вышеуказанных способов, где наночип стирается с использованием фермента, который лизирует заряженный полимер, например, дезоксирибонуклеазы (ДНК-азы) для гидролиза ДНК.
ПРИМЕРЫ
Пример 1. Иммобилизация одного конца ДНК рядом с нанопорой и контролируемое движение ДНК с одну и в другую сторону с помощью электрического тока
[0132] Разработаны экспериментальные методики для демонстрации того, что ДНК перемещается в одну и в другую стороны между двумя камерами, разделенными нанопорой, при помощи электрического тока в условиях, в которых соответствующий белок не перемещается между камерами.
[0133] Наночип, содержащий две камеры, изготавливают из нитрида кремния. Нанопоры размером <4 нм (для дцДНК или оцДНК) и 2 нм (только для оцДНК) получают, как описано в Briggs K, et al. Automated fabrication of 2-nm solid-state nanopores for nucleic acid analysis, Small (2014)10(10):2077-86. Эти две камеры обозначены как "ближняя" и "дальняя" камера, причем дальняя камера представляет собой камеру, в которой 3'-конец ДНК конъюгирован.
[0134] Показано, что оцДНК (пора размером 2 нм) и оцДНК+дцДНК (пора размером 4 нм), но не белок проходят через нанопору. Прохождение через нанопоры детектируют по прекращению электрического тока.
[0135] Конъюгация ДНК с поверхностью поры: Присоединение 5'-амино-модифицированной ДНК к покрытым карбоксигруппами полистироловым гранулам (карбоксилатные микросферы Fluoresbrite® BB, 0,05мкм, от Polysciences, Inc.) посредством карбодиимид-опосредуемого присоединения. 3'-конец ДНК метят биотином. ДНК имеет заданную длину.
[0136] Конъюгация со стрептавидином: Конъюгацию проводили на проводили на "дальней" стороне нанопоры из нитрида кремния, конъюгируя стрептавидин с поверхностью, как описано в Arafat, A. Covalent Biofunctionalization of Silicon Nitride Surfaces. Langmuir (2007) 23 (11): 6233-6244.
[0137] Иммобилизация ДНК вблизи нанопоры: Конъюгированные с ДНК полистироловые гранулы в буфере добавляют в "ближнюю" камеру и буфер добавляют в "дальнюю" камеру (стандартный буфер: 10 мМ Tris pH 8, 1 мМ EDTA, 150 мМ KCl). Напряжение (~100 мВ) применяют до тех пор, пока не наблюдают прерывистого тока (с использованием усилителя с фиксацией потенциала Axon Nanopatch200B). 50-нм гранулы не могут проходить через нанопору, так что, когда ДНК прошла и гранула надавила на конец нанопоры, ток становится в высокой степени прерывистым. Ток сохраняется 1-2 мин до тех пор, пока ДНК не свяжется необратимо с иммобилизованным стрептавидином на дальней стороне посредством связывания биотина. Для подтверждения иммобилизации ДНК ток изменяют на обратный. Различный ток наблюдают, если ДНК находится внутри и вне поры. Если оказывается, что ДНК не иммобилизована, тогда процесс повторяют.
[0138] Высвобождение гранулы эндонуклеазой: Фермент рестрикции в буфере для фермента рестрикции добавляют в камеру, где связана ДНК. В одном варианте осуществления ДНК является одноцепочечной и содержит участок рестрикции, расщепляемый ферментом, который расщепляет одноцепочечную ДНК. См., например, Nishigaki, K., Type II restriction endonuclease cleave single-stranded DNAs in general. Nucleic Acids Res. (1985) 13(16): 5747-5760. В альтернативном варианте осуществления комплементарный олигонуклеотид добавляют в камеру, где связана ДНК, и позволяют ему гибридизоваться в течение 30 минут для образования дцДНК, а затем добавляют фермент рестрикции. После высвобождения гранулы, она смывается. Ток переключают между прямым и обратным для подтверждения того, что ДНК проходит в/через и из поры.
[0139] Демонстрация контролируемого движения вперед и назад: С использованием стандартного буфера применяют ток в прямом направлении до тех пор, пока не наблюдают прерывание сигнала, а затем возвращается к "нормальному" после прохождения ДНК. Обратный ток применяют до тех пор, пока не наблюдают прерывание сигнала. Наблюдают, что сигнал не возвращается к нормальному, пока ДНК остается в поре. Применение тока в прямом и в обратном направлении повторяют на протяжении нескольких циклов для подтверждения того, что ДНК проходит через нанопору вперед и назад.
Пример 1a: Иммобилизация цепи ДНК рядом с нанопорой в чипе из диоксида кремния
[0140] Внутреннюю стенку наночипа изготавливают из диоксида кремния. Обе стороны силанизируют, однако олигонуклеотид конъюгируют только с одной стороной стенки чипа, а затем формируют нанопору.
[0141] Силанирование: Поверхность стенки чипа очищают раствором Piranha (различные коммерчески доступные торговые марки, обычно содержащие смесь серной кислоты (H2SO4) и пероксида водорода (H2O2), которая удаляет органические остатки с поверхности) при 30°C, и промывают дважды дистиллированной водой. Получают исходный раствор (3-аминопропил)триэтоксисилана (APTES) с 50% метанолом (MeOH), 47,5% APTES, 2,5% наночистой H2O, и выдерживают >1 часа при 4°C. Затем исходный раствор APTES разбавляют 1:500 в MeOH и наносят на стенку чипа и инкубируют при комнатной температуре. Затем стенку чипа ополаскивают MeOH и сушат при 110°C в течение 30 минут.
[0142] Конъюгация: Затем стенку чипа инкубируют в течение 5 часов при комнатной температуре в 0,5% масс./об. растворе 1,4-фенилендиизотиоцианата (PDC) в диметилсульфоксиде (DMSO). Его два раза кратковременно промывают DMSO и два раза кратковременно промывают дважды дистиллированной водой. Затем стенку чипа инкубируют со 100 нМ амин-модифицированными одноцепочечными ДНК-олигомерами (приблизительно 50-меры) в дважды дистиллированной воде (pH 8) в течение ночи при 37°C. Затем стенку чипа промывают два раза 28% раствором аммиака для инактивации любого не вступившего в реакцию материала и промывают два раза дважды дистиллированной водой. Затем в стенке формируют одну или более нанопор.
[0143] После завершения изготовления наночипа внутреннюю стенку покрывают ДНК-олигомерами длиной приблизительно 50 п.о. Это позволяет одноцепочечной ДНК, имеющей концевую последовательность, комплементарную связанной с поверхностью ДНК, локализоваться у нанопоры путем связывания оцДНК с относительно объемной структурой (например гранула, белок или структура ДНК-оригами, имеющая слишком большой диаметр для вхождения в нанопору), где последовательность, комплементарная связанной с поверхностью ДНК, является дистальной относительно объемной структуры, втягивания заряженного полимера через нанопору с использованием электрического тока, что позволяет оцДНК связаться с комплементарным связанным с поверхностью ДНК-олигомером рядом с нанопорой, и отщепления объемной структуры.
Пример 2. Синтез ДНК- присоединение единичного нуклеотида
[0144] ДНК перемещается в "резервную" камеру посредством применения соответствующего тока и детектирования движения ДНК.
[0145] В камеру "для присоединения" добавляют фермент концевую трансферазу (TdT, New England Biolabs) в подходящем буфере (50 мМ ацетат калия, 20 мМ Tris-ацетат, 10 мМ ацетат магния, pH 7,9 при 25°C), и обратимо блокированный dATP*. Также в "резервную" камеру добавляют буфер.
[0146] Для присоединения нуклеотидов к ДНК используют dNTP, которые имеют обратимые блоки на 3'-OH. При присоединении к цепи ДНК следующий dNTP не может быть добавлен до тех пор, пока блокированный dNTP не будет разблокирован.
[0147] Деблокирование может быть химическим или ферментативным. Используют различные подходы:
[0148] a. 3'-O-аллил: аллил удаляют посредством Pd-катализируемого деаллилирования в водном буферном растворе, как описано в Ju J, Four-color DNA sequencing by synthesis using cleavable fluorescent nucleotide reversible terminators. Proc Natl Acad Sci U S A. (2006);103(52):19635-40; или с использованием йода (10 моль.%) в полиэтиленгликоле-400, как описано в Shankaraiah G., et al., Rapid and selective deallylation of allyl ethers and esters using iodine in polyethylene glycol-400. Green Chem. (2011)13: 2354-2358.
[0149] b. 3' O-NH2: амин удаляют в забуференном NaNO2, как описано в US 8034923.
[0150] c. 3'-фосфат. фосфат гидролизуют эндонуклеазой IV (New England Biolabs). Другие возможные 3'-модификации, которые также можно удалять эндонуклеазой IV, включают фосфогликоальдегид и дезоксирибоза-5-фосфат.
[0151] d. 3'-O-Ac: ацетат удаляют ферментативным гидролизом, как описано в Ud-Dean, A theoretical model for template-free synthesis of long DNA sequence. Syst Synth Biol (2008) 2:67-73.
[0152] Затем ДНК перемещается в "дальнюю" камеру с использованием соответствующего тока и детектирования движения ДНК. Защитную группу ДНК удаляют путем смены буферов и добавления деблокирующего буфера/раствора, как описано в a.- d. выше.
[0153] Способ при желании повторяют для получения представляющей интерес последовательности.
Пример 3. Синтез ДНК: присоединение блокированного олигонуклеотида
[0154] 3'-конец двухцепочечной ДНК связывают рядом с нанопорой с отверстием размером 4 нм. 5'-конец ДНК имеет выступающий конец CG (считывание от 5' к 3').
[0155] Олигонуклеотидные кассеты A и B получают следующим образом:
5' CGAAGGG <КОД A ИЛИ B> GTCGACNNNNN
3' GCTTCCC <КОМПЛЕМЕНТАРНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ> CAGCTGNNNNN
[0156] Каждый из кода A и кода B соответствует информационной последовательности. N обозначает любой нуклеотид. 5'-последовательность содержит участок распознавания топоизомеразой и 3'-последовательность содержит участок рестрикции Acc1. Олигонуклеотид подвергается воздействию топоизомеразы и топоизомераза связывается с 3'-тимидином:
5' CGAAGGG <КОД A ИЛИ B> GTCGACNNNNN
3' *TTCCC <КОМПЛЕМЕНТАРНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ> CAGCTGNNNNN (*=топоизомераза)
[0157] ДНК перемещается в "ближнюю" камеру посредством применения соответствующего тока и детектирования движения ДНК. Нагруженный топоизомеразой олигонуклеотид "код A" предоставляется в камере "для присоединения". ДНК перемещается в камеру для присоединения посредством применения соответствующего тока и детектирования движения ДНК, в результате чего олигонуклеотид кода A связывается с ДНК, в "резервную" камеру добавляют Acc1, где он осуществляет расщепление в участке рестрикции, обеспечивая участок лигирования топоизомеразы.
[0158] Этот процесс повторяется до тех пор, пока не будет достигнута желаемая последовательность, присоединяя другой "код A" или "код B". Следует отметить, что не требуется непрерывное добавление нового Acc1 в "резервную камеру"; требуется только вымывание олигонуклеотидов кода A или кода B в камере для "присоединения" при переключении с кода A или кода B.
[0159] Для секвенирования в поре, которая позволяет прохождение только оцДНК, необходимы некоторые модификации протокола, описанного выше. Уже известно, что, когда дцДНК встречает небольшую пору (2 нм), только оцДНК пройдет и комплементарная последовательность "отделится". Таким образом, при проведении этого синтеза с использованием поры размером 2 нм необходимо убедиться, что надлежащая дцДНК способна "вновь образоваться" на другой стороне. Для этого можно добавлять ʺCGAAGGG <КОД A ИЛИ B> GTCGACNNNNNʺ в ближнюю камеру (чтобы гарантировать внесение участка рестрикции) и ʺCGAAGGG <КОД A ИЛИ B> GTʺ в дальнюю камеру (чтобы гарантировать образование участка, совместимого с топоизомеразой).
[0160] Описывая вышеуказанный способ, авторы изобретения демонстрируют последовательное "присоединение" закодированной в ДНК информации в растущую цепь ДНК с ≥2 последовательных присоединений (соответствующих 2 битам данных), каждое из которых включает стадию "присоединения" и "удаления защитной группы". Первоначальные эксперименты для оптимизации и подтверждения концепции проводят в микропробирках.
[0161] В подходе, описанном в этом примере, один бит информации кодируется в нити нуклеотидов. "Присоединяемый" ДНК-бит представляет собой короткую последовательность дцДНК, конъюгированную с топоизомеразой I вируса коровьей оспы (topo). В присутствии подходящей "акцепторной" ДНК "с удаленной защитной группой", нагруженный топоизомеразой ДНК-"бит" ферментативно и ковалентно связывается ("присоединяется") с акцептором посредством топоизомеразы, которая в этом процессе удаляется из ДНК. Затем фермент рестрикции может отщеплять добавленный бит для "удаления защитной группы" и создания подходящей "акцепторной" последовательности для присоединения следующего бита.
[0162] Нагрузка топоизомеразой: Общая схема нагрузки является следующей, как схематично представлено на фиг.22 и ниже, где N обозначает любой нуклеотид, и A, T, G и C соответствуют нуклеотиды с адениновым, тиминовым, гуаниновым и цитозиновым основаниями, соответственно. N друг на другом являются комплементарными. В то время как в этом примере используется фермент рестрикции HpyCH4III, основная стратегия будет работать с другими ферментами рестрикции, например, как продемонстрировано в примере 4.
+
Топоизомераза *
=
+
(нагруженный топоизомеразой)
(N друг над другом являются комплементарными)
Присоединение
Общая реакция "присоединения":
(акцептор)
+
(нагруженный топоизомеразой)
=
+
*
(свободная топоизомераза)
Удаление защитной группы
Общая реакция "удаления защитной группы":
+
HpyCH4III
(фермент рестрикции)
=
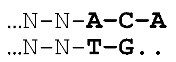
(с удаленной защитной группой)
+
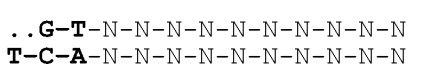
(побочный продукт)
[0163] Следующие олигонуклеотиды заказаны от Integrated DNA Technologies (IDT). ʺBʺ на конце некоторых из олигонуклеотидов указывает на биотин:
BAB: CGATAGTCTAGGCACTGTTTGCTGCGCCCTTGTCCGTGTCGCCCTTATCTACTTAAGAGATCATACAGCATTGCGAGTACG
B1: b-CACGTACTCGCAATGCTGTATGATCTCTTAAGTAGATA
B2: ATCTACTTAAGAGATCATACAGCATTGCGAGTACG
TA1: b-CACACTCATGCCGCTGTAGTCACTATCGGAAT
TA2: AGGGCGACACGGACAGTTTGAATCATACCG
TA3b: AACTTAGTATGACGGTATGATTCAAACTGTCCGTGTCGCCCTTATTCCGATAGTGACTACAGCGGCATGAG
TB1: b-CACACTCATGCCGCTGTAGTCACTATCGGAAT
TB2: AGGGCGCAGCAAACAGTGCCTAGACTATCG
TB3b:
AACTTAGTATGACGATAGTCTAGGCACTGTTTGCTGCGCCCTTATTCCGATAGTGACTACAGCGGCATGAG
FP1: CACGTACTCGCAATGCT
FP2: CGGTATGATTCAAACTGTCCG
FP3: GCCCTTGTCCGTGTC
[0164] Олигонуклеотиды растворяют до концентрации 100 мкМ в буфере TE и хранят при -20°C.
[0165] Гибридизованные олигонуклеотиды получают смешением олигонуклеотидов, как описано ниже, нагреванием до 95°C в течение 5 минут, а затем снижением температуры на 5°C каждые 3 минуты до достижения температурой 20°C. Гибридизованные олигонуклеотиды хранят при 4°C или -20°C. Комбинации олигонуклеотидов являются следующими:
B1/2
48 мкл B1
48 мкл B2
4 мкл 5 M NaCl
A5
20 мкл TA1
20 мкл TA2
5 мкл TA3b
4 мкл 5 M NaCl
51 мкл TE
B5
20 мкл TB1
20 мкл TB2
5 мкл TB3b
4 мкл 5 M NaCl
51 мкл TE
[0166] В этом ферменте используют следующие буферы и ферменты:
TE: 10 M Tris pH 8,0, 1 мМ EDTA, pH 8,0
WB: 1 M NaCl, 10 мМ Tris pH8,0, 1 мМ EDTA, pH 8,0
1x Topo: 20 мМ Tris, pH7,5, 100 мМ NaCl, 2 мМ DTT, 5 мМ MgCl2
1x RE: 50 мМ ацетат K, 20 мМ Tris-ацетат, 10 мМ ацетат Mg, 100мкг/мл BSA, pH 7,9, при 25°C.
ДНК-топоизомеразу I вируса коровьей оспы (topo) приобретают от Monserate Biotech (10000 Е/мл)
HypCH4III приобретают от NEB
Покрытые стрептавидином магнитные гранулы (s-MagBeads) приобретают от ThermoFisher.
[0167] Акцептор получают следующим образом: 5 мкл s-magbeads промывают один раз 200 мкл WB (время связывания 1 минута). К гранулам добавляют 5 мкл B1/2+195 мкл WB и инкубируют в течение 15 минут при комнатной температуре, а затем промывают один раз посредством 200 мкл WB, затем промывают один раз посредством 200 мкл 1× Topo, и ресуспендируют в 150мкл 1× Topo.
[0168] Нагруженный топоизомеразой A5 (см. фиг.20) получают следующим образом: 4 мкл 10× буфера для топоизомеразы+23 мкл воды+8мкл A5+5 мкл топоизомеразы инкубируют при 37°C в течение 30 минут, добавляют к 5 мкл s-magbeads (промытым 1× посредством 200 мкл WB, 1× посредством 200 мкл 1× Topo, ресуспендированные в 150мкл 1× topo), и позволяют связываться в течение 15 минут при комнатной температуре.
[0169] "Присоединение" нагруженного A5 к акцептору: s-magbeads извлекают из нагруженного топоизомеразой A5, добавляют к акцептору и инкубируют при 37°C в течение 60 минут. Аликвоту извлекают, разбавляют 1/200 в TE и хранят при -20°C.
[0170] Удаление защитной группы: материал промывают один раз посредством 200 мкл WB, затем промывают один раз посредством 200 мкл 1× RE и ресуспендируют в 15 мкл 10× RE и 120 мкл воды. Добавляют 15 мкл HypCH4III. Смесь инкубируют при 37°C в течение 60 минут, затем промывают один раз посредством 200 мкл WB, промывают один раз посредством 200 мкл 1× topo, с получением продукта, который назвали "акцептором-A5".
[0171] Нагруженный топоизомеразой B5 (см. фиг.21) получают следующим образом: 4 мкл 10× буфера для топоизомеразы+23 мкл воды+8мкл B5+5 мкл топоизомеразы комбинируют и инкубируют при 37°C в течение 30 минут. Продукт добавляют к 5 мкл s-magbeads (промытым 1× посредством 200 мкл WB, 1× посредством 200 мкл 1× Topo, ресуспендированные в 15 0мкл 1× topo), и позволяют связываться в течение 15 минут при комнатной температуре.
[0172] "Присоединение" нагруженного B5 к акцептору-A5: s-magbeads извлекают из нагруженного топоизомеразой B5, добавляют к акцептору-A5 и инкубируют при 37°C в течение 60 минут. Аликвоту извлекают, разбавляют 1/200 в TE и хранят при -20°C.
[0173] Deprotection: материал промывают один раз посредством 200 мкл WB, затем промывают один раз посредством 200 мкл 1× RE и ресуспендируют в 15 мкл 10× RE и 120 мкл воды. Добавляют 15 мкл HypCH4III. Смесь инкубируют при 37°C в течение 60 минут.
[0174] Подтверждение того, что описанные выше реакции работают, предоставляют посредством амплификации способом ПЦР аликвот A5 (акцептор с присоединенным A5: стадия iii, на схеме "присоединенный A5") и B5 (акцептор-A5 с присоединенным B5: стадия vi, на схеме "присоединенный B5"). "Без матрицы" используют в качестве отрицательного контроля для A5, A5 используют в качестве отрицательного контроля для B5, олигонуклеотид BAB используют в качестве положительного контроля для B5. Ожидаемый размер продукта для ПЦР A5 составляет 68 п.н., ожидаемый размер продукта для ПЦР B5 составляет 57 п.н. (Также на геле разделяют B1/2, ожидаемый размер составляет ~47 п.н., однако это может быть приблизительным, поскольку существуют выступающие концы и он биотинилирован). Реакции ПЦР (30 циклов 95/55/68 (по 1 минуте в каждом случае) проводят следующим образом:
[0175] SDS-PAGE с использованием 4-20% гелей с Tris-глицином используют для подтверждения того, что продуцируются олигонуклеотиды ожидаемого размера. Загрузку проводят, как описано выше, однако сразу после загрузки (стадия инкубации при 37°C), примешивают буфер для загрузки и образцы нагревают до 70°C на 2 минуты и позволяют им остыть перед разделением на геле. Гель окрашивают Кумасси. Для отрицательного контроля в реакционную смесь вместо топоизомеразы добавляют воду. На фиг.30 представлены результаты, отчетливо демонстрирующие полосы, соответствующие ожидаемым размерам продуктов для ПЦР A5 и для ПЦР B5.
[0176] Таким образом показано, что присоединение ДНК-бита посредством нагруженных топоизомеразой кассет ДНК и удаление защитной группы, проводимое ферментом рестрикции, является осуществимым. В этих экспериментах для подтверждения концепции ДНК иммобилизуют посредством конъюгированных со стрептавидином магнитных гранул и помещают последовательно в различные реакционные смеси, однако в формате чипа с нанопорами авторы изобретения создали отдельные реакционные камеры и использовали электрический ток для перемещения ДНК в эти различные реакционные камеры.
[0177] Наконец, ПЦР демонстрирует, что ожидаемые последовательности ДНК создаются при проведения последовательного присоединения последовательностей ДНК, соответствующих "битам" информации. Эти реакции работали намеченным образом даже с минимальной оптимизацией.
[0178] ДНК, полученную, как описано в примерах 2 и 3, выделяют и секвенируют с использованием коммерческого секвенатора с нанопорами (MinION от Oxford Nanopore), подтверждая получение желаемой последовательности.
Пример 4. Синтез ДНК: присоединение блокированного олигонуклеотида с использованием другого фермента рестрикции
[0179] Следующий синтез проводят аналогично примеру 3, но с использованием фермента рестрикции MluI, который разрезает "ACGCGT" с образованием:
В этом примере топоизомеразу загружают для формирования комплекса с комплементарностью последовательности, которая обеспечит загруженной топоизомеразе переносить ДНК в ДНК, расщепленную посредством MluI:
+
топоизомераза
=

("*" указывает на топоизомеразу, связанную с 3'-фосфатом)
+
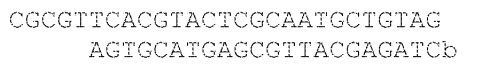
(b=биотин. Он может быть удален стрептавидином)
[0180] Посредством процесса, аналогичного предшествующему примеру, затем загруженную топоизомеразу используют для присоединения олигомера к 5'-концу синтезируемой цепи, имеющей комплементарную акцепторную последовательность, тем самым высвобождая топоизомеразу, а затем из цепи ʺудаляют защитную группуʺ с использованием MluI, и цикл повторяют до тех пор, пока не получают желаемую последовательность олигомеров.
Пример 5. Присоединение единичного основания с использованием стратегии с топоизомеразой
[0181] Авторы изобретения обнаружили, что система топоизомеразы также может быть предназначена для присоединения единичных оснований к одноцепочечной цепи ДНК (по сравнению с примером 3, в котором описано присоединение "кассет"). ДНК-бит, подлежащий "присоединению" находится в короткой последовательности ДНК, конъюгированной с топоизомеразой I вируса коровьей оспы (topo). В присутствии подходящей "акцепторной" ДНК "с удаленной защитной группой", нагруженная топоизомеразой ДНК ферментативно и ковалентно лигируется ("присоединяется") с акцептором посредством топоизомеразы, которая в этом процессе удаляется из ДНК. Затем фермент рестрикции типа IIS может отщеплять добавленную ДНК, за исключением одного основания (основания, которое "присоединяется"). Этот процесс удаления защитной группы-присоединения повторяют для присоединения дополнительных оснований (бит).
[0182] Нагрузка топоизомеразой: Общий протокол нагрузки является следующим, аналогично примеру 3:
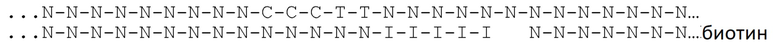
+
Топоизомераза (*)
=
N-N-N-N-N-N-N-N-N-N-N-N-N-N-N
N-N-N-N-N-N-N-N-N…биотин
(побочный продукт)
+
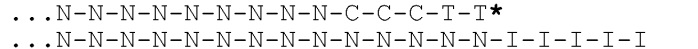
(нагруженный топоизомеразой)
Как и в примере 3, N друг на другом являются комплементарными. I представляет собой инозин. Биотин используется для удаления не вступившего в реакцию продукта и побочного продукта. Присоединение единичного основания проводят следующим образом.
N-N-N-N-N-N-N-N-N-N…
(нуклеотиды акцепторной последовательности указаны курсивом)
+
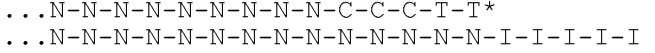
(нагруженный топоизомеразой)
=
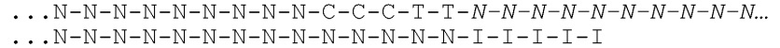
+
*
(свободная топоизомераза)
Ниже иллюстрируется удаление защитной группы с использованием фермента рестрикции BciVI (участок выделен полужирным шрифтом):
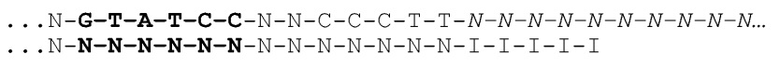
...N-G-T-A-T-C-C-N-N-C-C-C-T-T-N-N-N-N-N-N-N-N-N-N…
...N-N-N-N-N-N-N-N-N-N-N-N-N-N-I-I-I-I-I
+
BciVI
(фермент рестрикции)
=
T-N-N-N-N-N-N-N-N-N-N…
(следует отметить, что "T" добавлен к 5'-концу акцепторной ДНК)
+
N-N-I-I-I-I-I
(диссоциированный*)
+
...N-G-T-A-T-C-C-N-N-C-C-C-T
...N-N-N-N-N-N-N-N-N-N-N-N
---------------------------------------------------------
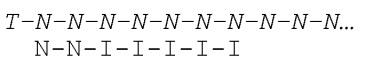
=
T-N-N-N-N-N-N-N-N-N-N…+N-N-I-I-I-I-I
(NNIIIII диссоциирует от единой цепи с присоединенным основанием)
[0183] Следующие олигонуклеотиды синтезированы коммерчески (B=биотин, P-фосфат, I=инозин):
NAT1 CACGTCAGGCGTATCCATCCCTTCACGTACTCGCAATGCTGTATGGCGAT
NAT1b P-CACGTCAGGCGTATCCATCCCTTCACGTACTCGCAATGCTGTATGGCGAT-B
NAT9cI P-IIIIIAAGGGATGGATACGCCTGACGTG
NAT9x P-ATCGCCATACAGCATTGCGAG
NAT9 ACGTGAAGGGATGGATACGCCTGACGTG
Nat9Acc CACGTAGCAGCAAACAGTGCCTAGACTATCG
Nat1P CACGTCAGGCGTATCCATCC
FP4 CGATAGTCTAGGCACTGTTTG
[0184] Олигонуклеотиды растворяют до 100 мкМ в буфере TE и хранят при -20°C.
[0185] Гибридизация: следующие гибридизованные олигонуклеотиды получают путем смешения олигонуклеотидов, как описано, нагревания до 95°C в течение 5 минут, а затем снижения температуры на 5°C каждые 3', пока температура не достигнет 20°C. Гибридизованные олигонуклеотиды хранят при 4°C или -20°C.
NAT1b/NAT9cI/NAT9x
8 мкл NAT1B
10 мкл NAT9cI
10 мкл NAT9x
48 мкл TE
4 мкл 5M NaCl
NAT1/NAT9cI
10 мкл NAT1
10 мкл NAT9cI
80 мкл PBS
NAT1/NAT9
10 мкл NAT1
10 мкл NAT9
80 мкл PBS
[0186] Буферы и ферменты: используют следующие буферы:
TE: 10 M Tris, pH 8,0, 1 мМ EDTA, pH 8,0
PBS: фосфатно-солевой буфер (137 мМ NaCl, 2,7 мМ KCl, 10 мМ Na2HPO4, 1,8 мМ KH2PO4)(pH 7,4)
10× Cutsmart: 500 мМ KAc, 200 мМ Tris-Ac, 100 мМ Mg-Ac, 1 мг/мл BSA pH 7,9.
BciVI приобретают от NEB и покрытые стрептавидином магнитные гранулы (s-MagBeads) приобретают от ThermoFisher
[0187] Реакцию присоединения проводят следующим образом.
[0188] 1. Нагрузка топоизомеразой: реагенты объединяют согласно таблице:
Затем реагенты инкубируют при 37°C в течение 30 минут. Побочные продукты удаляют с использованием магнитных гранул со стрептавидином (5 мкл) в 1× буфере для топоизомеразы после 10 минут при комнатной температуре для обеспечения связывания.
[0189] 2. Реакция: реагенты собирают в соответствии с таблицей:
Затем реагенты инкубируют при 37°C в течение 30 минут. Ожидается, что реакция присоединения будет протекать следующим образом:
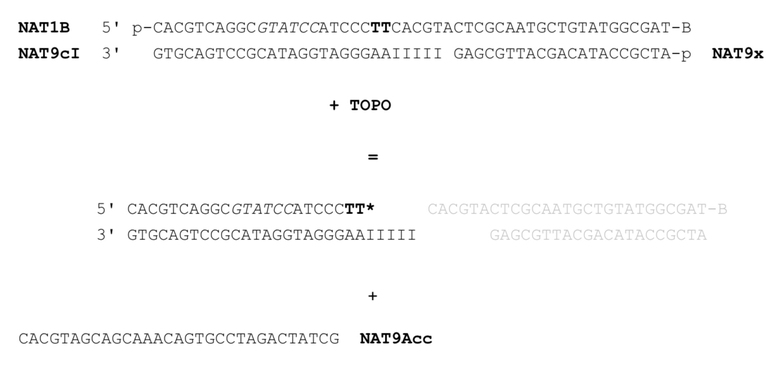
[0190] Звездочкой (*) обозначена топоизомераза. Следует отметить, что NAT9cI является фосфорилированным, однако это не показано для целей иллюстрации.
[0191] Когда с загруженной топоизомеразой присутствует акцепторная последовательность, она подвергается следующей реакции:
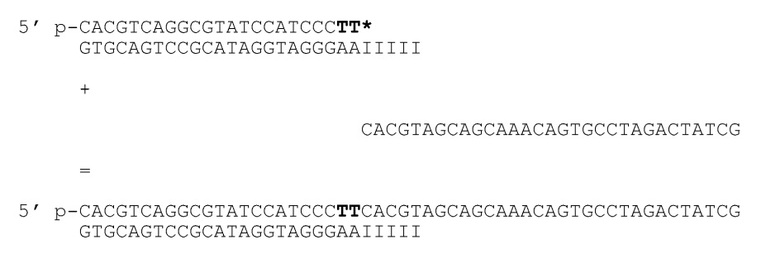
[0192] ПЦР-амплификация и измерение молекулярной массы продукта на агарозном геле подтверждает образование ожидаемого продукта. См. фиг.30, на которой представлена полоса правильногоразмера на дорожке 1 (эксперимент), и отсутствие полос в отрицательных контролях.
[0193] B. Реакция удаления защитной группы: реагенты объединяют согласно таблице:
Реагенты инкубируют при 37°C в течение 90 минут. Для реакции удаления защитной группы репрезентативный продукт реакции присоединения получают с использованием приобретенных олигонуклеотидов и исследуют в отношении расщепления ферментом рестрикции BciVI:
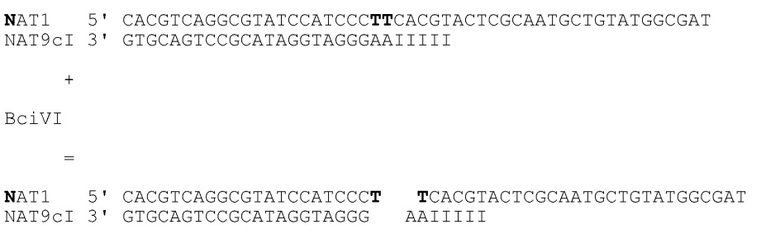
Не было известно, будет ли фермент рестрикции разрезать ДНК намеченным образом, учитывая, что 3'-конец участка разрезания представляет собой серию остатков инозина в противоположность "обычным" основаниям. В качестве положительного контроля получают эквивалент NAT1/NAT9cI с "соответствующим образом" спаренными основаниями (NAT1/NAT9c):
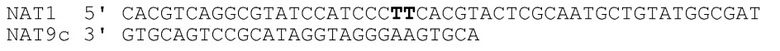
Амплификация продукта способом ПЦР с последующим измерением молекулярной массы на агарозных гелях (фиг.31) демонстрирует, что фермент работает предполагаемым образом. Для положительного контроля в нерасщепленном состоянии наблюдают более крупную полосу (дорожка 1), однако при распщеплении наблюдают меньшую полосу/ы. Ту же картину наблюдают для NAT1/NAT9cI, демонстрируя, что присутствие остатков инозина не подавляет или не препятствует расщеплению. Как оказалось, небольшое количество нерасщепленного продукта остается в случае NAT1/NAT9cI, что указывает на то, что расщепление не является настолько же эффективным, по меньшей мере в этих условиях, как в случае NAT1/NAT9c. Эффективность расщепления может быть увеличена пуетм изменения условий буфера и/или добавления большего количества остатков инозина на 5'-конец NAT9cI.
[0194] Вышеуказанный пример демонстрирует, что можно использовать комбинацию топоизомераза/фермент рестрикции типа IIS для присоединения одного нуклеотида к 5'-концу одноцепочечной ДНК-мишени. Сходную топоизомеразу, SVF, которая распознает последовательность CCCTG (https://www.ncbi.nlm.nih.gov/pubmed/8661446), используют для присоединения "G" вместо "T", с использованием аналогичного процесса, таким образом, позволяя конструирование последовательности, кодирующей двоичную информацию, с T и G.
[0195] Как отмечалось выше, когда дцДНК получают с использованием стратегий топоизомеразы, разрывы ДНК на противоположной цепи можно репарировать с использованием лигазы вместе с ATP. Однако при присоединении одного нуклеотида, как в этом примере, авторы изобретения конструировали одноцепочечную ДНК, так что отсутствовали разрывы, которые необходимо было репарировать, и отсутствовала необходимость использовать лигазу.
Пример 6. Присоединением единичного основания с использованием стратегии с топоизомеразой в сочетании со связыванием 5'-фосфата
[0196] В другом подходе для присоединения одного основания авторы изобретения используют 5'-фосфат в качестве блокирующей группы для обеспечения присоединения одной пары оснований в направлении от 3' к 5'. Реакция нагрузки нагружает топоизомеразу одним T (или G, или при желании другим нуклеотидом), имеющим 5'-фосфатную группу. Когда нагруженная топоизомераза "видит" свободную 5'-неблокированную (нефосфорилированную) цепь одноцепочечной ДНК, она добавляет к этой цепи T, обеспечивая ДНК с T, добавленным на 5'-конец. Это присоединение облегчается присутствием адаптерной ДНК, имеющей последовательности, с которыми может связываться топоизомераза и одноцепочечная акцепторная ДНК. (Следует отметить, что адаптерная ДНК является каталитической, ее можно вновь использовать в качестве матрицы в повторных реакциях). Добавленный нуклеотид имеет 5'-фосфат на нем, так что он не будет субстратом для дальнейшего присоединения до тех пор, пока он не будет подвергнут воздействию фосфатазы, которая удаляет 5'-фосфат. Этот процесс повтояют с использованием топоизомеразы для присоединения одного ʺTʺ к 5'-концу одноцепочечной ДНК-мишени и топоизомеразы SVF для присоединения одного "G", таким образом, позволяя конструирование последовательности, кодирующей двоичную информацию, с T и G. Этот процесс схематично представлен далее:
В общем:
Нагрузка:

АЛЬТЕРНАТИВНЫЙ МЕХАНИЗМ ПЕРЕНОСА

Пример 7. Использование ДНК-оригами для облегчения присоединения ДНК рядом с нанопорой
[0197] Цепь ДНК с крупной структурой оригами на одном конце улавливают в нанопоре и иммобилизуют на конъюгированном с поверхностью стрептавидине через концевую часть биотина на ДНК. После отщепления ферментом рестрикции структуры оригами иммобилизованная ДНК может перемещаться в одну и в другую сторону через пору, что подтверждается прерыванием тока. Иммобилизация позволяет контролируемое перемещение одной молекулы ДНК через пору, что в свою очередь обеспечивает как "считывание", так и "записывание" информации на ДНК.
[0198] Как представлено на фиг.35, образуется объемный двухцепочечный элемент ДНК, который является слишком крупным, чтобы вместиться в нанопору, с одноцепочечной областью, связанной с объемной частью двумя короткими двухцепочечными областями, которые служат для заякоривания ДНК, подлежащей присоединению в ходе синтеза. Затем одноцепочечная область может отделяться и заякориваться на поверхности, соседней с нанопорой, и структура оригами может высвобождаться. См. фиг.33.
[0199] Нанопоры формируют в 3-мм чипах с 20 нм SiO2, с окнами 50 * 50 мкм. Чипы предоставляются Nanopore Solutions. Держатели кассет с нанопорами и проочные ячейки предоставляются Nanopore Solutions. Усилитель представляет собой усилитель Tecella Pico 2. Этот усилитель представляет собой усилитель с питанием через usb, который использует usb-компьютерный интерфейс для контроля. Tecella предоставляет программное обеспечение (Windows) для контроля усилителя. Мультиметр представляет собой цифровой мультиметр FLUKE 17B+, способный детектировать электрический ток всего 0,1 мкА. Для скрининга радиочастотных помех авторы изобретения используют Concentric Technology Solutions TC-5916A Shield Box (Faraday Cage) с USB-интерфейсом. Олигонуклеотиды получают от IDT.com. ʺPSʺ представляет собой пропаргилсилан - O-(ПРОПАРГИЛ)-N-(ТРИЭТОКСИСИЛИЛПРОПИЛ)КАРБАМАТ от http://www.gelest.com/product/o-пропаргил-n-триэтоксисилилпропилкарбамат-90/.
[0200] Структура оригами основана на одноцепочечной m13 со структурой оригами в виде "соты", сторона которой равна ~20 нм. Рядом с сотой существуют двухцепочечные области, каждая из которых содержит уникальный участок рестрикции. Один из этих участков используется для прикрепления модифицированной ДНК для обеспечения прикрепления вблизи нанопоры, а другой используется для отщепления структуры ДНК.
[0201] Формирование нанопор: Нанопоры формируют в чипах с использованием диэлектрического пробоя следующим образом:
1. Чипы осторожно устанавливают в кассеты
2. Смачивание: 100% этанол осторожно наносят пипеткой на чип. Пузырьки должны быть удалены. Однако прямого пипетирования раствора на чипе следует избегать или чип может треснуть (SiO2 составляет только 20 нм).
3. Обработка поверхности: этанол удаляют и свежеприготовленный раствор Piranah (75% серная acid, 25% пероксид водорода (30%)) наносят пипеткой на чип. (раствору Piranha позволяют достигнуть комнатной температуры). Оставляют на 30 минут.
4. Ополоснуть 4 раза дистиллированной водой.
5. Ополоснуть 2 раза буфером HK (10 мМ HEPES, pH 8, 1 M KCl)
6. Собрать кассету в проточную ячейку.
7. Добавить 700 мкл буфера HK в каждую камеру проточной ячейки.
8. Вставить серебряные электроды, подсоединенные к амплификатору, и закрыть клетку Фарадея.
9. Протестировать сопротивление при 300 мВ. Ток не должен детектироваться. Если он детектируется, чип, вероятно, треснул и необходимо начать сначала.
10. Подсоединить электроды к току DC 6В и тестировать ток с использованием мультиметра. Ток должен быть низким и не должен меняться. Увеличить напряжение на 1,5 В и удерживать напряжение до тех пор, пока сопротивление не возрастет. Если сопротивление не возрастает через 5-10 минут, увеличить напряжение еще на 1,5 В и попробовать вновь. Повторять до тех пор, пока сопротивление не возрастет, и в этот момент применяемое напряжение следует сразу остановить (при достаточном напряжении происходит диэлектрический пробой и в мембране из SiO2 образуется "отверстие". Когда изначально созданное отверстие является малым, его размер увеличиться при поддержании напряжения).
11. Протестировать пору с использованием усилителя. При 300 мВ должен определяться ток от малого количества до нескольких нА. Чем больше ток, тем крупнее пора.
[0202] На фиг.34 представлена основа функционировния нанопоры. На каждой панели ось y представляет собой ток (нА) и ось x представляет собой время (с). На левой панели "скрининг шума RF" иллюстрирует пригодность клетки Фарадея. Чип без нанопоры помещают в проточную ячейку и применяют 300 мВ. Когда крышка клетки Фарадея закрыта (первая стрелка), можно наблюдать снижение шума. Небольшой импульс возникает, когда защелка закрыта (вторая стрелка). Следует отметить, что ток составляет ~0 нМ. После формирования поры (средняя панель), применение 300 мВ (стрелка) приводит к току ~3,5 нМ. Когда ДНК помещают в камеру заземления и применяют +300 мВ можно наблюдать перемещение ДНК (правая панель) в качестве временного снижения тока (следует отметить, что в этом случае используют буфер TS: 50 мМ Tris, pH 8, 1 M NaCl). Для этого эксперимента по перемещению ДНК используют лямбда-ДНК.
[0203] Электроды из хлорида серебра:
1. Серебряную проволоку припаивают к изолированной медной проволоке.
2. Медную проволоку заземляют, а серебряную погружают в свежий 30% гипохлорит натрия на 30 минут.
3. Серебро должно приобретать темно-серое покрытие (хлорид серебра).
4. Серебряную проволоку ополаскивают тщательно дистилированной водой и сушат.
5. Теперь она готова для применения.
[0204] Силанирование гранул: Способ силанирования первоначально разрабатывают/тестируют на покрытых SiO2 магнитных гранулах (GBioscience). Используют следующий протокол:
1. Предварительно нагреть гранулы в свежем растворе Pirannah в течение 30 минут.
2. Промыть 3× дистиллированной водой.
3. Промыть 2× метанолом.
4. Разбавить исходный раствор APTES 1:500 в метаноле.
5. Добавить разбавленный APTES к гранулам, инкубировать при к.т. в течение 45 минут.
6. Ополоснуть метанолом.
7. 100°C в течение 30 минут.
8. Хранить в вакууме.
[0205] Силанирование кремниевого чипа
1. Установить чип с нанопорой в кассету.
2. Ополоснуть метанолом, осторожно удаляя какие-либо пузырьки воздуха.
3. Добавить свежий раствор Pirannah (уравновешенный до комнатной температуры) и инкубировать в течение 30 минут.
4. Промыть 4× дистиллированной водой.
5. Промыть 3× метанолом.
6. Разбавить исходный раствор APTES 1:500 в метаноле и использовать для промывания чипа 2×. Инкубировать при к.т. в течение 45 минут.
7. Ополоснуть 2× метанолом.
8. Сушить в потоке воздуха.
9. Хранить в вакууме в течение ночи.
[0206] Конъюгация гранул со стрептавидином: конъюгация со стрептавидином конъюгация первоначально разработана/протестирована на силанированных гранулах, полученных, как описано выше.
1. Промыть силанированные гранулы модифицированным фосфатно-солевым буфером (MPBS)
2. Получить свежий раствор 1,25% глутаральдегида в MPBS (с использованием 50% исходного раствора глутаральдегида, хранящегося замороженным).
3. Добавить к гранулам 1,25% глутаральдегид и оставить стоять на 60' при осторожном пипетировании вверх и вниз каждые 15 минут.
4. Промыть 2× посредством MPBS.
5. Промыть 2× водой.
6. Оставить сушиться в вакууме.
7. Добавить к гранулам стрептавидин (500 мкг/мл в MPBS) и инкубировать в течение 60 минут (для гранул отрицательного контроля использовать бычий сывороточный альбумин (BSA) (2 мг/мл в MPBS) вместо стрептавидина).
8. Удалить стрептавидин и добавить BSA (2 мг/мл в MPBS). Инкубировать в течение 60 минут.
9. Промыть 2× посредством MPBS.
10. Хранить при 4°C.
[0207] Конъюгация кремниевого чичпа со стрептавидином
1. Ополоснуть силанированный чип этанолом 2×
2. Ополоснуть силанированный чип MPBS 2×
3. Получить свежий раствор 1,25% глутаральдегида в MPBS (с использованием 50% исходного раствора глутаральдегида, хранящегося замороженным).
4. Ополоснуть чип 1,25% глутаральдегидом 2×, оставить стоять на 60' при осторожном пипетировании вверх и вниз каждые 15 минут.
5. Промыть 2× посредством MPBS
6. Промыть 2× водой.
7. Оставить сушиться в вакууме.
8. На одну половину чипа добавить BSA (2 мг/мл в MPBS), а на другую добавить стрептавидин (500 мкг/мл в MPBS). Инкубировать в течение 60 минут. Сделать маркировку на кассете, указав какая половина чипа модифицирована стрептавидином.
9. Ополоснуть обе половины чипа BSA (2 мг/мл в MPBS). Инкубировать в течение 60 минут.
10. Промыть MPBS.
[0208] Буферы, используемые в этом примере, получают следующим образом:
MPBS: 8 г/л NaCl, 0,2 г/л KCl, 1,44 г/л динатрий фосфата, 0,240 г/л фосфата калия, 0,2 г/л полисорбата-20 (pH 7,2)
PBS: 8 г/л NaCl, 0,2 г/л KCl, 1,44 г/л динатрий-фосфата, 0,240 г/л фосфата калия
TS: 50 мМ Tris, pH 8,0, 1 M NaCl
HK: 10 мМ HEPES, pH 8,0, 1M KCl
TE: 10 мМ Tris, 1 мМ EDTA, pH 8,0
Раствор Pirannah: 75% пероксид водорода (30%)+25% серная кислота
Исходный раствор APTES: 50% метанол, 47,5% APTES, 2,5% наночистая вода. Выдерживать при 4°C в течение по меньшей мере 1 часа. Хранить при 4°C.
Исходный раствор PDC: 0,5% масс./об. 1,4-фенилен диизотиоцианат в DMSO.
[0209] Заказывают олигонуклеотиды (от 5' до 3'):
o1 CTGGAACGGTAAATTCAGAGACTGCGCTTTCCATTCTGGCTTTAATG
o3 GGAAAGCGCAGTCTCTGAATTTAC
N1 CTTACTGGAACGGCTATCGATATCGCAGCAGGACAGA
BN1 Биотин-CTTACTGGAACGGCTATCGATATCGCAGCAGGACAGA
N2 GTCCTGCTGCGATATCGATAGCCGTTCCAGTAAG
[0210] Гибридизацию пар олигонуклеотидов проводят следующим образом:
1. Получить исходные растворы олигонуклеотидов в концентрации 100 мкМ в буфере TE
2. Разбавить олигонуклеотиды до 10 мкМ в PBS
3. Нагревать до 85°C в течение 5' в термоциклере
4. Постепенно снижать температуру на 5°C каждые 3' до 25°C
5. Хранить при 4°C или -20°C.
[0211] Конъюгация со стрептавидином: конъюгация стрептавидина с SiO2 разработана и протестирована с использованием покрытых SiO2 магнитных гранул, а затем протоколы были адаптированы для чипов SiO2. Тестируют связывание биотинилированных олигонуклеотидов с гранулами, конъюгированными как со стрептавидином, так и с BSA. Как и ожидалось в случае конъюгированных с BSA гранул наблюдают ничтожно малое связывание, в то время как в случае конъюгированных со стрептавидином гранул наблюдают высокое связывание. См. фиг.38. Поскольку более удобно проводить конъюгацию в высокой концентрации соли (движение ДНК осуществляют в высокой концентрации соли), также тестируют способность гранул связываться в буфере HK. Связывание в буфере HK сравнимо со связыванием в буфере MPBS (фиг.39).
[0212] Получают конструкции оригами и подтверждают их функционирование, как описано выше на фиг.35-37. Биотинилирование структуры оригами тестируют с использованием олигонуклеотидов. Уже известно из результатов для "оригами", описанных выше для фиг.37 что участок AlwNI является активным. Ниже используется пара олигонуклеотидов, которая воспроизводит сегмент точной последовательности ДНК-оригами (o1/o3). Молекулу оригами изображают следующим образом:
[0213] Пара олигонуклеотидов o1/o3 представляет собой
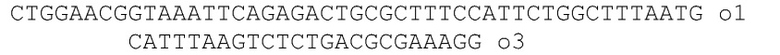
[0214] ДНК расщепляют посредством AlwNI в присутствии ДНК-лигазы T4 и биотинилированного олигонуклеотида, комплементарного выступающему концу на 3'-стороне последовательности оригами (которая сама по себе связана с длинной последовательностью оцДНК, которая сама по себе связывана с другой стороной оригами), в соответствии со следующей реакцией:

В этой стратегии AlwN1 расщепляет ДНК-мишень. При добавлении лигазы возможно религирование этой ДНК, однако фермент рестрикции вновь осуществит расщепление. Однако, если/когда (правильный) фрагмент (o1/o3) связывается с BN1/N2, участок рестрикции не воссоздается, таким образом, этот продукт не будет расщеплен. Специфическое присоединение подтверждают путем тестирования с ферментом рестрикции и без него:
Добавляют все реагенты за исключением лигазы и раствор инкубируют при 37°C в течение 60 минут. Добавляют лигазу и раствор инкубируют в течение ночи при 16°C. 10× буфер для лигирования относится к 10× буферу ДНК-лигазы T4 NEB. Лигаза представляет собой ДНК-лигазу T4NEB. o1/o3 и n1/n2 относятся к гибридизованной паре олигонуклеотидов, как изображено выше. Единицы представляют собой микролитры. Анализ в агарозном геле подтверждает, что в присутствии AlwNI образуется более крупный продукт, соответствующий биотинилированному олигонуклеотиду, связанному с длинным плечом оцДНК, связанным со структурой оригами. Сходную стратегию используют для 3'-биотинилирования, когда это желательно.
[0215] Выше продемонстрирована способность формировать и использовать нанопору для детектирования индуцированного напряжением перемещения ДНК через пору, создание молекулы оригами, длинная оц-область которой связана ее дальним концом с биотином, и конъюгация стрептавидина с диоксидом кремния и применение его для улавливания биотинилированной ДНК. Эти инструменты используют для присоединения и контроля движения одной молекулы ДНК рядом с нанопорой.
[0216] Первая стадия представляет собой конъюгацию стрептавидина с одной поверхностью нанопоры из SiO2 (и BSA с другой стороны). Это осуществляют в соответствии с описанным выше протоколом. Полученные поры имеют тенденцию к более низкому току, чем они имеют первоначально. После нескольких кратковременных импульсов величиной 6 В, ток возвращается практически к исходному току. Функционирование нанопоры в этот момент представлено на фиг.40.
[0217] Далее добавляют ДНК-оригами. Когда ДНК-оригами добавляется в подходящую камеру и ток включается, оригами входит в камеру. Иллюстрация этого показана на фиг.41. Результаты эксперимента, когда оригами подают в конечной концентрации 50 пМ, подтверждают, что ДНК с орагами входит в пору относительно быстро (как правило, в течение секунд), что детектируется результирующим снижением тока через нанопору (например, в этих экспериментах ток до вхождения оригами составляет ~3 нА и ~2,5 нА после вхождения). Если току позволить проходить в течение более длительного периода времени, можно наблюдать двойное вхождение. Если использовать более высокие концентрации вхождение происходит слишком быстро, чтобы его можно было наблюдать.
[0218] Связывание вошедшей ДНК с чипом. После вхождения оригами в нанопору напряжение вновь применяют через 15 минут после этого. Конец области в виде оц-ДНК в оригами содержит биотин, и с поверхностью нанопоры конъюгирован стрептавидин. Стрептавидин связывается с авидином с константой аффинности, близкой к ковалентной связи. Период 15 минут позволяет ДНК диффундировать и концу с биотином найти и связаться с стрептавидином. Если ДНК в действительности связалась с поверхностью, то, когда напряжение изменяют на обратное, наблюдаемый ток должен быть несколько меньшим, чем наблюдался ранее. Также переключение тока в одну и в другую сторону должно приводить к току в обоих направлениях, который является более низким, чем в случае свободной поры. В примере, представленном здесь, свободная пора демонстрирует ток ~3 нА. На фиг.42 представлено изображение присоединенной ДНК, и на фиг.43 представлены результаты эксперимента по переключению напряжения для прикрепленной ДНК-оригами. Следует отметить, что ток, наблюдаемый в обоих направлениях, составляет ~+/-2,5 нА, что ниже, чем ~+/-3 нА, наблюдаемые для свободной поры. Если ДНК не связалась с поверхностью, исходный ток восстановится при переключении напряжения (фиг.44).
[0219] Для удаления структуры оригами буфер в камере с проточной ячейкой, содержащей структур оригами, удаляли и заменяли 1× буфером Swa1 с 1 мкл Swa1/20 °L. Буфер в другой проточной камере заменяют на 1× буфер Swa1 без Swa1. Затем проводят инкубацию при комнатной температуре в течение 60 минут, а затем промывают буфером HK, и применяют напряжение. Движение ДНК вперед и назад, как представлено на фиг.45, подтверждается экспериментальными данными, представленными на фиг.46, демонстрирующими контролируемое движение иммобилизованной ДНК через нанопору из SiO2.
Пример 8. Альтернативный способ присоединения полимера к поверхности рядом с нанопорой
[0220] В приведенных выше примерах описано прикрепление ДНК к поверхности рядом с нанопорой посредством биотинилирования ДНК и покрытия поверхности для прикрепления стрептавидином. Некоторые альтернативные способы прикрепления полимера представлены на фиг.47.
[0221] a) Гибридизация ДНК: в одном способе ДНК, которая удлиняется в способах по изобретению, гибридизуют с коротким олигонуклеотидом, прикрепленным вблизи нанопоры. После завершения синтеза синтезированная ДНК может быть без труда удалена без необходимости в ферментах рестрикции, или альтернативно двойная цепь может быть образована связанным олигонуклеотидом и синтезированная ДНК может предоставлять субстрат для фермента рестрикции. В этом примере короткие олигомеры конъюгируют с поверхностью с использованием биотина-стрептавидина, или лигируют с использованием 1,4-фенилендиизотиоцианата следующим образом:
Конъюгация биотинилированной ДНК с SiO2:
A. СИЛАНИРОВАНИЕ:
1. Предварительная обработка: раствор nha в течение 30 минут, промывание дважды дистиллированной H2O (ddH2O)
2. Получение исходного раствора APTES: 50% MeOH, 47,5% APTES, 2,5% наночистая H2O: выдерживание >1 ч при 4°C
3. Разбавление исходного раствора APTES 1:500 в MeOH
4. Инкубация чипов при комнатной температуре
5. Ополаскивание MeOH
6. Сушка
7. Нагревание при 110°C в течение 30 минут
КОНЪЮГАЦИЯ:
1. Обработка чипа исходным раствором PDC в течение 5 ч (комнатная температура) (исходный раствор PDC: 0,5% масс./об. 1,4-фенилендиизотиоцианат в DMSO)
2. 2 промывания в DMSO (кратковременно!)
3. 2 промывания в ddH2O (кратковременно!)
4. 100 нМ амино-модифицированная ДНК в ddH2O (pH 8) O/N 37°C
5. 2 промывания 28% раствором аммиака (деактивация)
6. 2 промывания ddH2O.
[0222] Одноцепочечную ДНК, имеющую концевую последовательность, комплементарную присоединенным олигонуклеотидам, вносят, как описано выше, и позволяют ей гибридизоваться с присоединенными олигонуклеотидами.
[0223] b) Клик-химия: клик-химия является общим термином для реакций, которые являются простыми и термодинамически эффективными, не образуют токсических или высокореакционноспособных побочных продуктов, и осуществляются в воде или биосовместимых растворителях и часто используются для связывания выбранных субстратов с определенными биомолекулами. В клик-конъюгации в этом случае используется химия, сходная с химией, используемой в a) для прикрепления олигонуклеотидов, только в данном случае она используется для прикрепления полимера, который удлиняется в ходе синтеза в способах по изобретению. В то время как в этом примере полимером является ДНК, эта химия может прикреплять другие полимеры, функционализированные посредством присоединения совместимой азидной группы.
A. СИЛАНИРОВАНИЕ:
1. Предварительная обработка: раствор Piranha в течение 30э, промывание ddH2O
2. Получение исходного раствора PS (пропаргилсилан): 50% MeOH, 47,5% PS, 2,5% наночистая H2O: выдерживание >1 ч при 4C
3. Разбавление исходного раствора APTES 1:500 в MeOH
4. Инкубация чипов при комнатной температуре
5. Ополаскивание MeOH
6. Сушка
7. Нагревание при 110°C в течение 30 минут
[0224] ДНК, терминированная азидной функциональной группой, ковалентно связывается с этой поверхностью (как показано на фиг.47). Терминированные азидом олигонуклеотидов приобретают на заказ и связывают с более длинной ДНК-оригами, как описано выше для присоединения биотина к ДНК.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛИНУКЛЕОТИДЫ, СПОСОБ ИХ ПОЛУЧЕНИЯ, ГИБРИДИЗАЦИОННЫЙ АНАЛИЗ НУКЛЕИНОВОЙ КИСЛОТЫ | 1991 |
|
RU2142467C1 |
| СПОСОБ КОНЪЮГИРОВАНИЯ ПОЛИМЕРНОЙ ЧАСТИЦЫ | 2013 |
|
RU2636465C2 |
| НАБОРЫ ДЛЯ ГЕНОТИПИРОВАНИЯ | 2021 |
|
RU2835967C2 |
| СПОСОБЫ ИНКАПСУЛИРОВАНИЯ ОДИНОЧНЫХ КЛЕТОК, ИНКАПСУЛИРОВАННЫЕ КЛЕТКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2750567C2 |
| ВЫСОКОПРОИЗВОДИТЕЛЬНЫЙ АНАЛИЗ ТРАНСГЕННЫХ ГРАНИЦ | 2012 |
|
RU2603265C2 |
| ЭЛЕКТРОННАЯ СЕТЬ, СОДЕРЖАЩАЯ НУКЛЕОТИДНЫЕ ВОЛОКНА, СПОСОБ ЕЁ ИЗГОТОВЛЕНИЯ И ЭЛЕКТРОННАЯ СХЕМА, ВКЛЮЧАЮЩАЯ УКАЗАННУЮ СЕТЬ | 1998 |
|
RU2213393C2 |
| КОНЪЮГАТЫ ОЛИГОНУКЛЕОТИДОВ | 2013 |
|
RU2653438C2 |
| МОДУЛЯЦИЯ ПОЛИМЕРНЫХ ГРАНУЛ ДЛЯ ОБРАБОТКИ ДНК | 2019 |
|
RU2822154C2 |
| Новые способы получения олигонуклеотидов | 2017 |
|
RU2719641C1 |
| СПОСОБ АНАЛИЗА НЕИЗВЕСТНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ ОДНОЦЕПОЧЕЧНЫХ НУКЛЕИНОВЫХ КИСЛОТ | 2006 |
|
RU2322508C1 |
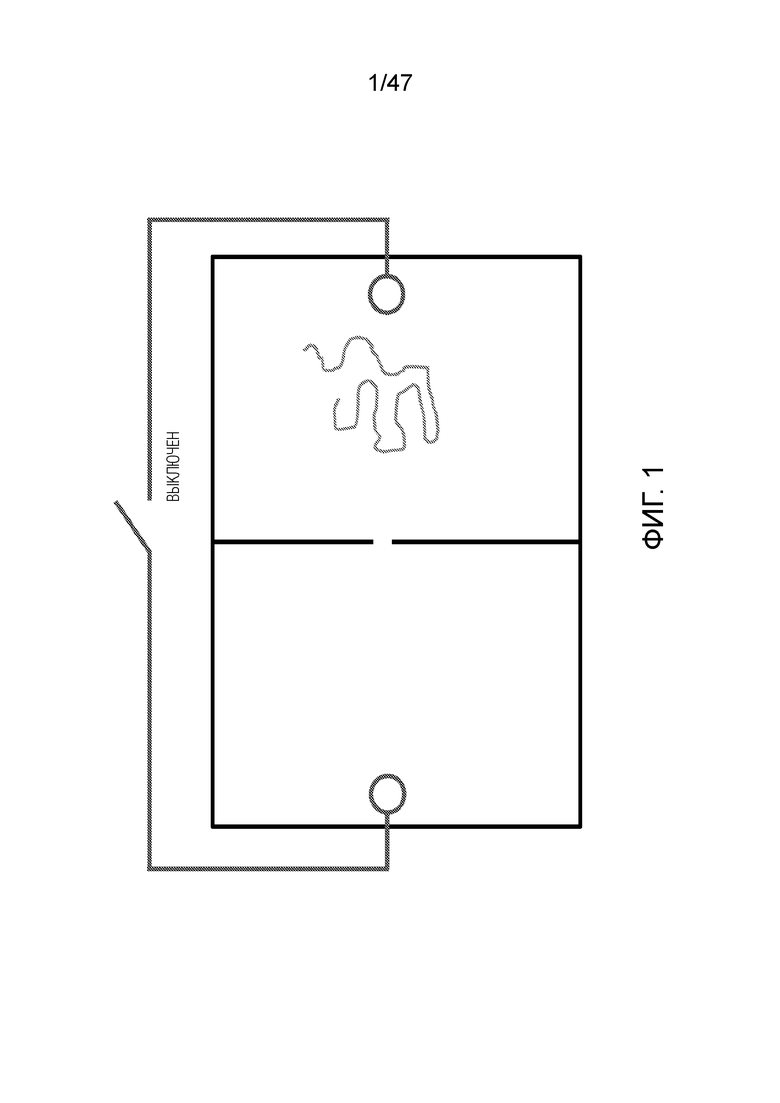
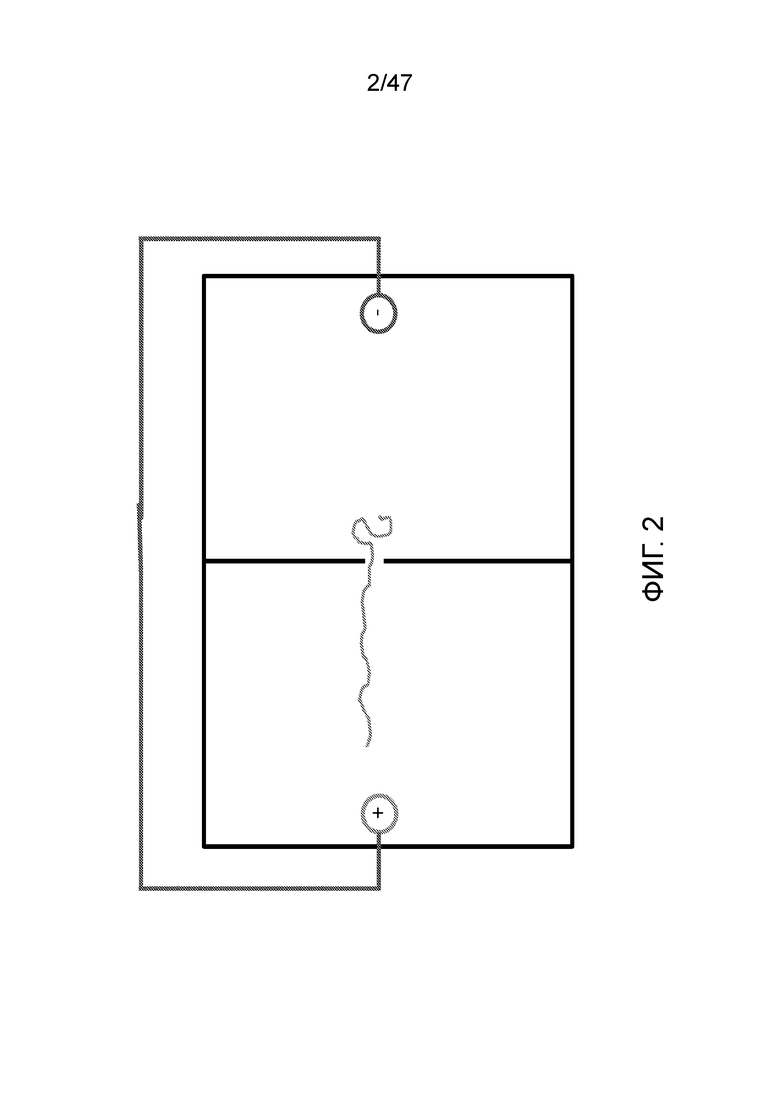
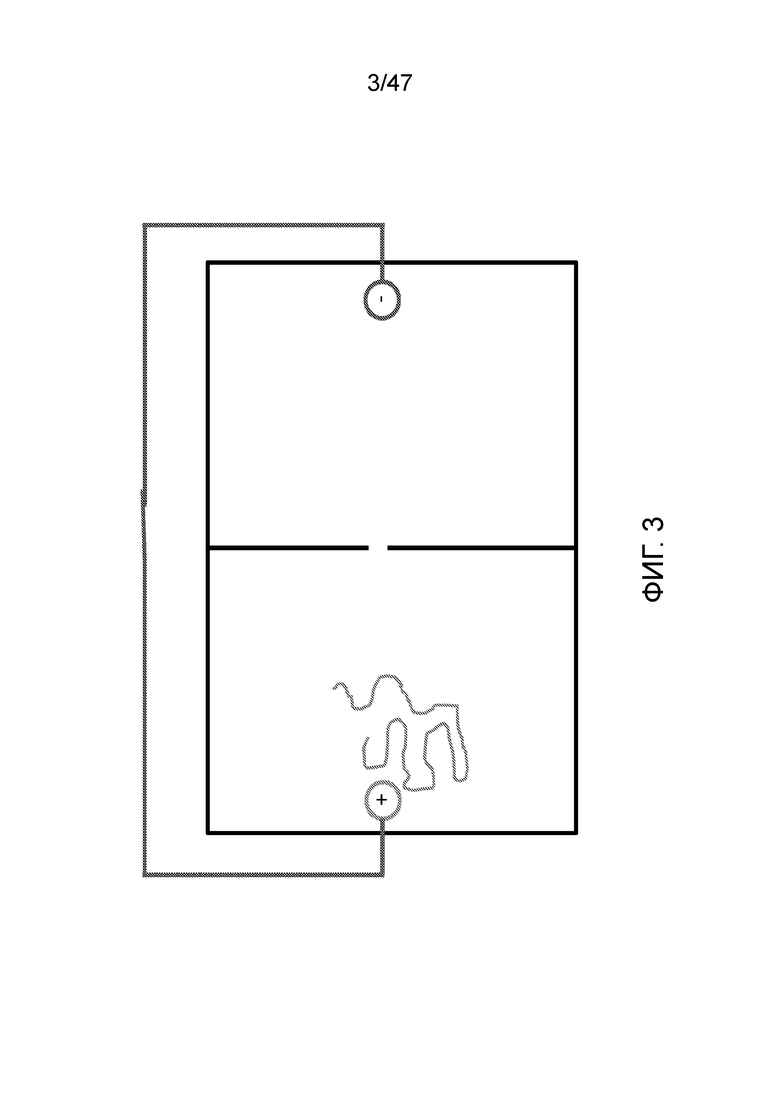
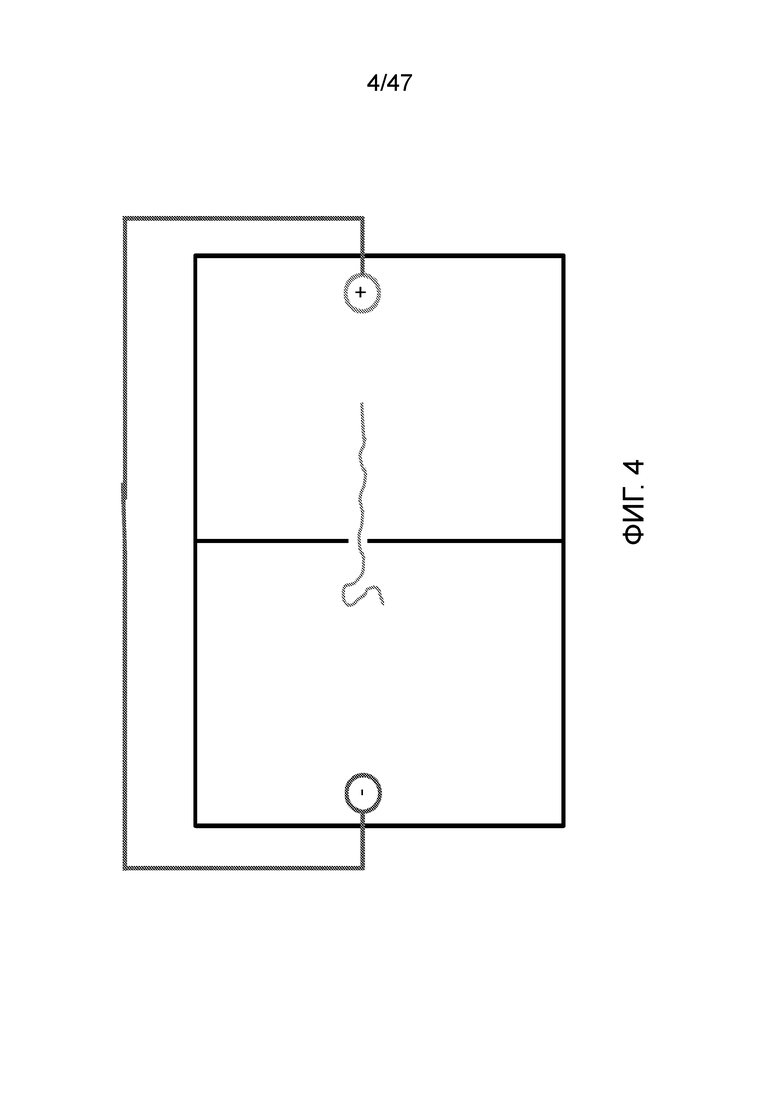
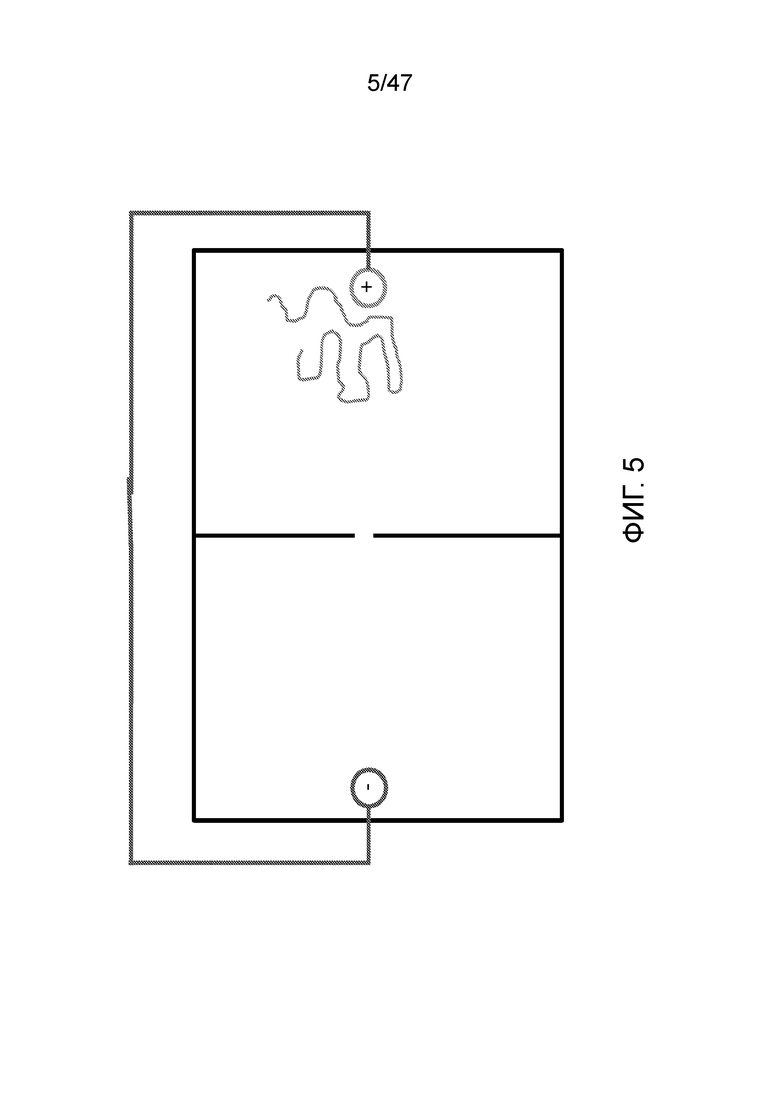
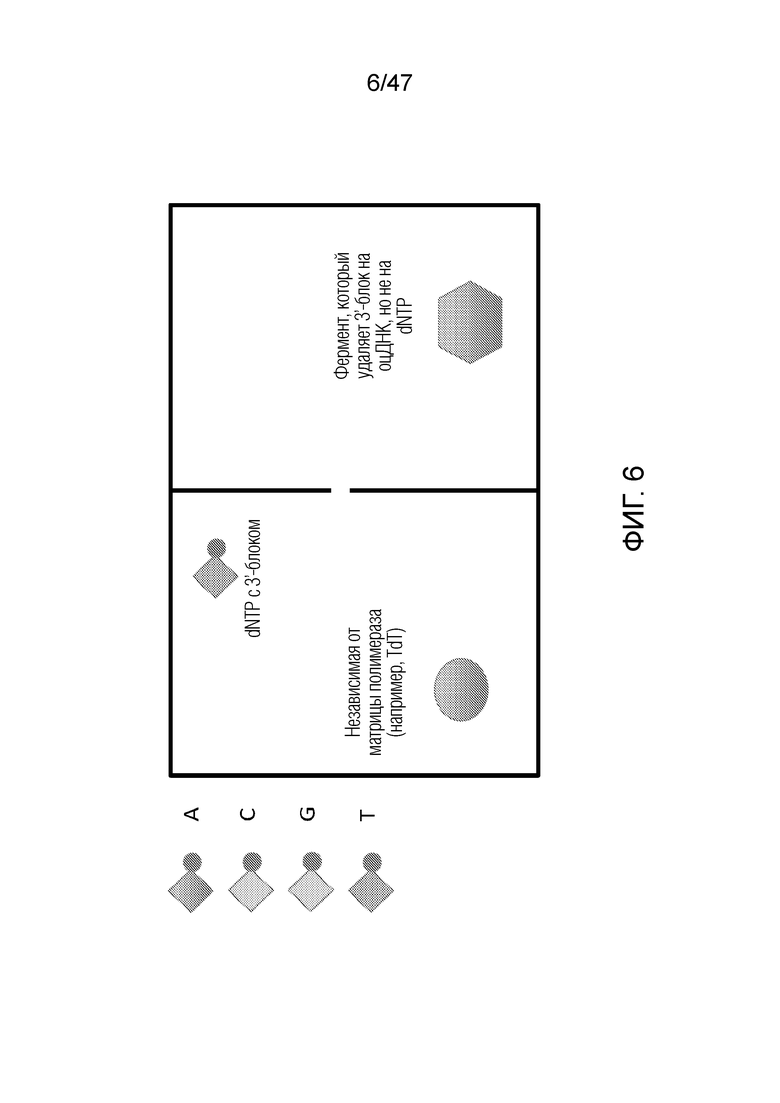
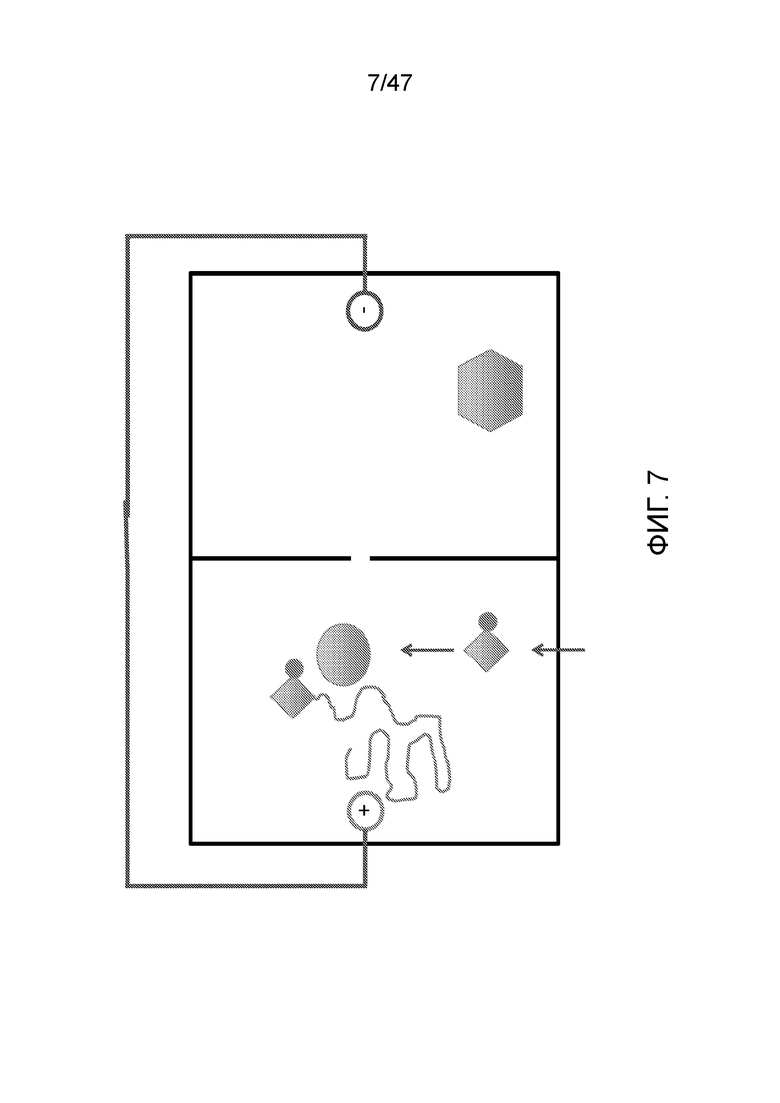
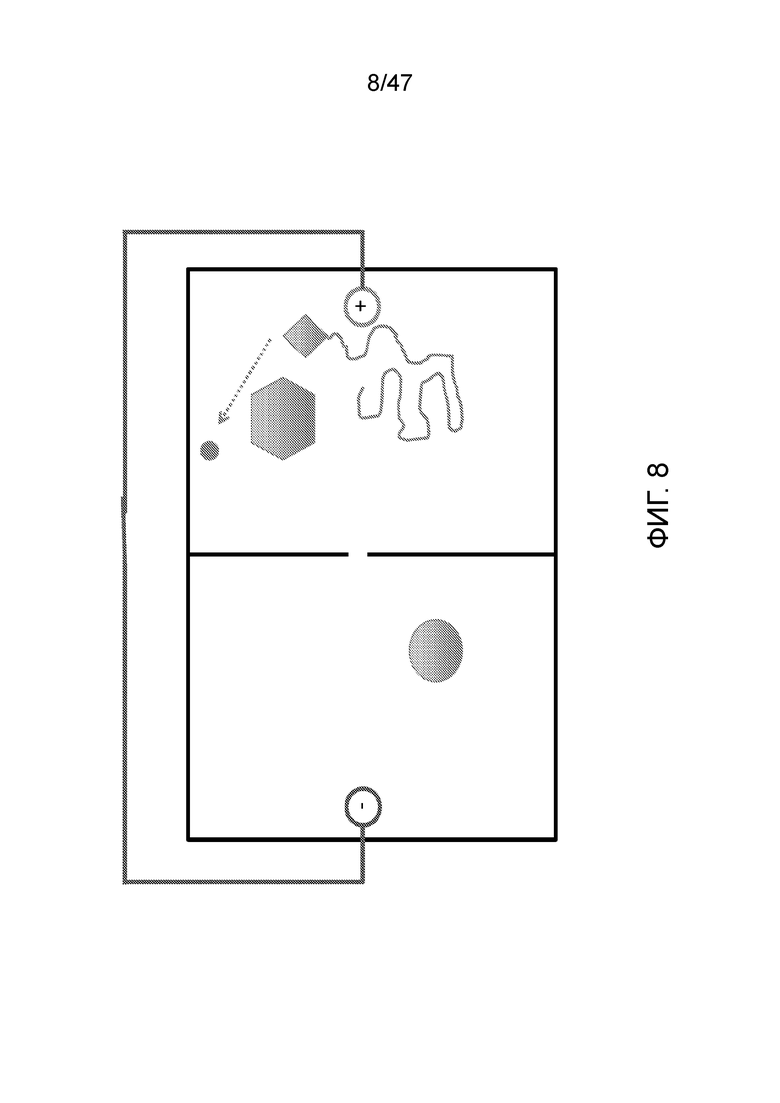
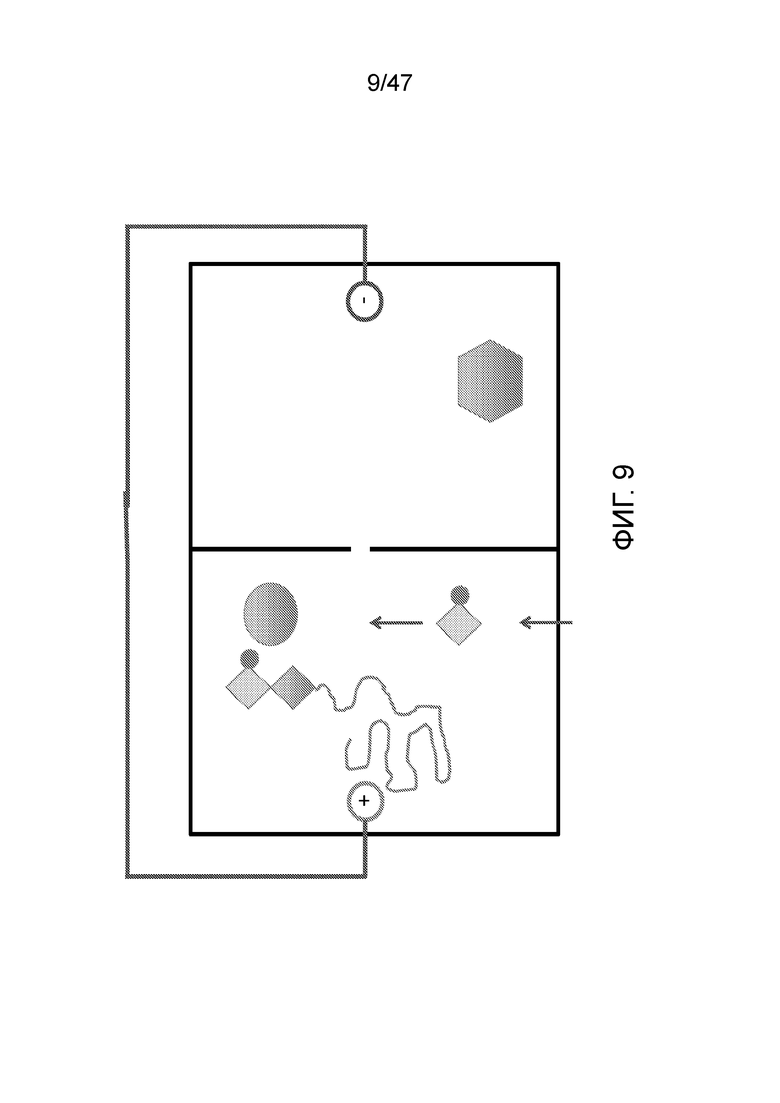
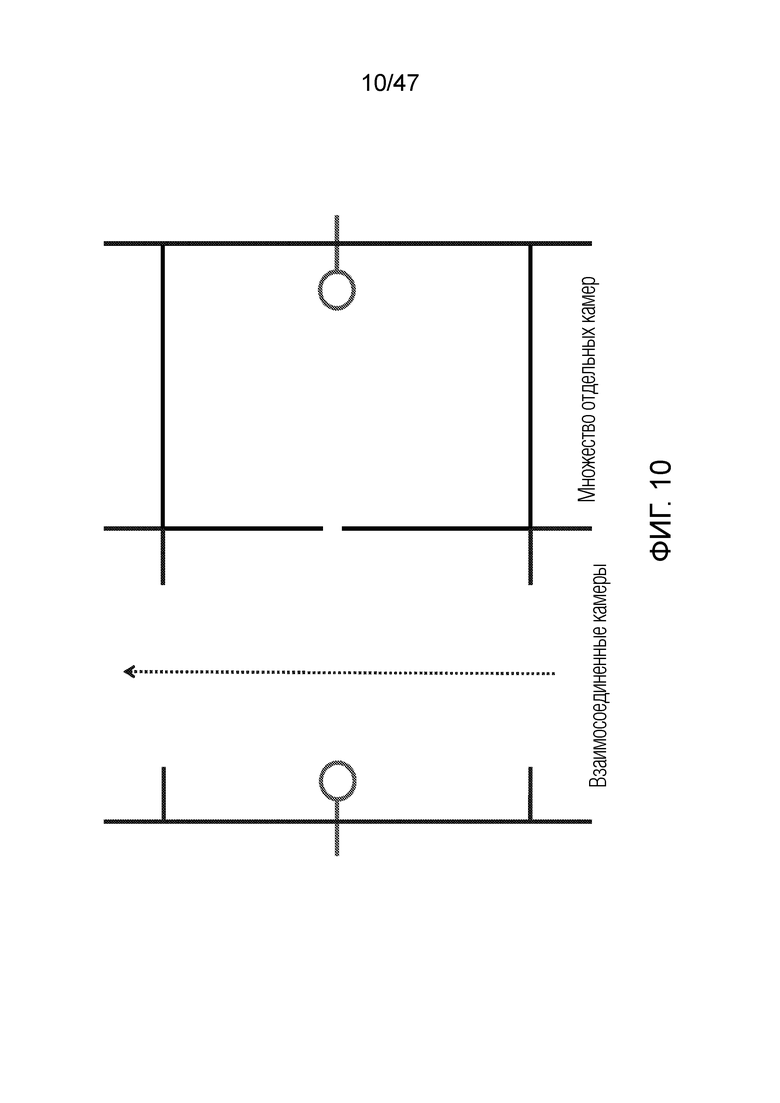
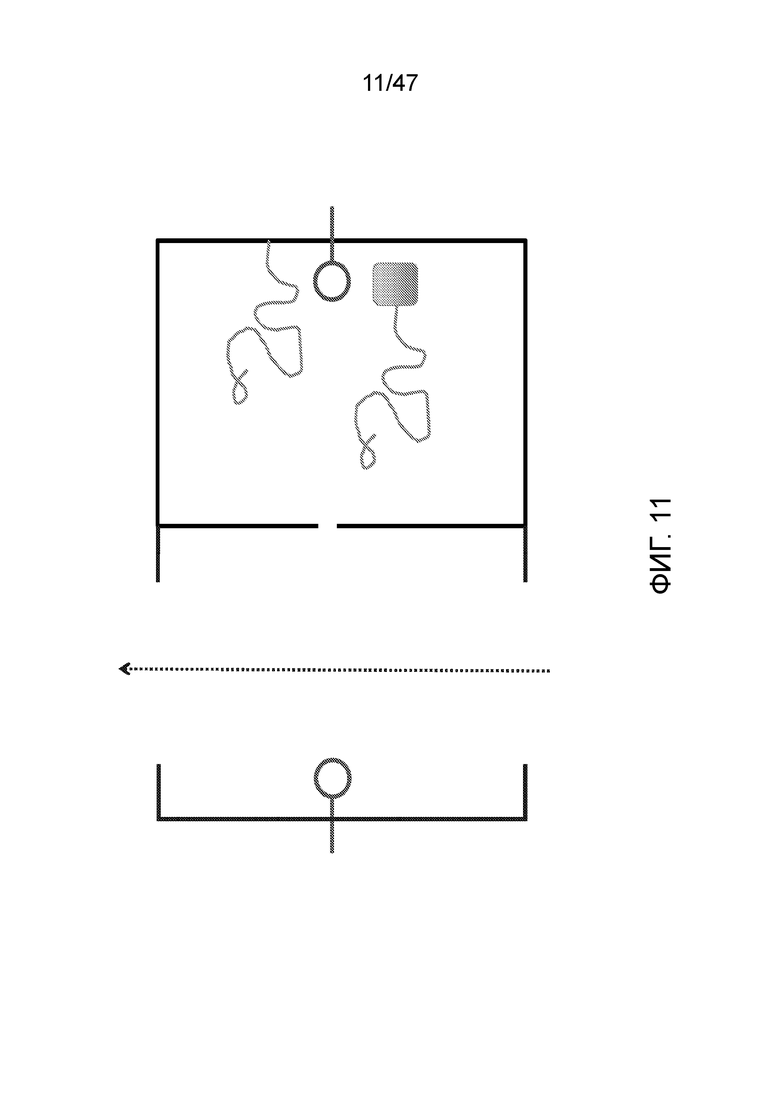
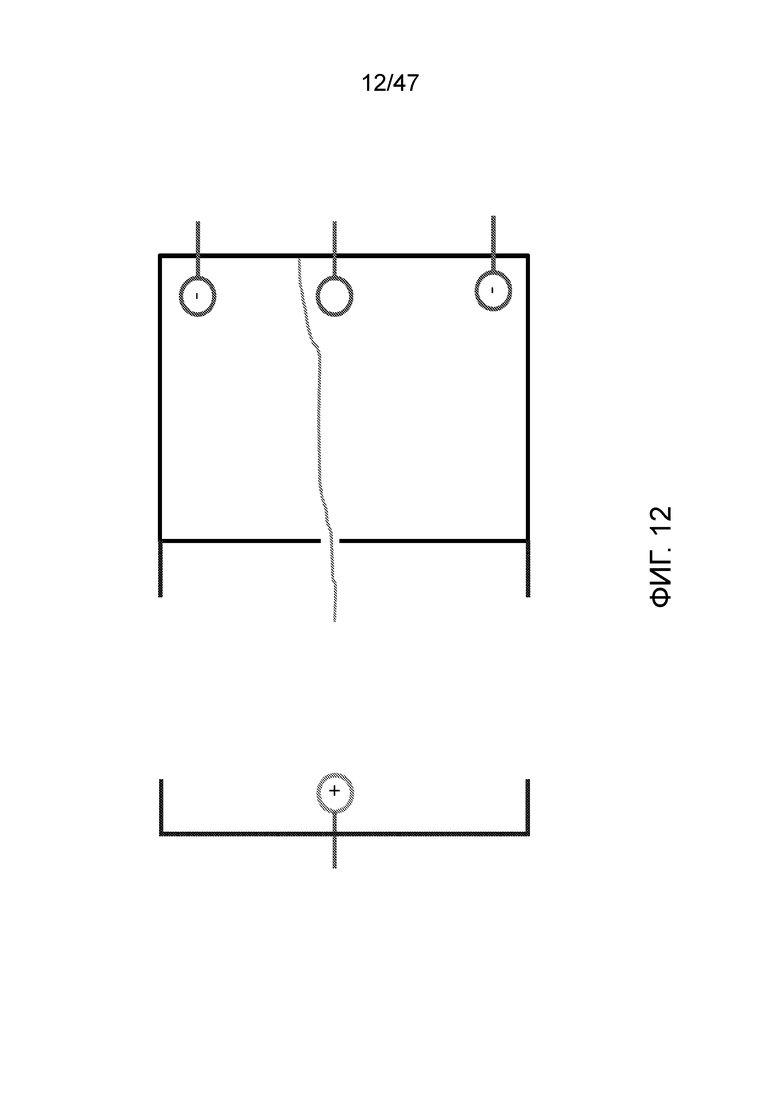
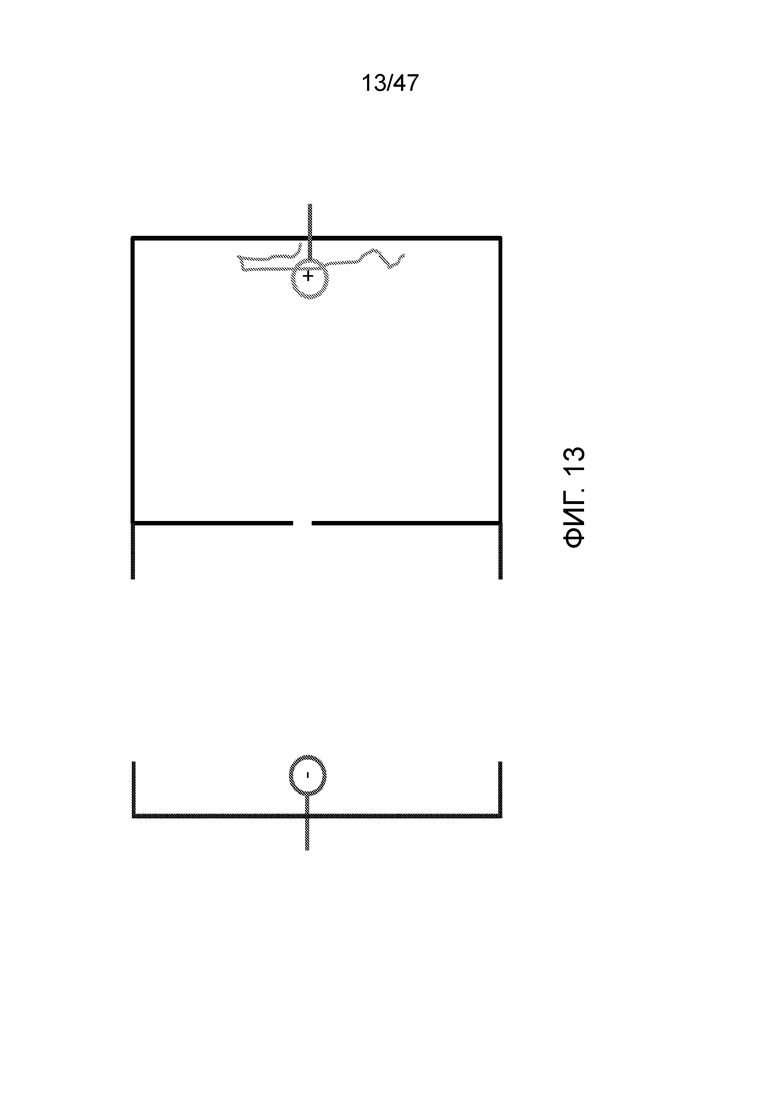
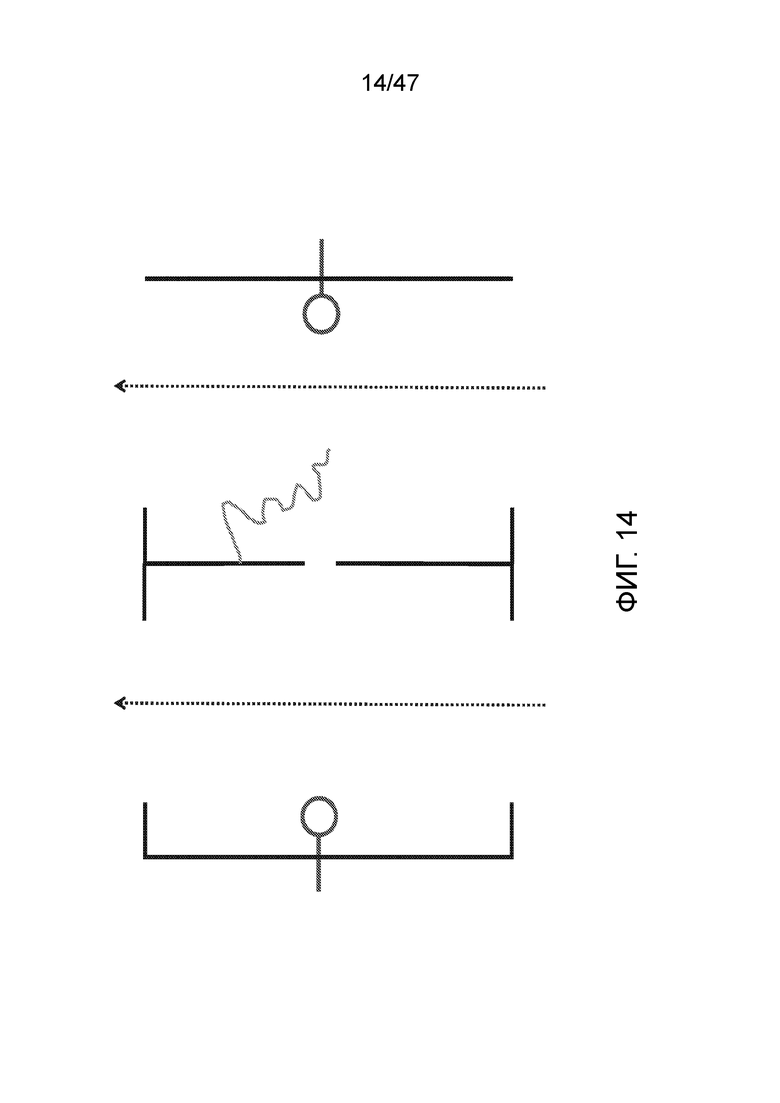
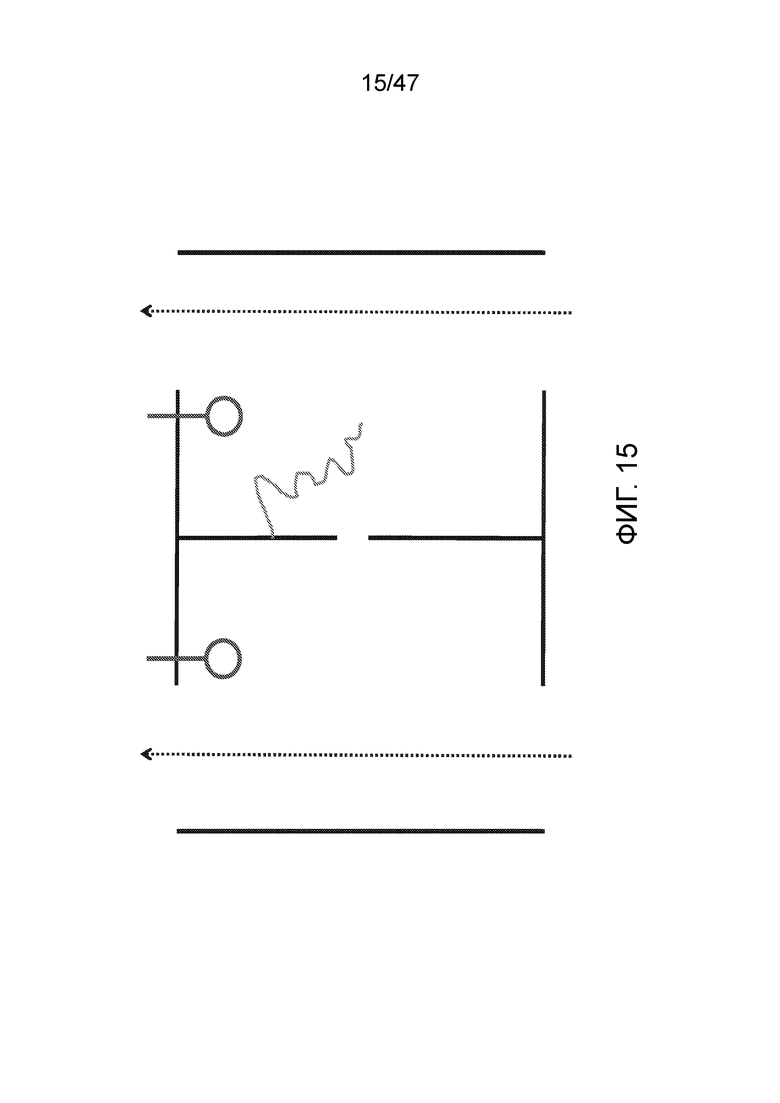
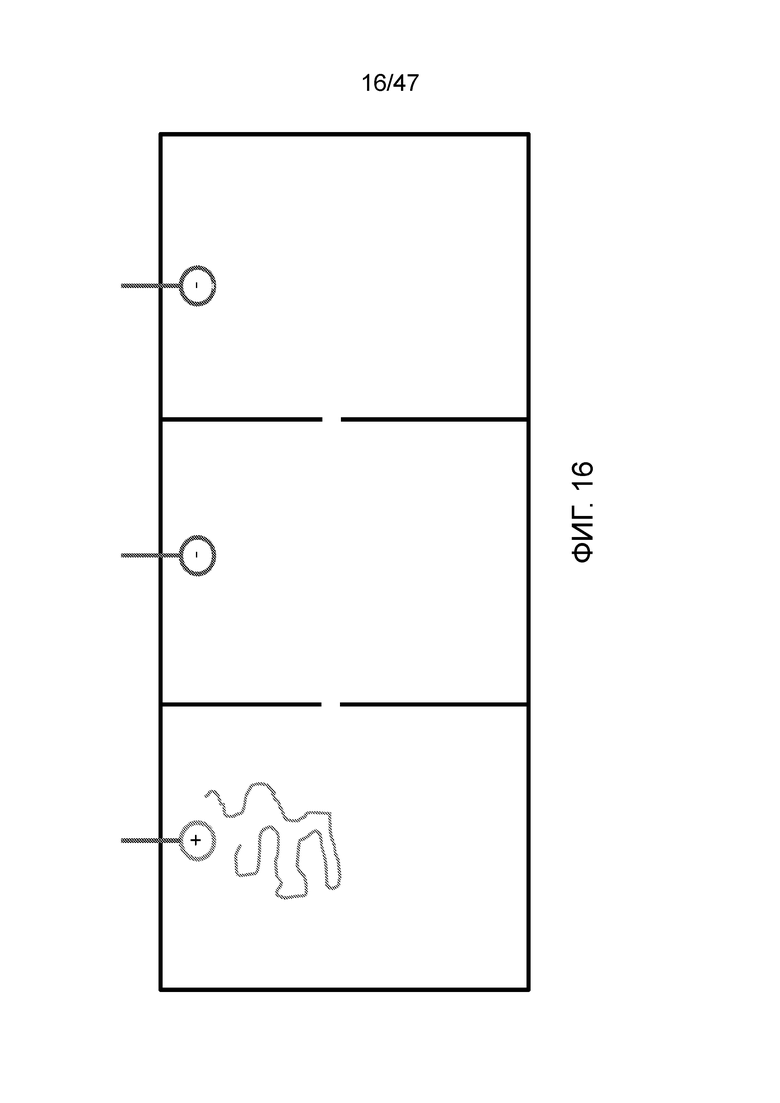
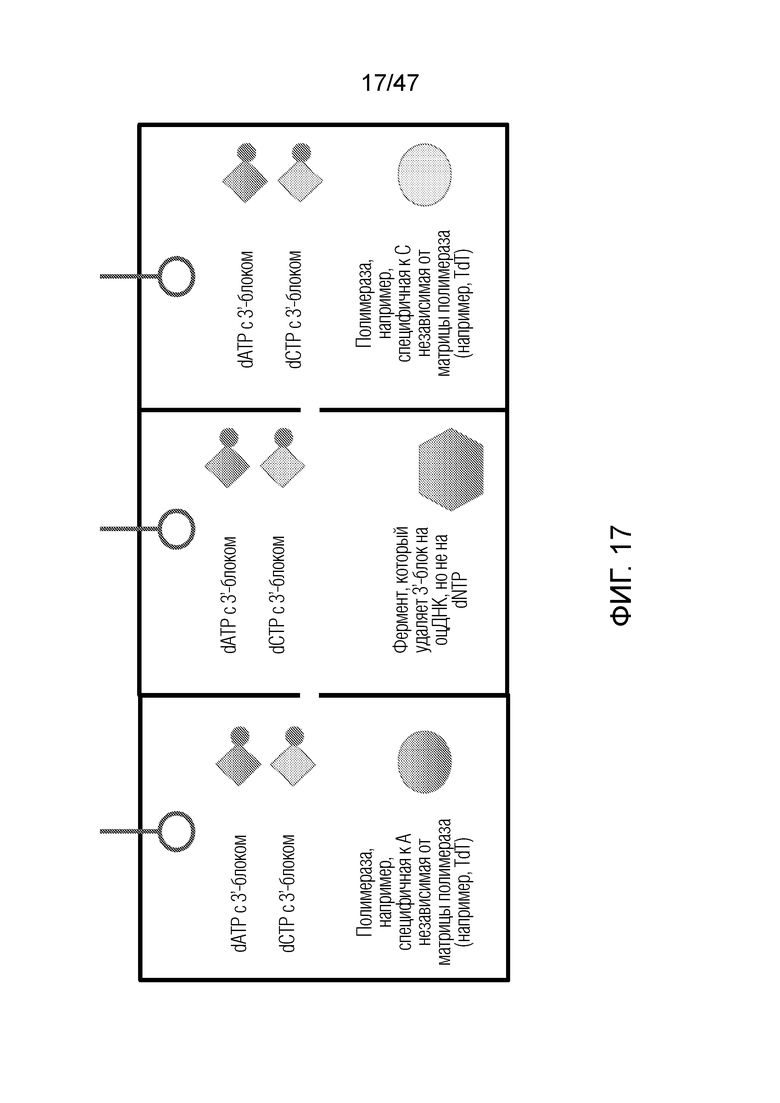
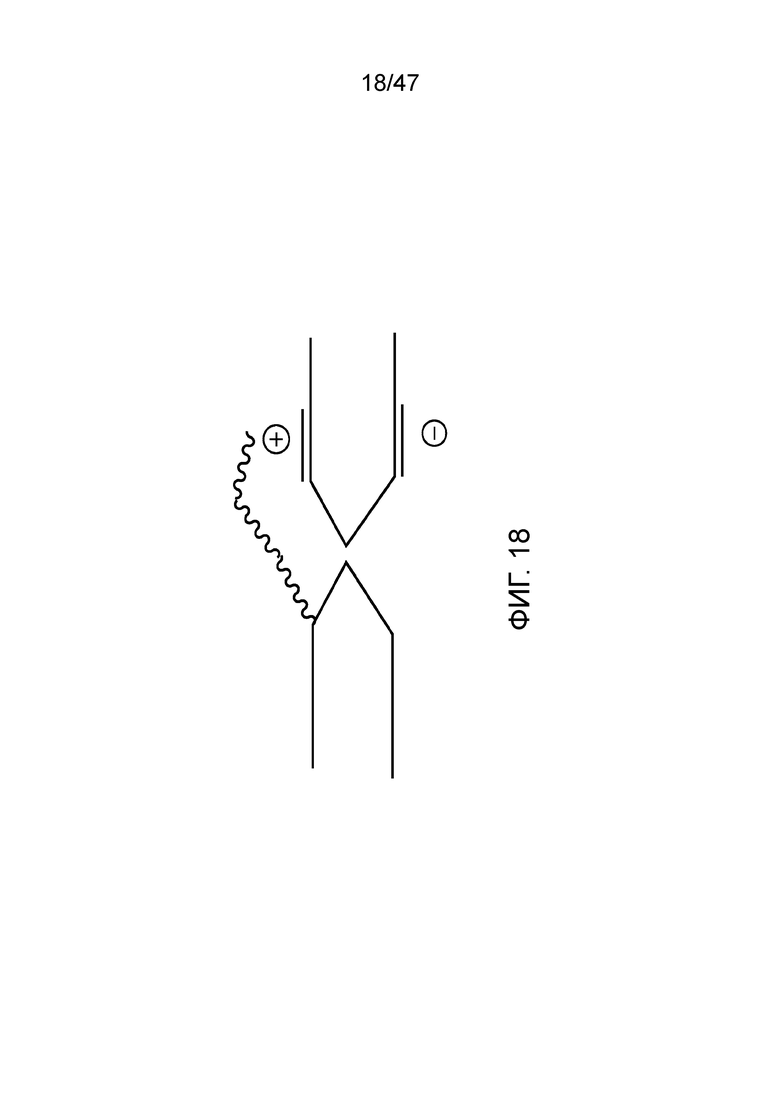
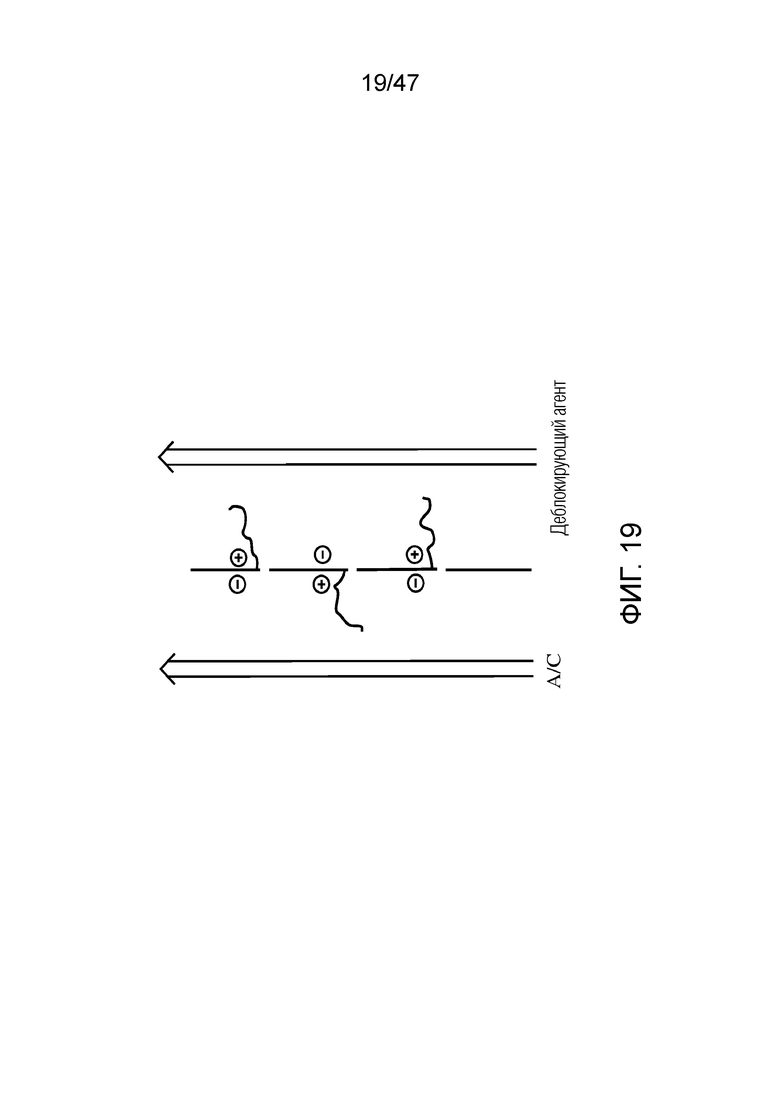
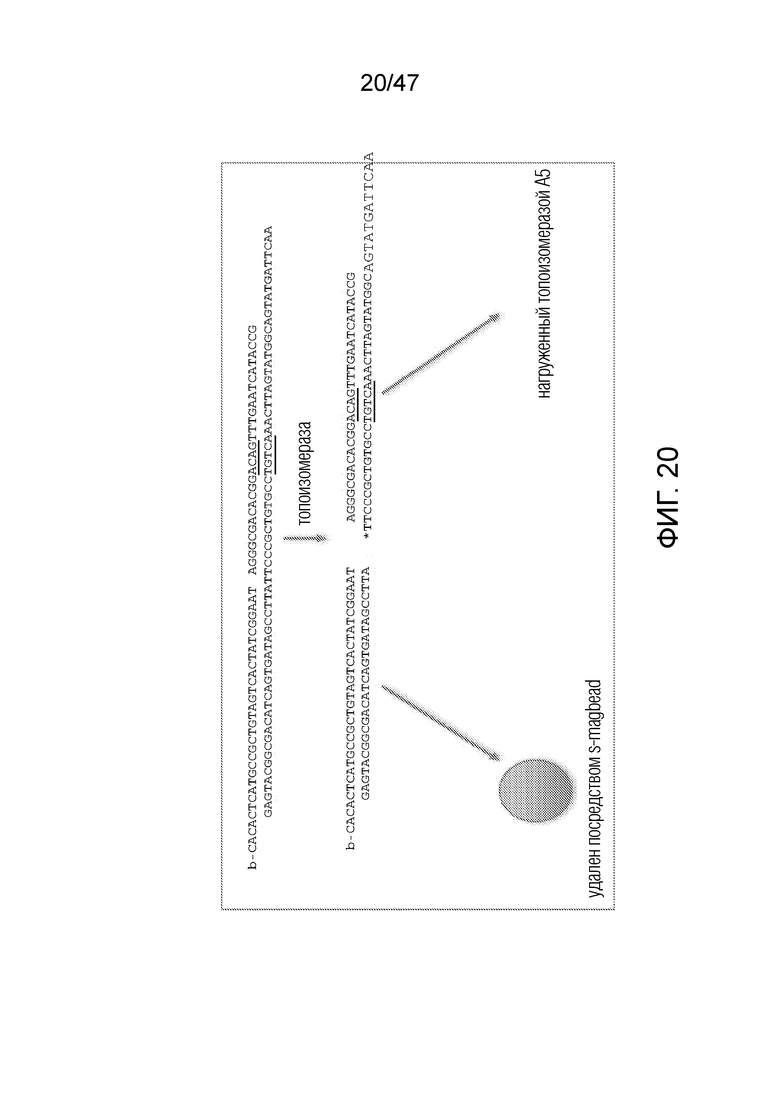
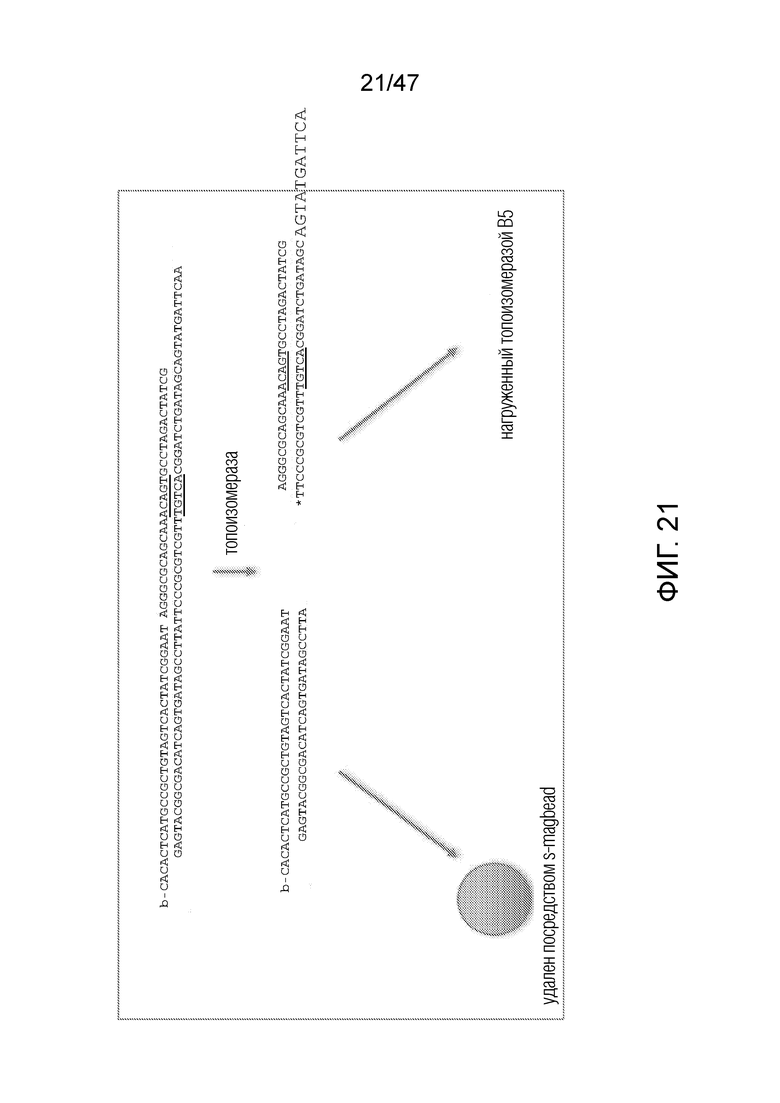
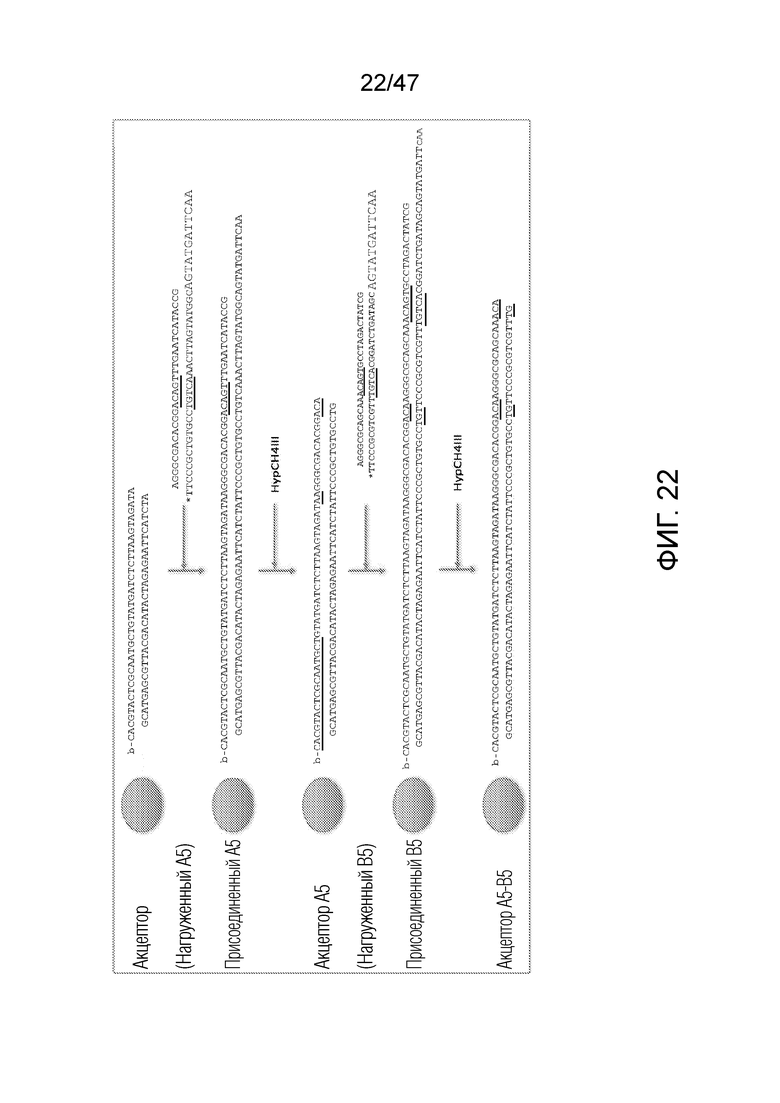
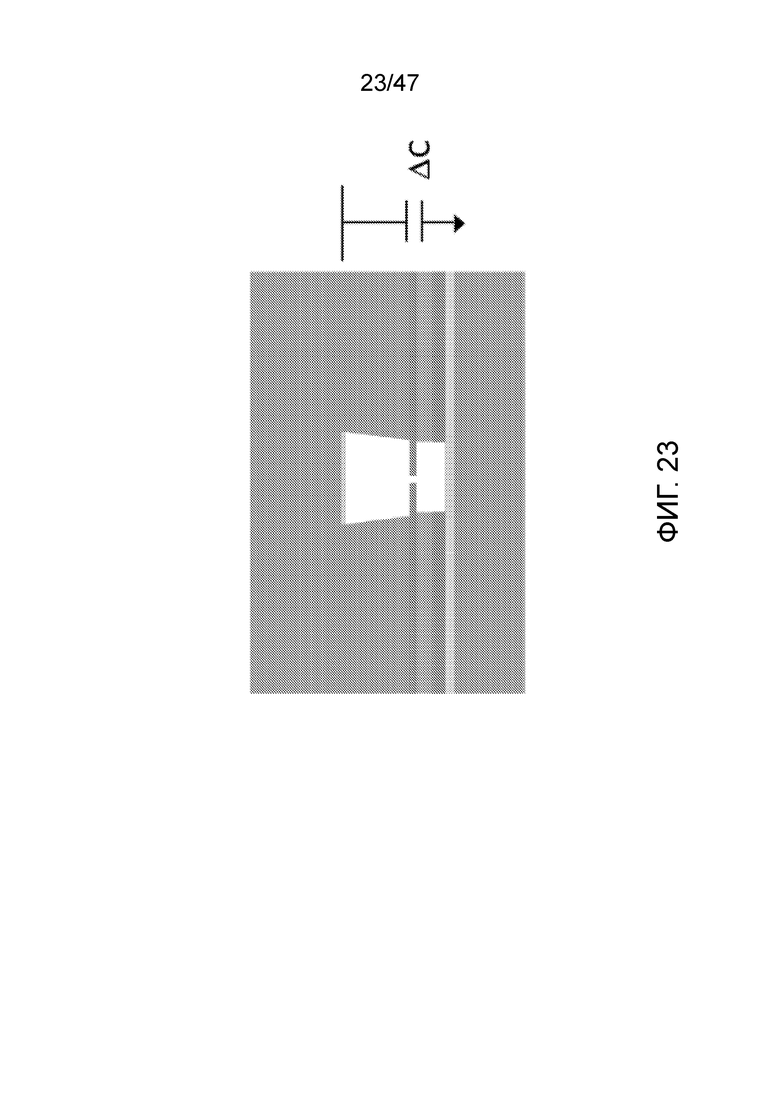
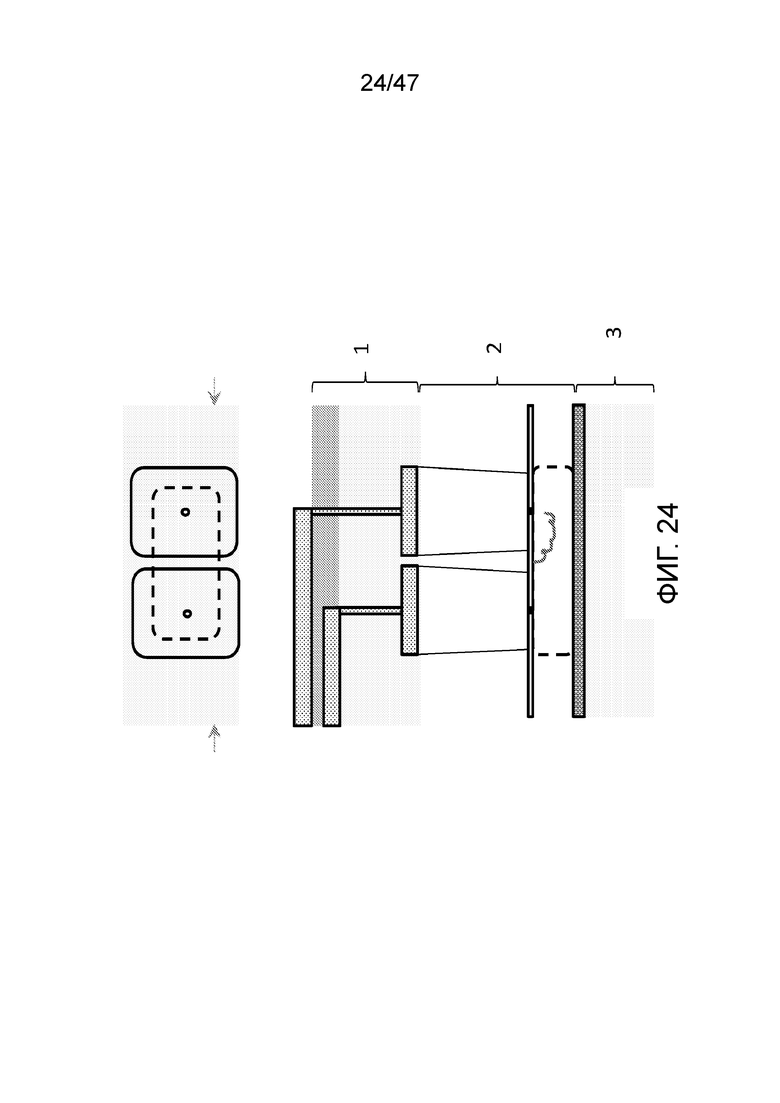
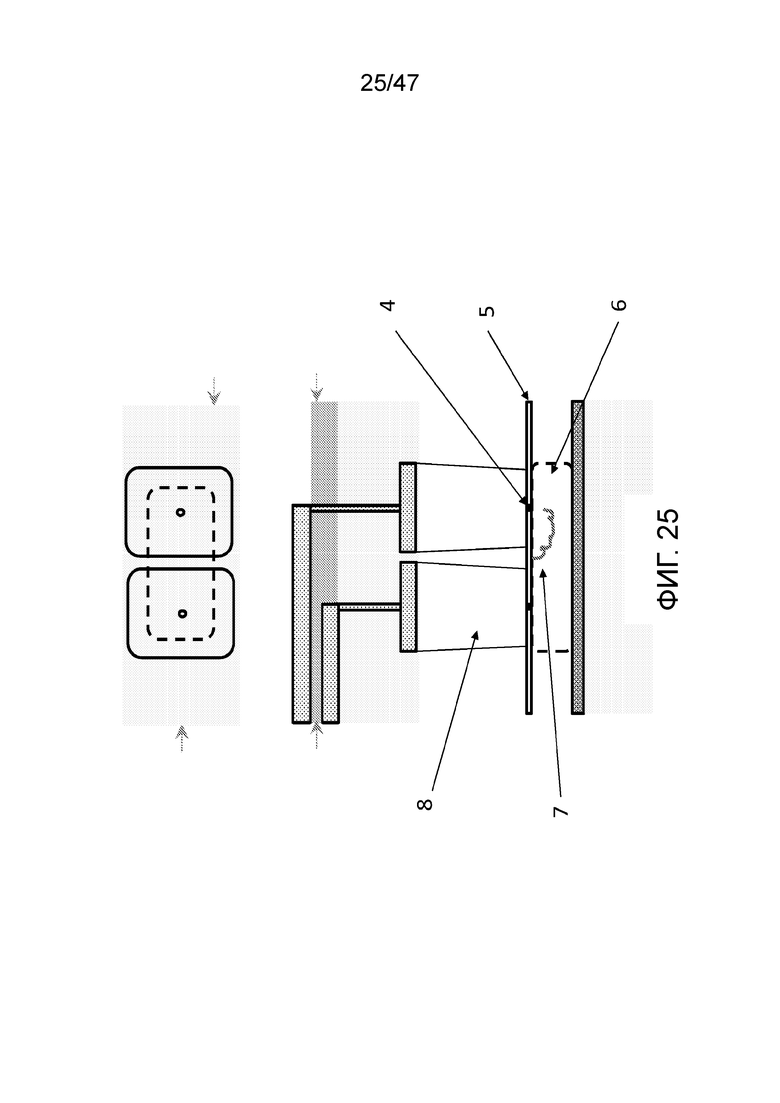
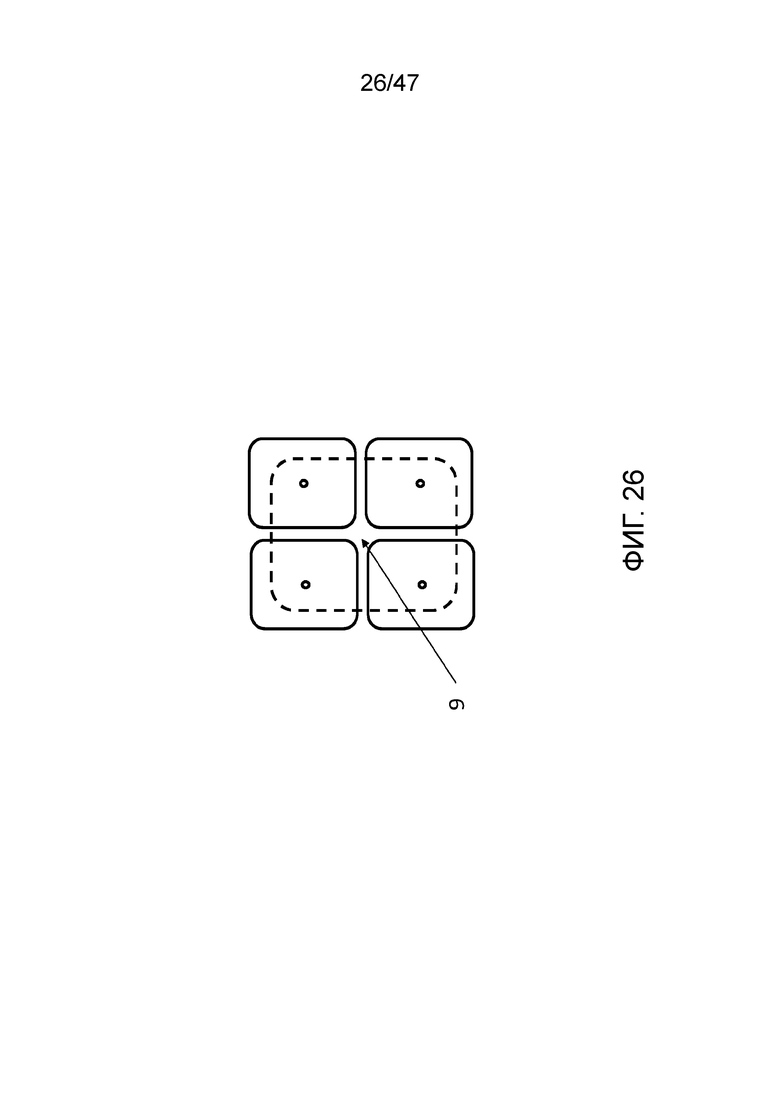
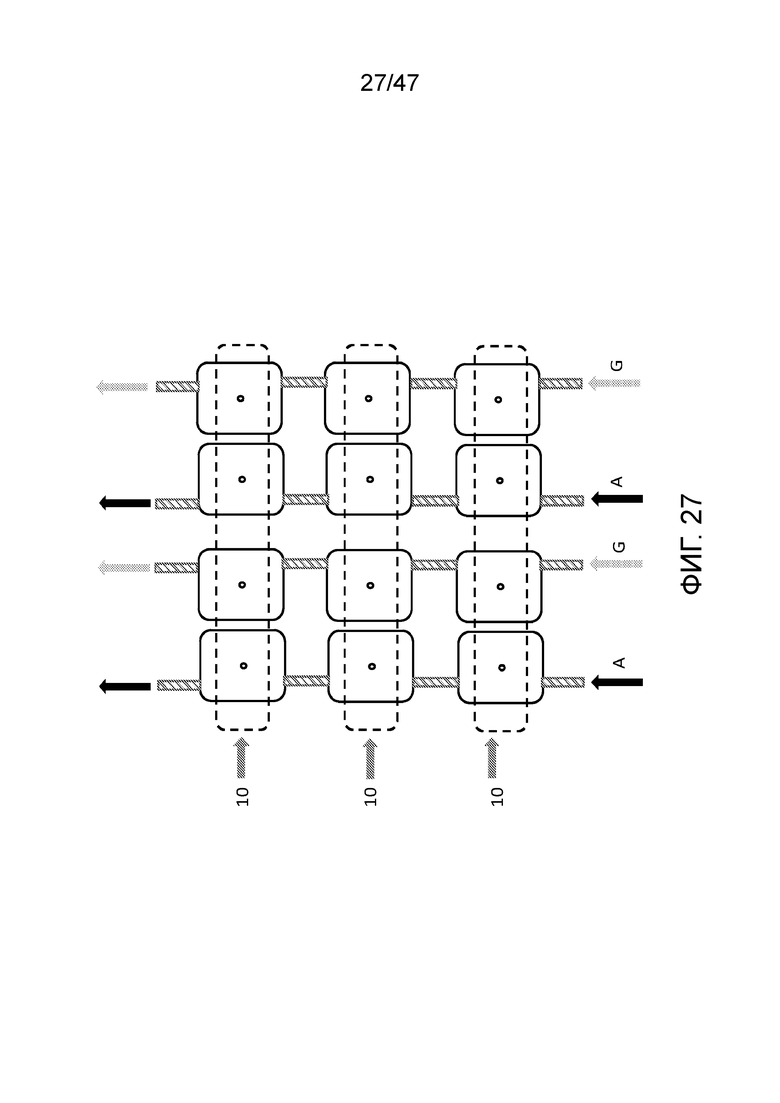
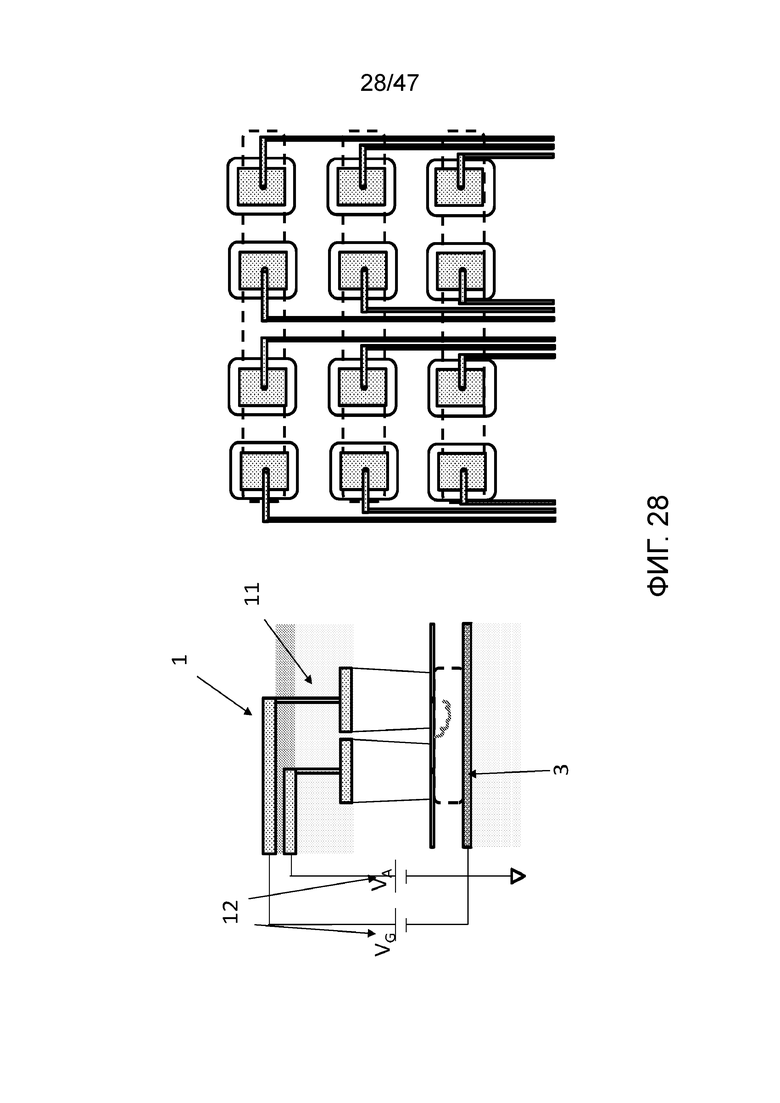
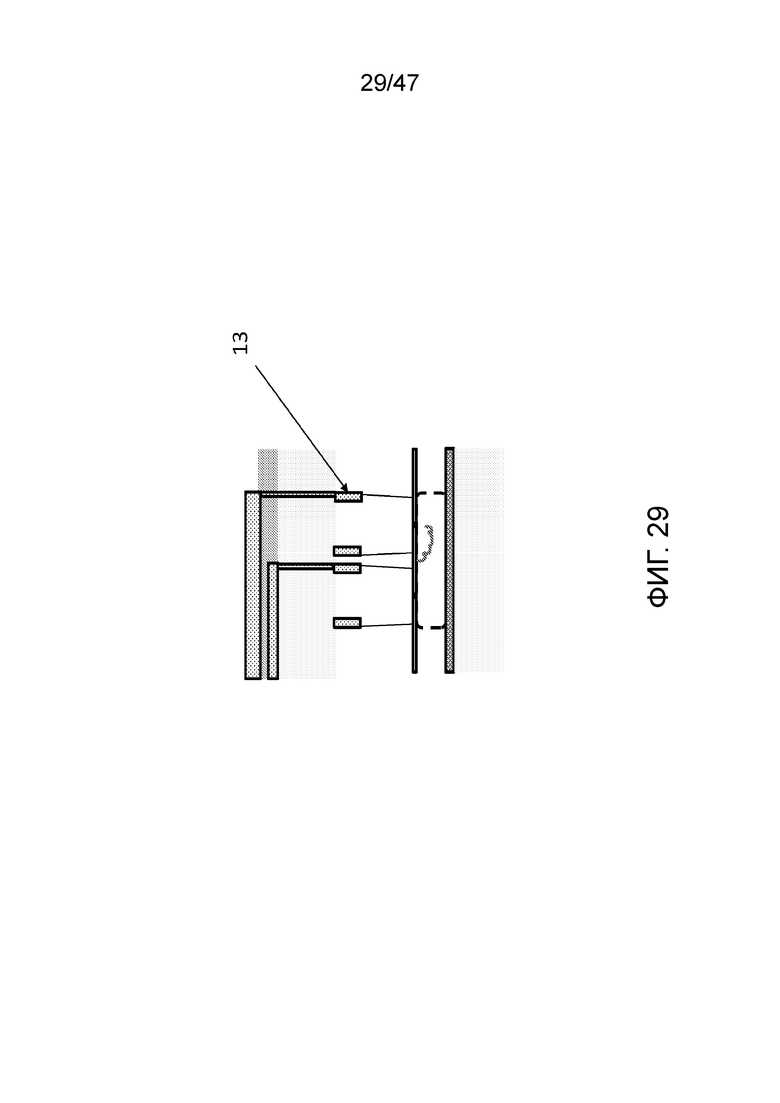
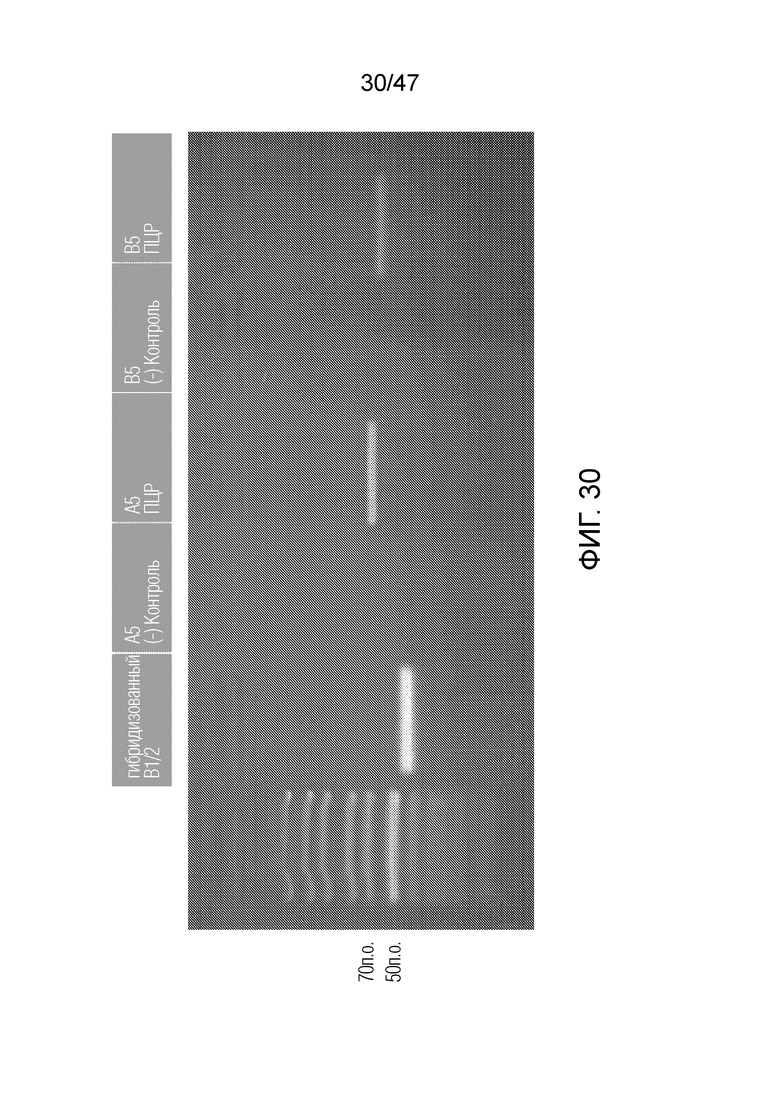
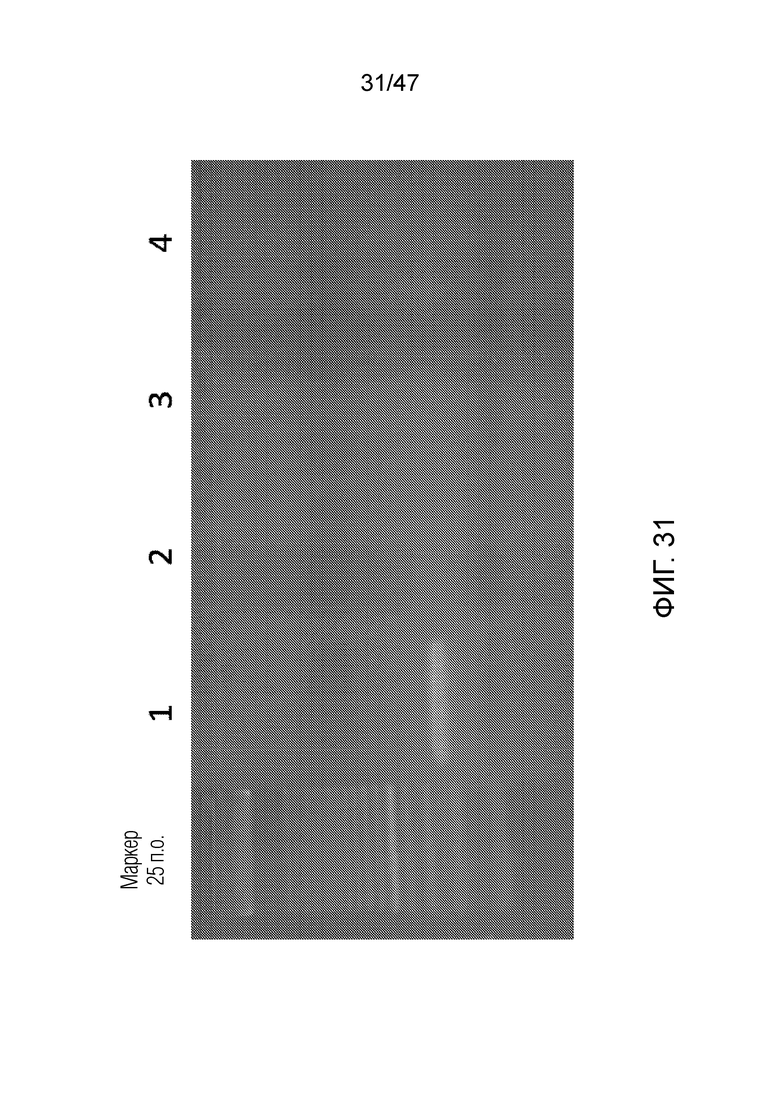
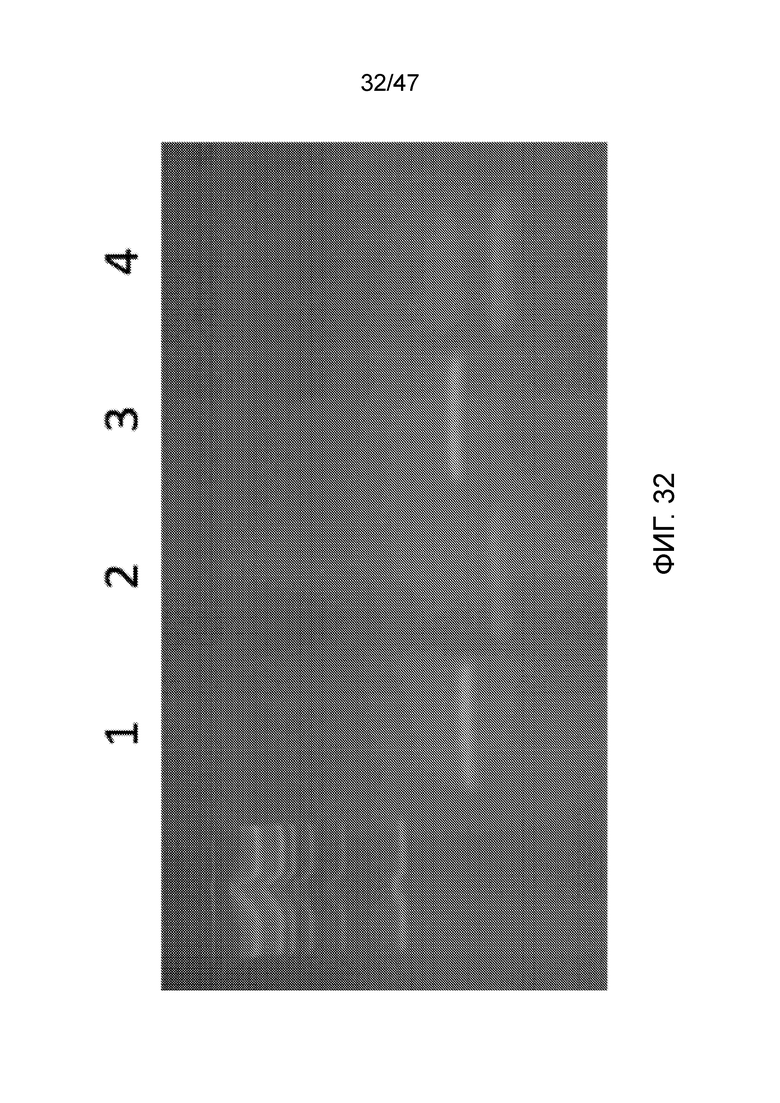
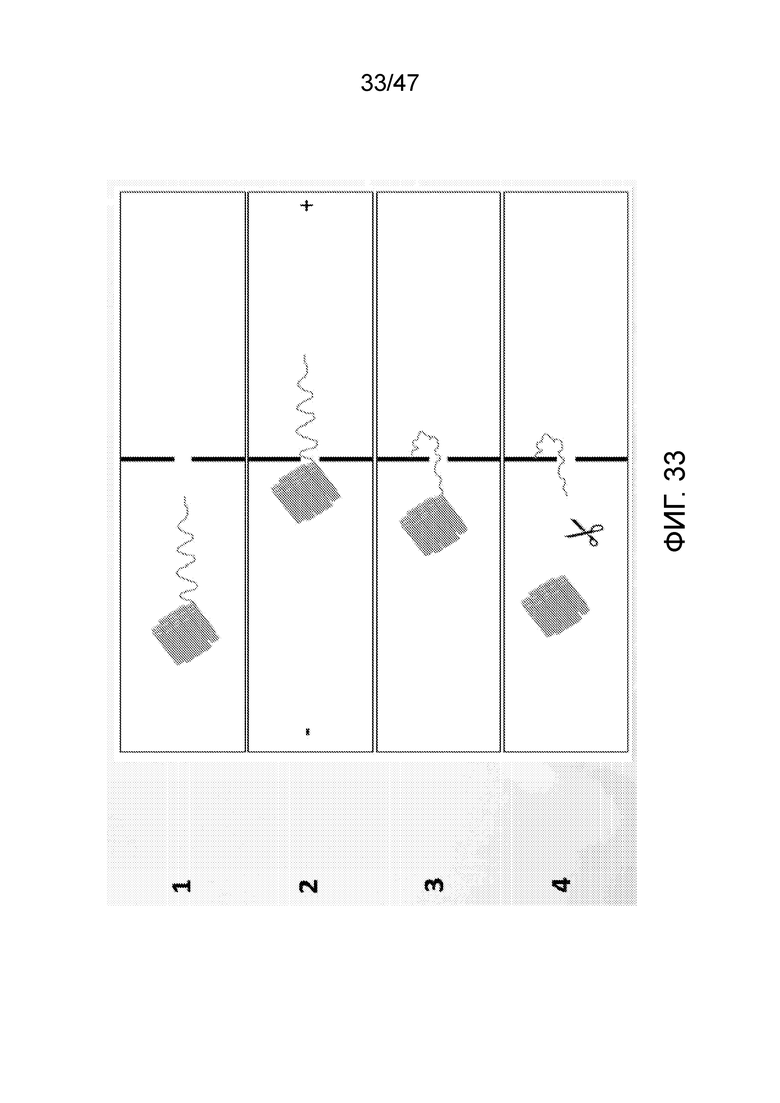
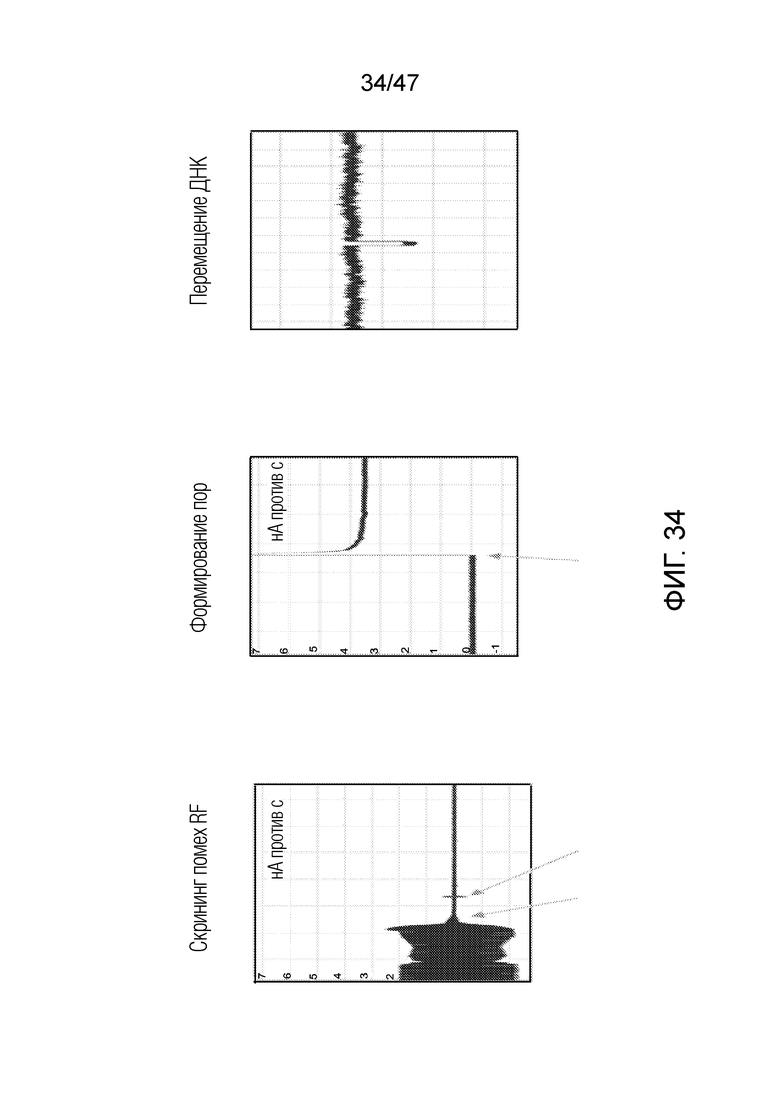
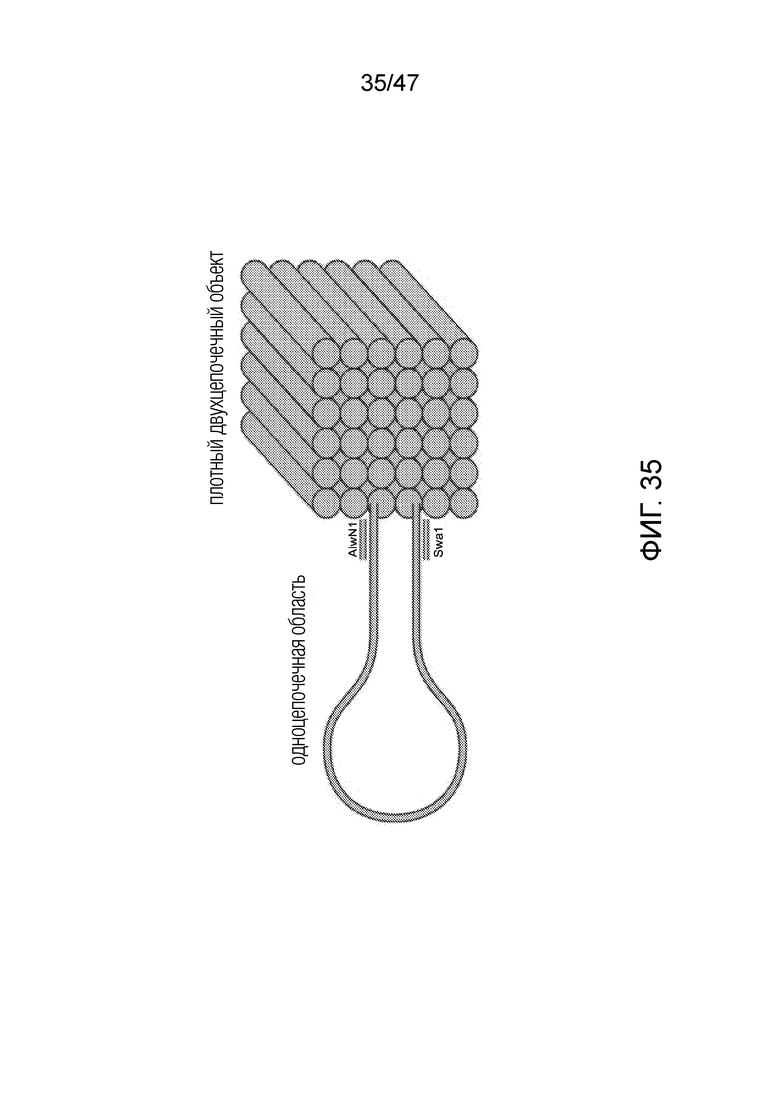
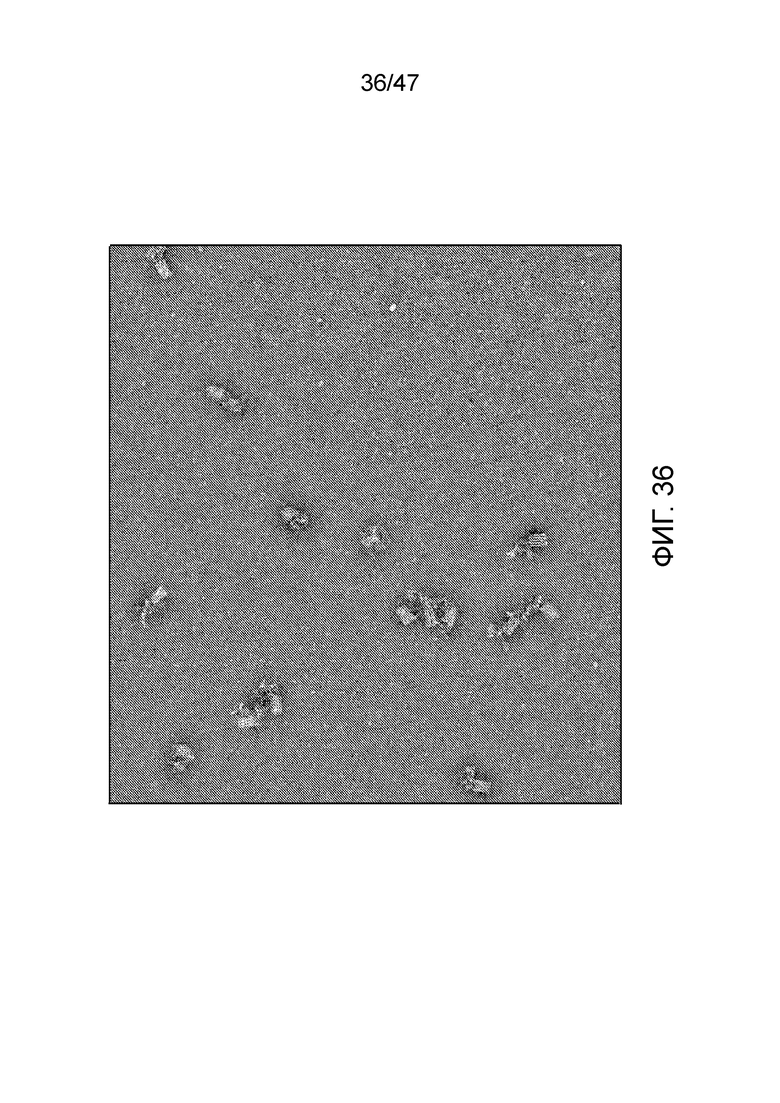
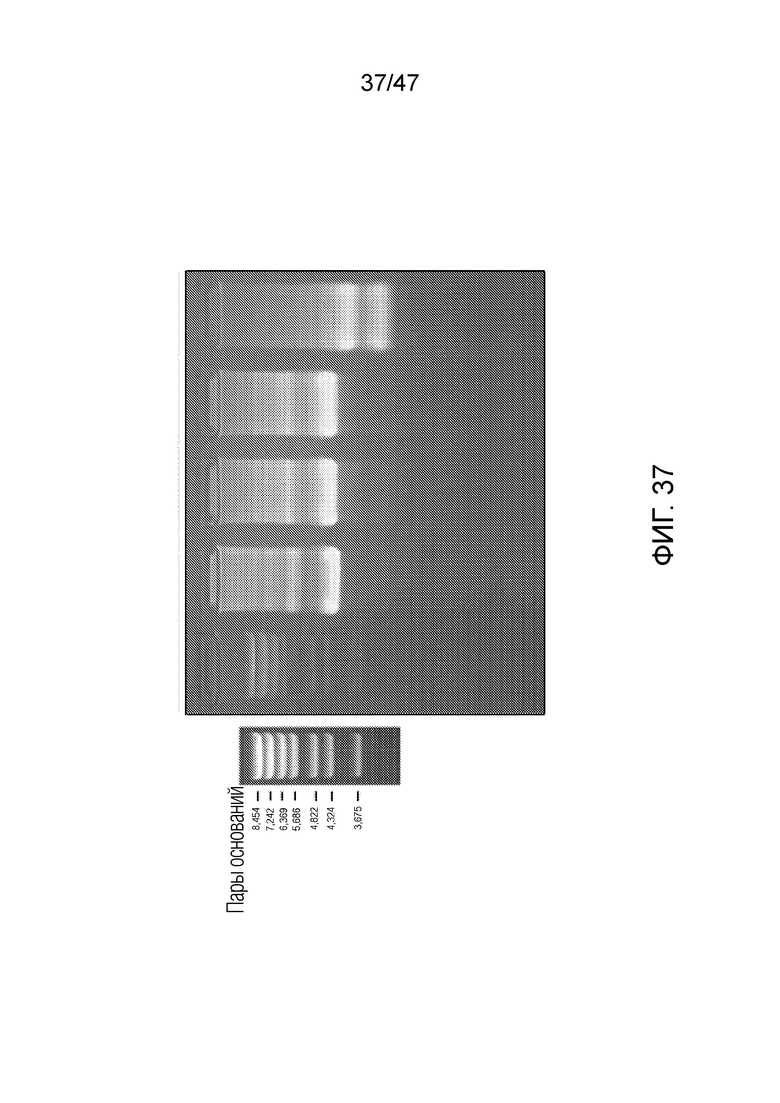
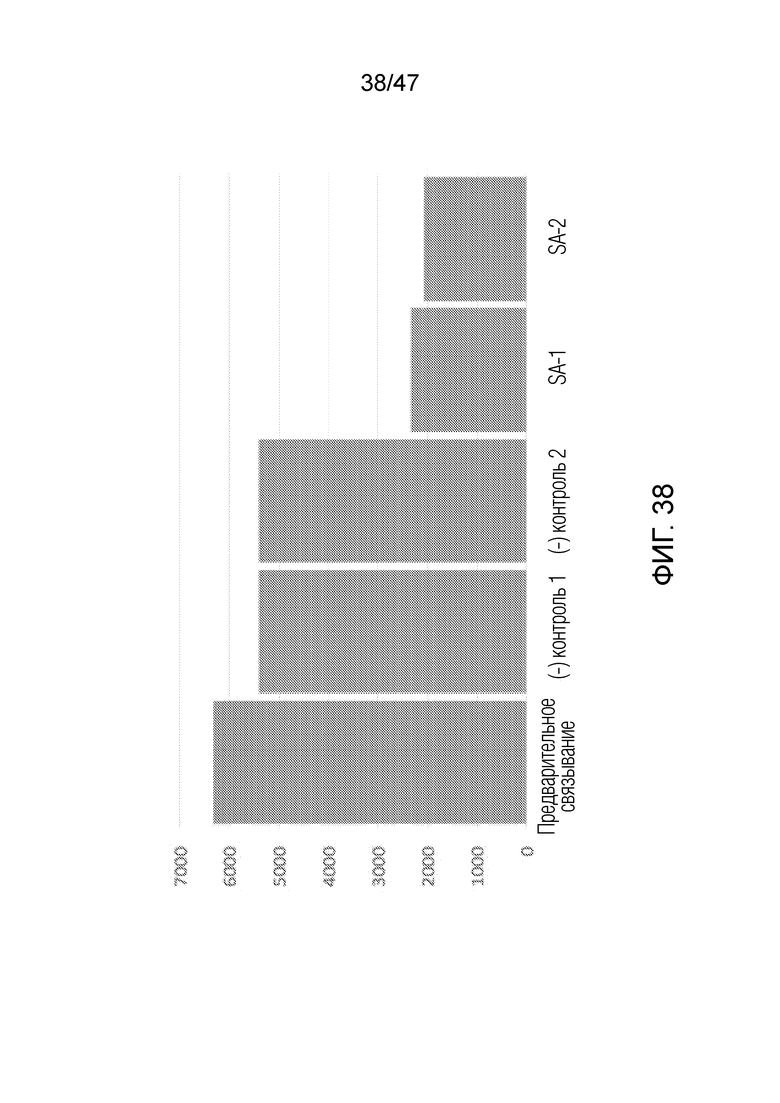
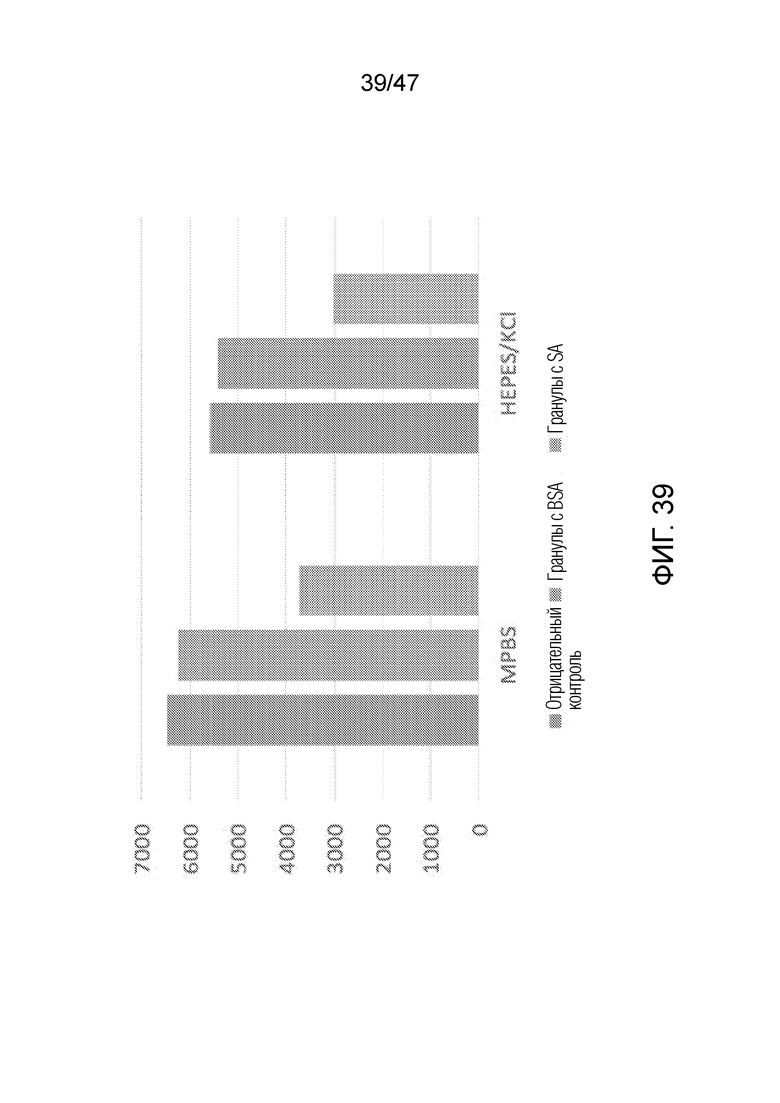
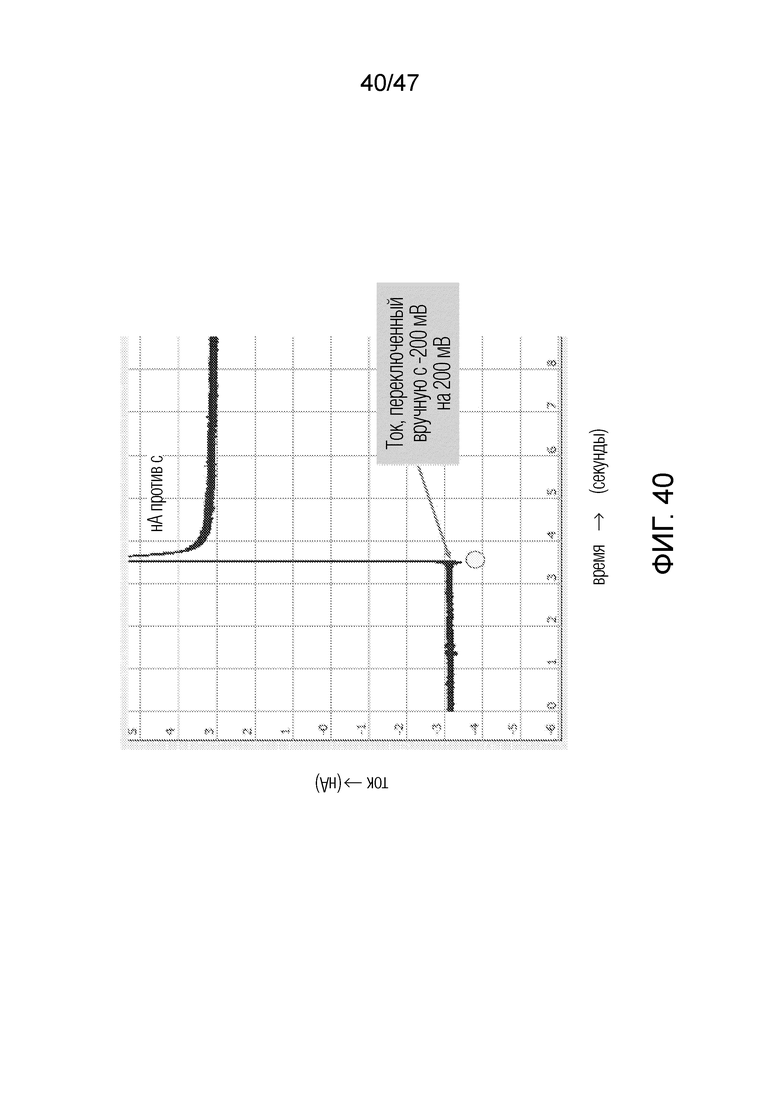
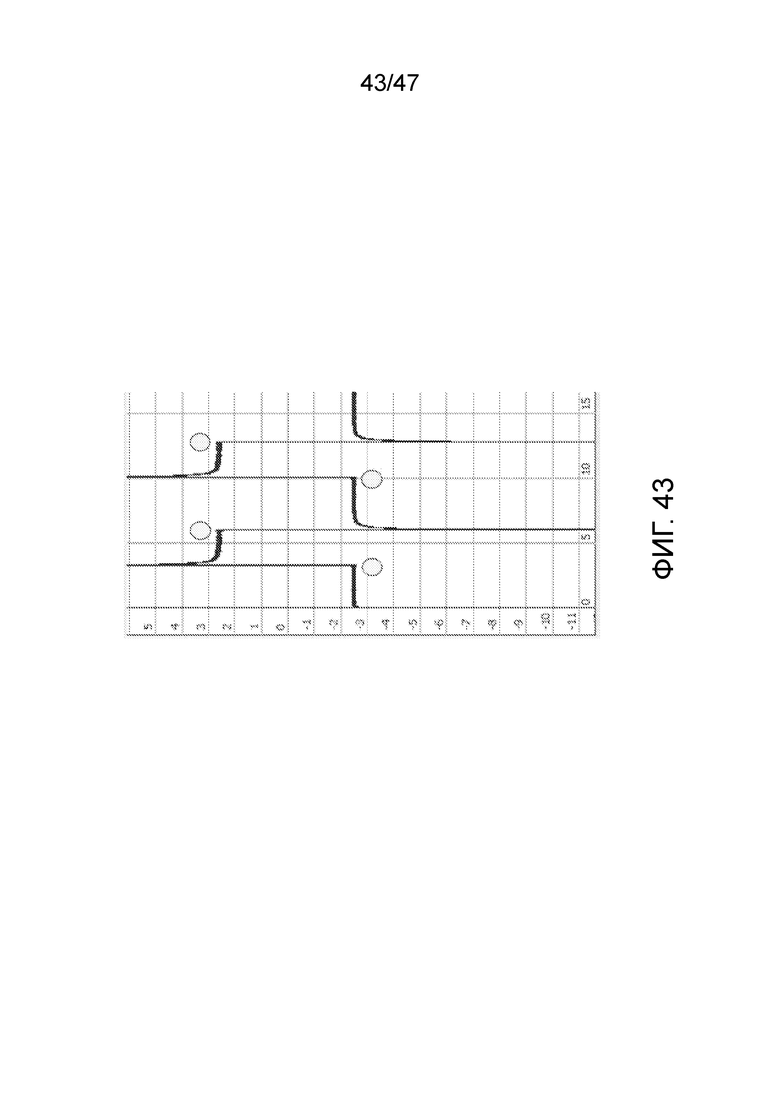
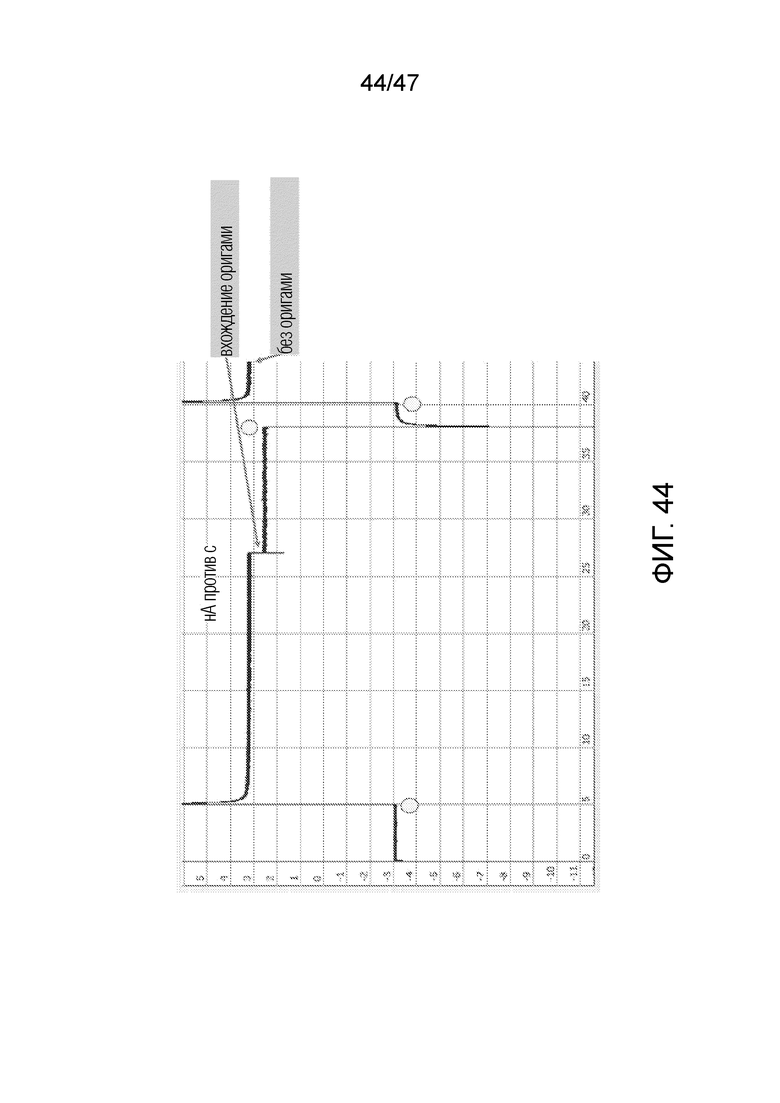
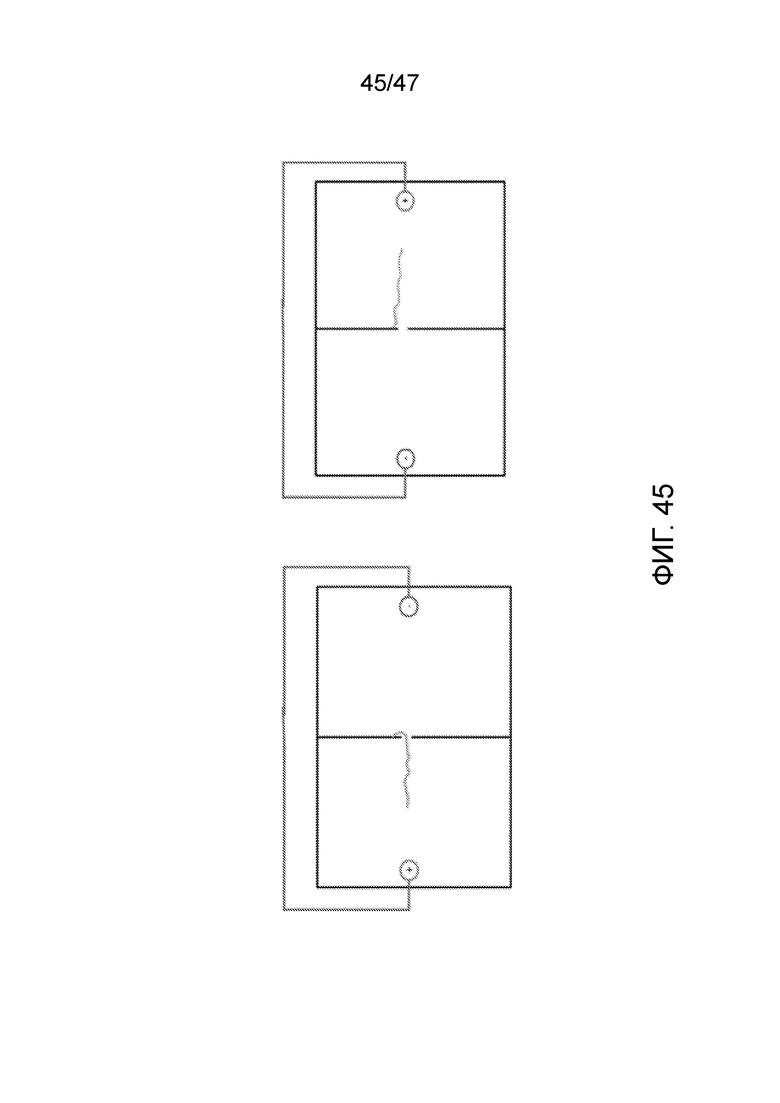
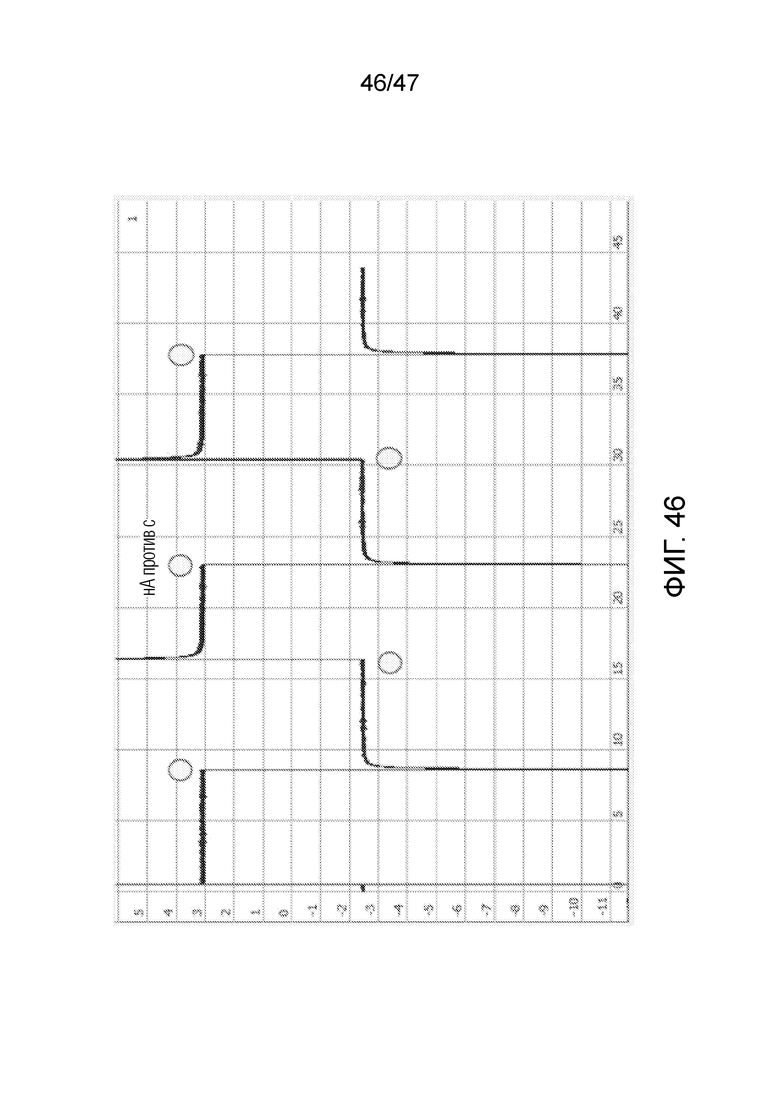
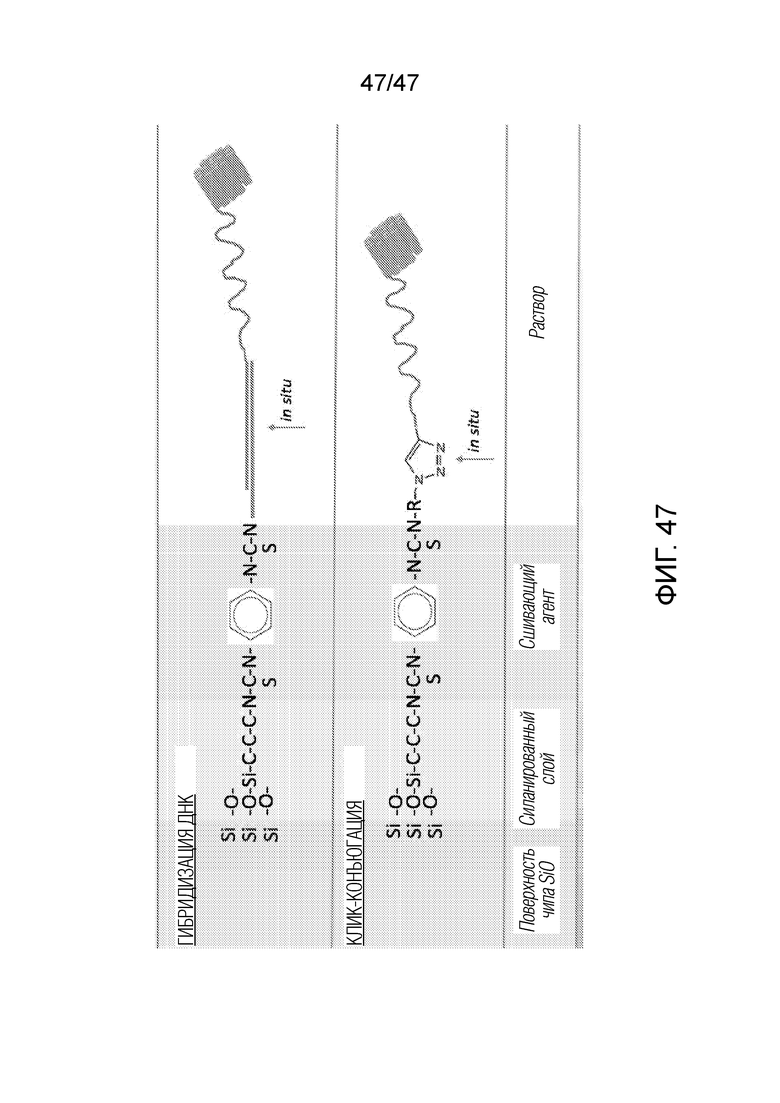
Изобретение относится к биотехнологии и представляет собой способ синтеза заряженного полимера, содержащего по меньшей мере два различных мономера, в наночипе, причем наночип содержит одну или более камер для присоединения, содержащих реагенты для присоединения одного или более мономеров или олигомеров к заряженному полимеру в буферном растворе в форме с защищенными концами, чтобы только один мономер или олигомер мог присоединяться за один цикл реакции; и одну или более резервных камер, содержащих буферный раствор, но не все реагенты, необходимые для присоединения одного или более мономеров или олигомеров, где камеры разделены одной или более мембранами, содержащими одну или более нанопор, и где заряженный полимер может проходить через нанопору и по меньшей мере один из реагентов для присоединения одного или более мономеров или олигомеров не может, при этом способ включает a) перемещение первого конца заряженного полимера, имеющего первый конец и второй конец, под действием электрического притяжения в камеру для присоединения, причем мономеры и олигомеры присоединяются к указанному первому концу в блокированной форме, b) перемещение первого конца заряженного полимера с добавленным мономером или олигомером в блокированной форме в резервную камеру, c) деблокирование присоединенного мономера или олигомера, и d) повторение стадий a-c, где мономеры или олигомеры, присоединенные на стадии a), являются такими же или отличаются, до тех пор пока не получат желаемую последовательность полимера, где заряженный полимер представляет собой ДНК, мономеры представляют собой нуклеотиды, где олигомеры представляют собой олигонуклеотиды и где нанопора имеет диаметр 2-20 нм. 2 н. и 6 з.п. ф-лы, 47 ил., 3 табл., 7 пр.
1. Способ синтеза заряженного полимера, содержащего по меньшей мере два различных мономера, в наночипе, причем наночип содержит
одну или более камер для присоединения, содержащих реагенты для присоединения одного или более мономеров или олигомеров к заряженному полимеру в буферном растворе в форме с защищенными концами, чтобы только один мономер или олигомер мог присоединяться за один цикл реакции; и
одну или более резервных камер, содержащих буферный раствор, но не все реагенты, необходимые для присоединения одного или более мономеров или олигомеров,
где камеры разделены одной или более мембранами, содержащими одну или более нанопор, и
где заряженный полимер может проходить через нанопору, и по меньшей мере один из реагентов для присоединения одного или более мономеров или олигомеров не может,
при этом способ включает
a) перемещение первого конца заряженного полимера, имеющего первый конец и второй конец, под действием электрического притяжения в камеру для присоединения, причем мономеры и олигомеры присоединяются к указанному первому концу в блокированной форме,
b) перемещение первого конца заряженного полимера с добавленным мономером или олигомером в блокированной форме в резервную камеру,
c) деблокирование присоединенного мономера или олигомера, и
d) повторение стадий a-c, где мономеры или олигомеры, присоединенные на стадии a, являются такими же или отличаются, до тех пор пока не получат желаемую последовательность полимера,
где заряженный полимер представляет собой ДНК, мономеры представляют собой нуклеотиды, где олигомеры представляют собой олигонуклеотиды и где нанопора имеет диаметр 2-20 нм.
2. Способ по п.1, где нанопора имеет диаметр 2-5 нм.
3. Способ по п.1 или 2, где реагенты для присоединения одного или более мономеров или олигомеров к заряженному полимеру включают полимеразу.
4. Способ по любому из пп.1-3, где нуклеотид присоединяется в 3'-защищенной форме и где резервная камера содержит реагенты, способные удалять защитную группу из присоединенного нуклеотида.
5. Способ по п.1 или 2, где одна или более камер для присоединения содержат нагруженный топоизомеразой нуклеотид или олигонуклеотид и одну или более резервных камер, содержащих фермент рестрикции или фосфатазу, где нанопора является достаточно малой, так что топоизомераза и фермент рестрикции или фосфатаза неспособны пройти через нанопору.
6. Способ по п.5, где на каждом цикле присоединяется один нуклеотид.
7. Наночип для синтеза электрически заряженного полимера, содержащего по меньшей мере два различных мономера, причем наночип содержит по меньшей мере первую и вторую реакционные камеры, разделенные мембраной, содержащей одну или более нанопор, где каждая реакционная камера содержит один или более электродов для втягивания электрически заряженного полимера в камеру и где указанная первая реакционная камера дополнительно содержит электролитную среду и реагенты для присоединения мономеров или олигомеров к полимеру, где указанные мономеры или олигомеры являются защищенными, так что только один может присоединяться за один раз, и где указанная вторая реакционная камера дополнительно содержит электролитную среду и реагенты для удаления защитной группы из мономера или олигомера, присоединенного таким образом, где нанопора является достаточно крупной, чтобы позволить прохождение полимера, но слишком малой, чтобы позволить прохождение по меньшей мере одного из реагентов для присоединения мономеров или олигомеров к полимеру и по меньшей мере одного из реагентов для удаления защитной группы из мономера или олигомера, присоединенного таким образом, где заряженный полимер представляет собой ДНК, мономеры представляют собой нуклеотиды и олигомеры представляют собой олигонуклеотиды, и где нанопора имеет диаметр 2-20 нм.
8. Способ по п.7, где нанопора имеет диаметр 2-5 нм.
| US 20060286024 A1, 21.12.2006 | |||
| US 20120058468 A1, 08.03.2012 | |||
| US 20120322109 A1, 20.12.2012 | |||
| УСТРОЙСТВО И СПОСОБЫ ВЫЯВЛЕНИЯ НУКЛЕИНОВОЙ КИСЛОТЫ В БИОЛОГИЧЕСКИХ ОБРАЗЦАХ | 2004 |
|
RU2359038C2 |

