Изобретение относится к сельскому хозяйству и биотехнологии и может быть использовано для получения константных (не расщепляющихся при самоопылении) безостых форм мягкой пшеницы.
С момента введения мягкой пшеницы в культуру было очевидно, что ее продуктивность во многом связана с признаками морфологии колоса. Одним из важных признаков является остистость колоса. Исследования показали, что ости колоса эффективно повышают транспирацию соцветия пшеницы и играют важную роль в процессах фотосинтеза и дыхания растений [1]. Было отмечено достоверное преимущество по продуктивности остистых форм в условиях недостатка влаги [2]. В целом, данный признак повышает стабильность зерновой продуктивности пшеницы за счет повышения адаптивности к неблагоприятным климатическим факторам. Однако с появлением комбайновых способов уборки безостые пшеницы получили технологическое преимущество перед остистыми, так как ости последних засоряют технику и снижают эффективность обмолачивания. Для гексаплоидной мягкой пшеницы известно, что различные количественные проявления остистости по длине остей и их распределению по длине колоса определяются тремя генетическими локусами: Hooded (Hd), Tipped1 (В1) и Tipped2 (B2), расположенными на хромосомах 4А, 5А и 6В, соответственно. [3-5]. Однако, наиболее сильное фенотипическое проявление имеет локус В1 на хромосоме 5А. Доминантный аллель В1 контролирует безостость, а рецессивный аллель b1 обуславливает проявление остистости у пшеницы. Несмотря на то, что аллель безостости В1 очень распространен среди отечественных яровых сортов мягкой пшеницы, в определенных условиях, особенно в засушливых зонах, возникает необходимость введения рецессивного аллеля b1 в селекционный процесс из-за его положительного влияния на засухоустойчивость. В зависимости от условий возникает необходимость скрещивания остистых и безостых форм и отбора тех или иных форм по признаку остистости. Однако, если остистые формы, контролируемые рецессивным аллелем b1, выявляются фенотипически и не расщепляются далее на остистые и безостые, то безостые формы, контролируемые доминантным аллелем В1, при дальнейшем самоопылении растений (в случае гетерозиготности) расщепляются на остистые и безостые формы, что существенно затягивает процесс создания константных безостых сортов мягкой пшеницы.
Применение молекулярных маркеров позволяет сократить время и снизить трудозатраты по созданию константных безостых сортов мягкой пшеницы. Молекулярные или ДНК-маркеры, основанные на методе полимеразной цепной реакции (ПЦР-маркеры) оказались широко востребованными в генетических исследованиях и селекции пшеницы для решения практических задач. Молекулярные ДНК-маркеры позволяют отобрать константные формы с целевым геном уже в ранних поколениях при гибридизации контрастных форм по изучаемому признаку.
Следует отметить, что вплоть до недавнего времени ген В1 не был идентифицирован, и его хромосомная локализация была установлена лишь приблизительно, что являлось основным препятствием для разработки эффективного маркера, пригодного для генотипирования линий с различным проявлением остистости и использование его для отбора константных безостых форм. Лишь недавно китайскими исследователями было проведено тонкое генетическое картирование этого гена [6].
Известен способ отбора безостых форм озимой пшеницы [7], заключающийся в том, что для создания сортов озимой пшеницы проводят скрещивания между остистыми и безостыми формами. Во втором поколении осуществляют отбор форм, которые делят на фракции по степени развития остевидных образований: полностью безостые, с короткими (1-4 мм) и со средними (5-8 мм) остевидными образованиями. Каждую фракцию высевают отдельно, а для последующего пересева отбирают фракции, в которых количество гомозиготных форм составляет 80% и более. Однако данный способ является трудоемким, так как для отбора гомозиготных растений фенотипическую оценку требуется проводить в нескольких поколениях на большой по численности популяции.
Наиболее ближайшим к заявляемому способу - прототипом, является способ создания линий озимой мягкой пшеницы с комплексной устойчивостью к бурой и стеблевой ржавчине и мучнистой росе, заключающийся в том, что коммерческий сорт яровой мягкой пшеницы содержащий комплекс генов Lr-Sr-Pm устойчивости к бурой и стеблевой ржавчине и мучнистой росе от вида Agropyron intermedium скрещивают с коммерческим сортом озимой мягкой пшеницы, и растения поколения F1 самоопыляют до поколения F2. Из поколения F2 с помощью молекулярного маркера Xicg6Ai#2, отбирают растения, содержащие комплекс генов Lr-Sr-Рт. Отобранные молекулярным маркером Xicg6Ai#2 растения F2 анализируют с помощью маркеров к рецессивным аллелям генов озимого образа жизни с помощью комбинаций маркеров VRN1AF/VRN1-INT1R, Intr1BF/Intr1BR4 и Intr1DF/Intr1DR4, для выявления растений с озимым образом жизни [8].
Недостатками данного способа являются:
1. Отсутствие возможности отобрать указанными маркерами константные безостые линии мягкой пшеницы.
2. Необходимость определения образа жизни растений, так как гибридная популяция F2 состоит как из озимых, так и из яровых растений.
Задачей предлагаемого изобретения является разработка способа, позволяющего быстро отбирать гомозиготные (константные) безостые растения мягкой пшеницы, которые не будут расщепляться по этому признаку в следующих поколениях.
Технический результат: сокращение срока создания константных безостых форм мягкой пшеницы и расширение функциональных возможностей известного способа.
Поставленная задача достигается предлагаемым способом, заключающимся в следующем.
Коммерческий остистый сорт яровой мягкой пшеницы «Любава 5», содержащий рецессивные аллели b1b1, определяющие развитие остей на колосьях, скрещивают с безостой линией яровой мягкой пшеницы «476-10» и растения поколения F1 самоопыляют до поколения F2. В потомстве F2 растения анализируют с помощью молекулярных маркеров b1for / Znfrev и B1for / Znfrev и отбирают растения с генотипом В1В1. Растения поколения F2, отобранные с помощью маркеров b1for / Znfrev и B1for / Znfrev, самоопыляют до поколения F3 и в полевых условиях проводят фенотипическую оценку для подтверждения безостости.
Основными определяющими отличиями предлагаемого способа от прототипа являются:
- в качестве исходного сорта для гибридизации используют коммерческий остистый сорт яровой мягкой пшеницы «Любава 5», содержащий в гомозиготном рецессивном состоянии аллель b1;
- из поколения F2 с помощью молекулярных маркеров b1for / Znfrev и B1for / Znfrev отбирают растения, содержащие в гомозиготном состоянии аллель В1, определяющий безостость, что позволяет на ранних стадиях создания линий мягкой пшеницы отбирать растения с безостыми колосьями.
Изобретение иллюстрируется следующим примером.
Пример.
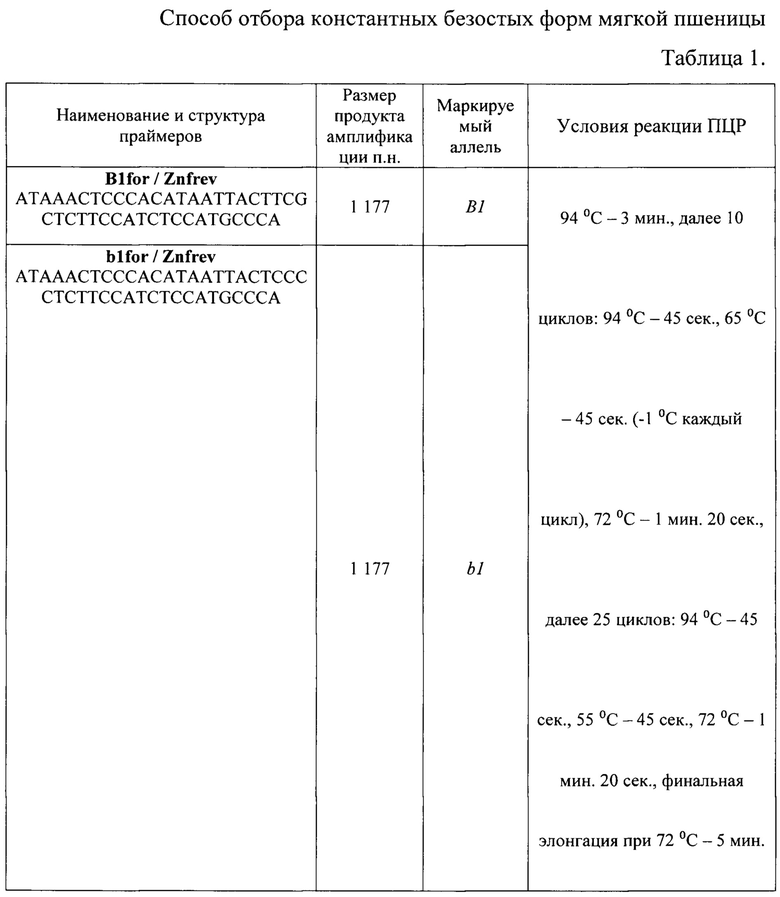
Коммерческий остистый сорт яровой мягкой пшеницы «Любава 5», содержащий в гомозиготном состоянии рецессивный аллель b1 (генотип b1b1), определяющий признак остистости колосьев, скрестили с безостой линией яровой мягкой пшеницы «476-10». Полученные от скрещивания гибридные растения поколения F1 самоопыляли до поколения F2. В поколении F2 с помощью молекулярных ПЦР-маркеров b1for / Znfrev, выявляющих рецессивный аллель b1, и B1for / Znfrev, выявляющих доминантный аллель В1, отбирали гомозиготные растения с генотипом В1В1. Структура использованных молекулярных маркеров и условия полимеразной цепной реакции (ПЦР) приведены в таблице 1.
Результаты анализа для ПЦР-маркеров b1for / Znfrev и B1for / Znfrev с геномной ДНК родительского сорта «Любава 5», линии «476-10» и безостых растений поколения F2, гомозиготных, содержащих аллели В1В1 (линия «476-10» и растения 8, 10, 12, 16, 19, 21), и гетерозиготных, содержащих аллели B1b1 (растения 1, 2, 3, 4, 5, 6, 7, 9, 11, 13, 14, 15, 17, 18, 20), приведены на фиг.1. Линии поколения F2, гомозиготные по рецессивным аллелям, имеющие остистый колос, в анализ не включали. Сорт «Любава 5», содержащий рецессивные аллели b1b1, использовался в качестве контроля. Обе пары праймеров b1for / Znfrev и B1for / Znfrev амплифицируют фрагмент ДНК длиной 1177 п. н.
Растения поколения F2, разделенные на группы гомозиготных (аллели В1В1) и гетерозиготных (аллели B1b1), самоопыляли и в полевых условиях проводили оценку расщепления в поколении F3 по признаку остистость/безостость. Результаты представлены в таблице 2, где константные безостые формы выделены жирным шрифтом.
Из таблицы 2 видно, что у родительского остистого сорта «Любава 5» были рецессивные аллели b1b1, у взятой в скрещивание безостой линии «476-10» были доминантные аллели В1В1.
Потомство F3 от безостых гомозиготных растений 8, 10, 12, 16, 19, 21, несущих доминантные аллели В1В1, было только безостое. Потомство F3 от гетерозиготных растений 1, 2, 3, 4, 5, 6, 7, 9, 11, 13, 14, 15, 17, 18, 20, несущих аллели B1b1, расщеплялось на остистые и безостые растения.
Таким образом, предлагаемый способ позволяет отбирать константные безостые линии мягкой пшеницы, и может быть использован для расширения генетического разнообразия мягкой пшеницы. Использование молекулярных маркеров b1for/Znfrev и B1for/Znfrev позволяет эффективно проводить отбор гомозиготных (константных), как безостых, так и остистых форм мягкой пшеницы без проведения полевых испытаний, что позволяет сократить срок создания новых линий, представляющих интерес для селекции этой культуры.
Источники информации
1. McDonough W.T., Gauch H.G. The contribution of the awns to the development of the kernels of bearded wheat. Maryland: Agric. Exp. Sta. Bull, 1959. P. 1-16.
2. Olugbemi R.В., Bingham J. Effects of awns on the photosynthesis and yield of wheat. Annals of Applied Biology, 1976, V.84(2), P. 241-250.
3. Watkins AE, Ellerton S. Variation and genetics of the awn in Triticum. J Genet. 1940; 40: 243-270.
4. Mcintosh RA, Hart GE, Devos KM, Gale MD, Rogers WJ. Catalogue of gene symbols for wheat. Proc. 9th Int. Wheat Genet. Symp. Saskatoon 1998; 5: 235.
5. Sears E.R. The Aneuploids of Common Wheat. Missouri Agr. Exp. Stn. Res. Bull. 1954; 572.
6. Wang D., Yu K., Jin D., Sun L., Chu J., Wu W., Xin P., Li X., Sun J, Yang W., Zhan K., Zhang Α., Liu D. ALI-1, candidate gene of B1 locus, is associated with awn length and grain weight in common wheat. BioRxiv, 2019; DOI: 10.1101/688085.
7. Патент SU 1606041 A1, опубл. 15.11.1990.
8. Патент RU 2598275 C1, опубл. 20.09.2016.

| название | год | авторы | номер документа |
|---|---|---|---|
| Способ отбора линий яровой мягкой пшеницы с повышенным содержанием антоцианов в зерне | 2021 |
|
RU2762804C1 |
| Способ получения линий яровой мягкой пшеницы с укороченным сроком колошения | 2019 |
|
RU2710729C1 |
| СПОСОБ СОЗДАНИЯ ЛИНИЙ ОЗИМОЙ МЯГКОЙ ПШЕНИЦЫ С КОМПЛЕКСНОЙ УСТОЙЧИВОСТЬЮ К БУРОЙ И СТЕБЛЕВОЙ РЖАВЧИНЕ И МУЧНИСТОЙ РОСЕ | 2015 |
|
RU2598275C1 |
| Внутригенный ДНК-маркер для отбора пшеницы с повышенным содержанием антоцианов в перикарпе зерновки | 2021 |
|
RU2774444C1 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ ПШЕНИЦЫ С НУКЛЕОТИДНОЙ ДЕЛЕЦИЕЙ В ПРОМОТОРНОЙ ОБЛАСТИ ГЕНА VRN-A1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2020 |
|
RU2772577C2 |
| Способ селекции мягкой яровой пшеницы с высокой обеспеченностью колоса метаболитами | 1988 |
|
SU1598929A1 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ ПШЕНИЦЫ С БИАЛЛЕЛЬНЫМИ МУТАЦИЯМИ В ПРОТОМОТОРНОЙ ОБЛАСТИ ГЕНА VRN-A1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2020 |
|
RU2772575C2 |
| Способ отбора гибридов мягкой пшеницы с голубой окраской зерновки | 2022 |
|
RU2790186C1 |
| СПОСОБ СОЗДАНИЯ ИСХОДНОГО МАТЕРИАЛА ДЛЯ СЕЛЕКЦИИ ОЗИМЫХ МЯГКИХ ПШЕНИЦ | 1999 |
|
RU2150822C1 |
| Способ получения озимых форм мягкой пшеницы из яровых | 1988 |
|
SU1581240A1 |

Изобретение относится к области биотехнологии. Коммерческий сорт яровой мягкой пшеницы «Любава 5», содержащий рецессивные аллели b1b1, определяющие остистость колоса, скрещивают с безостой линией яровой мягкой пшеницы «476-10» и растения поколения F1 самоопыляют до поколения F2. В потомстве F2 остистые растения удаляют, а оставшиеся безостые - анализируют с помощью молекулярных маркеров b1for/Znfrev и B1for/Znfrev и отбирают растения с генотипом В1В1, определяющим отсутствие остей у мягкой пшеницы. Изобретение позволяет сократить срок создания константных безостых форм мягкой пшеницы и расширить функциональные возможности известного способа. 1 ил., 2 табл.
Способ отбора константных безостых форм мягкой пшеницы, включающий скрещивание родительских образцов мягкой пшеницы, самоопыление гибридов первого поколения F1 для получения гибридов второго поколения F2, среди которых с помощью молекулярных ПЦР-маркеров отбирают растения с генами, определяющими безостость колосьев, повторное самоопыление отобранных растений для получения поколения F3 и тестирование последних в полевых условиях на оценку безостости, отличающийся тем, что сорт яровой мягкой пшеницы «Любава 5», содержащий рецессивные аллели b1b1, определяющие развитие остей, скрещивают с линией «476-10», содержащей доминантные аллели B1B1, определяющие безостость колосьев, затем из поколения F2 с помощью молекулярных ПЦР-маркеров b1for/Znfrev с нуклеотидными последовательностями АТАААСТСССАСАТААТТАСТССС, CTCTTCCATCTCCATGCCCA и B1for/Znfrev с нуклеотидными последовательностями ATAAACTCCCACATAATTACTTCG, CTCTTCCATCTCCATGCCCA, отбирают растения, содержащие в гомозиготном состоянии аллели В1В1, определяющие развитие безостых колосьев.
| СПОСОБ СОЗДАНИЯ ЛИНИЙ ОЗИМОЙ МЯГКОЙ ПШЕНИЦЫ С КОМПЛЕКСНОЙ УСТОЙЧИВОСТЬЮ К БУРОЙ И СТЕБЛЕВОЙ РЖАВЧИНЕ И МУЧНИСТОЙ РОСЕ | 2015 |
|
RU2598275C1 |
| Способ селекции озимой пшеницы | 1988 |
|
SU1606041A1 |
| WANG D., ALI-1, candidate gene of B1 locus, is associated with awn length and grain weight in common wheat | |||
| BioRxiv, 2019; DOI: 10.1101/688085. | |||

