Область техники, к которой относится изобретение
Изобретение относится к области биотехнологии и генной инженерии растений, а именно к способу получения однодольного растения при помощи технологии редактирования генома растений CRISPR/Cas9, путем целенаправленного воздействия на нуклеотидную последовательность консервативного участка промоторной области гена VRN-1A однодольных зерновых культур системами редактирования генома CRISPR/Cas9, в которых используются молекулы РНК-проводника или сконструированный на его основе экспрессионный ДНК-вектор.
Предшествующий уровень техники
Время от посева до колошения имеет важное практическое значение для производителей пшеницы, поэтому значительные усилия селекционеров направлены на сокращение вегетационного периода у культурных сортов пшеницы.
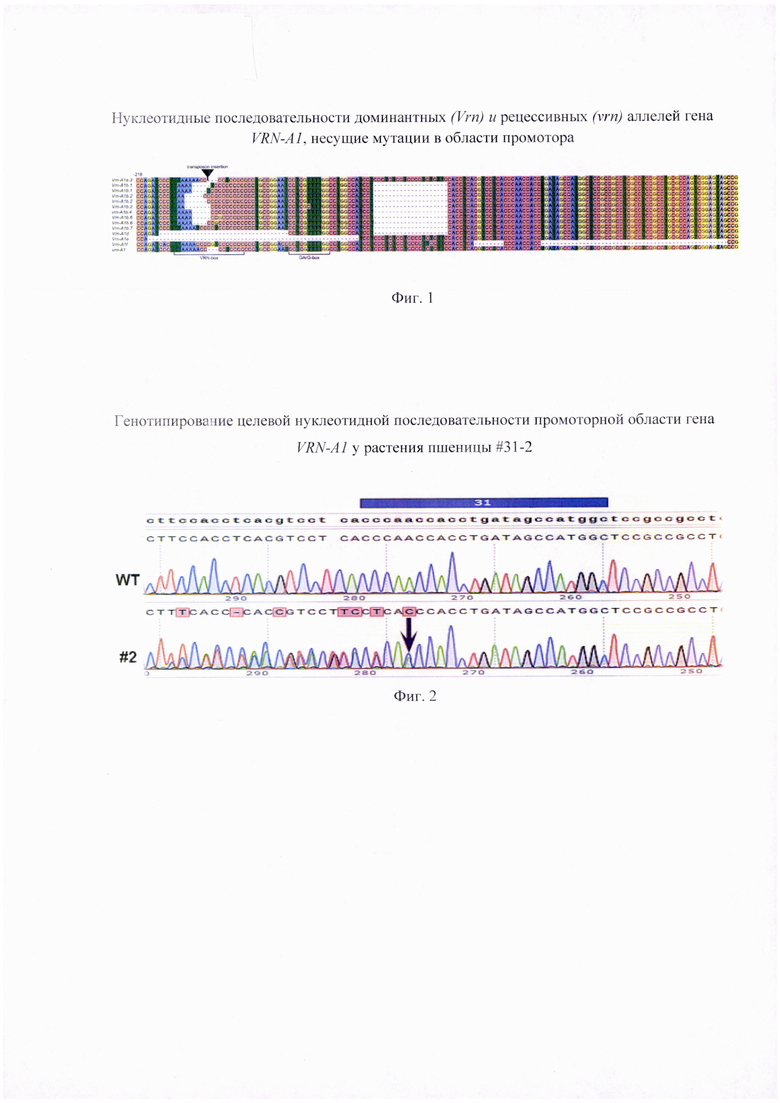
Способность растений пшеницы переходить к репродуктивному развитию зависит от функционирования двух основных систем генов развития - VRN (vernalization - ответ на яровизацию) и PPD (photoperiodic - чувствительность к фотопериоду). У представителей Triticeae чувствительность к яровизации контролируется группой скоординировано функционирующих генов VRN1, VRN2 и VRN3. Важнейшая роль в регуляции ответа на яровизацию принадлежит гену VRN1, кодирующему MADS-box транскрипционный фактор (Trevaskis et al., 2007), который необходим для закладки и поддержания флоральной меристемы в точке роста побега пшеницы. У озимых форм потребность в яровизации связана с тем, что VRN1 гены находятся в рецессивном состоянии и их экспрессия блокирована до наступления периода воздействия пониженных температур. Для перехода озимой пшеницы к репродуктивному развитию необходимо, чтобы уровень транскрипта VRN1 достиг определенного порогового значения (Loukoianov et al., 2005). Отсутствие потребности в яровизации у яровых сортов связано с существенными изменениями нуклеотидной последовательности промотора или первого интрона гена VRN-A1 (Yan et al., 2004; Fu et al., 2005). Показано, что промотор генов VRN1 содержит консервативные цис-элементы (CArG-. VRN- и G-боксы), представляющие собой мишени для транскрипционных факторов, участвующих в ответе на яровизацию. Известен ряд нуклеотидных модификаций промотора VRN-A1 (Konopatskaia et al., 2016; Muterko et al., 2016) (Фиг. 1). Наиболее распространенный доминантный аллель Vrn-A1a отличается от рецессивного аллеля vrn-A1 транспозонными вставками повторяющихся последовательностей в VRN-box участке промотора (Фиг. 1). Другие доминантные аллели содержат делеционные фрагменты разной протяженности в различных участках, таких как VRN-box (аллели Vrn-A1b, Vrn-A1d, Vrn-A1e), CArG-box (аллели Vrn-A1e), G-box (аллели Vrn-A1f), а также делении размером 20 п.н. в участке между -136 и -157 п.н. (аллели Vrn-A1b, Vrn-A1d) или крупную делецию размером 50 п. н. в участке между -62 и -112 п.н. (аллели Vrn-A1f) (Фиг. 1). Помимо делеций, доминантные аллельные варианты Vrn-A1b, Vrn-A1d, Vrn-A1e также могут содержать единичные нуклеотидные замены в VRN-box участке промотора (Фиг. 1). Крупная делеция размером 50 п.н. в участке между -62 и -112 п.н. а также 8 п.н. нуклеотидная делеция в G-box была обнаружена только у аллеля Vrn-A1f яровых диких видов пшеницы секции Timopheevii (Фиг. 1), и не присутствует у представителей современных культурных видов мягкой и твердой пшеницы (Muterko et al., 2016).
Возможность контролировать сроки колошения путем точечного изменения аллелей VRN1 позволит упростить процесс получения новых высокопродуктивных сортов, соответствующих современным требованиям. Примечательно, что в настоящий момент не обнаружены какие либо природные яровые и озимые формы пшеницы с измененной последовательностью промотора VRN1 в области между -113 и -122 п.н., а также отсутствуют искусственные мутантные формы зерновых с модификациями промотора VRN1 в областях между -62 и -120 п.н. Получение растений, несущих новые (или аналогичные диким представителям) аллели генов VRN1, имеющих мутации в регуляторных областях гена на неизменном генетическом фоне, представляет собой техническую проблему, для решения которой необходимо расширение арсенала средств, доступных селекционерам. В качестве решения этой проблемы настоящее изобретение предлагает способ получения однодольного растения с измененной при помощи технологии редактирования генома растений CRISPR/Cas9 последовательностью промотора гена VRN1.
Сущность изобретения
Задачей изобретения является разработка способа получения однодольного растения с нуклеотидной делецией в промоторной области гена VRN-A1.
Для решения данной задачи предлагается способ, согласно которому получают конструкцию pgRNA-VRNA1#31, смешивают эту конструкцию с вектором, кодирующим последовательность Cas9, переносят эту смесь в клетки пшеницы с помощью генной пушки для внесения мутаций в промоторную область гена VRN-1A и проводят отбор растений-регенерантов с отредактированным геномом.
В другом аспекте настоящее изобретение также относится к однодольному растению #31-2, полученному этим способом.
Использование РНК-проводника, в составе экспрессионного вектора pgRNA-VRNA1#31 в описываемом примере, позволило эффективно внести мутацию путем делеции нуклеотидов в целевой последовательности консервативного участка промоторной области гена VRN-A1 мягкой пшеницы, не затрагивая гомеологичные варианты гена VRN-1 других элементарных геномов, и в дальнейшем закрепить приобретенную мутацию в последующем поколении нетрансгенных растений, несущих гомозиготные аллели.
Краткое описание чертежей
Фигура 1. Сравнительная характеристика нуклеотидной последовательности доминантных (Vrn) и рецессивных (vrn) аллелей гена VRN-A1, несущих мутации в области промотора, идентифицированных у видов и сортов полиплоидной пшеницы (согласно Konopatskaia et al., 2016; с модификациями).
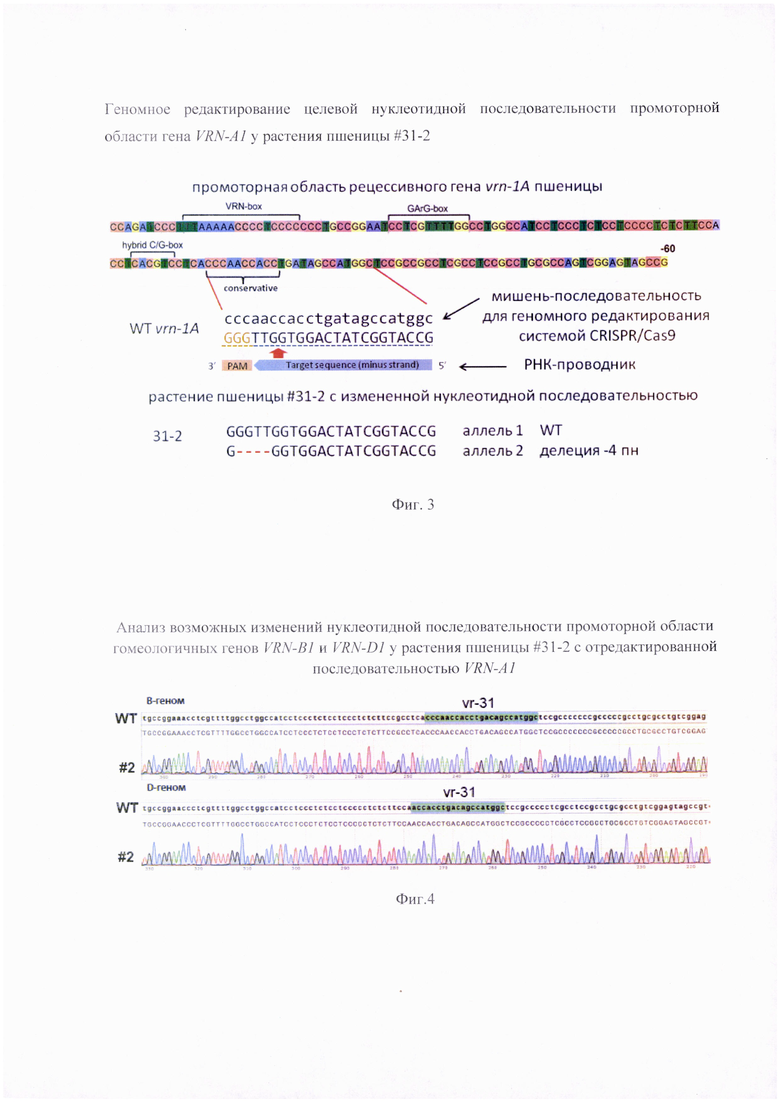
Фигура 2. Генотипирование целевой нуклеотидной последовательности промоторной области VRN-A1 методом секвенирования по Сэнгеру. Представлены нуклеотидные последовательности аллелей в целевом фрагменте контрольного растения (WT) и растения пшеницы #31 -2 с изменениями, которые отмечены стрелкой, а именно делецией четырех нуклеотидов в сравнении с WT.
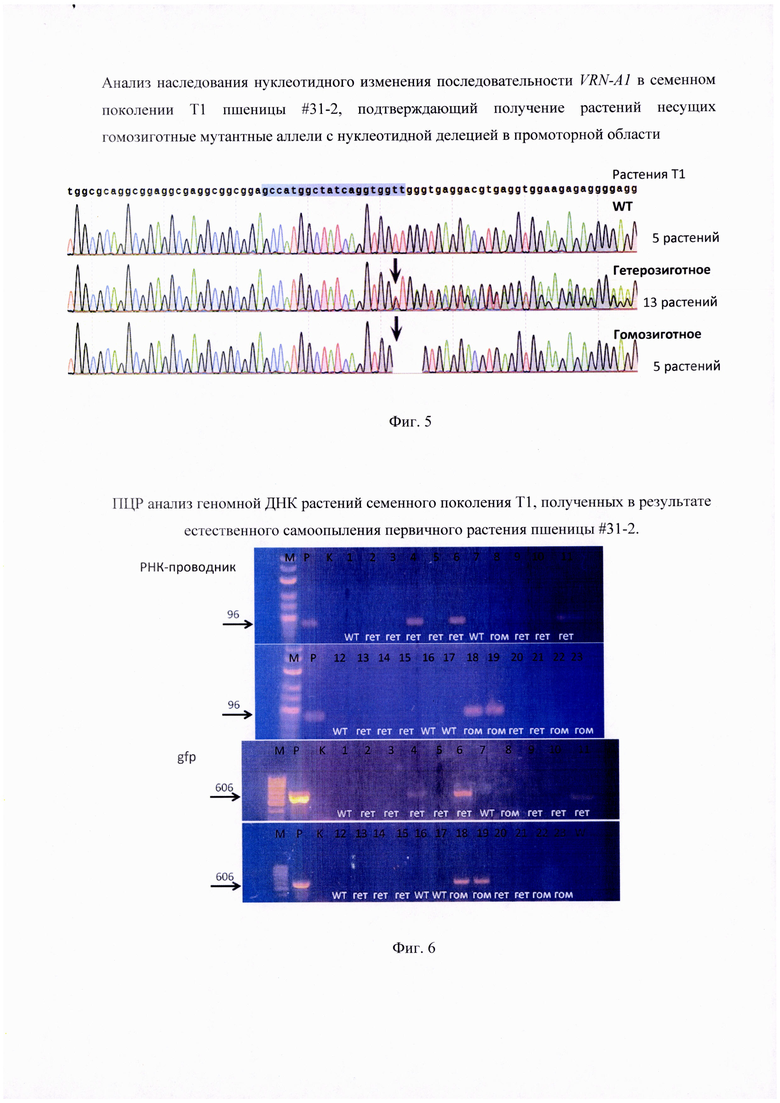
Фигура 3. Генотипирование мутантного аллеля относительно целевой последовательности в промоторной области рецессивного гена VRN-A1, являющейся мишенью для РНК-проводника vr-31. Представлена последовательность целевой мишени, которой комплементарна последовательность РНК-проводника (заглавные буквы) в которой указана РАМ последовательность и стрелкой указан предпочтительный сайт разрезания ДНК. При этом отсутствующие в редактированном растении #31-2 последовательности (делетированные нуклеотиды) изображены пунктирной линией.
Фигура 4. Анализ возможных изменений нуклеотидной последовательности промоторной области гомеологичных генов VRN-1B и VRN-1D у растения пшеницы #31-2 с измененной последовательностью VRN-A1. Представлены результаты секвенирования по Сэнгеру в сравнении с референсными последовательностями (WT) промоторной области генов VRN-1B и VRN-1D, включая фрагмент имеющий сходство с последовательностью РНК-проводника vr-31, использующегося для редактирования генома.
Фигура 5. Характерные примеры генотипирования целевой нуклеотидной последовательности промоторной области VRN-A1 методом секвенирования по Сэнгеру в семенном поколении Т1 растения #31-2. Представлены нуклеотидные последовательности аллелей у растения Т1 не наследующем мутантный аллель (WT), у гетерозиготного растения Т1 с делениями четырех нуклеотидов в одном из аллелей сравнении с WT (моноаллель), и гомозиготного растения несущего две аллели VRN-A1 с делециями четырех нуклеотидов (биалель). Изменения отмечены стрелкой.
Фигура 6. ПЦР анализ геномной ДНК растений семенного поколения Т1, полученных в результате естественного самоопыления первичного растения пшеницы #31-2. Наличие искомых фрагментов последовательности РНК-проводника размером 96 п.н. (находится в составе кассеты экспрессии pgRNA-VRNAl#31) и размером 606 п.н. репортерного гена gfp (находится в составе кассеты экспрессии одновременно несущей последовательности bar, Cas9, и gfp) подтверждена у пяти образцов. На Фигуре 6. представлены агарозные гели, содержащие ПЦР-продукты gfp и РНК-проводника где М - маркер молекулярного веса: Р - ДНК плазмиды pGCB (положительный контроль для gfp) или pgRNA-VRNA1#31 (положительный контроль для РНК-проводника); К - нетрансгенное растение 'Chinese Spring' (отрицательный контроль). W - смесь без добавления ДНК, 1-23 -индивидуальные растения Т1. Результат ПЦР сопоставлен с результатами секвенирования фрагмента нуклеотидной последовательности промоторной области VRN-A1, где WT - растения, не наследующие нуклеотидные делеции, гет-моноаллельные гетерозиготные растения, гом - растения несущие биаллели делеций, привнесенных геномным редактированием.
Пример 1
Получение гетерозиготного трансгенного растения мягкой пшеницы #31-2 содержащего нуклеотидную делению в одном из аллелей промоторной области гена VRN-А1 однодольных зерновых путем геномного редактирования с помощью РНК-проводника.
Для получения индивидуального растений с отредактированным геномом требуется взаимодействие комплексов Cas9-РНК-проводник с геномной ДНК. Векторную конструкцию pgRNA-VRNA1#31, полученную на основе вектора pU6-gRNA (Addgene plasmid #53062; http://n2t.net/addgene:53062) и содержащую первую нуклеотидную последовательность SEQ ID NO: 1 и вторую нуклеотидную последовательность SEQ ID NO: 2. фланкирующую первую последовательность с 3'-конца, смешивали с вектором, кодирующим последовательность нуклеазы Cas9 содержащим ген-селективный маркер bar, и репортерный ген gfp, и переносили в клетки пшеницы с помощью генной пушки для внесения мутаций в промоторную область гена VRN-A1. В качестве эксплантов использовали эмбриогенные каллусы, инициированные из тканей незрелых зиготических зародышей пшеницы 'Chinese Spring' (Triticum aestivum L.). В результате генетической трансформации отобрали растение #31-2, демонстрирующее одновременно устойчивость к селективному гербициду и флуоресценцию репортерного гена GFP (оба гена содержатся в экспрессионной кассете совместно с последовательностью Cas9). Наличие вставок последовательностей «инструментов» геномного редактирования, а именно РНК-проводника, находящегося в составе вектора pgRNA-VRNA1#31, и последовательности нуклеазы Cas9 подтвердили ПЦР анализом тотальной ДНК, выделенной из #31-2 (наблюдали амплификацию характерных фрагментов).
Наличие нуклеотидной вставки в промотерной области гена VRN-A1 подтвердили путем генотипирования фрагмента включающего нуклеотидную последовательность, являющеюся мишенью РНК-проводника. Для этого фрагмент гена VRN-A1 амплифицировали с помощью праймеров CTGAATTCTGAAAGGAAAAATTCTGCTCG/ACTGGTACCGAAGGCGTATTGGGGAAC. ПЦР-продукты очищали набором GeneJET PCR Purification Kit (Thermo) и секвенировали с применением секвенирующего праймера TTACCATGACTCGGTGGAG. Дополнительно к этому, для подтверждения избирательности и точности разработанного РНК-проводника проводили анализ аналогичного участка промотора гомеологичнных VRN-1 генов присутствующих в элементарных геномах В и D пшеницы. Для этого предварительно секвенировали фрагмент промоторной области генов Vrn-B1 и Vrn-D1, полученный путем его амплификации с геномной ДНК пшеницы 'Chinese Spring' с использованием праймеров
CTGAATTCATAGTAGTATAAAAAGGACAATTG/CTGGTACCACCGAATCAACCAAAC AGTG и CTGAATTCGTATAAAAGGAAAATTGTGCTCT/ACTGGTACCATCAACCAAACAGCCC CG, соответственно. Полученные фрагменты очищали набором GeneJET PCR Purification Kit (Thermo) и секвенировали с применением секвенирующего праймера TTACCATGACTCGGTGGAG. Сиквенсы фрагментов Vrn-A1, Vm-B1 и Vrn-D1 трансгенных растений расшифровывали; последовательности раскладывали на аллели. В качестве контроля (референсная последовательность) использовали геномную ДНК пшеницы 'Chinese Spring'.
Генотиприрование нуклеотидной последовательности показало наличие делеции четырех нуклеотидов у растения #31-2 в сравнении с немодифицированной последовательностью рецессивного гена VRN-A1, присутствующего в геноме 'Chinese Spring' (Фиг. 2). Эти изменения возникли в результате функциональной активности РНК-проводника vr-31, поскольку делеция обнаружена в ожидаемом сайте расщепления молекулы ДНК комплексом Cas9-РНК-проводник (Фиг. 3). Анализ аналогичной нуклеотидной последовательности гомеологичных генов Vrn-B1 и Vrn-D1 подтвердил отсутствие каких-либо изменений (Фиг. 4). Таким образом, получено растение пшеницы, несущее аллель с новой укороченной нуклеотидной последовательностью промотерной области гена VRN-A1. Эта новая нуклеотидная последовательность представлена здесь как SEQ ID NO: 3 в перечне последовательностей.
Пример 2
Получение трансгенного растения мягкой пшеницы #31-2 содержащего гомозиготные мутантные аллели с делецией нуклеотидов в промоторной области гена VRN-А1 однодольных зерновых.
В результате самоопыления первичного гетерозиготного растения #31-2, получение которого описано в Примере 1 (содержит одну мутантную аллель вследствие геномного редактирования с помощью РНК-проводника vr-31), получили семенное поколение Т1. Индивидуальные растения Т1 проанализировали на наследование мутантной аллели и закрепление мутации. Наследование нуклеотидной вставки в промоторной области гена VRN-A1 подтверждали путем секвенирования фрагмента нуклеотидной последовательности, являющеюся мишенью РНК-проводника, аналогично тому, как описано в Примере 1. Анализ потомства от самоопыления (двадцать три растения Т1) показал наличие трех типов наследования нуклеотидного изменения промоторной области гена VRN-A1. У пяти из десяти проанализированных растений Т1 сопоставление нуклеотидных последовательностей не выявило полиморфизм в структуре промоторной области гена VRN-A1 относительно референсной последовательности, указывая на отсутствие наследования привнесенной мутации в результате расщепления (Фиг. 5). Тринадцать из двадцати трех проанализированных растений Т1 наследовали мутацию в одной из аллелей, сохраняя гетерозиготный статус мутантного варианта VRN-A1 (Фиг. 5) аналогично первичному трансгенному растению #31-2. Пять растений Т1 содержали одинаковые нуклеотидные изменения в обеих аллелях. Это подтверждено характерным хроматографическим распределением нуклеотидных пиков (Фиг. 5). Таким образом, получили гомозиготное растения пшеницы, содержащие мутантные аллели с нуклеотидной делецией в промоторной области гена VRN-A1 однодольных зерновых, привнесенные геномным редактированием.
Пример 3
Получение нетрансгенного растения мягкой пшеницы #31-2 содержащего гомозиготные мутантные аллели с нуклеотидной вставкой в промоторной области гена VRN-А1 однодольных зерновых.
Успешное применение технологий геномного редактирования в селекции пшеницы ограничивается тем, что в геноме первичного растения, полученного в результате переноса векторов, остаются последовательности кодирующие компоненты системы редактирования CRISPR/Cas9, из-за чего мутантные растения являются ГМО (Miroshnichenko et al., 2019). Пшеница является преимущественно самоопыляемым видом растений, размножающимся исключительно семенами. Благодаря этому, существует возможность «очищения» генома в последующих поколениях от функциональных трансгенных вставок, кодирующих компоненты систем редактирования при условии, что трансгенная вставка (кодирующая гидовую РНК и белок Cas9) и целевой ген-мишень находятся в различных хромосомах первичных мутантных растений. В дальнейшем, в результате самоопыления или скрещивания с нетрансгенным образцом в последующих поколениях (T1-Т3 или F1-F2) отбираются сеянцы, у которых при сохранении отредактированной целевой мишени, чужеродная последовательность не наследуется. Трансгенная вставка в первичных трансгенных растения пшеницы проявляет себя как доминантный признак. Являясь гетерозиготной, при самоопылении она наследуется по классической модели однолокусного наследования генов. В результате, ряд растений последующего поколения не наследует трансгенную вставку.
Поиск растений, не наследующих трансгенную вставку подтверждали ПЦР анализом тотальной ДНК, выделенной из растений Т1, проанализированных на наличие мутации в промоторной области VRN-A1. На Фиг. 6 представлен образец агарозных гелей после проведения ПЦР анализа. Подтвердили, что геном пяти растений Т1, полученных от самоопыления трансформанта #31-2, содержит переносимые последовательности, т.к. наблюдали амплификацию фрагментов размером 606 п. н. характерного для репортерного гена gfp (находится в составе кассеты экспрессии одновременно несущей последовательности bar, Cas9, и gfp) и размером 96 п. н. характерного для РНК-проводника (находится в составе кассеты экспрессии pgRNA-VRNA1#31). Сопоставление результатов ПЦР с результатами секвенирования фрагмента нуклеотидной последовательности промоторной области VRN-A1 подтвердило получение нетрансгенных растений пшеницы несущих привнесенную мутацию (делеция нуклеотидов) в виде гетерозиготного аллеля (10 шт. растений), а также получение нетрансгенных гомозиготных растений (3 шт.) содержащих оба мутантных аллеля с нуклеотидной делецией (Фиг. 6).
Финансирование работ по созданию настоящего изобретения проводилось из средств Соглашения № 075-15-2019-1667 от «31» октября 2019 г. о предоставлении из федерального бюджета грантов в форме субсидий в соответствии с пунктом 4 статьи 78.1 Бюджетного кодекса Российской Федерации на осуществление государственной поддержки создания и развития центра геномных исследований мирового уровня «Курчатовский геномный центр» в рамках реализации федерального проекта «Развитие научной и научно-производственной кооперации» национального проекта «Наука».
Источники информации
Fu D., Szucs Р., Yan L., et al., (2005). Large deletions within the first intron in VRN-1 are associated with spring growth habit in barley and wheat. Mol. Genet. Genomics. 273, 54-65.
Konopatskaia I., Vavilova V., Kondratenko E.Y., et al., (2016). VRN1 genes variability in tetraploid wheat species with a spring growth habit. BMC Plant Biol. 16 (Suppl 3):244.
Loukoianov A., Yan L., Blechl A., et al., (2005). Regulation of VRN-1 vernalization genes in normal and transgenic polyploid wheat. Plant Physiol. 138(4), 2364-2373.
Miroshnichenko D.N., Shulga O.A., Timerbaev V.R., Dolgov S.V. (2019). Achievements, Challenges, and Prospects in the Production of Nontransgenic. Genome-Edited Plants. Appl Biochem Micro+(2019), 55 (9) 825-845.
Muterko A., Kalendar R., Salina E. (2016). Novel alleles of the VERNALIZATION1 genes in wheat are associated with modulation of DNA curvature and flexibility in the promoter region. BMC Plant Biol, 16(Suppl 1), 9.
Trevaskis B., Hemming M.N., Dennis E.S., et al., (2007). The molecular basis of vernalization-induced flowering in cereals. Trends Plant Sci. 12. 352-357.
Yan L., Helguera M., Kato K. et al., (2004). Allelic variation at the VRN-1 promoter region in polyploid wheat. Theor. Appl. Genet. 109:1677-1186.
Перечень последовательностей
--->
<110> ФГБНУ ВНИИСБ
<120> СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ ПШЕНИЦЫ С НУКЛЕОТИДНОЙ ДЕЛЕЦИЕЙ В ПРОМОТОРНОЙ
ОБЛАСТИ ГЕНА VRN-А1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ
CRISPR/CAS9
<160> 3
<210> 1
<211> 20
<212> РНК
<213> Искусственная последовательность
<400> 20
GCCAUGGCUAUCAGGUGGUU
<210> 2
<211> 76
<212> РНК
<213> Искусственная последовательность
<400> 76
GUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGC
<210> 3
<211> 695
<212> ДНК
<213> Последовательность промоторной области гена VRN-А1 с нуклеотидной
делецией
<400> 695
TGAAAGGAAAAATTCTGCTCGTTTTTTTGCTCTGTGGTGTGTGTTTGTGGCGAGAGAAAATGATTTGGGGAAAGCAA
AATCCGGAGATTCGCACGTACGATCGTTCGACACGTCGACGCCCGGCGGGCCCGGGGTGGGGCATCGTGTGGCTGCA
GGACCGCGGGGCCCCGCAAAGCGGGCCGGGCCAATGGGTGCTCGACAGCGGCTATGCTCCAGACCAGCCCGGTATTG
CATACCGCGCTCGGGGCCAGATCCCTTTAAAAACCCCTCCCCCCCTGCCGGAATCCTCGTTTTGGCCTGGCCATCCT
CCCTCTCCTCCCCTCTCTTCCACCTCACGTCCTCACCCACCTGATAGCCATGGCTCCGCCGCCTCGCCTCCGCCTGC
GCCAGTCGGAGTAGCCGTCGCGGTCTGCCGGTGTTGGAGGGTAGGGGCGTAGGGTTGGCCCGGTTCTCGAGCGGAGA
TGGGGCGGGGGAAGGTGCAGCTGAAGCGGATCGAGAACAAGATCAACCGGCAGGTGACCTTCTCCAAGCGCCGCTCG
GGGCTTCTCAAGAAGGCGCACGAGATCTCCGTGCTCTGCGACGCCGAGGTCGGCCTCATCATCTTCTCCACCAAGGG
AAAGCTCTACGAGTTCTCCACCGAGTCATGGTAAATTAAGCACGCGCTGTCTTTAAATTTGTTCCCCAATACGCCTT
CG
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ ПШЕНИЦЫ С НУКЛЕОТИДНОЙ ВСТАВКОЙ В ПРОМОТОРНОЙ ОБЛАСТИ ГЕНА VRN-A1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2020 |
|
RU2772578C2 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ ПШЕНИЦЫ С БИАЛЛЕЛЬНЫМИ МУТАЦИЯМИ В ПРОТОМОТОРНОЙ ОБЛАСТИ ГЕНА VRN-A1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2020 |
|
RU2772575C2 |
| МОЛЕКУЛА РНК-ПРОВОДНИКА ДЛЯ ГЕНОМНОГО РЕДАКТИРОВАНИЯ ПРОТОМОТОРНОЙ ОБЛАСТИ ГЕНА VRN-A1 ОДНОДОЛЬНЫХ ЗЕРНОВЫХ С ПРИМЕНЕНИЕМ СИСТЕМЫ CRISPR/CAS9 | 2020 |
|
RU2762831C1 |
| Молекула РНК-проводника sgRNA для внесения мутаций в консервативный участок промоторной области гена PPD-D1 мягкой пшеницы с применением системы редактирования генома CRISPR/Cas9 | 2024 |
|
RU2822358C1 |
| ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ НА ОСНОВЕ СИСТЕМЫ РЕДАКТИРОВАНИЯ ГЕНОМА CRISPR/CAS9, КОДИРУЮЩАЯ НУКЛЕАЗУ CAS9, ДЛЯ ГЕНОМНОГО РЕДАКТИРОВАНИЯ ОДНОДОЛЬНЫХ ЗЕРНОВЫХ КУЛЬТУР НА ОСНОВЕ ДВОЙНОГО ОТБОРА РАСТЕНИЙ | 2020 |
|
RU2762830C1 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ КАРТОФЕЛЯ С ТЕТРААЛЛЕЛЬНЫМИ МУТАЦИЯМИ В КОДИРУЮЩЕЙ ОБЛАСТИ ГЕНА EDR1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2022 |
|
RU2817383C2 |
| ГЕТЕРОЗИГОТНЫЕ ПО CENH3 ОДНОДОЛЬНЫЕ РАСТЕНИЯ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ ДЛЯ ИНДУКЦИИ ГАПЛОИДОВ И ОДНОВРЕМЕННОГО РЕДАКТИРОВАНИЯ ГЕНОМА | 2021 |
|
RU2832578C1 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ КАРТОФЕЛЯ С ТЕТРААЛЛЕЛЬНЫМИ МУТАЦИЯМИ В КОДИРУЮЩЕЙ ОБЛАСТИ ГЕНА EDR1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2022 |
|
RU2817384C2 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ КАРТОФЕЛЯ С ТРИАЛЛЕЛЬНЫМИ МУТАЦИЯМИ В КОДИРУЮЩЕЙ ОБЛАСТИ ГЕНА EDR1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2022 |
|
RU2817374C2 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ КАРТОФЕЛЯ С БИАЛЛЕЛЬНЫМИ МУТАЦИЯМИ В КОДИРУЮЩЕЙ ОБЛАСТИ ГЕНА EDR1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2022 |
|
RU2817379C2 |
Изобретение относится к области биохимии, в частности к способу получения трансгенного растения пшеницы, с нуклеотидной делецией в промоторной области гена VRN-A1. Также раскрыто трансгенное растение пшеницы, полученное указанным способом. Изобретение позволяет эффективно получать трансгенное растение пшеницы. 2 н.п. ф-лы, 6 ил., 3 пр.
1. Способ получения трансгенного растения пшеницы с нуклеотидной делецией, указанной в SEQ ID NO: 3, в промоторной области гена VRN-A1, включающий в себя следующие этапы:
получение конструкции pgRNA-VRNAl#31, содержащей SEQ ID NO: 1 и SEQ IDN0:2,
смешивание указанной конструкции pgRNA-VRNAl#31 с вектором, кодирующим последовательность CaS9, нацеленным на участок GGGTTGGTGGACTATCGGTACCG гена-мишени VRN-A1,
перенесение этой смеси в клетки пшеницы с помощью генной пушки для внесения мутаций в промоторную область гена VRN-A1 и отбор полученного трансгенного растения – регенеранта с нуклеотидной делецией в промоторной области гена VRN-A1, где промоторная область гена VRN-A1 имеет нуклеотидную последовательность, представленную в SEQ ID N0: 3.
2. Трансгенное растение пшеницы, полученное способом по п.1, с нуклеотидной делецией, представленной в SEQ ID N0: 3, в промоторной области гена VRN-A1, для применения в селекционном процессе.
| KIPPES K | |||
| and et.al., Identification of theVERNALIZATION 4gene revealsthe origin of spring growth habit in ancient wheatsfrom South Asia, PNAS, 2015, pp | |||
| Нефтяная топка для комнатных печей | 1922 |
|
SU401A1 |
| IVANICOVA Z | |||
| et.al., Characterization of new allele influencing flowering time in bread wheat introgressed from Triticum militinae, RESEARCH PAPER New Biotechnology, v | |||
| Способ сопряжения брусьев в срубах | 1921 |
|
SU33A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |

