Изобретение относится к сельскому хозяйству и может быть использовано в селекции зерновых культур, в частности озимой мягкой пшеницы.
Цель изобретения - возможность использования всего генофонда яровых сортов и лучших селекционных линий для получения озимых форм и увеличения количества получаемых оэимых форм.
Способ заключается в том, что, скрещивая между собой яровые сорта, в генотипе которых присутствуют неаллельные доминантные гены- ингибиторы потребности в яровизации (система генов 1-3 Vb ver- nalization яровизации), во втором поколении в зависимости от количества генов, контролирующих
яровой тип развития у скрещиваемых родительских форм, происходит расщепление на яровые и озимые растения в о тношении 15:1 или 63:1. Если у скрещиваемых сортов в генотипе имеются один или два общих доминантных гена-ингибитора отзывчивости на яровизацию, то в FЈ и в последующих поколениях расщепления по типу развития не наблюдается.
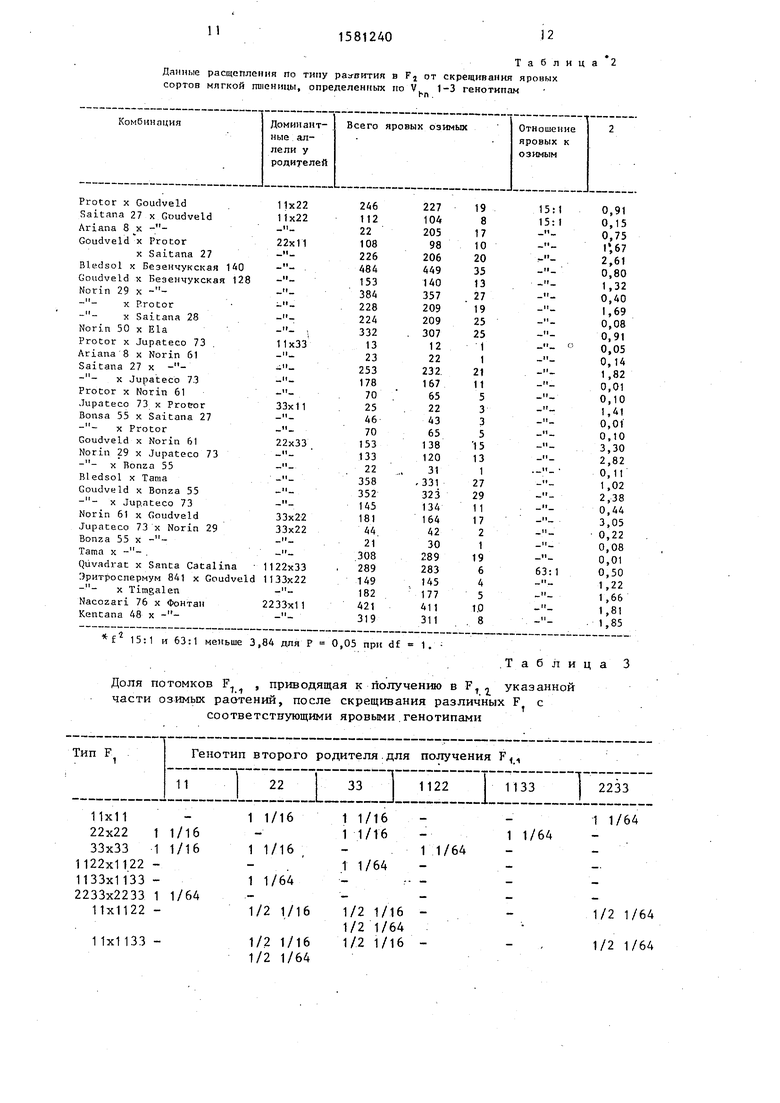
Теоретическое обоснование и практическое подтверждение указанных фактов приведены в табл.1 и 2.
Представленные в табл.1 данные четко указывают на то, что выщепление с частотой 1/16 или 1/64 озимых растений во втором поколении возможно только в случае, когда яровость ).
СП
00
1C
их родительских компонентов контролируется нсаллельными доминантными генами V 1-3. В иных случаях в Fz расщепления по типу развития не про- исходит (0). Все это подтверждено исследованиями по получению и изучению озимых форм согласно предлагаемому способу (табл.2). Абсолютно во всех гибридных комбинациях выявле- но расщепление по типу развития: для комбинаций с участием двух моноген- но доминантных по разным локусам сортов 15:1 и для комбинации, у которых в скрещиваниях использованы ди- генно и монотенно доминантные сорта - 63:1. О неслучайности такого отношения расщепления свидетельствуют величины критерия существенности, который во всех случаях оказался ниже табличного - 3,84 для Р 0,05 при dЈ 1 (табл.2).
Из сказанного вытекае0, что в большей части теоретически возможных гибридных комбинаций между яровыми генотипами в F практически невозможно получить озимые растения. Так, из 49 возможных гибридных комбинаций между двумя яровыми «сортами (табл.1) в 37 случаях не наблюдает- ся в F2 расщепления по типу развития. Таким образом, для получения озимых линий безвозвратно теряется значительная часть гибридов яровых сортов и селекционных форм, возможно обладающих уникальными хозяйственно-биологическими свойствами и особенно такими как засухоустойчивость, жаростойкость, устойчивость к грибным заболеваниям. А при скрещивании трех- генно доминантных сортов с любыми другими яровыми сортами или селекционными линиями вообще невозможно по0 лучить в F озимые выщепенцы.
Способ включает дополнительный
этап скрещивания полученных в данных комбинациях гибридов F г с яровыми генотипами доминантными лишь по локусам, которые у Ft были в рецессивном и/или гетерозиготном со- стоянии.
Условно обозначено получаемое на втором этапе скрещивания гибридное поколение символом F . При соблюдении указанного требования характе- ристики избираемого для второго этапа скрещивания генотипа ярового родителя все или часть (1/2 или 1/4) потомков такого F, 1 поколения оказывается носителями рецессивных аллелей (хотя бы в гетерозиготном состоянии) по всем локусам vrn. В результате этого в самоопыленном F-, i потомстве таких растений с соответствующей вероятностью ожидается выщепле- ние полностью рецессивных по всем локусам vrn генотипов, которые вследствие отсутствия доминантных ингибиторов потребности в яровизации оказываются чувствительными к яровизации и соответственно озимыми по фенотипу.
Поясним представленную общую схему рассмотрением конкретных комбинаций скрещивания. Если рассматривать 6 гибридных комбинаций скрещивания, расположенных в табл.1 но диагонали слева сверху, вниз направо, когда скрещиваются между собой два яровых сорта полностью идентичного мо- ногеиного или дигенно доминантного по локусам Vrnгенотипа, то F1 в данном случае оказывается тем же самым по Vbn генотипу (ив F не ожидается выщепления озимых потомков). Такое F1 на втором этапе должно быть скрещено с яровыми генотипами, доминантными по Vгп ингибиторам, которые у F1 рецессивны, т.е. с неал- лельным по доминантным генам V п генотипом. Оставшиеся гибридные комбинации из табл.1, для которых в F не ожидается выщепления озимых потомков, могут быть распределены на три группы: комбинации скрещиваний моногенно доминантных по Vrn локусам яровых сортов с дигенно доминантными яровыми сортами, у которых 1 доминантный ген аллелен таковому первого родителя; комбинации скрещиваний между собой дигенно доминантных по Vrn локусам яровых сортов, у которых один ген идентичен для обоих родителей; комбинации скрещиваний любого ярового сорта с доминантным по всем трем локусам Vrn генотипом.
К первой группе относятся 6 ре- ципрокных комбинаций типа (приняв обозначение генотипов только номером присутствующего соответствующего доминантного гена V,, и дефис - для соответствующих рецессивных генов) :
1А 11х 11 22 - F, 11 2;
2Б 1 1 х 11 -33 F, 113 К второй группе относятся 3 ре- ципрокных комбинации типа:
2А 11 22 - х 11 - 33 F, 11 2- 3-; 2Б 1 1 22 -- х 22 33 F, 1- 22 3- 2В 11 - 33 х - 22 33 F, 1- 2- 33
К третьей группе относятся 6 ре- ципрокных комбин-аций типа:
ЗА 11х
ЗБ - 22 -- х 3В33 х
ЗГ 11 ЗД 11
22 - х - 33 х
ЗЕ - 22 33 х 11
10
812406
Во всех указанных комбинациях генотип получаемого F., характеризуется присутствием в гомозиготе доминантных генов по 1 или даже по 2 локусам Vrft. В связи с этим во втором поколении не ожидается выщепления полностью рецессивных озимых потомков. Чтобы получить на основе данных F генотипов озимые растения, такие генотипы должны быть скрещены на втором этапе с яровыми генотипами, обязательно рецессивными по vrn локусам,которые у Гл находились в гомозиготном доминантном состоянии. Соответственно яровость таких компонентов скрещивания может определяться доминантными генами по локусам, которые у F были рецессивными и/или гетерозиготными. 20Например, для комбинации 1Г F1
может быть скрещено со следующими яровыми генотипами:
15






| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения озимых форм мягкой пшеницы из яровых | 1990 |
|
SU1734601A2 |
| Способ получения озимых форм мягкой пшеницы из яровых | 1985 |
|
SU1340676A1 |
| Способ отбора константных безостых форм мягкой пшеницы | 2021 |
|
RU2768393C1 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ ПШЕНИЦЫ С НУКЛЕОТИДНОЙ ДЕЛЕЦИЕЙ В ПРОМОТОРНОЙ ОБЛАСТИ ГЕНА VRN-A1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2020 |
|
RU2772577C2 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ ПШЕНИЦЫ С БИАЛЛЕЛЬНЫМИ МУТАЦИЯМИ В ПРОТОМОТОРНОЙ ОБЛАСТИ ГЕНА VRN-A1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2020 |
|
RU2772575C2 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ ПШЕНИЦЫ С НУКЛЕОТИДНОЙ ВСТАВКОЙ В ПРОМОТОРНОЙ ОБЛАСТИ ГЕНА VRN-A1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2020 |
|
RU2772578C2 |
| СПОСОБ СОЗДАНИЯ ЛИНИЙ ОЗИМОЙ МЯГКОЙ ПШЕНИЦЫ С КОМПЛЕКСНОЙ УСТОЙЧИВОСТЬЮ К БУРОЙ И СТЕБЛЕВОЙ РЖАВЧИНЕ И МУЧНИСТОЙ РОСЕ | 2015 |
|
RU2598275C1 |
| Способ создания гомозиготных гомостильных линий у гречихи | 1990 |
|
SU1738167A1 |
| КРАСНЫЙ САЛАТ ЛАТУК | 2006 |
|
RU2460282C2 |
| Способ получения линий яровой мягкой пшеницы с укороченным сроком колошения | 2019 |
|
RU2710729C1 |
Изобретение относится к сельскому хозяйству и может быть использовано в селекции зерновых культур, в частности озимой мягкой пшеницы. Цель изобретения - возможность использования всего генофонда яровых сортов и лучших селекционных линий для получения озимых форм и увеличения количества получаемых озимых форм. Скрещивают формы яровой мягкой пшеницы, в генотипах которых присутствуют один или два аллельных гена - ингибитора отзывчивости на яровизацию. Полученные растения F 1 скрещивают с яровыми формами, в генотипах которых присутствуют один или два доминантных гена-ингибитора, которые у растений F 1 находятся в гетерозиготном или рецессивном состоянии. Полученное потомство самоопыляют. Выделение форм с озимым типом развития осуществляют в следующем после самоопыления поколении. 6 табл.
а/ б/ в/
х 11 - х - - х П -33
-33
Только такие комбинации скрещивания обеспечивают получение в F 1,, всех или части потомков, у которых присутствуют рецессивные аллели по всем трем локусам vrn (хотя бы в гетерозиготном состоянии). В результате этого после самоопыления в F,
30
35
40
таких растений и ожидается выщепле- ние определенной части полностью рецессивных по всем трем локусам vrn озимых потомков. И такие озимые растения являются результатом последова- тельного скрещивания (яровой х яро- вой) хяровой сортов мягкой пшеницы, подобранных на основании знания их Vpn генотипов, и они выделяются в третьей генерации . Так, в комби- ,с нации 1 Г.а/ половина тригетерозигот- ных потомков F , 1 после самоопыления выщепляет 1/64 часть полностью рецессивных озимых растений, а вторая половина дигетерозиготных потомков F ,j ,, - 1/16 часть озимых растений. Соответственно в комбинациях 1Г.6/ и 1Г в/ половина гомозиготно доминант1.1
после озиV j,n потомков F е выщепляет в F а вторая половина в б/ выщепляет 1/16 и в в/ - 1/64 часть озимых
/2 1 /2 - /2
30
35
40
,с 50
55
-2- 3- + 1/2 1- 2;
-2- 33 + 1/2 - 2- 3- и
-2- 33 + 1/2 1- 2- 3Аналогично могут быть рассмотрены схемы скрещиваний всех типов указанных F., комбинаций. В обобщенном виде эти схемы приведены в табл.3. В первой колонке данной таблицы указывается тип скрещивания Vpn генотипов для получения F,, в результате самоопыления которого в F.J по табл.1 не выцепляются озимые потомки. В ГОЛОВКР табл.3 приведены V. генотипы третьего ярового компонента скрещивания (второй этап) для получения F 1 , , после самоопыления которого в F f ожидается выщепление части озимых растений. Первая цифра в соответствующих клетках табл.3 показывает долю или часть (1 - все, 1/2 - половина, 1/4 - четверть) растений, F1 , в потомстве F., Ј которых ожидается выщепление указанной второй цифрой (1/16 или 1/64 соответственно) части рецессивных по всем трем локусам Vrn оз.имых потомков.
Таким образом, здесь указаны все возможные типы скрещиваний соответствующий V генотипов яровых сортов для получения на их основе озимых форм. И указанные комбинации практически заполняют все клетки табл.1, по которым в F не ожидалось выщепления озимых растений. Остается незаполненной единственная угловая справа внизу клетка, в которой исходно скрещиваются между собой два гомозиготно доминантных по всем трем локусам генотипа. Однако вероятность такого скрещивания очень незначительна вследствие исключительно редкой встречаемости подобного генотипа среди существующих в мире яровых сор- тов. Если все же возникает потребность проведения подобного скрещивания для последующего получения озимых потомков, то данньй генотип полученного F1 может быть использован как родительская форма в указанных комбинациях ЗА-ЗЕ, и дальнейшая работа проводится по уже указанной схеме.
Итак, предлагаемый способ получе- ния озимых растений мягкой пшеницы при скрещивании специально подобранных на основании знания их Vh генотипов яровых сортов расширяет возможности и показывает, что практически на основании любой комбинации их скрещивания можно получить озимые растения путем привлечения для второго этапа скрещивания определенных Vh генотипов. Основным требованием для второго этапа скрещивания является то, что используемый для скрещивания на втором этапе яровой сорт должен быть рецессивным по vrn генам, которые гомозиготно доминантны у
полученного F . Яровость этого сор- , та может определяться одной или двумя парами доминантных V Пп генов, по которым F рецессивно и/или гетерозиготно.
Экспериментальная проверка предлагаемого способа проведена по комбинациям скрещивания между собой яро-1 вых сортов всех трех указанных типов: 1А-1Е, 2А-2В и ЗА-ЗЕ. Для этого
по каталогу были отобраны 7 яровых сортов, принадлежащие ко всем 7 возможным по локурам VPr генотипам (табл.4) .
Зимой 1985/86 годов в оранжерее фитотрона получены семена F (первый этап скрещиваний) по гибридным комбинациям, родительские формы -у которых содержат одну или две пары общих аллельных доминантных генов Vpn (фиг.5).
В полевых условиях 1986 года полуденные растения F1 были вновь скрещены (второй этап) с яровыми сортами, рецессивными по vrn генам, которые у полученного F были в гомозиготном доминантном состоянии. Соответственно, яровость данных сортов опрделялась доминантными V генами по которым F1 было рецессивно или гетерозиготно (табл.6).Полученные семена F 1 t были высеяны в оранжерее фитотрона зимой 1986/87 годов (без яровизации) для самоопыления растений. Созревшие семена F. убирались раздельно с каждого растения F1 1 и после прохождения периода покоя высевались по семьям ручной сеялкой (площадь питания 20x5 см 2 ) веной 1987 года на полевом участке в сроки, исключающие воздействия низких яротзизационных температур. После обычных агротехнических мероприятии по уходу за растениями через 2,5-3 месяца после всходов проводили анализ расщепления внутри семей по типу развития на яровые и озимые растения.
К этому времени озимые потомки ,находились в фазе кущения, а яровые практически все выколосились или находились в стадии позднего труб- кования (единичные растения). Озимые растения осторожно выкапывают, пересаживают в сосуды с почвой, помещают в яровизационную камеру, а после прохождения яровизации выращивают в оранжерее для получения семян озимых растений. 1
Данные по семьям, в которых не было выявлено расщепления поу типу развития, исключают из дальнейшего анализа. Результаты учета количества яровых и озимых растений в оставшихся семьях суммируют для каждой гибридной комбинации и анализируют на соответствие выявленных отношении расщепления теоретически ожи 1даемым отношениям 15:1 или 63:1 по критерию (хг. В большинстве комбинаций выявлено достоверное соответствие расщепления по суммарным данным теоретических ожидаемым отношениям. Однако в части комбинаций,по которым ожидалось выщепление двух типов семей с разными ожидаемыми отношениями, расщепления такого соответствия по суммарным данным не наблюдалось. Распределение таких семей на две группы с разными ожидаемыми отношениями расщепления на яровые и озимые растения привело к выявлению статистического соответствия фактически наблюдаемых результатов теоретически ожидаемым.
Таким образом, получено обоснованное фактами экспериментальное подтверждение предлагаемого способа, который расширяет возможности получения озимых форм мягкой пшеницы из яровых на основе последовательного скрещивания специально подобранных Vrn 1-3 генотипов яровых сор- тов. Указанные родительские сорта могут быть отобраны путем использования перечня генотипов сортов из каталога или на основании гибридологического анализа генотипа неизвест- ного сорта по локусам системы генов V 1-3. Получение озимых форм из гибридных популяций при скрещивании только яровых сортов мягкой пшеницы позволяет: практически полностью ис- пользовать весь генофонд яровых сортов и селекционных форм при выделении озимых генотипов, получив в ходе селекционной работы принципиально новый исходный материал для озимых пшениц, а т акже использовать более сложные схемы гибридизации, что дает возможность объединения в создаВероятность выщепления озимых растений в Гг от скрещивания двух яровых родительских форм, обладающих разными по Vh 1-3
генотипами
В целях удобстна запись генотипов яровых сортов проводят только номерами присутствующих соответствующих доминантных аллелей генов-ингибиторов потребности в яровизации.
ваемом озимом генотипе наиболее цен ные генотипы.
Формула изобретения
Способ получения озимых форм мягкой пшеницы из яровых, включающий скрещивание между собой яровых форм, в генотипах которых присутствуют доминантные гены: ингибиторы отзывчивости на яровизацию, и выделение в FI форм с озимым типом развития, отличающийся тем, что, с целью обеспечения возможности использования всего генофонда яровых сортов и лучших селекционных линий для получения озимых форм и увеличения количества получаемых озимых форм, предварительно скрещивают между собой формы, в генотипах которых присутствуют один или два аллельных гена-ингибитора отзывчивости на яровизацию, полученные растения скрещивают с яровыми формами, в генотипах которых присутствуют один или два доминантных гена-ингибитора,которые у растений 1 находятся в гетерозиготном или рецессивном состоянии, полученное потомство самоопыляют, а выделение форм с озимым типом развития осуществляют в следующем после самоопыления поколении.
Таблица 1
11
1581240
Данные расщепления по типу раз-вития в Ft от скрещивания яровых сортов мягкой пшеницы, опрелеленных по V 1-3 генотипам
12 Табл
f4 15:1 и 63:1 меньше 3,84 для Р 0,05 при df « 1.
Доля потомков F71 , приводящая к получению в F, г указанной части озимых растений, после скрещивания различных Ff с соответствующими яровыми генотипами
12 Таблица 2
Таблица 3
22x1122 22x2233
33x1133 33x2233 1 122x1133 1122x2233 1133x2233 11x112233 22x112233 33x112233
122x112233 133x112233 233x112233
1/21/16
1/21/16
1/21/64
1/21/64
1/21/16
1/21/64
1/41/16
1/41/64
1/41/16
1/41/64
1/41/16
1/41/64
1/41/16
1/41/64
1/2 1/64
1/21/16
1/21/64
1/21/16
1/41/16
1/41/64
1/41/16
1/41/64
1/41/16
1/41/64
1/4 1/16 1/4 1/64
1/2 1/64
Таблица 4
1
Генотип 1
Vj.h1Vrn1 vrn2vrn2 vrn3vrn3
vrn1vrn1Vrn2Vrn2 vrn3vrn3
vrn1vrn1vrn2vrn2Vrti3Vrt,3 VrJV IV V vrnSvrnS
841 Vrn1Vrn1vrn2vrn2Vr 3vro3
Dirk 123
x Quvadrat
x Эритроспермум 841
x Kentana 48
x Фонтан
Продолжение табл.З
1/16 1/16- 1/16
1/2 1/64 1/2 1/64
1/2 1/64 1/2 1/64
1/4 1/64
1/4 1/64
1/4 1/64
1/4 1/64
1/4 1/64
1/4 1/64
1/2 1/64 Продолжение табл.4
1
Kenta- na 48 Линия Triple
Dirk 123
vrn1vrn1Vhn2Vpn2V,,n3Vrn3
V 1V 1V 2V 2V 3V 3 vrn vrn vrr.hti m nn
Таблица 5
112233 x 1122 x 1133 x 2233 x 11
15
1581240-16
Продолжение табл.5
Для удобства- запись в табл.5 генотипа-сортов проводили только цифрами имеющихся тех или иных
доминантных аллелей Vh генов Vh
rn fn ,.а
| Способ получения озимых форм мягкой пшеницы из яровых | 1985 |
|
SU1340676A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
