Изобретение относится к сельскому хозяйству и может быть использовано в генетике и селекции мягкой пшеницы на повышенное содержание антоцианов в зерне.
Антоцианы - окрашенные природные соединения с ярко выраженными антиоксидантными свойствами, благодаря которым они не только защищают растения в условиях стресса, но также положительно влияют на здоровье человека (1). У мягкой пшеницы Triticum aestivum L. (2n-6х=42) антоцианы могут накапливаться в алейроновом слое зерна, придавая ему голубую окраску, которая обусловлена преимущественно дельфинидин-3-глюкозидами, обладающими среди антоциановых соединений наибольшей антиоксидантной активностью (2). В настоящее время пшеница с окрашенным зерном рассматривается как один из потенциальных источников антоцианов для производства функциональных продуктов питания и нужд медицины (1).
Насыщение современных сортов мягкой пшеницы антоцианами возможно за счет переноса генов, контролирующих биосинтез антоцианов, от донорных линий/сортов в реципиентные сорта, с помощью гибридизации. Биосинтез антоцианов в алейроновом слое зерновки мягкой пшеницы контролируется генами Ва (Blue aleurone). У дикорастущего дальнего родственника пшеницы - пырея удлиненного Thinopyrum ponticum (Podp.) Barkworth & DR Dewey (2n=10x=70, StStStStEeEeEbEbEbExEx; синонимы Agropyron elongatum (Host) Beauv., Elytrigia pontica (Podp.) Holub., E. elongata (Host) Nevski) был идентифицирован ген Ba1, который был картирован на длинном плече хромосомы 4Е (3). С помощью сравнительного транскриптомного анализа голубозерного и белозерного образцов пшеницы в качестве гена-кандидата для Ba1 был идентифицирован ген ThMyc4E. Показано, что этот ген кодирует транскрипционный фактор с доменом bHLH и регулирует синтез антоцианов в алейроновом слое зерновки (4). Ген ThMyc4E был выявлен в геноме пшенично-пырейных дополненных, замещенных и почти-изогенных линий с голубой окраской зерновки, но не был обнаружен в геноме образцов T.urartu, Т.monococcum, T.turgidum, Aegilops tauschii или T.aestivum, зерно которых не окрашено антоцианами. Известность нуклеотидной последовательности гена ThMyc4E позволяет разработать внутригенные праймеры для отбора голубозерных образцов пшеницы. Передача гена пырея ThMyc4E в геном пшеницы происходит в составе целой хромосомы 4Е, которая может заместить одну или две гомологичные хромосомы 4В или 4D пшеницы (5). При этом замена пары гомологичных хромосом пшеницы парой гомологичных хромосом пырея происходит случайным образом. Однако, как показали исследования, хромосомы 4В и 4D неидентичны по генному составу (6-12) и, следовательно, их замены могут по-разному влиять на признаки гибридов. В связи с этим, при получении пшенично-пырейных замещенных линий с голубой окраской зерна, необходимо контролировать, какая именно пара гомологичных хромосом пшеницы была замещена парой хромосом пырея.
Наиболее близким к заявляемому способу - прототипом, является способ отбора гибридов мягкой пшеницы с голубой окраской зерновки, основанный на фенотипической оценке наличия голубого пигмента в зрелом зерне пшеницы после его обмолота, с последующим цитологическим анализом митотических хромосом отобранных гибридов с помощью дифференциального С-окрашивания, флуоресцентной in situ гибридизации (FISH) с меченными зондами, специфичными к геному пырея и пшеницы, и подтверждением замещения хромосом с помощью микросателлитного анализа (13).
Недостатками такого отбора являются длительность, связанная с необходимостью получения зрелого зерна для его фенотипической оценки и последующего цитологического и микросателлитного анализа отобранных голубозерных гибридов, а также трудоемкий цитологический анализ, для проведения которого необходимо специализированное оборудование и реактивы - микроскоп высокого разрешения и меченные специфичные зонды, а также отсутствие возможности отбирать гибриды, полученные на основе фиолетовозерных образцов, накапливающих антоцианы в перикарпе зерна под контролем генов Рр, которые маскируют наличие голубых пигментов, накапливаемых в подлежащем алейроновом слое зерна.
В отличие от цитологических методов, методы, основанные на ПЦР-анализе ДНК с помощью маркеров (ПЦР-маркеров), являются рутинными и доступными во всех молекулярно-биологических лабораториях. ПЦР-маркеры могут использоваться для определения наличия аллельных вариантов генов уже на ранних стадиях развития растений, что существенно ускоряет и удешевляет селекционный процесс, экономит материальные и трудовые ресурсы.
Задачей предлагаемого изобретения является разработка более точного и менее трудоемкого способа отбора растений мягкой пшеницы, несущих доминантные аллели гена ThMyc4E в гомозиготном состоянии, с диагностикой замещения пары гомологичных хромосом 4В или 4D пшеницы парой гомологичных хромосом 4Е пырея на ранних стадиях развития растений, полученных на основе сортов/линий мягкой пшеницы с белой или красной, или фиолетовой окраской зерна.
Техническим результатом изобретения является расширение технических возможностей известного способа, повышение точности и сокращение времени отбора голубозерных образцов пшеницы.
Поставленная задача решается предлагаемым способом, заключающимся в следующем.
ДНК гибридов, полученных в результате скрещивания белозерных или краснозерных, или фиолетовозерных сортов/линий мягкой пшеницы с голубозерными сортами/линиями, несущими ген пырея ThMyc4E, анализируют с помощь набора ПЦР-маркеров, состоящего из внутригенного маркера ThMyc4E, специфически амплифицирующего нуклеотидную последовательность гена ThMyc4E пырея, но ни его ортологов из генома пшеницы, а также из микросателлитных маркеров Xgwm0251/Xgwm0375 и Xgwm0624, картированных, соответственно, на хромосомах 4В и 4D мягкой пшеницы и специфически амплифицирующих фрагменты ДНК этих хромосом. В таблице 1 представлены последовательности используемых ПЦР-маркеров для отбора голубозерных гибридов с доминантными аллелями гена ThMyc4E и для контроля замещаемых хромосом пшеницы хромосомами 4Е пырея.
Использование на первом этапе отбора разработанного маркера ThMyc4E позволяет идентифицировать наличие в геноме гибрида доминантного аллеля гена ThMyc4E, который может находиться как в гомозиготном, так и в гетерозиготном состоянии. На втором этапе отбора для контроля замещения хромосом пшеницы хромосомами пырея используют микросателлитные маркеры Xgwm0251 или Xgwm0375 и Xgwm0624, локализованные на хромосомах 4В и 4D, соответственно. Наличие у гибридов одновременно продуктов амплификации с маркерами, локализованными и на хромосоме 4В, и на хромосоме 4D, свидетельствует о неполном замещении одной их хромосом пшеницы хромосомой 4Е пырея. В этом случае делается вывод о том, что ген ThMyc4E и хромосома, на которой он локализуется, находятся у гибрида в гетерозиготном состоянии. Отсутствие у гибридов продуктов амплификации с одним из маркеров свидетельствует о полном замещении хромосомы, на которой локализуется неамплифицирующий ДНК маркер, двумя парами гомологичных хромосом 4Е пырея.
Принципиальным отличием заявляемого способа от прототипа является то, что гибриды, несущие доминантный аллель гена ThMyc4E в гомозиготном состоянии и полное замещение хромосом 4В или 4D мягкой пшеницы хромосомами 4Е пырея, отбирают не после созревания и обмолота зерна на основе его фенотипической оценки и трудоемкого цитологического анализа, а на ранних стадиях развития растений с использованием рутинного метода молекулярной биологии - ПЦР-анализа с помощью маркеров ТаМус4Е, Xgwm0251/Xgwm0375 и Xgwm0624.
Изобретение иллюстрируется следующими примерами.
Пример 1. Отбор голубозерных линий яровой мягкой пшеницы, полученных на основе краснозерного сорта «Саратовская 29».
После проведения серии скрещиваний растений краснозерного сорта «Саратовская 29» с сортом-донором гена ThMyc4E «Meropa», голубозерные растения поколения F1 самоопыляли, а полученные гибриды подвергали восьмикратному возвратному скрещиванию с исходным сортом «Саратовская 29», отбирая в каждом поколении зерно с голубой окраской. С помощью анализа ДНК голубозерных гибридов поколения BC8F2 был проведен отбор растений с полным замещением хромосом пшеницы хромосомами 4Е пырея. Для этого из молодых листьев проростков, высаженных в ванну гидропонной теплицы с автоматическим поливом ИЦиГ СО РАН (Новосибирск, Россия), выделяли ДНК. Для этого 2-3 кусочка листа (примерно 0,5×2 см) помещали в пробирку Eppendorf объемом 1,5 мл. Добавляли 200 мкл свежего буфера для экстракции (100 mM Tris-HCl, рН 7,5-8,0, 500 mM NaCl, 50 mM EDTA, 1,25% SDS, 0,38% Na2S2O5), нагретого до 60°С. Листья тщательно измельчали на гомогенизаторе. Далее добавляли 500 мкл буфера для экстракции, перемешивали и инкубировали на водяной бане при 60°С в течение 30-40 минут. После чего добавляли 700 мкл смеси хлороформ-изоамилового спирта (24:1) и после перемешивания центрифугировали 25 минут при 10000-12000 об/мин при комнатной температуре. К отобранной верхней фракции (~500 мкл) добавляли 1,4 мл 96% холодного этанола, слегка перемешивали и центрифугировали 10000-12000 об/мин. Осадок промывали 70% этанолом не менее трех раз, просушивали и растворяли в 20-25 мкл бидистиллированной стерильной воды. Оценку качества и количества экстрагированной ДНК производили путем проведения аналитического электрофореза, которую далее стандартизовали путем разведения в стерильной бидистиллированной воде до 100 мкг/мкл.
Выделенную ДНК гибридов BC8F2, а также неокрашенного сорта «Саратовской 29» и голубозерного сорта «Meropa», использовали в качестве матрицы в ПЦР с набором маркеров ТаМус4Е, Xgwm0375 и Xgwm0624. ПЦР проводили в реакционной смеси объемом 20 мкл, содержащей 5 мкл ДНК, 1,8 мМ MgCl2, 6 мкл Н2О, по 0,2 мМ каждого дНТФ, по 1 мкМ прямого и обратного специфичных праймеров, 1 ед. ДНК-полимеразы Taq, в амплификаторе в режиме Tdwn в случае маркера ТаМус4Е (2 мин при 94°; 13 циклов: 15 сек при 94°С, 30 сек при 65°С, опускаясь на 0,7°С каждый цикл до 56°С, 45 сек при 72°С; 24 цикла: 15 сек при 94°С, 30 сек при 56°С, 30 сек при 72°С; 5 мин при 72°С), либо 55GRAD - в случае микросателлитных маркеров Xgwm0375 и Xgwm0624 (2 мин при 94°С; 40 циклов: 15 сек при 94°С, 30 сек при 55°С, 45 сек при 72°С, 5 мин при 72°С). Продукты реакции разделяли в камере для горизонтального электрофореза в течение 1,5 часа в 2% агарозном геле с добавлением бромистого этидия в 1 χ ТАЕ буфере при напряженности электрического поля 80 V, визуализировали и фотографировали с помощью гель-документирующей системы Gel Doc (Bio-Rad Laboratories).
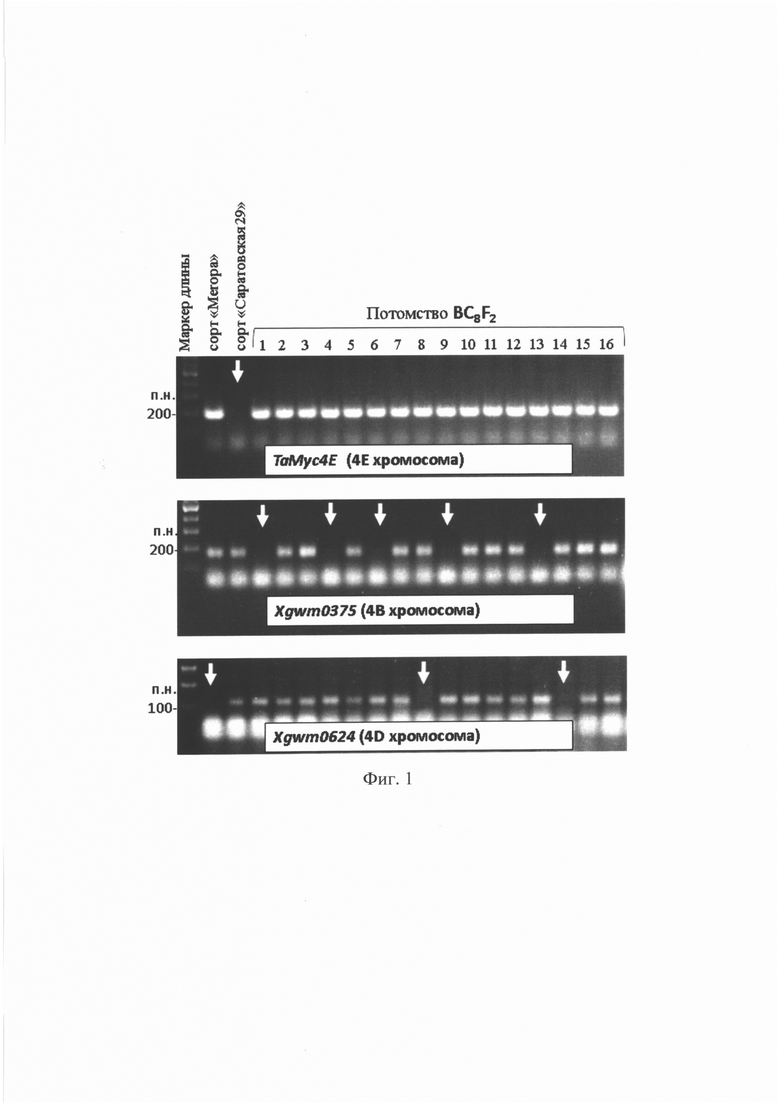
На фиг. 1 представлены электрофореграммы продуктов амплификации геномной ДНК голубозерного сорта «Meropa», неокрашенного сорта «Саратовская 29» и гибридов поколения BC8F2, полученных с помощью ПЦР-маркеров ТаМус4Е, Xgwm0375, Xgwm0624. Для контроля длины ПЦР-продуктов использовали маркер 100 п.н. Видно, что у краснозерного сорта «Саратовская 29» отсутствует ПЦР-продукт внутригенного маркера ТаМус4Е, локализованного на хромосоме 4Е пырея, в то время как у голубозерных сорта «Meropa» и шестнадцати гибридов BC8F2, такой ПЦР-фрагмент присутствует, что свидетельствует о наличие доминантного аллеля гена ТаМус4Е в гомозиготном или гетерозиготном состоянии, а также замещения хромосом 4В или 4D хромосомой 4Е пырея у «Meropa» и гибридов. Для контроля замещение хромосом 4В или 4D мягкой пшеницы хромосомами 4Е пырея, был проведен анализ амплификации геномной ДНК изучаемых образцов с помощью микросателлитных маркеров. Так, после применения маркера Xgwm0375, локализованного на хромосоме 4В, были выявлены растения №1, 4, 6, 9 и 13, отмеченные на электрофореграмме белыми стрелками, у которых отсутствовали продукты амплификации, что вместе с присутствием ПЦР-продукта маркера ТаМус4Е свидетельствует о полном замещении пары хромосом 4В парой хромосом 4Е у этих растений. В случае применения маркера Xgwm0624, были выявлены растения №8, 14, а также сорта «Meropa» у которых отсутствовали продуты амплификации этого меркера, что вместе с присутствием ПЦР-продуктов маркера ТаМус4Е, свидетельствует полном замещении пары хромосом 4D парой хромосом 4Е у этих растений. У образцов №2, 3, 5, 7, 10, 11, 12, 15, 16 присутствуют фрагменты всех маркеров ТаМус4Е, Xgwm0375, Xgwm0624, что указывает на то, что в их геноме хромосома 4Е и, следовательно, ген ТаМус4Е находятся в гетерозиготном состоянии. Эти образцы были исключены из дальнейшей работы.
Отобранных с помощью набора ПЦР-маркеров растения №1, 4, 6, 9 и 13, а также №8 и 14, представляют собой гибриды мягкой пшеницы с полным замещением пары хромосом 4В и 4D гомологичными хромосомами 4Е пырея. Отобранные растения были высажены для размножения в ванну гидропонной теплицы с автоматическим поливом в ИЦиГ СО РАН, в следующем поколении они дали однородное гомозиготное голубозерное потомство.
Пример 2. Отбор голубозерных линий яровой мягкой пшеницы, полученных на основе филетовозерной линии сорта «Саратовская 29».
После скрещивания растений пшенично-пырейной голубозерной линии с замещением пары гомологичных хромосом 4D парой хромосом 4Е пырея, унаследованных от сорта «Meropa», с фиолетовозерной почти-изогенной линией яровой мягкой пшеницы, несущей доминантные аллели генов Pp-D1 и Рр3, контролирующих фиолетовую окраску зерновок, полученные гибриды поколения F1 самоопыляли, а из растений второго поколения F2 были отобраны гомозиготные растения по генам Pp-D1 и Рр3, используя подход, описанный в патенте (16). ДНК отобранных образцов далее анализировали с помощью набора ПЦР-маркеров ТаМус4Е, Xgwm0375 и Xgwm0624. ПЦР и электрофорез проводили также, как описано в примере 1.
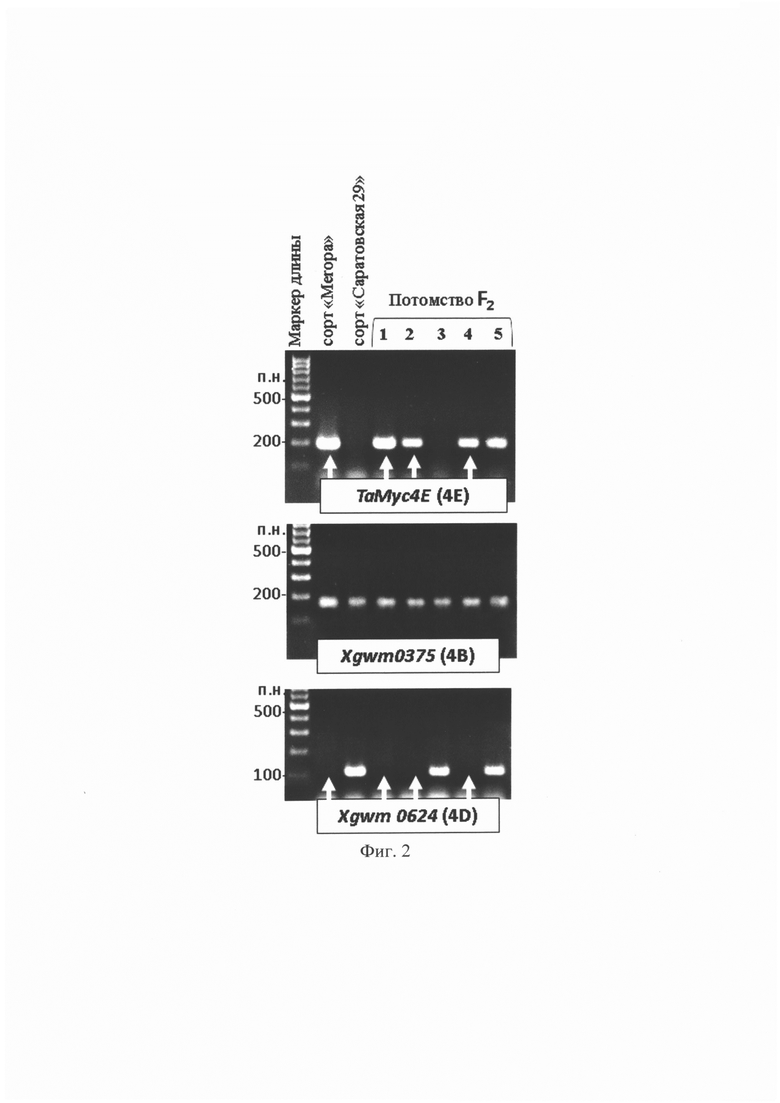
Результаты электрофоретического анализа полученных продуктов ПЦР представлены на фиг. 2. Из пяти изучаемых растений у четырех из них, а именно, у образцов №1, 2, 4, 5, а также у голубозерного сорта «Meropa», были обнаружены ПЦР-фрагменты маркера ТаМус4Е, тогда как у «Саратовской 29» и у гибрида №3 таких ПЦР-фрагментов обнаружено не было. При применении микросателлитного маркера Xgwm0375, локализованного на хромосоме 4В, показано, что все изучаемые образцы имели продукты амплификации этого маркера, что указывает на то, что в геноме этих образцов сохраняется хромосома 4В. Отсутствие ПЦР-фрагмента маркера Xgwm0624, локализованного на хромосоме 4D, у образцов №1, 2 и 4, отмеченных на электрофореграмме белыми стрелками, свидетельствует о полном замещении пары хромосом 4D парой хромосом 4Е. Следует отметить, что у образца №5 присутствуют ПЦР-фрагменты, соответствующие маркерам ТаМус4Е, Xgwm0375, Xgwm0624, что указывает на то, что хромосома 4Е и, следовательно, ген ТаМус4Е у этого образца находятся в гетерозиготном состоянии.
Отобранные с помощью набора ПЦР-маркеров растения №1, 2 и 4 представляют собой гибриды мягкой пшеницы с полным замещением пары гомологичных хромосом 4D парой гомологичных хромосомами 4Е и генотипом, объединяющем в одном геноме одновременно доминантные аллели генов Рр, регулирующих биосинтез антоцианов в перикарпе зерновки, и гена ThMyc4E, регулирующего синтез антоцианов в алейроновом слое. Зерна отобранных растений накапливали фиолетовые антоциановые пигменты не только в перикарпе, как исходная линия, но и голубые антоциановые соединения в алейроновом слое зерна, благодаря чему зерно таких гибридов имело насыщенный иссиня-черный фенотип. Отобранные растения были высажены для размножения в ванну гидропонной теплицы с автоматическим поливом в ИЦиГ СО РАН. В третьем поколении F3 анализируемые растения дали однородное гомозиготное потомство.
Таким образом, предлагаемый способ отбора голубозерных гибридов мягкой пшеницы с использование набора ПЦР-маркеров, состоящего из внутригенного ПЦР-маркера ТаМус4Е и микросателлитных маркеров, локализованных на хромосомах мягкой пшеницы 4В (Xgwm0251, Xgwm0375) и 4D (Xgwm0624), позволяет отбирать голубозерные растения и точно идентифицировать замещение пары гомологичных хромосом 4В или 4D парой гомологичных хромосом 4Е на ранней стадии развития растения, что позволяет уже в начале вегетационного периода выбраковать нежелательный для селекционера материал, не проводя уборку и обмолот зерна. Тем самым экономится время и средства, затрачиваемые на уход за посевами, уборку и послеуборочную обработку селекционного материала.
Источники информации
1. Юдина Р.С., Гордеева Е.И., Шоева О.Ю., Тихонова М.А., Хлесткина Е.К. Антоцианы как компоненты функционального питания. Вавиловский журнал генетики и селекции. 2021;25(2):178-189.
2. Ficco D.B.M., De Simone V., Colecchia S.A., Pecorella I., Platani C., Nigrom F., Finocchiaro F., Papa R., De Vita P. Genetic variability in anthocyanin composition and nutritional properties of blue, purple, and red bread (Triticum aestivum L.) and durum (Triticum turgidum L. ssp.turgidum convar. durum) wheats. J. Agric. Food Chem. 2014;62:8686-8695.
3. Zheng Q., Li В., Mu S., Zhou H., Li Z. Physical mapping of the blue-grained gene(s) from Thinopyrum ponticum by GISH and FISH in a set of translocation lines with different seed colors in wheat. Genome. 2006;49:1109-1114.
4. Li N., Li S., Zhang K., Chen W., Zhang В., Wang D., Liu D., Liu В., Zhang H. ThMYC4E, candidate Blue aleurone 1 gene controlling the associated trait in Triticum aestivum. PLoS ONE. 2017;12, e0181116.
5. Arbuzova V.S., Badaeva E.D., Efremova T.T., Osadchaya T.S., Trubacheeva N.V., Dobrovolskaya O.B. A cytogenetic study of the blue-grain line of the common wheat cultivar Saratovskaya 29. Rus. J. Genet: Applied Res. 2012;4:785-791.
6. Guan P., Lu L., Jia L., Kabir M. R., Zhang J., Lan Т., Peng H. Global QTL analysis identifies genomic regions on chromosomes 4A and 4B harboring stable loci for yield-related traits across different environments in wheat (Triticum aestivum L.). Front. Plant Sci. 2018;9:529.
7. Zhang J., Yao Q., Li R., Lu Y., Zhou S., Han H., Li L. Identification of Genetic loci on chromosome 4B for improving the grain number per spike in pre-breeding lines of wheat. Agronomy. 2022; 12(1): 171.
8. Yuan В., Yang Y., Fan P., Liu J., Xing H., Liu Y., Feng D. Genome-wide identification and characterization of germin and germin-like proteins (GLPs) and their response under powdery mildew stress in wheat (Triticum aestivum L.). Plant Mol. Biol. Rep. 2021;39:821-832.
9. Rahman M.M., Davies P., Bansal U., Pasam R., Hayden M., Trethowan R. Relationship between resistance and tolerance of crown rot in bread wheat. Field Crops Res. 2021;265:108106.
10. Shen X., Yuan Y., Zhang H., Guo Y., Zhao Y., Li S., Kong F. The hot QTL locations for potassium, calcium, and magnesium nutrition and agronomic traits at seedling and maturity stages of wheat under different potassium treatments. Genes. 2019;10(8):607.
11. Gupta P.K., Mir R.R., Mohan A., Kumar J. Wheat genomics: present status and future prospects. Int. J. Plant Genomics. 2008; 2008: 896451.
12. Helguera M., Rivarola M., Clavijo В., Martis M.M., Vanzetti L.S., Gonzalez S., Garbus I., Leroy P., Simkova H., Valarik M., Caccamo M., Dolezel J., Mayer K.F.X., Feuillet C, Tranquilli G., Paniego N., Echenique V. New insights into the wheat chromosome 4D structure and virtual gene order, revealed by survey pyrosequencing. Plant Sci. 2015;233:200-212.
13. Gordeeva E., Badaeva E., Yudina R., Shchukina L., Shoeva O., Khlestkina E. Marker-assisted development of a blue-grained substitution line carrying the Thinopyrum ponticum chromosome 4E(4D) in the spring bread wheat Saratovskaya 29 background. Agronomy. 2019;9(11):723.
14. Korzun V, Borner A, Worland AJ, Law CN, Roder MS. Application of microsatellite markers to distinguish inter-varietal chromosome substitution lines of wheat (Triticum aestivum L.). Euphytica. 1997;95(2):149-55.
15. Roder M.S., Korzun V., Wendehake K., Plaschke J., Tixier M.H., Leroy P., Ganal M.W. A microsatellite map of wheat. Genetics. 1998;149:2007-2023.
16. Хлесткина E.K., Гордеева Е.И., Шоева О.Ю., Кукоева Т.В., Шаманин В.П., Моргунов А.И. Способ отбора линий яровой мягкой пшеницы с повышенным содержанием антоцианов в зерне. Патент RU 2762804 С1, опубл. 23.12.2021.
Способ отбора гибридов мягкой пшеницы с голубой окраской зерновки

| название | год | авторы | номер документа |
|---|---|---|---|
| Внутригенный ДНК-маркер для отбора пшеницы с повышенным содержанием антоцианов в перикарпе зерновки | 2021 |
|
RU2774444C1 |
| Способ отбора линий яровой мягкой пшеницы с повышенным содержанием антоцианов в зерне | 2021 |
|
RU2762804C1 |
| СПОСОБ СОЗДАНИЯ ЛИНИЙ МЯГКОЙ ПШЕНИЦЫ, УСТОЙЧИВЫХ К БУРОЙ ЛИСТОВОЙ РЖАВЧИНЕ | 2009 |
|
RU2407283C1 |
| Набор ДНК-маркеров для отбора ячменя с повышенным содержанием антоцианов в перикарпе зерновки | 2022 |
|
RU2797678C1 |
| СПОСОБ СОЗДАНИЯ ЛИНИЙ МЯГКОЙ ПШЕНИЦЫ, УСТОЙЧИВЫХ К БУРОЙ РЖАВЧИНЕ | 2012 |
|
RU2484621C1 |
| Способ отбора константных безостых форм мягкой пшеницы | 2021 |
|
RU2768393C1 |
| Способ получения линий яровой мягкой пшеницы с укороченным сроком колошения | 2019 |
|
RU2710729C1 |
| ПОЛУЧЕНИЕ ГИБРИДНЫХ СЕМЯН ЗЕРНОВЫХ КУЛЬТУР | 2015 |
|
RU2685907C2 |
| СПОСОБ СОЗДАНИЯ ЛИНИЙ ЯРОВОЙ МЯГКОЙ ПШЕНИЦЫ С УДЛИНЕННЫМ СРОКОМ КОЛОШЕНИЯ И С КОМПЛЕКСНОЙ УСТОЙЧИВОСТЬЮ К ГРИБНЫМ БОЛЕЗНЯМ | 2013 |
|
RU2535985C1 |
| СПОСОБ СОЗДАНИЯ ЛИНИЙ ОЗИМОЙ МЯГКОЙ ПШЕНИЦЫ С КОМПЛЕКСНОЙ УСТОЙЧИВОСТЬЮ К БУРОЙ И СТЕБЛЕВОЙ РЖАВЧИНЕ И МУЧНИСТОЙ РОСЕ | 2015 |
|
RU2598275C1 |
Изобретение относится к области биотехнологии. Белозерные или краснозерные, или фиолетовозерные сорта мягкой пшеницы скрещивают с голубозерными сортами/линиями, несущими ген пырея удлиненного ThMyc4E, контролирующий голубую окраску алейронового слоя зерна, затем из зеленых листьев полученных гибридов второго поколения F2 выделяют ДНК и анализируют ее с помощью набора ПЦР-маркеров, состоящего из внутригенного ПЦР-маркера ТаМус4Е и микросателлитных маркеров Xgwm0251/Xgwm0375 и Xgwm0624, картированных на хромосомах 4В и 4D мягкой пшеницы, соответственно. Применение набора ПЦР-маркеров позволяет идентифицировать присутствие гена ThMyc4E, а также хромосом 4В и 4D в геноме полученных гибридов. По наличию ПЦР-фрагментов каждого из маркеров делают вывод о гомозиготном или гетерозиготном состоянии доминантного аллеля гена ThMyc4E у гибридов, а также о полном замещении хромосом 4В или 4D. 2 ил., 1 табл., 2 пр.
Способ отбора гибридов мягкой пшеницы с голубой окраской зерновки, включающий скрещивания исходных сортов мягкой пшеницы с голубозерными сортами или линиями, несущими ген пырея удлиненного ThMyc4E, контролирующий голубую окраску алейронового слоя зерна, с последующим анализом зерна полученных гибридов второго поколения F2 на наличие голубой окраски, отличающийся тем, что белозерные, или краснозерные, или фиолетовозерные сорта мягкой пшеницы скрещивают с голубозерными сортами или линиями, несущими ген пырея удлиненного ThMyc4E, из полученных проростков гибридов второго поколения F2 выделяют ДНК и анализируют ее сначала с помощью внутригенного маркера ТаМус4Е, представляющего собой пару праймеров, со структурой: 5'-GAAACAACAGGACCGAGCAG-3' (прямой), 5'-CTTGATGGCGTCAAACACTT-3' (обратный), на наличие доминантных аллелей гена ТаМус4Е, а затем с помощью микросателлитного маркера Xgwm0251, представленного парой праймеров: 5'-CAACTGGTTGCTACACAAGCA-3' (прямой), 5'-GGGATGTCTGTTCCATCTTAG-3' (обратный); или маркера Xgwm0375, представленного парой праймеров: 5'-ATTGGCGACTCTAGCATATACG-3' (прямой), 5'-GGGATGTCTGTTCCATCTTAGC-3' (обратный); и маркера Xgwm0624, представленного парой праймеров: 5'-ТТОАТАТТАААТСТСТСТАТСТС-3' (прямой), 5'-AATTTTATTTGAGCTATGCG-3' (обратный), точно идентифицирующих замещение хромосом 4В или 4D пшеницы парой хромосом 4Е пырея.
| GORDEEVA E., et al, Marker-assisted development of a blue-grained substitution line carrying the Thinopyrum ponticum chromosome 4E(4D) in the spring bread wheat Saratovskaya 29 background | |||
| Agronomy | |||
| Станок для придания концам круглых радиаторных трубок шестигранного сечения | 1924 |
|
SU2019A1 |
| LI N., et al, ThMYC4E, candidate Blue aleurone 1 gene controlling the associated trait in Triticum aestivum | |||
| PLoS ONE | |||
| Автомобиль-сани, движущиеся на полозьях посредством устанавливающихся по высоте колес с шинами | 1924 |
|
SU2017A1 |

