Изобретение относится к биотехнологии, генетической и белковой инженерии, и касается нового рекомбинантного штамма бактерий Escherichia coli (E.coli), который может быть использован для получения рекомбинантного белка метилцитозин-специфической ДНК-гликозилазы ROS1, состоящей из гликозилазного домена С и C-концевого RRM-домена и способной направленно удалять 5-метилцитозин из ДНК.
Важную роль в регуляции экспрессии генов и, как следствие, в их фенотипическом проявлении играет эпигенетическое состояние генома. Эпигеном представляет собой сложную систему из взаимосвязанных эпигенетических меток ДНК и белков, которые организуют хроматин в пространстве и времени. Метилирование цитозина по положению C5 считается одной из наиболее изученных эпигенетических меток ДНК.
Правильный уровень метилирования различных участков ДНК в клетке крайне важен, поскольку от него зависит транскрипционная активность генов. Ошибки в процессе метилирования ДНК могут приводить к тяжелым последствиям, в частности, у человека глобальное деметилирование ДНК или гиперметилирование генов-онкосупрессоров служат маркерами онкозаболеваний. Поддержание статуса метилирования ДНК в клетках млекопитающих, так и растений основано на балансе процессов метилирования и активного и пассивного деметилирования. Активное деметилирование mC у всех организмов основано на системе эксцизионной репарации оснований ДНК. У млекопитающих оно инициируется регулируемым повреждением mC, которое может происходить двумя способами: либо с дезаминированием mC до тимина дезаминазами семейства AID/APOBEC, либо с его окислением до hmC и дальнейших производных (5-формилцитозина и 5-карбоксилцитозина) диоксигеназами семейства TET.
В отличие от млекопитающих, у растений обнаружены два уникальных фермента с mC специфичной ДНК-гликозилазной активностью. Эти ферменты, DEMETER (DME) и REPRESSOR OF SILENCING 1 (ROS1), принимают участие в регуляции статуса метилирования отдельных участков ДНК растений, от которого зависит импринтинг генов при наследовании по отцовской или материнской линии и сайленсинг или активация промоторов отдельных генов в жизненном цикле растений [1].
Белок ROS1, относящийся к семейству DME, был впервые идентифицирован в качестве репрессора метилзависимого подавления трансгенов [2], насчитывает 1393 а.к.о., и его молекулярная масса составляет 156,5 кДа. Ген ROS1 содержит 20 экзонов и 19 интронов. В структуре белка ROS1 выделяется несколько доменов [3]. N-концевой домен содержит лизин-богатую область, которая вовлечена в неспецифическое связывание белка с ДНК и в скольжение вдоль неё в поисках субстратных оснований [4]. За этим доменом следует центральный ДНК-гликозилазный домен, содержащий мотивы HhH и GPD,а также FeS-домен с четырьмя консервативными остатками Cys, ответственными за удерживание железо-серного кластера. Помимо этого, центральный гликозилазный домен имеет нетипичную вставку в 230 аминокислотных остатков (а. к. о.), последовательность и длина которой варьирует среди числа белков семейства DME. Такой последовательности нет в ранее охарактеризованных ДНК-гликозилазах, имеющих домен HhH-GPD. Также в составе белков семейства DME есть уникальная C-концевая область, содержащая мотив, узнающий РНК, и мотив пермутированного цинкового пальца. Было показано, что наличие С-концевого домена необходимо для проявления активности фермента [3].
Таким образом, создание новых рекомбинантных белков для редактирования эпигенома, открывает новые возможности для перепрограммирования клеток, компенсации генетических дефектов, лечения патологий.
В частности, метилцитозин-специфическая-ДНК-гликозилаза ROS1 представляет собой идеальный инструмент для целей эпигенетического деметилирования ДНК, так как он способен направленно удалять 5-метилцитозин из ДНК за один шаг в отличие от аналогов у млекопитающих, которые задействованы только в одной из двух стадий активного деметилирования.
На сегодняшний день аналогов штаммов-продуцентов метил-специфических ДНК-гликозилаз ROS1 и DEMETER, а также их аналогов деметилирования ДНК у млекопитающих AID/APOBEC и TET не обнаружено.
Наиболее ближайшим к заявляемому штамму – прототипом, является бактериальный штамм-продуцент Kocuria rosea 307, выделенный из почвы и обеспечивающий получение сайт-специфической эндонуклеазы KroI, узнающей и расщепляющей обе цепи нуклеотидной последовательности ДНК 5'-GCCGGC-3'/3'-CGGCCG-5', в которых два центральных цитозина метилированы в положении С5 с образованием четырехнуклеотидных 5'-выступающих концов (патент RU 2394099 C1, опуб. 10.07.2010).
Недостатком известного штамма является его неспособность продуцировать рекомбинантную метил-специфическую ДНК-гликозилазу ROS1.
Задачей изобретения является создание рекомбинантного штамма - продуцента метил-специфической ДНК-гликозилазы ROS1
Технический результат изобретения заключается в получении рекомбинантного штамма-продуцента, обеспечивающего получение рекомбинантной метилцитозин-специфической ДНК-гликозилазы ROS1 с чистотой не менее 90%.
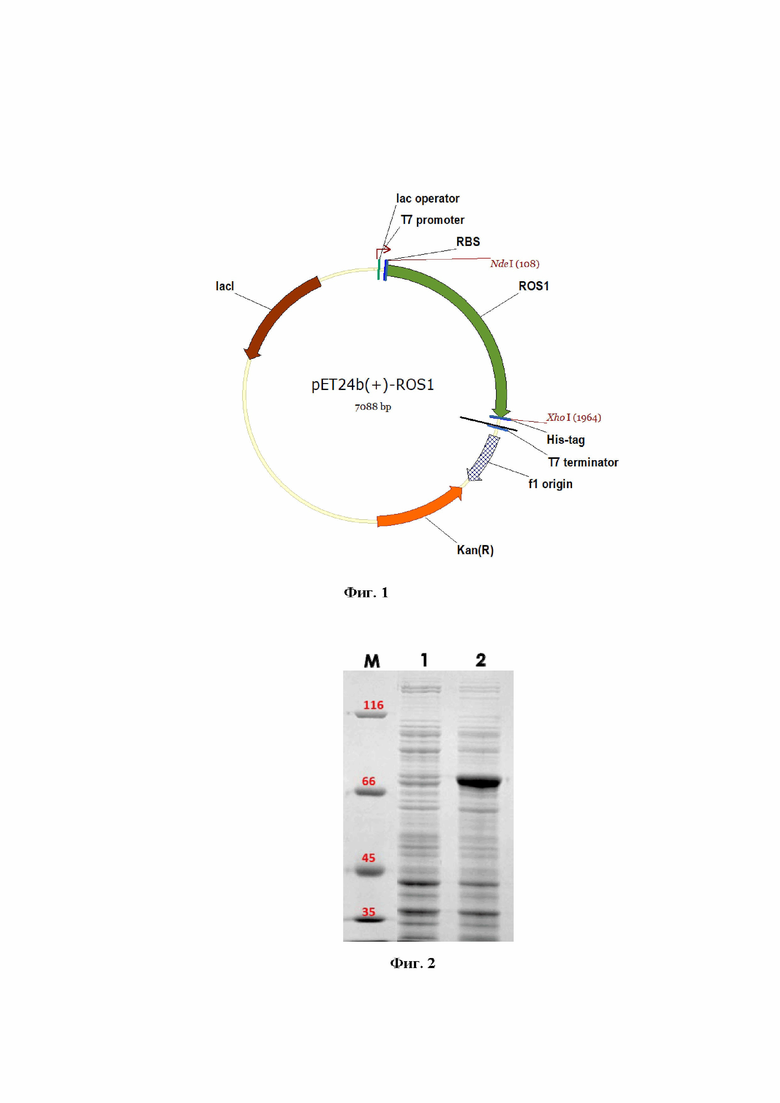
Поставленная задача достигается созданием рекомбинантного штамма E. coli Rosetta 2(DE3)/pET24b(+)-ROS1-продуцента метилцитозин-специфической ДНК-гликозилазы ROS1, полученного трансформацией культуры клеток E. coli Rosetta 2 (DE3) (генотип F– omp Thsd SB(rB–mB–) gal dcm (DE3) pRARE2 (CamR)) рекомбинантной плазмидной ДНК pET24b(+)-ROS1, сконструированной на основе вектора pET24b(+), несущего фрагмент гена, кодирующего ДНК-гликозилазу ROS1, из табака Nicotiana tabacum. На фиг. 1 представлена физическая карта плазмиды pET24b(+)-ROS1.
Культурально-морфологические особенности штамма E. coli Rosetta 2(DE3)/pET24b(+)-ROS1: грамотрицательные прямые палочки, размером 1,1-1,5×2,0-3,0 мкм, одиночные, спор и капсул не образуют. Каталазоположительные. Оксидазоотрицательные. Факультативные анаэробы. Клетки хорошо растут на простых питательных средах, содержащих и не содержащих хлорамфеникол и канамицин, например, на среде LB (питательная среда Lisogeny Broth). На агаризованной среде - колонии гладкие, круглые, слабовыпуклые, с ровным краем. В жидких средах образуют равномерную светорассеивающую суспензию, при хранении без перемешивания оседают на дно. Клетки растут в интервале температур от 8°C до 43°C, интервал для наиболее эффективного культивирования - 28-38°C, оптимум роста при 37°C. Оптимальный интервал pH для культивирования pH 5-7. Изначальный штамм E. coli Rosetta 2 (DE3) проявляет устойчивость к хлорамфениколу (34 мкг/мл), обусловленную наличием гена устойчивости в ДНК рекомбинантной плазмиды pRARE2.
Характеристики полезного вещества, синтезируемого штаммом: рекомбинантный белок ROS1 длиной 626 аминокислотных остатков, состоящий из гликозилазного домена С и C-концевого домена RRM (618 аминокислот), небольшого линкера (2 аминокислоты) и гистидиновой метки (6 аминоксилот).
Продуктивность созданного штамма - рекомбинантный белок ROS1, составляет не менее 20 % белка клеточного лизата при культивировании в жидкой питательной среде LB при 37оС, 250 об/мин, в условиях индукции 1 мМ ИПТГ (изопропил-β-D-1-тиогалактопиранозид) в течение 3 часов с добавлением антибиотиков хлорамфеникола (34 мкг/мл) и канамицина (100 мкг/мл).
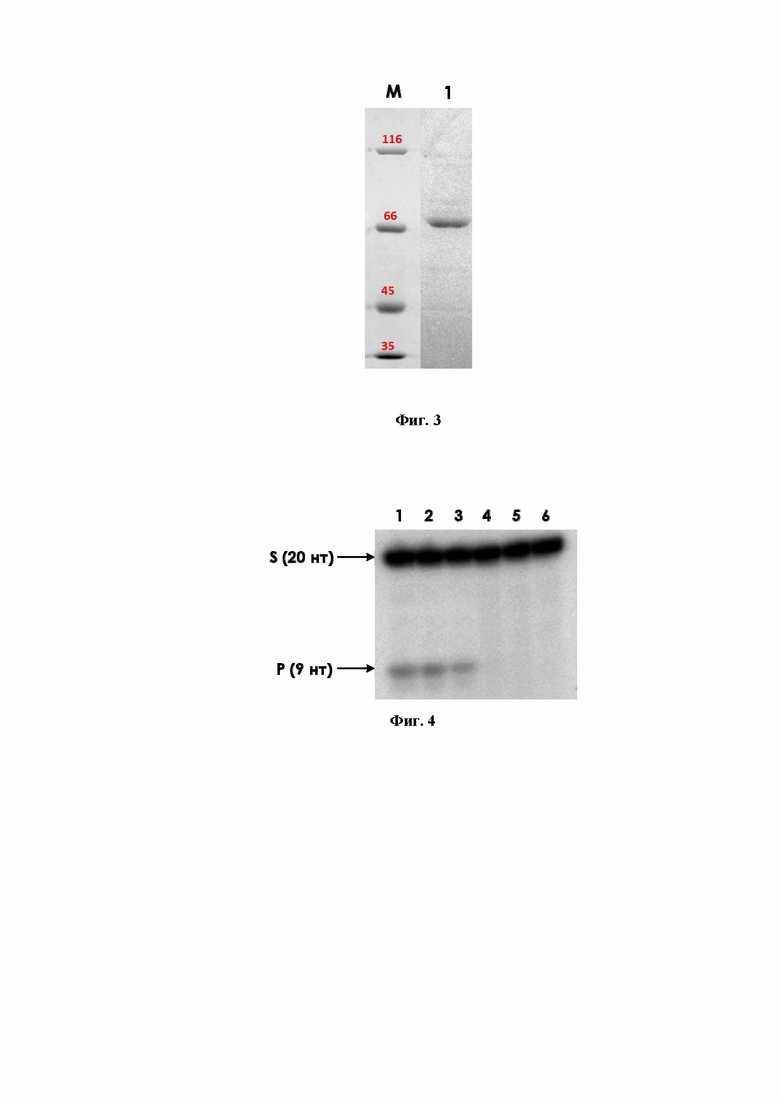
На фиг. 2 приведена электрофореграмма после разделения клеточных лизатов клеток E. coli Rosetta 2(DE3)/pET24b(+)-ROS1 при культивировании в условиях индукции синтеза белка при помощи ИПТГ и без индукции; где М – белковый маркер подвижности 14,4-116 кДа (ThermoScientific); 1 – контрольная неиндуцированная культура, 2 – 3 ч после индукции 1 мМ ИПТГ.
Сущность изобретения поясняется следующими конкретными примерами получения и использования штамма Е. coli Rosetta 2(DE3)/pET24b(+)-ROS1.
Пример 1. Создание генетической конструкции, обеспечивающей синтез рекомбинантной метилцитозин-специфической ДНК-гликозилазы ROS1 в клетках Е. coli.
In silico была проведена оптимизация кодонов для E. Coli, синтез полученной оптимизированной последовательности фрагмента гена ROS1 был заказан в компании Gene Universal (США). Клонирование в экспрессионный вектор осуществлялось по сайтам рестрикции Nde I и Xho I: фрагмент ДНК и вектор подвергались обработке обозначенными эндонуклеазами рестрикции, рестрикционные смеси очищались. Затем проводилось лигирование фрагментов при +4°С в течении 16 ч. После чего полученной лигазной смесью трансформировались клетки штамма Escherichia coli DH5α. Позитивные по данным метода ПЦР колонии были пересеяны в ночные культуры, из ночных культур выделялась плазмидная ДНК для подтверждения последовательности секвенированием по Сэнгеру. Последовательность нуклеотидов синтезированного фрагмента гена соотносится с необходимой аминокислотной последовательностью и совпадает с последовательностью ROS1 Nicotiana tabacum, имеющейся в GenBank (BAF52855.1).
На фиг. 1 представлена физическая карта плазмиды pET24b(+)-ROS1, содержащая ориджин репликации f1, промотор фага T7, RBS-сайт посадки рибосомы, сайты рестрикции Nde I и Xho I, по которым встроен фрагмент гена, кодирующего ДНК-гликозилазу ROS1 из табака Nicotiana tabacum, гистидиновую метку His-tag, терминатор фага T7, ген устойчивости к канамицину Kan (R), а также лактозный репрессор lacI и лактозный оператор.
Полученная плазмида pET24b(+)-ROS1 обеспечивает синтез в клетках Escherichia coli метилцитозин-специфической ДНК-гликозилазы ROS1, содержащей гликозилазный домен С, C-концевой домен RRM, а также C-концевую гистидиновую метку, предназначенную для последующей очистки рекомбинантного белка ROS1 c помощью металлохелатной хроматографии.
Нуклеотидная последовательность рекомбинантной плазмиды pЕТ24b(+)-ROS1 представлена в перечне последовательностей.
Пример 2. Получение штамма-продуцента рекомбинантной ДНК-гликозилазы ROS1 и исследование его продуктивности.
Полученной плазмидой pЕТ24b(+)-ROS1 были трансформированы клетки E. coli штамма Rosetta 2(DE3), содержащие в своем геноме ген, кодирующий ДНК-зависимую РНК-полимеразу фага Т7, под контролем промотора lacUV5, индуцируемого лактозой или ИПТГ. Кроме того, клетки E. coli Rosetta 2(DE3) дефектны по генам протеаз Lon и ompT, что снижает деградацию гетерологичных белков.
В результате был получен штамм E. coli Rosetta 2(DE3)/pET24b(+)-ROS1-продуцент бактериального белка ROS1.
Для поддержания полученного штамма-продуцента белка ROS1 использовали плотную агаризованную LB-среду, содержащую 100 мкг/мл канамицина и 34 мкг/мл хлорамфеникола.
Продуктивность полученного штамма-продуцента изучали путем культивирования клеток в среде LB, в термостатированном шейкере роторного типа при температуре 37°С, скорости вращения платформы 250 об/мин до оптической плотности А600 = 0,5-0,6. После этого индуцировали синтез целевого белка добавлением ИПТГ до конечной концентрации 1 мМ. В качестве контроля использовали культуру без добавления ИПТГ. Образцы собранной центрифугированием биомассы клеток лизировали при помощи ультразвукового разрушения и анализировали методом электрофореза в 8%-ном полиакриламидном геле в системе Лэммли в денатурирующих условиях. Результат представлен на фиг. 2, где М – белковый маркер подвижности 14,4-116 кДа (ThermoScientific); 1 – контрольная неиндуцированная культура, 2 – 3 ч после индукции 1 мМ ИПТГ. В результате выявлено, что индукция 1 мМ ИПТГ культуры клеток E. coli приводит к синтезу белка с молекулярной массой 70 кДа, что соответствует ожидаемой молекулярной массе для рекомбинантного белка ROS1 с гистидиновой меткой. Количество рекомбинантного белка составило не менее 20% от клеточного лизата при культивировании в условиях индукции.
Пример 3. Очистка рекомбинантного белка ROS1 и изучение его активности.
Очистку рекомбинантного белка проводили в две стадии. Первая стадия включала в себя ионообменную хроматографию на сорбентах SP- и Q-сефарозе. Вторая стадия очистки представляла собой аффинную хроматографию на металлхелатном агарозном сорбенте, модифицированном иминодиуксусной кислотой, заряженной ионами Ni2. После этого белок диализовали против буфера, содержащего 50%-ный глицерин, и хранили при −20°C. Результат двухстадийной очистки ROS1 представлен на фиг. 3, где М – маркер подвижности 14,4-116 кДа (ThermoScientific); 1 – очищенный рекомбинантный белок ROS1. Как видно из фиг. 3 препарат белка ROS1 является практически гомогенным, и его чистота составляет не менее 90%.
Для исследования активности очищенного белка ROS1 in vitro в качестве субстрата использовали двуцепочечный олигодезоксирибонуклеотид длиной 20 пар оснований, содержащий полуметилированный CpG-динуклеотид и радиоактивную метку на 5’-конце в конечной концентрации 50 нМ. Реакцию проводили при 37°C в течение 30 мин. Продукты реакции анализировали электрофорезом в 20%-полиакриламидном геле в денатурирующих условиях с последующим радиолюминесцентным сканированием. Результат эксперимента представлен на фиг. 4, где 6 – субстрат без фермента; 1-5 – конечная концентрация очищенного белка ROS1 900 нМ, 600 нМ, 300 нМ, 30 нМ и 3 нМ соответственно; S–субстрат, P – продукт реакции. Из фиг. 4 видно, что очищенный в рекомбинантном виде белок ROS1 проявлял активность в отношении субстрата с полуметилированным CpG-динуклеотидом, однако наблюдаемая активность не превышала 20 %. Снижение активности белка могло произойти на стадиях его очистки.
Таким образом, создан штамм бактерий Escherichia сoli Rosetta 2(DE3)/pET24b(+)-ROS1, являющийся продуцентом рекомбинантной ДНК-гликозилазы ROS1, состоящей из гликозилазного домена С и C-концевого RRM-домена и способной направленно удалять эпигенетическую метку 5-метилцитозин из ДНК. Рекомбинантный белок ROS1 может служить эффективным инструментом для редактирования уровня метилирования ДНК клеток человека.
Источники информации
1. Zhang, M., Kimatu, J. N., Xu, K., Liu, B. DNA cytosine methylation in plant development// Journal of Genetics and Genomics. – 2010. – V. 37. – No. 1. – P. 1-12.
2. Gong, Z., Morales-Ruiz, T., Ariza, R. R., Roldán-Arjona, T., David, L., Zhu, J.-K. ROS1, a Repressor of Transcriptional Gene Silencing in Arabidopsis, Encodes a DNA Glycosylase/Lyase// Cell –2002. – V. 111. – No. 6. – P. 803-814.
3. Hong, S., Hashimoto, H., Kow, Y. W., Zhang, X., Cheng, X. The carboxy-terminal domain of ROS1 is essential for 5-methylcytosine DNA glycosylase activity// Journal of molecular biology. – 2014. – V. 426. – No. 22. – P. 3703-3712.
4. Ponferrada-Marín, M. I., Roldán-Arjona, T., Ariza, R. R. Demethylation initiated by ROS1 glycosylase involves random sliding along DNA// Nucleic Acids Research –2012. – V. 40. – No. 22. – P. 11554-11562.
--->
Перечень последовательностей
<110> Федеральное государственное бюджетное учреждение науки
Институт химической биологии и фундаментальной медицины
Сибирского отделения Российской академии наук (ИХБФМ СО
РАН)
<120> Рекомбинантный штамм бактерий Escherichia coli
Rosetta 2(DE3)/pET24b(+)-ROS1 - продуцент метилцитозин-
специфической ДНК-гликозилазы ROS1
<160> 1
<210> 1
<211> 7088
<212> DNA
<213> Искусственная последовательность
<400> 1
agatctcgat cccgcgaaat taatacgact cactataggg gaattgtgag cggataacaa 60
ttcccctcta gaaataattt tgtttaactt taagaaggag atatacatat ggaagcccct 120
acctttagtg aagcaattgt ggatgttcgt gaagaagtga gtgtggtggt tgatagttgt 180
aaaagtgaac atattgcact gaaaagcaat agtaataaca agaaacacca cgccgatagc 240
accctggatc gcgccaatga taataccaaa gccaaaaaag aacgtccggg caaagaaaaa 300
cagaatgtgg attgggatag tctgcgtctg caggcacaga ataatggtaa aaaacgcgaa 360
gaaattgcac ataccattcg cgaacgcggc atgaataata tgctggccga acgcattaag 480
gattttctga atcgcatttt tcgtgaacat ggcagtattg atctggaatg gctgcgcgat 540
gtgccgccgg ataaagcaaa agaatatctg ctgagtattc gcggtctggg cctgaaaagc 600
gttgaatgcg ttcgcctgct gaccctgcat catctggcat ttccggtgga taccaatgtg 660
ggccgcattg cagttcgcct gggttgggtg ccgctgcagc cgttaccgga aagcctgcag 720
ctgcatctgc tggaactgta tccggttctg gaaagcattc agaaatatct gtggccgcgc 780
ctgtgtaaac tggatcagcg taccctgtat gaactgcatt atcacatgat tacctttggc 840
aaagttttct gtaccaaaag caaaccgaat tgtaatgcat gtccgctgcg tggtgaatgc 900
cgccattttg ccagtgcatt tgcaagcgca cgtctggcac tgccggcccc tgaagaaaaa 960
agtattgtga gtgcaaccga aaataaggcc gccggccaga atccgtttca gaattttagt 1020
cagctgctgc tgccgctgcc gcaggcagat cagaccccgc tggaacatag taaactgatt 1080
aatagtgcac cgattattga agttccggca accccggaac cgattgttga agaaccggca 1140
agtccggaac cggaacagaa tgccccggaa gtggatattg aagatgcata ttttgaagat 1200
ccgaatgaaa ttccgaccat taccctgaat atggcagaat tcactcagaa tgtgaaaaaa 1260
ttcatggaaa acaacatgga actgcagcag gttgaaatga gtaaagccct ggttgcactg 1320
accccggaag ccgcaagtat tccggtgccg aaactgaaac atattagtcg tctgcgtacc 1380
gaacatcagg tttatgaact gagtgatagt catccgctgc tggaaggctt tgataaacgc 1440
gaaccggatg atccgtgcag ttatctgctg gcaatttgga ccccgggtga aaccgcagat 1500
agtattcatc cgccggccat taagtgtaat agtcaggaag ccggtcgtct gtgcgatgat 1560
gaaacctgct ttgcatgtaa tagcctgcgc gaagcacata gccagaccgt gcgtggcacc 1620
attctgattc cgtgccgtac cgccatgcgt ggcagttttc cgctgaatgg tacctatttt 1680
caggttaatg aagtttttgc cgatcatgat agcagtctga atccgattga tgtgccgcgc 1740
gattggctgt ggaatctgcc gcgccgtacc gtgtattttg gtaccagtat tccgaccatc 1800
tttaaaggcc tgaccaccga aagtattcag cattgttttt ggcgcggctt tgtttgtgtt 1860
cgtggttttg ataaaaagac ccgcgcaccg cgcccgctga tggctagact gcattttccg 1920
gccagccgcc tgagccgtac caaaggtaaa ccggatgaaa atctcgagca ccaccaccac 1980
caccactgag atccggctgc taacaaagcc cgaaaggaag ctgagttggc tgctgccacc 2040
gctgagcaat aactagcata accccttggg gcctctaaac gggtcttgag gggttttttg 2100
ctgaaaggag gaactatatc cggattggcg aatgggacgc gccctgtagc ggcgcattaa 2160
gcgcggcggg tgtggtggtt acgcgcagcg tgaccgctac acttgccagc gccctagcgc 2220
ccgctccttt cgctttcttc ccttcctttc tcgccacgtt cgccggcttt ccccgtcaag 2280
ctctaaatcg ggggctccct ttagggttcc gatttagtgc tttacggcac ctcgacccca 2340
aaaaacttga ttagggtgat ggttcacgta gtgggccatc gccctgatag acggtttttc 2400
gccctttgac gttggagtcc acgttcttta atagtggact cttgttccaa actggaacaa 2460
cactcaaccc tatctcggtc tattcttttg atttataagg gattttgccg atttcggcct 2520
attggttaaa aaatgagctg atttaacaaa aatttaacgc gaattttaac aaaatattaa 2580
cgtttacaat ttcaggtggc acttttcggg gaaatgtgcg cggaacccct atttgtttat 2640
ttttctaaat acattcaaat atgtatccgc tcatgaatta attcttagaa aaactcatcg 2700
agcatcaaat gaaactgcaa tttattcata tcaggattat caataccata tttttgaaaa 2760
agccgtttct gtaatgaagg agaaaactca ccgaggcagt tccataggat ggcaagatcc 2820
tggtatcggt ctgcgattcc gactcgtcca acatcaatac aacctattaa tttcccctcg 2880
tcaaaaataa ggttatcaag tgagaaatca ccatgagtga cgactgaatc cggtgagaat 2940
ggcaaaagtt tatgcatttc tttccagact tgttcaacag gccagccatt acgctcgtca 3000
tcaaaatcac tcgcatcaac caaaccgtta ttcattcgtg attgcgcctg agcgagacga 3060
aatacgcgat cgctgttaaa aggacaatta caaacaggaa tcgaatgcaa ccggcgcagg 3120
aacactgcca gcgcatcaac aatattttca cctgaatcag gatattcttc taatacctgg 3180
aatgctgttt tcccggggat cgcagtggtg agtaaccatg catcatcagg agtacggata 3240
aaatgcttga tggtcggaag aggcataaat tccgtcagcc agtttagtct gaccatctca 3300
tctgtaacat cattggcaac gctacctttg ccatgtttca gaaacaactc tggcgcatcg 3360
ggcttcccat acaatcgata gattgtcgca cctgattgcc cgacattatc gcgagcccat 3420
ttatacccat ataaatcagc atccatgttg gaatttaatc gcggcctaga gcaagacgtt 3480
tcccgttgaa tatggctcat aacacccctt gtattactgt ttatgtaagc agacagtttt 3540
attgttcatg accaaaatcc cttaacgtga gttttcgttc cactgagcgt cagaccccgt 3600
agaaaagatc aaaggatctt cttgagatcc tttttttctg cgcgtaatct gctgcttgca 3660
aacaaaaaaa ccaccgctac cagcggtggt ttgtttgccg gatcaagagc taccaactct 3720
ttttccgaag gtaactggct tcagcagagc gcagatacca aatactgtcc ttctagtgta 3780
gccgtagtta ggccaccact tcaagaactc tgtagcaccg cctacatacc tcgctctgct 3840
aatcctgtta ccagtggctg ctgccagtgg cgataagtcg tgtcttaccg ggttggactc 3900
aagacgatag ttaccggata aggcgcagcg gtcgggctga acggggggtt cgtgcacaca 3960
gcccagcttg gagcgaacga cctacaccga actgagatac ctacagcgtg agctatgaga 4020
aagcgccacg cttcccgaag ggagaaaggc ggacaggtat ccggtaagcg gcagggtcgg 4080
aacaggagag cgcacgaggg agcttccagg gggaaacgcc tggtatcttt atagtcctgt 4140
cgggtttcgc cacctctgac ttgagcgtcg atttttgtga tgctcgtcag gggggcggag 4200
cctatggaaa aacgccagca acgcggcctt tttacggttc ctggcctttt gctggccttt 4260
tgctcacatg ttctttcctg cgttatcccc tgattctgtg gataaccgta ttaccgcctt 4320
tgagtgagct gataccgctc gccgcagccg aacgaccgag cgcagcgagt cagtgagcga 4380
ggaagcggaa gagcgcctga tgcggtattt tctccttacg catctgtgcg gtatttcaca 4440
ccgcatatat ggtgcactct cagtacaatc tgctctgatg ccgcatagtt aagccagtat 4500
acactccgct atcgctacgt gactgggtca tggctgcgcc ccgacacccg ccaacacccg 4560
ctgacgcgcc ctgacgggct tgtctgctcc cggcatccgc ttacagacaa gctgtgaccg 4620
tctccgggag ctgcatgtgt cagaggtttt caccgtcatc accgaaacgc gcgaggcagc 4680
tgcggtaaag ctcatcagcg tggtcgtgaa gcgattcaca gatgtctgcc tgttcatccg 4740
cgtccagctc gttgagtttc tccagaagcg ttaatgtctg gcttctgata aagcgggcca 4800
tgttaagggc ggttttttcc tgtttggtca ctgatgcctc cgtgtaaggg ggatttctgt 4860
tcatgggggt aatgataccg atgaaacgag agaggatgct cacgatacgg gttactgatg 4920
atgaacatgc ccggttactg gaacgttgtg agggtaaaca actggcggta tggatgcggc 4980
gggaccagag aaaaatcact cagggtcaat gccagcgctt cgttaataca gatgtaggtg 5040
ttccacaggg tagccagcag catcctgcga tgcagatccg gaacataatg gtgcagggcg 5100
ctgacttccg cgtttccaga ctttacgaaa cacggaaacc gaagaccatt catgttgttg 5160
ctcaggtcgc agacgttttg cagcagcagt cgcttcacgt tcgctcgcgt atcggtgatt 5220
cattctgcta accagtaagg caaccccgcc agcctagccg ggtcctcaac gacaggagca 5280
cgatcatgcg cacccgtggg gccgccatgc cggcgataat ggcctgcttc tcgccgaaac 5340
gtttggtggc gggaccagtg acgaaggctt gagcgagggc gtgcaagatt ccgaataccg 5400
caagcgacag gccgatcatc gtcgcgctcc agcgaaagcg gtcctcgccg aaaatgaccc 5460
agagcgctgc cggcacctgt cctacgagtt gcatgataaa gaagacagtc ataagtgcgg 5520
cgacgatagt catgccccgc gcccaccgga aggagctgac tgggttgaag gctctcaagg 5580
gcatcggtcg agatcccggt gcctaatgag tgagctaact tacattaatt gcgttgcgct 5640
cactgcccgc tttccagtcg ggaaacctgt cgtgccagct gcattaatga atcggccaac 5700
gcgcggggag aggcggtttg cgtattgggc gccagggtgg tttttctttt caccagtgag 5760
acgggcaaca gctgattgcc cttcaccgcc tggccctgag agagttgcag caagcggtcc 5820
acgctggttt gccccagcag gcgaaaatcc tgtttgatgg tggttaacgg cgggatataa 5880
catgagctgt cttcggtatc gtcgtatccc actaccgaga tatccgcacc aacgcgcagc 5940
ccggactcgg taatggcgcg cattgcgccc agcgccatct gatcgttggc aaccagcatc 6000
gcagtgggaa cgatgccctc attcagcatt tgcatggttt gttgaaaacc ggacatggca 6060
ctccagtcgc cttcccgttc cgctatcggc tgaatttgat tgcgagtgag atatttatgc 6120
cagccagcca gacgcagacg cgccgagaca gaacttaatg ggcccgctaa cagcgcgatt 6180
tgctggtgac ccaatgcgac cagatgctcc acgcccagtc gcgtaccgtc ttcatgggag 6240
aaaataatac tgttgatggg tgtctggtca gagacatcaa gaaataacgc cggaacatta 6300
gtgcaggcag cttccacagc aatggcatcc tggtcatcca gcggatagtt aatgatcagc 6360
ccactgacgc gttgcgcgag aagattgtgc accgccgctt tacaggcttc gacgccgctt 6420
cgttctacca tcgacaccac cacgctggca cccagttgat cggcgcgaga tttaatcgcc 6480
gcgacaattt gcgacggcgc gtgcagggcc agactggagg tggcaacgcc aatcagcaac 6540
gactgtttgc ccgccagttg ttgtgccacg cggttgggaa tgtaattcag ctccgccatc 6600
gccgcttcca ctttttcccg cgttttcgca gaaacgtggc tggcctggtt caccacgcgg 6660
gaaacggtct gataagagac accggcatac tctgcgacat cgtataacgt tactggtttc 6720
acattcacca ccctgaattg actctcttcc gggcgctatc atgccatacc gcgaaaggtt 6780
ttgcgccatt cgatggtgtc cgggatctcg acgctctccc ttatgcgact cctgcattag 6840
gaagcagccc agtagtaggt tgaggccgtt gagcaccgcc gccgcaagga atggtgcatg 6900
caaggagatg gcgcccaaca gtcccccggc cacggggcct gccaccatac ccacgccgaa 6960
acaagcgctc atgagcccga agtggcgagc ccgatcttcc ccatcggtga tgtcggcgat 7020
ataggcgcca gcaaccgcac ctgtggcgcc ggtgatgccg gccacgatgc gtccggcgta 7080
gaggatcg 7088
<---
Изобретение относится к рекомбинантному штамму бактерий Escherichia coli – продуценту метилцитозин-специфической ДНК-гликозилазы ROS1. Предложен штамм бактерий Escherichia coli Rosetta 2(DE3)pET24b(+)-ROS1, являющийся продуцентом рекомбинантной ДНК-гликозилазы ROS1. Указанный штамм получают путем трансформации клеток Escherichia coli штамма Rosetta 2(DE3) плазмидой pET24b(+)-ROS1, имеющей нуклеотидную последовательность SEQ ID NО: 1, сконструированной на основе вектора pET24b(+), содержащего ориджин репликации f1, промотор фага T7, RBS-сайт посадки рибосомы, сайты рестрикции NdeI и XhoI, по которым встроен фрагмент гена, кодирующего ДНК-гликозилазу ROS1 из табака Nicotiana tabacum, гистидиновую метку His-tag, терминатор фага T7, ген устойчивости к канамицину Kan (R), а также лактозный репрессор lacI и лактозный оператор. Изобретение обеспечивает получение рекомбинантного белка ROS1 с чистотой не менее 90%. 4 ил., 3 пр.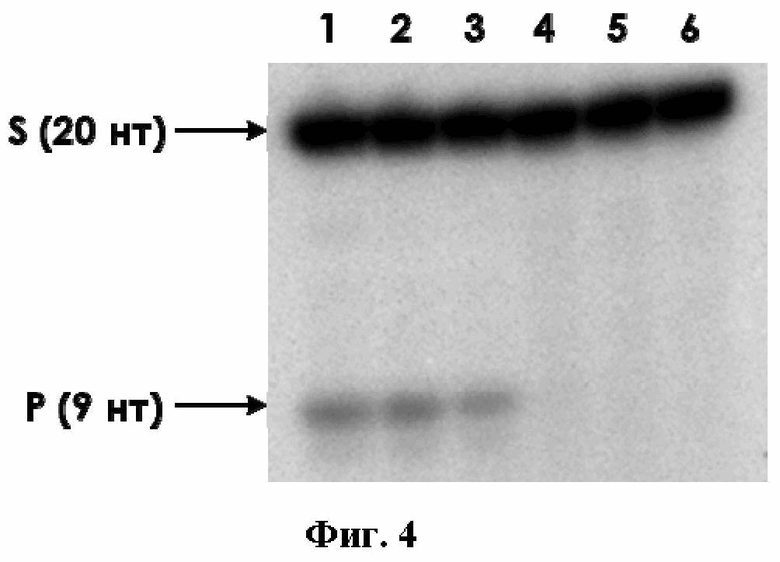
Рекомбинантный штамм бактерий Escherichia coli Rosetta 2(DE3)/pET24b(+)-ROS1–продуцент метилцитозин-специфической ДНК-гликозилазы ROS1, полученный путем трансформации клеток Escherichia coli штамма Rosetta 2(DE3) плазмидой pET24b(+)-ROS1, имеющей нуклеотидную последовательность SEQ ID NО: 1, сконструированной на основе вектора pET24b(+), содержащего ориджин репликации f1, промотор фага T7, RBS-сайт посадки рибосомы, сайты рестрикции NdeI и XhoI, по которым встроен фрагмент гена, кодирующего ДНК-гликозилазу ROS1 из табака Nicotiana tabacum, гистидиновую метку His-tag, терминатор фага T7, ген устойчивости к канамицину Kan (R), а также лактозный репрессор lacI и лактозный оператор.
| PONFERRADA-MARIN M.I | |||
| ET AL | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Nucleic Acids Res | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| PARRILLA-DOBLAS J.T | |||
| ET AL | |||

