Область техники
Изобретение относится к генетической конструкции (рекомбинантной плазмиде) pETZ, обеспечивающей синтез и секрецию инженерного рекомбинантного белка А в периплазматическое пространство клеток E. coli в растворимой форме, а также к рекомбинантному белку, синтез которого этой плазмидой обеспечивается и может быть использовано в биотехнологии и генетической инженерии.
Белок А - это многодоменный полипептид, одним из свойств которого, как природного, так и модифицированного, является способность связываться с Fc-фрагментом иммуноглобулинов, в первую очередь класса G (IgG). Благодаря этому свойству белок А находит широкое применение в биологических и биохимических исследованиях, таких как аффинная хроматография, блоттинг и иммуноферментный анализ.
Уровень техники
Природный белок А был обнаружен на клеточной стенке Staphylococcus aureus. Этот белок выполняет функцию связывания антител хозяина в неправильной ориентации, тем самым нарушая опсонизацию и фагоцитоз, что повышает вирулентность и выживаемость бактерии [2]. Белок А представляет собой многодомный полипептид клеточной поверхности с двумя функциональными участками. С-концевой участок выполняет функцию прикрепления белка к пептидогликану клеточной стенки бактерии и содержит 15-22 гидрофобных аминокислот, за которым следует хвост из 5-12 в основном заряженных остатков [5]. N-концевая часть состоит из 5 доменов (E-D-A-B-C), каждый из которых схож наличием пучка из трех альфа-спиралей и содержит примерно 58 аминокислотных остатков. Все пять доменов обладают одновалентной связывающей активностью по отношению к Fc-фрагментам иммуноглобулинов, особенно класса G. [1].
Природный белок А имеет некоторые недостатки, которые затрудняют его получение и использование. Во-первых - это наличие С-концевого участка, отвечающего за прикрепление к клеточной стенке бактерии, что значительно усложняет процесс выделения и очистки белка А из биомассы. Во-вторых - это наличие 5 отличающихся друг от друга функциональных доменов (идентичность относительно домена А от 74 до 91%), за счет чего связывающая активность доменов различна, особенно домена Е, который проявляет наименьшее сродство с иммуноглобулинами [3].
Логичным устранением первого недостатка является удаление С-концевой последовательности. Вторая проблема решается использованием одного, наиболее оптимального домена для построения белка. В качестве такого домена был использован инженерный аналог домена В - домен Z, полученный путем введения мутации для замены остатка Gly в положении 29 на Ala и остатка Ala в положении 1 на Val. Замена Ala 1 на Val предназначена для облегчения генно-инженерной подготовки гена, кодирующего несколько связанных доменов, и не влияет на функции домена. Благодаря мутациям модифицированный Z домен более устойчив к щелочам, чем домен B, и его можно использовать повторно с помощью щелочной промывки. Также остаток Asn заменен другой аминокислотой, чтобы придать дальнейшую более высокую щелочную стойкость. Еще одной особенностью домена Z является его пониженная связывающая способность к Fab-области иммуноглобулинов. Эта особенность выгодно облегчает диссоциацию антитела, связывающегося с доменом Z, с помощью кислоты [4].
Известна рекомбинантная плазмидная ДНК рММ 9, кодирующая синтез белка A STAPHYLOCOCCUS AUREUS, способ ее конструирования и штамм бактерий BACILLUS SUBTILIS - продуцент белка A STAPHYLOCOCCUS AUREUS (патент SU №1524485, МПК С12N15/31, опубл. 1991 г.). Целью изобретения является повышение уровня синтеза белка A St. aureus. Цель достигается за счет плазмиды pMM9, несущей ген белка A и стабильно поддерживающейся в различных грам-положительных микроорганизмах, способа получения этой плазмиды, включающего в качестве промежуточного этапа клонирование гена белка A в клетках E. coli, а также штамм B. subtilis IPM-12, секретирующего белок A во внеклеточную среду в количестве не менее, чем 100 мг/л культуральной среды. Однако белок, описанный в патенте SU1524485, идентичен природному, стафилококковому, из-за чего он существенно уступает рекомбинантному белку А в устойчивости к щелочам, поскольку не имеет аминокислотных замен, обуславливающих данную устойчивость. При использовании в хроматографии белка А проявляется его ограниченная стабильность в сильно щелочных условиях, которые обычно используются для процедур очистки хроматографических колонок на месте в биотехнологической промышленности. Наиболее часто для санации сорбентов используют щелочные растворы, в особенности, раствор NaOH с концентрацией от 0,1 до 1 М, в зависимости от степени и характера загрязнения. Эта процедура подразумевает воздействие на матрицу растворов со значениями рН выше 13. Для многих матриц, содержащих белковые лиганды, это приводит к снижению емкости из-за нестабильности лиганда в условиях высоких рН.
Известно изобретение, раскрывающее получение и применение рекомбинантного слитого белка и относится к технической области биоинженерии (патент Китая № CN 103184229, МПК С12N 15/62, опубл. 03.07.2013 г.). Способ включает стадии, на которых гены IgG-связывающего домена белка A Staphylococcus aureus и белка G-стрептококка соединяются последовательно и вставляются в носитель Escherichia coli типа термоиндукции pBV 220, так что конструируются носитель рекомбинантной экспрессии Escherichia coli, содержащий гены, и преобразованный рекомбинационный штамм Escherichia coli DH5alpha. Рекомбинантный белок A/G получают с использованием штамма. Растворимый рекомбинантный белок A/G, продуцируемый штаммом, превышает 40% от общего количества бактериального растворимого белка. Чистота рекомбинантного белка A/G может быть выше 95% за счет хелатирующей хроматографической очистки ионов никеля и анионообменной хроматографии. Белок обладает такими преимуществами, как высокое количество экспрессии, низкая стоимость, простота очистки и тому подобное. Кроме того, совместимая добавка, полученная рекомбинантным белком в качестве генина, обладает характеристиками комбинации белка А, белка G и IgG, может превосходно сочетаться с IgG большего количества видов млекопитающих и может широко использоваться для очистки IgG различных видов млекопитающих.
Однако и такой белок A/G существенно уступает рекомбинантному белку А в устойчивости к щелочам, поскольку не имеет аминокислотных замен, обуславливающих данную устойчивость. При использовании в хроматографии белка А проявляется его ограниченная стабильность в сильно щелочных условиях, которые обычно используются для процедур очистки хроматографических колонок на месте в биотехнологической промышленности.
Наиболее близким аналогом (прототипом) является рекомбинантный белок А-mer и его применение (патент Китая № CN 105481954, МПК С12N 15/31, опубл. 13.04.2016 г.). Рекомбинантный белок A-mer представляет собой мультимер двух или более Z-фрагментов белка S. aureus A, и, по меньшей мере, один из каждого Z-фрагмента представляет собой Z-фрагмент мутантов со следующими аминокислотными мутациями: Asn3Asp, Asn6Leu и Asn23Thr. Изобретение включает полинуклеотид, кодирующий белок А-mer, плазмидный вектор pET30a-4Z, содержащий указанный полинуклеотид, и клетку-хозяина E. coli BL21 (DE3), в которую интегрирован вектор с полинуклеотидом. Плазмиду pET30a-4Z' трансформировали в E. coli BL21 (DE3) методом CaCl2. Запатентован также способ получения полимера рекомбинантного белка А-mer, включающий стадии: (a) культивирование клетки-хозяина для экспрессии полимера рекомбинантного белка А по.1; (b) выделение или очистка рекомбинантного белка A-mer из культуральной системы. Способ очистки, включает стадии: аффинную хроматографию с использованием аффинной хроматографической среды, содержащий рекомбинантного белка A-mer.
Недостатком прототипа является то, что в его описании не содержится информации о продуктивности штамма E.coli по белку А, полученного с использованием описываемой плазмидной конструкции. Продуктивность штамма является важной характеристикой, определяющей его технологическую и экономическую эффективность. Так же плазмидная конструкция, описываемая в прототипе, позволяет получать целевой белок А, содержащий на С-конце остаток цистеина. Это может осложнить очистку и сохранность препарата белка А, так как свободные остатки цистеина склонны при хранении к окислительному образованию дисульфидных связей. Такая архитектура белка требует использования специальных методов очистки.
Таким образом разработка стабильного штамма-продуцента, обеспечивающего продукцию белка А, сохраняющего характерные для вирусного белка свойства, остается актуальной задачей.
Раскрытие изобретения
Задачей заявляемого изобретения является разработка получения экспрессионной генетической конструкции - рекомбинантной плазмиды pETZ, обеспечивающей синтез и секрецию рекомбинантного белка А в периплазматическое пространство клеток E. coli в растворимой форме без недостатков, содержащихся в прототипе.
Техническим результатом заявляемого изобретения является получение высокопродуктивного плазмидного вектора, обеспечивающего синтез и секрецию белка А в клетках E.coli до 100 мг/л культуральной среды и упрощение технологии очистки указанного белка А за счет синтеза плазмидным вектором белка А, содержащего на С-конце остатков шести гистидинов, что позволяет использовать одностадийную металл-хелатную хроматографию.
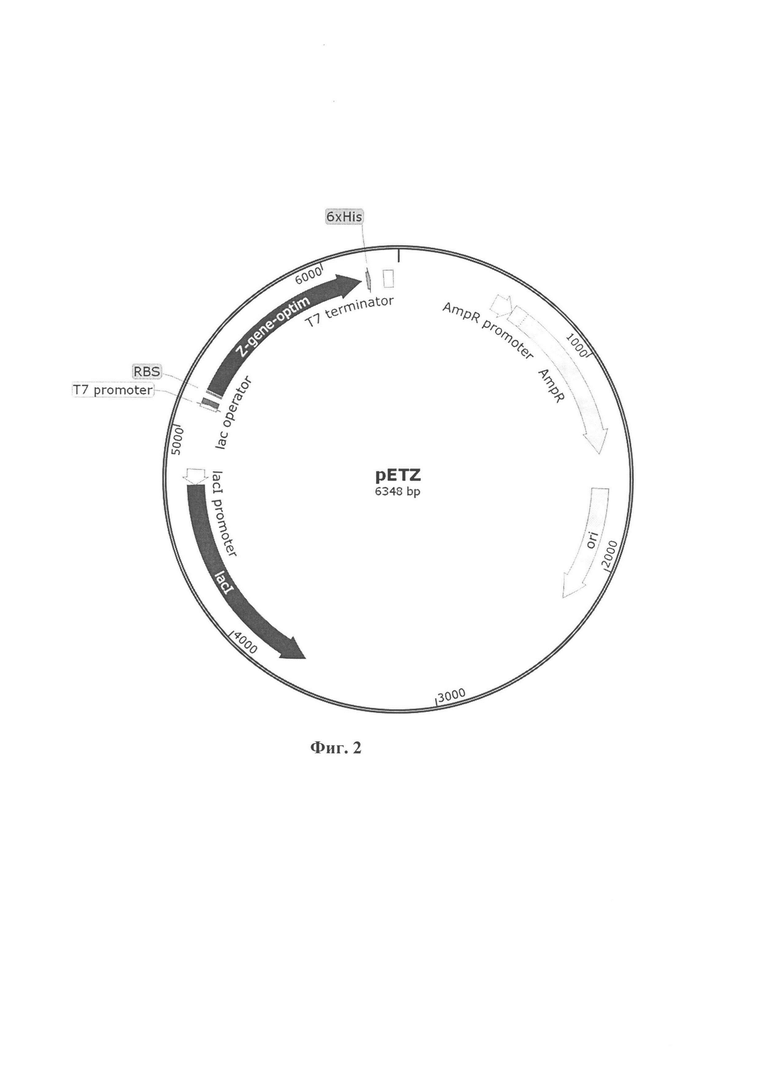
Указанный технический результат достигается созданием рекомбинантной плазмиды pETZ, имеющей нуклеотидную последовательность SEQ ID NO:1, размер 6348 пар нуклеотидов и содержащей в своем составе в соответствии с физической и генетической картой, представленной на фиг. 2, следующие элементы:
- AmpR promoter - участок связывания РНК-полимеразы (494-598 п. н.);
- участок, обеспечивающий экспрессию β-лактамазы - фактора резистентности к ампициллину (599-1459 п. н.);
- участок начала репликации ori (1630-2218 п. н.);
- lacI - участок, кодирующий белок, связывающийся с ДНК и ингибирующий экспрессию целевого белка до добавления изопропил-β-D-тиогалактопиранозида (ИПТГ) (3648-4730 п. н.);
- lacI promoter - участок связывания РНК-полимеразы (4731-4808 п. н.);
- T7 promoter - участок связывания РНК-полимеразы бактериофага Т7 (5117-5135 п. н.);
- lac operator - участок связывания белка lacI (5136-5160 п. н.);
- RBS - сайт связывания рибосомы (5191-5196);
- z-gene-optim - ген, кодирующий рекомбинантный белок А, имеющий нуклеотидную последовательность SEQ ID NO:2 и координаты с 5205 по 6164 п. н.;
- T7 terminator - терминатор транскрипции (6276-6323 п. н.);
- 6×His-метка, предназначенный для хроматографической очистки белка А, имеющий координаты с 6147 по 6164 п.н.
Указанный технический результат достигается также тем, что получен рекомбинантный белок А, продуцируемый плазмидой pETZ по п. 1, трансформированной в клетки E. coli, и используемый для иммунохимических анализов, имеющий аминокислотную последовательность SEQ ID NO:3 и молекулярную массу 35,8 кДа.
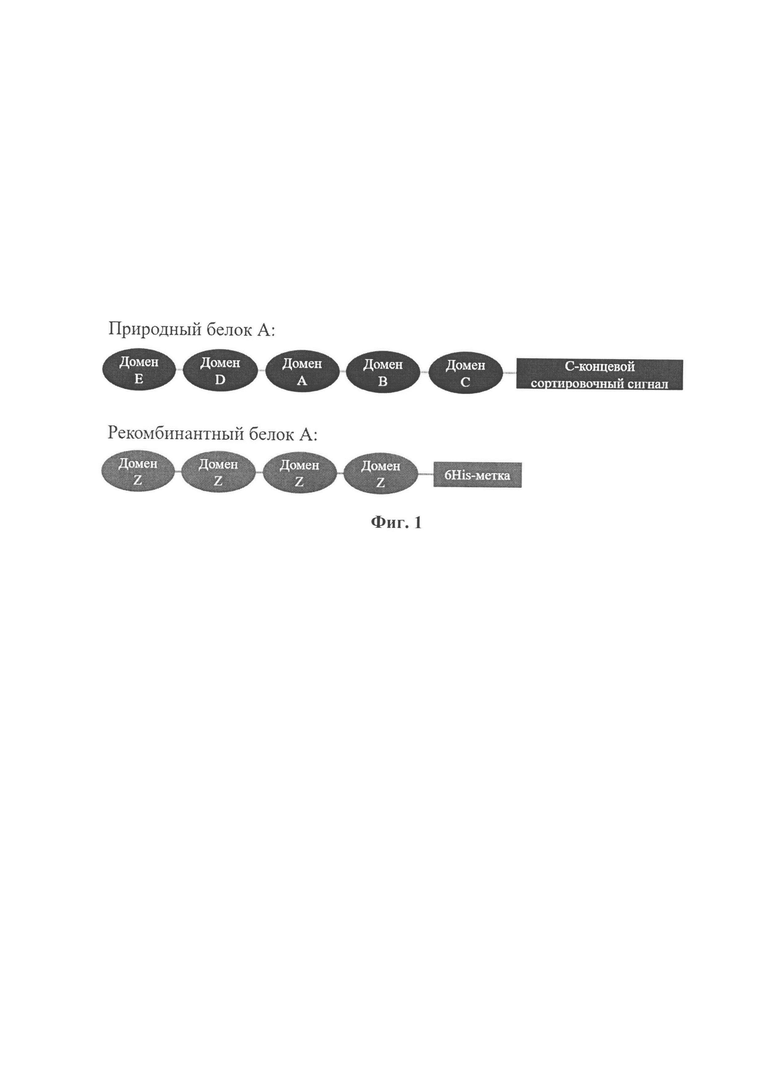
Конечный продукт - рекомбинантный белок А, состоящий из 4-х Z-доменов (в отличие от природного белка А, состоящего из E-D-A-B-C доменов), также в белке имеется 6×His-метка. Z-домен - это инженерный аналог домена В, полученный путем замены трех аминокислотных остатков.
Настоящее изобретение относится к рекомбинантному белку А и его применениям, и, в частности, раскрывает новый рекомбинантный белок Staphylococcus aureus A, который имеет структуру, определенную в описании, где функция включает, но не ограничивается применением в качестве лиганд-специфического связывающего иммуноглобулина, а рекомбинантный белок А может быть использован в качестве специфического аффинного лиганда и обладает отличной стойкостью к щелочам. Изобретение также относится к способу конструирования мутантного белка.
Рекомбинантный инженерный белок А содержит четыре Z-домена, а также последовательность 6×His, обеспечивающая очистку целевого белка при помощи металл-хелатной хроматографии. Сравнение структур природного и рекомбинантного белков представлены на фигуре 1.
В качестве вектора экспрессии была выбрана плазмида pET21. Карта плазмиды представлена на фиг. 2. Данная плазмида содержит лактозный оперон (lac operon), который, будучи связанным с лактозным репрессором, ингибирует транскрипцию, в данном случае Z-gene-optim. Ингибирование может быть остановлено добавлением изопропил-1-тио-β-D-галактозида (ИПТГ). Таким образом появляется возможность контролировать экспрессию гена. В плазмиде также имеется фактор резистентности к ампициллину (AmpR), продуктом которой является β-лактамаза - фермент, направленный на борьбу с β-лактамазными антибиотиками. Бактерии, обладающие устойчивостью, могут выращиваться на питательных средах с добавлением антибиотиков. Нуклеотидная последовательность гена Z-gene-optim приведена на фигуре 3.
В качестве продуцента был выбран штамм Escherichia coli SHuffle express. При культивировании в лабораторных условиях штамм обеспечивает синтез на уровне 100 мг рекомбинантного белка А на 1 л питательной среды LB.
Осуществление изобретения
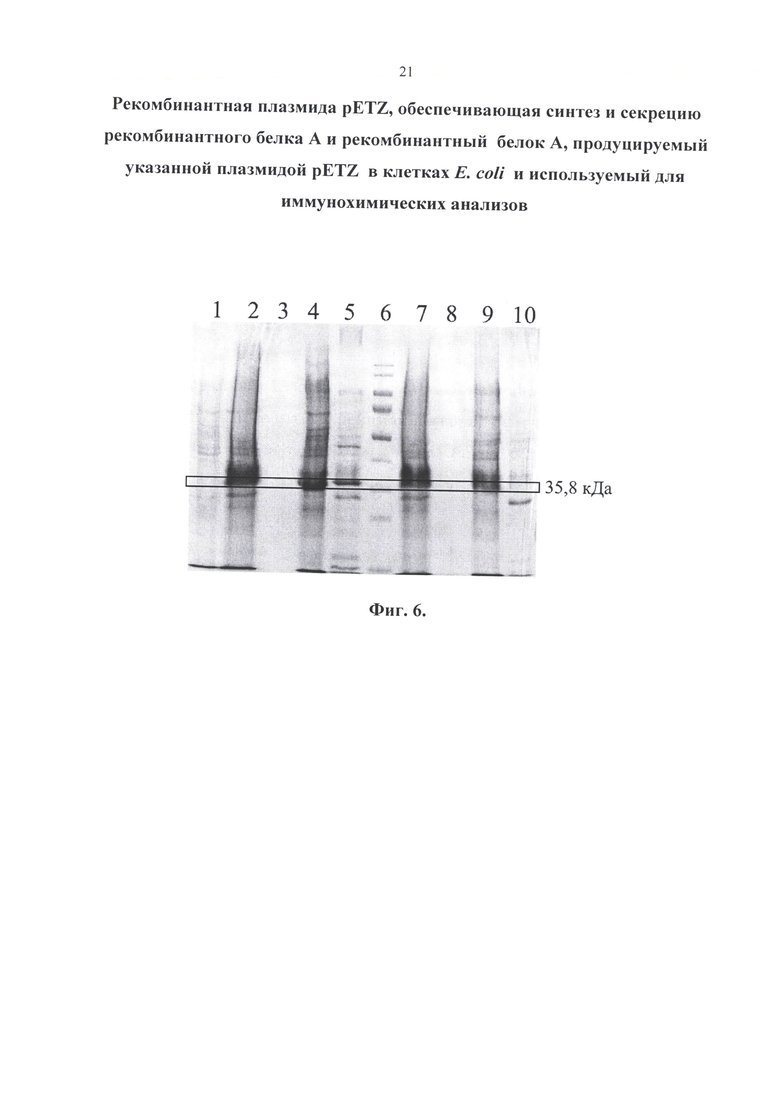
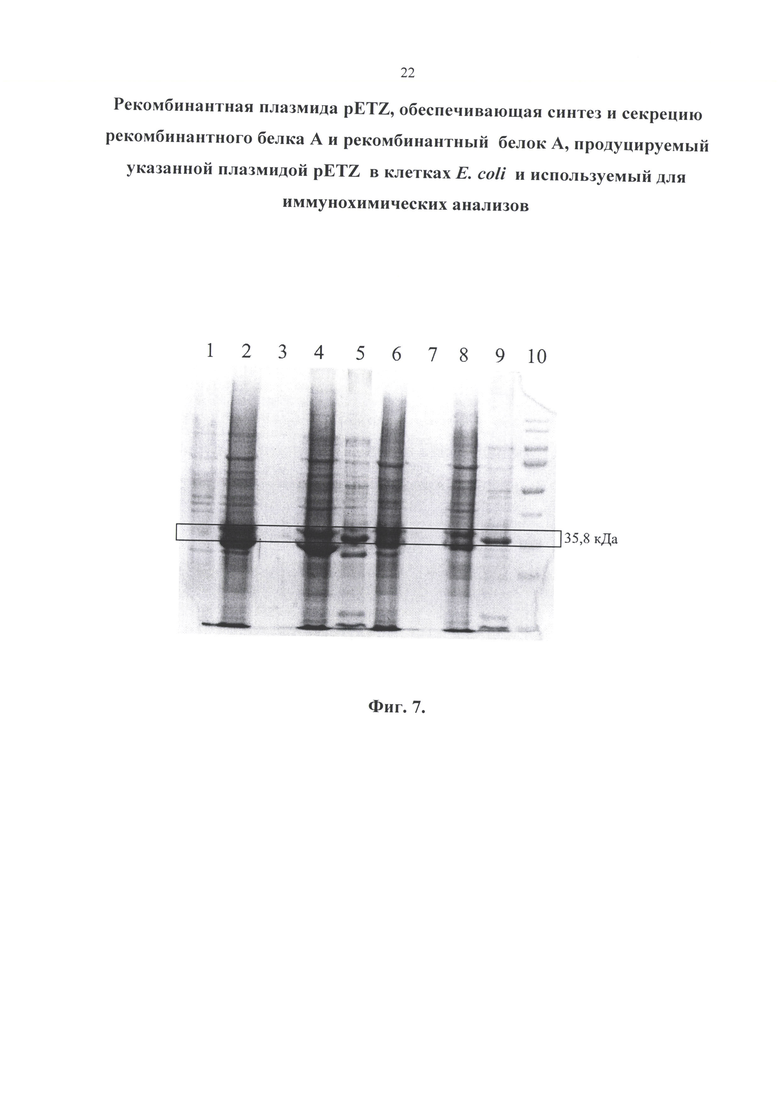
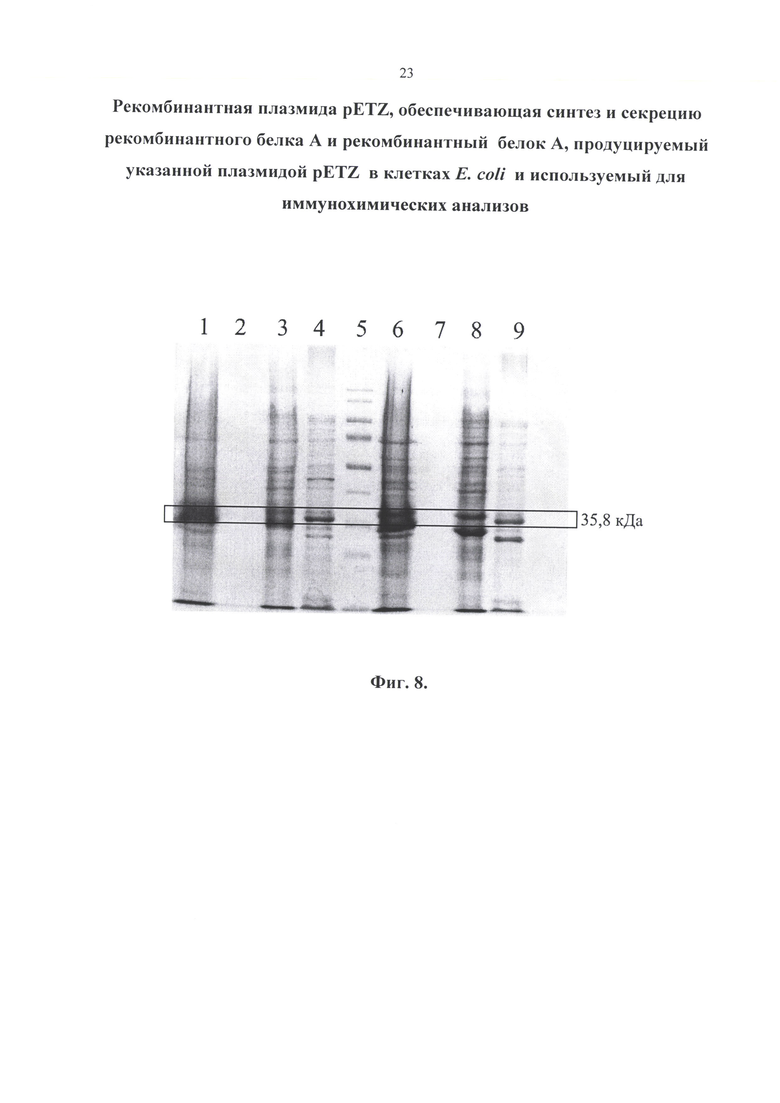
Изобретение иллюстрируется следующими графическими материалами. На фиг. 1 приводится сравнение структур природного и рекомбинантного белка А. На фигуре 2 приведена физическая и генетическая карта плазмиды pETZ. На фигуре 3 представлена нуклеотидная последовательность рекомбинантной плазмиды pETZ. На фигуре 4 представлена нуклеотидная последовательность гена (z-gene-optim), кодирующего белок А. На фигуре 5 приведена аминокислотная последовательность рекомбинантного белка А. На фиг. 6 приведен результат белкового электрофореза. Бактерии выращивались на питательной среде LB. 2-5 - 0,1 мМоль ИПТГ; 7-10 - 0,5 мМоль ИПТГ. 1 - до ИПТГ; 2, 7 - биомасса; 3, 8 - культуральная жидкость; 4, 9 - растворимая фракция (периплазма); 5, 10 - тельца включения; 6 - маркер. На фиг. 7 представлен результат белкового электрофореза. Бактерии выращивались на питательной среде ТВ. 2-5 - 0,1 мМоль ИПТГ; 6-9 - 0,5 мМоль ИПТГ. 1 - до ИПТГ; 2, 6 - биомасса; 3, 7 - культуральная жидкость; 4, 8 - растворимая фракция (периплазма); 5, 9 - тельца включения; 10 - маркер. На фиг. 8 приведен результат белкового электрофореза. Концентрация ИПТГ - 1,0 мМоль. 1-4 - бактерии выращивались на питательной среде LB; 6-9 - ТВ. 1, 6 - биомасса; 2, 7 - культуральная жидкость; 3,8 - растворимая фракция (периплазма); 4, 9 - тельца включения; 5 - маркер.
Для лучшего понимания сущности изобретения ниже приведены примеры его осуществления.
Пример 1. Получение рекомбинантного штамма E. coli.
Для получения штамма-продуцента рекомбинантного белка А была проведена трансформация клеток E. coli стандартным методом с использованием хлористого кальция. Ночную культуру E. coli SHuffle express (50 мкл) доращивали при 37°C и 200 об/мин до оптической плотности 0,6-0,8 в 5 мл питательной среды LB. Данную суспензию охлаждали на льду в течение 15 минут, далее центрифугировали 2 минуты (6000 g., 4°C). После удаления супернатанта осадок суспендировали в 700 мкл охлажденного CaCl2 и оставили на 15 минут во льду. Центрифугировали 2 минуты (6000 g, 4°C), удалили супернатант и суспендировали осадок в 100 мкл CaCl2. Добавили ДНК - 1 мкл (плазмида pETZ), перемешали и оставили во льду на 30 минут. Поместили в термостат на 5 минут при 37°C. Далее добавили 1 мл LB и культивировали 1 час при 37°C в термостате. Посеяли на чашку с нижним агаром и селективным антибиотиком (ампициллин). Культивировали в термостате в течение ночи при 37°C.
Пример 2. Наработка биомассы и выделение рекомбинантного белка А
Ночную культуру трансформированных бактерий выращивали на питательной среде Lysogeny medium broth (LB) при 37°С с добавлением ампициллина. Полученную ночную культуру в количестве 3 мл вносили в 6 колб Эрленмейера, содержащих по 300 мл питательной среды двух видов: Lysogeny medium broth (LB) и Terrific medium broth (TB) и растили при 37°С и 170 об./мин до плотности ОDλ600 = 0,6-0,8 о.е.. Транскрипцию индуцировали добавлением различных концентрации ИПТГ: 0,1 мМ, 0,5 мМ, 1,0 мМ. Образцы продолжали культивировать при 16°С и 170 об./мин в течение ночи. Биомасса была отделена от культуральной жидкости центрифугированием при 6000 G, 4°С в течении 15 мин.
Для разрушения клеточных стенок был использован метод ультразвукового лизиса, как наиболее простой и эффективный способ. Последующим центрифугированием раствор был разделен на супернатант (растворимая фракция) и осадок, который был растворен в буфере А с мочевиной и вновь центрифугировали. Во втором случае в супернатанте содержались тельца включения. В течении всего этого процесса отбирали аликвоты различных фракций, с которыми был проведен белковый электрофорез на полиакриламидном геле (фиг. 5-7).
На электрофореграмме обнаруживаются две полосы близких к значению 35,8 кДа. Они обе являются рекомбинантным белком А. Небольшое отличие объясняется наличием лидерной последовательности, которая влияет на длину белка в зависимости от того, секретируется он или нет.
С помощью программы анализа электрофореграмм GelAnalyzer 19.1 удалось определить, что концентрация белка при культивировании в питательной среде ТВ примерно в 1,25 раз выше, чем при культивировании в LB. Влияние значения концентрации ИПТГ на наработку белка с помощью данного метода не выявлено, то есть целесообразно использовать наименьшую концентрацию (0,1 мМ).
Результаты данного исследования показали, что до добавления ИПТГ синтез белка практически отсутствует. В культуральную жидкость секреция белков не происходит - весь белок остается в биомассе. Рекомбинантный белок А локализуется в периплазме и тельцах включения. Причем в периплазме сосредоточена основная масса белка А, поэтому, для экономии и упрощения процесса рекомендуется получать белок именно из этой фракции.
Надосадочная жидкость была трижды диализована против фосфатного буфера (K2HPO4 8,1 мM; KH2PO4 1,9 мM; pH 7,5). Диализат с рекомбинантным белком А в фосфатном буфере был профильтрован для предотвращения засорения хроматографа. Также в диализат добавили NaCl до конечной концентрации 500 мМоль. Он был очищен с помощью металл-хелатной хроматографии, основанной на сродстве переходных металлов к His-метке, имеющейся в рекомбинантном белке А.
При культивировании в лабораторных условиях плазмидный вектор и штамм продуцент обеспечивают накопление целевого белка А до 100 мг/л культуральной среды, что обеспечивает превосходное соотношение выход продукта/затраты.
Источники научно-технической и патентной информации
1. Deis L. N., Pemble C.W.T., Qi Y., Hagarman A., Richardson D.C., Richardson J.S., Oas T.G. Multiscale conformational heterogeneity in staphylococcal protein a: possible determinant of functional plasticity // Structure, 2014. - Vol. 22 - P. 1467-1477.
2. Grov A, Myklestad B, Oeding P. Immunochemical studies on antigen preparations from Staphylococcusaureus. Isolation and chemical characterization of antigen A // Acta Pathol Microbiol Scand, 1964. - Vol. 61 - P. 588-596.
3. Moks T., Abrahmsen L., Nilsson B., Hellman U., Sjoquist J., Uhlen M. Staphylococcal protein A consists of five IgG-binding domains // Eur J Biochem, 1986. - Vol. 156 - P. 637-643.
4. Nilsson B.A synthetic IgG-binding domain based on staphylococcal. protein A // Protein Eng, 1987. - Vol. 1 - P. 107-113.
5. Schneewind, O., Model P., Fischetti V.A. Sorting of protein A to the staphylococcal cell wall // Cell, 1992. - Vol. 70 - P. 267-281.
6. Патент SU №1524485, МПК С12N15/31, опубл. 1991 г.
7. Патент Китая №CN103184229, опубл. 03.07.2013 г.
8. Патент Китая №CN105481954, опубл. 13.04.2016 г. (прототип).
ПРИЛОЖЕНИЕ
--->
Перечень последовательностей, представленных по стандарту ВОИС ST26
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing SYSTEM "ST26SequenceListing_V1_3.dtd"
PUBLIC "-//WIPO//DTD Sequence Listing 1.3//EN">
<ST26SequenceListing productionDate="2023-09-22"
softwareVersion="2.2.0" softwareName="WIPO Sequence"
fileName="Рекомбинантная плазмида pETZ, обеспечивающая синтез
рекомбинантного инженерного белка А.xml"
dtdVersion="V1_3"><ApplicationIdentification><IPOfficeCode>RU</IPOffic
eCode><ApplicationNumberText/><FilingDate>2023-09-22</FilingDate></App
licationIdentification><ApplicantFileReference>RU-pETZ</ApplicantFileR
eference><ApplicantName languageCode="ru">ООО
Алтай-Энзим</ApplicantName><ApplicantNameLatin>OOO
"Altai-Enzim"</ApplicantNameLatin><InventionTitle
languageCode="ru">pETZ</InventionTitle><SequenceTotalQuantity>3</Seque
nceTotalQuantity><SequenceData sequenceIDNumber="1"
(SEQIDNO:1)><INSDSeq><INSDSeq_length>6348</INSDSeq_length><INSDSeq_mol
type>DNA</INSDSeq_moltype><INSDSeq_division>PAT</INSDSeq_division><INS
DSeq_feature-table><INSDFeature><INSDFeature_key>source</INSDFeature_k
ey><INSDFeature_location>1..6348</INSDFeature_location><INSDFeature_qu
als><INSDQualifier><INSDQualifier_name>mol_type</INSDQualifier_name><I
NSDQualifier_value>other
DNA</INSDQualifier_value></INSDQualifier><INSDQualifier
id="q2"><INSDQualifier_name>organism</INSDQualifier_name><INSDQualifie
r_value>synthetic
construct</INSDQualifier_value></INSDQualifier></INSDFeature_quals></I
NSDFeature></INSDSeq_feature-table><INSDSeq_sequence>tggcgaatgggacgcgc
cctgtagcggcgcattaagcgcggcgggtgtggtggttacgcgcagcgtgaccgctacacttgccagcgc
cctagcgcccgctcctttcgctttcttcccttcctttctcgccacgttcgccggctttccccgtcaagct
ctaaatcgggggctccctttagggttccgatttagtgctttacggcacctcgaccccaaaaaacttgatt
agggtgatggttcacgtagtgggccatcgccctgatagacggtttttcgccctttgacgttggagtccac
gttctttaatagtggactcttgttccaaactggaacaacactcaaccctatctcggtctattcttttgat
ttataagggattttgccgatttcggcctattggttaaaaaatgagctgatttaacaaaaatttaacgcga
attttaacaaaatattaacgtttacaatttcaggtggcacttttcggggaaatgtgcgcggaacccctat
ttgtttatttttctaaatacattcaaatatgtatccgctcatgagacaataaccctgataaatgcttcaa
taatattgaaaaaggaagagtatgagtattcaacatttccgtgtcgcccttattcccttttttgcggcat
tttgccttcctgtttttgctcacccagaaacgctggtgaaagtaaaagatgctgaagatcagttgggtgc
acgagtgggttacatcgaactggatctcaacagcggtaagatccttgagagttttcgccccgaagaacgt
tttccaatgatgagcacttttaaagttctgctatgtggcgcggtattatcccgtattgacgccgggcaag
agcaactcggtcgccgcatacactattctcagaatgacttggttgagtactcaccagtcacagaaaagca
tcttacggatggcatgacagtaagagaattatgcagtgctgccataaccatgagtgataacactgcggcc
aacttacttctgacaacgatcggaggaccgaaggagctaaccgcttttttgcacaacatgggggatcatg
taactcgccttgatcgttgggaaccggagctgaatgaagccataccaaacgacgagcgtgacaccacgat
gcctgcagcaatggcaacaacgttgcgcaaactattaactggcgaactacttactctagcttcccggcaa
caattaatagactggatggaggcggataaagttgcaggaccacttctgcgctcggcccttccggctggct
ggtttattgctgataaatctggagccggtgagcgtgggtctcgcggtatcattgcagcactggggccaga
tggtaagccctcccgtatcgtagttatctacacgacggggagtcaggcaactatggatgaacgaaataga
cagatcgctgagataggtgcctcactgattaagcattggtaactgtcagaccaagtttactcatatatac
tttagattgatttaaaacttcatttttaatttaaaaggatctaggtgaagatcctttttgataatctcat
gaccaaaatcccttaacgtgagttttcgttccactgagcgtcagaccccgtagaaaagatcaaaggatct
tcttgagatcctttttttctgcgcgtaatctgctgcttgcaaacaaaaaaaccaccgctaccagcggtgg
tttgtttgccggatcaagagctaccaactctttttccgaaggtaactggcttcagcagagcgcagatacc
aaatactgtccttctagtgtagccgtagttaggccaccacttcaagaactctgtagcaccgcctacatac
ctcgctctgctaatcctgttaccagtggctgctgccagtggcgataagtcgtgtcttaccgggttggact
caagacgatagttaccggataaggcgcagcggtcgggctgaacggggggttcgtgcacacagcccagctt
ggagcgaacgacctacaccgaactgagatacctacagcgtgagctatgagaaagcgccacgcttcccgaa
gggagaaaggcggacaggtatccggtaagcggcagggtcggaacaggagagcgcacgagggagcttccag
ggggaaacgcctggtatctttatagtcctgtcgggtttcgccacctctgacttgagcgtcgatttttgtg
atgctcgtcaggggggcggagcctatggaaaaacgccagcaacgcggcctttttacggttcctggccttt
tgctggccttttgctcacatgttctttcctgcgttatcccctgattctgtggataaccgtattaccgcct
ttgagtgagctgataccgctcgccgcagccgaacgaccgagcgcagcgagtcagtgagcgaggaagcgga
agagcgcctgatgcggtattttctccttacgcatctgtgcggtatttcacaccgcatatatggtgcactc
tcagtacaatctgctctgatgccgcatagttaagccagtatacactccgctatcgctacgtgactgggtc
atggctgcgccccgacacccgccaacacccgctgacgcgccctgacgggcttgtctgctcccggcatccg
cttacagacaagctgtgaccgtctccgggagctgcatgtgtcagaggttttcaccgtcatcaccgaaacg
cgcgaggcagctgcggtaaagctcatcagcgtggtcgtgaagcgattcacagatgtctgcctgttcatcc
gcgtccagctcgttgagtttctccagaagcgttaatgtctggcttctgataaagcgggccatgttaaggg
cggttttttcctgtttggtcactgatgcctccgtgtaagggggatttctgttcatgggggtaatgatacc
gatgaaacgagagaggatgctcacgatacgggttactgatgatgaacatgcccggttactggaacgttgt
gagggtaaacaactggcggtatggatgcggcgggaccagagaaaaatcactcagggtcaatgccagcgct
tcgttaatacagatgtaggtgttccacagggtagccagcagcatcctgcgatgcagatccggaacataat
ggtgcagggcgctgacttccgcgtttccagactttacgaaacacggaaaccgaagaccattcatgttgtt
gctcaggtcgcagacgttttgcagcagcagtcgcttcacgttcgctcgcgtatcggtgattcattctgct
aaccagtaaggcaaccccgccagcctagccgggtcctcaacgacaggagcacgatcatgcgcacccgtgg
ggccgccatgccggcgataatggcctgcttctcgccgaaacgtttggtggcgggaccagtgacgaaggct
tgagcgagggcgtgcaagattccgaataccgcaagcgacaggccgatcatcgtcgcgctccagcgaaagc
ggtcctcgccgaaaatgacccagagcgctgccggcacctgtcctacgagttgcatgataaagaagacagt
cataagtgcggcgacgatagtcatgccccgcgcccaccggaaggagctgactgggttgaaggctctcaag
ggcatcggtcgagatcccggtgcctaatgagtgagctaacttacattaattgcgttgcgctcactgcccg
ctttccagtcgggaaacctgtcgtgccagctgcattaatgaatcggccaacgcgcggggagaggcggttt
gcgtattgggcgccagggtggtttttcttttcaccagtgagacgggcaacagctgattgcccttcaccgc
ctggccctgagagagttgcagcaagcggtccacgctggtttgccccagcaggcgaaaatcctgtttgatg
gtggttaacggcgggatataacatgagctgtcttcggtatcgtcgtatcccactaccgagatatccgcac
caacgcgcagcccggactcggtaatggcgcgcattgcgcccagcgccatctgatcgttggcaaccagcat
cgcagtgggaacgatgccctcattcagcatttgcatggtttgttgaaaaccggacatggcactccagtcg
ccttcccgttccgctatcggctgaatttgattgcgagtgagatatttatgccagccagccagacgcagac
gcgccgagacagaacttaatgggcccgctaacagcgcgatttgctggtgacccaatgcgaccagatgctc
cacgcccagtcgcgtaccgtcttcatgggagaaaataatactgttgatgggtgtctggtcagagacatca
agaaataacgccggaacattagtgcaggcagcttccacagcaatggcatcctggtcatccagcggatagt
taatgatcagcccactgacgcgttgcgcgagaagattgtgcaccgccgctttacaggcttcgacgccgct
tcgttctaccatcgacaccaccacgctggcacccagttgatcggcgcgagatttaatcgccgcgacaatt
tgcgacggcgcgtgcagggccagactggaggtggcaacgccaatcagcaacgactgtttgcccgccagtt
gttgtgccacgcggttgggaatgtaattcagctccgccatcgccgcttccactttttcccgcgttttcgc
agaaacgtggctggcctggttcaccacgcgggaaacggtctgataagagacaccggcatactctgcgaca
tcgtataacgttactggtttcacattcaccaccctgaattgactctcttccgggcgctatcatgccatac
cgcgaaaggttttgcgccattcgatggtgtccgggatctcgacgctctcccttatgcgactcctgcatta
ggaagcagcccagtagtaggttgaggccgttgagcaccgccgccgcaaggaatggtgcatgcaaggagat
ggcgcccaacagtcccccggccacggggcctgccaccatacccacgccgaaacaagcgctcatgagcccg
aagtggcgagcccgatcttccccatcggtgatgtcggcgatataggcgccagcaaccgcacctgtggcgc
cggtgatgccggccacgatgcgtccggcgtagaggatcgagatctcgatcccgcgaaattaatacgactc
actataggggaattgtgagcggataacaattcccctctagaaataattttgtttaactttaagaaggaga
tatacatatgaaaaagaaaaacatctacagcattcgcaaacttggtgttggtattgcaagcgttaccctg
ggcaccctgctgattagcggtggtgttacaccggcagcaaatgcagcacagcatgatgaagcacagcaga
atgcagttgatgccaaatttgatgtggatgccaagttcgataaagaacagcagaacgccttttatgaaat
tctgcatctgccgaatctgaccgaagaacagcgtaatgcatttattcagagcctgaaagatgatccgagc
cagagcgcaaatctgctggccgaagctaaaaagctgaatgatgcacaggctccgaaacaggcaccgaaag
tggacgcaaaattcgatgtagacgcgaagtttgacaaagagcaacaaaacgcattctacgagattctgca
tttacctaacctgacagaggaacagcgcaacgcgtttatccagtcactgaaagacgacccgtcacagtca
gccaacctgctggcagaggccaaaaagttaaacgatgcccaggcacctaaacaagcccctaaagtagatg
ctaagtttgatgttgacgctaaatttgataaagaacaacagaatgctttctacgagatcttacatctgcc
gaacttaactgaagaacaacgcaatgccttcatccaatctttaaaggatgatccgtcacagagtgcaaat
ttactggcggaggcgaagaaactgaacgacgcgcaagcacctaagcaggctccaaaagtcgatgcgaaat
tcgacgttgatgcaaaattcgacaaagaacagcaaaatgcgttctatgaaatccttcacttacccaacct
tacggaagaacaaagaaacgcgttcatacagagcttaaaagatgacccttctcagtctgcgaatctgctt
gctgaggctaagaaacttaatgatgcccaagcgccaaaacatcatcaccatcaccattaatgaaagcttg
cggccgcactcgagcaccaccaccaccaccactgagatccggctgctaacaaagcccgaaaggaagctga
gttggctgctgccaccgctgagcaataactagcataaccccttggggcctctaaacgggtcttgaggggt
tttttgctgaaaggaggaactatatccggat</INSDSeq_sequence></INSDSeq></Sequence
Data><SequenceData
SequenceIDNumber="2"(SEQIDNO:2><INSDSeq><INSDSeq_length>960</INSDSeq_l
ength><INSDSeq_moltype>DNA</INSDSeq_moltype><INSDSeq_division>PAT</INS
DSeq_division><INSDSeq_feature-table><INSDFeature><INSDFeature_key>sou
rce</INSDFeature_key><INSDFeature_location>1..960</INSDFeature_locatio
n><INSDFeature_quals><INSDQualifier><INSDQualifier_name>mol_type</INSD
Qualifier_name><INSDQualifier_value>other
DNA</INSDQualifier_value></INSDQualifier><INSDQualifier
id="q4"><INSDQualifier_name>organism</INSDQualifier_name><INSDQualifie
r_value>synthetic
construct</INSDQualifier_value></INSDQualifier></INSDFeature_quals></I
NSDFeature></INSDSeq_feature-table><INSDSeq_sequence>atgaaaaagaaaaacat
ctacagcattcgcaaacttggtgttggtattgcaagcgttaccctgggcaccctgctgattagcggtggt
gttacaccggcagcaaatgcagcacagcatgatgaagcacagcagaatgcagttgatgccaaatttgatg
tggatgccaagttcgataaagaacagcagaacgccttttatgaaattctgcatctgccgaatctgaccga
agaacagcgtaatgcatttattcagagcctgaaagatgatccgagccagagcgcaaatctgctggccgaa
gctaaaaagctgaatgatgcacaggctccgaaacaggcaccgaaagtggacgcaaaattcgatgtagacg
cgaagtttgacaaagagcaacaaaacgcattctacgagattctgcatttacctaacctgacagaggaaca
gcgcaacgcgtttatccagtcactgaaagacgacccgtcacagtcagccaacctgctggcagaggccaaa
aagttaaacgatgcccaggcacctaaacaagcccctaaagtagatgctaagtttgatgttgacgctaaat
ttgataaagaacaacagaatgctttctacgagatcttacatctgccgaacttaactgaagaacaacgcaa
tgccttcatccaatctttaaaggatgatccgtcacagagtgcaaatttactggcggaggcgaagaaactg
aacgacgcgcaagcacctaagcaggctccaaaagtcgatgcgaaattcgacgttgatgcaaaattcgaca
aagaacagcaaaatgcgttctatgaaatccttcacttacccaaccttacggaagaacaaagaaacgcgtt
catacagagcttaaaagatgacccttctcagtctgcgaatctgcttgctgaggctaagaaacttaatgat
gcccaagcgccaaaacatcatcaccatcaccat</INSDSeq_sequence></INSDSeq></Sequen
ceData><SequenceData
SequenceIDNumber="3"(SEQIDNO:3)><INSDSeq><INSDSeq_length>320</INSDSeq_
length><INSDSeq_moltype>AA</INSDSeq_moltype><INSDSeq_division>PAT</INS
DSeq_division><INSDSeq_feature-table><INSDFeature><INSDFeature_key>sou
rce</INSDFeature_key><INSDFeature_location>1..320</INSDFeature_locatio
n><INSDFeature_quals><INSDQualifier><INSDQualifier_name>mol_type</INSD
Qualifier_name><INSDQualifier_value>protein</INSDQualifier_value></INS
DQualifier><INSDQualifier
id="q6"><INSDQualifier_name>organism</INSDQualifier_name><INSDQualifie
r_value>synthetic
construct</INSDQualifier_value></INSDQualifier></INSDFeature_quals></I
NSDFeature></INSDSeq_feature-table><INSDSeq_sequence>MKKKNIYSIRKLGVGIA
SVTLGTLLISGGVTPAANAAQHDEAQQNAVDAKFDVDAKFDKEQQNAFYEILHLPNLTEEQRNAFIQSLK
DDPSQSANLLAEAKKLNDAQAPKQAPKVDAKFDVDAKFDKEQQNAFYEILHLPNLTEEQRNAFIQSLKDD
PSQSANLLAEAKKLNDAQAPKQAPKVDAKFDVDAKFDKEQQNAFYEILHLPNLTEEQRNAFIQSLKDDPS
QSANLLAEAKKLNDAQAPKQAPKVDAKFDVDAKFDKEQQNAFYEILHLPNLTEEQRNAFIQSLKDDPSQS
ANLLAEAKKLNDAQAPKHHHHHH</INSDSeq_sequence></INSDSeq></SequenceData></S
T26SequenceListing>
<---




Группа изобретений относится к биотехнологии. Предложена рекомбинантная плазмида pETZ, имеющая нуклеотидную последовательность SEQ ID NO:1, размер 6348 пар нуклеотидов и содержащая в своем составе элементы в соответствии с физической и генетической картой, представленной на фиг.2. Рекомбинантная плазмида pETZ обеспечивает синтез и секрецию рекомбинантного белка А в периплазматическое пространство клеток E. coli в растворимой форме. Рекомбинантный белок А имеет аминокислотную последовательность SEQ ID NO:3 и молекулярную массу 35,8 кДа. Рекомбинантный белок А может быть использован для иммунохимических анализов. Изобретения обеспечивают синтез и секрецию белка А в клетках E. coli до 100 мг/л культуральной среды и упрощение технологии очистки указанного белка А. 2 н.п. ф-лы, 8 ил., 2 пр.
1. Рекомбинантная плазмида pETZ, обеспечивающая синтез и секрецию рекомбинантного белка А в периплазматическое пространство клеток E. coli в растворимой форме, имеющая нуклеотидную последовательность SEQ ID NO:1, размер 6348 пар нуклеотидов и содержащая в своем составе в соответствии с физической и генетической картой, представленной на фиг. 2, следующие элементы:
- AmpR promoter - участок связывания РНК-полимеразы (494-598 п. н.);
- участок, обеспечивающий экспрессию β-лактамазы - фактора резистентности к ампициллину (599-1459 п. н.);
- участок начала репликации ori (1630-2218 п. н.);
- lacI - участок, кодирующий белок, связывающийся с ДНК и ингибирующий экспрессию целевого белка до добавления изопропил-β-D-тиогалактопиранозида (ИПТГ) (3648-4730 п. н.);
- lacI promoter - участок связывания РНК-полимеразы (4731-4808 п. н.);
- T7 promoter - участок связывания РНК-полимеразы бактериофага Т7 (5117-5135 п. н.);
- lac operator - участок связывания белка lacI (5136-5160 п. н.);
- RBS - сайт связывания рибосомы (5191-5196);
- z-gene-optim - ген, кодирующий рекомбинантный белок А, имеющий нуклеотидную последовательность SEQ ID NO:2 и координаты с 5205 по 6164 п. н.;
- T7 terminator - терминатор транскрипции (6276-6323 п. н.);
- 6×His-метка, предназначенная для хроматографической очистки белка А и имеющая координаты с 6147 по 6164 п.н.
2. Рекомбинантный белок А, продуцируемый плазмидой pETZ по п. 1, трансформированной в клетки E. coli, и используемый для иммунохимических анализов, имеющий аминокислотную последовательность SEQ ID NO:3 и молекулярную массу 35,8 кДа.
| CN 105481954 А, 13.04.2016 | |||
| SU 1524485 A1, 20.11.1995 | |||
| Механизм автоматических отделочных форм в стеклодельных машинах конвейерной системы | 1930 |
|
SU37818A1 |
| WO 2019039602 A1, 28.02.2019 | |||
| US 9657055 B2, 23.05.2017, формула, Seq ID No.4 | |||
| Способ получения лигатуры,содержащей никель | 1974 |
|
SU496307A1 |
| Машина для добывания торфа и т.п. | 1922 |
|
SU22A1 |
| Прибор для промывания газов | 1922 |
|
SU20A1 |
