Изобретение относится к биотехнологии, генетической и белковой инженерии, и касается нового рекомбинантного штамма бактерий Escherichia coli (Е. coli), который может быть использован для получения рекомбинантного белка ДНК-зависимой ДНК-полимеразы А.
ДНК-полимеразы - это семейство ферментов, ответственных за точное дублирование ДНК. Полимеразы различаются по размеру, структуре, потребности в вспомогательных белках и роли в репликации ДНК, также различаются и их специфические свойства, включая процессивность, точность синтеза, селективность к включаемым нуклеотидам. ДНК-полимеразы играют решающую роль не только в репликации и репарации ДНК in vivo, но и в методах, используемых в молекулярной биологии, особенно в полимеразной цепной реакции (ПЦР). ПЦР представляет собой быстрый, специфичный и чувствительный метод амплификации последовательностей нуклеиновых кислот, который широко используется в исследовательских и клинических лабораториях. Непрерывно ведется поиск новых ферментов для генетических технологий, которые могут иметь улучшенные свойства по сравнению с известными на сегодняшний день.
На данный момент в ПЦР активно используются ДНК-полимеразы, принадлежащие к семействам А и В. Наиболее известными представителями этих семейств являются полимеразы Taq из штамма бактерий Thermus aquaticus и Pfu из штамма бактерий Pyrococcus furiosus. С помощью методов генной инженерии для улучшения каталитических свойств были получены различные мутантные формы данных ферментов. Несмотря на многообещающие результаты, ограничениями для ПЦР являются выход продукта, длина ДНК, которая может быть амплифицирована, скорость полимеразы и точность процесса. В конечном счете, время выполнения ПЦР зависит исключительно от кинетических свойств фермента.
Поиск новых термостабильных ДНК-полимераз представляет собой нетривиальную задачу. В настоящее время нет полного понимания причин термостабильности фермента, поэтому поиск термостабильных ДНК-полимераз выполняют в термофильных организмах, оптимум жизни которых находится не ниже 50-60°С. Однако, известны организмы, живущие при повышенных вплоть до 80°С температурах, полимеразы которых хоть и работают при таких температурах, но их стабильность значительно снижается при более высоких температурах. С другой стороны, можно предположить, что фермент определенного организма может оставаться стабильным при температурах, превышающих оптимальную температуру роста данного организма.
Были проанализированы структурные особенности ДНК-полимераз семейства А, на основе филогенетического анализа ферментов был установлен консенсусный «слепок», включающий 62 высококонсервативных аминокислотных остатка, распределенных в структуре фермента и непосредственно контактирующих с субстратами и кофакторами. Набор этих консервативных остатков может выступать функциональным слепком, характеризующим ДНК-полимеразы семейства А в целом. Были проанализированы последовательности полимераз всех микроорганизмов Коллекции экстремофильных микроорганизмов и типовых культур (КЭМТК) ИХБФМ СО РАН (http://www.niboch.nsc.ru/doku.php/emtc_collection). Из 4000 микроорганизмов было отобрано 44 с оптимумом температуры среды выше 50°С. Проведение сравнительного анализа остатков, входящих в функциональный слепок, у малоизученных ДНК-полимераз с ферментами из рода Thermus позволило выявить ряд ферментов, потенциально обладающих свойствами, необходимыми для использования в биотехнологии (Булыгин А.А., Кузнецова А.А., Федорова О.С., Кузнецов Н.А. Анализ функциональных особенностей ДНК-полимераз семейства А как инструмент поиска ферментов с новыми свойствами. Молекулярная биология, 2023, Т. 57 (2), С.185-196).
Одним из организмов с высокой схожестью функционального слепка является Massilia aurea (78,9%). Данный микроорганизм был выделен из пробы термальной воды (74°С), Камчатка, Кальдера вулкана Узон.
На сегодняшний день аналогов штаммов-продуцентов ДНК-полимеразы А из Massilia aurea не обнаружено.
Наиболее близким к заявляемому штамму- прототипом, является штамм бактерий Thermus thermophiles ВКПМ В-4892, выделенный из горячих вод Камчатки (Долина Гейзеров) и обеспечивающий получение термостабильной ДНК полимеразы (патент SU 1839189 А1, опубл. 20.12.1993). Недостатком известного штамма является его неспособность продуцировать рекомбинантную ДНК-полимеразу А из Massilia aurea.
Задачей изобретения является расширение ассортимента штаммов -продуцентов ДНК-полимеразы А.
Технический результат изобретения заключается в получении рекомбинантного штамма-продуцента, обеспечивающего получение рекомбинантной ДНК-полимеразы А из Massilia aurea с чистотой не менее 90%.
Поставленная задача достигается созданием рекомбинантного штамма Е. coli Rosetta 2(DE3)/рЕТ28с-Mau-продуцента, полученного трансформацией культуры клеток Е. coli Rosetta 2(DE3) (генотип F- omp Thsd SB(rB-mB-) gal dcm (DE3) pRARE2 (CamR)) рекомбинантной плазмидной ДНК pET228c-Mau, сконструированной на основе вектора рЕТ28с, несущего ген, кодирующего ДНК-полимеразу А из Massilia aurea.
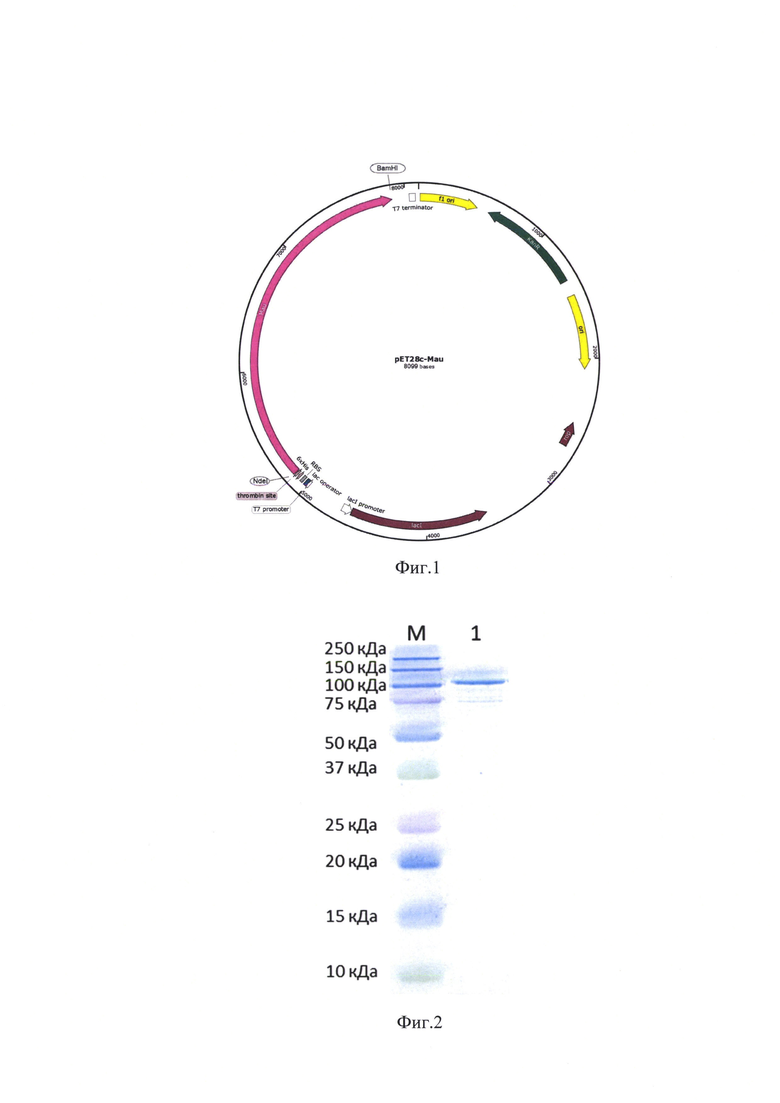
На фиг.1 представлена физическая карта плазмиды рЕТ28с-Mau, содержащей фрагмент ДНК, кодирующий ДНК-полимеразу А из Massilia aurea, с 1 по 913 аминокислотный остаток и N-концевой гистидиновый тракт. Указаны сайты для эндонуклеаз рестрикции Not I, ВатН I; сайт узнавания для тромбина; промотор фага Т7, промотор lac I; фрагмент ДНК, кодирующий ДНК-полимеразу А из Massilia aurea, с 1 по 913 аминокислотный остаток и N-концевой гистидиновый тракт; ori репликации плазмиды, f1 ori репликации для включения одноцепочечной репликации; KanR - ген устойчивости к канамицину.
Культурально-морфологические особенности штамма Е. coli Rosetta 2(DE3)/pET28c-Mau: грамотрицательные прямые палочки, размером 1,1-1,5×2,0-3,0 мкм, одиночные, спор и капсул не образуют. Каталазоположительные. Оксидазоотрицательные. Факультативные анаэробы. Клетки хорошо растут на простых питательных средах, содержащих и не содержащих хлорамфеникол и канамицин, например, на среде LB (питательная среда Lisogeny Broth). На агаризованной среде - колонии гладкие, круглые, слабовыпуклые, с ровным краем. В жидких средах образуют равномерную светорассеивающую суспензию, при хранении без перемешивания оседают на дно. Клетки растут в интервале температур от 8°С до 43°С, интервал для наиболее эффективного культивирования - 28-38°С, оптимум роста при 37°С. Оптимальный интервал рН для культивирования рН 5-7. Изначальный штамм Е. coli Rosetta 2 (DE3) проявляет устойчивость к хлорамфениколу (34 мкг/мл), обусловленную наличием гена устойчивости в ДНК рекомбинантной плазмиды pRARE2.
Характеристики полезного вещества, синтезируемого штаммом: рекомбинантная ДНК-полимераза Mau длиной 941 аминокислотных остатков, состоящая из 3'-5'-экзонуклеазного и полимеразного доменов, линкера (10 аминокислот), содержащего сайт узнавания тромбина, гистидиновой метки (6 аминокислот).
Продуктивность созданного штамма: рекомбинантная ДНК-полимераза Mau составляет не менее 20% белка клеточного лизата при культивировании в жидкой питательной среде LB при 37°С, 250 об/мин, в условиях индукции 0,2 мМ ИПТГ (изопропил-β-D-1-тиогалактопиранозид) в течение 3 часов с добавлением антибиотика канамицина (50 мкг/мл).
Нуклеотидная последовательность синтетического гена, кодирующего ДНК-полимеразу А из Massilia aurea (SEQ ID NO 1), а также аминокислотная последовательность самой ДНК-полимеразы A (SEQ ID NO 2) приведены в перечне последовательностей.
Сущность изобретения поясняется следующими конкретными примерами получения и использования штамма Е. coli Rosetta 2(DE3)/pET28c-Mau.
Пример 1. Создание генетической конструкции, обеспечивающей синтез рекомбинантной ДНК-полимеразы А в клетках Е. coli.
Из штамма клеток Massilia aurea была выделена суммарная ДНК. На основании последовательности гена ДНК-полимеразы А, имеющейся в базе данных Uniprot, были синтезированы праймеры, соответствующие началу и концу гена и содержащие нуклеотидные последовательности с сайтами рестрикции для Nde I и BamH I, Наработка гена ДНК-полимеразы А была произведена методом ПЦР.
Клонирование в экспрессионный вектор осуществлялось по сайтам рестрикции Nde I и BamH I: фрагмент ДНК и вектор подвергались обработке обозначенными эндонуклеазами рестрикции, рестрикционные смеси очищались. Затем проводилось лигирование фрагментов при +4°С в течении 16 ч. После чего полученной лигазной смесью трансформировались клетки штамма Escherichia coli DH5α. Позитивные по данным метода ПЦР колонии были пересеяны в ночные культуры, из ночных культур выделялась плазмидная ДНК для подтверждения последовательности секвенированием по Сэнгеру. Последовательность нуклеотидов синтезированного фрагмента гена соотносится с необходимой аминокислотной последовательностью.
На фиг.1 представлена физическая карта плазмиды рЕТ28 с-Mau, содержащая ориджин репликации f1, промотор фага Т7, RBS-сайт посадки рибосомы, сайты рестрикции Nde I и BamH I, по которым встроен фрагмент гена, кодирующего кодирующий ДНК-полимеразу А из Massilia aurea, гистидиновую метку 6*His (His-tag), терминатор фага Т7, ген устойчивости к канамицину KanR, а также лактозный репрессор lacI и лактозный оператор.
Полученная плазмида рЕТ28с-Mau обеспечивает синтез в клетках Escherichia coli ДНК-зависимой ДНК-полимеразы А из Massilia aurea, содержащей 3'-5'-экзонуклеазный и полимеразный домены, а также N-концевую гистидиновую метку, предназначенную для последующей очистки рекомбинантного белка Mau с помощью металлохелатной хроматографии.
Нуклеотидная последовательность рекомбинантной плазмиды рЕТ28с-Mau в SEQ ID NO 1 представлена в перечне последовательностей.
Пример 2. Получение штамма-продуцента рекомбинантной ДНК-полимеразы А и исследование его продуктивности.
Полученной плазмидой рЕТ28с-Mau были трансформированы клетки Е. coli штамма Rosetta 2(DE3), содержащие в своем геноме ген, кодирующий ДНК-зависимую РНК-полимеразу фага Т7, под контролем промотора lacUV5, индуцируемого лактозой или ИПТГ. Кроме того, клетки Е. coli Rosetta 2(DE3) дефектны по генам протеаз Lon и ompT, что снижает деградацию гетерологичных белков.
В результате был получен штамм Е. coli Rosetta 2(DE3)/ pET28c-Mau - продуцент рекомбинантной ДНК-полимеразы Mau из Massilia aurea. Для поддержания полученного штамма-продуцента белка Mau использовали плотную агаризованную LB-среду, содержащую 50 мкг/мл канамицина.
Продуктивность полученного штамма-продуцента изучали путем культивирования клеток в среде LB, в термостатированном шейкере роторного типа при температуре 37°С, скорости вращения платформы 250 об/мин до оптической плотности А600=0,5-0,6. После этого индуцировали синтез целевого белка добавлением ИПТГ до конечной концентрации 0,2 мМ. В качестве контроля использовали культуру без добавления ИПТГ. Образцы собранной центрифугированием биомассы клеток лизировали, используя пресс Френча, и анализировали методом электрофореза в 8%-ном полиакриламидном геле в системе Лэммли в денатурирующих условиях. В результате выявлено, что индукция 0.2 мМ ИПТГ культуры клеток Е. coli приводит к синтезу белка с молекулярной массой порядка 100 кДа, что соответствует ожидаемой молекулярной массе для рекомбинантного белка Mau с гистидиновой меткой (102,3 кДа). Количество рекомбинантного белка составило не менее 20% от клеточного лизата при культивировании в условиях индукции.
Пример 3. Очистка рекомбинантной ДНК-полимеразы А и изучение ее активности.
Очистка ДНК-полимеразы А представляла собой двухэтапную методику. На первой стадии проводили металл-хелатную аффинную хроматографию с помощью сорбента Ni SepharoseTM High Performance, который связывает His-taq содержащие полипептиды. На второй стадии проводили дополнительную хроматографическую очистку на колонке, содержащей гепарин.
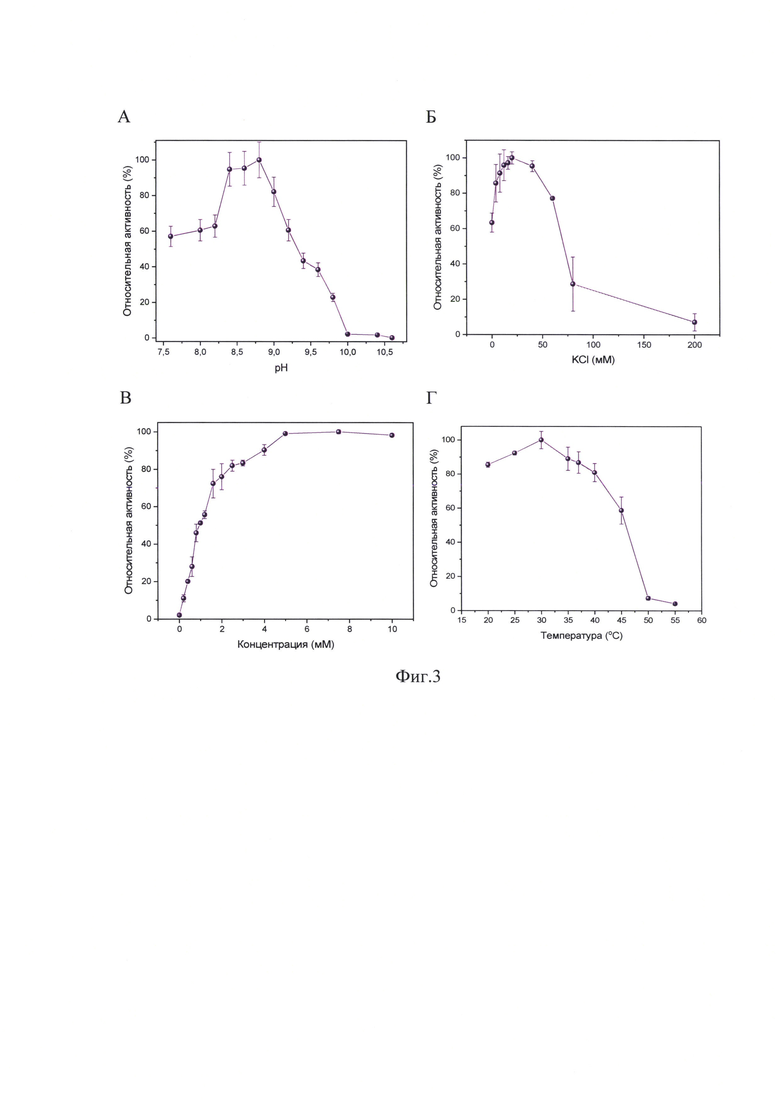
Результат двухстадийной очистки ДНК-полимеразы А представлен на фиг.2, где М - маркер подвижности 10-250 кДа (Kaleidoscope™); l - очищенная фракция рекомбинантной ДНК-полимеразы А. Как видно на фиг.2, препарат ДНК-полимеразы А является практически гомогенным, и его чистота составляет не менее 90%.
Для исследования активности ДНК-полимеразы A in vitro в качестве субстрата использовали ДНК-дуплекс, состоящий из 44-звенной ДНК-матрицы (Temp44-F, 5'-GGAGACATTTTGCCTTGATAGCTGCTCGACTCATCTGGGGGCCG-3') и ДНК-праймера длиной 24 пар оснований, содержащего FAM метку на 5'-конце (Pr_pol44-FAM, FAM-5'-CGGCCCCCAGATGAGTCGAGCAGC-3'), в конечной концентрации 50 нМ. Реакцию проводили при 30°С. Продукты реакции анализировали электрофорезом в 15%-полиакриламидном геле в денатурирующих условиях с последующей визуализацией с помощью гель-документирующей системы Е-Вох СХ.5 TS (Vilber Lourman, France).
Были определены оптимальные условия для протекания полимеразной реакции под действием ДНК-полимеразы А. Влияние рН (А), KCl (Б), ионов Mg2+ (В) и температуры (Г) на ферментативную активность ДНК-полимеразы А продемонстрировано на фиг.3. Показано, что оптимальными буферными условиями являются: 20 мМ KCl, 2 мМ Mg2+ при рН 8,8 (50 мМ Tris/HCl) и температура 30°С.
Таким образом, создан штамм бактерий Escherichia coli Rosetta 2(DE3)/pET28c-Mau, являющийся продуцентом рекомбинантной ДНК-полимеразы А, состоящей из 3'-5'-экзонуклеазного и полимеразного доменов и способной амплифицировать ДНК in vitro.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing originalFreeTextLanguageCode="ru"
dtdVersion="V1_3" fileName="pET28с-Mau-1.xml" softwareName="WIPO
Sequence" softwareVersion="2.2.0" productionDate="2023-04-27">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>1</ApplicationNumberText>
<FilingDate>2023-04-27</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>1</ApplicantFileReference>
<EarliestPriorityApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>1</ApplicationNumberText>
<FilingDate>2023-04-27</FilingDate>
</EarliestPriorityApplicationIdentification>
<ApplicantName languageCode="ru">Федеральное государственное
бюджетное учреждение науки Институт химической биологии и
фундаментальной медицины Сибирского отделения Российской академии
наук (ИХБФМ СО РАН)</ApplicantName>
<ApplicantNameLatin>The Institute of Chemical Biology and
Fundamental Medicine of the Siberian Branch of the Russian Academy of
Sciences</ApplicantNameLatin>
<InventionTitle languageCode="ru">Рекомбинантный штамм бактерий
Escherichia coli Rosetta 2(DE3)/pET28с-Mau – продуцент ДНК-полимеразы
А</InventionTitle>
<SequenceTotalQuantity>2</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>8099</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..8099</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tggcgaatgggacgcgccctgtagcggcgcattaagcgcggcgggtgtg
gtggttacgcgcagcgtgaccgctacacttgccagcgccctagcgcccgctcctttcgctttcttccctt
cctttctcgccacgttcgccggctttccccgtcaagctctaaatcgggggctccctttagggttccgatt
tagtgctttacggcacctcgaccccaaaaaacttgattagggtgatggttcacgtagtgggccatcgccc
tgatagacggtttttcgccctttgacgttggagtccacgttctttaatagtggactcttgttccaaactg
gaacaacactcaaccctatctcggtctattcttttgatttataagggattttgccgatttcggcctattg
gttaaaaaatgagctgatttaacaaaaatttaacgcgaattttaacaaaatattaacgtttacaatttca
ggtggcacttttcggggaaatgtgcgcggaacccctatttgtttatttttctaaatacattcaaatatgt
atccgctcatgaattaattcttagaaaaactcatcgagcatcaaatgaaactgcaatttattcatatcag
gattatcaataccatatttttgaaaaagccgtttctgtaatgaaggagaaaactcaccgaggcagttcca
taggatggcaagatcctggtatcggtctgcgattccgactcgtccaacatcaatacaacctattaatttc
ccctcgtcaaaaataaggttatcaagtgagaaatcaccatgagtgacgactgaatccggtgagaatggca
aaagtttatgcatttctttccagacttgttcaacaggccagccattacgctcgtcatcaaaatcactcgc
atcaaccaaaccgttattcattcgtgattgcgcctgagcgagacgaaatacgcgatcgctgttaaaagga
caattacaaacaggaatcgaatgcaaccggcgcaggaacactgccagcgcatcaacaatattttcacctg
aatcaggatattcttctaatacctggaatgctgttttcccggggatcgcagtggtgagtaaccatgcatc
atcaggagtacggataaaatgcttgatggtcggaagaggcataaattccgtcagccagtttagtctgacc
atctcatctgtaacatcattggcaacgctacctttgccatgtttcagaaacaactctggcgcatcgggct
tcccatacaatcgatagattgtcgcacctgattgcccgacattatcgcgagcccatttatacccatataa
atcagcatccatgttggaatttaatcgcggcctagagcaagacgtttcccgttgaatatggctcataaca
ccccttgtattactgtttatgtaagcagacagttttattgttcatgaccaaaatcccttaacgtgagttt
tcgttccactgagcgtcagaccccgtagaaaagatcaaaggatcttcttgagatcctttttttctgcgcg
taatctgctgcttgcaaacaaaaaaaccaccgctaccagcggtggtttgtttgccggatcaagagctacc
aactctttttccgaaggtaactggcttcagcagagcgcagataccaaatactgtccttctagtgtagccg
tagttaggccaccacttcaagaactctgtagcaccgcctacatacctcgctctgctaatcctgttaccag
tggctgctgccagtggcgataagtcgtgtcttaccgggttggactcaagacgatagttaccggataaggc
gcagcggtcgggctgaacggggggttcgtgcacacagcccagcttggagcgaacgacctacaccgaactg
agatacctacagcgtgagctatgagaaagcgccacgcttcccgaagggagaaaggcggacaggtatccgg
taagcggcagggtcggaacaggagagcgcacgagggagcttccagggggaaacgcctggtatctttatag
tcctgtcgggtttcgccacctctgacttgagcgtcgatttttgtgatgctcgtcaggggggcggagccta
tggaaaaacgccagcaacgcggcctttttacggttcctggccttttgctggccttttgctcacatgttct
ttcctgcgttatcccctgattctgtggataaccgtattaccgcctttgagtgagctgataccgctcgccg
cagccgaacgaccgagcgcagcgagtcagtgagcgaggaagcggaagagcgcctgatgcggtattttctc
cttacgcatctgtgcggtatttcacaccgcatatatggtgcactctcagtacaatctgctctgatgccgc
atagttaagccagtatacactccgctatcgctacgtgactgggtcatggctgcgccccgacacccgccaa
cacccgctgacgcgccctgacgggcttgtctgctcccggcatccgcttacagacaagctgtgaccgtctc
cgggagctgcatgtgtcagaggttttcaccgtcatcaccgaaacgcgcgaggcagctgcggtaaagctca
tcagcgtggtcgtgaagcgattcacagatgtctgcctgttcatccgcgtccagctcgttgagtttctcca
gaagcgttaatgtctggcttctgataaagcgggccatgttaagggcggttttttcctgtttggtcactga
tgcctccgtgtaagggggatttctgttcatgggggtaatgataccgatgaaacgagagaggatgctcacg
atacgggttactgatgatgaacatgcccggttactggaacgttgtgagggtaaacaactggcggtatgga
tgcggcgggaccagagaaaaatcactcagggtcaatgccagcgcttcgttaatacagatgtaggtgttcc
acagggtagccagcagcatcctgcgatgcagatccggaacataatggtgcagggcgctgacttccgcgtt
tccagactttacgaaacacggaaaccgaagaccattcatgttgttgctcaggtcgcagacgttttgcagc
agcagtcgcttcacgttcgctcgcgtatcggtgattcattctgctaaccagtaaggcaaccccgccagcc
tagccgggtcctcaacgacaggagcacgatcatgcgcacccgtggggccgccatgccggcgataatggcc
tgcttctcgccgaaacgtttggtggcgggaccagtgacgaaggcttgagcgagggcgtgcaagattccga
ataccgcaagcgacaggccgatcatcgtcgcgctccagcgaaagcggtcctcgccgaaaatgacccagag
cgctgccggcacctgtcctacgagttgcatgataaagaagacagtcataagtgcggcgacgatagtcatg
ccccgcgcccaccggaaggagctgactgggttgaaggctctcaagggcatcggtcgagatcccggtgcct
aatgagtgagctaacttacattaattgcgttgcgctcactgcccgctttccagtcgggaaacctgtcgtg
ccagctgcattaatgaatcggccaacgcgcggggagaggcggtttgcgtattgggcgccagggtggtttt
tcttttcaccagtgagacgggcaacagctgattgcccttcaccgcctggccctgagagagttgcagcaag
cggtccacgctggtttgccccagcaggcgaaaatcctgtttgatggtggttaacggcgggatataacatg
agctgtcttcggtatcgtcgtatcccactaccgagatatccgcaccaacgcgcagcccggactcggtaat
ggcgcgcattgcgcccagcgccatctgatcgttggcaaccagcatcgcagtgggaacgatgccctcattc
agcatttgcatggtttgttgaaaaccggacatggcactccagtcgccttcccgttccgctatcggctgaa
tttgattgcgagtgagatatttatgccagccagccagacgcagacgcgccgagacagaacttaatgggcc
cgctaacagcgcgatttgctggtgacccaatgcgaccagatgctccacgcccagtcgcgtaccgtcttca
tgggagaaaataatactgttgatgggtgtctggtcagagacatcaagaaataacgccggaacattagtgc
aggcagcttccacagcaatggcatcctggtcatccagcggatagttaatgatcagcccactgacgcgttg
cgcgagaagattgtgcaccgccgctttacaggcttcgacgccgcttcgttctaccatcgacaccaccacg
ctggcacccagttgatcggcgcgagatttaatcgccgcgacaatttgcgacggcgcgtgcagggccagac
tggaggtggcaacgccaatcagcaacgactgtttgcccgccagttgttgtgccacgcggttgggaatgta
attcagctccgccatcgccgcttccactttttcccgcgttttcgcagaaacgtggctggcctggttcacc
acgcgggaaacggtctgataagagacaccggcatactctgcgacatcgtataacgttactggtttcacat
tcaccaccctgaattgactctcttccgggcgctatcatgccataccgcgaaaggttttgcgccattcgat
ggtgtccgggatctcgacgctctcccttatgcgactcctgcattaggaagcagcccagtagtaggttgag
gccgttgagcaccgccgccgcaaggaatggtgcatgcaaggagatggcgcccaacagtcccccggccacg
gggcctgccaccatacccacgccgaaacaagcgctcatgagcccgaagtggcgagcccgatcttccccat
cggtgatgtcggcgatataggcgccagcaaccgcacctgtggcgccggtgatgccggccacgatgcgtcc
ggcgtagaggatcgagatctcgatcccgcgaaattaatacgactcactataggggaattgtgagcggata
acaattcccctctagaaataattttgtttaactttaagaaggagatataccatgggcagcagccatcatc
atcatcatcacagcagcggcctggtgccgcgcggcagccatatgaccctgctgctcgttgacggttccag
ttatctctatcgcgcctaccacgcgctgcccgacctgcgcagccccgatggcttcccgaccggcgccatg
catggcatggtcaatatgttgcgccgcctgcgcgctgatttccctgcggcatacatcgcctgcgtcttcg
acgccaagggcaagaccttccgcgacgacctgtatcccgaatacaaggccacccgcgcctcgatgccgga
agacctgggcaagcagatcgaaccgatccacgaagtggtgcgccacatgggctggccgatcctgatggtg
gagggcgtggaggccgatgacgtgatcggcaccctggcggtgcaggcgacggcgcgcggcatgaagaccg
tggtctcgaccggcgacaaagacctggcgcagctggtcaacgacaaggtcatgctgatcaataccatgac
caacgagcgcatggacgaagccggcgtgctggccaagttcggagtggcgccgaaccgcatcatcgactac
ctgaccctggtcggagacaccgtcgacaacgtgccgggcgtggccaagtgcggccccaagacggccgtca
aatggctgaccctgcacggctcgctcgatggcgtgatccagaatgcgagcagcatcggcggcgccgtcgg
cgccaacctgcaggcggcgctggagtggctgccaaaggggcgcgaactgatcaccgtcaagaccgactgc
gacctggtcaagcacgtgatctcgttcgaggaaaccctggtcggccgcccggaagatgccgaggcgctgc
gcgatttctttgcgcgctacggcttcaagaccatgttgcgcgacctgggcggcgccaaggccggcgacgg
cagcgtgcgtccggccgccggcagcgcgccgggcggcggaccgctcaattcgccggagggcgctgccacc
ctggccggcatgccggtcatcaagggtgagtacgaaacgatcctgaccgacgaacagctcgacaaatggc
tggcgctggtcgacgcggccgagttgacctcggtcgacaccgaaaccacctcgctcgacccaatgacggc
cgagatggtcggcatttcgctgtcggtcgaggtgggcaagggcgcctacatcccggtggcgcaccgctat
gccggcgcgccggaccagctgtcgcgcgaacacgtgctggaaaaaatgcgcgcctggctcgagaacccgg
ccaagccgaaggtgggccagaacctcaaatacgatatgcacatcttcgccaaccatggcgtaagcctgaa
aggcatcgtgcacgacacgctgctgcaatcgtatgtgttcgagtcgcacaagccgcacgatatggacacg
atggcgatgcgccatctcggctacaccacgattccctacgtcgacgtgtgcggcaagggcgccaagcaga
tctgcttcgatcaggtcgagctgggacgcgccaccgagtacgcgggcgaggattcggacatcaccctgcg
cctgcatcagagcatgctgccggaagtcgagaaagatgcaggcctgactcgcatctaccgcgagatcgag
atgccgaccatggaagtgctgcacaagatcgagcgtaacggcgtgctgatcgacagcgcgctgctcgacg
tgcaatcgggcgagcttggcaagcgcatgatggagctggaacagcaggcctatgaagcggccggcggccc
gttcaacctgggctcgcccaagcagatcggcgagatcttcttcggcaagctcggcctgccggtgatcaag
aagaccgcgaccggcgcgccgtcgaccgacgaagaagtgctgcaaaaactggccgaggattatccgctgc
cgaagatcctgctcgagcaccgtggcatgtcgaaactcaagtcgacctataccgacaagctgccgaagat
ggtcaacccgaacacgggccgcgtgcacacgaactacgcgcaggcggtggccatcaccggccgcctgtcg
tcgaacgagccgaacctgcagaacatcccggtgcgtaatgccgaaggccgccgcatccgcgaagccttca
tcgccgcgccgggccacgtgatcgtctcagccgactattcgcagatcgagctgcgcatcatggcgcatat
ctcgggcgacgccgcgatgctcaaggccttcgccgacggcgaagacatccaccgcgccaccgccgccgag
atcttcggcattgcacctgatgaagtgcagagcgagcagcgccgctacgccaaggtcatcaacttcggcc
tgatctacggcatgagcgccttcggcctggccggtaacctgggcatcgaacgcgccgcggccgccaacta
catcgagcgctatttcgcgcgcttctcgggcgtgaagcagtatatggacaacacgcgcctggaagcgaag
gcgcgcggctatgtcgagaccgtgttcggccggcgcctgtggctgcccgagatcaattcgccgaacggcc
cgcgccgcgccggcgccgaacgcgcggcgatcaatgcgccgatgcagggcacggcggccgacctgatcaa
aatggcgatgatcgcggtgcaggggtggatcgagggcgaaaagctcggcacccgcatgatcatgcaggtg
cacgacgaactggtgctggaggtgccggaaggcgagctggaactggtgcgggtcaagctgcctgaattga
tggcgggcgtggccaagctggatgtgccgctgctggcggagaccggtgttggcaagaactgggaagaggc
gcactgaggatccgaattcgagctccgtcgacaagcttgcggccgcactcgagcaccaccaccaccacca
ctgagatccggctgctaacaaagcccgaaaggaagctgagttggctgctgccaccgctgagcaataacta
gcataaccccttggggcctctaaacgggtcttgaggggttttttgctgaaaggaggaactatatccggat
</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>941</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..941</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Massilia aurea</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MGSSHHHHHHSSGLVPRGSHMTLLLVDGSSYLYRAYHALPDLRSPDGFP
TGAMHGMVNMLRRLRADFPAAYIACVFDAKGKTFRDDLYPEYKATRASMPEDLGKQIEPIHEVVRHMGWP
ILMVEGVEADDVIGTLAVQATARGMKTVVSTGDKDLAQLVNDKVMLINTMTNERMDEAGVLAKFGVAPNR
IIDYLTLVGDTVDNVPGVAKCGPKTAVKWLTLHGSLDGVIQNASSIGGAVGANLQAALEWLPKGRELITV
KTDCDLVKHVISFEETLVGRPEDAEALRDFFARYGFKTMLRDLGGAKAGDGSVRPAAGSAPGGGPLNSPE
GAATLAGMPVIKGEYETILTDEQLDKWLALVDAAELTSVDTETTSLDPMTAEMVGISLSVEVGKGAYIPV
AHRYAGAPDQLSREHVLEKMRAWLENPAKPKVGQNLKYDMHIFANHGVSLKGIVHDTLLQSYVFESHKPH
DMDTMAMRHLGYTTIPYVDVCGKGAKQICFDQVELGRATEYAGEDSDITLRLHQSMLPEVEKDAGLTRIY
REIEMPTMEVLHKIERNGVLIDSALLDVQSGELGKRMMELEQQAYEAAGGPFNLGSPKQIGEIFFGKLGL
PVIKKTATGAPSTDEEVLQKLAEDYPLPKILLEHRGMSKLKSTYTDKLPKMVNPNTGRVHTNYAQAVAIT
GRLSSNEPNLQNIPVRNAEGRRIREAFIAAPGHVIVSADYSQIELRIMAHISGDAAMLKAFADGEDIHRA
TAAEIFGIAPDEVQSEQRRYAKVINFGLIYGMSAFGLAGNLGIERAAAANYIERYFARFSGVKQYMDNTR
LEAKARGYVETVFGRRLWLPEINSPNGPRRAGAERAAINAPMQGTAADLIKMAMIAVQGWIEGEKLGTRM
IMQVHDELVLEVPEGELELVRVKLPELMAGVAKLDVPLLAETGVGKNWEEAH</INSDSeq_sequence
>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
Изобретение относится к биотехнологии, генетической и белковой инженерии. Предложен штамм бактерий Escherichia coli Rosetta 2(DE3)/pET28c-Mau, являющийся продуцентом рекомбинантной ДНК-полимеразы А из Massilia aurea. Указанный штамм получают путем трансформации клеток Escherichia coli штамма Rosetta 2(DE3) плазмидой pET28c-Mau, сконструированной на основе вектора рЕТ28с и имеющей нуклеотидную последовательность SEQ ID NO 1, содержащую ген, кодирующий ДНК-полимеразу А из Massilia aurea, имеющую аминокислотную последовательность SEQ ID NO 2, слитую с гистидиновым трактом на N-конце. Изобретение обеспечивает получение рекомбинантной ДНК-полимеразы А из Massilia aurea с чистотой не менее 90%. 3 ил., 3 пр.
Рекомбинантный штамм бактерий Escherichia coli Rosetta 2(DE3)/pET28c-Mau - продуцент ДНК-полимеразы А из Massilia aurea, полученный путем трансформации штамма Е. coli Rosetta 2(DE3) плазмидой ДНК pET28c-Mau, сконструированной на основе вектора рЕТ28с и имеющей нуклеотидную последовательность SEQ ID NO 1, содержащую ген, кодирующий ДНК-полимеразу А из Massilia aurea, имеющую аминокислотную последовательность SEQ ID NO 2, слитую с гистидиновым трактом на N-конце.
| Lawyer FC, et al | |||
| Isolation, characterization, and expression in Escherichia coli of the DNA polymerase gene from Thermus aquaticus | |||
| J Biol Chem | |||
| Механизм для сообщения поршню рабочего цилиндра возвратно-поступательного движения | 1918 |
|
SU1989A1 |
| ТЕРМОСТАБИЛЬНАЯ ДНК-ПОЛИМЕРАЗА, СПОСОБ ЕЕ ПОЛУЧЕНИЯ, ФРАГМЕНТ ВЫДЕЛЕННОЙ ДНК, СПОСОБ АМПЛИФИКАЦИИ ДНК, СПОСОБ ВВЕДЕНИЯ МЕТКИ В ДНК И СПОСОБ ОБРАТНОЙ ТРАНСКРИПЦИИ | 1997 |
|
RU2197524C2 |
| CN 104480081 A, 01.04.2015 | |||
| Bulygin, A.A., Kuznetsova, A.A., Fedorova, O.S | |||
| et al | |||
| Comparative Analysis of Family A DNA-Polymerases as a | |||

