Изобретение относится к биотехнологии, генетической и белковой инженерии, конкретно - к получению генетической конструкции, обеспечивающей синтез в клетках Escherichia coli рекомбинантного векторного pH-зависимого встраивающегося пептида pHLIP, обладающего способностью встраиваться в мембрану клеток при pH окружающей среды 7.0, предназначенного для диагностики и целевой доставки к зонам локального ацидоза.
Получение векторных молекул с целью создания систем для ранней диагностики и адресной терапии социально-значимых заболеваний крайне востребовано в современной биомедицине. Пептиды (молекулы белковой природы размером от 3 до 50 а.к.о.), обладающие меньшей молекулярной массой и более низкой иммуногенностью, по сравнению с белками, являются перспективными объектами современной молекулярной медицины [1].
Группа pH-зависимых встраивающихся пептидов (pHLIP) представляет собой группу высоко-гомологичных пептидов, содержащих специфический регион способный встраиваться в мембрану клетки при снижении pH среды 7.0 за счет протонирования остатков аспарагиновой кислоты и, соответственно, повышения гидрофобности пептида. При pH 7-10 пептид существует в виде неструктурированного мономера или связывается с поверхностью липидного бислоя. В кислом окружении пептид формирует трансмембранную альфа-спираль, в результате чего гидрофобный участок пептида пронизывает мембрану - его С-конец находится в клетке, а N-конец остается снаружи. Процесс встраивания сопровождается выделением энергии 1,8 ккал/моль, рассчитанная pKa для пептида составляет 6.0 [2]. Процесс встраивания в мембрану является обратимым - повышение pH вызывает конформационные изменения и пептид выходит из мембраны.
Известно, что для солидных опухолей характерен гликолитический путь метаболизма, в связи с чем, в результате экскреции кислых продуктов обмена наблюдается закисление межклеточной среды. Учитывая, что закисление межклеточного пространства, помимо опухолевого роста сопровождает так же такие процессы как воспаление, в том числе артрит и ишемию, диагностический и терапевтический потенциал векторного pH-зависимого встраивающегося пептида очень велик. Использование данного пептида позволяет селективно нацеливать контрастные метки в кислые ткани и переносить лекарственные молекулы через мембрану внутрь клетки при низком межклеточном pH [3].
Впервые рН-зависимый встраивающийся пептид был выделен из С-цепи бактериородопсина [4]. Пептид из группы pH-зависимых встраивающихся пептидов имеющий аминокислотную последовательность: NH2-ACEQNPIYWARYADWLFTTPLLLLDLALLVDADEGT-COOH имеет кодировку WT-pHLIP (wild type).
Использование pHLIP для нацеливания к опухоли предложено группой американских исследователей [5]. Описаны системы для диагностики и терапии, полученные за счет конъюгирования WT-pHLIP с флуоресцентными метками, в том числе для оптического имиджинга in vivo (NERF) [6] [7] [8] [9], метками для позитронной эмиссионной спектроскопии [10], радиоактивной меткой (99mTc) для однофотонной эмиссионной компьютерной томографии (SPECT-CT) [11].
Известны системы, полученные с использованием пептида pHLIP для нацеливания, визуализации и доставки терапевтической молекулы к солидным опухолям, таким как: карцинома легких мышей LLC и опухоли груди 4Т1 мышей, аденокарциномы простаты человека РС-3, LNCaP, АТСС CRL-1435, цервикальная аденокарцинома человека (HeLa), аденокарционома молочной железы человека MDA-MB-231, аденокарцинома поджелудочной железы человека [7] [10] [6] [8] [12] [9].
Существует система для нацеливания пептида к зонам ишемического поражения миокарда (на изолированном сердце) [13] [14].
Продемонстрировано использование пептида для доставки токсинов, таких как монометил ауристатин Е (ММАЕ) и α-аматин, подавляющих пролиферацию раковых клеток, а так же фаллоидина в раковые клетки [15] [14] [16]. Для этого биологически-активное вещество было конъюгировано с остатком цистеина на С-конце пептида за счет формирования дисульфидной связи, которая гидролизовалась в цитозоле клетки при встраивании пептида в мембрану. Показана успешная доставка анти-миРНК (пептидно-нуклеиновой кислоты), конъюгированной с pHLIP-пептидом, в опухоль для подавления роста [17].
Известен способ получения конъюгата pH-зависимого пептида с наночастицами золота, покрытыми люминесцентным европием [12]. Продемонстрирована успешная доставка нацеленных pHLIP наночастиц золота в опухоль и обсуждаются перспективы использования данной конструкции для диагностики и терапии (за счет активации золотых наночастиц «мягким» рентгеновским облучением).
Однако, не смотря на показанную эффективность использования pHLIP для выявления различных солидных опухолей и доставки лекарственных молекул в раковые клетки, данный пептид (состоящий из 37 а.к.о.) используется достаточно редко, что в первую очередь обусловлено трудностями получения длинного гидрофобного пептида.
В настоящее время пептид WT-pHLIP, равно как и все описанные соединения из группы pH-зависимых встраивающихся пептидов, получают методом твердофазного синтеза [18] [19]. Твердофазный синтез начинается со связывания карбоксильной группы защищенной аминокислоты пептида с соответствующей смолой, обычно через сложноэфирную или амидную связь. Проводят защиту групп боковых цепей аминокислот с помощью различных химических реагентов. Далее следуют повторяющиеся циклы наращивания аминокислотной последовательности, включающие: деблокирование аминогруппы промежуточно-образующейся N-терминально защищенной пептидной смолы, активация карбоксильной группы следующей за ней NH2-защищенной аминокислоты, сочетание аминокислот с формированием пептидной связи. В качестве сочетающего реагента применяют различные используемые в синтезе пептидов активирующие реагенты, например, карбодиимиды. Активированное аминокислотное производное обычно используется в 1,5 - 4-х кратном избытке. В случаях если происходит неполное сочетание, реакцию повторяют. После получения полноразмерной последовательности пептида проводят финальное деблокирование, снятие защиты с боковых групп аминокислот и отщепление готового пептида от смолы, например, с помощью трифторуксусной кислоты.
Данный метод рационален на этапе выбора пептида с оптимальными характеристиками, поиска пептида с высокой специфичностью к определенной мишени (патологической клетке, инфекционному агенту, очагу развития патологического процесса), в процессе которого может потребоваться перебор различных аминокислотных последовательностей. Однако, получение необходимого количества пептида, особенно гидрофобного и/или с длиной последовательности 20 и более а.к.о., методом твердофазного синтеза имеет ряд недостатков [20] [21], а именно: возникновение ошибочных последовательностей, пропуск аминокислот из-за неполноты протекания стадий деблокирования и аминообразования; самоассоциация пептидных цепей за счет формирования водородных связей, вызывающая экранирование N-концевой аминогруппы; а также высокая стоимость синтеза. Дополнительным негативным фактором химического синтеза является применение в работе токсичных растворителей и реагентов. Таким образом, получение pH-зависимого встраивающегося пептида в достаточных количествах для клинического применения методом твердофазного синтеза является высоко затратным.
Выгодной альтернативой для масштабирования процесса наработки пептида является получение его с использованием технологии рекомбинантных ДНК методом микробного синтеза. Это обуславливает необходимость получения рекомбинантного аналога векторного pH-зависимого встраивающегося пептида (pHLIP). В Российской Федерации и за рубежом рекомбинантные аналоги векторного pH-зависимого встраивающегося пептида (pHLIP) отсутствуют.
В настоящее время с использованием вектора pET31b получен ряд рекомбинантных пептидов, в том числе пептид rP11-4 (QQRFEWEFEQQ) [22], способный к полимеризации с образованием геля при снижении pH до 5.0. Данный пептид не используется для диагностики и/или терапии, предлагается его использование в тканевой инженерии.
На основе вектора pET31b получены четыре рекомбинантные плазмиды, обеспечивающие синтез пептидов антогонистов αvβ6 интегрина: A20fmdv2 (NAVPNLRGDLQVLAQKVART), A20lap2 (GFTTGRRGDLATIHGLNRPF), DBD1 (EKCPNLRGDLQVLAQKVCRT), TGFb3 (NEDDHQRGDLGRLKKQKDHH), предназначенные для диагностики и целевой доставки к опухолям поджелудочной железы и толстой кишки [23]. Нацеливание к опухоли в данном случае реализуется за счет высоко-аффинного взаимодействия и реализуется только в отношении опухолевых клеток, на поверхности которых сверхэкспрессирован интегрин. Тогда как нацеливание с использованием pHLIP, основано на феномене закисления межклеточного пространства - физиологическом маркере, характерном для всех солидных опухолей.
Новый технический результат - создание рекомбинантной плазмиды, обеспечивающей синтез рекомбинантного векторного pH-зависимого встраивающегося в мембрану клетки при закислении pH межклеточного пространства пептида pHLIP и наиболее приемлемого и экономически выгодного способа его синтеза.
Новый технический результат достигают рекомбинантной плазмидной ДНК pET31b-pHLIP, обеспечивающей синтез рекомбинантного векторного pH-зависимого встраивающегося пептида (pHLIP), в клетках Escherichia coli, имеющего следующую аминокислотную последовательность: NH2-ACEQNPIYWARYADWLFTTPLLLLDLALLVDADEGT-COOH и молекулярную массу 4,1 кДа, обладающего способностью встраиваться в мембрану клеток при pH окружающей среды 7.0, и представляющая собой плазмидный вектор pET31b+ гидролизованный по сайту AlwNI (PstNI), содержащий сайт инициации репликации плазмиды pBR322, промотор фага Т7, стартовый кодон, ген кодирующий кетостероидизомеразу, фрагмент размером 114 п.о. - сегмент, кодирующий pHLIP пептид, ограниченный с обеих концов тринуклеотидами, кодирующими метиониновые остатки, последовательность из 6-ти гистидиновых остатков, стоп-кодон, генетический маркер: AMPr - ген ампициллин резистентности, ген β-лактамазы, определяющий устойчивость к ампициллину при трансформации Escherichia coli.
Также, новый технический результат достигают способом получения рекомбинантного векторного pH-зависимого встраивающегося пептида (pHLIP), предусматривающем трансформацию клеток E. coli BL21 Rosetta DE3 pLysS, несущих ген РНК-полимеразы фага Т7 под индуцибельным lacUV5 промотором, сконструированной плазмидой pET31b-pHLIP по п. 1; выращивание трансформированных клеток в среде с ампициллином до среднелогарифмической стадии роста; индукцию экспрессии 0,5-1 мМ изопропилтиогалактазидом и последующим культивированием в течение 4-16 ч при 22-37°С; отделением клеток центрифугированием, отмывкой телец включения, содержащих рекомбинантный векторный рН-зависимый встраивающийся пептид (pHLIP) в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом, растворения их в буфере с добавлением мочевины, последующим отделением пептида от кетостероидизмеразы и 6 гистидиновых остатков с использованием бромциана и хроматографической очисткой рекомбинантного пептида.
Сущность изобретения заключается в в том, что конструируют плазмиду pET31b-pHLIP, путем встраивания в плазмидный вектор pET31b+ фрагмента ДНК, кодирующего векторный рН-зависимый встраивающийся пептид (pHLIP), трансформацией полученной плазмидой клеток Escherichia coli BL21 Rosetta DE3 pLysS, обеспечивающей синтез рекомбинантного векторного pH-зависимого встраивающегося пептида pHLIP в составе белка-слияния с кетостероидизомеразой и остатком из 6 гистидинов, последующего отделения пептида от белка-слияния с использованием бромциана и очистки пептида pHLIP методом ВЭЖХ.
Способ осуществляют следующим образом.
Генно-инженерными методами [24] получают плазмиду pET31b-pHLIP, содержащую последовательность, кодирующую рекомбинантный векторный рН-зависимый встраивающийся пептид (pHLIP), полученную методом отжига двух синтетических олигонуклеотидных последовательностей с последующей амплификацией методом полимеразной цепной реакции (ПЦР).
Клетки E. coli BL21 Rosetta DE3 pLysS, содержащие ген РНК-полимеразы фага Т7 под индуцибельным lacUV5 промотором, трансформируют сконструированной плазмидой pET31b-pHLIP и выращивают в течение ночи. Ночную культуру (1/100) засевают в свежую среду LB с добавлением ампициллина (50 мкг/мл). Синтез РНК-полимеразы индуцируют добавлением изопропилтиогалактазида (ИПТГ) в концентрации 0,5 мМ в тот момент, когда культура достигает среднелогарифмической фазы роста. Индуцированные клетки растят 4 часа при 28°С, после чего собирают центрифугированием при 5000 g. Индуцированные клетки E. coli BL21 Rosetta DE3 pLysS/pET31b-pHLIP используют для очистки векторного pH-зависимого встраивающегося пептида pHLIP в составе белка слияния с кетостероидизомеразой и остатком из шести гистидинов. Для получения векторного рН-зависимого встраивающегося пептида (pHLIP) выделенный химерный белок обрабатывают бромцианом. В результате получают рекомбинантный векторный рН-зависимый встраивающийся пептид (pHLIP), имеющий следующую аминокислотную последовательность: NH2-ACEQNPIYWARYADWLFTTPLLLLDLALLVDADEGT-COOH и молекулярную массу 4,1 кДа, кодируемый нуклеотидной последовательностью 5'-GCGTGCGAACAGAATCCGATTTATTGGGCGCGTTATGCGGATTGGCTG TTTACCACCCCGCTGCTGCTGCTGGATCTGGCGCTGCTGGTGGATGCGGATGAAGGCACC-3'.
Исходным генетическим материалом для конструирования рекомбинантной плазмиды pET31b-pHLIP являются:
а) плазмидный вектор pET31b+ (Novagen), обеспечивающий встройку фрагмента ДНК, кодирующего векторный рН-зависимый встраивающийся пептид (pHLIP) и его экспрессию в составе белка-слияния с кетостероидизомеразой и остатком из шести гистидинов под контролем позднего промотора Т7 ДНК-полимеразы;
б) фрагмент ДНК, кодирующий векторный рН-зависимый встраивающийся пептид pHLIP, который получают методом полимеразной цепной реакции с использованием синтетических олигонуклеотидов.
Полученная в результате плазмида pET31b-pHLIP (Фиг. 1) характеризуется следующими признаками:
- имеет молекулярную массу 1,8 МДа и размер 5856 п.н.;
- кодирует рекомбинантный векторный рН-зависимый встраивающийся пептид (pHLIP) в составе белка слияния с кетостероидизомеразой и остатком из 6 гистидинов;
- состоит из следующих элементов:
а) фрагмента ДНК, размером 114 п.о., кодирующего векторный рН-зависимый встраивающийся пептид (pHLIP)
б) плазмиды pET31b+, обеспечивающей эффективную транскрипцию ДНК, кодирующей векторный рН-зависимый встраивающийся пептид (pHLIP), и его экспрессию.
- содержит:
а) сайт инициации репликации плазмиды pBR322;
б) промотор бактериофага Т7;
в) генетические маркеры: AMPr - ген ампициллин резистентности (ген β-лактамазы), определяющий устойчивость к ампициллину при трансформации Е. coli и lacI - репрессор лактозного оперона Е. coli;
г) гены, кодирующие стартовый кодон, кетостероидизомеразу, рН-зависимый пептид (pHLIP), ограниченный с обоих концов остатками метионина, последовательность из 6-ти гистидиновых остатков, стоп-кадон;
д) уникальные сайты узнавания рестрикционной эндонукелазой PstNI (прототип AlwNI);
Таким образом, впервые получена плазмидная ДНК, обеспечивающая продукцию в клетках Е. coli рекомбинантного векторного pH-зависимого встраивающегося пептида (pHLIP), обладающего способностью встраиваться в мембрану клеток при pH окружающей среды 7.0.
Изобретение иллюстрируется Фиг. 1-9:
Фиг. 1. Нуклеотидная последовательность и кодируемая ею аминокислотная последовательность векторного pH-зависимого встраивающегося пептида (pHLIP).
Фиг. 2. Общая схема структурной организации плазмиды pET31b-pHLIP. pHLIP-ген, кодирующий векторный рН-зависимый встраивающийся пептид (pHLIP), KSI-ген кетостероидизомеразы, RBS - сайт посадки рибосом, Т7 - промотор фага Т7, AMPr-ген устойчивости к ампициллину, 6xHis - ген, кодирующий 6 остатков гистидина, lacI - ген, кодирующий репрессор лактозного оперона Е. coli;
Фиг. 3. Секвенирование кодирующего pHLIP фрагмента в составе рекомбинатной плазмиды pET31b-pHLIP. Сиквенс фрагмента, полученного в результате ПЦР с использованием в качестве матрицы рекомбинатной плазмиды pET31b-pHLIP и праймеров на Т7 промоторную и Т7 терминирующую области плазмиды (Т7 forward: 5'-taatacgactcactataggg-3', Т7 reverse: 5'-ctagttattgctcagcggtg-3').
Фиг. 4. Оптимизация условий экспрессии пептида pHLIP в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом в клетках E. coli BL21 Rosetta DE3 pLysS/pET31b-pHLIP. Электрофоретический анализ в 16% SDS-ПААГ. М - маркер молекулярного веса (Prestained Protein Molecular Weight Marker 20-120 kDa, Thermo Scientific), лизаты клеток: (дорожка 1) до индукции E. coli BL21 Rosetta DE3 pLysS/pET31b и после индукции экспрессии (дорожка 2) - конечная концентрация ИПТГ 1 мМ, культивирование при 37°С, (дорожка 3) - конечная концентрация ИПТГ 1 мМ, культивирование при 28°С, (дорожка 4) - конечная концентрация ИПТГ 1 мМ, культивирование при 22°С. (дорожка 5) - конечная концентрация ИПТГ 0,5 мМ, культивирование при 28°С, (дорожка 6) - конечная концентрация ИПТГ 0,5 мМ, культивирование при 22°С.
Фиг. 5. Электрофореграмма в 16% SDS-ПААГ белковых фракций и очищенного рекомбинантного пептида pHLIP в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом. М - маркер молекулярного веса белков (Spectra Multicolor Low Range Protein Ladder, Thermo Scientific), (дорожка 1) - лизат клеток E. coli BL21 Rosetta DE3 pLysS, содержащих плазмиду pET31b-pHLIP в буфере 50 mM Tris, 1% Triton-Х100, 100 mM NaCl, 10 mM DTT, pH 8.0 (дорожка 2) - солюбилизация телец включения, выделенных из клеток E. coli BL21 Rosetta DE3 pLysS, содержащих плазмиду pET31b-pHLIP, (дорожка 3) - фракции, не взаимодействующие с хроматографическим сорбентом, (дорожка 4, 5) - фракции, эллюируемые буфером 40 mM Tris, 500 mM NaCl, 15 mM имидазол, 8М мочевина, pH 8.0, (дорожка 6) - фракции, эллюируемые буфером 40 mM Tris, 500 mM NaCl, 300 mM имидазол, 8М мочевина, pH 8.0, содержащая векторный рН-зависимый встраивающийся пептид (pHLIP) в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом.
Фиг. 6. Анализ успешности отщепления пептида pHLIP, от белка-слияния KSI-pHLIP с использованием бромциана. Электрофореграмма в 16,5% трицин-ПААГ (дорожка 1) - полученного методом твердофазного синтеза (Bachem) пептида pHLIP, (дорожка 2) -растворенного лиофилизата, содержащего продукты расщепления белка-слияния KSI-pHLIP бромцианом, М - маркер молекулярного веса, кДа (Spectra Scientific Spectra Multicolor Low Range Protein Ladder, Thermo Scientific).
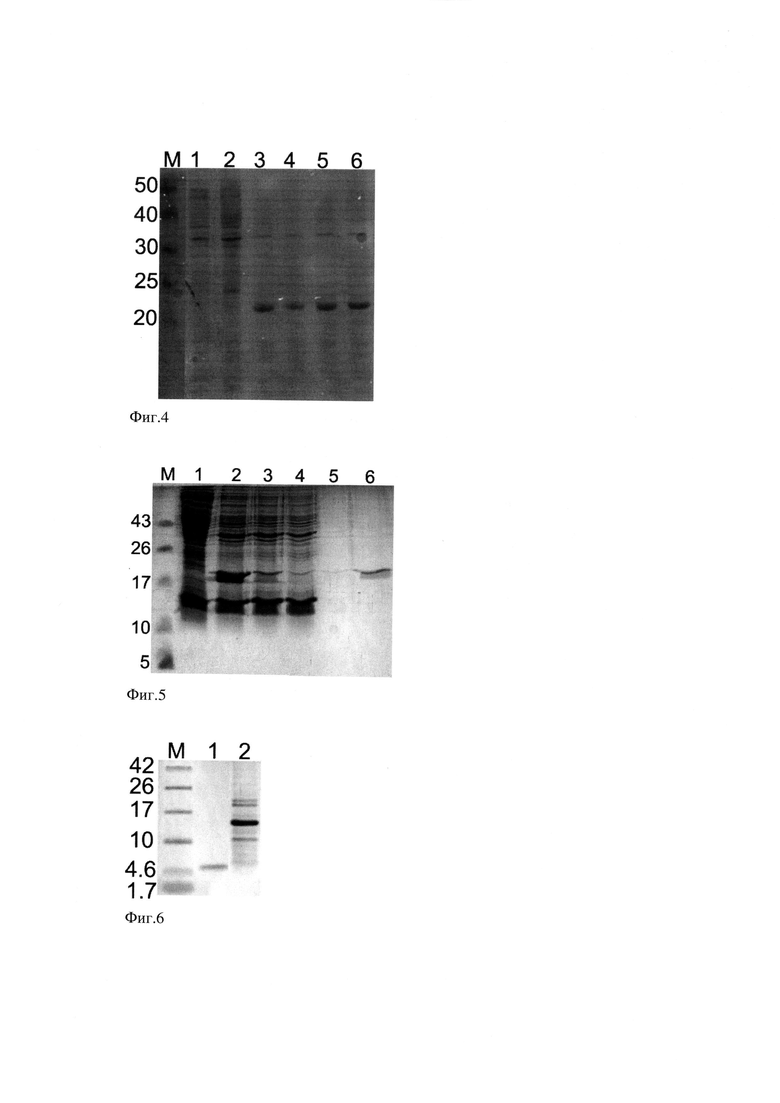
Фиг. 7. МАЛДИ масс-спектр (вверху) рекомбинантного пептида pHLIP выделенного из полиакриламидного геля и (внизу) пептида pHLIP, полученного методом твердофазного синтеза (Bachem).
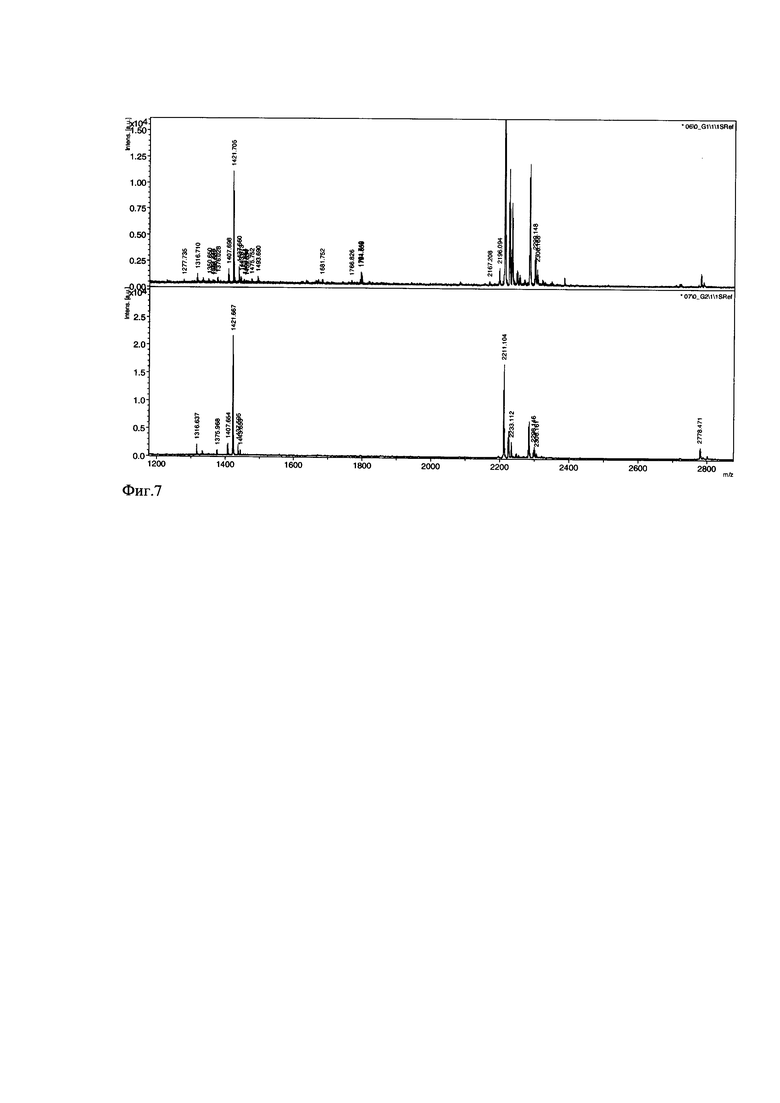
Фиг. 8. Хроматографическая очистка рекомбинантного пептида pHLIP. Профиль элюции супернатанта, содержащего пептид pHLIP на колонке Phenomex Luna 2 C18 (2) в градиенте концентрации ацетонитрила (5-50%) в 0,05% трифторуксусной кислоте. Скорость элюции 1 мл/мин.
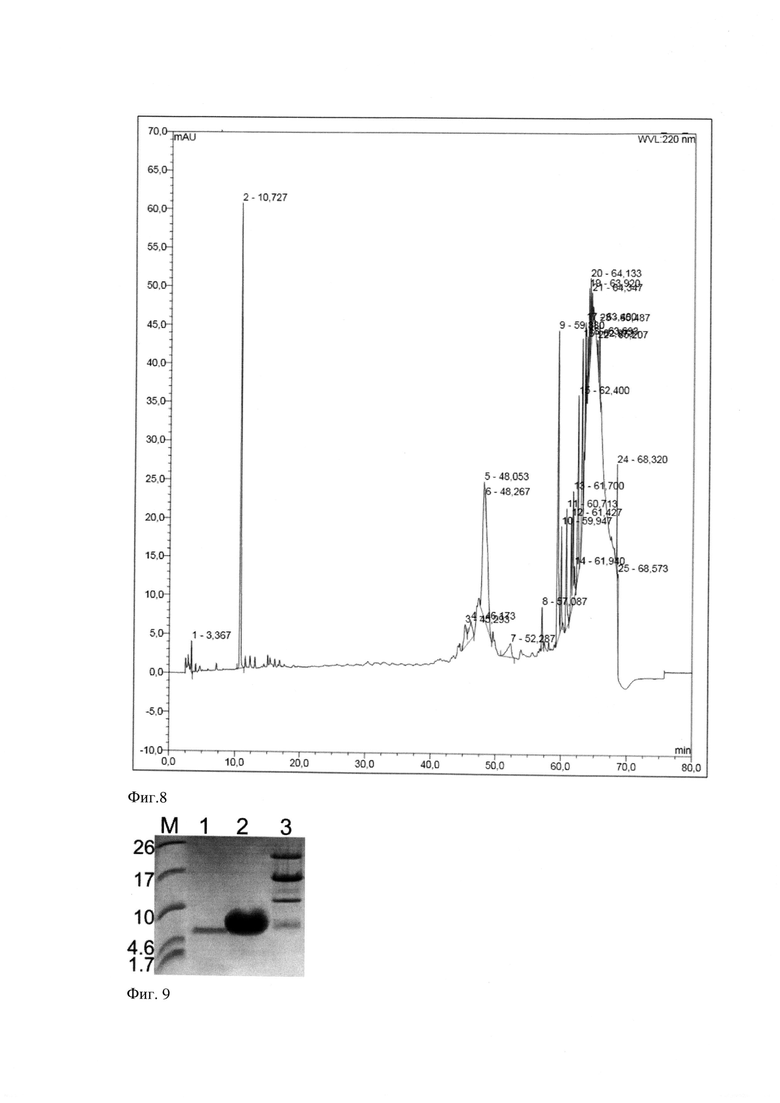
Фиг. 9. Анализ хроматографической фракции, содержащей очищенный рекомбинантный пептид pHLIP. Электрофореграмма в 16,5% трицин-ПААГ (дорожка 1) - полученного методом твердофазного синтеза (Bachem) пептида pHLIP, (дорожка 2) -рекомбинантного пептида pHLIP после хроматографической очистки, (дорожка 3) растворенного лиофилизата, содержащего продукты расщепления белка-слияния KSI-pHLIP бромцианом, М - маркер молекулярного веса, кДа (Spectra Scientific Spectra Multicolor Low Range Protein Ladder, Thermo Scientific).
Электрофореграмма в 16,5% трицин-ПААГ (дорожка 2).
Пример 1. Конструирование плазмиды pET31b-pHLIP.
Для получения последовательности кодирующей рекомбинантный pHLIP, используют два синтетических олигонуклеотидных праймера длинной 80 н.о.
Forward 5'-TCACCAGATGCTGATGGCGTGCGAACAGAATCCGATTTATTGGGCGCGTTATGCGGATTG GCTGTTTACCACCCCGCTGC-3'
Reverse 5'-GACACAGCATCTGCATGGTGCCTTCATCCGCATCCACCAGCAGCGCCAGATCCAGCAG CAGCAGCGGGGTGGTAAACAGC-3'
(подчеркиванием выделены последовательности обеспечивающих наличие в амплифицированном фрагменте сайтов рестрикции PstN1).
Готовят реакционную смесь содержащую: по 0,4 мкМ каждого праймера, смесь dNTP по 0,2 мМ каждого, 0,5 е.а. Encyclo полимеразы и Encyclo ПЦР буфер (Евроген, Россия).
Реакцию проводят в амплификаторе (ThermoCycler, Applied Biosystems) по следующей программе: начальная денатурация 95°С - 1 мин и далее 15 циклов - денатурация 95°С - 15 с, отжиг 60°С - 3 мин, элонгация 72°С - 30 с, финальная элонгация 72°С - 1 мин.
Продукт ПЦР разделяют в 1,2% агарозном геле с использованием 1×ТАЕ буфера с добавлением красителя бромистого этидия. Электрофорез проводят при постоянном электрическом токе 200 мА, напряжении 150 Вольт, мощности 1 Вт/см2.
Выделяют из геля фрагмент массой 140 п.н., соответствующий нуклеотидной последовательности:
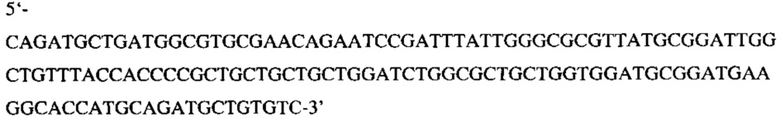
с использованием набора Cleanup Standard (Евроген, Россия).
Выделенный фрагмент и плазмидный вектор pET31b+ в отдельных пробирках гидролизуют рестриктазой PstN1 (Сибэнзим, Россия). Рестрикционные смеси разделяют в 1,2% агарозном геле и выделяют из геля линеаризованный в результате гидролиза плазмидный вектор и фрагмент массой 123 п.н., соответствующий нуклеотидной последовательности кодирующей векторный рН-зависимый встраивающийся пептид (pHLIP), ограниченный с обоих концов остатками метионина.
Выделенный линеаризованный плазмидный вектор pET31b+ дефосфорилируют с использованием термостабильной щелочной фосфатазы (СибЭнзим, Россия). Очищают линеаризованный вектор с использованием набора Cleanup Standard (Евроген, Россия), смешивают с выделенным фрагментом в соотношении вектор: вставка равным 1:10 и лигируют с использованием Т4 ДНК-лигазы (СибЭнзим, Россия). Полученной лигазной смесью трансформируют клетки E. coli XL blue. Трансформированные колонии отбирают на селективной агаризованной среде LB с добавлением ампициллина (50 мг/мл) и выделяют плазмиду, используя набор MiniPrep (Евроген, Россия). Успешность клонирования фрагмента подтверждают методом ПЦР с праймерами на Т7 промоторную и Т7 терминирующую области плазмиды. Подтверждение соответствия клонированной последовательности ожидаемой проводят секвенированием на автоматическом секвенаторе (ABI 3130XL, Applied Biosystems) (Фиг. 3).
Пример 2. Получение штамма-продуцента векторного pH-зависимого встраивающегося пептида (pHLIP) - продукта плазмиды pET31b-pHLIP.
Клетки E. coli BL21 Rosetta DE3 pLysS, несущие ген РНК-полимеразы фага Т7 под индуцибельным lacUV5 промотором, трансформируют [24] сконструированной плазмидой pET31b-pHLIP. Клетки E. coli BL21 Rosetta DE3 pLysS, трансформированные плазмидой pET31b-pHLIP, выращивают в течение ночи. Ночную культуру (1/100) засевают в свежую среду LB с добавлением ампициллина (50 мкг/мл). Синтез РНК-полимеразы индуцируют добавлением ИПТГ в концентрации 0,5-1 мМ, предпочтительно 0,5 мМ, в тот момент, когда культура достигает среднелогарифмической фазы роста. Индуцированные клетки растят в течение 4 часов при температуре 22-37°С, предпочтительно 28°С, после чего собирают центрифугированием при 5000 g и анализируют методом электрофореза по Лэммли [25] в 16% SDS-полиакриламидном геле (ПААГ) для подтверждения успешности индукции синтеза pHLIP в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом (Фиг. 4).
Результаты данного анализа, показывают наличие в индуцированной культуре клеток E. coli BL21 Rosetta DE3 pLysS/pET31b-pHLIP дополнительного белка с молекулярной массой около 20 кДа (Фиг. 4, дорожки 3-7), который отсутствует в контрольном лизате клеток E. coli BL21 Rosetta DE3 pLysS/pET31b (Фиг. 4, дорожка 2).
Пример 3. Очистка рекомбинантного векторного pH-зависимого встраивающегося пептида (pHLIP) в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом из клеток E. coli BL21 Rosetta DE3 pLysS методом металл-аффинной хроматографии
Клетки E. coli BL21 Rosetta DE3 pLysS, трансформированные плазмидой pET31b-pHLIP, выращивают в течение ночи. Ночную культуру (1/100) засевают в свежую среду LB с добавлением ампициллина (50 мкг/мл), экспрессию пептида индуцируют добавлением ИПТГ до конечной концентрации 0,5 мМ в тот момент, когда культура достигает среднелогарифмической фазы роста. Индуцированные клетки растят в течение 4-16 часов при 28°С, после чего собирают центрифугированием при 5000 g 4°С в течение 20 минут. Осадок замораживают -80°С для повышения эффективности лизиса клеток. Осадок растворяют в лизирующем буфере (50 mM Tris, 1% Triton-Х100, 100 mM NaCl, 10 mM DTT, pH 8.0.) из расчета 5 мл буфера на 1 г осадка и добавляют лизоцим до конечной концентрации 1 мг/мл. Клетки ресуспендируют и обрабатывают ультразвуком импульсами по 30 секунд с перерывом 30 сек во льду, 5 циклов. Инкубируют во льду 30 минут, центрифугируют 20 мин 10000 g при 4°С. Осадок растворяют в промывочном буфере 50 mM Tris, 0,5% (w/v) Triton-X100, 100 mM NaCl, 1 mM DDT, 1 mM EDTA pH 8.0 из расчета 10 мл на 1 г осадка и центрифугируют 20 мин 10000 g при 4°С. Отбирают супернатант, содержащий рекомбинантный белок и очищают с помощью аффинной хроматографии на Ni-NTA агарозе (Qiagen, США), согласно инструкции производителя. На хроматографическую колонку, упакованную 5 мл Ni-NTA агарозы и уравновешенную буфером 40 mM Tris, 500 mM NaCl, 15 mM имидазол, 8М мочевина, pH 8.0, наносят 10 мл супернатанта со скоростью потока 1 мл/мин. Для удаления неспецифически сорбирующихся белков E. coli проводят предварительную элюцию 40 мл буфера, содержащего 40 mM Tris, 500 mM NaCl, 15 mM имидазол, 8М мочевина, pH 8.0. Пептид в составе белка-слияния элюируют 5 мл буфера, содержащего 40 mM Tris, 500 mM NaCl, 300 mM имидазол, 8М мочевина, pH 8.0.
Полученные белковые фракции, содержащие рекомбинантный pHLIP в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом, диализируют против дистиллированной воды (две смены по 24 ч при комнатной температуре), анализируют электрофорезом в ПААГ с SDS по Лэммли и лиофилизируют в течение суток (Freezone Labconco). На Фиг. 5 приведены результаты электрофоретического анализа очищенного методом металл-аффиннной хроматографии пептида pHLIP в составе белка-слияния с кетостероидизомеразой и последовательностью из шести гистидиновых аминокислотных остатков на С-конце.
Пример 4. Отделение рекомбинантного векторного pH-зависимого встраивающегося пептида (pHLIP) от белка партнера кетостероидизомеразы и 6-гистидинового тага.
Водный раствор белка-слияния KSI-pHLIP полученный после диализа, предварительно лиофилизируют. Сухой белок растворяют в минимальном количестве 75% муравьиной кислоты и добавляют 100 молярный (в расчете на количество содержащихся в белке остатков метионина) избыток бромциана. Реакцию проводят при комнатной температуре в течение 12-24 часов в темноте. Реакцию прекращают добавлением воды в 10-кратном избытке по отношению к исходному объему. Затем пробу диализируют против 1000 кратного избытка дистиллированной воды дважды в течение 24 часов, замораживают при -20°С и лиофилизируют. Эффективность отщепления пептида от белка-слияния оценивают методом SDS-ПААГ-трицин [26] электрофореза (Фиг. 6).
Соответствие аминокислотной последовательности рекомбинантного пептида pHLIP заявленной подтверждают методом MALDI-TOF-MS спектрометрии (Фиг. 7). Для этого фрагмент геля, содержавший пептид, вырезают после проведения SDS-ПААГ-трицин электрофореза, промывают 50% ацетонитрилом в 50 мМ (NH4)2CO3, инкубируют 15 минут и дегидратируют 15 минут в ацетонитриле. Жидкость удаляют, добавляют 5 мкл модифицированного трипсина до концентрации 20 мкг/мл и инкубируют при 37°С в течение ночи. Реакцию останавливают добавлением 7 мкл 0,7% трифторуксусной кислоты и экстрагируют пептид в течение 1 часа. Полученный раствор сокристаллизуют с матрицей 2,5-дигидроксибензойной кислотой в 50% ацетонитриле с добавлением 0,3% трифторуксусной кислоты. Масс-спектр получают на приборе MALDI-TOF Bruker Ultraflex II mass spectrometer (Bruker, Germany) с YAG:Nd. Внутреннюю калибровку проводят, используя продукты саморащепления трипсина. На масс-спектре присутствуют ожидаемые фрагменты с массами иона 1350,6 m/z (соответствующий пептиду pHLIP) и 1421,7 m/z (соответствующий пропионамид-модифицированному по остаткам цистеина пептиду pHLIP в результате взаимодействия с акриловой кислотой). Масс-спектр рекомбинантного пептида идентичен масс-спектру пептида полученного методом твердофазного синтеза (Bachem, США).
Пример 5. Получение препарата рекомбинантного векторного рН-зависимого встраивающегося пептида pHLIP из клеток E. coli BL21 Rosetta DE3 pLysS
Проводят очистку белка по методу, описанному в примере 3. Далее проводят отделение пептида от кетостероидизомеразы и последовательности из 6 гистидиновых аминокислотных остатков по методу, описанному в примере 4.
Очистку пептида pHLIP проводят методом обращенно-фазовой высокоэффективной жидкостной хроматографии на хроматографе (UltiMate 3000 Standard HPLC Systems, Dionex, Thermo Scientific). Для этого полученный после обработки бромцианом и диализа лиофилизат, содержащий кетостероидизомеразу, белок-слияния кетостероидизомераза-pHLIP и рН-зависимый встраивающийся пептид (pHLIP), растворяют в водном растворе 5% ацетонитрила (MeCN), содержащего 0,05% трифторуксусной кислоты (Буфер А), пробу ресуспендируют и центрифугируют при 10000 g в течение часа при 4°С. Супернатант отбирают и наносят на колонку Phenomex Luna 2 C18 (2), с размером пор 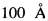 , уравновешенную буфером А. Пептид элюируют градиентом ацетонитрила (5-50%), с добавлением 0,05% трифторуксусной кислоты, при комнатной температуре, скорость элюции 1 мл/мин, в течение 45 минут. Детектирование проводят методом спектрофотометрии при 220 нм. Фракцию, соответствующую пику при τ=10,7 минут, содержащую пептид собирают.
, уравновешенную буфером А. Пептид элюируют градиентом ацетонитрила (5-50%), с добавлением 0,05% трифторуксусной кислоты, при комнатной температуре, скорость элюции 1 мл/мин, в течение 45 минут. Детектирование проводят методом спектрофотометрии при 220 нм. Фракцию, соответствующую пику при τ=10,7 минут, содержащую пептид собирают.
На Фиг. 8 приведены данные анализа собранной на 10,7 минуте хроматографирования фракции методом трицин-ПААГ электрофореза. На электрофореграмме в пробе присутствует полоса, соответствующая пептиду pHLIP, массой 4,1 кДа (Фиг. 9). Кетостероидизомераза, белок-слияния кетостероидизомераза-pHLIP и прочие небелковые примеси могут быть обнаружены во фракциях, соответствующих пикам, наблюдаемым на 47-49 минутах хроматографирования.
Обоснование режима экспрессии рекомбинантного векторного pH-зависимого встраивающегося пептида (pHLIP) в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом в клетоках E. coli BL21 Rosetta DE3 pLysS
Технические параметры способа основаны на анализе данных экспериментальных исследований (Фиг. 4), которые показали, что:
- использование для индукции экспрессии ИПТГ в конечной концентрации 0,5 мМ обеспечивает оптимальное количество синтезируемого E. coli BL21 Rosetta DE3 pLysS/реТ 31b+-pHLIP рекомбинантного векторного рН-зависимого встраивающегося пептида (pHLIP) в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом, тогда как увеличение конечной концентрации ИПТГ до 1 мМ не приводит к заметному увеличению количества синтезируемого белка.
- оптимальное количество рекомбинантного векторного pH-зависимого встраивающегося пептида (pHLIP) в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом нарабатывается при температуре культивирования E. coli BL21 Rosetta DE3 pLysS/реТ31b+-pHLIP - 28°С. При температуре 22°С выход целевого продукта несколько ниже, тогда как увеличение температуры до 37°С приводит к значительному снижению выхода целевого продукта.
Таким образом, впервые получена плазмидная ДНК pET31b-pHLIP, содержащая последовательность, кодирующую рекомбинантный рН-зависимый встраивающийся пептид (pHLIP), в составе белка слияния с кетостероидизомеразой на N-конце пептида и последовательность из 6 гистидиновых остатков на С-конце, что обеспечивает продукцию в клетках Escherichia coli рекомбинантного пептида pHLIP, обладающего способностью встраиваться в мембрану клеток при pH окружающей среды 7.0, предназначенного для диагностики и целевой доставки к зонам локального ацидоза.
ИСТОЧНИКИ ИНФОРМАЦИИ
1. Akhtar M.S., (2012). Antimicrobial Peptides as Infection Imaging Agents: Better Than Radiolabeled Antibiotics. International Journal of Peptides. 2012, 1-19.
2. Andreev O.A., (2010). pH (low) insertion peptide (pHLIP) inserts across a lipid bilayer as a helix and exits by a different path. PNAS. 107, 4081-4086.
3. Weerakkody D., (2013). Family of pH (low) insertion peptides for tumor targeting. PNAS. 110, 5834-5839.
4. Hunt J.F., (1997). pH-Dependent Membrane Insertion of a Transbilayer α-Helix. Biochemistry. 36, 15177-15192.
5. Andreev O.A., (2010). pH-sensitive membrane peptides (pHLIPs) as a novel class of delivery agents. Mol Membr Biol. 27, 341-352.
6. Segala J., (2009). Accurate Analysis of Tumor Margins Using a Fluorescent pH Low Insertion Peptide (pHLIP). 10, 3478-3487.
7. Yao L., (2013). pHLIP-mediated delivery of PEGylated liposomes to cancer cells. J Control Release. 167, 228-237.
8. Adochite R.C., (2014). Targeting breast tumors with pH (low) insertion peptides. Mol Pharm. 11, 2896-2905.
9. Cruz-Monserrate Z., (2014). Targeting Pancreatic Ductal Adenocarcinoma Acidic Microenvironment. Scientific Reports. 4, 4410.
10.  A.L., (2009). A novel technology for the imaging of acidic prostate tumors by positron emission tomography. Cancer Res. 69, 4510-4516.
A.L., (2009). A novel technology for the imaging of acidic prostate tumors by positron emission tomography. Cancer Res. 69, 4510-4516.
11. Macholl S., (2012). In vivo pH imaging with (99m)Tc-pHLIP. Mol Imaging Biol. 14, 725-734.
12. Yao L., (2013). pHLIP peptide targets nanogold particles to tumors. PNAS. 110, 465-470.
13. Sosunov E.A., (2013). pH (low) insertion peptide (pHLIP) targets ischemic myocardium. PNAS. 110, 82-86.
14. Moshnikova A., (2013). Antiproliferative effect of pHLIP-amanitin. Biochemistry. 52, 1171-1178.
15. Burns K.E., (2015). Inhibition of cancer cell proliferation and breast tumor targeting of pHLIP-monomethyl auristatin E conjugates. Mol Pharm. 12, 1250-1258.
16. An M., (2010). pH-(low)-insertion-peptide (pHLIP) translocation of membrane impermeable phalloidin toxin inhibits cancer cell proliferation. PNAS. 107, 20246-20250.
17. Cheng C.J., (2015). MicroRNA silencing for cancer therapy targeted to the tumour microenvironment. Nature. 518, 107-110.
18. Engelman D.M., Hunt J.F. Transmembrane polypeptide and methods of use. Patent for invention № US 5739273 A dated 9.09.1994.
19. Reshetnyak Y.K., Lehnert O.U., Engelman D.M. Selective delivery of molecules into cells or marking of cells in diseased tissue regions using environmentally sensitive transmembrane peptide. Patent for invention № US 8703909 B2 dated 12.12.2011.
20. Amblard M., (2006). Methods and protocols of modern solid phase Peptide synthesis. Mol Biotechnol. 33, 239-254.
21.  F.S., (2007). Peptide synthesis: chemical or enzymatic. Electronic Journal of Biotechnology. 10, 279-314.
F.S., (2007). Peptide synthesis: chemical or enzymatic. Electronic Journal of Biotechnology. 10, 279-314.
22. Riley, J.M., (2009). Bioproduction and characterization of a pH responsive self-assembling peptide. Biotechnol Bioeng. 103, 241-251.
23. Wagstaff, J.L., (2010). Production of recombinant isotopically labelled peptide by fusion to an insoluble partner protein: generation of integrin αvβ6 binding peptides for NMR. Mol Biosyst. 6, 2380-2385.
24. Маниатис, Т.Э., (1984). Молекулярное клонирование. Мир. Москва.
25. Laemmli U.K., (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 227,680-685.
26. 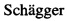 H., (2006). Tricine-SDS-PAGE. Nat Protoc. 1, 16-22.
H., (2006). Tricine-SDS-PAGE. Nat Protoc. 1, 16-22.
ПРИЛОЖЕНИЕ
Фиг. 1. Нуклеотидная последовательность и кодируемая ею аминокислотная последовательность векторного рН-зависимого встраивающегося пептида pHLIP
Фиг. 2. Общая схема структурной организации плазмиды pET31b-pHLIP
Фиг. 3. Секвенирование кодирующего pHLIP фрагмента в составе рекомбинатной плазмиды pET31b-pHLIP
Фиг. 4. Оптимизация условий экспрессии пептида pHLIP в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом в клетках E. coli BL21 Rosetta DE3 pLysS/pET31b-pHLIP
Фиг. 5. Электрофореграмма в 16% SDS-ПААГ белковых фракций и очищенного рекомбинантного пептида pHLIP в составе белка-слияния с кетостероидизомеразой и 6-гистидиновым тагом.
Фиг. 6. Анализ успешности отщепления пептида pHLIP, от белка-слияния KSI-pHLIP с использованием бромциана
Фиг. 7. МАЛДИ масс-спектр рекомбинантного пептида pHLIP
Фиг. 8. Хроматографическая очистка рекомбинантного пептида pHLIP
Фиг. 9. Анализ хроматографической фракции, содержащей очищенный рекомбинантный пептид pHLIP.

Изобретение относится к области биохимии, генной инженерии и биотехнологии, в частности к рекомбинантной плазмидной ДНК pET31b-pHLIP. Указанная плазмидная ДНК кодирует аминокислотную последовательность рекомбинантного рН-зависимого встраивающегося пептида и обеспечивает его синтез в составе белка-слияния с кетостероидизомеразой. Изобретение также относится к способу получения указанного пептида. Настоящий способ предусматривает трансформацию клеток Escherichia coli плазмидной ДНК pET31b-pHLIP, выращивание трансформированных клеток в среде с ампициллином, индукцию экспрессии, последующее культивирование, отделение клеток центрифугированием, отмывку телец включения, растворение их в буфере с добавлением мочевины, очистку методом металл-хелатной хроматографии, последующее отделение пептида от кетостероидизмеразы и 6 гистидиновых остатков с использованием бромциана. Настоящее изобретение позволяет получать рекомбинантный рН-зависимый встраивающийся пептид. 2 н.п. ф-лы, 9 ил., 5 пр.
1. Рекомбинантная плазмидная ДНК pET31b-pHLIP, кодирующая аминокислотную последовательность рекомбинантного векторного рН-зависимого встраивающегося пептида (pHLIP), кодируемого нуклеотидной последовательностью 5'-GCGTGCGAACAGAATCCGATTTATTGGGCGCGTTATGCGGATTGGCTGTTTACCACCCCG CTGCTGCTGCTGGATCTGGCGCTGCTGGTGGATGCGGATGAAGGCACC-3', имеющая молекулярную массу 1,8 МДа, обеспечивающая синтез pHLIP в составе белка-слияния с кетостероидизомеразой и состоящая из фрагмента ДНК плазмиды pET31b+, гидролизованной по сайту AlwNI (PstNI) и лигированной по указанному сайту с фрагментом, кодирующим pHLIP, содержащего сайт инициации репликации плазмиды pBR322, промотор и терминатор транскрипции Т7-РНК полимеразы, стартовый кодон, ген, кодирующий кетостероидизомеразу, последовательность, кодирующую 6 гистидиновых остатков, стоп-кодон, генетический маркер AMPr - ген β-лактамазы, определяющий устойчивость трансформированных плазмидой pET31b-pHLIP клеток Escherichia coli к ампициллину.
2. Способ получения рекомбинантного векторного рН-зависимого встраивающегося пептида (pHLIP), предусматривающий трансформацию клеток E.coli BL21 Rosetta DE3 pLysS плазмидной ДНК, выращивание трансформированных клеток в среде с ампициллином до среднелогарифмической стадии роста; индукцию экспрессии 0,5-1 мМ изопропилтиогалактазидом и последующим культивированием в течение 4-16 ч при 22-37°С; отделением клеток центрифугированием, отмывкой телец включения, растворения их в буфере с добавлением мочевины, очистку методом металл-хелатной хроматографии, последующим отделением пептида от кетостероидизмеразы и 6 гистидиновых остатков с использованием бромциана, отличающийся тем, что в качестве плазмидной ДНК используют рекомбинантную плазмидную ДНК pET31b-pHLIP, полученную по п. 1, а хроматографическую очистку пептида от кетостероидизомеразы и прочих примесей проводят методом высокоэффективной жидкостной хроматографии.
| АЩЕУЛОВА (БУРЛАКОВА) Д.О | |||
| и др | |||
| Получение pH-зависимого пептида pHLIP в бактериальной системе | |||
| Белки и пептиды: материалы VII Российского симпозиума, Новосибирск, 2015 | |||
| BURLAKOVA (ASHCHEULOVA) D.O | |||
| pH-dependent peptide (pHLIP) recombinant production | |||
| Приспособление в центрифугах для регулирования количества жидкости или газа, оставляемых в обрабатываемом в формах материале, в особенности при пробеливании рафинада | 0 |
|
SU74A1 |
| Н.И | |||
| Пирогова: сборник материалов, Томск, 2015 | |||
| BURLAKOVA (ASHCHEULOVA) D.O | |||
| et al | |||
| E | |||
| Способ использования делительного аппарата ровничных (чесальных) машин, предназначенных для мериносовой шерсти, с целью переработки на них грубых шерстей | 1921 |
|
SU18A1 |
| Постгеномные методы анализа в биологии, лабораторной и клинической медицине: cборник трудов IV международной научно-практической конференции, Казань, 2014 | |||
| АЩЕУЛОВА (БУРЛАКОВА) Д.О | |||
| Получение рекомбинантного векторного рН-зависимого пептида для диагностики | |||
| Функциональные материалы: разработка, исследование, применение: cборник тезисов докладов III Всероссийского конкурса научных докладов студентов, Томск, 2015 | |||
| US 8846081 B2, 30.09.2014 | |||
| RILEY J.M | |||
| et al | |||
| Bioproduction and characterization of a pH responsive self-assembling peptide | |||
| Biotechnology and bioengineering, 2009 | |||
| ZHAO D | |||
| et al | |||
| Устройство непрерывного автоматического тормоза с сжатым воздухом | 1921 |
|
SU191A1 |
| Protein expression and purification, 2007 | |||
| GALLOWAY C.A | |||
| et al | |||
| Increasing the yield of soluble recombinant protein expressed in E | |||
| coli by induction during late log phase | |||
| Biotechniques, 2003 | |||
| PRANCHEVICIUS M.S | |||
| et al | |||
| Characterization and optimization of ArtinM lectin expression in Escherichia coli | |||
| BMC biotechnology, 2012 | |||
| Коренев Ю.М | |||
| и др | |||
| Общая и неорганическая химия, Часть 1 | |||
| М.: Школа им | |||
| А.Н | |||
| Колмогорова, Изд-во МГУ, 2000 | |||
| US 2014080187 A1, 20.03.2014 | |||
| US 2009203081 A1, 13.08.2009 | |||
| Van der Velden A.W | |||
| et al | |||
| Vector design for optimal protein expression | |||
| Biotechniques, 2001 | |||
| ANDREEV O.A | |||
| et al | |||
| pH-sensitive membrane peptides (pHLIPs) as a novel class of delivery agents | |||
| Molecular membrane biology, 2010 | |||
| ERCIYES K | |||
| Distributed and Sequential Algorithms for Bioinformatics | |||
| Cham: Springer, 2015. | |||

