Область техники
Настоящее описание в целом относится к композициям, содержащим аминокислоты. Более конкретно, описание относится к композициям, содержащим аминокислоты, предназначенным для применения в медицинских целях, в частности в лечении заболеваний, ассоциированных с митохондриальными дисфункциями.
Уровень техники
Митохондрии представляют собой клеточные органеллы, основной функцией которых является окислительное фосфорилирование - процесс, посредством которого энергия, получаемая в результате метаболизма глюкозы и жирных кислот, конвертируется в аденозинтрифосфат (АТФ). После этого АТФ используется для поддержания различных энергопотребляющих реакций биосинтеза и других метаболических процессов.
Митохондриальные дисфункции могут оказывать влияние на любую ткань, приводя к огромному разнообразию симптомов, характер которых зависит от степени вовлеченности различных тканей.
К заболеваниям, развивающимся из-за митохондриальных дисфункций, относятся, например, митохондриальные отеки, обусловленные патологическими изменениями трансмембранного потенциала митохондрий, функциональные расстройства, вызываемые окислительным стрессом, в том числе действием активных форм кислорода (АФК) или свободных радикалов, а также функциональные расстройства, причиной которых являются генетические мутации, и болезни, вызванные функциональной недостаточностью механизмов окислительного фосфорилирования, необходимых для продукции энергии.
По мере старения организма происходит разрушение митохондрий, проявляющаяся угасанием респираторной активности, накоплением повреждений в митохондриальной ДНК (мтДНК) и продукцией избыточного количества активных форм кислорода (АФК).
Полученные в последнее время данные свидетельствуют о вовлеченности митохондриальных дисфункций в развитие ряда болезней, в том числе возрастных метаболических и сердечно-сосудистых заболеваний (атеросклероз), помимо серьезных нейродегенеративных заболеваний, таких как болезнь Альцгеймера, болезнь Паркинсона и болезнь Гентингтона, а также хронической обструктивной болезни легких (ХОБЛ).
Митохондриальные дисфункции также выявляются при ожирении и ассоциированных с ним заболеваниях, в том числе сахарном диабете 2 типа, гипертонической болезни, дислипидемии, сердечной недостаточности, заболеваниях почек и остеопорозе.
Примечателен тот факт, что саркопения, признанная одной из наиболее важных причин возрастного функционального угасания и утраты независимости из-за непроизвольного снижения мышечной массы и силы, а также ключевым компонентом так называемого синдрома дряхлости, обусловлена снижением массы и функциональности митохондрий. Известны также другие заболевания и расстройства, проявляющиеся атрофией мышечной ткани, например, такие как кахексия. Кахексия или синдром истощения, определяется как непреднамеренное снижение массы тела, атрофия мышечной ткани, утомляемость и общая слабость. Кахексия наблюдается у людей, страдающих раком, СПИДом, глютеновой энтеропатией, ХОБЛ, рассеянным склерозом, ревматоидным артритом, застойной сердечной недостаточностью, туберкулезом и нервной анорексией. На сегодняшний день лекарственные средства или нутриенты, способные предотвратить развитие данного состояния и/или излечить его, не известны.
Таким образом, учитывая всемирную эпидемию ожирения и усугубляющееся старение населения, в ближайшем будущем за медицинской помощью наиболее часто будут обращаться тучные пожилые люди с саркопенией.
На сегодняшний день, когда в мире проживает более 1,4 миллиарда взрослых людей с избыточной массой тела, ожирение и ассоциированые с ним заболевания, такие как резистентность к инсулину и сахарный диабет 2 типа, являются серьезной медицинской проблемой (World Health Organization. Obesity and overweight. WHO Media Centre). Примерно 500 миллионов из этих людей страдают ожирением, также 400 миллионов детей в возрасте до 5 лет в настоящее время классифицируются как имеющие избыточную массу тела или страдающие ожирением (World Health Organization. Obesity and overweight. WHO Media Centre).
С точки зрения метаболизма, ожирение развивается в случаях, когда из-за сниженной митохондриальной функции поступление энергии превышает расход энергии. Поэтому, помимо коррекции питания с целью снижение поступления энергии, существует еще один подход, заключающийся в повышении расхода энергии за счет, например, физической нагрузки. Однако это не является простой задачей, в особенности при работе с людьми с ожирением, которые часто страдают от связанной с ожирением мышечной дисфункции (Wells et al., 2008).
Поэтому существует растущая потребность в новых терапевтических подходах, направленных на снижение массы тела, а также уменьшение социальных и медицинских последствий этого заболевания.
Краткое описание изобретения
Целью настоящего описания является обеспечение новых композиций, содержащих аминокислоты, в частности эффективных в отношении повышения митохондриальной функции и, соответственно, в лечении ожирения и связанных с ним заболеваний.
Согласно настоящему описанию, указанная выше цель достигается благодаря объекту изобретения, конкретно упомянутому в последующих пунктах, рассматривающихся как неотъемлемая часть настоящего описания.
Согласно одному варианту реализации настоящего описания, предложена композиция для стимуляции митохондриального биогенеза и улучшения митохондриальной функции у субъекта. Данная композиция включает в себя действующее вещество, содержащее аминокислоты: лейцин, изолейцин, валин, треонин и лизин, а также лимонную, янтарную и яблочную кислоты.
В одном или более вариантах реализации действующее вещество композиции дополнительно содержит одну или более аминокислот, выбранных из группы, состоящей из гистидина, фенилаланина, метионина, триптофана, цистеина и тирозина.
В одном или более вариантах реализации раскрытые в настоящем описании композиции могут использоваться в медицине.
В предпочтительном варианте реализации композиции могут применяться в лечении и/или профилактике заболевания, ассоциированного с митохондриальной дисфункцией, каковым является ожирение.
Краткое описание чертежей
Ниже изобретение будет описано исключительно в качестве примера, со ссылками на прилагаемые фигуры, где:
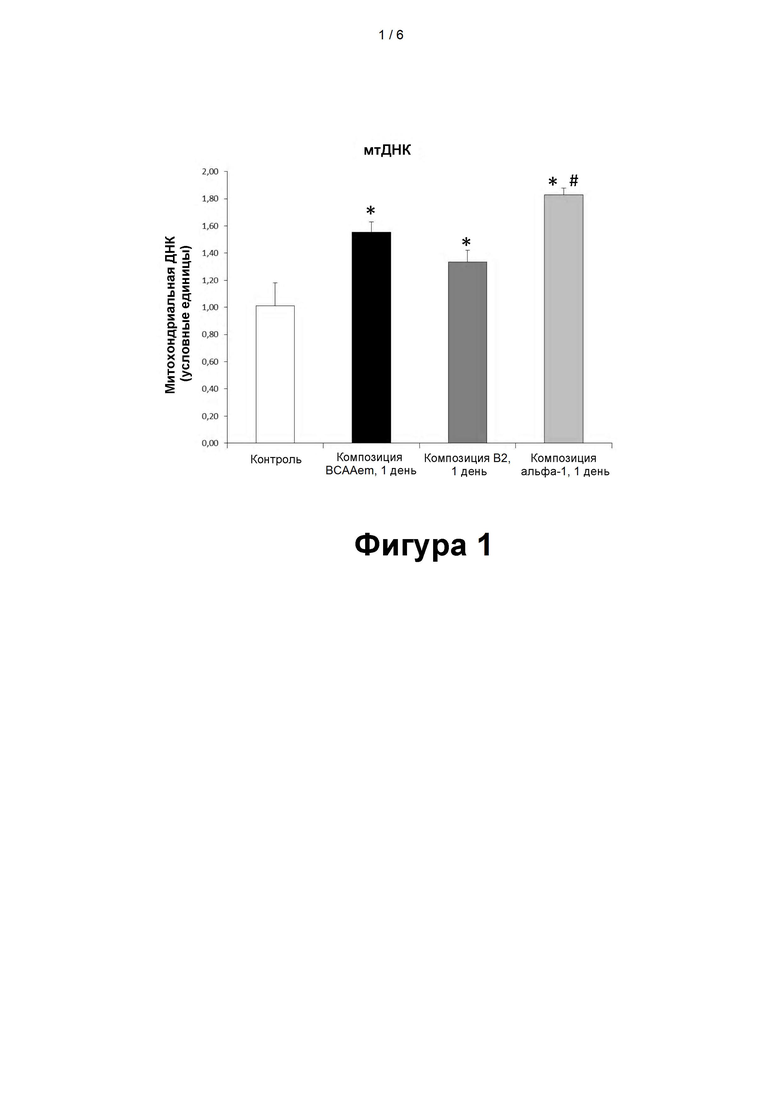
- на фигуре 1 показаны полученные с помощью метода количественной ПЦР данные по содержанию митохондриальной ДНК (мтДНК) в кардиомиоцитах линии HL-1, подвергавшихся воздействию различных композиций, содержащих аминокислоты, в течение 48 часов (48 ч, 2 сут). Количественную ПЦР выполняли в трех повторах, с использованием в качестве эндогенного контроля геномной ДНК, кодирующей глицеральдегид-3-фосфатдегидрогеназу (glyceraldehyde 3-phosphate dehydrogenase, GAPDH) (n=3, средняя величина ± стандартная погрешность средней величины). Значение для не подвергавшихся воздействию клеток (КК) принимается равным 1,0 (*p<0,05 при сравнении со значением для не подвергавшихся воздействию клеток, p<0,05 при сравнении со значениями для клеток, подвергавшихся воздействию композиций BCAAem и B2).
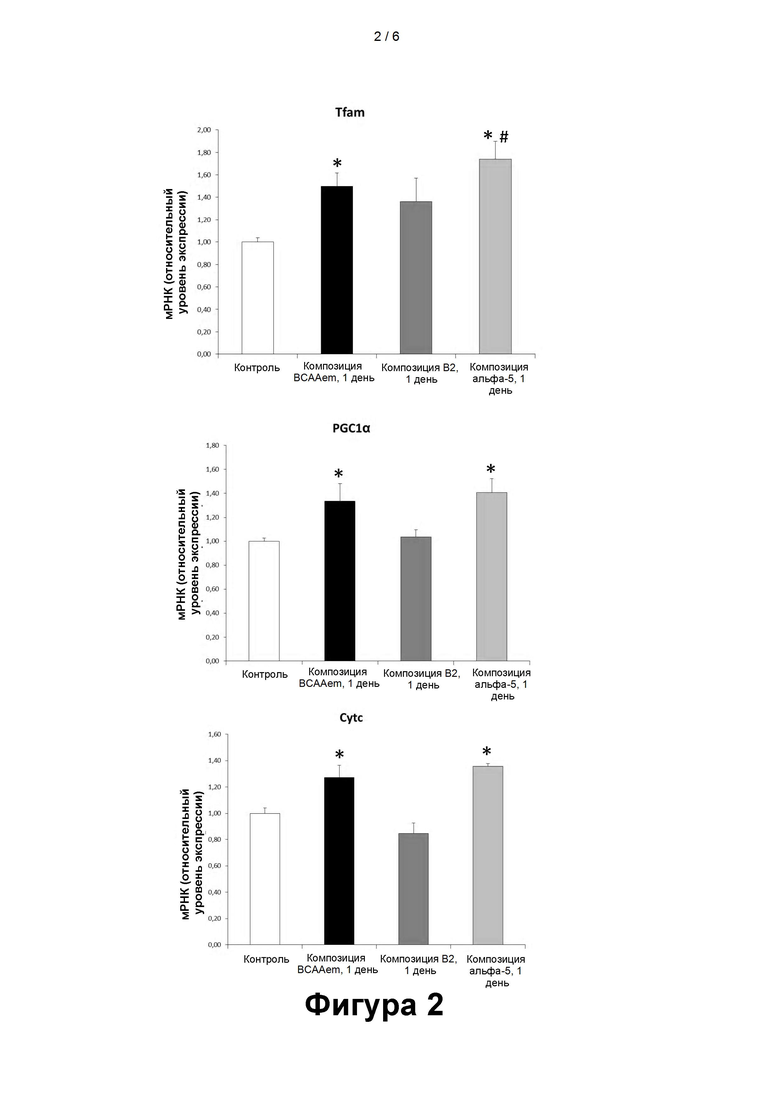
- На фигуре 2 представлены уровни мРНК маркеров митохондриального биогенеза (Tfam, PGC-1α, Cyt c), измеренные с помощью количественной ПЦР в кардиомиоцитах линии HL-1, подвергавшихся воздействию композиций, содержащих аминокислоты, в течение 24 часов (1 сутки). Количественную ПЦР выполняли в трех повторах, а полученные результаты нормализовали относительно экспрессии GAPDH (n=3, средняя величина ± стандартная погрешность средней величины). *Р<0,05 при сравнении со значением для не подвергавшихся воздействию клеток, принятому за 1,0. #p<0,05 при сравнении со значениями для клеток, подвергавшихся воздействию композиции BCAAem.
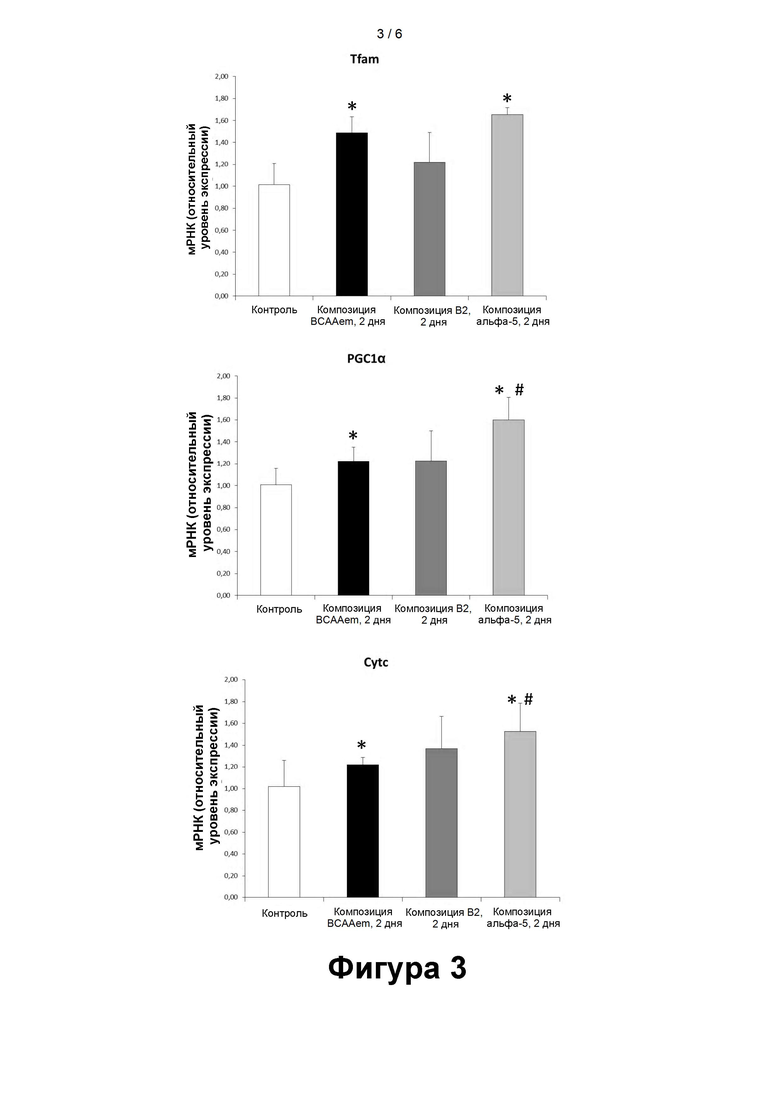
- На рисунке 3 представлены уровни маркеров митохондриального биогенеза (Tfam, PGC-1α, Cyt c), измеренные методом количественной ПЦР в кардиомиоцитах линии HL-1, подвергавшихся воздействию композиций, содержащих аминокислоты, в течение 48 часов (2 суток). Количественную ПЦР выполняли в трех повторах, а полученные результаты нормализовали относительно экспрессии GAPDH (n=3, средняя величина ± стандартная погрешность средней величины). * Р<0,05 при сравнении со значением для не подвергавшихся воздействию клеток, принятому за 1,0. #p<0,05 при сравнении со значениями для клеток, подвергавшихся воздействию композиции BCAAem.
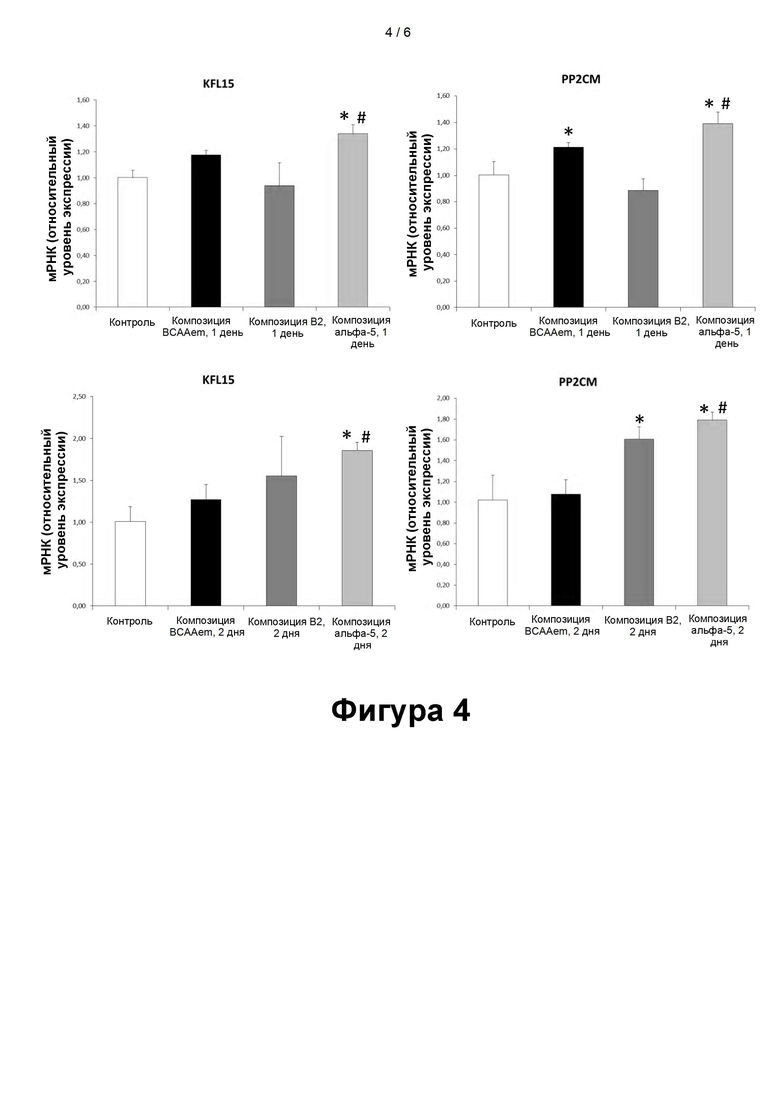
- На фигуре 4 представлены уровни мРНК Круппель-подобного транскрипционного фактора-15 (KFL15) и ассоциированной с митохондриальным матриксом протеинфосфатазы семейства 2C (PP2CM), относящихся к белкам, регулирующими катаболизм аминокислот с разветвленной цепью. Уровни PP2CM и KFL15 были измерены методом количественной ПЦР в кардиомиоцитах линии HL-1, подвергавшихся воздействию композиций, содержащих аминокислоты, в течение 24 часов (1 сутки) или 48 часов (2 суток). Количественную ПЦР выполняли в трех повторах, а полученные результаты нормализовали относительно экспрессии GAPDH (n=3, средняя величина ± стандартая погрешность средней величины). *Р<0,05 при сравнении со значением для не подвергавшихся воздействию клеток, принятому за 1,0. #p<0,05 при сравнении со значениями для клеток, подвергавшихся воздействию композиций BCAAem и B2.
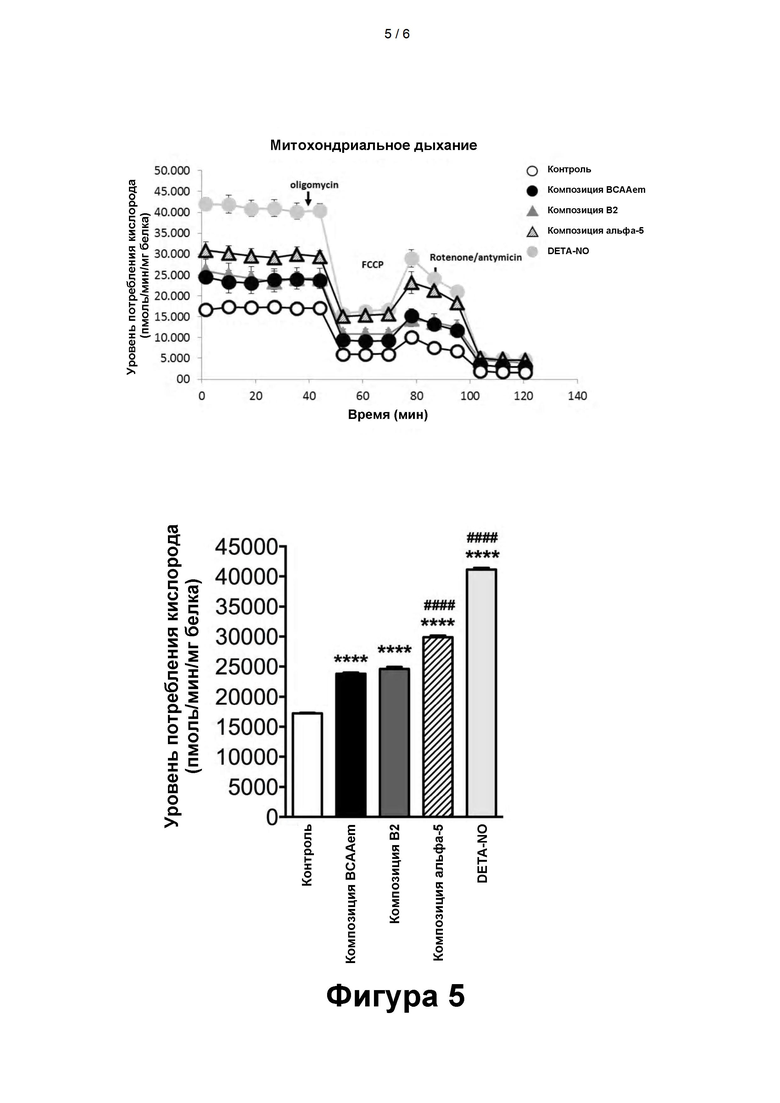
- На фигуре 5 представлены результаты оценки уровня потребления кислорода. Потребление кислорода кардиомиоцитами линии HL-1, подвергавшимися воздействию композиций, содержащих аминокислоты, или NO-аддукта диэтилентриамина (Diethylenetriamine/Nitric Oxide Adduct, DETA-NO) в течение 48 ч. ****p <0,001 при сравнении со значением для не подвергавшихся воздействию клеток; #### p<0,001 при сравнении со значениями для клеток, подвергавшихся воздействию композиций BCAAem и B2.
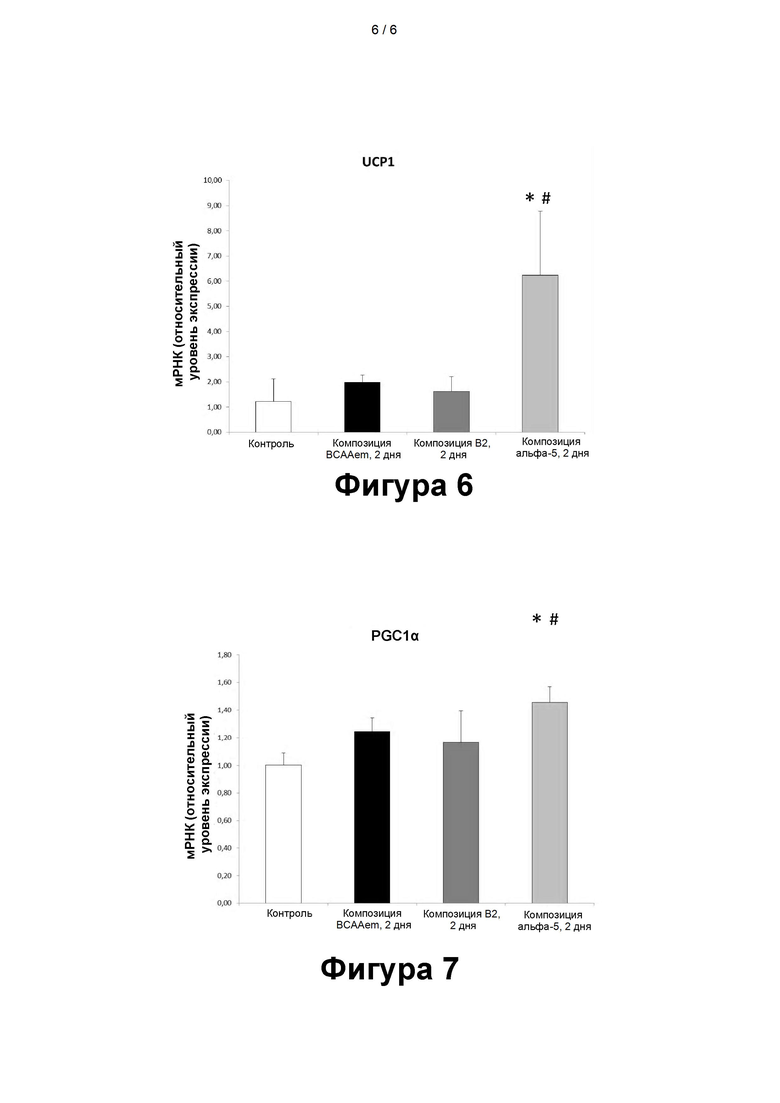
- На фигуре 6 представлена экспрессия разобщающего белка-1 (UCP1) в иммортализованных бурых адипоцитах (Seale et al., 2007) после воздействия в течение 48 часов (2 суток). КК - контрольные клетки, не подвергавшиеся воздействию. Значение для контрольных клеток (КК) принято за 1,0. *Значимо по сравнению с КК. #Значимо по сравнению с клетками, подвергавшимися воздействию композиции BCAAem; ПЦР в режиме реального времени выполняли в трех повторах, а полученные результаты нормализовали относительно экспрессии GAPDH (n=3, средняя величина ± стандартная погрешность средней величины).
- На фигуре 7 представлена экспрессия PGC-1α в иммортализованных бурых адипоцитах после воздействия в течение 48 часов (2 суток). КК - контрольные клетки, не подвергавшиеся воздействию. Значение для контрольных клеток (КК) принято за 1,0. *Значимо по сравнению с КК. # Значимо по сравнению с клетками, подвергавшимися воздействию композиции BCAAem. ПЦР выполняли в трех повторах, а полученные результаты нормализовали относительно экспрессии GAPDH (n=3, средняя величина ± стандартная погрешность средней величины).
Подробное описание предпочтительных вариантов реализации
В приведенном ниже описании представлены многочисленные специфичные детали, необходимые для полного понимания вариантов реализации. Указанные варианты реализации могут применяться без одной или более специфичных деталей или в комбинации с другими способами, компонентами, материалами и т.д. В ряде случаев, во избежание отвлечения внимания от аспектов вариантов реализации, хорошо известные структуры, материалы или операции представлены или описаны без подробностей.
Встречающиеся в настоящем описании ссылки на «один вариант реализации» или «вариант реализации» означают, что определенный признак, структура или характеристика, описанная в контексте варианта реализации, относится по крайней мере к одному из вариантов реализации. Следовательно, фразы «в одном варианте реализации» или «в варианте реализации», встречающиеся в настоящем описании, не обязательно относятся к одному и тому же варианту реализации. Кроме того, определенные признаки, структуры или характеристики могут быть любым подходящим способом скомбинированы в одном или более вариантах реализации. Расставленные в документе заголовки предназначены исключительно для удобства и не отражают объем или значимость вариантов реализации.
В одном или более вариантах реализации раскрытая в настоящем описании композиция содержит действующее вещество, содержащее аминокислоты: лейцин, изолейцин, валин, треонин, лизин, а также кислоты: лимонную, янтарную и яблочную.
Композицию, содержащую аминокислоты - согласно EP 2196203 B1 - вводили млекопитающим в качестве альтернативного решения, обеспечивающего преимущества низкокалорийной диеты (НД). Было показано, что данная композиция, содержащая аминокислоты (в настоящем описании упоминаемая как «BCAAem»), обеспечивает усиление митохондриального биогенеза как в сердечной, так и в скелетной мускулатуре (D'Antona et al., 2010). Описанные эффекты были опосредованы повышением экспрессии эндотелиальной синтазы оксида азота (eNOS) и последующей продукцией оксида азота (NO) (D'Antona et al., 2010).
Автор настоящей заявки с удивлением обнаружил, что при добавлении определенных кислот в композицию, содержащую схожую комбинацию лейцина, изолейцина, валина, треонина и лизина, можно добиться значительного усиления митохондриальной функции клетки и, соответственно, расхода энергии.
Автор настоящей заявки исследовал ряд композиций, отличающихся по содержанию различных кислот, и обнаружил, что композиции, содержащие в качестве действующего вещества комбинацию лимонной янтарной и яблочной кислот в сочетании с лейцином, изолейцином, валином, треонином и лизином, демонстрируют высокую эффективность при применении для достижения описанных выше целей. И действительно, композиции, содержащие описанное выше действующее вещество, а также композиции, содержащие описанное выше действующее вещество в комбинации с дополнительными аминокислотами (перечисленными в представленной ниже таблице 1), значительно более эффективны в отношении стимуляции биогенеза митохондрий и их функций, чем протестированная ранее композиция на основе аминокислот (BCAAm).
Тестирование композиций проводили на кардиомиоцитах линии HL-1, то есть на клетках, представляющих собой модель функционирования сердца in vitro. Результаты изучения этих кардиомиоцитов могут использоваться для подтверждения эффективности применения новых композиций в профилактике сердечной недостаточности.
Кроме того, раскрытые в настоящем описании композиции протестировали на иммортализованных бурых адипоцитах - модель бурого жира in vitro . Недавно было получено подтверждение присутствия бурого жира в организме взрослого человека (Trayhurn, 2016).
В отличие от белой жировой ткани, являющейся преимущественно органом запасания энергии в форме жира (триглицеридов), бурая жировая ткань использует энергию пищи для продукции тепла (термогенез), увеличивая таким образом расход энергии.
Этот процесс активируется как в ответ на факторы окружающей среды (например, воздействие холода), так и на пищевые (рацион питания) факторы путем повышения экспрессии и активности разобщающего белка-1 (UCP1; Cannon and Nedergaard, 2004).
Данные, представленные в настоящей заявке, свидетельствуют об эффективности раскрытых в настоящем описании композиций в отношении повышения экспрессии UCP1, подтверждая таким образом целесообразность их применения в медицине, в частности в лечении ожирения и связанных с ним заболеваний, таких как резистентность к инсулину и сахарный диабет 2 типа.
В одном или более вариантах реализации описанная в настоящем документе композиция содержит действующее вещество, в состав которого входят: лимонная, янтарная и яблочная кислоты в комбинации с лейцином, изолейцином, валином, треонином и лизином, причем массовое соотношение между общим количеством лимонной, янтарной и яблочной кислот и общим количеством аминокислот: лейцина, изолейцина, валина, треонина, лизина составляет от 0,05 до 0,3, предпочтительно от 0,1 до 0,25.
В одном или более вариантах реализации действующее вещество может дополнительно содержать одну или более аминокислот, выбранных из группы, включающей гистидин, фенилаланин, метионин, триптофан, цистеин и тирозин.
В другом варианте реализации действующее вещество композиции, раскрытой в данном описании, также может включать аспарагиновую кислоту и/или L-орнитин-альфа-кетоглутарат (OKG).
Согласно варианту реализации, композиция содержит действующее вещество, состоящее из лейцина, изолейцина, валина, треонина, лизина, гистидина, фенилаланина, метионина, триптофана, цистеина и необязательно тирозина, а также лимонной, янтарной и яблочной кислот, при этом в композиции содержаться только перечисленные выше аминокислоты.
В другом варианте реализации композиция может содержать аминокислоты: изолейцин, лейцин и валин в количестве от 35% до 65% по массе, предпочтительно от 42% до 56% по массе, по отношению к массе действующего вещества.
В одном или более вариантах реализации соотношение массы лейцина и массы лимонной кислоты варьирует от 5 до 1, предпочтительно от 2,5 до 3,5.
В другом варианте реализации масса или количество вещества (в молях) лимонной кислоты выше, чем масса или количество вещества (в молях) яблочной и янтарной кислот по-отдельности. Предпочтительно также, чтобы масса или количество вещества (в молях) лимонной кислоты превышала общую массу или количество вещества (в молях) яблочной и янтарной кислот. В следующем варианте реализации массовое отношение для лимонной кислоты и комбинации яблочной и янтарной кислот варьирует от 1,0 до 4,0, предпочтительно от 1,5 до 2,5. В рекомендованном варианте реализации массовое соотношение лимонная кислота:яблочная кислота:янтарная кислота варьирует от 10:1:1 до 2:1,5:1,5, предпочтительно от 7:1:1 до 1,5:1:1, и наиболее предпочтительно от 5:1:1 до 3:1:1.
Согласно некоторым вариантам реализации настоящего описания, предпочтительное молярное соотношение изолейцин:лейцин попадает в диапазон от 0,2 до 0,7, предпочтительно от 0,30 до 0,60, при этом (и/или) предпочтительное массовое соотношение валин:лейцин должно попадать в диапазон от 0,2 до 0,70, предпочтительно от 0,30 до 0,65.
В другом варианте реализации молярное соотношение треонин:лейцин находится в диапазоне от 0,10 до 0,90, предпочтительно от 0,20 до 0,70, при этом (и/или) массовое соотношение лизин:лейцин должно попадать в диапазон от 0,20 до 1,00, предпочтительно от 0,40 до 0,90.
В рекомендованном варианте реализации соотношение между общим количеством вещества (в молях) лимонной, яблочной и янтарной кислот и общим количеством вещества (в молях) метионина, фенилаланина, гистидина и триптофана превышает 1,35.
В одном или более вариантах реализации массовое соотношение между комбинацией лимонной, яблочной и янтарной кислот и комбинацией аминокислот с разветвленной цепью: лейцина, изолейцина и валина составляет от 0,1 до 0,4, предпочтительно от 0,15 до 0,35.
В другом варианте реализации общая масса аминокислот с разветвленной цепью: лейцина, изолейцина, валина, а также треонина и лизина выше, чем общая масса трех кислот: лимонной, яблочной и янтарной. Предпочтительно, чтобы масса отдельных кислот (лимонной, янтарной или яблочной) была меньше массы каждой из аминокислот: лейцина, изолейцина, валина, треонина и лизина.
В предпочтительном варианте реализации соотношение между суммарной массой лимонной, яблочной и янтарной кислот и суммарной массой аминокислот с разветвленной цепью: лейцина, изолейцина, валина, а также лизина и треонина составляет от 0,05 до 0,3, предпочтительно от 0,10 до 0,25.
В другом варианте реализации общее количество вещества (в молях) лизина и треонина выше, чем общее количество вещества (в молях) трех кислот: лимонной, янтарной и яблочной. Предпочтительно, чтобы соотношение между общим количеством вещества (в молях) трех кислот: лимонной, янтарной и яблочной и общим количеством вещества (в молях) лизина и треонина составляло от 0,10 до 0,70, предпочтительно от 0,15 до 0,55.
В одном или более вариантах реализации описанная в настоящем документе композиция дополнительно содержит витамины, предпочтительно витамины группы В, такие как витамин В1 и/или витамин В6.
В другом варианте реализации настоящего описания композиция может содержать углеводы, пищевые добавки и/или ароматизаторы.
Более того, при изготовлении композиций, в частности действующего вещества, в соответствии с настоящим описанием, следует избегать аминокислоту аргинин.
Также в состав раскрытой в настоящем описании композиции предпочтительно не включать следующие аминокислоты: серин, пролин, аланин.
В определенных концентрациях или стехиометрических соотношениях с другими компонентами композиции данные аминокислоты могут снижать ее эффективность или даже причинять вред.
Перечисленные в настоящем описании аминокислоты можно заменять соответствующими фармацевтически приемлемыми производными, а именно солями.
Как будет четко объяснено далее, применение композиций в соответствии с настоящим описанием особенно эффективно в отношении стимуляции митохондриального биогенеза и митохондриальной функции.
В предпочтительном варианте реализации раскрытые в настоящем описании композиции могут применяться в лечении и/или профилактике заболевания, ассоциированного с митохондриальной дисфункцией, каковым является ожирение.
В дополнительном варианте реализации раскрытые в настоящем описании композиции могут применяться в лечении и/или профилактике заболевания, ассоциированного с митохондриальной дисфункцией, каковым является ассоциированное с ожирением заболевание, выбранное из резистентности к инсулину и сахарного диабета 2 типа.
Согласно дополнительному варианту реализации, аминокислотные композиции могут содержать фармацевтически приемлемые вспомогательные вещества, такие как, например, белки, витамины, углеводы, натуральные и искусственные подсластители и/или ароматизаторы. В предпочтительном варианте реализации в качестве фармацевтически приемлемых вспомогательных веществ могут использоваться сывороточные белки, мальтодекстрины, фруктоза, казеинат кальция, рыбий жир, лимонная кислота или ее соли, сукралоза, сложные эфиры сахарозы, витамин D3 и витамины группы В.
Согласно настоящему описанию, предназначенные для перорального применения композиции могут выпускаться в форме таблеток, капсул, гранул, геля, гелеобразующего порошка, порошка.
Дальнейшие описания, касающиеся количества различных аминокислот, входящих в состав композиций, и их соотношений, представлены в следующих ниже пунктах, являющихся неотъемлемой частью технических рекомендаций, представленных в настоящем документе касательно изобретения.
ПРИМЕРЫ
В таблице 1 приведены составы различных композиций, содержащих аминокислоты, протестированные на клетках линии HL-1 и на иммортализованных бурых адипоцитах, как описано ниже.
В частности, композиция BCAAem - это композиция, описанная в EP 2196203 B1.
Действующим веществом композиции «B2» является комбинация аминокислот, аналогичная композиции аминокислот, входящих в состав композиции BCAAem, с добавлением лимонной кислоты. Данная композиция также содержит витамины В1 и В6.
Действующее вещество композиций, получивших названия альфа-5 (α5), альфа-6 (α6) и альфа-7 (α7), помимо аминокислот содержит лимонную, янтарную и яблочную кислоты. Кроме этого в состав действующего вещества композиции альфа-7 также входят OKG (L-орнитин-альфа-кетоглутарат) и аминокислота аспартат (L-аспарагиновая кислота).
Таблица 1
Композиции, составы которых приведены в таблице 1, можно готовить, предварительно просеяв все компоненты через сито с диаметром отверстий 0,8 мм. Для получения предварительной смеси каждый ингредиент (в количестве <10% по массе от общего количества) поместили в полиэтиленовый пакет вместе с навеской L-лизина гидрохлорида, таким образом, чтобы получить 10% от общей массы композиции. После этого пакет встряхивали вручную в течение 5 минут. Затем предварительно подготовленную смесь загружали в смеситель (Planetaria) вместе с остальными ингредиентами и перемешивали в течение 15 минут при 120 об/мин до получения однородной конечной композиции.
Кардиомиоциты линии HL-1 и иммортализованные бурые адипоциты подвергали воздействию композиций, состав которых приведен в таблице 1, после чего их митохондриальную функцию оценивали с помощью описанного ниже способа.
СПОСОБЫ
Клетки и воздействие
Кардиомиоциты линии HL-1
Кардиомиоциты линии HL-1 (безвозмездно предоставленные В.С. Клейкомб, Новый Орлеан, школа медицины (W. C. Claycomb, New Orleans, School of Medicine)) посеяли в покрытые фибронектином/желатином флаконы и культивировали до достижения 70-80% конфлюентности в среде Клейкомба (JRH Biosciences), содержащей 100 мкМ норэпинефрина (из 10 мМ стокового раствора норэпинефрина [Sigma-Aldrich], растворенного в 30 мМ растворе L-аскорбиновой кислоты [Sigma-Aldrich]), 2 мМ L-глютамина, 100 ед/мл пенициллина, 100 мкг/мл стрептомицина и 10% эмбриональой телячьей сыворотки (ЭТС) (Jrh Biosciences), как описано в Claycomb et al., 1998.
Клетки подвергали воздействию 1% (масса/объем) растворов композиций (растворенных в среде Клейкомба), составы которых приведены в таблице 1, в течение 24 или 48 ч.
По окончании этих периодов из клеток экстрагировали мРНК и ДНК или использовали их для оценки потребления кислорода. Контрольные клетки культивировали в среде Клейкомба без добавления композиций.
Иммортализованные бурые адипоциты
Иммортализованные бурые адипоциты были приобретены у профессора Патрика Сила (Patrick Seale) (Университет Пенсильвании, штат Филадельфия, США).
Представляют собой иммортализованную линию клеток, выделенных из бурой жировой ткани, SV40, которые, при использовании соответствующего протокола стимуляции, способны дифференцироваться в зрелые бурые адипоциты.
Клетки культивировали по стандартной методике в среде F12/DMEM (Gibco by Life technologies) с добавление 10% ЭТС; для дифференцировки достигшие конфлюэнтности клетки культивировали в среде с добавлением 10% ЭТС, 0,5 мМ изобутилметилксантина, 125 нМ индометацина, 1 мкМ дексаметазона, 20 нМ инсулина и 1 нМ трийодтиронина (Т3).
Через 48 часов среду культивирования клеток сменяли на среду с добавлением 10% ЭТС, 20 нМ инсулина и 1 нМ Т3.
Предварительные эксперименты подтвердили, что инкубации в такой среде на протяжении 6-7 дней достаточно для индукции экспрессии UCP1 и других маркеров бурых адипоцитов. В частности, в дифференцированных клетках была повышена экспрессия PRDM16 - белка с молекулярной массой 140 кДа, содержащего PR (PRD1-BF1-RIZ1 гомологичный) домен, являющегося маркером дифференцировки бурых адипоцитов (Seale et al., 2007). Поэтому через 6 дней клетки подвергли воздействию различных представленных в таблице 1 композиций (BCAAem, B2, α5), разведенных в среде F12/DMEM в концентрации 1% (масса/объем). Контрольные клетки культивировали в среде без добавок. Через 48 часов клетки выделили, а их ДНК изолировали для анализа генной экспрессии.
Способы оценки экспрессии генов и митохондриального биогенеза
Суммарную РНК выделяли из кардиомиоцитов линии HL-1 и иммортализованных бурых адипоцитов с помощью набора Rneasy Mini Kit (Qiagen). Один микрограмм суммарной РНК обратно транскрибировали в кДНК с помощью набора «iScript cDNA Synthesis Kit» (Bio-Rad Laboratories), как описано в D'Antona et al. (2010).
Относительный уровень экспрессии генов вычисляли как 2-ДДCt, где ДДCt соответствует разнице между ДCt для одного из воздействий и ДCt для контрольной группы с использованием GAPDH в качестве внутреннего контроля. Алгоритм Дельта-Дельта-Ct (ДДCt) представляет собой метод аппроксимации для определения относительной экспрессии генов при использовании метода количественной ПЦР в режиме реального времени (qRT-PCR) (см. Livak and Schmittgen, 2001).
Праймеры (последовательности приведены в таблице 2, ниже) были разработаны с помощью программного обеспечения «Beacon Designer 2.6» (Premier Biosoft International). Полученные значения нормализовали относительно экспрессии глицеральдегид-3-фосфатдегидрогеназы (GAPDH).
Таблица 2
Обратный 5’-3’
№ 2
Обратный 5’-3’
№ 4
Обратный 5’-3’
TTCATCCCTCTTGAGCCTTTCG
№ 6
Обратный 5’-3’
TTTTCCAAGCCTCATTTACAAGC
№ 8
Обратный 5’-3’
TGAGATCGCCGGTGCCTTGA
№ 10
Обратный 5’-3’
TGGCTCATCAATGCGGTTATCC
№ 12
(12SrRNA)
Обратный 5’-3’
TCACTGCTGAGTCCCGTGGG
№ 14
(GAPDH)
Обратный 5’-3’
CGTGATCGTAGCGTCTGGTT
№ 16
Ta температура отжига (°C); Учетный номер последовательности GAPDH: NM_008084.3; Учетный номер последовательности Cyt c: NM_007808; Учетный номер последовательности PGC-1α: AF049330; Учетный номер последовательности Tfam: NM_009360. 4; Учетный номер последовательности KFL15: NM_023184. 4; Учетный номер последовательности PP2CM: NM_175523. 4; митохондриальный геном домовой мыши (Mus musculus), полный геном: NC_005089.1; геномная ДНК (GAPDH): NC_000072.6; код праймера для 12S митохондриальной рРНК (NC_005098.1). GAPDH использовался для нормализации митохондриальной ДНК.
Для проведения анализа митохондриальной ДНК (мтДНК) общая ДНК выделялась с помощью набора «qiaamp DNA extraction kit» (QIAGEN).
мтДНК амплифицировали с использованием праймеров, специфичных к гену митохондриальной ДНК (мтДНК), и нормализовали к геномной ДНК путем амплификации ДНК гена GAPDH. Последовательности праймеров, разработанных с помощью программного обеспечения «Beacon Designer 2.6» (Premier Biosoft International; Palo Alto, CA), приведены в таблице 2 для геномной ДНК.
Статистический анализ
При обработке всех данных по экспрессии генов сравнение значений для контрольных и подвергавшихся воздействию клеток проводили с помощью двухстороннего t-критерия для парных выборок. Значение Р<0,05 считалось статистически значимым.
Потребление кислорода
1 мл кардиомиоцитов линии HL-1, подвергавшихся воздействию композиций, представленных в таблице 1, повторно ресуспендировали в сбалансированном солевом растворе Хенкса (Sigma) и центрифугировали для осаждения клеток. В некоторые образцы клеток, выступавшие в качестве положительного контроля, ввели донор оксида азота (NO), а именно NO-аддукт диэтилентриамина или DETA-NO (Sigma-Aldrich, Милан, Италия).
После этого клетки повторно ресуспендировали в буфере для оценки клеточного дыхания (0,3 М маннитола, 10 мМ KCl, 5 мМ MgCl2, 10 мМ K2PO4, рН=7,4) плотностью 3,0×106/мл.
Образцы анализировали при температуре 37°С в газонепроницаемом сосуде, оборудованном электродом для измерения содержания растворенного кислорода (электродом Кларка) (Rank Brothers Ltd.) и подключенном к регистрирующему устройству.
Электрод для измерения содержания растворенного кислорода откалибровали, приняв концентрацию кислорода в инкубационной среде равной 200 мкмоль/л при температуре 37°С.
Потребление кислорода оценивали при непрерывном перемешивании в течение примерно десяти минут. После этого полученную с помощью самописца кривую использовали для расчета потребления кислорода. Содержание кислорода может варьировать в зависимости от количества клеток. Поэтому для нормализации значений потребления кислорода образцами клеток использовали содержание белка, непосредственно коррелирующее с количеством клеток. Для количественного определения содержания белка использовали метод с бицинхониновой кислотой (БЦК).
РЕЗУЛЬТАТЫ
Митохондриальная ДНК (мтДНК) кардиомиоцитов линии HL-1
Для оценки влияния митохондриальной ДНК (мтДНК) на массу митохондрий ее содержание сначала оценивали в клетках, подвергавшихся воздействию различных аминокислотных композиций.
Как показано на фигуре 1, наиболее сильное увеличение количества мтДНК было зарегистрировано в кардиомиоцитах линии HL-1, подвергавшихся воздействию композиции α5, по сравнению с увеличением мтДНК в контрольных клетках (КК), а также в клетках, подвергавшихся воздействию композиции B2, и, что очень интересно, в клетках, подвергавшихся воздействию композиции BCAAem.
Экспрессия PGC-1α, Tfam и Cyt c в кардиомиоцитах линии HL-1
Помимо этого было проанализировано влияние различных аминокислотных композиций на митохондриальный биогенез. В частности, оценивали экспрессию кардиомиоцитами линии HL-1 следующих маркеров:
- коактиватор 1-альфа-рецептора, активируемого пролифераторами пероксисом, гамма (PGC-1α), являющийся мастер-регулятором митохондриального биогенеза,
- транскрипционный фактор А митохондрий (Tfam), фактор транскрипции мтДНК, регулирующий репликацию мтДНК,
- цитохромный комплекс (Cyt c) - белок респираторного комплекса.
Сравнение результатов, полученных после добавления в культуральную среду композиций BCAAem, B2 (композиции, аналогичной BCAAem, но дополнительно содержащей лимонную кислоту), α5 (композиции, в состав действующего вещества которой помимо аминокислот входят лимонная, янтарная и яблочная кислоты) показало, что композиция α5 наиболее эффективно стимулировала экспрессию перечисленных выше маркеров в клетках кардиомиоцитов линии HL-1
Также при этом наблюдалась зависимость время-эффект: 48-часовое воздействие композиции α5, содержащей перечисленные в таблице 1 аминокислоты в комбинации с тремя указанными органическими кислотами, вызывало более значительное увеличение показателей по сравнению с исходными значениями.
Применительно к композиции BCAAem увеличение было статистически значимым для Tfam, при сравнении с 24-часовым воздействием (фиг. 2), а также для PGC-1α и Cyt с после 48-часового воздействия (фиг. 3).
Экспрессия KFL15 и PP2CM в кардиомиоцитах линии HL-1
Круппель-подобный транскрипционный фактор-15 (KFL15) и ассоциированная с митохондриальным матриксом протеинфосфатаза семейства 2C (PP2CM) относятся к белкам, регулирующим катаболизм аминокислот с разветвленной цепью.
Начальные этапы катаболизма аминокислот с разветвленной цепью являются общими для трех аминокислот данной группы. Для их протекания необходимы митохондриальные ферменты: аминотрансфераза аминокислот с разветвленной цепью и комплекс дегидрогеназ альфа-кетокислот с разветвленной цепью.
На первой и полностью обратимой стадии деградации митохондриальная аминотрансфераза аминокислот с разветвленной цепью переносит аминогруппы с молекул аминокислот с разветвленной цепью на молекулы α-кетоглутарата с формированием альфа-кетокислот с разветвленной цепью α и глутамата.
После этого комплекс дегидрогеназ альфа-кетокислот с разветвленной цепью катализирует декарбоксилирование карбоксильных групп альфа-кетокислот с разветвленной цепью с образованием соответствующих ацил-КоА-эфиров с разветвленной цепью.
Данная реакция необратима и, соответственно, приводит к деградации аминокислот с разветвленной цепью.
Активность комплекса дегидрогеназ альфа-кетокислот с разветвленной цепью регулируется аллостерическим ингибированием конечного продукта под действием НАДН, α- кетоизокапроата и ацил-КоА-эфиров с разветвленной цепью.
Уровень активности комплекса дегидрогеназ альфа-кетокислот с разветвленной цепью определяется статусом фосфорилирования его регуляторной субъединицы Е1а.
При низких концентрациях аминокислот с разветвленной цепью происходит гиперфосфорилирование субъединицы E1a под действием киназы дегидрогеназы альфа-кетокислот с разветвленной цепью, что приводит к ингибированию активности комплекса дегидрогеназ альфа-кетокислот с разветвленной цепью и сохранению свободных аминокислот с разветвленной цепью.
При высоких концентрациях аминокислот с разветвленной цепью происходит дефосфорилирование субъединицы Е1а под действием митохондриальной серин/треониновой фосфатазы типа 2С (PP2CM) или Mg2+/Mn2+-зависимой протеинфосфатазы 1К (PPM1K), что приводит к активации комплекса дегидрогеназ альфа-кетокислот с разветвленной цепью и снижению общего количества аминокислот с разветвленной цепью (Bifari и Nisoli, 2016).
Помимо этого, было установлено, что KLF15 увеличивает экспрессию генов аминотрансферазы аминокислот с разветвленной цепью, комплекса дегидрогеназ альфа-кетокислот с разветвленной цепью и PP2CM в клетках сердца (Sun et al., 2016).
Результаты оценки уровней мРНК PP2CM и KFL13 показали, что воздействие композиции α5 обеспечивает повышение уровней мРНК PP2CM и KFL15 в кардиомиоцитах линии HL-1, по сравнению с исходными значениями. Это повышение сохраняло статистическую значимость даже при сравнении с результатами воздействия композиции BCAAem (фиг. 4).
Данные результаты демонстрируют, что композиции, содержащие действующее вещество, которое, в свою очередь, содержит лейцин, изолейцин, валин, треонин, лизин, а также лимонную, янтарную и яблочную кислоты, очень эффективны в отношении повышения митохондриальной функции и более эффективно стимулируют митохондриальный биогенез в метаболически активных клетках, даже при сравнении с композицией BCAAem.
Потребление кислорода кардиомиоцитами линии HL-1
Был проведен анализ потребления кислорода клетками линии HL-1, в среду культивирования которых вносили различные композиции. Также, в качестве положительного контроля, были проанализированы клетки, в среду культивирования которых добавили NO-аддукт диэтилентриамина (DETA-NO). Ранее уже было продемонстрировано влияние, оказываемое DETA-NO на увеличение потребления кислорода клетками (Nisoli et al., 2003). Оказалось, что NO запускает митохондриальный биогенез в очень отличающихся друг от друга клетках, начиная от адипоцитов бурого жира и заканчивая клетками линий 3T3-L1, U937 и HeLa. Оказываемый оксидом азота эффект был зависим от циклического гуанозина-3', 5'-монофосфата (цГМФ) и опосредован индукцией PGC-1α - мастер-регулятора митохондриального биогенеза (Nisoli et al., 2003).
Как и ожидалось, после 48-часового воздействия DETA-NO было зарегистрировано увеличение потребления кислорода.
Наиболее примечателен тот факт, что при 48-часовом культивировании клеток линии HL-1 в присутствии композиции α5 наблюдалось заметное увеличение потребления кислорода, что свидетельствовало о повышении активности митохондрий (фиг. 5).
Данное увеличение значительно превышает эффект, наблюдаемый после воздействия композиций B2 и BCAAem.
Экспрессия UCP1 в иммортализованных бурых адипоцитах
Как показано на фиг. 6, воздействие раскрытых композиций продолжительностью 48 часов увеличивает, хотя и в разной степени, экспрессию UCP1 в иммортализованных бурых адипоцитах.
Среди различных композиций, наиболее эффективную индукцию экспрессии UCP1 по сравнению с контрольными клетками обеспечила композиция α5 (p = 0,04 по сравнению с КК); композиция α5 также более эффективно повышала экспрессию мРНК UCP1 по сравнению с композицией BCAAem (p=0,05).
Экспрессия UCP1 α в иммортализованных бурых адипоцитах
В соответствии с данными, полученными для UCP1, композиция α5 также значительно увеличивает экспрессию PGC-1α (p=0,01), что подтверждает способность этой композиции активировать митохондриальный биогенез (фиг. 7). Следует отметить, что композиция α5 также более эффективно повышает экспрессию PGC-1α, чем композиция BCAAem.
В совокупности полученные результаты свидетельствуют о том, что композиции, содержащие в качестве действующего вещества комбинацию лейцина, изолейцина, валина, треонина, лизина, а также лимонной, янтарной и яблочной кислот, обладают значительно более высокой активностью в отношении стимуляции митохондриального биогенеза, митохондриальной функции и катаболизма аминокислот с разветвленной цепью.
Предположительно, композиции альфа-6 (α6) и альфа-7 (α7), действующее вещество которых включает аминокислоты с разветвленной цепью, треонин и лизин, а также лимонную, янтарную и яблочную кислоты, обладают схожими преимуществами.
Следует отметить, что катаболизм аминокислот с разветвленной цепью, которыми обогащена смесь, обеспечивает продукцию не только ацетил-КоА, но и сукцинил-КоА. Последний может стимулировать активность сукцинил-КоА-синтетазы, которая, в свою очередь, продуцирует сукцинат, являющийся субстратом для последующей активности сукцинатдегидрогеназы.
Таким образом, введение сукцината в смесь аминокислот с разветвленной цепью может дополнительно стимулировать активность сукцинатдегидрогеназы, что способствует еще большей активизации цикла. Следует отметить, что сукцинатдегидрогеназа, непосредственно продуцирующая восстановленную форму флавинадениндинуклеотида (ФАЛH2), также является компонентом митохондриальной цепи переноса электронов (комплекс II). Его стимуляция сукцинатом может непосредственно активировать окислительно-восстановительные переносчики в митохондриях и повышать мембранный потенциал, тем самым увеличивая градиент концентрации протонов, потребление кислорода и синтез АТФ.
В то же время добавление яблочной кислоты может активировать малатдегидрогеназу, что обеспечит повышение уровня НАДН. Это, в свою очередь, обеспечит субстратом комплекс I и, соответственно, повысит уровень АТФ, аналогично действию ФАДH2, получаемого из сукцината. С другой стороны, яблочная кислота может стимулировать активность малат-аспартатного челночного механизма. Это, в свою очередь, может способствовать дополнительному поступлению в митохондрии цитозольного НАДН, который при других условиях не проникает через митохондриальную мембрану, что делает его недоступным для митохондриального окисления. Запуск описанного механизма может еще больше повысить митохондриальную активность и потребление кислорода.
Кроме того, энергия, получаемая при расщеплении углеводородов или жиров, выделяется в результате процесса окислительного фосфорилирования в митохондриях. Катаболизм глюкозы и жирных кислот в цикле Кребса (цикле трикарбоновых кислот) обеспечивает синтез молекул НАДН и ФАДH2. После этого электроны переносятся с молекул НАДН и ФАДH2 на молекулы кислорода с помощью белковых комплексов внутренней митохондриальной мембраны; этот процесс создает на митохондриальной мембране протонный градиент. Протоны повторно проникают внутрь митохондрий через комплекс АТФ-синтазы, использующий энергию градиента для синтеза аденозинтрифосфата (АТФ). Таким образом перенос электронов взаимосвязан с синтезом АТФ.
С другой стороны, UCP1, являющийся митохондриальным трансмембранным белком, активирующимся в бурой жировой ткани после воздействия холода или избытка питательных веществ, распределяет протонный градиент по внутренней митохондриальной мембране, что приводит к разобщению с окислительным фосфорилированием. Энергия протонного градиента больше недоступна для синтеза АТФ и из-за того, что UCP1 не синтезирует АТФ, она высвобождается в форме тепла.
Повышенный митохондриальный биогенез и количество митохондрий также лежат в основе термогенной программы, запускающейся в бурой жировой ткани в ответ на указанные выше стимулы; при этом ключевая роль в этом процессе принадлежит 1-альфа-рецептору, активируемому пролифераторами пероксисом, гамма (PGC-1α) - индуцируемому холодом коактиватору транскрипции, способному регулировать экспрессию как UCP1, так и других факторов, вовлеченных в митохондриальный биогенез (Scarpulla, 2011). Соответственно, стимуляция активности UCP1 и продукция тепла в бурой жировой ткани может представлять собой потенциальный терапевтический подход к устранению проявлений ожирения; поэтому существует сильная заинтересованность в поиске агентов и/или стратегий питания, которые стимулировали бы активность этой ткани.
Также следует отметить, что совсем недавно в некоторых отложениях белой жировой ткани был идентифицирован третий тип жировой ткани, отличный от белой и бурой жировой ткани и получивший название бежевая жировая ткань. В ответ на воздействие стимулов, индуцирующих термогенез в бурой жировой ткани, в этой ткани за счет усиления экспрессии термогенных белков, таких как UCP1, а также индукции типичных для бурой жировой ткани маркеров, запускается процесс «побурения», результатом которого является усиление расхода энергии (EE, Song et al., 2017). Эти данные ставят под сомнение традиционное видение, согласно которому белая жировая ткань является инертным с точки зрения метаболизма органом, и подтверждают гипотезу более пластичной и поддающейся воздействию лекарственных средств ткани-мишени, делая бежевую жировую ткань еще одной потенциальной мишенью для предназначенного для лечения ожирения лекарственного препарата или диетологического подхода.
Все представленные результаты свидетельствуют о том, что раскрытые в данном описании композиции способны повышать митохондриальный биогенез и митохондриальную функцию, усиливать катаболизм аминокислот с разветвленной цепью в кардиомиоцитах линии HL-1 и стимулировать экспрессию маркеров бурых адипоцитов.
Все представленные выше данные четко демонстрируют пользу композиций, описанных в настоящем документе, с точки зрения лечения патологических состояний, характеризующихся недостаточностью митохондриальной функции у человека и у животных.
СПИСОК ЛИТЕРАТУРЫ
Bifari F, Nisoli E. Branched-chain amino acids differently modulate catabolic or anabolic states in mammals: a pharmacological point of view. Br J Pharmacol. 2016 Sep 17. doi: 10.1111/bph.13624. [Epub ahead of print].
Cannon B, Nedergaard J. Brown adipose tissue: function and physiological significance. Physiol Rev. 2004 Jan;84(1):277-359. Review
Claycomb WC, Lanson NA Jr, Stallworth BS, Egeland DB, Delcarpio JB, Bahinski A, Izzo NJ Jr. HL-1 cells: a cardiac muscle cell line that contracts and retains phenotypic characteristics of the adult cardiomyocyte. Proc Natl Acad Sci USA 95: 2979-2984, 1998.
D'Antona G, Ragni M, Cardile A, Tedesco L, Dossena M, Bruttini F, Caliaro F, Corsetti G, Bottinelli R, Carruba MO, Valerio A, Nisoli E. Branched-chain amino acid supplementation promotes survival and supports cardiac and skeletal muscle mitochondrial biogenesis in middle-aged mice. Cell Metab. 12: 362-372, 2010.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25: 402-408, 2001.
Nisoli E, Clementi E, Paolucci C, Cozzi V, Tonello C, Sciorati C, Bracale R, Valerio A, Francolini M, Moncada S, Carruba MO. Mitochondrial biogenesis in mammals: the role of endogenous nitric oxide. Science: 299(5608):896-9, 2003.
Nisoli E, Tonello C, Cardile A, Cozzi V, Bracale R, Tedesco L, Falcone S, Valerio A, Cantoni O, Clementi E, Moncada S, Carruba MO. Calorie restriction promotes mitochondrial biogenesis by inducing the expression of eNOS. Science 310: 314-317, 2005.
Seale P, Kajimura S, Yang W, Chin S, Rohas LM, Uldry M, Tavernier G, Langin D,Spiegelman BM. Transcriptional control of brown fat determination by PRDM16. Cell Metab. 6: 38-54, 2007
Song NJ et al. Induction of thermogenic adipocytes: molecular targets and thermogenic small molecules. Exp Mol Med. 2017 Jul 7;49(7):e353. doi: 10.1038/emm.2017.70. Review. PubMed PMID: 28684864.
Sun H, Olson KC, Gao C, Prosdocimo DA, Zhou M, Wang Z, Jeyaraj D, Youn JY, Ren S, Liu Y, Rau CD, Shah S, Ilkayeva O, Gui WJ, William NS, Wynn RM, Newgard CB, Cai H, Xiao X, Chuang DT, Schulze PC, Lynch C, Jain MK, Wang Y. Catabolic Defect of Branched-Chain Amino Acids Promotes Heart Failure. Circulation 133: 2038-2049, 2016.
Scarpulla RC. Metabolic control of mitochondrial biogenesis through the PGC-1 family regulatory network. Biochim Biophys Acta. 2011;1813(7):1269-78
Trayhurn P. Recruiting Brown Adipose Tissue in Human Obesity. Diabetes. 2016;65(5):1158-60.
Wells GD, Noseworthy MD, Hamilton J, Tarnopolski M, Tein I. Skeletal muscle metabolic dysfunction in obesity and metabolic syndrome. Can J Neurol Sci. 2008;35 (1):31-40. Review
World Health Organization. Obesity and overweight. WHO Media Centre. Updated August 2014. Available from http://www.who.int/mediacentre/factsheets/fs311/en/.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Professional Dietetics S.P.A.
<120> Композиции, содержащие аминокислоты, для применения в
лечении заболеваний, ассоциированных с митохондриальными
дисфункциями
<130> BWO20564-RC
<160> 16
<170> BiSSAP 1.3.6
<210> 1
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер 5'-3' GAPDH
<400> 1
aactttggca ttgtggaagg 20
<210> 2
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер 5'-3' GAPDH
<400> 2
acacattggg ggtaggaaca 20
<210> 3
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер 5'-3' Cyt c
<400> 3
ataggggcat gtcacctcaa ac 22
<210> 4
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер 5'-3' Cyt c
<400> 4
gtggttagcc atgacctgaa ag 22
<210> 5
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер 5'-3' PGC-1a
<400> 5
actatgaatc aagccactac agac 24
<210> 6
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер 5'-3' PGC-1a
<400> 6
ttcatccctc ttgagccttt cg 22
<210> 7
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер 5'-3' Tfam
<400> 7
aagacctcgt tcagcatata acatt 25
<210> 8
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер 5'-3' Tfam
<400> 8
ttttccaagc ctcatttaca agc 23
<210> 9
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер 5'-3' KFL15
<400> 9
acaccaagag cagccacctc a 21
<210> 10
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер 5'-3' KFL15
<400> 10
tgagatcgcc ggtgccttga 20
<210> 11
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер 5'-3' PP2CM
<400> 11
accacaggca ggcgactc 18
<210> 12
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер 5'-3' PP2CM
<400> 12
tggctcatca atgcggttat cc 22
<210> 13
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер 5'-3' 5'-3' mtDNA
<400> 13
acatgcaaac ctccatagac cgg 23
<210> 14
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер 5'-3' mtDNA
<400> 14
tcactgctga gtcccgtggg 20
<210> 15
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер 5'-3' gDNA
<400> 15
ggtcgcggtg tgggcatttg 20
<210> 16
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер 5'-3' gDNA
<400> 16
cgtgatcgta gcgtctggtt 20
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ, СОДЕРЖАЩИЕ АМИНОКИСЛОТЫ, ДЛЯ ПРИМЕНЕНИЯ В ЛЕЧЕНИИ ЗАБОЛЕВАНИЙ, АССОЦИИРОВАННЫХ С МИТОХОНДРИАЛЬНЫМИ ДИСФУНКЦИЯМИ | 2018 |
|
RU2770660C2 |
| КОМПОЗИЦИЯ ДЛЯ ПОВЫШЕНИЯ ЭКСПРЕССИИ PGC-1α | 2017 |
|
RU2814560C2 |
| КОМПОЗИЦИИ И СПОСОБЫ УСИЛЕНИЯ ЭНЕРГЕТИЧЕСКОГО ОБМЕНА | 2013 |
|
RU2655794C2 |
| ВЕЩЕСТВО, СТИМУЛИРУЮЩЕЕ ЭКСПРЕССИЮ ГЕНА КОАКТИВАТОРА PGC-1α | 2014 |
|
RU2559779C1 |
| СРЕДСТВО, СТИМУЛИРУЮЩЕЕ ЭКСПРЕССИЮ ГЕНА КОАКТИВАТОРА PGC-1A | 2011 |
|
RU2460524C1 |
| КОМПОЗИЦИИ И СПОСОБЫ МОДУЛИРОВАНИЯ МЕТАБОЛИЧЕСКИХ ПУТЕЙ | 2012 |
|
RU2631597C2 |
| СПОСОБНЫЕ К ТРАНСДУКЦИИ ПОЛИПЕПТИДЫ ДЛЯ МОДИФИКАЦИИ МИТОХОНДРИАЛЬНОГО МЕТАБОЛИЗМА | 2009 |
|
RU2529950C2 |
| САМОИНДУЦИРУЕМАЯ ЭКСПРЕССИОННАЯ СИСТЕМА И ЕЕ ПРИМЕНЕНИЕ ДЛЯ ПОЛУЧЕНИЯ ПОЛЕЗНЫХ МЕТАБОЛИТОВ С ПОМОЩЬЮ БАКТЕРИИ СЕМЕЙСТВА Enterobacteriaceae | 2013 |
|
RU2549708C2 |
| ВАРИАНТ ПРЕФЕНАТДЕГИДРАТАЗЫ И СПОСОБ ПОЛУЧЕНИЯ АМИНОКИСЛОТ С РАЗВЕТВЛЕННОЙ ЦЕПЬЮ С ЕГО ИСПОЛЬЗОВАНИЕМ | 2022 |
|
RU2827315C1 |
| УЛУЧШЕННАЯ НИТРИЛГИДРАТАЗА | 2015 |
|
RU2689606C2 |
Настоящее изобретение относится к медицине и клеточной биологии, в частности к композиции для стимуляции митохондриального биогенеза и улучшения митохондриальной функции у субъекта. Указанная композиция содержит действующее вещество, содержащее аминокислоты: лейцин, изолейцин, валин, треонин, лизин, а также лимонную кислоту, янтарную кислоту и яблочную кислоту, и фармацевтически приемлемые вспомогательные вещества. Настоящее изобретение позволяет повысить арсенал средств, направленных на снижение массы тела, а также уменьшение социальных и медицинских последствий этого заболевания. 14 з.п. ф-лы, 7 ил., 2 табл., 1 пр.
1. Композиция для стимуляции митохондриального биогенеза и улучшения митохондриальной функции у субъекта, содержащая действующее вещество, указанное действующее вещество содержит аминокислоты: лейцин, изолейцин, валин, треонин, лизин, а также лимонную кислоту, янтарную кислоту и яблочную кислоту, и фармацевтически приемлемые вспомогательные вещества.
2. Композиция по п. 1, в которой соотношение между суммарной массой лимонной, яблочной и янтарной кислот и суммарной массой аминокислот с разветвленной цепью: лейцина, изолейцина, валина, а также лизина и треонина составляет от 0,05 до 0,3, предпочтительно от 0,1 до 0,25.
3. Композиция по любому из пп. 1, 2, в которой соотношение между суммарной массой лимонной, яблочной и янтарной кислот и суммарной массой аминокислот с разветвленной цепью: лейцина, изолейцина и валина составляет от 0,1 до 0,4, предпочтительно от 0,15 до 0,35.
4. Композиция по любому из предыдущих пунктов, в которой соотношение между массой лимонной кислоты и суммарной массой яблочной и янтарной кислот составляет от 1,0 до 4,0, предпочтительно от 1,5 до 2,5.
5. Композиция по любому из предыдущих пунктов, в которой массовое соотношение лимонная кислота : яблочная кислота : янтарная кислота составляет от 10:1:1 до 2:1,5:1,5, предпочтительно от 7:1:1 до 1,5:1:1 и наиболее предпочтительно от 5:1:1 до 3:1:1.
6. Композиция по любому из предыдущих пунктов, в которой указанное действующее вещество дополнительно содержит по меньшей мере одну аминокислоту, выбранную из группы, состоящей из гистидина, фенилаланина, метионина, триптофана, тирозина, цистеина.
7. Композиция по любому из предыдущих пунктов, в которой указанное действующее вещество дополнительно содержит гистидин, фенилаланин, метионин, триптофан, цистеин и необязательно тирозин.
8. Композиция по любому из предыдущих пунктов, где соотношение между общим количеством вещества (в молях) лимонной, яблочной и янтарной кислот и общим количеством вещества (в молях) метионина, фенилаланина, гистидина и триптофана превышает 1,35.
9. Композиция по любому из предыдущих пунктов, в которой соотношение между общим количеством вещества (в молях) трех кислот: лимонной, янтарной и яблочной и общим количеством вещества (в молях) лизина и треонина составляет от 0,1 до 0,7, предпочтительно от 0,15 до 0,55.
10. Композиция по любому из предыдущих пунктов, в которой масса или количество вещества (в молях) лимонной кислоты выше массы или количества вещества (в молях) яблочной и янтарной кислот.
11. Композиция по любому из предыдущих пунктов, в которой массовое соотношение между лейцином и лимонной кислотой составляет от 5 до 1, предпочтительно от 2,50 до 3,50.
12. Композиция по любому из предыдущих пунктов, в которой указанное действующее вещество не содержит аргинин.
13. Композиция по любому из предыдущих пунктов, отличающаяся тем, что указанная композиция дополнительно содержит один или более витаминов, предпочтительно выбранных из витаминов группы В, более предпочтительно витамин В1 и/или витамин В6.
14. Композиция по любому из пп. 1-13, для применения в медицине.
15. Композиция по любому из пп. 1-14, для применения в лечении и/или профилактике заболевания, ассоциированного с митохондриальными дисфункциями, где ассоциированным с митохондриальными дисфункциями заболеванием является ожирение.
| СПОСОБ МОНТАЖА НАВЕСНОГО ОБОРУДОВАНИЯ БУРИЛЬНО-СВАЕБОЙНОЙ МАШИНЫ "БЕРКУТ" | 2001 |
|
RU2196203C1 |
| WO2016179657A1,17.11.2016 | |||
| JUAN C | |||
| BOURNAT et al., Mitochondrial Dysfunction in Obesity, Curr Opin Endocrinol Diabetes Obes., 2010, 17(5), pp | |||
| Способ пропитывания дерева | 1921 |
|
SU446A1 |
| В.К | |||
| ГОРШИНОВА и др., Дисфункция митохондрий как один из механизмов нарушения репродуктивной функции при ожирении, Акушерство и | |||

