2420-176692RU/061
СПОСОБНЫЕ К ТРАНСДУКЦИИ ПОЛИПЕПТИДЫ ДЛЯ МОДИФИКАЦИИ МИТОХОНДРИАЛЬНОГО МЕТАБОЛИЗМА
УКАЗАНИЕ О ФИНАНСИРОВАНИИ ИССЛЕДОВАНИЯ ИЛИ
РАЗРАБОТКИ ИЗ ФЕДЕРАЛЬНОГО БЮДЖЕТА
Аспекты настоящего изобретения частично поддержаны грантом номер AG023443-02, присужденным Национальными институтами здравоохранения. Таким образом, Соединенные Штаты имеют определенные права на изобретение.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится главным образом к композициям и способам доставки ДНК-связывающих белков, способных к трансдукции, в органеллы, а более конкретно относится к композициям и способам модификации митохондрий, например, индукции митохондриального биогенеза и окислительного метаболизма.
УРОВЕНЬ ТЕХНИКИ
В ходе эволюции многие организмы сталкивались с задачей введения макромолекул в живые клетки. Помимо специфичных для клеток, как правило, опосредованных рецепторами механизмов захвата или механизмов активного захвата, общее решение, независимо появлявшееся во многих линиях, основано на пептидах, специально образующихся для взаимодействия с двуслойными липидными мембранами и встраивания в них. Таким образом, бактериальные колицины, порины человека и домены трансдукции белка (PTD) различных видов содержат мотив положительно заряженной альфа-спирали, часто с амфипатической структурой, способной к встраиванию в липидные мембраны и транспортировке крупных соединений в клетку. Публикации о недавних исследованиях подтверждают успешное применение слитых с белками PTD для транспортировки белков через биологические барьеры, включая гематоэнцефалический барьер и плаценту.
Митохондрии представляют собой продуцирующие энергию и контролирующие гибель клеток б-протеобактериальные эндосимбионты, которые присутствуют во всех эукариотических клетках (Fitzpatrick, et al., Mol Biol Evol., 23(l):74-85 (2006)). Липидный состав наружной митохондриальной мембраны сходен с липидным составом эукариотических плазматических мембран, тогда как внутренняя митохондриальная мембрана содержит уникальный липид, кардиолипин, и больше напоминает бактериальные мембраны. Митохондрии содержат собственную ДНК и аппарат транскрипции и трансляции, участвующий в синтезе белков, необходимых для осуществления окислительного фосфорилирования. Многие заболевания связаны со снижением числа митохондрий, ухудшением функций митохондрий или, более конкретно, окислительного фосфорилирования. Кроме того, увеличение числа митохондрий, улучшение их функций и/или окислительного фосфорилирования в базальном или нормальном состоянии может повысить работоспособность клеток и тканей, сильно зависящих от митохондриального метаболизма, таких как головной мозг и мышцы.
Таким образом, целью изобретения является предоставление композиций и способов увеличения числа митохондрий, улучшения функций митохондрий и/или окислительного фосфорилирования у индивидуума.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Предоставлены композиции и способы повышения биологической активности митохондрий, а также увеличения числа митохондрий. В одном из вариантов осуществления предоставлены модифицированные полипептиды, содержащие связывающий полинуклеотид домен, нацеливающий домен, например, специфичный для органелл нацеливающий сигнал и домен трансдукции белка. В конкретных вариантах осуществления модифицированный полипептид содержит, по меньшей мере, один домен группы HMG, а в более типичных случаях, по меньшей мере, два домена группы HMG. Полипептид предназначен для проникновения в клетку и нацеливания на конкретные органеллы, такие как митохондрии. Нацеливающий сигнал помогает направить полипептид к месту назначения, например, в органеллу, и, таким образом, модифицирует функции клеточной органеллы.
В предпочтительном варианте осуществления последовательность рекомбинантного полипептида, по меньшей мере, на 80, 85, 90, 95, 97, 99 или 100% идентична

В другом варианте осуществления рекомбинантный полипептид кодируется нуклеиновой кислотой с последовательностью, по меньшей мере, на 80, 85, 90, 95, 97, 99 или 100% идентичной

В другом варианте осуществления предоставлены композиции и способы индукции митохондриального биогенеза и повышения митохондриального окислительного метаболизма посредством введения в клетку эффективного количества рекомбинантного полипептида, содержащего связывающий полинуклеотид домен, нацеливающий домен и домен трансдукции белка, для повышения митохондриального биогенеза по сравнению с контролем. В одном из вариантов осуществления предоставлена фармацевтическая композиция, по существу состоящая из рекомбинантного полипептида, содержащего связывающий полинуклеотид домен, нацеливающий домен, а также домен трансдукции белка и фармацевтически приемлемый носитель или эксципиент. Предпочтительно связывающий полинуклеотид домен включает TFAM или его фрагмент, способный к связыванию полинуклеотида. В другом варианте осуществления предоставлен способ снижения массы тела посредством введения индивидууму эффективного количества указанных полипептидов для снижения митохондриального окислительного метаболизма индивидуума.
В предпочтительном варианте осуществления предоставлен слитый белок, содержащий три полипептидных области или домена. Первая область представляет собой N-концевую область и включает PTD. PTD функционально связан со второй областью, которая содержит нацеливающий сигнал или домен. Вторая область функционально связана с третьей областью, включающей полипептид, связывающийся с митохондриальной ДНК или конденсирующий ее. Предпочтительно, связывающий полинуклеотид полипептид представляет собой митохондриальный фактор транскрипции. Предпочтительный полипептид, связывающий митохондриальную ДНК или конденсирующий ее, включает в качестве неограничивающих примеров TFAM или его фрагмент, способный к связыванию или конденсации митохондриальной ДНК или активации или индукции транскрипции митохондриальной ДНК.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1A-C представлены гистограммы, иллюстрирующие воздействие рекомбинантного TFAM на основное дыхание гибридных клеток LHON. На фиг. 1A представлено относительное основное дыхание в контроле в зависимости от числа суток, прошедших после обработки рекомбинантным TFAM. На фиг. 1B представлено относительное среднее число копий генов мтДНК в контроле в зависимости от числа суток, прошедших после обработки рекомбинантным TFAM. На фиг. 1C представлен процент среднего числа копий мтРНК (кДНК) в контроле.
На фиг. 2 представлена гистограмма процентов митофилина/b-актина, CI, CII, CIII, CIV и CV относительно контрольного буфера в зависимости от числа суток, прошедших после обработки рекомбинантным TFAM.
На фиг. 3A представлена гистограмма процентов окислительного метаболизма в головном мозге, сердце, печени и скелетной мышце относительно контрольного буфера. На фиг. 3B представлена гистограмма TFAM/контроль по сравнению с кДНК генов мтДНК.
На фиг. 4 представлена гистограмма массы тела (в граммах) у мышей, обработанных носителем, рекомбинантным TFAM, носителем в течение 3 месяцев и рекомбинантным TFAM в течение 3 месяца.
На фиг. 5 представлена гистограмма, иллюстрирующая относительное потребление пищи у мышей, инъецированных рекомбинантным TFAM или носителем в течение 3 месяцев.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
1. Определения
В описании и формуле изобретения настоящего изобретения применяют следующую терминологию в соответствии с определениями, изложенными ниже.
Термин "полипептиды" включает белки и их фрагменты. Полипептиды описывают в настоящем документе как последовательности аминокислотных остатков. Эти последовательности записывают слева направо в направлении от N-конца к C-концу. В соответствии со стандартной номенклатурой аминокислотные остатки последовательности обозначают тремя буквами или однобуквенным кодом, как указано далее: аланин (Ala, A), аргинин (Arg, R), аспарагин (Asn, N), аспарагиновая кислота (Asp, D), цистеин (Cys, C), глутамин (Gln, Q), глутаминовая кислота (Glu, E), глицин (Gly, G), гистидин (His, H), изолейцин (He, I), лейцин (Leu, L), лизин (Ly s, K), метионин (Met, M), фенилаланин (Phe, F), пролин (Pro, P), серин (Ser, S), треонин (Thr, T), триптофан (Trp, W), тирозин (Tyr, Y) и валин (Val, V).
"Вариант" относится к полипептиду или полинуклеотиду, который отличается от исходного полипептида или полинуклеотида, но сохраняет его основные свойства. Типичный вариант полипептида отличается по аминокислотной последовательности от другого, исходного полипептида. Как правило, степень различий ограничена таким образом, что последовательности исходного полипептида и варианта в целом очень сходны и во многих областях идентичны. Аминокислотные последовательности варианта и исходного полипептида могут различаться на одну или несколько модификаций (например, замен, вставок и/или делеций). Замененный или вставленный аминокислотный остаток может представлять собой аминокислотный остаток, который кодируется или не кодируется генетическим кодом. Вариант полипептида может быть природным, таким как аллельный вариант, или может представлять собой вариант, не известный в природе.
В структуру полипептидов по настоящему изобретению можно вносить модификации и изменения и, тем не менее, получать молекулу с такими же характеристиками, что и полипептид (например, консервативная аминокислотная замена). Например, определенные аминокислоты в последовательности можно заменять другими аминокислотами без заметного снижения активности. Поскольку биологическая функциональная активность полипептида определяется его способностью к взаимодействиям и его природой, в аминокислотную последовательность полипептида можно вносить определенные замены и, тем не менее, получать полипептид с аналогичными свойствами.
При получении таких изменений можно учитывать индекс гидрофобности аминокислот. Важность индекса гидрофобности аминокислоты при оценке биологической функции взаимодействия полипептида в данной области в основном понятна. Известно, что определенные аминокислоты можно заменять другими аминокислотами с таким же индексом или значением гидрофобности и в результате получать полипептид со сходной биологической активностью. Каждой аминокислоте присвоен индекс гидрофобности на основании ее гидрофобности и характеристик заряда. Эти индексы следующие: изолейцин (+4,5); валин (+4,2); лейцин (+3,8); фенилаланин (+2,8); цистеин/цистеин (+2,5); метионин (+1,9); аланин (+1,8); глицин (-0,4); треонин (-0,7); серин (-0,8); триптофан (-0,9); тирозин (-1,3); пролин (-1,6); гистидин (-3,2); глутамат (-3,5); глутамин (-3,5); аспартат (-3,5); аспарагин (-3,5); лизин (-3,9); и аргинин (-4,5).
Полагают, что относительные характеристики гидрофобности аминокислот определяют вторичную структуру конечного полипептида, что, в свою очередь, определяет взаимодействие полипептида с другими молекулами, такими как ферменты, субстраты, рецепторы, антитела, антигены и т.п. Известно в данной области, что аминокислоту можно заменять другой аминокислотой с таким же индексом гидрофобности и, тем не менее, получать функционально эквивалентный полипептид. При таких изменениях предпочтительны замены аминокислот, индексы гидрофобности которых находятся в пределах ±2, более предпочтительны - в пределах ±1 и наиболее предпочтительны - в пределах ±0,5.
Замены сходных аминокислот также можно проводить на основе гидрофильности, в частности, тогда, когда биологический и функциональный эквивалент полипептида или пептида, полученный таким образом, предназначается для применения в иммунологических вариантах осуществления. Аминокислотным остаткам присвоены следующие значения гидрофильности: аргинин (+3,0); лизин (+3,0); аспартат (+3,0±1); глутамат (+3,0±1); серин (+0,3); аспарагин (+0,2); глутамин (+0,2); глицин (0); пролин (-0,5±1); треонин (-0,4); аланин (-0,5); гистидин (-0,5); цистеин (-1,0); метионин (-1,3); валин (-1,5); лейцин (-1,8); изолейцин (-1,8); тирозин (-2,3); фенилаланин (-2,5); триптофан (-3,4). Следует понимать, что аминокислоту можно заменять другой аминокислотой с таким же значением гидрофильности и, тем не менее, получать биологически эквивалентный и, в частности, иммунологически эквивалентный полипептид. При таких изменениях предпочтительны замены аминокислот, индексы гидрофильности которых находятся в пределах ±2, более предпочтительны - в пределах ±1 и наиболее предпочтительны - в пределах ±0,5.
Как отмечено выше, замены аминокислот, как правило, основаны на относительном сходстве заместителей боковой цепи аминокислоты, например, их гидрофобности, гидрофильности, заряде, размере и т.п. Примеры замен, которые учитывают приведенные выше характеристики, хорошо известны специалистам в данной области и включают (исходный остаток: пример замены): (Ala: Gly, Ser), (Arg: Lys), (Asn: Gln, His), (Asp: Glu, Cys, Ser), (Gln: Asn), (Glu: Asp), (Gly: Ala). (His: Asn, Gln), (Ile: Leu, Val), (Leu: Ile, Val), (Lys: Arg), (Met: Leu, Tyr), (Ser: Thr), (Thr: Ser), (Tip: Tyr), (Tyr: Trp, Phe) и (Val: Ile, Leu). Варианты осуществления настоящего изобретения, таким образом, рассматривают функциональные или биологические эквиваленты полипептида, как указано выше. В частности, варианты осуществления полипептидов могут включать варианты с последовательностями, приблизительно на 50%, 60%, 70%, 80%, 90% и 95% совпадающими с полипептидом, представляющим интерес.
Как известно в данной области, "идентичность" представляет собой отношения между двумя или более полипептидными последовательностями, которые определяют посредством сравнения последовательностей. В данной области "идентичность" также означает степень родства последовательностей полипептидов, которую определяют посредством сравнения цепочек таких последовательностей. "Идентичность" и "сходство" можно легко подсчитать известными способами, включающими в качестве неограничивающих примеров способы, описанные в (Computational Molecular Biology, Lesk, A. M., Ed., Oxford University Press, New York, 1988; Biocomputing: Informatics and Genome Projects, Smith, D. W., Ed., Academic Press, New York, 1993; Computer Analysis of Sequence Data, Part I, Griffin, A. M., and Griffin, H. G., Eds., Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987; и Sequence Analysis Primer, Gribskov, M. and Devereux, J., Eds., M Stockton Press, New York, 1991; и Carillo, K., and Lipman, D., SIAM J Applied Math., 48: 1073 (1988).
Для выявления наибольшего сходства между изучаемыми последовательностями разработаны предпочтительные способы определения идентичности. Способы определения идентичности и сходства содержатся в общедоступных компьютерных программах. Идентичность двух последовательностей в процентах можно определять посредством программного обеспечения для анализа (т.е., Sequence Analysis Software Package of the Genetics Computer Group, Madison Wis.), которое включает алгоритм Needelman and Wunsch (J. Mol. Biol, 48: 443-453, 1970) (например, NBLAST и XBLAST). В настоящем изобретении для определения идентичности полипептидов применяют параметры по умолчанию.
В качестве примера, последовательность полипептида может являться идентичной исходной последовательности, то есть являться идентичной на 100%, или может отличаться от исходной последовательности аминокислотными заменами, число которых не превышает определенное целое число, то есть являться идентичной менее чем на 100%. Такие замены выбраны из следующих: по меньшей мере, одна аминокислотная делеция, замена, включая консервативную и неконсервативную замену, или вставка, где указанные замены могут возникать на N-конце или C-конце исходной последовательности полипептида или в любом месте между этими терминальными областями, и могут быть вставлены либо по отдельности между аминокислотами исходной последовательности, либо в составе одной или нескольких смежных групп в составе исходной последовательности. Число аминокислотных замен для данного процента идентичности определяют посредством умножения общего числа аминокислот в исходном полипептиде на численно выраженный процент соответствующей процентной идентичности (деленный на 100) и затем вычитания этого результата из указанного общего числа аминокислот в исходном полипептиде.
В настоящем документе, термин "низкая строгость" относится к условиям, позволяющим полинуклеотиду или полипептиду связываться с другим субстратом, имеющим менее специфичную или неспецифичную последовательность.
В настоящем документе, термин "очищенный" и сходные термины относятся к выделению молекулы или соединения в форме, которая по существу является свободной (по меньшей мере, на 60% свободной, предпочтительно на 75% свободной и наиболее предпочтительно на 90% свободной) от других компонентов, в норме ассоциированных с молекулой или соединением в природном окружении.
В настоящем документе, термин "фармацевтически приемлемый носитель" описывает любые стандартные фармацевтические носители, такие как раствор с фосфатно-солевым буфером, вода и эмульсии, такие как эмульсии масло-в-воде или эмульсии вода-в-масле, и различные виды увлажнителей.
В настоящем документе, термин "лечение" включает смягчение симптомов, ассоциированных со специфическим нарушением или состоянием, и/или предупреждение или устранение указанных симптомов.
"Функционально связанный" относится к смежному положению, где компоненты имеют конфигурацию, в которой они выполняют свои обычные функции. Например, контрольные последовательности или промоторы, функционально связанные с кодирующей последовательностью, способны влиять на экспрессию кодирующей последовательности, а последовательность локализации органелл, функционально связанная с белком, помогает локализовать связанный белок на конкретной органелле.
Термины "сигнал или последовательность или домен локализации" или "нацеливающий сигнал или последовательность или домен" применяют взаимозаменяемо для обозначения сигнала, который направляет молекулу к конкретной клетке, ткани, органелле, внутриклеточной области или группе клеток. Сигнал может представлять собой полинуклеотид, полипептид или молекулу углевода или может являться органическим или неорганическим соединением, которое направляет связанную с ним молекулу к необходимой области. Примеры нацеливающих сигналов включают нацеливающиеся на клетку сигналы, известные в данной области, такие как приведенные в таблице 1 и описанные в Wagner et al., Adv Gen, 53:333-354 (2005), описания которых включены в настоящий документ в качестве ссылки в полном объеме. Следует понимать, что полную последовательность, приведенную в таблице 1, не обязательно включают, и модификации, включая сокращение этих последовательностей, находятся в пределах объема изобретения, предоставляющего последовательности, способствующие направлению связанной молекулы к клеткам конкретного типа. Нацеливающие сигналы по настоящему изобретению могут быть идентичны последовательностям в таблице 1 на от 80 до 100%. Один класс подходящих нацеливающих сигналов включает сигналы, которые взаимодействуют с клеткой-мишенью посредством механизма, отличного от лиганд-рецепторного механизма. Например, нацеливающие сигналы включают сигналы, имеющие или приобретающие суммарный заряд, например, положительный заряд. Положительно заряженные сигналы можно применять для нацеливания на отрицательно заряженные типы клеток, такие как нейроны и мышечные клетки. Отрицательно заряженные сигналы можно применять для нацеливания на положительно заряженные клетки.
"Тропизм" относится к склонности молекулы притягиваться к определенным клеткам, типам клеток или состояниям клеток. В данной области тропизм может относиться к способу, выработанному различными вирусами и патогенными организмами для преимущественного нацеливания на определенные виды-паразиты или определенные типы клеток, принадлежащие этим видам. Склонность молекулы притягиваться к определенным клеткам, типам клеток или состояниям клеток можно получать посредством применения нацеливающего сигнала.
"Тип клеток" представляет собой способ распределения по группам или классификации клеток в данной области. Термин "тип клеток" относится к классификации клеток на основе их биологических характеристик, которые определяют частично посредством оценки их типичных биологических функций, локализации, морфологии, структуры и экспрессии полипептидов, нуклеотидов или метаболитов.
"Состояние клетки" относится к состоянию клетки определенного типа. Клетки динамичны на всем протяжении своей жизни и могут достигать разных состояний дифференциации и иметь разные функции, морфологию и структуру. Как применяют в настоящем документе, состояние клетки относится к определенному типу клеток на всем протяжении их жизни.
В настоящем документе, термин "поверхностный клеточный маркер" относится к любым молекулам, таким как группа, пептид, белок, углевод, нуклеиновая кислота, антитело, антиген и/или метаболит, которые представлены на поверхности клетки или вблизи нее и достаточны для идентификации клетки как уникальной по типу или состоянию.
"Домен трансдукции белков" или PTD относится к полипептидным, полинуклеотидным, углеводным или органическим или неорганическим соединениям, которые облегчают транспорт через липидные бислои, мицеллы, клеточные мембраны, мембраны органелл или мембраны везикул. PTD, прикрепленный к другой молекуле, облегчает транспорт этой молекулы через мембраны, например, транспорт из внеклеточного пространства во внутриклеточное пространство или транспорт из цитозоля в органеллу. Типичные PTD включают в качестве неограничивающих примеров ВИЧ TAT YGRKKRRQRRR (SEQ. ID NO. 1) или RKKRRQRRR (SEQ. ID NO. 2); 11 остатков аргинина, или положительно заряженные полипептиды или полинуклеотиды, состоящие из 8-15 остатков, предпочтительно 9-11 остатков.
2. Модифицированные связывающие полинуклеотид или упаковывающие полинуклеотид полипептиды
A. Связывающий полинуклеотид домен
Композиции и способы доставки полипептида, например, ДНК-связывающего белка, предоставленные в настоящем документе, включают связывающие полинуклеотид полипептиды или упаковывающие полинуклеотид полипептиды, необязательно имеющие PTD и необязательно имеющие нацеливающий сигнал или домен. Модифицированный или рекомбинантный полипептид может представлять собой любой полипептид, который связывает или упаковывает полинуклеотид, или его вариант. Рекомбинантный полипептид можно применять в качестве терапевтического средства как в чистом виде, так и в сочетании с полинуклеотидом. В одном из вариантов осуществления связывающий полинуклеотид полипептид включает, по меньшей мере, связывающий полинуклеотиды участок представителя группы белков с высокой подвижностью (HMG), в частности, по меньшей мере, один домен группы HMG.
Как правило, домен HMG содержит глобулярную складку из трех спиралей, стабилизированных в L-образной конфигурации двумя гидрофобными ядрами. Хромосомные белки высокой подвижности HMG1 или HMG2, общие для всех эукариот, неспецифически связываются с ДНК, например, для активации функций хроматина и регуляции генов. Они могут взаимодействовать непосредственно с нуклеосомами и, как полагают, представляют собой модуляторы структуры хроматина. Они также важны для активации ряда регуляторов экспрессии генов, включая p53, факторы транскрипции Hox и рецепторы стероидных гормонов, путем повышения их аффинности к ДНК. Белки HMG включают HMG-1/2, HMG-I(Y) и HMG-14/17.
Белки HMG-l/2 можно дополнительно разделить на три подсемейства в соответствии с числом доменов HMG, присутствующих в белке, их особенностями распознавания последовательностей и их эволюционными отношениями. Первая группа содержит хромосомные белки, неспецифично связывающиеся с ДНК (класс I, HMG1 и HMG2), вторая содержит рибосомные и митохондриальные факторы транскрипции, которые специфично связываются с ДНК в присутствии другого связывающего фактора (класс II, дрожжевой ARS-связывающий белок ABF-2, UBF и митохондриальный фактор транскрипции mtTF-1), и третья содержит ген-специфичные факторы транскрипции, которые специфично связываются с ДНК (класс III, лимфоидные энхансер-связывающие факторы LEF-1 и TCF-1; фактор дифференциации пола млекопитающих SRY и близкородственные белки SOX; и регуляторные белки грибов Mat-MC, Mat-a1, Ste11 и Rox1). ДНК-связывающий домен HMG1/2 имеет от приблизительно 75 до приблизительно 80 аминокислот и содержит высоко консервативные остатки пролина, ароматические и основные остатки. Типичные свойства белков с доменом HMG включают взаимодействие с малой бороздкой спирали ДНК, связывание с нерегулярными структурами в ДНК и способность модулировать структуру ДНК посредством ее изгибания.
Белки SOX (SRY-подобные HMG) выполняют важнейшие функции в ряде процессов развития, включая детерминацию пола, формирование скелета, развитие пре-B и T-клеток и нейрональную индукцию. SOX9 играет прямую роль в хондрогенезе, связывая и активируя хондроцит-специфичный энхансер гена Col2a1. Нарушение функций гена SOX9 приводит к генетическому заболеванию, известному как кампомелическая дисплазия (CD), - форме карликовости, которая характеризуется чрезвычайно сильной деформацией скелета, и к заболеванию, при котором три четверти XY индивидуумов либо являются интерсексами, либо демонстрируют реверсию пола от мужского к женскому. Клонировано более 20 членов семейства SOX. Все они содержат домен HMG, который может специфически связываться с двухнитевыми мотивами ДНК и имеет >50% идентичность с доменом HMG SRY, фактора развития семенников человека. Предпочтительный ДНК-связывающий участок SOX9, определенный как AGAACAATGG (SEQ ID NO: 3), содержит коровый связывающий элемент SOX (SCBE), AACAAT, и фланкирующие 5' AG и 3' GG нуклеотиды, усиливающие связывание с геном SOX9.
В одном из вариантов осуществления рекомбинантный связывающий полинуклеотиды белок имеет, по меньшей мере, один домен HMG, как правило, по меньшей мере, два, и более конкретно 2-5 доменов HMG. Домены HMG могут связываться с AT-богатыми последовательностями ДНК, используя, например, обширную поверхность на вогнутой стороне белка для связывания с малой бороздкой ДНК. Это связывание изгибает ось спирали ДНК в сторону от места контакта. Первая и вторая спирали контактируют с ДНК, а их N-конец встраивается в малую бороздку, тогда как спираль 3 преимущественно обращена к растворителю. В малой бороздке возникает частичная интеркаляция алифатических и ароматических остатков спирали 2.
В других вариантах осуществления связывающий полинуклеотиды полипептид может иметь, по меньшей мере, один связывающий полинуклеотид домен, как правило, два или более связывающих полинуклеотиды домены. Связывающие полинуклеотиды домены могут быть одинаковыми или разными. Например, связывающий полинуклеотиды полипептид может включать, по меньшей мере, один домен HMG в сочетании с одним или несколькими ДНК-связывающими доменами, выбранными из группы, состоящей из домена HMG, гомеодомена и домена POU; домен цинкового пальца, такой как C2H2 и C2C2; домен, образующий амфипатическую спираль, такой как домены типа лейциновая молния и спираль-петля-спираль; и домены сворачивания гистона. Связывающий полинуклеотиды домен может быть специфичным к определенной полинуклеотидной последовательности или предпочтительно может неспецифично связываться с полинуклеотидом. Альтернативно, связывающий полинуклеотиды полипептид может иметь дополнительную комбинацию из, по меньшей мере, одного связывающего полинуклеотиды домена, который специфично связывается с ДНК, и, по меньшей мере, одного связывающего полинуклеотиды домена, который неспецифично связывается с ДНК.
Конкретные варианты осуществления предоставляют модифицированные связывающие полинуклеотиды полипептиды, содержащие мотив спираль-петля-спираль или, по меньшей мере, связывающую полинуклеотиды область белка типа спираль-петля-спираль. Структура белков типа спираль-петля-спираль сходна со структурой бактериальных регуляторных белков, таких как l-репрессор и белки cro, lac-репрессор и т.д., которые образуют димеры и имеют палиндромные участки связывания. Они содержат 3 a-спиральных участка, разделенных короткими петлями, в результате чего их называют белками типа спираль-петля-спираль. Одна спираль белка (спираль 3) в каждой субъединице димера занимает большую бороздку двух соседних оборотов спирали ДНК. Таким образом, в другом варианте осуществления указанные связывающие полинуклеотиды полипептиды могут формировать димеры или другие многокомпонентные комплексы и имеют от 1 до 3 спиралей.
В еще одном варианте осуществления связывающий полинуклеотиды полипептид включает гомеодомен или участок содержащего гомеодомен белка. Содержащие гомеодомен белки связываются с последовательностью в 180 пар оснований, которую первоначально обнаружили в группе генов, названных гомеотическими генами. Соответственно, последовательность назвали гомеобоксом. 180 п.н. соответствуют 60 аминокислотам в соответствующем белке. Этот белковый домен называют гомеодоменом. Содержащие гомеодомен белки с тех пор обнаружили у широкого спектра организмов, включая позвоночных и растения. Гомеодомен демонстрирует высокую степень консервативности последовательности. Гомеодомен содержит 4 б-спиральных участка. Спирали II и III соединены друг с другом посредством 3 аминокислот, образующих петлю. Структура этого участка очень сходна со структурой спиралей II и III бактериальных ДНК-связывающих белков.
Еще один вариант осуществления предоставляет модифицированный связывающий полинуклеотиды полипептид, содержащий домен цинкового пальца или, по меньшей мере, участок белка цинкового пальца. Белки цинковых пальцев содержат домен с универсальной структурой: Phe (иногда Tyr) - Cys - от 2 до 4 аминокислот - Cys - 3 аминокислоты - Phe (иногда Tyr) - 5 аминокислот - Leu - 2 аминокислоты - His - 3 аминокислоты - His. Остатки фенилаланина или тирозина, которые находятся в консервативных позициях, необходимы для связывания ДНК. Сходные последовательности обнаружили в ряде других ДНК-связывающих белков, хотя число пальцев варьирует. Например, фактор транскрипции SP1, который связывается с GC-боксом, находящимся в области проксимального промотора ряда генов, содержит 3 пальца. Цинковые пальцы такого типа, которые содержат 2 цистеина и 2 гистидина, называют цинковыми пальцами C2H2.
Другой тип цинковых пальцев, в которых цинк связан между 2 парами цистеина, обнаружили в ряде ДНК-связывающих белков. Типичная структура цинковых пальцев этого типа следующая: Cys - 2 аминокислоты - Cys - 13 аминокислот - Cys - 2 аминокислоты - Cys. Их называют цинковыми пальцами C2C2. Их обнаружили в группе белков, известных как суперсемейство стероидных рецепторов, каждый из которых содержит 2 цинковых пальца C2C2.
В другом варианте осуществления предоставлен модифицированный связывающий полинуклеотиды полипептид, содержащий лейциновую молнию или, по меньшей мере, участок белка с лейциновой молнией. Первый белок с лейциновой молнией обнаружили в экстрактах клеток печени и назвали его C/EBP, поскольку он представляет собой энхансер-связывающий белок и, как первоначально полагали, связывается с последовательность проксимального промотора CAAT. C/EBP связывается с ДНК только в виде димера. Область белка, в которой два мономера соединяются, образуя димер, называют доменом димеризации. Он находится около C-конца белка. Изучение аминокислотной последовательности показало, что на протяжении участка в 35 аминокислот каждая седьмая аминокислота является остатком лейцина. Когда этот участок формирует альфа-спираль, все эти остатки лейцина выстраиваются в линию на одной поверхности спирали.
Поскольку лейцин содержит гидрофобную боковую цепь, одна поверхность спирали является сильно гидрофобной. На противоположной поверхности есть аминокислоты с заряженными гидрофильными боковыми цепями. Сочетание гидрофобных и гидрофильных характеристик придает молекуле ее амфипатические свойства. К области лейциновой молнии прилегает область, состоящая из 20-30 аминокислот и богатая основными (положительно заряженными) аминокислотами лизином и аргинином. Это ДНК-связывающий домен - часто обозначаемый как домен bZIP - основная область лейциновой молнии. C/EBP, как полагают, связывает ДНК посредством этих областей bZIP, которые наматываются на спираль ДНК.
Лейциновую молнию - структуру bZIP - обнаружили в ряде других белков, включая продукты онкогенов jun и fos. Хотя C/EBP связывается с ДНК как гомодимер, состоящий из идентичных субъединиц, fos совсем не может формировать гомодимеры, а гомодимеры jun/jun обычно нестабильны. Однако гетеродимеры fos/jun гораздо более стабильны. Эти гетеродимеры fos/jun формируют базальный фактор транскрипции под названием AP1, который связывается с различными промоторами и энхансерами и активирует транскрипцию. Консенсусный связывающий AP1 участок имеет последовательность TGACTCA и представляет собой палиндором.
В другом варианте осуществления предоставлен модифицированный связывающий полинуклеотиды полипептид, содержащий домен типа спираль-петля-спираль или связывающий полинуклеотиды участок белка типа спираль-петля-спираль. Белки типа спираль-петля-спираль сходны с лейциновыми молниями в том, что они формируют димеры посредством объединения амфипатических спиралей. Их впервые открыли как класс белков после обнаружения участка сходства между двумя энхансер-связывающими белками под названием E47 и E12. Эта консервативная область может формировать две амфипатических спирали, разделенных петлей, образуя, таким образом, структуру спираль-петля-спираль. Рядом с доменом димеризации находится ДНК-связывающий домен, также богатый основными аминокислотами, который называют доменом bHLH. Эти структуры также обнаружили в ряде генов, которые необходимы для развития нервной системы Drosophila - комплексе Achaete-scute, и в белке под названием MyoD, который необходим для дифференциации мышц млекопитающих.
В другом варианте осуществления модифицированный связывающий полинуклеотиды полипептид включает гистоновый белок, фрагмент гистонового белка, или, по меньшей мере, один домен сворачивания гистона. Домены сворачивания гистона находятся в мономерах гистоновых белков, объединенных в димеры. Гистоновые белки включают H2A, H2B, H3 и H4, которые могут формировать гетеродимеры H2A-2B и H3-H4. Следует понимать, что гистоноподобные полипептиды также можно применять в композициях и способах изобретения. Гистоноподобные полипептиды включают в качестве неограничивающих примеров HMf или гистон Methanothermous fervidus, другие гистоны простейших, известные в данной области, и полипептиды, содержащие домен сворачивания гистона, такие как MJ1647, CBF, TAFII или фактор транскрипции IID, SPT3 и Dr1-DRAP (Sanderman, K., et al., Cell. Mol. Life Sci. 54:1350-1364 (1998), в полном объеме включен в качестве ссылки).
Один из вариантов осуществления наряду с прочими предоставляет негистоновый связывающий полинуклеотиды полипептид, например, связывающий полинуклеотиды полипептид, включающий митохондриальный фактор транскрипции A (TFAM), его вариант или его фрагмент, достаточный для связывания полинуклеотидов. Последовательность варианта TFAM может являться на 80%, 85%, 90%, 95%, 99% или более идентичной последовательности исходного TFAM, например, природного TFAM.
TFAM относят к группе белков высокой подвижности (HMG), имеющих два домена HMG. TFAM, как и другие белки HMG, связывает, обматывает, изгибает и разматывает ДНК. Таким образом, варианты осуществления настоящего изобретения включают связывающие полинуклеотиды полипептиды, которые содержат одну или несколько связывающих полинуклеотиды областей белков семейства HMG и необязательно вызывают структурные изменения полинуклеотида при связывании или соединении полипептида с полинуклеотидом. Полипептид упаковывает полинуклеотид посредством изменения его конформации. Сообщается, что TFAM связывается с митохондриальной ДНК в соотношении 900:1 (Alam, T. I., et al., Nucleic Acid Res. 31(6):1640-1645 (2003)). Следует понимать, что количество связывающего полинуклеотиды полипептида, которое применяют в композициях и способах, раскрытых в настоящем документе, может варьировать в зависимости от размера и количества полинуклеотида, подлежащего доставке. Подходящее соотношение связывающего полинуклеотиды полипептида и пар оснований полинуклеотида, подлежащего доставке, включает в качестве неограничивающих примеров приблизительно от 1:1 до 1:1,000; более предпочтительно 1:100; и наиболее предпочтительно 1: от приблизительно 10 до приблизительно 20 пар оснований полинуклеотида, подлежащего доставке. Также следует понимать, что к полинуклеотиду можно добавлять TFAM, другой связывающий полинуклеотиды полипептид или комбинацию из двух или более связывающих полинуклеотиды полипептидов, чтобы обернуть или обмотать полинуклеотид, и, таким образом, упаковать его и защитить от деградации.
TFAM можно модифицировать таким образом, чтобы он включал PTD и, необязательно, включал нацеливающий сигнал. Нацеливающий сигнал может включать последовательность мономеров, которые облегают локализацию молекулы в конкретной ткани, клетке или органелле. Мономеры могут представлять собой аминокислоты, нуклеотиды или нуклеозиды или сахарные группы, такие как глюкоза, галактоза и т.п., которые формируют углеводные нацеливающие сигналы.
B. Домен трансдукции белков
Связывающий полинуклеотиды полипептид можно модифицировать, включив в него домены трансдукции белков (PTD), также известные как пептиды, проникающие в клетку (CPPS). PTD известны в данной области и включают в качестве неограничивающих примеров небольшие участки белков, способные проникать через клеточную мембрану посредством рецептор-независимого механизма (Kabouridis, P., Trends in Biotechnology (11):498-503 (2003)). Хотя известно несколько PTD, два наиболее часто применяемых PTD получают из белка TAT (Frankel and Pabo, Cell, 55(6):1189-93(1988)) ВИЧ и транскрипционного фактора Antennapedia Drosophila, чей PTD известен как пенетратин (Derossi et al., J Biol Chem., 269(14): 10444-50 (1994)).
Гомеодомен Antennapedia состоит из 68 аминокислотных остатков и содержит четыре альфа-спирали. Пенетратин представляет собой активный домен этого белка и состоит из последовательности в 16 аминокислот, производной от третьей спирали Antennapedia. Белок TAT состоит из 86 аминокислот и вовлечен в репликацию ВИЧ-1. TAT PTD состоит из домена длиной в 11 аминокислот (остатки от 47 до 57; YGRKKRRQRRR (SEQ. ID. NO. 1)) основного белка, который, как полагают, крайне необходим для поглощения. Кроме того, доказано, что основной домен Tat(49-57) или RKKRRQRRR (SEQ. ID NO. 2) представляет собой PTD. В современной литературе TAT приписывают функцию слияния с белками, представляющими интерес, для осуществления импорта в клетки. Несколько модификаций TAT, включая замену глутамина на аланин, т.е., Q→A, привели к усилению клеточного поглощения приблизительно от 90% (Wender et al., Proc Natl Acad Sci USA, 97(24): 13003-8 (2000)) до 33-кратного усиления в клетках млекопитающих (Ho et al., Cancer Res., 61(2):474-7 (2001)). Самое эффективное поглощение модифицированных белов выявили в экспериментах по мутагенезу TAT-PTD, где показали, что участок из 11 остатков аргинина на несколько порядков более эффективен, чем внутриклеточный переносчик. Таким образом, некоторые варианты осуществления включают PTD, которые могут являться катионными или амфипатическими. Дополнительные примеры PTD включают в качестве неограничивающих примеров поли-Arg - RRRRRRR (SEQ. ID. NO.: 4); PTD-5 - RRQRRTSKLMKR (SEQ. ID. NO.: 5); транспортан GWTLNSAGYLLGKINLKALAALAKKIL (SEQ. ID. NO.: 6); KALA - WEAKLAKALAKALAKHLAKALAKALKCEA (SEQ. ID. NO.: 7); и RQIKIWFQNRRMKWKK (SEQ. ID. NO.: 8).
C. Нацеливающий сигнал или домен
В других вариантах осуществления модифицированный связывающий полинуклеотиды полипептид необязательно модифицируют до включения нацеливающего сигнала или домена. Нацеливающий сигнал или последовательность может быть специфичной к хозяину, ткани, органу, клетке, органелле, неядерной органелле или клеточному компартменту. Например, композиции, предоставленные в настоящем документе, можно модифицировать посредством макромолекул с концевыми галактозильными остатками для направления композиций к печени или клеткам печени. Модифицированные композиции избирательно проникают в гепатоциты после взаимодействия носителя галактозильных остатков с асиалогликопротеиновыми рецепторами, которые присутствуют в больших количествах только на этих клетках и характеризуются высокой аффинностью к ним. Кроме того, композиции, предоставленные в настоящем документе, можно направлять на другие внутриклеточные области, компартменты или типы клеток.
В одном из вариантов осуществления нацеливающий сигнал связывается со своим лигандом или рецептором, расположенным на поверхности клетки-мишени, и сближает вектор с клеточной мембраной достаточно для того, чтобы разрешить проникновение вектора в клетку. Дополнительные варианты осуществления настоящего изобретения направлены на специфическую доставку полинуклеотидов в специфичные ткани или типы клеток, где полинуклеотиды могут кодировать полипептид или влиять на экспрессию различных полинуклеотидов. Доставленные в клетку полинуклеотиды могут кодировать полипептиды, способные усиливать функционирование клетки или участвовать в нем.
В предпочтительном варианте осуществления нацеливающую молекулу выбирают из группы, состоящей из антитела или его антигенсвязывающего фрагмента, домена антитела, антигена, рецептора T-клетки, рецептора клеточной поверхности, молекулы адгезии клеточной поверхности, белка локуса главного комплекса гистосовместимости, белка оболочки вируса и пептида, выбранного посредством фагового дисплея, который специфично связывается с определенной клеткой.
Нацеливание полинуклеотидов на конкретные клетки можно дополнять модификацией композиций по настоящему изобретению для экспрессии клеточных и тканевых нацеливающих сигналов. Эти последовательности воздействуют на конкретные клетки и ткани, но в некоторых вариантах осуществления взаимодействие нацеливающего сигнала с клеткой происходит посредством механизма, отличного от обычного лиганд-рецепторного механизма. Эукариотическая клетка содержит ряд различных молекул клеточной поверхности. Структура и функции каждой молекулы могут быть специфичными для клеток с разным происхождением, экспрессией, свойствами и структурой. Определение уникального комплемента к молекулам клеточной поверхности конкретного типа клеток можно проводить способами, хорошо известными в данной области.
Специалисты в данной области признают, что тропизм описанной векторной композиции можно легко изменить посредством модификации нацеливающего сигнала. В одном из конкретных вариантов осуществления предоставлены композиции, позволяющие добавлять к вектору антитела, специфичные к антигенам клеточной поверхности, для воздействия на доставку полинуклеотидов. Примеры антигенов клеточной поверхности приведены в таблице 1 и описаны в настоящем документе.
Специалистам в данной области известно, что почти все типы клеток тканей млекопитающих имеют какой-нибудь уникальный рецептор или антиген клеточной поверхности. Таким образом, в рецептор или антиген клеточной поверхности можно встроить почти любой лиганд, действующий в качестве нацеливающего сигнала. Например, пептидильные гормоны можно применять в качестве нацеливающих молекул для направленной доставки к клеткам, имеющим рецепторы для этих гормонов. Хемокины и цитокины также можно применять в качестве нацеливающих сигналов для направленной доставки комплекса к их клеткам-мишеням. Для идентификации генов, которые избирательно экспрессируются в конкретных клетках или состояниях клетки, разработан ряд способов, и специалисты в данной области могут применять данные способы для идентификации нацеливающих сигналов, которые избирательно или уникально экспрессируются в ткани-мишени, представляющей интерес.
i. Направленное воздействие на головной мозг
В одном из вариантов осуществления нацеливающий сигнал воздействует на клетки нервной системы, включая головной мозг и периферическую нервную систему. Клетки головного мозга включают клетки нескольких типов и состояний и имеют уникальные молекулы клеточной поверхности, специфичные для каждого типа. Кроме того, типы и состояния клеток можно далее характеризовать и группировать на основе типичных молекул клеточной поверхности, представленных на этих клетках.
В одном из вариантов осуществления нацеливающий сигнал воздействует на специфические рецепторы нейромедиаторов, находящиеся на поверхности клеток нервной системы. Распределение рецепторов нейромедиаторов хорошо известно специалистам в данной области, и квалифицированные специалисты могут направлять описанные композиции посредством применения антител, специфичных к рецепторам нейромедиаторов, в качестве нацеливающих сигналов. Кроме того, с учетом тропизма нейромедиаторов к их рецепторам, в одном из вариантов осуществления нацеливающий сигнал состоит из нейромедиатора или лиганда, способного к специфическому связыванию с рецептором нейромедиатора.
В одном из вариантов осуществления нацеливающий сигнал специфичен к клеткам нервной системы, которые могут включать астроциты, микроглию, нейроны олигодендриты и Шванновские клетки. Эти клетки можно далее классифицировать на основе их функций, месторасположения, формы, класса нейромедиатора и патологического состояния. Клетки нервной системы, например, стволовые клетки, также можно идентифицировать по их стадии дифференциации. Примеры маркеров, специфичных для этих типов клеток и состояний, хорошо известны в данной области и включают в качестве неограничивающих примеров CD133 и маркеры нейросферы.
ii. Направленное воздействие на мышцы
В одном из вариантов осуществления нацеливающий сигнал воздействует на клетки костно-мышечной системы. Мышечные клетки включают несколько типов и имеют уникальные молекулы клеточной поверхности, специфичные для типа и состояния клеток. Кроме того, типы и состояния клеток можно далее характеризовать и группировать на основе типичных молекул клеточной поверхности этих клеток.
В одном из вариантов осуществления нацеливающий сигнал воздействует на специфические рецепторы нейромедиаторов, находящиеся на поверхности мышечных клеток. Распределение рецепторов нейромедиаторов хорошо известно специалистам в данной области, и квалифицированные специалисты могут направлять описанные композиции посредством применения антител, специфичных к рецепторам нейромедиаторов, в качестве нацеливающих сигналов. Кроме того, с учетом тропизма нейромедиаторов к их рецепторам, в одном из вариантов осуществления нацеливающий сигнал состоит из нейромедиатора. Примеры нейромедиаторов, экспрессирующихся в тех мышечных клетках, на которые может быть направлено воздействие, включают в качестве неограничивающих примеров ацетилхолин и норадреналин.
В одном из вариантов осуществления нацеливающий сигнал специфичен к мышечным клеткам нервной системы, которые образуют две основные группы, тип I и тип II. Эти клетки можно далее классифицировать на основе их функций, месторасположения, формы, содержания миоглобина и патологического состояния. Мышечные клетки нервной системы, например, стволовые мышечные клетки, также можно идентифицировать по их стадии дифференциации. Примеры маркеров, специфичных для этих типов клеток и состояний, хорошо известны в данной области и включают в качестве неограничивающих примеров MyoD, Pax7 и MR4.
iii. Направленное воздействие на опухоли
В одном из вариантов осуществления нацеливающий сигнал применяют для избирательного воздействия на опухолевые клетки. Опухолевые клетки экспрессируют поверхностные клеточные маркеры, которые могут являться специфичными или неспецифичными для опухолевых клеток, но которые преимущественно представлены в опухолевых клетках. Примеры опухолеспецифичных поверхностных клеточных маркеров включают в качестве неограничивающих примеров альфа-фетопротеин (AFP), C-реактивный белок (CRP), антиген злокачественной опухоли-50 (CA-50), антиген злокачественной опухоли-125 (CA-125), ассоциированный с раком яичников, антиген злокачественной опухоли 15-3 (CA15-3), ассоциированный с раком молочной железы, антиген злокачественной опухоли-19 (CA-19) и антиген злокачественной опухоли-242, ассоциированные с раком желудочно-кишечного тракта, раково-эмбриональный антиген (CEA), карцинома-ассоциированный антиген (CAA), хромогранин A, антиген эпителиальный муцин (MC5), человеческий эпителиальный антиген (HEA), антиген Льюиса, меланомный антиген, меланома-ассоциированные антигены 100, 25 и 150, муциноподобный карцинома-ассоциированный антиген, белок множественной лекарственной устойчивости (MRPm6), белок множественной лекарственной устойчивости (MRP41), белок онкогена Neu (C-erbB-2), нейрон-специфичная энолаза (NSE), P-гликопротеин (продукт гена mdr1), антиген множественной лекарственной устойчивости, p170, антиген множественной лекарственной устойчивости, простата-специфичный антиген (PSA), CD56 и NCAM. В одном из вариантов осуществления нацеливающий сигнал состоит из антитела, специфичного к некоторым опухолевым поверхностным клеточным маркерам.
iv. Антитела
В другом варианте осуществления предоставлено антитело или его антигенсвязывающий фрагмент, связанный с раскрытыми рекомбинантными полипептидами и действующий в качестве нацеливающего сигнала. Антитела или их связывающие антиген фрагменты эффективны для направления вектора к типу клеток или состоянию клеток. В одном из вариантов осуществления рекомбинантный полипептид содержит связывающий антитела домен, полученный, например, из белков, связывающих антитела, таких как Белок A и Белок G Staphylococcus aureus. Вместо них можно применять другие связывающие антитела домены, известные в данной области. В определенных вариантах осуществления антитело может быть поликлональным, моноклональным, линейным, гуманизированным, химерным или представлять собой фрагмент антитела. Типичные фрагменты антител представляют собой фрагменты, которые связывают связывающий антитело участок невирусного вектора, и включают Fab, Fab1, F(ab'), диатела Fv, линейные антитела, одноцепочечные антитела и биспецифические антитела, известные в данной области.
В некоторых вариантах осуществления нацеливающий домен включает целое антитело или его часть, которая направляет вектор к желаемому типу клеток или желаемому состоянию клеток. Антитела могут являться моноклональными или поликлональными, но предпочтительно моноклональными. Для генотерапии человека антитела получают из генов человека; они специфичны к поверхностным клеточным маркерам и продуцируются для снижения потенциальной иммуногенности к паразитам человека, как известно в данной области. Например, можно применять трансгенных мышей, имеющих полный кластер генов иммуноглобулина человека и способных продуцировать "человеческие" антитела. В одном из вариантов осуществления фрагменты таких антител человека применяют в качестве нацеливающих сигналов. В предпочтительном варианте осуществления одноцепочечные антитела, смоделированные на антителах человека, подготавливают в прокариотической культуре.
Дополнительные варианты осуществления настоящего изобретения направлены на специфическую доставку полипептида к внутриклеточным компартментам или органеллам. Эукариотические клетки содержат мембраносвязанные структуры или органеллы. Органеллы могут иметь однослойные или многослойные мембраны и присутствовать как в растительных, так и в животных клетках. В зависимости от функций органеллы могут включать специфические компоненты, такие как белки и кофакторы. Полипептиды, доставляемые к органелле, могут улучшать функционирование органеллы или влиять на него. Некоторые органеллы, такие как митохондрии и хлоропласты, содержат свой собственный геном. В таких органеллах происходит репликация, транскрипция и трансляция нуклеиновых кислот. Белки импортируются, а метаболиты экспортируются. Таким образом, через мембраны органелл происходит обмен веществами. В некоторых вариантах осуществления митохондриальные полипептиды специфически доставляются к митохондриям.
Примеры органелл включают ядро, митохондрии, хлоропласты, лизосомы, пероксисомы, аппарат Гольджи, эндоплазматический ретикулум и ядрышко. Искусственные органеллы, сформированные из липидов, могут содержать специфические белки внутри липидных мембран. Кроме того, содержимое искусственных органелл можно изменять и добавлять компоненты для трансляции нуклеиновых кислот.
v. Сигналы ядерной организации
Композиции, раскрытые в настоящем документе, могут включать один или несколько сигналов ядерной организации. Сигналы или домены ядерной организации (NLS) известны в данной области и включают, например, антиген SV 40 T или его фрагмент, такой как PKKKRKV (SEQ. ID. NO.: 9). NLS могут представлять собой простые катионные последовательности, состоящие из от приблизительно 4 до приблизительно 8 аминокислот, или могут являться двойными и содержать два независимых положительно заряженных кластера, разделенных устойчивым к мутациям линкерным участком, состоящим приблизительно из 10-12 аминокислот. Дополнительные типичные NLS включают в качестве неограничивающих примеров GKKRSKV (SEQ. ID. NO.: 10); KSRKRKL (SEQ. ID. NO.: 11); KRPAATKKAGQAKKKKLDK (SEQ. ID. NO.: 12); RKKRKTEEESPLKDKAKKSK (SEQ. ID. NO.: 13); KDCVMNKHHRNRCQYCRLQR (SEQ. ID. NO.: 14); PAAKRVKLD (SEQ. ID. NO.: 15); и KKYENVVIKRSPRKRGRPRK (SEQ. ID. NO.: 16).
vi. Направленное воздействие на митохондрии
В других вариантах осуществления настоящего изобретения раскрыты модифицированные связывающие полинуклеотиды полипептиды, направленно воздействующие на митохондрии. Митохондрия содержит молекулярный аппарат конвертирования энергии в аденозин трифосфат (АТФ) посредством расщепления глюкозы. Энергия, хранящаяся в макроэргических фосфатных связях АТФ, становится доступной для снабжения энергией клеточных функций. Митохондрии в основном состоят из белка, но содержат также некоторое количество липидов, ДНК и РНК. Эти, как правило, сферические органеллы имеют наружную мембрану, окружающую внутреннюю мембрану, которая образует складки (кристы) и несет ферменты окислительного фосфорилирования и переноса электронов. В большинстве митохондрий кристы имеют вид плоских гребней, но митохондрии клеток, секретирующих стероиды, могут содержать трубчатые кристы. Митохондриальный матрикс содержит ферменты цикла Кребса, окисления жирных кислот и митохондриальные нуклеиновые кислоты.
Митохондриальная ДНК является двухцепочечной и замкнутой в кольцо. Митохондриальная РНК имеет три стандартных разновидности; рибосомная, информационная и транспортная, но все они специфичны для митохондрий. Некоторые белки синтезируются в митохондриях на митохондриальных рибосомах, которые отличаются от цитоплазматических рибосом. Другие митохондриальные белки синтезируются на цитоплазматических рибосомах и содержат сигнальный пептид, который направляет их к митохондриям. Метаболическая активность клетки связана с количеством крист и числом митохондрий в клетке. Клетки с высокой метаболической активностью, такие как сердечная мышца, содержат много хорошо развитых митохондрий. Новые митохондрии образуются из предшествующих митохондрий путем их роста и деления.
Внутренние мембраны митохондрий содержат семейство белков с родственными последовательностями и структурами, которые переносят различные метаболиты через мембрану. Их аминокислотные последовательности имеют трехчленную структуру и состоят из трех родственных последовательностей приблизительно в 100 аминокислот длиной. Повторы одного переносчика родственны повторам других, и несколько типичных характеристик последовательностей свойственны всем членам семейства.
Нацеливание специфических полипептидов на органеллы можно дополнять такой модификацией раскрытых композиций, при которой они содержат специфические сигналы нацеливания на органеллы. Эти последовательности направленно воздействуют на конкретные органеллы, но в некоторых вариантах осуществления взаимодействие нацеливающего сигнала с органеллами происходит посредством механизма, отличного от обычного лиганд-рецепторного механизма. Эукариотическая клетка содержит ряд обособленных мембраносвязанных компартментов, или органелл. Структура и функции каждой органеллы определяются в основном уникальным составом входящих в нее полипептидов. Однако синтез подавляющего большинства этих полипептидов начинается в цитоплазме. Таким образом, для биогенеза и обслуживания органелл необходимо, чтобы новые синтезированные белки могли безошибочно доставляться к соответствующим компартментам. Это часто происходит посредством аминоконцевых сигнальных последовательностей, а также посттрансляционных модификаций и вторичной структуры. Для митохондрий установлено несколько аминоконцевых нацеливающих сигналов, известных в данной области.
В одном из вариантов осуществления сигнал нацеливания на органеллы может содержать, по меньшей мере, два, предпочтительно 5-15, наиболее предпочтительно приблизительно 11 заряженных групп, которые заставляют нацеливающий сигнал притягиваться к органеллам, имеющим полный противоположный заряд. В другом варианте осуществления нацеливающий сигнал может содержать ряд заряженных групп, которые заставляют нацеливающий сигнал транспортироваться в органеллу либо по градиенту электромагнитного потенциала, либо против него. Подходящие заряженные группы представляют собой группы, заряженные при внутриклеточных условиях, такие как аминокислоты с заряженными функциональными группами, аминогруппы, нуклеиновые кислоты и т.п. Сигналы митохондриальной локализации/сигналы нацеливания на митохондрии, как правило, состоят из лидерной последовательности аминокислот с большими положительными зарядами. Это позволяет белку нацеливаться на митохондрии с большим отрицательным зарядом. В отличие от лиганд-рецепторного механизма, где приближение лиганда к рецептору зависит от стохастического Броуновского движения, сигнал митохондриальной локализации в некоторых вариантах осуществления притягивается к митохондрии вследствие заряда.
Чтобы проникнуть в митохондрию, белок, как правило, должен взаимодействовать с аппаратом митохондриального импорта, состоящим из комплексов Tim и Tom (Translocase of the Inner/Outer Mitochondrial Membrane). С учетом сигнала митохондриальной локализации, положительный заряд притягивает соответствующий белок к комплексам и продолжает втягивать белок внутрь митохондрии. Комплексы Tim и Tom позволяют белкам проходить сквозь мембраны. Таким образом, один из вариантов осуществления настоящего изобретения доставляет композиции настоящего изобретения к внутреннему пространству митохондрий посредством положительно заряженного нацеливающего сигнала и аппарата митохондриального импорта. В другом варианте осуществления полипептиды, связанные с PTD и содержащие сигналы митохондриальной локализации, по-видимому, не используют комплексы TOM/TIM для проникновения в митохондриальный матрикс, см. Del Gaizo et al. Mol Genet Metab. 80(1-2): 170-80 (2003).
С учетом важности митохондрий в заболеваниях человека, клеточной пролиферации, гибели клеток и старении, варианты осуществления настоящего изобретения также включают управление митохондриальными функциями для обеспечения способов, которыми можно лечить известные митохондриальные заболевания (LHON, MELAS и т.д.) и предполагаемые митохондриальные заболевания (старение, болезнь Альцгеймера, болезнь Паркинсона, диабет, сердечные заболевания).
vii. Направленное воздействие на хлоропласты
В другом варианте осуществления модифицированные композиции, раскрытые в настоящем документе, специфически доставляют полипептиды к хлоропластам посредством включения сигнала или домена хлоропластной локализации. Для хлоропластов обнаружили несколько аминоконцевых нацеливающих сигналов, известных в данной области. Хлоропласт представляет собой фотосинтетическую органеллу эукариот, окруженную двойной мембраной. Жидкость внутри двойной мембраны называют стромой. Хлоропласт содержит нуклеоид, в котором хранится кольцевая оголенная ДНК. В строме также проходит цикл Кальвина. Цикл Кальвина представляет собой серию катализируемых ферментами химических реакций, посредством которых из углекислого газа образуются углеводы и другие соединения.
В строме находятся маленькие мембранные мешочки, которые называют тилакоидами. Мешочки уложены в стопки. Каждую стопку называют граной. В каждом хлоропласте находится много гран. В мембранах тилакоидов происходят реакции световой фазы фотосинтеза. Тилакоиды содержат внешние и внутренние мембранные белки, некоторые из которых несут специальные простетические группы, позволяющие электронам двигаться от одного белкового комплекса к другому. Эти белки образуют систему транспорта электронов, которую иногда называют Z-схемой.
Простетическая группа двух основных мембранных белков (P680 и P700) представляет собой молекулу пигмента хлорофилла. Эти связывающие хлорофилл белки придают тилакоидам интенсивный зеленый цвет. Множество тилакоидов в хлоропласте придают ему зеленый цвет. Множество хлоропластов в клетках мезофилла листьев придают этим клеткам зеленый цвет. Множество клеток мезофилла в листе придают ему зеленый цвет. Молекула хлорофилла улавливает энергию света, и в электронном облаке резонирующей химической структуры, окружающей ион магния, электрон приобретает дополнительную энергию. Этот возбужденный электрон передается по белкам-переносчикам электронов в мембране. Перемещение этих электронов и сопровождающих их протонов в итоге приводит к захвату энергии в фосфатной связи АТФ. Тилакоид, таким образом, представляет собой место улавливания света и синтеза АТФ. В строме АТФ используется для сохранения захваченной энергии в углерод-углеродных связях углеводов. Некоторые хлоропласты накапливают крахмальные зерна. Они представляют собой сложные полимеры углеводов, предназначенные для долговременного хранения.
С учетом биоэнергетических функций хлоропластов, возможность введения экзогенных полипептидов может помочь получить растения с повышенной жизнеспособностью в неблагоприятных условиях и повышенной эффективностью фотосинтеза.
Таким образом, другие варианты осуществления направлены на модификацию хлоропластов для более эффективных способов биосинтеза для коммерческих композиций.
Нацеливающие сигналы для типов клеток и состояний клеток
3. Усиление митохондриального биогенеза и OXPHOS
В вариантах осуществления настоящего изобретения предоставлены композиции и способы, подходящие для усиления митохондриального биогенеза и окислительного метаболизма. Дисфункцию клетки, возникающую вследствие сниженного митохондриального биогенеза и/или окислительного метаболизма, можно лечить или сокращать посредством раскрытых композиций и способов. Например, гибридные клетки нейробластомы SH-SY5Y, несущие мутацию G11778A гена мтДНК ND4 у пациента, пораженного наследственной оптической нейропатией Лебера (LHON), обрабатывали нацеленным на митохондрии способным к трансдукции TFAM (MTD-TFAM) или контрольным буфером. Одновременно проводили эксперименты "с высоким разрешением", оценивая дыхание посредством оксиметрии с применением интактных клеток, метаболизирующих глюкозу. Показатели основного дыхания представляли как функцию от неизменного числа живых клеток, выраженную в процентах от соответствующих значений для клеток контрольного буфера в каждый момент времени. Обработка MTD-TFAM вызывала изменяющееся во времени, обратимое увеличение значений основного дыхания, которые достигли максимального ~3-кратного увеличения по сравнению с контрольными образцами приблизительно через 2 недели (фиг. 1A-C).
Поскольку TFAM представляет собой признанный фактор, необходимый для репликации и транскрипции митохондриального генома, воздействие MTD-TFAM может усиливать репликацию, транскрипцию и трансляцию митохондриальных генов с образованием дыхательных белков. Мультиплексная qPCR нескольких митохондриальных генов показывает существенное увеличение числа копий митохондриальных генов как в ДНК, так и в РНК (кДНК) после обработки MTD-TFAM. Уровни множественных индивидуальных белков ETC в рассматриваемых клетках оценивали посредством Вестерн-блоттинга. Анализ результатов Вестерн-блоттинга показал, что относительная митохондриальная масса, выраженная как отношение белка внешней митохондриальной мембраны митофилина к цитозольному белку бета-актину, в клетках, обработанных MTD-TFAM, удвоилась (в 1,9 раз) (фиг. 2). Уровни кодируемых мтДНК (CIV, субъединица 2) и множественных белков ETC из нескольких комплексов, кодируемых ядерным геномом, также существенно и обратимо увеличились в клетках, обработанных MTD-TFAM, с самым большим увеличением, наблюдавшимся в комплексе I на 11 сутки (фиг. 2). Усиление дыхания в клетках отображали на животных. Нормальных взрослых мышей подвергали интраперитонеальным инъекциям MTD-TFAM или контрольного буфера. В препаратах митохондрий головного мозга, сердца, скелетной мышцы, почки и печени оценивали дыхание. Обработанные мыши продемонстрировали увеличение показателей дыхания во всех тканях в условной стадии 3 (+АДФ) для индивидуальных комплексов (фиг. 3A-3B). Таким образом, в вариантах осуществления настоящего изобретения предоставлены способы усиления митохондриального биогенеза и/или митохондриального окислительного метаболизма.
4. Генетические заболевания или синдромы
В вариантах осуществления настоящего изобретения предоставлены композиции и способы, подходящие для терапевтических протоколов и лечения генных заболеваний или нарушений, где митохондриальная дисфункция представляет собой первичное заболевание или осложнение заболевания. Посредством раскрытых композиций и способов также можно лечить или сокращать клеточную митохондриальную дисфункцию. В частности, на вызывающую заболевания митохондриальную дисфункцию оказывают направленное воздействие. Также раскрыты заболевания, при которых улучшение митохондриальных функций можно доказать терапевтически. Заболевание может наблюдаться у детей, например, у индивидуумов младше 18 лет, как правило, младше 12 лет, или взрослых, например, у индивидуумов 18 лет или старше. Таким образом, варианты осуществления настоящего изобретения направлены на лечение носителя с диагнозом заболевания, в частности, генетического заболевания, посредством лечения носителя способным к трансдукции белком, способным сократить митохондриальную дисфункцию или улучшить митохондриальные функции.
Подходящие генетические заболевания, которые можно вылечить посредством композиций, раскрытых в настоящем документе, включают в качестве неограничивающих примеров:
Митохондриальные заболевания: синдром Альперса; синдром Барта; дефекты в-окисления; дефицит карнитин-ацил-карнитина; дефицит карнитина; дефицит кофермента Q10; дефицит комплекса I; дефицит комплекса II; дефицит комплекса III; дефицит комплекса IV; дефицит комплекса V; дефицит цитохром c оксидазы (COX), LHON - наследственная оптическая нейропатия Лебера; MM - митохондриальная миопатия; LIMM - летальная детская митохондриальная миопатия; MMC - материнская миопатия и кардиомиопатия; NARP - нейрогенная мышечная слабость, атаксия и пигментная дистрофия сетчатки; болезнь Ли; FICP - смертельная детская кардиомиопатия плюс, a MELAS-ассоциированная кардиомиопатия; MELAS - митохондриальная энцефаломиопатия с лактацидозом и инсультоподобными эпизодами; LDYT - наследственная оптическая нейропатия Лебера и дистония; MERRF - миоклоническая эпилепсия и с рваными мышечными волокнами; MHCM - наследуемая по материнской линии гипертрофическая кардиомиопатия; CPEO - хроническая прогрессирующая внешняя офтальмоплегия; KSS - синдром Кирнса-Сейра; DM - сахарный диабет; DMDF - сахарный диабет + тугоухость; CIPO - хроническая псевдонепроходимость кишечника с миопатией и офтальмоплегией; DEAF - наследуемая по материнской линии тугоухость или аминогликозид-индуцированная тугоухость; PEM - прогрессирующая энцефалопатия; SNHL - нейросенсорная тугоухость; энцефаломиопатия; митохондриальная цитопатия; дилатационная кардиомиопатия; GER - желудочно-кишечный рефлюкс; DEMCHO - деменция и хорея; AMDF - атаксия, миоклонус; непереносимость физической нагрузки; ESOC - эпилепсия, инсульт, атрофия зрительного нерва и снижение когнитивных способностей; FBSN - семейный двусторонний некроз полосатого тела; FSGS - фокально-сегментарный гломерулосклероз; LIMM - смертельная детская митохондриальная миопатия; MDM - миопатия и сахарный диабет; MEPR - миоклоническая эпилепсия и психомоторная регрессия; MERME MERRF/MELAS перекрывающиеся заболевания; MHCM - наследуемая по материнской линии гипертрофическая кардиомиопатия; MICM - наследуемая по материнской линии кардиомиопатия; MILS - наследуемый по материнской линии синдром Ли; митохондриальная энцефалокардиомиопатия; мультисистемное митохондриальное заболевание (миопатия, энцефалопатия, слепота, тугоухость, периферическая нейропатия); NAION - не связанная с артериитом передняя ишемическая оптическая нейропатия; NIDDM - инсулиннезависимый сахарный диабет; PEM - прогрессирующая энцефалопатия; PME - прогрессирующая миоклоническая эпилепсия; RTT - синдром Ретта; SIDS - синдром внезапной детской смерти; MIDD - наследуемый по материнской линии синдром диабета и тугоухости; и MODY - сахарный диабет взрослого типа у молодых.
Ядерные заболевания: мышечная дистрофия, синдром Эллиса-Ван-Кревельда, синдром Марфана, миотоническая дистрофия, спинальная мышечная атрофия, ахондроплазия, боковой амиотрофический склероз, синдром Шарко-Мари-Тута, синдром Коккейна, диастрофическая дисплазия, мышечная дистрофия Дюшенна, синдром Эллиса-Ван-Кревельда, фибродисплазия оссифицирующая прогрессирующая, болезнь Альцгеймера, синдром Ангельмана, эпилепсия, эссенциальный тремор, синдром ломкой X-хромосомы, атаксия Фридрейха, болезнь Хантингтона, болезнь Ниманна-Пика, болезнь Паркинсона, синдром Прадера-Вилли, синдром Ретта, спиноцеребеллярная атрофия, синдром Вильямса, телеангиоэктатическая атаксия, серповидно-клеточная анемия, лимфома Беркитта, болезнь Гоше, гемофилия, лейкоз, пароксизмальная ночная гемоглобинурия, порфирия, талассемия, болезнь Крона, дефицит альфа-1-антитрипсина, кистозный фиброз, тугоухость, синдром Пендреда, глаукома, гиратная атрофия хориоидеи и сетчатки, гиперплазия надпочечников, адренолейкодистрофия, синдром Коккейна, синдром удлинения интервала QT, иммунодефицит с гиперпродукцией IgM, синдром Альпорта, синдром Эллиса-Ван-Кревельда, фибродисплазия оссифицирующая прогрессирующая, синдром Ваарденбурга, синдром Вернера.
Инфекционные заболевания: вирусные - СПИД, СПИД-ассоциированный комплекс, ветряная оспа (Varicella), вирусная инфекция верхних дыхательных путей, цитомегаловирусная инфекция, колорадская клещевая лихорадка, лихорадка Денге, геморрагическая лихорадка Эбола, эпидемический паротит, грипп, вирусная пузырчатка полости рта и конечностей, герпесовирусный гепатит, опоясывающий лишай, папилломавирус человека, грипп, лихорадка Ласса, корь, геморрагическая лихорадка Марбург, инфекционный мононуклеоз, эпидемический паротит, полиомиелит, прогрессирующая многоочаговая лейкоэнцефалопатия, бешенство, краснуха, атипичная пневмония, оспа (Variola), вирусный энцефалит, вирусный гастроэнтерит, вирусный менингит, вирусная пневмония, болезнь Западного Нила - желтая лихорадка; Бактериальные - сибирская язва, бактериальный менингит, бруцеллез, бубонная чума, кампилобактериоз, болезнь кошачьих царапин, холера, дифтерия, эпидемический сыпной тиф, гонорея, болезнь Хансена, легионеллез, лепра, лептоспироз, листериоз, болезнь Лайма, мелиоидоз, MRSA-инфекция, нокардиоз, коклюш, пневмококковая пневмония, пситтакоз, Q-лихорадка, пятнистая лихорадка Скалистых гор или RMSF, сальмонеллез, скарлатина, шигеллез, сифилис,столбняк, трахома, туберкулез, туляремия, брюшной тиф, сыпной тиф, коклюш; паразитарные - африканский трипаносомоз, амебиаз, аскаридоз, бабезтоз, болезнь Шагаса, клонорхоз, криптоспоридиоз, цистицеркоз, дифиллоботриоз, дракункулез, эхинококкоз, энтеробиоз, фасциолез, фасциолопсидоз, филариоз, инфекции, вызываемые свободноживущими амебами, лямблиоз, гнатостомоз, гименолепидоз, изоспороз, висцеральный лейшманиоз, лейшманиоз, малярия, метагонимоз, миаз, онхоцеркоз, педикулез, энтеробиоз, чесотка, шистосомоз, тениоз, токсокароз, токсоплазмоз, трихинеллез, трихиноз, трихоцефалез, трипаносомоз.
Злокачественные новообразования: рак молочной железы и яичников, лимфома Беркитта, хронический миелолейкоз, рак толстого кишечника, рак легких, злокачественная меланома, множественные эндокринные неоплазии, нейрофиброматоз, синдром Ли-Фраумени, рак поджелудочной железы, рак предстательной железы, ретинобластома, синдром Гиппеля-Линдау, поликистозная болезнь почек, туберозный склероз.
Нарушения обмена веществ: адренолейкодистрофия, атеросклероз, болезнь Беста, болезнь Гоше, глюкозо-галактозная мальабсорбция, гиратная атрофия, юношеский диабет, ожирение, пароксизмальная ночная гемоглобинурия, фенилкетонурия, болезнь Рефсума, танжерская болезнь, болезнь Тея-Сакса, адренолейкодистрофия, диабет 2 типа, болезнь Гоше, наследственный гемохроматоз, синдром Леша-Найхана, болезнь мочи с запахом кленового сиропа, синдром Менкеса, болезнь Ниманна-Пика, рак поджелудочной железы, синдром Прадера-Вилли, порфирия, болезнь Рефсума, танжерская болезнь, болезнь Вильсона, синдром Цельвегера, прогерия, тяжелый комбинированный иммунодефицит.
Аутоиммунные нарушения: аутоимунный полигландулярный синдром, волчанка, диабет 1 типа, склеродермия, рассеянный склероз, болезнь Крона, хронический активный гепатит, ревматоидный артрит, болезнь Грейвса, миастения гравис, миозит, антифосфолипидный синдром (APS), увеит, полимиозит, феномен Рейно, и демиелинизирующие нейропатии, и редкие нарушения, такие как ревматическая полимиалгия, височный артериит, синдром Шегрена, болезнь Бехчета, синдром Черджа-Стросс и артериит Такаясу.
Воспалительные нарушения: алопеция, диастрофическая дисплазия, синдром Эллиса-Ван-Кревельда, астма, артрит, включая остеоартрит, ревматоидный артрит и спондилоартропатии.
Возрастные нарушения: болезнь Альцгеймера, болезнь Паркинсона, атеросклероз, возрастная дегенерация желтого пятна, возрастной остеопороз.
Раскрытые способы и композиции также можно применять для лечения, контроля или облегчения симптомов, ассоциированных со старением, для регенерации тканей/регенеративной медицины, трансплантации стволовых клеток, улучшения когнитивных функций, повышения работоспособности и косметических вмешательств у человека или животных, не относящихся к человеку.
9. Ожирение и потребление пищи
В вариантах осуществления настоящего изобретения предоставлены композиции и способы, подходящие для уменьшения ожирения и/или повышенного потребления пищи. Например, для 50 мышей (25 контрольных мышей, получавших растворитель, и 25 мышей, обработанных MTD-TFAM) наблюдали за потреблением пищи и массой в течение 3 месяцев, во время чего каждый месяц интраперитонеально инъецировали физиологический раствор или MTD-TFAM, проведя всего по 3 инъекции каждой. В начале эксперимента масса мышей была одинаковой. Потребление пищи измеряли каждую неделю. По окончании эксперимента мыши, обработанные MTD-TFAM, имели статистически достоверную (p=.0002) потерю массы, несмотря на то, что на протяжении эксперимента они потребили на 43% больше пищи, чем контрольные животные (p=.037) (фиг. 4 и 5). На фиг. 5 для 50 мышей (25 контрольных мышей, получавших растворитель, и 25 мышей, обработанных MTD-TFAM) наблюдали за потреблением пищи и массой в течение 3 месяцев, во время чего каждый месяц интраперитонеально инъецировали физиологический раствор или MTD-TFAM, проведя всего по 3 инъекции каждой. В начале эксперимента масса мышей была одинаковой. Потребление пищи измеряли каждую неделю. По окончании эксперимента мыши, обработанные MTD-TFAM, демонстрировали статистически достоверное увеличение потребленной пищи на протяжении эксперимента на 43% больше пищи, чем контрольные животные (p=.037). Таким образом, раскрытые способы можно применять для лечения ожирения и/или повышенного потребления пищи.
10. Введение
Композиции, предоставленные в настоящем документе, можно вводить реципиенту в физиологически приемлемом носителе. Предпочтительные способы введения включают системное или прямое введение в клетку. Композиции можно вводить в клетку или в организм пациента способами, как правило, известными в данной области для протеинотерапий. В одном из вариантов осуществления предоставлена фармацевтическая композиция, состоящая в основном из рекомбинантного полипептида, содержащего связывающий полинуклеотиды домен, нацеливающий домен, домен трансдукции белка и фармацевтически приемлемый носитель или эксципиент. Предпочтительно связывающий полинуклеотиды домен включает TFAM или его фрагмент, способный связывать полинуклеотид. Композиция включает эффективное количество рекомбинантного полипептида для усиления митохондриального метаболизма.
Модифицированные комплексные композиции можно комбинировать в смеси с фармацевтически приемлемым носителем. Терапевтические составы готовят для хранения посредством смешивания активного ингредиента, имеющего желаемую степень очистки, с выбранными физиологически приемлемыми носителями, эксципиентами или стабилизаторами (Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980)) в форме лиофилизированных составов или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы не токсичны для реципиентов в применяемых дозах и концентрациях и включают буферы, такие как фосфатный, цитратный и буферы на основе других органических кислот; антиоксиданты, включая аскорбиновую кислоту; полипептиды с низкой молекулярной массой (менее чем приблизительно 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие средства, такие как ЭДТА; сахарные спирты, такие как маннит или сорбит; солеобразующие противоионы, такие как натрий; и/или неионногенные поверхностно-активные вещества, такие как Tween®, Pluronics® или PEG.
Композиции настоящего изобретения можно вводить парентерально. В настоящем документе, "парентеральное введение" характеризуется введением фармацевтической композиции посредством физического нарушения целостности ткани индивидуума. Парентеральное введение включает введение посредством инъекции, хирургического разреза или проникающих в ткани безоперационных вмешательств и т.п. В частности, парентеральное введение включает подкожные, интраперитонеальные, внутривенные, внутриартериальные, внутримышечные, интрастернальные инъекции и способы инфузии посредством диализа почек.
Парентеральные составы могут включать активный ингредиент, комбинированный с фармацевтически приемлемым носителем, таким как стерильная вода или стерильный изотонический физиологический раствор. Такие составы можно получать, фасовать или продавать в форме, подходящей для болюсного введения или для непрерывного введения. Инъекционные составы можно получать, фасовать или продавать в стандартной лекарственной форме, такой как ампулы или мультидозированные контейнеры, содержащие консервант. Составы для парентерального введения включают суспензии, растворы, эмульсии на масляной или водной основе, пасты, восстанавливаемые сухие (т.е. порошковые или гранулярные) составы и имплантируемые или биодеградируемые составы пролонгированного высвобождения. Такие составы также могут содержать один или несколько дополнительных ингредиентов, включая суспендирующие, стабилизирующие или диспергирующие средства. Парентеральные составы можно получать, фасовать или продавать в форме стерильной инъецируемой водной или масляной суспензии или раствора. Парентеральные составы также могут включать диспергирующие средства, увлажнители или суспендирующие средства, описываемые в настоящем документе. Способы приготовления этих типов составов известны. Стерильные инъецируемые составы можно получать посредством применения нетоксических парентерально приемлемых разбавителей или растворителей, таких как вода, 1,3-бутандиол, раствор Рингера, изотонический раствор хлорида натрия и нелетучие масла, такие как синтетические моноглицериды или диглицериды. Другие парентерально вводимые составы включают микрокристаллические формы, липосомальные препараты и биодеградируемые полимерные системы. Композиции для пролонгированного высвобождения или имплантации могут включать фармацевтически приемлемые полимерные или гидрофобные вещества, такие как эмульсии, ионобменные смолы, умеренно растворимые полимеры и умеренно растворимые соли.
Фармацевтические композиции можно получать, фасовать или продавать в форме буккальных составов. Такие составы могут быть в форме таблеток, порошков, аэрозолей, мелкодисперсных растворов, суспензий или таблеток-леденцов, полученных известными способами, и могут содержать приблизительно от 0,1% до приблизительно 20% (масс./масс.) активного ингредиента с балансом состава, содержащего перорально растворимую или деградируемую композицию и/или один или несколько дополнительных ингредиентов, как описано в настоящем документе. Предпочтительно, средний размер частиц или капель в порошковых или аэрозольных составах находится в диапазоне от приблизительно 0,1 нанометров до приблизительно 200 нанометров при распылении.
В настоящем документе "дополнительные ингредиенты" включают один или несколько следующих: эксципиенты, поверхностно-активные средства, диспергирующие средства, инертные разбавители, гранулирующие агенты, дезинтегрирующие средства, связывающие средства, смазки, подсластители, ароматизаторы, красители, консерванты, физиологически деградируемые композиции (например, желатин), водные носители, водные растворители, масляные носители и масляные растворители, суспендирующие средства, диспергирующие средства, увлажнители, эмульгаторы, смягчающие средства, буферы, соли, загустители, наполнители, эмульгаторы, антиоксиданты, антибиотики, антигрибковые агенты, стабилизирующие агенты и фармацевтически приемлемые полимерные или гидрофобные вещества. Другие "дополнительные ингредиенты", которые можно включать в фармацевтические композиции, известны. Подходящие дополнительные ингредиенты описаны Remington's Pharmaceutical Sciences, Mack Publishing Co., Genaro, ed., Easton, Pa. (1985).
Дозы и желаемые концентрации раскрытых в настоящем документе модифицированных векторов в фармацевтических композициях настоящего изобретения могут варьировать в зависимости от конкретного предполагаемого применения. Определение подходящей дозы или способа введения находится в компетенции обычного врача. Эксперименты на животных предоставляют надежное руководство для определения эффективных доз для лечения человека. Межвидовой перенос данных об эффективных дозах можно проводить на основе принципов, сформулированных в Mordenti, J. and Chappell, W. "The use of interspecies scaling in toxicokinetics" в Toxicokinetics and New Drug Development, Yacobi et al., Eds., Pergamon Press, New York 1989, pp.42-96.
ПРИМЕРЫ
Пример 1: Рекомбинантные конструкции
В предпочтительном варианте осуществления последовательность рекомбинантного полипептида, по меньшей мере, на 80, 85, 90, 95, 97, 99 или 100% идентична

В другом варианте осуществления рекомбинантный полипептид кодируется нуклеиновой кислотой с последовательностью, по меньшей мере, на 80, 85, 90, 95, 97, 99 или 100% идентичной


Пример 2: Последовательности конструкций
Домен трансдукции белков (PTD), состоящий из 11 остатков аргинина, клонировали внутри рамки в связывающий антитело участок белка A Staphylococcus aureus, домен В. Кодирующую последовательность домена В PTD клонировали тандемно выше кодирующей последовательности TFAM и встраивали в бактериальный экспрессирующий вектор. Рекомбинантный белок экспрессировался в бактериях, после чего его изолировали. Очищенный белок концентрировали, а концентрацию белка оценивали посредством анализа Бредфорда (Biorad). Очищенный белок анализировали посредством SDS-Page для подтверждения его чистоты.
Участок пептида PTD-PA-TFAM (PTD подчеркнут жирной линией; тандемный домен связывающего антитело домена В белка А подчеркнут двойной линией; TFAM подчеркнут штриховой линией) (332):

Выбранные сходства белков модельных организмов, которые можно применять в композициях и способах, раскрытых в настоящем документе:
Организм, белок и процентная идентичность и длина выровненного участка
(см. ProtEST)
(см. ProtEST)
(см. ProtEST)
(см. ProtEST)
(см. ProtEST)
Последовательности, указанные в настоящем документе, известны в данной области, например, в GenBank, и в качестве ссылки включены в настоящий документ в полном объеме.
Следует отметить, что описанные выше варианты осуществления настоящего изобретения, конкретно, любые "предпочтительные" варианты осуществления, представляют собой только примеры осуществления, описанные только для ясного понимания принципов изобретения. С описанным выше вариантом(ами) осуществления изобретения можно проводить множество вариаций и модификаций без отклонения от сущности и принципов изобретения. Все такие модификации и вариации предназначены для включения в настоящий документ в объеме настоящего описания и защищены приведенной ниже формулой изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ТЕРАПЕВТИЧЕСКИЕ АГЕНТЫ, СОДЕРЖАЩИЕ ПРОАПОПТОЗНЫЕ БЕЛКИ | 2002 |
|
RU2319709C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ МОДИФИКАЦИИ ЗАДАННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ НУКЛЕИНОВОЙ КИСЛОТЫ-МИШЕНИ | 2012 |
|
RU2663354C2 |
| МОДИФИКАЦИЯ B-КЛЕТОК | 2018 |
|
RU2783116C2 |
| МОДИФИЦИРОВАННЫЕ БЕЛКИ, СКОНСТРУИРОВАННЫЕ ТОКСИНЫ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2002 |
|
RU2305684C2 |
| КОНСТРУКЦИИ ГОМОДИМЕРНЫХ БЕЛКОВ | 2011 |
|
RU2624041C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, ОБЛАДАЮЩАЯ ПРОТИВООПУХОЛЕВЫМ ЭФФЕКТОМ, И СПОСОБ СНИЖЕНИЯ ИНГИБИРУЮЩЕГО ЭФФЕКТА PD-L1 НА Т-КЛЕТКИ ЧЕЛОВЕКА | 2016 |
|
RU2688692C2 |
| ПРОФИЛАКТИЧЕСКАЯ ПРОТИВОРАКОВАЯ ВАКЦИНА | 2004 |
|
RU2556128C2 |
| ПРОФИЛАКТИЧЕСКАЯ ПРОТИВОРАКОВАЯ ВАКЦИНА | 2004 |
|
RU2385163C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ИНТЕРНАЛИЗАЦИИ ФЕРМЕНТОВ | 2018 |
|
RU2806021C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ LIGHT И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2542394C2 |
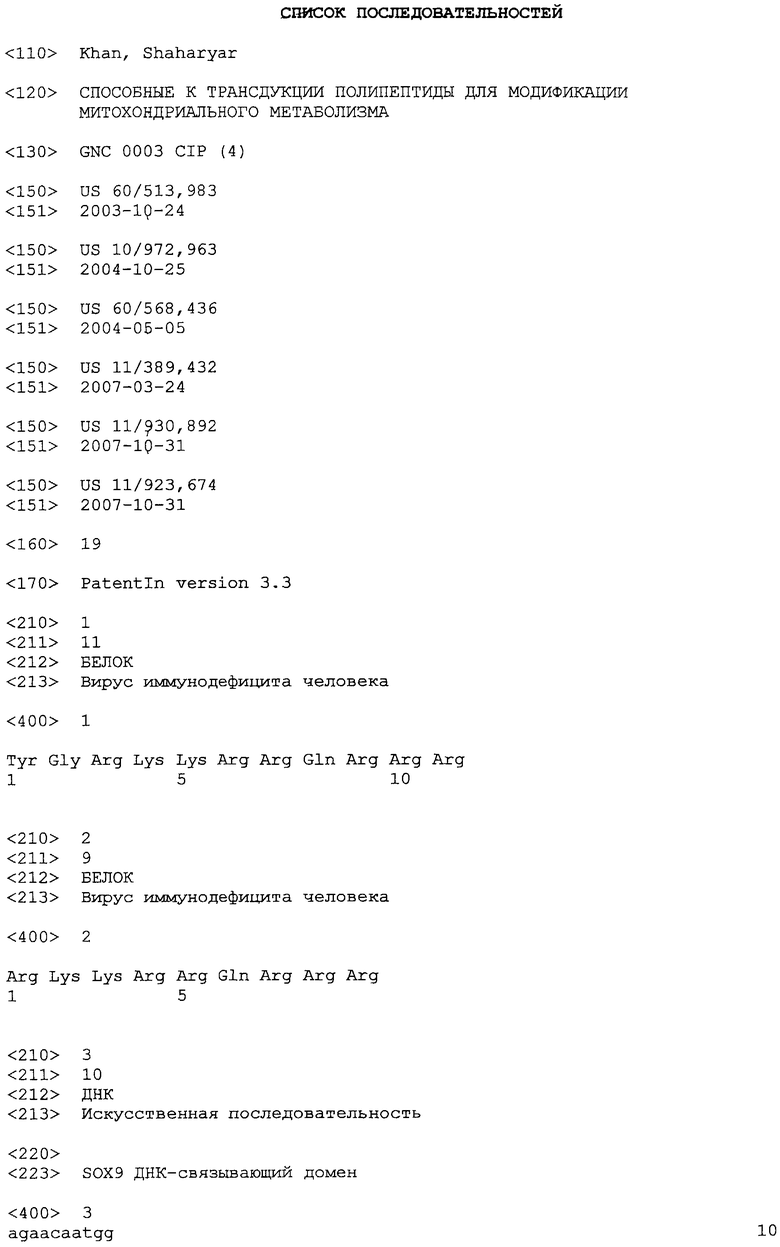







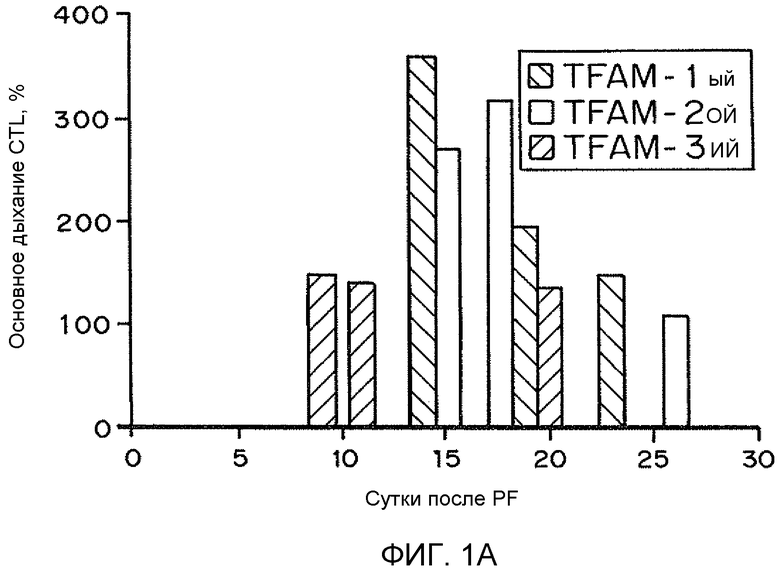
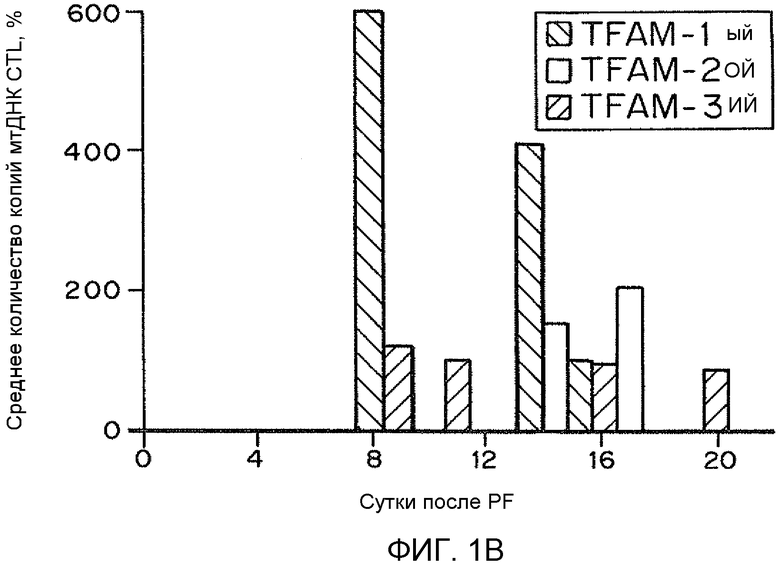
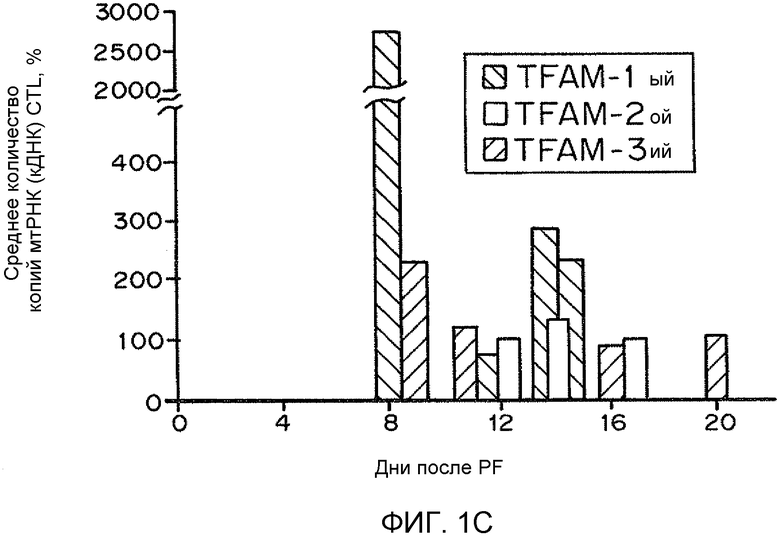
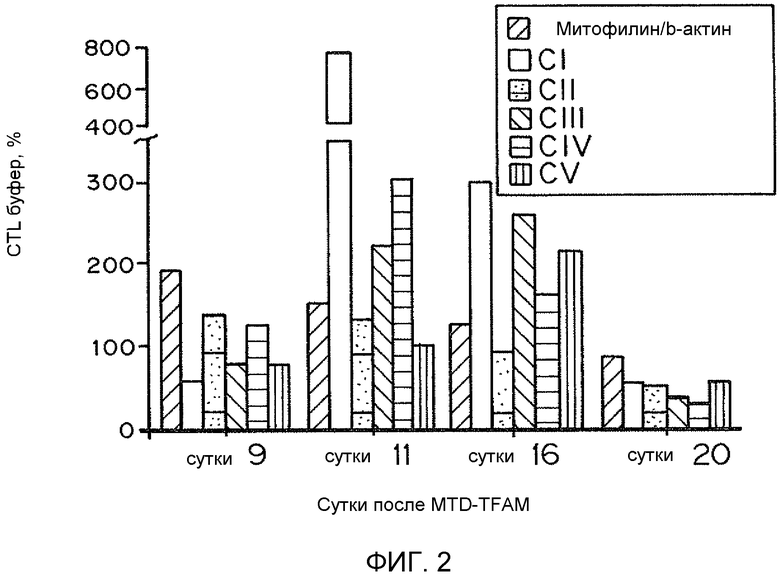
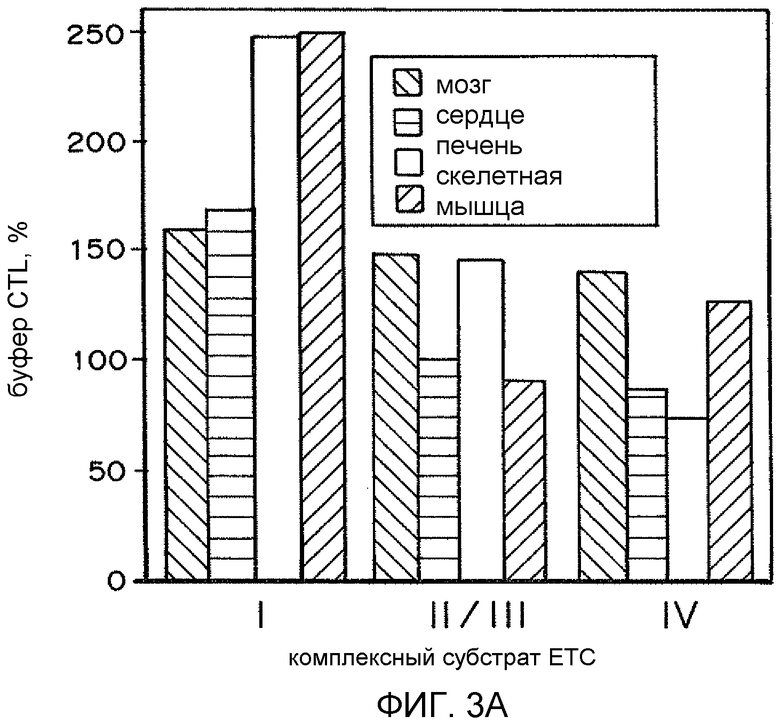
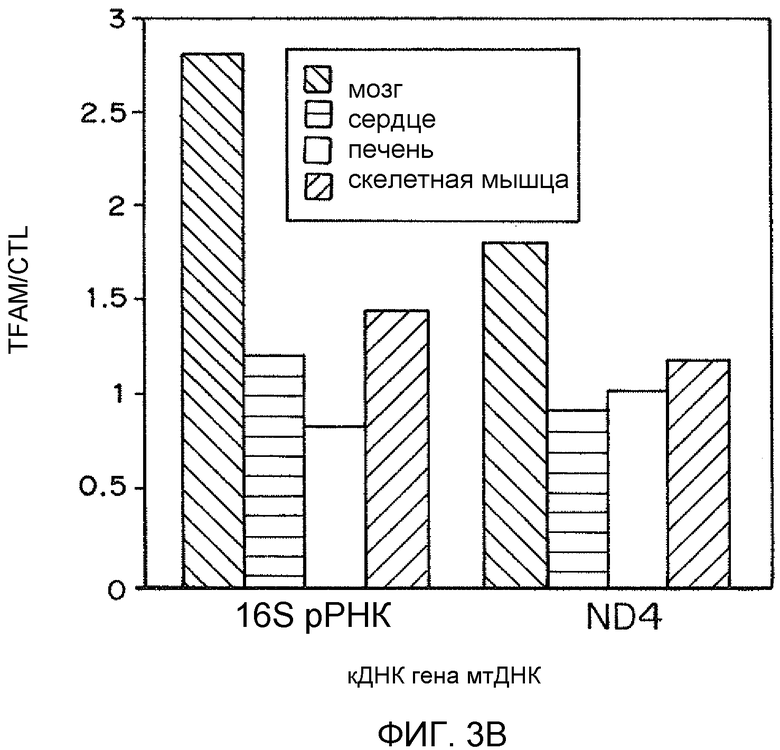
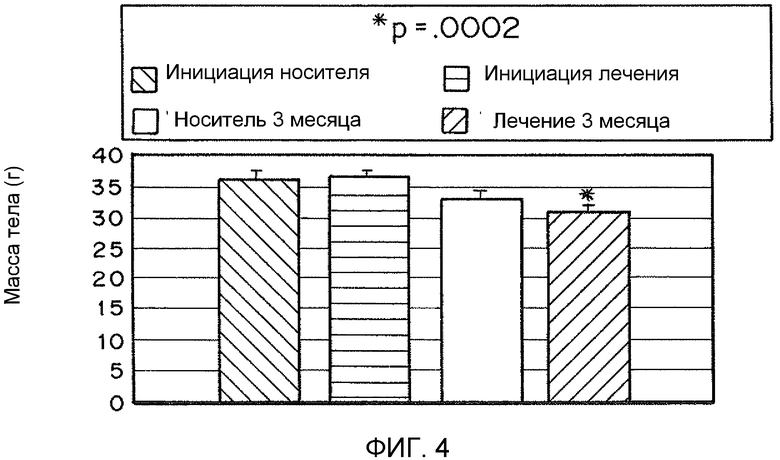
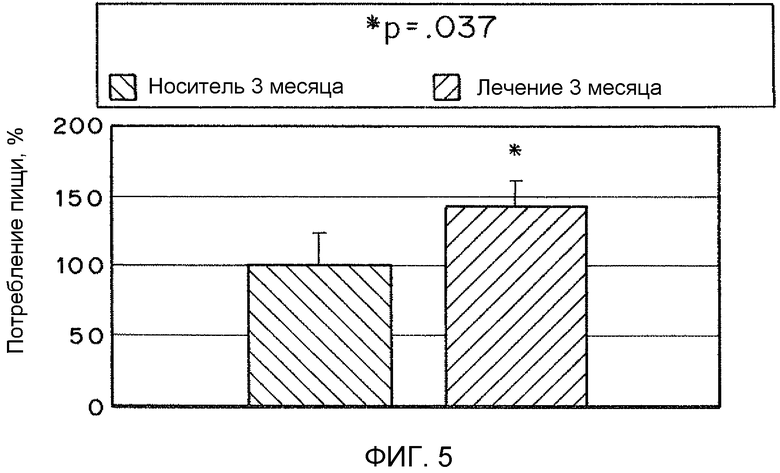
Изобретение относится к биохимии. Описано применение слитого белка для получения композиции для снижения массы тела. Слитый белок содержит домен трансдукции, сигнал митохондриальной локализации и домен митохондриального фактора транскрипции А, связывающий полинуклеотид (TFAM), содержащий группу с высокой подвижностью. Описаны способы лечения ожирения с помощью указанного белка. Изобретение расширяет арсенал средств для лечения ожирения. 3 н. и 6 з.п. ф-лы, 5 ил., 2 пр.
1. Применение слитого белка для получения композиции для снижения массы тела у индивида, где слитая конструкция содержит белок домена трансдукции белка, сигнал митохондриальной локализации и домен митохондриального фактора транскрипции А, связывающий полинуклеотид (TFAM), содержащий группу с высокой подвижностью.
2. Применение по п.1, где домен трансдукции белка содержит 11 остатков аргинина.
3. Применение по п.1, где сигнал митохондриальной локализации содержит сигнал митохондриальной локализации из последовательности SEQ ID NO:18.
4. Применение по п.1, где слитый белок повышает окислительный метаболизм при введении в митохондрию.
5. Применение по п.1, где слитый белок состоит из аминокислотной последовательности
(a) SEQ ID NO:18 или
(b) варианта SEQ ID NO:18, имеющего аминокислотные замену, делецию или вставку относительно SEQ ID NO:18.
6. Применение по п.5, где аминокислотная делеция представляет собой N-концевой метионин SEQ ID NO:18.
7. Применение по п.1, где слитый белок кодируется SEQ ID NO:19.
8. Способ лечения ожирения, включающий введение индивиду, нуждающемуся в этом, композиции с от 0,1% до 20% масс./масс. последовательности SEQ ID NO:18 для снижения массы тела у указанного индивида.
9. Способ лечения ожирения, включающий введение композиции, содержащей слитый белок, кодируемый SEQ ID NO:19, индивиду, нуждающемуся в этом.
| СПОСОБ СНИЖЕНИЯ МАССЫ ТЕЛА В ЭКСПЕРИМЕНТЕ НА ЖИВОТНЫХ | 2004 |
|
RU2262945C1 |

