Область техники
В соответствии с настоящим изобретением предложен экзополисахарид, происходящий из штамма Pseudoalteromonas sp. CY01, который представляет собой новый штамм, обитающий в полярных областях, и более конкретно экзополисахарид, происходящий из штамма Pseudoalteromonas sp. CY01, и композиция для криозащиты клеток, которая содержит указанный экзополисахарид.
Уровень техники
Многие типы бактерий, которые живут в морской окружающей среде, секретируют вязкие внеклеточные углеводородные полимеры, известные как экзополисахариды (ЭПС). Большинство ЭПС, продуцируемых морскими бактериями, представляют собой гетерополисахариды, состоящие из 3 или 4 различных моносахаридов, которые могут представлять собой пентозы, гексозы, аминосахара или уроновые кислоты, и объединены в группы по 10 или более с образованием повторяющихся звеньев. ЭПС функционируют, защищая микроорганизмы от холодных условий окружающей среды, и ЭПС, секретируемые бактериями в холодных полярных условиях окружающей среды, во многих случаях обладают новыми структурами и способностями обеспечивать криозащиту.
Обнаружено, что ЭПС, секретируемые Pseudoalteromonas sp. SM20310 (бактерии Арктики) и Pseudoalteromonas arctica KOPRI 21653 (бактерии Антарктики), усиливают жизнеспособность Е. coli (бактерии неполярных областей) на фоне циклов замораживания и оттаивания. Таким образом, была предложена возможность применения ЭПС из данных бактерий в качестве криозащитных агентов (КЗА) (Liu, S.В. et al. Applied and Environmental Microbiology, 79:224, 2013; Kim, S.J. & Yim, J.H., J. Microbiology, 45:510, 2007).
Образование и рост льда приводит к физическому повреждению на клеточном уровне и также уменьшает объем воды в растворе, вызывая осмотический шок из-за повышенной концентрации внеклеточного раствора. Это становится проблемой при хранении биологических веществ при температурах ниже, чем температуры замерзания.
Так как регенеративная медицина и трансплантация органов быстро развиваются, увеличивается потребность в криоконсервации клеток, тканей, органов и эритроцитов (ККТ - красных кровяных телец) доноров. Обычно запакованную кровь хранят в добавляемом растворе, таком как ADSOL, при 4°С в течение 42 суток без криоконсервации, и при этом показатель гемолиза указанной крови является высоким. Данная проблема может быть решена путем применения глицерина в качестве криозащитного агента. Однако, возможность сохранения высокого процента эритроцитов может сохраняться только тогда, когда к клеткам добавляют высокую концентрацию (40 масс./об. % или более) глицерина, и клетки охлаждают с очень низкой скоростью охлаждения 1°С/мин и хранят при -80°С.
Однако цитотоксичность глицерина устраняется только при разбавлении глицерина до 1% или менее путем промывки после оттаивания. Для преодоления данного недостатка глицерина осуществляли непрерывные попытки применения других криозащитных агентов, включая низкомолекулярные сахара, такие как трегалоза, сахароза, глюкоза, раффиноза, мальтоза и подобные им. Однако данный способ может влиять на финальное замораживание эритроцитов из-за таких проблем как осмотическое давление или окисление. Для упрощения сложного способа промывки после оттаивания осуществляли попытки применить непроникающие добавки, такие как ГЭК (гидроксиэтилкрахмал), поливинилпирролидон и декстран, вместо глицерина (Scott, K.L. et al., Transfus. Med. Rev., 19:127, 2005; Stolzing, A. et al., Transfusion Apheresis Sci., 46:137, 2012; E.P. Horn et al., Anesth. Analg., 85:739,1997). Такие полимеры не проникают в клеточную мембрану и находятся только снаружи от клеточной мембраны, и, таким образом, не требуют применения сложного способа промывки во время оттаивания после замораживания эритроцитов. Однако, для криоконсервации эритроцитов требуется высокая концентрация (20 масс./об. % или более) раствора ГЭК, и данная высокая концентрация приводит к высокой вязкости, что затрудняет осуществление процесса замораживания.
Соответственно, авторы настоящего изобретения предприняли масштабные попытки обнаружить криозащитный агент, который являлся бы непроникающим, являлся бы менее цитотоксичным и имел бы превосходные криозащитные свойства. В результате авторы настоящего изобретения обнаружили, что при добавлении экзополисахарида (ЭПС), продуцируемого штаммом Pseudoalteromonas sp. CY01 (KCTC 12867ВР), который является новым штаммом, обнаруженным в Антарктическом океане, во время криоконсервации эритроцитов, указанный экзополисахарид проявляет превосходные криозащитные свойства при относительно низкой концентрации, по сравнению с другими криозащитными агентами (КЗА), и не проявляет цитотоксичность, что и лежит в основе настоящего изобретения.
Краткое описание изобретения
Объектом настоящего изобретения является обеспечение экзополисахарид а (р-CY01), происходящего из штамма Pseudoalteromonas sp. CY01 (KCTC 12867ВР), имеющего способность обеспечивать криозащиту клеток, и способа его получения.
Другим объектом настоящего изобретения является обеспечение экзополисахарида, имеющего молекулярную массу от 1,0×105 до 4,3×105 Да, который получен посредством гидролиза экзополисахарида, происходящего из штамма CY01, и способа его получения.
Еще одним объектом настоящего изобретения является обеспечение композиции для криоконсервации клеток, которая содержит описанный выше экзополисахарид (р-CY01 или p-CY01_LM).
Еще одним объектом настоящего изобретения является обеспечение способа криоконсервации клеток с применением описанного выше экзополисахарида (р-CY01 или p-CY01_LM).
Другим объектом настоящего изобретения является обеспечение штамма Pseudoalteromonas sp. CY01 (KCTC 12867ВР), который обитает в полярных областях и обладает способностью продуцировать экзополисахарид.
Для достижения указанной выше цели согласно настоящему изобретению предложен экзополисахарид (р-CY01), который продуцируется штаммом Pseudoalteromonas sp. CY01 (KCTC 12867ВР) и состоит из глюкозы и галактозы.
Согласно настоящему изобретению также предложен экзополисахарид (р-CY01_LM), имеющий молекулярную массу от 1,0×105 до 4,3×105 Да, который получают посредством гидролиза описанного выше экзополисахарида (p-CY01).
Согласно настоящему изобретению также предложена композиция для криозащиты клеток, которая содержит описанный выше экзополисахарид (р-CY01 или p-CY01_LM).
Согласно настоящему изобретению также предложен способ получения экзополисахарида (р-CY01), который продуцируется штаммом Pseudoalteromonas sp. CY01 (KCTC 12867ВР) и состоит из глюкозы и галактозы, причем указанный способ включает следующие стадии: (а) культивирование штамма Pseudoalteromonas sp CY01 (KCTC 12867ВР) с получением экзополисахарида (р-CY01); и (b) выделение полученного экзополисахарида (p-CY01).
Согласно настоящему изобретению также предложен способ получения экзополисахарида (p-CY01_LM), имеющего молекулярную массу от 1,0×105 до 4,3×105 Да, причем указанный способ включает стадию гидролиза экзополисахарида (p-CY01), который продуцируется штаммом Pseudoalteromonas sp. CY01 (KCTC 12867ВР) и который состоит из глюкозы и галактозы.
Согласно настоящему изобретению также предложен способ криоконсервации клеток с применением описанного выше экзополисахарида (р-CY01 или p-CY01_LM).
Согласно настоящему изобретению также предложен штамм Pseudoalteromonas sp. CY01 (KCTC 12867ВР), который обладает способностью продуцировать экзополисахарид слизи.
Краткое описание графических материалов
На Фиг. 1 показана продукция ЭПС и криозащитные эффекты (свойства) 10 выделенных штаммов. На Фиг. 1 белые столбцы обозначают продуцируемый ЭПС, а черные столбцы обозначают криозащитный эффект (%) ЭПС в отношении Е. coli. Стандартное отклонение (±SD) рассчитывали на основании трех независимых экспериментов.
На Фиг. 2 показаны результаты проведения филогенетического анализа с применением последовательности 16S рРНК (рибосомальная РНК) штамма CY01.
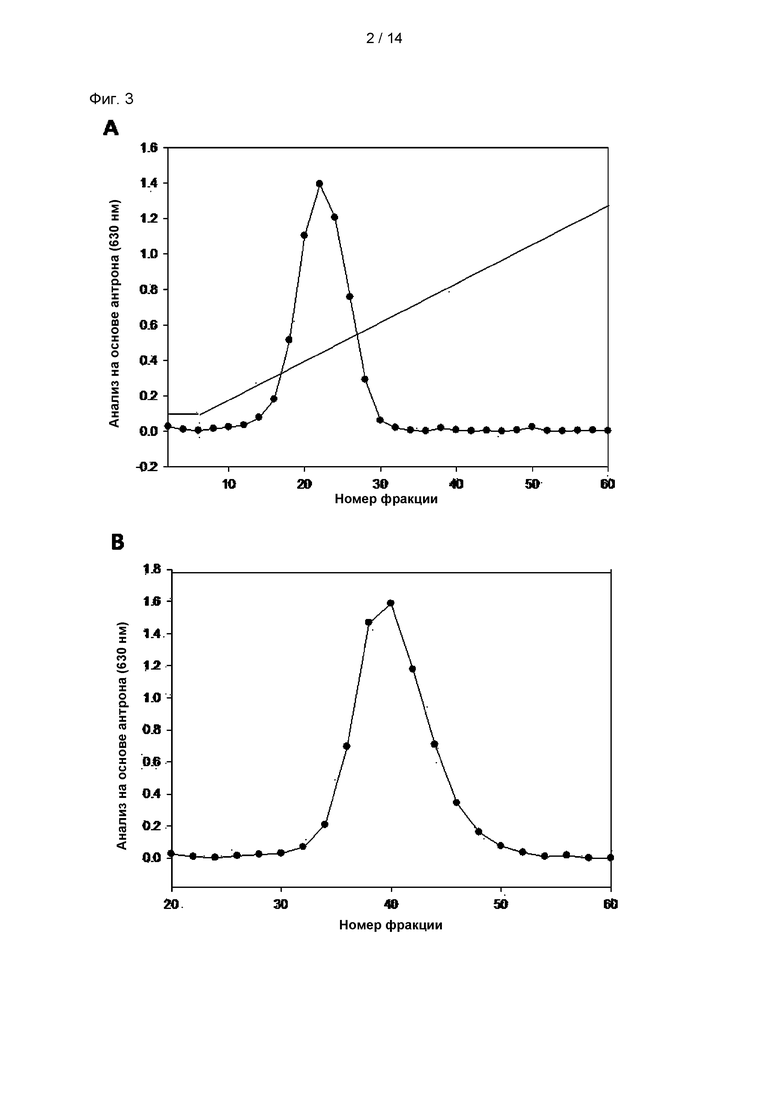
На Фиг. 3 показаны хроматографические пики ЭПС, происходящего из CY01. В частности, на Фиг. 3А показаны результаты анионной хроматографии, и на Фиг. 3В показаны результаты гель-фильтрационной хроматографии.
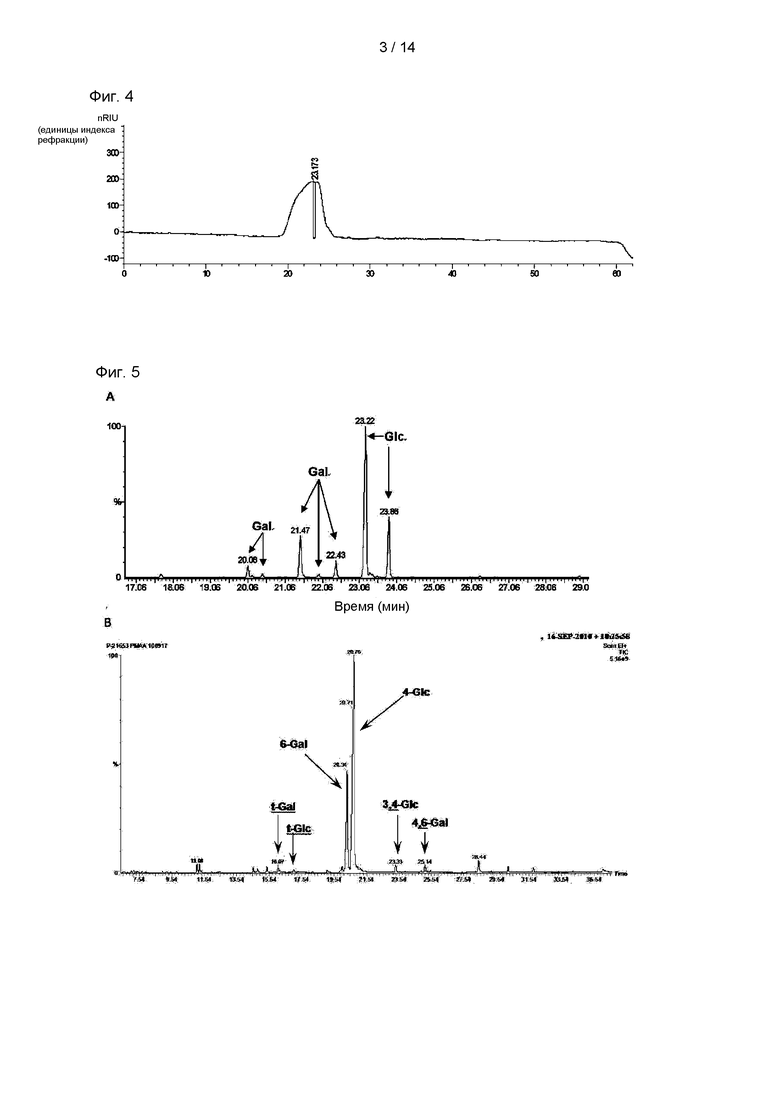
На Фиг. 4 показаны результаты ВЭЖХ (Высокоэффективная жидкостная хроматография), проводимой для анализа чистоты фракции ЭПС, происходящего из CY01, полученной посредством гель-фильтрационной хроматографии.
На Фиг. 5А показаны результаты анализа сахаров-составных частей ЭПС посредством ГХ/МС (газовая хроматография/масс-спектрометрия), и на Фиг. 5В показаны результаты анализа паттерна образования связей сахаров-составных частей ЭПС посредством ГХ/МС.
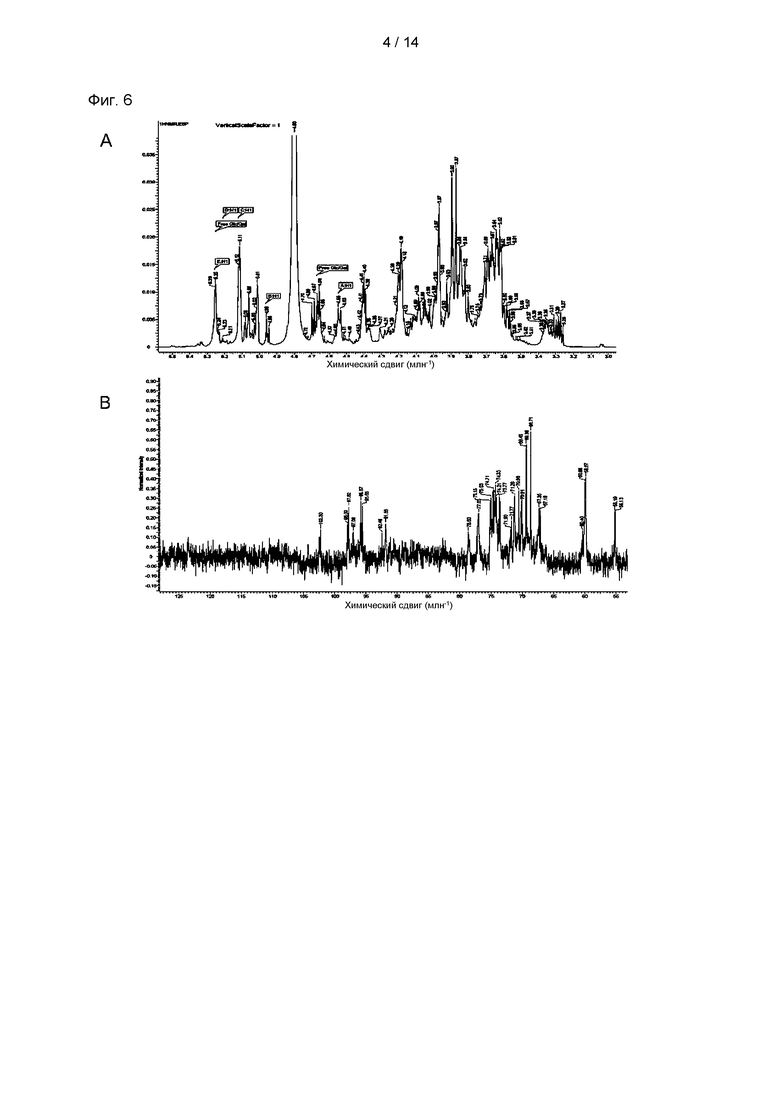
На Фиг. 6А и 6В показаны 1Н и 13С ЯМР (ядерный магнитный резонанс) спектры р-CY01_LM, полученного посредством низкой молекуляризации p-CY01.
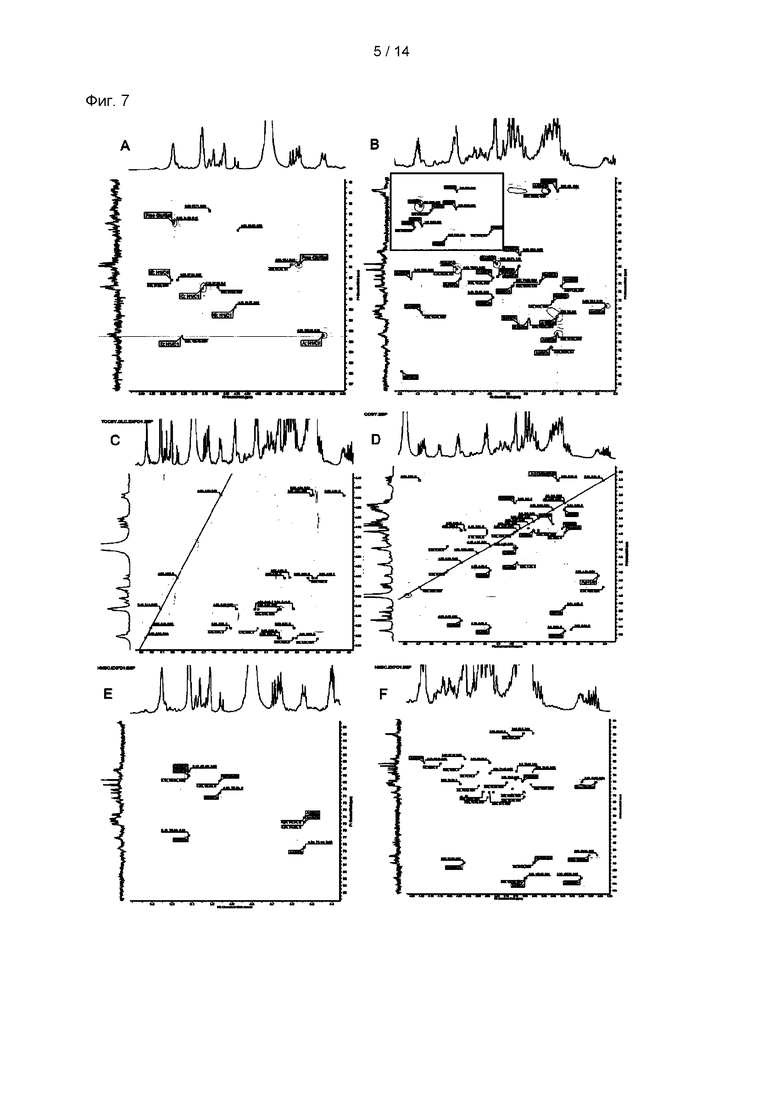
На Фиг. 7А и 7В показан спектр 2D-HSQC (гетероядерная одноквантовая корреляция) p-CY01_LM, полученного посредством низкой молекуляризации, и показаны результаты экспериментов по корреляции углеродов с протонами, и на Фиг. 7С, 7D, 7Е и 7F показаны спектры 2D-TOCSY (полная корреляционная спектроскопия), 2D-COSY (корреляционная спектроскопия), 2D-HMBC (гетероядерная многосвязная корреляционная спектроскопия) и 2D-ЯМР (600 МГц).
На Фиг. 8 показаны результаты анализа главных структур р-CY01 на основе результатов анализа ГХ/МС и ЯМР.
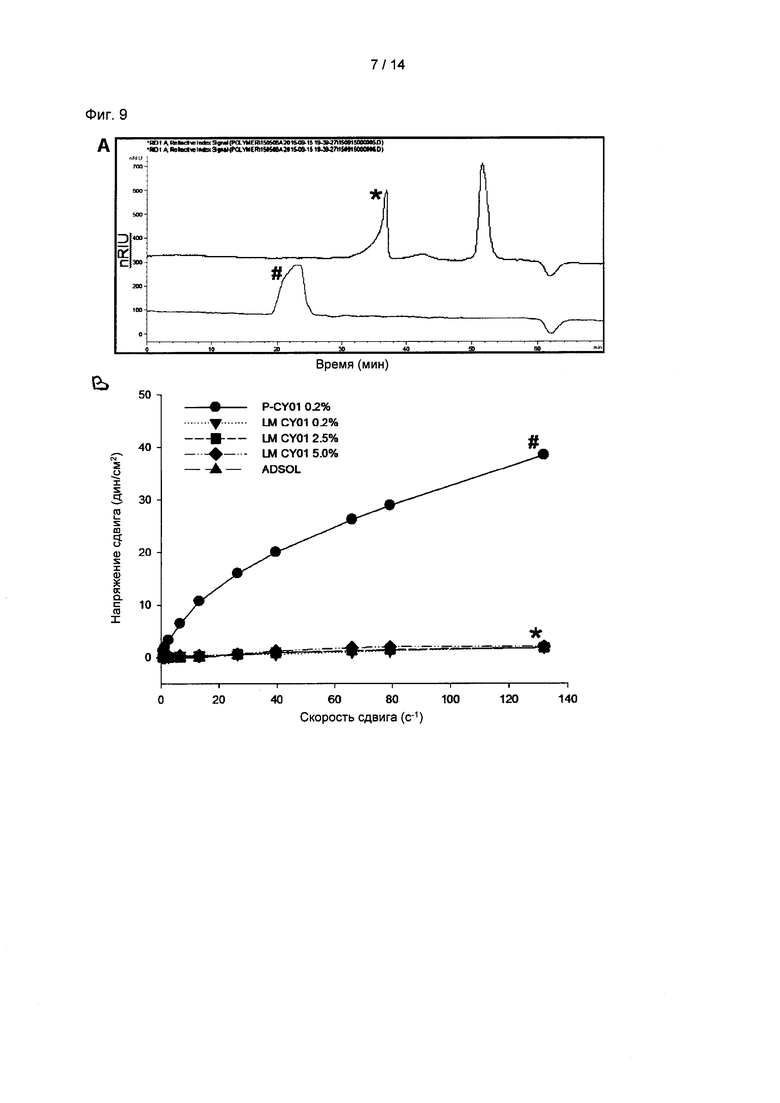
На Фиг. 9А показаны результаты эксклюзионной хроматографии р-CY01 и р-CY01_LM, полученные в результате обработки на основе кислотного раасщепления, и на Фиг. 9В показаны изменения в реологических свойствах раствора p-CY01_LM. # обозначает р-CY01, и * обозначает p-CY01_LM.
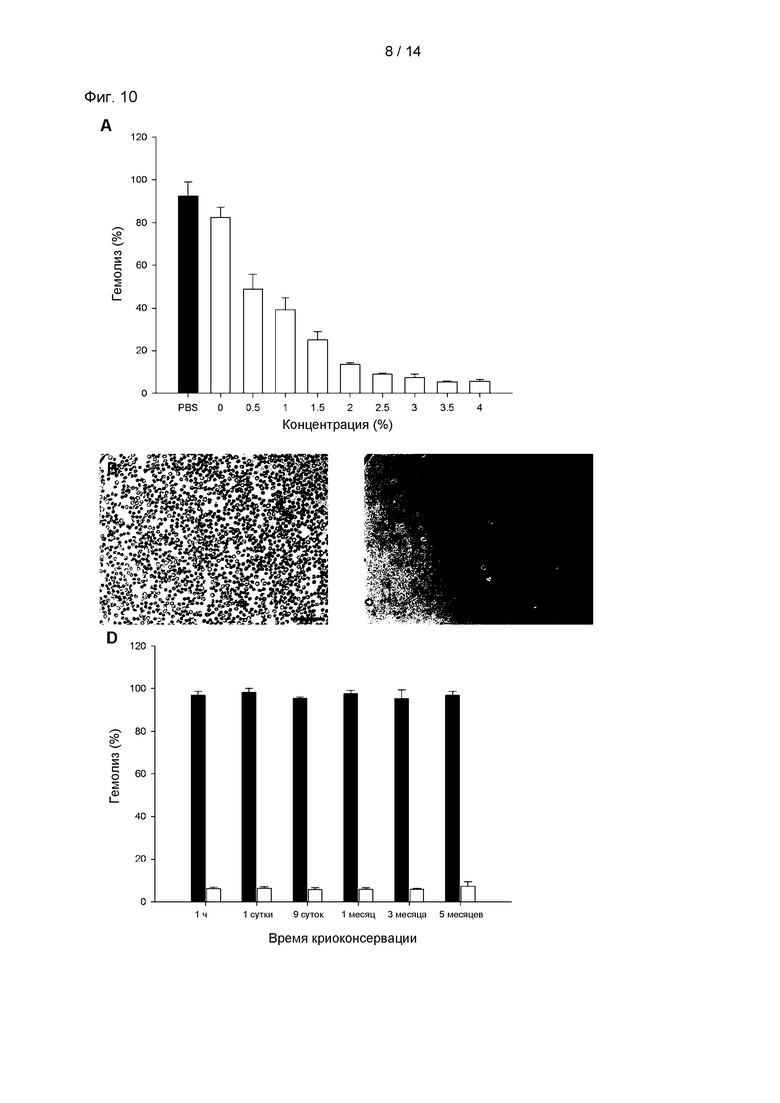
На Фиг. 10А показан гемолиз (%) эритроцитов в зависимости от концентрации р-CY01_LM в среде для криоконсервации; на Фиг. 10В показаны результаты наблюдения под микроскопом, проводимого для подтверждения целостности эритроцитов, оттаявших после криоконсервации в 2,5% растворе p-CY01_LM; на Фиг. 10С показаны результаты анализа целостности эритроцитов, оттаявших после криоконсервации в растворе ФБР (фосфатно-солевой буферный раствор); и на Фиг. 10D показан гемолиз (%) эритроцитов, оттаявших после длительной криоконсервации в растворе p-CY01_LM. На Фиг. 10 черные столбцы обозначают гемолиз (%) эритроцитов, оттаявших после криоконсервации в 2,5% растворе p-CY01_LM, и белые столбцы обозначают гемолиз (%) эритроцитов, оттаявших после криоконсервации в растворе ФБР.
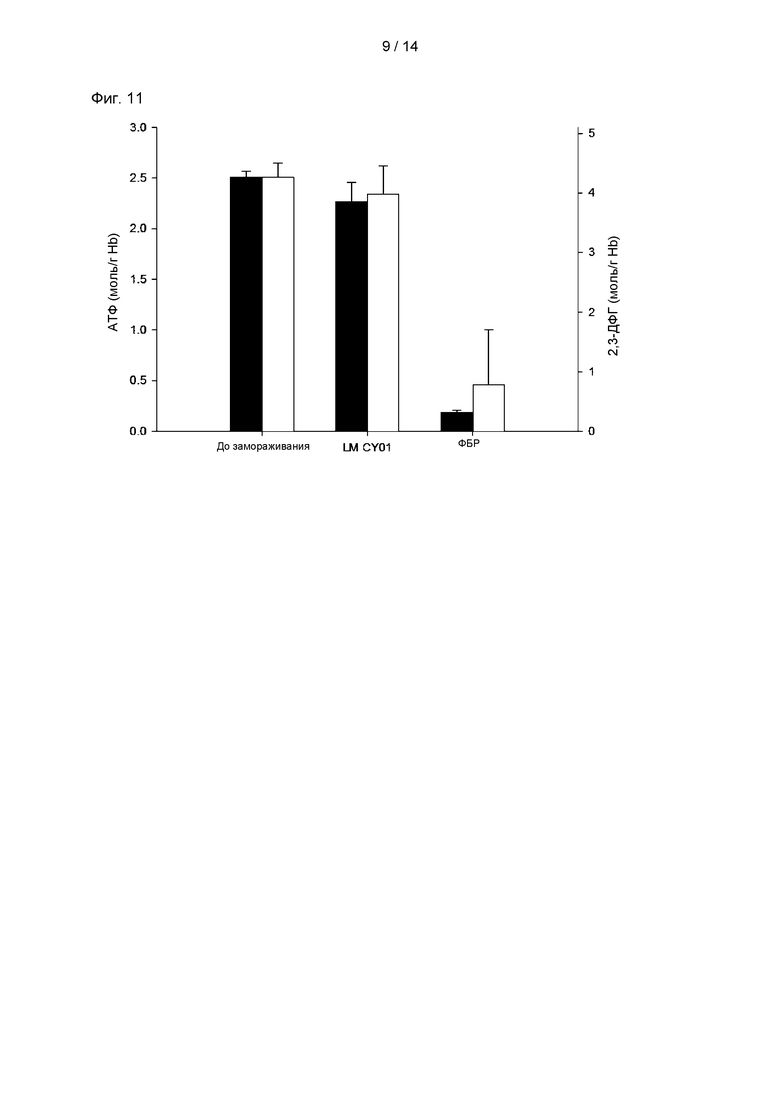
На Фиг. 11 показаны результаты измерения активностей АТФ и 2,3-ДФГ (2,3-дифосфоглицерат) после гемолиза эритроцитов, криоконсервированных в растворе р-CY01_LM.
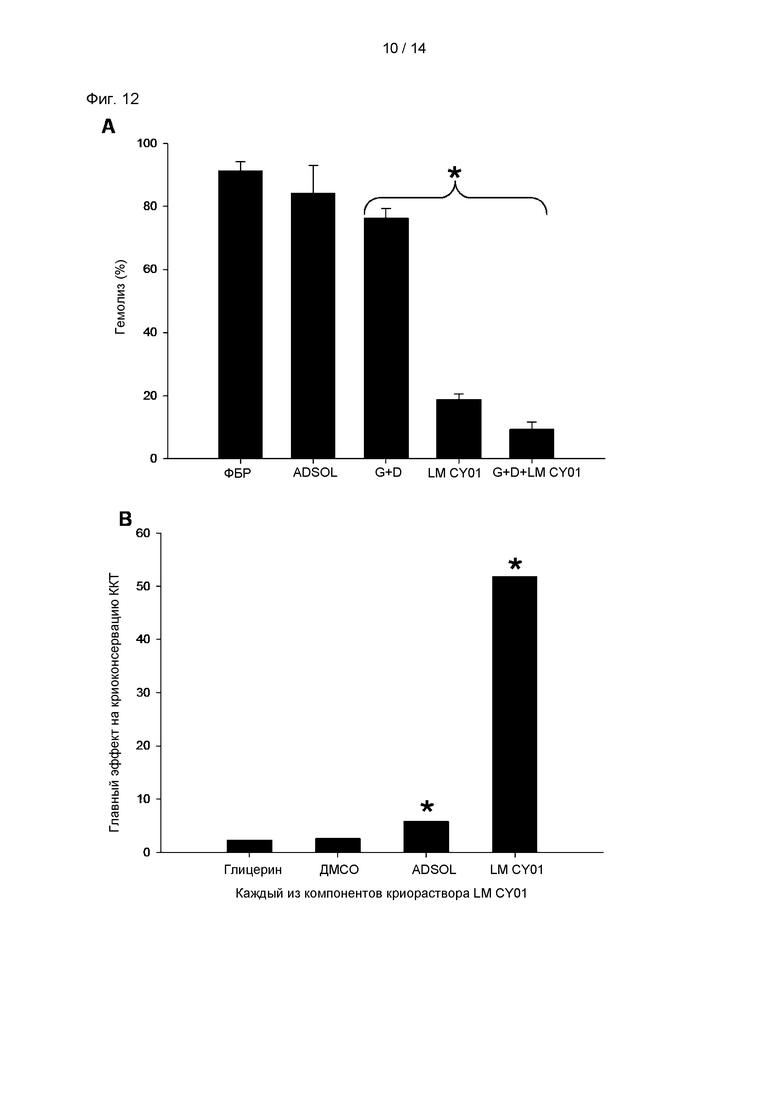
На Фиг. 12А показаны результаты анализа способности каждого компонента, добавленного к среде для криоконсервации, к консервации эритроцитов. На Фиг. 12А ADSOL обозначает раствор ADSOL; «G+D» обозначает 1% (масс./об.) глицерин и ДМСО (диметилсульфоксид); «p-CY01_LM» обозначает 2,5% (масс./об.) раствор p-CY01_LM; и «G+D+p-CY01_LM» обозначает раствор, содержащий 1% (масс./об.) глицерин, 1% (масс./об.) ДМСО и 2,5% (масс./об.) p-CY01_LM. * обозначает раствор каждого компонента в ADSOL. На Фиг. 12В показаны результаты статистического анализа Плакетта-Бермана, проводимого для анализа главного эффекта каждого компонента, который добавляют к среде для криоконсервации, на криоконсервацию эритроцитов. * указывает на то, что статистически различные значения равны или меньше чем 0,05.
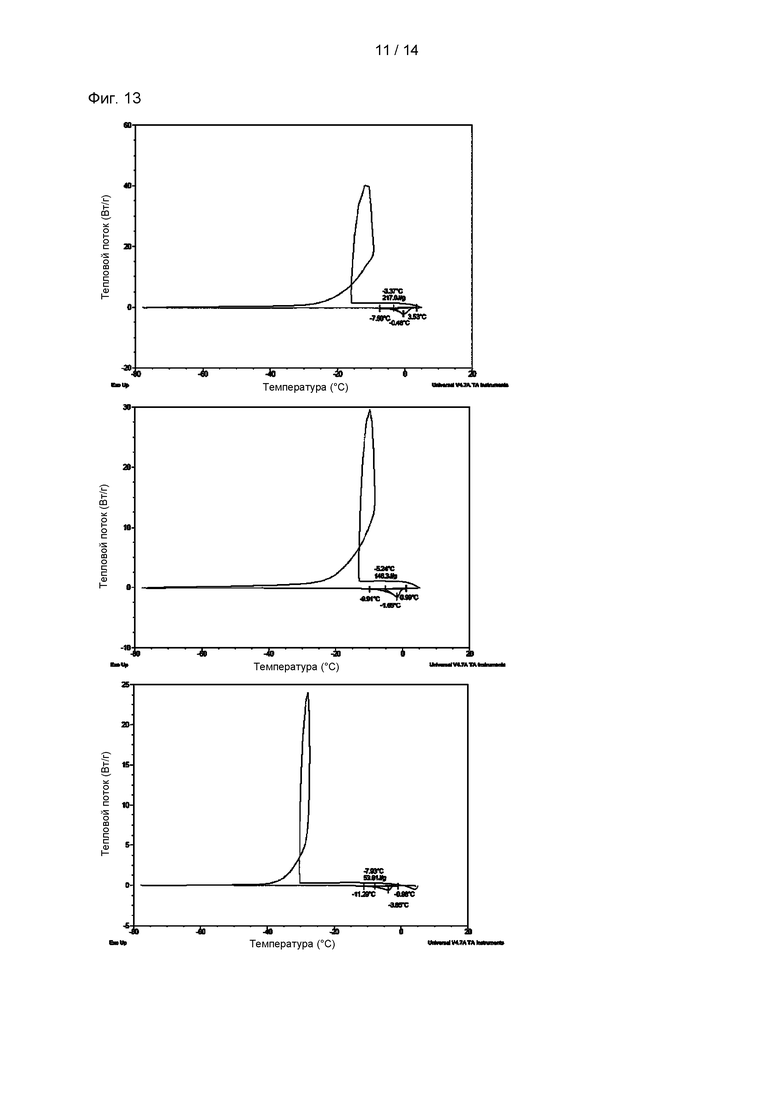
На Фиг. 13 показаны результаты анализа ДСК (дифференциальная сканирующая калориметрия) раствора p-CY01_LM. В частности, на Фиг. 13А показаны результаты анализа ДСК для раствора, содержащего 1% глицерин и 1% ДМСО, который представляет собой отрицательный контроль; на Фиг. 13В показаны результаты анализа ДСК для раствора, содержащего 1% глицерин, 1% ДМСО и 0,5% p-CY01_LM; и на Фиг. 13С показаны результаты анализа ДСК для раствора, содержащего 1% глицерин, 1% ДМСО и 2,5% p-CY01_LM.
На Фиг. 14 показаны результаты анализа способности раствора p-CY01_LM к предотвращению замерзания посредством анализа формы кристаллов льда. В частности, на Фиг. 14А показаны результаты анализа формы кристаллов льда в растворе, содержащем 1% глицерин и 1% ДМСО; на Фиг. 14В показаны результаты анализа формы кристаллов льда в растворе ADSOL (раствор p-CY01_LM), содержащем 1% глицерин, 1% ДМСО и 2,5% p-CY01_LM; и на Фиг. 14С показаны результаты анализа формы кристаллов льда в растворе ADSOL, содержащем 1% глицерин, ДМСО и 2,5% ГЭК. От 1 до 2 в каждой экспериментальной группе означает, что форма кристаллов льда становится больше при охлаждении зародышей кристаллов льда. Масштаб представляет собой 10 мкм.
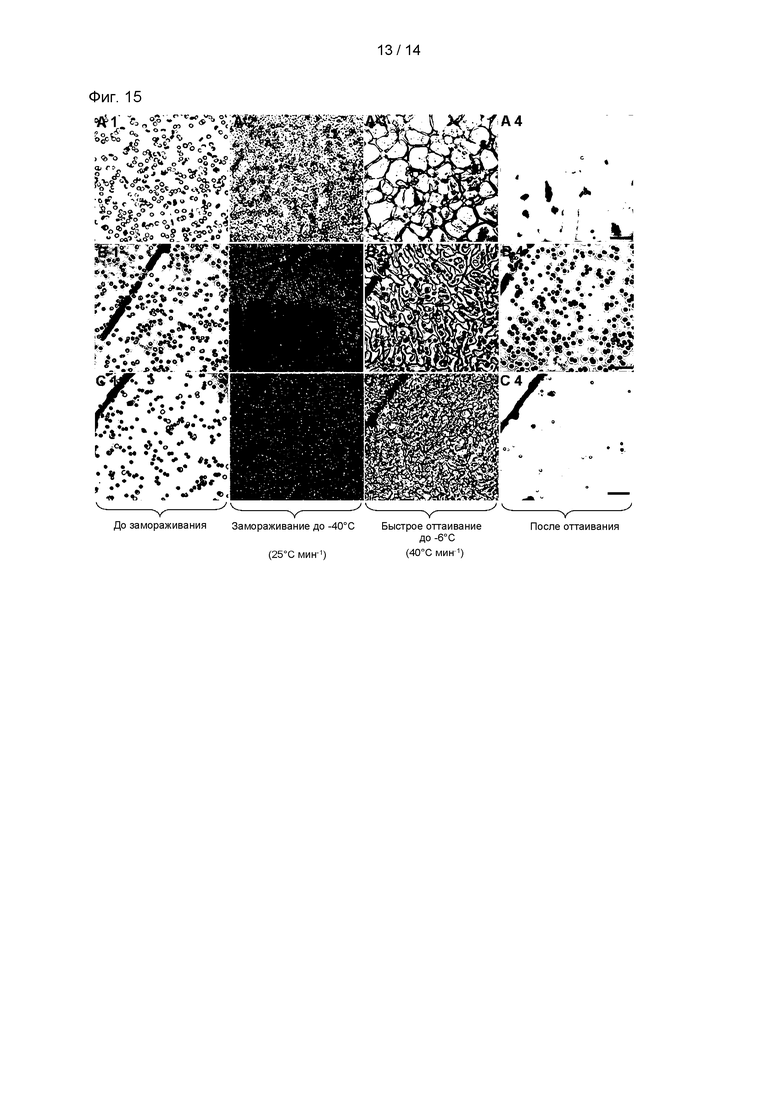
На Фиг. 15 показаны результаты анализа способности p-CY01_LM к предотвращению замерзания посредством анализа остаточной формы эритроцитов и формы кристаллов льда при наблюдении под микроскопом процессов замерзания и оттаивания. В частности, на Фиг. 15А показана форма эритроцитов в ФБР; на Фиг. 15В показана форма эритроцитов в растворе p-CY01_LM; и на Фиг. 15С показана форма эритроцитов в растворе ADSOL, содержащем 1% глицерин, ДМСО и 2,5% ГЭК. Изображения гемолиза эритроцитов получали после 5 минут на каждой стадии, и масштаб представляет 20 мкм.
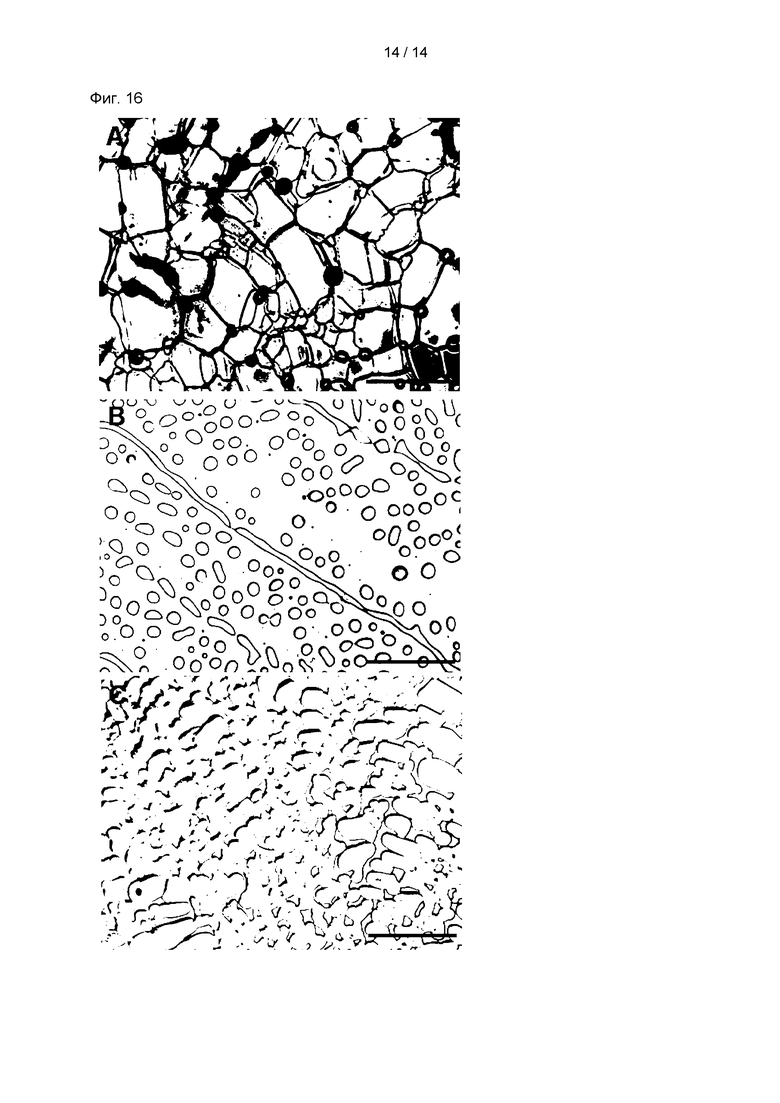
На Фиг. 16 показаны результаты наблюдения за активностью p-CY01_LM в отношении ингибирования перекристаллизации льда (ИПЛ). В частности, на Фиг. 16А показаны результаты для ФБР; на Фиг. 16В показаны результаты для раствора р-CY01_LM; и на Фиг. 16С показаны результаты для раствора ADSOL, содержащего 1% глицерин, ДМСО и 2,5% ГЭК. Масштаб представляет собой 100 мкм.
Подробное описание предпочтительного варианта реализации
Согласно настоящему изобретению новый штамм Pseudoalteromonas sp. CY01, который продуцирует экзополисахарид, выделяли из образцов антарктических морских вод. Было обнаружено, что экзополисахарид, продуцируемый штаммом CY01, оказывал сильный криозащитный эффект на E. coli во время исходного скрининга. На основе данного результата исследовали, будет ли экзополисахарид, продуцируемый штаммом CY01, пригодным к применению в качестве криозащитного агента во время криоконсервации клеток. Для обеспечения физических свойств, подходящих для применения в криоконсервации, экзополисахарид из CY01 частично расщепляли под действием кислоты с образованием низкомолекулярного экзополисахарида, и данный низкомолекулярный экзополисахарид добавляли во время криоконсервации эритроцитов. В результате было показано, что низкомолекулярный экзополисахарид демонстрировал высокую способность к предотвращению замерзания и не обладал цитотоксичностью.
Таким образом, согласно одному аспекту настоящее изобретение относится к экзополисахариду, который продуцируется штаммом Pseudoalteromonas sp. CY01 (KCTC 12867ВР) и который состоит из глюкозы и галактозы.
Согласно настоящему изобретению среди 2980 штаммов, выделенных из образцов антарктических морских вод, выделяли 73 штамма, которые продуцируют экзополисахариды слизи, и из них отбирали 10 штаммов, имеющих превосходную способность продуцировать экзополисахариды. Клетки Е. coli подвергали действию циклов замораживания и оттаивания в 0,2% (масс./масс.) неочищенном растворе экзополисахаридов, полученном из каждого штамма, и измеряли долю выживших клеток Е. coli. В результате неочищенный экзополисахарид, продуцируемый штаммом CY01 среди данных 10 штаммов, демонстрировал наибольший криозащитный эффект.
В примере настоящего изобретения доля выживших клеток Е. coli в растворе, содержащем 0,2% неочищенного экзополисахарида, происходящего из штамма CY01, составляла 88,16±2,92%, и доли выживших клеток Е. coli в растворах, содержащих 0,2% неочищенных экзополисахаридов, происходящих из других штаммов, составляли от 42,01±1,93% до 67,28±4,32%.
Согласно настоящему изобретению молярное отношение глюкозы к галактозе в экзополисахариде из CY01 может составлять приблизительно 3,4:1.
Согласно настоящему изобретению анализ гликозильных связей и ЯМР анализ экзополисахарида из CY01 указывает на то, что экзополисахарид имеет повторяющуюся структуру, состоящую главным образом из 4-связанной глюкопиранозы и 6-связанной галактопиранозы, и данные компоненты могут представлять собой компоненты, образующие основную цепь.
Согласно другому аспекту настоящее изобретение относится к экзополисахариду, имеющему молекулярную массу от 1,0×105 до 4,3×105 Да, который получают посредством гидролиза описанного выше экзополисахарида.
Согласно настоящему изобретению кислотное расщепление проводили для увеличения растворимости экзополисахарида и для уменьшения высокой вязкости экзополисахарида, которая вызывала сложность при обращении и промывке во время применения экзополисахарида в качестве криозащитного агента.
В примере настоящего изобретения экзополисахарид, происходящий из CY01 (р-CY01), нагревали вместе с 0,1 М ТФУ (трифторуксусная кислота) при 121°С в течение 1 часа с получением расщепленного под действием кислоты p-CY01 (p-CY01_LM). В результате средняя молекулярная масса р-CY01 составляла приблизительно 1,1×107 Да, тогда как молекулярная масса p-CY01_LM, частично расщепленного под действием кислоты, составляла от 1,0×105 до 4,3×105 Да. Подтверждали уменьшение вязкости и повышение растворимости экзополисахарида в зависимости от изменений в реологических свойствах раствора низкомолекулярного p-CY01_LM.
Согласно еще одному аспекту настоящее изобретение относится к композиции для криозащиты клеток, которая содержит экзополисахарид.
В примере настоящего изобретения образец эритроцитов, содержащий p-CY01_LM и криозащитный агент, замораживали без контролирования скорости охлаждения (охлаждали сразу до -80°С) и консервировали при -80°С. Образец эритроцитов подвергали быстрому оттаиванию на водяной бане при 40°С. После оттаивания анализировали функцию p-CY01_LM в качестве криозащитного агента посредством анализа гемолиза эритроцитов и анализа на основе оптической микроскопии. Гемолиз (%) уменьшался при возрастании концентрации p-CY01_LM. При концентрации p-CY01_LM 2,5% - 4,0% наблюдался выраженный в процентах гемолиз от 9,08±0,37% до 5,64±0,96%, и применение 3,5% p-CY01_LM демонстрировало самый низкий уровень гемолиза (5,40%). Иными словами, было показано, что при концентрации p-CY01_LM 2,5% - 4,0% 90% ККТ после оттаивания обладали целостностью (Фиг. 7А).
Согласно примеру настоящего изобретения материалы, подлежащие криоконсервации, могут представлять собой бактерии, грибы, клетки животных, клетки растений, эритроциты, тромбоциты, сперматоциты, ооциты, ткани, органы или тому подобное.
Композиция согласно настоящему изобретению может защищать клетки, образующие ткани и органы, от замораживания.
Композиция для криозащиты клеток согласно настоящему изобретению может содержать глицерин и/или ДМСО.
Согласно другому примеру настоящего изобретения исследовали, заменит ли р-CY01_LM глицерин в качестве криозащитного агента при длительной криоконсервации, и выраженный в процентах гемолиз эритроцитов, которые быстро охлаждали до -80°С и консервировали в течение 1 часа в ADSOL, содержащем 2,5% (масс./об.) p-CY01_LM, 1% (об./об.) глицерин и 1% (об./об.) ДМСО (далее в настоящей заявке, называемый раствором p-CY01_LM), составлял 6,09±0,64%, и выраженный в процентах гемолиз после 5 месяцев консервации составлял 7,24±2,15%, указывая на то, что в течение 5 месяцев консервации происходило небольшое изменение или не происходило изменения в выраженном в процентах гемолизе (Фиг. 7D).
Согласно еще одному аспекту настоящее изобретение относится к способу получения экзополисахарида, который продуцируется штаммом Pseudoalteromonas sp. CY01 (KCTC 12867ВР) и который состоит из глюкозы и галактозы, причем способ включает следующие стадии: (а) культивирование штамма Pseudoalteromonas sp. CY01 (KCTC 12867ВР) с продуцированием экзополисахарида; и (b) выделение полученного экзополисахарида.
Согласно другому аспекту настоящее изобретение относится к способу получения экзополисахарида, имеющего молекулярную массу от 1,0×105 до 4,3×105 Да, причем способ включает стадию гидролиза экзополисахарида, который продуцируется штаммом Pseudoalteromonas sp. CY01 (KCTC 12867ВР) и который состоит из глюкозы и галактозы.
Согласно настоящему изобретению гидролиз может представлять собой расщепление под действием слабой кислоты.
Согласно настоящему изобретению стадия гидролиза может дополнительно включать проведение тепловой обработки.
Согласно еще одному другому аспекту настоящее изобретение также относится к способу криоконсервации клеток с применением описанного выше экзополисахарида.
Согласно настоящему изобретению замораживание может представлять собой быстрое замораживание, которое имеет скорость замораживания, находящуюся в диапазоне от 10°С/мин до 196°С/мин.
Согласно еще одному другому аспекту настоящее изобретение также относится к штамму Pseudoalteromonas sp. CY01 (KCTC 12867ВР), который обладает способностью продуцировать экзополисахарид слизи. Штамм Pseudoalteromonas sp. CY01 (KCTC 12867ВР) может обитать в Антарктике.
В результате филогенетического анализа с применением последовательности 16S рРНК штамма CY01 штамм CY01 согласно настоящему изобретению относили к штамму Pseudoalteromonas sp., который распространен в Антарктике (Фиг. 2). Штамм CY01 демонстрировал высокую гомологию с Pseudoalteromonas paragorgicola KMM 3548Т (99,52%), P. nigrifaciens NCIIMB 8614Т (99,51%) и P. agarivorans KMM 255Т (99,38%).
Примеры
Далее в настоящей заявке настоящее изобретение будет описано более подробно со ссылкой на примеры. Обычному специалисту в данной области техники будет понятно, что данные примеры служат только в иллюстративных целях и не предназначены для ограничения объема настоящего изобретения.
Пример 1: Скрининг и идентификация штамма, продуцирующего экзополисахарид, обладающий криозащитной способностью
2980 штаммов, выделенных из образцов антарктических морских вод, культивировали с применением чашек с агаризованной средой S-ZoBell (рН 7,0) (изготавливаемых авторами настоящего изобретения) при 15°С в течение 3 суток, и выделяли 73 штамма, продуцирующих экзополисахариды слизи. Для измерения количеств экзополисахаридов, продуцированных выделенными штаммами, штаммы культивировали с применением жидких сред S-ZoBell (рН 7,0) при 15°С в течение 3 суток, и экзополисахариды выделяли и лиофилизировали, и затем измеряли их сухие массы. Криозащитную способность измеряли путем подвергания клеток Е. coli действию циклов замораживания и оттаивания в 0,2% (масс./масс.) растворе неочищенного экзополисахарида, полученном из каждого штамма, и затем измерения доли выживших клеток Е. coli.
Среди выделенных штаммов 10 штаммов продуцировали 1 г/л или больше неочищенных экзополисахаридов (ЭПС), и количества неочищенных экзополисахаридов, продуцированных данными штаммами, находились в диапазоне от 1,04±0,18 г/л до 2,04±0,13 г/л. Среди данных 10 штаммов штамм RosPo13 демонстрировал наивысшую продукцию экзополисахарида (2,04 г/л), и неочищенный экзополисахарид, продуцируемый штаммом CY01, демонстрировал наивысшую криозащитную способность (Фиг. 1).
Доля выживших клеток Е. coli в растворе, содержащем 0,2% неочищенного экзополисахарида, происходящего из штамма CY01, составляла 88,16±2,92%, и доли выживших клеток Е. coli в растворах, содержащих 0,2% неочищенного экзополисахарида, происходящего из других штаммов, составляли от 42,01±1,93% до 67,28±4,32%.
В результате филогенетического анализа с применением последовательности 16S рРНК штамма CY01, штамм CY01 относили к штамму Pseudoalteromonas sp., который в большом количестве присутствует в Антарктике (Фиг. 2). Штамм CY01 демонстрировал высокую гомологию с Pseudoalteromonas paragorgicola KMM 3548Т (99,52%), Р. nigrifaciens NCIIMB 8614Т (99,51%) и Р. agarivorans KMM 255Т (99,38%).
Пример 2: Очистка экзополисахарида, происходящего из штамма CY01, и характеристика очищенного экзополисахарида
Экзополисахарид отделяли от культуры штамма CY01 посредством осаждения этанолом и обрабатывали протеазой для удаления белка. Полученный неочищенный экзополисахарид подвергали анионной хроматографии с применением колонки DEAE-Sepharose, получая, таким образом, фракции, содержащие экзополисахарид (Фиг. ЗА). Полученные фракции, содержащие экзополисахарид, очищали посредством колонки для гель-фильтрационной хроматографии Sepharose 4В (Фиг. 3В) с получением единственной фракции. Концентрацию экзополисахарида в данной фракции измеряли при OD630 (оптическая плотность) с помощью анализа с антроном и серной кислотой.
Полученную фракцию подвергали ВЭЖХ (Agilent, США). ВЭЖХ проводили с применением 5 мкл раствора фракции экзополисахарида в дистиллированной воде (0,1% масс./масс.) при скорости потока 0,4 мл/мин, и выявление проводили с применением RI детектора (рефрактометрический детектор) (Индекс рефракции, Agilent, США).
В результате, как показано на Фиг. 4, выявляли единственный пик. Молекулярную массу экзополисахарида, очищенного посредством эксклюзионной хроматографии, измеряли, применяя RI, и в результате было показано, что экзополисахарид имел среднюю молекулярную массу приблизительно 1,1×107 Да. Очищенный ЭПС из штамма CY01 называли «p-CY01».
Применяя ГХ/МС, анализировали сахара-составные части экзополисахарида. Анализ проводили в режиме ионизации электронным ударом с применением Claras 500 (Perkin-Elmer, США) и масс-селективного детектора.
В результате, как показано на Фиг. 5А, р-CY01 состоял из глюкозы и галактозы, и глюкоза представляла собой преобладающий сахар в составе р-CY01. Кроме того, молярное соотношение глюкоза : галактоза в р-CY01 составляло приблизительно 3,4:1.
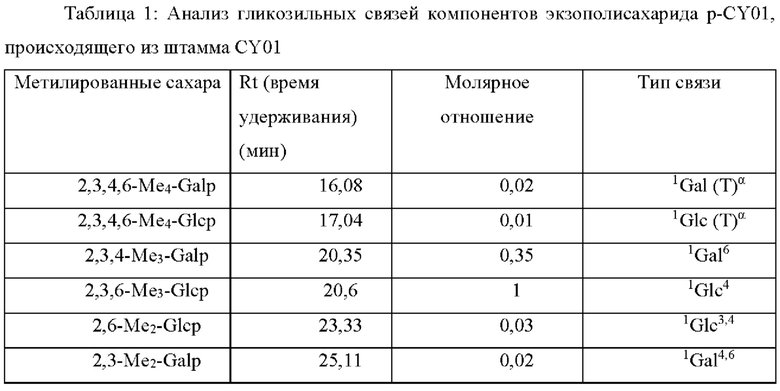
Как показано на Фиг. 5В и ниже в Таблице 1, результаты анализа гликозильных связей р-CY01 указывали на то, что р-CY01 главным образом состоял из 4-связанной глюкопиранозы и 6-связанной галактопиранозы. В виде минорных пиков выявляли соединенную через концевые группы галактопиранозу, соединенную через концевые группы глюкопиранозу, 3,4-связанную глюкопиранозу и 4,6-связанную галактопиранозу.
Для р-CY01, ID и 2D ЯМР эксперименты для определения структур проводили, применяя спектрометр Bruker AVANCE (600 МГц). Образцы измеряли при 25°С, 1Н ЯМР измеряли при 600 МГц, и 13С ЯМР измеряли при 150 МГц.
В результате ЯМР анализа типичные картины пиков полиглюкопиранозы и полигалактопиранозы можно было наблюдать из 1Н и 13С ЯМР спектров р-CY01 (Фиг. 6А и 6В). В частности, наблюдали, что р-CY01 главным образом состоял из повторяющихся звеньев →4)-β-D-Glcp-(1→ и →6)-α-D-Galp-(1→, и пики других боковых цепей были относительно небольшими и перекрывались с пиками главных повторяющихся звеньев. Кроме того, из спектров 2D-HSQC (Фиг. 7А и В) наблюдали корреляцию углеродов с протонами, а из 2D-TOCSY, 2D-COSY, 2D-HMBC и 2D-ЯМР (600 МГц) едва могли наблюдать необычную информацию, отличную от корреляции глюкопиранозы и галактопиранозы, которые являются главными компонентами (Фиг. 7С-7F). По совокупности результатов анализа гликозильных связей и результатов анализа ЯМР-спектра определяли главные структуры р-CY01 (Фиг. 8). Как видно из результатов анализа, р-CY01 имеет повторяющуюся структуру, состоящую главным образом из 4-связанной глюкопиранозы и 6-связанной галактопиранозы, и связывающая структура [→4)-β-D-Glcp-(1→4)-β-D-Glcp-(1→]n III и связывающая структура [→6)-α-D-Galp-(1→6)-α-D-Galp-(1→]n II повторно соединены друг с другом с образованием основной цепи. Кроме того, было показано, что р-CY01 имеет небольшое количество связывающей структуры T-α-D-Glcp-(1→6)-α-D-Glcp-(1,4→ I в виде структуры боковой цепи основной цепи.
Пример 3: Анализ изменений в молекулярной массе и реологических свойствах экзополисахарида после гидролиза
Поскольку высокая вязкость и низкая растворимость экзополисахарида могут мешать применению экзополисахарида в биологической промышленности, уменьшение средней молекулярной массы полимера в результате расщепления может понижать вязкость и повышать растворимость. В недавних экспериментах, когда ооциты и эритроциты криоконсервировали с применением высокой концентрации ПВС (поливиниловый спирт), недостатки заключались в том, что осуществление методики не является простым вследствие высокой вязкости раствора, и необходимо осуществлять промывку (Deller, RC et al., Nat. Commun., 5:3244, doi: 10.1038/ncomms4244, 2014).
В данном примере для преодоления таких недостатков полимера молекулярную массу р-CY01 согласно настоящему изобретению регулировали посредством кислотного расщепления. р-CY01 нагревали с 0,1 М ТФУ (трифторуксусная кислота) при 121°С в течение 1 часа с получением расщепленного под действие кислоты р-CY01, и осуществляли гель-проникающую хроматографию (ГПХ). В результате р-CY01, не обработанный ТФУ, элюировали ТФУ из колонки для ГПХ между 19 мин и 25 мин, и в случае р-CY01, подверженного тепловой обработке с ТФУ, главный пик элюировали на 33 мин и 37 мин и также элюировали на 51 мин (Фиг. 9А). Высокомолекулярные пики на 33 мин и 37 мин представляли собой фракции, полученные в результате частичного расщепления р-CY01, и пик, появляющийся на 51 мин, представлял собой моносахарид, полученный в результате расщепления р-CY01. Средняя молекулярная масса p-CY01 составляла приблизительно 1,1×107 Да, тогда как молекулярная масса p-CY01_LM, частично расщепленного под действием кислоты, составляла от 1,0×105 до 4,3×105 Да.
Измеряли изменения в реологических свойствах раствора низкомолекулярного р-CY01 (p-CY01_LM).
Реологические свойства измеряли посредством вискозиметра Брукфильда с применением оси S18, и измеряли напряжения сдвига растворов р-CY01, ФБР и р-CY01_LM (0,2%, 2,5% и 5,0%, масс./об.) при разных скоростях сдвига.
В результате, как показано на Фиг. 9В, в случае 0,2% раствора р-CY01, скорость сдвига повышалась при повышении скорости сдвига, а в случае раствора p-CY01_LM напряжение сдвига не повышалось при высокой скорости сдвига. Это указывает на то, что вязкость p-CY01_LM значимо уменьшалась. Данное уменьшение вязкости объясняется изменением молекулярной массы р-CY01. Когда экзополисахарид разлагается и деполимеризуется, вязкость уменьшается и растворимость повышается.
Пример 4: Криоконсервация человеческих эритроцитов (ККТ) в p-CY01_LM
Эритроциты можно консервировать в ADSOL при 4°С в течение 32 суток. Когда эритроциты подвергают деглицеролизации в Haemonetics АСР215 и консервируют в ADSOL, данные клетки можно консервировать при 4°С в течение 3 суток, при этом имея степень гемолиза меньше чем 1%. Глицерин представляет собой внутриклеточный криозащитный агент, и для предотвращения гемолиза эритроцитов конечная концентрация глицерина должна быть уменьшена до 1% посредством промывки после оттаивания.
ДМСО был одобрен FDA (Управление по санитарному надзору за качеством пищевых продуктов и медикаментов) для применения в качестве криозащитного агента в отношении тромбоцитов и эритроцитов человека. Таким образом, в следующем эксперименте по криоконсервации с применением p-CY01_LM, глицерин и ДМСО применяли вместе, и ADSOL применяли в качестве криоконсервирующего буфера вместо ФБР.
В клинической практике при охлаждении эритроцитов с высокой концентрацией (40%) глицерина с низкой скоростью охлаждения (1°С/мин) и консервации при -80°С или в жидком азоте, может быть достигнут высокий уровень восстановления эритроцитов. Кроме того, криозащитные агенты - экзополисахариды (ПВП (поливинилпирролидон), ГЭК) требуют физического замораживания и консервации в жидком азоте.
В данном Примере образец эритроцитов замораживали без контроля скорости охлаждения (охлаждали сразу же до -80°С) и консервировали при -80°С. Образец эритроцитов подвергали быстрому оттаиванию на водяной бане при 40°С. После оттаивания гемолиз образца количественно оценивали посредством анализа Драбкина.
Функцию p-CY01_LM в качестве криозащитного агента анализировали посредством анализа гемолиза эритроцитов и анализа на основе оптической микроскопии.
Как можно видеть на Фиг. 10А, гемолиз (%) уменьшался при возрастании концентрации p-CY01_LM. При концентрации p-CY01_LM от 2,5% до 4,0% наблюдали выраженный в процентах гемолиз от 9,08±0,37% до 5,64±0,96%, и применение 3,5% р-CY01_LM демонстрировало самый низкий уровень гемолиза (5,40%). Иными словами, было показано, что при концентрации p-CY01_LM от 2,5% до 4,0% 90% ККТ после оттаивания демонстрировали целостность. Согласно стандартам Американской ассоциации банков крови требуется непосредственный показатель выживаемости 80% (гемолиз меньше чем 20%) и требуется, чтобы 70% клеток крови оставались живыми в течение 24 часов.
Раствор ADSOL, содержащий 2,5% (масс./об.) p-CY01_LM, 1% (об./об.) глицерин и 1% (об./об.) ДМСО, демонстрировал выраженный в процентах гемолиз 9,08±0,37% (целостность эритроцитов после оттаивания 90,92%) (Фиг. 10А и 10В), и ФБР демонстрировал выраженный в процентах гемолиз 92,48±6,49% (целостность эритроцитов после оттаивания 7,52%) (Фиг. 10А и 10С).
Таким образом, 2,5% p-CY01_LM отбирали в качестве оптимальной концентрации для криоконсервации эритроцитов.
Исследовали, заменит ли p-CY01_LM глицерин в качестве криозащитного агента для длительной криоконсервации. Как показано на Фиг. 10D, выраженный в процентах гемолиз эритроцитов, которых быстро охлаждали до -80°С и консервировали в течение 1 часа в ADSOL, содержащем 2,5% (масс./об.) p-CY01_LM, 1% (об./об.) глицерин и 1% (об./об.) ДМСО (далее в настоящей заявке называемый раствором p-CY01_LM), составлял 6,09±0,64%, и выраженный в процентах гемолиз после 5 месяцев консервации составлял 7,24±2,15%, указывая на то, что имелось небольшое изменение или отсутствовало изменение в выраженном в процентах гемолизе за 5 месяцев консервации.
Кроме того, выраженный в процентах гемолиз эритроцитов, консервируемых в растворе p-CY01_LM при комнатной температуре в течение 1 часа, сохранялся на уровне меньше чем 2% (целостность эритроцитов больше чем 98%), указывая на то, что во время криоконсервации не имела место цитотоксичность или гемолиз.
Пример 5: Измерение биохимических свойств эритроцитов, замороженных с р-CY01_LM
Уровень АТФ в эритроцитах позволяет определять, будет ли сохраняться вогнутая форма мембраны эритроцитов и будет ли возрастать динамика мембраны эритроцитов. Измеряли уровень АТФ и 2,3-ДФГ (2,3-дифосфоглицерат), который контролирует аффинность гемоглобина к кислороду, для определения клеточной функции и терапевтической применимости эритроцитов.
В качестве группы положительного контроля измеряли активности АТФ и 2,3-ДФГ гемолизата свежих эритроцитов, и в качестве опытной группы измеряли активности АТР и 2,3-ДФГ гемолизата эритроцитов, криоконсервированных в растворе p-CY01_LM при -80°С в течение 1 часа.
В результате, как показано на Фиг. 11, эритроциты, криоконсервированные в растворе p-CY01_LM, демонстрировали небольшое различие или не демонстрировали различия в активности АТФ по сравнению с группой положительного контроля. Активность АТФ эритроцитов, криоконсервированных в растворе p-CY01_LM, составляла от 2,50±0,06 мкмоль/г Hb (гемоглобин) до 2,26±0,19 мкмоль/г Hb, и активность АТФ эритроцитов, криоконсервированных в ФБР в качестве группы отрицательного контроля, значительно уменьшалась до 0,18±0,02 мкмоль/г Hb. Концентрация 2,3-ДФГ составляла 4,26±0,24 мкмоль/г Hb в свежих эритроцитах, 3,98±0,47 мкмоль/г Hb в эритроцитах, криоконсервированных в растворе p-CY01_LM, и 0,78±0,92 мкмоль/г Hb в эритроцитах, криоконсервированных в ФБР.
Отсутствовало значимое различие в активности АТФ и концентрации 2,3-ДФГ у свежих эритроцитов и эритроцитов, криоконсервированных в растворе p-CY01_LM.
Пример 6: Исследование основных криозащитных добавок и анализ главных эффектов, оказываемых на криоконсервацию эритроцитов
Для анализа эффекта каждого криозащитного агента, оказываемого на криоконсервацию эритроцитов, способность к консервации эритроцитов каждого из следующих растворов: отдельно ADSOL, раствор, содержащий 1% (масс./об.) глицерин и 1% (масс./об.) ДМСО (G+D), 2,5% (масс./об.) раствор p-CY01_LM и раствор, содержащий 1% (масс./об.) глицерин, 1% (масс./об.) ДМСО и 2,5% (масс./об.) p-CY01_LM (G+D+p-CY01_LM), измеряли на основе выраженного в процентах гемолиза эритроцитов.
В результате, как показано на Фиг. 12А, выраженный в процентах гемолиз эритроцитов в растворе, содержащем 1% (об./об.) глицерин и ДМСО, составлял 76,29±2,95%, который значительно не отличался от выраженного в процентах гемолиза в ФБР в качестве отрицательного контроля или ADSOL. Кроме того, раствор, содержащий только 2,5% (масс./об.) p-CY01_LM, демонстрировал значимо низкий уровень гемолиза эритроцитов 18,73±1,86%. Кроме того, раствор, содержащий 1% (масс./об.) глицерин, 1% (масс./об.) ДМСО и 2,5% (масс./об.) p-CY01_LM, демонстрировал выраженный в процентах гемолиз 9,43±2,16%, указывая на то, что данный раствор демонстрировал наилучший криозащитный эффект.
Применяя метод Плакетта-Бермана, рассчитывали значение главного эффекта каждого криозащитного компонента на криоконсервацию эритроцитов.
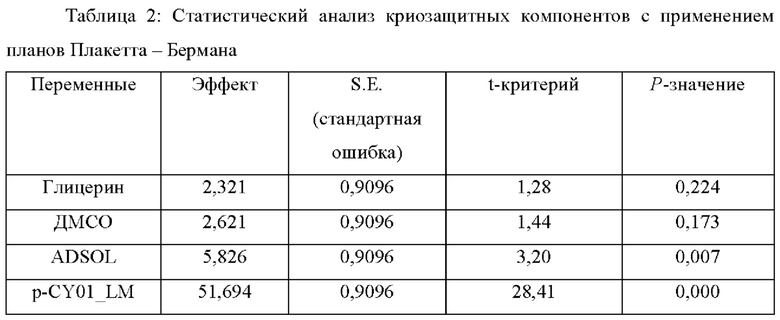
Как показано на Фиг. 12В и ниже в Таблице 2, 1% глицерин, 1% ДМСО, ADSOL и 2,5% (масс./об.) p-CY01_LM демонстрировали положительные значения главного эффекта на криоконсервацию эритроцитов. Значения главного эффекта составляли 2,32 для глицерина, 2,62 для ДМСО, 5,82 для ADSOL и 51,69 для p-CY01_LM, указывая на то, что даже несмотря на то, что концентрация p-CY01_LM, применяемая для криоконсервации эритроцитов, была лишь в 2,5 раза больше концентрации глицерина или ДМСО, главный эффект p-CY01_LM на криоконсервацию эритроцитов был по меньшей мере в 19 раз больше главного эффекта глицерина или ДМСО. Р-значения ADSOL и p-CY01_LM составляли 0,05 или ниже, что было значимо по сравнению с р-значением глицерина или ДМСО.
Пример 7: Анализ на основе дифференциальной сканирующей калориметрии (ДСК) р-CY01_LM
Проводили ДСК анализ раствора p-CY01_LM при охлаждении и оттаивании (Фиг. 13). Каждый образец охлаждали от +5°С до -78°С со скоростью 40°С/мин и подвергали оттаиванию со скоростью 2°С/мин.
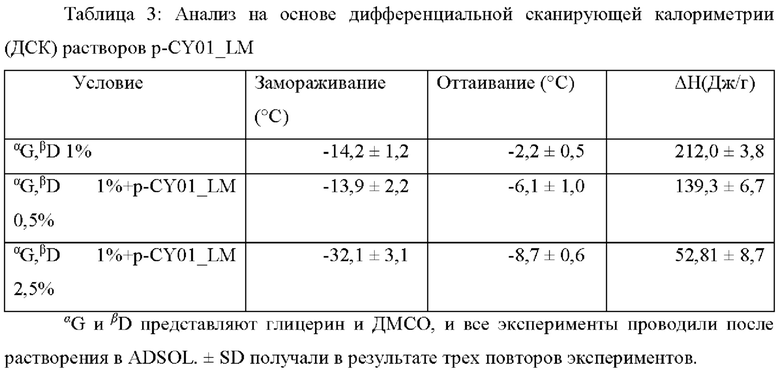
ДСК анализ проводили для раствора, содержащего 1% глицерин и 1% ДМСО (Фиг. 13А), и растворов, содержащих 1% глицерин, 1% ДМСО и от 0,5 до 2,5% p-CY01_LM (Фиг. 13В и 13С). Когда применяли раствор, содержащий низкие концентрации (меньше чем 1%) глицерина и ДМСО, который представляет собой отрицательный контроль, изменение в энтальпии (ΔН(Дж/г)) составляло 212,0±3,8 (Дж/г), которое было значимо низким подобно криозащитному механизму, который наблюдался при применении высоких концентраций (от 10 до 40%) глицерина и ДМСО в предшествующем уровне техники, что указывало на то, что раствор не способствовал уменьшению количества льда, образованного во время замерзания (Таблица 3). Температура переохлаждения раствора, содержащего 1% глицерин и 1% ДМСО, составляла -14,2°С, и температура переохлаждения раствора, содержащего 1% глицерин, 1% ДМСО и 0,5% p-CY01_LM, составляла -13,9°С, и температура переохлаждения раствора, содержащего 1% глицерин, 1% ДМСО и 2,5% p-CY01_LM, составляла -32,1°С. Ниже в Таблице 3 показаны изменения в энтальпии каждой опытной группы в зависимости от температур замораживания и оттаивания. Температуры замораживания и оттаивания в случае раствора, содержащего 1% глицерин, 1% ДМСО и 2,5% p-CY01_LM, были значительно снижены, и изменение в энтальпии ΔН(Дж/г) значимо уменьшалось до 52,81±8,7 (Дж/г) (Таблица 3). Данное явление означает, что общее количество воды, которое может образовывать лед во время циклов замораживания и оттаивания, уменьшено. Кроме того, данное явление представляет собой механизм предотвращения замерзания, обеспечиваемый p-CY01_LM.
Пример 8: Ингибирование образования льда раствора p-CY01_LM
Посредством анализа ингибирования роста кристаллов льда во время замораживания анализировали способность p-CY01_LM к предотвращению замерзания.
Как показано на Фиг. 14, рост зародышей кристаллов льда (Фиг. 14 В1) в растворе p-CY01_LM (ADSOL, содержащий 1% глицерин, ДМСО и 2,5% p-CY01_LM) ингибировался с уменьшением температур, и, таким образом, зародыши кристаллов льда росли в форме шестиконечных звезд (Фиг. 14 В2). Данное явление часто возникает в белках-антифризах (предотвращающих замерзание) и представляет собой механизм предотвращения замерзания, обеспечиваемый p-CY01_LM. Однако в контроле (ADSOL, содержащий 1% глицерин и ДМСО) и растворе ГЭК (ADSOL, содержащий 1% глицерин, ДМСО и 2,5% ГЭК) рост зародышей кристаллов льда не ингибировался, и, таким образом, зародыши кристаллов льда вырастали в округлые или плоские кристаллы льда (Фиг. 14 А1-А2 и С1-С2), указывая на то, что эффекта предотвращения замерзания за счет ингибирования роста кристаллов льда не наблюдали. Кроме того, температура переохлаждения воды в качестве отрицательного контроля составляет -15,9°С, тогда как температура переохлаждения раствора p-CY01_LM составляет -30,5°С, как показано выше в Таблице 3. Такие результаты указывают на то, что p-CY01_LM имеет эффект предотвращения замерзания.
Пример 9: Криомикроскопическое наблюдение в режиме реального времени за эритроцитами во время замораживания и оттаивания
Применяя анализ ингибирования перекристаллизации льда (ИПЛ), анализировали размер кристаллов льда во время оттаивания. В присутствии ингибитора перекристаллизации льда размер кристаллов льда, которые перекристаллизуются во время оттаивания, не будет значительно расти. Таким образом, в данном Примере, получали изображения эритроцитов и кристаллов льда, применяя криомикроскоп, во время замораживания и оттаивания эритроцитов. В частности, эритроциты охлаждали до -40°С со скоростью 25°С/мин в каждом из следующих растворов: раствор ADSOL, содержащий 2,5% p-CY01_LM, и раствор ADSOL, содержащий 2,5% (об./об.) ГЭК. Температуру образцов повышали до -6°С со скоростью 25°С/мин, и затем образцы фотографировали, пока им давали постоять в течение 5 минут.
На Фиг. 15 показаны изображения эритроцитов и кристаллов льда после перекристаллизации в присутствии или в отсутствии 2,5% p-CY01_LM. Было показано, что в присутствии 2,5% p-CY01_LM (Фиг. 15В) размер перекристаллизованных кристаллов льда был значимо меньше, чем в ФБР (Фиг. 15А) или 2,5% ГЭК-содержащем растворе (Фиг. 15С). В частности, после полного оттаивания эритроцитов наблюдали, что форма эритроцитов сохранялась интактной (Фиг. 15 А4, В4 и С4).
Такие результаты опять же подтверждали измерением активности ИПЛ р-CY01_LM. В частности, многоядерные пластины льда, имеющие диаметр меньше чем 10 мкм, вырастали при -6°С за 30 минут, и затем размер кристаллов льда сравнивали с размером кристаллов льда в контроле (ФБР).
Как показано на Фиг. 16, результаты измерения для ФБР (Фиг. 16А), 2,5% (масс./об.) p-CY01_LM (Фиг. 16В) и 2,5% (об./об.) ГЭК (Фиг. 16С) указывали на то, что раствор, содержащий p-CY01_LM, демонстрировал четко выраженную активность ИПЛ при одной и той же концентрации и, таким образом, ингибировал перекристаллизацию льда, а ГЭК не демонстрировал особого ингибирующего эффекта на перекристаллизацию льда.
Депонирование микроорганизмов
Учреждение-депозитарий: Корейский Научный Институт Бионауки и Биотехнологии
Учетный номер: KCTC 12867ВР
Дата депонирования: Июль 15, 2015 года.
Промышленная применимость
Экзополисахарид согласно настоящему изобретению обладает превосходной способностью к криозащите клеток во время криоконсервации и не демонстрирует цитотоксичности. Таким образом, экзополисахарид согласно настоящему изобретению может заменять общепринятые криозащитные агенты, которые демонстрируют цитотоксичность и требуют сложных способов оттаивания при применении в высоких концентрациях для криоконсервации эритроцитов. Соответственно, экзополисахарид согласно настоящему изобретению является эффективным для длительной криоконсервации крови.
Хотя настоящее изобретение и было подробно описано со ссылкой на конкретные признаки, специалистам в данной области техники будет понятно, что данное описание предназначено только для описания предпочтительного варианта реализации и не ограничивает объем настоящего изобретения. Таким образом, по существу объем настоящего изобретения будет определен прилагаемой формулой изобретения и ее эквивалентами.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ криоконсервации аутологичных вагинальных лактобацилл | 2022 |
|
RU2802074C1 |
| Способ сохранения бактерий в фекальной микробиоте и бактериальных культур, выращенных на плотных агаризованных питательных или дифференциальных средах в условиях низких температур с использованием составной среды для заморозки | 2019 |
|
RU2737321C1 |
| Способ определения криозащитных свойств консерванта для длительного хранения эритроцитов | 1989 |
|
SU1738198A1 |
| ШТАММ БАКТЕРИЙ PSEUDOALTEROMONAS SP. - ПРОДУЦЕНТ АЛЬФА-ГАЛАКТОЗИДАЗЫ | 1997 |
|
RU2113479C1 |
| Штамм гибридных культивируемых клеток животных мUS мUSсULUS - продуцент моноклональных антител против эритроцитарного антигена V F-системы крупного рогатого скота и родственных ему видов | 1989 |
|
SU1744107A1 |
| СПОСОБ ПОДБОРА КРИОПРОТЕКТОРОВ ДЛЯ КРИОКОНСЕРВАЦИИ БИОЛОГИЧЕСКИХ ОБРАЗЦОВ (ВАРИАНТЫ) | 2002 |
|
RU2236466C2 |
| СПОСОБ КРИОКОНСЕРВАЦИИ МУЛЬТИПОТЕНТНЫХ МЕЗЕНХИМАЛЬНЫХ СТРОМАЛЬНЫХ КЛЕТОК | 2010 |
|
RU2433173C1 |
| КОМБИНИРОВАННЫЙ КРИОПРОТЕКТОР "ДИМЕТИЛСУЛЬФОКСИД/РЕОПОЛИГЛЮКИН" ДЛЯ КРИОКОНСЕРВАЦИИ СТВОЛОВЫХ КЛЕТОК И СПОСОБ ИХ КРИОКОНСЕРВАЦИИ ДЛЯ КЛИНИЧЕСКОГО ПРИМЕНЕНИЯ | 2014 |
|
RU2563117C1 |
| КОНЪЮГАТЫ АНТИТЕЛ С ЛЕКАРСТВЕННЫМИ АГЕНТАМИ, ОБЛАДАЮЩИЕ УЛУЧШЕННОЙ СТАБИЛЬНОСТЬЮ, И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2670748C9 |
| СПОСОБ КРИОКОНСЕРВАЦИИ КЛЕТОК ФОТОТРОФНЫХ МИКРООРГАНИЗМОВ | 2012 |
|
RU2508397C1 |


Изобретение относится к экзополисахариду, имеющему криозащитные свойства. Предложен экзополисахарид, имеющий криозащитные свойства, который продуцирован штаммом Pseudoalteromonas sp. CY01, депонированным под номером KCTC 12867BP. Указанный экзополисахарид состоит из глюкозы и галактозы, при этом молярное соотношение глюкозы и галактозы составляет 3,4:1. Путем гидролиза указанного экзополисахарида получают экзополисахарид, имеющий криозащитные свойства, имеющий молекулярную массу от 1,0 x 105 до 4,3 x 105 Да, который состоит из глюкозы и галактозы. Также предложены способы получения указанных экзополисахаридов, композиция для криозащиты по меньшей мере одной клетки, содержащая эффективное количество одного из указанных экзополисахаридов, применение указанных экзополисахаридов для криоконсервации клеток. Экзополисахарид согласно настоящему изобретению проявляет превосходные криозащитные свойства при относительно низкой концентрации по сравнению с другими криозащитными агентами, а также не проявляет цитотоксичность. 7 н.п. ф-лы, 16 ил., 3 табл., 9 пр.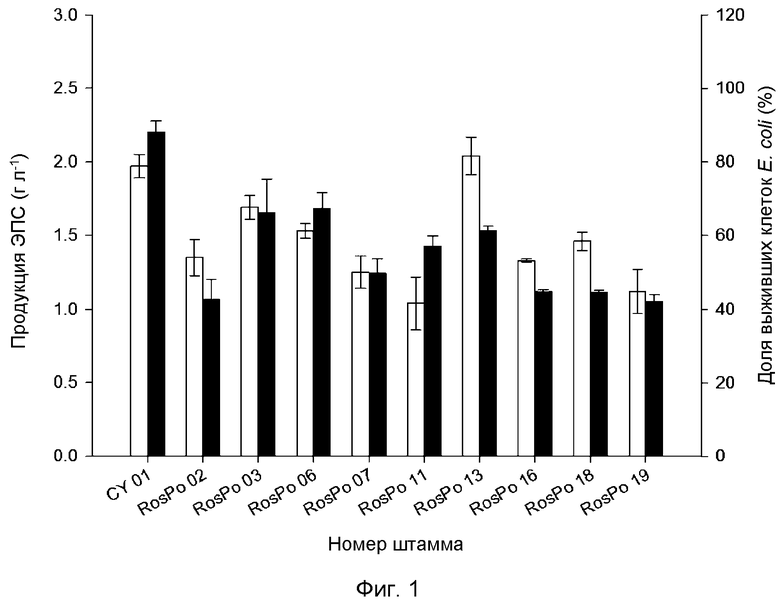
1. Экзополисахарид, имеющий криозащитные свойства, который продуцирован штаммом Pseudoalteromonas sp. CY01, имеющим учетный номер KCTC 12867BP, и который состоит из глюкозы и галактозы, при этом молярное соотношение глюкозы и галактозы составляет 3,4:1.
2. Экзополисахарид, имеющий криозащитные свойства, имеющий молекулярную массу от 1,0 x 105 до 4,3 x 105 Да, который состоит из глюкозы и галактозы и получен посредством гидролиза экзополисахарида по п. 1.
3. Композиция для криозащиты по меньшей мере одной клетки, при этом указанная композиция содержит:
эффективное количество экзополисахарида по п. 1 или 2 и глицерин или ДМСО,
при этом указанная по меньшей мере одна клетка выбрана из группы, состоящей из клеток бактерий, клеток грибов, клеток животных, клеток растений, эритроцитов, тромбоцитов, сперматоцитов, ооцитов, клеток, составляющих ткани, и клеток, составляющих органы, и
при этом указанная по меньшей мере одна клетка не является клеткой штамма Pseudoalteromonas sp. CY01, который имеет учетный номер KCTC 12867BP.
4. Способ получения экзополисахарида по п. 1, включающий следующие стадии:
(а) культивирование штамма Pseudoalteromonas sp. CY01, который имеет учетный номер KCTC 12867BP, с получением указанного экзополисахарида; и
(b) выделение полученного экзополисахарида.
5. Способ получения экзополисахарида, имеющего молекулярную массу от 1,0 x 105 до 4,3 x 105 Да по п. 2, включающий стадию гидролиза экзополисахарида по п.1.
6. Применение экзополисахарида по п. 1 или 2 для криоконсервации клеток.
7. Штамм Pseudoalteromonas sp. CY01, который имеет учетный номер KCTC 12867BP, который обладает способностью продуцировать экзополисахарид слизи по п. 1.
| NICHOLS C.M | |||
| ET AL | |||
| Chemical Characterization of Exopolysaccharides from Antarctic Marine Bacteria | |||
| Способ смешанной растительной и животной проклейки бумаги | 1922 |
|
SU49A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| KIM S.J | |||
| ET AL | |||
| Cryoprotective properties and preliminary | |||

