Перекрестная ссылка на родственные заявки
Настоящая заявка заявляет приоритет по датам более ранней подачи предварительных заявок на патент США № 62/430838, поданной 6 декабря 2016 г., и 62/479087, поданной 30 марта 2017 г., каждая из которых включена здесь посредством ссылки в полном объеме.
Область техники, к которой относится изобретение
Настоящее изобретение относится к биосинтезу антибиотиков, в частности, к композициям и способам для повышенной продукции эндурацидина.
Уровень техники
Глобальное появление бактериальных инфекций с множественной лекарственной устойчивостью привело к огромным затратам в здравоохранении и стало серьезной угрозой для здравоохранения. Для того чтобы опережать развитие резистентности к антибактериальным лекарственным средствам, необходимо проводить поиск новых антибиотиков, а также способов продукции таких антибиотиков более экономически эффективным образом.
Сущность изобретения
Настоящее изобретение преодолевает проблемы, связанные с ограниченной продукцией эндурацидина (энрамицина) штаммами дикого типа Streptomyces fungicidicus, а также ограничениями продукции промышленными штаммами, разработанными с использованием обычного радиационного и химического мутагенеза хромосомы и последовательных многочисленных раундов селекции мутантов для продукции повышенных уровней требуемых пептидных антибиотиков, в частности, эндурацидина. Настоящее изобретение раскрывает генетическое манипулирование регуляторными генами orf24 и orf18, связанными с кластером генов биосинтеза эндурацидина (энрамицина) из Streptomyces fungicidicus, для получения рекомбинантных векторов и штаммов, продуцирующих данный пептидный антибиотик с большими выходами. Рекомбинантные штаммы были сконструированы на основе продуцента дикого типа, Streptomyces fungicidicus B-5477 (ATCC 21013) и Streptomyces fungicidicus BM38-2 (ATCC PTA-122342), который был получен из штамма дикого типа и в настоящее время используется для промышленного получения эндурацидина. В микроорганизме дикого типа сайт-специфическая интеграция плазмиды pXY152-endorf24, которая направляет сверхэкспрессию второй копии orf24, обеспечила получение штамма SfpXY152endorf24. Интеграция мутагенизированной фосмиды pXYF24D3 в хромосому штамма дикого типа заменила нативный orf18 «разрушенной» копией гена и создала мутант SfpXYF24D3. Используя коммерческий продуцент Streptomyces fungicidicus BM38-2 (ATCC PTA-122342), интеграция плазмиды pXY152-endorf24 позволила получить рекомбинантный штамм Streptomyces fungicidicus BM38-2.24/16. Для создания штамма, происходящего из BM38-2 (ATCC PTA-122342), с отсутствием функционального orf18, была сконструирована плазмида pKS-T-orf18pfrd-AmR для делеции orf18 и его фланкирующих областей, заменив эту область маркером устойчивости к апрамицину и получив рекомбинантный штамм Streptomyces fungicidicus BM38-2.18pfrd-AmR. Было показано, что штаммы, подвергшиеся генетическому манипулированию, дают выходы эндурацидина выше в диапазоне от 1,2 до 4,6 раз, чем у соответствующих родительских штаммов. Повышенные выходы эндурацидина из рекомбинантных штаммов обеспечивают более рентабельное получение эндурацидина.
Вышеизложенные и другие признаки и преимущества изобретения станут более очевидными из последующего подробного описания, которое далее описывается со ссылкой на прилагаемые фигуры.
Краткое описание фигур
На фиг. 1 представлена химическая структура эндурацидинов А и В.
На фиг. 2 представлена карта интегративной экспрессионной плазмиды pXY152-endorf24.
На фиг. 3 представлена карта плазмиды с делецией гена pXY300-orf18ifd.
На фиг. 4 представлена карта плазмиды с делецией гена pKS-T-orf18ifd.
На фиг. 5 представлена карта плазмиды с делецией гена pKS-T-orf18pfrd-AmR.
На фиг.6 представлена карта плазмиды с делецией гена pKS-orf18ifd-T-AmR (NS).
На фиг. 7 представлена карта интегративной экспрессионной плазмиды pXY152-endorf24-camtsr.
На фиг. 8 представлена карта интегративной экспрессионной плазмиды pXY152-endorf24-blatsr.
На фиг. 9 показано выравнивание белка-активатора биосинтеза стрептомицина StrR (SEQ ID NO: 25) с Orf24 (SEQ ID NO: 26).
На фиг. 10А и 10В представлены карты вставок плазмид pKS-T-orf18pfrd-AmR (a) и pXY300-orf18ifd (b). В конструкции pXY30-orf18ifd внутренняя последовательность orf18 из нуклеотидного положения с 25795 по 26450 (идентификационный номер GenBank DQ403252) была делетирована и заменена сайтом рестрикции PacI (TTAATTAA, фиг.10B). Полученный делецией внутри рамки считывания orf18 (GTGTTTAATTAATGA (SEQ ID NO: 27)) может транслироваться в пептид из трех аминокислот (VFN). Как правило, любая делеция внутри рамки считывания по длине orf18 должна приводить к инактивированной функции Orf18 за счет его неполноты.
Фиг. 11. Выравнивание Orf24 с шестью функционально охарактеризованными ортологами StrR-подобных специфических для пути белков-активаторов из актиномицетов. Orf24 (идентификационный номер GenBank № DQ403252; SEQ ID NO: 26) из кластера генов биосинтеза эндурацидина S. fungicidicus; StrR (идентификационный номер GenBank Y00459; SEQ ID NO: 25) из кластера генов биосинтеза стрептомицина из S. griseus streptomycin; Tei15* (идентификационный номер GenBank AJ632270; SEQ ID NO: 32) из кластера генов биосинтеза тейкопланина из Actinoplanes teichomyceticus; Bbr (идентификационный номер GenBank Y16952; SEQ ID NO: 28) из кластера генов биосинтеза балгимицина из штамма Amycolatopsis DSM 5908; KasT (идентификационный номер GenBank BAF79690; SEQ ID NO: 29) из кластера генов биосинтеза казугамицина из S. kasugaensis; NovG (идентификационный номер GenBank AF170880; SEQ ID NO: 30) из кластера генов биосинтеза новобиоцина из штамма S. niveus NCIMB 9219; SgcR1 (идентификационный номер GenBank AY048670; SEQ ID NO: 31) из кластера генов биосинтеза C-1027 из S. globisporus. Идентичные аминокислоты(*), консервативные аминокислоты(.) и высококонсервативные аминокислотные замены(:). Подчеркнуты консервативные мотивы спираль-поворот-спираль (HTH), характерные для ДНК-связывающих белков, таких как StrR.
Фиг. 12. Выравнивание Orf18 (SEQ ID NO: 36) с другими функционально охарактеризованными ортологами регулятора ответа. SCO1745/AbrA2: двухкомпонентный регулятор ответа S. coelicolor A3(2) (идентификационный номер GenBank CAB50960; SEQ ID NO: 33). SCO3226/AbsA2: двухкомпонентный регулятор ответа S. coelicolor A3(2) (идентификационный номер GenBank AAB08053; SEQ ID NO: 34). SCO3818: двухкомпонентный транскрипционный регулятор ответа S. coelicolor A3(2) (идентификационный номер GenBank CAB46941; SEQ ID NO: 35).
Список последовательностей
Последовательности нуклеиновых кислот и аминокислотные последовательности, приведенные здесь и в прилагаемом списке последовательностей, показаны с использованием стандартных буквенных сокращений для нуклеотидных оснований и трехбуквенного кода для аминокислот, как определено в 37 C.F.R. 1.822. Показана только одна цепь последовательности нуклеиновой кислоты, но комплементарная цепь понимается как включенная посредством ссылки на приведенную цепь. В прилагаемом списке последовательностей:
SEQ ID NO: 1 и 2 представляют олигонуклеотидные праймеры, использованные для получения вставки плазмиды pXY152-endorf24.
SEQ ID NO: 3 представляет последовательность нуклеиновой кислоты плазмиды pXY152-endorf24.
SEQ ID NO: 4-7 представляют олигонуклеотидные праймеры, использованные для получения вставки плазмиды pXY300-orf18ifd.
SEQ ID NO: 8 представляет последовательность нуклеиновой кислоты плазмиды pXY300-orf18ifd.
SEQ ID NO: 9 и 10 представляют олигонуклеотидные праймеры, использованные для получения фрагмента oriT плазмиды pKS-T-orf18pfrd.
SEQ ID NO: 11 представляет последовательность нуклеиновой кислоты плазмиды pKS-T-orf18pfrd.
SEQ ID NO: 12 и 13 представляют олигонуклеотидные праймеры, использованные для получения фрагмента amR плазмиды pKS-T-orf18pfrd-AmR.
SEQ ID NO: 14 представляет последовательность нуклеиновой кислоты плазмиды pKS-T-orf18pfrd-AmR.
SEQ ID NO: 15-18 представляют олигонуклеотидные праймеры, использованные для получения фрагментов oriT и amR плазмиды pKS-orf18ifd-T-AmR (NS).
SEQ ID NO: 19 представляет последовательность нуклеиновой кислоты плазмиды pKS-orf18ifd-T-AmR (NS).
SEQ ID NO: 20 представляет последовательность нуклеиновой кислоты плазмиды pXY152-endorf24-camtsr.
SEQ ID NO: 21 и 22 представляют олигонуклеотидные праймеры, использованные для получения фрагмента bla плазмиды pXY152-endorf24-blatsr.
SEQ ID NO: 23 представляет последовательность нуклеиновой кислоты плазмиды pXY152-endorf24-blatsr.
SEQ ID NO: 24 представляет олигонуклеотидный праймер, который соответствует области гена устойчивости к апрамицину.
SEQ ID NO: 25 представляет аминокислотную последовательность белка-активатора биосинтеза стрептомицина StrR.
SEQ ID NO: 26 представляет аминокислотную последовательность, кодируемую ORF24.
SEQ ID NO: 27 представляет последовательность нуклеиновой кислоты, показывающую делецию внутри рамки считывания в orf18.
SEQ ID NO: 28 является аминокислотной последовательностью вставки Bbr.
SEQ ID NO: 29 является аминокислотной последовательностью вставки KasT.
SEQ ID NO: 30 является аминокислотной последовательностью вставки NovG.
SEQ ID NO: 31 является аминокислотной последовательностью вставки SgcR1.
SEQ ID NO: 32 является аминокислотной последовательностью вставки Teil5*.
SEQ ID NO: 33 является аминокислотной последовательностью ортолога регулятора ответа SCO1745/AbrA2 из S. coelicolor A3(2) (идентификационный номер GenBank CAB50960).
SEQ ID NO: 34 представляет аминокислотную последовательность ортолога регулятора ответа SCO/3226/AbsA2 из S. coelicolor A3(2) (идентификационный номер GenBank AAB08053).
SEQ ID NO: 35 представляет аминокислотную последовательность ортолога регулятора ответа SCO3818 из S. coelicolor A3(2) (идентификационный номер GenBank CAB46941).
SEQ ID NO: 36 является аминокислотной последовательностью, кодируемой ORF18.
SEQ ID NO: 37 представляет последовательность нуклеиновой кислоты orf18.
SEQ ID NO: 38 представляет последовательность нуклеиновой кислоты orf24.
SEQ ID NO: 39 представляет последовательность нуклеиновой кислоты фосмиды pXYF148 с orf24, расположенным в нуклеотидных положениях с 23109 по 24044.
SEQ ID NO: 40 представляет последовательность нуклеиновой кислоты фосмиды pXYF24 с orf18, расположенным в нуклеотидных положениях с 31091 по 31753.
Подробное описание нескольких вариантов осуществления изобретения
I. Введение
Эндурацидин (фиг. 1), также называемый энрамицином, представляет собой липодепсипептидный антибиотик из 17 аминокислот, продуцируемый почвенной бактерией S. fungicidicus B-5477 (ATCC 21013). Пептид выделен из ферментационного бульона и мицелия в основном в виде смеси эндурацидинов А и В, которые различаются на один атом углерода в длине присоединенной липидной цепи. Структурно эндурацидины различаются по группе разветвленной жирной кислоты C12 или C13 2Z, 4E, присоединенной амидной связью к остатку аспарагиновой кислоты, и наличию многочисленных непротеиногенных аминокислотных остатков, таких как эндурацидидин (End), 4-гидроксифенилглицин (Hpg), 3,5-дихлор-4-гидроксифенилглицин (Dpg), цитруллин (Cit) и орнитин (Orn) (см. фиг. 1). Семь из 17 аминокислот имеют D-конфигурацию, и шесть из остатков представляют Hpg или хлорированное производное Dpg.
Эндурацидин (для упрощения, пептиды будут упоминаться в единственном числе) проявляет высокую антибактериальную активность in vitro и in vivo в отношении широкого ряда грамположительных микроорганизмов, включая метициллин-резистентные Staphylococcus aureus (MRSA) и ванкомицин-резистентные Enterococcus (VRE). Минимальные ингибирующие концентрации (MIC) составляют всего лишь 0,05 мкг/мл, и эффект является бактерицидным. На основании результатов исследования с включением 100 штаммов S. aureus, отобранных из различных патологических продуктов, и включающих 40% MRSA, были установлены значения MIC в диапазоне от 0,09 до 0,56 мкг/мл при отсутствии штамма, способного выдержать воздействие MIC на уровне 1 мкг/мл. Для сравнения, типичные значения MIC для ванкомицина в отношении чувствительных штаммов S. aureus находятся в диапазоне от 0,5 до 2 мкг/мл. Кроме того, эндурацидин обладает хорошим токсикологическим профилем. В исследовании на мышах, кроликах, собаках и обезьянах значения LD50 по результатам оценки острой токсичности составляли: при внутривенном введении 30-125 мг/кг; внутрибрюшинном введении 750-910 мг/кг; подкожном, внутримышечном (в/м) или пероральном введении > 5-10 г/кг. В том же исследовании у обезьян, получавших эндурацидин в/м в течение 6 месяцев и у крыс, которым аналогично препарат вводили в течение 12 месяцев, было обнаружено только локализованное воспаление в месте инъекции. Эндурацидин вводили людям, 20 госпитализированным взрослым пациентам, инфицированным MRSA, внутримышечно (в дозе 100 мг каждые 12 ч). Сообщалось, что пептид не проявлял побочных эффектов, и также был высокоэффективен в лечении инфекций мочевыводящих путей и кожи, вызванных MRSA, но не хронических инфекций костной системы (Peromet et al., Chemotherapy, 19: 53-61, 1973).
Эндурацидин ингибирует биосинтез пептидогликана в клеточной стенке бактерий, посредством образования комплекса с внеклеточным липидом II, предшественником структуры бактериальной клеточной стенки. Сайт комплексообразования с липидом II отличается от такового, который распознается ванкомицином, что объясняет действие эндурацидина против микроорганизмов, устойчивых к ванкомицину. На сегодняшний день отсутствуют документированные данные о наличии перекрестной резистентности эндурацидина с каким-либо используемым в клинике антибиотиком, и отсутствуют доказательства наличия развившейся, приобретенной или передаваемой резистентности. Отсутствие какого-либо известного механизма передаваемой резистентности, отсутствие пероральной биодоступности, его низкая токсичность и высокая активность в отношении Clostridium spp. сделали эндурацидин ключевым коммерческим пептидным антибиотиком, используемым в качестве кормовой добавки в птицеводстве для борьбы с клостридиальным некротическим энтеритом.
Для получения штамма продуцирующего микроорганизма, который мог бы обеспечить количества пептидов, необходимые для промышленного применения, Japan Takeda Animal Health (ныне подразделение Intervet/Merck Animal Health) S. fungicidicus B-5477 подвергли различным традиционным способам улучшения штамма и отобрали мутантов, которые продуцировали более высокие выходы эндурацидина. Растущий мировой рынок эндурацидина стимулирует усилия для дальнейшего повышения выхода этого антибиотика в BM38-2 (ATCC PTA-122342). С использованием доступной генетической последовательности кластера генов биосинтеза эндурацидина (идентификационный номер GenBank DQ403252, который включен в настоящее описание посредством ссылки, доступной в интернете 3 октября 2006 г.), BM38-2 (ATCC PTA-122342)) служил в качестве исходного штамма для направленной генетической манипуляции с регуляторными генами, связанными с кластером генов, и составил основу настоящего изобретения. Настоящее изобретение раскрывает, что продукт гена orf18 оказывает негативное влияние на продукцию эндурацидина, и продукт гена orf24 оказывает позитивное влияние на продукцию эндурацидина, и что рекомбинантные штаммы, полученные из микроорганизмов S. fungicidicus дикого типа и из BM38-2 (ATCC PTA-122342), которые используют эти регуляторные эффекты, приводят к повышенным выходам эндурацидина. Кроме того, в настоящем документе раскрываются новые замены генов и интегративные экспрессионные векторы на основе на pBluescript II KS и pSET152 соответственно.
II. Сокращения и термины
а. Сокращения
b. Термины
Если не указано иное, то технические термины используются в соответствии с их обычным использованием. Определения общих терминов в молекулярной биологии можно найти в монографии Benjamin Lewin Genes V, опубликованной Oxford University Press, 1994 (ISBN 0-19-854287-9); Kendrew et al. (eds.) The Encyclopedia of Molecular Biology, опубликованной Blackwell Science Ltd., 1994 (ISBN 0-632-02182-9); и монографии Robert A. Meyers (ed.) Molecular Biology and Biotechnology: a Comprehensive Desk Reference, опубликованной VCH Publishers, Inc., 1995 (ISBN 1-56081-569-8).
Для облегчения рассмотрения различных вариантов осуществления настоящего раскрытия, предоставляются следующие пояснения конкретных терминов:
Введение: введение животному любым путем. Как здесь используется, введение обычно относится к пероральному введению.
Аллельный вариант: альтернативная форма полипептида, которая характеризуется заменой, делецией или добавлением одной или более аминокислот. В одном примере вариант не изменяет биологическую функцию полипептида.
Амплификация: когда используется по отношению к нуклеиновой кислоте, то она относится к методике, с помощью которой увеличивают количество копий молекулы нуклеиновой кислоты в образце или препарате. Примером амплификации является полимеразная цепная реакция, в которой биологический образец, отобранный у субъекта, приводят в контакт с парой олигонуклеотидных праймеров в условиях, которые обеспечивают гибридизацию этих праймеров c матрицей нуклеиновой кислоты в образце. Праймеры удлиняются в подходящих условиях, диссоциируют от матрицы, затем снова отжигаются, удлиняются и диссоциируют с увеличением количества копий нуклеиновой кислоты. Продукт амплификации in vitro можно характеризовать посредством электрофореза, профилей расщепления рестрикционными эндонуклеазами, гибридизации или лигирования олигонуклеотидов и/или секвенирования нуклеиновой кислоты стандартными методами. Другие примеры методов амплификации in vitro включают амплификацию с замещением цепей (см. патент США № 5744311); изотермическую амплификацию без транскрипции (см. патент США № 6033881); амплификацию посредством реакции репарации цепи (см. WO 90/01069); амплификацию посредством лигазной цепной реакции (см. EP-A-320308); амплификацию посредством лигазной цепной реакции с заполнением разрыва (см. патент США № 5427930); объединенные лигазную детекцию и ПЦР (см. патент США № 6027889) и амплификацию без транскрипции РНК NASBA™ (см. патент США № 6025134).
Аналог, производное или миметик: аналог представляет молекулу, которая отличается по химической структуре от исходного соединения, например, гомолога (отличающегося увеличением химической структуры, например, разницей в длине алкильной цепи), молекулярный фрагмент, структура, которая отличается одной или более функциональными группами и/или изменением ионизации. Структурные аналоги часто можно найти, используя количественные взаимосвязи между структурой и активностью (QSAR), с помощью методов, подобных тем, которые описаны в Remington (The Science and Practice of Pharmacology, 19th Edition (1995), глава 28). Когда изменения в исходном соединении являются существенными или объединяются многие дополнительные изменения, то соединение больше не является аналогом. Например, рамопланин не рассматривается здесь в качестве аналога эндурацидина: рамопланин не содержит ни аминокислоты эндурацидидина, а содержит другие аминокислоты, и хотя он имеет липидную боковую цепь, длина цепи существенно короче. Аналоги эндурацидина могут быть получены добавлением или удалением функциональных групп в аминокислотах, которые составляют липодепсипептиды, заменой одной аминокислоты на другую (за исключением аминокислоты эндурацидидин) или комбинацией модификации функциональной группы и замены аминокислоты. Типичные аналоги эндурацидина включают тетрагидроэндурацидин A, тетрагидроэндурацидин B, дехлорэндурацидин A и дехлорэндурацидин B.
Производное представляет биологически активную молекулу, полученную из основной структуры. Миметик представляет молекулу, которая имитирует активность другой молекулы, имитируя структуру такой молекулы, в качестве биологически активной молекулы. Таким образом, термин «миметик» указывает на определенную структуру, связанную с активностью.
Антибиотик: соединение, например, эндурацидин, пенициллин или стрептомицин, обычно продуцированное или полученное из некоторых грибов, бактерий и других микроорганизмов, которое может уничтожать или ингибировать рост других микроорганизмов.
Антисмысловая молекула, смысловая молекула и антиген: двухцепочечная ДНК (дцДНК) имеет две цепи: цепь 5' → 3', обозначаемая как плюс-цепь, и цепь 3' → 5' (обратная комплементарная цепь), обозначаемая как минус-цепь. Поскольку РНК-полимераза добавляет нуклеотиды в направлении 5' → 3', то минус-цепь ДНК служит матрицей для РНК во время транскрипции. Таким образом, образованная РНК будет иметь последовательность, комплементарную минус-цепи и идентичную плюс-цепи (за исключением того, что U заменяет Т). Антисмысловые молекулы представляют собой молекулы, которые специфически гибридизуются или специфически комплементарны РНК или плюс-цепи ДНК. Смысловые молекулы представляют молекулы, которые специфически гибридизуются или специфически комплементарны минус-цепи ДНК. Антигенные молекулы представляют антисмысловые или смысловые молекулы, комплементарные дцДНК-мишени. В одном варианте осуществления антисмысловая молекула специфически гибридизуется с мРНК-мишенью и ингибирует транскрипцию мРНК-мишени.
Связывание или стабильное связывание: молекула, такая как олигонуклеотид или белок, связывается или стабильно связывается с молекулой-мишенью, такой как нуклеиновая кислота-мишень или белок-мишень, если связывание можно детектировать. В одном примере олигонуклеотид связывается или стабильно связывается с нуклеиновой кислотой-мишенью, если достаточное количество олигонуклеотида образует пары оснований или гибридизуется с нуклеиновой кислотой-мишенью, чтобы можно было детектировать это связывание. Связывание может детектироваться по физическим или функциональным свойствам комплекса мишень:олигонуклеотид. Связывание между мишенью и олигонуклеотидом может быть детектировано с использованием любой процедуры, известной специалисту в данной области, включая функциональные и физические анализы связывания. Связывание можно детектировать функционально посредством определения того, оказывает ли связывание наблюдаемый эффект на процесс биосинтеза, такой как экспрессия гена, репликация ДНК, транскрипция, трансляция и тому подобное.
Физические методы детектирования связывания комплементарных цепей ДНК или РНК хорошо известны в данной области и включают такие методы, в которых используется ДНКаза I или химический футпринтинг, анализы сдвига геля и аффинного расщепления, нозерн-блоттинг, дот-блоттинг и детектирование по поглощению света. Например, один метод, который широко используется за счет того, что он очень прост и надежен, включает мониторинг изменения поглощения света раствором, содержащим олигонуклеотид (или аналог) и нуклеиновую кислоту-мишень при 220-300 нм, по мере медленного повышения температуры. Если олигонуклеотид или аналог связан с его мишенью, то происходит внезапное увеличение поглощения при определенной температуре, когда олигонуклеотид (или аналог) и мишень диссоциируют друг от друга или плавятся.
Связывание между олигомером и его нуклеиновой кислотой-мишенью часто характеризуют определением температуры (Tm), при которой 50% олигомера плавится с его отделением от мишени. Более высокое значение Tm означает наличие более сильного или более стабильного комплекса по сравнению с комплексом с более низким значением Tm.
Связывание между белком и его белком-мишенью, таким как антитело к антигену, часто характеризуют определением аффинности связывания. В одном варианте осуществления аффинность рассчитывают с использованием метода Скэтчарда в модификации, описанного Frankel et al., Mol. Immunol., 16: 101-106, 1979. В еще одном варианте осуществления аффинность связывания измеряют по скорости диссоциации рецептора и специфического связывающего агента. В еще одном варианте осуществления высокую аффинность связывания измеряют с помощью конкурентного радиоиммуноанализа. В нескольких примерах высокая аффинность связывания представляет, по меньшей мере, примерно 1×10-8 М. В еще одних вариантах осуществления высокая аффинность связывания представляет, по меньшей мере, примерно 1,5×10-8, по меньшей мере, примерно 2,0×10-8, по меньшей мере, примерно 2,5×10-8, по меньшей мере, примерно 3,0×10-8, по меньшей мере, примерно 3,5×10-8, по меньшей мере, примерно 4,0×10-8, по меньшей мере, примерно 4,5×10-8, или, по меньшей мере, примерно 5,0×10-8 М.
Биологическая функция: функция(и) полипептида в клетках, в которых он встречается в природе. Полипептид может обладать более одной биологической функцией.
кДНК (комплементарная ДНК): фрагмент ДНК, в котором отсутствуют внутренние некодирующие участки (интроны) и последовательности регуляции транскрипции. Также кДНК может содержать нетранслируемые области (UTR), которые в соответствующей молекуле РНК отвечают за контроль трансляции. кДНК синтезируют в лаборатории посредством обратной транскрипции с информационной РНК, выделенной из клеток.
Консервативная замена: аминокислотные замены, которые существенно не изменяют активность (специфичность или аффинность связывания) молекулы. Обычно консервативные аминокислотные замены включают замены одной аминокислоты другой аминокислотой со сходными химическими свойствами (например, зарядом или гидрофобностью). В следующей таблице приведены примеры консервативных аминокислотных замен:
Контрольный штамм Streptomyces fungicidicus: встречающийся в природе штамм дикого типа, Streptomyces fungicidicus ATCC21013.
ДНК (дезоксирибонуклеиновая кислота): длинноцепочечный полимер, который содержит генетический материал большинства живых организмов (некоторые вирусы имеют гены, содержащие рибонуклеиновую кислоту (РНК)). Повторяющиеся звенья в ДНК-полимерах представляют собой четыре различных нуклеотида, каждый из которых содержит одно из четырех оснований: аденин, гуанин, цитозин и тимин, связанные с сахаром дезоксирибозой, к которому присоединена фосфатная группа. Триплеты нуклеотидов (называемые кодонами) кодируют каждую аминокислоту в полипептиде. Термин «кодон» также используется для соответствующих (и комплементарных) последовательностей из трех нуклеотидов в иРНК, в которую транскрибируется последовательность ДНК.
Если не указано иное, то любая ссылка на молекулу ДНК предназначена для включения молекулы, обратно комплементарной данной молекуле ДНК. За исключением случаев, когда одноцепочечность требуется по тексту настоящего документа, молекулы ДНК, несмотря на то, что записаны только в виде одной цепи, включают обе цепи двухцепочечной молекулы ДНК. Таким образом, ссылка на молекулу нуклеиновой кислоты, которая кодирует определенный белок или его фрагмент, охватывает как смысловую цепь, так и ее обратную комплементарную цепь. Таким образом, например, целесообразно генерировать зонды или праймеры из обратной комплементарной последовательности раскрытых молекул нуклеиновой кислоты.
Домен: участок молекулы, такой как белки или нуклеиновые кислоты, который структурно и/или функционально отличается от другой части молекулы.
Кодировать: говорят, что полинуклеотид кодирует полипептид, если в своем нативном состоянии или при манипуляции способами, известным специалистам в данной области, молекула полинуклеотида может транскрибироваться и/или транслироваться с получением иРНК для полипептида или его фрагмента. Антисмысловая цепь является комплементарной такой нуклеиновой кислоте, и кодирующую последовательность можно вывести из нее.
Эндурацидин: эндурацидины A и B представляют липодепсипептиды из 17 аминокислот, обнаруженные в конце 1960-х годов при ферментации почвенной бактерии Streptomyces fungicidicus B-5477 (ATCC 21013). Пептиды A и B представляют гомологи, которые различаются на один атом углерода длины присоединенной липидной цепи. Структурно эндурацидины различаются по группе разветвленной жирной кислоты C12 или C13 2Z, 4E и наличию многочисленных непротеиногенных аминокислотных остатков, таких как эндурацидидин (End), 4-гидроксифенилглицин (Hpg), 3,5-дихлор-4-гидроксифенилглицин (Dpg), цитруллин (Cit) и орнитин (Orn). Семь из 17 аминокислот имеют D-конфигурацию, и шесть остатков являются Hpg или хлорированным аналогом Dpg.
Функциональные фрагменты и варианты полипептида: включают такие фрагменты и варианты, которые сохраняют одну или более функций родительского полипептида. Признано, что ген или кДНК, кодирующие полипептид, могут быть в значительно степени мутированы без существенного изменения одной или более функций полипептида. Во-первых, генетический код является вырожденным, и таким образом, разные кодоны кодируют одни и те же аминокислоты. Во-вторых, даже при введении аминокислотной замены мутация может быть консервативной и не оказывать существенного влияния на основную функцию(и) белка. См. Stryer, Biochemistry, 3rd Ed., (с) 1988. В-третьих, участок полипептидной цепи может быть делецирован без нарушения или элиминации всех ее функций. В-четвертых, в полипептидной цепи могут быть сделаны инсерции или добавления, например, с добавлением эпитопных меток, без нарушения или элиминации ее функций (Ausubel et al. J. Immunol., 159 (5): 2502-12, 1997). Другие модификации, которые могут быть сделаны без существенного нарушения одной или более функций полипептида, включают, например, химические и биохимические модификации in vivo или in vitro или включение необычных аминокислот. Такие модификации включают, например, ацетилирование, карбоксилирование, фосфорилирование, гликозилирование, убиквитинирование, мечение, например, радионуклеидами, и различные ферментативные модификации, что будет легко понятно специалистам в данной области. Разнообразные способы мечения полипептидов и метки, пригодные для таких целей, включают радиоактивные изотопы, такие как 32P, лиганды, которые связываются или связаны с мечеными партнерами по специфическому связыванию (например, антителами), флуорофоры, хемилюминесцентные агенты, ферменты и антилиганды. Функциональные фрагменты и варианты могут быть различной длины. Например, некоторые фрагменты имеют, по меньшей мере, 10, 25, 50, 75, 100, 200 или даже больше аминокислотных остатков.
Эффективное количество: количество или концентрация указанного соединения или композиции, достаточные для достижения желаемого эффекта у субъекта. Эффективное количество может зависеть, по меньшей мере, частично от вида животного, подвергаемого лечению, размера животного и/или природы желаемого эффекта.
Кластер генов: группа генетических элементов, сгруппированных вместе в хромосоме, белковые продукты которых имеют связанную функцию, такую как формирование пути биосинтеза природного продукта.
Гетерологичный: когда термин относится к последовательностям нуклеиновых кислот, таким как кодирующие последовательности и регуляторные последовательности, то «гетерологичный» обозначает последовательности, которые обычно не связаны с областью рекомбинантной конструкции и/или обычно не связаны с конкретной клеткой. Таким образом, «гетерологичная» область нуклеиновокислотной конструкции представляет собой определяемый сегмент нуклеиновой кислоты внутри или присоединенный к другой молекуле нуклеиновой кислоты, которая не встречается в ассоциации с другой молекулой в природе. Например, гетерологичная область конструкции может включать кодирующую последовательность, фланкированную последовательностями, не встречающими в природе в ассоциации с кодирующей последовательностью. Другим примером гетерологичной кодирующей последовательности является конструкция, в которой сама кодирующая последовательность не встречается в природе (например, синтетические последовательности, имеющие кодоны, отличные от нативного гена). Аналогично, клетка-хозяин, трансформированная конструкцией, которая обычно не присутствует в клетке-хозяине, будет считаться гетерологичной для целей данного раскрытия.
Гомологичная аминокислотная последовательность: любой полипептид, который полностью или частично кодируется последовательностью нуклеиновой кислоты, которая гибридизуется с любым участком последовательности нуклеиновой кислоты кодирующей области. Гомологичная аминокислотная последовательность представляет последовательность, которая отличается от аминокислотной последовательности, приведенной в списке последовательностей, одной или несколькими консервативными аминокислотными заменами. Такая последовательность также охватывает аллельные варианты (определенные выше), а также последовательности, содержащие делеции или инсерции, которые сохраняют функциональные характеристики полипептида. Предпочтительно, такая последовательность представляет, по меньшей мере, на 75%, более предпочтительно, на 80%, более предпочтительно, на 85%, более предпочтительно, на 90%, более предпочтительно, на 95% и, наиболее предпочтительно, на 98%, идентичную любой из аминокислотных последовательностей.
Гомологичные аминокислотные последовательности включают последовательности, которые идентичны или по существу идентичны аминокислотным последовательностям, приведенным в списке последовательностей. Под «аминокислотной последовательностью, по существу идентичной» подразумевается последовательность, которая, по меньшей мере, на 90%, предпочтительно на 95%, более предпочтительно на 97% и наиболее предпочтительно на 99% идентична референсной аминокислотной последовательности и которая предпочтительно отличается от референсной последовательности большинством консервативных аминокислотных замен. В соответствии с этим аспектом изобретения полипептиды, имеющие последовательность, гомологичную любой из аминокислотных последовательностей, приведенных в списке последовательностей, включают встречающиеся в природе аллельные варианты, а также мутанты или любые другие, не встречающиеся в природе варианты, которые сохраняют присущие характеристики любого полипептида из последовательностей, раскрытых здесь. Гомологию можно измерить с помощью программного обеспечения для анализа последовательностей, такого как пакет программного обеспечения для анализа последовательностей, представленный Genetics Computer Group, University of Wisconsin Biotechnology Center, 1710 University Avenue, Madison, Wl 53705. Аминокислотные последовательности можно выравнивать для максимизации идентичности. Для достижения оптимального выравнивания в последовательность также могут быть искусственно введены гэпы. После того, как было получено оптимальное выравнивание, степень гомологии устанавливается посредством регистрации всех положений, в которых аминокислоты обеих последовательностей являются идентичными, относительно общего количества положений. Гомологичные полинуклеотидные последовательности определяются аналогичным образом. Предпочтительно, гомологичная последовательность представляет собой последовательность, которая, по меньшей мере, на 45%, 50%, 60%, 70%, 75%, 80%, 85%, 86%, 87%, 88%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентична любой из кодирующих последовательностей.
Гибридизация: олигонуклеотиды и другие нуклеиновые кислоты гибридизуются посредством образования водородных связей, что включает образование уотсон-криковских, хугстиновских или обращенных хугстиновских водородных связей между комплементарными основаниями. Как правило, нуклеиновая кислота состоит из азотистых оснований, которые представляют пиримидины (цитозин (C), урацил (U) и тимин (T)), или пурины (аденин (A) и гуанин (G)). Эти азотистые основания образуют водородные связи между пиримидином и пурином, и связывание пиримидина с пурином называется спариванием оснований. Более конкретно, A связывается водородной связью с T или U, и G связывается с C. В молекулах РНК, G также связывается с U. Комплементарные относится к спариванию оснований, которое происходит между двумя отдельными последовательностями нуклеиновой кислоты или двумя отдельными областями одной и той же последовательности нуклеиновой кислоты.
Специфически гибридизируемые и специфически комплементарные представляют термины, которые указывают на достаточную степень комплементарности, когда происходит стабильное и специфическое связывание между первой нуклеиновой кислотой (такой как олигонуклеотид) и ДНК-мишенью или РНК-мишенью. Первая нуклеиновая кислота (такая как олигонуклеотид) не обязательно должна быть на 100% комплементарной ее последовательности-мишени для того, чтобы специфически гибридизоваться. Первая нуклеиновая кислота (такая как олигонуклеотид) специфически гибридизуется, когда существует достаточная степень комплементарности, чтобы избежать неспецифического связывания первой нуклеиновой кислоты (такой как олигонуклеотид) с немишеневыми последовательностями в условиях, где желательно специфическое связывание. Такое связывание называется специфической гибридизацией.
Условия гибридизации, обеспечивающие определенные степени жесткости, будут варьироваться в зависимости от природы выбранного способа гибридизации, и состава и длины гибридизующихся последовательностей нуклеиновых кислот. Как правило, температура гибридизации и ионная сила (особенно концентрация Na+) буфера для гибридизации будут определять жесткость гибридизации, хотя время отмывки также влияет на жесткость. Расчеты, касающиеся условий гибридизации, необходимых для достижения конкретных степеней жесткости, обсуждаются Sambrook et al. (ed.) Molecular Cloning: A Laboratory Manual, 2nd ed., vol. 1-3, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989, главы 9 и 11.
Ниже приведены примерные наборы условий гибридизации, и они не предназначены для ограничения.
Очень высокая жесткость (детектируются последовательности, которые имеют 90% идентичность последовательностей)
Гибридизация: 5× SSC при 65°C в течение 16 ч
Двукратная отмывка: 2× SSC при комнатной температуре (RT) в течение 15 мин каждый раз
Двукратная отмывка: 0,5× SSC при 65°C в течение 20 мин каждый раз
Высокая жесткость (детектируются последовательности, которые имеют 80% идентичность последовательностей)
Гибридизация: 5×-6× SSC при 65°C-70°C в течение 16-20 ч
Двукратная отмывка: 2× SSC при комнатной температуре в течение 5-20 мин каждый раз
Двукратная отмывка: 1× SSC при 55°C-70°C в течение 30 мин каждый раз
Низкая жесткость (детектируются последовательности, которые имеют 50% идентичность последовательностей)
Гибридизация: 6× SSC при температуре в диапазоне от комнатной до 55°C в течение 16-20 ч
Отмывка, по меньшей мере, дважды: 2×-3× SSC при температуре в диапазоне комнатной до 55°C в течение 20-30 мин каждый раз.
Выделенный: выделенный биологический компонент (такой как молекула нуклеиновой кислоты или белок) представляет собой компонент, который был в значительной степени отделен или очищен от других биологических компонентов, присутствующих в клетке организма, в которой этот компонент встречается в природе, например, другие хромосомные и экстрахромосомные ДНК и РНК, белки и органеллы. По отношению к нуклеиновым кислотам и/или полипептидам, этот термин может относиться к нуклеиновым кислотам или полипептидам, которые больше не фланкированы последовательностями, которые обычно фланкируют их в природе. Нуклеиновые кислоты и белки, которые были выделены, включают нуклеиновые кислоты и белки, очищенные стандартными методами очистки. Термин также охватывает нуклеиновые кислоты и белки, полученные посредством рекомбинантной экспрессии в клетке-хозяине, а также химически синтезированные нуклеиновые кислоты.
Мутировать: процесс, вызывающий изменение в последовательности генетического материала (обычно ДНК или РНК) клетки или организма. Мутации могут быть преднамеренно введены в генетический материал с использованием молекулярных методов, хорошо известных в данной области (например, сайт-направленный мутагенез, ПЦР-опосредованный мутагенез и другие).
Нерибосомальные пептиды (NRP): класс вторичных метаболитов, обычно продуцируемых микроорганизмами, такими как бактерии и грибы. В отличие от полипептидов, синтезируемых на рибосоме, эти пептиды синтезируются при участии нерибосомальных пептидсинтетаз (NRPS) из аминокислот.
Сборка нерибосомального пептидного скелета: вторая стадия биосинтеза нерибосомального пептида, которая включает образование амидной связи (конденсацию) пептидной последовательности.
Нерибосомальная пептидсинтетаза (NRPS): крупный многофункциональный белок, который катализирует синтез полипептидов по нерибосомному механизму, часто называемому синтезом с тиоматрицы (Kleinkauf and von Doehren Ann. Rev. Microbiol., 41: 259-289, 1987). Такие нерибосомные полипептиды могут иметь линейную, циклическую или разветвленную циклическую структуру и часто содержат аминокислоты, отсутствующие в белках, или аминокислоты, модифицированные в результате метилирования или эпимеризации. В конкретных примерах NRPS продуцируют дипептиды.
Оптимизация нерибосомальных пептидов: третья стадия биосинтеза нерибосомальных пептидов. Существует множество новых аминокислот-предшественников, обнаруженных в нерибосомальных пептидах, и многие из этих строительных блоков образуются или модифицируются при присоединении к доменам PCP специализированных белков или NRPS. Эта постсинтетическая модификация может иметь место после образования амидной связи пептидного скелета. Типичные модификации включают эпимеризацию α-углерода, N-метилирование, гетероциклизацию остатков Cys или Ser/Thr до тиазолинов и оксазолинов и галогенирование или гидроксилирование боковых цепей. Другие модификации, такие как окисление, алкилирование, ацилирование и гликозилирование, могут иметь место после высвобождения зарождающегося пептида из комплекса NRPS и часто необходимы для обеспечения полной биологической активности.
Биосинтез нерибосомальных аминокислот-предшественников: первая стадия в биосинтезе нерибосомальных пептидов. Нерибосомальные пептиды часто содержат аминокислоты, которые отсутствуют в пептидах и белках, которые собираются на рибосомах. Эти непротеиногенные аминокислоты вносят свой вклад в разнообразие этих пептидов и часто играют роль в их биологической активности. Биосинтез этих аминокислот может происходить через белок-связанные промежуточные соединения или в виде свободных растворимых молекул.
Нуклеиновая кислота: дезоксирибонуклеотидный или рибонуклеотидный полимер в одноцепочечной или двухцепочечной форме и, если не ограничивается, то термин охватывает известные аналоги природных нуклеотидов, которые гибридизуются с нуклеиновыми кислотами путем, сходным с природными нуклеотидами.
Нуклеотид: данный термин включает, не ограничиваясь этим, мономер, который содержит основание, связанное с сахаром, такое как пиримидин, пурин или его синтетические аналоги, или основание, связанное с аминокислотой, как в пептидной нуклеиновой кислоте. Нуклеотид представляет один мономер в полинуклеотиде. Нуклеотидная последовательность относится к последовательности оснований в полинуклеотиде.
Олигонуклеотид: множество соединенных нуклеотидов, связанных нативными фосфодиэфирными связями, длиной примерно от 6 до примерно 300 нуклеотидов. Аналог олигонуклеотида относится к молекулам, которые функционируют аналогично олигонуклеотидам, но имеют не встречающиеся в природе участки. Например, аналоги олигонуклеотидов могут содержать не встречающиеся в природе участки, такие как измененные сахарные группы или связи между сахарами, такие как фосфоротиоатный олигодезоксинуклеотид. Функциональные аналоги встречающихся в природе полинуклеотидов могут связываться с РНК или ДНК и включать молекулы пептидной нуклеиновой кислоты.
Конкретные олигонуклеотиды и аналоги олигонуклеотидов могут включать линейные последовательности длиной примерно до 200 нуклеотидов, например, последовательность (такую как ДНК или РНК), которая составляет, по меньшей мере, 6 оснований, например, по меньшей мере, 8, 10, 15, 20, 25, 30 35, 40, 45, 50, 100 или даже 200 оснований длиной или примерно от 6 до примерно 50 оснований, например, примерно 10-25 оснований, например, 12, 15 или 20 оснований.
Открытая рамка считывания (ORF): последовательность нуклеотидных триплетов (кодонов), кодирующих аминокислоты без каких-либо внутренних кодонов терминации. Эти последовательности обычно транслируются в пептид. Например, ORF, открытая рамка считывания, и ORF эндурацидина относятся к открытой рамке считывания в кластере генов биосинтеза эндурацидина, выделенной из Streptomyces fungicidicus. Термин также охватывает аналогичные ORF, которые присутствуют в других микроорганизмах, синтезирующих эндурацидин. Термин включает аллельные варианты и однонуклеотидные полиморфизмы (SNP). В некоторых случаях термин ORF эндурацидина используется синонимично с полипептидом, кодируемым ORF эндурацидина, и может включать консервативные замены в этом полипептиде. Конкретное использование данного термина будет ясно из контекста.
Открытая рамка считывания, которая была инактивирована, является открытой рамкой считывания, которая стала нефункциональной в результате делеции, инсерции или мутации одного или более нуклеотидов в кодирующей последовательности.
Streptomyces fungicidicus, включающий ослабленную открытую рамку считывания-18 (orf 18), представляет микроорганизм, у которого наблюдается снижение, например, снижение в 2 раза, или даже полная потеря биологической функции продукта гена orf18 по сравнению с Streptomyces fungicidicus дикого типа, например, в результате генетической модификации orf18, включая инактивирование orf18, как показано в примерах ниже, и/или посредством регуляторной манипуляции, например, модификации, инсерции, делеции и/или замены некодирующих областей гена, кодирующего ORF18, которые приводят к снижению экспрессии продукта гена orf18. Например, промотор дикого типа для orf18 может быть модифицирован таким образом, чтобы существенно снизить транскрипцию orf18.
Streptomyces fungicidicus, включающий усиленную открытую рамку считывания-24 (orf24), представляет собой микроорганизм, у которого наблюдается повышение, например, повышение в 2 раза или более, биологической функции продукта гена orf24, по сравнению с Streptomyces fungicidicus дикого типа, например, в результате генетической модификации orf24 для усиления биологической функции продукта гена orf24 и/или посредством регуляторной манипуляции, например, модификации, инсерции, делеции и/или замены некодирующих областей гена, кодирующего ORF24, которые приводят к повышению экспрессии продукта гена orf24. Например, промотор дикого типа для orf24 был заменен сильным конститутивным промотором, который усиливал транскрипцию orf24, как показано в примерах ниже.
Модифицированный ген: последовательность гена, которая содержит модификацию по сравнению с последовательностью, найденной во встречающемся в природе гене (дикого типа).
Операбельно связанная: первая последовательность нуклеиновой кислоты операбельно связана со второй последовательностью нуклеиновой кислоты, когда первая последовательность нуклеиновой кислоты находится в функциональной взаимосвязи со второй последовательностью нуклеиновой кислоты. Например, промотор операбельно связан с кодирующей последовательностью, если промотор оказывает влияние на транскрипцию или экспрессию кодирующей последовательности. Как правило, операбельно связанные последовательности ДНК являются непрерывными и, если необходимо, связывают две кодирующие белок области в одной рамке считывания.
Ортолог: две нуклеиновокислотные или аминокислотные последовательности являются ортологами друг для друга, если они имеют общую родовую последовательность и расходятся, когда вид, несущий эту родовую последовательность, разделяется на два вида. Ортологичные последовательности также являются гомологичными последовательностями.
Полипептид: полимер, в котором мономеры представляют собой аминокислотные остатки, которые связаны вместе посредством амидных связей. Когда аминокислоты представляют альфа-аминокислоты, то можно использовать оптический L-изомер или оптический D-изомер, где L-изомеры являются предпочтительными в некоторых случаях. Как здесь используется, термин «полипептид или белок» охватывает любую аминокислотную последовательность и включает модифицированные последовательности, такие как гликопротеины. Термин «полипептид» конкретно предназначен для того, чтобы охватывать встречающиеся в природе белки (независимо от того, продуцируются они рибосомными или нерибосомными механизмами), а также те, которые продуцируются рекомбинантным или синтетическим путем.
Термин «фрагмент полипептида» относится к фрагменту полипептида, который обладает, по меньшей мере, одним пригодным эпитопом. Выражение «функциональный фрагмент полипептида» относится ко всем фрагментам полипептида, которые сохраняют активность (такую как биологическая активность) или измеряемую часть активности полипептида, из которого получен фрагмент. Например, фрагменты могут отличаться по размеру от полипептидного фрагмента, такого небольшого, как эпитоп, способного связывать молекулу антитела с крупным полипептидом, способным участвовать в специфической индукции или программировании фенотипических изменений в клетке.
Термин «по существу очищенный полипептид» в контексте настоящего описания относится к полипептиду, который по существу не содержит других белков, липидов, углеводов или других веществ, с которыми он ассоциирован в природе. В одном варианте осуществления полипептид, по меньшей мере, на 50%, например, по меньшей мере, на 80% свободен от других белков, липидов, углеводов или других веществ, с которыми он ассоциирован в природе. В еще одном варианте осуществления полипептид, по меньшей мере, на 90% свободен от других белков, липидов, углеводов или других веществ, с которыми он ассоциирован в природе. В еще одном варианте осуществления полипептид, по меньшей мере, на 95% свободен от других белков, липидов, углеводов или других веществ, с которыми он ассоциирован в природе.
Зонды и праймеры: нуклеиновокислотные зонды и праймеры можно легко получить на основе молекул нуклеиновой кислоты, обеспеченных в данном описании. Зонд содержит выделенную нуклеиновую кислоту, присоединенную к детектируемой метке или репортерной молекуле. Типичные метки включают радиоактивные изотопы, субстраты ферментов, кофакторы, лиганды, хемилюминесцентные или флуоресцентные агенты, гаптены и ферменты. Способы мечения и рекомендации по выбору меток, подходящих для различных целей, обсуждаются, например, в монографии Sambrook et al. (в Molecular Cloning: A Laboratory Manual, CSHL, New York, 1989) and Ausubel et al. (In Current Protocols in Molecular Biology, Greene Publ. Assoc. and Wiley-Intersciences, 1992)).
Праймеры представляют собой короткие молекулы нуклеиновых кислот, предпочтительно ДНК-олигонуклеотиды, длиной 10 нуклеотидов или более. Более предпочтительно, более длинные ДНК-олигонуклеотиды могут иметь длину примерно 15, 17, 20 или 23 нуклеотидов или более. Праймеры могут быть подвергнуты отжигу с комплементарной цепью ДНК-мишени посредством гибридизации нуклеиновой кислоты с образованием гибрида между праймером и цепью ДНК-мишени, и затем праймер удлиняется вдоль цепи ДНК-мишени с участием фермента ДНК-полимеразы. Пары праймеров можно использовать для амплификации последовательности нуклеиновой кислоты, например, с помощью полимеразной цепной реакции (ПЦР) или других способов амплификации нуклеиновой кислоты, известных в данной области.
Способы получения и применения зондов и праймеров описаны, например, в монографии Sambrook et al. (в Molecular Cloning: A Laboratory Manual, CSHL, New York, 1989), Ausubel et al. (в Current Protocols in Molecular Biology, Greene Publ. Assoc. and Wiley-Intersciences, 1998) и Innis et al. (PCR Protocols, A Guide to Methods and Applications, Academic Press, Inc., San Diego, CA, 1990). Пары ПЦР-праймеров могут быть получены из известной последовательности, например, с использованием компьютерных программ, предназначенных для этой цели, таких как Primer (версия 0.5, © 1991, Whitehead Institute for Biomedical Research, Cambridge, MA). Специфичность конкретного зонда или праймера повышается с увеличением его длины. Таким образом, для получения более высокой специфичности можно выбрать зонды и праймеры, которые содержат, по меньшей мере, 17, 20, 23, 25, 30, 35, 40, 45, 50 или более последовательных нуклеотидов требуемой нуклеотидной последовательности.
Белок: биологическая молекула, экспрессируемая геном и состоящая из аминокислот.
Очищенный: термин «очищенный» не требует абсолютной чистоты; скорее он является относительным термином. Таким образом, например, препарат очищенного белка представляет собой препарат, в котором указанный белок является более чистым, чем белок в его природной среде внутри клетки.
Рекомбинантная: нуклеиновая кислота, которая имеет последовательность, которая не встречается в природе, или имеет последовательность, которая получена посредством искусственной комбинации двух иначе разделенных сегментов последовательности. Эта искусственная комбинация может быть достигнута химическим синтезом или, чаще, искусственным манипулированием выделенными сегментами нуклеиновых кислот, например, методами генной инженерии. «Рекомбинантная» также используется для описания молекул нуклеиновой кислоты, которые подвергались искусственному манипулированию, но содержат такие же регуляторные последовательности и кодирующие области, которые обнаружены в организме, из которого был выделен ген.
Регулирование продукцией антибиотиков: означает вызывать изменения, такие как увеличение или уменьшение количества, изменение типа или качества продукции антибиотиков. Настоящее изобретение раскрывает рекомбинантные штаммы Streptomyces fungicidicus с повышенной продукцией эндурацидина.
Идентичность последовательностей: сходство между двумя последовательностями нуклеиновых кислот или между двумя аминокислотными последовательностями выражается через степень идентичности последовательностей, общих для последовательностей. Идентичность последовательностей обычно выражается в процентах идентичности; чем выше процент, тем больше сходство двух последовательностей.
Способы выравнивания последовательностей для сравнения хорошо известны в данной области. Различные программы и алгоритмы выравнивания описаны, например в публикациях: Smith and Waterman, Adv. Appl. Math., 2: 482, 1981; Needleman and Wunsch, J. Mol. Biol. 48: 443, 1970; Pearson and Lipman, Proc. Natl. Acad. Sci. США 85: 2444, 1988; Higgins and Sharp, Gene 73: 237 244, 1988; Higgins and Sharp, Computer Applications, CABIOS 5: 151-153, 1989; Corpet et al., Nucleic Acids Research 16: 10881-10890, 1988; Huang et al., Computer Applications in Biosciences 8: 155-165, 1992; Pearson et al., Methods in Molecular Biology, 24: 307-331, 1994; Tatians et al., (1999), FEMS Microbiol. Lett., 174: 247 250, 1999. Подробное рассмотрение способов выравнивания последовательностей и расчетов гомологии можно найти, например, в публикации Altschul et al. (J. Mol. Biol., 215: 403 410, 1990).
Базовый инструмент поиска локального выравнивания (BLAST™; Altschul et al. (1990)) Национального центра биотехнологической информации (NCBI) доступен из нескольких источников, включая Национальный центр биотехнологической информации (NCBI, Bethesda, MD), и в интернете для применения с программами для анализа последовательностей blastp, blastp, blastn, tblastn и tblastx. Описание того, как определить идентичность последовательностей с применением этой программы доступно в интернете в разделе «помощь» для BLAST™.
Для сравнения аминокислотных последовательностей, содержащих более 30 аминокислот, используется функция «Последовательности Blast 2» программы BLAST™ (Blastp) с применением матрицы BLOSUM62 по умолчанию, в которой установлены параметры по умолчанию (цена открытия гэпа [по умолчанию=5]; цена расширения гэпа [по умолчанию=2]; штраф за несовпадение [по умолчанию=3]; балл за совпадение [по умолчанию=1]; значение ожидания (E) [по умолчанию=10.0]; длина слова [по умолчанию=3]; количество однострочных описаний (V) [по умолчанию=100]; количество отображаемых выравниваний (B) [по умолчанию=100]). При выравнивании коротких пептидов (длиной менее чем примерно 30 аминокислот) выравнивание следует выполнять с использованием функции «Последовательности Blast 2, применяя матрицу PAM30, установленную на параметры по умолчанию (штраф за открытие гэпа 9, штраф за расширения гэпа 1). Белки (или нуклеиновые кислоты) с еще большим сходством с референсными последовательностями будут демонстрировать увеличение процентной идентичности при оценке этим методом, например, по меньшей мере, 50%, по меньшей мере, 60%, по меньшей мере, 70%, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90% или, по меньшей мере, 95% идентичность последовательности.
Для сравнения последовательностей нуклеиновых кислот используется функция «Последовательности Blast 2» программы BLAST™ (Blastn) с применением матрицы BLOSUM62 по умолчанию, для которой установлены параметры по умолчанию (цена за открытие гэпа [по умолчанию=11]; цена за расширение гэпа [по умолчанию=1]; ожидаемое значение (E) [по умолчанию=10.0]; длина слова [по умолчанию=11]; количество однострочных описаний (V) [по умолчанию=100]; количество отображаемых выравниваний (B) [по умолчанию=100]). Последовательности нуклеиновых кислот с еще большим сходством с референсными последовательностями будут демонстрировать увеличение процентной идентичности при оценке этим методом, например, по меньшей мере, 60%, по меньшей мере, 70%, по меньшей мере, 75%, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95% или, по меньшей мере, 98% идентичность последовательности.
Альтернативным показателем того, что две молекулы нуклеиновой кислоты тесно связаны, является то, что две молекулы гибридизуются друг с другом в жестких условиях (см. «Гибридизация» выше).
Последовательности нуклеиновых кислот, которые не проявляют высокой степени идентичности, могут, тем не менее, кодировать сходные аминокислотные последовательности за счет вырожденности генетического кода. Понятно, что изменения в последовательности нуклеиновой кислоты могут быть сделаны с использованием этой вырожденности с получением множества молекул нуклеиновой кислоты, которые все кодируют по существу один и тот же белок.
Трансфицированная: процесс, посредством которого молекула нуклеиновой кислоты вводится в клетку, например, методами молекулярной биологии, в результате чего клетка трансфицируется (или трансформируется). Как здесь используется, термин «трансфекция» включает все методы, с помощью которых молекула нуклеиновой кислоты может быть введена в такую клетку, включая трансдукцию вирусными векторами, трансфекцию плазмидными векторами и введение ДНК посредством электропорации, липофекции и «обстрела частицами».
Трансформированная: трансформированная клетка представляет клетку, в которую была введена молекула нуклеиновой кислоты методами молекулярной биологии. Термин охватывает все методы, с помощью которых молекула нуклеиновой кислоты может быть введена в такую клетку, включая трансфекцию вирусными векторами, трансформацию плазмидными векторами и введение «голой» ДНК посредством электропорации, липофекции и «обстрела частицами».
Транспозон: мобильный генетический элемент, имеющий практически идентичные повторяющиеся последовательности на обоих концах и содержащий, по меньшей мере, ген, кодирующий транспозазу (фермент, необходимый для встраивания транспозона в последовательность ДНК). Транспозоны могут быть встроены в разные положения в геноме клетки или за выделенной ДНК-матрицы плазмиды, космиды или фосмиды in vitro. Транспозоны могут также содержать гены, отличные от тех, которые необходимы для инсерции.
Вектор: молекула нуклеиновой кислоты, введенная в клетку-хозяина, с получением тем самым трансфицированной клетки-хозяина. Векторы рекомбинантной ДНК представляют собой векторы, содержащие рекомбинантную ДНК. Вектор может включать последовательности нуклеиновых кислот, которые позволяют ему реплицироваться в клетке-хозяине, такие как ориджин репликации. Вектор также может включать один или несколько селектируемых маркерных генов и другие генетические элементы, известные в данной области. Вирусные векторы представляют рекомбинантные ДНК-векторы, имеющие, по меньшей мере, несколько последовательностей нуклеиновых кислот, полученных из одного или более вирусов. Плазмида представляет вектор.
Если не указано иное, то все технические и научные термины, используемые в данном документе, имеют то же значение, которое обычно понимается специалистами обычной квалификации в области техники, к которой относится настоящее изобретение. Артикли «а», «an» и «the» в единственном числе включают множественные ссылки, если контекст явно не указывает иное. Также слово «или» предназначено для включения «и», если контекст явно не указывает иное. Кроме того, следует понимать, что все размеры оснований или размеры аминокислот и все значения молекулярного веса или молекулярной массы, приведенные для нуклеиновых кислот или полипептидов, являются приблизительными и представлены для описания. Несмотря на то, что способы и материалы, сходные или эквивалентные тем, которые здесь описаны, могут использоваться в практике или тестировании настоящего раскрытия, подходящие способы и материалы описаны ниже. Термин «содержит» означает «включает». В случае конфликта настоящее описание, включая пояснения терминов, будет иметь преимущественную силу. Кроме того, материалы, способы и примеры являются только иллюстративными и не предназначены для ограничения.
Подходящие способы и материалы для практики раскрытых вариантов осуществления описаны ниже. Кроме того, любой подходящий способ или методика, хорошо известные специалисту в данной области техники, может использоваться при выполнении раскрытых вариантов осуществления. Некоторые традиционные способы и методики, применимые к настоящему раскрытию, описаны, например, в Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd ed., Cold Spring Harbor Laboratory Press, 1989; Sambrook et al., Molecular Biology, 4th ed., Cold Spring Harbor Press, 2001; Ausubel et al., Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology, 4th ed., Wiley & Sons, 1999; Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, 1990; Harlow and Lane, Using Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, 1999; and Kieser, T., Bibb, M.J., Buttner, M.J., Chater, K.F., and Hopwood, D.A.: Practical Streptomyces genetics, John Innes Centre, Norwich Research Park, Colney, Norwich NR4 &UH, England, 2000.
Все публикации, заявки на патенты, патенты и другие ссылки, упомянутые здесь, включены в качестве ссылки во всей их полноте. В случае конфликта настоящая спецификация, включая пояснения терминов, будет иметь преимущественную силу. Кроме того, материалы, методы и примеры являются только иллюстративными и не предназначены для ограничения.
III. Сконструированные рекомбинантные экспрессионные векторы Streptomyces fungicidicus
Настоящее изобретение раскрывает сконструированные рекомбинантные экспрессионные плазмидные векторы Streptomyces fungicidicus. В некоторых вариантах осуществления сконструированный рекомбинантный вектор Streptomyces fungicidicus содержит, по меньшей мере, одну выбранную открытую рамку считывания Streptomyces fungicidicus. В некоторых вариантах осуществления сконструированный рекомбинантный вектор Streptomyces fungicidicus содержит, по меньшей мере, одну выбранную открытую рамку считывания Streptomyces fungicidicus, экспрессированную под контролем промотора. В некоторых примерах промотор является сильным конститутивным промотором Streptomyces, который приводит к повышенной продукции эндурацидина, когда вектор экспрессируется в штамме Streptomyces fungicidicus. В некоторых вариантах осуществления открытая рамка считывания операбельно связана с гетерологичным промотором вместо ее собственного нативного промотора. Например, она может быть операбельно связана с конститутивным промотором, таким как сильный конститутивный промотор экспрессии или индуцибельный промотор. В некоторых примерах сильный конститутивный промотор представляет собой ermE*p из продуцента эритромицина. В некоторых примерах индуцибельным промотором является tipA. В некоторых примерах используется система P(nitA)-NitR (Herai S., Hashimoto Y., Higashibata H., Maseda H., Ikeda H., Omura S., Kobayashi M., Proc. Natl. Acad. Sci. USA, 2004, 101 (39): 14031-5) или промотор стрептомицетов SF14. В некоторых примерах используется нативный промотор гена устойчивости к апрамицину (amRp). В некоторых примерах используются PhrdB, Ptcp830, PSF14, PermE* и/или Pneos.
В некоторых вариантах осуществления сконструированный рекомбинантный вектор содержит открытую рамку считывания orf24 (SEQ ID NO: 38) и/или открытую рамку считывания orf18 (SEQ ID NO: 37), которая была инактивирована. В некоторых примерах открытая рамка считывания orf18 (SEQ ID NO: 37) была инактивирована посредством делеции внутри рамки считывания, сдвига рамки считывания и/или точечной мутации.
В некоторых вариантах осуществления сконструированный рекомбинантный вектор содержит открытую рамку считывания orf24 из кластера генов биосинтеза эндурацидина из Streptomyces fungicidicus. В некоторых примерах открытая рамка считывания orf24 (SEQ ID NO: 38) операбельно связана с гетерологичным промотором. Например, она связана с сильным конститутивным промотором, таким как ermE*p. В еще одних примерах открытая рамка считывания orf24 операбельно связана с промотором tipA, SF14, amRp, PhrdB, Ptcp830, PSF14, PermE* и/или Pneos.
В еще одном варианте осуществления сконструированный рекомбинантный вектор содержит открытую рамку считывания orf18, которая находится в области апстрим кластера генов биосинтеза эндурацидина. Открытая рамка считывания orf18 (SEQ ID NO: 37) инактивируется посредством инсерционного разрушения, делеции внутри рамки считывания, сдвига рамки считывания и/или точечной мутации. В некоторых примерах открытая рамка считывания orf18 инактивируется делецией внутри рамки считывания, такой как делеция внутри рамки считывания, показанная на фиг. 9В. В одном примере открытая рамка считывания orf18 (SEQ ID NO: 37) инактивируется делецией внутри рамки считывания. Например, открытая рамка считывания orf18 (SEQ ID NO: 37) инактивируется делецией внутри рамки считывания нуклеотидов с 5 по 660 из orf18 (SEQ ID NO: 37). В общем, любая внутренняя делеция внутри рамки считывания за orf18 приводит к инактивированной функции Orf18 за счет ее неполноты. В некоторых примерах делеция внутри рамки считывания включает делецию, по меньшей мере, 3 нуклеотидов в orf18 (SEQ ID NO: 37), например, по меньшей мере, 3 нуклеотидов, включая 3, 6, 9, 12, 15, 18, 21, 24, 27, 30, 33, 36, 39, 42, 45, 48, 51, 54, 57, 60, 63, 66, 69, 72, 75, 78, 81, 84, 87, 90, 93, 96, 99, 102, 105, 108, 111, 114, 117, 120, 123, 126, 129, 132, 135, 138, 141, 144, 147, 150, 153, 156, 159, 162, 165, 168, 171, 174, 177, 180, 183, 186, 189, 192, 195, 198, 201, 204, 207, 210, 213, 216, 219, 221, 224, 227, 230, 233, 236, 239, 242, 245, 248, 251, 254, 257, 260, 263, 266, 269, 272, 275, 278, 281, 284, 287, 290, 293, 296, 299, 302, 305, 308, 311, 314, 317, 320, 323, 326, 329, 332, 335, 338, 341, 344, 347, 350, 353, 356, 359, 362, 365, 368, 371, 374, 377, 380, 383, 386, 389, 392, 395, 398, 401, 404, 407, 410, 413, 416, 419, 421, 424, 427, 430, 433, 436, 439, 442, 445, 448, 451, 454, 457, 460, 463, 466, 469, 472, 475, 478, 481, 484, 487, 490, 493, 496, 499, 502, 505, 508, 511, 514, 517, 520, 523, 526, 529, 532, 535, 538, 541, 544, 547, 550, 553, 556, 559, 562, 565, 568, 571, 574, 577, 580, 583, 586, 589, 592, 595, 598, 601, 604, 607, 610, 613, 616, 619, 621, 624, 627, 630, 633, 636, 639, 642, 645, 645, 648, 651 или 654 нуклеотидов между нуклеотидами с 5 по 660 orf18 (SEQ ID NO: 37).
В связанных вариантах осуществления сконструированный рекомбинантный плазмидный вектор включает две или более открытых рамок считывания из кластера генов эндурацидина и/или областей, фланкирующих кластер генов, или из других штаммов актиномицетов. Две или более открытых рамки считывания могут быть связаны с одним промотором. Альтернативно, они могут быть операбельно связаны с двумя разными промоторами. Два промотора могут быть промоторами одного типа. Альтернативно, они могут представлять промоторы двух различных типов.
В еще одних вариантах осуществления дополнительные или альтернативные открытые рамки считывания, которые могут усиливать продукцию эндурацидина, могут быть введены или инактивированы в сконструированном штамме Streptomyces fungicidicus.
В некоторых примерах рекомбинантная плазмида представляет собой pXY152-endorf24 (SEQ ID NO: 3). В некоторых примерах рекомбинантная плазмида представляет pXY300-orf18ifd (SEQ ID NO: 8). В некоторых примерах рекомбинантная плазмида представляет pKS-T-orf18ifd (SEQ ID NO: 11). В некоторых примерах рекомбинантная плазмида представляет pKS-T-orf18pfrd-AmR (SEQ ID NO: 14). В некоторых примерах рекомбинантная плазмида представляет pKS-orf18ifd-T-AmR(NS) (SEQ ID NO: 19). В некоторых примерах рекомбинантная плазмида представляет pXY152-endorf24-camtsr (SEQ ID NO: 20). В некоторых примерах рекомбинантная плазмида представляет pXY152-endorf24-blatsr (SEQ ID NO: 23).
IV. Сконструированные рекомбинантные штаммы Streptomyces fungicidicus
Настоящее изобретение раскрывает сконструированные рекомбинантные штаммы Streptomyces fungicidicus, способные продуцировать повышенные уровни эндурацидина по сравнению с контрольным штаммом (таким как штамм Streptomyces fungicidicus дикого типа или промышленный родительский штамм). В некоторых вариантах осуществления сконструированный рекомбинантный штамм Streptomyces fungicidicus содержит, по меньшей мере, одну выбранную открытую рамку считывания из Streptomyces fungicidicus, введенную в хромосому и экспрессируемую под контролем промотора, такого как сильный конститутивный промотор Streptomyces, который приводит к повышенной продукции эндурацидина в сконструированном штамме. В некоторых вариантах осуществления экспрессия введенной открытой рамки считывания в Streptomyces fungicidicus регулируется гетерологичным промотором вместо ее собственного нативного промотора. Например, она может быть операбельно связана с конститутивным промотором, таким как сильный конститутивный промотор экспрессии или индуцибельный промотор. В некоторых примерах сильный конститутивный промотор представляет ermE*p из продуцента эритромицина. В некоторых примерах индуцибельным промотором является tipA. В некоторых примерах используется система P(nitA)-itR (см. Herai S., Hashimoto Y., Higashibata H., Maseda H., Ikeda H., Omura S., Kobayashi M., Proc. Natl. Acad. Sci USA, 2004, 101 (39): 14031-5) или промотор стрептомицетов SF14. В некоторых примерах конститутивный промотор экспрессии представляет собой amRp. В некоторых примерах используются промоторы PhrdB, Ptcp830, PSF14, PermE* и/или Pneos.
В некоторых вариантах осуществления сконструированный штамм содержит открытую рамку считывания orf24 из кластера генов биосинтеза эндурацидина Streptomyces fungicidicus. В некоторых примерах открытая рамка считывания orf24 операбельно связана с гетерологичным промотором. Например, она связана с сильным конститутивным промотором, таким как ermE*p. В еще одних примерах открытая рамка считывания orf24 операбельно связана с промотором tipA, SF14, amRp, PhrdB, Ptcp830, PSF14, PermE* и/или Pneos.
В еще одном варианте осуществления сконструированный штамм связан с открытой рамкой считывания orf18, которая находится в области апстрим кластера генов биосинтеза эндурацидина. Открытая рамка считывания orf18 инактивируется посредством инсерционного разрушения, делеции внутри рамки считывания, сдвига рамки считывания и/или точечной мутации. В некоторых примерах открытая рамка считывания orf18 инактивируется делецией внутри рамки считывания, такой как делеция внутри рамки считывания, показанная на фиг. 9В. В одном примере открытая рамка считывания orf18 (SEQ ID NO: 37) инактивируется делецией внутри рамки считывания. Например, открытая рамка считывания orf18 (SEQ ID NO: 37) инактивируется делецией нуклеотидов с 5 по 660 (SEQ ID NO: 37). Как правило, любая внутренняя делеция внутри рамки считывания за orf18 должна приводить к инактивированной функции Orf18 за счет ее неполноты.
В связанных вариантах осуществления сконструированный штамм включает две или более открытых рамки считывания из кластера генов биосинтеза эндурацидина и/или областей, фланкирующих кластер генов, или из других штаммов актиномицетов. Две или более открытых рамки считывания могут быть связаны с одним промотором. Альтернативно, они могут быть операбельно связаны с двумя разными промоторами. Два промотора могут быть промоторами одного типа. Альтернативно, они могут представлять промоторы двух различных типов.
В еще одних вариантах осуществления дополнительные или альтернативные открытые рамки считывания, которые могут усиливать продукцию эндурацидина, могут быть введены или инактивированы в сконструированном штамме Streptomyces fungicidicus.
В некоторых вариантах осуществления сконструированный штамм Streptomyces fungicidicus получен из родительского штамма дикого типа, такого как, не ограничиваясь этим, Streptomyces fungicidicus из Американской коллекции типовых культур (ATCC) 21013. В еще одних вариантах осуществления сконструированный штамм Streptomyces fungicidicus получен из промышленного родительского штамма, такого как, не ограничиваясь этим, BM38-2 (ATCC PTA-122342). В еще одних вариантах осуществления сконструированный штамм Streptomyces fungicidicus получен из обычных мутантных штаммов, таких как, не ограничиваясь этим, Streptomyces fungicidicus ATCC 31729, Streptomyces fungicidicus ATCC 31730 и Streptomyces fungicidicus ATCC 31731.
В некоторых вариантах осуществления повышенная продукция эндурацидина представляет увеличение, по меньшей мере, в 1,2 раза, например, увеличение, по меньшей мере, в 1,5 раза, по меньшей мере, в 2 раза, по меньшей мере, в 2,5 раза, по меньшей мере, в 3 раза, по меньшей мере, в 3,5 раза, по меньшей мере, в 4 раза, увеличение, по меньшей мере, в 4,5 раза, включая, не ограничиваясь этим, увеличение в 1,2-10 раз, увеличение в 1,2-4,6 раза, увеличение в 2-5 раз, например, увеличение в 1,2, 1,3, 1,4, 1,5, 1,6, 1,7, 1,8, 1,9, 2,0, 2,1, 2,2, 2,3, 2,4, 2,5, 2,6, 2,7, 2,8, 2,9, 3,0, 3,1, 3,2, 3,3, 3,4, 3,5, 3,6, 3,7, 3,8, 3,9, 4,0, 4,1, 4,2, 4,3 4,4, 4,5, 4,6, 4,7, 4,8, 4,9, 5,0, 5,5, 6,0, 6,5, 7,0, 7,5, 8,0, 8,5, 9,0, 9,5 и 10 раз продукции эндурацидина по сравнению с контрольным штаммом Streptomyces fungicidicus. В некоторых вариантах осуществления контрольный штамм Streptomyces fungicidicus представляет штамм Streptomyces fungicidicus дикого типа, включая, не ограничиваясь этим, Streptomyces fungicidicus из Американской коллекции типовых культур (ATCC) 21013 или промышленный родительский штамм, такой как, не ограничиваясь этим, BM38-2 (ATCC PTA-122342) или обычный мутантный штамм, такой как, не ограничиваясь этим, Streptomyces fungicidicus ATCC 31729, Streptomyces fungicidicus ATCC 31730 и Streptomyces fungicidicus ATCC 31731. В одном примере, контроль представляет Streptomyces fungicidicus ATCC 21013, и увеличение усиленной продукции эндурацидина составляет увеличение, по меньшей мере, 1,2 раза, например, увеличение в 1,2-4,6 раза. В одном примере контролем является Streptomyces fungicidicus BM38-2 (ATCC PTA-122342), и увеличение усиленной продукции эндурацидина составляет увеличение, по меньшей мере, в 1,2 раза, такое как увеличение в 1,2-4,6 раза.
V. Получение сконструированных рекомбинантных штаммов Streptomyces fungicidicus
В вариантах осуществления рекомбинантные штаммы Streptomyces fungicidicus можно сконструировать интеграцией рекомбинантной плазмиды, содержащей, по меньшей мере, одну открытую рамку считывания, усиливающую продукцию эндурацидина, в хромосому родительского штамма Streptomyces fungicidicus. Интегративный конъюгативный вектор может иметь, или может быть сконструирован таким образом, чтобы иметь сильный конститутивный промотор Streptomyces. В некоторых вариантах осуществления плазмида может не иметь репликона стрептомицетов, и может быть интегрирована в хромосому сайт-специфической гомологичной рекомбинацией с одинарным кроссинговером. В еще одних вариантах осуществления плазмида может находиться в виде свободной плазмиды. В некоторых вариантах осуществления можно сконструировать конъюгативный вектор, в котором плазмидная вставка несет частично или полностью делецированный представляющий интерес ген и его фланкирующие области, которые могут быть интегрированы в хромосому после гомологичной рекомбинации с двойным кроссинговером для генерации мутанта с делецией внутри рамки считывания.
VI. Продукция эндурацидина из сконструированных рекомбинантных штаммов Streptomyces fungicidicus
Сконструированные рекомбинантные штаммы Streptomyces fungicidicus, обеспеченные настоящим изобретением, обеспечивают способы получения повышенных уровней эндурацидина. Такое техническое преимущество в данной области позволяет значительно сэкономить средства, связанные с производством эндурацидина. В некоторых примерах способы получения эндурацидина включают культивирование раскрытого рекомбинантного штамма Streptomyces fungicidicus в условиях, достаточных для продукции эндурацидина. В некоторых примерах способ дополнительно включает выделение эндурацидина из культуральной среды после культивирования. В некоторых примерах способ дополнительно включает определение антибактериальной активности продуцированного эндурацидина, например, с помощью анализа ВЭЖХ или биоанализа с использованием S. aureus ATCC 29213 или Bacillis subtilis ATCC 6633 в качестве индикаторных микроорганизмов.
В некоторых примерах эндурацидин продуцируется раскрытым штаммом Streptomyces fungicidicus с использованием условий ферментации, описанных ранее для получения эндурацидина (Higashide et al. J. Antibiot., 21: 126-137, 1968). После получения соединения можно очистить и/или анализировать, включая анализ ВЭЖХ, как описано в примере 1. Способы получения эндурацидина и сбора этого соединения из культуральной среды можно найти в патенте США № 4465771, который полностью включен здесь посредством ссылки.
В некоторых примерах раскрытый штамм Streptomyces fungicidicus культивируют в триптическом соевом бульоне (TSB) на встряхивателе (например, при 225 об/мин и 30°C в течение 48 ч) и затем переносят в среду для продукции эндурацидина (EPM, таблица 1 ниже) на период времени для непрерывной ферментации, такой как, по меньшей мере, пять суток и до одиннадцати суток, включая 5, 6, 7, 8, 9, 10 или 11 суток непрерывной ферментации. В некоторых примерах продукция эндурацидина дикими типами и производными штаммами проводится в автоматических ферментерах.
Состав среды для продукции эндурацидина (EPM) (pH 6,7)
В некоторых примерах биомассу Streptomyces fungicidicus получают процессом ферментации в промышленных ферментерах с глубоким танком гигиенического дизайна с системами мониторинга и контроля pH, температуры, кислорода, аэрации, перемешивания. Например, каждую ферментированную партию S. fungicidicus получают из охарактеризованного и контролируемого рабочего посевного фонда, происходящего из производственного посевного материала, хранящегося в безопасном месте и содержащегося в низкотемпературных условиях.
В некоторых примерах процесс ферментации протекает в одну или несколько стадий, таких как следующие три стадии, и может дополнительно сопровождаться дальнейшей последовательной обработкой:
Стадия I:
Охарактеризованные стабилизированные рабочие посевные культуры используются для начала ферментации. От одного до пяти замороженных флаконов с посевным материалом извлекают из хранилища при низких температурах и размораживают естественным путем или помещают на водяную баню при 28-32°С до размораживания содержимого. Размороженную культуру(ы) в асептических условиях переносят в стерильную воду, которую выдерживают при комнатной температуре, и осторожно перемешивают для ресуспендирования культуры.
Стадия II:
Ресуспендированную культуру в асептических условиях переносят в 0,005 м3-0,05 м3 среды для посева. Среда для посева состоит из глюкозы (0,1-1,0 г/л), декстрина (0,1-3 г/л), жидкого кукурузного экстракта (0-5,0 мл/л), соевого порошка (1-5,0 г/л), сульфата аммония (0,1-0-0,5 г/л), фосфата монокалия (0,13-0,54 г/л), сульфата железа (0,00-0,5 г/л), гидроксида калия (0,13 мл/л), карбоната кальция (1-2 г/л), силиконового пеногасителя (0,1 мл/л), воды, q. s. Среду стерилизуют при температуре от 125°С до 128°С в течение 30-45 мин, и затем охлаждают до температуры от 28°С до 32°С. Объем среды доводят стерильной водой до требуемого рабочего объема. Значение рН доводят до 6,5-7,0.
Рабочие параметры цикла наращения посевного материала включают: температуру инкубации 28°C±2°C, внутреннее давление 1,0±0,5 кг/см2, скорость аэрации 3±2 N м3/мин и скорость перемешивания примерно 80 об/мин, в зависимости от размера и конфигурации резервуара. Проводится мониторинг pH, потребления кислорода и вязкости, но эти параметры не регулируются. Культуру выращивают в течение 40-80 ч перед переносом в основной производственный ферментер. Вязкость на время переноса должна составлять 200-600 сП, и значение рН должно быть ≤6,0, и должно иметь место увеличение потребления кислорода. Культуру посевного материала в асептических условиях переносят в основную ферментационную среду для завершения цикла ферментации.
Стадия III:
Состав производственной среды в ферментере (10 м3-250 м3) включает природные и химические компоненты, такие как кукурузная мука (13,0-15,0% мас./об.), кукурузная глютеновая мука (3,0-6,0% мас./об.), мука из семян хлопчатника (0,1-0,3% мас./об.), жидкий кукурузный экстракт (0,1-0,6% об./об.), хлорид натрия (0,3% мас./об.), сульфат аммония (0,25-0,6% мас./об.), молочная кислота (0-0,5% об./об.), хлорид цинка (0,01% мас./об.%), сульфат железа (0,0-0,02% мас./об.), гидроксид калия (0,20-0,5% мас./об.), сульфат кальция (0,0-0,5% мас./об.), карбонат кальция (0,5% мас./об.), амилаза (0,02-0,06% мас./об.), гидроксид калия (0,05% об./об.), растительное масло (0,5-2,0% об./об.), пеногаситель и вода, q.s. Ингредиенты добавляют в указанном порядке. Добавляют воду к ингредиентам, затем нагревают до 70-90°C, чтобы фермент расщепил сложные углеводы в течение 15 мин при температуре. Добавляют оставшиеся ингредиенты, доводят рН до 6,6-6,8 и добавляют воду, q.s., стерилизуют при 125-128°C в течение 25-50 мин для стерилизации среды. Среду охлаждают до 25°C-32°C и добавляют воду, q.s., до рабочего объема.
Содержимое переносят из посевного ферментера в основную ферментационную среду и ферментер устанавливают следующие условия: температура 28°C±3°C, скорость аэрации 20-60 N м3/мин, внутреннее давление 0,1-1,0 кг/см2, скорость перемешивания равна примерно 1,85 кВт/м3. Скорость аэрации, внутреннее давление и скорость перемешивания доводят таким образом, чтобы гарантировать, что растворенный кислород не является ограничивающим фактором для скорости. Тщательно контролируют пенообразование на протяжении всего цикла, чтобы предотвратить загрязнение или вытекание. Значение pH начинают контролировать после увеличения потребности в кислороде. Следующие параметры контролируются и/или подвергаются мониторингу в течение всего цикла ферментации: pH, аэрация, растворенный кислород, CO2, вязкость, чистота, скорость перемешивания, внутреннее давление и остаточный сахар. Значение pH поддерживают на уровне 6,8, пока не прекратится рост бактерий, затем значению pH дают измениться естественным образом до сбора. Типичный цикл ферментации составляет 210-300 ч. Культура готова для сбора, когда активность превышает 5000 мкл/л, pH повышается до 7,5 или выше, вязкость снижается, и потребность в кислороде прекращается.
Ферментацию собирают нагреванием культуры до 70°С в течение 30 мин для инактивации бактерий, и затем охлаждают для сбора жидкостей до 25°С-32°С.
В некоторых примерах дальнейшая последовательная обработка включает удаление воды из биомассы, сушку биомассы и формуляцию высушенной биомассы в премикс.
Депонирование биологического материала
Следующие биологические материалы были сданы на депонирование в соответствии с условиями Будапештского договора в Американскую коллекцию типовых культур и получили следующие идентификационные номера:
Вышеуказанные штаммы были депонированы в условиях, которые гарантируют, что доступ к культуре будет обеспечен в течение нахождения настоящей патентной заявки на рассмотрении, определенного Комиссаром по патентам и товарным знакам и имеющую право на нее в соответствии с 37 C.F.R. 1.14 и 35 США.C. 122. Депонированный материал представляет собой по существу чистую культуру депонированных штаммов. Депонированный материал доступен в соответствии с требованиями иностранных патентных законов в странах, в которых поданы аналоги данной заявки или ее производных. Однако следует понимать, что доступность депонированного материала не представляет собой лицензию на практическое применение предмета изобретения при нарушении патентных прав, предоставленных правительственными действиями.
Следующие неограничивающие примеры приводятся для иллюстрации определенных конкретных признаков и/или вариантов осуществления. Эти примеры не следует истолковывать как ограничение раскрытия конкретными описанными признаками или вариантами осуществления.
Примеры
Пример 1
Материалы и методы для повышения продукции эндурацидина
В данном примере описываются репрезентативные способы для повышения продукции эндурацидина.
Бактериальные штаммы, плазмиды, фосмиды и условия культивирования.
Streptomyces fungicidicus B-5477 (ATCC 21013) и Escherichia coli S17-1 (ATCC 47055) были приобретены в ATCC. Штамм S. fungicidicus BM38-2 (ATCC PTA-122342) и стандарты эндурацидинов A и B были предоставлены Intervet/Merck Animal Health (MAH). Штаммы E. coli DH5α (Life Technologies, Inc.), EPI300 (Epicenter) и XL10-Gold (Stratagene) использовали в качестве хозяев для E. coli плазмид, фосмид и челночных векторов E. coli-Streptomyces. Плазмиды pSET152 (Bierman et al., Gene, 116: 43-49, 1992, публикация полностью включена здесь посредством ссылки) и pIJ773 были предоставлены профессором Кейт Чатер (JIC, Norwich, UK). Плазмиду pWHM860, несущую промотор ermE*p, предоставил профессор Брэдли Мур (UCSD, San Diego). ISP2 (DifcoTM ISP среда 2), ISP4 и TSB (триптический соевый бульон BactoTM) были приобретены у VWR. Праймеры, использованные для ПЦР и секвенирования ДНК, были синтезированы в Fisher и Sigma-Aldrich. Среды и условия культивирования для выращивания S. fungicidicus описаны Higashide et al. (Journal of Antibiotics, 21: 126-137, 1968). Все процедуры c E. coli выполненяли в соответствии со стандартными протоколами.
Выделение и манипуляции с ДНК.
Для получения геномной ДНК из S. fungicidicus B-5477, BM38-2 (ATCC PTA-122342) и производных рекомбинантных и мутантных штаммов для секвенирования, конструирования фосмидных библиотек, субклонирования и ПЦР свежесобранные споры из отдельных штаммов высевали и культивировали в 100 мл жидкой среды TSB с добавлением 5 мМ MgCl2 и 0,5% глицина. Репрезентативное культивирование проводили в колбах Эрленмейера емкостью 500 мл в роторном инкубаторе-встряхивателе при 225 об/мин и 30°С в течение от 48 до 72 ч. Клетки мицелия собирали центрифугированием при 4000 об/мин и 4°С в течение 15 мин. Супернатант отбрасывали и осадок последовательно промывали один раз 10,3% раствором сахарозы и дважды 10 мМ Трис-HCl и 1 мМ этилендиаминтетраацетатом динатрия (ЭДТА), рН 8,0 (ТЕ-буфер). Влажные клетки, эквивалентные объему 80 мкл воды, распределяли в стерильные микроцентрифужные пробирки емкостью 1,5 мл. После добавления 300 мкл лизирующего раствора, содержащего 200 мкл 10 мМ Трис-HCl и 1 мМ ЭДТА, рН 8,0 и 0,3 М сахарозы (буфер TES), 50 мкл 0,5 М ЭДТА, 50 мкл лизоцима (50 мг/мл) пробирки инкубировали при 37°С в течение 30-60 мин, пока раствор не становился вязким. Затем в каждую пробирку добавляли 5 мкл протеиназы К (20 мг/мл) и 180 мкл 10% додецилсульфата натрия (SDS). После осторожного, но тщательного перемешивания растворы инкубировали при 37°С в течение 90 мин. Затем добавляли 80 мкл 10% раствора цетилтриметиламмония бромида (CTAB). После тщательного перемешивания пробирки инкубировали при 65°С в течение 10 мин. Растворы дважды экстрагировали 600 мкл смеси фенола/хлороформа/изоамилового спирта (25/24/1). Геномную ДНК в верхней водной фазе извлекали и осаждали 0,6 объема изопропанола. Собранную геномную ДНК дважды промывали 70% этанолом. После высушивания при комнатной температуре в течение 10 мин геномную ДНК растворяли в 50-100 мкл стерильной воды. Высокое качество препарата геномной ДНК подтверждали расщеплением HindIII и Sau3AI, которое показало полное расщепление и отсутствие деградации нерасщепленной геномной ДНК с помощью электрофореза в 0,8% агарозном геле. Затем объединенную геномную ДНК расщепляли РНКазой для удаления загрязнения РНК. Чистоту и количество геномной ДНК определяли на спектрофотометре Nanodrop. Проводили общие манипуляции со стрептомицетовой ДНК, включая электрофорез в агарозном геле, и использовали наборы QIAprep Spin Miniprep (Qiagen) для получения плазмид и фосмид из штаммов E.coli. Рестрикционные эндонуклеазы, ДНК-лигаза, ДНК-полимераза, транспозаза, фермент Кленова, щелочная фосфатаза и лигаза были приобретены в Biolabs, Invitrogen, Epicenter и Roche, и их использовали в соответствии с рекомендациями производителей. Фрагменты ДНК очищали с использованием наборов для экстракции геля QIAquick.
ПЦР.
ПЦР колоний проводили следующим образом: споры из независимых мутантных колоний-кандидатов инокулировали в жидкую культуру TSB. После культивирования в течение 48-72 ч мицелий собирали центрифугированием и дважды промывали ТЕ-буфером (10 мМ Трис, 1 мМ ЭДТА), рН 8,0. Мицелий ресуспендировали в стерильной H2O и использовали в качестве матрицы в реакционной смеси для ПЦР в конечном объеме 100 мкл, содержащем 60 мкл мицелия, 150 пмоль каждого праймера, 20 мкл 5X буфера A для амплификации GC-богатых участков ДНК AccuPrime (Invitrogen) и 1 мкл Polymix (добавляли при 80°C) из набора реагентов Expand long template PCR system (Roche). ПЦР проводили следующим образом: 1 цикл при 95°С в течение 3 мин, 30 циклов при 95°С в течение 1 мин, при 55°С в течение 1 мин и при 72°С в течение 2 мин. Реакцию останавливали проведением одного цикла удлинения при 72°С в течение 10 мин. Продукты ПЦР очищали в геле и секвенировали. Обычную ПЦР проводили аналогично тому, как описано выше, за исключением того, что выделенную геномную ДНК, плазмидную/фосмидную ДНК использовали в качестве матрицы вместо непосредственного использования ДНК, выделенной из мицелиальных колоний без предварительной очистки.
Конструирование интегративной экспрессионной плазмиды pXY152-endorf24
Для эктопической экспрессии предполагаемого регуляторного гена orf24 из кластера генов биосинтеза эндурацидина в штаммах S. fungicidicus дикого типа и BM38-2 (ATCC PTA-122342), orf24 клонировали в интегративную плазмиду pXY152, полученную из pXY152aR20 (публикация Yin et al., J. Natural Products, 73: 583-589, 2010, которая в полном объеме включена здесь посредством ссылки) и orf24 амплифицировали с помощью ПЦР из геномной ДНК S. fungicidicus с использованием прямого праймера (End24Ndpf: 5'- CCACCACATATGGAAATAAGTTCGCTCTCCA-3' (SEQ ID NO:1, сайт NdeI выделен жирным шрифтом) и обратного праймера (End24ERpr:5'-GTGTGTGAATTCCTCGTTCACCCGGCCAGATG-3' (SEQ ID NO: 2, сайт EcoRI выделен жирным шрифтом). Продукт ПЦР расщепляли NdeI и EcoRI. Очищенный в геле фрагмент orf24 затем лигировали с аналогично рестриктированным вектором pXY152. Полученную плазмиду обозначали как pXY152-endorf24 (фиг. 2; SEQ ID NO: 3). Секвенирование подтверждало безошибочную вставку orf24.
Конструирование плазмиды pXY300-orf18ifd для делеции внутри рамки orf18
pXY300-orf18ifd конструировали клонированием двух фрагментов, которые фланкируют orf18 и предназначены для делеции в pXY300, чувствительный к температуре, конъюгативный челночный вектор E.coli-Streptomyces, содержащий ген устойчивости к тиострептону (tsr) для отбора в S. fungicidicus. Фланкирующие последовательности «апстрим» 2 т.п.н. и «даунстрим» 2 т.п.н., обозначенные как orf18ifdNP и orf18ifdPH соответственно, которые фланкируют orf18, получали с помощью ПЦР с использованием геномной ДНК S. fungicidicus в качестве матрицы и двух наборов праймеров. Фрагмент orf18ifdPH амплифицировали с использованием прямого и обратного праймеров (Ifdenorf18pf1, 5'-TTATTGAAGCTTGCCGGGGCCGACGCGGCGGGCGGCCT-3'
(SEQ ID NO: 4), Ifdendorf18pr1, 5'-GTTGTTTTAATTAAACACCAGGCCTCCTGGGGTG-3'
(SEQ ID NO: 5), сайты HindIII и PacI выделены жирным шрифтом). Фрагмент orf18ifdNP амплифицировали с использованием прямого и обратного праймеров (Ifdendorf18pf2, 5'-TTTATATTAATTAATGACCCTTCCGTCCCGCCCCCGAT-3'
(SEQ ID NO: 6), Ifdendorf18pr2, 5'-TTTGGTGCTAGCTGGTCGTGGCGCTGTTCC-3' (SEQ ID NO: 7), сайты Pacl и Nhel выделены жирным шрифтом. Эти два ПЦР-фрагмента были соответствующим образом вырезаны и одновременно лигированы с вектором pXY300, полученным расщеплением NheI и HindIII, с получением плазмиды pXY300-orf18ifd (фиг. 3; SEQ ID NO: 8). Безошибочная вставка делеции внутри рамки считывания pXY300-orf18ifd была подтверждена секвенированием.
Конструирование плазмиды pKS-T-orf18pfrd-AmR для делеции orf18 и его фланкирующих областей.
Фрагмент oriT амплифицировали с помощью ПЦР из плазмиды pIJ773 с использованием прямого праймера (Oritnhexbahd3f, 5'-AGCACAGCTAGCTTCTAGAAGCTTCATTCAAAGGCCGGCA-3' (SEQ ID NO: 9) сайт HindIII выделен жирным шрифтом) и обратного праймера (Oriter1pstxhor, 5'-GCCAGTGAATTCTGCAGCTCGAGCAGAGCAGGATTCCCGTTGA-3' (SEQ ID NO: 10), сайт XhoI выделен жирным шрифтом). Фрагмент oriT расщепляли с помощью HindIII и XhoI, очищали в геле и затем лигировали в аналогично рестриктированный производный вектор pBluescript II KS, с получением плазмиды pKS-T (Alting-Mees and Short, Nucleic Research Research, 17: 9494, 1989). Вставку плазмиды pXY300-orf18ifd вырезали расщеплением NheI и HindIII, очищали в геле и затем лигировали с плазмидой pKS-T, линеаризованной NheI и HindIII, с получением плазмиды pKS-T-orf18ifd (фиг. 4; SEQ ID NO: 11). Фрагмент размером 1 т.п.н., несущий aac(3)IV, ген устойчивости к апрамицину (amR), амплифицировали из pIJ773 с использованием прямого праймера (ApraNcoIpf, 5'-GAATGGCCATGGTTCATGTGCAGCTCCAT-3' (SEQ ID NO: 12), сайт NcoI выделен жирным шрифтом) и обратного праймера (ApraBamHIpr, 5'- TCTCGAGGATCCGAATAGGAACTTCGGAAT-3' (SEQ ID NO: 13), сайт BamHI выделен жирным шрифтом). Расщепление фрагмента AmR и плазмиды pKS-T-orf18ifd с помощью NcoI и BamHI подготовило вставку и вектор для лигирования. Полученную плазмиду обозначали как pKS-T-orf18pfrd-AmR (фигура 5; SEQ ID NO: 14).
Конструирование плазмиды pKS-T-orf18ifd-AmR (NS) для делеции внутри рамки считывания orf18.
Вставку pXY300-orf18ifd вырезали расщеплением NheI и HindIII, очищали в геле и затем лигировали с вектором pBluescript II KS, линеаризованным SpeI и HindIII, с получением плазмиды pKS-orf18ifd. Фрагмент oriT амплифицировали с помощью ПЦР с использованием прямого праймера (Oritnhexbahd3f, 5'-AGCACAGCTAGCTTCTAGAAGCTTCATTCAAAGGCCGGCA-3' (SEQ ID NO: 15), сайт HindIII выделен жирным шрифтом) и обратного праймера (oriTXhNdSpr, 5'-AGGCAGCTCGAGCATATGACTAGTCAGAGCAGGATTCCCGTTGA-3'(SEQ ID NO: 16), XhoI, сайты NdeI and SpeI выделены жирным шрифтом). Фрагмент oriT расщепляли с использованием XhoI и HindIII, очищали в геле и затем лигировали с аналогично рестриктированной плазмидой pKS-orf18ifd с получением плазмиды pKS-orf18ifd-T. Фрагмент размером 1 т.п.н., несущий ген aac(3)IV, придающий устойчивость к апрамицину (AmR), амплифицировали из pIJ773 с помощью ПЦР с использованием прямого праймера (ApraNdepf, 5'-GAATGGCATATGGTTCATGTGCAGCTCCAT-3' (SEQ ID NO: 17), сайт NdeI выделен жирным шрифтом) и обратного праймера (ApraSpeIpr, 5'-TCTAGAACTAGTGAATAGGAACTTCGGAAT-3' (SEQ ID NO: 18), сайт SpeI выделен жирным шрифтом). Плазмиду pKS-orf18ifd-T линеаризовали расщеплением NdeI и SpeI, и затем лигировали с аналогично вырезанным фрагментом AmR с получением плазмиды pKS-orf18ifd-T-AmR(NS) (фиг. 6; SEQ ID NO: 19).
Межродовая конъюгация, процедуры разрушения генов на основе pXY300 и pKS.
Плазмиды для разрушения генов индивидуально вводили в E.coli S17-1 посредством трансформации, и затем переносили в S. fungicidicus или его производные конъюгацией. Вкратце, свежесобранные споры S. fungicidicus предварительно проращивали, и клетки E. coli S17-1 культивировали в течение ночи при 37°С в бульоне Террифик. Готовили серийные разведения суспензии пророщенных спор, и 100 мл каждого разведения смешивали с равным объемом E. coli S17-1, содержащего плазмиды для разрушения на основе pXY300. Растворы высевали на чашки с агаром ISP4 с добавлением 10 мМ MgCl2 и инкубировали в течение 22 ч при 30°С или 37°C. Каждую чашку покрывали 3 мл мягкого питательного агара, содержащего налидиксат натрия и апрамицин (0,5 мг/мл), и затем инкубировали при 30°С в течение примерно одной недели. Выделенные экзоконъюгаты, которые выживали при отборе на антибиотике, очищали посевом на чашки с агаром ISP4, с добавлением налидиксата натрия и апрамицина (по 50 мкг/мл каждого).
Для проведения исследований по разрушению генов с использованием плазмид на основе pXY300 экзоконъюгаты сначала культивировали в жидкой среде TSB, содержащей апрамицин (5 мкг/мл), при 30°C в течение 24 ч, после чего мицелий собирали, гомогенизировали и использовали для инокуляции жидкой среды TSB с добавлением апрамицина (5 мкг/мл). Через 3-6 суток инкубации при 40°С мицелий гомогенизировали и высевали на чашки с агаром ISP4, содержащим апрамицин (50 мкг/мл), и инкубировали при 30°С в течение одной недели. Геномную ДНК выделяли из произвольно отобранных отдельных выживших колоний и анализировали с использованием ПЦР или саузерн-блоттинга для подтверждения того, что произошло разрушение одним или двойным кроссинговером. Для плазмид для разрушения гена, на основе pKS, и плазмид для делеции внутри рамки считывания, экзоконъюгаты пропускали через три последовательных цикла инкубации на чашках с агаром ISP4 для споруляции без дополнительного какого-либо отбора на антибиотике, чтобы стимулировать превращение в рекомбинанты двойного кроссинговера. Экзоконъюгаты на основе pKS не подвергали температурному отбору при 40°C. Правильную конструкцию всех мутантов подтверждали с использованием ПЦР и/или саузерн-блоттинга.
Конструирование интегративных экспрессионных плазмид pXY152-endorf24-camtsr и pXY152-endorf24-blatsr.
Для эктопической экспрессии orf24 в мутанте, устойчивом к апрамицину, несущем делецию orf18 и его фланкирующих областей, конструировали интегративную экспрессионную плазмиду pXY152-endorf24-blatsr. Для конструирования этой плазмиды вырезали кассету (camtsr), несущую ген устойчивости к хлорамфениколу и ген устойчивости к тиострептону (tsr), из производной плазмиды pUC57 расщеплением SacI и NheI. Затем кассету camtsr лигировали с плазмидой pXY152-endorf24, линеаризованной SacI и NheI, с получением новой конструкции pXY152-endorf24-camtsr (фиг. 7; SEQ ID NO: 20). Ген устойчивости к ампициллину (bla) амплифицировали с помощью ПЦР из pBluescript KS с использованием прямого праймера (amp2956SwaIpf, 5'-GTGGCAATTTAAATGGAAATGTGCGCGGAA-3' (SEQ ID NO: 21), сайт SwaI выделен жирным шрифтом) и обратного праймера (amp1973SacIpr, 5'-TATATAGAGCTCAACTTGGTCTGACAGTTAC-3' (SEQ ID NO: 22), сайт SacI выделен жирным шрифтом). Затем bla клонировали в сайты SacI и SwaI pXY152-endorf24-camtsr, с заменой кассеты camtsr на blatsr. Полученную экспрессионную конъюгативную плазмиду обозначали как pXY152-endorf24-blatsr (фиг. 8; SEQ ID NO: 23).
Конструирование кассеты Tn5AT для мутации транспозона in vitro
Кассета Tn5AT была сконструирована для объединения трех генетических элементов: транспозона Tn5, oriT и aac3(IV). Tn5 специфически и уникально распознается транспозазой Tn5 (Epicenter) и легко встраивается в ДНК с высоким содержанием G+C Streptomyces, клонированную в плазмиды и фосмиды E.coli (также упоминаемые в патенте США № 8188245, который включен здесь посредством ссылки). oriT необходим для конъюгативного переноса ДНК из E.coli S17-1 в Streptomyces, и aac(3)IV является бифункциональным селектируемым маркером E. coli-Streptomyces, обеспечивающим устойчивость к апрамицину. Оба oriT и aac3(IV) вырезали из плазмиды pIJ773 в виде фрагмента XbaI и затем клонировали в донорную плазмиду с транспозоном pMODTM-2 (MCS) (Epicenter), предварительно линеаризованную с помощью XbaI. Полученные плазмиды pXYTn5ATa и pXYTn5ATb различаются только ориентацией фрагмента XbaI, и их использовали для получения кассеты Tn5AT расщеплением PvuII в соответствии с инструкцией производителя.
Мутация транспозона in vitro и отбор мутагенизированных фосмид pXYF24D3 и pXYF148D12
Для создания библиотеки случайных мутагенизированных фосмид, несущих сегменты кластера биосинтеза эндурацидина, для исследований по замещению генов, проводили опыты с инсерционной мутацией транспозона фосмид pXYF24 и pXYF148 in vitro. Два предполагаемых регуляторных гена биосинтеза эндурацидина, orf18 и orf24, находятся во вставках фосмид pXYF24 и pXYF148 соответственно (идентификационный номер GenBank DQ403252). Реакцию с транспозоном in vitro проводили при 37°С в течение 2 ч после смешивания 10 мкл (0,5 мкг) фосмидной ДНК-матрицы, 2 мкл (20 нг) ДНК-кассеты Tn5AT, 2 мкл 10× реакционного буфера, 1 мкл транспозазы Tn5 и 5 мкл стерильной воды. Трансформацию компетентных клеток E.coli EPI300TM-T1R (Epicenter) с использованием реакционной смеси с транспозоном проводили электропорацией. Мутагенизированные фосмиды отбирали на чашках с агаром LB, с добавлением 100 мкг/мл апрамицина. Чашки инкубировали в течение ночи при 37°С и выжившие колонии отбирали произвольно и выращивали в жидкой культуре LB с добавлением 100 мкг/мл апрамицина. Мутагенизированную фосмидную ДНК из этих колоний и контрольные фосмиды pXYF24 или pXYF148 расщепляли HindIII и анализировали с помощью электрофореза на 1% агарозных гелях. Кассета Tn5AT содержит один сайт HindIII, который пригоден для скрининга наличия одного или более событий разрушения в фосмидной вставке. В фосмидных вставках pXYF24 или pXYF148 отсутствуют сайты HindIII, и в фосмидном векторе присутствует только один сайт HindIII. Следовательно, расщепление с помощью HindIII легко идентифицирует фосмиды с одной вставкой Tn5AT по наличию двух полос в геле. Колонии, несущие мутагенизированные фосмиды с одной вставкой транспозона, отбирали произвольно и культивировали в жидкой культуре LB, чтобы обеспечить выделение фосмиды и идентификацию разрушенного гена. Скрининг проводили анализом последовательности с использованием праймера 5'-AAGGAGAAGAGCCTTCAGAAGGAA-3' (SEQ ID NO: 24), который соответствует области гена устойчивости к апрамицину. Таким образом, было установлено, что фосмиды pXYF24D3 и pXYF148D12 имеют Tn5AT, вставленный в orf18 в положении нуклеотида 26386 и orf24 в положении нуклеотида 34333 (идентификационный номер в GenBank № DQ403252), соответственно.
Инсерционное разрушение orf18 и orf24 в S. fungicidicus дикого типа ATCC 21013.
Фозмиды для замещения генов pXYF24D3 и pXYF148D12 по отдельности трансформировали в E.coli S17-1 электропорацией, и затем вводили в S. fungicidicus методом межродовой конъюгации (публикация Mazodier et al., J. Bacteriology, 171: 3583-3585, 1989, которая в полном объеме включена здесь посредством ссылки). Колонии экзоконъюгатов, выживших после отбора на апрамицине, подвергали трем последовательным раундам споруляции без отбора на антибиотике на чашках с агаром ISP2 с получением стабильного мутантного штамма посредством гомологичной рекомбинации с двойным кроссинговером. Полученные споры объединяли, разбавляли и высевали на чашки с агаром ISP2 с добавлением 50 мкг/мл апрамицина для подтверждения наличия устойчивости к апрамицину и для использования в посевной культуре и ферментации для продукции эндурацидина. Мутантный штамм с инсерционным разрушением orf18 в S. fungicidicus дикого типа обозначали как SfpXYF24D3, и мутантный штамм с инсерционным разрушением orf24 в S. fungicidicus дикого типа обозначали как SfpXYF148D12.
Продукция эндурацидина в лабораторном масштабе и в ферментере емкостью 10 л.
Лабораторные условия ферментации во встряхиваемых колбах для продукции эндурацидина штаммом S. fungicidicus дикого типа, BM38-2 (ATCC PTA-122342) и производными штаммами были такими, как описано Higashide et al. (J. Antibiotics, 21: 126-137, 1968), за исключением среды для продукции эндурацидина, которая была раскрыта в патенте (патент США № 4465771). Для ферментации в лабораторном масштабе 5 мл TSB использовали для инокуляции посевной культуры свежесобранными спорами стрептомицетов. Обычно 5-10 мл посевной культуры инкубировали на роторном встряхивателе при 225 об/мин и 30°С в течение 48 ч, и затем переносили в 50 мл среды для продукции эндурацидина на 10 суток непрерывной ферментации. Продукцию эндурацидина дикими типами и производными штаммами в строго контролируемых условиях также проводили в автоматических ферментерах емкостью 10 л.
Сравнение выхода эндурацидина (энрамицина) в штаммах дикого типа, мутантных и генно-инженерных штаммах Streptomyces fungicidicus
Экстракция эндурацидина из продуктов ферментации для анализа ВЭЖХ
Для экстракции метаболитов для анализа продукции эндурацидина методом ВЭЖХ свежий мицелий собирали центрифугированием, промывали деионизированной водой и ресуспендировали в 5× объеме (соотношение водного метанола (мл) к массе влажного мицелия (г)) 70% водного метанола (рН доводили до 3,5 с помощью 1 Н HCl). Суспензию встряхивали при 200 об/мин при комнатной температуре в течение ночи и затем центрифугировали при 4000 об/мин и 4°С в течение 20 мин. Затем 1,4 мл супернатанта из каждого образца переносили в отдельные микроцентрифужные пробирки емкостью 1,5 мл и центрифугировали при 13000 об/мин при комнатной температуре в течение 10 мин. Фильтрат пропускали через шприцевой фильтр 0,22 мкм и затем анализировали ВЭЖХ. Экстракцию метаболита из мицелия, продуцированного в ферментерах емкостью 10 л, проводили в небольших масштабах, аналогичных лабораторным ферментациям.
Анализ ВЭЖХ и определение выхода эндурацидина.
Образец 50 мкл для ВЭЖХ, полученный, как описано выше, инжектировали на колонку Gemini C18 (5 мкм, 4,6 × 150 мм, Phenomenex, Torrance, CA), присоединенную к жидкостному хроматографу Shimadzu HPLC. Разделение проводили с использованием 18-мин ступенчатых линейных градиентов с растворителем A: вода+0,1% ТФК и растворителем B: ацетонитрил. Скорость потока составляла 1 мл/мин, начиная с 10% B, увеличивая до 40% B в течение 10 мин, и затем дополнительно увеличивая до 95% B в течение 8 мин. УФ-область в диапазоне от 200 до 300 нм сканировали с помощью фотодиодно-матричного детектора SPD M20A. Выходы эндурацидинов рассчитывали сравнением со стандартной кривой, построенной с использованием стокового раствора стандартов эндурацидина в 70% метаноле. Серию инжектированных растворов, содержащих 2, 4, 6, 8, 10 и 12 мкг эндурацидина, использовали для построения стандартной кривой с использованием суммарных площадей поглощения для эндурацидинов А и В при 230 нм. Уравнение регрессии получали из стандартной кривой и использовали для расчета выхода эндурацидина.
Оценка антибактериальной активности.
Staphylococcus aureus (ATCC 29213) использовали в качестве индикаторного микроорганизма в биоанализе. Клетки использовали для инокуляции бульона LB, культивировали при 37°С в течение ночи, и затем 100 мкл культуры смешивали с 5 мл верхнего слоя агара (смесь равных объемов питательного агара и питательного бульона). Верхний слой агар накладывали на чашку с питательным агаром, в которой были сделаны соответствующие интервалы между лунками агара. Стандарты эндурацидина и аликвотные порции экстрактов культур растворяли или разбавляли 50% МеОН в концентрации 20 мкг/мл, и 100 мкл каждого раствора вносили в лунки. После инкубации чашек при 37°С в течение 16 ч, измеряли и сравнивали зоны ингибирования роста, и чашки фотографировали или хранили при 4°С.
Пример 2
Разрушение orf18 и orf24 в S. fungicidicus дикого типа и влияние на продукцию эндурацидина
В данном примере описывается разрушение orf18 и orf24 в S. fungicidicus дикого типа и влияние на продукцию эндурацидина.
Последовательность ДНК размером 116000 п.н. из S. fungicidicus ATCC 21013 дикого типа, которая содержит кластер гена биосинтеза эндурацидина и его фланкирующие области (патент США № 8188245, который во всей полноте включен здесь посредством ссылки), была ранее идентифицирована и доступна в GenBank (идентификационный номер DQ403252). Среди 48 аннотированных orfs имеется восемь предполагаемых регуляторных генов: orf5, orf12, orf18, orf22, orf24, orf41, orf42 и orf43. Для расшифровки роли каждого из генных продуктов в продукции эндурацидина, фосмидные вставки, несущие сегменты кластера эндурацидина, включающие эти предполагаемые регуляторные гены, произвольно мутировали с использованием транспозон-опосредованной вставки маркера устойчивости к апрамицину, как описано в примере 1.
Последующий скрининг на устойчивость к апрамицину и положение вставки в колониях E.coli, несущих мутагенизированные фосмиды, идентифицировали pXYF24D3 для переноса разрушенного orf18 и pXYF148D12 для удержания разрушенного orf24. Наличие одной инсерционной мутации в каждой из этих фосмид и сайт инсерции подтверждали секвенированием. Затем эти две мутагенизированные фосмиды по отдельности вводили конъюгацией в штамм S. fungicidicus дикого типа. Экзоконъюгаты, демонстрирующие устойчивость к апрамицину, затем подвергали трем раундам споруляции на агаре ISP2 без дополнительного какого-либо отбора на антибиотике для стимуляции превращения гомологичной рекомбинации с одинарным кроссинговером в мутацию с двойным кроссинговером. Полученные в результате стабильные мутантные штаммы SfpXYF24D3 и SfpXYF148D12 ферментировали в среде для продукции эндурацидина (EPM) в лабораторном масштабе во встряхиваемых колбах. Анализ ВЭЖХ экстракта мицелия 70% метанолом после 10-дневной ферментации выявил увеличение выхода эндурацидина в 1,3 раза у штамма SfpXYF24D3, с разрушенным orf18, и полную потерю продукции эндурацидина штаммом SfpXYF148D12, имеющим разрушенный orf24. Экстракты мицелия также оценивали на активность в отношении S. aureus. SfpXYF24D3 с разрушенным orf18 сохранял активность, тогда как SfpXYF148D12 с разрушенным orf24 терял активность в отношении S. aureus.
Пример 3
Конструирование рекомбинантного штамма SfpXY152-endorf24 и влияние на продукцию эндурацидина
В данном примере описывается конструирование рекомбинантного штамма SfpXY152-endorf24 и оценка способности этого штамма продуцировать эндурацидин.
Потеря способности продуцировать эндурацидин у мутантного штамма SfpXYF148D12 указывает на возможную регуляторную роль orf24. Поиск BLAST в базе данных GenBank с использованием последовательности белка Orf24 выявил высокое сходство последовательности со специфическим для пути регуляторным белком StrR, участвующим в биосинтезе стрептомицина. Выравнивание последовательностей Orf24 и StrR показало, что белки имеют значительное сходство (54% идентичность, фиг. 9). Потеря способности продуцировать эндурацидин в результате разрушения orf24 и сходство с StrR указывают на то, что Orf24 может функционировать в качестве специфического для пути активатора в продукции эндурацидина.
Для исследования роли orf24 в качестве позитивной регуляторной мишени для улучшения штамма, была сконструирована интегративная экспрессионная плазмида pXY152-endorf24 (фиг. 2) (пример 1). Плазмиду pXY152-endorf24 вводили в S. fungicidicus дикого типа посредством конъюгации, и экзоконъюгаты подвергали скринингу на фенотип устойчивости к апрамицину, что привело к идентификации нового рекомбинантного штамма SfpXY152-endorf24. По меньшей мере, десять колоний независимых экзоконъюгатов из этого штамма были отобраны произвольно и очищены. Данные штаммы колоний несут плазмиду pXY152-endorf24, интегрированную в сайт attB в хромосоме S. fungicidicus гомологичной рекомбинацией с одинарным кроссинговером с сайтом attP в плазмиде.
Для исследования метаболитов, продуцированных рекомбинантными штаммами, споры из колоний двух штаммов инокулировали в посевную культуру TSB, и затем переносили в среду для продукции эндурацидина в ферментации в лабораторных условиях. Анализ ВЭЖХ 70% метанольных экстрактов собранного мицелия выявил 2-кратное увеличение (60 мг/л) продукции эндурацидина обоими рекомбинантными штаммами по сравнению со штаммом дикого типа (30 мг/л). Повышенные выходы эндурацидина, наблюдаемые в колониях этих штаммов, которые способны к сверхэкспрессии orf24, являются еще одним доказательством позитивной регуляторной роли, которую данный ген играет в продукции эндурацидина, и результаты согласуются с результатами, полученными в опытах с разрушением orf24, которое привело к потере продукции эндурацидина.
Пример 4
Конструирование штамма BM38-2.24/16, сверхэкспрессирующего orf24 на основе S. fungicidicus BM38-2 (ATCC PTA-122342), и влияние на продукцию эндурацидина
В данном примере описывается конструирование штамма BM38-2.24/16 (депонирование ATCC № PTA-124006), сверхэкспрессирующего orf24 на основе S. fungicidicus BM38-2 (ATCC PTA-122342), и влияние на продукцию эндурацидина.
Для дальнейшего изучения позитивной регуляторной роли Orf24 плазмиду pXY152-endorf24 включали в хромосому коммерческого производственного штамма S. fungicidicus BM38-2 (ATCC PTA-122342), как описано выше для организма дикого типа. Отбор экзоконъюгантов, обладающих фенотипом устойчивости к апрамицину, позволил получить ряд колоний рекомбинантных штаммов, в том числе S. fungicidicus BM38-2.24/16, способных продуцировать повышенные уровни эндурацидина на уровне до 200 мг/л (увеличение в 3,3 раза по сравнению с BM38-2 (ATCC PTA-122342)) во встряхиваемых лабораторных культуральных колбах. S. fungicidicus BM38-2-24/16 был отобран для дальнейшей оценки способности продуцировать эндурацидин на основе выходов, установленных в предварительном скрининге.
Продукция эндурацидина рекомбинантным штаммом S. fungicidicus BM38-2.24/16 во встряхиваемых лабораторных культуральных колбах показала явный потенциал для значительного улучшения по сравнению с BM38-2 (ATCC PTA-122342), и, кроме того, наблюдали значительную вариабельность выходов. Для более тщательного контроля условий культивирования в течение 10-дневного периода культивирования, включая pH и растворенный кислород, которые трудно регулировать во встряхиваемых колбах, продукцию оценивали в нескольких циклах в 10 л ферментерах. В этих более строго контролируемых условиях выходы были более стабильными, и ферментация по 10 л в трех повторах в среднем составляла 375 мг/мл (в 4,6 раза выше, чем с BM38-2 (ATCC PTA-122342)). Повышенные выходы эндурацидина в рекомбинантном штамме S. fungicidicus BM38-2.24/16 (депонирование ATCC № PTA-124006) также подтверждают положительную роль Orf24 в повышающей регуляции продукции эндурацидина.
Пример 6
Конструирование делеционного мутантного штамма BM38-1.18pfrd-AmR и влияние на продукцию эндурацидина
В данном примере описывается конструирование делеционного мутантного штамма BM38-2.18pfrd-AmR (депонирование ATCC № PTA-124007) и влияние на продукцию эндурацидина.
orf18 находится в области апстрим кластера генов биосинтеза эндурацидина (идентификационный номер GenBank № DQ403252). Вероятно, Orf18 играет негативную роль в продукции эндурацидина, поскольку инсерционное разрушение гена в мутантном штамме SfpXYF24D3 повышало выход эндурацидина. На основании этого наблюдения были приготовлены конструкции для делеции одного orf18, и orf18 и участков его фланкирующих областей. Для этой цели конструировали плазмиду pKS-T-orf18pfrd-AmR (фиг. 5). Эта плазмида, полученная из вектора pKS, не имеет ни репликона стрептомицетов, ни элемента для интеграции в хромосому стрептомицетов. Она может только обменивать свою вставку с определенным сегментом ДНК в хромосоме хозяина посредством гомологичной рекомбинации с двойным кроссинговером. Карта вставки этой плазмиды показана на фиг. 10. orf18 и его фланкирующие области, содержащие полный orf19, и область, кодирующую N-концевой участок orf17, делецировали в плазмиде pKS-T-orf18pfrd-AmR. Левое плечо размером 1 т.п.н. содержит область, кодирующую C-концевой участок orf17 и его область даунстрим, и правое плечо размером 1 т.п.н. содержит неполный сегмент orf20, кодирующий N-концевую область. Следовательно, делеция после гомологичной рекомбинации с двойным кроссинговером приводила к получению рекомбинантного штамма, в котором полный orf18 плюс orf19 и область, кодирующая N-концевой участок orf17, были делецированы и заменены геном устойчивости к апрамицину.
Плазмиду pKS-T-orf18pfrd-AmR вводили конъюгацией в S. fungicidicus BM38-2 (ATCC PTA-122342), и гомологичная рекомбинация с одинарным и двойным кроссинговером стимулировалась на чашках с агаром ISP4 без добавления апрамицина. Экзоконъюгаты, которые были способны выжить после последующего отбора на апрамицине, очищали, и этот новый рекомбинантный штамм обозначали как BM38-2.18pfrd-AmR (депонирование ATCC № PTA-124007). Споры из этого штамма инокулировали в среду TSB для получения посевной культуры и затем переносили в среду для продукции эндурацидина. После 10 суток ферментации мицелий собирали, обрабатывали и анализировали с помощью ВЭЖХ. По сравнению с родительским штаммом BM38-2 (ATCC PTA-122342), наблюдали 1,2-кратное повышение продукции эндурацидина в этих лабораторных ферментациях. Относительное увеличение выхода было аналогичным тому, которое наблюдали со штаммом, полученным из SfpXYF24D3 дикого типа, и результаты показывают, что orf19 и orf17, которые фланкируют orf18 и которые были затронуты при конструировании BM38-2.18pfrd-AmR, оказывают незначительное влияние или вовсе не оказывают влияния на продукцию эндурацидина. Следовательно, повышенная продукция эндурацидина в рекомбинантном штамме BM38-2.18pfrd-AmR обусловлена элиминацией негативной регуляторной роли Orf18.
В отношении делеции одного orf18 с использованием плазмиды pXY300-orf18ifd в BM38-2 (ATCC PTA-122342), то возникли трудности с положительным отбором экзоконъюгатов и одиночных/двойных мутантов с использованием маркера устойчивости к тиострептону. Следовательно, альтернативный вектор pBluescript KS II использовали для конструирования безмаркерных плазмид для замещения генов, таких как pKS-T-orf18ifd (фиг. 4) или pKS-orf18ifd-T, pKS-orf18ifd-T-AmR (NS) (ген устойчивости к апрамицину переносится на векторе вместо вставки в orf18, см. фиг.6).
Пример 6
Разработка ряда pKS-производных векторов для инактивации гена pKS-T-orf18pfrd-AmR
В данном примере описывается разработка ряда pKS-производных векторов для инактивации гена pKS-T-orf18pfrd-AmR.
Был разработан ряд векторов для инактивации гена, полученных из pKS (фиг. 4, 5 и 6), которые обладают конъюгативной функцией и не требуют прохождения трансформантов через высокотемпературный отбор для элиминации плазмиды, как это требуется для некоторых других векторов для разрушения генов. Эти pKS-производные векторы несут нестрептомицетовый репликон, обеспечивающий репликацию в E.coli, и могут поддерживаться и отбираться по маркеру устойчивости к апрамицину в Streptomyces и E. coli или ампициллину в E. coli. Они генерировали обильные стабильные копии рекомбинантных плазмид в E.coli для конъюгации, и они были сконструированы с несколькими редкими и уникальными сайтами рестрикции, обнаруженными в ДНК стрептомицетов, такими как PacI, HindIII, NheI и XbaI, которые можно соответственно использовать для сборки ДНК-мишени в плазмиду для исследования инсерционного разрушения гена и делеции внутри рамки считывания.
Пример 7
Разработка pSET152-производных интегративных векторов экспрессии генов pXY152-endorf24-camtsr и pXY152-endorf24-blatsr
В данном примере описывается разработка pSET152-производных интегративных векторов экспрессии генов pXY152-endorf24-camtsr (SEQ ID NO: 20) и pXY152-endorf24-blatsr (SEQ ID NO: 23).
Были разработаны два новых вектора: pXY152-endorf24-camtsr (фиг.7) и pXY152-endorf24-blatsr (фиг.8). Они обладают конъюгативными и интегративными функциями, такими же, как вектор pSET152, который представляет наиболее широко используемый интегративный вектор для экспрессии и комплементации генов стрептомицетов. Оба этих вектора несут несколько сайтов рестрикции, которые редко имеют место в ДНК Streptomyces, для соответствующего клонирования и сборки экспрессионной конструкции. Вектор pXY152-endorf24-camtsr можно поддерживать и отбирать в E.coli с хлорамфениколом в концентрации 12,5 мкг/мл и в Streptomyces с тиострептоном в концентрации 50 мкг/мл. Вектор pXY152-endorf24-blatsr можно поддерживать и отбирать в E.coli с ампициллином и в Streptomyces с тиострептоном.
Заключение по примерам 1-6:
Генетические манипуляции с регуляторными генами и генами биосинтеза Streptomyces для улучшения штамма
Среди многочисленных микробных продуцентов природных продуктов примерно 75% известных микробных антибиотиков продуцируются актиномицетами. Streptomyces, грамположительные нитчатые почвенные бактерии, являются членами семейства актиномицетов и известны своей высокой способностью продуцировать широкий ряд структурно разнообразных, фармакологически и биологически активных вторичных метаболитов. Поликетиды, продуцируемые при участии поликетидсинтаз (PKS), и природные пептидные продукты, полученные с помощью нрибосомальных пептидсинтетаз (NRPS), являются типичными представителями.
Исследование биосинтеза антибиотиков, представляющих собой природные продукты, имеет ряд общих проблем: во-первых, как преодолеть типичную низкую продукцию исходных или структурно модифицированных соединений, продуцируемых штаммами дикого типа или генно-инженерными штаммами; во-вторых, как активировать множество непонятных или «бесхозных» путей биосинтеза вторичных метаболитов, идентифицированных по геномным последовательностям, чтобы можно было изучить биологическую функцию продуктов. Достижения в изучении биосинтеза антибиотиков, представляющих природные продукты, за последние десятилетия показали, что продукция вторичных метаболитов регулируется многими путями. Например, гены биосинтеза предшественника и структурной сборки (такие как PKS и NRPS), регуляторные гены и гены саморезистентности могут быть кластеризованы в бактериальной хромосоме. Продукция антибиотиков может регулироваться посредством специфических регуляторных генов, включая активаторы и/или репрессоры, плейотропные эктопические регуляторные гены и двухкомпонентные регуляторные системы. Мутации, имеющие место в любом из этих регуляторных генов или систем, могут приводить к увеличению, снижению или полной отмене продукции антибиотиков. Криптический путь биосинтеза может быть активирован непредполагаемой мутацией, приводящей к образованию ранее неизвестного продукта.
Улучшение штамма может играть важную роль в экономически эффективном производстве антибиотиков или других вторичных метаболитов в промышленном масштабе. Мутантные штаммы, способные продуцировать определенные метаболиты с повышенным выходом, могут быть получены посредством случайных мутаций или целенаправленного разрушения определенных генов или введения гена(ов), которые элимируют узкие места на пути биосинтеза. Генетические манипуляции с позитивными и негативными регуляторными генами, а также с биосинтетическими генами для генерации гиперпродукции целевых вторичных метаболитов, как было доказано, являются эффективной и весьма успешной стратегией улучшения штамма актиномицетов.
В настоящем описании была показана позитивная регуляторная роль orf24 и негативная регуляторная роль orf18 в продукции эндурацидина. Направленная инсерционная инактивация orf24 приводила к полной потере продукции эндурацидина в рекомбинантном штамме SfpXYF148D12. Последующая сверхэкспрессия orf24 под контролем сильного конститутивного промотора ermE*p в рекомбинантных штаммах SfpXY152-endorf24 и BM38-2.24/16 приводила к повышению выхода эндурацидина примерно в 2-4,6 раза. Делеция orf18 и его фланкирующих областей, включая делецию полного orf19 и части orf17, увеличивала выход эндурацидина в 1,2 раза. Эти результаты предоставили убедительные, генетические доказательства, свидетельствующие о роли orf24 и orf18 в качестве позитивного активатора и негативного репрессора, соответственно, в биосинтезе эндурацидина.
Наличие ортологов Orf24 из других путей биосинтеза антибиотиков было функционально подтверждено.
Запрос с использованием BLAST в отношении последовательности белка Orf24 в базе данных GenBank выявил сотни попаданий (идентификационный номер GenBank DQ403252). Многие показывают очень высокое сходство аминокислот (идентичность на уровне от 60% до 99%), и они аннотируются в качестве регуляторов транскрипции в биосинтезе аминогликозидного антибиотика стрептомицина. Однако ни для одного гена из этой группы функция не была подтверждена экспериментально. Анализ результатов BLAST выявил несколько родственных белков, которые имеют более низкое сходство (идентичность выше 40%, но ниже 60%) с Orf24, которые были функционально охарактеризованы. К ним относится хорошо охарактеризованный белок StrR, который имеет более низкое, но значимое сходство (54% идентичность в совпадении 311 аминокислот) с Orf24. Генетически и биохимически было показано, что StrR функционирует в качестве специфического для пути позитивного активатора экспрессии генов биосинтеза стрептомицина в Streptomyces griseus. StrR представляет собой семейство специфических активаторов пути, несколько из которых были охарактеризованы либо генетическими манипуляциями, либо биохимическими исследованиями. На фиг. 11 показано выравнивание Orf24 с шестью функционально подтвержденными StrR-подобными белками актиномицетов. Типичный и высококонсервативный ДНК-связывающий домен спираль-поворот-спираль (HTH) присутствует во всех семи белках, он подчеркнут на фиг. 11. Orf24 также имеет значительное сходство последовательностей (54% идентичность) с Teil15*, специфическим для пути активатором, регулирующим биосинтез нерибосомально синтезируемого гликопептидного антибиотика тейкопланина. Tei15* положительно регулирует транскрипцию, по меньшей мере, 17 генов в кластере тейкопланина. Actinoplanes teichomyceticus дикого типа продуцирует примерно 100 мг/л тейкопланина, тогда как генетические рекомбинантные штаммы, полученные из родительского A. teichomyceticus и несущие tei15*, с экспрессией под контролем различных промоторов, увеличивали выход тейкопланина до 1 г/л в случае промотора ermE*p и до 4 г/л в случае нативного промотора гена устойчивости к апрамицину.
Как показано на фиг. 11, Orf24 также имеет значительное сходство последовательностей (54% идентичность) с Bbr из кластера биосинтеза гликопептидного антибиотика бальхимицина; KasT (50% идентичности), регулирующего экспрессию генов биосинтеза аминогликозидного антибиотика касугамицина; и NovG (45% идентичность), специфический для пути активатор, участвующий в биосинтезе новобиоцина. Мутант ΔnovG продуцировал только на 2% больше новобиоцина по сравнению с диким типом, и сверхэкспрессия novG из мультикопийной плазмиды в рекомбинантном штамме приводила к трехкратному увеличению продукции новобиоцина. Orf24 также обладает 42% идентичностью с SgcR1, одним из четырех регуляторных генов (sgcR1, sgcR2, sgcR3 и sgcR), экспериментально подтвержденных в качестве участвующих в продукции противоопухолевого антибиотика C-1027 в S. globisporus. Сверхэкспрессия sgcR1 в S. globisporus SB1022 увеличивала выход С-1027 примерно в семь раз по сравнению со штаммом дикого типа. Сверхэкспрессия позитивного регулятора sgcR3 в рекомбинантном штамме приводила к увеличению продукции C-1027 на 30-40%. Напротив, инактивация негативного регулятора sgcR приводила к повышению продукции как C-1027, так и гептаена. Кроме того, сверхэкспрессия sgcR1 в мутантном штамме ΔsgcR приводила примерно к семикратному увеличению продукции C-1027. sgcR3 обладает регуляцией более высокого уровня за счет контроля sgcR1 и sgcR2 в иерархической регуляции продукции C-1027. В заключение следует отметить, что эффекты разрушения и экспрессии orf24 и сравнение Orf24 с другими функционально охарактеризованными ортологами указывают на то, что Orf24 функционирует в качестве специфического для пути позитивного регулятора/активатора в продукции эндурацидина.
Orf18 является предполагаемым атипичным орфановым регулятором ответа и соответствует функционально подтвержденным ортологам
Продукция антибиотиков видами Streptomyces жестко регулируется сложными генетическими сетями, которые ограничивают способность многих продуцентов антибиотиков дикого типа генерировать выход, необходимый для крупномасштабного и экономически эффективного промышленного производства. Одним из важных регуляторных механизмов являются двухкомпонентные системы передачи сигналов. Двухкомпонентные системы включают сенсорную киназу и когнатный регулятор ответа. Сенсорная киназа отвечает на определенные внешние стимулы/сигналы окружающей среды, такие как стресс, питательные и химические вещества, и т.д., и затем передает сигнал цитоплазматическому регулятору ответа, который запускает и активирует транскрипцию генов-мишеней. Регулятор ответа, который не спарен с сенсорной киназой, обозначается как орфановый регулятор ответа.
Двухкомпонентные системы и орфановые регуляторы ответа присутствуют в геномах стрептомицетов и могут функционировать для подавления продукции вторичных метаболитов. В кластере генов биосинтеза эндурацидина из S. fungicidicus orf18 кодирует предполагаемый орфановый регулятор ответа, который имеет сходство последовательности в диапазоне от низкой до средней с тремя другими охарактеризованными регуляторами ответа Streptomyces, включая один орфановый регулятор ответа, SCO3818 из S. coelicolor (фиг. 12). Orf18 имеет более длинную N-концевую последовательность по сравнению с другими выравненными белками и, по-видимому, является атипичным орфановым регулятором ответа, поскольку высококонсервативный лизин в положении 118 (по сравнению с обычным положением 105) отсутствует в Orf18 и замещен треонином. Предполагается, что лизин необходим для формирования «кармана» фосфорилирования.
Только несколько регуляторов ответа стрептомицетов были функционально охарактеризованы. Геном S. coelicolor содержит в общей сложности пять атипичных и семь типичных орфановых регуляторов ответа. Orf18 имеет 26% идентичность с совпадением 191 аминокислоты с AbsA2. Делеция AbsA2 в S. coelicolor приводила к повышению продукции двух антибиотиков, актинородина и ундецилпродигиозина. Orf18 показывает 32% идентичность с совпадением 176 аминокислот с SCO3818. Делеция sco3818 приводила к повышению продукции актинородина. Orf18 имеет 29% идентичность с совпадением 166 аминокислот с SCO1745 (AbrA2). Делеция оперона регулятора ответа, содержащего AbrA2, приводила к 100% увеличению продукции противоопухолевого антибиотика овидомицина в рекомбинантном штамме S. coelicolor M145 по сравнению с продуцентом дикого типа. Наблюдаемая негативная регуляторная роль Orf18 в продукции эндурацидина согласуется с продемонстрированной активностью родственных негативных регуляторов (фиг. 12). Кроме того, отмечено, что Orf18 имеет наибольшее сходство белковой последовательности с членами семейства LuxR транскрипционных регуляторов в поисковоц системе BLAST.
Отсутствие полярных эффектов у мутанта BM38-1.orf18pfrd-AmR
Делецированная область в мутантном штамме BM38-1.18pfrd-AmR включает три гена, orf18, область, кодирующую N-концевой участок or17, расположенный ниже orf18, и полный orf19, расположенный выше orf18 (фиг. 5 и 10). Предполагается, что orf17 кодирует рибонуклеазу, вероятно, не имеющую функции, связанной с биосинтезом или регуляцией биосинтеза эндурацидина. Также ген устойчивости к апрамицину, заменяющий orf18 и его фланкирующую область, транскрибируется дивергентно с orf17 и не должен создавать каких-либо событий считывания от промотора гена устойчивости к апрамицину. Следовательно, не должно быть никаких полярных эффектов, возникающих в результате частичной делеции orf17.
orf19 транскрибируется и транслируется в том же направлении, что и orf18. Данный ген аннотирован для кодирования белка с неизвестной функцией. Мутантный штамм SfpXYF24D3, несущий один разрушенный orf18, и мутант BM38-1.18pfrd-AmR, несущий делецию обоих orf18 и orf19, оказывают сходное усиливающее влияние на продукцию эндурацидина, что подразумевает, что orf19 не играет никакой роли или играет незначительную роль в продукции эндурацидина. Ген orf20 расположен апстрим от orf19 и транскрибируется и транслируется в том же направлении, что и встроенный маркер устойчивости к апрамицину (фиг.10). Orf20 остается интактным в BM38-1.18pfrd-AmR, и продукт, вероятно, не играет роли в продукции эндурацидина. Следовательно, полагается, что любые полярные эффекты на экспрессию orf20 не ответственны за повышение продукции эндурацидина в BM38-1.18pfrd-AmR.
Пример 7
Дальнейшие применения и манипуляции с orf24 и/или orf18 для создания штаммов с повышенной продукцией эндурацидина
В дополнение к вышеприведенным примерам существуют и другие возможные способы применения регуляторной роли orf24 и orf18 для повышения продукции эндурацидина.
I. Экспрессия orf24 с альтернативным, конститутивным или индуцибельным промотором сверхэкспрессии
pXY152-endorf24 (показанный на фиг.2) был сконструирован для интегративной эктопической экспрессии orf24 под контролем ermE*p, широко используемого сильного конститутивного стрептомицетового промотора экспрессии. Сверхэкспрессия orf24 также может регулироваться другими конститутивными или индуцибельными промоторами. Промотор tipA представляет собой индуцибельный стрептомицетовый промотор сверхэкспрессии тиострептона. Была разработана мультикопийная, содержащая промотор tipA, челночная плазмида E.coli-Streptomyces, pXY200, которая была успешно использована для сверхэкспрессии генов стрептомицетов. Для применений, относящихся к настоящему раскрытию, промотор tipA можно вырезать из pXY200 и клонировать в pXY152 для замены ermE*p и регуляции экспрессии orf24. Аналогично, orf24 может быть легко перенесен из pXY152-endorf24 в pXY200 для экспрессии на основе плазмиды. Другие варианты промотора включают, не ограничиваясь этим, систему P(nitA)-NitR и промотор стрептомицетов SF14. Недавно интегративная плазмида pKC1139 и нативный промотор гена устойчивости к апрамицину были успешно использованы для экспрессии регуляторных генов для гиперпродукции пептидного антибиотика тейкопланина. Регуляторный ген sanG кодирует специфический для пути активатор продукции никкомицина. Экспрессия дополнительной копии sanG под контролем пяти различных промоторов (PhrdB, Ptcp830, PSF14, PermE* и Pneos) приводила к увеличению выхода никкомицина на 69%, 51%, 26%, 22% и 13% соответственно (см. Du et al., Applied Microbiology and Biotechnology, 97: 6383-6396, 2013).
ii. Двойные мутантные штаммы S. fungicidicus с делецией orf18 и сверхэкспрессией orf24
Как с делеционным мутантом orf18, так и со штаммами сверхэкспрессии orf24, демонстрирующими повышенную продукцию эндурацидина, можно получить двойной мутант, содержащий оба, и возникает вопрос о том, наблюдается ли аддитивный эффект на выход этого пептидного антибиотика. Двойной мутант может быть сконструирован введением плазмиды для сверхэкспрессии pXY152-endorf24-blatsr (фиг. 8) в мутант BM38-2.18pfrd-AmR. pXY152-endorf24-blatsr представляет конъюгативную интегративную плазмиду, несущую ген устойчивости к тиострептону (tsr) для отбора в стрептомицетах и ген устойчивости к ампициллину (bla) для отбора в E.coli. Поскольку штамм E. coli S17-1, используемый для конъюгации, естественно устойчив к хлорамфениколу (cam), то маркер устойчивости к хлорамфениколу в pXY152-endorf24-camtsr (см. выше) заменяли устойчивостью к ампициллину (bla), для отбора трансформанта S17-1. Альтернативно, pXY152-endorf24-camtsr и ее производные могут быть введены в стрептомицеты с использованием другого конъюгативного штамма E.coli, ET12567/pUZ8002.
Используя плазмиду pXY152-endorf24-blatsr или pXY152-endorf24-camtsr для введения второй копии orf24 в дефицитный по orf18 мутант, можно отобрать двойной мутант по устойчивости к тиострептону. Для генерации инактивированного мутанта с делецией внутри рамки считывания в BM38-2 (ATCC PTA-122342) были сконструированы плазмиды pXY300-orf18ifd (фиг.3) и pKS-orf18ifd-T-AmR (NS) (фиг.6) для этой цели. pXY300-orf18ifd позволяет провести отбор делеционного мутанта orf18 с тиострептоном, в то время как pKS-orf18ifd-T-AmR (NS) использует апрамицин для отбора делеционных мутантов внутри рамки считывания. Несмотря на то, что мутантные штаммы S. fungicidicus дикого типа легко селектируются с использованием маркера устойчивости к тиострептону, при использовании этого маркера устойчивости в штамме BM38-2 (ATCC PTA-122342) возникли трудности. Таким образом, две плазмиды, pXY300-orf18ifd и pKS-orf18ifd-T-AmR (NS), были сконструированы для одной и той же цели.
Ввиду многих возможных вариантов осуществления, к которым могут применяться принципы раскрытого изобретения, следует понимать, что описанные варианты осуществления являются только предпочтительными примерами изобретения и не должны рассматриваться в качестве ограничивающих объем изобретения. Скорее объем изобретения определяется следующей формулой изобретения. Следовательно, заявители заявляют в качестве их изобретения все, что входит в объем и сущность формулы настоящего изобретения.
--->
Список последовательностей
<110> Oregon State Unversity
Zabriskie, T. Mark
Yin, Xihou
<120> КОМПОЗИЦИИ И СПОСОБЫ ПОВЫШЕНИЯ ПРОДУКЦИИ ЭНДУРАЦИДИНА В ГЕННО-
ИНЖЕНЕРНОМ ШТАММЕ STREPTOMYCES FUNGICIDICUS
<130> 127789-236560-P005PCT
<150> 62/430,838
<151> 2016-12-06
<150> 62/479,087
<151> 2017-03-30
<160> 40
<170> PatentIn version 3.5
<210> 1
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотидный праймер
<400> 1
ccaccacata tggaaataag ttcgctctcc a 31
<210> 2
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотидный праймер
<400> 2
gtgtgtgaat tcctcgttca cccggccaga tg 32
<210> 3
<211> 6210
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая плазмида pXY152-endorf24
<400> 3
gaattcgtaa tcatgtcata gctgtttcct gtgtgaaatt gttatccgct cacaattcca 60
cacaacatac gagccggaag cataaagtgt aaagcctggg gtgcctaatg agtgagctaa 120
ctcacattaa ttgcgttgcg ctcactgccc gctttccagt cgggaaacct gtcgtgccag 180
ctgcattaat gaatcggcca acgcgcgggg agaggcggtt tgcgtattgg gcgctcttcc 240
gcttcctcgc tcactgactc gctgcgctcg gtcgttcggc tgcggcgagc ggtatcagct 300
cactcaaagg cggtaatacg gttatccaca gaatcagggg ataacgcagg aaagaacatg 360
tgagcaaaag gccagcaaaa ggccaggaac cgtaaaaagg ccgcgttgct ggcgtttttc 420
cataggctcc gcccccctga cgagcatcac aaaaatcgac gctcaagtca gaggtggcga 480
aacccgacag gactataaag ataccaggcg tttccccctg gaagctccct cgtgcgctct 540
cctgttccga ccctgccgct taccggatac ctgtccgcct ttctcccttc gggaagcgtg 600
gcgctttctc atagctcacg ctgtaggtat ctcagttcgg tgtaggtcgt tcgctccaag 660
ctgggctgtg tgcacgaacc ccccgttcag cccgaccgct gcgccttatc cggtaactat 720
cgtcttgagt ccaacccggt aagacacgac ttatcgccac tggcagcagc cactggtaac 780
aggattagca gagcgaggta tgtaggcggt gctacagagt tcttgaagtg gtggcctaac 840
tacggctaca ctagaagaac agtatttggt atctgcgctc tgctgaagcc agttaccttc 900
ggaaaaagag ttggtagctc ttgatccggc aaacaaacca ccgctggtag cggtggtttt 960
tttgtttgca agcagcagat tacgcgcaga aaaaaaggat ctcaagaaga tcctttgatc 1020
ttttctacgg ggtctgacgc tcagtggaac gaaaactcac gttaagggat tttggtcatg 1080
agattatcaa aaaggatctt cacctagatc cttttggttc atgtgcagct ccatcagcaa 1140
aaggggatga taagtttatc accaccgact atttgcaaca gtgccgttga tcgtgctatg 1200
atcgactgat gtcatcagcg gtggagtgca atgtcgtgca atacgaatgg cgaaaagccg 1260
agctcatcgg tcagcttctc aaccttgggg ttacccccgg cggtgtgctg ctggtccaca 1320
gctccttccg tagcgtccgg cccctcgaag atgggccact tggactgatc gaggccctgc 1380
gtgctgcgct gggtccggga gggacgctcg tcatgccctc gtggtcaggt ctggacgacg 1440
agccgttcga tcctgccacg tcgcccgtta caccggacct tggagttgtc tctgacacat 1500
tctggcgcct gccaaatgta aagcgcagcg cccatccatt tgcctttgcg gcagcggggc 1560
cacaggcaga gcagatcatc tctgatccat tgcccctgcc acctcactcg cctgcaagcc 1620
cggtcgcccg tgtccatgaa ctcgatgggc aggtacttct cctcggcgtg ggacacgatg 1680
ccaacacgac gctgcatctt gccgagttga tggcaaaggt tccctatggg gtgccgagac 1740
actgcaccat tcttcaggat ggcaagttgg tacgcgtcga ttatctcgag aatgaccact 1800
gctgtgagcg ctttgccttg gcggacaggt ggctcaagga gaagagcctt cagaaggaag 1860
gtccagtcgg tcatgccttt gctcggttga tccgctcccg cgacattgtg gcgacagccc 1920
tgggtcaact gggccgagat ccgttgatct tcctgcatcc gccagaggcg ggatgcgaag 1980
aatgcgatgc cgctcgccag tcgattggct gagctcatga gcggagaacg agatgacgtt 2040
ggaggggcaa ggtcgcgctg attgctgggg caacacgtgg agcggatcgg ggattgtctt 2100
tcttcagctc gctgatgata tgctgacgct caatgccgtt tggcctccga ctaacgaaaa 2160
tcccgcattt ggacggctga tccgattggc acggcggacg gcgaatggcg gagcagacgc 2220
tcgtccgggg gcaatgagat atgaaaaagc ctgaactcac cgcgacgtat cgggccctgg 2280
ccagctagct agagtcgacc tgcaggtccc cggggatcgg tcttgccttg ctcgtcggtg 2340
atgtacttca ccagctccgc gaagtcgctc ttcttgatgg agcgcatggg gacgtgcttg 2400
gcaatcacgc gcaccccccg gccgttttag cggctaaaaa agtcatggct ctgccctcgg 2460
gcggaccacg cccatcatga ccttgccaag ctcgtcctgc ttctcttcga tcttcgccag 2520
cagggcgagg atcgtggcat caccgaaccg cgccgtgcgc gggtcgtcgg tgagccagag 2580
tttcagcagg ccgcccaggc ggcccaggtc gccattgatg cgggccagct cgcggacgtg 2640
ctcatagtcc acgacgcccg tgattttgta gccctggccg acggccagca ggtaggccga 2700
caggctcatg ccggccgccg ccgccttttc ctcaatcgct cttcgttcgt ctggaaggca 2760
gtacaccttg ataggtgggc tgcccttcct ggttggcttg gtttcatcag ccatccgctt 2820
gccctcatct gttacgccgg cggtagccgg ccagcctcgc agagcaggat tcccgttgag 2880
caccgccagg tgcgaataag ggacagtgaa gaaggaacac ccgctcgcgg gtgggcctac 2940
ttcacctatc ctgcccggct gacgccgttg gatacaccaa ggaaagtcta cacgaaccct 3000
ttggcaaaat cctgtatatc gtgcgaaaaa ggatggatat accgaaaaaa tcgctataat 3060
gaccccgaag cagggttatg cagcggaaaa gatccgtcga cctgcaggca tgcaagctct 3120
agcgattcca gacgtcccga aggcgtggcg cggcttcccc gtgccggagc aatcgccctg 3180
ggtgggttac acgacgcccc tctatggccc gtactgacgg acacaccgaa gccccggcgg 3240
caaccctcag cggatgcccc ggggcttcac gttttcccag gtcagaagcg gttttcggga 3300
gtagtgcccc aactggggta acctttgagt tctctcagtt gggggcgtag ggtcgccgac 3360
atgacacaag gggttgtgac cggggtggac acgtacgcgg gtgcttacga ccgtcagtcg 3420
cgcgagcgcg agaactcgag cgcagcaagc ccagcgacac agcgtagcgc caacgaagac 3480
aaggcggccg accttcagcg cgaagtcgag cgcgacgggg gccggttcag gttcgtcggg 3540
catttcagcg aagcgccggg cacgtcggcg ttcgggacgg cggagcgccc ggagttcgaa 3600
cgcatcctga acgaatgccg cgccgggcgg ctcaacatga tcattgtcta tgacgtgtcg 3660
cgcttctcgc gcctgaaggt catggacgcg attccgattg tctcggaatt gctcgccctg 3720
ggcgtgacga ttgtttccac tcaggaaggc gtcttccggc agggaaacgt catggacctg 3780
attcacctga ttatgcggct cgacgcgtcg cacaaagaat cttcgctgaa gtcggcgaag 3840
attctcgaca cgaagaacct tcagcgcgaa ttgggcgggt acgtcggcgg gaaggcgcct 3900
tacggcttcg agcttgtttc ggagacgaag gagatcacgc gcaacggccg aatggtcaat 3960
gtcgtcatca acaagcttgc gcactcgacc actcccctta ccggaccctt cgagttcgag 4020
cccgacgtaa tccggtggtg gtggcgtgag atcaagacgc acaaacacct tcccttcaag 4080
ccgggcagtc aagccgccat tcacccgggc agcatcacgg ggctttgtaa gcgcatggac 4140
gctgacgccg tgccgacccg gggcgagacg attgggaaga agaccgcttc aagcgcctgg 4200
gacccggcaa ccgttatgcg aatccttcgg gacccgcgta ttgcgggctt cgccgctgag 4260
gtgatctaca agaagaagcc ggacggcacg ccgaccacga agattgaggg ttaccgcatt 4320
cagcgcgacc cgatcacgct ccggccggtc gagcttgatt gcggaccgat catcgagccc 4380
gctgagtggt atgagcttca ggcgtggttg gacggcaggg ggcgcggcaa ggggctttcc 4440
cgggggcaag ccattctgtc cgccatggac aagctgtact gcgagtgtgg cgccgtcatg 4500
acttcgaagc gcggggaaga atcgatcaag gactcttacc gctgccgtcg ccggaaggtg 4560
gtcgacccgt ccgcacctgg gcagcacgaa ggcacgtgca acgtcagcat ggcggcactc 4620
gacaagttcg ttgcggaacg catcttcaac aagatcaggc acgccgaagg cgacgaagag 4680
acgttggcgc ttctgtggga agccgcccga cgcttcggca agctcactga ggcgcctgag 4740
aagagcggcg aacgggcgaa ccttgttgcg gagcgcgccg acgccctgaa cgcccttgaa 4800
gagctgtacg aagaccgcgc ggcaggcgcg tacgacggac ccgttggcag gaagcacttc 4860
cggaagcaac aggcagcgct gacgctccgg cagcaagggg cggaagagcg gcttgccgaa 4920
cttgaagccg ccgaagcccc gaagcttccc cttgaccaat ggttccccga agacgccgac 4980
gctgacccga ccggccctaa gtcgtggtgg gggcgcgcgt cagtagacga caagcgcgtg 5040
ttcgtcgggc tcttcgtaga caagatcgtt gtcacgaagt cgactacggg cagggggcag 5100
ggaacgccca tcgagaagcg cgcttcgatc acgtgggcga agccgccgac cgacgacgac 5160
gaagacgacg cccaggacgg cacggaagac gtagcggcgt aggtgagtga agatctagac 5220
gtggcaccgc gatgctgttg tgggcacaat cgtgccggtt ggtaggatcc acatatggaa 5280
ataagttcgc tctccaccga cggctccccg cggatcgacg gggagagtcc cgagcacgtg 5340
gaaatgctgg ccgccgccga caccgcgctt ccaccgatca tggtgcaccg ccgcaccggg 5400
cgggtcatcg acggcatgca ccggctgcgc gccgcgatgc tgacgggccg tacgacgatc 5460
gcggtgaggt tcttcgacgg caccgaggag gacgccttcg tcctcgccgt gaagtcgaac 5520
atcgcgcacg gactgccgct gtccgccgcc gaccgccggc gggccgccgg gcgcatcatg 5580
gccacccatc cccggtggtc ggaccggatg atcgcctcgg tggtcggcac ctccgccagg 5640
acggtcgccg agatccgccg cgacgccggc gccgccgggg cgggggagcc cacccgcatc 5700
ggccgggacg gcagggtacg gcccgtcgac gtgagcgagg gccgcagact ggcccacgac 5760
atgatcgtcc gcgacccggg cctgtcgctg cgccaggtcg cccgcgccgc cgggatctcg 5820
ccggagaccg tcagggacgt cagacaccgg atgctccgcg gtgaggaccc ggtgcccgcg 5880
ccgcggccgc ggaccctggt ggagcgcggc gcggaccgcc gggcggagcc ggccgggaag 5940
gccgccgcgc cgtgcgggac ggagccgccg cccgccgtcg tgatgaagcg gctgagggcc 6000
gatccggcgc tgcgtctcaa cgagaacgga cgcgacctgc tgcggcttct ggatatccac 6060
acggtccggc tggaggactg gaaccgcatt atcgaaagcg tgccgccgca ccgtctggag 6120
acggtggcgc agctggcacg ctcctgcgcc gacaaatggt ccgagatcgc gtcacgcatc 6180
gaaagcaacg catcacatct ggccgggtga 6210
<210> 4
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотидный праймер
<400> 4
ttattgaagc ttgccggggc cgacgcggcg ggcggcct 38
<210> 5
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотидный праймер
<400> 5
gttgttttaa ttaaacacca ggcctcctgg ggtg 34
<210> 6
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотидный праймер
<400> 6
tttatattaa ttaatgaccc ttccgtcccg cccccgat 38
<210> 7
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотидный праймер
<400> 7
tttggtgcta gctggtcgtg gcgctgttcc 30
<210> 8
<211> 10670
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая плазмида pXY300-orf18ifd
<400> 8
ccgcggctcc tgcccgccga acgcgtcgtc gtcgacggcc tggtgctcat cgacgagcac 60
ccggagccag gtgaaaagcg ccggcggacg ctcggactgg gcgcgggatt ccagcagtaa 120
cccaggtccg ccggccacct cacggcaggc agaccctcgg ctttcgcgcc gggaccgcca 180
tgagaccgcc acccggatgt ccggggtggc ggtctcatgg cggtccctca gcggccctac 240
gcgaccgctg tgtcgacgcg gaggcagtct ccggggagcc ggccccaggg ctggagtacc 300
ggggcgagct tgcccttgca gcggcggcac acgggcgacg cggcgccggg cgggggagtc 360
ttgacgtcga cggtgaccgg cttcggcttc aggcgcttcc gaaggctgat cgtcgggcgg 420
atcgcctcct tcttcggctg acgcactcgg tcgagcgaag cggccagctc ctccgtgcgg 480
gcctcgttgg ccttccggcg cgcctcgcgg aaagcagctt cctcctcgga catgacctcg 540
aacctcatct ggtcagcgtc aaggtcgccc ggcgcggccg gcgcttccgg ggcgggcggg 600
tccaggacgt ccttgcccca caccaggccc caggactcga cgagccgccg gacgcccggt 660
aggccgtacg tctcggcgac cttgatgagg tcgaggtcga ggcgacgtcc ggcgacgcgg 720
gcgatgtatc ggtaccagat gtaggccggg atgaccgcga tggcgaccag gccctcggtg 780
tcgtcggtga tctcctcctc ggtgcggacg tcctgctgga tgccgagttc cttgatcagc 840
cggttcaggt tctgcgaccg gtagtgcttg cggacctgga agacgccgaa ctcgcgctcg 900
cggtacttct cgacgaacgg gccgggccga cgaagccgct gcagctcggc ggccgccgcg 960
tcgcccaggt cgagcggtcc catgcggtcg tcgccgcgac cggccttgaa gttctgtccg 1020
gccagctcca ggccgatctt ggcgacgccg cccttggtct tgtcgccgtc cttgtagagg 1080
tagcgggcct gcttgcccgc atcgccgtca gcggcgtccg cgccgttgag tgggcgcacg 1140
tcggtgccgt ggcccttgcc ctcacaggag caaccgggcc ggtcgcacgt ctcgctgacg 1200
gtgtagccgc ccgcggattc gaccccggcg gcccaggctc cggcgagtgc gtcgcggaac 1260
gcctgggcgt ccgggccgag cacctcgcgg gtgacccaga gcgtgtgcca gtgcaggtgc 1320
cagccggagc cccagccgaa ggtgtcctcg aaggcccgct cgtagccgat gatcccgaag 1380
tcgtcgcgca tcgtgcgcac gcggcggccg gacgagccgt acgcgccctt ccagccgtcg 1440
tgcaagaccg cgaccaggcc gtgccgcatt cccttgcgga cggtgccgaa cgccatgcgc 1500
tcgaagtggc gcaacgtgtt cgtgccaagg tgcagcccgt acccggcgtc cgcgagaccg 1560
tcggcggcga gctgcacgtt cgagccccgt acggccagga tgcggctcat gcaccacggg 1620
caggtgtgga cgttgttgca gcggcacgtg ttgccccacg tcgcctcgcc cggcttccac 1680
atcagctcgc ggtccggcag tgagccgggt cccgcagccc ttgaacgcct cgttcagcga 1740
caccgtctgg tgccggtccc gccgggcgaa ccgctcgtcg cggcgggtcc tcccgcccgc 1800
tgtcgcggca ccctcgtttg gggtagaacc cgttccagtt acagcgctct gacctgcagt 1860
ggacggagat tttcccttac tactaaagcc cgcgtccgga ttacccgctg tagtcgtgct 1920
tgctacgctg cgtgactggt ccgcaatgag acgctttgcg cgctttcggc aggcgtccga 1980
gcagtagatt ttggggcgct tcccggggat gtggacgatc ggggtgccgc agtggcactt 2040
cggtccggcg gggcgcggtg gtgtcgacgc gctgttctct cgtacgcctc gtcacagagc 2100
aaacgtcctc actcggcatg ctgcgccggt tcgggggcgg cgagccggga ggccaatccc 2160
gggctcgtgc catttctggg tcctgttgat catcactgac gaatcgaggt cgaggaaccg 2220
agcgtccgag gaacagaggc gcttatcggt tggccgcgag attcctgtcg atcctctcgt 2280
gcagcgcgat tccgagggaa acggaaacgt tgagagactc ggtctggctc atcatgggga 2340
tggaaaccga ggcggaagac gcctcctcga acaggtcgga aggcccaccc ttttcgctgc 2400
cgaacagcaa ggccagccga tccggattgt ccccgagttc cttcacggaa atgtcgccat 2460
ccgccttgag cgtcatcagc tgcataccgc tgtcccgaat gaaggcgatg gcctcctcgc 2520
gaccggagag aacgacggga agggagaaga cgtaacctcg gctggccctt tggagacgcc 2580
ggtccgcgat gctggtgatg tcactgtcga ccaggatgat ccccgacgct ccgagcgcga 2640
gcgacgtgcg tactatcgcg ccgatgttcc cgacgatctt caccccgtcg agaacgacga 2700
cgtccccacg ccggctcgcg atatcgccga acctggccgg gcgagggacg cgggcgatgc 2760
cgaatgtctt ggccttccgc tcccccttga acaactggtt gacgatcgag gagtcgatga 2820
ggcggaccgg tatgttctgc cgcccgcaca gatccagcaa ctcagatgga aaaggactgc 2880
tgtcgctgcc gtagacctcg atgaactcca ccccggccgc gatgctgtgc atgaggggct 2940
cgacgtcctc gatcaacgtt gtctttatgt tggatcgcga cggcttggtg acatcgatga 3000
tccgctgcac cgcgggatcg gacggatttg cgatggtgtc caactcagtc atggtcgtcc 3060
taccggctgc tgtgttcagt gacgcgattc ctggggtgtg acaccctacg cgacgatggc 3120
ggatggctgc cctgaccggc aatcaccaac gcaaggggaa gtcgtcgctc tctggcaaag 3180
ctccccgctc ttccccgtcc gggacccgcg cggtcgatcc ccgcatatgg tgcactctca 3240
gtacaatctg ctctgatgcc gcatagttaa gccagccccg acacccgcca acacccgctg 3300
acgcgccctg acgggcttgt ctgctcccgg catccgctta cagacaagct gtgaccgtct 3360
ccgggagctg catgtgtcag aggttttcac cgtcatcacc gaaacgcgcg agacgaaagg 3420
gcctcgtgat acgcctattt ttataggtta atgtcatgat aataatggtt tcttagacgt 3480
caggtggcac ttttcgggga aatgtgcgcg gaacccctat ttgtttattt ttctaaatac 3540
attcaaatat gtatccgctc atgagacaat aaccctgata aatgcttcaa taatattgaa 3600
aaaggaagag tatgagtatt caacatttcc gtgtcgccct tattcccttt tttgcggcat 3660
tttgccttcc tgtttttgct cacccagaaa cgctggtgaa agtaaaagat gctgaagatc 3720
agttgggtgc acgagtgggt tacatcgaac tggatctcaa cagcggtaag atccttgaga 3780
gttttcgccc cgaagaacgt tttccaatga tgagcacttt taaagttctg ctatgtggcg 3840
cggtattatc ccgtattgac gccgggcaag agcaactcgg tcgccgcata cactattctc 3900
agaatgactt ggttgagtac tcaccagtca cagaaaagca tcttacggat ggcatgacag 3960
taagagaatt atgcagtgct gccataacca tgagtgataa cactgcggcc aacttacttc 4020
tgacaacgat cggaggaccg aaggagctaa ccgctttttt gcacaacatg ggggatcatg 4080
taactcgcct tgatcgttgg gaaccggagc tgaatgaagc cataccaaac gacgagcgtg 4140
acaccacgat gcctgtagca atggcaacaa cgttgcgcaa actattaact ggcgaactac 4200
ttactctagc ttcccggcaa caattaatag actggatgga ggcggataaa gttgcaggac 4260
cacttctgcg ctcggccctt ccggctggct ggtttattgc tgataaatct ggagccggtg 4320
agcgtgggtc tcgcggtatc attgcagcac tggggccaga tggtaagccc tcccgtatcg 4380
tagttatcta cacgacgggg agtcaggcaa ctatggatga acgaaataga cagatcgctg 4440
agataggtgc ctcactgatt aagcattggt aactgtcaga ccaagtttac tcatatatac 4500
tttagattga tttaaaactt catttttaat ttaaaaggat ctaggtgaag atcctttttg 4560
ataatctcat gaccaaaatc ccttaacgtg agttttcgtt ccactgagcg tcagaccccg 4620
tagaaaagat caaaggatct tcttgagatc ctttttttct gcgcgtaatc tgctgcttgc 4680
aaacaaaaaa accaccgcta ccagcggtgg tttgtttgcc ggatcaagag ctaccaactc 4740
tttttccgaa ggtaactggc ttcagcagag cgcagatacc aaatactgtc cttctagtgt 4800
agccgtagtt aggccaccac ttcaagaact ctgtagcacc gcctacatac ctcgctctgc 4860
taatcctgtt accagtggct gctgccagtg gcgataagtc gtgtcttacc gggttggact 4920
caagacgata gttaccggat aaggcgcagc ggtcgggctg aacggggggt tcgtgcacac 4980
agcccagctt ggagcgaacg acctacaccg aactgagata cctacagcgt gagctatgag 5040
aaagcgccac gcttcccgaa gggagaaagg cggacaggta tccggtaagc ggcagggtcg 5100
gaacaggaga gcgcacgagg gagcttccag ggggaaacgc ctggtatctt tatagtcctg 5160
tcgggtttcg ccacctctga cttgagcgtc gatttttgtg atgctcgtca ggggggcgga 5220
gcctatggaa aaacgccagc aacgcggcct ttttacggtt cctggccttt tgctggcctt 5280
ttgctcacat gttctttcct gcgttatccc ctgattctgt ggataaccgt attaccgcct 5340
ttgagtgagc tgataccgct cgccgcagcc gaacgaccga gcgcagcgag tcagtgagcg 5400
aggaagcgga agagcgccca atacgcaaac cgcctctccc cgcgcgttgg ccgattcatt 5460
aatgcagctg gcacgacagg tttcccgact ggaaagcggg cagtgagcgc aacgcaatta 5520
atgtgagtta gctcactcat taggcacccc aggctttaca ctttatgctt ccggctcgta 5580
tgttgtgtgg aattgtgagc ggataacaat ttcacacagg aaacagctat gaccatgatt 5640
acgccaagct tgccggggcc gacgcggcgg gcggcctcgt ccatcgggac gcgccagtcg 5700
acgccgacga tgtccgcgcc ggcctcgccc atgagcccca gcagctcgcc ggtgccgacg 5760
ccgaagtgga tgcgcgggac gccgtggccg gccaccgcgc ggaacacctt cgccgaggcg 5820
ggcagcaccg aacgccggta gtcggagggg gcgagcgcgc cggcccagga gtcgaagagc 5880
tgcacggccg aggcgccggc ccggatctgg acgtcgagga aggccgccgt gatgtcggcg 5940
aggcggtcga gcaggtcggc ccagagctcg gggtcgccgt acatcatcgc cttggcgttc 6000
tcgtacgtac gggacgggcc gccctcgacg aggtaactcg caagggtgaa cggggcgccc 6060
gcgaaaccga tcagcggggt ggacccgagc tcacgggtca gcatgccgat ggcctcggtg 6120
acgtaggaga cgtcctccgg ggtcaggtcg cgcagccggg cgaggtcggc gcgggtgcgc 6180
accggctgct cgacgaccgg gccgatgccg ggcttgatgt cgaggtcgat gccgatggcc 6240
ttgagcggga cgacgatgtc gctgaagtag atcgccgcgt cgacgtggtg gcggcgcacc 6300
ggctggaggg tgatctcggt gaccagctcg ggccgcatgc aggagtcgag catcccgatg 6360
ccctcgcgca ccttgcggta ctccggcagt gagcgcccgg cctgccgcat gaaccacacc 6420
ggcgtgtgcg gcaccggctc gcgccggcac gccttgagga aggctgattc ccggacggcg 6480
tcgttcttga cggggtcggg ggtcgcggta ggcgtctggc ccttcgggct cgtgttggca 6540
ctcacaccgg ccagtctcgc acgacccgtc cgccgccccg gaccccaggg gcgtccggcg 6600
ggcggggccg cggtgcagga tccggacagc gccgggcggc gcgggtgtcc ctccctgcgc 6660
cgggggcccg ttccgcttaa tcttcccggc atggctgcgg ctcagggacg actgtcggac 6720
ggcgctggcg gaatggacga accgaaggag ggcggggggg atcccgggca cggaggtgcg 6780
cctccgccgc ccttccgggc cgctgtcgag gcgctgcaga gcgcccggct gcggccgcag 6840
atcgaggtgg agacggtgcc cgcgccgaaa cggctcgccc cgtacgcgca cgcgctggag 6900
gcggcggtcg tcgacggcga ggaggatctg gccgacggcc ggctggtgct gctgtgcgac 6960
ccggccggac acgacgcctg gcgggggacc ttccgtctgg tgacgctggt gcgcgccgag 7020
ctggagccgg agatggcggc ggatccgctg ctgccggacg tgtgctggtc ctggctgacc 7080
ggcgcgctgg cggcgcgcgg cctgtcgtac ggcgagccga gcggcacggt gacgcgggcg 7140
agttcgcact acttcggcgg gctgtccgcg cggcccgccg cctcccagat cgagatccgt 7200
gcctcgtgga cgccgcgtga gggtctgggc ggggttccgg acacggccgc ccatctggtc 7260
gcgtggtccg atctgctggc gcaggtcgcg gggctgccgc cggccgctcc gggggacgcg 7320
tccgtggtga cgctgccgca gcggaggggg ccgcagtcgc gctgagcctc ccgctgcggg 7380
gtcccgacga ccggcctcct gcatctctct ttgtcgatac ggccactttc ggaagcgttg 7440
cgacgcagac cgaaaccgtt cgatcttcga atgatcgatc gtgcgtccga attgcccgga 7500
ttgttactca tcacttcgtg atcattcgtt aaaggacacc aggtttgctg ccgaagacga 7560
ctgtgacctt gaaagcacgg ttcgtcccgc cttcaccccc acgagccggc ccgtcccgca 7620
ccccaggagg cctggtgttt aattaatgac ccttccgtcc cgcccccgat ccgcccgtcc 7680
ccgatccgcc cgcccacggc caagcgaaca cgttctttca ctcttctgac cggaatacga 7740
cccaccggcg cccgtcacgg agcacaaccg tcgacgggcg ccttcgcggc acggataccc 7800
ttgacaggtg accgacgccc acgacaccgc agcagacagt tcactgcgca ccaccggagg 7860
cgctcctccg gacgacggcg gatcttctgt tacggaggcg ccgaccccct tgctggaacc 7920
ccgcgagggc attccgcccg tgatagcgga cgaggccgcc ctcgccgagg cggtcgccgc 7980
cttcgcggcc ggcagcggac ccgtcgccgt ggacgccgag cgcgcctccg ggtaccgcta 8040
cggccagcgc gcctacctcg tccagctgcg ccgcgagggt gcgggtaccg cgctgatcga 8100
ccccgtggcc tgccccgacc tgtccgccct cggcgaggcg ctgtccggcg tcgagtgggt 8160
gctgcacgcc gccacccagg acctgccctg tctgcgcgag ataggcatgg tgccctcccg 8220
cctcttcgac accgagctgg ccggccgcct tgccgggttc ccccgcgtcg ggctcggcgc 8280
gatggtcgag aacgtgctcg gcttcgtcct ggagaagggc cactccgccg tcgactggtc 8340
cacccgtccg ctgcccgagc cctggctgcg gtacgccgcc ctcgacgtcg aactgctggt 8400
cgatctgcgg gacgccctgg agaaggagct ggaccgccag ggcaagctgg actgggcccg 8460
gcaggagttc gacgcgatcg cctcggcccc gccgccggag ccccgcaagg acccctggcg 8520
ccgcacctcc ggcatgcaca aggtgcgccg gcgccgccag atggcggtgg tgcgggagct 8580
gtgggagacc cgcgaccgga tcgcccggcg ccgtgacgtc tcccccggca aggtgctttc 8640
cgacgcggcg atcgtggagg ccgcgctcgc gctgcccgcc aacctgcacg ccatggccgc 8700
gctcaacggg ttcgggcagc gggtggggcg gcgccagctg gagcagtggc aggcggccgt 8760
cgaccgcgcg aaggcgctga gcgaggccca gctgccgcag cccggccagc cggtgaccgg 8820
ccctccgccg ccgcgcgcct gggcggacaa ggaccccgtt gccgcggccc ggctgtcggc 8880
ggcccgcgcg ggggtcgccg aactcgccga gcggctgaac atgccgccgg agaacctgat 8940
caccccggac acggtgcgca gggtctgctg ggagccgccg gggcccgacg agcggtccgt 9000
cgccgcggcg ctgacggcgc acggggcacg cgcgtggcag gtcgaccagg tcactcccgt 9060
gctggtggcc gcgctggcta cttcgtcgcc ccccgcatga acccgttgta gatgaagcgc 9120
tggaggaaca ggaagacgat caaggtgggc aggatgacca ggaccgcgcc cgccgagatc 9180
gtctcccagt gcgcgccgaa ggggcccttg aagcggaaca gggacgtcga gatgaccccc 9240
aggtcctcgg agggcatgta gaggaagggg atgtagaagt cgttgtagac gttgatcccc 9300
tttacgatca ccaccgtcgc gatcgccggc ttgagcagcg ggaagatcac cttgcggtag 9360
acggtgaacg cgttggcgcc gtccaggcgc gccgcctcgt ccagggagac ggggatggag 9420
cggatgaact gcaggaagac gtagatcgag acgatgtccg tgcccatgta gagggcgatc 9480
ggcgcccaca ggctgtcgaa catgccgaag ctgttgacga tctggaaggt cgccacctgg 9540
gtggtcaccc cggggaccag cgcggccagc aggaacagcg ccacgaccag ctagcgaatt 9600
cctgcaggtc cccggggatc ggtcttgcct tgctcgtcgg tgatgtactt caccagctcc 9660
gcgaagtcgc tcttcttgat ggagcgcatg gggacgtgct tggcaatcac gcgcaccccc 9720
cggccgtttt agcggctaaa aaagtcatgg ctctgccctc gggcggacca cgcccatcat 9780
gaccttgcca agctcgtcct gcttctcttc gatcttcgcc agcagggcga ggatcgtggc 9840
atcaccgaac cgcgccgtgc gcgggtcgtc ggtgagccag agtttcagca ggccgcccag 9900
gcggcccagg tcgccattga tgcgggccag ctcgcggacg tgctcatagt ccacgacgcc 9960
cgtgattttg tagccctggc cgacggccag caggtaggcc gacaggctca tgccggccgc 10020
cgccgccttt tcctcaatcg ctcttcgttc gtctggaagg cagtacacct tgataggtgg 10080
gctgcccttc ctggttggct tggtttcatc agccatccgc ttgccctcat ctgttacgcc 10140
ggcggtagcc ggccagcctc gcagagcagg attcccgttg agcaccgcca ggtgcgaata 10200
agggacagtg aagaaggaac acccgctcgc gggtgggcct acttcaccta tcctgcccgg 10260
ctgacgccgt tggatacacc aaggaaagtc tacacgaacc ctttggcaaa atcctgtata 10320
tcgtgcgaaa aaggatggat ataccgaaaa aatcgctata atgaccccga agcagggtta 10380
tgcagcggaa aagatccgtc gacctgcagc ccgggggatc cccgggtacc gagctcgggc 10440
tggggctcgg ggccgccggt gagctggtag acgaaggcgc ccgagtctcc ttcgttcact 10500
gcgtgccact cgtggtgcgg gtacttccgg cgcaacgtgc tgtcgtccat gggcggcatc 10560
atggcagagg cggagacgcc gttccgcgcc tttcgtcggg gcccgtaggg tttcggacat 10620
tcttgtgcgg ggtggggggg cgccggcgga cccggtgcgc ccggcgtcgc 10670
<210> 9
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотидный праймер
<400> 9
agcacagcta gcttctagaa gcttcattca aaggccggca 40
<210> 10
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотидный праймер
<400> 10
gccagtgaat tctgcagctc gagcagagca ggattcccgt tga 43
<210> 11
<211> 7173
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая плазмида pKS-T-orf18pfrd
<400> 11
gtggcacttt tcggggaaat gtgcgcggaa cccctatttg tttatttttc taaatacatt 60
caaatatgta tccgctcatg agacaataac cctgataaat gcttcaataa tattgaaaaa 120
ggaagagtat gagtattcaa catttccgtg tcgcccttat tccctttttt gcggcatttt 180
gccttcctgt ttttgctcac ccagaaacgc tggtgaaagt aaaagatgct gaagatcagt 240
tgggtgcacg agtgggttac atcgaactgg atctcaacag cggtaagatc cttgagagtt 300
ttcgccccga agaacgtttt ccaatgatga gcacttttaa agttctgcta tgtggcgcgg 360
tattatcccg tattgacgcc gggcaagagc aactcggtcg ccgcatacac tattctcaga 420
atgacttggt tgagtactca ccagtcacag aaaagcatct tacggatggc atgacagtaa 480
gagaattatg cagtgctgcc ataaccatga gtgataacac tgcggccaac ttacttctga 540
caacgatcgg aggaccgaag gagctaaccg cttttttgca caacatgggg gatcatgtaa 600
ctcgccttga tcgttgggaa ccggagctga atgaagccat accaaacgac gagcgtgaca 660
ccacgatgcc tgtagcaatg gcaacaacgt tgcgcaaact attaactggc gaactactta 720
ctctagcttc ccggcaacaa ttaatagact ggatggaggc ggataaagtt gcaggaccac 780
ttctgcgctc ggcccttccg gctggctggt ttattgctga taaatctgga gccggtgagc 840
gtgggtctcg cggtatcatt gcagcactgg ggccagatgg taagccctcc cgtatcgtag 900
ttatctacac gacggggagt caggcaacta tggatgaacg aaatagacag atcgctgaga 960
taggtgcctc actgattaag cattggtaac tgtcagacca agtttactca tatatacttt 1020
agattgattt aaaacttcat ttttaattta aaaggatcta ggtgaagatc ctttttgata 1080
atctcatgac caaaatccct taacgtgagt tttcgttcca ctgagcgtca gaccccgtag 1140
aaaagatcaa aggatcttct tgagatcctt tttttctgcg cgtaatctgc tgcttgcaaa 1200
caaaaaaacc accgctacca gcggtggttt gtttgccgga tcaagagcta ccaactcttt 1260
ttccgaaggt aactggcttc agcagagcgc agataccaaa tactgtcctt ctagtgtagc 1320
cgtagttagg ccaccacttc aagaactctg tagcaccgcc tacatacctc gctctgctaa 1380
tcctgttacc agtggctgct gccagtggcg ataagtcgtg tcttaccggg ttggactcaa 1440
gacgatagtt accggataag gcgcagcggt cgggctgaac ggggggttcg tgcacacagc 1500
ccagcttgga gcgaacgacc tacaccgaac tgagatacct acagcgtgag ctatgagaaa 1560
gcgccacgct tcccgaaggg agaaaggcgg acaggtatcc ggtaagcggc agggtcggaa 1620
caggagagcg cacgagggag cttccagggg gaaacgcctg gtatctttat agtcctgtcg 1680
ggtttcgcca cctctgactt gagcgtcgat ttttgtgatg ctcgtcaggg gggcggagcc 1740
tatggaaaaa cgccagcaac gcggcctttt tacggttcct ggccttttgc tggccttttg 1800
ctcacatgtt ctttcctgcg ttatcccctg attctgtgga taaccgtatt accgcctttg 1860
agtgagctga taccgctcgc cgcagccgaa cgaccgagcg cagcgagtca gtgagcgagg 1920
aagcggaaga gcgcccaata cgcaaaccgc ctctccccgc gcgttggccg attcattaat 1980
gcagctggca cgacaggttt cccgactgga aagcgggcag tgagcgcaac gcaattaatg 2040
tgagttagct cactcattag gcaccccagg ctttacactt tatgcttccg gctcgtatgt 2100
tgtgtggaat tgtgagcgga taacaatttc acacaggaaa cagctatgac catgattacg 2160
ccaagcgcgc aattaaccct cactaaaggg aacaaaagct gggtaccggg ccccccctcg 2220
agcagagcag gattcccgtt gagcaccgcc aggtgcgaat aagggacagt gaagaaggaa 2280
cacccgctcg cgggtgggcc tacttcacct atcctgcccg gctgacgccg ttggatacac 2340
caaggaaagt ctacacgaac cctttggcaa aatcctgtat atcgtgcgaa aaaggatgga 2400
tataccgaaa aaatcgctat aatgaccccg aagcagggtt atgcagcgga aaatgcagct 2460
cacggtaact gatgccgtat ttgcagtacc agcgtacggc ccacagaatg atgtcacgct 2520
gaaaatgccg gcctttgaat gaagcttgcc ggggccgacg cggcgggcgg cctcgtccat 2580
cgggacgcgc cagtcgacgc cgacgatgtc cgcgccggcc tcgcccatga gccccagcag 2640
ctcgccggtg ccgacgccga agtggatgcg cgggacgccg tggccggcca ccgcgcggaa 2700
caccttcgcc gaggcgggca gcaccgaacg ccggtagtcg gagggggcga gcgcgccggc 2760
ccaggagtcg aagagctgca cggccgaggc gccggcccgg atctggacgt cgaggaaggc 2820
cgccgtgatg tcggcgaggc ggtcgagcag gtcggcccag agctcggggt cgccgtacat 2880
catcgccttg gcgttctcgt acgtacggga cgggccgccc tcgacgaggt aactcgcaag 2940
ggtgaacggg gcgcccgcga aaccgatcag cggggtggac ccgagctcac gggtcagcat 3000
gccgatggcc tcggtgacgt aggagacgtc ctccggggtc aggtcgcgca gccgggcgag 3060
gtcggcgcgg gtgcgcaccg gctgctcgac gaccgggccg atgccgggct tgatgtcgag 3120
gtcgatgccg atggccttga gcgggacgac gatgtcgctg aagtagatcg ccgcgtcgac 3180
gtggtggcgg cgcaccggct ggagggtgat ctcggtgacc agctcgggcc gcatgcagga 3240
gtcgagcatc ccgatgccct cgcgcacctt gcggtactcc ggcagtgagc gcccggcctg 3300
ccgcatgaac cacaccggcg tgtgcggcac cggctcgcgc cggcacgcct tgaggaaggc 3360
tgattcccgg acggcgtcgt tcttgacggg gtcgggggtc gcggtaggcg tctggccctt 3420
cgggctcgtg ttggcactca caccggccag tctcgcacga cccgtccgcc gccccggacc 3480
ccaggggcgt ccggcgggcg gggccgcggt gcaggatccg gacagcgccg ggcggcgcgg 3540
gtgtccctcc ctgcgccggg ggcccgttcc gcttaatctt cccggcatgg ctgcggctca 3600
gggacgactg tcggacggcg ctggcggaat ggacgaaccg aaggagggcg ggggggatcc 3660
cgggcacgga ggtgcgcctc cgccgccctt ccgggccgct gtcgaggcgc tgcagagcgc 3720
ccggctgcgg ccgcagatcg aggtggagac ggtgcccgcg ccgaaacggc tcgccccgta 3780
cgcgcacgcg ctggaggcgg cggtcgtcga cggcgaggag gatctggccg acggccggct 3840
ggtgctgctg tgcgacccgg ccggacacga cgcctggcgg gggaccttcc gtctggtgac 3900
gctggtgcgc gccgagctgg agccggagat ggcggcggat ccgctgctgc cggacgtgtg 3960
ctggtcctgg ctgaccggcg cgctggcggc gcgcggcctg tcgtacggcg agccgagcgg 4020
cacggtgacg cgggcgagtt cgcactactt cggcgggctg tccgcgcggc ccgccgcctc 4080
ccagatcgag atccgtgcct cgtggacgcc gcgtgagggt ctgggcgggg ttccggacac 4140
ggccgcccat ctggtcgcgt ggtccgatct gctggcgcag gtcgcggggc tgccgccggc 4200
cgctccgggg gacgcgtccg tggtgacgct gccgcagcgg agggggccgc agtcgcgctg 4260
agcctcccgc tgcggggtcc cgacgaccgg cctcctgcat ctctctttgt cgatacggcc 4320
actttcggaa gcgttgcgac gcagaccgaa accgttcgat cttcgaatga tcgatcgtgc 4380
gtccgaattg cccggattgt tactcatcac ttcgtgatca ttcgttaaag gacaccaggt 4440
ttgctgccga agacgactgt gaccttgaaa gcacggttcg tcccgccttc acccccacga 4500
gccggcccgt cccgcacccc aggaggcctg gtgtttaatt aatgaccctt ccgtcccgcc 4560
cccgatccgc ccgtccccga tccgcccgcc cacggccaag cgaacacgtt ctttcactct 4620
tctgaccgga atacgaccca ccggcgcccg tcacggagca caaccgtcga cgggcgcctt 4680
cgcggcacgg atacccttga caggtgaccg acgcccacga caccgcagca gacagttcac 4740
tgcgcaccac cggaggcgct cctccggacg acggcggatc ttctgttacg gaggcgccga 4800
cccccttgct ggaaccccgc gagggcattc cgcccgtgat agcggacgag gccgccctcg 4860
ccgaggcggt cgccgccttc gcggccggca gcggacccgt cgccgtggac gccgagcgcg 4920
cctccgggta ccgctacggc cagcgcgcct acctcgtcca gctgcgccgc gagggtgcgg 4980
gtaccgcgct gatcgacccc gtggcctgcc ccgacctgtc cgccctcggc gaggcgctgt 5040
ccggcgtcga gtgggtgctg cacgccgcca cccaggacct gccctgtctg cgcgagatag 5100
gcatggtgcc ctcccgcctc ttcgacaccg agctggccgg ccgccttgcc gggttccccc 5160
gcgtcgggct cggcgcgatg gtcgagaacg tgctcggctt cgtcctggag aagggccact 5220
ccgccgtcga ctggtccacc cgtccgctgc ccgagccctg gctgcggtac gccgccctcg 5280
acgtcgaact gctggtcgat ctgcgggacg ccctggagaa ggagctggac cgccagggca 5340
agctggactg ggcccggcag gagttcgacg cgatcgcctc ggccccgccg ccggagcccc 5400
gcaaggaccc ctggcgccgc acctccggca tgcacaaggt gcgccggcgc cgccagatgg 5460
cggtggtgcg ggagctgtgg gagacccgcg accggatcgc ccggcgccgt gacgtctccc 5520
ccggcaaggt gctttccgac gcggcgatcg tggaggccgc gctcgcgctg cccgccaacc 5580
tgcacgccat ggccgcgctc aacgggttcg ggcagcgggt ggggcggcgc cagctggagc 5640
agtggcaggc ggccgtcgac cgcgcgaagg cgctgagcga ggcccagctg ccgcagcccg 5700
gccagccggt gaccggccct ccgccgccgc gcgcctgggc ggacaaggac cccgttgccg 5760
cggcccggct gtcggcggcc cgcgcggggg tcgccgaact cgccgagcgg ctgaacatgc 5820
cgccggagaa cctgatcacc ccggacacgg tgcgcagggt ctgctgggag ccgccggggc 5880
ccgacgagcg gtccgtcgcc gcggcgctga cggcgcacgg ggcacgcgcg tggcaggtcg 5940
accaggtcac tcccgtgctg gtggccgcgc tggctacttc gtcgcccccc gcatgaaccc 6000
gttgtagatg aagcgctgga ggaacaggaa gacgatcaag gtgggcagga tgaccaggac 6060
cgcgcccgcc gagatcgtct cccagtgcgc gccgaagggg cccttgaagc ggaacaggga 6120
cgtcgagatg acccccaggt cctcggaggg catgtagagg aaggggatgt agaagtcgtt 6180
gtagacgttg atccccttta cgatcaccac cgtcgcgatc gccggcttga gcagcgggaa 6240
gatcaccttg cggtagacgg tgaacgcgtt ggcgccgtcc aggcgcgccg cctcgtccag 6300
ggagacgggg atggagcgga tgaactgcag gaagacgtag atcgagacga tgtccgtgcc 6360
catgtagagg gcgatcggcg cccacaggct gtcgaacatg ccgaagctgt tgacgatctg 6420
gaaggtcgcc acctgggtgg tcaccccggg gaccagcgcg gccagcagga acagcgccac 6480
gaccagctag ttctagagcg gccgccaccg cggtggagct ccaattcgcc ctatagtgag 6540
tcgtattacg cgcgctcact ggccgtcgtt ttacaacgtc gtgactggga aaaccctggc 6600
gttacccaac ttaatcgcct tgcagcacat ccccctttcg ccagctggcg taatagcgaa 6660
gaggcccgca ccgatcgccc ttcccaacag ttgcgcagcc tgaatggcga atggaaattg 6720
taagcgttaa tattttgtta aaattcgcgt taaatttttg ttaaatcagc tcatttttta 6780
accaataggc cgaaatcggc aaaatccctt ataaatcaaa agaatagacc gagatagggt 6840
tgagtgttgt tccagtttgg aacaagagtc cactattaaa gaacgtggac tccaacgtca 6900
aagggcgaaa aaccgtctat cagggcgatg gcccactacg tgaaccatca ccctaatcaa 6960
gttttttggg gtcgaggtgc cgtaaagcac taaatcggaa ccctaaaggg agcccccgat 7020
ttagagcttg acggggaaag ccggcgaacg tggcgagaaa ggaagggaag aaagcgaaag 7080
gagcgggcgc tagggcgctg gcaagtgtag cggtcacgct gcgcgtaacc accacacccg 7140
ccgcgcttaa tgcgccgcta cagggcgcgt cag 7173
<210> 12
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотидный праймер
<400> 12
gaatggccat ggttcatgtg cagctccat 29
<210> 13
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотидный праймер
<400> 13
tctcgaggat ccgaatagga acttcggaat 30
<210> 14
<211> 6034
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая плазмида pKS-T-orf18pfrd-AmR
<400> 14
gtggcacttt tcggggaaat gtgcgcggaa cccctatttg tttatttttc taaatacatt 60
caaatatgta tccgctcatg agacaataac cctgataaat gcttcaataa tattgaaaaa 120
ggaagagtat gagtattcaa catttccgtg tcgcccttat tccctttttt gcggcatttt 180
gccttcctgt ttttgctcac ccagaaacgc tggtgaaagt aaaagatgct gaagatcagt 240
tgggtgcacg agtgggttac atcgaactgg atctcaacag cggtaagatc cttgagagtt 300
ttcgccccga agaacgtttt ccaatgatga gcacttttaa agttctgcta tgtggcgcgg 360
tattatcccg tattgacgcc gggcaagagc aactcggtcg ccgcatacac tattctcaga 420
atgacttggt tgagtactca ccagtcacag aaaagcatct tacggatggc atgacagtaa 480
gagaattatg cagtgctgcc ataaccatga gtgataacac tgcggccaac ttacttctga 540
caacgatcgg aggaccgaag gagctaaccg cttttttgca caacatgggg gatcatgtaa 600
ctcgccttga tcgttgggaa ccggagctga atgaagccat accaaacgac gagcgtgaca 660
ccacgatgcc tgtagcaatg gcaacaacgt tgcgcaaact attaactggc gaactactta 720
ctctagcttc ccggcaacaa ttaatagact ggatggaggc ggataaagtt gcaggaccac 780
ttctgcgctc ggcccttccg gctggctggt ttattgctga taaatctgga gccggtgagc 840
gtgggtctcg cggtatcatt gcagcactgg ggccagatgg taagccctcc cgtatcgtag 900
ttatctacac gacggggagt caggcaacta tggatgaacg aaatagacag atcgctgaga 960
taggtgcctc actgattaag cattggtaac tgtcagacca agtttactca tatatacttt 1020
agattgattt aaaacttcat ttttaattta aaaggatcta ggtgaagatc ctttttgata 1080
atctcatgac caaaatccct taacgtgagt tttcgttcca ctgagcgtca gaccccgtag 1140
aaaagatcaa aggatcttct tgagatcctt tttttctgcg cgtaatctgc tgcttgcaaa 1200
caaaaaaacc accgctacca gcggtggttt gtttgccgga tcaagagcta ccaactcttt 1260
ttccgaaggt aactggcttc agcagagcgc agataccaaa tactgtcctt ctagtgtagc 1320
cgtagttagg ccaccacttc aagaactctg tagcaccgcc tacatacctc gctctgctaa 1380
tcctgttacc agtggctgct gccagtggcg ataagtcgtg tcttaccggg ttggactcaa 1440
gacgatagtt accggataag gcgcagcggt cgggctgaac ggggggttcg tgcacacagc 1500
ccagcttgga gcgaacgacc tacaccgaac tgagatacct acagcgtgag ctatgagaaa 1560
gcgccacgct tcccgaaggg agaaaggcgg acaggtatcc ggtaagcggc agggtcggaa 1620
caggagagcg cacgagggag cttccagggg gaaacgcctg gtatctttat agtcctgtcg 1680
ggtttcgcca cctctgactt gagcgtcgat ttttgtgatg ctcgtcaggg gggcggagcc 1740
tatggaaaaa cgccagcaac gcggcctttt tacggttcct ggccttttgc tggccttttg 1800
ctcacatgtt ctttcctgcg ttatcccctg attctgtgga taaccgtatt accgcctttg 1860
agtgagctga taccgctcgc cgcagccgaa cgaccgagcg cagcgagtca gtgagcgagg 1920
aagcggaaga gcgcccaata cgcaaaccgc ctctccccgc gcgttggccg attcattaat 1980
gcagctggca cgacaggttt cccgactgga aagcgggcag tgagcgcaac gcaattaatg 2040
tgagttagct cactcattag gcaccccagg ctttacactt tatgcttccg gctcgtatgt 2100
tgtgtggaat tgtgagcgga taacaatttc acacaggaaa cagctatgac catgattacg 2160
ccaagcgcgc aattaaccct cactaaaggg aacaaaagct gggtaccggg ccccccctcg 2220
agcagagcag gattcccgtt gagcaccgcc aggtgcgaat aagggacagt gaagaaggaa 2280
cacccgctcg cgggtgggcc tacttcacct atcctgcccg gctgacgccg ttggatacac 2340
caaggaaagt ctacacgaac cctttggcaa aatcctgtat atcgtgcgaa aaaggatgga 2400
tataccgaaa aaatcgctat aatgaccccg aagcagggtt atgcagcgga aaatgcagct 2460
cacggtaact gatgccgtat ttgcagtacc agcgtacggc ccacagaatg atgtcacgct 2520
gaaaatgccg gcctttgaat gaagcttgcc ggggccgacg cggcgggcgg cctcgtccat 2580
cgggacgcgc cagtcgacgc cgacgatgtc cgcgccggcc tcgcccatga gccccagcag 2640
ctcgccggtg ccgacgccga agtggatgcg cgggacgccg tggccggcca ccgcgcggaa 2700
caccttcgcc gaggcgggca gcaccgaacg ccggtagtcg gagggggcga gcgcgccggc 2760
ccaggagtcg aagagctgca cggccgaggc gccggcccgg atctggacgt cgaggaaggc 2820
cgccgtgatg tcggcgaggc ggtcgagcag gtcggcccag agctcggggt cgccgtacat 2880
catcgccttg gcgttctcgt acgtacggga cgggccgccc tcgacgaggt aactcgcaag 2940
ggtgaacggg gcgcccgcga aaccgatcag cggggtggac ccgagctcac gggtcagcat 3000
gccgatggcc tcggtgacgt aggagacgtc ctccggggtc aggtcgcgca gccgggcgag 3060
gtcggcgcgg gtgcgcaccg gctgctcgac gaccgggccg atgccgggct tgatgtcgag 3120
gtcgatgccg atggccttga gcgggacgac gatgtcgctg aagtagatcg ccgcgtcgac 3180
gtggtggcgg cgcaccggct ggagggtgat ctcggtgacc agctcgggcc gcatgcagga 3240
gtcgagcatc ccgatgccct cgcgcacctt gcggtactcc ggcagtgagc gcccggcctg 3300
ccgcatgaac cacaccggcg tgtgcggcac cggctcgcgc cggcacgcct tgaggaaggc 3360
tgattcccgg acggcgtcgt tcttgacggg gtcgggggtc gcggtaggcg tctggccctt 3420
cgggctcgtg ttggcactca caccggccag tctcgcacga cccgtccgcc gccccggacc 3480
ccaggggcgt ccggcgggcg gggccgcggt gcaggatccg aataggaact tcggaatagg 3540
aacttcatga gctcagccaa tcgactggcg agcggcatcg cattcttcgc atcccgcctc 3600
tggcggatgc aggaagatca acggatctcg gcccagttga cccagggctg tcgccacaat 3660
gtcgcgggag cggatcaacc gagcaaaggc atgaccgact ggaccttcct tctgaaggct 3720
cttctccttg agccacctgt ccgccaaggc aaagcgctca cagcagtggt cattctcgag 3780
ataatcgacg cgtaccaact tgccatcctg aagaatggtg cagtgtctcg gcaccccata 3840
gggaaccttt gccatcaact cggcaagatg cagcgtcgtg ttggcatcgt gtcccacgcc 3900
gaggagaagt acctgcccat cgagttcatg gacacgggcg accgggcttg caggcgagtg 3960
aggtggcagg ggcaatggat cagagatgat ctgctctgcc tgtggccccg ctgccgcaaa 4020
ggcaaatgga tgggcgctgc gctttacatt tggcaggcgc cagaatgtgt cagagacaac 4080
tccaaggtcc ggtgtaacgg gcgacgtggc aggatcgaac ggctcgtcgt ccagacctga 4140
ccacgagggc atgacgagcg tccctcccgg acccagcgca gcacgcaggg cctcgatcag 4200
tccaagtggc ccatcttcga ggggccggac gctacggaag gagctgtgga ccagcagcac 4260
accgccgggg gtaaccccaa ggttgagaag ctgaccgatg agctcggctt ttcgccattc 4320
gtattgcacg acattgcact ccaccgctga tgacatcagt cgatcatagc acgatcaacg 4380
gcactgttgc aaatagtcgg tggtgataaa cttatcatcc ccttttgctg atggagctgc 4440
acatgaacca tggccgcgct caacgggttc gggcagcggg tggggcggcg ccagctggag 4500
cagtggcagg cggccgtcga ccgcgcgaag gcgctgagcg aggcccagct gccgcagccc 4560
ggccagccgg tgaccggccc tccgccgccg cgcgcctggg cggacaagga ccccgttgcc 4620
gcggcccggc tgtcggcggc ccgcgcgggg gtcgccgaac tcgccgagcg gctgaacatg 4680
ccgccggaga acctgatcac cccggacacg gtgcgcaggg tctgctggga gccgccgggg 4740
cccgacgagc ggtccgtcgc cgcggcgctg acggcgcacg gggcacgcgc gtggcaggtc 4800
gaccaggtca ctcccgtgct ggtggccgcg ctggctactt cgtcgccccc cgcatgaacc 4860
cgttgtagat gaagcgctgg aggaacagga agacgatcaa ggtgggcagg atgaccagga 4920
ccgcgcccgc cgagatcgtc tcccagtgcg cgccgaaggg gcccttgaag cggaacaggg 4980
acgtcgagat gacccccagg tcctcggagg gcatgtagag gaaggggatg tagaagtcgt 5040
tgtagacgtt gatccccttt acgatcacca ccgtcgcgat cgccggcttg agcagcggga 5100
agatcacctt gcggtagacg gtgaacgcgt tggcgccgtc caggcgcgcc gcctcgtcca 5160
gggagacggg gatggagcgg atgaactgca ggaagacgta gatcgagacg atgtccgtgc 5220
ccatgtagag ggcgatcggc gcccacaggc tgtcgaacat gccgaagctg ttgacgatct 5280
ggaaggtcgc cacctgggtg gtcaccccgg ggaccagcgc ggccagcagg aacagcgcca 5340
cgaccagcta gttctagagc ggccgccacc gcggtggagc tccaattcgc cctatagtga 5400
gtcgtattac gcgcgctcac tggccgtcgt tttacaacgt cgtgactggg aaaaccctgg 5460
cgttacccaa cttaatcgcc ttgcagcaca tccccctttc gccagctggc gtaatagcga 5520
agaggcccgc accgatcgcc cttcccaaca gttgcgcagc ctgaatggcg aatggaaatt 5580
gtaagcgtta atattttgtt aaaattcgcg ttaaattttt gttaaatcag ctcatttttt 5640
aaccaatagg ccgaaatcgg caaaatccct tataaatcaa aagaatagac cgagataggg 5700
ttgagtgttg ttccagtttg gaacaagagt ccactattaa agaacgtgga ctccaacgtc 5760
aaagggcgaa aaaccgtcta tcagggcgat ggcccactac gtgaaccatc accctaatca 5820
agttttttgg ggtcgaggtg ccgtaaagca ctaaatcgga accctaaagg gagcccccga 5880
tttagagctt gacggggaaa gccggcgaac gtggcgagaa aggaagggaa gaaagcgaaa 5940
ggagcgggcg ctagggcgct ggcaagtgta gcggtcacgc tgcgcgtaac caccacaccc 6000
gccgcgctta atgcgccgct acagggcgcg tcag 6034
<210> 15
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотидный праймер
<400> 15
agcacagcta gcttctagaa gcttcattca aaggccggca 40
<210> 16
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотидный праймер
<400> 16
aggcagctcg agcatatgac tagtcagagc aggattcccg ttga 44
<210> 17
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотидный праймер
<400> 17
gaatggcata tggttcatgt gcagctccat 30
<210> 18
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотидный праймер
<400> 18
tctagaacta gtgaatagga acttcggaat 30
<210> 19
<211> 8113
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая плазмида pKS-orf18ifd-T-AmR(NS)
<400> 19
gtggcacttt tcggggaaat gtgcgcggaa cccctatttg tttatttttc taaatacatt 60
caaatatgta tccgctcatg agacaataac cctgataaat gcttcaataa tattgaaaaa 120
ggaagagtat gagtattcaa catttccgtg tcgcccttat tccctttttt gcggcatttt 180
gccttcctgt ttttgctcac ccagaaacgc tggtgaaagt aaaagatgct gaagatcagt 240
tgggtgcacg agtgggttac atcgaactgg atctcaacag cggtaagatc cttgagagtt 300
ttcgccccga agaacgtttt ccaatgatga gcacttttaa agttctgcta tgtggcgcgg 360
tattatcccg tattgacgcc gggcaagagc aactcggtcg ccgcatacac tattctcaga 420
atgacttggt tgagtactca ccagtcacag aaaagcatct tacggatggc atgacagtaa 480
gagaattatg cagtgctgcc ataaccatga gtgataacac tgcggccaac ttacttctga 540
caacgatcgg aggaccgaag gagctaaccg cttttttgca caacatgggg gatcatgtaa 600
ctcgccttga tcgttgggaa ccggagctga atgaagccat accaaacgac gagcgtgaca 660
ccacgatgcc tgtagcaatg gcaacaacgt tgcgcaaact attaactggc gaactactta 720
ctctagcttc ccggcaacaa ttaatagact ggatggaggc ggataaagtt gcaggaccac 780
ttctgcgctc ggcccttccg gctggctggt ttattgctga taaatctgga gccggtgagc 840
gtgggtctcg cggtatcatt gcagcactgg ggccagatgg taagccctcc cgtatcgtag 900
ttatctacac gacggggagt caggcaacta tggatgaacg aaatagacag atcgctgaga 960
taggtgcctc actgattaag cattggtaac tgtcagacca agtttactca tatatacttt 1020
agattgattt aaaacttcat ttttaattta aaaggatcta ggtgaagatc ctttttgata 1080
atctcatgac caaaatccct taacgtgagt tttcgttcca ctgagcgtca gaccccgtag 1140
aaaagatcaa aggatcttct tgagatcctt tttttctgcg cgtaatctgc tgcttgcaaa 1200
caaaaaaacc accgctacca gcggtggttt gtttgccgga tcaagagcta ccaactcttt 1260
ttccgaaggt aactggcttc agcagagcgc agataccaaa tactgtcctt ctagtgtagc 1320
cgtagttagg ccaccacttc aagaactctg tagcaccgcc tacatacctc gctctgctaa 1380
tcctgttacc agtggctgct gccagtggcg ataagtcgtg tcttaccggg ttggactcaa 1440
gacgatagtt accggataag gcgcagcggt cgggctgaac ggggggttcg tgcacacagc 1500
ccagcttgga gcgaacgacc tacaccgaac tgagatacct acagcgtgag ctatgagaaa 1560
gcgccacgct tcccgaaggg agaaaggcgg acaggtatcc ggtaagcggc agggtcggaa 1620
caggagagcg cacgagggag cttccagggg gaaacgcctg gtatctttat agtcctgtcg 1680
ggtttcgcca cctctgactt gagcgtcgat ttttgtgatg ctcgtcaggg gggcggagcc 1740
tatggaaaaa cgccagcaac gcggcctttt tacggttcct ggccttttgc tggccttttg 1800
ctcacatgtt ctttcctgcg ttatcccctg attctgtgga taaccgtatt accgcctttg 1860
agtgagctga taccgctcgc cgcagccgaa cgaccgagcg cagcgagtca gtgagcgagg 1920
aagcggaaga gcgcccaata cgcaaaccgc ctctccccgc gcgttggccg attcattaat 1980
gcagctggca cgacaggttt cccgactgga aagcgggcag tgagcgcaac gcaattaatg 2040
tgagttagct cactcattag gcaccccagg ctttacactt tatgcttccg gctcgtatgt 2100
tgtgtggaat tgtgagcgga taacaatttc acacaggaaa cagctatgac catgattacg 2160
ccaagcgcgc aattaaccct cactaaaggg aacaaaagct gggtaccggg ccccccctcg 2220
agcatatgtt catgtgcagc tccatcagca aaaggggatg ataagtttat caccaccgac 2280
tatttgcaac agtgccgttg atcgtgctat gatcgactga tgtcatcagc ggtggagtgc 2340
aatgtcgtgc aatacgaatg gcgaaaagcc gagctcatcg gtcagcttct caaccttggg 2400
gttacccccg gcggtgtgct gctggtccac agctccttcc gtagcgtccg gcccctcgaa 2460
gatgggccac ttggactgat cgaggccctg cgtgctgcgc tgggtccggg agggacgctc 2520
gtcatgccct cgtggtcagg tctggacgac gagccgttcg atcctgccac gtcgcccgtt 2580
acaccggacc ttggagttgt ctctgacaca ttctggcgcc tgccaaatgt aaagcgcagc 2640
gcccatccat ttgcctttgc ggcagcgggg ccacaggcag agcagatcat ctctgatcca 2700
ttgcccctgc cacctcactc gcctgcaagc ccggtcgccc gtgtccatga actcgatggg 2760
caggtacttc tcctcggcgt gggacacgat gccaacacga cgctgcatct tgccgagttg 2820
atggcaaagg ttccctatgg ggtgccgaga cactgcacca ttcttcagga tggcaagttg 2880
gtacgcgtcg attatctcga gaatgaccac tgctgtgagc gctttgcctt ggcggacagg 2940
tggctcaagg agaagagcct tcagaaggaa ggtccagtcg gtcatgcctt tgctcggttg 3000
atccgctccc gcgacattgt ggcgacagcc ctgggtcaac tgggccgaga tccgttgatc 3060
ttcctgcatc cgccagaggc gggatgcgaa gaatgcgatg ccgctcgcca gtcgattggc 3120
tgagctcatg aagttcctat tccgaagttc ctattcacta gtcagagcag gattcccgtt 3180
gagcaccgcc aggtgcgaat aagggacagt gaagaaggaa cacccgctcg cgggtgggcc 3240
tacttcacct atcctgcccg gctgacgccg ttggatacac caaggaaagt ctacacgaac 3300
cctttggcaa aatcctgtat atcgtgcgaa aaaggatgga tataccgaaa aaatcgctat 3360
aatgaccccg aagcagggtt atgcagcgga aaatgcagct cacggtaact gatgccgtat 3420
ttgcagtacc agcgtacggc ccacagaatg atgtcacgct gaaaatgccg gcctttgaat 3480
gaagcttgcc ggggccgacg cggcgggcgg cctcgtccat cgggacgcgc cagtcgacgc 3540
cgacgatgtc cgcgccggcc tcgcccatga gccccagcag ctcgccggtg ccgacgccga 3600
agtggatgcg cgggacgccg tggccggcca ccgcgcggaa caccttcgcc gaggcgggca 3660
gcaccgaacg ccggtagtcg gagggggcga gcgcgccggc ccaggagtcg aagagctgca 3720
cggccgaggc gccggcccgg atctggacgt cgaggaaggc cgccgtgatg tcggcgaggc 3780
ggtcgagcag gtcggcccag agctcggggt cgccgtacat catcgccttg gcgttctcgt 3840
acgtacggga cgggccgccc tcgacgaggt aactcgcaag ggtgaacggg gcgcccgcga 3900
aaccgatcag cggggtggac ccgagctcac gggtcagcat gccgatggcc tcggtgacgt 3960
aggagacgtc ctccggggtc aggtcgcgca gccgggcgag gtcggcgcgg gtgcgcaccg 4020
gctgctcgac gaccgggccg atgccgggct tgatgtcgag gtcgatgccg atggccttga 4080
gcgggacgac gatgtcgctg aagtagatcg ccgcgtcgac gtggtggcgg cgcaccggct 4140
ggagggtgat ctcggtgacc agctcgggcc gcatgcagga gtcgagcatc ccgatgccct 4200
cgcgcacctt gcggtactcc ggcagtgagc gcccggcctg ccgcatgaac cacaccggcg 4260
tgtgcggcac cggctcgcgc cggcacgcct tgaggaaggc tgattcccgg acggcgtcgt 4320
tcttgacggg gtcgggggtc gcggtaggcg tctggccctt cgggctcgtg ttggcactca 4380
caccggccag tctcgcacga cccgtccgcc gccccggacc ccaggggcgt ccggcgggcg 4440
gggccgcggt gcaggatccg gacagcgccg ggcggcgcgg gtgtccctcc ctgcgccggg 4500
ggcccgttcc gcttaatctt cccggcatgg ctgcggctca gggacgactg tcggacggcg 4560
ctggcggaat ggacgaaccg aaggagggcg ggggggatcc cgggcacgga ggtgcgcctc 4620
cgccgccctt ccgggccgct gtcgaggcgc tgcagagcgc ccggctgcgg ccgcagatcg 4680
aggtggagac ggtgcccgcg ccgaaacggc tcgccccgta cgcgcacgcg ctggaggcgg 4740
cggtcgtcga cggcgaggag gatctggccg acggccggct ggtgctgctg tgcgacccgg 4800
ccggacacga cgcctggcgg gggaccttcc gtctggtgac gctggtgcgc gccgagctgg 4860
agccggagat ggcggcggat ccgctgctgc cggacgtgtg ctggtcctgg ctgaccggcg 4920
cgctggcggc gcgcggcctg tcgtacggcg agccgagcgg cacggtgacg cgggcgagtt 4980
cgcactactt cggcgggctg tccgcgcggc ccgccgcctc ccagatcgag atccgtgcct 5040
cgtggacgcc gcgtgagggt ctgggcgggg ttccggacac ggccgcccat ctggtcgcgt 5100
ggtccgatct gctggcgcag gtcgcggggc tgccgccggc cgctccgggg gacgcgtccg 5160
tggtgacgct gccgcagcgg agggggccgc agtcgcgctg agcctcccgc tgcggggtcc 5220
cgacgaccgg cctcctgcat ctctctttgt cgatacggcc actttcggaa gcgttgcgac 5280
gcagaccgaa accgttcgat cttcgaatga tcgatcgtgc gtccgaattg cccggattgt 5340
tactcatcac ttcgtgatca ttcgttaaag gacaccaggt ttgctgccga agacgactgt 5400
gaccttgaaa gcacggttcg tcccgccttc acccccacga gccggcccgt cccgcacccc 5460
aggaggcctg gtgtttaatt aatgaccctt ccgtcccgcc cccgatccgc ccgtccccga 5520
tccgcccgcc cacggccaag cgaacacgtt ctttcactct tctgaccgga atacgaccca 5580
ccggcgcccg tcacggagca caaccgtcga cgggcgcctt cgcggcacgg atacccttga 5640
caggtgaccg acgcccacga caccgcagca gacagttcac tgcgcaccac cggaggcgct 5700
cctccggacg acggcggatc ttctgttacg gaggcgccga cccccttgct ggaaccccgc 5760
gagggcattc cgcccgtgat agcggacgag gccgccctcg ccgaggcggt cgccgccttc 5820
gcggccggca gcggacccgt cgccgtggac gccgagcgcg cctccgggta ccgctacggc 5880
cagcgcgcct acctcgtcca gctgcgccgc gagggtgcgg gtaccgcgct gatcgacccc 5940
gtggcctgcc ccgacctgtc cgccctcggc gaggcgctgt ccggcgtcga gtgggtgctg 6000
cacgccgcca cccaggacct gccctgtctg cgcgagatag gcatggtgcc ctcccgcctc 6060
ttcgacaccg agctggccgg ccgccttgcc gggttccccc gcgtcgggct cggcgcgatg 6120
gtcgagaacg tgctcggctt cgtcctggag aagggccact ccgccgtcga ctggtccacc 6180
cgtccgctgc ccgagccctg gctgcggtac gccgccctcg acgtcgaact gctggtcgat 6240
ctgcgggacg ccctggagaa ggagctggac cgccagggca agctggactg ggcccggcag 6300
gagttcgacg cgatcgcctc ggccccgccg ccggagcccc gcaaggaccc ctggcgccgc 6360
acctccggca tgcacaaggt gcgccggcgc cgccagatgg cggtggtgcg ggagctgtgg 6420
gagacccgcg accggatcgc ccggcgccgt gacgtctccc ccggcaaggt gctttccgac 6480
gcggcgatcg tggaggccgc gctcgcgctg cccgccaacc tgcacgccat ggccgcgctc 6540
aacgggttcg ggcagcgggt ggggcggcgc cagctggagc agtggcaggc ggccgtcgac 6600
cgcgcgaagg cgctgagcga ggcccagctg ccgcagcccg gccagccggt gaccggccct 6660
ccgccgccgc gcgcctgggc ggacaaggac cccgttgccg cggcccggct gtcggcggcc 6720
cgcgcggggg tcgccgaact cgccgagcgg ctgaacatgc cgccggagaa cctgatcacc 6780
ccggacacgg tgcgcagggt ctgctgggag ccgccggggc ccgacgagcg gtccgtcgcc 6840
gcggcgctga cggcgcacgg ggcacgcgcg tggcaggtcg accaggtcac tcccgtgctg 6900
gtggccgcgc tggctacttc gtcgcccccc gcatgaaccc gttgtagatg aagcgctgga 6960
ggaacaggaa gacgatcaag gtgggcagga tgaccaggac cgcgcccgcc gagatcgtct 7020
cccagtgcgc gccgaagggg cccttgaagc ggaacaggga cgtcgagatg acccccaggt 7080
cctcggaggg catgtagagg aaggggatgt agaagtcgtt gtagacgttg atccccttta 7140
cgatcaccac cgtcgcgatc gccggcttga gcagcgggaa gatcaccttg cggtagacgg 7200
tgaacgcgtt ggcgccgtcc aggcgcgccg cctcgtccag ggagacgggg atggagcgga 7260
tgaactgcag gaagacgtag atcgagacga tgtccgtgcc catgtagagg gcgatcggcg 7320
cccacaggct gtcgaacatg ccgaagctgt tgacgatctg gaaggtcgcc acctgggtgg 7380
tcaccccggg gaccagcgcg gccagcagga acagcgccac gaccagctag ttctagagcg 7440
gccgccaccg cggtggagct ccaattcgcc ctatagtgag tcgtattacg cgcgctcact 7500
ggccgtcgtt ttacaacgtc gtgactggga aaaccctggc gttacccaac ttaatcgcct 7560
tgcagcacat ccccctttcg ccagctggcg taatagcgaa gaggcccgca ccgatcgccc 7620
ttcccaacag ttgcgcagcc tgaatggcga atggaaattg taagcgttaa tattttgtta 7680
aaattcgcgt taaatttttg ttaaatcagc tcatttttta accaataggc cgaaatcggc 7740
aaaatccctt ataaatcaaa agaatagacc gagatagggt tgagtgttgt tccagtttgg 7800
aacaagagtc cactattaaa gaacgtggac tccaacgtca aagggcgaaa aaccgtctat 7860
cagggcgatg gcccactacg tgaaccatca ccctaatcaa gttttttggg gtcgaggtgc 7920
cgtaaagcac taaatcggaa ccctaaaggg agcccccgat ttagagcttg acggggaaag 7980
ccggcgaacg tggcgagaaa ggaagggaag aaagcgaaag gagcgggcgc tagggcgctg 8040
gcaagtgtag cggtcacgct gcgcgtaacc accacacccg ccgcgcttaa tgcgccgcta 8100
cagggcgcgt cag 8113
<210> 20
<211> 7120
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая плазмида pXY152-endorf24-camtsr
<400> 20
gaattcgtaa tcatgtcata gctgtttcct gtgtgaaatt gttatccgct cacaattcca 60
cacaacatac gagccggaag cataaagtgt aaagcctggg gtgcctaatg agtgagctaa 120
ctcacattaa ttgcgttgcg ctcactgccc gctttccagt cgggaaacct gtcgtgccag 180
ctgcattaat gaatcggcca acgcgcgggg agaggcggtt tgcgtattgg gcgctcttcc 240
gcttcctcgc tcactgactc gctgcgctcg gtcgttcggc tgcggcgagc ggtatcagct 300
cactcaaagg cggtaatacg gttatccaca gaatcagggg ataacgcagg aaagaacatg 360
tgagcaaaag gccagcaaaa ggccaggaac cgtaaaaagg ccgcgttgct ggcgtttttc 420
cataggctcc gcccccctga cgagcatcac aaaaatcgac gctcaagtca gaggtggcga 480
aacccgacag gactataaag ataccaggcg tttccccctg gaagctccct cgtgcgctct 540
cctgttccga ccctgccgct taccggatac ctgtccgcct ttctcccttc gggaagcgtg 600
gcgctttctc atagctcacg ctgtaggtat ctcagttcgg tgtaggtcgt tcgctccaag 660
ctgggctgtg tgcacgaacc ccccgttcag cccgaccgct gcgccttatc cggtaactat 720
cgtcttgagt ccaacccggt aagacacgac ttatcgccac tggcagcagc cactggtaac 780
aggattagca gagcgaggta tgtaggcggt gctacagagt tcttgaagtg gtggcctaac 840
tacggctaca ctagaagaac agtatttggt atctgcgctc tgctgaagcc agttaccttc 900
ggaaaaagag ttggtagctc ttgatccggc aaacaaacca ccgctggtag cggtggtttt 960
tttgtttgca agcagcagat tacgcgcaga aaaaaaggat ctcaagaaga tcctttgatc 1020
ttttctacgg ggtctgacgc tcagtggaac gaaaactcac gttaagggat tttggtcatg 1080
agattatcaa aaaggatctt cacctagatc cttttggttc atgtgcagct ccatcagcaa 1140
aaggggatga taagtttatc accaccgact atttgcaaca gtgccgttga tcgtgctatg 1200
atcgactgat gtcatcagcg gtggagtgca atgtcgtgca atacgaatgg cgaaaagccg 1260
agctcaataa ctgccttaaa aaaattacgc cccgccctgc cactcatcgc agtactgttg 1320
taattcatta agcattctgc cgacatggaa gccatcacaa acggcatgat gaacctgaat 1380
cgccagcggc atcagcacct tgtcgccttg cgtataatat ttgcccatcg tgaaaacggg 1440
ggcgaagaag ttgtccatat tggccacgtt taagtcaaaa ctggtgaaac tcacccaggg 1500
attggctgag acgaaaaaca tattctcaat aaacccttta gggaaatagg ccaggttttc 1560
accgtaacac gccacatctt gcgaatatat gtgtagaaac tgccggaaat cgtcgtggta 1620
ttcactccag agcgatgaaa acgtttcagt ttgctcatgg aaaacggtgt aacaagggtg 1680
aacactatcc catatcacca gctcaccgtc tttcattgcc atacgaaatt ccggatgagc 1740
attcatcagg cgggcaagaa tgtgaataaa ggccggataa aacttgtgct tatttttctt 1800
tacggtcttt aagaaggccg taatatccag ttgaacggtc tggttatagg tacattgagc 1860
aactgactga aatgcctcaa aatgttcttt acgatgccat tgggatatat caacggtggt 1920
atatccagtg atttttttct ccattttagc ttccttagct cctgaaaatc tcgataactc 1980
aaaaaatacg cccggtagtg atcttatttc attatggtga aagttggaac ctcttacgtg 2040
ccgatcaacg tctcattttc gccaaaagtt ggcccagggc ttcccggtat caacagggac 2100
accaggattt atttattctg cgaagtgatc ttccgtcaca ggtatttatt cgcgataagc 2160
tcatggagcg gcgtaaccgt cgcacaggaa ggacagagaa agcgcggaag cttatttaaa 2220
taccgcgcgg gtcccggacg gggaagagcg gggagctttg ccagagagcg acgacttccc 2280
cttgcgttgg tgattgccgg tcagggcagc catccgccat cgtcgcgtag ggtgtcacac 2340
cccaggaatc gcgtcactga acacagcagc cggtaggacg accatgactg agttggacac 2400
catcgcaaat ccgtccgatc cggcggtgca gcggatcatc gatgtcacca agccgtcgcg 2460
atccaacata aagacaacgt tgatcgagga cgtcgagccc ctcatgcaca gcatcgcggc 2520
cggggtggag ttcatcgagg tctacggcag cgacagcagt ccttttccat ctgagttgct 2580
ggatctgtgc gggcggcaga acataccggt ccgcctcatc gactcctcga tcgtcaacca 2640
gttgttcaag ggggagcgga aggccaagac attcggcatc gcccgcgtcc ctcgcccggc 2700
caggttcggc gacatcgcga gccggcgtgg ggacgtcgtc gttctcgacg gggtgaagat 2760
cgtcgggaac atcggcgcga tagtacgcac gtcgctcgcg ctcggagcgt cggggatcat 2820
cctggtcgac agtgacatca ccagcatcgc ggaccggcgt ctccaaaggg ccagccgagg 2880
ttacgtcttc tcccttcccg tcgttctctc cggtcgcgag gaggccatcg ccttcattcg 2940
ggacagcggt atgcaactga tgacgctcaa ggcggatggc gacatttccg tgaaggaact 3000
cggggacaat ccggatcggc tggccttgct gttcggcagc gaaaagggtg ggccttccga 3060
cctgttcgag gaggcgtctt ccgcctcggt ttccatcccc atgatgagcc agaccgagtc 3120
tctcaacgtt tccgtttccc tcggaatcgc gctgcacgag aggatcgaca ggaatctcgc 3180
ggccaaccga taagctagct agagtcgacc tgcaggtccc cggggatcgg tcttgccttg 3240
ctcgtcggtg atgtacttca ccagctccgc gaagtcgctc ttcttgatgg agcgcatggg 3300
gacgtgcttg gcaatcacgc gcaccccccg gccgttttag cggctaaaaa agtcatggct 3360
ctgccctcgg gcggaccacg cccatcatga ccttgccaag ctcgtcctgc ttctcttcga 3420
tcttcgccag cagggcgagg atcgtggcat caccgaaccg cgccgtgcgc gggtcgtcgg 3480
tgagccagag tttcagcagg ccgcccaggc ggcccaggtc gccattgatg cgggccagct 3540
cgcggacgtg ctcatagtcc acgacgcccg tgattttgta gccctggccg acggccagca 3600
ggtaggccga caggctcatg ccggccgccg ccgccttttc ctcaatcgct cttcgttcgt 3660
ctggaaggca gtacaccttg ataggtgggc tgcccttcct ggttggcttg gtttcatcag 3720
ccatccgctt gccctcatct gttacgccgg cggtagccgg ccagcctcgc agagcaggat 3780
tcccgttgag caccgccagg tgcgaataag ggacagtgaa gaaggaacac ccgctcgcgg 3840
gtgggcctac ttcacctatc ctgcccggct gacgccgttg gatacaccaa ggaaagtcta 3900
cacgaaccct ttggcaaaat cctgtatatc gtgcgaaaaa ggatggatat accgaaaaaa 3960
tcgctataat gaccccgaag cagggttatg cagcggaaaa gatccgtcga cctgcaggca 4020
tgcaagctct agcgattcca gacgtcccga aggcgtggcg cggcttcccc gtgccggagc 4080
aatcgccctg ggtgggttac acgacgcccc tctatggccc gtactgacgg acacaccgaa 4140
gccccggcgg caaccctcag cggatgcccc ggggcttcac gttttcccag gtcagaagcg 4200
gttttcggga gtagtgcccc aactggggta acctttgagt tctctcagtt gggggcgtag 4260
ggtcgccgac atgacacaag gggttgtgac cggggtggac acgtacgcgg gtgcttacga 4320
ccgtcagtcg cgcgagcgcg agaactcgag cgcagcaagc ccagcgacac agcgtagcgc 4380
caacgaagac aaggcggccg accttcagcg cgaagtcgag cgcgacgggg gccggttcag 4440
gttcgtcggg catttcagcg aagcgccggg cacgtcggcg ttcgggacgg cggagcgccc 4500
ggagttcgaa cgcatcctga acgaatgccg cgccgggcgg ctcaacatga tcattgtcta 4560
tgacgtgtcg cgcttctcgc gcctgaaggt catggacgcg attccgattg tctcggaatt 4620
gctcgccctg ggcgtgacga ttgtttccac tcaggaaggc gtcttccggc agggaaacgt 4680
catggacctg attcacctga ttatgcggct cgacgcgtcg cacaaagaat cttcgctgaa 4740
gtcggcgaag attctcgaca cgaagaacct tcagcgcgaa ttgggcgggt acgtcggcgg 4800
gaaggcgcct tacggcttcg agcttgtttc ggagacgaag gagatcacgc gcaacggccg 4860
aatggtcaat gtcgtcatca acaagcttgc gcactcgacc actcccctta ccggaccctt 4920
cgagttcgag cccgacgtaa tccggtggtg gtggcgtgag atcaagacgc acaaacacct 4980
tcccttcaag ccgggcagtc aagccgccat tcacccgggc agcatcacgg ggctttgtaa 5040
gcgcatggac gctgacgccg tgccgacccg gggcgagacg attgggaaga agaccgcttc 5100
aagcgcctgg gacccggcaa ccgttatgcg aatccttcgg gacccgcgta ttgcgggctt 5160
cgccgctgag gtgatctaca agaagaagcc ggacggcacg ccgaccacga agattgaggg 5220
ttaccgcatt cagcgcgacc cgatcacgct ccggccggtc gagcttgatt gcggaccgat 5280
catcgagccc gctgagtggt atgagcttca ggcgtggttg gacggcaggg ggcgcggcaa 5340
ggggctttcc cgggggcaag ccattctgtc cgccatggac aagctgtact gcgagtgtgg 5400
cgccgtcatg acttcgaagc gcggggaaga atcgatcaag gactcttacc gctgccgtcg 5460
ccggaaggtg gtcgacccgt ccgcacctgg gcagcacgaa ggcacgtgca acgtcagcat 5520
ggcggcactc gacaagttcg ttgcggaacg catcttcaac aagatcaggc acgccgaagg 5580
cgacgaagag acgttggcgc ttctgtggga agccgcccga cgcttcggca agctcactga 5640
ggcgcctgag aagagcggcg aacgggcgaa ccttgttgcg gagcgcgccg acgccctgaa 5700
cgcccttgaa gagctgtacg aagaccgcgc ggcaggcgcg tacgacggac ccgttggcag 5760
gaagcacttc cggaagcaac aggcagcgct gacgctccgg cagcaagggg cggaagagcg 5820
gcttgccgaa cttgaagccg ccgaagcccc gaagcttccc cttgaccaat ggttccccga 5880
agacgccgac gctgacccga ccggccctaa gtcgtggtgg gggcgcgcgt cagtagacga 5940
caagcgcgtg ttcgtcgggc tcttcgtaga caagatcgtt gtcacgaagt cgactacggg 6000
cagggggcag ggaacgccca tcgagaagcg cgcttcgatc acgtgggcga agccgccgac 6060
cgacgacgac gaagacgacg cccaggacgg cacggaagac gtagcggcgt aggtgagtga 6120
agatctagac gtggcaccgc gatgctgttg tgggcacaat cgtgccggtt ggtaggatcc 6180
acatatggaa ataagttcgc tctccaccga cggctccccg cggatcgacg gggagagtcc 6240
cgagcacgtg gaaatgctgg ccgccgccga caccgcgctt ccaccgatca tggtgcaccg 6300
ccgcaccggg cgggtcatcg acggcatgca ccggctgcgc gccgcgatgc tgacgggccg 6360
tacgacgatc gcggtgaggt tcttcgacgg caccgaggag gacgccttcg tcctcgccgt 6420
gaagtcgaac atcgcgcacg gactgccgct gtccgccgcc gaccgccggc gggccgccgg 6480
gcgcatcatg gccacccatc cccggtggtc ggaccggatg atcgcctcgg tggtcggcac 6540
ctccgccagg acggtcgccg agatccgccg cgacgccggc gccgccgggg cgggggagcc 6600
cacccgcatc ggccgggacg gcagggtacg gcccgtcgac gtgagcgagg gccgcagact 6660
ggcccacgac atgatcgtcc gcgacccggg cctgtcgctg cgccaggtcg cccgcgccgc 6720
cgggatctcg ccggagaccg tcagggacgt cagacaccgg atgctccgcg gtgaggaccc 6780
ggtgcccgcg ccgcggccgc ggaccctggt ggagcgcggc gcggaccgcc gggcggagcc 6840
ggccgggaag gccgccgcgc cgtgcgggac ggagccgccg cccgccgtcg tgatgaagcg 6900
gctgagggcc gatccggcgc tgcgtctcaa cgagaacgga cgcgacctgc tgcggcttct 6960
ggatatccac acggtccggc tggaggactg gaaccgcatt atcgaaagcg tgccgccgca 7020
ccgtctggag acggtggcgc agctggcacg ctcctgcgcc gacaaatggt ccgagatcgc 7080
gtcacgcatc gaaagcaacg catcacatct ggccgggtga 7120
<210> 21
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотидный праймер
<400> 21
gtggcaattt aaatggaaat gtgcgcggaa 30
<210> 22
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотидный праймер
<400> 22
tatatagagc tcaacttggt ctgacagtta c 31
<210> 23
<211> 7162
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая плазмида pXY152-endorf24-blatsr
<400> 23
gaattcgtaa tcatgtcata gctgtttcct gtgtgaaatt gttatccgct cacaattcca 60
cacaacatac gagccggaag cataaagtgt aaagcctggg gtgcctaatg agtgagctaa 120
ctcacattaa ttgcgttgcg ctcactgccc gctttccagt cgggaaacct gtcgtgccag 180
ctgcattaat gaatcggcca acgcgcgggg agaggcggtt tgcgtattgg gcgctcttcc 240
gcttcctcgc tcactgactc gctgcgctcg gtcgttcggc tgcggcgagc ggtatcagct 300
cactcaaagg cggtaatacg gttatccaca gaatcagggg ataacgcagg aaagaacatg 360
tgagcaaaag gccagcaaaa ggccaggaac cgtaaaaagg ccgcgttgct ggcgtttttc 420
cataggctcc gcccccctga cgagcatcac aaaaatcgac gctcaagtca gaggtggcga 480
aacccgacag gactataaag ataccaggcg tttccccctg gaagctccct cgtgcgctct 540
cctgttccga ccctgccgct taccggatac ctgtccgcct ttctcccttc gggaagcgtg 600
gcgctttctc atagctcacg ctgtaggtat ctcagttcgg tgtaggtcgt tcgctccaag 660
ctgggctgtg tgcacgaacc ccccgttcag cccgaccgct gcgccttatc cggtaactat 720
cgtcttgagt ccaacccggt aagacacgac ttatcgccac tggcagcagc cactggtaac 780
aggattagca gagcgaggta tgtaggcggt gctacagagt tcttgaagtg gtggcctaac 840
tacggctaca ctagaagaac agtatttggt atctgcgctc tgctgaagcc agttaccttc 900
ggaaaaagag ttggtagctc ttgatccggc aaacaaacca ccgctggtag cggtggtttt 960
tttgtttgca agcagcagat tacgcgcaga aaaaaaggat ctcaagaaga tcctttgatc 1020
ttttctacgg ggtctgacgc tcagtggaac gaaaactcac gttaagggat tttggtcatg 1080
agattatcaa aaaggatctt cacctagatc cttttggttc atgtgcagct ccatcagcaa 1140
aaggggatga taagtttatc accaccgact atttgcaaca gtgccgttga tcgtgctatg 1200
atcgactgat gtcatcagcg gtggagtgca atgtcgtgca atacgaatgg cgaaaagccg 1260
agctcaactt ggtctgacag ttaccaatgc ttaatcagtg aggcacctat ctcagcgatc 1320
tgtctatttc gttcatccat agttgcctga ctccccgtcg tgtagataac tacgatacgg 1380
gagggcttac catctggccc cagtgctgca atgataccgc gagacccacg ctcaccggct 1440
ccagatttat cagcaataaa ccagccagcc ggaagggccg agcgcagaag tggtcctgca 1500
actttatccg cctccatcca gtctattaat tgttgccggg aagctagagt aagtagttcg 1560
ccagttaata gtttgcgcaa cgttgttgcc attgctacag gcatcgtggt gtcacgctcg 1620
tcgtttggta tggcttcatt cagctccggt tcccaacgat caaggcgagt tacatgatcc 1680
cccatgttgt gcaaaaaagc ggttagctcc ttcggtcctc cgatcgttgt cagaagtaag 1740
ttggccgcag tgttatcact catggttatg gcagcactgc ataattctct tactgtcatg 1800
ccatccgtaa gatgcttttc tgtgactggt gagtactcaa ccaagtcatt ctgagaatag 1860
tgtatgcggc gaccgagttg ctcttgcccg gcgtcaatac gggataatac cgcgccacat 1920
agcagaactt taaaagtgct catcattgga aaacgttctt cggggcgaaa actctcaagg 1980
atcttaccgc tgttgagatc cagttcgatg taacccactc gtgcacccaa ctgatcttca 2040
gcatctttta ctttcaccag cgtttctggg tgagcaaaaa caggaaggca aaatgccgca 2100
aaaaagggaa taagggcgac acggaaatgt tgaatactca tactcttcct ttttcaatat 2160
tattgaagca tttatcaggg ttattgtctc atgagcggat acatatttga atgtatttag 2220
aaaaataaac aaataggggt tccgcgcaca tttccattta aataccgcgc gggtcccgga 2280
cggggaagag cggggagctt tgccagagag cgacgacttc cccttgcgtt ggtgattgcc 2340
ggtcagggca gccatccgcc atcgtcgcgt agggtgtcac accccaggaa tcgcgtcact 2400
gaacacagca gccggtagga cgaccatgac tgagttggac accatcgcaa atccgtccga 2460
tccggcggtg cagcggatca tcgatgtcac caagccgtcg cgatccaaca taaagacaac 2520
gttgatcgag gacgtcgagc ccctcatgca cagcatcgcg gccggggtgg agttcatcga 2580
ggtctacggc agcgacagca gtccttttcc atctgagttg ctggatctgt gcgggcggca 2640
gaacataccg gtccgcctca tcgactcctc gatcgtcaac cagttgttca agggggagcg 2700
gaaggccaag acattcggca tcgcccgcgt ccctcgcccg gccaggttcg gcgacatcgc 2760
gagccggcgt ggggacgtcg tcgttctcga cggggtgaag atcgtcggga acatcggcgc 2820
gatagtacgc acgtcgctcg cgctcggagc gtcggggatc atcctggtcg acagtgacat 2880
caccagcatc gcggaccggc gtctccaaag ggccagccga ggttacgtct tctcccttcc 2940
cgtcgttctc tccggtcgcg aggaggccat cgccttcatt cgggacagcg gtatgcaact 3000
gatgacgctc aaggcggatg gcgacatttc cgtgaaggaa ctcggggaca atccggatcg 3060
gctggccttg ctgttcggca gcgaaaaggg tgggccttcc gacctgttcg aggaggcgtc 3120
ttccgcctcg gtttccatcc ccatgatgag ccagaccgag tctctcaacg tttccgtttc 3180
cctcggaatc gcgctgcacg agaggatcga caggaatctc gcggccaacc gataagctag 3240
ctagagtcga cctgcaggtc cccggggatc ggtcttgcct tgctcgtcgg tgatgtactt 3300
caccagctcc gcgaagtcgc tcttcttgat ggagcgcatg gggacgtgct tggcaatcac 3360
gcgcaccccc cggccgtttt agcggctaaa aaagtcatgg ctctgccctc gggcggacca 3420
cgcccatcat gaccttgcca agctcgtcct gcttctcttc gatcttcgcc agcagggcga 3480
ggatcgtggc atcaccgaac cgcgccgtgc gcgggtcgtc ggtgagccag agtttcagca 3540
ggccgcccag gcggcccagg tcgccattga tgcgggccag ctcgcggacg tgctcatagt 3600
ccacgacgcc cgtgattttg tagccctggc cgacggccag caggtaggcc gacaggctca 3660
tgccggccgc cgccgccttt tcctcaatcg ctcttcgttc gtctggaagg cagtacacct 3720
tgataggtgg gctgcccttc ctggttggct tggtttcatc agccatccgc ttgccctcat 3780
ctgttacgcc ggcggtagcc ggccagcctc gcagagcagg attcccgttg agcaccgcca 3840
ggtgcgaata agggacagtg aagaaggaac acccgctcgc gggtgggcct acttcaccta 3900
tcctgcccgg ctgacgccgt tggatacacc aaggaaagtc tacacgaacc ctttggcaaa 3960
atcctgtata tcgtgcgaaa aaggatggat ataccgaaaa aatcgctata atgaccccga 4020
agcagggtta tgcagcggaa aagatccgtc gacctgcagg catgcaagct ctagcgattc 4080
cagacgtccc gaaggcgtgg cgcggcttcc ccgtgccgga gcaatcgccc tgggtgggtt 4140
acacgacgcc cctctatggc ccgtactgac ggacacaccg aagccccggc ggcaaccctc 4200
agcggatgcc ccggggcttc acgttttccc aggtcagaag cggttttcgg gagtagtgcc 4260
ccaactgggg taacctttga gttctctcag ttgggggcgt agggtcgccg acatgacaca 4320
aggggttgtg accggggtgg acacgtacgc gggtgcttac gaccgtcagt cgcgcgagcg 4380
cgagaactcg agcgcagcaa gcccagcgac acagcgtagc gccaacgaag acaaggcggc 4440
cgaccttcag cgcgaagtcg agcgcgacgg gggccggttc aggttcgtcg ggcatttcag 4500
cgaagcgccg ggcacgtcgg cgttcgggac ggcggagcgc ccggagttcg aacgcatcct 4560
gaacgaatgc cgcgccgggc ggctcaacat gatcattgtc tatgacgtgt cgcgcttctc 4620
gcgcctgaag gtcatggacg cgattccgat tgtctcggaa ttgctcgccc tgggcgtgac 4680
gattgtttcc actcaggaag gcgtcttccg gcagggaaac gtcatggacc tgattcacct 4740
gattatgcgg ctcgacgcgt cgcacaaaga atcttcgctg aagtcggcga agattctcga 4800
cacgaagaac cttcagcgcg aattgggcgg gtacgtcggc gggaaggcgc cttacggctt 4860
cgagcttgtt tcggagacga aggagatcac gcgcaacggc cgaatggtca atgtcgtcat 4920
caacaagctt gcgcactcga ccactcccct taccggaccc ttcgagttcg agcccgacgt 4980
aatccggtgg tggtggcgtg agatcaagac gcacaaacac cttcccttca agccgggcag 5040
tcaagccgcc attcacccgg gcagcatcac ggggctttgt aagcgcatgg acgctgacgc 5100
cgtgccgacc cggggcgaga cgattgggaa gaagaccgct tcaagcgcct gggacccggc 5160
aaccgttatg cgaatccttc gggacccgcg tattgcgggc ttcgccgctg aggtgatcta 5220
caagaagaag ccggacggca cgccgaccac gaagattgag ggttaccgca ttcagcgcga 5280
cccgatcacg ctccggccgg tcgagcttga ttgcggaccg atcatcgagc ccgctgagtg 5340
gtatgagctt caggcgtggt tggacggcag ggggcgcggc aaggggcttt cccgggggca 5400
agccattctg tccgccatgg acaagctgta ctgcgagtgt ggcgccgtca tgacttcgaa 5460
gcgcggggaa gaatcgatca aggactctta ccgctgccgt cgccggaagg tggtcgaccc 5520
gtccgcacct gggcagcacg aaggcacgtg caacgtcagc atggcggcac tcgacaagtt 5580
cgttgcggaa cgcatcttca acaagatcag gcacgccgaa ggcgacgaag agacgttggc 5640
gcttctgtgg gaagccgccc gacgcttcgg caagctcact gaggcgcctg agaagagcgg 5700
cgaacgggcg aaccttgttg cggagcgcgc cgacgccctg aacgcccttg aagagctgta 5760
cgaagaccgc gcggcaggcg cgtacgacgg acccgttggc aggaagcact tccggaagca 5820
acaggcagcg ctgacgctcc ggcagcaagg ggcggaagag cggcttgccg aacttgaagc 5880
cgccgaagcc ccgaagcttc cccttgacca atggttcccc gaagacgccg acgctgaccc 5940
gaccggccct aagtcgtggt gggggcgcgc gtcagtagac gacaagcgcg tgttcgtcgg 6000
gctcttcgta gacaagatcg ttgtcacgaa gtcgactacg ggcagggggc agggaacgcc 6060
catcgagaag cgcgcttcga tcacgtgggc gaagccgccg accgacgacg acgaagacga 6120
cgcccaggac ggcacggaag acgtagcggc gtaggtgagt gaagatctag acgtggcacc 6180
gcgatgctgt tgtgggcaca atcgtgccgg ttggtaggat ccacatatgg aaataagttc 6240
gctctccacc gacggctccc cgcggatcga cggggagagt cccgagcacg tggaaatgct 6300
ggccgccgcc gacaccgcgc ttccaccgat catggtgcac cgccgcaccg ggcgggtcat 6360
cgacggcatg caccggctgc gcgccgcgat gctgacgggc cgtacgacga tcgcggtgag 6420
gttcttcgac ggcaccgagg aggacgcctt cgtcctcgcc gtgaagtcga acatcgcgca 6480
cggactgccg ctgtccgccg ccgaccgccg gcgggccgcc gggcgcatca tggccaccca 6540
tccccggtgg tcggaccgga tgatcgcctc ggtggtcggc acctccgcca ggacggtcgc 6600
cgagatccgc cgcgacgccg gcgccgccgg ggcgggggag cccacccgca tcggccggga 6660
cggcagggta cggcccgtcg acgtgagcga gggccgcaga ctggcccacg acatgatcgt 6720
ccgcgacccg ggcctgtcgc tgcgccaggt cgcccgcgcc gccgggatct cgccggagac 6780
cgtcagggac gtcagacacc ggatgctccg cggtgaggac ccggtgcccg cgccgcggcc 6840
gcggaccctg gtggagcgcg gcgcggaccg ccgggcggag ccggccggga aggccgccgc 6900
gccgtgcggg acggagccgc cgcccgccgt cgtgatgaag cggctgaggg ccgatccggc 6960
gctgcgtctc aacgagaacg gacgcgacct gctgcggctt ctggatatcc acacggtccg 7020
gctggaggac tggaaccgca ttatcgaaag cgtgccgccg caccgtctgg agacggtggc 7080
gcagctggca cgctcctgcg ccgacaaatg gtccgagatc gcgtcacgca tcgaaagcaa 7140
cgcatcacat ctggccgggt ga 7162
<210> 24
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотидный праймер
<400> 24
aaggagaaga gccttcagaa ggaa 24
<210> 25
<211> 321
<212> БЕЛОК
<213> S. griseus
<400> 25
Met Asp Pro Thr Arg Val Asp Ile Phe Ala Leu Pro Ala Val Glu Ile
1 5 10 15
Glu Leu Ser Arg Leu Ser Ser Ala Ser Ser Pro Arg Thr Ser Gly Glu
20 25 30
Asp Pro Glu His Val Glu Thr Leu Leu Ser Ala Glu Gly Glu Leu Pro
35 40 45
Pro Ile Leu Val His Arg Pro Thr Met Gln Val Leu Asp Gly Leu His
50 55 60
Arg Leu Lys Val Ala Arg Val Arg Gly Asp Thr Lys Ile Leu Ala Arg
65 70 75 80
Leu Val Asp Ala Thr Glu Ser Asp Ala Phe Val Leu Ala Val Glu Ala
85 90 95
Asn Ile Arg His Gly Leu Pro Leu Ser Leu Ala Asp Arg Lys Arg Ala
100 105 110
Ala Val Gln Ile Ile Gly Thr His Pro Gln Trp Ser Asp Arg Arg Val
115 120 125
Ala Ser Ala Thr Gly Ile Ser Ala Gly Thr Val Ala Asp Leu Arg Arg
130 135 140
Arg Ala Gly Glu Asp Gly Thr Glu Ala Arg Ile Gly Arg Asp Gly Arg
145 150 155 160
Val Arg Pro Ser Asp Gly Ser Glu Arg Arg Arg Leu Ala Ala Glu Leu
165 170 175
Ile Arg Ser Asp Pro Gly Leu Ser Leu Arg Gln Val Ala Lys Gln Val
180 185 190
Gly Ile Ser Pro Glu Thr Val Arg Asp Val Arg Gly Arg Leu Glu Arg
195 200 205
Gly Glu Ser Pro Thr Pro Asp Gly Thr Arg Arg Leu Pro Ala Lys Pro
210 215 220
His Pro Leu Arg Leu Ser Glu Pro Asp Phe Gly Arg Ala Val Asp Gln
225 230 235 240
Asp Arg Leu Ala Leu Leu Glu Arg Leu Lys Ser Asp Pro Ala Leu Arg
245 250 255
Leu Asn Glu Val Gly Arg Ile Leu Leu Arg Met Leu Thr Met His Ser
260 265 270
Met Asp Gly Gln Glu Trp Glu Arg Ile Leu Gln Gly Val Pro Pro His
275 280 285
Leu His Gly Val Ile Ala Gly Phe Ala Arg Asp His Ala Arg Val Trp
290 295 300
Ala Glu Phe Ala Asp His Leu Glu Ser Arg Ala Thr Glu Leu Ala Ala
305 310 315 320
Gly
<210> 26
<211> 311
<212> БЕЛОК
<213> Streptomyces fungicidicus
<400> 26
Val Glu Ile Ser Ser Leu Ser Thr Asp Gly Ser Pro Arg Ile Asp Gly
1 5 10 15
Glu Ser Pro Glu His Val Glu Met Leu Ala Ala Ala Asp Thr Ala Leu
20 25 30
Pro Pro Ile Met Val His Arg Arg Thr Gly Arg Val Ile Asp Gly Met
35 40 45
His Arg Leu Arg Ala Ala Met Leu Thr Gly Arg Thr Thr Ile Ala Val
50 55 60
Arg Phe Phe Asp Gly Thr Glu Glu Asp Ala Phe Val Leu Ala Val Lys
65 70 75 80
Ser Asn Ile Ala His Gly Leu Pro Leu Ser Ala Ala Asp Arg Arg Arg
85 90 95
Ala Ala Gly Arg Ile Met Ala Thr His Pro Arg Trp Ser Asp Arg Met
100 105 110
Ile Ala Ser Val Val Gly Thr Ser Ala Arg Thr Val Ala Glu Ile Arg
115 120 125
Arg Asp Ala Gly Ala Ala Gly Ala Gly Glu Pro Thr Arg Ile Gly Arg
130 135 140
Asp Gly Arg Val Arg Pro Val Asp Val Ser Glu Gly Arg Arg Leu Ala
145 150 155 160
His Asp Met Ile Val Arg Asp Pro Gly Leu Ser Leu Arg Gln Val Ala
165 170 175
Arg Ala Ala Gly Ile Ser Pro Glu Thr Val Arg Asp Val Arg His Arg
180 185 190
Met Leu Arg Gly Glu Asp Pro Val Pro Ala Pro Arg Pro Arg Thr Leu
195 200 205
Val Glu Arg Gly Ala Asp Arg Arg Ala Glu Pro Ala Gly Lys Ala Ala
210 215 220
Ala Pro Cys Gly Thr Glu Pro Pro Pro Ala Val Val Met Lys Arg Leu
225 230 235 240
Arg Ala Asp Pro Ala Leu Arg Leu Asn Glu Asn Gly Arg Asp Leu Leu
245 250 255
Arg Leu Leu Asp Ile His Thr Val Arg Leu Glu Asp Trp Asn Arg Ile
260 265 270
Ile Glu Ser Val Pro Pro His Arg Leu Glu Thr Val Ala Gln Leu Ala
275 280 285
Arg Ser Cys Ala Asp Lys Trp Ser Glu Ile Ala Ser Arg Ile Glu Ser
290 295 300
Asn Ala Ser His Leu Ala Gly
305 310
<210> 27
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотидный праймер
<400> 27
gtgtttaatt aatga 15
<210> 28
<211> 321
<212> БЕЛОК
<213> Amycolatopsis
<400> 28
Met Asp Pro Thr Arg Val Asp Ile Phe Ala Leu Pro Ala Val Glu Ile
1 5 10 15
Glu Leu Ser Arg Leu Ser Ser Ala Ser Ser Pro Arg Thr Ser Gly Glu
20 25 30
Asp Pro Glu His Val Glu Thr Leu Leu Ser Ala Glu Gly Glu Leu Pro
35 40 45
Pro Ile Leu Val His Arg Pro Thr Met Gln Val Leu Asp Gly Leu His
50 55 60
Arg Leu Lys Val Ala Arg Val Arg Gly Asp Thr Lys Ile Leu Ala Arg
65 70 75 80
Leu Val Asp Ala Thr Glu Ser Asp Ala Phe Val Leu Ala Val Glu Ala
85 90 95
Asn Ile Arg His Gly Leu Pro Leu Ser Leu Ala Asp Arg Lys Arg Ala
100 105 110
Ala Val Gln Ile Ile Gly Thr His Pro Gln Trp Ser Asp Arg Arg Val
115 120 125
Ala Ser Ala Thr Gly Ile Ser Ala Gly Thr Val Ala Asp Leu Arg Arg
130 135 140
Arg Ala Gly Glu Asp Gly Thr Glu Ala Arg Ile Gly Arg Asp Gly Arg
145 150 155 160
Val Arg Pro Ser Asp Gly Ser Glu Arg Arg Arg Leu Ala Ala Glu Leu
165 170 175
Ile Arg Ser Asp Pro Gly Leu Ser Leu Arg Gln Val Ala Lys Gln Val
180 185 190
Gly Ile Ser Pro Glu Thr Val Arg Asp Val Arg Gly Arg Leu Glu Arg
195 200 205
Gly Glu Ser Pro Thr Pro Asp Gly Thr Arg Arg Leu Pro Ala Lys Pro
210 215 220
His Pro Leu Arg Leu Ser Glu Pro Asp Phe Gly Arg Ala Val Asp Gln
225 230 235 240
Asp Arg Leu Ala Leu Leu Glu Arg Leu Lys Ser Asp Pro Ala Leu Arg
245 250 255
Leu Asn Glu Val Gly Arg Ile Leu Leu Arg Met Leu Thr Met His Ser
260 265 270
Met Asp Gly Gln Glu Trp Glu Arg Ile Leu Gln Gly Val Pro Pro His
275 280 285
Leu His Gly Val Ile Ala Gly Phe Ala Arg Asp His Ala Arg Val Trp
290 295 300
Ala Glu Phe Ala Asp His Leu Glu Ser Arg Ala Thr Glu Leu Ala Ala
305 310 315 320
Gly
<210> 29
<211> 395
<212> БЕЛОК
<213> S. kasugaensis
<400> 29
Met Ala Glu Thr Val Arg Ala Asp Ser Pro Leu Lys Ser Ser Tyr Arg
1 5 10 15
Asn Val Pro Ala Ala Glu Val Gln Gly Ser Gly Leu Ser Val Gly Gln
20 25 30
Arg Thr Thr Arg Ile Ala Ile Ser Ser Leu Leu Ala Ala Asp Ser Pro
35 40 45
Arg Ser Ala Gly Glu Asn Ala Glu His Ile Arg Leu Leu Ala Asp Ser
50 55 60
Gly Ala Arg Leu Pro Pro Ile Val Val Gln Arg Ser Thr Met Arg Val
65 70 75 80
Ile Asp Gly Met His Arg Leu Arg Ala Ala Ala Leu Arg Gly Glu Thr
85 90 95
Glu Ile Glu Val Arg Phe Phe Asp Gly Ala Glu Glu Asp Ser Phe Leu
100 105 110
Leu Ala Val Arg Ser Asn Ile Ala His Gly Leu Pro Leu Ser Gln Glu
115 120 125
Glu Arg Ala Ala Ala Ala Gln Arg Ile Ile Arg Ser His Ala Gln Trp
130 135 140
Ser Asn Gln Ala Ile Gly Glu Val Thr Gly Leu Asp Ala Lys Thr Ile
145 150 155 160
Ala Ala Leu Arg Arg Asp Ala Lys Asp Val Pro Gln Leu Asp Ala Arg
165 170 175
Ile Gly Arg Asp Gly Arg Val Arg Pro Val Asp Gly Ala Gln Gly Arg
180 185 190
Arg Leu Ala Gly Glu Leu Met Ala Glu Gln Pro Asp Ala Pro Leu Arg
195 200 205
Lys Ile Ala His Ala Ala Gly Val Ser Leu Gly Thr Ala Ser Asp Val
210 215 220
Arg Arg Arg Ile Arg Asn Gly Gln Asp Pro Val Pro Ala Gly Arg Gln
225 230 235 240
Lys Ala Asp Pro Gln Pro Pro Ala Arg Tyr Ala Ala Ser Glu Asp Arg
245 250 255
Ser Gly Thr Thr Ala Pro Arg Thr Gly Glu Gln Asn Arg Arg Val Leu
260 265 270
Leu Gln Lys Leu Arg Lys Asp Pro Ser Leu Arg Cys Asn Glu Ala Gly
275 280 285
Arg Ala Leu Leu Arg Trp Leu Glu Val Gln Ala Val Glu Gly Glu Asp
290 295 300
Trp Glu Arg Leu Leu Asp Ser Val Pro Met His Cys Ala Ala Thr Ile
305 310 315 320
Val Glu Leu Ala Arg Arg Lys Asp Pro Ser Leu Arg Cys Asn Glu Ala
325 330 335
Gly Arg Ala Leu Leu Arg Trp Leu Glu Val Gln Ala Val Glu Gly Glu
340 345 350
Asp Trp Glu Arg Leu Leu Asp Ser Val Pro Met His Cys Ala Ala Thr
355 360 365
Ile Val Glu Leu Ala Arg Gly Cys Ser Gly Val Trp Gln Asp Phe Ala
370 375 380
Ala Gln Leu Glu Arg Arg Gly Arg Ala Ser Ala
385 390 395
<210> 30
<211> 367
<212> БЕЛОК
<213> S. niveus
<400> 30
Met Thr Asn Ser Gly Asp Glu Glu Ile Thr Pro Ala Ser Leu Lys Ala
1 5 10 15
Thr Arg Lys Gly Glu Arg Val Ser Ile Gly Ser Leu Leu Pro Pro Ser
20 25 30
Glu Leu Val Arg Ser Gly Glu Ser Thr Glu His Ile Arg Val Leu Ala
35 40 45
Glu Thr Asp Glu Asp Leu Pro Pro Ile Val Val His Arg Gly Thr Arg
50 55 60
Arg Val Val Asp Gly Met His Arg Leu Trp Ala Ala Arg Phe Arg Gly
65 70 75 80
Asp Glu Ser Ile Glu Val Val Phe Val Asp Gly Ser Pro Ala Asp Val
85 90 95
Phe Val Leu Ala Val Glu Leu Asn Arg Ala His Gly Leu Pro Leu Thr
100 105 110
Leu Asp Glu Arg Lys Ser Ala Ala Ala Gln Ile Met Asp Ser His Pro
115 120 125
His Trp Ser Asp Arg Lys Ile Ala Arg Thr Thr Gly Leu Ala Ala Ser
130 135 140
Thr Val Ala Ser Leu Arg Ser Ser Ser Thr Ala Gly Thr Val Gly Arg
145 150 155 160
Arg Thr Gly Gln Asp Gly Arg Ser Arg Pro Asn Asp Gly Thr Asp Gly
165 170 175
Arg Gln Arg Ala Ala Ala Leu Leu Ala Arg Asn Pro Asn Ala Ser Leu
180 185 190
Arg Glu Val Thr Arg Ala Ala Gly Ile Ser Val Gly Thr Ala Ser Asp
195 200 205
Val Arg Ala Arg Leu Arg Arg Gly Glu Pro Ala Leu Thr Ala Arg Gln
210 215 220
Gln Ala Val Met Lys Leu Arg Pro Ala Ala Arg Asn Pro Asn Ala Ser
225 230 235 240
Leu Arg Glu Val Thr Arg Ala Ala Gly Ile Ser Val Gly Thr Ala Ser
245 250 255
Asp Val Arg Ala Arg Leu Arg Arg Gly Glu Pro Ala Leu Thr Ala Arg
260 265 270
Gln Gln Ala Val Met Lys Leu Arg Pro Ala Ala Ala Gln Arg Ser Gly
275 280 285
Pro Asp Tyr Gly Arg Val Leu Glu Asn Leu Arg Lys Asp Pro Ser Leu
290 295 300
Arg Phe Thr Asp Leu Gly Arg Arg Leu Leu Arg Leu Leu Asp Gly Ser
305 310 315 320
Val Pro Gly Ser Val Glu Gln Ile Ala Gln Ile Ala Asp Gly Val Pro
325 330 335
Glu His Cys Arg Thr Val Val Val Asp Met Ala Arg Glu Cys Ala Ala
340 345 350
Ala Trp Gln His Leu Ala Asp Gln Leu Ala Asp Arg Asp Thr Ala
355 360 365
<210> 31
<211> 359
<212> БЕЛОК
<213> S. globisporus
<400> 31
Met Lys Ser Asp Ser Ala Gln Arg Ala Val Glu Arg Ser Arg Arg Val
1 5 10 15
Val Arg Ile Asp Glu Leu Ile Pro Ala Asp Ser Pro Arg Leu Asn Gly
20 25 30
Ile Asp Arg Ser His Val Gln Arg Leu Ala Thr Val Tyr Ala Ser Leu
35 40 45
Pro Pro Val Leu Val His Arg Pro Thr Met Arg Val Val Asp Gly Met
50 55 60
His Arg Ile Gly Ala Ala Arg Leu Lys Gly Leu Asp Thr Val Glu Val
65 70 75 80
Thr Phe Phe Glu Gly Ala Glu Glu Gln Val Phe Leu Arg Ser Val Ala
85 90 95
Ala Asn Ile Thr Asn Gly Leu Pro Leu Ser Val Ala Asp Arg Lys Thr
100 105 110
Ala Ala Ala Arg Ile Leu Ala Ser His Pro Thr Leu Ser Asp Arg Ala
115 120 125
Val Ala Ala His Val Gly Leu Asp Ala Lys Thr Val Ala Gly Val Arg
130 135 140
Thr Cys Ser Ala Ala Gly Ser Pro Leu Leu Asn Met Arg Thr Gly Ala
145 150 155 160
Asp Gly Arg Val His Pro Leu Asp Arg Thr Ala Glu Arg Leu His Ala
165 170 175
Ala Ala Leu Leu Thr Gln Asp Pro Gly Leu Pro Leu Arg Ser Val Val
180 185 190
Glu Gln Thr Gly Leu Ser Leu Gly Thr Ala His Asp Val Arg Arg Arg
195 200 205
Leu Leu Arg Gly Glu Asp Pro Val Pro Gln Asn Arg Gln Ser Ala Met
210 215 220
Leu Glu Pro Gly Leu Ala Pro Gln Lys Lys Ala Thr Ala Lys Pro Pro
225 230 235 240
Val Gly Pro Ala Ala Arg Pro Val Pro Lys Val Pro Pro Ala Val Ala
245 250 255
Gly Arg Pro Pro Val Ser Pro Arg Ser Arg Ala Pro Leu Glu Ala Leu
260 265 270
Arg Lys Leu Ser Asn Asp Pro Ser Leu Arg His Ser Asp Gln Gly Arg
275 280 285
Glu Leu Met Arg Trp Leu His Asn Arg Phe Val Val Asp Glu Ala Trp
290 295 300
Arg Arg Arg Ala Asp Ala Val Pro Ala His Cys Val Asp Ser Met Ala
305 310 315 320
Glu Leu Ala Gln His Cys Ser Asp Ala Trp His Arg Phe Ala Glu Glu
325 330 335
Met Val Arg Arg Arg His Ser Ala Ala Ala Asp Gly Ser Gly Leu Arg
340 345 350
Thr Thr Gln Pro Thr Arg Arg
355
<210> 32
<211> 329
<212> БЕЛОК
<213> Actinoplanes teichomyceticus
<400> 32
Met Thr Pro Asp Glu Glu Ala Leu Asn Arg Gln Pro Ile Met Glu Met
1 5 10 15
Glu Ile Ser Ser Leu Ser Leu Gly Gly Ser Pro Arg Leu Ala Gly Gly
20 25 30
Asp Pro Val His Leu Glu Ala Met Val Ala Ala Gln Gly Glu Leu Pro
35 40 45
Pro Ile Val Val His Arg Pro Thr Met Arg Val Ile Asp Gly Ser His
50 55 60
Arg Ile Gln Ala Ala Leu Arg Arg Gly Glu Thr Thr Ile Ala Gly Arg
65 70 75 80
Phe Phe Asp Gly Ser Asp Asp Glu Ala Phe Val Met Ser Val Trp Leu
85 90 95
Asn Val Ser His Gly Leu Pro Leu Ala Leu Ala Asp Arg Lys Arg Ala
100 105 110
Ala Glu Arg Ile Ala Val Ser His Pro Gln Trp Ser Asp Arg Arg Val
115 120 125
Ala Ala Val Thr Gly Ile Ser Pro Ser Thr Val Ala Asp Ile Arg Arg
130 135 140
Arg Val Ala Gly Thr Ser Ala Pro Glu Ala Ser Arg Ile Gly Gln Asp
145 150 155 160
Gly Arg Val Arg Pro Leu Asp Cys Ser Ala Gly Arg Leu Leu Ala Gly
165 170 175
Arg Leu Met Ala Glu Asn Pro Ala Leu Ser Leu Arg Gln Val Ala Lys
180 185 190
Ala Ala Ala Ile Ser Pro Glu Thr Ala Arg Asp Val Arg Asn Arg Leu
195 200 205
Leu Ser Gly Ala Glu Leu Val Pro Asn Arg Arg Pro Arg Asp Ala Ala
210 215 220
Pro Val Gly Val Lys Gly Gly Arg Asp Arg Arg Pro Leu Asn Leu Ile
225 230 235 240
Arg Ser Gly Asp Arg Pro Glu Pro Val Pro Asp His Ala Val Val Ile
245 250 255
Asn Arg Leu Met Ser Asp Pro Ala Leu Arg Tyr Thr Asp Thr Gly Arg
260 265 270
Asn Leu Leu Arg Leu Leu Ser Leu His Thr Arg Trp Ala Lys Glu Trp
275 280 285
Glu Ala Ile Val Asp Asn Leu Pro Pro His Cys Ala Asp Ala Val Ala
290 295 300
Asp Leu Ala Arg Gln Phe Ala Asp Leu Trp Ala Asp Phe Ala Ser Arg
305 310 315 320
Val Gly Pro Glu Glu Arg Met Ala Ser
325
<210> 33
<211> 221
<212> БЕЛОК
<213> S. coelicolor
<400> 33
Met Thr Ile Arg Leu Leu Ile Val Asp Asp Gln Glu Leu Ile Arg Thr
1 5 10 15
Gly Phe Arg Leu Phe Leu Gln Thr Gln Asn Asp Leu Glu Val Val Gly
20 25 30
Glu Ala Asp Asp Gly His Gly Ala Leu Ala Gln Ala Ala Ala Leu Arg
35 40 45
Pro Asp Val Val Leu Met Asp Ile Arg Met Pro Arg Met Asp Gly Val
50 55 60
Glu Ala Thr Ser Arg Leu Thr Ala Ser Asp Ser Pro Pro Arg Val Leu
65 70 75 80
Ile Leu Thr Thr Tyr Asp Leu Asp Glu Tyr Val Phe Gly Ala Leu Arg
85 90 95
Ala Gly Ala Ser Gly Phe Leu Leu Lys Asp Ala Ser Arg Asp Arg Leu
100 105 110
Leu Glu Ala Ile Arg Val Val His Ala Gly Glu Ala Leu Leu Ser Pro
115 120 125
Ser Ile Thr Arg Arg Leu Ile Glu Asp Tyr Ala Thr Arg Ala Ala Pro
130 135 140
Val Arg Pro Arg Glu Ala Val Leu Ala Gly Leu Thr Pro Arg Glu Arg
145 150 155 160
Glu Ile Leu Leu Leu Val Ala Arg Gly Leu Ser Asn Pro Glu Ile Ala
165 170 175
Ala Arg Leu Val Val Thr Glu Ala Thr Val Lys Ser His Val Gly Ser
180 185 190
Met Phe Ala Lys Leu His Leu Arg Asp Arg Ala Gln Ala Val Val Phe
195 200 205
Ala Tyr Glu Asn Ala Ile Val Leu Pro Gly Gly Thr Gly
210 215 220
<210> 34
<211> 222
<212> БЕЛОК
<213> S. coelicolor
<400> 34
Met Ile Arg Val Leu Leu Ala Asp Asp Glu Thr Ile Ile Arg Ala Gly
1 5 10 15
Val Arg Ser Ile Leu Thr Thr Glu Pro Gly Ile Glu Val Val Ala Glu
20 25 30
Ala Ser Asp Gly Arg Glu Ala Val Glu Leu Ala Arg Lys His Arg Pro
35 40 45
Asp Val Ala Leu Leu Asp Ile Arg Met Pro Glu Met Asp Gly Leu Thr
50 55 60
Ala Ala Gly Glu Met Arg Thr Thr Asn Pro Asp Thr Ala Val Val Val
65 70 75 80
Leu Thr Thr Phe Gly Glu Asp Arg Tyr Ile Glu Arg Ala Leu Asp Gln
85 90 95
Gly Val Ala Gly Phe Leu Leu Lys Ala Ser Asp Pro Arg Asp Leu Ile
100 105 110
Ser Gly Val Arg Ala Val Ala Ser Gly Gly Ser Cys Leu Ser Pro Leu
115 120 125
Val Ala Arg Arg Leu Met Thr Glu Leu Arg Arg Ala Pro Ser Pro Arg
130 135 140
Ser Glu Val Ser Gly Glu Arg Thr Thr Leu Leu Thr Lys Arg Glu Gln
145 150 155 160
Glu Val Leu Gly Met Leu Gly Ala Gly Leu Ser Asn Ala Glu Ile Ala
165 170 175
Gln Arg Leu His Leu Val Glu Gly Thr Ile Lys Thr Tyr Val Ser Ala
180 185 190
Ile Phe Thr Gln Leu Glu Val Arg Asn Arg Val Gln Ala Ala Ile Ile
195 200 205
Ala Tyr Glu Ala Gly Leu Val Lys Asp Ala Asp Leu Asn Arg
210 215 220
<210> 35
<211> 218
<212> БЕЛОК
<213> S. coelicolor
<400> 35
Met Arg Glu Asp Gly Lys Ile Arg Val Phe Leu Leu Asp Asp His Glu
1 5 10 15
Val Val Arg Arg Gly Val His Asp Leu Leu Ser Gly Glu Ala Asp Ile
20 25 30
Glu Val Val Gly Glu Ala Gly Thr Ala Ala Glu Ala Gln Ala Arg Val
35 40 45
Thr Ala Thr Arg Pro Asp Val Ala Val Leu Asp Val Arg Leu Pro Asp
50 55 60
Gly Ser Gly Val Glu Val Cys Arg Asp Ile Arg Ser Arg Asp Glu Ser
65 70 75 80
Val Arg Cys Leu Met Leu Thr Ser Phe Ala Asp Asp Glu Ala Leu Phe
85 90 95
Asp Ala Ile Met Ala Gly Ala Ser Gly Tyr Val Leu Lys Asp Ile Arg
100 105 110
Gly Ala Glu Leu Leu Gly Ala Val Arg Glu Val Ala Ala Gly Lys Ser
115 120 125
Leu Leu Asp Pro Ala Ala Thr Ala Arg Val Leu Glu Arg Leu Arg Gly
130 135 140
Gly Gly Ala Arg Pro Asp Asp Arg Leu Ala Arg Leu Thr Glu Gln Glu
145 150 155 160
Arg Arg Ile Leu Glu Leu Ile Gly Glu Gly Leu Thr Asn Arg Ala Ile
165 170 175
Gly Glu Arg Leu His Leu Ala Glu Lys Thr Ile Lys Asn Tyr Val Ser
180 185 190
Ser Leu Leu Gly Lys Leu Gly Met Gln Arg Arg Ser Gln Ala Ala Ala
195 200 205
Phe Val Ala Arg Leu Glu Ala Glu Asn Arg
210 215
<210> 36
<211> 220
<212> БЕЛОК
<213> Streptomyces fungicidicus
<400> 36
Val Ser Val Leu Leu Glu Gln Pro Ala Ser Leu Val Ala Tyr Arg Pro
1 5 10 15
Asn Lys Pro Thr Ala Met Val Val Val Ala Asp Pro Arg Val Arg Ser
20 25 30
Thr Val Thr Arg His Leu Trp Ala Leu Gly Val Arg Asp Val Ile Glu
35 40 45
Ala Ser Ser Val Ala Glu Ala Arg Pro Arg Ile Gly Asn Pro Arg Asp
50 55 60
Ile Cys Val Ala Glu Val His Leu Pro Asp Gly Ser Gly Leu Thr Leu
65 70 75 80
Leu Ser Glu Thr Arg Ala Ala Gly Trp Pro Asn Gly Leu Ala Leu Ser
85 90 95
Ala Ala Asp Asp Ile Gly Ala Val Arg Asn Ala Leu Ala Gly Gly Val
100 105 110
Lys Gly Tyr Val Val Thr Gly Thr Arg Thr Asn Leu Gly Leu Pro Thr
115 120 125
Arg Pro Gly Ala Ala Pro Ile Gly Ala Ala Ala Ala Arg Leu His Arg
130 135 140
Arg Pro Pro Gly Ala Pro Ser His Pro Gly Gly Tyr Arg Glu Leu Ser
145 150 155 160
Gly Arg Glu Val Glu Val Leu Arg Leu Val Ala Glu Gly Gln Ser Asn
165 170 175
Lys Ala Ile Gly Val Ser Met Gly Leu Ser Ala Leu Thr Val Lys Ser
180 185 190
His Leu Ala Arg Ile Ala Arg Lys Leu Gly Thr Gly Asp Arg Ala Gly
195 200 205
Met Val Ala Val Ala Leu Arg Thr Gly Ile Ile His
210 215 220
<210> 37
<211> 663
<212> ДНК
<213> Streptomyces fungicidicus
<400> 37
gtgtccgttc tcctcgagca gcccgcaagc ctggtcgcct accgcccgaa caagccgacc 60
gccatggtgg tcgtggccga cccccgcgtc cgttcgaccg tcacccgcca cctgtgggcg 120
ctcggcgtac gcgacgtcat cgaggcctcg tccgtcgcgg aggctcgtcc ccgcatcggc 180
aacccccgcg acatctgcgt cgccgaagtc catctgccgg atggttccgg cctcaccctc 240
ctctccgaga cccgcgccgc gggctggccc aacggcctcg ccctctccgc ggcggacgac 300
atcggcgccg tgcgcaacgc cctcgcgggc ggagtcaagg gctacgtcgt caccggcacc 360
cgcaccaacc tcgggctccc cacccggccg ggtgccgctc ccatcggcgc cgccgccgcg 420
cgcctgcacc gccgcccccc gggtgccccg agccacccgg gcggctaccg cgagctgtcc 480
ggccgcgagg tggaggtgct gcggctggtg gcggaaggcc agtcgaacaa ggcgatcggc 540
gtctcgatgg gcctgtccgc actgaccgtc aagagccacc tggcccggat cgcccgcaag 600
ctcggcacgg gcgaccgcgc cggcatggtg gccgtggccc tgcgcaccgg catcatccac 660
tga 663
<210> 38
<211> 936
<212> ДНК
<213> Streptomyces fungicidicus
<400> 38
gtggaaataa gttcgctctc caccgacggc tccccgcgga tcgacgggga gagtcccgag 60
cacgtggaaa tgctggccgc cgccgacacc gcgcttccac cgatcatggt gcaccgccgc 120
accgggcggg tcatcgacgg catgcaccgg ctgcgcgccg cgatgctgac gggccgtacg 180
acgatcgcgg tgaggttctt cgacggcacc gaggaggacg ccttcgtcct cgccgtgaag 240
tcgaacatcg cgcacggact gccgctgtcc gccgccgacc gccggcgggc cgccgggcgc 300
atcatggcca cccatccccg gtggtcggac cggatgatcg cctcggtggt cggcacctcc 360
gccaggacgg tcgccgagat ccgccgcgac gccggcgccg ccggggcggg ggagcccacc 420
cgcatcggcc gggacggcag ggtacggccc gtcgacgtga gcgagggccg cagactggcc 480
cacgacatga tcgtccgcga cccgggcctg tcgctgcgcc aggtcgcccg cgccgccggg 540
atctcgccgg agaccgtcag ggacgtcaga caccggatgc tccgcggtga ggacccggtg 600
cccgcgccgc ggccgcggac cctggtggag cgcggcgcgg accgccgggc ggagccggcc 660
gggaaggccg ccgcgccgtg cgggacggag ccgccgcccg ccgtcgtgat gaagcggctg 720
agggccgatc cggcgctgcg tctcaacgag aacggacgcg acctgctgcg gcttctggat 780
atccacacgg tccggctgga ggactggaac cgcattatcg aaagcgtgcc gccgcaccgt 840
ctggagacgg tggcgcagct ggcacgctcc tgcgccgaca aatggtccga gatcgcgtca 900
cgcatcgaaa gcaacgcatc acatctggcc gggtga 936
<210> 39
<211> 39331
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая фосмида pXYF148
<400> 39
atcgaatata acttcgtata atgtatgcta tacgaagtta ttagcgatga gctcggactt 60
ccattgttca ttccacggac aaaaacagag aaaggaaacg acagaggcca aaaagctcgc 120
tttcagcacc tgtcgtttcc tttcttttca gagggtattt taaataaaaa cattaagtta 180
tgacgaagaa gaacggaaac gccttaaacc ggaaaatttt cataaatagc gaaaacccgc 240
gaggtcgccg ccccgtaacc tgtcggatca ccggaaagga cccgtaaagt gataatgatt 300
atcatctaca tatcacaacg tgcgtggagg ccatcaaacc acgtcaaata atcaattatg 360
acgcaggtat cgtattaatt gatctgcatc aacttaacgt aaaaacaact tcagacaata 420
caaatcagcg acactgaata cggggcaacc tcatgtccga gctcgcgagc tcgtcgacag 480
cgacacactt gcatcggatg cagcccggtt aacgtgccgg cacggcctgg gtaaccaggt 540
attttgtcca cataaccgtg cgcaaaatgt tgtggataag caggacacag cagcaatcca 600
cagcaggcat acaaccgcac accgaggtta ctccgttcta caggttacga cgacatgtca 660
atacttgccc ttgacaggca ttgatggaat cgtagtctca cgctgatagt ctgatcgaca 720
atacaagtgg gaccgtggtc ccagaccgat aatcagaccg acaacacgag tgggatcgtg 780
gtcccagact aataatcaga ccgacgatac gagtgggacc gtggtcccag actaataatc 840
agaccgacga tacgagtggg accgtggttc cagactaata atcagaccga cgatacgagt 900
gggaccgtgg tcccagacta ataatcagac cgacgatacg agtgggacca tggtcccaga 960
ctaataatca gaccgacgat acgagtggga ccgtggtccc agtctgatta tcagaccgac 1020
gatacgagtg ggaccgtggt cccagactaa taatcagacc gacgatacga gtgggaccgt 1080
ggtcccagac taataatcag accgacgata cgagtgggac cgtggtccca gtctgattat 1140
cagaccgacg atacaagtgg aacagtgggc ccagagagaa tattcaggcc agttatgctt 1200
tctggcctgt aacaaaggac attaagtaaa gacagataaa cgtagactaa aacgtggtcg 1260
catcagggtg ctggcttttc aagttcctta agaatggcct caattttctc tatacactca 1320
gttggaacac gagacctgtc caggttaagc accattttat cgcccttata caatactgtc 1380
gctccaggag caaactgatg tcgtgagctt aaactagttc ttgatgcaga tgacgtttta 1440
agcacagaag ttaaaagagt gataacttct tcagcttcaa atatcacccc agcttttttc 1500
tgctcatgaa ggttagatgc ctgctgctta agtaattcct ctttatctgt aaaggctttt 1560
tgaagtgcat cacctgaccg ggcagatagt tcaccggggt gagaaaaaag agcaacaact 1620
gatttaggca atttggcggt gttgatacag cgggtaataa tcttacgtga aatattttcc 1680
gcatcagcca gcgcagaaat atttccagca aattcattct gcaatcggct tgcataacgc 1740
tgaccacgtt cataagcact tgttgggcga taatcgttac ccaatctgga taatgcagcc 1800
atctgctcat catccagctc gccaaccaga acacgataat cactttcggt aagtgcagca 1860
gctttacgac ggcgactccc atcggcaatt tctatgacac cagatactct tcgaccgaac 1920
gccggtgtct gttgaccagt cagtagaaaa gaagggatga gatcatccag tgcgtcctca 1980
gtaagcagct cctggtcacg ttcattacct gaccataccc gagaggtctt ctcaacacta 2040
tcaccccgga gcacttcaag agtaaacttc acatcccgac cacatacagg caaagtaatg 2100
gcattaccgc gagccattac tcctacgcgc gcaattaacg aatccaccat cggggcagct 2160
ggtgtcgata acgaagtatc ttcaaccggt tgagtattga gcgtatgttt tggaataaca 2220
ggcgcacgct tcattatcta atctcccagc gtggtttaat cagacgatcg aaaatttcat 2280
tgcagacagg ttcccaaata gaaagagcat ttctccaggc accagttgaa gagcgttgat 2340
caatggcctg ttcaaaaaca gttctcatcc ggatctgacc tttaccaact tcatccgttt 2400
cacgtacaac attttttaga accatgcttc cccaggcatc ccgaatttgc tcctccatcc 2460
acggggactg agagccatta ctattgctgt atttggtaag caaaatacgt acatcaggct 2520
cgaacccttt aagatcaacg ttcttgagca gatcacgaag catatcgaaa aactgcagtg 2580
cggaggtgta gtcaaacaac tcagcaggcg tgggaacaat cagcacatca gcagcacata 2640
cgacattaat cgtgccgata cccaggttag gcgcgctgtc aataactatg acatcatagt 2700
catgagcaac agtttcaatg gccagtcgga gcatcaggtg tggatcggtg ggcagtttac 2760
cttcatcaaa tttgcccatt aactcagttt caatacggtg cagagccaga caggaaggaa 2820
taatgtcaag ccccggccag caagtgggct ttattgcata agtgacatcg tccttttccc 2880
caagatagaa aggcaggaga gtgtcttctg catgaatatg aagatctggt acccatccgt 2940
gatacattga ggctgttccc tgggggtcgt taccttccac gagcaaaaca cgtagcccct 3000
tcagagccag atcctgagca agatgaacag aaactgaggt tttgtaaacg ccacctttat 3060
gggcagcaac cccgatcacc ggtggaaata cgtcttcagc acgtcgcaat cgcgtaccaa 3120
acacatcacg catatgatta atttgttcaa ttgtataacc aacacgttgc tcaacccgtc 3180
ctcgaatttc catatccggg tgcggtagtc gccctgcttt ctcggcatct ctgatagcct 3240
gagaagaaac cccaactaaa tccgctgctt cacctattct ccagcgccgg gttattttcc 3300
tcgcttccgg gctgtcatca ttaaactgtg caatggcgat agccttcgtc atttcatgac 3360
cagcgtttat gcactggtta agtgtttcca tgagtttcat tctgaacatc ctttaatcat 3420
tgctttgcgt ttttttatta aatcttgcaa tttactgcaa agcaacaaca aaatcgcaaa 3480
gtcatcaaaa aaccgcaaag ttgtttaaaa taagagcaac actacaaaag gagataagaa 3540
gagcacatac ctcagtcact tattatcact agcgctcgcc gcagccgtgt aaccgagcat 3600
agcgagcgaa ctggcgagga agcaaagaag aactgttctg tcagatagct cttacgctca 3660
gcgcaagaag aaatatccac cgtgggaaaa actccaggta gaggtacaca cgcggatagc 3720
caattcagag taataaactg tgataatcaa ccctcatcaa tgatgacgaa ctaacccccg 3780
atatcaggtc acatgacgaa gggaaagaga aggaaatcaa ctgtgacaaa ctgccctcaa 3840
atttggcttc cttaaaaatt acagttcaaa aagtatgaga aaatccatgc aggctgaagg 3900
aaacagcaaa actgtgacaa attaccctca gtaggtcaga acaaatgtga cgaaccaccc 3960
tcaaatctgt gacagataac cctcagacta tcctgtcgtc atggaagtga tatcgcggaa 4020
ggaaaatacg atatgagtcg tctggcggcc tttctttttc tcaatgtatg agaggcgcat 4080
tggagttctg ctgttgatct cattaacaca gacctgcagg aagcggcggc ggaagtcagg 4140
catacgctgg taactttgag gcagctggta acgctctatg atccagtcga ttttcagaga 4200
gacgatgcct gagccatccg gcttacgata ctgacacagg gattcgtata aacgcatggc 4260
atacggattg gtgatttctt ttgtttcact aagccgaaac tgcgtaaacc ggttctgtaa 4320
cccgataaag aagggaatga gatatgggtt gatatgtaca ctgtaaagcc ctctggatgg 4380
actgtgcgca cgtttgataa accaaggaaa agattcatag cctttttcat cgccggcatc 4440
ctcttcaggg cgataaaaaa ccacttcctt ccccgcgaaa ctcttcaatg cctgccgtat 4500
atccttactg gcttccgcag aggtcaatcc gaatatttca gcatatttag caacatggat 4560
ctcgcagata ccgtcatgtt cctgtagggt gccatcagat tttctgatct ggtcaacgaa 4620
cagatacagc atacgttttt gatcccggga gagactatat gccgcctcag tgaggtcgtt 4680
tgactggacg attcgcgggc tatttttacg tttcttgtga ttgataaccg ctgtttccgc 4740
catgacagat ccatgtgaag tgtgacaagt ttttagattg tcacactaaa taaaaaagag 4800
tcaataagca gggataactt tgtgaaaaaa cagcttcttc tgagggcaat ttgtcacagg 4860
gttaagggca atttgtcaca gacaggactg tcatttgagg gtgatttgtc acactgaaag 4920
ggcaatttgt cacaacacct tctctagaac cagcatggat aaaggcctac aaggcgctct 4980
aaaaaagaag atctaaaaac tataaaaaaa ataattataa aaatatcccc gtggataagt 5040
ggataacccc aagggaagtt ttttcaggca tcgtgtgtaa gcagaatata taagtgctgt 5100
tccctggtgc ttcctcgctc actcgaccgg gagggttcga gaaggggggg cacccccctt 5160
cggcgtgcgc ggtcacgcgc acagggcgca gccctggtta aaaacaaggt ttataaatat 5220
tggtttaaaa gcaggttaaa agacaggtta gcggtggccg aaaaacgggc ggaaaccctt 5280
gcaaatgctg gattttctgc ctgtggacag cccctcaaat gtcaataggt gcgcccctca 5340
tctgtcagca ctctgcccct caagtgtcaa ggatcgcgcc cctcatctgt cagtagtcgc 5400
gcccctcaag tgtcaatacc gcagggcact tatccccagg cttgtccaca tcatctgtgg 5460
gaaactcgcg taaaatcagg cgttttcgcc gatttgcgag gctggccagc tccacgtcgc 5520
cggccgaaat cgagcctgcc cctcatctgt caacgccgcg ccgggtgagt cggcccctca 5580
agtgtcaacg tccgcccctc atctgtcagt gagggccaag ttttccgcga ggtatccaca 5640
acgccggcgg ccggccgcgg tgtctcgcac acggcttcga cggcgtttct ggcgcgtttg 5700
cagggccata gacggccgcc agcccagcgg cgagggcaac cagccgaggg cttcgccctg 5760
tcgctcgact gcggcgagca ctactggctg taaaaggaca gaccacatca tggttctgtg 5820
ttcattaggt tgttctgtcc attgctgaca taatccgctc cacttcaacg taacaccgca 5880
cgaagatttc tattgttcct gaaggcatat tcaaatcgtt ttcgttaccg cttgcaggca 5940
tcatgacaga acactacttc ctataaacgc tacacaggct cctgagatta ataatgcgga 6000
tctctacgat aatgggagat tttcccgact gtttcgttcg cttctcagtg gataacagcc 6060
agcttctctg tttaacagac aaaaacagca tatccactca gttccacatt tccatataaa 6120
ggccaaggca tttattctca ggataattgt ttcagcatcg caaccgcatc agactccggc 6180
atcgcaaact gcacccggtg ccgggcagcc acatccagcg caaaaacctt cgtgtagact 6240
tccgttgaac tgatggactt atgtcccatc aggctttgca gaactttcag cggtataccg 6300
gcatacagca tgtgcatcgc ataggaatgg cggaacgtat gtggtgtgac cggaacagag 6360
aacgtcacac cgtcagcagc agcggcggca accgcctccc caatccaggt cctgaccgtt 6420
ctgtccgtca cttcccagat ccgcgctttc tctgtccttc ctgtgcgacg gttacgccgc 6480
tccatgagct tatcgcgaat aaatacctgt gacggaagat cacttcgcag aataaataaa 6540
tcctggtgtc cctgttgata ccgggaagcc ctgggccaac ttttggcgaa aatgagacgt 6600
tgatcggcac gtaagaggtt ccaactttca ccataatgaa ataagatcac taccgggcgt 6660
attttttgag ttatcgagat tttcaggagc taaggaagct aaaatggaga aaaaaatcac 6720
tggatatacc accgttgata tatcccaatg gcatcgtaaa gaacattttg aggcatttca 6780
gtcagttgct caatgtacct ataaccagac cgttcagctg gatattacgg cctttttaaa 6840
gaccgtaaag aaaaataagc acaagtttta tccggccttt attcacattc ttgcccgcct 6900
gatgaatgct catccggaat ttcgtatggc aatgaaagac ggtgagctgg tgatatggga 6960
tagtgttcac ccttgttaca ccgttttcca tgagcaaact gaaacgtttt catcgctctg 7020
gagtgaatac cacgacgatt tccggcagtt tctacacata tattcgcaag atgtggcgtg 7080
ttacggtgaa aacctggcct atttccctaa agggtttatt gagaatatgt ttttcgtctc 7140
agccaatccc tgggtgagtt tcaccagttt tgatttaaac gtggccaata tggacaactt 7200
cttcgccccc gttttcacca tgggcaaata ttatacgcaa ggcgacaagg tgctgatgcc 7260
gctggcgatt caggttcatc atgccgtttg tgatggcttc catgtcggca gaatgcttaa 7320
tgaattacaa cagtactgcg atgagtggca gggcggggcg taattttttt aaggcagtta 7380
ttggtgccct taaacgcctg gttgctacgc ctgaataagt gataataagc ggatgaatgg 7440
cagaaattcg atgataagct gtcaaacatg agaattggtc gacggcccgg gcggccgcaa 7500
ggggttcgcg ttggccgatt cattaatgca gctggcacga caggtttccc gactggaaag 7560
cgggcagtga gcgcaacgca attaatgtga gttagctcac tcattaggca ccccaggctt 7620
tacactttat gcttccggct cgtatgttgt gtggaattgt gagcggataa caatttcaca 7680
caggaaacag ctatgaccat gattacgcca agctatttag gtgagactat agaatactca 7740
agcttgcatg cctgcaggtc gactctagag gatcccacca ccgccggacg ggggtcgccg 7800
gcccggaagt ccaccaggat cgagccgccg atctgccgcc cgcgccacca cgactccatc 7860
agcccgacct cctcctccgg gctgcgcacc agccgcagca gcagcgagca ggaccgctcc 7920
accaggacgc tctccacccc ggagatgaac tccatgtaga acggctccag gccgagcagc 7980
cgggcgggcc ggcagaccgc gagacccacc acgtccaccc gcgacccggc cagcgtgcgc 8040
gcggttcggc tcggcgccca ccccagctcc cgcgccgccc ggaagatgcg gtcccgggtc 8100
gcctccgaca gcccgggctt ccggttgaag gcgagggaca cggcgccctt ggacacgccg 8160
gcgcgcgcgg cgacgtccct gatggtgacg cgaggggtcg gcgttgccgt catcgagtgg 8220
gctccacgca gtacagggcg gaacgggcgg tgtccgggtc cggagtcttc cacccccgca 8280
ccccgatggt cacctgttcc ccggggagca gggtcaccag cccccggtcg gcccgcgccc 8340
cggggtccag ccggtcggcc tggagcagca ggtcccgtac gagggtgcgg gccgtgaccg 8400
tgatcccgtc cggcgcgagg gcgacctcga actccggcgg ggggtagggg atctcccggt 8460
ccggcgccgg gaagtgccac gcccgcaccc cgtccgcgtc ggcgaccagg aactccccgg 8520
ggccgtccgg cagcagttcg accgggacct cgaccacggc caccgtccgc cccccggcgt 8580
ccagcgccgg ggccgcctcc gcgatcgggg cgccgtcgac ggacatccgg cgcagccgca 8640
gcgttccccg ccagtcctcc gcggactggt tgaccgccgc caccaccaga ccgtcaccgt 8700
ccgcgcgcac ggtcagcagc cggtccgcgt acagccggcg cagctcgtgg tagagcggct 8760
tctcccgccc gtccccgtcg atcgcggccc acgacgtcac cggccagcag tcgttgagct 8820
gccagaccac cgtgcccgcg cacaccggcc agtgcgagcg ccagtgctcg acaccggccg 8880
ccaccgcacg cgcctggttg acctgcgtca gatagtgcca gcggtcgaag tcgccctccg 8940
gcacggcgaa gtggcgggcg aggccgcgct ccagcttgcc gttgccgtcc tccgccttct 9000
ggtggtgcag catgccgggg gagtccggcg cggggtcctc cccgggcagc gcccgccgca 9060
gcgtggcgtg cgcgggaggc gcctgccagc cgaactcggc cacgaagcgc gggacgtcgc 9120
gccggtagtc ggcgtagtcg gcgcggttcc acacctccca ggagtggtgg gtgccgtgcg 9180
ccggatcgtt ggggtggtgc cgccaggaac cggaccaggg actgcccgcc gtgtacggcc 9240
gcgtcgggtc cagctccgcg accacccgcg gcaggacgcc gaggtagtag ccctcgcccc 9300
aggagtcccc ggcgagcccc tgctcccagt cccagtcccg gaacccccac aggttctcgt 9360
tgttgccgtt ccacagcacc agggaggggt gcggcatcag ccgtacgacg ttctcccggg 9420
cctccgcctc cacctccccg cgcagcggct gctcctcggg gtaggcggcg cacgcgaacg 9480
ggaagtcctg ccagaccagc agccccaact cgtcgcaggc gtcgtagaag tcctcgtcct 9540
cgtagatccc gccgccccag acccggacca ggtccacccc cgcgccggcc gcctgctcca 9600
gccggtgccg gtagcgctcc cgggtgatcc gggacgggaa cacgtcgtcc gggatccagt 9660
tgacgccccg cgcgaacagc cgctcaccgt tgacgaccag ggtgaacccg gtgccgtgcg 9720
cgtcggccga ggtgtccagc tcaaccgtcc ggaacccggt cctgcgccgc caggcgtcca 9780
gcgcctcgtc accgtgggac aacgtcagct cgacgtcgta cagcggctgt tcgccgtatc 9840
cgcgcggcca ccacaggcgg acgtccggca cccggagccg cacggtcccg gccgtcccat 9900
cgacccgcgc ccgggcgcgc acgcccccgg cgctcgcctc cagggtgagc ggtgcctcga 9960
cccgggagcg ctccacgtcg accgccagct cgatctgccc caccccgtcc tcgacggtga 10020
ccagcgggcg cacccgggcg atccgcgccg tcgaccagcg ctccagccgc accggccgcc 10080
agatcccggc cgtcaccagc gtcggccccc agtcccagcc gaacgagcag gccatcttcc 10140
gcaggtactg gtacggctcg gcgtacgctc cggggcgctc gcccagcctg ccgcgcaccg 10200
cctccgcctc ggcgtacgcg gaggcgaacc gcaccgtgag ccggccgctc agtcccgtca 10260
cgtcgaagcg gtacgagcgg tgcatgttcc gcgtccggcc cagtggccgg ccgtcgagca 10320
ggatctcggc gacggtgtcg agaccgtcga agacgaggtc cgtctgctcg tgcgggcccg 10380
tcccggcggt cagctccgtc tcgtacgtcc actcccgccg gcccacccag gccacctcgg 10440
tctcgttgcg gccgaggaac ggatcgggga tcagcccggc cgccagcaga tcggtgtgca 10500
cacaccccgg caccgaggcg gggagggcgt cccccgtgcc gtccgggtgt cgcaggatcc 10560
atccctcggt gagcggtgtg acctgacgca tgcacactcc ctaaaccggt tgagccttct 10620
ctgaagagtg gtctggcatc gttggcgcga ttgcgacttt accggttcag ttcagggctg 10680
ccagagtgcc gaatcagcca tcccactcgt gctcgtccgt cccgtgaacg gagccgtgat 10740
gcatctgaac cgccgtacga cactcaccgg atcgctcgcc ctgctcgccc tcctggcctc 10800
cgcctgcacg ggcacggggg gttcctcgaa gggcgcggac gccaaggctc ccgacgaccc 10860
gtcaaaggtc aaggggtccc tcacggtcct cacccaccgg accgatctgg tgcaggacgg 10920
gacgatgaag aagtacgccg ccgagttcaa cgagacctat cccggggtga aggtggagtt 10980
cgacggcctc accgactacg agggcgaggt caagatccgt atgaacacgg agaactacgg 11040
cgacgtcctc atgatcccgg cggtcgtcga gaagaaggac tacccgaagt tcttcgcctc 11100
cctgggcacc aaggccgaac gcgccgccaa gtaccggttc accgactact ccaccgtcga 11160
cggcaaggtc tacgggcaga gccccgtcgg cgtcgtcccc gggttcatct acaacaagcg 11220
ggtgtggagc gaggccggcg tcaccgactg gcccaccacc cccgccgagt tcctggacga 11280
cctgaaggcg atccggtcga agaccgacgc ggtgccgtac tacaccaact tcaaggacat 11340
gtggccgctg acccagtgga ccaacgtcaa cggctccgtc ggctgcgacc cgcacgccac 11400
cacgaagctc gccgagggcg acccgtgggc cgagggggcc gacctgcgcg tgggcgacac 11460
cctgctccac gacatcgtgc gcggcggact cgccgagaag gacccgacca ccaccaactg 11520
ggagggctcc aagcccaagc tggccaaggg cgagatcgcc accatgtggc tgggctcctg 11580
ggccgtcgtg cagatgcggg acgcggcgaa gcaggccggc gccgaccccg ccgacatcgg 11640
cttcatgccc ttccccgcac agcgggacgg cacgttctgc gcggtgacct ccccggacta 11700
ccagcaggcg gtcaacgtca actccgacaa caaggaggcc gcccgcgcct ggatcgactg 11760
gttcaccgac aagtccggct acgccgaggc caacctcgcc ctatcccccc tgaaggacgc 11820
cccgctgccc gccgtcctcg agccctacga gaaggccggc gtgaagctcc tggacctcga 11880
ggacagcaag ggcgccgagg tgaagtccct cgacaaccgc tccgaggtcg gcatctacaa 11940
gcccgactac cgccaggaac tcgtcgacct cgcccgcggc gcccgcaagg gcggcctgga 12000
cgactacctc ggcggcctcg gcgagcgctg ggccgaggcg cgcagcgcgc tgggggcctg 12060
atgacggaca ccacccgcaa ggcggcgcgg ccggttcccc cggccgcgcc cgccgggccg 12120
ggccgcgcgg cgccggcccc gcgccgcacc cggctgtcgc gccgcctcac cccgtggctg 12180
ttcctggccg caccgctggc cctgctcctg accttcacct acgcgcccga tcgccaacat 12240
ggtcgcgtac agcttcaccg actgggacgg cgtgagcccg gagctgaact ggacgggcac 12300
cgggaactac accgaactcc tcacccgctc cgagctgttc gaggtcttct tcgtcagcgg 12360
ctactacctc gtcgcctccg cggtgcagat cgtgctcgcc ctctacttcg ccacggtcct 12420
cagcttcgac gtccgcttcc ggaacttctt caagggcgtg ctgttcttcc cgtacctcat 12480
caacggggtg gccatcggct tcgtcttcct ctacttcttc caggacggcg gcaccctcga 12540
ctccgtactg ggcctgctcg gcgtcgagac cgaccacgcc tggctgggca cgccgttctc 12600
cgcgaacacc tcgctggccg gcgtctccgt ctggcgctac ctcggactga acttcgtcct 12660
cttcctcggc gcgatccagt ccatcccggg cgagctgtac gaggcggccg agatcgacgg 12720
cgcgaaccgc tggcagcagt tccggcacat catcgcgccc ggcatcagac ccgtgctgag 12780
cctgagcgtg atcctctcgg tctccggctc gctgtcggtc ttcgagatcc cgtacatcat 12840
gaccggcggc gccaccggca cggagacctt cgtgatccag accgtgaagc tggcgttcca 12900
gttcaacaag acgggactcg cctcggccgc cgccgtcgtc ctgctgctga tcgtcctggc 12960
ggtcacctgg gtgcagcggc gcatcgtccc cgacgagaag gtggacctcg tatgacccgc 13020
cgtaccgcgg cacgcgccct ggtcctgacg tccctgatcc tggcgacgct ggtggtgctg 13080
ctgccgctcg ccgtggtctt cctgacctcg ctgaagtcct ccgaggagat ggcgaacggc 13140
agcggagcgc tgacgccgcc cgacgacccg ctgaacttcg gcaactacgt gacggcgttc 13200
cgggacggcc agatgctgtc cgcgttcggg aacacggccg tcatcctggt cgtggccgtc 13260
ggcggaacga tcctgatcgg ctcgatgacg gcgtacgcga tcgaccgctt ccggttccgc 13320
ttcaagaagc tggtcgtggc gctgttcctg ctggccgcgc tggtccccgg ggtgaccacc 13380
caggtggcga ccttccagat cgtcaacagc ttcggcatgt tcgacagcct gtgggcgccg 13440
atcgccctct acatgggcac ggacatcgtc tcgatctacg tcttcctgca gttcatccgc 13500
tccatccccg tctccctgga cgaggcggcg cgcctggacg gcgccaacgc gttcaccgtc 13560
taccgcaagg tgatcttccc gctgctcaag ccggcgatcg cgacggtggt gatcgtaaag 13620
gggatcaacg tctacaacga cttctacatc cccttcctct acatgccctc cgaggacctg 13680
ggggtcatct cgacgtccct gttccgcttc aagggcccct tcggcgcgca ctgggagacg 13740
atctcggcgg gcgcggtcct ggtcatcctg cccaccttga tcgtcttcct gttcctccag 13800
cgcttcatct acaacgggtt catgcggggg gcgacgaagt agccagcgcg gccaccagca 13860
cgggagtgac ctggtcgacc tgccacgcgc gtgccccgtg cgccgtcagc gccgcggcga 13920
cggaccgctc gtcgggcccc ggcggctccc agcagaccct gcgcaccgtg tccggggtga 13980
tcaggttctc cggcggcatg ttcagccgct cggcgagttc ggcgaccccc gcgcgggccg 14040
ccgacagccg ggccgcggca acggggtcct tgtccgccca ggcgcgcggc ggcggagggc 14100
cggtcaccgg ctggccgggc tgcggcagct gggcctcgct cagcgccttc gcgcggtcga 14160
cggccgcctg ccactgctcc agctggcgcc gccccacccg ctgcccgaac ccgttgagcg 14220
cggccatggc gtgcaggttg gcgggcagcg cgagcgcggc ctccacgatc gccgcgtcgg 14280
aaagcacctt gccgggggag acgtcacggc gccgggcgat ccggtcgcgg gtctcccaca 14340
gctcccgcac caccgccatc tggcggcgcc ggcgcacctt gtgcatgccg gaggtgcggc 14400
gccaggggtc cttgcggggc tccggcggcg gggccgaggc gatcgcgtcg aactcctgcc 14460
gggcccagtc cagcttgccc tggcggtcca gctccttctc cagggcgtcc cgcagatcga 14520
ccagcagttc gacgtcgagg gcggcgtacc gcagccaggg ctcgggcagc ggacgggtgg 14580
accagtcgac ggcggagtgg cccttctcca ggacgaagcc gagcacgttc tcgaccatcg 14640
cgccgagccc gacgcggggg aacccggcaa ggcggccggc cagctcggtg tcgaagaggc 14700
gggagggcac catgcctatc tcgcgcagac agggcaggtc ctgggtggcg gcgtgcagca 14760
cccactcgac gccggacagc gcctcgccga gggcggacag gtcggggcag gccacggggt 14820
cgatcagcgc ggtacccgca ccctcgcggc gcagctggac gaggtaggcg cgctggccgt 14880
agcggtaccc ggaggcgcgc tcggcgtcca cggcgacggg tccgctgccg gccgcgaagg 14940
cggcgaccgc ctcggcgagg gcggcctcgt ccgctatcac gggcggaatg ccctcgcggg 15000
gttccagcaa gggggtcggc gcctccgtaa cagaagatcc gccgtcgtcc ggaggagcgc 15060
ctccggtggt gcgcagtgaa ctgtctgctg cggtgtcgtg ggcgtcggtc acctgtcaag 15120
ggtatccgtg ccgcgaaggc gcccgtcgac ggttgtgctc cgtgacgggc gccggtgggt 15180
cgtattccgg tcagaagagt gaaagaacgt gttcgcttgg ccgtgggcgg gcggatcggg 15240
gacgggcgga tcgggggcgg gacggaaggg tcagtggatg atgccggtgc gcagggccac 15300
ggccaccatg ccggcgcggt cgcccgtgcc gagcttgcgg gcgatccggg ccaggtggct 15360
cttgacggtc agtgcggaca ggcccatcga gacgccgatc gccttgttcg actggccttc 15420
cgccaccagc cgcagcacct ccacctcgcg gccggacagc tcgcggtagc cgcccgggtg 15480
gctcggggca cccggggggc ggcggtgcag gcgcgcggcg gcggcgccga tgggagcggc 15540
acccggccgg gtggggagcc cgaggttggt gcgggtgccg gtgacgacgt agcccttgac 15600
tccgcccgcg agggcgttgc gcacggcgcc gatgtcgtcc gccgcggaga gggcgaggcc 15660
gttgggccag cccgcggcgc gggtctcgga gaggagggtg aggccggaac catccggcag 15720
atggacttcg gcgacgcaga tgtcgcgggg gttgccgatg cggggacgag cctccgcgac 15780
ggacgaggcc tcgatgacgt cgcgtacgcc gagcgcccac aggtggcggg tgacggtcga 15840
acggacgcgg gggtcggcca cgaccaccat ggcggtcggc ttgttcgggc ggtaggcgac 15900
caggcttgcg ggctgctcga ggagaacgga caccaggcct cctggggtgc gggacgggcc 15960
ggctcgtggg ggtgaaggcg ggacgaaccg tgctttcaag gtcacagtcg tcttcggcag 16020
caaacctggt gtcctttaac gaatgatcac gaagtgatga gtaacaatcc gggcaattcg 16080
gacgcacgat cgatcattcg aagatcgaac ggtttcggtc tgcgtcgcaa cgcttccgaa 16140
agtggccgta tcgacaaaga gagatgcagg aggccggtcg tcgggacccc gcagcgggag 16200
gctcagcgcg actgcggccc cctccgctgc ggcagcgtca ccacggacgc gtcccccgga 16260
gcggccggcg gcagccccgc gacctgcgcc agcagatcgg accacgcgac cagatgggcg 16320
gccgtgtccg gaaccccgcc cagaccctca cgcggcgtcc acgaggcacg gatctcgatc 16380
tgggaggcgg cgggccgcgc ggacagcccg ccgaagtagt gcgaactcgc ccgcgtcacc 16440
gtgccgctcg gctcgccgta cgacaggccg cgcgccgcca gcgcgccggt cagccaggac 16500
cagcacacgt ccggcagcag cggatccgcc gccatctccg gctccagctc ggcgcgcacc 16560
agcgtcacca gacggaaggt cccccgccag gcgtcgtgtc cggccgggtc gcacagcagc 16620
accagccggc cgtcggccag atcctcctcg ccgtcgacga ccgccgcctc cagcgcgtgc 16680
gcgtacgggg cgagccgttt cggcgcgggc accgtctcca cctcgatctg cggccgcagc 16740
cgggcgctct gcagcgcctc gacagcggcc cggaagggcg gcggaggcgc acctccgtgc 16800
ccgggatccc ccccgccctc cttcggttcg tccattccgc cagcgccgtc cgacagtcgt 16860
ccctgagccg cagccatgcc gggaagatta agcggaacgg gcccccggcg cagggaggga 16920
cacccgcgcc gcccggcgct gtccggatcc tgcaccgcgg ccccgcccgc cggacgcccc 16980
tggggtccgg ggcggcggac gggtcgtgcg agactggccg gtgtgagtgc caacacgagc 17040
ccgaagggcc agacgcctac cgcgaccccc gaccccgtca agaacgacgc cgtccgggaa 17100
tcagccttcc tcaaggcgtg ccggcgcgag ccggtgccgc acacgccggt gtggttcatg 17160
cggcaggccg ggcgctcact gccggagtac cgcaaggtgc gcgagggcat cgggatgctc 17220
gactcctgca tgcggcccga gctggtcacc gagatcaccc tccagccggt gcgccgccac 17280
cacgtcgacg cggcgatcta cttcagcgac atcgtcgtcc cgctcaaggc catcggcatc 17340
gacctcgaca tcaagcccgg catcggcccg gtcgtcgagc agccggtgcg cacccgcgcc 17400
gacctcgccc ggctgcgcga cctgaccccg gaggacgtct cctacgtcac cgaggccatc 17460
ggcatgctga cccgtgagct cgggtccacc ccgctgatcg gtttcgcggg cgccccgttc 17520
acccttgcga gttacctcgt cgagggcggc ccgtcccgta cgtacgagaa cgccaaggcg 17580
atgatgtacg gcgaccccga gctctgggcc gacctgctcg accgcctcgc cgacatcacg 17640
gcggccttcc tcgacgtcca gatccgggcc ggcgcctcgg ccgtgcagct cttcgactcc 17700
tgggccggcg cgctcgcccc ctccgactac cggcgttcgg tgctgcccgc ctcggcgaag 17760
gtgttccgcg cggtggccgg ccacggcgtc ccgcgcatcc acttcggcgt cggcaccggc 17820
gagctgctgg ggctcatggg cgaggccggc gcggacatcg tcggcgtcga ctggcgcgtc 17880
ccgatggacg aggccgcccg ccgcgtcggc cccggcaagg cgctccaggg caacctggac 17940
ccgaccgtgc tgttcgccgg ccgggaggcc gtcgagacga aggcgcgcga ggtcctggac 18000
accgccgcgg gcctggaggg ccacatcttc aacctcggtc acggagtgat gccctccacc 18060
gacccggacg ccctcacccg tctcgtggag tacgtccaca cgcagacggc gcgctgaccc 18120
accgctcacg cgccggacgc gagtcggaat ccgggggcgg ggtactgggc acgggtgccc 18180
accacgttca cccccgggta cgggcaggtg gaggccccat gaggctcgag atgttcgacc 18240
ccgccccgat cggcgtcgtg ttcacccagg ggccggagca ccggctcgcg tacaccaacg 18300
ccgtctaccg ggagaccttc ggcgaccgcc cgctggggcg gacgatccgc gaggccttcc 18360
ccgacctcgc gcagtccggc tacttcgaca tcttcgaccg ggtcctcacc acgggcgcgg 18420
ccgaggtggt caccgcggtg cccctcgacc tgatctaccc cggctccacg ggcgagggca 18480
ggcgctactt cacgttcagc atctcccgcg ccacgatgag cgacggccgg ccgggagtgc 18540
tcggcgtgat cgtggaggtg accgcgcagg tgaccgccgc ggaacggatc cgtgtgctgg 18600
ccgaggagcg ccgccgcgcg ctgcagcgct accgcagcct ggtgaacgcc ggaacgcaga 18660
tggtgtgggt ggcggacgcc aagggccgga tcaccgagcc gagccccggc tgggaacgcg 18720
tgaccgggca gacctgggag gagttccgcg gcgagggctg gatgaacgcc gtccaccccg 18780
acgaccgcgc cgcctcggtc gaggcgtggc ggcgggcgac gaccgaacag gtgccgcgct 18840
ggatccacac ctaccggctg cggctggccg ccggcgggta ccggcacttc gtcgtcgacg 18900
ccgcgcccgt gcgcgacggg aacacggtga tcgaatgggt gggcacctgc acggacatcg 18960
agcgggaatg gcaggagggc cgccgtacgg aactgctggc gcgggccgcc accgccacgt 19020
ccggcatcgc gcggctggac gagatgctcg ccgccctggc cgatgtgatc gtgcccgaca 19080
tcgccgacaa ctgcaccatc cacctcctgc cgcaggccct gcaccgtctg ccgggcaccc 19140
cgctgaccac cgaacgcgtc gccgcggtca cccgcccggg gctcccggac ctgcccccgc 19200
accacgagga gcacctgcgg cccggcagcc cgctggcccg cgccgccgac cgccgcagcc 19260
cgctccactt cgtcttcccg cccggcgagc cgccggccga cctcgctccg ctcgacggcg 19320
agccctggat ggccgaggac gtcaacagcg tcgtgctgct gcccgtcgtc gtcgacggca 19380
ccaccgccgc cctggtcgcc gtctccacca gcggcgcccg cccgcccctc ggccaggcgg 19440
agatcggcct gctgcagaca ctcctggaac gcgcccacac ccccctcagc aacgccctgg 19500
agtaccagcg cacccggcag gtggccctgg ccctgcagaa cagcctgctc accgacccgc 19560
cggacgcgcc cggcctggac atcgccgtcc gctaccggcc cagcaccgcc gccgccgagg 19620
tcggcgggga ctggtacgac gcgttcgtgc tgcgcgacgg cgccaccgtc ctcaccatcg 19680
gcgacgtctc cggccacgac ctgccggccg ccgtcaccat gagccagctg cgcaacatgc 19740
tgcgcgggct cacgctggac cgccaggaac cgaccggcac catcctgcgc cggctggaca 19800
tcgccgtgca gaccctctat acggagtgca ccgccacctg cgtgctggcc cgggtggaac 19860
gcccggactc cggcggcgtc cggctgcact actccgtcgc cggtcacccg ccgccgctgc 19920
tcgtcgaggc ggacggctcc gcgcgcttcc tgaccggggc gcggtccccg atgctcgggc 19980
tcgtccccgc gccggagtac tcgagcgcca tggaaccgct gccgcccggc tccaccctgc 20040
tgctgtacac cgacgggctg gtggagcgcc gcgacgagga tctcaccgtg ggcctggagc 20100
ggctgcggca ccacgcctcg gaggcggtca gccgcccgct gcaggacttc tgcgacacac 20160
tgctcaccgg ccagctcacc gtcgacaacg acgacgacgt ggcgatgctg gtcctgcgcc 20220
ggtaggagcg tgccgaggag cgccactctg gccgatttta cccttgcttt tccatcggga 20280
ttcgttctcc ggatttcccg atccggcgcc gacggcgaga ccgttgggat caccaatacc 20340
ccggaattcc cgcctccgcc accgttgggc agcgacggat cctgtgatat ttcgactacg 20400
cgcggtgatg aattggctcg gtgccggtcg cgcccggctg tagcagttct ggagcgcgtc 20460
tggacatcgt cacgagcgct tgtgattctt ggtcctgtac acgcaagccg gcgcaacgtc 20520
cacgttgccc atcagcggtt atcggcggtc caccggcgcg acggtgaccg cgggcgggta 20580
ctcatagggg gaactgcaat gaattactca aaagcagcga gaggaatgcc gacagccgga 20640
caaggtgccg ttcgggcggc gcgcgtcgtc cgtgaaagtc cggcggaatc agaaacggtc 20700
acagttcaga tagcgtcgtt attaccgggt gagtcgctgc gctcgaaagg gatcgagcag 20760
aaccacgtcg cggcactcgc ggaggtagac gcgccgcttc cgcccatact ggtggaccgg 20820
aagacgatgc gggtcgtcga cgggatgcac cggctcctcg cggctctgct caacggacgg 20880
cagacgatcg aggccgaact gttcgacgga accgcggatg agggattcct gcgcgccgtc 20940
cgggagaacg tggtgcacgg actcccgctg tcgcaggcgg accgccgggc cgccgctgcg 21000
cgcatcatcg tgtcccaccc gcatctgtcg gacagggcga tcgcccgggc gtccgggctc 21060
ggggcgaaga ccgtcgcggc cgtgcggcgc agttcaactg ccgtcgtgcc gcagttgaac 21120
acccgggtgg gccaggacgg cagggtccgg ccgctgaacg ggggcgaggg gcggcgcagg 21180
gccatggcgg tactggccga acaccccgac gcgtccctgc gcgaggtcgc ccgtctgtcc 21240
ggggtgtcgc ccgcgacggt cagcgacgta cgccggcggc tggccgccgg cgagtcgccc 21300
ctgccgtcga gacgggaacc ggccgaaccg cggacgggcg ccgactccca ccgcaaccag 21360
agcttcgtgg atcccgtccc ggtgctggag aagctgctgc gcgacccctc tctgcggcac 21420
aaggagggcg gccgccagct gctccagctg ctccgccaga acgcggtcgg cgtgcaggac 21480
ctgatggagc tgtccgacgc cgtgccgtcc cactgcaggt ccctggtgat ccatctcgcg 21540
cagcagtacc gggacgcctg gcagtccttc gcggagaagc tggacgagcc cgcctgcgcc 21600
tgtcccgggt gacgaacggg cggcacggac ccgttcaccg gacatgaccg gcgccgcgcc 21660
gcgttcacgg cgcgccgccg gcactcccac ggcacccgga ccaccgccgc gtatccggcg 21720
gacccgggcc cgggcgggcc ggattcagcg ggcgggggcc caggtgccac ccgattccag 21780
ccaccgggag agctccgccg ccgagtcctt gcgcacgacc agttcgacga ggccgcgggt 21840
ctggtcctga ccgtgctcga tgcggacgtc ctcgatgttg acgcccaagt cgccgatcga 21900
cgtgaacagt tcggccaggg cgccgggctt gtcggagatg gtcaccgaga cggtcgcgag 21960
ctccgtccgg cgcgtaccgg gtttgcgcac gatcctggcg cacccccggt tcccctcccg 22020
caacagctcc tcgagctcct cctgcgcgcg gcggcggacc agcgggtcgg cgtcggagac 22080
ggcgcgcagc gcgccgacgg cccggcccag gccggcggcg agggagtcga gaacgtccgc 22140
cacggccgtg gcgttggaac gcaggatgtc cccccagagc cgggcgtcac cggccgcgat 22200
ccgggtgacg tcggcgacgc cctgccccgc cagccggacg ctgtcctccg ccgcgtgctc 22260
cagccgcgcg gcgagcaggg aggagagccg atggggcgcg tgcgagacga gggccaccgc 22320
gtggtcgtgc acaccggcgt ccatgaccac cggcatgccg tcgcacaacg acaccatctc 22380
cagggcggtg ttcagcacgt cctgcccggt cagctccgac ggggtgagca cccaggggcg 22440
cccctcgaag aggtccgccc gggcggcgag cggcccggaa cgctcggtgc cggccagcgg 22500
atggcttcct atgtagctgg ccgggtcggc ccgcatcgcg cgcacgtcgt cgtgcgggac 22560
cttcttgacg ctggcgacat cgaggtaggc tcgggccagc ccgctctcct gtgcgcgcgc 22620
gagcacgcgt ccgacctgtg ccgggggcac ggccagcacc gccaggtcga cctgacggtc 22680
cggtctctcc agggatcccg cgcccatcgc ctccgccgtc ctggcggcgt tccggtcgac 22740
gtcctccagg tgcacgccga ccccgcggcg ggtcagcgcg agagcgacgg acgtgccgat 22800
ggccccggtg ccgatgactg tggtggtcct caacgcgcgc ccccaggtgc ggtgatccga 22860
aatcggctcg gacaagtgcc gtgcccggca cgggaaaagg gaattcccat ggcgccgtgc 22920
gccgccaatt taacgcttcg gcgcgcatgt tcaactgcgg cgtcgcagcg gtcgaacaca 22980
gtagcggtac accggaccat tgaggcatcg tgctcagttg gcgacaccgg gtcggataaa 23040
cgccggaatc cgaggagttg acgttgcagt cagcgctgag acacgacgac ctgcatccga 23100
tagaagaagt ggaaataagt tcgctctcca ccgacggctc cccgcggatc gacggggaga 23160
gtcccgagca cgtggaaatg ctggccgccg ccgacaccgc gcttccaccg atcatggtgc 23220
accgccgcac cgggcgggtc atcgacggca tgcaccggct gcgcgccgcg atgctgacgg 23280
gccgtacgac gatcgcggtg aggttcttcg acggcaccga ggaggacgcc ttcgtcctcg 23340
ccgtgaagtc gaacatcgcg cacggactgc cgctgtccgc cgccgaccgc cggcgggccg 23400
ccgggcgcat catggccacc catccccggt ggtcggaccg gatgatcgcc tcggtggtcg 23460
gcacctccgc caggacggtc gccgagatcc gccgcgacgc cggcgccgcc ggggcggggg 23520
agcccacccg catcggccgg gacggcaggg tacggcccgt cgacgtgagc gagggccgca 23580
gactggccca cgacatgatc gtccgcgacc cgggcctgtc gctgcgccag gtcgcccgcg 23640
ccgccgggat ctcgccggag accgtcaggg acgtcagaca ccggatgctc cgcggtgagg 23700
acccggtgcc cgcgccgcgg ccgcggaccc tggtggagcg cggcgcggac cgccgggcgg 23760
agccggccgg gaaggccgcc gcgccgtgcg ggacggagcc gccgcccgcc gtcgtgatga 23820
agcggctgag ggccgatccg gcgctgcgtc tcaacgagaa cggacgcgac ctgctgcggc 23880
ttctggatat ccacacggtc cggctggagg actggaaccg cattatcgaa agcgtgccgc 23940
cgcaccgtct ggagacggtg gcgcagctgg cacgctcctg cgccgacaaa tggtccgaga 24000
tcgcgtcacg catcgaaagc aacgcatcac atctggccgg gtgaacgagg aaacacacga 24060
atccttcgag gagccgtcgg agaaagcggg acggcccgtc ggaacaccct tgtggagggg 24120
caatggagat acggtcgatc gatcacgtcg aattgttcgt cgaggacgcc caggacacgg 24180
ccggcaggct gtgcgactcc ttcggcttcg tccgcgtggg ccgcggcgcc gggaccaccg 24240
gactgcgcgg ctgcgagtcc gtcctgctgc gccagaacga catcgccctg ctgctgacca 24300
cggccaccga cgccgaccac cgtgccgccg agtacgtgaa gcagcacggg gacggggtcg 24360
cggtgatcgg catcggggtg gacgacgcgc gcgccgccta cgccgaggcc gtgcggcgcg 24420
gagccgtccc ggtcgccgcg cccgaggagt tcgggcccgc cggcgcccgt gtcgtcttcg 24480
cctcggtggc gggattcggc gacgtggagc accgcttcgt ctcccgggag gaccccggag 24540
cgccgttcgc gcccttcatc gaggagaccg gcgcccacgg ctccgggggc atgctgaagc 24600
gggtcgacca cttcgcggtc tgcgtcccgg ccggcgaact cgacgggacc gtccgccgct 24660
accaggaggt gttcggcctc agccagacct tcgaggagcg gatcgtcgtc ggctcgcagg 24720
ccatggactc caaggtcgtg cagagcgacc gcggcgcggt gacgttcacc gtcatcgagc 24780
cggacaccac ccgcgcaccc ggccagatcg acgcgttcgt ggcctcccac ggcggggccg 24840
gtgtgcagca cgtcgcgttc ctcactgagg acatcaccac cgcggtgcgc acctgcaccg 24900
ggcgcggggt ccgcttcctc accacgccgc cgagctacta cgagatgctg ccggggcggc 24960
tgggcccggt cggcgtaccc gtggaggagc tcagcgcgct caacatcctg gccgaccgcg 25020
acccgtccgg gatcatgctg cagatcttca ccgagtcgac gcacccgagg cggaccctgt 25080
tctgggaact gatcgaccgc cgcggcgcgc agaccttcgg cagcaacaac atccaggccc 25140
tgtacgaggc cgtggagcgc cagcaggcgg cggaggcggc cgaccaggaa tgaggaagct 25200
ccccgcagac gcgtgtggac ccggaggaca cgccctccgg gtccacacgc gtctgcgggg 25260
ccagcgtcgg ctacgccccg aggagccggc cgccgtgcag cgtctccccg tacgcgaaca 25320
catggccggc ccccacctcc acgggcgcga gcagtgccag atccgtgccc ggacggccgg 25380
agagggcgag ccggccggga ccgctccaga ccggtgagaa ctcgacgccg ctcacctccg 25440
aggcgaccag gcggggccgt ggcggggcgt cggacggcac ccaccgcgga aggacgagac 25500
tgtgcgccag cgggacgtcg tgcagcgcgg gcggccgctc ggagcggcgc tcgaccgtga 25560
ccgacgcctc ggcggtgagc cggccgtgga cggagagcgc gccgtcgaag cggcccccgg 25620
gagcgagccg tgagcccgcc cggccgacgg tcaccggcct ggtctggtgg atggcgccga 25680
actgcttggg catgccctgg acccagccgc gcaccatcgg cacgggctgg tcgacccagg 25740
cgaacgggca gcgcgccatc ggccggccct cgaacgcgca cccgaggagg atcaggaact 25800
ccgagaaccg gcagacggcc gggtcggcca gctccgcgcc gtcctcggag caccactgcc 25860
aggtggcgaa cacggcggcc gccgcacccg gatccgctcc cgcgtccagg cccggcggca 25920
ggaaacgccg tgcggcgtcg gggtcgacac ggtagtcgac catgaggatc tcgccggaga 25980
agtgccacgg cggaggcgtg agcatcgacg cctgccccga aggggacagg ggaaggctgt 26040
agccgatggg cccggcggcc ccggccgcgt ccggatccgt cggatgtgtg tgcccggtgg 26100
tggccgtcat gggttccctc cgatctgccg gtccggcggg ccgccggacc atgcctgggt 26160
cagccgtcga gcggcgcgtt cgagcagcgg gggcggattg aagctgtagg ccaggcgcac 26220
gctcggctcc cgtgtgccgc cgaagcgcga acccgcggtg acgcgcacac ccgcccgctc 26280
cgcacgggcg agcagttcgt cctcgccgag cccggtgccg caccggagcc agaggaagaa 26340
cccgccctcc ggacggctga tccgcaccgg gaggtccgcc gcctcccgca gcgcgtcgag 26400
gagggcgtcg cgccgcgccc tcagacccgc ccgcaacatt tccagatgcc ggtcgtagcc 26460
gccgtcggac agcagccctg cgacggcgag cgaggtgatg tggttgagcg acccgccgct 26520
gcggaacagc ccgtgcgacg cgatccgttc ggccagtgcc ggctccgtca ccagccagcc 26580
cagccggagc cccggcccca gggtcttgga gaagctgccc agccgcacca cgccccggtg 26640
tccggcgagg gccgccagtg gcggcggggc cgggggaccg tccgtcaggc ccagttcgcc 26700
gtaggcgtcg tcctcgacga ccaggacgcc gtgctccgcc gccgcctcca gcagccgcag 26760
ccggcgctcc agcggcatgg tggcgcccgt cggattgtgg tgggtcgggg tgaggtacac 26820
gaacgcggtg cggccggtgc cgccttcgcc gccccgcgcg gtcccggcga gggcgcgccg 26880
gagcgcctcc ggcaccatgc ccgacgcgtc gagggcgacc cgcctcaggc gcagcgcgca 26940
gtccccgagg atgcgctgcc cgaggtcgta gccgaggccc tccacgagca ccgtgtcgcc 27000
gggcctcgcg agggtggtcg ccagcaggtg gagcgcctgg gacgtgcccg ccgtgacgac 27060
cacgtgctcc ggcccgcacg gggaccgccc ccgcacggtg gcccgggcgg ccagctcggc 27120
gcgcaggggc agggcgcccg gatcgtgtcc gtagcccagt gccgccgctc cgtactcctc 27180
cagcgcgcgt gcgtaggcgt cccgcaccag ccccaccggc agcagcgccg gttcgaggta 27240
gccgggcccc aggtcgagga cgcccgcggg ggcgacctcc tgcaccacac cgcgacgcca 27300
ccgccgcgtg tgcgacaacg ggcgggccgt gccgtacggc agggtcccct cgccggcagc 27360
ggtcatcagc ggggtgtcag cacatggcgc aacgcccgta cgcactgggc cagcggggcg 27420
gacgcccggg cgagcgccag gcgcagcgtg cggtcgccgc gggcggggtc agcccagtag 27480
aaggcacggc agggcagggc gtacacatgg tgctcgcgca gcgcctccca gacctcggtc 27540
ccggtcagat gcctgatcag cacccgctcc acactggccc ggctgtccgg gtcgggcacc 27600
ccggtggtcg acaggtccgc cagcccggcg cgcaccaccg accgctgggc ggcgatgaac 27660
tcgtgcagct ccgtcagccc gccggcggcg gcgtcctcgg agaagcggcg gaccatcccg 27720
aggatcagcg gggagacgcc cagcaggatg tcggagtaga tcttctccac cggcaggccc 27780
aggttctcgg agtggaccag catgccgacc ttgaggtcga gggtcggcca gagcttgccc 27840
gtgtcctcga tgacgaccca gcgcacatcg ctggcgtcga ggatctcgta gtggtcgtac 27900
tgggcgcggg tgtcgaagcc gcggaaggac gtgtcgaggg cgaggatcac gccgtgccgt 27960
gcgcactgcc cggccagccg gcgcagccgc tccgccgaca cgacccggcc cgtcgggttg 28020
ttcggcgtgg tgacgaagac acagcccacg gactcgagca gctccgcggg caggtcgtcg 28080
gcgtgcagcg gatcctcctc caggggcacc agacccaggc ggttgccgcg caacaggtcg 28140
gcgatgttgt cgaaggtggg gtggaccagc gccacggagt ccgtgaccga cgccagggcg 28200
cgggagagga tctccatggc caccgacgag gagtagcagc tcagcacacg gccgggtgcg 28260
gacgggtagc ggtgctggcc gagggccttg aagaaggccg cgtgggcctc gcgttcgagc 28320
tgctcgacgg ggcgcttctc gccgtcctcg aaaagcagcg ggagatcatt gacgatcttg 28380
ctctggccgg gagtgagcgg ctgccgggca tgcccgtcgg cgatgttgaa ctcgctgttg 28440
agtgcgagga attccagttg ggtgaggttc tccgcgcctg atccggcgtg cgcagcgtgg 28500
gccttgcttt gcagtgttcc ggacacaggt atgcctctct gggatgtgag ggtttccaga 28560
agcggagcgg acgtaaatga gcggcccact ctacggcctt cgccctccgg ctgaaatgcc 28620
tcttcttttc ggcaccgtgt tcaactgcgg tggtgcggca gtcgaacgag ccgtctcgcc 28680
cgccgtatcg gccggacatc gcgttccgac ggtgacgcgc gtgcggttcc cgtgtccaac 28740
tgacctgagg gcgcagttgg acgggccacc ggcacacggc cgcccgatcc ttgtcggacg 28800
ggcccgggca cgcgaaagtg gacgtgcggg atctgtgttc cgccgccggt gtctcttcgt 28860
aagccgtgaa gtggggcctt gatggaattg tcgctcgatg aattcgcgtc gctcgcccgg 28920
gaacggctgg acccggccgt ctgggatttc atcgaaggcg gcgccggaga ggaacgcacg 28980
ctcgccgcga acaccgcggc attcgaccgc gtcccgctgc ggccgtcggt gctgcgcggc 29040
gcgggcagcc cgcacaccgg caccacgatc ctcgggcgga cgtgggacgc gcccctcgcg 29100
gtcgccccgg tggcctacca cacgctcgcg gacccggccg gtgaggtcgc caccgtccgg 29160
ggaacggcgg ccgccgccgg actcccggtc gtcgtcagca ccttcgcggg ccgcacgttc 29220
gaggacatcg ccgccgaggc caccgtcccg ctctggctcc aggtgtactg cctgcgggac 29280
cgctccctca cccgaggcct catcgaacgc gccgagaacg cgggcttcga ggccctggtc 29340
ctcacggtcg acgcgccgca cctcggccgc cggctgcggg acctgcgcaa cggcttccgg 29400
ctgcccgccg gcacggtccc cgccaacctc ccggtggacg gattcgcgga ccccgcggcg 29460
cactcccgcg ccgacttcga ccccggcctg gactggtcgg tggtggagtg gctgcgctcg 29520
gtctccgaac tgccgttgct cgtcaagggg atcctcaccg gcgccgacgc ggtgcgcgcg 29580
gccgaggccg gggtggacgg cgtcatggtc tccaaccacg ggggccgcca gctcgacgga 29640
gtgccggcca ccctcgacgt cctgcccgag gtcgccgagg cggtcggcgg acgcctcccc 29700
gtcctcctcg acggcggggt ccgccggggg cgggacatcc tggcggcgct cgcgctcggc 29760
gccgacgcgg ccctcgtcgg ccgcccggtg ctgcacggcc tcgcggccgg cggggccggc 29820
ggggtgaccg gcgtcctctc cgtcctcctg gaggagctga cggacgcgat gtcccttgcg 29880
ggcctgagga ccctcgccga catcggcccc tcactcgtcg gccgggctcc tgaccacccc 29940
cgccgaagca ccgtggacgc cgggaagggc gcggggagcg accggcgcac cgccgccggg 30000
ggaggggccg ggctgcgcct cgcggacctg cacccgagtg tcgccgaccc ggtcatggac 30060
accatgaact tcctcaacga ggtgacactg cgctaccccg aggcggtgtc cttcgccccc 30120
ggacggccct acgcggagtt cttcgagacc gagcaggtct tccgccatct gcgccgctac 30180
ctcgaccacc tggccgagca gggccgttcg cccgcgcagg tgcgcgacgc gctgttccag 30240
tacggtccgt ccgccggtgt gatccgcgag ctgatcgccc actcgctgcg ggtggacgag 30300
ggcatcgacg tgtcgcccga gtcgatcgtg gtgacggtcg gctgccagga ggcgatgttc 30360
ctgacgctgc gcgcgctcat gtccggcccg gacgacgtgc tgctcgtctc cagcccctgc 30420
tacgtgggga tcaccggggc cgcccggctg ctggacgtcg cggtgaccgc cgtcgaggag 30480
ggcgaggacg gcctgtcgtg cgacgccctc gaggccgccg tctcggcgga gcgggcgcgc 30540
ggcaggcggc cgcgggccgt ctacgtggtc ccggaccact cgaacccgtc cggcgcgacc 30600
atgccgctcg aggcccggaa gtccctcctg gagctggcgc agcggctcga cgtcctcgtc 30660
ctggaggaca gcccgtaccg gcacgtcagc ccgggcacgc aggtggcgtc cctgaaggcc 30720
ctcgaccgga cacggcgagt gatccacctc ggttcctacg ccaagaccgt cttccccggg 30780
gcacgcctcg ggttcgcggt cgccgaccag ccggtgctgg cgccggacgg cggcacgagt 30840
ctgctggcgg acgaactcgc caagatcaag agcatggtca cggtcaacac ctcgccgctc 30900
agccaggccg cggtggcggg cgcgctgctg gagtcgggcg gccgtgtctc ggagctcaac 30960
gcccgcaacg ccgcccacta cggggaggcc atgcgcttca ccctgcagtg cctggagcgg 31020
gagttcccgg ccgcgcggcg gacccggctc ggcgtccgct ggaacgcgcc cagcggcggg 31080
ttcttcctca ccctccaggt gccgttccgc gcggacaact ccgcgctggc ccggtccgcg 31140
caggacttcg gggtcatctg gacgccgatg tcgtacttct atccgcaggg cggcggcctg 31200
cacaccctca ggctctccac cagctacctg acccacgccg acatcgagaa gggcatctcc 31260
cggctggccg ggttcatcga gttcgagtgc ggggacccgg tggcctgaac cgccgcgacg 31320
acgaagggcc ccggccgcgc cggcggggcc cttcgtcggt cgcgacgctc aggacggatg 31380
cggctcctcc cagaacatgc tgtcggacga ggcgacgaga ccgccgtcga agagcgggac 31440
gtcctcgccg gcgtactctc ccagctgcgc ccgggtctgc acctgcgagc cctcctgcat 31500
cgcccggctc accgccgacg accggaacag cggggccatg ttctcctcct gccgcccggc 31560
catgtcgtcg acggcgtcgg cgaactccgc cgactgctgc ttgagccgca ggacgctcga 31620
ctcggcgtcg gagaggtcga agtccgtgct ggacatgccc gccaccagct ccacgaacga 31680
ctccagctcg gcgtggctgc tccgggtgac cttcttcgcc gtccagaagt aggagtcctc 31740
gtcgacgtgc atgtcgtaga acgaggtcag gaactcgtag aagacgccgt actcccgccg 31800
gtaccgggcc tcgaactcgt cgaaggcccg ccgctcgtcg atccgccccg ccagcacgct 31860
gttgagggag cgggcggcca gcagcgcgct gtaggtggcc agatggaccc cggaggagaa 31920
gacggggtcg acgaagcacg ccgcgtcacc gacgagcacc atccccggcc gccagaaggt 31980
cgtgtggtgg tacgagtagt ccttgcgcac ccgcagctgc ccgtactgac cggtcgtgac 32040
ccgggtggcg tccgccaggt actccttgat catcgggcat tcgtcgatga ggccgcgcag 32100
cgcgctctcc gggtcgccct gcaccttggc cgcgtcctcc cggcggacga ccgcgccgac 32160
gctggtcagc gtggagctga gcgggatgta ccagaaccag ccgctgccga acgccacaca 32220
gaggatgttg ccggcgtagg gcgccggcat ccgcttgccg ttctcgaagt agccgaacag 32280
cgccaggctc ttgaagaagt ccgagtacgt gcgcgagccg ccgacccgct tgtggatgcg 32340
gctggtgttg cccgaggcgt ccaccacgta ccgggccgac acctcgtgct ccgtgccgtc 32400
cgggtcggtg taccgcagcc cgcgggcccg cccgtcggcg tcgtccacga catcggtgac 32460
cgtgcggtcc tggcggacca cgacaccctt gcgggccgcg ttgtccagca ggatcttgtc 32520
gaatttgctc cgctccacct gataggcgaa cgaggtcggt ccggagacct tggacgagac 32580
ggagaaggag aaattccacg gcttggggct cgcaccccac cggaacgtcc caccgcgctt 32640
gtgcggaaaa ccggcggcgg cgagttcgtc ggtgacaccg agcagatggc agatgccgtg 32700
aatggtcgac ggcagaagcg actcgcctat ctggtacctg gggaaggtct ccttttcgag 32760
cagcagcaca ctgtgcccct gcatggccac cagggtcgag agcgtcgacc ccgaagggcc 32820
gccgcccacg accacgacgt cgaattcctc gtgctgtcct gtactcattc ggcctcccgg 32880
cacgcactga tgcggtcatc gcgctggtga ctcttttgtc agggttccac aggactcaaa 32940
ggggccaaag ggtggacgac attccgtgaa tggacagccg ggcccgggcg cgcgcacgca 33000
aaggcgccgc cggggaattc ccggcggcgc ccggaccggc gtcgggtgag acgggggtca 33060
actcacccgg cgccggatcc gccacacccc gaacaccgtg aggattccgc tgagtgccac 33120
atagatcgcg gtctccagcc actggaacgc ccagtagcgg ctgctggggt ggtacaggac 33180
gtcgacgtgc aggtcgtgtt cggcgaggca caccgcggtg tcgccgaacg tgccccccgc 33240
gccggtcttg ggcgggtcgt cgaggcagcc gttgaactcg ctggaggcga gggtcctgcc 33300
gtccgcggtg cgcagcggac tggtctcggc gatccacgcg tccggcgcgt ccgggatccg 33360
caccccgccg atgacggatc cgccgccgat actgcccagg ttctgcgccg agttgatcgc 33420
ctcggccgtc atcgccagcg tcgtcctgtc cggcggcatc aggctgggcc gcaccacgtt 33480
cgggaagaag aactggaagg cgatgaagac caccagcgtc accgccatcg cgggcagggt 33540
ccgccgcagc aggagcccga cgacggtgcc gaacgtgaag gccagcgcgg cgtagccgat 33600
cggggcgatg ttgcgcgcac cgaacacgaa ggtgtcgaac tgctccttga cgacgtcgtc 33660
gaagggccgg gccgcccagg tgagcagggc cgcggccgca ccggtcacga tcaccgaggc 33720
cgcgccgatg agcaggatct tgctgagcag ccagcgcggc cgggtgacgc tctggttcca 33780
caccagccga tgggtgccgt tctcgagttc cctggcgatc aggggagcgc cccagaaggt 33840
gccgatgagc gcggggatca gggccaggcc ggtcgccagg aacagcaggg tgttctggaa 33900
ggtgctgcgg aactggctcc tggcctgggc gcagttggcc gagttgtcgc agttggcctg 33960
gtagacgtca tgggcgtcac ggatgtcccc gcccaggtag agcaggtaga cggcgatcac 34020
ggccagcgcg ccggcgccga acagggcctg gacgcggaac tgccgccagc tgagccacat 34080
catcgggtgg ccccccaggc tgcggcctcg gtgcgggcgg cgggaacggc ggccgcccgg 34140
gtcatgtagg cgagcacgag ttcctcgagg gtgaccggct cggaccggta gggcagtgcc 34200
tcggtcgcgg cgccggtgcg gacgaccgcg ctgctgtgct tgccgctgtg ctcgaccgag 34260
atcacctcga tcccggcggg cggctggtcg aactcgccgc gggccgcgac cagccgggcg 34320
tgcccggcca gcagctcccg ggtgtcgccg gcgacctgca cccgggcgtc gcacagcacg 34380
atgagatagt cgcagacctg ctccacgtca ccgaggaggt gcgaggagag gacggcgctg 34440
gcgccgagct ccagcacgaa ctccatcagg ttctgcagga acccccggcg cgccaggggg 34500
tccagggccg ccgccggctc gtcgaagatc agcagctccg gccgcttggc cgccgcgatg 34560
gtcagcgcaa gctgcgcgcg ctggccaccc gagagctgcc cggccttctg cccggcgctg 34620
agccccacct ggctgatgcg ccgctctgcc aggaccgggt cccagcccgg gttcatcttc 34680
gcgccgaact tcaggtgctc cgccacggtg aacgcgccgt acaccggcgt gttctgcgcg 34740
acgaacccca cccgggccag gtgcgacgcg ttggccgccg gacgcgagcc gaggacgctc 34800
agtgagccgg acgtcggttc ggtcagcccg caggccaggt gcaggagggt cgatttgccg 34860
gccccgttcg ggccgaccag tccgatgaca cggccggcgg ggacgctgag gtgcacgtcg 34920
ctcagggcga gcttgccgcg gcggccgtac ttcttcgtca agccctccgc gtgaagcacg 34980
ggaggggagt ctgcgtgtgg catgactcca tcctcgaatt ccgccccgtt cacggcatca 35040
gtccaaagca cggttcccgt tgccggcggc cgtactttcg gccggtcggc cacggccctg 35100
ctggtgggcg aactgggtgc ggtacagctc ggagtagaga ccgccgccgg ccagcagctg 35160
gtcgtgggtg ccccgctcct ggatccgccc gtcgtcgatg acgaggatct ggtcggcgtc 35220
ctggatggtg gacagccggt gcgcgatgac gagcgaggtg cgcccggtca gggcggtctt 35280
gagggcccgc tggatggcca gctcggactc ggagtccagg tgcgccgtcg cctcgtccag 35340
gacgacgatc ggaggcgact tgagcaggag ccgggcgatg gccagccgct gcttctcacc 35400
gccggacagc cggtagccgc ggtcgccgac gaccgtgtcg agaccgtccg ggagctggga 35460
gatcgtcggc cagatccgcg ccgcctcgca cgcctggacg atctcgggct cggaggcgtc 35520
cgggcgggcg tacagcaggt tggcccggat ggtgtcgtgg aacaggtgcg cgtcctgggt 35580
gaccacgccg accgtgttct gcagcgagcc gagggtcagg tcgcggacgt cgtggccgcc 35640
gatccgcacc gttcccgagg tggcgtcgta gagccgtggc accaggtggg tgatcgtggt 35700
cttgcccgcg ccggacgggc cgaccagcgc cgtgagccgg ccggccgggg cgtggaagct 35760
cacgtcgttg aggaccagcg cgccggggcc ctgctcgctc ttgcgctgcg gcatcaactc 35820
cagtgagggc agggacactt cctcggcgct ggggtagcgg aaggcgacct ggtcgaactc 35880
gacgggggga gcggtgccgt cgccgttcgc cgaggcgcgg gccggcaggg ggcgggcgcc 35940
gggacgctcg gtgatcagcg gcttcaggtc cagcacctcg aagacgcggt cgaagctgac 36000
cagcgcggtc atgacgtcgc tctggatgtt cgtcagctgg ttgacggggc cgtacagcat 36060
cagcagcagg gcgaccatgg ccaccagcgt gccgatctgc agcgagccgt cgatgacgaa 36120
ccagccgccg aagccgtaca ccatcgccgt ggtgacggtg gtgagcaggg tgacgaggat 36180
gaacagcagc cgtgcgtgca cgtccatcga gatcgcgatg tcccggacga ggcccgcctt 36240
cttggagaac tcggcggact cgtcctccgg acggccgtag agcttgacga gcatcgcgcc 36300
ggagatgttg aaccgctcgg tcatcatcga gcccagcttg gcgtcgttct gcatgccggc 36360
gcgggccagc ttctccagcc gctgggcgat gatcttcccg gggatgaaga acagcgggat 36420
catgatcagc gccgccacgg tgatcggcca cgagaggtag agcatcgccg cgagcaccag 36480
gaccagcgtc agcagcgtcg acagcgactg cgacagcagc gaggtgaggg cctgttgggc 36540
gcccacgatg tcggtgttga tccggctgac cagcgacccg gtctgggtgc gggtgaagaa 36600
cgccaccggc tgccgctgga tgtgggagaa caccgcggtc cgcaggtcga agatgaggcc 36660
ctggccgacc cttccggaga accacgtctg cgtgtagacc gccacgacgt tcagcagggc 36720
cagtccggcg acgagcccgg cgaggccgaa cacgacggac gtcttcccgg ggatgatgcc 36780
gtcatcgatg atcattttga gggtcagcgg gatcgacacc gtgatcaggg agtcgacgat 36840
cgtcgccacc atgaccatcg ccatggcccg gcggtagcgc atggcgtagg gaatgatccg 36900
cttgaacgtg ccggacctga ccggctgcgg gtccaccagt ccttcgaccc gcagtccgat 36960
cgtgcccatc gtcgggtcgt gtcccacggt cacggagact ctcctcagtg tgtgtcgcgt 37020
cgctatgtgt cgcgtcgctg tgtggcgcgt cgctgtgtgt cgtcggggcg tcgcgtcagt 37080
gcttgtcgag gaactcgacg atcgccgcgt tgaccgcgtc gggccgctcg aagtacccga 37140
ggtgcccgca gtccgggatc tccacgagat cgcagtcggg aacggcctcg gcgacctcca 37200
cgcccaggtg cggcggggtg atgaggtcgt cggcgaaggt cacgacgcga cagggggcgg 37260
ccacccggcg cagcgccggg cggcggtcgt ccatgatgtc ggcccaggcg tgccgggcct 37320
gcgcctcccc gcccccggac agctcgaaga cgtccagcca ggcggtcacc gcctggtcgt 37380
cgttgagcgt cgcgggcgag aacatccgga acaccgtcga cgcggcgtcg tacgcggccg 37440
gcagccgcac cccgctctcc accagtgccg tctccgcccg cgtctgggcc cgccgcgcgg 37500
cgtccgcacg ggcccgggtg gcgatgagca ccgcgcaccg cacgagttcg ggatgcccga 37560
tcgccagctc ctgcgcgatc atcgccccca gggaggtgcc cacgatccgg cacggcgcca 37620
gatccagggc ctcgatcagg cccttggcgt cggcggtcat gtccagcagc gagtacctgc 37680
ccggcggcgc gtcggacggc gggacacccc ggtggtcgaa gacgaccgtg gagtagcccg 37740
ccgtgtgcag cgccggcgtc tggtgcaggg tccaggcatg gccggccgag cccgagccca 37800
tgatcatgag caccggttcg ccccggcccg cacgctggta ggcgatgcgg acgcccccca 37860
cggtgacgaa gtgcggggcg cgccggcccg cggtgtggtc catgccgcct ctccctcctc 37920
gtcgtcgcgg gggccgcccg gtggtaccgg ccggccccgg gggcggctca ctatcgcacg 37980
cggccacggg gcggggcagt gtgcgcgggg cacgtccatg gacacccccc ggcccgcgtc 38040
caactgcggt gatgccctca gttggacacc ggccggccgc gtccaagcag gcccggccga 38100
cggttgatcc gctgtgtgga gctgagccat attgggccgc cgtgagccac tgacgcccac 38160
caagtccccg cgcttcttcc gcaccggccg cctggcgcgc ccgtcgcgcc gagggaggga 38220
ccaccttgtc agcttccgac cttccagcca cccggctgac acccgagaag atccggtcct 38280
ggctcgtcga ccgggtcgcc tactacgcca ggctgcccgc cgaggagatc ggcgccgacg 38340
tcccgctcgc gcactacgga ctggactcgg tgtacgcctt cgccctgtgc ggagacatcg 38400
aggacggcct cggcctcgtc gtcgagcccg tcctgctctg ggacgtcgac accatcaccg 38460
agctcaccga ccatctcgcc gaactgacag ccgactgagg gccttcgagg gggaggacga 38520
tgcgtcgaaa ggacctggag aggctgacgt ccggtcagct cggcgtctgg tacgcgcagc 38580
agctcgaacc cctgagcccc gtgtacaaca tcgccgagta cgtggagatc cgcggcgacg 38640
tggacgtcgg gcttctggtg tcggcgctgc ggtctgccct cgacgaggcc cagacctacc 38700
ggctccgctt ccggcaggag gacgccggcc ccggacagta cgtcgacgac tcgctggagc 38760
ttcccgtcca cgtcgccgac ctcggctccg caggggaccc gcgcgccgcg gccgtggagt 38820
ggatgaccgc cgacctggac cgccccgcgg accccctcac cggcccgctg gccgcccacg 38880
ccgtgttccg gctgggaccc ggccatgtcc tctggtacca gcgtgcccac cacctcgtcc 38940
tcgacgggac cagcctctcc gtgttcgccg gtgggatccc cgggtaccga gctcgaattc 39000
gccctatagt gagtcgtatt acaattcact ggccgtcgtt ttacaacgtc gtgactggga 39060
aaaccctggc gttacccaac ttaatcgcct tgcagcacat ccccctttcg ccagctggcg 39120
taatagcgaa gaggcccgca ccgatcgccc ttcccaacag ttgcgcagct gaatggcgaa 39180
tggcgcctga tgcggtattt tctccttacg catctgtgcg gtatttcaca ccgcatatgg 39240
tgcactctca gtacaatctg ctctgatgcc gcatagttaa gccagccccg acacccgcca 39300
acacccgctg acgcgaaccc cttgcggccg c 39331
<210> 40
<211> 40551
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая фосмида pXYF24
<400> 40
atcgaatata acttcgtata atgtatgcta tacgaagtta ttagcgatga gctcggactt 60
ccattgttca ttccacggac aaaaacagag aaaggaaacg acagaggcca aaaagctcgc 120
tttcagcacc tgtcgtttcc tttcttttca gagggtattt taaataaaaa cattaagtta 180
tgacgaagaa gaacggaaac gccttaaacc ggaaaatttt cataaatagc gaaaacccgc 240
gaggtcgccg ccccgtaacc tgtcggatca ccggaaagga cccgtaaagt gataatgatt 300
atcatctaca tatcacaacg tgcgtggagg ccatcaaacc acgtcaaata atcaattatg 360
acgcaggtat cgtattaatt gatctgcatc aacttaacgt aaaaacaact tcagacaata 420
caaatcagcg acactgaata cggggcaacc tcatgtccga gctcgcgagc tcgtcgacag 480
cgacacactt gcatcggatg cagcccggtt aacgtgccgg cacggcctgg gtaaccaggt 540
attttgtcca cataaccgtg cgcaaaatgt tgtggataag caggacacag cagcaatcca 600
cagcaggcat acaaccgcac accgaggtta ctccgttcta caggttacga cgacatgtca 660
atacttgccc ttgacaggca ttgatggaat cgtagtctca cgctgatagt ctgatcgaca 720
atacaagtgg gaccgtggtc ccagaccgat aatcagaccg acaacacgag tgggatcgtg 780
gtcccagact aataatcaga ccgacgatac gagtgggacc gtggtcccag actaataatc 840
agaccgacga tacgagtggg accgtggttc cagactaata atcagaccga cgatacgagt 900
gggaccgtgg tcccagacta ataatcagac cgacgatacg agtgggacca tggtcccaga 960
ctaataatca gaccgacgat acgagtggga ccgtggtccc agtctgatta tcagaccgac 1020
gatacgagtg ggaccgtggt cccagactaa taatcagacc gacgatacga gtgggaccgt 1080
ggtcccagac taataatcag accgacgata cgagtgggac cgtggtccca gtctgattat 1140
cagaccgacg atacaagtgg aacagtgggc ccagagagaa tattcaggcc agttatgctt 1200
tctggcctgt aacaaaggac attaagtaaa gacagataaa cgtagactaa aacgtggtcg 1260
catcagggtg ctggcttttc aagttcctta agaatggcct caattttctc tatacactca 1320
gttggaacac gagacctgtc caggttaagc accattttat cgcccttata caatactgtc 1380
gctccaggag caaactgatg tcgtgagctt aaactagttc ttgatgcaga tgacgtttta 1440
agcacagaag ttaaaagagt gataacttct tcagcttcaa atatcacccc agcttttttc 1500
tgctcatgaa ggttagatgc ctgctgctta agtaattcct ctttatctgt aaaggctttt 1560
tgaagtgcat cacctgaccg ggcagatagt tcaccggggt gagaaaaaag agcaacaact 1620
gatttaggca atttggcggt gttgatacag cgggtaataa tcttacgtga aatattttcc 1680
gcatcagcca gcgcagaaat atttccagca aattcattct gcaatcggct tgcataacgc 1740
tgaccacgtt cataagcact tgttgggcga taatcgttac ccaatctgga taatgcagcc 1800
atctgctcat catccagctc gccaaccaga acacgataat cactttcggt aagtgcagca 1860
gctttacgac ggcgactccc atcggcaatt tctatgacac cagatactct tcgaccgaac 1920
gccggtgtct gttgaccagt cagtagaaaa gaagggatga gatcatccag tgcgtcctca 1980
gtaagcagct cctggtcacg ttcattacct gaccataccc gagaggtctt ctcaacacta 2040
tcaccccgga gcacttcaag agtaaacttc acatcccgac cacatacagg caaagtaatg 2100
gcattaccgc gagccattac tcctacgcgc gcaattaacg aatccaccat cggggcagct 2160
ggtgtcgata acgaagtatc ttcaaccggt tgagtattga gcgtatgttt tggaataaca 2220
ggcgcacgct tcattatcta atctcccagc gtggtttaat cagacgatcg aaaatttcat 2280
tgcagacagg ttcccaaata gaaagagcat ttctccaggc accagttgaa gagcgttgat 2340
caatggcctg ttcaaaaaca gttctcatcc ggatctgacc tttaccaact tcatccgttt 2400
cacgtacaac attttttaga accatgcttc cccaggcatc ccgaatttgc tcctccatcc 2460
acggggactg agagccatta ctattgctgt atttggtaag caaaatacgt acatcaggct 2520
cgaacccttt aagatcaacg ttcttgagca gatcacgaag catatcgaaa aactgcagtg 2580
cggaggtgta gtcaaacaac tcagcaggcg tgggaacaat cagcacatca gcagcacata 2640
cgacattaat cgtgccgata cccaggttag gcgcgctgtc aataactatg acatcatagt 2700
catgagcaac agtttcaatg gccagtcgga gcatcaggtg tggatcggtg ggcagtttac 2760
cttcatcaaa tttgcccatt aactcagttt caatacggtg cagagccaga caggaaggaa 2820
taatgtcaag ccccggccag caagtgggct ttattgcata agtgacatcg tccttttccc 2880
caagatagaa aggcaggaga gtgtcttctg catgaatatg aagatctggt acccatccgt 2940
gatacattga ggctgttccc tgggggtcgt taccttccac gagcaaaaca cgtagcccct 3000
tcagagccag atcctgagca agatgaacag aaactgaggt tttgtaaacg ccacctttat 3060
gggcagcaac cccgatcacc ggtggaaata cgtcttcagc acgtcgcaat cgcgtaccaa 3120
acacatcacg catatgatta atttgttcaa ttgtataacc aacacgttgc tcaacccgtc 3180
ctcgaatttc catatccggg tgcggtagtc gccctgcttt ctcggcatct ctgatagcct 3240
gagaagaaac cccaactaaa tccgctgctt cacctattct ccagcgccgg gttattttcc 3300
tcgcttccgg gctgtcatca ttaaactgtg caatggcgat agccttcgtc atttcatgac 3360
cagcgtttat gcactggtta agtgtttcca tgagtttcat tctgaacatc ctttaatcat 3420
tgctttgcgt ttttttatta aatcttgcaa tttactgcaa agcaacaaca aaatcgcaaa 3480
gtcatcaaaa aaccgcaaag ttgtttaaaa taagagcaac actacaaaag gagataagaa 3540
gagcacatac ctcagtcact tattatcact agcgctcgcc gcagccgtgt aaccgagcat 3600
agcgagcgaa ctggcgagga agcaaagaag aactgttctg tcagatagct cttacgctca 3660
gcgcaagaag aaatatccac cgtgggaaaa actccaggta gaggtacaca cgcggatagc 3720
caattcagag taataaactg tgataatcaa ccctcatcaa tgatgacgaa ctaacccccg 3780
atatcaggtc acatgacgaa gggaaagaga aggaaatcaa ctgtgacaaa ctgccctcaa 3840
atttggcttc cttaaaaatt acagttcaaa aagtatgaga aaatccatgc aggctgaagg 3900
aaacagcaaa actgtgacaa attaccctca gtaggtcaga acaaatgtga cgaaccaccc 3960
tcaaatctgt gacagataac cctcagacta tcctgtcgtc atggaagtga tatcgcggaa 4020
ggaaaatacg atatgagtcg tctggcggcc tttctttttc tcaatgtatg agaggcgcat 4080
tggagttctg ctgttgatct cattaacaca gacctgcagg aagcggcggc ggaagtcagg 4140
catacgctgg taactttgag gcagctggta acgctctatg atccagtcga ttttcagaga 4200
gacgatgcct gagccatccg gcttacgata ctgacacagg gattcgtata aacgcatggc 4260
atacggattg gtgatttctt ttgtttcact aagccgaaac tgcgtaaacc ggttctgtaa 4320
cccgataaag aagggaatga gatatgggtt gatatgtaca ctgtaaagcc ctctggatgg 4380
actgtgcgca cgtttgataa accaaggaaa agattcatag cctttttcat cgccggcatc 4440
ctcttcaggg cgataaaaaa ccacttcctt ccccgcgaaa ctcttcaatg cctgccgtat 4500
atccttactg gcttccgcag aggtcaatcc gaatatttca gcatatttag caacatggat 4560
ctcgcagata ccgtcatgtt cctgtagggt gccatcagat tttctgatct ggtcaacgaa 4620
cagatacagc atacgttttt gatcccggga gagactatat gccgcctcag tgaggtcgtt 4680
tgactggacg attcgcgggc tatttttacg tttcttgtga ttgataaccg ctgtttccgc 4740
catgacagat ccatgtgaag tgtgacaagt ttttagattg tcacactaaa taaaaaagag 4800
tcaataagca gggataactt tgtgaaaaaa cagcttcttc tgagggcaat ttgtcacagg 4860
gttaagggca atttgtcaca gacaggactg tcatttgagg gtgatttgtc acactgaaag 4920
ggcaatttgt cacaacacct tctctagaac cagcatggat aaaggcctac aaggcgctct 4980
aaaaaagaag atctaaaaac tataaaaaaa ataattataa aaatatcccc gtggataagt 5040
ggataacccc aagggaagtt ttttcaggca tcgtgtgtaa gcagaatata taagtgctgt 5100
tccctggtgc ttcctcgctc actcgaccgg gagggttcga gaaggggggg cacccccctt 5160
cggcgtgcgc ggtcacgcgc acagggcgca gccctggtta aaaacaaggt ttataaatat 5220
tggtttaaaa gcaggttaaa agacaggtta gcggtggccg aaaaacgggc ggaaaccctt 5280
gcaaatgctg gattttctgc ctgtggacag cccctcaaat gtcaataggt gcgcccctca 5340
tctgtcagca ctctgcccct caagtgtcaa ggatcgcgcc cctcatctgt cagtagtcgc 5400
gcccctcaag tgtcaatacc gcagggcact tatccccagg cttgtccaca tcatctgtgg 5460
gaaactcgcg taaaatcagg cgttttcgcc gatttgcgag gctggccagc tccacgtcgc 5520
cggccgaaat cgagcctgcc cctcatctgt caacgccgcg ccgggtgagt cggcccctca 5580
agtgtcaacg tccgcccctc atctgtcagt gagggccaag ttttccgcga ggtatccaca 5640
acgccggcgg ccggccgcgg tgtctcgcac acggcttcga cggcgtttct ggcgcgtttg 5700
cagggccata gacggccgcc agcccagcgg cgagggcaac cagccgaggg cttcgccctg 5760
tcgctcgact gcggcgagca ctactggctg taaaaggaca gaccacatca tggttctgtg 5820
ttcattaggt tgttctgtcc attgctgaca taatccgctc cacttcaacg taacaccgca 5880
cgaagatttc tattgttcct gaaggcatat tcaaatcgtt ttcgttaccg cttgcaggca 5940
tcatgacaga acactacttc ctataaacgc tacacaggct cctgagatta ataatgcgga 6000
tctctacgat aatgggagat tttcccgact gtttcgttcg cttctcagtg gataacagcc 6060
agcttctctg tttaacagac aaaaacagca tatccactca gttccacatt tccatataaa 6120
ggccaaggca tttattctca ggataattgt ttcagcatcg caaccgcatc agactccggc 6180
atcgcaaact gcacccggtg ccgggcagcc acatccagcg caaaaacctt cgtgtagact 6240
tccgttgaac tgatggactt atgtcccatc aggctttgca gaactttcag cggtataccg 6300
gcatacagca tgtgcatcgc ataggaatgg cggaacgtat gtggtgtgac cggaacagag 6360
aacgtcacac cgtcagcagc agcggcggca accgcctccc caatccaggt cctgaccgtt 6420
ctgtccgtca cttcccagat ccgcgctttc tctgtccttc ctgtgcgacg gttacgccgc 6480
tccatgagct tatcgcgaat aaatacctgt gacggaagat cacttcgcag aataaataaa 6540
tcctggtgtc cctgttgata ccgggaagcc ctgggccaac ttttggcgaa aatgagacgt 6600
tgatcggcac gtaagaggtt ccaactttca ccataatgaa ataagatcac taccgggcgt 6660
attttttgag ttatcgagat tttcaggagc taaggaagct aaaatggaga aaaaaatcac 6720
tggatatacc accgttgata tatcccaatg gcatcgtaaa gaacattttg aggcatttca 6780
gtcagttgct caatgtacct ataaccagac cgttcagctg gatattacgg cctttttaaa 6840
gaccgtaaag aaaaataagc acaagtttta tccggccttt attcacattc ttgcccgcct 6900
gatgaatgct catccggaat ttcgtatggc aatgaaagac ggtgagctgg tgatatggga 6960
tagtgttcac ccttgttaca ccgttttcca tgagcaaact gaaacgtttt catcgctctg 7020
gagtgaatac cacgacgatt tccggcagtt tctacacata tattcgcaag atgtggcgtg 7080
ttacggtgaa aacctggcct atttccctaa agggtttatt gagaatatgt ttttcgtctc 7140
agccaatccc tgggtgagtt tcaccagttt tgatttaaac gtggccaata tggacaactt 7200
cttcgccccc gttttcacca tgggcaaata ttatacgcaa ggcgacaagg tgctgatgcc 7260
gctggcgatt caggttcatc atgccgtttg tgatggcttc catgtcggca gaatgcttaa 7320
tgaattacaa cagtactgcg atgagtggca gggcggggcg taattttttt aaggcagtta 7380
ttggtgccct taaacgcctg gttgctacgc ctgaataagt gataataagc ggatgaatgg 7440
cagaaattcg atgataagct gtcaaacatg agaattggtc gacggcccgg gcggccgcaa 7500
ggggttcgcg ttggccgatt cattaatgca gctggcacga caggtttccc gactggaaag 7560
cgggcagtga gcgcaacgca attaatgtga gttagctcac tcattaggca ccccaggctt 7620
tacactttat gcttccggct cgtatgttgt gtggaattgt gagcggataa caatttcaca 7680
caggaaacag ctatgaccat gattacgcca agctatttag gtgagactat agaatactca 7740
agcttgcatg cctgcaggtc gactctagag gatcccacgg ttcttctccc gggtccaccc 7800
ccggttccgg acgccgtacc ggccgacgat cctgctcggc gtcgccatcg cgatcctcgc 7860
cggcttcacc ccgctgaacg agctcgcggc gctggtgaac atcggcaccc tgttcgcctt 7920
cgtgatcgtc gcgatcagcg tgatcatcct ccgcaggacc cggcccgacc tgccccgcgc 7980
cttccgcacg ccctgggtgc ccgtgctgcc gatcgtctcg gtcgccgcgt ccctgtggct 8040
gatgctgaac ctgccggccg agacctgggt ccgcttcggc atctggatgg cggtcggcgt 8100
cgtcgtctac ttcctgtaca gccgcaaaca cagccgtctg gccgaggagc gcggcgggga 8160
acggacgtcg tcctgaggcc ggcgcctccc ggcgctaccc gtcccgccgc ggcgcccagg 8220
cgccgcccgc gggccgcacc gcgcgcgggc cggtgacgtc agcgcccagg gacgtcaccc 8280
ggcggcgcag ctcgcggtcg gccgtgacca ccaggaccgg gcggtcgccc gcctccgcca 8340
ccaggtcgac catgcggtcg tcgccgctgc ccggggccgg gtccacccgg acaccgggga 8400
ccgactccac cccgcgggcc gccccctcgg tcaccaggac gatctccacc gggcccgggt 8460
gacccggcac gccctccgcg gccagccggt cgcgcagccg ttccgcggcg ccccggcggt 8520
cccgccacca tccgtcgggc accgacccga ccacgttggc ggcgtcgacg atcacgagca 8580
ggccggtgtc atccatggcg tcagggtccc acgcggcgcc ccggccgtcc gggtcgcggg 8640
ggtgcgaccg cgcgcggggg aggacgcccg ggaacacatg aacgatcgac atatgatgtg 8700
tcgagtttca gcgttcccgc gtcggtgacg cggcgggccg caggaagggg cacctggcga 8760
tgcggagaag gtcggcgggc cccgtgggcg cctccgtcaa ggacggacgt gccgccgggg 8820
agcaccggga ggccgcgtcc ggggccgccc acggtgactg gctcacccgc ggcaaggacg 8880
gccggctgac gctgtacgtc cccaccgacg gcggtctgct gcgctggacg gagaccgccg 8940
tgggtggccc cggctggagc ggtccgcact tcgtcccggt ggccgggctg acgcacctgg 9000
cggtggctca gggagccaac ggctacgtcc acttcctcgg ccgcagggag cgcgagggcg 9060
ccgactccac gccgggcgtg gacatcgtgc acgcgatcca gtaccagacc ggactcgcct 9120
tcagcgactg gcggtccctc ggcaacccgc accgggtccc ggaggagccc ggaccgctcg 9180
ccgtgccggt cggggcggtc gcccgggacg gcaccgtgca cgtgttcgtg cggggcgcgc 9240
acggagggct gatgctgcgg cgcgaggccc cgaacggcaa gtggaaggcg tgggaggacc 9300
tgggcggcgg cggcgccggc gcccagcccg cggcgctcgc gctcaccgac gggcggatcg 9360
aggtctgcgt ggccgccgag acgggcgtgc tggcgtggag ccagtccaag cccggcggcg 9420
acttcaccgg gccccggggt ttctcgctgc gccccgtgcc gggcacggtc gcggccctgg 9480
agaccggtcc gggccgggcc acgttcttct ggacggacgc cgagagcggc ggtgcggcgg 9540
cctggcgggc gggggcgtgg cccgtcgcgc tgggcggtac cccggccgag cggccgtgcg 9600
cggtgctccg cacgtcgctg gacggctacg actgtgtcgt cctcgcctac cgtgaccagg 9660
acggcacggc cgtgctcggc atgggcggca cggagaacga ggccgccggc ttctggtggt 9720
acgcgctgac cgagtcctgc cagggcgctc cggccctggc cctggacggg cggggccgcg 9780
tggtgatggc gctgatcggc gccgacggca ggcccagggt cgcccgccag gaggacggcg 9840
acggcctctc gctcacccgg tgggacgtcc tcgggggctg agcgcgccgt cgccgtgccg 9900
gaggtcagcc gcccgtcgcg ggcgacttct tcgcggactc ggggatctcc gcgtcggagc 9960
ggatcgcctt ccacagctgg tcggcctgcg ggtgcgcggc caccacccgg ttcgggtcgg 10020
tcttgtcgta cgccacgggg agcatgacgg tctccatgga cgccgggtcg accccgttca 10080
tgctgcgcgc gaagtccgcc aggctggtca gcgaggccag ctcggagtcg gtggtcaggg 10140
ccgaggtcag ggtgtcggcg atcttgtacg tcttggtggg gctgccgaac aggtcctgct 10200
tcttcacctc gctcagcagg gcgatcatga actgctgctg gaggcctatg cggcccaggt 10260
cgctgccgtc gccgacgccg tgccgggtcc ggacgaacgc cagggagtcg gtgccgtcca 10320
gcttgtgcgt cccggcgctc aggtccaggc cgctcgtgct gtccttgatg ggctcgtcga 10380
cggtgaccgt gacgccgccg atcgcgtcga ccagcccctt gaagccggcg aagtcgatct 10440
ccatgtagtg gtccatgcgg acgccggaca tcttctccac ggtcttgacc acgcaggccg 10500
ggccggccgt cgagtacacg gagttgaaca tgacgcgctc ggcctgcgga agggcggagc 10560
cgtccgcctt ggtgcactcc gggcgggtga cgagggtgtc gcgcgggatg ctcacggcga 10620
cggcctgccg gcggccctcc gggatgtgca tgaccagcgc ggtgtccgac cgggcgcccg 10680
ccaccttgcc ggtgcccagg ccggcgttgt ctccggcacg cgagtccgag ccgaggacca 10740
gcacgttctg tccggaggtc ggcagcttct cggggcggtc ctcgccgagg gcctcgtcca 10800
ggtcgacccc gtcgatgttc ccgttcaggt cgctgtagag ccagtagccg gtgccccccg 10860
cggcgagcac gacggccagc agggacagca ggacgatgcg tccccagcgc cggcgcttcg 10920
gccggcggcg tccgccgccg gactcgggct gcgcacgccg ggactccgtg gtggcgctgt 10980
gcgtcatggt tcttgggttc ctctcctcac gggctccgcg cggcggagtg gggcccgggt 11040
ggggggagac cggtccacag atgcgttcac gtcagtggga gaactatagg cagcggtcca 11100
cggcacatcc tgcacgggtg gaattagcac ggctcagcga tgaatgccac atgaaccgac 11160
cttgaccact cttaaggctg gaataagatg tgtcgggcct gtgaccgtcg tggagggggg 11220
ctttcacgag gtcggaggtg ctggtggggc ggagcggccg gaagccgcga agcctcacat 11280
aaggggtggg cgtccggtgt ggacgatttg gtgtcccggc gggcgaaatg tcttggtagg 11340
agtacgagtt ggtggttcag ccgcgtctga gtattgtcgt gcccttccag gacgtcgagg 11400
tgtacctcgc cgagtgtctg gaatcgatcg cgcggcagtc gttccgcgac ttcgaggtca 11460
tcctggtcga cgacggctcc accgacgggt ccgtgcggat cgcggcggac ttctgcgccg 11520
ccgaccgccg tttccggctg gtccgccagc acgcccacgg accgggccac gcgcgcaaca 11580
ccggactgcg gaacacgcac cccgcggcgg agttcctcgc cttcgtggac ggtgacgacg 11640
tcatccccga gtacgcctac gaactcctgg tgcgcacgct cgaggagtcc gagtcggact 11700
tcgtctcggg caacgtgcag atgatgaact ccaccaagaa gtggcagtca ccgctgcaca 11760
agggccccat gcagaagaac cggcgcggga cgcacatcac gaagttcgac gcgctgatct 11820
acgaccgcac cgtctggaac aaactcttcc ggcgctcctt ctggaaccag aactccatca 11880
ggttccccga aggcgtgctg tacgaggact cgtgggtcaa catgtacgcc cacttccgcg 11940
ccgccaaggt cgacgtcatc acggacgtcg tctatttctg gcgccgccgg gacggcggag 12000
cggcgccctc catcacccag cgccactccg aactgtcgaa cctccgggac cgggtcgcgg 12060
ccgtgcagtc ggtgagccgc ttcctcggcg accggcgctc gcgtgagtac gcggacagca 12120
agcggaagta cgatctcgcc tgcctgaagt ccgacctcct gctccatctg aaggtgctcc 12180
cggacgcgga cgaggagtac cagcacgcct tcatgaagtg ggccaacgag ttcctcgacg 12240
agacggatct caccatcatc gacgagctgc ccgcggactc ccgcgtcaag tggctcctgg 12300
tgcgcgagga gcggctggcc gaactgctcg aggtcatcga gttcgagcgc cgcggcggtc 12360
ccatgcccgt gcagcggcgt ttccggcgct acctgaacta cccgtacctc ggggaccggg 12420
gggtgggcct cgacaagaag gcctaccggc tggacaagga gctctcgctg cacggctcgc 12480
tgtccggagc ccgctggagc accggctccg acctgctcac cctcaccgga acggcgtacg 12540
tccgcttcat caacgtgcac aagaagcaca tgtcggtgaa ggcgatcgcc ctgcggaaca 12600
agaagcaggg gcgcatgcag atcacgacgg cgaagaccgt ctacgcgccg caggcgaccg 12660
aggacagtaa gcagaatcgt tactgctatg actgggccgg cttcgaggcg cgcatcgaca 12720
ccacccgcct caagcgcaag ggccagtggg tcgagggcac ctgggacgtg gccgccggtg 12780
tcctcagccg gggactgttc cgctaccggg gcatcgaccg gggcggcgcg ggcagcgccg 12840
ccaacccgcc ctaccgctac gtcgacaaga acacccgcat cctcccggtc ttcctccagg 12900
gcaaactcaa gctgcgcgtc gagatcgtgc gctgccggat caccaagcac cgtgtcgtcg 12960
gcgaccagct ggagctgcgc ggcgtctacc tcggccccaa ggtcccggag tggggcaagc 13020
tccgcgtcac cagcatgagc ggcgcgggac gccacgacgc acgcgtccac ttcaccccgg 13080
gcggtgaggg ctggtgcacc ttctccgcca agctccccct gagccgtctg gtgcccaagt 13140
cccgcgtcca ggcgggaacc gacgcggacg tcccgcagtc ctggggcatg ggcagcaacg 13200
gctggaagac caccttccac gtcgagggcc gcaagtcggc catctatccc gtgatggcgg 13260
aggagacccc ggacgggcac tactccatgc cgtcctccct gcagaccccg gagcgcgacc 13320
gggagatcgt cgtgcaccgc aacggctccg gctatctcgt gctcttcgaa cgagcgaccc 13380
tgcccctggc gacccggtgc gactggcagg aggacggctc gctgtggatc cagggccgtt 13440
acctggccgc ggaccagctg accccggagc agtaccgctc cgcccacctc gtggtgcgct 13500
cgcgcgccca cggcgcggaa cgctccgtac cgctcacctg ggacgggcac gagttccgct 13560
gcgtcctggc ccccgccgcg atgcggaccc tggccgggga catcccgctg gcggccggac 13620
ggtgggactt cttcctgcgc cgccaggacc tgtcggccgt ggcccgcgag gaccggctcg 13680
aagacctcat ggtgaagatc gagcaggatc tcatcgaggc gttcccgcag gagtacgaga 13740
gaaacgaacg ccgctacgag acgcaggccg aggcctacga ccggctgtcg ctgctcgtcc 13800
actcggcgat gcccgaccac gcccgcggcc cctaccggca gaagctcctg aggaccaagg 13860
cctaccccga cgcccggcgc cggccggtgc gtgacgccgt gctgttcgac gccttcaagg 13920
gcacccagta ctcggacagc ccccgcgccc tgcacgagga actcgtgcgc cgccgcaccg 13980
gcctggaaca cctctgggtg gtgcgcgacg accaggtgca ggtgccgccc acggcgacgc 14040
ccgtccgcat gtggtcgccg gagtggtacg aggccctcgc caccagccgc tacgtcgtcg 14100
ccaacaacca cctcccggac tggttcaaga agcgggacgg acaggtcgtc gtgcagacct 14160
ggcacggcac gccgctgaag aagatcggcc acgacatcga gtccatccac ttcgccgacc 14220
agcgctatct ggaacgcgtc gagaaggagg tgcagaactg ggacatgctg gtgtcgccca 14280
acagcttctc caccccgatc ctcaagcgcg ccttcggctt ccccggcgag atggtggaga 14340
gcggctaccc gcgcaacgac atcctgcgcc ggccggacac cggggcccgg gagcaggaga 14400
tccgccgcag catcgggctg ccggagggca agcgggtggt gctgtacgcg ccgacctggc 14460
gcgacgacca gttctacgcg cccggcaagt acaagctgga cttccggatc gacctggccg 14520
ccgcgcgtgc gcagctcggc cccgaccacg tcctcatggt gcgccgccac cccaacgtcg 14580
tggacccggt gccgggcgcc ggcgacggat tcgtcttcga cgtgtccgac tacccggaca 14640
tggccgacct ctcgctgatc accgacgtga tgatcaccga ctactcctcc ctgatgttcg 14700
actacgtgaa caccgggcgg cccatcctgt tcttcaccta cgacctggac cactaccggg 14760
acaccctgcg cgggttctac ttcgacttcg agggcagcgc gccgggcccc ctcctctaca 14820
cgtccgagga actggtggcg gcgatccgtg acatcgacgc catccaggac ctctacgccg 14880
agcggtaccg ctggttccag cgggagttct gcgacctgga cgacggttac gccgcggccc 14940
ggctcgccga ccggatgctg gtcgcgggcg gcgacctcgc ccccgggcag gcgcacgcgc 15000
cggccgtcgg cgcggtcgac acccggcaca ccggaaggcc gatgaccccc ctccagtggg 15060
ggaactcgga gtggttcgcc ggcccccgcc cgccggcggg tctcgtcgac gccgtgcccg 15120
cccagcccgc cccggcgtac gacgccgtac cgcagcacca ggcgggtccg ttcggccata 15180
ccccgcccgc cggcgaccgc agctacgaag gcgtgatcgc gtgacgccgc cgaccccgcc 15240
cggcgcgagg tccccggcgt gccgcgcggt gccggaaggc cccggccccc gatgaccgcc 15300
gtgccgttcg ccacgcgggc gccggccgcc gctgccggtc cgtcgtccgg cggccgcccg 15360
ggcgcgtcct cagccggtgc cggtgtccac ccggatcagc agcgcgcggt ggtcggagac 15420
cccggtgtcg ctcacccggc acccgagcac gggcagcccg gtgaagaggt agtccagctt 15480
gtggtgcgag acgtgcgtcg gccgggccgg ccggagggga ccgggcgtcc cgtcgcactc 15540
ccggtgcgtg ccgtaaggct ggtcgggcca gacccgggag agcgggttgc gctctcccgg 15600
cggatccacg ttgaggtcgc cgccgtagac ggtgcgccgc tcgggcaccg cgtccaccag 15660
ggccttcagc tgtccggcgc ggaactcgcg gtccggatgc gccagatcgc cgccgcgcgg 15720
ggtcagatgc gcggtgcaca ccgtgaggtc gtgcgccgcg acgaacgcgc agagtattcc 15780
gcgctgcacc ccgaccgcgg gctggggcgc gggcaccgcg cgcacggacg acagcggata 15840
cgccgacagc agggcgtagc ccgcggagcc ccggccgggc gccccgcagc gcaccgcggt 15900
gcggcgcccg tcacggccgc gccaggtgta ggccctgaac tccgcgtgcc acgacgcccc 15960
gagggaggcg cgcaccgcct cgacgtccgc cgcgcaggtc tcctgcagga acagcacccg 16020
ggccccggat tccgcggcga ggcgttccgt ccggccgcgc ttggcgtcct cgccgcccgt 16080
gccctcgcag ttccattccc tgaccccgca catgttccag gtcgccacgg tcagtgtccg 16140
gtccccggtg gcggagcggt ccgctccatt gccgctccgc tcgtgggtca ccacggcggc 16200
gagccccgcg agggcggcgg ccgccgtgac ggcggcgagc aggcgccgtc cgcgcgggcg 16260
gggcgaccgg gttcggttcc tgagcacccg gccatcatga ccgatccgcc gctgaccagc 16320
caaaaccacc cggcgccagg cgccttccgg caggtgtcct ccccctgccc gcgcacccgc 16380
ccgtcgtcca agtacctctg agagtggagt ccgtacatgt ccaaggcacc ctcgaacggg 16440
cggcagctgc tcaacggcat cgaagcctcg ggaacgttcc cggtggagta ccggttcacc 16500
cacgccaaga gcggcaaccg gcacctcgtg gtggtcttcg ccaacttctc ggcacccgag 16560
gactacggct ggtcgaacgg cgtcttcgac aacgtccgtg ccaacatcct gtggatccgt 16620
gaccggttcg acgggatgaa cgcctactac ctgtgccgga acatggactt cggtctggcg 16680
gactcggtgc agaccctgat cgcgaacgtc accggggcgc tcgggctgac gccggaccaa 16740
gtcacgctct ggggcggctc caagggcggc agcgccgcgc tgtacttcgg cctgcggtac 16800
ggctaccgga acatcgtcgc catcgtcccg cagttcctca tcggcgacgc cctggagaag 16860
cggcacccga aggtctccgc gtacatgctc ggcgaagggg cgcaggcgca caacgcgcgg 16920
atcctggacg cgctcctgcc cgacctggtg cgcgccaagg ccaacccggg cgccaacatc 16980
tacgtgctct cctccccgca ggacgagcat tacgccgtgc aggtcgagcc gttcctcggc 17040
atgttccacg gctacgagaa cttcaatttc ctgtacagcg agtcgccgac catcacgggg 17100
cacgccacgg cgacccggcg gaacgtcccg gcgctggtcg gcctgctcaa cctgctcgcc 17160
gacggctacg ccccccggct gggcttcacc cgccacgccg ccgaggactt cgaccacgac 17220
cggtcggaca tcaacgccta cctcgcctcg acctccaagg tccagggcgc cgacgcgttc 17280
gcgccgccgg tggtgaccac cccgggcttc aacagcgagg tcccgcgcac cggaccgtgg 17340
ttcaccggga cggcccacgg agcggtgcgg gtgagcatgt ggcgcaacgg caagttcgtg 17400
gcgtcgcccc aggtcgcggc cgacggcacc tggtcctggc agccgaccgg gccgtgggag 17460
gccgggaagc acatcgtcaa gatcttcgcg gtggacccgg cgggcttcca ctccgcccgg 17520
gtcgagatcc ccttcaccgt ggtcgaccgg gatcccgtcc ctgccccgcc ggtcgtctcc 17580
gcaccggtgt ccgggcagca gaccggagcg gcggtcgggt tccacggcag cgcgccggga 17640
gcgtcacagg tcggcttccg ggagaacggc gtgctcctcg gcgcggtggc cgtcgcgccc 17700
gacggcacct ggggctggga ccccggccgg ccctggcccg aggggcagca cctggtcgag 17760
atcgtcgcgg tcgacgcgta cggcatggag tccgcgcccg ccgccgccgg cttcaccgtg 17820
ctcggccacg cggtgcccgc cggacacttc acgccgcggt actgaccgac ggcccaggac 17880
gacgacacac cacgagtggg aagcagacat gccgaaagaa gcgccgacaa cacgcgagct 17940
gatcaccggg atcgacacct ccggcgcgta tcccgtcgag taccggttca cgcacgccaa 18000
ggggggcaac cggcacctcg tcgtcgtgtt cgccaacttc gcggtcaagg acgactacgg 18060
ctggtccaac ggcgtcctca acccggtgcg ggccaacatc ctgtggatcc gtgaccggtt 18120
ccgcgacatg aacagctact acctgtgcga ggggatggac ttctccctgg agcagtccgt 18180
gatcgggctc atctccaagg tgatgaacgc cctggaactc accccggagc aggtcacgat 18240
gtggggcggc tcgaagggcg gcagcgccgc gctctacttc ggcatgcgct acggcttcgg 18300
caacatcgtc tccatcgtgc cgcagttcct cgtcggcacc tatgtgaagc gggtgcaccc 18360
caaggttgcc cggttcatgc tgggcgaggc ggtgccggag gagaacgtcc gcgcggtcga 18420
cgcgctcatc ccggacctgg cccgttcggg cgtcgcccgg cactccaaca tctatctgct 18480
ctcctcgccg caggacgagc agtaccagga gcaggtcgag cctttcctcg gactgttcca 18540
ggggtacgac aacttcaact tcgtgttcag cgagtccccc cacatcaccc gtcactcgga 18600
cgtcacccgg cgcaacgtcc ccttcctgat gggcctcgtg aacatgctcg ccgacgggat 18660
gtccccgcgg ctgggcctgg tgcgcaacgg gtacgaggag ccggaccgcg acaggtccgc 18720
catcgagggc ttcctggcgg ccacttcggc ggagcggccc agcgccatcc cgatgcccgt 18780
ggtgacgcat ccgcttccgc acatggaact gcccacggac ggcgtgtact tcacagggac 18840
ggcccccggc gcggtgcggg tgagcctgtg ggagcacggc aagttcctgg gttcgccgtc 18900
ggtggcgccg gacggcacct ggtcctggaa gcgggacaag ccgtggagca agggcgacca 18960
tctggtcaag gccgtcggct gggacgcgga gaagcgccgc accaagggca ccgtggtccc 19020
gttcaccacg gtcgccggcg cgaacgccgc cgcgcccggg gcaccggccg ccgcgcccct 19080
ggcgcccggg cagccgctgc cggcgccgac ggtccacacg ccgggggcgt acgagcagat 19140
caccggcacg gccgtgcgct tcagcggctt cgccccgggc gccgcccagg tgggattcag 19200
ggcggggggc accctccttg gcacgagccg ggtcgcggcc gacggaacgt gggcctggga 19260
ctccggctgg ccctggcagg cgggcatgca caccgtggag gtgttcgccg tggacgccgc 19320
gggatccgag tcgcccgtgg cgccggtgcc cttcgacgtc atgcacgcca cggcgggcgc 19380
ctcgccgttc gcctactgac cggtcgccac gcgcggaagg gctcctggag atcgccggcg 19440
atctccagga gcccttccgc ggtctacggg gactaggcgg gcacgctcgc cgtgcccggc 19500
tccaggaacc gcttcccgtt cacccgctcg gagacgccct cgcggtccag gtacggcgtg 19560
atgccgccca ggtggaaggg ccagccggcg ccggtgatca ggcacaggtc gatgtcctgg 19620
gcctcggcga cgacgccctc gtcgagcatg agcccgatct cctgggccac cgcgtccagg 19680
acgcgggcgc ggacctgctc ctcggtgagg acggtgtcgc cctgcttcag gagcgcggcg 19740
acctccgggt ccagctcggg cttgccgctg tcgtagaggt agaagccacg cttgccggcc 19800
tcgacgaccg ccctgaggtt cggggagacc gtgaagcgct ccgggaacgc cctgttgagg 19860
gtctcggaca cgtgcagacc gatcgcgggg ccgaccagct ccagcagcac cagcggggac 19920
atcggcaggc cgagcggctc gacggccttc tccgcgacct cgaccggggt gccctcgtcg 19980
atgacgttct ggatctcgcc catgaagcgg gtcaggatgc ggttcacgac gaacgccggg 20040
gcgtccttga ccaggaccgc ggtcttcttc agcttcttgg cgacaccgaa cgccgtggcc 20100
agcgccgcgt cgtcggtccg ctcgccgcgg acgatctcca ggagcggcag gatcgcgacc 20160
gggttgaaga agtggaagcc gacgacccgc tcggggtgct tcagcttcga cgccatctcg 20220
gagacggaga gcgaggaggt gttggtggcg aggatcgcgt gcgccggggc gaccgcctcg 20280
acctccgcga acacctgctg cttgacgccc atctcctcga agacggcctc gatcacgaag 20340
tcggcgtccg cgaagccctc ggccttgtcc agcacaccgg tgaccagggc cttgaggcgg 20400
ttggccttgt cctggttgat ccggcccttg ccgagcagct tgtcgatctc ggcgtggacg 20460
tagcccacac ccttgtcgat gcgcgcctgg tcgatgtcgg tcagcacgac cggcacctcg 20520
aggcggcgca ggaacagcag cgcgagctgg gaggccatca gaccggcgcc gaccacgccc 20580
accttggtga ccgggcgggc cagcgacttg tccggggcgc cggccggccg cttgccgcgc 20640
ttctgcacca ggttgaacgc gtagatgccg gagcgcagtt caccgcccat gatcaggtcg 20700
gcgagcgcct ggtcctcggc gtcgtagccc tgctggaggt cgccgttctt ggcggcggcg 20760
atgatgtcca gggcgcggta ggcggccggg gcggcgccgt gcaccttgga gtcggcgatg 20820
aagcggccct tggcgacggc ctggtcccag gcctcgccgc ggtcgatcac cgggcgctcg 20880
atccggatct cgtccttgag gacggccgcg gtccagatca gcgactgctc caggaagtcc 20940
gcgccctcga agatcgcgtc cgcgatgccg agttcgaaga cctgcgcgcc cttgagctgc 21000
ttgttctggt tgaggctgtt ctcgatgatg accgagacgg ccttctcggc gccgatcagg 21060
ttcggcagca gcgtgcagcc gccccagccg gggacgagac cgaggaagac ctcggggagc 21120
gagaacgccg ggagggcggc cgacaccgtg cggtaggtgc agtgcagacc gacctcgacg 21180
ccaccgccca tcgccgcgcc gttgtagtac gcgaaggtcg gcacggccag cgtcgacagc 21240
cgcttgaaga cgtcgtggcc gcccttgccg atggccagcg cgtcctcgtg ccgcttcagc 21300
agctcgacgc ccttgaggtc ggcgccgacg gcgaagatga acggcttgcc ggtgacgccg 21360
acgccgacga tctcgccgtc cgcggcctcc ttctcgaccc ggtcgatcgc ggcgtcgatg 21420
ttcgccagcg actgcgggcc gagcgtggtc ggcttggtgt ggtcgtggcc gttgtccagg 21480
gtgatgagcg cgaagcgccc ggcgcccagg gggaggtcga agtggcgcac gtgcgcgctg 21540
gtgacgacct cgccggggaa cagctcggcc gcacccttca gaagctctgc ggtggtgctc 21600
acttgtcccc ctcgaagtgc gggttctccc agatgaccgt cgcgcccatg ccgaagccga 21660
cgcacatggt ggtgaggccg taacggacct gcggctgctc ctcgaactgg cgggccagct 21720
gcgtcatcag acggacgccg gaggaggcca gcgggtggcc gaacgcgatg gcgccgccgt 21780
actggttgac gcgcgcgtcg tcgtcggcga tgccgtagtg ctccaggaag gccagcacct 21840
ggacggcgaa ggcctcgttg atctcgaaca gaccgatgtc ggagatggac agccccgcct 21900
gggcgagggc cttctccgtg gccgggatcg ggccgtagcc catgacctcg ggctccacgc 21960
cggcgaagga gtaggagacc aggcgcatct tgaccgggag gccgttctcg cgggcgaagt 22020
cctcgctcgc gatgaccgag gcggtggcgc cgtcgttcag accggccgcg ttgccggcgg 22080
tgacccggcc gtggacgcgg aagggggtct tcaggccggc caggttctcc agggtggtgc 22140
ccgggcgcat cggctcgtcg gcggtgacca ggccccagcc ggtctcaccg gcctcctcgt 22200
tggtgcggcg caccgagacc gggaccaggt cggcctggat cttgccgttg gcgtaggcct 22260
tggcggcctt ctcctgggag cgcacggcgt actcgtcggc gcgctgcttg gtgatcgagg 22320
ggtagcggtc gtgcaggttc tccgcggtca tgcccatgaa cagggcggac tcgtcgacca 22380
gcttctcgga gacgaagcgc gggttggggt cgacgccctc gcccatcggg tggcggccca 22440
tgtgctcgac gccgcccgcg atggcgacgt cgtacgcgcc gaaggcgacg gagccggcga 22500
ccgtggtgac ggcggtcagg gcgccggcgc acatgcggtc gatggagtag cccgggaccg 22560
aggtgggcag gcccgcgagg atgccggccg tgcggccgat ggtcaggccc tggtcgccga 22620
tctgcgtggt cgcggcgacg gcgacctcgt cgatcttctt cgggtcgaga ccggggttgc 22680
ggcgcagcag ctcccggatc gccttcacga ccaggtcgtc ggcgcgggtc tcgtggtaga 22740
tgcccttcgg gcccgccttg ccgaacgggg tacggacgcc gtcgacgaag acgacgtccc 22800
tgacggtacg aggcacgatg gctctcctcc cagggtgcgg gacgctgagc gcttgcttac 22860
gccatgctac ttatgagtaa cgtgactgcc cagtcccggc cccccgagcg gcgaacatca 22920
cacgtacggc ggcgcccgcc aaacgccgga ggggctggaa tcagccccct ccggcgtttg 22980
aggagcggga acccctcacg gtggagccgt ggagcctcgc ggggtcagga ccggggtcgg 23040
taccgggtgc gaccccggct gctcctcccc ggtgatgacg ccgaacagcg tgcgcgccac 23100
ctccgccccg aacccgtgca catcgtgact catcgcggac agcgtcggat gcgtcagccg 23160
gcacagctgc gagtcgtccc acgccagcag cgacacgtcg tccggcaccc gcagccccat 23220
ctccgccgcc accgacagcc ccgccaccgc catgatgtcg ttgtcgtaca cgatcgccgt 23280
gggccgttcc cccggcgcgg ccgccagcag cgaacgcgtc gcccgcgccc ccgcgtcccc 23340
cgagaagtcc gtggccgtct gccacgcccg cgcgggcggc tccagcgccc ggaccgcctc 23400
gtcgaacgcc gccgtgcgga tcgaggtgtg cccgagcgcc gccgcgccgc ccacccgggc 23460
gatccgccgg tgcccgagcg ccgccagata ccgcacggcc tccgtcacgg ccgtggcgtc 23520
gtccgtccac acggaggtga gcccgcccgt cagcgccggg tgcccgacgg ccaccgccgg 23580
cagcccgagc cgctccgcca ccgccggacg ggggtcgccg gcccggaagt ccaccaggat 23640
cgagccgccg atctgccgcc cgcgccacca cgactccatc agcccgacct cctcctccgg 23700
gctgcgcacc agccgcagca gcagcgagca ggaccgctcc accaggacgc tctccacccc 23760
ggagatgaac tccatgtaga acggctccag gccgagcagc cgggcgggcc ggcagaccgc 23820
gagacccacc acgtccaccc gcgacccggc cagcgtgcgc gcggttcggc tcggcgccca 23880
ccccagctcc cgcgccgccc ggaagatgcg gtcccgggtc gcctccgaca gcccgggctt 23940
ccggttgaag gcgagggaca cggcgccctt ggacacgccg gcgcgcgcgg cgacgtccct 24000
gatggtgacg cgaggggtcg gcgttgccgt catcgagtgg gctccacgca gtacagggcg 24060
gaacgggcgg tgtccgggtc cggagtcttc cacccccgca ccccgatggt cacctgttcc 24120
ccggggagca gggtcaccag cccccggtcg gcccgcgccc cggggtccag ccggtcggcc 24180
tggagcagca ggtcccgtac gagggtgcgg gccgtgaccg tgatcccgtc cggcgcgagg 24240
gcgacctcga actccggcgg ggggtagggg atctcccggt ccggcgccgg gaagtgccac 24300
gcccgcaccc cgtccgcgtc ggcgaccagg aactccccgg ggccgtccgg cagcagttcg 24360
accgggacct cgaccacggc caccgtccgc cccccggcgt ccagcgccgg ggccgcctcc 24420
gcgatcgggg cgccgtcgac ggacatccgg cgcagccgca gcgttccccg ccagtcctcc 24480
gcggactggt tgaccgccgc caccaccaga ccgtcaccgt ccgcgcgcac ggtcagcagc 24540
cggtccgcgt acagccggcg cagctcgtgg tagagcggct tctcccgccc gtccccgtcg 24600
atcgcggccc acgacgtcac cggccagcag tcgttgagct gccagaccac cgtgcccgcg 24660
cacaccggcc agtgcgagcg ccagtgctcg acaccggccg ccaccgcacg cgcctggttg 24720
acctgcgtca gatagtgcca gcggtcgaag tcgccctccg gcacggcgaa gtggcgggcg 24780
aggccgcgct ccagcttgcc gttgccgtcc tccgccttct ggtggtgcag catgccgggg 24840
gagtccggcg cggggtcctc cccgggcagc gcccgccgca gcgtggcgtg cgcgggaggc 24900
gcctgccagc cgaactcggc cacgaagcgc gggacgtcgc gccggtagtc ggcgtagtcg 24960
gcgcggttcc acacctccca ggagtggtgg gtgccgtgcg ccggatcgtt ggggtggtgc 25020
cgccaggaac cggaccaggg actgcccgcc gtgtacggcc gcgtcgggtc cagctccgcg 25080
accacccgcg gcaggacgcc gaggtagtag ccctcgcccc aggagtcccc ggcgagcccc 25140
tgctcccagt cccagtcccg gaacccccac aggttctcgt tgttgccgtt ccacagcacc 25200
agggaggggt gcggcatcag ccgtacgacg ttctcccggg cctccgcctc cacctccccg 25260
cgcagcggct gctcctcggg gtaggcggcg cacgcgaacg ggaagtcctg ccagaccagc 25320
agccccaact cgtcgcaggc gtcgtagaag tcctcgtcct cgtagatccc gccgccccag 25380
acccggacca ggtccacccc cgcgccggcc gcctgctcca gccggtgccg gtagcgctcc 25440
cgggtgatcc gggacgggaa cacgtcgtcc gggatccagt tgacgccccg cgcgaacagc 25500
cgctcaccgt tgacgaccag ggtgaacccg gtgccgtgcg cgtcggccga ggtgtccagc 25560
tcaaccgtcc ggaacccggt cctgcgccgc caggcgtcca gcgcctcgtc accgtgggac 25620
aacgtcagct cgacgtcgta cagcggctgt tcgccgtatc cgcgcggcca ccacaggcgg 25680
acgtccggca cccggagccg cacggtcccg gccgtcccat cgacccgcgc ccgggcgcgc 25740
acgcccccgg cgctcgcctc cagggtgagc ggtgcctcga cccgggagcg ctccacgtcg 25800
accgccagct cgatctgccc caccccgtcc tcgacggtga ccagcgggcg cacccgggcg 25860
atccgcgccg tcgaccagcg ctccagccgc accggccgcc agatcccggc cgtcaccagc 25920
gtcggccccc agtcccagcc gaacgagcag gccatcttcc gcaggtactg gtacggctcg 25980
gcgtacgctc cggggcgctc gcccagcctg ccgcgcaccg cctccgcctc ggcgtacgcg 26040
gaggcgaacc gcaccgtgag ccggccgctc agtcccgtca cgtcgaagcg gtacgagcgg 26100
tgcatgttcc gcgtccggcc cagtggccgg ccgtcgagca ggatctcggc gacggtgtcg 26160
agaccgtcga agacgaggtc cgtctgctcg tgcgggcccg tcccggcggt cagctccgtc 26220
tcgtacgtcc actcccgccg gcccacccag gccacctcgg tctcgttgcg gccgaggaac 26280
ggatcgggga tcagcccggc cgccagcaga tcggtgtgca cacaccccgg caccgaggcg 26340
gggagggcgt cccccgtgcc gtccgggtgt cgcaggatcc atccctcggt gagcggtgtg 26400
acctgacgca tgcacactcc ctaaaccggt tgagccttct ctgaagagtg gtctggcatc 26460
gttggcgcga ttgcgacttt accggttcag ttcagggctg ccagagtgcc gaatcagcca 26520
tcccactcgt gctcgtccgt cccgtgaacg gagccgtgat gcatctgaac cgccgtacga 26580
cactcaccgg atcgctcgcc ctgctcgccc tcctggcctc cgcctgcacg ggcacggggg 26640
gttcctcgaa gggcgcggac gccaaggctc ccgacgaccc gtcaaaggtc aaggggtccc 26700
tcacggtcct cacccaccgg accgatctgg tgcaggacgg gacgatgaag aagtacgccg 26760
ccgagttcaa cgagacctat cccggggtga aggtggagtt cgacggcctc accgactacg 26820
agggcgaggt caagatccgt atgaacacgg agaactacgg cgacgtcctc atgatcccgg 26880
cggtcgtcga gaagaaggac tacccgaagt tcttcgcctc cctgggcacc aaggccgaac 26940
gcgccgccaa gtaccggttc accgactact ccaccgtcga cggcaaggtc tacgggcaga 27000
gccccgtcgg cgtcgtcccc gggttcatct acaacaagcg ggtgtggagc gaggccggcg 27060
tcaccgactg gcccaccacc cccgccgagt tcctggacga cctgaaggcg atccggtcga 27120
agaccgacgc ggtgccgtac tacaccaact tcaaggacat gtggccgctg acccagtgga 27180
ccaacgtcaa cggctccgtc ggctgcgacc cgcacgccac cacgaagctc gccgagggcg 27240
acccgtgggc cgagggggcc gacctgcgcg tgggcgacac cctgctccac gacatcgtgc 27300
gcggcggact cgccgagaag gacccgacca ccaccaactg ggagggctcc aagcccaagc 27360
tggccaaggg cgagatcgcc accatgtggc tgggctcctg ggccgtcgtg cagatgcggg 27420
acgcggcgaa gcaggccggc gccgaccccg ccgacatcgg cttcatgccc ttccccgcac 27480
agcgggacgg cacgttctgc gcggtgacct ccccggacta ccagcaggcg gtcaacgtca 27540
actccgacaa caaggaggcc gcccgcgcct ggatcgactg gttcaccgac aagtccggct 27600
acgccgaggc caacctcgcc ctatcccccc tgaaggacgc cccgctgccc gccgtcctcg 27660
agccctacga gaaggccggc gtgaagctcc tggacctcga ggacagcaag ggcgccgagg 27720
tgaagtccct cgacaaccgc tccgaggtcg gcatctacaa gcccgactac cgccaggaac 27780
tcgtcgacct cgcccgcggc gcccgcaagg gcggcctgga cgactacctc ggcggcctcg 27840
gcgagcgctg ggccgaggcg cgcagcgcgc tgggggcctg atgacggaca ccacccgcaa 27900
ggcggcgcgg ccggttcccc cggccgcgcc cgccgggccg ggccgcgcgg cgccggcccc 27960
gcgccgcacc cggctgtcgc gccgcctcac cccgtggctg ttcctggccg caccgctggc 28020
cctgctcctg accttcacct acgcgcccga tcgccaacat ggtcgcgtac agcttcaccg 28080
actgggacgg cgtgagcccg gagctgaact ggacgggcac cgggaactac accgaactcc 28140
tcacccgctc cgagctgttc gaggtcttct tcgtcagcgg ctactacctc gtcgcctccg 28200
cggtgcagat cgtgctcgcc ctctacttcg ccacggtcct cagcttcgac gtccgcttcc 28260
ggaacttctt caagggcgtg ctgttcttcc cgtacctcat caacggggtg gccatcggct 28320
tcgtcttcct ctacttcttc caggacggcg gcaccctcga ctccgtactg ggcctgctcg 28380
gcgtcgagac cgaccacgcc tggctgggca cgccgttctc cgcgaacacc tcgctggccg 28440
gcgtctccgt ctggcgctac ctcggactga acttcgtcct cttcctcggc gcgatccagt 28500
ccatcccggg cgagctgtac gaggcggccg agatcgacgg cgcgaaccgc tggcagcagt 28560
tccggcacat catcgcgccc ggcatcagac ccgtgctgag cctgagcgtg atcctctcgg 28620
tctccggctc gctgtcggtc ttcgagatcc cgtacatcat gaccggcggc gccaccggca 28680
cggagacctt cgtgatccag accgtgaagc tggcgttcca gttcaacaag acgggactcg 28740
cctcggccgc cgccgtcgtc ctgctgctga tcgtcctggc ggtcacctgg gtgcagcggc 28800
gcatcgtccc cgacgagaag gtggacctcg tatgacccgc cgtaccgcgg cacgcgccct 28860
ggtcctgacg tccctgatcc tggcgacgct ggtggtgctg ctgccgctcg ccgtggtctt 28920
cctgacctcg ctgaagtcct ccgaggagat ggcgaacggc agcggagcgc tgacgccgcc 28980
cgacgacccg ctgaacttcg gcaactacgt gacggcgttc cgggacggcc agatgctgtc 29040
cgcgttcggg aacacggccg tcatcctggt cgtggccgtc ggcggaacga tcctgatcgg 29100
ctcgatgacg gcgtacgcga tcgaccgctt ccggttccgc ttcaagaagc tggtcgtggc 29160
gctgttcctg ctggccgcgc tggtccccgg ggtgaccacc caggtggcga ccttccagat 29220
cgtcaacagc ttcggcatgt tcgacagcct gtgggcgccg atcgccctct acatgggcac 29280
ggacatcgtc tcgatctacg tcttcctgca gttcatccgc tccatccccg tctccctgga 29340
cgaggcggcg cgcctggacg gcgccaacgc gttcaccgtc taccgcaagg tgatcttccc 29400
gctgctcaag ccggcgatcg cgacggtggt gatcgtaaag gggatcaacg tctacaacga 29460
cttctacatc cccttcctct acatgccctc cgaggacctg ggggtcatct cgacgtccct 29520
gttccgcttc aagggcccct tcggcgcgca ctgggagacg atctcggcgg gcgcggtcct 29580
ggtcatcctg cccaccttga tcgtcttcct gttcctccag cgcttcatct acaacgggtt 29640
catgcggggg gcgacgaagt agccagcgcg gccaccagca cgggagtgac ctggtcgacc 29700
tgccacgcgc gtgccccgtg cgccgtcagc gccgcggcga cggaccgctc gtcgggcccc 29760
ggcggctccc agcagaccct gcgcaccgtg tccggggtga tcaggttctc cggcggcatg 29820
ttcagccgct cggcgagttc ggcgaccccc gcgcgggccg ccgacagccg ggccgcggca 29880
acggggtcct tgtccgccca ggcgcgcggc ggcggagggc cggtcaccgg ctggccgggc 29940
tgcggcagct gggcctcgct cagcgccttc gcgcggtcga cggccgcctg ccactgctcc 30000
agctggcgcc gccccacccg ctgcccgaac ccgttgagcg cggccatggc gtgcaggttg 30060
gcgggcagcg cgagcgcggc ctccacgatc gccgcgtcgg aaagcacctt gccgggggag 30120
acgtcacggc gccgggcgat ccggtcgcgg gtctcccaca gctcccgcac caccgccatc 30180
tggcggcgcc ggcgcacctt gtgcatgccg gaggtgcggc gccaggggtc cttgcggggc 30240
tccggcggcg gggccgaggc gatcgcgtcg aactcctgcc gggcccagtc cagcttgccc 30300
tggcggtcca gctccttctc cagggcgtcc cgcagatcga ccagcagttc gacgtcgagg 30360
gcggcgtacc gcagccaggg ctcgggcagc ggacgggtgg accagtcgac ggcggagtgg 30420
cccttctcca ggacgaagcc gagcacgttc tcgaccatcg cgccgagccc gacgcggggg 30480
aacccggcaa ggcggccggc cagctcggtg tcgaagaggc gggagggcac catgcctatc 30540
tcgcgcagac agggcaggtc ctgggtggcg gcgtgcagca cccactcgac gccggacagc 30600
gcctcgccga gggcggacag gtcggggcag gccacggggt cgatcagcgc ggtacccgca 30660
ccctcgcggc gcagctggac gaggtaggcg cgctggccgt agcggtaccc ggaggcgcgc 30720
tcggcgtcca cggcgacggg tccgctgccg gccgcgaagg cggcgaccgc ctcggcgagg 30780
gcggcctcgt ccgctatcac gggcggaatg ccctcgcggg gttccagcaa gggggtcggc 30840
gcctccgtaa cagaagatcc gccgtcgtcc ggaggagcgc ctccggtggt gcgcagtgaa 30900
ctgtctgctg cggtgtcgtg ggcgtcggtc acctgtcaag ggtatccgtg ccgcgaaggc 30960
gcccgtcgac ggttgtgctc cgtgacgggc gccggtgggt cgtattccgg tcagaagagt 31020
gaaagaacgt gttcgcttgg ccgtgggcgg gcggatcggg gacgggcgga tcgggggcgg 31080
gacggaaggg tcagtggatg atgccggtgc gcagggccac ggccaccatg ccggcgcggt 31140
cgcccgtgcc gagcttgcgg gcgatccggg ccaggtggct cttgacggtc agtgcggaca 31200
ggcccatcga gacgccgatc gccttgttcg actggccttc cgccaccagc cgcagcacct 31260
ccacctcgcg gccggacagc tcgcggtagc cgcccgggtg gctcggggca cccggggggc 31320
ggcggtgcag gcgcgcggcg gcggcgccga tgggagcggc acccggccgg gtggggagcc 31380
cgaggttggt gcgggtgccg gtgacgacgt agcccttgac tccgcccgcg agggcgttgc 31440
gcacggcgcc gatgtcgtcc gccgcggaga gggcgaggcc gttgggccag cccgcggcgc 31500
gggtctcgga gaggagggtg aggccggaac catccggcag atggacttcg gcgacgcaga 31560
tgtcgcgggg gttgccgatg cggggacgag cctccgcgac ggacgaggcc tcgatgacgt 31620
cgcgtacgcc gagcgcccac aggtggcggg tgacggtcga acggacgcgg gggtcggcca 31680
cgaccaccat ggcggtcggc ttgttcgggc ggtaggcgac caggcttgcg ggctgctcga 31740
ggagaacgga caccaggcct cctggggtgc gggacgggcc ggctcgtggg ggtgaaggcg 31800
ggacgaaccg tgctttcaag gtcacagtcg tcttcggcag caaacctggt gtcctttaac 31860
gaatgatcac gaagtgatga gtaacaatcc gggcaattcg gacgcacgat cgatcattcg 31920
aagatcgaac ggtttcggtc tgcgtcgcaa cgcttccgaa agtggccgta tcgacaaaga 31980
gagatgcagg aggccggtcg tcgggacccc gcagcgggag gctcagcgcg actgcggccc 32040
cctccgctgc ggcagcgtca ccacggacgc gtcccccgga gcggccggcg gcagccccgc 32100
gacctgcgcc agcagatcgg accacgcgac cagatgggcg gccgtgtccg gaaccccgcc 32160
cagaccctca cgcggcgtcc acgaggcacg gatctcgatc tgggaggcgg cgggccgcgc 32220
ggacagcccg ccgaagtagt gcgaactcgc ccgcgtcacc gtgccgctcg gctcgccgta 32280
cgacaggccg cgcgccgcca gcgcgccggt cagccaggac cagcacacgt ccggcagcag 32340
cggatccgcc gccatctccg gctccagctc ggcgcgcacc agcgtcacca gacggaaggt 32400
cccccgccag gcgtcgtgtc cggccgggtc gcacagcagc accagccggc cgtcggccag 32460
atcctcctcg ccgtcgacga ccgccgcctc cagcgcgtgc gcgtacgggg cgagccgttt 32520
cggcgcgggc accgtctcca cctcgatctg cggccgcagc cgggcgctct gcagcgcctc 32580
gacagcggcc cggaagggcg gcggaggcgc acctccgtgc ccgggatccc ccccgccctc 32640
cttcggttcg tccattccgc cagcgccgtc cgacagtcgt ccctgagccg cagccatgcc 32700
gggaagatta agcggaacgg gcccccggcg cagggaggga cacccgcgcc gcccggcgct 32760
gtccggatcc tgcaccgcgg ccccgcccgc cggacgcccc tggggtccgg ggcggcggac 32820
gggtcgtgcg agactggccg gtgtgagtgc caacacgagc ccgaagggcc agacgcctac 32880
cgcgaccccc gaccccgtca agaacgacgc cgtccgggaa tcagccttcc tcaaggcgtg 32940
ccggcgcgag ccggtgccgc acacgccggt gtggttcatg cggcaggccg ggcgctcact 33000
gccggagtac cgcaaggtgc gcgagggcat cgggatgctc gactcctgca tgcggcccga 33060
gctggtcacc gagatcaccc tccagccggt gcgccgccac cacgtcgacg cggcgatcta 33120
cttcagcgac atcgtcgtcc cgctcaaggc catcggcatc gacctcgaca tcaagcccgg 33180
catcggcccg gtcgtcgagc agccggtgcg cacccgcgcc gacctcgccc ggctgcgcga 33240
cctgaccccg gaggacgtct cctacgtcac cgaggccatc ggcatgctga cccgtgagct 33300
cgggtccacc ccgctgatcg gtttcgcggg cgccccgttc acccttgcga gttacctcgt 33360
cgagggcggc ccgtcccgta cgtacgagaa cgccaaggcg atgatgtacg gcgaccccga 33420
gctctgggcc gacctgctcg accgcctcgc cgacatcacg gcggccttcc tcgacgtcca 33480
gatccgggcc ggcgcctcgg ccgtgcagct cttcgactcc tgggccggcg cgctcgcccc 33540
ctccgactac cggcgttcgg tgctgcccgc ctcggcgaag gtgttccgcg cggtggccgg 33600
ccacggcgtc ccgcgcatcc acttcggcgt cggcaccggc gagctgctgg ggctcatggg 33660
cgaggccggc gcggacatcg tcggcgtcga ctggcgcgtc ccgatggacg aggccgcccg 33720
ccgcgtcggc cccggcaagg cgctccaggg caacctggac ccgaccgtgc tgttcgccgg 33780
ccgggaggcc gtcgagacga aggcgcgcga ggtcctggac accgccgcgg gcctggaggg 33840
ccacatcttc aacctcggtc acggagtgat gccctccacc gacccggacg ccctcacccg 33900
tctcgtggag tacgtccaca cgcagacggc gcgctgaccc accgctcacg cgccggacgc 33960
gagtcggaat ccgggggcgg ggtactgggc acgggtgccc accacgttca cccccgggta 34020
cgggcaggtg gaggccccat gaggctcgag atgttcgacc ccgccccgat cggcgtcgtg 34080
ttcacccagg ggccggagca ccggctcgcg tacaccaacg ccgtctaccg ggagaccttc 34140
ggcgaccgcc cgctggggcg gacgatccgc gaggccttcc ccgacctcgc gcagtccggc 34200
tacttcgaca tcttcgaccg ggtcctcacc acgggcgcgg ccgaggtggt caccgcggtg 34260
cccctcgacc tgatctaccc cggctccacg ggcgagggca ggcgctactt cacgttcagc 34320
atctcccgcg ccacgatgag cgacggccgg ccgggagtgc tcggcgtgat cgtggaggtg 34380
accgcgcagg tgaccgccgc ggaacggatc cgtgtgctgg ccgaggagcg ccgccgcgcg 34440
ctgcagcgct accgcagcct ggtgaacgcc ggaacgcaga tggtgtgggt ggcggacgcc 34500
aagggccgga tcaccgagcc gagccccggc tgggaacgcg tgaccgggca gacctgggag 34560
gagttccgcg gcgagggctg gatgaacgcc gtccaccccg acgaccgcgc cgcctcggtc 34620
gaggcgtggc ggcgggcgac gaccgaacag gtgccgcgct ggatccacac ctaccggctg 34680
cggctggccg ccggcgggta ccggcacttc gtcgtcgacg ccgcgcccgt gcgcgacggg 34740
aacacggtga tcgaatgggt gggcacctgc acggacatcg agcgggaatg gcaggagggc 34800
cgccgtacgg aactgctggc gcgggccgcc accgccacgt ccggcatcgc gcggctggac 34860
gagatgctcg ccgccctggc cgatgtgatc gtgcccgaca tcgccgacaa ctgcaccatc 34920
cacctcctgc cgcaggccct gcaccgtctg ccgggcaccc cgctgaccac cgaacgcgtc 34980
gccgcggtca cccgcccggg gctcccggac ctgcccccgc accacgagga gcacctgcgg 35040
cccggcagcc cgctggcccg cgccgccgac cgccgcagcc cgctccactt cgtcttcccg 35100
cccggcgagc cgccggccga cctcgctccg ctcgacggcg agccctggat ggccgaggac 35160
gtcaacagcg tcgtgctgct gcccgtcgtc gtcgacggca ccaccgccgc cctggtcgcc 35220
gtctccacca gcggcgcccg cccgcccctc ggccaggcgg agatcggcct gctgcagaca 35280
ctcctggaac gcgcccacac ccccctcagc aacgccctgg agtaccagcg cacccggcag 35340
gtggccctgg ccctgcagaa cagcctgctc accgacccgc cggacgcgcc cggcctggac 35400
atcgccgtcc gctaccggcc cagcaccgcc gccgccgagg tcggcgggga ctggtacgac 35460
gcgttcgtgc tgcgcgacgg cgccaccgtc ctcaccatcg gcgacgtctc cggccacgac 35520
ctgccggccg ccgtcaccat gagccagctg cgcaacatgc tgcgcgggct cacgctggac 35580
cgccaggaac cgaccggcac catcctgcgc cggctggaca tcgccgtgca gaccctctat 35640
acggagtgca ccgccacctg cgtgctggcc cgggtggaac gcccggactc cggcggcgtc 35700
cggctgcact actccgtcgc cggtcacccg ccgccgctgc tcgtcgaggc ggacggctcc 35760
gcgcgcttcc tgaccggggc gcggtccccg atgctcgggc tcgtccccgc gccggagtac 35820
tcgagcgcca tggaaccgct gccgcccggc tccaccctgc tgctgtacac cgacgggctg 35880
gtggagcgcc gcgacgagga tctcaccgtg ggcctggagc ggctgcggca ccacgcctcg 35940
gaggcggtca gccgcccgct gcaggacttc tgcgacacac tgctcaccgg ccagctcacc 36000
gtcgacaacg acgacgacgt ggcgatgctg gtcctgcgcc ggtaggagcg tgccgaggag 36060
cgccactctg gccgatttta cccttgcttt tccatcggga ttcgttctcc ggatttcccg 36120
atccggcgcc gacggcgaga ccgttgggat caccaatacc ccggaattcc cgcctccgcc 36180
accgttgggc agcgacggat cctgtgatat ttcgactacg cgcggtgatg aattggctcg 36240
gtgccggtcg cgcccggctg tagcagttct ggagcgcgtc tggacatcgt cacgagcgct 36300
tgtgattctt ggtcctgtac acgcaagccg gcgcaacgtc cacgttgccc atcagcggtt 36360
atcggcggtc caccggcgcg acggtgaccg cgggcgggta ctcatagggg gaactgcaat 36420
gaattactca aaagcagcga gaggaatgcc gacagccgga caaggtgccg ttcgggcggc 36480
gcgcgtcgtc cgtgaaagtc cggcggaatc agaaacggtc acagttcaga tagcgtcgtt 36540
attaccgggt gagtcgctgc gctcgaaagg gatcgagcag aaccacgtcg cggcactcgc 36600
ggaggtagac gcgccgcttc cgcccatact ggtggaccgg aagacgatgc gggtcgtcga 36660
cgggatgcac cggctcctcg cggctctgct caacggacgg cagacgatcg aggccgaact 36720
gttcgacgga accgcggatg agggattcct gcgcgccgtc cgggagaacg tggtgcacgg 36780
actcccgctg tcgcaggcgg accgccgggc cgccgctgcg cgcatcatcg tgtcccaccc 36840
gcatctgtcg gacagggcga tcgcccgggc gtccgggctc ggggcgaaga ccgtcgcggc 36900
cgtgcggcgc agttcaactg ccgtcgtgcc gcagttgaac acccgggtgg gccaggacgg 36960
cagggtccgg ccgctgaacg ggggcgaggg gcggcgcagg gccatggcgg tactggccga 37020
acaccccgac gcgtccctgc gcgaggtcgc ccgtctgtcc ggggtgtcgc ccgcgacggt 37080
cagcgacgta cgccggcggc tggccgccgg cgagtcgccc ctgccgtcga gacgggaacc 37140
ggccgaaccg cggacgggcg ccgactccca ccgcaaccag agcttcgtgg atcccgtccc 37200
ggtgctggag aagctgctgc gcgacccctc tctgcggcac aaggagggcg gccgccagct 37260
gctccagctg ctccgccaga acgcggtcgg cgtgcaggac ctgatggagc tgtccgacgc 37320
cgtgccgtcc cactgcaggt ccctggtgat ccatctcgcg cagcagtacc gggacgcctg 37380
gcagtccttc gcggagaagc tggacgagcc cgcctgcgcc tgtcccgggt gacgaacggg 37440
cggcacggac ccgttcaccg gacatgaccg gcgccgcgcc gcgttcacgg cgcgccgccg 37500
gcactcccac ggcacccgga ccaccgccgc gtatccggcg gacccgggcc cgggcgggcc 37560
ggattcagcg ggcgggggcc caggtgccac ccgattccag ccaccgggag agctccgccg 37620
ccgagtcctt gcgcacgacc agttcgacga ggccgcgggt ctggtcctga ccgtgctcga 37680
tgcggacgtc ctcgatgttg acgcccaagt cgccgatcga cgtgaacagt tcggccaggg 37740
cgccgggctt gtcggagatg gtcaccgaga cggtcgcgag ctccgtccgg cgcgtaccgg 37800
gtttgcgcac gatcctggcg cacccccggt tcccctcccg caacagctcc tcgagctcct 37860
cctgcgcgcg gcggcggacc agcgggtcgg cgtcggagac ggcgcgcagc gcgccgacgg 37920
cccggcccag gccggcggcg agggagtcga gaacgtccgc cacggccgtg gcgttggaac 37980
gcaggatgtc cccccagagc cgggcgtcac cggccgcgat ccgggtgacg tcggcgacgc 38040
cctgccccgc cagccggacg ctgtcctccg ccgcgtgctc cagccgcgcg gcgagcaggg 38100
aggagagccg atggggcgcg tgcgagacga gggccaccgc gtggtcgtgc acaccggcgt 38160
ccatgaccac cggcatgccg tcgcacaacg acaccatctc cagggcggtg ttcagcacgt 38220
cctgcccggt cagctccgac ggggtgagca cccaggggcg cccctcgaag aggtccgccc 38280
gggcggcgag cggcccggaa cgctcggtgc cggccagcgg atggcttcct atgtagctgg 38340
ccgggtcggc ccgcatcgcg cgcacgtcgt cgtgcgggac cttcttgacg ctggcgacat 38400
cgaggtaggc tcgggccagc ccgctctcct gtgcgcgcgc gagcacgcgt ccgacctgtg 38460
ccgggggcac ggccagcacc gccaggtcga cctgacggtc cggtctctcc agggatcccg 38520
cgcccatcgc ctccgccgtc ctggcggcgt tccggtcgac gtcctccagg tgcacgccga 38580
ccccgcggcg ggtcagcgcg agagcgacgg acgtgccgat ggccccggtg ccgatgactg 38640
tggtggtcct caacgcgcgc ccccaggtgc ggtgatccga aatcggctcg gacaagtgcc 38700
gtgcccggca cgggaaaagg gaattcccat ggcgccgtgc gccgccaatt taacgcttcg 38760
gcgcgcatgt tcaactgcgg cgtcgcagcg gtcgaacaca gtagcggtac accggaccat 38820
tgaggcatcg tgctcagttg gcgacaccgg gtcggataaa cgccggaatc cgaggagttg 38880
acgttgcagt cagcgctgag acacgacgac ctgcatccga tagaagaagt ggaaataagt 38940
tcgctctcca ccgacggctc cccgcggatc gacggggaga gtcccgagca cgtggaaatg 39000
ctggccgccg ccgacaccgc gcttccaccg atcatggtgc accgccgcac cgggcgggtc 39060
atcgacggca tgcaccggct gcgcgccgcg atgctgacgg gccgtacgac gatcgcggtg 39120
aggttcttcg acggcaccga ggaggacgcc ttcgtcctcg ccgtgaagtc gaacatcgcg 39180
cacggactgc cgctgtccgc cgccgaccgc cggcgggccg ccgggcgcat catggccacc 39240
catccccggt ggtcggaccg gatgatcgcc tcggtggtcg gcacctccgc caggacggtc 39300
gccgagatcc gccgcgacgc cggcgccgcc ggggcggggg agcccacccg catcggccgg 39360
gacggcaggg tacggcccgt cgacgtgagc gagggccgca gactggccca cgacatgatc 39420
gtccgcgacc cgggcctgtc gctgcgccag gtcgcccgcg ccgccgggat ctcgccggag 39480
accgtcaggg acgtcagaca ccggatgctc cgcggtgagg acccggtgcc cgcgccgcgg 39540
ccgcggaccc tggtggagcg cggcgcggac cgccgggcgg agccggccgg gaaggccgcc 39600
gcgccgtgcg ggacggagcc gccgcccgcc gtcgtgatga agcggctgag ggccgatccg 39660
gcgctgcgtc tcaacgagaa cggacgcgac ctgctgcggc ttctggatat ccacacggtc 39720
cggctggagg actggaaccg cattatcgaa agcgtgccgc cgcaccgtct ggagacggtg 39780
gcgcagctgg cacgctcctg cgccgacaaa tggtccgaga tcgcgtcacg catcgaaagc 39840
aacgcatcac atctggccgg gtgaacgagg aaacacacga atccttcgag gagccgtcgg 39900
agaaagcggg acggcccgtc ggaacaccct tgtggagggg caatggagat acggtcgatc 39960
gatcacgtcg aattgttcgt cgaggacgcc caggacacgg ccggcaggct gtgcgactcc 40020
ttcggcttcg tccgcgtggg ccgcggcgcc gggaccaccg gactgcgcgg ctgcgagtcc 40080
gtcctgctgc gccagaacga catcgccctg ctgctgacca cggccaccga cgccgaccac 40140
cgtgccgccg agtacgtgaa gcagcacggg gacggggtcg cggtgatcgg gtgggatccc 40200
cgggtaccga gctcgaattc gccctatagt gagtcgtatt acaattcact ggccgtcgtt 40260
ttacaacgtc gtgactggga aaaccctggc gttacccaac ttaatcgcct tgcagcacat 40320
ccccctttcg ccagctggcg taatagcgaa gaggcccgca ccgatcgccc ttcccaacag 40380
ttgcgcagct gaatggcgaa tggcgcctga tgcggtattt tctccttacg catctgtgcg 40440
gtatttcaca ccgcatatgg tgcactctca gtacaatctg ctctgatgcc gcatagttaa 40500
gccagccccg acacccgcca acacccgctg acgcgaaccc cttgcggccg c 40551
<---
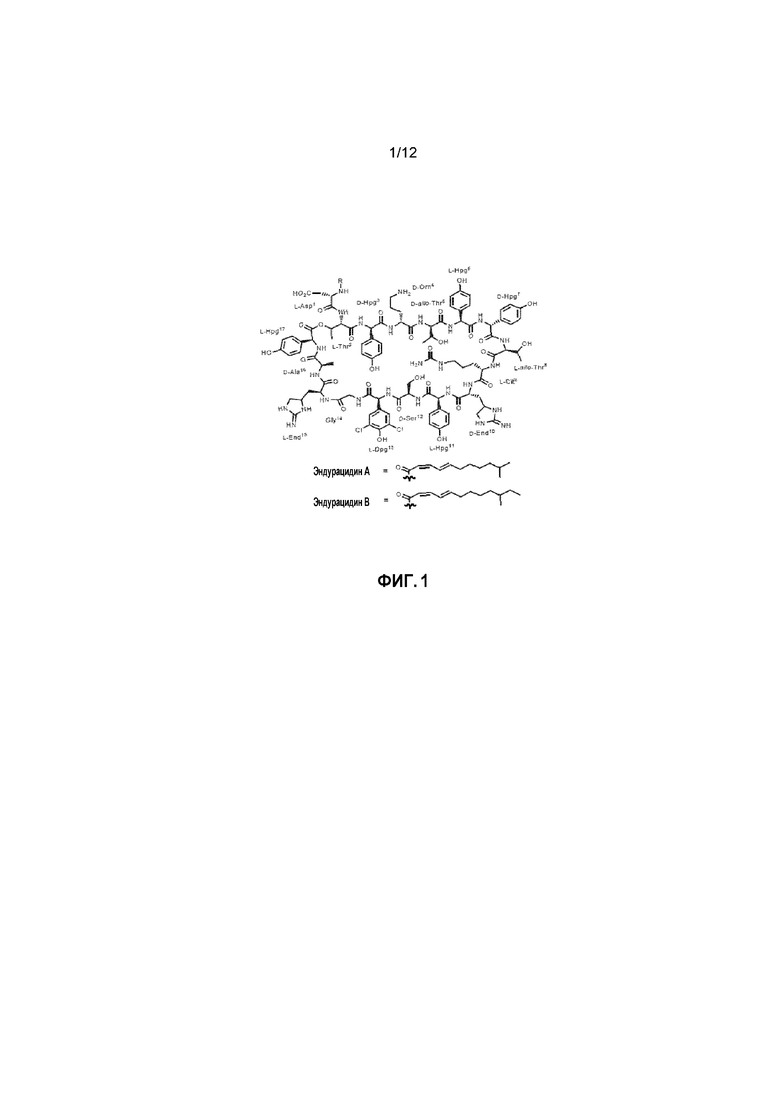

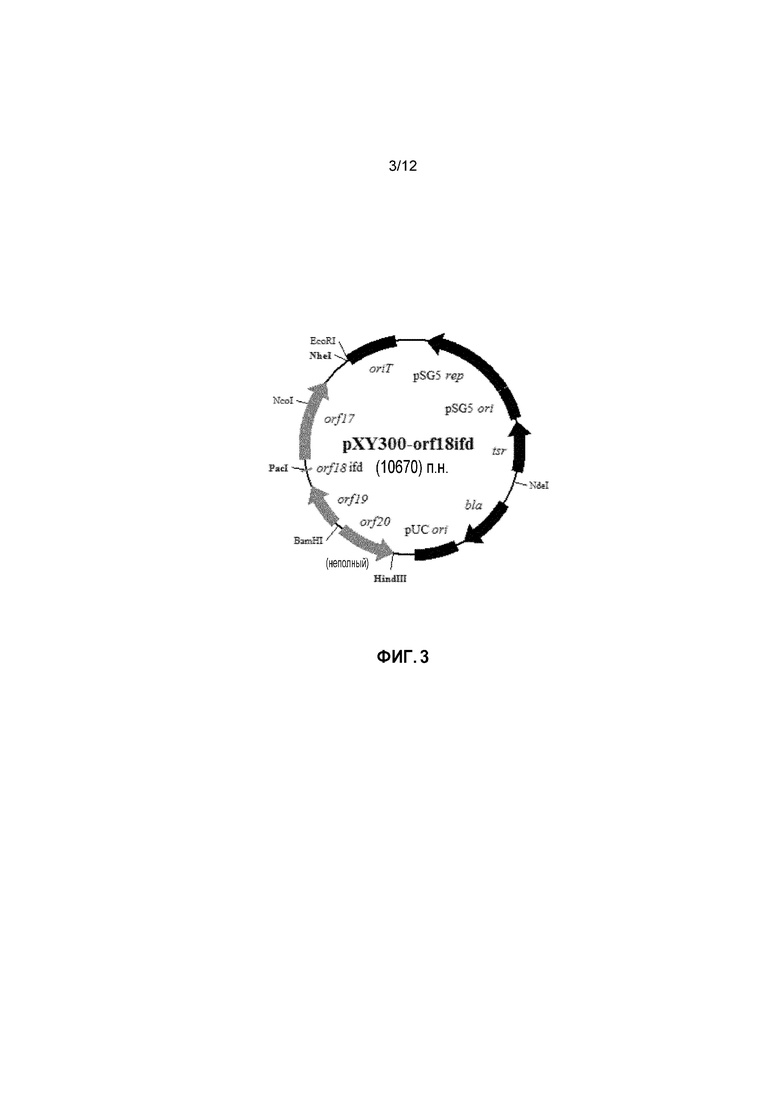




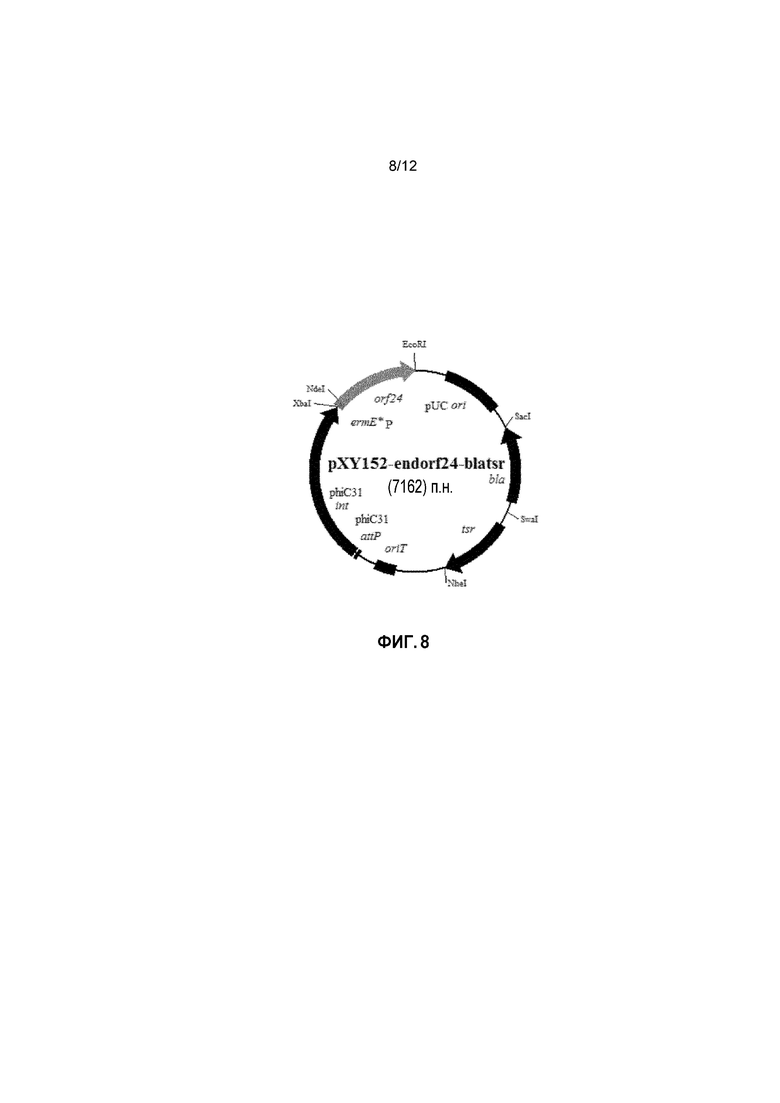




Изобретение относится к биотехнологии и представляет собой бактерию Streptomyces fungicidicus, содержащую один или более модифицированных генов, выбранных из группы, состоящей из усиленной открытой рамки считывания-24 (orf24) c SEQ ID NO: 38, ослабленной открытой рамки считывания-18 (orf18) c SEQ ID NO: 37, и также усиленной orf24 c SEQ ID NO: 38 и ослабленной orf18 c SEQ ID NO: 37. Изобретение касается также способа получения эндурацидина, который включает культивирование такой бактерии Streptomyces fungicidicus условиях, достаточных для продукции эндурацидина. Изобретение позволяет получить высокий выход эндурацидина. 2 н. и 15 з.п.ф-лы, 12 ил., 2 табл., 7 пр.
1. Бактерия Streptomyces fungicidicus для повышенной продукции эндурацидина, содержащий один или более модифицированных генов, выбранных из группы, состоящей из усиленной открытой рамки считывания-24 (orf24) c SEQ ID NO: 38, ослабленной открытой рамки считывания-18 (orf18) c SEQ ID NO: 37, и также усиленной orf24 c SEQ ID NO: 38 и ослабленной orf18 c SEQ ID NO: 37; где повышенная продукция эндурацидина достигается с бактерией Streptomyces fungicidicus по сравнению с таковой, полученной с контрольным штаммом Streptomyces fungicidicus.
2. Бактерия по п.1, где ослабленная orf18 ослаблена, поскольку она была инактивирована.
3. Бактерия по п.2, где ослабленная orf18 была инактивирована способом, выбранным из группы, состоящей из делеции внутри рамки считывания, мутации сдвига рамки считывания, точечной мутации и любой их комбинации.
4. Бактерия по п.3, где ослабленная orf18 была инактивирована посредством делеции внутри рамки считывания.
5. Бактерия по п.4, где делеция внутри рамки считывания представляет делецию нуклеотидов с 5 по 660 из orf18 (SEQ ID NO: 37).
6. Бактерия по любому из пп.1-5, где усиленная orf24 операбельно связана с гетерологичным промотором.
7. Бактерия по п.6, где гетерологичный промотор является сильным конститутивным промотором.
8. Бактерия по п.7, где сильный конститутивный промотор представляет ermE*p.
9. Бактерия по любому из пп.1-8, где усиленная orf24 усилена, поскольку она была сверхэкспрессирована.
10. Бактерия по любому из пп.1-9, где Streptomyces fungicidicus представляет Streptomyces fungicidicus ATCC 21013.
11. Бактерия по любому из пп.1-9, где Streptomyces fungicidicus представляет Streptomyces fungicidicus ATCC PTA-122342.
12. Бактерия по любому из пп.1-11, где продукция эндурацидина бактерией, по меньшей мере, в 1,2 раза выше, чем продукция эндурацидина контрольным Streptomyces fungicidicus.
13. Бактерия по любому из пп.1-12, где продукция эндурацидина рекомбинантным штаммом в 1,2-4,6 раза выше, чем продукция эндурацидина контрольным Streptomyces fungicidicus.
14. Бактерия по п.1, которая представляет собой BM38-2.24/16, депонированную под номером ATCC № PTA-124006.
15. Бактерия по п.1, которая представляет собой BM38-2.18pfrd-AmR, депонированную под номером ATCC № PTA-124007.
16. Способ получения эндурацидина, включающий культивирование бактерии Streptomyces fungicidicus по любому из пп.1-15 в условиях, достаточных для продукции эндурацидина.
17. Способ по п.16, дополнительно включающий выделение эндурацидина из культуральной среды.
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| MARK ZABRISKIE, The enduracidin biosynthetic gene cluster from Streptomyces fungicidicus Microbiology (Reading) 2006 Oct;152(Pt 10):2969-2983 | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Орлова Т.И | |||
| и др | |||
| Биологически активные нерибосомальные пептиды.I Нерибосомальны антибиотики полипептиды | |||
| Антибиотики и | |||

